Ease on down, ease on down
Propagation is the way…
Today's Topics
- Equilibrium potential and driving forces
- The action potential
- Synaptic communication
Party On
- Annie (A-) was having a party.
- Used to date Nate (\(Na^+\)), but now sees Karl (\(K^+\))
- Hired bouncers called
- "The Channels"
- Let Karl and friends in or out, keep Nate out
- Annie's friends (A-) and Karl's (\(K^+\)) mostly inside
- Nate and friends (\(Na^+\)) mostly outside
- Claudia (Cl-) tagging along
Party On

The \(K^+\) story
- \(Na^+\)/\(K^+\) pump pulls \(K^+\) in
- \([K^+]_{in}\) (~150 mM) >> \([K^+]_{out}\) (~4 mM)
- Outward flow of \(K^+\) through passive/leak channels
- Outflow stops when membrane potential, \(V_m\) = equilibrium potential for \(K^+\)
Equilibrium potential
- Voltage (\(V_{K}\)) that keeps system in equilibrium
- \([K^+]_{in}\) >> \([K^+]_{out}\)
- Nernst equation
- \(V_{K}\) = \(\frac{RT}{(+1)F}ln(\frac{[K^+]_{out}}{[K^+]_{in}})\)
- \(V_{K}\) = ~ -90 mV
- Negative in/positive out keeps in/out concentration gradient
Equilibrium potential
- \(K^+\) flows out through passive/leak channels; most remains near membrane
- Separation from \(A^-\) creates charge \(\frac{K+K+K+K+K+}{A-A-A-A-A-}\) along capacitor-like membrane
- \(V_m\) –> \(V_{K}\)
Equilibrium potentials calculated under typical conditions
| Ion | [inside] | [outside] | Voltage |
|---|---|---|---|
| \(K^+\) | ~150 mM | ~4 mM | ~ -90 mV |
| \(Na^+\) | ~10 mM | ~140 mM | ~ +55-60 mV |
| Cl- | ~10 mM | ~110 mM | ~ - 65-80 mV |

The \(Na^+\) story
- \(Na^+\)/\(K^+\) pump pushes \(Na^+\) out
- \([Na^+]_{in}\) (~10 mM) << \([Na^+]_{out}\) (~140 mM)
- Equilibrium potential for \(Na^+\), \(V_{Na}\) = ~ +55 mV
- Inside positive/outside negative to maintain outside > inside concentration gradient
- If \(Na^+\) alone, \(V_m\) –> \(V_{Na}\) (~ +55 mV)
"Resting" potential
- Sum of outward \(K^+\) and inward \(Na^+\)
- Membrane more permeable to \(K^+\) than \(Na^+\), \(p_{\)K+}\(>\)p_{Na+}$
- Outward flow of \(K^+\) > inward flow of \(Na^+\)
- Resting potential (~-70 mV) closer to \(V_{K}\) (-90 mV) than \(V_{Na}\) (+55 mV)
- Goldman-Hodgkin-Katz equation
- \(V_m = \frac{RT}{F}ln(\frac{p_{K^+}[K^+]_{out}+p_{Na^+}[Na^+]_{out}}{p_{K^+}[K^+]_{in}+p_{Na^+}[Na^+]_{in}})\)
"Driving force" and equilibrium potential
- "Driving Force" on a given ion depends on difference between
- Equilibrium potential for given ion
- Membrane potential = effects of all ions
- Anthropomorphic metaphor
- \(K^+\) "wants" to flow out (pull neuron toward \(V_{K}\))
- \(Na^+\) "wants" to flow in (pull neuron toward \(V_{Na}\))
- Strength of that "desire" depends on distance from equilibrium potential
Action potential
Action potential
- Rapid rise, fall of membrane potential
- Threshold of excitation
- Increase (rising phase/depolarization)
- Peak
- at positive voltage
- Decline (falling phase/repolarization)
- Return to resting potential (refractory period)
Action potential components
| Phase | Neuron State |
|---|---|
| Rise to threshold | + input makes membrane potential more + |
| Rising phase | Voltage-gated \(Na^+\) channels open, \(Na^+\) enters |
| Peak | Voltage-gated \(Na^+\) channels close and deactivate; voltage-gated \(K^+\) channels open |
| Falling phase | \(K^+\) exits |
| Refractory period | \(Na^+\)/\(K^+\) pump restores [\(Na^+\)], [\(K^+\)]; voltage-gated \(K^+\) channels close |
Action potentials and driving forces

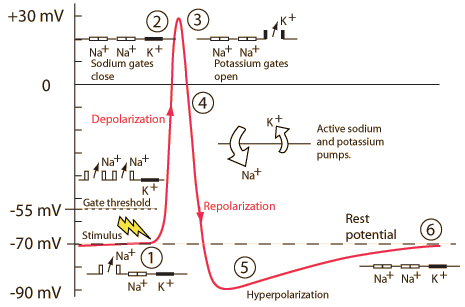
Neuron at rest
- Driving force on \(K^+\) weakly out
- -70 mV - (-90 mV) = +20 mV
- Driving force on \(Na^+\) strongly in
- -70 mV - (+55 mV) = -125 mV
- \(Na^+\)/\(K^+\) pump maintains concentrations
Action potential rising phase
- Voltage-gated \(Na^+\) channels open
- Membrane permeability to \(Na^+\) increases
- \(Na^+\) inflow through passive + voltage-gated channels
- continued \(K^+\) outflow through passive channels
Peak
- Membrane permeability to \(Na^+\) reverts to resting state
- Voltage-gated \(Na^+\) channels close & inactivate
- Slow inflow due to small driving force (+30 mV - 55mV = -25 mv)
Peak
- Membrane permeability to \(K^+\) increases
- Voltage-gated \(K^+\) channels open
- Fast outflow due to strong driving force (+30 mv - (-90 mv) = +120 mV)
Falling phase
- \(K^+\) outflow
- Through voltage-gated \(K^+\) and passive \(K^+\) channels
- \(Na^+\) inflow
- Through passive channels only
Absolute refractory phase
- Cannot generate action potential (AP) no matter the size of the stimulus
- Membrane potential more negative (-90 mV) than at rest (-70 mV)
- Voltage-gated \(Na^+\) channels still inactivated
- Driving force on \(Na^+\) high (-90 mv - 55 mV = -145 mV), but too bad
Absolute refractory phase
- Voltage-gated \(K^+\) channels closing
- Driving force on \(K^+\) tiny or absent
- \(Na^+\)/\(K^+\) pump restoring concentration balance
Relative refractory period
- Can generate AP with larg(er) stimulus
- Some voltage-gated \(Na^+\) 'de-inactivate', can open if
- Larger input
- Membrane potential is more negative than resting potential
Review AP phases and driving forces
Neuron at rest
- Voltage-gated \(Na^+\) closed, but ready to open
- Voltage-gated \(K^+\) channels closed, but ready to open
- Membrane potential at rest
- \(Na^+\)/\(K^+\) pump still working…
| Phase | Ion | Driving force | Flow direction | Flow magnitude |
|---|---|---|---|---|
| Rest | \(K^+\) | 20 mV | out | small |
| \(Na^+\) | 125 mV | in | small |
| Phase | Ion | Driving force | Flow direction | Flow magnitude |
|---|---|---|---|---|
| Rising | \(K^+\) | growing | out | growing |
| \(Na^+\) | shrinking | in | high |
| Phase | Ion | Driving force | Flow direction | Flow magnitude |
|---|---|---|---|---|
| Peak | \(K^+\) | 120 mV | out | high |
| \(Na^+\) | 20 mV | in | small |
| Phase | Ion | Driving force | Flow direction | Flow magnitude |
|---|---|---|---|---|
| Falling | K | shrinking | out | high |
| \(Na^+\) | growing | in | small |
| Phase | Ion | Driving force | Flow direction | Flow magnitude |
|---|---|---|---|---|
| Refractory | K | ~0 mV | out | small |
| \(Na^+\) | 145 mV | in | small |
Animation
APs and Information Processing
Information processing
- AP amplitudes don't vary (much)
- All or none
- \(V_{K}\) and \(V_{Na}\) don't vary much b/c \(Na^+\)/\(K^+\) pump always working
- AP frequency and timing vary
- Rate vs. timing codes
- Same rates, but different timing
- "Grandmother" cells and single spikes
Information processing

Generating APs
- Axon hillock
- Portion of soma adjacent to axon
- Integrates/sums input to soma
- Axon initial segment
- Umyelinated portion of axon adjacent to soma
- Voltage-gated \(Na^+\) and \(K^+\) channels exposed
- If sum of input to soma > threshold, voltage-gated \(Na^+\) channels open
Axon hillock, axon initial segment

Axon Hillock" by M.aljar3i - Own work. Licensed under CC BY-SA 3.0 via Commons
{kind=link}
AP propagation
- Propagation
- move down axon, away from soma, toward axon terminals.
- Unmyelinated axon
- Each segment "excites" the next
AP propagation is like
AP propagation
- Myelinated axon
- AP "jumps" between Nodes of Ranvier –> saltatory conduction
- Nodes of Ranvier == unmyelinated sections of axon
- voltage-gated \(Na^+\), \(K^+\) channels exposed
- Current flows through myelinated segments
Question
- Why does AP flow in one direction, away from soma?
- Soma does not have (many) voltage-gated \(Na^+\) channels.
- Soma is not myelinated.
- Refractory periods mean polarization only in one direction.
Question
- Why does AP flow in one direction, away from soma?
- Soma does not have (many) voltage-gated \(Na^+\) channels.
- Soma is not myelinated.
- Refractory periods mean polarization only in one direction.
Conduction velocities
Hodgkin-Huxley Equations
More on HH
- Measuring APs in actual neurons. https://www.youtube.com/embed/k48jXzFGMc8
- Interview with Sir Alan Hodgkin, https://www.youtube.com/embed/vSIXbfunHYg
- Simulations
Synaptic transmission
What happens when AP runs out of axon?
- Rapid change in voltage triggers neurotransmitter (NT) release
- Voltage-gated calcium Ca++ channels open
- Ca++ causes synaptic vesicles to bind with presynaptic membrane, merge
- NTs diffuse across synaptic cleft
- NTs bind with receptors on postsynaptic membrane
- NTs unbind, are inactivated
Receptor/channel types
- Leak/passive
- Vary in selectivity, permeability
- Transporters/exchangers
- Ionic
- \(Na^+\)/\(K^+\)
- Chemical
- e.g., Dopamine transporter (DAT)
- Ionic
Receptor/channel types
- Ionotropic receptors (receptor + ion channel)
- Ligand-gated
- Open/close channel
- Metabotropic receptors (receptor only)
- Triggers 2nd messengers
- G-proteins
- Open/close adjacent channels, change metabolism
Gap junctions
- Cytosol flows through adjacent neurons

Chemical synapse

Receptor types
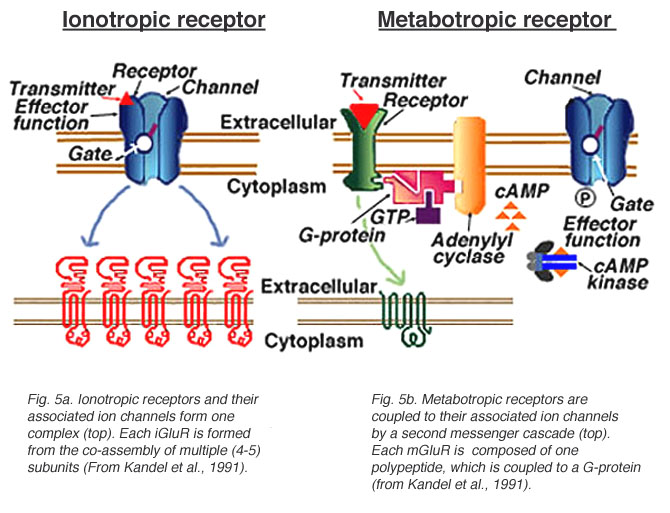
Receptors generate postsynaptic potentials (PSPs)
- Small voltage changes
- Amplitude scales with # of receptors activated
- Excitatory PSPs (EPSPs)
- Depolarize neuron (make more +)
- Inhibitory (IPSPs)
- Hyperpolarize neuron (make more -)
NTs inactivated
- Buffering
- e.g., glutamate into astrocytes
- Reuptake via transporters
- e.g., serotonin via serotonin transporter (SERT)
- Enzymatic degradation
- e.g., AChE degrades ACh
Questions to ponder
- Why must NTs be inactivated?
- What sort of PSP would the following induce:
- Open \(Na^+\) channel
- Open \(K^+\) channel
- Open Cl- channel, \([Cl^-]_{out}\)>>\([Cl^-]_{in}\)
Types of synapses

References
Eyherabide, H. G., Rokem, A., Herz, A. V. M., Samengo, I., Eyherabide, H. G., Rokem, A., … Samengo, I. (2009). Bursts generate a non-reducible spike-pattern code. Frontiers in Neuroscience, 3, 1. https://doi.org/10.3389/neuro.01.002.2009