Cellular neuroanatomy
How many neurons and glia?
- Old “lore”: ~100 billion neurons
- New estimate (Azevedo et al., 2009):
- ~86 +/- 8 billion neurons
- 85 +/- 9 billion glia
“These findings challenge the common view that humans stand out from other primates in their brain composition and indicate that, with regard to numbers of neuronal and nonneuronal cells, the human brain is an isometrically scaled-up primate brain.”
![[[@azevedo2009equal]](http://doi.org/10.1002/cne.21974)](data:image/jpeg;base64,/9j/4AAQSkZJRgABAQAASABIAAD/4QB0RXhpZgAATU0AKgAAAAgABAEaAAUAAAABAAAAPgEbAAUAAAABAAAARgEoAAMAAAABAAIAAIdpAAQAAAABAAAATgAAAAAAAABIAAAAAQAAAEgAAAABAAKgAgAEAAAAAQAAArugAwAEAAAAAQAAAhwAAAAA/+0AOFBob3Rvc2hvcCAzLjAAOEJJTQQEAAAAAAAAOEJJTQQlAAAAAAAQ1B2M2Y8AsgTpgAmY7PhCfv/iGyhJQ0NfUFJPRklMRQABAQAAGxhhcHBsAhAAAG1udHJSR0IgWFlaIAfgAAQACAAMADMAA2Fjc3BBUFBMAAAAAEFQUEwAAAAAAAAAAAAAAAAAAAAAAAD21gABAAAAANMtYXBwbAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAEWRlc2MAAAFQAAAAYmRzY20AAAG0AAAEGmNwcnQAAAXQAAAAI3d0cHQAAAX0AAAAFHJYWVoAAAYIAAAAFGdYWVoAAAYcAAAAFGJYWVoAAAYwAAAAFHJUUkMAAAZEAAAIDGFhcmcAAA5QAAAAIHZjZ3QAAA5wAAAGEm5kaW4AABSEAAAGPmNoYWQAABrEAAAALG1tb2QAABrwAAAAKGJUUkMAAAZEAAAIDGdUUkMAAAZEAAAIDGFhYmcAAA5QAAAAIGFhZ2cAAA5QAAAAIGRlc2MAAAAAAAAACERpc3BsYXkAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABtbHVjAAAAAAAAACIAAAAMaHJIUgAAABQAAAGoa29LUgAAAAwAAAG8bmJOTwAAABIAAAHIaWQAAAAAABIAAAHaaHVIVQAAABQAAAHsY3NDWgAAABYAAAIAZGFESwAAABwAAAIWdWtVQQAAABwAAAIyYXIAAAAAABQAAAJOaXRJVAAAABQAAAJicm9STwAAABIAAAJ2bmxOTAAAABYAAAKIaGVJTAAAABYAAAKeZXNFUwAAABIAAAJ2ZmlGSQAAABAAAAK0emhUVwAAAAwAAALEdmlWTgAAAA4AAALQc2tTSwAAABYAAALeemhDTgAAAAwAAALEcnVSVQAAACQAAAL0ZnJGUgAAABYAAAMYbXMAAAAAABIAAAMuY2FFUwAAABgAAANAdGhUSAAAAAwAAANYZXNYTAAAABIAAAJ2ZGVERQAAABAAAANkZW5VUwAAABIAAAN0cHRCUgAAABgAAAOGcGxQTAAAABIAAAOeZWxHUgAAACIAAAOwc3ZTRQAAABAAAAPSdHJUUgAAABQAAAPiamFKUAAAAA4AAAP2cHRQVAAAABYAAAQEAEwAQwBEACAAdQAgAGIAbwBqAGnO7LfsACAATABDAEQARgBhAHIAZwBlAC0ATABDAEQATABDAEQAIABXAGEAcgBuAGEAUwB6AO0AbgBlAHMAIABMAEMARABCAGEAcgBlAHYAbgD9ACAATABDAEQATABDAEQALQBmAGEAcgB2AGUAcwBrAOYAcgBtBBoEPgQ7BEwEPgRABD4EMgQ4BDkAIABMAEMARCAPAEwAQwBEACAGRQZEBkgGRgYpAEwAQwBEACAAYwBvAGwAbwByAGkATABDAEQAIABjAG8AbABvAHIASwBsAGUAdQByAGUAbgAtAEwAQwBEIA8ATABDAEQAIAXmBdEF4gXVBeAF2QBWAOQAcgBpAC0ATABDAERfaYJyACAATABDAEQATABDAEQAIABNAOAAdQBGAGEAcgBlAGIAbgDpACAATABDAEQEJgQyBDUEQgQ9BD4EOQAgBBYEGgAtBDQEOARBBD8EOwQ1BDkATABDAEQAIABjAG8AdQBsAGUAdQByAFcAYQByAG4AYQAgAEwAQwBEAEwAQwBEACAAZQBuACAAYwBvAGwAbwByAEwAQwBEACAOKg41AEYAYQByAGIALQBMAEMARABDAG8AbABvAHIAIABMAEMARABMAEMARAAgAEMAbwBsAG8AcgBpAGQAbwBLAG8AbABvAHIAIABMAEMARAOIA7MDxwPBA8kDvAO3ACADvwO4A8wDvQO3ACAATABDAEQARgDkAHIAZwAtAEwAQwBEAFIAZQBuAGsAbABpACAATABDAEQwqzDpMPwAIABMAEMARABMAEMARAAgAGEAIABDAG8AcgBlAHMAAHRleHQAAAAAQ29weXJpZ2h0IEFwcGxlIEluYy4sIDIwMTYAAFhZWiAAAAAAAADzUgABAAAAARbPWFlaIAAAAAAAAGFsAAA4MgAACkJYWVogAAAAAAAAbxwAAK6VAAAWmVhZWiAAAAAAAAAmTgAAGTkAALJSY3VydgAAAAAAAAQAAAAABQAKAA8AFAAZAB4AIwAoAC0AMgA2ADsAQABFAEoATwBUAFkAXgBjAGgAbQByAHcAfACBAIYAiwCQAJUAmgCfAKMAqACtALIAtwC8AMEAxgDLANAA1QDbAOAA5QDrAPAA9gD7AQEBBwENARMBGQEfASUBKwEyATgBPgFFAUwBUgFZAWABZwFuAXUBfAGDAYsBkgGaAaEBqQGxAbkBwQHJAdEB2QHhAekB8gH6AgMCDAIUAh0CJgIvAjgCQQJLAlQCXQJnAnECegKEAo4CmAKiAqwCtgLBAssC1QLgAusC9QMAAwsDFgMhAy0DOANDA08DWgNmA3IDfgOKA5YDogOuA7oDxwPTA+AD7AP5BAYEEwQgBC0EOwRIBFUEYwRxBH4EjASaBKgEtgTEBNME4QTwBP4FDQUcBSsFOgVJBVgFZwV3BYYFlgWmBbUFxQXVBeUF9gYGBhYGJwY3BkgGWQZqBnsGjAadBq8GwAbRBuMG9QcHBxkHKwc9B08HYQd0B4YHmQesB78H0gflB/gICwgfCDIIRghaCG4IggiWCKoIvgjSCOcI+wkQCSUJOglPCWQJeQmPCaQJugnPCeUJ+woRCicKPQpUCmoKgQqYCq4KxQrcCvMLCwsiCzkLUQtpC4ALmAuwC8gL4Qv5DBIMKgxDDFwMdQyODKcMwAzZDPMNDQ0mDUANWg10DY4NqQ3DDd4N+A4TDi4OSQ5kDn8Omw62DtIO7g8JDyUPQQ9eD3oPlg+zD88P7BAJECYQQxBhEH4QmxC5ENcQ9RETETERTxFtEYwRqhHJEegSBxImEkUSZBKEEqMSwxLjEwMTIxNDE2MTgxOkE8UT5RQGFCcUSRRqFIsUrRTOFPAVEhU0FVYVeBWbFb0V4BYDFiYWSRZsFo8WshbWFvoXHRdBF2UXiReuF9IX9xgbGEAYZRiKGK8Y1Rj6GSAZRRlrGZEZtxndGgQaKhpRGncanhrFGuwbFBs7G2MbihuyG9ocAhwqHFIcexyjHMwc9R0eHUcdcB2ZHcMd7B4WHkAeah6UHr4e6R8THz4faR+UH78f6iAVIEEgbCCYIMQg8CEcIUghdSGhIc4h+yInIlUigiKvIt0jCiM4I2YjlCPCI/AkHyRNJHwkqyTaJQklOCVoJZclxyX3JicmVyaHJrcm6CcYJ0kneierJ9woDSg/KHEooijUKQYpOClrKZ0p0CoCKjUqaCqbKs8rAis2K2krnSvRLAUsOSxuLKIs1y0MLUEtdi2rLeEuFi5MLoIuty7uLyQvWi+RL8cv/jA1MGwwpDDbMRIxSjGCMbox8jIqMmMymzLUMw0zRjN/M7gz8TQrNGU0njTYNRM1TTWHNcI1/TY3NnI2rjbpNyQ3YDecN9c4FDhQOIw4yDkFOUI5fzm8Ofk6Njp0OrI67zstO2s7qjvoPCc8ZTykPOM9Ij1hPaE94D4gPmA+oD7gPyE/YT+iP+JAI0BkQKZA50EpQWpBrEHuQjBCckK1QvdDOkN9Q8BEA0RHRIpEzkUSRVVFmkXeRiJGZ0arRvBHNUd7R8BIBUhLSJFI10kdSWNJqUnwSjdKfUrESwxLU0uaS+JMKkxyTLpNAk1KTZNN3E4lTm5Ot08AT0lPk0/dUCdQcVC7UQZRUFGbUeZSMVJ8UsdTE1NfU6pT9lRCVI9U21UoVXVVwlYPVlxWqVb3V0RXklfgWC9YfVjLWRpZaVm4WgdaVlqmWvVbRVuVW+VcNVyGXNZdJ114XcleGl5sXr1fD19hX7NgBWBXYKpg/GFPYaJh9WJJYpxi8GNDY5dj62RAZJRk6WU9ZZJl52Y9ZpJm6Gc9Z5Nn6Wg/aJZo7GlDaZpp8WpIap9q92tPa6dr/2xXbK9tCG1gbbluEm5rbsRvHm94b9FwK3CGcOBxOnGVcfByS3KmcwFzXXO4dBR0cHTMdSh1hXXhdj52m3b4d1Z3s3gReG54zHkqeYl553pGeqV7BHtje8J8IXyBfOF9QX2hfgF+Yn7CfyN/hH/lgEeAqIEKgWuBzYIwgpKC9INXg7qEHYSAhOOFR4Wrhg6GcobXhzuHn4gEiGmIzokziZmJ/opkisqLMIuWi/yMY4zKjTGNmI3/jmaOzo82j56QBpBukNaRP5GokhGSepLjk02TtpQglIqU9JVflcmWNJaflwqXdZfgmEyYuJkkmZCZ/JpomtWbQpuvnByciZz3nWSd0p5Anq6fHZ+Ln/qgaaDYoUehtqImopajBqN2o+akVqTHpTilqaYapoum/adup+CoUqjEqTepqaocqo+rAqt1q+msXKzQrUStuK4trqGvFq+LsACwdbDqsWCx1rJLssKzOLOutCW0nLUTtYq2AbZ5tvC3aLfguFm40blKucK6O7q1uy67p7whvJu9Fb2Pvgq+hL7/v3q/9cBwwOzBZ8Hjwl/C28NYw9TEUcTOxUvFyMZGxsPHQce/yD3IvMk6ybnKOMq3yzbLtsw1zLXNNc21zjbOts83z7jQOdC60TzRvtI/0sHTRNPG1EnUy9VO1dHWVdbY11zX4Nhk2OjZbNnx2nba+9uA3AXcit0Q3ZbeHN6i3ynfr+A24L3hROHM4lPi2+Nj4+vkc+T85YTmDeaW5x/nqegy6LzpRunQ6lvq5etw6/vshu0R7ZzuKO6070DvzPBY8OXxcvH/8ozzGfOn9DT0wvVQ9d72bfb794r4Gfio+Tj5x/pX+uf7d/wH/Jj9Kf26/kv+3P9t//9wYXJhAAAAAAADAAAAAmZmAADypwAADVkAABPQAAAKDnZjZ3QAAAAAAAAAAAADAQAAAgAAAFYBLgHrAsQDgwRTBUAGJwcaCBgJHwo9C04Maw2QDrkP7BEtEmUToxTjFioXeRi/Gg8bVxykHfMfPCCEIc0jESRSJY0myygAKTMqZiuXLMot9y8oMFExgDKrM9U1AjYxN104ijm2OuE8DT04PmA/iUCxQdxDCEQ2RWNGkEe4SN5KAUskTEdNak6PT7RQ2FH4UxZUM1VPVm1XjViuWcta4VvwXPdd+18AYAphGWItY0FkUmVfZmVnYWhTaTxqIGsBa+Bsv22gboNvaHBQcThyIHMIc+901nW+dqh3k3h+eWx6WXtJfD19N342fzmAQYFLgleDZYR0hYWGlIekiLOJwIrMi9aM343nju2P8pD1kfaS9ZP1lPqWBJcVmCyZR5pjm3+cmJ2unr+fy6DUodqi4KPppPSmA6cVqCqpQqpcq3eslK2yrtGv8bERsjOzU7RztZC2qbe6uMG5w7rAu7u8tb2vvqm/o8CdwZXCisN8xGrFU8Y2xxjH+cjgyc/KxsvDzMHNvM6vz5jQc9FD0g3S09Ob1GbVNdYF1tfXqNh22UDaBtrH24DcNdzn3ZHeO97j343gO+Dr4Z/iUuMD46/kVeT55ZTmLOa/51Ln5OiF6S/p5Oqo627sM+z17a/uY+8R77XwUPDm8XzyF/LG84/0bvVh9mb3ivjk+sX9Lv//AAAAVgEjAbACagMyA+gEsQWABlgHPQgtCScKJwsiDDANOg5RD2MQfhGfEr4T6RUQFjgXYRiIGbQa3hwLHTMeWx+AIKEhvyLbI/UlCyYfJzUoSSlgKnkrkyyxLcwu6DAEMSEyPDNaNHQ1izakN7k4zTniOvQ8Bj0bPjE/TUBsQY9CsEPQRO1GCEcdSDJJRUpVS2NMbk14Tn5Pg1CFUYZSiFOKVIxVilaEV3lYaFlUWkRbP1xGXVped1+VYLBhwmLKY8VktWWcZntnV2gwaQxp7GrQa7psqW2bbo9vgnBycVxyQHMfc/t01XWtdoZ3YnhFeTB6JHsdfBl9FX4QfwZ/9oDigcqCsYOVhHeFXYZKh0SITIlhioGLpYzGjd6O7o/1kPeR9ZLzk/CU7ZXslu6X+JkQmjmbbpymnc6e45/poOWh36LZo9GkyKW+prynyqjtqiGrXaycrdmvFLBOsYiywrP5tS62ZLebuNK6Bbs0vF+9hb6mv7zAvsGqwpDDfcR3xYDGj8edyKPJnsqOy3XMU80tzgTO2s+y0IvRatJR00XURdVR1mLXddiE2Yfafdtm3EPdGt3s3rnfh+BY4S3iCOLm48PkneVx5kDnDOfU6KPpf+pp62TsaO1t7nDvcPBw8XLydvN59Hf1afZH9w/3s/hF+Mv5Wvnt+tD8DP3a//8AAAArAMQBSwHYAmUDBgOPBCsEzwV6Bi8G6QelCGgJLwn5CsoLnwx3DVIOMg8OD/YQ2RHAEqkTkxR9FWcWUhc9GCkZFRn8Gugb0hy+Ha4eox+bIJYhlCKPI4YkdiVhJkUnICf4KMwpnypzK0gsIS0CLeku1y/IMLoxqjKYM4A0YDU5NgY2zzeVOFg5GTnZOp07ZDwyPQY93j62P49AaEFEQiZDEEQARPNF50bWR7tIlkllSiVK4EuYTE9NB03DToNPSVAYUOpRvlKTU2hUO1UNVd5WrVd8WEpZFVnfWqhbb1w1XPldvV6AX0NgCGDRYZxia2M8ZAxk2mWnZnFnOGf8aL9pgGpAawBrwGyBbUNuBG7Fb4VwQXD4catyWXMBc6l0T3T0dZh2PHbed4B4HXi9eWB6Fnrae6t8g31Yfih+9n/DgJOBZ4JEgyqEGIULhgCG9IfjiM2Jr4qMi2GMNI0HjdqOsI+KkGuRR5ISksaTW5PslHqVQZZCl4iZCJqKm+SdGZ4ynzmgOaE1oi+jKKQhpRmmEacKqAmpD6ohqz2sYK2Hrq2v07D8siizWrSStdG3FLhZuZy617wFvSm+R79iwH/BnsK+w9/E/8Yhx0PIZ8mMyrHL1sz9zinPW9CX0d7TMtSN1e7XU9i52iHbld0v3w7hd+SL6HntTvL2+VH//wAAbmRpbgAAAAAAAAY2AACTlQAAVo0AAFbpAACRtwAAJrsAABcKAABQDQAAVDkAAoeuAAJHrgABa4UAAwEAAAIAAAABAAQACAAPABYAHwApADQAQABMAFoAaQB4AIkAmgCsAL4A0gDmAPwBEgEpAUABWQFzAY0BqQHFAeMCAQIhAkICZAKHAqwC0gL6AyMDTQN6A6gD1wQIBDoEbQSjBNkFEgVLBYYFwwYBBkAGgAbCBwUHSgeQB9gIIQhsCLkJBwlXCagJ+gpNCqEK9wtOC6gMBAxiDMMNJQ2JDe4OVA68DyUPkRAAEHEQ5BFZEc8SRhK9EzgTthQ6FMMVUhXjFnUXBheWGCYYuBlOGegaiBsxG+Mcnx1iHise+B/GIJUhZCI0IwYj2ySzJY8mbCdLKCspDSnwKtUruyyeLX4uXC85MBQw8DHNMqoziTRoNUo2LjcVN/847TneOtI7yjzFPcQ+xz/OQNpB6EL2RABFBkYIRwhICEkJSgxLFEwhTTROTk9uUJRRvFLlVA1VNFZZV31YoFnEWudcC10wXlZffGCkYc5i+GQkZVNmhWe8aPpqQ2uZbPluY2/ScUZyvHQ0da93LXixejx70H1vfxyA1oKUhEyF9oeUiSyKxYxljhWP35HNk92V/5ggmjmcTJ5foHmioKTapy2poKwwruCxobRetwu5rrxdvyvCH8VGyKLMDM8o0fbUkdco2dHcnt+X4tLmYOn57RLvrPIA9DD2Off/+Yr6uPvU/Lv9o/5w/zf//wAAAAEABQAKABIAGwAmADIAPwBNAFwAbQB+AJEApAC5AM8A5QD9ARYBLwFKAWYBgwGiAcEB4gIEAicCTAJyApoCwwLvAxwDSwN8A68D5AQaBFIEiwTFBQEFPQV7BbsF/AY/BoMGyQcQB1kHpQfyCEEIkwjmCTsJkwnsCkYKoQr8C1gLtAwSDHIM1A05DaAOCw54DucPWQ/OEEYQwBE+Eb8SQxLKE1MT3RRqFPoVjxYqFssXcRgWGLYZURnmGnkbDRukHEAc5B2RHkofDh/dILMhjiJpI0IkGCTrJb0mkCdmKEIpJSoSKwgsBS0ILg4vEjARMQgx+TLoM9c0yjXBNsE3yjjbOfI7DjwwPVE+aT90QHJBaEJYQ0dEOUU0RjhHR0hfSX5KoUvHTPBOG09FUGtRhlKUU5dUlVWUVqBXv1jxWjVbgFzNXh5fdGDPYiNjZmSVZbVmzmfkaPtqFmszbFRtdm6ab8Bw6nIXc0Z0dnWndtp4EHlNeo972H0mfn1/44FngxCEyoZ0iAaJh4sEjIiOG4/DkYKTWJVDlzuZO5s7nTmfKaEEosakdqYbp72pYqsVrN2uxbDPsvW1Mbd8ucu8D75EwHLCpsTpx0LJsswxzpzQ3dL01PHW59jj2ujc9N8B4QrjFOUk50jpluw374rzpvds+c77hPzD/fT++f//AAAAAgAIABEAHgAtAD4AUQBmAH0AlgCwAMwA6gEJASsBTQFyAZgBwAHqAhYCRAJ0AqYC2gMRA0kDhAPAA/0EOwR6BLoE/AVBBYoF1wYpBoEG3Qc+B6IICAhvCNQJOQmdCgIKagrVC0QLuww5DMENUA3mDoIPIQ/AEFwQ9hGQEi0SzRNuFA0UqBVCFdwWeRceF84YjhliGkIbKhwUHPsd3R64H5IgbSFMIjAjGCQGJPkl8SbvJ/Mo/SoOKyQsQC1gLoAvnTC4MdMy8zQaNUk2gjfDOQw6XDuwPQc+YD++QSJCk0QWRa5HWkkRStFMmk5vUFVSMlPaVVpWzFhIWdNbZlz4XoNgAWFwYtVkN2WcZwlogWoKa6NtTW8AcLdyb3QedcN3dXlke+5+pIB5gdaC/IQChQiGEIcziHOJ1ItVjOmOiJAukdmTiJU7lvGYqZpenAqdp583oL2iP6PApUSmyqhQqdSrVKzQrkavubEospe0CLV9tvy4ibomu9C9gr82wOnCnMRPxgTHu8lzyyvM5M6e0FjSFNPS1Y/XStj/2qzcT93o33Xg/eKC5APlgucC6IPqBOt/7O7uRu958J3xjfJ180Tz9/Sq9U712/Zo9vb3dvfp+Fz4z/lD+av6Dfpw+tL7NfuY+/H8Sfyi/Pv9VP2t/gP+WP6s/wH/Vf+q//8AAHNmMzIAAAAAAAEMQgAABd7///MmAAAHkgAA/ZH///ui///9owAAA9wAAMBsbW1vZAAAAAAAAAYQAACc3wAAAADKKtWAAAAAAAAAAAAAAAAAAAAAAP/AABEIAhwCuwMBIgACEQEDEQH/xAAfAAABBQEBAQEBAQAAAAAAAAAAAQIDBAUGBwgJCgv/xAC1EAACAQMDAgQDBQUEBAAAAX0BAgMABBEFEiExQQYTUWEHInEUMoGRoQgjQrHBFVLR8CQzYnKCCQoWFxgZGiUmJygpKjQ1Njc4OTpDREVGR0hJSlNUVVZXWFlaY2RlZmdoaWpzdHV2d3h5eoOEhYaHiImKkpOUlZaXmJmaoqOkpaanqKmqsrO0tba3uLm6wsPExcbHyMnK0tPU1dbX2Nna4eLj5OXm5+jp6vHy8/T19vf4+fr/xAAfAQADAQEBAQEBAQEBAAAAAAAAAQIDBAUGBwgJCgv/xAC1EQACAQIEBAMEBwUEBAABAncAAQIDEQQFITEGEkFRB2FxEyIygQgUQpGhscEJIzNS8BVictEKFiQ04SXxFxgZGiYnKCkqNTY3ODk6Q0RFRkdISUpTVFVWV1hZWmNkZWZnaGlqc3R1dnd4eXqCg4SFhoeIiYqSk5SVlpeYmZqio6Slpqeoqaqys7S1tre4ubrCw8TFxsfIycrS09TV1tfY2dri4+Tl5ufo6ery8/T19vf4+fr/2wBDAAICAgICAgMCAgMFAwMDBQYFBQUFBggGBgYGBggKCAgICAgICgoKCgoKCgoMDAwMDAwODg4ODg8PDw8PDw8PDw//2wBDAQICAgQEBAcEBAcQCwkLEBAQEBAQEBAQEBAQEBAQEBAQEBAQEBAQEBAQEBAQEBAQEBAQEBAQEBAQEBAQEBAQEBD/3QAEACz/2gAMAwEAAhEDEQA/AP38ooooAKKKKACiiigAooooAKKKKACiiigAooooAKKKKACiiigAooooAKKKKACiiigAooooAKKKKACiiigAooooAKKKKACiiigAooooAKKKKACiiigAooooAKKKKACiiigAooooAKKKKACiiigAooooAKKKKACiiigAooooAKKKKACiiigAooooAKKKKACiiigAooooAKKKKACiiigAooooAKKKKACiiigAooooAKKKKAP/0P38ooooAKKKKACiiigAooooAKKKKACiiigAooooAKKKKACiiigAooooAKKKKACiiigAooooAKKKKACiiigAooooAKKKKACiiigAooooAKKKKACiiigAooooAKKKKACiiigAooooAKKKKACiiigAooooAKKKKACiiigDm/Fej6xr2jtpmh63P4euJJYGa8to4ZZ1ijkV5EQTpJGDIgKbmRtoYkDOCPgU+K/jVafDOw+Jtr8QNT1K4Txq2gPpk8OmxQXMC+KU02IGaOzWSPNqjI5BOWfeMYwf0bkjWWNomJAcEHaSpwfQjBB9xzXkVt8B/hjaeEovA0NhdnRodVXW0ifVNQdxqK3P2wTec9wZf+Pj98U37C/zFc5NKKtJPzX5q/4J/Mbfutdf+A/1a+48stf2pk1OSbSNC8GajrPiDTXvl1CwsiZjELDUpNNcwy7Aj73ikkj8zyVKKdzI5CntP2lNf8TeHfhWdY8H6zPoWotq2h2q3VukEjiK/wBSt7SUbbiKWM5jmbGV4YA9uZ5f2Z/gxNcw3jaLcCeK4vrlnGp6gGmOpzC4vIrgi4/f288qh3t5d0JOfkwTnt/Gnwt8EfEK+0S/8XWMl6/h+cXFoi3NxDD5ivHKvnQxSJHOqyQxuqyq6hlBAzVQ+GF901f7/wCkErc0uXbW33aHlOoftGaZpNmmoR6Nc3mlPrGqeHYboyqsr6hpENzJKZI9uFika0lRJFZju2kooJK1L/8AaUZLu203R/Cs19d3sfhqWGNrqOEFPE32gQF2KkL5L2zCUDPykMu4/LXp4+CvwzGv3XiQ6MGury5nvZImnna0+2XUBtp7lbQyG3SeSEsjyrGHIZ8t877uQ0n9lz4K6IkK6bpV/E1vJp0qOda1V5N2klzYgu10WK2+9hGhJULhcbVUBU/7/wDd+5fH9/T9OpO2vL2lb1+z9y0ffe/bl9N/aXvJNal0bWvCElm8GrajobmLUIJD/aGnaWdWZQZRDGIngUoJHkUCTAYBPnGNF8dNQ8faj4PuPDbXOhxweMLbRdSgBjlgvYbrRX1IbZHiDMi748PGVVypKNJEys3qmo/s5/B/V57241TRp7o6jf3up3CSajfNHLd6jaGwuXaMz7MSWrGLZt2KpIVRUOjfs3fCPQNTttZ06w1H7ZaX1tqSSTa1qlxm8s7ZrOGZ1lunWRxbsYiXB3IArZCriote7foo39U05fJ6pL8tS58tmo9pL701H7tH69+nS+MfiS3hvxNZ+DdK0ttV1i80vUdYWMzJbx/ZtNaCN1DsGzK73CBF2hcbi7pgbvKPhb8QNb+M3xCm8V6XqN3pvhKy0rRNRsbJXiH2mPV7SaYi8iaB2DfMh/dzDY0S7SVeQN7X4x+Gng3x5d2Go+I7SVr3TEuIre5tbq4srhIrtAk8XnWskTmOQBdyFipZVbG5FYJ4Y+GXgnwZrd9r/hewbT7nUba0s5US4nNsLexUpbxxWzSGCJYlJVfLjXA46UoWS13/AOC/0t9z72M5+X9bf8E8Y0PU/GnxV+J/xM0RfF154X03wRdW2kWVrpcdr5pluLKG8a+uHuoJ97Zm2RR4EQCMXWQkFbVp8arnwlpq6Nr0E3iCTw1q+j+EtV1YNFbvPq+oQWu25W2XIEDS3cQchgylm2xsiAt6V4g+DngLxJ4ll8Y3Vtd2OtXMCWtzdaZqN7pkl1BGSY0uPsU0ImCZOwyBigJCkAkGtH8DfhZBrA1u00NbWXfaSmGCeeGzabT4/KtZmtI5Ft2mgQKI5TGXXZGQ2Yoyip6JJ+V/k9XfzV9O7XSKHJ3v+H3afdpr1tfds8KtP2ubmfwi/jabwTLFpsXh+58SvjUI2kWysrg21wm3yxmYkb4lB2Ov33ibCnsbz9oy803V7i31DwhMNMh12+8PJdR31vve8tLJ75G2S+UiRSpGU3vKuyT7w2fPXQr+zF8F00X/AIR1dHu/7O/sq40Qw/2tqW06ddTefNAT9pyQ8nJYndj5Q23iue8Lfs4eH/7R8XT/ABDsotbttc1K+urWJ7++uIo4L60WxkElvM/lC4MAZDcKPMKyOu4ZYuntZb2l9/MuVf8AgN/nd9r3eO9uq+7lf/t34aabrK0/9p2719dPtvDHhCS/v9Q1saEqPepBB9pbRxrKOszRfPEYSyeYqlWYboy8bK5o+Iv2pLmCLV7Hw74eR9T0+W3SNri632jxvrS6PN+/t0kjaSNmEhjjdyudkpjkBSvS9C/Zt+EnhzW7TxHpdjqI1Gxu4b+OWbW9VuP9KgtTZJM6TXTrI/2ZjCS4JaPCtkAAUz+y78FN18y6ReJ/aEUsMgXV9TVUjlu1vtsKi6xCEuVEsflBPLbJTbubNya5lbbX8tPx/AzS92z30/PX8NF5mBD8cIfCt34iuvE0N0dOsvGUPh+9ubi5ga10s3dlaSW8kbLDEfsryTxofM3OsspJby/uX9I/aEuNc1+z0DS/Bmp3UgfTU1Jo1JGnjVIvPjaQsioRDE0bTgurLvwiyFWx3sfwU+Gi6L4m8O3Gkvfab4wmW41aC9u7q8S6lSOOFWP2iWQqfLhjX5NvCL6VPqnwd+HeseOYfiNf6Y512JLdGeO6uYoJxaMz2xubWORbe4aB2LRNNG5jbDIQQCGmvdVtuW/yik/vab6OzKlZ3fr+en4fkvM8r/aE134y+Hte8H6l8HJ1vbm2N9d3+hTLF5Ws2lqiNJbxymNpIrnDEwOrqm8ASBlPHC+EP2mNO1BdR1vwtPqXjo+KNQuG0bTRCsb2MNhptrc3VrIIIWkSVLicxEShirsqsyopevrPU/Bug6x4l0jxdfJcHU9CWdLRo7u4iiUXACyb4I5FhlyFGPMRsY4xXC698AvhP4hEputFaznm1WXWzcadd3WnXKajPCLeaeOezlhljaWIbZAjBX5LAsSTnG6TXf8ADVfon6PybKvF2v8A1v8Ar+HojmbH9oCK81i30Obw1d2N5Pq9ho7RXEsayQz3+lHVV80LvAMagxSAE4flSy81h+D/ANozV/F9ho0tv4Me1vfEOj32v2NvJqEJ32FgYEfc4TCzPLcIsaYK7cs7xnCntrv9nT4QXfiKy8Uf2NPa32nSWk1utpqN9aW0cthA1tbyC1t544C8cDGIOYy2z5c4AqzH8APhZDoeg+H7bTbqC28Mw3Ftp0kep36XUFtdrtngF0s4uGhcAZjaQplUIAZEKuWqdvO33afjq99NPMLx006K/rd3/CyX3njPgr9oRdV8VzahbTXmqab45bw2PDtldLFALRtV064vnWR4kLKgjgZmJ81t/wAqnaRi7rf7W2l6XaGK28Nzy6xE2pQtYz3MduZ7nStSj0ya3s5MOLiRpZBIigKTGQSAx2V61P8AAD4TTvcyDRXhNw2nOghvbuFbV9JXbZNZpHMq2hhXKqbcR/KSpypIPk/jX9mSfVPE2n3vg7Uv7G0my06a0gWK/wBWs7y1u7u5e5u74z2N5A97JcOyNIty/LpvLks2abXtP7t391tPx/PcyjH3N9dPzd/wa+56ao9O0b4zxar8Qb74YTaQ1r4h02+EM0BnVv8AiXNaC5TUk+UFrdnItxxkTZU4wTXt1eP+Cvh9r2m+Odc+IHja70/UtWuLePSdOms7RoJY9Jt5ZJo1uZHdzJO7yZlKbIvlUpGuWz7BQ7WX9f15+d0HV/1/Wui8rMKKKKkZ/9H9/KKKKACioLo3ItpTZKj3ARvLWQlUL4+UMQCQM9SATjsa+KPh9cfGrULDxn4H8e+IdeXx5oUWnXt2mlto08Nxa3IuTH/Yks9paRwrcSwvG4v0eSNU2iTJE1C2k+yuUl/X9f1c+3qK/NTTv2h/inqPhLQkvI7+5tvB3hmbX/Gs9sLW01aWWzvbixls4llXysxPZ3JuGiEZkKJ5LxBsH9IbG7gv7G3vrUlobmNJEJ4JVwCCc89DVcu/rb8WvzTt6CmnF8r3/wCAnb7mr+qLVFFFSIKKKKACiiigAooooAKKKKACiiigAooooAKKKKACiiigAooooAKKKKACiiigAooooAKKKKACiiigAooooAKKKKACiiigAooooAKKKKACiiigAooooAKKKKACiiigAooooAKKKKACiiigAooooAKKKKACiiigAooooAKK+HvhX4n+KviPxpqul6jqXia9sNN8aarpq6gsejHSlsLDfIltcIiC8+ZV8rzBGvzsp8w9G639p34z+J/hlFpa+CMzXWk41/WIVtpLhptFs5USe1QojLFPcK7tCzlQRBJzwSDS0JN/FZ+iaTu/Ravro/K9+zlzyglqrr5ptW+bVl6o+tKK+f8A45/FG/8AC/wcj8b/AA/vbbzdcudHs7HUJV863gj1m7gtlvNmQJBGk3mKpIViBnjIPYeGvA3izw74lttSuvHOp69pK6fJbT2WoxWZ8y8MkTJdrLb28DI21ZFePmP5gUVMHc1F3aeltPmle35ff62zUk0rPfX5f1f7vS/qFFcL8SvGkHw98D6r4skjNxLaRhbaAKzGe7nYRW8ICBmJkmdE+UE89K+Z/h58VtW8U/BbVbr4g+NZPDOsfDPUri08UarBaxwSXNvYAyx3EcN1AwjS9gaGUEREkMyx4bDCW9G+i39Or+Wl/XS5ooN289Pn/wAG2h9pUV+c2o+KPjzpXw10j4i6t4m1y0ufGPivRIdN0Iw6SuoWuh3lwtuIZTNbpH9ruUfzH3sohYpHuXa5bu/GfiX4seCvAp+Kr6x4gtdP8Ka+l1rOj6rDpb3cnh5fLguiHsonUrHta7jaOQsyb4y2cBKUe+lnZ+T929/TmV+1n85a2trfbz3t99tPkfb1FfBnj34heP7nwmnxP8E+KdTl0bxL420jTdHtbBNObztIedLGf7O91Dsb7XKJZYZHlxsMTK6qSDra1rfxQtb7wLaSeJfEegf8JH4zl0q4tNRi0hrxNP8A7NuZVG+3glhKPNaebG6szbXZSx4CEU396XzfL+XOr9VqE1yq9+jf3c3/AMi7dHpqfbtFfnzrnxa+LGh/EeT9nb/hJmu5bnxBo9gnik21qt/b2Op2V5fPBJEsQszeKLPZHJ5ATZMjmIsPm9+8GeJPE+ifHjxB8IdT1iXxBpK6FZa9ZzXSxi7s2muZrWS2Z4o4xJE3lCSNnBkU7wWZdoUiruy8/wAFzP8AD+trk1yp36Wfybsvx/4J9EUV8M6p4s+Ks/xg+JHhbTtR8T3el6Tf6FFZS6XHorW2mJqUMbTG4S5QXMsSMxkfYkhEeQGUjcs89t8Xm+P6fCmH4p6qLafwlNrqzCz0vYL5dSWJQE+yFjAI28vZv3lefM3/AD0QXNKMV1v+C5vyT+4cotRlLtb8Wl+bR9v0V5X8ZfGV34L8B3d1o8wg1rU3j03THML3AS8vG8uOZoo1Z3jtwWnlAB/dxuegr5hHxn8ZeMfgl8PvENlrdxofidfFuk+GPEQtooQzXH21bK+Ux3ELmNZlPnRYVHVXQ8ciimuaSit24r5yaX4Nq/qtxuDS5ntq/uTf42dvR3PvKimK6bzEGBdQCRnkA5wSPfB/Kn0iAooooA//0v38ooooAz9W0u01vSrzRr/zPst/DJBL5UskEnlyqVbZLEySI2DwyMrKeQQcGvO4vgz4It9JGkwHVIyZ4Lh7tdZ1IahK9spSJZb4XP2mSNVZh5TyGPk5XJJr1SilYdzyPVvgV8LdatbWyvNGKQWtpPYFYLm4gFxZ3TiW4guvKkX7THNJ88iz7w7MxbJd93rSIkSLHGoREAAAGAAOgAp1FMmwUUUUDCiiigAooooAKKKKACiiigAooooAKKKKACiiigAooooAKKKKACiiigAooooAKKKKACiiigAooooAKKKKACiiigAooooAKKKKACiiigAooooAKKKKACiiigAooooAKKKKACiiigAooooAKKKKACiiigAooooAKOvFFFAHn3hH4W+C/AumazpHhm2ube18QXVze3olv7u5eS5u8meRZJ5pHjZycnYy88jmo9B+FPgrw2NSGlQXYOr2Fpply02o3twzWllE0MCK00zmMqjNl02szMzsS5LH0Wih6pp9Vb5LZeiK5ne/z+fc8a0j9n/4S6L4Z/4Qy20V7nQf7LGi/Yb29vL63+wLI0qxeXdTSr8rMdr43qoVVYKqgWbT4HfD60sJNOaPU7xJI4oVku9Z1O6uIYYZUmEcM81y8sSs8aGQRuvmBVEm4ACvXKKrmd+a+v8Aw/8Am/vZNv6+7/JfcjmfEPhDQfFVzpF1rcc0smhXiX9p5dzPAq3Easqs6wuiygBj8kgZOc7c4rzPxf8As4fB/wAd6zq+v+J9Jurm9197CS+MeqajbRztpbFrMtFBcpGPJYllwo55PPNe5UVK029fn3/BfcO+/mrfLt6Hhuq/s5/CzXZhdazBq97Os9ncLLN4g1h5El093ktmRzeFk8t5GcBSAWwxBIUjtdB+Gfg7w7Z6zYWdtcXcfiEn7e2o3t1qUlwpj8rY0l5LM4jCcCMMEGSQoLEnvaKTimnF7P8Ar9EPmd076o8v134NfDvxH4U0DwRqGnTQ6L4XktJdNt7O+u7H7M9gALYq9rNE58raCu5jggHrzXO3X7OXwpvru21G9ttVnvrO9/tGK6fXtXa5S6EBtlcTG78z5ImZEXdtQO+0De2fcqKpu7bfe/z019dF9yJ6W6bf197PJ1+B3wtXwlceCv7DU6dd3SX80hnna+kvoypS8a+Mn2s3SlV23Bl80bRhxgV0PhH4deEvBF1qeo6Dayf2hrLRte3t1cz3t5ceSCIlkuLl5JSkYJEce7YmTtUZOe3opJ/1/XovuBniOr/s7/CzXNR8Sapf2up+f4vZW1cQ65q1vHeBIxEqSRQ3aR7BGNmwKE2fLjbkV16/DDwWnxCj+KaWky+JYrD+y0nF5dCIWW7f5ItvN+z43gNny85Gc5rv6KabVrdP8rflp6Dcm9/6/qy+45jV/B2g67r+ieJ9Tjmk1Dw688liUuZ4oo3uIzDIzwxyLFKfLJUGRGKhm243HPmmrfs4/CTWtR1XVb7T78XOtapba3dNDrOp26vqNmoWC4VIbpFjeMKoGwKPlTj5Vx7lRS6p9v8AO/5pP1C/9f16s8y0P4QeB/DurRa9p0d/JqMV095591quoXkjyvAbbEjXNxIZI1iYhInzGjEuiq/zV6bRRQIKKKKAP//T/fyiiigCG4mFtbyXDKXESliFGWOBnAHrXzxrf7U3wl0D4HWH7QuoXV0PCWpeUISlu73TPLKYRH5A+berhgy/w7T6V9GN0r8mPBHh2HXvjl/wybqUJl0XwJ4j8R+KJoSPk/s3UrdX09MdNon1GYKP+mXTigD9FtZ+MXgnRNV8HaNPPJPc+OkuJdLEEZkDw2tv9qklc/woI8cnuwHU1vfDzx94e+KHgrSPH/hV5JNJ1uAXFsZUMchjJIG5DyDxX5u/srjVfGXj19L8QFppvgF4VvfCTs4/5iE99PDu57mxsYGz6Se9cN8GvA9t8PvhP+zJ8T/DurarHr/iPxDYaZfs9/cPbTafexXW61NqX8gRpsXZhAQRuySSaAP2N3DOO9cP4X+Inhzxf4g8U+GdGeRr3wdeRWOoB0KKs01vHcqEY/eHlyLkjvkV+T2s67ocvwq8YfGDVfG1/a/HnTPFF7a2lguqTLNBcw6kYLPTItMEnlvby2wTjyjvVi+7uGfGPxJ4z0fxd8StM0W7h0zSvEPxI0Wx1q5ub2fTLZbZ9AhdIp722VpbeKWdY0Z1x1ClgGJoA/ZzdnpXD6H8RPDfiDxt4m+H+nPIdX8JJYvfKyFY1GoRvJBsfo2VQ5x0r5A/Y5j17SvFnxB8MS+IdHv9EsBp0kGl6Pq17rkOm3Mqy+aBd3kalRKoRvJWRthBOF3c+M+ONF+DWt/tU/HYfFrxhP4bOn6FoN3ZRx6tLpQDJaS77pPKkjMssJ2hQdwXd935qAP1eJA603eM471+Vfwp0/xD8ZviZ8LE+MV/qsjxfDO21q4tI7u5slnv478JDczxwvGWk8s7iDwS3IOBXzdqHxDM2veAvin4T1CSzutf8c2US6hqHiya51y8tLjUDDPby6NAotbe3WMlTG+Ni7c/OeQD9xvDniSDxJa3V1BZ3dkLS7uLQreQNbu7W7mMyIrjLRORlHHDLgjiug3Z6V+RsKax46fwL4V1bX9UgstW+MPjCyuja308EstlAmoutsZUcOIsIF2gjaPu4IBr6u/Ze04+E/Gnxk+G+m3l3P4f8MeILVNMgvLmW7a1iu9Ot7iSJJJ2eTZ5jsQpY4zxQB9jUhOKUdKRulACbqN1Npe1AC7qdUdSUAITik3UN1ptADycUA5pD0FIOtAD6QnFLTD1oAXdSk4plObrQAoOaWmr1p1ADd1G6kPWgdaAHE4oBzTT1pVoAdTd1OqOgB4OaCcUi9aQ9aAHjmg8Ug6UHpQAm6lBzTKctABup1MPWnjpQAhOKTdSt0plADwc0m6lXpTKAJKQnFLTWoAN1OHNR08dKAAHNLTB1p9ACE4pN1IetJQBJSA5pT0pq9aAHU3dTqYetAC7qceKYOtOPSgABzS01adQA3dSg5plKOtADzxSA5oPShelACnim7qU9KZQA8HNBOKRaVulAC0hOKUdKRulACbqUHNMp69KAAnFLTWp1ACE4pN1DU2gCQc0hOKB0oPUUALSE4paa3WgA3U6o6koAKKKKAP/1P38ooooAQ9KxYdA0G31m48R2+m20WrXcSwTXiwoLiWJDlUeUDeyqeQpOB2raPSvkjwlq/xUsf2lPGejeL9dj1bTx4at9R03TLSIwW1qrXtxGg+d2MkzpGvmSEgZOFAUCgD6f0/w/oOlzX9zpmm21pNqsnnXjwwpG1zLt275ioBkbaMZbJxxUEfhPwtDY6fpkWjWSWekSLNZQrbxiK1lQEK8KBcRsu44KgEZOOtfllaeO/i/8E/AWoeIfigPFFh4713w7qNzpz6jrMOp6T9uTbJKotIiy27wK4eIEspRWBIOVr0v4ka745+BcXjbwjoHjvU783/gtNVsbrV53v7m31f7YtkBbtskYG7aZFjjCMiygFVAyKAPu+b4feA7jxOnjWfw3psniGIBU1JrOE3igDAAnK+YMDj73Sr114R8KX1vqdre6LZXEOtkNfpJbRst2QoQGcFSJCFUL8+eAB0FfKv7MOv+LP8AhMvHPgzxP/b2lw6fDpd3Y6T4ou01DVYUuUlWa4+1RPNG9vLIm1EEzsjo+QgZVrxPVfih4v8ACfjm41bxDrXiWPxr/wAJza6UujGC4Ognw/fX8dpAyDy/spV7eRZBMH83z/kzjK0Afot4Z8I+FPBemjR/B+jWWh2AYv8AZ7G3jtotx6tsiVVye5xWdrXw2+HXiW8OpeI/C2lardtJFKZrqygnkMkAKxOXdCSyAkKc5AOBXg1jqXxPtP2s4tF8SeIY7jw3qHhvUrqx0q1iaKGAQXlrGkszMzGadldsthVUfKo6k6f7RPxw8Q/CK20OPw/4X1TWJNR1TSreS5trWOe2EN3epBLBuaaMidkJ8sYxuK5PWgD6J/sbSF1Ma2LGAaisP2YXPlL54g3bvK8zG7Zu525xnnGa5GL4VfDCC9vdSh8IaOl3qUiy3Uy2FuJJ5EcSK8jbMuwcBgSSQwz1qp4hGt+PvhnOdH1C98A3mqQIzXFxDGL2xi3gzZQu0aSGMMFbcwQkNg4xXwsfEvxGj8D/ABB1f4UeIPEWt/DuO/0W2s9XmeTUdUNuLjZrl1pcjq808KQkeU+H+cSNDkBaAP0Rh8JeFLd4ZYNFso3trqW9iZbaMGO6nDCWdSF+WVwzbnHzNk5PJrQtNI0rT7q8vrCyhtrnUHWS5ljjVHndVCK0jAAuQoCgtkgADpX5cXHxU1241jVfA3hvxJ4sn+GGka1osuo60Yr19ag07UbK8Z0jleL7YbY3lvCGm2eYqyPtbywGH13+y746vfGXh3xZYTXmoalY+GfEF1pmn3eqxvHfTWawwzxGcSqkhZfOKhnUOyBS3zEmgD6gHSkbpXxavxB8XfDv4/ePNU+L3iyH/hD9J8JLrUNrbwtDa2Fsl9PGGYFnaado0G9+MsdqKBjPEfst/HrxD8W/jp46PiHxNYzWGp6JpGp6LoVrdwz/ANm27z3kbJJ5TtuuSixSXJH3GdU6KpIB+g1L2r4++MX7RPj7wReePpvAXhGy1/TPhjpkV/rL3moPZzyvcQtOsdqiwSq3lxKHdnZc52rzX1fo1+dV0ex1Nk8s3kEcxUHO3zFDYz3xmgDRqSo6koAa3Wm05utNoAcegpB1pT0FIOtAD6YetPph60AJTm602nN1oAF606mr1p1ADD1oHWg9aB1oAD1pVpD1pVoAdUdSVHQA5etIetKvWkPWgBw6UHpQOlB6UAMpy02nLQAh608dKYetPHSgBG6Uynt0plAD16Uynr0plAElNanU1qAG08dKZTx0oAaOtPpg60+gBh60lKetJQBIelNXrTj0pq9aAHUw9afTD1oAB1px6U0dacelACLTqatOoAjpR1pKUdaAHHpQvSg9KF6UAB6UynnpTKAHLSt0pFpW6UAKOlI3SlHSkbpQAynr0plPXpQAjU6mtTqAGtTac1NoAeOlB6igdKD1FAC01utOprdaAG1JUdSUAFFFFAH/1f38ooooAQ9K8+n8B+FZfG+o+OLos+p6jpCaTcRvIPKNlHLJKPkxkEtIwLZxjivQT0r4z8HeCYdH/ay8b22qape+IRr/AIStJ5/7RkEqJHNqF0gt4olVY44UjAQKFy3LOWYkkA6jw7+zd8IIlfSr7V77xVZJpt1pFjY6lqIuotP0+7UJNDaqoVxuRVTe7PIFAUMBnN3R/wBmr4YXOh6/pmr6pqXi9tctLfTZr3UL/wC0XVvbWLmW3ht5YgnlGGU+YGHzmQBmYkCvkHxb8NPAmlaB8b/i98KfC+n+Fk8E6ffeG9Jl0u2S1kaVI1OrXpeMKxZNxgj5+TypCOXNfQ3wx8NeB/h3+0PN4c+F1tbaX4evvBdrfX9vZ4W2Nwl2Y7W4cL8vmyxGQFz8zhQSTgGgD1fw18GvDPg7ULm/k8Uavf8AiLWJLLzdRv75Hvp7fT3MkVouI0QQZZt6ogLb2LMSc1Sj+Anw+X4gf8JFLq2ozldROux6FJfbtOj1IjH2xbYjfuDfOFLmNXO8IGwa+HPivpOr6p4u+NXxQ8U6Zomq2fgXWbCKNb2W4j1qKwt7S0uIk0q4idRZO0kjujBX8+RirYAqpFZ3k/xL/wCFzpZWK6G3xOjs/wC03dv+ErUmRdO+xAfc+xeeceVu3fZ8tsz81AH6f3HgbQ7nx7Z/EeTzf7XsdOn0uPD/ALr7PcyxzPlMctuiXBzwM8VyXxK+Dln8TruzuNR8T61pENkYnS20+eBIDNBJ5sc5WaCU+ajYKsCMYGORmuD/AGsfEnxX8MfB3xBqPwqtLaS7jsLlpLh72W1u7YqF2PapFbzea/XgsmOOTnj23wZd+MNT8NRz+NbOy0vV3Lho9Ou3vYVXojCWWCAlsckGPAPc0Acl4x+FGh+PfhVffB/xJruqXNhd26W95eLdImoyxhg5Ekqx7f3gG1/kAZSRjBritF/Z78JyeDNS8JS+MPEGv6TqC2ywNPqSM1g9k4khezeCOPynR1U9x8oBGMg+G+CPB3g3w5r/AO0L4U+JHia/udBurnRE1LU9RvTHcSC7sYtwaaPyxEjs2wJGERVO0ACut/Z20ePwB8Y/ih4Gk07TdDheHSNTtLLQ2P8AZcNpKJoEYowQx3crREy4UKyhCMnJIB6G37O3gFNNk0aXxBq//CS3mpRa0dbN+o1l7yCI28cgfYIzGkOYhF5XlbSflySa7nwd8NfCvge00/w54d1W8Sawu59Tut92JLnUbi6DiWW9LAtJuZ93AUBlXGFULX51/E+wvrz4l/EH4tW1np8mj+GfG2gW1xqVw7DxLZtZrZJLb6Z/AtrKzrhGZTIJJyFbcuX/AAj0zVtP1X4cfGDxnpei3mpeJ/Ft7ZTXVrJcJ4iS9uJLqI/aLjfsmt4wmx7MxhYo1VgxKUAfpFqng/wHB43uPHmvTRLqOo6UmkSRXUsf2eS0imacDynGCd7nJyRjAxVfw58Jfh54c8c6l8TfDOnQ2up61YWunyNbpGkH2e1eWRDGqKMMxmbeQfmAX0r5R/aP8NX3ir9pv4baZYeF9A8WuPDWvyGz8RuY7MBbiwHmArb3H7wZwPk6E8jv3/7HDvpfwy8TeHJ4Y7e98OeJdYtbiztH8zTrSQy+eLbT3PLW0aSKEyFIO4FVxtABwH7SP7OfxH+KninxHb+FoIbTSPGWm2+nXl5Bq89gMRb1LX9msUgu/KDZi8t4if8AVyEpX3ZZwWui6Va2TSBIbSOOFWc4HygIvJ7nivzxb4//ABZ8OfBqH9obVPFmkammv6PqOoW3hZ7RIZIJoUaSKO2mVxLKbcD/AEoSnkBipjxiuA8YfEX41av4Uv8Awt8SI57jTbt/DupwXN9HpdrdJL/blkhWKHTry5L2rq2Vd1BUrtLvu4AP1ZimhnUvBIsiglSVIIypwRx3B4NWa/I+z+Jfxi8G6HcWPw9u3t9Jh1XxjqF4dOtLTU9Qjli1q4WOS4s7iaKVrEKDva3Bk3cZHGf1K8F+IrTxf4P0PxXp9yl5bazY215FPGjRpKlxGsiuqP8AMqsGyA3IHB5oA6RutNpzdabQA49BSDrSnoKQdaAH0w9afTD1oASnN1ptObrQAL1p1NXrTqAGHrQOtB60DrQAHrSrSHrSrQA6o6kqOgBy9aQ9aVetIetADh0oPSgdKD0oAZTlptOWgBD1p46Uw9aeOlACN0plPbpTKAHr0plPXpTKAJKa1OprUANp46UynjpQA0dafTB1p9ADD1pKU9aSgCQ9KavWnHpTV60AOph60+mHrQADrTj0po6049KAEWnU1adQBHSjrSUo60AOPShelB6UL0oAD0plPPSmUAOWlbpSLSt0oAUdKRulKOlI3SgBlPXpTKevSgBGp1NanUANam05qbQA8dKD1FA6UHqKAFprdadTW60ANqSo6koAKKKKAP/W/fyiiigBD0rFTQNFTXZPFCWUS6tNbpaPdBR5zW6O0ixFuu0MxYD1JraPSmUAYWmeFPDWkaXd6JpumW9vp9/LcTXECRqI5ZLt2knZ1xhjKzMXz1JOa43QPgj8I/Cmhat4Z8OeEtO07S9dXZfW8MCqlygGAsgx8ygHAXoB0Ar1JeleZfGzWtU8OfBvx14h0O4a01HTNC1K6tplALRTQ20jxuAQRlWAPIxQA/xB8IPhd4r8TWnjPxJ4W0/UtcsfLMN5Pbo8q+Ud0eWI52Hlc52nkYpp+DvwsPjb/hY58K6d/wAJPv8AM/tD7Onn+Zt2eZux9/b8u/72OM4ryrxv4+8VaH+ynD4603UvJ8ST6Fp0kVyyLJJJeXaQriNCrIZpXfbGCpXzGXcNua+Ybb4u/GmbUE+E9tP4iOp3viKS3WK4GmJ4jg06LShehZJ9n2BVmn3FJQHIiDL/AKzGAD7/APHHwu+HnxLS0j8feH7PXlsC5txdxiTyjJjftz0ztGfpW14Q8G+FfAeix+HfBulwaPpkTu629smyMM5yxAHcnrXzDb/FvxTpX7JmvfEOHUJtR8TaDbalbGfULaKKeG9tbiS2xdRwMYWeBgPMaMhJNpZQA2B5N43+IPxe+HvjZ/gbo/iLV/FU+tXGhNb6iI7AarBHfw6lJdQxu0UVqu46cCjyIfLWRj821RQB96XXgjwfeDXPtmjWk48TIseqCSFXF6iR+UqzgghwI/lAPbiuZ0D4NfCrwtoV34Z8PeFdPsNL1CWOe4gigVVmkiYNG0ndihUbcnjAxiuH+BnjXW9Q+GOq3niSfUtb1fw3f6hZ3UFzbwDU0e1csts/2VvInlCFQske1ZAVOASRUnwO+M+p/Fc6/FqfhrUdFbSdT1C1jlurUwQtFa3BiRCxkf8AfgD94o4BzigDuNS+Dvws1nxjD8QtV8K6ddeJLdo2S/kt0acPEMRsWI5ZB91jkr2IpbL4QfC7TfGcvxEsPC2nW/iWYuzX6W6CcvINrvuxwzjhmHJHBJr5A+FHxY+Jms/GfSF1zVdSl8O+JNU8SWUUtzFarol3Hp7yGyj0sxRi6WZY42MjTnZIElKlvkNe7fGTxH8S/D/xE+GMWg6la2PhfV9ehsL+ERGS7uzLbXUmwu3yRxL5Sn5QXZuMqB8wB6Z45+EHww+JlzZXvxA8MWOv3GnJJHbSXcKyPEkpUuqE8gMVXI74FdN4Y8J+GPBOiQeHPCGlW2jaXbZ8u2tIlhiUscsQqgDJJyT1J5NfMUeqeP7L4/aX4b0rxtd+Jp5ru7utd0tLa3TStJ0V4pPsallj85Lov5QXdMxl/eNsCYxx/wAdPGPjyD48S+ENCv8AxaNMtvCMWpx2vhWKxkk+2Pe3EXmTfbI3yCqKqgHGRyKAPp3S/gr8JNF17UfFGleEdMttV1dJo7u4S1jDypcczK3GMSH74A+b+LNUtB+AnwX8MWV5p/h/wXpdjb6g8DzpFbIBIbWQSwZ46RSAMg6KRkCvN9E+Net6P+yto/xn1eKLxRrSaXZy3Mds32VLi8kkSCVRvX92wkYhgVADAjAFcX4l/az1nwXPfeFfFXhW2s/F1vq8WmxQR301xYNHLYjUBO9xFaNMAsR2sot2O/GDtO4AHvmv/AX4MeKYLe28Q+DNLv4rWa5uIhLbIdst5J5twc4z+9c7nHRjyQa9YtbW2sbaKysokgt7dFjjjjUKiIowqqo4AA4AFfKng79p5fFlnJM/hqawmttA1bWZkld0HmaVcC3aOPzIkZopc745SqnbjKA5AwV/ao8S3f2y50zwXFPaxHQrS3Lajsln1LxBBBNbw7fIKpFH5x82UtkBcqjE4AB9nN1ptfIb/tG+PX8RWvw4tvBFq/jdtWuNKuLdtUK6fH5WnjUo7hLn7MXeOSJgMeSHVsjGOa5Yftj3P2vweLnwpFpdl4jgtpJrjUL820QnluntZoLSYwG3mkgMZcrLLCXUrsBY4oA+5z0FIOtB6CgdaAH0w9afTD1oASnN1ptObrQAL1p1NXrTqAGHrQOtB60DrQAHrSrSHrSrQA6o6kqOgBy9aQ9aVetIetADh0oPSgdKD0oAZTlptOWgBD1p46Uw9aeOlACN0plPbpTKAHr0plPXpTKAJKa1OprUANp46UynjpQA0dafTB1p9ADD1pKU9aSgCQ9KavWsDxD4t8K+E4I7nxTrNno8MzbUe8uI7dWb0BkZQT9K2LS5tr2CO7s5UngmUMkkbBkZT0IIyCD6igCzTD1p9MPWgAHWnHpVFtRsEv49Le5iW9lRpUgLqJWjQgM4TOSoJAJxgEj1q8elACLTqatVU1HT5b6XS47mJ7yBFkkhDqZURyQrMmcgMQcEjBwaALFKOtJSjrQA49KF6UHpTI3SQEowYAkHBzgjqKAHnpTKeelMoActK3Sqv2y0S6SxedBcyI0ixFhvZEIDMF6kAsAT0GR61zw8d+CJNc/4RePxDp7ayDj7ELuI3Of+uW7f+lAHWDpSN0rK/t3RBq48PHULcaqYvOFp5qfaDFnG/wAvO7bnjOMUk2u6JFq8Xh+XULdNUnjMsdoZUE7xjOXWPO4qMHkDHFAGnT16UysceJPDo0V/Ef8Aalr/AGSgZmvPOT7OAjbWJlzsADAg88HigDaanVlXGtaPbxWc9xfQRR6g6R2zNKqrO8gyixkn5ywGQBkkdKNJ13RNeimm0PULfUY7eRoZGt5UlVJF+8jFCcMO4PIoA02ptYdt4p8M34lax1e0uBb3H2SQxzxvsuSQBC2GOJMnGw8+1blADx0oPUUDpQeooAWmt1p1NbrQA2pKjqSgAooooA//1/38ooooAQ9KZTz0plAD16V49+0P/wAkC+JP/Yt6v/6SS17CvSvHv2h/+SBfEn/sW9X/APSSWgDF0DRPB/iL9nHw9pXj8xLoEnh3TXu5JpTbrEkVtE4l84MpjMZUMHDAqQCCCK5bwr8KfgL4y8Jz6v4S1yXXI2v/ALc/iC21ma41BLyCHyN329ZTIuyAmPZuC7CQVwTnnfiBp+oar+xbZ6bptjJfy3Ph3R0KwxvNJFHstzJOkSZaVoFzKIwDv27cHOK+boPDfxh8X6H430f4fQSeNfDHiPXIZ9Z1m5b/AIR+61Wxi06GN7SCPyFQJvRYZZY413IGUfOSwAPuzQ/hv8Jf+Fex6ZpTLeeDbvTLuGQNeyT2d3a35824nmdnKyvISzGdiW+Zju5NcB4K+GX7PHjHwrqz+D9ebxLEbu3mudYi1ua8v7a4sEIttt95rSw+TGx2AMBtdiQd7Z8V0ez8X6t+wdpXw4/sefSfFMvhWHZp0cUlxI1navEsq7diKZJIcjyCQxzt55NdP8K/DWjfEC7+JekXT3Xi3wbf2WkhNTvrH+zLi6uYVnaWxbyYbXz4IAIiAUIHmPExYAqAD6c8C+CvAdh8PDovgW7ll0XW0muDqEF7LLc3T3mWkuRe7zI0jk5EgfI42kADDPAHwk8N/Di7vLvQL/Vrg3xdpY7/AFO6vYjJI295Ak8jqJGblnA3HnJrgPgal74Z/ZT8IpcSNoV1pvhqDc8tq8rWjRQfea2GHfZjJjGCcYrz79lDX9fv7zxXp+u+L/7eMup6lexW50iewIinui0dwJZSRtdTxD1TPPSgDtfCXgH9nwfEXULPwpraXfiLR3vZv7Ji1iSZdKmvtwupbezEpW2kcyMGZVBXeQNu4g9v4uuvg9Y6x4N8I+N9ftLfWdLu4LzRre91AJdy3Ecb20b4dw8xYSMvzZ3Me5r4v+G1n4ksfiv4S8K6cCNG8A654n1G/kk0q6t9QS0ukux/pk7Zhk82WZGi8kubhQJSFKEVT/aEg1LXviJ4m1DS3voBrmm+GzpelxaZLPH4nFtcvcCJ7kxF7Qxs2xtrRsgPmPkYwAfUXg74d/Ajw58WtVh8KeK7pfGM95Pq9/pCeILmTfPcDLyTWJnKkYYYDJhV24AAFRfGWx/Z4tvHttrnxL8bHwh4nn0tLVRDr82kSzaes0jruSKaPeglZ8MR1yM8V5tql1pGp/H7wbH4P0e8s9Q0TxBqLanpEmmNBEpubeZJtaN6i7XWRSoQNIVffgoJB8uR8d7vU9H/AGkLm/j8TTeELS/8EwWgvF0OTWY55ft90xiCqjqrqGDYPJBHFAH1vbfDT4dah8MbP4caZZxt4Q+zwLbxW8zFGhjdZo2WVWLNlgGLbiW5JJyay/FHwK+Hni3Ur7XdQt7m21a+vLe/+3Wd3Na3UNzbW/2WOSGWJlMf7nKMBwwJ3A14DZW/jfQf2HdH07Q7O/8ADHiGz0nT7WGO1EgvYFW4ii3qjhpEZovnKMCUyVI4NcT4nPjPwX4r1jwbquveJk+G9r4ksftuopNd3N9DZ3OkGYot2ga4W3a+CiRkPyZ25VSQAD6V1T9mb4WatYafp9xHqUYsba8s3li1S8jnu7a/cSXMV3MsoedZXAZg5PPTAyK3n+APwwfw/qnhk6bILLVvsBlK3MyypJpcUcNpJFKrB4pIliQq6EHIznOa+LdJtvit42t5oNR1/wAVWmj6f4c8S3mkTJLcWN1dfZr8LpU1yVVXebyOVVwGkXBdWya9f8ba78eNa+Eeh+bbnSE1JvC6tqWmXcw1ctdXVmt75lt9mVYRseXcRI2AOQBnAB7v4Z+Bnw98JX+n6xp9vc3GqafeXOoC9u7ua5uZrq7t/sskk8srM0h8kBFDZCgAKBiuYvf2X/hJfW9pYPa30VhbxrDLaRajdR213Ck73Kx3USyBZlWWRiNwPBKnK8V8fePtQ+LnhPVbzwXFrOrx+BtO8T6hA2o3tzqRmWE6ZY3FpFJfWaSXZgM8tztbO0uqxs+AFMl/8Tviv4S+H3iBfFOq63d6rrHw9uf7AmtrC+8ybVYp74RuqCIOlyIWt2ZpFRmADkDnAB+oeAAAOgoHWvmn4H6d4kvfGfxE8T+KNR1WdodXjsLC2up5RZx2Y02wmLQwNhCTO8uZME53LnqK+lh1oAfTD1p9MPWgBKc3Wm05utAAvWnUxetR/arX/nsn/fQoAkPWgdarm6tc/wCuT/voVOpBwQcg0AKetKtIetUNT1bStCsJ9V1u8h0+ytlLyz3EixRIo6lnchQPcmgDSqOvzh+Mn/BVD9lP4UTyaZpGrz+OdTjba0WiIJYVI9bmQpEf+AF6+TdW/wCC3ngtTt8O/C3Ubs9vtGoRQ/okUlAH7pL1pD1r+fC+/wCC0fxLvyw8L/B6AA/dMt3cT4+ojhTNOsP+C03xE09lXxX8H4ePvGG9ngJ+gkhfFAH9B46UHpX4Z6N/wW88AyAL4i+GGp2jd/s19DPj/vtIq9i0D/gsp+yvqm1NZ03xDozHqZLOKZAfrFMxP/fNAH6z05a+DfC3/BS/9i7xXNHbQ/EKLTZpOi6ha3Nqo9jI8flj/vqvrLwd8WPhf8QYhN4F8XaT4gVhn/Qb6C4P4iNyR+IoA789aeOlMPWnjpQAjdKZT26UygB69KZT16UygCSmtTqa1ADaeOlMp46UANHWn0wdafQAw9aSlPWkoA+EvhV4F8G/GL41fGrxL8WdHtPEmreHNej0LTrXUokuorDS47KCaPyYpQyp9oeV5HcDLdM8Vyvj7x34d/Zf8H2Xw0/Z8vptQufEXiy507yYbeTXf+EeZ7dr24t7ewtish2IoMVuWAXzCxOwYr6X8ffs4eE/Gni+bx/pWua34M8RXsCW17eaBfGza9hiyI1uEKvHIUBIRyu9RwGxxWa/7KPwj/4VzafDiygvbGKx1H+2YNUgvJV1ZNWOd199sJLtO24hmbIKnaRt4oA+V5/2mP2gtG+E/jq+m0+eTVvD1/oMOj6zrWgXOiR3yatex20sU1nK334cnLxvgqynAINaHxJ+OPxz8F+P7f4Iw6/dX2q6TpMer6lrek+EJ9XkuGvbiZLaBbO3mKQRokR3yM7M5wFCkE19K2/7KvgA+F9a8Oa5quta9c+Ir3T76/1LUL4z30z6ZMk9sgcqESNGQfIiKME9zmug+Jf7P/hf4j+JrPxvHrGseFfEtnatYf2lod4bO4msmbf9nmyrpIgcllyuVJJUjJoA+GNL8V/Hf4u/Gz4TarZiH4feLr3wn4lt9Qn1HTZWZILXUbVPPgsZ3jdTcbEZVlY7Ec53EDPdp8f/AI9eJI/Bvw80rUdK0rxbdeMtb8IavqgsmntXXSrWS4F3b27SgqzIqnYzkB8gkjivdNQ/Y8+GVw3he60XVNd0DUvCFld2Wn39hqUiXa/bpVmuZZZXDmWSVlO4vlTuPy5wR2fh79m34a+FofBkejpdq/gi/vNUtpZJzLNdX1/FJFcz3cjgtM8glZiSR82OwAoA+Kvjv+0v8YPhtc+Mv7D8daff33gK1ib+zdM8M3uqJdyQWyTXLapeRlYLAuxYiNXPlJtZmbJA+gfhHrD+If2qfGuvyRiJ9S8EeFLkoDkKZpL1yAfbNb3i/wDY5+GfjW/8WHUdX1+00XxtNJdatotnqT2+m3N3LGI3naJRu3MFVmXfsZgCynpXr3gz4PeFvA/ii48X6VLdTajdaPpuiSNPIHVrbS/M8g4Cr+8PmNvboeOBQB4BrXjL9oD4k/Ej4h6F8IfEGj+F9O+G8trZJFqVg12dUv5rVLxxPKJUNvbhZFQNGrPnc3bbWZL4+/aI+Jvi7xzpvw617Q/CVv8ADdLO3linsn1JdT1OayjvZQ0pkhMVookWNHRS7ctxjbXqfxE/Zf8AA/xD8Tap4pfWNc8PT+IbeO01mHR9Qezg1WCFdiLdIAckISm9Cr7Pl3YxjO8S/skfDXXNQkvND1DWfCEN9YW2l6jbaFfvZW+oWVpH5UMdwgDZKRfuw6FX2fLuxigDxjwr8cfjX8dfGfgbSPAGraf4L0zxB4JtPFN6Z7H+0pUuHujA0EWZYx5bf3jkgDjk8fOnw4+M/wAXPhN8JNA8J2mryaxrPjvxh4sjjvrTQZtTnsbfTb64a7eOzglLTyTSEFAzBYlJB3BRn9P/AA38FvAng/xZp/i/w1avYTaVoMXhy1to2xbRafDL5qKqYzuDfxbun515Uf2QPht/ZN1pVtqut2mNbufEGmTwXxiuNGvrxpHuDYyqgKRytK++N96tnBFAHjug/HH9oS8+Dvj3xPcWht9T+HVzDfwXuuaPNoMWvaVHH59zA0Fyx+zTBQ8YlVim/Y2ACQPb/wBl34teK/j34T1D4xX4gsfCmv3LL4e05QjXdvZ2xMTyXkiMw86WRWPljiNQASSTUg/ZR+Hc/hO68I67qWs67batqltqusTahfNPPq0lrjyoLtioDWw2r+5QIpCgHgnPp3gX4U+FvhxrfifWPCfnWcPiu7W/ubEOPscd3sCSTQRbR5bTYBkwcMw3YBzkA+F/iJ448VL8OP2mvjPo1xJb67ol/J4U02dPv6dp2nLAk0sX90+ZcTTsR1wufuitr40fCD9nr4YfBS2sLX4T6jr+mpp8l7J4h8NwWTapYNbRrN/aD31xNFN5xOZFcF8kHIxwfpnwz8LLjw/8RPiHaXNjDqPgf4iqmpTxS7WEWomJbS8heNvvR3MSRyAgEBhIGxlc8LL+xj8PpNNPhRvFPiv/AIQwgofDg1qcab5J4+z4H7/yMceV5u3HGMcUAfEnjfxJbJ+1r4Z/aR0Z7n7DoPhPwfd3L3OBMdG164urS4acIdu5BPHK+MgFDjiun8OXM3jL9vzw78ZlVriDVG8W6HpShsK9h4ftYIcpnj95dvcHP0r7y1b9mj4W6xc+IZprSaGDxJ4atvCk9tDIEgi0y0MhhWFcZR08w4bJxgccUvhH9m74c+B7j4e3WhC7V/hrZX9jpnmTB96akFFw9wduZJGKbt3HzEk9aAPnrwZ8dfj3qn7SfiTwtqfww1WHS00nRGawfVdNZNOE9xdpJekrMQ4kVRlEJf8Ad9ORnxO60aG7sda/YouVzban8TJFNuDj/imrqP8A4SOTA/uffg7DnH1/VmLRdIg1a416Gyhj1K7ijgmuVjUTSRQljGjPjcVQuxUE4GTjqa8v/wCFEeAW+OK/tCGKf/hKk0r+yB+8/wBG8ndu8zysf67B2b8/c4xQB8J/Bi7v/H/jP4JfB/WWM0/waPiG41VW5zPosh0fTmYerpKZVz6ZFeN/st/EIfs6eGPiF4guAzWHjGy8Ra/YRk5Ems6PqtxZSQoP70ySWuAOuDX6qeDfgb4D8B/Evxl8V9AhmTXfHRtzfl5N0K/ZlKjyUwNm8nc/J3NzXnN3+x18Hb/w74S8MXsV7NZ+Ddcn1+yLTje1zc3DXUscxC/PA0jZMeBnavPFAHyB+yt8GTp3wj/aC+FOrTsNUh1iOWW73HzI9XOjWN2bpG6h0uz5qnsQK/Qr4C+Or74m/BXwR8QNUQJe6/pFnd3AAwPOkiUyED0LZI9q47xf8IbzQvCvxJT4VeYPEfxQuxJdT3Eq+VaS3EEVjJcoMA7YYI/MCDJZxjvx7L4L8J6T4D8IaL4J0FDHp2g2cFjbqeSIreMRrk9zgc+9AHUjpQeooHSg9RQAtNbrTqa3WgBtSVHUlABRRRQB/9D9/KKKKAEPSmU89KZQA9elePftD/8AJAviT/2Ler/+kktewr0rx79of/kgXxJ/7FvV/wD0kloA5rTPHtp8M/2bfDXjK7tXvzZ6HpEcNtEQr3FxcRQwwRKzcKXkdVyeBnNdN8NviXqXiy48S6D4t0VPD3iHwnPFHfWsNz9tgMdxAs8M0M3lxFlZSQQY1IZWGCME8WvgK4+Jf7MfhrwnY3a2F9Loui3NpPIpeOO6s0guYGdQQWTzI13AHOM45rW+HvgT4iaDruv+NvFE2ly654vvrQ38VoZzbW1hZWpgijgaQBpJS+XYuqr8xH8IJAPMYv2ubew8Njxt4s8LSaboWr6Rf61oTw3kdxc31vYFMxyw7UFvLIsiMg3uoBO9lIIr2z4bfErWfFGuaz4L8ZaFF4e8SaJb2V7LbW94L6B7TUPNELrMI4juDQyI6lBgrkFlINfM+h/sh6zHqeq6hrE2iQSx6XrOnac9vbzzwzSauRumubO4cwQxqq4a3t8RuzOxIOAPXvgB8Crz4W6r4i8V64tlDqmvwWFn5FhNdXMMVtp4l8sme9Zp5Hdpn+9gIgRFGFyQDutH+Pvwd8Qa5P4a0nxTaTX9ut2zoSyIPsDFboCR1CFoSD5ihiVHJGOaz7P9o34I3ujX/iGDxbaLp+mNbC4lk3x7BeP5du4V1DNHM/yxyKCjHoTXkOr/ALLOo694H0bwTqGsQwRWlx4se5mhRt5i8SJeInlg4+aL7UpbPB2nHaqt9+z18TPGmqWXiLx3qejQX+kpoVnbRabFMIJbbStTh1CaWXzBkPL5IWOMArHk/M2SQAfUngrx94P+Imjy674N1FNRs4J5LaVlVkeKeLG+OSOQK6OAQdrAHBB6EGuJ0L9ob4K+JZryHRvFtlN9gtJr+V2LRxC1tyFmlWSQKjLExAk2k7CQGwayNJ+B8aeJvFHiHV9WvYv7X8QHWbWPTr25s0Cmxs7Xy7pImRZvmtS2GBXa2O5rxjxB+yRq3iT4ceFvAVxrkFo2heH9Y0mWeJJPmn1Ca2nikUAo2xWg/eDcrEHgg80AfSXh/wCN/wAK/E0KTaN4ghlL3lvYCN0khl+03QJgQxSKrjzQCUYrtYA4Jp2ofGv4W6fq1loF1r8P9o6je3OnW9uiySSSXVm6R3EYVFJzE0iBz0XOScZI+YtN/Zh8e2MM3ia1k0ez8UW2o6Pe2qNearqEEy6VJK5jnub2WSVVk859qxx4jPXfk103hj9n/wCJWha5L4zuda0w688fi+RDDHMII7rxHNaTW5G7LbIDbEOerZBA6gAHa6b+1N8KVtJLjxdq0GgO9xdfZ45WeVpLCC7ls471yiYjhleFiGY7VGCW5FeizfGX4YQeMF8Ay+ILca48iw+T8xXz5EEiQmUL5QlZCGEZbeQQQORXiN7+zNcxeG/EfhnRNQt4rfVfAVv4PtmlViyTQC5BuJMdVczBjjnIJrjrL9kzW7L4gS6pJdWV/oV1r0XiBpLi81MXEMqMkrQpZRzrZviZMxzMMqpAKMVBIB9Q3fxi+GNholr4kvPENtFpl7Y3upwzsW2PZ6cFN1MDj7sQdd31rBP7R3wTWXToZfFNvE2qqjweYkqAJJKYY3kLIBEkkilY2k2hz90mvma+/ZX+KuseE18CalrmjJpWj+G/E/h7TZYo7jz5f7d8ryp7gN8q+WI8Oibs9QecD0v4jfATx54j1PxnYeF9T0uDQPiRZWNlqzXkUrXdmtpGYGe02fJJviPyq5URv8+WzigD1uH49fB+68VSeCovE9qdXhmubZ4W3qq3ForPNCZCojEiIrOU3btoLYxzWz4D+K3w9+Jwuz4G1mLVTYiNpQqujBJgTHIBIqlo5ADsdcq2DgmvFD+ztq402y04ajaMLbxlqXiVjJE0ivb31vdQLCynAZ189d2TtIBGal/Z4+CPjb4S6rqs+uanappFxZ21rbaZY3F3c2qSQM5a4jF6zNbK6sFFvGxjQDgmgD6rPQUDrUN1dW1lbSXd5KkEEKl3kkYKiqBklmOAAO5NfDfxB/4KSfsdfDm5uLC/8eRaxe2rFGh0mCa++ZeCBLGphP8A38oA+7ahlkWMruz8xAGATz746fWvxR8a/wDBbH4VWTPB8O/h/q+uS9Ee+mhskJ9cR+e36CvjTx3+1b+3n+1q01j4cD/D3wjcZBSy32StGTj57p/9Ilz0Pl4U/wB0UAfuJ8f/ANt/9nX9nGCa38c+JY7vW41JXSNOxdXzHsGRTtiz6yMor8ffHX/BVf8Aan+LOpXVt+zx4Ot/DejRnalzNAL+7HPDPLLttkJH8Oxsf3j1ryL4c/sVeG9NuRq3jq4k8S6izbnWTKwbzzkrnc5z/eOD6V9kxeENN0TSV07T9Pht7ZFCqkcYQLt/ubRx/wDWIoA+Btevv26PjUZP+FkfE+/tbOXiS3W+aCEg9vs9kEhP41gr+xl43aNHl8eyrIy7inlyFhk+nn557V+j/hbwpHeM0O1d2WZQRxjj0zz/AJ9a7+90K206dobiSGO4b5W8tllcAc5cJlcDsD39jQB+Sd5+x941tU8w+OJyD0BhkB69/wB9xXbfD347ftt/siOi+GNdk8VeFLdudPvd1/ahB2EbETwf9snUZ65r9DtU0yJw0TICmSwfG0k+vqD6jpXz74v0zUrm4/snRbQ3N1cDIVcAAf3mZsKo9yaAN7xN/wAFqPGWteEbXSPh58NI7Pxrcgxyy3Nw93aRP0DQwxqkkhPXDsAvfdXxf4j8NftVftR6qNe+Ovi69NtI26O0kb93FnoIrOIpBF9cZ9cmvo7wH8JL3Qbk+Lryxslv52+QmMyMAMj/AFqrsyecnd7DvX1B4a0GH7Gtyc+a43NlAME84A9O1AHxb4P/AGUvAnhMR3EumtqlyuMy3f7wE/7K4CA/8Br1geB9J04Lb2unw2+RglI1TGPTaB9a+rP7Ot1QFwFxxkDHP/168z8T2kdpNHMh8wBs+w5GMY7UAcdp/gW2lVZZFLOOMnJIHoPSt4/D+1lhQ+SG3Ejjjt09yf5V6h4aXTWiQlm5BJZEPAHuRjOT9eePbrY7W0e3WVFATPAA3OB3BI60AfI3iH4S6VeMYZ9Nhkz13wq3Ue4NeU6x+zd4Oc/aBotoZR2FunXr0xg1+iC21leeZN5QU5wABgDA4A9s9azrrwpHdhnT5VACsQOCW4GB/KgD8kPEH7N/hJJHNzpbxqct5lozR4H+6Sy/pXm837N+nectx4T127tZlIKllVyD7MhjIxX7Dan8PbXymSSLzPUlM4NfOfiPwK2jamb7S49rLyyhcK3PQgdDx1oA+S/D3iP9t74T7T4B+Jmrtb22Nlu97NLFgdB5F0JIce2MV9GeEv8Agqp+2X8N/Lg+KPhTT/FlpFw80lq1nOwH/Ta1Pkj/AL9Gvf8AwNZ+H/F0cVnbyRwX4GJIJQqZIyeGYhT1z1yQePWtHxD8N9Ovkkt5bVFlORkqASASCR7ZoA7j4d/8FovgTr3lWvxI8K6x4TuG4eSHy9QtlPuy+XL+URr7V8Kft6/sgeMY430r4o6RbtKMhL6VrFx7EXKx4Nfhd8S/gr4dtZpL2/0eC5ijz5gaMKQqjJKsMNkd+envXip/Zu8FeJdkuiWU9ik33SJyc+4Vwxx+tAH9YPh34n/DfxZGJPC/irStXUjINpewzcf8Ac12isrqGQhgehHIr+Slf2Ergwpc6X4kntJiNw3RBgCPdCp/Gu38E+P/ANtz9iu6Op+EtUm8YeD0fdcWVx5l5a7epLRMfNgOP44mx6k9KAP6paa1fmJ+zN/wVN+A/wAb2s/DXjuQfD7xZORGIL6TNjPJj/ljdkBVyeAsuwk4Aya/ThJY5o0lhYOjjKspyCD0IIoAWnjpTKeOlADR1p9MHWuG+IPxQ+HXwp0OTxJ8SPEdj4c02MH99ezpCGI/hQMdzt6KoJPYUAdyetJX4tfGf/gsH4Pg1BvCH7M/hO78da1O3lQXdzFJFbNIeB5VugNxN9CI6+yP2HPGf7VXjfwJretftT+Hzoeo3V8J9K3Rx2zmzlQZia2Ql4xGw48z5iG56ZoA9Y8S/GHxbpPx48I/Cu28LvDoGum9SfWLl1Cyy29oblY7SNWLkDpJI6hc/KoJyRyGufFn41eDvG/h3/hK9H0aDw/4r8QtodlpUMksmteQfM2X3mhzA67Y/NkiCDy4zkyFgRXsXjP4f3/ib4i+APGltdRw2/hCbUZZ4nBLyi9tTbqEI4G0nJz2rwTwz8Ivj9YfGbUfid4mvPDOt/bbqSG2uJvtpu9M0d3H+jWMZHkRuUAMj4zI/wB8lQAAC54b+OXxWT4sp8N/Ful6Jd6hqGm6hqS6PpVy76jpS2u02yX0rsYX+1BwFZRGFbgB1yw2/AvxQ+L8PxW0f4afFWx0Q3XiHRbnWBFozTGbSPs0kSeTdmVmWVXMu1JkEYZ0YBMcjLi+EHxw1XxX4e8SeMNd0O8uvAy6jJpd/BayxXmo3N1byW0P28DCRRIrhpEi3B2VSNoAFZ3wT+E3x9+G8+s6j4nufDGta3rkMkt9rOb57+9vlU/ZxKXARLaMnCwxgKi8IM5JALHxM+PXxB8O+KfH6+E7TRk0H4V6bbalqyanJKl3qCzwvcMloyMqQhYkIWSRZA8nyYUAtTPib8ffiH4K1rRdatLTRI/C2uXWlWumWN1NIdZ1r+0GiE0loI22RfZxKCVdH3bWJKDBND45fs0+KPjD4luryceHJrLWdLh02a7vbFn1HSx84uHsnTAm3hyYhOf3Mg3rnJWt/wCJ3wT+Knjaw1P4c2Ot6QfAWrpZxRtd2znU9Iht1jWRLMoBHIz+WHjkkIaNyT8wCigD2/4p/FXwf8H/AAbqXjPxhfwWsNjbTzxQyzxwy3TQRmQxQ+YyhpGAwAK1fDvjzQvGnhFvFnge5g1+Mwl0jtLiKQNMIw4gMisUV8kA5PGeeK4D47fD/wAT/EbwHceCvDdno92mpQT2ly+sebuihniMZe3aJJCsoz1Ira+H/h3xx4Z+Hs2iXtpoena3BFIlmNNWX7CWWILC825EcksPnwOR0oA8f8N/Fn436b4/sfAPxB0bRL/Vtd0G91q3sdHllSfTpbQxhbW7lnZo3WVpPLSdRGpdW+Qjmsjwf8cPjNqviLxh8PpNM8PeJPFuhaRb36Q6TdSxW1je3ErRf2ffyzF/nQAyeYm0sqt+7U7ct+DXwZ+PHgltdfxfq+gT6x4lt5jf+I7RbqfV5LwoRbuFuR5CwQsfkgUBFHAHJJuQ/Bj466v4kl+IGt+JtE0TxbpuhS6Np11pVnJJHcyTXEFxJcXyzYJVjbhBEmfLEkhV9xGADuPgH8XPEfxH1Dxj4f8AEU+katN4Su7e1/tXQjKLC4kmi8ySEJM8jLLAflkw7A5B4OVHj93+1Zr998V7zwr4ZufDi6ZpuvroH9nX1xLFqt88biK5ninyLaAo5cQwyAvOI22lSyivVvA3wz+KGh+KPFXxO1qbQ4fE3iODTbFbKwSddOjt7GR2eWR2CySzusr4O0BQEXkAmuJ1/wDZ08c302v+CdH1LSYPAninxGniS7lkgk/taCU3EV3PBEV/dNvmj+SViGjVtu1tqmgDF8G/tSeK/Efxstfh7cxaM0N7req6PJosDTf27psGnRyumoXZZjCYJvLXChFwJo9rucivob4j/HPwD8LdRttI8Svez3s9tLevDYWNxfPb2UJCyXU4t0cxwqTgu2MnpnBx4X4W/Zs8c6LrvhrSL/UdIbwt4R8Q3XiG1voYJRrl3JcNOwguHP7sDM5EsgYmRVC7VyTXPftL6l4o8D/EtvFfg5biO98Q+GJtGleXRb3VbSQJM0kSQNYbnS6VpGIjmVY5FI+cFTQB9peC/F2h+PvCekeNvDMxuNJ1y1ivLWRkKF4ZlDoxVgCMg9CM10zdK8d/Z68L6v4J+BfgHwj4giMGp6Roen211GcZjmjgUOpwSMq2QcGvYm6UAKOlI3SlHSkbpQAynr0plPXpQAjU6mtTqAGtTac1NoAeOlB6igdKD1FAC01utOprdaAG1JUdSUAFFFFAH//R/fyiiigBD0plPPSmUAPXpVLU9N0/WdNutI1a3jvLK9ieCeGVQ8csUilXR1PBVgSCD1FXV6Up6UAUrKytNNs4NO0+FLe1tY1iiijAVI40G1VUDgAAAAVZoooAKeOlMp46UAB6UynnpTKAF7UlL2pKAJB0pG6Uo6UjdKAGUvakpe1ACVz3iTxp4P8ABsUM/i/XbHQ4rltkTX1zFbK7ddqmVlBPsK/Mr/goz+3N47/Zq1Dw38Lvg9Y2tz4u8VW7ztc3CGdrSNpPJh8qHhWkkcNjflRt+6c8fko37NXxw+POrzeOf2i/GV7dapcLlYmk+1TR7uQvJ8mJR/cjBA6cHigD+pCL4jfD66s5dStvE+lzWkCGSSZL2Bo0RRkszB8AAdSTX5sftDf8FZ/gL8Kjc6D8MEb4i+IY8oDaN5WnRv0+a5IPmYPaJWB/vCvyfl/4J+WmALLxbeIh4KtbISRnBAxItfQPgD9k34eeBI4ZtP083l+gB+13ZEspbuVGNqf8BA9yaAPnH4j/ABZ/bA/bMumf4hazJ4Z8HStuXTbcNa2mzqP3AO+Y46NKxHoRXZ/D/wDZp+CmixQrrGnnWrvIG+8kPLfL/wAskYLtySRweODzX2WfAsdvGpihUdeBz+uK8u8YeFbyO2abTG2XNvh1IGA5Bzg89D/+qgDv/Dnwv8FaE0b+H9DsNLjiKkmKBEbnvuC54HPXgda950jwTJcwPLLi2woYearxswB525XBxx6H0zXjfwt1ay8QSWHnzR3JlOJVGVTgnerFuAOzMegyee/p/ib4h6Et2nm300cDQqQifOQ20HAB2n2LY569xgA7fTvCukksklzDFngEOeG2sRkEZ28YJ6DNc/r+kNHYr5MYJnYrtQ5OQu4dcAgg8YJ6HOK8nufizo9pL+4kafPYwtkEqeqkFfXHPvXUeF/FsHiSWWS6LRjY8m+RdzEqPlGOqgn0P6UAVYZ7zw+TeREb+iZG4bSPmDdeSP6GuU8GalN4i13U7x5DtSYIGP8ACqgDt1ya9H8dEyaPb3yqvkmMD5QibQMoBtDszZKltzYJJIwAFr50+H++8truJn/cz3kgcjqzKAFHHYZzx/8AWoA+i9Rn0dZBZtdRCVs4QOM47ZA5BwemOPXrXjutRRaXerPa6qlpNcICH3Lszz8h8zBLHphcnOOK37nTVtXa3sI1VowodVXBG4ZXJ/iJHcZx35NeZeIPCmpaxqVldXUjLFZyCZV28FhwOc5HXPFAHYpbapauWOoww35AMUMkm3epwGCIO7c9v516/o1pPJZJuUr5e0MSNpORwR7e/wCHauE0XwxZaiitd26zN13MNzAjPOTyDz27Gt3TVvvCuqKLmPz7d1Kqj5wQx5UHPynup7GgDvFg8xMO3APHcY6cA9D6H868+8ZabGbJ3xyBkY5/CvbG0+2nghvbKRbi2uQDFJtPIJ5Ug4wy8gj1B+tcnrujrNp0reUcbT2xgd/880AeG6JOb3T7aHO3LFWUHj933x/wL69a9Q04G8RYbVWwvBJGPavnSzu7qw8aLpaoTbzQyOT3yhAAA/Efhn0r7R8DWI/siEW0C+awyZDznI7DqSM9qAMSOwkiiiMuSh554zg+9dna24kICsvBx05Hfnnpj61a1m1uLWIRum4yZKybQuGHJHbPelsLCeCOASsgeQHCq+Wz6Fe3t698UAQXOmxiLM0uI0UsWZsDA7jPsOAOT25rxzUdAW9jedxyxOO/y9u1ew69HPKsMESkrcZYk9NoI/mcDrUQ0tBbrG8Y3ZGSSOw459wc9O35gHxX4l8CzSX6vp5aCccpKpwfxA6j2rc8L/25oNuP7ble6miYrlAcPEx44JAUr37Ed6+idV0GRZIruKBXWP75cMVwT2xg+v5mnSeFoja3F3OY4YU2qS7AP84+6F6ttBwf8mgDxz4gaFZ6hpnm5EySKQe2cjODnnOK8N+E1gkd/eaEVaSXTbiSFAoyxA5B6/3ea9+1iCz+ztbWshMSYCBju3Mw55J428ZPfoK8X8M3KaR8RdZaFQjSx2ku4ErtfDLnPJ5A9PwoA+vtM8MrNaq1tCXCpvLdAozgg46Y9DjrVqTwo6o14QBBtVz8wBIfIzt4OAQRnpx1qM+PL0aHPDaxCQXCktIyhGxhQo2AFc/KeQR1Oa5T4b+OH12Q30KtJ9jklh2Nn5Tlgdvpgk/5zgA+NP2iv2WfAHjnWYrvTLJNG1O4hMst1aIFDOxO0yxjCNnHLcE+tfMlvoX7ZHwz0z/hG/CPxW1Gz0XTTsghttUu4o40POBGMhOvKjpX6jX2naw11qEwmR/tUj7pD2KnYAMegHYYx0rxDXfD3mXdpodsTM08itM+MkqBuY8+poA+OYPiV+398PpLbxzp/wAR9X1v+y3ExtnvZ7yN1HUSW842yKRwVwT6c1+pPwY/4LCfBPW/h5cX/wAbLW58M+LtKQCWzs4HuYr9+m62P8BJ6pKw29masO08NeGvD+mRTX08AmdgrJKGCxx4zvLIHbPUbQh69a+KPjr+zD4L+KNzdaz4JgbQ9bRS3nlAsF02M4ljUnaT0Dglu7A5woB714n/AOCkn7Vf7S+sT+CP2Nvh1cWEDsYzqUkIvLpFPRndgLW24/vlvY1ufD7/AIJRfFb4t65H4/8A2yfiPeajfTYd7G0uGu7nB52NdS5SMdtsSEDsa8f/AGPf+Cgnin9li8svgD+0hoQi8L2rCG01O1gRLizQnAaRYwFuYO+8fvFH9/7o/o00bWdJ8RaTZ69oV3Ff6dqEST29xCweOWKQBldGHBBByDQB4r8Gv2ZPgZ+z/YLZfCvwjZ6RNtCyXmzzr2X/AH7iTdIc+m7HoK94pT1pKAPjP4h6F8SV+NHhzwt4P+KWuxXGvXMmq3en+TpzWNjo9m6+cMm0Mx812SGPMmcsWJOwgpq2hfEmP9oDQvCHhz4pa7dQkvr+sWU0OnGztdKWUpDagraCXdcy5jQmTIjjkYksBn6Tsfh7pdl8SdX+KH2iabUtV0200vyn2+VBBaSTS/u8KGBkeYl8sQdq4Axy3QPh9pmhePPFPxBiuZrjUPFUdhDKsm3y4ItPR1jSLChgpaR3bcT8zHGBxQB5X+1h4n+Kvg74KeJvE/wpubLTrzStPvLy4vbsNJJbxW8JcfZ4QNjyuRtBchU+9hvu149+0h8T/Hug+JfDXh/w3qut29vL4ev9Vni8MWUF/qZu4DELd7mOeORUtDlwSoBZ8DOBX1T4q+G0fjb4Xa58L/E+s3d7Dr9nd2VxfbYUuRFdbhlQkaxAorbV+ToBnJyTyPjj4H/8JL4isvF/hjxZqfg7XLfTTo891p6WspurDf5gjkS7gmQMj5ZHQKylj1BxQBpeEviUyfBLw58SvFRbVZ7vTLG5um0W2mvxLNcIgdoIYFeRl3sTwvAyTwDXP/s6fHaH45+DE1ybTLzTb+NphMs1hdWluVW4liTyZLhFWQ7YwXCsdrHBx0ruLL4axeG/hnp/wz8Aazd+GINKtoLW0vYFgubmJISMk/ao5Y3ZwCGLIepIwcEZvwj+FWo/CnS5tEfxdfeIdN5Nvb3VtY26WzPI8khT7HbwEmRnJO8t7Y5oA8Q1rQviVH8f9B8I+G/ijrtzFI0mv6vZSw6cbK00lZSkVsCtoJibiXMceZNwjjkYksoz6B+0mutaN4Pm8caZ431rwwNKjEMVlo8dk76jeXUiRWsObu3nw8krLGu3AG7J6V6joPw+0zQ/H3ij4hx3M1xqPiiKwt5Uk2mOCHT0kWOOLChgpaV3bcT8zHGBxWV4i+FkHi2DQrfxLrd5froOvJrsW5YFEjwNI0FvIEjAMUJdSvG8lFLMTkkA+dfHGmfFP4f/AAw0vWviP8YbvR10TSiLm4sbS0e/1LXJ3JjjCvA6yRjIiihijV5Dyxrd+L/j74p+FP2L9W+IepynQfH1n4dgurl7ZFZra+YR+bsRw65BJG0ggHjmuy+Kv7P1z8TfH2hfEK28c6t4bvPDlvJDZQWkFhc28UkzZkuFS9trgLOy4TeMEKMDGWz2Hjn4SxfEf4O6j8IPFmv310mrWIsrnVAtul5IQQTNtWIQByRkgRhfQUAfNfwd+JPiab472XgaPxH4l1PQdR0S7u5ovF+mx6dcG6gliEZ09hbWzTAIz+cu1go2HIzXt3jr41+I9D8aX/gb4f8Agq48Z3+g6fBqmqiK7itPJgunkWGOESA+dcSeS5CfKuAMuCQKteGfgdcab44034heNfGur+NNU0O3uLfTVv47K3gtBdhVmdY7K2twzuqhdzlsDOAM5pvj74EL4w8U3vi/w94v1fwbf6zYR6Vqp0s23+m2kLO0QJuIZTDLH5sgWWIo4DHnhcAHi1l+3P4Gv/HUHhtLKCPSp9YTQTM+p2w1NL15hbbm0zPnCBZzsZ924AF9mwbq8v0T9p3xt4QvrvxJ4tuLrxBpWm2Xi+eSziSNC7WHiRdPtGZwnyJBA21nPAQFmBIzX0t4b/Zh03wdrUL+GfGeuWPhmLUv7WOhq9u0L3RfzXDXbQm8MMkuZJIvP2sxIPykrVaL9lPwvaG4m0zxLrOm3csGsRR3NrNFFNC2samNVkdGEeCY5lCqrAo0eVdWyaAPQvgf8Wj8YfDF14gOn29l9luTbh7LUYNUs7hdiSCSC5gwCvz7SHVGDAgjGCfZ26V4p8HvgppXwlm8RapHqk+tax4quIbi/u5oLW1Dtbx+VGEgs4YIVwucsE3MTlmPGPa26UAKOlI3SlHSkbpQAynr0plPXpQAjU6mtTqAGtTac1NoAeOlB6igdKD1FAC01utOprdaAG1JUdSUAFFFFAH/0v38ooooAQ9KZTz0plAD16Up6Ui9KU9KAI6KKKACnjpTKeOlAAelMp56UygBe1JS9qSgCQdK+av2if2svgn+zHoR1P4ma4keoSoWtdLtsTX9ye2yEHKrn+NyqD1zxXxV+3N/wUn0P4KPd/CP4HtF4i+I0ubeadB51tpcjfLggZEtwD0jHCn7/Pyn8oPhx+zH4r+JmuSfE74+311rOs6rKbmWC5lZncnnNxITn6IpAA49gAe2eMf+CmP7ZHxp8U3F78AtJi8LeHLbKxQ+RBcySDP3pp7kbC/+zGAB79axZP2uP+Cn8KK8mpxKGOB/oumcnr6V9NaT4CsPD1hHY6bYx20EC4SNBsUBeqqFwOBzSXVlZ3msW8dpIskaKGbthmGNrKcjcO5HFAHxf4W8E/tIfHH9ovRfjB+0VD9p/s+Nc3P+jbcWqHyI1hgYAYdtx4HOSTmv1F03QrcWqNE4bIB6Yx3PHfB/Pr7VDpGjRwLD02kD5uo44OOmcH613kE82kWUxkEyQMrMfLUgFlXcnPHHpzQBiab4Zlvpkt7RQs7M6hZfkBIwBsbo3XpjrW/N4bstNjQTXMay43MhwDh/ujjcN3XOemMZBNN8SeLrO9tordLKGVbW1ihC3LSNghD80JDjb94HGOvOSOD47rvxEs9Nj8u9vCWZmIiLk/M3JCgk8k8knJ96APZrSXwXBZXT6gzy3MWFiRdgyD1LBlJJz15GAa8W8a6EbBFuHKMk4LYjbcFB6KTgdsc9OcfXkLXWvEetzs9qn2O2kYna33m7hueB+Na15pLRQu7DDAdCD8xPLH6f40AfKujatdaL40vdD0t3iW6wYtvGN/DBew6kD0r3jSNBaRPNnG+ToTncOPVq8bltdN0/4qaeNVt/NW83QKrHCF5V2qTjnHPX15r6j0HwvdRfvbC6WOByAluzbm3HgBOd7AcDJyeeSaAMiHwbZMvmyx5BHP8AwH65rpdP8NpagCKMyYwDhN2Dz0A9RXVaBqPhyK8lt/E94LB7bcpDHBdlB6YyNpGcEdTgVe8UfFn4e21hJp+llYGO3Y7TMzx7TkhBnI+v3c5OM80AeQ+ONdl0/wAPyabMymFXaUEjMgJQIRuP8J29OxrgvhBbC48OWlyx2tcXE0h5xjfyvr/DXA/GD4i2uo2rWWmzLI03ykjrhupJ65r2D4YFYtKt1iA27Rg4zgDpz6UAe42+gvtR5eQ4OwcncOmR1z79+9T2/h6CcmGSMF25GByR68jp+VeqaLrXhe+0KC3uYHW+RP3k4BfCBCwZCMgAMMEDH+E2lav4Ygkd9TYtHsCgwk5Ziuc7c7QM4BA4z2PYA4PSfDD2sm6OPBGSqkfe+mOo/wDr1U8V6fZLbWc+qRfLNMI90RCEsOSBuzjrxXY+I/iVbadMl1oES20sTnEs3zErtAC+WBsOGycn1HfJr5G8f/E4X+oSXEspvbmP5WRWVTwflRRwF4yenHU0AfVHh3xTpf8AZsuhyKysGDgKgaMysOdnJIJPXPU9AM4rqdduPD0vgm5SAfZ7uYDD8HnqvD5I/ukAdT27/Bel/EDxralrxNIVtxLEedhtpOeCRj6e9Tax8ZvEscqunh65jnRsoyybirdvnU5OPbr0NAGD47stR03xjp0FsVIuHjWQowbZnkZ5BHXBxkcngnGPrHw7NfpYq+mai9tC2Y5TGA+7A4KFhww65wM9D0r89Nf1Txj4luYWg03ZOknmGWVtzDuAoP3eeeOT9K+pfhl438WWajTdZ0/bHcoglZGyshXI3bSPlIz3zg5xwcUAeuatLcO/mX15NeTICPMmkJIA4wFG1BnHULmvILv4g3vhbxHYJOxktLyUQFupjlPTJOMg9MnvivZdVhivrcXUC4wASM5/HsK8J8f+G0v7WReQ5+ZW7q6/MrY9mGaAPrbw3fWWr2D39zeHzIY08qNsEux5Zc+h6jrjkela8ur+FtOXz7+5efg+XFCNxJ425JGFBOc+lfE/gv4nQRwDRtdmW3vbUbX3jAJzywzjIPrXUXfi2TxFJ9j0aci1GPMmT/loSNpVT2yOpHNAHqHiT40pA0+kaXbKsjjGIU8yQA54aVztBx3Cj6CvMft/i3X3fzWW0ikJxty0uOvMh4z64H412Gg+ELWKFFhXaqcADnH/ANfnr1NehL4Zu7azF9NA8MQyVkZcBscgDkDjHagDx46EbbT3uXYs4VuCOh9v/wBVfNvh24u9Z8Xa59mSMpBf+WkvO9liQIQW/ug5x7knvX0r8Y/HGmeGvDlzd8QXCwmOAMdzGZlO0gDBOCcgHPHtXmfwV8AaloPh2C51dHOoXhNzMkg+Zd/O0jsR39zQB6N9v1Sz0pbdFTy2IYkoNwK5HDdcHPIrjfhnqkOmWWualgiS8vJkVD0HzYbgEddvX36816bqdqZ7ZlxhWGOM8+9c9ofhZdOeZHz5Mz+Y6sTjJwSc9RnGaANZbkatatMlzHbb0ZsyZViccHHccDnniuKt9Om/4TC4iEZBsQIZCTu+ZsMQOwwpAzj1rrvidGNBm0PUUlW7vo4ypgjcMSrnKrnnDKPmOOg681X8E6FfW1s11eu011dyPLM7dWllbc7egyaANGfwQjyPqck0rEnLKSGQk54XI7Htmm6E1nrmkmVCEnV5RtO1U8uIqmAFG4uT156V3msQHTLKO0eYq02yR0VyVxztO0EAkHgk5wcYNfMXwg8V3c9pO0j+X/pV1sbplWc8g9RkHrmgDB+OPwh0r4l+Hrnw7rMKs0e6Szu0VfNtpfY9WGeq5wQa5z/gn9+2Nr/7NHjn/hlz4/3Jh8L3E/l6VfTMSmnzSt8o3n/l1mJ4PSNjngFsfVt6lrJEvl5aPbzxjDex6dMV8dftFfAez+KmhO1siw65YgtaXGMFj3jc90b8wefXIB/ScHWRQ6EMrAEEcgg9xS1+J/8AwS6/bC8UeIb2b9k/4yPI3iPw7BJ/Y93cNmWW3tsB7SUnlniX5o25zGCD90E/thQBIelNXrTj0pq9aAPHfgl8S9R+KPh7XNZ1O0is5NK1/V9IRYixDR6ddPAjnd/EwXJ7Z6V5fqf7Tx0Sf4oT674XudKsfh/aafcWxvJUhl1Fr+SeCNtuSIImliAVnOdp3soGBU/7IX/IjeMP+x18T/8Apxlro/G3wFg8b6r8QL2+1MQxeNNN0iziUQhzaz6RLPPFMwY7ZAZJVJQgDCkE88AFz4F/F3U/ipBrn9pRaU7aPPFELnRr83ttJ5se8owkSKaN4+h3IFcFWQkHjH8PfG7xLN8WZfhz4y0Sx0mKazv9Qi8nUFuLyztLKVY0m1CMKI4VuVbfEVdsAEHkHGt4G+Gnj/QPE2t+PPEGuabd+IfELabb3QtLGWCzXT9O8zCxxtO7+e/mufMZiB8q7SF54bxB+zZr3xH1hLf4u+JYtd8O2KanDbi0tn0/U7iDU1MZhu7uGUBo4om2ARom8hWbkcgHb/A7472Xxu1PxoNI0uWy0nw5fW9tY3Uxw2o29xbpOl0qEArFJuzFnlkw/G7FdXp/xs+GmqfES8+GFlr1nLrdnbwTlFuoGDtNJNH5KASFjMhhYum3Kgj1rjPgx+z1pHwZ8XeMfEWj6xqOoW3iY2Iht729ubv7NHZ26Q4LTyvuYlflbAKphBwBWdofwa8ZaZ8Y7v4nXEnhgwXqx28kcOkzR3S20Es0kbJKbllW4JmIkk2YbA+UUAWvil8W/iF8OtWk1JfC9nL4RtLvTbJrie+Md9fTajKkRFjbpG6sYi4ysjKzkMFGAGPWfHX4p3/wd+Gur+O9M8PXPiW5063mmW2tyqIqwxPM8k8rECOJVQ5PLE4VVLECvPfGfwf+KviH4y23xJs/EmjXGk6SkSaVpmp6dcXAsHxi4uIzFdRIbiXJUSshKJ8q4Bbd6p498Ea58Qvhn4y8AapqNvBL4ls7+wt7iKBgsEN1G0cZdGcl3QNliCobsBQB5r8Wfij8YvBlrpeseD/D2h6rY6x9itrSC71C4gvbi/uzxDHHHbyJtA+YsXGFDM2AtbPx1+L+s/CXSPCEtnHpMV14n1iLSpJ9Wu2tbC132s9w0jyqpOMw7RkDO6uv1H4cS6n4x8EeJLq+VrPwbbXapamP/W3dxFHAk+7Py+XF5qgYP+s6jHOD8bvhZrfxNt/Cc3h6+sLO98K6ymrIupWjXtrPttp7cxvEkkR/5bbgd3UdKAH/AAO+LF58V9A1rUNQtbNJND1ObTTd6Zcm80298lEfzrScqhdBv2NkfK6suTjNZuh/tPfBXxClxcWWuyQ2sFjNqQuLqyurWCa0tmCTSQSzRIs3luyq4jLEMQMZrS+D3wn1D4cDxTqWsX9pc6l4tv1vriLTbQ2On27JBHbqsFuZJSCyxhpGLEuxJPavGdc/ZEi8RfDzwx4A1LxAFi8P6BqGjNMltnzZLya2njm2F8bUe3G5CTuBxkUAe46F8evhnrrRwQX9xaXct7a6eLW9sbqyuhPeh2tt0FxEkgSUI2yQrsO0jOQRTb/4/fC2w1qz8PSanNLqF9eXtlHFDZ3M2JdNkSK7LlI2EccLSLvkchADndgE14Ho/wCyhr2j2M2raTf+H9I8TQalpOoWjWGlSx2R/sppSEuA9y88gl85+kqhONo+8W6zwn+zTqWn6rNq3jDxDFqj6hD4phvBbWrWwf8A4Sa4t5m8vMkmwQiEqMlicg545AO0tf2pPgncaTqGuNrk1tY6daR35luLG7gE9lLIsKXVsJIVNxCXdR5kQZfmU5wQTK/7THwjGnNe/bb4zrftpv2AaXff2j9pWD7UV+xeR9ox5H73ds27Oc15Hd/sueOPE2kWWl+NvGFldnw9o8Oh6S9ppzW/+jpdWlxJNdBp3DyutnGmI9iKdzAcgDd+J/7M99451bxJrNvf6ZOdd1Wz1JbbUrGSaOP7Np62PyTQTQ3EUoK+YksMiEcoQQTQB9R6DruleJ9EsfEWhzi60/UoUuLeUArvikUMpwwBGQehAI7itlelcH8M/CepeBfAWh+ENY1mbxBe6VbJBLfXBYyTsvcl2d8DoNzM2ANzE5J7xelACNTqa1OoAa1NpzU2gB46UHqKB0oPUUALTW606mt1oAbUlR1JQAUUUUAf/9P9/KKKKAEPSmU89KZQA9elKelIvSlPSgCOiiigAp46UynjpQAHpTKeelMoAXtX5S/8FHv260+Anh9/g98K7oT/ABJ8QRbXkh+dtKtpeBJgZ/0iTpEvUffP8O76g/bQ/am0D9lP4O3vjG5aO58R6lutNEsWPM92V++w6+VEDvkP0XOWFfz5/s4/DnWfiB4nvvj58WJpdX17Wp3ubc3BLOZJDk3LA/lGOAByO2ADrP2cP2eV8NiPxp41iN54ovj5x83Lm38w5Iyc5lPJduueAe5/Rvwtof21Y0tEEjKjuzgBQPLQuxLNtACgZJ/E8VyNto2LeOZTlOgGT27cgf5961LbWL3RQwtxGkjDaZfm3bW445wGH3gcZDYz0FAHY+I7PQ7K0lguZjHMN/lKkW3zAMlT8x6sRxge4JNef+FvC0koad4y9w+ZOwz0ByTwFGcc/rWBq2rJJ4mS1mZdrJvyCTk5Xp2/i/SvZdPbTrLw612jxz3LMAqptLxgj5nYZDYBAC4HrnHcA7jRbqLTl+y6mqyRrvJ8wKrqVGxI1UYYZOdqkfwkjqau6hqnh+Pw4xeWOSaYlmtQ8mGZcZMqkk9QSu4YIPriuDt9euDbNa3MFqQ0iy7xBiXOQANwOMHbk8DnPrzzfjLxZc2ltLFdGHgNIPIJVhuw3LHO4AdBnrnmgDzL4ieLk0ex877S5eFV27skBcYCknoFzwDXJ/D7wrqXiHZ4i8SROqzMfLUADjGVVSR94gE88L1PHXD8OaVd/EzxjJd3bk6XpznJYcSSD+LOcFVPvya+pbVbPTExbRYEYEcZz93uxPP3u9AFdZVgItLWwj0+NVwwU73PAxlmyxJ7jgdx1rS1W4+36eTJGoEKAZUZbC+pwcDnpmvnvVbu7uJbvVIb5fNVNQeOPzLkTySQS3AtvJVc24jKpF8jsGk2zHZJuiJivfE8sFrZ6jrkMltqdtd2wMougbf7Nd2mpYiSCP5QN0COXd3djj7gG2gDxz4lSRT+ONGSE/OJ0ZgucjnAORzX1X4W8VW1zo01pqvmGcqoiIj3q2Su5m4OOBk8jkdDk18j+G4I/GniXXNRK27TWQtY7J71bh7cSzNJujCWoeV5mRNyfIyqiyE7W2EesXkLeHre91K0leWGF5UsfKe5eRVP2prZbcxkrJLKWs3TzS26J0JJUzGgD0fVfJuJg11Cb6LAUq43KFUYAG7AGMnHp2rm7axsZluI49GmtXQAnG0QjcT91sHeMdQpyD1x0q74DNprXiZ7rVPMku9Qb7KkqzrLDIyNNvZIhIWVPMikhikKKjrDuB3Pz1ep6le2OqXen6YkaW+nuI3kaLzQOUUySMJojGm99vyLK/yOSuAoYA8V8W+FrGa0f7Xbqycj5l5Ax16du3evPvBfxIXwxNJoF/vma3cRrgZLgHoQPUCvS7S1vPGL6JquJ4bxra/utWiP+qE5urizhiRxK+Y41gb5fLUEgSE7iQOCh8I6Fpuq6ZFpk7jV9VtRJNB9mLRPcfbLmKJJpzMGRpjGqqVidV+XcQM4APTJPiffz3CTWemzAEAbVYbOOOFydvGcAEelaz/FnUkjSKfTrpFySQu3JOOOSf8A6+M155Zaxq9t/YcNlBJqVvqenW17JJa6YZGhlurK1vREq/axuCJcfPLIYkXA5LELU3inTtauNZ0y/tr1IW06KCfaI1ZJ1vhDsEiwzyIrRFyHCyOGxgMMnABV1rxP8QvE8v2bSrJ7MOdvnO43pngkYJOa9B8DfDWS3iWfUlMsrEtuYBiWPUjOTknqTya6Lw8txbawun3EiXscs/kCQWv2UOpW4KTxD7ROSjfZ2BVwjDKkZ5A6OPW9bttaks4ru0S2kuDbqjWTuIAXSKN55RcodksjYBjicrgkggDcAdYvheBIQEiVSB93HPsP/r/lmufv/Cts+MRJuB6gAj6c8c1zlz8Y9fs5bDTksGv5NVEro9vZmV4vIjglbdDHdOrKUuY28xpUVMOHAIUNPL8QdaeaDSbe1iuL+W0jv4o4IoZrm4jkfy/Lit1vNjSRsH80JPIdoUhclggBdg8GRLdJJsGJABwAM49vfP14r0mx8ORREhogpj9EB2n614tZfFrX9X0+4OmaLcm50i5mt7qyTT5Li+QoqMLiaz8+O5hicNtULHMVKSA9F3fQ3hy//tvRbHWUZIvtkEEwQOH2+dGH25BOSucHGen4UARS2tvHCBI2/Yfu46enA/LJ49K8+1W286OaFAPLUHggbhnrx146ivXpNOkKqEmh4y2N38OPyJAxkZ6++agm8K6PYzRz+JdUhihl2sEhZZpASQCGCMx4B9O3GaAPh7xb4FgvZfNuLfdnOGxhlB9D1r0j4YeHvsKCFwcxjoR+X+e9et+KNM8L3atDorvO0cgjEhxtcc5OOuM45x61s6Nouj6FceVcXInhV13PEDkoQcgI4BOSBzjA980AdfpGkkK9zIViSBWZ5XVWROPk3A5z15/Lk8V5Z8W/jGXlttG0ZftV6o/0eyBZIlQHZ5rcBUXAGBwT0Azmrfxa+L0WmeHoNA8MxlZZnMNvE8geSWSUhiGwMbEK7vl9846V4b4G0GFbh7zVZS99cOHlY5BaQ/eZd2OP7q+mKAOKvvAuvareJ4k8R3EmpXcDrJHCc+QhiIfbGhOBnA5OSe/HFeoWkPi+W9g1O2uo7qyuXV23IVmWNsAg54JHXHU46CvY7Lwpd3KRxrH/AK0O0ZciMSlV3BUJ+8xOFUAHJIHvWDLo+oaTKrWq+UUbeY2+5v8AUr74wfWgB6TqW8pxu3cbh8vOcAdhnPpnio/EusWvhXSHvp3/AH4+VUK44PXB6deOOPWul0+xbWWnvLW1EBiieaYMz+WhBydpVTgEDgHGOcnGK+PvifrGs+MfFQ8N6PC0VnYyomRudFZsNy2DkYOR75+lAHZeFbm+8V602s3wLMBiLuIU64X3/vH146V7XDp+u2EIvIBJeQwFUkV2aPIOeUcYjL45y/3uxLYrmfAfhW38P6Vb2NtuaRsPJI5O5nPJJ69+n5VueOPGs/hPwwbe4kEttBJiKIYBa4nOFXgksC5HXgHkUAef/FDxzqd9MfDdgrQahqEOXePLJZx7gu0Ek9s7eTzz2rlfC/hmLQ7SG2tHZEjGMbck+5zzzXQ+HtPG17y9USX103mzsP4nbn04A6D2r0mLTI5YykwA7DHVePzP1oAx4ftRKiRMkngsRjGB05rTFnBc2u8Nu5w4zyG9+nfr2/CtGGMadYrqOtwyw6YqT+XdRoT5jRrkRjOFZyQdoJA6AkZzXO+JfH/h/QrDzdNma2juYwrJdsnzk5BIQcEAcqR909+KAPzg/aFGt/AH47+Df2jfBY8q4sb2FrgIcK0tv/CwH8M0OYznsD61/Uf8PvHGhfErwPoXj/wzMJ9L8QWcN7bsDn5JkDYPuucEdiDX83Px01bTvih4Z1LwRp2LySb51nAHkxyp8ynzCQCQRjjjBPJ6V0f7Cv8AwUGuv2YrW1/Z9+PunSp4Ut53NlqcQMktgJnLMHQZ8233EsCnzLk8MOAAf0rnpTV61h+GfFHh3xpoFl4o8JalBq+kajGJbe6tpFlhlRuhVlyD7+h4Nbi9aAPkH9ly7uNP+F3j6/tI/NntvFviySNP7zpfzED8SK+QrDx78UfB/wAO7bxtq2oX9rrPj3wTc6rFdxa3LqcTTlrV57ma0miWOzkgS4LQfZmMarmN8lVNfZ/7IeP+EG8YZ6f8Jr4o/wDTjLVr4Yx/sveJPEXiHT/hxpenyajqdtOLwfY5I47yxkmKTm3MyCOa2abIcwZjLkZ5IoAf8G4JfCfxU8d/DjTtTvtT8P6ZY6Jf25v7ya/kgub1bhZ0Wadnfa4hjk2lsAsSAAa+SfCPxW8W+BfFVr8WfE39r6h4b1l/FckEsN9PqM2uCzleSztxpHKWJijjZY2j3Z24k2biK+oYfEn7Mnwn1qP4aWukyaNcNq1mVSPSdQaB9ScolsftQhaNyMqqkyFVHy8AYra8B3/7NDfFa/XwLZ2cPjG5a9BuI7SaKOeSOQfbhazuggdxJ/rxCxO77/INAHlX7HHxY8QfEXxd8UYvF2uzapqf2zTr1LU211b2unQ3FlGTaW4uI4/lifKliA0jAyYwwr1PQfin8Vr/APaB1jwBqHg6S38OWunWM6TG8s28rzri7jN0QreYyyrEgEX3kxkgZr6E0/QNE0vU9T1nT7OO3vtZeKS8mUYed4YxFGXPcqihR7CuEsPgj8KtM8TjxlYeHoIdaWd7kXIeQuJpCSz4L7ckk9sUAeIfGX4f6X4q+LnhXw/pGra3Ya3r7tqF9Jaa1f21vbaXpXliTZbRTLCHnkeKHlOjO/LCup+MsE3ib4l/Dr4bX2pXuneH9Zi1q7uzY3c1jLPPYQw/Z4TPAySBR50kpUMNxjGcgGvoA6Doza8vic2cZ1ZLY2YucfvBbs4kMYP90sA2PUVyPiT4SfDbxhpjaL4m0C21C0e8k1DZKCSt3ICHlVgQyswZgdpGQSDwSKAPgiK/8Q+Ofh34Q+w+Ktd1b4n6xYC20S3tb+W2trKKyvZYjrd8IiqPHJGqNIbgOJSBHGuWavf/ANrCTXzZ/DDR9Jhu9QfVfFUNtc2dlqMmlPeR/wBn3khjNzE6FV3IHxnBKivU9a/Zw+Bev30Gp6r4M0+W6tLWCyidUMfl2tsCIYVCFQEQE7RjAzXa+Mfht4F+IelWei+NdGg1ay0+Zbi3jmBxFMiNGroQQQwVmGc9CaAPB/2XdV19bXx/4T8S/bLS88O660cWmX16dUfT7We2hnhiW/Z3adXDGQbjmPd5fRQT5FaftU/FbRfCGl+OvF+laLd2fibw7qGr6fa2QuIHtp7Ke3gVbmaWSRXik+0B3ZUUxhSPn619geB9L+GPgm/1H4beArC20q4s4odRvLa2iKjF4zpHLLJjDO/ksPmYthRnjFXl+GHw8WwstKPh+yay06zuNOt4WiDRx2l3t8+AKcjZJsXcD1xQB8va58dPjT4Y8TH4YXsXh7UPE899oEcN/BDdR2C22uNdofMtzO8glga1YjEuJEZeEJOPQPBPx61K40K60/xxp80viWy1LVdLM2k6Xfz6ZJJpszRLLJKqzJaq4AJWabjn5iOa9Q8OfBb4VeE7ZLTw94Ys7KOK8h1BdqZb7VbqUhlLsSxMakqmThRwABXZ2HhvQNItb6y02xitoNSnnublEGBLNcktM7erOTknvQB8EN+1f8W/DHgzR/E3jDR9I1C48YeF9M17S4NMiul+yz6jdWlp5NxvlkMyA3iPlPLPylOchq9B8DftCfE2+8T6B4O8beH4rKfUvEA0t7uSzm0/zraTSry/V47WWeZ4pEktgjbndWRty4Jwv0lc/Cn4bXul2+i3vhyynsLbSxosUEkSvGmnDYRbBTkeWDGhA7FR6Vz3/CgPgwvht/CI8J2R0uS7S/aMqS5uo12JMZSfM8xV+UNuyF+XpxQB498Pfjj8SPiP8QNI8NadDpFlpjR63dX0rxTyyvBpervpsccGJlVWkQB2dtwDA4Ug8fYa9K4Twx8NfAPguS3k8J6DaaS1pBNbQ/Z4xGI4bib7RLGoHAVpTvI9a7telACNTqa1OoAa1NpzU2gB46UHqKB0oPUUALTW606mt1oAbUlR1JQAUUUUAf/U/fyiiigBD0plPPSmUAPXpSnpSL0pT0oAjooooAKeOlMp46UAB6VzHjHxd4e8A+FdV8aeLLxNP0fRbeS6up5DhY4ol3Mfc9gByTgDmunPSv58P+Cov7TOrfFfx5Y/sd/CS48+C3uYzr0sTfJLdj5ktmK/8s7cfPL/ALWAeUNAHyb4z8aeLv8AgoL+0dffEPxDHLa+BPD7eRp9m33YrVWzHDxx5sx/eTMPp0C1936f4fGlRQ2+nx/Z1hAUFVAAAGAAOcADgY/OqfwP+Eel/Dvwhp/hbSohIlvGHllH35pW5kkYAc5xkegGOwr3abQ2MSKiZDMMe4/DnAx+FAC6NNouuQySpAlrPIDmBdxWLymJIViQCGTkZ6kHmszx/oH9keQUUIjI4UgY37DtJwSTnpk9D1GOlUYozoetPDM+3zd1vKEZcjPyg+mQTxz681P4z16TUtMtHuNq/ZYjEGBLFwxJ3lyTvYDjPHAoA+bdKgurjxpcru/0aCNFAPTe5ycfgK+i7GIWlqix4Rm+ZsqDnAIH6f59PmrSPE1ifEWqQLIN8flyjBwdoGDjqOO9fTGjS22taas9vKCAMx5/jz1HTr9eKAHiTILN9wY5A6/TPb1rwv4s3mrR2kb2RVo5mWJRg/uyeMn1AOa9weC4tpZILldkiHBU8YH4j9e9c34n0yC+08h03CKVH3Y4wpDcUAHgLQ/+Eb8NQ2drybiMb/lwcJzzzzk8nJ5rsGZnh8y4O1eTknof8a1LbwxrtvDalYhKktsLlBG4Y+UAGJYcZ2jkj69etYOuTadp0El1fSKzICpVclcdScnA/T8aAPPf+ELiurkrbTmG3Z5OCrGWJZCzEIQ4i6yNsZ4nkQHEboFQLmeKvhrFrN0YorkwWUnkO0AjDEvBHcxxEOWG0AXTkjac4Xkc56bTfGl9q2n22pWulhtEt7n7NJqQAhXfNKIdsYPNz5cnD7DiPoWDYVhfGtpNdxedEn2aYx7ZC8gfynUOsxHleUE8siYp5pmEXztEFyQAeK/Cz4XX2g+K5PDMV6d17LBPG7LP9+PMashS4SdXTe+CJwDvIYEYA+qdU+HEMnh220jTNX+ypZ3ZmmnjiDG4CReTGEA2LEYlA8kqCE2R4Hy15v8AEUweGfGKNpN1FKLAo5uneVYGE4YLGPIimmZn2OQEjIwjMxVRmpLr4ny2sTRW0bOdPU+fE0qg52SyGOAoJEmzFC8iklVZNmGLSIrAHe6P8PvD+g6tb38F8sKWhBht1iGVyZcIz5wYkad2VQobkAuwRArLvwUNb8TvLbXf2eW6k8+NUj84eaqx5ZAHVTny0YLKs0e5ciMNuJ+SvGvxr8S2+qW48P2yz2t5EtzGpYyNJEXKqzIiq0e/aShDcphsAMpPU+EP2hrrw/etd6/ob28zIpbc3mhYy3UgsrxqSMZBBPTNAH0foHw5g8MS37QXXnWjqkUEBQhreNJp5mDyFiZGL3DZbA6DjOa8X1rwdqS69aR3U1tNBGJoILlLUxXcFvPLLMVEvmlGKvK4WR42dM7oyjc11F3+0DA40e9n01raw8RG4ktnxOWdLYsjyqvlGMRGRCi5mMhPzbCh3VlQftAaVDrD32q6LHE0sC/YBGWZRHLI8UcrbgoWRnjbCMWznHPQAHoHh74IQzaLLox1BLqe1SK0DGArE9vaw20EUbJHLG/yiyhcSJKjbgTjaStLJ8C9V1XUX36+Ykjiit44INPjG2O3x5TnEgztKpkkZYLz8xLni7b9obRbaKG40baZZflEMzoDLlVP7tlcEnBU4APBHOCKi1v9qyTTWsFi0yVZrxpQ7G4Hlp5O3KgqjDcuct5jx8cLluCAe7W/wmNve6Je/wBoeYmmRKkyPCIxcSxrMok3BiUA89yVAb+EZrjfEng3RNLvNX1CfxS0C3ktvJBttFKwNBKJ8MDLmQE4HVSOeteW3fxx8V+K3iXQ47UmSLcAbvdkKxyQqHJCk4Y84OAa4dn0/wAR6g8HjvxLC13ZtuktUn8mKNT13n7xx3JIHTOO4BS8QX/9ta/Jb+FkTULK3thafZ5IitsytHaKfNkSSOT71lDIvlsjK4PzFSQXy+AfE+uX0Woa3/ZdxFDYpYxWp09hCsMUryrl45opt6lyPMWRXYffLlnLew6VY+ENIvY9IgvrKCWVwot1kjVtzjco29mcEbQRk9QDXRT+I/A9tYPqv9tWqWMcxh85pVKCVV37CRtGdmWz6c8DmgD54k8FeI7PUX1ySfT9QvfNinhe4snBtngjjii8hop1YKkcMShJTKnyKzKWLFtzRfiV4i8P2g0rUdDaWOD5PtEE6yNIc5LlWCHLEknjrnivQr/xN4LuNLjmttdsZLi4kZIYhcorMArMzBPvH7u0dfmwMc150t74Xlkge81S2D3Y3QoJFIflvu4+8cqwwPQjtQBc1P48WkcT/YLK7kmjIQx+Sy7WHqxOOv19q8yufjfq9zMGn0+a2ijOcAjJIGOuB069K9DvI/CohEv261USp5inzV2+WTguxBwFyCM5xkEZzXB614Tl8Qa5F4d0sHbuVppF5Ko3QLgcue1AG/ovxW1bU1ij0TTy7dS5OAvvnH9P0runn8farERNqi2ynnCRZyMdCS3Ir0Hwr8N9L0WwWOCLYjbR0AAGMADjnFdPb+HvtTHTtiI0W0F1yGZecbu2DzyBk4HTnIB8n65pXiOy8Z2njTUXS8tYCRLFGhTykZQDIq5bceORwcdO9ezW2saL4ljtJ/BzlGSON2cgEpcLzuVuMhuGwR8udozjJ9K1DwPAEKyFm4GOAd3JB6fpXk2m6ePh34jlnkjebQr1z5yJy8e7gMg4A6DjtQBV1C6+IOjXEep2t79reCcXCxsmx968khwxCnIBzt616p4T+KOheIpLa21mx+zzWbD7XCvyvKrEZByevU5B54A21uappdpLbRXcDxlJ081Nr7tysOmQBk8+g6dq8R8a+CheQyXOnzPZXkQPlzR43Ke3+8PXPb86APUfjN8QPDOiTx6N4HQXU0kHlAruiadmO7c687Y1OABgEnPXjHz/APD7wp4oTWb288QQqr3biaOeGQSAnpsdOCMD7p+ucVg/CnT7zUA97rU3n3qyyI7McnKNjqffp9a+lfDOi23hyDzIWLfM0pDkkSsxyVYjgdccDigD0XwtILO2l1HULGG5S2jAdNzxCNm+VT1bd9On0r5j+J5bxB4o0fQ1PGnSfbJyB1ZQUjBHThmLY9RXsvjf4h6JZWCTWMBhlZgqQIxZ5JpDycE8AcDjC8EgAHFeI6P4f1iO9u9a1S4E11dFWm7oCP4U9Bjp1NAHfaTaOtuVjJZ2QhWxnaR0JXIzj079K6bT9btLGeNfEDulinLSQqCZGUDKYdl2k55574FT6TAv2VZ4/lUgYz1z2rzbxvqsVlZXd0xj8mBWOF5BA6g+p/rQBznxw+Lpi05ktwlqqxtFYwKD5af7RBznruP5V4P4Q+H2seNFTXPFk8829QqBzwUxx/8AsjAHrnNSeD9Ib4heLG1TUAz2enunkxPyhPUKwPUDqR3yB0r7Q0vS1t4ooVTacDj9B1A6UAeP6X8MdKsoBAkO8DsRwM/T8q4n4i/Abwp450eTTNZshvIbyZkXEkTf3kYe/Y8HvX2PY6RZm3a6muYkEecxZbzOmRjHYkcHJ+lU7nSDeKRAm5SA2Bgtt/lj0oA+J/8Agnr+0trn7Kfxgu/2YvjBdGLwl4guh/Z9zKSIrS8lOI5VJ6Q3HAfsr4PHzV/SWpB5HINfzdftR/s/J8TPDQutHiCeKNMRpbJhwZlHzNCx/wBsDKZP3vYmvuj/AIJh/tjy/GnwU/wT+JN0V+IHguHy0ac4lv7CIhA5zyZoThJR1I2t3bAB9Yfsi5/4QPxltAY/8Jp4owDwD/xMZeK5PwfL4quPjLH8SP8AhC9c0a207w/eWWrWF5HHLb2ssckLQW+ilNvmeaY3aRoxscBC2GAA+n/h58OtC+Gml6jpPh95nh1PU77VZTOwdhcahM08oUgDChmO0dh3Nd0etAHhuqaFrvi3426ReanZyx+GfCmkte2jOMRTavfu8AbHd7W3RwM9PPz1xj5i+Anw58d+CfF3gfw+th4ggn8ORalb+I7nU5Wl0eeIhhbPp6sxjDySFXUworbN4mJY8/oeOtOPSgD4L+L3wk8T+I/GPxI8a6fpV/c6taHwu/hyeKaVRHJbyk3jWyK4QMF4lOPmXg5HFchD4D+IH/CYo1v4b1uHxymv6zc6rrzyP/Z93oU0d19mgSTzCkilHt0igCBo3QthcFj+kS06gD4D+EHwj+I/gG58KyeC4G0LUNQ8C2KaxcastxfWz61DLBv+0x/aI2acIZVBDjA45AAqpr3wg8eXviPxZ48v7C5u/F1r4w8Jy6Ze2rzww/YI10uPU5LWAyuEgZRciVSWyoIYttFfoHSjrQB+TPhb4SfHKe91iK/XVoPFlxp/iKHUrhdPa3tr2S5gnS1D6jJfulwrSNG8HlwBosAYjUEHu9e8P/Eb4q3kaN4U8Qado7ab4J065jvFe0eVrPV5JNTwEk3bUgb94/G5ehK4NfpaelC9KAPgOf4Jan4h8TaFc6toFzDoeoeNLkXdmrvEkWg6Xo13YWCzbGBMEkqo4QkhjKNw5NeDeIvhP8YktdM0nWtP1eXw9p1rrNnpEENhNq09pOur3ZtSNt9atAzWZt/s9w5YKq43J/F+up6UygD86L/SPjBpP9seCbzRNe1jUtZ17wZqUepxopt/slkumx6g0sqSFI3D28xkjXOQ24blJNZ1t8IdQ0HTfDOteOPCGr69o15qviSbxDYWpmuLqaeW7k/smeaASBpII4S6oq5CF0bb8uV/SpaVulAH5a/8KN+JuueDPEN54u0jVrnXNM8ERDw8sl5LJPbagl/qM1pFvSXa97b2zW0byEsc5+YgknuPBvg74i6J+0+niEaHqd7bahf3TX13qEMsQtrSW0O14b+G58i4t/MVFjtJbfzI2JbI27q/RMdKRulADKevSmU9elACNTqa1OoAa1NpzU2gB46UHqKB0oPUUALTW606mt1oAbUlR1JQAUUUUAf/1f38ooooAQ9KZTz0plAD16Up6Ui9KU9KAI6KKKACnjpTKeOBk0AcB8WNX1Hw/wDC3xhr2kTG3vtN0e/ubeQAEpLDbu6Ng5HDAHmv5c/2HPD1rr0/iX4k6y73uuzXnktczNvf96BLI+487pHb5ieTX6ift6f8FHPCXgK01r4AfB+zj8Y+L9Xt59Nv5kZntLD7ShiaMeXzNPhj8qkBD94k5WvzL/ZcsfE/wk8P3Nr4vsXsk1S6SZRncYwVVP3gXO08d+g64oA/YP4ePZ2sFzPL5abI42UlSTlWO4AqwZTll2t749K1LnxjaaZqd1cw2/mrJGNjOohIP8I3kb+B1AxkjmvFvDHiGym2ykhtoy3ltyU65Ug8jr069q6PU7/TLpXjTzBOQJAjjDOrufmZuv8Au0AeK+MvFMFr50gmAfpyAnJJY8DAxnt0/DFeNp4zu/F9sdNf93at8joOshHXOOg4/KvWvEfg2bXblpJyfLYklP4efUfzrL0b4Y6dplws9tbhX3ZY7BkZ4x+tAHGR/Dmxu0DLCI3GGDLlcD2IORW9o+leMfBN4txod8Ly1kIMlncjII9UkHzLn8Qe9fRmkeG1ls0iiU+c7oi/KrId/ChSCTknjOMDPUGrXinw1a+G1mstVZIpY0V4/LUSRyKTtYF0JXeGzwxHTnBIFAHI6D8YLPX7i08K6tJ9mvULLDa3jIMbwARE55A44AOM545rt59d8Of2ZJ/aiWNtC53BrmSVfMUfKUXB4APXGOe+QK+IvGOlzeNtbFloK5W2kDNcEEeUe2COc8dAc/zr1jRfg9ez6XBc+JNQuLsOTIjXMzO8nPJ25+UE55bGexPWgCz42+OWm6bMwe6bUnjMaKRgNtUABgTkds8DPvnNcDLd+JfipFGX36fp0+GVflErRjpjA4B9+vpXd6r8DvDwsnubeyiaBiI+UUS7m9F6n0yOp4612vwvt7XSIWhv7cXBgR4UBPIwPlIweoxnHt6UAeCj4Y2Hg+5bS9MS4hn1iCONJFMjvvj1G1uCMgEJgCR8cA8mqdjqnijw3p8ln4kS0vpU06S0Ajju/tm+W0+xAtC0n2dWgiYgzhAJUQbSZHNfdU95ZGyVorOMKNx+UdeRkZ4714R4k05PEGqG0cG2kJXBAxCsbFixYsN2VGKAPLbH4k2fibUZNaso0tJpjBsiu/ljkaOO6ikTzEV0Rj9pRl5BwHCncADn6lpc0EiJPfzLa61Da2KxW9qGnlh06wFk0sbTRs8ElzEsqRFdjDz13kEfL73YfD6xnjhgSZ7KW1fgqA29D1LLxkMDkAHrXsXg/wAB2cUL3Burq9v8EvcThXKqOm3HQbj2/rmgD5X8N+BNSOrQNqekeVeiXzLzyY7gBVX7Quz5j5ItktxapAByu0Y5EtaWpWGpaFrP2e0abT4nvvtGR54DqfJ/eKkSEXDbEaHyjzgcjZKxr6x8RarpdrGHs18t+F+Rflb+8wJ557dq4iLUV1SWRt7qwIDzLgyLg8HryeQAfy5oA+avAHhqfUILTR/7Om8zQrV7WKBkcpZ+ff38pjQn5eITESUyCu3npW6dM16SLT9PRby7hsIbi0msdl1EgBu7p57cxlBDItzHIoMmSqhgXKhFJ+u9Mh0yyhi+yn7TcONrvJkSKcfdyeNuByQeea7uG/W70pYJCWBOEGVbt1254yfbrj6UAfmxbaDf6jo2j6tevqAuNJ0mx0pJVnu7WMmDTdMjmtXKFSsXmJcM33A8ibTIAxzV8Q39u+pFPnmlFjpAkaK3vmj32jQ+aA93vlYokJ5dySACpwVr9Db3Q7Jyv2aJmjJ4XO1cH0zgYP1rnF0a4hugTbpKmwupceYpdevyj0BHzdB9aAPkXzNZ1bVrN7m4vLmbWrqHUUaYzSKTsvhI43ZEfyvGpHHG1cYUAaUH/CUWt4TLHfwWPh+9jvJ/LjnWFRcXcZkkOwYkDW6/7Q2FhwGIP1DZwzrPNDujtmidW+eHcFZ+QQuQTwPb3rbm1DSrOGSeayk1W4RHZC5PfODtUbVA64A4x15oA/PF4vENn4W0vTtMs0g1NdIltGs76fUY1knnWUFjbxOInMnmI6tsKo4UyshjBHXa2l3Y+EbvRdTV0ul0549oinkd7uPRP7NEQCxkfvJELpISEZSADuJUeoeF7K+8R+JZNZvgYrexZreBGHPykbmJ57jAA7c17jLpbXTR2um2w8yXCCNBk889QPzOaAPifWbvdqH/AAkMt5NBc6rDFZxzTPPAWt7aVc2gcjMKSogIOFRvJ2M2HFdf4YXUI9fNpqjXc1tri6Z9okQTr9sWztrsTO0h2Oy+b5YJkw0hAkIO6ul8dRN4h8bafp1qBJ9mkJHlg8+UNuVHXq3J+te76Xo93IUtFGzDDcxxtUDAy248DJGe3IoA+KbPUtQP27UNemmttMu5rC8a6W5vNNv1um06zhvbhZ1WSOVvNjk3RyQSP5gJTImkx3/ww8MjTtHtYHaR24Ku+VmKKcRFuchggUYB+U9+K6/4maqmr+Kh4XaFBBZESTgL8vyMQmef42BY8103hvS2F0jNhViAwykN83YAr1PcemKAPS7HUHjJTVpNrzAlZyAm44/5aHpuOOH/AIuh+YisLxR4p8JeA5heaxMkkzKu5XkCQRNhj83OTIB0xwATnnFaOrX9rpVrF5sRvL684tbfIXzQActJuIwO4zwcZzivmdvB1p4gvpNZ1W+TUrlZCvXMNuwHMcangccZ6n9KAPatC+J+h+L9ME9mohaLJ2cFCCSAQTkjAHcnPbvVLV47e9jlt5kQiQHbkDkMo9Ox/Wuc0jwzDp9wLq0gEbqQWCjAYHsw7g/SuulsIV2vBKyJ8x2qcBG68r/ex1HQ5oA4HwhrQ0K7l8F6pJM1tIjSWLK4AZyeUZm5wF4ODnbhuxrtdTgiltnMnygrzjGCB05HA6VQ1Lw9Z6wY96YmVt6FRhg2DyuP5j8a3/FgvdD8FXniLWIBDMzJDAmNvnzAMGIUfdO5MP2BPHBoA8Q+HOjWzzXOqrbpKXu7gLuHDANgYYcjHsee9eo6/fW+j+HZ7+Rnt4IgWeNxwoH8Q/hxk9vxrH8D2kdhZW8TRhSQGYtkFmJ55Hufxr0vXNHiv7Bra+hVopFKtG+HUq3UYPYjqMdKAPmaTxHH42g0yaz0vbLYAo1zMVLzoCQioVGdiA4GeT+tdRpWv2q3troGqp9iiunVGuWPynJwiFRwBnnjknAHpXbWXhKw0yBbXTrZI4YhtVI1+RVHYAdB7Gl1XwjFqytp9zAFYfLIrDlccHdnpg+uKAOzWz0xJ1tYriOU7MxuWUIQ5GC3OOew68j1r5r/AGhUXR9FuLESZlluYIZBgghnBfafwH0r0yw8La9o7+fYa1coYUKxBjnaMFcZYE9Mjn/CvCfj7d63HoL3N2Y5pftls4dEWMfJnAKrx04z370AdR8HNCls/D1sjKGdsszdzvY49/8A9Ve2sJIQPLYhmGCuPw69MfSuM+G32O+0C3u4UMYdFbBbkFhkgAdMZI/Wu/vI4go81tg7tg8c4yAMk+nr6CgDlLy11O9nuHiu5m+0DEwV3HmRqc4cA/MoI4DA11OneHLyOETbDOqggBiW6DoDyenpjpSaTuVpgwPkRgrOysEYoCN6qzHYScZA5yfUdNnQvjJp5upLVLSAHT7n50P+sZUBxHIehHckcnjBxQByfiWJ9FvY74XDzieNQpIZfJKnO3J64PIx2r88vjz4C8X/AA18e2/7UHwN1EWHiLS7r7TdQ2vMh6hroRgbWjflJkxgg5x8xr9ULzX/AAnr1tcT3uj7Zny8AgfMaPj5chucAkk8ZA46Yr548QaPDAZdO1BIbsEblIVWUhurRt1we49Rg8gigD7s/Yn/AG5/An7WHhaPT7h4tG8f6dCDqOlM2PM28G4tc8vETyR95CcN2J+7j1r+Qj4pfDPWvhfrrfGb4N6hPoes+Hbgzv8AZSUZdjf66Pb04P7xCNpGeMZB/o2/Yf8A2mbT9qT4E6V41upI18Tad/oGuQJhdl7EBmQKOiTLiRewyV/hNAH2COtOPSmjrTj0oARadTVp1AEdKOtJSjrQA49KF6UHpQvSgAPSmU89KZQA5aVulItK3SgBR0pG6Uo6UjdKAGU9elMp69KAEanU1qdQA1qbTmptADx0oPUUDpQeooAWmt1p1NbrQA2pKjqSgAooooA//9b9/KKKKAEPSmU89KZQA9elKelIvSlPSgCOiiigAr8Kv+Cin7dvj0eM9W/ZJ+AdrPbasAtrrOqJxMfOiWRoLUg/IoRx5kpwRyBjBY/urX8n3h3Uv+Fh/tv/ABc8agiaP+1NVMLDnKG88qLH/bNfyoAufBz4G2/w0SPxBqiJqOvTrmSZhuEW/qsW7jn+Jup9hX1PY3emasjxKqjdnfCXAcKW4BLYDHoeO3tmuxg0ATW+ZQSOPlwM/wD1/wDPFed+I9Av9LmOqaTmOUKwIBxvQjBUk9NwPBxxQAsC6n4Vuhc2W97HeGlt1IYYIxmPB4Oeo6Gvovwjrmk6ppMtyZMykKqgkglWIIyO2CMEHHX8K+c9F1mXVY0tViDThShjY4YMuSQRwOAMk+2arWlprtrfm70S78kMPnXG6NznIOD6Hoe9AH37p2jeG9YS+uLi4MFwxLQxKgG5C5VURQTyBjAABPbNV7XTNBsblRe5ntoJVjkSMjzGU4JztJxjkcE88HAINfL+k+MPiLo/797e3u5CCAVlKMO3dSDkcYzV27+K+prbynUtPmimIIOApTJ5P3MfyAoA9s8WeK9G0y3uRCkVgIyJI1WfziAOSHc7euAcDAUjqTxXzveeNdb+IM11Z6bOw0qb5LieQmR5iD0icjjHQkdPxr5o+Kfiq/1i+lhtJHe2uERUJBXcMdMdvm7V9S/Cazs7bw/ZQxJtgt0iQ4AO1mBODzxk7vqRQB6n4E8G6RpUCvcWizpCEVYt+0Mx7sRyVGOQMknjPevRpGkvpnlvCXklGWYYABA5wegGQcVWIis7aMlfLbZlCSCJIyzZbgZGMYIz29aaLt0jUHDDPQc5PXGT/n+dAFa8RIYWj+WUMF3KSFDEHqT25+ntmuF1HTbPTrqHULSUNFMCjBSCTIuCDjrg57gdK1dWGrbftMaAwGUKxzj5vY9TwfTH41zuqXq2yRWVttYktnzGBAlKMg2sAucDJ+uB70AdPbSSXkUdtaqZGkzjHLk8HAH4dKwNNtLW48Zz6dfMYFRU3nAJVRnecZGeen+RXqWjaho/hySW9soElZFCqu4qXAjUNtfLbctuLcDgjBGMV86ahe65qXjKXxFpPlRWSo0TRsMCU7yeG7BM4BPU5J9aAPZ9Kma1WaaVQSzMcZ9Ccf8A6qjv/FdyijTrSVo4T80i7sF3Pc9OgPA96zk1TUZbJ9RvtLX7PbYaR43yxwTklFxkk9gAOPrXVTRaRb25ultgZJFO0gYyDjlskkLyOnU45xQBxdzf+Yy+YjzSSA7Yk45xj5v7q/mfaqt7Za5p2mS384iht4wHZgn3SOxJ+Y59uv612dtoyWyz3s04guAPvFlCoBz65PsB1/DBxfEnjXwJpOnI2sa7FJKoAji6HI5JI2gdegx16560ALoNrrGpWInguI5UwGO04IHXBwSQQOvUjvXVWniq10P99eSi1kRQWWRs7wT2GM5P/wBbmvne0+MHh+01OYabBcy2cw3ARwyktLtCYBXGc4z+Q7c2x468m4ikuPCOp6re3L/6OrxSxjdj5dgEXLA9CSRjtn5qAPtuyv7fV0jmG/ZIpwjY+bkHkckEfTPqMZrKuYrOe4kUSeXAqZ+V8gNnue56YHBzzjivka41v45mNY9F8CalY8xoJJ7iJVDdM4J3KN3I6kdDkV1kt1+0ydtvd+GrHcMci5iX3+YKvJ96APbmkj01XaJwJS/zEoGJOARjOMjHB569qqXM06wzwyxJL9oOdqpufJYnGBnGD24weorwMt+0neyy28Gg21mYWBMhuFVeeAQBGTx9K07bwN8f4ZLq+fXbOVpsOLfyihjkDbj++A3Od3JY9SOgHFAHc+H1FiNQgiXBFwSynAZTKQcDdgE856+9X9Qu7tbOaz0ksJJgfPnRmZIEUENllGNzc7eemRjJFfM95N+0BpV1cae/hxr1mzukhuImVy3OfnKMSe+R06eoj/4Sz472On3Nk3he8EWoczJC0Thjg4yFcscZyPfrQB23w6m0y78UX2s6iJ0Fvm1t2TAYeWd29c4yrN1AOcAe9e5eJPEtvp+j39/buZ7uUpFAwJi28fMw/vEDAP8AdzkjJr498H+IfiB4WEn2zwreiNwxbfBJhTnJYbQOg+vGfrXV3n7QHhvVNObw1rFvHZSIXGX3ebFJLj5wkmOdqgYOMjv6gGd4Guvt66jr1zIz3epXkjtjpGIyIkUdOFRRn3JNfR2gSafp1tNfybktbdS5CFUZueVHqM45HIHOK+bvhNrvhc3uoaVc3aJE9wZIHkwF+fBYEYPqcHvjJ68e2alp9wNNvJtGmAS4ibbGhBVgeSoLfTPUcetAHk2r2+u/E7xRJr+p3qnT4pJIYljYojLkAnjgrkADnGB6dfZdK0uPR7KICCI28Iz5KgIcAckNyCT3B4Jx3rhPh9Y2uk+E7C2u3ELRRbi8n3Bj1J55bge/WvTnuEIWPrsQMcE5I68DgfhmgCh/bPhSUPqA1uGxWHLEXAYFf9hgoO1unU471xev/Fbwbpbu8N3LrMv3QtmjyBmXoQQOAOR2J69K8W1vwnZ+IPHF5qlxGfKjVRjJ2kjPJ5wT9c16NYaJY2cESgKUIXaNuMducfy4oAltfjpBBcefa+H7mFocMjyQKfm49X655GQAK8wuviI/jnxpaaK4ntIYC8zrchQ885HByGOQOpJ5JxxxXqc+kRSRO0SZQKWIIHQHoOv0rhNc8PW9zbCSEtFdQjfBIiAyIw4BUEjPuM8jg+tAH0HBpf2JIkVQrRqMjkHJ68dvx5HpzWfqOvSaNB9qmZkS35Cj+FepwDwTjseDR4G8YN4j06fTdXuo4tZ05lR02grdAjCyK4IALDAII/h4Oc07xBo73lrcRbQVY7JASAq9c5Zjxx74oA4C0+OVksRXS7Oe8WVFYyx2zny2B3BA0vlksD94rlT6kVCvxymuLrzNesLuDeR5jSRBwUAxj90W+bHQnjP3iBzV7QfBREARDGh3EBS6D5cE53bsYAHGDz2p/iDQtE0h7n7RfW7CBSzLvUttU4YZyBuGOVUt7ZoA6IeN9JvrhrjSPNFk5/dM+x5AnXc4iZhn6V4Z8c/st94Lu54Jlk+620I6uhRx94OByc5+UsMd88V6Lo9s+k2080VtHNaSxoT1D27Eg5Ug4+YfKwZSPTB5rjPjb/Y8HgS5ktrsXDz2m+SNo2iMUnmBfLOeCe4ZSQRQBt/A26ludFNoInlZYg0ZHJUIMsSADxg9c/yNes6jcWkcW2R9szqRhlO3K579iTwK8B+EEMk2hW8s7sI3i4CsEPyjH3jn0r228txcQkxynCtjD4Dtj/lp06H055oAs2NuUiuJZN5LjLqg7DuxH3VB7EcnHua+UvE3h+4v/HVze6ZPJbymOI/u2wu75uWAznivqDTdTn0oXIacR2728y9SA7bSBknccg846HkcdR5LoJtdR8RX8wkDrlVJA6GMfj1zQBT0jwvrZPnavqtxKZNqjYxjUHPyjAzk+/6V0svh/XrON2tNRnUFWVgz78r/ABA8dPWvRY7BTAFQcjHI4wf8j2ro7SWPVLmOy1tWhuJWRFlhQEPnC525HOR0yMk9aAPkafwnrkqalNqVutzZSO6NsG9nUggfIOqsBg+xzivB/wBjr4tX/wCxr+1unhXVZXj8D+OpI7G4Vj8kYmfFtMe26CUlGP8AcLHuK+8tdtG0O5nMM5YRSh44mT5ZEzjd3X5WBDKTX54ftj+B7bVPCVr8QNJ82LUtIvMSRAbs28w4kVxghkZUDA5Jzuz1yAf1dg5wRTz0r8xf2P8A/go58C/i34N8NeDfG/iNPDvjy2s7e1uotTxbw3lzGgRpIJyfLbzCMhSytk/dr9N0kjmiWWJg6OAVZTkEHoQRQA5adTVp1AEdKOtJSjrQA49KF6UHpQvSgAPSmU89KZQA5aVulItK3SgBR0pG6Uo6UjdKAGU9elMp69KAEanU1qdQA1qbTmptADx0oPUUDpQeooAWmt1p1NbrQA2pKjqSgAooooA//9f9/KKKKAEPSmU89KZQA9elKelIvSlPSgCOiiigDn/FmtReG/C2s+Ip+I9Lsri6b/dgjZz+gr+Tb9i1ZNQ1jxZ4rvsvJe3MSs/cs5eR8e5LCv6Wv2wfEP8Awi37LXxU1sMUaLw7qMaMOoeeFoUP/fTiv54/2K9CkT4YXOrJGryT6pLIodQwYRpGoyp4IyDweKAP058HeF49fgeO2kXzgMqhYBmAUsxB4A2gGuV8Q+HluIMCPcH4U7cArjOQeM8c9K8rh1rUtEWUS2zXEAzhY87wOmADy3Fe4eEPiJpXiCxNnrDreWOf9YijzY3UYxj2B6YByAe1AHzFrvhm70m8k1TRyyTlSrDorg46474BAPoTXX+EbhNVghEERErtt2kbQO3JPQA9TXtHiHSNGtpZpVuRcwKQolA2K5wCFQAnOBgnvnIxkGvPtL0G2tNbvNV0oxeXMkcR2sSDMTu3LnnOOCPXtxQB3Nt4aLC4R54mlQElOR9wc9RkHPGOc9hXEeK9KtYLd/Nwm0AnBHAPQ8dj/wDrruzDdW1vHtcrtXIOSGxnPDDBH1B/Gvmrx349l0C4utGnQOJV3RqxyP3h569y3ftn6UAZ3gnwWPEWvSapsElvGBGqsoP70NliM9scD8a+tLDwHpuhRQz2V6Yb5CGwgUoj8MFwR82VPT1z1rA+Gugww2MMcFqCQvIQHAJGTwPxPP1969CjZ7eNomBJjJB3EFiMn1oA5bUdfu9GDT6vGHgaQiQoOULH74UdOT82OPb19g8O2dlPGsqwIxGCC5yMA8HB+XnGDweOmK8w8U30l/biTUQrmb93kIB0XBDYHJwOefwrrfhTfNcfD2y1C8ctcRTz2zMfvOEbA3N0+7z1474oA7TxHaaVZxWFosaxPdKWlYjcgdWUeoCH5umMYFfPWsSJqIGrRSBYpJGgCZ8yQJFlmkIIPy5wcg5xwCK9a8VarZWcyXEyrMtpKsuwYxIQeR/dCnGOme45GK+fodU0PRdWj1fXozJp9224xK2S6lsuAVOVO7tjGOOKAO1ns5bjTE2hzBcqQjupVn2kbsJnjrjnPGMVC8elaQjvr9yttB5QCBANwcYC8c7gRyTkHr1rzjV/jNr2rX0mheCoZru6uwLeOONQ85jHKqzDJ+XnbzwCRzk16H4S/Zx1LVgus/FC4ecXOwJZ2z7Ioe/72TguRkAqpAFAHnr/ABK0ue6t9P8ADdjeanfsBGEjXOXYYPyrnA3E4yenNegvovx21uBI7Ky0vTLZUC/vZ3mkWPPO5IwTxgDl/bpX2N4X8F6L4c0GHSdAsI7K0RlEQhjA2kt3A5LZ5B+970t54du4832n70vU3EtGu8DccYYHp3yO/OKAPm/Sf2dNb16xjn+IXie4uZXy5h01vs8OOgAyGkJHcZHNe3+E/gj8LvDLB9G0mKWRFGHuY1uHyed258tu4IJz6Cuhs7+5td1re2j2TMFJcBmicrx/F0+939e4rro724aFDLZqpQbYnU5XJGSOBgD/AGSW9RjGKAMzUfDltdtDHpiLDHGB90IoHI3feHHHQdRWsfCsUtpCY5RJJGiuSepYjJz1xg5HHQ11Om2d3cSGaWHaAq4CqQc7cHGeo55/P0rTg8MGSMxC3zC7FlQHIQNwSD6EE9Pr1zQBwTaVcQQTfarVJFKrhlbk5xhieMbsjPHNcvqMMX2oC2bYQpYq65bAHygcj8q9uh0R9M0saZLcFZLcMsLPyGjQDYC2BllHXPJwTXP6zoehWl3Pq+vapbQ2NsjZa4cQrGcjJc8ZJ4XJPTIxyMAHk5uTOgRF+ZUZmbvwfTv17+1Yk17HDGytG8jE5PHAAzyfxBx9K8j+In7X/wCyp8P/ALb9p8dWMt4rLiGzZ7xgwyCAsAkxjGSGIxnAr4/17/gqj8EdHlFn4d8K6x4lADB5pPKsVdixIKbmlcAZ4yAf5UAfoux8q6mE8QjQKrFhjO49Fx16Vlwx273IW9tlEW8ohK5Y4P3j6cDAGT1r8rNW/wCCouqajaXA8H/CKWRmJPm3V9LMoJPJZYYE698MK8hm/wCCgf7WEt9Fq9r4V06DT4CS1t9hmYOp7FmkMnTuuKAP3CltdP8AMe0W3BO3cp2DkZ4I+h/SvPPFXw08K+I3jN1ZQG6nbDSPEku5RjHDqT0Pavzu8Af8FRfCDzmy+LHgi70l22q0umyCZE2858qXy3Az23tX3L8PP2s/2b/iXgeHPGdjHfMSYbS9ZrK4L4wF2z7Vcn/ZJ9qAMfUf2XPhlr/+kaX9q8N3+SDJZS5RscEmNwy8nPAAx0rkrv4JfFrwpfCXwn4ls9Vs0ddsd9DIh2qQwEm0sjDIwRtAr680qRLq2UuuLh4dykH5F3E7V/3sYzz1zirGoHyYWm2YBAVeRtz6g9O2SaAPiDUPCvxxtdOa01DwzYXVkgKbrS6LMFbkYExBUc8Y+p55rjbKbxb4OvILzXtG1e2tYWXDShpYkBPLEDcp+Untg46c195x6gftc1uFEqfNx9/O0YUZPqe5qjLB9rjjgabbKRhEzu2j0wee3WgD450HxX4OuleDzik0uYy4QMMlm+fLFSCysOMcY6Zxj0S80XyNO2m4WS8uFEtvFFhz9kdmBnkAzsIAwATkEj2NbPjv4IaT43luryDbo2sQoggvIc7ZDgY86POHGcjJ5HY9a+aPD3jLxP8ADHxXP4T8R2/2eRJTHOTEn+kwnKM8EhAby5BngHHY8igD1uTrHHEWikyCCc8IOB7gkDPqKt3Nu7qqeXufadu1dxIxkEHP/j3/AOqtiLTlb/SZnVvtKLLuDFlCuMjliT35JJJPXNQ6xdQafpstw8fmWwjcOQS3lzH5k2sDhTkc9jgjGSDQB8z6hYavqnj+a80G/ksYrOBLeVoSAXfJcqwIKnaGHJGea7G/0zxeLGYx6rJO20/I+0Ru3Vd5A5APtxml+FHhrUJby51u9UT/AG55GZ0PyhyVO0kfLx6+g5r3S+0Q2E7Wtztjkt+Co2kg43Hr/Fgggd6APBdN8Pa/q2mLNq9xNayl8mOOTAHYfMBkn2ziqKfDJTcRyG9uSQVKZfgEe3TJ+mTXtt1LZx3Zgi3W0SnB3H7zYGHPoWHTH8sVPFBHOhjCFUGSc/KCScEA9B68+maAMPS9BmWNbe5JkCL8zFslgO5HX6n1xXlHxo0byPCt5FNMXBjUorEnBDjG0ntx0r6RMWnWGny3+uSCyto4/MWdmyp29w2Rg+p4AGWNfEvxL+I+rfFrxAdA0CM3draH57gjBmER+U8cCMZyDjLHk9qAPY/hDdWNjp9kdS3vaoqeYEOGKE/MAexxyD617/a2Vjql55OnTqUk+eLzWG/ZjuCcA8c/nXxnoug+PdOtPLs71VQLwvlhgB7E81pWbfEPw3ZeZK0F2nmMduxkGyTB6YwQDk/iO1AH2F458LJ4Y8ORR3t1bvfSPKI44XEjR7cgozL97JB5PcccdfmvwDbPFq8kUcDebJdyDywOpdhgD15PArP1XV/iDdxfKI7Ziqq5VCcrjp8xyMVW8DS6hZao1hrQRvMcyJKPlXLHJBHXI4oA+rNOhfzishRGtwXZW7qvJB9CQMZIODXnXiPx9Breuyab4fsmsLaJWEZADne2ChdzycnqV2hew4NdXr1leRadKJI5FjYKw28E5Xcp3Dggg8j39q8r+Hej3klve3V9EZZ1mZ5GVduEBCITgYxxgZ9OlAHU213quveZDqcIicSM/kRvIyoTxgKSc445xnFeU+NbCwi0gzyss0c7vEVbqjE/KcDgjtzxXtt/YzmHz7V3hKRMCUZs+XjkdQMY4I6Yrw34ia9ca1fLZ6qhF9dSozyKxAkCj77A+vB574PWgD5U+Iv7NfgrxNZf2rYW39kag0ZcPbYVHJHyl4+nXrjBxmvv/wD4I8/H3VdY0HxX+zp40vZZtU8MSG+01bhy7i1ZhFPCu4k7YpApA7BzjgV4BHpl3psYuYQXi/iQ85+ma+ZoPGs/7LP7XPg7456XmPRdSnVr9EOQ1vN+5vFPrlDvUH+Ie1AH9aa06qGl6lY6xp1rq+mTpc2d7Ek8MqHckkcihlZSOoIIINX6AI6UdaSlHWgBx6UL0oPShelAAelMp56UygBy0rdKRaVulACjpSN0pR0pG6UAMp69KZT16UAI1OprU6gBrU2nNTaAHjpQeooHSg9RQAtNbrTqa3WgBtSVHUlABRRRQB//0P38ooooAQ9KZTz0plAD16Up6Ui9KU9KAI6KKKAPz5/4Ki+If+Ef/Yr8cIr7JNUfT7Jcd/Nu4mYfiqtX5X/sdiLT/hD4f0+9iMLT+dMjnlX82dyPocfpX6B/8FhbDWrz9kcT6Yu61ste0+W9x2hKyxqfp5rpXwh8Dtf01fgh4UXRrcXFw9hEm6QfLE8PyNtI5yXVs+3FAH2FP4PXV7Eahp6qy+XIzhSFaPy1JO5TjqBx69gcYryfUfCt1pNwbyy3WU4bkqPkk2/wt2OR+PpWbY+L/GNtGUMMLwkgsoLA5AwDnnnHH0rotM+Jdk0/2HxNaPAj4UljgKD94q3OTgnAYAZ7jrQB0HhfWdK1eKbSNVAW6QBdhOGVnYHcCeCuF59z6iuqvW0L+zk8GWM8QuZZ/PeYDJjkUFIlDbQNxySRnjAz1ryPxd4S02+a01TRbwxiRd0ciyKzkOWzgKDwAoDBu5GBXjek6d4i8DeJv7U1WT7bpszP5r7WdwG6H/gP6enFAH2Akt5aGTTdTBdoxgZPybs8PweN3AbuK+bPjp4IGqWjX8MZW4tgcMMblGemc/yJr0/TPGdtf6U0vm+YhdjGW++qljgMe4x7V6j400TRx8N7HVi5jupYo4pxMHTY7IzGTlSpjOBtIO7ORjtQBmfD2UyeHodSCjypI484k25JAYZ56ZAPfkdO47S6YXE7XgQRiUDKgHaT079jjvz7V5T4FulTw/ZLYyFIJEAXAIyh6Z6cH0NeoOXlihmRPmKsDt6cH/PbtQBw/jNWbTUaE7WLkgjswyPp7Vw3g74n2eh+GG8M30/lRWE80kcag5zN8xCqASc9/wBa9B8URn+xpJGHKP8AMO3Ttx/+qvj+e2SXxBqF3HjICJn3C84/PmgD1TW/iS2oXEst/Gws4clIvMA8xx91pGwTtB5wDk98Vd8G/Bn4g/FdoNTulk0vRZ/mErg+bKuf4E6gHsTjNYXwM+HjfEb4gSf2pF5+haCFnu1J4kkYkQx/99DcR3Ax3r9TLPbZxolgiIFbajbcKA2AVwpJAA6defxoA4j4f/Bbwn8NrO3XRbRPtAjJuJJeJpTuHJc8Zwemc4IHUiut1IG5nSHRykM8bfvEYlUxnlQc4U7cFu46Edao39zPc6tFCYJI0kBLTFgUznjOOgzng89PfDtOc6hJc2V7azGC3IhWRWKXJYZcNGRkFCTnu2SOMYoA7vRSYUIaxkZTjIUjjPBwWIwe2OvritjykeB7kx74fkVnLMhjyQB8xA55GOwPPFZWlXd1aXYtLwrJHcOFTLKHUMcAOM5UjoSPr2r5o/a4/bY8DfsseG20TUkl8QeJtdikk07T4FCxRxg7SZJzwIg2RnBcjgDHNAH2BqaeGvCHho6l401K30rTbeIG5mu50jjVcEsHkchQD3PccV+Znxg/4Kjfs2/Ci6udC+FunTfEW+iwkbWh+yWKY/h+0uGZwO3lxMD/AHq/H3x543+OX7V/iAeKPi5rM9toysWtLCLKQRqeghhJx9ZHyx969M8B/AfTbHdNp2nrbNCqs09yC8xVsjcoI6cHJGAKAPo6/wD+CwPxzvUmg+H3wo0vTEmJI85ry9Ks3U5j+zqSTyeOteeXX/BSP9vzVV/4ldnpejp/dg01F/8ASiSQ17Pp/wAAtBuJNEuI5r67sL1US4eNUGyf5i4GOQgAHJx14zwT7s/7Nnwg0mERTRXV3cEDc8szBFycHAUqcgcjP4j0APzd1v8Aaq/b/wDHYFnq3jl9Jgdt2+GO0tjGe2Ht4jMMdtprx/W/h38Q/H179p+IvxB1LxJdSHJ8ySe7LH2e4fdnt9z/AAr9rx+z98L7M24tfDEc5ZwhJeWTDMPlyGY/U5IwK+n/AIb/AAu8IaE6ahpvh+0sY1DmNoYEVwwAyXIBYrjIA7/rQB+DXgD9hTxd4l8uTRfA+qakjcie+zbw4653P5KY/Ovtzwd/wT5+M2gR2cGmajoPhWe8QvHDHcPHcMEwWz5EJLFdwzhjjIr9hLe/IFzqN5i0s4GX95IAgb1wOp7AAD/CsfXviBo+j2y397PHpqShgvnbGupCeMxxgkBRjJY9O4FAH53Qf8E//iXqNzb2fxA+Ikc9gjgvDai5u5QD3VZvLUZ/vEceh6V9L2P7Hv7PPhzRrfw3c+H11r7Shd7ueeX+0JCBgMpjK+WoPJChRxzmsPxB+1Xo2kO1t4B0Wa+lRgr3V6TDHKinOYsB3OTwWcKB1GeK+Qvih43+KXxItfsus6wlpptyRMbSwcgMC2QJXRt7MBx85H+6KAK3xl/Y4+AF1rN1YeFfGENtFHGSI9ViW7hEwJzEtxGqlcDo2G6EE5HPwT47/YbltY1vPDklvqCTEhJNHuhdDI/vQSbZQP8AgIHoa+mB4LFrYPdMC8hlESAHe53qSGCnryvJB46HGRnHW1v9PtCbuKaGYnKsgGFRch9yfeznGAPegD4x0ST9qf4C3Pk+BPFN9Bbwf8uMzN5RAOcG1ug0X5c1+gf7M37dNx8VreX4bfE+OHTPHMbCOyaONooL5sgBdoJZJx02jhuoxgioNP1Bta0e6027vBf2DwKDDdkoxYAD90CjkMOCMfNnqAOa/Ov9oT4fy+GpF8YadHJZ6hY3CfvojtbAPyPuXGHRscjn9KAP6FjJNplkbtF2wRAKxwdzyMevHJGR1/8A11yq3F5c3n9pNK0TEBdqZGR2G/ueeg/rWd8BvG9t8Vv2ffB3ju3uxeXNzp0FvqLAgzC8tVEMwkx3Zwze4PvXbp4e1KWHy1g+xDILFvmdhkYAGeOuRzQAzTb9r6eOB7h1cDdISpcbeRhj79u/HFcb8YvhTp3xQ8NLFZGODxFpClrK6YfMQMkwE9Qj/pwea6UJLpUUljbwuXmO0ucFxzyST/kV0tlYzG9WOB9ixkYPbjqQeRQB8P8Awx8YQy20vg3xbF9g1LS5WQLMrM29W+aL+6OmQxIGM89K9S8TaZYT+FdUlklEKlY4vKHKXAZgdoOc5AG4EcADHfnif2lfCEPg/V9I+JFoStzq90bO6RD8m5Yy6MR2J2kA59Kx/CtjqPjKwY312qW4aOMq4wzDBbCkZAUYGQdvbtQBb8A22o/2DbWYuDbIA7gIdrScgYJGC2B0HQZPrXRXhNijWgiE63QZX5G9TnOWUeue+M54zXb2nhu20+wk04XKI1v++jmYhQUIQgdSCck5GONvU1yGu6dqtxc6Pe/b0ntFjuswxkFiYtxJdVywTklSepJ5wKAJrSeWZQl02CBtDY5IHTAHXrxivRNKsYLWyvNe1QBbPT4xJI8pIQnICqx4GPTGQO/y5I880GP7TeLbLmCNoXuWlLqY/KiHzEHOMjjj88dvEPj38WtR8UyxfCLwWrQafHIrXWwqDLJIB+6JBPILfPzgfdxjNAHLfEb4o+Jvjlr9x4Z0GT7L4dt3IAhLCGRCy7QsZx5cW5dwQdSNx5xj1rwV8PNO8IaSsMK73cbpJCMM7D+99O1Q/Df4YXHhOzin2xTmA77zDrIjZGFxjB4JI/uk4wc8V7FLBE8aRBtpU9+3HX268/8A1qAMaHQJZAGjQYBUEEFWG/lSVbBwQchhkHt3q14o8OR+HQmn6mYmuLiIMI4HErKjH5XdkO1dwzgAk8Z+6Qa898ZX1/pl3ZwWNy227mSJyvIZRyAGYcAEHGMfSu30i0gtreNidzFQXZufxJNAGZp1hc6ho8EoTe1qxind3BctwV3rngBSApAwQCexxzmteGl1WIquY5YiJFdM7lKnIbPHP611epXS+GdQg11GCwuDHKRht0bjg46HBIPXpmvStCfwzqtuk2pzpa/Z/JMuyRJAXlXcoDM6IST/AA7uOp6YoA4fw9LqEFnDZa6st8IgVUyHgkj+IoQ20dcEdepNc/4UnktfHGs6EQrSTQrJ85O04TOI0wNp4yuRkk49K6v4g+NPB+jW8un6Av2y7mJQ+S6TSjA5J8tvKQA4BO9iecA4zXzFaSeO7XxIni/TdkN/bKVjyrNmPnCueMn3oA+prlWaKSJ2kQRkA7SQoI7+uQR3B/kK8C8e+FLuaT+2tMPmPYkTMoBLBAO+R06Ajse2K72z+INveeR/aNlLZ3KxpHKxUuDIMBpA65JyBk8Zya7OSfw9qWnNJb6jHNGRnyJh5czGXAIH3gNueRzwM8dgDxTSxHqegxXTyB2lVmwOSCDhgQOh5Br5v+NngGLxn4PvfDzJ/pMBNzZSZ+5OOikcja/3T/dJzX018ONQ0aOTWtHupVBeWNolIYkKc7ym1T0xk57ZrJ8aaO9neShGVgpKHYQyMPUEZBBBz+VAH0p/wSP/AGlpPiN8Kbv4EeLbgnxL8PQFtBIf3k2lO2EHPJNu+Yz6KY/ev19r+Q7SvG+pfsk/tPeFvjb4dWQaXPcFdRgB4lgkbZdRY/2omDrn+MZ7V/Wz4f17SfFOhad4l0G5W803VbeK6tpkOVkhmUOjA+hBBoA1KUdaSlHWgBx6UL0oPShelAAelMp56UygBy0rdKRaVulACjpSN0pR0pG6UAMp69KZT16UAI1OprU6gBrU2nNTaAHjpQeooHSg9RQAtNbrTqa3WgBtSVHUlABRRRQB/9H9/KKKKAEPSmU89KZQA9elKelIvSlPSgCOiiigD52/a1+G4+LX7NnxE8BpF51xf6PcvbLjJN1bL58GPfzI1r+dD9jbxRb3vw2vfD17IqPot82N3/PK4Xeo/wC+w9f1ay7PLfzBlcHIPcd6/kI/Zo0KLWvGnxLe2HlWkVwFRI8gASTzbAAOmAMCgD9XPDvhxLyzRj8xYDPBzyPbj8f8nO8R+AUnRvMj3dRwOcY574r0b4K6vpUGhw2ettKii2jhlZcl1KAfMAT8xYLgejHPbB9XWPwdPp91Fq7NHM2/yxEcsJBygKHsc8nPB7UAfBOn2t34Q1dYLl2Omz4Rd3/LBxnHXPysT+B/T1m40211TTyhUSBvvgHgj6f4GnfEzR7EaFKY0WSXDKVUchjjYc+59OmD6itbwTpU8WkwLcpiRUXceT2Gfr/OgD5e1Z2+HniS2W7WW40m8kAjiRd7g5BZBz82edvNd7f+MNd+I7DTdItG062mJV5rokFPT90CSx4wOVrvPHQ0/RtQ0/U74iNYp18uQqpEcj8K5yCMA8n0+ldJ4V8FxWlstxcnc7kyF3wC2W+Zs4x8p754AoAwPATSWuk29hc5EtmghbnGGQ7enuea9hurjyNOtnA3E5HXkszEdenbNeQa1cw+EPEonvBEmk6s8aQXCMcfaTndGwJ43BfkYZBwcjNeo20NnfwR3M0rgw/PEAxxn7pBHfrwMdfyoAwdQv7bVbeWxllCSSwnyyflDbTjcM9eQQeR2xXyXpVhcTyXss3y4uJkIz90h2/oK+qPFFjBPYmzYm12gIHxsYbjgFScgc9x+VfMuoaZL4a106c4Z4LiNZY5HHzMGALbvcHPPfrQB7H+zTNe22v+ItIjPkm9kt7lJGJUbIt0THg/NgtxwRn8a/SXR9RsI0W3u2X5wcSj7pOeAVHAOep/SvyO0DxBdeBPFtr4otE8yKFHikiyfnhfp+R59ua/QfTPEtlqehQ6jYXKPBcw74nB3LkjJUtzyOnt+dAH0PLYwz23m2JGyVm3FcshK4wep5zwM5qlotneWQFxczGGFy2UwrKA3y5BwDk7QOD1x0xXhmkapd3epSRWuoS2jRoEiaByB5igEttPyNknGWUj2r3DRblp9HtdShk3SGMiZipZyRjcSg/iyDyMcY4oATxdZXWq+HL218KXsNlq8yO0ErRl5gq5VCQ5yecA4av53v8Agonp+saXrHg6fXrKaDVNMmvobmaUEifLRPHIr8ja21iFB+XkV/RFqWoW0sMVwpS1njjDK0o/eO/cAKAeRwMYyOfevnT43+ANM+Lvha68KeKrSxuFnLFLaZPkUE/89A25Co5UoRtOOoyaAPxw+GHi/wAOXMtpqgMM9pdPGrXDKXMCbhvIA5V1GecE9sc161N8QdAs9RiurW2Nw1szmEh2d5GLYG4bVXayfKQfr14r5e+InwX+In7Inip9atdO/wCEv+H2qMSXAYDav8MjoC0EqZ+WTG1scgjK12fgLxh4O8e3C3Hw51pbDVwMjStTeOC6J/uwSNiG49gCrn+5QB+lXwl8Y+L/ABOFs/8AhEpdMhtvLVJS+VIROARtHLnIyegUA9CT9IxaLdi1ZtUhaOR3DCPAJLKdzPknAAHJI/AV8q/B39phLHUF+H3xY07+wJW2pHexI6t5m1UUTpISQGAJ3g4yew5r7c1S2iUHTbJS0kq7ZGc/NtbA+QDPPpzgn1HFAFXw/YXev6kijK28CbFHzYbIIbp1OPU8V7xAum6Lpk96EbZZK80pV/mYquQnHKoMZPOex4rlvAWmiDRFZlHyyGNxllY+WdrEDvk/4cV4T+0x4yvNM0u38K6RdSRTakS195I8sC3kVgCxXqSwUYA+7uYn5aAON+K/7UWkaLcR6P4evoNV1+eLzln2sYLLc37tY1I2yPtySx4UgDBJOPAbqXxV4+uGvLiSQGcnzbq5JeaU5ySF+6insAP1rjbbQINQ1bSrxWhuWtVEUbbeAgJzz1AySQGz68Z5+odD0XTpIUSS5d9iHcVG1M87OvDDIyeenpQB51afDrT5lEV1qEgkIGfKAkYj/bJwqjPsatf8IDppSJdJ1SSKXBOHCuhxjOflAP4H8K9EvtLghiKQE+WCA4TO49sEsM/j0rEmjt4ZlaRjlT68DHoM4HFAHlOuaXd6Kf8Aia2m6E5XzYz8gJPXB5U8f/XrhNctlNur2jtIqndtbnOTlvxP19a+p7O/s7yRrTWYzLbXSkKpUyuzDgsc4G3B5OOw4NeO/EvwLc+DYE17RMz6PPksjf8ALNgcDjqFPJFAHzqb/SVlklvD5SCQqA6mMKewDLkqfc8d8YNcp438KjxTod34bv4DPHdR4Hzg8uOG3bevPPf3r2Oz8M6Trls8+qoYvPUyQxxgZnkQbgshJAWInALL85GQMVTj0q/0uKYaurSWN8DJE6JsWN1yBjZjCY4Cr1OKAPgr4A/tCfEP9jPxTdaVqFnJrfgnU5leaEfwsCB5sJOVWXaMMp4bHUYBr+gj4X/Erwf8afAln8QPCV8LrTNRXMLD5JYXTh4pFOcOjD5ge54OMGvxc8feGLe0ZzeQJPp+tJKkkDAlcHhwM+mQVPVT9K6r/gmF8S7Xwz4v8Z/ALxNefZ1v3F9pqu23dc2x8ueNeRzJHtfA6iM++QD9dtXsEeZzZfN533nOedvGSBngHrz+dbdtaHTLRL6BlkaFSxUZOQMk/gfb+dVHZdOuRAZd8cYYktwwXJPI79cZ9h15NYmseJ0t4lWHCybWAK5BIJJJxnkkdAfp70AfNf7XmpXEujeH9IUD7PqOoLMSBz+4BkODknliAfTmsfwteW2laJZItvFBO0Sb3TBLHkbjkYLMGxnt15IrA/as8V+DdF0jw/eeK9XhsrqGfy7XznILhyu8FcgKNuSWx1ABPNT/AA+8Y6DrOlifSLq11BJIGQPE6TocAcrgnYc98Z6jrQB2+p6isv2JzGkCECFAzBzCdyETMADuXrgHgZPXtJdadpcUDrFLJLO+7E+QmQ2QrhBkgDrtzz7Cquo655DXc7Ja3NnIsEG25h2uPPDFWPOYmAQ4YZBUduh4o+Krdbb7PbN5zJlMkjkZxksO35ZHagDkPiH4hsvCOn3+laK8c2qvO0EO08TlZf8AWhc9Mjcy49s9q88+DPw/mvNWF5Nt8+eQQxSSngSs2C7Mc5JPH1NcDq/iW01/x1LdW8clzDZs8KbcfOzOWdl9FLEhSewBNfQHhWDxYLD7NZWlvbQupf5w8p65yQm3HPPWgD29bQ2rfZptsrQ5QDGCpIIIG04I4HP0GM81JLDM8TSuAI+EBAwoIHI47+vqP14iD/hJdJjd71WyVDo8R3R7wMAlW+fb34fI/luWfiya1cm/s8YAKyqAYMBcn73IfjvQBleJdFe4it541MhtpY5emAdpBIHHv/nNegzLbwxNfw2qvEzIkZLBoUdhkqY2G9wM4AyO2WNRQfETw3f+HI7aSb/R7KOdkZUEhPmL1I27wwZQD8zZHAAwc7fhzV/Bmo2FpqV+ZY/MjDpKiHbLGcMGbkYJ4HAHQE56UAeC+L7LVPEmqLazgRWC7flwAwxg4AQIqnj+73xU2mfDpb50sNMsnm8wMyRxZbLbM/nsBPrjNe8a94w+EXhzT21CMtd38u0MJU8wsoI3BFHzAnlR8vfuQK8P1X4p+KNU1H/il9Hl0yzdt7STH7O4jXJX92hLjBY7fmDkEqSVJAANmy+Hz2lpFNFZFElww+UjKkkZ9xlTn6VsP4Snij8xgwPRVxgkkkZbONoGOp69h1rzG1134rrAs1xLB5kjfujJ5qssRB+UKsmQnHGeeuc1euvHXxXjjjZ4rC8WMjKuZELL7lg+cdBkbcUAd63h+AwuAri4U7UjChg6EdVbIA54wc5pp8NlJPKJjZ+NkYJDknGOWAXBzkE4JHauds/i1dxQ/Z/EPh/C7QQwxKqnrkNC0b9ezDHfHauug+Lmg3GkXN7c6Juhs42Z5pY540Bz8rsTkMV47kHuM0AfOWq2LaZ44E1uu3z2ZcKcYOcgZHHtXd6yr3+nrmUSGMBWXA3IP4TwBlTz83Y8HtXK3WvaL498Syvp00DPI/yxp8qKw5JOc4UDJAznOPpXpPi3womkWumz2c6H7RZpMssRwcFmDIw4H3geCOetAHxt8YPh5B448Kalo7RgXhXzrSQ/wzxDjn0YfKfY1+hn/BID4/3njr4Sat8EPE0xfV/h9Iv2TefnbTrhm2pzz+5kDL7KVFfK+veehjeSFGDvtLIGBAPrz+VeBfADx0P2W/27/D3iCSb7P4Y8bOLO7OcIINSYRlm7YiuQrn0UUAf1T0o60nXkUo60AOPShelB6UL0oAD0plPPSmUAOWlbpSLSt0oAUdKRulKOlI3SgBlPXpTKevSgBGp1NanUANam05qbQA8dKD1FA6UHqKAFprdadTW60ANqSo6koAKKKKAP/9L9/KKKKAEPSmU89KZQA9elKelIvSlPSgCOiiigClqUnladdS/3Inb8lJr+TD9h/T5dbu/HlzzmaW1BOSOXM57fXr2r+svWLKTUtIvtOhk8mS6gliVyM7S6lQ2O+M5r+Ur9mr+1/gB8c/Gf7PvxAthYa412IlkOQHltNxUJ6pNG/mRnuMeooA+77XTvGmiOLiwlSYLyvmq24gnpkfe59ea2Lnx74wtrZlksYmKj5gr8H2HHUfpXpmnXTXFmqyKBFu3sSgdlIOQORkL6gdTzWTqWim/yIIs5BJK9QQCenpwaAPAofiLe654t07RtRgksEkWQjIAV5FGVUMOvGTj2HpX2V4E1DSLeyvFv1AeVAqHG89jwB0z9M9O1fL2v+BItRiTylbzRh0Zc5UjlSp6g+h69apQeMfE/hK3W31+1kvbeEBVuYiRNs9COpPpkEfSgD1z41vo114Ys1gi/fgAzec25Xdm5G0Haq5zjocYyMis/wtJ4pvLK1S2v5JVRQEQ/MOBkjaeCPXjGOtfI3jH4ieJ/FN0NK0RJpGOH8vyyDgEEE7guBn86+sfgR4xuJtJTQPGdl/ZuoySJCm8JsuIgufllztjO7rGxG4YCknigDP8AG+hXGrxQaBr9ol1DeSRJHFMpAHmMNjhePmJP3+TjIzzWR4Y8Kw/D3VZtGsJpHjuJ38sySyeWI0wB5YZtuQ27J9OtfT+p2S6ldpqr3Md0ybCnmRghCMLGc9EYY46YrzPxFpcSyRQPbJLeK7ySkN+7877oJIG3b/eIP50AdlJo97qFuEltJLjCv5csSl4mHIYKwyvVSSPQc14xrfgPVNUtLVbW3lubou1xGVQ+Y0cnzKyoOSDnPy5yOelddp3jyPS7geEbjVmWO5BcRvIfIlcjazqhOzPUfMQSOcV6pZeXdyR313I090pQB3OW+XheeoKgADA47UAfJMOizi2EUg/ejGSODuB//V/+uqmmXniDwbN/xIV3WpyZId2FZj/EB0B/Svue207SLzN7e6fDO12snnedFvdnc58xX5IYE/NjaO/PSub1T4beEL2ZFQPaXEj7k8tfMjA7l8kYH0J56dcUAeKeCPjLY6Iqxa3oUxSPqPmIOTknKkgHtzj869Q0X4seDNOs3n8KXd7pd7NMszQyQu9oy5O+NlALJuznK9MDjtXM33wjkS+EenTx6iHchWj3Kc8YUq4XDc8DmseXwTcWauLu0OI3dGO0H5kOGG4ZB28UAe56p8WvBySGTS4Ly9kbGyQxyICmOAQ+0HHbJxjrxisK++JOqX9pJFpFmLXzGyPNlZlVNxKr5aHZwCQOfavK4NGEc4tgvlurFWVlwQwPIx2OePaumtdE1WwjW4urKaOBkLxsybcqHSMkFsEruYfnxmgDjPFr+O9Z0+WwudelW2uN7NbwRRQxOG+Yh0RQGB9GJr86Pip+yh4d8STyaho0K6DqZOfNtI8W7MO8kAxtOe6Y9wa/VSbRriZTKkRfO7OFJZcA8kAdMDI+hxnBrlbzQ4pmCSwkkgEFlI4Ye/0oA/F+fxZ8Svhdrmm/D743TG90h1IsdSbM0kKHgFZjhniDY3xvlkHK46N+ov7LvxJur6N/hvLPDHqgy9rcTyFoprdVJ8sYJztyWXZ94E+mam+Nf7O2hfF7wDdeHrmOOC8jHnWVyq/NBMF4I/2T0de6+4GPyf8Ahx408X/CjxsPh74teTTdb0C5C2c5PMbxnKKG7oeqHoQcdMCgD90vFE2oGyS28Q69dWt7oxuUjisZfKivpbx18pQFZf41CISeAxzk8185ePvCHiXQ7LQfF2oaubaLVbGbLF5jFt8vzYpMSM7K0wjZyARhdq43KSfcfhv4Vj+PngCfxoLtdJ1Ke5VGfyvPEPlo6OsSllKksd6nPy4BIOMHU/aK+H02tWGladqWsMqaHavHJaojxrdwGMpDMdrqgZeUJKtjqu0nNAHz9Y2d94fDNpMpWZP3KQpLcS3SsYZVcXCzboWmS48qMmLcqznZlUYKcDVPEOph9S0SxuXFnOGMiw3c0aG58y6kVvtF1KrIIbFEubgmRVUqiOFaZhXuvw48IWPiCeO71PVVk1K1dppYWSRPOLN53mHdNJEB5ih3SGOBS4ViDhQdSf4NxXQuofGGoGTVLh7hkaS2YQG3u0eORHRZlZxJG4jMiSIwSOLaVMYNAEuhfEi3s/B1zbSB/NstloiXLMSkiEIxncF9yRKrMzpu3BSVySDXmWveK9c/tWy0vxP5ltp08V7cXDwQCG5LW1k91AhjS8IMchGdyThjsZGVB8x9sk+FsE2gf2faTh75ZftIukVBmbf5ozG4KhAfl2MGGz5WyCc8LP8ABy61Owkk/tIQfYIpkaVLaaaCFbpPsrDM9zNM0jRHag8zCYwkeC+4A5K88F614f8ABvhpfEWsPFqSR31zM8bSmRLiKzuLiIMqS5cB1UKhkKnABJAzXQ3/AI+8TQ3M3hLUI7MI9tJLM08BmeSbz7WARKgvRFG/+lo+7z2QqRnawIHp3iTwhL4itLGOw1GG3MXnIRd2xuY5YpraW2nUp5kZ4SXIYHIP3eeniWt/D+38P3UN6uoiUWy+VNbi3lEbQ/aIbgiOSaeWZWZ4FLPM0hIwvCqoUA8R0S/8S6f4X1bxJDbPatHYSXklvcWHl22x2RJ4rS5kuPNmZN3DLA0ZKkBypDNX8L65qXhbw3e2+u68lzb2E627LJZNdynz7Zbx5YhLdxJGEUgEht7MeAQSB6Jp/gi51fwv4g15dS03QdM+yjR11G9tZZrya2ijSGEFVnMSyIkUah4okLFfnLgtu8jv4PDpstQj1DxVbanBdRS4iis5IJluvs62yDbIzHYyoDuxjJJyPugA2PFenm8jvNJuzGZbGe6t2KfdLxTGN2UnDbXKBhnkDGa/OX41+GNN0xf+E20+6kstTjmjjUxHb5jA8MGBBVlA6j0r72utce9jvLu+mSe6vJJ7mV408tN80hkYKrMxABbj5jXwB8b55dV1vQPCcJ+a5l3n6yuI0P4fNQB+tP7O3xv8cn4UeE7fxjbz6rdGxQi6mPmvIhzsyxJIbbjOcE+9YXxl/bG8FfDnT2tNe0ObUtUkSUWtmZBHnGVQyEMSqKwznGe3WtXwlrOh2Xh6w0628owWMUcKxk7crCoULkcjIHNfHX7K3hbRv2nP2ztZ8VeMdPiv9C0t7q+SykG+3CRMY7WMqeCi8HaeCRyOTQBa8CfswftD/tseIl+LPxLxoHhq5XbZGTMKeQOFW1gwzeX3LsP3h5LEkmvVfE//AATFTwrKs/w5+L8On63CoJhvB9nbcfSSCQuoPuhr9Pdc8R6vfa9d+GLeSSzskkFtAkChUcD5dm9fmQnoCRtHSvJPFWhPoviGx11DHa6dcsUdJVG4s/EQLnns3bjqxoA/L3xB8Jv28fh2v2VEHjiyto9qPZyxai/lA5wq/Lc44/u+teT6l+0p4208/wDCK/EXQrjQLuOIQOzRPFKsYzwY5ArdyM5JAwBwBj9ptWjvtQjtJtJvZbdrdkLPasG81FPHzL6HB4OeKseKJfhd8VYrf4d/HPw5B4gSdG8m9e32tFgAH94u2SN8n70Z+tAH5m/ALWfDmq3sl9aXMd1AdgYIw3ADnHPKn8K++LPWdMurO2s2tbe3ktowm+JAGkUEffycNn72SM9ecYFfKHxX/wCCY/iPw9dN41/Ze8UvIBl00+8mEcwHXbDdLhXHbbIB7sa+Zrb48fGD4Ka4vhH4+eGLuyuIzjzmi8qVlH8QH+rlH+0jAfWgD9X7XxpBH4gbTLtWDQqDGowokR+OPbtn+9irRt7SRY5J7ZGiyx2jIb2JPQduRwe4FfKfw3+KPw/+Kfny2eqxI0kKxJMTKslo24NveOMNJzjaRsYYJ47j2gav4j8IXllYeLIl+y3ybrO9ik822uoweHRl74P3Gwy91oA7K68J+HdYgNrq9ojxygB1blcH1HJxjriuIv8A4FeEtTjS5stOsx1cMYgFBQ/dYcDHYk8V6ba3MN+qPA28E9gOvfJPXiteFb3cNknU4KumUYE5B29OOvtQBxGgeBdJtkW3tEjt0b5AFQRkDO0ZA4HrjqB1r0Wz+H2nXmnTrJqENrMybI4mZRLLM4zGjISGB6nr2544NG6tr+/1WK7uJjDPcOTM6ZZXLNkFVxgbFGMDAOPXmp9U8O6q9lCLSWIxTweYsziPz9k5WPYdoLRhmYEISc7SzfKpoAw9N8KJeAQ2KLOUkZX2EMoWPAaQkfw5z8x49O9ad54HYt5EpjWWbnyhkOVK7t5U4JXb+OOeARWbY6V4i0iC3eJZInth5MskYYFmQljvZcdAQPQCq80/ijTEzpeo3Fs7E/cZl8sn37E9T2PfOKAI5/CGnx+WkwZ1ZQyKsYlYBvlUtn5QTycDofQ1zur/AAs1DxBrJivNPmtkLsytJKwErKmY225VASQAMD9ajvNR8Spci7NyxulVdrsgJACjbj1IGB+HFef31z431aX7HPqUkFsoBRof3ZTnGAPTHGM9ABjigBniX4dpol7HFZFDLDiQNDKrlGbqCyM2Dn1P4VJa+JL7T2XSvEZEtu33bkDGw4Aw2PX1H49q5+78M6xBau0eqXe5eVPmt2/H9MVytrPd3Mc4v7o3kdspBJwSAOoIHf296APStXs7OW2kkyrIFO7kEBcdc9h6c18O/tdfDy8uvC8Pi+15l0LyWbafnSKbhwSO4cg+1fTHhWx1PW9TltIroNpsRDxqRhShG4bgeWwCAB1OMAV7L4m8CWeseE9Q0vWYo5be9hltdpQNneCjlgfulQc+obHcUAfpJ+xR8a1+Pv7NXgzx7cTCbVVtRYanzlhfWf7qUt6F8CQezCvqwda/nV/4JTfFfX/hF8cvFX7KniN4/susXEtzbGaQptubSNt5hAVgxniCMASo2pnOcA/0VDrQA49KF6UHpQvSgAPSmU89KZQA5aVulItK3SgBR0pG6Uo6UjdKAGU9elMp69KAEanU1qdQA1qbTmptADx0oPUUDpQeooAWmt1p1NbrQA2pKjqSgAooooA//9P9/KKKKAEPSmU89KZQA9elKelIvSlPSgCOiiigAr8Rv+Ct/wCzVfXWl6T+1h8PIWh1zwqYYNYMIw7Wqv8A6PdHHUwOdrH+4wzwlftzWR4i8P6R4s8P6j4Y1+2S803VreW1uYXGVkhmUo6kehBNAH4Y/s5/FK0+Lfw+sfEUbqlyqiG7i3f6u4iA3jGeM8MvqCK9m123uVhaWx/dyKMqc49eDjsR1Ffm78LbHVf2P/2u/FX7P3iaVl0i8u/9Bkk+7IP9ZZy88fvIjsb/AGuO1friyaV4ptRcWQVnjVfNII6c/LgD26++O9AHkPhDUF8QFbC9QQXFuTDIo9VG4EHuCMYbn0rp7zwxHeMVZMY69enTjjFaEnhK80u8XV9OwjjBV4+cDOBuGcdj/kivZBYWsqxhyoaREZmY7VDP95QOQBxkDkjrzmgD5vfwFZWzrMtuAQMcEED6EdR79K6M+CoptLdybaTy0B8nzUMsgYHPlxjJJABLDqMH2z6wJ9HsjJPNdLJ5Q+aAuEMoA2lUZUIBfke/SvlLxn8RXt/iRZGELE2o3MsgTIYB1y6jkDIGMdOcdO1AHZ6f4kuPBkL2M8b3gmkQJPI5IRc5xIQM8YwD34DcV1f9hPc2ZumncRpIQ0iAMMlcplSynnB5yOR0zxXEahfx6mp8kBvNUlwfuKWzlVzyVA9c1laL421vwNOpebNhGTtkKLI1vkbScENuX0PVfcUAT614as1vZVm8m6AIHB3DJAP/ANbg4yCM96xLLSrnRpfP8P3s1mW5MW/dEc9cKxI/Kvo3Q4vCevwrqltPDausRceYxnW4wNxyV8vaHPHHIGcEVU1H4fSsLm5vLY2l1PHPcqImQwRbJNkcQOcEP0yDlegBNAHmei/ETxZokpOsWaahb53brf8AdyjHbYSVY+vzDvXpWk/GPwPOE/tIXWnSZ+ZZ4WQjPfK71A79fpmuDuvC+o2sksLgSCCTyGkQ5TzR1jXONxB4/Csg6ZcF3ChfNAZWy3QqfTPrQB6Zqnxi8FRIU0nUvtSxEJ+5OZN0nC9cE59QDjnOK9JsvFum6npMaTQ/MFzv3AON3X17/wBa+cbPw1IW37A8kuFBUZYtnGBjmuyt7PUIS1jahZLpWEZjDgEMGK+WSSF3Z7Z6ep4oAqapJd6dqravZSx7ZGdvMnDrDI55dTgEMMHBGQR2OcVz/gz4rweKvEXk3mZ7bTmCeQzFkEYZiRHyCy7sMD3IBIxU/iHQvEWqmd9b8w21qVT7OMrFAMDaDtwu8jG4kZY8nNcRd+Fl0q2t/EGlhUuIQxXaADhWwUOMBgw6emaAPpS/8Stp1g9vGY5FZdzNFuO0M2QNxAJAGOMDnOcDg89peo2PiG2kWd08448pm3ZJHbqPz/SvnjUfigNTtVsNDVbi5nGSN3yxjGCzZ9CcfXtU/g3RNeneJ9Q1WWdW+ZY0URxke2Mk4PfP4UAfVhsPItGieMlAPl5BOOoyDjB6joemc1+I3/BRPw9Z2fjvwhPaWgh1DUY5Ud1XDsI2jEYyOoG449K/bPw7Z36xxwzDdIw43MckHt69BX49/wDBQc28nx9+HOljEkKshJGUVw10iMcdVzt59O1AH3d+w747tvD3hbXtN1m6SGztLmN2U7jIDLGSGUKGOAYyDx3r2X4reJbfxtqT6hYiQWUdutvG0oCyPtJZ2IGcKeAoPPt6fB/wHvng1bxBHAuIp2iycnhQ78decg4Oc8V9ewQSXTCCMrz7nGO/egDmtItnsLgS25bKAKCM5A7kY/CtTxH8RfG9hNHotjq9xbwJFH5VuhhEUjyG4dzM0sMz8JCduzac45xmu2TT4rNMRFWYjH3uRj2/GuN1zwpY6nrVpqd0ZWntkKhUlIjbIkT5lBwxUSNtPvQB59qfxO8Zaay3Whtca1LLJbx2ltdtbZlM89rbFDst40WQPdKY3Y7CUPmja5CZlz8ZNZTTbHV9Z1xDb3t69lKYTJdpZotutzHJGY7CNTkZEjQxzoI/3qj5drddc/CnQmsrG3+13heycyRvHduJN4kilRjIuGzHJDGyEEFSoOaXTvg9qGqxQ3OoanfRw6bcfaLaJJ2jUTsrATxxw7EjfuWG0scFtxAIAPI5viJ4qkv20zW9VhtNPu7RpbC7h1AXFpc3cNxFG8Ml1a2rPaxrHMCEuIkfzSis6rIuen0C58Ra1pd1BqqhJY5MIFuob3fBLGkscgubcLDKGVsqyAAjGQGyK+gtN+C3hYW0UN68kl08bolwkhXbCx3SQiJcQ7HbDOuwq7AMwLKpHbxfDTwr4ftFiiJf7Q4Jkkdnd2VAoG4sOFRQoA4UKFUADFAHkXgeKaXwdqvw/kcQieOQxMwyBuJLHJBPcj2GPSsnXf2ZfCWo6bLL4ea4g1AIqo7yEgyEfL5i7TkZxkccele4z+CLGNJGidSdoHDFdu4ZwfmySR1HTBqT7b4i0Af2pc3Bv4VQKZHGXQLjBcAksmOCRyO3SgD8xvGfws8beF4p4bvTJp9v/LW2VpI8A8ngZA9cjFfDvxj8EXuo2f8Awm+lysLjSEUOiZ3BAxbeCORtJJz/ACr+kiy1zw/4ktoksVWGVkbMQfs33sN3HXkevI4r87P2lvgXJ4T1K78aeEoRcaDfKRfQRjItnl4fI4zE/rjAPHAxQB8B/D74jajr3hFNRNwVvYI2inKnBLoPvEf7Qwa+lf8AgkdZLN418e6s4y0dlbx5/wCukpJ/lX51aFeN8O/FWveF75/LtbiOTymbgZVS0Z/4EvH1xX6lf8Ek9Lax8K/ErxdMMQrNaW4Pr5Uckj8+2VoA+qfB/wAQV8V6nrmnXbRSX1pfzqYx8s7QtKdjbTyVA4yM4POMV6RH/YPiN0gnnxLah0SPf5gLcMyu5xjjvtPp614Zrnwus/HOmNq+myPYajFvdJ0G141fIOSCCVyRnnjqMVz3w28bf2Pqlr8NdVtCNS05DELon93Msa8yBuuSOSPmOe/WgD6d8KAWzi2uVRG06ZoxESiyrC3EbqoYlkLHAbB7g8gV5F4+i1+z1qC5toBZf6QyCW4dl278BWbcAEXP3jk8c+tew77SGa21jZHNdKrREgjequVb24ygODn/AB4P4yaPf654eu3iuxbDUEB8wyBnCMCc4IOQX+Xb8vB4NAHonhnxNrUnh67jtdltqKHzPPikU27naeVB4KM+TkcHt1rA8RahB4x8GDSPi94e0vxfZSqfNjMLKucdY/MBZWzwCNp78VF8LdKu4NOsvC0+qHUTbxCGIuEjZIlGFj+RR93GCd3vVbxlrcmn39rbxxF5buVFjSFdy7EYNuJ6DaOWyefTmgD86fix+wtFHLH8Rv2UtXk0+/2+d/YNzcjzOmStpdE7X9PLlOT2Y9K8K8FftD32kapL4A+NdlP4f1S3k8ieaaF0Mb55E0LYMZ7kgY78V+xd3Y3Y0+W70/y4Zr0kKxzsDLgMCgwfrjFeb/Er4N/BP4zaCPDvxmih07U7WIRWesQMUu7Zh0VpG37kJ5KSEoP4QtAHzx4V+Ib6TDGVulu7LjbNG+9dpPB3cZUjBBx+FfQGk/E7R5oFkM6leMFiv+Pf61+cPjX9lX9qH9nO9lvPh/A3j7wdKPNhubBDcoYzyC9upMkbY6lMqfU15FY/tBWMOorZeMNEuNHuISFlA3MAw67o2CsMntg4oA/Z+38eaLqLCO2nR+dpO9doI687s4966XTvGMptL/TLeVXV5Y2Ds2QqruI8sDpyevpn1r4d+F/xZ+E3ieO3txPZ3yBsmAFYp2z0BDYbAwBjH416fqslyt9Lq/gW3aztY1BktVk+0R4LYXDM26Pk8A7s+2aAPqOzv4PJIBLFWd+TjDSAbmzu5PGKvyyaXfWax6mrreJnYdwEYBIzuUAknAPPqRjPNfINh8ZRYyLZ6zbvaydwfmBHrxXdW/xJ0/ULeW5sRJcJHtEuwHC5J2k5xnODigD2K40iSS5eONg21wAwbDHPAHBJ6c+3Wreo+D9E0FYrzxBqlrbxyKrYjlWSRjt3mONMDdJgEbSRycdq8gs9a8QzzLJFpjJ5n7xPMmUNICu4MEBLEbcE1laloepeOdfstJ1mH+zEtpFkRzIzbFI+ZjgKcDqeuaAO51bUvBWu6w3h7wnHcyEKRLPJHmXEzIOYo2KoQMgZbLZ6CvGPG2jWnh2+lt4lWMWrspK87tpxwRkHP196+k/CPgLR/D8V3dW8qS+UqzMwO0yOOAQx5zkk89cdq+ePiIt1qWtR2yOHeV2mbdyNoYDB5H6HnFAFXwF4dljgk1XzltmJ3xmbcBI4JKqoVWJPIyeAuMEg9PcYPFj63pE2hTWptr21J85ZGOQy54QbOR0G4MQ+MhjmvMdFNxavDbrN5cNswwGJI3HqVOQQWbk16JOsF81rfyRRxTQxyLvQ4lcE4RiRtDFDyMjJPBOMCgD8z/jr/aXgX9qz4beKvhzJ5fi6a7smRF+60yXCxRZx1WQEofVRX9Y8JkMaGUAPgbgOmcc1/MN8GPD7fGX/AIKaeHNNlxc2Pg+UXUuB8o/s2Ezf+lDAV/T4OtADj0oXpQelC9KAA9KZTz0plADlpW6Ui0rdKAFHSkbpSjpSN0oAZT16Uynr0oARqdTWp1ADWptOam0APHSg9RQOlB6igBaa3WnU1utADakqOpKACiiigD//1P38ooooAQ9KZTz0plAD16Up6Ui9KU9KAI6KKKACnjpTKeOlAH4m/wDBYb4A3Wp+E/D37TPhCEpq/g6WO01KSIfP9jkkzbzEj/njMcZ9JPQVyH7O/wAWT4x8G6T4y08oY7+Hy7y33FQs8YxKgIOQQw3L+HHNftv478GaD8RPBmt+BPE9uLrSdfs5rK5jP8UU6FGx6EZyD2PNfzSfsqWer/AH9pzxb+yX8QpDHFdXcsdm7g7XniG+J49w4F1Bggj2FAH6W6z4subWwWaxkfzlC/aIGdd7P0+THQbcHkHnn3rzxvjXpEt2NIE94l82cWzRurbV4J/ukDPDA17/AKp4C0S30/xBciN0OnrEIXDAx5kxksT/AAnoOfWvhH4gXc9p4usm0VlN3akyMWGVCHIaNv8AeyeO1AHtmpan4i1JRa6aotI3wgdgZJBnjdgHb7968a1/4KateXia42qzy6nE6zRSuR8jqwIwAAAMjnHavV/DPxR0GCeNtaP2CdMn/SNpTg4wW+7z79a9Ev8A4kaF4ouoW0y2tbowrECtrkRyKuB+8ZSAoKDtgHGeTQBjeBV0zxHpUVneQeTdzP8AZ/nBXyZ9pGD0GN+35j/CSax/EfhPUdPuJbSRdssR2MDkgkeh6Ee9ZOvReJbXVG8Q6BZRRsSvm2gdgrwrk7AzE4cc7SfoeK978G/Ffwj400c6P4iLjULRFBjlI86CMgKN0WcnGDhhlT1zQB8eRzeLvB1zNe+GpWRW62k6tLak88gKQyE56qce1dd4a/aGtrWzksvGWn6jpuHMrvGrXkGSxY4ZBlV6dVOMDuM19W6p8NtLu7KO+029tmW4ywjlnjR9gzglSQQxx05/nXkGrfDi+sIjdvbOIMgCUYeJyem2RSVOR0weaAO38JfEbwl4w3XOj61Z6hvyTDKqOxZgCxdHGd5Kgsw+YkZzkmvRUSGdGTZaMJDywtIlfPXGVUbRkc4xgdMDr8MeJPhxpF1IH1KzLXJ4R48+YPxXrWVp2p/EnwQCmg65P5AOPIu1Djk/9NARnge9AH3XcaWZJ91lILWTaVDxKYwrBcKUAKjJJwSDnr3rhZ9Aks7W4iuYxNbo6u4Eroc9VXj5CcDpjOMkHJrxHSf2hNd0yJ4vF2kkqQMyW+I9xzzwxCkY9D/OuuHxz8G3lsGnlmslYE7ZoiqgkDJDDIzjjOecUAeuWF3qcaXnhyzd76LV0SGRWQ+YBC2c4DEbkHyhskFQB2GPJviX4p8OeGfCUsujQT3VysBMxlH7seXwCuPuKPvMc5PQcE1Z8P8AxZ0pLuNPDGsrO0ytGSjCSELglhICGUj0B6npzXkXxg1jVLnw3q9hpVk8gdFiE8WzaqAhZOASTuXcCR0HzUAYnwd8LRxeHba9vWEt1qJa4ZsfwuflUewH45NfXreEdR0y20qKO5tknKOyRTEqscbYJDMBx7bycHjpXi2m694A8BaZbL4n1eLS0toI3WEB5J5QoUBI405yc5ycDjrXmnij9oz4gfEjVrqx+E3h9raAnaswiEksSNgDLY2R5x1YnqfU0AfYWu6peeBdAOoeNNSsNMsm3FXa4Uu4YZUAAncxXptHPpX5F/tJ6xofxf8AjLofxJtbx4Y9DigiiV40jE9zDMXVgD/CflGMBjzmvfP+FRyfvPFHxi8QtI6KZZIxNuKooyS8r9FGOQo+hr84PFPiPSvi58cIdR8F6e2n+E9CeKKMITueGJjmZiT/AKyUkkdwMelAH6Wfsq6R/bWqeIDJ1SOFiT0539a+z7DQI9IjkadkaQk7fYdsj8f89vjP9mN5tGOq63IwhtrpkiRQ2GJQMxIz/CM4z68V952ms2N3Av2qaOK5eQIyuVUAMMAnPI/Hjg/WgDi5riNJ5CSFji5JwefXGOw/nXNaemqazdfYbFzJLLKRkjBUM33fwGP8muT+M3xo8M/DfWItB0u2XVbmVCbhUkwkSEDa2cHLFuccZHXtntP2fPiT8NvEcDBtZitdbRk8q0uWEE0rklmZWb5W57KSTn6UAekTeFk0CeIXLA5cl2dWXgDHTONpBz79fSremeIPDupTJplpcRuEJRgN2VYDIPXPbkV6L4+vrWfwzqOp60iWrWKNIWLANIE5AAHO4jn5eoBzzXxB4F19dW0+2nL/AOmNKZE2/LJ579RlcYP8gCBQB9d6z4l8PaNFHDcbzKSBDFGhLseQB2xuPGTwB7VW1G41WXT11O/jFjD92MFSNrY+YAt95iOMgfTivKL74tvpjrp8FtDJfWxYSXUxaMKFQM6qY7efb5ayRtJI/lom9cvk8cvc+OdMsYotavrO6u7m7RoVhm2QxQOqkSBjlmRI9rmRxk/K+MnhgD1W91G8KM0JD4yh2oFGcY5Pdu54qWw1CzuLJJby5YXltkuU+WNgP7uecc8g5r5xufiqmoWMt5M1rE4RpIjGZlg8mFJXlc5t0lXyhE25TCXYlNobeDVvTtcn8QapYaTo2mi51DzmtrllnCLbsu8syEI3mgIpfnZgPF0aRRQB63f2Wm2d7DqPhqV4VuCWaLBRS3rGf4GYZz2Y4Ar1Cxu7TW9LlgvolnhnRklUrnejAAgrjHIPzAcd68K1YzeD7pNN19THDckRImM+cV5KRrjc7dwqgkYzVXUPi3Jo9neX2gWsmv6bpywi7ki85vJmncpFF5sMM2JGZSWEuxU2ney7lyAfnL4u/ZY8NfED4v3/AMNNZ1d9A1GB5Y9PvUiE6Sj/AFkUciFlJDIeCGBDevSv0J8EfCvQf2QP2br/AMDwXkuoaprkk7yzKu12mnQIWCj7qxxqM8nnvzXmXx3uf+Ef+IHhT4s25WS40q9iiuyAAXWBxIjNj+IruU/hX1f8YdGh1fUtK1cTl7O8tXjUBvlJX94pX3YHsOcYoA4fw7quhWGkmae/eT7TF5Q2j5QJMZYkD5jjnA64rlNb+HdncFr7T7oIrbpEvIxidHxnzFI6HbyQQQemK8j06x1Hw340e4vIJY9DeLCRP9y0mccy+URgKRgMR0yTwK93sPFOjaVALee5hlF3hREHXdgdSEBLZGcnHTmgDg/h343kiit9I8ckw61NvLRyx+WLmNW2h4mjGzGMFk4Knk8GvTfiVo1p4i0UpFctFZXkTRq0DFGbgAqcZ4B6+5644rjvGGlJJHHqnhnM0sUiOkUrY5PDqzdlYZxgcGtTRboa3o8mnXkLWQZWZYGIJXDEOpdRhgeuR1oA6H4ZRWbGwhF60rxW+wyOAHYrHhiSoGWznn8+9aVva32s63fyX80MVtEobzsHaAoAb5RwAT0x96uQ+GGkL4VvhpksqiCN7l7cbfmjiZjxk5yQec9x7V1eueJArQWUNvI1uFMgMYCeWygY3liMlskeoHI4oAt+INVk0tilnbvILZHAiA2PJgZyXxhCTj26V4v4r8QzG/Gj2lnI11erj94uQq/7ZDEHGemDn2r1m3jtIUn1fVLkG0eQFY5DkIOjiMDlR3Yjk+5rxA61p1/8QYbkP9ssoVZlltI90SuzhQrbPlB4OCcHGSaAPR/hfquv+FIv7Ajv1S5hkAkiiDPEBgEHD5ChgR+Oa6bxzB8H/ifL/Y/xU8B2OuAoQ1w0KmeI9gGwsq/VXqEaZdaTrzXsDbrSdI3BJyU2Ej5ceu7r61r3lvNNdiG1UNFHkJnYQUydvI/P8aAPhvxx/wAE8P2cvGMl1e/CzxffeCb6FyPIvQbi0Vh0CmQpIBz1ErfSvn/xD+y/+2/8DoG1Lwrcp470G2+dTYTfbfkXkH7PKFmH/bPP1r9MvFNjFaSW2nXFphrybbJl9uDsZjkDPOBjGR1zWfeap4l0WbyPBd9JbXEce6QK26NFGM7k559scdT7gH42aT+0FDfa/wDZviHZSaJfRN5UuUfYhGchkb50OeoIPueK+4vh34r8M39iZ9KmiuYph8kkZDqSOgzXpvjvUfgr8YIf7I/aG8Gx3Goq3k/2zp8X2e8iGDtk8xMFwSDxyP8AZPb5d8TfsBfEXw3aD4jfsneMl8V6RKPMW0eRbe9A67CGxDKR3Vwh9qAPtjS/F+m30kZvPLnaOJU3LHHvCAcLkr27jOTWx9kgmmOq6K6iR5QGtyCqbMHJRsnDZx8uSCeeowfyEtvjl468F+JX8OfFXSbjw/rdsQkrSwvCw7AyQuOh7MvHpxX234G+MU97axuI/tCSKCkkRyrDthh/SgD63utai0q1capG9tNtDhXDHdnpsZchuPSvGdWmivtQN1ArIyhVBb7x5JLN1x1wB16Vzvif4vXtvapCqzKsx2hYgN7+oD9vfHb8Kw9P1zUtTQNBCIVPJH3iD7/T3oA9N0yx2Jv8wM5JxuHHPP8A9bNUvHPxX8J/DrwzcaxrtxHZRWi4XB3FpeSqRqMEs3Zc9OuBzXK+JPE2ieBPCF74q8Y3DfY7CPfhSv7yTOECpkZYthQO35183/An9nr4u/8ABQ74pJ4r8TxXHh34Y6bM4E6IRbxxow3W1qSArzMMB3AOO4wAAAfUf/BHXwZqvi34o/FH9oHWrWXZcJ9itriRCFea9nNxOEboSgRA2Om4V/QKOtcn4H8D+FPhv4V03wT4J02HSNF0mJYbe2gUIiqo68dWPVmPJOSea6wdaAHHpQvSvlXWPjx8RPEnjDxF4S+BPgWDxZD4PnFnqmo6hqY0y0+3bBI1pbEQzvLJGrLvJCopO3Oc49A8E/G/w5rnwz/4WT46gf4fQ2c01rqMGuulp9jureQxSIZpCsciFh+7kQ7XBBHoAD2o9KZXEQfFD4bXfhaDxva+KtLl8PXTrFFqK3sJtHkY7Qgm3bCxPGM5zxWDf/HT4K6XouleJNS8eaFaaVrpIsLqXUbdIbra21vJdnCvtbhtpODwcUAerrSt0r538TftG+B/BPxXTwB4w1Cw0TR5PDya4msXl9FBbsZbv7MkC78KSw+cMH5HbvXq2vfETwB4Z8MR+NfEXiTTtN8PzKjR6hcXcUVpIJBlCkzMEbcOVwTntQB2Y6UjdK8M8bfHPQPD1h4B1jwubbxNpfjvXrTRYLu0ukeBVulkbzkdA6yBTHjaCOe4xUnwY+Nmi/FrwppmtTiDSNV1WTUxDprXCyTvDpt49pJKikKzLlVLELhSwBPTIB7ZT16Vwd98Tvhvpdj/AGnqXinS7Sz8+4tfOlvYY4/tFoGaeLczAb4gjF1zlQDnGDWNbfHT4LXQ0U23jzQpf+EkYppm3UbY/bWVthWDD/vCH+Uhc/Nx14oA9UanU1qdQA1qbTmptADx0oPUUDpQeooAWmt1p1NbrQA2pKjqSgAooooA/9X9/KKKKAEPSmU89KZQA9elKelIvSlPSgCOiiigAp46UynjpQAHpX4b/wDBXH4Jazolz4R/a6+HcbW+s+F7iC11KWFcsqxv5lncNjskmY2J7MoPAr9yD0rC8R+HdD8XaDf+GPEtlFqWlapC9vc20yho5YpBtZWB7EUAfk58OPjf4f8Ajn8KLPxj4cvEmudTSNNUtgHE1jdRgGWJuMbd3MZycrg4Feeaz4OtxczXMUPzPklgMliepPXnjrXyP8X/AIa+Pv8AgmZ+0APEXhpbjVvhJ4ykKqMkq0Ibe1rKeguIPvRsf9Yn/Awv6beGNT8CfF3RdO8V+Cr6KRNYjR4zv2xlmAyCoB24PXnIPBAxkgHyvF4Dh1ucRTQqMD94OnJP64r2r4ffDmy8FKixwi3s52SWR9vdvlBwM7uSBx0/OvWPD/hc2N/DdrDuMP7xwy7SvY9c4I9+M1qeNNda+8Lx3IR41urSGeNZRslRd6ybZE5KyAAqy5BB546UAR6t4aFss9pLCEeLIwSMdOm7p2r5w8d/D2a8uoNT0wrp+p26ny7hWZJo8kk7WQHcGB5UkA4HvX13pnia11vSoo70jz9i4f5SrRhT1PGSe1Y2paVp1+U2PveQHKlgCcJnrj+Y7cUAfMXh74jeO/CaR2OoWj3NqxBkmtWCliuMM8DfK3ruBz6CvcPDXxt8M6tJPZzSWV5cyxiMxzoLaZVCleAyjccY74yOtZer+ELuygtbi5hO2dAyFQdpwcfgf5+vWvM9U8IafKGa5t0cHJzxxnPtQB7td6boGrXHmFWszkfONsikHq2QM+/v254rJ1v4bLMBLYtDqVrKSiSIMO2OxQ9PQDP618kvb+JfCl+8vhq6ZIwxIikYtFj/AHc4+mMV7N4Q+MU8bxR65u069QbvMi4jbHoSeD6g0AV9R+GMM8bxpa4jYHI2b1/LtjvXleqfCvTJZTFbR+UrcN5ZKqTn0969J8XftL/DfQbSezdJNa1IhmX7E4RVlJJPmudyMCNowOVx0rwywg+PHx5uDDodsPD2hvkmVi0ERU+smPMkJ6fKNvrigDZu9b+G/wAMrE2ryxS3YzmC2AkmJ9GPAHPOWNeQah448fePBLaeGbBrKwl+V5AMlgeDukbgfRRn3NfTug/sx+CvA8FvfeLGk17UbqNmi3bVtxMOg8sFiw7nd1GCOuK7bVPBwktVtraEQInARV2hO2AvG0e39aAPmv4dfBHw7qVnH4n8YXsmply26IMyR70x8jMMyOxJGAMD3wCR9Ii703w5YRaVpFhHp1tIpeOOJQibfu7ht+U52+vOPXr5BZeENY8Oag1pbzvLZXDmQxMT8rseo7EDnryAT2Ncv+0F47i+CvwxvfFH2oi/nPkafBn5WunBAO0k8KNzH2HrigD5O/bK+L9xr2rW/wAEvAjfadQvnT+0XibJBb7lvn3+9J6DA/vAP+AnwI1XXr6z8AeD7c3d1tM99cKOBgDzHJPRR91AevHc14f8DPBWs6pd/wDCVXMcmp+JfEsjCDILShZWO5sn+OQ8k9l78mv2I+EvxR+CX7LngebT9QnbX/G15KzahFYBZVyhwkQuM+Wsac55JJJOOlAHWeC/gZdeDreO+12CArZgeXZqN0ROMK0rHljzkAAgnrxXJfFbx5a+GLbUtZ8wSS2x8uKBieZT8qj8+T7CvD/Ef7SHxB+OPjW1tNPvR4Y0N3bctrJuZIFOWZ5iOWAGFKhRk4r5c/aC+LNjoVgsK3C3RhL+TGrl/MmPTLHlti/fbkZJwTxQB5R8TfjTq3h7UZP7H/07xjqsqSLLIiTpEjNyPKdWDvJ9xVIwFzxyMcd4U+NEN1fy2HjOI6PqqyZDhfJiWTOSNuB5eD0HAGOD0FepfsufA3WfF+s/8LZ8bwl3uyXsEkGN244M5HZQOEH49MGvqD46fsw+GvGfh+8u0tUj1yG1L2tzH8reYCdqSHA3A8degPGKAPbPhP8AHO5+Jml2nw98c36pLBFsimVQXvo+6O2cbivBxy4/GtDSdEg8M/ErTLtIRNpwM05iJ3Ru5UqEORzktycc96/J74F6/KdG1DwxfO1rrHhyYPEM7XMTPtYDvuikx+De1ftP8GLGbxJ4TtfH1+fOu5LdYrcEAl5/uSNzjCqQzEqCQQOKAOH8X3VnZeKNa0tY1tzfxXcLBo7hwG1G2s4mdPJgfmNrQsYsAsHUKThtsN3N4s1BrvTYvD19YXGnXkupm5eJpFJuTNc2u0KrlxbzyxtIiBwdjja4yrTeKEv7bxpca1rTZsbZmgDI1vbsl6YopYVaW5mijcMGceWvzEJkd69LtP2hNDs9Mht/ENhdRBrz+zorhRD814Ngkijt5JluGaNpFWQGLCdWIAJoA5DQPCEer6NrWv6rp9wdP0mxmWER2MRmkUDMVvFbPapDLnzbvzAYGjdZ0Vi7KdnnsfgbxRoTwa1HBcyXN0DNbTLbzi6aSX7S93InlRjyLie4+zNEzCLZDHGhIRJFPp3jH4/ajpHhp5vh5bfb79pZTCJpLS5SSOFPMuAJLW6eIGJSpZC6uFYEKdwy6/8Aizca1Hbyz2M9hHLDFdrJK9sHa3lUMtwIUuHn8k55cR7VGS+0KSADR+IGo6r4hvNK/tiwe6NlA1rcmFZH88tJE7RuIkkYLIIzk7NpKhXwrE14vpPhmW58Wiy1nTZZV1V9PZ3ntZfLnOn2l2okkaVA2ImeKOMyhXYIrgDNespq+tWl6s9ujbw65IXBPrj5hkn3r2kzNq9hF4hNszSqnkzJhdvzfdfqWG3kH6/kAfEXjpbrVrDW9D1lvNltoZZvMbJkkZVLozsfvEFcA+gwa+t/hpr0fjn9nTw9rM6G5utGjS3k2sA6taN5JbJ7+XhiO9eC/GrRbuxujqdpa5e7heJoifmPmLx7ZyCBz0Ndf+xTfS3/AMK/F2hLtdre7aRFYkDM8AHJHIGU7UAdh4m8LaNrelRz2kBY3O+GeGQnygu0KyoxHyqV3EjnB4Ga+Y/GHhNvAd+mveGtI+1wR2twlxaWc4smaFU37fNKuyoNhD+Xtcg/I6n5h9TGx8SW9wjmSA2zDfIcMFAHJ5Lvz6HHP6Vzeu+Gk8Ti4sTOot76CWF9hwUV1wWViDyBxgjn+YB4LJe6RLqeseH7Bbe00/TtZ1C0jtXgSe1tlmvrpwFhuI5Yn8uOFY0EkbqokzjcAVm0+Sawsb66ttSGi2t5Ha/vSzG0t2nt4jJ5aBhsDl22BCoQMgXhQK1LjwlJBeah59wtz58k7zvNbxs081zN9plDqVMYUOEZMKrqRwwBOc6DwZFd6q+r2uqXErhFWe2YrDCdsZiQKIQqxFUKhFVV27E2kYoAb4furv7LbXFtGsYv7lUuIrG3Fm0tgxd+DHHF55uI4mMUm+aVNpVp2Morlb25F5E+iavHby2Mfy3CmH+z7TzI2UXdwkMsUPlRxJNBAgEShrqYyoN0TMffNO8N6dod+l7fSTpdACUt5VuIvOIYbj5KoS212C5G1dzEKCTXGXHhTRNN0xprnUJtSuIgY5JXhgnaSHeXMTQ+WUcNKTMflOZDvyDjABpeLviDqENvFLJbGG3aK7aR5UWQ2Q0+KWW4jWJHXzGRYXRRuVS+PmC/NXjekamuq6p9v1WBprfUrPV2jjlt7bzU1C0t4DGJo3Wa3mgWO4jkTcn3zhkOzLe522k2WqWK211cyx/Z5HmG/aZGklRxK0gZWR/M8xg4IKMHKsjA4GEngPT7nT7Se3u5NNuLbzVQCCBIRHLkSRrGiAZkH3nxvJVRuCqoABf1jQrS00PwzY3lwqX+j6TC0MwRZNt9DqGmwqV3tEEWRmKFvl2o7HBHymrfeOvG73ssEE8P22ws0e1jMCDzpzNPB5cjtLtjCG3ZhIPvKV+VDkL6ZrHhG+1y1t7i3uzAskU9rLtgguIpbed4pGR1mR1GWhTkDI7EHkM074SeHfNstUuL+cvGipcwoIQksfzmNNqouxQZnfCYJLMWLMSxAPIL34l+Jb7wfrl/4mSK2EWmK0ltN5MLt9qaK23pAl1NcIqSTo6tIrK64yIyyirWg+Ide8H6BceH5taEtlpN/qVoy/YIwfIspbdnuAEmiQsWucCFUVdowrpjnudV+EsviS11bw/p2pXVpFqTCKYmC0aWSOOWOVTLL5OQd8asWBG48tuYlqiufhTpOlprFpP4gutV1C+F7LNiOKNIpdSMQlkiwo3ZMSHDAKuDwc0Ac8/hK3v9N+16zBDJdXJkjnZSNolikeJiityU3IdvoAAeevm2na/4r+FniISeC7l7WVpAjIceRMDg/NH0YDkE9e4INd6NU8U6VM/h5NS86xzNOWu1UOJriUvMAFAUKSd2BjHOK8t8YW87zZ0u9W5nB+eeLcyIp7IxXHXPrjBzQB9K+ONN+AP7WOlr8PfizYx6f4ot4QYLuPEc9vI/AMFxyMMSD5UmQ3oSOPyj+LX7Pfx1/Yv1l9Xty3iXwJNJhL6AMIgrH7s6DcYJCOO6N6noPrzQ9DsNOVJL++jOXMpa4lHMhAy20ZbPQAtiu+1n4xahoKX3huy1H+09Ouf9G8t0S5tpflCsPKkLIUc5xkDIIyM0AfIXw78feF/iRaHVLa586+tCHa2nAWSAkbcqo4284yOOlem3Pjjwh4WDz69qtpp4GMh5VRzx6A5P4A1zFx+xHovxn1WLXfglr0HgjW3Y/wBo6bcGVbcI2d0tm0eXA/vQngZ4YDAr67+Fv/BHH4SpbJrvxY8f6jr2NpeOzSOwhLYyys7maTg8dVJ7UAfn54Q8G+P/ANvv42Wfwy8BebZ+CNGkWfUL9lPlwwA7WuHzjMjjKwRnnnPA3Ef1OfDf4e+FPhP4F0X4d+CrNbHRdBtktreMddqDlmPd3OWZu7EmvFPhH8Pvg1+zf4UbwN8FdAWys5ZTNM+5nkuJiMb5ZnJkkIHA7AcDAr0sWnjHxN8105s7Zuxygx/u/eP40AdfqfivR9Myjy+dKP4I/mP4noPzq1oGsrrlobtYjEVcrtPPToc/SsjTPBOkWOHuAbuUd3+7+C/45rsI0SMBEUKo6ADAoA+BvA3jmL9l7xR8RfBnxI0LWX0zX/EV94i0bVNN0y61O3vIdSCO8DG1jkMc8MgZdrgbl2kHFeWfHKf40/E7QPhn8VvFmh6h4N8P6V4g1Sea1s9Li1rULGwliMWmX15p06TK0o+bzFVGaLzARhgcfqexHTvSKyn5cjPpQB+O0Xwyk8RfDnVri0tNe8TaT4o+I3hK4nbVNDh0iG8ht7qBLu5i0+3jjKQlBiaSSJN+0scjJrq/jt4P1jwl+0V4j8R61fap4f8ACut6FY2ukT6T4VtfEdufJMpu7FkktrhrdnkfzMAKsu75iSor9XyR0phZQQCcZoA/HrwRoHh/4Q/Ez4ba78TfBHiHWPD+k/Dyazhub7RjqFxp9xc6nJJHDPb2aTJE5gby1VAfLQhCFBIrnLr4ZfErwx4c+FHi7V4dX8J+DdP1DxTcxW9rosOtXGgxard+dphm06aKfYBBujykZaDdtyoJr9qgyg7SRk9qc3SgD8lPBngHV4NA8Gavoltr+o2OpfFiy1Yy6lpMWlboFtGilvIrC2RBbW8jrkmSOMs5LkfNk1/hPJrHwc1j4S/EPxr4d1hNFt7TxvpVw9rptzdzW9zfawlzaiWCFGlVZkibY23BOOec1+ug6U1iOnegD8kvAngbVvGH/CrZvFPg+8Syf4q+K9WuLPUbNs28Esd7Jby3CMCqjeU2k/LuxgnipPGXwYtYfhF+09NpXgvbq154wFxpZisD9okjjSwlSS12puKrKZWBj4DFz1zX6zBgehzinBlHykjJ7UAQwbvs0O7Odi5z1zirNMZlyFzz6U+gBrU2nNTaAHjpQeooHSg9RQAtNbrTqa3WgBtSVHUlABRRRQB//9b9/KKKKAEPSmU89KZQA9elKelIvSlPSgCOiiigAp46UynjpQAHpTKeelMoA8s+NHwc8EfHn4cav8MviBZi70rVY8bhjzIJhzHPE38MkbcqfwOQSK/BDXf+Ce37c37NU09/8AvEsXi7RI5GlW0tpRFKRnq1nc/ui2OpRyTX9IfakoA/mr8Gft9fEv4aeKV8GftO+Dbrw9fOnlPdtbywyBR8oaS2mB3p2LIxHoDX1be/Fvwp4u0xdTt/EdnaaNfRlkvRLEwK4P3UZ0GSfl5I28kjjB/UD44fs9/Cb9onwlJ4Q+KuhRapbYJgnHyXVrIRjzIJh8yN+h6MCOK/KW2/4IifDZL2d774navLZtIxhiisoI5EjJ+VWkZnDMBwWCKCedo6UAZsfx7+EvgeyWzTxxp0kEagBJ76GWUDOcAx54Hpyfeq+n/tc/Be+uTBc+LbCJRjaxmG3Pc87c/QivbdD/4IzfssaaVfVdY8Tasw6iW8t4kP0EVsrD/vqsT4if8ABK39ifwdZR67r+v6h4X0qHPnNcagC8noImkHDe2xyfSgDQ0T9on4Qak8MV7420m9s9oTyjqUEZHzA7kO4EEAEDPqa9d1LTPB97oq6vba/Y2yOis63DtBCpYE71lbcCrcdSMe9fjZ8S/2af2U7vWF8N/AAeMvEeoSyBUmuZ7byXOekVulmJ3z6lkx6GpPjD8OvjX+zZ4C0J/GPgvWl8KnBhZ5zLbW248rId0vkSN1Cuq+3pQB9f8Aj/41eCNKuZNN8MxHXb1WMe6I4tQ3T5Xxuf2wMehri7X4U/Gv4mmOfxBbtoOlvhljljMWVPQrD99vYuRWp+zb8T/2eNb0ZdW8KXFlp+sQJuuF1W4SO/i2j5iPM4K+8Xy+uDkV7vrX7ZPwS0G6uBrvjaze7hjYcs115m44IDIHBx94ehAxQBL8Mv2f/Cng68h1E2o1TU7ZgyTXSCRAwHaIjaPUcE+9fQ82bWN7s3y6e6l2dgpU7eCfug9+AOg/Dj89ta/4KL/BLTo3SxvbzUXAyPs1k53Hjgm4aIHGMDgDFeJ61/wUp8ONLPNpPhXUr9WAA+0TQWy9T2jWUAc+hoA/WzS9FTVbh9dnuXvZbMJbfvkdRlFG0RkbcleO2Prit+SyspFFxfTpbi5J3ySuNhkBBPON2CT79fy+A/hx+19feLvh9pWraVojw3V2ZZJI5LnzER/MK8sEVmxtHXHsKk8R/Fbx1rto0ImTSoT2s1KMfbzHLMMD0xxQB9ZeIZ/DH9oyRafZxMPIjQNK7l1kBO6QEMg+fPOQcADA7n8N/wBp/wCIN78d/wBoS38Bb2h0XwzK1jDEV2GaVMefIq92kKhU/wBkA96+kfEGta1Elw0GsXkcl0himxcy/vYyQSjfNyMgEjuRzXyD8RvBWvXms2XjTwTLNB4jsJY5RLC5SXfG2UlVwQQ6EAg5z+QoA+g/EuiwfCLR4NT+I+oL4RSWH/RrFsnU7iIrgeXZqRIqEcbpTGhHQnpXw748+KXiHxJiPRYJ9K0a5LRxZz5k+3AbMgAHcZVeBnBzXsfhz4M6l4g8QL4g+Jt/NrWr6lMGme5lZwXc5LSyOS8jfjjPHNfVf7VP7NkurfDSw8W+BLYJe+CoTutrcYV7PhmKIvG+MjdxyVznkCgDxuXxVpPwj+H1lpk9yI557WJZthBldQoxEmPX7zn3AOMGsP8AZ7+BniL9pbxpJ8QfGFu9v4M0iTaqsCFuHXlbeP1UdZWH06nj2L9l39h7xJ8ZNXsviT+0PNPY6E53Qaa5MV1dqgyPMAwYIT7AM3bAIJ/b9Ph74V8N2NnoejabHpunWMSolpbFIY1gXlfLAUAP2x3zzk80AfP/AIJ8BabHcQWjutpEuEBGFAGPkCj7oAxjAx+tdL4w0XSLi2trACKKEI4uJQSSH3lNmAQMHZuz6N1PfU1nQlsZftGkXEsCvu/d3EecN/CpPToQcnHUV4zr+ua9p/mLcpDPGnB27g27njB/75PUcUAfj1+0n4Q1D4L/ABjt/Humx7dL1x3WXaQVeQBfPX/gSsrj3r9IP2ffipeaN8PpNLRnZtOuG+zhQDlZwXPLA7cEt065NfFP7ed9Nf8AhzwuphRFtbydXkUctK8SllPsu3AxjOM19DfsmaB/wm2sxaBIX8qaGC4kKMQRGiFn/McfjQB9FWfgz4n/ABTg1LXdMs7qwjnuXMeoR/ZHga2lgjhlh8ueaJt25MiQZAwRtfJA6Ff2dvFN8ur2+t6bDfWouRdRI80b290JpIZJIMMMsUWMoTKqLIRz8rHHt+u/EzVPDOsTeGrDQbO50nR4Q+DdzxSKkUSyu5gt7G4/dxo43OWBBOAMsM8nqHx0e71axtdJ0maVJbiSz+yPLKqm7hm8iaKJobWWOXEvyCSaSJSw6KgL0AeDar8LfiaNM02xstIm1IWR1dnkzp9nGo1C2it0SO3gl2oEeLc2GbIbPBAWsO58LfETRdFXTr/RZBLHD9m8sSQlif7FbTfOB342ea27Gd23nGeK9q/4X7farp0+oaDpduVu3hGnCS5cGeGe/ubHzJyIcxMGtmYqocYYDIPTy1viz4x8SWOkx/2baQJrlmt1vinuJBaLIkTIs7CzGZXMqKkcPmMWZBtIYkAHoNv4htbidjd74JVbgkAYz0II6V6z4J1+G3v102edT58bKEcg5DcHbx1P+FfJ8t9q8dw+nNDaefZwNcSPPPcKrQi5jtAqhLNpmm85yhQxJ90kZUq9ap8aTW0c872lvPb6PHby3EsgupoYxd+b5MkZis3uECm3lSUyRIEdQoLlk3AHtnxs0R7jQ7iEMZZIFOyZcgnIBX6EdPwryD9hC+Yap4x0ORh5c0VvL5eOpVnQ8+mGxivWNNFv4v8AA8Hi/QgZLbV4ys8UkryhJEJVWiaQksjEErgAYwQBmvnH9lTX7Lwh8b9Y0jU2+zrqAmtk3A8OsuVH09z9elAH0jq3gjxlo1xjwlqUOo+GbOXa9pJGZ7iGNGImjhmLKQxGQocNtPAOK898aXVhYXlna2Vzd2Fj5bSxiJ57VldpoIozMU2uyoZSXQN8w4BBwR9x33h/RGu5LuZ5LKe45M0LCJyVB7gYbGPuvuB9OteP/ETwvos9vaT6p5OoHTpRLH5gkhOVYEBhbSRK67lVirIVJAJBxQB8VeItYntrHU9R0S4mMlnYXMvmPcXEqC5tYLudWVHcq0MotcFZvMlG9gJQFUVDDNq2g6ZqurXcgiv9JurSC5isHu791815IZo5PtCi3NyCuYvKhRNynlo812FxbXOqXN14d8N6Bp1zDdSSXtwktzebfuvG5Zmn3CIrMy+SPkO8jadxz7HeeBNJudEtINctm1Gcx5aNb+9ksl8ohvLFrJO2Y1KhTGwKfLnbjFAHy5feLtQ+2wsNQW78OXU1zb3Mlo2pzXcE/wBnluIFurW4X7VCFETNO0EhHlhwAjKM9x8Om1HVrh7GWS1v7GSNLnT5NPnuLlGBae3d43ulS4Qh4yDFLuwQcMVK7dvVv7Lub2WzuIrhriKVRBJLd3MstosZXJtZJpGeIMQDlSO2OlJaajHpd9KNLW5nunZTJd3lw13dMkTHyg0khJb5f4TlckkDFAHo1xoWqoLUTGP7Ha7sYQK5BGW3P1LfUH2AFNltmhRZ5Gki46Egx7fXp9ecf41yl14l8QXFzuuL6Z0GWwXbgt0I244Iz/SqTave3Mg+3ILjb8gZmw232b+L2zmgDuZ/EkmnQsIEMb2o4AP/AD0HGRjvjGR6Ypg8eNcaeZ2h8yd1LFM7Czhjk52kHP0HtXE3FlIyi5spCySYCMhzIhPGMAELg9D3/GsTVdG8QwadJb2twD1wJCj7gxyylxhgD0BByO1AHvVt4utFsQtrcmM3Bw0Tcv5mduAy9c9BVZ5o5p1urlijhW25HIViOvqT+Iz16Cvn/TtdnbWNPtLq3nsYbp1+aQqymQcrGCmcH06ZIA617XcTWS2T3VzckrDwWTBbaeOwPQ44x35oA8F8Z6jqFp4kdl0yPUdOiRTIXfAVnPsCeBgA8jOfWqt54xaSzOnrDJZxOQkcK7HQyMSQA6hQOSeoxzXV6bremC8uW1eE5v5GS1lQ/wAIyoLRlXLE/KTgggkDIGa0bvw3LrGn3N3dW6QMwC44wHXqecgEDBJJ6n0oA8G1axmaUxQKrzSt8xLfdGehGAPr+lSRfDaHWLZmv7lLW4Td8saHzQRxkkkAjPTAB969v8JeHNGfU911N9oW/jZInuQJIYWkZkIZSMcEYBPANNu9G0nTrh7K6VhEirJDMByGPID98dQSD19sigD588LHxB4I8TW5t5bi3dZ1Md2g+XYD9/cScHGcg5yK/ZT4XarD4+sbCczbo5mZGkj6O0YwzKD0yRX5ma1EiWscrKqRqVJCcnrg4x9cfj9K/Qj9lNZB4P0DzAd3mXXUY4EkgH6UAfY2meH9J0sZtYBvH8bfM35n+lbdNWnUAR0o60lKOtAHyB4i0LWNJ/a+8Fa7f+Ir3ULXVdG8QeTYSMkdlZxwCywI0QDc7MzM0jlm5AGAMV85fDKxPwF1vw78TvH+iaN4nufGkuu3CazoF1d3+omVkudRIVCfKnjeFGiURKChCL82c194Xni/4K6h8VdP0G71zS5fH+kw3ENraG6T7bFFdKjzIIt2fnWNWIIJwAeleafDe3/ZI034p3lj8NbzQT44je8jeztLsSz2z7y12sNvvZYCXBMojRckfNQB8x/sx/FzU/H/AO1RrGt+LdR1Ean4q8LRXEOjzWV7b22kpFeymO1XzoY1LrCA0sv3XlZlUkACus+NGhaP4x8V/HjXPF80kd98PPDmn3Hh2ZZ5Im052tbi6N3AEYASPcIFL4yQgQ8ZB+uviBrXwW+H+rr4z+I+qaboGoazaHR0ur65W3ae2VmlMKlmGQrOWJHIzyRXmfxB8NfslR6z4UtPiPJocOpfZLa10mO8vPLe5s4WX7PGwMg+0RB8FBLvXccjknIB4J8QvCGj+PNX0zT/AAxFe6x8ZvEcOjalNqRuJFj8J2qRxF5dyFUgR9khS3wXnd23Apkr9VftJ+K/iL4K+D3iXXPhxpaajqVppd/K07Xa2z2flWzutxGrRyCVlYZCcZ9aNd/Zh+BfifxRf+NNa8LpPrOqSRzXNytzcxNK8ahELCOVV+VVAHHQV6B46+Gvgv4ladbaZ4xsWv7W0cyRKtxNBhiu05MLoTx2JIoAp/CjXPGviLwRp+p+PNHj0XU5Io/3cV2LwSoY0IlLiOMKzEnKY49a+TdL8OeM9B+I/wC0Dpdr4zubzXbrwtpdza6lqUiRQ2VxMmpCMoqhY4IYtqngcYLMSSTX0V8Ptb+BPgnXbj4OeBNd0+DW45JJ5dJGoG5u1kCLvyssjyAhQCVzwOcCtLxrrnwV+G2q3PiXx/qel6DfeLIYrCaXUJ0i+2w2wfZFtkbDKglfOBgBvmoA+PP2d5v+Ed+MXg/w14a8H6l4Qs9V8I3lzrQvbuO5XULy2mtFjvAY55xKSZJMXBKmUP3C8dD8efAXhfxV8Rbrw74ES91r4tazPp93HqX2iQReE7CAoonDoVSFHEbskHL3EjtuBTJX1Kfw9+yT8FbOXQb+40XwvF4wtkgC3F8Y3ubMH5I4nkl3pbqW+VYysak8AVd+JHwT/ZhsJtY+MXxL06z0/wC0tBNf6rcXtxbRMwCQQtIyyqnQIi8egoA+Q/iydc8K+JvH/jKXQb3XPGNp4p0kaJ4ltL2JrWxtJJbOJNOdRPvic73SS28k+d5oY8HI+2fjB408e2fjXwR8MPh7fWei6h4t/tCeTUb22N2kMOmxo5ijhEkYaSUyDktwqscE4xkeEtG/Za+LPjmb4h+CX0bxL4k02SCeea0uPOaOaNfLgnlgV9hlVRtSVkLADAbitj9oH4Oaz8XtN0W20i60pTpNw87W2sWL3dvNvQoGWS3mt7mCRMnDRSgMCyurA8AEn7NXxE8V/FD4WReKfGz2cmrpqer2Mr6erLasLC/mtUaMMzHBWMHJJzXvleTfBD4U2HwU+GelfDywnjuVsWuZ5ZYoRbRNPdzvcS+VCCwjjDyEIm5iqgAkkZPVeH/H/gnxXcpZ+GtctNTnkt/tapbzLIxtxK0HmgA/c81GTPTcpHUUAdkOlB6igdKD1FAC01utOprdaAG1JUdSUAFFFFAH/9f9/KKKKAEPSmU89KZQA9elfE/7d2rNpPwu8JmW61K2sLzxn4etb8aRJcR3s1lNc7Z4YvspEzM6ZAVPmJwBzivtheleQ/GL4Vf8LXsPC1l/aP8AZv8AwjXiTSPEG7yvN87+y5xP5ONy7fMxjdzjrg0AfCPwl1Tw/eftD+H7D9nufxja6VoMd2/jW38TT6h9ljtJrdjZgQao7Tee0wVkaNQNgbJra0z/AIKK+GdQ1+wvxHoB8JalqselxRx67E/iFVmn+zR3b6aI8CMuQxjEpkWM7iOCK+u/G/wWj8RfFfwp8YPDup/2JrehxzWGogQ+bHquk3Ay9pMAyYKSYkifnY2eCCa8a8J/sqeNvBU1h4U8PfEJLT4f6XqP223s00mD+1lgExnFj/aJc/uNx258nzNny7u9AGd4K/a81/xv8Ybr4faZ4b0w2NnrV1o1xbtrCR6/ai2LqL6bTpY0zbSMmV8uR22srYPSuo/ZE8bfFnxr/wALSf4nxW4XS/GOqWNm0V21yYlgKq1soaKMLFCAvlt1fcxIB68/rv7I3iTxb4803WvF3ja31fQ9F12PXLJptIiOvQmGf7RHZrqok3i3VvlwI9xj+TPevcvhB8Jtb+Fuv+PZ312LU9D8X65ca7a232UxT2dxe4NwjzeYwlQlV2fIpUZznNAHlfi39oz4mHWvHMvwt8BW/iTw18NZWt9YurnUfslzc3EMK3FxDYQiKRXaGNxkyOgZvlHrXkvw58TeL/jL+2M/i2WBn8F6Z4V0bVtHC6tdQrFBqy3TRzyWKIsMs02NrrIWEYRSpJOB694v/Zr8eXGt+Nv+FafEP/hFPDnxIkM+t2T6cl5PHPLCtvcTWM5kTyXmiUBt6SAN8wGeK9I+HPwK0n4Z+O73xToN639my+HdF8O21kyZaCHRTP5bmUsd5dZgCNoxtzk54APCvih40+Lui/thaX4c+F2kx+IZLzwLNO9pfag9lp0DJqQBuJNqSkyEYjTahPzckKDUv/DWnjPxDa+AtJ8B+BobjxX4xvdb0q6sb+/8iDTL/QTi6WSeOKTzI8htrKuW+Xjk49I+JnwM8f8AiL4wWPxp+G3jmPwtq9joT6F9nuNOF/bTxS3P2hnkUyxHg427SCGGSSCVNLwV+y9Z+CtX+HetQ+IJr+78GXGu399NPCvmapfa+ubiZirBYgJCWCgMAML2zQB5L8Zv2zfGXwZutXvvEXhfQbPSvD0tvFPa3niGKHWdQDhPNm0+yEbF41LkR+YyNJtJ2rWj4+/aQb4a+NfiNqXh/wALXGu6jZXPguyWBtRkVLo68zQxiKJw0Vu0e7krxIcFiMZrF8c/sLaz4uX4haLZeN7PTtE+IF7dajPMdDgn1mOW6IfyDqDybmtldQAgQME+QMBzXp/ij9lN/EviHXPEM3iURSa3f+D75kFrkI3hSUS7QfN5FwRjP8H+1QB2fwo+L/jrxJ8R/E3wn+J/hi08Pa/oNjZapE+n3zX1rPZ3zyxr87xQssiPEwYbcHqDXnnxt/Y10D45fFK08d+IfEF3Z6bHapDPYw/M0kkZOGjdyViDLgMAvJGepJr3jTfhgNP+NeufGD+0N/8AbOi2Oj/Y/Lx5f2KeebzfM3c7vOxt2jGM5Oa9a7UAeW/Db4L/AAy+ElgLHwFoNvprEYefbvuZf9+ZsufpnHtXo2qaXpmt6fcaRrNpFfWN2hjmgnRZIpEbgq6MCCD3BFW6koA/H341f8Eefgx8QPGK+Jvhvr1z4AtLpmN7p8EAurclu9uHdDFnuuWX0A6VpeDf+CNv7MGg+XJ4p1PXfE0q43CS5S1iYj/ZhQNj/gdfrc3Wm0AfIXhH9gf9kDwTGg0j4YaVcOoH7y+R75zjvm4aTmviv/gqJ44+DXwI+Acnwh8G+E9Hs/E3xCAgiitLKCF7eygkV5bg+UqnJZVSP1JJ52mv2SbpxX8m3xJ1rxP8b/28vFVx8Z0+xahoN5cQWemSEtHHFp7Yt4UJ4KhP3xOPnOT3oA9K+AvhG+8BfC7S7DXSUvJg9y0ZGDEJW3BD7gdfyrv7vU7e8tVisd73W/awCnYAuQWZjwCeMBffPIrQvoZ9VuPJRtkQI3MM8+1dhoHhhCYwVCB+5HA/CgDzey+G91r0m66dzkdvlH4V3WjfCy10yVreMAbACxxuIJ9e/Pb1r2e2sRbBbWz4ZlO5+6r3x6H+ldzpugAQD90SSCS7ZJY+pNAHzbr/AMNbe4sBJDEReRYkjdsjleVBAz1x0NfUfw31Kx8QaZa3CARs43FGGCjqfnU+mCMY6djSTaSs0DRFRnH1zjrXl5XxD4G1xde0UNd2MrBrm1U/dH8UkfqcdR0P1oA+v0vLaxlBObhm/eOqkFh2U4z6kZ/Wuyt9Th1ENfqzW4xsXgElScueDyDgADtk8V5rout2fijRba+0aVHju41Zd3yZAHQZA5wMfNgj0rt4ZdOg0ddNEa7g29pHH3jtILPjngHA4xyO+KAOc1rWYnkujFuMSqdmF3AqxOckA5Gc8Z9CBXzh41vba2t5TGhWPJYKXV1IBJXgnIIHYjr1Ar2fxHq0SQSx2kahYTjManb83cDoPXnmvnLxndPcWbiPAIHJAxnjqeOp9f6UAfm9+2tAsngTSrlOfL1FAc9Rvhfv+FfXP/BPCU3HxGtTFavcGXQzIkiuEWEhYysjjOWHO3aO5HbNfK/7XL2958JQyIBJb39s545AZXU5/MV79/wTi1ua2+IHht4GQfatHNs29towBGpA9ztwKAP0U17wBeXOveINcTVbaKy1HSbzTNm1jJHc3UcMZdzkL5a+SDj7xzXm1x+z9qN5o/nXGrwOL24u55U/fRhHvbh55Fi2sGaPexIw0UmS3z7SFX6v1hbHSjfpqix2y7iyoOg4JDZx168c++OM1NJiYwCGX50Kh1d8ZIb0UE4zx2/+uAfCcfwv1DwdBp2g+FEjuLXSEWKBb4zMSkc01wol2H53SWeWRGBXG4BkcKDWt4P+E2sWHhO10u61C1e+szAsTxLLhoLeGCJQ2DuEnmwRyggkK6LkMuVb6x8Q6ro2nIsHlmcqvMSqXYnJOTtHBzXkc58X6zKx02IWaLnDyDe+CMZAXJB9/wAxQBzlp8GtWTU9O1x72EyxEC63CQkxLdx3o2MDzI0qtuYqowwCqoUCvPpvgN4km8RXni6e4+zzo4htJraWeznhh86eYMsytkF/OIkUKwbYhBVlFfWHhbSru2iefVJPPlDbmLMDjjAAxwFOfTPp6Va8Ra1Z2sT28k6xK20E4OAvXaBzuJPTjk0AYXgfR7TSdKn0m7uBfSXsk11I3llYjLPIZCVGXICs2Mklj3JJJr8/f2iPDlx4B+Jdt410tQEmlHm7F2p5qdQAegK5XHtX6D+GzdK7bAZIy6BQSP3OTk5AyM44PPGPXiuW/aI+HEfjzwPcyw4SSJWW3z0LROTkHH9/PpwSBQBt/DT4jad8SNBGh3lyFv5YA9vLnJlTaPnXP8a55HX9a+SfiVr3i7SfEsvhXV0b7ckgSMINwl8wgIyZ7N/9Y9DXzj4H+IOr+DkSwd3tr/RLjzod2Qysh+ZMexyD6gkV90fG7TdL8d6Z4A+MWnyyQ6eXjN3LAwSRInUyw/N2KzqEJ7FqAOa+H3hT+xZpr/W5o5759sh25Koi9A2RwV3dcYJJrp9c8UaRolzZpPOkP2qcjyXPmSykg/LFGvzysSQFUA9eBTLPVLa5sJJ7SIW6oecliZu7AknBIAz6E9a8e8WeLfDXijWdPtbRI9TuLJL7YIMrM8NxbfZpolYJJtYrMZGIVyEhcmNhwQDsb3V9NvtcgSSa58kl4TBBA/nLPjcITAFE24J8xUL05I2mo2uPD9/5iWc5byFfzmZQhikB5WZeCjDurcjvg15XHbwnxF9gNhKbE6XAr22TIfs8eo/vgxWKIMkm1ZRiJAIDtC+WgY9HpkeivZ6loXjHUP7JS+tY4vJkYLN+7gyCN67t0dmIVww4xgrnIABoJr+n28CTGQ2kbo0sE00MkMM0KjLvHJIqpIAuGJUkAc9Ko3+u6RYwzXd7PLElvkSJJbypKvy+ZzEU8wDZlwdv3fm+7zXG6pp9ymu3Okw77e8kvLa4c6ihjkluYC8sfnDAEk8rt8yjHlxbozhnKR5M9tfLcSqILoxx291DHDE/9o3Eltb3F1dz7WKRB5bm4klgX5OIIJsht+SAe0aZqklrLazWUpiilIYHbtzn1yB1HY/j3qxLqWkXD+RMxtrqOQL5CI8rzuf4Yo0DMWOD8gB7+lcZqLahJax2bA3BS8ubNjEWh+0vavLHHGpU7o/tMqJF8r7l8z5WBAauQuA+ia1YaxDbqXm07xDC0F1BN5EiDTC6zfZ7hmKMrmSLcMBhuUgsrkgHp9/rXhm5sbZBiGz1JjFG7ROzSEttKsm0lBuOHL4APBwaz5/COuRQLpfh2/8AsukQgmSG4RvIXnIEZYZRF+YsE4Pakj0FNM0zTPDelWb6pbWekTLbxfPu2wxxneBAY3Jjx5gVCCxGO9cfq99qEhu7OeWSZ49OY26ztdLHGZtRhgW48kyhv3kG5RGx2fxqoVwKANnw0L+cz6t4omS5MQCO9uh2wQRuzhAT2GWYnHfnpXqdt4o0u+0Y3Gjl5IrlWij3K+SysQwCMoYN8uDkCvnnwzoup6T4N1N74SyyWGi2d2LqK2uVhb7SbUZmu57tvOeaGZt6LCYw27yyPL3VJYaFoGhadrVrpGlwwnSLzVGt1drkxC3s4rWXyBHHKigO8xbfjcuPlIJJoA938G6lPod2fEi232iySZTMgABZHPDgfXkg9ehrqdWPn6X9rgdZJ7ciSNVHMaudsoYMNvzdcA8d8VV8JWxk0/UfDoUH7PdXMNv5jgma3trp4RvYjcXAQBuSzdcc1kNfW+kXl/o9luNpOztE7ff2K3CuO/bjtQB5JrdsYWS40wM63UiqYiMs55IwOeSVHTvX6/fBXw2ui6ZaWYXjTrWOI/8AXRh8x/PP51+Z3w78O/8ACU/EjSrQh3g026+1TDnaAFEgz+Kgf8Cr9ffA9l9l0NZmGGuWL/h0H8qAOxWnU1adQBHSjrSUo60AfkWL2MfE77QNdtBcj4pNc/8ACD+Uh1cSF/s/9oeeD9o8ry/9M27PK8r5N+OK9D+AmuaZ4N8XeAvDPw6+IQ8cw67daydc0y4sbW3vNLT/AEi6lupVjUXNs63RETrcO28yfLjAr9Jjpunfbf7T+yxfbNuzzti+bt/u78Zx7ZpLfTdOtrie8t7WKK4ucebIqKryY6bmAyfxoA+LvHXjv4ZaT8f9P8dePda01PB2seB7uDS767ljNlNKLsSXkcUrfIzyQ+UdoO51U4Bwa+B2tbyz+HuqaX4k8RW3gi6Hw+0O2/snVYVkvPEdqvnzQ21qZSJIXRXFtiEPIHYEqCq5/ca40nSbu2jtbmzgmghYNGjxqyIy9CoIwCO2KLjT9NvZYbm7toZ5bY7ondFZo29VJBKn6UAeS+PfEt5ovwmttbj8Q6b4Ankgts3GuqJbeAyIMwPunt8yH7oPmdR0NeS/sVa/rGq/BrSNO1nxpo/iq602ytYmg00KZ7BirZju5Fubje7Y4OE6Hg9vru4s7O/hMF7BHcxEg7JFDrkdDg5FRWem6ZpyuNOtYbUORu8pFTOPXaBQB+T3wbu5fD958MtJ1HxNp/iHxWPFeoJq3haSytxeabO8t011qRkA+1pLHne80jGKRHCqACle4/HPxv4b8OfFnw78U9H8Q+G57fWPCuoWUY1+eSGwe0WeOV57K6jjljlmyQsluMNKm3aRtNfeUenact2+pR20QupVCPMEXzGUdAWxkj2zTLjSdKuYYbe4s4ZYrdg0aPGrKjDoVBGAR6igD8mYvHvgzwX8I/Bnwb1+40TQPiV4x8IWtrq+q6/JFaxaXojmQRb453DSShZGENsv8QzIQq8/R/7U1romhfsYfY/C+rxSaRp8fhuOy1GV1niNvFfWYiuHfcFddoDk7gCO4619oXei6Lfzedf2FvcS4xuliR2wO2WBNXJLKxltPsM8Eb22AvlMoKYHQbTxgdqAPgD4O6+2u/tUtqMnjLRviXLN4UnhbUfD8CW1vpaRXkTCG5EUtwsj3JbMZaQMojbCYJNO8ZeOrRfi343Tx78TdT8E6n4c1XSoPDmkWci4vbO4ht3LpYbS199oneWFiA2wLxtKk199Wun6dpqmPT7aK1VzkiJFQE+4UCln0/TZ7qG/ubaKS5gyI5XRS6Z67WIyM+1AH5S/DD4q/FzX/ilol5rXi6ys9evvE95p2o6DPrU0s62cUsytbDREsf3HlwqsiXHmgHAZ5Cr4ry/w1rGt6F4TGoaB4kj8KXI8JWiNfStJFEIn8U3iyRvcRJI1ssqkobjaRFnccAZH7UDTdNW9bVEtYRduu1pwi+YV9C+Mke2aT+zdO8tovssWx0MbLsXBQkkqRjpkk46UAfL37IXji98YeB9estQvLzUZtB1eWz+0XGow6xAVaGKUJa6jCqC6iTfjc48xWyjcrX1ieoqtZWVnp9slpYQR20EfCxxKERfoqgAVZPUUALTW606mt1oAbUlR1JQAUUUUAf/Q/fyiiigBD0plPPSmUAPXpXIeO/HnhX4beGrrxb4yvlsNOtcKWIZ3kkc4SKKNAXkkc/KiICzHgAmuvXpVDVbVbvT54jEJX2OUBAOH2kAjPQ89aAPnXUv2rfhXpHgLwL47v5LpB8RE0+TStOWNXvmj1F4o0eRFcoiRmZfMcvtBIAJYqDc+Lf7QN38Ip72bUPhz4k1vR7COJ31PTxpxtWaXAEaC4vYZmfcQgUR5ZiAuSRXj8fwt8bD9jn4a+Azokn/CUaND4Rju7X5POhFheWktypbOMRrGzMAT04zXufxF8MeIPGXxX+HmmyWbyeEdDe91u/lyPKfULVY4tPhcZycNNJOOMBolPUCgDq/HXxR0j4feEbHxRrOn3s1zqstta2el28aSX9xe3X+rtkQuI/M67izhFCszMFBNefn9pPQ4PDi6he+FNetfEUmrHQ4vD0lvB/aU1+IvtG2JhObR4vI/emYXHlBAcuGBWuZ/aO8Hal8SrHQpV8M6jq1l4G8SW99d2MEq28uqWT2csMv2VhKm8J9o+ZGZC+x0GcjPzg3wP8QXkdj4ru/A+sP4A0fxXc3tl4RNyBqEWlXWlraSssfn4EbXm6X7L5v+rZuOdlAH6DfDn4gWnxH8PSa1BpV/odxbXM1ndWOpRLFc29xA2HRtjPG46FXjd0YEEMa6DS/Eeh61qGq6Vpd4lxd6JOltexrndBM8STKje5jdW47Gvn/4A+FvEXhn4ZeJrPW9K1HSNJvdR1C40XR5bjzNQsdLkRRFbh1kby3Zw7xoJD5YdVDDHHjv7OvgQ+Gfi34m1ybw14406LXbwXNpNrN9PLaRxJZQwMt2r3swllLxtsZkYgFQCMDAB9P/ABi+N/gP4H6RpeqeNriQPrmoW2mWNtbqJLi4uLmQINqEqNqBt0jEgKo9SAee8UfHtNE+I9x8M9A8E694sv8AT4bOe9uNMWy+zWi3zOIhK1zdQOW2xs5CI2Fwe4FfM/7UP7Ovxt8aa9r/AMQvCOuafrDXJ0az0zTLjTpJZ9Otbe+t57jyJRdRxjzJI/OnYpvdEWMEACpPjT8LNV1zxF4rXTfhldXXxB16HTv7G8Y6fcbLaC7hiSMXMpeYPZ/ZnUkxor+bGAMuSRQB9/69r2jeFtDvvEniG7jsNM0yB7i5uJTtSKKNdzMx9ABXzppX7VHhrxZ4P8LeK/h94T1/xVJ4vivrmzsbO3gjuEtdPl8maadrqeGCIbyoRWl8xtwATIYBvxo+Gvj/AON2naTomiapF4dsvDmrLNfW2rWBu7TWvsyI8DFYbiJjbrKd+1mG5kAZdo5+XvDkP7UPwy+Bvhj4Vjw5qM1zq99rEmqanoNnD9r0vT3undI4I7i52G5uS7MkpO2JCPkZwMgH6HfD3x7oHxM8G6Z458MtJ/Z2qRs6LOhimjaNjHJHIh+68bqyMMnBBwSOa1dK8WeFteme20PWLPUJozIrJb3EcrKYiFkBCMSCpYBvQkZ614PNoNrbfsqax4Z8JWFz8P4ovDuo2trHqzrFPYEQyoJrmVHkAJP71pN5PO4nOa+Yf2eotAu/2l/DVx4Y8Ap4Ki0/4eyW90UNni5f7XahJF+xySK8fDeXK+DIM44WgD9NakqOpKAGt1ptObrTaAHHoK/ny/4K3/BbUPhz8SPCX7WfgiDyhdzRWOqlBwLyAE28j47TRBo2/wBwDvX9Bp6CvGv2gPg5ofx++D3if4Ta/tSHXrRo4piN32e5T54JgPWORVb3Ax3oA/Ff4f3+k+KfCmj+J9NcSWuowLPwM4ZuWUn1Vsg+4r2rQLBWw5UZbnA7+gH86/M7wZ4o8d/se/EO9/Z6+PFibOxtpy0F0uWjjWU5E0T/AMdvJ97IGVOcjOQP1M0W9sZtOjv7KVZYXjWWGVSDHIpG5SpHBBHIwaAOks9ImmceQnzoTnjIORn0/Tqa9Ait72OAW5hEc3C9SenP3QOf6VV8ManZxwZfAAO8k9eRwPbk/hXVabq8S6h9rnZC0gyGbjYq5yGxjkfhnp2oAr2dg4It7yEhnwcqvAB4OM8A45/Os+98NxLHI0gbaQWXC7uQOQMcn6YzjtXfC7W8jZ7OVljZl255wvB2jcSDznPp+VaMsluLdo9TAZCyMCuAVY9JAc8ADt3/AEoA+fH0+78G3zXenqFtp2zcQjk7j/GuQMEfxetdnbeIrbUbIywNvDZViDtZWX1A9O39e3nvjzxwmnXKWNyWnZ02wxRrud9uMgAnJz6+/pXlQ8M+Lr++OsWt9JpHmAHyosMQBz82RgnGfagD1LxTqNraNIEBXcOuSxJz1bkY9Mjt2rxXxJepNCyxktxtzjuO/wBO34Vtan4Q8WXefN1W4lDjgBI1zjjPCZ5PFefN4F1GeWS3upZ7ksSBl2yRySOCBjr9KAPkL9p1Vm+FWsqxUNG9s4XvxKue/vVn9iXxLYeHPE/w61jV4zJZfalhcdMbpmjVsnj5Xwfwr2r4tfCD+2vA+t+G/L+zSXlu3lyPxtlTEiAk54LKAee9fAfwN8T3s2jTeEbt/KudAkeSFcYdUkfLc9fkk/mKAP6cPiO9y2oPLa2fmyqGOZOIweoOP73pyc+1czp+qRXdrbSyl1kUHzMIWK/gGHXHAz9as+DPF8fxO+EvhjxqnzT31qsdyVGdtxEDHKDjp86E+wI9a5O/e5sbOVYvkJblySCxGRjI6e54/WgDe12JL6HdE0ccXVy4IbAORhQR16df/r8ldeJbe2ytrHuPIzGO4IOFAP8AP6e9czY6Nq+szwrNcu0eHIjRS8jpkkhFXn15JHua7ez0uztIDbQ2ptJo/wDWQIwaUbugklORGCPTkdjQBm6drt2zol7tVG5Csf3v444A/Wob21tLmd55pDEVDl3B45IOFz0BPU/QL1OMW6traC/WPG2McmCIsBgd2diXbHrkZ9K3ZLKHXjHFBEzw2+G3Nnyw44xgEZP14oA0tO1K1dVSWJUtIUKZXqX7njGeOOT2OK9D0/VrKWy8uQCdvmCrgMhD9cKeSQOpyAD2r511RzpFz9muHxFBuMhLDy9xIODnPABH4/Wuw8P6nrmuTQ4kOlWUablfaFkkXuVByFViD1HPt1oA+Hv2qfhinh3xG/i7RojDHOUF3DgbonYYVyBkfN0PPXB717D+yXr0PxC+FHib4QarIPOsldrfdziG5yRgHtHKM/8AAhX1n8UPA+meO/h3faVcuGCxyGLH3mdgdx4568nJPT6V+ff7GlheaL8ctT0m7GyW30+6ikHT5kljH5elAHW6Tqc+rWV5psp+w3Fi0lnJGABJbzQqVYMuRx3DE454zyK8l+Evh7QNX0i+n8TahYpHZaibdBcuqR70XcrqWZSd7E4x3U+lXPG0VwnxX8ZXlqQwGr3okTo23zWw3ykMVzwffFZba5DoNncapcraRXk1vqHkSzIo/fPZTRoqqwKsxYgrgEhsEcmgD3ka78LI4bzR7f7BssroLJKyI8JmRRvkWX7p5yu/cCcY5HJxr6HwHqOlQyL9knktrWc2sDRwZeOZCZViAJ3ltzBiAc5J9j5ZY3VteXVxqF5cG/1iz09rb7TFfR2csVyL8sA93PHN5cot/MSNpcOUHlhwSoqbTob+fVrvVNRsIrKzguLCe0nN9bT/AGuSzjdZ3+0Fba3aRmljaRFOHIkO52DmgD2fw2/gCH7C2oXWm3Os24RkU3MZU3AUhyoLDkcgccE+1aVvpPw0TQGisE09ba1l827jQxqsZI/du5B+VhyDnpjFeARNomtQ+JPB9rcw36X1tppm/s+YKZlt9IUSqskZUlPNJjYZ2F8BsnFdbql5pN29g1pLbPFpuoWsjmOeKSKK2W7yzLErsbS2EGFaKRYvm8vKF0ZgAeoahF4Tm0UtaahYz6TaBFeGJ4niWEfeWRFyAuDx6is7WND8D3Whx6ha2NneQGUMDGiujupKAZXIJwxUAnGDjpXkuv32pQWvha9gWC8ur6XTNQjguZTbwi2uYNUFyY2VH8iBZIrYgqnlRSJFIVG0k+ifD2PzrnUJWurq7W/gQSx3l3baibeWJyI1+12gRZy4ckF/3iKoB+UpQB6RqfhDR/EOmWA1DToCtpeFUFyqyBVY/wCzkA4PBBODjriuU8Z+DfDdjawC00+1FxAWEcoVUKSOQWkDHnJ2g568D2r1pnicPow23AdCzMGO0SEZ/A9vX9K8P+Iohm1KGz1R/Ot1DSEnI+QHDK3+8cYx6HvigDgtJ0Xw5JrDab4l0aC9tX/1n7sSgZJLEBsqm8sM4BBx09e9tPAvhMQ6lNpOn20N2JRB5kMY8gpJtaSJ8D7shHzAjkYzSeErSO61S41fUSZHuxsJRTIFjA/djaT8wjGFLHk+/Ndnos+keFoZ9MtwIoriUzKzNnzXY7SYgRyMAcc49ulAHZ6lbeFtGmsbfSbCfSLOIyIsKbECzsHxIoX5fL8zAJ3EsATjJrxTxJOVuJpGyXinDDAAyXboR0wc/wBK6vVtXt7rT4ot0stzbmNgzc+jg4PQYxj9aXwP4OuPGvje00263Jb2bLc3THkvGhDKjdhv46c0AfUH7OXw4urXT11G/jC6hrj+a5xgxW/Yc+vX8hX35DDHbQJbwrtSMBVHoBXD+BNFXT9OF66hZLgDaP7sY6D8etd6elACLTqatOoAjpR1pKUdaAHHpSfwmlPSk/hNAH5+/DLXvFelfsC+I/EPheaV/EFlpvimazkDFpRPFdXhjZTydykArwegrxO80qLwtp/iPwfpdxpE1heWvhjVLpdG1Ga3sNRtJb8xPY3EtxK6i7u1+X7UzxrcLxIIxzX2D+xe0afs4aG0pAQXesli3QD+07nOc9qXQviV+zM/gPxfqHhDStPufDNtqi6TfwafpkbQ6pfzrFsjhhjTF0ZDMiq2CGOcHaM0Acl8BPEOsw+A/ipb6Jpo0K007V7+Dw3pv2mO58jytOgke3ieJ3iOy5Mm5IndEbcqsQteJ+BIvhhpsXh+A67JYeHfFXgCO98XXdteSh3vZbu0SK4uZEZmjlmeW4jkc4bbuBIC8fc3w21n4ceIfDKX3h7w+vh+28J3E9t9iurBLKXTJkT96FjA2x7o3zujJVlbqQTXGfCvxr8EPGl7q+jeDvCyaKdftP7UlFzpSWUes2MrGP7Wp2gXEbE8l/mwwJXDAkA5r9mBW0G6+JngmBbSOPRdbSa2t9NneXSLWG7s4ZI7e2dxuQgqZJ024V5Cyjawqx8HfH/xj8Q/FLxro/jWz0KLRbe8gFubLWZ7ySJfsED7bSN9PgE0bSFjI5dSjllAbbk+n/BXxV8I/GXg+8k+DVtZxeGtP1C804rY2yW1o9xbttmMaIqqyMTw4GG6jI5rR8LeAvg1omv3tz4I8NaFp2t6SwguZLCxtoLm3aeNZNjvEiuu+N1bGeVI7UAfBnjqXVv+Fw+KvH3iFtPuZvDXjLQ9OsbY3FxHri2dwtmsQsCrrHHE7zSNJGYpBcL5isy7Rj0/9pazv/HHxQ8OeFGvNEHh6Pw/rV8BrckxsJNQtZ7aNvkglh3TRRs21jJ+53M4VmHHsXib4j/Bux+KFuNd8NPda1pV3a6V/wAJAdKWWCxu77b5Fsb4rvRpDKo+XKguAxBbFbPxm1r4DfDnwSt/8WdM0ttGF4ZrezmsobgzXszlmaG3KndKzMzuwGRlmYgZNAHwLql1b6tb6P8AHG/m0jV08MeGPC7w+GtZe8udYzJJI2be4EkKedcM4EUhin81kVWVCa+l/wBrjTjrniv4PaU2laTrMV1rGob7LXbhrXT5QumzsPOdIZzlSMqPLPzAdOtd58UfiP8AC/wZ4r0+/wDF/wAOtS1W80l7aGx1eDQ0u44ZLplSKO3uT8ysXcLtTndwK6r42+JPhtpY8NaH8QPBz+Np9eu5YtNsE06DUX8+GBppGCXBCpiNWy2c9qAPL/2WNQvNM+GXjfSXMKXPhrX9XhWGzna70q2G1LmO30+dwrPbxLIq4ZVKyb02qAAPDbj9pD45eDfAGi+LNcvrHxFdeL/COn67BHFYx2qabcXV5ZWjtmS4RJIlS88w+bLGoZOXRCdv3J8KfE/gDxX4OceBNNXRtM0y5uNPuNNe0WyayubdsTQSW4AVCCcnGVIIIJByegt9N+H99btZ2trplxBY2n2Jo0SB0hs3APkFQMLCwUHZwpAHHFAHxx4J+PvxVt/EmjeG/Hj21vaS+JbfTJrq6k0z7U1tc6ZdXXlzppt5dwwuJoY9jFkLo4GzILHFg/aJ+K2uJfa/oGo2dzp2g6N4u1028FoszamND1SW0tLdJAx2LLEoDOoLE4I7ivsSHwZ8Fl8GnQIND0AeFZpDKbVba1+wPKnzF/LC+UWG3JOMjHtXVaVoXg3TLeLUNC06wtIFhkVJbaGKNBDK3myBWQAbHf52wcE/MeeaAPhHWfjr8XfDwi0DTPGOjeKrzWbLw9fw6nb2CiGwfVdUgsZInijmIkikjlZ4Nzq/yNlm6iv4w+Nnx48OtqfhOz1m01K+8Pa/f6fcXdva2Salc2sGnWd9E8Vjd3UEMgia6K3HlyeYUVSi5LEfbOh+FfhNpVlqEPhzSNEtLQXCXN4trb20cf2iIiRJZggA8xCAys3IIyDU2qeEvhh4zshFq+jaPrlpqU/2sCe3t7mOe4RAnnYZWDyBFC7uSFAGcCgCz8OvFMHjjwF4e8Y2063Mes2FtdiVYngV/OjDFhFIS6Ak8KxJHTJ6121YtvqGh24ttOtbm2i3qEghR0XKqCAI1B6AKeAOx9K2qACiiigD/9H9/KKKKAEPSmU89KZQA9eleH/Hv4rah8JPC+l6rplrZyT6xqttpYutSna102x+0Bz9ovJlVikQ2bBx80jouRnNe4L0rwT48fDXxH4+g8Kar4Zi0/Urnwnqw1JtJ1Z3TT9RX7PLBsldI5SjxmUSxMY3AdRle4ANX4F/E+8+LXgMeKr+zgtZ4b29sWks5jcWN0bOZoftNpMyoZIJduUYj1HOMnyTTf2pbrxd8SvHPgT4deFZfEUHhTRmvbK5SZYl1a/juWtpYLdn+Xyo5BsaYkjcrgA7edz4b/CDxn4c8G634P1s2Wm2XjS+1W8v4tHuZoV0aO+iWOKDTWMQ3YZS7uViAdmZU5xXkujfsnePvhL4svPFPwX8WzXI0/widC0W11y63W8V357vH56wW3NvGrb1AyxkHPBzQB7v8M/ih491X4ja18K/iVpGm2ms6Zplpqyz6PdS3NsIbuSSIQzedHG6TK0ZI6h1+YYxiu48RfGDwL4W8a6H4C1fUYYtS10XRTM0SpB9kjWRhNucFCysNnHNeU/s4/Dr4m/DmPU7Xx9pOjLcapi6vdXtNUudR1HUr44UyXJntLZVULwiodqDCqoFcf8AFL4G+KvF3xe0/wAf2Hw48GavbaQ0+2bUb14ri/FzDHGWu4xpc4LQlf3f71uADkdAAfUHxB8YXHgnwpdeIdP0S88R3SbVgsrBA0s0j8KCzFUjTu8jkKo5PpWH8IfiDN8SPhH4W+Jmp2qabJr+l2+oywIxdITNGJGUMcEhc9cV6Jc2zz6XJaKqxvJCUAH3VJXGBx0H0rxj4U+BfGnw8+FHgX4ZTmwnOjaOmn6ldLI7bJYYAkbW8bR4lUvncHKfL2PSgDwT4Oftc6j8Q9H1nxxrsvhiDw7pekXestZ6dqc1zrUVtBlozNbvCkYyg+fa52sQpr1P4Y/GP4ha14v0Xwr8TfDthoreLtGk1zSXsLuS5KxQNEJba58yKMCZFnjbchKt8w4xk+dN+z/8TPiHJpWlfFaPQdJ07w3omr6PFcaEZPN1FtWthaNI8DQwpaxKmXMCPKDJtwwCDPb/AAw+GXxaTxt4c8V/Fd9IhXwToU2iWC6VNNOb17loPOu5hNFF5OUt0CxKZMFmy54oA5Lx/wDtHeOtA+JXjzwdoR8KWFj4GsrC7La9qM1lPefbIHmZYtqso27NucHkjivojQvinol38ItM+L/ixT4X0y70uDU7pL07WtEljDlH4ySpOBgZJxgZOK+a/G/wE8e3/wAXvHnjjT/B3g7xhpvjG00y3gPiCeVZ7NrKB4nxGtjcBlcuDgSKTjt1Hod/8BfEE/7MOm/BA68t3rmlWmneVqFwjtDJd6ZcRXcQkUkv5LSQhCMlvL9TQBvp+0p8FtR8O3+rT6rKIbWeKynsbiwuo755bpS0MS2MkQuJPOQMybYyGUMegOPHJvib8KPhjpGi6r+z94Y0i0tfFdzq0N8YLI2EsVxpOn3F75dxAEikWQNFsKSAFQ2QORmt8S/gf8b/AIw/2b4o8YQ6BZ6j4d1OC7sdGsr++iguIVt5re4FxqkMMNyrv526LbCVj24O7e2M+0/ZY8Y/2HpXlw6Ro18NU1zULy3gvL68QDUdIl023BursPLPIpKGRysQ2jCp8oyAe0eE/wBqH4Z6vD4Z0zXNQ+xa9rttpjTRRW9xLaWt5qkKSwW0t0sZhjkk8xdiu4ZsrxyM/S9fnB/wyt8W9O17w/eeG7jStHu7KDQIp9bsNRvrS4VdLjgjuYrjT1ja11ESCJliklMbKrAEfKM/o/QA1utNpzdabQA49BSDrSnoKQdaAPmD9qT9k34X/tV+CJfDfjW0W21i3Rv7N1eJB9rspTyMN1eMn78ZOGHo2CP55rHxJ8W/2FPiHP8AAz482ct14aLM9jexhpE+zsSPOtHP3oW/jj+8jdgcg/1d15Z8VPgr8KfjbpEWg/FXwxZeJbK2YvCt1HuaF2GC0bjDoSOpUjNAH4peAf2o/hBrIWyh8W2ZDYEf2ljA4z0DCQL09fzr6QtvEseq2UFzo063Nlt6qyukpPHBHQcZ56n6Vu+Pf+CQX7J3ivzZvDSav4RnfJX7FeGaIH/cullOPYMK+SvEX/BHP4t+D5nvvgl8X0DrykV7HPYN7Ay2zyg/98CgD7Q07xLcS27x3bII92BnKsMDt/iKZrvjCDTNPa8+0ybVbOzzCyneeFXIyeTgAk/zr83dS+C//BU34KBnXSW8a6fB/HbNbaoWA9FBW7/8drz2b9tb4meBtVis/jt8KLiwuYmyQyXGnvkcA+VdIwbB7bgDQB+nvhbwvb6zezeJvENwtvJKHS3XAZ4wgyETg43HBY9+gr1ORtK0RItJe3jl8xBIkwGJs88uefVsL9DxX5y6J/wUQ+CviVAmrSXvh2YgDE9rmEY7D7MX4HGMqPevctJ+OPwt8d2oXwx4x028uH5jjFyiS7znAMTlX5OAfloA+j9Qv9H8ryoC8T4OPKOVOcDJUgrx24/rXF6hHazozuFVkPDA/N8vPzdznJzjPHHaua0nxVaX8ypJMLa4jyGiZgC3umSA6+hXOO/NdDcvA0UjMWaVjlSeFC9MH1P+RQBwXiWO0v4WRsM0md/fJY88fp/Ovxy+Lehn4K/HO18UwoV0XWy3nADjbJ8lwPqARIPev2O8Q6l4f0y0eXWb63tbeEEl5pViVBjksXYdMf4V+Uv7V/xa+EPjPRn8N+Hr99a1O3mEsM9umbeJ84YGRsBgVyPkyOnPFAH6k/sT/EVtN1DWfgP4huMQ6gHvtLcEYMwTMiIfSWLDr24OOSK+x9e0+H7R9gt49nGNpOVQY+8Rxk+lfhV8E/EfjTVfAfhjxXpthfw+JvBBCiZbWQrLYWrK1vc7gOVi3CJyeMbeTzj9m/hv+038HPjekel61KPDfiKeILJBeMFinlIwwgkHysG7KcN2APWgClFqP2dHttGl8mxgEkVxe7iskjKSCI88HA6nGwdACwJrFg8Sz6rN9ltU2WDsJAE4aXDclmYE/ieTXuPij4ZC70yPTrNkksQQSkXAVUXAX0AGAAPwHFcHfeE9trLGqNbsThW/iJX7uOmBjn+maAJNF0m3m3TrHuhGQ28jrkcKAMn8APaun1CO28P6f9ruiIUhXcxY9mBI47ZwT16dazvBwhsLWeV08qOxYBncZLFRghc/KzbiB1xmvnL4ga14p+JfiGPRri2lt7JZBHbW5Qh9xP8ArmyNvC9M/wA8YAPR9Qsob23j8STK6i5UtFFKAqx4Jy64/ibHVycDGMd+Fi8TvY3ijTF+13MgJAbPlqccnbgb+RgZ47gVwvxX+KmlfDjSo/CS3D6hd28YVYxMS0pOSGkb7yoM/Vj06ZHnvg/4L/tC/GTTk1zUtXbw9o1+S8STvLH5kbdGSGMZKnsXYbuuT1oA+1YfiboWk6b5vjbxJY6GNv3PMRJSueyEu+cdlH0r4h8O/GH4d+A/2gm8Z6ZM+oaLfwvZ3NwiMmzcwAnCkAsMKu4ADuR6V7j4c/Yh+H1g8dx4s8QXutucMy26rBG4B5H8b+3DZz1roviD+zb8LbHQZE8IaHHZXC7gvmO8kkgH3smQsw+Uk8dMEnpQBx/xW+Ey6T4jX4x+CmGraNq0hu7lC3mJG85DmVCP+WbnnjoT6Hh+j6La6xoZkurUW94ybl5UKjOeGIxuwevTrXH/AAr+N9z8Kpbfwv4x33nhWaZreWVgZDYlzgOfWHP3/TOexB+r/Enhl7S1/tHw0Y7jSblfOUpIqgCXuJB95GBGMHGKAPn/AFbwZHotpG00zraXCs7NFJ5bhpPmabb1Llsnce/qa2LDxglvbSWDyyooyikkI20thDncCuf4lzjrgVvXC+QbhBOBHC80UsoU3O98kfu8cAgDCtnryOKw9Y8HXlzbT6rqVjc2kbIkm0gHYi/dYnpkkjJHAB64oA0YrOyvoPMuI0t7eVFMkgwGAY4JQEFmYnjjk579a5bSPC0Md7e6jMogSfCx7VB2CLO3PGMkNyfoM8Ct/Ro7qE2dxqccn2K5UmOU7o9rZCK2z0Y/QE4IHAr0aGK1e7+zQ7pkhJJt498e0sCCHZsYfgZPUjGBigDxZ7O802ePdcG2t443dYRIzAFt5KMpBwC3zBOnOf4hV4arrE9lbQ6QkL3UHM8s+TMXByPkG0KvOOhPHWvRYvDnmajM11C8txGjbUQBYol7eYZGUxxg5y3XOdxHWuF8arZyTJb2oLPauC86LiPd2EZHDr2JHBHTPBoA4TUfjf4d8M32oeGtYs7m51W3YLmEIUaRkH7sPnAVWOMgZHeuC1G38ReKdQTVdRTybW4BzFEGaNVjwUU7sk9OOnPPUCs/X/B1vL47ttUt4QscRSWUhmxAu5V3sRxgEjBPU++K+ltA8JQ6fMkV25db2GORH3qSGcEkEE7shuD/ACyQKAOL8PXE1vfS6YY1aWaNPKEe1FUR5ZnYkYI/vYx07mrkvjeCxu7bX0SNpISxSF4g8cRb/nirA7Nxy2BgAtXbeLorObRraO2CWd5aM8UpiIj85MgpuAwG/iByOe/t5PHZte3cUenIZLtVKxKh+Z5jjYAOOpbB7gCgCvqGqLOt1qFyRHdSzN5yA5EbAgIme4AxgA9MYr6+/Zt8KG80tdaZGV9ckGzdyRbQ5AP4/MR7YrsPhD8DdPsdDisbu1trrUEBlurqWJX/AHshyVQkZwOg9hmvqDwx4Ph8OyNKJFkOwIiqmxUHfAyaAOxjRIkWOMYVAAB6AVKelNHWnHpQAi06mrTqAI6UdaSlHWgBx6Un8JpT0pP4TQB8p/sXKr/s46EjgMrXmsgg8gg6nc1wXjL4FeKtWsfG7R6Mbq1fxvpviGz0+3ultGv9Ps7G0t5YY5EdREx2SBVdkBKgEhWzVj9njxivw8/YtuvHzwfaR4cg8S6j5WceZ9kvbyXbntnbivTPhR40+JK+PJ/h78S9QstYubrQbPX7e4srQ2iw+dK0M9symWXeqNtMb5BIJ3ZIzQBg/CzwJ4v0rwR4q8IXmg3ehaL4vvtQXTLCW6huH0Kwms1jAlYSuMSTrI6xwtII/MA4AOPG/wDhRvxp+JukaP4c1BW+GreGPCa+F5b5jDfDUTLPatc+QlvOjrbyRWuzczxyASnABGR7TP45+MGhfGXQvCGoappGq/8ACRz6i50O0tpPM03SbeKQ21/NeF8/PKsUbhogrNKVjzsJNz4S/EL4lXfxT174ZfEa+stQubDTLbUC9vYzaesVxLK6SwWpnbN5bxjZ/pCgAMdrfMdqgCfsyfC/4gfCzw14y0LxvLZSw3mu3tzp0Wm2osYxauqqrIizyhFcKNiZBjAwSeo4j4FfDzXvC/xY8S+I9S8L+LtMtddnSeCbVtfW+to1is4oCtxENRuTNIzRkxu0ZKKVXIAFelz/ALSmh2vjS68L3PhnV49NsddTw5ca0VtvsMeoywRzxpt8/wC0FGWRV3iLarHBx1rnZv2uvCNloVx4p1jwxrenaLNpN/rWk3csdsV1e005BJL9mVJ2dHaMiSNJ1iLp8w6EAAyfG9v8VvF3xrs7PxH4F1G++Hvh28tLjT/sN3p6QXl6m1/t18JblJzHaucxQLHyy+Ydx2KvpXxv8L+IPid8FfFOg6XoZj127intrOCd4d7bZgodZA5VVlRd4ywO0gMA2QNPwT8a9G8UajrWi+IdKu/Bmo6J9keSDWJLVDJBfK7W8qPBPKnziNwULB1KkFRXMT/tLaDaeMbrw1P4b1ZNLsNdTw5ca2Ra/YI9QmgjuIl2+f8AaGRllVd4iKqxwSOtAHV+KPCWveJviv4LvLy33eFvDFre6gxLrh9Xfy4LXKZ3HyoXnYHGAxBzkCvN/wBprwB4j8Za18ONX0fQtU1+y8O6neXF9Do+orpd6sc1lLAjRzm5tTjzGG4LIMjOQRXOaf8AtzfCTUrXU7+O3ufstppd9rFm0dxYXEl/a6enmS7IYLp5YHKfOiXKRFlz0IIHqXjr9obQfBGuXPhlNB1PWtWhl0iCO3s1twZ5daNwtuqNNNGowbdvMLFQoIOTzgA574BfD3xZ4M+HXivRNY0+bS4NW1PUbvSrC8ukvb+C2uo1O28u1Z/OmefzH3NJIwVlUu22vmk/ss+MNI8B+GNI8J+GbSwu08EWWn69bwyw241K+tb2xuZrO4kQ/vDPFFcRea25QHILYavf3/aM1qz8b6low8L6hriX2pHRtHsLAWom+16fYLe6i0881xHCFRpPJyHIDxMASWrH1r9uL4TaNp+m6i1rdyJc6ZHq15HJPYWs9hayPImHiubqJ55Q0UmY7YStheM7kDAHAad8ANZ8T+ObDXrz4fW3hvwVN4mstQfw7cG0eOKO00m7tpbuS3gd7cGaeWFfLQsSIw7DJOO90H4L/FDSfg/rXgzTNTXR7PZ4mt7fQY7e3kinhvbm8ayVbgtmFfKliAVcBAMHHNevw/tAeDbi2nu7W3u54IfEVn4aEiKm2S5voYJoply/+p23CZJw3X5TxnhdH/aot/EtjpVx4a+H/iHUbzXIru8sLNDp6TT6fYskc92S92qJH5jrGiuyySMcqhXJAB8+eNv2W/GUGkR2Hw60KLSLI6B4Zi1K1sDZRNqNzpl28t3CyXCSW8spjYENOjRyEbWOCca/gT4S/En4dX/h3xzpXhTVNQgsfEOo3kmkT3OkQXccF7pgsxNHBZJbWMKmZdzxo7MQTISWYqPZbj9r7wYyNqWi+G9a1bRLWx0rUr7UoY7aOGytdXd44jKk06TF0aNhKiRsy4PBxXZeAv2hvDfxF+IWseAdD0i+B0ae8tZb1ntWiE9jJ5ciywxzvcwBjkxNPCglAJQkYyAeOfs7/AzxJ4Y8W6T4r+Ivhy2hvdL8M21lbyu8Ny9penUb+edInUkqfKmjy64yDjPUV9zVHUlABRRRQB//0v38ooooAQ9KZTz0plAD16Up6Ui9KU9KAPHPiD8S7vwZ4++HHg63sUuYvHOpXljLKzlWt1trGa7DKAMMWMQUg44Oa5X4rfFrxr4Z8US+Efh5ollq99peh3HiHUPt9zJbr9kik8uOGExxyZmmZZMFsKuznOa5z46/8lw/Z+/7GDVf/TNd11XxT+D/AIp8X+JG8V+BfEkHh2/1DRrjQNRNzYterLYzyeYrwhZofLnhYvsZt6/OcqcCgCp8SP2gdM8I/DPwn460dLdpfHMtnDpv2+VoLeP7Xbtdb52RXfCQxsQiKWdtqLy1ef6V+0j458T2fhnTPC+k6FNrWs2+tXstxLqT/wBltaaLPHAxgkSIy+ZMZVPlyIrQ4fzBlcH0zxB8F9Tu/D/hvRvDesWunSeArixufDjz2TXCwG0tHs3S7Xzk89ZY5HAKGIpkEEkZPmOtfsqa5q+g28U2v6Rf6vcapqerai2paGl3YPdansBktLczrJbtAqKsf75w+WMoctkAHvvhz4weD9a+FGi/F3VL6DRdG1mygvA93PHEiGdQwj8xyqFs5Uc89qPBHxi8C/EH4cRfE/w1qEV1pRsVvpljljlltlMInMUwiZgkqqfmXOQaon4XXXh74Q6V8LfAVzYQLo9pbWUMur2TX8LRQqFZngjmtyXbqCHABOcGqfwn+Feo+Bvhovw08TXWmajZQWi6fEdNsJLANaJAIMTCS4uTJIQDl9wz6d6AOT+E3xq8Y+MPEWiaN430Kz0iPxjoLeI9Ga0uZJ3W1jkhWSC6DxoBMq3MLZQlTlh/CC3Tar8W9YsPjjoXwmj8NTrpuq2d/ctq0zqkcktmkT+VbxglnwJRvdgqg8DJzjj/AAB8CvHPg68sNU1HxdZ6reeFtCbw74ec6a8SwWjyQvJLeKLkm4mdbeFSUMS/KSF+bA9Z8ReAJtc+InhTx9HfiBvDFtqcAgMW4THUEiUNu3DaE8rOMHdnqMUAed+Cfix8Rrv4laf4A+IXh3T9Jm1vS7rVoIbK8a6urCK2ljjEd8NixhpBINrxsV3K6jIG6vJfiv8AtW6/8P8Axz4+0C2PhqO28Cw2UyWWo6hJbapqpurVbgxWkaqylyT5cYwcuQD1r0z4JfBn4m/DXxDqmueMfFuleKZtdd5tQvE0ia11K5lz+5U3D306LDCp2pCkSoo6YJJOD49/Zy8YeJ/FvxA1fRvEejWulfESC1t7yG/0Nr+6tlgtBaMbeb7ZFGCygsN0TANjOcUAe7eMPid4Y8AeGdO8UeMDcWMGpzW1rFClvLc3Burv/VQiKBXYuW+XAB5rhz+0p8LG0ay1e2n1C6kvpbyEWMGmXkuoRPp7iO6860SIzRCFiA5dQASoGcjO9r/wph1Xw34J8NWmpSRQ+DNQ0u9SSZfPluF01doR23L8zjq/PPavn7xf+x5Hr3iGTxdZ6npd5qUmoa1ceTrOkm/sxBrE0U+0RpcQuJYXiG2QOAwJDJ0IAPoEfHX4Xvo2q+IYNZE+naNZWOo3M0cUjqtrqO77O64X5t+08DJXHIFYl5+0r8LLS61Ky8/ULifTNQbSSkGmXkxuNQQtvtrbZEfPkQIWcR7tq/McDmvHtX/ZO8Rf2PfeFfCXi6y0nRdd0fStL1KI6OpctpUjyJJaiGeGKBZfMIdDG+B90g813t1+z9rVnFb6p4V8SQ2fiHTPE+p+I7Ke5sjPaj+1FkjltpoVmjZ18uQgOsiEMAcYyCAauo/tTfBzT7Gxvxf3t2L+0u75Y7XTby4mit7CYQXbTxRxM8JgkO2QSBSp6iuh0/4/fDLV/F1v4M0m/nvby4aGNZ4bO4eyE1xbi6iie6EZhWR4SHCswOCO5Arzrwx+zTcaDc6hqt/4kF9qetaPr1jfSraeUj3evXaXcs8aeY2yOMpsWIliRgl85J5nTf2UdV0vx74Y8W2nia1t4/D66arz22nPa6ncR6fbpA1s9zDcrHLbTFNzLPDK6glVf7pUA+0z0FIOtKegpB1oAfTD1p9MPWgBKc3Wm05utAAvWqOp6RpWt2j2Gs2UF/aycNFcRrLGw91cEGry9adQB8ieOf2EP2RfiF5r6/8ADHSYZpSWM2nxtp8m49Tm1aLJ+oNfGHjv/gjB+zzrvmTeBvEmt+F5nyVR3ivoF9trqkmP+2lfsOetA60AfzleMP8Agkh+0x4Itp3+FvxLi1qzQAC2SW40+4cEgYVC5hyOvMi8Drniuh0v/gj18eNftrefxn8Y44DIis8Wy6unjJGSpLSqpK9Dg49DX9Ch60q0AfhDpn/BELQnlEniX4s3lzz83kaYiEj/AHpJ3/lX3t8H/wDgnN+yf8HbRRZ+DofEupYw1/rmL6YnjlUYCGPkcFEB96+6qjoA4bT/AABoXhfQptF+HllZeGTI25WgtEZFLOrSZjBUHeBg8+/avmb43fsO/Cf4qm41jQYR4T8QSZb7RZoPs8rnvNb8KcnqybT3Oa+1F60h60Afis15+1j+x3M0GvWZ8VeDVb/W5e4ttvQESgebAcAYDjb6A19EfD/9sD4U+P4YLaW3XRdXJ/1F5IFBY/3Jdu0gfgcdu1fo/JDDcQtBOiyRyAqysAVIPUEHqK+KPi7+wd8G/iVctq2hI/g/VJG3SSWCKbeTJ53W7YUE+qFeeuaAPHfjn8XvAieF57bTPENja3Blixb2sqPkiQbycfN8o7Y9+1fL3iD49614iu7TwF8A9LuLvVHd92oCPdNKZAqny0x8qjbku/ryAK+ytB/4Js/BmwCtrms6xqrjqBLFAh/BIyw/76qOf4TaN+zN4vgbwnYvH4b1dVWO8lId1uSW3QSyghsEYaPcMHkZyBkA8n+DP7KGl+HL9/G3xnvI9a8SIftD2TkzRwMed0rNhZZPXnYv+0en1dJrAvoHaE/ZbUny1YkYbcMjHOctkYHpjOK7mK5s/EukC9jjXMC5kGAFJHOCOTgnnnPbPfPCaVDJfpNqOsQi2VWyVYY2QjlCoP8AE4G5iBwTg9KANWC0hW1DGYkICZC2MbevOOgx2HT361kSXcuoRvHY2qeVEFPmMABycFgDySVzXQ2VuuvXSam8DRWaErHCpUK2ONzgZBPcjp78V1V9a/6HILONE2AZyNqFT6EDggE/jigD8t/jh8OLrTNTuLrTov8ARLkCR4u4Y5zjnoV/DivTP2cNQ1DTdOk+EniO0aTSLm2Z7Z2mMvlmXJaAj/lmrLkxgE8hhkHivqHxJ4Wi1KylXUELGIF9ydQU6dQeMHr37YridF8B+Hr3T5NZt3M2q2h3x3Fu/lsGyGV24Ikx02kcD6mgDwK9ufEXhnxDN4G1OGKeCw4tyARm2/gPzdWK9Tnjp1Ga9E02S48SQj7XcmaWR0SKHBCeWVz0HAOQAc8nknGK6n4reFbzxN4dg8X6TD/xONKhl3RqDmVMEOg7na3IH1HWvDvBmreIL6wmux5dvaRKzeZKfIk8wLgFTub5A3ryR6UAeoWlzc6zoR0XTla4ubckTbH+6MnZgMCdxXd04X73JxWHDd6cU/sLTg0+oiHzTFBjDH5dpdmwB1w3G4Y6HNc7Y2epPpOjzrdKsupKxM0TlPNVN3mHco5G4AZ4OOgJIq8k9v4Mv45DbmMXIjdWcH96mTtJHUqSueefWgD0LTrveBY+J91qwwfJiVWVvmKgKAFLgAHryTXN+IZNNjjktoE8vYw2oAH3MTj5iPu4HRQCSeuOtZ/jDXrnUtGS7fZG0pIicZDY43CNMEg54Jc8/wAIxmuesnisbcy37SS3DoPMPDtCuQpVRhAGJyQTk4P40AYWjWlpLqOr3d/a77hvs6R8/JGmQXV1I+Yn09TXSXuqraXMy+XiLTXVkA6rDOcBVHQBfb0rnYb8WEP2S9bM5ka4kVMseW3Kmep6Dk9B9azlj1DVLmRZ7Vp5bx4gIYVYs6o3yDOeOT1x0H40AWdc1OK6hkH7x9vzKIuWY8hVGOc5PHB6V9K/BD4N3unNa6rqyeZq8sY8uL+C2BUBmJx949/TOBV/4S/B69tr7+1dZhjm1KbHlQqMx2q+pJyN3qR07Zr7i0HQrXQ7byYfmlbl5D1Y/wCHoKALOjaVDo1hHZQ8kcu395j1NaR60+mHrQADrTj0po6049KAEWnU1adQBHSjrSUo60AOPSk/hNKelIvSgD5D/ZT8M2ev/stWfhLxLaM9lqr67aXUEgKF4LjULpHU9CNyN1969B8E/A0+DJ7nVD4w1bVtZmtrHTor+7FoZoNOsJDIlqipAkZD7mEjspkbOdwIBHveAFwBgU2gD5w8J/s8TeFPGuveMLfx5rd0vie7lu9RtZo7ArcCRSiQm4W1F0sUCkLCqzDYBx1Oei8A/BK38FeIofFOqeJ9V8VX+n6e2lae+qPCxs7F3WR4w0UUbSu5jTfLKXdtg5zkn29aVulAHit38C/Ct99vFxdXTLqPieDxVKu5Mfa4IoohEPl/1RWFcj73J+avN3/ZI8K3Wiy+GNY8T6zqWi2ulX2jaRaTPb7NKtNQURy+SyQq8jrGBHG0xcqnHOST9ZjpSN0oA8oufg14E1TxRq/ijxHpsGutrEFhA9tqEENzbx/2cJhFJGkiHDkTsCfyxznHufgN4RvI9QhluLkRaj4mt/FEiAoFF1bRQwrCo2cQlYFyPvcnDDjHtlPXpQB8paV+yZ4T07QNQ8HXXiLVb3w3daVd6NbWDfZYltLS8Ty2AlhgSWZ40+WJpnfaPU5NbOk/s16Zb68PFXiLxZq/iHWPtekXjXF19ljy2i+f9nQJBBGioRO28AZJGcg5z9JtTqAPGNE+CXhrQb3w1qNveXU1z4avdX1BHkZCbi51oyG5ebCDPMrFduMcDkDFeXr+yN4VsEt18NeJdV0U/YV068eFbSR7u2jmlmjy09vIYZUMzqJIdjbT6gEfWrU2gD5vu/2atEuvFj+IYvE2rW2my61ZeIG0lGtzatqVlFFCsjO0JnKskKbk8zG4bhg1I37OOladpHhay8H+KNV8N6l4UsbrTLfUbX7NJPNZXjrJLDMk8MkTfOiMjBAysuQcEg/R46UHqKAPnC0/Zf8Ah/p3hbXfB+m3N7b6dr2maVpUo8xXkSLSWkeN1dkJMkjSMZGbOTyAKsaX+zroOn/Fe3+LF7rl/qd9p8l5JZQ3K2xNub4Mskf2pYVupYVDHy4ZJmROMD5V2/RFNbrQA2pKjqSgAooooA//0/38ooooAQ9KZTz0plAD16Up6Ui9KU9KAPk/46/8lw/Z+/7GDVf/AEzXdQ/tNeDNI8TP4c022utStfFPiq8i0TT5LPVL2yjt49slzc3TQ200aO0EEcrqWU5fYpODU3x1/wCS4fs/f9jBqv8A6ZruvYviH4q+GXgiTQ/E/wAQ7m0s7mO8Sz0uWdQ0/wBrvyLcJbgAuWcPtbaOFyThcmgDlfjDongnRPg5eT+LpdSl0bwfZfax9l1G6tbyY2kJVEaeCRJZHk6YZjuYgnJxXgOpfB/VNF+D+k+Ej4iv7zxyLHUdXj0iTxFdWayXc/lmRxclpLl4dO3qkSlihJBk+Zgw+ltb1H4O3PjeT4Ya0LC68SeLLf7dPp7xiWW5g0/YFlnXBAWMlQhfGcfLnBxyvx21r9n2AaVpHxw0621bas15BHNp8t+ba3j2pPcyGKOQwQDeqySPtjwcMcUAaPhTx14j1P4A+GvHHhSzl8WanfaTZzf6RJHYyzM0I8yZy29A2csVGQexI5qx+zz4x8eeO/hJ4a8TfEPSf7M1a+06ymd/Ojk+1GW3R2nKRqoi3MT+7xlax/GPjP4E63qlp8DPEem/22JEsXSwh0q4vLGBJyRaGSSGJ4IVwpK72UBRngV6n4O8A+BvhxYT6b4I0W00G0uZPOlitY1iR5MBdxA6nAAzQB4f8D08V2HxZ+L2i+K/Elz4jktLvR5InnCxxQLcWXmGKCFPljjBPA5J6szMSa+fvEvx/wBe1n9rPwVoUs+r6D4Z0zW77RU086ZfRpqkxsZt15LN5PlNCswVIQGICh5nwuCv31eeDfB962r/AG7SbWVvEQiXUN8ak3YgXZGJf7+xRgZ6CtC90TQtTv8ATtQ1CzgubzSJHms5HRWkt5HRonaMnlSUZlJHYkUAfDH7PY8TeFviJok3je4g1+8+ItvrV/aalp+vXuo26rFcico1rKRbJGI5USN4VGzbsOd2T5t8bbfx5qXxF+O+qeHdNvr9vDNlpktlqEPiK400aQ39mLM0sVorrFMVIMu1iocjaxAOa/Qvwz8LPhl4I1u/8S+FfDWnaPquqbvtNzbW6RSyeY+9wWAGAzncwGAzcnJ5rE8T/An4KeMfEU3ijxb4L0jV9ZuxGJbm7tIppZRCoWMOXB3BVAAznA4oA4/x98S/FGlfDnwLrfgCW1ur7xhqGj2EdzqcEnliLUVyZ3hjeNg4HzbNwGeOOteUaH8b/jJ4p1uD4ZadLoOn+JrKfxELzU7m2uGsp4tCuIYEEFsLhXV5fPVnzMwjCkgNkY+yNR0LQtWjsotTsoLqPTp47m2WRFYQzwf6uRAfusnYjpXlPjPwL8CtZ1DSfBfjLw3pmoXOvXl9f2sEtosu+5CB7ucsFO0spAkZiA+QpzkCgD5fi/ai+Lev+HNT8f6DZaNZaL4c0fQdWvLSeKeee6OpSyx3EUM6yosagRlopDG+cjK45r3H4g/GrxO2seHvCXw2sH07U9X17+x5rrXtLvIrVU+y3U5lt8mAXGDb8FH2kEcjINezyfD74fS2N9ZPoNgbPUbe3tLmPyIxHLb2efIidQMFIsnYvRe1dHqGiaJq9xp93qdnDdTaVN9ptHkUM0E2xo/MjJ+62x2XI7MR3oA/Pqf9qf4z6i9p4X8PaFbXXiDTbTVbnUprawku7ac6bqdxpqiKM3cDQRubYu7s8vl71GD1PpEX7Sfiufwv4h1+bS7OwuNM1PwhZw20j+aVTxD9h+0K7o+13j+0yCNk+U7QcEZz734g+CPwa8U29tZ+IvB+k6hFaS3NxEs1rG2yS8kMtyw46TSEtIOjnlgafqvwY+D2ua5b+K9Y8IaTd6papbrDdSWsTOi2bB7fa2OPJZQYz1XtgUAYPwM8Y+O/iBous+KfF0mnpZHWNW0/Trezgljkjh0vULmy3TySSuHeQQhvlVAvPXPHuI61k6Ro+ieHbIaZodpDYWzyzz+VCoRTLcytPM+B/FJI7Ox7sxJ5Naw60APph60+mHrQAlObrTac3WgAXrTqavWnUAMPWgdaD1oHWgAPWlWkPWlWgB1R1JUdADl60h60q9aQ9aAHDpQelA6UHpQAysvXNC0nxNpF3oOuWy3djeoY5YnHDKf1BHUEcg8jmtSnLQB8evo+vfAjUYft7za54QuHkJvRCzzafFHlgt2VJ3x7TgS4GNvzcnNep31tpfiaKzm0ydfs93iZZ4iGR4nGflbkfNnJwf15r2uSNJUaKVQ6OCCCMgg9QRXyn4i+H3in4WTXeq+BPP1DwfPcC6uNKgDS3liSf3n2BOhh/i8kZIyRGMYUAHoJjtLFWkkRbSOLYjBiNgjHXCkgDI49s45qw5Mg+zRtksC20gMEBznjvjuenGBXMWWt+HvGOj2fiCwnF/ahy0TqXQmQEqFkQ4ZSGGWUjOR7CsKz1nWTqF3CksENsjARlhtlxn7qrnbgdvlIHXHegDstasYTAt0TuKoEfJwShzwB07n/APVXm+k6PZ6VqM1hGphjYZRwNoXqcEdCCOBjpXQm78Sazb4hgcRFirNMJIZQysykhZNrFeCVYDDA5HUGqMxlRl+1qWuLRuN3BIBPAI6n8f5UACafNazv9m3McBvLfksvfaRxnnIzXyp8S/CNv4b1SDxJptvHc+G9WfMwxuS2lk5Dgf3HPPXAP4Cvt+zEF8Y5UjBlQcHy+u7pyOMeo7HpxXlviHSra0vrvQtZRJ9C135UjIwIpXHzqSOgc/MpzkOTzyMAHzhp9/aXRt7a0t4UtLfKwkhii7uC6jO4E9gOM44rD8X31jdxTXEjrNOxLM6qFXJARQAo4wB689etUdR+F3izwdq15bWtvfaxp4bdbyhi4KHkBtvIK9D+dcTrdxrT20emXVvPYW8T+YyeXJEGI6ZU7c+mcc+poA1hf6cLKOGR3M9umY3YEM0jddoB25XAwTn3Bqs2r3FzshRlhiHzNk5zJ3ZzySfYUuj/AA48eeLVkfSNNe1ikBxPN+6Qn2JOdvsM/SvpTwf+zbps0kQ1gy6pKoX9zEzpEDjB3MCCR+IoA+c/B/h/VfE3il7G0t5Jhb7SZgmUjlYg7pH6cL/DnPPTNfdPwv8AhDFpe54CZ53wJ7uQdMfwoO30/M17D4X+Fml6LZw2jRR2ttCMJbWyhI1+pGPxx+depwW8NrEsFugjjQYCgYAoAq6Vo9lo9t9ns0x/eY/eY+pNaS9acelNXrQA6mHrT6YetAAOtOPSmjrTj0oARadTVp1AEdKOtJSjrQA49KF6UHpQvSgAPSmU89KZQA5aVulItK3SgBR0pG6Uo6UjdKAGU9elMp69KAEanU1qdQA1qbTmptADx0oPUUDpQeooAWmt1p1NbrQA2pKjqSgAooooA//U/fyiiigBD0plPPSmUAPXpSnpSL0pT0oA+T/jr/yXD9n7/sYNV/8ATNd11X7R3gOXxr4Htm0fRk1TXLHVNHe3cRo08MCapaTXJR25VfLi3Pg8he/FegeLfhvo/jHxX4N8X6jPNFdeCby4vbRIyoSSS5tZLVhJkEkBJSRgjnHbivQqAPn3xL4DlP7Qfgbxzo2jItvFZa2mqX0UaKxlmjtEtxM3DMSsbKuc4C44rjPj9da7408G3HhiLwP4huLPxFp11Clzpbwx3sV2j7I7W7jZgFtpx85YuYyo2yAZGfrWnjpQB+dmp/Dzx/a+LoL7TvC2tad8RL+48NSPrenag/8AwjzQWcdvHfiWASRxIixpNH5LxOz5Uxnn5fc/jD8KU+JfxS8KnX9IfVvDNlomupcKXZYBeTPZ/Zg6qw3NhZCmQcEZ4IBr6gPSmUAflZJ8MPizL4WjtPFXhPWtZ8c3fhzw7beGdWjnBTRL+2tlS6+0SmVTA6XIaaZ8N56EJlyu2vWNE+C3jDS/FGk+OrDS5rbxTJ461yW51CR3kC6Nc296IC6eYM2plMLCIEfNg8HJH352pKAPhf4kfBXx1498T3c/xFt4fFLWng3WIbSewglsbQapJPE1oFha5lPnqFJRy3HUbTXiMnwn+N994/tdS8TW2sNrEk+gS6bqNtZw3BtbaC3tRcxtfSXcf2YJKk/nxGJvODHAkL4H6tDpSN0oA/NK+8MfF+90yx8Cw+FNZS40K+8cyz3zNGLSdNVi1BtO8mQS7pN/nxgfKNjcNg4q/r/wH8TTeCNU0bQfD1xZLpPhbSdMsoopCk1xqGoXUVzrEwk3794WCFWctksHwa/Rul7UAfmT8Tfgh470fW9a0jwRot1F8Ol8RWV7Np1rbjUI5oH0ny3ljspJ4vORLzDSxhsl/wB4EYrVnQvCfxb8BaLZyv4c8Q69BqnhjxHpNnAscCT2U95eJNZRzRC5dYIvKBEZMjeUuEYggCv0sqSgD8y/+FH+O38PeI9f1Pw3e3uqB/CULwpPtu7vR7Syshq1pbMZFUGRo5FkXcvmldpJyK1/DXwV1TxT4z0ePUfCF9pnwyfxBqF3baLfOUW2tG0lYczW6yN5cM14GdITkAncVXNfo23Wm0AfkxefCT4vaSfAmoQ+GdW1bWNGsbW1toLtftNpH9k1K4dFW7juoZ9PkWAxl5T5iSxhVKsVKn9Z0zxu4PenHoKQdaAH0w9afTD1oASnN1ptObrQAL1p1NXrTqAGHrQOtB60DrQAHrSrSHrSrQA6o6kqOgBy9aQ9aVetIetADh0oPSgdKD0oAZTlptOWgBD1pw6U09aeOlAHzh8Sfhrqela1J8S/h9EJLxvm1XTiWKXsKJgtBGAVFyABjoH6E5wa5y28P2HjjTU1XRrgLEcqUYFZYJF5KuGwysDkYYAg8Eda+sW6V414n+EFnqWo3HiLwfq914U1u5YSSz2m2SCd9yktPbyZVyVXblSpwTk0AchF4a8XabD/AKReQCBOS7hlxjk5BGOncHH9Of8AEzOmnNeHU7YyW43xrE+8nJJ5UAs27jgA+1auoeKviN4Ie8j+IWj/AGvRrSOJk1fTQZ4X/hbzrXmaJsjJ2+YgBHzDBFZ03xO+GmpNZLo+rWt/qmqofstvYKLu4fO3GI4wSFXcDlto55I5oAh8PeINOgs/7bup0SO3QmVpDhIhjMgJbAXb/F2yDXcado1z8XPDGnarJ9s8NaVdM8jW8kNsbm6t3j/dMHzL5I3Nu6bzjB25rnfC/wAK9f8AHTx6x8UoBp+jARNb6BGyssm0BxJfsqjMgJ5hRjGCuSz5r6e2qqhVGAOAB2FAHg0Xwf8AEFrcym38XSS2mR5UdxZxSOigAYaRGj3HPOcCt6X4aXDQwp9sjuHLjzC8exQnOSq5bJzjgn8a9ap46UAcHZ+ANIhCfane4dSrddq/Kc4wOx6EE9K7a2tba0iENrEsSLwAowKkHWn0AMPWkpT1pKAJD0pq9acelNXrQA6mHrT6YetAAOtOPSmjrTj0oARadTVp1AEdKOtJSjrQA49KF6UHpQvSgAPSmU89KZQA5aVulItK3SgBR0pG6Uo6UjdKAGU9elMp69KAEanU1qdQA1qbTmptADx0oPUUDpQeooAWmt1p1NbrQA2pKjqSgAooooA//9X9/KKKKADrSbRS0UAIBiloooATaKNopaKAE2il6UUUAHWk2ilooATaKNopaKACkIzS0UAJtFG0UtFACbRS0UUAIRmjaKWigBMZowBS0UAFJgGlooATaKCM0tFACAYpaKKAE2ijaKWigBMA0AYpaKACk2ilooAQDFGAaWigA6UdaKKAE2igDFLRQAm0UtFFACEZo2ilooAaVVlKMMg8EHoRWBp/hLwtpOoz6vpWj2dnfXKqstxDbxxyyKgwoZ1UMQBwMmuhooAKQjNLRQAm0UvSiigBMAUtFFACYBo2ilooAKQDFLRQAUm0UtFACbRS9aKKAEAxS0UUAJtFGAKWigA60gGKWigA60m0UtFACAYoIzS0UAFIRmlooATaKAMUtFACEZpaKKAEIzRtFLRQAdKTGaWigApCM0tFACbRS0UUAFFFFAH/1v38ooooA5rxne3WneD9d1CxkMNza2F1LE4wSrpEzKwzkcEZ5r4G+GXxB8d+Hfhd4Z+KXxD1Lx1dDVhockg1KbQDZ3Ju4zPK9lHp8clwYTtwY5/LkKMoBD7iP0YuLe3u7eS1uo1mgmVkkjdQyOjDBVgeCCOCD1r55034vfALxJ4W1u10eWC40j4fazbaBNDBaMosNQWaO0txbIqDaEkkVY5YhtXBKsApIhKScnF6+7b0Td/vul/SNNHFRa7r5tafdZs8l0P9sTV7jXNC0XxF4Ns7U+I4PD17ay2GuLqCx2fiK9FnC1xi1jCSxk7mVS6NwElOciPVf2zdRtb7xJ/ZXgdb7S/B9prmoapdf2oEZbTRdQn0/MEa2z+bLO0G9VLIiAkNJ8oLemeAP2Svg74Et9btryyXxVB4gggtLmPWLXT5Imgt5WnRGit7WCOZzK5kaaZZJnYKWkJUY9XtfAvwr+Huh3txo3hbStF06ysZ0mjstPghUWWWmli2RIMxsxZimMMxJwSTWlSpCK5nolzP8Pdv/h6vrd9hKPM+Vbvl/wDtreutu2nmch8G/i9rPxQ07X49Z8Onw5rGgTRxSRvJcPbSCeBZ4nR7q2s7jGGw++3TkZQuhVj5B+zN4y+IGu61qeo+J/Gh8YeGrmxs4zqUqQW9g/iHzpkvIdFK29vLPYAbFjlkDBmA8t3Jfb9IeANB+Gnh3wPBcfDzQrDQPDeqQDURBY2UdnC6XEav5jwxoo3MmM5GcDB6YrlvAHwu/Z5n0XSvGXw68D+HbSw1qKy1OzubTR7a0aVBtubSYAQo4ZCVkTcAyNg8MK0ceWo7rayflvv+LW2q7JoxjLmp6Pfr9z0/rZvvde3UUhIGMnr0pNy4ByMHgVkWOopMjJXPI7UAhgCDkGgBaKTIzjPNBIAJJwBQAtFc34n8W6J4Qs7W/wBclaKG8vrLToyiNITc386W8CkKCQGkkUEngA5PFdFuXAORhunvTS0v8vnp/mvvAdRTSyg4JA6fr0pSQoyxwPekAtFcZ/wnvh8+P2+GkX2mXW49PTVJdltK1vDbSyvDEZLgL5SvI8cgRC25gjEDAJrssjOM8ijpcHvYWikBDAEHINIWVfvEDjPPpQA6ikyB170AgkgHkdaAFopCQMZOM8CjIzjvQAtFcppnjXw/q/izWvBNhOz6t4fgs7i7QxsqJHf+b5BVyNrbvJfIUnGOeorqsg5x2oG1YWikJA6nGa5u68W6HZ+K9N8FzzMNU1a0vL63QIxRoLGSCKYlwNoIa5jABOTk46GjyEdLRSZGdueeuKTcuduRnpigB1FISAQCcE9K47w1470DxbrHiHRdE+0SS+GbpbK8kkt5YoPtBQSNHFK6qspQMN+wkKSATzQtXb+v61X3g9NX/X9WOyopCygFiRgd6CQMAnr0oAWikyMkZ6UAggEHINAC0U0uq/eIHfk0uQMZ70ALRXNeK/FuieCtFk1/X5Wis457W3ZkRpGEl5PHbxDaoJ5kkUE9ADk8A10hIGATjPSgduotFc14w8W6H4E8Lav4y8STGDS9DtpLu7kRGkZIYVLOwRQWOAOgGTR4W8XaH4y06bVdBlaW3t72909i6NGftGn3ElrOoDAEhZYnAPQgZHBBp2E+50tFZZ1aEa0uheRcea1ubnzvIk+zbQ4TYZ8eX5mTkJncVycYFVPFHijQvBfhrUvF/iW6FnpGj28l3dT7HkEcESlnfbGGZsAZwoJ9BQle1g62N+iuT8MeNfD/AIvudctNDmeWTw7qEml3geN49t1FFHKyruA3qFlT5lyMnGcg11YIPIOaXRPvZ/J6r70D3sLRSBlOSDnFcfrfjvw9oWoy6NcNcXOpxWR1D7JaWs1zM9sJVhLosSNuIdgNoO7HOMAmhb2/ra/5K/oD0V3/AF0/M7GimllBwSB0/XpSkhRljge9AC0Vy3i7xloPgfTbfVfEUzQ291e2WnxlEaQm41C4S1gUhQSA0sigseBnJOKl8WeK9F8E6I/iDxBK0Nkk1tblkRpD5l3PHbRDaoJ5kkUE9AOTwDQnfYdtbHSUV5tqXxb8CaZ8P9U+KE1+8vh3Rnu47meOCVmV7G4e1uFEW3zGKTRsnCkHGRkYJ9IVgwypyKbQNC0UhZRwTjv+VG5eBkc9PekIWikyM4zyK5zwt4s0Txlp9xqegytLBbXt7YOXRoyLjT7mS0nGGAJCyxOA3RgMjIIoA6SimhlJwCCef061maJq8Gu6TaaxbwXFrHeRiRYruCS1uFB7SQyhXRvVWAI7igDVorkfGPjjw94Ds9P1DxLM9vbalqFnpkcixvIq3N/KIIBIUB2K0jKm9sKCwyRml8PeN/DvinVfEOj6LO08/hi9XTr5ijLGt00EdwY1cjDlY5ULbcgE4PIIAtdv62/zX3ruNq2/9b/5P7jraK5zw94q0bxQ+qpo8jSHRb6XTrncjJtuIVVnUbgNwAcfMODXRZGdueeuKFsn31+T1X4C7rtp92jFopAQwypyPak3LtLZGB37cUAOopAQc4OccGgso4JA6fr0oAWikyM4zz6Um5du7Ix69qAHUUEgDJ4Aqnp2o6fq+n22raVcxXljexJPBPC4kilikUMjo6kqyspBBBwRyKALlFfM0H7W/wAGJY9RupLy/t7Gx0+/1WC8m026jtdQstLYJdy2MrR7bgREjIQ5IIZQVO6u/wDh78a/A/xK1bUPD2iDUNP1nTIYrqbT9W0670u7+zTlljnSG7jjZ4mZWXeuQCMHBIy0m9v6/qz+4Jab/wBf1c9aopAQSQDyOtUtS1K00nTrnVb19ttaRPNIwBYhI1LMQBkk4B4HNROSinKWyHGLbsi9RWD4Y8S6R4w8MaT4v0KUy6XrVpBfW0jq0bNBcxiWNirAMpKsCQQCO9UfG/jbQPh74XvvF/iWSVNP0/yxJ5EMlzMzzSLFGkcMSs7u7uqqqgkkirqJwbUtLBBc1uXqdZRVa0ukvLaK5RXjEqK+yRSkihxkB1PKsO4PINZWs+J9D8Pz6Vbavc+RJrd4LCzGx3825aOSUJlFIX5InOWwvGM5IBGrOwlqrr1+W5vUUgIOcHOODXnOn/FjwLq3iaz8IabftPqV8+qxxoIJVXfos0cF6pZlAHlySooPRs5UkDNJauwPRXe239fcej0UmRnbnnriloAKKKKAP//X/fyiiigDnPGEXiebwnrMPgl7aPxDJZzrpz3jOtsl20ZELTFFdtgfBbapOOgr8/8AUv2LfH3h7wzd+GvA/ie28Q2es+H7HS9St9eK2kUlxpV/Fe2zK2n2gdkkD3aSvIWm/eKd7EcfoH4s/tf/AIRfV/7AuI7TUvsk/wBmmmjaaOOXYdjNGkkTMAeoV1J7EV+bHhD9rj4jeC/h/wCBIdZgtPGNq/hzR9fvtZhFwz/2LbK9vrjysZZTJf2UwhDgkeY0jZRSCAopWlK+1l96nr8km39/QvlbS0vv+cbL5uyX/BJ/F37G3xL1/S5H0XS/C+iltTvr7T9DgvJG0nRPtFraQI1sJtLlik3S28k0oS1tpVaQ+RPExdn9DP7L3xGl8cXuu3LaF9qub/Ur+XxLHNdJrN9bX1hJbR6TPCYmVbOGSRSv+kyLshjIhEhLVU1H9qP4raNqOkWniG10LRdWmsNBvF8O3EN0dT1qTWpissGmyLMFVrJMLIRFcZkDFxDGVauz/aI8WfErwl8WNG1j4ealZ2raR4J8VatcW2pQ3FzaXIsJtPdV8qC4t8SnOxZSW2Kz4Rs4qKsEouE9veT8uVXf/pPr3vsVRu2lDfRrz5np/wClenVa6nC6X+y18U45tM8Ha4nh/UvBsOtrrd3LNd3MtxKH8P8A9kS2f2NrPyWj80s4dphujIBQNmubsP2OPiZbyfD4tLo1n/wjGk6NprtZXbJ/Zk+l3TzXN3YM+mtMz36sGmEctoxfKSyTx1Brn7bXxKs9a8RNp+haXHb6Zp95dQaXcsv9oCK30Y6nHfuVvBM9u8n7raLNI9pyLnzB5Z9O0T43/HKPxidE8TTeHrrT7DWND067+yadeW8s8ev2guEMLveyrG1sx2ksr+cDnERHPVOtNPme94ffrGK+52/PW5zWj7O3Tlf3L3m/v1/LQ6D47fsx3/xe8Za54rVNLkml0fRLHSZbwyefY3Vjqct3dyxlYm8rzLd1jV423k5U7VOT458WP2NvH2vaTd+Fvh5a+GrPQvtOvTaZbSObVdKk1Nbf7NNbodPvI0MTRysVgWCRWYGO4XLCs34e/tceMLPVPCXhGWHSItMvPD1hNbWMfn3mo+dJoov98slxfm8MQcFfMNrNEUGZLtZSVrnD+0b8VPi3P4B0bw94k8P6heXeqeGNSOqaJBeLp1hcait4s2lajAL0yTvGIw+wywFsDfGnBrOlRvJRj35f/A5f8C/4Xu0b1HyqU59Fzf8AgMbfdrb73ayZ2Ev7IfxZv/EfjDW7qPw/CfGGmahb37tqF1PNeS30MKiE3AsIryKNXjwWa6ni2gGO0j3Mg+g/2e/AnxR+Dtjpnw91nStMbQ7ybVr6SbT5I9mnLm2+ywHybTT4ZXmd53YxWkKrgDaTudvnvUf2y/itp2paNpM2laMlyrXNvMJQIE1y8tNaudKkg07z76OSJwtuspSOO+kVpkVkCje1vxb+1T8bvDmlW/iO8/4RvTNG1nXfEOmWd3c2zLDZx6BcT26x3Ut3qdlC094Y8oFlTaschVJmYIj9rpz9LNfJP/N6dfIXs237Pqnf5ta/gtfzOn8W/sc614gm8Qarp0+lafr3iXWfE813qi+abt9H1rTLq0trV38rc4inlikaEsIxsLKxfGVj/Zk+JPjz4h2Hiz4w2HhyXRIrnS/tGkRXlzqUE1vpulajZ5cXFnAju9xdxSbCu0IpyxZRut/Dr9qjxx4o+OWjfDvxLZ6XptprVtbsunW6mfUbaaXS0v3M7tdJOqK5dA504W5Xb/pPmkx1B8TvjN8QPhj8bPHVoPEmlR6dLp3hb7HDqMNybbSba8vbq2utTmRbpQ8UJwLh0ESsXtw8kSoXefZe6oPZpJf9u6L/AIfbq7bjcnJyl11b/wC3nd/1vpZdnzEX7GPivSfAdtoem2Xhy+1GXS/CkOqw3MtxFaatf6FqbXly11Kls7us0BESyMjOQNrKEArn9e/Yt+KWsWtu1vNo1javNq72mhW94yaf4bOoSwPBLpcsmmSsZIvKZ8xRWciu5EM0a5zs+I/2wfiLpnhd9YivvDOnW9lZ69c22s3Vpey6Z4ml0iYRwwaWqXCGNp03P/rrg8ZiEqBnEE37Vnxh8W+JNT8D6VZaVo9zdWc8MNu6ltRgH9kC/XU1VL9Lh4DIfLRRaogBVvtRkBiObqOMXUWtk38nv+VrPWK1Vk7mnK3JJ9bL/wABSX5Ppo9tdiz40/Yk1jVvC12LGw0PVvFGoeJtX1e6vb24ZGmtbt7gWQd7mw1CF2gjmw0T2rKrM7QyRyfvG9/+JvwY8Y+MvBvw7069s9D8Y3HhLadV0fV3nttH1SU2TW/mZEV2QYZT5kSywyjrna4V1+WvhT8Y/ippGo3vjiDxHp3izw/dT+AdOvY3juppZ5dYt4LSSSzmN48UBjeXfKGSUyyh9xjJrS8Nftk/GTxPpF61hpegDVp59GhjhlBP9jXOq6tFp32LUYre+nmaRFkLbpFtJN8bj7PtwR0VKL5nQWuqXzilby2f9OyOWnWvGNbpa/ylq/PfT/gH0d8Bv2dLn4U+K38X66dO1DVpvDOk6PLfQo7XbXFpPdPcDzJVMjQ+U9rDGzSM7LAocDYteG6t+xb4yfwy1p4Z1DStG1zVLHxHba1ewPMj6p/aOqwX1jDcSeQWkiW3ie3kLq3lLIRGkikqfsu68Spqnw48UQxazbanrfh+zurHU57BHt0i1GK1EkgWNpJXiPzq6qZHKhh8x61+cH7LPxS0TQ/FnhbxBqfiS3/sUeCJ5/EE1r4q1HxFDFeq9iyXOqw3aJFps5zMqJGZMszoDtXJiLdSo1fZXv5SU392n4rc2d4RUn3WnnHlS/P8D1Cw/ZY+NHhbRbiTweugx3eqab4l0k6ZPqUsNhpkOupZCOW1ez0uGM7JLRpHhjtLdC0pIO7c7aMH7Ger2emzazqGmeHfFPiI69aXs1vqUkwtNU0e306K1/s66nNvK6RrdKbtU8mWJpI4y6k8p7H+1B41v/A2q/C3VbPxZpvg2GfxBcwT6hrPmNpio+k320XMaXFqHBcLsDTKok2sTkAV8u3/AMa/FnxGi0nXLq1fQr27i0meWWyurtIbuOWy8SIkkUTSlEgl+ypcRYXeyunmM5jjK5yqunCVVa2svTW6/F+tm+hrTpOo1G/SUvws/wAFbTb8+yP7F3i280DVP7Ul0WXXItBitPD0hkuJI9B1BNWvdQRbOSSEvHDawzw28EqgSFYsbEU7T03hD4J/GP4OfETxt8T/AAvpOi6p/bT3TJax3TNc38l7qUcyO8hsoZYvKhaUstxd3ah8LE0EQIrjdG+P/iTwfYWukJquj+Fbe9ubOG68SeJDeXtgskPhnS7xIpFN3CqXFy0zBNssSERO2ySZiX6bwd+1D8YPEl1p/iTVdI0rR9B/tvwzol3pcltdHUg/iGxtbhpVuXmRIxBJcjCNbEugIJRhk9E6fJW5Funb/wADf6t219XpqccZupSU39pc3/gK/wAl+dt7HU/tafs9/Er4331hL4LOjeXZaXeW1tNqEpgubHUJpI3juoHaxvugjAzD9mnUjKzYyo4m/wD2O/G17ptzqemX+leH/F+t3ni6TVNVtJJ3uJbLXo7j7FbGcQxySJBK8LMhKBdpaM7+v354d8Q6J4s0Ox8S+G7yPUdL1KJZ7a4iO6OWJxlXU9wR0rZrDkSi4td197u/6+Rs6jbUu2v4W/rz13Pkj4EfArXPhxrPj3Wbjw94d8H2vi200yC30vw7NLNawy2MVxFJK7Pa2ozL5iNlUzwd2WBZvGNJ/Y98deC9L8LXHgf+w7bU9E0rws2owLcT2sGra1oN+LmeaeZLV2IlhLok7I0mSAybBx+j1FWptVFUW6s/uvb87+qXYn7Lg9n/AMD/AC/Pufmbe/sX/EDxZZeKNT8Z/wDCPtr+rWeqtpZS4ubmPStS1HWptSSaCaS2jdTHFIE85EWTcCAApJrc8Vfsg63B4lubnwt4R8H694W+1a29loWqTT2VnZR6rZ6VFHJDFDZXEayR3FlPIVCgfvAysJCSv6LUVKVoqC2s18m7/f57mntXzOb3bv8AO1j81/8Ahjr4rR+MdO1m78S2utS21lpcJ1u5uWi1KL7Bpq2c1qq/Ypp3t7mZWlcC/jQ+dIZIZHG52w/sV+JfDOq/DbUvAlh4dsb3w1Y6Omp3zu5dtQtZlkv7pY5LKZ5nuFGFlinspmIVZnkjCxr+lVFaRqtS5lvdP7r/AOeveyuZNJx5elrfl/kfGP7Qn7PXjH4peMLjXdCt9Fv1vtFi0uyu9VnuIbvwzeR3Ekx1TS1hhlD3Dh0yN9u26CIeaULAdND+z7Po/wALfjB4K8Lw6ZpmpfESTWJLSaBTDGGv7UQwtctHFuBWQszbVfAJK5JIr6oorHlXK49Gmvk2m/xRfO+ZS7NP5pW/I/PvV/2RdY03V5JPCOkeHL7wmtzo17J4VvZJrbS9SurSxu7S7kugltOodnlgmR2im8x4QZFDBWGR4d/Yz8YabaSalq95pF74ks5PDUmj3pkuXfSU0zVJLy9trWaSNpEi+yutrCRzLGirKEXgfo3RWjnq35p/NEKPu8vS1vl/X39T81Jv2JvF9l4Ls9G8MXmkaTql3ol5Z6/NA8iDWrttWtL6BLp2t3MsLW8M9u7yo5jWUqIpIyyGI/spfG7RfC2p6d4UfQjN4i0HxHoM1hdanPFZaTDrMlu8L2jWulxxusflOzQpa2yBpPkGMk/phRUx0Vl2a+Tvf8/l0sae0d+brdP5q1vy/wAz89H/AGPNStbebX59H8O+Ktdn8UXOrXtnqksy2eq6XJA8VvZ3M5tpmUW8jLOkfkSReagYjcd651n+xn4wGkXE+q3mkXviKz07QYtEu3kuHOkXOnavdX86WsskTyRxR280dtA4+d0j2uqKcH9HKKtVGreVvwVkZKK18/8AO/8AXkfmTp37JHxqh8c6j4hnk8O20GrSyNfy2t1JG16w1u01SGeSJNNSRpFigkjC3F1csjOdkqoSte8/Hj4EeKfiR48tPFGmaXoHiK0GlDTYY9envIW0W6Fz5x1Gw+xqXaZ0wrbJraUGKPZOoLY+vaKiHu8lvs7fj/m/0NXUb5r/AGv+B/kvXrufmxdfsd/E251L4iSHVtPMnirT/EVtHqLXTLPftrMhe1S/ijsUk2WinYpkurraFHkpEpK1b8YfskfErVfE+ieJNFfQ5L3TNW1W9iuru4kf7HHfa9NqqMlvLZTh38mRVY28tnKHXabh4uB+jlFXCo48lvs7Eyd1KL2bufK/x1+BPiH4s+JLK7tZ7I6OselQ3dtdSSr50dnrNtf3C7UjZSHt4nQZIyxCnCksMa8/Zru7j9n34kfBKK30kW3iG91m40C1ZW/s/T4ryQzWSbBF+58mU7sRIwjPKZwK+waKin7keWPn+LUvzWnbXuDl73N5p/cnH8nqfnr/AMMe+I/tF34l0IaJ4O8Vajr11d/2jpTStPY6Rc6C2mrZ283kQM6wXRWRIsRxlUD/ACvhR337OX7PHjH4T+GvFekarc2ehy67ZWtpBHpFwJ4Ip7eGSJ7/AGpY6cizSF1JzE8jCNfMnkIG37MookrxcOjSXySt/XnruPmd0+zv+N/68lbbf8zfAn7E/i/T9O03RfGP9jz6TBqmh3GpWC3BuLTVI9MjuVubmaJNPtEae5MyblmEzuFxNcSYWuht/wBkHxZpQf8Asi18PLcPp2s6ZDd+ZNFPZWs+stf6dHAVtWJjitSLdk3IIwNqb0FfojRVKbvfzb+bjyv8Pu6GdWPOmn5fg7r8fv6n5reLv2Jtdv8ASbe80u20TUNYuNd8Q6jq0F1KEg1GLU7id9OkkmuNPvx5tjDJtSM22EMkpikRvnf134yfs/8Aj3xz8L/h/wCALCXS/EP/AAjUaR6kdblybmRLJrZLhJLmz1GN3V2LnzbdnYHKyRuNx+zKKzcE4OHTT8Db2j5lLrr+O5+ZWjfsh/FcP4am8W2fhjXtZ0298J3j+Ibq9u5tUsIvD4tY7mysy9md8Vx9neYEyRDzJnV0YgS1vWv7L/xoul0Hw/rM2hDRvCVlPp9pcC/up7m+jl16w1QSXELWaJCRb2ZTaskoMh67TkfovRWrnd3fdv71b8v83qZx0tbpb8Hf8/6sfGFr+zZrVl+zR48+BOl2OiaRP4gvdZexe0eRLWS2v7tp4GuQtujRyRxMsBVBIAsa7XxhV56X9mvx/wCBPHN14q+D1noEGjw6jeS6doct1PptlDbajpFrZTvi3tJwkgurcy7EQq6uzF1c194UVnUXPe/VWfmv6/4BSlqn2bfzZ+bfg39h/UbKz0BvG9voGsX+m6h4cNzM4knMmk6XokWnXtkDLACY7idGbyjiORCDJyNgyfEX7InxmF54Wk8Nf8I5IPCaacNOne7ltp7JbDVZb14InbTbqVY5bdo4QYJrbaAyyLMgUH9OqK0c25KT6O/ztb9Pvv3IcU00+3L8r3/rysfG/wACP2adT+D3iPwz4hjXTIJ4tD1Wx8QS2Rk87U766voLi0nkZokMwhjSVA0hDJv2oCpOPFfFP7HvxM1+bxVDpNp4Z0G61a58XXCa7bXd1/aWpweIFuha2N+gs1CQwPcJKxWWYBoUKR7iXr9MqKztol2TXybu/wCu2hp7R3b80/mlb8v6ufnXr37I3i/Tfi14f8VfDTTfDukaD4bv9FuLNo5Ta3kVpZsTfWxP9n3Ex+0tLNIzx3cSSFsTROxZzJ4T/Y68VaMbfxBqs2j3PizT7/wlNY6mJJ3ntbTRmhGpRRStCHT7TGsihVwJQwEpA6fohRVRbVvJ3+en+S/pkdGvK3y1/wAzxvx74E8R/FT4Maz4H8V/YtN8QatZyKktnJJNa2t9G3mWk8byRxyHypUjk5QEMOM8GvjD4i/sgfF7xT8OdC8Ow3Ogapr8lvr15q95dStAIvEGuTLOby0aTT7zMcB3JHsS3uFVY9k6fMK/TOipa1v/AF/X+S7Fqey/r+v833Pz51j9nb46INQ17Q5NCutb17TvEem3UV1qV3FBbjXodPAuEnSykeV4ZLR8oY03hlIdTkVzMP7G/wAT08V3+q3GsWFybqzniGpG8liu2jl0caeunSCGzS6+yrMPMBW/CAAOLcTDfX6WUU+lvJL5LZfe7/8AA0Ji7WS7t/N2v+S/4fU+Mvg78CPiT8O/gv47+HpbRdNvdcW5Gjw2zebbWzTWawBrmW1s9NVt0q7iYrZHC4LPLISx8T+HX7E/i618U2MfxEsPD0ngi31RNTfRoJ/tMG8aVJYkfZo9MsLViZxDOSYhu5373QM/6c0VTk7t90l923zvrcUdEkul/wAdz4o/Zg/Zx8b/AAX8S6lqvii9tJkmsBZST2dxvk1e488ynULyJbK02zkZ5llu5cyOPP2gbvNL39h25v8AQr6C7s/D8+r3+ieLLea6kWRml1jU74T6NePIYC5ayhLoJT88O4rCCpJr9IqKTfwvtf8AG/8Anf1t2KjK17dWn9x+b2tfsj/Fufx34y8WaVq2ltd62NZkttSuLh1uJ49Sszbwafdxw2Szm1t2K7Sb6WMCNHS3V+mX4A/Ys8Wp4jSL4laV4bufBlvdX97BoyT/AGuCOa8063tFAto9MsLQFJYPO3LCvzOWwZF3v+mtFS1eLi+1vkHNqn2d/m9z5xX4WeLm+Ang/wCHGsx6f4i8QaHYadbXj3l7eQ2009rbiKWVZ4EE7EtkruQbgcsAa0f2f/hXqfwm8Hnwpe2NhZJbQ21vC9leXV6Zlt4vK8yb7Skao5wOI1Cn0HAHvtFaTm25t/a3ISsorsfmtP8AspfGy58Pa94P0660Tw7pV7oWuaabe01TU7nTb+61FCttLFpt3DJHo6IzNJKtpJICTswygGvp/wAB+AfibdfFE/FT4npo+mT6fojaFYWGj3NxfK0c08dxNPPcXFvanduiRUjWHCjcS7EgL9FUUlN3UuqVvvuvyk16bDeqa/rp+qTPy0uP2NPjLqfinVb7UG8ORafr8zDVWt7qWI6hGdctNTEksUemxys/2eGWELcXVyVMhCSqhIrs/En7HviRrzU7PSNG8L674UlutdXSvD+pSTW+naTDq8VmIru2iitZUW4glhnIjVFGJiY5o2LZ/RiisfZR5eXpZr5O1/y07dLGntXzOXW9/mv6+fU+TtY/Z0l8V+Bvgt4F8Yx6fq+m+AHtDrVtOZHgvRbaTNZgRoYwJV+0PG+2UKrIDuH8J+cdB/YZ8Wrb6tpvixtF1SO4e333E0gl/td49WhvXu72FdPhKzi2SWNTLNePmZ0WVIiwb9P6K3nUcpuo922/mzOn7sVBbK34an5l6n+xh8TLu88erZ6tp0J8R22tQw34umimvV1O5WW2ivYoLBJFS0jHlxl7q627V8lIVJUfT/xB/Z98O6jonw80fwH4a0O2tPAWuwalb2dxEIII7crKs/kFIZtsvmSLOMph5EBZgTuH0rRWbXuxj2t+DuiuZ+953/FWf4Hxh+z7+zz4x+FnjC21vWrfRdPisNHuNMvLvSp7iW78TXctxHMmp6oksMSpcIqPgF7ht08gEgQAHhD+xZa6h4mvDrXh3wxN4dWfxfcW0IRmaSbX2tXspZYDbLGj2oiePId9qhGTnhP0KopVIcys+zXyer/4HboONRp/NP5pWX4M/PXwB+yt8VfDvxW8N+PPFGuWeqS6cNNlutUW7b7fi00uOymsEVrHzpbaWdGl+a9jjO8s1sZRvP6FUUVtOo5b/wBf1/WtzJRS1+QUUUVmUf/Q/fyiiigAooooAQqpIJGSOntS0UUAJtXduxz0z3xS0UUAUtR06y1WxudN1CIS295E8Eq5I3RyAqy5GCMg9jTNJ0rT9D0y00bSoRBZ2MMcEMYJbbHEoRFyxJOFAGSSfWtCihaXt1/T/h394Pp/X9bL7hCASCRkjpQQGGGGR70tFACbV3bsc9M98UpAIwaKKAPMfH3wd+HvxOltZ/GWnzXL2sM1qPs97d2QltrgqZbecWs0QngfYu6GXfGccr1r0mKCC3RIoI1jSNQihQAFUdAMdAPSpaKForA9dWFIABkgYz1paKACsvW9E0jxJpN3oOv2cWoabfRmK4t5lDxTRt95HU8MrDgqeCODxWpRQAhVSpUjIPGO2KWiigBGVWGGGR70tFFABRRRQAUUUUAFFFFABRRRQAUUUUAFFFFABRRRQAUUUUAFFFFABRRRQAUUUUAFFFFABRRRQAUUUUAFFFFABRRRQAUUUUAFFFFABRRRQAUUUUAFFFFABRRRQAUUUUAFFFFABRRRQAUUUUAFFFFABRRRQAUUUUAFFFFABRRRQAUUUUAf/9k=)



Glia (neuroglia)
- Functions
- Structural support
- Metabolic support
- Brain development
Astrocytes
- “Star-shaped”
- Probably most numerous cell type in CNS
- Physical and metabolic support
- Support blood/brain barrier
- Regulate local blood flow


- Interact with neurons
- Ion (Ca++/K+) buffering
- Neurotransmitter (e.g., glutamate) buffering

- Shape brain development, contribute to synaptic plasticity
- Disruption linked to cognitive impairment, disease (Chung, Welsh, Barres, & Stevens, 2015)
Myelinating cells
Oligodendrocytes
- In brain and spinal cord (CNS)
- 1:many neurons


Schwann cells
- In PNS
- 1:1 neuron
- Facilitate neuro-regeneration


- Mnemonics: COPS/SPOC
Microglia
- Phagocytosis
- Clean-up damaged, dead tissue
- Role in ‘pruning’ of synapses in normal development

Neurons
What makes neurons “special”
- Long-lived (most generated b/w 3-25 weeks gestational age)
- Extended branching (dendrites and axons)
- Electrically excitable
- Connect to small #s of other cells via synapses
- Release neurotransmitters
Macrostructure
- Dendrites
- Soma
- Axons
- Terminal buttons (boutons)

Dendrites
- Majority of input to neuron
- Passive (do not generate current flows) vs. active (generate current flows like axons)
Dendritic Spines

- Concentrate effects of local current flows, biochemical reactions
Soma (cell body)
- Varied shapes
- Nucleus
- Chromosomes
- Organelles
- Mitochondria
- Smooth and Rough Endoplasmic reticulum (ER)
Axons
Axon hillock
- Transitional zone between soma and axon
Initial segment
- Action potential generated
Nodes of Ranvier
- Gaps in myelin sheath
- Neuronal membrane exposed to extracellular space
- Action potential regenerates
Axon terminals

Synaptic bouton (terminal button)
- Synapse (~5-10K per neuron)
- Pre- (sending side) and postsynaptic (receiving side) membranes
- Synaptic cleft
- Synaptic vesicles
- Store/release neurotransmitters
- Autoreceptors & transporters

Classifying neurons
- Functional role
- Input (sensory), output (motor/secretory), interneurons
- Anatomy
- Unipolar
- Bipolar
- Multipolar

- By specific anatomy
- Pyramidal cells
- Stellate cells
- Purkinje cells
- Granule cells

Pyramidal cell (Wikipedia)

Pyramidal cell (left) | Stellate cell (right)
Morphology, physiology, gene transcription
](data:image/jpeg;base64,/9j/4AAQSkZJRgABAQAAAQABAAD/2wBDAAMCAgMCAgMDAwMEAwMEBQgFBQQEBQoHBwYIDAoMDAsKCwsNDhIQDQ4RDgsLEBYQERMUFRUVDA8XGBYUGBIUFRT/2wBDAQMEBAUEBQkFBQkUDQsNFBQUFBQUFBQUFBQUFBQUFBQUFBQUFBQUFBQUFBQUFBQUFBQUFBQUFBQUFBQUFBQUFBT/wgARCAQSA7IDASIAAhEBAxEB/8QAHAABAQACAwEBAAAAAAAAAAAAAAUEBgIDBwEI/8QAGgEBAAMBAQEAAAAAAAAAAAAAAAECAwQFBv/aAAwDAQACEAMQAAABo+6Uc87AAAAAAAAAAAAAAAAAAAAAAAACGi48fTHsDx8ewPHx7A8fHsDx/wBITYESAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA8w9P8wmv47PnRzehaF7VjVt5xG9qxDx/r9n8XmOX7P/ABh+z6X9DGW4AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADSN3I/IT9er0/IT9ej8dZH6gun5Cfr0fkL9H7ciQrcAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAawbOii0ii0ii0ii0ii0ii0ii0ii0ii0ii0ii0ii0ii0ii0ii0ijhd0u0WkUWkUWkUWkUWkUWkUWkUWkUWkUWkUWkUWkUWkUWkUWkUWkUWkUWkUWkUWkC+AAAAAAAAAAAAAAAAAADrOxPTFBPFBPFCNkTjYk8UE8UE8UHR3xIAAAAAAAABhcEUE9KgnjCu63UKCeKCeKDCzYAkAAAAAAAAAAAwuExQTxQTxQTxi2tdpFBPFBPFBPFBg50ASAAAAAAAAAAAAAwM/ARqkS15/rl6A8rpw9A++cj0Zo3XLfXmffD0V550S9J+6BtZu1mNZz0CLAAAAAAAAahCu6npjsPRoPw9FeZ0DfHl+QekPPsQ9MaFs8tj27UdupcK3AAAAAAAAAAA1DXdi03XGrR06XDfcjyqmneuWny0elffNLktuea5UPQHm3aeiPOqZvW56ZudbhW4AAAAAAAAAAAADDzCNT+batXUm2jUm2jSee5xiW20ak20al92wTKZWwJAAAAAAAA1vG21ampNtGpNtGl99C6ak20ak20a7sRFgiQAAAAAAAAAANbxttWpqfDbxpWRto1Jto0zup2jUm2jUm2jUm2jWdmIsESAAAAAAAAAAAAAAAAAjWYxZAAAAAAAAAAAAAAABCuwroAAAAAAAAAAAAAAAAAABFtRbQAAAAAAAAAAAAAAAAAAAAAAjWYxZAAAAAAAAYksvoQuoQuoQuoQuoQuoQuoQXdJsl1CF1CF1CF1CF1CF1CF1rOzAAAAAAAAAAAAEW1FtAAAAAAAAAAAAAAAhGRV4cwAAAh4l7bO1hM7O1gbPGwcM3NrA2drA2drHbEbEKVANf6L22drCZ2drAqZuq9xtTWBs7WBs7WMqIuilTEh2ts7WFp2drAyrulZxs7WBs7WBs7Fys6Gs/NL7O1gbO1gUK+m5Zs7WBs7WBs7WNhrXuFYAAAEqZqtYXts7WBs7WBs7WBm2tLzTZ2sDZ2sDZ2sDZ2sUIiuKVAAAAAAAkzfuKXs3xHaD0eTBzjawAAAaR5x6PqfZ0w703Hmaqb0FrsmpVOmRgQ2DumcS1d1TapeiDh5QPP9Q2/Xe3qnZs0U0zvM3pxOoo9+tVzPmfekzN0839IiN5HHzStI3fSOrfQb07AtbasTC6Jm3l6dtpxweHUdmzaNvMzvVOZT4uXzTWNn1Xt6aPOFkQqcIdo4ZEvBNrx8Lrl2dXXgQ3n0Lz30LPPLHNiAAA1XatV0v5700IPTvV5zuRXwMTrLvLUO82PlMgF+nF5FfslZUs3Wu/rhs+1aFvsRtw4+YAAAAAAAABEt4ZmI9gAAAhYe0r21ZtKZ1bF3OUSujbp5G7NgyDVm0jVu7YwGdANd6NpXtqzaUzqzaRqXPYO81ltI1ZtI1bLvIgKVxIO0rW1b7tC06tibpGJ+HukUwW0jVvu0DFyjOmscdpaX1ZtI1ZtI0vLo2DVm0jVm0jVtj7lahWAAAEmsmdWbSvbVm0jVm0jV/m0jSs3MuGrNpGrNpGrNpGrULKIClQAAAAAAAAAAAAAAAEqrKKU+hPMvIx8gAAAAAAAxe/o7zmAAAYhlNVzi4hi5H6+4rRbUUtgAfPupm1oYuIYuIY7LGnVS4hi4hi4hzzbQAAAPjUTbkMXEMXEMXEOSV7cHkXEMXEOUbihi4hi591umUQAAAAAAAADEMtHsAAAAA+H2VUllKfQnmXkY2QfXz6AAAAAAYvfj952AAAYeZhnVRm0gAD5FtRS2ABEtRS2AACPYjWQABHsRywAAAD5DuQy6AABDuQy39+fQBFtRS0+fQDHxMrFKYAAAAAAAAES3ELYAAAAPmnbjEPny8HDmIP26IWLs8MuAAAAAAxO/o7zsBoGrbR55w+RsXbrcqKbz06FWTtXPSOJvXRqUs9Ht+Y+mzb0GLai93r24tqLTPz2bg9nF5FfC0nqtp6R26Bu9c/QtF3rRNN/s3UsquW64GrUYbJ6D5b6jpto0qrpGeG29nn1NO3ctLxzfNo829ItfdePLj2+t5NiYmh8Hjeo9GozD0jC1asj1Ha9U2vr9PyXr7PPuPy9x7PPsm1926tX3GK9e8eZ+l31h69sGiVz2DePNPSrabf5N6z5NfaZ26X35c29+q+Tes69HeOn0AAAAAAAES3ELYAAAAPkS3ELgAAEO5DLgAAAAAMTv5/QDQNY9k+c/F433evIr413ev4h5TO9hop8c4ezEeQbBv/ANm32Lai9Hbbi2vla+R4fszDi8g6fZIx5789dEDRPXON9fH+n2ZTLx/r9XzTyP0C5yvr5PP9mUy8X5ezR0+a9nrxHjm47l9tflx5N+zyDh6+5uDwur6/gTPlXd68iNX2vjy36/JInubHl8h6/Yo8R5rw9lHkG47d9tfzKd67hRXy7b9v+zb55L67wtp4199cxs8PNfV+znpuGvQAAABqOy6HONvy9FyjcKPkGym9T5HabaAAAAD5EtxC4AABDuQy4AAAAAAAAABh5mGdVGdRAAPkW1FLYAES3ELYAAMLNws0AAR7EcsAAAATqM8oAAARLcQtgAYOd0ncADHxMvEKYAAAAAAAAES3ELYAAAAPkS3ELgAAEO5DLgAAAAAAAHHl1muZvbVIvXeEDttCBfh3AD5FtRS2ABEtxC2AADDzMPMAAEexHLAAAAE6jOKIAAES3ELYEqrEPqz9Iq0Is/aopw4bAIq0Iq0IFHvwioAAAAABEtxC2AAADQNa2Hz/AIvI2j7rMqtN87PPN8PXB3+0Ah3IZcAAAAAAA13Ytbzx0jGh8uTy6nZplybek7ppO7dPfp2n7d5lhx3cjzjNrT2Lc9K3Xp9D5FtRdei2ABEtxC2AADDzMLNAAEexHLAAAAGBn4JnAAARLcQteV+qeT8/D09UPW8eTesnVNniNm27Ttw37fN8ON8w4tgzfHfW5tuem7h5zbSt16bh0y3j5BwTaezTs+W07hpO6Tptw7fVAAARLcUtMHOAAAPPNY9l48/F41keh/Yr5nS9S+neOnvAQ7kMuAAAAAAAa3snCtPIsX2Phhx+Tc/RaiNP3fr7NuvTNL9j4Z4eQdnoVmtNQ3br7Nur5FtRb62wAIluIWwAAYeZh5gAAj2I5YAAAAwc7BM4AACJbiFryb1roy5/Iev2LBzw8y7PVyND3HN569PkWH7JhZc3lud6iTrulet9dtPHOHsyufjnz17CPK+32IeS7ns/O1+wb9gAAAGLlAAAABD5ceRaAAAh3IZcAAAAAAAA6+zrJ1WVVAAIluJbAPkW1FLYAES3ELYAAOnuAABHsRywAAAB0d4AAARLcQtgT6GAZ4AGBn4JnAAA6MLNwioAAAAAAAAAAACHy48i0AABDuQy4AAAAAAAB19nWTqsqqAARLcSlSclrLyuzZYuH0G3NYGzolv0OZEtxOjO2AAAAABHsRywAAAAAAABEtxC2BhZuEZoAGHmYRmgAA6MLNwioAAAAABNpRDnYAAAACHy48i0ABrWy6n43d84eN7b4/ZvXf5T6fFttH2XhgAAAAAOvs6ydVlVQACJn4Gfz6eWydwwvkfZwevOzCq48ubehsmt7J9T46Jbietx2wAAAAAI9iOWAAAAAAAAIluIWwMLNwjNAAw8zDMwAAHRhZuEVAAAAAAIluIWwAAazlcrRDXBD76ohrgh4uzQi5qm14/n9Pn/AB9BeZ1ed7Dsf2zJH0PmAAAAAAOvs6ydVlVQACJn4Gfz6eVzd0xfkPa1rtzaVq6ZnbBkmw7Jreye/wCeiW4nrcdsAAAAACPYjlgAAAAAAACJbiFsDBzp5QAAws3AM8AAHRhZuEVAAAADXDYwIluIWwAARbUW0AAAIV2EXPvz6AAAAAAAAAOvExjnV1euUQHVPOm1qVmFF87az1uzrCYKnOQtFeJ2SJbclCqlCqlCq4cw+SyqlCrH5yTa0oVUoVUoVUqgdoAADqllmJ9jm5IotdcmucwAOvsAAwzMRPhVwsLENsQxcRK52AAecej4x5X3bRsBH6dkmlsAAEW1FtAAACFdhFz78+gAAAAAAAAEP52Do+/fpc+/PpF6MngdDv8Ah9txbQi2opw+vpwfRz6+3gfOHf1n3h39Z8493A+24lswI1mSd3Z87DH7PnYdPP52mPy+8zqcx87PnIsAAAh24lskfPvwAqzaU0rAAAAQb0EuxrUU+2Y1kj/PvwsR7MYsgAAARLcQtgI/Hg6bSKgtatnlpF+Ftx5d/MTOji6LULng1nZ/sT6WkXO0pmDsxAAAAAAErC/WwXNtk/cIbT9l1OrGPww/nLtm/MPiULWr7Nvnyi5kalsv7hM7ZbDFDhifDP68XiUevE4lDhicTOtws7bNJ54mOlHsncjK7J3Iy+2dyMrngfTMYQz+U/kbMx8jt5wvAEO3rOZ5/Tz+YIzmCNjm9Py9bo7cAAAEGvrvLttMVhY3zrOsZpy+YI2eNxwzZ3U7eftF4AARLcQtg1DTtx0v4n3udjoxIjN4xuNmx09Y2fDTcefDn918/rOm7lD+O9vjjTuWVs3nD+XrtWz6ZufTnYH1njgAAAAANW2nVvG7tVxOzD8Pu3LWcmfS+9bPrOzfQ+b5LxzsLwPQ2STMzaWyPR/NfSva4ZHmvpPnPJt2cOrK5dNn13NmZaZ/RxWrk9kzmnNnZ0Wa1Ni1nvpfdOXHl6nLrsvJl+Z1dey6zTmv3MkdSczH7exF7T6+BEyqXHD2z3lIocnRtdCfQ+z8IOrIDTfPfQtJ+I97r3nRtiJPLvn3pl7zpm56NjH2HiAAAYXnXovn3zfqYmy672eV1YG0QeyU25M+2j5sETZM7bQPsfGyx04gAIV3rOf2RXI/C28/piLYiLYiLaHHkehzS+i24uiItqzEWxEzs1pQOzEAAAAABKqsLxFtzbRFsS6h1Yxflty7RFsRLPJvng4VtnaItqWiLYiLYiLYiLYiLYn/AHPbZxFtjpEWxEWxEWxEWxEWxEWxj5B284XgCL8tvP6Yi2Ii2ImTSWqHdgAAB0S7bl2iLbG8RbERbERbHS7nbzheAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPEPYfIt7NwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAec65NfaXiyXtLxYe0vFh7S8+9BrIJAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA/JPmnpfnu/NSg7DtEtJxtzy4eadHqnmEx6/+qvyr+qstgpoAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB4Pof60Wp+S360TH5Byv0vdPyW/Wg8P9wK3CJAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAEgroYuIYuIYuIYuIYuIYuIYuIYuIYuIYuIYuIY+XdayS4hi4hi4hi4hi4hi4hi4hi4hi4hi4hi4h4pswAAAAAAAAAAAAAAAAAAAAAAAEepLKvMAAAAABPlQRVrWkUWkUWoqbDbET7K0ii0ii0ii0+faVAAAAAAAAAAAAAAIq1rSKlaRRaRRaipsNsRPsrSKLSKLSKLSLlxGeKwAAAAAAAAAAAAAAA+fRoedj4RsWb5/cNp6uXEqAAAAAAajt2o6303uwJvTvtWNqWbDYPunfTd/mkdB6Fw1mfLdeWjcYbx90gbx26vtFp9L5ceXByAAAAAAAAAAAAAAeaSa2id3VsWXqPOF/nq3w3r5pXA3rokwpbl3aZMh6Hz1/Wj0nr0jmbo0PLNz2fQN/rXahyc4AAAAAAAAAAAAAAAAAHXMrxiy4cwAAAABr+wJnSW7NL6S3YaS3YaTjb/FIbdhpLdhpLdhpLdh8+mWYAAAAAAAAAAAAAGl8d2a6aS3YaS3YaT074NK4bxFIbdhpLdhpLdhpLdhpNi8iAzoAAAAAAAAAAAAAAAAAAAAAAAAAAAAAi2opaAAAAAAAAAAAAAAAAAAAAAAAi2opaAAAAAAAAAAAAAAAAAAAAAAAAAAANfNgRRaRRaRRaRRaRRaarQLSKLUV0GwootIotIuCbQii0ii01fOLSKLSLwLoAABLKiKLSKLSKLSKLSKLSKLTVs8tIotIotIotIotRXQbCii0ii0ii0izja0UWkUWkUWkUWkUWkUWkewAAAAAAAAAAAAAHzWjZnDmDFiMqGl1z3BDqTbIFrgAGDg1p9t6hTitxw530BIAwYjOkdU+tNsQ68z2oX2K9ljUqJcYWLNq8T5Miu3offM1RbQRYraifJlabehfS4h15t2i1wBOitFD+VpztahTLiHbm/0WsAAAAATsSuduJ8mQ3BDIuPn2+oAiRW2hq0ufInw+3NPqFxDFxDFxgZ9tAmQAAAAAAAAOnBzsEqAQb0GmXmHZNx+Lyr/AKV5T6tr0Wh1+kABqXn/AKB5rx+X1X4lvPD1TJxsnv8AZCbANS23Us8POpWRy5PNt+o+Z+ma9HmUivruPJztw9hTuMC/A16Nd4y5OeO1bX5T6tNvRB3eu8T9s8Gw5HR1/Ofis4ERLZ/X/JPW9OigOr0AGkbvpGXNonX1x+Xz69CJs0Rz9b8k9b36+Q6e8AAAADSdH3jTOLycHGmZkVu5nz7TL1vnw5+h7gTLxT2vw3n4vs753YcfDOhdszSyYm2RWPP5zJtsNjr7KZbVuembn2eqGvQAAAAAAAAB04OdglQCTWRXzvq9Jh5c2sbhTW0DTcADX9b9EZc/mnftVqKdHea9QTIDXthVp551ekYefPo+8Zi2vnb0RXLzft3bNNfk7stp529Ewq56RT3EsG3S0LfVMvPMf0uJTHVnog883fMW1DTYBq+0K087eiM+fzTI2m0eeeg8l9QvsAAAABrEP0Nlzed9HpUSK6x89EHHkbdYDz70FTLzx6Gpj5589E+HmuRs9w88ehjzx6GNa2U06AtcAAAABruxaobHF0uUeyyNC7T03l4vs56Dg9fYVAAIdyGXAAAAARbUW0AAAAMPMxDLABh5mHmADCzcEzgAAIluIWwAAPk3FLiKLSKPtqB2FtEFtEFtquznMAAAAHyJbiFwAAAAD59+ES5DuAAAAAABhdx3gAAAAA6cHOwSoABDuQy4AAAACLai2gAAABiZeIZYAMTLw8wAYGfgGeAABEtxC2A1+bnhuTTURcqaJlG5tNG5NNsTa01fFRuTTURuTTRdo6PkG6NNG5NNG5NN2u1+8X1A+RLcQuAAAAAfPvwiXIdwAAAAAAQb0csAAAAAA6cHOwSoABDuQy4AAAACLai2gAAABiZeGZgAMPMws0AYGfgGeAABEtxC2DRPM/TPNeLymdg8Yp2503siKMz5yKfoHnnod9IvnXoujVrldmBwiuxaNtUwr40a0ce7A6zN49HcUfX/ABz2PbpzR0+gB8iW4hcAAAAA+ffhEuQ7gAAAAAAj2I5YAAAAAB81rZhw5gAh3IZcAAAABFtRbQAAAAws3CM0AGFm4WaAMDPwDPAAAiW4hbBCnbczx1FtyK6jg73FmZjbkRqNawtbW8XbkRqLbkRqLbhoedStzOpNuRGotuGo7P3raDqvp2sMZcTPimzsMZjDGYxcoAAfPvwiXIdwAAAAAAR7EcsAAAAAAAAAQ7kMuAAHA5td7i4hjla0yuXEMXGubCcgAMLNwjNABhZuDnADBzsIzQAAIluIWwAE3FLcX5gG1ofItAHE5Nc7y4hhb0ywXEMXGubGANf2CIZagJ6gJ6gJ6gNX2eJbPoAHz78IlyHcAAAAAAEexHLAAAAAABr5sDhzAEO5DLgBhZ2zcfq6c7fKkGnLLY/VMYVrW69ZzWLytGFQiZ9Zoujv2oOuY7MJ0YaVGHmaVOrHrPzOl5UTlHHbPlhOjDSowszSv0aVARLcQtgMXhhpi14FKGawsu9eUbMk0tsbCy715dfHFiemtBqQynXjSwLevVazmsLtmJlqLa3p9Yfzn0zYmdHNlYQzWFlXrzYfyls1hDAt63YM1hDNYXfaO759+bUiXIdwAAAAAAR7EcsAAAAAA6p+fglQACHchlwHzQd+818H0cvpjTPH7L2zaPvKc/WNn836crtXS4/Np6x39Hf1Z65wgxea/sV2Fd+k81gZ+B0Za0Piff7dv1Db/ovLl6/sGv8AB0svEy+Pfaenu6ftPB1EfCfRNy03cve87kPofMARLcQtg1aHc0L4v3bfDW6dY2Ld/OvRfU5tB5S+Pkdl7cfNPS/W48HUdu8yw0vbB5j6dz6VNN3LzXrxqd+tZHn9FfZfIvXenPvtRbX13jafqe2eUfFe76Zk+T5cxvdLzGPMey7Rp249uOt6htfl/B0ekYWkzrV9UyPNOis+hUPKeyY9Tt+c+jdOdj59+fV+REuQ7gAAAAAAj2I5YAAAAAB04OdglQACHchlwHzV9pcW+rNpce+rduyJiPO2lpXVm0srwO203z1ZtLDSRXPS5WHmL11ZtLyuzWtlO3nnxtpY6atk7Ayu6u16vHqzaXkdurbPydmAduACJbiFsGv420vJ7dWbSpOrbJ2uvHVm0uTbVti73VjMk7SpbVsu8ztgxNpa01ZtLn11bOttKxLUW16vHrPHaXj92m5O0oas2katfynVlrPDaXNrqzaUNQ57YNWbSNWrU2+b59+elyxLkO4AAAAAAI9iOWHHXDZQAAGlUDZWrdxdwfn0qAAQ7mrG0Igtogtogtogtogtogt/YfQbGAAAAAAAAAARSyiC3EcS6iC2iC39h4JtQAAAAAAAIlqLaPoAAAAAAPjVc8togtog+XIf0togtogtok0258+gADWdmjmXC2oAAARNZ9BHlVjfRj4edglRGwvM69mayzts0PFxjb2sjZmsjZms5l62iD143msuHo2ZrI2aHi4xt7WRszWRsyVV9DlDegAAAABiw+Do2aJiY3Prt33WRszWdj6seaNhY6bM1lnbZoWNjG3tZGzNZGzIlvu5g6cwHz7xhHah0fJ+xuzz/laNwztGvVtbRVbWqOqbF381IfQeaMHK2c1l5vXszWRszWRnWNRyDZmsjZmsjZmNI6sdgay5dtmayNmh4uMbd91kbM1kbM1kbNHysX0uSwNagAAAAdODnYJgeV+p6R8j7GL1ZFDG+JlYNiloUja14j8Kv1GD6Dq+0XbJq206t7HF5tTz+rw+6Va6OUTi/eFSYn4dHITD7szMmu5XIdz6Pzg9HlAAAAAwvJvWdD+c9PV8zN4ed0SlrIT3ei6Dv3qcuv6Fvuued061n5nVnOB3ZfWar92fnpWdW7MvDS7sutbL9D5wevxAOPLjWdChWdY+J97P6eyVrnU2rRMultwaP31tuOxaDv3fz0h9T5CZTmce3l8zd5Hy3r4HfndERi/M7X7PtXM6YjW7WZ3zMm7j52d921/YNf8AW4/Osmx88zpwYtXpvXPjbfOzv0dvPgY/bhZ1onXOWRnbcurt6vrPHsDsxAAAAA6cGprZZ6s3nz6T1BWZ6gJ6gJ6gJ/blrQws1pWeoMbz1AT1AT1AT1AY2Sb5heAAAAAOvDoMbz8W1iZ26mf9J+f9a1xOqgpM9QVmeoCeoCeoDDzDagaVAceSHm8/1V4Po+dR/XUx5o9LQ816PUR5T6Pn8urL6PV5HT3KzPUHPpPUBPUBr1HHsE9QE9QHXi5zWs9QZWnqAnqAnqAnqAnqA6ZtiP0Z2BaAAAAAAAAAAAAAAAAAAAAAAAAAAGJl4hlffn0AAAAAAAAAAAAAAAAAHwl9fXdIyyNRqWhF76ccsAAAAAAAAR7EcsAAAAAAE0pOHMAAAAAAAAAAAAAAAAAAAYeZhGZ9+fQAAAAAAAAAAAAAAAAB8+/CHdhXQABHsRywAAAAAAABHsRywAB0d8ApdnmWGexPHb56G8nyD02N28yoAAAAAAAAAAAAAAAAAAACLNnxuL6DcKfnm2Wz2kdfhgAAAAAAAAAAAAAAATZnX59ye1ulLzaxn1ep8uHPu+bQ7nnuXdewdH6ef1fTHk+yHqfbiZfb88E1AAAAAR7EcsAAdHeJvymJ2LbGs188dODT1o2Zw5gAAAAAAAAAAAAAAAAAAGhxtxwOD6TXdsxbVsrI7fAAAAAAAAAAAAAAAAA1by/2CLx+75zsmxM+vcOfHl3/AC7z30LV8fQ8sxfUnP6uh9m8IttGX0d/f80E1AAAAARszGKwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAIGsWdCPWJ/j/Ue89HifaepYk/ST2jE8jonrXPw7APeePjnpxj1JehY+d7H98o+zb1Hu8z5J9Gw9M12K+xffNeibepZfjvsFtuQv0AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAOoO0AAAAAAH//xAA4EAABBAECBAUDBAEDBAMBAQADAAIEBQEUFRMgMzUGEjAyNBARQBclMVBgISQ2BxYiRSNBkEKA/9oACAEBAAEFAvHfiEjbXw3NJY0H+anrIkkwxtCz+xubcFHA/VKnX6pU6/VKnX6pU6/VKnX6pU6/VKnX6pU6/VKnVTaBua//ACz/AKkf8UWVM8J6fHDd5GUx2ysDfnGBuyz6f9P/APiP+Wf9SP8Aiiz/ABbeJYtqA/iGLmmH4jAS5p/EgIipLWthVuP4X/T/AP4j/lnjGpPdUX6Z3a/TO7X6aXa/TO7X6d3Go/TO7X6aXa/TO7X6Z3a8J1pqeg/zHHev82x3r/FYeulxOBOXAnLgTlwJy4E5cCcuBOXAnLgTlwJy4E5cCcuBOXAnLgTlwJy4E5cCcuBOXAnLgTlwJy4E5cCcuBOXAnLgTlwJy4E5cCcuBOXAnLAZm78CcuBOXAnLgTlwJy4E5cCcuBOXAnLgTlwJy4E5cCcuBOXAnLgTlwJy4E5cCcuBOXAnLgTlwJy4E5cCcuBOXAnLgTlwJy4E5cCcuBOXAnLgTlwJy4E5cCcuBOXAnJj5IZ39bTdq/oMd6/pz91/pSEwJuvCteFa8K14VrwrXhWvCq6QyNA14VrwrXhWvCteFa8KEZpsfhvmCY7XhWvCteFa8K14VgzNy14VrwrXhWvCteFMljI78l8wTHa8K14VrwrXhWvCteFa8KKdjp+vCteFa8K14VrwrXhWvCmzROd+bO+Mq+wdYs+n3/wBOb+V9/wDRVnT/AA5fyLGa2ug5/wBMsMwr/wCeX+V9/pD+T+TL+TOn5iEgzcTgoJuKz+FmQPBRE4ovp/8AXLH6/wCbO+Mmwn7jGqTtr31HkeKKdRaghJMKKQdtZVTpUiRWSTypseV5SVBcxpdcSQ2RV/8Ay1fF26s6f4cv5HiCOSVS2NUUqlVZfMWpcSJLpWvlnrJJyWEWVwT1JXjnVxJA5VV/uKni4r4fyfyZfybaE6ZLPShzYCgHw8UU0gQ67IHYgYZcggPGFtNIK2fENLqpVXIw8UMkMkOqIeQKCfBIdUUDaKKSGaP1/wA2WzJAaIy0JloTLQmWhMtCZBqMx8hA+QLQmWhMtCZaEy0JlojKCJwmfhyIhXm0RloTLRGWhMtEZcB/G0JloTLQmWhMtEZRopRn/JkRCvNoTLRGTq8r2x6nMUOhMtCZaEycB7S6Ey0JloTLQmWhMtCZaEyDEK0v9LTdq/oMd6/pz91/rabtX9BjvX9Ofuv9bTdq/oMd6/pz91/rabtX9BjvX9Ofuv8AW03avwJZtPFY6xc39xX7iv3FfuK/cV+4r9xX7iv3FfuK/cV+4r9xX7isa/d/3FfuK/cV+4r9xX7iv3FfuK/cV+4r9xX7iv3FST2EWP8Akn7r+cy0ilN61N2r8C07aHo+tjvXq2/avyT91/Ntv/kGxuGN55Vlpjb0t6W9LelvS3pQ7HSRN6W9LelvS3pCtuITkNa8Eu9LelvS3pb0pVnqYzLfyM3pb0t6W9LelGs9Qb6ST6cO9LelvS3pb0tw/wB7vS3pb0t6W9KMbUBWbn7Z3pb0t6W9LelMstVF3pb0t6W9LelvSE/ii9GZO0jt6W9LelvS3pb0t6W9Ilj55W9LelvS3pb0t6W9LelDn6t/r2lkOqhksZsUEScyTF4rOIMrC4sep6Fp8yys3xp8e9+xz2caMRlqB0vFvEyLNtFwwtnGDl1hHYdttEeUF1GMzNtEaIdiApIvyeSb8uVaNh2sO5GVr7iGwTLBvmbbxHgdbRG4xcxnSj2guGGa1tZuUfhmtPvKgWjDRaz5v0s/h4/mDcmkHFbh4ObSLwm2PFsd3iYFrQ6Xco/CJZeaXWWzTwFW/CTvdPs3xJ0i2iRCNs4727nG4TraK1MtRNbukbItWHEXc4/CfZeeXWW7TwFE+N6Nz77iY+EGDaeaMe6jAbm0EFzbKO8W5cafKso0NxbGOF2baLg2bSK08CyIdobOKckW1izHFsI4TYsI+ZMC3MZ8aUOWyn6/r+IK4thClSJNnCsquynVhq02L3wvEkQwyoTASPQtPmYhZ3WXVPkI1SfLD0zjZnVEmzZYVbpE8NcWAbY/93Dhnmxn1JnNLUmknlV5DWMX5PJN+W6G7NpmoK046kurk1knLcQTiNAqHw5MGoNByKhkZI6uISqm15rBNpnNKymO0Nb836Wfw8fzHqJDGMhHr5kepPDKOjOBO8PGI20ibhFnQD2Qx1DhlZRyGQlW/CTvceiFMGWnkSI0yrOaQCkLGI2KZ1hGqCBPsD2PLVYJUzoB7IY6lwzNo5DIKifG9G598+HmYiUT9PmpfgJKgj5Bqo+TQKUsIt594yl0cmY01SQgjNcIoKYjHR6k7WRKl8fDYsiPNBScCVFpjxnVUB8DFP1/xLLqehKrdSbZsrZsrZsrZsqVW5jRsU2c4k1mnjjqMvZs2Vs2Vs2Vs2UKpyMvIaqyU2zZWzZWzZWzZWzZRKh7R4ps5xs2Vs2Vs2Vs2Vs2VGrMxz/SUDUB2bK2bK2bK2bKPXcEmzZUWv1ONmytmytmytmyowdOFZps5zs2Vs2Vs2Vs2Vs2U+u8krZsrZsrZsrZsrZsrZsoLOEL0ZsHV52bK2bK2bK2bK2bK2bK2fK2bKzX/aZs2Vs2Vs2Vs2Vs2Vs2Vs2Vs2VDgaUn4kuM2WAGJzH+pa9rb7bPt0foevKwTMZn38nqz/kKq9nqG7r+S7vP9Xa9rb7bPt0foevKa98Zn3wz1Z/yFVez1Dd0/Jd3n+rte1t9tn26P0PXkjcWMzH2bzy8/aNBp4hIWyQlskJbJCWyQkGpiAKqr2csevBMkbJCWyQlskJbJCWyQlskJFp4eLLZIS2SEtkhLZIS2SEtkhKZWRorPRh10eXnZIS2SEtkhLZIS2SEtkhLZISsKmKJuxQfvskJbJCWyQlskJWdREFE2SEtkhLZIS2SEj0sNoa3P3r/AOgte1t9tn26P0PXlCyeMzHlZzzPi13wOaq9nLXdXmN3Xmtfj+jVezntfby2/wALkkdCs7d+FKktiBC6c8nqWva2+2z7dH6HryxceKzHlZzzPi13b+aq9nLXdXmN3XmtPj+jVezntPby2/wuSR0Kzt34Vl1PTrKqLJhbHCWxwc/R7MEZscFbHCWxwlHiih23qSxtNFZjys+lz8080MXIjNOP7r75RTMFjzYZj7r75QjtkD+6p8/75VXsVv8AA++UOzjFJxm8XcIozuM1j/uh9k+6+6dZxmFR7AEZzCYI2l+ETP8A5mso0d7TseTP/kvP/wCX3yvuqHqrP8ffK1YuB9/umFZ5/vlMngIah9idnP3a/wA2JE8ETJJQhBDNDIaKzjHdHmhlqhz/AOU7P+8bZxXlBPBKzQ5/83e375TLSKV5ZwI5M5ygdD8Cx6np0vbOfPefUlsYSKzGMM+l182Dnh3RZWTT4lhKsCCt5sxlvkj4TphNSCxkuFGsjvUGY8dVWz5NlIp/nKq9iuPgX7SvpSTghjnO7Eifl2B23GlHAZkkI+yKLPmljRJwoHhaVYSRKnzgcisl66vpfhP6tMfEWmxMNxNxLJJMlPjHPaSoI64spxqHqrP8Wch0eGfG3wDzT8Uko48zLYo5XkdItqD2LPurZZpDcTZsk0TgCkXsrjMyUT1Aa48qg9075tJg3kiMcewoPe/2t9wCA/7Wk5kVwcZ+7QdD8Cx6np0vbOfPefUl4G6Kz7eT6XXzTRxSMPihKPyN+74oCNINhWMAIbYVYGFjEcTTZhx8po2MVP8AOVV7FcfATYcdpeGzh6UHEw3DUxrRtH2RYExqdEA8uRMyjRQyVjGG4pfhE6hYgDvPH4jWxAti4ALGPI37hjij4oeqs/xnGHZcNhEOpjslZExyzGC4mBtw+g9id7i1YDSvI3GcgFkYQCj4FEABzBsGqD3T/mtY1iaNo1Qe9/tTIgBPfGCQiB0PwLHqenS9s58959T+eTy4yvI1eRq8jV5GqW3Glrm4zA8jV5GryNXkavLjH0qvYvt915GryNXkaq/H3L5GryNX2/08jV5GryNT/NiV5GryNWMfZeTC8jV5Gq0bjT+Rq8jV5GrGMY+vkavI1eRqiRsBx5GryNWMYx9PI1eRq8jVbf8Axw/I1eRq8jVjGMLy4UyNk4PJhYxjH08jUduMBrv/ADgeRvpxbc0601IvtT3YrMU22DHBGtQki6sGojygy22PU9Ol7Zz57z+LM+LXfA5qr2ctd1eZ7iYl81r8f0YYhibz3PweWYDMkHJI6FZ2707esNZzc1E5RaSVhkKokhp7CmtptWarkbv4dhyoY5VeCNI9Ol7Zz57z+LM+LXfA5qr2ctd1eZ+S6vmtfj+jDwHDee5+DylC07OSR0Kzt34Vj1PTpe2c+e8/guz9mwwypMPRSVopKfXyCMFXHCPRSUDJw2HJVezlrurzP4uq5rX4/ow3Cc3nufg/W0M8EHRyVopK0UlaKStFJVgOXFhOgyHNFXHCLRSVopK0UlaKSiRJLWQSuPC9Sx6np0vbOfPefwX+yq7Xy57zyVXs5a7q8z+Lqua1+P6MMrCt57n4P1uu28t12nmN0avtvqWPU9C3e7E488cZNPl7cPdjHEcmEd5+XPefXu85xGybLWgnjlIdnHe4FkKS6jdlx1euy0hpjYzdYPh5PnDqN2XG+lV7OWu6vM/BdVzWvx/RiGyfHPc/BTyO85ZeI7NaN4MW0fLAzWyW1D3ZnWP+kLJXYQp4paxLxlgSO4167LQ8Ry4jkSRwmBnMkpk4bivsgiKeeyMmSOI2idl3o2PU9C5+dV4/cXSHkmhtZU7G7zZSDn755c959e9+Lc/ZWGP3Goab/t+kwbFdQ9dX3UD3+eEZzRCvZcUPW+lV7OWu6vM9pMyua1+P6MUhSY57n4KJ73f8gvcY+9w2RiGHBMMp/nWPwbz41rjHFJ+3VUb333RspWYMCZZyKxXZzDiWnlxMDx6s1YccXwtW/wDlbD8nkoPRsep6Fx888MErJIMYzOCPzPgxiofU5c959e9+K5rXtjw48TLQjYNrWsbQ9dX3UPGDKa2MFgsjZklD1vpVezlrurzPYTMrmtfj+jF43257n4KJ7zgFJY2IBoHsYXCp/nWPwc4w7EeDHiuzHE4IOtfdH+UGHHj4ZAjDECJHi5HBjBKSvimIeICUmMYJlB6No1/DBNBJbz5Ex2eANcJm8WAmNJwBrgD5s959dzcPXAGngH5KsLHVvAGmjaz6OG164A1kTN34A00bWfWq9nLXdXmeJ+ZXNa/H9GKMo8c9z8FcAa4A1MZHHG4A1wBrAmNznGM44A1NZHHE4A1wRpzGvXAGuANcAaMEfCrAsdXcAa4A1wBprGs9LSh4vo/+6serz57z+C/2VXa+XPeeSq9nLXdXmyLGS81r8f0RBwHHPc/B+s8owxeWeRgYXMbo1fbPx/8A3Vj1efPefwX+yq7Xy57zyVXs5a7q+ra/H/Bufg/WafTxuWcbTQ+Y3Rq+2fj/APurHq8+e8/gv9lV2vlz3nkqvZy13V9W1+P+Dc/B+swrwx+WYV4YvMbo1fbPx/8A3Vj1efPefwX+yq7Xy57zIdlgdYZawq1hUIrgY1hVrCqLIIQv0rur6tr8f8G5+D9Zjitj8sxxWROY3Rq+2epPkujBBAcx/o/+6serymMTBtS/78R3nzJJhBOTJfXf7KrtfLnvMv4/iZ7mVVfYYixy3umdVmMS7+kHr/Su6vq2vx/wbn4P1mYNmPyzMFzF5jdGr7Z6lj1PS/8AdWPV5ZHWjt89kS7MwdnMPPih6/rv9lV2vlz3mX8efGFLAeljyCOpAlbErmxTtdh+FB6/0rur6tr8f8G5+D9ZgyFj8swZCxeY3Rq+2epY9TmiyZ8uN+4r9xX7io0eRrZ8ch8fuK/cV+4qTKnRBKR131UUgHVMV5300QhAdb13+yq7Xy57zL+P4m823TLEmDbxLaJtzI8lZMfGq666lWhoPX+ld1fVtfj/AINz8H6zQ6iNyzQ6iJzG6NX2z1LHqc1L2n0LvtqzHG7OmEtMJaYSxHHjPrv9lV2vlz3mX8eTGFME2sisZBowxybZFxJzTwsqPXxomYPX+ld1fVtfj/g3PwfrOEw8XlniYeHzG6NX2z1LHqc1L2n0Lvtv4r/ZVdr5c95+33XDauG1cNq4bVw2rhtWG4x9a7q+ra/H/Bufg/V7GkbyvY0jOY3Rq+2eh/3FW4P9bHqc1L2n0Lvtv4hZ8YD32sLy1lpDbXMsYpX/AEIVgWbrCWbKJu+6wkIrDs+j3tEzd4K3eCt3grd4K3eCq+zhtLusJbrCW6wlusJbrCW6wkx7SMWc/bG7QlusJbrCW6wlZ2cNwN1hLdYS3WEt1hLdYS3WEt1hIZWGZzEKwI95greoCtbaESHvUDC3qBhbzATH4I30ZEsMTG9QFvcDKNdQOHXW8IdfvcDK3uAt6gIZGlZyWDTv8Tn8TyZdXY3k2tJSnmEUmbiRI5qXtPoXfbfxIjcZmjY3d8txvVm3GHfS1x948wI9bkLN4iBHrarH2D9Lrtdp07DH/wAk3H++k91fj93w3GbkLG7tEZjcITG8asZhVTG6amZjbaXtSn/BmMb/ANvXDG7bZsbp7RmFNY3jy2Y3AzG7rlmN6Yxu8RmY3SExutr2N41T/pD5rrtiq+gLu075Vx2knsqe1ejM7iqj4U35iq/jz+uqbtXIQP3y/wAPMJUjgRhAjxhRB2PV5qXtPoXfbfxIfzR94z3u0/n6WnQmfOz3mH86r6X0ue12vSsOpN+fJ7q/u7e8h7tD7jB61Wqr4tN22l7Up/wZf/Hrnttn8a1U3rzO4G7rnvTO8Ru6Qvm1/Wqvic112xVfQF3ad8q47ST2VPavRmdxVR8Kb8xVfx5/XVN2r0LHqc1L2n0Lvtv4kP5o+8Z73afz9LToTPnZ7zD+dV9L6XPa7XpWHUm/Pk91f3dveQ92h9xg9arVV8Wm7bS9qU/4Mv8A49c9ts/jWqm9eZ3A3dc96Z3iN3SF82v61V8Tmuu2Kr6Au7TvlXHaSeyp7V6MzuKqPhTfmKr+PP66pu1ehY9T6On+V24rcVuKgycxIe4rcVuKxn74RpnBJuK3FTT6yPuK3Fbio8jj49cs3hEDJ4RmyfLM4/8AvpR9TkMzikUuTqGGkcU+o/3gZGRSIknMZjc/fEiTwFNkauLKkalsiRx3GPxjlPxJeZP3mYkfaayT5JYT8KQCTwXxZGmUSTphQpGkjVWeHHkSeBk8zjgOXi102Rq40qTqByz6lHk8YhpHFkPk+aXx/wDfYkfaaORw5QJHBPHkcB8WTpRALxh8s0+rj7ioknTDbJ8ss8njFmSdXEdYfdtYTyA9AxOEMx+LJ3FRJGlCeTxjbiosnTDkSeO/cVDk6SI1/mbz2PU+herm1KzxHR2pbMo7KOWQG4iSHR7mHKUWxBMcm+1TfkXk99eKKduYTbiI4ObmG0O7xeHGlDlsrvb68r5EOwMezjWzy3Kn3GlsYXyFZ2roc+9szVoWuw9thMfHJWTczow/ZY/zS2j55ItsQ1zDky7cOP4lTDEsymLXwayeeVXU059lW2s/FXXgDOa8VsR12adIj2ar+pY++XJbDjUs00yNaWhocmMTJY1vKPGiQJbbCESwMO5NNlhtlVWbpk6luHTI8Es6fD+kHocv/wDUHxAc9bm7kDoVf2parGbQufElHautRwOv6Ez497MJX1JZMmulpluQUYWH4FFuC7zVSXzKzxBIlQoIWEGMXS57HqfQvVsqN8/EKu0M2HXHiCFSyMsf4dyaLV1WYZE32qb8iVqUCjw2nkQJc6FIpTzSTvD7pkithaIVd7fXl/IZDlss8+HGBFnFn5JlAOTIhfIVvGPNjNoxvNEhWFdClxTSx1Fdirgj9liiV0yTKz4dYAceDLrWfadxdBL45oB7BRKt1bMqoUyugGgyrKIPck7w41kUMKUy3+1pw6/q2PvsYW4MZRDHNl0XEeMU4ONtLNNEoAiHIhyCW+YJsT27h5318mXMqa+XCiEppmad2v4oOJwYPQ5f/uL4eyGS+lk48PG1+XGriziwKIkPIoJn2sDr+hM+PcwX2VZmFJmSsYs+HJqOPcfa04R6bUAixJ8KFYQZFjXZQulz2mfJ9XQPM7blty25bctuW3Lblj/TCND4pNuW3Lblty25bco0fgY9csLiE25bctuW3IMPhEW3Lblty25bcm4+2JEbjrblty25bctuW3Lblty25bctuW3LblHi8B0iNx87ctuW3Lblty25bctuW3Lblty25bctuQBcEfLty25bctuW3LbltyBE4L/QMPij25bctuW3Lblty25bctuTGeVvOQbSjBX5ju//AE4leMdn8bwZoLKJ/mttR3EfxN4So3eHKD//ABBfeOYXh6d+q1Wv1Wq1+q1Wv1Wq1+q1Wv1Wq1+q1WvD3jWH4kmf5d/1P/5LXQC2k6bCqY+HQJLIsXw9ZSzbNOzK0Z+GQbhE/wClXf8A/Lv+p/8AyWls3U1rODTOUzxDBK03iUefE8O7gxxV3iqPCmymjbI/6Vd//wAu8aeCbC/uP0uuF+l1wv0uuF+m1rqf0uuF+l1wv0uuF4H8Gz/D1p/mWO9f5tjvX+O2BSjWnnLTTlppy005aactNOWmnLTTlppy005aactNOWmnLTTlppy005aactNOWmnLTTlppy005aactNOW3S9Tppy005aactNOWmnLTTlppy005aactNOWmnLTTlppy005aactNOWmnLTTlppy005aactNOR8zIn9i6yxxW58zfxLDq/19r0f68mfKyoZhlX6siaOM7dgLdgLdgLdgLdgLdgLdgKXPGYm7AW7AW7AW7AW7AW7AW7AW7AX8/nbsDGd2At2At2At2At2At2At2At2Ap08cge7AW7AW7AW7AW7AW7AW7AW7AW7AUeYOVn+hrZVheQ41vnErf4PCZZxyS5MgQ2VXbPVuetZTNvgBedOI1jXymseOaAsd0wLZOCsc9phvd5sYw0rCJpWPbkjWrjj++Ssw/BWZem/wAfmu9wZ2Cy5dg2OLMnH2rrFs8bbEWFxGcTjDy9zvs1slnC1YdVgzHLUCw1xWDb5sLisw5pWPTSsemEaVtN7/6GFWT6iPKoZrz/APap2V7aWQ23lRAFbVds9W5615GJMp3xGmgsgSRNfAcaWOJJbBra08V8KtOPG3Zq/Ds2tK7w7Z05jkJWE8jqw8qRNrCubLqCkECKUNwm/wAfmu90+rNJJLqSFlurDYmRawzFHqzAxtp1tHCpYgSErAU5PKCtOw2KU2azNY3cgU8kLM1x4ij0xfuStMYL6j/zqRnECm9/9KTHmZUPw+r9Wxhkkk2o62o62o62o62o62o62o6NTOJnajrajrajrajrajrajrajrajrH8fm5qz5ztR1tR1tR1tR1tR1tR0ajfIZtUhOrTNW1HW1HW1HW1HW1HW1HW1HW1HW1HVdEJGd/TOrfsVmMtb+JY9X+vtej/g1h1f6+16P+DWHV/r7Xo/4NYdX+vtej/bGyY1jo5C0chaOQtHIWjkLRyFo5C0chaOQtHIRRymTNHIWjkLRyFo5CJWGLnRSFo5C0chaOQtHIQBSil0chaOQtHIWjkKGKVIHo5C0chaOQtHIUXJhz+e0M+NX6OQtHIWjkLRyFo5C0chaOQtHIWjkLRyFo5C0chQxypDdHIWjkLRyFo5C0chaOQtHIWjkLRyFo5CNWGO3RSFo5C0chaOQtHIWjkLRyFXjlS4ejkLRyFo5C0chaOQtHIWjkLRyFo5C0chaOQtHIVYUhA/0Le8+lI7r6UL5PJWdHkZ3nnu+0+pV9P8ACpO2epVez8rOftiLGzZCY3DG/SRIZGHvMZYsQ7lvMZAO2QPmkzBxFvMZFsgvnbzGTHYezmkzBxFvMZRrMAzbzGWCYyLeYy3mMoVkAA95jKNLHLwW0AEm8xk2yDix3mMgWQJBPq+2jsdvMZWdkGTA3mMt5jLeYyETBh8smeKK7eYy3mMoNkADN5jLeY3rSZwojt5jLeYyrLEMaDvMZbzGWM/fHJm4jYzvMZbzGW8xlvMZQLEMdu8xlvMZbzGW8xlvMZRpopefxDdKr7Z9br4UyXiG2XKHBjANiQCn+BzX/wDEuXpcgsJBpqjfG5r/APiVMeGS66Jph5dkY+3p85z5UuxkRBDy/LKD2WXzo0vUGBN1L6+ZifDqPn/WxnEHbQLVlgSRdjjxCS8DmybJ4ZALAMh9f8LlvfkSZuRSSzntkS554pQy2FKm+31L7rzZmILJ9liDiRbOC8MwJypns5LSXoIkqZkBg2El5oVoSQSPNcWTEtGypbJmoissmmijtyyFCsmyxKh9/wCIbpVfbPrdfCuBEKCbHJZgrBPj11P8Dmv/AOLEDzPjgIy1Ub43Nf8A8WEHMuwnwv8AaIfb/wD6YMkCfYDNNhqg9ll86LksSwh1hIxqqvIHFR8/62ddiX4gtYRvs+rycwBzI5JkTVTYQyDjV/wuW9+RaBFJa6G4lbpJWQ4bj7pvt9S+68uM2bF0Us9G2qaey+2Pumezku475dcWpGyYOIRpqqrJVkZHkDlsqSQj4x9lX1xI1gGme5DG0I1Q+/8AEN0qvtn1swPkxtnlLbD8TZ5Srgujxea2iFlY2eUh1xzZ2eUgNywPNbRCysbPKTaw73bPKTBuxE2eUtnlIdYcuNnlKpiEitm1kg0rZ5S2iUtnlKvrjx5f1PVSXm2eUj154wdnkrZ5K2iUog8ijctrBNKNs8pbPKQq458bPKWzylj+PUtYRpRdnlLZ5SBXnki2eUtnlJv+jeR1RJy7Z5S2eUtnlLaJSDXnPjZ5S2eUtnlLZ5S2eUqmEWK78Q3Sq+2cre8+lXdb0gG4heSKbjs5MMLrOe77T6lX0/wqTtnqVXs/DsbR0SWEjnDdcjZbEMwSi2jZEh0gTR8VnEN0qvtnK3vPpV3W9IJXEJyRSuMzkwJ+s57vtPqVfT/CpO2epVez8O+AKWGugWNaeuppEFRKc8BTIU5h9jljrRQpQ/EEyC0+KrtnK3vPpV3W9ILyOJyRnkezkwH7Tee77TzWZHCrsVn+m2LbFti2tDphDxtQ1tQ1tQ1tQ1tQ1YV+AQGf6t9Ok7Z6lV7PVkymxcBMyQL0zdKr7Zyt7z6Vd1vSDxeJyRuL5OTAmYnc932nmt+1Y/j0rbtbPZ6dJ2z1Kr2esweltfTN0qvtnK3vPpV3W9ILSYJyRmkazkwwWu57vtP0n2eYZt/ct/ct/cpVvmVF35y39y39y39yr5+ZqlXGY8jf3Lf3Lf3Lf3Lf3KVb5kxsXrsY39y39y39y39y39yATjB5aTtnqVXs9Y3dfTN0qvtnK3vPpV3W9II3sJyRhvEzkxwtdz3fafpe/LFaPZbVVkU8PNxFw11yxwSWsYRX2UdhiX4RgHL4sygVn84Nsbi4tQiCa2igfIJwo5LuU3w8awDHxIuI4E2za9wrWKZEuY7I+LgWTQj6gUL4fLSds9Sq9nrG7r6ZulV9s5W959Ku63pBFwy8kUOQs5MOFrue77T9L35cuk1aNUkdmDTOhkPUkKGTUuOI1SQjT1TzBHFJr6BWfzpNNqYGaswk6k/25q0RBlo8PNOoSzGbd92Zqykwek1Ay1hpmcQTcWviZhAhfD5aTtnqVXs9Y3dfTz/qo5DVg2O87ORvefSrut6QBNGXkiCaFnJgo9dz3fafpOrNaXYFsC2BbAoVTrIuwLYFsCgV+hUmn1B9gWwLYFsC2BTanSRtgWwLYFsC2BbAgj4IeWk7Z6lV7PWN3X8JvefSrut6QGDaXkiMGxnJg2Mzee77Tz0va/Suu3eg8jRY1gFrALWAVNLDis1gFrALWAWsAmyRPdz1Xs9Y3dfwm959Ku63pA4fF5InCwPkwZ2ZnPd9p56XtfpXXbvQsBMLN2+Ktvirb4q2+Ktvirb4q2+KtviqwiAC7nqvZ6xu6/hN7zyvd5GhnTTh1E5aictROUE83BdROWonLUTkedMjhbnzN5QZHkvJEyPI+TBCaznu+08k2U+OtROWonKDr4kTUTkGYfVfXOftgE+ZJBqJy1E5aicrY83MDUTlqJy1E5Gny4zOSZ8/0LT0Kr2esbuv4Te88p+jV9t+td1vra9rF0+UD2OLyRCMIPkxkus57vtPJM+VyH7r9Seyp7V9brt31ue3ckz5/oWnoVXs9Y3dfUHJlTk37+Xkb3n66wS1gkaWLIa2UNlfiUJ2SGYJawSgSRtLrBJskb3EOwWbOUN9aOWLAxmYX6vfgbdYJDmD8+JQs5RCNFjWCQZg/K2SN7lnPlxrBLUM42sFz3fafo6SNjtYJSjsfI1glrBLGfvjWCRpI82esEmuw7BCtEnyxZbWnYGuZIGRzyNHjWCVoZh4OrEtYJMkMJm57ctWLC1glrBKVJHmdrBLWCWsEmvw9uZYsLWCWsEtYJWUgb1rBLWCWsEtYJDK0v1qvZ6xu6+mXp1XbOVvefpn+JcnEOJmTjEWNYClC3oWBB61h7wTWnlAsRyZKifIsOtFmtlFjz2SiV3u+k34/wBBdVWHS+kX5CN0fq328t32n6SvkQprZw41mOTIzaMUeQyXHH0bCa2uhTZo4EcJOKyN0LH24mtzYSLEEWTC+RP6Eqa2ISXYDiFky2xmxp7Jj4HXue3J/vFfNI4BuOxEmtHP+kT47/eHxHHNSxjYkx3WIxzOPjjKxmYroOP9cKu9v0qvZ6xu6+mbpVfbOVvefpn+LgTj1DbN4qtkXOau+G4yF1rD3gK6FccDIrmpFK3WJ8iw6sArodlUQDQz13u+k34/0F1VYdL6RfkI3R+rfby3fafpK+RTGdFUOSTGCQxxKaoaZlUPo+IgvkUcgsexiUmJDYkboWPtO90TxBOrJb7OF8if0LvD8FtOFMjxByJE6mgGhmgde57cne+LBK12RZGcMQrVHBMjFfCmOjR4wog4nxz/AOih08lhYLJMNuaySWBOE5gGxTkDNgvPFtmSpUiogcKVXe36VXs9Y3dfTN0qvtnK3vP0ytKVaQq0pVpCoUYmCTQvI7SFWlKtIVRo5GGmBeQukKtKVQgvHn6SmZeHSlWlKhxi4Ipg3EHpSrSlUeORpkXH3HpSrSlWlKm/xy3fafpIjkebSFWkKtKVaQqZj7D0pVpCrSlQMZaKaJxG6Qq0hVFARhpjHEFpCrSFWlKtIVQwPGW57cnRS5dpCp9ZxDaQq0hVpCrSlUZuWBdFLl2lKtIVaUqNXukCZCeNmkKtIVaUqhDcNv0qvZ6xu6+mbpVfbOVvef6e77T+Nc9u/FqvZ6xu6uz9sQ7Uhp3om6VX2zlx3n+nuu1fjXPbvxar2etOhAlWseECIqaHwpPL4oNKhwotoBgI/iSHKfFvokyRLnxgYqu2cpoYZdzssNbLDWyw1ssNbLDWyw1ssNbLDWyw1ssNbLDWyw1ssNVwWR534t12vZYa2WGtlhrNHBdjZYa2WGtlhrZYa2WGtGKHa/gXPbvXh1seU3ZYa2WGtlhrZYa2WGsUcLC2WGtlhrZYa2WGtlhrZYamV4Ijuc3dee4rt1hNr8SfFlDTSpFdT+GG1JTNxw6vtnK3vPqxe6fi3Xa/Rk919PP+mNxW4rcVuKmyNXG3FbitxW4oBuMz0qvoepa/xzm7r6ZulV9slyHiLrDLWlWtKtaVcV+o1pVrSrWmWtKtaVRZJCFUiURhtYZa0q1pVrSrWlTCvGbWlWtKtaVa0yhneXPqyX5GHWGWtKpBXyg60q1pVrSpv+rZch4i60q1hlrSrWlTyPefWlWtKtaVa0qjSSENzO9tqVwarXti1hL4Qcb4LGBXUfLcZ++PrA6H1llcIetKtaVa0q1pVrSrWlQTPjt1pVrDLWlWtKtaVR35eEssrS6wy1pVrSrWlWtKjFedawq1pVrSrWlWtKtaVRCOKM3dfTN0qvtlh1q2Vtr8XuSAZeuaHF+LOLO1xWkzevjHsbzVwI0/MVM8QllOZ4mASZB66l/InkzX2ce1O6S3xOxw5li+DDdfcEpLt2p31m5xvEJZeMeIQqPIkE8R13u9WZ8e+dnWV9o6LWi8SDOqa1x9meKMHBWyCS4DPbP69g90m5tw7aHF19gg8Q6zEa+1qqr/ABFrD2+Cvl+JgQ5EOxfMlwvkczvbKj4lxdle+NmjaV06qa11HFKPlgdD62HR8QtkkTfEI2vZ4jG5Sr7ADFvXBZLv+Awcxwpbb1ubLw/YFbWjv8yC4v3vfWztwjRPjyOswmYFvFuc/aPe6zA7xpZdbeMsTx7QkEjrzDc4umZGK94zgXozAi+KBzVSzC2FXX9E3dfTN0qvtlhj7lLT/dkCjw2IOmjixtOHWc+BiwE+ljvcap1MR1GLJmUYRMDW4jmg9dS/kSqlszMikDJWxhcW7hEnR5tV9oAKoUdwaEACRqcEVM8Ow2BjVbQS673erM+PNr2zSNogjfsAdEyijDMCraAEONo4rPbP68yAyZl1U0qJSBeeLSgiEHRhjoVEGMOVSBlrNY3EiNDZEdC+RzO9s+VoYEm+NBFixluPnxAL748SPGG0m2EOHItnQTvvChLXTiSTQOh9bDomjMOQlBEKYtXqY7aMPEk0oZL7eoLJy6mC6RGogxSjpAgDFqI8QrKcDMQ4Y4I4nx5HXdVNIYFOCOQtU2Cq3w60cCJEzFT6MJCZpAuJsQeKajcEwKEIHw6xsJsCHiviV/RN3X0zdKq7YSMwrtCJaES0IloRLQiWhEtCJaES0IloRIcZgnJ8QZHaES0IloRLQiWhEtCJaES0IloRLQiQgMD6z2YI3QiWhEpcL7RdCJaES0Ilj/TBIzCu0IloRLQiWhEtCJaES0IloRLQiQ4rBu5ne20jumVjKOPkEivFJe2obuDamPwH1QSxyVMYuZ1YIraeASG6B0PqQTSt0IloRLQiWhEtCJaESggacWhEtCJaES0IloRJjMDa6GNztCJaES0IloRLQiWhEtCJaES0IloRLQiWhEhiaJpu6+oJkqtazOXM/qJeCOi4/G8jV5GryNXkavI1eRq8jV5GryNWMfb06vofiG7r/bSxvLFx/TEtIgSbzBW8wVvMFV1tDYHeYK3mCg2cSQT8E3df7aWLMiLj+mrOpzTPl/gm7r/bTBYPEx/H9LWdTmmfL/BN3X1JFjHivY/BGf1E1gyQ8fx/S1nU5pny/wAE3deQ5xxhRJoJwzSRR88js+VtOLDYP9RJs2RSntQyA72NQ5rZn9HMmthrexqHZNju3saxn74+h7Vkcu9jR7NhTb2Nb2NDfxB+sbuvJdxRTYGL6fHLaTjT5s2fOKTFvLHaQbqc6PQ3FlYvmTxAxVds/qLf5v0o/wCP6K8/jWZ3U1hkVimez6WXza6XmYME4jpxLjLUzOcsi/G9Y3deQwRyB4rYjYu2RODmui5iirs4s6ihj1IRwIwpBulV9s/qLf5v0o/4/orz+JIJLLWUGSawTPZ9LL5teKVDfFDOCN0RseDVRHQK6L8b1i5+9t6kcpKxjHedv9PNrHyT7IVbIVV0J0P+jsYTpmNkKtkKtkKm4+2PpLqiHkbIVbIVbIVbIVBZwxerJKUWIcR43/8A6U31pslPG8ZhmwcWURxa7xBX2gHTo7Fm1hYx4kus0FXTeIXT5w7SGZlhaaR+vjalkwBMPs4glNm6U1z4kHSzwWrSzmTAEG6UFqzPjYw6UFhszQ+cFmN0Is2OBGlgjp8sIi/494sgGtPDlh4QlM8S1/h24JfB8L2MWgP4Ys7HN/4PkEtfFdIez8Nu8HTotYPwYadXu8MzyQIPhKaG6keCbeMWz8KS4Yrb5dnU5sbbHhWybEfQy8RX0M6WrWiNiXimmx7SnqpEFuPDU4UdvhonHBSy4jrDw7KkWOMfbH+VPG1+fV//xAAzEQACAQMCBQIFBAICAwEAAAAAAQIDERIUUQQQEyExIEEiMDIzQEJQYGFScCNxRICB4f/aAAgBAwEBPwH8rJGSMkZIun+7vxy7FuUPP7xgjBGCMEKNv5FcuXLly5f5dy5cuXL/ADrly5cuXLl/wHytzsW5Lx8r3525x+b78rc7FuUfwMTExMTExMfl4mJiYmJiJW+biYmJiYmJiYiVv9EYswZgzBmDMGYvnizBmDMGYMwZi+Si2YMwZgzBmD5YMwZgzBmDMGNNem1zBmDMGYMwZgzBmDMWvwF4O53O53O/o9judzvy7j8coeDudzudxD88u53O/LuVPTDzy7ncvy78u5L6fwMmZszZkzNmbMnzyZmzNmbM2ZsyfJSaM2ZszZmzN8s2ZszZmzNmbG2/SnYzZmzNmbMmZszZmzJv/S0/rkYmPfuYMwZBWmuVT6GJdhQGrIrfp/6MDHtdlDtUEjBmDKXaouUY3FAx3KP20JXMC1il9xD8spfcRxPsY9zhvL/Mlw95XuaZ7mme5pnuaZ7keHxd78pLJWNN/Zpnuab+ydHO3c0z3NM9ynR6bvc039mme5pv7IUMZZX5ab+zTvc0z3IRwjY0zXuab+zTPchQwllcfDXfkhQwllcq0uoaZ7lKl0/f/Q0/qkYss18ur9DLDVrlD6Cr9bMSh9H4vE+wlcfi5R+0xIasV/uCiyxbyUfuL5UuHble5pnuaZ7/AC5xyjY0z3NM9ynDCNidByllc0z3KcMI2/Fq0+oaZ7mme5CnjFxNM9zTPcqUM5Xuad7mne5p3uQoYSyv/GEsnY0kzSTNJMqU3Tdn+1cIk5O4pQflHwtPt8yn9aJReV0WldX5cV9z9qoVVSd2amnsPiYW7L5lP60Si2+wlMUZ7nFfc/ebsuy7/kGnqbGmq7Gnq7Gmq7clQqNXSNNV2NNV2JUZwV2vkQhKbtE09XY09XYnCUHaRp6uxp6uxOnKH1EaNSSukaersaersaersaersaersaersdKeWFu5p6uxp6uxp6uxp6uxp6uxp6uxp6uw1i7Pnp6uxp6uxp6uw6FSKu16YUpzV4o01XY09XY09XY09XYs/U8sViLJIynsQba7jIfaReVvBlPYr36Pf5HCfWz4yTk18Jxf1jy7pDbaOJvjG5Sv042Pi/USbPiv28F3j/Z8X6RN+5/5I8suwsk+5eV7nxEm34FlYTKv3Hznl+ki2vI879id+k7+nh79J2ItoV15Fkn3EviuS8+rUVNzUVdzUVdzUVd+Sr1ErJmoq7moq7kq05qzfyITlB3iairuairuTnKbvI1FXc1FXcnUlU+ojWnFWTNRV3NRV3NRV3NRV3NRV3NRV3OrPLP3NRV3NRV3NRV3NRV3NRV3NRV3NRV3G8nd89RV3NRV3NRV3HXqSVm/TCrOCtFmoq7moq7moq7mpq7l3/6F3RkjJGSPP7w/PLtyh4/eMUYIwRghK38mxZizFmLMWYsxZi/wMWYsxZizFmLMWYsxZZ/lw887l+Vy/wCJflcuXLkvp/LTsZszZmzNmbM2Zszf4GbM2ZszZmzNmbM2Zscm/wDR3g6kNxTjLw/Q5xXZs6kN/Q2l3Z1IbilF+GdSG51IbmcbXudSG51Iv35OSj5OpDc6kNxSjLw+baXk6kNzqQ3FJS8etziuzZ1IbnUhvzclHydSG51IbnUhudSG51IbiafdfNqfQxJHD/W/RW+47kv6F451/tsWPY4f3IjtYn9mJZCSVRW5cRbtc+HyWRw/1PnX+gSQ7HC+/rq/cdy0V59HEfpPhGo+w7ex8I/6KH2182SyVjTQIUowd16JUYzd2aaHolHJWZpoEKUYeDTQNNAdKLjiaaAqEU78p01U8mmgaaBClGn3XOcVNWZpoGmgQpqn49cqEZO7NNA00Oc6aqeTTQNNA00DTQNNAjFQVl/IHWhF2Z16e516e516e5GrCTsh1oLsdenudenudenudenudenuRqRn4/cZ2zdz4fJ8IsSl9xDtk7lokcfctE+E+E4f6n+4ulBu7R0Kex0Kex0KewqUIu6Q6MH3sdCnsdCnsdCnsdCnsdCnsRpxh4/jKi5eEdOex057ChJ+EdOexhK9rHTnsNNdmeTpz2HFx8ihJ+EdOe3Lpz2HGUfK9PTnsdOew4yj5R057DjKPlChJ90jpzXsKEn4R057DhNeVy6c9jpz2OnPYcJR8o6c9jpz2OnPY6c9hwku7X4vCeJGexd90yk7U4im/cf31/0ZvscT9xlL7i5cZ5Rwv2yp9DI+Vy4z29MXaMTJtHFd4xG+7RxLvTTKLtSRNtwlc4d2pGTuSblSd+Um1awqj2Oq17HFfQiTatY6jOo9jqMru9K/4tKs6Xg1c9h8XN+xDiZQjijVz2NRLPM1c9ic3UlkyLxdzVz2KlV1fJT4iVNYofFSatYXY1c9irWdXz6VxUkrWNXPYqVnV8mrnsVK8qqsyHESpxxRLipSVrFOvKmrI1c9iXEykrctXPY1ctjVz2KleVVWZq57GrnsauWxq57E+IlUji/4bHhpyV0aSZpJmkmaSZLhpxV2U6bqOyNJM0kzSTNJM0kycHCWL9dOhKoro0kzSTKlGVLyR4acldGkmaSZpJmkmVKbpuz9HRpJ2sdKgKhSfsaelscRRhCF4rlGOTxRpJmkmaSZpJmkmaSZGjKcnFexpJmkmaSZpJmkmaSZKOLs/kQTwi0JVD47MtMUZ+CeXTlkcJ9bLSXg+OzufH4LTFGfucT9x+vh1en/APRKoi1TwcV9MbkFekjGS8FqhjO1hKd+5xf180Tx9x4WsRnFI6kTiXenyo/cRKMr3R/yEs0JTQozXYip37lD7sjGXsfG3YtOyP8AkF1LkVL9RW+4/kKpNdkzq1Nzq1Nzq1Nzq1Nx1Jvs2KTj4OrU3OrU3OrU3OrU3OrU3G3Lu/Wpyj4Z1am51am5KcpeWKpNdkzq1Nzq1Nzq1Nzq1NxycvPofExbu4j4mL/SamH+JqY/4lSspQwS5J27o6tTc6tTc6tTc6tTc6tTc6tTcU5J3TOrU3OrU3OrU3OrU3OrU3OrU3G793/pSjwdKdOMmjieFpU6TlFfhcPw1KdJSmLg6D9uXB0o1ptTNJw/bsLhOHl4RJWk1+x0eJpRpxTkcVxFKdJxi/wuHrUlRUZSsxcRQV/i5cFUjTqNzZ1qH+YuIoJ3yJd5P/ec51ozdldf/h16n+B1qidsR1qtr4nWqf4lOcptqSt+/f/EADwRAAECAgcFBQYGAgIDAAAAAAEAAgMRBBITFCFRUhAVMTKxBSBBYfAwM0BQcZEiIzRCYHCB4UVTgKHR/9oACAECAQE/AfipFSKkVIqXzccdmKnsfw+cVyq5VcquUXT/AJFJSUlJSUlL2klJSUlL20lJSUlJSUlL4g/AH4V3wE1NTU1NTU/ZzU1NTU1P201NTU1NTU1P+iajirNys3KzcrNys3Ko7bUcrNys3KzcrNys3Ko7YGk8FZuVm5WblZuVm7ZZuVm5WblZuVm5WbkWkce6BNWblZuVm5WblZuVm5WblZuRYR8AOULFYrFTKx7ngsVisVisUeXZC4LFYrFYoJ3Eo+SxWKxWKxUXh3YfMsVisVMqZWKxWKdy/AV3BWjlaOVo5WjlaOVd22u5WjlaOVo5WjlaOVd2wOI4K0crRytHK0crR2y0crRytHK0crRytHIuJ490GStHK0crRytHK0crRytHK0ci8n+lncxUlJSVVNEnDY7lOySkn+CkpKHzbJKSZzbAJqSkofKpKWxnMj4pnMo3gpKF8YYUzOasvNWXmrLzVl5oQpGc9hExJWXmrLzVj5p0OasvNWXmmw6pmrLzVl5qy802HIz2WXmrLzVl5pokJKy81ZeasvNNhyM1ZeabDkZp7KysvNMZV/oY8SpFcPZv5dhUPlT+YqSh8vwsVAbGch2cFE5lLbD5vZGHMzVkc1Zefs3CYkrLzVl5poqiSdDmZqy800VRL4V7aysjmrLzTWyElZeasvNOh1jNWRzVkc1ZHNNhyM/4w91RpcfBbzg5Fbzg5Fbzg5FQI7Y7a7flXabi1jZFOEUcHIGI1zZu8faR/dP+hTHtqVXeuCmwNMvXDZ2b7n/PyqmUd1IaA0rd0bWh2dFrAl3D2kf3T/oUyI0NkUXQ8vXrxTokMzMsV2b7n/Pzmq3JVW5Kq3JAS4fx806jj93VX+j6uqv9H1dVf6Pq67HU2Aw1S5X+j6uqv9H1dVDpcGK6qw4+wixmQRN5V/o+rqr/AEfV1UKKyMKzCr/R9XVX+j6uqhUiHG5Cn0uDDdVccVf6Pq6q/wBH1dVf6Pq6q/0fV1V/o+rqr/R9XVXmFZ2s8Ff6Pq6q/wBH1dVf6Pq6q/0fV1V/o+rqr/R9XVX+j6uqa4PAcNt/o+rqr/R9XVX+j6uqbTIDzVa7uxaTCgmq8q/0fV1V/o+rqr/R9XVX+j6uqDgce8Ktd1ZOqEkhVYWaeGg/hQ4KLK8OnmVJk+KDYXifWCoUryJew7U90Pqvy0wMB/GuzPcn6oVMC5BrQ7jguzZWj6vBUuVu8H1gjZ/sTQBxX4auPFSbXn4TQq/vTgDKS/4/1mhUqyPFOqFokpMqy8V+H/0mgCc0ahcZegiJKi+5Z9NrKmNZPDTy5lCzq48cVBleG1e72hK8tnww6pwBknVTwTqhGCJFQBM5R3jQaOcaquFH09VcKPp6q4UfT12OoUB5rFquFH09VcKPp6qHRIMJ1Zgx9hFgsjCTwrhR9PVXCj6eqhQmQRVYFcKPp6q4UfT1UKjw4HIE+iQYjq7hirhR9PVXCj6equFH09VcKPp6q4UfT1Vwo+nqrtCs7KWCuFH09VcKPp6q4UfT1Vwo+nqrhR9PVXCj6equFH09U1oY0NG24UfT1Vwo+nqrhR9PVNoUBhDmjuxaNCjGs8K4UfT1Vwo+nqrhR9PVXCj6eqDQMP8AwLkVVKqlVT85HDuO4/OKxVYqsVWKJn/JZTVR2SqOyVR2SqOyVR2SqOyVR2SqOy+AquVR2SqOyVR2SquyVR2SqOyVR2SqOyVUj4uHzbJqZU1NTUyh8DMqZUypqZUypqafy/FgyxVoVaFWhVoVaFWhVoVaH4C0KtCrQq0KtCrQq0KtCrQovJw/o+sM1MHuVgFWbn3OCrDNTBVZuarDNTCrDNVhsJA4qs3NVm5oEHhtnJVhmqwzQIPDv1gqwzVZue0kDiqwzVYZqsM1WbmqwzXH2r+U7IfN3H86KG2JyrBQvFDYeQKSHMNkRYKQULjticqwRUHx77+ZYdyL4LBYIyWGyHy+1IngrIJrA3uGGCZqyHcImJKyCawNVkFZBVBKSsghDA2OaHcVZBWQTWBu0isJKyCsgmtDeHfMMEzVkFZDa5odxVkFZBWQVkFZBAVRL+QF7QrRqtGq0ag8HBWjVaNVo1WjVaNVo1Bwdw+YulWM1gsFgmcywmVghJYLBYKFxPzEsaVZtVm1WbUGAKo1WbVZtVm1WbVZtQaG8P4y6IxnMZK8QdY+6t4WofdGNDaZFwVvC1j7q1hyrVhJXiDrH3TXB4m0okATKvEHWPumva/lM06LDYZOcEI8I4Bw++y8QdY+6bEY/lM+6Y8IYFw+6vEHWPumxGP5TNW8LWPumxGP5TNOjQ2mTnBCPCJkHBOiw2GTnSV4g6x90I0JxkHDZeIWsfdXiFrH3V4g6x902Kx+DXTV4hax91eIOsfdXiFrH3V4g6x902NDcZNcPhe0+aHP1wVnmgBNpCpTa0eJ6yRhg4gpv6J31VmMfXguz/04VJ9y/wCh2dl8rl2j79QPet+oUTkOzsri/wDx3YrZvf8AVFjQV2aJRHhBoIB812cJRnBU0VqQ5QmtEWHI5dVThOkyOSqCUyoLQ2kslsa1pnNGCM/WKsQfH1Ndme+P0TWtM6ysmzxKsQRxViDLFUIVaSAPhaRRW0mVY8FuuFmUOzIQM5lRez4cV5eScVuuFmUKCwQrKeC3XCzKgwhAZUanstGlp8VuuFmVR6M2jghqjUFkd9dxTezYbHB0zgiJiS3XCzKo9FbRp1Tx7ruzYb3FxJxW64WZVHojKOSWnit1wsyqPQ2Uc1mlRqDDjPrkpnZ0Njg4E4KPQmR313FbrhZlQ+z4cN4eCcNm7IWZW7IeordcLMqj0NlHdWaUezIR8St1wsyt2QtRW64WZUGgw4L64J/hsTtCFDcWEHBbzg5Fbzg5Fbzg5Fbzg5FM7QhRHBoBxUeO2jtrOW84ORW84ORW84ORW84ORW84ORUGKIzK7e/HpkOjuquW84ORW84ORUClMpE6vgonaEKG4sIOC3nByK3nByK3nByK3nByKgR20htZu0oUykkTrdFeqXn0RptJbxd0V/pGroqFSosWLVedkR4htLz4LecHIrecHIrecHIrecHIrecHIrecHIqJSmQmNiHxW84ORW84ORW84ORW84ORW84ORW84ORUN4itDx4+wjEWzw5Ewgvy5hB0IEGWSrQuPrgoNW3ZVXanuh9VWYR+L1wX5ZcAF+Vx9epqcLL1j/pF0PGquz/0479PIbSATl/8AUTCOKLoRM5LsyVd8lHIFIfNVmESciYXgPWP+lXhVp+uKJhywC7M9yfrtPBQw8j8KAiz9evBOhvc7FWTl2eKtIkctlL9w5NeyUnesVOEmWRIRdDPFOdDJJl69STzDl+EKm/poXrwRewjFflAT9eP+kHQ5nJflI2UvXn/pPMOX4QqJ7hvsDR4TjMtCu0HQFdoOgK7QdAV2g6AhR4TTMNCfDbEweJq7QdAV2g6ArtB0BXaDoCu0HQE1rWCTR33wYcQze2au0HQFdoOgJkJkPkEkaPCcZloV2g6ArtB0BXaDoCu0HQExjYYk0S2lDs14Eg9Ds6IP3rd0T/sQ7OiD96gUN0KLaF2xzQ4SKu0HQFdoOgK7QdAV2g6ArtB0BXaDoCdCY4BrhgrtB0BXaDoCu0HQFdoOgK7QdAV2g6AmtDRIf0o6I4GShxHOdI/BPe4OkFav2RXFowVo9Wjwhw+RuY4uOChscHTPwT2urTAVR+WyKC4YKq/JVH5IcP7zJcCqxyVY5KsclWOSBJ+ff//EAFIQAAECAwMCEgYJAgUEAgIDAQECAwAEERIhMRMyECAiMDM0QVFhcXOBkZKTobGyBRRAcqLRI0JQUnTBwuHiRGIkU2CCg0Oz8PEVlGOQBoDSw//aAAgBAQAGPwL0gzLzb8o7JMJIrOZFJUb9SimrPHHo+ZeNp11hClEbpp/rbKuyrLrtLNtbYJpvQlCEhCE3BKRQD7SXNzAWWkkA2BU3xmTXZfvGZNdl+8Zk12X7xmTXZfvGZNdl+8Zk12X7xmTXZfvGZNdl+8Zk12X7w1OMWg05WlsUONP9WzHvo82jNobnkPTMqwJhxnJlOoNMDzxbsqsYWqXRLszNJIPIyiVvA0pS7CKhCiKVqBBWEqsDFVLtGQ4leY/6tmPfR5tH0jIethlpTTamH0ppaISKtqurSsBLDrCU+pBgyrhdtW+BI1ON9qPQc496StMNMWHW1lRKHLBBURw78f8A8daVOFuXZS6JpF9LyaV34aQqcACmHkusuKczzgAnNpwwNCQ4leY/6telZayXVKSRbNBcYzZftf2jNl+1/aM2X7X9ozZftf2gs2WLYTb2Xc6IzZftf2jNl+1/aM2X7X9ozZftf2iVlJiyHm61smozj/rJf4dPmP8Ardf4dPmP+lmnvWWk5RIVTI4fFG22+w/lG22+w/lG22+w/lG22+w/lG22+w/lG22+w/lG22+w/lG22+w/lG22+w/lG22+w/lG22+w/lG22+w/lG22+w/lG22+w/lG22+w/lG22+w/lG22+w/lG22+w/lG22+w/lG22+w/lG22+w/lG22+w/lG22+w/lG22+w/lG22+w/lG22+w/lG22+w/lG22+w/lG22+w/lG22+w/lG22+w/lG22+w/lC/8S3ayAvyP9x/ujbbfYfyjbbfYfyjbbfYfyjbbfYfyjbbfYfyjbbfYfyjbbfYfyjbbfYfyjbbfYfyjbbfYfyjbbfYfyjbbfYfyjbbfYfyjbbfYfyjbbfYfyjbbfYfyjbbfYfyjbbfYfyjbbfYfyjbbfYfyjbbfYfyjbbfYfyjbbfYfyjbbfYfyjbbfYfyjbbfYfyjbbfYfyjbbfYfyjbbfYfyjbbfYfyjbbfYfyjbbfYfyjbbfYfyjbbfYfyjbbfYfyjbbfYfyjbbfYfyjbbfYfyjbbfYfyhpp11DqHEKNzdmlKcPD9nSnJJ8PsFf4dPmP2RKcm5+n7GtKwjO7ozu6M7ujO7ozu6M7ujO7ol2lnVoQAaRnd0Z3dGd3Rnd0Z3dGd3RVJr7IUk3jgjO7ozu6M7ujO7ozu6FPWtQWgjnqYzu6M7ujO7ozu6M7ugJBvPB7UUk3jgjO7ozu6M7ujO7ozu6M7ujO7ol3QdQhKweenyjO7ozu6M7ujO7ozu6M7ujO7oACrzwe3K0EPJYsyywShzKA9I3NGu5rFdzQVx+yOccPTKklaWxWyOPQcQhYUps0UBuHHWUe1OccSzaWss5MLKEi1ZpRNfyhS7BbUham1oV9VQx0CqwtuiiPpBTDd4tBbdqrqU2y2Maf+CELsqRaANlYoRwHWW/e9uVoJekJd/0etYX6yFijZNnUmmBVapeIdFiYTNCScaWnIpSFrI+9XVmt4MWEsONsOej8m76vnFdodJxiVMxKZWTacdq22xZtVAsOFrrXc8SPrMuTLI9ZKW3L8mCU5NJhyTp/hJdfraDX7+COY2z0R6XX6vlCqUSlgn79F4cOEPl/L1cQ1knGmgsookV1RzDaqY9IsJl3lqenUPIUkamxVFb+Y3R6TcTL/wCIdnFK3luM2wSkHhFYm/VpVbMo4qXozZsVIc1arO5qfCPSKA041LKdZcayDQWmoTebG6N8RL5ZpLDtm9tAoB8oVx+yOccTbTSC44tFAkbt4j006mXKphb7apdf1rgi9O9ux6eTLMZN+YIU06lNLSaJtAK37jDwZbesuOsVZLAZTQLvIA4MYn7MonI+ohtgBOpCqrw4cIPrIfVblmkNqbaDikKCdVeTqDW+selmEsPPLmHm3G1pFxGorf8A7TdHplxtn/FOzKi2qtFLa1NQDuVAMT3qco4xLOIZSGbNi0oOVJCdzU3R6SQlpxqVWWFI9XbChaFamxu7lRDIeaSy4BSwhNkY3XbnFCPanOOPRlAvJtvKUtTarNkZMjEcMejkCVC5NpLxWDeLRs0rvnGGR6usekEzmUXOUuLdr728U3WYYXRS5ducmsogN5W+2bJsnHd6YlfWGH5yUSwtKW1thSm1FVb0+7cN6HnvVCA5JpS2uzWyoWqgncNCIlhOyTk02JBtpLaU2sm59YcBN2q4If8AXGvWHv8A41DQWb6u6utOHC+PR7SkKcWHZcujgFLVYmUtNKRJ+uodySEAhSMnfRO7qr6RIPJRMOMofdUpGSoUBSKCiBuV8YkvW5QlkOza1odvAtKqisND1dYn0zmUVOU1Jbtfe3rOpsxIO+rlMwmfWtxX1smSvE72bD7YaUJeyCHHmrDlqp1JP1/ehv3vblJTeYzO+MzvjM74zO+MzvjM74cLbNkuKtKNcTCHEJqhYqDGZ3xmd8ZnfGZ3xmd8ZnfCgoUv9kWQmoMZnfGZ3xmd8ZnfGZ3xkqfSBNqnBGZ3xmd8ZnfGZ3xmd8JUpNB7UshNx4YzO+MzvggouPDCWmmQhtOCQYzO+MzvjM74S2Rq1AkDi/8AcZnfGZ3xmd8ZnfGZ3xmd8ZnfCCU3A7/2NKcknw+wV/h0+Y/ZEpybn6fs6U5JPh9gr/Dp8x+yJTk3P0/Z0pySfD7BX+HT5j9kSnJufp+zpTkk+H2Cv8OnzH7IlOTc/T9nSnJJ8PYXnQK2EFVOaAf8NeP7o/pvij+m+KP6b4o/pvij+m+KP6b4o/pvij+m+KP6b4o/pvij+m+KP6b4o/pvij+m+KF7XtZAfe+8Y/pvij+m+KP6b4o/pvij+m+KP6b4o/pvij+m+KP6b4o/pvij+m+KP6b4o/pvihx5QliEJtEC17VKcm5+n28NIeStZu1N+vynJJ8PYZrkleEI4hr6/wAOnzHXpvkleHtUpybn6fbmWa6h50IXTdTiR3QEpFkC4AawUZO1zxsXxRsXxRsXxRsXxRsXxRsXxQyzYtZNITWsbF8UbF8UbF8UbF8UbF8UJTkqVNMdKpGTrQ0xjYvijYvijYvijYvijYvih1rJ2baSmtYAyWA+9GxfFGxfFGxfFGxfFGxfFARk7Nd2uiV0tU3I2L4o2L4o2L4o2L4o2L4oMxYxbCLNeGsbF8UbF8UbF8UbF8UbF8UJXSldzQ2L4o2L4o2L4o2L4o2L4o2L4odZydm2kprWNi+KNi+KNi+KNi+KNi+KNi+KErwqK60kWLVeGNi+KNi+KNi+KNi+KNi+KNi+KNi+KNi+KGnrGxpUKVxrT5RsXxRsXxRsXxRsXxRsXxRsXxRsXxRsXxQU2LNBXH2BUw4FKvCUoRnLUTQAc8OvTEkhLaGVOfRv2qECtDcIlXVUbL6AsIKt8VjJ2hbpWzW+KoUFjDUmsSXLjyq1lXEIdZ9aZlkIlssMqmtpVSKY8G5DwnP8OlLDLtmyTYtA2qwUOOhJFCq4kJrhU7kTMubSDLgFalJonCuMOuZayltOUVbSU0T96/chKratVUgZNVaDE0phwwkF2tU29SCrU7925GRLmrqEm40BOAJwrGTD19vJZppb+7XCsTK6qbQwsoUpxJEKcU9YSlYQq0kghRwFMYShKlFSgDSwq6uFbrueGveGld96GmHXG2mVMKcqu7VWgIm3HHEFpEyWWii+1cKcZxhLhe1KlFA1JraGKaY1iYtEFLa0pGTSoqFU11QpDbqXbaHahFlJJVTGgxhr6a3lUlbYQCoqAxpSJZlBU56w3lELQklNLvnD4bX9Mhpa0hSSK2d7fhmamFhALSVqVwkCLdsjVZOyUG1a3rOMS4YUlTTjL6zduopEnlVf4h5lDiglJuqO6+Ec+ivm0JZOXZdW7MLaVLpTRSEgq1Va8A6YaLriMo6VhAZClWrJ3LoacDtpLtbFgFRVTG7GJdDa0rlnJZb1ocCgPzhxxT1hLYClW0lNAcDfuQZgrsMi8qWCnxguFZSAoJopCgqpwFMYkEsqBbeW4ldReLKa04L4klvq/wAQ80FkIQeKt2A0G9AxLoCQZeoEwv7lu5HfC0OvWFIAK7jqAcCd6HSF7FS2LJqK4XcMZTKXW8nSybVv7tnGsNAu3u3oSEKqb6G6kDLOItKdW2gNBSrVncwxhtwOWg5UICEkqNMbsbo9ZyqchZtZSt1ILhWUgKCaKQQqpwFMYkEsqBbeccQuovFlNacESS31f4h5oLIQg79K3YDQa90a01xGGChxDWUfQ2paxUJBrD7sw4gtNu5ND6Bc6LsBxml0NK1aso7kaBBqlXCIfyzraEIcQ2KVrVQqAeGFLSokJXkymwbQVvWcYkUMLC2HkOkml9U0/eFB1yhSm2qiSqynfNMISlblLVNVZNm/C/C+C1ldWlzJG40C94mMkXNVlA1ck0tfdrhWGLamquPvN0NQSEk0p0RYQ6CTUi40VTGh3eaEJZdt2020XEWhwb8ZJTmrurRJNmuFd6MhlNXbyeBpa+7XCvBEllSgIcTMKWaUzFXRbbJKeFJT4wv3fYEZCz6ww6iYbCjcopNaHjiYlkSDzRcZWkqeKQAbNwxvgMepKSsSLaGbKEWrYxClHClBSkOuNSzi2XrWXygTTY6VbXiDgKeEPtONlEuCMipxCUOkU+tZxpvxJKC3lHLjPdUoZqt86yriEKmyUlJZDQFLwQomvfHpSjiR64wlpP8AbRJF/TE60043kpxICyutUaiwab90T7dsBibZQ3X6ySkU54eLy2kOmWMuixUi8gknowgTSLC1ZLJFDi1J3ag1TClSmRCVtIbKFWgEWd7gvwh9RQ06089ljlFLqnC6guOEPMqsIl/XnHCb7dztad2MTKQtAtTXrjSr86oNFDeuhUw4ttDqnWFWU1ICWzXHfvhqYRYaKSmrqVKC1J3UkYEeENe8NK770ImrQspYU1Z3b1A/lCn0OIygmzMoCq0oUWSDDcytxGU9ZVMLCa02OwAI9IBh5CDNuJNbwQiyARx3RKvtJYQthtTORBNixdShpdhEs6XAqwl23QUqpawq7gj0cbaF+rtLZXiKhSgajohtTzyVrSy60p20pSnLYpavw4olpcqQl9jJqBvKbSPyuiXccyWUYcKkthSrJBFKVxiXWkNN0DwcSm19el4ribokBVpt6XQhv1htSgqgxHCDvGEc+ivm0Jdla2sizM+s2k1tnVFQHBjHottFhxaRMk1qE3kH84amW1trmPpcolVQk21Wrt6hENlqYSlzIvIU5TNUtVq0niMTFVtoU6y23nLXeldokk78ONBVhRUFpJvvCqjwhou5JLjLodQgKVZNxBBOO7Eo6hLTZbeW44lJUa2k2cTuxJshTSHpdNhMyhSgtN/fUbh0G9AxN+sVU5ME6pKiLIwT0XRPpceRlpqXbaKhWlpINT3xNOtPBGVDIpUioSTUVGFa7kZZCmsqmZU+lFVWaKRZKa488MTTikVQyttSUV3VA/lEs4XEnJTD7xpuheAhpyqHVoW8bKlKSCFqtYjdEeposMkUULANkKCrXRWGi7kkuNPB1CEqVZNxBBOO7Eo6hLTam3lOOJSVGtpNnE7sSbIU0h6XTYTMoUoLTf31G4dBr3RrTXEYlqEDJPodNd2kGXS4nINPpfl0Krqd9BpuXmm9DZaQy08mZTMUClEKoKXk34Q65lEi3NszHMhIFIfWh0WXZhLqm7Sk202LNkkcN8Si7baskp+0BXBw1u4o9J5Nxq1Ny9MkutqtkpFnfrvQpJcTZIas21K+is0qKC6+mMTScon6adE0OAWkmnwwzJNONugekA9ZvygFu2ajg34l7S02Gn33Lq1KXK998SLLjjeSkgbCk1tL1JSK71xj0VVxJ9TZU2r+6qQLuiJh1gtlEwUqVlK1SQKbmN0ElDTrXrBmApal2hfWlMMd2JXVNKyOXTfW9KzUf+odBsoaURYYQoqS3v0rv70L932WS5ceCtZK7dnmjZfhjZfhjZfhjZfhh57KWsmgqpTejZe6HXcpWwkqpSEqyuIrhGy90bL8MbL8MbL8MJVlK0NcNKteUpU70bL8MbL8MbL8MbL8MbL8MKKV21AXJpjA+kpzRsvwxsvwxsvwxsvwxsvwwleUrTcpolFaV3Y2X4Y2X4Y2XujZfhhhNuuVXYwwuJ/KNl7ocNuzZcUjDeMbL8MbL8MbL8MbL8MJRWtNDZe6Nl+GNl+GNl+GNl+GNl+GGmbeelSq03qfONl+GNl+GNl+GNl+GNl+GNl+GEoxoKa0k27NOCNl+GNl+GNl+GNl+GNl+GNl+GNm7o2X4YSxbxQV1pwgRs3dGy/DGy/DGy/DGy/DGy/DGy/DGy/DBVbtVFMPZS2qo3QpOKTuEQlLpZcRurFUno12c5FfhAia5JXhDfuj2B0NXO2TZ44FcdekOX/QvQmPxDnjrsryTn6Pam+QV5h9mTnIr8IETXJK8Ib90ewOpaNlwpISd4wK469Icv8AoXoTH4hzx12V5Nz9HtTfIK8w+zJzkV+ECJrkleEN+6PYHUIVZWpJAVvGADedYdIxsnwhhSmQSW01NTvRsA6TGwDpMbAOkxsA6TCXEMgLTgd7QmPxDnjpp1byLag9SpJ+6mNgHSY2AdJjYB0mNgHSY2AdJjYB0mJYZEULbm6f7Y2AdJjYB0mNgHSY2AdJjYB0mNgHSYacabsLDzd4J++NamXHm7a8usVJO/GwDpMbAOkxsA6TGwDpMbAOkxsA6TGwDpMMWWqVeQDed+K+rprv3xsA6TGwDpMbAOkxsA6TFpLVk20CoJ+8I2AdJjYB0mNgHSY2AdJhZDIzTumJYm85JPh9gznIr8IETXJK8Ib90ewOthVkqSU2t6AMdYe9w+ES3Jp8NPMfiHPHTT3Lnyp08ryTn6NOjlmvONamPxDnm1iW/EN+Om/3t+caVz3TErySfD2MuLqQNxOJO4BCS4hlpvdTaKlD8tdnORX4QImuSV4Q37o9gebKrAWgptb10AY0GsPe4fCJbk0+GnmPxDnjpp7lz5U6eV5Jz9GnRyzXnGtTH4hzzaxLfiG/HTH32/ONK57piV5JPh7HJcuPA642440FrVUkknfjYE98bXSdApUKpIoQY2unvjYE98bAnviyymwks1IB/u115ClWEqQQVb10AC+7R/2iAHXggnAE3mAttYWhWCkmoOiMouyFGwK7pO5CRWyMBfohba7aDgoHQTxHQmPxDnjoL5vHQDaJlClm4AKx4oydsZSlqzW+m/Cmsu2l21QprfWEJUsBS7kgnHQ/4zo5MzKAutmlrd3tCy6+lCqVoTuQFJVaSbwQcY/3GFccFDkwhChiCcOPehSErBWmhIBwrhF9+7FLV+9XRd4tA6CnsqnIprVdbhTGMYcbQoW03rSNyugWUvpU7fqQemHuMaBioVUb4MAPPpaKhUBRgOrdSlrcWVXQotvJWE51+HHFluZQo0rnbm/ByLyXKY2TD3ND3vRkhMIU5WzZruwQy+lwi/UmHuIQdCwmabUomlLWJgNuPJQs7hOg37o9hkuXHlVrjPEfHWE8gfNrryXFWWyghSt4UgUvFNH/AGiPSWUNHVWC3Xdbs7n+6sMyzL6WWlNqcyqaGpBpQbnCYlk5UMW5ZbhKEA1IXZBFdw4wxYohRk0zFRZAKjX725dub8SBUQ06qZYrTVBJhDKyl9Lc802HFIG6mp5xvw0+X0rtThl8iEjNtlPHUC+JOYLgOXmciZayNSKkcdRSpiVZaWpLpS85qQnALOJVDNXQ236q0+pCUDVFVa3710J4joTH4hzx0F83jE4GaleTwTjTd7qxJFiw4y66203ZNwB3oHpbJKySHbOVqKZDNPDjqo9NkuN5ATCCtr6y9Si4H9ofmGWFOKkyMkoEZ41S+7Uw262atrSFJPBH/GdD0evLBbk405qbAASoJJTSG3UJCsi0LTZNNX9YHhrHpB0PCzLTKGkt2BeDYrU/7o9JIWaTPrSiquJR9Q8VIYmLAbyia2RuR/uMH3omCunrDSnVTCVG8qqTfxikT82wnVhmWeLWNU0JUOiJfJOhEtMvKS26AM1Kdz3jXoioW26tEk+5lbArVJEFVoTJMiZmhQBRVRvbl/dC0vELbsBQUVItV/27kO+6NFwt3vK1DY31G4R6Wkskpltcsp9kKIO5ReHDQ88ekLM0llMqwhxAKQa6km/guj0tOoXkVtsMO5IpBvydaGKtKJbRMNsLSoJCdVSv9xxihISzKALQkU1S1A3nm8Ye4xoc8ejGUuolkOyq3Dk204hVLok5hgNqmvVH6Ai5yjgF3HHoUoXbkwwtLS13fS3Y7xpaial7FW2sg48ReS2VaodAhAtoVbzBUGvFEzNOEVqWEJSa2UJPiYe5oe96Jj6RGQ9Zf1Fi/PO7WJiYWRRqsu0hJwTcSef8oe4hBgQ4h4pUDlQEVvKraqUG/WHJ1uYbU8hlHrcs59chO/uHxiuF27Dfuj2GS5ceVWuM8R8dYTyB82uvB02WrBtHgpAphTR/2iAHW0OgYW01gNrabU2MElIoIBsioFK03IQlTLakozQUi6ChaUrSdxQqISlLaEpSagAYGKgJW5aWrKFItao1p3wXQ2gOnFdm/pi9ho0VazBjvxqUpTdZuG5CeI6Ex+Ic8dBfN46GUDDQc+8ECsWLKbFKWaXUjKZFvKffsCsXADigJSAlIwAj/jOgmiEixm3YRlFMtqc++UCsKqhJtGpuxMDKtNu0wtpBigoBvCP9xhXHAU4y24obqkgwstFLTyk2MrYqQIRL5NKmUJCQlQrhAAbRQCzSyMN6AbIqBZrTcghptDdcbCaQ77o0RUA0NRGqSlW5eIdeKEKtWLKSgURZFLoXVCTbzqjO44yhabLn3rIrClhKQtWKqXmHuMaBht1SUFCGy2GigWbyD+UA2RUCgNMBBRk0FCrymzcYo02hsf2CkFTbLbajupSBBsJSmprqRS+HuaHvejUgDduhVlKU2jaNBiYe4hB0LaGWkr+8lABhLi2m1OJwUUio0G/dHsMly48qtcZ4j46wnkD5vYcIwEYCMBGAh676h8Ilrv8App8IwEYCMBGAjDQmPxDnjo4CMBGAidrf9OfKmMBGAikYCMBGAhpIQMkUqKjTA3U/OMBGA0MBGAjAQi7/AKzXnEYCMBGAi4aOAjARgIc1QctOKVxVOEYCMBGGhgIwEYCKp1JtovHviMBGAjARcIwEWEEIVaSa84MYCMNDAQu76piWUq8ltNSeKMBrcyw0lgIlnMm4hbhDuFbVN6+K5RGdYzvrb0apbTcxlHE5EL1VErKa05omy2604+w2pZatitw3YlFvOtMuzDaVpbUsX1G5GQyzeXpXJ2hapxQVMPIeSDSraqxJcuPKrXGeI+OsJ5A+b2Z73D4RLcmnw08x+Ic8dNPcufKnTtJA+iKVWjw3U/PTo5ZrzjWncmu3VxSjwGt41g8o35xprAVZNpJrxEHSue6YleST4a4ypMgGJpl9Jb9IJcFzYN/DeKizhDcuJbUJ9K+uZW2KZMrKuOt8SssZUNFqfVNqmbSaFNtSrt2pBpCvRy5NJeDTyPWypNFFVb9++t8KlcgU1km2kJSpsUWM62cd6lIdW0wr1d616wha05N36OlU/WSrAb0PNPBaZcWQwl4pLgFLwSnEb27EkttJCsuMVk/VVrjPEfHWE8gfN7M97h8IluTT4aeY/EOeOmnuXPlTp2gB9DZVa47qfnp0cs15xrTuRNRlFWverfrB5RvzjTWVioqDpXPdMSvJJ8PY5Llx5Va4zxHx1hPIHzexEww6Z5YK0BRo2jdHFG33OzR8o2+52aPlBSZ92hFMxHyhKEz7tlIoNQj5Rt9zs0fKMit8vILVrVJAoa8GlmPxDnjpp7lz5U6dqzsNlVrjup+enRyzXnGtO5FNn6RQV71b9YPKN+caR1bZsrAuNK0jb7nUR8o2+52aPlG33OzR8o2+52aPlG33OzR8ofeTPLKkIKhVtHyggz7lDdmI+UIbTPu2UiyNQj5Rt9zs0fKNvudmj5Rt9zs0fKNvudmj5Qo+vuXD/LR8oYcVnLbSo9GuyXLjyq1xniPjrCeQPm9iVxRJ8ijw0yeQPmGlmPxDnjpp7lz5U6dqzsNlVrjup+enRyzXnGtO2EWKOKSeE1x1g8o35xpHubxGmnOSVp18RiU5JHhrsly48qtZVQkXDdgZaYDVcLa6QFJcKgcCFRQEgccZyumE6pWO/pk8gfN7AmhpqoJLhAGJKoORmA7TGwusWETbZI+qlzeijM0l040Q5WHKknU7ug1QkXHCKuv5MH7y6QHsunJ4ZS3d0wE5Q2jgLUO1JOp0Zj8Q546ae5c+VOnaIP0VlVocN1Pz06OWa841pyqLFlxSeOhx1g8o35xoK1Ssd+LTr2TTvqXSC7l0qZGK7ephS/XGylOKsrcIKmnw6kXVQusJqom47sPe7GeemCluYS9vhK6wpYfqhNaqt3CmMN6o5w3YboaarcjOV0xnK6YK1u2EC8qUq4QclMJdpjYXWCyiYSXE/wDTC7xGTXNIQ591Tl8DLTAarhbXSApDlpJwKVVBh6pJwx1mS5ceVWsq4hHpS3tjLDjydkWebGPVZZxthCGMrbshQN9OgUvgKQpEuPUhM0sWtVU910H1dq9Ms29SgIUVCt5JFBuQ2SKG67TJ5A+b2BHvxJZTa3rCcrXClDSvBapHouxtjLc+TobfNhBKi1kck7QBJtfWiVyhas5BFnJpIObuw57ugzxGJnKZ+RRka/dvtU5/yj0g0EhUqt2WDiRhlLWq7rMS0m+Sp6Xbdos/9RGpsq/I8Ih33dGY/EOeOmnuXPlTp2lA/RBKrQ4bqfnp0cs15xrTmVTZo4oDirdrB5RvzjQVxmEZTD1f6Cv3rWqpw0pHpbJUs+pjLUwt2tTz0/KGrZZt+ts2bCTTP3Y+lsFf/wCMUEJ4jD3uwzb2vl28tvWK7vBWkejrFPWfWUWKY2fr81mJt4bWmsshz+1y0oJVz4HmhnjTDXvRMTATbLSCqzDoWpEz/hsuFBNmmqAv/tvrzGPSMo6tL/8AhMqFhNmmqpT5R6OLdPWvWBSmNj6/NT8ol2UKbmZN9aksq+u2o1V/uHDGXU3lShtS30qxLn1rXDWPSRXs9UWN/JWbqc9Y+js2f7cIe5tZkuXHlVrKuIQC80hwjAqEIQthtSEZoKbhBVYTUpsE0+rvQi2w0qwKJqnAb0J49MnkD5vYEe/BSoBSTcQYORZbaJxsiMmlCUt4WQLoCUgJSBQAbkOe7oM8RgJebQ6BeLQwgNpbQG03hIFwgOFKcoBQK3QId93RmPxDnjpp7lz5U6dpYV9GEqCk75up+enRyzXnGtOZbHKKs+7W7WDyjfnGgrjiw6hLid5QrBYSy2GjiizcYFsBQBtX7+gniMPe7BBoRBLLDbSjuoTCmi2gtKxRS4w37whr3tBQaZbbCsQkYwtpLDQbXnJs3Kg5FltonGwmkFxthtDh+slN8Fxcu0tZxJTjAyrSHKYWhhAQhKUJGCU3AQ9zay06hJXkXA4UjEjd8YBaeQuu8rWKlIJ4RGxp6IpZFMhhT+6JKiAKvjc/tVGxp6IzE9GmTyB83sF4B442NPRB1CcN6JQlCScknc4I2NPRGpSBxDQ1SQeMRsaeiEiyKZA3U/uEbGnojUpA4hozH4hzx009y58qdO0sKo2lKgU75up+enRyzXnGtOZVVqriinirdrB5RvzjQzE9EbGnohanWxY3aCNjT0RsaeiKhAB4BFDeI2NPRDynWxkwklVBuRsaeiMxPRGqAPHGxp6I2NPRGxp6IXqE4HciVJQknJJ3OCNjT0RsaeiNjT0RqUgcWtZTJIyn37N+tf8AB+qJLlx5VawnkD5vYlcUSfIo8NMnkD5hpZj8Q546ae5c+VOnS5U1SCKVuv8A/WnRyzXnGtKoSbSiq811g8o35xpFqdRlEDFPPpn1uoyjaUEqTvjTr4jEpySPD2j/AIP1RJcuPKrWE8gfN7EriiT5FHhpk8gfMNLMfiHPHTT3Lnyp15HLNecexHlG/ONItyxlKfV0zztjKWEk2d/Tr4jEpySPD2j/AIP1RJcuPKrWE8gfN7EriiT5FHhpk8gfMNLMfiHPHTT3Lnyp15HLNecexHlG/ONIpbaMosYJ0zq20ZRxKSUp3zp18RiU5JHh7R/wfqiS5ceVWsJ5A+b2JXFEnyKPDTJ5A+YQpQxjO7ozu6M7uhVg0tKKjxmM7ujO7ooo1GjPcufKnXkcs15x7EeUb840iywLTm4NM6WBaeCTYHDp18RiU5JHhroyYCnVqCEA4VMJccmnnV7t9Enm1r/g/VEly48qtMsBZximUNeOLddVSlY2Q9MIFs4+wK4ok+RR4aZPIHzCFwbNq95oag0J+kTdE/NBLvq2XDTMstVVheaRfhVUOtPS5TNJLYS2hdQ5bNE0PGDHpQPCxZQxRAXaGCsNHm0Z7lz5U68jlmvOPYjyjfnGkVkLndzTO5C56ybHHp18RiU5JHhrsly48qtb/wCD9USXLjyq0znHARkUslXpN1SZyt5smuT5x+cPTIaR6mzMZBQqcodVZKt7E4QhwJbTLJ9INtDG3qXgK72IwhHvewK4ok+RR4aZPIHzCFxk3jRFtCsaXhQI74mVKC/8RZthKqC0nBfArh4Ieyq3XXXLNXlK1Ys3ppTChh57KOOuvWQtTh3sPGKpIUN8aHNoz3Lnyp15HLNecexHlG/ONIpLS7CzgdM6hpVhxSSEq3jp18RiU5JHhrsly48qtO28lMulLibQBKowlulUYS3SqMJbpVBffLex2AG679YZLRTbbct6vA3H5xhLdKowlulUYS3SqC6tMupIIqElVcdBfHDjRa1C3MsRU59a16YLpb1RXlCLRslX3rOFYKy0b3A7ZtmzbH1qYVhHH7AriiT5FHhpk8gfMIXDdilr1pilrDZBCpaYQ28tqalqLTVIos71cRSHZkpZyDc56tkwDaIt2K1378Ibm7Lfqqpv1bJ0NvPsWq8e5vQw0yRl3H5ghOSLhIDiq3DmiTDaWWUOSqZly0Co51CBHNoz3Lnyp15HLNecexHlG/ONItAXk6/W0zzYXkypJFre06+IxKckjw12S5ceVWnk+ST4ay7/ALfMNCpQKxmCMwRmCKhA9gVxRJ8ijw0yeQPmELgtPIDiDfQxZDSaZQO33m2MDWHXHQlxxUwt9JvoKm67fG/GXyKcrat1/u+9TCvDA+gTctSxQkXqzunehJZZS3ZbyQpuJrWkc2jPcufKnXkcs15x7EeUb840i0OLyaT9bTPNuLyaFIIKt7Tr4jEpySPDXZLlx5VaeT5JPhrLv+3zD2ZXFEnyKPDTJ5A+YaGaOiM0dEZo6IzR0RmjojNHRFwGjPcufKnXkcs15x7EeUb840llSQpO8dMUqAUk3EHTr4jEpySPDWcj661la0sVv3tJJcuPKrTyfJJ8NZd/2+Yey2HJhptf3VLAMK/xbGH+YIlEmbYCg0m4uDegIRMsrWcEpcFTolbighAxUo0EbbY7QQk+tM0yBvyg+8I22x2ggLbWFoP1kmo0StaglIvJO5G3GO0EbcY7QRtxjtBG3GO0EbcY7QRO1mmRV8kfSD7qY24x2ojbjHaiNuMdqI24x2ojbjHaiNuMdqICkkKSbwRu6FTcI24x2ojbjHaiNuMdqI24x2ohFJpg/TNf9QffEbcY7URtxjtRG3GO1EbcY7URtxjtRG3GO1EbcY7QQFtqC0HBSTUadS3FBCE3lSjcI22z1xG3GOuIKUzTKlW2zQLH3xG3GO0EbcY64jbjPXEBSSFJN4I1oF51DQNwtGlY22z1o24z14UPW2akH68SyVTTQUGkggq4I24z1424z1422z1oStCgpChUKG7pXBKrDcz/APGKyalCotZQUhU5LKTLITkGlW01suqcAcH+0XROy6HRM2FS4EwQkFvKEg13LqXe8ImUzQqG10QoqSV0pgqzdWJJOReR9OL1tkDNVp5Pkk+Gsu/7fMPZZ+7/AKifIImNSNgb3OFcJuG1z5hEncNsJ/PRbBvGWa84iR1Cc9W5/YYTqE7Adz+4RP6hOenc/sEPUu+nc8x0Zn3IY/EN+aJLlx5VR6P99XkMSXuufphnkF+ZMOXDYE+YxNakbE14rieuGKPLE/qRs29/YmJq4bYXCrhszv8A3DDNw3fGJX3BoTHJq8IeuG1v0w7cNzxEJuGzNecRKXDbCIkLhs36FRI3DFfliVuGxOeKITcNrnzCHbhsCfMqJy4Zrf5x6QuGyJ8iYnrhs/6Ux/yOec6d/i/PQd5d3zmJnkW/Fcej+WP/AG1ROcirwhXFEnyKPDWpDjX5dD/kc86o9H8qf+2rQXyzvnMSPL/oVoSnJJ0pWgID9myHCmsCTU7eXkvuOWc9Vu2buGFMoYaQyrObSgWTzRYZaQ0j7qE0ESXLjyq08nySfDWXf9vmHss/yifIImOQb8Vwn8OfMIk/xCfz0W+Xa84iR99XkMJ5A+YRP++nyCHuXd8x0Zn3IY/EN+aJLlx5VR6P99XkMSXuufphnkF+ZMOcgnzGJrkmvFcT/GjyxP8AL/oTE1+IXCuWd/7hhnn8YlfcGhMcmrwh78MfLDvN4iEcs1/3BEp+IRHo/lv0KiQ41+WJXknPFEJ/DnzCHeQT5lROe63+cekOUT5ExPcv+lMf8jnnVp3+L89B3l3fOYmeRb8Vx6P5Y/8AbVE5yKvCFcUSfIo8NakONfl0P+Rzzqj0fyp/7atBfLO+cxI8v+hWhKcknWZLlx5VaeT5JPhrLv8At8w9ln+UT5BExyDfiuE/hz5hEn+IT+ei3y7XnESPvq8hhPIHzCJ/30+QQ9y7vmOjM+5DH4hvzRJcuPKqPR/vq8hiS91z9MM8gvzJhzkE+YxNck14rif40eWJ/l/0Jia/ELhXLO/9wwzz+MSvuDQmOTV4Q9+GPlh3m8RCOWa/7giU/EIj0fy36FRIca/LEryTniiE/hz5hDvIJ8yonPdb/OPSHKJ8iYnuX/SmP+Rzzq07/F+eg7y7vnMTPIt+K49H8sf+2qJzkVeEK4ok+RR4a1Ica/Lof8jnnVHo/lT/ANtWgvlnfOYkeX/QrQlOSTrMly48qtEixhwxmd8ZnfGZ3wyyU2i2gJqDGZ3xmd8ZnfolNmvPGZ3xmd8KapZrS/njM74zO+Mzvg3Up7AU2K04Yfcs1yqgab1wH5Q4/ZzkJRTiJ+cCYp/07FOesM3Wcm4FwE2ac+glNmzRaV9CqxLuWaZIk037qQH7OCLFOeJhyzXKkHHC6kLBTW04peO+awDCdTWsOM2bNsUrCBSzZcSvoNYYNmmTXb47j84l3KUyRJpv3Uhl6lMmFCm/WnyhD9nNQUU4yPlCn6YthFOcmHXrOelKacVfnD7tK5Wl29QUh9VmuVXb4rgPyh261bcK+mCizaqtS+lRP5whqlqzuwhjHJppa34As1rDjdmlpJTWsLlQKWmsna5qQpqzZrS/ngJs0otKuhQMM3WbDiV9ES6rNMku3x6kj84l3aUyRN2/UUhp6zmIUmnHT5QJimDdinPWFP2cWwinOfnDz1nZAkU4qxMOUrlVBVN7UgflD5s1yjlviuA/KLFm1qlK6ST+cWqU0y2aWbW7GZ3wpNmtXFrx31Vhx6znoSinFX5xLrs0yS7fHqSPzh5mzZyiCmu9BFjvhmXpsbYTa36ayVUrSGHaUyVbt+opGZ3xk7NrVKVXjJP5xLrs0ySyrj1JH5xmd8KTZtVWpfSomGFWaZJdvjuI/OMzvhpmzayaQmsA7+sSXLjyq0V8cCTVZ9VW2AlW7lL1eAPRE9lAAhDgyNB/0yKisFlKypVooqEmzaGItYViiHq1SVpJSQFgYlJ3eaBk3rlN5VJUkpCk74JijSyTZt0UkpqnfFcRoDQVDK7RaYLlHnwi3k00xpx7sIeMw28izay6bkkb8LdylltCQoqWgpuOBvxhxxbuTS2oJWFpIKScKjG+HF5SmTUEKSpJCgo4CzjfBU2a0NkgihB3iIX7AuJyWcZDQZShSdVUkG1j0Q5LqSBLKBDC/vKRsnj3HQk5VtvKF1wIcV/lgg05zSE6EtLhTDQdQtRcmDQXEXd8ILDYdWPpFp//ABJz4Ckm0k3g78SjLQTlZhywCvBNElRPdClLSEOIcW0sJwtJNITxQ3CkuzCBMJrlZIt2VtmveOHdhyWUkCXUD6uv7ykGi/HuMLmWHkS7JWpLKS1btAGlVX7tNyL8YRIS5S2cllnHVJtUFaAAb+MPOPrD6k5llFmu8OOsWnGgZ1pRadaSaDKA0PNuwzMOISha61Sk3XEj8ofmim3kxcn7xwA6YQp6ZbX/AJjaWqD/AGmvjCpYoAlVBSGnN9xOeO/4TEswpLSm5hSkpCa20gJraP8A5ujQVxQjih19ea2kq44UmaSETjKy08hGFeDmIiZQizRuQcmRUfWBuhpZxUgE9Ef4VKVzbhsMpVgVf+gYYmW62XUBVN6GJQsgMuNrUHLV5KabnPErLUl1tvlRutWkoSMfAc+g4y9MIamEKUFSSm6KArcQd3jgTD88ySErUuXQ2KpAJ4eCGZwvtS4dAcDBbqAk75rjTRGm54nVrQn1ptdGkjBQVsf/AJwRIzoZQ6p0N5Q1sgVIFw59CUW2EqbLn01f8sC8iG5JIT6tk1WlbuUuPge+C4pTAx+ibJK03karojm1lUTEwzTKIApaFRiBEsh9aJhh9zI2giwpCtzjF2h6Ry4CpiUcKQEilsHY+moEIyhBcpqiMKxNSr6Uhq2US6xukJBKTw316YlX3KW3GkrNOKFzMu8lFimpU3arVQH5xZdcyq65wTZ7oRxaxJcuPKrRXxxNlL2SccU0ptYF6LIp+ZiZcQRknEtJSj7tgU+ULlLTapMqcNq/KUVU03t3GJNp51rJybakNlANV6iwCd66JJhx0WWZNcqsgY2kpFR0RbcbYC0t5MLbKiTv44YYaA0FQky+SqM5Ltb+cYRNSS1BPrOUKskKJRb+6N6Mk8tlC0FtbamwSLSTWp4LsIeedcbQ84pjUoqUhLa7XOTfEy9lE1U808hKq01CaUPHWF/RttrWq0ckSe84wv2BcTsyksAPNJQi81BTWhPTEmZdVmZllpWFrUohX37uGpg0Mpasrpcqlquo5qYww8klCxMB936RWq1JG/xQnQdl2gzYdQpClO11NeDdgqfJdSlpDLYtKGpAvrTfMJlmXJdaWm1JbLoVXO1Ff9t0LS63LvUdCmRaUigpjUfWxwgM1BVaUtVnCpNYTxQjniXmHTLJelgvJuN2tUSml+8N2l8SapZVmZllpWFrUohX37uGphxmULCpcqUtvLVBbqa0uxFeKM6XLdtO4qtimq564QxO2mPXQ2WnUiobWmtRwinzhpM2Wg0lzKFtoq3BqRXjviadlyMk8lKsmtR2QbteEU6Il5esuShzVnVXoJJNOG+Fy06WMk41RRYrULtXEV5ueEhZlbsXBa1fNud8S+RXSdZWl0OqUqhVXVXcNVdMPzTmRWldEI1RtNt72GNb4xk8pY3l0tWvCz3wrihHFDTSj9DlApwbqgMB006IfUAFSsw2A60sk1Wk3K6PAQ7kChptUkuVSneKjWsIQky+TTkkioVWzTV/tDTk8UHJJVZSwVJ1ROPVoOmHWHUIelg8XGEmuoBxHTWJSZTksiyhaKEm0bVPlE1OBSC4WQ0wFVondNeM+ECplbNpFaBVaU1ffhEm5M+r0lXcolxqttVxFOAX33wiSe9WclgFAkFVogk7nPCvRYcZUxZyaXl1thG5Ubp54NDLWLaqXKrZs6nntd0N5azlrIt2M2vBA0/o51Tu127LiRg4Rek81VRL+jm1tFaLNVqrTUrtQcn6sBRNAq1jXVd2EI9aLRbGWRZRW9ChQc9I9HKW+HXGMoXVkXuWhT8hCZ1/JIKGi0lLVTWpBqTzYRzayqH5ZtSUrWBQqwxBhl2bU0lDCsohpmpqqlASTvVi8yduwncVS1XVc1MOGGJwOWW0j6Rv75Teg81TGMnlMnvLpbteFnvidQXLK3X8u04nFtVBQ90MS7a5Y5JltFVBWcDquamETMutTSVLXqCmtLIUCK8N2gji1iVcNyEPptHexH56JNvHgjP7oz+6M/ujP7oz+6M/ujP7tEqtU5oz+6M/ujP7oz+6M/ujP7oN9a+wFVuleCM/ujP7oz+6M/ugKtV5tDP7oz+6M/ujP7oz+6AITqqUjP7oz+6M/ujP7oz+6M/ujP7oz+6M/ujP7oz+6M/ujP7oJtVgG1SkZ/dGf3Rn90Z/dGf3Rn90Z/dGf3Rn90Z/dGf3Rn90Z/dGf3RZrXTZ/dGf3Rn90Z/dGf3Rn90Z/dFq1Xm1kprSsZ/dGf3Rn90Z/dGf3Rn90Z/dGf3Rn90Ab2sKQtIUhQoQd2E2Zl/Jj/pqUCO8V7//ANnPpCXnplY9HtyyFIaS3aoq6puFd+GpmWcDrDgtJWnd/wBbT/pGQlpebbnJX1ejjtjJ8JuviVkFuZVxsG0oYVJr/wD0h9VmGn1rsBdWwKeMbXm+qn5xteb6qfnG15vqp+cbXm+qn5xteb6qfnG15vqp+cbXm+qn5wuXl2n0LSi3VwCnj/q8cgjxMMSjOyuqsisPttT0w5MNXAlgZNw7wvqOeBMql3Uy5wdKDZ6YabRJPAumylSkEJ38eKHZZMm+t9vOQlskjhi3kXLFvJ2rJpa+7xwpC0lC0mhSrERM/hj5h/q8cgjxMS06lNstKrZ3xuxMvMzkwq3UtS+QoUk/eVWlOKJ6ZQ46pyakhKiRKNQ2aAVrhQUuiQnQ48qUYDVU37iaKuielTMWg5MpmETLkspdf7bNa3bkTsy6p18zj2qSlFiwn/M9+HA06XmrWpcUKFQiZ/DHzD/V4mpZTIbySUfSLod3gjPle0PyjPle0PyjPle0PygsW5a2EW8879N6M+V7Q/KM+V7Q/KM+V7Q/KHZiaUyUKZsDJqqa1HB/rNf4dPmP+t1/h0+Y/wCnmEsqCFOOWLSk1pcT+UbcR2H7xtxHYfvG3Edh+8bcR2H7xtxHYfvG3Edh+8bcR2H7xtxHYfvG3Edh+8bcR2H7xtxHYfvG3Edh+8bcR2H7xtxHYfvG3Edh+8bcR2H7xtxHYfvG3Edh+8bcR2H7xtxHYfvG3Edh+8bcR2H7xtxHYfvG3Edh+8F/1xNsosbDw13424jsP3jbiOw/eNuI7D9424jsP3jbiOw/eNuI7D9424jsP3jbiOw/eNuI7D9424jsP3jbiOw/eNuI7D9424jsP3jbiOw/eNuI7D9424jsP3jbiOw/eNuI7D9424jsP3jbiOw/eNuI7D9424jsP3hpaphDiS6hBTkqYmm/9oqbZZdmSi5RbpQHeqTAJBTXcO57LI8v+lX2gzy7XnH2go7wiVputgnjN514Bdb94R9boj63RH1uiPrdEfW6I+t0R9boiWKa/Ru2jduUPzj63RH1uiPrdEfW6I+t0R9boj63RH1uj2/63RH1uiPrdEfW6I+t0R9boj63RH1uiPrdENhNbnULvG4FAx9boj63RH1uiPrdEfW6I+t0R9boj63RH1uiCEVu3/sIT7L7csy4SWWi1aqmtxVfu8EelRMH6KWmEtIsIJN6EncxvMNryxVbKkhKUKKqpztTSt0GWSoqdTjRBoDStK4VpC0qcQk0wKolOSR4a8j3YmJmxlMki1ZrSsH1hDbX3bLla9IEEqUlIGJJwgV2KwVl60LIpCH0vIyK7wsqoITLl1IeUm0EV3IUgLSVJxSDeIolxCjvBQgmooMYNlaVUuNDWkWkrSpJuqFVEGqkim+YQMoiq706oarigJK0hRwTavgotptjFNb9Ae3HjibYIs5CxqicbQrDLiLLqXHkM1CrhU0hotjLJWsIqhQoOGCdShdtabFq/UqIr3Q0l05Bx0qCELUKmkZO2nKY2K39EWcoi1920Kwo5xAwEIW4QyVAGytQurBlsonLhNqxW+kKotJs51FYQVZVuyN22IClLSlJwJVQRiLoAtpqrAWsYVZWlVnOorCFWVpVZxorCLSFJWnfSaw7xD7CVKSapdcsCcip61abBNaUGdTjETqmn0BD8w2+U21ItgICVIJGGAMLZGQU6p919DgWtCmSrApVjd3wzNBxCLNMq6gkKfFilFJzcb6wtS2W1qs4qSDEpyKPDXke7E4w0m2442UpTvw/Lt+jDL5Sg1ZSB71x3MYlco16x6s+pSzdWYqLnL/rcHRDS0ywZZ9VebKDS4qIphHo1n1UoQ23YdCUNlVuyBu3UN98eiVOsVU3KFhahQlCqih7o9HNmXyTssVF2YqPpLjzm1UG+JN1LKETkkEOUFNUcFJrwg0iZlEjKTDjarX9yzeYnBLIS225LtoomibRS5Up6ILzbT9r1hl0tuWAVBG8E3V8aQtbktRtfpBt+ysg6gN0qeePSLSZYOKmbOQfFAGrgBwiyRW6PSpS0lTz0w2ttd1SBYv4MDDq221Jl3FLU5lLJvOBQcb946A9uPHE/wDRBaHnpZQqReE0tRMWWU5Bc1LO0uoQnOuhSktgNf8AyDT4pTNCKKPTDDXq2TcROmYVMamlm2TccbwaR6MU5LBzIvPlaRQmiiqhjJZH6b1zL+uXZtuvHWmppEzVCGJpLrkyl264hZUk14odWtFmYmkqcUjeKhcnmFBByrCVH/4xEsLVDq76iEkt2XF+j0MZYUOTcFbz04w82GXkviSVLgKLYQoncux4zHo5Yl28gy24FakUBNnc5jEnlG1qQ2y40Wm7Cimq6i5V1KXQ6ltlx5LkgJdNVpJChazj/uxiZJQlt1Ukyy09cShQQQe+AGpT1QoknGFCqfpFFNALsRW+phA9X+g9QLDiWiEkmqbvGFpeH1zYKkpSsp/uCbqw7xD7GUN8RK0/ykju15JRSgG/GCemME9MYJ6YwT0xgnpjBPTGCemG8qhpRCqotH63BGCemME9MYJ6YwT0xgnpjBPTGCemME9Pt5uT0xgnpjBPTGCemME9MYJ6YwT0xYdbbcTjRV8YJ6YvsDcvVGCemME9MYJ6YwT0xgnpjBPTGCemME9MYJ6YWV0v4fsda2H3Ja2aqCKFJO/QiACbR3zu+yyPL/pV9oM8u15x/oeR5f8ASr7QZ5drzj/Q8jy/6VfaDPLtecf6HkeX/Sr7QZ5drzj7XLKH1MoDQVqUg1NTvxt93qI+Ubfd6iPlG33eoj5Rt93qI+Ubfd6iPlG33eoj5Rt93qI+Ubfd6iPlG33eoj5Rt93qI+UMNevOWVpWTqEblODhjb7vUR8o2+71EfKNvu9RHyjb7vUR8obKp53UKtDUIx6OGNvu9RHyjb7vUR8o2+71EfKNvu9RHyjb7vUR8omUmeco2uyPo0fdB3uGNvu9RHyjb7vUR8o2+71EfKNvu9RHyhajPOXOLRchG4ojejb7vUR8o2+71EfKNvu9RHyjb7vUR8odYceLycmlYtJAped7i1iYcbNFpQSDG33eoj5Rt93qI+Ubfd6iPlG33eoj5Rt93qI+Ubfd6iPlG33eoj5Rt93qI+Ubfd6iPlG33eoj5Rt93qI+Ubfd6iPlDhM85qXFouQjcPFG33eoj5Rt93qI+Ubfd6iPlG33eoj5Rt93qI+Ubfd6iPlG33eoj5Rt93qI+Ubfd6iPlG33eoj5QAuedoFBWYjEGu9G33eoj5Rt93qI+Ubfd6iPlG33eoj5Rt93qI+Ubfd6iPlG33eoj5Q06qecClDcQj5Rt93qI+Ubfd6iPlG33eoj5Rt93qI+Ubfd6iPlG33eoj5Rt93qI+Ubfd6iPlG33eoj5Rt93qI+Ubfd6iPlG33eoj5Q4HF5RSHVotUpcD9hL5BPmOtynuOfp1uf5YeROld5d3znSu8gjzK1ia5M66/y7nm9jY4vz12Y/EOeb2xMzMLc+lFpDSHCkITuYYmAkYC686NteEZyurCnqmwWgnDdqYzldWAtGadOnKEivBGcrqxLugmwhKwbt+nyjOV1YChgb9OnKEiu8IzldWJpRJo45aF39oH5RnK6sZT6tKxnK6sZyurCwompdWrDcKiYzldWFZMk04IUhRNocEZyurC3qmwWkpw3an5xnK6sBCCbR4NIUkmoNM2M5XVh9pBJWpBAujOV1YzldWM5XVhK05pv0wS4SCb8IzldWM5XVh0KJ1Tq1C7cJjOV1YzldXXgHCQTwRnK6sZyurDTayQtIvujOV1YzldXTZx6sZyurGcrqxnK6sZyurD1snVPLULtwmM5XVjOV1YzldWM5XVjOV1YIbJNOD2VfFEpySPDSf7hDRKbVt1DXWNIcfdNEIFTSG3U5q0hQrwwjn8dOxzxLizayzoaxwrX5Q5LmVSjJhJWrLVuVXg4NBr3Rp2OeJdhtrKuPWqVXZAsjihp5EraQtwM0U7QpXas72/uwCtIQvdSFVhPJ/loOS8u0HVtgFxSl2UprgOOG1mUGrUlFMsLlE03oBcSEL3UhVYe4xD3HE02E0yDljj1IP5wqw2stJJGVuoSO+GXwLOUQF2K1pWG+fw0jks0xll2VO3rs3Wqb0BKUKTaZQ+i19ZJ+RhMwpCi2p0oFPug3r4t2GJazUupWq1XCzT5w+03L5XIsh5Zylm6/g4IQlKtUtsOhJ3jDPu6Zv3YZl20BbziVKAUuyKD/wBw3LoZCn1t5WypyzdhThhhAlkrDy7CTlqX0rvcEFmoDyUgqQDWmgNdb92G3FJqguJQo/drdWHfoy5kmVPqodwbnPf0RNWZfKNyzaXXFW6GhFbhzQptC6qSASOPQHFpZqYs28kkrs78SjaUhRfUU3nDUk/lE2hbLIEtnFLir9TauuiVS6ylHrLGXRYVWmFxu4YUwthTKwjKCqgbq03MDE0xZKclmq++MCRzgiHHWEFahasoVqakRJvNC16yUhKePHoviVDTCbb7jqNWo0FjmgEpKF2lIKRVQqDTHm0HuIeyr4olOSR4aT/cIZybanSiYbcKU40Bvi0MrLqbC7LTiE6o2ccYlW3dkS2kG7C7CEc/jp2OeJEoTaycylauAUMTjpTRtbbQSd+lqug17o07HPEiopJabDtohVKVAphEszLNAJbfaVZTuJCqnQTyf5aE25klvMzJS5Vu8pUBQinMIZoypC8u2soJFQAvQe4xD3HE9alXlodeStK0AUpZSN/giVQlgoU066XJjccQbVBw4i7cpHotPqpl1yyTl3CALWppThvv5ob5/DSKcdbKmMitFbRF9vC7giWckkhLrdWqYANqFD0XHmjJ2izKtM5BAshVoHOx5hHo5bsu68Zdt1lRboScLKsd0CH3XZB55DsslCRcCFVVcb7tyGMuQuYDaUrWN0wz7umb92EtvyS5pvG03ik9NRxxLMT8o7OOpRsrZBWlXHUUOF8ehw79K8y4lTy/9ihWK0vO7oDXW/dh1heY4kpMT2XRWefaKLIO8KD588Tbkw1abUhkJqq40Brd84JpecToDi0s6w2KuOIUlIiQdl5dtvJOEuFIAusEeMelFUufpYvx+jp4xLlDSbLjCUPiuYtIxHB/7iZmJeW9XttZi1D6R2tysdyJFxl1b2R+iWF0H0ZxOG/QxddDpVT1ZCiqXG9bvV/5wxIpfaSpDbswtYJ+9WzCUNpCEJuCU3AaD3EPZV8USnJI8NJYQKqqIzB1oydE26WrNrcjMHWhKFiihXTtZMVpWt8Zg60OBKRVCrKr93/wxmDrQ2k4hIGnayYrSu7GYOtCwAklBorVYRmDrQEfWsU7ozB1ozB1oJSEqAJGduiMwdaHMoKVO/Di0pFknfjMHWjMHWjMHWhC1pASK7ukcUEChUTnRmDrQt1xICEipvjMHWjMHWjMHWhtCs4DTIU2moApjGYOtGYOtCihINlRSb90RmDrRmDra8gtpqAN+MwdaMwdaEuNpBSrC+MwdaMwdaBpTqB1ozB1ozB1ozB1ozB1oVYSDZUUG/dEZg60Zg60Zg60Zg60Zg60OZQUrTd9lXxRKckjw0y+QT5jrc9y/wClOtzCbNnJrs139SD+elUbNii1J6DTSqVa+hsAAcNT+2sTXJnXX+Xc83sbHF+euzH4hzzeySsq02hb8xaKcouwnU489+EIyiQ24RUotVpHqarITkMtlSq7Os0hNtaU2rhU4x6QbUkNplHA2VlWOoCq98BZcSEHBRNxjJ2026Vs1vhfFEpySPDTL5BPmOtz3L/pTrb6SmgQuyk7+pB/PSqKk2SFqTzA00qnLf0ZQE2eGp1ia5M66/y7nm9jY4vz12Y/EOeb2RLMz6NV6QlzfqKVQrc3R0iPR78w07NkSjkuqiwpaarBQFHiFK8Eej1zfowzQR6O9XLYsqou1Wl574Y9ekz6UHqTbApRVhQraGq36i/+2PSdiVttvzbLtqylf0eTANkG60Cndg/4d/1hEy+uXFltaaKwC04UPBhDUwiXIylPWbaUlsfR0q2rOG9SFrLjyTZwQ8pIiU5FHhpl8gnzHW57l/0p1t8LFEpXRHCKD866VWVTZNtQHFW7u0qnbeLYTY5zfrE1yZ080tBsqS0ogjcujbUz2sbame1jbUz2sbame1jbUz2sGjszeSo/TqxjZpn/AOwv5xs0z/8AYX842aZ/+wv5xs0z/wDYX842aZ/+wv5xMuIfmQtDalA+sLxpxwNcY4vz12Y/EOebXklSHFA7qEFVOiEuNqC0KwUnXF8USnJI8NMvkE+Y63Pcv+lOtv28239HxUH510qsrnW1U92t3dpVuW9WWwmxwVN+sTXJnTznIq8NcnORX4QOLXGOL89dmPxDnm18hFzcwgrKf7gRfz17tcXxRKckjw0y+QT5jrc9y/6U62/bNUleo4qD866VWVNTbVTird3aVarX02TAKeCp/fWJrkzohAbtXVxjYR1o2EdaNhHWh1nJhOUQU1rGwjrRsI60bCOtGwjrQuqLNnhhbeTBs7tY2EdaNhHWjYR1o2EdaNhHWh5nJhOUQU1rvwBkR1o2EdaNhHWjYR1o2EdaNhHWhC6UtCumY4vz12Y/EOebX5XknPFGuL4olOSR4aZfIJ8x1ue5f9KdbfKlVStdUjeFBpVBarRK1HmrdpV/5+TFfdqafnrE1yZ0U+5E6y/QS6E2mzT7oFvzCHHJlJyoeKLDaKncoKDjho21KytqwlKCVGmN3BE4UVaMuuwVPoVZ3PnDjalkFtYQs2DRBOFTzwWyu8KyZVZNkK3icKxOu5J6kqqyRYN+HzjJggDJZSwpJC86kP8AND3HE829ZFMqZZQGNjFJ4RjDZeWbeSS6uygmyCMTTAQ4lTh+jSFLKUEhKTgSd6HXPuoKu6C7qP8A5BNUKuuqBaJp7vjCS4VJTQErsEpTXfO5EyPpHFS4JcCGyaUFaVhlVQ22tlTpStJC7qfOLnKDJ5UFaSkFH3hWHnNWck3lbFghRTviu5Eo3knqzCSofRm6lPnBVaQui1JqgGlxpuwz7g0zHF+euzH4hzza/K8k54o1xfFEpySPDTL5BPmOtz3L/pTrb6rVq2u1Te1IH5aVQKrdVqV0mulWLP02TFT/AG1P76xNcmdFPuQbTlmszlrh9UgAp5wIdKXU2VzOXLaqhKhZpZNOmJdWUSckX7kppnmvdHpBnKICJlzKg0NUnU//AOY9IIyoHrTyXRdm0s//AOYmGMogSr72WVcbYqbRG9iMY9Js5RIRNqKwql6SQPlHrTik2iwGilO/arD/ADQ9xw+xlLDinVutugZhMO5B1sZZlDS7YN1lNmo5tyJ5lDlEvy6GE1GbZTZqYesttofcbLeWs33ikTKw5QPS+Rs0uCrNm10UhaC8hQU0hAK7X0ZTjQYXx6SSV3TalG4ZtUBMMlbiApEq5L6kGl4Ar3Q02t3UplFSxoL77N/wwtUy43a9XXLpyQP1sVX8QuiQdKm8pLpKFC+hBAF3RCmyq1Vxa6+8omGfcGmY4vz12Y/EOebX5XknPFGuiXcYcebRc240LWp3KjfgKoRXcOlXyCfMdbnuX/SnW5hQVaK11I3tSB+WlUEqt1cUrpVpVt2PpQ2CVcFTrE1yZ0QvKWbqYRs3wxs3wxs3wxs3ww29lbNsVpSNm+GNm+GNm+GF6u3a4IW5laWtyzGzfDGzfDGzfDGzfDGzfDCncrapS6nDGzfDGzfDGzfDGzfDGzfDGzfDCEVrZFK6Zji/PXZj8Q55tfleSc8Uexr5BPmOtz3L/pTrcwUKqpS6rG8bI/KmlVk1WhlFk8dq/SrasXhsKt85u7tYmuTOsS/u627zeI1mq1BI4TGzN9YRszfWEbM31hDFXkYfeG/GzN9YRszfWEbM31hGzN9YRRLqCd4K1iY/EOebX5XknPFHsa+QT5jrc9y/6U63MWM63q+OyPyppVZK9OUXXjtX9+lU1Z1AQFWuc6xNcmdYl/d1t3m8RrMilaQtNpdyhX6sbXa6gja7XUEbXa6gja7XUEbXa6gja7XUEbXa6gja7XUESqkMtoV6wi9KQNYmPxDnm1+V5JzxR7GvkE+Y6Yq3oQ4mTRZWkKFX/wCMbTb7f+MbTb7f+MbTb7f+MTtJRs/T/wCf/angjabfb/xjabfb/wAY2m32/wDGHHVSaLKElRo//GAdNMWBRQXq+E2R+VNKrJCgyi68dq/v0qkWfobAIVw1P7axNcmdK0G28qtxdkAqs7hP5RtNvt/4xtNvt/4w2yZVpVgUrl/4xtNvt/4whl6XDdtJUCly1hTg4dJWG3UyaLK0hQq//GNpt9v/ABjabfb/AMY2m32/8YdtSrYF3/X4R/bG02+3/jG02+3/AIxtNvt/4xlHJRAQKVo9Xd4tLI+8vy6zK/iEaxMfiHPNr8ryTnij2NfIJ8x0znumJTkk+GknuX/SnSTnIr8ITxaaYCU2VJXRR3zZH5U0qy2mwMosHjtGulV/kWBT3qmv5axNcmdLIcqfIrSyvJufp0iuKJPkUeGkd5vEaR3m8w0sj7y/LrMr+IRrEx+Ic82vyvJOeKNdK5fJtS+CVuAqK+Gl10C0QVbpGlXyCfMdJnxnwvVbhiVBVQhpO5wRQKvjVGkZ8TtTSr9139qYz4oFXxRSqGJoBVSWlbnBCdVuRqTXRqq4Rnw5dY1WP3rsf/N6KWtCqjSM+DUZPVG7nxigVfoVMZ8FeW1NmlinfGfp5rkzo0Kr4z4k1A1CHCVcGoVGfGfFdyM+JZVbg25fT3Yz4BGEao0g6qJVCzZWlpII3jSKJVUxVRoIz4cQg2lGl3OIz4z4olVTDvN5hoZ0Z8Z8SRBuClVu/tjPjPjPiowjOjPjPjPiWoa0fRuRnxnxnxnxqTXRmPxDnm1+V5JzxRriuKJTkUeGmXyCfMdI9MKBKWkFZA4BWA/ZWoFIVZSKqvh1YtN5JRQ4lwUKCMawy6pp9DbgQbZRcm1m1hHHCOKJpgAhUuUhRO7UVh1ltDpySihS7GpBG5XQRA4omUJBBYcyRrumgP5woNocU2CRlrOoJGNDC9FWijj0Bx6KNBfFpBpprkzorhxaQUhDq2r99JpBZsOtOBGUCXUWbSa0qIdsMvPZNwtfRorUgVMNvNG024kKSeCE8UPzKwVIaTaITjBedtWagakVNSaCAqwtv+1wUMI4oRxwqUobYaDtdyhNPyiWl3F0dmFFLad+6ExzxLIUCS+7kk03DQn8oZaKXHHHbVlLSLRuxhoqCiXFpbSkYkmDkkOFsEjLWdQSMaGOaHebzDQPHCCZdxDK3zLJdqCLdSMOaCoIWm8ii00N2gxKlJtOoWsHcFmnz0UQrjh70kEOBtq1abI1VRuc93TDTyc1xIUOcQ9Lr1GSZD6nFHU0qR+UJbsrNpNu2E6np0H5paSpLKCsgYnRXx6Mx+Ic82vyvJOeKNcXxRKckjw0y+QT5jpJ5tCSta2FpSkbpsmCWpOaW+0ymjSmSm0qlKQ3JJDylzLv+JeW2UVrqnFc+aIZSwh5U02tCm0hJyWd9bcuEI96EcUekiuXmFIeU0ULbaKgdRSGjJJmEAurXNWq5JQpw7taYRLPql1S9tt31hKWikBVRQFROq3b4RA4o9Ipcl5ij00FoWlolJFhIx5okGktOodZU6Jlw1sLRfZ4DfQ8F8L0VaKOPQHHoo0F8WkGmmuTOiuH2HJeYSpU08oKyJs0KiQaxPekHJV/1xafo2VNKFlAzEcZN54+CESjxmFVFFqlgq0pWKsN81iVTMpCHw2AtKRQCE8UT7bSC44pohKU4mFpfl5oNpKVXtLSa1uIpfC8vlaZVeSy+fk66m1COKEccLfLD7jSpRKApporvtk7kSc0hYctTKVXsmrSAhVBjhf0mExzx6NdS046lqZtrySbRAsKGHPCH8jOpmUpWlktIWhYP/lMYljNC+UYTaNLlPKF5HEK9aPR7SWnW3GQ4maWa2Fj6vAb6G7C+OaHebzDQPHDNhiZE0ifW5VdckGy4am+7NO5fHo1E206pBmZtRboVVF9m7dESxmWHV+jQ68QwUlRQDTJ2k40zrtyoiTd9XdXZbmg0hWKQSC2hR3LhE8GWHUpdkkiy2ypr6S3hea1puxk2UBtFa0EIhyl5viXayZEq4wh52u46lFmnPqT/tiULrD9/oxLNlKSaODcO9DrLku4pxz0O22LQ/6ia3cd4guMNLZl0+i3xmlNlV3fjCT6MbdlnPUVJcUsFNpwgWMcTjfE2mQlHmWlSK0LbWkptuXWccTnXwpbUstLiFMFlaWVWlJqLRtV1O6KR6QmHGiHlTLlhav8u6lOCF8ejMfiHPNr8ryTnijXF8USnJI8NMvkE+Y6TMjMjMjMhJKN2E2U1ujMjMjMhJKaCAUprdGZGZCrQpokJFTGYYzDCTY3dABIrfGYYzDCSU0GgoDejMjMMZkDTTXJnRUQmojMjMjMjMhI3aRmRmRmQgHGkJsisZkZkAlNBFEiprGZGZGYYzIqpNBSHebzDQOojMhp1TNXGq2Fb1cYzIzIzIzISDcYOo3YzIzIzIW041aQtJSob4hKUt0SBQCMyMyMyFWhTRmPxDnm1+V5JzxRri+KJTkkeGmXyCfMfsia5M+zu83mHs0x+Ic82vyvJOeKIJpXgEeqTEt6u8WsskBwL1NaX7x1pfFEpySPDTL5BPmP2RNcmfZ3ebzD2aY/EOebX5XKtJc+iczhwphWRaS1axsiPSMwtDmWdmDq3QKlAzQP7f30wmJabWwQ423ZCEkG04ATeOGHAqZW8pl/1dZWiirZIoKAcIhoJLgQ6stIdU2QhSxXU15jCG21K+kqG1lBCHKY2TuwtDkw02uzmqWAYlORR4aZWVRbowKdYxsA6TGwDpMbAOkxsA6TGwDpMbAOkxsA6TGwDpMbAOkxsA6TGwDpMbAOkxsA6TE422LKNQbNeA+zTO5qDhGwDpMbAOkxsA6TFDLpI3jGwDpMbAOkxsA6TGwDpMbAOkxK5FNi0hytDjh7C7zeYewPOOt21l5y8k/fMbAOkxsA6TGwDpMbAOkxsA6TF0ukbu7GwDpMbAOkxsA6TGwDpMbAOkxsA6TEstpuwrLoFQTv6xK8k54o1jIW8n9I2utK5qwr8ocmEWsi2hJdSU0Sp4VCTzJJ+GJFMyoNS7Ewp/J5MhwkOKsg8G7DQCZZbTFcm5kfpeCp/Pdhd25EpySPDTL5BPmOvTvE34H2aZ9zWpL3XP067md8ZnfGZ3xmd8KapZrS/njM74zO+MzvjM74tUprbvLu+c67LfiG/HWJXknPFGuL4olOSR4RRJoKRnd0Z3dGd3Rnd0F61qymxzRnd0Z3dGd3Rnd0Z3dFFGo0FAG7ijO7ozu6M7ujO7ozu6HHQrVLpXmjO7ozu6M7ujO7oVaNdeUoYxnd0Z3dC2lq1KhQ0jO7ozu6M7ugRRJupGd3Rnd0Z3dGd3Q26VapAIHPGd3Rnd0Z3dGd3QAo3acxOOIVZWhhakkbhsmJd960tSktgBN6lqVS6Fhxl5LyHG2yzcVavNONCIWlTTqX0vBjIXFRURUU3ML4ey5EmWXMkoPrA1VAbjxGKi8aTn0gKbjWM7ujO7ozu6M7ujO7ozu6ClKrioqv4TWM7ujO7ozu6M7ujO7oSo4mFAKuB3ozu6M7ujO7ozu6M7uhFs5qgsc0Z3dGd3Rnd0Z3dGd3Rnd0EqvNYleSc8Ua4viiU5JHhHNBZnmXm57JKVlcoVtzNm8kcPBDRdl1y6ZlhTrSkrCjcm1TgNISG5dcxYk0TSlrcANkg990P/RLqhDbjad10Lzac90S6CgfSki2tdhA4K753I9LGZaCZWSpqkqqo6muHPDzaPo3kKYVVp22KF0DEeEelVqUlVJ9SE5V2wkXJ3THo0y0ulSJlxxtdpeBRXDoxjJAAoy/q9q2LVrCtneruxzaC4cmJ1p1cmVIycy04aMblFJ3q7vDHpQOsoS1KKolWUArqQb68cOWWcq6h5tmy06FJJXhRUMurYq644hvJJWLio0xgtvsFtaHW23KKtBIXmqrvVuhDDEqXlLU4lJKwkailo9JpzQmTKU1WpaUlDoUapFbxuRK2JE1mmS81aeGApWt12MCbOXDJkRM5O6mdTD725AS83kP8HWwHLQ2TxhevKj0Wiy64hbq7TbSrJV9GeEQcooFfrDjdiZeslu+5JJxNKb8SFhuymbRbSXVhG7SyN9XBE8hRU5kJh9Tqybmk2jT/wBQ4pppOU9XU+2nKA4DBVM0ww88gNrWgKok13IEc0SkiVqRLlpby7JpboQAmu9fWJT1bK6qdZ+jyp6L8IfttoadZeDKg48AmtAa2uI70SYYlytyZyoAKwAkoxviWTLy5W86lSyhSwmwEqsm/jiRaXq5h1Ljn0ztkABZGPdE5NoW8hn/AOMS8Ak0UnVq37qw83ZtBiyHDbAVf91P1sYmW0y9lphxTRdKxeoUwHPA05h5hRIS6goJHCKQlh2cW4G7BaOTSkoUnAwpx55Tj6nWnCsJAzDqU03oemUl4vKfQ+nJJCihQTZw3RSJ518K+nmMonKJANLKRhuYaXn0g44kESjxZeL9UmtxIQo0PBdExOrtpaRKotM7qXMopJTx1uhQLYK0utNnJOBadWaA1h1pMup1xEw3LgBQFStNYmFmVJRKAesFLg1JpU0+9QcUTTrbGWl5ZQQ45bpfdmjdzhE0FOO3+lUsiwr+xN1+5CZOwhS1laUWXgo1SK6obkSSciXpyaSp42nsQMVE7mIFIl2mJVS3XQ5aSpYTk7CglQPTDdmTJS5MLlkKLoz017tSYylgtqC1NqSTWikmhhEL44/xzTv0z9GJtLhLd+ahQ3N7eiXabbcmHH3n0JLqwKWFHG7CJdDEuVTDoWS2pdAiyqyanjhieXbal/UHnVt1rSy4BBZshK8nlRYcC7t40wMelHHW1OSzc6Uqct3oBCMBvXwpzIH1NL/q5ftfWtWa2d6t0NqyStXOGTpXdtEV7olVJlz6tMvZBt23ecb7O9qTEg7k1JE2tSBU5tAo/phOSbByraltDKCpoK6ofViWmXm0trdbC6JNReIPHEryTnijXF8USnJI8Ip/bClF56ZdS0tDWWUNTVNN7vMSwmXHHHG5fIhKlCjdU0VT5woC3qpZMqb/AKgr33xIqyaktSTVlLhVn4WRzY8cFpTriG1JKFoRSigeMRNVtZKZQEONVuNBQHfrSHGHpqYeC7OqURVNDUUuhToddQ5l/WARTUrs2TubohgIcdSpl1byV1qaqra8YUpp55DRWXCwDqLRx4eaObQXCw4++WHDVbFrUH86cETlpbliboXEAjOFKKHQIU646664t1t5SlEZyMNyGENVqmYaWSDQgBV5idQlK5yYmxZUtZA3KJO5QDgiTUCq1KoLaTv1pUnohlQcdssrW422SLKSqtfExJ2Lf+FaLLdTuGmPRAaIWtv1b1WilfUrXpj1lT7z72TyVpwjNrXcEL15UMOZRxpxhRUhTdN0U3YacbdeQ8hS15a0CpRVnVu4BDUnlXvVUADJVBtUNb7vCC6m1bUpwrvzws1KTviCwX3nWMnkg24q4J/834bYC1OBAsgrpWkCOaGlEqbdaNW3WzRSd+EZZ955SXUvAqVupwuwgvJW429l/WAtJFyrFjd4IYWguFTJcUm0queaqrDJZcdZW1bAWkipClWiDdvwwGHXmVspUhLiVAqKVGpBqL74cyjjpykv6svVZyfnDjrbzzGUslxLarl0/wDNyHyiv0zpdVXfNPlA05iYmLNvItqcs79BWFLfl0VMquZbDblc0CqTdwi+G5YMMesrbL17hsJRWg3MawzNrbdSkSj7qkW8LCgCKbpruw8t2VOpbC05O1SpIFglQF94hKrEulwzDTdQokEKUAdyJ85IuOtmXTZyxsVXddvfnD0sthBmkvNsoCF6hVsVF+5gYm2XUISuXWE1bJoqqQrd4459IOOGFqrVlVtNN+hH5xOOFKrU2lIcoaZuBG8flCm3pl902krCyRVJSail1ILinHXHFPomCpSvrJFBzQ/VbqG5imXaQdS5uX8110TTMuh5tMytClELTktypIxwGAxhbtpzVTKZqzW4LApEupLjpTLrWtpskWU2q148YlEMuOsrlklDbqSLVDiDUXwy43btNIWkVVWtogqJ4aiGaW/ophUym/6xrXzGFobrRTinDXfUamEQvjhKnH33W0uZVLK1VSFbm5XmrDC026srdWmp3V50MKlkTBWguatlabVFG0RqsRXoiVRM1LiGFsuIBqlSVmpB+cUMy8+KBIDpFw/83Ydq47knXcstiosqVdwV3BdCtW5kVO5csV1BXWtd/G+kJVlXrCZj1pLVoWQutd6JD1Za8gzN5bIlQsoFFVpznCJejjpbl1qW00SLKa1ruX4mEtoeeVLpTZQypWpSP/N+G5dC1rbbFlNvEDcEHjiV5JzxRri+KJTkUeEVVjxxgemMD0xgemMD0xgemMD0xgemMD0xgemMD0xaTjx6BURfxxgemMD0xgemMD0xgemMD0xgemMD0xgemMD0wbOvWThGB6YwPTDuRH01g2L93cjA9MYHpjA9OhVWMYHpjA9MYHpjA9MYHpjA9MYHpjA9MYHpi0Bfx6cxNsIpbdZUhNcKkQpD1t0rYyBtuE0Tugb0NrJW242LKXGl2VU3oboyhEizLLYCK1t2iK3c3fDjKy480tNiy66VUTvCFtOLecCilVpbpKgRhQw6VpKi4WyslWNjNh9xLOUedKFH6QovTgQdwiJx13OmHAuyV2yKJAvPNHPpKKjA9MYHpjA9MYHpjA9MYHphZViHVp5goiMD0xgemMD0xgemMD0wEjCCSLzwxgemMD0xgemMD0xgemMD0xgemMD0xgemMD0xgemMD0xROESvJOeKNdyLbQmWBsdF2VJG8awCRZO9vfZLwZNHSg2Dw+z4CMBGAjARgIwEYCMBGA1x3l3fOfZZXknPFH2u6htVhxSSEq3j9jqQuZbStOIKsI2011o2011o2011ocCpppP0zhvV/eY22z1422z14Dbcy0tZwSFX+xSvJOeKPtd5sKsFaCm1vfY87+IV4DTyHKnyK9ileSc8Ufa7zalWErQUlW99jzv4hXgNPIcqfIr2KV5JzxRrthx0BeNnEwFDAio+yX0uqsNlBClbw+x538QrwGnkOVPkV7FK8k54o0qnXVpbbTeVKNAItsOJdSDQ2dww3lFhGUVYTXdO9pSd6G3Te6+A64rfJ+ybBQonghxpba7K0lJvjY1wqykps7/2Gm0kqtb0bGqJglCvpHSscVBGxq0imyhRIjY1RLLCFfRLKj1SPzjY1RsaoSrfFdfleSc8UaUsuTHqpUpNh2o1KwapxxvGEeqvllTzU6wwt9pOpcQvg+qqEtZVKch6YbabUBm/Q18SYaYE4pOR9KIl8uhI+kBRW/cuN0OejFEGZU8hbK7OLBvUeaik84j0dNrmEveszapdUuEAXWlC7dqKViTfcRRmYt20KKAEUrSzfaJFKGvdC0KylbP1WlK8BEpyKPD7JPENF7m+w2eeBK2RZyGVtbudSJeVyKqPWvpa3XCugNF2HlKSE2HnGrv7TSDKushteTyosrtXVpfCbLSVByZMu2VLsg0BqekGBaACqXgGsNe6NfleSc8UaUtuoS42rFKxUGDLCWaEucWrAs9EFn1VnJE1KMmKR6sZdr1f/KsCz0QuccWFGxkmkpTSwmtTx1u6IQAhDj6bX09gBV6ifzhT6GG0vKxcCBaPPC+KJTkkeH2SeIaL3N9hs88JmmWkvJyBaILlm+1XeiQeSymwzaK/pPvJpoDRdh1tTCFNLfccygd3FGuFInHFMI9deqbYdBA3EJ4h84Zl0SyZtCBZsuEDnvhiXWq0pCaGmHFDXujX5YboZcPenXRLusuuNIubdbTa1O4CN+Aq8V3xT7IK0qSBTdjPRGeiF2iDa3vsNFkgWd+M9EZ6Iz0QNFbgWkA78Z6Iz0RnojPRCEncFNeGSYLyj/cEgQt99QXMOY2cEjcSP/2VTc9k8tkEW7FaVj0VMssk+uTIlVoKqFlVDWvRDjQmWS42KrRlBVPHGWl5ptTdvJ1JpfWkKtPtJsqsmqxcd6Gj62xR7YzlBq+LfgzaWfWFW0ICCqznKpjE/JTUsJWak7JXZdyiCFYX0EBbc0wtBVYCkuAgq3uOGUNoEwtTyGlpDiUlsK3b/CDL+sNZcCuSti1TihsoebUHLkEKGq4oTbmmU21WE1cF53ok0WbXrDuSrXDUqV+mJNl5v6J+tp2ux/8AlYn2FANplbByhVcbQrCVpebUhRspUFChMGryBQ2TVQx3obPrDVHMw2xquKEtKdQl1WagqvPNC20uIW8kVyQWLUMTEwUSmVGatwY71Y+kfbb3dUsCE5V5tu1m2lAVhDa3UJcXmoKrz/p/0hKyybb7rRSlNaVMejp6S2qp9D02zUCi0gi2OmPR8zOyoAaefL6gWg2UqBzUi+/drDcsj0an1mT9IJmahaB6wgLUbjwA7sTi3pFCBMelGJktFxKvowNVE2WJRT8jMS6WUNyy2mw1Tc1QuG7qYRIsIy6wtmoWvEJUK1MenvREo2hMrMnLS8xaAJvvaVu8FY9KW2H5OZdQ2Wcs40fpEZpo2ABvV3o9HOutoc9Ir9JtT05QjUgbg4hQQDMMvOhM+qaE2h5sJpw3W67lMImFSSkhEjMesejG7QobRqsHwj0en0dKl6bYYsesKW2WiortLtoVw1NRfHoeuPrX/wDyciXU42Fynq7rTl/3qR6QZUvLfSslolQ+lbRuf+96Ms1LO5UTrcyWXXUVUEj+3Ugw8p2USkO+km5gtlYP0YF8T2QkG5pmZlwy1elIYx7t26JRbTRWtIaQ9MLUhSFhIvNDqgrij1d30chbgLp9etJ1Va89dyPR1tlboblCwtlpbdUqr/ddfBty4cbT6N9XRlVBZC77v3iVce9Hp9Ij1FEuWlLT9GoY47hibKmHHWZnJlJacQnJ03DUVu4P9WIKkhRSapqMDr3/xAAuEAACAQIEBAcAAwEBAQEAAAABEQAhMUFRYfBxkaHxECAwQIGxwVBg0eGAkHD/2gAIAQEAAT8hwCgMkEZlBRBLhATQzkwGf7sDDQwc+7ENQdeKIGQA/kgphC+EKOdlR2VHZUdlR2VHZUdlR2VHZUCVFkNgKofH9t23L4EgTB0ix2ATKSCQlJqkuasnaEBuIjIkUJqraw6ADCSFnwhMIiAacTb+4V9ty+FZcI/CMgUZWEydjpDjsjBi+Bb4ipYZZaFUQEZMQwd6NCEDARnCRqNyGo79KmXMRdD+31xdg0owSa+UoqooFoSkLGJH08pVRRQbQAjfCX4H/wAf48Z23VmasP4FZZZZZZZZZZZZZZZZZZZZZZZZZZZZZZZYtb2w/iFlllllllllllllllllllllllllllllllllllllll//AA4XFZv/AMdYx9kpEE1HNNRzTUc01HNNRzTUc01HNFQweBIYE1HNNRzTUc01HNNRzTUc0NkgKNF7QjMUDVNRzTUc01HNNRzTUc0DRKlC0NbgZqOaajmmo5pqOaajmh6ZsVe6IzFA1TUc01HNNRzTUc01HNNRzTUc0q4S0W6P2mo5pqOaajmmo5pqOaajmmo5oTISIVe++v8AcAZAzh6ZISiiqNx4lYqO+l9Jd6RLg1OkNL04+FvqYjW3hisYCgIq8olwanTw2+ntUemHKiaB+ywxa4wdDh5kwAA/BB+YMleEYTYUstbeGIGJgKUrwiJsLPw68/XuwM/QBBgySdIDItpDJQMUPHWYrG8ZQKNupfJgcRDmpxgAhewEmitSQHCa/wDjYWMDpGvrwBBZ0z8LR1WN46rG8d9J0T331/uWhtQ6lxXBqDJbgG0IApqLVOpLIal6dcAKFxojHcMR5EACdJLVEzCZYarrnwLuAChVWwjPuNQD0t0g30vzo+CBNfCPZBabYglBg2LxtLqDd7JVUVa6GNVkDcUVAOinWD1s60cQNRQDZwuqMwAyrlQrTjBAAwt4JSDNiKdGpt9PaoJ8S7k78gEcBhYwTUMsdQNeEgsEIhVLvWWtOayJle0sULwGHKEqRDhVXrCzYOGZAHNudFAaKpXQJlkCCzki7tWJL3oE4VgvI8XjWyysaNYpwAzD5gcoNKStIBN4JADNWagjVRqdefr3YP5kUkQIsBFQFJiOCj0TmoWZdYaeaC6TdVHLOFHAYTmagC2ooxVSlAlqAUklYqqBMQKhUg0LkAXjnHYMEQAuD6CxCUDa28FRrP7QpMjmrG9Tg8gKSTBnKAKWNGhi758V2BRTVxgEKgtACCg0WLXhR5o6ZN1SXVwo4ISpv7WQaOFIojBl1BGwF8SZnRPfBwakPmbwTbCbITbCbITaCAgXDTnEk9omtLwDBm2E2Qm0E2Qm2E3gjcUsK6e0c3DBYm8E2wm8E2Qm8EYdsG+Ip8xNkJtBNkJthN4IsSmpYy90xuRgpNoJvBKEURS9QY3GiDEs43ZJc2Qm2E2QifrRCoR/SbQTbCbITbCbYTbCbYRPUEkp/wDBNjH2xj7Yx9sY+2CiQxZFNEYj4oN/X/8A/wD/AP8A/wD/AJ3l61//AP8A/wD/AM4lxgCQKr+Z7OTRA4PiKfwDG355u2Xvce6Zv5jsCTIMYig/Io4EwNgJAgBkvQwRAC6fybeybeybeybeybeybeycMdNoXtNvZNvZNvZNvZNvZMHybD08uu1MfSbeybeybeybeybeycUz9MJ2l0yA+ybeybeybeybeybeyZ3jFw4eOVcuWM29k29k29k29k29kCfSiEz+Zt7Jt7Jt7Jt7Jt7JzF5rwYDCdrTb2Tb2Tb2Tb2Tb2TicbphO029k29k29k29k29k29kplRJl6WFTJwKbeybeybeybeybeybeybeybeyWP/8AD1sZt7Jt7Jt7Jt7Jt7Jt7Jt7Jt7Jqtir2ANSAkwINmSAh7sssXJXsw4WofqdBBnecq4UZqEY8LEAP48PAt6G0ZRNGUuawDgpVWLwwnhnNAoAUGUoCGRCDcEEJwZEK4TDIJEaFQDH4cLAB5SRbIDOoSiCAQXiETQoCkQxWArGQKLMw6SCtWHKhYAnEZxYReyRAqz8DhMCtlYIDFMSUr6Vg3wJXrAwhOFKyhMIpW4qiglpt2fl6lGwzkxAQA8CaQHkyil4ABkhNkA/AbBTE0RQVk4aQdAjBbA1wwTrGDhoeQDLNQggKINZgAmjjEoGgAhcDHkVVAaeGIossABBTUMA1VBgA4mghA4IqAmjVGKra0Ob2A1qQVqESWDKghE4JVKBAlk5stPHrvtAYDMw4gDSBnABzFYEK9SOUEBUSKPV4QAApDJIEWY0pjA3TsTIIg5IpXI/nikkGxxgouZRwAqwA/7K6IOEEGGJwQlW5nQQtA1IXAyqbSgBmoANSjPh0J+z4dQYOvji7Cr4V0UEeDZwuCAg1MNdxkfCDORCs3V2sfAofMKJAGAxAImrtLnP+Ai0qUCuGUqt8gbZUYFaUheA/tKVUmcBIMMTghBh5hKDFoGpAioMqG0oQNQANRBnw2rL0t4zEWsVaohqQMIAkpKQohA2WXmRSHcRh1uAphZJ1pCXWsUQQKudFgsYgLhfDKqowgHKC/DArUIkCIVIluFEKUNTlMFhaFa6pkZhPChf0kogSwqwSuQJInYlTQ4n5T4DpFGgbUCg2NQG4YJcaoDiRyKDuTIARhq0ptCCEABJoCbBwBzBxqzZzOWVR1AKLSl5YKlXwIYIQMEVc3mvsMJo3C7WABh4OHWdwWQAiuNxTWAkUsivXbiYrbdhCJukKLjxDNLnMQDfwYlIsUABQT1K1wFw4EEFvQ2jKMIDMAQnfIgamWiVTL80hAFGCgKRzl0chJWdLBY3GOcFv1K69QeFYK1MCs0GpUipclg30gen6gCQCrJQqj83hBUecA1zVwSCbYtCF+IN6wpQSqQuthFPZIZfoQBx0jbAXJxGAJJqoC0JKhoC8WWBMbdn5epRDzMIWPCDDlbJCNYFNEWgwYsHAWPQIkm9YTw3MgQsQKEyODzgQ+GphK4ECOFRDPdcRiUwBEVgiHgugCXGAx4aFIQIUJki/LWyQJ2Lcj0g4EKxG2QoDVggfBrHLv6QpqmIEbT0golmZuKCQo0CTeb7Q+PXfaEgORgwUkH8XIWATWnGOrXngGhgFHgwMAxxKhbNwygwqITSFhA0aZYBw1hMoZc4STBViEkAzhiGDjo4hwWAg2RApS8AY8KDVVCRCugVhAGrgmB6ABBcpPOG5wnQn7Ph1BjXUJchIDRUFUXcLDh4QA4JLQkdrHmp6wCjqg5hAM0wm1GOiD36RBikTkFgGICAWz8gdYADTemdihAzBrpBm4lAhYSyitXWOpkBwAINkQKUvWBW6lBqqhIhXQKwiw1maHYACC5SecNScJtWXpbxmJhX1WjsdYUZUCVDaqrYFW0hwgp2oA0zXWApBJCAhGhBxFUj/wDqggG4xQKKFIxJAUhpAqZaAVvnACJQ2A6CBjLIa2MMDldLfWsRbPhAHAWAFBaa1c4ltiV4qIkxvRCxg2PwTIo0A9MKCiBWKWNEqJs5QE0kQNTH/SNWzoQs1gEBSiMGXCwLNtLkWdEIS7wpOghXBobFjDXkFXQu1gFXAFN5r7FReJIpnC3oGS2ACqml24zS7cZpduM0u3GJYwHNGoAFK74wpwMPLQJUQwFJw8Zpd+M0u3GaXbjNLtxi10k7/KtGqWHrNLtxml24zS7cZpduM0u3GDfFTQOqBukNRElUYes0u3GaXbjNLtxml24zS7cYoKtg4eOR+seM0u3GaXbjNLvxml24wqveuJ98ZSs34woTWk6Dml24zS7cZpduM0u3GZzuqWPgQFKne80u3GaXbjNLtxml24zS7cYVBOmW/wAppduM0u3GaXbjNLtxml24zS7cZVKib0iBqEjFNLtxml24zS7cZpduM0u3GaXbjAPb+zS7cYW4+GyHWUdv3NLtxml24zS7cZpduM0u3GaXbjNLtxhzQWCj2pd3oghCyMwQDAgfYmjmD+fV3DPOmE2nPN4y9gRVgzyWnWIycPj63UpG3rE7jn/RivcM86YTac83jL2FXC8qhQ84AE2IDOvouOOOOFzUjbzkuOOOOOOb3nDjjjjjj9Jxxxxxxxx+Atxxxxxxxxx/wW4Z50wm055vGXsK2KeTBQwp4gAE+gQ8IA4I4oWQ4RxKazcn7Nyfs3J+zcn7GJ+LyTQqM5Ew2PmJNozjI0IWLzcn7Nyfs3J+zcn7Nyfs3J+wZXEeJr1m5P2bk/ZuT9m5P2bk/ZuT9lIqt1UJxyPomxj2FhiCgXm5P2bk/ZuT9m5P2bk/ZuT9m5P2HkdPqwNQvAhaVUyyvNyfs3J+zcn7NyfsMg8kGRnHIzcH7Nyfs3J+zcn7ASRHIuMIeEhSSWTT/A7hnnTCbTnm8ZewQK+zCCHDkiwAD9Dac03vL5jY+Yk29CRuOfsD82PpVIiw8t/yzvGU2nL7On8AAmUUBmSQIdbKhAOKDbr6bjhbDHOmE2nPC3GEccfqhYgWoQcr3QB5+htOaHs8PmNj5iTb0JG45+wfzY+lViLDzaX5N4ym05fZgwGoni3pGOPEcJLazu3+oRERsQSSD18ATjVgIMAgAAAUv/qd2/1O7f6lc2yBEgEb6+q0M3YCQJgyRAAB8TIKp7kritMahAFYGaz5iI2Z5zUPOIEBs5AfKUIUB0cAI+Z5xliecHHcqQKKPUGNmecMax/5Q2PiTTxYah5wnjTdEXBWPxCpXF5yqMnSME2BatAQOpC6Q4ViiEwGVnSsbM84Rqxk2Z5x8zzhYFHFZmGwOkZdy5i/9j1ECw1MCfHqYBmDCYwJfN+OsCTcHG9v+kGtVmELnyjBAqkwhzFQYiXoYuLNTUPONmecImth+/Dpow3POEVPJY+JjojAYNiDrDXAgjUqwTxvNQ84AlENbSzVY5QyWbB4XEbnGBG92YBhD1EEkC54Bw4eI1A7I4vBTCXCXPdb5laDlFwXB3GomCRkLANjwOcMWl0/cNfN+MqPyQuUuOIUE4Rg2meo1FIYsOwzpzLFTzg6cQR0iGZ0gROAisRKHCucQNTzm4Zey8C3pGbpm9Dasvq1A1RcJnlBCaAR8b+68Umc4kQotAqrExaOcr0nYGcTrFrmN4eRAs1lROh8gFtbddmFIVRkRAgarAh2lTXP0g4KUB6IjWGNpkWDZCLJVZC2MFUnXkUPEil6CkMro0IGshAAkUFTpCGTHkaU1RoTesobHxJ6n6IEY0s3Qo1gMEKIaiAeAC0Fmunw2fIYQkgWgslqqEoAVR6XguAVKJdUo1mqGpCDE23GYzEV1EIIVuKvpBBsN+qA4lJLq4bsyia4HUkvmCN1nIhfRjChgBQCZmKjTpLcMTZvuOBKSA/CUByUAEKHrPOtSOYhcoSo1rajAjwoi82laRBiwzAoSIaYmBEYafLR+UYMKwwl0NOKg8bzfM/C/wAIDOnHv3Z8Rpk0cKG+P+nABgpLETHNSRAUV7x/qEANQ6quKIys1DCUAZYhLFONYRZ0RsCkFwAyzJGqmwZeFR7XiwqGKEFNAESUo0HopSIgDRACMKqCZqbSFRkogGBYhK2JmyXDIclwpRQ2JVUxCywlOu3AI3IxKnKgnQ/udbhpHJF3SHSBFShAgTBYkqUQDObBrOjM6kRLIrhMRuWEqxgM0ggFYKnZVxRSJ0LMrhNwy9l4FvSM3TN6G1ZfVaTAiwczyggE2pHTxv7Lw+OGAF4OXrKu4AwgGKFShjJw0l1tjhwhUl1GQhPgxB43QMwDA6wwobKAFraUQUUEEH+0Q1AF0DvZc5w6ziUQosOGk3rKGx8Sep+iOERyqRHMoScEtwFZaSlpyI+BqnBaEslAqm5lEHgYAHATbcfAcIJkAj45fExZaw51A4AkZHAqczQcoaI5cQ84MAACAEBLcOp/cox6B5zIlNDhGU4fGDi0gCAgpSBKzRBAO/BpAMQKBDGThpAYcmAH5TfM/C/wh4IVgWOYhwE4AQUaG4+YADiNWIHV0lqQSJT7fMfSFWai1dIDgAAgQLM4qbBl4ZjEwgt+hGmi+0A2ClAeQMhQUhVkRLBhqSRYwrMZZAW5S8Sg8PKCQAkMAYrniZ0P7nU4IICkkgCZNzDRGCphqHM6zYNZ0Z8L1rPWABy/xT3CMNjNwy9l4FvSM3TN6G1ZfVIAEGo8hIZAnUTsU7FOxTsUHIAGjqhaIE5emdinYp2KdigKYAHhDY+JJAEQxrOxTsUKPyglFAg8IdinYolgLKdinYp2KGS7BzCp/PJOxTsUACAAGk7NOxTsUDWA0p9inYp2KWUHAePYp2KUPygfNQ6zHJOxTsUsgDh4dinYp2KVNt7BRCZ2KdinYpZQcBCcsm+IVWQDAGCmYBE7NLKA4Dw7FDOABCCBpAAQ91BJWdilvSpDSwCJW6x3zFpQ0RWUxL5vCLFAMnUoqG+YDgvVWFAKiDnKAEgFNU1KiipHVxiUb0gFEHKnh4FvSM3TN6G1ZfbbTmm95fMbHzEm3oSHwzarAb3z7A/NoIEz2lVvgPogx5TzeiTTy7xlNpy+pQjEwyRojuAXXgxAEoq/QAdCghYXMBknJEIFLgTYCDraCOSVAIN3VUBjUyiRPIyhWVQTVwMrH8jUFHEBBMgOkMoIBfWggFAjZeJcQfwQTBb0jN0zehtWX2205pveXzGx8xJt6EgaqhzVm9hn5tHomOzpYc/RBjy4UiGqggjqB5d4ym05faeBb0jN0zehtWX2SouA4KPfIAgPiEiTDhxZRkN4GI2gEPCJeXqtgCMAz8hsfMSbehIPpi4uP7C/MdgAeY3HP0QY8Ue8zQQGjxjvK0SJEiQu5BBgQnZCUo0QdloBDyxIkSJB1pD8EmHBMICqQJ9bwLekZumb0Nqy+y65Nwy+leNj5iTb0JAtFwXF7DvzD4MRwKgh+Xogx49BEPLtGXn3bKb1l9bwLegBQ1oEMJVTVhVdYICTbAjMFwSBGAITv2aWuPPzbVl9gdyEtisDB5mZAADMmFBLWKLrFjMaVgXUBwUJhaLjWdDLcouePhqygWIg4Mcgaw5CsxrECWZSqz6wjQAJKTKvyYluEXLx8DY+Yk29CQcpCN129hn5hbpxGRHN6QMXOuPOXpdUx5XnH7VBZlqAQZACBhsy8YaGDIAcqGCUFaJHCESgUWqIEJIwKkk6Q0flkvg7QPQgBcQF0WMPqYOJzjBKlywnfs79jegaJBmTE3BCj4oynEA2Zo8OktigKiq2M5mKXCsIfR2lGhBgrjCefpeBb0Nuyjl2gHwTtblLagHE1AYAuAzFoZkGDyiqm/5owGklLnHSxmoY8FCPCJlmjSnm2rL7DpH0ZlDbuNi0tBA4AU6cWdpqOA6sm8tZ4Q9G6UBiP1Oj/fhuGYiGg2Y70tRdONRJR43tVjBRWrovOUOoZzon34Gx8xJt6EhdMW+5G18c3sH5hmboNManyPSB7dnFIVq5GyN09YJTCLzG+V4rggCPNoO1YSz8RXJw1Oj5Jm/ZTrUX5i8V1HSCGAENwvPC2kEvSBcPiTRrPN40nXfqJfilYrPT8j9MEkzFiJqpG6gCmOLlQMgCQjctDeACBwtbNXaeaUqN2gFY2aDgIxcIRF7kmRbqaQluor2EPwX1jKQUJIFOKttXLN+PpeBb0N2ygpOpgQMnlBMsQROQyGkZgGAz+FbQcIUQwcJppaFi4fvzbVl9h0j6MEAlGCCMiIVLwhgn5/JSchSBDcL5POCcCoQAWAnR/vw3DMQ6EkCLZjKEO8AUGBYKzdYXwKNAYuAcihyhcp9+BsfMSbehISyfC4XLRc3sH5mFcAXI6ekDIXxf9yq32sDz4x+A0hrMYwYByAEFAWDxEeoh77CdaggCAiDUEQGaEQAVlw0jogOGsWWNTEuD/oJ136hQEFEG4MpvDEgBkdNLQFlUMEZHMaQ63NAI/Mc0xgCq/OVmQBieLP5lY3eYeAwao0AANAJbvx9FqqKbBYQYlEVioOQ2KJ5ehrJ4Bnbc4j1BRQIgUFYBP452P5tqy+wDIbkDnbcAzdhhKAOSQZpnbcJMnwPA0KHqTtuYj9wIdtwkSTOh4Gx8xJt6Eip1EjULvhc3sH5lYnUTR0PgeiDE7HnbcPiCsVcZTsudtzWSwBCoAJgZ23C5l+OiqU7LgEWBnhgAAKMjnbc7bnbcCMOHcoSwCkkK0ztudtztuPKjdF6Wot0Ou/qMwLefasvsuuTcMvpXjY+Yk29CQZwQEKlFxtf2L8QMgtCS6aaeiDHjQOJirRn5rcQBtVFfPu2U3LL7pmBbz7Vl9l1ybhl9K8bHzEm38FIfgx4uy14qgeZoI4Li3n3bKbll90zAt59qy+y65Nwy+leNj5iTb+CkPwY8aPoUCXUZeah+ESaVBTz7tlNyy+s/UZgW8+1ZfZdcm4ZfPeOygsZsBF/8JvhBwhQVyMzfCb4S26E28Db+CkPwY8VcVUniPx+Zw0SG0p592ym5ZfVzyTNVzoKk6CAwarD8SEt19RmBbyjQQDQBgOQGDC+Yp+bV1LgPCsEIUEWCfYdcm4ZfPe6b9hjJyYXiAgWLgq+MxUC9cHsawUJoK4wgOEY40kKgLFFjHBdAO6xIX4C3jZ4vA2/gpD8GPEBmKDlZh30fmBZgrhVFpfz7tlNyy+t4FvUZgW8vXYxbrwmqA24VKIGJCGGWJQAE0awgXwg4dISxzxicGOE6R9+w65Nwy+e9037A4tUItYvgFMYDYUq4DFkIfBlGRuYQFOzAVBoLktxX1gQtSEAAAutKtIIYxS/hZ4vA2/gpD8GPGn+1VKo81R3oKhQ+fdspuWX1vAt5gbYGIgH4j2X6j2X6j2X6hYdxU1C4OtGANYFSHsv1Hsv1Hsv1LHMwQgKMa+HU4AOadBoLLYKA0hy5Cpz2JmoFVgIU8JEAsAircAqqzo/sOuTcMvnvdN+wS6pZTagKozGoTgMUZBpHHDjodgLKoDRWGEW8ohxqJq6eJwY0Eb4QEFGWJOOcT1CKEnWhlc21lni8Db+CkPwY8SsUvBUHzGWluuL+fdspuWX1vAt66HUQCGhAqkqdrna52uDwEioK9h1ybhl897pv2HoUCzgWDoQQ3HOmJMkuJFkhC+UYFbDBzcSCqIFkGIFUK5ZyOWQWEmARtisOUMIxZNQOjLlni8Db+CkPwY8RiUmSpUZ+YLgk6pIvXz7tlNyy+t4FvXQ6iAe165Nwy+e8QBEMZHzqUpSlF2EdB4G38FIfgx4lRSuFg+Ya90bBHn3bKbll9EvHyhPBxp5fAt66HUQD2haHKkMfBMIEHtT/ALwTJMgyKdYbSKIeAH42mOAD5M2f+zDttjzJsv8AZZIYAT5HiMQ7NAMyYRXDvzisF77xSxd95UVXL/vDYq5f95cBgBYuXldVduM3r+wH2vWP2vWA9trrAcMF25wC92bAZg+ADEAKkmUXRz/6wjqdrrCPa9Yva9YEyjEMM5xym5f2V9rzlfa85X2vOb1/ZvX9j9v1lpjoE+R5xJYogGZPh8CWNKujAgrJ+DCQiIm2JCAyIDEyVGBfEEmCMCPSP/Fg2XpO1YBmCjMBCqaIBK0g5CoQQVgti8AhGmF0DCs7VgOiDrAGxB8pp8owXAZOHzksiah5nyOkoRV7FNrgBZYmo1UJRCEkFICdGCKQZy2fKQYLeuh1EA9o/AOYPD0K6fiSYBkjSBYeAhgEcDMIfxOCrStrI4VrBwEIAAUx43VoItdnBWRIBaiCLQ/5wC1BcRNKfTIQN5cydIs2rhGonQdR+izXBXhW4ZpqC4SJ0LLpOpi0C09TAM+4TF5Na50VMFWlkyQPi4Eh+J0kjkTIjMuExCAEMPz/AEz6QeQGuW7zmnVPqbtl9Le8/kOmrx/04TaMvKBmrRTFGxIdUxDrCguiJTYiC0EYrEgDbtEZVmbSdwEGMC3rodRAPbw41LacoFh6PereKWusTYcvj4Cb3l5iFX4HT2mo8DPRfb4y3DN5NRhOy1M67Pas/kSdkm2ZeH91md/z90j6QeQGmW7zmnVPqbtl9Le8/num/wCnCbRl6XgW9dDqIB7eHGpbTlAsPR71bxS11ibDl8fATe8vMQq/A6e01HgZ6L7fGW4ZvJqMJ2WpnXZ7Vn8iTsk2zLw/uszv+fukfSDyA0y3ec06p9Tdsvpb3n8903/ThNoy9LwLeF/7EeRthPCBQFDybZQc/DDGVfFt8r9aQH88i2db5XfsPnMRqUCwSLgL5sy98ITKVA6+Yt0hyQLOy2nTrMc7HwEx05u0nyodYZ2xH+oWxJq5gW/iGUABgMAfxADLTsenWYGMOO5lipi0tdqcZ621pD2h7sAFB81vYj/U1bmxDawbdh7Ywlh1Hkx1hquv+zm98IW/Q6oBMmE9G2xhPlrSyQhi4zWEwxb65OpP7Mmz/wAEw+Tupyg4GFCNgQDPyKEDAV4XYD+Q4+65LwnpC0Bu0t5IQEFSD0PbCESQJLQiU8K3u576wmUqJXzFukOqKtciODJhABrJ/qERQENghHJ2aHoEcZgamkFwSE/M1qRKWq/88EUkBuBmk6y8a4bOb3wmJDI8Sf3+JqCjdQTliLCi7a5MABr0fh0TGqHV4BvnwCiCYGS8Sf2+PDGxUZrCfhwmTKejbYeC1rUpoSn6QFP0fAt4dZ+4L4EQoVBORkOE/BRLbqN/mBhnMGKKhiHNa4QBAEI4ofVJApVMQAQGGrQ+UICY1lAzMQ2nTDw6AfUALEqqEaBQMwHKuNibtYSAFrHPjEo2hAhwV49aWfIAlCVBUgKcYpZoqAxYJCsJLA5WPuQiDxnUj2HXfkOnqw/QeAaQ7we/d3MAA/ILwiMw7KBLxs0yBOU+z9eBq8FQkUAlU/WNHMcCUkkNUQBqYP8AhgOwGxhe458REACppQNYTqZzYkngU/mdEn2vyCcvkEiJL+QGooIXaG4gJ+QUB5cSETeIXFREMXGSiiqtAs3dh4dMRUbAWrG/dZl0DBkMSpnaG+A5lIJsVA0MqpEkQNEJg9OtCU0hjBkBBL7ISlqih1PSYqqALngKmJhFZ8JkmgRhcPhvdZ1CM4JgBdMBqbfMF53Cgag6CRBZ3w5pm00il6bWZAwzA34XCa6AiDwSJXYVB1BY+IQrLXwiFhrrSUsAwg+E6q540DDCL4hM9DhYkIoxXCBAq+CS9VABlyI4uwCx1AgCbHx6o+ZNM/1DEqzoqj+TQ8UPU7r2fuE1JDSphCJGsPlCVshzWBg/BjQhSagCwcgONHxKIiGzDWn2fR+v9xMkwdQNp8xFE50QS4BIIwwIKv4CVNHCJUf9AGAQTEColqhk4QSMT9G+RmBkllZjQZYqU3VkcDYNhUhaIiYrVpQTOk/Xo+Bbw6j9waGuvJi5gPzBhyMQqzX8dEvkyAt8SBO62DhrBoZEgJoh1AbOUZVXJ8gz3guIQGphq6ClD4w2nTDw6AfUAAaRDQAOlx8Fxk8V6CJIJgfzXOMn+BBxCVUlFs4iNlZmWUSDUgqTOK7gQyiqZUWpBgldoI0ADqkhenCdSPYdV+RvQjLRMSqawMsYSPhIoAWIC2RczWLzVKv2nakPOfAEKQhYc2QU+z9eBCtprAYEKgmhIqo+F5qMZgzc4Cad/DgJgKmLAlWySYdxxDXYDGDIsBDck6oNVrSdEl60KhXjYMeFDmsBYgQtvCxQAsQFsi5j1iwcxIBURIqVVrCGi65EfFrkFefDJOlDUhWNbhBSkKj0CSqxpYCGNXjSHioykDBbmkhYWRAnQOlIMeLGDWrEgAVaplBAHxE3glS1OUTXEoV3WurxlkAIKCK2XpINVFDVC60ynFzyNj+nBN3rOoRIIFnIqACLXjB8DJ+4RJxI1gmobptDghA94pJNLoPtCp/AGGJsGziHRG6mbEUFzreHGLZlGLiRRU53hAGfeRD+ShrAIH4ZZbXoXhLgKEWgU3sw1A6t39C4LJeKSueEIhAhAATRxhcRmomGZhgrLsOoPZrVVPOdUfM0+RgGhCfLI/KG+RDoJeigASBOAEYzLOCRGfGHWvCA54dFPeo8MoeApsYMSOA5MHgd5IJwoCyLUaz7Po/X+4Iwo2hFYuENMikmkJRiAF6uK8VVzW+1ipKAAhd4flenlKgXhcCbnwSyVWUQ+MD8wVBcsJXFDp+p3pKPfuRWhkqVHCZJ1nSfr0B30GABATzCC3genmJh5PJ5PJ5PJ5CgZeB77KkPJ5PJ5PJ5It87L2Hx2IeTyeTyPbbDwEiZPJ5PJ5aGBR3MsHHk8nk8nk8nk8nk8nk8nljahJKYfJWceTyeTyeTyeTyeTyeTyeTyQWBJfmJEmTyeTyeTyeWntL0Xz6I8nk8nk8nk8nk8qvtAH6BciB7AG4MYXLRR5Oh/wDpyIaItuNoRiTSYIXaB/dhnEi6JAKNClhAGCa8GEtKr/xCKm1h0JOYZeg1atWrVq1JOWYZGBgWf9w8jZBCwQzJ0AZjztgwUokCF65Jb+2o+RQD6LGiLiCVzyl1AR8YIrHDObWiW+l4TNAYiC4I/uMHzVBHZOBAciYbLQnW8wCMD5GFWUTGJC1QgbMBLNeCtD8eMATEPQgGpIlqmpTNr4QYQyMKANK1tAOsTPOCDY5/3CCJlUCMBbFn5EKFCC0CCtIl9PkQoUJFBnNRYhl/5Ix4xw3ZUFgGI7SnsKewp7CnsKewp7CnsKewp7CnsKewp7CnsKewp7CnsKewp7CnsKewp7CnsKewpAaDFlIF9s7CnsKewp7CnsKewp7CnsKewp7CnsKewp7CnsKewp7CnsKewp7CnsKewp7CnEhwUBlXi/kTTgkvBgD0DWMOWQGVPQV7Xpv9gtpQtyGCkLuzAz5JJ9aulBip6F3d3d0NoH5H+PRu7u7uwUfviQBtp6O7u7u7BW5qhUHQeld3d3d3XEiyi/gTKKPBVIEoWjSJi8pzRSELkxJsIAQMMkggjMxSENwhJvkqMBRLRjQZ0QNps2X1ttrMfQ6YeEBx8A06CzNOwOI4RKqqcZNXVtilImhAEEYYvjpeE1WOqIEB9RxloAUniDCFAhuUOdoTgK4JoOMEGJNo2ZFWg+fiDEOTEFENoJCFjWAoBgJqBnVX4hctIgMeEA0gGEUGZF/Dovfde+4AiJja9Kyh8FkdyFjLKG1oRrMzqBpWGgvT5Sir1qhYVf1pZoSLB/KvSaQLGnPFAokyugcXQZFwZo186yiTMclgunwgPSQPGCQPo0l7/SB+eXzCcBEIJoBF6vCLh4AxMkTCAkIIMh2GcJzHSMD4BjLtakj88vmFgLYMfnlBUsWHA/Im8Zn+CIVGIAsaBAxWAgc4As+SaqlAqEPIhQYZoiQw2xAwe5IGyzGNi8IARrYg6W1E2zL6221lZEoANSjMAqDQ4vNnhVaUCFO8SAUJKwmpY8EseMROwIoqiaWj4hhBAEivFQraXS1E3kJuACqJYMGaJGUQagjAs1CVz6IF1oXc4iIEVdwBm6dSeUERHEAY+RSyFVQkrCdTgWVgQqpsaQZghX1oGRZaF3kSEgQc0AlXnCaQMgl2jU3xoqSHKrSGnh0XvuvfcXYAqB9nRHjGywykKr6OMLfA4gCqHcY7PhelhYgIs3DFAZhL5XFQdHBROnfrjW+FV8Sur3TEcDCzgZUOY2Eu7FKMZbEI+6K2l2sRATEvgodoFoQQIUEQRYYsPMmDiUaBLCOLEhLdiMlJUYtFQhhHVo9kwQilFGUFKuIEGhuEQqIGaikAINqAGgvqYS5k6uxDmgp/2GFAClYgjiB2uAHN4zP8MUJcghJNonQhCPggj1g3EiWmPoJSlKUpQCNEVgOq/opSlKUpSKD3x6CCT56UpSlNSEhsAxangEKCqSAoVNh6SUpSlKUpGaAAAJv4d01rfBkjmk8YUyIIqeqntei/3e29N/u9t6b/AHe29N/s9tr8VNEFgOA9PPnz58+fPnz5wtYlWwet5c+fPnO1ml4AjpLzJ8+fPnGVBqqC+4y8ufPnzhBiMwQjp8ufPnzhY8wSIjNgyegEoeINHh6ufPnz58+fPnz584XQhMEUD0+fPnz58+fPnzlWwtAoA5x50+fPnz51w7IwCErn1OfPnz58+fPnz5yuHUiAICJIDVP6W7PR4zJB6ZtN7z+yM6v9vUNjN9ze7AYmgEVXsByoVigmTjZRriQDCeZ8WJIcCgc74g9F3L2G+iJ3xDdEsTC84oqhSad8R1GS96P2nfEsqIDziiqFVJ3xEP4Sdv3FO+IHAasPBOd8TviDPgNGaA6Gd8QsAKS0g88qKOd8Qll9L2HP0nfEK1sTMeRvmiqnfEB6kyYZif8ARO+J3xKsAUdPMDgCIjO+If8AqIBHKUZqSJ3xO6ICw/VDMBhEZ3xG/wCiJ3gAGcTO+J3xEBFj5SgFoKunfE74nfEKf0QNBFCM3InfE74ndE74nfEOIBBLIe16r9TdsvkvwMVWACkaj4hyhdRk5ADEmwEFoQGrkDH3Op85135gChmHSMBPbGGLQsEBQIX3eG75efrvzBv2dgAnFnKwxoBgzFzACww+MCxR8oOdDgWEZRUFAbESSFUqCGPgXMRV8TFi+ky2KKPlBzZMps9BAHUwGSW5ZHRAVODkDCKDoYIaTBg/hkElrTpvs8goZDyklIVVg66YkL0GmBAjxEZlAbJIHAAHg4xzAFCj/RCKIyhUNIZVruRKW+QRwvmdH8251lrvpqWiizRQDWH/AFSHAAQQFFmcKYwZkRgI8mHpVWFNcZgdHjbw6D1erfcMTHKUpoejIHzDqwEgCIhxF1I/UFAsVhVAsRAqtOwGPDpXlQuORUYOBQrykGHfpE0ecCyqvjWOjYQ6gDICGVEUgSxRMTKrMC3WA5SyQsPPAviIhZwBNWCaxI6xyEeilc/AG4QwuiCSZDoRqobYxAwzQhEO7w3jM+16r9TdsvkvwMajTZ9GRLs8jNIC5mQDgyYephtAwAWZW+J1PnOu/MPiBPwWnqIc8y24egx4bvl5+u/Ma749wVRA1IMyUHoCyfvh0OBZB0XogMmMKEEBGriDgcwGJJRVg14bJlNnoIXiTM1oSwIqWEHQOiFzDbGuEQ9BsAOSSDejYR032eQbhkAK7swd2kIRrQ/wKjEGgAGk3qyUHMi0TPSxrBiYUaFsgBDQ4qqiBLDOwFfhudH8251ltv6Tp6CLBHJJivGrxFmhUYFWRGY1VIgBqAgDiIFvs8/DoPV6t9y/e1kxf4v8QpYWBi6e9azrV8M9QQKKKsgEOAXqK3h0ryj9GUKBJ1jXsbr/ACqhC2G5pY3hC3yeDAo2JVBAxWaEHMg5mBEAAdcXakq/8apNBBUDU1gBozUlFfoNWBowRBA0KYGTwQZFS9xwgonJEMgPDeMz7Xqv1N2y+QpCZIlTsaBkVXIdSfSdhQdohgC8fONBvqJlOwoYmqoQkB+gnYUoklHLzrTezTKdhRedkAqQK5Ec52FCvFFN4wED6U7GhIYUgKERHwROwoDxmIhNCCzQKjCdjSt1knY0HFBiBOHkKSBBSznYUOhEcCaTs6dvTtKC9QYIfmEAcmAxnY0P/DQQEYhFRGdhTsKChGnqjOKSwGM7CnYUGVJkRE7CnYUFA3A8prCCSbJ2FOwp2FO0oVQDAIqIzsKdhTsKdhTsKE8YiATZ+16r9TdsvsXfU/T8CI5lfT48og9p82t8p+UQuq82rDyPoiD0zab3n9kZ1f7eobGb7m9phrHuACDRJoIBmcIhAoVViHjxgmIsRFFStjCQESSINkJTPLCfgmH4iV0gCGWRlPUbNRmp1H6m7ZfYu+p+nrt9kNRzIfHlTfmFakgfkB+UctgZzQCE8iOXokHnYzjGcYzjGcJCvN7z+yM6v9vUNjN9ze0dN8q4lwBIDepSEY2hlKgQ0Iq66nEtfGnrh0ACKYIaBQqv51Qp9mwgGOUG6FCWBUC1Q4rmkUTHEXuC6oxKw2EjLlYwMMhlC5fCgYIZAqbZl9i76n6egjhFiz5g+PKa0IxJiYP5Ty3zcDIn26eiQeYhG6WIEjGh9r/Jul/k3S/ybpf5Nkv8gYGeANkWTQ+a5cuXLjMkSEBEQRLNyB6ZnV/t6hsZvub1joiL4tEVQefmQwfRUXh1X6m7ZfYu+p+nlRbWDPeUMk1k2Lh4PKMIEyooBB1Hl6JB5thzy3w9PaM86V6ZnV/t6hsZvub1xUVPWWHiAXwep1X6m7ZfYu+p+nm5SEXZbygpdnJ4zXo8ogNZotSHmeT0SDwHgE1imM2D/JsH+TYP8h5CXHTCdoAADY+JsH+TYP8AJsH+SmylZm4HIpKy6cJsH+TYP8mwf5Ng/wAmwf5D8ER3SJ2ltgFspsH+TYP8mwf5Ng/ybB/kKXYFMn5TOr/b1DYzfc3u4nVfqbtl9i76n6esvDxtS5gn58q7c5ZKIyHwCB5RcBNiquQX9EQeHQPswpRqGhD4E40JFGjyhCq8EoGYIxCCcGhQMHE4Nar6pddA+G+No+zJlUiEGnOFTAOqPZVKhTUQzjlrTjAUhxcZaLaHVMXQClr/ABLtuM6J9CDgiFTQWGYQNQOksODZwjXHfLSPRK2GoQIaomF9URgFhsa3wzI/hCNoOBFASFVnLDHkjqCJoiUtQbx5IokOaigxm11lEAMBhwA3lM8OgBXVhipVON/LKc3zaFP0jpUpqCrg7TdMvKZ1f7eobGb7m93E6r9TdsvsXfU/TzmlCD4V9vny3np8mp8NeVUkpU6Ac/REHh0D7MBeRCN41/gQnoW+kBFJ3Gizh1DEEgCsAGckGoHF4kAixGDjAqZASosnP/UGkyuoSDkOoAPCVfLcVJBFiOesoshHKAE4JwqpdtxnRPoTOKzR5XGhIOYJiIAcc6C71Xc7xIu28SzqLw1GLcSibyj8EdSZ8xH4gPVOEQAwjguteMem7cD8hZwqKy4SqvnGso8HusNjwwQLkdI1rs3wK1gqZG38MTpvSsCxNOCwH4am6ZeUzq/29Q2M33N7uICI1EokLowWOYAUyKeksEQKURx9g76n6egGWe0XID8+VJ8SORISPgleUwBENFSABzB5+iQeGCjSriZ9uMTPtxiZ9uMTPtxgqs4jXWJn24xM+3GJn24zgEyJQLm53F1iZ9uMTPtxiZ9uMTPtxiZ9uM5FM3AZ6xM+3GJn24xM+3GJn24xM+3GJn24zDEdZeUzq/29Q2M33N7yIvYu+p+nocSxhK5C+fKB3ZB2MiHwWPLTK1GYNmfokHmM3+p9RYehXpyYhN9fs31+xmy6wWgl+c1m+v2b6/Zvr9m+v2DxjsAT5zYzfc38bEd9T9PAXyTCt0pfKAxm4Hjft5aN4PXUkBHQc/RIPMZv9T6iw9ByuSAX5zd/5N3/AJN3/k3f+Td/5N3/AJN3/k3f+S6KAS5yEFh5jYzfc38REdhLVAE0mMsgCIfkbbbPICTsR8l222xeXIQDMJCxD8wLMFBnyqN3WAdZ+ryklUly5sD4Dm9Eg8iH6NEKwtHxbLAVI0F3g2KzltgwNHkO8gcFG3zEQw/I222G2KoaS74ttsUlEBvAEBQcfl2zP6Nu7UwWHmNjN9zfxER3vGU2vL5Op+T3DPOn/XmX/HVS+YPjyqgoLMQF8lnyq5GL07B6Ig9Hft+fk9Um7ZfKsPHrJB5Nsz+jbu1MFh5jYzfc3vIh2rhQwogFMjNb0giNAoECeHoOzNoGbQMEocnCOURWSQSpTBlkihQwoA8bUhT/AIMIyRixVENoGCzxWCMDaAaUROWACqXgoNaYHKFCHheniSGRjNoGFtAACXK/4rAUDklCh8AhISptAwtIfyg2DO/zBx4rBHwGSgAGZtAwDekwCyCzdqwPibQMBYYt5yDwPFguEZtAxiJUBYh8yJtAzaBgRhcGJtAwAnEFiFZ2gYRplUGBwVDaAMVkHAxsUvFgQRAhwwRl5wU2gZaBIA1ov/gzaBlrwaU6yQQnEGsaGbQM2gY5K1AqVzaBm0DNoGCR2VjCRBrGhm0DNoGbQMCX5FFQMwJH4M2gZtAzaBgYmkv4Gxm+5vdxDRxd/qAl7B9F3cl7U/JBCA5RPJBuoQAGNZauKiDD4EFjAzPaO/prxJFmmGp0H7nXPuJxwAqOviEALXAUCeYIp9768BZ9m6ijSmW08tMIGVRYaVDWdEPHqB9+PSfvw2+nj1v54dZ+vJ0nnIPDrvyGhwBnJnhSVJiAL7SwKNqS+EUtQTVINVxCDlCKipkGJ0v6hq870AycoEK5zBAA4kS7odj8ROjzqn1KtfucV4uFaGEMkGJ0FLzoz9T6MZd0ainRHAo7I4QCCWlxAcTnqf4GToDBiHkSnEDK1eCoVLnHOskE6l9w0rclArQLAJNGAEGH3WJo4G4OIlYZpwqhCQeXlEcjLGdN+zrn3FYCAK1IM2TggeCAcboBH3LVnoAH8JOYwdpAmlgI2MugxDlpQ88GAYBygKvWI5Ton14Gxm+5vdxOq/U3bL6Lu5L0IkCAAEF5UpdAB3rfRxEc5uzgKAB8gGEG4fFrM44ZeooqqYayfc659wB4QPASqLIxSDysg6i0lTOEF0hy0MpYhgfBZllPvfXgt1RAfUlBUuU0jGGxEmoQlgsnQDx6gffj0n78Nvp49b+eHWfrydJ5yDw678lZ7iuXhRED2mh2VMSYpQHGzkwxkdrLqVgdO4CALIUGFqTpf1B9JyZcgIXIeC2a4EQDS0GBkRmMeoXvVJ1nR51T6g+9FhTgcFCIAvFaQomUSwF3SdGfqfRgxAwFrKStw5xKFmg4FMgyMmkqmICilnEWRjGJbmkm4QMCpS5xzrJBOv8A3H6NQujVX0sQjWExppxJERwCCq8IzGFiVEZAKKrrSvw+yag4qSzagbgXy4QWoAQhI5zrE70gtsmpnTfsIsBFIGN4VYvtFOmOKEtSGrSrQuWJpS8Ewcylukzg9yVRVMErgEG1Bc4qWMjeaxL3p3hRkWBqALEBhuxUwi7IB1YAIzK6VbpeSXLJHgCWE6J9eBsZvub3cTqv1N2y+i7qBmv5ianpNdzE1fSF8AACTSEYwipTU9Jr+Ymp6QhcDZ+IfAFE1PSa/pDOdwE/F3wVPmd8E74IdRAACajwcw1ITvgnfBC/wGp8CDjJICa7mJ3wTX8xAQjl5yDwOfIaGk1PSanpNfzE1XMQxQIBC+Jruk1PSa/pAsIEIgtpzVTU9Jq+kK+A6/EskCk1PSanpO+CavpCUxgOdZIIYQSJOImp6Q95pDVSSuImr5ianpNT0mu5iBPQKiGMEiWImv6TU9Jr+kbY5TQgRF8oWKKENgAgJqek1PSa/pB7XkLwNjN9ze7idV+pu2X+PdkHtuskHtDYzfc3sIicIwHiGcciYCCAIJGYuiV6XVfqbtl8/txxxxxxxxxxxx+4ccfhwGOOOOP2PWSD13HHHHCaGb7mjjjjjjj9B3+m5tcEhpROKqjVwABEmgNWuSr5iJAQnVMC1mifTLJhAGUQLVhrYq8AIEL85UhQkEbBt2UR4okNR/uDgxkTNsy+YSDu4mk96fs3p+zen7N6fs3p+zen7N6fs3p+zen7N6fs3p+zen7N6fsS8QJhAJZ9e2IhZNEyRjdp1m9P2b0/YZn+hISD1m1P2b0/ZvT9m9P2b0/ZZrQkSUuvseskHrgggZcUAMchN6fs3p+zen7N6fs3p+xnUiSRqOJvN6fs3p+zen7N6fs3p+zen7A7stIUIvBYevEOnjnkEtafmIvnMoDE3LRU+EA8BmSSQSqiLGafKnwSWS7lMAkIPhDDECXw0m7Zf4p2I69B6O95eoJhy8u23y1x2A/nlW21tpN+5BsCw9nE6r9TdssDL0Wm4M0XJNFyTRckALUSoEhJ/ZouSaLkmwE0XJNFyQTpQm3gDeANI3Bmi5JouSaLkmi5JQNYuArl9zRck0XJNHyTYCAekAVF6xIUC+47/jNFyTCJYAGaLkmi5JouSEwbkSjKg2mi5JuDNFyTRckspNQFY/oTRck0XJNFyTRckGlJA0Xn6CG1XyQCA847AWpCAAMyTAOzsHiQiGFavAzKrNh8QNBuOgBcOqy4sFCjQgBEBAwRj5Ps+QxVhaaLkmi5JouSaLkmi5JouSVpT0FzF1M0XJNwJouSaLkmi5Ib13IJYCAKJuDNHyTRck0XJNFyQSBbYAFSpNNyTRck0XJNFyTRck0XJDlWFvVidV+pu2WMaCjaYYYh4oG7QxMAQDSkvRE81FRMMXFPiFzR+9QLVr4AcIZwy4QjHAx8iIelUBkBXQRUoKaNY6LkVwABqJ+HWDVFbinQpVuDqxBidhSgTTqIXoAeEAyBBggMCSIbZhlAQ8kIUo6mkqtUqyzxeHUfkPk8sRBBEVVFS6DaJW9mcVQHisOssIjgWtACzpRQ4NwtRFYAIGcAbcoYSbYMgagIMc7e102F0ZxQBlUBGs14wCqk5gQhiAcF0TGUK70h5MugEKhiqzKErg9dylaAUWtwJnQD1vr/AHKPi14BRwDW8IgGKOOCLcRYGIVVYzIEXMYiIFU1Bizi5GiaCDWpJAoGGUPzFV1DqKwQVXENiHo3lVQL62wnRifRgOm+czViuKAvSVELHAGpBBqDiLRreZ1gAIVCKVaQZbQokp0LDsQPiUkweKjIXhCvekQSVk1ljbLIAaYARZfrjqpAYKU0bQ5wfJyIOYgAm2jhXlHfQrLWYzoT9efoJf4JwAQlzlHOhK0EZAraxoawaWTCnAwVHWph50JYBTV4xepUqRjM2LpHUQuk/L9nydFjgHUAZc8yh4x9VJTdABoKDfNoOeIQacUNFsJ84BdLEWSzYDGMm8YpRiroE8jjE0OTJ1hBoqEqIXRECRrEOdgIwNsARSAC8PPUCPS8HghFAbgAEIrgZopKtGhX+YNBWYMlZCodyp0gTLVYw1hcMXnTfs6vKuLSYC1TbYwxq3Cb79Dww4AQJSvEQAiEfpLwhCukTY8QAihQmhXGUgfQNSJAosU5ExcUgUAG3A1VF6OYa1KxUYi3m8UotogRQfg8kv8AjgXaa7MdqSiqyF8E5385Wfu6sQQvFRfKhlf0SYgvS9p1X1YnVfqbtllaMApQo4xLa+QDcqAZNBUK+Y99EhDSCpomdIU1HrCgMM2sI2M4lWgQL0liBYpn+3AMCzmKiAUzwVIEKQAFjg7wDuAH6IiI2ASSC8Ydv282MgEqXAXoodX8xs4WEQWguq88wtQTWpOWeLw6j8gLIEVFUshIMCAfmFxEoQKKgYOFalpcIpKbaAAFbCI6Asg4gJxAim4GdiCS5ofEw0obRwdUJLfJhmde0I4WdYlSp+LnjLKmmCKDNmgzYwCCHTQAqxfGdAPW+v8AcJI0IYEptCDQmFwPAQ24QrgAJBSmmNIQU2LN4oYvrgAMBVdLAwzvHAOHSmlQAkqjIlQYh/ZgEBQCwnRifRhIvHAkK5ggi4IIMDEDcwTkAAAK1QrnDDcBAOGACEc8NpXKw4QrW0rl0aqZiBFwowpXXuEAwDEtSFjwLxLGCSRYSa6w5cMGAggAlgkFAAlGBK09zukBWlE6E/Xn6CaxiFUI/iNt0gBcLIIqkpeAuDAUIAJqJkqBBOFADCcFgfrAP9QrZTwTFUE1YMJnCD7joHoGBquFRlBzMGbBLZFrvBGSIGEISDABlDal4T2/NKQ0MYJ9nydFlUpWKNW/BQW9QPMnqlF6IMrMXw4UAFwdq4wMew1W3gAAmAhRogEJGQwwBMGBGRyGxBdKTRsINZkVBLmWaITGmEZHoOdK7lyVCgBAM0AAaXGAlVRF8zpJj1gcK4nH1LYHCFAN7XW/AzOm/Z1OBEmEABZOoAagkFMof+7Jqtqa0hA/shBtignyQQ2tWYk0qoqhWgZKGSAuADOqsMJg1MW44FapUhoNu/wIoQVE6qWBIioRHEQSSUTjSEMkAmjYoZwWKZULrcJoQDEGcSoRiIWJuFmVYMUIPKijIdGABoZ1nVfVidR+ptmWK0SipHcE7gncE7gncE7gncE7gncE7giaEWV8BCRK8dwTuCdwTuCdwTuCdwTuCdwTuCFCAh3ZfrHdUp3BO4IAk5BA5ObWBFWcU7gncEBAFhFaJRUM7gncE7gncE7gncE7gncE7ggOUAz87oIWMXNTEB6VlfjG1wG8BI40GUPEKQCNNhgUOTgwSxamNbAETZZMBClTGwWYacb0FaRpRJAQTwEQ/twvTwrM2XwecGqqumDbUaPOHJIpwwDmk1UoLCfZ8gkIkAuhncE7gncE7gncE7gg5yQNFUJ0E7gncE7gncE7glIkLQmRjMdwTuCdwTuCdwTuCdwTuCdwTuCdwTuCKUQzqfViGYwJgYGKggWBBsmMS0WgEmW2X8SvJliS06xkHf23ap2qdqnap2qdqnap2qdqgBQCGn9XBxKm8UTBQygB/wAMeJCGMsjOx52POx4bAEIBFCQHlO3p29GkmRklen9JiLL5Z5CcBAD+FNj4U1FF/Te8TS/xMhOAg/hTY/w9PvEHikKLTmgyoYdjESp/E8BI9FTLC/hTY+/p94gXa0cNTCN8kXwHIwAHwjNPBIDWh5eXHlHNUSRHfRoZAD+JMRkAWkoMeIAohGAIXWECWkfF/BgDqRUYQq/cQptRBFwPyd7EUHPxxyIhTvYg2gABVXRR3sTvYgxYQEAPtIj3QxgIUoFFBVeN+qcBJNxOAwzcK8C8YS5So8ziIBPKdskbiZlQq0TewAJMDOqZ5sWxlUFaIWIk6J0F4PYWsTQKwAAMSbFA71xo1syCbZl/jDdd+/4PrvxCGpJaxV4QjjiQgAmiAufDoR49cPoQuACZwkeMOzj9PZ0hHgwa5R+rENKOUaJ/EJJlYAHjjN+y9pEG9RDeIBhhEsDh1RQlphVhIoCQr0FYBEgqCQq9F6wCjjhGglliGQIKQ+DKxkKXyfEYN9UZd3TqP1N2y/xhuu/f8H134inNjmEvUpfECCwaAUquvh0I8euH0IPyNHYNDW+ZRqFMIUAVmvHVBoZJIoXIbm/Ewdp8MlSUeAaGgm/ZeuIIWQDIEPqEMTGqLLgCoAUsinoAhgAAU4OIw/iDdQATObYzbGAhp5cP8GCX4vjm2M2xm2MQsh4kRTQM7TbGbYzbGbYw50yMiOHrPORdYE/gMU4gYHxGFSXiTwA9Rf8A0dvhFyfg0VL9JzlBoqQbKNgwwEwxRi5BsfMq9qAJFwBGtSCs8IboQWG2KtDpCnFSMvBj6IOSBVZIMI55QGTRxARJCksWCBNJ6uMJB5IF0zJArBVtZU4RFwTQc1TUM9ckYGL6rG0vUxFWUQrUg4Qg8alQxzXB8wu00jShAZCswrhBJTJQNVoo2ec7JAupmT65LO10hbjy651fiV8thBwrjHO2LoB0h6cAguBkAUfiZTQwQbGpgQwtJfGWay1Fwxwxj/XwxTbB8g0iqHmEGgtQQKg+dTz0Igwwqkky2f3xCHUQFukGWBmU7YHhycc1rYgs3vJqrbpClFgmcmROgPGCj0s5jiVAQJhGsY7CXRIUQcRkjaI/mKDAZiFgGqhxFmORJBBySt2GMoDlMIPIwAweucDvouVIrIOZRj21W0qtFiGJQr4Guks9AUXEbYBSsTyTATQbgkICnbSHQUuOhbPhEw1rqMhsxzGIjeCo6gSqjdjDu8uayIzmCa4w0NCwRF0lQKJFaSmjpWRuIWBVAgIE6EucwhmkOEAByQGHFgwb4gDJWJ/tbOEplIhjIokfPrf/2gAMAwEAAgADAAAAEGPPPPPPPPPPPPPPPPPPPPPPPPOcssstPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPK73a9g/PPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPP/AP8AX/t88888888888888888888888888888888888888880wwwwwwwwwwwwwwwwYwwwwwwwwwwwwwwwwwww88888888888888888884wwwMwww0888888888ywwYww/888888888888wwwwQwww388888888888888qN/8A/c4cPfPPPPPPPKAz6972KfPPPPPPPPPPKBV+m6X+y/vPPPPPPPPPPPPPP3ffcffbf/PPPPPPPLHffdffTPPPPPPPPPPPLHTXfdfffXPPPPPPPPPPPPPPPPPPAPPPPPPPPPPPPPPPPFPPPPPPPPPPPPPPPPPPPFPPPPPPPPPPPPPPPPPPPPPPAPPPPPPPPOPPPPPPPHPPPPPPOPPPPPPPPPPPPFPPPPPPPPPPPPPPLBPPPP8Azwjzz/zjfzwTzz/z7zxTzzvzzxTzz3zzzzjzzzxzzzznzzzzzzzyxRhTzzxMxe3jKXyhMWtufOLwHcKtu6sHPea0Cnzzzyr9Z+jTh0gDzzzzzzzywwhjzzx/zywTyzyz/wA8Y88v8esso8svc88U88v8888s8888c88898888888888888888888U8888888o888888oo88w888088808884888488488o888888888408888848c4888888s8ww0Ywwo4wIwwwYwwgQw8w8w0ww4wgY4c088888888sU88884cMsc8888888879mSTmo+3q/wB8WJ1HsPfCA6dc/aSQQPvSfPPPPPPPPFPPPPKPPPPPPPPPPGPE042owqOgvwAM68Exo9eEhI3UEg56BYn4PPPPPFKJJFPPPPKPPPPPPPPPPPPPPPFPPKPPAPPPPPPKFPPPEPPPKFPOFPFPPPPPPPPPPFPPPMKMPPPPPPPPPPPJKFCGNKPPAPPPFPPKFPPPFPPPKEICBAMCAKMNPPPPPFPPPHS7FPPPPPPPPPL8YNfQVKPPAPPPHPPKFPPPFPPPKKTZRtZP86/WvPPPPFPPPMNHHPPPPPPPPPKICdwAN6PPAPPPFPPKFPPPFPPPKNxMZIdokpohfPPPLHPPPPAFPPPPPPPPPPPPPPPFPKPPAPPPHPPKFPPPDPPPKFLFPLFPPPPPPPPPPNPPPPAFPPPPPPPPPPPPPPPEc7//AEDzzzzzyhTzzzzzzyhThTyxTzzzzzzzzywTzzzwBTwJ2zzzzzzzzzzzxZLHb8DzzzzzyhTzzzzzzyhShTyhTzzzzzzzzzxTygQwwwzw5zzzzzzzzzzzxJJf/cDzzzzzyhTzzzzzzyhTxTyxTzzzzzzzDTxTyhTzzzzzzzzzzzzzzDzTw7f/AP8ARMMMNMMIEMNPPPPOLEPHPPPPPNPOPPLPEFPKFPPPPPPPPPPPPPPMDPJJPKPcNINNHFFBGOBEPPPHBFFPPPPPNBHIPPPPPEOIGNPsMOM/PPPPPPNytvWnsoNgkgnls0FnookgN/PNlnFfPPPOEEOiM/PPPAKMUI/qQqYfPPPPPPFwdNVpvXIVo/b+qpF/C+gff/PACiVPPPPEIw+G/vPPOKHvPPv/ADzz3zzzzzzw/wA8tc8se888888P8888888f88c88v8APPPPPPPPP/PPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPNPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPLPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPMsMMN/PPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPB+Ex1fPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPLjZTfPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPOPPPPPPPPPPPPFPPPPPPPPPPPNPPPPPPPPPPPPPPPPPPPPPPPBPPPPPPPcMMNmMMMPPPPPPPPPPPPPPPOsMMImMMMNfPPPPPPPPPPPPPPPOBDKPPPPPPPdSxKlbtoPPPPPPPPPPPPPPKO3NJpVPn/i/PPPPPPPPPPPPPPPPPPAPPPPPPPvvvutvvvnPPPPPPPPPPPPPLPvvritvvvvvPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPFPPPPPPPPPPPPPPPPPPPPPPKFPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPPHPPPPPPOPPPOPPPPPPNPPPPKHPPPPPPPPOPPPPPPPPPPPPPPIHP98vPPOM9fPPss6+8s9PPI28/POZ29vPPPPPOas9POM92889vPPPPPPPPPKPKbgvPPPCFfPKG1NQab6/BjlYvPKVpgvPPPPPFJVl/F1xKOQ/vPPPPPPPPPKPLPTfPPG9X/PP2buTbabvHSdTvPLOZXfPPPPPDaXXPDDXRTTfvPPPPPPBAGAPPPPPPPKFPPPPOPOPPKPPPKFPPHMIBDDFPPPPPKPPPPPPFPPPPPPNPPLDDDKPPPPPPPKFPPPPKPKHPKPPPKFO8nstuMslsss/PKPPPPPPFPPPPPPCPPPPPPKPPPPPPPKFPPPPLPKPPOPPPKFIvH+9Ao18p3yfPKPPPPPPFPPPPPPKPPPPPPOFPPPPPPKFPPPPPPKFPLPPPKFPv+P8A13/+z/8A+8woww8888U888888o88888888888840oc8w8848oc84888oU80k488088848oMMMM0888U888888o888888w888806wtW6r09r+mw/v988oU0X/AKu/t9s+1NrvefvUvfvFPPPPPPKPPPPPPGPPPPAQGONqHfP/AKr6kBYDzygDjuKMJMOsmlTyAil0ts9XxTzzzzzyjzzzzzyjzzzxtEOIFIHT7GfqI5EXzygQBESOekMlEJSAIHQAGIHXxTzzzzzyjTzzzzyjzyzzzzzzyjzzzzzzzzzyjzxzyzzzzzzzzxTzzzzzzzTyzTzzzzzzhjzzwiyjTzzzzrTzyzzT/wA88888+88208o88388pHDBMO87888c88y48s088j888888oPf3SvOX1KZwFU888888t3Y2+NNsSW88qmIO0+8rhvy/4gou9i88PB888888k1//AP8A+3z/AP8A/wD/AP8APPPPPHa37Pf/AP8A/wDvPK4oMw8vLP8A/wD3/wD/AD//AP8A/wD8jzzzzzzzzzzzzzzzzzzzzzzzzzzzjzzzzzzzzzzzzzzzzzyyABCDzzzzzzzyjzzzzzzjTzzzzzzzzzzzzzzzzzzyxzzzzzzzzzzzzzzzzzzxTzyjzzzzzzzyjzywxDTBzzzzzzzzzzzzzzzzzzzzpDzzzzzzzzzzzzzzzzzsPeEwH3zzzzzyjzzzyyyzTzzzzzzzzzzzzzzzzzzyofzzzzzzzzzzzzzzzzzqBcfIP3zzzzzyDzzzzzyxzzzzzzzzzzzzzzzzzzzzzzzzzzzzzzzzzzzzzzzzzzzzzzzzzzzzzzzzzzzzzzzzzzzzzzzzzzzzzzzzzzzzzzzzzzzzzzzzzzzzzzzzzzzzzzzzzzzzzzzzzzzzzzzzzzzzzzzzzzzzzzzzzzzzzzzzzzzzzzzzzzzzzzzzzzzzzzzzzzzzzzzzzzzzzzzzzzzzzzzzzzzzzzzzzzzzzzzzzzzzwSnzxTixtitdj/zzzzzzzzzzzzzzzzzzzzzzzzzzzzzzzzzzzzzzzzzzzzzzzyCDxxxzzzzzzzz/xAAtEQACAQIDBwMFAQEBAAAAAAAAAREhYTHR8BBBUXGRoeGBscEgMEBQ8WBwgP/aAAgBAwEBPxD8luKlwuFwuCoE/wBvjbGpUZDiNQv3E1Kj6QBDyv8ARQ4kOJDiQ4kOJDiQ4mP2ocSHEhxIcSHEhxE08Puw4kOJDiQ4kOJDiQ4kOImnh9/AxKTmIXEg5thowvtMQuJCOYgaoYPu3ELYhELaNGD77UqCBAgQIECAlCj7TRkCBAgQIEfdaMgQIECBAh/wkJ6lIslkslkske7an1gslkslksjWpa2IJSLJZLJZLI1DhlkslkslksmKL6UzQiyWSyWSyWSyWRpS1+BJrBQSJEiQ5hztUwgkSJCbGxKc7eUxJCewlFTGHMUJ4BMSJZIwL6cEczQkSJDbQhL2MSfwEtQmXS6Xy6XSfftS6SXS6XS6XRvUN7EEIul0ul0ujcuWXS6XS6XS6Yp9LGlF0ul0ul8ul0ujUhv/AIssse7Mm661QmkTFCZKGOq5DV3xWzsWSyENw3/ZVO5CMw8gkqOrmPdfBRobsxWxPgySWxodNUkTIh4iQp1GzEzreMe/h3jMoU8BqcxqtzquQ0Sq617TlqBqkvfkmKkpaoKm18RElLVDH6/gjJXg9r+Yxyxm3k1V5NVeSg17fJoryMTXi3nZK8aE6w7fJqryT/nyPRKIUYEHj5IvHyPxYlMrt8lCPb5J/wA+RqmlFtk/58jdj2+TVXkjJzAmJXb5J/z5NFeR6GlFhjHv28j0NKLFDWIN57fJLbqn/gyS57n8iZWRsqX9zKSVcZ7CsTBwVr6lT+HgdEp6iTD9fxXhev4JyU4iJQMbu+xIlXElpp7/AGEbgt8DTVaMU1KY48njMd9T2+0xLGdj4v28lPEhUGYuU7kY0hOxsU7n8VqpJxBRg6qGzuM+xDQqSSxyw+j/ALkNKP8AMPQuLoXV3yLq75F1d8jePt+qSiTQUSleg1GrRcPud+vcds8xPdgXnxs9r9UwomSb+SWTVPL7nfr3Ja2I0qvWtwk0sCg9r9zeLxeG28f8+nqV3LM0WszVazNFrMwFqgd1maDWZotZj5GXNfYhRLNVrM1WsyBEM1WszVazE6aRPIS5yd0arWZqtZmq1marWZqtZmq1meCDVazNVrM1WszVazNVrM1WszVazHsxFt1WszVazNVrMcKBXWf01UF6Gi1marWZqtZmq1mNTh/VWbB1E1x15E2umF+YiNYZiYjcHBEaRudxwytK+B2Z4qe6+x2nyiYKNzfrXIoBwMPl8sq7xv4GDWmXrX4Im1Y+4lOJRbXPsMKEnfkYDlueRRlDo7jglB0q59haYPEeDl8DwNKDstXBgyo9Zjk3E49hkmjERJT8km4717UkmGVLwXXeSUqSig8D+m9pfsiQlONw73U4Hxhqo6dUQYvP6ktQuxZGolkaiWRqpZDqLVArLI1EsjUSyHycuS+xKiGaiWRqJZEqJZqJZGolkJkmmBOjJWRqJZGolkaiWRqJZGolkaiWRvIaiWRqJZGolkaiWRqJZGolkaiWQ9mI9uolkaiWRqJZDxSOy+migvQ1EsjUSyNRLI0Ushucv/wW1UbLpdLomklfuIzkhEcQ0lh+5W5y/oAQkL/SpNuEWSyWSyWSyWSz+BZLJZLJZLJZLJZGpS1+YtuSWSJcCXwJcCXATn8BykoJYmyXAlkuA2W4lwJcB5b8tjSv0P8Af/8Af+kh/wDDm0kvAteo4hT+hvATLXqJzXahkhFr1MNGWvUteo0IyQy16iahJ12Y1gtepa9R5CntSy0Itepa9THs/W2gJlr1LXrtxrBa9S16lr1LXqWvUQySvu9qMJNvmIkpcH7r6ET1MCE8i9jC2+w9ygjWOPV35GPlETmSgpU7jMxq7l7V7kwKT87KHMEnhsNeO4lutf7z4EVFw+duP6DKTeu+REoWqnw/P1w4SRIPFHjyONwsNkZlhOQoL0+chhvT3z4kbkJLOtz8eoybHvPf7qWPgy49ehVR9Dy5yy49eglCjap+Ey49ehIuVS49ehcevQSHbhFx69BeRumuGxUlulx69C49eg4cu3cmLj16Fx69Ce4/rZ3OWXHr0Lj16GGyO4S49ehcevQuPXoXHr0Lj16Czhf6BwZVWZqJmomaiYlOq+Y2bOqszUTNRM1EzUTNRMdNPP7Gjlp8Xp6ZkPEvVPIlMvVPJGhqg8fj+SrV6pbmMohSbc6Xkh8WQq1wp3I08P2L1UPmaDZoNmg2LFd6jJtU7s0GzQbNBvM0GzQbHUrH+Zx6fIv+jKEz6MTy1rkybf6M3qnhBe9GMYIYk2hYl/0ZhVrmI5c1yGhS36MSmiL/AKMxyXNfSmKU/Rl/0ZXHLmi96MWS5c0J5DXJjQlvHJiuXNci/wCjEktS5PZf9GQb/Rl/0Yoly5ov+jL/AKMvejL/AKMTyEuX4rNPVvkbSsVO5IqyF8D+F/ok4SXOWZpLsqouNep7L2O9Xvs7NmDzZ2L9jvlswev4+mMvBpeyFTHrcM2mPSD3U7jmXEe7f5ZCKKOOgyRcchzkqzEDxx7F+jcNU0wzHiNIJubj8MTIu7+DalCqQOKiKW1T+jXsow9/xZKKc8S075iBqFeeYiIULnmWnfMbEgpSgtO+Y4Iqxq13VLTvmOk1SjgQEUD8yV55jSTW4tO+ZBRSjh9K8qUpvzLTvmLkkSjgWnfMUEJQLCFCGV0rTfmQuUFp3zH90q889iQoh3zEnBO+Zad8xRWuIkKId8y075lWYd8y075jwhQ9cf8AGoblD1wLq75F1d8i6u+RdXfIa3KFzyG7c8S6u+RdXfIurvkXV3yLq75DliL62N6jCpdXfIurvkKE3VeAhuUPnkXV3yLq75F1d8i6u+RvP7VVoc43ZxeZa7swfueZqt5lDRzfY5eIy6u+RdXfIurvkXV3yLq75F1d8hwCkXV3yLq75F1d8i6u+RdXfIurvkOfjL7DOMpDFZFKpXX8G9NTx+fAikjp5ESwZ2nyic3gnPtgJKxq7hJ3VqciXFxpeRlq+T2Xsvrc9LSggSQlKoVpGKvwORXBEhulPcUKvVPJTM8MjEVDB5fL24kVS02DXkoOXFiLHWoETrjs78aZuooNQhPVfBCcPVfBRtwqUtU8iFWh7/3El7i/g0pwdN3kbmTVDSKj1kVCesPIq02O9+wlTKXMvepe9S96l71EkmOY7l4Ze9S96l71L3qXvUeSy/rXw5Ivepe9TEZiWSlzL3qXvUvepe9R1Ly9qcOSqB6+BlL7vBuPf4G1Rp2KKFeOxjpkMvepe9S96l71L3qXvUYmE2XvUvepe9S96l71L3qMeSX/AMUqQmk8WV1ije+KX4SpqttYvi0SyTUpix0YmqUlPdCYmlXCrHjTKXYpBub/AEaqyaSFOW3Huvwotw2+7sPWWP8AnAdWyLIo+UJSS3MNQQWlFHp0HTC4v/ubcZK4puWHk3EPimrFOJTyd7U5DeM+KVE72rxxhJ1rRpIrzwh0qrViXhR+0OMV6uOinoR2SdMa4/316/vf/8QALREAAgECBAQFBQEBAQAAAAAAAAERIWExUdHwQXGx8RAggZGhMEBQweFgcID/2gAIAQIBAT8Q+5xLRaLRaGyxX5fC8E4VJZCc/mBOHPlAGJD/ANFLIlkSyJZEsiWRLL6cPIlkSyJZEsiWQ01j9WWRLIlkSyJZEsiWRLIaj66xQySSSSSTF9JYEkkkkmP6vAkklkkkkmL66oTJkyZMmTG5c/STkyZMmTG5+rMmTJkyZMmNz/whISkWCwWCwWBqrHinKUiwWCwWCwNSlrwXyhYLBYLBYGmnDLBYLBYLBYKynlY8IsFgsFgsFgsFgRS19grcBKkrwJFTAkuYc+KmMWE8MEid/QTxQNhnKcvDF5iaKu8Ccgm00GxOKnyiUCcgmJy7oN5CZzvgYHlwBwcokzKIHWhDQJoJwqS++RNvP2CQhMuFwuFwuDdSfFKUSXC4XC4XBuUN+C+GLhcLhcLg225ZcLhcLhcLhSW8rHlFwuFwuFwuFwuCKG/+LJITdSaaTJkoGIPw+IJSpFnY4JM6RTiUKWLDJiUyxuJ3gJCrwgCd8d0KFLMAUpZSNQpEhVvARNsyhY8CMwY3945gTExITDkeBMZiYmJChrgTExjSSeIlgJhq5eEw2eImIYJlgJiYh5DZtsQ0ihrEExU1n/g+k4jTar+niCVBImpheAwTO4ML7XCiRpSOimTrOgqoabKpKlCZtVIlSmNRNhnH6TWI/CT+mkhMT8GGPRk/tmKOvhJiXniTCcOqPIAGLl/mE4CRv2O0LU7QtTtC1IGtKYrt/imU5V4OOA1Se5v6Ljx7k4FYRL4NXv8AU3jJiMLjPp+1HX04jWVq1HHNqWo8a/ox+bovxSVihzUScE93oKpTTLPT6m8ZMQky02+nH0ai4u2lL04ZY6rCBFRt1ji3/DH5ui/MNTRlp7Fp7Fp7CEhI/wA+0afw0G0tBtLQbS0CcqUMMJpw6PQ2loNpaBAmtZ/tfQR4ybjBvobS0G2tA1ylMcVX1NpaDaWgZtTRjRrqh+jpZ8+CNpaDaWg2loNpaDaWg2loPYOYfLCJNpaDaWg2loNpaDaWg2loNpaB9MpqV47S0G0tBtLQLchuio9PKmw3E4N9EbS0G0tBtLQbS0CBJg/NTMLlcm3j6YiUwUKF6V9nw45jW4nubcae44p5Rh8h0akr9j5/NMydponlTOI7DThq3Rhd1sKi8Nej+htrMiTniklZxV+/OklUJUPDO+HofKdEOqWwmVjOnLJGAjq/j/cHopHuyWm4Q+zjaqxwkaEwmiqspnNVo/cdSywpzvvGJoYyhx2yjq/ao58l+qdCduSrRQqK9IwcXghjJUU8/wCYP3Fi5hZWVV/nV+1REDqeI0zOjil8Y14PhxE1SdNUtL9P0FSOqrHL3XpnxnEdAySnGE+K3K9yEnKZ8J08XRcekektr1w9UYkS6FI/foQqoQXucx7R8rInfQm1TKuE8eflgyaXKQgg1PHLHTGOoiU3Sbxtb3TfJoReaTp0q9eUkQxMvhwhRX3Pjrp5mjbFd6m89RvPUbz1CUKENMpty6vU3nqN56hAipd/t/QR5yTnFrobz1G89Q1wk3PF9Taeo3nqHzcU41b6sfpbXepvPUbz1G89RvPUbz1G89R0Bl88Zk3nqN56jeeo3nqN56jeeo3nqFwwlReO09RvPUbz1ETFqqq9fKny2lGLVPRm89RvPUbz1G89QgSKi/8ABadVIsFgsDTVH+YwYJZLyE3x/MyQo8lTGl/6VM0L7gAAAC19EAQABpS193hDbTJPgWieMDZcCeRYGn7BtpKCpgKhQsDbgi0NHAlkSyGl39294fgf/wD/AP8A/Ty/4c2kpZbCchPyNqGy08jaSWWwsIy0LYalM0LYTaJ+GIC0LQxR4tEllsWxiifO0uGy2LTxxRBbFsWxaFsJpJX1gkmlJS0eSMp3QhNDAvHFFgkxeA4jw0BEoM/34RhSJYxSV3UhOPHGEoKSCX0CE5ITHeByFh4QmQqN5jTOhBgJb+o4mhh/VQjY5o0leSbDmiUU8VTC6yQgususbl4UXWNk14JYHNLrGErxXILrOaSvnOzHNLr8YUXWXWXWXWXWLgf6BpDZdLpdGcGNDhsul0vl0uj6G/JAaepG/oJcZGMDdVmQWXEjOQYMKX5FpLRZLJZGcpDc5aLJZLJZLJXk/wAzExyzaXU7A1E9wpuTUfFzXBtDXoNROi4CZUTzOJ8DUhkazVRiZCR2BqL21wyafQhEPJtIZJk3ZqNpKWdgakylwyafTysmias1OwNR+0uGTT6E+g1H8LazT6EGh5NpCEybutSEJrtI7A1Fhs3k1r4Q0+BqdkanYGo7hNZp9CHH2Gp2BqdkanYGpJoeSaf2qJowVCSHhr8Yv49+HEVp0vD1Ud9KozSiNrSJqBRNcePWKc1mfE9EOFpiia+T+VHNwdX1ZtGXh89GLyRsmaPgvp4fC/byof4purkgbT4LPnzj5EIsEv2OAm6pjGKasUqeC/aEp8YUeyHPM25c9v2L3Cjj5ElaETP6i2C5iceVP7a+Ynk/By+fX+Z8hCqz/DSlaunyUySq/qWdKEKObqhm7xDnHFVlevASmweHvXjwQkDp58OdafIkkUXWZ4YRKm7fIWyTR9Pta0lVhHGM08jZLQUkpctBCc8kcsjZLQZFaTmaT0sbJaC65tKcbuRWCka9zZLQZi3OcfpIdGE7RoKhMyfDh6C3vxNktB8znRjHDkl5ZoJm+HH0NktBiB3RojZLQY2G2orGiGp5N5RwpkT9TJ8OHoS4JxFI0NktCVCaeGng3NvpaDfCdC60NktBqcbaisWssho2/i0NktClFDmtDZLQWnjWccaZf42RkaMFqdoWp2RanaFqdoWpPWNGC1FxLabindHYFqdoWp2RanaFqdoWonKaTnGzjzoC221NI5ZrI7YtTtC1H6K1RjHH1ZISNGC1OwLU7YtTtC1O0LUYVtKYrt+LQmxMTE4w0G6rLYVVcKmlGg21oK3Kh8FbJeGCzVQ7QtTtC1O0LU7ItTsC1O0LUXy4jERNVNanYFqdoWp2RanaFqdoWp2hamGTVX6C+HN+vbOeGFRk1E+8Y85w+Rxp0ePTh7+sGOAruETTnM+kDZUpfrmXWH7+rvDhSuu7G2syMWKEuNK8E4xM8HS44khTXHJfEzcbRUZ0rljjOi40qkqXx9Xq9YEVBwar8f2bQdX1fn3iXBfI3kqt3v8AyM+OY6Vt13Sl6jM3BTqxrZn6OKP0Egqtca2+PnoVKzM+Sa7SOcqxb/bDl/TOaL40n958D5TovHGERZ7Y2n5HEePKPbk+gpMlLhVVv4TqUsuKyTzutyPuD5h+HxyLXhl7l7YZeopW2vStuVxEjUYTVxwnjjjEcI9EcpWPmNY+fXHybed444ahiED9b3yg+EhQmXF4wccqxKVMhNxeMq8AgqKozzrF+dJJZYdcNc+AtZXz5Jv7MfVhlFVRjhX+Hx/oOj5vjCO2I7YjtiO2IVFzVkIkgl1J2xHbEdsR2xHbELayWS88AjXUnbEdsQzbTLJQOj5uyO2I7YjtiO2IhmS1PFJUFLHjh/RbC+Lp88/crTzYPUaJ9Ltey9kRLOkREZXeXgzLlPgdsR2xHbEdsR2xHbEJ4tME1gdsR2xHbEdsR2xHbEIy4S4f8UeUeAgMp9k9Moho4iwENjGrgPGMaUf4N4IIaKfZPPIQ3QuAWAspJqYxbFASEX/c2yFK/mvhqsDBe2HgnDlR+e//xAAtEAEAAQMCAwgDAQEBAQEAAAABEQAhMUFRYXHwECAwgZGhwfFAsdFQYOGAkP/aAAgBAQABPxBgDBYEiiEDAkslBFFgKoCxLLa17f8AbBDakAQGSCiTF3egpNJAkAAAbH+kaAmU4xITdJv4OPHjx48ePHNQwbiLMYgWLvXEf9fd4GCx5U4ERvVQjEbF0rUjC+ht3CZoWMY8AwA2yEkFAaXlwjCwtC0rTihoqSXcAkHgvYZ/7B1dCMyoDyooEev864orAbwCmxNeGIeVyDBzLS/lUTODTPnUWJpc6VL7WsqgsJ0aJRbJ2IYROGwwzBQpQJneL0Z/651k60xCoMWHTteevzVWYtVEjTAQW+qVaoaoM7KeJK6M6OiQYgTY0/8Aj7hwaUa0CkMwW6Jia+90+90+90+90+90+90+90+90+90+90+90+90+90+90+90+90+90+90+90+90+90+90+90+90+90+90+90+90+90+90+90+90g2IrNCWTczea+90+90+90+90+90+90+90+90+90+90+90+90+90+90+90+90+90+90+90+90+90+90+90+90+90+90+90+90+90+90+90+90+90+90+90+90+90+90+90v4MW7heNEmI0L/AOa4rrWz/ieHUd/8YlyPKC5Y0rpr4rpr4rpr4rpr4rpr4rpr4rpr4qB7yCIMMXvrXTXxXTXxXTXxXTXxXTXxXTXxS2pIVBidfxEUlAJD6V018V018V018V018V018Uy94QlwxBeumviumviumviumviumvil7sgQm05jh+UqFoBIfSumviumviumviumviumviumviumvip0qYAI4kX9iDeumviumviumviumviumviumviumviieAKCVxp+d7v9dIBlBejJGcNyAmAsSQiS1QUSSoG7MOzwoZAuqEGUdnZrhkm1ZvOIpRWwXcI51NKQlCWEsS7HHhSgCgyg5YzG9TQyNTAZeRRMQ4KkeUZq6sErFYZvipkr2L9Pw3FdZwKAXYUNhi2LlvtQEaXBfIzG9RxCvsi0WI2FIAoKwKkWphcEBkhXAcWlEGyoDldjd4dk+gDV5GtACwrAqRfKrm0zMiCM34a7VMwlyuubqMfkOK6HgUGmPPgztpCAbpTVscZP2QmEDCDDJQjeLLJuExMbcaB1kzuWIlwTJcG00zLmGi7gvrwow/nZUeVE1VRIwdlOYksOGTcc0iEoSgliVwc+FF2Al2KhEMVgiRvOI40Mg6JImpvSgVYC6rAVDIQLJvG8bcahkwsm8bxtxoCgRVCDh2dnhXXd/wA73f66VChKXCYpHA8WvAEnDXLbIUxMIwfhiCuGagIKMEyLY3KhkySzDrUOwAIggUGEJGhdKiSfSAtUhElJppI28RKxmIQ2lHFOvH1c8k3TXCLBzRpPCfAGo1kGUoaQtZ8Q2zC9mBGaZEJkSUtmRIKoxdQCso8B4N9rpmGJXFvMMYybWcxLJAM0pwwBlAuUlJivYv0/DcV1nAoM1CESMShhPlQJmeN3GzCUkmEZKHae+QspNA3FMMASy7oP7wlYgiYzRzMOIVY5LABIQl6XPwz+LiGXggTlFNjIx9UobCyIRszVCQib1kOEKvGahcwU0oECOBVlXixO+WAsLKNwpJHwZY4hcZWSblauubqMfkOK6HgUyqHTxQA3BeYbURz80bgJDvJCrMNKdBETl9aDhYLChTJa+OLRDiBMCySV7i6cAwUBUVSLSzp6pwVcgltgUUPLPIugF5CUSlq5GEQUrAzhYwVhBLS29XXokSQjJe+agJgH0hMdKbCEM0Dl/E3M8NbYLB/LbBFNpIiwTRCrKMqEqtYQhYFhTC6R6ELclybF1BvTJM4+lYgRCJyZTru/50lMXATAdaeq/NAY6POg+j+66H9atxht/wCtEhtx9tNj4KYgSBsABgWAKnzYkBSMLJbelOj+6lZhO/8A610f61CR0fOlM9HnUXV/dW/akGSBp+G4rBWUUkHGpur+6Und05q9Oe//AK1Ho6eNdX+tBeWACzTLGQiZtV+cunWsm/pzUJGHTrSk7unNRHV96nMEyUXGjRj8hxWKtoJINFpTq/uur/WnHq4xIkMATmI7Vg7cIKIqlSCVVVVasxh0610P61LkdPGhFy2mZzMEamZtXR/rSk6HpzVuMNv/AFrof1q5OW//AK10P60BiHTvV+3AEA31/wAVxXWtn/E8Oo7/AOacV1rZ/wATw6jv/mnFda2f8Tw6jv8A5pxXWtn/ABPDqO/+acV1rZ+DFxYIZApiYihQKBwSDtxqdnrpOz10nZ66Ts9dJ2euk7PXSdnrpOz10nZ66Ts9dJ2euk7PXSdnrpOz10wBmb6Z4zmp2euk7PXSdnrpOz10nZ66Ts9dJ2euk7PXSdnrpOz10nZ66Ts9dJ2euguXSyyCllBv+V1Hf85NNkjIMCpEzIZvpRc8VxXWtn4RfqWz/X4TOo7/AJwW24lwQQgZ4pDhuGGIALAFgPA8xSJJOJd9ERESbJlRgxZKJjE95EREbj5vcxMdw2rUPsOzWO8RERPUSQe1lEzFXz6JJgCfR3kRERcHVoITiG3braBqJBmHfvIiImfnCHRrliNO8iIiaJpdgUzHDsymkJ4HvERET1XeGsQmJxPfRERE45eZuJifC3wo0yGzv4KIiIiJzWxLl84Rw32R4KIiIiJ6ziVwiIN/wAIjYRHkE0lAmVAWpuEMEQcJgluIzaFnC8oW6ULLLHlUvwZ3lmOMRT4ahQMipubdmzBy8G4aIyTbMgRYGVmlERNBKKsh7CgzrB9BwyoaGciOcXo9lHrJTAXCgcwC00e5AEAVLYJKTLMRLhFjmLxeQtIHlG3g6OxEMMTDB036AMSEKkIuCybnD7RBNETEHJ0mSVFq4srW6CDexoTDw90rHMiXZCJoTV5sCjICLyDK2Kv6CzuOK6RwqIvqaFC4nWaTNqcKEd2A0iShiy0hi2jiQQSljguVCR4+3ASt5BBSEilWixeaTUIsiBGcTKesOAKpISQJeSzSPkKWrLMssSTSCpvMrtWMNKxJozQ2KQRWQJZYwlVAFoWYKWcCJ3ALVgWkknUjNxBRTDJDEVfX3PoluEpgwg2r2f7e3rm2gewA+bUMQ6rUhMsCLBDFAvVZ4DCSGIwSJpWqB+JRTwt4mRBtQ/uQ8FQiSRkvCRSdFvM3aqyCIo3tVtW6GaKAJxbIiZJCYmRRCKhshJOTBke9OXAgwgTFovNDAR9ZZATQIE2nMNmMR2BmujbtC+BNgVrBfZ6t1MkmzzRFSiSBJeKi30YP0uZLVQ0YayiJpygu6XL5ovi9LwPkZCCgxAamxNPktZA4biwQuGSXodWdlTgXKQKLIYYl6U3fqe2kZm0TaiXENcEkIrYi+TEy/XYWCDUAY0hmiLImvUAFkAlicwkKV1zZ4agzouodnIEIVgmnedFVLyYAQTaS2DixluHAM5IQS9MHCiwYLUtTJSSRRNYI6QRWbEDQji9HQpDJFgCrAmSGIqUMp5e9FITKJhpLEDRpNmaqABKm5JT/AJA2F8BEqBmlOzQr4ANKhKQoMNEMTADVIoqbCCl7VopEtboLeQzi9WTBE2RuMgWRNKr4tsSuLIlJzi9PLHghM9QHoJtSie3ACZmwm7om1LrqCMAIpAAkRzXV8P4AKk+I3QDAWTIMMU8lQ9LD2kYJLoKgVnC3LBGgi4HZQRc5luCy26Zx0Cey4gm3ygoRkYKNbGIVuQ4xJpWDl4NwpwORiBSAQFryNOIOWVOLKQ2vA0YChiMVkRqbiUMUE1ianrMnEkpYQ3FwToNNyAogNyUbQiVAYTRcAFARFF4yxMsvyAUREHOhL65CGQL4QkwKEMKAyAr8nJMJhrDbea28MkSmAwNyoRoodMHvgWaKq24fBQudhMyUJieobO44rpHCiQ6kGCxcQEbylH45oDVhYDEyiyLS8SRSmwBUCRLAsU/OA8jmRgIgbgUAJotmnFW4iTMNCzmKHbMyCCnDmaWdMEMkaUBbGckUj+HHqg417egxAPbmdusSLlWm4TCT/wAcTEWgVBBIQVQOh10mU0GNSUsCgjmCAlujqyNYCxYCCIG3bdc21JV4GOTUh+MQigF6KZBA0PE40ppYiAKhbGo9JBreVQhJV6YYiIDtecXJBgYbGbinZ+xmNiMCQBCkzdosbw92NTCAMwsXihXKjCIQwPcpIm1I9pFGiNhyLBYEgIRmNSAMDKm4pCgQVQ24dgZq7R2ubQ256SgElJIkzYij0YjtWEmyAuXmo2oPwJjkIQzZaDuXiKw5kUFvmCgjZU2kW9SQWJWQC1CwoJBIcndZi1NNFlfxXsFsCG4ayaALVnMtGQTZosy2IgCIh5JkqnkRx2MRsORYMJSVBmYWIGRzUKQoIKobG1dc2eGoAAp9xZUxLGJtZmmQRkEiwBeLACEI31n3FlxaRIQWE1dwgFmH3GrEJN6WJQKn9ElugUWNR+GSkhGDsVuLtxdiWbJkQuDfXKJ2SzIG+1WDMEQQr/FSiS1iElpFGZvOQgaOb0gCk3G5AZXCGo3SAjLAHpPWVBoUJcIiFq0HLurSYBTqhKN1lMUREREm8ujFFZGAg0ZB1i0S0lnS4Q9l0AlSSkrUEEAOqQBAISyjLXV8P4IDAFQ2KgmYvQAWIolEAUvEj3QrBy8AZEnNqCMzX3qn3qn3qn3qjsBcW63pYmImok0Bie1CyML3OJtMRPGgSGUNiQx6qVynN0+9U+9U+9URZI2yYZie4b0SQtfPJNPvVPvVPvVPvVPvVDWI+yplQkwS4maWGQrOUXJp96p96p96p96p96oyQkQUpRmePa5IaUrEBxPCvvVPvVISLGpOKfeqNqRExttE30RbVpSKdPfQNFlOZa1piY03a+9U+9U+9UJ/lUVUsCNyU48+yCXI56tPvVPvVPvVPvVPvVHqzBt4RE3nVNuavvVPvVPvVPvVPvVPvVFWmjJEwRMeEfAwDclHc2r71T71T71T71T71T71QaABsIp96pBQ6EzGJ1vmdIilYQjESin3qn3qn3qn3qn3qn3qn3qhbWyWLjOXb8UVK4vmijABIloRFKZjcQprOLKJiGYDFHiu+rbdl3o+38CQjpoSOctjDNElyCe8L+/j2svKuk7/APlK0V31bbsu9H2/gahySFk0MQhmKj2SmmYEvr4MePo1Hj6NR4+jUePo1Hj6NSQDxirLyrpO/vIN/So8fRqPH0ajx9Go8fRqPH0ajx9GlKAwabVHj6NR4+jUePo1Hj6NR4+jUePo1Hj6PhIN/So8fRqPH0ajx9Go8fRqPH0ajx9Go8fRqPH0ad6GNlpjx9Go8fRqPH0ajx9Go8fRqPH0ajx9Go8fRqPH0ajx9Gj/AAnfVtuy70fb+BemeSUkhchRte1XuZjMoAvgLAWCESBGm+kyoZVzVVnut27duziUFikyAYE7LvXtq6Tv7uTlWM/tRiAAAlsbvfbt27du3O7JM6Re9699u3bt27c0RUqARuFBGyLR4HtKKQxTkgEQAAA8Bu3bt27du0FUHsqaxLJSUMI7IkWV8SDG4d1u3bt7BoomCJ3oPBall1+527duhmgSUZ1XKIZDKq3Xj/hO+rbdl3o+38DPUsrGWI2WbOlSrTNyAT46zz7auk7+7k5fge6zs8D21de3eB0HZXtu9bMdzr+/8S6HzEO/a0MC126EtOHOzqMEbhEgIXhy8OPH0ajx9GjcnKa+rbdl2KUeW7ajx9Go8fRolv5nihgQAkLFyYmc6UI0QA2AT2yGtSblSblSblSblIyVhZamTcqTcqTcqTcoR1r21dJ39kxUm5Um5SJXMUwv1cm5Um5U2nSpNypNypNymZaaTcqTcqZqTcqTcqTcpmerE3Kk3Kk3Kkde2TcqTcpmYxTMv3Kk3Kk3KEdeyTcqTcqTcpjcpMzGKk3Kk3Kmak3plVgjNCOvZJuUjL9+mZa2Tc/CI4IyOJHPRB8isHLwsPMoi7opp1ewUaONho2RgnBoAIMFHt86RIRNRFKJgAAiAwdooUhQTMiSCiQRPHxSLqIAcGbWFb2tRXRh1AAe0yAMYpQBHqYBqJBaWINWrY9GhMSBhuJ5Nffq+7UZhPqE4e7cBrWXeVBWIFOYLBtavt1CAWrASvTTLCSkieAHiNffqAVDCq09tXSd/YlAol8eCvu1LjGEkrUEhsls7UFEhycB3JgiW7FJ6xBbbBiRZcNKk2LpaMrKAoMBNE5+9SFJ1E3w0z/Or7dRT/W86BqChJGUImk5MCSVZN6LoVzjKLwyDgLN7UDIcv0kAwiYSkaqudeJQ/WluptcBZGREgciy18VGPX6NW4BDcGjJRkughzAgjohSSQMbfCCJSgpfFyvu1ffqlNO+rq7LeY/VMSOrBdS5iFzFlBgkDsjQgAJGVxpvOCgUk7kZa192psAoLPE0WgK6gqRSxmZ1divX5u0Coswc3EYfKnug6TgRuCXBJUPEYuezLiF0mlGnDZOMEG0WwtfFI4AqgNKIAl1EC6xSwUCCsiww2GBIwwtSIQzM61OQQWxb0rKWgDstoEUk2igKfXKagdxEJJa1IoxlZ1p1LahkDrBAqiVXYQVTQSCQVkSKaOg8EhYsGwYC2JoQxoJEqu6Cz8LZg5eFh5n7/G5gbVC2EYmbEJb7VE9RNMgEPp3CBDxKxQK3bDwBcjS2ET9tE7tgVYkJUNVV+I0mSwyAHWhMqKM4QwcQUISBJUkDirKsJMmzZvTG03FoYGliASBmWCFFxsABpkswaP6osGbQ4xRqGwlRrnybEBTGBk0ErFU5Iq6qBGRaHM1V7auk7+14Ce8lulBvKIi97UPGiqOMSWhAEuMXp/qKZ8ZNNNnZY4vTeRDhxtjPAScRMjH+5eKguRIOBmaPqc7MMnonvWTqtQJBu0ugODLOsREiMzEKM4w6GMIpaQq7UaCReCxQNvaSEJpZTtWMCllAEwRszQ7jRPwAQJOA1rp+5UCwyJ2nSChEWxZ3QTBFwkIoe+HB4IxJIgWd4pmky6oUKp5DForQWBw69MNyzrggkD+6GIvAsGkUwuatsx6kCZACibCcq6Ru7Pev1SY0LASsyhZVywDWxSQwYWBCLBCZvVo9maz0QCYwlBVxRBaSJgJ1RebRRgdaS0tAQiBEQJU4rg4g42cCF4QV07d2CFu6Eb8y4sDGUCmFtdpmiAcimbK1ZDBkgVkIIsMvDIDvkUXU6aCJlFJZdoZe5rGF9dO5w5KWZn9vCQylJiNshno29et70cXWKpgdtN402402Mb2U9MFg7MiUvTd6dC2roW5ToqPyycy0pBhIiazR6xPGRwSFORmmtPo0SEwm5MPEa6Bs/C2YOXhYeZ+/wAbmBYYmaFgJhiJNN6IG1gQ+nbuCtA1EablgYXhmkE6MYxBKRZawWtUDSOCQMWLSCy1i1q9+GU/QwwWqPixt4SCUamZ+yoQi42Xu3pnKKkDAyxJlcPKoRVhTsQglYNdDagCAHboabIicnWaPBxlUkbJhLGBLBemZyl7auk7+yyvIGGHeaREymq5YSl1cutKWymWCjHgtZEVEySQhIAjSAAzYIKYsKQBSUDVVV1VWjWGAX2CAOVZOq1JByetBKQFRSEiZlmyS1HSxLlDDOSmi3KLoZVCsNLFhZbNipDpiYW6CFBdMUF9wgBgAsHArp+5SMhzeKiSAD+GCQwbYppAjURuLyiVCwgw4UlDn5hNEKRMxm9qHokxgAiBwTgxcahy1UjiSxaw2WLWqEW6E92BNLWn+js96/VThABLElzDCkl7u9NBblQ4JSwCEwlmaQEl2E5oxMGAIhFGAsBgAgJFoWCVqUGwriMb5NwRe0EUWUGyViBIkwLaWKWv9zsUWXO4tWQKcMGJCUADV5vm6YIiWCdAWsbFIPiBcsGRbqkrfNQm5F1uwJeLQb3QtplFAxN4xQvW8DZcAlLq3XNLZ/8AaqMpz6A59MKlAyrKuXLTQVFFwGMuQSr2K6bvToW1CWZNNaUtSkmuWJJlm95qFqtmW5KSQ3L20pmYw610DZ+FswcvCw8z9/jcwJACEIkiUAABBt28fOAtfVa+q19Vr6rSMYvgKEAyZQrT9Vr6rX1WvqtccOAGvbV0nf2RBJoJK+q19VqU2ttDogiJhNBsXa+q19VoPQsWW9K+q19Vr6rVz6g0A0hJa15K+q19VrgpIQV9Jr6rX1WmRZGQFr9fVa+q19VpZUXMB2JNfVa+q0sj2NYjsJmajweWCvqtfVakbzmAdn1WvqtfVaXkSaCSaIo7i0IcbbX1WvqtONxzATSxA3VFIAVzHFg9yXtR/wCZqUvOYCkmvqtJP4AER3oUA6hJFVyrU38tAAAgNPCPEuVCYDCBpEWgjszBZEGbxJqkSKXQg0IpMkQu0GqL0lx7xJ0kWAlLTUclErK2Q4Re1LoUhbYTnwaxFPA8Y+SpaCSWpTm5dGDl4WHmfv8AxuYFZ59tXSd/dycvA95JQLsjdaSzrHDwHZ4AlTiKLUDCpKw5F77+Clw83uizaRlsoWTIzzoI7nX9/i3ZnuwKCSWKtlO0moOv/SPnCmQXJIFLyzYIEiMxrlAFBwhxzdCyGAljExUIzZQYY1yWZlDz+IUSzRjRKMsqG3bE3aUCFgZNqsCJBwyXZ7Vg5eFh5n7/AMbmBWefbV0nf3cnLwPcgKAJhgTkkcdr4PAdngQlOKAEiWpVFIWJYtt4KXDze63c8E7mpEbI8qO51/f+Ld2YOXhYeZ+/y+YFIZJE8CjTe2MAJbAtpXtdOgECYagjfgaR4TigpHKwdjomffiwrOExZwdz21dJ393Jy8D2AxfLizeNmi2+ngOzwMGcU8BeFIkFK2ZPweClw83tNqAcSaIFJWbVMUMlg4/dOnTp0rS+9yAQNvOnSQAOyI+6hgCqUNI5MB3XTp06byDzDIkpC6QEXgNLrbxtmDl4E1NK3mfvspzU/jgZp9Bo0poOe5PYxnt9tXSd/dycvA9phAEzdvF8aLb6eA7PAsGhQr4J6y3XvffwUuHm1NTSihFbzf3U1PZNMOivobVPZNTU0+i1UqrmpqfD2YOXgY2IUeQafIwyGTKDJDVwUG4zN64BCcRqyjgMHIGumPmnmoNl2HGj8UC7rErLyqYN4ua6hgDdoAujKS2FCQOjhoYeUXEXQAKdgdqlydmCZglrl+JRUWMCg872EJLOLLkNSjceEU3LmLwTUxVEOCEC1eJE6DNBLdAOEwmUuNpN6KiyEeF+72e2rpO/u5OXge7D5gEsiIlgMd+J4Ds8BQLUoRDDcBcGBPy+ClMefzR2BFk3HGnBIE1OwFuXYvVj7KoKicBDEzi1I+NSVQ2IoQnMWrEyC+BkoDCMcaRALKPItK6KBITGtF+ZBgF1VYANaFZpnObEoUptMJU/yx6pBYJCBxF64yYQRPGkttJVlftXTHzXTHzSJg0BGUMBxaSWTbUW1iIOJSKCjSCShJGyESBomKvphAJwI2LoMLpQLElkI5gyY1jGtFxQ0QwoCcqthrCWFtz4WzBy8CVnNKaAMSkW3XN0Wz3qIPjlBjJSbiSHJe1FlBHI2AlrBEaMXfqUmA5Q1uVyPDeMJlMizDJJmKPxgKJQNu7XEa2r1LrU2AGxDsGvm7O9FUCGJbUqe6Bpxp3ShhTVMNowLzp3GdhAKLRXvzvc0XWirVmSjQIsNWbkjlR63xw6fWPGBorou/s9tXSd/dycvA94bZJvjZeC7NubwHZ4FgxmhEnccsL6zYZ14eClMefzXSN1QtRQb3m9PsrUByFAMORLenMqRguMbYkbm4txq+HJ2HWZO9+wLrHCmbWZcc/NNrptNEEECBNYkdE3pvPO9mLQRWxMtcl/anQN9AQcoii0i4JlTQ0pP8Kc4TMgWKXIr+oNZhhxZAtk0ahWEKaY3bEz6BTHswAigqwKISWL0SI8pOiDhRiwbIq82GGUndvaSN7xUMJILZFAxHJxmbzXvf08LswcvARBIOq0TDgGtkZS1Jh2pwep7RCIAgAiyFxrGVQRBUbGKtl2nMHTgRARjByC1Bgc3go/FAoJkKJHwUDrKLImjRDkAZBkHKBxKDaoGBfQixbQshrLehfWaCgAYAADhTGNHYeywJQ32jHSmZIkyotIlENSbEICAAlmSc0g1fJkc4IiYYbUTCjxePs9tXSd/dycvA934PRJJaQwvacz4Ds8CUMZpOEPMy/QzffwUpjz+a9xDxVZj4gAwJwL3Ib1C4ZdojYwlBVlYKELihgnOEJHI3Kh/wClEkIu7Rx1GKSsY8QsiNkS0NSVAUcpglyV4QcKAHBWgwqQyKyXVaQMgDgakEqG48dGK6SAiNkTWg7xCZCAagUMAsBNY8TvRALgLCsFgKEOADaMgglJvCxUSyzmlNxiWYidZp4RCeDDpZBCHFS2cYoDkyAwSDDqVwyDNggAUhhRvg8PBiiXVmH0kgXQC6UGybUDGGUiaiCQyeA9TLLT1SugvimPnuoI3iImhoISKEkMFyx6VCbHVagUQkZkse34wE6ZsgRPnXQXxTWE5OA8KQ8+cjKql2ugvinCUQoJPI7ERWwQhymugvii3WRBdniImugvijApCgn0Oz21dJ393Jy8D3aXB9JIMMLzcwy+A7PAkjGaCSegpY4yLBHu+Clw82lVVlZlue1dBfFFhuGNyRFjlNakl1HP8K6C+KOAmEnqFEzCEZE4jXQXxQtaGJiUkGY2RobovahwQyJIPpToUkDA8JroL4roL4roL4o9JGEBLuFJXHOUyqpeugviugviugviuTjOXp4KTUxbCBRmIm26LZ8Jw5/07PeDl+SB6hs+GDI+2rpO/u5OXge3UHgATXApCHSdz4Ds8BJEo9+udSQlgOMAt4KXDze0kpIpEwNllHyox3R4PaIBOiuWvQz3upbvyTbhz/p2e8HL8kD1DZ8MGR9tXSd/dycvwPbs/CS4eb2kUEOe/aOJnGlDJ3RCWV4kOlzjD3+pbvyTbhz/AKdnvBy/JA9Q2fDBkfbV0nf3cnL8D27PwkuHm9raxwLiTa6wr5UMm3dZsEkAlBcy7Xo73Ut3jm4TEk7eE4c/6dnvBy/JA9Q2fABkYdXGAYuFTME3Y/lSqGKaJfFKiQ6OFJJm8G4N+LjSuG6OFcN0cKFnlhEuRsdmTl+B7dn4SXDze0WgpAlabKepRMX7pzEBANQhQb8SjvdS3eMbTH7NFAIbxEK6oXSn83E5RCBhdYu2JVd8Fw5/07PeDl3HFGp4gQNqHDEpkDdMlLGzoT15S2kGKBElQMMqwHOgx8WFJx+B1DZ8AGR9u/ShbWaB7EEzBuQqRBwy7ICoNAbQlMVI7zBRFo19sSYN6EpZbQIEyk3GHt63w7MnL8D27PwkuHm9rKSSADhCUExfultQUCRNgmdzv9S3eOb2YOXhOHP+nZ7wcu447AEkdUWkYBCy3QQkMb1Gy3AIYpMFKOFYtdEcAQhJBZP4KjqGz4AMj7d+lQXmRbIRZkfEgzUsjaoAKEwWDclGjsussKKVw5UXEoNttMcUchAjGszj0IcyQSWkRHZE7Ot8OzJy/A9uz8JLh5vbCsV3ts3L4E86MZ7t/fj5yJC5Dqd/qW7xzezBy7rRi4TSyCkFjucOHCEBi6zNITgAG9Bu3uIJTeMSGHFAgdsOHAAtpBJ5QTlfapkaM0jZ/mQcSwYCiItZSg+hkwwKhbDlKXQRqR2LFWRNN6F5lroe/wCB1DZ8AGR9u/SiUPEF0Uuidr7UHWS20uGrSRS0hSspT6iyxC6IBuKQmAAyGyC3mGFoXWOYS9H4BDhAJQUyVUhIrIEaTNkmjTdcdmTl+B7dn4SXDze2LpiX46RMxGdaCDu5YprchijbOSi3e6lu8c3swcu611PZ4QHDzf3SnNkXL3Pe8jhwFwmH8DqGz4AMi45b9KRW2LAY6IgAERBGik0odRWnoFMAMEUPfCioa5YhImiWq9/JMNyC0VkObQGo0pwyDDAleuypPVmUmLhYNDdpmY+vsycvwPbs/CS4eb2m0cvBFjZdA86CDuqdZICgtssXvagjvdS3eOb2YOXda6ns8IDh5v7/ABeobPgAyLMyZCRr6J/K+ifyvon8r6J/K+ifyvon8qzQokB7MnL8D27PwkuHm9sdqYWAiSPEHyqI7qZxiXrIjZHbv9S3eCbaYAlNY02E8bE2rPc2YOXda6ns8IDh5v7/ABCWCAMSRQJJT5WInuMVTkGwAURkI2qelBCQsAlYFttWewhsjJiwSgF0L70lR8I5FkW3OqhqfJGi1gwwiNxO1bHx5pKiwBq1BXRAkJXBRQTMQTceFFQScBkdbUSCJYUjGYoNb4KXEb020+GZCbiRJLWamViJTDUZQVMhZrKQsMOjUmQsMMZaiykmLUxQJwlMw8xLhICyJqdh6RUIALqtJz284Mb1EQQ1aYsokxemxJGWDVa3n7iMIIMAK7AtJoMrjVraFxMU8S5ExTxLkTFMmS4ZKYy6GaYxBIYYrmNiQxmJERuJ32ANAEZRYOLUPWe9BSBMKNcyZo5IZDJlgzEJOw7NBEQgbq9i97C+VLBNKEBus0kVELs/0oIBSliUFkREfCf08UQKJZYF5FdB/NXk0kuSMOu4nlRNzg9zYCbsGCjOwEEKI4RtFESaKSskYTOjakQo4ByiYL3YG3B7IydaDNSBZERnuxHWav8AXzYHgtJNj5xg5rRJfVsqJc7cR6nRxBkkNAOIpUMyjAhJUN3ShSdWME1g5d1rqezwgOHm/v8AEQ38oNMHs7Ct5KGOO9TI7PYKfbdh8QxiJrjRMETQ/X15PsHo0zFII/r6OqSCAcI7RMgJr+VTuEz4rBGHBoBbcusXoHREIcurD2dw2w0kGHa/porUMSdlTRh6P00HNXbmgSBpGht5NJVnCFtqSCAAYBGnYZr5TLuWPwpkDmLbetTrrIxr1MBY5pjc5wmsrDG2Ggg23T/prgQCNKrCwxraCGBNZmuFqP6qmB4jtoUgCIEB3++nRbzf32TBNSmsTSQ6jVRjw9+CUZoAp2HH1oU2KfFmfWuqbe63xBtlYQMCEZIyN6uPWhzuEEoSyBeLstFhS2FcEDJeKCmSMyMsAS70BABWYOXda6ns8IDh5v7/ACLpNJt9t4OP8Rz4uv8ADwC9Q/M27SPUNle1U4dpCtHpXHufsPfdng4fgkzjPpEfWBXN33Dzf33dm3jS6Ru8Tvwcd4A6bPMK6pt8LZg5d1rqezwgOHm/v8i6TSbfbeDj/Ec+Lr/DwC9Q/M27SPUNle1U4dpCtHpXHufsPfdng4fgkzjPpEfWBXN33Dzf33dm3jS6Ru8Tvwcd4A6bPMK6pt8LZg5dj6gLbMMbVLr/AJUuv+VLXr8qS3TwUhQSYtUuv+VLr/lLh6/arHxAY27HwmWDJOIqXX/Kl1/yp8ZahwQwZs86Ged62qXX/Kl1/wArSrIZpJ28dsUuWAYJsOI41BksmjhsxebtMxTWI879ovNsabptFyvrlbgjEQjjUMEEsJKLWXVwpsT3WiCcR2AwKeXtrcGdJoohUJoPBi0M/Kjng7v6KckRrTiauCHxteWfJpHKMsQAVshBat7ZQ2kpzYa2ojhxpPNjH1iCfWmKNytpoC0xE6UBRLxnDQVtU8KMRKinj8xaL9cRSBRM3WADMWjgvNRkBq+rujS2I9lRluqMb4jW2I0qAIiW3MkXn0bppjEjW7HKLzE6U6gYgjDar655NKvAwaLVDovEZoBFhbRYEaWTrE0hSgE5mJDGirT8GLGJ2erS5jJYoeVSjFmC7lEXicUOGUSBb0HnE1a3KaDQQ5POgrvJ0IWGUjOkzWXJOFSotZZzRlVkNgwFrXLPyouMGtZAhi0LNHgqGVhIxaJW4NqjhXtytyRiIRxqSYZnkb+jUBHCm8FzFjFWLz+tNIphiIhYvLLzij6BwRgzlrt7PGpUXZ/aSIcWTrE1z5wMMZ7xvJK8oFxHRoYdf6oJaFAAHhkIzwpeGjetkXmyItumycVG9YItuX0a0/n5fdiCYmYkpMyUtMkbUigbIh5NJic+DtGs4mUM+dLFp/evcotEzrSnr+KX4DlsQ4s4xNqgImF/WiLb3JrUuv8AlKgKKWsAjgzrE2oSAS1OG1W1z8qHAaeulMlcHbyYhidpr0klJJ8HZg5U10HdVozJLzTYY8Rlb1bZpAilN4XlBpSpjSERBdgOQQmRKGEWASmHVFWI3Ea33hGLOzhNxNqkzTWjBnKsTGNycnKurbduUt10XNqLAHAMk1JWzlGZhgQXSFltgZb05lElJspyQCUlEpfP4hkSLGbNmDrb1xUZKYpdabxCX35LAQ5GAWRLI11Lbx3FX8l+lQIufVABLmJAEsyASwSRurCFl0XsJqzDhAZDmhSzJy/D93ZOiknocC6iXlaQiTwRHl80+xT4JpkIQdkR86Mru7UIi2hSW5AaUBQE1N3Wgm4QbldM2KAk2EL60I2+cVDEYaYBWFA5WpETMIKjo1TXWJAqAFSZBKVs7UEjCYvE6TTQTnk4lRLGQySIlT+k3BVAUoCCxmgO45ton3CGbaCwMRxtocoFMpQzgqFx8ucxINoK3inUQbr1ZLEJsBskg0w2GNdcWRDXYp3bwDBo3xCBKJFK0rruCukblGbol4k3kgG4pTNSLeY3jp4tHUEjQfMJi31blYIV2JMg0JWhasMqEsbzByN6JmBhX5CkBuqjCeOA0FGHKkqAIXPaXFBVNjAAeBVzJhJLtvRYYcppskuS2hGnh00bTBXgkvMWmjGF5jz1LYtgABdy7cezpe/e9APWkm6EUtxM5HNa3qhJrbCUqsALQHhYj0YolIwssINhk3sGtIsg3JImQJUi6NqbvlwTUowAsu610vE8HpuCmAi9dJGRNnadquq5OZ0im2QViwMgxJmKahWQKqRuWQ14Ou56E5GFKCcRS7ryDcyQqXIgs1JQiNp0CWCXE0ibN5EOtBGAuhVqPUUShAEFpm9dN2eDswcqa6zupw6jFIXdt7S+mGJOAhlhGxG6aBIMXAeAYEmkFt1Bgj7HpIUq7FkAUSVN+d9bBKjKQ3qefVHQkxOcRMwCsnKurbduUV+JuSC+Qb3IMWzR9ZwmBBCiBykhYYJcOboZQZEKjQrVaMwWEAVgIXupWUdjK6QBCsqUMIpAm84m9FMYCAABfqW3juK9i/SjyhQ0uIoCosWsrXdldcVF42CAcDSDtu8dThKEol2nR/HO4JFC+CBGiUfZ/u7F+zGoDPMKMCuiaDmRMzxKqciD3piNvsgXsl7DcsYmh/jgPEBDOEBa1aBCi6TMplQSUBWVrpmxRUlJAxMNqYq6yKEsmixBIFxGV+y6i8YhAOBo07StM4FowEigUBASyCLAQcW5EyTekAw0TdE5cxAMwhEbsV4CBkzBIY4YVvU6zwXarmIlBKfiKEKYYALiCW7BO9Jt4s0R1kKKkUCzucxsoiAZEc6Nd25NEIEOCjMcl0eTqVCCjg0yYOAPGbVppxaaLH8K6RuUoczTqtIiDJLQLtPcrh8QezE7DDU0tfc3EW3kmb1eKuAikhu4Ni9Xs6/MjUpETEvDRM4PtoVuJLaLcqG8hgjGDLHAu4wvdJTQRoXRXLGzV26yAU0ZMTjNsTvaF8RVhBwSQBjABkGyxdFuCZvbJSp8f+U6KbxjAUGRJLtw0uwh1Uy6gQVqBKit2r0ZRE10vfvPmz6NWku+g3OI9N3hVbg7A4yoo2YiVJiFXeUixLAOJwOdlqSpYbhxsrkySh7bxNiggr6VYFPxQB4hEIKykK6XieD03BRqXkWsAYbbb0KmuoGnpBRLpKAoIwJLL8zKcYUEdF1fMk4FR3mpjluLjE3nQTiUU+aRR65ysoWSstBeWnSICIYYYRoLmdh7eZLOFkEWqDrCn1a6bs8CRcjtkQOhHXSZpiL9kINi3xLO9cD151wPXnXA9edcD151wPXnXA9edKCRv11q78wCewkEkZsEZmuB6864HrzrgevOuB6864HrzrgevOkbMSRYgjfx29JGJByRYMzwrgevOuB6864HrzrgevOlUSXSmSMzTekDG7PWa4HrzrgevOuB686UEjfrrV3JilvBFO4BI3pjjwrgevOuB6864HrzrgevOuB6864HrzrgevOuB6864HrzrgevOuB6864HrzrgevOlSOHWrzvTiEJDNLzrgevOuB6864HrzrgevOuB6864HrzrgevOuB6864HrzrgevOuB6864HrzrgevOuB686d3TBGWe8gRur1muB6864HrzrgevOuB6864HrzrgevOnli7gZjjw8FAsLCYhHHlXA9edcD151wPXnXA9edcD151wPXnXA9edcD151wPXnSQ3BUTBHgEqUKKgDZEUika0btCBsQ0mVi8Wo/8AzBkmNf8At3HQDzRKAJggeVCCcsz7awiIiIIiIJ/20eYWIuWGbJcdNZREFxIzvBgWFiYJj/4hNCxivQlk36beALFixYsWLWCJrF4LJk6b/wDYaUh7TScVRgCRoNXt7Sz5MywoRbktBIJa2CHkRBmGLUqWoWDKEBD5la62ALCCRaUhBImakN+j9sHWtT63GtoQuI2Rx/2MPSWsNeQF0Uw6MURzEKIlE1FEERDRkRwqvNyQgmzF6n1R4IJoDdWnqo9+jiCErBlSGwJRJcGPOjqUsValhJ2hkjI8qzqby6/9hDShgkbrAItvO/cHDhxakmx4E6pemIv3Bw4cf70PIMlhDvO3/wAkcOEQS8lXJmWEzq2aGPyKlSpUqVKlSpUqVKlSpUqVKlSpUqVN0gTFgxqlmYiLa/jlSpUqVKlSpUqVKlSpUqVKlSpUrbRNFq4LhCYw4ouH+esUu8ZGKJTsySilhR5Wppk1CSYspsv42Uwf6Aowf58SMBncFKhZKZklLqrHVXxg+lbUiY3rh9XOuH1c64fVzrh9XOuH1c64fVzrh9XOhhxjmFre7NjntQQW6udcPq51w+rnXD6udcPq51w+rnXD6udcLq50YjCT+cQUkqNjG9cPq51w+rnXD6udcPq51w+rnXD6udcPq51w+rnWPN9Fmb5kg1aCC3Vzrh9XOuH1c64fVzrh9XOuH1c64fVzrh9XOuH1c6IwA0Mzx4f4Kglq6LX/AHRgN8hKUKmwAWIeMrGgmENJoixdmiNTsxLyYkkDrHQCBTUDIDFyhkN2BlRZZpTFfxr92r6WEChgdWYaZg7drFIbJxATMtRbzujxEiGRmMm9FwxUmaxzQQBgpRQW2wwwQSiEJdA1Ci9rTHgPMgzJgQYCnwiy4TZ8wVFnKFdoSFS1+V8UgAUFFMymDJnemkzBxEqJkF4Yq/oq6MQRFm0DNP1KFTixLZi8Ol8VAAkW2BceMio5tpYUwNlYRgJvQWvMbsBYDunZ7J+vzWuibqSmhAIxsxZGpmfKltFnNXMLm+pKKJHnGQtEQQk7rDeE6hj2ONDYQTEtJpzslXBAwJdGpTqlNuWcYirc7mSzTeyJjR2oZyJCYQuQGFpBecXqbjGcyRo1KSUijC0Irf8ARQI1VsLwTEXrjRoLtFv4Qo6ImLgBsCkE0kmJp7toUtwgGdIb0n5kxG5cTYs3bWdqScwEuihNKQUiZCayxi4YyRb7NoYaZAMFNROK4XvFPtdIBswiPr/hKEkqToSQQcQJyouCRjQtWhSslpjMKeoW14uVSRAEBQkothv/AOjy4khEgJqWegkgoJRoRAQYDxv3auUzAVcgBIN1Cry1VpgNCYgsgOSjnwreOoqOvICqA1lm7m9E5LcLLNTlZq4WgQQyy4BaNsWycuAgJOlgAaGeGyCW5HiCd8ij6XKJoH1Xl5GolKpDPjAXILoDSncNqucRBLfoTSSUixi2VrBsDkASZcabQCZAGa86WdMDXkoJOxhhNR/Xetgd0KxI5jJK9LaczXxHK0AiQDXsn6/Na6JuqWAuxeSnAsPJNLJNu2d7C0BEmBmKCzjtiEpDBkiS8OaAjZFkcBB6WAHAEuASluFMAXAzciRrhJP1CnBrVVlltabTICFsohRq4PDKyGl5dzrTgigUJimYJ3Te1NwiGKeZvwXUiSCiHIhg7qTbLk0CPPFJqQugoDi7co2k0ifBRbLBBuQwn/2HnLAQEZkFpYMoIrynRsnRYoeY+QmqQVzDaYTHKaxSTAghbDJOp/1XKM7NmCgBU/x1ERcRndEKJyCJk55ooDRE8ZjcL0m6vrn8r65/Kn/k/lfXf5X1z+V9c/lfXP5ULgmwQsgxBEXsulT/AMn8r65/KD/k/lfXP5V74n8r65/K+ufyvpv8pDuQD81qYyIswrwr65/K+ufyvrn8r65/KX/k/lfXP5Wdl+FpSiFG9KMsmZj/ACk5QgQpATqqAarX1z+V9c/lfXP5U58T+V9c/lfXP5X1z+V9c/lB/wAn8rWwDkFn9/4zepKWBI9GXyry4W9AXuMEC6AAubAbH4onhUpg/wA8TYneqMH/AA+Uwf6Aowf8PlMH+gKMH/D5TB/oCjB/rRRzqw1W2BgIy+HMmTJkyZMmTJiXaQjOJsyWcGO7MmTJhMrxmBPdRgcqAO6SZMmTH9RpVFuowrRaO5MmTGKmBy5BixMq8FeM2O7MmTJhwR7NsJkTIN5vfwBFh1JlmVmMw1MsQy+GZkyZMmTJkyZMTFph19kYuTOUF32PDmTJkyZMmTJkycRH0KPBBJrQB3iTJkyZMmCqSSFFmIjeUVhTgNXxJkyZMmTJkyZMmDToVF9J7yCCSiBQJjH5k/4Dn5O48bIi6LtWHm+Hk5fhlcPM/fisfbfmSGDBKroGaEmJfzKko0luQAK0C9IgglFXiquvaoO5rqXFiug/il3kCknIicicV0H8UuJAnVmGzy74IhidOInGM10H8U1wjATBkXnQxHGoR6Pypl1aUhhJLeffFEMTJxE4xkroP4oOZvUpfJFnB5OtdB/FPIRFlPAcqHB6PlUh0fpR95G+M4zBbSug/inTCS0VJM8qgoi1EwOQ410H8VDYB5EeiJxc48K6D+KA86BFiW7wO4L+QiEijeNyug/iglmGELEtir4mvn/zroP4q50fpSVoahSPDvQaZhsTGldB/FKHo/KjJMkgl4tIltK6D+Km6P2oiGEnxVLzK2zGldB/FMYiuZ3+VLyiGBmyENkroP4roP4oMSBO7JshRlGNq6D+K6D+K6D+KUjIddqGJrTCFsWkcZNa6D+K6D+K6j+K6D+K6D+Ks+xHRmM8vxeg7u7r6Hu0JTFOyDkuZJrWGU4Hw11QDKlAW10AUmFJgTdrqe7v9K2pDkAQgjkZC2PhRiz6IJxVl4k0vTh5V1rb3+hbUCTrIrJmVWIAaXpQ1EmoIFugDIaUmpFwt7GkRoZ7JfbFSxjWNQRpmIIK3CoVuGCbYpWCjDgiUyiDcPLjSIcGa6Xuq2oDwt6ICAtYwZuTrR3OnFY71CbyBYlh1f0YkIQ+aE9wo0QsC5ZWlSjeDdqQ0swmyaApS4MNL0MwTDIszGYF7KUhSJBdEiWzCNo1mi8NoEZCBA9hcvS5poYjITSDKaV1Hbve/wD70eHaaITJuwWAKQKLViImHj6wASQSnqo9UOtAjgssMXtDP5geJEsVLBk3revaP141p0O3iNBLhoIi/RpvRnCxORsAbCM0LyDJV5GJRGawVGoKCJIL8MmihThromx3GmHEc451wMTvDSBbqEeoFRQthW8QlCnTT8AxCkapNOfpD9AqMXkkSzFQOmz6vrBUS4oYmQmzhuBMPS8odSr/AELCpTSCCIMQm4lXrNdiFlDnxGY2mlSTYENNhM4i05aP/hk3tGoiizcH8fZ0Hd3dfQ92hv565jAChpJUPwKyvNAITK6JII8VPEokEjZcN7ta6nu7/StqNwJkEfWW4JWlvilcmMQIITJugzanDyrrW3v9C2pI14rmbYoQMWvVuDUwwAXASsrdutOHz7JRPIKB+Ie9SRmBQwBZqUcuJzySJgVMXSnLzrpe6r6g4RYbLRCJN1ib1GZe6hEZDDcE2tN9xTWQ8l5lgGyh3DjTiJoEsRQQoZhUsLwhQgYhAOEzR+dvCEjlq6yppaM1J1kMsDK8jMlOb6opqEieSGdErT4MNDFgm6wXVgmuo7d73/8AelkcRC9hvUmHgwxV206EAABYlplPFTBAFEMm2HULrMQ0O4F7IQXYsbS3revaP141tncSZLAeKgcRWat+ATJi8a9lzilQnqDmWilqG1pqNgEC8SS3gUNpacNdE2O40LN8c2AVYOLRWsZCFxDoRz0oO086yb0L88UEjd5cAbmXRAi9GfMui/JFCUiYFihEMD5kMSwKlcLtyQAFAYBWV9VebTbXSdTegqNOmmtQWpCmRDmocoQoGAWD8fZ0Hd3ddmmibBvdpTKVpjFAiJDkVHl2Fr0YBALS5we+jGZCjJDOcNMG1W9xdoCOLkyOLtTC7VYIAhGYQDfvkyzI0ZIZ5NdF/NBGilrlXGXyFWer9aAeGcEYonnrQSvQVrZTnSraCcR55A8TsbOYKlMBHHOpau0xIGHlUzNym9L8Tg/uomaEiRCASwscXtaVhQhlNLefYWtR4yDMBdpfsFG91frR2C2kCcTvKiqkKycPClspSglSl+BdAJO+kjfXtbbZrIB9PFdNNo1u14djbBpLS7wiJSUuNy49jaolWtnBH07jSZYRcKr3G220hauyHHAgJOSTOvebbbbbYrEnNGWOZ+L0Hd+Dr8uHxBwClLFzdD04n6na2ozIjrk9if0Zc90KToXKxRqCZ0x4HRdqw83w8nL8Mrh5n78Vj7b8WQZ4S48xcXAUGgUopHUwoAEBYsJ4UhNi4YCNpW45YihQBZq4kSXgXoWKYIptIBMSuqbxSDfTYwIwzpDekmhEKbOMsxOsRS6nV+DrcuHxB1glAJKq/OhW5p7thFTjI3sNl7W7qPNFMScXHOdWngdF2rDze8sVwnrXCetcJ61wnrUpZjekZaWTcqTcqTcqTcqTcqTc8TDzP34rH234shg0wygSEwxmMvAzQCTQR8NFEiZZAgbYnzdAQ2FylNbca+7NEIx3wsiTSmlaJ+DsBROCrNva2hVASGKdkrR9wx05jDnhqJaFmxlhXcPLjF6EBeDJ+BuXD4g7F7A0zpTfIW0adxoEhybiw4FOs6Y7qw0bw3+1txzbd/ou1Yeb3oN/FSUEiSINTO+g+VSNcjXI1xDXF00hLUIQlZsHelSpUqUsNVSDEMGELNqUCUl3YPDw8z9+Kx9t48gQnEEQmEQcYajxoyeYsnER2RG/dQck0g6VA6Ug5KQ6Uh0KgmYvXQd34Oty4fEHMMggguz0X59/KO40lFAEBq/2d792cqfpbzEKW3q8Dou1Yeb3yntX68Q31zY8PDzP34rH23jyEHJQPkkiBtMEkarZVfD6Du/B1uXD4g4LozBvYxax3nPkdrRcRGDDdgMIxw1z3WJMf0C8cagz4Dou1Yeb2AslOolERDtX26n26n26jbRYSn2ITEzE1j4gZU+3U+3U+3UfMPjJmcCMUUlISUg45q+3U+3U+3U+3U+3UXChQitrKJmKNChSVeCKfbqfbqfbqfbqfbqEAbyyQTE93DzP34rH235khp0Hd+DrcuHxB1z5AED4tjHFssr3Gt4Njh94IwRBbu4In4w3F52OHgdF2rDzeyyk+OB0sh5A7ElTx73pDggNuCqUmXxXMtqDABEM4oj6ggAu6WA2IlQcI7YTvRlmJExUnvsULnSgk5BhQbbsHCXJkGykCVkkJEQSKgxMEIvJyK9tTbXUTqcoZDVcU3XQwDZC4qINZdCcJFXQcMKCQC3WL4qbqT9S2vEtQgCTK75RiYmhQQmSRWCVF3FgvShg4YC7DSC+JcDT3B34UjENTJMlMmlTUysYgBdAxJOShzKpjGHFEiNpTBUSFkAAJAzujYBJESr4h1ieF2Ii4LZFdR2d3DzP34rH235khpd0N34Oty4fEHZ5xlaajurI/buN6lQjjNh164DyYMd1RZN9CjmnQqI18Dou1Yeb2GRo5+qkk3x2JBxi1qjqbQl0mBmLkASiGuWmaAhZus2pe85EiPcRCOstdvExIIEpuZ3ILNqk9julbjICFcCFELee2cMCKERII0mpxkFrGi1gIJMle2pvrqeCrjLbhbAXgLWpOup10tZJhJByDLiwgp0DmFgjDvUfg4djFxEWnAFTZRQOpPdSmYd70Syd6gEoQSINrIiA2dkmCEWETsgZCi9abSRWIBJNYDa4zvCMKSdgFKUzITE069SEQQKmTALQuKRHBvslTJAiQMhNhoBG1YASK5EzrE11HZ3cPM/fisfbfmSGgqAhCOGim5w6REYSDACKiOtdp5MDRNTx3Lh8QcZkXUtkcb3C/LHccUHkbU0T9xute/dBhcdJWc2XGPV4HRdqw83sFNso7sKzMm9fVafVafVaJ5zbaIRSMR3pEwnG1fVafVafVaEmKE31zizmnIkYBgDPJX1Wn1Wn1Wn1Wn1WhBpMDidRlxeVDc7bT6rT6rT6rT6rT6rQgstCLETHdw8z9+Kx9t+bIaIcg0EeO5cPiDnCuUGElhbAM9w1P/pQ4KwywcO6ZSuSJJ+GknPJv3+i7Vh5vew8z917V4Z7mth5v78A8xwGV2lS/aMGMYGVCONQumtId1DBgwYszYTnIGXv+2/zZDRy4fEHPD9mxBAm0RdrXdZ7jRisR3hqL6EJptbusa0xyeYiw2Z9HgdF2rDze9h5n7r2rwz3NbDzf34CN8ieSNgJPc+fPiz3efPnz4I8CFMgwg6te273tv8AJkNHJOKaZIC/FE/mYgGBJXhJu9oADA0DA0JdgTXaGbZ7gAADVYCanQCBYGLlGSQQHNye8cElGJISXEjaMPcabQIKikbi2Q/O0d2KtLPZN9FkRHu8Dou1Yeb3JLdDoSTaMgMp2ALRMEjRJopcFKyDxdgCpsLypQWZijfD3DaFEUcCaK3MGE0AQMJN3uAAASEbKY0rQqJAhLfm7QABE7jSgoZS6JMZoZ8b2A9t3vbf5Mho56xv7kdw94dj6bs71nEXBOW3OoeyO41YxEIYI9ncve/dSHC0MmxTn2r218Dou1Yeb4sqr0DZ7vf3NbDzf33MmHm/vxvYD23e9t+ZIaLFWk/PdKPmRkiMAkliZE4y6JUJ0ljd8BygFbReuC6uFcN1cKKMCAvM+FNUI8KJRIs0EwIMy+VD9LIUyeRUtZjqtSQ0MC1yWuSN+DXDdXCiX6cJ11KGM0EjbyKYc18UEAi7QiQGmKzhUOiiBEDzO23lUwXLGlcN1cKOSAOUNsSQzrckUfoYMy407LNTCi38q4bq4U/RjCTFBa0xHOq80C18QE+3Y10skYCuG6uFOBW2xoiEyIGLd6t4dXCjJJRI97ou1Yeb2EtfEhHoVw3Vwp6bEsKFWxC864bq4Vw3Vwpp5Am4k1Do6uFPZAqQYBY1hjk1w3VwqFlbCJKms2FCy+VD1Qg1oeFIuRblAIMiJR1RMQJg5VjakkW7yrhurhSimT+WEZNhfKg0Qy9UVw3VwpuIlAi3mdmTDzf3RFRETEnlXDdXCuG6uFMpcwgok2tdiuG6uFcN1cK4bq4Vmn+BJoSoiJiTyrhurhXDdXCuG6uFSp3nEEi2wb0WQY6rVw3VwrhurhXDdXCg8lwgSHz7PbfmSGiPECic1EMAEAeDufaP6qOzj1/AQlFEtHNXvilkVk2LqgLTgDGG8RKQTIiCNKw0UxiWEyIN2qCC2Q08nZpeu3wjaLNlDMXxT7I5MpFuxiIlilGROZXvP2UoXh/bVgf9IwZdAXhkeFE/iu4+kkECRAUNdH3e3rOzt6bsor279u33X9qMV0XdRjscPKvZ/wBd7ou1YebTiruT/SoeYuyiKFuUmsZJo/gjsCgK2jDATISVLBQl84UBdVEsCq/dtkKSG5Zw3K6vsqHbjuqEgTfVqTYbIZduqbzp3kMYAUyMYtfHaQgtm7YmWF5uHEQl6mZI2xOjCQlaUMtOYMP7VYunNFbFExky6BaWUp+cHBBAgAzN5tQ4l4sQCFAgvrKvFX7mHC5SgBNxkMV1Dc7MmHm/uhJ7l7qF3duMET8QckxM07ZanKshLFoEEs1EkhkyRQxTCgAA3ltRunSrkXMRDUMCI8SK92/arwmNd0uq5UWRiiMMhfcSoQfWwAmLTAxQ0oQBeSrNyLaEoVMUiBoqiq3BGKRyE5kVDqh4DKkCedJNUAw4TSBKgdUt20e2/MkNOg7vC1ufaP6oIxYyaNqqgc6urWAQgLC9kwVe1M4cQEKguAwDURO5w0ugijunlUjkYE7Ydmi04kkummCzigzZ0OaKy2+HQo9shrzbEyEsWQMPefsq/l/207cgNFnoAquJNLnhKUlijaZuMh6fu9vWdnb03ZRXt37dvuv7UYrou6jHY4eVez/rvdF2rDzacV7H+lIyw8GCguInMMtFIhEKgclxTdaTq3M6VWKsu1IDag2X2mkcMASwiForq+ypK/l5iNQu1RjYizxq5x0hbUdtBkWwm9sjCjk7SEEKFFo0jKZfervSZkJxiR3GZgoeB/VWLpzW7X+LaUQbUiyOGCqAhi26JwNNvkuBqsQDpsFmlPtwUQTKSxCkBkHqG52ZMPN/dW0AnPgthRcsmLsRdTrFZyu1SCgIRsLPsJ0vlGKKGEEFEjevxctNBTUQFLYSMSJaQKQCZhNAZBYZBFVVbqt1r3b9qWdkBSrAObWUuXp9yJnF0AkS8UtgdER3CEUa0KwpjsCARK2mGEeeBOoBLRpJgQxTlWlZuDobFgDLkB/3z5K6k4mEKVIDupFiQhznigBQSOd7YRNRSQAKL20e2/MkNOg7vC1uQiLsNdSfNR/3/qosH0b1J/X+qSRcGwHnTvBCQQzxaVfl/qr2z0Zr7X+qf0chLXcaEcykTMu7V/5f6q98v9UBEIkMwuz2ogcgReyuqPmuqPmkJXDYBONFNLEpAgh3rqj5rqj5plNlEtZ40YqHA4NVKIus9a6o+aYnpPOkMQgE8u90XasPNpxQSLIN1jjwq/N3n/VAth5P9V1J80oJ1nOp9QC0YUtMuc/6oHA8n+qC/r/VKWLRo0iBOhBFuNLt15v9V9j/AFRm2CktPNQp81BC196VLrzf6qzFnaf6rqj5r7H+quAlCZk2ezJh5v7pKNIS4K8aJZu8/wCqm0tES45CwDI8KA4PRmvtf6rL+3+qOrP3Sc1hItdprNAcgrxr7n+qs+Tn+q+5/qlBmHS5UShSWdaPMciDAX0AK+1/qvtf6oL+v9UqBOCjJHDs9t+ZIadB3eHrcxUVFRUdkVFRUdkVHdioqO2Kio7YqKjv9F2rDzeyKioqKjsioqO2Kio7IqKio7cmHm/uoqKioqKio7IqKioqKioqKjt9t+BIy9CkRR0ILJMEoS4ulT3hBCwzgIE2pPC6Du7+twt+VRDj6NQ4+jUOPo1Dj6NQ4+jUOPo1Dj6NQ4+jUOPo1Dj6NQ4+jUOPo1Dj6NDO/p+NgqHH0ahx9GocfRpyQPJ4UAa5dKhx9GocfRqHH0ahx9GiW/p+Fkw839+PA39Khx9GocfRqHH0ahx9Gi1sbNKJB5VEOPo1Dj6NQ4+jUOPo1Dj6NQ4+jUOPp4CgcInEFD1XrROSDFnJiYzEtKp1Vqi3+vyVBUvdSPybEkYHiASCjeR3/SKSapHgFkoloq3SlZcMwNJBLapp8/g5IkC2wiCQO7QbCoYDDSFBEyHezEcMS6TCbHp4zhw4cOHDhw4cOHDhGZdSIgKxMJjMfjSvzIIDAwiJZ0oUudfU7ThwYoYVGyMEqdld37ocOHDgNR1ioHAoYVjaX8LJh5v78d+zYyAnAABoB3nDhw4clgwwSRlIuVuuvfcOHDhw4tWnKZCNQiiNe08FpEd5wVO9ubhxE2uvEUeBUo4PER37rpsgWbuCY4yoRhYloYjoJcAWZdlDOUweJyh1f5etzV6XuVh5v7/FmWwmCxWHX+q6frXT9a6frSAzNTtaDBwedEfP1xXT9a6frXT9amtR4R+TsR9t+A0Dt6Du7NZQkMEt5d6iJQDhbntUPVe1ddfFdNfFGpTJiFtGZd9orpr4qXW5fyqDT1cK6a+K6a+KBtZgS5G1OKEO4EGLFTCgQy2D2rpr4rpr4rpr4rpr4pgBKAgBAi2U1018V018VzyrgY3/APKi4hkDCV28bCS5QcgxQyweSfFc26tKf6utYdmLYpHQ1irpr4phdrqtTpJAr5UNFaiW8u9XYGdj+VRKIEyMh7V018V018U7q2AgTIi+l50y3Lh/5VCTa3/8q6a+K6a+KaawgmDh3/eP1TZoDgGHEAnKoMJAppSDryBdUBaU6h8XwgQQsSQSrCLiESctqmSwIvjwxBFERUWZzJaj8iSkCSI7R3M3TjuFzQpQ2h35V018V018V018V018V018V018UhAkZNqbYlg0K6a+KSiQTib3tXTXxXTXxXTXxUCJKgjVKJ0ESQDUxMIxNj9dmXTXxVmbXVpXTXxTKRr/ACStcnJQYGLqxXTXxXTXxXTXxXTXxXTXxR8wYQFoNvFadB3dmsgJsiEw3hjXlSrJZ7mg4YS9CQrEOGaMyDm70pRJMVuVqSSLiWSrcxQsB5AYzqUgzcYSkTdrlMzmlTAQDDHiMb4gkQLhIGb0RZTQAm0CixgIyKiKCo4rAEocVYyqCR207tSAQiABMjIx2E1IkZ2ZqoX9finFezfpQxMMyFLiaQRBAUMayEEcAXhmgwMwoBO/+kOIFwoUokiYk/vA4SAIsC2lDnMTcHgKpFcS7Hx/G5CCINAqtoRF48WQQeRcwEBDRQhiN0ArDwL17WrvNLisgJJgb4ltaNJQmZGUACV6BXT93xum4KvHUiEJQ0AinGrRRPyfHTEBMhpCr1/oooZIFhACkxQJaOpLtyGhgCrYMOD7UR8YFhAAlTfLM8aGCBuycJc10rasPRdpZS8/IKAKUGBWJGzIAwUub2YySiY0YZPucbLIqF2TCaAlsd3FyauSkMJYlYFt55xpTFgpBS224LGHmgAlysikqWKQIIBkBZkimDDagXgUjVySQSrVWhAcCRBuEJhm3f8AveP1UR9E50JWkFE2rCyy6QyZERgQ5qYiFxonJMrdS04Axb1aZUSL+yMwKObFYMFxJsyo1C0EEGm3czdOO51bZpAUtIKuYIDpLIUBN5ix6AM1QQCbKjBGiEQwhCMIIkC6DkAmnhrSvlJxTmmXBlsWbeUxIJRjshv1A0KoMqEw0CFFsnA7iiuxkS8rUkHEoiiUFUJCLVOd2odhcrD3hgBQNv1qOSkuhSBgkOKPlkGXSJeCBORTzBejJCC8kCRLDavfv2rrm9NApP4YPRhgURAqseNB2YpEhEAZS0kF6b/JgWT7lAGpo1RfWoLLUTALS1JCgQLBMESXiMwGFElVoIkoRBAJEGmE8LQgEhJot4GJq4PaTs6eNuqEpZPUdgwd0aBbUoTTdB4q/mYJYNm1RUyNNAMFZxNlNikhIoZgREzVeqRda6NseK06Du7NZuwJLAMLJceJco13B0JhE0qyG90go/fF1msIqwQWWXO4CVkE4Euq1rU3eAWZ60iVhFclR31KMYBwWCCKRh2VFgRpYhHMMiEJp5cFIu4b+AjotWPuP5ts4AA3qfEBXrKuCeJLIiUOBaNKt6JixOli6V1fhTihPLfpSFmW90vWpmMWJTFc7IQYLCDJXlc1ExycxEYQqSB0inns3dKkgSEMpEOKjUOy0SAsMlxKCLVmrukhDEiZzmSZrEa6iUQizEyQDFqvp2sSMuxoLbW16ES9yI41QJlCkQQSVAEbXLJIHWZyZm0dP3fG6bgoRQZQAgYmiONEcbkCW2EkKCUQhhK53dHQOuyAE60Xj7xbO+A6ruYuk8vMw1mi1sgInK4WN1uUkYAmJYlVrpW1Yei7QdhiQECLDDAJJBABgikq8ZoA6rCjymMEFbSQgbqiMUL3M6ORQwbIlzRUu4xEiYeoQjmXRY0AQ4rCskCOZlY2zM9qszikzkYIkLyC+uSZCAOJp9EbCAotIhAX1v4H3vH6phyhnw+TE2TFprUZzZJxAAKsgQl/iuQhzPVEEpNhZzNtfIFoTSCYIcMykgF3VLIkhBYw7mPIoBAyrN2YKa9xUxYYKSBYCglCbORl/wBy7xjcwBXQ4QgAgJBnEzWbpx3OrbNO6T+BJIRcwrXh0oqqc6WnXCyt1YkZKfv6YNiEpc5uLEpsknniQkG0UCVzC75G6DBg70CdHWP8RhVwViiyJituEaAkKSIuZjYQ5EBAQZxZIBihpjpTiZECiysQl8rxbG6kk4rMiIiIl+JDLsamNZGq14gU9uAhgiRBoate/ftXRt6WoOevmF5AhZAB2eAcK4uQKhaONWDPK05S9RLiGtDOX2zLJKwVKnDFLcDraMYzAyoCXdKpavYE1KhMmSJGIhY4YJzMZE5lkMBYWgbuNXUFWWg3firDayAkCyBhq07UL8EGIZKBitMLPyShMcFRT2BlERYoABCtVro2x4rQSRl/apSlGYLBJCx9+G0010000000eDBkjZ7GAXKEG2PDaaaaaaaaaadkEJOHPxilYcgw2Zz2tNIv5mBnhCjRm1MMKBPZNW6ocCAKsmxJSx9+E000000000GKkFEvm3f94/VI+FIyXkMSEsNqKuxETmmRCpdcbALRz20T15uEYQISanSeVE5CQGmUd2ryRaxFwRYQZ6BCR1q/7zeQIXZbjUSK2MjkSEJYjdSrGhm+DEkQELaxaKa1fLuQJxAATBTldOO4SIkEL9PfaaaaaaXsqrAVtsBOue800000OCOAsuZzTLKEEXfFaaaaaaaaaaaaZKEgiz58vFaCSKGpZJek5QsEYCQoaeWYkLyLMYkt/kiTcUSmOUYiF4aAJTC/4qSQ4r6fX0+vp9fT6+n19Pr6fX0+vp9Qx5oEf8vsaXVX2lpKXIUZL2oIlKF3/FWKj7YqiDBowjHE7kEEC6q7DEh0UI6iNdZ/NdZ/NFoFQ+pgGWAV4H/EtMt0iuItRtM5MVcyYIn/ABfbUJomHH1qHH1qHH1qOfrUVH/Fbmh2VSiEU5gsM3dKAgyAE/4vtv8AHE7mkXCoRaWYSElIpfTwKSJLIJbR/wAmMPyoTsmRCCW5pQAFJBD/AIvtvzxO5pFwK3cEsZPWkGxmqAKeQEYQYR1KJbctb4FDRxVIk9w1hCsNYJq70ilutXYAsAEBRY/yJwgUhfm8KeoIppoGbMOaFCAI6WmWQCUZuxDw/wAM3AgGIgZl40lGU6b0csk/gFN7Mu3KrfS+tAeIAYe1rWgvBkG0vGutfmlIsRIil2ZLfQa61+a61+aRtn0Akw+v4jRBt54I4LFEvNaGX3jHA4ACCJ1AKdGnjlGJgx7FBGIbHZiHIwkkQMJCIMN2odEKuzd4wYe1JTLPSspQkkUJ5VZXAfNBUC4YnLLdW/qk21pSpr/kn1vZ7eqbf4br+1JZN3GzYcWM7zV8oQ+berFlgBQuzG9dW2Oxx2OhwbCUVTOoJTE4ry9CRGp2ElAA3Tkk+hIcgAgMwdaLKxWWLhAhxgnaut7PxGkPvBGmYEiW1KVHAVnCtwZBlJkGlk9saxYpABkQXtTSNFJiBIQTJEzfNQiYmojhJUSAZKVecJPGDMsAM7g5Vo8+cqwJTBN7wTMUJ6m7/K19b2e3qm3+G6/tQN/SDgCAkEaXrCchMgjr7zaGIp1rq2x2OOx0dwIC8QlAQT1VB0BwFIijbnMkTUaW8oKKEWRsm9fC8H6aRItyYZvDjFdb2eODTYqGfNH0dvEJhBHI60flFeT4V02UAIrQDkawSTCXW43P8jW6YwJnBxr6v+a+r/mpPKdy0HM8/wDDTSpuXgREcq+r/mvq/wCa+r/mkdZQFOB2NB6MgggF4OFfV/zX1f8ANfV/zTF8f80HzH4UAp40/QoGyVsw8RbFFKN4SZckhQoWKB4oQ5BrH/6OILA/MoN0TmHlVwgmxnUrV0FpMUbogwUhwTVAFOrTEUKc1VBJISh1wu8aZVyXFdNKR9iBqMhZptle1R604kS2YJcsVLIQERVEExCzcorJlQMIB4F+FAzAnIg22SOBmGlxFr4wSu1heYiKDoUtBQxhgiSiHanyuNzvOMIi42Sh/qosCNIchC2qbURaXGAPPTeJEL3o6db5C1IBhEqzm1ElwhZg0LDYvWGG3QiSRsi8rxVnECCkxfZfbKifBA/3Z0HBpR4nNqAm7lK12C96dgASW5PmadRkYRLRYLLYdWl+GC1OCJK5jevci9W7PkH/AJ8uoK1EgkBjKlXv3pNpJKd3qDDMkYUw6smbKgUy0QgCh9kSQItSmEVFzePtPGQJHXKQXZ24vJpXwCUbiYob+5mKgqnKQqYJYp+opkT7cgCSArpRYjj8xKLilKiF2xIEtIhgJgzdKaT1dj+k3RlhAIp/9oJ/xQGFCZoY0pIADcWMYNCozeSwFuRVkesl4MmSjALIbMUFhJg9MECzeQTN1S2JMkB+7AZCqNqhLAD4dTCMOTeJLo4HOA4S3KMy6CYa1VgL03gBkSlXaZcla3szMEgABZFqDWbUxSrEWnCFLVLCzqdrskERINKlfxoSUSlzBDNCRQ3Af+sWLBeIFaZ9IDUeLx/6GO7EdmvbCXCZXJu4BeEavjf/2Q==)
![[[@Boldog2018-dj]](http://doi.org/10.1038/s41593-018-0205-2)](data:image/jpeg;base64,/9j/4AAQSkZJRgABAQEASABIAAD/4QAYRXhpZgAASUkqAAgAAAAAAAAAAAAAAP/hA3NodHRwOi8vbnMuYWRvYmUuY29tL3hhcC8xLjAvADw/eHBhY2tldCBiZWdpbj0i77u/IiBpZD0iVzVNME1wQ2VoaUh6cmVTek5UY3prYzlkIj8+IDx4OnhtcG1ldGEgeG1sbnM6eD0iYWRvYmU6bnM6bWV0YS8iIHg6eG1wdGs9IkFkb2JlIFhNUCBDb3JlIDUuNi1jMTQwIDc5LjE2MDQ1MSwgMjAxNy8wNS8wNi0wMTowODoyMSAgICAgICAgIj4gPHJkZjpSREYgeG1sbnM6cmRmPSJodHRwOi8vd3d3LnczLm9yZy8xOTk5LzAyLzIyLXJkZi1zeW50YXgtbnMjIj4gPHJkZjpEZXNjcmlwdGlvbiByZGY6YWJvdXQ9IiIgeG1sbnM6eG1wTU09Imh0dHA6Ly9ucy5hZG9iZS5jb20veGFwLzEuMC9tbS8iIHhtbG5zOnN0UmVmPSJodHRwOi8vbnMuYWRvYmUuY29tL3hhcC8xLjAvc1R5cGUvUmVzb3VyY2VSZWYjIiB4bWxuczp4bXA9Imh0dHA6Ly9ucy5hZG9iZS5jb20veGFwLzEuMC8iIHhtcE1NOk9yaWdpbmFsRG9jdW1lbnRJRD0iQzhDQTUxRTNGNDc4QUNFRDQ4NzU2MTdGNTVBOEMzRjkiIHhtcE1NOkRvY3VtZW50SUQ9InhtcC5kaWQ6Q0RGQjhCQkJBMDA2MTFFOEE0RkFBRjdEQ0ZBM0E2MUEiIHhtcE1NOkluc3RhbmNlSUQ9InhtcC5paWQ6Q0RGQjhCQkFBMDA2MTFFOEE0RkFBRjdEQ0ZBM0E2MUEiIHhtcDpDcmVhdG9yVG9vbD0iQWRvYmUgUGhvdG9zaG9wIENDIDIwMTggTWFjaW50b3NoIj4gPHhtcE1NOkRlcml2ZWRGcm9tIHN0UmVmOmluc3RhbmNlSUQ9InhtcC5paWQ6ZTY2NzhhNTItNWE2ZS00NjUxLTlkYjQtNTJmNjYwMzFhZWEwIiBzdFJlZjpkb2N1bWVudElEPSJ4bXAuZGlkOmU2Njc4YTUyLTVhNmUtNDY1MS05ZGI0LTUyZjY2MDMxYWVhMCIvPiA8L3JkZjpEZXNjcmlwdGlvbj4gPC9yZGY6UkRGPiA8L3g6eG1wbWV0YT4gPD94cGFja2V0IGVuZD0iciI/Pv/bAEMABgQFBgUEBgYFBgcHBggKEAoKCQkKFA4PDBAXFBgYFxQWFhodJR8aGyMcFhYgLCAjJicpKikZHy0wLSgwJSgpKP/bAEMBBwcHCggKEwoKEygaFhooKCgoKCgoKCgoKCgoKCgoKCgoKCgoKCgoKCgoKCgoKCgoKCgoKCgoKCgoKCgoKCgoKP/AABEIAYkCuwMBEQACEQEDEQH/xAAcAAEAAgMBAQEAAAAAAAAAAAAABQYDBAcBAgj/xABSEAABAwMDAwMCBAMEBAkKBAcBAgMEAAURBhIhEzFBByJRFGEVMnGBI0KRFlJioQgkM7ElNDVDcnOCwfA2N1NjdHWS0eHxFyais0Vkg7K0wtL/xAAbAQEAAwEBAQEAAAAAAAAAAAAAAgMEBQEGB//EAEIRAAEEAQIEBAQEBAUDBAICAwEAAgMRBCExBRJBURMiYXEygZGhFLHB8CNCUtEGM2Lh8RUkcjSCkrI1ohZDU8LS/9oADAMBAAIRAxEAPwD9U0RKIlESiJREoiURKIlESiJREoiURKIlEXhNEVU1BJlXO6/2ftrqou5jrzJaU5Uy0olKUo8BayFYJ/KEk98VW8knkC1QBkcf4h+utAdz3PoPurJDitQ4zMeMgIZaQltCB2SkDAH9BVgFLM5xcS52pK2KLxKIlETNEXm4fNKXgNrwuJxmi9te5FEXtESiL5WcDNF4TS5DqZ1rXmrLXFt78VMCLJfiKlpAccLoaDh2YI2gbQAonk54I745B47hynQL6bFJ4XjvdIDzvDSBsKuhe932VkHp5HEGaym96gTIlLLjkkT1BW44ydowjx5H/dU/w7QC2zr6rA7jMjpGv8JlNFAcor+6vDCOm0hG5StqQNyjkn7k/NaBpouU48xtZK9XiURKIlESiJRFjkvIjx3HnVBLbaSpSj4AGSaE1qvWtLyGt3KrGktd2PVUlTFpkOLeQ31VIW0UFKdxT585H+Y/aqKeOWww2uhm8Jy8AB2Q2gTW4Ou/RWrcPmrVzl6DmiJmiKgavhTtQ6rt1paQgWmGpqdMWJC2nFZ6gSlO3uMp5/bkVmlaZXhg2GpXa4fKzDxn5B+N1tboCBVWTfXsrLaNOWy1yFSIUUIfWjplxTi1nbnOBuJwM47fAq5rA3ULn5OdPkgNldYHoB+QCmqmsq8Xyk4ovCobTdjFmNwKZUqQZkpcpXXXu2FWPanjtxUGR8l63a1ZOUcnkLmhvKANPTqpqprMlESiJREoi+VjcMUC8IsKsOaPiszJcy0y59rky19V8xXRscX/AHihYUnP7VV4LQSW6Wt54hI9jY5gHhooX0HYEarNp+fNauUq1XlbbkppAdYkNp2iQ0SRnb/KpJwFAccpIxnAm0keV2/5queOMsE0WjTpXY9r6g9Ou6sQOe1SWW1ilLW2y4ptHUWASE5xuOOBk9q816IBZAVB0PMVeLu5Pv8A9XAv6ApLdredKEsMKAwUI46gOMlZHCgRxiqInlxt2juy62fAIIxHjkPj/qA3PY9q6DsrheLPBvEdLVwYDoQre2oEpW2odlJUCCk/cGrnND9CudDO/HdzRmj++h0UFH03cbJEab09dXOkyNqIc9IdZI+6gA4DnJ3ZPJPFRDHN0afqtDsqKVxM7LJ6t0I+W329ltaQ1I7dlTI1ziIt10iuqQuGXd6ijPtcGQMpVzgjjikcnN8Qopm4bcctdE7nYRvVa9vcKzBQParFhBte0XqURKIlESiJREoiURKIlESiJREoiURKIlESiJREoiURKIlESiJREoiURKIlESiJREoiURKIlESiJREoiURKIlESiLSvMwW+1zJq/wAkdlbyv0Skn/uqL3cjS4qyGEzSNibu4gfVVP0ntsSPppq5xlSXH7sBMkrkAhRcPcAHskHOO/HOTmq8dgDOYddV0OMZD5ckxOqo/KK7D9VeauXLQnHeiLDIlMx2lOvuIbaQMqWtQSlI+ST2ptqUaC93K3UqqXH1EsMSVEZYeeniQvplyC0X0NnaSAopzyccJGT5xjmqHZDAQ0a+y6UPCcmVjnkBvKL8xAJ9gVge1RqF9UVy26SmfTuvhtRmSEMuJTgkrKBu2jtyf6dq9Mj9KYvW4WKC4SZAsC9ASL7Xpqt5tnUdzWFyZTNmY2kBqKUyHVH5K1p2pwPAB58+KlUjvQKlsmJCBQ8R3r5R9AbP1Hsou46fbsYmXd7VV7Yc2DLrz6HEe0HA6ZSEqz/dAz8YzUHR8lvLitDMx2QG4zYGEegN+/Nd6f8AKgrZ6jSrPcp0bVkecmM2y3JbeXDDbqAtQTsWhBUkDuQSrIwQcniqRlljiJRXVdF/AW5MTJMJ4LiS0i9DQuwTRv5b7aK233XNts17t8CWHA3LjrkiSCOmhI7Zyckk8ADnJHzWh8zWODT1XJx+GTzwvmYPhIaR1s/26r50RreLqhT7X0cy3TGgF/Ty0hK1oJIC0/I4544PFeQ5DZdtPdT4jwqXA5S5wcD1abAPYnoVbFnI4q+lyyuaaEMa6a1vT7FumW1mC5vREkNhv+O5uQ48AP7yUAY5Ayo/zVlhcHPcAKpd3ibHw48PPIHlw3BugNmn2Nq5aweusfTc12wIS7c20bmWykK3kEZTg9yRkVfIXBpLRZXLw2wvna3JNMJ1PZQGiNVXK9Xe7ousdi3sRiy2iKsnrtuK3ZCycA5xkYHYjGaphldITzaenVbuI4EOJHH4Ti8m/N/KQO3X3vqr2DmtC5K9oiURKIlESiLxQyMUSrUDcNLxZUsS47si3zQnZ9RCX01lOSdqhgpUMkn3A4JOO9QdG0nmGhWmLLkjaYzTmnoRdHuOqgtRM6osFuXMgXdNwhRlh95uTFSqSWU8rSlSdqVHGcZAPHeq5DIwW02At2KcLJf4crC1ztAQfLZ2JBs/f5KQ01rzT97gxn2LjHZdfTkRpDqEOpOcYKc1OOZsjeYLPl8MycWV0Tm83L1Go91YWJ8aSHvpn23ukrY501hWxWM4OOxwRxUwQdlicxzK5gRaq+g7hFvU7UV1iL6iXJv0qVYI9jLaQOD8lSz+9VxODyXD90t3EIXY7Y4HCjy3/wDLX8qVzHarVzwlESiJREoiURKIlESiJRENEULfbMufIiSo0x2FKi9QIeaShR2rACkkKBBBwk/qBUHt5tjRWiDIEAcHsDgehvp10UZY9UstzGbJfXERr+VFsNHITJATnqt/4FDJ+xBT3FeCUE8rtD+atkwX8hngBdGNb/pvofUK143DntVixKm+olrT+Hm9wYcl6+W5BMJcPBdBJ5GDwpJBOU4JxnHNUTsBHOB5hsunwvId4n4WR4ETz5gdvfuD2r5qJTqHUsDSQv0hmBcWJDbUlDSFlpUcOLSCgkAhSEpVnfwRg5B714JXtZzuCt/B402T+HY4tokXuDV69wT22XQ2Vh5sKBBBGQQc1o0XFFrRvNgtt5bQi5wo8npnLalp9yD/AIVd0/sai5jXaELRBlT45uJ1fvtsoNxF/wBNtH6ZKr9bm+Q2pzbMQn4BPtdx4ztUfknvA87BpqPutQ/D5TvOfDcev8v03b9x7Kw2a7RLvCTJguFbZUUkKSUqQod0qSeUqB7g81Nrg7ULHLE6J3K8f29x6KQByMipKtKIlESiJREoiURKIlESiJREoiURKIlESiJREoiURKIlESiJREoiURKIlESiJREoiURKIlESiJREoiURKIlESiKu69mxoekrsZb7TIcjONI6iwnctSDhIz3J+KqmcBGbW/hTHPzI6F04E+wItTcQf6s1nvsH+6rAsT/iKzk4FeqKqGr9Q3Fh9q2aZhCfd1qbU4FpUWY7SlfndUCMZAOByeCcVVI9w0YNVuwsaKQl+U7lYL23J7Afms0XSwlvolallfi76fc2y40lMZk/KGvJ/wASipX6V6I7Pn1XhzSxvJjDkB3o6n3P6CgrMlpKRhKQkfYYqxYTqbKxSYjUhhxl5tK2nElKkkcEHgg17voV60ljg5pohUuLbf7MaxhJYnzV22ew5GbiSHittlbeFoS3nke3eAOeEn4qgN5HgXoV1JJhlYjy5o52EGwNSDYN+xr6rPrKDHn6k0qieyHoyJLziUrztDyWtzZ+54VgeefivZGB7mhyqxJ5IIZ3RGiQNt6J1UPrmBGe11YmEJmofurL8SWqE6EqUwACCsHI2gk5OOx78AVVOAXtb3sFbuGSPbiSv8oEZa4c39XpXdS2n9FW216jNxkTpdxuSWEtsfWrSpTDY4OwADGeBnH+81NmO1r+cmz6rPlcXnng8BrAxl2eUVZ9e6kddW76qwSJEeH9TcoiC7DUhfSdQ4OxQvwft2PY8Gpyi22BZCz8PkDZmskfyscadpYr1H7rdUTSmq9V6itaZMN62Ov29xpcqGy3/GktKTnAUTsSogngY9ycZHas0MsszbFWNwu1xDh2DhzFjg7keDyuOwINE9yB9ffdT2mLu9dvUCVIYQ0m2v2xsjaSXEuNuEFDo/lWkrWMfbvVrHF8hI2r7rn5WMyDCa13xh59iCNC3uDurlepyLdaZk13bsjMrfVnthKSf+6r3O5QSuTDH4z2x9yAqUj06gTbRGlPjF+UtuY5NeCllToVvwtG4BSRkpA8DGDxWf8ADNcLO/ddr/rM0T3RtP8AC1FChp6GjV7nupyyagfRdxZdQJjMXVaOqwphSunKRzkoCvclSccpP6gkZxa2Sncj91glxB4P4nHssGhvdp9fQ9CrP1Bx96tpYrK1rjcoltiLlT5DMaMjlTrywlKf3NQc4NFuNBWxRPmeI4mlzj0ChomuNPy7i1BZuTQlOp3ttuJU2VjIAxuAznPHzzjtUBPGaFrZLwrMiYZHRktG5FGvoSs8vV1ji3ZNtfukNE0oU50y4OAnvk9gcZOCc4B+K9M0Ydyk6qpnD8p8XjMjJZdXSi7x6hWmBClSY7c64tx9oUuFFW43k4OOqBs4CgTz/nUXzsa3mGvsrsbhWRNKIXUwn+o19jr9lZbZc4lzhtSrfIakRnBlLjagQft+v28Va1wcLCwyxSQvMcgojcLaLqQsIJG45IGeePt+4/rXuirVd1bq626cbaEtSnpDrqG0xmMLdwo43be+B3/yHJqqSVrN1uw+HzZnNyaAAmzoNOl9/RTEm3RZrBZmRmH2j3Q62FpP7EVYQ1wohY43vjNscR8yq5P0TDjNuSdMMxbRdgrqNvtNYQVYwUrQOCggnjweRyKqMLRZj0K6DeJSyEMy3GSPtetehWfR8u6IckWq+pYXPiNtuKlMcIkBZV7tuAUqyk58HuO/CMuvlduFHNjhLRk49hjiRR3FVpfUa6K0jtVywJREoiURKIlESiJREoiURKIvlZp1Rc81jdHZV0abiWOfMZtE1l1+ZGSham8ALUhtOdyiQUhQHhR79qokeeYANuuq6+FjNbEXOmDS8GmmxfYk1XtasMHWNmeJbfnMQ3wkLLUpxLaiggFKhk8ggg/5HBBqzxGjQ6LF+DmItjS4emqn9wKe2fIqayGiaKpmkmbdd9L3CxKWzLgwZDtuUArOW0nKQceQkjn5H61SwtkaWHYGl1MwS4s7Mhnlc4NePQ/8hbfpfIjvaJtTcV1lxEdr6Y9M5wUHbg/fABP6580gc0xil5xiJ0ebIHdTf11Vtq5c1eKTu70XhFqs6gtEll127WAhu6Jwtxr/AJuaEjGxwf3scJX3BxnI4qDm1q3da4Zmmo59Wfceo/spey3Ji6WyPNjK3MvJ3JyMFPykjwQcgjwQamDzCws8kboXmN+4UhRRSiJREoiURKIlESiJREoiURKIlESiJREoiURKIlESiJREoiURKIlESiJREoiURKIlESiJREoiURKIlESiJRFSPVW3/XWOIer0OhcIrvX6aV9AdQJKyFcYAP8A8+M1TPHztHoQV1OEZBx5nEC+ZrhXewrj1A2jK1BP3PFXLlA60FVIl2vd+kzHbI7bWbUy50WX32VvCUQBuWkpUkBIOUjvkgnNVBz3k8uy6D4seBjWzAl51IGnL6HQ69VtaO09Ks4mvXW5KudxmPdV2QpvZgAYShIycJHOP1NexRlgPMbJUc7KjyXNEUfI1oqt/mfVWYDFTrVYl7XqLwnFEVG13eLdI+p08hDky9vMpcjRmUHelRJ2OhfZGwjduzxj7gGmZ4I8MakrpYOJM0ty3U2MGiTtpqRXW9q62qFHRqdqx2Sbdb/bREjOnpJmydiXnW1EIyQ3vVgoJ/Nkj4zWRplEbS4jRd6ZmC7KmiggdbgPhF0CAT1rrXopXRLDGuL1crtfm3rjEgj6SI5KioZaOSVOKSkcgggYCslIxzknFkFTuLn7DT+6o4k6ThEMcGOQ1z/M7lcSR/SCdvoNSo68X+32Ke7ctI2R1+LAQlS7lHWpxt0rO1TDilclGNpBBO1QScYJzGWYREmNt11VuBw5+ZG2HKmDS4/AegAvmHS/TS+6un9ptSIes0iVYmVWuYkre+jW5KdYTsBSTtTg5J8ZH381f4kltsaFcf8AB4ZjlDZTzsOlgAO16a36rT9EgXbBc31tvN75ziAlxBTwj2jv3+CPBGKhhm2F1dVo/wAQsEeQxlg+Vp02sj9lZnmFO+o90btk9m3XUQ2HQgtpdRKbyoFTicggg4GQUnBHfPEw3+KeU0aCq56wozLGXR8ztdqOnwn23sUtu6afu2opNtZ1EzbxBhyPqlKiyHf45CSEoLZAATk5OSrtgdzUiwvID+ipiy4sUPdjE8zgRqBpfUEde2ivOMp5q5cxRN90/bry0BOjJW6gYafT7XWTkHKFjlJyAeKiWNdurYciWD/LNA7jofcdVBxdNX9h6QTq+etpbm5pKocdSkDAGCSnnt4AqLY3A6uJ+i1SZmM4DlxgCN6Ltfuts6LtcsE3vr3lw/zT170p7flQMIT28DP3rzwmn4tfdBxGZmkH8Mf6dPqdz8ypuZaoM1pTcyIxIbUjplLraVDb8cjtwOKmWh24WSOWSI80biD6FaitMWNUAwlWmAYhzln6dG3nGeMfYc/avDGwiiNFaM3JEniiR3N3s2tq22qDa4aIluitRYyM7W2UhKRnvwK9Y0MFNFKqeaTJcXzOLnHqd1RPUCE7YZEW+acLzd1ekJjmE0UpZnLX26oyBkAK93fkc1nyAWU9m/buuvwuVuS12LlaxgE8xslgH9Nd+2y+rPZEatmm96mC27hH6kEwo7q20xgFe5KlpOXCrg5yAQRgea9bH4p55Nx0TIy/wIOPiG2GncxAN6aUCNO1dwtmal+y3iJaNF2m3B+Q2p2Y+6FZYRyELWRyvKt2ATk7SOB7hYbaQGBZ2Obkxuny5DpXKO/cDoK70rxES4lhtLywtwJAUoJ27jjk48fpVq5nXQUFkUcd+1earwqqaYmM3LVWpZMeS2+2w4zBAQc7S2gqUP8A4nCP1SaqY4FziOmi6OVFJDjwsd/MC76mv0v2VtHYVcuelESiJREoiURKIlESiJREoi+Hc7fbjPjPavF4VXPT3pu6ThO5KnnS45IUe6nitXUyf+lkfoBUIzbQVt4gAMh7W6Abe3T7KdkRWH2ymQ2h1GCMOJChz+tWEA6ELK17mG2miub3OVbbUzK/s9I1K2uIFNIagNuSGFqQcFtHUSpvKexxjGCPGKyvcxjSW3p2XbxmT5T2tnbHR3LiGkA9SQQSeut2sGmLnd9I6Tiv3Oxb4ryFSHlxGum8wokqPWR/MTkAKGAPISBmvI3PijBc3RW5sMHEMtzYZfMKABOhAFeU/p9LVo0FIhqVeI9tdjuwmppU0WCkpT1G0uKTlPHClL/8CrYnNNhuwXO4jHKzwnTWHObrd9CWjf0CuFXLnJRF8qSFd6IotjT9tj3Ry4xo/QlO56qmlqQl0nytIO1R+5GfvUQ0A8yvfkyvjET3WBtetex3ClgMAAVJUJREoiURKIlESiJREoiURKIlESiJREoiURKIlESiJREoiURKIlESiJREoiURKIlESiJREoiURKIlESiJREoiwTIjE2M9HlNhxh5BbWg9lJIwR/Q14QCKKkxxY4PaaI1HyVaOgrAbixKdiOvlptTSW5Ehx5vBI52rJBPH/g1V+Gi3I/NdA8YzOQxtcAD1Aa0/UAK0MsNstpbaQlDaQEpSkYCQOwA8CraXNs3ayYFeolESiKsau1O3aVm3wU/V3+QyVw4KUkl0+CSOAngkkkcA1VJLy+Uau6BbsLBM58WQ8sQI5ndv7nsFWrFPeRpqcvTbUO7asbebTdAt3/nyfeCrgEJ9wATwMYGcc1MeQ0lmruq3TwM8djcolkBBLNOnQ13PW9V86gtbNk0ZOnXpUL6luebmwgpKmWXisFLSMjOFEYJwMlajgCvJGBkZLu9/NMPIdlZjI4LALeQ60S2jZPt+i0n9arv9stNvctEq3pvryY4cS824FNHPW2lJyFADacgEbs9xivBk+I1oLa5loPBhhSyPbIHmEEnQjUaN397sdlYNY35iLarlbba1LXIbj7Fux2FLaiBQxlShwnanKsfAHbIrQ99NoLlYOLzzMfKaBN67n+9nRY7/AKlt0W03Oy6ffbXcIdtW40htO9DaEJSMbvylQSc7eTUS8fC06gK2DCmc5mRMzyucB2sn7/NWLSVktun7K3CszZRDGXEkrKyvdzuJPzxU42NjbTdljzMyfLlMuQfNt9NlH2Z2NJ1xqBQdZXKYajMEJUCptG1SsHyMqUc/tUWG5HD2V87HMxITWhLj+QWbV0mZ1LXAtUxcWbJlJO5CEq/gI9zpIUCMYwM/KkjzXshOgadVXhtYOeWZttaPudtvr7AqzjsKsWRDyK8ItExXqUlESiJREoijrtZrfdBHNxityDGdD7PUGdjg7KFRcwPrmGythyJcfm8JxHMKNdQei5ZqOTe7Vd9WfQpkfSPyYLi3lTEt7EuEIX0yB7CQkJ3HsOfisb3PY94Hovo8WLFnx8YyuFgSD4T01F661d/ZdC0XbWYNnbdTa122ZIAcktOu9ZzqDj3uEkr/AFz5rVEOUbUuDmyuklIL+YDYgUPkOi+7pqWFbdQW20yi6h+elZacKD09wxhJV23HwP0+RmTnhrg3uoxYskkL527Nqx117e3VTiiCk7q9OizGiqvpZhr+0eqJTCAhC5bbJATt3KbZTuUfuSsjP2FVsFFxC3ZL7ihYTdAn2sn9Fax2FWrElESiJREoi+VK20XhNKrNa+0+47JbE1wLjvLjuAxneFpOFAYTzzVYlaeq3O4bkta1xbo4WNRsfmpqzXmBeYxftktqS2lRQooPKFf3VA8pP2ODUmuDtlmlifEeV4pSIqSrSiJRF8OdqLwqoaHhx7Pc79aIKUIityES20pVuIDyMqB/RSFYHxj7VVEGttjf3a6OdLJkNinkNkiv/j/sQrNcmHZNvfYYkuRXXEFKX2wCpsnsoAgjI+9TokVawtcGODiLHbuo3SNkXYbR9I9OfmvKcU86+6AkrWo5UQB2BOT55J5ryNpYKJtXZWQMmTnawNGwHoNt1N9NOKms6qGtYf4WI9/tjLiJkZ5lEhTCCepGLgDgWhP5wlJUocZSeR5qmW2+do10+i6GC9sgdjynykEi/wCqjWvSzp2KnbLfrdeer+HTG3yyQHEjIUgntlJwRkduOatDgdljlgkiID21alRXqrSiJREoiURKIlESiJREoiURKIlESiJREoiURKIlESiJREoiURKIlESiJREoiURKIlESiJREoiURKIlESiJREoiURKIlESiJREoiUReK7cUQrn+oYF0kayg/U3FEOJJYkxI70SP/ABElWxWxSlEjcUoUQQBjZ96pkaeYEGl1MSWFuO8cnMQQdSa0vWh2JG/dbku0v6bdTP07FQ9FDLbMi3ISApaEE4W2fLgCjkHO4ADIOK9ot1YFUJm5fkyXG9SHb16Edj3GyxagvBuStNs2uBJmRp8puQZTSSG2EtrSo7wRkHvwcYI+RioSyE8oYLsq/CxxEJnzuDC1pFdSXAjT+/ZV31ms0K229u8sIbacMkKebPDbig2tQXke5LmUpTuSQTkbs4FV5jQGc/Zb/wDD2TLLkDGNusUPQWOmxAHQ7dKWnpyPP/E2bQu4SLnari5+ITZcNsIUlTqApLTi0/lSrGSpOCcgcA8+RB9hhNg63+isznwuYcprAyRnlaD1DT8QvqNqI6dStnTF0tto9NdTM25SlxrS9IYbdWkslZXyn83Ygr2/fbnHOKQvZHE4DYEqHEcfJy+IQvlbb5A00PT29r+y6BoiHGt+mIUSHcl3NhpGxMpboc3474I4wOwHgACtMTQ1gANrh58rpch73s5CTdVVKBvVztWm9fJdmrZjfiUE5UhBK1uNLyNwSMqJSsgefaR8VAvbHLZ6hbI8efMweVjSRG6/Qcw/2191n07JnXXWNxnP2mTEtiIqI8R+Uktrc95Ushs+5IJI74/KP29YXOkLq0rRQyo4oMRkIkDnlxLgNa0AGo0Kuw7CrlzEoiURKIlESiJRF4rtRFTLm9Hia2bhyU9WNeovRcQ4yS2lSN20FWNvvSpY2nykY71USGyUf5v0XSibJNhl7DRiNjv5t/XSgVHaavy9NNrtOpmFW2I1JcZt0pZKmXmdxKElfO1QScAKxkD7Gqo5DGC2XTXRacvFZmOE+E7nJaC8AUQdLPrZ7d1v+ol5gI00Y+RLcuaehGbZb6+5Suzm1OcpSSFZH2xyRVk72hld9Fm4XiyyTh40DNTZrbpZ7/vS1G2X1As1sstqiXZFzhTChMfpPxX3VlaQAcLKcryex7nPIqsZUbWtDrHRa5uCZM80r8flc0a2HNAo/PSlr6A1JpqA1NL9xjxLnOlvSZLb7i0DJcUlJT1Me3AH/fg8UhkiBIB1K84hhZ0oY8x2xgABFHprtuV0uO+2+ylxpaHG1DhSFAg/vWq7XEc0sPK4UfVZFEAZovFAK1TCTrBnTqQtc1cZUhSkYKUYIG1XwSMnn7fIqHit5/D6rX+Cl/DHL/kBA+ZF/RSrtyhtSUx3JUdEhWAltTqQo/oM5qRIG5WcRPc3nDSR3pbDqwhBWohKUgkknAAoTW6hROypOoda2lVlmraYus6HsLSn4EZ3aoHglDwATxn8wP6GqZJmNabv6LqYfDMiSVgBa07jmcPfUFVbTd701arvZGrLdbkiCt2RGdM155La3AhKgVJXhCSSoc4ByaoifEwtawmte66mXjZszJn5MbeemkUG7WRpW+3cq7X6HJYcN+0+lLs8NjrRgr2zmhkhOfCxk7V/seDxrcDXOzf81w8d0bj+GyNGnr/Se/t3H01VgtU9i5QGJkRRUw+kLQSMHB+R3BHYjwRXrXcwtZpI3ROLHjULcqSglEXw6kLRtV+VXB/eibKo+lUCJA0dEREZS3vceUvByVK6ihkk8ngAc+AKqhYGMFLocVnfNkkyG9B9KCuNWrnpiiJREIzRFWtYaYRfG47seY7bbjHcCm50dI6qU/zIz/dOe3bOODVUsXiDQ0e624eb+FJ52h7TflO1nr7ra0rcHZsFxqapJnxHVRpO0YBWnsrHgKSUrH2VUmO5hruq8qJsbwWfC4Aj2/2Nj5KbzU1mSiJREoiURKIlESiJREoiURKIlESiJREoiURKIlESiJREoiURKIlESiJREoiURKIlESiJREoiURKIlESiJREoiURKIlESiJREoiURVnWxD7VshNAKlvz2FNYOFIDaw4tY/RKVf1x5qqUXQ9QtmE4Nc97tg038xQ+9H5KyJGE4q1Y1RrhbdQWS4F3S7UOZb5UtUmTDfUW1JUpJKihzOBuVzjHCj8E4oc2SM3HVLrQy4eTHWYXNeG01w19rHoNN9vVUXVGuZWotMT7a9ZC3LZfUuY2mQlRjx2XAd5JBG4qTtxgjjPYjOSXJ8WMtLff0C72DwVuHksmbN5SBynlPmc4fD3qjZULZLpdYHpzIlxunJkMqE1E6HKQVQl4SkNvozggt9gAR4IyM1VG97ILbuNfb0PyXQzcbHyOKsikBDT5eVzTThqSWnfR2/XrdFZkwrncdPXJ27SGghtllUVht5SxJmPo7LKslxwoUn8pG3ICcYNWtjkkaef5e/qsD8jGxpmNxr3NuI2a06cu1C79+q6rH0zMtDTDmm3YsNwNobkRVoUqM8UpCd4AO5Khj8wPuAAVngja2Pk+DT8l81JmtyS78SC7UkH+YenYj8uirDd0vj2v7c7qOC3BbtzgjoDKypD/XCmw8CRkDclAwTxux3rO17zL5xVfquo/ExmYBOM/mL9TpVctHlPyJPyXWW8bRgYravm19URKIlEXhIFEXoOe1ESiJREPaiFU/1BQxddOz4CL1Dtqmy24+67tX0UbtwJBPtJ25Se+RxVM1EEc1fouhw0mKdkhiMg1oa66fu1yzWFwtV/dj29epJNxeYUOi4bepBbC1BCnFON4yUpyrJTggfesE72S0wusj0X1XC8fKwuadkAa1w1HMDZGoFOs6nQ1r9FKMJtdpsF9t9kb+q6LKXWLlFjILvsw4lD4SAoYUke4gApV3yDWhhbGwtZr2P91z52S5WUyXJpvMfM0kga6W2ybNdOnZVKz3a9x7/bkSG35l+NxCm5LzjjzCW3k8lLe3btOSoKSoHCeOBWOJ8geOYEuvc7Uu/l4uFLjvfE4Mi5PhAAcS01RN3d7iuq6PofTf4Dru+Q33XJqX4aX3X5ASrrdRfAAx7QFB3I7HKfjNboYhHI4b3qvl+JcQ/FYULwA3lNAC9OX8ybbr6Kdv9os9ihP3eHLRp9bR3rkMD+EonAw40PasHgdt3wQate1rAXDRc/EnnyXjHc3xL6Hf5O3H5eihkPX3VMtLdqvr6bceXpsKMGGUpwPY2V7lOLOT7gdqfueKr88nwu07j9FteMbAH8SHzdGuNk+9UAB7WVo361fhtoOl1IipnSNjlqnsM9J3eFDqrWrnC0J9ylggqST2NRkZTPDG/QqzEyDJKMwAlo+Nt2K6AehOgFaFeSoEKyyLcYtuf1ezdm1fULcQiRvWjaS8HFZAJTkbMgHCQMY5crY6oF1/P5qTZpclsnM8QeHsBbRrs0gak+pHurZC0xpm72xh+NEWYT7YPTDzqErQf5Vo3AEcYKSPkVe2KNwsDdczIz8yN/JK7zN9B9lbUBLbYSgAJAwEjgAfAq07aLmk2SSqPap0addtV3S4x/prUwUxC4+E9F9LW7e6SeDySnPwkDvkClhBLi7YLqzwujZBHC65DrQOouqHpY1UXF1HbbVcA3o9Ma5Rrm0XGIUZ0NoZkAnuTw0had2QcHLfAJUagJWt/wArUHotTuHzzNd+Ntjo93HUlvy3IJ0PY76K96XguW2zxo0hYckJClOqAwN6lFasfbKjj7Yq9jOVtLj5WR48peP2AKH2CmKkqUoijNQXRiz2t+fJJLbKc7QMlaicJSPuVEAfc1FzuVpKshhdkSCNm5/f23VFs1ve0fdtPqk3BTUW4o+klRC4CgzVJ3hxOfKiFA7cc4Pk1QxhiIJOn6rr5M7M1kobGOZpsHqGDTl9hpS6Yj8orSuIvaIlESiLwjNEVZ1RpGHfG5CkuyYEx9rorlQ17HFo/uq8KH68jnBGTVT4Q8EXVrdh8Qfiva7lDwDdOFi+/usWjHnbck6euCUplwG0dJaSoiQxjCXBuJIIIKVDJwceCKR8zfIen5KObyyO/EsOjySR2PbT7bK1hQxnNWrGvRzREoiURKIlESiJREoiURKIlESiJREoiURKIlESiJREoiURKIlESiJREoiURKIlESiJREoiURKIlESiJREoiURKIlESiJREoi+XDgcd6LwqiWq/WuVrO7vypzHWivN2qOhSvylSStQHjctYIP8A1YHiqWyNLzZ20XUlxJo8eMBhoguJ71pfsL+6sU+c9cNPyJGmJMSRKW0r6Vwr3tFeOMkeM1MklhMaxMjbHO1mUCBeo2NfNamiNQpv1mSt9bP4jGPRmstKyGnkkhQ/Q4JH2qEMnO2zv1WniWF+Em5QDyO1aT1adR/v6rmKZVglen93MaHKXIdc3TZ7MRTiS6h3qe5WDwO/bbz96xczDC6vma9V9KYslnEYzI4BorlaTRoitvX3tV7UGn4suxr1W5PQ1ZHJLZTGUtCnpKd2FLK0pT/E7kN4O3BqqaJrh438unzXQ4fnzwzf9Oa0mUAi9SGmroAkiu79F2zTGkrBa0tS7Vb2krWC6h9YKnPfyTlXIyMfHxXTjhjZq0L4nL4jl5Q5J32B06aK0JGM5q1YAuZep0x2FOnoZgomvzLc0wxHUso6h65SrBHO4dRBAHOTnwayzEtJIF2P1X0HDGtljY17+UNeST28ul+hohdFtiHW4EdElwuPpbSlxZ/mUByf3NaW3Wq4cjmueXMFAk0Ow7LZr1QSiLxXbiiKoavnXm23G2T40iMixMvJTcEKby5tUdu4HOMAlPA57nkDFVyFzSCNuq24bMeVr43g89eU9LGpv3CtyDx9xViwj1X1RepRFFaovDFhsE+5yv8AZRmlOEdtx8J/c4H71CR4jYXnotOHjPy52QM3ca/3+Q1XLY9us06/aWuGomWJz98Q6+t0x1FK3wlAbbwRkNpQVgZHJSFGsvIxxa54suXeORlQxTRYri1kNNHmF8pOp03JNXWwXWokKNCa2wo7MdH91psIH9AK2NAGwXzT3ulNvJJ9Ta5T6h6SXJ1hbpUcT5xlNvJkto6S3W2UkEdHqAJThSwOSTzWPIhL5QbOtr6XhHEhBhSMdTeUtLdwC438Va7BQGh2oOj/AFLet96fkxoyWimCZwKUKV/KdxO3ISpxGR7c5xjNZ4AIJyxx06WupxQy8W4Y3IhYC6/NydL3FDXcA66qV9VZLSL3a7xa7q4pElHRcVBWrPSRuP8AtEHG0rUByMgjIPBq3LIDg9p3007LHwKNxhlgmYPLZHNW5roeoANfRSbOnndH6SgSbc2JTzgZanwpD2+PJLitpUcggFKlJwrsUjnPermxGGMcupWCXPHEst/jHlbrykDUVsKFb1r6ldJnTWLXaX503c1HjtF1zCdxSkDJ4Hx9q0lwaLK4UcTpXiNm5ND5rkcu9o9SdV6fjwoUpdjjS3FSC9FJacCW+CV9hkKI2H5B5ziueZhkvbQ8oJvT0X1rMF3BMed8sgEzmt5adqPNrp8t1dYDL2jXVwo1vkSNPqBdYERBdXFWTlTZQPcpBPuBGSCVD4raB4WgHl/JfPvcOIfxHvAl68xrm7G9r6Hvuq1cL9edO2o/2fQ5JitvLc2XC1uxg00pZUR1FKG4jdgJSknA7cVne+Rg8n3FLqY2Ph5b7yyWuI3a4OJNf0hu3ckrZuepr/dtL364WdyDBagNqbcCkqcdDgbSXNqwoJG0qOFAHOAcc1J8znxuczoqsfAxoMmFmQHO59a20sgWPWtdlcdGPwpelLWIrSW2TGSnonnbj2kHPfkK58960RkFgpcvPa9mS/nNnmOqgPUSwWuZptqwR47cV2bLQIzcRsI/iZytZSnGQlG8nP255FUzxNezwxot3CM+XHyTlPN0Dd62K2+Zqld4rbUOMyw3lLTaUtoCiScAYAyeSf8AOtOy4xdzEuO6yKmR0vpZU82HVDKUFQCiPkDvUSQF6Gki60UXd9TWu2u9B6QXJh5TFjoLzyv0QjJ8jk4FRc9o0KvixJpm87RTR1Og+pUA7p5/WSVOaujdOAlW6NbEuEbOeHHlJPuWRkbR7UgkcnkQMfiD+INFpbmfgXkYbvNsXVve4APT16q5KiMrDW9ltXSIKNyQdp+RntVtDZc7md0K2AMDFeoEoiURKIlESiKJvligXnpGeypamc9Nbbq2loyMHCkEHnA4z4HxXhaDurY5pIr5Duq19ff9PXZq3rg3C/WxbSlty0BHWZKf+bcJUlK/GFHBP3OTVQL2u5asd1tMeNkwmUOEbwa5daPqO3srXaLtEusFEqEtS2ySkgpIUhQ4UhSTylQPBB5FWtIcLCwzROgdyP3/AHt3W+CDXqrXtESiJREoiURKIlESiJREoiURKIlESiJREoiURKIlESiJREoiURKIlESiJREoiURKIlESiJREoiURKIlESiJREoiURYpUhqMyt59aW2kAqWtRwEgDJJPgUPcoAXENaLJWuzcYsqG1JYfacjvBKm3UrG1YPYg+c14CCOYHRSfG9jjG4GxuOq5LqqdZpms7Tc5RjN2Zbqo6uu0Si4FKF7nMEYKUEpSlXdRWQOBzilLDI1x229/+F9RgxZLcOWGMEyAc2m7NRp/7tSR0rVb9pZ1Na1D+ydnQxpwyOuIs5aGnyg53pbT/ACJPBSFncDwcDgTAkYQIh5Vne7ByWufnSkzVVtFj0s9fWvzVW1rq9LEof2S/4MTemx9U/IbSyEqC8FxIzg+1SklYyDt4JIyM889aRiube9F1uE8I8UeJnHn8LVoBLt9hpsOoG/el1zS+nrZadMt2u3JachONnetP/P7x7l8f3h/lit8cbWMDWjRfI5edPkZLp5D5wfpWw+SjtIx4N90la27lFZlqtzqmEl1CTtdYUpvePg+37d+1RYA9gsbfotGY+TFyHGN5HOLu9SHC9f1V0R2q7quYF6eKL1U27QGbl6j2tUpx0GBDXKitpXtSVlexZOO+AUD/ALXmqTHzShzum3uunFkeFgSMjrzuAd7AWK+dq4oGEgVcuYvqiJRF4rtRFynXtpnDUN1kFEpq13G3JjqmsdSQWHkLCkksj8qcD8wB+eCTnJK1xc6tARv6r6Dh2RD4EbHeZ7X3ymgCCKOqvWh7i5dtJ2q4PqUp2TGQ4sqIJKiOc7QB3+AKvidzsDiuTmwDHyZIhsCQpK6z2bZAkzJO8MR21Or2JKlbQMnAHJNTc4NHMVRFGZXiNu5NKHtesrHcbQ3cmp7TUVa0t7nz0ilSvypIVjBOQR8jkcVBsrHN5wdFpn4dkwTHHewlwvQa7b7b0or1HlzZFvi2qyxmJNxnOBaEPubW+m0pK1nPk/lGO/JPio5HMW8rdbV/CPCZMZ5yQxo1oG7IICt7YSQlTiEFwc8c445xV+41XLIDSa2VU15rJFiggWxUGXcQ8hC4rsgIKUd1KP8AdwPJwBnP2OaefwtBqfddbhnDPxjrltrKPmDSbPQDv8lz65vXfWlqXd4ERNwVH3rb3SS1FaCTy02jAU+SB7lqKRuxtxiqHl0zQ9gsj6f7rs40cHDpvwmQ/kDqsAW89iTdN9ALNE2pzSr9ttTdpVfbfbtt4SyuDcGWVLC3FJB6RCypSCM+05wRngHObGPDQ3xANdv7LFmRyTGT8K538O+ZpNUAa5hVX6irCr7WmxG9WpM6+2yNGgKkFSVMvqDYW5kMuKH+IoUkg4G44xyM0NhJyC5w0/dLqScR5uDCHHkJdWtgWa+IX6WD7bdasV8skjS1hRa2rgF6cmPIYW9Odw5b96xyhWMFOBgAgbVYOcEgaSPCbyg+Un6Lkw5A4hN47mfxWtJAaPjodfX8xoui3O6Q7fbVy5rqURxgbgCrcVHACQMlRJIAAznNXlwbqVw2ROkdyM3+n/H6KAtU6Lp3Skm735f4e0665MeDwwWgpXsRtH8wSEDaOc8d6i2mt5naLVk3kTCKHzcoDR123+V2uRxvUzUV/wBUXubp2O8m1txS00qQ4kMRsKyHnArCQojPt3Z584xWIZD3vLmDT7L6Q8Ix4MaOLJI8QuB0B5iP6RX3WOfpjUkyZHu+oJ0abDeQZLLdxnIjuIyAPclSSgJAwcADG8ecijoHOIdKbHurIuJxQskx8NvK7QW1pOnuNbPW+yqN31HftPt3C1QFQHrZKZIdTGbD7YZ9wA3pSkHAVjcOMYGeMVnL3t5mA2Cus3GxcgR5UrS1zTuSQSaHQkkf8rN6caqc+rkLevz1ru7cZpmLIcQHWFMIIyyUbSc7R7cdzxwTk+Qu681H7V7KPFow8tBgEkZJc4CweY9ebsu1aX05cdRlV11hOelJO5uGy0r6dC2CchbiWz+ZRAJTuONozXQijc7zyn29l8ln52PABjYDQ3YuO55uwJ6D/hTk/wBN9PTEtjoS2NjgdBZmvJ947K/MRkc4PcVY6Bjt1mg4vkwk8pBsVq0HT6KQj6Ns6OouZFFxkOEFUif/AB3TjgDcrkAeAMDk/JqXhM/mFqj8dOCDG7lroNApeBbIVuZDUCKxGbH8rSAkf5VMADZZpJHym3klbaUgdq9UF7REoiURKIlESiJREoi+SgEmiKq6l0VFvK1vsS5trmOKQtyRAeLZcKD7d6fyqI+SM9v0qmWEPGhr2XQw+IvxtHMa9oug4XV9juF8MXGZYLw3Ev8AcPqYMwJEWa40lra9kgsr2gJGRtKSe53Dvip/CacqjGyZhfC2iNx6d9furaFfY/0qayL3d+tF5a9BzRepREoiURKIlESiJREoiURKIlESiJREoiURKIlESiJREoiURKIlESiJREoiURKIlESiJREoiURKIlESiJREoixSY7UlhbL6EuNLBStCxkKB4II8ihFiivWuLSC00QqZK9PrWtoRnlzH7My99U1atyeihz4HAVt74QVbcqNUHHYRynbsum3jGQxxlZQkIou61/f13VIQv+08WEdQKeQuLdkpGnltpYLbRVsbKQkBZ2pUD32narxWehNQk6HZdcg8Oe52LrzM/wAwWdasg9NdvTSl1PU8sQdN3GQVJBRHWU7hkKUU4SPvkkD963PIDSV8xjNLpmMvqPzXNrVbEa+tcEOW+Pbm7YyiI5KfZbefL7aeW0g8dMdzke7JAA5NYWMblMGm3fXVfTZGTJwaaQiQu8Q81AkN5T1PXmPTstSwOat0A+3GuMOTOtKnEIbZipL6Et4UVqSskFspwDtICSMgYxmoxmXG0eLHpqr8xvD+MeeBwY+rPN5ddKFdb77rXYvlm1fery04bt+Fygw+1b7cglctwD3uL2ZKSNqeMpGACea8bMzIe4a1pQHVez4GTwqCN9NMjeYFztgOgAO97g0e2y6do7VBvLsyJLt8i2S4yyEx5WQ4tnOEOc+DyD3wRjJrZFL4lgiiOi+czsA4oa9rw9rh8Tdr/p91t6i1VaLBJhx7rNajOS9/S3g4wkZJJHYfc+ak+VkZAf1VeNw/Jy2udAwuDd69Vz7TWqJtwv8AedUrjLl2BsJjMOIcR1IbQCFrUpA7g5CjglQ24xxVEcxe4yfyrr5+DHjwx4V1Nu6wdSdAAfTv1tdK09frffoKpdpltyo6XFNFbecbk9xz+oP3BBrSx7ZBzN2XDyMWbEk8KdvK6rpS1SVKURCM0RYnAB5wabrzqqZa7vb7BrCTp1UltsTimXDa59q17uo2OMDJTvA/xmqhI1r/AA++y6JxpsjFGW0WG6OP5H9PkrqpAXzmrhoucVxXUdrh6TmXiRPkiTb1bJbMaWyl5p8dX3sgADC0rVlJHYOc8ZrBI1sRc5x07Hb2X12FPJn+FHC3lfq22kgjTRxN7Eb+3squyq4xJUG5W2woksn6m6QGn563EQ2wR/FI9oSCUkgH82QO4rM0PBDmNvcjXZdZzcd7ZMfIn5XDlY6mgFx1063Vi+vXZX3VKX3/AE8fvF61FKSqVGQWm4CkssBa8bEj27iNx5JPYHtWyQExcz3EX2Xz+GWR5/g40APKT8epobk60NPTdW6w6KslsLDyIiX5qPcqVIJdccX5WSrPJIzkYq5kLGdLPdcrJ4nk5JNupvYaCu1Dop9yI0Iq2AAhtaSnCQBjOc4/rVhAOixB5a4POpFLlmpHX7f6PSG4vRLtncTEUXE7tyWXggKHwop2nI7HOMVjlcW4/l/l/Qr6XAjZJxdhkBqXzaf6hf09OvVZ9BPPagg6rava4Ey7vOBC2WXAWyx0gWSnv7CVEhXf55Fe473SB3Pv+6VXFIocJ+McUObGBdkWQ4O81juKFjZa7l0eToFFj1gxHh3mOhtLTUtQ6cwNKBSUrOUlSgnB5yCcnvUQ7+FyS6H81c6EfjzlcPJdGb1F22xrYGoAvTRSMqUdUKtt6Q5Itul4bDr6VOutsCQop2tqHOW9oKyCrGDtI+au/wA0h901YW/9gHwOaHSkgHc8oGpHrelj3VO0xYH/AFC043ZrrdlxbdbXMoYYfS/JfXvV/FWpWQG8EhG3IJyc8AVVGwyt5HnQfVdDOyhgZJycdtl43qmjTZo79719FJaX0zZ7tfZ9ggw1DTMFTEgrWyT9U4hJQAVnukq3qJ7qycYHfxjGyOMdeUUp5OXPiwR5Zf8AxnhwPcAm7/Qdl16VZ7fLW0uVDjPraBDanWkrKAe4GRx2H9K2FgO4Xy8c0kbS1jiAd6O/utgxWiSShJJTsOQOU/3f0+1e0FXr3XLdY+kdm6b1101a2G7w04JDcZZzHfIOS2WydqQrHjHP2rLNih3mZuF3+G8clhPg5LiYyOX1bfUFVi06qVou5O3d2JLi2GQnozrOwwvbb5YPKm8pCAkpwopBzlfY1Bs/Ked3zHYrZkcJE7RDE4Odu11jzNq9Rd302XbLPfrZd0ZttwiyiEJcIadClJSoZSSO4z962hwdsvl5YpIjUjSPdSC3QgZUpKR8k4r1V77Kuv640+zdo1uXdohkyCoICXApIKe4UocJPwCeaqM0Ydy3qt7OGZj4jOIjyj96DcqwRpLUhsLZcQ4g9lIUCD4PIqywdlhIINEUs1eolESiJREJAoi0btdYdpgPTbjIajRGUlbjrisBIH/jt3rwkAWV6xjpHcrBZKirJrSwXu4OwbbdYz01v8zGSlZ4B4CgCcZ5xnHmotkY800rRNhz47Q+VhAPVT6HULTuSpJHPIPHHeprMvrePkf1oi+u9EWvMhx5sV2NMZQ/HdSUrbcTuSoHuCKHXRSY90bg9pojqoE6G04eDamMfqr/AOdQ8NvZaPx2QDYcvl3TLNoYkydMMpizygYRvV03tpyELBJAz+XcBuAJwaBgbZajst8xa2c20e1rGjVr2xKTpu//AFJ4U0IowFeR1CoII++cGvC81sVIYY5q8Vtd7/Sr+ym7Nd4t2hokRHQdwyW1e1aOSMKT3SQUqGD5B+KmCDss8sToncrx/b6qRBzXqrSiJREoiURKIlESiJREoiURKIlESiJREoiURKIlESiJREoiURKIlESiJREoiURKIlESiJREoiURKIlESiLTuk6NboL8ua6GozKStxZBO0D9P91ebar1rS9wY3crnOqpOkL5drHcpL8F5bElUV4Sj09jZbcUQtK8FOCAQSP071mk8N7muO67uGM3GilgbzNBaCKF2QRWotQeohp57Vun7Rp5yCu2TXtlyjw5RDboBTsSpCVYPJz84HPAqqUsfI1rDp1W/DGVFiTZWQ13MAOQubtZ1okdlZ9VaJsdo05ebpY4DEG5stLmtSEA5acQN+UDOEj24wBjk+OKvdA1jSYxRXMx+LZORPFHlvL2WGkHsdCLV2tiHHrVGE5SHnVMp6qgnalZKRk48A/FaBdarjy8viHk0FmvTsqFr+xupvOnRpFuPDvLTq3EAJ6bBYSBv6gSORlYwO/vV81lmjNt8LQrt8NzY+Sf8fbmOFHqb6EWtO4a1gOxY95jh5F4t0kwZcNopWNvKnUHJAKcNqUhY5ykfcUdO13mG40pTbwuSM+BKajeOYE7dgdNjZojpauWm7bOW9Nud9ZjInS9qEsNHeGWkg7UFR7q9yirHHPGe9WsadXP3P2XMy5oyGwwXyt6nSydzXbtetKHautu0lebwxd/9RjzJTK4hSwotKQWm2gAUp2pIUhWQcYGD2NeeIxh82lq38NNmsYYfMWg3rqKJPX0KmrFBVar7dYsVnp215LcpoJRhCHVFSXUjxztSrA8knzUmN5SQNlRO/xomSOdbhY9aG350rMOwqxZEoiHtRFR5343ddbzrUq5CDaGYjL6fpUhMhzepSSN5B2jKFZxzjGCM1Qed0hbdCl04zjRYbZg3mkLiNfhAFHbrutOTp29afvbL+mWY8+3KCnXmLhIJdS7tCctuqBUMpAHJI48A1Eskjdceo9d1ezKxMmAsyrbJoAWjSv9Q29VYrBeZytLpuWqIbdpkIQtyQ2V5S2lJPuzzjgZxVsbnFnM8UsOXBC3IMWI4vbpRqifRcQ1t/w0/epUNVwhyGri1IaiPpW2G21RytTgTjAWvYokHnAH941zpo/FLuXoQftuvseFZAwmxCUBzXMcLAskh9BpO9CxqPlspjTTl+sdmXfIMNL0GLDeidHrtKbX0n1KS4peRxhbn5O5HwRU4jJEzxK0qq9lRnRYWVkDEeac5wddOvztFgD3A32B7hWnTF8ieot7jSBBmptcGNvKX8paXJUtJ8HCyjZx+pOO1XwzDJdYboPzXLz+HS8EiLHSDxHmtKvlo69xZK6elIT2rWvnKWGawiTHcZdSVNuJKFDcU5BGCMjkV4pAlpBHRcjuabPafRG4W94tx8daN0VPe76oOE7c85IIBz2IGe1YncrMctPt87X1MDp8jjLJmAnY7fy1qfp91NelsG4puV+nXNEVtTpjsdOMgJQ0ttshaEkd0p3JGcnnI5xVuMHBxc7rSwcYmheyKKG6HMbJNkE6E++9K8XSBb5bA/Eo8d9ppXVHXQFJQRn3c8DHPNXuAd8S5EU0kRuNxBOmhpc1szEZ29R5twQmVpVch1u0mSgBtp1at/U29ilRK0trIyAlOOF5NQ0Pm+FdSVxkYS3SUAXruKr6jS+/yVr1bAtl50w7cmJSmVx46pEW5QiC60kDcS2oEZBAIxnB/wA6lIwPbYNeqzYU0mPOGFtgmi07G9Nf79FGei8ZSdNOS3XXHVSnlKQtZGemn2p7ADPfPHf7AVVhg+HzHqtv+IJWuyhG0VyiiPXr/suhFQFalw14VYovLWIrSrdhWSDggeKBDpqqPohluVF1HarzKYurv1zhkbgkpdaWlO0qSOMYBTj5SR4qiAHzBxvUrr8Uc24JYWFjeUAb7gm9fv8ANUbVPpZd7XriJePT5tliK/xJjiSYyGhxuTlGFdNQAJAyQRx9qnY5bIHR6d1tx+MxyYxgzRzEfCas/fdW202GzX24txNRQLg3d4AEhUKdcXJLSkqynqIyohaM8ZwCDwQM82mFjz5hssAz8jHB8BwAdpYAB0+Wivi7Rb1wvozDimJjHRLKdmPjbjFXcoqqXPM0vP4nMebvZv6qst6Vk2BlxGjJjcFpSy6IUpvqxt5POMYWjP2JH2qoQlg/hmvRb38RblODs1vMf6ho759D7lWHTVzVdrLFmOt9J5aSHW//AEbiSUrT+ygRVoII0WLIi8KQsG3T2O32UpXqpSiLDIkNx2lOPOIbbSMqUshIH6k0QWTQFlctuvqDfdQSHInprZ/xFpJKF3WSNkYK/wAGSN2Oef6A1mdK9xqIX69F2IeHwQDnz38vZo+I+/ZRbWmUyb9Eiep98GoJTyVrjQm1bY8c7CpZWkbT8hJIxhNeBo5wJXWfsrXZB8Bz8GPkaKs3bjZ6f7Kdk+kGkZ838RiImx3lhSw5GmLAKlJICwcnGM8YIHg5FTONGTzBZm8Zy2N8Emx2IX1o70wc0g1JRY9SXNrrlJcS60y4g4B52lPCue4P65r1kHh/C4rzJ4n+LcHTRN07afktljQl6Qy4HNd6hVIW7vU4npBO3GNoSUkD9v6UEJG7youzozoIGV87WWMv1EgJejFnT92Q3/sZbrzkVbg/xoSlQ3cckYHNejxBvSi78G8cw5mntQI+R7KIjettgbnuQr3FuVoltvdFxEhkKCPG5RSeBn7duarGUwHldoVqPAsp0QmhpzSL0P6LpdtuEa5xGZcCQzJiPJ3NvMrCkrHyCK0AgiwuO5pYeVworaIBr1eL5LacYxRecoVLuuln7debjf8AShYau8pv+MzIRuakEdhngoJ+QcE9x5ql0RBMke5XUjz2SxsxcyzG3YjcX6dfnspHTGoHZiUQbyz9FfW2wp+MU4C/lbZyQtGfgnGcHFSieXCniis+XjNjPiQO5oydD+h7H9hWYdqsWRKIlESiJREoiURKIlESiJREoiURKIlESiJREoiURKIlESiJREoiURKIlESiJREoiURKIlESiKMvV8t1m+lN0mMxRJdDDRcVgLWew/8Ar2qLnNbXMd1bDjzT34LboWfZSKTmpKkG1r3OYxAhOypbyWY7KStxxR4Skck14SALOymxjpHBjBZOyp8HTR1FJiXvUhfMht8SIkQLKER2hyhC05wpfZSj/ewBwMGkRiQh7910X5rsUOx8aqIpxoWT1rrXQfXqtq8tRFa+s31LTHEOQsLWlO5a97SUpyfjerA+Tx2qwgF4KohkezEkDL3G3zP79lE630nY4UOXqGDHat94if6wzKYQMl3PtBR+VW5RCe383eqpMdgPO0eZbsLiuU7lxJXl0R0IJ6enYjf5KC1Rrm4zLdebGmxOqlotzjUxxtaXWmXSNpBKCeCN2AcKJwNveqpMl9OYG60tmJweFro8gzDkLwAKpxG96/8AHqumaYlfW6ftslSmlF2M2sqaJKCSkZxnnH61radAvnshoZM9o2BNfVRWrXmbddbHdpKg3FYfcYfeVwlpt1sjKj2Cd6W8k/bkVF+ha4q/F88ckQFkgEe4PT1q1XtaaQY1VquG39fJgpEFbpXFCMuZcSDk45yD9/H75pYRLKNa06Lq8N4i/AwnP5A+3AU7WqBP5rNpnU0qxPSrNrKahbkeQGWbnghp3ekLS2s/yrAUO/B+SQatjkLAWyn5rLlYjMupsBu4st6itCR6fkrVqe2JutpcQha25TKhIjOIAKm3kcoUAeDzxg9wSKuc0OFBc7GmMcgPQ6H2O6jNOa2s90T9O5Mbj3JlDSZEd8FkpcUnJSkLwTg5HFVsyGP0vVa8rhM8Hna0lhuiNbAO+l0tPUOrJMi5W23aMk2i4TnFLckNOPEhLSUnncnO33YHk9uO5EXykuDYiCVbjcPYyN82eHsYKo1ue2vpqvNL6tnoegWzVzDUa7TFuJjORiHGJG0nICkk7VJ4yDj/AH4RyuFNlFEqebw+Lzy4JLo2gXejhfU3V30pWTU1/gaftpmXJ3Y2VBDaUjKnFnshI+T/AE+ate9rBZXNxsWTLf4UQs7+wG5+SrsGyahnX+33y4T2retsqQ7b4wC0lggkIUsj3K3YJI4HOPmqhHIXiRxr0W+TLxWQPxYW8wNeZ2hsbkDoK011V6wCOeav3XJVS9SrS/c9MupgJeclRnG5TbDbmwPFtYVsV8ggHA+cVVO1zmeXcarocLyGQZI8SuV3lJIugeo9QuQ3GJJft0DVCILy0rxIuci5vlbEheFBOG2lHCcnb2AT7QR+asT2EtEoHvey+rgnjie/hz36CwwMA5gCQbtw9L9dfRT9ssSdV3FFomGXDtMF9Ul+zKUECOD/ALJnjlWTvXkHalJAT34tEbZf4Z0A1I/Rc+XLfgNdlR06R45RJrrW7uwoeXuTqrzPtY03Oi3KwwXfpAenNhRPyqb2YS4lrOCtJCfy+4pJ74FaeXkotGnouE2Y5dxzv13Bd3va+3vpal7bqmz3GUI0a4M/VkkfSu5ae4/9WsBX+VSa9pNWqZcOeJvM5hruNR9Ropse4ZqazA2uQeoenZlqamCxQoKLLOLb0lKlAKbl9dIS4gKBAKtwSfGO+Mc4p4nDRgFH819PwvOgkLTkvd4jbAr+nlOhqtjr3K0NNX+ZbNGS1x77bITlsDxRa+iiQoIQ4sJG8LClAlJG7GT388whl5Y7JGl6eyvzcBs2YGmNzg/lt+rdwL0qhXa9NlV9Q3q/eoepWoLLF2bsTDDE25wk7RtTtyoIHBWlSSdqSSVZzjsKiJnzkUCB1CvdgYvDIiHuY59kNdrvel9L+w9V16XPterfTm5rs8VUqN9M603FDRQoOIB2oCRykghOMfbFay5s0JLRYXBiim4dxGNszuVwIN7ijub66KEt2o7Vb9KWvSunZBm3p+AG2Gm9p2LLeSpwr9qcEklJyeMbfFQbIxjREw6rTLiZGRO/PyW8sYdZPz2Fb/s2oK0a9s3pdZTp+9B6XeY76y8IWFhe/CgsqUQBwQnaecp7YwarZI3GYI3brTl4c3GZ3ZeOKjI05ugGlK56PRM1PcBqi6pnRYv/APCoDjhQENFGC64hJwpaiVYzkBOMDNaoyXHmOnouJlckA/DsIJG5HU9h6BSU2Bc75PkNXF1UGytq2Jjx3cOy8d1LWOUIP91JCiM5I7ULXOOp0Rk0OOwOjHNIep2b7DqfU/Jcy1xcLXEuS7Pol6HGfV/BuEb3BmSnaVDJTkEjaUqVkH3gE4zjFPI0HkhIvr6r6nheHNIwZXEGOLf5SKsG9TR6dR7XWylIE+23axMdLTjsa6w2R1pNsZabXEUMe5tsK6i0g4O3aQR8+bGOa9ujaPp0WCeGXHnLXy2xx0DyTzA9Ca5Qa31FK3enOo/xO3iDcLpHn3Zkne6yUhL6M5C0AeACARgFJGCBxm6B5I5XGyuTxPEbFKZIWFsZ2B3Hof06EbKW1Nptu8SrfOYlvW+5QVqUxLjpSpYSoYW2QoEKSoYyCO4B8VcW3qsUcxYC2rB3CpMLUGsNFvTRraO5erIjK2rpAZSp1OVDCVtgjjB7gcfJHajmkjJ5xY9F0nY+JksacZ3K/q136H9FLW/1e0TOQ+oXpthLIGTIbU3uzn8oI9x47CvW5MTuqhNwXOhrmju+2v5KGsnqdpOBdZbDV8Q5bZT4eYcUhY6C17i6lWQClAUArceBvx2HEWTRg1atyeHZb42yOjIcNDfUDb51p8guqsPIeTubWlST5SQR/lWlcbbdaGob7bdPW1c+8zG4kNBCS45nuewAHJP2FRcQ0W4qyKKSZ4jjFkrgvqe7K9UNW2CDpYvz7GhsF2Q2haWW1qX7yskAAhKeAeeeO9Y5gZnNDNQvpeFuj4bDK/I0l2AO/wAl0PUc9+VcJ2h9OW2OkN29BD5fLDcYKVgHCcHgYICe578Zq17yXGFo6LDi47Yom8RyJP5iK3JoX10+qtUTSVkbtzMZdqt5SkpcVtjpAUsfzH55+atbE2qIXPOfkF5cHnW+vQqBiR7jpS+3iTGsjTtkkqQtCLav+K1tB3KLJACiScnYc4xwTVbWuY9xA0K1yTQ5ePFGZCJG3fNsbOmv91d7dMj3GEzMhupejPJC23E9lJPY1oBvZcx7Cxxa4ahbOKKK8wKIoO/6TsV/W25eLXElutJUlDjrYKkgjBwf/GKg+Nr/AIgr4cmbHvwnEX2XJn7tM9EGG7YuI5dtPyZZdjyuptcaSU/xGyPy7t2FDsFZV5rOXfhhVWF2WQjjkhkDg2QDUd/b9V1rR+prdqyzN3O0OLXGUooIcQUKQsd0qHgj/wC1aWPa8czdlxsnGlxZDFKKIU5UlQmOc0RV7Wds+vszrkdA/EYgMmG52U28kEpwfg/lI8gkVB4sX2WnElMcoafhdoe1FSdpuDVxtkOax/spLKHkZ+FJBFSbqLVErTG8sPQ0t6vVFKIlESiJREoiURKIlESiJREoiURKIlESiJREoiURKIlESiJREoiURKIlESiJREoiURKIlEWrcYEWewWpsZiQ132PNhY7Y7H7E/1rwgHdSa9zDbDR9FVNGQ1acvMjT5nSpkYsCZGMhW5TSd5QpvP90ezA+5qqJnhEsBtdDOyBmNbkcoa7Y1pdDRx9TrakNZlDv4JGUneHroyCk8g7dznP/wAH9QKlILoeqqwnBviOP9Lvvp+qsTWNoNWLDsqX6jxbY5N07KvgZNvjzsLLy9qUKUhRQon7LSnvxyapmawlpf0K6vDZchrJmY+5b8yLGg+qi7/qRrU6Zds0mUTpEBxuTIkgnptltW8JRj/aKUUlIx7RySe1QdMJSWRnUbrRBw9/DwzIzgWtfYA6mxWvar6qpWORB0TpyPfIL8wQb03JDqEq6ikOJ3Fkt5BwruCVZHPNZ4nMgZ4g2da7GeyXiuV+De0F0XLR0GmnNeuvpS6l6eQzA0hamVyVylGOlzqrTtKtw3dsD588/NbIRUbQvl+IyiXLkfy8uu3tp+isT6EraUlQBSoYIIyCKt91j16aKm6Ptdvh6ovyrS0ExGUMxkhC1Fppz3rcQgE4T+ZskJwM4HiqY2gPJbt+7XTzJ3vxYmyHzGyfUaBp9euqwTbexPnawt82EmWqQhqUzHVtHVSGAgFJPAIWgjPg4PmjgHBwItQifJE2GWN3KASCR01uj8vqtS2a8Ra4dptWorZdWL46wEhgMdTrLSlIG1YO0lXxng5CscZg3IDeVjxTitknBnTulmxXtdE03d1Q9Qdf79Fh1vcL6m0vy0aXtn0rbTi5Dk99t0hAbUc4A55CfJz2wO9eTF/LYYPmveGxYviCPx3WSKDAddVU9MQAlOj46rzbYIfU6lhNoWnc5/CUla1uKBUVqUdvGAk8DnGM8bP8vUC+y7OZOf8AuiInP5QL8S9PMKFDSgNfXrorxcrVaNHXS13qNETEgMpcjSltJUUoSpI2uKHPOUAFeMndznxrLGRHn+S+dhnnzo3YzjzONFu2pvUX+m17KN9RdXxXrZbV2NX1+y5MOrcRFdda2oWVHC0jBI2nIBJwDVc845QWa6j1W3hnCpBJI3I8nldVuAN9AQTa6NarlDucZMm3SmZMdYBS40sKByM+PsRWprg4W1cKWGSB5ZK0g9it4nFeqtYTIZ63R6iOtt39PcN23OM474z5pfRe8prmrRcwuEaRdtK6qasEnGLutf8AqqUuFaEhpS0oH94nJ47njzWN4c+NwjPVd/GkbjZkL8tt+XrY3Bon06LPp/TQkIn321alcnXqQ4pxuclKQCjaAGXm+xTlPbAKfGMVKOMG3tdZPVQzM/lLcSaHljbu2zv1cD3+ZCs+g71OvdmU9d4SIU9mS7GeaQvckKQrBxV0TnvbbxRXO4jjw40/Jjv5mEAg7br49Smm16MurpRHU+0wVsKdRu2OD8hT5Cs4wR2JFeTA+G6t6U+FvDcyMOPlsX7db9KUtpxFwbtEVN6dZeuIbAfWyMJK/JA/8ftU2BwYA/dZskwmZ5xwQy9Ad6Va9Y4UaZoa4OS1uBERIkhtLhQl0pPCFY7g9sfOCOQKpy2h0JtdL/D874uIM5NzpdWQDuR6qp6fs8f0+mW5NziQBbJEBTkya5hzpyGklw7CU7gnaDx2yOMHg0wxCAtFaEan1XSz85/FmSSNe7ma6mt2tp01rS7Wtp2ejVepNSar06l11UWTCYYQyvpOvsIGXdyCQFbkqUE7/wC6MYIqxjvEcZG9FkniOHFHhz6cwJN6gE/Dr0r0Vl1LddLRrO9qeZImQ1H/AFdz6V1yNIecScBlSEkFSwc/m7DnOKm9zK8Q6LNisymyjEYA7tYDh7gm9PUbrnnpxY9T6ojynbdNOnrBIkurcdbO+c/uVuALh54Ck85APJwSSaywRveDymm3813eLZWPjuYMhvPIGtHL/IK02FduxV3tvpJA06BOsEgLvTThcbfuLSXkEEDKVJGCPJCgQrJ8jir24obqw+b1XLm467KAjyWfwx0Zofcb37HRTti1PJit3NvWiYdslQyXgttz+E7HzgLSSSScggjv245qxsrgD4oAI+ixZOFC8sOAS9rtNRqHdQf3W6p+oHpGt5ino1vlbo4WxbWlMr6S1rAC3JCwdgSBx0zzzg7uUjPITMbG/T/ddbGbHw5gY9wp2r9rAGwaCLs9/pW6vWgdJRdO2qOpyLCF2WjMqQw3grWfzYJ5A4HAwOOwq+GJsY9eq5HEs+TNlJ5jyD4QTdDovvV8C2NSIWoLi9JZVa1KWj6dWC7uwkNkDleTgBPknHmpyNaKkd0UMSWdwdhxgESaa7Cuvp7qkyz9NqVT+vGIFtttyZT9K606tKmJCCcBTgxsd2KIKwQCE45ArM5xD/4+gO1LsQxtfjAcMLnyMJ5g6jbT2HVtjbfVXOPaLmtlaGNWy3EI9rSgxHWpPkb1bTvOCPjPfzWjlJ2cuOZ4mH+JANe/MPpqF8RtQvWZxMbV5Yiudm7g2CmK/wDuc9Nf+FRwfBPgH8v+Zv3UpMRs55sKyP6T8Q+m49Rr3Wa76e0pKCLvcrfaloaaViS4hAR01DnJ7EEE8nPfjvXrmx/E4KmGfKvwYi6ydhe4UDoDSFhRKuF6tlojRoctH08RKmyd7HO5whWcBwk4HHsCfmoRRsBLmjRauIZeQ4MhmeS5u/oe3yofO1BStF6z07qi83nRUq0NQpiwRa1BSW1cBO4g8BY5OQRnt24qJjlDi5h3VzM3CmhZFlNdzD+Ye/Zadx9KNQ6qehyta6pU8tDyluRo6MtNt4HDXACVcHKsdsVE4z5P8xyvi43j4RcMSL2J3+as6ro4i0rsHphGjOrtpSyt1ah9MxzkoyTlazzyAR3JOak55osg3CojxWtlbkcVJDX66fEfX0Hurdpu1PQutKubwk3WVgvupBCEgZ2ttg9kJyceSSSeTV7Glos7rm5MzZCGRimDYHf3Pqev0U7UlnXytIIonuqjcbFKtMw3LSrLQccWTMgrcUhqQCclSf5UO5zg4wcnd8iosczWNb4Z45h4WW40BQIGo9+4+/ZStjv8a6OPR+m/FnMAF6LJRsdbz2OOyknwpJIPzUmvDtFmnx3wUbBadiNQf32OqmxU1SlEWGZGZlxnGJDTbrLgKVocSFJUD3BB714Rei9BLTY3WvZbXDs1uYgWyO3GhsJ2ttNjCUigFbL18jpHczzZW9XqilEWtPj/AFcN9jquNFxCkdRs4UjIxkH5FeEWCEa7lcHVdKnNaDaZtLMf8SnSpsNhLcCTIc/4qpI9qkpSAByBkkEkcZxxVX4cBu9noum7irnSEhjWtcfMAPi97+wGl6rJYtcQ3owTfW12Wal5TCm5mUpKk+Qsjbg4OMnxRmQ0/GKK9yOESxn/ALf+I0iwQb36VvffRb41naXAVRRPltZ4diwH3W1fotKMH9jU/FaVldgTN0fQ9yP7r1Ws7KgbpEh+IAcK+qius7f1KkgAfevPFZdEqX/T8gi2tB9iD+qmrfc4VxbLkCXHlIH8zDqXB/kamHA7LPJDJEeWRpB9RShbxq+Fb7ou2MMTbhdENh5USGzvWlvBO4k4SBx85JIAHNQdK1p5dytEeBLJGJjTWE1zE6X+f2UdFn6k1Ehl+AyLFbnEJWVzGQ5KVnn2oztQPGV5J+KiDJJRboPur5IcTE5mSHxHj+k035nc/KvdSA0q3sUpy731b6uVO/XrQc/O1OED9AnH2r3wRe5+qrHEXAUI2Af+Av6mz91pvLuunbjb1OT1XK2SHm4i0ygkPtLWralSVJAChkjIUM8ZB8UpzXCjYQOgnjdbOV41FbH3s6HtWiuKFZAq1c8L6ovUoiURKIlESiJREoiURKIlESiJREoiURKIlESiJREoiURKIlESiLDMfRGiuvOkhttBWogE4AGTwOTXhNC161peQ1u5VEs+oIiYFw1pd1fS214IZiqUglaY6SQCpIBIKlqUcDwE1Q2UcpldoP0XXmwZfFZw6HzPF3tXN1r0A+9qp6xf1FcdU6UkOmDFYlTCu3xXmlrW0QnKVulJHKgclIPHA55rPM6Rz2EEUToutw2PDZh5LacXBvmIIo67AHt36+i6fY5V9W+GLzbYjSQjP1MSSVoWr42KSFJzye5x81ubzfzL5edsANwOJHYiiPpotbUIB1Vp8Sv+Jfxwj4MgowgH/wDp9b968dXMLVsF+DJyb6e9X0+dWoVzTdpd12sRYiWAIHVeciLUwpDhcIQdyCD7gXMjztH2qvwm89AV9lqGdkNxQXPsXQ5vMKrXQ36fVV3VPp/FsVvjyoU5TFujSm3nSthBeRlxI3peACvaTuO7OcVQ/Ea0DlPX96rqY3HZMh7mStsuaQKJrbblutRp6L40xerrpiWE3mEn6B1Qcd/Dll6Oy2tStkhIydiDsXuAOMbSACCCZI6N1SD6be6ZmHDms5sd2o25qDiW1bOlnUUd97tdbdmMCEZAcSpkI6u9JBBTjOQe2Mea3AjdfL8hL/DrXb9FQvRti7mBc7nd1OssXKUuVHiOthJbC1FRX8+7cOPtnzWTEa9oJf1K7v8AiCbGdJFDj0eRoaXDrWn2TU2oHIetZTNphSJ93btiUNRUtq2rUp3IyrGAkAZJyB4zmkj+WU8os1t81LExGzYLfGdyRl+rvZp6b7/ulHtRdQOa20+9q4wTDdkyHIkVDYV9MsNDYkuYwVY3EfcEgngDz+L4rTLVWa9FIyYQwpW4IcHANDif5tdTXbZWn1Iudsg6Sns3OWxG+rYcYYL2SFOFB2jABJ5wauyHMZGec1ei5vCcfJyMpgxmkuab06ALl2h75bHLjCdlsXORs/5AgqbSVAkqSsA4AUeM71HCc4ySkmsWPKwkEj/xC+o4pgZLY3tY5o//AMpBPoRe/tQGtdirjohcLW7t2l3+KiVMiy1xxGdSVsR2+yQjPsKiASVjJ58DFaYXCXmLxquJxGKThgjZju8rgHWNCT69RXQH81qzTY9Fahu9qZkGAxdLUXWWA4Q226gLSdufancMY+Sk/bPlxwOLRpzK2MZfE4GzObzmJ2przEGjr1ICzenVlb0WmxRmHFrTeY5+o3pwRISjqpIHgbN6T/0U17jw+AA0dVVxfiL+KvklfQ5Dp/47V+v1Vt1NfVxZMW12kx3b3Oyllt1R2tpAJLq8c7Bjt5OBkdxdI+vI34iubi4viAzS34bdyPXYD1P2Gqx2XSUW33P8YluOzr2potOzXjypJOSEoHtQBjAA7D5OSTIg0851K9nzpJYvw7fLHdho9q1O5PdafpTFjwtK/TstsNvNyX0SA1tOXA6oZJHBO3bz8YqvHaGsob62r+NyvlyjI4mi1pF9q0UtctLWS4PqkSLdHMlf5nmwW3FZ/vKRgn9zVxY3elkjzZ4xysca+o+hVAsls1XEvd3YtFwZTDt1yDv0clRcXLQtpJwp48gkY7jhRyScc5GMmDjynQHbqu7k5PDXQRmRhL3MrmFANIJ/l/322V6kv2rUcGVZ5chn6kpCZMVmSOqyobVEZSc5SSnkcVrLmvtt6rhtZPiFuQGmuhI0Krib/ftHwpq9VRzPtrKldG5MKSSEZAbS6kAHcTwVgEDIzVHPJECZdQOv5WukcTF4jIxuCeV7qtp79S09uwKrF/1/YdS+mTj19THE0rClQmtzpaUlz2qP90EA9z5+9Z35Uc0NnddWDgeXw/ifJH8Lb8x0BFaqvervqJbdT6cjxNOdRRV1VLU6UN7WwnCgBuznkYB7jOM1HInErQGBXcJ4W/Akech4/l2s63fals+l8tXp76QXTUU+OsSri+DDb2Al3KMNE+dudyufAJ81dB/BgL3LDxUHifEm48RuqB/VSHproJ/UtnuN/wBbtLkXS5K6kX6kE9EbRhwN5A5wnCcflQB2NIYTK0ul3K84hxNuDK2HBNNYKNbnuL+qvPprAlacbkafuchuRKbCZaH0IKQ6hftPB8pKdvHGCmroI3RDkJtc7i2QzNeMqIFrTpR1qvX11P1W7ri8y7e9EjxZceA26hx16U6wXlBKShIQ2gH3OKKxgc9uxr2Z7mkAGlVw/GZK1z3tLqIAANam9SegFarl2rrFOWqNd9QXNtxuS8qK1DuyC79P1GztUtLZHTUopB9g9mR+bk1inhN8znb9Cvp+F8QY1hxseM20cxczQmjrRO9C99/RdI0vLuliscO23HTjvUYbSgKtXTWwofIBUkpPyMcnJBOa2Mc5jQ1zfpsvncmGHKmdLFMACb89h32BB9/ss9y9QbLbWVi5fWRJSc4ivRlpcUR2A42nd2B3YPz3o7Jjb8WhUIuD5M7qgpw7gjbv0P2tQtpXab7eLbMvOoBIu7L5eYtzMpJZYWQohACR/EKU91ZPIPYHFRZySEOLtVdMcjFjfHDFTCAC4g2frsCfbRTXqn0V6IuLLkYyHXkpZjoS3vIfWoJbUB8hRB/b71PIAdGQRuqODOLM5jgaDdSfQb/ZfOh4ce03e+W2E0yiM2tiR/CRtCXHG/en/wDSFY8BYHbFSjYGkge6hmSGaOKR5s0R8gdPzVvkBC0bVhJSe4I4I+9WGtisAJBtq4XP1Lam7s7peY4q56ZjKEgdFAUpSRkhjAI3oSsghScH2gc4zXPdO3nMLhbV9hDw6UY7eIxnkld5df8A7ehI0++i6TE1hZEMMs2RuVcYyGx/yZEU8hlHZIO0ecEADPY5xWxr2nRuy+efgzhzjOQ11/zGiT++q2LnqR1ouR7fZ7rLnYAbSYqmmVEjI3OqG1IGeT9iME8V65/YKiPH5jzPeGj310/v0WjE0DDedMu9yZ1ylvNoElL0pZYcUnn/AGYwNoJOEngfFQELTq4kra/i0ob4ULWtaDoQ0c31/VWy32+LAZ6UKMxHbznYy2lCc/oAKtAA0AXMe98h5nuJPqtuvVFKIlETFEUBqGwquTjcuHMkQLlHQtLEhkjA3YO1aTwtGUpJSf2wag9pdsaK042QISWvaHNO4P6dj6rURer1Hjj67TM1braf4ioj7K0KI7lAKwog+AQD4oHOrUKbseAklkorpYN/Ogp203GPdIDMyIsqZdGUkpKSMHBBB5BBBBB5BBFTBBFhZZI3RPLH7hbtFFKIlESiJREoixPModQUOpC0HukjIP7UOu6AlpsaL6bSEgAAgDxReBerTu45xRCoO4aTsc94PSbXGLw/51COmvtjlScE/wBagY2k3S1MzciMcrXmu3T7rYs1ht1oDpt8RLCnSC4rcVKVjsCpRJwOeO3Jr1oDdlVNNJPXOdlLAcVJVoe1EVJ9RkzXhY2LfKjsuLucclDjW9Tm1YVlPPG0JKj9ge1UzhxA5T1C6XCzE0yumaSAx3sCRQv5q6Njj7eKuXMpfVF6lESiJREoiURKIlESiJREoiURKIlESiJREoiURKIlESiJREoiURYZsdMqK8wvOx1CkHacHBGOD4rwixSk1xa4OHRVVvRz4hR4Ui/XF+2s9MfSrbYCVoQQUoUoNhRHtGeeR37moiOtLsLY7NBkMwjAcb1BO53O6y3RSZ+rLVbN7ZEZK7i6gq5OPY3x/wBJRV/2K8dReG/NeQAx4z5a3po/M/YV81aUDAGasWGlRdbOSLtqqyWC3PPx3WnUXKRIQ1vShtsnagn+UqOeTxx5zWeUkyNY33XYwAyDHlypRYILAL6kb/JSN2s96GpoVxskiEzHdLbdwbeQSpxtBUQUkecKI/pzgYqT2vLw5u3VUY82L+GfFktJdqWkHY6b+is60bkYUAauIB3XO1XK9PWC5WK+XBu1HryIriymPKeO1+E8rcjY5g7FpcQvIIIPPyDWSKMxOIb+wvoc7JhzY2PlHKHAC29Ht3JHWwRrahnX7raG9T2a4MoZt7qUSI8SM4FtxW1rJKVK4KW17Vp7BAOc7QQagPEZzNd8h2W6sKfwJorc4WHE6FxA0oXuND3rayr8rX+nBFJZmh2SGkOphsoKnl705QlKB3UR48ZGcVo/ER9Df5riDg2ZzBr2couuY6NFGib7Kr6Tv4l61duOpJiIMxTRgx4DqC2pvctKsYIzjhKd6jhS9wTjABqifzSFz9DtS25uKWYgix2czb5i4G9gQdtPWhsNyVLesyQrTsOQwGVTYs5mTGQ5uO4oO5WAPhIJOeAATUssUwEbghVf4fp+Q5jz5HNLT6A7feq9VHTLFLiS7ne79tkCY4EwbOqQZKPqVpLaTuUAOxV7QMJGT4GImI257uvT1VrMyMtjxsckcvxPrlJbdkaH097UtoLSFiVoS1NLtsZxMlhqS6spJUtwpzu3d/JxzwDirIYWNjDaWTiPE8qbMfLzneh7fl/urrAgRbdEaiwY7UeM0nahppISlI+wFX7CguY97nuL3GyVSfVVp2SzaGrfCan3QTQ5HivICm3UhJ6m/PASEqzye4Hc1nyWlwHKLPqutwblY+R8zi1nLqQdQTt76jZVWZZdRqu8C2w212stb5FrdFx6zUY7cOoIKdy04I2pI9u487eBQ9khIaBR6a37rp4+RhNjdkOPiDQPby8tj+Ug3ob3PX31XRtN6XatrqZ0903G9qbDbtwfGXFDylPhCM/yj/PJrVHGGizqe6+fys185LGANjuw0bD37n1KsC+2DVqxbLluhJEPS941Fabal+7MuS0yWhbWEqQ1vSQW1kYQgpKPnkYwM5rHFUTnAakm9Oi+l4gyTNigmlAjpvL5jV1sa1JBvsrNNVcb/LZYLM+02ppKnJS1qQ25IPG1tKkKJSnuVEYPAAPJrRrJo4EBcn+HiW9jmvf0qyB6mx9FVLFB0u9rVuJYbgqW1NjSJUkN3B1X8RLiAk5ChnguDnPAB+5zRsgL6afuuxlZPEm43iTsDeUgC2N2IJ7fsq2zdJadhW+S+5FTDKVGS5PQ4pL7agD/ABOqTu4H3x9jWjwo2AmlyW8Ry5HBvNfQNrQ+nLt+q47B0nq31DuUq6xtSTWdNy1ORm3pDhS49HT7QeigJSQog98dyT98phkmN83lK73/AFHE4bGGCIeKNa6An1Nm0vPp0/Y2kNawvURVkbZWmI62gR2lP8KCZOBuO4JIBBUcj5xnz8NyDzHRSZxn8Sf4LHc5IvUu07t9uypmtLPoc6Zi3HS0yd+LSXkt/ha3A70jglWeN2OQEnJz484okELmc0Z1XXx5eJRZRgy2gsGt1v2+fosP9jtXaWtLVyk2lTkGcgNuMcqdSCQdpSPe2ohOMgZwSD5FPClazXYqB4hg5c38J3nZZBrTtfatV+gvSr1CtGqoAgRW3INxioCTCfd6iygAAKSs8rHyTz8+CejDM2QVsV8fxHh0uI/mdTmu/mGy0L8/qNy5Xm9x025UWyPERiXVNOpbQhKnkHCSFpWD2URghJHaqXOktzhVDb9V0IIcMRwQP5g6UW7qNbDSOuh39FVp2qJZ1YrUlziuq+kt7y4MNwJ/1cqQChzAV7woKGV8AZIHKaoM3n8Z3QaLqQ8PjGN/06J2r3gOcLN6mxtoR0HXfYqS0terNNuuy+T3egyGZqi4yUtPS1clx5YyBtIAbQSEgBJGTjF0MjXu5T7/ADWHPwsiGIPgA15m6EWGjSgNzfU72T8+1I2qGU4IravlvRFtBSSCkEfB5FerwEollCQnCEjAwMDtXm2y9JJ3WvdbdGucB+HNZS9HeQULbUSAoft2/XxXjmhwoqccj4niSM04bLmrGjtXaf1RcbpZLo1PiyGSEs3KS5necAbsA7tuODwSDisQgljkLmGwe6+kk4nw/LxI8eePkc06loG3Xr17d9VpsRNV6xvD9j1BIcZs8dKVXBLTKGd6/wAwaCklRUCNqtySPb3GTQNkmcWPOg3/ALKfj4PD4BlYzP4pvls8wA25iNKO4rXX2XUbdaINtbCIEKNFGMfwWwjP9P0H9K2Na1ujRovmpciac3K8uPqbUNEabs+q3ILSCiLcmlym0hR2JeQQHAkdk7gpKsDuUqOO9eDyuodVe/mngD3allD5Hb6bfQKzpSNvapLEvsDAr1epmiJREoiURKIlEXhHFEVTsspFkvMmzzMtNypC5NvUQAhwLwpbYP8AfSsrO08kHI7HFbTyuIK2zN8aJszNSBTh2rQH2qte6tiVA1YsS9oiURKIlESiJREoiURKIlESiL5cOEHHfxReO0CqmlGBdZ0m/wAxQfcU67HhHA2sx0rKfZ91lOSryNvgYqtg5iXlb8txhaMZooAAn1JF/a6roraOKsWFKIlESiJREoiURKIlESiJREoiURKIlESiJREoiURKIlESiJREoiURKIvF9qIVVLRGZd9Qb1Lditty2IkeO24U+5bZ3qKgfgn247+z9KqDR4pd1W6SR4wo4gfLzOPz0FfIa/NWtRwKtWFU7TF1iXPWepxDUpaoxjsO5QU4WlKwRyOaoicHPfXoupmwPixMcvG/MR7GqVxT2q9ctFdqIqlOkKtWuITmWnG7s0mGpsH+KhTfUWlwDyjBIV8HafJqo+WS+/6LdHc2I5lfAS6+lOoV76KSvmnoV12vvt7JaAelJaUW3WyfhSecfIPB+Km5geKVEGVLBfIdDuDRH0VJ0iV6Oslygai+m+ohMCQH4LW0usKKkoTwAorCkkc8+4c9zWaK4Iy1+4XY4iRxTJZLiXTjVOOgduT10r8l7puwuI9P4TFsg/UfiTe64G4PLbeIUgj2kpJyFEbcjAAyOTmpQxlsQDRZO9pnZUcnEHvndQYabyCxp6Ai7G6hpDV+lvIjXmbDeu/1bNpfaQjaW4rg3rcbVySXAkZXgY2KA7GqiJHinHXY+y0iXEgPiY7SGUXNJ6vGwPo09PXdS+vbqLpd7TGt1waVa4jhcub0RwLcjpUekk7hwk+9Q77hyr+XNWSOL3N5DoN1k4fC2GKUzMPOQAwHr1OnyHoduq6XDYZiRmY0ZtLTLSA2htI4SkDAArSBS4Tn8xs7lZ1HCSa9RUb6xx7W9xucxUZFjs8VUYPqWU7XlbFu58EABIz4PA5JqkWZCegXR5GsxWRMB8V5uvTZv3sr6s71xvWsUXZEVTNhYhrYjqkoU0664paSVhB5CcJABIB7/NG8z5OYfCvZWxY+L4PNcpdZrYADQX1NlS9+1jYNPzWYl4u0OFIeQXEIeXtynnknsBwcZ744qxz2tPKSskWLPMzniYSBvS55rb1WRHXc4tukwGGm/axODvXW8QnK9jaM4wSBuUQBgkbsYrNJkUS1v13+y7eFwVrvDfOCb1LaoAdLcep7AE9NFa/T9qzWfRzT0e6wZLG0OSJqHUhrdgDkk+0DsAefnkmroQGs3+a5nEpHz5LiWlvYHceimJmobeh24w0SGkz4jHWLLx2bklOUqTuwFJJ4yCQDwam51ArLHC5xbexO6p/pLZ2bVp5WoZqkNGUwlxKnQEBpoJypZJxt3qys/bbntVGMwNbz912uO5j8if8ADAk8p+96D/2jQfNUGNrqxa/1hJRrZ5iFp6G0Vw4jkhaUOuhQypxSSErOBkD4PHnNbZmSvLXbLTLwyfAx2yY4t7tzW3oP7roUL1AYmyFWfQFiduaIjQR104iw2Dj2pJUM8DwB9h5Iu8Wzyxi1yXYBY0TZTw2/m4/L+6q2odDXKY9GvPqhq6I9b4zmfom0KZYJVwEJVwQCeCcFWAee5qmSM/FM7Tsunh5zWgwcNiPiEVzbmuunT6rnnqVYlRbfpy/u2u320T3lJEOF/BSGk7Sg4xnJBUSrGeU8DtWWePyiRwq13uFZBdM/CheXclGzvzdRvVDatt1f7JdVL1Doi0We5uXK2IZbdeQgbsOpKytZUvCgBkYHxwB4qxjiXxsYbH9lkyIAzGzMnIi8N9mvY0AABopL/SA0v+IW+3XW0J6d+julLYjoIekJCCogKTzlASVZPYAjyAdOVHYDm/EuNwHM8J7opv8ALI1vYdB9Vg9HdR2rWekZmnNRLZdubjzin2nXClcsEhfUHY7hjBx22g15A5szCx+/VT4rBLgZLcnGPl/lI6LHdYbjOvrTpdFujG3NS0vRS64l1LcYhK3G8EZB3N5AznBI5B4pkH8VsVafot8Dw7AlzzIeeqNCjzbNNjfRxvuddwr24+NP61ZbRbHforuhqOl2OlIbS8gOElY4wdm0ZwchJ+K1nySUBoVwGxtyMMkuHPGSTZ1INAV31v6q7N9jirrXMC+qL1KIlEXy5+U0XhNBV7SriZN01FIRjYZ3SGDnPTabST/XI/aq493H1/Rbss1HCwiiG/mSVY6sWJVnWp+mRbLkGnV/RTW1rLSFKUlteW1nCQSQArJGD2qD+h7LVikHniP8ze9ajUfkpe0XOJdYaJNvkNyGFdloOefII7g/Y81IOB1CzvjfGeV4orf8V6oqu3i+/Q6osVs6sdIn9fclZ952IBTt5+c1Fzw1waeq0xYzpIXzAGm1r01VhBzUlmXtESiJREoiGiKG1RaE3exyomB1lIKmFngtOjltYPghWDmoubzCldjTeBIH3psfUdQsem7yq5JcYlxnolyjBIkx3APaSDhSVDhSCQrCh8HsRijXWNd17kQeDTmm2G6Pt09CLFqaK8frUlnWhc73brUWvxKdFhh3IQZDyWwrAycZNeOc1u5V0UEs1+Ewmt6F0qzfvU/TNnkx46pyZjzygAISkuhAPYqVnAH75qiTKijIaTZXTxOA5uUx0rW8rR1dpft3WS3+pOnpiy2X5DL2TtbVHUsrA7lJQFBQHnB4r0ZEZNWqZOEZUbBJy+U9bH3vb5rZi+oGmZD7zKbxFadZVsWmSosEHGf+cAqQmjJIvZQl4XlxND3RkgixVHT5KRZ1VYZC1IjXq2PKSkrUG5baiEjuTg8Dkc1MPYeqznFnaLLD9CsM3WFhhXFUGVdojUtKQpTRXlSQfnHaoOmjaeUnVXw8My52eJHGS3vS3LDfbffoX1dpkJkxt6m96QQNyTgjn/xyKmxzXjmabVGVjS4knhTtLXVeqkd4qaz2vQsGvEte7uKBenRVnUl1kLkN2WzLWLtISlXVS3vTEaKsF1eeBwFBIP5leMA1B7v5W7rVjwAt8eb4B9z0A/XsFMWS2MWe2xoMQKEeOgIRuOVH7k+STkn7mpgACgqJJHSvL3blb9FBKIlESiJREoiURKIlESiJREoiURKIlESiJREoiURKIlESiJREoiURKIsTrwQgqJAA7kngfrSu6jYOgVX04+/edQG9NQzHtqoqmGXlOe6UnqAoc2eE43FOeSF581U23O5+i6MwbBD+HJt1gn/ToQR77X7K2KGRVqwLWhW+NBS4mIyhkOOKdXsSBuWo5Kj8knzXgAGyk5731zm60+S2hxXqihoipWtRb7Pd7PqecltpENxUeRJ8oacSpIyPKQspz5GSe2apl5WubIen6rqcPMs0cuHHrziwPVps/a1bwtLrCVIIUlQBBByCKvC5R7ELk19s120tGutxnTHLzCdWytT7rm16K0h/qbdp4UnnuCCPAxWGSN0bXOJsafLVfT4+Zj5kkUMUfhuAcKAsOJbV6bH92urx3W3mEutOJW0sbkrSchQPIINbbsWvmSxzCWuFEfsrlvqnFusu9Le0+XTMhwUg9BCFOBp1a0ulOedwSE4xz3x3NYckOcf4Z1r7FfT8Ekxooh+JHlc7S7rmaAW36am/la1bLHt2l4Ny0zZYt3uj1yjJkstyGShhQcRsO4n8g4KlFQB7J5IApGGwgwtski17lyz8ScziOSWtDHcprfQ3tuew9Fv+kF2uUZDVkvs6PJdcgMz4W1alO9FX5gsq5zkg+e5xwBU8YvH8OQ61YVHHIcaQnLxGkN5i11gAX0qvRdLmS48WOXpT7bDIIy44oJSOcDk8d61k1qV881jpPKwWfRcpjKlyfSvT9qgJbclXd0suFxRB6e9bjrmfnCe58nycCsrS7wm1uV9FMIm8RmklPlZdVrrVNH77KY1h6oWuxOraZeivpbJDznWIAURkNo2JUVL89sDjJyQKlJkNbo3VZcXg80g5pWubew5dfc2RQ7dT91yrR1gaOlZeqtcPSnotweDaI8hHVfmJB/hp6p9zYKvIxkJ7gGsoYAPFmOhXdkmcZ24PD205g+K6ruaujptfVdf9PPTqz6ahuvKiR3LnKQUyXAnKAD3bQDnCPHyrufitkUDYh6lfO8Q4pLmOADiGDbXX3Pqfso++ejWlpkG7N26KYEqckbXEKUptlQIIUG87cZHb4JxivDjR0QBupR8aymvY57ubl7gLnPqfpy/WjR1vs05H4rFtoW7BnxSELQhKOUOtKzwkDO9JPAxis07HhjWHVdnhOTDLkSTsby81Ag7amtCoa5+ose5+lf8AZWM083LdeaaSTgIDW4KUCcnACsgZ7p58Gqzkh0fhhb28GfFmjNebbqTW97V6k7+66r6M+nMDTEJVxfkxLncpacCUwd7SG/7rZPfkZKu5xjxWvHgbE29185xrisubNyEFoHQ9/VX7U0W5yLNKa0/KaiXJYHSedRuSk5Gc/txnBq94cWnk0K5mI+FkzXZLS5nUA0VoWzSMQ9KTfFKu9yABW9KJWgL4JLbZ9rYyOMDOB3NQbEP59SrpM95JbB5Gdhoa9Tufqqr/AKQmm/xnQT0xn/jNqX9an/EgDDg/+Hn/ALIqvKj54z6LZwHM/C5jb2dQVB0BfLlcLtpZ2dAjzo1piOFn6YIW+tBSkbgkqBKkZA4BJKVAAnNYoHP52cw0AX0vFYMQQ5DoHkOkcLBsNB10uiNd7NLsNoms3/VKLjBcDsKHC6SV4Iw66oKUnHhQShGQeRuxxzXTb5nWvkJozjQeG8eZxv5Db5EnQ9Vo+omhEX1TN4sriIGp4JDkSZj2kg52uDsRyeSCRnyMioSw8/mG4XuFnmAGKTzRu3H6j1G6oGjbb/ajWjrdxjNR59tmKm3NCXSpP1AXhIb57HYCT4GU98VkjYJJTzDUGz7r6DMyHYeAwxutr28jdB8O5J+ZXXdZ2+TcNPyW4CUquDYS/F3YA6zagtHPgFQAPI4JrfILaa3Xy2G9rJmmX4TofY6FbGnb3GvEZSmSW5DRCJEZwFLjC8cpWk8j7HsRyMijXBwXmRA6BwB1B2PQj0UwDkVJUpREoi8WMpNF4VVdBw48J7UjURltlsXVwhKE4GS22o/5k1XG0Nuu6350r5fDdIbPKB9LCtdWLCvlad3kj9KLylUbJAjW/wBQdRfRt7BKixZT3JO50qeST9uEp4/fzVTGhr3V1XQyJpJcWIPN8tge2h/VXAdqtWBVzUunV3SXDnQ7g9AuMNK0supaQ6kBeN2ULHPCcZBGKrfHzkOuiFqxsrwWOic0Oa6rFkbbbEKO05c75bpoturUNOOyH1phz4wAacGMhtSe6V4CiO4PIzkc+RueNJPqtGXFjSDxMKwAPM07j1B6hXNKtwBq1cy17RepREoiURY3FAJ5Ir0KLiB1VHudzGndcLfW2Rbp0Iuy5C3EpQwpkhIVyc8haQUjzggZJrO9/hya7Eb+y6+Njfi8UtYfO1wDW1ZdzCz+W/ZQ90v93nXovQby5b4TjJEWAxA+plyOeHi2R/DSrnBURwB81U5zy62uoH0srXj4+PHBUkXM8HVxcWtHpY3I6gLDM0Zc705GejsJtMhh5LxuFwdTMmufbGChsYOcA4yAMCj4HPojT1OpU4OKw4vMxx8RpBHK22sv8yuiWaxW+129MWJGQGwSSVjcpaj3UpR5Uo+Sa1NYGiguDJkSzHmeTp9vQLX1RYWLzZH4W1LbuwiM8BhUdzHtWgjlJBx2qMrA5tKzEyn48okB06juOxHX5r5004xcrLHdWwEucIfbdAWpLqPYsKJHKgUkZPcYqTNQFDIBjlcAfatqOui2bxpqz3iGYtzt0WQwTnapsAg/II5B+4NeOja8U4KUGXPjvEkTyCPVZbbYrZax/wAGwIsT27CWGkoJTnOCQMnkk/vXrWNZ8IUJp5ZxUrifcrTmaRsst5x5UMNSHMFT0ZxTCyR2VuQRz96iYWE3SvZxDIY3k5rHY0fztRN00ZJelQHIGpb5EDDoW6DJLodTg8YVwD9yCO/FQMOoIcVqj4o0Nc18DDYoeWq9dFhv2jLjPRG+m1ZeWnI7weT1ShaFKHbcEBBIz4zSSEvohxFKOJxKKDmEkDXBwrrY9t/yX1dJ2r4VolNptsSXK6KujKhLxtXjgqZXyeecJJ+MV650jWk1ZXkEWDLK23lrb1Dh09CNPrQW76exkM6ejSTKE6ZLAclTCFBTzo9pJCuRjG3bgYxjA7VKFtM136qjiEvPMWtHK0aNHYf7731VrqxY0oiURKIlESiJREoiURKIlESiJREoiURKIlESiJREoiURKIlESiJREoi8Xnacd6Lwrj2pXr7btQ3Bu4SXLm+6+2qxQlgsR1rWSDlQ4UpsDO1R+VY+MUnO0kk3ewX02I3FniYGN5AAfEddmgL0HS9rCuugnnIcMWCbhMy1toaTyMPMYw26n7HBSfhST9q0xkgcjtwuPnASP/ExfA/X2PUfL8irYDmrFiXtESiJRFUvUBxh22xLa4povzpsZtppZB37XkLVx5ASkk1XLRAB6rbgNk8R0jdA1pJPbSvutvUeqbdptIXdUzW4+wuF9qG680gA8hSkJO0/ripucGjVZooHzHlZv7r6ZnWXV9jkJgyotyt7yC070lhY5HY/B5zzzUNJGkbgqwePhzB2rXNNhUvRes3mNBx31WZ0xbc39K6pMloKy37ANqiFbjgYSQCScCs8Ex8IeXbT6Lt8V4a38e9plFvNjQ1Ttd9qHdaml1/g8yfq6TN6zNwk9G6suEgwOQG87gFewkBQIHtUCMhPKMBlzE77+n/ClmE5LGcPayuQEtP9d7nSxrWnqNd1PX5YGoZzM+FLmwLrb0sMmCdy0pQVFzIBBAJcThQz+1WyENceYXY6LBitL8dro3hpjdfm0Fmq16/DstW2X62Wtk2i1WmXFu0ZDcOAzcG9rkgK5GFEkltJyVc4AH6CoskY3yNFEaaq3Jw55T+KlkDmvJc4t1DT6gbE9Pdbl10DFn2BbE2dKenLSlbst91brbjiSFb1MlWzaDnCewBxUn47ZG0SqcbjEmLP4jGDlHQAA1toQLB9d1yG1TmkOyP7R3p6dZLXFeZbebUvplRwlKEJGCpG4pA5OSlPZOcc+K7Ieba219bnsDWB2NF4c0pB5TRI9b6Hr873W16JNNJssRFossCfqF5x1S7m6kKTb2+Ep6nGdx2qIQCM8c8mrsVwoNYLPX0XO43G50rpciQtYKAHV/evbqdV06TIvYhXGJrSDEVbJRWyiRb0rd6KSAEqWj82M8hQyUkc4GDWol4vxRp6LitixnPY7Ae4PGpDqF9wDt6Ue/yWtafU62xIsaFeDLN0bKWX+jGW4lagDlwHAykhJURjI544NVNzWAU7dapf8OZL3Okg5Qw6i3Ab/wAu570O+6vGnL5C1DaGLlbHFORXs7VKQUnIJBGD9wa1RyNkbzN2XGy8SXDmdBOKc3dcx9QrnJja36U8Smreu2PR7ekJDjb8laFAnGOMZCTnOOOwNYslzhNR2o17lfScHgjkwv4dF3O0v3BDAQfv/dcm1YLDpT1CYftDKHrXKtyVqaSVBLRfZUkqSfzdiF44POBiqHckbxQ0I/NdXGORl4xEhp7X6evKe32Xd9LaK04/om3C2K64WwHGLmhKmnd5yQ6kZBScnOP2NbooGMYA36r5fO4xlz5LjNsTq3p7Kb03eLn+Iqst7gvGXHjpcVcGk/6tIGcZBwNqj5RjjBwcYNWMcbLHDbr3WTJxohEMmB/lJI5T8Qrv6dirUkYFWLCql6rwLjc/T6+Q7MnfNejlKUYBK05G5Iz5Kcgfeq5QSwgLXgSMiymPk2BX5q9InXJN7k2BcCFIlymloiuTxgQ1pJWtQTwdxweBghQHYZrm4wtxZWq+44w4MhbktkIjuyG/zdiu+aSseqdPwVdN22SVySXnozzi0IZdJ5LawlRUFDBO7nOT5NbY45YxuCvl83Lwct4sOaGgAEUSR6gkAV0pbeotXzbVZZYuNvct1xW0URHStL0Zbx9qB1BgJwSD7wOM98VJ0xaKcKP2VOLw9k8rfCdztvUbOoanTrp2tUy8RV6UnQ9WaSkx7u60wi0zITSgoyV+5aldROffu93Yk9vOKrcPDqRmvRb43ty2vxMppYSeYON6DYCj0XQdKa8sWobGzcmp8aOFYQ6y+6lC2XMZKFZxyPnyOavbKxzeYFcibh2TFKYeQkjsCb9dFHaudZjXjTd7t8mCy39V0JUsqB3x1JPt3A8p3Y+dpIPAzUZSQWvBFdVdgtBZNjSscTXlGujr7e1q9R3EuMoW2oKQsBSVA5BB8g+au3XMILTRCyjtREoi+XVBKCSQAPJ8UXhF7Kt6IUZDNzuDf/FbhNckxyRjc3tQhKv0OwqH2IqDNbK15gLXtjO7QAffevlas1TWVKIqRNujGldRXiZe0qZt85bbrc4DchG1oILS8cpOUkp4IO7A571OeIiS/buuhHjPzWMZj6vbY5eps7jv69qVst06PcYTMuG6l6M8gONuJ7KSRwRU2uDxYWKSN8TzHIKI0IWG5XeBbEJVcJsaKkgqBfdSjIHfGTzivXOa34ivYsebINQsLvYE/ktOU1a9V2AbXESoEpIcbeaV5BylaFeFJIyD3BFeGnt02UgZMWWiKcNwfyIUG1qSXpt6PbtUJkSC4FdG5xoylIdSBkl1KQS2scZxlJ7jHIFRkMRDX6+v91ubhNzGulxaFbtJAI/8Sdx91ITddWCG7AQ5cW3PrhuYLCVOgpwTk7QcD2kfqD8GpPmY0gE7qiLhuVMHFrPh32H519lmTrTT5ZdcXd4bIaIStL7nSUM4x7VYVzkY45rwTsOxR/DMxh1jPuBY+o0Wk1rFy5OSWrFZrnMWyraXHWxGZzjIO5zBwc+Ek/bkZeMTYY0n7BWHhzYqORK1oPY8x+jbH1K0169/BbclzWUBy0yulvIQtLza1YJ2pUkkgnHZQHPYmvDOGC5RSsbwo5MnJgv8QexB+h/utCRdFvsW+dqfUEaHDkoKo8K0urSp9eBx1EkqcKc4CUYBJ/QV454Ornb7AKyLHLS+PFhLnNOpcAQB7HQX3Kr970Vc72yu73V6Y/Bt6C5CtdxIW462PcpLhRyCQAASVKBAKviqX47n+dx0HRdPG4xDij8NA1odJo57bAB2BF9utACtl1DS9vtMa2tO2SLHjxJKEPJLKQAsFI2nI78YxWxga0eUUvmsqWaaQ+O4kjTVTQSAMVJUL0DFEQjNEVO0pETE1hq5Lbjqm3Xo8gpWskJWpr3Y+Ow/yHYCqWDle6l0cqYyY8AIGgI29equNXLnJREoiYoi82ii8pfCgPPmvU2Ko9pvsO1XTULCo9wcH4ipxP0sF59Ay02VDKEkA7t2RnuTnvVDZKLr7rqZGN4scL2EXya24A/E7ue1Kci6ts76JCnJgimMEqebmIVGW2lRwlRS4AdpJwD2zU/Eaeqyuw5mkANu9q1H2tTyVbsEYxU1mX1REoiURKIlESiJREoiURKIlESiJREoiURKIlESiJREoiURKIlESiLxZwCT4oi5h6iXBibqrStut11Y/EWbmhbkJKkq7DO5wdxgBQH3X2rHO8Oexodrey+h4VBJFjZGQ+I8pZQd8607r41fe7fpjXSbk6xH+ukxGorKnipsKCnFlSisJUQBtSDgE+4eKlJI2KUE7nRQwsOfNweSMktaXOIGtUANr62foVK6Zt13s+q3XrtInzhcmEBTiHCuOw8kqUpOw8oTggJPbgg4JAqxjHNeSTdrJl5MM+O1kQDSwnpRI0o31O9q+5wDVy5ihl6nsqbs7a1XOIi4N7dzCnQlQKuw57n7d+3zUPEbzct6rScLJ8Lx/DPIetaKQiTY8ouiO+06ppXTcCFhWxXfacdjyODUrvZZyxzQOYVfdVzU6I6tRaWUqO0/JTNXsSUgrSnor3LHwEkIJ+5HnFQkALmnst2JK9sM7WmgWj/7DRWlaEqRhQBBGCMdxU1g2NrmEvT0fQWsI19sbTEKwTv9VuzKSUoaOT03+cgAKO09gM5PGcVeH4bw5u3VdL8U7MgMMxt41aevqPpqFp+n9pk//iBqRq5RUrjxJipjS05Q2X3cEL2HJUQgcK7D3Y5PGeBpErwdgb+q6nFZo3YOPJE7zObyn2b0v57eyntcafgtT3r/ACkOPRlttxrlFJJacjAkFZSOSpG4KB7gI4q+SJush+awYGbKWtxWaOBJYRuHaaX2O1barZ9PoUKTFau0S9S7wx0vpoy5CEthhsEZSlISnklKck5PtHNShDSOdptVcTdMyR0EkYjN24Dqe51PrQ9V5e2Yupb8iGHFfRWpK3ZDzStnTkKGGwlY7LQNyjjtlOaOAlOvReQPkw4TI3QyaAHW2jfTsTQ9dVTtN2q4a5u8W6Sb3cXbLa3XmGZTbpZVccLGPa2QAlOCCscq7DAFUxtMlEk0PutuZLHh8zI42hzwDW/L169T26LBon0qt896eL/OkXe1Q5T0aFEdUpCWyF5U4ClWCT24+9eRYzaI3Csz+NTEtdGOR7gOY9T/AGC6/Z7LbbOyWbVBiw2jglDDQQDgYGcfatLWNboAuHNkSz14ri6trN17LfWn471K60VFWuXxdExJ2tdQOLmSm2Q+2pcQBKkONuNoUtJyMhK1JwcYJAIJIzWZkIMjj7L6CXibo8OFnKLo69QQSBt2G1/JTMlUXS1tVYtJQmzdHQtxiKjJCCru66SfagHyTzgAA1YeWIckQ17LE18mdN+KznHkFAu6n0A7/wDKgL9pW5DSpn3x56+3ltCQ6ylYbSpogpdaQUhOEkKKirvlI8AVU+MiPmf5nLfj58JyvCxx4UV6aXqNWkjW6qq9VxrSUK3StN6iNznKbZmzIsRMjoOLU02hzJWcAp8pAG7OAf3wxeGIyHnsF9TnHJflxyQR6gONWBZPzF1rrSvnpFqiXo68f2R1Q/i2OlSrdNeSppA8lI3gEIPPf8qsjyK148pjPhu26FfPcXwPxTBmwDzHRzQQT76Erq0nXemY6XFG7Rl9MElLWXCcD+XAOf2rUZ2AWSuHHwvKc4NEZBOmun5qHV6hu3CfaY+nrHNlJntrdS9L/wBVb2JHJSog5wSM8fpnIqr8QS5oY2weq3HgwhjlfkzNaWUKHmNn0C19Qi/N3JF0vGooumoCGvpwww6JIdWSVFQ3oSN2BgYCjxxXknMHc7n8o+q9xDjmI48WOZn3dm20PWidO+oXDLh9VH9YoD6npyC9Mb6MqRGbL5WrA3qbGAVBSh7TyBgEAjFYtRMDa+ntknDnCm1XwtOgr3s/TRfoGTpW+ImIujus5Ykx21D+JGbTG28Z3NjAxxyc58gjFbvCeHc/P/ZfKDiOMYzAMUUTvZ5vqo+TPu109N9RvXcRXRNS4xbjGbUnqoc/htEpOSNyiCOScHNHc74nB9a7KyJuPj58JgvyUX30I1d9AtfXekLbA0auZbtPsvXCOpqQ4mIotKUUqSpZ9v5uArxnnivJoWtYS1uvpop8O4lNLlNZLOQ1wLbd5gLGm/c0qNa7Yz6c64XqJwRV6Ont7gpDjbym0u4KQlJO9QQvA3JzlJz84qDPBeXn4St0mQ7iGM3FbfjNNdga9ehV31PP0GdIyrrDt9hnLkM7mmPY046SRkcDck574AORU5nQCMvoG/us+BDxV2YzGc97eU6nUhvr2pRekJ1lYsNuTYr2xa77HjNOymJTyksPkjBQtJO0dvzI5Twee1RifHyjldRHS1fnY2W6eT8REZIy4gOAHMCDuK1+R0OyvFt15CnREmJEny5KEkSWITXX6Ck8EFYwk8jjByoYIHNaGztcNNfZcaXhUsT+WQhgOxd5bHQ1uPnspzTeobbqOI5JtEnrtNOqZXlJQUrHcEEAjuKmyRsgtqx5OJNiuDZhRIsexX3qhFvcsFwReVhFtUwsSVFRSA3j3cjntXr65TeyYplbMwwDz2K66qt2OTqlmyQXVwIcpIaSC2t4sSVJA9qiNpQFkYJTkAEnmq2GWhoCFsyGYLpC1r3A3vQIHf1IvYqbsmpIdzlPQv4kW5sAKehSMJeQD2VgEhST4UkkVY14dp1WSfFkhAk0LDsRsVnv17iWhlBkqcU86drMdpJW68r4Qgcn7nsO5wKOcGbqMGO+c0zbqToB7lc4vrqNW29hjWk632CKyrrPQWpyXJCl4ISFnskDOduFEn4rHKWzCpSG106r6DFD+HvMmAx0pOgdykNA613PTpovjS1w1A1dpEewQY8yL7VSHn1KhsuE7il9DZBKN/Y7U7SUkjvkSjdKHUwWPomXBhGEOyHlrugFOPTykjTTpZujrsp/RXVlTJ72rYSWtQPPFgNupCmyykZSlgnOU/mJwSc5J4xVsPM+zKKP72XP4iI4S0YLiY636315h0Pbp2X1dLdp23XBMO1x50a5OJ3GPZHFNrCOfetKSEAZ4yrk+M145rGaNv5L2HIy8gc0rgWjS36/IEgn6Kv33Sd5ucVb2m4Rs8pKkuJfmznXJL5Cs84UpKPnncT2wOarkie9v8PQ+p1W3Dz8THmH4w+K2iCGtAaPsL+3dYI3phqeDcHbhb9W9KdKeR9U4lnbvbxkq7EFW7d4AIPJ5NVjDlaS5smp3WuT/EWDNEMeXEBa0Hl12P8AburPG0FMjsPts6ru+XlFS3VtMLdJIxnqFG7IAAGDxjjFXeCQK5z9lyncYY4gnHZp0HMB9AQP79Vu2v09scS3xY8xpdweaSAt2S64rqq7lRSVbe/jFWNgZy8p191lm4rkPmMrCGE/0iq9lNRNLWKIoqj2e3NrPBUmMjP9cZqTYmN+EAKmXOyZjckhPuSiNMWNueJzdot6JgX1A8lhIUFf3gccH708NnNzVqvTn5RjMJkPKel6KWUlO3HGKsWQ6qtaWbat17vdpZShqO2tuXHaRwENupwQB4HUQ4cD5qtg5SQFsynGVkczjZNg9yR3+RCtGasWNKIlEVQs0eaz6g6hzIbNveZjyEs7fcHCkoJ3fGGu36duc0MB8V/bRdPJMTsGBwaecFwJ7i7/AP8AZW+r1zEoiURKImaIqpr1lUmNa4ynnExZFwZYkNIO3qtqz7SocgZAPBGex4JqqYkctd1v4cxrnSOO4Y4j36KwW23xLbDRGgRmY0dOcNtJCUjJyeP1qwANFBY5ZHSvL5DZ7lReq9K2jVERLF6htyEoOUKyUrR87VDkZ8jsahJEyUU4LTg8QnwH+Jjuo/b5jYqOVZbzZn+pp+4CRCxj8NuC1FKeP+bewVJ7diFD9K8EbmfCdOx/urDlQTtIyGU7+pun1boPmNV5c75qaLDLsbS31DqSP4YuDeSMjOOO+Ki6SQCwz7q2PEwXuDXZFevKd1NWG/Qb006qC6S40vpvMuDa4yvylaTyk96tY8PFhYZsaTHNSDfUHoR3B7KXHapKhKIlESiJREoiURKIlESiJREoiURKIlESiJREoiURKIlESiKM1Jck2qzS5uwOrabPTazjqLPCEf8AaUQP3rxzuUEq2CPxZGsJq/y6n5DVcyhenqbVf25F0mm5P3hxaHSsdNTToR1AtC0ncCFoUMgj2kDxWJuM1j7OvN/yvo5+NyZWN4cTQxsQFeoJ5aI21B1WpqBUm9WLTiZMWa+v8LJExOA2qYVNpQlS8jaStsgKx/On74lJUgaCL039V5iM/ByyuDmg89cut8tG9Oo127Aq5Wu5aiesrb0CPbbmlbJ6MhchcdZUBj+KgoOFZzuAPcEcVcHScoLRa5ckGH4pbI9za30Dh8jY07eizhrVN2gtRZiYtn3oT9TJivl509twaG0BBPI3EqxnjJGaDxHb0Puou/BQPL4yZOwIofPU37KVh6XssWIuOzaofSc/2oWylZd85WVZKjyeTk1MRsGlLO/OyHkOc82NtdvYbD5KsQrbE056gR49ijw4sWdEefmtIeUCShSdqw32GCrGRjgq+BVTWCOUNjFWNVvknfmYLpMpxc5jmhpI01uxfy6+iyWt9uX6u3Isz0SUx7YhpTKUg/TKLgynd8nG7/f2FGm5yAb0+iTRlnDIyY+XmcSHf1Cu3ZX88JrRS46q+qprspz8Btu766Y3/Ed6QWiMwolKnFbvaScKCUnOT4wDUHn+UbrTjMaP48vwjp3PQf39Fy67tuelepIszdc5FkX0wHxl3a0FbVMuZ4O0EKQRgjaAB3zjcTjvBOxX0WMDxjFLABzt/lFDpdge/wAXcLoOsdQNPaEfuFmcVNjSwlgSYixhpKyEqcJyMbQT9wcZxyRqlfcdt1vsuNgYp/FiOamluuvcagfNaC7rH0ZJuloYeZXHRBeucKOFHcwEjlrnjYVcox29wxgCoF7YiWDtYVjcWTNY3KeP5g1x7k637gb/ACKkJuln39CR9PW+SiKl9CG5jykErU2rl8pH/pFkq5P94nvVnJ5OVZXZI/FGc61denb6afRaen4atCS5EWXJfk2WY6X2ZBZwiI4VBPTVt4SkgpIUcDIOcZ5riZ4BLSbBWvLnHFGiRjQ17RRH9QHUXufTU7dlZNDWp2x6Zg2+Sppb7QWXFtZ2qUpalkjPzuzVsbORoCx5+Q3JyHStFA7D0pWCprIholqsTtIWuXqCRd5Jl/UvMNsKDcpxpO1BJB9igf5vnHHbJOanQNc7m6+5W6HiU8MYhZVDXVrT+YKz2TTdosk6XLtzBakyglL61urWpe3JGdxJzyea9bE2MktG6ryM+fLDWSusNuhQFX6BVCSqdrq9vRGJrcbTkRxsrQhG9VwbUVgndnGzc2QBg55PxVJ5p38oPlH3XTYI+FwiVzeaV1gdOSq+p1+S5RoSfJsV1kaXnTozNliXhT0uS8dqVhg7ikJ+VFKDgc8D4NY2SFknhE+W7K+lyMVmXijNiaTLy8rQBr5tL+Wv1XW9bydH6z0qmPJmsz1vDMRMT+LJS4UnBQ2Pdn5BwMd63yGOZndfJYsGdw/IPKC0t3vQV6nZVv8A0fdRSGZF20jdJXWcgK6kRThWlRb7KQEq5AScHb3TuI7CqcSS7Z2W3j+IGcmWG8ofvWovuDtqrN6zapj6Vt1qkJjPv3Ay0riIZITyn8wV52qSooOAfzVfPIIwCsHCcR2VI9tgN5df0++vyXCNX+p2rLuwpi5RGI0dUkPxwYnuZKOAlClDPChyrvnI4HFc6bJe9tEL6/A4LjYr+djySBRAO9/oei2NA3J3UutrfcL+6/IuEaWiTHajpbbW8VOZWcHG5IIBITyBkjPOJQuL5AXlQ4jAzHxnQ47aBbRJ7Duenuv0BPhQ9Q6xeiXR1t+LbmULbhKV7HFuA7luIz78AAAEYGc+a3kB8haei+Sa92LiNli0c8mz2A0oe6tE+1xJ1tXBksJXFUAnYMpwBjGCMEYwMEYIwMVa4A6Fc+OR8b/EafN3VUh6UvzLsmM7qqW5alvJWhJQfqUNgDDaXt2QDjBOCTjuCTVLYnjTmsLpTZuNIGubAA8DUg6E96/TZYdS6K0e1pSbFubEeHCcTl2WpeHUqzkK6isknPjJB7Y5r18UTWEuFDuo42fnzZDRCS542aPvoNFQfT29G/usyXLhY4f0aTHjww2EKlyW0q2PuBICwnBB2gnkcCs8MnObsAdB+q7WfhnEjLTG9xJtztaa01bR0v1Vy9Lr/AXpNxF2W1Hk2pZjzHJKQ2lJKlFABV4wcAeO2KnjTB0fn3G6x8awZI8sfh7c2TVtak99vVYpeopca5zndLQUzItxKEx5D+5mN9UElOASPduASMjAKk4zzXpkcCfCF39LRuHFIxjc1/K5l2Bq7lv7Ub01Ot1ovZen9WSrmq6oEC3SHGEIULdKKHN4JKivc2UuA7uys42jBGTk6OUnm0B9EizMGNgicXPaCT5hYroNDbfcH3BWC4ztS3GbD0veLfaOu6hMsrVMUES0NLBUjaEHBJ25HbGcZHbwvlcRG8D67r1kGFDG7MgkcQPL8Pwk9bvb7qzTLvqlMtiLG04xueCiZSpwWy1jH58JCvngDnjnviznkJADfuueMbCDS8zHTpykE+2pH1VP1PY73CtSrldL1C/EGW1pekw4oalLilfvSlZWAQlKgcbRjGM85NMkbw3me4A9wNaXUw83GM4hihPhmqa423mrQmh12/PRZrLoZm/sQ5Tku4t2ttLYaUt5RfntpH5nFE5Q2Rja2kjAOTyRj1sAkp1mvufdeT8VdiOfGGtL9bFeVhP9I2Lh1JvsNF023WqDb2EtQocdhsHIS00lIz88DvWsNAFAL5988khLnuJJ7lVnWv1NomM322LDsohENcJSSfqwXMpSkjkLTlZB5GCrPHIrl8vnG62YPJOw4sujb5ub+nvfcHQV32U1fNO2i+uR3LrBalLj7ukpefbu74wfsK9exjviCzQZc0Ac2F1A711XumollixnE2BEFLG8oWYu0grTwQojuoffmvWBgFMTJkne4HIJJ6X29FNYHxU1QmKIlESiJRFWtU6qj2J6BH+kmzpUxzY2zDbDi0j+8oZGE5IGfvVT5Wx0Ku+y2YmC7LDnBwa1vVxoe3utZxzUl43phNpsMUcB6ShL8hZ+UoCtiB91FRPwKW9+o0U+XFgPnPiH0sN+pFn5ALRe0NNVMbnDVl8FwbbLaXglgAg84UkNgKTnkA5x471X+HJPMHm1qbxaNrPD/DM5SdvN9jeh9Qvi26qvMR2bb7vZJEu4wvdugqQQ+2c7XAhSgUhWCON2CCO4xU2yvFh7dR26qEuDjP5ZIJQ1rv6rtvpYBuvWtFP6c1TCuzTbbu6DciP4lvlEIfbOAcbTyRgggjgg/tUmStfpse3VZMnCkxyT8TR/MNW/VT4UD2qxZFWrO4XNaah3HIbbiNp+w2rV/vUag0eYla5T/wBtG31cfy/srNU1kSiJRF4pQT3OKLwkDdV266iQ1cFQLVGcudyTjeyyoJQx93XDwjt25UfAqHia03UrVHiFzfFldyN7nc+w6/vVa8OzXK43RifqFxhKYp3RoMValNJXjHVWpQG9XcJGAE9+TyAaS7mKkciOOIxQDV27jvXYdh37q1jsBU1kSiJRF4ocdq8RVK96ecjTnL5p1CWLtjc+yAA3PSBwhz/F/dX3BPORkVWYw087d/zW+HLDmDHyNWdD1b6j07jqFP2W5xrpbmZcVZLTg7KGFJIOClQ8KByCPBBqwODhYWOSJ0TzG7cLfr1QSiJREoiURKIlESiJREoiURKIlESiJREoiURKIlESiIe1EVCvlwfuPqPZ7ElsKhRm/wASkq2rzvTuDScjjG73YPfA/Q53PLpQwbDVdbHgEWBJluPmceQe38x/T5rQ9a3ZDFns7keUmMj8QQ06tSVY2LSpJClJIKU4KskY4PcVDMJAaQa1Wv8Aw6xkkkrHt5rYaHqCDY6WKUl9XZ9T6Vuto0rJhuLjtqioaQChDSx+XAx+UHGFAEccVaJGSsLYzssEkGTgZEc2WCL19wf19FTNA6d1BeIU2U3qC8WiOu6SHC3htfWSchSkrHc788kY4OBnmskMMj2kl5GpXe4rxDDgkYxkDJKY0X5tNO1/nquywWlR4jLTrqnnEICVOrxuWQMbjjye9dDpRXyL3Bzi4CgeizkivVG6XLfUvr27VFonQ2X0Gc2bWZURpK321KWFJSjcQApWCAo8AFVZcglrgQN9LXf4Q1uRDLE94pnn5XE8prSzWunbqt3Tc6Hom0sRLxBmxHVkJMxUZJMtwq7qLal5XySdx55I80jcImgPFevf6KrNhfnSukheHDflBIDR6Bwbp7KfvuudPWZlSpVzZWtKwgtRz1nAc45SnJHPzVj8iNgsn9VmxuEZmUf4cZrudB9TQVHu11VbbbfvUOIlrrLKI0ZExakoVGbUEnakc71qCiAe3HbnNRdQM43XQZEJHxcLmPlGp5RZ5iPuBptooaXqC460uD6L3PTp3T8dpK5FmXzMmtEA/AOFZCRt557ZINR8UzOIdoB06lWnCbgxtfADK9x0ePhafl1HW1Ho0Dp9GmpTE/SupGpSC4tMlgFIUgKJQpSSsoSrbhJyk+ftXggaGEFp6q6TiU78pr4pWUa0rboRt3/NUaxw2n4NmVNu15g5dG2e0v6hiOhKiEjHdCgrHGcYKT8VijfzBvNfvvS+jzYhDJM2ANIA+HYuJGpHQivnuFPRL7KgT79ZNY64uY+mCVQpUSTuEhZUM7c43DGO6glJ3fGa0Fx8zXv9qXHEDB4M2NjDzWHB47fverVrv2tm7xYkRb3dWYUF+MpLxs7rc96QQQkhQGemkpJV58jcMYNz5DJHyv0Hpraw4uI3EyPEx2+I8GwHAtA9Rep101r2UD6S6zslt1JBbuU67Mt/Srix3pbw+nGXSUpKR+Ubcck43Z7DFZ8WRjX0XH5ldTj+HlTQOlbE2iQTyt1+Hv117Luum9TRr1Pu8RsdN+3vhtSd4X1G1JCkOpI42qGcfBSQea6Yda+HkhMQBPX90vrVOq7RpqO27dZaWi6oJbbSN61Z8hI5I+TUZJWRi3FXYnD8jNJbA263J0A9ydFS9U3uZcp0Fh+6P6SipV1lrc5fdQcpG/aChlJO7BWrkjtxVMjnOI15f39l08KCKFr3cgndWnRoPpdFxHYD5qIvcN11x6Nou6XS5216O6zKix3VPNs78ZcDyjhSuCdgUVEk44JFVStJsRONdf7rfgzMaGv4hGxrgba4iia2aWjp60B3Uvp6627Ty1twHZc+PJitMwkqQoyFusAtqYUkgFKvclXIAAUewFXMe2MADXsublY0+US6SmkEk7UA7UHtX56BU8QdR6Y1Pqa/29mFcX8pcu1oHO5txIXlo7dxCcqB4GSCcHsKgx8bnP37hbZZ8XKghx9WtGjHWasb2Ol7itr7Kalesujbbptm4WmIldydb2pgNshtbeONriwMBPbtnI7CpnKia3mG6zR8CzZpjFIdB/MTY/3XJtW+o8S/6lsd8jWQQp8VX+ulD/EpOQNuQAr8oIyeeftWOSdj3BxHuvosLg+RjRSQeJYPw9QD31BV+m2y5XNuJq222tFttx6LMaI9KW5IdCn2ilawTtKSoDCCRwM5zjF7mO0kjFD7rnxTwtkdi5b+Z4vUAUBRsWBYPqAfYqJuSLrc5F4/ERDuMKBKTcZjp2hLAX7FJT01KH5UkkA5O0Hkjmh3iPJ5hYBtdaE4uO2M45cx72ljRrrWoJ5g06nS9v0mbtpK16mt2n7uzI/C5F0mSEsSWsjprLji2eMjwjbxg5KfIq8wseGuB3J191x28RycV0uO5nN4bW2N7DQA7+/VQNuu7OnPUNDD6RIv1tW6H7g2tbwuZVsyCk7ihWwrGQcAjt3FVWyOa71G57rY6PIzOHkbROotboOSr1vTQmvcLuMbXtgcZtjzk36di4pdVHckILaVdM+4En8p+M8HxW4Ts0JOh2Xyx4RltdIwMssq6o77bb/Jad5v869TIMPREph7a+lc2WWuqw01tzt3ZAUs5B2JOfkpr173OoRH/hIMaLHDznNOxpux5vUdB6r1eirZPth/FpMq9uhKlF2ZJW40XOfcGkqCE45GEgYHHzXjoWubTtV7FxXIgk5sf+H00A29yLXKv9GUxLlD1DZZ8RqVHUtmUeskFKsEpAKT90g1nwiPM3suz/iQSRuhyWuokV6/vVdLvdhjQ9eaXlW+BbG23FPMvIEcBRCWytKxjgFJBAPf3/HbQWASNLaXKhypJcKdsrnEiiDegs0QetH6Kyart0mfZFCA2hyfHcalR0uL2pU42sLCSfGcEZ8Zq2S+XyrBiSNjk8/wmwfmKWa3XqLMspuDhMZlAX1g/hJYKCQtKznAKSCCc44oHAjmVckD45PCGpO1dbFhQWn7Db7zbH7jcYCFPXKQqWFkFDqUbj0ffwoYRg4yMbjUGxhwJd1W3Iy5IJQyF3wCvTbXQ6ei1hCuOipM25CXMudjeUlTzEh1bz0NIGCtBOd6R/MDg4GcnHMOR0ZLuax+SsdPHnxxwlgZILFigHX32o9llusC0aj1vaV9FiY5a21PPuhW5KN2Omg44JJysA9gkHyK9c1skg9P1XsM8+FhyNstElAd9Nzr6afNSnpwP/ydblY9qw4tP3SpxRSf6EVZH8IWTPv8Q69/7BWaprIqfrlc1cqyMWUMKuf1anWw/nYhAaWlbiseE9QceSQPNUzXbeXe10uH+G1kpmvlLQNN75ga+dFZWNEWgtBV1jC6SykB2VN/iOOEDGT4A+wAFS8MVrqqH50vMTH5W9hsP1WO0xY8LXctm2stMR/w5ovoYSEoDnUUEZA4zsCgPsP0r0ABxAXsj3Pxw6Q2eY1ep9fv91b6msaURKIlEXilADk0pFS/T+1x3PqNSKK13G7je8oqJSlAUrYhH+EDA++M1RC0V4nVy6nE53AtwxXJHtXqBZPfVXQADsKvXLXpoihdQafjXjpuKW/GmMghiXGc2OtZ7gHsQcDKSCDjkVBzA4ditGPlPgNUHNO4OoP77jVVG/aIvd4Mdbt5hNToiVCPdG4akysEEYUQvbjnnAHyMZNUSwOkA81EdaXVxOKY2MCBES127C7y/lenqfe1m0hqVdrNwtWr7nEZlQFIQh2S+lDjySCrd43J7YUACexGUnPsUnLbJDqFVm4fils+Iwlr9gAaB7eh+Z7jRTekFonSrzdWUK+mmyU/TuKSUlxtDaEbgDzt3BePkc+atj1LiseZTWxRdWjX3JJ/IhWerFjSiLxSgkZJovLpVS+yZF8uRstrU8yw0pBuM1Ci2Wkkbg02e/UUMZI/Kk5zkiq3W48o+a3Y4ZA38Q+if5QdbPc+g6dz6KetNrh2qGmLb4zUdhPIQ0kJGfJPyfueTUg0DZZZJnzO5pDZW8BipKCURKIlESiLxQz4zQixSKh3y0yLBqReqYcp/wDDtmLhASohGDwp8AcFSQAojGSATnwaTGWyeIDp2/VdSPJbk434J7RzA+V3X/xPp2V6acStCVJIIIzkVdd7Ll7br7oiURKIlESiJREoiURKIlESiJREoiURKIlESiJREPaiKvzdS/TvuMt2e9SVoUU/wontVjyFKISR+/Piq/E1oArazCLmhxkaL7u/QaqL0nNauWrNRS2GX2WtkRhaX2i0oOpS4pQIPnatvn4I+K8jdzOdSnmQGCKJj3A3zEUb00H6FfV8gs3zUL8KfuMOJALnTTnlbxWjf+qUIUB8FZo4B7uU7UkL3YsAmj+Iu3/8aNfMnX2Wj6aWdaYkC7KjQ4rJgJZitxkBJW2pQWXHOOFKwk7QSE88nPEYWVTiK0V/E8oOLoWuLrdZJ6Hah6DXXS9NBS3bdYdQWBgxbNcIMuGVrWhE9lSVtblFWN7Z9wyo9wD96k1r26A37rPLPj5DueRha7T4djQ7H+6zz7lqqGyhabHb5SitKFJYnrJAJwVYLY4Hc9zj5rwulH8o+v8AsrGQcPcaMrh7sH//AEq4nVusnri6iDp+FcIbEwxn3Iz5BTtI3pSFkbinON2ACfFVeNNflaCFrHDuHCO5Zyx5bYBG/a6ur7LR1oLpJ1BfgqLGkNw7ch6GPq8LaTuUVOJQU43kpT3I4SMHk0le8udQ2FhW4LMaPHieXkczyHeXQmhQJu+XXoDruNFadbTI8r08nvSw0gS4gDYcwUpccACOT8KUOe/mrZXN8Lmd1XO4fDKM5scV20nb03Oi8u9p0npmzyLzJs1rYRCT1itEdCVFQ/Ljj8xVjH3IqRjijHNQ0VceTm5j2wCRx5tBZPVULQXp9I1Fc4Oq9SusuQZBdnN2gtr2NOuq3AkKOMeeByQD2rNDj2edx0OtLr8R4qI2/hIWgOaA0uG5AH7tdjdtEB6WJT0KMuSNpDqmkqXlOdvJGeMnHxmtlC7rVfOtlkY3ka4gdrNKL11JVbtHXqQyB1W4bqke8Iwdp5z4xnNRlfysLuyv4fC2bKihOluA+6r3pGWrjpGU8htrpyJTwCU4KNqQG0gcAEBKEjtziqcV3PGT3K6HHITi5bY7NtaPezZPf81semlktUXTiYf4TFZmxFLhzUlCVlToxuO7HKVApUB8EDAxipwsYG0BSzcSyp5p/EfIXA0QrfHgxIiNkWMwwnBGG2wkYP6CrgK2WB7nPNuJJVE9SNMJcSLpBsVquiWWHUyITyNipGSghSVAfmTtJGeeSB3rPPH/ADhtrr8Ny6PgSzOZZFGyQDruPVc8kelembnajN0rqQxVvsh/6Z+QlTZazkpV2WBuH5iDjHY1nGNG4c0bqXVPGcuJ4iy4OajV1179r9lp6JSNCG33u7MOr05eWui4t5SJCWm1ZLeFYCgQCQpGMEFSgSRtqUIEQDzsVHiD35z3Y0Rp7DoBYsjQ6bexJ/uruj1M9PtPuvQ7Mh6S46sJWIMZTgcITgDco+4eAATUxPCzyt1WGThfEsoeNNpXc1X0RXqhdorUZb+gLu3Glq2wwlxO9wcn8m3KTgZx8c9ql47h/KVFvCI3uIbO0kb9h8+q5td9YRnJku43vTF8bZfnCQ08HzHMfICg2lW3nfjKufjH5QaxveHHmkB309P31X0UGMY42wYsrSeWnCru+p/8f5fne6+r7LvHqxNZTo21SYcO3wzHefkzD7wcHpqWTz2IxyT3VgVJ95Lrj0pU4/hcEidHmODucg0BdV1Cp+g/Tu8a0m3CPBXHYEJP8R11e5vfnASCnOeyjkZHH3FUw4zpSR2XTz+Mw4TWveCebstrU/pjedLNsLmzbUqS6peyO3I95SgElfuAGOPJ7kCvJscxVzEar3A4yzNLjCxxDRrp1JAA99V9W31M1NEjSIt4lSJ9ulxVxQ0+AAjvtcQQPzJUR+wxxgYk3Kfy0dQqpeA43iAx2x4Nncn1XUPSPWNgZcu/4xMtdufvD6Hmo7Q2tD2lJTyNqTkdieSqr8SVp5uagT0XI/xHgStMQgDnMYKDj11v6dPkq+1qRy23KJb2S6bZHfkuN/XQypTTi1lAWptPcpB3AAD3LI4BGKBPyP5OgJXWdw5uVjOyDRkc1gNOqwG2RfrsbOwB1Krmr48zROp7XfbLDkRWkgKhyZDIR9RgHepaBjbu3fk9pCCngV5JzQPDgNOnr3VuOY+JYr4nvt104DZorygXvVb3qb1WkzrC4XaxSrZKcdXGcCUMQ0NKWlte4KHTOCQSQUhHkEjIAqLpJJWlu4KtixcTAljnA5XNsl10HCqqr+/zV/j+k2sLzpC0Jk39MByOVOsW1xsoTG3HPK0c7/JJBIyee9a2Ysnhgc1V0Xz+Rx3DOXJJ4XNzHV139Adgq96f+osjQFrvlgnJU46088mMAkqDD4BHIJGUFQyQOQSTzmq4sgwWx2q15/B4+JuZkwjlBonoK/v9lP8ApLe3dPWeNPXpt+FFlub7jfnELfS+gKXkBCBlvkgA4xwSRzmrsd5AHl+a53FMYTzPYZbLdGt2I+u+i6FrW86ZnQbLfU3mAp6FLafiOJkpG8KUkLSRnONuSfjbV0zo3APvULm8NiyWvfAGGntIOnbUfel0KFMjzoTUiG82/HdTubdbUFJUPkEVeCHCwuS9jmOLXCiOiokm2uL1QdPuILlpmSFXd1JUFJCR+dpQ/uqeKFYPcFQ8Yqkg8/IdjquoyUDHGS009o5PrsR7Cwfkug8Np+B4FX+y45Nalc9v70LX81Njjonhm2ziqc+AplKSgEBsH+YqKsfYBR49pOV/LOeTsdV3scTcJZ+JdXM9vlGh36+lV9a9VC6ht0jTi42mdMz1IiXZfSVGyN8BClf7RCxhWD7kgLJz4PBqEjTEBHGdD9lqxJmZrnZmW23Ri/8Azro4enoum6btybRY4NvQ+7IRGaS0l10+5QAwM1rYzkaG3suBkz/iJnS0BzEmhtqpM9qkqFV7H/rmp79OcIWGXEQGCPyhCEha8H53rIP/AEAPFQbuSVqnJZDHHtuT7nQfYKSv99hWSF15zhBWdrTSE7nHl+EISOVE/A/fAr0uAFlUxQvmNM+foO5UdoO0z7baVrvb6ZF0lOqkyHB3BOAEE9jtSAnjA44+TCFrmt8x1WnPyI5pQIRTGgAD8z8zqrPVqxJRF4TiiLzeCCRk/pReWqdebm9qFb9n064enktTbik+xgHhSG1DhTuD44R3VzgGBJdo1boY24wE8w13De/v2H59FbIcZuLHaYYQltlpIQhCeyUgYAH7VPYUFjcS4lx3Kz0XiURM0RRGo7wzaLa9IcWnq4KWW8ZU66R7UJA5UScDArxzuUK2CB07w0bda6DqfT5qps2C+sut6gkNW2TfnEhEqKlO1tbYSn2NrVnasFJIJ9uVEcDBrOGuB8Q/F1XUfmY72fg2lwiGxJ1B11IHQ3RA1oK3WO7x7rFW6wHm1tr6TzLyChxlYAJSseDyD5BBBBIq9hDhYXLnhdA7lefYjY+ylAakqbRSsd6ITSpes5T97X/Z+wTJLE/qtqlS4xGIbYIUd6v7ygOEDk9zgd6JbeORv/C6WCWY7hk5DQWi6B/mP9h1KsdjtMe0QExowWRuK1uOLK1uLPKlqUeSSfP/AHVc1vKKWGSZ0zud2noNh6D0UkM+a9UAlESiJREoiURKItafGRLiux3clt1BbXg44Iwef0NeEWCF615Y4PG41Vf9OYogaTiQUqUtENT0UKWoqKum6tOcn5x28dqrhoModNFu4qS/KdI7+YB2nqAf1Vpq1YEoiURKIlESiJREoiURKIlESiJREoiURKIvCcUS18OupabUtxSUpSMlSjgAfJogs6BY4MxifFakxHmn47o3IcbUFJUPkEd6AgiwpPa6Nxa8EEdCvt3CUKJCexPJxXmqiAL0XM9B6imPWd9MKzGTc5VwkvKWlw/S8uKyoyNu0gYCAE7icDx2zxSlzbA1+31Xc4jhsjyAHyeQNb08223Lenzpeaivl6s1+YuD9qSJs1kQY0aO4qUhZSouKKylKSDg7UgJONxJIAo+RzHC26nReYmNDkwOYH6M8xJoHXTSzR7nUdletJQHbZp+3w5BHVZZCVhJyAe5A+wzj9qujbytDSubmTtyJ3ytFAlTBNTWZYVqSoqAUN2ORQbrw7UqdBuR0uqTCvTDrdvD7jsaehsraKFq37XCnOxQKiMkAEAHPeqWkx+V216LqTR/ji2WE26gCLo2BWl7igPW1W2CjWerrs7brY2i3bGYci4qWkF9tLnUIQByoLASnnjb38CqQfFlcANqFroyA8OwohK+3EucGjoSOUX7Gz3tRHqxHU5fHZGpQlURUcxbRAi9R5chxSgFOFAGErAUMZ4zt54qGTGXOJk26BX8Hy2QQtZi2Hk29x6AdB6d186k07e73bLZK1Y+3ZrTHlRWItniO/kSp1CN7rhOCtKCcY7Y8c1OVjnN8+gsaLLg5UcD3HHbzvLXEuI7AnQe/wBdl2u3xWYURqNFbS1HaSENtoGAlI4AArYAAKC+fc90jy95slbNF4qd6m2CfqOwog2x9plwSWnVdZSghSEnJB29/Bx9qpyIjKzlaV0uE50WDkGWVtiiOmhOxX1/aSPaY4jtWe4qEb/jH0kFaGGQD71JUoJCkjkjbkkDtUucN0pUDHfKeYvFnazqew6/dQnptqW2M6fvKFvhDFrlvuOPrGAtpbinEucZJyDgnyR96ogmZyHXZdLi3D8g5ER5dZAKA7gVXotiVMuutWoLunZEq0WcOh5VwyEuyUDI2ttkEhJP8ysdvykVO3TNBjNDuqmsh4a97cpokfVcutNPqf0H1UxpNTs20ToVyeVcWWJL0MSX0j/WWxjJUAADjcpBwMHbnzVrLohywZJa2RskQo0DQ6H96qqa6h6T0zFs1u+ggxA5NbfbK2UdNIQtJWXFq7Dbu798VTIIogGALp4j83OfLO55cA3XU3roKA9VTdTXtR06q3WDWtr1Mm5uIiCFLjNl9rqHaFthGNoTkHlBx3FQe4kEMddq7HhY2USZMLmcupIO/Wtf7qTtHooWtFNNPSI0TVjEgyY9wjBR6ZBG1ClHlQ4zkAEHGAcHJuLysA/m7pkcfMmUXgHwyKLT2X0nXsm0aniI9SIX4dItzLrYlw0qdYeU7tIyRnaSlHA8n4xUhMWu/iaUq3cMjlg/7E83OfhJoij991GXu/QvV3Xlhs1nC3bRFZfkSnHkFJBUkpztPxlGD8q+1VOeMl4aPhC1RY8vBcZ80pqRxAAB6XZVgl2WN6V+nb9rtAeuV8vL30jSwkJU88tJSCE9glKcn9e/ereTwI+Vu5WRuQ7iuYJZtGN1PoB/dJ2mIttb9PrA9E6a2sNyX0sBbSwEEraU5wfcskgec57gVF7NY2UtGHluLMvK5umgvW7ABA60N/oqzO0m07rSVZdOS0SFqYdwt9P8OGrrIWUhairepODkJGecH+aszsf+KWxn/ZddnFXfgW5GWyhzCxer/KRZAAoHTe/yUb6uaXuNh0gr8TXaX0LuDaI8ltBacQ2ELIbS3jahJVuUduSSSTnwnjfHGeajr+9F5wnMx83NHhFzHcuoJsE+rt9qGo0ApcbWtxiO0ODkqyk+4EVz2gFxIX2bi+CJre9+vVbEO7TLapt+A+7EkpOUuR3FNkD9Qa9bbXcwKoyWxz44ilYCLuq0XY/T+8O+qjDem9XPNuKhtuPsvgYefO3bz4JTu3E+cDPk10Ynfih4cnRfH5sP/Qv+8xL85qjtW/0P2Vm9NrG7DhXzTtmm2eLOadcjTpDUd0StoUpKHAd4Ge+Mds/Pe2Bh5XRtdqFg4pOwzRZ0sTuR1EAkcvSwN9L6UofSWpLlJvatNaBuz7jLKCuTOvgDiW0tHaVsoCs7VEgbTzwDxkmvIXG/DjdfclS4hjsdGMzLiDeb4Ws0u+/9/spi0egdsRdEzL5d37kgnquMBkNJdWTlW45J25J4GP1qxuGOfmcbWab/ABHIYRDCwN0ruaVp1hBft97trmjYbKb8+5vdAcLTKmEp2qLyRwRygA43A4xnBFSmDmkGIarPw+SKSOQZzj4QGmlnm6AHptfboq5d/TS76gnzX7oxpiIiWztCokRSlsuZ3bkk7cqUeFLOTjsBVT8aWQnmrX7Lowcaw8ONghEji02bIAIqtd9B0HdaunNI3OHPnSNISJVgmQpJZkWyQ8Hob+5tJy2OSkHIIKgTyPuBKOJ4JLdCOnRUZmZDyNbPUrXCwQKcPQ9Ce6w2r1TTp+XcJWqLFcUTpL3T67aB01BsbUoQT7dvCzwo+7eag3LLLMgPuFqdwJuWWQ4srQABod9RZOm52Gw0AXTNfXYNaAu0yPIMSQqCtxglYQ4FbMjHOcjPitMj/wCGSD0XD4ZCH5sbHDmAdR0sLE1qHTunIce3x3WipCEgRISC84nPkpTkjJ8nk/evTJHG0UonFysp5kcN+p0Hyvp+ShYU3T2opepFzZsREaS4w2gPOhl5KmkfnAVhSSFH2n5SagHRvLgStToMzGbGWMJIBOgJGvQ9CCN1O2bUjduKbbqCawH08MTlKCWpiBxu3flDg7KTnvyODxNr68r/APlZpcN0v8THadd29W/Leux+XRWWROYYjF+Q600wACXFrCUAHsdx4q0kN1JWBrXvdyMaSey5nY4F41JAjqt9xetNoM2RKdkR3Muzt7yydmD7Ec9zyc5wMDOSMSSgEGhZ+eq+gyn42DI9j2c7+VoAOzPKPqfTb16K6xtLx2NVuX4ypTslUVMUNur3IQkEElPkE4Gccdz5q8RjxDIuS/Ne7FGLQA5uawNSrDjaKsWRRN91DbLEw27dprEVDhKUdRWCsgZIA7k/b7j5FRe9rK5itGPiT5RIgaXEb1091Govt/ktIXD0wpAWncPrZyGsA9gQkLIPyMcVEOef5furzj4zQLmv2af1oL7Q1qeXudckWu35OEsBlcnAx5XuRk/on+tefxDsQPuvObCZoWud62G/anfmsDWio8lSndQXCfeHVnJQ+6W2Ec5wlpBCQO3fPanhA/GbUjxBzKEDQyuoFn5k/pSssOGxCjtx4jLbLDY2obbSEpSPgAcCrGgNFBYHudI4uebJ3vqtivV4vFHAoiibxqO02ZxDdzuEWK44kqQhxfuUPsnuai57W/EVfFjTTf5bSVBO+oNt+vdYbh3Z2O2x1lymYDymxzyn8uc45zjH3qvx2k0L+i2nhE7Yw9xaCSBRcL166HbotexaXReZVu1NqCS9LnpHXix0rH08QKwUhAT+YgbcqJOSM9q8bCHEPdqQj89+PE/DgAa06ONaurue3YK8cBOM9q0LlWFXr9pqNcFLmxFmFeAkdKa0SlSVJ5TvAIC0+ClWeCRxVb4w831WvHzJIRyO8zOrTtr27H2XkDVkAWREy6yI8B5BLUhl1wAtvJ/OgDuTntjuCD5p4jasleyYUol8ONpcDqCOoOxWCZqz6mP07JbLlMmOcM9SE6yznHCluLSAEjufJ7AEmvPE/pXv4It1mcABvqCfkBupbTVpRZ7SxECg44lO513bguunlaz91KJNTDeUUFmml8eQvI9vQdlLivVBKIlESiJREoiURKIsT4UUnYoA+CRkV4dAvCqz6bty2tMIbnvtyJSJMlLjiEFIWoPrBOCT3OTVUHwUe5/NdLivKcm2ChTa/wDiFa6uXOSiJREoiURKIlESiJREoiURKIlESiJREoigNaLkR7SmdEU6FwXkSVIbJ/iNpP8AESQO42FXHyAfFQeaFrTiNEkvhu05gRr36fdRuopCNQpTYbe624zNZ6kx5KQ4G4yhgY8b15wnPYBSscCouAd5R1VkAON/3Eg1afKO5/s3r9Oq0bVIj6GnSrZN/wBUsCyHLa6QpaEe3+I0pQyQd2VAHuCeTjA8YBD5apvRX5JfxGpweaT+b17EfLRVr1C1a1qW33K2WG5Q/pIjYdm7num5KbHuW215wEg7jjk4SPNZ8iUSAsYdt11eFcOdhyRz5UZ89hmlgHoSOuuwv12XRNHTLJLsbCdMvsO22OAy2GuzeADtPkHBBOeeea1xOYW/w9lwM+HJind+LBDzrqoPW1zdsWoLDcGbbMuHU68NTcVKVLwtKVjAJHOW+/bGckcVXkP8NzXVfRa+G4v4qCaIva2qdZ9NKv5/M0rvGX1GkLKVIKkglCsZT9jjzV5XK06KC13dJNp07JkW8BU/2ojt9PqF1wqACAnIyTyPtyfFVSuLGFzd1s4fAzIyGxymm9TtQ6lUG36evTOpp0qJdnYN7utsElYdbDjQc6h3pCMnHTy2E8nGSTnOKobC8PLmmnEfJdqTOxjitifEHxMeQCDTqrf562rDdJ+t40KK3Fs9vdfBSJEhuakgowdxCVhO055HcD4NWPdMAA0Wudjw8Ne9xkkLRryir66AkdK9ioH0qkahFleuJMG+NzHcKcRL2OpLY2e4qThRISDnIPY854qxjJyFx81ro8ajwfGbjs5ouUdRY1101vqsVu0zcNaa0l3HVM1Bi2iQGo0OIvIbUrY4UqcGMlOEBXkkHtxUvCfK+5DoDpSgeI4+BjBmEzzvaQ5x69NB+Vq6eoMONLt1rZkstSFG6ROm04kEKPUG7j/obz+37VdMA4AeoXJ4fIYXvcw15XD7K2N9uDn71asNL6ovV8FQ3Y80peX0WtNkxocZ2RMebYjtJKluOK2pQB5JPahNCyvWRl7uRgslUT1EtlmjQY+qHojDkaEC9KCP4apLRSSlPGN537CEq45NUStYB4jgurgZOTZxWONu0HWj1Ou2ljRQdh9SNUantSXtL6KHSLojiTJmAMIPngBJUAAQSOAcD7VFkz5BbGq7J4bjYkpZkT2fQEn/AJUVrD1J1A7BuNptkNhEmOpH1l2tDq5jUNoqwTjak7xjt8Z7eIyTOILR9Qrsbh0AIlkO9013ls/2WjAsnp/NsgucC5t6guTLzDsg3STh51CV5U2lC8AFWDgHOe2eaiGxFtg37q8zcQEphczw2kEANGg9dL0HU9F1C33rR1vbelRXrTBWy3l7+Elh1Ce2FJwFDnjGK0CSIC7AXGdg8QkIaWudZruCfcaLydrpAtEmdabXNltNMl7quo6DO0AncVKO7GAeyTk8d848M1i2C1ZHwzlfyTyBpuqFl30H917owOXOTfZl4gpizH3W2noSlB1LaENJKMnsokLJP9PFexkvsu37KOcGwiNkR5mgEh21660OgG3yvqoufqaBatSO6f0Zp9mffA2C+IyEMMRv7vWcA475wMn9zXjpGh3I0aryPGfJF+IyH0zpe59l9aX0XentVM6m1tdWpdzjpW3EiQwUxowUMEjPJURn/wCZwMGRu5ud5XuRmReF+Hx2U07k7n/ZW3Vlha1BZHre6+9H6hSUvM43tkEEFOex4/zNTewPaW3Xss+FlHDnbO1ocR0O3zCr8izsaWh6aVGU4u22ha2nVr5UlLqSnqKx4ClAnjgEnxUA0RhtbBa3ZL8+WUuHnfqK206D9P8Adc5/0qZjP4bYYIfH1HWcfUxu527doUR8ZyAf1qjPcOUBdT/CcZMz3gaV91+fMJLbbKgoKAyCPvzXIsgly/QwAWtiO4/VfTzCt6N3DYSAVfFA4KUkDuYA7VqVkt9xkwbnElwVFt+M4lxnA7KScj9fvU2OLDzBZMqNuU0wuFtql3f8Rva9L3HU7cWZBXckl+X0kIkw32tuzacK3tK7+7AIB5PGa2l72sMzRqfovl/wuM/Ij4fM4OEflFktcDvYsUfY/JVX0pscWfp++oflQ4wmlMN5+VLS0mO2lQXkIzucUfbgHCcYyTyKjCwPjNmr6rRxLIfj5UZYC4x6hoFl3TXoNPcrrFptem2bm30Nczlzmo7LCCm4ISkhvsAMbFf9E58+TmtTWR2CJD9VwZ8rLc10bsRobZcRyG9e53HuKtSFt/HoOqLyX5lplpRHjhEuWeiotZcI3bBtBCt3bA7V63xBI7UbBVSfg34kR5Xt1doKIuh31qvfqt7T/qLaJ1pdk3KTGt8lh1TDsdx0KVuT3KAOVpPggVZHkRvbroVnzOCZUDwI2lzSAQ6tKPft6qta7u94jRX7/YYcy1KU2llcqWW0pdaBUUqLJyoEE4SpW0+/BGDxVkOe1viR6LdwfFxpJRi5RD9bAbZN9uYaV1O+261IzcC+aQctOodS25TAYW+uHDKBuUf4hUXVg5UFK7IA25xzio6Oj5XPG3RTeHxZfiQQOJsCzeg2Gg29yTfZVnTSmV6ed059DBk3KOFde6FDWWYpQhSFFxZAC8rUjdk7dh4JwKqGrOXr1Pot8hcMoTuc4MB8rDzG3WdgNfWut71asFr0rJv9lt8G3W6Omx28KLCp0kqRJfCv9qUtJBcRjKRkgd8ZGMzbF4jQ0DQdz99Fnn4i3FnfNJIfEfuGtogEfD5ieX1oEq8s6WuojJaVd4ZSW9hbVamVNt57hsDGE9+FZrX4TqGv2XF/Hw2f4bv/AJkH56EH5UtV/wBL7KuI7035zE5w5XKZdCCr7FoAN7f8O2qnYbHagm1czj+S2mFrSwfykWPqfNfzXOLnpPU0S/yLG0I14hraVLaYDiWMIyhClISfa2s5KeOANxGDWR0M7Xlg8wX0UXEOGz4wyXc0TweUnV2tEjXcgbjW9hqFbdB6xESRMtLlhukZaZi9sZhpLrcMEoCkkpOQnesqzjACuOBWjHyNeTlI/RcfivDLAyBOx1gGySC7fXUb0K3V1ja505ILyWrvD3slSVpWooOUnB4UBnB+K0maPuuR/wBLzKBbETe1a/l+q1oWuGLtCafsNrutxLgyAiP0Up5x7luFKR28E1FkoeAW2VZNwuXGcW5Lmsr1s/QWVltOnlTLp+NajZZdueMMMpUXGoSP7qMjlZPKl4BPAGABUwyzzO3VL8oxsMGOSGHfoXe9dOwVp2jiprFVKMvN8hWZLP1rqkrfXsaQ22pxaz5wlIJwByT2FRe8NFlXwY0uQeWMbeoA+p09ljteo7Xc5C48Oa0uUj87CsodSPnYoBWPvivGSNfspT4k0A5nt077j6ix91LFfx5qdLMSBooW76lg2uQmK6tx6csbkxYzSnniPkoSCQPucD71F0jW6FaYMWSYFzdB3Og+q0XbjqG6YattrRa2u6pNzIWcf4Wm1Zz59xT+hqPM8/CK91cYsaGxI/nPZu31I/IfNSOnrCxaEPOdV2TOkq3yZb+C48r747JHhI4T4qTWBqonyHTkWKA0AGw/ufU6qYKM55NSWci1V3dC2oPOOQXrlbQ4dy2rfNcYbJ+diTtB/QCqTA0mxY9iulHxOZopwa71c1pP1ItYxpN+ES5adQXlp84z9U/9W2ofBQ5/vSQfvQQlurXH56o/iAl8s0LSPQBp+RH6grILLfiSpeqHk5/kagMhA/QKCj/UmphrurlSZoBoyLT1cSf0/JRcCyotWvobr0lc+dPhvreffQgEdJTQRsCQAgYWQcd+PiqxGGyc25K0PynS4fhAcrWEUBfW7vXXb+yviRwOavXLX0Bii9SiJREoiURKIlESiJRFEajvLVnhdVSS7IWdkeOj877h/KhI+T5PYDJPAqLzQV2PCZ38o0HU9h1P732Xul7WbRYokNawtxtJLih2K1EqVj7blHH2o0ECimRI2SUuboOnsNB9lLVJUpREoiURKIlESiJREoiURKIlESiJREoiURfK05FF4RarumtH2rTtwuMu1NONLnqSp1JdKkjGcBIPYcn/AHduKgyFkZJaN1uy+I5GYxjJzYZtoFYto7jvUyVhrqoS+6Ztd0iSkSbdFeW8Q4rdlBcWkEJ3KTz9v0JFVviY8EELXj5uRjua6N5FbdavegdPX3UboGyTLdHlS7q1EjTZhRuiQkhLDCUDakJx3UeSVfoOyRXkTCBbtCreIzxzPDYSXNb1dufdZNRPRYuqtMrdfbbfeefjoQtYG9KmiSAD53JRyPnHmkhp7Pn+S8xYy/Hn0ugD/wDsB+RKtLeMDHxVqwAqo6neknXOlo7UYOx8SXnFleOnhAQFAeSN/jnk/eqXE+I0Aaaro44i/Bzue6neUAVvrr8lsawQuGwxfYzalv2sLcUhP5nGCP4qB98AKH3QKnIK84GyrwgJHnHcaD6F9Aeh9uh9CojV0xN79J50yVEnxhJhdUsI/wBsjsRnHjsT/hznzVMx54C4jot3DY/w3FI42OB5XVfQ/Vcdfvr+g7hIuOlEMliUyhpUFsuSmWnFAFAccwEh3sdgJPJGcEVj5TjuDo/hPRfRl8fFo/AziS9hJ5tAa7AdvX2K7B6X3OEbBIQVhFwbeU7PU46D1H3AFqc5wQDnG0gFONpAxW+GVrm0Tr1XynFMWWOUENJafh06A6d/rZtY7Pel6q1uiRFbWuyWxLyEPBHtU/gJ3lR+xWEhOeMk43JqLH+LJbdgrsjFOBi8s3xvo16b9Pld12GxUpqnXlosFrjTQ6ma09IEdIiuoWrvhSgM5O3HIHOcCkmQyNvMT9FVg8IyMyYwtHKQL1BA+vRRf9uZVzlCLBgu2gOOhhEy8NLaSVYyQlGMFXIwCpOf8qiMgvNAV6laXcJZjsD5HiQ1Zawg6ep6euhUs3pF1TyJi7/e/wASTx9QH0hOPI6O3p4/7OfvVnhC75jaxniJLfD8Jgb2o/nd3869F8T9CwrqjbqKZcLwkghKJDoQhBJ7pQ2EjPgE5I8UdA1/x6qUXFZsc/8AbBsZ7gan5kn7LmnrBfomqGG9N2GQzJVEmMI+jaJ3SXMkbOPyNISCVOHjOB4JrPM8SDkYV1uGQPwXfislpBcCQT07H1JPTetV5JMN5q5w/S+ApsNQ1sXC5CQtcRhGNy22txwtw9gpOAM5+9ehrWgiHTueiePLM+M8TdduBDaAcelmgKC90XrOBpHST9st0m13KQ242IDcULaLxczuCwRuynGST/eAqqHJbGwtBBPT5rXncDkzMkSva6NtHmLiDQGxB03vQDstO6E6jnP6sstsVCtEJtMK5NsJBemJ6qCtDaUDIIB/PxkDB44qQcJyXgUBofVV+GeHMGJLJzSOPM2/haKNE2SL9Fs6ab0BfdQx29NPfQXhhIQ3HvEdT7T/ABkgJUvO8fZQ+QD3pHDjucOQa+q9y8/i8OO5uUeZjju0gfcDY+y+tV2uxaWbkyr5qBgXyY60tBht/ToMcPI3Atp3JOBuPPOUjvgk+OjZGeZ7vMey9xs7KzQ2OCH+E0H4jfmo68xor1zX1vtTlwm6fuN4vM+8pbZjtSYCwGdmQXCQlJcwFEhKeTgAkdxYJWt8wcTfoskmDLkARyRtjEdkkO3vpqTS3vTfVnp7pGO9bWbzMEx9zqypdxjuNqcdxghRIwnHwf6mpQyxR+UHX1VGfg8Qy6ndH5QNOWqAXZoUuPMZS9EebfZV+VxpYWk/uOK1gg7LguaWmnCisF/tbN6s8u3SVvNsyWy2pbKyhaQfKVDsa8cOYUVKN5jcHjouN6l9ItUulp+2ayelORWehHZlpU3hodkFSSQrgkEqTz5rI/HednlfQYvGMVliTHAvWxv/ALfJcqu/p9rFFnnXS+RHUR7WEthuQ9klG7BLXJykZz3AOeM1idjSBhJ6L6bH43hvyGRR6l56AD60qWtSEPPLbwpff5FYacRRX1ZkiY5z49T9liS4+4sYRkHxt4NS5WAbqkSTyEaaFbtuYZ6q1yumhplBcdbDgClgcBKc+Sogcc4yfFTYA467LPmSugYOSg4+5+vZX/R9pbv9ustv1ZNucK3vuOiAIrKUNKUSEqcXlPPu43HtgDgHJ1xM0Eb7AOy+bzZiJZMzF5XubXNe+nRov9Payut+nlnt+nbyLOuHFACnEfxGkrKZKAhe5tZG7attW/aSSkpUBwK3Rjk8pC+azpn5ERna49OtWDYojbykVfYi10+XBizIyo8qMw/HV+Zp1sKSf2IxV/KDoQuI2R7Hc7SQe9qgR9I2n/8AEeW9HiR3IqYKUyY7kdBZaWSOmEDHCiEqUc+CPms7YI/GJA6LtycVyjgNY55vmNGzZ73rttXr81qy5F4smsPo7YxZZEy6KSG20lTKWGWgv8wGSFbVIG4dwkgDih8SN9NANqcbcXJxeaUvaGbnQ241te4sH6qH1bBt9lvMWTqq2SrjBU0uRcLkltxSFvLyhLaU8hCEhRwMj+U5JHNMzGRu/iNsdStnDp8nJiLcCTkdYDWWNhqST1uvU79Fprht37UMTSVwZZetyYzbUaYhwZ6LalO7kJA4UtPTSo+Ck/IoWtkd4BGndWtnfhwnicbiHlxJbRqyK19AbIHt6qDvceIrX1ptTD7EV+a+/a5qNpAaZS6hTATnj8raCADkk896ODfEDBudD8l7CZhiuyni2tAc093OB5vkbJ/4X6QaAx2xXQXxt2oXWF9Vp+2NSWoMict19thDMcp3lSjgYz3/AG/3ZIrlk8NvNVrZg4gy5fDLwwUTZ20Wi3EvV0Lkq4TpNqZGenCiKbKwB/M44QoFR+E+0fJPNC1ztSa9kjmghIbG0PPUuuvkAR91B6QhQ7Oy5qm9TVLnXVlBQXVlxaG8FYbTxlaj3ISAOBhIAyaoWgDxSdStnEZpJn/gIxTYydAKF7E/8k+6ipM+82nU0vU/4GxGs0oIjuIdX0n3QB7HXO4Qc+0bsDkbsdxAl7JDJy6H6rY1uLkYjMMSl0jbIrVovcDYk+1+iszULSOsip9MeO/Lb2reSlRafaUodnNhBzx2OeR9qt8OKY3Wq534nOwGhnMQ07Xq011Fgj6KPhW25WfUMq22CcpMeO01Ki2+W6VsKaUVpcSVbStGFAFPJ/cZx4Iyw0w/JWS5MeRCJMhgskguA1sVWlgV30UzFlazU7KS9bLElCVgNH61wZTgfDZzz9h+nmph0tmwFnezBDW8j33WvlG/pqviIdcLS+3LTp9tS3ldN9K3V9JvPA2YG8487k/pRvi9aXr/APpwcCwvIrUEAWffXT5FSdk083b5js+TJkTrm8gNuSXz/JnO1CB7UJyM4A5wMk1JjOXUmys8+T4w8NrQ1o6D8ydyf2AFCa1LF6lizRLQzd5bOHX1OPlgREkZSeqkFSVk4ISnnGSeMZhMOfyAX+n+618PJxwZ3ycg2FDm5v8A27V3taTlhTIvkO2aknT50NcbqQ2XZG1PVSf4iStAQpZSkpKd3ONx7ivDDZ5XnTorm5zmRGfHY1r+bXTodtDYGt3XoFc7HYLbY2HGbXGSwhxe9Z3KWpZxjJUoknjtk8VYyNrBTVzcnJlync8ps/IfYaKSCAnsBU1nX1RepREoiEZoi8IFEK5/6k2lL9zsV6kolOQLQtx98RFhDqPyqDhz+ZCdp3JHuweM8g0ys1a/tqurw3KdHHLjsrmkAAsX8h69uivcZxLzSHG1pW2sbkqSchQPYj7Vddi1yuUtNHdZqIlESiJREoi8UcChRYnX22tnUWlG5QQncQMqPYDPc/amiAF2wWhd77bbS3vuU6PGBSVJDiwFK8e1PdXJAwAai5zW7lThgknNRNJ9lBrvOoXozlxiWuM1BbBUI851TUh1AGSrsUt8dkq5+dviJc/etFtEOI0iNzyXd2gFoPbufcLTMtvUOsdNy7eC43FhPSX1FOOkl9COkCf7xAJx8DJ7jPhHNI0jYKTP+2xZmP3cQB68pN/L9VfEjCR+lWrmr2iJREoiURKIlESiJREoiURKIlESiJREoiURKIlESiJRF4QMUXhVK9Q3Ex5emZX0K5JZuzQLjaQSyFpUjPzyVDt8fpVMzuXlNdV0+HNMjZow/l8hNd61pXNvsP0q5cwKl+oBgpvmlF3QPdATyG1NBXteKf4ZO3nGf2+azT0HMLtrXa4YJHQZDYt+Udtr13VxWhLjakqAIUCCD2IrTW64u2oVHh6JuKIkm1zdSS37EvLbcRDSUOJY27Q0XvzbccEgAnHcZNUNids52n76rqy8RhJbIyECQbmyQT3ra1cW7bDEVlgxGC0ypK20FAIQodlD7j571dyjZc4SvsuB1O/z3VW1roaxX9uXPnRB+I/TLbRJSVZT7TglI4VjxwapmxmSWSNV0uG8Zy8EiKJ55SQa7+notLR2r7G5oKJJTMK0QY7UeS26SXG3NoGxXHOTkZ7YGfBryGaPwgQdAp8R4bmNzXRllF5JFbEXuFzL0RjWeRryQX2GXZTLSn4ZaSS02d3u8kZSCAk/r9qwYTY/Gd6bL6z/ABPNls4dHRprqDr32/Xqv0JPhRp8J6JNZbejup2ONrGUqHwRXXNOFFfnkb3ROD4zykbELnN2v1z9PXWxc3vxXTzbQaZDSU/WIVn27wVDeAgKGQOdoJ5yazPe6E27Vv3XahxoOJMJj8kt2b+A+3Yk9PoodnWWq9coktW2yKsunVDbJuElZ63S/n6fGN+3OMBWD5qDZnzfCPL3V7+HY3D6M0vNL/K0a69L/VU/Rml7Nb9a3yPFdl6isce2tzekwsJD5KwUNqwoBYAOSCQkke4cYquFjGPPKbFLbn5eRkQRmVvhycxGt9t/7FdltmqbG7Haj6VhruLRyotW6OEtt987lHahJJ8E5PxjmtLZmEfwxfsuHPw7IZITmHkP+q7PyFk++y+U/jKWz+EaTtdvkYGx2RJb2jx2aSSeM+RQh4+BoBXoOO41kTuI9ATf/wAiK+hVstzb6I6BL6HXxlwspKUlR7kA5P8AWrhtruua/lLjybdL3VT116cae1dJbl3Jp5ia2jaJUVzprx4zwQceMjiq5IWSfEt2HxTJwrbCdD0ItV3Q1h0jpg3+dHRDRa2loZRNkrS4FBLe5ZC1d/cvB28e0DxVULGMJcNlr4jk5WS2Jj75yCaqqs0NPZfPpXdLtq7WN81DMQ8xZWE/R26O81sLYJSVFPxkJTu+5A4xXsL3SOLjt0UOJ48OHDHC0286u/srXqX070vqSeJ13tLT0zABeStbalgdgopI3fvVr4Y36uCx4vEcnFFQvICrJ9HottuJmaQv91sDhVv6TSw60D/0Vd+M8Emq/wAMG6sNLb/1mSRpZksD76kUfqFuzbFr+3NCZb9Xt3h5lQWYEuA0wh9PlO9HKTjsfnvXvJINQ61SJ8OTyvi5QeoJNfJfVx1Pq1FrckvWCFY2o7XUkS7jMD7aOOQhtnKlfuU/pXrpHNbZFLyHEx5JRGxxe4nQNFX8zsucazuGtr1pm0PuR4c+Ku6gxH4yiyZg7tAsk8AkHBJzwMjyckxklY33+q+h4azFwcqUWeYMPMCL5e+veu2i1dW+mU68agjTJrllsEi7havpWVrcShxKd6itRIGTwCUDGSOD3pNjczg40LXvDOMtgx5GRh0gbWvoTXqqtefSm72BKn7rdLY3Ey2lLjTq3VKQtYRvDaU7toKgT/QZJqiTE5NXEUupif4ibknw4GOLqPUDYK2+jFhsVp1vKtd/iJdvCVhy3uyW1ozgKPDZ4BKfcCSft2q3F5GymN416Ln8c8ebAZl4zvIdHgHY+/2pdy1jp5F/t/SDgaeb3bFqRvSQpJQtC05G5KkkggEHsQQQK3vj59LXyOJlHHk5iLH0PcEHuDt9DouUxJs6zasVp2Zb5EqT9TFnpWy6H1BlpOMI3ELP5eN2VYJBz+Y4w4sk8Ei9ja+odAzJwhnxPAADmkEVq42NtL+166bLqTGsLKvciTMEF5P5mp6VRlD9nAM/tmtvjMuia918yeGZWpYwvHdvmH2VW0bdhc/U7UK7ddo8u0ux2XEoSke5YGzKDnJCQDk4IORzxVEUnNM6jYoLo52J4HDofEiLX24HfbfUfNNePhd/Zj2t1i2uxVouEy6OMo6DakJUG23lEg8g+DnBHBB48m+Km6db/Qpw1vLC6TI87T5Wss81HdzRtQ+lrWTqO0amvljjXUhlLQcU5Gknaw69x01JKsdUcLKTjbhWfzYFPHbM5od66fvdWDh+RgxSSQ2TpRGrgL1uvhOwPXtoqDr+Qxoz1KbuWlDBaV9P1FNoV1WkqXlKsoT+QHIPf7j4rDlO8CYPj2r5L6rgkbuLcMdjZ1nXQ7HTXc7/AL91LyItnuWllSbi4iVHCZIkhDwRMZ6q23EPoSvGTvb3BIxkLGM4xWimcviH199SuK78R43gNtp8gBrynl5mkEjbQ1t017qx6CuWqdTyF26/yTBahRWZH1FuWkKlh0HYVKJJTwnOEjv3Pg6GGSQU/T2XKy24mEQ/HHMSSCCCWiug2v3U/e7c3pyVbrxIuFwlRmJID/1j5dQ0laFI6uMe0pyOe2Cr5zXvL4dOJJ1VcUxzeeBrGtcRpQokijXrfZTup7kmFp6Q+0s/UOt9OOlob1OOr4QEjPuOSO3jntVz3crSufiQ+LMAdhv6Ab32Vb9NNOsIjRdQSZLsuc9FbYQlwANxggbFJaHgHb+buQPuapgiAHOdz9B7LfxPPMl47RTASf8AU6zdu9ddla7xfLNawGrnPhRy6Q2G3nUgqJ4xg/rVzntb8RXOhxZ5dYWE+3oqxL9PItvjzHtHy5djuLqD7mXlKaWocgKQrIx445Gf2rOcUNsxHlJXVZxyWRzG5zRKxvQjUD0IpSHp7HabhSXHRIN2LxauC5L3VcU6gfPYJwoKSAAMK7CrYWgA9+qx8RldJIAK5K8tChX9+5Nm+quGB8VasCcURV3WGpYem7c5IkqQp0jDLHUShTqsgcZ7AZyVdgATUJJGsGq1YWFJmShjBp1Pb/np3KzaXt7sOAtyW80/MluqkyFtDDZWrHCM87QAlIJ5IGfNGNLQb6qvJlErxyimtFC99O/rayaisjV6gpYWtbLrTiXmJDfCmXU8pWPnHweCCQe9eubzCiowzOhfzN2O/svjR9yduun4UyUpsyHEHq9MYTvBIOBk45HbPHavI387Q5XZ2O3GyHRNOg/tamXVhCCpXAHJJ8VNZdei0LJeIN6hJl2yQiRHJKdycggjuCDyD9jzUWva4W0q2eCTHfySiipGpKpKIlESiLUu0CPc7bJhS2+pHkNqacTnGUkYPIrxzQ4cpU4pHQvEjNCDYVZ0dGTp+6StOoefXEYZbfg9dW5QaOUqQFeQlSc/bePGKrjHJ/D6DZbMxxyQMwjzOJDqFC9x8zqrjmrVgTNEXhUBRFq3G4xLdHL86SzGZTwXHnAhI/c0sDdSax8hpgsqCVrCC+VItbFwua8hOYcVakZP/rDhH/6qrErXfDqth4bMwXMQz3Iv6Cz9lHx7ZqK9T5b14uUq2WtwJTHt8RxAcSMe4uOgE5J8JPHzUAx7yeY0O3+6udkYuOxjIGB7xu4g17AE/mEkememnktEw3kPtuJdTIElwu70nIJWokk5+aHFjOvX3UmcezW23mBB0rlFV2qlN2jTUG3zHph6sua6U7pMtfVcAT+UJJHtAyeBjkk1NrA31Kxz5kkwDaDWjoBQW9erXGu9slW+a3viyW1NOJBKcg/cVJzQ8cp2VMMz4JGyxmnN1C1tM2CFp22iFbkuBncVkuLK1EnA5UeTgAAfAAFeMjbGKap5WVJlymWY2T9P3191MDtU1QlESiJREoiURKIlEQ9qIqTqb1N0rpy4uQblc8S2iA40y0twoyMjdtGAft35qp88bNHFdDG4Tl5TeeJhrvsFrxvVnRkhnqovaA2BlSlsOpCOM4UduArjtnJoJ4yLteO4VlsdylmvoQVZLZqix3K2ifDu0F2GTt6oeSAD8HOMH7HmpCRpFgrO/FmjdyPYQfZbkC7264PPNQJ8WS6zt6iWXUrKMjIzg8ZqQcDsVB8T4wC9pFrezXqrSiJREoiURKIlESiIe1EVN9Q7a6qNCvUREuRLsz31TURgkiR2SpJT5O0nB8c98kVVONA8akLocNmAc/HdQbKOUuPT1VistyjXO1xJsRWWX2krSDjIyOxx2I7EeCKm082o6rHLE6F5ifuFAa/ulsRbJER24MMXNCEyozedzhcQsFvCByrKwlOB3yRUXvaNL1WnCgle8SNaSzYnoL319l9o1TIiISu92K5W9hSQovbUyEIOOQrpFSk4+SMfpXgkNW4UvTgtJLYJGvPzH/2q1YrdOi3CG3JhyGn47gyhxtQUlQ+xFWAh2oWN7HsJa8EEd17cJ0W3Qnpc59uPFZQVuOuK2pSkdyTQ6CyvGNL3BrRZKpM71X0bFuCYbl5bUpQSrrNNrWyMjIy4kbf/AJVUciNvVb4+EZcoJaz5dfuqFrLUen7o3Oa0df0NSryUMSmyjZHdO5KDlakhSFlC+Sk4KUnIzg1mne0giI6uXd4XjzRPa7Pj5mxWRrbhoToL1Fj5FXGNZ41p9WLbIhxozKJdqdQtLLKRhbake7OcgEEDOPAHmrREGTDlHRYDlyZPCntkeTyvFWejgV0GbJZiQ35MlxLbLLanFrV2SkDJJ/atJIGq4TWF55QLJ0XDNPWWL6p369X5T8hEB1z6cOhOxxlCAUobb8ZUDvUSOAoJ7lRGIxNySXHZfTjMl4K1jABzjWjrv1P5AfO1dL/ARoqxgWSY8WHMRWbRICpCZLigQG2jnehSuc4JSBkkAA1cWGMUw/JYI8z8dLzZLfNvzjQj1I2P5+qh/TvS1zls3dV3fes9wKkQJMeElr+IyhAKCVYJ3lKyCsEZGDgEVXFEXXeh60tXEcyONzCz+IPiaTzaE71rqLHULqVptcO0QGYVtjNx4jXCGkDAT5P/AH81qa0NFN2XAlmknkMsptx6rfxUlWnaiKq6rlm4SUaegvJEiUgmUtKdxjxyCFKIIwCr8ic/JODtqt2p5B1WzFaI/wDupBbW7eru3y3P0XCFWm5al/sdZbTDYFsYlKmrMIrUGGFugJW9uJwohDhGecHtWFsRIa0DQE/mvqp82KGWeaR5MhaBr3rXl9Ngv1Antk10l8SvaIvFcJNF4udax1w/HvRstgUw9MyhlRCC6tDzmdidoISMAEkqOBx35rNJOAeRp1XaweFmRgyJweSid6sDcg0eumgUBK0feb1PtsDVWoJZmKtsjqLi7UtqytCSjgDd3SpWfzYAGBmqTA+Smyu1o7LpN4pi4ofNg445Q9tF1k6XqddL1qtll15p5iw6dhyFXSWbk0tliNcpakq+kCAV+xKQAM9PHYkkgZIqU8LWR3zaitT0VXC892RkuaYwWO5iWNvzF2m++m/YVYWgzB1Dr23Q/wAVtjDzTzSY70op6TbDe4KW41k5W4dqeQNgxgZBJqAEk7QHgHuf7LS6XF4TNI7GkI5TbW7lx2AOlACzoTZWHVGmbXYHvqdJNTG7pZv4zrqit9tPbZHAVkFS1KGAnO3uecVCSBkR5ortv7pW4vFMnNHg5xBZLpQAaetv76Ub77KwazkF/T0HVKI6LZqeA2JLUSQ4grW3n3NKGQVJIyoYwR24ORV07vIJdnDofyXO4XGPxL8DmL4HnlLgDXo4fOvkrBpvXtquOlmbpcpcOG7ktPNB4Lw6O4RjJVkcgDJwaujna9geSsGXwjIgyXY8bS6ttCLHdVXVUyG9Mdut00S/IgSo4ZbklwJlOqSdzYDWQpHk5HuAGTgDFVSlt8zo7HfquhgNlDPBhyuVzTdV5B3JOo9KOnqomFMfEaI9eJUa/mK3mL9S1JTHjrKTwohkpWoA7d6jxnGAeara48tupw9br8lrkgYZCyBroid+Utt3qPOKB/pGnuouC03qa49aFZotl1QqP9WgKW+wpxaMoy0eEIyRnB3Zwc+TVYDZdWt5Xb9fsts0smAwMlmMsF8p+EgdfMNSSOuoroVt3KMvSNmTM1UluTqJSTMhzDILoD4IKm1Nk8eElaQQRxlOBXpHgN5ptXd1RFJ/1PI8LAHLEdC2q8ve/vymte6lvULWelbo7are7IZlsuErlSo6A6WGSg7kpPJClZCcjlIz2OKnkZUDuVjjYO6o4RwPiMIknjBaW6NBNczgevcCvmorTtqVqfUD0Rq2/g+nZiUyBHWAr6qM2pIISRynKy2c5x8ZqEcRkcRXK38wtGXmjBibIZfEmbYv+lxHr2Fj8+intS6DYsG66WW5TmS1HRFiQcJeKnN4KG0KXyElROQCCMk5A4q5+MGedh26LBi8ZkzP+3yGNNuLnO1bpWpIGhoeh6aKK1SNR+mMOLfYb8e5RVoiwZiFRdmxtoKCACCeDuUCo452fJqT+eEB2/dVYn4Tib3QuBaTZbruT3+ivcT1Bs16iRPwFRuk2WgqRBaKUuJA/N1dxwgDsc/tmrROxwAbqT0XPdwnIiJdkeRoNWfyHc/sqE0fZIq7jAd1BLeRebElQYgrUG2o7OSlKh/6UYwOpnHYEAjFVxsBIL929P3v7rdnZT2MeMUDw5at3Ukakf6T/p6dyFNylzdI+m+beIz8yEyV7ZbgbQobipXIOMgE4GeeOatcTHGS3UrnxCLNzAJSQ0nWhZUnpK0R27a3OfZZduE4fVSX+mMrWvCsAnnaBhIHwkV7G0VZGpVWXO4yGNhIY3QCzsNPqdz6qykcGrFkVTurg07flXh3KbXNShmYs52sOJ4bdPwkglKj49pPGSIE8p5jstsTTkx+C34m6j1HUfqPmrSw6h1tK21pWhQBCknIIPkGprFRBo9F9KovCqhAgxka71AVtpddejRnVOL9xShW9BbGeyfZuwOMqOag0DxDY7LdLK78LGAaou+uhv36Wtr076bOmWYgAQuI68wtrt0iHFYTg8gYKcD4IqEApgCnxU82SXdwD76DX63fqrG88000tx1xKG0pKlKUcAAdyTVt1queBzHlCp2iJkObf9RPWh1t62uusPJcZ5bU6ps9Qg9s+1Ocee/OahG4OJLV0M2KWGONk7SHAEa71eiuivy1YucVT75BmWy+m/WW3IlrcYUzNjpd6a3wCFIWngpUpOFAA4J3YzxVT2lpL2i1vhkZLGMeZ/LroasDvfYdVKaX1FFvsQrQh2LLQB14clOx5knkBST4Pg9jUmSB400PZV5eI7Gfykgt6OGoPsp3IqaypREoiHtRFCahsbV2Ed1LrkafFWXIspv8zKiMHg8KSRwUngj9jUHM5tVox8h0BIq2u3B6j9PQ9FGMXWfYprcTUbqX476gI9wajFtAUT/s3QCQg8jaonB7HBHMeYtNP+qu/Dx5DObHFOG7Sdfcd/Ubj2X09qV+4uFnSsT8RVnYqYslERo9id//ADhHwjP6ivfEvRg/sojDEY58l3L6buPy6fNa8jTN6mXVu4StRyY7zbam22oLCUMjODlSV7t3I5z9gMc5j4TyQ4u+iuGZjRxGGOAEE3bjbvlVALX0jAReXDer2UTrgw67FZStooTGShZSf4RJCVqxuUe+CAOKRs5/O/Uj9/VMyc4w/D49tYQCdbJv16tGwHzKvYAwKuXNXuKIlESiJREoiURKIlESiJREoiURKIvFdjRFxnRVnsOo/VbWVzXb0z/opbRYlOtnYh4DCwnnaSkoBBxnn9KzMax0jnUu3lT5EOFDCX0CCaH2tdebhsIbW2hhpLaySpCUABRPckec1poLiWd+qr9v9P8ASlulGTD09bWn8qIX0QSM8EDOcD7DiqxEwagLU/OyZBTnn6qBlaJmWPWcnU+kvo0mRH6Uu2ujpofIIIKFpHsVx5BGf1NeeFyvL2q78b4+O3Gn1o2D1Hv3CkLH6hW+TKTAvkaVp+6k7RGuKQhLis9m3fyOdx2Oee1SEgJo6FVSYT2jnjIcPT9R0V1Cwamsh01QuJFEXwuSyhaULdQlauySoAn9BXhIG6DXZePSWWAC86hsHsVqCc/1r26XoBOyqUn1O0axcW4S9QwS+450hsUVICvusDaB9ycVUZmA1a2N4blOZ4gjNKwWO+2y+xfqbROjzGM4K2VhW0/BHcH9amHBwsFZ5oJYHckrS0+opSYORxUlUta5To9thPTJroajMoLjiyCQlI7nivCQBZU443SvEbBZOyr7usIrzGbfAvExahubDcB1IX8EKWEpweOc+ah4o6C1pGC+y17mt93BUxLGoNPSp8+9XBiyWqbJL4VASh5qO6pKc9YLTkhWMZSQN/P8wrO3xGEueaBXXe7FzGMhxWGSRorzbkdOWu3Y617Ldvkmw6TUzqbUs5d4uziQzbylpHUUDnCWG0cc7jlXPfv4q5wYw+I7U9FjjkmygcOAeGz+bU1Y/qJ7dAvYOqNeXB9t1jQzcaBnJ+ruKW3lJ+AMe0+eQRXgklOvKoPw8KMUZ7PoDQ9+/wAlVp1/1zZXLvNsejJUSK4pyRJamuNutNuHG51vYUkghJJGSPPFVudKwktat8cODkNYzIm8w0BAINa6H17H6r7d0v6ieoNohwtWy7ZAs6nUylmMnL7gwNqcJO3ySMnggZzgV5ySzNp+gQZWDw6UyYoLnDQXsD3XWI+mbUxp1qxmEy7a22QwGHRvBSPnPc+c/Nagxoby1ouC6eR0pmLjzE3aqeu9MWSU9Yrbc4zDNgPXaLaMNJQ90gW1AjHIS24P6cGqpY2mg4aLoYGXPG2V8R/ieU9yRev3q/RUbTEDUGmbpZJhZlXxH0y3QylQ+pZhISpKE4UcZIcSoI4OQU9+2eNkkbheu/0XYz5cTJilEVRgcorcOeTZo+n5aqyax9TtL3P08v30c9P1a4zsVMN8Fp4uKG3G3v8A93HirXTsLDqubjcKyo8qMlliwbGooeqgfTG+6Y0HZroTqIyoq3mksxW8OOOO9FBcUhAGeVkpGeMIHNVwPjiB82nZb+K4uZnyRgw06iSfQk1Z2FD1Unebpd37enWVwgosz6JMeNDZlBTjjUVTg6i9oHtW5kAgj8icDkgmcj3cgkIrULJhY0Lp/wAIHc4LXE1QHMASNT0HT1XRrA4n8c1ERnH1TQ5H/qG6uYDzvPr+i52W4GDHA/pP/wBnKxA1YsKV6i+Hc44BNF4dVxqZrGXYvTS86jcjBq63C4ONRlKWhYWrPTbWnBIKUIR4yCUZ7GsplcyMvI1Xf/AxT5keMx9sAFmqrSz91N/6P9sehen7b0tSly5kt997eBuSvdsIJHc+z/OpYo/h33VPHXf94WN2AA+VWPzXTR2rQFx0oi8WMpIovCuG3++Cwu2xyBbl/Wyrw9MlS1M4aWlLj6dm/gFWwqwM8dya50k3Lymtb1Pta+zxeHtn8Zr3+VsYAbetuDTYHa91I6s9RW2lWy9WiIl5hsribpUhttKlOFBIASpSjtCfccADIOTUpskN5ZWjvvpuqOH8EdL4mHM6iad5QXaNv0A1vbdRc6RK1VcLK7qeUfoDIcMuxskqUkoGWihDaVKdSUqSSvOCD3HAqJDpy1z9uo/JWxuh4cyWPEHnocsh00OjrsgAjat7XVLJqWFcrm/b4rM9txhlLpMiI4ynCiQANwHPHb+mcHG5sgc4tA2XzEuG+GMSuIokj4gT9FDR4UmXNvlpvNnluwZ8tx4TUuo6RRtTs7KC0KTsSAQM5Gc1ENJ5muGhWh8oa2KaCQczRtRv19CDZX1pizQod4u1umRo8qaC3K+sdbCnZDS8hJWT/MkoUk4wDwcAk0YwNJBFle5OTK+Nj43FrdRygmgRuB6a39lrX70+R+MovOl3xabjlX1HTJSmSlWMgkZ2HjuB+1Vuxhzc7ND+a04/Gn+CcbLHiM6XqWnuO/taqOqXnbHIjXOfMvUS629ZK98hEtLrSwUdRoEJSoJJAUnCVYVz4NVTnwyJCSCP3+9l0eHRtzQ7GjawtfoDXKQRrRIsixsTzCxp1VpsOsZ+p4q4VlYQ3cmNrc2S8MMxyc+9LaiFrJCSQkgDkZPerWT+KaZuPsublcJZg1JknyOvlANk1uC4aCuvXsFWblHXB1izLWxLVEs0gS7pPddbWt3cAA4QM4SEpBCBtKUgnHIzU8VJdaN1JXQxpGuxHMsc0o5WNo6UdgTXf4jdk1ehWjqN3Tlit7osUF+8l58vSpz69zTo9wLPXOAf9r+VPyc8kkVSmONtsHNrd9PqtmDFl5klZTxHQ5Q0DzD15R7bn5LN6e2mFfpxtOorPHW7CglTzhUlxMwuOAodStPcJSkpBB8kZqUDQ88kjdQPqo8WndjM/F4UxDXuoDUctDUEHra6M+uJF19aI8dxtL34c+2pjcMoaC2ykgeBkEfoPtWwkCRrR2XzcbHPwpZHXXM3XuSDeq+dW/W3C9W2DaURlyYmbioydwbBTlDaSU8jJUs/9jz2r15JNNUMURsY6SW6Pl0+pI9h+azPyNQyWHGJWm7e6y4koWhdy3JUCOQQWsEV7bjuFHw4BRbKQf8Ax/3XEbDJc9LvV1ca6MR4Fkug3hCV9VEZCioIIWQD7VApP+E89qxNPgS04aFfSZEY4rw4PjcXPj+VrueqrCm/RmVsqa6zYUEdTOxxC07VoURzhQxgjsQDzjFbnN5gCN181i5JgLmnZ1e4rqP7ddlSfTy0u6iui7xqW2LfSI0f6VyQ4l1kOJCkLU2n+UnaCcjjP3rNCxznF8g+67XE8qLHhbjYb6ouugQ6jqAT1rZdbQkJSABgDgCta+bXtEWOQ31Gyn54NEVJtMa86MhMxHEu3u0oylC2GgmTHBJITszhaBnjbgj4I7UNa+JtXzfmupO+DPkMjajca3PlPrfQ/u1ZrReoN3il+3SUPISdqwOFNq/uqSeUq+xANWtcHagrBPBJAalFfvp3UVpf/W7jfLqDuYlSUtMK/vNsoCCR9ivqYPkftXke5KuynOayOE/yi/Yu1/Klp+n0aQpy9XO4SEPzZE12MtTbQbTsYcW2j2jue/J5xgeK8iB1LlbxB8f8OOFvK0NB1N6kAnXt2C0NSIk33XsOxOrlrsKYSnZrTB2tqWokIS6oEKwQD7QefjBJqmS3yhn8tarXiGPGwHZTQBLzgNJ3obkDa76q7223xbXCYhwWEx4rCAhttHZIFaQA0UNlx5ZHzPMjzZO5WK73q22hlLlzmsRUqOE9RYBUfgDuT+lC4N+IqUMEs5qJpd7dPft81XLjrVbEd6XCsN3nwGiEl5loJKjnB2IUQtQB4KsY/oaqdLQsAkLbDwznf4MkzWPPQnT5uGgPpajrrd414u9nk6QUxMvbSiHM5SluMeHEPqwSjkDCSN24cDGa8c+6MepU4YDC18eXbYz6XbhsW9/U7V12U0rU0i3AG+2WdBYGN8lsokso+6ig7gPuU4HnFSMhGrhQWcYbX6QSBxrbUH2F7/X2UknVVgwM3u19s/8AG2+39a98WP8AqH1URw/LO0Lv/if7Lbj3m2yW+pHuER1GcbkPpUP8jUudp6hVvxpozT2Ee4IULL19puJdhbXrqx9SUb/Zlae+Nu5ORu/w96rM8Ydyk6rVHwnMki8ZsZ5bqzp+dfVbCtX2YD/jTnH/APLO/wD/ADQZEff80/6Tlf0//s3+6h745J1nDdtMGDLYtcjamVNlNlg7MglLbahuUTgDJASAe57V6/8AiDlrQr3H5cCQTvcC5uwBvXoSRtW/VbUG7TrItmFqSM020VIbZnw2z9Mc4AStJOWjnjn28jB8UDuSmuH7/RVux2Th0kDtdy0nX5f1fmrZvSR55q6lg6Ks6UbS/eNQXKOAIsuSltBH/OKaR01r/dQKc+dn6VWzclbMo0yOJ24H56gfvurTU1kSiJREoiURKIlESiJREoiURKIlESiLHIWhtpS3FJShIypSjgAfJNE9lTPSaO43pyRKefZfXOuEuUVtfl9zyhgHz+XvVUIpt2t3EHgyhoFUAPsrvVqwpRF4RmiKA1jpS16usrlsvTHVYUd6VJOFtrwQFJPg8n7HPNQewSNpy0YuVJiyeJEaP5+6q1s0hrS3xm4LeukmA0ChCl2xC5ARzgb1KIJAwMkGq2skGnNotUmVivPP4Ov/AJGvota0emsm6XSVdfUKei8ylDpR47W5tmOgHhScEEKOAeMYJPc8jwQW4uebUn8RDImw4reQddiSfft6KXd9KdIvqfcl2sy5DyipciTJdcd/ZZVkftUvAjO4VY4rlNADXUB2AH6KDHoTpNSsyHLrKTztS9L4TnyMAGofhI+q1f8A8gzAKbQ9a1VwsWhNN2OG5GttoiNodbLLqlI3qcQRghSlZJz5q1sTGaNC58+dkZDg6V5JG2qql+9K4cAJuegyLHfI5Cm1NurDLwHPTcTk8H5/qKrfCG+aPQrXFxN7/wCFl+dnruPUHdTlv9RLXlMe9x59jnBOVsz4y0pzxkIcAKFgE9weata+99CskmI5mrCHD0K8vPqfpK2R+rIugcB4AYZccJPxwnA/civHytYLKlBw/IyDUbfuB+a0YXq5o2Sy+t64uRFMY6jcqO4hSSc4GMHJ47CojIjq7VjuD5QcGBtk9iCoibrGZr9YtGjbT9RaneJtzukZSY6UZGQhBwVq8jt2HHciD5fFHK0aLS3DGAfGyH8rxsGnW/0V00rou1aeT1I7a5EwoQ2Zck73difyoSeyEDwlIAFXMYGCgudkZUmS4ued9dFZggCprPS07xAFwtU2Hv6f1LC2SsDO3ckjOPtmvCLFKcb+R4d2NqH0ne2Xo7NrlYj3eK0G3orgKVez2laM/mQcZChkYIzUI3g+XqtWXjvafGaLY46EevQ9iNtfkrGpQx96sCw7rmvqe1H1BcbRY1Noc2ymXnVbuU7lbQkY8lAcUfsj75rNkNEtNPdd7g8kmIyTIb1a4a7EdTfoaHuVPR5TEvXim4bjTyosFbcvYc9FSnEltJ+5wvjxj71YCDLodgsTmyMwQHiuZwI9aBB36bKjXcxxr7V7kKNHcvyI7DcV9TSdkNvpbnHnCeOCrk8qPtABAql5Be7lHmr6eq6GNE9uLC+UkRFxsXq4gimgd/t1taMARoNxsLdyaj2WQYfUt8ti2oefeJ/OpYwrYsjB2gE8klWcioNcIyGu0PTRbJ4XZLZZIP4gBpwL6aOwGwcBtem2gpb0dqdqG73VmXquYIltKH1SQyqIho4JaHTIAUQfeon4SB34AGQkF+2u1Kt7o8FkZZjtt45a5uYnXXUaixoB6rb0ToSb+HN3M3y6xZrkpyczvUFAhadqVOtngqUn3EZBAVtzkZqUOPWvMbu1DiXGmSPMTYW8vKGn0o3p2o6etWr9p26y335FvvDLDNzjpQtXQWVNutqyEuI3cgZSoEHsR3PBrU1xJ5XbrhTwta0SxElrr33BHQ/Kip7dUlmtfK1DsTigT0XFfV2GjVV8ZtJc6FqsiUvyek2FOuyXgQyw2k8FRA5HgK5rJOzxDyjYfmu9wmQ4zDIBb32Ndg0bk/krRY7oz6f6QjwL4+ZlzQlyUuPBZK14WsqOEjskFWNxwD4r1hGOwNduozwv4tlukxxTDQBcQBoO566bbqe/t1p4x+q3cWnVcAMNhSnlE9glsDcSfsP91XeK3usA4dkk/Aa79B7nZY5WvrFDQwZr0uKt5wNJbehPJVvPZOCnv9hUHZEbfiP2Kvi4RlzX4bQQBejm7fVYrlr+0RXGGGkT5cp5wIQxHiOFzsTkhQHHGM/J/WjshgqrPySPg+S++blaALsuFfYn6LnF1f1C7KFtfbjxbBFfcfnqBVJQ11VlxLEjZgnaT7thxhYyQOazPMgpo0aDr136Fd2FmF4bpyS6ZwAbdC+UVzNv7XreoBWLWOnH1xZ066R9Pply8pachP8ASVKSnbt2tqQslauQChQPIBJ4ryXH5gXOqz26qXD+LeG5jIHPLW781kNJ3stI0H+q/QLds+nNQ+n1ruF6trFvKHG0Kehr3vOsNIBPC8pClcnKRgeR2xXkcUuM0vZ9F5l8RxONSsxp3OBF07QNJJ6iiQOx1Pdbtqvbmpr3alzL3emW57K22GrfEchx9yRuO5xSlFSuCPaSOP63CTxHtJcaPb9Vjmw/wMEgETOZhBJLg52u1AVornG/HrIv6f6dd9g8hp1LqESWx4S5vISv43gg8cgnmtA526HUfdcZxxp/PfI7qK8vyrb227Kv3/VLlkvabs/Ybyw0m3uollbaCjCFgt+8KI4UpQz8L844qknMZLi00unh8KZlR+A2dnMXDlFm9d+nb8lvWj1Ebdg21d8tU22rmNqd6oQXoyUjGFdVOeCCO4GD3x3r1s91YItZ8jgzo/E8GRrww0daP0OqocWZbdYXP+y13mOzkC6vORpjjoC3GC05whQHBCgOMYI48YrIC2d3gvN6nX0/5X0L4puFQjiMDA0ljQWgaB3MN/dq10dWyvSZUDUb0VUO5/hMuXNSFLLRPsKSR/E2p/lVnG3cCBnPlcmodsaJP2XlMzCGSwXzM52tadLG/wD42R0362VbLLHlxbNLh35KbtDVIcYu7QjHrBa8EPgpJLiSNh4AISRj8pFamghpa/UdVxZ5I3ztlxR4btCzWxp01Gmt77m+6nX5ekDoZ9DbcOdY4CEgxUYVgp9yUbVEHeT2B5JP61O4hGQKoLMI+IfjA82JX7HbfrY6eu1KiaDduWoNeRLm5Ecs6ULlqVsVt67KOkgMFGONhKQew74wayQOfLIJCKq/mOy7/FYsbh+E7GY4SEhnycbJdf8AqHz76K03VvT17vOoZepfp9tpSlhDayUustgbi7x7sqUrCcZztGOTWlwje4uf/L9lyIXZmNDFFjWBLr6E7V8gNe1qV9NlPLZmKu6ib+C2iYF90pSnDQHPI25JP98rHGMVKAkg83xdf0WfijWtc3wf8rWiO/8AN99v9NK8mr1y1yP/AEirZb3dLxblOhpkLircbSoOFBTvbVgZHgrCOCOTgcZNZcpreTmPRdzgEkonMUTq5h+o/RWH0xlPx/SmzSpTyp7jULq5ZG5SkjJSgfKgAE/qKthJ8MErHxFjPx0jWChzUpvQUX6XR9oR1EuEx0rKkHIO73cHyPdjNTZo1UZrued5qtVYKksyUReKVtHNCaXhNLUuM2LCiLkTnmmI6BlTjqglKf3NCeXUqccbpXBjBZ9NVyDX9/hLv1qkwW7wqLccxZkeNDcZVcG0glAQ4oDOFHacYJSs84rBkSAOBF0dDpuvq+EYUhikjk5Q6MczSXAhhvXQXv8AYjupRjVN1dbtNutMGLpkNKDS2bwlQSoBPDLWAM/Y5BOOB3qYmc6mtHL7rK7hsMZklmcZrsgsr6m/voply5N6UsrltbmsT9RPuOPIYCcFx95wnJQkkpbClHufyg8+avc4MFA2VzWwOzH+I4csY6+jR9zQ+qs2n7Mi0xncvLky319aTJcGFPOEAFWBwBgABI4AAFTa3lWSeYzEaUAKA7Dt+trV1DqARelCtgbk3iSdjDG7ITzhTjmOUoT5P7Dk1454B5RqVZBimUF79GDc/oL6notfS+k27VIcnT5S7reHSd0+SkdRKCc9NA52IHwPmoxRcnmJsq3N4gcj+HG0MjH8o29z3PqVZijJ+9XWufS+W2ENqUpCEpUo7lEDG4/J+TXi9JJ0J2X0pGTS14ogaXs4vH4n+HRfrQ10A70x+TOcY7fvjPioeGwnmIFrUM3J8PwfEPLd1fVfErR+npj6n5lktj76sbnHIqCo4GBk4+Ki6CNxstCsi4lmQt5Y5nAe5UlCtsWA0hqDHZjNIASlDTYQAB2HAqYAaKAWaWWSU80jiT6m1tbfuf61JVIlODmiLWukBi5QZESY2l2O+2ppxB/mSRgivCLFKbHujeJGGiNlWXv7QafZcU2pu825gb/4yyiWEBPuA2o2uEYJGcE5wT5qvzs0Go+62H8Nkb2x57fD9zax+mN0iSrO/CjvJdkQpDvWCew6ji3EkHyClQIP9cGkUjXAgdFLiWPJFI2R40eAR8hR+YIV0ByKtXOte0XqURKIlESiJREoiURKIlESiJRF4s4SSaIuD+o+s4vqFGVpLR8mauY7LQy842wosvM5KVncDnppJBOQAQPNY5ZBKeRh1X0WDgvway8tnko179AuraA0vH0hpqLaIrzj6WipSnHOCtajlRA7JGewHb+prRGzw2hoXHzMx2bM6d4olWOrFlSiJREoiURKIlESiJRF4Rmi8pebeMAkfpQoBS8U2FDCuR9+aFK1ta0+2Q7hHWxOisSWFkFbbzYWlRHIJB74rwgHQqbXOYeZpIPotlDYQkJHAAwABgV6or7FESiJRFXdV2B+8KtrsK4qgSoMjrodDKXCQUlJTz4IPODyKrkYXUQaIWrEyGwczXs52uFEWR63p17KtauVc4em59w1RcI8eFFZOI1ucW0JThGEhbh9yQokDYn91KqEpdykvND0W3DbG/IZHiMLnk7urQeg2PufoFH+l2lG7PqG5YdaeahpbQ26FKK3XXG0rW4oE4ztIAIHYkc81TjQCN7q1r9Vv43xN2XBESCC+ydABQJAArXcWbJ1pTNydtWktaIlhCIqLtHeMgNNFSn3m1N7NqUgkrIcXnA57n5q8hkT77rmMM+bjeHv4ZFf6QdwT2sdVo3SBK1nclsmxybLESnaq6PK6cl5tQwptKEnsex3kgDnbnGK3tMxIoj16rRBMzhrGuEgkP8ARuAbsEk/p9aS3Ws2D1AskWU/dLkpdufbZmzV70h3KVFKf7p2IVnyQRyea9a3kka02TW5XuRkHKw5ZIw1g5mktbpprr6i/uttdsGvk3Bu6SXG7IxNXHTCZASX1Mq2lbq+SRuBwlOOAMknsLfxGjtgfqqo5jwktdEB4haDZ1oOGwG111V+Q2EoCRwAMVpGi5B1Vf1HpVm8z4U1E+db5MYKQXYTnTW42r+RR+M4I/f5qt8fOQbqltxc047HxlocHVuLojqPXotX6bVbSTFTLtT7ecCe62sOgecsp9qlfcKA+3in8QaCl452G48xa4f6RRH1OoHyUHd4upHdUW62xb7KUytCn5jraGWQy0DtAQnYoqUT8k4x4zxS4SeIGh3uuhE/EbjyTmEbgAG3WdzZsVp23XK/SC3XBfqRerDe7pc48uMXZR6boKlvghBcJIIJKFnnzms8LHGUtcTouxxLKjZgxzRRtIcA3UbAa0Krr69F+hrHYINnQ79GhfWeUFvPurLjrygMArWTk8dvA8VvYwMul8jkZMmSR4h0GwAAA9gNFvSC2w2p5woHTBO9ZAwP1PYVL1KpF7BcqPqUufpm5yzYnHJ9vWVNhpPXYaVkpbcUo4PcHsM8E8ZrG3M52EhuoX0zv8PCHJjjMoDHjXodrcKF+la9VYXbTqq52hkXe+ohrTtU+1a4xClgfmQFqVnJHwBz81dyyuaOZ1ewXO/EYMMrjBDzDpznb1oCtPVatosk24o/Dvw5dg0olj+HHZd6cp9Sj7g4RkoB5JAOTkZPcCLYyfJVN+/zVk2VHE7xy/xJ71J+EVtyjr8xQ6BfWj7TbtKXkWmTGSmSoKbts1RUsvM5Ky1k8IWjyBgKGFfIHsUTYjX0TPzZuIMM9+XdzQAADoLobg9LvqF0Dp7gQScVoB1tcWt1z+aiMjUM/TcpExhdxfNwt0hhBUlpaUIKlBX8pDiScHj3DPCqz3T/AAz11C7LA4wNzI6PIOV4J3smh7ctD0r0Uozqg2lKouqunElNpUW5I9jEwAZygk4Sv5bUcjwSOas8Tl0eaWY4fj07E8wP8u7h7jqOx270oO6QJepdFT7nOflH6mE4/GtjLobaQFNktpcxjev8pO47cnAGBmqXtMsRcdiNltxZ48DNZFGB5HAF1WTR1roB2oX1tRlp1SuxW6GiyB6920sNJJcdDLUBeBlBfUMFJzwnJKdpHkCqxP4bByeYfl81pm4YMmVxyCI32ehJeO4aDd9b0BCqrlsguaeut3vanLPfSfxO3bjjpoOSlDSuCtJUr8vBSVJ48mhzG+G6STR24/2XZjy5hkxY2J/Ei+F/qepcDtQ69aKwWyVamRb7dfHHLymMEzXRFWp9Kl84YKE/w0EOFZUtX5hjn3HBjmABj/NWun5LzJiyXGTJx2+EHeUXTdKHmBOtEVQG2qsX9qLtIlXmdMtblphz2PpY0jrLZSvZuwvqBsndhXBATkds4zV4lkcXEtodNVzncOxGNijZNzuBsjluttKsaehvXtaiGLFILzt3buUG4Pxg0/NeiOHbJeWSltDS0j2O8DLh92XM8DvSIzfiA3X0J9PXuV0JctnL+EcwsDiQ0HdrRqS4HdvVrdgARqrRpR38MlaVZRCdZWZdwhy3Qd7TS1KUrpFZJ3KK0owrztPkkDRGeXkFdTa42e38T48vMDQYQNiQABYHTuR09lD6ynWy6awbmx4LyYcCcwzOu0da0qU4lSv4O0DKyCEYKcnIH2quVzHycwGgIs/ot+DDPjYZhkd5ntcWsIGgIHmvp1v0VitkB6xzokm2RlC/XKQ9LkW9TpI+nWrJ6ijkI2nbhXPvUpIyCSLms8M2PiOtLlyzjLZySu/hMDQHBv8AMOw0u9j2FE7BXFF8uzSymbp2VtxlKokht4foclJB/YirQ9/8zfp+wsJxcd3+XOP/AHAj8uZU31JkM63siNOwi7FnPOB55mSypt1pDaVLztI5yRjIyPGeRVUxEo8MdVs4e13DnnLdRAGlG7vTp23o0fRSvorMblemVreZbS0hHVSGAonpBLisIJPOQMZz81ZA64wVl4rEWZjmk66X7kAqf9PmPp9F2dG7dujpc7dt3ux+27FShNsBVXERWVI3sSPporDVixJRFpXqX9BapcspChHZW6QTjO1JOM/tXjncoJU4o/FkbHdWQPqVXLLpSHKRGuV7Crjc3AiStUhSi2heMjY3nakJyQnjI75yc1W1jXU52pW6bMliLoITyt1Gm5+e+vuvn1JXMt1ojXe1xRKlWyQJHSKsBSClTa8n4wvP2xnxXk7nNaHN1IXnCYYppzBM7la8EfPcfXZa/qNJkuafYtkW3Rp8y6LEf6d1zIQCnKnBxzsODnjHB+BSdx5AALtT4VE0TGZzywMF2B16D5qw6b09AsEBMe3R2WAQC4W0beooAAqV8k481NjGximiljysqbLfzzuLiNrOy+r5fItmbZ+oS8668ooZjx2y468oDOEpH+84A8kV65waNVGCB0xpugHU6AKJ9PtNNWG2OOrjJYuc5xUmYQred6lFWzd5Cc4/XJ85quGJsd11WrP4hJmOa158rAABsKHoOpVtAxVywJREoiURKIlESiJREoiURfDrYcSUq7Hg0Rc/kaWuun3uvo2UpTK0pbfiS1hwlKBhvpKUOCBxhRxjAyMCqHQubrGV148+DIby5rLomiNN97r9Fs6fv2orhaGp0WJa7gx7kqw8uM8VJJSpOwpUgKBBH59p8HGDXrXuc3mAtQyMbGil8KQuafYEe+lGvlanbVqm2XBxtgSExpylFswpJ6b6VAZKSg8njnIyCOQTUxI06XqssuHNEOYttv8AUNW/X9lTgOe3aprKvaIlESiJREoiURKIlESiJRFz/wBYL8/AsKbPa2nn71fN0KG21wQSMKWVdgEg/wCf2JFMz6bQ3Oi6HDccSy+I8+VnmP8Ab5rS9IvTBjRDbsuW63LvDwKFPNghDbfHsQDzzgEn9uw5jj44iHqr+K8XfxEgDysGw/uum4rQuQlESiJREoiURKIlESiJREoiURKIlESiJREoiURfK1YFF4TSoa7rG1zJetUFC3LZEk7bk660UpWW1ZDKc99yhknwkfKhVHO2Ylo2G66/4aXhgbPLo5wtlHWj1Pb0/wBlEw7za7JrLWDsOPJl3RxTCExWUOKdfWEKKgkHjAPOewH6jMOdjHvIFnTRaJMbIyMTGErg2Mc1ONaC9br7KXZs18u8mHf5spu23Rhs/TRA11G2ULwVtu5OVKICQSkp2lPHnM2sc6nv0PRZpMnHgDsaMF0ZOrtia2I6AehtTdhVqL8SmJvbdsENIR9OuIV7lHnduCu3j/61YzxNeevksmT+F5WnH5rO/NXyqlqepXTj6UnXLcpuXb2zKiup7tupBCSPsc7SPIURUMk8sZcNxsr+EN8TMjgcPK8hrh/pvX6b+lKQ0TEai6WtaGm1Ngx0OKCklKitQ3KKgedxUSTnnJqcQ5WAKjOfz5Ehu9T9AdPsp2prKlEXm2i8pav0McTlTA2PqVN9IuHvsBzj+pzXhFm1Pndy8l6Xdeq4z6hRP7I+sen9Uxwl78UC4jrC1hA3hARu3ngZBT34yn71km/hSteOuhX0XDg3OwZMR5rk8wNX76DVXK7X/WkBKX29LxZLDgIDbEze60okBO7gA9yTtzx5GMmckkzfhZfzWPFxOHTeWTILSOpboe9dvmobUzGttURlWSRbI0W1zAESJO1O9A3gnCeqeMA8/tjzVbxPL5CKB6/srdiu4ZgO/FRv5ntOjb0P/wCgWM6ZmaPSJERxl+FJlx7dIYfdUpC4agGUlSdoAc3KJJTx7jx3oITCPIdDV+yh/wBRbxC2yghwDnNIGvPfN8xQr0oK0aJNytUhdhvbjLhQFOwHW1KVvjhWNiioklSMp7k8KHJwa0R8zCWO+XsubnCGdoyMex0cD/V39j+auRSAKtGi5ypnqJdWIcaIYzgdvcd5EmHCSrLj5zsUnaMkgoUsZ8ftVE7w2iNxsF0uGQOmLmu0jcKc47AHUE+xAVnskp2bbI8qRFdiOuoC1R3sb2yR+U481a0ktBIpYZmCORzGu5gDuOqgPUKDcHrQJtiZccvENW+P03ktqIPC05Ukggp/lI5IB7gVCbm5bbutPDzF43JkGmO0Ol+3b/hck9QLpbZkK2sqvcmS+l1Lk9ifIWCyQgkDo7AhW1ZwU7cnAHGSRz8mRjqBPqQvrOC42VE97mxUC2mlrRrrrrd6jrem/or7aNJ6cjaeN5vMIsMSY7UqTFedWY7HsBKQ3nG0H+Ug44A7AVsbDGG8zhv9Fwp+J5jp/AgfZaSA4Acx9earv1VKs6NKSL3DkSOu1HbcU+44t11mP/Ddc6HSSlOFKA4KSRgdud1ZIxCTYvf5ei7uS7PbE5jKJIqqDn6tHNZJsD119Vb/AFWtls1NoldyZloeS02VwVoWNq3VKSkAYBJUcbdvye2e2nLjbNFY+S43+Hsubh2e2MigTTru6r7VvahvTtDtun3Jy1xYTF7Q4li4WYqS0pSEoThbaj2O7er+6oHwQDUMYBpJaNeoWvjL3TMjbMS6KiWP33Ox+w7ilb7petUMvRI8a3WluZLOGmFynHFISnG9aylIASnPJHkgDJNaHSSdtVyIcXDdbnvdyNGpAA16AAk7/lquU6ls2oNHofgsXCEiPOW/LcEcFIISAsL2AZbKSDtIV/KODg1zpIZYfKDof+V9bh5+BxAtmlY7maGtFkE6mquxYI3BHU91e79L+g0PdbTMjRrTKgQxLiLZdLja9qwQ42cBQWHNv5hkKWDzmt0ruWIjaha+dwoTPnxy/wCYHO5SDofUH0r5UKUBLlBOjkvOXBuPFiTGGoqorCipbyyhYlu4yVK2qJ2jgqycnKQKS4cl9ARX910Y4XnM5Gtsua4mzsBY8MHoLFXuB21vrtotEOC9Jlxw4uRMVvdeeUVLUOdqcnslIOAngD+tbQ0A33XykuQ+UNY7QN2A2Hf5nqVqTtW2GBdnrdNucaPMZbS4pt1W3hXbGeCfsOcVF0rGnlJoq6DhuXPGJYoyWk1Y7qna3vlkubmn5Me+x4Cm5Djzclw7dpbBG1YyFbSvAKcg81RO9pLTzUurwvEnjE0ZhLrABHudx61sVQdG68RpiBqCyMPx98h1Um1r5WgLcXtcbz52nKk/I881THkNa0tGvZdPK4PJkTRvkBFaOvfQEgn3Gh+nRfoi1Q27dbIsJkqLUdpLKSo5JCRgE/fit7WhoDR0XyEsrppHSv3JJPzW1UlWlEVU9R7zIsum3XYsFcpb60xcjbta6ntC1gnkZIGPJI7Dmqpn+GwmrXQ4XijKyQwu5a1961oepU/aIibfbosNta1ojsoZSpZyohIABJ8nirAKFLC95kcXnrqss6M1LivR5CA4y6hTa0HspJGCD+xoddF40lpDhuFz6DbrzD1VMjRbgxK+ghtpiC4oWtSGnCrPuSRk5bCSogqwkffNDWyB+h0C7Uk+I/HaXsILieblIqxtuPUmh1WG9+qBtra4jVllSL4y4hEiE2oLCM8kpWnJVx24884wRUJcsMsAW4dFbicAdkVI6QMiIJDjpdfkpGx3dN91ImXNQ5b3IjKxFgS21NPqQsp3PKB4x7cAJzgE5POBZFJ4j9dPQ7rJl4rsTH5YzzhxFuFFtjZvpv1q+g6q8tqCxlJBHg1dfZclZK9XqURKIlESiJREoiURKIlESiLzaKJS+emkDAHFLTraqesNOJmTrZeocJiTdbW6FtJcO3qo53I3dgRnckngKHgE1VIwFwfVkLdh5roo34rnERv3rXUag1+forHapJmQmXzHfjFacll9IStHPYgEj+hNWg2Fje3lcRd+o2W5RRSiJREoiURKIlESiJREPaiLjl8urd69etPWyFcZLa7SlxySw4NrSlFsnDflSyleD4wOPNZXOBmDQdl3IITFw2SV7dHEAHrv+S7EAK1Lhr2iJREoiURKIlESiJREoiURKIlESiJREoiURKIlEVX1akTplotC9/SmPqckBBIyy0kqIyOwKy2D8gkVB+pDe624Z8ISZHVooe5P9rPyW049ZNL29lhbsG1xOUtpUtLQJ84yeT/U05mRjXQKoMyM2Qua0vd16/8AAVM9OTL1FqSfqF9hMeGJEhMdW4qMgFLTYIyB7Uhkc45Kvtk0Y5dIS8ih+a6/F2x4kceI13M4NF+mpNe+v29V1AJHwK1LgL3A+KIql6o26Tc9FXKPBa60nCHG2+oEbihaVck8Y4J5qjIYXxOAXS4NkMxs2OR5oagmr3FLS0p6k6f1BB6qJiIclO/fGkrCVpCUlSlDwpISCcj4NeRZUco0Kt4hwLMwH8r28w7jbU6ex9F5p2VrG8QxcZDtttiHCejEcjLe3N90uKVvSUqIx7fgZIBOB6wyubzGgvMlnD8eTwmczwBqbA16jY6LPfpOr7XBVMZXZp6GiFOshpcchsDKilanFDIA8ivHGVnm0IXkTOHzu8Ih7SdjzA69NOUK12+QmXDYkN7wh5tLiQsYUAoZGR4PPatA1FrluaWOLD0WzReKq+pkD63R1zWzGQ/NisqlRAUBRS837kEA/cdvIyPNQlFsK04buWdtuoHQn0Ken2q4OsdOR7jDcQXSAmQyO7LuMlJGTj5HyCKMeJGhwTNxH4UzoX/L2VoCU44Aqay0ojVNrcu+np8CO62w9IZUhDq0bwg+DjI7f5d/FRe3maQtGLP4EzJSLAINd+4+YVNgOzLrf7DHmTnRNsypS5zrSQhLqk7EIyD2StK93HgH7EUtJcWgnUbrpPEcMMz4meSTlDb1rcmvaqUvdL7cLnc49v0lIiOFCiZ0txtTrTCNvABBAU4Sc7c9hzgGpOeXECKvVUQ4kcTHSZoI/pGxJ767D9hTOnrI1bDIfdecl3CVtMiW9je6RwBgcJSMnCRwMnuSSZtZy63qsc+QZ6FANGwG331J7kqbAA7DFTVK8UkHgiiKv3q26djOSLveIltQQhKXpUhtPZJ9uSfg4x+1Qe1g87gFqx5sotGNC5xHQAlVZ+LA9SJdyQ1cZirTDKIyFw3FNocczvdz4cGAgcgjByPmqPLkcwB8o7LosdLwYRuLB4hskOGoFU0+nU/JbzFztenJv9l9M2h5+chH1K2WSG22wo/mW4o8E8ccntgVNrmxnwoxsqJIZ82MZ+W8AE0CdyR0AVHulnvc7UwhxIDFquLqzMQqHIWuM0Ao/wAd4DjqK5SnCQSSo5zjGaSORz+UDlPpt7+67mNl4cOOZHvMjAKpwHMf9Lf9I3OvYLd0bYJqNaXe5REJuy4rqWjOuR6ZceCVIcLDiAr2gAJIIPxkEcooiJXOaL9T+ihn50T8KKCQ+GDZ5WagCwRzAkanUgivbVZY0zVS3tRXx25WJK4CFRWVuNK+n2hRUvC8gjaoBJOCFFGOSKmDP5nuI0VL4+HVDjsY8hxs0RzXsNNR61dgH1VZeD2rbBedTakcS9Ehlzb9ItbJXtQUoZSk8hO5zcVK5yrGMZrPZnjdLJsPddTlHDsqHCwhTnV8Qa7c3zE9wBQA2GqldRSbncdDWKIxb5yLm6GYkW5IUCw8h1IR/EUr3ALAGUqT3AOTwTa/ndC0NBs9f7rHhtxoeISzSOb4Y5iW9bB0qtNDsQdlM+pNr/BbBbZNvuDcCUl2HGLjq9sfLWSlaspOCNp5+OOeKtyW8kYLTWyycGmOXlObKwvbTzQ+Lzb9e5UtpPUt71jajNtbMG1spUppf1e6QvqDGQEpKMJGe5OT8ealDOZ28zRSy8Q4ZBwuUwzOc87jlporpe9+v5qxWTTMK3SzPfzPuyx/FnyQFOqPwnwhPwlIAFWiNodzEa91z5s2aRghBpg/lG3v6n3UFa7Nb7vq+ZLaQ0YFpBgtxklKmVPL/iOr2YwCCpKfnIOfFVhjJJC7oNK/NbZMibGxWxnQyHmvrQ0aL7b/AKLkX+kFpr8DnRJ8BC2YanNzAb2hplZ5WkDuCVALA7HKu2OceWzwyHN2X0f+Hsn8WxzZTbgKd3cOn02+i6lon1FVc7bp9d5gPxVXRva1LSpKo63E5BCjwUKKgcJI8jBNa2ZF8ocN189lcJ5DI6F4cGbj+aj1rr69l0gKynNaVxlT9QXK/TLsmBpRURCojiFTX5gJbwoEhoAc7sYUT4BHzVMhkJqP52uhix4rGmTM5qcKaG7339lVNbX28TS3plUKBNuD8phRVBl+1oIWHNroWBtUQg4GeRk+OaZpHnyAWfTp7rp8OxII7zXPLWtBrmGpJFDl7jXfoust9z9+a2FfNgUta4XGHCKBLlMMKczsDriUbsd8ZPOK8JaPiNKxkUkn+W0muwtc+t78zUurLq5brgqFb32GQ3KYTudkMoUtJ6alDalO8r92CTxjjmqG295o6LrysZi48ZezmeLsHYE0aPcgdNu6u1h07a7IHVWuCzHceA6riAd7uM4KlHlR5PJ+TVrI2x/CKXNyMufKrxnl1bdh7DovjUunbfqGH9PcGiQkKDbraihxoqGCULHIyOD4I4INHxtkFOCYmZNhP54T7g7Gu46/ulBWme5pCLFtd/ITb2UJZjXNO7pkDgJeznpq7e4naf8AD2qAPhDlcdO61TM/HvdNAPOdS3r7t7j03Hrurq2sKAI7GrR6LmX3X3Xq9SiJREoiURKIlESiJREoiURKIvCkEYIoiBIA4AFEXtESiJREoiURKIlESiJRF4rtRFwrX1qf0n6v23XRYcVZFFCZ74G4RyUloqI7hO0oOfnI81kkYWSiQbbL6LDyGZPD34Tj5xq386913NtYWkFJyD2IrWvnOq+6L1KIlESiJREoiURKIlESiJREoiURKIvCoCiL0EHtREoiEgURc7uL9wvXqO1Etcn6eFboa/qJbKUuKDrih/B9wKQfYCeMgZzjIrM4ufLTDoBuu3C2HG4eXzN5nPcKabGgHxaUa1pRXqRaYtmtMa4S50qVNcnMIckPtJfecb3hRabTgJQMpCvaBnGCTUckBjbJvULRweZ+RM6JrQGhrqAJaAa0JO57ak+yvGhbdNtemLZDubqHZTDCUKKUBOwY/Jx3I7bvOM1ojBawNK4ubJHNkOliFA+t/P59lYamsyURRGqbWbzY5tvDqmi+3sCwMgHOcEeUnGCPIJFRcLFK2CTwZBJ2XOoXpipWoLfMnw4aWGUuGStEpbq5JJylKgUJ4BJ57lI2nIrI3FHOHUF9FLx6sV8MTnG6oEABvevMf+dd11OXJjwYjkmU62yw0kqccWralIHck+K2EgCyvmmMdI4NYLJ7bqmBF11fOc3qVC0mtCQltTWyRMwed2eUNKHjhRHwDVA55CejfuV0iIMKMD4p7/8Aa3+7vsFeW07QAK0BctfdEWN9pLzS23BlC0lJGccHg151XoNahcY0bY4Ppz6sN2C3uTZMe8wS4gvOJwxsUsgEAe7hJG48j75OM0bRDJyDqu7mZE/EcHx5K/hkD6rtY7CtS4K8WPacURc4vmmW4t3u9/vV3krgSEttfQxmg2XEDASwVD3K3KIGBgq4B44rO6KnF73aFdiDM8SCLFx46cCTzXfua2FD6bqc9NGWW9KxXG2ERlSFuyFx0I2hha3FFTYTxjafb27ipwfAKCzcUJ/EuBN1QB3sAb/Pf5q2AAdqtWBebh80S1CalujluaZbhNh+4S19GM0okIK8ElSiOyUgFRPwMDkivHP5aHUq/Hg8Ul7j5W6n2209SdlSNZaZvEjTt1N01G5JamdJJjIioS02orSkbN25SUgkHg5OO+ayzxu8M87tCuzwzNgZkRHHhDXNs2SSToTrsrHA9P8AT0KGzHYhqSW0BBdQ8tpxzHlZQU7j+tWsx42igFz5+LZWQ9zpH3euwoe1rNNZs2lbO+I7sO2OSFFLbz4LhceIO3OTucP2znAqZ5YxY0UIzPnSta4F9dBpp19Bp1VV0S7e7xa0yYrim5s1KHZt3kRsDgYS2w0cBQSP5j7MlR9xJxTFzub69T/st3EGYsMpBb5W/CwGz6lzh+Q19lf7LbmrLbGIbS1rS2D71kFS1ElSlH7lRJ/U1paKFLjTSGRxeRS5BYrczJtF5v8AIhLTOMgwIMeYd7aX+upKVqRkpO1x0DnIBSrHesDGh3NJWuwX1WRM6B8WCJAWVzOIFGiASL32H3C6FerA4zohNss7LT70fpuBp44EpSFhakrV8uEcn781qMZbHyt1XDiyhJmePKeW71H8t6aewWRqRC1FoNiZeenCYdjpeeWh3YIy0n8yXONpSpOQrxivebmZbtFWYnY2UYoPNRIGm46aeo6Lmc2K3fpU2FNl6jvQ9htCJATHS+4oLUpQ3IAKEpCSVkcAkAe4A4i1sri1xJ7L6WPIlwWMlhayMm+etSANADroT26nXoux6YtotNkgwQ20gsMoSvpJwlS8Dcr9zk57nNbmMDGBq+UypzkTul7kn7/2Uqvt9/FTVKpXpk01Fc1LEZguQkNXZ0pQsAZCkoUCME8HOf0IrPAAOYAVqV1eKPdJ4L3v5rYPlROnuOq1fUbT7esp7ViecDbbcRyWkkEgOlaUNkgEe3HUGPOeO1JoxKQw+6s4blHAYclu5IaR/pok/oue+nMm6MaFuVvmWF6+xbY+q3SIIkI2pWhZVltAQVKPvBKs9wCBxVEZcIiC2wNKXTzWQyZsb2SiNzvMHEHqNATt+9Ve7XcLlaYSbo0/Ju2lnWApDO0uTIffO493ADlJ5KhjztJNzXOYObdvbqFzZ4YZ3fhyAya6JumO9f8ASTuDsfotPSlzm6gXcW9PS2oEd19Ux2Q9FK3gHTlsJSSE52Jzk7vGOBUYZDLfJsruJY8eCWeO3mcAG0DQtoomxvrdjTZSWpoFqsunTFt02HGucJxFwbLzyA884g7lKWSQVKWnekk/3j4qcnK1tWAd1mwjkZU/O9he13l0GgB2rtRorPZLxqDVEMSrcxDtEFal9KS6TJceRuIStKBtSkEDOSTnwMc16x75RzAUoZOPi4UnhvJkcKsDygHqCdbordGhrXKlszL91r1Na3bXZ6gtKc9wGgAgD7bf3r047XHmdqVWOKTxsMUB8Np6N0v3O5+qzXJsQdVWKQ0npsOIdgK2cDlIW2MfA6asEds47GpO8rm/ReQ/xceVrtXCnfo78wrOn8oqxYAvaIsMtlp+M4082hxtaSlSFAEKB8EHuKUDuvQ4tNgqtaDT0485qIsqtDcpbcEKOSlCeFgHygOBYT8AY7YquPY1t0WvONuYXfGQC79PnVX6q11YsaURKIlESiJREoiURKIlESiJREoiURKIlESiJREoiURKIlESiJRFzr16miF6ZXf+MhlT/TjpUrdzuWMgY84B/pVGQajK6XB4/EzI9LrX7K46aStFhtqHZapjiYzYVJUOXjsHv5+e/wC9Wt2C58n+Y41WuylKkopREoiURKIlESiJREoiURKIlESiJRFS/VJLJssQiQtmemW2qFh4tgug91Y/MhIyoj4Saz5Fcos0ei6vCHFsrvLbCCHaWaPa9iToCpPQd2mXzTEG5XBhlh6S31NrKlFJT4IyARnvjn9TVkTzIwOPVZuIYzMXJfDGSQ01qKKsNWLGo6/3Jm0WuTOkfkZTu2jus9koH3USAPuai48otWQxOmkEbdyoL08scOx2l5qOWjNeeL07puFYTIUAVI7nG3IAHxg+ahDG2MED5+61cRzJcqQGT4QAG/8AiNPvutHXsVUnUWj9+1cNc5TL6SfzZR1Ecefc0M/p968msOb7q3h72thn78um/ev1V6QMAd6uXLApfVF6lESiJRFRNcOsXm8WzSUhSyzOC5EtCElJUw2CQAvsMrCQcc4+M1RIWvcIj1XTwWy48T8+PdhAHufT0Fq3BUW22/cShiJGayfCW0JT/uAFXaNC5tOe7uT+qyxJjEuM1IjOpcYdQlxC09lJUMg/uDXo11RwLCWu0IWcKB7Glrxe0Rcc9YbvKs3qHoR+FFipcdkFhUtbYUsoUtKFNg+BtWT+uMec5pnlj2Uu3wzGbPi5HMToLr1XYQQB3rSuItefPiwIjsqbIaYjtjct1xW1KR8k145waLcpMY6VwYwWT0CqWuI6nbvpe5IdaUxGuCELbU3v39XCEqTzgEEjnuMkj4NcjSXNd0W7Cka2OaIjzObp0qtT9RuvvTTLlq1hf4v1Li4LiG56UOYCY6nFObgjH8pKCo9ufnk0Y0se4dDqmTK2bHicQA8W01/MBVE+vQK0wLlCuKHVwJbElDay2tTSwoJUADg484I/rUw4OFgrHJE+Kg8EXqL7KB1I7KtMxu9NSHXYLaUtTIhI2hsq/wBqj/GknkfzJyO4FeOJb5ui0YwZO38PXmJtp9ex9D9j6LBqOM1J1jpUqLvXackuhKHFJTtDWCSBwfcpA5+T4zUXt87D2v8AJW4kvJizgV5g0f8A7X+QK+vURlu4WJNkJw/dnExWgFYIP51L/RKUqVjzgDzSYB7OTuocMldjTjJA0js+/SvnanrRAatdrjQo6nVNR20tILqytRAGBknuamBTQAscknivdI7c9tlSQ8b/AKnu12srn1sixtqhRoindjS5JG5ajkcY4Tkd8HnFUh3iPc5hvl0+a6xiOJjxwZA5RLTieob0A/Mhbfp9MusSJbrFcbDNi/SwkpXLWUKaLiTgpGD2I5B/+59he8gNc2lDiUMHO6aKYO5naDW67nSvkpXXwcdszMRt96MJktiI46ydq0occCVbT4OOM1KbVtd9FVwwN8YvIB5WucL2sCxfoqhd2LHYb9o/T8mQ9Ct8BLs1Dz7xbQ44n8oUskJUSStRH6D+bBpeY4yyMmgF0cc5eZFlZbG8znUCALIB6gdtgr1qm6G0aYuFyjI67kaOp5CAkqClAZGQOcZxn7ZNaHv5Gl3ZcXDxxk5DILrmIH3XPpEdifbPT+1R4Ux9DikTnN/sSlpG0uF1GcHJWMDsM8Z7GhwDwxtb6rtMe/Hky5ucAttvckk0OU9Nt1bLnAjNeoNguCkK+ocjyIu/cSOAlYGM4HAWc/8A0q17R4rXe650Eh/BSw9LafzH9lb8irVzisb+SnCTgnsfivQlgLlEK8TtH6uetlykG7P3JhUlAZyp991JCE4bxhvKQRjO0BvII5zj5zFJyO1Ll9EcVvEcTx4m8jYyG+gB1Nnc0fmSa7K66V0+u3yJ9znuqcu1zKVytrhLTe3O1tsH+VIOM+eTx2q5kfIS47lcrMyhM1sDBUbLrub3J9/tsqNp1bmmvXS92dDyfoL2x+IhDgCSHueEf3v5+3gfaqmu5JizodV0choyeGMnI8zDy/LcKWvn4podq9XtiUxNtch8vGG8OimMV4AUlQzn3n3DHOcjnOfXOMAdJdjt2UMZsXFHRYnLyvArmGpd1+taD6bKI0nbFyJ8C6Pretb17YMiObe6Uhgp9xZwoFKkrSpTgBHBCsVCFpdTjpzdvy/VauIZDYw6CP8AieEaPNu69ObTUVQbd66eqlLF6ax/9YXqV1yetc1b6EOOhwKRuBR1F7ApR4GRnb4x3zJmKATz6qjK448lv4RojoVoCPehZA/P1XR2UNR20toCUNoSEpSBgADwBWqqFBcEus8zioSRq+zMvvtfUuuKYWW3CzFddSlQ7p3JSRkHg88GqjMxuhK3s4ZkyNDwBR7uaPsSCoy0S2dWXs3ONJLtotznTihtXsef2e9w+SEhYSkfO447UY5sp5hqB+asmjkwGeE9tPeNb/pvQD3q1c0/lFW7rmhe5FEUZqC5RbbaJMmZKEZpKCOpjJBPA2p/mVnGB5OBXjiALKtgidNIGMFnt++nqo/QECZbdJWuLclqXKaZwoqSEkAkkJIHkAgH7571CFrmxgO3WjiU0c+U+SEU0nRWOrFiSiJREoiUReL/AC0RQ+mbk/cY0gTG0NyY8l6M4lAIT7FnaRn5QUn96i13Naunh8JwrYgEfP8A3UzUlSlESiJREoiURKIlESiJREoiURKIlESiJREoi5T/AKR/17np041AiLkMrktGUpA3FttJ3bsd/wAwSM+M1mygTHTV2eAGJua10pqrVu9MrbNtGgrHAui3FTWYyQ6FnJQTzs/7IIT/ANmrowWtAK5+bK2bIfIzYlWiprMlESiJREoiURKIlESiJREoiURKIlEUFqzTVv1LEjsXNouIYd6zYyQArBGSP5hyeDwfNVyRNlFO6LXiZ0+E4ugdVij7fvqoT00vyZUaXY5CFNzrIpMNeUFHVQkbUOBJ5GdvI5GexORUYZOa2dWrTxTCMPJkg22UXvdG9QfXVXcqGKuXLXMvVG8S/wAatlptzpafQkT9qmeol9SXEpab8fzqCjz/ACjucCsmRK4ODGmuq73CMVngvyZW8w+HQ0RYtx/+IIXlghP6Gl3WMlt28SpUVVydLDe1x51Lm0gJJI928HA7EKx3AqUTPABO53+ajl5A4qWGxG0EMF7Btaa+i0tBzJN9i3LUGohIuC7fIcXCiIQhXRwAvCEgAl0fkye2CB3NV4zjKC+TvoOy08WiZhPZiYvl5gOY2db09g07/ddStk1m426LMjEqYktJebJHdKgCP8jWwGxYXzsjDG8sduDX0W1XqglESiJRFWtW2a43By2SLNLjw5sSTvLzzRc/hKSUrSADzkEcHjIHbvVUjXOotK24U8MPO2dpc1w2BrXoVjOl3pwIvt3nTW1ZBYaV9MyU/BSjlX3yo/pTw7+In8lL8a2N148Yb6nzH76fZaydCx48ll633a8wgwhbbLLUvc22FYzhKwrjjscivPAF2HH6qw8Ue5rmyRscTqSW0fsQpLSjF/jNym9RyosopeIjOsp2qU143jAG79BUo+ejzrNluxnOa7HBGmoO1+nWvdTkh9uOy46+4htptJUtazgJA5JJ8CrFlAJ0C/P2vfpfUj1T0sxpu4R7hCit9WWpknDCEuhSiVfKgAAB5/WsUo8aRvIdl9NhPdw7Dm8dvKXaC+q7teEz12x8WhcdE4p/gqkJKmwf8QHJrY660XzcfJzgyXXoqHrWJqXUmm5sNyzoYQlslUdExClS3MjaEqx7Wwfec7VKKQngZNZ5g+VnKQPquxw6bGwcpk7ZCaO/LsOul7nbsN1H3mBfrZo5ybqC9QLci3KD8SPFjJ6aVoI6SFKOc88BKQO/c1E+IyImQ1S1QSY0+c2LEiL+fQlxNm/iIA2/ei0laebvGsrHcbzcfxGBfIC/9XfSqPkJSlaEpCFYz7skEnso/pWY/Eka55sEey1NzzjYU0OMwMdE4ainbkg6kbab+oW9boZ0rcLtYLXc7bZIz5XKjLlRlKW4gtgqKXC4kHpkKGMEgAE1YxjYbjaaWPIyH8RazLnY6RzaBo6aHSxWnN+ajtaIl6R05Ch2+5TrwxeEvR3Pqn0uBxS2yQpvjcFFRyMZHODyQarlJgaOUk33WnBazic8j5GNiMdEcoIqjWtmvcmj91JXLWMVGoLbdFf6tb7RuhXBMxpTT7S3kIKShP8ANjYMgcgZPIwasfOA4OOgG9+qzw8Je6F0A8z5KLOUgimk3Z6X0v566KwXKS3rbTkSdpW5MtuMTEutPPMkjc2SCg5wU57E/BPg1bfis5oisDYjw3JdBnRk6UQD366dvzXzZdaSb+l1FktSJCo2G5fUmJbQ075QkhKt44PvAAI7ZOQPGzl9+GNvkpS8Mbi1+JkLQ7VtNux3Ooo+m/cBSWhrdcYjNxl3mPFjT58xUhxmMvc2gBKUJwfJISCT5J8dqnE1wBLtz2WfPlie5jYHEtaKt2/U/IdlaMDNWLCq7rePIcgRJkRtT7ltkpm/TJ7vhAUCgf4sKJHjckZqDxpY6LVhvaHljjQcCL7X+nf0W2tuDqCyocKW5MKWyFoUpAUChScggEd8GpHlkbr1VTXSY0vlNOB7n9FzGTcHNP6JvOnr+481IYQ4qCZcjqfXx0rBCQUjPYbVJHISrPbtjc7woix+429QvomRDNzY8rHaCHVzUK5HVV/XzA99FbNATDqNbmpFRlRYzjf0cFhWMpaSolS+P7ysDHgIFXQPEo8SqHRc3ieO7h7vwRdzOHmcR3Ow+mvzW9qm3sv6j0pNc6nWjzVoRtcUBhTLhOQODykf7vJpIBzMJ7/ooYUr2wZEY2c0fZzVk1Xfp1sm22Fbrd9Q/PcU0iQ65sjsqAyN55OSM4AHOMVN73NIDRv9FRiY0c7XySvoMo0NSfb9e26irtqm9WOYzFuNnZlGSlQivRHyEvO8bWilQylRyfJGAT4OK3SvYQHD6d+y1w8Px8tpfFIQG1YIsgdXaaUFSXrCdRXO9yprjkl2DDdUZDKthXJOfyHulCemUpH91OTyok0Oh8VxJ3A+660PEPwcccbNA5wsHUcvqOpN2T8ugXSfT+6KlWFmM+71pUNDba3ck9ZJbStt3nn3IIPPY5HitcJJaAdwuDxFg8d0jBTXEkDtrqPl/ZQnrLo5epdP/W2hhP8AaKApL0N5PtcODkoCvvyR9wPmoTxlzbG4VnDMsY0vLL8DtCP1UJH9TLDqX0+nLnvW9m6IjKD9vnHCS6BxgEe4EjIxnBwDgio/iI3R27fstjOEZUGa0QBxaCKc3t7/AJqu2pdxvTsGx6SnuRbSVtOxpEghb0ZTbf8AFU3hR9hKgCM4Cl7QACQMrS+QBkRpvruF2ZhjYvNk5zOaTzAgXymz5QTWhGtdwL9+mptGs1XFDy9RW9thtrphlEAqS6rIO9YK8gkce08eK2hsvNfNp7L5vx8ARFghJcTd82w7DTX5reTZL1Le6lzv7iAlOEN21kR058qVvKyo/HIA+M1Isc7c/RUMyIovgiB/8rP5Upm021i2W9iHERsYZTtSM5J+ST5JOST5JNTA5RQWWV5leXv3Krtwei6f1jFfcU3HjXdP0yjkJSqSj3IJ8ZUncnPf2pHPGInyuvutkbX5OO5rQS5hv/29foaKsUa6RJEuVEYfbclRCkPtJOVNbhlO4eMjmpNIJq9Qsbo3sY17mkB23rS0r3e/w96NGixHp06QFKRGZUhKticblkrIASMgcnuQKOdWinFDzNLnHlA66/TRQ8Oz3e7agYud+LDMCMnMW2pId2O/+lWvABUASABkDOc55qnle91v27fqtsk0EEJix7Lzu/bT+kDXTv3VxSnaAKuC5q+q9RKIlESiJRF4rtRFXLIenqvULaSdivp3ikHIC1NlJP2yEJ/pVTf8xw9lvyGgYkLzueYfIEV+ZVjBz2q1YF7REoiURKIlESiJREoiURKIlESiJREoiURKIudetF/at2nUWduM/Lud7V9JEYZxlRyMkk8Acgfqf1NVyPDaHdbcHGMzi+6DBZK6C3yM4xznFWLDSyUXqURKIlESiJREoiURKIlESiJREoiUReEZoiqmvIkhEBNxs8aQ9e4xCYpjgZO4gFK8nBb8qB+MjBANVyg1zN3C3YD2B/hTGozv/cdj2P10VXv+ur3bLUtu6WGVaZKhgTErS83xyemBnKyM7Uq4z3OAaodkvaP4jaXSi4PjzvBxZhIO2oI97rQdSPotW3CVdNW2N15N6MZ9JecNzS02tQaAcQEoQAdoWpO5RHJIHOOPW297XVovJeSDFliHLzNOnKTXm0Ju9bGle6s+uHXrRMtl6hux0PbjBc+qJDPTX796iOU7VIB475I+CLpCW+YLnYTGTh0LwSDqK3sf3tRfp5pNNuZkaklSXVy7rF6r7KGikI3/AMRXtycqyft+nJqqGENJkJ1ct3E+Ivn5cMNpsbqF6nTSr00U96W3Fq46Hs6mA502WExsuIKSemNmef0/31PHfzxgrJxfFOLmyRk9b3vfVW2rlzkoiURKIlESiJRFH366xrJaZdynr2RIrSnnVAZISBngeT9q8JABJUo4zK8Rt3K4zZdV3n1euU+zxG1WfS6B/rTyEBx95CuAypRO1JV7ido4Ax+uVkpyCQNGr6DKwI+DsbI93NKdh0Hr6rsVjsduscP6W0wo0OPndsYbCQT8n5P3NaWsa0U0UuFNNJO7nlcXH1UoBgVJVLwpGK8Ras6DFuMRyLOjsyI7nC23UhSVeeQaOAIohSjlfE8PjdRHUKlazZkWO+W/UCUTLjb4+I34ZHRwyteUpeQB3PO0g+FcVnm8jhJuBpX6rrcODcmF+GOVjna8xNWBqWn33+SnX27Pq/TwU6y3cLXKb3oyj8wweRnBSocjwQaucGSs7grAyTI4fkWwlj2n9+4XGrXGYVre1zrI+bY01Ict8G3XhKl9J0Nq37EhWUjJb8/mc+1c1oHihzDVGgCvtHSPODLBkt57DXufGRZBOgOm+/yCm2NFajtCpNw22+VKZaLpkuvOyVP53FxrpKTykjgDIOTncckVoED2W7c/u1y38WxcgNi8zWE0WgBoA0AdfVw7nT0VbtJYQvSydQ28N2SWx9WUxwsgBsqCFuoxwkJXtKvduQlJOMGs7TrHzjQ66fqurNG5oynY8hMrTy+atebcNPckaAVWq6/cdOvsXJu92GV05TEVMdMEhKYzzYVu2nAyknPChwD4xkHe6Ih3Oz6dF8nDmtdCcfJFgm+bXmBqr9fUdfdb2i9Tt6jiSCqKuFOivKYkxHFhS2VD5I8Edq9hl8QGxRHRQ4jgHCkADuZrhYcNAQrJVqwKua0k3FLESBZHY7FxnLW22++nelrahSydvk+3HPAznnsa5HOHlbuVtw4YXF0s4trKJA0JsgVfzW7pe2rtNggQHnQ87HZShbiU7QpQHJA8DOeK9jaWtDSqsucZE75gKDjddlG+oyWE6Kvjspht9DUN1aUrQFYVsOCM9iDjmoz14biR0K0cI5znQhriLc381UvTqFd7DpK2ybXI/HrdJYS99IpxLbjClAFQaWTtUN24FKsc+e9Z8Vj44gW62urxyaDLzZRK3w3NJFgWCAa1HTStRalNSyYOpounn0OzWWG7whpxDS1MPNu7Vp2K7FJCsZ+3Y81a+peV3YrBjiTDdMxwBLmGjuCLGo+Q0PdS+oyHtQ6ahrHsXKcklWectNKKR+mVf5ferXk8zQsmM0eFLJWtAfU/20W7qqws3+zrhPLDK/zsvpTlTLg7LT9x+vIJHmksQkbyleYWY/DlErNtiO47Fc202f7LQrhcZ0kCEu4uxb1HdUp5Ec4IQttR92CC2CDuyFjymskY8EFzjpev6LvZl8RfHBE238gLCNCdrB9jdVWxUrAj3yyriXTTsaJfbSqGGUJbkFl5xgHcyTuBSooBUkKBBIVyMirT4jTzsHMFj/7WcmDJcYnA9RY5tnbai9O9LcR6nRXNMSLumx3nayFEoMfCSBj3dT8oTk475+1RGUHML+Uo/gL2ZDcZ0rNeoddfLdUqRoi+ah1ObvGtR0vcF7pAmhba0JQRtDSmUjClkDcVHH5jnOAKrMb5H83LynutozsXEgMJl8Vu3LRHzDr26BSelNSHSF/Xa9fxI1uuMtJdbvCXAWJSU/y5P+zA8JACR8AnJujk8I8svXqseVj/AI2IS4Z5mt3b1b6+vvuuwxpTUlvew4h1Hbc2oKH9RWiwdlwyC00RSy7hmhUbXilhOScADknNe6ItS4QIt0iuR5rDMiO4MKbdQFJP7GvCAdCpxyPjdzxkg+iq0/02sD7zbsRuRbVpWhxf0D6mOqUnKd2O+OcHuM96pdjRuPMND6LpQ8Zy4meG4h4ogcwBq+ynrPp23WlS1w46UvOf7R5ZLjrn/SWolR/c1axrWfCFhmyZp6EjtB02H0Gil8VJUr2iJREoiURKIlEXiuRRFUdZxIjbsK5N/wAG7okx2WXmllC1JW8lJbVg+9JBVwrI896qkbQDm7rbguJJif8ABTj6WGn6FWxocd81aVgavuiklESiJREoiURKIlESiJREoiURKIlESiITgURcU1A6i/f6QdmtdwuLaY1rZTLiRm0Hct/buIUrHBwkK5OMJAHJNZHHmnDT0Xdgb4PC3ysbZeaJ7Dp/ZdqArWuEvaIlESiJREoiURKIlESiJREoiURKIlESiLxSd3eiL4WCkEilLxxNLnkeVfbVr1+Rf4EddsmqESLNjrKhHTu/hoWkjIK1KGVf3tvgCs9yNlJcPKdiu29mJPgtZjOPiNsuaRv3IPoFh9aXXJVhatcd2KlT6XpLrb4zvaYbKyByOdwR/wDQA15lguZyXv8AopcBLYZjlUTy6AjoXGux6WsbV5v93s8+26RszLcJtlMSHcnJHTaICdq1JGAo7ew25GR3qLZXvBEQ06FWHCxcZ7Js6Q2SS5gFn0s7C+oOoC6JaIog22HFASkMtIawj8o2pA4+3FahoKK4Mr/EkL+5JW7XqilESiJREoiE4FEXwV14fReE1uvz1/pH65ucWY7pWK001CfjodfdKdy3gon2jPCRlPccn7Vhy5i0+GF9b/h3hkcw/FvOrToPkum+nws+lvTe2uqk25iII4fektO5aWtXKlBSuTz88+PFa2BrIx2Xz+W+TLy36EuJ26rWZ9XNOO3GBF23JCLg6G4clyIpLL+VbQpKjyU7uM4+/bmo+OywO6sHC8gxvkFHl31Git6r7BTe02hUlv8AEVMmQGBkkNg43HwP3qfOOfkO6yjGmMP4jl8l1fr2Vdd10ifcWYOmbe7eVOdTdJQrpRmynGQXSCD3H5c96r8cOIEYv16Lc7hboY3SZTxGRVNOrj61/dS2kY18ixHf7STY8qS4+pbYYRtS0ggYRnA3Y55xU4w/+crLmPxXPAxWloArXcnqfRWBQyKsWVUG+aVuFvE6fYL5cm3tr7zUJbzYj9Va+oc5Qfbkng/pkCs8kbgC9jj7aLq42ZE8shyI2kEgF1HmrbSivv0zscN3T8G9TI7T94mf649LcSFrK1Z5Sf5U4PAHGDTHZbA9w1OqlxbJP4h+PC6o2+UAaaDv3Pe1m9RLldoEB9m3299yG9EfDs5g7lxV7cIPTHKhnuR2HPipyvc3pYVHD8eKV3mkAdYoHrZ112FfdVdtnT9x0odT6dkCDJiW9MR0N7n222w2MsONkgKA3DkYPAPzVDBHJH4kemlLpTOy8XK/B5YJBdzdiT/UD60o6134Wm5W3RF1mz7MmJHDbrzcgOF91RG1KXiCUNlJyDgEZ25TtGYeKGPEBNUtcvD35MMnFoWCQOcdCKoDqRYBP1WeRdIvpdqmQZEN8QLj1HElDqVBzaEbDlSt24ZcSrPcqSc96PkbiONj4l5FjTf4ggaGPHNEAKqtyQRoKravmum6V1LbtUWwT7S8XWNxQoKSUqQoAEpIPnkVrjlZK3mYdF89nYM+BMYMgU4fu1qX3/y103/1cv8A/sRXp+ILyP8AyJPl+qsmcCpWsq5j6oSrpd5g0za1RUw5qEMS3HEq3sqcXlO3BAJ2IWrb8Ak4yM5cjmf/AA27HdfQcGEGN/30wJLDbQKo1vd9rH6K0xLLNtmkoVrs02My/HZS0JDkXKe3Kg2lQAOee5++avDHMYGMOy5j8tmRluyMlpcHG6Br5Wb06Kk3S42m0uRWpl5+tvCbwwucstFKlKaQNxS2gcJCSnsDz3JxxS5zI6D3Wb/ei6kMGRl8zoIeWMNIAvQWerj1vosWqb9+I3BV8YtdxWxp9JVFcWPp2y8tCVKLxUQoICCj2492fviovkLj4gB8vfRTw8URN/Bukb/FI5q8xAB0Aqxd39ll/txfJyYiYP08hbriGl/hMRUpDalIKgFOuKQjPByBnHkjyGQ81Wvt/deP4RBHzCQ8lWRzmrANfCAT87F9lu6ebuV5a1dY7w2I9xeCHFPqYSWsONbEgJBwogI5I4znHbmTAX+JG7f+6pyHx44xcuE2wWBqbsGz0FXeg7K36Gcfk6Osz0lxDj64ralKSgJBOPAHAq6H/LbfZc/ikbY8yVjNg4/mqDfzIYt940al6JGjsR3pbRc3KdejcuJQhONvBygqKs8HA81S83cV0f0XUxQGOZxINLtQNKADttdb1Gu266naJBm2uJKUkIL7KHSkcgbkg4/zrSDYXBkZyPc3sStbUFgteoIgjXmFHmMA7kpeRnafkHuD+leOa1wpwtShnlx3c8LuU+i5216KWS3ynn7NeL9ayvBSIsoJ2cknnGSMcc9qoGK0G2khdU8dneA2ZrXe4VXd1NqTQN6nphzpGr9MwkJTIW84FOQ1qJ4W6AeQR9xgjO2qjK6Nxo8wW9mDj50TOdohkdt2cPQH7ahdBsMGDrq2oud3nR7tEeH8KHHKhGY+yk/mWseSvGPCRV7WNkFuNjsuVLkP4fIY8dpY4dSPMf0A9B9VeocZuJGbYZG1ptISlOewAwBV/suY5xcS5xslZqLxKIlESiJREoiURKIlERXai8K57qfTdnuPqJZn7oxvdejOBpZcUj+KytC0JGDz7SskHwKzyRtdIHFdjEz8mDDkjhdQJF6DYghdAQMCtC44X1RepREoiURKIlESiJREoiURKIlESiJREoijNRPT2LTIXaI4kzyNjLalhCdxOApSvCU53HAJwOATXh9F60AnzGgucel3pW9pm9v36/3IXG8uFYStsnaN/wCZZKuVLPP2ArPDj8ji9xsrs8R4v+JiGPC3kjHTqV1oVpXFSiJREoiURKIlESiJREoiURKIlESiJREoiURCM0RRmobS1erRKgPKW2l9G0ONnCmznKVJPyCAR+lQe3naWlXY8xglbKBdH6+nzXKrNYGbrG6up9TylyG3F2ZjKkIcadGUuI34JXuIyDwdvfuayRxBw8776D0X0eRnOhcBiwAA1IasgjcGtgB19VdPS5bw0wmHJk/VOQJL8AOFISShpZQnIHY7QKuxr8PlJujX0XK4zyfivFjZyh4a6vVws/dXQCtC5aURKIlESiJREPaiKneocZbcOJd2H5CJVsfQ6wyhwpRIUpQR0lAd927APgnzyKqlaNHdQuhw9/mdDVh4IPp1seypfrvpS7atkWWFY7Q086hS1uXBxSUBpIwA2Vd8Endjn8vHmqsqIy01oW3gefFgF8sjtNNO6idJ+iE14Mo11c/rIERBbiQor69jeVZPJAwD8Dnnk8VCPEO0h2WjK4+wEuwmcpduTVro3qTYLleNLoh6cVGjzmX2XGnHFbC0EKBy2rB2q4ABx2yK0ytJADd1xMGZkUvPMLBBse65xrXWdluV0jQZsEWW7NOGLNl3FtQUy1tO9La2jhzPI2nhW4cYzjLM8OeBsep9F2+G4ssULnAlzSLa1tG3WALBuvfcLbh64ski6W5iH6hvsYQpsNqtaWo2TgJSQUgJx4yT+o8+iRvMOWT7JJiStheZMT1vmPN+f5BXea7qGwTItxnXdq4WNB2zGzES042FEJDiSnulJIJ+2e/GLzztN3ouaw4s8bo2xlsn8pux6ghXhJzVy5ihtZrYRpe6/VyREYMV1Kn1HAbykgH+pH61CQ0wrTh3+Ij5RZsfmvNEOrf0jZnnGeityGypTW3Gw7BxjApFqwey9zmhuVKAb8ztfmo/1AkSFWtNstYjOXK5L+nbbkJJQUYy6VYIIARnkeSPJFRlJ5eVu5VvD2RmQyz3yM1Nb30q+pKomm7BpqDrubYC68281GacREQ44ht3/nFpV/6QJISRuPZRBzt4yxRxMlLB0Xezs3OnwG5cgBDnGyQLFaCu1jQ0NKvqbuGqPTixajujtynIkolPNhp1TL20LAxjIIODwBxir340cjuZw1XIw+N5mFGIoXeUGwCAdVEafsMOx6lfgXiRKkNBC12luY91GEM7QHEp3fzjcQQf5SMcZxCOIRuo/K+y0ZedLlQNliaGnQPLRRLrsE107eu6r2r/AKHQ93jTNL6jj2WLcHNkiG22JDQUEnDobGSBxg4+2PIqqYDHIMbuUHfS10uHOm4sx0WXAZXMHlN8p9r6nqAVka1NqFjV9mfmMx9SQ5DC3Ir9rbKC2hwtpJKScDBTj3H+ZXIxgBLKJG2OYHsouwMGTDlDHGGRp1D+pF6Cu99O3qpbV+trqrTUu8WXEC2RXkMuPyGQ48tfU2uJQ3naAk5BJJyQccYVVs0zvDLmaAfsrFw7hcX4tmNk+Z7tQAaG1tsgXr9tLVc0z/wpPlTNMzNSTvpglLUzLK8yFpHUcWl4gYICU45wEnBHaqoiH26Mn3/5W7O5sXlizGRtvduugBNC2+5Prdqyl71ObtCIjMW2OSUoKTNkLSFn4O0KKd3YdsVYTk8tAC+6ytj4IZzJI93L/SAfzIC0NJaX1c3d5twuKYTFxlJA/FZDgkutA90tND2JBzwCeP8AF2qEEEwJc8Cz1/sreI8T4a6JkGOSY268gHK09iT8RP70XxJ0/b3/AEkkXeTBl3K6TIf1TriXC46XF91JBOAAecAdh2r18LRES4cxXkHEZxxFsTHiNgNDoAB0Nan3XTdMdJen7a4yw1HaXGbUlpoAJQCkHAA4xWxhHKCvnMkOEzw42bOvfVUf1SkOWkT3EuuJZnQU9QIzu2sPJK0ApGQFoeUnPg8kgZNUZDuQFw/dLq8IZ+JdHCRfK7S9rcDqfYgLoNjfiyrRDft6dkN1lC2U7NmEFI2jb44xxV7K5QRsuTkRvikdHL8QJB9+qqPqvZLbcbXGentBvEphhyYg7XGmVuBK8K+DnGORzVORG17fMulwbNnxZT4Otg6HUEgXt6L50wxqK42aO8xfBFhAKRFCoSFuraCiELdJONxAHCcDyeTxJjXFu6qyJMZkhBjs9dSAD1A9PdSEqz6sW7FLOp46UMuFS825OXQQRtV78YB+MV45kpOjtPZGZGCAeaAk/wDnVfb818ztGG7xFx77e7tNbdGHWkupYaV8YShIxg84ye3OakYQ4crySoxcQMEgkgYGkbac353r8lYoNrhwLczAiRmmojTfTS0lPtCcYxjz9/mrA0NFBYpZXyvMjzZK4jAtjvpN6opeddV/Zi/uqYb6fPTcJCkBaeANpKgCP5SfisgHgSf6T+a+gfL/ANWwuUD+LGN+7f8AZd7QcithXzYNr6ovVXrzqZiBOTb47Emfc1o6iYsVAKgnOApSiQlCc+VEfbNQfIGkNqytUGG+VhmJDWDqfyA3PyWrZ7zdzqEW69x4cbrRTJYSw4pZOF7VIKiAFFIKScAY3Dv3oHO5qIXssEQhEkJJo0b+xrpfS1agoE4qayL2iJREoiZoiUReKIA5oiqUtH4vruEmOo7LIlbr6scFx5spS2P+ySon7pHnisi3iun6rfG/wcVwP/8AYR9Gm7+eytieKsXPGy+qL1KIlESiJREoiURKIlESiJREoiURKIlEQgHuKImBREoiURKIlESiJREoiURKIlESiJREoiURKIlESiJRExRFTJfp1p56S5JaiORpK5H1fVYeUkh7Od4SSU5HOOPJqg40ZN16rqN4zmBvIX2K5aIHw9r3pQ9ztLHp3AevtoRLfSlwu3JnO5UtKlcueEpUgnIIAG3cD4Ii5gx287fmr4siTi8rcacgaU07AUNBfW9vddGYdDjSF8jckHB7itA2XEOhpZa9RKIlESiJRF4ojHNLrVFUrtIYn62s1uQ6H1ROrLfZSdwbwgJbUsdgdyvaDznkds1U88z2tXRx2+HiSyPFXQae+vmA+W6tiUJHYCrVzV9YA8UperzAoipGuGYSbzaJGoUsv2NRXGLT6QW25CxhC1gjkFO5HPCSrPnIpk5Q4F+36rpYXiGKRmP8dXpuWjcD8z6LW1B6TaPvSGt1qbgrbACVQT0CU5zgge0+fGee9H40btwvYONZuOSY5LvvqvmD6fybXpC92GHfZMhiXzDMxO/6Qce3OfcOB4GPivPBIYWtP1T/AKgx2SzIljGm4Gl/2UmmRqyHFUw/Bg3CQlGES2ZPRStXyttQ9vPcJJHxjtXvNI0VVlQEeFI7nDywdiLPyI/ULyxWC4SFMTtYSmZ9yaO5plkERmCCcKSk/mX/AIlcjxjuTGOIuXf7L3KyomkswgWMOln4j6E9vQfNZrnqFT04WvTamJdyUNzi925mKnOCpwpPfIICAck/ABNSdJZ5GalQjw+Rn4jJsM6d3HsB+Z2HqdFu2OxCDJdmzpTlwujyQhyU6kJwkdkISOEJzzgdzySeKNjINk2qZ8rxGiNo5WDoO/cnqfX6KPvelXJ12M+Dd5dsWstF5MdCD1emTjJUMjIO044IAyDjFHx8xsGlZjZrYYyx8YeNau9L/srDcpzFut78qW4lphlBWta1BIAH3PAqwmhZWWON0jgxgslUeBDX6hQmZ19guw7SEkxYbi8rWs5AfUR2wOEj7qJzkVnaBOAXDTp/ddWVx4RI6PHkDn/zEbV/SP1PyWaXoRTESUxbJbCo8tstPx5sVC0rBGCQtsIWk88ckA/FSMI2Coj4jy09wpwNgtJ0PsbBXM47tj0dqW9W7Vkh+WVx2lgW/ewyVJ3bWShChlW3acqPcnPPJwNczGlcJD9PyX1pizOLYkUmFGGgE/FRdrVuBOtXYIHorrG0yrUz9vnQ7fbLRZFxwFFlDbz76SABscSP4ftG3IOeT+o0iHxCHUAPuuJJxD8G10RkdI8GxdhoPqD8WuvZXSe9F0zZGGoMIKCVtxYsRjCNy1HCUgngDnJJ7AEmtQqNtNXEBflSF0jtTqSdfcrWRbtQXFpKrjd24La8qVHtzQ3JHhPWXnP3ISM+MVAMedyr/HxYx/Cj5j3cdP8A4ivzKrWr7BA0npu63th66vzWSX0uKuTiVhSyEkA9sHPbBJwOe2KpmNhjc+z9VvwcqbiGTFiODQ00NGDp17rFDhu2D0/t02Lqp+3sqitIBlpRLYSpYATtwAUjJ7pOMdwcV5QZEHc1bb67qbnDL4g+IwB55jo22mh/sP8AdaenpGp7HqBWnmbnFu6bXCQ6IYY6BcbIACUuYIC0kAjPBCu4OcRYZWv8Mu5qHstGY3CycYZQiMfO4jmvmAI127H7V9bNdY39totpuFhuyYjCeoh9YaCnek4gBbWDwhfAyCMgj7c3vHigFh/fZcqF44e58eTHzHprWo1B9R+auMCO1BhMx2UhDLKA2hI/lSBgD+gFXDaguYXOcS525VK9Sr/bHLJdrMhJuM52M4Fxo+VdIAZK3FJyGwn83PJxgCs80jCDHuey6/DcOZsjMonkYD8R0v2H83bTTurTpRG3TFpCkhJERnIBzj2DzVsY8gBWHNcHZEjgb8x/NS1TWZMURKIqR6taL/txpn6BMr6V9h36hlak7kFQSRhQ+CFHt2479qqniEreVdDhmecCfxQL6KB/0db5PvOhXEXJ5byoMpUVtxasqLYQlQBPnG7AJ8YqGK9zmebotHHseODL/hCg4A17q96iv0SxMMrmKdUt90MsMstlxx5Z7JSkefucAeSKuc4M3XNgx35Di1laaknQAd1qaStcllp243dxS7rPS2uQngIZ2g7WkAeE7iMnJJySaMaRqd17lPY53JF8DdvX1+akL3Zol3ZaTLQ4FMr6rTjTim3G14IylSSCOCR9/NeuaHaKMM74XczevQ6g+4KrrOlLxDmyp0XVdyVIcCUIRKbbdaCEjgKTgZOc5UnaSD+9ViItNhxW12dDIxrHQNAHUWCfW/0Nrebu9/YATL067IUMgqhTGlpJ+QHCggH781Lnd1aqnY+MRcc3yLSK+lrx6+3xLTjydLyg2gZKHJjIcIHfCUlQz3wM8/aomRwOjdPdesxccjWYX/4mvqtO3azk6ggpk6Ys8qTHcOxMuUtDDSVA4VkZKzj/AApPPHioNn8QczAr8jhn4J/h5UgB3oan09PqVqzNS3OfPTYfpDZJrqAVyJMhP5P5zHIBDigO2cYzkjivTMebkqvf9FJuBEyE5XPztGgDR16c39P3vori7cYrFsXOU+kxG2i8XUq3AoAzuBHfir7oWVymsc5/hjcqrMaim6qdmx9Mux2YjQbQ5OcUrqoUtOTta28KSMj3nhXcVTz+IPJst5xGYlHKB5v6RVfM3t7KwWCzpta5S/qZMp6U4HHXZCgpSiEhA7AAe1I7CrAzl62sk+QZyPKGgCgBf6kqYqSpSiJREoiURKIlESiJREoiURKIlESiJREoiURKIlESiJREoiURKIlESiJREoiURKIlESiJREoiURKIlESiL4eaQ60pDiQtCgQpJGQQfBFeHVAa2VNk6Mmm2G3Q9T3eJCTgMobDW5pAVlKAvbvwAMDJzjvmoOjLhQcQt0eaxsniuha4+t/Wu6uTSSlIBJJAxk9zU9VhX3XqJREoi0rtcYlpt7824PojxWE73HFnhI/8eK8JAFlSijdM8Rxi3HYDdQurNTx7HYU3Fth24JecQyymMN4WtfCcqHAGcc/cYySKrklDG8wFrXh4DsqYwkhtAk32G+nf0WTSdhTalS5knou3aevrTJDTewLV2CUjwlI4Hk9zyakyPl16ndRyct04bGNI2aNHbufc/wCysdTWRCcURQOo7w7AMaPb2UyrlLX02GVL2pGBlS1nuEJHJwCTkAckVF7qoDdXwQh9ukNNG/r6D1Kp+v5d0a0/Kj3y1R5aHAlcV2E244A+lQUlC0kEp3YwFDjkg+M0ZBd4Za4X2pdjhUeOcpr4H8oGh5iBodLHQ11G6v1lkyZttjyJkJyC+4gKXHcUFKbJ8Ejg1e13MNRS4c0bY5HMYeYDrtakKkoLBMkNR47jrziG220lalLVgAAZJJ+KWALOyAFxDWiydlRGr3N1nKlxLIlpGniGkqurbqgtaVJCnG0JxwvBCc59uT5wBQ2Qykhvw911pcRnD2tfOf4oJ8lbVsSf06+ytun7LbLBBMO0RWYkbcV9Nv8AvHuTnk1axjYxTRQXPyMqbJf4s7uZ3cqEvGp5qriq36Xtguspl1CZTing2xHBBOCvnKuAdoBwDz3ANb5CCAwX+S14+FEWGTKk5B0FEud7Dp7khfMmJd/p1yLzqdm2NkAFMNhtDaCePzu7iTzwfb+lSIcBbnUoiTH5w2GEu66kkn5Nr9VCux9OaYEJq73KTfb0HgmO1Lkl55bij7cN7tiSAeFED9cmqaijoOdZW3nzc1r348QZGBZ5WgADr5t9e1/JdHChjk85rWuFY6LmmvfUNq2W+5fQu2qWy3ui7G5ikyQ7jaohAT2STkncOAeQaxz5TY2kggld/hXBJsyVjXtc0HW+Xy17+vRVfSukLgix2a3v3EJ/FlCa59O2Q420kBZJcVkgjfgBIHucJJPY1Q47vDDHHfVdLiXFIjlyZEUekflF7E6jYaEGr16Cl0aD6e6ahx0sxraUpbUVJV13d6TnOQrdkftWtsETRTQuBLxbLmk8SR9n2FfSqWxE0ZCYvUS6OyrlKkREqRHEmQXEtBQIOOMk4OMqJP8ASnggODibIUZOJSOgdjtY1ocQTQokj99FaE4CQM1aSsGygNcWVeoNL3C2MvpjuSW9gcUjeByD2/bFVytL2Fo6rVgZIxMpmQRfKdtlTo91VdbhE0fO2v3KHKS5OKI+xhcdslSTxwCr+F7Rxzj5FVBwcfCO43XSdjHHjPEI9GPBDdbPMdx3011UvruenT0uDeYaBJuKkqifQIyXJjZ92EgAnKFDdnGMFQ8ipyu8PzgWeyz8Mg/GXjPPKzfmOzffbcaV7LbdtD01mJe7dGTab8pCFupWNyVggbmngnG8fCu4IBHkGQbzeYaFU+O2MmCUl8QOlfm29v1CretNR6ggvxbRItcl4SFJedk2VxZcSwlY3AJI3JJ7Zz2PHPbPLM9pDS2/ZdXh/DcXIa+eOQN5dAJKomu919lJKdusDTElNo0fHNtS0rEJ6WUyH0HO4qSEqG4gk4UoqPOeattzWktbp2vVYQ2GXIAnyDzf1BttHtqNPYKR07q+BKcg236ddvkqbUhcZ8hBjrb2DpHJ5JCgU47pGak2VpAB0PZUZGDKwve087QdxeoPX97HRXEHNWrClESiL4dAUkg9j3ovFxf/AEf1GyXnV2kpSgJEKaX2/laD7M/0DZ/7QrJjnlc6P1X0PGv48cOWP5m18x0XadqVEEgZHY1rXz6+gAO1ESiJRExREIBGKIsbTLbLYQ0hKEDJwkADnk8URaF5stuvLTTd0hMykMudVtLqdwSrGMj9iai5jXaOCuhyJscl0Li0kVp2UFF0NAauMh51+a7BW4lxm3KeUmNHUDk7UAgEE87TkDwKr8AXvp26LW7ikxjaygHCwXUOYg9z+u/qrWyw0ypZbbSkuK3LKUgbj8n5PA5q5c67WWiJREoiURKIlESiJREoiURKIlESiJREoiURKIlESiJREoiURKIlESiJREoiURKIlESiJREoiURKIlESiJREoiURKIlESiJREoi1bhCj3CI7GmsNvx3U7VtuJCkqHwQai5ocKKkx743B8Zpw2K5nrjSbluem3HTlneeD0MpLEN8M9OQhaVtPJb4BI2+ASSBxyazSxFlujHTZd/h+e2ZrYMuWqd8RFktIIcCd+vXbe1ftL3ePerPGmRnCrejDiVDapDg4UlQ8KByCK0scHNBXEyYHY8zondD9uhHcKYqSpXysZ/SvV4Vz5TEPT3qoHEwpDj2oGNofTlaWlt5K85/KCNnbgY7VmNMm/wDL9F2bkyuGlpcAIjdHc821d6N/VX/gpHJ/rWhcXYL7BAFFJRupZr9vsM+ZEQw4+wypxKX17EHHPuV4FRc4tBIV0EYllbG66JrTUqr27ra5hMz7g3KhWB5sKahbyhyTkcqdKf5PAQD7hyeCBVbf4zeY7LdO0cMldEwh0jT8Q6V/T69z8gvu3XqXHkTLRZNKSWmoDnQaUtSI8ZSO+5KvjJ7JBPzzwIiQ2WNZoPovJMWMxtyJ8gEu1I1Lh7j+5CjLihy3W6dP9QNQLcZQoOKg29RaabbWralJCQHF85HJwcH7144cgL5nLTA4Tysh4ZD5jpzHUkjWxflGnRSli1ZpiPbHURFItcSIjqJZfY+m3NH8riEEAqSrwQMk8d6kyaOraK+yzZPD8wSDxPOXGrBDhfYkWBXqoO4PXHXNydYZtLZs8PYpUe6OqZ6zigFJLjaUqVgAghJI7knwKiSZ9C3y+q1xCPhbecS/xXXq3WhsQDYHvV+hUffJY0+qPDaRo22XZDiBDbbYceWhaztCidoKOCe4PHNVyExkAcoK04kf4wOe8SvYBbtQBQ97tWyVY9TXBURT2qkw+ksLWmDACNxxgpJWtWR3xkfcg1e6OQ15q+S5UWXiRc48DmvQcxOn0rVVKTpaHIusC5tNoMO33ZuMFFCS5MUpzDzrqse4lwgAdhsV8gCh8TC4PrYrqQ8QyWRPx71eyzv5Q0WANdDy/mrRqa5SI2s7fFiwrg++5bpIS5GaSpLZKkYVlZCSfZ2J8p+aukeRK2hehXPwcdr8F7pHtaA9u5OtB3YE9VJ+nmozqKwfUPJUmSw8uM9llTQK0HBISrkZGCRk4ORnipRSiRt9lRxDBOFL4dgggEUb0PqrOVgdzVqwWFAXnU0aDLRAiIXcLqs+2HGKSsDypZJw2kfKsfAyardIGmhqVphw3yNMjjysHU/p3+Sgn7zqKVFuEmZGa05b4Kllx5wfVPOoSAcoSPaB9/dnsB5qPM+iXCgPmtjcfGD2MY7xXO6fCAfUn/Zbfp3bEtW528PF12ZdF/UKekYLymsYbSrAAB2gEgAAEkeK9haAObqVXxKdxeIBQbHpQ2vqfmevVZrE09cNW3e6lttEVtAtrW7l0ltaitQP8qSVYxyTtBOOBRmri75KGS3woI4TufN8nAUPlX3U9d7lEtFuem3B5LEZlOVrV2HgD7knAA7kkCrCaBJWOOJ0rhGwWSojTTEyXNlXq5RTFelIQ0xHUcrZZTkgL8BalKJIHbgZOK8Zr5itGQ5rWiBmtXZ7k9vQUrKBxUllUJdtLWW7zW5dytcWVJbwEOuoBUAO3P2yf0qDo2ONuC04+bkY7SyJ5aDuAVOAYqazJREoi8P6UXhXE/Wu0SNNXuD6hWFYamRHEMzWj2ebPtyePj2n7YPdNZMhpafFZuF3+FTsyI3cPn+F3wnsVcNC+p2ntUwG3Ey2YExS+l9JKfQHCrGfbz7knwfODxV0c7JBYKw5nC58Nxa4EjuNj81fARVq569oiURKIlESiJREoiURKIlESiJREoiURKIlESiJREoiURKIlESiJREoiURKIlESiJREoiURKIlESiJREoiURKIlESiJREoiURKIlESiJREoiURKIlEXhVilLwmlglS2Y0dx6Q4lppAKluLUEpSPkk8AUJDdSvWAyO5WiyqrGul11LucsS2IVqCilE2SyXVyceW28pAbz2UTzjgY5qsPc/Vug7roSY8OJ5cgEv6tGle57+lfNRN0gXzTEm6aoVdIb7RZSZMBEdTbbpTgdQHedrm0Y3c8d8gDFXhyNcXl3yWrx8XKhZiRxkOvRxIJ16bCwujN8/tWgLiBfVer1RF/s5ugjKaluw5EZ4PNPNJSpQO1SSMKBBBCiKi5tq/HmETiXtDgRRBsflqoO4I1lboaxBctN3WnGzroVHcVzyDtJRwPPt/Sqz4rR5KK1RHh8sg8bmjHp5q+uv5qDV6hzrczaJl6gxW7ZOfUw4+w4vqRlgEFCmincVBSSOP/AL1uySwNLxoVtj4K3JdJHjvt7BdECiO4N1trqsevZ8nWUFFr0c9HnAfx5gK09IoBIS2pR5CyoFQGP5cn4LI5pG1HqnChHgTGXOtnQVvff2/urBf47kmBaNPi3rESUUplqaSVMsMtgKLZVgcKwEA4HBJ4q14oBgCwY7wHyZPP5m6i9ySd/luttrSNsZ3i3ql25lwe5mBJWy0T8hKTgE/IxmvTC07GvZRbxCWqeGu9XAE/VaEbT2j4t8kO9CA5dIjaVvqkOl11tJyQpe8nHHO4142KIOsbq2XPznQt5nERk6UKF7VoFE3zXlokXBxm1Jt06TD5Dz+V7VnsllCEqW4rjPtwO3PNUuyGE0KJH70WzG4Nktj55A5rXDYdR/qJIAHvfsq7Dj39y4xLxqoOu2iY4I0knfDW2ncek4tpGDsCllI3k4BycDBqtokLg+TVp+X2W6Z+E2N0GH5ZW6iqeCa8wDjptroAPddQRpayi0rtybZEREWreptLeMr/AL+4c7v8Wc/etvhMI5SF81+Mn8Txg8837+VemyjJlo1BaEJTpicw/HLiMxropbvSTuG7Y5ndjGfarOPB8VWWPbow/Va458WYk5bSDR1bQvTSxt8xXqq3p6TOm6cYs62GIk9Zcl26QXd7L7rT5WUlOAoYJGRySnJHaq47cwNdvrX1XQzuWLLdMwktoNcK1AcwDvX+6uujr81qeyInNxZEUhRacakIKSlaThWPkZ8/99XRPD281UuPn4f4SXw+YO2IINij+vcL417eJGnNKz7nAifVPR0b9ngDPK1cgkAZJxXksnhMLgLU+G4jc3KZjvdyh2l/p81GK0/f7zGhOS9UyYzW5L5RCioYc5H5SrcoEDPwf3qJY94B5q9lfHlY+MXAQBx28xsD1rRWWy2WFZ4oYgR22k91qAypxXlS1d1KPck8mrmtDdlz5ppJ3c0hv8h7DopDaOa9VVL5OAKCyl0NlXdBqUuw/VukFcx96UpKRwjc4r2j9MVVALbfe10eKeTI8LowBv0CwS5LF91fGgsLQ6zaMypQByEvkFLST9wCtWD2wk17Yc6gdt1ERyY+MZXCvE0HqBvX2HzVrQMDFWLCF9URKIlESiJREoi+VoStJCgCkjBBGQRRFy/1G9HrLqeOXrY1HtNzTyHWmR03f8LiBgf9ocj79qzzYzJddiuxw7jWRhnlceZh3BVL0z6l3b08mvaZ9Q2JEn6cJLEphQdWGyOO+Ooj4PcYIOccUtndD5JfqujNwmPiTTlYBA7tK7lpu/23UlsbuFmltyoq+NyDyk45Sod0qGexrY1wcLC+bmhkgeWSCipSpKpKIlESiJREoiURKIlESiJREoiURKIlESiJREoiURKIlESiJREoiURKIlESiJREoiURKIlESiJREoiURKIlESiJREoiURKIlESiJREoiURKIlEUBrF19u3MNRX3oy5MxiOXmQNzaVOAKIJBAOMjOD3quR1Ae604cTXyHm1oOP0BKqUyxagl3mLaL84i9aYdWtxbh/hLSEo9iHgkAL9xB4xkpycYxVPhyF/I/VpXSblYsUJnxrjmFeoPctPT7ro0VhuOy20ylKG0JCUpSMBIAwAK1AVoFxCSSSdyvZLKH462XkJW2sFK0qGQoHggj4ogJabaaKqGkjLgamu9lYS89ZYTbJZeff6imVqGSyCRkgJwQCSQCOcKAFTba4t6LflFksLJyfO4mwBQobH3JtXMnFWrAvN4ovCa3UNdLq11F2+HKjC7PMLdjNKWMqwMbsfAV3/Q/FRL2/CDqtEcD6Ez2HwwQCR+X0VWXbF2q/aNgtx25bDaZJeW87lQe2pUp8AjKlZ3e7/H4zVRaWuY0C910W5DciHKkLy0nlAA2IuuX2GmnorRfL7DsiG+sh16Q+ohqNGaLjzxHfakcnHkngeTVjpAwWR9Fz8fFfkuppAA3JNAe5/TdULWEl+99F24aeuS4TzyLdHgPvCO4XHASt/gkHaEgJ+PeTgVnmPO0czbBNVt812OHsGM8iOdrSAXFwHNsaDduv8AZY2YF3ablWm2WO/RWo8ZKY7q7zlvKgoDjcAQMcgHP6ZFQEbxbWgivVa35GM8iaWZhJOo8PWveuvsoXTulJt9mm13K3QoX0Kv+EJrUNLipD21JDZcUpW7IXk8Y4GfivGwmTyvAHfRSyuJxYnnx5HPafgHMQANelDWx9FfIXplZYroksv3BE5O4olNyOmtsqGCUhACf8ufNXDEjBsaHuuZJx7LkZ4b6LTuCLvtvrp6LXh2G8adtcqKSzqSDIW47KbeHSkOFw+/aSShWefadv6162N8beW+YKM2XBmTNkA8JwoCtWitvUe+qyen17gOT7hZ4Uh4JY6bzUSYpQkRgpPvaUFkqISoZ7nG8DOMV7DIHEsvZQ4jhyxRsneL5rFiqNbEV3V7cUAkkkDHPNXarlA2uU2eGq76quVojXFS7LDm/WITHUAY/sCkpQ6nkb1urPfs2R2JFZWjneWg6D7L6GeYwY7J3RVI5vLZ1Dtd6PYCvnfRSkty86At76IEGdqSG84pbPvUuQ04o5KXDzlvuQoDIPBBzmvXF2OzyguVcTYOLTAyvbC6henlNdh0Pp13WprLWUWdo+5NybbdoDi4ilD6yC4horA3dJSyMHdjb8HJryScOjOhGnUKeFwmVmYzkcx4DgPK4E0dOavTddHt76XYbC0hIC20qASeBkZ4+1am6gFcKRpa8t7FbJVgV6oWoPWF3ds2nps2K0l6WhISwyrJ6jilBKU4HJySOBUJHcrbG604cDZ52xvNNJ1PYdSqhftRaotLUIS27IzIdZJUywpyRIdd9oCWWsp3cn5IHJJ45ofJI0AGgT8z9F08fEwZy4x87gDuaa0DuXeavpqdFGWu1XWyyrNpy6apkOG6OPPLYjtpacSkILi9ro9wy4SOMZB4xUA17HNjc+7WqefHy2S5cOOG+GGgEkkE2ALGg2v9V0ux2O3WVpxFsitsB1W9xQypbiv7ylEkqP3JNa2ta0mguBPky5FGVxNaD0HYDYKUAxUlQlESiJREoiURKIlESiKB1ZpOzaqhpj3yC1JQglSFHKVoOMZSoYI71B7GyCnBX4+XNiO54HcpVL9FLQ5YJmsLW2xIat8e6Yi9ZJypOwDO4j3cBPNVwMEdtG1rocVyTleFK824tFrqVXrkJREoiURKIlESiJREoiURKIlESiJREoiURKIlESiJREoiURKIlESiJREoiURKIlESiJREoiURKIlESiJREoiURKIlESiJREoiURDRF8KVj9aIVqXG5xLbGXIuEpmMynut1YSP058/avHODdSVZHDJK4NjBJ9FU39XXt2VEkWzTE6TZlqWHHFYbkKGAUqQ0oj2k+VHJ+B5oMr7BDCQunHw7H5XNlyGtkGw3b7FwvX2Vrstzj3e2MToitzLydyc8EeCCPBBBBHgg1e0hwsLmTQvhkdG/cH9keh6KA1q7KmvQbRaS2m4OOJlh5xO5uOhpQO5aQQTlWEhPGTnkYqqUF1Nbv8A2W3ALYQ+eX4aLaG5LhWh6UNT/ut/UttmXC3R0xLs9bJDLqHlPMpBSsJ5KFAnlJ+M+Oc1N7S4aGlnxJ44JCZIxICCKJr6eqw6Evjl9tCnnnI7zrL7kZT8bPRe2n86PsQR+hyOcV5G7mbYNqWbjHFl5CCLANHcX3/eqn5T7cdhx15xDTaAVKWtQSlIHcknsKssDdZQ0uIa3cqhaQvdvTO1Zd13uK7bHJyem4VoSlG1pAPPc5IwPnbkd6oie08z+axa62diztEGOYSH8vrZsn6KOv3qAu9LctmjGrhKfDim3Z0NhKw2AgKBQVnYSo+3KiMYUeeM0uyRIeWIE+oWuLgbsb+NnlrBQIa4nXXrWvr66LHcrdqzTenV3RF7kXJSYf8Ar0OY2X9yyRktbcFO0KP2O0Z75Hrmywt5+a9NR/Zesnwc+cY/hCO3eVwNaf6ru/8AdRyLfp6zaWsF20vLiwJiT9SyuW11pElBSUrb2g5Oc4wOAe2DzUQxgY18Wh313Vz58ubJmx88F7fhIB5WtI2dtX9wrFYNJahM2Re7rqBxm7SwAWWGUrZYbIB6aUuA4wQORjOOc1bHC+y97tSufncQxnMbjY8Xkb16k9yR0VssdgZtjjslx5+ZcXkhL0ySrc4sDskYACU5/lSAP35q9rAz1XOnynTAMADWjYDQe/qfUrS1AQjVmmexC3ZLeD2BLBVuH3G0j9FH5rxw8wKnAf4EzR/p/Pb7/ZWQj28Adqn1WOgFQtIajiHUd+tyGJz0x27O9ZTcZRaZAQhKSpXZIKUAfcg+MVRHKHOcK6rs52A6OGKXmFFgOp11JvT3+y6AntV64wXhSKL1UH1BtJj3G06hhynILsN3oyX20JVhhftysEe5CVbSe2ElRBBFUTRguEg0IXW4blERSYb28zXUQPUduxI+9aFamv7bfjp64yXNSDoJhutGIxBSkSVrASlJJUruSEgj+9UJY5OQku+y0cNycIZLB4HUGy4mgNSdh0WlGsR9PLdYJlsiFxpnKLxIUsJPSUAVKKexCFcjn2gHvuJrxsX4dreT5qybN/6zNN45pxH8MV1B0A9xv336LqTa0OpSpCgpJGQUnII+RWv2XzdEaHdV71CITpO4BZ/hqCEuZOAUFxIUD9iCQf1qqYfwzZXS4S28yOh3+tGvotB3T15t0lr+yl1Yi25CnFG3yo/UaBPISgpwpKc54zx4+K8MbmkeGdOyk3Nx5WkZUdu0pzTR+d2Cfolygauuym4zk+32yCpC0yHIO9b/ACONilgBPnn7149sxNWAPTdTim4dCC9rHPeNg6g35gEk/ZRP9l1/jdiskq+3maxGZcmKLjwQcIKUIG5ACs5X3JzhNRMOoaXH6q8cRbyPyGRMa4kDRumu+hJH2VthaVtNvnpuDUdTk9CShMmS8t5xKSMFIUskgfYfJ+atbExputVzpc7IlYY7pp6AAD5gAArRfdbuutYTEUtuC0JcekOJwrY44nYlrPg4KlEfAT80IDng9lZHzwYr+bQSUAPQG7+oA+qtiasXPC9ovUoiURKIlESiJREoiURKIvAMURe0RKIlESiJREoiURKIlESiJREoiURKIlESiJREoiURKIlESiJREoiURKIlESiJREoiURKIlESiJREoiURKIlESiJREoiURKIlESiIe1EKgdSWJd6VC/wCEp8Jth3qLTDe6ZeG0jaojkD9P/tW9hdsaWnHyRBzHkDr2sXXqBsvq2aXslucQ9FtcRD6MkPFvc4D871ZV/nQRsBsBeyZuRK3ke8126fTZS56e4jjJGasWQhUnUqHNPOS5dnuAjvTVpKLeWkuB+QohIKASCncSndgEcFXfOangt+E0univbNTZmFwH81kUPXQ/Jc+fus636hu7svUtzmXFgR4zse3Rm0KfcUVDDaVDOxvvwCVFR7ZJrGXFrjbyT2AC+iix454Y/Cx2NY7mIc9xIAH9RBAsn6aKxmXqiVouRD1lEfSxIBacuFvKes20oA71sgdhnCscgZ44ybR4pjLZh8xv9FznNwGZjZsFwJGvK7ax0Dr+l+i09Gyplovcq26ajxZcJ5pC477yVwmXlNj+LtSEqBX705UAAcfakIdGSxg0+mqnxLwsmFk+S4tcCbGjiAdrN3Qo6HVbGpNZyJWoLVZrhp+4NYJmPxXH2B1QgEo92/aUhSSogkZ2DjuK8kyCXhhYe/RMThDG48mWzIbpTQadpe96XddlJs2Oc/GXIhaU07bFupUULfWFvslecrIQ2Uk852hWPGeauEZIsNAWB+YwEMdkPeBV9Aa2As38yFr+lt/t/WXZLXanbbaGUkRHpR2OS3Eq2ukpPc5Kc85HbHgV48jTbGCmjb1WnjeHMKyciUPldq4DXlB+H5Up/XuoXrbb5cazpS9chFckKUeURmkpJ6i8c8kYSPJ+wJq2RxHlZuufg4rJHtkn+CwPUnsP1PT3IWxoPRtr0nbEt25ta3nUhTsl1QU4s4zjI7Dk8DjnznNewwsibTU4nxKfPl5pjoNh0A/ffVWoADxVq5+y8XgJJoEOqpdluCdR6zlOoZfbj2Tqw8vI29R9ZTuKfOAhI7/36pa8SPOnwrp5GO7DxmhzgfFp1DoBtfrauu0Yq5cyl8hpAJISBk5P617aL7rxEoiwTGGpEV5l9tLjTiChaFjIUkjBBHwRXhAOhQEtPM00QuPR7wxbLvdWr3Nmq05p13bEitRXVblJwpBccAwoIztSFKA4ST2zWAS05/OfK1fWnBdLjwuxGjxZgbPM0abEAE3r1IHdWi4a5jS3IMXT1vevapeQ4hA6YZRtzlZWnGDnHPHf7A6XTg1yC7XHi4W4eI7JkEfJ87PYUkHT93sFqYl2191ctnLjlsQ+fpVoJyWGkq/JtBwlXHI5GDgT5C0ab9lU7KiyJeWUANNDmrUeprf1S6Jf13H/AAmTaJ8C1na5LcnNdNa8HKUNAE85HKjwB2BJ4g4eOORwoLTC4cJkGRHI1zxty6gepsfQde+ivEKM3EiMx2k7WmkBtIJJwkDAGTyeKuAA0C4xcXEuO5UdqlE9ywXBuySURbiWVdB1QBCVeM54HxnxnNePDi0hpoq/EdEydhmbzMsWPRVLTd9s0CI1crpf/rbtLjth0uAFxIGTsDTQJSASrjGSfNUxOaG8zjZXRy8fImeYoYuRjTp2168zjrp12pZbzeNR6ggfR2C13azrfcSkXGU20Etoz7iW9+8HGccZ7du48c98g5WAj1U8eDExH+LkvbIBfkBO/TWq0PqrfYrXHtFuYhxW0pQ2nuB+ZX8yj3JJOSSSSSe9aAOUUFyJZXzPL39f3XyUlXqrSiJREoiURKIlESiJREoiURKIlESiJREoiURKIlESiJREoiURKIlESiJREoiURKIlESiJREoiURKIlESiJREoiURKIlESiJREoiURKIlESiJREoiURKIlESiJREoiURfKzhNF4dFAWfVFrvyZf4JKRMciqUhxtHtO4Z492OCRgHtVcczX/CtWTg5GKW+O3l5tQfRco01PvVw1PKdjMFGrDDeS+Li+rpxEl7LY2DxtKQEgeNxPPOKN8rnENHmrr01X1WZj4kOM3mdcHM2i0C3eXXX3s/YLo8WzRoM93U+ppERVzaa6YkZLbMVrGNqQo8ZJJKjydxHbitgjAPPJuvnJMp0zBiYgPITdblx7mh9uiri7Va7/AH6cdIXOfbJjqQ/MuEBRWy6TwG1ZOAvndxjjP7UlrZXExmieoXR8fJw4WjOjD2jRrXaEeorWumqs7Wkg2w2U3y+iWkf8Z+tUok+TsVlv9tuKv8MAblcs5/M4nw2cp6V+u/ztU3WqE2HU+n7zq6eubCiuOBl1oKbWlYbKkhTafaokgDcMZ4BGMms05Eb2PlOgXa4aHZmPPjYLKc4CxuKsa8x10/ZVpsunU3yyS3dWQUuSrpnrMOHPRaCj0m04J24GFHB/MVHNXMj523KNSuXPmHGma3CcQ1mx2s/zE97O19KX1J0wbHEZkabfltuQwdkWRKeeYcbwct7MnH2IBIIHBHFSEYYPJ0Vf4w5DyMkA825AAIPfp8/+FSNCWu7at0115qYxaTcnZkZ6eyS628VklSUIKRgKJOFZyrPAAFZcYPlYHO79V3eKvxuH5JjhJ1YAQ06EVsSbOvpWldV0RemI7WmbnbWXnutNZcS/LePUddWpBG9R8n7dgOBgVrEYa0gdV867Le+Vsp2bsBoB6D96nVb2jrn+M6XtdwU0GTIjocLYOdvHbNexu5mhy8zIDjzviJuipdZwkmprKVTFaqVeZJh6RLMx1ta0yZLyF9CMU8YOMFSiewB7Ak8YzV4weai1XSPDXYoD84FoNUBXMb/Qd/kpLRNrn263uu3t5l+7y3S/LdZzsKsBKQkYGAEJSP2r2Jrg3z79VVnywSyn8MCIxo0Hf1v5qx1YsaURKIhoir+sb23abLLDMltFzcYWITIwtx10jCAlHdXuI8Y+eKrkfyg1v0WrCg8aVoePJY5j2HX7LPpq1twbBFiONqKi3l8OncpbiuXCo+SVFRNSbtSqnf4kpf8AT26fZTGxOMY4+KkqkwKIvAlIOQBReUq3ri6y4luMKy7lXyYC3DSEBQQRjc4rPAQkEZJ+RwTxVcrnBtN3K3YEMb5Oef8Ay26u/sPU9FA3m1ux7RFN5mPXu6vuIaZiPL6cV6Qe2W0j8iQCo5zwknviqnRlrRzmz26Wt2PktklIx2eHGLJcBbg33PU7A6alWfSNjbsNqEVKkLdW4t51xtpLQUtaio4SOyRnAHOABzVzGBgoLmZWS7KkMjr+ZJP1KnAkDxU1nXuKIlESiJREoiURKIlESiJREoiURKIlESiJREoiURKIlESiJREoiURKIlESiJREoiURKIlESiJREoiURKIlESiJREoiURKIlESiJREoiURKIlESiJREoiURKIlESiJREoi8JA80RV/Xd1kWrTcp62pZcuatrcRlxWC46pQCQB5POcfbnA5qqZ5Y2xv0W3h+KzKnaybRm7j2A1K2dN21m02WJDbbS0UIHUAOdzhGVqJwNxKsknzVjG8rQAs+RMZ5HPJu9vboqnrO4ORNVW5/TwYm3RhK25cBttSnXmlbSElaQQjGAQV4798Eg0SyOa4Fmp7dV0+H40c0L25NtYfhddAEb6de2mq2oUO9agvsG43u3R4NqipWpqBIV1XutwEuq2+wEDcAOSMk9zwjD3O5nCh26quZ2PjROigeXPJHmGgrqNdfdfeudOOS1tXe3qmGdFQUuMRpCmFS2eco3JIO8ZJRk4zweDU5mWQ8XY7KPD8xsYdA8Dld1IvlPf2OzvRe6H1KJxXbZMj6t9prrszEowmSwTgKPw4k+1afCh4zgeMk5vKd91HOwvCHjNFNJqux7ex3aeoXP71fI+u51rlS2xAs7D70dIlxlEZU2tIeU9jYgBYQkDJ55PYVlLxkuDjoB+a70WO/hDZIh5pHNadD0sEtrc2NT6LrOi7r+MaZts9YKXH2QVg/3hwo/pkHH2rdG7naHL5nNg/DZD4r2JU2ojHepLOVW9AAi0zgpW5Yuk4KV23H6hfOPH6VCPb6rXmfG3/xb+QVkUnNSKyKhrkTtF3CFBjwRMsc+b0mlNfw1wVOKJ2nPCkbicHg9xzxmkkxEACwT9F1mRxZ7HyF3LIxtm/566+hW1qi5XaVfoNhsLjLSnm1Ozpe3cuI12QUgkDco5A79s4xzXsjncwY35+iqw4YGwvycjoQGjo49QfQBWSy2yLabezCgtBqO0MJT3P6k9yT5J5NWNby6BYZJXzOL3mypCpKCURKIlEWjd5Ai22W/vQgtNLc3L4AwknJ+1eEaaqUbed4aOq5boaJbr7edO3OK8uTOjxVTbnIbcyDJcSkJQvj8w3OYSDhKU4xzWOINlLXjcan3K+j4gZ8Fk8D28rXENaOvKOw7ba9e67AOBW1fNJRF8rUEjKiAPvRKtU6+a5j24zXGLbcLhCghX1UqKlHTZUnuklShuIHfbnB4PNUyTiME1YHZdPF4U7Ic1hkDHOOgN2b66A6HparNjkar1ZcP7TW9qFAibgxDZmoUsrjHClqBSeCshHOP5eD3zTE+aU+IBQ6X2XQzIOH4A/CSEvdqXFpAAdsBqL06+62tT6T1BOn2e8pmhV2izW/bGTtajx1HasIC/zq5ySruBgAdq9mhkcWvB1B+yjw/ieLCyXFcz+G9pu9y4aiyNhewHzXTkDArWvnl7REoiURKIlESiJREoiURKIlESiJREoiURKIlESiJREoiURKIlESiJREoiURKIlESiJREoiURKIlESiJREoiURKIlESiJREoiURKIlESiJREoiURKIlESiJREoiURKIlESiKv6znvQbKoxnAy4+63GEhX5Y/UWEdQ/8ARzx98VCXRhC1YUPizai6BNd61r5qPs+gbDCgpZeiG4O4wZM49Z4/HvPKf2xUW47Git/dXzcWyZX+Ix3IBsG6AfL+9rInTlzhIej2a9LjwnPyNyWjJWwfPTWpeceQFBQB+3FeFjxo11BR/Fwy0/Iitw7HlB9wB9SKtTVktEazwBFhoUEbitalrK1uLPKlKUeVKJ8mrAxrRQWWed87ud+/psPQDoFIpGBUlUhFEXI/VODbLdebNKkMvxYNwfXFujzBU22ttW3BcUnsdwSc+QnBJAxWPJDWFp6HQ+y+m4C+aeKVjaL2DmYDqbH9IO+l326LJaLjH076QT8NvzIEYyI8VxLaiJLalkIXjwn34zwPaSOCK9byxY5rUD7quaKTiHFmhxDXuon/AEkDUe/Ybra03p+bAgwWNJaoQYMQFL0R9pMlBd2+5JUkhSBu5Kc8E8fFexxlrQInaDpuqcriEc0j358NvfsQeXTvRFHT6/dfUb1PixL07BvybfFjsoVumxpgfaU4lQGwAJ3A4OcHkV4MxodT6HrdqZ/w/LJCJMUOcSfhLSCARvZ0I9lY/TBl9jRNuTLbcbkrC3XA6CFkqcUrcrPOSCDzzzV0F+GLWDirmOyneGdBQ020AH5q1Vaueq7r16NH0jd3pzanGG4yllKVbTuHKSD4IVtIPjGajI4NYXFasBkkmUyOPckD9/JV70ogsKbu11XIbuM+VLU27ckABMpKAnaUge0JBJHHxzzVOOyrfdk9Vt4xMXPZCG8jGDRv9JPfqSd9V0OtC5CURKIlESiKlXhyDedcRbLLW2+iJFVNXEV+VS96UoUsdlYBUQO2SCfFVFwdJ4fpa6MTJIMN2S3TmcGg+lWR+Ss9rtcK2NuIt8ViKh1xTy0soCApau6jjyasDQ3QBYZJZJTcjifdb1eqC8JwOaIqRqwsaukjTjDji4aV7rm6zkdMJ5Szv7BalbcgZISDkDIqmRok/h3p1XSw5H4FZlDm/lv1612HQ9193O3RBcNOaeZjhFuSHJC2Uk7ShlICUKHkFbgUc9ynnOakWjRg2CjHNIRLlOPnOl+p/WhorkygIbSlIASBgAcAVZ7LnnfVfdESiJREoiURKIlESiJREoiURKIlESiJREoiURKIlESiJREoiURKIlESiJREoiURKIlESiJREoiURKIlESiJREoiURKIlESiJREoiURKIlESiJREoiURKIlESiJREoiURKIlEQ0RVH1FisXKysWiS0hxFzlsxRuGQn3byr9QlCsffFVTAOZynqt/DJHQzeM00WAn7UB87VqYQlptCEJCUJGAB2A8CraXP3WSi9SiJREoiofrQ+hjQUwusKeQt1lCkjG0AuJ5Vkjjj+pFZ8qvDJcLXY4Ax7uIM8N1EWfoOiwvQm34tr0hbpIQwzFCpzsYp3IQgJCE9iAXF5PI5CVcc5qQZbRGD7qps/I9+c4ak+UHubv3ofelXo2hbpYrnLX9HY7zCkFyS49LbEctqwnCAEjCQfdzynz7fOYYzmEig4b66LsnjMOQxvnfG4ANAb5gd9dTZ/dLNprTcLWbFwduVjTBtj3EIxHkJZAGElbYSASslJO9ScbcAcZzJsQmB520PsqMjNl4W9n4efneN7BB9jZIr0HXddUtsNECG3GbW6tDYwC64XFH9VK5P71sa3lFL52WQyvLzWvYUPoFs16oKn+q7S3vT+9obksxssHct1G4EeUgfKuwPPJHBqjJBdC4DsunwR4ZxCFxF04fv5bqX0oiQ3p22pmxG4ckR2w5HbA2tKxykYq1gpgCyZjmuyJCw8ws0T1CmAfFSWde0RKIlEUZqOYbdY7hMSptCo8dx1KnPyghJIz9s4qDzytLldjRePMyL+ogfUqH0BCQuyxLw48ZdxuUdp6TJcUFFSikHanHCUJJICRwOe5yajCAQH9SFfnveJXQDRrCQB21qz691as4FWrCsbz7bLalvLShCRkqUoAD9SaHRSAJNAKq6yusWfZvw+2XBtcyc61HQiNIG8grBXyk5SNgWSfAqt5BFA7rZixvjk8V7dG66jT0+6sFtgRbbCaiwWG2IzYw222naB/48mrAA0UFie90z+eQ2T1Va0nKOo77cryHMR4jjlsjNdJSDtSpKlrUVcklQAAwAAPkmqYnh7i7toulmRvxImY/9XK89ehr7FXQVcualESiJREoiURKIlESiJREoiURKIlESiJREoiURKIlESiJREoiURKIlESiJREoiURKIlESiJREoiURKIlESiJREoiURKIlESiJREoiURKIlESiJREoiURKIlESiJREoiURKIlESiKrax/5Q0v/AO9kf/svVXJ091pxNpP/AB/UKzo7CrFlC+qL1KIlESiLnPr3/wCbS5f9Yz/+4ms2Z/kOXd/wx/8AlYf30KhP9H3/AJBl/wDWI/8A96hw/wCAq3/Ff/rf32Ctnql/5A37/wBmP+9NaJdiubwj/wBcz3Cs1n/5Pif9Sj/cKm3ZY5/85/ufzW/XqrSiKmeqv/koP/eEL/8AyG6qyP8ALP76rp8E/wDV/J3/ANSrensf1q5ckbL5H/GB+lOih/8A2LNXiuSiJRFU/VL/AM32of8A2NdUZP8AkuXS4N/66L/yC+fSX/zc2D/2VP8AvNe4/wDlN9gvOMf/AJCf/wA3fmVbHPymrguY7Zcd/wBIv/kG0f8Ato/3VlzdgvoP8O/5r/b9VEekn/lk3/1Dv+6kHxBWcU/9M799V3Xx/wCPitZ6r50/zKG0d/xSf/7xl/8A7yqqi/m91szfjb/4s/8AqFP1YsaURKIlESiJREoiURKIlESiJREoiURKIlESiJREoiURKIlESiJREoiURKIlESiJREoiURKIlESiJREoiURKIv/Z)
Neurophysiology
Why animals need brains
- Escherichia Coli (E. Coli)
- Tiny, single-celled bacterium
- Feeds on glucose
- Chemo (“taste”) receptors on surface membrane
- Flagellum for movement
- Food concentration regulates duration of “move” phase
- ~4 ms for chemical signal to diffuse from anterior/posterior
- Paramecium
- 300K larger than E. Coli
- Propulsion through coordinated beating of cilia
- Diffusion from head to tail ~40 s!
- Use electrical signaling instead
- \(Na^+\) channel opens (e.g., when stretched)
- Voltage-gated \(Ca^{++}\) channels open, \(Ca^{++}\) enters, triggers cilia
- Signal across cell within ms
C Elegans swimming.
- Caenorhabditis Elegans (C. Elegans)
- ~10x larger than paramecium
- 302 neurons + 56 glial cells (out of 959)
- Swim, forage, mate
Neural communication types
- Electrical
- Fast(er)
- Within neurons
- Chemical
- Diffusion slow(er)
- Within & between neurons
- or other cells
Electrical communication
- Electrical potential (== voltage)
- Think of potential energy
- Voltage ~ pressure
- Energy that will be released if something changes
Basic principles
\[E = IR\]
- Current flow (\(I\)) across membrane
- Membrane varies in resistance (\(R\)) or permeability (\(1/R\)) to ion flow
- Product \(IR\) is electrical voltage \(E\)

- Membrane stores (& releases) charge like capacitor


Resting potential
- Measurement
- Electrode on inside
- Electrode on outside (reference)
- Inside - Outside = potential

- Neuron (and other cells) have potential energy
- Inside is -60-70 mV, with respect to outside
- ~1/20th typical \(1.5V\) AAA battery
- Like charges repel, opposites attract, so
- Positively charged particles pulled in
- Negatively charged particles pushed out
Contributors to
- Ions
- Potassium, \(K^+\)
- Sodium, \(Na^+\)
- Chloride, \(Cl^-\)
- Calcium, \(Ca^{++}\)
- Organic anions, \(A^-\)
- Ion channels
- Separation between charges
- A balance of forces
Party metaphor
- Annie (\(A^-\)) was having a party.
- Used to date Nate (\(Na^+\)), but now sees Karl (\(K^+\))
- Hired bouncers called
- “The Channels”
- Let Karl and friends in or out, keep Nate out
- Annie’s friends (\(A^-\)) and Karl’s (\(K^+\)) mostly inside
- Nate and friends (\(Na^+\)) mostly outside
- Claude/Claudia (\(Cl^-\)) tagging along

Ion channels
- Macromolecules that form openings in membrane
- Different types of subunits

- Selective
- Vary in permeability
- Types
- Passive/leak
- Voltage-gated
- Ligand-gated (chemically-gated)
- Transporters

Conditions
Neuron at rest permeable to \(K^+\)
- Passive \(K^+\) channels open
- \(K^+\) flows out
- \(K^+\) outflow creates charge separation from A-
- Charge separation creates voltage
- Voltage prevents \(K^+\) concentration from equalizing b/w inside and out


- Force of diffusion
- \(K^+\) moves from high concentration (~140 mM inside) to low (~4 mM outside)
- Movement of charged particles == current
- Electrostatic pressure
- Voltage build-up stops \(K^+\) outflow
- Voltage called “reversal potential”
- \(K^+\) positive, so reversal potential negative (w/ respect to outside)
- Reversal potential close to resting potential
Equilibrium potential and the Nernst equation

Neuron at resting potential has low \(Na^+\) permeability
\(Na^+\) concentrated outside neuron (~145 mM) vs. inside (~12 mM)
Equilibrium potential is positive (with respect to outside)
Some \(Na^+\) flows in
Calculate net effects of ion flow across membrane via
Goldman-Hodgkin-Katz equation


Ions contributing to the resting potential
Summary of forces
| Ion | Concentration gradient | Electrostatic force |
|---|---|---|
| \(K^+\) | Inside >> Outside, outward | - (pulls \(K^+\) in) |
| \(Na^+\) | Outside >> Inside, inward | - (pulls \(Na^+\) in) |
- “Driving Force” on a given ion depends on its equilibrium potential AND current membrane potential.
- Driving force >> if membrane potential far from equilibrium potential for ion.
- Equilibrium potential
- Voltage that keeps current (inside/outside) concentrations the same
- Voltage membrane potential will approach if only that ion flows
Under typical conditions
| Ion | [inside] | [outside] | Voltage |
|---|---|---|---|
| \(K^+\) | ~150 mM | ~4 mM | ~ -90 mV |
| \(Na^+\) | ~10 mM | ~140 mM | ~ +55-60 mV |
| \(Cl-\) | ~10 mM | ~110 mM | - 65-80 mV |
Action potential

- Rapid rise, fall of membrane potential
- Threshold of excitation
- Increase (rising phase/depolarization)
- Peak
- at positive voltage
- Decline (falling phase/repolarization)
- Return to resting potential (refractory period)
Components
| Phase | Neuron State |
|---|---|
| Resting potential | Passive \(K^+\) allow outward flow; passive \(Na^+\) allow inward flow; \(Na^+\)/\(K^+\) moves \(K^+\) in and \(Na^+\) out |
| Rise to threshold | + input makes membrane potential more + |
| Rising phase | Voltage-gated \(Na^+\) channels open, \(Na^+\) enters |
| Peak | Voltage-gated \(Na^+\) channels close and deactivate; voltage-gated \(K^+\) channels open |
| Falling phase | \(K^+\) exits |
| Refractory period | \(Na^+\)/\(K^+\) pump restores [\(Na^+\)], [\(K^+\)]; voltage-gated \(K^+\) channels close |
| Resting potential | Passive \(K^+\) allow outward flow; passive \(Na^+\) allow inward flow; \(Na^+\)/\(K^+\) moves \(K^+\) in and \(Na^+\) out |

Neuron at rest
- Driving force on \(K^+\) weakly out
- -70 mV - (-90 mV) = +20 mV
- Driving force on \(Na^+\) strongly in
- -70 mV - (+55 mV) = -125 mV
- \(Na^+\)/\(K^+\) pump maintains concentrations
| Phase | Ion | Driving force | Flow direction | Flow magnitude |
|---|---|---|---|---|
| Rest | \(K^+\) | 20 mV | out | small |
| \(Na^+\) | 125 mV | in | small |
Rising phase
- Voltage-gated \(Na^+\) channels open
- Membrane permeability to \(Na^+\) increases
- \(Na^+\) inflow through passive + voltage-gated channels
- continued \(K^+\) outflow through passive channels
| Phase | Ion | Driving force | Flow direction | Flow magnitude |
|---|---|---|---|---|
| Rising | \(K^+\) | growing | out | growing |
| \(Na^+\) | shrinking | in | high |
Peak
- Membrane permeability to \(Na^+\) reverts to resting state
- Voltage-gated \(Na^+\) channels close & inactivate
- Slow inflow due to small driving force (+30 mV - 55mV = -25 mv)
- Membrane permeability to \(K^+\) increases
- Voltage-gated \(K^+\) channels open
- Fast outflow due to strong driving force (+30 mv - (-90 mv) = +120 mV)
| Phase | Ion | Driving force | Flow direction | Flow magnitude |
|---|---|---|---|---|
| Peak | \(K^+\) | 120 mV | out | high |
| \(Na^+\) | 20 mV | in | small |
Falling phase
- \(K^+\) outflow
- Through voltage-gated \(K^+\) and passive \(K^+\) channels
- \(Na^+\) inflow
- Through passive channels only
| Phase | Ion | Driving force | Flow direction | Flow magnitude |
|---|---|---|---|---|
| Falling | K | shrinking | out | high |
| \(Na^+\) | growing | in | small |
Refractory phase
Absolute
- Cannot generate action potential (AP) no matter the size of the stimulus
- Membrane potential more negative (-90 mV) than at rest (-70 mV)
- Voltage-gated \(Na^+\) channels still inactivated
- Driving force on \(Na^+\) high (-90 mv - 55 mV = -145 mV), but too bad
- Voltage-gated \(K^+\) channels closing
- Driving force on \(K^+\) tiny or absent
- \(Na^+\)/\(K^+\) pump restoring concentration balance
Relative
- Can generate AP with larg(er) stimulus
- Some voltage-gated \(Na^+\) ‘de-inactivate’, can open if
- Larger input
- Membrane potential is more negative than resting potential
| Phase | Ion | Driving force | Flow direction | Flow magnitude |
|---|---|---|---|---|
| Refractory | K | ~0 mV | out | small |
| \(Na^+\) | 145 mV | in | small |
APs and Information Processing
- AP amplitudes don’t vary (much)
- All or none
- \(V_{K}\) and \(V_{Na}\) don’t vary much b/c \(Na^+\)/\(K^+\) pump always working
- AP frequency and timing vary
- Rate vs. timing codes
- Same rates, but different timing
- “Grandmother” cells and single spikes
![[[@eyherabide_bursts_2009]](http://dx.doi.org/10.3389/neuro.01.002.2009)](data:image/gif;base64,R0lGODlh7gKYAfcAAAAAAP////7+/kxDRVFJS/79/vr5+vTz9OTj5NbV1tTT1MzLzMvKy8fGx8C/wLy7vLW0tQYDBw0KDhgVGSglKS4rLzMxNDg2OURCRUtJTEpIS1NRVFFPUk9NUE5MTxIQFB0bHyIgJEA+QkZESEhGSl9eYFtaXFhXWXx7fXt6fG9ucGtqbKemqKSjpaOipKCfoZ6dn5uanJmYmpiXmZSTlZKRk5GQkpCPkYyLjYuKjIeGiIaFh4WEhoSDhYOChIB/gX9+gL69v7u6vKqpqzw7PkxLTl1cX1xbXllYW1VUV2loa2hnamVkZ2RjZmJhZGFgY2BfYn18f3l4e3h3end2eXV0d3RzdnFwc3Bvcm1sb2xrboiHioGAg2ZmaXt7fXJydGdnaaSkppycnpubnZeXmZaWmJWVl5SUlpOTlY6OkI2Nj4yMjomJi4eHiYWFh4ODhYCAgsPDxbu7va2tr/39/vn5+vf3+PX19vPz9PHx8unp6ujo6efn6OTk5ePj5ODg4d/f4Nzc3dvb3NjY2dfX2NTU1dPT1NLS09DQ0c/P0M3NzszMzcnJysjIycXFxsTExcPDxMHBwsDAwb+/wL6+v729vry8vbq6u7m5uri4uba2t7S0tbOztLKys7GxsrCwsa+vsKysraioqaWlpqGhoqCgoZ+foIaGh4SEhVdZXNfY2cfIyb/Awb2+v0NHSkxQU1tiZaWtrs/R0f7///3+/vv8/Pn6+vf4+PX29vP09PHy8u/w8Ovs7OPk5N3e3tna2vT49/3+/fv8+/Hy8f39+v7+/fz8+/r6+fT08/z799rVz728u/349Pz07/rn4Pzt6KegnvDk4d7Auvrd17qdmMWOhvfPyfGlnfW9t+LZ2MlmXe19dKKIhulXTap3dJJubJlLSP79/fr5+fb19fTz8/39/fz8/Pv7+/r6+vn5+fj4+Pf39/b29vX19fT09PPz8/Ly8vHx8fDw8O/v7+7u7u3t7ezs7Ovr6+jo6Obm5uPj4+Li4uDg4Nzc3NPT0////yH5BAEAAP8ALAAAAADuApgBAAj/AAMIHEiwoMGDCBMqXMiwocOHBAVIZCiA4CyIGA0KmCWAFseNGR1OFDgypMmTKFOKTFhx4EWDL1UqlLhxFkeZCEEG2NgSp8+fQIMKHUq0qNGjSI1OtOWuqbtc79qVW/funEF079YlTVrRnLt25kpuHcs1QLp3Tt29Y1euHZ5aNweeXReTKE2SPYWewyN1J9m/gAMLHky4sOGZAiN10IABw4gLS+hRueDIoJwLVPIKbUkoE7+iF/kceULvsOmUc0gwbnzhh70jI375HRimwo2tiTDt2VkXKKEiVNAFsHm6uPHjyH+KTV685RwJEkJQoDDBg70fIxoZnCQCiGaiVSLI/zj6R/ru5B/vMg9wI8KH6dSVzFuyIZDBIUTMbD0hYdNRBhKMUMdw62kUjx7qFKjggoFVFMgKJ2jCYHGeSHCBLwfiY485+fzSzjn5zCMAIYG4I0g+tMzDB4iQHJJgRbs0wAA78eSTzk4j3dKHiAkIEkA5gWTyACAVufMEAEDAU45ZhwjhS0W9BcBHJIzMQ1IA7SASyR+0zOYHJb7sU4EFeghETySO7HJlYb90UYIJJiDhBBq84GicGQBwYE89evCxyzl/BJKOHX1UdYgf8fxyjwD34HMOIJL8Ys5AerRoQD171IJjS+0gcMA5h+wTwDmEaDJJPgLtMQIAMrBT0R0MWP/SB0np7fTHJIm8Q1A8CzyCql8CBFKJHoqA4MGAUlLCgDsC2fSdYYlUcQEFRdBQD4ETZqstUDYAAMAJmm5LWIVE6DrQLUtUkMggFSihwwcpTFKBFQFQUUEVGYAAghdWJULCBBSYUAQRCfA2UQMVrNCDBFLgUkUIFUx3gy5MTGBxEfLkU4K+IeQgHEG1jGGBviJkItACHYAg3Q9WBsBCBR9UkEUFF9QJygUkR9JsYY5AJ10IEgCQxbOG4XlCQXtwQEQfQVCAAhUSkBEKBTaUY8QFVRAxQQg1CHTJBRNU8IQIHuxW0hwURGGFBDLUAwXEFFQwhB4eWDyBEwEYkjIIFcD/cGVFuWxBgb4ZLOC1CCpXQMON5dwQwgcXqAACB0uOUYG+HRjSLNGDaUIBABWI8AEAT8gj7umoQ1QRHhlIAPAiqQfmydYrfKHFFbJpEIEjh0wQsxGWzBHB0U0AcEEoYoDwAQMBeAAAFa08EUEIiWz3QcxQtOKCBPU9Qh0jDJAQwRGW3JPEB2UwYEQELxQ0RAQWgDLGBxP4Uo8FH+BwyQlIBoAICBPIQQswEAEi3KERE8AAK+YAMUAcphExY8A74lEGCVTgD8gxgwWxcIUVpKAe9ajABP7ACddZQAkMQAMApGAOEgCABBBggwQo0IddWEACOcgEB4z3K4KQwoQrYAQb/yKghHyA4gMW+AUmLiCBFTRiHxcIgSggQYIPPABbAUgDAEaQiRyAzh0J0NcYOCGCCIghAJqYoQxicIHhBWAIEjhBA2AwARKYbjmDqYg+LvA8PdSiARYAABtiR8hCGuQSH+jACiIwSENuZXYT0FckJREADkigAYUAYCgEAgoJGCEAKwBAGgRSAgCAIgEfEEE8AvAHC4SAEPBgASlIEQp0NMBinxCIL7Zki0fgbxIB+AIAyBCATEigA4PgBRxFwI6BmCMJEQCFQEahBEGMAgBfEIgg4jaPG/QvAA74ABHooQIA1EAP+1gCAM7wQMj1QiCQkIAIShOlwpiBfvqKWSDwEP/FfRiTAo8QSA0iAIRyFCECm9wFBiSgCEwA4JMBYAAIapYPUrTgBf6BIwUaIBBCxIEP6fiEdHy0gQhAIAAxAIAS/qCHGgAACleqB9i0cw4zfIEPPBAlPD9AgnRkAQBnDMAoIoAEPHhAAqK4xy+c5wnkmGKLLQsAJzbQNUdaNXXChEEj5LlKPF7VJxWywCICkYAEqMMWlmxEISCnpgB0sgSgBAApBNIFAMyhEhI4QkXW4QEQAGIRE4AOBXyhCHGaCx4ySIIIiBACEEAiAFcwZwA0SAELWBYEIuDDQPhgAQoMoiBWAEAMBPKODEzgEFQAwBAEoo8KEMEXG0CiZUXohgf/bg0LPOAC4po6m9OYIQJFGMQvCiEbPvQzExFIwkAG+gM6ZCACwGSH7hjxggikQCCctUA7nvOBAubiE9/abA44IAKwVcA+Je1EAKZgQctaYAInuMVADgECItijINCUUAD2YYHzbuCSAlHEBJAQCLC593JzPU5or9tbcyzpqxBm0B8qEAJflGMEEpCQTiL8kwpdwFwCWUda1/phTkoArqFMcPE+cQh5mo4fFCZELobAglHM4RwNEGczAyCFbzXgF0X4gM4iW4YAkIKoDYDEIyYBCa0IxB5ag10AEmGKPmwBAGsQCC+IAIJA+AAA+gnAIEKwNBNIAA2LkIQjHkCIBwIw/wRwjqQNrMI5wZwhvATJRz81gecAMNe5EaAElsS3iE089CISrRkfwsACF0hoCAAwgUBscQShHUIRnUVvBCTEgwhcgQFoskQjNFWRMVdAVAGARAv4sIQIrDYAhAjBBfbghAjwNhISQEIftDYKBih5SL01TRMiMErecPjYC3JBBC5AyyIIbU3IlomH4bEzEUtArYY1MYrlKpDirZaAVKhECWZ4CINAQpx3CMA6CIiJACRAhI/9qQ/cPdFZHaIJfiNIZH8QgHsQkAGMkKeoWBABEqAxAiIQRDCAAAAi4IIGAOCCQEjhhEIEOzAQrMAC3kEPR9wwl8fBk3KNrecQ+BMAI//3cwSa+1xBt0N8DKgHBXCoCQ8gvIcDuaakA9AHEWpuEnzzRQA6AAC/WUICHGhHACJRAvX65RwbAGoAfjE4BED6BKtcAwC6EAAyfIsX76grEiAbAb+dQw0reGedA1MFADC4IoCAQSZuFO26HyYd/LEY/SRgAaGv3e4NCQXo8mBsXGAAAI84BOio/UYAbCAARxptAJAgWv8VAWAlYCwigk0J0Ck9AD5Y9goy0F3/lCECud6FGN6zAZUFoSC/EIEEMHBDBkfBghoIGyXZoYTpXaACFpQHLkoqAgImQU1/PwoELaCPgTABADxAjhoAoAG88JwCHwBEJ6g/EC9SgQ4iAED/u3NBBABcsRJaq4ATaIYq9ZQCAB0QSDnqKgIlLHQCm08BAD6ABXb8gO8dgESaMxurADMZgH3s5A5dYCEYkEoWpwc7RAG/V3D7RQQSQAJEEAFYYBX1RBjXRAQ9xAZ9BngkCBiMoDIy8AlzwAJllG8lGBKlxgamQHcCcQ5DkAMIcA84AAN0RwhbkEuZ4APlJlVcoACw5ggN8Af0MAIg0GZ58Qds8AIfswtmYAJO8AmfwAOaE0tXwAZKdwlfcAQoMIQFoQ82AAVLEAoJYhadgAVQwAOyYSYx4AQp8AimMAbMogdl0AROEANdlXxGgUAVkAmC8AuZEEguYBwVsQA+sEl2/wIPYlAD8hAIPeCIEcUDkUALLLADDmQLLKAD+nAOhvAIjaAHE1YzwxEXCuAD0iQQvcAGSLACmMACOYBBejAGWjADozIHWWAEOiB0BjEIPGAEKoAJD/YO1PQENzArAtEHawAFOtAAZyAKmgIIOAAFTCAKNGgcenB4T3AI9jAEIRABLRAY9TAHLtACLkAKnfArHzEcNTEL5eBgEkEL5VAOLQElI1ERNMGPBkMrBHEPosAJWtESN7GPE/GOm9KP/PiEpcBRBdGPEdETHOEsLSEPQ6AJa2gnHVgUUwAAcDUQVzYC6faCGKEeFwdtPDEQPVES8ZAvoIAOMBABHDAgzyIRdP9wkxOpERGhEVHCOXFhEV5FFj0TARYDAtDRBPR0GjHxERa5JunxHSUBCJb1COqQU1jgEjtBByxpJwVBHDMxlNA2kTcJiITREozAR1sDHShgC4FxCDN3PdBxATqTkgFQChugX57gAYmIRQu5JmIhkQMBWBdwD5vjj2QZkSiJI2AZAJ5ARD7JkoK5kD1BCJiFfIOBDjKgAgF1E4SQAj5wHib5ELTAkJpRKxORjx1hmgIBASNDYRpQboHpLF6JIzl5F5O5mOrRkF8plTSRk5K5mIEBCECQAlRQBVaAAqCgK2bJFaa5j1EpEe8oAAUgnaspEKUwOJdjAs0XE6ZJm3hhnS//IZ2S2Rt5UZolMZ06EZ3j2Zxj0RJ/YANIkAFK8Alu6Z44YQjUcQMQMASlNE8DcY8EIUxVhQMAoAI7EwBdUpsHARJBeTITYF+zERe0ABdrIqDWp6C0chGz0CWdEAErgBClmaG9NRKTIgghoEqSSRjO8hLyiJ8vKJGB2ZBQYn08kZogoQebQAqR8If1hJvoKaMggaMPGhcUyZDg+Z0esSlMWhhEI5xnKRH4GJV/Y526mYpx8QfouAo7lqF38Y614pU4Co+pOJEXoY9gyqFVSqOTqYg9YQ63oI+BoZ8gUDABAAgwox0LIAUbcAI+4CNncAEfUARmIAYaIAFEoAOG2QAq/7ABVlA9AQABPEAIQ7AEheAJOLAKsGgFilCmg4lZnhCGYvB5fGADJtABxbgk92AGR8ABV6Ad8JQFGzAFZLgAX7ABZeACHxCiSFMDJnAEZVAnAYAOmwAhKQCp++UDHBAFnqOiemADJ3AEo+Bko8kgVsqQRYGbD0oW10oT21qtGZGPZAmjjtSiHQmuD0GndpoA1DEIjEA/WmBmI8APWgBAFAAFSiBCIdAB+bAJIEABJ/CvzNNqJPCvcVBpF5AE5UcEmlUQizA4FtABFLByAaBQjncFITABlRAAXTA8V2Ax2uECfBOwGhcAilABy0YE+IOgV6IPRRAgZbQBq6QDFrQBW/9jMvfgbKEDNj2Vsx+gNBLAbwyKrm56mmKpEru5YdzqrQEAnORJtBkRpBOJj+i6ESMKtSahnxOABWzQA4d3AuXQCSbgH7xwQ5VxBRFgAwJxZdnUCxVQAUJ3dEeTAtMDA32gC6WEoHomAZcQABOUB4SnCI8zCgHACI01CPvQBThwI/qnA3nwODrTAFawAHxQWX4QAJsgAU8QAOXEBbNgPgBwBWuSUwiKC1AwAY+ACJAjdJkrArTwVCcwD+WAAhGQAbagAxGgi7gQgJVxtFgrGF6lkJtxtUr7nhXppef6uxpRADlBrsoLtYYAM9fzASBQAqogEIAACmxgBPoSUFMQAcT/xB7PEwCdgERY8AVdQL1/8GVTIH9IIAFz8CPvewn7UAQXcAEYEAiWKQLXEgDDlojz0Ao0oAJtNG+LFAJP0AKoEgoWcgVf0ATkhgGnJRDvkwVXggsjIGQCEQ+CQAfeJLTtMAITwABUEAEsIBCGAAIZ0AdBVgJfYAXl1wbPexwtwQA9sARUMAdq8q0m0ZLmMCkB0AgyQIaA0RK9UAo68whnMIAz3BAX4QvpOAqjEAYvkAluCRH3+GCAMY/Oy5IOBk8yYKdN7BB0ygKJAAlm1Zr9BQVAIEIcVcJFFgDT174swHdQgAQlgAVRcA+0q4sBYA78wQk/YmaW0AsbIAIiUASA/2CZRYALAhEepEAPGxACHhAF/BMF6vYJXTBzFnAIcHQBJYAEUIAFbkAIOOMj5CsBWnAl9DAyQygAk5JT0RcAt8ABHzAJVyABThcIIaABgCA6G/AmWaAF6uW7YzwWZoCUyhNH73SmPUkrNXolwmsn+jDK14IFWLaiEeGdtFkSbXqjwenMXhMBm/tTaNCg5jmWUDsEHyABEfDODcd44XmlRsYBghzOxRtsSAqQLEmlA6EDG8BRSnuaTXolC7ABW3ARSsAqGoGjqqnO4EqnbUYQ6HB4m3QHRPABHBUeYWagVBAAC/ABGeDItiAJARUe4yG/EtBU5vC+6nUL7LAOWiG4FP/QZruAYY0wBgCwBALRA9C3Cy2webUQWVJQWBtwI+kwCatwC9DkiF+WlSxpAq4mUBcACeDVBAIxYSDQC1qHyQHwHBkQD07AtwIRCJ0wK415zINBXxMwB3sACXzEYAH60ICpmk15JQowQ2USWWpLIGLxElQbmOEpEAuqmHhxpkuCV3gzCJ3gQASxoDpR2HYJrgxcAVtAAzegA6OwkZD9pkAsEMJ0G9qMEC+xoBUpzj8C2VpJEKsCci4REz9MEly5E1rsmABQBBVhCBAgKj95ntKcvIDXOx9gcSBDdDswCFwgAR/wWD2mAfF7Z0TQAvXQalgwCVPwAW/AuQBwzn+8Q/H/aw7ebRASRZOfUE4YUA4wkCeHEAoVAAA+oAeNBQq/UGn6IdVWMAlV8AEyLAozRAM+EAIqVRCdYEGicE8hAAjvIHspAAEnEAEf/T8fwAVlwEciIAAPYCGhIAoWcAGOrdaGUUIWwIyQ8AVh4Jg74AhbkARRAAiQYAQmEAOmEwDzMAZGgAQ48E6pkgYmYATB6m5LQDuJgAIAsAWjgARKcEUG0QhY4KrMg7koAAkxcAJKsLECkQh82gWc8DH2IANHcARpwIwBcAhW0AFpwAIfgNWSsAWGwA41IANx8AUn0APNFwD6oAMnMAUPsAUwUNtQKzxF8DEE0QgqwAFYYDgCsQhT/8ABSQAEbSYGgjoCMfALN3AGhMcHbFAGt8AIP6AIQgAFldEAWeCqyEoQfZADwSwKwlEJVnDC7uDfDEADMMMBoZAAQPAJiLAEJ8wAVaDoPfBZf6wJXZAEU9Cpj3A+FTAFg+AIazCEiiAFJ7AEmrAk+rADLGAJS2ACNlAazzsI5YXKBDEJOGMBJbABFvB6jcCEReAOgMABIGABCJAHSwBnIZAF99UDIUC4f5wFFNC35qDvfVsQh0AESfAE1DEC1ZMPR/CvHLDJ0ccJRBA3FDAFarIPTvAzX3AtwnAD1EEEV2ABQGAQLeBKIUACAZU3JwBnFAAH2g4K72UBKiACJuDILv8g8kQgB/zs4YIRCO0tAmwACbowEAhL7ss2AhwwjmpLD2YmAq2TcAHQDxhGAgu1AbrgCFozUZ8ABPBzAiMwPZ1KEAxUAUgAZwGlThWQBAREAdXzAI+zAROrAwHAC0kweyQQIA5UCBawbCKAP0oQAM8nA/MwcyJwAp/DBAHADlFHBBzQ3kRgFcr7HETQAAmQCIhAbSJFAWEfAsDECr6TBeMmAoEwBfaKBXIAMO+UCO8hDy4lAtinCZggHUjwr3U5EH/wLxtggW6fCIElBynFAYTQBPaqA3MAOkx0A+c2ASpwBLN3LWrAetRLCey9NRjQANhcZJ2gL0kAMzsQUdczAgH/G3EQXa3nYA/3wPjWNyUMgAvrsAefBw+/sA9c2Q6C4AuTgg6B8Ai/wIHwoAfpJhHyABB61gUQIE/PrQAJFaK75w5dIQa7FLZbEGnPOT3yBATY1cBSIHMBZgVIF+iRoJAbA/xhhO/cvTwjRaq0x0CRO5Ej06lq4CehTD4LAJmrR4/mggUSBahU2NTpU6hRpU6lWtXqVaxZtT7VJEICgA8XcsQLsAKAkgCDKEhoEaBFhBMBygBYci7ADgBXAqQAAIcgFgBrAhyaUMFXACsAkrgrdwJAmYQb81WgECgApA8eAkgBcALXuSQAXgQgEYFFAEMWiMSrASDLRiAApASoInsW/7wSZwMAJrXrgoQxlwvT6xRhw7sAUQCQsLvV+XPo0QPMmVB9wocPj9ZRFhTg0YcMASBA6RSAnogIQWhHUBMgQQgL+VCHuKArhoQJa/gkqGA4wKQPOrBFpISuiICUAHQZ4YNFppuAAhAsGCQAdDqIYIgAMPlAgiwIkScUKDIJQI8LIvAHkAcLCUCUCZYIgJMIOMAlABUiGMUWIj4QERALPiBkkBAmEJGFCEbAgyDpklRySSabdPJJKKOUckoqt1IJHkyiwOArK8qKYI4A7iEChEMCWOUDE8yBIoJNEjJkAiIAKWICBhKyJMAAApnAAj4C+AIAGxJKLI2mMvmgAhW+6P/iAxD0ia2GAgGA4ZcJLiBLAD56eWcDCR5ISJEPROCjgwkaSWgTCZjYDYDeLAghgQD2oWwf5WZIaJEPRmiuSl6ZnEOCCqxAgYoq8oEEWCywWBQEWAEBRQcjQvhgks0ioME9+uQzhD5dxgDAiHLEQ/aKZQlBsp4cofgCCwsAMCMhFCKYAMyEOIigvEwiuKAPhQIJRQcTQJjAkCHySkgdQHzSBIANEqoxlETAmzE5AEohBAQR7gmAAYx5IbBXkEMWeWSSSzb5ZK0amSGRhM4Z4wMKflFBgkpGHJNlRz4o4ZYTOk2IkEoNwSAEQxI6E4MAfAGhAvn+DBQxAG5oao4PLCj/wYQnrniDHs7eDeAKVhWRAAN1EgomgHgy+ACShA75IM4iQGD5v1RXbTXmWCnrRbkzEmJAAl1RFhyqOSIgYeKEQAHWCROguIILPSixoIISgLjgA0mqRQPbC/p874I8YogghYSGWBzrL37YR6F8LpjghKuzsEKOhKqQNxSF7P0kgHw5qCMhT/pzAogeEYkBAC4UKqdsCBjeqMY5GoggiZACyCECMxLA2J6NMa7n48HDF3988ss3/8m5MNDj1glCEAQLCS4Z8QK5A8jZBKi9hiCCDm4xIgJQJOQFAOhCWva0B6g9LTFP2wioRECWdESCWonZ3NckpYcQ4C0eTihCH1QA/wAxJKRwcWkCAEyRkDXQxW67sAAFzCUrCuSjcB34Xm2Yc76TFW4EuWhKxESgiwDUARKQuIMHACCKALxjBBFwwHo2954KACIAlHibfUaXkAV8AAM4SYckWjGglnVAAtRyjycQyAKYLQ1WAfBABDiRIf4hxA4YAIAnAjAPIkggAQ/ojF0QIYIs8E4xDouAKPoQIX0EoBz/y8QvtrexCYjge0vBYSUteUlMZrJXf7gAAEbgA8sZ7AkA0MSIKvABRVwmRgFoBAhCQAYWWEACGPoEsFwghiCpRxAgkEAKAAG2HCQkCwAIpvKUEIEVVCIFH0ABjQDQngAoAVAU20AmhhmXSP+0bwwuOKUdPSGBEKhhCxQAgBPKAsJ5QAhWf4CQL2oRmgtoQJYkQIcmQVYwIsCDICrRQgSW0IplRiEAjuHCIHoggQhYIgBeAAAGOvEHEQCgCp3oAAAskAczAOALCaEFGJDZCikwMzKlkwARQFEKCqwGECEYIw0A0IGBOCYJjPiEJxFyjiIAYAuDiIIEJLAAc2BgdJugY3ssEYEQ3KAPwwzOn07ACSmM7RyHkMAFPLaKqnJPJvbkale9+lVNjiQBTZAWzKIwjwBMoQJsq8cGiACrRVhACyOBgAhCQJ8wbEQAo7jAXUVQngCcYwsVAIELbAACGCRkByA4YVP4kIW7hmD/BRrzAQgQFIDKIkgXXqDAXZWAwAB8ggh3vQCGAkCLMlAATlewwGx6AIJQpE0Eh+kDCTBA2xqUIAeemAAG6gnWKGWCAkbAySxmsRE9QLazS1hfJMZkASecoAJCCMACMgCCDOyCE4S9gBQysAE8sCAExQwAH1aQwRBkQWNL2QgtxOCqEGCgAQF4Awh8EIA8HIECGMqECCaQhUtQoAvpSMgD6AfdDVggEgEgBBLuWoEb2IUeKqAABSihBgogUR5S6GwIngCrQ1zgBGgN8YhzAlwUp1jFK54SJc/hi0cwok8Jgcc9CFyOedSjnujgRTxUogtEKIIeTqmHIhKRh6aUAx+//4BHO/Rwh4TgQQ/teEotfNGAXzTnHXpgR0K2DGWC+AHLtfiYPBKRCLQqRAD6UAQfzGEPfW55HeWgRz3sQpTvJWIFpggXKCJQQBYraSTquIc8aBEZSprjD1gOyUj0AIlFtGMd96ByANoRiH1spA+M+IMA4lEPWrCjIfsMgDkAweiZsDch9lhAIvRpDj7ooTnuyAf30PaLPaSj0EzBByQUcQd2cDkh6SDEKvKhVwrtQxDraMc9BqLIfTDiF79Fhz12Ea5z2GMe4WJKoL39bXCHW83d/olUthoVZDuFklA5t1PaLYBzb3UpW50FLdatEOO2Oyt/sEAEmkCFuzpC3M8xbv9T8j1vdd8b3/p+yrqPC2935/u4DW9Kt8ltFYWfGCoZ1zj4jsvwgYdc5CMnuch9wQUSjOAKDSp5ky7upJe3XOYzp3nNbU4ydIDx5jvnec99/nOgc9XidIh50I1+dKQnXelLf0q+f8Jxpkdd6lOnetWtfnWsZ13rW+d6173+dbCHXexjJ3vZzX52tKdd7Wtne9vd/na4x13uc6d73e1+d7znXe9753vf/f53wAde8IMnfOENf3jEJ17xi2d84x3/eMhHXvJxn/jkp4LwKKna8pcvOpXY23nJsxfkm0c66KGjedKTD+qpR7TPz5EHXcRe9rOnfe1tf3vc5173u+d9733//3vgB7/370iJlCiZjngIX/nLZ37znf986M8+Hu2oHskEwI7kR1/72+d+971f+3iwY/QyT24JlgCGLqRf/etnf/vd/374x1/+86d//e1/f/znf/5IIO+UBvEEJ9A/ARxAAixAAzxABGQ/E8CdkoEAJEhACIxACZxACny/JniCVfA5f7iAUYAES/hAEAxBERxBEixBEzxBFExBFVxBFmxBF3zBFGwEHxgBAjO959gICLiATJAEGOxBH/xBIAxCIRxCEHSEJmimkuEBJHiESiBCJ3xCKIxCKRRBSpiEJLCVnjuEIpgx1lOIS+CAgbBB58DBDfitLkwINEiekskBHTjD/6ZIASzkuUPIAPlwwwD4wjDMPPEowzNUiTT4AZNhAx6ww4SgAhnwuTmUDzFsPDxEEighQzM8QzUAxJIRREIMACqIw507BA1QRDdsxEXUCkgkxD8MxEEkRENExE50xC4ERT2EAD60w1KsxFO0w1TMwlUMxcVzxUfcw0jswlkkGUtExUPERU88Q158klGURUoUxlp0w1vEinq4hEuwB1djReDixGNsRTDERpfzRVJsxpEZRlssxqtghCzAghJIADiggiEruGzMxU/sRl3EimV0w2Acx2c8w2isCj1YgQdYhyjoh+SADG/sKm08yM1LRpgDR2Y0xUvsR6qAgC2wtCj4hf8AQIQUsAMVS8h6TDyG/EZY/EXWy0eRIUdoNMeq0ITkwYUo6I4GQIEB+Ugl+bx90iucXD2F8Mh5zMNeHMlwhEhilEYtCIU8+IFAqAclOI2TIbd5Ywqd3El5REZ6fMVYxEdxPMl97EKJpIoEUIEsIAEtaAIbsIuopBJz8IN9iMRbAIR8CBeiowqepEqfVEaHxEqhLEetQIdDEIJKQABEG78pyYceoAIUiILDCIBf+IEfkII14B7Qm0turEuXu8czNMmQQUl+VMmoQIdMeAEXGIJP8ARO8IRPGIKigTiQOT4VgIN5eAccCKQ7UIIxuAN7+IH7UreRapupvAqcbD3pgLf/c6PJ8bHJ0yLOJQnJmoQ4WNyVrKg844yOdYvOrxI94DQZzDS+btPMrFCJh0NONXs64USxexMAWhBMKunKp7iFFvCBGviCC1gCNeCCEeCAMjlLJyk4THgCfQqAehgDdoAAKpAJPViCMnEKomMKybSKygvP4FTNciNP/AwZ5ZTObmvOMXQKe3sSeAQrzLtOkslO42sK7nSO7wxOddtQ4FK4CZUS9aw4OkiIdsABTFCIe8iB+WpRJ/GBQJGEIAAtHgghlXgDpoRRVVvQqtiIdIgBJqCXJFEJT2CCMkCyFJu4PFCDJfAUlKlQCxWAGO2EDXDOrJiHLVCCBWMSdCiFJmCB/98BT/HJBxTIgjpBGRGNkuOig5HYgq3Eil+wAiroDlKTDj3QAS04UzcNn0XIgh9IpEOFjhdVM5lghX10hB4Il5JJARQ4AxzYgRWoExQopXIYiRu4lorbiD9QFCxIAhGAzK34ASIAAgyQmiWJJSwoghIgC/IMAHVQAgyQAhEIoZPh0uhggCVQgisYgQ34SHeAghGogtJikhywACoQASo4G+8Eq3tIAg64AhEopZOp0yiZgybQAhUgAhx4Dl8gARNYARJAhECFDjwoAQyAz7ZYMUYgAiWAAg4AVJN51KcAhXNtikNIgWcLGZVQARFw1wxpgl7wAflRiBoQDHcLAHuYA/8W6AQ2UB+FnIohwACWgYQLCKAkgQQiwJ17yIDZ2FhNUgMMkCIIIIIFIAj0pAov7biqEFbo2AcWYIFNqIINqL6sAAIS4BcXuIA5VdmoqDwWIIJHCAB/IAKDJAY6KAaw+oINAKIzEAF+0dGGoyR4U9GqAFcoSYQWGAJQMAI2EEVdNQInEAbE8AB5kE5B2YD1CYULiIMVPTR60AAgSIguQAKgvUHUwwp/dQpAUIKjLQcuIAOSkYkuUIGmkAJPwIHdkQkcMMhxOzckpQp7GIHLCgAZIAGliI4uqIKdFIEa5Vry8QURsKOEcIMTQI6alFmswNkMVQigvIqJKwQiyJyESIH/EjC47twId+iAGMBdIpCFACiGuPSqRxCBuTmHJggkpKVZ4/o8MRTbJmnQALiBPb28ABgFDKjDXOiAtIWOkTAEIjjaHkgCMAOuNOCALguAPsCAxHrS6oWKwnWKTTACNwgDGVgBKmBVg00ILmhDhfABUIiBvlGIFAAsfFO4zZ2KMuCAIxmJPOCAUpAOSNCAA02IKDCn/MUhIHCCQ0MuEgCTRg1F2xVFm8xdBk2IKUALAvmFEaiZ2yWFDEAgqqUFFagGkQBbrnoC0kESQxCBOiHOd8MK7f1GlcgBVNgKd0iC40USUCABjYkOK1iBkcoHk6rOjdgHDKjchCgFD8BVktlf/6cIhFKwgTN4gLJRXSbRBCWoND9YAl9gADCQiMFoAn6hOKncxqqohwwwLZUghQ3g4+dYgSlwCkLAAGpp1JHxBQxoonXrATBQEj7IhMSs3apkEoQbiU0IU61IAPkKVCCAhROe2fDEhRMwSAEIVWzohmlY3kPTJJWABAywDHjsAi+IUHTNhD4phuzNSl6RiRKlijmgoZE6BySooOdIBBFgG0fkgRMgM7C6Aer5GD3QAAiOjkQIgkojXM6cCnT4g3y4BwTgg3jIh+8Bn5BJByuwAgZ4hCt4l1nYASpgAAdYgnqVy96UCkrihAx4R4WIhw5AolCkpF8gAZZriigANKFLiP8x2ACObApBIAFqhg5EKAIRIAGBC2LQa+EbvMurOAP8iQwyywZwkIYAMAZLzQpKKII6nIVQZYZu2IbjOptMEgBs9gIXIZCRcABwiIZSg45OwAAR4ADLiGmqaOIoSeapWIH+Kx0P+B7uvQodEOGm8IUMKKWHw+UAsIUk8JqmwIEScGp7VIkbEAEiwAKOFEM1Tgh6oAIo6AIlYAINMIEklmStuAcY4IIdyITmCC8u4AFMSDepmOBSTQgsyM1uAwIssJKNgIETqMGmaAASSKSJRgcoCI6n0AIEfo512IAssAMgKIIhG1yoIOkMhWGraAcTuF8vPYdDy+lZOAe1toof4GL/gigGYgiXa+gGZpgFqu3pcEEAD/DWj5uFZuiGa5gFYniOPiACNWiHEoAFbLYKqIYSqY6KQnDop8gHDMgc1VWH2YaKL6ACjmLlwdmIB+CAP24KfxgBjIRQjCswC3gAiAoUuS5nqagFPeCDWMsHTKACd81qkaGDGKVdReI202PsinMPEkhip3gED8DI7tRVE6hip1iHI/DwnnYTDpgQchuCDaBS5ygDDVifTWngs3RtKzHpqsgEElidjaADcwjV4X4GWrhlrBgVb12KcgjVaegGbBjh8KGkWHiF1Ya3jZiFbegGYnBvp6CCEqinQGhpJjZmkPluqFgDKBDThPgC031X/6t4AA3gbHwLAE0YWiUfH1TYKOEVgCMQDAUH3zsogoDlBBGQoquY66h4ABiHCowIhH7QA5IUtHGDcqjEb6iQcKeIgScI3IRAhyOAlA2fBA3AyA5NCBm4bHuSiRyg3qfggyIQEefQAxIYBYVggatF89b+ZLuE7arIASRsL2Iwh1lwBuj+dKtoASRAMogrhlANgG3QaStHGZX4Bm5w0I2whm6wBrXNSCJgG+Oa8uMOWy/vFTB3iltIAlfXzQcgASniWhU48wlPhyQAVq6qh+WmODHYgIK1xwBwAQwYMoI4gfawQUGHigSQAiqzSXNAhDJIAStYgi7AghSwAe2YifGR9P+myIKAhQoXSIL5DVq9iIp9yAARYW3zGQlaKIHEAvYA4ILJVnKVCAUNIPYRKQIwQU8ZV1tbnwpm0AaXjopkX/YAgAVCiQppr2VcpoVnQHJ2E4Cdt0dz2IgcQIKECBdpp3Y6KIdth4rufpJvP9IO0PCFI4kSwMJF5IPyBmQZMAHKvCRIyADAhIp8KAIt3Yon2IGR0uH+rApBp4V0UAd1SIdaSAc2yE1S+4UcAAIYkARf0AM9CARIIAUugIOQ9mvpkPiE6AMPePun+IUiUJHnfIL7jQoV6AEQPZ+N+IUOYBkWvQwO6BMbZIopKGKVYAMaHmlab8ial4qgB7lZfgYrj4b/bzjap0gG6LYneJtlZ9i4WRju6fbNkKgFJyjG8wwG3KaFcui8q3eSbyc1GchyQHaDoEbPmo2EDRDkgntkFYF8KsEBLZgKFUBCew8EEmCEpoD3N7IKQZcHN7gCKqACKVACJQCIQAEEEAwQQE6lcgYXLjTHqNM6hhInUqxo8SLDQxryDcRocMiJdhbRlTDjUSKiDoUqCgggKskug7NO0qxpkyGpI7UCzGq5sCW9DZ8GzsQoYCagDJEGFkzE4RdPjJc4RPR582RLCBvQ1dy2bVbRic66YbuJrduzsBO3dSN29S3ciWC9spw1rZu1mrTMBWBV5A/TcrOuoQ1gzirFND/i/zI2yIaHx56GS8igWBCTBgRRjRrsocJyy3NHzjQufTWdCVIdKbrg8M5mSzNH+K7mUoUmlco0zykKMklSpUbxFoKVbI4BJj0BGESRkuknQcSmGWvkKL2ilS0W6QSQgWRnTTNMLLa8V8TT6unqK2rRTpShTylSYAeYs6FqwXRGhpycWnU9fAFoxdVFBTXTzTWzKFSRV2GpxVBYDT7IkDV4AXihRAdeU1cxCG5mlELUfGMQQYKNVdZhFymGIU2PYRWAAhwkkB6JBqWzwQs0tcTOBvxJR9BMo3CAB4ssJqLBjNcZpIcHS93EBA3EzZQJB7yclJtRRc2TCG0G7cGIQgUZ9v9DElT8MIQWMbRQwlBKFllTdTRWxMcGk5AXwB8dSFIjRulAkSNLBukwnpxvktiSOpQ04KZEgRTBCEstZZIBYBNO5NMXKFDExRcFtuRfodNltVWB5YQzy1nT0OImqt1MM0s44XBHkTkmkjXLrHIVQ1dPjLLYUjyZIBJgRWc5wxJ3hDWjoK9EMeMhiebM0mGCtBRzkRqLGWqRi5EFYIoTYbppRhPnvDgVHxj14UEn20YawD4Q6MOURbQEkAMUfFKkgrbNLkRIBgtMlA8JkNBr0RS63elOH5ksgQgggvySzxlRTBRJExxZIsImBjUwhTvuvhWnv49scI9HWqihr0WKZAD/iEVFraLBPqEWKSYORFygm6+SJCFPXQHgcUIoH8YcAD4bUHKwQZNsoNydmFBlc2mjElgRrEcRRoy1ls3y7IbFiF1ROYIpS0s4lil0jTfABECLpTcHUAsVFxCBHnEVXfOVpQIIdpc1sNpb4CwVTgNfMAEQFoxkFt3AhcgUdXuSFGzIFLdGhNSEAhUnWXFb5AH+kcQFHgxis09WWI4RJxvEJCZGMZQAnkRX7MA0RVLMwFkhKpDuxBJLdNGEEXtGZ1ALMBhUTxanB0APCjCHDudGVC9UxgoFGlRDE/4aFAMUC1pm0DsnqOa9egVdckEjkFygidET3ZB90AFEYYVNTu8R/wAdiPGRhJPGFwAzkCAd1mOM1TByqllI6FqXmomEBODAidCCDgz8CtooUoyd+OIVS5veQsaAAUCwgAias9kFwVKRWtCCFgiaRTCaBRbCHIMhwVBIqhonEZ9UAQggXMjkMBIPE3ACdxLJRRGKdpJ2IIE/8ftJAD6hAaj9UAtOiAcVkhCyQNHjBM+Jm0HygYFH8MR7VbjdDgMghieIqyLv2IAYOHMOdzDiBwiQRz3scY+QRacgLnAiHn4gCPKhACo/9AjJ+gSFMDxRIgnIwLA8UgsjVMZNRzFIGU5wi+m1BB4nqIFBcnACA7ppHSZwAXl8QokO9KEmP8BfoargHgGuQP8EfEHfVRIYs5acSEETZMjYjIUrS86il7WqCG2oQL/QteQXGBhKAJTwGdzNwl7GJFvitkaLxGGkQ9sAZjlaoqFqWqQcSbDYIYN4kUskwUo6pAgPlnm0BxTBOpYMAC44EINDdmIEAqlHBlQDmgBs4mQHVAHkwMgQPmiAjGkchIzeQxFfUEB5BaLFTtaxjy4NRFwCsNcoRGEQeERhkAEgqUkPeZFEDvQQTznJJBU20BjNSKEG4YMH4LfDluCSPpXQQLoCwIcM6LQig+hAJDEyjyRA0yPlKIFAKdKCJ5hrfOYggQjC9CsBkSpm1zpLWgrwy4UUIxy0MGau4GM2tPTqXYj/0MAhjNarntakJaZIgi0MwgANKCMAZWNIOfgizLFOpEJ5GdxFnkEWi0jIInqwgA/TCZmT7AB0HrFEPY0oER5cwSM+EYMWQ2XTXBZlBej81gnUodkAsEELLINQS16SC9QVZBId2B9iWpKOJoiUUZYAQVQtso41zIEdargCFZJLBS1Y4lDIE6nQgGBSPEhXpRhh6aVa0oJyNVIiMjhCVSlSlBzQz3vKlMhM5nrAtwBhmgaBA6EqAj4DngQInTtJAoqgCIswYAMcEW8ABhECDGgVQ7qMWQpxxc0dmqMWCRYffObyzWoS0yAl0IFc2oqhYzgBB1GphTe+2TUSGaMldFlV/zeh5RFh6q0bzLhIJT4w2R+qsyLoMMIoaHKHE7TgtQyphwcg4NmW5KMIl5AodABUEEBooBL8C0CR21WoN+YYyWnchwcaQDWfsCELF5FCCqi2BQmwwCPnOIQfzIGIRiygzQtoQCuPF4AXnGANOeACBqpg5yhwADDWtQh20xgAKcwYkRpw3vhmAYU4wu4iixCBlu8EoHx44AE1YkQRpEeRL3iYJp1Iwms88gI2WuQeSXiORUgRgg2Yi6515erVKkKLZHiIDn+9VC2MEYBlmIAehvHaWPJChwJY0iefGMG8QrcKV2gusIPpRjNoIT4JzoLW1fIXYS6JkcZSxLBg7MEHcv+g0hpTJBAdSIQrp8koSXiglUYZnA9KAOE3FQQGSVBt6mCZ2wD4YgSak/OlDMKElTU6QKbs8YOKUgoT2MEi5aMAdC9SDF74QhB9yAfGMd4HeDDFJ/0QgwxiMAZSiCEGMZhBKNjxZ0BX7567SEIRvWcLI5ThTojIQFxxGQUkQLgciwhFI1oNoFAYYYsGwcUJADWRdhCtuxLxhQYCeJEsoNEiXogsra6wASPge6sDIpxhAxCMcLiJDnzJxyuOXADxtiot4TAHYseHjiNMQSLqiMMcEOHqoxmkGt9YVTAseKIRG6QYdsGLguJeLFd5BGzvZAgxVDwRejhhBGsYd6EtMoT/I4jEswGgBM0wwgX3ev4PGKjyQuiRCU/ULH2hKcE+w8IJD6BsQpxAAtAKDp+WoAEJPBVvSx65Ejf9wQPoVpIjNoAEVGKEHWwoQRei3wUlBO8IRVThan8y2h8GeiKZQEJMtt+dEwidIjtoAn36IIIZJ0AJIzACBrjQag03BgsYlkganrDTADyiA4BBXxXAEhj5QpZhxOapnLYtRAJAQRqYwH8YGKxdxExsjU1EQWdhTQpdzkkwggVEFSQcAQkYgQiQwXSAxalY29swRIPE3dpA27QQ1kRoCHmkTQ4piYRZBCU4QRTMEgiRm3MFgBaggU2owwaYQqDwggd0zE2MwQXs/5dhjIIHbMAJZIAD+Fgu2csg+Ft6xAMHFM11UMFpnQTUOVSgpEG+2Iw5NIFFwVYA7MAX+MA+GcU87IEe7BE7wMM7rMM8hIyGLUByAQEKdMEVSMESYMEYEMnKUUT3SYQadEr2SUSeLM3jnVQSOJGOtAQkWIAKPAAcEAETzAgiYICm8MQkXsXMWeJCLMKhEYsZuNZ6GcQDcEC6lGIACEES6MKd/AIHHN+DaEIXgMIJPOCFHBjb0YVNOAIJINpEZAM45MVVDEEF+MAlXMEFVEHNWMIFkIZpAEkvQRhhPANiFYeE0MGCBYqI5NWlmANfEIbhKUlLGEtFiAEVoIEPYB55zP8EPXCApTWLmJALV+RWS3gCB/jaTdDBFFQAKYBCCRCBDOhCObgABhjM3gUcJpjAPPwgFZDeQsBDEhQVTWhBD/BdFliO7i2EGniZoGUBDWxBHKZSRpgBEPgAKAxHH7WELwzBJ7RAETRBC4CCDoyAECaiIracRbRDCRQNLVJEFkBOGUmEfRDkJepVEhDBE1wCYiUAETBfAsYFImzAjEhEHZjAGnZEOSgBz3iePWzAc5RkAJykR5jDETDfRKSBFFjCfbziWxAjRQCO+BnEOSTBWErE89GOjhiEJhSBCGgBAzDEJFyAwZRGMaxVWqSVM2iDNIjFYknbvAWcLLiCAqgN3DT/SDDAIE+cSEVQwQ3IAFPSWObtXgBITbrwY0EYAgbkHIQYRBe05m6ggQhgABy0nkHMAAmgjHpswX310Wtmllo8AgcAJ024QDBeBD0kASY8Iis8TRp1kRy4gUyBRlEMARSYASiwABVcgXL0UViEgtIFACHkQD0I5UQs4kIoQAfADPq0xBxwQO1JxDlAgTbWRHEYhDrMA4FgHymIwP9J2kmYgRJURA40KBT5gwf8m7+kjmVRxC2ckvYEgBfMx0Tgggl0QgNMzUR63tdll0GcxbLcxA2UQKwtBE6BQgCkTV35BDvMA200jg8ggcpdRFpZBtmlEOEZxBcU2juy1djxYwCc/wMlUUQFFdOtHJOTBoOHvBMCcAADjMFq9qBu8skWXKjnCYARhGQagZ4v4OVEwEM8+EQ4BYAtJEGYCRBeukMSSBkUvdEcTMToXUUgYECkyUUAaAIHnOdF7MMIQEqAFNQ8AEF3XsQvdMFXhpINvBY7oMBKIBYOeCR8BoB8GgQMHMGLcoY7wNHl1BYGCMR6tEQ5QMEFXocuxMAGVMG8HEWjickSgBJFJAIJ5NwlgUISIOBJSIYDZEDN/EgA3Jzm/ChxSEoS+BpiJIAsQgKJel1XSUQ4CEYANMhV/AEJOBlTFEULdEAeoClN0EMRXB50+MQe6MAG6ADQlNMsyGA5jNUceP8AFd3gVbjAEZCSRDAOWM0oS7Qg19RCDEnKBqRDGWzp9PhgR9hCCSBcXxKUBuDW7wXAF7QqgDQAETgJTzWOP1jBBpQCd/jKAhTBmVIEHGBB3gRAF9TcVWjBKDKLRACBI3rECqyMlATAlw5ao1oEKOCsCiVACvQoWEqBI8ysEGxqRhClZTjBf0alGHQAAkrGLDhB3SlZSyyCY8aPPZiABsgAB2jArP6ITW6AE95mAIRnelSBh7IlhMwEO5gPjRTFFkAoq8yEHyTB0SIGDOQLJUwrBJ6otV6LYm3IZnrEDywBiUjGPHSAy0as53kCEQxST4BFSwgCCSQBGYiAEXCcZej/mjC1aY0sFaq1BNjcREv8gQYEATVdEE+QnQYJAy0YDi0IQ/8YhBfchg0obOgwbEswZ4LaxDokAa4eTyOIAKTcLoCkQBJwlE9sggV8ARtYQKddxBo4gUU0AAkUa/B5wCq8BSZsgJUAXADcQwYUEWeQAfZKxB2YAH9Qwe7QBCTc30I4wClYRChkwcmugylgwbgqrUF06i8UwSKgbkvgwwhEnGGKQJIomUG0QQe8J0O4gxF0AEfIwwk8gb2UJHPu58rawBPsREv4QREsxVa6ZADQQAk8SEvUwhH0GFuKiQqkwUQoASgJAeAOYwRORDAIVjc4w7TQRyIQgUP5xAwUwXuW/2igACETKERb+QEJdEHI/AERhGG/Kk43mAuK8UkVjKJBGA7qGsQUeIFMkBWUlkUt1II7lgMtjJMxpIgdnICMrohkvQsbKC65GgQpFEH4+cQSTJPyrkcfAOUPWoIF4OolVICMXkQPWUQ8aECZScYMGEHF1gTSMRL2LYQl6CdNGEIRJFUAb28AYAlNqEMacIJPAIIPHC3ZxIATUIEUrAAWKKPSdioNhI8BG4QpYMDJBkA7dAAaKTHW+HIR8GAATAEJBBWejECV7Vt3dI+cXG4GNMJCtEASEMl9XhJtxtVEKIIGDB8Ku4ARhFcA8AIHVCeoCPOdCC4whSbfkKbobcD/wv+LCDCSgbXEPoiAE7WEHZjAEaiWQTwCEVuGO7/NkBrEpCizVxieTajQYVLR4n7jjAYyfNAGN3hDAESmQRRCEcwIHf9QDlQdRaiDhuZxAKQDEngxQRFBpLptXHjCBUCFT/wCEYjbQsgAEhelCeSpRaiB/vmlCURJxPpEFKAf7tBsTZSDE3RnC4DqKMfvSZxDDlAlCnwBBmBAFExBFThhOchDeAHCJgBd55UDgTodCGHl8aGXQTzBDSCQEXBdAKDDCmyAs74JJrzPQuCABQxLP25A1+VNOTSBSVzEF5yWEeAsXKxAv1jFKciToTVwALAAXb9mDjtwJohAbMIHMcSrh9T/wuF6xC6QADqZ2grUwbbEABE44TmswAhwRONMgbpZxVEoVllYxNDoRuGm4FXIAwcE5tsYo6e9QhKHkhK0xEeD0A7Y7XUsQAf0Q15mrQWUWQAwwgW0JAQGwAqcwEUKFQY4djokwWAXXBx4gKalUQKQgMAEQAOcqpXRxAKMALqpl3lo6p1EAUrOBDqYAA2Pcs9WBDs4giRggiZkgiRAAicMQSj4GUHdwAJ0HkOcwyDIgAvwq1D6AgiQqWsmgAYIDOQyxCCQwAnEwAmIgEwT25vIQAWYAj/4QAVUp0QRTMfsWx9swN66SSRgwCBVAgbQ8k1MwgiczvHsAQeg2m6orUGU/wMTdBqVCKMDR8IH9NiHwFCrHItZwYUkEIESzMAIIAFUGgoQXEAm+IISsPShtMQCYABfy3YAgPFFwIAGDMc3BvFbwAAHvOtChF1N3AMHoG8A1EMHQNdxLywIUOhcOjZ9DJAFcMEaWIBK3/PzbAAHKAIjjMAJeC5DlEER3EGhxEB8WYQKXCAWXOBVzEoA1khLkIJrHJAKTQIJAIa9PMIIGBIpv4guXBzG+UGoNeU7aIJ9vQEMdIImhEINgBkLoIxLT48vhMAFlDe92MAKm3QZ8wMQrMAW+NlHqXOglMMobEAGANCyLkQOGAFHLUQoNGv2lcMSlAAjZADvoq4J3J8KQf+h0dGENb+rImhhACw5tL/II0gAB1g6hERQiR7CctHAcG8LO9RABxRBCXzlqmCfFhynlLCFWwQKkKVAKwwAf0jpVfg5KgZANWgD3NgEFyzTHGiAPRjE46jUGUQA1kkEF348bPhEJoR6KHCF6F5IW+XDFBQBCUyBBEOR+Uo3YsyCEkzqnVA3C8CABaT1zLdEIxABkJOICbS198yEOjSpQSzXQsz6SegrE0TfElhfGcGOOiSCC9zADwABDsBAJAzHDyZifmVA5s0EF+bYsaMwFOV9XIjJLvwCTfLU8QDCCNjJRESBtmDE6FRAFrwOY4SCCAxSS7RDElRvTdQDCUDXG6z/bwCk81Y9wAh4gFwyBOAwBTwD8Gbcwy+IRK8UBSaOQErJhAttyLsEgDdD1iYZg+LZRBp0AE3iCQHIaE8lwgjw9ROcVqCHjhlwwC5SBCgUQcrDRSkCidzMRDnsgy8QJv4dQYEZxCCjG9+Ngt0QfYerqhFIvCbgeHsHyhB4wHD0AQnwNH/XRCI8gXDkkR5p+r7/ipwBhAABAQgWNBjg0AkXJHwFGCgwQKUiewgOPHgRY0aNGzl27JhFSkEBswh2geExAB9E6hyixDhwVpclBUuN6OPS4I0MvCYRgVTwEod1LXG6HKgJiowN7QySvKYtWVGpU6lqRJfEzMVn3bChBGTI/1xVg/E6tCkIJNUtoi6/lLAFQwSggml+iLWLQ40UKwRJkgygoq5dwYM1DkwgYpFDgSQdbcjnsVyhhoMXEblE8FySwBZRztOwJgAWE7YKUpHhMhSOl3QItxb4kC/HQxz2MeFS0aIWFBVb9279oAi/giQHcUjU0e9gWoYv5AjQwIILqQPdOcFAJI3BoEM5+x4IwUQfDi8OxjMBzXd6wjCSuDuIrZuz5IVdB8BUYUiAIRZ+QsTZiwMSLgjFILrUQ4kNNBohAhHFBkpkBEXWOpBCjmaxSIkUeCNoDRWK8s+uLS4oJAAvMMAngAuLqoSIDDAwxCDTXDpkDXYOEoCO7iqsSv9HjPzJAJ5GRCBxpAAiedEh1nZcciNbTEDjIDKamC+jxVoqcir/NCHCAyLcCGbCjizS45MHmAJKqDDrg2CDAFjI4B6DXBiBDybt3IiPIkA5aJttUBppFiqzJEgGCzggYgzccBroj1AcOYcoA+88KIct/vKQqC8w7XHSCouUg4RAFAugnSQIBHEjVKkayI4vWsSgkVGlSkSUBDaU0aUhmHBhk046mWMQWTsNIJ9HGnAEkj8KSgSGMET16Ec9AuhiBYOO0FDRYZkkiT07WkKnhDL+TC7QwX5ZI5Ns/+SUoO3UJOy7NtPxQI2C9MDgNEG1XRKILgxyhqtZCvAoUHapYiD/jcQU07ew7vyTVNstegjAEMsKaoCIxAzetzeIBEDCuZaE0EAfhxi+8eRFA6iDkzSEe21jnHD1KB0piogCjh9+oMKBdz21yIcucuABBTkIYmGJF8zowhGPDsngJkKIsDQAHOLieNKB9Bkh3YEKIYFErCl0N2YeA2ATUk0s6CQAc1Y4wcayxSaMkREICcAYWuB7ZhYw5/YN4mHZ4IGgNC44hFgSNvt7yTM4YImkH2ZiPL2ZO7oECxsNUtXOgeiZ4tlZaAkgASjuDkCTLt7p6OnH7LNACy0ueIDyHYvcwoR0CCrlBN1rb43sA+NFh1ALpDgCA1s5/701FXZLsU9aVGTe/67AOx0cM1d9GMGI1cul3rd7RmiBoHRMSDRH8MWynKNMbMCo4E4HIoQHd8wJi6AyqA4AnS8oYb0IrgvAApagBQmpzzsDyccI5hAAXXDAFD5DoEeCRyE2+e5sR4iCXAIgvQm2ZhUiQNw0AmaOcHxQKtabVBoIR5A7iMEENNiFg+SGQo7IoAOrm4MGpJU+G7qEfRuhxwxstbnXTMovldBCKXyQArYFAAiiGFUOYpARdzQiEot4AZ1GBZHp/VAwWxDBL7bAgRmCEScV7I09IBGJRGxhA5ASiUWOiMbplCQJf6DCN4gRgHIoyY4bUeGO/iCJRzBACTrY0HBStLxAaoQk9//AABd8QQJxqeuRFwmiRsxBBQt0wQc74EEUmracrI2uBUagRCAsgQQpouAyFqnBDW40EEEgYQQcuAAR6iHBTEoFDyuwAAZ+8sVfXkSNrXFEETTAAQtsoIbHTJXWkmCBV6hCmoIMzJ1IgYEidKAC2cnmYB6BAQuooA7jzMgmM9IOT8RgBjBwQShIobFoDgYifkBAQRqhBF74ABNEqUENahmAc+QhHuVoAAnqdE91XgQeD/gFJtWZTMEIBB3xkEcAQhHHh2YJInvIxD4+epFBCk8A7JDHO8zRA0WW9I4BGMQD4AHTGJ2GKoHIAQSExSSBmMN3A4HHFRKAA/IVhA2lgF//cg6hgcc4FKacuZAjj2lRu6BKExsgnk3XRdWPnvRA81FDC7m6LnKVlZ14CoUSNjCFu0HVLiQ5hw0YURB9OGEPo7gNQdyhgqa9pCBNfWpZ10VDuH7Qqlc9Ipu2SthpThVLXAWrer4XAOw51kKLeU3KpJnWivhFHY+IQhIqAIQ47csva1ACU8qhgygEwA9Q+EkAXKAClsjGqb7E7G7blabDiulsWuXtcLXJscsSF7kH8WxB9jAEJSCBC4pYgyawZhF59OAKZuCCDqQVgEwowQZtUEERcTvY5CY3sfgMbmPPi9zJ7ui47UXucgNwCyQQYQ7eCgAaPqFbCh3xHImAQCJ0/2eRfWSCEvboKUYE61/5fjS9F13vg927TcGRlcK8pW85XLCEH3wCRTRg22+vGr+CNqVsDSZxhsEY4atOmMW7fW+F4htjwi7XIrrwxBeMsAUmcGKOnP2gim1MWBeLZXhFduyMKVRjJcMUx5wpxyFuwAEkvICkC7YjkZ8M0yObjbFdlqyFr4dhMX90uQX7Ih4goIUM9Deyj+TymSvqW+HBmM4PZfKBnJznzuKUIzBLTjkcwYBxztnPVbWzepKc6HHuWT19dvQj6btbRE86kF9eFZ4xnUlIp0fSnQZjpTF7aVH/UNODCvOpH/lp34Sa1R8ktWNNHesJpvqOq7Y1GF3dG/9Y7xp8syZsrYFNPVx/iNPFnmCvW/NrZddO2GUl9rMpd2yV6Zra6mM2YZyd7blFm6vT9rbYrG2UZI+7dtseTLfRvS9w27TB7WZeuf90bnnP7QZknhQb3HBv5kkB0A8+BAYGUQd5xAPhCVf4whnecIc/HOIRlzjEV7oOhE4c4xnX+MYxHoA5eIA7d4ZAEezBDo6fHOUpj7g83LEOlb8c5inXRS14AASObQEF54BHzHne85WvQ6U+F/rQEf6OdqhgBiwGxAiuwAMuPB3qUZf61KledatfHetZx3oTtQAEH2gd7GEX+9jBroMSlECOFYqECFDwdbK/He5xv7oPqKACOMj/He95f/sPuMCDIoRMW2UggdP1XnjDbx0LX/AB3w/feMdzwQcakGKG9RGFRfxBEJnX/OY533nPfx70oRf96EXvh0xo4RCYJ/3qWd9615MeAZzYAqRWTJ9EAIEQvnj97nnfe9HnYxRWCITufV9847v+F7/ggw0CfqeBhGELfQjE8alffdHzgQtsyIf1ud/9zP+iFz3wBIv3oQN6JJeM7kHuIXJQ4LAGQLr4I64j3pdcUVRxXy2IYHLFMHnk0mBPjml5hMwgAOEHeokAH4oQfkD9iCsRUAGDKIQBeED+hisS2AC5BqIFzkBbSMIUki65yoAUUsS9COSRVEUAlsNh4gwj//qgBubBwR7KF87gTIgrAWaAeBJQKkbHH2SgAnmLAcQgAwNgE1iAY+bA/4hLFOBsuEjiBdLFjgCFJNRhHxTMjwbiHPwARfzrHHiB9ogrHeqhHJJrHejBhxgtANahl5KLHeih9tBoIPBgo/YlD84IufKgpt4QjeghFwIJSxpAClDgCsbgtnzBB6SACmpgoxLQq0qqEQnLSu5MD09wEm1IdCoR2eQLUDARgTyGE30qAH7hCD7hDgShC6oIHlSADOJhD1BgB1DiQioLXpridx4RuKRCFvdls97PFrmqF1NEBy8iF7VlMYKxY+qoPpinR35xnDTLD0lCDWyOIBqACVDnC/+UJB+6gLz4Yhj9jXoiEV4+0RszyRjH0c8gIhQmoSUYoVqoiCh8IAlNhhnNUX7mMdDskR4/ahbGMB+Zpxsn6Igs4hy+gAwCIAo2oUAI6kYCYB+0AAnO7gpq4AxqAA0q0iIvEiMr0gxyIAokMiMrsgzYIAXQgAY+0iRJ0gZSIAfOwCRR4BRY8iLPAAjawAxOsiLPIAreoAxs0iTNQA2i4AZYkgaioAd28iLNwLXQACbRgCLN4A2AgAYGiien0gzYQAoo8iRRIAdqEg3MwAdiQBwXAQqO4Oyo4AyWkie1ywe4kifLgGjQEiM38irhcirr0i7vkgZQgAfKgAaqEghskgb/biAF1KAkaYDvzAArM/IMfmAH2BIjzyAl08AxLXKgolIK2KAmE9Mif4C6JmUOTuDsnIAL2FK7RvMu0aAMfCAKYHIoeaAmy2AHUoAGStIMfgAOJjMjNzIFbIAkz8DpapIlowAz7zIK3AA4b0AK1sAxywAIfgA3FRM512AizWALouAuvdIHjBINtLImzQAHpOAGNFI1n9MiITMKCLMrucA5L3IoG8API4seqEALmCIFYokg0KBejCgA5uH+RgEIKiAUHqESBpRAC9RACbQBtgADIAASDrQSLKEB2mBBJcFBK7QSIAEUMEANGuBAJeEBSAAFGKFAPZQEokBELZQSHIEE/5RgASzUQh0BBoigBR6BEuJgBLSgRQu0AYCABC4hEgp0EhogC5JAEhzARY+UQBmBDSxAExr0QCNhEzBABxphQBlhe26rN0jCD0phDERhBS7AR5G0ElKUCUogDibhSINgFb6gAySBQh20AWxABObAScXUTu/UTidhEjzAClaBEhohBzCgE37UQSFhCESgDBpAEiDhCJiGEhx0EiDBBJSgAR71QB+BBYggBji0QiEBAixgC060QFO0CbDlThAhBmCgBTjACCp1TB0BClrVUsV0EZaABCpVTzfgCziUEVCABB5AElLUCKDAVS20AdYAAzyhQRegCTKgUiNBE6SUU48UTf85gAo49BFCwQJkYFop4RGQgAmK1UIfYQgqYFMroQF0gAQ2oU5ddAGOgAMYgRIgwRNEYEPRtQxEIBQg4U+vgAMkAU079RNEwAY4lAFOIAkYgEAnQRI24JLg0D8cwQl8AAahaPzKARoLsryoYhKYoAYxohWYABekYh6U4BGqRAUa6CBoIQtUliO6QCGlYhBKQFkIogke1iA4YQV+kCDGQN+qwhI6QHMwYh264DIKAhM6IOQsSLhcogYwECVaoAo4ohGc4Pw+6AuMkCAkgQn6UCP04AmOgyDcgAM3wjg3Ih+gAHE2gh04wBI0ogZ+1k5s4KUKQg3q1iVsYHIIIgWkgyD/5iALOIMNAG8jIEEJ5rBqqoUg2KELgkBm/I8PNKBBDuINYpYj+oAE1jYiVqABO4IL9oIg5KEJ3JMgEOEJKGJ3qKAj5sEJ6oogomAKLiIFQBBi/9YIeKogcEBcLAIF+mtjoUoOnOBjkakJhtcj6IEJZusi3MYEDaIWlMBlNwJmqWIQTGAyAuBmMQIUlKAWMEIGnmcwLIEDjLcg3KEJAgpNljY9Gg0l0uAVUeIFvoAjGqAEeAGFsOCoAmASniAe0NYIDigAfMByMYILBvgg/OAINLed3FYj1E0wamwLzMwjamBvA4AKyONvlUD+eGB/NiISmAAGBwIHFDcA2OF8XWIW/75gFAoicifXIICAljyiDzRAcy8BDPDAJXwAdANAHp5AeRXBCLYwAExBfjmCHkrgrwwyJJSr+WyoSCQhC7KsIChhBWqKdJpAijVC3DIieMkXKIpXKpBXeQ+CeS/ieaNXI6Z3Kqr3erP3Ira3ey/ie1tDfL04AMwXfXtLfb3D3hr4fT0ifue3fu83f/e3fzUiH/63IASYIwp4IxBYgTGiHRg4IxzYLiBYgjuCgmMEgz1OgwuCgz3ig0M4cQvChPMYOVSYhSX3ImAYJWa4hm84h3e4h384iAuCiDviiJM4Cpb4pvyQILTgCYIgFEKBBSIhAAoACFDgEBxBCU6CdXILqv8eAIw14hKaYGSLQowzoowP4hyg1yPUWGaPoI1xtiDg2Hu9gI7HVyPwWDsWbX37uJLxliMCeSPo134/CH8LwpD9F4AZ+SI4w5E1wg+MIJIvYpLftpLllkkg+I8TcJNLo5Pn4JMJggegliMgAYRbYoRN+YRh0QpWmCBa+CJ+IIY7ApaBQpZRQocLooeRmSCAWIhhoIiFCAp42ZdLo4lRaCBqIRNkQAxiIAbKYBMgJR5cgAd6oBPGMGYQzYuqJAAuQXjDpEiEwAnc457qoQlgeiQ4Ax26oHkL4qvDOo0NeJoGonqfJQCcgAN1BBS6IO0MggywxT/KkSCCtgaNqTqgUI//YzDXmhYlbKDf4LemM6IBjOC0wMciVGAECWISSuCMAoVK8sEEFCYAuCCGV5AgfqD+HqI7EHhyjaklJrkS3sWSxQKCJ8YhTKkjbqCCqSB/Q2EFRseiCVcjHqELELCUCWIdmAAKPbswvgBySeA4usMHxKkj8iFzC6ISliAPq+SIuKCm5aEEkHkgFsEE6oQgYOAKdNkI4qAgYDdb3m1ukFE/DQp/WFuLpfkgEiGJhTEAKGED5uC5DyIQgKCaccIeSqABfoF0A4AQfoIOmECKuqMcVoAKJoojnMAMBMGyNYIOhEBUSCIBTuB6S4AGMCIU4Log5kETNsoMVHcaxZYqLqEI/9DBEFahp96hBFCZ3jyCfTtCFTqgg+u5sDGiAY4AFBS8Nw7hZAfDEBoBCzr5AYzAHc4BE7rLIA7BE0oAgKNAnHTEHh4gHeBAIe8BE3DYIPbhBAjBD4wmoElFAxqIU067KmqMBwgHAm7iEc1ABBIbC1SDIDwBA04ngDfgDjpiDpDgkAOgCrSgIAYBA47WSMJGI1agBNbwHprjIAZhBOqvIxBBBMjLFIxgZA2GJOLBEVRAxH2hySUhDSGhE47gMfKhEWgAUzaiHkwAElZBLqYgBfxgjMlbbMxbJITF1i/i0nDAVMP8qDeARATlBpyAAZgaJ9RBEt4BB7aJpgnCEURFR/8MwQlqHCMcwQ+cLR2cgMwDgB4moQEhQXkOAhAaIDkqvCEGwXUDAAUweir6wHHzcpH4BxK0u6/FMcY5Qg06gMc7wp414h4aYAUSxTfM4MarAg2oABHUGgF+gg+QYAEu4g18YBEQdxFgpEcSwQj8IAGCJQAS4QiyOADeQR1LwQTiHUOgRE3MnCpqLBEGwR1IoJiETNA787L3lg84QKlI4hEsgMQ1Qg1KoCDUwQSUiiBCIQMESAWIPswDQAiIQB0J4gZI4CYKIgeuxiNa4AS0PACm4HkwPQD+IANGgeNfYAUcgSKOGBQWwT0YAAoiAYA1IrTyIQv2hAEGwRSw4JftaKr/5oNzIOt3DyIaO2IfTABGMCIH9ooZ/WILfKAgXmCHNUIHMhkjeICeMSIduoAJz1tHLKLcrZeRoMi2qQInY2MjXhy4sI0j1sAsXKLfN8IK8K83ZODxxSLE4z0lKvsi4OCkI0tVEKEEsjgRoADkC0IMjMD2CWItG5ihl6TP5CEDTvYfDcIWugDICEJvDUIK9i8A6iED0l0j1oCE24EJ/udoUj0AviB/w7wdnEAICkIVjKBmCUIHNqUjSKGwrVPLGGkfRoC8AALGlQAE55WIRDDAoiP4Ejp8mNCWljkJY3xxSEUGxI0cO3r8CDKkSI6HNOQLIOBhGiAg9x0xxHENl5Ec/9n0SEiKCsgtOkTuaAMyXRdPNDkSMuELIhc1RTmaYRnyEod1KJuKTAlhA7qRanqOfHERZJUxVj/G0Fn2YRkpHPkYUQTRh42Rh57sc4joiT6OMEoQTAkRiJmOaX6kPUyQDY+N8jw4GpmOyaaENpZgLJWwXgYGINesSMiuCaaEQ7QADmAlzMd2Th4kNHTkbkIdKka2qJ0wSg2R+jQkSPhiIEF5TyAlXFSioUh0WUAlFGMFo0bE1KtXL3nyNMGVLV/GnHnYJk60HnmK1LEl6FDqhI4kfbi0+lORUqlqt/43QNatItXsoAlWSGPhVxF5iK3V1ltxzSVSXbIRlNdeG/WVX/9gg3FUGIFFKcZYEY+JFNlkBFV2WWabdfYZQaGNRtAQK9CSUGqrtfZabA7RZhtuBOnGm28JlRJWALtAYRxBixihXEjMOUcQdNJpCGWUJJlUlUPcfbSPCTBtJBNi4hGU005egYSeekQh1t57DsVH3XxRTVUllFhpxZUbAAbp0YAanmUdghu5BddDctFlF1568eVXnA4JRphhUoLEIUTyeAgZExBQZllCVGBGkGacfeQZaKKR9mKMqnnUThOW1PhgADiK1AIWDqHAIEi9/UZQKcIJSeRxSI605HPRaTrdo8ZqiJ2iAawRRUta1uToR7P8Na1DW/gA3LAe6bBYSD3Y+VH/ZJ9QlwASaib0Qw7VnYFCR9USVJ+y+M3JX0en4dCtSAGKRZaGM2iL2BlVtHVEoGveMBIiJbSqcKsoTTuGCRXCd2FCp2V4bEeRPjSPBo9A1sSl2zHhUBWmJGQPCYyAhEOKAaRaSUKiKFFOQle08NE7TkiS0CFItLpDFiORsmsAXiAc0j4jFJIQDDoaVKRCJiQZ1AoUEWTRkxlvTV2y96nR7EZ2cNJLAH8gAZN2QRRCA1QdsePJXSm148legkiiw00EBTjIJfcldMobIE37xn8f2dIEk4clcEJS1epxyRe1grQJ0xCkDRgNXGRyiEN8hPLOQ/H+bR29IDGCUBqCf4Un/0eMEPGCSIpcslEitDskw8DyGehQykDY8tAbTImEiBNhJJJQIiVI6FA+kNDghL1NnJFQH5+4kxDGXD+0cTEQEBKAPBkYR8u7HQlFBa5mPJFJHwRJ4UMr7QRATwY8yE16AGoYkVA6JsQgMyWc5gqksFcA1oEEGPQMCX+wFhYY0DmQDIEIriFICpDmkZT8oQiJsIT7SkGFRyBkFyWYBCQEEQAGOCEBmghdRxQRs3SogBSbqEcAYjCFQLAoI9vroVW89pBfRFBSJXBNOyCxiwDQwSFKwAEvhsgRehiBElWphxEmEQBOOOEQv6gKIJAnkI2kxBBM+0hKCgG+j5SjEe5DDP88HBE6AUxLAUjohNxCYgIxBOB/AZhFtQQRCCMAMHlJcNjopGS6j7DBMIIoo75at5E2bOAkISGDjhxSBlk5RBBQPIwQOyIDDTgsAbgKSTwUYYVa7YIR7IDIIqDQgC09ZFrtyMC4UsIAE5RtO9HyYQA2lo4nUOQcj9ADSvLnkFkcogvpCUAfLKGBBhDkF6Zwgj0CUAtIlMAFE3uIJ46QRAtqISF/WER+EnGuhwBGCk+Lw/wS8gtFXIGbIMmDEpSQvDSCxB0M0IcReLYPX0RhIOlYQD6yMIQA8MIQjkACHzbixwDIADeFcMQIBhGAQAiiE0xIRwB46MuQggSIFkNmPJj/sKqSOkQL6gLJPJqAxZTQowk8y0QT7COA05hCd+pEZk9FUi3zHSanKUkAUuTFkRIosASDJCpBmOqQQxzBD6KDk0+Hqh86fUQN+crpI0GSAxyMJAYAS0juVFodr3okAU5I51+uWiUu7OYjiYDCXgRAC+3MrQOuSYkinoCA7PXSh8BkwpmOKUeRtOE/09JMIxICCWsmpAqx84gjlmDDaa1hnA4h6lW9ygah9TElEU1IObDAApGMAgvTgutoEyIPKCCEIFzQ1jlUEIo+BmABDBFjSsQQJECQoIsE4YQSPApSkSp3IyR9a/7y0IUgPGwWaiVIFtR11XkwgWcybQJCMsEE/6o87EdouU91zXhejsgxvU2RI3W9moAjNJC9G4HCUslCWsCUQI89MwJVT3PIKCXSIzcw3DG/+pEcNDMkYuCpQ2LgYNdeJSX5K8QTAAERp5qRtnPtZvIOpdvOBgAXHFgVjBTRBEoGQHu+3Bg61kMtCQdgB2OiRxEemxJIMMGGBNnURyLRhXlUpWUQ8eN77ZUSNuCGqKUlCC2+IApYadK9IKFyPIpTldrq9ra5NZIRjCnGAAA3IYDQAAoJsokVILdYy20zQZqL5ADEowtU/GkAWBoS7c42AHoOgCaa0EqIgNC39E0Lac0HmEQL1bMHtlgA4jtfkdg3AE/Ab2KfqsCeTf+1qvZBZFbrRRhwaRgk+/JIDtgwkgY7F2u6yyuBGL0RC/Ojp5cOiVxBUjyqdqQdHIgZQRThBBWzmLD5QgcYziTj2Sy4HjfWrY55/NECQqQciY1EE+ihWxxwttZvdde0tnBJMX4htRuOlYeRnJJ4yDbLA0sJOnDrqz14BAbaKvOZA5Bm4CXXzW2Gs3rTTecMA+a6ed6ubvsMXvE+ZNAZbnTpCCJUOlAY4p01shLN+2gTRDokk640xfWb6Tdv2iEBltOnQZIGcB05JKXuCA4WDBJVd/uGuqOuhgodAAtjuLNUlhaHQZKIJ+iaI7xe1bSALezB9tDFSyBKspWdmWZPCxL/TYA2FaRN6wBEggnY1uy2k9naq4LbjFcgdwYDcJtVozfd604JF3h626spxAjy7gi9yWzmhOT7o2zmt3L9DdFpyaMJdRYxQVaAapcyASHTmul3u3CLjbzAwSUVKnWaXNLTmNfmF1dnxjEsYaVSepBxMgJ/32yCocPLqp7eD0i4ilaQmKJoGIL5R8YwrOqSleIECipHCgGFnSck7FX+OV1TjKoOGD0Af23jipW+PRd3gRMOH0kOatxsgjRgxzGqbNYlce2EENmAEE3IFjibpyiHxAWiHcm74nEQdNk2C13e7RHqzpEX1DvvaF6BvvvudyEFeH/yC0MiBxuhB8izAgbm/xHzAAVoIA8EIUVUpAkk4FZipkkbMQgYRSD1kAi1IGLpgAjQNksBMAgodF7q8AuQ0DhItREJFAAYoEen8QdZQnoBkAiFxGkuSB0DpjGGUQ6KcA9jhX4coQdHkC8gYUkBsA9QVAa48QfEhR+AoE8PwQ4t0IIPoQoXyBG/wAQd1hGIUATS9DYaUAMutAAm4HzDtnTF1gRsMIRWcQVj4gcYsDIE0QJJIGQEoQIhBxHtEAibAAU2lA99oANgEADlgAjYNhyH4FE+9Q4m4DIcoQ5OYHYdcQ6AcAOixQuIcA4hsQfIEw9I0AD58BsoICuGQA+zsARzQA8wsQBJ8FAdsQXtZ/9veud/fBeA/UYlrjUEVbAOmoBhQnUJTWAOkCBNV4UOn8ABTLIOc4BhfUAEZbARiMAilsctGvIIGxAPEKEHTfBYHeEDYgURe9AFEJAJEciDDsEJgKAJRFBKBFEGPbAJiOAQ+CAKEQhgrCdgJ/cRq4AQeQAFPDM4CpEJH1EDJGAwIMEAmhAAuEdxicAiMcAWBOInEAEJHtACuaAUYPgRV6AFZWR5CQEPJxA2HFEOQ2AE0pYPn4AHgqVcGzMLQgAFaWAV8jACctcAHbBztWACFUkQQvAbrvYQg0B4mdBKQ2AFj7AqqZJSAQAJXcCI6hUAZXAByZg/e4ABhdcR8aAEaED/hg/QBS70EZkAeebgCeywBTrBCjGDBZ3gZ3zQAgNxD3PwkijJBDZJELfYf/+3i8s1gBAxl4BBbQ+BCVCAPXg1krO0AlGWWIChBGvAc4nGXjugN/gBCSSwhw6xByVAhp4XAOmyEflQAisjABIXEoAxMzDiEDSQAh52HyV3c/5IfvRzBA4QErRQmB+xBYmXmmYVFniFO5AUMJSXEJQABfCgLHCgl1W2AuRGX4BRA+3iWwSBAmDIeWwYfd0CGNdiFfaQAeEIldwXACsyc+k1LYVwAqAXAKKwBDUzYlBwkARBCU6wmWGmA9u2mHygAZ20EXlQAtJFEOBFlh5hlthDED8Q/yTnoAUJ1SQZ6BHpoAVEsUR8iW+5uG9/6UOB+RAs8AWECZ8J8QBNMD+Y1xF00KDjBRgrMDxqN2rmNybWEQceoI758Y13yBHj+CdP8Cmn+ZsBEApaEKIEYQbQFwB5tY+d1o+ud3YEMSR7lkG7GWYBgAO+WXwEMQa6k1PvAmEaQgZA+RCTAAXdeB9vMDkZpALqF53TUgMnaWeMglbayTWR0p2XSRPMhqOP0ATXdEBMMJ+cl15FZQLsWRqsaZ4EIQlMEE4cMS1Kdm4JwQdF4J8QAQ9OgEXTIgRLcJdlqQQIGgA+wFPvVn8wQJwQYQvNoUQBYKF7l6Eauj0c6hAeOnH3If8EJEotIFEOKcpkh9eiTLaYBGEe5WcVkFAENVoVeuAEd0g6OgoRfOAEnOFVOHerobACNfMubiJw7yKbr0abZ7cLTwCl1FGlvJdBgJGlHnauIUYdX8oRDvAE3QgRZgpUKnCJHtGm/5agFzJqcro1kVItbMCAI8Fs4hkHe0oQ6sAEDqlX5QMRaVIVLgKfhxoAktAFi1p+jqquzSqpIFGphXcJS1CgHZEJS/BOAaBlBBF3TROcHmELKqA4vsB/F+qXrupLsJoQo/ChHVGMnhoSueqgW3p4NimtDxGsRTa0G/EINIpYBLEHTWCay0qOD8EHTQCt7pVoVAmkWhAM4xUAbnL/Vds6L936EcRBkKmZP4DxcjHGbTyHpcbJardKaCLhJ+c1CU+QBxsxKNW3EVlQrx1hA27KtgEAp+fFrxmzMYkRsCIxsAmhp7xwsAnbaNEqYvGlJi5iDgSRKrczsRULcS+aGOH2J/2ptw5RqVhEEJgABnGEc5mgBCULqgmRsilhCsGJTO/mHNNyiynBqgBIsxljswQRBlfAmhtxCU/As7iqBD+rVk0Et2SShIFbFJCQAeoItHvwBDgGtX+SrG+1clg7BDTzWl0LHiABtg/HpCGhM6ZrtljLBjHavXa2extxVqMLvR5RBlPgZNpRt3e7IDPXEfQqEvdanQEAB9RDrs8X/5PPuwWnYBX04AHi2QBd4LgBgLAKK7ncJqhqIgorcLkvwz4JMQlNsIfvpVcBUIsgwQckYI9cS6lQIF0pgQlMEDom+hCZsAK4kBBvVxUtOy4EAQPlhUzpkAVEMS0wi4JZlAWf2Kq9eyy/GwBhIKQiewQcORK00AWhEA+QcLmAsQQ7kADyJmE+ML6edwjIQxORoAHzoAh7UZh7cASPsbYEwawPkQ9HsABah3+GsJANJwpMUDOXhgYmoJzky48md76UQwRRYy+TsHEN5yqCYw+QEHk+VQ6VQA9OwxFlcBGDcMeug1GuVQZa0ABJVF37uxFRYANfdMCO9g4n0LexpgWA2/+5BGEIJEAGCXEHk8CnhXsshxsAPGACCjcSgIABnxIAnnACjFjBuiUIH0I6CZAEDSQH5jAEJOMHjOAOJSAEE5sL2oQHuxAJdqC3vnAEEMoR+TACC4AI2TGSumAEQpAI7oMJXbAPcUDFIpsEcfipXQC7XRByplAEYOwR59AEKoCgf3AB4zItm6ABgaCLTPyqvRgSv1AJQ/oQ+4AJn0gTlJAPluAEwuwIk7AECAFXuGTMG+EGdhoSCJAJ7tAFzpFY64AJ2ZGjUesQd5AJu2APpZkQNECdHvELlCBUkzACkwoR5ZtWYusRSdAEOAwSSaB+H7EI0tQAR+C4lLwBndAH4Zr/EIgQBxJVVg6BBX74EYMwByVgTmFqtxvRAILArh9RCBmwxxyhAlYgSyUouBzg0ASxD0gQQbxsLL4cCRZw0iLRABvwIDYgMY/rkC3ynmE2D5xwC/TQn/kgXS6SDnKwB+dQQu8wGpE1lQ8BzGntEe6QCYCgKhnrEOogB4PABNKlD47ACGr4EQkgAvrEAiUQaAGgAjyVACNwWB4xB0SgrEzQfpF6MksM0VLixMaiCU+w2xLoBIpMHTyQuCFRB0zQwyMxx6NZAp1clV5QFvngBHNNvIU8m4f8EScwCiIB1TThCFa9jgKwAfXnETIQ1gmhAt4HiiZQ2AFgyhyBBvj7EcXj/3wesQIO6hEoQI3khAT+wEsJ/BDiQ90h8QhSmRA3kCkUDLkEEQpdsMUecQ8kYMwzs0Tl+cEEEQRNYKyLYtM5UwK+5hHwUELJQ9seoQ8kUEoPsAKe+gPaYg7w5lIl8DE+DDBSoBHKvdxQ0tyPYlPRTT9MUOGHUSZNUQcwxt0vLrVOAN4zIMsi0QdOYMYfgdQ9qNQdgQSvvBEnoOAh0QAlcE0+JQBJIHcewSccQUA0sQdGAN4gvNYccQav+REOEhIp+hFwytdHoAARLlK+PClFThN6yqcBoOEJscwezsHreA8ZAN4ucuISOwlM0OIJAQctChI6I6AzbqldbQQqxhG3kv8QrFuyOoyyKlDnHUEPJfAhAQADDjYFAKTkS04gTS4lT/4Q9CDlBFLlRXHlwR0S3d2sXJ4QXl4WYT7mHlHmiOGDH5HmIsHmNPHmcZ5BdB4Sd74ReT4Se97nBBHghhfoI1UoIKEFUd0RiM6Eis7oIeXoRQDpIyHpGb7hlw6kme5Tm97pK/Dpo8Luos4RpT4SqM6xqw4hrf4RsD6grZvDcGfrIZHruy5zPQbsvCvsyCLRIQUYNiXM9NMEU54WOhCjI9HsRcGsI+ms4C0DblMU1v4mSmrIWhUS3R4S3z4S4U6/f0HuIGHuEIHuBanuENHujta1gu4ReeEwHJEF9A6a9v7/B/iOwI3+vLvA703x7wRR6YydEHNA8CLO6aRy4qnCIgCuqPLyA6aeM05QZ8hUuqzu6hth8QEg6xkPuxsPEvQABbve6w7x6w898o/S3Of1q1cxLcbuEPWweEj1+BxR5UfbP1huZxUC7XQs7Vj65fhKEH3wBMhzVZiA3tyq3h4B9CBxAlj/EW9+1chEC0f/EfN77ljnc3xQAnBhPpTQBCHrEO9e4FAgG3HsZPO+tIYHBAasQSYQQTRgpP36vPNAKYf2qIxrsClR9hze2AMPgh9xD0Vwx9MSpB3MGqMxLazAuX/DJh5hc5UqXZxXgvHQBI8xLYlwBACRL4CAAAUNGtSn/yGBwUxa2BnkUsXgORVzDl4sOCtAvRKOAmgUY+XglBgBpsjAmFLlSpYtXb6EqfKQBoEEWc7yVW/gQT38YqokyMlEu4E5A9g7QunjQXiEzv00+INLAED2fqYzAiodoIPlAuk8aDPFjgCBdqXMh6RRwXVYqLhcN2jdzoJ+kCyge9GcIHkBMHGYaxPqS4IQNqCLyYEUTA6j8qqEJyjAoyT3HmOk5WHISzQqVl5ZTDjAniSRME46YTmsoHkB0nxxqejEvgCC5gLtQmrPu4GCD0pJo4deQUAbEBWc8mPwcpVseFych0HSvXg73QlKt3LcCyO8Cr6j0sSgoQ2cDLJwYm7pSv9EFxYZrPGkXMF2JjQttWRkuMaLUnS4nIUgeE7I5LKL3kkitAAW2cAPAw3aBwO8CtLEhOEIimiiiloiqJ4TLslHnQDMwCIe2gK4wgZ7VhCDORdfhNGlmWpqyZ0sWlmvIBamiDGAQ2B4yhwsPgmgnTIIwYiBDYYbbBPzVPDkJ3PGGGSIKA6yJYtNLrJpjgJXKBAjeGLwRccLiGyJkCdoEyyeGLjyzSB3lFDqr8B6TKmww2I6o4EHLzJDkT8NaoSJO/4wgzcOyRDUpUo2Uwk0mDTCJYkWMDKkg0PyQucITALIJJSMVvqjDDsE6eJElcJoxQlJeguroE4wmQKUguiZwUH/PSwAAs/lnDtQhkNK8JMgRsBQLaUXMmiBqAB0gMK8AOZJQgXJjA2NP5V8wIDJc5JIwSB1xFiIoF/GcCdHg4bY0iWC1iGjkIG0TcmKJG4paB8yWmOJnRNKKsgfEiYMIMOCKLKIJYLYcSETEwIJQJJNMvniqVA6UUEEx3zluOOXZhy0IHmasETdFlSI86dZVh61nC5EWU8AehsZwSrm0GEiYahkeEtcJ9A0SAChCbLDiU5cMsOLj0/4w8CUC3KnhALtDPlFPRGLKcCnMZLZpQZMSJZljzGS9CcsXMBoDyMILigdD9r9iN6LtE3ECFUx0midDAqUW+sAglkCZt/24HVs/5iAxWieER7Z6ZESksUoDfEMqqLFguwZYa15CyJI7ou4yMGgdpy4BCjOheZya65Rh9V0IZiAh66hWYrCBoPiKeHVggwOAGEOUfcFg0EM6qSLh6aFgoiNDWd+bJBVLyieLpS6aBTYOC5nBVFXYqQIsJZDZwlbBxsjXHG7OHqlW7qAeyUZenVJgSPufsmdJkoXArCqXby6ea9L8I7/yKYgmGDhUhfZQwkaZZB0bOA+UDnEE/TREnZwQAgcWgGkeFI4ATbnORiRRxE8UpAGNCGAKanBEg5ChRcYpB5FYMRy3oADg7CjCRfsYEzksARFxQQFNDCIPJ4ACYhI5GAbggkgNP/wC4NsYgW32UUTiBCGHFbRRc9rifSodxDrdSx721NJ9743mPCNDyrlO4g60MeS9bVPJTK4UvzmBxX74U9/0LNaAAyDNSsepAEA7COKCPgSA2KED0ZYYNscOJgITrBfFsSgBg2yK/gFMgCIu0gIRxiAEp4QIylcYQsL8sIYDuYNoSuIDT9lyZUIAQx4gEoKgCgyKBBxd0bsHRJfokQmFoQTTyxIFKfISmKuBIss0SJGuog97bFEjC4q43LQeL70qaSNLoHjS+RHP5fUMQD5u9PY+kfMP3oyh2WLSSEvcshEBqCBD/xJIykISYVlECOUJCYmD6JJg3RyJaA0CAtdCEP/GaIyAKosZkpcCcufyDKItSyihnTmEl4a5JdQlCIVE7rRghxzJdKrhLqspy2xuagcWgDj3ALwTObYQglAUxi9plkQNVYzJenogrTwZpBsqvQi24RKO+7nlzsabpweK+lKGgAFneCxY+iEyRUOeJAEthMdi4Tgmub5AHVxyZ4I5KAl9bkUeXhgkyWE3EUAWhCBjpKgg+GBQRHqVBgllSVCUEK6GnqGIDohDhE9ohkpukSLAjMAwtQoRxPqUZXgrkALI0opVhAAdCiKrhgxxy6eUo4mlOIxrKsEBmx2kHPkYT4tSccTUroSAWS2DgWZaQBuUYKJYoQdToiSQcqRB1sU/6SyAUADLoMWAFzoNQCI2ABXMHKLHl7EHfYhajg9dlSWqCN2MXkHH1niiCTQ6CXtMO5KdIELlihhDJP6GxNgcLqC/GIDDLjIOt4WE5s8AglNM107SHA0zxlkHiVgAUbyUYFKBnKsBdEHEYhIEEiYYA8+DYAPoGAQXKwAYAHwhQj8NItciOgltaACWQZyDxO0qx3HI8ws4oHildzBWe4C1RFOWA55aDclVEiDQXZhhC2iQAkGscVXnVpRXxoWsYrdKGNTcoct4IUgmrhBAC5hu0zkeH9okUKZ6ICGN6jBw5wriA1S8IO+/FQFZW6JEqbyknr8YFMBiO05dGCalRQic/8HsccPDFGQQKAgD5hASUqGYAaDAIILfEjJHMigknW4AS9Uu+xgqJunAESiCuqBSRQm8ZJAiGC9MXmBKFmSg9yqpAZuXIlGHmGB0oE5ASr4Qj9y5IsRLNgltChIC3iAgwebLgBJyAJL7CEFJvjpIrsYQYH7eOAApIAIsiaIIXzAL4PwZw5FsF0A3GEFMJQsAIX4QhdkjQ4d/NUl6NiCE8YXCh6kYUI2aBddaZEDeLpvqi8xhAhgphE8RKGdGAECFgxyCyN8OgBiuEAvA9AFH8SEyAG4aDAziuTF0mR/7HhKQSQ7i3I85AVawHWkD+KLJMirIKAwAov544T/nPYgjsD/wBjdJ5KX7MEEjIOz+TjXjt6uZBIaqI5B+qCWgG2gTF++iA16FgBasKMcrDPIDXikEoxH98rLmTTXAgCKDNQiMZfq70V+0PCY+KAHLrEwS9aRHacSxAUdyDjqHnACgZR0WMqFcbNDpzrUyQAJYMbIHzZgbIxsQTms1CdB0gFgMJ+jHXTgUkF8oPN6YMDblT6CgwKwDiPYdCW4OAIYdyCe+QggC6b4iQBWMMiUTIEN6EXRGnYiDyRcXiWm8IxBpFCDneghCQ4wCBBgE/aLPDzih504xYmp5FQXxHqCCcMXCCLyQh9hIQXJRBNuA3glRDlljSgCk1ryAinARA9NMHZs/22iOkl4oLl8cAJ8A5AAI/gE8Acpg7Jnd5AzKDsllygqcdKjPXEJTtgATHuJE4CZl1CD14sJNjColfgCUaurABgFExiuAKAEJ8iD1vERJ+AmX+MCNGiJAAkAGCgBltgHI9gzz7GBwxOrDzqIdFgCCPBAqKMLHfiPW/GAtdAISGgCsFgHJrBBl2AHJgiTAMCByTIIK0CbrLGC5VmJH4gy0SgY3isIeHiCTWMJUriCg/ACINIIeXACWwqAH1i6wVK44zsy5WMl5muJUbACbYk+6jsIQDgCJME+Jti+g+g+lQA/8WMJGJg6lzg/zYmtl4gE9zsI+JuQBDgC/FIJMoijlv8wAxhcCQCUro7JusgzQAR0iROQpFHbgp+AQJeYQDwhiAs8jSfoQN+IIEmECS7AQpdAwfs7iH04An9QiTTAxGWTQYNAhzXikILQQRfKAM0JACAEC3VggnpbiXVogiRcwoOwgnsDkC+QQpUAgirMuyioRS0Evi78QoNwqGCCqIJAQ4cjrCLDqGFyQ0uCQ5aQQzqUPhfBQz0EFT5MiT9MiUB8CUI0vyZARJ2DiUV8Pyd4xEhkCUp8iUt0CU28OkkbQBvTuk+MCVGEiRwoxQeMQJVIxR5ZRQy8iElwxceIRaigRZi4xQfRRV5MCV/Mp2D0LWJUGGPcwQCghyJQRmakqWf/fAlppEYmLIhrVBltdIluhAmbAEeDEMeW8MKDOMfDSsczTMOWMD4jS7547KN5XIl6NIg6dBF+MILr28c+NAh/xIhGyABBXAmBfIlDhC2DVERGNAhHLAhIlEWMcEik+cX/C8DpqshixEiY0MiX4MifWIOPTImQjJFV/LuSPElYfIK9dAku4KuATEGX3MVe/MsqOrBh9DxKe5ac3MmeDMKfhEZGm0aDqEbK2UYOsYIFZIkfqMVMC8cnGMeViEpznCWqNMN1TKJ2DABNMCx5YAJ45Eor8spy2AN8OYiP44Wge4EVsEODcIQMMMtOMAG0LIgS6MhdGCNHEIE90AUbk4cx/5qBcnSJPUACnDOD60kJejiLi0DIg5GHPzABYzsEDiiTg6AHNFO6btKB8msJibxO0dijYpwDDSgHeKjPlvAAbLyIctCD13oDsoMJKniDlLADPTgtJQi0xwyAF+AAjJi7vvCNRECCQnixl/ACB2SJdDAHM0gCXDQIwTuOmPzMHBordMgHI6it2vyBdXgIe8AASmAH9ZAEI/CFDryFI+AEdEAz1kxCNpicgsiCCzMHPcgOllgBEuyqkcjJlngHy6gCGkK0dzgB20uJGFACdQCLL7gBARiOeTgBSsCFh5gCgRstlvgFDNBHU4AC6CQx5VnOrrQ41dmDK9DHghACF/ABgP+xhDLAugC4gxLYAKsgCAZYA+2yCRoYnyHoSI0gBB8wBCooExMMADLITF+yHJfYhRy4PgiIVYxAgwsziPssCyo4hBoYHqp4Az1IuljtBNRjiXsAgjYIsAMNTE4czJsIgEZ4DhggNJfoATNMiV2wAkFhgVFkCUJQzoNYACuozzJYpRjBNUnoyFE5hC3gDd/wBRmAAQMtRiU8Aq8jvgTwgRbINpW4hx3gCs+RScSjSUqQAhyQv5sMAFEYhTcAvnfogU1IAdowBBkYgoajBRtgADRYLYwQynXJgGSRAqK0ByYwuZV4A+8ZU4PIASPIuJb4BDgIAFLYhHKwgkS4pB2lNEz/IIVHiAIRgQFWMIXXawc1SIQtKB0XGIJMELGWCAUSIFbiSoKlO4QO8IAnTNQc8so/SII3060AUIIPKgd0wEHRoIcSuCCtoYVa8I2hKYfTEgOB6w1c+4Uk0EP+4AINFQB0sMiUCBB0KAeNMIeYBbin3VW79IckKJPSK9s4QYF2LdyW8IMjgC88QlCjilbWmh026NGwOAc6OFuD4AUkIKJaOAdWbYlISI2UYF2bqQVQ5B+4DQsBKNuQ0xaCMIUbdReCuAQj0IUH0QhFMIGmebr+EoBzmL7I+VwBGqtQeALvzBOWaYcSABpCSAImmo8hcALEEABauAKDs1KDIISH2QkW/+gCTMMHCQEKgkCEJPAJ1bGEJohQljCFH4seEyiZWnC51ZkPCHCCXAgAr4uC3AuAOlgBmJmPGAg2bJLP0jWNTm2CJqDArfUfr9SHI9izDFSB11u/cBA5gqAHJohgv0md6SOIUlg61BEEE2Ai1tmCNsiI9QOQ08GjHmDMRbwuSIQT1okTLlADHFWJPnACncXcZ+WYTuyKrgkANShThRndy+mIWJHWR3CC+jUISCiBC1nex0yZlVm/s3UBkvydAJjf6wLcAEgEJ3AQWmDiPHk65p3Ji/iEJfjbuREajUgHJoCbXzgCaAuAUFCCmKUCrR3fgiAEI4CTABgCLTgtPciAf/8brgQogaahhbCjhCUIupZwAQOOByfgwigOgDgGlSV4MS4QCYLwnYIwBflkCTHAJXuAgrWQ4M5KPQu+okVdiX0wgQ2+iA7uDY2gPhEm4d6QmTiZPrFR4aAhiH44AoXDSWYW5YMQZqG5ZJboARq6iPbb4SPoYSHmAiujKyLW2YhEYl9RYpVQAzcgDKiTm3qAAo8YGrpyBKZKiUewZ2MWyf2z4WrGCBc4gnYOgAdoguZKiTV2kDduXzk2WDoGg54rY3fqgiIMAD+GGIIIhRVAQCpgvfhCQoNIABPAO1HIgkeO5JYwhDkS5Urogg50iZO5nS0U4hneiUxYAaIgCB+QCI3/OIcsAKMXcOW3pDmdhALCW4AJRlbCeDqom2dcZonnOWaMyGBfPghgnuaFHqVihmLfWGZ9bmEmYtVjNOarXp2mxggfyGEP6OYymeYfCOJqCws31pY+eAJJxojMFcAFjYk0eGIA4eeNgALGUV2XcIQSkLmCcBwmsWbiSz137hzicwEj6F2CEAInYKiVWGMEmOyUUYPm9Z+x+oQueK2ryCmD8GNAzmivY6uOPggbSkIeXhdHLgg9KAK7XgpDsJuWsIQu2GSW6GSnlOmbmB1MyKul4J1VDoBWfomQcCFiMYijtuWlVInBdmqMOKazneqUsOoQLqkRpjPW+gjB6GrB+OqL/9ABGZYdqBDdjKDuCItAjYiEDlAU+mOT2nxrZH5soSvilj0IvBZMvYYJvpY0HIRnnAvvlyjsw56MLaZmxs4jTg7o3h1oy24JRHACR3oJz7kBz26esfIEO4YKNYKnXzACiCmIOZDt1aYgJuCqvKy/dVEB/qDtcl6JlA7Bg7AEJTBolXAB9tRCLuRcm8iE4r4liTII5bZFXKKHojYIRWACCv6JebAEnQAFLJiDkKtuY1pUWhDcwPvalFiCqWjvmGI6xEjbLVqJcwDFMaBbdMi4uw1Wc9AILlizACjcYR5lszVhg4gCxC2IIBgBsPCHDXhhlkiBjuxb1S6IOz8IfTCBhf9lCf+GVgBHTA2FiXNY6oMoXSJSXrIuCEg4gT/d1RMACwGwhf59kXMw3IGgg1NfCTHg3TKuEDRWiUQogRuvNi4n5YMo2Bi8CO790phQBygAmkFIAkAeAibgIyy41ZRghxKAm0I4AbxDX0zjAwyA9MhLhCSwzIu4BMN+iRdggtOKB/3t3fnQhCaApflAAQO+hcDZCQZ+CRlgz3q4OYNYhCd4As+69AJWB0tQQC3YErvK8o6yuAC4BzcwcZ6wgkc1CB2wACeDCoJQhBwwh3aoghhSnVmwASAPZAd0gSfkBxXgCkEAgg6UAUstiAcYUaj4Azd4sJSpgZbvKCRAAYjpgw7/YAJYUp0bOK+C+AQK7IRm1wMg6IQbMKe7PmdV3Nxzg4J/fQl42AIFACEr8IdMiE2WSIQqwOKCWIAvKLM/2AKB6JFP+PmDEHuyxwhNMAJ75RAY0AEfYLGUGIUOcMuU8IUcmFoXqoEsaFcDC8Y/IAED/gkmYALV5gMRsDIzdg+DYIMhvYhwcIIlwLQ+IILFxwQiKOd3wAAUYIkhaAMpUHuVaA9tbolKuIBrBY9sp7Q92AIW8IG5eIBROANUKoVQWINPeQcagAPZa4lzMAFcYoAR4Fm/U4GkdolHwAIBBgIqCgIucHCnBpkA2IcNoHHd0gM7qN2jsBQXgQATIC8+IC9R/zYHKAgw1IEHsPgB+EEHPkCMRxABndgF8SMDwv+JBOCAa8GIelBwgDiUpFGAAJaKAAogoCDDhrzoMbRRpeGNiQ3L3Wm04U/DjgUvcVin0CPJkiYLLoSwAd3JjvOQWGrZsV6SSR7N7QnghovMhun01CK5jk+5gglO/OrZ0wYWjwJ9lYwSxZ3SAF1kpJOp5krPBSf0ddxnBAqOqmaVsuHR0FGHQ2cLdjrBi2EKJwznJenE0B48maBOQCw4hQlDmpoY0nCysGQKLrnOBaB1kg3hnlu6FCynR53SPyIYuVsYY4W8eQW/qEF3JwC9E55EtqSX5HBBT1De0Z3iJkbVOWkC2P+7EihAAintRr5NrrznIQ35CiI44rahgMXUZxW8wgL52UxMcKGsbr2juRWhFFZvuGWHxwZF6jEUgB3GFOWDTAwfrxA7ST5OGBhlREImydcQGVE0VAYQJB1iwj4tgSSSfst5lNJKVdHzBCRm0QPFhiXlsEVVBba0ECFPDEchQ2VI4VEhTwzokQ82mJXFdtyRVMMPPSXiRB8d6bNEEzmoSGFaDTXAxFxvNdDFPSPdsARD6ihB24QlxdEFRAutoQVD7DCByUhDqECHST+oUdAs/HmEHQsqKNVCUzjKpI8GCYxEyhcNpXDGXU/E0VOGjIzESRacFfTDD23MUNUhU7wzRBb/RcngBkpFYkphc88FEN108Uk2Xnq02EinUpg0wQ5yAkhGUnnnpacee+gt1Mh7tBZEn30mQKWfeONhx0cTABJnQkJX0rrYgQku6BEiRjx4UoSmZmohS0rtAkUkHHpYYUEhjigfsqxKdmKKRbLoIowlzVjjdrGWVAOCMvX4Y0NBDplpckcy1EATObGp1CNKQillQep0YSW8JkXChGkL4eBlQew0IWZBoqjQKkk+pImrty3AWWIAcoYX8El24ilZKXvS5Sd29JQQqEwwOzLSJivAFoCiW8hglgwlnLAtGibgSaK+Ryu1KXRGfBreQvyJp5AK7yaHqqrpLdxQLeYly9B6//HV6sHDseqa3CBH9BrwLFkH+59RR8RI0trWkdFsQWbY3RAiR/gBYUjUYmptVbs8sS2G3Xa0GLhKsU1dgYM8IUim6cYXwItxN9RuVaWiZ5K8PPoYFhj5Il0VvwX5q0cAJsv0SBNzLRQlQ7cwAcFIWZMECRNbBtAlQ7g0cQlDomihsUdc/LY67iONErJMIF9aFcojlcIVXWbchXhsJdRcECcr2MEQED/wXKJ15vzCRwDpLKB+1KXDb5LSAfjBtEnYzY1SqciWtFgmqSpvXAF4Fa7WFICv4cpWvGOIGOqzuuiRRDzYOVuv+NcR/xAqAIQoAVSqUoZ53S1vDNnbg9Z0pf9pWZBCglOKPJxgOKV06EPU+ZaI3rKQAr2vICeSHKYoV5SFGGJdJHmDn5QyiywMQU2sY8jnFFIOZPWobw3ZxxKYQKT4yeR0AUhdBA1YEtctKQA2UMKUmEAbwHUkElpiCA5W8LuKPZAFWhBVrNCklObFKWQpBNKdGLIyPhUxAPOAgswIxBoodC8AnNBC+AoCBCDooGctOQckQvGJTVhCCJ3wxAM0MYjOYTGUDJnfHzaQCJOUIx6QWdM60tEFUuCoHPAwh0w2YYLjrC0AtsDDReABmVo84UYDnGVBuNAsAbiDM4/AgD1wJAPrCUAXWSGJLGlZoARsABFVMQc6eoGEBhT/RBEb6KBSakCFhtjgnA3BDiKS4At01MKLHUFh/FYoKCPYpCr1OEE+ZxgAHvAEjSQ5xztoiYvQBCwBSEhKD1tUkGQGABEmKIQSGxIFtVSlCzDoCQ7q4w5rlkQSJgBLQ/5QAieURZQm0eIjjpCPcxTlYO/In0dYEokjPOk4OHiCTEugF4LuMQBBMIIe6lCUHRAmFwFohxE2EYCgtKAJMY2eqlKgA5ksxAVKWKJHymAwAeABMj2ZXkH+yBAr3IAhrZEEGsuBjnucYEOc+YQTVKVUKUyBDY1qyS3MoAIqUGEKUhisFKywIVoEVaWack5B9JAChpLkHT5QBEpwYIkyHGY8//gAwrnahJ1G/EBVI2kAF6YZAD6gACrmaENMoJMCjgTgBRstCBvmYLkryANHnEhrAOLBhVOSZB8p6NtiAOEDHySxJ4DggiG2gKcAUCEDfdkjdkRRhoYMAQ0k+QVzdZCUJdITfvaUSQKKsACzxCMKwPVnC8QAwZ4YAg7xCAApXuARX0RBiuhyaABwcBhA9IAL58HRDDAA2558oQRBseAQYrCLKPCQJIkogRSa6RAdrKCGivWIFhGhA0P84MCU0MEPSUIJGyzAB7oIgBg+8QkivWMKibjBhhLBhV30RBE5OAQXnuMCGUCCSOdgwyZSQIgAZELDlxJAGihBChcoRQosa/+JC0xw3QDg4QfrbQlZA2DWgtBAFAx5h5Zb0gcrHIIHhghAJNAQCTZkBQ2ZaIEMeCDJk/BvH3ji6oYzNT8BxONaJJFHCSwmACZAWVUBA8QGmtaRXKJDHmbKJSeOINoABGIDz3VHHUYiCE1P7NJPKEst8lDiKfUlAHo4ggw7QogNJKVA5jAHCnqgFENsIBDBoGUATNAxPlcOD6thyDpwU1Nb7AMJ5y1JeEs33pYwAgmcUgotVOmtANyBl8tjGGAC4AOMNiTSU1UR5QKwhNneQgpsIAmjl92TTphguvxbhzn2YUqTQOAE02bILOiQg4D2uSFaTEctfrGBIxckFCXo9BL/h+CEdhxHZz7ANnAwAE6RTAKnPTlHMXANldWMogmSqcU+RAAgdhib3wGgwwpgmbOT0EILLejJFCZeEHmc4AFK6fKXlxpxhTwGjb4QQQLoQMtPNAEesMGCdm0xhTuPVQw4qMENbgAFWAbcs+VIRzGsw45B+CIozGHsHvPQBUqoiXMe+cMRntudJuTMF0f45H1N8En9LIG3MuFFE8C5Xfx4pAdXlAkh0NYQJ0C9dL14wpbn+bfE2jAAKhG0TBhQgidlnSSPKAGOtzD4+JUbCzMvSA/03hA9lICySrFEE7Qtk/o5uiHeuXRH0rCjzNMW3AwJxBEg+4klUL4joFiC2AOg/wMN16MIBCmI7uCjlMLzgyFDWIGZVJ0B1ZvkC6OoCi2+kNyWRIFGBYHHE/rJ5T6WdcrKAYQGIHuznE1hr1RI/EnksQIdRMEELPiCEX4E+fglQBmYVgJMQQpQgQ6oDrAFgNKU3dmlnTB1BNu5nVn8T9zNHUn4gt3RyRKI3971XUn8AuB1hOA9n+ExBOKFEgIwnkk0G9I820lYXk7gXkdsXud9HvyE3ugFQOmRRD4YAfbJxAM4geu1BOyZxP8Mm0fY3gwGgBYVBO/5HvCZxPAV3/EVhvIxRPNVBfRJH/U11vW1hPaNyBeImUyEH0OQn/mdDPp5mfolB/u5H/gwRPwVxP/8VYUloEAA7IMaqAoM2BYTFoAebBAU8Ac9LIELEJQb5KFMMKBMmB3arY7aRWDbJUcFMoTc0V1HZODddQQHKgXf+Z1HhGBnkd4NmkThkVMAoCAWqWDjNUQLHs0LmkQMMmFD1OABmSLS5CBD8KBH+CAQtoQQEuFJGGFJIGFJLOEMOiGm9R5D/F7wNQQVMoQVFkTyLV8AaGEJRl9BTF/16QEYnoQYMg4Z9sQZjl/57Rwb9twbth9D3EwjBQAdQhf9mcQc1EAA7MIbcIQiAIFpZZ46jIIUEEEWMEQoWISqLUHsOQXZyUQ8dEEQMMTUlIQEViLcXeIRIJzKYeIGdqAFgSL/CIpgQ5BgT6Di4dGjvrQiCz5eKE1eVVie6hTJ/2leCZjGFqQUFu1iQbiB6THEL96hE+DGHumD/RhjE4AHSSQj7uGA7j1hMxbEM07hEohVAOxAG1zhNcZBEzgfScKN9GlBTH0jMFbOLIhjTwjAF0CgSQDBPRZEPDwBW40VG+rJo50FHDIEBMihYMjfSZKEInxBbvmAmDQAFSAK7gmASETCFNhCQfjARvEHD2zfOs1NORQFIjBkBKVHPDSBzhXECoyeyQDCCVAUKPWEJT7hCUzgAwnCCRzZwjRBxwTAE5nEPSBSSQzCCUjOePjATbbEUfRKQRgB9mwb44wHYtGPEZyX/8ksBCyqiHhQpiJdiFIwgAmoD1atysIMZ09EgglABA8oGfzQgBUwRBZgXQC8wRqQRB8oW9dEUAAIQQn0hTw5TkHswwloU0lowhNcjRLe3obNAmUuxBagwgytJieCQhN02pWEQhMU33HtxQgQBHZEwhNYmMiE4G8OgRLwmh6MgLsljtTMXHaihHb0hBfQCHbIg7bIRAEEgD6MgAL40Zzc0K+Ex0kAwghkpCYoQRJOAfbMo1mwwQrUAyVoQSg8AZQ9EBMm5jShwGHwhw1o1zotRCAYQQecgAgQgfMhghn4Y5I6whlowSMEgDyYgQmQQgv0TcDogxOMZgB8AigohWnqYf8TZOKlAMIK3IAQeIQKNAog4ECqkYQ9wMEGiOUTNsFvMgQPWI9M/EITcMRirIApkKZZ5AMaqI5+AIIMFIIWuIV12IEZwChzFokjbMAGIMEFbADkRQEJrJh1CoIN5JZ8BsA90IB+nUUjNIFppAFvwo8M2A0V3IgfYAANqMnpdYEixABlDSdebABnGE1H8EIN2IM9NAEgJMJedQQkFAEO5BZ/Zl0LFEESJEEFwKZReMETQMU5sIAPTAGvkQQnSAEofEJBqAEOsIA2zUMJ2EByNYIKlEEh9c9CdEIGTNsPVMas2mf/FEQUiEEMpOIpEsEftgQKLGo7HEFamoQZWEC0zML/E5BRbXwfOozBeQ3nA4jAgW0BFMRUHTyBe/1oVdgBI6hOJqTAENBSLjGhJExBp5UDCugpQ6BB8vDby3BCKGiCGmAATGbCCQCqmiwEKTDBIKjKH3BAJvxCEmjTeKTDLyhVQfBACsSpRR6MIPzcpagDHzSGRwRCTjyCCMDHlQxCB2wCNBbELfyCSARMGDjBYsoEOwiCsz7hpcpkQRzCBhxLRyRCEvyCPigaQ+QCEhyGqKqIH8zBHGCCFGyAu8oEB+Riez7CBgCMyQxCEqRmVeDBICxmHxxY/ORDtAQAIMAkJWhAfnTEOfxBPWiAXgynZRoasuwDB2hTUsRAEgwUCySB/60yRFJumCEMAShoQgl85xygJmSkwxPEAOaVxDzowfgUxB4AQgl4wgDpQQww6FL5wgq4rEzgABQ0RBYEEjr8Qtn2xyFgwCL0hBxwgOuexBh8bEGglVJ8AX/JwwaI2UKkgPOgAxOImfF4RAx0wVRhAfIhQT7ZYUuUgyC8lC8AAiEkgC/8giBsiQKKks7ybRTY1mLgALbKD2MFQCU8QdMm6f4thlj4gj2YwJpdCQrbnEzIaQohEEkokEkIwhGQotx0BCcoQZcqbOdgh4iehCEcgf6OUhRPqjsETwBIrnIsTEsqxQlQjcg0QgnApEcIAhTUqQ0tBs46m9OqXBB0wWpcSf86bEBm8Q8iNMGPAJsfSAdDwEAJlESP9AJS9qdiBcwNMCVUrk8XcMKkegQ1BoA6mNFIzIESFB8VlIJStFFDVAHGBix24EMGuCJJVMIKUIVMjMKcOBJbygQKCKcgOcE1csFBnoMK2BawiYE6FYQU9Ax21EMJLF8Fn8Q6tMEQrMMWrIAKYMEVfMESiIkIY9FCOMAUTBMORKlgOFVLzE8lAGVJyDB9ErE9HMGatQQO7HBLyKlM/LBH2MrDkIQgmECEVQUiZ54CULFT1HNHWLHwCMFKipfkRWdPIAEnm0QYP8mElPEZ12JyTMIT5AG1yPEZyc8TSDFJ6HGnBkAf/3HoeKv/Mh4yGHRaIuuFTDhyOkRyQczBClRyeRrSGrjRWSGpWeCDBoSyR1RCF8BwSUAPQ0SBKrdEChDrzWlPLDPELEusScCAeM4htvay3wEzKtHDHezCIejBPfBCVd+D/GbeQlACFVzLI3TBkiRCE1TnSWTzNpNEN+vhN4ez+pbzSZxzS6RzR6wztbgzPCuFPGcdPU/0At5zQ+Sze/LzGnMxQAt0SRC0qRx0QqvIQjd0HM8xI0r06+3xQmC0hGl07Q1y1jnhJ3h0QbhSSLfESJd0AJx0SlsnSzeEFbx0VcT0THdETd80SeT0wvL0Sfg0Q6SoDAl1QRC1TBx1Q8hjACx1HfJl/40SBxUEhuMMbqZgByVUwUc34RdAgneUITazsDanXEeg9T6otTifBDmH7cu1BBvMChAXgWmURF2/BV4HnF4zSF8zxF/vs4TUkz/TbUkEtFkctkE/AUIrtlkwtkk8NGTvdUdUNB/7sWXbC2Yz4WZLIUj3hGifUWkzBBWo9Emgtku/RWv3RCUsQWx7xGwHwE4rxW0DtW7LMi37NlLrZWGUAFMXNyidwxYscuI4cSjxgSKUmDqAAhusASSm0PxYwm18aACcckwlWyDYAxJ4t0nkQGYfoROI92QwJZJoQHKL4kK9BSh0wX2HEnYUwhGs7ig5yNA+FBRkAmDTdz8Ptkx4cf+SykQjmMClqu0TZOR/vwUllMB8eUs6cEBmkXVkF6EJZC2LHUFJKEIJLHhDKK9mMyUoaKhnN8G8ovNVeXYXXDNpozRDVIF99UQOGMxpXHhP8AEJuHZDWIISlHJLkAJB0gVPmoQUqDKhwWUAAAGLo0ORysQYLGo8+qhwG0H3NHVJnAM9VAe4SoFswUAZnNKz4vlZzM8DGAEML0YLXMFC2EMMjBMvnMCaOcAp4c/6XMKDbAHAtYQmSPnmmLtcewAgTAKWM8QvIAUarUMmLPocOEGCBoAvXMK4NUQiJNJTYUL0jcc9QEB22zibdUCaElscXMITFEIkNNNCxEMl/AEcYUL/YLugPwsAImRQS2zAQYpMAyBBTkzIJszucoC5GibHInx8STjClmm1E/hCEHRr5aiDB9iOBR2CoJ8Edy8CJSBAbCW4R0wY8hZEowecE24CBjyXOjRBpp/EFVi6cD+2SW96QXS6TOzDImBYQSACJFCBfZWDHMRIAkBkSfABB8BAoXZEK2zAvpmEC1yBH1gCZEiBGSTAhuRwHWoXJtCDOxxBA9CD8PzAnjBCzayAbVVCERdEITSBA5VDHCQBbxQEL5iAmL7sSejBGeyCOrTADeRAGpC+DhDEjT87g7BwPkhC8KXHLxBLJWwAJ6iDOkwCjimBiERNh8REIpz6pehDHPi7/0ngHKyQRD00gD8ggX/rQis0tknsQbCXFCRM1YJOZUeYAS4HwDpAAWgzhD+4kyEFwBCYAO0FQB88wQMcgiqk3khwNyMowo9k/JoL9gaIEZSTRAxkQIXKBEDombQuAK0ABxHqaGILYUOHDwMIEHBwhgqIFzH62ILx4I8cDSfqUbTAiC+HE81Z0hdxFsZDT/ZxPJjLUZ8OlwIAcnSRXiQ8ENP8kDmUKEY2PBzy4TDmoDlI+Yamg9IC4S8OiBDOWWEOIaNfQyl1caToIJsVhQQFYFdCU8QAo7pMhHgrkRUsQ3+JOEQ00CFMTeAFWJBvh0WMLRkIcmfiUQBJdjA5OVeIQf+AKF8C1AwAZsjFHxtUHUy3YkovhOso6TlIRcbQfD/4YJRYlHZt2xwPaYA62y1CiXInCnHSrncALToizp7XhNLt2vaYNAjQ8uIvJF8v8r6ox4l03wflgupy7qKMFA3tNOH0sJAJQLIPDtEiF2EfKGR73Xerr0Qit5g4IIg+5ziaCIINAqDBB6IYKGEPogSgRa5yBjwoh48IlCsGK8AjMIA31JDJhxB9k8uQEkwCabbfcINJplkmwuWmF5N7KCgPcSwLKYe8YArGotTpIhOEBilpugC0qmU67TiSpIl63MphBYTaeWLIiVhYoRyIZtsii6H4yGCvoVq6pIt3EOrhigL/5ZKnhEgQ0mSJXBD6gcOD0MlijouioMKtdLQApaCJBmQNTDdUyy7HRRkNILfdgHtoFuoCeMAJd4rLgg2WJtqFiUloI5TSoephorFRHernCOy4ZNIhPZpoBKTpqANlCfIgksEL9LqA4KEEjvhjqCG0dKiPJ/zrwwmyJtIH2f8CLK7R7w4M4IweGCwhtqEEoJCjHDbN8KAxqljUDRIx4uGGEmcz5In3VASuQodeiokodjgQQlqQDHI1gBunte2oh1KA4SBUMVKHiU2qMiKQrJRQ8kiiImmCHoTU0ILKJjBBiIX5FGXDMI7CxIrbSpdAsyUeMLsIxtnieQIShDJRAtOD/7jw86BzVNgTIiCmcMuWLD5JLtIADJWpHk1JgcHpp8vwz9+Aqb7o0X05ssQJchzStMYAPAX1oRXjnRc8bxGqp4nGOBJkVYdejhDhhmCVlV25Prm1PBQODmAdJnx1KAEjhJWJ2C1r7MUJ/3p59iAEHA/gkmjNZtRABM3YUaYFHCxqaoRwCJdFmX7jLYZyH6Lwc4d2SEMmHlxnN4BCnID3u9Ubqhfrh/DVl01/Aa6aqIEdkkIM24JkeKJBjnj4oCRti4SJiyfCYcqD7uAYIVFA5jIAkbObLcy9Sm81ACGW0MUtHu4s/2ABqIsHCjgnymQJ4ibiAvUAePb5ISCk8KcsCP/KaPRJGkfqsQIgoOEGDXTgFmSFO+FV7WqVg4jWMEUfr1EqbK2qkAS/FgC1sQ0jbmMVp2hlwQDUTUV0wJvecnUet/wtcA0ZnO0wcjhjLS4AjfPP4yI3OQFN8HIByBxROPcgbqkwAKEr0RIJdZDTmY+JB2nd62IHHvjN7l1w2x1EdHevfHELbQ0J3gQ5QryGpIAptUlHF5QXgF+Y4GETgV5tJNGFebjFehvDyUREkYXKTQR8HgxAPjRgsg55Tw5LCIwVWxZCLR5EHlCY2UFqdrMA6A8h/bsIAP+kAk8EwIXSOiBG+IAo2WwRja1ESAVpI4QnZBAhK0BOQ+bhhOZIal7/INSOPWQmk1+c4ITwa4n7IKIHKHhHRc9jAq4eQoYAzlA9v3LPsJaAOIT4oT8B4CZZDsLNH0qOckQMQLXQwAUGHWFbpGMiG275xZMcTQaRbGZRrsgRH6yhRNQ5hJFmBcIAIKIEhWMiO4owpBcBTyiuNIrmEJKCGNgmHUxYz0Ss04+sjMc2TtpFlJRAJSdYAiGj+FLIRtaQlx0kHyNQpAVbIgQmoOkgPljTkVj5vpbI4wiSiBMTiHOQy3RyBaG4CAr81JJ0ZGGUhIoi0lojk9e003wObSUsT1YJJTziAQ1RQYg4EYeD7II5GPHEmPr2i1BAsyGLYBgumTAzCwbibQhp/0kuhhCTuSHkHt2x4Ry0+QmOQiQGbxiEJ7aUnhoipAwigIqiAkAsucgBTnxYVgD28QSsTGQf3TyIEOWZoyKiAaIXcccVirBHotRDFPcwSg3uMYSgkikA7RCFvcZwp4cAQhQEGUo+MfKGLLrFHJ2wVYoQ4g4WrIQohyjBPQjRGZm0YwNB4Mgv5sAVoDTUqg9R40F8MQJS2OYTFiBpAHzRBiTASxFZuII2i9KAC4z3IGiowA/PUYInJAoUFmiL9wopqQDwARSD6ABWEjHKw0xCDVkITDw2cQW+yREUbD2SIi6hCyg0JhP+iMQSbmGOIcSDDTrjn54uwgUM0CkA5nhCFP869ICOrSaqHNlDGhLVXR3nTjehnSQvfgEDFUTxEPwIABOwFQB0JEI1rqJDE0bxkE0cwbcOEUNKW4wI11oQF4t4JN1McEmO1CERF0NIKKCADoTwIRGV20c+QuEErpQjEfiQlgn25z1vonUKHzlHmWuriMBMpB2LUN9nyym8y5UDECe8iCIwsJ6iDDMBGBnEPRqQBNfShhcnYNsfxlQhSZwASr8drhcTgFyEoMMDLPCFJlmahMoQRRcLCIApklAg/i2g1GaDwBHucJEz7vh7pfVEBoxcmytkgR3PS0ID0nGQHnQBh0WpwxScIJd8bCBcAWgABnx3hyOEFMBYdggikiD/iEN8lBRd4Ig5VFCDQJBHHyP4BAIOookn/OQhYlhBLRARjwBYAQ228I88kPCIPQziSOVABFUbAmlmmqEE6kAIFSJ5yougYw9qJjaxsfq7g4zivSH0mnZcVY4VSLchmWhClRtSijzb9XNkc0hfd8KmhwhWzbiTyxyWoF2jNcQJE7VgKXsThXVpkT6TWiQ5h6joc27A40xkxBPs4eOD9KMEhIBsA4zAi+eUQDpyWWlDIFECM8sEuGOrHDo4IGmH5OMI4FzoQV5gApFLJBwXycRwhM3dj383AKDQgsRoQwUXHEkr2uTBRn5UmzloQeg6wJBamNCxieRAY+XGzRFW0pIW/5z0IuXAgijqowEF/KcLLHZIKe4S0RogZBeWXJIkHzKPYNZvBc0+CAqCRmPS7fXjrgz5Kkv6hS1pRwUYcqqTV/4Ql8McIaQwcbyyM6l53aMJzMyOhAaUt54/dWzPU4J240Z0o4fvaEBYegElov0BgbaK4jpQz4fSoAfVPxBQ8Lr5GqAExI427IHsDqYlgqFCIAEK1o4j2u4kJgVh0GEDfCWnDskI7O4weKMUjGCVvAWZWq4Jgm27ik9HHOITwIAhaoMKXoDx/g0h3GAjnEOHaurycEF7LIT0TuJ7zK0hDAH0SqoHEaIcrqAzJqIPMmD1JuISlgDWqu+mDsILaAAh5v+hBGYGVfaKHqBgJyZiE1agyqJgmqCKRgSqBBvlaiBkIlpiFKzA7KhDBbrNqVIn+hxi+iDC+tyO+H4DYbiPC4FjVGZjVDyhC8av5g4CFJQAmoxJ/bDmmFTqINyP6coHRkaF/szp/rSOAYwgNvjP/8LH2wRw7KQj/frmIBawATFCB9ClNiZwsRAiHzCQKMAvAEihA1cJBOtQBAPPDAkvb1SQNqagBZ9nCRAvBgmEBgPABy6PHXAwAHJAB2fFSzjiB1diIkZBCA+CCFmuD1QPITABDJzwIEgBCgMgBaawJbTwkp4PI+qhBEjICwmiJcLw4mrMDO2Rx3Zj6CAi8t7iC+b/pfn0ESLKQQtYjmaaoNkqBA+npQ+HrubwhhCJolYS8YuKbmKe6JOWLv6ID9Girmoux+Pyr3OKoh+ewOtG531AUeyqaCJ4wQCZpHzSDhUvQhU9hzdaESJgMQPhQxxtkf2GwuVG0EYEj9gIzxOWoA5sQwqEEUkiBiF2oNtsAxmVESGYccacsfPKL8B28BCOICYmogVUgPi0MfVW7yC+kaa+oxTI0RzdovbEbEUqhx7c0S0goPfcApSE7x718iCwyiHdYhT88SG0QAZXR+VQzyE04SCfLgBkrlFY0glORQ39MgAE6xd1DhG5Yq8q8oMssCGiIHYkhCgsUeowMVuajHQC/0AQPBHpngjsVPJkeMEIym6eECLtSu11+Kkm8YQCcdIIxgk1A+AFjkDnflIXSbAECc9WLGwoUMBgzA/xckAGb0Mql7EZOU9RpNF7DsEE9CoARk8si5AsvfFMHGJLTIEcpWAKD2IelimEPkcuxeogOEEJhigKgm8M91Iv0XAPGgEkL4IRTKIUBOkhloANzoERlrMhyoEJWEDKSsAW7IER4EsMYm9RQsIILqkcViEfIYIOwOYQwoAJ/HMn46wBNg0ioODyDkIPHGBEi+fyBOE3L2I0PXLqXPQiGGADQEHgiEIQkGAMqKdC3OEQNMEEThQ1eSHh9qUlfsE/JAEJbvMi2v9BEawAB3pUkZSMNx+CD05ABrKOWxqhHUjhBDiiJRIArfzOOIWSF0vrE6rL4ooiHZ7AOZkSmrBgB46xWGqQKq0TK3ewB5bAfA4hCYSlJUphCcBzG7sx354ALfuGFEygEaKtHFcgLQIgDzZgC/olAPShEYTuIezhCDzBECzOE5QAEcYEBZRAAYhD4/Kz+NDQEkrAUSFiBRbPLyAiEwghEqBgtvyu0hxCENbj72AOVy0nAN4BFB6roopmkAIgAUoAEyYBvobCF0DBCf4LInzg+gKAEoqATionEpjpDHZFJmiUaj6yKOjBFDiAhGQCDzoBAwQFYfrBCeRADoLyXjqhDyz/siHIgEP6wBPgFCMIwQlcYBGI4gzy7CYfAh3ctCAPQwlAYR/eiiPUAMY4Aih3ETlLyw9sIN1oAxA0AE7MD1fkYQSMaiMvgjr51Cqvcx8DgBEU6iLuARTkwS0MwSoFMjwPghvLMjU3AfH6Jr1MYFsSQQlMzAwswHkCYBO6wPcgoh0uoROgQDV8IRLO4E4UoRQ64GFc9VVBrscCwBKggFYfYgWiTCYw4Ql89TY0wQmor5XQgQn8x9KQoI5s4xaq6SLGAGMRohKKAH+IAg2AYCjONWDStSjcxAFqIwkaFCJ+oQQYjmrqiTagVdU4wgyuj2EhQguogii0IG2H4gZkCCM0//Y4i4/wAiAQTGByieIXmgdiJKYeNIAR8lSbpvIgqhIhXra7xrJnF3UoEsEEHis4R2YfRgBYA0BOcEEmGMAEcmwMIim8vLYewfbj0LASnMBsu+ZxOeISmqBtbSMTmCBu0QgdlqBoZGIQTMAk6q8hgsQVG0IG/PYgKKEDBHcozGAoIeJwpyVxiaL2SLYoTgBiEUI1XXdapqgoEuAJMBcjykAMs1R+EUIFwqAossB7ZcIG6jdX1dQhhm3HVJeuHO26jkCjnvMg6qEIbHcG9TQZq9Nl/dShfjcAfLYoEmETnxAh9kED/m957ZIjSCLHxGB/RLZ6r9cMs3d7ZSKDDTd8cf+EfM13gtBXfTmCfd3XNuwAcDCCfh3ifvNXJvbXcBOtRkszgOenNgr4IhC4ahaYKBr4gS8ighuCcx/CgjFYgzmCg2XidNeUYx+ChGkDdlGYKWe3COxmOl84d9WiT3eshm+YKHK4ndSSh304ToIYI4YYIYoYIY4YP5MYe8VWe7m3IZzYXKF4MYtCE8rXNlSWI6p4KJgni2vDDnqlizv4fmlJf4dyXvzXMW10YlRIgCHkINYYYX4BCkp4Ud54KOK4KOh41bQUIvCYKE55KGqgg6Uvlf84dUtrdevqdU9YdlUYkQkkFLRAm97ABrVn82bYlSA5eGUiEbQFIUohknr4hzX/IZMvgnNyDAaMWAOQOJRF+bHI9suaaRZW4HPLzywBD/eI4m2n2GXMBvxG5yTRoQv2ZEB4Y3mQIC3kpZdGRW87geYaogzKFSGCwAMw5ZUDIJsdghZQ5ZePFRMNIiAdogp96mQCIAlQT6YdYhCcoJBDRZURQgbaR2WhlWnFeHOnWTAZuolFVybQQJsn6S/C8SBCWMdUFxCOIIGFaZxTWITM+Z7gIxRe8CDc4PKqZGbNYh8v2u0IxYW2qDMRAt4adCIQgARWT/s4QhHq+SBMQbf+gARMcnm1YIp/ZBGMQDVaQgwiCRBIYKAJesfQUA5m1SHOYRKYSwBsdTsyweIyoQTQ/+EQbE3rHAICnmCiS08SkMseMgEhJ+IWKKF4V40JBIWj0Wtan/VuD2IBpGZAFkAnA8AOnGA92sEScuwgyKB0DyIIimCXZYJcH4IRsJSmLTSYA2AQuBAjduEIdok4N2AFbqFfc4IDBriYva0QupgKGACsL2JwHob4DKEyNLeOn9ohFpoofAEDDBgi1gEKUhoicOEKTsAONtabH6IGMKDaTLiQ56ApyTqR+aASWLshqAAKaC8JrPQg2gEKZnYLrkfA7OES0MRsBCASWGUQ0vshzKEKqmBM+gADzOA0WdxqRICkWoIUPCBR+uACvBcTMABNH9EP1AB6DyIBVOA+xcACYv/iayv7qsTWDyTBwvLgCXDiIBpgmQNgETRANfbBp9igcM17KML8U4libo3qIBpBA6BkNlpSruijHCChGh9iDpwg2uiBEm4GCNqgFCHxzx3CHCQhJmCxhati1tbsAfBPlhEdIaRgFbFbtLTbFE7q7SiBX3n6E0TA+1atBKwXOK+gjQaESRPhCuiUI/JgEj7KgmrATxJA0SfYZSDBUmXCBYgA4qxmBCrBfBBBBA4TdQevtOCBCOJJnCFcwlc4kSMhFIeCHkTgeExxBGy3JdBhEvi1JQ4h58pvUIXlg1pMCy54IkghUK/LCFpGHTjBBDY6AGBgxDU7CfbnEDAgPo1jxPX/gAPIgCNIAQkmAU6lYAUmdx46gAsmAsqjfIKOryHigQlIiokSgQSoKgf4tzZcuiHOAQzWPADaPEgDAOdQ+yA+QUQfgguksyG4QEWfjg+e4NG/w4tW0mxQ4P2gLuRvI3FdABp3skwP4spb/OLaqCioA9Xh+iCuYCkjkvjKgFt3k4L5cSheoCfn2Qlse1YUgep3Z6u7i/DmIQPcFS/E2pDLOZEh4UmIQg8ywNZaYhK64GZpxa5Q0xC4c18mghawIMq+UudnRU2GUAWCnRSw0QciKR6cYIBJwZ6iQD0vQgx0C2nKwC0KsIURPuEpqMcqRxe6oNeFNwOU6CDWQJ10rBaU/8B/GiG1eoMXus82BnE5e0Dl9xSVnOC0HSoKbKAhJh1HcL5CpyUdL6IKgr42rCDVIeILkD5Hotkh7JhAOLAoEOEJ8A0jFOEJNF3YibK05KEIvj6sk/2QyZ4JohQj7iHtEUISmEC1nKMQulI2vsB7r1HXWGYIsYDlXED3HcIHdEseggnv7CkFzAAgAggcSBCGFYJTZgysV6KBQCoyCEqcSLGixYsYM2rUeEhDvgACKurqUkljogx7CK7hsrElxlpK5gxsVGQeSIG8mjh06anLuYk9clD0IdQiHycLXCqVGMUGwUsc1t1cqjEkhA3oBLrAQhUjPSiQKlYZQ9UKDIxfXv90pVhGCkV0GyCsHVjKyEZETxBcVPSkj8U0P+YKZsNDorwij1z+OtJv4BwltQTWK9JoICQm9TTey5BUoCQmNrsWOrLPooAvLAaOUnExJI8vA8thGTJwq0UfBwXKexJW4AvYA1OYuWgQoULJDR9GFMy8efOOH0NSHFky48mUA1c63whTpkCaNqXn3Nmy50+JQYcWrXi083aLTZ9GnfpeoFWsWrnW/9p74tiyZ12U1nZtvRUXc3Xdlddeff0VWH0tEWYYYoox5hhkklFmGWaacTbQZ6FRNVppFZ2WmkCrtRbAa7HNVpt+Q+UWwG79/UaQcMTJGEBCCyUXAEQQBimkRND/0SdRHiSZpAE+BOHA0lqzKNXdTBnQM1VOjhiJkSdgZIXeegQRdVF7ULoUBRnySSUdhPd56QJrE0W5Fn8VTUHWUlWYgtEVajlHRgoGylWRnBm9YMJAa1KUSBMfBUAoQXz5VRFgQ2YkIUGHJdZSIEYIMtAnK0QWwGSVCXRZZhlt5h4kXViZqKOPThTSiBSFRMsVJwYQBpwlrnhFi7TlZxEXMsoD1kA2DiTFcBbBANxDaCK3E5CVVvteka8OJM8RmViUzyACMSACH44KxAOgIIXERwIlhtQHIfZF+YcvBAkQkj6/wBrAOVAEG4AjItiTbgB6IBGJloMGEEoJ55UbgBc+/1CEQmEC4RuAH54GkM8J5AlUTgICb+RLoxhVwWsAUKkpZJsCidGFfQTNEuW3Ls0SSAImSKLlO0kct5F0S8Rg0TuDMHFnczV0QK5E6ZDwycAD7fMHwhTFcMI7v5RTUQLkJoJBtw5LxIAJgwgi1USUWlvRpQM5YkGWLRmShKchteCEqKRuiCpGCGDAwECZlGClQIPccxM+8FoEzxAbTG1vvQGUswQpAezDiwsvX3RPF1kIZA4iR4gyUAyZDxQlO/lcASc9jZxAyb76bNF5AHkEkoVTFsmwRCDyCGQFFXoIZE8SMLDz43JqJ78WthWlU4oCFrGQGz9mvDOwJps4Kp30pv8F0AIV8Qaghg4UleLWTbWQgkjUZsAzsDsw5Eu1RFEuQsqa0nkCdqKaXDI6EAGAAQoEAg8YBEIit1gC2DTiA2ZhhAVEAJdAUjY/5rAsAI/wF0VaAJxsVaQJpJhDY2KFiQqU6mcBiMcGQiGrkDSACTI4IXMeYYEyQA4dZigE1ATChhq0xBFzkMQS3EEf6biIHiZAQh0oEiVfiOIRXZCURNKmPIKwLQDsyEAJ9qaRqxhOIDtYwUI0ZCoOZaQBGqBbAGiABK0FQB0rwES5hqACDwrkExuIQTxqFQA6jGIRAcABEAihwYmUAwVQ8F8A+NABL8gPg6JD1L2OoIYHCIQFWXD/Qb7ikYUtCEoRS5ABIy6yCBmAYScPKMGT3EEKJKSGWlWMZUuYZxo7tgALWpuFvQSgS+nQ4XEBIMWz6CcQYcIsAKd40jGLoz03xmmX0rFjC5+ZKPyFRDplAFQMzhe+gbDDCdnbiBfWoJE+OEERA6GgNAXDMunQApjHfMHsnImRJIiulxIJxQbWWRFelKAyjxIALQIQiSO4ajs8IKdAerlLkDwKCGnQiMwmWAJd0CdKAlDCiS7xBDw4NGbSWUQJ/EARKsryivMgQVhidZE5rMCZbtjCGE94Ko1Iogu7uEkOtDCQdkBhgaNQAh0sMooVXBOe9rnmimaHVIKYQwWpiZIf/0ZwCJD88pgw28cIdBgSGMCIHkc4WEgWYQJy2TEk9TBBHEY3zCooBJayjOtFaLmUUXxhoDEj1KNaUIVeBVNHAdgCxRYapVKAT18Jg1pT2xUnQvHyIjEYoBi8cBF2MEETLQHCDTTSCyckIp3z4edaWCanbEmHFL8KG0aQUEiCgCIJ9MxIlOyRnFiFBBJQCJFzdhDRmBEWqT7AXUse4ATrxUkLkbxEE4i4WIEowglSJIhJY3lFeXggS6IdCCiU0DAdyDRDNDUjRiLRqpvggKcCwUUT5CgQUWhBmpgsbTWjGQAd6Eea5biCv/qQAehlRB8ZYFdISvGsXRxLIIswAnYuQv+PEmQpSmLo60CmIDS4yvXCRPJIBTFiV5e0ALATcYGEB3JFuhwWwxmJQRQCIIYBWmQdl23JDzabEQQ84bMTDO3KAnAVL2UEtUo5QWu1m4TsSoS2HZPIIwS3YaXwoLcbCa5SiOtRWWUhkphYbgUTAd2SPiiu1aWQS0CxBFHV97ujImMAapqRSIDGvGIUCDvWOxD3wjcLLJVVfWFkkfzuVwP+xYg+NMAugRB4IAbuT4KZ5BUHDyTCBKFChZGH4krTVSkdbsmHMSJiiZTY0CeutEVUzGIXVwTGmN3IjDViYxyjTMdBuuCPU9sSIVvktUYmCJItsuSDOufJLpGyS6hcqyv/CyTLzLUIl6M7kOlWMcyaagmZzezdmepNI24OTyDjjEU6t/e90cNzRkJi34z4eSB9ALRGBl3oABxaIIkeyKIz0mC4sXjEP5q0qEV9aZdkeiObvogLQh2AT7ub4PuWCKlbfBFUy5jGfLsxaFUWax7jRyNAdomtK4LrpXwlyQTpdX2A3RJhD5cJdyg2lrtAcUV1GW1fPulgZyRmaZd5INUG77XbDIbehQQH3LZsdQIgipNNpOh5lgi5+VwiK/w50BdhN10KfOAAzNsrUDihGKaAEDEcL+Eo7rdGbGELGBhVIvRIR0VggF6LvEAFj/LBihElkBgYHSPtCFEe3NcVW/Ai/7YVQUNfy5AbddADrwGogzvucARPZOQcNpGCUNI+EXjkQSD7OIEiBEBEde64xxiR0xiWcBFz0MPMAehACwKADnvEdghFUMc7spULiyo9AHs4gb0HUgcrSSIJmVnH4Cayjpy2JApoHkg6XD8U4W4EExxgtn2c4AKBaIIDpZkoRS5xApLC/MLQVsoQSsALUcEhCujQmj0wIAQ3TsIIvmiYRSaxAXoJZAsn8CYSqi+QFiDh8hURVEbmA233Yk4wCgNxCERQVUk3EFJnaFSnaEbAaBdRD0igM1EiA0zgTFaAJhYGdrEkdirCCG/gCZQWAHgQBaNUTQHwAD7UUkXgcyHBAv99slACwQnLkV1zsB4zcIIuEQhVQDIX4QlC0wlCEwCMsALtEADhEAAMoAJ5kAbRZhGCQAX24AKc8A5SoEMS0QLMcg86sA8xsHpCAGtsYnE+5leZ8IIVoQ9SQC/S4QOWEACHYIUSEQk5kAgp4GsCEQoKBTmJ4AFctSaKIAXoQAg64D6ZMEwEoQkgdhGkoEFRsgBSsEdGYnIbMQhEgG8EcQNzGACa+IgDsQAngAJc1GwxR10zlylK0QBb4AVrFQBD4AZAoA8B8A46sAY4MFCF0AYoEE4XEQgjwHUC4QkVcCIC8ANEgGOLcAFsYBE9MDsaoQURgxGXQASaog5ggBI7RBH/D+huEShvE1hL+4IEz2h9REAeUgA+UuCDIKg8Itg9mWAEu6BU8FYClsSC6JBytRISC6ABwZMu6XA24XMLt9BkAyEDMAIH1UgVCTA345YOaocOahcAljAC7hMSF/kOwZCGFYEISTA1AdALSPA3EnED50MLakcFONCCZlgfslYiUYIO6tBQEyEISGAIH2UHtvAvJxCQA2EOa/aTEyED0kh3kGAEAgNPkYAEAnMLITEKHFARLTCVVZF4S0R3D2ACMhgmzrcRaJAEepYOPxESaJB/VAMBJsALk5KKz7aKNdcS8FACjicQh7AB+RKVTSAq66ACXpcRptAEzkQFOzAQ9nAC/6kWAEAABhYBBC2pEU8VSRdRCl3gRvKABKn2TlFHaFOHaFV3df0YJSgAPlHiDkzgHQEgAweRAu74jmoTj70iB02whPAkD13wOgHVXEkVAAtQBIZjj74lHbq0ETBAjGe2FoNgAvbnEiHhAB1gPc35nFhVEQpwBCKJAOc0EWeQilHgQ2XYcjCJhhhhk7/kQYHwBOwiMxgVAI1gBEA5MA1QAl9UEJx4E48ABZnxKPeZnyExBGh5dIeyESwVEpPwBJcXK5i4EaRgF0p3VCHxAlAwnYDTBCmXLc6mPOGnFOnABNkTEovRGCHxGGZGBaWgEaGwAkIJRo/Zbd0SEjlwlBLxA/9qsBHl8AWSaREu8Czx4ARipX1tyJkQ6JkSSIH9GAA/wHUziQVPIxDFGQCs6ZpxBZtMFABCMJsTIQ9MMAktpJvD2ZtLKTOaKUkOSpzGqQNtgJxHsJwXITPCGQCS4AEZGQDOeQAD81gUUZ0kYk7oJBFnAEAEgQI+9AAv+R4xWaTD2VyBAAUCZi/D2QjyCVIBEJ9sKRHMBDmOUAJLGXJbNDD+WRGi8J/jGRKORaBPsEd2KhAJuqbFxKD18jjSAQMlcBGZYKVeBn5wuXsboQ5dwAnlshgHFKIr0DBUQDkZMQQvNRA+sKLt4G0B8KIWAQd/iBE1eqNUySvw4ARaqpvl8o3/7zYjvDEQilACRDqlQMBNdZAFS8pi4DMFrQmllXIIGeAXuVYuQuAERGQYTKAz0ARNGdGbvylQummTGdGkYJR8VfE4g3AEB9QSDSUJ0hklDsAB95ouYUp3h0AaAqGn6SIdZuCnA9GdVDqo21GordGA51loAlAAj+Wo80kQLKtaYiCKl5oZ8LSfNtipFCEKSLAU/BoABbpHCPqV7SIAQ7WgGgGrskqhFXEDbomhcJkY85oOXQCMH1ouoYAhD0Gs2joEWuBMYiIQy8pezQqjBCGjNIoaGdEC1voEWjqelcOZA0Zr83As/jiOGfEDbhES55quYsCO7equQuIPRSCELlGv/8mmLfmaVPjTr0WQqd24FgQbWAYbqtKRsAuLQiHhsMYlpx1Ap3RXEf6AsRrTBOgETx3LFE4hqOBJqOKJQhaRqO3GSy5kBC07EA3gBHzIYjK7qTWLnwSBs0e3szy7JpRgoM1Ho6fFqsQRqxYxq/w4EUxrq5hyXaqVEbfQBWL7C/B3E6GQBYhHBau3YcbqTG+wHnMmtjs1LDOqEbRgo2m7tm2rIn4QYOXyArQmD04QFiGBCCWwNBiBt+WSDirAqy0DKO0IuFV0CB6wD/WgDkuRCUiQCwSzN/NgApY0MPHAaPwkLomgNe/QB4A3ELzwnhghA7yCAg15leUAwoTAARnzM//54D6WgAGXVwe6AAEYsLnwJoTqQFKHwAGAIBCM4AGAVJofkQYEVwXPqLoI2RUkaxG40AcpOhG/kARcCBL50DuOkAQkLBCOUAQvPBBmwG13eAKCoG0CMQnARxAt4AEVQQpu7BLq0Ac9+TgPgASDQJteiULkIgYboBEyIJbNq7QT0Q5RoEwyRxDzgAGhEMIXkQ5IYEMC8QskcDACwTj+mwVAMFDSdA430AQpGg8qUJgBsA6+gATA2ANNUBH0oAWTSxHqkA9LwH8XobYDAQ9suxGOcAGFZg46wG2FoAGWrDAdMK4VIQVOMBC24AQBEgAzcAS4gAJ/i8D1cZeKUAXtJlH/AVAIaCAVPxAtAdAONcCAjuED2dUHUsAE6EQJVeDAFWEGbJgRrVCiAZALJ3A02owOVBAJ8pAGXXwR6mAFc2gIOiAVfIACLIADBHmDUkAog6AF9qAH/rwvTJAEKXFbq4wJuYIinRCyqzuyrasRiKACkzoR+WADvUAQWtDRgVADvTMR+0ACNDARl0DPE5EAMCAGZ0oQCXAD9xoljhCtBAEJULYRv3AFpVG5ZUAFdbnHLYECQcAIEHcRkHAGSfu8BGEGFQAmqkgQ6sAGJZCuLcEDF5A4u2ACJbCEAaAAIvBlpHABiUmVJOACIZEOXrAEiqQJVVADOikQoOCO6iAFSvA6/2c0BTUwShhhywSUyxlhDxngBC8dAxrAQgJxBRrgfYIwAihQkRxmAe1WBU2AVw1gAWagA9NMzdtxl4xQAuvTErNAC9oHBjNXDhUbAG/XEgyhpZtQAguNHog8npp5D08AchmRDk8gE/MaAHZAlxurMUYgQwMxBIKZgCEZOSGRC1AgKCGRCSSACwFgDolSDloDfR/tHE9cEa3jv4YETxyAgAJQDnYkWIb0TomifTPAdLNQDsOpPWfFqLOknLsZAEpg08gqtBcxywHeGgJVEc47FCCwooksEVVw2hPBCyOAXQRFfgOBA6s8EFjQzGvDBD1lBJgVJaPQBUMlu7BNEe7ALf8bwVEvndjwqxF/gAFcFQAowCvooAT+wgBJEB30ZgL6GwCcAAZSESVSIAVbQOGozRx3yQBMUFXMuSZZcI6QMxABpxH00ARhkQlM0NsEsQWkLFswMx5LUQdd4HjaShF2wARyoVT5gBQUcaLORCs3sawtGhKCyo95Rt5NTBXnTREMUAILxkRrYmvrhAOv/LgDMQb1GQC0sOLVCzlrLhGF8ARBbJNYEAYTkaoXkQW5Ilr+TRENPhFbEAIQ3tUSkQJ/6RL0oCFD5QhMIDBRcgNKEGnEahFq0HbnW2daMFA+ShHLOocNOBCV0AV8J+O3zNgYsQ/0GyVcMGLooAKoKVIXXez/AVBvA7EJWkCQQAAEbcDkTT4XTx7lSmGTVd6q8JTlGVEPTNDlX24RYv4z/n0POnHmaf7nA9Hmb/44cW6S+XSsAkErV3UHTZDnLqnH9TJUfp7cDBvSGTHohd6gwITo0pQDVt6qFuHoW0pf/SiwGXHpma59m97pBm4RoJ7gtWRHpS4Rp57qbzkRUgDiZJ43IREHTZCptk4QVZDrFbEGQXfwcyTwulTbA+EOTSAHG3Hstqfsix2/UUe/IeEDfRUl50Dt4mjMrOxoUbIJK6AyQPAD4T7u1VLuUh7sQysnWQAmaO8CMqKeFkEP7x4AXg7mA7EFbkDm9GXvxN09Uut4+52r/xyaLjMj5xNB5wNBK49j8AgvqMYT7I/T8J93cRoh8RYR7IiO9hu+6MFOKBxPP1EyUQ34WLkm8hNR8hIhbJo/ESlfQbqE7XRPyC4fAlwd86ve6rhdBOTxCDl/Ezs/ECSKEbvuTXQWJVw7VCpPEO3gBEqvEcee7ALHK/HA2HYUJQAmYAFA9Tdx9ZQdAOGq9RSh7SHh9UeumOAu7mRPFU/eBVUF8isvECpQ+7sZcJU+D11wMHUv7xTj/ooLELMCDNTTpMFAhAkRCmCIUN0STwFmCVSIcFZDhOu6QFDIx8kChQJDaSmH0J+RfRIFBLjTJNPAlQ84sKuIEROHdQFWVuTZ0/+nzgAQNqD7qZBBiT06d/48IUopxYpqthTlOcYK1aIXYU7s2bCQE18Vr7So6MPG1qICsrCgSmvpz0xN7vDcEiIHVrx5EbLhUVHKGL0D6xVphPARE14Ib6xISIUUzJ5qsiBk1wQTwiErSkLt2c6JHLyWwOSBKYBzwhZXEMZzAknlT30ZEkgM4IMKUHQqQtFeVAIf0KL1oDgCymlFzoFAfrSREdj5c+jRER7awKgEo9JU1aVLqGXqzxcqltbi3jMfEkkBNpVAztMHkAC2yl3kbCsdw5XnuOs54jotrVnSuSUAdaCYQ6fTBprllpLeWqeET+Irjx8TCqtIlCZKGgiRJP7/QFCAXYzQhMCBMiGBJp6Iuimnt6SrKaih8mrkiKQEKAcXDXvigC1bgCvrDefKwEI6twJIpw6lugongEOQCKQiJV4o6yydbmxRISZcoEoAOtQxp6i45qqoBxBwcFE6vhSq5Qoy9KolAHswIE6dACIx4p4AkMzBiQDKIQoLwH7KgbGBKrtsoFGYyPEnz0DDChMo5DHnTYwqKkWL1UwQQqIEE/pDBEWAQgHTgdBZgq2BGEmCDx99qucE17gDpYl1zjknACmkYKO5M3v1VbpDODgEji4igayoG8haqZYTaOiJK062WMqSGq5MjojZIomiPJ5kgCGAHIw1DbXHgBqllADi/0EhVLw+KSOAc6KYREGe0tmisLfUQcFYU8IIYIgb3iCEp0z6QqgfK/JBaIwWtlhAiBgGaiSL9hSSwxQhNmDxV4VWEooovAzxIJGB9PEhJZ+kqCSMCHvyRYSIAwuFDel2OoOT13qyRYdRuECg1TZaTsgshALx4TefeHiJqnSiAAkuuaq64C6O9UozoyigWBovSdKgJwtEAiAFlAS8wEUdKvYJxQcmt3gnjU5yrkiGE0A2FKFNDKaqUbwc4aEeHUCydiAIMEDZliNUwEqeDTbAM4BSRhCj1DcsQUgVKvDsVCF5pgBklBkCgISHXGxwrQwzeIi5atZbr+iQDPhYB4rHNv/Hku2B5Hnikq4GuoOecQOAYTKe6HjCjELp0YonedYx3qkWufghoR/gGIieip8dqIwvBMteIXWMAKVVAejhLgUdAojCB/lCCqCdeBI6x55bB9L1zRqmGEidegZvoQpMJGFjrfNYjPBiDxPMKwCCOIECniWAd9QCC2fwiSM24AfntIMevVLC6nyCjgwMoRz1S0g83DElhCTiBIDwySzoYQesrGMEW+tJmHhiizdwwXVYuVruMBARvQzhCXWIB1F+wLYN2kMEB4nfJI7AC1ywaHOVgMIGA3C3gbhDHkli1Gfwwg52vAMJh/LJPjBwCMyU4B2t4gkiLlAIhKiBUOXjTn7/6kEpqpijHgGYwpDSAQ91MGE37mCHFHi1Q0RW7RAaSIoKRpEXFdBsIPEoVmD+15NyrGA3eTGHEg5UkS2kDyE6+E5gxiAFvKijCzjzCRfSEAAuqCE6P7jBQMwAH6qIAgWXECAbf1VAkGGFNekJgC+gMJuiUMGDCmnAE6yYyJ5coVw/OccGWIkXog3kEE9AmYvWwYFNQU1MFbnB9KDZkx4GQB5FeERgQLGENw1kB6IMwGAsJDom7JEqkOiCFbEYGL7l5R1OCEJR+qABNA7EEkpYI1X0oQFkBuAFqvkVEPQ3EFtkQWhUOOQ5PQqdRTbykXiJ5Gq6oMC8XJInmdwkXjr5/0mFhDIhpHzOKVO5yp+4sjayhA4tbYnLoujyEhrzpa+AmZddQMFYxTwmVZTpkwaUIDEfVYg0qVLNTeglm0ziZq++GU6f2JAnaTAnVfeiN3Wy053wHGUp7YkQSOQTK5FgwjwKZZnnBBQvAy3oTw6a0ABcAgx4wMpDI1qK7lX0ovHRKEI4albI5iWkAXAkJCWZrpNasgqY1KReXkoXegaAps6xKVZUeU2e6NQHPH2OTwNwS6wIlaiDO9NR8ZLUpRozoj55ak+iOtXIWrUoWNUqlbjaTel8FUxRG2tZIZvOdbZTL++Mp2jdShi4ypUqdLXrFfGaQS8KlKAGRShCBEtYh/9CFCGI5ZhFEZLRjXY0svPlyWQrS9uElHSSmdWLCzZbvM5iRSC1WAFME9KG0J6iDTX1wk+gclp6VUS1PLWdT4BQy9cC9SdC7SV+awajYFIFHk0gZiCamsxlJqQBUNijh10nXNpWE7VUeUMNptPVMymXUzzBBBPGqRCy0he6ap3uEkg42npidyCQaII9sBJXf34XoOHdqxPmFbyK5EMD/kCIEMCwRg/vQ70DKQVFIxwdIKASo40dyGPp++bXZSApXTBFXrrANoGECKx4GV7H3GGjJqCqY3jo0ULgBYVvtYoLOkxHTn6gw7zsRAa38VE53PGlpdihBOOjjzneQcIULBj/BaJsUTne8SWemIYKVLsBpeuAh821wAoBHCDrbIuVe5yAEgP5xQkM4ZN00EkFFOyJI5KgMBdzbCdKmFxRDBBCpawkHbjwSRQgHQAVhqUn5sADqovCjhEc6CJvaYgmSnDCKwV5vum8Bwb8kxch7kJDcIDPSuJEnJU48XE8GXcAMlECffLCBBx5H4r65A48VqQdJqDhT1aShxI8oKi8FkHYBjIHKMBj4kD51K9zEYAYMMYcc6lwUdrBR9WUYx136MJu0FGLKcgXzm+GHatw8JLl/UQNWsriGxYRGEwYdyDl0MGut2C5isCDCz83NK4w8KS3kEJKLvhWC9AV6ZW0gAM//64nF2azFHTooJ0N4UUUACsGf5XCXzzRAxAGxpNERMEGnwwFrxqgAjrxeAwZq3XVbk0VFxDh7b2AAz98QokoBIAM4+tJIESQaKrupAZAtN052PCDNUAGAonvySiI8ElB/IBVPREEEBR21azFjXwD2UITVpIgdUc2naGwgCACAwkuREG6LHgDD/oQAHhMAY4rQQQP4tcTWkiEDDxIg8YdwIU3YEfxUhrIOoCAnU6pwwf3TEt8oNCcwZ1DCyQ4/ShOUIYB0XYle/gCH+aRBXtQ4pWO2EIdoXMDTYwCebsAgiLOYKwYkIEdkLmZmy/YURg8qCMXaweaYAhagIdCy4t0+P+4hKiFJnikd6CT07CHI0CpJAEEWGmVW8gJONAhdTA4rGgISCCBXfARfEACfEuId+iRhsgHEziIlWCHk1sHaquUgegFJJC+isCEEbgHDVGHE/K3EUDBhMgJIcCJjXORvyuKLVgsc4CHRVGIUCgCP/mehAACIDErhpgFXKAJruC3lRCDDYCMF1jDrvgCM4EXXcjChFAhw+u+c7ACDCO3gUgBKkk35zKrdLIBIzgzrKgHIxCaRNiAXwgAWnCHc9CKc8ADOmw6FQC/ABiFJsgDVEuBHUCIhRuRroAHbnG4nUABWfLB1TgBIAqAH1iC0tgc0yiHj/sUOFqjTXgCCoQOLLD/MWqzhxDkjimYgl0pwNZJB0Dog0qsrww4PS7akuzolXLQgiH4iXpognfbMXrogrHjCR0ILed4BA9owaXQAyfgvo4ZCI/ADlWkDYXIBydgl4qohA44OR+0hJn4iUuAwmR7jinUuVKiClBIgmVMCKmAs35LtZUYBRNIiDBoSJ9wAxuLwgBABBxzuIH4AWepCIr4gTbpidh7LrSiASY4FrxIBybIKoH4hSOAumhbiMF5vSrguX/RAjpACB+Iw/dpAt7plZ1wrZ4YMXEJAB5ILA/DCMNCiExQgpOLjikIHcEogYPISCDQAQI0RucQiJX4BS9IgSpggxYrikVCtpnLpGr0/wl6wEaesIcmkC5vBMfAeIQiiJ9ydAIh9AmPYAC9gEd5VIggqMd0lAkmVIh97DtlA7G8UINPbBeCpIocCEisrAiGhIwWgMieqDG82CbkKoofILaeAALkAclApKp0IsnnQMms4rWWlI6ZxAzNwEmdrIyerK3kwLCeGKhdG4geMLO8eKi38zcloDanhMp6kkqEAAIgYI7IlMLcWQFTaAd7kB6sGEuKPCez/Im0zEaEYEu3rIhvlA65pEuEMMe77Im83Mt47Im/bEpDE0x95EfX+UefUMy8+ITGLIrHXM7Og8iss0yewMzpvEiq6MyfAE2fCElBHMmSdA7URAiWfJLoaP/NgciMHMlJyuBJo6rNn8BNhChK5/BNpQxO6XhKhKgH40yO5LxK/dSLOaC0ANCDJQAsn6DOAlSLs+yJbezOEu0C4uiJLbis5whPhdADJtDLotiDJngavOCDJiAZnlhPnnBPnyjM6pSOj8mLG4DMn7DPgkSINaCaFU2IyUSIyvwJN0CDzGwCfcgLLngXn4CDj+QJGxjNj8IBN0gI02TQLtiaQCgBCIUOKiCLCVUBqOiBzCsUJkC6X+ECoauId2iCpSLKxNILEB0ITNCCwdSLFDgkeniCe1KOYgxT59iJHZicnegBp0gIihCAO9A4RhgBQaiFd5hVWq1VW71VXM1VXaX/1XVgDXTJ1XTYByO4DFs9hj84At651ROKgih4xFltB3XYVVsNAEzAgD5AB1pFB18wAUqAIF1FB0FIIAGAB2md1XMIV0jw1loNAE4gAT1Ih2n1BHeFV1wNAE+Az18RCGF4B42bAw6Yh2iV1nYoBx6IgnNoh3LNxLkMWFxtB3PwgR8wB4QtV4qlWHi4Azuo2FkNgDHYADBaoxjQmDCy1XZAh1CzNGkVgAZAAkI4B2mFh3ZIh1VD2VqFWbTBAZql1T/bAuuhKpd1h3LoASAoBndYoxwwgnKY2IrlHyOoxncoh0U8BFPT2FmFB3ZwhywInTV6gSaIBx00BylgG3dIB/6I/xCNhYd1WIdcRTgqSB9ywNV04AMkyKo1QgElCICirVhzIAQRWAAICoA5eAI9yFiqfdZ0UIFXeodj2IcTkLg1ogIq8IEUE1VoVB+O2IkbmEiYDIA/KAES4AAMCIHP7QAPKF3TPV3UTV3VXV3WLV3SrQAi2IDV5YAMqAAM4ADU7QANqAAR2ADSNd0O6IANuIAL4ADjJd0i+N3WLV0OEAEKKAIO+F3dtd3oXd0O4IDdxQDfVV7WxV7qTd3mrQDoPV3hdd7kXd0NsIAOwDSOEYgG6IAM2AAieF7u7V7ird7l3QARqAAPqF/U5YD79d/lHeDWRd7gJWAPkN8KOF4FPuDcBf/g4h1gDhiBCvjcAr5eCDZe8sXg+8Vd4DXeCtDSRBIFEvAADrAAC/Dg4V1gAe7eDuDdDTBeEqgADdheBO5fDLaA2OUA+bUAExbe++3fIuBd/L1gDiiC5C3i/uVhCyheDc7dIb5dIE7hG75eEqCAEcDd8IXeFl7eEy7e641iLUZhC1g7yo20ADCHFOCdhqgB1lIQgYCHR5gERigFIgCFRogEPd5jPu5jP/5jQA7kPmaABpAEQJYESFiARzDkPoYEBljkPpYESW6ERnAAQ14EM7CATWgAQd5jSXiERYgEB+BjR4ZkQU5kSGZkQUZkRVZlTwblSHBlSXCAR1gAQV6EHOD/APszqpXQg1lWhBzAAEpwhE7WY0loBEaQ5GJ2gAZYAFf+42NO5mcuZmoO5ERAASKAhEeoZkloZkme5QZggGmOZQeg5HHuY1peAEg4Z0/WY0ZohGlm5Hce54nBnY/yBUqIhEbQgiVoBEiI5UZgAEuuZj4mZEN2AEdeZ2Um6D026FhuAEbYYweY54Z2hFEuZkNuBCNYAnWO5EmuZEFeAEeQ5IlmhIuuZoRehHWOZUdggFhmaD6WBEZoAEs2ZAYY6VheAEUwAsg7Y734AU4TiDUgzoSolAP06dZpgCLoLqTmGCrtRwHzQU0woKb+FRY4gaquGjToWY96iyzNao5xA9sE/+szQQGiJuuewAEK2gkpUL2iVghEGIHfK7lzwo8WascUrAhHIIFnwouJgGoEsYhkS8hIO0O4KALDdBGuaAhO4IAQSwu8rpnI3qFRSILn+OvKjep+XArMfms45ok10LC63gkdsGeLyDm8UMXJxsjPpg3TUEWt1AsUMBMfhOrYjo7VHlXTIGyFsAI3RWufiIR+2pAm+L2iKIQiMG7gjo4G8ACmXu7n4Dto2gSqhm7nuGrrjo4bEG3SNO3szusAAII3/u686C3yXgmCrYIGsIQuENSieIdHsEfyzot6aIAInG+s4IMG8DYCCgA+eIQuxW+F4IepFPDuEwQZ/SiBGIQEN//wmliJQmhEB6eKRNC277ZrdxAFVNCBS1iJZLPrCU+1EMe6HQLxEe++E+8YwK4a1E7xhCiHnYBxF9/c3FZVG7eIAmzx1jbFFb/wGe+KMSzxHsdvHT/xMaRr19HKIV9uqDDxGffwzbYWJwfuxf5xK/8Jt1jyLZnyK+9y/EByW+PyLr9yAcjywEgHdjAHVOvsMW9z/A5yAtJyN5/zDKVzN69xUAwDFVABQtgCgsNzyq2DRIiEOCh0Qz90RE90RV90Rm90R390SI90SZ90Sq/0SI8ECZdzjHyHiLZ0T/90UA91UR91Ukd0SNhMquIDSICEUm91V391WI/1RF/1BujroqD/BTdIgVbIgl+QhAR6RuDuBwyogi3YAR44dmRPdmVfdmZvdmd/dmiPdmmfdmqvdmu/dmjPASgwgVvRdJ9YCUgQgR/QAWwvd3M/d3RPd3Vf92PfATbgAJ2ELDMogi1gd3u/d3zPd31fdmPPgFTFCgagAnWwhR8YhKAwJ2+HM0SwVju/8kzQgMTO0E4ogsdu+BPHARSgLze4Not3cSsITayYg1fKhShoxASQAvaEbhrt+BR/6vhETJZPcQQlTbSK+RCPOcnSgj0wBy6wvRj4joSnOQ0gS5ufcJfv7yst+hCf+Y9KJ6U3cDfDixhYAhcwAlKwgRL4uuymzqB/+sg8eluD/3mvF3Cm9yinz3HP7pXNKfK0qJW0zwsw56R18LCo752GKIdMQIElUAEc0Laupy+uH3v8Bnu/E3vBJ++yP6erWQpWaHCq2IMaQPWsXAlBkAF98sfgcQdPAK7S2Ik/yITq+vYA+AU1eAMgWBq2T8d9QAM9AHZ/HIhIaAMecAMj7Ym6V8gs8oUGKQl3AISb/PvICvzD/27CP8ykH/7vTnxouppZKAlfKAKhyYscAAEnPZNZUAELUO7XLwlP8IDN9HAY94EmgMY+YIIY8IVKEJ8dy4qBQAEKsL24Lwq6sgRAGAIjqP6KuP0X+RJDAIgo8wSUExaAU48AAQQobOjwIcSIEv8nUqwI8ZCGfAstcuzo8SPIkCJHkgx5icO6jSUnMoSwAd3KmDJn0qxps2GaHzc9suHRUFAZJhTkgKykwoiimaG0NPEV01GaERj0OJw1K0AdUFwsKOlYQ0pDCCpsfdyU5QkhlSvLqQDV8AabiVRkVGSYwAqSC0tUZMmiZcOQAFd3EpaIUSPDwooXM24c8WTKxDdbvnRs+TJmmjkzN+zZcBehX184cbQaYN+XR1UYrbyaiAoDFYHUhpzFJxAkJ/seXjXn6w8pKeU4lsq0MTY82hBNE/rS4Eoh5SPfkdKocA0NuXQpMpyXiIWTIAwaNYA0iKFkzjMPS1fv/n1myO1lUob/Cf8+/sub1Xt+6KMTR4m5MUoAWTQiWHodCcBQO1NUAs8S/czURxf8UFRJCuZYNIsd5zTkQw4KJejQggGYE0UntiihwHwgJTaPIzC4wYd2dTUUDyHp6ZPWiPl9xF6PPgo55EjyBRlTfUQqueRI+3HWn0M/ANjRJ1HQcY4WBwowmEclBhADDgHc08VsXJJEoYUTXZLCcNxJdk4aT9DoZUSJheETPEok0GKXiQGSQxJK0KiQmQHMVdcsJSIQAxs13IBDCTAsVCiTEwFZKaaZQmQkYUlq+qmmTmYGZUNSdiSIFocEgEuWCx0pUYkNrEAjH10IMmGFFK3ZJncK+cLXbiKy/xTAIVo4Rc8ShriKpJd0cJFCjR3FowIqU5zgwgolzAmqpRnxyS246nG6k6fhmosZpQqJihmpCgHhSUcyjJCDDj+I8MQDIiXmAwdqbCEFEVgwIBOaFAkhhYYcUWKEGHcgSKdEa2SwRhsoiNAFJDP5Eko6PzmBj3SHdiQHCqetwU4AY4Ry7kXevsoyzIqNO1kALtkXM86TJTriupe1GwAQU1rExwINMCDJCTEM+vJDDM2yT9GKaLJBKPYQ3AUgBktRC0eJLJEIibQcyVAfCzSywCVJuECVTIRscKtCkGSRi0QicxSKDQHM88YfASwCRB3fZnppzoUvNrNN5Rq+eEkL8v+sE38+PVTFynWlt44TjyzbZTlbOpTPEbemC5I+Jsw2kVgJV6SFEoZAIkklhwxnWp3p0WOEqhCTdIoKg9TjCBSV6263RYtYIU8APhjXCBXq5Ew449HPhHhNikt/fUXpwBNkz5b9/AkiGzrOEDqjzOb4R7OILVk8awtu0Tyl3ENRIXPwOtE5MUSxxQ47ADEE19yEngC0wwV9eB9H3FEKKXhBCvDanEOIZxE1ZGEelVjCEKBACsEsByJMew/0sCdCkFCPJtaryOhGyC2GRCIKHYtI9xzzsz4JS0RW+SCJYCWTFNqwI6PjYWJuGESnJWokXnoHH27hKqsUSoIVecckDhj/iizAIHCeawj6PJgY3YHQZSr8YkdKOJMTChCMjSPXy65CihEk53HumWFtRLQlHppRhzXcYuMSVKIEOXEiv4iBHWgzx4ikYw+EIAQfUiLHIYWwjo5UiBjpU7PK9AmHj3wIIipnwtM4RYekIALIKBXDxsARJFcZnyUvSQf0jA+BtZNMESGokD5KRA8rAFvTIFaHBZiBCli4Aha0cIU1UOIdNcxPIy9pxkgiaZI3s4gpkpJKZQZABxzYCRu4ACuGkKICThEl5J4kuR1apXPUtMiCaHHKQXJRQQEoBx/awUE76OF+s9xOR0Khggf84pCF4AUECdGGH4SBAX/Qgx58sYBQ//AACBlTDB0jksxzjpCZKyFjRMyRgRgo8xyBQBntKlICDZDFJl9gQkXMMAGwuTFyMkFEJRziCFXALKILOcc0GWIOGGgpAHzYgkZC2qUAvOMKktjIIZgwqAji0yK6uIEJTHAFYHbBOAsaDCso8cKHoAMdiYAASC+6kHa80EvtUGIAxFaRiVIUexY9o8064g4S5M2RDNmHE3LHEXlcYARbpQkTTrDIhgymDB84KgzDOapxCqudEwHCNRWSjg2YISbnyMUcBQCPjk3TRQRhWh+uMAiRzEMEYmgIKyqgLIg5Nof2wACBGCIJC4w2loZqakXOoYd67OIdvNADPvAQgAJc1f9LQ3hDG4mqhr65KpWayAEa0ICDBShkHzbwgakUwosy/OAHLODsWr3YVjC+lSQYhQgeRqBNuwagEBjoqUUSAIIMPHMmSPCAhoSaGDRI4BMSGSVj2kXEj3ThAlYLgB4s0IZjhsQRXQBZAGqBhTUwuCaxjOgjPpAGkfiCAmhoiCgoQN1YCoAO7uRDBSp7FU1UYBG0ocIMPDKYdfhDkbMohzoFQAuH4IAIU0CAQuJRhV/I0ZK7yIIY5KAJT8wmHlq4ASAY0QRRBCAdVPBBPw6hgjJwhK3jjV55jejMjujhAld4JEMWUIEgTMoiDQBBB5BXPam4QzBmSswbIrDBxL6RsQr/GcUkPmIODoBgTwEQRAjO7EqI1EACByKgBS5gzCvSrCOaiMASTBmAX4AACBykwQQisZFy4LS1WGSIL0AQzhaAgM0MUSstIzKYfERhqV4aDEPM0AkWlIA18EAB3CzJEEIAQZ7CGkUVGtKALqADEipQpCCY0EnCtgwxX/5imEkSV47gwwJI+EhnmXSJCbB5I7CEiCcq8AQ/oFMh6HjHy9RBhAvUg4M5DIAbPlBXiAB4MTnwcy084GmPyAMDFBB1ABjwgSc0BB0KMCaumbaCD1xCIfmwQAX0sWh308OeEpHDwCwSig+sQNajs8sEvtCQHkgAAg0BRQ+G80GGZFgFDfnC/wcssZE5cFkKuOUIH7iwrYnYgDSaOIEncAEHuNHwATXoAyUSkDAgUJkhdrBCIspABpUAYQ65TI+Xr224bCuITlOr70T+QIEReLxOAVCHHjRk6kzNQQJC81JmHRKKCnThgGV8gBXAKxF6YPyAtlUJGygQhf8qljHpSUNCHCKPCqDUI384gQfcEoBMhOAIxgyAPzDA0lZSJB1PqACVA0AIElRAVQiWgXU4kgkPBIsivNDAvilyBgpQwdiHIIXzKN20ALw5CWj1ggRK0ZAkjIAsqByWGECABa7hQwNE8LpC/hKAFPzcIvjQwR74hAPsH6IJPvjCtH3IkBgkAS4q6AE70P9BhVU4ZAdDwAGASnSDrZOIIfnIARfkgBKIwPyEm9hxC9l5RCH4gA+sARIkQdtFxCCEgAjoQoAQSxYMHcu8gARo0lXcQwvgUnqEggU0AXNxhBgQwS60iB8QQQWMlh4tBBBggBYwHmZQwg+4ARtoQJg4BKJ9m0f4whMggfIFAAyIgAkQWQC0wAcEmo7ZlDwgAQlkB/GZQAcg3AJQgNBYxBZIAC5RBCZIwBZwRBqYGT0oBBqAgFO0U4lAAgaIAJGdQxRUgOTMgwVgwOehh1pNxBYQgRXMQwAMwgYcgQtIVgZowIlwFEigQz6gHUS4QEwpBD6ogAjEnjsFACmkgXDRQxb/zEA6fAFLKQQPMErFlQgabFj/BUA+qAEPpEEWEODGHSC4JGC7JcAp7IANHAEEdoQhVAAGsNuGbN4FrKFNMYkNREARNgQLAIAbRIQpnEATuF6vBIANXACQ0U8SWIAjuF05AEEGZIENXgYl+AAbqEEGhIhMgcAJBE5HHMISKEENKMQNMEEJIBwVRAC8zF1D3AMSnAC0BEAkOAEUaJ4lTIAicoQSTEAkUgQnRABYWAQbHMEKWAcKSEADtAhDbMITmEBMxcMXIIEXKIQ9UIAFUIVkBEli9AA97sYidEETUGEfVMAF5MEPdJ9EoIMv+MIvCMIv6OQvfB6hNIQ5NcQ6CMJf/11gghxCFuwDCgxMYuwAC+QAaSTGDZyBRYSdLMYMLbZbWFCSRQQBEZxA+HTEHFQAj0jEOTRA+GEKHEjAQSoEDUjAej0EDDQBE3yhRMSSD4SAoU1EAxgBCWACg+nUFBiBCjgPvzXeYphJGvhZKFCAE9RbRzjCFVCB5GyBDlyBJhDQCVjABrmGC3gIRQBCE1SBygXAJ6iAyAxBBPhAQ7DDGUpEPHDABfhXRYjBB/xjRfDACjhB1gQAzj3Qq8BAFliB1+EDFAABFXRMH1gABfxmLsBA+tWJFGwB7viNCnQB5CzABVxAPnCBTUaEO0iBBWxAFZTABXQBFpQA9g2bGZiBGP+IwRjIwAzU5yco0lDdwSFERgDoQxP0gQ6sDEPYAhUsABrQxb4ERnhZW1ZKz1Z+xLaxBENkgggYwUV2xAuEQFKoU0TsQgY8UKVUQQXEo0rUAAUoZgDIwBMwwVhO4xVIwDbqyhM4AfZFBDs8QRVYgXAlpnv820PIAAYsQZpwhCNUQRTYXACggAxIQWDQQxJsABUGwADWGUUQwgqYwRfgQgAMQRVwwQ0ohAx8gBXsWACMwhIcZUPoQQdwwJ5RxA1QwBfs6ETMAgpswQawyDlUAQXkTZAMxhYYaWDswwbIgAqcYQJkAAYkBbFQgG1OxDpcgQtowTZGwhVEweJtHgd0wC//+EB4QoQ5uAEq+MIuIEANpIA+QEIJZAxDIIIO6IAapMAIRIG/9EAZAOVHtMMKLCQEdIULPGQAJEIX3MIlqIBo/gETKCFFYGWDnsuDXmCEVsQcJMER5IuNBEANTICqSkQfXICkVEo5NMEIxIVKVGPJQcQaeEGqWgSZIsEHbCZFjEIJSEG3Dt8dHIEVWIHD8KhLOUQNQMESMJ1FZMIU7AA4sgVDmUIA2MMRnEDJBIA7RFpsTkQiKMEQKAFVlAEV4ADkoMEIdME1dgEIACNECEIJlEBlVQQblOx0QgQ6VMEZiCVRfUEG6CZFeAEZuAFdJEAJQMARWAginEASGAdATgCJ/07EPHRBJ2RBxnDCFIwBFSgEC9ilInCqRzBCD/zVDcTUEKAjnfBBG7DNYCnIVbiACTSAPQSkYPICGIyBHiSAEkgKO3zBGuCDL1QBhV2leC3r4jRruz2rrClEGFiBCaQeR/hABGzm6PxCBYgrk8RDEyDBeqXkCFTeQ7ABDpiAu/bKO5wAEbBARYyBCjRKUGJRAOSBCQDBFwCivorTQ+gAFTwBi5BlCuBAybFDCchBDyhfICzBFZymL1iABdSeRDhbHDyBU+RACsxAuebAFEDBReKBCICAND6EUi5Bw1KED/BAulKEO2hBKChBxuiBE6hASLZIOmDBJmxBdizCEijCBv+oCiUogRLsWSR8QLLpEAJsAANQAYBoAhXok5aawgqowCT0QKc+xCegoufAQCE+gg6EbT6gAtjKElJ+whSkAA881Ka5gRRQAQskDALkABVQwRhQ6YLGot5iCt/aiN92UAC8QBWYgIJyhBZ0YIUphCJMADhWCh8AU6Y5RBQkwRI4n0PwwOUKba/8QRMYQZRKxBnoQBk0o1oU0T40gSlkgcg6RL8pRrv8QBogwYWi4BSQQqbJAwcwAg8o3yBoARt8AdcsQAdcQItKxCRkQQJsQPjowBbIQLm+wRaogHHsAwmIgBBIxCNYARWcJktQQXBo0uCVQCZkASUEQAI8gQ0EXov/3E4isEFCSMIS+EITiJom9C6XoSYFlEC+SoQveEACTAG8iAIVQEITWE0OqIAUZIIBe8QgqMAJygMWZIwanOxD5AMPTPBKrIMesGNDmIM9JAeJ2MMKCo6ypjACogQKO6tXVkQN6MAKFGJHmEAIEO69WYIECFal+IFqNoGw1MIAX0E8QAQVwAAUuJyNIEIWTMH9El0amMJppgdD6MP8ZsE1PsQWFwapmMMXjMEJxKiEKgQc3MADdIU+nIAv5MCGJcAKzEAWJAckHAEHINZEzIEK6MEJiJoPxMAclKsUiEEVkAYhNMERlDJEUAIV3AAW3LBCqEMWzAEQeC5F9EEHLEAV/7iFJVzBFNENRSBAEeRDGljB5lXQFbjFKKAACgycKHhAEQAsRCCCCejDFBCIGADBIHBA3yinDwwBDxwwXToBC0ACKDSBNrkARUcEMRuzTRQj3jIoNefMCvdKC0vEDawBeHaELnQBCXQqCRoBQDFJAqgAGSxBnZGPEUzBFRyYQ1QBC3iw5QSAJXzBDUBBRfhAKYhCCUyELyxBGGiBxq3uYjmEOiiBKIRHu7FzJlxCyR2CE8xDGpxCAKyCCnzCCgBUJ3TBCtSoRIQCFtiBCsDLD4iBJoDjMSgBJxAU8fXu/UIMJ1DBEKyAlo6OOzBBAR/wIJxAIKAAlZlCCjQAE9Tbkf/oAwnwQRok23PDA+V8yRvswNNS4xM0AWtMBAM4gTwAwdaZAhDwAQeoShSwARu8wA6o9UNIQg8AwQ98Qi3QghCkRUT8lF0bEUr6cyvN3TTvtab09bD8dUT4gBmcdEf4wQo0wd0OX8qUABNEB5MogAqUwgqsIEO4QwmsARYIr0KoAAtEQRhYxHCMwgoMwRNEoEOHASY4gYf486+uACs0QV9qMYrKEGPFQxJYQhVsoQ6Z8SFYQqY1AhO0Aw7ERStggSOYYAAsBSN/rs1pNhXMQfzOApc3ABe0QADQ9BmowFUcHgukQBwwgdUEST2cQCLgQO5FBAMolXYHwA6kgC+UgCX/SiAH2IMMnNkoWEEtfMHK8EAPuMBp+oAWWEHFTcQiQMEuAEE8lkEKvAMUHIgUkEIbyIAOOLhDsMMvKMIg7EMCsI1kBAILiMIckMGgzoEohAEn4Ke+fNDhdVneijjMkLgOmTjpbt8LrEHRVgRGp8Cl9ggOeMEKPNqSNIAWQEAXxF6tlEIW/GbDIQEERAGB2MgMSAEkMIGcRoRxqve+O0QirICUzTFOZDkpMZYZSwIVNGpF0IMG+IMiNEE5OEAXlEMOoMKbY8EvIMGenMGEoeJE4MCcF/kXdELEr0MfbEAgbEE8fgL/roCtNsQoTMEhlEDfCFVD8EEGCEIOcG26XIIS/7hDCkgKELDBPiABkEOEIhhBOwzBmbVAsk0BR/1AD2CCCjAEEGxBCjgyREwCFNDCswQAFyxeFxRyCrBANu3xR4zCCWCBaV5BVTWXQhxCFKQAVds9CkQBFeSAM2cRfoQ4tVeKtbsdto+uFrSAGEixRTDCCqRBucI4FaiBEkwyk0yCFjTAEmSN/3UBKCiBXknWBmCCDrwlRaTBDbR5HrBEFmhCIziBBY4IIyiBmdM4QRt8gDHWHmzAIrzBCygEh07EPiSBLyBCF5xDJ3RFDYAjDaiAO2TAwKjBFNTAizfNVahBDcJBdizBJJCsPBgCgOMAhYnCFJj5nDAmFfCCB7BIkP/8QQfwwQ04NW2oj0JMUfJ8WBTAgDwUwZVHRCIAxJF2LKwEkHElABAzAVLEaLCEXQAqLbjACHARY8ZNKwLkqJIwSoAsngJ8AYUmCo8YGVm2TGCkUrx69uzdaxeAjoBZs8zxWdcyY7o/P4EWNXoUKdBDGvIFEJAUalSpU6lWtXoVa9VLHH4+zWr0KYQN6JDOuojkEwwdUTEtaYGkXEuvR1owCfUVL9JOTA45IeQ0AKEukJYoYrkuyaQzN6L2iEHISR+j56AI+fVEMsunQlb8MsEIaJofea+y4ZFRkIl9abZE/WVkT6ImtjIpCVBjSQA2KASciBSARw00o4GaDbDloxv/GwKOMMJ3go+DFbN0tCZDZd8JQYAz6rCCjoMj7i0NmXCnJstF4y0PBvixEEiMeCQUII3kxByo9Ds+pigTwAtSCoFiFwGyyKSNhYzShCNTPvohpC9aCECLIEChYgoxpJpDDaDK8eqiTNzgBIG4MqInEjZaIGs90rJaqikQXZyRxhptvBGjrbq6MayxygoglxIeIQWIqEJpwpIS4hkPo3ZMyEQFFnDMqwUo9IBikQDMMgSMBZwQLyM9jDjkhhqimsKFQJ7Yxyh2OnAEECd6kSsAUJaQZwMhQiNuSqNMy8gXI/goA4uLZNQsgECO6MOXI+YJpYsAYrBtizQCWCITiUCJ/4Gjo3zIIQAfalAng0Pa0cAXCLToSAXdvmCnCAaYDGCHkJq4RMuiEjGCnTFadUpGAZ7aodAfcAjAChbY2SDLo2oLgJLcdigIiNOqYKEPDvJRx4hV3DDTqE84CgUhFMycYiUVLFlAiRUsiqqBHEzMSNj1ylkAhy+mwMEMGcx44woeHrBj1j6TgrFggxVemGEXdUw4rx7JOsosPqAgZA6EoHKBikWc2INJs/Z4QpEf3o2qjwUaJmWJekpoBCNGmPjDBE3GyweKBNLoEKos5tjjiASMcoeDRvooIZBcDX0KhibO2cCSPRtm6U+MDDFCHlGaiOoQI+7Zx4h5ZMhtDo62OP/2irvAgCSG3I7qYY3jgKiHA0HOSeKQTDhKg6MdRoMiap0y4oKxLO5qMSNWmpgllEKVbumH0XqgIgAsPgnAiUmMMutZOWzjokguKFdBE3ky8KWeIvbZIoWjXihI73OikIGhY5WgpI8TkHBBKltycCEXdtxx5510Es5HkhbWyEGMTAKpZWqpED40+uqtvz6jh6knTWKknvoFCj6G+CKqMnz4xQlf6BTkiX52UDCqHTKw5fEpyeDtBJvNmqSLepDY5HH6OMIvYBCSpJyjBJnAQwkQYZR8mCAB8zhCAxEXAEKVYwMPoNdFhnO9ql2kAU5YhwO6kIv6AeURTXBHH5KwBzH/5AYUqwLCGCTCAgweggVuExZGdhiAN4AqDVXYRwma0oUGsGBv6ekBDQLABEyoJyM/YKIVRnEUS0RKXOc4ShTMpIaCXOETAoACK5DSglY5ognt4AITJxcAFUAAHRMUxAbcgZyjwIB8UErHFCwCBCAI4AkLYIcJMDChqLhDCxVAQhaUoIQSQOCE2MPK9CRZSUsyTHs8CoBYJoaURRgBD3OIFFTYkIZ8NKEQBUvEE3hRA1Al5Sm3wAAFJLO9GpVhNEngBGAs0QR4ZEFK6ynEE/RgCilAxU2SsMUJGmAUsAmCHmMqyg18EIAmgGKDEuFT9D4YAEg0QR0MaMI9IIaRBvjy/x0ZGEQMCvUAJbgjCryLAgze0YFATGIJRGGJcdAVgDVQYZj1cOMlZrAqF6RHVCIZAlCqwLsqlCKSF5mDbSgBBnjMyjhAsEEAWvCFdixBDgHoQieQQgryHaIE9XiPpKiACyf8RgmRaMQTdPMRo+QAdlmQxxfmEAAU/MAcJkiEMJhABENCBR1/CAQhmEoIBZCzh0uDIg9tWUlKXhKrWaVRJm3UPYoFQAgbCEAmkgC9pESBDHh4Asz2CS0ToKMMXIDKUx4hAhFAgmE5OBYTFnoRUZQAWUe9iCGawAsxtC4pejhBljqQqaIkoAQISMcGfkM9HcRNBaJoyR8ssM2pddMTkf8yxBHYVNWntKIJ7GhHBxSBhrU4on8p4J0baLCPI+gBEVAA2aGMQwXexYAKkeiCOyTiiRmMCwuzmMKErCBYjHxBSkBgjFFY0CpJNGEXTqkgsiAKJT4sAa9X6KtRaFCoruljClIqRQryUQShqeABmkgPDApilChQ7pz5sAJJtvADPRThFxSigJSQUo5f9KEc+/AFIBgMCF9kN6qb2+4lr0qVQ1VVKhHWqlVmASKzeGXCc0WUVLhaI68iZQ4mCIAmNmAOqKjABbQogeYqCIokBIAUNoUKy7Jwsq/iJTgBuALvLuICIzBkJUpbABTa0QK3bS4AfzDCIAKwAUjq6gkg88D/E4EShZVo4S4sKUQI5Gq9bsaApr4wQYA1jBGz2CkA53CCJMzw2iXkAwsk1UEODtGEXOSDA0K7cABo8YUwBCAUKQhFFuLiAzHYoHVDUAEdpKDZ5hoKI+hYweW4YCmjyCA9iDBCZhBVjitotgFdSMARVIYF50KOfH/gQCEUHQBRUCERG9BHST4xCvKZcYshWQATBKEEzYkBCIpAAjmvMOCkrEMHQ1gHG7JwBWtfoQtPbPOGkVJhqHwYxMLadlTEnROpCo7bsKRFD3Wybgx7TyfiHjdQSkyjEx+FFAixRBIikhQkLPQEPQWKC2zDAttEBQdpMINnA/Chpc17KiigHRU0/3QRGZCPDdPFCCYAO4QTYNooCYACm7ow3pY8ogTzCEASbAYUKxza1S1pAAh64MHTYCQNqwIEEv5SVbN8QglPMQkONloIJyjACVFLwxQIU4t2cIDGQGkHFGw2hymQwnE8WIylMqGFnfZUoyAPwDtmHIAeFEkuw0rP1eYU4aecQwUL/cURIDFrhjAxxD74yD6KsIgvADATVdBEF+TBEBbcoCCDv0lRpsBEBjCBEEwQzxh8gAkl0A8LzqZYOebBDnogghf0qMfoeXETc6eba0wpJ1BM+26wyIgW2US9VSBOsQ5fpd4zurdRajCaT4IMKerIXwCMcGigkKF1c7hxVJQwB/8IHIG3t+dhXrBw6DdoXA2nQQPadziKI4zVCHfQrlEYQEw3EpjeTlBHAFZgcoyUQwnYVEhLNCGBtZj55hfZQs3p0QG2giUAXGCUvMwHIOoXmKABkkBllq5sLkILFgpxzMIePKCZKoEKfMBxcoAL1iBuIGEF+mAF9MQHeCYj9sADsmQL0qMljKMHpiAAyqO0WsQs3KEJ9CQfNmAOBigAuAgpooANAuA5rigOAuASHuoKtCgFxgAHjkkRTqApikIK/oMQTuARTuAQcIwLYqC+qkACNOsoeogQqECgWKIcaCHEZi8jvO2ABuEeCCEGXCAGYgBXvqIdEmABOCEUnKAF4EH/HeYFDYvCHHCBHQChBbZgC9QglbBCcMpBE/4vKnLPRXavKN7AUhigBPQgKeyhBJrpCyquJXKgDVbsBNYvKe4gCSBhCp+QqjyMNJSgitwgbi4CCMxkDFTQK2TANh7hCegBWIyCEvwsAPqpKDYBCqBnBbzwMJIgU37gP1iCBSLgB/EvI4AADQLAdMAEAG9glPbsDSyCfTqhCbbjBaSgBjrl6lgvUTbgLyKBCqTAcfjGBywCEZoAMjTnDcosIwrhBAAhAHLuKH4gbvgulWTk9uTBCH5jHrrADJZAMvwIKbzgDAKgHVTAFKwwABhABXzgmNzjB3Jgo/hOaIriCyCKH3Bw/x8DwBOiYAoQawoiABmLAt3OgQ0Ejl5c7w8PIQNipCg+jCXCQAQ4oANOoAm6IAkuYA0SYWLWI8S8gg8wYQVEQAMuoAKuAAmSoARQIBHmZSfaikdsMiPWw/Ke4AmSAApU4AqOYARsQBXFDiYzoh52oAJGYAc0QYtypaogkXs2yUeSAgtoRxGOYE6QQhBKgMqioBq/8iKkoBobIKWgIlB+QReSABsxbRZIoQfIKS8mi6RqAAUw4gogqgVUTGnSQK4Q4Qn8AGKeAhRKwMW44DCBohQAC1l8LFfcoQMkISEikiViAABaQxoNxdAk8gQ0aClvgwku4gayIAUW6h5OAA1UAP9k5gAF2OBXHmqqMI0RiCgAEmEFlGAHLG4FvEBKVOEJIMEiqU3sKIEJsqsFVJArUYCJ+IAEGmjQ+KAIEiEA1uELgCALyIkHHMcopGAhaAEFoqAE+DERVGAFOpNWpMANaCceyg4o6iALsIkXuiAHVsAeVowKruD+pMAl4e0paCEMiuALysAMaAAHZAXd/tAocFInWU/cLmIdzEADLoERFKEO0KEM5wAKOIAJOuEdenHe0GENSOAEvsAR9sEfOsEc7IESNqEKRCAaiRTcZlTe8CJLA8AcgiXeEBMliUANLEEIDoEUV+wINMAMdMFQ3A0my8E4ysACFFAKMCADkoDLtpT/JfAyYvSyk9AxAEyAdwbzL5DCPFWODUIRKJ6AdxZBO5OCEk6ATU0A/TKCDzwgBDYqL/ZBqAKADDayHI7gLjihBNbPK7hgowihBLYDKWDgyIDjWIqiBmwKDnSTJfLhBPCzmy7CBiSgSrkp/9ShCXYJHZZgl5LCIy4CBp5gBTLFHk6AC6zAhEABC37AN6WLLUNkuALgF5rgCZoRUpaApPIhCUpBCXaNUx4uAJCIDmwtC2IPUWjBCngnHzSggejkD9yLS6PACawADwJADDqlKMohSi5CB5LgCgTqF0rgCMoMBrCgChbKFiq0KNBBBXoKHaTACVoqAFKtC95FB0KUYrzC/x04gRRoAA1SNgdAo0VdVCly8gvRTQZIIANkpSj2oQaCksrq5ShsQAQqod+KAhIuoLYCwXhEVEuNIx02YQlSQAfEgA/UNSPYQQwsgDYz4hwgoAg4AAYw0wzbMgAgCwNAYUj5UA9s4AJ2AGR2Ynv4VEv9NCns4AggSU1C8iiG4PtuA+1uFQmaaTDt9ihcQDZRIBZZIgiuaQP4ES8gwQR4IQBg4ODcIQk0CBKeQKC84gpeIAD6wAgoKMRsgHIC4AYMCCh8YLpcEygAIWhEd28xwg1CIFar54PiwQMcgEsfMFtZog3uDwJOgAIDIA+WAArqKxKywAo0lQeiMVhsbQViD/8fOiADqmisGJYSAiAemkAKrOCiQmEFoEf6cGBVVmwJFm+DaiHuAoAekGAIV/AFVeMieGAEpiAuLoEJ8sAozDfMbIACquAn9KEDkoB21nUFVGCXaOEKjK8l4IEJNAg4LAAIYm+VoIDI0EACKrUoaAEP2EEAasHF0s5ljaIQPOAMyGBDZfQizIAIRGG36kVPcUEFkmBItcSWXOACmomqeBIjOOEENMACWoAdIIEfCEEUEmEfHKEBSjh3q8IrHOAEimAH+DMJSEDgNCwGKsAM6lLeBOBrA2APdkADTOAJQ8wchoAIvoDUMmIORAAJ2CSi3PYrJLElAKFTOVdlkKIMEAv/j4BCEpKAnOTBBCoBKragmiTlyTKCFC4PCUgKL0LzIlrg4wIgH4xAZRIBClDzKWrBCHqqHk5x/BhPN83APVtCCwwJB/AxIz5p19IgdDNCCiyABIEVI7LFMILxasH0B3hmASwAAzLDB/b3IhhhCT72ffNvg2zAbeCBBCyAyxohCUQAZmiBCzAgdB+gC9hUabzjIiaBCVSOIAMAHo5Ag96hCRgYKD7pCdMABBCrEDhAcYFiYgUuFCTgVxSLBEghRIzACNZXCmagKOgBCtaXBiKgzARhBDzgykbhA16tJdShB1gAD1KgCbqzkR4piT84USrgAoiAT1rEK26ACHBTaYzj/ymkLwDmwQQ4oAYANyPmAQYuQJEBQ8OM4x18QRRGoKA3oAOegAREIANEAAtoIAoe4U/PMClI4QJyABMvIh2MOgWONnsuIGqusyjYYQUwYAtULmE84QJIoarqQQuIAICmL0e4YvWyAo5ZwgGSgBfT4QQuB2zN5SJGgaZa4gW25iKMQHqRogneBRNOIIYzInklonCzAgcMaAhMIC4GoQQCjFBrkwNwRR08wGbG7RyQQOA+4QjONCNuYQNazgZaFyM+oQSMZwxkEyPOYQlE4JU/K/9eQn0YIoCRIkMuQhBCwAPoJwDQIAI87ReQoAMELgdC2c2Ow3Ge5gNsNgEswAL4gf9XJaDMQgiqjONTLoIBFsXNzMIs8iED8HMWvsAU1ANxKoEJliS3AQBc9KADQKOTMSIeoEBzvOkDNtIcugAE+ooSkqADbDYFwKUlogmv6mS3L6IeRqACABklJ4DIjqIWFGAfzsEfDqEQFCEREmEBotYrGWaHxI2oA+MCHsEQRCBL2mwBPlqq21IPWqAJMIDLMKIRooADNqCvLjwALAEICOEXBqEcQOEG/GAQkoAIVKAIsmAtnaIcImELYKAK3oCNvUKLo8oQiACbWmIQRmCVAyAdzOACahlQ6yEUMOCVYHqH0OEEPM2EA+AFLkAHxK/DesiNzVovjWfeYGCUmsgTNcP/K4zAkBwhCaKWJVLgy6WAiZAiMrnMMxKxD7DaCXinaYxii6WKqi5CCd7FEp7goiyxKfiA1QADaGTFHEqgp6qKD5KArRLBBJj70cnVZmuAfFqCBg4OAkqAuDBiHZpY42D5l52Az93glTpsJxBHAGIuAOyBAo7zIj5BAt6lHkSgAn4jAMzg4ICiB9wAI5QABJImAO6BAjCAYAIwAt7lEEirk8MuAHzBA3oOSwPgEE7gDwzWpjQcI4RgCWL4BT6AJMbuCUKqXlqkD1jrIlQBBMjcByTgvxPhAizA2qPgmCrIF4qgPgJAES5gxVXgAxJxEkAgwY2CHuag8AIAD8bw4Ta8/2GChaSLwh8yAB/c6FcO5RcywAUJzfVIWgaIAHshABOsAANKIBS64vTwAgHWfR+6gAS4oBP6oB4SAAiSAAO8tQqUQARq2dzk4QTqy9xmdOWh4Ac04Q9S3MDnais7VgRUIBJM1TjaAQs6YMi/UvoaQAMovSXaXBHhVrZ9EzgGuyYDYB84wIb9571P2wTCLABswICqahI6YNcCgB2SgCR+QQSchheSoJkawQNe2yra9fAdCxJKYEMzAQkuCg8yCDAOwQSsHQvyuigWwX0DgBc24BHaCqUUlwVKgHrgIFYVoQRQEyMCoQScIHZtPQAmoQmwOgda3lBGPj+fQJHLYQVi2/8QLGAOV04CrrBOlKAORgwF4AcUTIAXrdwIbBUSQoDLfCHQWIIK7DnYO8CG2XaHKuH3L2IIwAC3Z/QiuHdiPAEE2OocfObRWRwJ+NwdAMIHpAACAlDCMChAgHoULMRTyKOKQgEFFQaoZISewnWGchEM0MhLPYWJKJSyiDJlvij5FHKSkZJixZQ0a9q8iZMmnkODxBGcafNQkZaKiDT4aNESESnzCM4CilPArADoFoHiciKDlEgon07NSZPi0wCzxlrE02IKEg0kNGiRwfXcVE5EUuya+DOAuiYn+EwsKJOkGiBFLijxizQnncABEF0RsaKpwkhJnOi7KTMwrxNJWAD/qgchyowZSdQlBosaZUEIJ3z5uHRT3YlQFmVItFnwk4l1FlWUSZmIwy+yAVoY+WuTSQqUX8wEwOJkw5BGSTSeMwEj5zkXVRTELMiogy+Ff5IoClBmiUJ0GzgRH4KkjkIpMXK6MIJOobkStC1SDODJCR4FwIARfaQUDBMsKLRPEomg1MIKOPCQWoUWskGhQp10cYtCNDQRQDq8AAbVHkkw8tE98hDHTiUjFTTHCfZM5sSMNJmjwoIFsZNPORPx4Y5FfKDRUgD3FHGUVFRp0Z8tSgyh2n8tLEGLQg08oYd/gbWggkV94FBkAFWQEkA8+QHlwgp0fGSOjwWhA0iHCsVQ/0ZFpGBhkw1d5kUcQeX85wsF2eXEBw/4KBTKDV2JZWGjFX7VmBVATPHDgZAGlUGROnRgo0L+EAHTaTmVRRZQ9CBQ0aWOWlTWV18BtcshiwiSH00JmJABVwp9FcMIe5xWkZIK9ZHIAf6hBtiuCgFihAZrcMLGBTqsqKpqNO2jwwlFcHBCFFlQ4IE5oq6aEqkFZbLBCkSIYMmxFkFSxGUFhYKEjzd9AQRKL5iAC0o1MIFXEEX8ehMmIgxX0Rlf5MNBAqCUwMMKFsVQhEYo1eNCJucEEAMGTYzwiLIV6SCxQuWUAGUTYlj0xSkW8YCCRXCkYS1K8CSxskVSRMFqRfhaxP9LEkehZE8RkyjkDgcPoKTEGGNwQW7UKGFokQsgKsQCwGZckIlNQmSU2KvHpvPQVIWYMB5N7EChSVh92pQOE1D+R08SAymUwg51cMEDO3jpcIVFvyRBCBlHUIJcDFoom5IUNshGZko9WIHXrmapqqQlTTxEExU6sFp5Su0kUR+hPBAMChpR/id168fewwRt6uQwBVRkVWnRIRkcGMAtJjQxUgCQiOAD464fjzzo6dRwwYIWNUJE28lbGI8ZXZTABOJRs7NAJDbakEE640pNi0xTRRLCCH2IcgEmPSukxG0FRZLp6gptQsSD/92TwaAB4LEBF+BlHSZQQQI29pOKvIP/A2pADiOQ8IYSBMAeHKiAKCwCj9/x5n4dKAIJUPCIESyNebApVQD0MYJOoEQHPOADBxbwkU8U4UUmcF4AyGCCO3wnALyAgglWpKRKEKESKfkFBhqBFCeMoXKm4EDwzNEEGxIiA4GQwQ+ml5PMKIRqCqnB4hSCiCSYAgkr6ADBUmKDkjmKD0U4j7V44QEkrioKPAuAjwSxAYShpwttIAIRBGgRKqhOIfcwwgxEcIHd5cUHMcsTFKYAAgpsISUpwIGjCmKIDfwhAOVL1RVaUCEvhAonfHBDlgLQiTPUxHZYRE35AjAEL1ikHitwo39Ilbv6MagJHeDCFzCABla2cpir/9KECN7Qi8YAk5gWOseapncJDvBGmORinUIcEQElKMQFIoCBZBSSAxIAAiW7WMIJfDCHMClkFCKwoUU+gQFSvCMAMuCAxSyyCCdwgAmYAMoeUnACeNBECSCAUgB+0YAgWYSCKfARJohACgHoQQUhmIJFWjACLuhCIXboghbspRBMnEAHUKhVAHJhAizgYhVFGI5C5HGCL3iCEsNAiTxK0IR71kYEVEiAfpQQOJTAAAM3KFIjRPAJlLihA5fAQwBs0IUAoOGKzHSbRXCwA4tMwZIWqcIHnLMBUNKECzkgVzpMINaU3GqcqyqFE2YCiZxaRBAZuEAiLqHLdXRhExahxf8KQFCDACzBDRZRhxP6UxNFUCB/kBgBISxSjhyRKx8eeFBKbnq01KRglDfRgxZGMQlHRKELkLjEJSAQiKo26j89MAVSeDCKmJSlFu4IEiNIIAhzvAMe59iFKJ7wBkWUQx3weEdtj4vc5Cp3ucxtrnOfC113sOMY7sDDO97ht0cYQQQnIIIP0pGO6xo3uuSFrnjbsQ52jLe8yBVvLt7h1E9Ic3xRK8g5ihsACFAgEwJQhwBC0YERsKEQimDCBQYiXnjYIhAvmAIHMLCGQehjDke4wBAEsI4EswMdGE1CCi6QVHcY9x25IFsigEAEKMxhAZTQAhE2ELLiipcOmcABPDL/LK51yFighxiBE9pwAZiISw9RaIAA3sGbRBTBA6GYwwYy8IcjXzcdumgCCCLqDnK8wxyHQAIityCAdlxXeEZwggdIIAMGQGIIJ9gALwIg3uuu4xwNaMIFcJAILIwgEFJGMi9C4QEPWGIOF+CBOTKcC1scggojSIIPLnDBLUBNtaq5BTzaQQseAMGhRJCDANyRC3MkAAUJGQIGHGCO46KjDx04STvY64523JADfLDFcdlRCx0koR7pgLU7BLCIETAizOXgQhNwwY5ctIMdmAAFOgRghBUECXoDccc4bAGDLPgBlhbgBG9CgYFf6Fa57JhHKBxhxyNcYdnmQMQIjgZr/yS/wwhSOIeYaysAR2AAEeWALjtuYYXS4YQdaLhCClJAhSocPAVXICI1KV2zFLRNSTcIbLkC4IsVGKELG6CACZjwhCc4oQsqyELJm+CElKt85SxvuctfDvOYy3zmKvehyEWe8iwwwQISqIASlhBynNN86EQv+tCf0AURdGBjDyfXKkpQgi5koAJdaMITmqCCI1DgAxewwAcwoAInCP0JSsACFrQ+gQuIIAQfGMEVmMDyJzABCycAgQQssASU3zzlXciCFUbwgRBYYOsWKMEKxK7ykC9BCyCPO+JVwIEJfMACK4B7CZ6QhS4E3QlY2MAEQACCD3QgC0J3ghI6YAEmWP9d5VlwQggmgAQwiF3kS6j6BkIAAgtUIPQeOHzcsa4EC4CgAhMggRZWjnQrlCAEFGA+EzQ/e6Amwe4UQPoFJglxi8DHCUzwIxPAQIQKqB7xi6+6EipwgS6onAkd34D6jT5yE1QgCXBPeROgQIH0w98J969AEVQPBRUgAu/HfyYHBUygASGQBEqAAR5Xf06wBJnXBCbAfCWgBESQejCHdYfXBRgQAifwfCRAATi1fx1YASSYckyAARVgBCgnc1ZnAYCEGQVRDjVIETWIg8mSfZgRAOaQAkvzHzSgKDWjC5DwAA0AA0QwBHEQBE0YBJQwCZTghFNIhVVohVeIhVmohVv/aIWrUAMW4AlMOIWR0AgL0AgOUAlcqIZryIZt6ISMsAUcID5Ntyp8YAmW0Ag6gAEPEAlNSAmQwACMAAlxsAANcIWU8IeMwAiRMAkNUIhSeIVxwABmOAlYSAmOUIiQ8AiBCAlXOAmVqIWbyACNUIkMkAIiIAl96ISVQIgN4IiPkIZTyIqM4ABUWAmRMImREIt+mIqOwACrEAdxUIawiIWS0AiNEAeMYIhVWAmA6AgNwACSsIt+uIkLwAiSEAcr8AY7qBC+gAlB4AhK0AWPEATJCIp+WImVMAnIOI2TsIjTuIa3eIZV6Ipu6ITrqItNWI9TOAlxcARdkAiNQAm3SIu7//iJQXCLDOAICBkHq6CFiNiEjkCKCEmGaGiPlfCMtkiG55iFn3gE/mMTuHQTi8GNYAEEc5AXOSBwFkEHz6QQh6AB6lSSM4kSDeAB30STORkA0TRN08MY+bUBJqWTNCkKJjCUQ4kGk5Z9QJEGn3OUOdkDQ/iUM7lZWRQsmcEYdAhx/6EGivIVVCA9WpISPWYpU8mNjkACOmWWEJcJRdCTyUMic8EBQrmWlMYCSVCX3JgDdbSDrNMGSpmXzFQRUcBVgUlpVvAbhrkqFdEITIAYjNAEp4QT/lAEvKOYzGSTOHmZrSQEG9AhWrlaxAEBSUCXmzk9RWmazFQDgElpIslFqf8JlwrxAzQDm61EBStZm6hxA1iQCXPABCgJFvDQADqUm8ljD44gPsWJPHqwAG5CTFPBB40AUsrZOggAQ9QpNYDyWDk5CD6FnfWlEIaAMKD5nTVxCGxVnjaBDpqAAzcwNFr5k+lZIUpCnvJZE15BTNZkn9UkLPsJFoyik8Hin4/yKvo5oDcRDsZzoOVSLTTxFAa6oA76oBGaGvGJRRZKoTwIoRmqEMUAoDm5oRyaFxgqok7RoPsJFSdqLWZRoi1qISLpoiUZoi1anxcao24jFSx6ozvKoz3qoz8KpEEqpENKpEVqpEeKpEmqpEvKpE3qpE8KpVEqn0Chov45FfB5oDP/0Z+VQ6XHs6UwGqHwWaOX2aCX8h9fWqWLyUpjmpsC+hGYs6Csw6Yl6ibpMKe1SaL/UQ62cKW0kKaw+aBXKhWvtKd44SPlkJxeake2oBB+SqFiYTtKYg6MKgCLcadrqUVaFADncCbJYg614qhwSRFrUguUSqL2uaGvVAudOqXBYi8yYS/nIC58cg4IdKOFkAMowAaGIKIFEQ+kQAMycAal8GYBkAhsQJgJMaN4GixCsA99Ygk+EAVlcCgK0Qg7gAI3kDau0wA8EAU2gJ4cmg8xYAYyQANDMCCSIK00gAChY5/rkAkzMhXn8AlAEAWkkAcW8QBcIBpnFJsBYAtzAARA/+ACAnWpqZkINyADMXAD7RIA7CAKAysKG2SfJBkAgBAEycILYwAEcMBfCvEOLgAEPzAHtlqihdAEYfALYdAE3kGhY+EIRsACoTAKoKAR+RQKgwADTCAIb1OeX8EAlWURnuAEl5AAOPAFKwIJTuAJg0AGSrBtrSMJTrAJhIAGSiCTBzoVc9AEczAEYZAJfpMJTYAJhKAGKjAjy1qcncABlikGK9AAh+AFPiAuvkkJCaADVjBPPqkQMpAFjIAIU/AGwUBf+8kFU+AJQ+ACQ3MDV7AAivAFDYSqrAMEWWAR6UAFPlAIkKAgCqEDVIAIjOAbMfoGzqEQM8CaEToKrkVJL/9gEWlQVv4JhyRgAS6VB0ogCRZhBbRhBQYVADxAA61TDlcAnAHgA2RQomigQijBDivQNXgDSAc7ldF6ASMQPIDQBNspD0vQCLUwjibzBYiFPAUhCE2QWgvRBUOTofAABAmBEojQBEXSB01wCP7ZDi5ABSHwBe+kAgjEAE2wDoqwBMHTD00QrhlqD1rgU1NBCF2QttJbl19BAxCQDwxgI/qwAuNUEIuwAntrn3ygAIywApbVACvAG1PxCTrgCyrgF1MBCVnAqo5CCFrwZgUxCSowqxR6DmzQAH+gCJyzACowTwXBCbexn/tACEKwAskUAHMgBShBBmOwCFYgHwUhCo3/lDxTMQRYHAA0MEgZ6gs98AeHkADJCQMZAk4g+Z1vQgiCQAZUFQA7wLqbSgULUAo0UxE+kFQlSghKkLYTZECF+51TMbxZkAY/sAQqlAhaECRToQ9awA/7+UruUMcKUa94ERKXUAVnclBZ4BdzOglXkMOEkAX3wKH14ARSsAYpoAQhswlUoCyKgAX5+qe1uQ9f8Kz0ZAN4AQpbMAeACQlU3EpnYLoFMQR8SaGPcAI4sAVWQAXPigODUhAv4DKtWhGaAAcKcQ4pkLsWkQMsYANQMhZngJsUighKIFBTEQ9fcB5gOqUBsAtRAAriQxmDgAgrYBoFoQdXsJ3y+Sdl8gXX/ykKbaAsgevEyvIHWDBO7owamkA5xOELVxC1FAoIVBAJ5jAL9CIPENBIBVEIV2DKKHqxKpDLNbCSmtADLpBVH7EAViBQWHQD0oxKsMyhlpAC48EOPgDLOXBBpDIK+WLNGgLU7VAFclQQOFAGaaBC/xEDX8yhCbAC9zQPWFC/tVycYnEOM1EGM1AIKiBrBZEPKrCt6fkfefAFi6AQc1A8FmGKmUAFObzCZbkqlfAFM/ELKuCvcWqyPjBTy0EciIAFKzKggEDSCkEGqmQRnbAFQ7DWVmIFdtBKaBAqW+vXGWoOORwPKkAIZiCDAUAKqICqKNEJQJ0OUnA3U4EDpFADKP9JKjPAWRmaDyswHgXRD12QtSLNCxMdAJrAA76QBS1REIegBHcxoGaN1gEQCTD8EZngA/6gAk1RECXsN1GDCFlgsMKTBQYgroihEKYQA5A7hwGQCfvrs+lJ2LnMArJkEaVgBpBABVfaxE/cSi5AVQUxBmvAoeUACEFCg1TACGGAfV2UmAM62hbBBbFVESgQB02dF4vtolLgu0Mg32qLnY2gBZwTAFwQvF/AVwrxAqkrn8atEHzQBfWrEKhAJiqQWQFgBmyAHI6CBytwN1JVmBTqCbVjMirwCemgBMcdAFtguladmuf9kl2AGOXwBUIgD13Qz1zguq20CGAQ0ufAmyL/qgP+4wsl0AeKsAIvnQtaEDIHWuDbJN8BIAhMUA+SoALJWQ9KYOIlOglQ4Aju0AAlMBAQnJcF0Q5Z4AP5QA8s0AXjoQlPsAjtMAlGwAALur0oMicrEAjwIApNcBmhwASH4A6aUAKFIMhgMQRMYAjusAkl4J2wDQVlYA/3cAbPHQAvoAS/8A6hAAVpg+dP6QtMsEkKsQNekA/zQANZME9mINbw0AJNwN1eWhFcAAR7QA+7qVAZKglJcAnxEAhVEFgCMAVucOptMAUmu5+hQNMBQA9gYArxcMu7nA5XkAb0sAc/8AO406JufbjPm6FgnQOZmwMuFQCfQAWHqz0HCrHo/5kOpkApUYAIjToEVAAHKYBurlMOLIDwKaC+HPoLXJACXGADRYIOLkApKGBLA3oPLNApu1ADXhAFb8BW7BADA++yPjkV9XADKRAFO5DLIhoEUzCyLJCce5Crulrs/pkIezwVgwAHcDAFMjDd+qADKOAFadApLuoOCODBdKoHfMDt76APsjbvgTEL9dAHjIoSeYAA04088aAP/bKlOswHejCdATAPCADZWUqfyVIOU5/DCnEq+SzkjTL3fLD2EUoqt9ALxP0f5oAPeuDuA8pK6ZAPI4IS55D2s26aE/riUkqdYgEpkBolrtOlkF+XoJn3qamf8UkiXfH5LxqjMJoZIv9J4cUpqKNS+tYsFasf2qEf+49aEE+xGK1yppb/Ssau+4cfp4CBOZpqpU6RGDkaGJZfFhZ7ocRPowmU+2Uh+2169sSPlT76+j/7o61v/BeHPJfP+Ya5oQxd/N5fM3zynNlfnruvpeq/mdXCSuRP+fNP//Vv//eP//mv//vP/wARQOBAggUNHkSYUOFChg0dPoQYUeJEihUtXsSYUeNGjh09fgQZUuRIkiVNnkSZUuVKli1dvoQZU+ZMmjVt3sSZU+dOnj19/gQaVOhQokWNHkWaVOlSpk2dPoUaVepUqlWtXsWaVStCAQIEehU4C+zFsSNnbd0JVmyAtV/Pauwa8i3/WqFz57I927Zm3LNdBdylG3iiX8IBvMYlG/ZjXMSCZc7qa5gwYbFlLTb2iNlxzq50/Eqm5bUy4IefDZezTBZsV1qkN7+mmFpiV3S0QJo7Bzuo5onlarvGWOucgHC6c0Lm+lV2w8/n0PFOjK6cZOPVEWZac4ZGDe03fhWKpJEBppC4QuWz7tJrnVBqzqCpsZ1GPgaKNLZyFFJPqHhs09fcYwzu4DsjjSF24aQX6ipCBxRfDOvoLEQ0wes/CxsRRZMtMGDhkyH0GCINg4BTKB0f8lMoNdKgg8EUC1c665xJRMmECg8+mWOOXcxowaK37kHhFwhVMyiNTl6kqZ5OQuHE/wQtNBlCknq+sI9EvQ7yqoE31CkIsL8Igu4gPaIQZEEkrWNAC3YGYgeXgc7hxc0A3LknN7DWuScdw+5qBAU9DaOnvwBuyXPIdvRAhzo43VlwHT3WESgQKfQ4syVPpCConVsGWocXO+mZRzSB3NHDTtM+YWOgcup5Z9Q69wwAnnvMoS6deyAdEg9eaA2gkR/+rFSmG0ZRFY9EBcqlnunKsSePBeepR7TDBMoBlIFs5TIAaKctB9SvBHpn1wXliVagUsYI9j8GVpAzgEUgCSCISUgB4otFEmGDCh/4EMgRKaL4AUWCaiAlgAUsAYUHFTzxxQYUqCgkgHJA+QGIKAwR6P8XLlDw4pOBHkjhhykqEQgOCtNNyRMqCIIEETswYaWGkPfZ5I0rxjgWgiqi4EGBgcDigjxIJBkFDiwi8SeHKYBQkB0XuEihh30EYgAFIFCQRCB2SEkBBTgGCYCdKhJBOaYcXLgWFD3qyaQSNaxQgw8WuMAilFFloAKFGuphCyw+sJhakzjKmCJIS974ogw9+7iBCypiyBaUKX5wQ8gA7EEDhRRyQA+RKuAp27h1242BhwCuKKIBecIQYYd+9LDCdEWcyIQeTZpIgKBzpggPBhFAqSeSEZRYJB4zugggEi0I2QWGEuCRhwkx7EmkCwofOGKSeTQpAl4Yzgj9JJUJcoP/FHQ6UGEQe1DAoBQ9CjGBkwBub4SeFpbgNy49rhDSBw0skcccROAFQtQDCJhiQRX2cY8tZCEAgijBJ+hBCSfYxwZKIAQvbHCCewQgB0MIn0vO5p93PEEB+RABD/7ghyXcaB6QOAHZcEAFQPTBBzv4VgAkMYUApCMJK1DAPNwgAhnkIxBQYEEAolAGeviCCTQIwByakIh6wEALoEtBCvbBBxR0QQDuuELulhNCrIxuIKVYQwCUsLLLbeBkjlCCOdogioG8II0DyYcKAhEA5x3rCVwQyD6YwIchqEAg6dDELligg4E8ggq5qILHBOICjz3gB2QkyfgGggMWnKMDkpRE/wb8IBAziMgKDhhIDl7wLURgwVk+sIJAzFGEggWAAUq4BQ56IJBdaGIdN4jBQIbABkB04UEBsAMODhGAMNwAkysZ4ahWkIB8kMA+AXBBEuxQslD4QQuUCsA7qEQQF+xyHUjoUQAQQQKwBUAGQAhACY4UgAQEwR0qaMBAeLAJRaxAFwLRgw74xYUHPFMwZhQIKdJIhVII5B5fIIRAGoGFe6xAB3NgASi4oILpCGQQXeAFH7cwECqAMAD4+AIg4pEFFZSiAblJgQo6wYJQnKEJkrgCPqjjFUhkITcG/YgmBYIDUdxhBcsMACRU8NNSmIEPRjhDKDr0BUD6ZxIq8AobYP8gEHUoAUWG+EI7AuGEKYwCEQLpAhBAkVEeVIEFKHBLXIYAV6CeJJoBaMcKCLEPJexBIKKIQio94QAkhEIUohiCESR5FhrgIJxLWIBADvEFegikBZfsxAnYMIc/PDAJMZCqJ5RABlKkai1g2QId67oVhAZAoUq0WwD2F9FefSEfS8hBKEbRAlE4oqMB+Kg9+OhMgUhhDgL5m5DgUQkZdEEK6aCCCj7Rghaw4BHrqiyEapFULPx0tRsRagCICsZ2RsKnAjGFDPZhBDIMIQxhGMI1vSIJrHqQWAFYhxYaIdkr8M0emqBBCdKwDifAYQ5hcAELEBEGTEFIANMRBV2/O5L/u7ZDCQn4wxf4FgAWBJZam9BEYXcbhjkAQjFmqEGslEA2dX5hHpOEZwB8EQodIEEIgjhBDEQxCh77ggZq2KlX3GDSCV+lta9FgUn1gIXc9UoFd5gCBAaSAPIMpA9fGCUMgFzc4wbgb4FoQJPrsIRPiMEMA+FDJ/ChgkcM5BPW2sQli6yR8I4XC7Q1709hYIa8stiWbV5LIqiwphykcx0rWIVkv6AHS3RWtkaAxBa6DFxK3HLD5TDDIvhogzmLpMIX/gMWhMthDweADZ9AhAraIZBybGKPX2pBDsK5YoEggrKT5EEuNLGmpDbhEFkoU78OoYkv2CYA8UjD1FJgiSF1/1oqjXDCqgUiBkZW4b57UELEAuCIJkSxBAgIAC+a4KJrTSGfMhipQK5g0tv6gRRK6M8utCAEXyAhn+f4QQpc2wR+qeIEBZUB+Jx9kVAYciA7cIE7LiwQSjDhWDJIVQ2y0B9BmCATBOHDFx70hlXi1wltDkAiVgCPHqQgUYDA3SOM0I9jrwB8UthBbuZwAvTwgMgD5wgPtoosJyjAF0oYtQuqMBAfoG0FZRDIJkwwNbBUAq7vgIKmA9DP7MLgB+gwQhIDcIkVsOMGVVh1IU7QgHJ0QQwCsWk5dLFwnEuFEDmA1Fk0kUQZkExbNzAxPXFgDkxnQQcqqMGmCHKDHm1C6/8BKMMkBMKLNPDhHArLwRdiQCtQKIEHeePXLST+9/vC4eJtn01SzzyQUVyCHTbIeyLS8FNOJDEPO7gCD7SANv8IBAgka8Hn1bEGfwhEEDZQhz2kYIUtYMGkpljBDrDABkb54WE+yAK82lGFs4J+I2H4fABwkQNA5MMG8hDIJYJp2YsPwgpe4MIK8omYPlghH+bIgeV+YYN/zg9djFjCD3zwBQYEoB5w+AI3UALVKoQvsJovyJ1EqAJkaDbrWwpzYIdpsQU9UYdEEQA6WIdaiBF2mA4BCARI+AVaKYsGgAPpSIfKqMDTWAdaOYcEiANA+C0+cIRD2BSvCAdBiAN98D3/KgAnB2QO2sgWrkqHcmAHDQyAc1AH1BCAdPgTYyAESHC0rngLUBARdKgDscDA4RCAc1gHr1CHQ3CEPpiLP4CEQdguL0SEBsiuBogCYPHBipgFdbCFWQiNclhBOzQHr0iHLuQqWxiVRWgEvjENDzqSFRQILkSNHsqWemgEBtgFCEmHQoCEqRmId1iABWAUPpoBM3lDpRCA0HiL1RiLv+iLxhjEAFAHHdivn1GOUgQTJSyIC7SMFji7TvxBhaiMw2AMy/ALzzgLPfiBPVIOyRjFsYAMzPiL1KiBebLF2IgrUTGM0FAOtSgLv4gMR9ABP1wQOlSLu5BCgsjFgqCHHwCbYzFqxt2oPXA8Rr5AjgZMjUSQg2GskL6RFrtwixx6C3aYA3A7R4ZoxxEJC7swRtEIxYGABIFpwH88iLxYkQC4h1Bwln4cDHNMyFgkxYI4h07IO1bEx8MgjcrokgAohCojEZ0ICAA7)
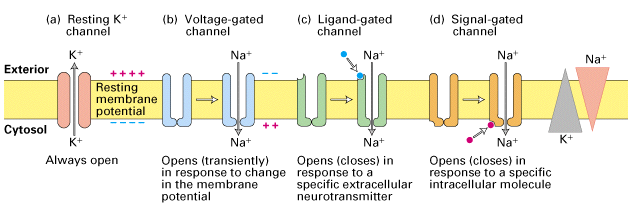{kind=link}
Generating action potentials
- Axon hillock
- Portion of soma adjacent to axon
- Integrates/sums input to soma
- Axon initial segment
- Umyelinated portion of axon adjacent to soma
- Voltage-gated \(Na^+\) and \(K^+\) channels exposed
- If sum of input to soma > threshold, voltage-gated \(Na^+\) channels open

Wikipedia
Propagation of action potentials
- Propagation
- move down axon, away from soma, toward axon terminals.
- Unmyelinated axon
- Each segment “excites” the next
- Myelinated axon
- AP “jumps” between Nodes of Ranvier –> saltatory conduction
- Nodes of Ranvier == unmyelinated sections of axon
- voltage-gated \(Na^+\), \(K^+\) channels exposed
- Current flows through myelinated segments
- Why does AP flow in one direction, away from soma?
- Soma does not have (many) voltage-gated \(Na^+\) channels.
- Soma is not myelinated.
- Refractory periods mean polarization only in one direction.
Conduction velocities
Hodgkin-Huxley Equations
- Measuring APs in actual neurons. https://www.youtube.com/embed/k48jXzFGMc8
- Interview with Sir Alan Hodgkin, https://www.youtube.com/embed/vSIXbfunHYg
- Simulations
Chemical communication: Synaptic transmission
- Synapse permits neuron to pass electrical or chemical messages to another neuron or target cell (muscle, gland, etc.)

- Gap junctions
- Cytosol flows through adjacent neurons
- Bidirectional ion flow (current) between neurons
What happens when AP runs out of axon?
- Rapid change in voltage triggers neurotransmitter (NT) release
- Voltage-gated calcium Ca++ channels open
- Ca++ causes synaptic vesicles to bind with presynaptic membrane, merge
- NTs diffuse across synaptic cleft
- NTs bind with receptors on postsynaptic membrane
- NTs unbind, are inactivated
Receptor/channel types
Leak/passive
- Vary in selectivity, permeability
Transporters/exchangers
- Ionic
- \(Na^+\)/\(K^+\)
- Chemical
- e.g., Dopamine transporter (DAT)
Ionotropic receptors (receptor + ion channel)
- Ligand-gated
- Open/close channel
Metabotropic receptors (receptor only)
- Triggers 2nd messengers
- G-proteins
- Open/close adjacent channels, change metabolism


Receptors generate postsynaptic potentials (PSPs)
- Small voltage changes
- Amplitude scales with # of receptors activated
- Excitatory PSPs (EPSPs)
- Depolarize neuron (make more +)
- Inhibitory (IPSPs)
- Hyperpolarize neuron (make more -)
NTs inactivated
- Buffering
- e.g., glutamate into astrocytes
- Reuptake via transporters
- e.g., serotonin via serotonin transporter (SERT)
- Enzymatic degradation
- e.g., acetylcholine esterase (AChE) degrades acetylcholine (ACh)
References
Azevedo, F. A., Carvalho, L. R., Grinberg, L. T., Farfel, J. M., Ferretti, R. E., Leite, R. E., … others. (2009). Equal numbers of neuronal and nonneuronal cells make the human brain an isometrically scaled-up primate brain. Journal of Comparative Neurology, 513(5), 532–541. https://doi.org/10.1002/cne.21974
Boldog, E., Bakken, T. E., Hodge, R. D., Novotny, M., Aevermann, B. D., Baka, J., … Tamás, G. (2018). Transcriptomic and morphophysiological evidence for a specialized human cortical GABAergic cell type. Nature Neuroscience, 21(9), 1185–1195. https://doi.org/10.1038/s41593-018-0205-2
Chung, W.-S., Welsh, C. A., Barres, B. A., & Stevens, B. (2015). Do glia drive synaptic and cognitive impairment in disease? Nature Neuroscience, 18(11), 1539–1545. https://doi.org/10.1038/nn.4142
Eyherabide, H. G., Rokem, A., Herz, A. V. M., Samengo, I., Eyherabide, H. G., Rokem, A., … Samengo, I. (2009). Bursts generate a non-reducible spike-pattern code. Frontiers in Neuroscience, 3, 1. https://doi.org/10.3389/neuro.01.002.2009