511-fear-stress-reward
Rick Gilmore
2020-11-12 09:19:00
Fun
Fear
Animal models

http://www.cns.nyu.edu/labs/ledouxlab/images/image_research/fear_conditioning.jpg
![Adapted from [[@davis1992role]](http://dx.doi.org/10.1016/0165-6147(92)90014-W)](data:image/jpeg;base64,/9j/4AAQSkZJRgABAQEASABIAAD/4QB0RXhpZgAATU0AKgAAAAgABAEaAAUAAAABAAAAPgEbAAUAAAABAAAARgEoAAMAAAABAAIAAIdpAAQAAAABAAAATgAAAAAAAABIAAAAAQAAAEgAAAABAAKgAgAEAAAAAQAAAo+gAwAEAAAAAQAAAaUAAAAA/+0AOFBob3Rvc2hvcCAzLjAAOEJJTQQEAAAAAAAAOEJJTQQlAAAAAAAQ1B2M2Y8AsgTpgAmY7PhCfv/iGyhJQ0NfUFJPRklMRQABAQAAGxhhcHBsAhAAAG1udHJSR0IgWFlaIAffAAYAFwAUAAIAFGFjc3BBUFBMAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAD21gABAAAAANMtYXBwbAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAEWRlc2MAAAFQAAAAYmRzY20AAAG0AAAEGmNwcnQAAAXQAAAAI3d0cHQAAAX0AAAAFHJYWVoAAAYIAAAAFGdYWVoAAAYcAAAAFGJYWVoAAAYwAAAAFHJUUkMAAAZEAAAIDGFhcmcAAA5QAAAAIHZjZ3QAAA5wAAAGEm5kaW4AABSEAAAGPmNoYWQAABrEAAAALG1tb2QAABrwAAAAKGJUUkMAAAZEAAAIDGdUUkMAAAZEAAAIDGFhYmcAAA5QAAAAIGFhZ2cAAA5QAAAAIGRlc2MAAAAAAAAACERpc3BsYXkAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABtbHVjAAAAAAAAACIAAAAMaHJIUgAAABQAAAGoa29LUgAAAAwAAAG8bmJOTwAAABIAAAHIaWQAAAAAABIAAAHaaHVIVQAAABQAAAHsY3NDWgAAABYAAAIAZGFESwAAABwAAAIWdWtVQQAAABwAAAIyYXIAAAAAABQAAAJOaXRJVAAAABQAAAJicm9STwAAABIAAAJ2ZXNFUwAAABIAAAJ2aGVJTAAAABYAAAKIbmxOTAAAABYAAAKeZmlGSQAAABAAAAK0emhUVwAAAAwAAALEdmlWTgAAAA4AAALQc2tTSwAAABYAAALeemhDTgAAAAwAAALEcnVSVQAAACQAAAL0ZnJGUgAAABYAAAMYbXMAAAAAABIAAAMuY2FFUwAAABgAAANAdGhUSAAAAAwAAANYZXNYTAAAABIAAAJ2ZGVERQAAABAAAANkZW5VUwAAABIAAAN0cHRCUgAAABgAAAOGcGxQTAAAABIAAAOeZWxHUgAAACIAAAOwc3ZTRQAAABAAAAPSdHJUUgAAABQAAAPiamFKUAAAAA4AAAP2cHRQVAAAABYAAAQEAEwAQwBEACAAdQAgAGIAbwBqAGnO7LfsACAATABDAEQARgBhAHIAZwBlAC0ATABDAEQATABDAEQAIABXAGEAcgBuAGEAUwB6AO0AbgBlAHMAIABMAEMARABCAGEAcgBlAHYAbgD9ACAATABDAEQATABDAEQALQBmAGEAcgB2AGUAcwBrAOYAcgBtBBoEPgQ7BEwEPgRABD4EMgQ4BDkAIABMAEMARCAPAEwAQwBEACAGRQZEBkgGRgYpAEwAQwBEACAAYwBvAGwAbwByAGkATABDAEQAIABjAG8AbABvAHIgDwBMAEMARAAgBeYF0QXiBdUF4AXZAEsAbABlAHUAcgBlAG4ALQBMAEMARABWAOQAcgBpAC0ATABDAERfaYJyACAATABDAEQATABDAEQAIABNAOAAdQBGAGEAcgBlAGIAbgDpACAATABDAEQEJgQyBDUEQgQ9BD4EOQAgBBYEGgAtBDQEOARBBD8EOwQ1BDkATABDAEQAIABjAG8AdQBsAGUAdQByAFcAYQByAG4AYQAgAEwAQwBEAEwAQwBEACAAZQBuACAAYwBvAGwAbwByAEwAQwBEACAOKg41AEYAYQByAGIALQBMAEMARABDAG8AbABvAHIAIABMAEMARABMAEMARAAgAEMAbwBsAG8AcgBpAGQAbwBLAG8AbABvAHIAIABMAEMARAOIA7MDxwPBA8kDvAO3ACADvwO4A8wDvQO3ACAATABDAEQARgDkAHIAZwAtAEwAQwBEAFIAZQBuAGsAbABpACAATABDAEQwqzDpMPwAIABMAEMARABMAEMARAAgAGEAIABDAG8AcgBlAHMAAHRleHQAAAAAQ29weXJpZ2h0IEFwcGxlIEluYy4sIDIwMTUAAFhZWiAAAAAAAADzUgABAAAAARbPWFlaIAAAAAAAAGFsAAA4MgAACkJYWVogAAAAAAAAbxwAAK6VAAAWmVhZWiAAAAAAAAAmTgAAGTkAALJSY3VydgAAAAAAAAQAAAAABQAKAA8AFAAZAB4AIwAoAC0AMgA2ADsAQABFAEoATwBUAFkAXgBjAGgAbQByAHcAfACBAIYAiwCQAJUAmgCfAKMAqACtALIAtwC8AMEAxgDLANAA1QDbAOAA5QDrAPAA9gD7AQEBBwENARMBGQEfASUBKwEyATgBPgFFAUwBUgFZAWABZwFuAXUBfAGDAYsBkgGaAaEBqQGxAbkBwQHJAdEB2QHhAekB8gH6AgMCDAIUAh0CJgIvAjgCQQJLAlQCXQJnAnECegKEAo4CmAKiAqwCtgLBAssC1QLgAusC9QMAAwsDFgMhAy0DOANDA08DWgNmA3IDfgOKA5YDogOuA7oDxwPTA+AD7AP5BAYEEwQgBC0EOwRIBFUEYwRxBH4EjASaBKgEtgTEBNME4QTwBP4FDQUcBSsFOgVJBVgFZwV3BYYFlgWmBbUFxQXVBeUF9gYGBhYGJwY3BkgGWQZqBnsGjAadBq8GwAbRBuMG9QcHBxkHKwc9B08HYQd0B4YHmQesB78H0gflB/gICwgfCDIIRghaCG4IggiWCKoIvgjSCOcI+wkQCSUJOglPCWQJeQmPCaQJugnPCeUJ+woRCicKPQpUCmoKgQqYCq4KxQrcCvMLCwsiCzkLUQtpC4ALmAuwC8gL4Qv5DBIMKgxDDFwMdQyODKcMwAzZDPMNDQ0mDUANWg10DY4NqQ3DDd4N+A4TDi4OSQ5kDn8Omw62DtIO7g8JDyUPQQ9eD3oPlg+zD88P7BAJECYQQxBhEH4QmxC5ENcQ9RETETERTxFtEYwRqhHJEegSBxImEkUSZBKEEqMSwxLjEwMTIxNDE2MTgxOkE8UT5RQGFCcUSRRqFIsUrRTOFPAVEhU0FVYVeBWbFb0V4BYDFiYWSRZsFo8WshbWFvoXHRdBF2UXiReuF9IX9xgbGEAYZRiKGK8Y1Rj6GSAZRRlrGZEZtxndGgQaKhpRGncanhrFGuwbFBs7G2MbihuyG9ocAhwqHFIcexyjHMwc9R0eHUcdcB2ZHcMd7B4WHkAeah6UHr4e6R8THz4faR+UH78f6iAVIEEgbCCYIMQg8CEcIUghdSGhIc4h+yInIlUigiKvIt0jCiM4I2YjlCPCI/AkHyRNJHwkqyTaJQklOCVoJZclxyX3JicmVyaHJrcm6CcYJ0kneierJ9woDSg/KHEooijUKQYpOClrKZ0p0CoCKjUqaCqbKs8rAis2K2krnSvRLAUsOSxuLKIs1y0MLUEtdi2rLeEuFi5MLoIuty7uLyQvWi+RL8cv/jA1MGwwpDDbMRIxSjGCMbox8jIqMmMymzLUMw0zRjN/M7gz8TQrNGU0njTYNRM1TTWHNcI1/TY3NnI2rjbpNyQ3YDecN9c4FDhQOIw4yDkFOUI5fzm8Ofk6Njp0OrI67zstO2s7qjvoPCc8ZTykPOM9Ij1hPaE94D4gPmA+oD7gPyE/YT+iP+JAI0BkQKZA50EpQWpBrEHuQjBCckK1QvdDOkN9Q8BEA0RHRIpEzkUSRVVFmkXeRiJGZ0arRvBHNUd7R8BIBUhLSJFI10kdSWNJqUnwSjdKfUrESwxLU0uaS+JMKkxyTLpNAk1KTZNN3E4lTm5Ot08AT0lPk0/dUCdQcVC7UQZRUFGbUeZSMVJ8UsdTE1NfU6pT9lRCVI9U21UoVXVVwlYPVlxWqVb3V0RXklfgWC9YfVjLWRpZaVm4WgdaVlqmWvVbRVuVW+VcNVyGXNZdJ114XcleGl5sXr1fD19hX7NgBWBXYKpg/GFPYaJh9WJJYpxi8GNDY5dj62RAZJRk6WU9ZZJl52Y9ZpJm6Gc9Z5Nn6Wg/aJZo7GlDaZpp8WpIap9q92tPa6dr/2xXbK9tCG1gbbluEm5rbsRvHm94b9FwK3CGcOBxOnGVcfByS3KmcwFzXXO4dBR0cHTMdSh1hXXhdj52m3b4d1Z3s3gReG54zHkqeYl553pGeqV7BHtje8J8IXyBfOF9QX2hfgF+Yn7CfyN/hH/lgEeAqIEKgWuBzYIwgpKC9INXg7qEHYSAhOOFR4Wrhg6GcobXhzuHn4gEiGmIzokziZmJ/opkisqLMIuWi/yMY4zKjTGNmI3/jmaOzo82j56QBpBukNaRP5GokhGSepLjk02TtpQglIqU9JVflcmWNJaflwqXdZfgmEyYuJkkmZCZ/JpomtWbQpuvnByciZz3nWSd0p5Anq6fHZ+Ln/qgaaDYoUehtqImopajBqN2o+akVqTHpTilqaYapoum/adup+CoUqjEqTepqaocqo+rAqt1q+msXKzQrUStuK4trqGvFq+LsACwdbDqsWCx1rJLssKzOLOutCW0nLUTtYq2AbZ5tvC3aLfguFm40blKucK6O7q1uy67p7whvJu9Fb2Pvgq+hL7/v3q/9cBwwOzBZ8Hjwl/C28NYw9TEUcTOxUvFyMZGxsPHQce/yD3IvMk6ybnKOMq3yzbLtsw1zLXNNc21zjbOts83z7jQOdC60TzRvtI/0sHTRNPG1EnUy9VO1dHWVdbY11zX4Nhk2OjZbNnx2nba+9uA3AXcit0Q3ZbeHN6i3ynfr+A24L3hROHM4lPi2+Nj4+vkc+T85YTmDeaW5x/nqegy6LzpRunQ6lvq5etw6/vshu0R7ZzuKO6070DvzPBY8OXxcvH/8ozzGfOn9DT0wvVQ9d72bfb794r4Gfio+Tj5x/pX+uf7d/wH/Jj9Kf26/kv+3P9t//9wYXJhAAAAAAADAAAAAmZmAADypwAADVkAABPQAAAKDnZjZ3QAAAAAAAAAAAADAQAAAgAAAFYBLgHrAsQDgwRTBUAGJwcaCBgJHwo9C04Maw2QDrkP7BEtEmUToxTjFioXeRi/Gg8bVxykHfMfPCCEIc0jESRSJY0myygAKTMqZiuXLMot9y8oMFExgDKrM9U1AjYxN104ijm2OuE8DT04PmA/iUCxQdxDCEQ2RWNGkEe4SN5KAUskTEdNak6PT7RQ2FH4UxZUM1VPVm1XjViuWcta4VvwXPdd+18AYAphGWItY0FkUmVfZmVnYWhTaTxqIGsBa+Bsv22gboNvaHBQcThyIHMIc+901nW+dqh3k3h+eWx6WXtJfD19N342fzmAQYFLgleDZYR0hYWGlIekiLOJwIrMi9aM343nju2P8pD1kfaS9ZP1lPqWBJcVmCyZR5pjm3+cmJ2unr+fy6DUodqi4KPppPSmA6cVqCqpQqpcq3eslK2yrtGv8bERsjOzU7RztZC2qbe6uMG5w7rAu7u8tb2vvqm/o8CdwZXCisN8xGrFU8Y2xxjH+cjgyc/KxsvDzMHNvM6vz5jQc9FD0g3S09Ob1GbVNdYF1tfXqNh22UDaBtrH24DcNdzn3ZHeO97j343gO+Dr4Z/iUuMD46/kVeT55ZTmLOa/51Ln5OiF6S/p5Oqo627sM+z17a/uY+8R77XwUPDm8XzyF/LG84/0bvVh9mb3ivjk+sX9Lv//AAAAVgEjAbACagMyA+gEsQWABlgHPQgtCScKJwsiDDANOg5RD2MQfhGfEr4T6RUQFjgXYRiIGbQa3hwLHTMeWx+AIKEhvyLbI/UlCyYfJzUoSSlgKnkrkyyxLcwu6DAEMSEyPDNaNHQ1izakN7k4zTniOvQ8Bj0bPjE/TUBsQY9CsEPQRO1GCEcdSDJJRUpVS2NMbk14Tn5Pg1CFUYZSiFOKVIxVilaEV3lYaFlUWkRbP1xGXVped1+VYLBhwmLKY8VktWWcZntnV2gwaQxp7GrQa7psqW2bbo9vgnBycVxyQHMfc/t01XWtdoZ3YnhFeTB6JHsdfBl9FX4QfwZ/9oDigcqCsYOVhHeFXYZKh0SITIlhioGLpYzGjd6O7o/1kPeR9ZLzk/CU7ZXslu6X+JkQmjmbbpymnc6e45/poOWh36LZo9GkyKW+prynyqjtqiGrXaycrdmvFLBOsYiywrP5tS62ZLebuNK6Bbs0vF+9hb6mv7zAvsGqwpDDfcR3xYDGj8edyKPJnsqOy3XMU80tzgTO2s+y0IvRatJR00XURdVR1mLXddiE2Yfafdtm3EPdGt3s3rnfh+BY4S3iCOLm48PkneVx5kDnDOfU6KPpf+pp62TsaO1t7nDvcPBw8XLydvN59Hf1afZH9w/3s/hF+Mv5Wvnt+tD8DP3a//8AAAArAMQBSwHYAmUDBgOPBCsEzwV6Bi8G6QelCGgJLwn5CsoLnwx3DVIOMg8OD/YQ2RHAEqkTkxR9FWcWUhc9GCkZFRn8Gugb0hy+Ha4eox+bIJYhlCKPI4YkdiVhJkUnICf4KMwpnypzK0gsIS0CLeku1y/IMLoxqjKYM4A0YDU5NgY2zzeVOFg5GTnZOp07ZDwyPQY93j62P49AaEFEQiZDEEQARPNF50bWR7tIlkllSiVK4EuYTE9NB03DToNPSVAYUOpRvlKTU2hUO1UNVd5WrVd8WEpZFVnfWqhbb1w1XPldvV6AX0NgCGDRYZxia2M8ZAxk2mWnZnFnOGf8aL9pgGpAawBrwGyBbUNuBG7Fb4VwQXD4catyWXMBc6l0T3T0dZh2PHbed4B4HXi9eWB6Fnrae6t8g31Yfih+9n/DgJOBZ4JEgyqEGIULhgCG9IfjiM2Jr4qMi2GMNI0HjdqOsI+KkGuRR5ISksaTW5PslHqVQZZCl4iZCJqKm+SdGZ4ynzmgOaE1oi+jKKQhpRmmEacKqAmpD6ohqz2sYK2Hrq2v07D8siizWrSStdG3FLhZuZy617wFvSm+R79iwH/BnsK+w9/E/8Yhx0PIZ8mMyrHL1sz9zinPW9CX0d7TMtSN1e7XU9i52iHbld0v3w7hd+SL6HntTvL2+VH//wAAbmRpbgAAAAAAAAY2AACTlQAAVo0AAFbpAACRtwAAJrsAABcKAABQDQAAVDkAAoeuAAJHrgABa4UAAwEAAAIAAAABAAQACAAPABYAHwApADQAQABMAFoAaQB4AIkAmgCsAL4A0gDmAPwBEgEpAUABWQFzAY0BqQHFAeMCAQIhAkICZAKHAqwC0gL6AyMDTQN6A6gD1wQIBDoEbQSjBNkFEgVLBYYFwwYBBkAGgAbCBwUHSgeQB9gIIQhsCLkJBwlXCagJ+gpNCqEK9wtOC6gMBAxiDMMNJQ2JDe4OVA68DyUPkRAAEHEQ5BFZEc8SRhK9EzgTthQ6FMMVUhXjFnUXBheWGCYYuBlOGegaiBsxG+Mcnx1iHise+B/GIJUhZCI0IwYj2ySzJY8mbCdLKCspDSnwKtUruyyeLX4uXC85MBQw8DHNMqoziTRoNUo2LjcVN/847TneOtI7yjzFPcQ+xz/OQNpB6EL2RABFBkYIRwhICEkJSgxLFEwhTTROTk9uUJRRvFLlVA1VNFZZV31YoFnEWudcC10wXlZffGCkYc5i+GQkZVNmhWe8aPpqQ2uZbPluY2/ScUZyvHQ0da93LXixejx70H1vfxyA1oKUhEyF9oeUiSyKxYxljhWP35HNk92V/5ggmjmcTJ5foHmioKTapy2poKwwruCxobRetwu5rrxdvyvCH8VGyKLMDM8o0fbUkdco2dHcnt+X4tLmYOn57RLvrPIA9DD2Off/+Yr6uPvU/Lv9o/5w/zf//wAAAAEABQAKABIAGwAmADIAPwBNAFwAbQB+AJEApAC5AM8A5QD9ARYBLwFKAWYBgwGiAcEB4gIEAicCTAJyApoCwwLvAxwDSwN8A68D5AQaBFIEiwTFBQEFPQV7BbsF/AY/BoMGyQcQB1kHpQfyCEEIkwjmCTsJkwnsCkYKoQr8C1gLtAwSDHIM1A05DaAOCw54DucPWQ/OEEYQwBE+Eb8SQxLKE1MT3RRqFPoVjxYqFssXcRgWGLYZURnmGnkbDRukHEAc5B2RHkofDh/dILMhjiJpI0IkGCTrJb0mkCdmKEIpJSoSKwgsBS0ILg4vEjARMQgx+TLoM9c0yjXBNsE3yjjbOfI7DjwwPVE+aT90QHJBaEJYQ0dEOUU0RjhHR0hfSX5KoUvHTPBOG09FUGtRhlKUU5dUlVWUVqBXv1jxWjVbgFzNXh5fdGDPYiNjZmSVZbVmzmfkaPtqFmszbFRtdm6ab8Bw6nIXc0Z0dnWndtp4EHlNeo972H0mfn1/44FngxCEyoZ0iAaJh4sEjIiOG4/DkYKTWJVDlzuZO5s7nTmfKaEEosakdqYbp72pYqsVrN2uxbDPsvW1Mbd8ucu8D75EwHLCpsTpx0LJsswxzpzQ3dL01PHW59jj2ujc9N8B4QrjFOUk50jpluw374rzpvds+c77hPzD/fT++f//AAAAAgAIABEAHgAtAD4AUQBmAH0AlgCwAMwA6gEJASsBTQFyAZgBwAHqAhYCRAJ0AqYC2gMRA0kDhAPAA/0EOwR6BLoE/AVBBYoF1wYpBoEG3Qc+B6IICAhvCNQJOQmdCgIKagrVC0QLuww5DMENUA3mDoIPIQ/AEFwQ9hGQEi0SzRNuFA0UqBVCFdwWeRceF84YjhliGkIbKhwUHPsd3R64H5IgbSFMIjAjGCQGJPkl8SbvJ/Mo/SoOKyQsQC1gLoAvnTC4MdMy8zQaNUk2gjfDOQw6XDuwPQc+YD++QSJCk0QWRa5HWkkRStFMmk5vUFVSMlPaVVpWzFhIWdNbZlz4XoNgAWFwYtVkN2WcZwlogWoKa6NtTW8AcLdyb3QedcN3dXlke+5+pIB5gdaC/IQChQiGEIcziHOJ1ItVjOmOiJAukdmTiJU7lvGYqZpenAqdp583oL2iP6PApUSmyqhQqdSrVKzQrkavubEospe0CLV9tvy4ibomu9C9gr82wOnCnMRPxgTHu8lzyyvM5M6e0FjSFNPS1Y/XStj/2qzcT93o33Xg/eKC5APlgucC6IPqBOt/7O7uRu958J3xjfJ180Tz9/Sq9U712/Zo9vb3dvfp+Fz4z/lD+av6Dfpw+tL7NfuY+/H8Sfyi/Pv9VP2t/gP+WP6s/wH/Vf+q//8AAHNmMzIAAAAAAAEMQgAABd7///MmAAAHkgAA/ZH///ui///9owAAA9wAAMBsbW1vZAAAAAAAAAYQAACc3wAAAADKKtWAAAAAAAAAAAAAAAAAAAAAAP/AABEIAaUCjwMBIgACEQEDEQH/xAAfAAABBQEBAQEBAQAAAAAAAAAAAQIDBAUGBwgJCgv/xAC1EAACAQMDAgQDBQUEBAAAAX0BAgMABBEFEiExQQYTUWEHInEUMoGRoQgjQrHBFVLR8CQzYnKCCQoWFxgZGiUmJygpKjQ1Njc4OTpDREVGR0hJSlNUVVZXWFlaY2RlZmdoaWpzdHV2d3h5eoOEhYaHiImKkpOUlZaXmJmaoqOkpaanqKmqsrO0tba3uLm6wsPExcbHyMnK0tPU1dbX2Nna4eLj5OXm5+jp6vHy8/T19vf4+fr/xAAfAQADAQEBAQEBAQEBAAAAAAAAAQIDBAUGBwgJCgv/xAC1EQACAQIEBAMEBwUEBAABAncAAQIDEQQFITEGEkFRB2FxEyIygQgUQpGhscEJIzNS8BVictEKFiQ04SXxFxgZGiYnKCkqNTY3ODk6Q0RFRkdISUpTVFVWV1hZWmNkZWZnaGlqc3R1dnd4eXqCg4SFhoeIiYqSk5SVlpeYmZqio6Slpqeoqaqys7S1tre4ubrCw8TFxsfIycrS09TV1tfY2dri4+Tl5ufo6ery8/T19vf4+fr/2wBDAAICAgICAgMCAgMFAwMDBQYFBQUFBggGBgYGBggKCAgICAgICgoKCgoKCgoMDAwMDAwODg4ODg8PDw8PDw8PDw//2wBDAQICAgQEBAcEBAcQCwkLEBAQEBAQEBAQEBAQEBAQEBAQEBAQEBAQEBAQEBAQEBAQEBAQEBAQEBAQEBAQEBAQEBD/3QAEACn/2gAMAwEAAhEDEQA/AP3M8a/ELwD8NdKi134jeJdM8K6bPMttHdareQ2MDzsrOsSyTuilyqMwUHJCk4wDXmH/AA1j+yz/ANFk8Gf+FDp3/wAfr5v/AOCiGk6Vr+lfs/aFrtnDqOm6j8X/AApbXVrcxrNBPBMt2kkUsbgq6OpKsrAggkEYr6Q/4ZO/ZZ/6I34M/wDCe07/AOMUAH/DWP7LP/RZPBn/AIUOnf8Ax+j/AIax/ZZ/6LJ4M/8ACh07/wCP0f8ADJ37LP8A0RvwZ/4T2nf/ABij/hk79ln/AKI34M/8J7Tv/jFAB/w1j+yz/wBFk8Gf+FDp3/x+j/hrH9ln/osngz/wodO/+P0f8Mnfss/9Eb8Gf+E9p3/xij/hk79ln/ojfgz/AMJ7Tv8A4xQAf8NY/ss/9Fk8Gf8AhQ6d/wDH6P8AhrH9ln/osngz/wAKHTv/AI/R/wAMnfss/wDRG/Bn/hPad/8AGKP+GTv2Wf8Aojfgz/wntO/+MUAH/DWP7LP/AEWTwZ/4UOnf/H6P+Gsf2Wf+iyeDP/Ch07/4/R/wyd+yz/0RvwZ/4T2nf/GKP+GTv2Wf+iN+DP8AwntO/wDjFAB/w1j+yz/0WTwZ/wCFDp3/AMfo/wCGsf2Wf+iyeDP/AAodO/8Aj9H/AAyd+yz/ANEb8Gf+E9p3/wAYo/4ZO/ZZ/wCiN+DP/Ce07/4xQAf8NY/ss/8ARZPBn/hQ6d/8fo/4ax/ZZ/6LJ4M/8KHTv/j9H/DJ37LP/RG/Bn/hPad/8Yo/4ZO/ZZ/6I34M/wDCe07/AOMUAH/DWP7LP/RZPBn/AIUOnf8Ax+j/AIax/ZZ/6LJ4M/8ACh07/wCP0f8ADJ37LP8A0RvwZ/4T2nf/ABij/hk79ln/AKI34M/8J7Tv/jFAB/w1j+yz/wBFk8Gf+FDp3/x+j/hrH9ln/osngz/wodO/+P0f8Mnfss/9Eb8Gf+E9p3/xij/hk79ln/ojfgz/AMJ7Tv8A4xQAf8NY/ss/9Fk8Gf8AhQ6d/wDH6P8AhrH9ln/osngz/wAKHTv/AI/R/wAMnfss/wDRG/Bn/hPad/8AGKP+GTv2Wf8Aojfgz/wntO/+MUAH/DWP7LP/AEWTwZ/4UOnf/H6P+Gsf2Wf+iyeDP/Ch07/4/R/wyd+yz/0RvwZ/4T2nf/GKP+GTv2Wf+iN+DP8AwntO/wDjFAB/w1j+yz/0WTwZ/wCFDp3/AMfo/wCGsf2Wf+iyeDP/AAodO/8Aj9H/AAyd+yz/ANEb8Gf+E9p3/wAYo/4ZO/ZZ/wCiN+DP/Ce07/4xQAf8NY/ss/8ARZPBn/hQ6d/8fo/4ax/ZZ/6LJ4M/8KHTv/j9H/DJ37LP/RG/Bn/hPad/8Yo/4ZO/ZZ/6I34M/wDCe07/AOMUAH/DWP7LP/RZPBn/AIUOnf8Ax+j/AIax/ZZ/6LJ4M/8ACh07/wCP0f8ADJ37LP8A0RvwZ/4T2nf/ABij/hk79ln/AKI34M/8J7Tv/jFAB/w1j+yz/wBFk8Gf+FDp3/x+j/hrH9ln/osngz/wodO/+P0f8Mnfss/9Eb8Gf+E9p3/xij/hk79ln/ojfgz/AMJ7Tv8A4xQAf8NY/ss/9Fk8Gf8AhQ6d/wDH6P8AhrH9ln/osngz/wAKHTv/AI/R/wAMnfss/wDRG/Bn/hPad/8AGKP+GTv2Wf8Aojfgz/wntO/+MUAH/DWP7LP/AEWTwZ/4UOnf/H6P+Gsf2Wf+iyeDP/Ch07/4/R/wyd+yz/0RvwZ/4T2nf/GKP+GTv2Wf+iN+DP8AwntO/wDjFAB/w1j+yz/0WTwZ/wCFDp3/AMfo/wCGsf2Wf+iyeDP/AAodO/8Aj9H/AAyd+yz/ANEb8Gf+E9p3/wAYo/4ZO/ZZ/wCiN+DP/Ce07/4xQAf8NY/ss/8ARZPBn/hQ6d/8fo/4ax/ZZ/6LJ4M/8KHTv/j9H/DJ37LP/RG/Bn/hPad/8Yo/4ZO/ZZ/6I34M/wDCe07/AOMUAH/DWP7LP/RZPBn/AIUOnf8Ax+j/AIax/ZZ/6LJ4M/8ACh07/wCP0f8ADJ37LP8A0RvwZ/4T2nf/ABij/hk79ln/AKI34M/8J7Tv/jFAB/w1j+yz/wBFk8Gf+FDp3/x+j/hrH9ln/osngz/wodO/+P18P/8ABR79nv4BeBv2MfiH4p8FfDTwz4f1qy/snyL7T9GsrS6h8zVbSN/LmhiV13IzK2CMqSDwTX3B/wAMnfss/wDRG/Bn/hPad/8AGKAD/hrH9ln/AKLJ4M/8KHTv/j9H/DWP7LP/AEWTwZ/4UOnf/H6P+GTv2Wf+iN+DP/Ce07/4xR/wyd+yz/0RvwZ/4T2nf/GKAD/hrH9ln/osngz/AMKHTv8A4/R/w1j+yz/0WTwZ/wCFDp3/AMfo/wCGTv2Wf+iN+DP/AAntO/8AjFH/AAyd+yz/ANEb8Gf+E9p3/wAYoAP+Gsf2Wf8Aosngz/wodO/+P0f8NY/ss/8ARZPBn/hQ6d/8fo/4ZO/ZZ/6I34M/8J7Tv/jFH/DJ37LP/RG/Bn/hPad/8YoAP+Gsf2Wf+iyeDP8AwodO/wDj9H/DWP7LP/RZPBn/AIUOnf8Ax+j/AIZO/ZZ/6I34M/8ACe07/wCMUf8ADJ37LP8A0RvwZ/4T2nf/ABigA/4ax/ZZ/wCiyeDP/Ch07/4/R/w1j+yz/wBFk8Gf+FDp3/x+j/hk79ln/ojfgz/wntO/+MUf8Mnfss/9Eb8Gf+E9p3/xigA/4ax/ZZ/6LJ4M/wDCh07/AOP0f8NY/ss/9Fk8Gf8AhQ6d/wDH6P8Ahk79ln/ojfgz/wAJ7Tv/AIxR/wAMnfss/wDRG/Bn/hPad/8AGKAD/hrH9ln/AKLJ4M/8KHTv/j9H/DWP7LP/AEWTwZ/4UOnf/H6P+GTv2Wf+iN+DP/Ce07/4xR/wyd+yz/0RvwZ/4T2nf/GKAD/hrH9ln/osngz/AMKHTv8A4/R/w1j+yz/0WTwZ/wCFDp3/AMfo/wCGTv2Wf+iN+DP/AAntO/8AjFH/AAyd+yz/ANEb8Gf+E9p3/wAYoAP+Gsf2Wf8Aosngz/wodO/+P0f8NY/ss/8ARZPBn/hQ6d/8fo/4ZO/ZZ/6I34M/8J7Tv/jFH/DJ37LP/RG/Bn/hPad/8YoAP+Gsf2Wf+iyeDP8AwodO/wDj9H/DWP7LP/RZPBn/AIUOnf8Ax+j/AIZO/ZZ/6I34M/8ACe07/wCMUf8ADJ37LP8A0RvwZ/4T2nf/ABigA/4ax/ZZ/wCiyeDP/Ch07/4/R/w1j+yz/wBFk8Gf+FDp3/x+j/hk79ln/ojfgz/wntO/+MUf8Mnfss/9Eb8Gf+E9p3/xigA/4ax/ZZ/6LJ4M/wDCh07/AOP0f8NY/ss/9Fk8Gf8AhQ6d/wDH6P8Ahk79ln/ojfgz/wAJ7Tv/AIxR/wAMnfss/wDRG/Bn/hPad/8AGKAD/hrH9ln/AKLJ4M/8KHTv/j9H/DWP7LP/AEWTwZ/4UOnf/H6P+GTv2Wf+iN+DP/Ce07/4xR/wyd+yz/0RvwZ/4T2nf/GKAD/hrH9ln/osngz/AMKHTv8A4/R/w1j+yz/0WTwZ/wCFDp3/AMfo/wCGTv2Wf+iN+DP/AAntO/8AjFH/AAyd+yz/ANEb8Gf+E9p3/wAYoAP+Gsf2Wf8Aosngz/wodO/+P0f8NY/ss/8ARZPBn/hQ6d/8fo/4ZO/ZZ/6I34M/8J7Tv/jFH/DJ37LP/RG/Bn/hPad/8YoAP+Gsf2Wf+iyeDP8AwodO/wDj9H/DWP7LP/RZPBn/AIUOnf8Ax+j/AIZO/ZZ/6I34M/8ACe07/wCMUf8ADJ37LP8A0RvwZ/4T2nf/ABigA/4ax/ZZ/wCiyeDP/Ch07/4/R/w1j+yz/wBFk8Gf+FDp3/x+j/hk79ln/ojfgz/wntO/+MUf8Mnfss/9Eb8Gf+E9p3/xigA/4ax/ZZ/6LJ4M/wDCh07/AOP0f8NY/ss/9Fk8Gf8AhQ6d/wDH6P8Ahk79ln/ojfgz/wAJ7Tv/AIxR/wAMnfss/wDRG/Bn/hPad/8AGKAD/hrH9ln/AKLJ4M/8KHTv/j9H/DWP7LP/AEWTwZ/4UOnf/H6P+GTv2Wf+iN+DP/Ce07/4xR/wyd+yz/0RvwZ/4T2nf/GKAD/hrH9ln/osngz/AMKHTv8A4/Xt+k6tpWv6VZ67oV5DqOm6jDHc2t1bSLNBPBMoeOWKRCVdHUhlZSQQQQcV8QftN/syfs26B+zb8V9d0L4UeEtO1LTvCWu3NrdW2hWEM8E8NhM8csUiQhkdGAZWUgggEHNe4fsnf8ms/Bv/ALEzw9/6boKAP//Q+/f2+v8Am3L/ALLN4Q/9uq+/6+AP2+v+bcv+yzeEP/bqvv8AoAKKKKACiiigD5l/aT+Kvjf4XWXh248EW9vdTX89ybiKdC+6C1gM7hMEYbarY616b41+ItpoPwrvPiLo2LtZbGOfT1xkTzXYVbVMDrvkdBx61578aYIbr4kfCW2uEEkUurXqOrDIZWs5AQR6EV5L4AF5qfiLw9+z5e75E+HOsXt9ds+cyWFiVk0nJPBDtcxkD0h/GvraGBozw1GbjrG8pecby/KyS/xH5ti82xNLH4mkpu02oQ7Rnywat680pPyge4/Bz4oXfiD4O2vxB+JV5Z6dMkl2l3NkQW6fZ7mSEcsxA4UDryenXFeieEPiN4F8fJO3g3XLXVja481YJAXj3dCyHDAHsSMHtX5+Rx6g3wU+GMv2u3sdJi8W3zXU97E01nFKLu5+zNcorpmLzODuYKCRk17rp0eq/wDC8tI17X/Ful6rq+maRfPcWuiadMjzWDKNouJfPmUbZdrRBsEnIHUY1zDJKPNVmnZ3nZK9lyt6bP8ANWujmyTizFcmHpyjzLlpKTbV25xTcruS2vtyy5mmk0z3C++N3wj0zXW8NX/i3ToNRSTymiadfkkzjYzfdVs8EEg1t+MPiV4C+H/2ceM9dtdJa7z5KTSYdwvUqgyxUdzjA7mvgXxN4gutW/Zs1PUrCfw14X8I6hFO9npMSS32pSSvKSEeaSZQJi/zMwRio56Dj2PVYzN428N6z4L8XWeh+Nh4VtEktdYtjLZXtiW3gpMSrKwkLb/LYkjGQMEmanD9GLTbdryT9YpdVF231+JLuXQ41xVRSUYxbahKPTSbeylNOWiuruDf8vR/UaeOfB0ul6XrcOs2sun61PFa2U6Sq0dxPMSEjRhkFmIIx6gitS517RrPWLLw/dXkcWpaikslvAxxJKkGDIVHcLuGfrXwhc+KrDUPhv4Z1a40qw0C38N/Eezj1CXTnLabIYZiZryF26Qsz9TwCD9K9k8Q+JtC1v8Aaa8AadpF9Dey2Ol6u83kyLIE89I9gJUkAkITjrjB6EVx1ci5ZNa2Sm+n2Vdbaa6HqYbjDngn7t26SW6vzys7J66a29Op7HpPxU+HOvazb+HtG8RWd5qV1HJLFBHKGdkiZlcgf7JRsjrgE9Oar6R8X/hfr+vnwto3iiwvNV3MggjnVmdk+8qHo5HcKSRXyr4D8OvF+yR4m1PwtZqNdv01xzNEg+0ORczRNtYDdnyk2jBz6Vz+o2N7q3wl8HaWPHXh630pptMGjLp+lzyahHdo6GMRBLp380NkSnZnlt2DXZ/YOHc5xUnpJx76rrZJ6eWmz1PM/wBcccqVKo6cW5QVTsrO1o3lJWfeWqV0uXqfZvin4t/DPwTqcei+K/Ellpl9IocQzSgOqt0ZwPuA9i2Aa722uba9toryzlSe3nVXjkjYMjowyrKw4II5BHWvjnxHBNa/Efx7qPw58V6PDfXC2w17RfEluUt5NluER47jKyeU0WOgZAc84IFe7/A7X7HxR8J/DWuabpA0G1uLbEdkhLJCsbtHhCeSjbdyf7JFeVj8shToRqwv0vfu1fayfo9U1rdbH0WT59Vr4yphqtlbm5ba3UZct7ptPdXTUXF6We5ZsfGtvp174su/FGvad/Zmj3kEKCIsr2azIgEd0W43u7grjjBFXND+K/w28TeIJvC3h/xJY3+rQ7t1vDMrOdn3tuOG2452k4718Z/E9BJ4J/aHjbo+saSD+P2SvdPi7oukaJ4l+EA0izisxZeIY7WERIE8uCS2l3RrgcK20ZHfFd9TKKNo3bvK9rWsrU4y173bt07nj0eJcUnPlS5YNXvdt81apTsnfSyjfr2tbb6ZJAGTwBXy18L/AI36/wCL/iHJpmtQwReGvEqX0vhqdFKvMumTmGYOSTuLriVcAYUGu3/aD8U3/hz4a3lhoR/4nniaWLRdNUHBNzfnywQexRNzg+or528d+A/jT4G+H/hzW1Hh+W0+FjQX9uunpdpdyQWy7LhGaVmQrJEWaXgE4yKjJ8vpTpP2rSc3yxv5Ldf9vOK9LmvE+dYiliV9XUnGklOfKrp3ez/7cU3bvys+wfE3xI8B+DJpbfxVrtppcsMCXDJPKEbypHaNGAPJ3MrAAZJwa53xH8Q9K1P4aX/jTwD4l0uOGPYI9RumMllE3morCXZ8wODgDggkZryzS5PD3jP9p3S/FdvHHe28ngW3vbKR1DbfPv3KyLnOG2MRkcjJFeNfEK3gsfAX7Q9jZosNuus6dKI0G1Q832VnYAcZY8n1rXB5NSdSEHfm9xu9re9JRatbz/Sxz5nxRiY0a1WKi4fvIxte/uwlNSvfra2ivs79D7T8S/FH4eeDpbm38U+ILPTJrRYmkjnlCuBPu8vC9Tu2NgAHoazL/wCNfwk0xdNe+8XabCuroJLVjcIVkjZiofIJCruBG5sDIIzkGvONE0nTrz9qbxZqN3bpNcWnh/TkiZ1DFBNJJvxnpkKAcduK8j8LeGdAtP2cPjCbfT4ULXniTJCLnFsX8kZxnEe0FR2PI5rOjlOGajzOV/cvt9tenT8dtNzfE8R49Sn7NQsva2unf92+uv2tult9dj7E8XeP/BXgO1hvfGOtW2kxXBKxefIFaQjkhF+82O+Aa84+Ifxg063+DHiH4lfDTVLPV20yIGKVCJohLvRSrqCCCA2cHB5BryNPEt7P4y8GaH4XtNHtfE1r4RtLp9X1x5mVLWbarRW0EboGfcu533DjjOMivF7m8W7+HH7RG3U7XV8XGns9zYwi3tZJSFWRoow8gA3KRncd2N2ea68BkFO8JTvdON+zTmo7W036u77HmZxxlX5akKVknGok1upRpud7t6tNW0jZae9fQ/TS3dpII5G6uoJ+pFfLPwC+Nfirx/4u8TeFvGcdvG9s0txpbQIU821guZLWbfknLI6p0/vZ9K+pLP8A49IP9xf5V+e3hKeLwX4U8OfGFz5dt4d8V6xZak4BwNN1O6aF3fHJEUoicD6/j52UYWnVo1oSjeTso+T95q3q0l8z3OJcxrYfE4arCbUIqUpro4pwTb/wqTl8j3HxP8ZfEtp+0P4e+F+iRQHQpcQ6lK6Ey/apLeW5SONsgDbGqMeD9/6V614i+L3ww8Jayvh7xL4nsNO1E7SYZZlVkDfdMnZAe27FfKmi2F2fEnwg8X6pGYtS8Z67q+tzq3VEu7Nvs8fr+7t1iXHbBrufgzqngfTfB/jy0+IFxZwaoms6q2vpesod0aRjGXD/ADNEYSoj6qeQvOa78bllBQhJRb5Uk+Xdvmkm+umn4pHj5Vn+LdWpCU0ueTknO9ox9nCSja61tLvpaTs9j6V8Q+MPC3hPRv8AhIfEmq22naadoW4mlVY3LjKhCT8xYDIC5J7V5b4O+LNv43+K2o+HfDmoWmp+HINFtr6Ga3O9jcSXEkUis4PYKPlIBB6182fDL7RpafAm98fhrfSI7bWo7JrzKpFcySZsPMZ/lDNajEGcH+7zXR+IJrDUfir8YG+GjpNqr+DQsjWZDM2ofvR8pTrLs8scc7uvOaUckpQc6b1dn732VafL+l7362t1KnxZiKypV4pRjzRvDXnd6XtPu1ta3S9+h9O2Hxg+F2p+Iv8AhEtP8UafcauXMQt0nUs0i9UU52s47qCT7Vn+G/Ges6r8XPGngq68v+ztBtdLmttq4k3XiymTc2eRlBjgYr40ksrnVfgD4c0d/HHh+z0KZdPSxSy0yeXU4r0OhQRCO5ZzciXiQqmfvZGCa+mvAQkH7QnxKExDSDTtA3EDAJ8ufPFZ4vKaNGnVcXeye99GpQV9lvdq2tu+ptl3EmKxNbDxqJRTlF6NaqVOq7NKUtE4ppu3N2Vj6Jooor5M/RwooooA+AP+Co//ACYp8TP+4L/6eLKvv+vgD/gqP/yYp8TP+4L/AOniyr7/AKACiiigAr5n17xT8Xdd+MWtfD/wLqmmaVZaPp1peFr2ze4d2uCykApImMbfSvpivljT/EOgeH/2nPGUuvalbaak2h6YEa5mSEMQ7kgFyMke1exlEL+1lyptRurq+t10+Z8xxNUt9Xg5uMZTSbT5dOWT3VuqR0/hjx/8QNB+IVl8NPixb2Mk2uQTTaRqemrJHDctbKHmgkikZmSREO4HO0jpzWJpvi/4s/E7xH4pg8CarpXhzS/Cupy6Xsu7V726uZrfG+SQCWMRRMT8mASQCc1ma/4k0b4ofHrwFpvga9i1e18HC/1HVLu0cSwQ+fD5MEXmoSpd2zlM5289jjDm074G/FTxHrmo+InfwH450e7ms7qS31E6ffFYTiOcHKJKjoAyuUbjAzjFezHDQjapOnaTim7Rvyvmavyt7NJel7pao+WqY6rO9GnX5oRqNRvNxc0oRbSqJO7jJvtzWs37rOs8R/FX4leHvgb4q8X69o0Wj+KvDMzW3zRu1ndbJY0FzAGIYwyK52jcSCDk16N8HPHl945+H8Gt+JFjtdZsJbmy1SNPlSG6s5Gjk4JOAQA/XgGvkvxL4j13Wf2cvixpF7rb+KtH0G6itNM1mTBe8hEkLOGdeJTEx2+YPvV03j671LwV4u8WfDbRS0T/ABagsZNMZQdsV3cMtjqLZ/vCHbOT7fltUymE6bpKKU3Nu6vslC611slJyae1mctDiWtSrxxDnKVNU4pp21k5VUnpo23BQTWkuZM9J/Z9+MPin4reIfG8euQQ2+m6ZJZTaWqRlJDZ3omkiaUkncWiWNug6133x58ca18OfhZq/jDw6Ijf2L2ax+cpePE91FC2VBGflc45615J4K0nXNH+J3xi0P4cfYrS/sbfwtb2Iv1ke1SOKzK4dYirnEYIGD1xnjNZ/wAe4/iVH+zt4tHxLn0qe6NzpnkHSY544xH9ut87xOzEtnpjjFYSwVCeYU3BJQbp+71s1Fv89fmdtPNcXSyWuqjlKpGNf3+icZVEtejXKradj2vWPHGt2Pxt8PeAIPK/svU9KvLyXKEy+bA6qm1s8DBORisz4d/FCbVtM8f6141ubeysPCniPUtNSbHlolnaCPY0hJOW+c5PH0rC8R/8nR+Dv+xf1L/0alZPwQn0K20n4uT+JpLaLSk8ba2blrwotuI8Q5Mhk+QL654rmlhKX1ZS5deWO2+s5J/Ox3QzHEfXpQ9ppz1FrtZU4NX8k22fTtrdW99aw3tnIJYLhFkjdTkMjjKkexBzXz7qXjr4jeO/G+teC/hQ9hpen+GXSDUtXv4nuc3bru8i2gVkBMY++ztjJwAOCfoGzezks4JNPaNrVo1MJiIMZjI+UoV424xjHGOlfKHww8TaL8K/iR8QvAHjq8i0e417W7jX9NuLtlhhu7e+C5SOVyFLRMu0rnOc4zg44csopqrOMeaUVonr1V3brZf59D1s/wAVKMsPSnU5ITlaUk7fZbS5unM+qd9LJ6npfhTWvi1onjSHwb8Qba31vT7+3lmtNa022lgjjkg27obuMmRY2cHKMHAONoBOcdPrvxe+GHhnXF8Na/4nsLDUyVBglmVWQv8Ad3nome24ivnKy8Ra3pXxp8G+FdA+Jtz42gv5bx9UtFW1ljtoI4GMbSPbR/JmTA+Zgenrz5z8Pl8Rw/D3xta634q8O6Uj6hqn/CQW2qWMs14JWkcMZG+0xlwyYMWE9lywNezLJYTftKrS0Xwpq93JXtyu22yVnpqrny8OK61JOhh021KWs3GVlGMZW5lNJ/Fu5NpJ3Tsfd3iHx14N8JeV/wAJNrNrpYniknjNxKsYaKLaHYE8EAuo+pAHWk8JeO/B3jywk1TwdrFtq1tC2yR4HDeW2M4cdVOORkDivknQ/DVm/jH4A6XrE/8AbsVjo+ryQzXNu0RkVIYfIdoZcspVCuA3IIBp3j+z1SDxh8c7fwnG0d9eeF9PmKwDDySbZ1dgF5LmMEAjmuX+xKLtTUnzWvfp/E5Nt/Pfyseg+LMUr1pU1yJ8vKtZX9j7XdO2/u6LXe/Q+mdI+L/wv1/Xz4W0bxRYXmq7mQQRzqzOyfeVD0cjuFJIqzrXxT+HXhzVTomu+IbOx1ATRW5gllCyCWZQ8alevzKQc9ORnqK+LNRsb3VvhL4O0seOvD1vpTTaYNGXT9Lnk1CO7R0MYiCXTv5obIlOzPLbsGvdfAehaPf/ALRfxb1S/s4rm5tk0GOJ5EVzGrWhZtpI4yUUnH90elViMmw9NTnzO0U9Ot1KMd3FfzbWdrbkYLijHVpU6ShFSnKKvurOE57KbenJo20pJ3srHp2v/Gj4UeFtYbw/4h8VWFjqCEK8Mkw3Rk8gSYyEOP7xFUNf8c6nY/FbwP4R014ZdJ8R2ep3Ez43MxtUiaIxuDjB3nPByMV8q3Him4134Y+O9b0OXw54P8KXs+qfaLe6jlvdVu7jLIxlDzRok07D5Fw5XIxnv1Pw+lM3iD9nh2bcf+EY1EZ/3bW2H6Yrd5HTpxcne6U00+6g32smmtrv1ORcW1681TTVm6ck1p7rqxi1q7tNPdxjfXQ910j4teGNA8I6frHxF8W6T5l/cXkUV1bsY7aXyJmXYm/ndGu1X/2ga6rwp8U/h144up7Hwj4hs9WntY/OlSCQMUjBA3H0GSBXy78Ovihp/wANfgHo4iijv/EGsanq1tpVk7hBNOb+bLOzEBIY8hpHJAA4zkitDxJ4Xj0L4FePpdE1yDxH491ywkutWvLWVJJZQMCWOJUO5YIoiyRqAAB2ycVnXyalzyjK6bm4p6W+KzdraJeu+i622wnFOI9jTnBxklTU5Kz5l7l1Hm5tZSfTl0Wr3XN9F6H8Xvhh4m1xvDWgeJ7C/wBTBYCCKZWZyn3hGej477SaueKPid8PfBV/b6X4s8Q2WlXl0A0cVxMqOVJIDkHlUJBG44XPGa+ONWsb3VvBHgTTj468PJpy3mmnQ10zS52vVnjKmNYwl07KcArKSvHO7Br2D4f33hvSviN8WofHM9rbapPfRysb1lQSaObZBDgy8NEpDhsfKGyDWdfJqEE5xbaSenXRpbuK766OzVvM3wfFOLqONKajFya9525dYylaym3f3dG2rp3tpY9H+CvjbVviB4KbxDrJiaf7ff26mFdqGK3uHijPU87VGTnnrXOfHX4ieL/CMGj+H/hxBBdeJ9ZN3PFHcIZEFrp9u88xIUg7mISNMkDLVkfsnHTT8HoDooxp51LU/sw5GIftcnl9efu461xGmN8TPH/xl8VfEf4evpH2Dw/nw1atqq3DqTAVluniEDKOZTtLEnIAA701gqccdXbSUIN6PRXvZJ/Pp2TJlmteeUYSMZSdSqo6x1la3NKS+Ssn3aPpTw94/wDD2vfD+y+I7XKWukXNkt9JJI3ywps3SBz28sghvcGs+w+L/wAL9V8Rp4S07xPYXGsSHC2yTqXZsZ2jsWA6qDuHpXx/p0uueBvhx8ZPgj4sNul5pul32s6eLXeLY2OoRu0iQCT5xHDMSvzZOWPJ613/AMQNB0fRvgr8LxplnFbGw1bw3JCyIFZHd4w7ggZ3Nk7j1PeqnklBVOVt2lK0bWtZpNPz31X46EUuLcXKjzqKvCF5pp35lJxlFWemzabv00d9Pqey8WeGtQutWsrPUoJJ9CYLfpvAa2JUsPNBxtBUEgnggZzXIr42tta8ReEZPDOvafJo2uRX8nlNuNxeiBU2tasONsZJMhPUEYr5S/aItl134hXz+B7S7u10bTY/+E2FhKIvtOlmaORLTod1x5au/BU+VlckHA9W1i/0HVPi38FL/wAIPC2kT6ZrrWRiGIhD9lt/LAUdABgbeCMY6is45PCMIVLv3oydnbS0G9fXeO2mu+28+J6s61Sg0vcnBXTfvc1WMbryivdnv711ot/X9U+L/wAL9F8Qf8Irq3iiwtdV3rGYJJ1DI7fdVz0Rj2DEE1ynjL43+HfCHxS8N/Dm+u7WEavHO91LLIQ8DAKLZAvTMzMQCT/DgDuPkTwDFrg+A+vWHiHxb4e0uyd9RTXrbULCWbUUu2kkEvmkXKF5jwYiEz93aCRXtFsY/DvxC+Blvrep+cY9B1O3F1dK1s07eRbBNySncrsMZVvmz15rrnkeHpVJRbcrKa66tRbuvd/DXo7tHmUuLcbiaMJpKHM6Tvo7KVRRcWue+z+K0dmrJ6n1PpPijw9rukPr+kahDdadGZQ86MNimAlZAx7bCpBz0xV3SNX03X9Mtta0a4S7sbxBLDNGcpIjdGU9wexr4d8cvqvg/wAR+M/gZopaH/haF7aXWlOoyIU1JjFqx9hGsTPgdA+fr9yaXplloumWmj6bGIbSxhjghQdEjiUKqj6AAV4eY5fCjCM0782q/wANk9fm7eqZ9dkmdVcVUnSlFJ09Jf47tWXlZc2vSUS/RRRXkH0p4B+1j/yaz8ZP+xM8Q/8Apuno/ZO/5NZ+Df8A2Jnh7/03QUftY/8AJrPxk/7EzxD/AOm6ej9k7/k1n4N/9iZ4e/8ATdBQB//R+/f2+v8Am3L/ALLN4Q/9uq+/6/OD/gpNqOu6R4V+BereFtI/4SDWrL4s+GJ7HTftCWn266jjvGhtvtEgKRea4CeYwKpnceAa6D/hof8Abs/6NH/8v7R//jVAH3/RXwB/w0P+3Z/0aP8A+X9o/wD8ao/4aH/bs/6NH/8AL+0f/wCNUAff9FfAH/DQ/wC3Z/0aP/5f2j//ABqj/hof9uz/AKNH/wDL+0f/AONUAfd15o+kajdWd9qFjBdXOnuZLaWWJXeB2G0tGzAlCVOCVwccU2DRNGtdUutctrC3h1K9VEuLlIkWeZYxhFkkA3MFH3QScdq+E/8Ahof9uz/o0f8A8v7R/wD41R/w0P8At2f9Gj/+X9o//wAaq/aSta5m6ML81lff52tf7tPQ+4ovC3hmDRZPDcGkWcekzb99mtvGLZvMYs+6ILsO5iS2RyTk81U8NeCPBvgxJk8JaHZaMLkgy/Y7eODeR03bAM47Z6dq+KP+Gh/27P8Ao0f/AMv7R/8A41R/w0P+3Z/0aP8A+X9o/wD8aqvbzs48zs99dzNYOlzRnyK8dE7LRdl2PsS3+F3w1tb671O28KaVHd36SR3Eq2UIeVJgRIrnbyHBIcHhs85q3rXw98B+I7Cz0rXvDun6hZ6coS1hntYpI4EUBQsSspCKAAMLgYGOlfGH/DQ/7dn/AEaP/wCX9o//AMao/wCGh/27P+jR/wDy/tH/APjVX9bq3Uud3XmzJZbh+Vw9nGz3VlZn3H/wjHhs6F/wi/8AZNp/Y2zyvsXkR/ZvLznb5ONm3POMYrP0XwF4H8OfZT4f8P6fprWRlMDW9rFE0RnCiUoVUFTIFUOR94AZzgV8Wf8ADQ/7dn/Ro/8A5f2j/wDxqj/hof8Abs/6NH/8v7R//jVQq87Nczs/M0eDotqTgrrRaLReR93aVpGk6FZJpuiWUGn2kZdlht41ijVpGLuQiAAFmJY8ckknmucsPhv8PdL1xvE2m+GdNtNWcsxu4rSJJ9zfebeqhstnk5ye9fGn/DQ/7dn/AEaP/wCX9o//AMao/wCGh/27P+jR/wDy/tH/APjVEa81e0nrvrv6jlg6UuXmgny7aLT07fI+zvEPw78A+LbyHUfFPhzTtXuoBtSW7tYp3VRzt3OpOM9uldbDDFbxJb26LFFEoVEUBVVVGAABwAB0FfAn/DQ/7dn/AEaP/wCX9o//AMao/wCGh/27P+jR/wDy/tH/APjVKVWckot6LYqnhqcJSnGKTe7tq/Xufb1z4R8KXsOo215otlPFq7pJepJbRMt08eNjTAqRIy7VwWyRgY6CtC+0jSdUltJ9TsoLuSwlE9u00ayNDMAVEkZYEo4BI3DBwTzXwj/w0P8At2f9Gj/+X9o//wAao/4aH/bs/wCjR/8Ay/tH/wDjVHtZdw+r09fdWvl53/N39T7sv9F0bVLizu9TsLe8n06TzraSaJJHgkxjfGzAlGxxlcGrtxbwXcElrdRrNDMpR0cBldWGCrA8EEcEHrXwN/w0P+3Z/wBGj/8Al/aP/wDGqP8Ahof9uz/o0f8A8v7R/wD41U87012LVOOrtvv5n3BpfhTwtok8NzoujWVhNb24s43t7eOJktg5kEKlFBEe8ltg+Xcc4zTbrwj4UvYdRtr3RbK4h1h0kvUktonW6ePGxpgVxIV2jBbOMDHSviH/AIaH/bs/6NH/APL+0f8A+NUf8ND/ALdn/Ro//l/aP/8AGqv28783M7mX1Sly8nIrdrL0/LQ+7otI0mHU59ahsoI9RuY0iluVjUTSRxklEaQDcyqScAnAycVVj8NeHIdNvNGi0q0TT9QMrXNusEYhna4z5pkjA2uZMnfuB3d818N/8ND/ALdn/Ro//l/aP/8AGqP+Gh/27P8Ao0f/AMv7R/8A41U+1l3/AKRfsIfyrr077/f1Ps7XPh54C8TQ2dt4i8OadqUWnKEtkuLWKVYUAACxhlIVcADaOOOlWW8EeC3t720fQNPaDUYoYbqM2sRSeK3GIklXbh1jHCK2QvbFfFH/AA0P+3Z/0aP/AOX9o/8A8ao/4aH/AG7P+jR//L+0f/41Wn1mpZLmdvUxeAoczl7NXe+i7W/LT0Pv4AKAqjAHAArnz4S8KHR7jw8dFsv7Ku2d5rT7NH9nlaRt7s8W3YxZvmJIOTyea+If+Gh/27P+jR//AC/tH/8AjVH/AA0P+3Z/0aP/AOX9o/8A8arONSS2ZvOjCXxJPp8nufdU2haHcTWFzcadbyy6USbN2hRmtiy7CYSRmPK/KduOOOlYms/D3wF4j1SLW/EHhzTtS1CEAJcXNpFLKAvQb3UnA7c8dq+MP+Gh/wBuz/o0f/y/tH/+NUf8ND/t2f8ARo//AJf2j/8AxqqhXnF3jJr5mdXCUprlnBNb6pbn3VrGhaJ4h06TSNf0+31KxlxvguYllibHTKOCOO3FUNE8HeEfDLK/h3RLLS3SEW4a1to4WEIYuI8ooO3ezNjpuJPUmviT/hof9uz/AKNH/wDL+0f/AONUf8ND/t2f9Gj/APl/aP8A/GqSrTUeRN27DlhqbmqjiubvbX7z7Lt/hv8AD2018+KrXwzpsOsli5vEtIVn3nq/mBd2455bOT610cGkaTa6jdaxbWUEV/erGlxcJGqzTLFkRiRwNzBMnaCTjJxXwj/w0P8At2f9Gj/+X9o//wAao/4aH/bs/wCjR/8Ay/tH/wDjVOdecvik30+QqWDpQ+CCWt9Et+/r5n3/AEV8Af8ADQ/7dn/Ro/8A5f2j/wDxqj/hof8Abs/6NH/8v7R//jVZHQff9FfAH/DQ/wC3Z/0aP/5f2j//ABqj/hof9uz/AKNH/wDL+0f/AONUAH/BUf8A5MU+Jn/cF/8ATxZV9/1+IP7evxl/ax8V/sneOdA+Jn7Ov/CCeG7r+zPtetf8JdpuqfZdmpWrx/6JbxrLJ5kipH8p+XfuPCmvr/8A4aH/AG7P+jR//L+0f/41QB9/0V8Af8ND/t2f9Gj/APl/aP8A/GqP+Gh/27P+jR//AC/tH/8AjVAH3/XGa58OPh54nvzqniXwvpWrXpUIZ7uygnlKr0XfIjNgdhmvjP8A4aH/AG7P+jR//L+0f/41R/w0P+3Z/wBGj/8Al/aP/wDGq0pVZwd4Oz8jGvhqdWPLVipLzVz7o0Xw/oPhy0+weHtNttLtc7vKtYUgjz67UAGaxvEXw98BeLrhLvxV4c07V54wAsl3axTuFU5ADOpOPbOK+MP+Gh/27P8Ao0f/AMv7R/8A41R/w0P+3Z/0aP8A+X9o/wD8apxrzUudSd+99SJ4OjKn7KUE49rK33H3HN4Y8NXGhnwvPpNpJoxQRmyaCM22wHcF8krswCM4x15qxdaHol9f2Oq3un29xe6Xv+yTyRI8tv5q7X8pyCyblGG2kZHBr4U/4aH/AG7P+jR//L+0f/41R/w0P+3Z/wBGj/8Al/aP/wDGqXtZd/6e/wB5f1en/KunTtqvue3Y+7rbR9IstQvNWs7GCC+1Hy/tVxHEqyz+Su2PzXA3PsU4XcTgcCk1fRtI1/T5NJ16xg1KxmKl4LmJZonKMHXcjgqdrAEZHBAPWvhL/hof9uz/AKNH/wDL+0f/AONUf8ND/t2f9Gj/APl/aP8A/GqXtJXUr6lexhyuFlZ3uvXf7+p92yaNpE2qQ65NYwPqVtG0MV00SmeONzlkWQjcFYjkA4NUv+ET8LfYdR0v+xrL7Hq8sk97B9nj8q6llx5kkybdsjPgbmYEnAzXw/8A8ND/ALdn/Ro//l/aP/8AGqP+Gh/27P8Ao0f/AMv7R/8A41TVWS2ZLw9N3vFfd30f4H31b28FpBHa2sawwwqEREAVUVRgKoHAAHAArF8Q+FPDHi6zGn+KdJtNXtlO5Y7uBJ1VvUBwcH3FfD//AA0P+3Z/0aP/AOX9o/8A8ao/4aH/AG7P+jR//L+0f/41SjUlGXNF2ZVSjCcXCaTXbofbnh3wf4T8IW72nhTRrPR4ZMb1tII4A5HQtsA3H3NZ2s/Dj4feI9Vj13X/AA1pupajFt23FxaRSy/L9352Uk7e2Tx2r40/4aH/AG7P+jR//L+0f/41R/w0P+3Z/wBGj/8Al/aP/wDGqtYmopOak7976mMsDQdNUnBcq2Vlb7j7um0fSLjULXVrixglvrBZEt7holaWFZQBIsbkblDgAMARnAzRFpGkwalPrMFlBHqF0iRzXKxqJpI487FeQDcyrk4BOBk4r4R/4aH/AG7P+jR//L+0f/41R/w0P+3Z/wBGj/8Al/aP/wDGqz55bXNvYwvey7/O1r/dp6H2XYfDf4e6XrjeJtN8M6baas5ZjdxWkST7m+829VDZbPJzk966O20jSbO/vNVs7KCC91Hy/tM8carLP5K7Y/NcAM+xeF3E4HAr4R/4aH/bs/6NH/8AL+0f/wCNUf8ADQ/7dn/Ro/8A5f2j/wDxqrnXnL4pN9N+hnSwdKnpCCWt9Elr39fM+yx8OPh8uvSeKR4Z03+2JWLPefZIfPZm4LGTbu3EHk5ye9atn4T8Lac2nvp+jWVq2kpJFZGK3jQ2scuPMSHao8tXwNwXAPevh/8A4aH/AG7P+jR//L+0f/41R/w0P+3Z/wBGj/8Al/aP/wDGqcsTUejk/vJhgaEXeMEuuy33v96T+R9j33wy+G2p2ltYal4T0m7trIytBFLYW8kcRnbfKUVkIUu3zNgDceTk1Y0D4e+AfCl4+o+FvDWmaPdyRmJprOzht5GjYhihaNFJUlQSM4yB6V8Yf8ND/t2f9Gj/APl/aP8A/GqP+Gh/27P+jR//AC/tH/8AjVU8XVceVzdvVkLLsOpqoqceZdbK/wB59l6X8N/h7omtP4j0bwzptjqj7ibqC0ijmy/3jvVQQWzyc896ua/4I8GeKri3u/E+g2OrT2ZzC93bRTtHzn5S6kjnnA7818Uf8ND/ALdn/Ro//l/aP/8AGqP+Gh/27P8Ao0f/AMv7R/8A41SeKqc3PzO/e5SwFBQdNU1yvW1la/ex936bpOlaLbGz0ezhsbdneUxwRrEhkkYs7bVAG5mJLHqScmmaToujaBZ/2foVhb6da72k8q2iSGPe53M21ABljyTjk9a+E/8Ahof9uz/o0f8A8v7R/wD41R/w0P8At2f9Gj/+X9o//wAarN1JO93ubxowVrJabeR9wal4U8Lazdvf6xo1lfXMls9m0s9vHLI1tJy8JZlJMbZOUztPcVZutA0K+srbTb7Tra4tLNonghkhR44ngwYmRCCFKYG0gfL2xXwt/wAND/t2f9Gj/wDl/aP/APGqP+Gh/wBuz/o0f/y/tH/+NU/bT01ehP1anq+Va76b+p91adoWiaQ95JpOn29k2oTNcXJgiSMzzP8AekkKgb3PdmyT61Q0/wAG+ENINgdK0OxsjpfnfY/Jtoo/s32j/XeVtUbPM/j243d818Sf8ND/ALdn/Ro//l/aP/8AGqP+Gh/27P8Ao0f/AMv7R/8A41R7aeur1F9Vpae6tNtPO/5pP1PsrUvhv8PdZ1pPEer+GdNvdVQqRdTWkUk2V+6S7KWJXHBJ47Vsa54W8M+JxbL4k0m01UWcglg+1QJN5Ug6Om8Ha3HUYr4d/wCGh/27P+jR/wDy/tH/APjVH/DQ/wC3Z/0aP/5f2j//ABqq+sVNHzPTbXb0I+o0bSXIve30Wvr3PpPTPAvi/WfiqPiD48bT1tfD8N1aaHBZGR3CXTjfcXDSKu2QxKqbFyoy3Pr7ZXwB/wAND/t2f9Gj/wDl/aP/APGqP+Gh/wBuz/o0f/y/tH/+NVWIxMqrXN0Vl5L/AIfX1IwOAhQUlC7cndt7t7fkkl5JH3/RXwB/w0P+3Z/0aP8A+X9o/wD8ao/4aH/bs/6NH/8AL+0f/wCNVznafQH7WP8Ayaz8ZP8AsTPEP/puno/ZO/5NZ+Df/YmeHv8A03QV8P8A7Qvx2/bO1f4BfEvSfFP7L3/CP6Le+GdZgvtS/wCE20q7+w2sllKs1z9njiDy+UhL+WpDPjaOSK+4P2Tv+TWfg3/2Jnh7/wBN0FAH/9L79/b6/wCbcv8Ass3hD/26r7/r4A/b6/5ty/7LN4Q/9uq+/wCgAooooAKKKKACiiigAooooAKKK/K/xB+0f8LfgL+3v8SZPjH4tHh3SdQ8K6FHZLOtxLE0yvI0mxIUkCnbjJwM+tAH6oUV+T/iT9qT4O/Hn9s39mvTPgr4zXxBFp1x4qOpxW63MKBZdJJg8xZUjD5MbleuCO3FfR3iT9uj4WeHNe1qA+H/ABJqXhPwvqX9k6z4ss9OWXQNOvQ6xyRzT+aJWWF2CyvHC6oSOTkGgD7Sor5C+J37ZvgD4TeI9S07xN4W8US+G9CmtbfVPFFtpfmaFYy3gjaNZLkyK7gCVN7RRuqlgudxwOg+KH7VXgv4X/Ez/hT83h7X/Eni2bQRr9pZaLZpdveQG5e2MMQMqMJVMbyMXCxhB/rNxCEA+naK+C/Fv7cf9gfEDwT4J0f4S+M9afxXocmuyQQ6ckeowR4TZCLaeaJWeMuPtB8zEZ2qN7Ftn25qetW+kaBd+IryKRYLK1kupIwB5oSNC7LgnG7AxjOM96ANiivirwd+3N8PPHOj6d4s0HwZ4wl8LahpV1qn9tDR/M0+I2cLTS2jSwyyFrkbDGFjDoZf3QcuGVer+HH7WnhT4geI9Z8EX3g7xV4Q8VaPpDa8uj63pfkX15piyeV59rHDJMJP3hEezIYOduODgA+qqK+GfgV+2/ofxV+Hms/E7xl4J13wJ4Z0W21TUJda1CCI6S9rp949uI4p45XlkudijfGIQPMWRELYUt3Hwu/a68G/ErxnpHga/wDCfibwRf8Aii0mvtBfxFp6WcWsW9sokmNo0c0vzpGRIY5Aj+WQ2MGgD6uor55/ag8aaP4B+FEniHXdX1zQ7RdU0iA3Ph77P9v33F/DGiD7TiPypGYJN38stgE8HhPHP7ZfhHwd498cfDfT/A3i3xXrPw9jsbjVv7F06G5gjtr62+1LMsr3EYwkfDI22Rmz5aOquygH2DRXyx4r/bA+E3h3wF4H8c6NHqfi2T4lKD4c0nRbQ3OqajhBJLsgdowggU5maR1CdCc8Vn65+2H4J8G+AvDvj74ieEvE/hGLxD4ki8LrY6lp6R31veTQyTpLJEszb7crGdrwGVnJAVCcgAH1vRXx7fftm+EdH8Mpq/iDwH4x0nW77WG0TS/DtzpKprOq3KwrcF7OATFHhWJg7ytIqpyrENgG7pv7YfgbVPBet+IoPCvij/hIvDupQaRfeExpm/xDFe3KebAn2ZJGjKSRZlSXzfKKAncCCKAPrWivkrSf2zfhRd/C3x18UdfsdZ8Mx/DaVbfX9K1Oy8rVLOeQIYkMKO6N5vmKI2VypzkkDmug0v8Aah8Ir8OvE/xO+IXhzxB8OtH8JQRXV3/wkNisDyQz58prcwSTpOXYBAqOWDsqkAkCgD6Vor5g+FX7VHhf4meNY/h5qnhLxL4B8QX1i+p6da+JrBLJtRs4mVZJLZo5plLR71MkbFZFBztwCR9P0AFFFFABRRRQAUUUUAFFFFAHwB/wVH/5MU+Jn/cF/wDTxZV9/wBfAH/BUf8A5MU+Jn/cF/8ATxZV9/0AFFFFABRRRQAUUUUAFFFFABRRRQAUUV+V/iD9o/4W/AX9vf4kyfGPxaPDuk6h4V0KOyWdbiWJpleRpNiQpIFO3GTgZ9aAP1Qor8n/ABJ+1J8Hfjz+2b+zXpnwV8Zr4gi0648VHU4rdbmFAsukkweYsqRh8mNyvXBHbivo7xJ+3R8LPDmva1AfD/iTUvCfhfUv7J1nxZZ6csugadeh1jkjmn80SssLsFleOF1QkcnINAH2lRXyF8Tv2zfAHwm8R6lp3ibwt4ol8N6FNa2+qeKLbS/M0KxlvBG0ayXJkV3AEqb2ijdVLBc7jgdB8UP2qvBfwv8AiZ/wp+bw9r/iTxbNoI1+0stFs0u3vIDcvbGGIGVGEqmN5GLhYwg/1m4hCAfTtFfBfi39uP8AsD4geCfBOj/CXxnrT+K9Dk12SCHTkj1GCPCbIRbTzRKzxlx9oPmYjO1RvYts+0fEfiay8L+EdU8ZanDN9k0ixnv54kVTN5dvEZXUKWC78KQAWAz370AdHRXxL8PP27/hh46vfCsGseFPFngaw8cvDFoWp+IdKW30zUZrld0EUN3bzXEQeYcxhyu7oOSAeq8d/tf+BPB/jDWfBGgeFvFPj/UPDAT+2n8MaS2owaW8ieYsdxKXjUybPmMcW9wOoBBAAPrCivnW0/aj+FGrr8L7rwzc3Gu6f8XLm4tdGvLSNfJSS1geeX7SJXjki2iNkKhGdXG1lGCRmeNv2r/h14DuPi1baxp+qzN8GrHS9Q1jyIYGE8WrxvJALTdOu9lEZ3iTywDjBbsAfTtFfKngL9rjwT44+I2h/DW78MeJPCl74tsp7/w/c65p62ltrENqglm+zESvIHSNhIUmjjbZzjkZ+Uf2xP2uND8XfAT4veFfhh4X8Ua5YaTb3OlXPirTrSNdFtb+F1EqC5aeOZxEfleSKJlBIwSDmgD9WaK+J7H40eFfA/jfxTPrWqeIryTwb8NrHxHfaYotn0sWcAnkaW1DMsrXsnlsreYyxlQvzf3dLwL+2v8AD3xv4l8FaFN4V8UeG7H4iwq/h/VtY05LbT7+Yw+ebdHWZ3WTZnaWQJJgmN3UqzAH2NRXwz40/b4+GvhTX59G0nwX4v8AFlnBrf8Awjn9raTpsH9lPrG/yjZpd3l1bI0gk/d5HyluFY8V9wW8pnt452jaEyKrFHwHXIztbBIyOhwTQBNRRRQAUUUUAFFFFABRRRQAUUUUAeAftY/8ms/GT/sTPEP/AKbp6P2Tv+TWfg3/ANiZ4e/9N0FH7WP/ACaz8ZP+xM8Q/wDpuno/ZO/5NZ+Df/YmeHv/AE3QUAf/0/v39vr/AJty/wCyzeEP/bqvv+vgD9vr/m3L/ss3hD/26r7/AKACiiigAooooAKKKKACiiigAr5B8H/Cvxrp37Z/xG+LGqaUE8Ka74a0awsbwywv5t1au5mTyg5lXaCPmZAp7E19fUUAfIHxn+FXjTxT+09+z18RPDWkrceH/A8/iZ9ZuhLDH9lXUNNWC2Plu6ySb5AV/dq23q2BzXxbB+y78RvhvqHjT4d3fwOT4veHNe1jUL/S9WXxa2kWy2uoy+d9m1GykmBJhZmDSRxuZB0XPJ/ZGigD8i/2q/gv+1F8U9G+JHw6tPCV7rugz6VpUHhBNO123sNHs/Iiia9W7tpJ4Z7q4MyMsJnWSILtIMbDJ+pvC3w++Imqftg6d8ddf8LNouiSfDSLRHM1zaTy2uqHVjdPasIZXYsIcM0iBoj90OTX2dRQB8SfHvwh8YtE/aE8AfHz4W+DR49t9E0bVtEvtNj1G3064Q3zwyQzI90VjZA0ZD87gOQpr6k12313xF8N9QtbiwFprOp6TKj2azLKIrqaAgwib5FYK527/lB68Cu3ooA+Qfgl8P8A4t/C/wDYp0L4eaFYW2j/ABK0TwvLb2tvcPDLBDq/lOY/MeJniYecQWIZlOSSSM186fAn4Z/H7Qv2m/CvxZ8aeAtcjt77wZcaBrmpav4gsdSuE1N7uC7kuPKjuXWK2YxFIYbVAoyCYo/mNfqTRQB+UHg39nv49ap+zN4t/Yn8X+DotG0lodaNh4vXVbea0uZbnUn1GyAs483Khnk2y71Xaq9GLYrrfgL8LfiBB8Q/Bd749/Z8j8Maj4Y843Hiafxe2qQRyNavC0mn2fnSybrgsAyyKgRGYEsQM/plRQB8q/tl/Dbxp8V/ghL4Q8Aad/amrtrOh3Yg86KD9zZ6jBPO2+Z0T5I0ZsbsnGACcCs34a/C7xxoP7R37RPjvWtLEOgeO4vDKaPcGaFxdnT9Mkt7keWrl49kjBf3iru6rkc19eUUAfkn4C/Z3/aG+Evw5/Z0+Img+ELbX/Gnwn03WNK1jwxcahbQSy22rucyWl5ve1WaIKCcvh0YruBGK9w+Kfhv45fH/Q/g/rGu/DNvCl54T+J+j63e6dPqtjfSRaLY28+69keN1j3CWUL5EZkkGAwzk7fvyigD4Y/bM+BHjT4m6p8N/id4A0WLxZqnw5vb95dAm1F9J/tKy1SBYZ1ivUZfJmQxoyFiFI3AkjKt5RY+APj1ovw58ba/8Hvg+/wu8S6/e6ElzCPEkGqa7qmmWkkgvfLubiSW0tp0gk2WxeQkbnJ2sI8fp9RQB+V/wy+Ffxk8HX/x/F/8Fp9c0Tx+ukXVhpmu+IrS9bUYoYIbS7s7q7luLmT7S0Zll3yFog6hBIVKtXNaB+yl8c/HX7O3xh+CmoWVz4F8KavLpdz4E0PW9Vj1i50qWwkW6nglubdpAtpLNFGsSGSRo1LNjd979daKAPzp+AXw48Zr8UfD3iPxj+z4vgS90CC7WTXrrxc2tBHnhMTJp9sJpW2z/LvMoTaox8zAEfotRRQAUUUUAFFFFABRRRQAUUUUAfAH/BUf/kxT4mf9wX/08WVff9fAH/BUf/kxT4mf9wX/ANPFlX3/AEAFFFFABRRRQAUUUUAFFFFABRRRQAV8g+D/AIV+NdO/bP8AiN8WNU0oJ4U13w1o1hY3hlhfzbq1dzMnlBzKu0EfMyBT2Jr6+ooA+QPjP8KvGnin9p79nr4ieGtJW48P+B5/Ez6zdCWGP7KuoaasFsfLd1kk3yAr+7VtvVsDmvi2D9l34jfDfUPGnw7u/gcnxe8Oa9rGoX+l6svi1tItltdRl877NqNlJMCTCzMGkjjcyDoueT+yNFAH5F/tV/Bf9qL4p6N8SPh1aeEr3XdBn0rSoPCCadrtvYaPZ+RFE16t3bSTwz3VwZkZYTOskQXaQY2GT9TeFvh98RNU/bB07466/wCFm0XRJPhpFojma5tJ5bXVDqxuntWEMrsWEOGaRA0R+6HJr7OooA+JPj34Q+MWiftCeAPj58LfBo8e2+iaNq2iX2mx6jb6dcIb54ZIZke6KxsgaMh+dwHIU17z8WLjUbv4AeMrvWLMaffzeGNRe4tllEwgmaycvGJAFDhGyu4ABsZwK9irF8SaDYeKvDuqeF9VDGy1i1ns5wjbX8q4jMb7T2O1jg9qAPyG+FXhf48/tFfs5fAT4RnwB/wiPgnQ18N6vfeJrvU7O4F5Y6UiTRJY2sDNcJNcEKd0gTyxkHOa+hND0T9pL9mrxr8TLD4d/DGP4peG/iB4ivfFNhewazaaXPZXepInn2t6l4QTGjx/u5It+E4IyQq/b3w98DaF8MvAugfDvwwJF0jw3ZQafaCZ/MkEFsgjTe2BubAGTjmuxoA/KyH9mH43fCz4c/CLxXoOmWnjXxr4G8War4o1nRrK6SyhmGvLOLmCwmutsY+z+au0OVD4YjBIByPGXwP/AGj/AIl6V+1brmq+AV0PUfitonhm08P6eNVsrl5n02KeOaOWVZFjjkTcpbcRHlsI8gBY/rVRQB8b+O/hT481n40/s1eL9N0rztL+H/8AbY1ufzoVNoLzR/ssPys4eTfNhf3QfHVsLzXyF4n+Cv7T/gD9mnx/+yJ4O+GC+NtI1ibVf7G8R2Ws6daZttRujdoL22vZIJfOjLsjFWcHAAOACf2FooA/N7xr8BPizq/jH4uarp+heba+J/g9H4W05/tVsvn6ysd0ptsNKCnMifvHCx8/f4OOh8RfBP4mXnw4/ZS0W30YNf8Awz1Xw7c+IENxbj7DBY6VJbXLbjIFlCSkLiIuW6qCOa/QCigD8IPD/iP4ezePvEvxL8eWnjaL9nxfGZ8R6JNpNq0vg+W4WZGOoXzTKL5YzfF5CkaC23j5dy/e/d1HSRFkjYMrAEEHIIPQivkV/wBh34ASQHQzaawPCLTG4Phca7qQ8PGYzefu/s77R5O3zfn8rHlZ/gr67VQoCqMAcADtQAtFFFABRRRQAUUUUAFFFFABRRRQB4B+1j/yaz8ZP+xM8Q/+m6ej9k7/AJNZ+Df/AGJnh7/03QUftY/8ms/GT/sTPEP/AKbp6P2Tv+TWfg3/ANiZ4e/9N0FAH//U+3f+Ck2gf8JX4V+Bfhb+0r3R/wC2Piz4Ys/t2mzfZ761+0R3kfn202G8uaPdujfB2sAcHFdB/wAMC/8AVxvxm/8ACv8A/uWj9vr/AJty/wCyzeEP/bqvv+gD4A/4YF/6uN+M3/hX/wD3LR/wwL/1cb8Zv/Cv/wDuWvv+igD4A/4YF/6uN+M3/hX/AP3LR/wwL/1cb8Zv/Cv/APuWvv8AooA+AP8AhgX/AKuN+M3/AIV//wBy0f8ADAv/AFcb8Zv/AAr/AP7lr7/ooA+AP+GBf+rjfjN/4V//ANy0f8MC/wDVxvxm/wDCv/8AuWvv+igD4A/4YF/6uN+M3/hX/wD3LR/wwL/1cb8Zv/Cv/wDuWvv+vK/jF8ZvAfwK8HHxt8QLqWGzluYbK1gtoXubu9vbgkQ21tBGC0kshB2qOwJJABIAPlT/AIYF/wCrjfjN/wCFf/8ActH/AAwL/wBXG/Gb/wAK/wD+5a+iPgr+0J4S+OMuuafouja94b1fw79ma+03xDpc2l3kUd55v2eQJKNrJIYZACrHG05AyM+m+O/GOj/DvwR4h+IHiHzP7K8M6dd6nd+Su+T7PZQtPLsXI3NsQ4GRk96APir/AIYF/wCrjfjN/wCFf/8ActH/AAwL/wBXG/Gb/wAK/wD+5a+sfDHxh+H3iy+8OaPpmqKmr+KtDj8R2NhKCly+mSeWPOK8qMGVVI3ZznGQCa9OoA+AP+GBf+rjfjN/4V//ANy0f8MC/wDVxvxm/wDCv/8AuWvv+oLm5t7O2lvLqQRQQI0kjscBUUZJJ9AKAPgf/hgX/q434zf+Ff8A/ctH/DAv/Vxvxm/8K/8A+5a+2fBnjLwx8QvC2m+NvBeoR6toesQie0uos7JY24DDcARyMYIBFdNQB8Af8MC/9XG/Gb/wr/8A7lo/4YF/6uN+M3/hX/8A3LX3/RQB8Af8MC/9XG/Gb/wr/wD7lo/4YF/6uN+M3/hX/wD3LX1p47+K/hT4d+IPBvhnxD5/2zx1qR0rTvJjDp9pWF5z5pyNq7EPODz2r0ugD4A/4YF/6uN+M3/hX/8A3LR/wwL/ANXG/Gb/AMK//wC5a+/6KAPgD/hgX/q434zf+Ff/APctH/DAv/Vxvxm/8K//AO5a+/6KAPgD/hgX/q434zf+Ff8A/ctH/DAv/Vxvxm/8K/8A+5a+/wCigD4A/wCGBf8Aq434zf8AhX//AHLR/wAMC/8AVxvxm/8ACv8A/uWvv+igD4A/4YF/6uN+M3/hX/8A3LR/wwL/ANXG/Gb/AMK//wC5a+/6KAPgD/hgX/q434zf+Ff/APctH/DAv/Vxvxm/8K//AO5a+/6KAPgD/hgX/q434zf+Ff8A/ctH/DAv/Vxvxm/8K/8A+5a+/wCigD4A/wCGBf8Aq434zf8AhX//AHLR/wAMC/8AVxvxm/8ACv8A/uWvv+igD8Qf29f2Q/8AhV/7J3jnx1/wur4meLf7M/sz/iV+IfEf2/S7jztStYf39v8AZ037N+9PmG11Vu2K+v8A/hgX/q434zf+Ff8A/ctH/BUf/kxT4mf9wX/08WVff9AHwB/wwL/1cb8Zv/Cv/wDuWj/hgX/q434zf+Ff/wDctff9FAHwB/wwL/1cb8Zv/Cv/APuWj/hgX/q434zf+Ff/APctff8ARQB8Af8ADAv/AFcb8Zv/AAr/AP7lo/4YF/6uN+M3/hX/AP3LX3/RQB8Af8MC/wDVxvxm/wDCv/8AuWj/AIYF/wCrjfjN/wCFf/8Actff9FAHwB/wwL/1cb8Zv/Cv/wDuWj/hgX/q434zf+Ff/wDctff9FAHwB/wwL/1cb8Zv/Cv/APuWj/hgX/q434zf+Ff/APctff8ARQB8Af8ADAv/AFcb8Zv/AAr/AP7lo/4YF/6uN+M3/hX/AP3LX3/Xj3xx+N3g79n7wMfiB44hvbjTvtdtZLFp9v8Aabl57p9kSpEGUsS3HBz7UAfMP/DAv/Vxvxm/8K//AO5aP+GBf+rjfjN/4V//ANy11en/ALeXwWTVLHTPHeleKPh2mpzpbW934o0C80uyeaQ4RGuXQxJnP3nZVHcivtNWV1DKQQRkEdCKAPgH/hgX/q434zf+Ff8A/ctH/DAv/Vxvxm/8K/8A+5a+/wCigD4A/wCGBf8Aq434zf8AhX//AHLR/wAMC/8AVxvxm/8ACv8A/uWvv+igD4A/4YF/6uN+M3/hX/8A3LR/wwL/ANXG/Gb/AMK//wC5a+/6KAPgD/hgX/q434zf+Ff/APctH/DAv/Vxvxm/8K//AO5a+/680+MPxX8K/A/4caz8UvG/n/2JoSwtcfZoxLNieZIF2oSufnkXPPTJoA+S/wDhgX/q434zf+Ff/wDctH/DAv8A1cb8Zv8Awr//ALlr7/ooA+AP+GBf+rjfjN/4V/8A9y0f8MC/9XG/Gb/wr/8A7lr7/ooA+AP+GBf+rjfjN/4V/wD9y0f8MC/9XG/Gb/wr/wD7lr7/AKKAPgD/AIYF/wCrjfjN/wCFf/8ActH/AAwL/wBXG/Gb/wAK/wD+5a+/6KAPgD/hgX/q434zf+Ff/wDctH/DAv8A1cb8Zv8Awr//ALlr7/ooA+AP+GBf+rjfjN/4V/8A9y0f8MC/9XG/Gb/wr/8A7lr7/ooA+AP+GBf+rjfjN/4V/wD9y0f8MC/9XG/Gb/wr/wD7lr7/AKKAPyg/aF/Yk/4RT4BfEvxT/wAL8+LOsf2P4Z1m8+w6l4p+0WN19nspZPIuYfsy+ZDJt2yJkblJGRmvuD9k7/k1n4N/9iZ4e/8ATdBR+1j/AMms/GT/ALEzxD/6bp6P2Tv+TWfg3/2Jnh7/ANN0FAH/1fv39vr/AJty/wCyzeEP/bqvv+vgD9vr/m3L/ss3hD/26r7/AKACiiigAooooAKKKKACiiigAr5z/aZ+B9x8dPBWlaVoHiFvCnizwxq9rr/h/UxGs62+p2IfyzJC3EkTK7K6+hyQwG0/RleO/Gj4KeHvjfoGnaNrWr6x4eu9GvV1HT9T0K+aw1Czuljkh3xSqGU7o5XRldWUhjx3oA+TdB/aD+LuveEviN8Kvifqei/CD4rfDyLTptR8Quq3miyaTeS8alZRzsuWZFdVimOFlZA3JaNfHNN+MXxF8Z/Br9rj4W+OfEtx4zsfBXgqW60nWb7RjoN/dW+r6RfOwntDHCNqND+6kEa+Yp38gjH1DbfsJ/Ct/BXjTwx4k8QeJPEus+PZLCXU/Eep6gs+s7tLlWayEMvlCKNIJFBRBER2bcAANCz/AGMfCseo/EPVNX8d+KtZuPijoDeH9ea7urI/aIjBLbJcKI7NFSaKKZxGFAiGcmJjQB4d4C8ZfFK/8T/Cv4MeCPEY0C31f4JLqNpL9ktZzb6un2SC3uS00MjMqbvmiJMbDqpOCKPwn/aq+Kfx18R/B/4TeG70aL400oahe/E0fZ4He1XQnNi9oyOjpF/aF5hsxhXjj+4w5r7L8L/s7+CvCXjnwl8QNNvdQk1Hwb4UXwfZxyyQmGTT1eJxJMqxKxnzCvzKypgn5OmNTwR8Bvh18P8A4j+Pvin4csjDrnxHktJNUYkbM2kRjAiAAKCQlpJOTuc7j2oA+FPh/wDHL4xaB+0Z8OPAmrfFe3+Kmi+PLjWbDVvsvh9bHSdNv9PspLpY9L1GJQt0I3Qxyq0srKBl9rtxpfs1x/HzVPHXxy8aeIvi3eajpnhHxdrenSaM2mWf2a5a202H7O8crq81skRdCsMb7CY8tuLyFvWvDv7B3gXwrd+B5tF8e+Lobb4cX1xd6Bam8s2gsYrsOtxaqDZ72ikWRlLOzSheFkWvQE/ZP8IWXxN134k6B4r8TaHH4ounv9V0Sy1BY9Ivb2SD7O08sDRM5Ypyy+ZsZgpK/KBQB8pfD79oD47/ABL+HX7Pfwz8JeILPRPGXxN0K+1nV/Elzp8M32W00wqrLaWKCK2aeVnVcFQkaAttJIx0uu/Gb4+fBu/+Lnwa8d+K7bxdrei/DjVfHPhvxJDp0FlcxfYxJbmG8tED2zMk+x42CBGUMGBztX3S8/Yw+Fs/wv8AAfw2sdU1zSrr4Zoy+H/ENjeJba5ZeYCsu24jiETCVflkRoSjADK5ANWfDP7Hvw80Pw/490/WNc17xTr/AMSNJn0TV/EWsXqXWrfYJ4Xh8i3fylhhRBIzIqw7d2C4faAAD5p8K/E39pTwXL+zn8RPHXxCi8XaL8ZJNN0zU9HOk2llFaTalpzXcFxbTQgTb1ZD5u9ijEnaka7VX7Y/aS+LF18DPgT41+LNhZLqN54b06S4t4Hz5bzsRHF5m3B2B2BfBB2g4I61nah+zl4I1Lw58KfDE99qK2vwfu9OvNIZZYRJPJplo1nCLsmEh1aNiXEYjJbkEDivYPFXhbw/438Nap4P8V2Mep6NrVtLaXltLnZNBMpR0OMEZB6ggjqCDzQB+VPjLwd8b9J+KP7L/iv4vfE5/Gk2ueJ1mk05NMsbOysbmXTZ5D9lmtkErxopMY8x23/f4JrnD+0n+1f8RD4w+KfwvutZkXw/r97p+keEbPwqt7pV7aabceS8d9qhX7RHczhSx8tlERwMYY4+xvD37EfhDQPFngvxJJ8QvGmsWPw+uxeaLo+palbXen2rLC0CIoe0+0FFjYqoM2QMDNT67+xR4JvPF+v+KfB/jnxl4CtvFl099q+k+HdY+w6deXc2POnMRid4pJsfvHhdGb1HFAHAah4z/aA/aA+P3jX4W/DXxq3wk8P/AA107RZb2VNLtdR1G+1HW7Y3iI63gZIoYIxtYKoLOGG4g4RvjbxP+0pcftIeDv2dfDPj+DTVv/h7JqOsauNKtZGGo213HBJqFvbyAgPK3yLC0jRIshbazIM+wfEP9kfwZ418ZQfEXw14q8S/DzxOLCHS7vUPDWoLay6hZ2/EUd2s0U6StGMhJCoccZYhVA7fw38APDHhz4jeH/ikdZ1jVde8PeFz4Ujl1C6S5NzaNPHctcXLtEJZLpnjG6TeFIJymTkAHwr41+K/7SX/AAtxv2eLLx7q9rdeBPD1heatr2geErfVb3WNRvy7I7Wjh4La1jVQCEALvuXcAcL2F58Sv2iPEnwk+H2qePPHjfBTxXeC/tLvT7bw5/a2ta3qFrNsga006VJZBA8KieVY4yy7wpZEGa+lvi7+y34Q+K3jSw+Jlj4j8QeA/GNjaHTzq/hm+Wxubmy3+YLe5EkcscsatkqGTIJzngYxLv8AZC8Kyv4M1Kx8ceLbLxB4LXUYotaGpR3OpXsOrMr3cd3LdQTKwdlG0xpGY8DyyuBgA+V7j9of9pjx1+xn8L/jZ4ZNxZXeoX9xF4xvtA0uHUNStdOs7i5tHurOxut0bZeFXlG1igJKqFBxr/Ej9pT4g2Hwz+Cvhv4X+P8A/hLtT+LF/qSHxlpmhJc3AsNODzssGkjMX2zaUhZXGFaOVjGp4T3bw3+xboXgv4daR8OPBPxN8baBa+H729u9PurPUraO4hjv3Mktqw+ymKaAyEuBLG7hyTv6Aalx+xP8GJfhDovwgt5dXsovDuoyaxp+t21+0WuW2pzSNJLdx3gXiWQu275NmMfLwpABV/ZT8Z/HDWb7xt4R+L0Oq6lp2hT2cmheIdX0caJdanbXUbGaKa1QLF5lrIm0ugAdWViAc19i14n8HvgpF8I/7Wup/GviXxxqOsi3Wa78SagL1447XzPLjgSOOKKJcyuW2oCxI3E4GPbKACiiigAooooAKKKKACiiigD4A/4Kj/8AJinxM/7gv/p4sq+/6+AP+Co//JinxM/7gv8A6eLKvv8AoAKKKKACiiigAooooAKKKKACiiigAooooAK+B/8Agoz/AMkM8P8A/Y5+Gf8A0uWvvivFPj78CvC/7RPgH/hXvi3UtS0izW9tdQjudJmigu457N/MiKPLFMow3P3M+hFAHQ/GS18AXvwo8XW/xUWBvCLaXdHVPtOPLFqsbGRjnoVAypHIYAjnFflZ8G/iT+0f4f8AgP8Asf8AgXw5ra6Vqvj5tYsLx761juQdNghnksJisqhz9ntRHLGqOnmbVRm2sa+ul/YN8A6zfWz/ABT+IPjv4naTbSJMNH8Ta6brS5JIzuRpbaCG3WTacHD5U9CCMivojxf8GPCXjLxr8PfHN7Lc2d38NLi7uNLt7Roo7ZjeWptHSZDGxKLG3yBGTBxkkcUAfKvxQ8SftBfsweHPA3xH8dfERvHnhPSNfe08XyPpNlYt/Y2qMsFpdsIE3K1hKVLmNlEiv8y/LmvTf2UPiV8QPjda+NfjHrmoE+Bte1mW28Haf5EUYj0nTibc3hkVFmdryZXfbI7BAo2YU1iftwX/AIk8Q/DbS/gB4Itnm1/4xX6aB54hMsdjpjfvNSvJeCoWK3BUZIO5wV5FfV3g3wloPgHwlo3gjwvbCz0jQLOCxtIR/BBboI0BPc4HJ7nk0AfGXxJ8R/HTxh+1/P8AAf4f/EI+B/Do+H8GvySRaXZ39wt62qT2u+FrlDtLIqBg+9Nqnaiu3mL8r3fx1/a3tv2Zdb/aZvPiJZNcfC7WptKvdFg0W2jtddg07UFsrma5mcvLFLMrFgLfy0XaAFydy/qKnwg8NJ8b5fj4Lm7/AOEgm8Ox+GTBvj+x/Y47x70OE8vzPO8xyC3mbduBszzXlt3+yL8Nrz4F+L/2fpdS1YeHfGuoXupXk4mt/tsc19di8kWF/s/lqgkGFDRsQvBJPNAHnniLxr8cvjT8f/G3wc+EfjO3+GuifDOy0mXUNR/syDVb/UL3WYXuYUjjuj5MdvHEvzNtLl8gH+74prP7T/xy0bwBqnhfXNVs4/HHgP4k6B4T1LVrC0jFrqmm6jNC3m+ROJVgllhk2yqh/dsMoy5GPrj4n/sr+EviH4zh+JOh+J/EXw+8WizTT7nU/DN8lnNfWcZzHFdJLFNFLsJOxigdePmwFAor+xv8Gl+C2p/BEJqTWGs3o1a81Zr131qfV1kWYai94wJN0JEUhiu3jbt25FAHNePviH8UH/atuPg34X8RLo2j3Xw01DW4Q1nBc+RqsepxW0d0N4V2KROwEZk8snBZGxX52ata/FvWP+CVfib4kfEL4j3ni+28T6XY3EGn3lnbIbKdtciaST7XGouJy5znzWOM8V+oXw3/AGVvDPw9+KC/GO98X+JfGHiptCk8PS3Gu3sN0klnJcR3P+rjgiCMrRADy9qkFiys7F681h/YB+Gdv8OvEPwhi8aeMB4I1y2+yQaM2pxSWelxfbYr8/Y45LdsP5sWA8vmsEZ1UjcTQA3SPFHx0+Ff7UPgT4Z/ETx5H450L4n6VrVwITpdtYDTNQ0dYZm+ytAPMNu6S7VSZ5XGMlyck/dteVeKPhB4a8WfFTwP8XtRubuPWfAMOqwWEUTxi2lXV4o4pzOrRs7FViXZsdMEndu4A9VoAKKKKACiiigAooooAKKKKACiiigAooooA8A/ax/5NZ+Mn/YmeIf/AE3T0fsnf8ms/Bv/ALEzw9/6boKP2sf+TWfjJ/2JniH/ANN09H7J3/JrPwb/AOxM8Pf+m6CgD//W+3f+Ck3inQvA3hX4F+NfFN19i0Xw/wDFnwxqF9PseTybW0jvJppNkas7bUUnaqljjABOBXQf8PRv2E/+imf+UbWP/kKj9vr/AJty/wCyzeEP/bqvv+gD4A/4ejfsJ/8ARTP/ACjax/8AIVH/AA9G/YT/AOimf+UbWP8A5Cr7/ooA+AP+Ho37Cf8A0Uz/AMo2sf8AyFR/w9G/YT/6KZ/5RtY/+Qq+/wCigD4A/wCHo37Cf/RTP/KNrH/yFR/w9G/YT/6KZ/5RtY/+Qq+/6KAPgD/h6N+wn/0Uz/yjax/8hUf8PRv2E/8Aopn/AJRtY/8AkKvv+igD4A/4ejfsJ/8ARTP/ACjax/8AIVH/AA9G/YT/AOimf+UbWP8A5Cr7/ooA+AP+Ho37Cf8A0Uz/AMo2sf8AyFR/w9G/YT/6KZ/5RtY/+Qq+/wCigD4A/wCHo37Cf/RTP/KNrH/yFR/w9G/YT/6KZ/5RtY/+Qq+/6KAPgD/h6N+wn/0Uz/yjax/8hUf8PRv2E/8Aopn/AJRtY/8AkKvv+igD4A/4ejfsJ/8ARTP/ACjax/8AIVH/AA9G/YT/AOimf+UbWP8A5Cr7/ooA+AP+Ho37Cf8A0Uz/AMo2sf8AyFR/w9G/YT/6KZ/5RtY/+Qq+/wCigD4A/wCHo37Cf/RTP/KNrH/yFR/w9G/YT/6KZ/5RtY/+Qq+/6KAPgD/h6N+wn/0Uz/yjax/8hUf8PRv2E/8Aopn/AJRtY/8AkKvv+igD4A/4ejfsJ/8ARTP/ACjax/8AIVH/AA9G/YT/AOimf+UbWP8A5Cr7/ooA+AP+Ho37Cf8A0Uz/AMo2sf8AyFR/w9G/YT/6KZ/5RtY/+Qq+/wCigD4A/wCHo37Cf/RTP/KNrH/yFR/w9G/YT/6KZ/5RtY/+Qq+/6KAPgD/h6N+wn/0Uz/yjax/8hUf8PRv2E/8Aopn/AJRtY/8AkKvv+igD4A/4ejfsJ/8ARTP/ACjax/8AIVH/AA9G/YT/AOimf+UbWP8A5Cr7/ooA+AP+Ho37Cf8A0Uz/AMo2sf8AyFR/w9G/YT/6KZ/5RtY/+Qq+/wCigD4A/wCHo37Cf/RTP/KNrH/yFR/w9G/YT/6KZ/5RtY/+Qq+/6KAPxB/b1/b1/ZO+NP7J3jn4afDPxz/bHiTWP7M+yWn9malb+Z9n1K1uJP3lxaxxLtjjdvmcZxgZJAP1/wD8PRv2E/8Aopn/AJRtY/8AkKj/AIKj/wDJinxM/wC4L/6eLKvv+gD4A/4ejfsJ/wDRTP8Ayjax/wDIVH/D0b9hP/opn/lG1j/5Cr7/AKKAPgD/AIejfsJ/9FM/8o2sf/IVH/D0b9hP/opn/lG1j/5Cr7/ooA+AP+Ho37Cf/RTP/KNrH/yFR/w9G/YT/wCimf8AlG1j/wCQq+/6KAPgD/h6N+wn/wBFM/8AKNrH/wAhUf8AD0b9hP8A6KZ/5RtY/wDkKvv+igD4A/4ejfsJ/wDRTP8Ayjax/wDIVH/D0b9hP/opn/lG1j/5Cr7/AKKAPgD/AIejfsJ/9FM/8o2sf/IVH/D0b9hP/opn/lG1j/5Cr7/ooA+AP+Ho37Cf/RTP/KNrH/yFR/w9G/YT/wCimf8AlG1j/wCQq+/6KAPgD/h6N+wn/wBFM/8AKNrH/wAhUf8AD0b9hP8A6KZ/5RtY/wDkKvv+igD4A/4ejfsJ/wDRTP8Ayjax/wDIVH/D0b9hP/opn/lG1j/5Cr7/AKKAPgD/AIejfsJ/9FM/8o2sf/IVH/D0b9hP/opn/lG1j/5Cr7/ooA+AP+Ho37Cf/RTP/KNrH/yFR/w9G/YT/wCimf8AlG1j/wCQq+/6KAPgD/h6N+wn/wBFM/8AKNrH/wAhUf8AD0b9hP8A6KZ/5RtY/wDkKvv+igD4A/4ejfsJ/wDRTP8Ayjax/wDIVH/D0b9hP/opn/lG1j/5Cr7/AKKAPgD/AIejfsJ/9FM/8o2sf/IVH/D0b9hP/opn/lG1j/5Cr7/ooA+AP+Ho37Cf/RTP/KNrH/yFR/w9G/YT/wCimf8AlG1j/wCQq+/6KAPgD/h6N+wn/wBFM/8AKNrH/wAhUf8AD0b9hP8A6KZ/5RtY/wDkKvv+igD4A/4ejfsJ/wDRTP8Ayjax/wDIVH/D0b9hP/opn/lG1j/5Cr7/AKKAPgD/AIejfsJ/9FM/8o2sf/IVH/D0b9hP/opn/lG1j/5Cr7/ooA+AP+Ho37Cf/RTP/KNrH/yFR/w9G/YT/wCimf8AlG1j/wCQq+/6KAPyg/aF/wCCj37GPjn4BfEvwV4W+If23WvEHhnWdPsYP7J1WPzrq7spYYY98loqLudgNzMFGckgZNfcH7J3/JrPwb/7Ezw9/wCm6Cj9rH/k1n4yf9iZ4h/9N09H7J3/ACaz8G/+xM8Pf+m6CgD/1/v39vr/AJty/wCyzeEP/bqvv+vgD9vr/m3L/ss3hD/26r7/AKACiiigAooooAKKKKACiiigAooooAK+CPjt8VvjFqn7S3h39mj4YeMtK+GX9peHX15dY1LT11OfUbhbloBYWcEskURZEQyyfMZNpyoAU5+96/Pj9srxx+yDf3sfwm/auiudCC2SanouvmzuQIp3aRXFhfW0cjR3UJiV3iPDBo8rJ90AHtx+NVh8B/AvhKy/av8AGug6f4s1q8m09bu3b7FZ3ZWZ/KnCTt+5UW/lvcMzCNJCVU/MgPreo/FX4X6P4Us/Hmr+MNGsfDOohWttUn1C3isZ1cEqYrhnEThgCQVY5HSvx01HU9W1P4C/s1fGT9ozTpNS0/w/40kg1DWtY0/fdyeHCt5HY3WopsZljdfLLF8hjtkYlnyfVPFvjv8AZ3g/aK8M/GP4sWdrefBHUPCIsPBepXWlyvoNjqMV7Kt7GbdoSsE8yKpileJQ8a4RiMUAfqJcfET4f2nhEeP7rxPpcPhchCNVe9gWwIlkESH7SX8r5pGCL83LEKOTiudi+OXwVnt9eu4PH+gSQeFiF1eRdUtSmnktsAumEmIct8vz4+bjrxX4u/Eez0LVP2bf2p/Ffwr0qXSvgp4k1nwe/huFrd7OzubtNQs49WurK3kCFLeWTywCFCsVO3G0qPszx98IPhVY/t4fBzTrPwhpMFlJ4U17dbpYwLAxsmgFsWjCBCYg7bCR8ueMYFAH3NL8XfhTB4Hj+Js/jLRo/CEwBTWG1C3XT2yxUYuS/lE7gVxuznjrXQaF4w8JeKPD6eLPDWt2OraJIjSLf2lzHPasicswmjZkwMcnPHevxy0jxB8P/hL4H+KVjrHhHSNU0PTPjpfWOkjWYs6D4dkvbKIfb7iJVKpbW4llCoAqhpFAaPh18u0VH8Tfs0/tM/DT4c3S6+8XjqHWLzTfDlpJYNd+HpjYPePYWe92ihnjSQxoHbeisFLjqAfqf8W/2l/Az/An4oeM/gb450PxF4h8GaFfX6f2fe2uo/ZpoYmaNpY43cbdw/iGD0r0X4RfGrwH8QtJ0bQ4PF2kap4zGk2d5qem2t5bveQSSwo0rSW0bl4wHbkFRjIHcV8I/ET4q/sc+P8A9mj4w6f+zimjrrNn4B1MzrpukvYywWCxbRDNIYIgpWQr+6ZtwOSFxk1Z1jwV4P8AA3ij9i3WfB2iWWi3r3DWTy2kCQvJb3ehyNMjlAC4dvmO7J3Et1JJAP048ReJPDvhDRbrxJ4s1S10TSLFQ9xeX08dtbQqSFDSSylUUZIGSRyQKjvPFPhjTtX0nw/qGr2dtqmvCY6daS3EaXF6LZBJMbeJmDy+WhDPsB2qcnAr5m/bw8N654u/ZC+KOg+G7KXUNQm0oyRwQIXlkEEsczhFXJY7EYgDk9q+Y7n48/C344/ta/stXfwv1j+27fT7bxabmWOCZIoZJtIjxA0kiKvnLsO+MEsgKlgAy5APpv8AZf8Ajlf+Nf2fG+K/xh1ux0/7LqmtW9zfXDRWNrFb2Woz20JdmKxrhEUFiRk9eTXu/hr4s/C3xn4ZvPGnhHxhpGtaBp2/7VqFnfwT2lv5a7n82ZHKR7V5O4jA5PFfipotlYxfs3/BXxd8QtNuNX+FHhn4j+Jr3xXbw27XcUcf2+9Wzubq3RXMltDOcygoww2MEkA+7/tL658HfjH+zXq+qfsqwWeqeHNF8U6HqHjCLQ9IcR3enWzb5t9uEgW88tRG8iqW+WPaxXGQAfpn4Y+Lnwq8baBqPirwd4y0bXNF0gOb2+sdQt7i2tREpdzNLG7JGFUFiWIwvPTmo9O+MHwl1fVF0TSfGuiXuovp6aqtvBqNtJMdOkjEqXYRZCxgaMhxLjYVIbOCDX5P+D7r4L+KX+MHjv4VfEa28U39z8MNastQsNF8MSaFpqQwJvt5bo8R/bE3tHGjDzPL3ggKlfQPwu+DHwA039g7wxr/AI10GK0t9U+HlsmrazZ2Ym1eO21ewia7eOWON5225BVQGwiKu0qNtAH3b4L+J3w2+JEd1N8PPFeleJ0sX8u4bS76C9EL5IxJ5LvtJwcZxnFdxX5h/sA+NdIvPEni34beHzovjTSPCunabHYeOtE0v+zf7Qs/nWGw1ACNVa6thk5RmBBJID7s/p5QAUUUUAFFFFABRRRQAUUUUAFFFFAHwB/wVH/5MU+Jn/cF/wDTxZV9/wBfAH/BUf8A5MU+Jn/cF/8ATxZV9/0AFFFFABRRRQAUUUUAFFFFABRRRQAV8n/tb/Fnxp8I/D/w61DwTcR203iPxzoGhXhliWUNY6hI6zooYHaxCjDDkdq+sK+Jf29fB/i/xH8HtC8S+CdHuPEN/wDD/wAVaJ4ok06zTzLq6ttNmJmSBP45Ajlgo5IUgZOAQD6A+Pvi3WvAHwK+I3jvw1IsOr+HPDesalZu6CRUubOzlmiZkbhgHQEg8HpXnnw5/aO+Hdt8IPhl4g+MvjzQPDvibxf4Z0jVpItR1C006S4lvLWOSWSKGWRDsMhYDaMA8DpXy18dv24fgT8X/gV4v+GHwZvr7xl488d6PfaHYeH7PTbwaglxqMDW7G4SSJViWASFpGZtuFOCeteUeJYvgx8Dv2kPg94R/aXk0gaHoHwch0V21a3S8sjf2t3BFwHjkXJWKTaxAyOM5YAgH626r488D6F4W/4TjW/EOnWHhzy1l/tOe7hisvLf7r+ezCPa3Y7sHtVey+JPw71LwjJ8QNO8U6VdeF4UaSTVYr2B7BET7zNchzEAO5LcV+MmmeF/hdqP7O3jPUvFWr3Xwt+F+p/E3+1/hxfXOnSXNlZPBCZILmSyljKpptzKkpSKVUj+YA4LDLfHPiPxL8W/2SPC3jJfCtjpfh3wp8SoZPFE2g6PJLpOtabZAoddh01lie5tGkaJ5IWB3lCTwpAAP1+g+OPwcvPAWqfFHTvG2j3/AIS0VHe81S0vYbm0g8sAsryQs438jCfeJIABJANj4XfGL4Y/Gfw/H4k+GPiXT/ENqYoJJls7qGeW1NwnmJFcpE7GGXHVHwwIIxwa/KPw/wCGfhD8UvCP7QesfCHxfbfEfUfEfgC8hn07SPC76Vpi6hbW839nzAEGM6hmQqqqPMA5IBHP2t+xB8Q/gh4u+EOh+HvhQ1outeH9E0WPxDDa2L2jw3xtvLZLlzEivOrxSBxuZlxk8MpIB9J+LPit8L/AWpWOjeOfGGj+HdQ1P/j0t9Rv7e0mn5IzGkrqzDIIyAeeOtcd4i/aS+BPhH4jx/CfxT440nR/E0lql39mvLyG2wsrqkUe6V1UzSl9yRAlyoLY2kE/nL+274g+GPw8+Nt58SdK1DS9Z8eR6NZ6ZqfgbxFpMl7a+JtLeUvGunTeTIY7oF2X93uXIHmIACX6v4g+Ivgj8KP23I/iN8bNDtfD3h/xN8PLFrKa+0o3CtrMWpSySQgRQyk3qRFMhQXIAAPQUAfpF4p+Knww8DalaaP428X6P4fv78A21vqOoW9pNMCSB5aTOrPyCOAelXvEnj/wH4Nks4vGHiTTdCfUUuJLZb+8htTOlpH51w0Qldd6wxDfIVyEX5mwOa/J3Q/FP7MXwy8b/G7Rv21NHtX8ceIPEupXdtPrOlSX76r4elCrpkemSiKXMccQ8sRRMGRxg/MOOc+GngHW5NS/Yl8HfGPRJJUF544mttL1qMTz2+nx2MtxpcVyku795BEsJVWGY9qggMvAB+qtx+0F8B7Tw5p/jC6+I3hyHQtWma3s799WtFtbiZCA0cUpk2O65G5QSV74re8X/Fj4W/D59Oj8eeMNH8ONq5xZDUr+3tDcnj/UiZ139R93PUetfml4U+DnwqvviL+2jo914S0uS0tY7JbeM2kRFv8Aa9D+0TGD5f3RebEhKYJcK3VQR4+3jfwJc/Bj4QeCfGOiaBBrvib4U2+PFHiWwfU2lt7VTDFpGmQqUL3rtKZMiQFRtbZKQqgA/afxH408HeD/AA8/i3xbrthouhxKjNf3tzFb2qrJ9wmaRlTDZGOee1fKvxu/aTsNL0r4QeKvhB4r0nVvD/i7x9pmganf2s1ve2psJre6luE85WZI3UxIS2QyjrjNfnloPibwRffB/wDY9+JHxQDeIvhD4Is7+x8RsLd7+x0/WRZxQ6fJqEKrJujhbegbaRE5wTztPuvx01n9nD4weE/govwas9K1TwbffGjRrXUorTT/ALNY3V3/AGfcGbdG8UaTBo2jDOFZW+6SSGAAP018E/Ef4e/Eqwn1T4deJ9M8UWdrKYZptLvIb2OOUdUdoWcK2OcE5xz0rs6+CvhroujeEv2/viZo3hixg0qx1bwToV/cwW0axRyXUd3cwLKUQAbvL+XOK+9aACiiigAooooAKKKKACiiigAooooA8A/ax/5NZ+Mn/YmeIf8A03T0fsnf8ms/Bv8A7Ezw9/6boKP2sf8Ak1n4yf8AYmeIf/TdPR+yd/yaz8G/+xM8Pf8ApugoA//Q+/f2+v8Am3L/ALLN4Q/9uq+/6/OD/gpN/wAJV/wivwL/AOEF+xf8JJ/wtnwx/Zf9peZ9h+3eXefZ/tPk/vPJ8zb5mz5tudvOK6D/AI2m/wDVGf8Ay4qAPv8Aor4A/wCNpv8A1Rn/AMuKj/jab/1Rn/y4qAPv+ivgD/jab/1Rn/y4qP8Ajab/ANUZ/wDLioA+/wCivgD/AI2m/wDVGf8Ay4qP+Npv/VGf/LioA+/6K+AP+Npv/VGf/Lio/wCNpv8A1Rn/AMuKgD7/AKK+AP8Ajab/ANUZ/wDLio/42m/9UZ/8uKgD7/prokg2yKGGQcEZ5HI/I18Bf8bTf+qM/wDlxUf8bTf+qM/+XFQB9/MqspVhkHgg9CKR0SRSkihlbggjIIr4C/42m/8AVGf/AC4qP+Npv/VGf/LioA+/6K+AP+Npv/VGf/Lio/42m/8AVGf/AC4qAPv5lVgVYZB7GkCqpLAAFup9frXwF/xtN/6oz/5cVH/G03/qjP8A5cVAH36qImSihdxycDGT6mnV8Af8bTf+qM/+XFR/xtN/6oz/AOXFQB9/0iqqjCgAZJ49Tya+Af8Ajab/ANUZ/wDLio/42m/9UZ/8uKgD7/pqIkahI1CqOgAwBXwF/wAbTf8AqjP/AJcVH/G03/qjP/lxUAffqqqAhQACSePU9adXwB/xtN/6oz/5cVH/ABtN/wCqM/8AlxUAffqIkY2ooUcnAGOTyf1p1fAH/G03/qjP/lxUf8bTf+qM/wDlxUAff9FfAH/G03/qjP8A5cVH/G03/qjP/lxUAff9FfAH/G03/qjP/lxUf8bTf+qM/wDlxUAff9FfAH/G03/qjP8A5cVH/G03/qjP/lxUAff9FfAH/G03/qjP/lxUf8bTf+qM/wDlxUAff9FfAH/G03/qjP8A5cVH/G03/qjP/lxUAH/BUf8A5MU+Jn/cF/8ATxZV9/1+IP7ev/DfX/DJ3jn/AIXX/wAKz/4Qz/iWfb/+Ee/tr+1P+Qla+T5H2v8Ac/67y9+/+Ddj5sV9f/8AG03/AKoz/wCXFQB9/wBFfAH/ABtN/wCqM/8AlxUf8bTf+qM/+XFQB9/0V8Af8bTf+qM/+XFR/wAbTf8AqjP/AJcVAH3/AEV8Af8AG03/AKoz/wCXFR/xtN/6oz/5cVAH3/RXwB/xtN/6oz/5cVH/ABtN/wCqM/8AlxUAff8ARXwB/wAbTf8AqjP/AJcVH/G03/qjP/lxUAff9FfAH/G03/qjP/lxUf8AG03/AKoz/wCXFQB9+CNFdnVQGbGSBycdMmvny/8Agxqd7+1Lpvx8N9B/Zdl4RufDjWTKxmaae+juxKDjbsCptI65rwb/AI2m/wDVGf8Ay4qP+Npv/VGf/LioA+/WVXUo4DKwwQeQQacAAMDgCvgD/jab/wBUZ/8ALio/42m/9UZ/8uKgD79VVQbUAUcnA468mlCqv3RjPPHqa+Af+Npv/VGf/Lio/wCNpv8A1Rn/AMuKgD79KIzKzKCV5BI5GeOKUqpwSM45Hsa+Af8Ajab/ANUZ/wDLio/42m/9UZ/8uKgD79ZEYqzKCVOQSOh6ZH4U6vgD/jab/wBUZ/8ALio/42m/9UZ/8uKgD7/pCqkgkZK8j2r4B/42m/8AVGf/AC4qP+Npv/VGf/LioA+/VRFXYqgL6AcUKqooRAFVRgAcAAV8Bf8AG03/AKoz/wCXFR/xtN/6oz/5cVAH3/RXwB/xtN/6oz/5cVH/ABtN/wCqM/8AlxUAff8ARXwB/wAbTf8AqjP/AJcVH/G03/qjP/lxUAff9FfAH/G03/qjP/lxUf8AG03/AKoz/wCXFQB9/wBFfAH/ABtN/wCqM/8AlxUf8bTf+qM/+XFQB9/0V8Af8bTf+qM/+XFR/wAbTf8AqjP/AJcVAH3/AEV8Af8AG03/AKoz/wCXFR/xtN/6oz/5cVAH0B+1j/yaz8ZP+xM8Q/8Apuno/ZO/5NZ+Df8A2Jnh7/03QV8P/tC/8PI/+FBfEv8A4Tr/AIVN/wAI3/wjOs/2p/Zv9vfbvsP2KX7R9m879353l7vL3/Luxu4zX3B+yd/yaz8G/wDsTPD3/pugoA//0fv39vr/AJty/wCyzeEP/bqvv+vgD9vr/m3L/ss3hD/26r7/AKACiiigAooooAKKKKACiiigArnPEnjHwj4Ot4brxdrljocFwxSOS+uYrZHcDJVTKygnHOBXR1+Yn/BRm78MWPiP4AXnjPw3N4v0OLxbK13pMFgNTlvIhZvmNbMgiYnrtx2oA/SrSdY0jXrCLVdDvoNRsp8mOe2lWaJ8HB2uhKnn0NaNflv/AME+k8O+I/iV8afiX8I9FbwV8LdUvbCwsPDsrRwzQapYxMt9PJYRO4sS5KgRnBcYOAFArfT/AIKIW8qj4hReBkPwgOrDS/8AhIjrdqNRMZufsf28aPs877J53GfM8zZ+82Y+WgD9KaK+MYf2qPFuu/tC+JPgf4J+HqavB4NutPt9WuptatrK/WG+SOQ3ltp0se6e2hSTLuJVLdFUkgHjdT/bF+LS+MPiRYeEfgjP4q8LfDDU5NP1TUrLXrZL1hDClxJJDp88EZkIjbIRZiWPAOegB+gFFfFWlfGn4cfEb4+fB7VvDOmXOo/8Jn4R1fVdM1X7fNbx29pvtjJBLp4BikkYkAu7ZjKkDPUc38Gf2w/ih8bfCvh/4i+FfgpeS+Eb8alHqN7Fq9vJNbXFgZ8Ja2zxRSXayeUqbh5eJZNgDBC5APvmivjHwB+1B8RNQ+K9j8H/AIv/AAqm8Da54g0m81jRlg1i11X7VFZFRJBMY1iWCbDZGWZO2/vXjXw8/ac+NHxV+F/xf8R/E74XWtt4L8PQ+K7aW4stf8m4/wCJTE8baeEjh81XYK4N2knDHciABQAD9M6K+FdM/aYGgeBfhD4C+D3w/vfF3jDxf4R0/XLPQRqaxx6bpC28IEt9ql2pJVWcRLIY2eVwSQCQCl/+2rLoHw2+KWueMfAN14e8e/Ca0hvdT8NXV9EyT290f9HntdQhjkjlhkAI3iPKsCrKOCQD7ror4y8GftS+Nb34seCfhv8AE/4Zy+C7X4m2V5d+G70apDqDyvYQLdTQXkMUai2kEB3jEkgJ+UEndt9y+Ofxk8MfAL4W638VfF0c9zYaOkYW3tVD3FzPPIsMEESkgFpJHVRngZyeAaAPT9Q1HT9IsbjVNWuorKytEaWaed1jiijUZZndiFVQOSScCpre4t7u3iu7SVZ4JlV45EYMjowyrKw4II5BHWvzp+NXxE+PHjL9l74023xd+FCfD+yPgzVLm0uItct9W3uYGBt5o4oomjlUHdkbkPIByOdz4L/tHeONCuvg38M/iN8OJPDWg+PdIitdB1capDdyyXFjYLPsvLSOMC3M0SFo9s0pzhWwd2wA/QCiviTTf2rPiHpvxQ8IeC/ip8JbrwXoPxCv59P0LUpNVtrq6aaKN5o/tljGga281F4AkkKE4bBzjj/EX7afxKtB8VtV8J/B19d8PfB3Vr2x1m/Ouw2xktbGFJ5ZraFrdneVI2Z3iOFChdsrsxVQD9C6K+LvHP7W2tJqegeG/gV8O7r4k63rnhWHxmYH1CDR4YNHuW2QM00yy7ppHyoiVSRjJOOa87s/279W1X4e/DbWdI8BWtx4y+Kkd1eaVpJ16OGxt9Os1Uy3V7qU9tEIirNs8sQsWcFVJ4JAP0Vor55/Z2+P1r8eNB12S70Y+HPEfhPUpNK1jTftUV/HBcKiyo8F3CAk8MkbqySBVzyMcZP0NQAUUUUAFFFFABRRRQAUUUUAfAH/AAVH/wCTFPiZ/wBwX/08WVff9fAH/BUf/kxT4mf9wX/08WVff9ABRRRQAUUUUAFFFFABRRRQAVxel/En4d63qzaBovinSr/VEZka0t76CWcMmdwMaOWyuDkY4xXaV/MWdQ+FesfCbxB8M9P+HVxH8Y/E/wAQNStfDnjOSzi0y0trv+098X/E8dkYtEisPJUsckADJBoA/p0or5a+Ov7QU3wD03wT4Tt9KHjLx34zkey060ku4tLt52soVkvLq4u5gyQQxqQT8rMWZVVTkkeOXP7etnonwj8f+NfE/gt7XxX8NtR07TdV0a21OC6swdWZBZ3S6oiCIWsivlpGjBQqysmcEgH6EUV8QeNv2qviT8PvgpbfE7xD8MLd9a1HXNN0bTtOtfEVvcWOoJqZRYLuDUYrdx5Rd9uHhU/KT93DHT8L/tN67qviXWPhH8bPhxd/D3xc2h3ms2Nr/acOoWWqWNsNs4t7+1CFJUJG5CiuqkOKAPsyivzb8EftXWXgb4Zfs6eG/APw5vNRX4saVqC6Xp39sNc3FnJptuksMMlzdRZmWRnxJM7R+UgL7XwEr1DXP2gP2mtH8M2epj9n2R9Ut7e7vNYhk8SWcVpaQW0rogguvKb7TLMkZlVAibFKb2BYUAfalFfnV8SP2zPiBP4I+B3jb4F+BU1y3+Lt/bwiPUr6KyZGaN5WslfEu2RvKcNOUKKq/KGZwU7vT/itpdh+0lCnxN8Kt4U8TWfwwm1/VLwa3Le2VhZQ6mFmtPsyRpBKyMPN+1hQ5UbNu2gD7aor8+2/bc8Z2XhK0+NWu/BfV9O+Dd55Uy+ITqFrJfxafcMFiv5dIQGVbdtyuSJWcRnftIxnsfFX7UHxHk+MPif4N/B74Xp44vvDukadrIvZNch061lgvw5CEyQSFXO0CLbvV8ksYwoLAH2nRXj3wC+Mmi/tAfCHw58XdAs5tNtdfilLWtxgy289vM9vPExGAdk0bqGwNwAOBnA+WvFv7Q3hz4P+Mf2oPGum+D573VfhxZ+F7q/aTWJvJ1X7XZsYViheKSKx8lSVYxq/mn5mANAH6DUV8beDf2ofGt58XfB/wx+KPw0l8FQfEixvbzw3eDVIdQeZtPhW5ngvIYo1FtKIW3/LJKp+6CTkjJ+Hv7VHxQ+MWvxa18JvhHJrXwtbWjpH/CSz61b2k80UNwLa5vrfT3iZpLeFgx5lV3VThQwK0Afb9FfDOl/tY/E3xn428a+C/hh8IJvEr+APE50LVLg6zBZxC1xHtuo/OhG+U7nZrdc7EjyZcuingvEP/BQhdLuPEHjHSfAY1L4W+FNVk0u/1w61aw6i/kTCC4urTSGQyz28UhIz5is65YKNrAAH6R0V8P8Axr/a08efBefVPGWp/Ci5vfhToU9lBd+JP7WtobiVbwxL59ppxQyTQxtKFLGRCxyQNqk1qfFH9qbxZ4T+Oq/AX4e/D5PFmuRaPHrcgutattHkuoJJJI/L06O4jcXUimMl8vGq4IJ70AfZlFfFnxc/ay1vwl8ULv4QfCzwVb+MfEGh6dBqWsNqGuW2g2tml1uNvbpLcJJ51xKqlgoUKoKkv1A98+B3xf8ADnx5+FegfFfwrDNbWGuxO32e4AE1vNBK8E8MmMjdHLG6Ejg4yOCKAPV6KKKACiiigAooooAKKKKAPAP2sf8Ak1n4yf8AYmeIf/TdPR+yd/yaz8G/+xM8Pf8Apugo/ax/5NZ+Mn/YmeIf/TdPR+yd/wAms/Bv/sTPD3/pugoA/9L79/b6/wCbcv8Ass3hD/26r7/r4A/b6/5ty/7LN4Q/9uq+/wCgAooooAKKKKACiiigAooooAK+ZPjx8F/FPxQ+IXwc8WaBdWdvZ/D3xE2r6gl08iSSwG3eLbAEjcM+5gcOUGP4u1fTdFAHyTqvwE8W+Gv2nbD49/CO50+y0zxPa/2d440q7eWFdQjh/wCPS/t/KikU3kOSpEm1Xj+XcpZmr5l8HfsbfGr4Sx3Hw48A6R8MvEHglby4l07VvEelTT69YWlzM0phkSOIw3TRbyImaVBgfNhcKP1QooA/PL4y/s4/Hn4s/Gfw/wCLEbwjoln4W1u11DTPFNn9uh8TwabC4eXT5I1XyJ1lBZDvl8sqTmPcST4t8PdL/aX8Q/Ez9pzwv8EJPDNhpms+Lp7S61LWJr1b6wlmsIEaa2ht4mjlKxkFA7piQc5Wv12rkfDXgPwh4P1HXtW8M6XFp954nvDqGpSR7s3N0UWPzXyT821VHGBx0oA+UPh9+ydffC74j/BvU/DGo29z4X+GPhPUvD85nZ0vbm5vXhcTpGqNHtZkdnBkBXcAA3Wu2/Zq+CXjj4L/ALLekfBbVdWtIPFGm2uqxLqFgXnt4Zr27ubiCVPOjiZvLEykhkHzAjkcn6nooA/Ln4Sfsm/H7wJ8V/hZ8SNYtPB7XXhO11HT/EOoQX2oXGqa2b9YVl1Keee0DTXLCMlYpH2xk4EhU7U7LQv2cf2jPCmgfFj4Q6NqPha7+H/j+bxXfWt1cNfR6tDN4hgm8m3dEjaBUhndS8oLsybtse7GP0UooA/Psfs0fGv4fD4V/Ef4O6voTePvBXgmx8E61Yav9pOkanZWqRyZhuIY/PieO4RmjYxfOrAMFwQ3O+J/2RvjP8SvAnxn1/4ja7ocvxO+Kmj2uiWkFj9pi0TSrGycvFCs0kb3MpdmZ5JDF97AVQK/SaigD5Z8Y/BDxX4h+KnwD8cWV3ZJYfCz+1v7TSSSUTTfbtLNjH9mAiKtiTlt7R4XkZPy12P7SPwX/wCF+fCPVfh1b6q2halLLa3unagsYl+y31hOlzbyGM4DLvQBx3UnHNe60UAfDOveAf2xfiv8JfiF8MPi03gazHiHw1faVYT6PNqRaXUrlBGk1wZ4sQ2+0tuVEkcNjBwMHqvFPwB8Y65c/s9TWl5YIvwmvIrjVd8koMyR6a1mRa4iO8+YwI8zy/l5znivryigD8oF/ZA/aRg8X+EvGd0/hDXfEfhTxhNrs3iG+vL86rrFlN9oWKGc/ZXFulrHMqR20TtEdoKmPBL/AEToH7OHjjSvAn7SPhe4vtOa6+MOqa9e6QyyzGOCPVNOS0hF2TCCjK6kuIxIAvILHivtaigD8TviT8M/FPjLx/8ADr4G+GfCMfjXVfgb4J07S/GEVrrR8NG8W+tIkhgt9TUrdT2x8mV2iaJIssNzBiQPXI/hhp37TvgD4V/HH9nLQdC8O6p8N11Pw+PDHim2W90WS1jb7LPaSyWvm7vJlhElvPEWB3bm5JA+8/iP+z58D/i9f2+q/E3wNpHiO/tVEcd1eWkb3CxKSRH5uA5jySShYqcnI5Nej+HPDXhzwdolp4a8JaXa6LpFguy3s7KFLe3hQknbHFGFRRkk8AckmgDxn9n7wN8QfBmjas3xF0fwjoF7qFwjxWXg+0kt7SKJEwPOlmSN55SSTu8tQowADyT9A0UUAFFFFABRRRQAUUUUAFFFFAHwB/wVH/5MU+Jn/cF/9PFlX3/XwB/wVH/5MU+Jn/cF/wDTxZV9/wBABRRRQAUUUUAFFFFABRRRQAV8LfDX9j2FP2c/G3wC+MrWWpW3i3W9a1JJbBnk+zLf3BntZUaaOMrcQNtbhSoYYywzX3TRQB+bHiv9lP49+O/ht8L9Q8Y6x4X1r4rfCO5vIrO51OGfUNE1zTrqNYGGoxPCkqTSRpGXKCTEiF1YlgU9B8PfCr9o7Svh/wCJ4I/DXwt0nW9aks400ax0+6XRrqxjEouYr+fyklleQSgJiEpHtPyuHavuaigD8Tfix8CfiZ8Bf2Zl0e+udIt7zXPid4e1PRdCsJbubRNFM11GFtopZwJzC0w3yBUVVySijJr7J0X4DfHHx98VLj4zfHm+0Gz1LR/D2oaD4f0jQJLqezhfUhi4vLm4uoonMjgKgRI8BMEksOfrvxl4D8IfELTrXSfGmlxatZ2N5b6hDHLuwl1aOJIZRtI+ZGGR29q66gD8/wD4afsp/EPwbc/svzapqGlSj4KabrtnrPkzTsbiTVLFbaE2e6Bd6q4y5k8ohegJ4rt/2iPgh8Ufif8AEXwxr2gHQvEHhHT9PvLS60DxFLcpYx387q0OqLBBFKl1LAilEim2hcko6Mdw+yaKAPzI0P8AZP8Aj7o37O3wf8F21/4bi8d/BfX4tU08NNdzaXqFtDHNH5U8vkRzRPJ57HKxMFAA5JJHqcX7OfxM+IHxavPiH8bbjRPsfiL4a6h4I1e20SW6GJ77Umn3W4uYuYhasFMjuGMuSIwpGPuSigD8z7v9nL9sLX/hJb/ss+IfFfhJfhxHaQaNP4htobz/AISCfRbcKiwizdTapcPEgjeTzWG0lgC3NfR/w5+BWs+BP2iPGvxPimtB4Y1zw/oOjafbRySNdxHSRKrearRhApV12lZGJwcgd/qGigD5x/ZO+EHiX4E/AvRPhj4uubS71TTbrVZ5JbF5JLcrfajcXcYVpY4nJEcyhsoMMCBkYJ8C+LP7JnxG8eP+042kajpUI+NNh4ctdH8+adfs76RbGGc3m2BtgZjlPL8wkfeCniv0LooA+XfGnwT8VeI/jH8BviFZXVkmnfC4a2NTjkeQTzf2lpf2KL7MojKttk5feyYXkZPFeS/B74M/tWfs9R2fwj+HWo+D9a+FWn6nLPY3Orfb4tas9Murpria08q3UwTSJ5jiKQyKM8soGEH35RQB84/AH4QeJfhVr3xZ1TxDc2lxF488YXniCxFq8jtHaXFvbxKk++NAsoaJiQpdcEYY8gfI9h+x78Z/hlrfiTw/8LdH+GviXwfrmqXepWN34s0yebV9KF8/myQAQxNHcxROzeUGkjP95sHA/UaigD8rf2j/ANjX46fGK5+JFha3PhbW7HxPZ6bDoF9rEt3Hc+H1so4vPtbK0jt5YIUuZYyzTRur4YqyuMY7r9pb9nz9pD9oGystEns/A1oEitLiz1rztSj1rw3qSbWnl064iiUzoWUGPcYCT99SAAP0YooA/PD4qfsqfExfjHP8avhbB4O8YX3iHSrHTde07xvZO0Es+nr5cN9azW8UrRSMh2yIUKkDucbfsD4OeF/E3g/4eaZoXjG30S11mNp5LiHw5aNZaVEZpnkVLeJ/mwiMql2wXYFyBnA9PooAKKKKACiiigAooooAKKKKAPAP2sf+TWfjJ/2JniH/ANN09H7J3/JrPwb/AOxM8Pf+m6Cj9rH/AJNZ+Mn/AGJniH/03T0fsnf8ms/Bv/sTPD3/AKboKAP/0/t3/gpN4W0Lxz4V+BfgrxTa/bdF8QfFnwxp99BvePzrW7jvIZo98bK67kYjcrBhnIIODXQf8Ouf2E/+iZ/+VnWP/k2j9vr/AJty/wCyzeEP/bqvv+gD4A/4dc/sJ/8ARM//ACs6x/8AJtH/AA65/YT/AOiZ/wDlZ1j/AOTa+/6KAPgD/h1z+wn/ANEz/wDKzrH/AMm0f8Ouf2E/+iZ/+VnWP/k2vv8AooA+AP8Ah1z+wn/0TP8A8rOsf/JtH/Drn9hP/omf/lZ1j/5Nr7/ooA+AP+HXP7Cf/RM//KzrH/ybR/w65/YT/wCiZ/8AlZ1j/wCTa+/6KAPgD/h1z+wn/wBEz/8AKzrH/wAm0f8ADrn9hP8A6Jn/AOVnWP8A5Nr7/r4V1j9srxVc+JvFUPwr+DPiD4g+EvA2pXGkaxrWn3FpEwvbPH2qOyspnE935RO0lMbm4HGCQDK/4dc/sJ/9Ez/8rOsf/JtH/Drn9hP/AKJn/wCVnWP/AJNr7/ooA+AP+HXP7Cf/AETP/wArOsf/ACbR/wAOuf2E/wDomf8A5WdY/wDk2vv+igD4A/4dc/sJ/wDRM/8Ays6x/wDJtH/Drn9hP/omf/lZ1j/5Nr75uJltreS4ZWcRKzlUUu5CjOFUZJPoBya5rwN4utPHvhHSvGNhYX2mW+rQidLbUrZ7O8iDfwzQSfMjcdD25HBoA+K/+HXP7Cf/AETP/wArOsf/ACbR/wAOuf2E/wDomf8A5WdY/wDk2vv+igD4A/4dc/sJ/wDRM/8Ays6x/wDJtH/Drn9hP/omf/lZ1j/5Nr6b+Onxu8K/AHwMPG/im1vNSNzeW2m2Fhp0QmvL6/vH2QW8CMyKXc5PzMBgHqcA4PwL/aE0X42z+KNBbw/qng/xV4KuLe21jRdYjjS6tjdx+dbyBoXkjeOZASjBsnB4xgkA8C/4dc/sJ/8ARM//ACs6x/8AJtH/AA65/YT/AOiZ/wDlZ1j/AOTa+/6xPEuuweF/Dmq+Jbq2ubyHSbWe7eCzha4uZVgQyFIYU+aSRgMIg5ZsAdaAPhv/AIdc/sJ/9Ez/APKzrH/ybR/w65/YT/6Jn/5WdY/+Ta+6dA1iHxDoWm6/b29xaRanbQ3SQ3cTQXESzIHCTRPho5FBwyNypyDyKxNX8a2WjeMfD/gubTtQuLnxHHeSRXVvaSS2VuLJEdhdXCjZCZA+Igx+cggcigD4x/4dc/sJ/wDRM/8Ays6x/wDJtH/Drn9hP/omf/lZ1j/5Nr6Y+APxksvj18MrL4l6fpkmkQXl3qFqLeWQSup0+8ltGbcoUYcxFgMcA4r2egD4A/4dc/sJ/wDRM/8Ays6x/wDJtH/Drn9hP/omf/lZ1j/5Nr7/AKKAPgD/AIdc/sJ/9Ez/APKzrH/ybR/w65/YT/6Jn/5WdY/+Ta+/6KAPgD/h1z+wn/0TP/ys6x/8m0f8Ouf2E/8Aomf/AJWdY/8Ak2vv+igD4A/4dc/sJ/8ARM//ACs6x/8AJtH/AA65/YT/AOiZ/wDlZ1j/AOTa+/6KAPgD/h1z+wn/ANEz/wDKzrH/AMm0f8Ouf2E/+iZ/+VnWP/k2vv8AooA+AP8Ah1z+wn/0TP8A8rOsf/JtH/Drn9hP/omf/lZ1j/5Nr7/ooA/EH9vX9gr9k74LfsneOfiX8M/A39j+JNH/ALM+yXf9p6lceX9o1K1t5P3dxdSRNujkdfmQ4zkYIBH1/wD8Ouf2E/8Aomf/AJWdY/8Ak2j/AIKj/wDJinxM/wC4L/6eLKvv+gD4A/4dc/sJ/wDRM/8Ays6x/wDJtH/Drn9hP/omf/lZ1j/5Nr7/AKKAPgD/AIdc/sJ/9Ez/APKzrH/ybR/w65/YT/6Jn/5WdY/+Ta+/6KAPgD/h1z+wn/0TP/ys6x/8m0f8Ouf2E/8Aomf/AJWdY/8Ak2vv+igD4A/4dc/sJ/8ARM//ACs6x/8AJtH/AA65/YT/AOiZ/wDlZ1j/AOTa+/6KAPgD/h1z+wn/ANEz/wDKzrH/AMm0f8Ouf2E/+iZ/+VnWP/k2vv8Ar42+IP7VXinS/id4g+Ffwb+FOrfE7UfBcVpL4gmtLy00+CxN9H50EMbXTA3EzRfOUQcAgZJyAAcT/wAOuf2E/wDomf8A5WdY/wDk2j/h1z+wn/0TP/ys6x/8m19veEfEB8WeFNF8UnTrvSDrNlbXv2K/j8m8tftEayeTcR5OyWPdtdcnDAjNdDQB8Af8Ouf2E/8Aomf/AJWdY/8Ak2j/AIdc/sJ/9Ez/APKzrH/ybX3/AEUAfAH/AA65/YT/AOiZ/wDlZ1j/AOTaP+HXP7Cf/RM//KzrH/ybX3/XHeCfGtl460++1Gx07UNNSxv7vT2TUrSSzkkezkMTSxpIAXhcjMcg4Yc+ooA+Mf8Ah1z+wn/0TP8A8rOsf/JtH/Drn9hP/omf/lZ1j/5Nr7/ooA+AP+HXP7Cf/RM//KzrH/ybR/w65/YT/wCiZ/8AlZ1j/wCTa+vvit8TvCnwZ+HWvfFDxxM8GieHbY3NyYk8yVhkKiRrkZd3ZUUEgZIyQOa8b+CX7Uuj/F/xlf8Aw41fwfrvgLxXaaZDrkWn67DFG91pU8nlJcwtDJIuBJhHVsFWOOSGwAeS/wDDrn9hP/omf/lZ1j/5No/4dc/sJ/8ARM//ACs6x/8AJtff9B4GaAPgD/h1z+wn/wBEz/8AKzrH/wAm0f8ADrn9hP8A6Jn/AOVnWP8A5Nr7O8AeNLL4h+ENP8Zafp2oaVb6krsltqtpJY3sflyNGfNt5QHTJXK56qQw4Io8XeNbLwdNoMF5p2oagfEGpw6XEbC0kuhbyTq7ia5MYPk267CHlb5VJGetAHxj/wAOuf2E/wDomf8A5WdY/wDk2j/h1z+wn/0TP/ys6x/8m19MfCP4yWXxZ1T4gaZZ6ZJpzeAfElz4dlaSQSC5ktoYZTMoAG1WEwG05PHWvZ6APgD/AIdc/sJ/9Ez/APKzrH/ybR/w65/YT/6Jn/5WdY/+Ta+/6KAPgD/h1z+wn/0TP/ys6x/8m0f8Ouf2E/8Aomf/AJWdY/8Ak2vv+igD4A/4dc/sJ/8ARM//ACs6x/8AJtH/AA65/YT/AOiZ/wDlZ1j/AOTa+/6KAPgD/h1z+wn/ANEz/wDKzrH/AMm0f8Ouf2E/+iZ/+VnWP/k2vv8AooA+AP8Ah1z+wn/0TP8A8rOsf/JtH/Drn9hP/omf/lZ1j/5Nr7/ooA+AP+HXP7Cf/RM//KzrH/ybR/w65/YT/wCiZ/8AlZ1j/wCTa+/6KAPyg/aF/wCCcP7GPgb4BfEvxr4W+Hn2LWvD/hnWdQsZ/wC1tVk8m6tLKWaGTZJdsjbXUHaylTjBBGRX3B+yd/yaz8G/+xM8Pf8Apugo/ax/5NZ+Mn/YmeIf/TdPR+yd/wAms/Bv/sTPD3/pugoA/9T79/b6/wCbcv8Ass3hD/26r7/r4A/b6/5ty/7LN4Q/9uq+/wCgAooooAKKKKACiiigAooooAK/Lb4yaPr/AOzAfGf7X37Nniy21LwhPqUt94z8IXc6yafeXazi1vZ7G4GWtb4SjEiHO5xg52rEf1Jr5N1z9hn9lTxH46uPiNrPgG2n1i8vP7RuV+03SWVxebt3nzWKzC1kcsSWLRHcSS2Sc0AeAmLXv2qv2n/iH4G1n4geI/BfhLwBpGgT6Rpfh7UX0a6vn1q1+1y31y8Y8yQQsViVclFYDIDZ3cn+1bZ6h4asdc03wl8T/iDq/jr4feCl1K3ttG1CK0s7CLT7eXGqa27GGK5ku3icumXkdQwjhAO6vtv4tfst/An44a1Y+JfiR4YF9rOnQm2hvrW7utPu/s5JYwvNZSwu8WScI5KjccAEmsXW/wBjn9nLxCLFNS8JkR2GjReHljg1C/to5tKgBEVtcpDcItwseTsMwdgeQcgEAHyhrWs+PfjL8XP2b/Dmo+N9d8Oad438AXmsa1HoV8+n/bLj7PZy5Jj4Q75Dh0AkVcqjKCa8c8VePPi34Y/Z6+Kvw/sfiBrjXvgP4qad4d0nXJLx31ZNLubmzdYprnhpygmZWMmQ6nawKYWv0/8AB37Onwf8BXvhLU/DOiyw3ngawutM0eaa+vLl7azvCrTRfvpnEgYqoBk3FAAqFVGKj1T9mz4K6zYeIdM1Lw751t4r1uDxHqSfa7tfP1W1MZiuMrMCm0wp8iFYzjlTk5APlLSbPxd8E/2rbz4YaZ458Q+JPDPizwFqOtyQa9qMmoy22q2F0kQntpZMGFXjkOY48JnkAAKF+fPAXiH4mfE3Rf2LPC2o/ETxLpieOdI8Wvrt3Y6nNHeaiLO1ilj8+ZixdhyFkOZI9xaNkfDD9YtV+FngPW/HVv8AErU9M87xJaaXcaNFdedMu2wu3WSaHy1cRncyg7iu8YwGAzXKeHf2dvg54Uk8AS6B4f8AsrfC6C+tvDh+13T/AGGLUoxFdL88zed5iADM28r1Ug0AfnXfeMvjV8OPhh+0v8OPC3jTVdTj+Fut6QbLVtQuo7rWbPQtQit7q/SK6uyBJNBA0vkmUls8KS2wV6l+xfoHibVPix458e2fiH4h618OVstPg0GXxvf34a4uplZ74w2VyUV4k2xbJjCpDM6ozDOO8/au/Z81LW/A/iL/AIVF4cl1Cb4ja7pc3jy2tLmEahqui2cJhkSxOpObWG4VVi2bfKJCkhhJgnJ/ZZ8LfGXw98U9RNva+M9K+EH9iCKKz8e6ja3+pLrK3C7PsSwzXE0VstvuDCWTDMRtXAyACr+3fc3nxQk8Ffsk+DrKE+MviDctqlnq11O9vFocWiMk730ZjIkkuBysUadfmLEDr85fsWfF7xnpHxv1P9nF9OtdW+Ja6rqepfEnxTf6gt6uoW9kEisn03ynTnEiRCEoFtlU7k3FlX9MPjT+zl8GP2hbLTLH4u+G49cGjStNZSiee1uLdnwHEc9tJFKFfaNy7tpwCRkAjn9F/ZG/Zy8M3HhC88L+CLXQ7vwJNJcaRcadLcWdxFJMAsplmglSS5EigCRbhpFccMCCaAJP2q/DPxF8R/BHXp/hFq97o/jPQPL1nSjZTyQm6uNOYT/Y5VjZfOiuUVomifKMWBYcV8H6d8efGX7SnhH43ftJeB9b1bw74I8FfDq903RrW3up7VH8RTaY+oX12yoyZnsC0dvHJg4OXQg4Nfr1Xlel/BP4X6L8ONb+EmlaDFaeE/EQ1Nb6xieSNZl1hpGvBvRhIu/zXA2sCi4VNoVQAD8vfil498b3Gl/BfTPHfjLxzpnhbxN8N7S6sn8ESyS67qXikQxNMbowiS7kTyXRlJAjMrEyOO/oHwn/AOFyeAPiB+yV4K+IWv68ur+JNG8XXniSy1HVru+Nxdx2Vo8QufPlfcYSxZY2JELs4TFJ8afgX4rtfjNY3934V8Z3/gLwhoOn6R4Hm+H2o2dpqGlkJ5d5Fey3NxBeSBjFFsZpJIQo+cbixr6b/Z5+E3jO+8JeA/HP7SdnJffE3wS2sxaVd3N4Jry1sNTk2CO7azZbS4uGtkiWRwjDK5UhixIB8LfBeLRtV/Zf+F3hXUPGHi7Sb7Wdf8Yx6foPgq5Wx1TW7mPVbmQlrotGIoLSMO8m+WKLLqXbIVWSH4nfGfxR+yn8HLfUfHGtaV4kvPi9ZeE7vV7e5VNSksl1K6tdlw8eYZyEChw6vHIUBdXGc/ovc/sk/AC58MeHvCK+GpLbT/Cd5e3+lNbalqFvdWlxqUjyXZju4rhbjbOztvQyFCDtxgACPQP2RP2e/DGmWmi6J4Xe3sLDxDbeKraA6hfPHBq9m7yQTxK05CKjuzeSP3RJO5DQB8hal4t+IP7MPxX+NHgrwz4m1rxppGlfDCTx1plv4hvZdVmtdTt57i3ZUmmPmeS5jEjx7sf3cV61+zd8JNbsvA3hP9oW++Kfizx54l1jQxql7ZzawZ9Ev7i9tTJ5MNltEUSRSPiHywpUrz3FfXL/AAw8Cy/EO4+K0ulrJ4ou9HXQJbppJWV9MWdrgQGAv5OPNdmLbN5zgtt4ryHwD+xz+zj8L/HNt8RfAXhEaPrNjJcS2oivr02lrJdxvFM0Fk87W0W9JGUhIgAD8oFAH5u/Aq//AGqfif4J8G/tKeFNX8Rarr2qaqt5qsl34ksk8LS6cLtoruwXR2kH2cRRAojYEokBbPzDH7e18p3H7EX7Llz45l+Ij+BYE1i4vF1GVY7q7ispbxX8wTyWKTLau4f5stEeeetfVlABRRRQAUUUUAFFFFABRRRQB8Af8FR/+TFPiZ/3Bf8A08WVff8AXwB/wVH/AOTFPiZ/3Bf/AE8WVff9ABRRRQAUUUUAFFFFABRRRQAV+evx4+F2v6F4z8ZftQ/sxeOLbRfHfh+1jTxZot26z6RrEWm2wnjhvkyGtrlbVl8qYEEIV+4GZz+hVfLnxN/Yt/Zn+MHjC48e/EDwXHf65fLEt5PDd3lmLxYQBGLqO1miSfaFUDzFY4UDoMUAfJ8nxQ8Q/tffF34V+Ak8Uax8NvBXiP4fr41uLbRb5tP1HUb+e5+zGxF4iiRobZQzkR7S6ncRjBX0D4wfDyy8PReBPg2vxR+JGta79l1J9K03Q9VSHWb8rP5rXupajJ5KNDZrJHEnnyIpGOJXyK+nviX+zR8Dvi5oWheHPHHhS3uLTwsoTSGtZJrCfT0CKgS2ntHhliTaqgorBTtXIOBXNn9jz9nf+yPDOiQ+F5La38IfbP7OaDU9QhuI01B/Mu45biO4WaeOdzmRJndW5BGCQQD8/wDSfib8XPir8AP2SLnU/HWsaLrHjjxLPpms6jpdx9luru3t1vYSJCoKMzLCpyyth/nA3AGtfxn4g+Inwd0j9rX4VaL461/U9N8I+FtN13QL3UdSnutV02fULefz1jvmbz9u+IMmW+Tsclifvjwl+yf8BPA2meG9G8NeG3trLwhq8uuaRE2oX0q2d/KjRs8Yknb5NrsBEcxgksE3Emun8SfAD4SeLr3xpqPiHQftdx8QtOt9J11vtVzH9ssrUOsUWElURbRI3zRBGOeScDAB8O6JY+O/hD8X/wBnDW7b4i+JfEsPxVgu7HxFZ63qDXlpPIul/bIZ7e3IEds8cq/8sgMrwc5Yt45fePfirr3wm8I2dt4+17Sr7Wv2g7jw7JqFtfSG7TS3ubyMWqtIXVoUCrthdWiG1coQMV+sGo/CL4eatqHgvVNQ0nzbr4eMz6E/nzr9jZ7c2rHCyAS5hO396H9fvc1ykX7NnwVg07TdJi8O7bTSPE58Z2qfa7v93rxd5Dd5M2W+aRz5TExc42YAwAfF9vJ8YvhF8V/jf8Ffhj4w1XxM8fgCPxR4eHiW/OozWWrSSXFuVS5uzxG7oH2yt5YbGcLurg/2Q7fxX8R/jd4b8e+HPF/xM8ReB7Tw0bvVrjxRf3tvpcmvyuiJFaQApb3UZjaVmVEkiTCMr52g/Z37Svwd1/WfBvjvx58ENLgf4s69oUGgJcTzDbcaWl0JZ7VY7ovZq7RPMI3kjxvYb22jj5p/Z0+G3xX8BfFbwXpPwt0Xx94W+GGk217a+IrHxzqlheWYVbfbZJpUEFxcyK4nwWeMpDsBAHOKAPc/26fGVnZfCO2+D8WgweJNc+MV8nhXS7W7mNtZpcXalvtNxMvzKkG0OAmXZ9oUdSPz5+BfjX4l/s+ftHv+zh4iFr48+OGv3WlaZH4o1HUPP0+Hwda27TGFIBJHLDcQiN2+zn55pG8xmI2l/wBifi78F/hj8ePCTeB/ivoUWv6P5yXCRu8kTxTxghZYpYWSSNwGI3IwOCQeCRXk1h+xH+y1pXhiy8J6X4BtrK20/UYdWgube4u4dSW/gJMc/wDaMcy3hdcnGZiOelAHsvxc8G6v8Qfhl4m8F+Htbu/DeratYzQ2WpWNxLa3FpdbcwyrLCyuAsgUsFI3LlTwTX54fs4fG/4mftR/FfwJot7d6l4fh+Dej3DePLeGWW2S98UvJJp8VlcKhUSRILeW72ZZMsqsCRx+qYAUBR29ea4vwp8O/BfgjVPEmteFdKi0+98XX/8AaeqSR5zc3flJCZDknGVjGQuBuLNjczEgH46aB8VPiTefsofs3+IPHnjjxVa+EvEl3rdr4n1XQrqSfxLdXQnuBpcUcp827eMsjiTyVZwqqOF5HU+EV+M3w7+G/wAE9Q8S+JPGVveeMfjFZ2uPEWq3r6lc+HWW/wDskV9BLKRD50So01uAEYqrMm6vUPj1+zXc+H9S+G3g3wd4P8RXvwS8C2d7c2tj4Nv4Y9f0/XpZ2aO4FxfzpdvAYppQFgn3BzlgVCivXP2fPg/458V+GmX9pCw1a/svC3i1Nf8ABEfiLUY7nXrKK3t9kEl/Pp0nlSurySlY5GkIDYk3YXAB84+HrrQoNH+Ptr4i8Y694QtdR+M1zaIvhhjHrOrXE9jbJBptpIoLxvPLtYspU7UYF0Qsw88174n/ABi8Mfsb/tOaRP4m8SWOq/DrxBaWejXmpaismv2FpcPYSCCa/s5G3uokYF1lYkMV3EcV+n+ufsv/AAN8RaPr+h6r4bL23iXXh4nvGS9vIp/7aVVRb2CeOZZbeRVRQvksgHOByc8vH+xd+zcmjeL/AA8fC00mm+PEgTWoH1XUnW7Nu8ciOS1yWWUvEjPKhV5CPnZsnIB4c2l+NPgJ+1r8M/DWneOvEfjDSPibo/iNtVstbvjexte6NDBcR3FrGQsdq0hk2FIVSMLwFAAxgfsfeCvFHx78DeF/2pvHXxX8UT+J9Y1Ge/m0nTtWaDQbWK3u3jGmNpwXyzGI0Cy7h5hYk7s4Y/fmt/DbwX4j8b+GfiPrOnfaPEXg6O+i0q686VPsyakiR3Q8tHEb+Ysaj94rbcZXBJz4x/wxn+zYnxGX4q23g5bTxCuoxauWtr29t7RtRhkEqXTWUU62rSh1DbjEcnlsnmgD86NPv/2o/j1qHxS+IXgXVPE8HjDwr4r1PSdHhsvEVnp+gaUmlShYrW+0qVwLjzFG6eSRTvDDb905/aDw/d6hqGg6bf6tFHBfXNtDJPHC4kjSV0DOqOCQyhiQCCQRzmvnTx/+xf8Asz/E/wAZXfj/AMZ+Co7nXNSCi9mgu7yzW9CABRdRWs8Uc/CjPmK2cYORX0xZWVpptnb6dYQrb2trGsUUaDaiRoAqqoHQAAACgC1RRRQAUUUUAFFFFABRRRQB4B+1j/yaz8ZP+xM8Q/8Apuno/ZO/5NZ+Df8A2Jnh7/03QUftY/8AJrPxk/7EzxD/AOm6ej9k7/k1n4N/9iZ4e/8ATdBQB//V+3f+Ck2v/wDCKeFfgX4p/s291j+x/iz4YvPsOmw/aL66+zx3knkW0OV8yaTbtjTI3MQMjNdB/wAN9f8AVuXxm/8ACQ/+6qP2+v8Am3L/ALLN4Q/9uq+/6APgD/hvr/q3L4zf+Eh/91Uf8N9f9W5fGb/wkP8A7qr7/ooA+AP+G+v+rcvjN/4SH/3VR/w31/1bl8Zv/CQ/+6q+/wCigD4A/wCG+v8Aq3L4zf8AhIf/AHVR/wAN9f8AVuXxm/8ACQ/+6q+/6KAPgD/hvr/q3L4zf+Eh/wDdVH/DfX/VuXxm/wDCQ/8Auqvv+igD4A/4b6/6ty+M3/hIf/dVH/DfX/VuXxm/8JD/AO6q+/6KAPgD/hvr/q3L4zf+Eh/91Uf8N9f9W5fGb/wkP/uqvv8AooA+AP8Ahvr/AKty+M3/AISH/wB1Uf8ADfX/AFbl8Zv/AAkP/uqvv+igD4A/4b6/6ty+M3/hIf8A3VR/w31/1bl8Zv8AwkP/ALqr7/ooA+AP+G+v+rcvjN/4SH/3VR/w31/1bl8Zv/CQ/wDuqvv+igD4A/4b6/6ty+M3/hIf/dVH/DfX/VuXxm/8JD/7qr7/AKKAPgD/AIb6/wCrcvjN/wCEh/8AdVH/AA31/wBW5fGb/wAJD/7qr7/ooA+AP+G+v+rcvjN/4SH/AN1Uf8N9f9W5fGb/AMJD/wC6q+/6KAPgD/hvr/q3L4zf+Eh/91Uf8N9f9W5fGb/wkP8A7qr7/ooA+AP+G+v+rcvjN/4SH/3VR/w31/1bl8Zv/CQ/+6q+/wCigD4A/wCG+v8Aq3L4zf8AhIf/AHVR/wAN9f8AVuXxm/8ACQ/+6q+/6KAPgD/hvr/q3L4zf+Eh/wDdVH/DfX/VuXxm/wDCQ/8Auqvv+igD4A/4b6/6ty+M3/hIf/dVH/DfX/VuXxm/8JD/AO6q+/6KAPgD/hvr/q3L4zf+Eh/91Uf8N9f9W5fGb/wkP/uqvv8AooA+AP8Ahvr/AKty+M3/AISH/wB1Uf8ADfX/AFbl8Zv/AAkP/uqvv+igD8Qf29f2vP8AhaH7J3jnwL/wpX4meEv7T/sz/iaeIfDn2DS7fydStZv39x9ofZv2bE+U7nZV75r6/wD+G+v+rcvjN/4SH/3VR/wVH/5MU+Jn/cF/9PFlX3/QB8Af8N9f9W5fGb/wkP8A7qo/4b6/6ty+M3/hIf8A3VX3/RQB8Af8N9f9W5fGb/wkP/uqj/hvr/q3L4zf+Eh/91V9/wBFAHwB/wAN9f8AVuXxm/8ACQ/+6qP+G+v+rcvjN/4SH/3VX3/RQB8Af8N9f9W5fGb/AMJD/wC6qP8Ahvr/AKty+M3/AISH/wB1V9/0UAfAH/DfX/VuXxm/8JD/AO6qP+G+v+rcvjN/4SH/AN1V9/0UAfAH/DfX/VuXxm/8JD/7qo/4b6/6ty+M3/hIf/dVff8ARQB8Af8ADfX/AFbl8Zv/AAkP/uqj/hvr/q3L4zf+Eh/91V9/0UAfAH/DfX/VuXxm/wDCQ/8Auqj/AIb6/wCrcvjN/wCEh/8AdVff9FAHwB/w31/1bl8Zv/CQ/wDuqj/hvr/q3L4zf+Eh/wDdVff9FAHwB/w31/1bl8Zv/CQ/+6qP+G+v+rcvjN/4SH/3VX3/AEUAfAH/AA31/wBW5fGb/wAJD/7qo/4b6/6ty+M3/hIf/dVff9FAHwB/w31/1bl8Zv8AwkP/ALqo/wCG+v8Aq3L4zf8AhIf/AHVX3/RQB8Af8N9f9W5fGb/wkP8A7qo/4b6/6ty+M3/hIf8A3VX3/RQB8Af8N9f9W5fGb/wkP/uqj/hvr/q3L4zf+Eh/91V9/wBFAHwB/wAN9f8AVuXxm/8ACQ/+6qP+G+v+rcvjN/4SH/3VX3/RQB8Af8N9f9W5fGb/AMJD/wC6qP8Ahvr/AKty+M3/AISH/wB1V9/0UAfAH/DfX/VuXxm/8JD/AO6qP+G+v+rcvjN/4SH/AN1V9/0UAfAH/DfX/VuXxm/8JD/7qo/4b6/6ty+M3/hIf/dVff8ARQB8Af8ADfX/AFbl8Zv/AAkP/uqj/hvr/q3L4zf+Eh/91V9/0UAflB+0L+23/wAJX8AviX4W/wCFB/FnR/7Y8M6zZ/btS8LfZ7G1+0WUsfn3M32lvLhj3bpHwdqgnBxX3B+yd/yaz8G/+xM8Pf8Apugo/ax/5NZ+Mn/YmeIf/TdPR+yd/wAms/Bv/sTPD3/pugoA/9b79/b6/wCbcv8Ass3hD/26r7/r4A/b6/5ty/7LN4Q/9uq+/wCgAooooAKKKKACiiigD80v2pvG+uSftM+DvhF43+J+r/B34b65oMl1Y6vo8kVk+o+IFuihs5tQmjkSFY4NrrGQA5bBbcUFet+K/wBoLwb+x54R8DeBvjn401Dxnr2t3k1rFqQsGku5LQzSSRz3MVmj58mApESiF5nUsE++UT9p34iaJZXN58L/AIsfBDxF8S/AOtafHLFeaFpg1lDeb5Ukt5YEZZbaVAI2hnBGWc4ZCma+QtI8A/HLwH+zx+zx4n8YeFdc1qT4c+NJdUudJgRtS1uw8PSreR2iSxqd80lvFJGpRBuVdq7RtIAB9xeI/wBt79mDwlJpEXiLxn9iOs2cGoRb9Pv/ANzaXJIhlu8W/wDogcjAFx5Z74xzXonxA/aK+C/wuvdM0/x14nh02bWrGfUrHEU863NrbmMO0TwRurMTLGI0B3yFgI1Y8V+f/wC0h8Qvi18WdB8ceENF8AeMPDHhzxV4Lu5tOhtPDXnajrWqulxb/ZtVmaOdbKKJUiZYz5c7o3yyBisR6rwB4S1/xZ8Yf2U/Gd94Q1e2sfCngfVra6m1DTbi2/s/UEtra0VZ/NQeU8m2Tyg+C6/MoI5oA9p8Uf8ABQL9mrwyvgK7bXLq/wBP8f8A2uS1uLXTr2Vre3s1mDyzW6wG4GZ4vJCCIvuJYgIrMGfD34tX3iP9rHx1b/2/M3gWHwPoGuWUNyXt7eAXkk7vcGOYI0TNGFL7wpUDDAEGqH7VNn4k8K/GH4EfGrSPCuq+J9A8B6jrw1aDQrNr69iTV9Na1ilW2i/eOokOXKg46nkivG/FXw6+IXx2+I3x0u9F8O6v4Wh+Inwu03T9Ml1e0ezAvnN0fsk0nzxrINyrMiszIrcigD69+Gv7XH7PPxe8Wr4G+H/i5NR1iaOaW1jktbu1S9jtziV7Oa4hjiuVTBJMLPwC33RmvWPiT8SPBfwi8Eap8RviHqP9k+HdFRJLu68qWfylkkWJT5cCSSNl3UfKp656ZNfl38C9FHiDxF8I/D/jjwL8WLTxV8PpbfYmp2ttD4b0mWC1+zzPFerGiSWrJlI0jdnddoKjGR9bf8FAPDHiXxn+x/8AEjwz4P0m713WL61tFt7KwgkurmZlvYHIjiiVnYhQScA4AJ6CgDubX9rf9nW8k8YrH41to4vAMK3GsTyxTxW0MLyGJHjneNYrgNKDGvkM+5/lXJIBs/D79qn4CfE/SvEOs+E/FcZt/Cdqb7VFvre502a0swhk+0vDexQyeQUUsJApTHfPFeSftsfBfxH44/Z5svD3wq0t7m68Ialo2pxaVaMkUt5ZaRJk2kJkVk3LGS0SsCCygYJOD4V4FsvCXjfxP4q+IGr/AA3+LPiO+j8F6lo2oR+LbKCzF3YTyI8mk26Yha4nkbeUKNsUg/OCwoA+wPDP7Xv7PXi/w14k8X6F4oeTSvCWntquoyy6ff27R6eAxFzHHNbpJNEwUlWhVw3bNO+Ff7Xv7OXxp8Qp4S+HXjSG/wBbmiM8Vjc211p9zPEo3FoYr2GBpQFBY7A2FBPQZr83be8+Jmlfs6/tAeALbT/FUnwe0fwLeN4fuvGumtp2r2FyYnSTS4zLtkubaKNQVkZfk2iMEgAn0jSdU8SftITfs0eHPBHw/wDEOhr8Nb7Sdd1fxFrelyaZbwWdjZeW9paTy4Nx9tLKMRblKhWbgZUA+1fHv7Yn7NXwx8Wz+BvG3jq0sNZszGLqJIri4SzMv3BdTQRSRW5PXEzpwQehFdJ8RP2lPgh8KpvDkHjjxTFZv4vt57rR/IguL0X8VssbubdrWOUOSJUMag7pNw8sMa+H/hv4p8Tfsz+H/iF8GfHnwZ8UeOdX13Xta1K2v9G0r+0tN8SQatK8kRu7ofJA+xhFOs4wiKCNw4ql8HvgP8Ufhn42/ZA0DxfpVzfzeCtB8Yx6vdwRPc2mmS38UUltbS3Kho0KK3kRkth9hCFhzQB9u6b+1T8ANV+FV/8AGq18XwL4Q0q4NneXUsM8Utvdh1T7NJavGLhZizqBGY953AgEEGr2h/tLfBHX/h1rfxXtfE8dp4Y8NOYtTub+C4sHs5QqP5csF1HHMrsJE2rsy5ZQoJIFfDWtfDm0Rf2ndP8AiV8PPFGteE/FHi/S7q2XQLKX+0CBZwP/AGlp33TMbe5QMzRbzkEFW5Q8Jrvg79pz43fspa9o2s6dreuTeCfF2nan4fl1e0TSPEHiLRNOZJWS5tpQwFwu5vLeRAZiikpuyGAP0m+EP7Rvwb+Os2pWnwy1/wDtG80lY5Lq0uLW5sLqOKbmKXyLyKGQxOPuyKpU9M54qb4t+Nf+ES1z4d2P/CWweGP+Eh8RQ6eYJtOlv21XfBM/2KN4zi2diu8TP8o2YPXB+U/2cLfQ/F/xzHxJHhj4oW2vWfh+fS59T8c2kNhax273MUoskARGnk80FwU3Im1vm+YA+n/tV+GPEviLxn8Abrw/pN3qcOjeP7O8vntYJJltLVbO6Vp5yikRxhmUF2woJAzkigDW139uH9l3w1rGqaFrXjZbe70LVJtG1L/QL9o7C8gcRuLqVbcxwx7ztWZ2ETkNtdtrbe/+LX7SHwX+B8Wkt8R/EaWM+vbjp9rbQT393dqg3M8VvZxzSsgBGX27BkZPNfEWj/DPxk37Nf7ZOhXnhXUP7T8UeK/HF1pNq9jMJ9RintIvsctrGU3zpJIv7lowwZh8pJqGys/G/wACPjD8Pfjz4o+H/iDxfoN/8M9J8LzNo1jJf6noeoWzC4mSaxXEoimBwzgFldSrAA8gH2VfftY/s+aZofhTxJqPi6O203xxbahd6PNJa3Si6i0sKboYMO6ORCwURyBZHchEVn+WsW9/bQ/Zs0/wNoXxDuPFxOleJp7m202OOwvpL66ms2KXCpYpAbr9yw2yMYgqnGSMjPjHjG01T4y/tF/st/FGHwJrumaJpf8Awmlzdxatpr28umTPbRRWr3qLvS3aeSMvAJGDNwQNwIHLftVeBvGnhf8AaU8IfH7TdL8V6p4TTw5deH70+CYorjVtOuXuzdi5+zPG7PDMGKymMbhsBPZXAPqXWv2uP2d9C+HWh/FW78Ywz+HPEsr2+my2tvc3c91PHnzIktYInuA8e0iRWjBjPD7TWrN+098BoPhRb/G4+MLWTwXdzC2hvokmkaW5ZzGLdLdEM7TbgR5Qj38H5cV8O6De+Jvhf8P9P1b4ReB/HGh6b4z8a6hca3rGtaImq+J7CG9tYnnvrTS4YnaJbu4jWMmWFwpDSPE25BXnPw/8LWX/AAzt8RPAPxX+HXxDuIU+JF/qtrc6fpUsOuWS3UslxY6va+WiLLJC0QMptldUMg+QqcUAfpZa/tMfA65+F+o/GR/FMVp4R0iY215d3cFxavBcLs/cPbzRpOJiXQLH5e9iwCgkitL4RfH/AOEvx1g1OT4Za5/aU2ivHHfW01tcWN3bGYFozJbXccUyrIASj7NrYOCcHH5peKvCf7UHxk/Zf0HWPFuj61qur/Df4hWusWKyWsWleItf8O6chjiums5A6RX4ExZFcDcYgShYjd9Hfsw2WgeI/jHrPxNtPDfxKs9Xm0FNMutT8dWkVhE8aXKyx2sUQSN5ZY2MhMgBRRldxyMAH6B0UUUAFFFFABRRRQB8Af8ABUf/AJMU+Jn/AHBf/TxZV9/18Af8FR/+TFPiZ/3Bf/TxZV9/0AFFFFABRRRQAUUUUAFFFFAH58fsjftARH4XW0/xi8T3F3q/ifx5rnh3SJLlZrh5po7iU29sGjRgirFG20uVQAY3Zxn6o8SfHr4T+EdS8X6R4h137Jd+A9Jj1zWk+zXMn2TTpQ5SbKRMJM+W3yRl3GOV5GfyR+GuieKz+z/4I+JPhnw/f+KLb4ffGfVta1Ky0qE3N82nCe6gllt4FO6ZkaVTsXJIyegJHo/j6y+IPxZ179qLxlo3w88T6Zp/if4b2enaMmo6VPBc6jPEt0CkMGGfzCzYEJAlAwSgDLkA+1rP9uP9le+vZbCLx7BG62f26F5rW8givIchf9CkkhVLt9zBRHbmSQt8oUkEDe039rr9nvVPhX4g+NFr4qx4T8KXAtNWnksryO4sbgyJEI5rNoBdKxaRAP3XOc9ASPBPGPgDxPdeM/2Pru38OXk0PhR7gak6WkjJpqtonlj7SQuIAZVVBv2/OAOvFfLX7XXhTxDoPg79uHXb3Rruw0TXW8BzafcyW8kVrevElsly1vIyhJSknyyFCcNw3NAH6afCb9qT4CfHDV7rw98M/F0Op6xZRefLYTQXFjeCLODILe8ihlZASAWVSBkZIyM+d+Ef2pvhX4T+E3hnxn8UviXaazD4m1PU9PsdYi0m70+G7ntLm4H2cW2yR42hWIxBnwJWTK7i658jl1TxJ+0f+1L8IvHXhL4f+IfCOifDFNYudY1jxFpcmkTS/b7U20OnWyTYecFyXlIBjUAENk4PhPwq+FnxHsvh5+y1Yar4Q1aCfw98QvEV9qUU2nzq9jbS3OovDcXKsmYY33oyO+FO5SDyKAP0q+HP7RPwa+K2ga/4m8GeJIpbHwo7pq/2yGbTpdP8tDIWuYbyOGWJNgLB3UKQGwflOOf+Fn7WPwA+NHiN/CXw68VLqGrCBrqKGa0u7I3Vsh2tNatdQxLcRqeC0RYDr05r47+LfwH+IvxL+If7Wnh7wvplzpx8eeFfDEOmXssTwWeoXllFPvhS4ZRG5wqwyYY7A43YFZPwTtbfxb8Q/hne+JfAnxYh8V+A4roRjX7S3ttC0lpLFoLhI7kRRrNDMAsUSRklgUZlUKcAH278Pv2rfgD8UvEcvhXwL4sTUr2OO4lR/st1DazxWhxPJb3U0SW86RnhmikYD6VZ+F37UPwM+M/iCXwx8NvEv9r6hHBLdIPsd3bxXFvA6xyTW808McU8aO6qWiZgCa+F/wBnGfxx4M+KMvg74VeBfGdr8HbzTdVvNT8OeLtLNrFoGo7TJFa6RdTNieO6d2QwqzxqG8wOdxxc/Ze1b4g/Dnx9q3hf4Z+DvHB+DNhod9qK6N4r0trS90XVYmDx6dpNzOwa6jnBYCPc6IdrCU5YkA/VkkAZPAFfNHg79sP9m/x/46h+HPhLxpDf6zdyzW9qRb3MdpeTW+fNjtb2SJbW4dQM4ilYkcjPNbfh7xVH+0v+zcfEvhmK58Pf8LB8P3K2q3Y2T2j3kLxKzbcj5WO4MM5GCBzivzL+C3gm+j0b4V/Bn4qeBPisPEvw+1jSZIoYba2/4RW1u9NmCrqMWoCMJ9mC73YbzI4LIA2QxAP0e0D9r39nPxP8R/8AhU+ieMoZ/Ej3U1lFE1vcx29xd25Ilggu5IltppEKkFY5GOeBk8VoXn7U/wABrD4mH4Qz+KFPidLyDTnhjtLuW3jvrkqIbaS7SFrZJnLACNpQ2eMZ4r83fij4h+Mnizxf4H1XxR4J8V2qfDv4pWlyNC0fwzI2j2WhWklxHHexXUUDy3s9wrozNDL5S7mBiT5Wb0rxBceO/Cf7VC658AfBvjPS9Q8SeJbaDxhpmoaYz+EtU05WEUms298W8q3uFiVWDI292AVozht4B91R/tJfBST4a+JPi6fEyReFPCF1c2OrXUttcxPZ3VnIIpoJLeSJZxIrsqhBGWYsu0HIz63DrulS6EniVp/s+mvbC7MtwrW+yAp5m+RZQrR4Xlg4BXncAQa/Jv4hfBJvHH7etx8J/DGohvh7r0ek+P8AxzpKpmIajpbSwWkbH7gF8/kyTRfecRmQ8AV+rHi8xL4T1pp9KfXYxY3JbT4wpe8XymzbqGIUtKPkAJAyeaAPBfC/7Y/7OXjOx1vUvDvi37RbaBYTarcSSWN7bq9jA6xvPbmaBBcp5jqgMO/czADJIzxfgL9vn9m3xz8Or34lPrlzomnaYsD3Ud9p16kkf2uVoreOMrAUuZZCv3LZpSPoM14B+yra/EXTvF3iP4UeBdD8X2nwXbw9cmwi8b6a9heaDqkkgRdNs7iY+Zc2pjYtjDrHtUCQ87+X+F3xQ+MXw2/ZL8O/C7QvhD4mh8dfDyCytr65v9Aku7S0X7WIJ7rTlVwb+eK3laWNIcgjO5sAqwB98+HP2ofgV4s+GniH4u6F4oS48MeE1lbV5jbXKXFj5KB3We0aIXKsFOdvlZPYGuTn/bX/AGaY08Stb+Lxdt4T0w6veiGzuyPsqssZMLmIRzOJHWNkRiyudrAEMB+fMPh3x/qnhv8AbF0ODwz4z1e68d6JY32j3ur6DLaXGqtFpq28oWOC2hjE7TMdlukSyBB8yBlav0gl+Hl3P+yY/wAM/D2mCz1GTwO+jWloy+U0Usmm+QsJ34KfPgMGxgjnpQB8q/FT9sXw/wDFj9n3RPHvwN1rUtI1H/hIfCEWoxta3VlLbx6tdx+ZbebNFHHOCodHaBpEOMhsMpP1f8U/2sv2f/gv4kXwh8RPFa6fq4gW6mt4bS7vTa27nCzXRtIZVt4ycYaUqCOenNfAeqan46+IH7IXw5+D1n8MfGGmeIfh5d+BLPUVvtGmiikfTbiCG5e2YbmmjjEJkeRV2KjKS3UC18QfBfir4WftF/F7VvGOh/EPVvCXxSawurK/8C2kWpJKkVn9lmsdRhMMkiFCCsP8BRjkgk0AfrhpupadrOnWusaRcx3tjfRJPbzwuJIpoZVDJIjqSGVlIIIOCDkVdrxv9nvw1p/g74KeDvDGkaTqeg2Gm6fHDb2GsvHJqNtCCfLjuTESnmKuAQCdv3c8V7JQAUUUUAFFFFABRRRQB4B+1j/yaz8ZP+xM8Q/+m6ej9k7/AJNZ+Df/AGJnh7/03QUftY/8ms/GT/sTPEP/AKbp6P2Tv+TWfg3/ANiZ4e/9N0FAH//X+/f2+v8Am3L/ALLN4Q/9uq+/6/OD/gpNp2u6v4V+Bek+FtX/AOEf1q9+LPhiCx1L7Ol39hupI7xYbn7PIQkvlOQ/lsQr42ngmug/4Z4/bs/6O4/8sHR//jtAH3/RXwB/wzx+3Z/0dx/5YOj/APx2j/hnj9uz/o7j/wAsHR//AI7QB9/0V8Af8M8ft2f9Hcf+WDo//wAdo/4Z4/bs/wCjuP8AywdH/wDjtAH3/RXwB/wzx+3Z/wBHcf8Alg6P/wDHaP8Ahnj9uz/o7j/ywdH/APjtAH3/AEV8Af8ADPH7dn/R3H/lg6P/APHaP+GeP27P+juP/LB0f/47QB9/0V8Af8M8ft2f9Hcf+WDo/wD8do/4Z4/bs/6O4/8ALB0f/wCO0Aff9FfAH/DPH7dn/R3H/lg6P/8AHaP+GeP27P8Ao7j/AMsHR/8A47QB9/0V8Af8M8ft2f8AR3H/AJYOj/8Ax2j/AIZ4/bs/6O4/8sHR/wD47QB9/wBFfAH/AAzx+3Z/0dx/5YOj/wDx2j/hnj9uz/o7j/ywdH/+O0AfXXxd+Hdp8W/hd4r+GF9ePp9v4q02502S5jUO8K3MZjLqpwCRnIBrovB/h2Lwh4S0TwnBM1xFoljbWKysArSLbRLEGIHAJC5Ir4j/AOGeP27P+juP/LB0f/47R/wzx+3Z/wBHcf8Alg6P/wDHaAPv+ivgD/hnj9uz/o7j/wAsHR//AI7R/wAM8ft2f9Hcf+WDo/8A8doA+/6K+AP+GeP27P8Ao7j/AMsHR/8A47R/wzx+3Z/0dx/5YOj/APx2gD7/AKK+AP8Ahnj9uz/o7j/ywdH/APjtH/DPH7dn/R3H/lg6P/8AHaAPv+ivgD/hnj9uz/o7j/ywdH/+O0f8M8ft2f8AR3H/AJYOj/8Ax2gD7/or4A/4Z4/bs/6O4/8ALB0f/wCO0f8ADPH7dn/R3H/lg6P/APHaAPv+ivgD/hnj9uz/AKO4/wDLB0f/AOO0f8M8ft2f9Hcf+WDo/wD8doA+/wCivgD/AIZ4/bs/6O4/8sHR/wD47R/wzx+3Z/0dx/5YOj//AB2gD7/or4A/4Z4/bs/6O4/8sHR//jtH/DPH7dn/AEdx/wCWDo//AMdoA+/6K+AP+GeP27P+juP/ACwdH/8AjtH/AAzx+3Z/0dx/5YOj/wDx2gD7/or4A/4Z4/bs/wCjuP8AywdH/wDjtH/DPH7dn/R3H/lg6P8A/HaAD/gqP/yYp8TP+4L/AOniyr7/AK/EH9vX4NftY+FP2TvHOv8AxM/aK/4Tvw3a/wBmfa9F/wCER03S/tW/UrVI/wDS7eRpY/LkZJPlHzbNp4Y19f8A/DPH7dn/AEdx/wCWDo//AMdoA+/6K+AP+GeP27P+juP/ACwdH/8AjtH/AAzx+3Z/0dx/5YOj/wDx2gD7/or4A/4Z4/bs/wCjuP8AywdH/wDjtH/DPH7dn/R3H/lg6P8A/HaAPv8Aor4A/wCGeP27P+juP/LB0f8A+O0f8M8ft2f9Hcf+WDo//wAdoA+/64H4n+ENd8eeB9R8K+GvFN74L1G+8oR6tpyxtdW4SVXcRiUMmZEVkyRkBsjkCvj7/hnj9uz/AKO4/wDLB0f/AOO0f8M8ft2f9Hcf+WDo/wD8doA+pvgh8GvCPwC+Gmk/C/wU1xNp+mea7XF3J5t1c3E8jSzTzvgBpJJGLHAAHQAAAV6zXwB/wzx+3Z/0dx/5YOj/APx2j/hnj9uz/o7j/wAsHR//AI7QB9/14v8AtDfBnT/2g/g54j+D2q6lLpFr4iS3R7qFFkki8i4juAVViAcmMDk9DXzR/wAM8ft2f9Hcf+WDo/8A8do/4Z4/bs/6O4/8sHR//jtAH3/RXwB/wzx+3Z/0dx/5YOj/APx2j/hnj9uz/o7j/wAsHR//AI7QB9/0V8Af8M8ft2f9Hcf+WDo//wAdo/4Z4/bs/wCjuP8AywdH/wDjtAH3/UNxbwXdvLaXKCWGZWR0YZDKwwQfYivgb/hnj9uz/o7j/wAsHR//AI7R/wAM8ft2f9Hcf+WDo/8A8doA+9bKystMs4NO063jtbS1jWKGGJAkccaDaqIq4CqoGAAMAdKtV8Af8M8ft2f9Hcf+WDo//wAdo/4Z4/bs/wCjuP8AywdH/wDjtAH3/RXwB/wzx+3Z/wBHcf8Alg6P/wDHaP8Ahnj9uz/o7j/ywdH/APjtAH0d8JPghp3wv8TfEDxxPqkuu+IviJq/9pX15PGsbRwQxiKzsowpP7m1jyseSSdxJNe418Af8M8ft2f9Hcf+WDo//wAdo/4Z4/bs/wCjuP8AywdH/wDjtAH3/RXwB/wzx+3Z/wBHcf8Alg6P/wDHaP8Ahnj9uz/o7j/ywdH/APjtAH3/AEV8Af8ADPH7dn/R3H/lg6P/APHaP+GeP27P+juP/LB0f/47QB9/0V8Af8M8ft2f9Hcf+WDo/wD8do/4Z4/bs/6O4/8ALB0f/wCO0Aff9FfAH/DPH7dn/R3H/lg6P/8AHaP+GeP27P8Ao7j/AMsHR/8A47QB9/0V8Af8M8ft2f8AR3H/AJYOj/8Ax2j/AIZ4/bs/6O4/8sHR/wD47QB9/wBFfAH/AAzx+3Z/0dx/5YOj/wDx2j/hnj9uz/o7j/ywdH/+O0Aff9FfAH/DPH7dn/R3H/lg6P8A/HaP+GeP27P+juP/ACwdH/8AjtAH0B+1j/yaz8ZP+xM8Q/8Apuno/ZO/5NZ+Df8A2Jnh7/03QV8P/tC/An9s7SPgF8S9W8U/tQ/8JBotl4Z1me+03/hCdKtPt1rHZStNbfaI5S8XmoCnmKCyZ3DkCvuD9k7/AJNZ+Df/AGJnh7/03QUAf//Q+/f2+v8Am3L/ALLN4Q/9uq+/6+AP2+v+bcv+yzeEP/bqvv8AoAKKKKACiiigAooooAKKKKACiiigAooooAKKKKAOMl+IfguDx/B8LJdVjXxXdac+rR2GG81rFJfJaYHG3aJDt65z2o8ZfEPwX8PjoQ8ZarHpf/CTapbaJp3mBj9p1G83eRbrtBwz7GwTgccmvjfXCE/4KaeGC3Ak+F98q+5GrKSB+HNO/bxBkm/ZyiQbnPxi8KNgddqLdlj9AOtAH3zWfY6tpepyXcWm3kN29hMbe4WGRZDDMFDGOQKTscKynacHBBxyK+DdG+OH7VXxO+JPxR8D/CnR/CltZ/DfxHDp/wBs1kXyi7s5IY5TCgt5G/0hcuzSnaigxqI2LMy+b69+0t4u+Gngj44eMvh/4P8ADlvrWhfE+38PpGttLbx6kLs2UTXN68cu5rphLjzhhRhSY2wQQD9SqK+G/Dvxr/aK8P8Axr1j4H/EzRvDmtaxqPhO78TeHJdDa6s4JJ7SUQNp90128pyzuuJ1CgLyU52rzPwn/aW+M178e7j9n/4kXHgnxFrF1od9qdtceFbm68mwvrKSNGsL8TNM6n94GLgBgP8AlnzwAfoDe3tnptnPqOozx2tpaxtLNNKwSOONBuZ3ZiAqqASSTgDk0+3uLe8t4ru0lWeCdVeORGDI6MMqysOCCOQR1r8oPhj8Xv2m/Ff7NXxj+K3xcfwh4o8L6Vp/jIJpslhdtLLdaYZkS2mUzLC+nlYmQoFErIRukLFmPs9t8ffinrr/AA3+C/wA8N+H7fxTqXgrTfFOpXGp+fDoej6fOiQwQQW1sfOkZ5AyxxiRRHGoJJByAD7+or81fGn7Y3xc+HXwp+NNv408OaNZfFX4QW2nXm23e4udD1Oz1SVVguYVLxXCjG9XjZ8qwB3EHA9L8NfGz4/eGPj74G+Fvxr0zw62lfFHT9TutJk0L7X5+nXOlQx3E1vdSXDFbhTE42yJHFls/IBxQB9v0UUUAFFFFABRRRQAUUUUAFFFFABRRRQAUUUUAfAH/BUf/kxT4mf9wX/08WVff9fAH/BUf/kxT4mf9wX/ANPFlX3/AEAFFFFABRRRQAUUUUAFFFFABXMeM/GXhj4e+FtS8a+M9QTStE0eIz3d1IGKQxA4LEKCcc9ga6evkP8Ab4/5M4+LP/YEl/8AQ1oA6T4cftjfszfF3xbaeBPhv49std1++WV4LSFJ1d1hQySEF41X5UUk89q+mK+FPgT8Qv2kX8OeHYPGHwd0vw34atdCWRdZi8Qw3suIbLdATaJbxv8AvWVQw8z5QxOTiuB+E/7UH7SHin4I6P8AtS+O9E8Mab8N7fQtT1PV7O3+2rrMrWEUzLLZh3eBI5pYgiRyO7bCJC+W2KAfpTRX5Y/CD9tz4peIvHHw+g8cjwrqehfEi4S1Gn+Hlv21Xw/NdRGW1+2yzkwXCZAimeNYwrtlcrgM3xn+1z+09rvwk8cftIfBzQvCWn/DTwnNqEdl/bwv7rVNVi02Y28tzGlrJBDFG8isEVnYjacnoaAP1Qor5X+Hnxx8V+Lf2g9W+E+o2llHpFh4O0bxCksSSi5N1qMsiSozNIyeUoQbAEDA5yxr5hv/ANu3xtZ/A/wl4new0O28a+OvFWseH7GS7+0w6NZW2lXU8b3l0qSSTvsiiXckbAu7ZXaBtIB+o1FflRc/tz/E7RPgx8Z9bvLbw9rviv4X22m3thqmmwXo0HVbbUZli/1M0izpLAdyyoJsbipU7evuGj/G39orwt8cvB/wt+LWk+GZ7X4naXq13oZ0h7yNrC90qFLiS1vJpy4mjKOB50cUZzkiPHFAH3PRXwB+xR8Qv2n/AIpW2veLfivq3hy+8MJrOt2Kx2drdRajFdWV35CJG7SmD7KiowUGPzCMFmLbifcvjH8X/Enw8+Kfwd8D6LbWk9j8QtZvdOv3uEkaWKK3spLlWgKSIqsXQAl1cbc4APNAH0bVHU9T03RdPuNW1i7hsLG1QyTTzyLFFGi9Wd3IVQO5JxXwr8S/2pviB4N1X9pex0zT9Llj+DOgaJqukGaKdjcT6la3E8q3e2dd6K0ShBH5ZAJyScEcX8Uvil+0Wv7Nvjj4ufE/wn4LvvBN54Si1Ox0O5hvLq5FzM0LCDUo3dYZI2jdmZYmBjcBAz48xgD9KlZXUOhDKwyCOQQadXwNrXxo/aI1346v8CPgzZeFNOt7bwVpviL7brEN46wTXM8kBhWG2mQPGdqhB8nljLEvwlZ3h79pH47/ABL/AGf9C+JXhW38H+CdUtrzVLHxRqHie6nXR9Om0q4NqRAsbxyN9ocbkLyARgFWLkqSAfoVWfcatpVpf2ml3d5DDe6h5n2aB5FWWfyhuk8tCdz7AQW2g4HJr8q/G37XXxz8a/sfab8V/h6mi+HvE0niqPw1qdwjTXNrkaitmJLEEBvLm3KWZ23IhYJlyrr7zqvi/wCJHh/48fs9eD/jDo/hLWPE/iL/AITAy6rp1jcq2nR2dlHLH/Zz3MzyQmZGCXO7eHAwNooA+66K/OXSP2jv2qvib4D1L9oL4O+EPC8/w1tWvZtO03VLq8TXtXsNPkeOSeOWIfZrZ5vLcxRyI/bc2ME3rv8Aam+K/wATfiL8N/CH7PdnoMWmfEXwXL4pW78QR3TyWLJPHHtZLWVRLt3+W0QK/Od3m7V2sAfoZRXzN+y18ZfF/wAYvB/iZfiHptnpvivwN4l1Pwvqo04yGxnutNKEz2wlLSLHIkqkK7E9eew+maACiiigAooooAKKKKACiiigDwD9rH/k1n4yf9iZ4h/9N09H7J3/ACaz8G/+xM8Pf+m6Cj9rH/k1n4yf9iZ4h/8ATdPR+yd/yaz8G/8AsTPD3/pugoA//9H79/b6/wCbcv8Ass3hD/26r7/r4A/b6/5ty/7LN4Q/9uq+/wCgAooooAKKKKACiiigAooooAKKKKACiiigAooooA+ZPjz+zh/wtzX/AA18RfBvi29+H3xD8Heemma3ZQxXS+Rc4E1vdWs2I7iFsZCMRhuQeSDw/hf9ljx7rHxI8N/E79ov4ozfEi78FyPcaHp1tpUGi6Za3bqUN1LDFJM08yr/AKtmcBDnAr7TooA8N+EnwY/4Vb4t+Jfin+2P7T/4WJrv9t+T9n8n7H/o8Vv5O7zH83/V7t+E6428ZPhniP8AYw/4SDwx8QvDf/CY/Z/+E88dWnjXzf7P3fZfsr2r/Y9v2geZu+zY83KY3fcOOfuWigD5r+Iv7O4+IPxaX4oN4km0of8ACIat4UNvbQ4nX+1ZEf7XFc+YNjxbflXyzk4O4YwfFfhd+xx8Qfhz44+Gfi2T4k2F5a/DjSJ9Ajsrfw4lmlzpsxiJyy3j7bmRogZZiGDEkrGrZY/f1FAHwTY/sceOdG8M/EX4YaJ8VXg+HfjuHxEI9Il0WCWazuPEMcquxvPOV5YoJJfMSNVjLH5WfFdL4h/ZP8RW03gXxj8JfiDL4L8feC/Dlt4Wm1M6dHf2Wq6ZbKpEV1YyyrjEqmSNkl3IWYEvxj7RooA+BtY/YgufFHwp+Knhjxj8QbjXPHvxdSyj1bxNPp8aRxQ6e6tawW2nxSokcMahgFEu4sxZmPAH0B4x+CX/AAlvxh+FHxY/tr7L/wAKwj1qP7H9m3/bv7YtI7XPm+Yvk+Vs3fcfdnHy4yfeaKACiiigAooooAKKKKACiiigAooooAKKKKACiiigD4A/4Kj/APJinxM/7gv/AKeLKvv+vgD/AIKj/wDJinxM/wC4L/6eLKvv+gAooooAKKKKACiiigAooooAK8h+Pnwq/wCF4fBvxZ8Jf7U/sX/hKLJ7P7b5H2nyNxB3+Vvj34x03r9a9eooA5LSvC39meBLPwT9q8z7Jpsen/aNmN3lwiHzNm44zjO3d7Z715T8K/2fdE8Bfs36Z+zd4lvf+Em0i20efRbyfyTafa7e5EiS/uxJIY8pIRw5I6g19B0UAfF3wo/Zw+O3wxuPD3hl/jrd6v8AD3wu0KWmkyaHZx38lnbALDZz6kHLPEgVQSsSuw+XcBxXy/8AtU/s5fED4N/s0fGK1+GnxGWz+GV5b3+qv4ZvdJS7mtmu5fOuLezvxPG0UEkjMwV4pCmTgkktX641S1LTdO1ixn0vV7WK+s7lSksE6LLFIp6qyMCrD2IoA+NLn9njxrrPjDwv8cPhP8QT4H1m88LWGhatFLpcWqw3dnD+/heJZJIvJnjZ2Ac71IxlCAQ3Oab+wvFpfwQ8NfDO08f3tv4s8FeILzxJonimGziW4tr27uJZm821d3imRkmZJFJCyddqj5a++I444Y1hhUJGgCqqjAAHAAA6AU+gD4f8Xfsu/GP4p/Bvx58Lfi/8ZT4iufGNpa2lvcw6Bb2Nppy286ztItrFPvmklKgMzTgAAbVGDn2jxl8E/wDhLfjH8Kfi1/bP2T/hWMWtx/Yvs3mfbv7YtY7bPm+YvleV5e77j7s4+XGT7xRQB8l/Bn9nTx18FvGurSaF8R3uvh3qWqanrC+HZdKg85LnVHaR0OoeYXMUcjb0VY1YnhmI69b+0F8C9Q+Mlv4U1jwr4nfwd4u8DaoNW0jUvsiX0SSmJ4ZYp7Z3jEkckbkHDqQcEHqD9D0UAfBDfsX+KNW0z45Dxn8TG1zWfjdpGm6ZcXp0iO3TT206GeEPFBFcAPGRMNsZZWUL80kjEtXvfxX+B3/Cz/2ddW+AX9tf2b/amjw6T/aP2bztnlKi+b5HmJnOz7vmDGete+UUAfPXhf4D/wDCN/He6+Nn9ufaftPhSw8MfYPs2zb9huHn+0+d5pzv37fL8vjGd56V4Lb/ALEut6LoHg+18M+P4YdW8GeK9a8T20l7owvNPmfWnlJjmsjdJuktllPkzCUFW+YKCePv+igD8/8ATv2IdZPwS8a/BjxF8RpL2PxD4iPiXTNSt9LjtrjTr43ovy0iGZ47jMoAIAiXbkKFJyPSNJ/Z1+It/wDEX4YfFH4ofEhPFWt/DqbX5D5WjRafFdRa3Zw2qwosU7eUtuYy+5vNaQsQSoAr64ooA/Pz/hjL4meHdD1f4V/Cz41XnhL4U6zLdMdEGkW13fWFvfuz3NrYak8gaGJy77C0TtHnIJOTXsnhf9mHw94I+LHgj4geEtRNjo3gXwhL4RtNJMBkZoXmilWdrkyA7lEWGUxksSWLDofp+igDw74JfBj/AIU5N8Q5f7Y/tf8A4T3xbqXinH2f7P8AZP7QjgT7N/rJPM2eTnzPk3bvuDHPuNFFABRRRQAUUUUAFFFFABRRRQB4B+1j/wAms/GT/sTPEP8A6bp6P2Tv+TWfg3/2Jnh7/wBN0FH7WP8Ayaz8ZP8AsTPEP/puno/ZO/5NZ+Df/YmeHv8A03QUAf/S+3f+Ck3hbQvHPhX4F+CvFNr9t0XxB8WfDGn30G94/OtbuO8hmj3xsrruRiNysGGcgg4NdB/w65/YT/6Jn/5WdY/+TaP2+v8Am3L/ALLN4Q/9uq+/6APgD/h1z+wn/wBEz/8AKzrH/wAm0f8ADrn9hP8A6Jn/AOVnWP8A5Nr7/ooA+AP+HXP7Cf8A0TP/AMrOsf8AybR/w65/YT/6Jn/5WdY/+Ta+/wCigD4A/wCHXP7Cf/RM/wDys6x/8m0f8Ouf2E/+iZ/+VnWP/k2vv+igD4A/4dc/sJ/9Ez/8rOsf/JtH/Drn9hP/AKJn/wCVnWP/AJNr7/ooA+AP+HXP7Cf/AETP/wArOsf/ACbR/wAOuf2E/wDomf8A5WdY/wDk2vv+igD4A/4dc/sJ/wDRM/8Ays6x/wDJtH/Drn9hP/omf/lZ1j/5Nr7/AKKAPgD/AIdc/sJ/9Ez/APKzrH/ybR/w65/YT/6Jn/5WdY/+Ta+/6+f/ANpT47p+zv8ADhPH50CbxNLPqNlpkNjBOls8k19J5UeJJAVHzEdePcUAeAf8Ouf2E/8Aomf/AJWdY/8Ak2j/AIdc/sJ/9Ez/APKzrH/ybWnqH7Yvjv4fRR618efgX4l8C+GBIqXOswXNjrVrYq7BfOu1spWkjhXPzOFbHoa+3NJ1bS9e0u01vRLuK/0/UIknt7iB1kimhlUMkiOpIZWUggg4IoA+EP8Ah1z+wn/0TP8A8rOsf/JtH/Drn9hP/omf/lZ1j/5Nr6z+MfxV8NfBD4YeIvit4uWV9K8OWxuJY4FDTSszCOKKMEgb5JGVFyQMkZIGTR8JPGPjbx34Lt/Enj/wVceANVuJH/4ld1dw3kyRD/VyNJB8qlwclDhlPDDNAHyZ/wAOuf2E/wDomf8A5WdY/wDk2j/h1z+wn/0TP/ys6x/8m19/0UAfAH/Drn9hP/omf/lZ1j/5No/4dc/sJ/8ARM//ACs6x/8AJtffNxcQWkEl1dSLDDEpd3chVVVGSSTwAB1JrlNc8a2Wg+KvDfhK407ULmfxO90kNzbWkk1nbG0h85jdzqNkAcfLGXPzv8ooA+Mf+HXP7Cf/AETP/wArOsf/ACbR/wAOuf2E/wDomf8A5WdY/wDk2vv+igD4A/4dc/sJ/wDRM/8Ays6x/wDJtH/Drn9hP/omf/lZ1j/5Nr7/AKKAPgD/AIdc/sJ/9Ez/APKzrH/ybR/w65/YT/6Jn/5WdY/+Ta+/6KAPgD/h1z+wn/0TP/ys6x/8m0f8Ouf2E/8Aomf/AJWdY/8Ak2vv+igD4A/4dc/sJ/8ARM//ACs6x/8AJtH/AA65/YT/AOiZ/wDlZ1j/AOTa+/6KAPgD/h1z+wn/ANEz/wDKzrH/AMm0f8Ouf2E/+iZ/+VnWP/k2vv8AooA+AP8Ah1z+wn/0TP8A8rOsf/JtH/Drn9hP/omf/lZ1j/5Nr7/ooA+AP+HXP7Cf/RM//KzrH/ybR/w65/YT/wCiZ/8AlZ1j/wCTa+/6KAPxB/b1/YK/ZO+C37J3jn4l/DPwN/Y/iTR/7M+yXf8AaepXHl/aNStbeT93cXUkTbo5HX5kOM5GCAR9f/8ADrn9hP8A6Jn/AOVnWP8A5No/4Kj/APJinxM/7gv/AKeLKvv+gD4A/wCHXP7Cf/RM/wDys6x/8m0f8Ouf2E/+iZ/+VnWP/k2vv+igD4A/4dc/sJ/9Ez/8rOsf/JtH/Drn9hP/AKJn/wCVnWP/AJNr7/ooA+AP+HXP7Cf/AETP/wArOsf/ACbR/wAOuf2E/wDomf8A5WdY/wDk2vv+igD4A/4dc/sJ/wDRM/8Ays6x/wDJtH/Drn9hP/omf/lZ1j/5Nr7/AKKAPgD/AIdc/sJ/9Ez/APKzrH/ybR/w65/YT/6Jn/5WdY/+Ta+/6KAPgD/h1z+wn/0TP/ys6x/8m0f8Ouf2E/8Aomf/AJWdY/8Ak2vv+igD4A/4dc/sJ/8ARM//ACs6x/8AJtH/AA65/YT/AOiZ/wDlZ1j/AOTa+/6KAPgD/h1z+wn/ANEz/wDKzrH/AMm0f8Ouf2E/+iZ/+VnWP/k2vqn44/FO0+CXwk8UfFi+099Vt/DFm149rG4jeUKQNodgQDz1Ir5dj/bD+L2k+H4fG/jb9nLxRYeFXgS7e90y/wBN1eWO2dd4mNrBMspUKdzcZAySOMUAQf8ADrn9hP8A6Jn/AOVnWP8A5No/4dc/sJ/9Ez/8rOsf/JtfUFt8cfAer+CPC/xF8ISXXivQPF95Z2dlc6PayXoU3j+WJZ1QboYomBE7OB5RBDANxXr9AHwB/wAOuf2E/wDomf8A5WdY/wDk2j/h1z+wn/0TP/ys6x/8m19/0UAfAH/Drn9hP/omf/lZ1j/5No/4dc/sJ/8ARM//ACs6x/8AJtfTHw6+Mll8Q/iH8Sfh9baZJZTfDi/s7Cad5A63TXlql0HRQAUCh9pBJyRmvZ6APgD/AIdc/sJ/9Ez/APKzrH/ybR/w65/YT/6Jn/5WdY/+Ta+/6KAPgD/h1z+wn/0TP/ys6x/8m0f8Ouf2E/8Aomf/AJWdY/8Ak2vv+igD4A/4dc/sJ/8ARM//ACs6x/8AJtH/AA65/YT/AOiZ/wDlZ1j/AOTa+/6KAPgD/h1z+wn/ANEz/wDKzrH/AMm0f8Ouf2E/+iZ/+VnWP/k2vv8AooA+AP8Ah1z+wn/0TP8A8rOsf/JtH/Drn9hP/omf/lZ1j/5Nr7/ooA+AP+HXP7Cf/RM//KzrH/ybR/w65/YT/wCiZ/8AlZ1j/wCTa+/6KAPgD/h1z+wn/wBEz/8AKzrH/wAm0f8ADrn9hP8A6Jn/AOVnWP8A5Nr7/ooA+AP+HXP7Cf8A0TP/AMrOsf8AybR/w65/YT/6Jn/5WdY/+Ta+/wCigD8oP2hf+CcP7GPgb4BfEvxr4W+Hn2LWvD/hnWdQsZ/7W1WTybq0spZoZNkl2yNtdQdrKVOMEEZFfcH7J3/JrPwb/wCxM8Pf+m6Cj9rH/k1n4yf9iZ4h/wDTdPR+yd/yaz8G/wDsTPD3/pugoA//0/v39vr/AJty/wCyzeEP/bqvv+vgD9vr/m3L/ss3hD/26r7/AKACiiigAooooAKKKKACiiigAooooAKKKKACvgf/AIKM/wDJDPD/AP2Ofhn/ANLlr74rzP4tfB74c/HPwe/gL4paT/bWhSTxXLW4uLi1PnQHdGwktpIpBtPPDY9aAKXxy+IPw6+Gfwq8S+KvipdW0HhyGxuEuYrllAuleNl+zIjf6x5s7FQAlicYr8idAufjn4D+AH7Jv7Ol1PrOkN46j1ufVBpd/FpGrSxWxN5p+nR307AWxaKdd4ysjCPylwSVr9FPB37B37JfgXxLaeMNC+H0EurWDB7ebUby+1QROOQ6JfXE6BlPKsFyDggg817f8V/g38Mvjj4X/wCEN+K2gQeIdIEqzpFMXR4pkBCyRSxMkkbgEjcjKcEjOCaAPyD/AGjvh58adP8A2RPixoHxa1XXrfRPDms+H7/w0l9r0WoanJZ397BbT2upzWzN9phhlLSWxmywba2SYwF9O8eWfxS8T/tH6n+zjpGt+Nr/AMPfD3wzptxp8ek+KItJ1XUJb0v5uo3t9cOs12YmCxKpJRWALLyd33FpX7Hv7OujfD7X/hfZeE8+H/FU1pcaqkt/fS3N5LYyrPbtLdvO1yfLkQFQJAo5GMEg9B8YP2Zvgh8eL7TNW+KHhldT1LR1aO0vYbm5sbyGNzloxcWcsMuwnPyliBk4AyaAPhDU9c/aL14/s8/s1/GDxjdeDdT8XSeIh4j1jRr6GHUr9dCANnaw3luXWKeZHQ3OwhiQwB6qfZPit8M9A+Gngzwv8NNQ+KvxFvV1rWLn+zNO0zVTN4j1a4e3Vlsk1B/LdLW2SOSZvMlRAG/eSgBQfftX/ZT/AGfNc+F2mfBnU/BlrL4R0WY3NjarJOkttcF2kaeK6SQXKSszsWkEm9tx3E5NYdv+xp+zraeFLLwba+GriGy07VH1u3nTVdSS/j1GSJIJJ1vluRdBniRUYCUKQBxQB+Z/jzXPiF4+/YE8SQ+PvFeuC+8F/EP+wHmlvI1vJ7SDWoLVEv7iEETPCsp+ZX2l1V8ttU19meJNH134OftC/szfDHQfGniTV9G1i+8YvqH9r6vcX018ItIaaFLp3b98kMnzRK4IQ8rg17von7JX7Pfh/wAH+Lfh9p/hJG8N+OJxc6tp9xd3dzbzSq/mB0SaZ/JIk+cGHZ82G6hcW/BX7LHwK+H194a1Xwr4dkt77wjd39/ps8uoX1zLFdaparZXcrtPO5mMluix4l3BQAUCnmgD4d+HenfEH9oT4I+JP2oPEPxs8R+CPEcFxrcttp2nX0Vtonh9NJnmjhtb2xKMlwQkSvOZzuZX4wPmah4e8cfEf9pX4k/s+23iLxXr3gyx8b/D7UdX1mz0G/l05bu4ilgVXGwkxhyd6OmJFQ7VcBmz9l+L/wBib9mDx34wvfHPibwPDcanqk63N8kV1d29nezqdwlurOCZLadyeWMkbbiSWzXs7/Cr4fv440T4jjR0j8QeHNOm0nT545JI47exnKGSFYEcQ7SUXBKEqBhSBxQB82fsU694vm0b4o/D7xZ4gvfFCfDvxxqug6bf6nKbi+fTYooJ4EuJ2+aaRPOYF25IwOAAB9q1w3gz4beC/h9d+I77whp32Cfxbqkus6m3nSy/aL+dEjkmxK7hMrGo2ptQY4UEmu5oAKKKKACiiigAooooAKKKKACiiigAooooA+AP+Co//JinxM/7gv8A6eLKvv8Ar4A/4Kj/APJinxM/7gv/AKeLKvv+gAooooAKKKKACiiigAooooAKKKKACiiigAooooA+Q/2+P+TOPiz/ANgSX/0Na9k8H+JvD3g74J+HPE/ivUrfSNJ07QrGa5urqRYYYo1tkJZnYgAV1Pj3wH4U+J/g7VvAHjmx/tPQNcgNveW3myQ+bExBK+ZCySL06qwPvXy3pH/BO/8AY40bU7PVYfh1FeTWDK8KahqOo6jApXpmC7upYWHsyEe1AHwd8MdW8X+E/wBnzwp4x8IXWoeF/DnxE+PlpfaJbwSSWbHwxql+ES2ZEKn7PNscmNhtdTnBBBPd/Gbxjr/hD4oRfEX4W/EHxv4iutP+ImnaHrskt3EnhG1ivL1YpNEisXZTJLDDIiNLDG+GDM8qyHaP1L8Z/C/wJ8QLDQtL8WaUt5aeGtTstY06JJJbdbe+05t9rIogdMiM9EbKHoykcV4x4q/Yu/Zv8Z6nrOr694Xma41/UF1a7EGq6lawnUlZX+2RwwXKRRXDFBumjVXYcFiCcgHztc+EfHnxv/a2+Ovw1u/iV4m8LeEtC0zwtPb2+h6lJZzQXl1bTlXgfDiKMlXeZIwvmuIy5IXBytB+Hvj/AOMn7Sv7QPww1P4r+LtB8L+Dh4RTTl0nVXt7pLubR0dpfMKsEQsrSSRxqqTSOWkBKrj9APC3wp8A+C/FeueN/DWmG01vxJbafaahcGeeUzwaVG0VopSSRkXy0dhlVBbOXLHBq14f+G3grwt4y8VfEHQtO+y6/wCNmsn1e586V/tTadB9mtj5bu0ceyL5f3aru6tk80Afl5YfCa/8bfFb9rPULPx34l8Mz+HbrTpbI6NqL2GbyHRIpEuLloQr3BBRR5bt5eCx27iGGH4h+NHxk+MWmfsy+A2v9Zih+IXhSfWtWOganBoWo6zqFnDErQpeuY/KQZaeWOIguG2jAAx+qWmfCX4faPqfjPWNO0rybz4gukmuP587fa2jtxaqcNIRFiEBf3QT1+9zXBeJf2VfgD4v+GXh/wCD/iPwjDe+FfCixrpNu09yJ7Lyl2qYbsSi5VgOp83J4yTgUAfGKWHxN8NfCeXwB+0L428W6DA3i5LXw5Do2qQXni7WrO7t2e10qe+tiWDrKHdpmaPdEg3uqZNe2fsH+K/Huu+DPiP4Z8f6jqmpXHgjxvqmh2Ta3PFd6nFYQQWs0MF3cwM6TzRmZg0iuwPYkAV6Rb/sb/s5WfgpPAFp4TaDS49Uj1tZY9Qv1v11OFPLS6F+Lj7WJVQlQwm4XgccV2/wr/Z++E/wV1DXdT+GujyaTN4klW4vg17d3McswUKZBHcTSIrvtG91AZyAXJwMAHs1FFFABRRRQAUUUUAFFFFABRRRQAUUUUAFFFFAHgH7WP8Ayaz8ZP8AsTPEP/puno/ZO/5NZ+Df/YmeHv8A03QUftY/8ms/GT/sTPEP/puno/ZO/wCTWfg3/wBiZ4e/9N0FAH//1Pt3/gpNr/8AwinhX4F+Kf7NvdY/sf4s+GLz7DpsP2i+uvs8d5J5FtDlfMmk27Y0yNzEDIzXQf8ADfX/AFbl8Zv/AAkP/uqj9vr/AJty/wCyzeEP/bqvv+gD4A/4b6/6ty+M3/hIf/dVH/DfX/VuXxm/8JD/AO6q+/6KAPgD/hvr/q3L4zf+Eh/91Uf8N9f9W5fGb/wkP/uqvv8AooA+AP8Ahvr/AKty+M3/AISH/wB1Uf8ADfX/AFbl8Zv/AAkP/uqvv+igD4A/4b6/6ty+M3/hIf8A3VR/w31/1bl8Zv8AwkP/ALqr7/ooA+AP+G+v+rcvjN/4SH/3VR/w31/1bl8Zv/CQ/wDuqvv+igD4A/4b6/6ty+M3/hIf/dVH/DfX/VuXxm/8JD/7qr7/AKKAPgD/AIb6/wCrcvjN/wCEh/8AdVH/AA31/wBW5fGb/wAJD/7qr7/ooA+AP+G+v+rcvjN/4SH/AN1Uf8N9f9W5fGb/AMJD/wC6q+/6KAPgD/hvr/q3L4zf+Eh/91Uf8N9f9W5fGb/wkP8A7qr7/ooA+AP+G+v+rcvjN/4SH/3VR/w31/1bl8Zv/CQ/+6q+/wCigD4A/wCG+v8Aq3L4zf8AhIf/AHVR/wAN9f8AVuXxm/8ACQ/+6q+/6KAPgD/hvr/q3L4zf+Eh/wDdVH/DfX/VuXxm/wDCQ/8Auqvv+igD4A/4b6/6ty+M3/hIf/dVH/DfX/VuXxm/8JD/AO6q+/6KAPgD/hvr/q3L4zf+Eh/91Uf8N9f9W5fGb/wkP/uqvv8AooA+AP8Ahvr/AKty+M3/AISH/wB1Uf8ADfX/AFbl8Zv/AAkP/uqvv+igD4A/4b6/6ty+M3/hIf8A3VR/w31/1bl8Zv8AwkP/ALqr7/ooA+AP+G+v+rcvjN/4SH/3VR/w31/1bl8Zv/CQ/wDuqvv+igD4A/4b6/6ty+M3/hIf/dVH/DfX/VuXxm/8JD/7qr7/AKKAPgD/AIb6/wCrcvjN/wCEh/8AdVH/AA31/wBW5fGb/wAJD/7qr7/ooA/EH9vX9rz/AIWh+yd458C/8KV+JnhL+0/7M/4mniHw59g0u38nUrWb9/cfaH2b9mxPlO52Ve+a+v8A/hvr/q3L4zf+Eh/91Uf8FR/+TFPiZ/3Bf/TxZV9/0AfAH/DfX/VuXxm/8JD/AO6qP+G+v+rcvjN/4SH/AN1V9/0UAfAH/DfX/VuXxm/8JD/7qo/4b6/6ty+M3/hIf/dVff8ARQB8Af8ADfX/AFbl8Zv/AAkP/uqj/hvr/q3L4zf+Eh/91V9/0UAfAH/DfX/VuXxm/wDCQ/8Auqj/AIb6/wCrcvjN/wCEh/8AdVff9FAHwB/w31/1bl8Zv/CQ/wDuqj/hvr/q3L4zf+Eh/wDdVff9FAHwB/w31/1bl8Zv/CQ/+6qP+G+v+rcvjN/4SH/3VX3/AEUAfAH/AA31/wBW5fGb/wAJD/7qo/4b6/6ty+M3/hIf/dVff9FAHwB/w31/1bl8Zv8AwkP/ALqo/wCG+v8Aq3L4zf8AhIf/AHVX3/RQB8Af8N9f9W5fGb/wkP8A7qo/4b6/6ty+M3/hIf8A3VX3/RQB8Af8N9f9W5fGb/wkP/uqj/hvr/q3L4zf+Eh/91V9/wBFAHwB/wAN9f8AVuXxm/8ACQ/+6qP+G+v+rcvjN/4SH/3VX3/RQB8Af8N9f9W5fGb/AMJD/wC6qP8Ahvr/AKty+M3/AISH/wB1V9/0UAfAH/DfX/VuXxm/8JD/AO6qP+G+v+rcvjN/4SH/AN1V9/0UAfAH/DfX/VuXxm/8JD/7qo/4b6/6ty+M3/hIf/dVff8ARQB8Af8ADfX/AFbl8Zv/AAkP/uqj/hvr/q3L4zf+Eh/91V9/0UAfAH/DfX/VuXxm/wDCQ/8Auqj/AIb6/wCrcvjN/wCEh/8AdVff9FAHwB/w31/1bl8Zv/CQ/wDuqj/hvr/q3L4zf+Eh/wDdVff9FAHwB/w31/1bl8Zv/CQ/+6qP+G+v+rcvjN/4SH/3VX3/AEUAfAH/AA31/wBW5fGb/wAJD/7qo/4b6/6ty+M3/hIf/dVff9FAH5QftC/tt/8ACV/AL4l+Fv8AhQfxZ0f+2PDOs2f27UvC32extftFlLH59zN9pby4Y926R8HaoJwcV9wfsnf8ms/Bv/sTPD3/AKboKP2sf+TWfjJ/2JniH/03T0fsnf8AJrPwb/7Ezw9/6boKAP/V+/f2+v8Am3L/ALLN4Q/9uq+/6+AP2+v+bcv+yzeEP/bqvv8AoAKKKKACiiigAooooAKKKKACiiigAooooAKKKKAPzU+H03xj/bbu9e+I1v8AEXV/hl8J7TUrzS/D9l4aMFvqmppZSeTLf3V9KkxRXkVlSKNRgDnkbn+lvh54J8Wfs/aR4s174j/FjVfHHgzTrL7dEdbtYZb/AE2O0SSW7ke6tY1kuVZApCGPcuzC53Yr5W+FmofEb9iGDXPgr4q+HXiDxv8ADL+0L6+8Na34WszqctvZ30pnezvrSJhNE0Lu2JVBV88AYOPFvhB8KNThb9ozXfhf8MfF3gvwHr/gOfTdF07xGl2dQutTFvcCUQ211cXNyfMLLsHfgAAnFAH6M+Ev2t/2dvHPiCPw14Y8ZQXV5LpH9uh3guYLYacIkneV7maJIUMcciNKjOHjzh1UggV/AH7YX7NfxP8AFsHgXwR46tL/AFq88z7LC8VxbLeeV9/7LLcRRxXGBk/uXfgE9Aa8C134DeMPF/8AwTX0b4LeDdLOj+LJvBmip9guVNpKbuNLa7vLObzNrRPcOkkUgfbhnO8jBI4L4k+KvE/7T2m/Dj4T+A/g14o8Ear4e8Q6Nqt1qOt6V/Zmn+HbfSpFeb7Jct8k8hQeVCkAw6Nk7RxQB9jfE/8Aaz/Z4+DfiM+EfiL4zt9N1mOJZ5rWOC5vJLaJxlXuRaxS+QpBBBl2Agg9Dmtnxl+0n8DfAGi+GPEfinxda2+leNC66NdwrLdwXpSIzYikt0kUlk+4CRvYhEy5Cn498JeKPEn7LPxH+Mmm+OPhb4p8aL8QPE114h0rWfDelHV476zu4o1i064ZCGge12tGqy4QqSwwPmbzf4afAr4p+CV/ZltfEPha4s/sPjLxNrl5p9pG11a+HbLU7e5ltbaWWJTHGsZdVBJC+YdoOcUAfXk/7ef7Kdt4dl8Sy+Nv3FrPLb3NsNO1Br+1aDb5rXFiLc3UMab1zLJGqc4DE8V3+rfFPRr74lfCax8O+ObOLSvHdlq17aaclhJdnXoLe2hnSaC+Q+XbLbrIJDv/ANcHCryOPEfAPgLXIf2k/wBqvWNT8PXMek+KrDwvFY3Mtq62+o+VpM8NwkEjLtm2MQkgQnDEBua+fvgL8OfiRpeofsTy634X1azHhTQPGEGsNcWU8f8AZslxawJbx3ZdB5DSFSsaybS2MLnFAH13pf7c37LGs6lpel6d45R31e9OnQzPY30Vsl6JWhFvPcSW6wwSMynaszoWXDjKMrHovi7+17+zn8CdaPhz4o+MotK1RIknktobW7vpYYpTtjaZLOGYxBz93zNu7tmvz5n+E/xDb/gltr/gMeDtV/4S59auLqHSxp8/9osf+EoE6Srb7POP7geYGC/6v5vu81zn7S/ijwjaftLfFr4cJ8Q4vBHhD4jaLpUXjPUX0pteVbiGJ7T7NFJagjTpRaKNxuyQC3mIo4JAP2Z8J+KtF8b+HLDxZ4deaTTdTjEsDT281pKUJI+aC4SOVDx0dFPtXRVzPgr/AIR7/hDtC/4RK9XUdDFhaiwuklE6T2oiXyZFlGQ4dMEMPvZzXTUAFFFFABRRRQAUUUUAFFFFABRRRQAUUUUAfAH/AAVH/wCTFPiZ/wBwX/08WVff9fAH/BUf/kxT4mf9wX/08WVff9ABRRRQAUUUUAFFFFABRRRQAUUUUAFFFFABRRRQB8x/tneKvEfgf9lr4leLfCGozaTrOl6TJNa3du2yWGQMoDIw6Hmvmf8AZ11L4YvqWh+ItO/ar1bx14jfSZbqXw/e69p+pW/mmzZ5meyt4kuH+zfNJtD5Gzk8GvpD9tnw9r/iv9lH4neHPC+m3Osatf6RLFbWdnC9xcTyFlwscUYZ3b2UE1q+EvhF4I8O/CSxu9I8FadpniNPDoiMkGmww3wney2SISsYkDsSVYdSSQaAMXQv2k/hn4J+AXgz4o/FD4g22s2viGCGO21W3025tX1e5kzj7LpiJJc7mx/q1QkYyQB08V+A37UPhvxZ8Vf2j/Het+L5ofhp4Ufw09i+qxz2MWnJJpxF2ogukiliL3KkFSgLvjaDkZ8H8B/Dr4l/DjwR+yb8ZNW8B6zr9p8M9G1bT9a0O2s3bWdOl1SERxXcVhIY5JGQrslXG9UYMqnHHH/Ee+tfFXhj9r34q+MvDXiDwhoniZ/B114eXXbFtEu77UtGtF8qOGLUECTH7VAoaJQ5dDtUbmWgD9Hvhz+2R+zj8WNZ1PQvAnit7240W1kvb6SbTNRsra1t408xpJrm6tooEGz5hukGVBIyAasfDv8Aa/8A2c/iprdx4e8FeMY7m9t7Wa+Vbi1urFLi0t/9bPbSXUMSXEaYO5omYAAn7vNfn34M8PRftPfDr43Cz+J1r4m+NXjvwpa6fLpUelz+HFsbWx3tFE1td/vZRLNIY5bk5QBwo2rjPZfArTrfxZ4/+FSeLfA/xXs/FXgPeqHXLS3t/D+jubI29x5d2sUazW8igRxJGSzDaSq4OAD6C8Mfty/BX4v+E/Hw8AeI7rw9qXhrTtaulvr7Rb6aC3t9LjIbUCoiEUqI37wW4l851GCisSo7+7/av+Cnw/0XwhpvxC8area74h8PWut2r22l32/Vbd1RWuLa1hilcNIzbxbjMqLkldqMw+MfCL+PfCnwD+PP7NOqfDfxU/iPVF8f3VjfQaVJNpV1BqEV1NZmG7QlZJLgsI44ow0hcgFR29M+DngHxnp37Q3wR1zVvDmoWtho3wZg0y6up7OVIbTURNaZtZZGQLHcBQ+Y2IfAbjANAH1jD+058BpvhMnxyHjKzTwS7mEX7iRD54cx+R5DIJ/P3DHleX5n+zTPg3+038Fvj/d6pZfCbW7jWpNGJW7Z9M1CyjicEAp5l3bwozgkZRWLAEEjHNflt8StNvPhZ8FfiX4g1uzuvDviTTvjvfeIvBsVzb7DdSmWFY7qCzmXffW3lSzPtt1ZmALIRtJH21+xNfeBdej+IPjTSviHB498aeKNUt7/AMR+Rp8ujLYyLbLb20I024/fwqI4iPMkyZSCSSQaAPumiiigAooooAKKKKACiiigAooooAKKKKACiiigDwD9rH/k1n4yf9iZ4h/9N09H7J3/ACaz8G/+xM8Pf+m6Cj9rH/k1n4yf9iZ4h/8ATdPR+yd/yaz8G/8AsTPD3/pugoA//9b79/b6/wCbcv8Ass3hD/26r7/r84P+Ck2na7q/hX4F6T4W1f8A4R/Wr34s+GILHUvs6Xf2G6kjvFhufs8hCS+U5D+WxCvjaeCa6D/hnj9uz/o7j/ywdH/+O0Aff9FfAH/DPH7dn/R3H/lg6P8A/HaP+GeP27P+juP/ACwdH/8AjtAH3/RXwB/wzx+3Z/0dx/5YOj//AB2j/hnj9uz/AKO4/wDLB0f/AOO0Aff9FfAH/DPH7dn/AEdx/wCWDo//AMdo/wCGeP27P+juP/LB0f8A+O0Aff8ARXwB/wAM8ft2f9Hcf+WDo/8A8do/4Z4/bs/6O4/8sHR//jtAH3/RXwB/wzx+3Z/0dx/5YOj/APx2j/hnj9uz/o7j/wAsHR//AI7QB9/0V8Af8M8ft2f9Hcf+WDo//wAdo/4Z4/bs/wCjuP8AywdH/wDjtAH3/RXwB/wzx+3Z/wBHcf8Alg6P/wDHaP8Ahnj9uz/o7j/ywdH/APjtAH3/AEV8Af8ADPH7dn/R3H/lg6P/APHaP+GeP27P+juP/LB0f/47QB9/0V8Af8M8ft2f9Hcf+WDo/wD8do/4Z4/bs/6O4/8ALB0f/wCO0Aff9FfAH/DPH7dn/R3H/lg6P/8AHaP+GeP27P8Ao7j/AMsHR/8A47QB9/0V8Af8M8ft2f8AR3H/AJYOj/8Ax2j/AIZ4/bs/6O4/8sHR/wD47QB9/wBfH2sfsmXKax4wn+HnxO8QeCdE+Id9cajr2l2cOm3MU1zeRLFcS2015aTTWzSqo3bXYA8qF4xwP/DPH7dn/R3H/lg6P/8AHaP+GeP27P8Ao7j/AMsHR/8A47QB9lfDjwB4d+FfgLQPhx4Rjki0bw5Zw2NqJXMknlQqFBdzjLHqTgDJ4AHFdrXwB/wzx+3Z/wBHcf8Alg6P/wDHaP8Ahnj9uz/o7j/ywdH/APjtAH3/AEV8Af8ADPH7dn/R3H/lg6P/APHaP+GeP27P+juP/LB0f/47QB9/0V8Af8M8ft2f9Hcf+WDo/wD8do/4Z4/bs/6O4/8ALB0f/wCO0Aff9FfAH/DPH7dn/R3H/lg6P/8AHaP+GeP27P8Ao7j/AMsHR/8A47QB9/0V8Af8M8ft2f8AR3H/AJYOj/8Ax2j/AIZ4/bs/6O4/8sHR/wD47QB9/wBFfAH/AAzx+3Z/0dx/5YOj/wDx2j/hnj9uz/o7j/ywdH/+O0Aff9FfAH/DPH7dn/R3H/lg6P8A/HaP+GeP27P+juP/ACwdH/8AjtAB/wAFR/8AkxT4mf8AcF/9PFlX3/X4g/t6/Br9rHwp+yd451/4mftFf8J34btf7M+16L/wiOm6X9q36lapH/pdvI0sflyMknyj5tm08Ma+v/8Ahnj9uz/o7j/ywdH/APjtAH3/AEV8Af8ADPH7dn/R3H/lg6P/APHaP+GeP27P+juP/LB0f/47QB9/0V8Af8M8ft2f9Hcf+WDo/wD8do/4Z4/bs/6O4/8ALB0f/wCO0Aff9FfAH/DPH7dn/R3H/lg6P/8AHaP+GeP27P8Ao7j/AMsHR/8A47QB9/0V8Af8M8ft2f8AR3H/AJYOj/8Ax2j/AIZ4/bs/6O4/8sHR/wD47QB9/wBFfAH/AAzx+3Z/0dx/5YOj/wDx2j/hnj9uz/o7j/ywdH/+O0Aff9FfAH/DPH7dn/R3H/lg6P8A/HaP+GeP27P+juP/ACwdH/8AjtAH3/RXwB/wzx+3Z/0dx/5YOj//AB2j/hnj9uz/AKO4/wDLB0f/AOO0Aff9FfAH/DPH7dn/AEdx/wCWDo//AMdo/wCGeP27P+juP/LB0f8A+O0Aff8AXj3xq+C/h/43+HdK0TWtRvtGu/D+q2muaXqOmvGl1ZajYlvJmQTRyxOAHYMkkbKQTxnBHzD/AMM8ft2f9Hcf+WDo/wD8do/4Z4/bs/6O4/8ALB0f/wCO0Ae1fDz9nWfwz8TP+Fy/EDxzq3j/AMYw6XJo1rcX0NlZW1rYyzLO6RW1jBCm93UbncsTjA2jivpWvgD/AIZ4/bs/6O4/8sHR/wD47R/wzx+3Z/0dx/5YOj//AB2gD7/or4A/4Z4/bs/6O4/8sHR//jtH/DPH7dn/AEdx/wCWDo//AMdoA+jPjH8DNN+LeoeFfE9vr+o+E/FXgi4ubrRtW0z7O8lu93CYJ0khuop4JY5EOGVkzwMMOc5/wk+AMHw38YeIfiZ4j8V6n438a+Kba0s73UtQS1t0W2st5iht7azhhiiQFyTkMxPJbrnwP/hnj9uz/o7j/wAsHR//AI7R/wAM8ft2f9Hcf+WDo/8A8doA+/6K+AP+GeP27P8Ao7j/AMsHR/8A47R/wzx+3Z/0dx/5YOj/APx2gD7/AKK+AP8Ahnj9uz/o7j/ywdH/APjtH/DPH7dn/R3H/lg6P/8AHaAPv+ivgD/hnj9uz/o7j/ywdH/+O0f8M8ft2f8AR3H/AJYOj/8Ax2gD7/or4A/4Z4/bs/6O4/8ALB0f/wCO0f8ADPH7dn/R3H/lg6P/APHaAPv+ivgD/hnj9uz/AKO4/wDLB0f/AOO0f8M8ft2f9Hcf+WDo/wD8doA+/wCivgD/AIZ4/bs/6O4/8sHR/wD47R/wzx+3Z/0dx/5YOj//AB2gD7/or4A/4Z4/bs/6O4/8sHR//jtH/DPH7dn/AEdx/wCWDo//AMdoA+gP2sf+TWfjJ/2JniH/ANN09H7J3/JrPwb/AOxM8Pf+m6Cvh/8AaF+BP7Z2kfAL4l6t4p/ah/4SDRbLwzrM99pv/CE6VafbrWOylaa2+0Ryl4vNQFPMUFkzuHIFfcH7J3/JrPwb/wCxM8Pf+m6CgD//1/v39vr/AJty/wCyzeEP/bqvv+vgD9vr/m3L/ss3hD/26r7/AKACiiigAooooAKKKKACiiigAooooAKKKKACiiigCnqOoWWk6fdarqMogtLKJ5ppDnCRxqWZjjnAAJr5Z8K/t0fsmeN/Eul+D/CnxIsNR1nWbiO0s7aOO4DzTzMFRFLRAZYkDkgV7t8VP+SYeL/+wPqH/pO9fnJ+wh41/aMT4H/CXQbf4PabN4L+yW0R8QHxDCtyLMyEPc/YfsxfcoyRH5mTjrzQB+q9Ffnv8H/jv+1J8ZrrV9f0XSfCeleEvCHjLVND1WW9F8t3d6bp9zseWzVJHRZIoSCWkYrLLuUJGq5fwXw7/wAFEfiHrkek/FRbfwxL4E1fVo7QeG4BfSeKoNMlufsy3rTAm0aRf9c0CpgR8bwwJAB+wVFflp8WP23fiNZ/Er4g+GvhdP4S0zTPhpKLSWDxEL6S/wBdvo4hNPBZm0Kx26rnykeUPukwcBc43fjb+05+0d4a+EOi/tB/DWPwjaeF/ElhpZ03QtbstTufEFxq2oqoWwjFtcQxPJ5hKgBQQqsx4FAH6PS6tpcGpW+jT3kMeoXaSSw27SKJpY4iokdEJ3MqFl3EDAyM9RWhXwvpfjbx/b/tB/BPwj8WvDnhqXxlrvhTXL3UtQsrOUT6fPE1uWtbGaSeUrCd4SUkt5hXcuwHbXgPgv8AbH/ahu/gZ8Pv2n/GWh+Ej4G8QapbabqdhYLfJqSxXWoNYC7hklmeKPZIB+5YSZHPmDdtjAP1mor8uf2if2yPjf4H1b4rS/DK28H6bo/wiksIL6LxJLdSarqT3sMU/mWVtBNbqECyhYw7nzSDgjoP0l8JSeIZvC+kzeLZLeXWpLWJrxrSGS3tzOVBcxRTPJJGueiu7MOhJNAHQ0UUUAFFFFABRRRQAUUUUAFFFFABRRRQAUUUUAfAH/BUf/kxT4mf9wX/ANPFlX3/AF8Af8FR/wDkxT4mf9wX/wBPFlX3/QAUUUUAFFFFABRRRQBwvxD+JvgL4T6FF4n+I2t2+gaXNdQWSXFySEa4uW2xJwCcse/QAEkgAkd1X4r/ALdfxQ+A3xR/aKsP2dvjh4vTw14J8FaFfajeuI7iYy+IdUtzBYJst0c7rOCX7SpOFJba2QcV6t8Hv2ydevf+CeeufGPR57PWvG/wvspNLvftQlkt7m6sGSOK4kVWhlZbiBklJyhLlh2IoA/VKivgvwx8e/2ifAfjv4e+G/2jNG8M3WhfFCb7DpereGHvITZ6g1ubiKC7tr0yEiZVKq8chwwORg1414e/az/a3f4Va9+0hqfhvwfrnw78NajqcN9ptib+y1xbHTLt7aWeN5pJ7Z3VUMhU7cgEDBIAAP1Zor4F8QftDfHjxl8dtU+DXwE03w9LbP4Q0zxPY6rriXYhi+2yupWcW0m6TzF2CJFVNvzuzttCN5V4/wD2zfjJYeP9f+Gnh268E+HdX+H+n6d/bb63/aE8Oqazd2q3MtppotmRooIs7POnySxX5cBjQB+kN9498Iab400v4d32pxw+I9Ztri8s7IhvMmt7UqJpFIG3CF1zkg88V11fkjqfxQ+IPxn/AGgv2ePiV8KtLsdE1/xR4I8QStBrwne305vOtlud8cPlyzmN0KIoaLfkMWUZFeiXf7bfjbwR8CfHXiL4haHpc3xE8FeMD4IWGzlmh0m8v5jE1td5kLyxW5ilLuhZn+QjK7htAP0qor8yvAv7ZHxMN3478LeLpfDHinUdF8H6j4o0rV/DcF/Fpnn2CEyWF5FduZN4Yq6skih48/dbOLfg/wDae/aSsrP4FfEP4o6P4XPgv403Gl6WLfSRerqFheatZtcW05lnkaJ45CjFotmYwQnmSEF2AP0qrP0/VtL1YXDaVeQ3gtJntpjDIsnlTxHDxPtJ2up4ZTyO4r4P+Hn7SPxk8cftL+K/hFqA8LeHbXwzq89sNB1Fb6HxBfaLGMxatZzlvs06Sg7vKWP5FB3OON3nGk/tLeJvhv4Y8YfEfT/Cnh7SfBnh/wCMOteHfFbWNnLBMNNe5S2XWHdZyrXQmeM3LsjCRSNqpjNAH6j0V+bPxJ/bg8T+EB8WfFOg6Pp2peE/B2s6Z4O0CWUyxtqHia6x9tM84cotpZ7gGCRb2Kthx2639nH9p/x746+Ld38IPiJc+HfEbz6QdYsda8KxXsNkjRSrFPY3Ed6XcSrvDxurYZAcgNkAA++aK+Sv2tv2jNQ+AHh3wxa+GYNPk8S+NtVTSrCbV3lTTbNQjSz3d35H7144kX7keGYsMH18Z+Gf7Sn7QHxN0j4i+B/CcfhTXviD4Ngs9Q0rUbWC+TQNbtbjdvttk0yTW90hQoWMroGZWxtyaAP0bor4I+CH7WvjX9pr4h2GkfCzwwug+F/DFuf+E2udcglNzaaqxKDSLFUkiDTRsjNLM6sgTHyBiFbxbwr+13+1mPgtP+1F4l8N+D9a+HGm3l8l/p+mG/s9chsrG+kspZ4zPLPbSMmwyFCUyowCCeAD9YKK/NL4v/tn+L4/i7q/wu+EGqeE9Bg8L6Zp9/e6j4rW+lW9uNUh+1W1taxWTIUUQlGklcnG8AJxltGD9sX4nfE/w58H/D/wW8LWOn/ED4p2eo310uvmdtO0S30dzBeSSpBsmm3zApb4KhsqWwDQB+jlFfm1+0F8Rf2zfh54M+FmnG/8IWXijxT4607Qbm8s4r57KeK5ZpbZTBKfNjifymW6CylyPljIBJro/jn8X/2rvg14Ph8U6re/Diwi0fS3ur6TUJr63/trUkaZzY6VCZg8ZEKJgyvIXkfAUKCaAP0CrPj1bSpdTm0WK8hfUbeJJpLYSKZkikLKjtHncFYqwViMEg46Gvzb+JP7QH7RXjDxD+zLcfBCbQvD9n8XtPuNVeDWEuLtGmTSGvTb3PkGJvIjWTKGNwzyqpbCKVbr7z46r8K/jL8TtU+JugaJ9r8D/DvR9a1XVdJtHjv7243ziW1SWaVibfzVP2dHG5Sw3uewB+gtFfnNrf7Rv7VXwu8EaX8fvjH4Q8Lw/DW7eyl1LT9Lurx9d0ay1GRI455ZZR9muWhMqeckaJznacZYaXiP45ftSeJPjH8bPhx8HrHwlFp/wpj0aeCfWIr6S4u/7Q01b1rcrbzIm5n3BZcqEUAFHLF1AP0Horx/4AfFVPjh8F/B/wAWUsv7NbxPp8V3Jbbt4hlOVkRWwNyh1IUkAkYyK9goAKKKKACiiigDwD9rH/k1n4yf9iZ4h/8ATdPR+yd/yaz8G/8AsTPD3/pugo/ax/5NZ+Mn/YmeIf8A03T0fsnf8ms/Bv8A7Ezw9/6boKAP/9D79/b6/wCbcv8Ass3hD/26r7/r88P+CiGraVoGlfs/a7rt5Dp2m6d8X/ClzdXVzIsMEEEK3bySyyOQqIigszMQAASTivpD/hrH9ln/AKLJ4M/8KHTv/j9AHv8ARXgH/DWP7LP/AEWTwZ/4UOnf/H6P+Gsf2Wf+iyeDP/Ch07/4/QB7/RXgH/DWP7LP/RZPBn/hQ6d/8fo/4ax/ZZ/6LJ4M/wDCh07/AOP0Ae/0V4B/w1j+yz/0WTwZ/wCFDp3/AMfo/wCGsf2Wf+iyeDP/AAodO/8Aj9AHv9FeAf8ADWP7LP8A0WTwZ/4UOnf/AB+j/hrH9ln/AKLJ4M/8KHTv/j9AHv8ARXgH/DWP7LP/AEWTwZ/4UOnf/H6P+Gsf2Wf+iyeDP/Ch07/4/QB7/RXgH/DWP7LP/RZPBn/hQ6d/8fo/4ax/ZZ/6LJ4M/wDCh07/AOP0Ae/0V4B/w1j+yz/0WTwZ/wCFDp3/AMfo/wCGsf2Wf+iyeDP/AAodO/8Aj9AHsPirRP8AhJvDGseG/O+zf2rZ3Fp5u3f5fnxtHu25Xdt3ZxkZ9RXn/wAA/hV/wo/4N+E/hL/an9tf8IvZJZ/bfI+zeftJO/yt8mzOem9vrXPf8NY/ss/9Fk8Gf+FDp3/x+j/hrH9ln/osngz/AMKHTv8A4/QBc+AvwXHwR8NeIPDp1f8Atsa94h1bXvM+z/Z/K/tScz+Rt8yTd5edu/I3ddq9K8B8Gfsk/Fz4Vg+CvhL8brvw38MUvJbm30J9EtLy8sYZ5mmltbXUZX3JGzO20vE7IPunOTXuX/DWP7LP/RZPBn/hQ6d/8fo/4ax/ZZ/6LJ4M/wDCh07/AOP0AeXeKf2YPiTpvxR8TfE34AfFaT4dN44aCbXNPuNGt9atJ7uCPyluoFmliaCUoAGAJViMsDwBzHxT/ZK+NHxB8afDvxnp3xqSzuPhvZ7bJdR8OQ6n5+pyoUn1KZftdvCZmBAjAhCw4ynJJr3j/hrH9ln/AKLJ4M/8KHTv/j9H/DWP7LP/AEWTwZ/4UOnf/H6AOU8Ofs++OR8TvAHxb+I/j+LxTr/gvTNZ02d4dHTTkvhqsqPG4RLiRYfISMJgB/M+9lelcJY/sX/Yv2RfDn7K/wDwmO//AIR+8tLv+2P7Px532XVP7S2/ZftB27v9Xnzjj72D92vZv+Gsf2Wf+iyeDP8AwodO/wDj9H/DWP7LP/RZPBn/AIUOnf8Ax+gD8ofil8RbG9/aC+KH7QWt3Xw8ttR+HVy2jWPhbx7K0msMuh75EvNKRFjaFr1nD25EdxubBDjgD9q/h14ruPHfw/8ADXje60ybRJvEGm2eoPYXPE9q11CsphkyAd0ZbacgHI6DpXjl3+0n+x7f6hbatffFPwJcX1lnyJ5Nc0t5Ys9djmYsufYitb/hrH9ln/osngz/AMKHTv8A4/QB7/RXgH/DWP7LP/RZPBn/AIUOnf8Ax+j/AIax/ZZ/6LJ4M/8ACh07/wCP0Ae/0V4B/wANY/ss/wDRZPBn/hQ6d/8AH6P+Gsf2Wf8Aosngz/wodO/+P0Ae/wBFeAf8NY/ss/8ARZPBn/hQ6d/8fo/4ax/ZZ/6LJ4M/8KHTv/j9AHv9FeAf8NY/ss/9Fk8Gf+FDp3/x+j/hrH9ln/osngz/AMKHTv8A4/QB7/RXgH/DWP7LP/RZPBn/AIUOnf8Ax+j/AIax/ZZ/6LJ4M/8ACh07/wCP0Ae/0V4B/wANY/ss/wDRZPBn/hQ6d/8AH6P+Gsf2Wf8Aosngz/wodO/+P0Ae/wBFeAf8NY/ss/8ARZPBn/hQ6d/8fo/4ax/ZZ/6LJ4M/8KHTv/j9AHz/AP8ABUf/AJMU+Jn/AHBf/TxZV9/1+UH/AAUe/aE+AXjn9jH4h+FvBXxL8M+INavf7J8ix0/WbK7upvL1W0kfy4YZWdtqKzNgHCgk8A19wf8ADWP7LP8A0WTwZ/4UOnf/AB+gD3+ivAP+Gsf2Wf8Aosngz/wodO/+P0f8NY/ss/8ARZPBn/hQ6d/8foA9/orwD/hrH9ln/osngz/wodO/+P0f8NY/ss/9Fk8Gf+FDp3/x+gD3+ivAP+Gsf2Wf+iyeDP8AwodO/wDj9H/DWP7LP/RZPBn/AIUOnf8Ax+gA/Z/+Bn/ClNO8VXGra3/wk/iXxtrt7ruraobb7J501ywEUKRGWYpFBEqoi+YQOSMbsD48/ba+Bf8Awh/w6/aF+N/h7W/s2neOfCdrbavov2bKzalYTIkF+twJRsYQExNH5R3feL54r7D/AOGsf2Wf+iyeDP8AwodO/wDj9UNS/ad/ZG1qwm0vWPiz4Gv7K5XbLBPr2mSxSL6MjTFSPYigDyf4bfs1ePNf8UfD74ofHL4j/wDCbxeBbXz/AA/plppEWk21vc3VusZubopNM1xKkfyp9xVPzAZJFfJH7M/7PPxI+PPwB1TwjrPxNOkfC7WfE+ujUdCs9JiF/PFDqsrSQLqbTFkjmddzYhLAZUHaa/RuP9q39lWJFii+MXgtEQAKo8Q6aAAOgA8/pVHS/wBpn9kPRLX7DovxX8C2FsXeTyrfXdLiTfIxZ22pMBlmJJOMk8mgDT8MfAmz8K/HjWfjNp2pqlpqfhzTvDsOlLb7Vto9OlkkWQT+YdwIcKE8sYxncc4rzbx3+zL4/Pxa1z4yfAX4nSfDnV/GEFpDr9rPpMGs2V+9jGYbe4EU0kTRTRxELlWwwAyOufR/+Gsf2Wf+iyeDP/Ch07/4/R/w1j+yz/0WTwZ/4UOnf/H6AMHR/wBnvXLf4pfD34reJfG8/iHVfBWiappNy9zZRRy6lLqckcrXBaF0jgEZTasSxMNuBuGOeJ1j9jHw/wCJfC/xY8M6/wCI7hv+FkeJl8U2d3aQ/ZrnRr6FIBbtExkkErRPAGLEJuDFdo+9Xqn/AA1j+yz/ANFk8Gf+FDp3/wAfo/4ax/ZZ/wCiyeDP/Ch07/4/QB57o/wD+PepaB4n8OfFv42v4vsdb0LUNFt4YfD9ppscT30XlfbLjyZWaeWNeiK0Sctxkgi7qP7Ln2/4Y/Ar4cf8JN5f/CldV8Pan9q+xZ/tP+wbSS18vy/OHkedv3bt0mzGMNnI7X/hrH9ln/osngz/AMKHTv8A4/R/w1j+yz/0WTwZ/wCFDp3/AMfoA8i1v9lLx941+L3hfx/8Q/ieNb0PwRrsmu6LZR6JbWmpQFi5Syk1NJC8lqoYKyeUpkUDeSVDV3fg79mDQNE+H/xZ+G/inUv+Eg0j4r+IPEGtXK/Zxbm2j14gm3XMku9ocZWX5cthgikV0X/DWP7LP/RZPBn/AIUOnf8Ax+j/AIax/ZZ/6LJ4M/8ACh07/wCP0AeXaV+xT4Fsf2UdM/Zel1O4jXTViul1u1UQ3a6zHN9q/tBAS2G8/kIzN8nybuAa9A+Enwv+PfhTxD/a3xa+Mb+PrG3tXt7exh0K00iMu7KftFw8LyNLKFXAC7EG5jtJxjR/4ax/ZZ/6LJ4M/wDCh07/AOP0f8NY/ss/9Fk8Gf8AhQ6d/wDH6AH/ALQXwItPjr4d0a1tdeuvCfiXwrqcGs6HrVmiSy2N9AGUMYZPkmjdGZXjf5WB56V53H8Ef2mb7wV4m0LxF8e2utc162isrbULbw7b2cemwbj58kEEVyGN1Kp2iZpSIuqR5Ga9B/4ax/ZZ/wCiyeDP/Ch07/4/R/w1j+yz/wBFk8Gf+FDp3/x+gDzDwH+xl4W+Cvjrwt4y+AesP4Nt9Psk0zxDp8lub2DxFax5ZJbn97D5d8jszrdAMfmKlCmVPxX+yv8As5/EP49/sw6f4T8TfEs2Hwo1bW9Ye+8PWekxJfXEdtrNw7W51QzFlillj3tiHfglA22v0j/4ax/ZZ/6LJ4M/8KHTv/j9Z2lftM/sh6HZLp2ifFfwLp9ojOyw2+u6XFGGdizEKkwALMSScckknmgDivH/AOzD4wk+KFx8X/gD8RX+Geu6vp9rpmsQPpUOr6fqEFiCtq7QSyRGOWFGKqyPyoAwBu3ReNP2W/G3ifTvAniiw+Kt9Y/FjwH9uEHip9OtpY7yHUnL3Ntc6apjhaAjasSKwMYVSGLcn0v/AIax/ZZ/6LJ4M/8ACh07/wCP0f8ADWP7LP8A0WTwZ/4UOnf/AB+gDzfxp+zV8TPiX8LtI8NeP/io1/438N+JLTxRpOv2+i29tBaXdiCLeE2CyFZYV3PndLubdy2BiuN8R/sh/FfxX4jXxnrvxehvtf1Dwze+FtUupvDkDKbS8uJZjJp8S3KrZyBJRCzHzvMRFL5bmvev+Gsf2Wf+iyeDP/Ch07/4/R/w1j+yz/0WTwZ/4UOnf/H6APDn/Y/8bxfDf4P6JpPxLTTvG/wWDRaPraaMj2r2r2n2FoZrF7glt0ACs/nglssAMgDsLL9k9dY8U+MPEfxW8VN4yXx14Qs/Cmqx/YUsGmFu0zSXStDIVQyed8qKg8sqCGY16D/w1j+yz/0WTwZ/4UOnf/H6P+Gsf2Wf+iyeDP8AwodO/wDj9AHz5/wxn8TPEmiaR8Lviv8AGm88XfCvRZrVxoh0i2tL6/t7B1ktba/1FJGaaNCibysSNJjJIbBr6B8I/Ar/AIRX4rfGD4nf239q/wCFrjSB9k+zbPsH9lWH2H/W+a3neZ9/7ke37vzdad/w1j+yz/0WTwZ/4UOnf/H6P+Gsf2Wf+iyeDP8AwodO/wDj9AG3+z58JP8AhRHwY8KfCL+1f7c/4Ri0+y/bfI+zef8AOz7vK8yXZ97GN7dOteyV4B/w1j+yz/0WTwZ/4UOnf/H6P+Gsf2Wf+iyeDP8AwodO/wDj9AHv9FeAf8NY/ss/9Fk8Gf8AhQ6d/wDH6P8AhrH9ln/osngz/wAKHTv/AI/QB7/RXgH/AA1j+yz/ANFk8Gf+FDp3/wAfo/4ax/ZZ/wCiyeDP/Ch07/4/QAftY/8AJrPxk/7EzxD/AOm6ej9k7/k1n4N/9iZ4e/8ATdBXh/7Tf7Tf7Nuv/s2/FfQtC+K/hLUdS1Hwlrtta2ttrthNPPPNYTJHFFGkxZ3diFVVBJJAAzXuH7J3/JrPwb/7Ezw9/wCm6CgD/9H9zPGvw98A/ErSotC+I3hrTPFWmwTLcx2uq2cN9Ak6qyLKsc6OocK7KGAyAxGcE15h/wAMnfss/wDRG/Bn/hPad/8AGK9/ooA8A/4ZO/ZZ/wCiN+DP/Ce07/4xR/wyd+yz/wBEb8Gf+E9p3/xivf6KAPAP+GTv2Wf+iN+DP/Ce07/4xR/wyd+yz/0RvwZ/4T2nf/GK9/ooA8A/4ZO/ZZ/6I34M/wDCe07/AOMUf8Mnfss/9Eb8Gf8AhPad/wDGK9/ooA8A/wCGTv2Wf+iN+DP/AAntO/8AjFH/AAyd+yz/ANEb8Gf+E9p3/wAYr3+igDwD/hk79ln/AKI34M/8J7Tv/jFH/DJ37LP/AERvwZ/4T2nf/GK9/ooA8A/4ZO/ZZ/6I34M/8J7Tv/jFH/DJ37LP/RG/Bn/hPad/8Yr3+igDwD/hk79ln/ojfgz/AMJ7Tv8A4xR/wyd+yz/0RvwZ/wCE9p3/AMYr3+igDwD/AIZO/ZZ/6I34M/8ACe07/wCMUf8ADJ37LP8A0RvwZ/4T2nf/ABivf6KAPAP+GTv2Wf8Aojfgz/wntO/+MUf8Mnfss/8ARG/Bn/hPad/8Yr3+igDwD/hk79ln/ojfgz/wntO/+MUf8Mnfss/9Eb8Gf+E9p3/xivf6KAPAP+GTv2Wf+iN+DP8AwntO/wDjFH/DJ37LP/RG/Bn/AIT2nf8Axivf6KAPAP8Ahk79ln/ojfgz/wAJ7Tv/AIxR/wAMnfss/wDRG/Bn/hPad/8AGK9/ooA8A/4ZO/ZZ/wCiN+DP/Ce07/4xR/wyd+yz/wBEb8Gf+E9p3/xivf6KAPAP+GTv2Wf+iN+DP/Ce07/4xR/wyd+yz/0RvwZ/4T2nf/GK9/ooA8A/4ZO/ZZ/6I34M/wDCe07/AOMUf8Mnfss/9Eb8Gf8AhPad/wDGK9/ooA8A/wCGTv2Wf+iN+DP/AAntO/8AjFH/AAyd+yz/ANEb8Gf+E9p3/wAYr3+igDwD/hk79ln/AKI34M/8J7Tv/jFH/DJ37LP/AERvwZ/4T2nf/GK9/ooA8A/4ZO/ZZ/6I34M/8J7Tv/jFH/DJ37LP/RG/Bn/hPad/8Yr3+igDwD/hk79ln/ojfgz/AMJ7Tv8A4xR/wyd+yz/0RvwZ/wCE9p3/AMYr3+igDwD/AIZO/ZZ/6I34M/8ACe07/wCMUf8ADJ37LP8A0RvwZ/4T2nf/ABivf6KAPAP+GTv2Wf8Aojfgz/wntO/+MUf8Mnfss/8ARG/Bn/hPad/8Yr3+igDwD/hk79ln/ojfgz/wntO/+MUf8Mnfss/9Eb8Gf+E9p3/xivf6KAPAP+GTv2Wf+iN+DP8AwntO/wDjFH/DJ37LP/RG/Bn/AIT2nf8Axivf6KAPAP8Ahk79ln/ojfgz/wAJ7Tv/AIxR/wAMnfss/wDRG/Bn/hPad/8AGK9/ooA8A/4ZO/ZZ/wCiN+DP/Ce07/4xR/wyd+yz/wBEb8Gf+E9p3/xivf6KAPAP+GTv2Wf+iN+DP/Ce07/4xR/wyd+yz/0RvwZ/4T2nf/GK9/ooA8A/4ZO/ZZ/6I34M/wDCe07/AOMUf8Mnfss/9Eb8Gf8AhPad/wDGK9/ooA8A/wCGTv2Wf+iN+DP/AAntO/8AjFH/AAyd+yz/ANEb8Gf+E9p3/wAYr3+igDwD/hk79ln/AKI34M/8J7Tv/jFH/DJ37LP/AERvwZ/4T2nf/GK9/ooA8A/4ZO/ZZ/6I34M/8J7Tv/jFH/DJ37LP/RG/Bn/hPad/8Yr3+igDwD/hk79ln/ojfgz/AMJ7Tv8A4xR/wyd+yz/0RvwZ/wCE9p3/AMYr3+igDwD/AIZO/ZZ/6I34M/8ACe07/wCMUf8ADJ37LP8A0RvwZ/4T2nf/ABivf6KAPAP+GTv2Wf8Aojfgz/wntO/+MUf8Mnfss/8ARG/Bn/hPad/8Yr3+igDwD/hk79ln/ojfgz/wntO/+MUf8Mnfss/9Eb8Gf+E9p3/xivf6KAPAP+GTv2Wf+iN+DP8AwntO/wDjFH/DJ37LP/RG/Bn/AIT2nf8Axivf6KAPAP8Ahk79ln/ojfgz/wAJ7Tv/AIxR/wAMnfss/wDRG/Bn/hPad/8AGK9/ooA8A/4ZO/ZZ/wCiN+DP/Ce07/4xR/wyd+yz/wBEb8Gf+E9p3/xivf6KAPAP+GTv2Wf+iN+DP/Ce07/4xR/wyd+yz/0RvwZ/4T2nf/GK9/ooA8A/4ZO/ZZ/6I34M/wDCe07/AOMUf8Mnfss/9Eb8Gf8AhPad/wDGK9/ooA8A/wCGTv2Wf+iN+DP/AAntO/8AjFe36TpOlaBpVnoWhWcOnabp0Mdta2ttGsMEEEKhI4oo0AVERQFVVAAAAAxWhRQB/9k=)
Adapted from (Davis, 1992)
Amygdala circuits
![[[@medina2002parallels]](http://dx.doi.org/10.1038/nrn728)](data:image/gif;base64,R0lGODlhWAJNAsQAAMzisP///46Srv/xxWFpaewLCvOvjqiechITE6mrxHGyTMvM2AceZGFrjP/44NrsyHyCf7LBmtXW4/bRugltK+jq6kJJU+j121tVTb6/vOXl9DIzNPro29/h5b291vP29CH5BAAAAAAALAAAAABYAk0CAAX/YCCOZGmeaKqubOu+cCzPdG3feK7vfO//wKBwSCwaj8ikcslsOp/QqHSa7AggEEHmQ+16v+CweEwu6yQECyZt2VgS5rh8Tq/b7/PE+goh+NsZeIKDhIWGh4YdCQQCjVgNfmyBiJSVlpeYmTeLAgkRjXx9fxuTmqanqKmqZB9YGQsZCQmOj34YbRWruru8vb44CwQJHRKwEZ60ogQYGwS/z9DR0qkSEBIfxLCxs7SQy20C09ALcOLm5z0fGa4l6xAJXDPqBB0BHxXZGbGfod63pOh4ESgVo4K+EQZLuUuQK6BDVBA2IJiIAGAHCxQnhouR4N01Efc6LNAmK1mkWxbq/z2ECKFGhorxXiK4hyEjgo0rc1JaMNHCq5obAkBAQAAf0Bh9MnToUIHLh5Aj93HDooxZS52ZEmxYUCMBTBFeEQQQgABDhQpHsaolRHZDvA4SOxAgCjILDFcjJTB1iq+YvpLdTgJcu3ONA5cTY04UStSpXcKQ6wy9CpJLWKIC4rWo4CeBvlcSzvLNtw9UrT8WNEcWxBlDORpe3YJdLLOsgIarc5cZirME2Yy9U6AVJkvfAr1NRfSNCjhUJEC68XwgcADDYcRiRcjMdXki5ejgu0we8eHKUpECCEicuYIzo0bFQe+1h0+kMWTO/zgLP6fVgQMEzRDbCNvptUB666nG3/+CTYTFVQBhSSBRKYmtAIEFVMHnmXzJheTXNqZhoR+DcWRwgAEQXIfdg7F9INFrEuFG4oxIVEARAUPRNdcGjaSVwgcW4IiFAAd04lle83lIEj/OrUHjGNUYgIFKNnQw0QZYeMcYlulV+OSXRWSw3kRmBYCWTcGZsEAzQoJyjHHIOXVPMSTNQlUfToLpxXQG/KeiDb9RZEEu09n0nZ6I+lBBeoyUsAiOD6qQQZB+ZGgknEku9xeTVFmQZqJOQHAiBhLssIAowT0KQaSgttqFmJFYKsCbSHaoKYi0WFCqq09c0ecBvAbLy5prVDrkpa8ssJdTS0V1DJM4CttEBCf2KaP/tNhqgpIfDVga30hJYmPfNrNEm20S6Rmg7gF/nusuJVqpwa23Gx6XKWme+KHgu0F0gIG6UgbI78CEMINBsRkWSau9tmZDDoYEF1Gtugq0G/HFdcAlr7FZaAhaaHLWJ8DBVHnGFMY7JKAAwBiwivLLcoi5cbcdy1JvnPbck14GEsRyRSQ4agHyvjCnsMC/AFdc9NJxxFuskDXfPN+pw9D5V0nKAA21Fh08xbR2EAAsJZXifm22eBtvrfDNTS2ygLgfesJNP7ZY0MYGEm31tQRIUxyBme5s8NrZhDuhMaVbI3tcNRCMJkHc+NFd990Co/xB3+picAAEmv93aOGgKxHR/8yW0pqBa17fs1ydRIZSFTNFmU2k2AYoMPEBBFgc+u5GGIywtxnsLNpo+B4TIgTeqPc5zB1MTDvLRPMuvRBouRHrkB1dmMDj89HXbJ1zU5UGBp++3EHYz4utwADTt3/EdNZz7EcwyeLs/bgZPAtKep5GjzJa1QJA+jBQOfcZ8AdaccMa1BAOucBiceFa3dVAkYYCwkxU66oO5hTwtwN6cAhW6EMWVLIO4zAsdfUphgfyJ7c+XItpnDuAAkQFAOqoCwCa+6AOlXCFz9QqHkqqkx9euDS0QCACEzhMBghww//s8IlHwMK3lCUa5UhwEY1rn78Ahjv/QfGLOmDEFJFjq//l9NB9FcAgxTgAxjb+AC0akloVVVcNC4LuPwDLohv3aCryGSkCJrRfAOh3QA1MDAMA4KMic8CJItkMU/NxmwfViIEOLvKSNBChm+rFsDTqsQT76hooXRaALWRLDxQDFiZXKQMRdkxxyqIaCgSwK/J8rhqg/GSwttin5bHylyi4HM1A8UhYjBAFp1JQLEzgmRKcD1toSCUwp8mCi9wpC44Mnmt+FLsSQIBKI/hmOwbnqstlkJroTMGkCDBMN/WhlieAgMDUYYI0mkAL0pqdlHSXTnQKgFI06xiOvPgYZwoseCaAABERxcuW9fOhJLgQx66wzR8V1JvwFIE4SfBMYVX/q5L8hCgw0xCJbnUmBR0IXgFnYQJcOoqcoDJRnywp0n66aGO3gEcJKgALPrwNBeuIJzjtoUtQHU1KAAhpTVkpIbuNL6NgS8ACFiqCYOxLAmlCKA4+8Cg7SsOcAFqqSOPVBgJ4kQVoIKJSEjrUGcgFR+qBqjkwmCKxQjQieFOoDYLRVjN97lQ4OF8GelnRgJxubHa9a0VgGoNq9FUoAlvVDQA4gc1xzpfRQAtSE3tXUu01eI9laTvK94JWDKSyohgUOvwTVs4+lKotWIcAHisUxmamtFiQ0ekOMAETiWgw5jARBibg2uKqABsdoaUKLjoCWNxFDWXSKPkM4Ns/kNYX/63wrHGLW54sJKBIJaOtCK5wgunANpdYwABlIgKB3m7OD1g6B+e8ul2Irg5ksS2fPGEwnU49KCK87eWFBBfcHNb3wPztCNFE+wLOIA9PubhQe92hHr1NAy1TQrCG29OIAr5DBtOJ1TLU44cDfPdCZaVvDbABC25I9bxOqEAzYLzhlz3lxl6TwhJNGU/GsiDEyEvDBtbwL1EdoA0HIykpV2yFb9hNDU8OUhY+MzeGNMFuSiUBjs/K3xrnJoF4C/N1jXCq2wJ1zCpIAJSH3CkT4a4ZG6BZGt5xMhAHz8lP09rkosxnedKYBwLYwJ/xKubSLtlMqm0lZr2MCp6eCgHJEv8vCLN3aLAsmgViasadlmGA7wZJ05u+xRrCK5w7b6ukek61qtkQpNv0Tq4oWMNIwPWCo6HARlw+gR8YDQ0x7VQfDeFCBbZHAnJIGkICoOKN+xI8LETgvFfI9QkWNepr+sEdB8MQVQIqoudIWaot7oPdELbqcpv7G1j6c1d8HOtPSUCqWtZHPNQsSnVMwkYp5THgrDyCd+9q1yIQJa91oRXNjOwPXBmx8ohK5EsbhJg2u4JnYBw8VcrgQMu4prW1pnGaBVTO2R43udl57pKXWw0q7soFYKDenY5YvVw4nS0YUhMC9IzIpPoAUZaht4OnIRytmXMAdo0G7Q5cFQXXzgb/VGLrW3CBJ2MJigh4JIRW6FQGrdG41rd+TY+7EkfcfjAkskZyk5s9ElSXgnqejIHptH3qjUNAOV4iEgyIIE+NgrQIliGhXaHF7cLeAOPgguajWyLpY/mOWVpuppkczGZqCMI6Kr2Ct3L98nfqVkChJkKTproPJk1e2c9+bmaknAiPL46LSvFPrZDAU3wbLwJWJWwENKT1+xmvmErxFM41w/C7iIBsAlAkEpCqUWYSdFkjwW4byPbYKOCbkMLMecxr3evsFCHYRXQaoJGd9CZP+xPIp2ULQ4h8FjD+ASSQfu0wQ/A6b8iRO0ICE5tfo0RBAOWBf/jhL8L4l3MVuMZ4/6VEU4yUBbDWAgsgZRGBAAzAAAjAgNa3bZm3aQ8GVxgIV6AHfhwIXwQ0fsFBYNL1EpohOLYGISCRBjJ2exYQAXYXTmvyGtbAOUM3fPx3Cl4RDxKgdxDiDIyHa16RC1byKcnRYAeSBep2ApOSBVgAgRMBgdo2gdbGfXJ2gaEnCqLXgWb3NGW1f0JAgBq1dI0HB/HFGB+wJnCnUYNie2FoJXOXfhGRCy/hHvbwezd4CqdDAv+UXlzxYfbwdn1wcCgQH/XzOIYYFfrQYUmohFHYBw5IEVJIgQ/2CFSofdxXKRqYgVmoheZmN8olOuTkdulxFUeDBT4RANaUUtpGQJczb//OoGZZ4Cn24AcjEw6NUFUW4IV3eAlnwQL44EWqw1PkYmKeERpFCAzVhxFXUn2RSImTuH0YOHabiIncwomqRmRA0waXhgTYcC29SAJF+I0q8ItaJnAmIG27+C5GpHHX04xbJyRVaFLdZomjZ43llmfYuHDpuI/pQIvuKIWaR4nQeIHZN3pYaI8lVywKqY/82JA3MFH/uHVh13X0SJDaR40IuWpPs5BrYIcO+ZExMDJMGJGYN5DyOJAYuIH16A3TiJAbeRIkBZIy+WOu1AjgMCQk+QgSKD4UmIlBlokk930ZaQswOWJB0pGFN5MNyTlXwChsQFI46Y7qoV4fJ5CdZ4n/noeRQ6mR+qFwRCaGShmWIoAGoVBWQ6FeKLGTE7iTVQk1mZg8obeV96hnxXKUDCmWM7kOc1ETB7NzGMIHbBCJavCOmyaPY3eR2gcJLCmXMJmPHCloeKmUctETQzZkoraTUymRQaaNkvhxbnmRcJWVjJlq+KgfSRYkEBOZM7l2F9ITGGGKHINiHMc5UNaIzjiJF5ibJ1kpY5eBo+mYRPlypqeaMrmEV2A3WDI+vsdmP+NU6aFeEygkJkmPvSmUo0mXCmeXC5RoxOmQaoYjbCZlGKFtbSBlXFEk/tKMYeeTvFl2QQaXv6lwXblARnl63clrp/KUQOEGFfGX5Ecgc6aZ/2K3bdMJdphondcJNPmonYF5nzJ5D9qQUt80DxuxObhRPZH4mSdJjweqlWaHOOYGnAupnanhoHgpF5NQfKVkcyMQL8xIgVVZkW95kKnWknrGJme3kdh4Czyniya6i1bHBZ/AFdXwIP6SBmE3ka7Ulu+ZkiS3mGeHoyHqlVRKKai5jT+6j0skbFgwXhvxoh33jJcYZBeZlQgaolJqcjr6ckdpN4uYpcBHf2DBohbqSuzIjIaZm06KkWeqak6VkPKJOAyKlHCKl8nWbwTAFetQbV2HJbrZkxVZndWoNTYKNGmqpkSJjaj5DblXqDN5D1omWez3ogFKkRq6gTSaNZWqNf9/CqgvmalGyXNvugJbVqs35qk1hQ9gAZ3XdAuUKI+1MKahCZp9mmpDxoEjGqubClw6IAAP+Kx89mQNMK3UOq37Iwse4AH2smW4Ch5PcQEX8AAAMK7jGgEAEAHomq7quq7oykHo2AJ2eidqaZWgx6GqKmLXCZyN+XKc2gPO+qwAG7ACO7AEywBRVq3dAh/aao7dygsfEK7iqq4KMLEUW7EWe7EYW7HvSqu8ek0MmKfBGqmY2JuUmqCaqh+bepRg2awF27Iu+7IFazfWKgAegF8Niwgf8AAR264Z27M++7MUu7FpZpudUn0DWYkp6Q3KsKouSZr8Sp+pqQMegLBUO63/fAazWJu1FmCtUsWwN0sG4XquEQC0ZFu2P7sDl0OqjnqBjXiYPimp9XhurZqjRDmoy0Cf/wB9NqAgtjonIpGtzUGtT5a1WLu1DUCzH/G1UpCzYmu2ZTu260qu46qzOguulguuOZYyRGuKRAtq0rmh0cincatnc6uFJ4uaC7QGEahvY3BjGrAAHoA1Vku4BXu4UqUBaPujwWZmPxa2jeu4Ewu56kquDwCuIEEF6al1viqJvup11MmevjmpN9qBO6qg/EpS5WkwnUBFt3I1j/QKQlt1EOoBoCC4FkC7Bmu72jqr5KAc97lRkvUjv+u46Dq5FxC+UqAHWleqjcqT22dS/yMnvSV3qdT7tOOznQoacmzHkQejkP0jB2c4C7NLuFubbNJmDX51n/hUSmnCuGNrtvVbvIgQr4/gBhIYUFLGpLbwnio5usZawCJKUgt5rAipBgnICh0QuwLQABTcANvjP60Qc0nZkAeiHDhxAecKwgAgwpfAfuyYi6XEox4LnaCLJ8SqieZGwOBXmlC7duAno7Tpo2UgAeTbAOcLs1vLb8W2ERtMnEEsAuYKAI87rvjbuiPRITJnbSlBHvxDKbTJfbqZcVDJcUDTkmlDt9VrpbGKEmRXkVMIyNq3x5SQwzt8xi57uB4wbx1UVJH5iSrzs+b6AHVcBjKTZN+gcQfTVv/3sA5OhSHxKEJr0ABu0Hku7Kcgao91ya92w452Gr0jBsmBmAkfgFVmDLM0q1cdRZzhekTagbEhbAoi0RGba4FntShQ6chslzXFqjVaPKUnS5/KqkCmnMDtiMrV94GnoAESbDcFux/tG5kfkMQT684YELwAMMp1sCbTPIUNxjF5apZA2ZtMu3zWyIUK5wYZ8HCigDBaMFW3OkqxeSF6i7NWwMMAK3dgwQA+PNGG9wAfXLFlwhmivAvYMK+N2nwmsESPCl+hWXabqJiji2SkV70LaZRYsi+Z+wJ806tYigkV4AEX8oC3B7ANIMaM5tEZOxAahc+CEBFK+hzo/AJYJJ3/D7bLmyesVRFl/4AhcnHLdBvOSg1omiMq6cXUhMBVuUcAAvsGuLuL8nyx5hpUY2Gf8PKcy0BRntJhQjuKZGqKJqzV0cp2jABI5IAguThVEYHIRckMbwAEUPMfDvUMWAUSG1C7HsB/D9CzEfAA4fR0Q4wJrdAMD2wD9UqJGFIBcaMFyfJAU3WMATCqmoY3CV2eyBplrgYEeuA5uIPSqLDBwTO1tftTXobUcL3ECoJPb/wLZ3EcOACLX/cG+Js9qiY+2XsSgf1kszxxQ9BfrUMd0GBmGzXMCWDRAZvGZo1JxG2x5mo0bGzU00AMmnE+J3whdF1s2nacbdKUTIghQubH/4DpSsJtBFfwH5vz2bx4b8uDVZYMsBZw2dslx3DN2cd1FSZCGHwDXXA1mM7Y2DUwPncbK/rtCPvBU64NBbhTJLv9DAahHaTFVeRN1LyNTulNsZvtAhs1q79gBWn6xD2dAq5wOpSiXj/zHDHeBAMO2TecCvHQxingAQv+rA3g4DX1AR9dsffcYOe9CkuUbXlVgd0ii5M1FtoWNFYsN0kOBVhgYrgjDuoG3ALLu/0E4RW73okyMguJJ5yXJ8DA39c0W3GABmoe1ZEhAcXM4J9ITRdQ5Ry0cq2CFvz9BxiQPL66AxIiDOTRH5qTACjS4w/BVU+u0WfeRnJO41mOFedDUv/xw4B+zgNThQdYYADUYlYLUh6fLuWXROXqLeHSsihlIWvosESdVkIzogEv/oAGbkAfcLFXfi56sQ44bgpLhK59El0zkgAC2wChfkC4XrG67qkjcyLCle2rkUYCW+Ttk9lz3u389xT1sRRV5AI98+r6sDn1HRluDuXPbjaKru4Dp9CX6H13ciR6cR491QcAQl16me+QYe0MbusfpOiMfnQhVGL/EQHUdfETcPHU4jmNbGS9RV2oJSzsF7DmTjiKXurUZFoYNFjz/nU4YmIG8PGDNQE0T/MxnwG9RfMRYC7C8gHFfuxLg+4Uy+81Fgwn0lstjyPDcBb4QGGikgA5z/L/SG/zuBPWpxSwQP8yF2CxiTRw5VFJ7oUijMC69dRsL3/wg9VLOFLvT8Lwz1ryMCP0i35063AiuKPaAe4CqI2IPpS4BOPkAJv1BLP1Gkv3THT3HN0+Ph/4vLPtE4vy0yQqjKDwu0PsAAv3BFPlET9w6yDuO1QBL+7wXyP3Bqi4S/MBAev5/KL5pk84bm+wkA8mo775rf81HsD4ZmPltQ86//qAiY8to777oLP4D9gAX8Ptwg86EnD5SyPnpZ/8X9P77WdjH0300H/65C34ieL81x86FQCwv98q1d/9oZP9KEP4E0v75B/9ABv74BH86184y/+sov8uH/388b805M3p/60CAoqoPIF5oqm6sq37wrE807V947m+873/A4MmD6PYECKTSt9lpPgso9IpNfjpdBaLDLe7kHQq0Cq5bD6rPkWGZYx+w2XNUbxuv786mQSk3z8ABgb69QlkSLjhKS7aNawlMkZSPYxESF5iCukJ/B1MTAyEio6SfkYc+AlIZLK29gisrbrO9lCKWNLm6pp0cEIcRICSDhMTT6BCGELuMrfCFnk0S8PYKgBMY+PpZUT8CheDh49OdB9wyWan2xEVJai/V1+/z1NJ9AV/i+vvD5D3JSyjJzAJOwbuBkqLh3Dhjwz3+EGMGOoYgYMML/YoaBHjrGq4OIKM4RCCAYkmJf8eIJAhJMsZGlu2UghzpokKnPKdzKlvAgSVNH+eeAk0ksyhHD8kIHBAJ9OIPCF0MApTqFQ8RasOrJABUNOuEU9liIoVI9WxcTyaFbgAQjCvbiF2ExAwbbqydM9cvTtNAAGcb/+GS7lA7zy7hKvkPZxLglLAjvnxHKwYm+HJURJbZpWBQITHnndWzNyssmgkaEu3cuj3M2tSfSqgpkU69g/MtO/YW9p6N7hTAm6zmg18h+3hbzZ35q28GCrjkoQ7v1E8Opmkq5djP7CReh3o3Gec/n4mKfbyxfqI775me3ob09sjWWt+/rDm8M94v8/ivX4fqukDKAoq6PQnRX4FosD/H4I5/BKggxNBtWAUB0oYnoQ/CKDbgw7ydKESFN6XyFVzeSgDKhuiaF+JOyRSFon6RRBBCQHI9EAE8qxYAx8o8phMjjp4YMFBQtnDHoK2yAjACADYKMIFP9JQAQQ86uPAchAYCSUMH1jAQAMeaOSBIwxEs2IETqB5i5YydADBdSdFkMFn5BQjp2NsranjGnvu2UaOc6Q5wot5BlABZ38hsIE4VobjAKOjOHqAosM44OZjB/xGqAxd8rlnlgueGagCH2maxqFuOSApAhE82s8Ejk4AwACthsKBlZ88eusAESgKKyiOWmplBm+adABspb6QQKdr+PljNWk+iawKGQK2/8EBGGBwKwQYbIDAL91iMEAfVlbq5gbnYuBmquduQEC4A3B7LgCVygoBuxo21Y20LqixLAOZQikqqfsGsONbDkSAwK4IvDoBAktlkOgED2zAlgWzDrAqBgQM8AABCie8VMIbOEDABg/Am62bCXfG61tYEszCmJ2KBaWSaeJIsKHEmuQAtlY+7MAE2VpZsaMEEODzq5IOPauVF2/saMkkAxOKvZWSo1vEPEdEgGQxo/AMn0es+UGg0cbsyV8ORLwBtt0OAADHoVhwwNFJA+JA3RxMGQrWdZN7wMUZbHytBanG6m4fDL/F00pgn9DBsjVreXMlkCeAr1smA8BNyBMk/f+03SUnHYEFLjftKLwTWAABufZmPOW6WGNggTAIoNw4AcdCHoDYRmhqthM5S9tBX4A9ELTTJoMuCuCkF92uo0aLqzDs0yvKsKMWUA2A0Qgz/pe+vQcgAZ9f5xmqCINqyRZgqSo8SsSoqP480sBqD74FFmzMwQDnbmsDUKvY/h7mpgN4y10ImNtfIPC43s2MbJpS0I8MBhguDCMYsgrFsG4lq3VhDBQxGkBfrJQST9jphLsa1qxSwpljOKZD5AMRgoSnALRJSz6eaRW5MOa3XCHMWuRK1ye29UOpkUtqGGOUElXnmDiRLwBdalapzkS8Uu1sN7TymyiSKK7RzUputcP/wKt6yEQukmKLniEA5WJGBAmWqgk4RBa1WHNGMy7Rh04zIxPvmEc88rA14+udkAg2o319QHPv62Ia/7hHQCLRaY4EJBfVeKcHgs0DGohiqbihxUpO0od8pKTqSJlHSOkxhgRgHydbWaAPHO8xPQSlKUsZSj7u8ZanRKNjtOPKX+Yoc7JEZS0f+cgkIhGXyNQlLx/DkzYCM5r30eEOG0lKZeZylLYs5hlB+UQIsFKa4rwNSRwzyyVes5TqbGIytblOSeIxlZ45APrGqY4KJEAAmcvnPvWpHX/y85/9HKhACxrQgwI0oQRF6EIValCHMvShDR0o71hCzWqKEp3bXCZH/zW6TXh6MZ6OBMwEMAmTDAggpfnUp0pbytKXrjSmLeUnS2lq05ri9KY6zSlPdwpTl8r0p0JdqSGCsADOACOpp1iqUpvK1KXGKKpSnSpVq2rVq2I1q1F1Klef2lWvzcR95tSjHzfK0XaiNa3ZvOZujhHOgfSkAXKdK13rale6QqABhNhrIZLh15QCNrCCHSxhC/tXvhJCr3m9K2MZizSjQgAAkp0sZStrWco+QLKZfQBnO+vZB1wAtBcYLWlLa9rTova0oP3sZ5k02cxeNrayhUA9OXJRVDUSniB9Z1p7G8nffnSWlASMA3/CicIil7D8TABzm9tcMCUATNKdrnQX4P8BLWA3u9q9LnW7C13ngpe5yR2vYNEDhLXINr3phS1r29vZ1MIXvu6d72bVa1/K0rYlH7DUWNOp23X6NsBoPSY2c+sWnryVHskgL4MDG97nMte7ErYudyssYepGN8IPFmqDkatXyN73vvWlr2fja2LUina0JOasZkMc4vyyRJhrs+YkkSngG6s1uE4cLmB8ORMBNKDDyX1weC883e0iObtGli6RwSvk5Jr3B+h1sXpHTOLQYjnFJ47vakO7YhbDlsqxhXFI+DvjWvIWx2oWMDp3eU6vgC7B71jwkwUrUyJ7IMMT5q51tetnLVwX0EvO84ZpWmfBfvi8kRVzbDf75Sz/bznSKs4yiV1raUZXlsxHSc7BiKnRNq851O3UpR8vZVKQAPnQg13ug/O85D/DOskXznCTX6pqwEbZB1PG9GUdPV8sSzrYpF1tpVnM60zXliES4PSZ/2hjUUMbx6BmpCVPIsOW0PnWNW11hC8c628jedbfJbKtVZ1oKS/62K79cpe1LOwTt7vSl1a3pjFi5q68GZIejTa/IRnPajel3hxJtbZZ3WpXdxfQ4F44di08XQg7Wbza9gWI1d1iJtEX2O/eOLHlHWZeC3wht8WtWQnc75P/1tkjhTMEYJLtQ9uU23sGtwS+YPOag9vI5Da0uVuuaIuz98qidffGTTx0L893/97HDjlCBMDspgiXlrtFOcqNGXXPMH0hBL+1wZ2L8IQH+s8SGPvNyw5rPndXz4WeeK57sGt1+/rX7/7AaOku7KGvWOmMzjo9pITRdFI98B797y6bgilsc128eI6ud/usXZzj/Atjn3zNJx9rCdN657c+t67TjWkrZ/wCfZD0fuN0ALoTnctId+/F6Z3sgWyF5KJ0Z+D5ZqvAFzOGB2jJ1hncAHZtQKVd1/CE/XzdylM++WS3ufEpnPbMN7fch247D97++XVnPGIIAMCJP+CAD6QrAgQIAK8uYHfvG13F9G09pvk+j3IOM5v7jnYAJDW6qqfclG6JgJyx8fLxNgACWP+AXJ3L8DEedTWcFlieBCTABkje2GEB5UWg8kWekmHeuIVXz1Uc3GGce5GWu1hA0oTWApCWBIDWB9hIAlxAulzAAjwABKzKA6BgBCRABjhAaHEfF6SYZiXdx4Hc69FDm3QamuEesBiASuDep72Z+BAIRvTeeBUgS21AA4jXP9DaceVZSgWaBwDWF8CgAHjA2PETGNScSlmeSn0BdKVUn+UTlriaL+gTUelVYP0fclHfDlgflYHer0UMACTMvGTAyVwA6QRAIPbEuTgdZ1jAAj1AxKSLtQQABOzPuRyAimFABqwea63b0gHhPFQNvnka4J3cUh3h7RUh4YkUUxiAz4H/RB0i1yIKn0rBYAFBgJgI4Mdwj8lYgBbwjy4uAAYgAAYIgAaYjL044AK4jTFKgAcEYzF2ALeEYDAuQANiCwKAoQC4zSJWhAAlyhQCFgIEme+xYudZXAd64GgRwOFcQMWQnyAGwGO1iwkgkHZgQDuOlreYQMJcgAAsUMlYwGjJjfq11yZyIkbsF9dAhCgWIQdgQAQYAAD8QhJiE8DpxCqFxBNCWbcIEBXm0wKBycckhQVcFwxCAAMiQD5tgARoADPmmQNqAD8ugAZIQLskgAVogAYgY01iwE0GyQIsogf0AgL8YkpKALd4wBSCyRd2C5akYz4FYIfdoQ7koYs5mjmy/1ZoWctoSSLbbEAFDCLSJM8DfN8HBJ/4OUADfkD5kZa1IJD5AcD2JRIEZKIm+uDedaI6jFxOBFJIzR+/eUMwGAAGSEAR5t/KGd6pLYQrJlcDcE+iJEAAOpe3yBWY8CN3bUANMh4MMpcDdkAIyqQGJMBJeksYbsFJUthlhmAWsOQXquFJEprbCJB4WUBKCVCHcZ7beR6jBd05jswkbsAFBCJovaPXdGXdhV89Jsx+/SPdgV8y/OZotY7PRIBAsl5dUpn7pYPTNRvtBR5jkMN30o9EoiJgDJIThiODLdhKNeNlRtgU9gFlboBlqoLJcONmfkHFyGQWCCU2JgqWdCO7JP9AutQcSy7A7wlgRz7XJFYESiaDNULlOOIm0F2Z6J0OIJhO5/wmFGxMWH5Ah1pLUpzlBqTlb36fAwSfpNSdpAQi93VcdRYkQxwkblFbjVHdLwDAJ8TKMZiiROYeUyBYKwrZFMaUyfyec1XMZHJhfFrXZZoMaW7AHqRkZxKABnQAaCJAWOAkNsLgIWhBAvhkAwwoAiAcNgafa0aXANGkPu2PyQjZbVZfbuoh9rWXl2EAADBnImFA8iQA+IDTxtDdF0YAcuJOBbSLDKKovZQWMMoldWqi3l3nXWaDBCjSSeSKsxFmpLhJrNxorABCAPAo4fUYYg4ERr4iOH5jRRxoTV7/pl5RppguaU1aQBZ8jD5daQeEZgJogCE2YEzOpADUzgLY6i6m5jKKqVEW6EmGYJ7NKk0yFwya6oNu4PVdmemY1gLgzixuTMs9wCTWjnasUgeciwtyD/dcgySSVlpuH6Wtn3W+GKRigyfJaB9ZKsp5Z45+wqZ6zSmyE48xRXkyhGIW1iIiwMAKCUpqJBiy6gJgo2Vu5rm4iwMu5QdgozZS6SKOa82NqzVqgIBuQXweJbuAYQOCbAMuqD5FoW1C6JseW9zNqZep1iA2YorRXQYsgN3V3Q2eYAtGQAfYXYpF4j9OWg9aHHa+K0JWCbX10aV6w6Zu6sSYJWHuEcosYVP8/6h51lkhOJk+QVfC9dkyMhfZLV+V7kFMYsFK1iAWdAAD1uDyNVygaSGTCZ/ixWG7PFlU5sBUxumExtf3md/NjmWHdqj5Be4gDm5pAcA8Bu26Oup9Ee00tIVXAJIfflARrsUD3OvlWi5PWILgSU2BNsDtzWjANSFCACxyxRzEHSACZhcFRmDZUinaVimVTt4EWp6SCdrzORksPpmb4iGcUqVVXqWwMWffDm/h2uxppWXrlFaLDiS7Mq67SkMiyWibLYnJQRv54YPlctbEcC8AEA3uvWARCECpeYWPXQSpMliTeZufUSAEwu77pm3aKt/Z4e6D2ct5QuvPceCKdd+5Ev+v4BLv4B4vvCnuiyKEEEKd1M2K+ljD5PJbm3yC9k4ABzwABU8MB4Rnvw3ANM6MBYilLclTTlzbvxZcHB6c6mIX8rEu/Mav/LZvuGEYoUWc9IljtIoZywYv6dXd/wauzfowAc9lZxFk+0FvM8iYXqLRKA1AoNwIv3EGBXNAFEOxFHPABOMJv2UAp+zJ+AJY4Z0EBFRU05VuYRlg6qLw6rIuGLAw7c6v8aWdDDsXTl1tyvYu0AFviWlc9/HtnQbw+QVbsdFbBgDuIBNyIRvyISNyIivyIh/yJ34GwgjMB6sZ+UFAFUNxBUdxBWtyFRPAIIYaBwiAFvdJBwyAFJdyKVP/sSlTcD90DCuDwie4Ciw7ECPTci3b8i0n8hgPmQkX2Rkr4APO7uuqMdoOc/uyrQU+HBxHH1HtLh1Lpe+GGA5fJWgdbnwxZwSQYPEO7w5LGvO2Vn19HgSIyjiTczmb8zmjczqr8zoLzLzgGAf0xRSn8jxfQKeqWZv4Sz7r8z7zcz/78z8DdEDn82zCHC97XePp0+MlXwdwYTG/rzFX4O16V62pmt3iAN7+7q+hAnAKojW3TgDUnR8D7v9umTd71uKGmAIQADuzdEu79EvDtDm7s2/V31Js8gXMcyZH8QRwsYDZg0ADdVAL9VATtT+XsPrmmZD4JAFENDD3QpQ+dAu//3BTO9/W7hwzo6wNx+kdd1alHI5aapndBUCHposJMCdIQ8ENjnQAk9YNDhujNqrz2pc4x3Rd2/Vd4zWaTCdNG88EdEBO4zROU3EFB4OnBlgGzExRK/ZiM3Zjr0HwqVoZU9dJSsAAMvTNkV3ZYqNKyiRnx27a3mQW0K4CRh6YVLXmVbQz3y00R7OcflagXiavoAIBQIAMQqQfyGC6xGVP8B9KzfZW9MTpbQVpOd0FdANvBzGYyfVcj8qZOPeoQPdzS3d0U/d0W3d1Y/d1a3d2Qzd3b/d3czc5x4gk+5b7pHJg20oUC7YUO4SaIbYod8oA7g9jyvf+2Pd943d+67d9M/9mf8u3fwP4fwt4gBP4gBt4gSP4gRs4QUe2QXtdUixQKLcO0pSkwiJNRTy1rV44AYShXxFASvVERdhDGDIgbT2m4jCZMi+zBuqvtHI1Z6VjIzLiBdC23hAAZ4VgAAijvUhAC/4jDEaAlHhLaOUpio5WuryljCAQi7Le0NZs/7nCvQ2h0zDw8EwyIOi0enOA93G5o3DAeqdKy6nZB9gDfBdBB0jN7e2ol7N5ep+y/8D5KY/CAXgAFNj5WOP5net5nvP5nvt5nwP6nwt6oPP5cRU0nrVmLcJiS64FZcOgBIRyr46du0iANCJjfNqDmFI2pIsp/+SZR2JgHGO1kFn0DWD/tIi9OGhtC/hwn3FjQD3L4ArWY7pkwDUQueiJKPkhgPlF4h4A7QqW5MOMlgC8+BDbJULsF8ntq5WkSROHmgBMsBRXQBQ7wJaX6Ads+ZdTMVitWQD0Qqf0NF/uH8A0Hf4+meId3K0udeVxOHOlbWgqbEomgPymo7XiamdCgEwyYyAmgBjGJ/dc19tCn6izOLrZsTSL3qsH52gFKgqm4yLOuutsC/f85rZ4XwPWnQMJDmmlC/y0C5PTJUo/7wH3DVNs0TYxsIyIWvkcwBSbZYeOpQMUzAakNxV3Q7SBshZbAOh2E0VChmqrgy6TcdyCl6uFpgCsexhSYZmfy35ugK3W/w66WCuwPqMApC1LBl8ZxmcD9qdVZ+CokzrQmzpr2xd7cXXCB5FoJQXFNCSNR7zJJACTdLLFJ1JxVgo3/DoLhpbprEpyi/zIw96k9kyb5VIlkHeoOZAUp8r3wryGXim2r7e4rAS/PWY7FFgIS8QxnK+5n7uDd9vRf6kCcngxZgAzKuwu0qTCYsDUj50wXv1JXubYLex1PSao15rE1RnvPvPBe+C2GHdHi1/EPAlZYkBzbgAuzGTCXzzumB9bNGAJJI/FDOI6ymXzsp+YNe4uHLFOVGo2KYnK059DdAwFO4CvxnyHBgAy9oSXazsHbAbK1eTnKpFh6gT/kTDXeb7aWf9jZTM1CHhSk2yQpgnIImwZ8aGYtaySZEGd5CGeuYMgSoIbYeNJJBPMpiAhiEqn1KoAAglot9yu9xtYQADksvmMLj/Ij7b7cYkgIJEN/HIgPGaEvouAtdFHgHEAgeHwUQfnQPdgYdGXsxc5A3Dx9gawltaZBrEAJjpKWmp6yiVgMMDa6voKC+sw4FBLa3tbq7vL2+vwQDAxwUHscID4kZgYgEHQgZDhcEH8wDEQAeGrve2wYD1rO8sqHlseG/GBqr7Ovi7QYBVvxfTUZJ+0YWLRsOAhlgBhA4aBggTQsNCs2YYSIiRgKMLDR8BmFpBMrAjBwxJ7TqDI+/guSztRYjz/mTSzJpPKCAceZLgAcw0mQy8zsFEEoeWDBQ8kxHwZE84DQxFkxskpVCWbk0wBgBoJNSo7CBPMWXUFbty4rNy66gpAZ0C1ag4EYEiGNgyCDgEiBSAGtxoor3R30dJ6NW+rCBWk+v1LCgJIkB45NkmSRDATjQsSLGgMqF+CDBl4QCAhIUOSBTcYP3YsQQCgxZBJbDQcxePgeFgAbynZ1GTKTSrvwMR0G2ay2x94f+idO7jwC9Ju266tiVPsT6FcO/f7gare6eCq1/X6QtiEahyMIUuU7shjIR0cUONQNUOEANe53a2uldx0WOie2/f7bvVHeoaVKNEIoGeP3TAggTfs/3BDggM+tmCD/TAGoH/9qaZfFQ2I5Bpsy3mCnEoxDQdiiCIah0mHs23oyVP3rahOBRDMZw5X773Xni8fEBDBdsNw8EBZiCjjQAX5DInAAeadJwwE3dXYSy53PQmjOXyxSKU6glU4T2oTBsilgAx+CWaYYj7WZYQT0pMallS05pyGKJ4xm4lujEgnnUKVKOeJb5qhYpV+ftHBi1HGSCOT2gRgyADacWcMAew5oMUBaxVzhDRxbXfAAY8aalc44mQ1qCsRsPVnqV/kpyYV/J35H5cPjgkrrP6UGeFpHVGYakjPubmnGrSZ+KFtdQ6LW1ImbtIrc6YuG8YBoWL1pIzWGf8aRjATAKAjWQdsEEEC3daRRS0fZDAENcI8IAwBC3Cqyy3uQvmsqM0xu+yVuaq6Zatl9hNrvwzOWqaEZ967JoaA8ZrsUnnixjCxdRor51IJk9EnvVVm4Gy8r0jL7i50XAsyutVkMFDJzZT3QTFYxIUuthHA0LF17soYrwEJWLwsqgQ/oeWZtAbo75gA/2yrPTzjmuqFu44xscJ5ttGwwyHe+TSyyjVdMc4reqtxfDN6yi4HB4C8KFxL9gLXNEeiqx3ImW7K7tddi6qp1n/au3M9Pvv3M7/+BD00rQL3x3PeUbCZIdNNW/0rsFRLHdzjVes5cdZ2P3fAKnN7DR+7iB7/gO21oW/Ho9lmT2M6NS0Lgy0GD8Bd48xyyzfoBAJc7qfOhkORL9EA+i008L4jxmrhOyvdpuKLW/10sZCTWOzkEi9uOe6ACRBB16BOm0tXbfASQAcY5Jhj6CIPw13qcfHItnahA2CIL5l1cLZ74WzeygQGW28f3jvzziq+DW9WwiPg8AKItLwh7mAtmVhKnias5xELYse62p4eUD3+SUU6+OOYL8QCgAiIUAEKAEAvcjKB8rFOR6QrnQvPY7Y2kI0MBpgAATIAuw4wgAH7EEwGvLENGs0oXvrT4IquYDgpAJAjxPMdv/xlwAMGMInIyxCORIjFLGpxi1zsIhbVk0X1/1AmApQpoxltgsYzjlGMYfSiG9/4RnUZ0TkcfJa03NWGMmCRhHzkYwTA9wIDRMAAACDkCrOFvvVx53xtC50gJxC/XmRgh5Sk5D609MMewWtuRZzjcy7TgFCKcpSkLOWFBINKJKqJd0fTm9FYCUssoRIL8LiQKW+Jy+dcIAEH4F0vn/BLXvpymMBcIgI50ssmdMtbzATIICDAhG8dk1XJDKY1iSnMYhazL578Sx3nJqMI9HGc4/wjL/YgyHQWkgyHPF8bXChDtmFrniks5CAPIAHYdaOS/OSnBaLRuU0OCgLp6CZgMuDKaXIkTaxs5TCZkEyISlSYAKFkAyJ6TYpO1P+hDK3H0RSKGhwadAuQes4D+AgAMJTUFCsdqRGjA85o0QgA5Kxpj3YBlgMMcpDrPGTIWMhIFr5PGDlKJ0uy4QsB9LOfFpDA/QQaKv0V1KVUjcoCKHmzqmrVpCjdqldLEZ2qdLB7tDhpTf0oSQwYQJ2ENOQK0TUAa8i1FXJ9gKLk6b62GuCG+nRAA5ZqSacGFC/xggA3v4rYUXgAq4ltLDvMqoD1ONaxMNXYHbl3VhIShxeEGIRnIdAH0GJhtKP9LGlJCwjPenYg2tAhYBvQrjvOzbCTra0WFrvDrNp2t16AbEp569Ww4u9runiAOM96U17wCKiIdOcEJLAd8SFgugf/6ICOoOvc7NaPFxAALAMacNMhEvZZtAVuYnHLAN2al7e+Xa9WXRTTT+UihJlVQHK5wZ78Qmq/JNVCIxCQD5Gmg78E1q9XKvBX7+ogtlAd6GHdq1X0qhfCk20vhUcK3w5WB7KZNWHMehEAAgDYBX3tWAUs4N0dCuBd4w1VeS9MVQnDuLaQleyM55jheIHqFjQtp3HR2rFk6De/RwjwkMUVs0lSMjRMBWiDo/TiG3dTxlJOrIWrzD/hbq4WPy5nLY6rgM3WCC2H+q+RcbqLZHBKqTykxQISXEkB4IJ2DsbylBlrZ69eOc92q6xl7wLmPnrYAT0eNLuEXOAik5jAylAz/7v+agFHs7mSN8SFxqLMZ9xROdNU3TOn6aVlHQ+gx+M09C8ie2gg+SLEI8ZhVxxdow7sgxcZgDMlIZCBJ+sF05/G2aZ73c0aA9tifg7VLEgt6A9GQMxBrsWRFe1q/Toa1oZCqC84MGmLajKqBB223X7tbQ16Otx++iaMCB1oEhYltrq4QPfaM20kg0/EZ+4FtTlVDE/dYgEorqQFEqBrc3Sb3PQCN8EvN+6Dr8jc80k3CT1cqA/rgtpHFsKZ2UNmiWuDRg+w9Q4rHaVOKtxUBh+5xRJucjqKFUZd7qM5p6VxXmQcp/RetI1U/WHZ3SID/a4kBOw6H5GnvEolH7qphP9t9Psw/CrI5uO2ZxdzexM4IBdvNM6j7qRcZHuHNAj6/pJun6KDvUooHztUdDqfpqs7WlyJOpBm/pWau9rtG7+fpbvhcQbgWi8GuJ3Z7yP2v9+n7IJfBy/1ovYS3j0+MKe7moUM7Ylfne5DpB0Gm3wVlhT+OYHfvGsI73lTYOwqLfejTOXbObdnHNZyDwC1H491u9OMFjzv54qlJNLQ+6XzupcK0nvPjgVkrBwOL2E52t54uiM5xEOaO9xjP/tYzGLr3wVAOQyQe+CPhPfabwfou+8FCQgKFonPnvQ5927V45zV9VY+L8hq91gkoOe3Pkc+wd8O7uMfFd/fvxZy7Ar/ahcB1icLrcAx6ed4uhB5SBZvlLdjeUF9//YKU+J/6qB/FUgK/ed/oUZf5USAhAJ16id5cddqrud+nTI7AfcA88dPOLIXU4WBpXCBMQgGv0eDX8BBxfeBeYF8Ead8iLaAMud27yZeiJd3N3QND3aDYDCDS8gFGuh/ELAKDmd+eiEfyJeAINZ6I+h+p+c1elF7/EQHMOiEXtCEZRgAULh/vFRTO3gVqCdEJ+gLQSiHRPiAMBKBGoCGTIhne3gKagh+pbd2UQKHMuN+sMZ+i3ZvQ3hZARcLYVhJE+aHZ1iGNuiEcUBOAyhq6CeHnNV8ncg9d8ht9PddCUAqk9iHflgK/4Coe4qQiX+Wgp3oX5CyhSV1gnJThKECialYhpTohKzoeeW3ZYw3M+4XDY8SeVoAAOvigLKFPweQdw1wik7oi0sIjIKHiaWGdsN4eskncQRgAQfQFxZHYi9gc4zYiLPVASzITwJAhjFYjTdoif7HYXx0AQFweLBIjN4ocQFALvkAWkNyBRuAAASgfMXoiCHXbR8QgfNCg/FIg9cIdtnocgUlfmPFdlgYdRYAYCNGJAQ5d83ohXSmkFzwZv0kif4HkTEokUNHkX70OltQbJsYUHKIj0OCk/lQkAdZk8PFCm/TBQbBTySghPi3khjYkiZHhe8IFlUIi6inkTGHAR6Zk/8hyZPOeGn31wUfwI5xxpTAd5QVOI+6V48kFJNfkI/DmHoniI9UqZM/woiV55OsMHBfwJD91AB6uH9hSY9dpX0vqW73KAoXGVNyGZUSxwEcmZMboCm3CB+5+CxCBwZX5U8pqXt8CXxwsAVXhgnBaFNgNXzxJTs2SXUfWQEOSFYJCSNASQoIhpcOWXj9sAWbpgFaqYoBQFNFkYZd9WO/9XeuWGqokCNz+SmyR3cSQJA4aZCOuRW4qDEHAJuKlXfu6HkSwEOhQJnpFQBMZpll+AEuB56RJXiCWEKCeQoZsHKi1nY+qHylSZByFpf6pprTIVXrcJL+JgBFCXZstg+W1HP/+rmHZTlOZ5l0gBlZ5okKC+CU3HhZdNcBObld0CdeJDkoEeB37JAATNWdKacBKaZ3t7mV9aUAZiegBKoOhFmYIymHU0mQSBV7Oidfz5h96+CaQxmdRpd3lWSbIIqbHQZ2wFmRUBFqhSl77Clx5MiMXThYFFpnVpV3HvB32YmXPNoFF5BZX0lwieebUBEWDLqPJ5gBBLmcL0ozMTo3BvB17fAB0bijQ0eKvEilxWdjJkeeAICl6jB6GNmI/MguUwlQGgd/WDk3BzCjUeEB03mjCpejkUalIUpOJkpwdfocweClHmSMY0qmMjWX9ekcGtBdQ3mn3iallASljdoFDheq/5xmoMtmHwIQmvpIXMQVczyCjk4Sf0wKI9iwIuhlSaVqcmvqT6bqBVbqciOnpSsiBj7ZoAgIikxiqzGKq88yKizSAWyachkaZ8LqBYGGoN4GpH5EJUOqp7HYrIYShxM6XBTIIh7wphd6cBpAf3qprVtAanPqbcdaJWmpPZpqiOVqru9XpnN5AKkKHdQnjSM3aWkqrMRqX+F2AQL4AATrFwDIjZVnqdchfFiQKRtrCIQaodcRi5q6OQfgrlWSAnjpq+EGrzskr/O6BeJkr5/GsILGLK46l+SKby9DABwrSDq1saH1p14Rcc45n7s2jX9SrXjZpr2mVA3gsl5AU5DKZ//fqm4S6xwoWrG4eLHXdgw5sVaDlE5hC7Z58HOxs6dziaa+RooWkLLApkMKq62KYLUUpqVzq3LECXXpx6wOEAEYIIVGJUg7JbiBu1bHYCRBtJ7Rd7OEqjXAOpQt22sN0LZP63q9tqp2+xzqcbPQUojFyAt5MLgixFZ6BbZYtFfq0iQfFLCiSEQwcDldCaeZhrmUC2MC2q3EtqD7uj2jyQ0YRABfC7wpJLwDSEjkA7bpZLiqm4JFGyW9ZD0n60+JSrvTex+KkG4DmGVjs7lfuKe1kAeZI7rEG0Luc0hFFUKAmxPGmY6sqzGc+rwGO7nUK7+AIaB2akQZMH7DtWM12Qv/eQC8bSW815IjIENPNHS+O4WmLpq4crm5zjtHI4CXRzu/EzwSAji7fkKp22uYbde3hDu+A+hTBTxDRJVC6VQIqbe8zJsXNnTBfgK7KkbB4ZoZakTDNWzDN4zDOazDO3zDkyEBLWwK9TtS+jpWUNmNA5ATpVtDAzxUo9NIJFzCCIwBThawBtjAG4o70FtJDQDEplABC8DDYSzGY0zGYrwAAPon+AsIOcGxbezGbwzHcCxMvUTHc2zHdUzHrhrHe8zHbowFgFCofrGqt/tSS5e152oMmePBTBzCBDzCwxvFx7Akscq+I4vGGjSqPITFI1EBotEMBBBKoCzKDTDKuGRL/5dxWqmcyleQSq3Myqysyqp8yqYcyqRsy6XcWfm5LGJAByjyQLURNRIkzMGCHMiyIdiQQezwARCrVXmKtwwMDgswPqL7yObTXOQ7wghsAIkSfxrsCim0VSlAiiTQxTL5AoCQKsbERPrSJU/kzkHzID+DQLlyGQSQAJfsGlewOL/cPBEEOXcAQXlEPSWrzIlHyCPVpc9sxfdTCGtVvG5lPiJTOouEV2RzLYS7s2cbreS1tCPFlWsrvajgEPDwPwnVHwMUOCjdJYsxIa10L6RczqSQAATQNPy8MMH8z84zOZSzJ/YMFUKMWBJAABp8h977uyJUSI7cXKlTDduBV2RAPv/A+yKVfLO6mliOG2eQ2yIYQNL3sionPUBP9Dey4juDQzgJhCUwzSILQNMO5DTA8iE4ndMUVMzGPDFytA5Ue6COlSnejH47i8Dja83pMxbwRAxO3TbaEb4HMMlEO5cE0NFVxZUaOhKgnEQBhBh9Q0CxMtYpvdJFcys7AwgsgiNunUf9PMwiQtd1bUFv8jLrQJ5S61UaMNTb+4AOEEiPNMDku0jqsz7osTofjLx/1GLbawi2VaNbHNlgwHN5g0CZTStkAs//4jeCA9pGYzhNdR/4u89ssDB3ktrQQzURI9CVE8h2uZS7tY1+DQ5JvFOsgy3u5NtMvVxkUz49i1QLrcH/BIDPW8Vv/RS/YHAj/rNKSmQYS0A07zzdYCJFC8UfxwO3UdHLCcMJzSPX4R05eNIh05MsVj0KdSrbV/2qyhotNuTQ67TbLXQ608DiaWMu7eMyYYsBYrXRs3XejcWVHmcBukwKL7A7rJLgCr7g1O3ZCN7SaI0lGLDc7RAoy2PTHgI1/ozhJLLaSmHXvcLfoyCABz1ZNuvXtPAyhLvb8Z1IpoMW3YE6ZqMjoxPVdFDjgxrhlIWt/obFl1HS6szS0A00Yw0rEsAZf94vRGPWr3Q8BC0VCOPLzAPXUz4secI4lZOokgphN5Kem+u9OvVISl3YHEAcJJUWcMM+5BvVLGEk/1/OwhfmIraHpVjg3CZ9GOwMIX3OGX5O63/u52LyNwFz3Q1FMFV0MMrTK08O5ZADHMDR6AxD3le+J5aj16x6Yfj75Y0AvoFLNhINF6C1s2iBv6cFAMDtyPa0zdnw5WR0Y7zKdQF+566e59ENJrXu7gny7sGj67tuPKmyQH+B6MtR4Y0DzHiwbMQSAQsQQsY+QRpezK39JlkD1DOWE+ydxJnuyNxxATVgAgHhulNJCCXjDUGFwEf15QMwsFKWtEPZpqyOJRUBCR510rDuJWFC6/AO8wQC6GEizy1d70kT58EH7BfE78AMANMl5cHxGx/gtwmAAf5IALnxGw8TMcqOIv99ssyZGNNVVQEZrMEPv1NOXDoO0AclxQEmwAyOsguXIsJgO9Rv/iwka2fsipJBmdYABgkb8G+urue5/kQGYvSzfiAxjyAxj+uCM+geRc85b587vyFxAteEEAnFISxw0BsiRPQQMPF4sAIX0BsY41QfQgbEXEFYEwrkqZt2Jn6V/tjA28TpEwEWsCTpkAE30wyzaB4yVL7Im78NTJ129gGH6k+3b/KrIfcetQEkwARYICEecAWOYfwC8CXHfwMq4A8SsI4CkAF9zzvU/wR+PitP4DdXoBH+QfyrAuFLY9py8rAIEEII0CNh6vh9kIZxPwPYIxjTtQE8kQ8YQGKHIAT/CPDve/ASIPCIIwk8JpCqK9seWaTIc3QFN57rO9/7PzAozEAGxiMyqVwqIYeIIRKZACZW68MBwTgcH6/jhrFEEgdBhKMWXaeAqIHAnNOXE8JHqN/z+/yPAIPgoIVHgECDgOIiY+PGhkBCJESkwOOjgAfEJUKDxQYCwcICwYZF6ALoBoTG5imGhMTnKUFHqilkx9gjAsRCwiWkBzAvBDAGaO9iZ2PzIgSEH9ACRIv19YkJyfbDhffmxQOCgEPGRncAAUGABdcHBMJBAtfB+cUGxsVH/QW8hQSBfBcAEOjGbQSKbNcWqoAw4yEAaRInUgxyYEKdjHScwKHi0YQaLe44/3zJEwADglkbCKyxUqUKFCgHimisqURUxZw6KXZoMGhQA0rOhjZAkDJRAkkpk37yUCrBgqKiBCDwYGmBhARVgXnooDWBBg+9gEnQoDUDMqwCYJ1KIAEeqQ0LPJjy0G4YgXGgLKxtIKnoUGdBd+KgxvBwQm0Hu907cCPghwgbKlxwoO4CggwfPmBGgyEAsA8ZEDzYjDnCAdIOIiAAsA9CuMUiUCBmCIHAQwU2CPPu/eOtzeBGnMSE8hILB5EOSHrpcOADhnXL1aiZ8MDlxygzHQi3acw3+PA57P5kACnwUGRGLSSAlzTpuAYN5lL1MBfBewm/8ANboKGdBh1oYMlXZf8lIFYCHcTCngW13OcUfr8Yg98wwJTCniR8WaIhes9Ew5thtVlzAomydTOaOgFtcIE56KiTijc2YCDPZ6y9YwFn+mBAyYrerJLOAd6YSKKI1zg0Q0TiKembE91p5MABB3R0nHXWcSBZF8kFsAl00k3HwQMcVEkFHDLR5GQdMy25Jm8a+DTIeR02IgkinTz13koQNOCBVRvYR5cAb30CSlJySbCKBhJ0IBYpCOCTwGgWSNoWBg3oJxafp2ywJ1Xv2fUJAUlZoggzcioyGIjVFNlCibJ9YwE00FgAgDnenFQQaabBGgEBDoTGT44WRNnjPhuYA4CQQ9K2akMyRGASm9H/UvTOAWjWwWtxE0zhkZUcEABBcg50AOutAVAHppjXfaQtFBBEwJ21TEwAAbTS2itNAm8yYOoiFvgVSQKfNLDBe8DoCQGflvxJ8CaZiJUBWeMSkKhZmcUiQClUAdyeXRBcmtmB+ZrS6TCfriRJwA3GaSo0vYXIbAolKrZNjBgA0EUYtwGQWQAL9BLASjekRmMAqemzShis9dOjN7Q4EJtsMcO8ghP3Wj1Rk/EygQEcBlBpgpUTPIIBShB0gZKkuKShLkxdR4ABRlovQe/VdffR08pygqKIJKZohQGfeXkQVMIEz0UwgB3kBZ9/VC3wwaEECGBBB5vxBWAFC1igOQGx/2Aq6cX4bconYI8mBU+p/KJK2MtTt7rYBRFYwJg3LealYjSpBPQJGutIkBKtppQSwZYWxLhPZuEkG7VCMPtiN/RCfEBABHI3IaUUUXBbZZhXQvNuchlEOf5FbLehrQEzWd9EAtG7HwQ0/AqwiaOXTJLS3k4hvIDCh/c9xqDMAQkNlKIUG4jFIwjwiQUIsBSsgNXHhpGSU8xnQxQEhl9Qtjf5ze9DrFOV62YGuxTEKEYZ8EZ7TtgNBzyAEhfowAMqIAF9LOAAHQjAA2aimXAsIEYOsESMlNW8qT3vfUbUQQUIELf1rYYAcIDJuq7TvSxRpwtfmI75oBiTKMFrfQNQ0/8Rw5gDRHAQEQpEynsI4Bc+JYVPJRvFgbIiOf24xSoZEND8BCAgBeVxAR2wBcYCJQE+jeIXoxhcqNyIMckNg28A8xsHV7eT1oXQVbQrYQmtqA9v5AEMm/GCPgLAmU7mCJMAQJryoHaQxBBxAWIUYw29eAQMxER7VKoSus6lyzBJETttk8kS1wfGV4pRKGUsGDIrdCA3/sk+hVRULKD5RwUJSAMBmqY1AxSLP2ozmoU8ZCGZ6UZkigowZWxZqqYmtSFdEpPu5OQmRxnPUr7zeJKZXRDZuSznuZKYR8yaF2fyRO15RIq8RNdBwzQmbk2hlqiR5Rfp5s8w5rGMi0BZMpf/Kc5vjiKaHo3mNEMqUgV9VD8cPaQ4Q0bO04XCopLUCSVh9jqaiaCemMyRPDczyp3m9J01DeLyVjlEZhVxou77QHsgSgDsaS8Ct7yCQtHFPexkp5YEKIAsz1Avo7qPjBatxEpDJk5CnrSk21TUSKdpVm9+s5nMVCkyK/FVdH5QnQkZkk3ridOemgaeeXWnEPdJ1H5y1X3q8yJaHOrLgmJhqmOiknHgUI8CGGB9qClsGI15zI1ldKzO/OZat5lWaob2pChN6UrB+tWX5iSmzJopTf96PL/mlKfxlG0qTcRKfmL2fYcV5hPKpC2qcktdC4Us+mKygQIwN5hoelsFemtE/69+FaOdHatpS0pSao42tCYN51w8m1o6WZSukwShTEXIjdz+Faf6sK0ob4rboArVrikoqnTtNr+Ack2xw/0aVY8Dkzc49KrMZa7WeHXD/LpPsxy07nU3ylHvchet1iTpWk0rXmWS86un8uB57XtX2LUzr/KE52Ze0NP5Miaw6sQvg68WJesZgLlOdOi2uFUFHa/LCpFtFwQmcOACVAtNcVhwjKFHXYtC2FMalTBotStNbmYgUKT9KFY46ta3hhVgcwUxTNH7WvWuN5Ul3GsoOblTL3zgR++11ZvraRDdDnWwSYYeQNEkZOb2QrhNXewtG+pQ7chhADU+cGW7o+A7R//PwQ/mbDKxG2WsbJek2szmgDbwx7Kcla2xCOdY4Vowua4WzK0V85hhSzNMPiAgASFeq6NTkACgZi15WUVkorOjftArHBAo8ZyZZ1cYM1paonGXnoecvqUWp6kN/a+23vCG4dYyz4dmbqJtIo+tFvteS97sSkvm2Y4GbAyi6MDkzB0LY5DtFRkgYDsABxAPrHs+oEbteD1s3jDblUgmeqcWNgCADsDjAWnphwW25KgXrKSHq6jAaF4Qj5MYr4TBrm+di0TsbkcLINXrzpCxOoAJzKhMHXl2TCZQpkHvKAl7PrBzN+IYjtvN0fJrcsE8+1m6+EIrE/KFOfQIilA9RQL/k7MPMmSxgYoJ4LQp5fB7SO1SU1fEtUVS9XovKYqnXUByrfGCOKqMAFe2ORpEwIHs3tGL1gBWldzYLW9pbjVvFVnbykbCt0yevVoSuKEGKNMBaLmEayM4I3don9zr9u0H47xCOhfUNjHWIIoloHOhsKYl/sieD1hTGKFYyaXC+1aoj5q856Q6Rax+9ZiRmNUYoB2UNoBTDAgAWPeIRsBuow7jOQAlz7G429+e8VVtPPFrqr1NXl54JASeqYOu5crTt6OYHyHk1EfC2xYQBuNbzeb8olNYdd5Rspl0QXr8NAZSAZZMI1DWtxuQo0yqc9KLyvRTdxmqV0/mrAdRIJwE/wdnOICwABFnkEvgxYq7+EgvcB1Q6dPw1UbxcZ94vMOZ1AHhUV8EbEFwPV8U1NJMUE9GhBwdzER0SaDVLN6jhd+TuZEEjEH5aU6CoB9dJAi6/YPPfEtQGAMBOQq9bdk40R9G6RvqTYTqiUg24JWZUc8XPICwbADx4FBm2N6PZMA63EAGXgDZsEaQCAl9vZ1gxZ0JSosZXAQdEF62LQEAQAP5RAlqrOFM1J0F3p0S1FoYXo33fR+k5Zz4WcI2BQT5YV7nEMwfWUKA8AXntaBSTI6fiN7odZn9lRFrVV3+pRc7lRBrfAs+IA8mRgM46MgqYOEq5MUBSIo+HIvFsRORDP8bYdUhm4gGAQAAE5hhTfhYlOieE1yEFQSHA8jiEcwLBCAZK0oLCt4cznnKuDlQW9BFKOLH78Tg33iAOaiRKZiDW2jO5uhcWNWfh3UQ/okY1sVWjPDKjHASFc4IZ8BAEEGDr0XHCf1aOJ5Q8K2SiDXEKgbjmhAcHBqB8okcyCFaF3UHLx4A9dhj922jXKmg+DlFg7jFIEFAO1SjAGRANN1GgqzFjtCH/CFMqIla1End/aXTPI7YQSgP7G2SD/WVSXIS15WENzAg8MWj8IlYBBKkb7wDCFbfkFnLASCa1lgfCdKktyWCQRZjG60gIbEVVpxVUn4aWpVUR2nZ0wGhqBj/ZCTyxCSu3jeOgJnNF1fKmYvNo8cApbRQQ5AZmhw6yVkaWchlgFjei54MJViRk7iN22eVVZbdJV6elA9yWZd5JCQKQG9MT0iKpCV1pWE2ICp+IczgQVtKSwbuZE6iiQOkJZpYX2MKo1BuI1EWpfiZlmd+JnhhY8mM1yOWEQGwZW+YgTey3v7VlEEcJouRZGA9oIicwWUam/XFC2RSVrxMwBOEHLfdpm/ogkE6EkIapRvNBWguJzjNn1ROZXEyZm8kUUgeISqSJGzKGRcmJm0WiXQK55IQ3lLFC2XaRAae5g0M2cyBp3gMYwpmo3P+CXNqmHKKJkdqY2lyEHuER3tU/2dWuuZWZmfbcafU2Nd3sKeSKJ8BcEDlkWE/Yptw+CY0rKKCImh4VACHeNhmOp5zKud8glN9zl82aiNVfmdgOsFg/qdWYqeAYicqzkZ3IsYwWSh4AOcNVEB7LNX1McFuniEdaMtMJEAJ5sAHXCCN+kYGZKhm5mFRImdyElJd0icjhtpoOmJ+ys8/KMn0HMBqvuhrBihXfunFCVuMHoZADumR8gbhccAOeIUa5mIdlKfLzQQ0aIYPcEDIpWlvTM5QbuYyOelGyaeg7qV9OmJxTg5qaimKVtJ1vmaJtWiLwaQ8Tk0GBqeeUoQZ+oADZEWs5BicGsFk8qQ+XgGQqqEEWP+qDlTope5EwGRmdY3oXCZkfWpZc07pho0odEKiBSTqkohGB0kBsAarsAYrADiVsQ4msqZABhQrs0pBsQ4rtAKrGtrpqu6EGaIqkdpCBsAADNSiEwzZUo1PBGxrBowCMOpBplZrTuCoJ8iHfOhJULzru1aUnKBMXOIqvo6oanWIUFBCvP4rvMKrJxiDBFhNB0RAByGgwi4swzasw3YQxFKCxOZR/FSsw14sxl5sRJ6rulLEPrKpRGxGBVRAB1SAsgkItv6BjXZsRXwA/2RsDmaszM4szdaszUKDW6Qsy+4sV+0mZfFGj/oGniKazvKs0R4t0jKaqhLGkE0AeBhp0kb/rdRObX6FHMgSxrUZQHhYLdV2rdd+LfQUadP6xthu7ZBdLdimrdqurbWWbW8ErXjkKdvOLd3WLRCkK9nCnJLgrd32rd+q7dAemHhkrZZa5t8eLuJKreHWqN4mqNwmLuRGbrXuo5LA7d6ereRmruay5wc8rtk215pwANRuLumWbjB6bngQ7pp07pAVrem+Luxi1ujGbeOurnrGLu7mbn4pqOtShOWuCd/qrvAOrxGt7JK47er6LNoSL/M2r7SsabSobrRQrvNWr/WmrrJJC/KySfBer/d+bx+wruBKy+9GL+aCL/qm7w+I7YEt7/HWbrRsBuqqL/2mb/eGJ6JdzdLW/y//em/gEpnVbO/zCnD/FrDw/m8BuO+alK+9+KzTGjAEC+/8RgsBa+/5RjAGm+79cm/+2s21ZjAIa+4+9m5vVLAFt28IpzDkLq7VMPC9iC+RkbAKz3DU7m8Aw2/d2DAN7/DXKqgCDzC2vc8G8zARHy0MF4AM5y3oHtXsFrETG63PJrFvuPDVHPETXzHPDrGSiK7WpmfjqlwX53D2YjEZX+r/GoAU6wRkLui1WQFk/nD8hlwYlzEdt+VWsfC97GPIBXH0ILAOqEEdB3IYqhy0aDEH7/EFQ8/+/qwgN3Li1VgX6/ALI7AcH1G64ul6OrImM9r4AmcaqykiL7ERQe/4bv+yKccYokGvEIdyJr8PAr8cHJ+yLIsRJfNx8SLyHBuRHtvyLPcyMRFeBx/RLvsTMPOyLx+zEfmssqHxEe1xLh+VyoUyEiMzNR+VNJuwvezjM9tNLe9xLFczOG/xNWPbJ/OGFR8R+17zNoczOw/uNV9EGI1xGImuOrezPQOvNDNzGF3bN9fNB+xyKd+zQJcwLvvT0K6zJYdyPw80Q+uBHi+oUdVYOQOxPDe0RUsDME/ARK+JykmXb1b0RYc0ugYz26ZzAYg0SusBSc/tP6NwSr80D+ApRB8uJMO0TRPpQq8tIN80T/e0T/80UAe1UA81URe1Ue9B5pCrUm+rBKCp3Mn/0FIrdVNLIOQwUFSXa+Uctfci1RZwhJR8dfp0dQJs9PtQw46ANVpvgeQ49Z29BdmgdfqkT+DNkVY3bwXYmpT8nV7vtV4HHu0VW+WVHF8P9t9tgQCQdd0QgWATNl8fIFvX9evOAwQwNmX/nUDWI2YtANlUNmUHHq/2llvnNWc3Nu0hNmSD7VqMdmUHHmBK1zyodmXvCMdOlGaLNmzzNdlg9mlHruJwzW1TtjrMthi9g2//NmF7Nma9tnEzNgYg3m5Lrhoud2cfqD8RgW1LN1+HilG9xXVjd18P4XPX7RZ4d2V3jj8pN3kT9l8TUwt2d3obwL6Ft92ixWirnBUYwH2D/3FsP3b00Ddn2zeA4/doY8Bnv08FLDZjB3h+Dzh4yzfYvkNl27eAT7iE6/dgC6Rp74QTUXaF/x0Ydzhlw4IYLRWHC/iHmzh+W/hgN7eD2y00lDiAk+oVTHiIt3J/u7eHm7iMz7iEM3bgZXhFEMF/6zipUnhs63aLe61/D3aF53eRO/l9qzeSW82Bl3iKx7iM6zhjU8I/RTiW7ziPq3hfm2iSe20LJjiRg3mWczYGCLe9UAthf7mar3mIF/jV1F6Cy/mcn/hx23iZTy1HxPmV7/mcH3eDs8mS83WTEzqYb7n7VLmgDzqj8/iWA/mfN6YuoPmkN/p+R8+GM7mkb7qYN/82fwvjZIO6qO84jTd2wV461QoUqqe6mh/3lLPJmee5rHP6Xh9Ara9ucTN5rj+5mNumq0/tpwP7nIvJBCi7rl84QrMJniN7qFuBsjO7qvv4oSuJZse5nlO7txc6c1t6sdvjre/1ol/BuXg7dYB7Y9sNdEQ6mIuJvFf7nqu3uOtBoCP7jlfHsve7tV/7XhP4uCdttCv6vovPReiSbyY8nQ82mY/lr5t7vPs7tdP7mou5wFfxsev1jOs4vyt8s194tg/8keY7x3s4qaLNIxwAdWwCL8BNyBuAiMuYe3/5TMw7BzjkunNAdPz7cbe2wUa8ueeFURiFEq1Cus9ILzB7QET/OV+PPMlbKI7vODJcATwcgBbEw7KPBswv+GBDgJ2zicnnuIznPLicyyZUhxq0A7Oj+K5DfW8IucFbQdX7WC9swbkETGogwILS/QaMuszfe9Qb3z4IOqmmRgZUEdlMQDwsx7IbAAJ46qpbNtC7Zc1L+s0ve3Kk/bmQjbVPfmHXDawD++JT+yaYQ+c39yVcwRh0/F4TQKkPPo0WvqJLOttrvpgQwQboEgdIScP3tZ+LveGTqtnrEudT++0XOWE/PLR3N4B7fkhsAslpPuM/wd+HgpgsPnPntOzfJu3HutiwPM6fBAZQB52CvbA3NtzzhtCnuRuffUikBtnMP/bHvETd/0uU5HlexAoygMDhQAQ3btyBTBlycNg2zUZtE0ug73zv/8CgcEgsGo/IpHLJbDqf0KhU+TnYbLTZbAPhTDhgLwETHmMQJe3kajtApkkIG6v+QrphDkQ2WcwsZGpzNx9wRwJWbGoEaBgYBBlgGSgcXA4HMilkjmuDGBOGoaKjpKWmp6ippVWDBnWOYF8cERh7eWCcWq5zBwKqPB8QiVeddW5eYCe3j151g2+/O26tM7C3Xi4tE1WZExsHBHydVxgV0efo6err7O1Gw8SuWiqRYZwuDsgTaM27c8LphA2qYwdPGFuxOABqRmOQm0LnMshRJC/XDGQwHpFJwUcPgg2fWv9hgOiupMmTKFOq5CFwzho1MSJNgIAgwQgXXiahETTuRg50B+DVKKbGjRo9lGTBIsgL2rkJGKhBtVAnTAucKVZoOfOppw0IJFeKHUu2rFkkLeMRjGFhw7eDHy0guENAl9caZNIliOqyqgoEgNFA+JTwkSCRGdJV4Etxwhgt+ry4lYVJEIKQTc9q3sy5s8kMjNU6O5BBlukM8gjeNXBAHYxWQ1VXle2MDYYO6tISe0lQ35ffqofOwZDAs/HjyJNL+RBa9FHgzRjWdmhgnW7htCNfDN7Kzbq9UmmbFr/aQF7l6NOrXx/gOpbU5Gn7u0J6HWhq8OPLZ3OguOvmwuWnn13/87XhFHsIJqjgSve5JOCADa2GgQTrLDbQgxDCxhoH7LiXGobxFdhGYguWaOKJ6AAYIIR2lWcAAe1Mgx+LvIlYA1jsNBgejTbWcIA5KAYp5JBT6KgIbyxqiAGJ62ig4i40NiRSa+14mEWSPbKGI5FcduklEQc8CSWIWbw3HJXtGBkPlOKNWR6M7rw2I5ktwnbbl3jmiSdzGtYYXJmrhXmSh9ghydNLSv7kTgZCNfagPy6ah6aelFZ64gJiQuqmpnYqGicBfWK3aZZ4+WJSB5luimiNsIFj6auwKhiUhqqOqaR/J4VJ66qjwnZHSuDtKiqiGobJYazIJoscDI3CFikb/wCl9AEBE9EaoLWsERCWSboKS6onnior7rhlVdAttuhiMClKVaSK7hXCHKtSBgQ0+25ToJCr775inXuvSLiKBZq9/5q3pVgZFdwKCdvy6/DD9qmrMDlLNqxSB/VO3Ia6mvk7cS3yQizyyNFwMIbGtVgsVjASK/xIuGS1SzCtjjBJ8s04m9Kuu/Q5AvNZ9LacbsWeLTDGzLyoq3LOTDcNhQMJHG1Fo1M/ksDSQA82K9WsPfJjcjPVMjV1jkCAm9Nopz0FB6glYAAEBWBQgEAJRLIe2wa4HWbcBVhRd77p5cSaG3LLbUUGC4Ss9uKMN/GBAQVEXl2QkUse5AeVFwB44/+cd64E5JFvbiLomguZ+eSep666EKSjfuLpplcu+uq0r0767AseIHvsltfuu+2Vu24i7JTv/vvxnd8upPLF947882q3znsBwpeYOe7QZ08y8yjqHvr01WsvPsTSN0/99NiPrz653J/Y/vDBry+/w+WjSLz9xs+vv7jvl+h96eYL3/4GSKn6vS5+5ksfARfYpf4tyIEKuh8DJ/glA8LPeQf8HgU32MD8neh/CmSPBDlIQvchEH8YvCAAS8jCEkEwQS9E0AhbSEP1WNB6J8zgCmvIQ/TEkD0gBF8Ph6icGy5ohkf0IBGXSJYZ7IB5j1NcEeXFvGawxwDyIp4VmchFlHD/QHKPk10YCyBF5IzRAJiTHQdAV8bjnDEA11tD5NrYxTqiI3N4DB7WPDOBPOYRjezpox9Pt0c7GvIUpBtkCI0zyOslqJFKPKQkVZFIPypIkI28JCQLMMlOngOSAkQPJBd5nFF68pSpqKQjNTlIF2YSlbA0xSuTmEdSIkeRsczlKFR5Plfm0YS/1KUw4ZBGPBYyOcXMHDCROMxmLqGSoVwPNE+UzDk685pOwKQGTaRNMlLzf5HDpjiZ0LpjilKPKNJmNMfJTh9gko4IIh081zNGb7bznmCinjnRI0hABqmf+8QnNsNoS/Vss3gFFag4+8glyDWUkwqNqA8+ME8FVTRB/xeVqEY3ytGOevSjIA2pSEdK0pKa9KQoTalKV8rSlm4mAXe4gwACCgTEceYDAogpBEgkgZ8ZIQNAQkcGKLSDClzNpbH8wAaoFVOa/qAWnFmAYAaDhgDktAkWsFk0uMADqQYVqZ5kTig+4FR2SJUHLZCAd5iAAZ+iYgOm0oFXwYpKsfKgAwLoAARmKoCyQaSvGPCFXnVaAcR9IAOAxRHLlhTXdpyVBwjIAEx1ELXA7kACY4CEDhYrAAu49RTf4IEEEGCOoBGgOBXI6x0yQC8LQGABELAAAc7WWs1KoG5jwFVfNRsAjL2Mrqr7gAUEkIDiLkCqgTUaDmBrgcUslwACMP8qazdgAeaABQECOK5rK8AF7SKgrKKQqkxj+4arjgFxBKguphKwgAS4VrivXYAAEPBZU1jAtTkVACMOawH2ZsC1o00uSBAXEwnsIQDuHWqCW/DaFhTnZf8FywYSIAEBbICowOWcUh8xBgGMNjHMIYlrNwARCZBYriMJQGCntYOoWXgH8zUJch0hFxjltAOk3cGESbCDSbiXBxuobynaYgYMyKU9uMKUVEmUVcpiYLPffW2LBdACiFALxxBZwAYqEFm5fjXDiwtxV09cAUAAIgaMcISRvxsA7pIosBVwCkwH0+MTu0OqJOFyf3a6AR7Uglo7+MBcDtRWdcBVtFE+WwD/ptUCIN0JwXDicgUIEFS1/ncH1GqBmuVyWLfwFsyNE/MOtFyISR8XcfINMqqZ/GQdrBhOkI5ti+3s2BzvYDA51bKfSeCUDnAB1gHwrDpc29UoY3jSjXY1kPaiA+7GOajyvbQOMq3qUxfisIzQKqjRJmq5nli4QbWACs4mAQu0p887qEWcW4wD+k6bzXeG96K5AFNBkwjH8jW3DvZAaigLmRSHHjUCrOqUvo7W0WeLWrOjjKu9SjsABKA0Aog6iYfLdtuM63YA+t2ewFY2GMONWt3mUtwDYGymcoYRrvtKa7PORQAwT28HvNNZmD+izdCF6QYS02Gd/3sUwy32B8oN/4EEWHioti4HZSNN4gWEPLYdePivYgvz4cLX6E3G+OIUvVmu3/YAisIr2Ht76uNW4OxtDnQHKrCAD3SgAwlAtzuGvgDWLgBIaG9zBBKg6A9EgMokUSvfKwDeKKwdGEQ9LGmujeG+K5qoFfh7Bkqt6LfrYOZ8b/Hitc5DHPvn4pwP/c3i3hali/70IvvAcVHP+ta7/vWwj73sZ0/72tv+9riXpASiTgISAVW+bW6qDugl5UXr1z91b/HbO1Bhc8y58LmP/hJIf4edBzsGbQVJbJ9s4eq3XftVjTiUWVv6YHChLV+WvvpDgQkgYVa4G8ANoPeNqVazdrQ5WLf4501+Cv9NAkj7t34CaAhXlW4dEHTBJluPMGFatlQ5MC2X4R/7p1SsBQ00kVluAX0DuIE8UICLVg7EFmzjlV2LBlPYpQMGloE8xn9Gt29wJVMBw4EyuAQHoG+Lpi0BRxw9Ri9nU3PQQIEriGMVqAMvNnwHMoNIeAQJMBdnB2hcZVX0VVhbdl9r9wgOZlQTt4S7dwaS9YNMKF5JGIaHQC0RR2ntwSSD0WG9VYazZVUaURwsw1SnNmoRB1VieIdDcFU01TAaiId+2ANr9YeC6A55N4iGeIiImIiKuIiM2IiO+IiQGImSOImUWImWeImYmImauImc2Ime+ImgGIqiOIqkWIqmeIr/qJiKqriKrNiKrviKsBiLsjiLtFiLtniLuJiLuriLvNiLvviLwBiMwjiMxFiMxniMyJiMyriMzNiMzviM0BiN0jiN1FiN1ng8EnGNtWM0vTZ/QEBWUhA1bHiERHB8oTBfYREBCoAr6hgBiqEAB6ONybGENSFXHxEEDCYFNBEYgNFqRnAG5AgFFhYWBEABFEAiEEABCpAOHWCQ6SePnnEVbCZVcrdoJJGPREAS4OgDNLEkJthlm4U1t4VhG2kEZNUw6MgDCmCQFFAIK3kg+6SRfMiHgbYAAKCRP1CSgTZREKkO9FhVFEmEG/AR2vKTEwIIhRBbxeFaNDFTZ4AGXNce/3ORbvUYDIBhho4gV/ilg1J5GTkgcwjWZP81IeHwEfGICQTJkguZkAvpdwaJAwEAAASwkAEQATjQAQogACspAW6pkERllwapAGcDmAQAAAFwARCgAIUAmBRAIR8AjyupkNcWmbyFlwbpjj0ZDRIZWTjWZ/u4B3OhEw4IGObACG8QGHB1BmPwEVx3gaylE3zJFWewZdkGcWhgmrb5CICBKSc4lDCSZoxwfvUIY/IWACtZkBRQAS9ZlwoZmQ8QAQapAysZAQDAkoX5kpFpmIEpAJLJnQp5mQ2ZnNCpkAX5AB/wlpHpjscZmRcQnvCokJn5C0sYWIAxX33WXt5lbvkoVf9sBpABsJtQCAk6cSD7yI+JYZ/TNRc4pi27aZoBxlpcMV8WgGMfwWX0NZtFB1Qd2HIFGQELoJDcuZAGuQAVRgERkAHRGQwmKgEGeQF9SVbQqS0GKQAnqgEPYJC4AQA0Gp4oepALAJ1gYZDmuZKKSQFwgpcSQKQ/iqPxmQotYG5PeY9Ox4/6OZX8WQi4iQD6VqCBAWw0MVz6dY9cepX/WRP3aJr8yY9BBhg0MZT7uHFDCRhB1oHFSaTGqZYawJIsGQHjKZ0rCp9WBagfSgHMuaeD2gM3aqJ6ypLnmZwIRgEQUAGAugOLOqJNigpLaG6jJaeChgZGo6UBsJ+kGWxzIWj/cNKUEtBTGdCaU6kDQ6lXH5GqdUchUfoGAClVs6Wq3xcY80WmHUCiFtaqOoCWPFCQxcGiMxoAClkBD9ABANABfdoeB1mdMOKWcBKjzPqr2WmQhbAAChAB4TmeEtCezdqo5kCkkgoNeAkACRldFSABN3mpp9AC6LaPW1afKgaqT4p0JMem/9lqo3VhUxowNOEU2bap7CUXiVGgX1mqH7EAGDOV9kqhXSYXxbGEB5KSO2CndwqfKzl50JkB1fmnf6qYAZCoAICsCiCpCympLRmYlgkBF+CQgRl5Jmquj/oG4LmSS0oBD9Cj8noKmbpZQwml/Aiqm0pacsGP1wVrXFpd1ywhrDRRHFyaYleBbrj5kxUaqmQap0l5tDYzkMVKAf6RAIGpeotqDpUqska6A+P5lliqpws5qIwankOXtsrqqHb6tpDaZpFpkNoWtIZQWMdmU4dlXDYVABLAWiXIXlGHG0OFVsSlbY+7A5W7ccVlM6x1NpfbXoA3aqvaW5ErVwnQH+GisTrwAAvwAJcFABpweRngoRewWT/6rDepeqxrubHLlzpwAT9qWM32oz/xAQ8AecILEcUbvJwbuwswu4v2u84ruLWDutLbUmJbvS6lk9i7vdzbTCEAADs=)
- Direct (fast) pathways via thalamus
- Indirect (slower) pathways via cortex
- Input and output (behavior, physiology) specificity
Specificity of learning stimulus/response mappings
![[[@Pellman2016-xu]](https://doi.org/10.1016/j.tins.2016.04.001)](data:image/jpeg;base64,/9j/4AAQSkZJRgABAQEAcQBxAAD/2wBDAAMCAgICAgMCAgIDAwMDBAYEBAQEBAgGBgUGCQgKCgkICQkKDA8MCgsOCwkJDRENDg8QEBEQCgwSExIQEw8QEBD/2wBDAQMDAwQDBAgEBAgQCwkLEBAQEBAQEBAQEBAQEBAQEBAQEBAQEBAQEBAQEBAQEBAQEBAQEBAQEBAQEBAQEBAQEBD/wAARCAEEAjADASIAAhEBAxEB/8QAHQABAAICAwEBAAAAAAAAAAAAAAcIBQYDBAkBAv/EAFsQAAEDAwIDAgYKCw0HAwMFAAECAwQABQYHEQgSIRMxCRQiQVFxFRkyOFdhdpW01BcjM0JScoGRwdHTFhg3VVhigpahprK14SQlQ0RTkrFUg6M0NqJ0dZOz8f/EABwBAQACAwEBAQAAAAAAAAAAAAAEBgIDBQEHCP/EADsRAAIBAwIDBQcDBAECBwAAAAABAgMEEQUhEjFBBhNRYXEigZGhsdHwFDLBByNC8RWisiQlM3KCksL/2gAMAwEAAhEDEQA/AJ6vXEzm9l0NynO0ag2yy3/OdWZuHYlNva2UwLDDZm+Kdo4HNkcjceHJfVzb7rX8YFZbEuJjKs1044b9XmL+2p3IsvTg+Y2+IttUaXLejyWHHBy7gckqM283ynbkc84VUp4LwrYXjmQzbzkrbF/Zh5Ze8nxmO8hYatfsqhBltLa5i28e17coWpO6UukDY7kp3CvhrOaYxecU7OzWG1ZpL1AutpQHFonXlcNUZlxvdXKwhJWpxSEJ2UoJ6DckgThSlKAUpSgFKUoBSlKAUr4VJT3kCtduGo+ntqUpFzzrH4ikHZSXrkygpPoIKulM4CWeRsdKxlnyfG8hbLtgv9uuSEgEqhykPAevlJ2rJ0ApSlAKUrjffZisOSZLqGmmkla1rUEpSkDckk9wA89AcFzutrssNdwvFyiwYrXVb8l5LTafWpRAFRhlHE/pPj7yoVsuzuTTminto1kDb5ZCgditxS0NJ326Dn5iOoBHWoK1dy+26lJk5FMyh5uy3WLJbsLsKWeXxdLLhD0VQ8nt1tBbwKfLIG3UN9IRhYNeomYRLzgt0QrEVx2Apm7LkOSiv7YHVsLcKlhpaCyS2vlBcR2g2689G1LtZOjKpC3ilw5SlLOG14Y9+Pjtydv03s5SrqEq8m84bUcZSfjn5ly0cXGDq91iOUo+ItRPrFbFb+JPSOXEZkS8hftzrygjxeXCeCws9yN0JUhSviSo1VT2O+IVxv2ovMONIWptS0KSFp23SSNtx8Y76rVv271OEv70IyXo1/P8Fkr9jNNmv7U5Rfqn/H8lrLRxMaQXucYcS/TWmwstplSrXJjMqUDsdlOISeUbHyyAjbqFbVKSVJWkLQoKSRuCDuCK87MYxZuLlkKFfrg4trxUGFKM5TG62uXtkPNDlQ4FhRKQSpKAghKEbbm9WlUpqdpnis2O6HWJFmhusLG/lMqaSWz1678pTV47Pa3X1dz72KSWGms9c7PLe/w9OpS9c0qhpjjGlJ56p48t1hLb4m1UpSrMcAUpSgFKUoBSlKAUpSgFKUoBSlKAUpSgFUw4htfdVMQufELf8MyV63p0wteH2m0xUxW5LfjdxkpdlSi0sELc7F9ptIPTyT03O9XPqE8r4X8WzjO9QrrlUhU3FdS7NZod7sza3Y7xnW19a2JTchpaVI8hTaSkbHdpJ32JFAV+h8SWrzeA5Lfciyq4C5aY6o47CkxbhYxZ7pPsM9yOypE6HsUIDnjbq2lo25uxQrYEEG9tQHfeEHA12WDjuIvSoEWbl9oyfK5VzmyrrPvrduV2jEdyTJdW5t2jbAG6iEoSsJSCrep8oBSlKAUpSgFKUoBSlKAUpSgFKUoBSlKAUpSgFKUoBSlKAUpSgFKUoBSlKAUpSgFKUoBSlKAUpUL6sa/s449IxjA249xvLfM1JmOEqiwFjcEED7s6COrYI5SPKUk7Axru8oWFJ17iSjFfnvfkSLW1rXlRUaEeKTN01Y1Rs2k2JyMmubBnSE+TDtjL7bcme7tv2TAcUAtZAJCd/NVV8i101mynI5V3hZhNxvH5LTXi1hiMRVuMkJ8suSlM9qST12QU8vUcyq1+a3cLxd3cjyK5SrveH0lDk6WQpwIJBLaAAEtN7gENoATuN9t+tPFT+DXyzXO2lxePutPzTh4/5P7e7c+k6N2Tt7Rd7fYnPw/xX39+x08hmXXLmTGyq4S7yyoj7VcHlSUd+48lwkdD3VoOmeJy7VdcpyhlsQLfkc5l6Bb2k9mhphlkMh7lGwSp7l7QjYHYp3677SBc44ENSXGedpa0NvAjoGlLCXCfiCCon4hWV8S9AqofqK86c1Uk5cWzy2+TT6+eN/LzLV3dGE4uEVHh3WElzTXTyya8q1sKlsz1R2zKjnmZfKQXGz6Uq70n4xW+4trDqbiUhlUPJJE6Kjo5DuRMltxPm2Uo9ogjuHKoJ9KTsNsH4l8Rp4l8RrOzvbywkp21Rxa8Ht71yfvMLu2tL6LjcU1L1W/x5os5p/xBYxloEO+siw3HcAIeeC2He4eQ5sPOe5QSfRvUppWlaQtCgpJ6gg7g1Q19pmIw5KkvIZZYQp1xxxQSlCUjcqJPQAAEk+avmOa555DZLGnc+8uQkjkbmPJbEHlPUKaS/uXUn8JtGx8yutfQNJ7bVWuHUIZS/wAlt8U9vg/RFF1PshST4rGe7/xe/wAGt/j8S7WXZnjeDWlV5ye6NQ4/NyNhXVbzmxIbbSOq1kA7JHXpVRNXdXcg1YK7ZIbVbMYA2NqK0nxo7ghcpQ6LA26NjyAdyechJTp6p+oWRXORedR8obv89RUI8hKCylhokfakMjdtoeSnco25ykFW5G9fZdtZnRH4MpHOzJaWy6kKKSUKBBAI6joT1HUVy+0Xa+rfSdtYvhpdZLnL7L5vr4HT0DsvSs0rm8XFU6Lovu/kunifnTmzXq54nHkZPdF3KC9cZN2s0RxpIRDjPOOLZ83MtRQ5zeWTycwSkJ5dzufsePwa6mkcaSMJi2a5XCVOn2V123SZEuQp994oVzNuLWoklS2VNLO56c+3mrZ7vjz90t7sKLOkwH1bKZkxz5bTiSClW3csbgboUClQ3BBBIrh16fe1Xl7dPToTKVdUIYSw+vr1+Zg/Y8fg09jh+DUTaaXjiUtWRWuJrm7iLNtnXAxGXok1kPzklEgNvMMtpG7a3HoDfKVFYICiNlnawCreeU8qRzbdN+7evbnTnazUXJSz1i8r4nlvqiuIuWGsdHzNPuOO2+7wX7ZdITUqJJQW3mXUBSHEnvSoHvFWh0OvK73phZ3FtJb8R8YtaQkbAoivrjpO3xpaB6dOtUD0HdvzWoeQRtQNeGcquoaflLsjLbzKbSQ5yupeaClMshktrQBv9/uT7mvQHRi0PWbTm1svsrZVLVIuAbcQUrQmS+4+lKknqlQS4NweoO4q9dk7OpZXdWkpZhwp8mll8uaXnuVLtFe0763p1MYll+Gce43elKVfSoClKUApSlAKUpQClKUApSlAKUpQClKUApSlAKUpQClKUApSlAKUpQClKUApVQ9dvCM6c6eXi64FpTYZWoWYWp9yFOSysxbXbZCCUqRIlKB5lJIO6GkrPQglJqmOW8TfFblGVQNSpeqz0a6WKUifbcctKTEsZ5TuqO81uVyUrTzIKnVEgLJHLUG51K1tJKFWe76ffwLHpPZLWNbpSr2dFuCTeXsnjpHP7n02zvzwexVKj/QjWfFtftLrLqfialNx7m0USobit3oExB5X4zo8y0LBHcNxsodFCpAqdzK6008MVjMnedj41dn2HFNutQX1oWg7KSoNqIIPmO9ZOuKVFYmxXoclHOy+2ppxP4SVDYj8xoeFG+FrP87vngtL/nd6zS+3DJWMWy99q8yri87OQ4yZnZLD6lFwKRyp5TvunlG22wqDc31L1Xb0A4Ysr1HzHVJOkM6yyXM8vmJz3vZVcvmUmOqVJ3K+yB5D5R8oBzbdQRUvWXg74z9N9Ksi4WtMtT9MnNKr8udGYu12izPZ2Bb5hV27KUNjsFKIW51JPVaiCjcBMw3PR3id0gwjC9OeGHIdN7njGPY41Yp9tzmHKBfkIKuaYhyLuTzhXlNK8kbdN96AkDhWm4HcNGLTJ001evmpOOrekKh3u+TTKnhJcKvF3lqSlfM1vy7LSFAAb+apcqBeC/hpmcLmkkrCbxf4l2vN7vcrIbq7AYLMJqS+htBajIOxS0lDKANwNzudgNgJ6oBSlKAVjbJkmO5K1Kfxy/W66twZbsCUuFKQ+liS0dnGVlBPK4k9FIPUHvAqF+MvXC7aO6VC14InxjUTPJaMYw6Ik+UZ8jyTIJ7kpZQS4VHyeYIB2Ct6rhoHY5/AXrJjOAXvIZtx051gYjwZVymLUtMLNEo259z7hEtIIAO5Kkp3IS3XFvO0GnWGpW+k3FRKtXUnCPjwrL9PLxw8cjZGlOUHUS2R6E0pSu0axSlKAUpSgFKUoBSlatqblxwjCrjfGXGUzeVMaAHgShUt1QbZCgnc8vOpJUQOiQo9wNYznGnFzk8JbmUIuclGPNkc636ry47r2DYjNWw+ABc5zKtlMg/8BtQ6hwjbmUOqQocp5juiBWLczFZRHjNJaaaSEIQhOyUgdwArMIiubFT8h6Q8tSnHXnju464olS1qI2HMpRJOwHUmv14r8VfEdb1WtrFw6kn7C/avBfd9T6zo9jS0qgoR/c/3Pxf2XQxPipp4qay3ivxU8V+KuN3Z1u/MR4p6a+tOhqU1CkIID+4ZcA8nmA35Feg7bkHuOxHftvlvFfirrXGzRbpDcgy0KLbmx3SrlUhQIUlaVd6VJUApKh1BAI6isoQSftcjCpWco7czl8S+KniXxV3rCmRNtEaRM2VIAU08pIAC3G1FtagB3AqQogeYV3XIobbU4UKIQkqISkqJ29AHUn4hW50cPBGVzlZI0yO2s5Hf02KY0ty2WttuVJZP3KVIWT2Ta/wg2Ec5QehLjKvvay3i/wDNNdfGHXbm5fLk7HfjiVdS6iPIaU08yjxSMkJcQoDlV5O+w3Gyh133Azni382sLmLUu76L8ZItaqcOPq/xGL8X/mmni/8ANNZTxb+bTxb+bUbgJPfEX5rdslwTJ7HlmLwX5KjOSqbFLqURZzKW1Nlla1AJjv8AI66tp5aktHkU2sgrb3lxrINJ9dbNKwVN1efVcWPt0JK3ocpPLsoqacTylRQdvLaUpO46KI7+mI5B3A2NZ/Sqy23JdQ5llvuCMXq3+xCg/KloadjsJU4khK21c3MpakJCAQCAh89w623R6i1OpSsnHhnFPE0+i3WV1xy2aKlrFv8Ao41LyMsqT3i11ezw+njyIPv/AIOPS64Xux3az5fmNoVYBGRFSm5KfdZS06Vkodd3UgkciUhOyEcpISSomsJxC8V+sOh2WjE29FrtNsdsSx7I5g7b3Xo8ltSUlTrCWuzZSvY7FKlgBYUNgAKtrK4VrPCnvyMA1VzzCIEh5ySq02mXFfgtuuKKllpqbHf7JJJJ5EFKB3JAHSvzprw5yLheLxd831LzHL8eL0Y2Zi8vRUsvFsczjxjsMNsuN9pydmVoJ3bK0nYoUbpT0S4qVU7qUasY8s5T39F/LKfLUIQg+4Tg3zwaJwrfuj1RjW+551p+/aS9fZt2kRXoaoZchIbKYb0qOVqLbjqi052S1K3KFKG6QDVzaxOL4pjmFWVjHcUs0W122NzdlGjNhCElR3UfjJJJJPUmstVgsbKFlTcIdW3+ehza9eVxJSl0WBSlKmGgUpSgFKUoBSlKAUpSgFKUoBSlKAUpSgFKUoBSlKAUpSgFKUoBSlKAUpSgK28SHA1phrvJfzKyOrwnPyjycgtjKSmWR3JmsdESU/GdnBsNlgDavMaYu4Y9nl90zvk6yXW546vs37pj00TbZJ7vcOjqhwb7KaXspJBHmr3KICgQQCD0INUM4h/By26CLjqFwz3CBjb47SZcMTuL3Z2eVsOZa47p6wlkb9OrXd0QBvXI1bTY31JuEV3nR8vz3l57EdrKvZ69jCvVkrZ/uilxL1xnbza38maX4LrL7TjWo2oOCZVmbtuv2UPN3C0492Abt85plGy5Ud0qJXJ5dg63sk8qEq8sDdHpRXgTdcuTcMenZDZGZjF9xSZ2jTsCWkPW+ayv7sxJZKkrCCCrnbKgpI8+9e3Og12yu+6M4bes4yixZJfJ1njyJd3sYUIM4rTul5rmAOyklJPkpHMTslI2SM9JuJ1qPd1I8Moey17vz+DR220uhYagrq0q95Srp1Iv1bym8Ye+fPxSfPfaUpXUKaKUpQClKUArjffZjMuSZDqGmmklbji1BKUJA3JJPQADz1yVU7jlz2/ZInHOETTe5OxMq1VKxdpzKtlWjHGjvMkH43UpUykHordwbg7bxr28oafbVLu5lw04Jyk3ySSy38DKMXNqMebIHxTiJ0v1f4voWrue5O0xFmvycP0jtLp32YTzolXdY35WhKdSqO0o+Uvco2PISJi40rnoynReRiutGSIsUPJ5jVutNw5VKXCufVyPJHJ5SUtKRzLUOgTuD0V1rlqH4L++5bq4xm1o1YhY/jNudgM22BEiOIk2+BFQ2hKGnQrlS7shS+fb7ooqO5JrXcl0D1c8INbmsgybVHCbTLwi5TLVGj2i4Iusd2C4GlsPuBh5YbfVssKKiFKCUboQU9fyhrlXs/2t7Q23av8A5RwpU1F1cJqVF7ulGHs7+1tJpSXFmXKWF3KSq0KUqHBlvl5+OS+fBTrzc9adK12nOJDf2QcGkewOVNJUk9o+gHspaCnopuQ2A4lY8lR5+XoKsHXmtZMAyjgHm6b60y76LzY4kRvDtUHobK0tvW4vFFuuPZEk80YKaaUeqilISBso16SRZUabGamw5Db8eQhLrTragpDiFDdKkkdCCCCDX6V7K9pbLtVpyv7GopxTlFtbe1F4ez3Se0op78LWTj16MqE+GSOWlKVYzSKUpQClKUAqF+IdbsufilpS+pttpc26qSk/dFNNoYCT6RtMUr1pTU0VVnV/VBF/1vs2J2qMzKtdvZm2l2WgnmFwWgSHB6C2hEQNnbr2iyOnId+J2irKlp1RZw5LC88818MnU0alKpeQaWVHd+WOvxwY3xX4h+anivxD81ZfxU+j+ynip9H9lfIO7PovfmI8V+Ifmp4r8Q/NWX8VPo/sp4qfR/ZTux35iPFfiH5qxeTTnrJajIhxUSZ0h5qHBYUSEuyXlhtsKI6hAUoKWQCUoStW3k1tfip9H9lQ9hd8ves+Xuar4rb0O6faeXKRaIjzrrpXc7qpRjPzWWo7bqnWmWHHENpVyc5kKVuOUVP0/TKt7UfBHKisv7e/kQr3UoWtPMnhvZff3Ep4xiS8cgORHbzPujr7pfdfl8m/OUpCuRKEpCUkp5uXrsVHrtsB181vicUsyZTTCJFwmyG4FuiqUE+MSnD5KfxUpC3F7dQhtZ81ZODmLIvHsBk+PXPGpxZQ4hNyQkMuLPumUvoKmi4kKaVy825S6nbchxKNM1czDF4uUYPjci7QzJdyB3xmKpQDzIagyVpdWD1Q2HOyHMdgStA3IOxkysK1Oq1Vg8pN4x4LP8EKF9TnFKnLm0vizswrcuLGQ0672zu3M69y8pdcPVSyPNuok7ebfaufxc1lPF9/TTxY+g1wnHieWWKNRRWEYvxc1wyHYkRbDcuU0yqU6GGA4sJLrnKVBCd+9WyVHYddgazXi3rrTpthcybIIs1ySs3GI/IjY3biwpYRNAWyqWtvdJeICiAdwhDalKKgVFSd1vauvPgXy336fngaq12qMeJmSfRc3LlBttstkqcZSyHhBbS/KaR02WlhSkBae8FRWkIJSdlDfaxOnNkhWLG0xYmMzrKpxwuPN3BxhyXIc5UgvPLYWtBUQkDorYBKUgJSAkcmDYHZcFthjW9Dj0yTyrnTpCw5IlOgbcy1hKRsOuyEpShO55UpB2ruXmRDn3S24a7IWhy9JfcdQ2opWYjKU9sQobFO6nGUEghQ7XcbEbj6joej09Lhw7Ob5vHy9PhnwKDq2pz1CfE8qK5IxNvRP1RnKLCvF8KiuqadWpKg7fFp7w2enLEB6FfUvEKA2aAU9JqEJbSEISEpSNgANgB6K/MdhiKw3FjMoaZZQG220JCUoSBsAAO4AVyVZ0lFYRwm8ilKV6eClKUApSlAKV8NUpkaiX/WfNLpMsOoF8tEa3P9vzWu4qR4sguuIjx0N9WeflY7R5S0rJU5yA8vROqrVVGPEydp+n1NRq91SaT8y69KrjZ9T9W7lAuWOOXqAyYEtLCL8YyFzn21NocI7DkTHbWntAkObLSeU7tg1xw3MztchEq2anZQ2sJ2cRJfZmNvH0qS+2vl69ftfJ6OgrOMlNKS6kavRnb1JUp84vD9xZKlQ3ZNab7Y1MR9QrYzLhFAS7ebQwsBlflbqeiFS1pR0QOdtTh3USpCEAqEt265W+8QI90tU5iZDlNpdYkR3A426gjcKSpO4II84rI1HZpSlAKUpQClKUApSlAKUpQClKUApSlAKUrWdSdScJ0hwm56i6jX5qzY9Z0IcmzXG1uBsLWltICG0qWola0pASCSSKA2alVV9tE4Gvht/u9dfq1PbROBr4bf7vXX6tQFqq6l2tNsv1qmWO92+PPt1xjuRJcWQ2HGn2XElK21pPRSVJJBB6EE1WD20Tga+G3+711+rU9tE4Gvht/u9dfq1AQ3rt4Ny74oJWU8LUpD8DdTz+DXWTs2N+qhAlL6tk/9J0lHU7LT0Fap4P8A4m3dH9R3eFPU9ufYrddZixYIV5aVHk2O6LJUu3OIX3MvElbKh5POSkE842sf7aJwNfDb/d66/VqiPiB4nvBa8Stlbt+pGpINyhj/AHbfYFguke525e+4Uy+Iu+wPXkUFIJ6lO+xqNG0pwrOvT2b545P18/M69TW7q4sVp9y+OEd4Z5w/9r54a5xe3LGGj0GpVRMc8JfwRWLH7ZZJnELLu79vhsxXbhMx65mRLWhASXneWKE86yCpWwA3J2ArI+2icDXw2/3euv1apJyC1VKqr7aJwNfDb/d66/Vqe2icDXw2/wB3rr9WoC1VKqr7aJwNfDb/AHeuv1antonA18Nv93rr9WoCzWQXmPjlhuWQSo8qQxbIj0xxqKyXnnEtoKyltCeq1kDYJHUnYCvNXQbiJxaLm+d69ay4rn7OdZ1PLTEJvDrlITZbKwrliwkOJZ5SdgFrKeijyk9Qash7aJwNfDb/AHeuv1antonA18Nv93rr9WqvdqOzdt2t02elXlScKc2uLgaTaTzhtqW2cN+nhk20KzoT44rciTXDifwTU7SbJ9P8Smai41c7/AVBbuStPbo+GkLIDqeXsgfLb52+YHdPPzDcgCqy8E0WJwuZ5dsov2aZfOtN0hGHItFv09vSkSlBQU06pbjKQhaDzbbJV0Wsbjfer6+2icDXw2/3euv1antonA18Nv8Ad66/VqqGm/0m0fStIuNDtq1VUK7zNPu23yX7nSyuSx4PdYe5Inf1J1FVkllev3NIyziw0CzfGbph+T4fqHOtN5iOwpkdzBbps404kpUPuO4Ox6EdQdiK2Dwb2qGSX/Ty+6QZLb8lda06nCFj96u1okQvZKyLKvFAe2Qk9q0EFCk+ZPZ7b9TWX9tE4Gvht/u9dfq1PbROBr4bf7vXX6tXV7Ff090zsGqsdLqVHGpjMZyTWV1SUVh42fiseCMLm7ndY40ti1VKqr7aJwNfDb/d66/Vq5I/hPeByTIbjI1vbSp1YQFOWG5oQCTtuVKjgJHxk7Cr2RS01KUoBXwkAbk7AV9qG9bs+kNc+F2SQUFSP94OJ6KAUAQ0D5twd1H0ED01B1G/pabbyuK3JfFvwRKsrOpfVlQpc38l4mD1l1pkyg/iWCzlst7qZm3Jk7KUO4tsKHd5wXB1G3k9fKFeChrEJdvyO3Wd6W/bj2DDSHVbJC0qQPJJ2O5UE7gFZ5gBzHYGQrJid6yW4ItVgthlyVDnO6+RppP4Ti9jyp36dxJPcDsdpZtXCpp7NgxjqM05k09iUxOaJecjR4j7SkrQWENqB6KSDzLUpR6jcJPKPndG01TtVcq6qvgpJ7c8eiXV+L/0Xmrdad2etna0lxVGt/H1b6eS/wBmuQ2nXorS5AaD/KA8GVc6EuDotIPnAVuPT0rm8V9f5qkW9aV9pe1zsaeh2+NPHPNQtpSwh5LaENrbQFJGxS2lKk7juCh15t9Wk4Jqbb3VREWi0Xcg8yZjEpUNtaN+oLSw4pCxuABzqCtieZPRNb7nsxeUZPu1xLO2PscujrdGaXG8Mwfivr/NQRCTsAT+SuW/27KoEhuBKwPKktK2Uqdb248hpJ/B5UOLdI/9utTvGHYPa7i7lGTpyhQmNpguN3GPdPE0hSgTzoWgNJBKUgqX5IAAG253570i6p/+pTkv/i2b/wDk6T/ZJP34NR1Tx3W3WvDrjhnDS5aGFzu0g3DLbhO7KFDT1S6zHU2hxbr+26SpKOVvf3XOCE6vw+cOHHzwm4QnDMKkaQZnjzc164u26TMnRpq1uBPMlp8tBsb8oPljp/ZUyY9h2mdsd2xBi1wwpXahm0zOyZ5idyoNsqCASfPt1rdbDqVcbdqHjuBRFTLmbi6tueylh11MCN4tIcbkOO8pS2C5GDQClDm7Q9CQKteh3tpQ4bOlTknLm34+fl9DiajSrVc3E5ppcl9iB7Rxf66z7ldMK1V4C86WqFsiULI41dozg90Oi0IbUOnmWr8/StWw7iSwC9okW3P+CvVWy5RObXbFMxMfVMjulAW4ltkuLSEDdJWE9nypPnO259Dq+FKSQSBuO6rY6UHucbjZ59aM5s/Fv8/TS7YJnOK21sNyMYVl0BTEmQypvd6Kp0lSXnm3EurBCypba99h2aqmfsKmvVzSrHtYsMfw/IH5sT7a3MgzYT6mn4M1o8zEhspIBUheyglW6TtsQRVe8XmZZZbu/pnqgw2xllrZ7VExpHJFvkQK5RNjDfp96HWu9tagPcqQT8/7SaF3E3eUF7L5rwfj6P6ls0fVeOKt6r3XJ+PkY3VTI1YVp9ecjbgXOa6y23HZYtqUqlLefdQw12QUQkqC3UnqQOlSxojp1Kxpufl19sibZdL43HSITjqZD0NltJASpzchKlbgrbbUWgUAjmUVrVGuYaeP6g5PiuPXq93GPh0y4ITe4sBSWXlutntYa+3250I8YQ2lYQQoktbFIC+abPsKTrlFu9pyfVHLZtpuEh1UaHFmeJrjR1JSkNeNNgSV7cqjv2o6uKG2wAE3svpsHQ/Up+1lrrsttmvEi61eydXun+3C+O5+811b0209tV7u+WZlaYSMeguXG4MKmNmQ0wjbc9lzc+5KkpA26qUkDqRVJeCDjHzvio4zMhm3hhqDh9sxu4x8ct6Y6AqKHZURQ7R0DmW643GKiCdhyHlAAO9WvCPcFEzhmyqPnGA3G+XPB8rStD65shcl+DJStKiy+7t5baiWy2pe53SQSSATq/g0c/ueF8Q0aOysotktlMu5OJHlNMRlha3OnUgNF5JSO8L9VWuov0tN1FvjHPwys/BZZxIf358D258vHG3zPfSlY+DkFiuikott6gy1KBKUsSEOEgd5GxrIVLTT5EbGBSlK9ApSlAKUrXdRMonYTgeQZfa8dm3+ZZrbInMWuEnmfmONtlSWkDqSVEbdAT6AT0oCFuI/WrFsbYucbJsp9hMOx0NIyKakr5ZMl/ZLcI8iSsoCXEKcCfdFxtJPKHUnRH7dKiXFvKsBs8O6QbzaYcJhMeQhhlhLReWw96CypMgglAKwEo5ULBPLhIt7xfUDAWYea4zCvNuuykyroi/NMqQ9JW6HHFuMtpcbSrt1bhJ22PKB5qkXH3mHoY8Qm21+C0EsxkQW+VDKUjbk3CiOg2GwCdtu6sJwjUjwy5Ei1uqtnVVai8SX+j8Yvj6MbtQhqlLly3nFSJstY2VJkK92sj70dAEpHRKUpSOgFZelKzSxsjTKTnJyk8tiufBr/I0/yqKywVKx7IpqIsuLzeTDmvL5WpLQJ8kOOKDbiADzKWhwcpS4XOCtcz27221Yxc3Zl0aiPsQX58fdSe0Co6e07RCSpJVyKCVdCNunUd9DFLJaulQfC4pbILbFk3fEJqZTzCXXG4F0t77SVFIJSlxx9oqHXoeUAjr0rsxuK7TMMrVe4V+sz5QpbEZ+IiS5JIG5S2YjjySfWod4qPG7oT5TRJnZ3EP3QfwJnpUe6cXzUzNpjmXZTYlYjYFtqbtdhkJSu4vpKv8A6marbZhWwATHRuU7qK1qJCG5CqQRuQpSlAKUpQClKxGJ5bjWdY7By3D71Fu1nuTfaxJsVfM28ncgkH4iCCO8EEGgMvSlKAUpSgFVT8KP7xzUX8a0/wCZxatZVU/Cj+8c1F/GtP8AmcWgO9w08MnDbfuHPSy+Xzh700uNxuGF2SVMmS8TgPPyHnITSluOLU0VLWpRJKiSSSSakn96dwsfyadKv6m279jX64VPev6Q/ISw/QGalOgIq/encLH8mnSr+ptu/Y0/encLH8mnSr+ptu/Y1KtKAir96dwsfyadKv6m279jT96dwsfyadKv6m279jUq0oCKv3p3Cx/Jp0q/qbbv2NP3p3Cx/Jp0q/qbbv2NSrSgIq/encLH8mnSr+ptu/Y1Trwq2hOh+nfCwch0/wBG8Gxm6/ukt7Hj1nx2HCkdmpLvMjtGm0q5TsNxvsdhXo1VHvDD+9APyptv+F6gJV0M4X+Gi76J6fXW68O+mU2bNxa0yJMmRiVvcdfdXEaUta1qaJUpSiSSTuSSTW7/AL07hY/k06Vf1Nt37Gstw+fwCaa/JCzfQmq3+gIq/encLH8mnSr+ptu/Y0/encLH8mnSr+ptu/Y1KtKAir96dwsfyadKv6m279jT96dwsfyadKv6m279jUq0oCKv3p3Cx/Jp0q/qbbv2NP3p3Cx/Jp0q/qbbv2NSrSgIq/encLH8mnSr+ptu/Y1Snwteh2imm/DNaL9p3o/hOLXN3MYUVc2y4/EgvqZVEmKU2XGW0qKCUJJTvsSkHzCvSuqFeGe96hZPlxA+hzaAvrSlKA6N9ujNjslwvcgEtW+K7KWB50oQVH+wVWNiLOvchMh5lSps9ztHEJ7y84rmUB5uqlHv6dasLqXEk3DTjKoENBVIk2Scy0kd5WphYA/ORUUaetpuTtqyRI2Z5GZpbPQqCk8yfN5jsaqXaa1nfVra2X7ZN5+X0WTv6Jcxs4Vq3VJY+f8AOCXcKxKDh9lbt0ZtsyHNnJbyRt2zuwBV167dNgPMAPjNZ+tccyxfczCSPjUvf9FdVzJ7kv3PZI/FT+urVSpwowVOmsJbJHCnOVSTnN5bNtpWlLvd0c91MWPxdh/4rrrmzHPdy3letZrMxN87u+vyXGh7pafymtAUpSvdKJ9Zr5QG13vFMRyeOqFkWOWi6sL6qamxG30K9aVgg18xjDMQwqG5b8NxWz2KK852rjNsgtRW1r225iltIBOwA3PXpWq1+kuuI9w4pPqO1MAkGlaKi5XBv3E14fFzmudF+uqO6WT+MkH9FAbnWqaiaeWfUG1MR5o7C42x7xy1T0D7bDkgEBQPnSoEpWg9FJUQfi/Kcnuae/sVetH6jXIMqnedhg/kP66xlGM4uMllM9TcXlEPXKxP3q2rts96Tbn2n0JkeKrRztvMuBRSFFKhtzIHXbcpIPTepAs+ok63utov7wfircS2p4oAcb5lbBR5QAUgkb9Bsnc79NjhL3gGL3bOGdRolrYtV+7FyLOlQmkIVcmF9n9rk7g9pyllooX0UkJKQeVSgeu9p7DlXidc5eQXp6NPjsxXLaX20xEIbUpXkhKAsFfOoLPPupOyT0AAqtvo99YXH/hKi7vOcP6denU61W+oXNL+9H2+WTftXdM7DrJpjkul+SoBt+SW56C4vl5iypSfIdSPOpCwlY+NIrx4wPgh4peFPX6z5ZdNObzkmNQpC2XbxiLqJSzGdBbLiWfdlSQrmLTjfKrYjr317CqyG7HukAepCf1Vxqvd1V3zF/kAFWirSjXpypz5NYfvOVCbpyU480QyzjN9fesLmItKXOamMuMpdaWw60NwFuFXL/sxSgrUQ42QsAtFPl7VZaKmSiKyiY6hx9KEh1aEFKVL26kAk7Anzbnb01pvsvcv/WOfnr6LzdB3TXP7KgaXpdPS6bp05N5ed/sSry8nezU5rkbtStMTf7sn/myfWhP6q5U5LdE9621etH6q6ZDNupWqpyqePdMsH8hH6a5U5Y8Pdw0H1LIoDZaVr6ctR99BUPU5v+iuROVxPvozw9WxoDqu6YaavX4ZU7p7jS72HhJFyVaWDKDw7nO25OfmH4W+9QLqRw6616i3K5XB/K8XtT893fxy1OSoL6mkqHZtuFlIU6ORCUKLi1q5eYIU2SCLDfuqgf8AQkfmT+uvhyuF97HePr2H6a8ayZwqOm8rHvSf1IWsmnmtSkMQrhjeOMiO0lt6U/kDiy8sDbnQlEQ8wO2/lch691ZRzRnVV5a3Gs7xOIj7xpVhkyT6ivxtv8/LUnryxP8Aw4RPxqX/AKV1nMpnK+5tMo/ISf8AzXpgRbO0b1sTBkG35vhSpaWllgO2KXyLcCTyJVtL3SCdgTudgd9j3VGGOcNHEBk+f41e9Vp2NOWuyXBifIQ8Wp6Hm0c/aR22UMM8hcS7y87i3QnkUAD0UuzDl+urnfKKfxUgforhN1uR75z35FkVi4qXM2060qWeHrtyN4AA6AbUrRxdLkP+ef8A+81t1pcddtzDjyytak7knvPWsjUdulKUApSlAKhnX7iVtXDhcMeumf4beHMFvDpiT8qt48YasskqSGhLYA50srBOzqSrYp5eXcjeUMryKPiOM3TKJVvuM9i1RHZjka3RlSJTqG0lRS00nq4sgdEjqT0FaPphrboTxN4tP/cFk9myy2qbMa62yQz9taSsbFuTEeSFpB6jZaeU7HbegJBtd1tORWiLebLcY1wttyjokRZUZ0ONPsrSClaFpOyklJBBB6g1D/B1pHP0E0KtOkF4yq332fj825do9CdKm223przzSNleUk8i0kg9yioDcDcwVgeK2bW+Te8bsYlY7oPg95nWDH8VtE16MzfJLLyxLlyXUL7RUUPLdQ1GSoN7J3UkjZI3m68KuhsiKk4tg0LCbswk+J3vEkiz3GIvbotD8flKtu/lXzIP3ySOlfGO039cOz/ZnV5aRUhOo4PE5RSxF9Vu05NdcYxy3ex0aOm1a1PvE0vA2riw13zXTWFjmmOitkZvWquospcDHozyCtiAygDxi4yAO5plKknr0JIJCglQM4WCHcrfYrdAvV1VdLhGiMsy5ymktqlPJQAt0oQAlJUoFWyQAN9gAKrrw/66qTE1CsXEFcbQzmOjATFvGUKjNx03Cyvt9vGm8qdy2XEN/bGkeT2jfkjchKcho1xV5Vr9nKY+nOguSRtOGS4H82yJ72MRI5Uq5fE4im1OSApYA5ipASNyrYgJP16zu6N/b07u2lxU5xUotdU1lP3pnPlFxbi+aLE0rT87zxOOKasVmbalX+cgrYZc37KM3vsZD+3UIB7kggrV5I26qTrOkb09GZZdCm5Lc7wExbZJU5Me5gHnDJDhbQNkNJIbR5KQB5IrpxtJujKu9kvnul/JAnf043EbaO7ez8tm/jt9yVqqn4Uf3jmov41p/wAzi1veea0XRyBKyfTuQst4TdjGyW0TIgQ9IYUrsw4knyko5grY9D98rogoVG3hK7zAyHgDze/Wt7tYdwZsslhfpQu5RFD1HY91eVrSrQgqk1s9vR89/c015GVvf0bmpKlTe6w/JrdZXo00/BomfhU96/pD8hLD9AZqU6izhU96/pD8hLD9AZqU6jEwUpSgFK6aLxaXLo5Y0XSIq4tMiQuGH0l9LROwWUb8wST03223ruV4mnyApSlegVR7ww/vQD8qbb/heq8NUe8MP70A/Km2/wCF6gLRcPn8AmmvyQs30Jqt/rQOHz+ATTX5IWb6E1W/0ApSlAKUpQClKUAqhXhnveoWT5cQPoc2r61Qrwz3vULJ8uIH0ObQF9aUpQHwgEEHz1FuM6O3TCrtclWrLpdysUz7ZDts5psuW9xTrq1pbeQEksgLQhCFhRSlsAK795TpWE6UKjTkstbryMlJxTSfM1dvFZqvur7SPVuTXZbxNkfdZi1fipA/XWfpWZiYlvGbYj3QdX+Mv9Vc6LHakd0NJ9ZJ/wDNd+lAdZNtt6fcwmP/AOMV+xEiDuitD+gK5qUBwmHEV3xWT60CuNVrtq/dQWfyIArtUoDHLsFpX/yvL+Koj9NcC8XtyvcreR6lD9IrMUoDAqxNk+4mLHrSDXGcSPmn/wDxf61sVKA10YkfPP8A/i/1r9pxNke6mLPqQBWfpQGFTisAe6efP5QP0VyDGrYO9Lp9a6y1KAxf7nLV/wBJf/ea+HGrWe5Dg9S6ytKAw6sXtx7lvj1KH6q41YpFPuJTo9YBrOUoDXlYl+BO/O3/AK1xKxSSPcS2j6wRWzUoDVFYtcR3OMK/pH9VcZxu6DubbPqWK2+lAad+527f+nH/AHp/XX0Y5dT3soHrWK3ClAaojFrgr3bjKf6RP6K7LeJnvem/kSj9O9bFSgMO3i9uT7tbyz8agP8AwK5hjtpHfHJ9a1frrJUoDGnHrSf+VI/pq/XXfZZbjtIZaGyEDYD4q/dKAUpSgFKUoBUcnh20WRqnH1sh4BBt+bRw6k3e3OOw3JIcBDnjCWVJRJ3B/wCMlfcD3gbSNSgKP8BkFWG6SXrRq6yVryHTjK7vZbuh0EOFapTjzT/X3SHW3ApKu49du6rJVGHEjhR0ozePxb4i0WmobDVr1GhNjybhYgoBM/l878LftAe9TIcRvsAKyOqmoqMc00ym/YbOh3K/QYao1sjMOpdK7o+QzDZUBvsVyHWU7H8KvxJ/V3+nt1Z9roVLNcUL+eY+VSUkpJ+XFJNeTx0LJYXcZW7Uucfoapwn25eX8Suv2ssBp0Y+7LteGQX1jZEyTbmVJmrT6UodWGwobg7K694Eha557k+M5vbbPacsm2iG9aVylIjNR1Fboe5dyXW1+bzDat50L0otGh+kmMaW2dYdbsUBDMiTy7KlyleXIkK/nOOqcWfjVUBcZ1szaTmmOzMUxi73RAtbrTrkKC6+lB7XcAlCTsfXX7h7HaTbabC30yGO7pQUFxeEY4Wc9dig9prmrTsp16eeLK5Zzu/Lcxqs+bivSpj1yemTZrnaSpchQLrygNhvsAAAOgSkBIHcBXYxuBK1Gx7US7Wi7TIlzsEa13WA7GdUCX44mLCCAdlbjcDfuUUnzbGrF1v9+iTHrfdhKhyWFlDsd5tTTjah96pJAIPxGpo0C1MhYNpXqbOe7d643cQLRamGWytx6Y83LSgAAHu6qPpCdhuSAfo19Yu3tnOhvLMceH7l8vlg+b6fqaurtU7jaOJ58f2y+effkkzH8hj5nqTg+TKQhLOqWNzrJfkM7pC32EONvOJ9BKmmAPQkfH1iLiRvMy5+C9z+BNB7Sy3KHA5z3r/3rEd7vNt23L/RqTMQsRwvUzSnCJrqWnMBxu5ZLkaisKEPxkOOLSfSUqLSTt5lA1Euvcab7VvqJe5ieUXy8RbgyCNjyeysNrr/AEmlfk2quar3f6V8HL2cf/aoo/8ARj3YLXojqfro8f7sT4vXhpOX/Xn35LncKnvX9IfkJYfoDNSnUWcKnvX9IfkJYfoDNSnVWLuK1DU7U2x6U2BnI7/bbxNjvykQ0t2uEqU6FqSpQJSCNk7IPX0kDz1t9KxmpOLUHh/E8llrYpBZ+JXCYvFRftR3Mcy5VumYmzbER02dRlpcS+2oqU1zbhGyT5W/ftVudO8/tGpeMM5XZIF0hxX3HGktXKIY7wKFbElB83oNaTaNNsnh8UF81VeZj+wE/FGbSy4HgXTIS82sgo7wNkHr6qlyudp9GvTc3VltxS2xjrz58iPbwqRzxPq+gpSldMkiqPeGH96AflTbf8L1Xhqj3hh/egH5U23/AAvUBaLh8/gE01+SFm+hNVv9aBw+fwCaa/JCzfQmq3+gFa/n+bWbTfCr1nuQtzF2ywwnJ0pMOOp98toG5CG09VK+KtgpQFPvbTOGr+JNSP6qPfrrZtM/CFaD6r53Z9O8ZtWdNXS+PmPFXNxt5lgL5Sry3NyEjZJ6npVm6UApSlAKoV4Z73qFk+XED6HNq+tUK8M971CyfLiB9Dm0BfWlKUApSlAKUpQClKUApSlAKUpQClKUApSlAKh3UHjC4Y9Lb25jWc61YxbrswvkfhCWHnmFehxDfMWz8StjUIcYvFmifCvfDrw83SVc8/lBEa83a3rCImOxisB4OSt9kSVIC0oSgLUk7kgEAGAsN0nwnHceYtb+C2FD4B8YUUJmreXv1W4842lbild5JA6k1yNR1elpzUWuKXhlbHUsNLqXyck8LxwegeCcTHD5qc+iHgWs2H3mW50TEj3Znxg/+0VBf9lSXXk7lvDdoxmLCkTMHgQJBO6ZdsbER5B8xBQAD+UGshpXqtxC8H17hBzKrpqXpEuQ2i5wrkXJN0sscq2W9HI3K0oB5ilPQ7EcqfdVrs9etbuSg/Zk/H7md1otzbR417S8vseqVK0fSXW3SrXTHP3VaUZtbsit6VBDxjLKXY6yN+R5pYDjStuuy0g1vFds5ApSlAKUpQClKUApSsHmmcYdpzjkvLs8ya22CzQU8z824SEstI9A5lHqo9wSOpPQAmgM5Sq8tcaOL3weNac6I6z5zbVfcbnZsOcZhvj8Jt2atgLHxgVJelOqFw1OhT5dw0ozjBlwHUNBjKYceO5J5gTzNdi+6FJG2xJI6kd9Ab3SlKAUpSgFKUoBSlKAUpSgMDn2Px8swTI8VlobWxebTMt7qXGy4kodZUggpAJUNldwBJrzJ4NLdpTl3E8zEjydAHrhEkrkGJZbLf7JeVuxB2yOwizEpjEodZQ6pKfL2aUo7AGvVSuuIEFMsz0wmBKKeQvBsdpy+jm232+KgOxSlKArzxT8MjGrtsXlmHtNR8xhNbJBIQi4tp7mnD3BYHRCz+KemxTHvg+7PMiDUO05JZ3Y023zralyNMYKXGHkB8jdKhulQOx9PdVyK/CWm0KUtDaUqWd1EDqr1+murHV636GVjPeLxh9Vhp49PocOeg271KGpU/Zks5WNpZTWfJ7+/wCZW7VPRnKrNb73ZtMY1yu1/wBVruWL5kc5xBFqt2xUWvJKSG+UFAAT1Hkk79mK0/wi+L2vCfB5ZhiVlb5IVpjWWKzv3qCblFHMr0qJ3JPnJNXEqqfhR/eOai/jWn/M4tRK15Vr01TnyW/r039EsLwJ1tp9G1qyq0+bWPRZy8erbbb3b9CWeFT3r+kPyEsP0BmpTqLOFT3r+kPyEsP0BmpTqKThSlKAUpSgFKUoBVHvDD+9APyptv8Aheq8NUe8MP70A/Km2/4XqAtFw+fwCaa/JCzfQmq3+tA4fP4BNNfkhZvoTVb/AEApSlAKUpQClKUAqhXhnveoWT5cQPoc2r61Qrwz3vULJ8uIH0ObQF9aUpQClKUAryX11mcVejmvWTW7OeJfPMag3q7PXDC7ibkRZpsVSyoReRYLDbrQUkFtewOwAGxBr1orC5fheIZ/Y38ZzjGbZfrTJGzsK4xUSGV+g8qwRuPMe8VrqwdSDjF4fibKU1CSlJZXgUBxvij42sRip9nZ+A5vCbbC/GpVtXDlrT5yrxZfZ77bnyUbdPjrabVxtcTN8hGZA0u05QgLKApd6lqCiPddEtnuO47/ADVKF28Hbw/uLccwubm+C9oSeyxzJZDTCT/NZe7VtPqCQPiqJ8j8F3fXZzkzFuKHJX0L7mMqtTN3SPUQtoD8iRXMdHU0mlVi/cdBVdPe7pyXvPtz4z+KizxVXK4YPpeiOlSU8omT991HYAr5dh185+L01w434THOoU5DeoPD8mVAV7uTi17TLfbT51mO6hCiNu7lUSe70Vpdw8GrxDY+1Kdx/NtMspLzxfLU6JcbO53AcjbkZxQQOnQd25NV+jXDJ+HnL28S1PsWRW7JbPGLlytE8IltTogQVPXG3y0gcyG0NuOKQ4VdN0ghdap1dRtouU8S932ZthTsLiSjHMff9z1z0a100v18xb912l2Ts3aG2vsZTRSpqTDd2+5vsrAW2r4iOveNx1rfq8srrk83QnKbdxIYAtUdcB+OMpiR+jV7sy1pDodT3KcbSorbcPVJB7xXqPElR50VmbEdDjEhtLrax3KSobg/mNTtPvoahR72Kw+TXgyFfWcrGr3cnnqn4o5qUpU4hiqo8e/EXf8AS/F7No7pbL7PUbUtbkG3PoPlWuCkbSZx27ilJIQenXmUN+TarXV5VNZgNfeJjUvXZ4qetdomHC8WKlbpRDik9s6jzbOuHnB/nqFQtRu/0VtKt1XL1fImWFt+ruI0unX0NL05w3HcZzm5YZcJJYjYrDjXNpiQvZV3kOhReushR+7FKk8idyQ2Uk95BE4tOtPtoeZcS424kKQpJ3SpJ7iD5xUQcSU7DbPjsK63eb7G5ElbiLJcWZfi0iKQnmdXz96mwkDmb2IWShO26gRjNKeHzjc1ExpvJrHppAbs0lAetsy63mTj8qcwrql0xI0jkRzDr1QnfffrvvVNp6fW1aPfxbz5/Nr1+pa6l7R02Xcy/PL3HczzR/Mpt8XNxHJ7q7Fkw1MR2Zl9lJFqlEq2kNeUSR1QeXqR2ew8lZ22K3Xq+aZ2pkat6oWCe3yhKX3Yfikl0ge5SlKyHTv5koBPxmu9D4KuOrIgI8q24HYmVdFOXHM7zKIHxJbcUD+UVu+EeCvzmRc27lqfrvBgNEjt2MOsLUWUtPnSJ7wLo9fKa6MdEuq0VCs1heSz8d8fmSBLV7alJzpJ5fm8fDb85EUcGeW3e++ERj/YysEm0WF6xTU5cygFtMhtLThZfkNbANudsqMAD5Y367EqA9bqjTQ/h00h4d7C7YtLMUatxlqDk+e6svzZ7g+/ffXutZ3JO24SNzsBvUl1aaFJUKUaS6LBXK1V1qkqj6sUpStpqFKUoBSlKAx2R3kY7j9zv5tk+4i2w3pfidvYL0qR2aCrs2Wx1W4rbZKfOSBVSbrh2o97tjfE1rjpPcs6zJUhpnAtL42zkDGi+rlZdllX2syeoU/KcBDI6IA5RVx61rOc9suB21uXcUPypcpRbg2+KkKkzHQNylAJAAA6laiEpHVRFAQfZ+ENzUZo5XxWZxfM0yeb9t9jLTe51psdk37mYTEV1tSuXuLzpUtewJ27qsHjVggYpj1sxi1KlKh2mI1CjqlSXJDxbbQEpK3XCVuK2A3Uokk9SSag+4XTULM30zMlySVZIaXCtq0WKS5HSlO2w7WUnlfdV1J8ktoPTdB23PRfxRSwpyFmOZwZRIUmSxk05SwodxKXHVNr9S0qHxUNndsshSq9xtccq0qkMq1emMXjD3XAyrKGI3YyLUT0Sq4Mo+1qZJ2BktBAQSOdpKOZxNgGH2JTDcmM8h5l5AcbcbUFJWkjcEEdCCPPQwaa2ZyVomK6l+y+oGV6e3qCLfPsUlkweboJ0N1hDiHU7nr5Xap6bAlpYHVC9t7qEeJDCUuQmNTMVlsxs0tDBiw4rhPZXxntEueJOhPlAhY52nx1YWVL35C4FAibqVGejGoMnNnsltyX1XCDj01qEzcj1Drpb5nWOb78tbo3X5+0AO6kqJkygaw8ClKrprNxm2TTnVBGh+n+mGWan583CTcp9px5pHLbYygClch5Z2QSFIIGx6LTuRzJBHhYulVfyfjptFgxrBRG0Tz+VnuoMydAteDPxG4lybdhqKZCnlOK5ENjvSvrzJO+wAVyzTpBqJeNTsPGRZBptkeC3JqU7Dk2e+tIS+hbZHloUglLjSgRyrHQ9fRQG70pSgFKUoBSlKAVVPwo/vHNRfxrT/mcWrWVVPwo/vHNRfxrT/mcWgJZ4VPev6Q/ISw/QGalOvN3Q/wsfC/p3ovgWAX6150u541jNrtE1Ue0sLaL8eK204UKMgEp5kHYkDceYVu3tyvCT/FGoXzNH+s0BeylUT9uV4Sf4o1C+Zo/1mntyvCT/FGoXzNH+s0BeylUT9uV4Sf4o1C+Zo/1mntyvCT/ABRqF8zR/rNAXspVE/bleEn+KNQvmaP9Zp7crwk/xRqF8zR/rNAXsqj3hh/egH5U23/C9XU9uV4Sf4o1C+Zo/wBZqtnH/wCEP0F4m9AvsZ6cwMsZu/s3DuHNc7e0yz2TSXArykvLO/ljYbemgPUDh8/gE01+SFm+hNVv9edWlXhb+FrC9LsPw672vPFT7FYLfbJRZtLCmy8xHQ2vlJkAlPMk7HYdPNW0+3K8JP8AFGoXzNH+s0BeylUT9uV4Sf4o1C+Zo/1mntyvCT/FGoXzNH+s0BeylUT9uV4Sf4o1C+Zo/wBZp7crwk/xRqF8zR/rNAXspVE/bleEn+KNQvmaP9Zp7crwk/xRqF8zR/rNAXsqhXhnveoWT5cQPoc2uz7crwk/xRqF8zR/rNVf8If4QPQrik0Mt2nWmsHKmbrEySLdnFXS3tMM9g3HktqAUh1Z5uZ5Gw27t+tAexlKUoBSlKAUpUAa1ccGgui093GJF+k5Xl6d0oxrGWDcJ3OPvXOTyGf/AHFJPoBrxtJZZ6k28In+lebGZcfnGXf1Ov4NoNjOD2cglMq/yXLjLSjzKKGlNpSrb71SSAem/pjp/XXi7zWW0zmnEhc7XCfdCHUYza4cAIRuObleKCvpudz5tt+oNQqmp2tP/NP03+hMhp9zN/sa9dvqeqeU5liOD2ty+ZnlFqsVvaG65VxmNxmh/SWQK8t+NzXbDuKnUXEbbozAXPsuJtXWFcsrlMqjxpbU5lMdyPEKgFuAJ5/tmwSD5Q3SCThr/pxoXizyM41Sv71+mqIWi45VeHJ7rqvNyNqPKs+gJQa1HUDO7zqBcrTiGnWATm3JclluEpEPnvExokBRiW4bKCAnr2kjkRy+auZLWJXn9q2pvD2y/wCF1/NjoR0pWv8AcuJrK3wv5fT83MjqFkly1UsTuiOJwyi53l6Jah2B3LZfWG47Kkq6guLG59DSHVHYJBV7EWC1IsVittkacK0W+IzFSo/fBtASD/ZXmToi3w68GGQDUTXO/C46tvR1M23C7As3q420L5+dclbP2tcxwOEFayhCEkobG25O737j64sM4cWjSPhysmJW9ZKW7jmdxW48R+F4s0UKQfiPOKl2dOhpVHhqSUc77si3U62pVc04t422PQ6leZkvPeNrKx2mTcTkawpX7qJjWMxkJR8SXnQV/lIrV8kwTVW+xXXrxxKar3l4NLPi67/4u24vbdIAbSkJHeNvjHUbdcJ6/YxeFLPomZx0S8ksuOPej0h13zJWnmimd5y252btix24Tmleh1uOtSP/AMgK8meHeBlVs0qtePWSUnxqUwi4OKWoIDReV2q+YnytzzBPkpJHXuHKVRrrVp7boOAXO4KuuVz7q2mOw2Lnd33i4868hvZTaln8Ikd3cfdA+TYmz2WLpgqwWq3MSXm1sR4ThXylKSQEEjuIJKUdANvcjuAFc3V7+ndUIqn1b5+SX3OhpdlO2rSc/BcvP/RjsFxCdq9xIYbgeSIh3Cxt3WNarkpb6i4Vw2XLpJbS2UkKbcKYSFKKgftQGx3O3rilKUJCEJCUpGwAGwArxUw+53xevlnvmlGYz8WyxByG8zLiu3GUylCnm4rSUsyB2KwptKklSOvd13HS0tr4yeMDTx5IzHTnENULQ1tzybI8u0XMp86i04VtLPoSgDep2nXlra0o0KklGT3328vToQ7+1uLio60I8UeW2/n69T0HpVfdF+ObQHWe5NYoxf5OKZgrZK8ayaOYE0L/AAW+f7W959uRRO3eBVgq7iaayjjNNPDFKUr08FKUoBSlKAUpSgKi2jXTIsazbCM8zXLpxs+f3+42SXb3VJTCtMbs31wyhsDdKmzGbQ66pXUvOqOyQhKNkmZG0pN11Wz6UiFzNrU0HiSm228K3bYSAPdq8lS+UErcVsNwlAGi5jov4lqWsXC7h5nE5l3uNtguRQtmVFvDwkIKt1eStl1uSwOigWwD0KiBukyxt5Bb1229tB6I8n/6dYB5yNlIUfRsoAjz7gGsW9zfCK5kZ4XkGdWsacZzfcqm3eHqcytqXBUhPi8GTIjLnQ3GBygobbaYdjqSSedTjZJJBJnFhLiUDtFbn46jWNjb0Z7T3A4CyLfiT8ifIfO26whL6GI6U+ZI8ZG6t+gjpTseclMkvvdk2VDqdwPy1lkzRjcuEdWMXQSW0uNmI6FIUNwrdJGxHnB7j8VRnpDxGQNGcGiYfcsjxKRaYgcXbo94vLtulWyOVbiLuGXkvNIJUG1boKW+RBB5eY7Tq3cp/sHCw/HEtP5Nl8v2GskVxXKHX1NrW4tZ8zbTKHXVq7wlshO6ilJmzSXRDA9IcbgWiw2K3uXNlkCfeVQ20zLhIPVx51wDmJUrc7b7JGyRsEgDzfJhUa5Mhtniry3UBzxTS3C7/eF8hKlWmwSFsLUR5PZ3KUluIP6Y69O6u9bdFNZtU5SZ+qt+GIWR5A7a2W+YZd5lNnYlp+WPtUUbjqiP2gIO6VtkVZelemri8DGY3jVhw+xQ8axi1R7ba7e0Go0ZhPKhCe/8pJJJJ3JJJJJJNZOlKGIrzzwDU3CeGDwguvCdfL01jMfUWNbbpjV8uSVIiyI7SCFtJd22HVYT12G8cjfflB9DKwuT4ThmbR2omZYjZb8wwrnaaucBqUhtXpSHEkA/GKApnxM5fwWcTVrwzI5/EcvT/IrYu4TcJzKKt6DyqZeDMjkcdShDqA62noFpWeXdCtid978HNrDqprHotebjqheRkZsOSzLJZsnEXxf2dgspRyyeXYc3lFQ59gTtsrdaVE2JvGnOnuRWiJYMgwTHbna4G3isKZa2Ho8fbu7NtaSlH5AKzUC3wLVCZttrhR4cSMgNsx47aW220DuSlKQAAPQKA7FKUoBSlKAUpUPI4lLKtce9fuEycYVKvacfay4iJ4gqUqV4ohfZdv4yGFSdmg8WuUkhX3MhdATDVU/Cj+8c1F/GtP8AmkWrWVVPwo/vHNRfxrT/AJnFoDa+GHSDSa5cNmlFxuOl+IypcrCbG8++/ZIy3HXFQWSpalFG6lEkkk9STUm/YT0Z+CPC/mCJ+zrAcKnvX9IfkJYfoDNSnQGl/YT0Z+CPC/mCJ+zp9hPRn4I8L+YIn7Ot0pQGl/YT0Z+CPC/mCJ+zp9hPRn4I8L+YIn7Ot0pQGl/YT0Z+CPC/mCJ+zp9hPRn4I8L+YIn7Ot0pQGl/YT0Z+CPC/mCJ+zql/hadN9O8V4TjdMYwLHLRN/dNbm/GYFqYju8hS9unnQkHY7Dpv5q9BKo94Yf3oB+VNt/wvUBOWg2jukc7Q3TubN0sxCRIkYnaHXnnbHFWtxaobRUpSijckkkknvre/sJ6M/BHhfzBE/Z11OHz+ATTX5IWb6E1W/0Bpf2E9Gfgjwv5gifs6fYT0Z+CPC/mCJ+zrdKUBpf2E9Gfgjwv5gifs6fYT0Z+CPC/mCJ+zrdKUBpf2E9Gfgjwv5gifs6fYT0Z+CPC/mCJ+zrdKUBpf2E9Gfgjwv5gifs6o74X/TrT7E+F2zXPFsEx6zTF5nBZVIt9rYjuFsxJhKCpCQSklIO3d0Hor0WqhXhnveoWT5cQPoc2gL60pSgFde4XCDaoEi6XOWzFhxGlvyH3lhDbTaQSpalHoAACSTXYrz28JjrNf8jvln4SsFvC7cLrD9ncwmtvFsi2gqCIe4O/2zlUpSfOkI8xUDhUnGlFzlyRnCEqklCPNmm8QfG3nPEVfZ+mmgl+dw/TmNIMO4Zg2SideiFcq0Qv+kz6XPdEDfce4PzBdN8O05t3sfi1nbjqX1kSl/bJMlZ6qW66fKWSep3O2/cBUSwLJDs8CK1aEQ4kNpsIjtNI2CEDlSny0kFXXywB17wdwNk4rUbVXKbtMt+mNuXLUu4pJnG1JPsg812iG2o7Ck+SHH3FpaCgPJ8o+qmX1W41aoqcHiPh+c3+IttnSoaZTc5rMvH85Eg6la34pYrTerXZWZeQ3CLGebkotyQpiIrk/wCO+ohtvv7uYq8229RXieLcRmtltWnSfHblK8ZQhUdyy28rhtK7kl25SuzaHKQNw2hwjbvFXq4aPB0YZhtqhZDrlGYyW5h1ybCxVSy7YrCXHFOdklpXSW4kq2LroI6DlAABNzokOJb4zcKBFZjR2UhLbTSAhCEjuASOgFdqx0SlbR9rd/nu/OZyLzWKleXs7L89/wCcjzj018FrlN+bRL1lzODj7LhBeYsS13K8OpCgtIVcpQ7NkhYBIYjgHqNxUban6tYXZMivOgvCPBTiOLwXHIOYaisLMq8Xl1CCXY0WU4SvbdJSp3mPXYpCUgBdwvCQ64X7Rrh0lQMJkvM5ZnU5rGrQ4wvldZLoKnnUEdQQ0lSQR7lTiT5q8+8GxeDhWNW+xMeMlEFBD/IPKfd2JJIHUbr5jv37EjfyTW6/rxsafDSXtM1WVGV7U4qr9lEm6R2LSfHY64eEQkIuBUoypUw9pNkuffKW6dyokncgHbr3CpKqvcZbimkusyJEVTYKiVbpIQnvCdvSFJ2Pd0A22G46E7XrMmXZ7GKJiuQbRKYiXW83pe0C3qdeQ2lPMNlOuJ5wVJ3GyUqUdyfJptWyq3NRyi8vz+5aqd1ToQUWsLy+xZOsTkGTWvHGOec+A4pKlIaGxUQATuR5k9Ntz061XtEDVXVO7Qm8MvOrWo8ZUlYnqw2zG1WtLfZq5UIk7bH7Z2e6lOEcvN59q1DVTTBzCM6gYhetF4tzyq7w1x27FJzt65XCK8ogplzUsEhlCU7DkUtJIJPm3EqloU2uOo3jnyx/3OP0I9TWIJ8EMZ9c/wDan9Ta1NR9UdW8axFtpL7NrfTk98cSk7MdmN2WR0A+2OqSog9dk7jbrvYrI2bYLa9dLmyhabY2uWlSvvORPMfybDqK0Th/0Wh6MYd7GuvNS73cFB+5zEA7LWBsltJPXkQCQN+/cnz7VlNbb5AsundwYuEvxZF5Ui0dqAd0JfPK6vp18hntXCfQg1DuKka9xGjReYrZPx8X+dCVQhKjRlVq7Se78vBfnUinh5uEm8an3qXfUx2pFkx63WWJ2IUUK7VS5bwKlf8AECnE7j4/z2RqjWnOfexUmRGt0ASTk16kz4sSSp+LcfF+QpZUzJkKLcpPI0nlQVJVzq2BINW6xXUfE8ix5V3iT3IwhK8WmRZyS3LiPDp2TzZ8pK+nx794JHWt+r2tSFXjxtsvz1eTVplxCdPhzvv+e5Gnas2zBs5kv2LKsdRJRBQlSJ7Cw3OjuDmPMw4NyFII35VbenYgjffuF3jay7Q7MIGiHEHkErIMKnvoi4/mk4csmApQHJHmnchTe5ADhO6eu+46IjTILoLhcbjNtqUtxZICkIWyoEK6cw6EBKioFR2/CG5O/k65lFjh5bYp1gvLLbrUxvkWjsT5J2BCkbDcKHknfzH73cVO0y8nY4j/AI9V/KIeoWsLzL/y6M9oELQ4hLjagpKgClQO4IPnFfqqY+DL11u+c6aXjRHNZypeT6USUWsSVrKlS7WoHxR1RJPVISpv8VCOpJNXOq6RkpJSXJlRknF4YpSlengpSlAKUpQFb9Zsrn23Xu3YhDwy4SWb3jTUqTfGz/s8MR35JQ24OUjyisjcqSd1J5QvyyjVp2od5teqWK6fwdPLvdId+jypE+/NIcES1BptamwtQbUjdaklIC1oPlJ5ecnlr7xTaiXjAtYrPkF+wyTHwi2WJyDcMlbK1tNuTXkKZD3khCEpdh9mCVkgygVBCSCetH1Dn5q1CxvR5pnJMlnRUPtI7T/Ybchfc/Peb3DSB1PID2jm3K2n3Sk49TdB+ybflWU4Vgsb90WXXu3WhtX2ht6S8EKeUSNmm0+6cWTy7IQConYAGowz7LLvk+n8u+2nLnsPlOP9hjVpLSFXW8zAeplRllLsWIlJKlA8j3L5SuQJDbsvYtwvpwCx3TO0T0Z1rM/b3kRcov23LEkKSeVuEyoLbgsJUeiG0kn79S6hHTjhr1cu8hrHLlZGMbhJswkG4zIrq3Eqecd7WH2qXw4HFLS04FJ9wlIKgFFKE+tHneZNu4RsIvOcaq5Hrrk0m4zrRYGF4ziDs1YUl11S+a5S2htsQVIZjpWDseyfA6HrcOsHg+PHEsLsOLKEQKtFtjQleKMBlgqbbSkltsbBCSQSEjuFZyvVsa28vIpSlDwUpSgFKUoBSlKAUrD5JluP4nHaevc/slyV9lFjttqekSnPwGWUAuOq267JSSB1PTrWqq1IyeHc2kXbBAxEkNKfbhM3Rp+9IYSQFPrhJGykAnqGnXFjpslRVygCQqVjbBkdjyi3puuP3NidGKi2VtK6oWPdIWk9ULSehQoBQPQgGslQHxSgkFSiAANyT5qq9hum+a5Pg9p0YhX3Gl4BDn2/I03AqkovarSqf7IRIbkRTQaacJaDSnu2J5ElXZJUobS1rtmON4/g8yyXu/QoKr+WrU4HZojutxJDiWpMhB35gWmFPOAjzoArIaSdhcsfl5nHVzM5VOcuUQhPKnxEJSzDKB3hKozLLmx++cV6aA3iqp+FH945qL+Naf8AM4tWsqqfhR/eOai/jWn/ADOLQEs8KnvX9IfkJYfoDNSnUWcKnvX9IfkJYfoDNSnQClKUApSlAKUpQCqPeGH96AflTbf8L1Xhqj3hh/egH5U23/C9QFouHz+ATTX5IWb6E1W/1oHD5/AJpr8kLN9Carf6AUpSgFKw1rzDGL1f7vi9qvUaTdbD2IuURtW643ap5m+YfzgCRWZrxSUt0zxNPkKUpXp6KoV4Z73qFk+XED6HNq+tUK8M971CyfLiB9Dm0BfWlKUAryKyyM5qRxjax5NIHO7DyU2VL6juWI0NhtsBKT03JHnBHUnzEH11ryritKxri04gsKks8jy8kbvzRPepqW0Fjb4uqfz1ydblKFjOUfL6nT0eMZ3kVLz+huTFot0e3i1ojJMYNlspX5RUCOvMT1JO53J6kk1GmgenzSvCTYbHurfb2tmxyLzAbc6jtWGHktn4yhaioeoGpXqPM5zV3RjU7T3X63QH7g9hs19q6wYzXO/ItEpvs5JSO7mQPKTvt98fMaqWhXHc3seLlLKLPrNDvbR8PNbnqzStW001OwTWDDrfnunOSRL3ZLk3zsyY6t9j50LSfKQtJ6KQoAg94raa+hlFPObwpEucvWXQW2Ps9rbUvXaUELBKFPhtvv269AE/nqLcFxZi+ylTrlblogs+UyObZL/mSSU+SpPRShynbmIPdy1bvwjGhmY6r6SWjNNMLabhmmm13Rf7bDSjnXNY25ZEdKfviU8iuX77sykdVCq1aYalYnqdjTN7xeQ2kpSEzIJ2S9Ce++bcR3gg79dtjtuKqnaRVYcNWK2xjPh/ss2gSpzUqcnvnOPE5G9M8WSXA7HeebXtyoW6RyEb9QRsTvzHffffz1GOEcP+T6Z39q72iTheZRbatx+0wMwtUh9iI+talqeLbLyG3XTzAc7qFkBKeXl2qeaVWrfUbm1fFTl8d/qd6vY0LhYnH+CL9TNc+OfJIpxyXqpYcIsr20d1OMWZMXkjbALWJDilONbb7DlWnp13HdUfYHhMbAVKnW2K8/dlSSufdJbhVKccKEFbrqnNykeVzHnHnOwG5Jm7OZ8Buzu2t3kcmTU9nGa25lJWSEpc2BBASpSTuCDvsAdyK0rF8Mevz7U1+Kpm1pIKUl7dDqeUe5AA5zv99skDvHNsOXpz1Svd0v772+pAhptG2qf2Vv8AQ3S15vAltMInMvxpCj2b3M0UobWB1KgTzITv0HMAeo+MiK89gZJrtrXiuiWETWy3dJj1llqQ2pammSlKrlKCgdkdjH+0gkHdT7yRsRXZ1FymCLvE0k0NxdWU6lzFdlbrbACnUQFd5kSOvICgEqBX7k7KOwG9XZ4GuC1nhosMnLc5mRrxqNfmA1NlMqUti3Rubn8UjqV1O6yVuL6c6+vm3M3RdL45q6ksR6J9fP0ImrajwQdvF5fXHQm7LtCdKM904Y0ozDC7fdcbiQm4EaM+3uuO22gIQppweU2tISNlpIO4768htXNNLvofrFlWN3bJLhcZuE3G0wocp/kAuWOzSRGDqkpClyGlqCS4on3Gw6b17b145cTuUY3r5xqZVbMPIujltulltfjbBJajRLYFOTXlHflO8hxLSe8nkVt0PWxaio9w5y6fTr8jhWDl3yjHr9enzM7YcGyidZHHzIYYVIZPZ9tzlw+UroT96dgNlDfpy7e5Brtw8EyGS+uDN5YqE8qXJSPK507A7o6956b7jvCieiusn0r507ubbL2raCwR3wm3RenHhIIuJQ5vPHzXEH48pA3AW602t9ClDcjmAikDbuCvXXq1XjzwlzGc28IHiuXRo8gqm3O9vxJCkgtLtMO1vxG3G1A9y5Bd3H4tew1fRtO4lawU+aRQr/hdzNx5NilKVNIgpSlAKUpQHXuEC33aDItd1hR5kOW2pmRHkNpcadbUNlIWlW4Ukg7EEbHeuhi+IYlg9qTYsLxi0WC2oUVph2uE3FYCj3kIbATufOdqxupeWzcCxGXmMazG5x7StqRcmkKIdbt4Wnxl5tIBK1NNc7gR3q5CkdSKzy5hk2pVwsxZmF2OXonK4Oze3TujZY38k9Oo36HegO1uD3Gla9p1lrOf6f41nTEbxdvIrRDuqWebm7Lt2Uucm/Tcjm27h3VitN89VkukFj1Kyow4Jm2Vu6z1NApYYT2fO4RzEkJABPUnoKA3elavpjfr/lWn9hyjJ7e1AuN5hIuDkNtCk+LIe+2NsqCiT2iG1ISs77FaVEAAgDoafZpc89u2R3aJHipxOFLFrs0kJV2099hS0zJAVvy9h2uzLeydyWHF7qStGwG70pSgFKUoBSlKAVqmpl+u+PYw1JsbzLEybdrVakPOtdoGUy5zEZbgTuAVJS8pSd+nMBuCNwdrrRdY/wD7Xtfyrxr/ADiJQHewXGccgsryKD43cLpN52pV2uSg7Nf5F8qklWwS23zI3DTYQ0D7lCayuS4nj+XwRb8gtyZKG1Fxh1K1NPxnOUp7Rl5BDjLgBIC0KSob9CK6uBf/AGwx/wDqJX0hythoCLHdMcoi5M3cYV+UtbxUkX5kojXSOgJUUtymwgx7i10S2ntEJW2k8wKlkuD7gluybVDCLNlmaZrcGmb7a2ZDtqsW9sjoLgC9w8hSpYUNwN0vpSfwetSlWiaEfwL4R/8AsUL/APqTQGxWHDcVxcPHH8fgwXJKUpkPNMjtpHKNkl1w+W6R16rJPXvrD4ZaLjhk+RhqIy3McbR4xZJAUNobe4CoKh38qCQWiOnZnk6dkCvcagLi3wh7UVrSzEot0k2uVKzkvQ7jH354MxmyXV2NJG3f2b7bS9j0PLsehoCe+dG+3MN99u/z1VTwoxB4HNRgCDsq0g/F/vOLWi4xkOoGdznb4xa42O5cvXiHGmMXKE8/Eizo+Gsx5WyErbU6z2rTvZrCwFJKFb7HatU4uo+TR+DPikRmEu3yrqc/tJfft8RyNGc/2fH+UttuOOKSNtgd1q3IJ6dwAt/wqe9f0h+Qlh+gM1KdRZwqe9f0h+Qlh+gM1KdAKw2ZZA/ieI3rKI1guV8etFvkTm7ZbWu1lzVNtlYZZR984sjlSPOSKzNKAp9+/wDM7/kH8Qn9WlVn8A41MxzbNbLiM7g21wx2Pd5jcRy7XSwFqHCSs7dq8s7cqE95PmHp7qtFSgFKUoBVHvDD+9APyptv+F6rw1R7ww/vQD8qbb/heoC0XD5/AJpr8kLN9Carf60Dh8/gE01+SFm+hNVv9AK0XVPUe+adxbfIsmmWR5iua44hxuzNBZjBIBCl79wO+w9RreqVhUjKUWovD8f9nkk2sJ4KQaY625ZZ9eNV8li6EZrcZV7VajItcZgGTbezYUlPbjzc4O6fiFXFwzIJmVYxb8guGOXCwyJrZW5brgkJkRzzEcqwO49N/URWlYBpVeMR1i1I1Gm3GG9CzM24xGGubtWfF2ShfabgDqT02J6d+1SfUHT7etRi+9lneW2F1k3nbx5+8j29OcE+J9X9RSlK6JJFUK8M971CyfLiB9Dm1fWqFeGe96hZPlxA+hzaAvrSlKAV5m+EJsMjRPilwziEaT2OOZ3bRit9kDfZiU2ftbyvU32RHpDKxXplUX8SmguLcSej190qyltKRPaL1vl7EqgzkA9jITsR1So9Rv5SSpJ6E1qrUY16cqU+TWDZRqyo1FUjzR55TsNyhfZAz3ZqQ05yvJlFWxJHKrZRB7iegKubolSu9Stfu8K7tNpcyJ6WG2XlhJlOLQ2SW+7mKQAgk9FJO4JICU78iY+wHU/VfQq+XfQLWqwrFyxEqYblqStxxqKnYNuciUkvR9vKS6nfZOwVsNiJbx/J2tQo71lukyFNhXdt4sOR0haUABJACgOU7b77Hc7cu569aHXt7iyqONVLC6rwLrQr0buClTe76Ea4mvU3SjIE5lwz549Zr5KW29MtSRzWy8ggb9pHCS2FDmHMrZPLzjYkjtFW8058KTYLG+zi/FhpxdtN70rcC6RI65tpk9ehSW+ZxP5O0HT3Q7qr3bzPxC/sGZ2TfYOMsKbSkoR2J3TzJSeZQHMXSkJPKDufOQJVuFutd9t7kC6wI06FJTs4xIaDja0n0pUNjU2nrtWzajJcUPmQ6mjUrrMovhl8i9mnmtOkmrUQzdNNRsfyRoDdQt89t1aPxkA8yfygVB2uvg9tHtW8gf1Aw+4XTTfN5C1OvXvHHOyEpw+d9jfkXuepI5VK67k1SvIeErRO9yDcLfj0nHp/MFIlWWY5GU2f5qNy2n8iRXdgad8QuKNtMYFxjak26NH6MRZ0pUxpA8w5VLCdv6NdWn2hsayxUyvVZ+mTmz0O8pPMMP0f3wSNdeDfj4wVShi+o2nuokFvo2m5tOW+WoebcJRyb+tw1hHNJfCONHkHD5hD2336MlYAP5DIBrqIzfwhdsY7K28WltmJQPJEzEoBWfiKiwo/nNai7xKeEPbdWy5xG4+hTaSpe2M24bbKSlQ3VG23BUd+u2yFHcAb1gv+FuHlKPwa+xn/AOb0Vht/FM2eNwz+EVyG5Ldf0y04sDjx28buF3RIS2Ntttm1uHbb+aak3GPBza85isHX3iccYgKA7W1YZDEULG/VJkrSlRSRuNuT8tV/c1t468nns2y88W4gMPJWp12DYIUctJSvl3JQ02R3pPugQDsdj0rIztJtbctjGLn/ABf6nXqI790jMXFxhpYP83tFJ/sr2VzpFo08LPTZv+DFW+qXKay8eqRfvDsM4ReCrHFsW6ZiODNvpCZVwulxbE6Ye/7Y86ouubnry77A9wFRDqh4WThvxVarPpfGvupd+UstNRrPDWzG5+4cz7qQSCe4toXVWLHwgaG2l5My42CbfpgVzqkXWe66pavSpKSlCvypNSZbcFwuyWtdns2LWu3QlnmUzEioZTzeZXkgdR6e+tNbtPQjtSg367fc3UuztaW9SSXpuQ3qnxWcafEo85ZZVwGlOIPJUl6Dau0bkyGwCSFugF5Xk9FbFCBv1HXasjobaML0jtS8Xat5ZmOBDsy5uN8z8palKG7pTvsgHyQN+hCtx0UquteEKRPMSG9zRoRebbBZWEujZCtyrdRUByjbboFISVDYJFbVbsStsVuNKya9M29K2kvGMlwoW4AT0cKienKrl6ddiRv1Vzc7UL+d5DhqvCfRE+ysoWs+Kmt11ZJbMhiS2l2O8hxCwFJUhQIIPnqK+IPPbjYrBHwPD1BzLsxKrfbWwdiw2ro7JUfvUoST5R8/XzGtd1WvlnwHxFzFb4zKuNzmpft9rivFcyVIUoIACuY8rZUAkqO3UqG5KjWvaYaGarcSGsPsBYspceu8hhUTPMjhIS5b8ftjm3LbYqzuFP8AKlYJSdyVK338o1H03TO+qxnzXRNeHj5ePjyN9/qCo05Q5Pq15+Hn4fEn/wAFNpk9kmaZDry9aHItgxuyxsGxjtFhXMtAS5PdSR0Vu715x0JdWPSK9Mq1rTbTrEtJcEsunGC2tFvsdgiJiQ2E9TyjqVqP3y1KKlKUeqlKJPU1stX2nBU4qK6FJnNzk5MUpSszEUpSgFKUoD4pKVpKFpCkqGxBG4IqPNI7JesFVetMpUB/2AsLzb2MzSN2zbH+dSIe/wCHFWhxoJ/6Pix3JKqkSlAaVo9jN3wzAo2K3lhDSrZOuUeGlCwseICa/wCJncdx8WLO48x3HmrUZOlWUscPdg0VSqLKdcj22xXx9l3la9je0QJ5QVAKVzxw62npvu4DtsDUx0oDS9VYeZXbGmcUwbtYci/ykW2ZdmXEoVaIKkqU/JRudy7yILbXKDs662ojlSqtlsNitGMWSBjdggNQbZa4zUOHGaGyGWW0hKEJHoCQBXfpQClKUApSlAKUpQCtezvFn8vx8WqHcUQZTE+Dcoz7jBebS9FlNSGwtAUkqQVNAKAUk7E7EHrWw0oDR7Tm7Ngej47nFkbxiU852caQhztLZLdWrolmRskJWpSujbqULUSeULA5qy2UZi1jq24EO0T7xdZLanGIUNsbco33W68shplHT3S1DfYhIUryazkyFDuMR633CIzKiyW1NPMPNhbbiFDZSVJPQgg7EGtORo/iRUmHKcucyxtHnZsEqat22tK6f8FXu0DYcrSyppGwKEII3oDU7Pfs+za7t3exT2bghlR7B6K441j0PcLSVB0cr12cCVnonkj7pH3NxIVWcxjHtSdNcbt2NWsWTKrZaITcVhJ7S2zeVsAAbkutOqIHn7Iek+epGACQEpAAHQAeavtARVqtrXO04xKzZC7ijdvevF8Zsji8kuCIEG2FbbixIlyWQ+lDRLaW0qG4LjzSSU7kjc8Vg3WQo5VfpafHrnDipVBiTTJgwwlBKksLKEdrzLWs9qUJUpPINgEgVpWV5NlOZXAYZiMy3Wl2XcJcVtd0tAukebBjslEt5+MXmFpZEhaGEqSslSyk8pbVzV0bNpPk2m+GYjgOI3OfLtWPRkxW5sGSmJOjPKWeZ7sHCYsiMA4f9nWB2SWx2faK5QkCZNh6Kqp4Uf3jmov41p/zOLW33jiGzi1Y3kupkPA7VPwDCrlMtl2nruzjNzkogPqj3CZHihhTfI062/yoU8FLSyogp3SDqHhR/eOai/jWn/M4tASzwqe9f0h+Qlh+gM1KdRZwqe9f0h+Qlh+gM1KdAKUpQClKUApSlAKo94Yf3oB+VNt/wvVeGqPeGH96AflTbf8AC9QFouHz+ATTX5IWb6E1W/1oHD5/AJpr8kLN9Carf6AUpSgFKUoBSlKAVQrwz3vULJ8uIH0ObV9aoV4Z73qFk+XED6HNoC+tKUoBSlKAgrig4RtP+Jq1Q5V0lSsezCx7qsWT27YS4St9+RX/AFGie9B9JIIJJry1z/TPUfhKyOS3rJZrzj8l6Y7KgZ1jzZmWe7ulS1JEmMryG1nmKQlPZq2J6co3Pt/XSvNls+RWuTZMgtUS5W6a2WZMSWyl5l5B70rQoEKB9BFaK9tTuI8M/wA/PB7G+hcToSzA8jce1mytjFo2WZhiduv1oTGQ/JveJy0S2oo5QpYdjrV2qCj77lK9tiem1b5jGrOnOYOiLYstgrmFXIYT6jGlhXoLDoS4P+2pi1c8FDorlcu43zR/Jb5plc7g24h6PbHC7bnQtOxQphRBSg9d0pVt16AVAeqXBRxsMRYsLIsR0+1htVvkNyA5FeRCnvIRuShXbBATv03CCd9tuu9Vq57O5eafy2+Tyn8UWC313CxU+e/zX2ZJHd318qpN5jZNppeIse5aZ656UwW0vCWhlh+bCQ75HZhoOBTZT90JP4u1Zqxav5dc7o5bce1dZdhMxmnG5ORYi+0p11SlhTZU0EBPKEpPNtsebp3GuPV0SvDln3p/xxL5nUp6vRnt9Gv5w/kSTlWT3a9TTAiKkR4aX+UNtJU3IWgHYL5T5Q3UlWxIAHQkb9/z9whFsSvKb0xbWHxyBp0c4DqiNio8wTsAnuI69N9uUbanbNSNR27rdA2dKHHIpaQqe9PfhJmcyO03QSk82xUQf529a9Pz3UzJ7tc0OO6ahyA6iPucoAaO7SXPI5kbrR5ad+7qlQ9NbI2taPsrEUvP7mLuaUvaeW35EnXTTO8NwHmbdcEz4zrSk8nNs45s3ypJPQK327t9upB7+ZPWx+43vGr01AbadU2pxLL8c9ezRuTz7HbkTuVBJ71lB22AKRH2Oa46kwMWevi5GmniqW3n0RJmTqEhJbKgpKUBG/lFO6U7+cbbAgDgyDiIyVti0XCTnGkkJUt1CSph2RcH4KVNqUVrQNiNtgkgdd1D469VpcyzCaTX54ZMXdW8UpxbX55lpK45IWqM6EAFXZq23PTfbz1Tu9cSRlzodv8As8z5zTodEn9zGK9k6lQA5Ajxjcq3PMCfN0rmsmGakat39X7mNAdYdS4jjLCIZyaQ/Fgof5nO1WsjlZCCC1sOYbcqt++vKWg157v6P/8AXCe1NZoQ5fVfxxG/nUHDMZvsNvJb7AmPNdohu0WpPj0t9zlRskNMFZ90kdFBHlN79wTWpaiZ5k2UMuZ5LuVr09x2TDC7a/d3UybpNSEnYsRG1K5CVEg79w2OwPNvPukvg9OK6c5NRdL9h2jOP3aSiTJg2FInXFGzaUFKFjyEbhIPRw9So7datjoH4Pfh20HnMZLHsD2WZW0Q57O5CoSnkOjrztII5Gjv13A5h+FXft9GxLjns/j67cl/1HEr6snHghv8vnzfyKPcN3BlqdxDWdEqbjVx05wK8oYcvmQXrZ7I8hKdllMdKx9oZ5wOVZSE7bKTzdRXqNpHo/p5obhELT3TLHmbRZYW6g2klbjrivdOurVupa1HvUT6B0AArc6V3KVGFFYgjjVas6zzJilKVtNYpSlAKUpQClKUApSlAKUpQClKUApSlAKUpQClKUApSlAKUpQCog1Wn3u+at4BpQxlNzx+y3+2Xy8T5FsfEeVMchGEhmIh7YqbSoTHXVchSsiOADy84Mv1r2aafYVqJAj23NsahXdiI+JMbxhHlx3gkp7RpY2U2rlUpPMkg7KI7iaA0Th+n3OYvPLReLu7fl4llcjHLfe5aUKly4LcePIS286lI7Qsuy32ObbclolW6iomXKxWMYtjeF2SNjeJWOFaLXDCgxEhspaaRzKKlHYedSiVE95JJO5JrK0BB924cLpcYmRYRH1Icjad5Zd5V3uth9ikLlKMt8yJkZqZ2gDcd91TqlpLSlgPOBLid08uh+FH945qL+Naf8zi1ayqteE5t8+58EWo0a3Qn5TyU2x0tstlaghFyjKWrYddkpBJPmAJoCUuFT3r+kPyEsP0BmpTrzO0W8LPodpto7gund50v1MkXDF8btlmlPRbdDLLj0aM20tTZVJCikqQSNwDttuB3VuXtzfD58E2qvzbB+tUBf8ApVAPbm+Hz4JtVfm2D9ap7c3w+fBNqr82wfrVAX/pVAPbm+Hz4JtVfm2D9ap7c3w+fBNqr82wfrVAX/pVAPbm+Hz4JtVfm2D9ap7c3w+fBNqr82wfrVAX/qj3hh/egH5U23/C9WB9ub4fPgm1V+bYP1qq7cd/hDdLuKXQr7F2DafZ5bLn7MxLj292gx0Mdm0HAobtPrVzHnG3k7d/WgPUnh8/gE01+SFm+hNVv9ebWl/hc9CsI0zxLC7npbqc9MsFigWuQ4xboZaW6xHQ2pSCqSCUkpJG4B28wrZvbm+Hz4JtVfm2D9aoC/8ASqAe3N8PnwTaq/NsH61T25vh8+CbVX5tg/WqAv8A0qgHtzfD58E2qvzbB+tU9ub4fPgm1V+bYP1qgL/0qgHtzfD58E2qvzbB+tU9ub4fPgm1V+bYP1qgL/1Qrwz3vULJ8uIH0ObXV9ub4fPgm1V+bYP1qq3cevHxp1xd6N2rSrTjTnPoV4ayWJdAq529gNuIQxIa7NPYvOKKyp9Gw5eux6924HshSlKAVj7nd2rYUJcZWsuAkbbbdKyFYm/W2RcUM+LhJUgnfc7dD/8A5QHRcyx0/cYaR+Mvf9VdVzJrmv3Kmkfio/XvXM3iktX3WS0n1bn9VdpvE2B91luK/FSB+ugMOu83Rz3U1wfi7J/8VwLmS3PdynletZNbQjGrWj3SHF/jL/VtXOiy2tHdDQfXuf8AzQGllSld6ifWa+VvabdAT7mEwP8A2xX68UijujNf9goDQiARsRuDWIuuH4jfUlN7xa0XAHoRKgtOg/8Ack1KhhxFd8Vk+tAriXarav3UFn8iAP8AxQFd7vwucNt9WXLpoNgLziupcGPxULPrUlAJ/PXPZuGfhxsCkrtmgOnSFp7lrxiE4sf0ltk/21Pa8ftK/wDleX8VZH6a4F4vble5ceT6lA/ooDT7DbccxdAaxzDsetaB0CYVtaYH/wCAFZ8ZVNA2Mdj8gP667SsTZPuJix60A1+DiXon/wDxf60BwHKp/mYYH5D+uuNWTXM93ZJ9SP8AWu2MS9M//wCL/WuROJsD3cxw+pIFAYtWQ3ZX/MgepCf1Vxm9XRXfNX+TYVnU4tbx7p18/wBIfqrkGNWsd6XD610Brvsvc/8A1rv56+i9XQd01z+yti/c5av+iv8A7zXw41az3IcHqXQGBTf7sn/m9/WhP6q5U5LdE9621etH6qyysXtx7nH0/wBIfqriVikU+4lOj1gGgOmnKp490ywfyEfprlTljw93CQfUsj9FfpWJfgTvzt/61xKxSUPcSmj6wRQHOnLG/voKh6nN/wBFcgyuJ99GeHq2NdBWL3EdzjCvUo/qrjON3QdzbZ9SxQGV/dVA/wCg/wDmH66/Ksrh/exnj69h+msV+527f+nT/wB6f11+hjd1Pe0getYoDvLywf8ADg/lU5/pXVcyi4L6Ntso/okn+019Ri09Xu3WU/lJ/RXZbxMd700+pKP9aAxi77dXO+WofipA/RXEbpcj3zn/AMiyK2JvGLan3ZeX61bf+BXKMetI/wCWJ9a1froDWBdLiO6c/wDlWa3KApaoTC3FFSlNpJJ85IrqHHrSf+VI/pq/XWQbQltCW0DZKAEj1CgP1SlKAUpSgFKUoBSlKAUpSgFKUoBSlKAUpSgFKUoBSlKAUpSgFKUoBSlKAUpSgFKUoBSlKAUpSgFKUoBSlKAUpSgFKUoBSlKAUpSgFKUoBSlKAUpSgFKUoBSlKAUpSgFKUoBSlKAUpSgFKUoBSlKAUpSgFKUoBSlKAUpSgFKUoBSlKAUpSgFKUoBSlKAUpSgFKUoBSlQJlnGHiOD6rOaUZVprndteaQJbt8lMW9m0t28ym4xnqfXMCkxw66gElHP16JJ6UBPdKjmPxH8Pstb7cXW7BXlRYJubwRkEVXZxBsS+rZfRsAg83dsQe4iujkHFJw+Y9hsnOndX8RmWxlcthkw73FcVMlR2u0cisfbAFvcpT5G/36N9goGgJUpUeYZr9pJnzLbeM5xZ5V2VaG709ZE3CObjGYU0h3Z1kOHkUlLiArc7JKhudiCevgvEforqA7Y7XZdRscRf7/EalxrA5eoTtxAXHTI5C2y84FKDS0r8hSklJ5gSOtASXSoGxzjR0ayibJtlsN5E6Dnx07mRXWGUOR7h9v5ZCx2u3iq/FXuVwEqJQRyAhQEhY9rhozlymUYrqtiV4VIffitCDeY75cdZaLzqE8izuUtAuEDuQObu60Bu9KjR7iZ4dY7Lkh7XTAkNtMIlLUchi+SytLSkLPl+5UJDBSfOHUbb7isozrhozJvNvx6PqviDtzu0RM+BFReo6nZUdSFOJdbSF7qSpCFqBHQpSojoCaA3elRa1xN6ITpltbsmo+N3e3z/AB/trrBvsBcOB4mwHni+ovBQHIoHdCV7DylcqSFV2neJTh5Yg265v644I1Eu2/iLy8gipRJ2c7M8hK/K2X5J27iCDtQEkUrRpGuuisO4X60y9W8PZm4s0Xr3HcvcZLluQFBJU+kr3bAUpKTzbbKUkHqQK/MXXjRKdNsVuhauYe/KydCXLMy3eo6l3BKlqbSWQF7r3WhaBt3qSpI6gigN7pSlAKUpQClKUApSlAKUpQClKUApSlAKUpQClKUApSlAKUpQClKUApSlAKUpQClKUApSlAKUpQClKUApSlAKUpQClKUApSlAKUpQClKUApSlAKUpQClKUApSlAKotxx2iHc9TsgbmJK23tIzHcQdilSDktuJBBHn229VKUBW/iVx2yw+InW7StmG77E26x3LOIrhmP8AOzOmQWmXW0Nc/i6WuVxW2zQc7klwpATWS4s7zJ0l1Y1AzHEmYol5Pkk3BJjMphLsdu33DHbSX1tN7DkeBYRyr6jqoKSoHYKUBmuEq6SXeJ29YLG3g26x4JJuShEccbTcHZNvhgmQzzeLlSN+jrbSHVf8Rbld3hZsNvjYppk+2gl1GrOJkOEJ5tk4S2kJ32322J6ekmlKAZbovh2TtYLfJK7hEnX3WTMMZuL0OQGjJhPTrmtIX5JBW0pTpaV3p7Zzv5qgnEc/vGMaZWjXmxMtMZTaM7Y0/aW8/IlMuW42mTE7RaX3VqDobfWU8qktJITythI5aUoDf8UlDSThLn68YvEiu5Pp/luK3O2Gayl1h5ZxeDF7N9I2UtsIkuqACklKyFAjrvjYGMQJOrWmuiLMmbBtef4U1eLxcIEhUaa4ZUGc47H2b2juNcz6+TtWXHGwEpbWhI2pSgOhbchu3EVptCyXPXY8aRZ42SWyKizQmIIDVntEZyMVuIR2rilKbTz86yAN0thoHastp41BzfR3O9RrhASxc8w0kyq5z0IfekIS+bpCZKkLkrdeJ5Y7Z3W4s77nfu2UoDtZfdJ70nXDSNT6E2HQ3xvL8dWhhtE6TOkXmPOUmTMSkP8AZh0qSAytlRSU8ylKQ2pOOwa9uNaocOLlrii1ytUnol8u8yHKkpkNOi9yXVtJWp0+MNLVvzJl+MEbqUlSVqUsqUB//9k=)
- Specific stimulus/response, \(S \rightarrow R\), patterns
- Visual OR Auditory \(\rightarrow\) pain
- Taste \(\rightarrow\) nausea
Circuitry
![[[@Brandao2008-hw]](http://dx.doi.org/10.1016/j.bbr.2007.10.018)](data:image/jpeg;base64,/9j/4AAQSkZJRgABAQEAcQBxAAD//gAuSGFuZG1hZGUgU29mdHdhcmUsIEluYy4gSW1hZ2UgQWxjaGVteSB2MS4xMwr/7RrIUGhvdG9zaG9wIDMuMAA4QklNBAQAAAAAAAccAgAAAgACADhCSU0EJQAAAAAAEEYM8okmuFbasJwBobCnkHc4QklNBC8AAAAAAEoAAwAAAEgAAABIAAAAAAAAAAAAAALe//8CQAAA/+7////u//8DBgAAAlIAAANnBSgAAAP8//8AAX//AAEAAQAAAAAAAAAAAAAAADhCSU0D6QAAAAAAeAADAAAASABIAAAAAALeAkD/7v/uAwYCUgNnBSgD/AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAF//wABAAEAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADhCSU0D7QAAAAAAEABxAAAAAQABAHEAAAABAAE4QklNBCYAAAAAAA4AAAAAAAAAAAAAP4AAADhCSU0EDQAAAAAABAAAAB44QklNBBkAAAAAAAQAAAAeOEJJTQPzAAAAAAAJAAAAAAAAAAABADhCSU0ECgAAAAAAAQAAOEJJTScQAAAAAAAKAAEAAAAAAAAAAjhCSU0D9AAAAAAAEgA1AAAAAQAtAAAABgAAAAAAAThCSU0D9wAAAAAAHAAA/////////////////////////////wPoAAA4QklNBAgAAAAAABAAAAABAAACQAAAAkAAAAAAOEJJTQQeAAAAAAAEAAAAADhCSU0EGgAAAAADOwAAAAYAAAAAAAAAAAAABB4AAAamAAAAAwBnAHIAMQAAAAEAAAAAAAAAAAAAAAAAAAAAAAAAAQAAAAAAAAAAAAAGpgAABB4AAAAAAAAAAAAAAAAAAAAAAQAAAAAAAAAAAAAAAAAAAAAAAAAQAAAAAQAAAAAAAG51bGwAAAACAAAABmJvdW5kc09iamMAAAABAAAAAAAAUmN0MQAAAAQAAAAAVG9wIGxvbmcAAAAAAAAAAExlZnRsb25nAAAAAAAAAABCdG9tbG9uZwAABB4AAAAAUmdodGxvbmcAAAamAAAABnNsaWNlc1ZsTHMAAAABT2JqYwAAAAEAAAAAAAVzbGljZQAAABIAAAAHc2xpY2VJRGxvbmcAAAAAAAAAB2dyb3VwSURsb25nAAAAAAAAAAZvcmlnaW5lbnVtAAAADEVTbGljZU9yaWdpbgAAAA1hdXRvR2VuZXJhdGVkAAAAAFR5cGVlbnVtAAAACkVTbGljZVR5cGUAAAAASW1nIAAAAAZib3VuZHNPYmpjAAAAAQAAAAAAAFJjdDEAAAAEAAAAAFRvcCBsb25nAAAAAAAAAABMZWZ0bG9uZwAAAAAAAAAAQnRvbWxvbmcAAAQeAAAAAFJnaHRsb25nAAAGpgAAAAN1cmxURVhUAAAAAQAAAAAAAG51bGxURVhUAAAAAQAAAAAAAE1zZ2VURVhUAAAAAQAAAAAABmFsdFRhZ1RFWFQAAAABAAAAAAAOY2VsbFRleHRJc0hUTUxib29sAQAAAAhjZWxsVGV4dFRFWFQAAAABAAAAAAAJaG9yekFsaWduZW51bQAAAA9FU2xpY2VIb3J6QWxpZ24AAAAHZGVmYXVsdAAAAAl2ZXJ0QWxpZ25lbnVtAAAAD0VTbGljZVZlcnRBbGlnbgAAAAdkZWZhdWx0AAAAC2JnQ29sb3JUeXBlZW51bQAAABFFU2xpY2VCR0NvbG9yVHlwZQAAAABOb25lAAAACXRvcE91dHNldGxvbmcAAAAAAAAACmxlZnRPdXRzZXRsb25nAAAAAAAAAAxib3R0b21PdXRzZXRsb25nAAAAAAAAAAtyaWdodE91dHNldGxvbmcAAAAAADhCSU0EKAAAAAAADAAAAAE/8AAAAAAAADhCSU0EFAAAAAAABAAAAAk4QklNBAwAAAAAFLoAAAABAAAAoAAAAGMAAAHgAAC5oAAAFJ4AGAAB/9j/4AAQSkZJRgABAgAASABIAAD/7QAMQWRvYmVfQ00AAf/uAA5BZG9iZQBkgAAAAAH/2wCEAAwICAgJCAwJCQwRCwoLERUPDAwPFRgTExUTExgRDAwMDAwMEQwMDAwMDAwMDAwMDAwMDAwMDAwMDAwMDAwMDAwBDQsLDQ4NEA4OEBQODg4UFA4ODg4UEQwMDAwMEREMDAwMDAwRDAwMDAwMDAwMDAwMDAwMDAwMDAwMDAwMDAwMDP/AABEIAGMAoAMBIgACEQEDEQH/3QAEAAr/xAE/AAABBQEBAQEBAQAAAAAAAAADAAECBAUGBwgJCgsBAAEFAQEBAQEBAAAAAAAAAAEAAgMEBQYHCAkKCxAAAQQBAwIEAgUHBggFAwwzAQACEQMEIRIxBUFRYRMicYEyBhSRobFCIyQVUsFiMzRygtFDByWSU/Dh8WNzNRaisoMmRJNUZEXCo3Q2F9JV4mXys4TD03Xj80YnlKSFtJXE1OT0pbXF1eX1VmZ2hpamtsbW5vY3R1dnd4eXp7fH1+f3EQACAgECBAQDBAUGBwcGBTUBAAIRAyExEgRBUWFxIhMFMoGRFKGxQiPBUtHwMyRi4XKCkkNTFWNzNPElBhaisoMHJjXC0kSTVKMXZEVVNnRl4vKzhMPTdePzRpSkhbSVxNTk9KW1xdXl9VZmdoaWprbG1ub2JzdHV2d3h5ent8f/2gAMAwEAAhEDEQA/APUqqqqKmU0sbVVU0MrrYA1rWtG1jGMb7Wsa1TVS7qvS8e11ORmUU2tjdW+1jXCRuG5jnbvoqH7c6J/5YYv/AG9X/wCTSU3klR/bnRP/ACwxf+3q/wDyaX7c6J/5YYv/AG9X/wCTSU3klTr6x0i2xtVWdjvsedrGNtYXEn81rQ73KxkZFGNQ/IyHiumoFz3u0AASU0eq0UZGV06rIrZdWb3kssaHNkUZEe18qs0/VF2Tfi+jhsuxrGU2h9LWAW2jfTS2y2tldttjP8HU96VF3U77en29ToZi3OyrvTpY7ftq9G/0fUf+da5vuf8AQVjI+rfRcnKdmX43q5LnNd6j3PdBZu2tra55ZXV73epRW30bv8NXYkpr1O+plr2sq/Zzy/YGbRSQ42myuhlb/o2Ptfj3NZWz3/o0xs+prTcHtwGHGea8jeytvpuH5t29rfS/kb/5z/BotP1V6JVVVWaX2moMbvstsc5/pkln2iX/AKf6XvZb+jf6dFX81j47Kueysz6uOxC8dKzMd1PqWsZRFVrWYuS31/Q2XM21/aLqP6J/g8v9U/S03fZ0p6AV/VEv2BvTi/eatoFM+oB6jqY/0raxv9NQc76ltYLHHpjWODXNcfQALbA40uDv3bfSt9P9/wBN656v/m9V61NnQMu91T7oYPcw0+vk+njtZZZj1txGto+1U9P9H7NRken/ANrv066Hp/ROjZVbOoOw7WZFjQ132myyy0Guattj7Lrt+33fpN/6X+cSUp5+prK32EdPLao37W0uILmuurbsYHP321MdZVX9O1n82i4VGFV1bdhVV01XYjX/AKJgYHDeSxztrW/vfnIY+qHQCHCzHdaHO3APsfDRrtqrY17WMpa52/0/9L+n/nv0ibqD83DynnpOK3LyacSsU4rnisOaLmtsZ6z52O9Hfs3JKdtJDovqyKxZU4ObJaYIMOadllbo/wAJVY11djPzHoWT1Lp2JYKsrKposI3BlljWuLZ27tr3D27klNlJUf250X/ufjf9us/8kl+3Oi/9z8b/ALdZ/wCSSU3klR/bnRf+5+N/26z/AMkmPXuiNBJ6hjADUk3MgD/OSU//0O+w8ujGy+oi0mbcsRAnRuLhmyx38itv03qy7rXTGuDTeNxIG0BxIktbLhHtaz1K/Vc7+a9Wr1f52tBwKKL8jqTb62WtbmMc0PaHAOGNiQ4bvzkfLw62YpOJiUWX1DdRW5rQ3dPbjYkpY9b6UAC7IazcC73S0wAC7duHs2bmb9383ZZVU/8ASW1pn9c6WxrXm8em54rFoBLNxY+7+cjZ9Gl7P+P/AFf+f/RqnTj9TbuNuBiOZ9F1bGtD3slvtbuf6bfZ9Fln+i/we/8AR3MDDLsTbn4tDLnPLnsra0sMSyp3H0vS9iSkHUs7Dy8UNx7W2mrLww/brB+1UAf9Qi17+o5QvMjAxnfoG6t9W1pex9z2n+cxqv8AtJ+Zbd+t/wCDw7kLreDi/sp2NXWKasjIxmWej+jP6TIore5rqtjm2e76aL0zJtqf+y8wNZk0MmlzAG13UthgvoY32sdXuZXlY7f6PZs/wF2P6iUk6hRl2W4l2K2ux+Pa57mWvNYLXV20+17K7/dusb+Yq/2/6wf+VVf/ALFD/wBILVSSU5X2/wCsH/lVX/7FD/0ggV9V6lZlXV1dOx35dAY3Ia3LabGBwNlLbv0G9jX7nWVLcWbg4+PX1nqdtdTGW2+h6tjWgOfDCG+o9o3Wbf5aSkI6r1s3OoHTKzbW1r3t+0jRry9tbp+z7fc6m1T+3/WD/wAqq/8A2KH/AKQR6Qf23lnt9lxv/PmarySnK+3/AFg/8qq//Yof+kEXEq6hZnHMy6q6AaBW2tlhtdO4vdvmmlrdv8l1i0EklNHKrtxXuzsVpc3V+Vi1MaXXnayttrXTW77VTXUxjPf+lp/V/T/o76YYtjLOtZb2GWuxMWDx/hM/mVYzs1mHSHlpttsd6dFDI322EFzaap/ktc973fo6aWW5F3p0VWWLN6LiW4nVc4Xlpvvpx8i8VyK22WWZu9lDXf4P2fT2frFvqZNv6W5JTpZPUcLEeGZNoqLmlwLpAgHaTu+j3Qv230r1W1HJYHvgBpkauLWtaZHte7f9B3/Df6C707VlFNv85W15jb7gDoSHbdf5TVi1Y3WCXXPwMJl8i3VokvADv5xrnO3+t7vX/M/0fs32JTfPXekgP35LWGt7qntfLXB7S9m3Y4bvc6v9H/pP0ez+drT9c/5E6h/4Vu/89vQ8PEddbddn4VFZNgfj+1jngfvWObu/S/nfSROu/wDInUP/AArd/wCe3pKf/9Hvvs/XMbLy34bcaynJtbaDa57XAiqnHc2K2Pb/AIDcpb/rP/osH/ty3/0ktVJJTlb/AKz/AOiwv+3Lf/SSW/6z/wCiwv8Aty3/ANJLVSSU49tH1gy/SqyW4ldLbqbbHVusc6KbK8naxrmMb7/S2fSV/Owq8ykMc51dlbvUovZG+qwAtbdUXbvdtc5j2O/RXUvtx7/UottrVlJJTR6fn2Wvfh5jW1dQxwDY1shljDozKxt253oW/ufTx7f0Fn+ltDX1fKvY27GwXvos1qe6ythc382z0y/cxr/pM3/pP9J6f0EH6yMx3P6WX1X25Az6PQ+zODHNG4OvsyPzn4LKmfrVex/+D/m/5+mfSI/ZOHH+hYkpL+0eo/8Alef+3qv/ACSFXkZteRdkNwHl9+3eDfVA2Da3br5qv1HF65bm139Py6seiqp9ZpfuO99u7dkPG19W7D9PGfi7mWep+tV/olVtZ9bqQ5jLWZIFZfvY2lr3OBqa3HqdayutlljH5dnqWY9ldXoYf876t1SSm7Wx1XUruqM6UG5uRWym231qpLKy5zB9L+X7v+Lq/wBGrX7R6j/5Xn/t6r/ySxvsX1vtaS/NordcxjbADGyG3ep6LKqD6Vu66v8ASsyHv9XG9b1fQ/Va9TprOpsruHUra7XutLqPS4bTtZsre706dz/U9T83+2kpL+0eo/8AlcT5C6qf+qRh1TF/ZrOpv3Moexr2tIl5L4bXS2tm7ffZY9tNdVe/1bv0dakz6bfiFmdE6W2xvT+p2X2ObThsqpxCQaWWe7fmtYRu+1uqs+z+q3/tP6tf+FSU3en4mQ+49S6gIy7GllVMhwx6iQ70GOb7XXWbWPy7Wfztmyv+ZopUMvE6s3qL8vp9mO1ttNdVjb2PcZqddY1zDVZX/wByVppJKcrb9af9Jg/9t2/+lktv1p/0mD/23b/6WWqkkpytv1p/0mD/ANt2/wDpZBzMT6zZeJfivuwmsyK3VOcK7ZAeCwkfpv5S20klP//S9VSSVbJ6l07EeK8rKposI3BltjWEjjdte5vt0SU2UlR/bvRP/LDF/wC3q/8AyaX7d6J/5YYv/b1f/k0lN5JVaOq9LybRTj5lF1pBIrrsY5xA+l7WOLlX6oTmW19Ir4vG/NcPzcYHa6vT6L85/wCrV/8AA/a7av0mMkpbpn69e/rDtWWNNWAPDHkON/8AWzrGNu/8K14X+F9RVbqa8PqOH0ui7PrZlttdV6Ia+ikVBr/TsutqsfUx+/8AV27/APgf0f6JbgAAAAgDQALMsGU3rVBvex1bvW+zNYCCGbKNzbi5zt7/AFm2fQ/waSmGRh3U3Y1X2/Ld9psNU7qhtiu3I3f0f3fzGxH/AGVd/wCWOX99X/pBB61lMqzOlUBxZkZGRYMd3pvsbuGNkibPSG3279+19tP6P1P9HYtUTAnU90lND9lXf+WOX99X/pBL9lXf+WOX99X/AKQWgkkpx6sS63KyMb7dlt+z7Pduq928bv8AuP7dq1MfHpxcevHobtqpaGMbJMNaNrfc/c939Z6zun5uNb13quLW8m/GGObmbXDbvY51fvc0Vv3N/wBG5aqSlJJKrkdT6bi2elk5dFFkB2yyxjHQfou2vcHfmpKbSSo/t3on/lhi/wDb1f8A5NL9u9E/8sMX/t6v/wAmkpvJKtj9T6dlWeli5VN9gBcWV2Ne6BALtrHO9vuVlJT/AP/T9VWNk35eP1fMtxKPtLxj4m+sHaSwPz3P2E/4T/Rsd9N/+jWysezqXTsHr2UM3KpxTZi4xYLrG17oszd2z1HN3bdySmf7V6hMfs6wCYJJJEay/wBjHO/Ru9j/AKdm/wDo1eRjfrKizrHU3lrR0uxrpHqNc7VoO7b7tvovd7fzLv0f/F/Zn5NXLzeg5OUb/wBv1VNOz9FXlVho27txZ+k9nqtftf8A+lFGnL6DVdW8/WGuxlTg5tdmVU7WGtdue95e5vt/8EtSUmyuo2l+Jk5eJZisxsp7gD+ke5gw8u0uZXSHu3/8HX6n7n85+jT9M6lgVMsycq4fbMt3q3gBztgjbTite1vuZjVfo/8AhLvWyP8ADqd3Vul5vUOm1YeZRk2tyHvcym1ljg0Y+U0vLa3O9m57G7lspKaGH1zpmdm34GLdvysZjLLq9j2w2wTU7e9jWO3fyXLOxMS/I6Li5/rZF2ZbTXY79KebA117mMa6pvta5zvRrdX/AKNb656uz6y9J6U1hxcK2jAojf8AaLQ9zKW/S9P7K9rbHsZ9D1f7aSlg3PDN5oyHDRpLc4fSdvHs3nbt9RldX87/AIX/AAvvWhjdMc+lll9uVTa4S+r7Q923+Rva73LJZ0TqjWxX07ErDmNrJZnZAOwNNbR/Q/p+m+xvrfz36S307VpC/wCtffDwP/Ym3/3iSUxx8CyzpteQ2/KtvNQfs9dzd7o3beWMZvVQt6i2S6m0Ma/a956hADQdr7f5Oz2e1Wcb/nVj49dAxMBwraG7vtNomO+37G5VHdJ6u603jp2Ey10zYzMvBlzjY4n9T9/6Q7vf+5T/AKCpJToYOBkXVl+WcnFcHkMq+0ufLIb7nvY78525N07q+NV07EGbkE5DqWPcXAlzgfaHu2N+k7amZZ9aa2NY3CwdjAAB9quJgfynYTle6XjW4nTsbGuLTbVW1thZO3dHv2bvds3JKQu670prXON+jQXGGPOgG535ir9EzcXPyM3Pw3erj5Ipspsgt3NNQ2+14bY3+01bCxWdU6Zg9X6izNy6MV7zS5rbrGVkt9MN3NbY5u5u5qSkh6p1Wqms2dNfba+sPeKiAGv2se6o7i76HqbN/wDhPSu9Ov8AwSa3rWZVXbc7AsFNQJLnEg6Ocwez0zZ+Zvf6bLP0fp+l66z2W9Ca9pP1jYW1ya6xlVhrXe/a7+c3O/nPoP8A0f8A6D/q6X2jo/phv/OZu8Ajf9pp03Tuc33btzfZ6W91lf6P3121+pUkp1ch2/qvTXlpaXV3naeRLavaVpLEq6h03K6r0+nBzKsw01Xb/Ttba4Nipm+0tc93u/fettJT/9T1VQdVU8y9jXHxIBU0klI/s+P/AKJn+aEvs+P/AKJn+aERJJTBtNTDuYxrT4gAFTSSSU0T1vpYc5v2gEtc5pgOI3NJre3c1u32Pa5ip9W6piZPS8vGw76/tN1NldPqts9Pc5pa0WbA1zWfyksN/UWdIrPT62W2/aLi9tjto9Nt1z3taf8ASWtb6NX+j9T1ER2T9YmttLcOqwwPSBeGR7QXbvfdv/Sb2fTq/wAGkpnjdbwm49Qyb2HIDGi41ssDN8fpPS3hz/T3/Q3on7c6X/p/+i//AMihZF/1gOQ6vGx6m0uaA257gdji2XO2b/0zd5/dp/m/8J636uzcn6wnZuwqW7jDgLJ2aH3nje3e5n0f9H/wv6JKTftzpf8Ap/8Aov8A/Ipftzpf+n/6L/8AyKH9p685r3fY66y11Xps3h5c0z9pa526nY5n7/8A1zZkfzSg3J+scBz8OmXvaPTFn0Gx+le+yffz+j2V/mf8L+iSk/7c6X/p/wDov/8AIpftzpQ19f8A6L//ACCA7K+sW15bg1SAXNabAZ7sq3S33u3bX/4Nno/4X7R+raySmFVtd1bLanB9djQ5j2mQWuG5rmn+Uk6ml53PY1x8SASqnQ/+ROn/APhWn/z2xXklI/s+P/omf5oS+z4/+iZ/mhESSUwZVUwyxjWniQAFNJJJT//V9VSXyqkkp+qkl8qpJKfqpJfKqSSn31n7Hmz1Ptnqetdv+yftT0J9W3d6P2f9B/xnofovX9X01L/If/mz/wDhsvAEklPv/wDkP/zZ/wDw2S/yH/5s/wD4bLwBJJT7/wD5D/8ANn/8Nkv8h/8Amz/+Gy8ASSU+/wD+Q/8AzZ//AA2TO/Ye0/8AKfB/8t/BeApJKfqDpe39mYm309voVx6M+lGxv8x6nv8AR/0e/wDMVpfKqSSn6qSXyqkkp+qkl8qpJKf/2ThCSU0EIQAAAAAAVQAAAAEBAAAADwBBAGQAbwBiAGUAIABQAGgAbwB0AG8AcwBoAG8AcAAAABMAQQBkAG8AYgBlACAAUABoAG8AdABvAHMAaABvAHAAIABDAFMAMgAAAAEAOEJJTQQGAAAAAAAHAAgAAAABAQD/2wBDAAMCAgMCAgMDAwMEAwMEBQgFBQQEBQoHBwYIDAoMDAsKCwsNDhIQDQ4RDgsLEBYQERMUFRUVDA8XGBYUGBIUFRT/wAALCADuAYABAREA/8QAfQABAAICAwEBAAAAAAAAAAAAAAYHBAUCAwgBCRAAAQMDAgICCQkPEgQEBwAAAQIDBAAFBgcRCBITIRQXGCIxQZbT1BUyN1FXlJWh0iMzNDVER2F1doGSk7S14RY2OEJFRlJTVFVWcXORo7Kz0SRDcrEoYqTBCSUmY2aChP/aAAgBAQAAPwD9VaUpSlKUpSlKUqiOJ/FrTm+QaM2K+QmrnaJ2XqRJhvb8jqRa56gDsf4SUn71bQ8Hei5+t3Zz/wDov5VfO470W9zqzfi1/Kp3Hei3udWb8Wv5VO470W9zqzfi1/Kp3Hei3udWb8Wv5VO470W9zqzfi1/Kp3Hei3udWb8Wv5VO470W9zqzfi1/Kp3Hei3udWb8Wv5VO470W9zqzfi1/Kp3Hei3udWb8Wv5VO470W9zqzfi1/Kp3Hei3udWb8Wv5VO470W9zqzfi1/Kp3Hei3udWb8Wv5Vfe480XA6tO7P+Av5VceD5lEfhxw9lscrbaZSEJ9pIlvAD+4VctKUpSlKUpSlKUpSlKUpSlKUpSqd129kLQz7sXPzRcKuKoDj2vumuXZtKw6yZ5j11ymKVpdtES4tOSEqR68BAVuSnY8wG5Tsd9tqn1dUyWxAiPSZT7caMyguOPPKCENpA3KlE9QAHWSa+x5DUyO0+w6h5l1IW242oKStJG4II6iCPHWDkuTWjDbFLvV+ukOzWiGjpJE+e+llhlO4HMtaiAkbkDcnx0yHJrRiNt9UL3dIdogdK2x2TOfSy30ji0tto5lEDdS1JSB4yoAeGtlSlKUpWsybJ7PhlimXq/wB0h2Wzw0c8mfcH0ssMp3A3WtRASNyB1nx1sW3EPNIcbUlaFgKSpJ3BB8BFU/widfDvif8AXM/LHquKlKUpSlKUpSlKUpSlKUpSlKUpVO67eyFoZ92Ln5ouFXCslKCUjmUAdk+Dc1+ZWjb5xrU/S3GsOYvGQQ4WTSen06zTFC3ccLbc6YvTEXRCEpKUkjYKUsLDgAJO+3LTPU7V2RYMwnXnLb1GyWJhmSScntMyRcFqt8xttwxXGUKioYgrQtOyEtvKC0HmG5TudXNu+UaraP5harDluoeXW2ZpG7ccoRd+yR2PfEhpxllgqbTt0qenStprdC2hv178xkuU5xfLZYOxWc31ItVujYFbn9N3LI3LdF6u5S8Xm5JDSi+4HUx2wy9skNEnYA81WnxYv6y5PwsZ7bb9idm9S5GJxFrft0x125ruJEZTzXYqWygIDheAKXFdSUnxnaN6lp1CwS56x4za8jzG+2qFdMJl2uTcZDsp5CpE7lnpad236MpbSVIT3qAo7BIO1RuLqnl83WK9RrHmmaXLOYur7tohY0VSHLQbAl1HZQWOjLXIhC3CVKVzN8rfLy7jmx9I9VMxvmfYiLJmeb5Fm7moV4gXOwXEyHLOnH0PykqcJKeiCWylsJc5isL3b9akBObpBqFqrcb7hq7PkGa37VB+Le1Z5jOQR3k2q2uoYeMUNoWhLTH/ABAYQ2Gld+lR33G5rswzVmTatD8nurGUay5VmTGGx3cnt7gcZRbLouQ2h4Muvx1mK8grd7xlCkhlCl8pUlJrU4jlOp+RrGPRcuy1Nkd1RsUVi5QZ86Sv1JkW55UlLcuU0h12OXE7dItISFAKT1cpqX6YJ1Jx/OMImycvzm7x06o3zFnId2luvRzZGm5fQKdSU/NDzIQoPr3V1gBWwSBAuHTWXMrqzpddLVlmo2bZO9Dvz+X2aUl5+M3BZakdiOMdK2Wg6XG2EtKSVlalqSvfYpEIyXLc8z7SbVa2MzsryDG7xpuxcWoF0mzbs43cU3FlK0F16OykSA2shxphAbSQAB1V+rlhQUWK3JUCFCM2CCNiDyiqr4Q/2O+J/wBcv8seq4qUpSlKUpSlKUpSlKUpSlKUpSvH+qfGvktg4mMd0asWEm3y511iRpF3vLqXOkiuODmdYaaURsUcxClL3G3fIBBFW9rt7IWhn3Yufmi4VcVK1uS47bswxy62G7x+zLVdIjsGXHK1I6VlxBQ4nmSQobpURuCCN+oiueP2GBitgtlltbHYtstsZqHFY51L6NptAQhPMoknZKQNySerrNZ9RLPtXMI0qRBVmWW2XFhOWpEU3ec3G6Yp25uXnI3A3G58A3G+1dsjVPDIkW9yXstsbUexssyLo8u4NBEBp1HO048ebZtK0d8kq2Ch1jcVDsPzPRvAMxv+KWHK8dgZPd7hIv1ytPqwlyS7KcQlbzpQtwlJKEJUUJ2ASN9gOuojojr/AKD2nRl3JsayazYnhjt+nsOKu91QjaeuU4t0qLjquUukl9Kd/nbiVAJSeqysn1+0ywoQ/V/ULGLP2ZGZmxhNu7DRfjuqKWnkbr75tRSoBY6jynr6jW1w7VHDdRJl1iYtldmyOTaXAzPatU9qSqKs77BwIUeXfZQ6/GkjxGpRSo9p/p/YNLcPt2LYvAFrsVvC0xogdcd6MKWpau+cUpR3UtR6yfD7VSGleTMX1ZuehnAxas4tdnZv6rQ465Jt7rymi4wq4ONrKVgHlI5wrcgjYHqq1OGbiLh8TOCuZNb8au2PRmnuxybiEFp5wev6FaVbrSk9RUUp6+rwg7W/SlKUpSlKUpSqkyniAXZ9QLniFjwLJs0uNshx5kx6zGGlplL6nUtgl+Q2SSWV+AHwVjnXbKvcNz78bafTadvbKtvYNz78bafTadvbKj9Y3Pvxtp9Noddsq29g3Pvxtp9Ooddsq9w3Pvxtp9Noddsq29g3Pvxtp9Noddsq29g3Pvxtp9Op29sqP1jc+/G2n02h12yrb2Dc+/G2n06nb2yoj2Dc+/G2n02pZpNqnF1asNwuDFouNhkW+4v2uXbrqGunZfZVyrB6Ja0kb+MKPgqb1BNYtRpOneMxzaLYu+5Td5KbbZLWkEJkS1pUQXFD1jSEpU4tZ22Sg+PaunDtF8esaW7pc7Tb7jlkicLzOupbU4o3AtdGtxlTilLbQE7oQjm2SjZPt1HtdhvqFoZ92Ln5ouFXFSlKV5j4ueH/AC7VjIMfveFsMM3aDbZtuNyj5A9aZjQeLZCFbMPsyIxKO/Zcb3JCdlDrqmdR9IbrkvEJpLp96ss3efc8cgRdT24EJTMV2Pa3WZcZ4hKQhAed6RkJAHer2AA8FgN8Meotq4lBldgXbbHi8nKl324pTenZUWZHW1yu/wDy5+OoNTF9SS+0+hIH7XxVDIPB1qjYMd0/bhRbF2dg98yJxpmDelRDdIlycU4h/pVRHAy43uGygoXujm2Wnfati3wO5NCx68QGGLH0z2jr+DQt5jrqY9xdlSH1BKlo5uhAeQAs9fe7coFXPoroLddMtXbhf1s26NY3cKsePssw1nnTIhl/pd08oHJs4gA77nY9Qq/aUpSqH4bsVtWccKdlx++Q0XC0XJidGlRXCQHG1S3wRuCCP6wQRW6veOWrh0tN8yvC8PflwZb0Zy8WSzu9GhmO0goclRooBSp0I5OZCeUuJbHWVABVmY1klrzLH7fe7JOZudpnspkRpcdXMh1ChuCD/wC3i8BrZVA9a9Ue1BgUjIG7Wb3LTJixI9uEgMdM4/IaYSOkKVBIBdB32PgqMdsnWH3F4nlfH8zTtk6wn6y8Tyvj+Zp2ydYT9ZeJ5Xx/M07ZOsJHsLxPK+P5mnbJ1hI9heJ5Xx/M07ZOsJ+svE8r4/madsnWEj2F4nlfH8zTtk6wn6y8Tyvj+Zp2ydYSPYXieV8fzNDqTrD7i8Tyvj+ZrotuuOYwdQ8SxnL9OG8ZZyN2SxGns39qbyrZjreIKEtJPWEbb7+OtdpR18U+r32lsX+pcavulKUpSlKVSHDF6/VL7ubx/rmrTzfNrLp1i1wyLIZyLfaYKOd59SSo9ZASlKUgqUpSiEpSASSQACTUQ0uwm/s5HkGa5fOD19vCuxoduiuuGJbLc2tRZaQlQSVOK3Lji1JB5l8oACRVl1Tuu3shaGfdi5+aLhVxUpSlKUpSlKUpSqd4Q/2O+J/1y/yx6riqns6vbvDyi3XW0Y9BRpiHpDmQN22MoSra485zmelKTstkKK+lSE8yQrnG4SRVtw5ke4w2JUR9uTFfbS6080oKQ4hQ3SpJHUQQQQR7dUZxkdelcT7fWb86RavmlKUpSlKVSGt3s36FfbS5/m52sPSgf+KfV77S2L/UuNTGJqxdL1cZb1lxKRdsZhXddmfuTctKZCnm3+gfcaj8p52mnQpK1FaVfM3ClCgBzbFeuGCtm48+RxWkW9DjrzrqVobKG3UsurbWU8rqUOKShZQVBKlAK2JrAvvEFhNkxeReTdi8GkydoXYzyZIWwgLdC2ijnaCUqbKlrSlKQ4gkgKSTrVcRVkZjRi/HW0/JgvzmwCtbLCWoEeYsSHUoIZPLJQBvuCOvwnlqR3XWrCLJcLnDnZDHiu21t9yStxCw0joGi88gOcvIpxDQK1NpJWEpJI2Brra1wwh+3LmNXsOoTKRCSyiK+ZDry2i6hLbIR0jgU0lTiShJBQlSgSEkjCv+v+HWhFmTEujN2kXZ+2tRkROZSCibKRHZcU4ElCNypSkpUQVdGoJ3I6mX622rDdSIeLT2jHjm0vXibdZPSIZjtIcQ2hCNm1JcWtawnl5knwbcylJScudrrg1utLdxkXwNsK7I5m+xHy+yGOXp1OshHSNJbC2ytS0pCQ4gkjmG+9smfY/kt9uVmtVybnz7aEdloYQtSGSpCHEpLm3JzFDjauXfflWk7bEGqu4ZnENDVRa1JQhOcXgqUo7AAPncmueHuxOJK/Rcwn21f6icfuJexUuOqCLq8lJQq4OMkbFCVb9AT1+uXt3yNrupUL1P0ns+rMK0MXWXdYDtpm+qEKXZp7kKQy90TjXMlxBChuh1YI+zVYZZwzy0stiyag6jIWfXKcy+arb73PUXPDLl5Hslah+VMv5dO5ly8/XK1C8qZfy6dzLlxHslaheVMv5dO5ly7b2StQvKmX8uncy5efrlaheVMv5daLEtHbjnkSdJsGrud3RmBOftslbGVzCGpDKuVxs9/wCEH+8EEdRFb08MuXe6VqH5Uy/l1Hsz0dv2CtWdy46kakqTdbpGtDHQ5PKOzz6+VBVu4Nk7+E9Z+wakPcy5cfrlaheVMv5dDwy5dt7JWoXlTL+XQ8MuXbeyVqF5Uy/l07mXLvdK1C8qZfy6dzLlxHslaheVMv5dO5ly4/XK1C8qZfy6meOcMxXbwbtqBqOZPtozCakf3c9Wtp7gdq0xw224xZA+LZb0qSz2U8XXTzLUtRUs9aiVKJ3Pt1Iq4uNpdbUhaUrQoEFKhuCPGNqqm63rLdONU4oktSMi08yN5mGwIkUKex6XypbQlSW07qiukDdZ3Laz1nkV3se4yPYrifb6zfnSLV80qAT9Z7DY8kvlpu612826VFhMuBC3lTHn4630ttttpKirkbX1AEnlNc0a7YE7MtsVvJYrrtwaiPMdGhxSQiS4W4ylqCdmw44lSE85TusFI6+qtVlPEjg+M2S83BM+RdDbGlPKjwYby1yEJfRHWpg8oS8lDriErU2VBJUN9txUmtWqWLXzI1WGFd23boC4gMqacQlbjYBcbQtSQlTiAe/bSSpPXzAbHaMM8QVhTmGW2ac3IgMY/cGbQH1R5Dj02YuKiUpDDKGiXAlpfMSkqOyVKKQjlWrejWjCXJ9tiNZBHkruKYyo70ZC3WD2Qdo4U8lJbQXSQEBSgVEjbfcVtcKz2w6jWhV0xy4puluS4WhKbbWltSgATylQHMBzAEjcA7jwggSCqQ1u9nDQn7aXP83O1h6UdfFPq99pbF/qXGpUxpFfLROnQ7HmS7Pis+8LvL9vZt4M1DjrxfkNNSukAQ066VqUC0pY6VwJWndJTo2OHKUpm3wZmVJk2ezMvRrRGbtgbdaaclMPkPudIQ8pIjpbSUpbGylFQWdiMm+6ATJGY5Bk9kydu13e8uTUOKlWwSm2o8mHbo7qEp6VHzQKtjDiVklI5lJKFDY1hT+Gp16yS7dFydLAmRX4L63bd0m7LtpjW9QSA6nZYMRDoUdx3yk7eBVcMy4aHM1s92sTmVBjGpMm83CHGbtwVIjy7jGlsPKW8XNnG0mfJWlAQhW5QCshOx22o3D61nmQM3tF2ajT4siPIjtyoi3mAW48hhSXEtvNKUFJkkjlWkpUhPWoEg6m08Nr2PswoNmytmLbFyrTOukZdpQpUmRAkofCmSlxKWA4W0pWkpc2ABBCioqkOreg8PVqeuVKubkNQgNxWUJZ50pdanR5jTitlJKkhyMgKQCndJOyknYiFXfhPlXeyP21eWsMx7h2d6pRGLW4iI6qQ2wyh1DQk7l1puOlKFvrfHfq70DZItzTvAU4DDubImCa5PktSXFhnotlIiR4/g5jvuI4Ph6ubbxbnybhuRXG6aqXTT03OTimP3rPL1OlXpl1TLlwcYkhSLYw6nrbcc251ndJLaClG5KtvbjTSGGkNNIS22hISlCRsEgdQAHiFcqUpSlKVpsytVzvuJ3e22W7CxXWXFcYjXQsdMYi1JIDoRzJ5infcDcdYFeUv/h+6B3XRtzUB4Zku82dV7mWh22Owej5pER8tplJX0quUrTzBSNjvunvjyivY1U7xLdVt05PtZ3ZPykCripSlKUpSlCNxXhPWy8XLDZV60uTdH8tsFvudjusS4Puqek2UO3eNtAlOq359xutoklfICFDYJUfdlKr53SJl3Uc5abkorNzYuXYhYBAU3b5EMJCubxiRz77dXJt49xGsc4b42O2+5Rk3xcjs1NsSVLiABHYdwkTQQOf9sZBR9jlB699qjMvhMl3K4XGbcM27NuEi0XC1MzX7etyQrsiVFkpdkLXIV0hbVEQnkbDSOVSglKOraU4Xw7MYhn6ciNziy2W7jcLu0x6nqD6ZUxTqnj0qnVpDYVIf5QhtCgFhJWoA83Rm3DjHya/3K+tXtqNc5WQG+MpmxFux0b2uPAWytCHmlrBTFS6FBaCFdWxAPNqHeFJTkvHkJysuWmzSbNLYiPW8lTK4Ehp8pYCXkssoeU1urZkr3We/KQlIt3TfC29OsBsGMNSlTkWmG3ETJWjkLnKNubl3O2/tb1JKpDW4b636FfbS5/m52sPSjr4p9XvtLYv9S41fdKV42uOU57ktvn2+83jIH0QcgsM1Vwt8JLbUdpN5bS6hTa4Ta2VJb2WpsqfASyFFwpJ582x3nVB+2rTBu92x5iHIsLDUG32SIhpYm36VGmuKSqOo7ojdG53vKEkBatwo7yS9anZlg+OZJFuFzvdwuSbTfW7LLNmDrj8tia83GUehY5OcNFk7FISpKSvlICyNrodGu8HVbI2JNucj25b98fRIdhBKlLVeXSkB4p5ikoIUE83Kdwrbx16DpSvNek2nll1SxbV3Hr6wp2G/nN2Wh1lZbejPJkbtvMrHWhxCgFJUPAR/XUy0r1DvVhybtZahyEuZZHZU9ab2EdGzkUNHheSPAmQgbB1rxHv07pV1XFSlKUpSlU7w1fSzUQe1nd8/KSauKqc4meqz6en2s7sP5Wkf+9XHSlKUpSlKqrWHU26Wy5QcEwZDM3UO9NKcZLyedi0RQeVc+SP4CT1IR4XF7JHUFEVXrtpja9KNAoNotzj02S9kdol3C6TFc8q4y13SKXZDyv2y1H7wAAGwAFeqaUqkeJrLsqxCFZ5WNXOZDQGZjj8aDH5nZS0JbLSW3FRJKOk618rCg30pJ2cTyEGrX52bWjKc1dtT13RcVyMgkxn59paecgKWxblRQ0otetIK+9CilXRbHco6pVlM/VLFJt3Rbcnv+Qlu8TbVCYftcQhTP6n3ZrLyi3HSVLEwJbSoFKCOVBSVbkxvUXOL3qRl1tkWmLcrhYLc6iTDTKs6koS+5jt/S9uFtjnHOY7ageZIWQjwkg+rbJzmy2/pEhDnY7fMkJ5djyjcbDwf1Vm0qkNbhvrhoT9tLn+bnaw9KOvim1e+0ti/wBS41fdKUpSlKUqkOGL1+qX3c3j/XNT3VXS62asYyLZNdet86M6mXbLvDPLKtstHzt9lXiUD4R4FAlJ3BNRzSHVG53W6TcEzlpi36iWZoOPBkcse7Rd+VE6Lv4UKPUtHhbXuk9XKTatKUpSlKp3hr+gdSB7Wd3r/WB/96uKvKXHFr9i+lbmBWi/N3Jp8ZFa76h5mKVsuR4stC3wle/WtKQDy/8AmT7delcOyiNm+KWnIITEqNDucZuWw1Na6J4NrSFJKkbnlJBB2PWN+utxSlKUpSq71h1WVp5Bt9ss0EX/ADi+uKjWOxpXy9O4Bup10j1jDYPM4vxDqHWoA89H9KRpvbJ0y5zzf8zvbol3y/Oo5Vy3gNkoQP2jLY7xtsdSUj2ySYPxkDfSyJ9vbN+dItXzSlKUpSlKpDW72cNCftpc/wA3O1X6NXMf0Y4mNR5uUpu0eJdrRaG4b8GzS5qHFMuTS4nmYaWAQHW+o7euqbnjY0r2+jMj8k7p6PQ8bGle30ZkfkndPR6d2xpWfqzI/JS6ej07tjSs/VmR+Sl09HoeNjSvb6MyPyTuno9O7Y0r2+jMj8k7p6PX3u2NKz9WZH5KXT0evndsaVn6syPyUuno9O7Y0rI+jMj8lLp6PTu2NKz9WZH5KXT0evnCVcE3yyZ5eGGJbEK65dc58QzYrkZxxhx3mQvo3EpUAR7YFXzUA1e0oa1MtcGRCnLsOX2V0y7Hf2EAuwn9tiCP27Kx3rjZ6lJPtgEY2j+q7udt3Cw5DBRYM+sRS1eLMFEpG/rJMdR63I7oBKFeLrSrZSTVkUpSlKVCtL9PndPWspQ7MRM9Wcgm3pHIgp6JL6goNnrO5G3hqa1W+t+gWKcQFnstuyqKZDNpubFyZKQN1FChztK/+24jdKh9kHwgVY6UJQhKUgJSkbBIGwAr7SlKUpUN1V1QtulGMC5TGXrjcJTqYdss8Mc0m5S1/O2Gk+Mk9ZPgSkFR2ANR/R7S+5WOdcM2zV5m5ai3xtKJbrJKmLZGB5m4EXfwNIJ3UrwuL3UfEBaVURxjxpcjSXmhQZNyfjXS2yzFiI5nVoZnx3XOUbjchCFHwjwVr+7TsPufag/A7Xn6d2nYfc+1B+B2vP07tOw+59qD8Dtefp3adh9z7UH4Ha8/Tu07D7n2oPwO15+ndp2H3PtQfgdrz9Dxp2Hb2PtQfgdrz9O7TsPufag/A7Xn6HjTsPufag/A7Xn6d2nYfc+1B+B2vP1GJWriNbtb9KF2nEcotMaxzZ8iZKvUFDDSUuQnG0bFLityVEDweOvQ1006tF2mLkvtczi/DuKxe1TYtvnPxCnapsW3zn4hTtU2Lb5z8Qp2qbF/E/EKHSmxH/k/EKdqmxfxPxCnapsW3zn4hTtU2Lb5z8Qp2qbFt85+IU7VNi2+c/EKkFjsEXH4hjxE8qCd+utlSqz1h0plZg5bsnxWY1ZNRLCFKtVycSS0+g9bkOUB1rjubbEeFCtlp6x17HSTVaJqlZZRXDdsmS2p7sO9WCWoF+3SQNyk7euQod8hwd6tJBHjAj3ErkV4s+JY9a7JdH7JJyTIYVkeucTbsiMw6VKdUySCEuFLZSFEHl5tx1gVEhwu6eugKlQrxcJJ+eSpmRXFx11XjUpRf6ya+9y1prt9Jp/w7cPP0PC1pqf3Gn/Dtw8/Q8LWmv8AM0/4duHn6dy1pr/M0/4duHn6dy1pqR9Jp/w7cPP18PC1pqR9Jp/w7cPP197lrTXb6TT/AIduHn6dy1prt9Jp/wAO3Dz9O5a012+k0/4duHn6+HhZ01P7jT/h24efr6eFrTU/uNP+Hbh5+nctaakfSaf8O3Dz9fF8L+BspK7c1fbLNSPmU+3ZHcG32VeJST05G/2CCPbBqccNeWXjMNJoki/TTc7rBuNxtLs9aAhcoRJr0ZDq0p6gtSWklW2w3J22qYZ/ntl0yxKfkeQSxEtsNAKilJU44snZDbaB1rWtRCUpHWSQKr3SrAr1k+T9tDUCL2PkshlTNksKyFox6Evwo9oyXAAXXP6kJ71PXcdKwrrZol6YDUpsOIB3APirUnT6yn6lHxUOn9kI+hR8VO1/ZP5KPip2v7Jt9Cj4qdr6yfyUfFTtfWT+Sj4qHT+yEfQo+Kh0+sh+pR8VO1/Zf5KPip2v7Jt9Cj4q74OGWq3SUPsRgl1HgPtVvKUpSlKUpSlKUryVxv4XqehVuyvRewzRlnYj0C7Xm1S2m33IKknZgsK63VBR50LT37ZB5fXHbsud/u2T6B8PM6/xJsG/DJrRHuDFxaW1ITJaQ806VpWAoErQo9ft16BI3FKEb1QmivE2rVXUvJcfk2pu3WlDT03G7ihZPqrEYkuRn3evq3DiAQB+1Vv9k5kbjI0vnR2Hotxu8pEmIubELNhnK7LabOzpZ2Z+adH1lfLvygHfbY7STujdPjarrcvV4CDbLNHv77yozoCoL4PROtgp3XuUlPKnchWySATtWFf+KHTzGrrLt8+5TkOw1x2ZTzdqlOMR33+XomHHUtlCHSFg8hII699iCK0GovFVYscyG2WHHw5d7mvKYONzXXIUgQ2XHnUoebTJCQ0Xm0qB6Pm38PV1HaRP8RuIv3m4WOC/NVeWmZyofZdsksRZzsVBU+hl9aAhzk5e+5VH7G9aCx8V+Lw9PMRv2WGVbZl3ssa8TBb7bKlRYDb2yQt11CFJbRz7gFR36qvNKgtIKSCCNwR46+nwU2p4apLTfPO1dwj59liVIS9aLnlMpgLG4U8m5zOiSR9lfKPv1D+EfVCVxk35Gb5tCcju4mGkWiyMRHRbBJKNnbh0qhyuPE8yUNkksp3I3KuavZtKUpSlKUpSlKUpSlKUpSlKUpSqb4qdfZvDlpfIyqFiczKFpWGeZlaUR4ilEJQt9W/OElRAHKk7nYEp3BqlbFqhfdaeHTQnNMkVHN3u2eMuOJitdG0hKZMxtCUp3J2CEJHWSTtuSTXp4jehG4rQ55ZLlkuFXy02e5ps1znQ3Y0e4qaLnYylpKekCQoblO+46x1gVSOP8GVkwG64Dd8QvVwt12xpzo5Dk+VIlszIq2lIkNIZW7yslZIWCgbAj1p6tsnDuFh/FLZgEU5G1JOLY/dLIpYhFPZBllJDoHOeXl5fW9e+/hFV7kXDlNlZpoxhUaLdJVux21twcmvoiBm33GAw41IajklRJUZEdPeDcgOKJO1ajWDTTN5ea5vimM2y/Gx5RkdtvK0OWdtyG46lyOp54Tw98ybCWeYtqb5+ZCQDsTvZ1w4Wb87fFMQ8yiMYgM1ZzdFudtRXLEkPpddYL/SgdGSFFPecwKhuSBsdfZ+EO9xc+g5Lc8yiXZ+C5dgmW/bnVTpTU1l1tKXnlPqHzLpEhKUISnZJ6tzuNRlnBJesmw624yrOYr1ri41FsaUT7Y68IrzG5MmM2JCUIU51BRUlagE9SvBt6viMqYiMNLUlS0ISklI2BIGxIFdxG4oRvSq54aLBbMo0SuVsvFvi3W3PZTfy7Dmspdac5bzJUnmQoEHZSQeseECryjRmYcdtiO0hhltIQhptISlAHgAA6gK7KUpSlKUpSlKUpSlKUpSlKUpSlUZcx29NcmrWn5thGnklEqafC3PvhSFMs/ZTGQoOKH8Y43v1oNY/Evb7XhuI6fSI8aLZrDa82t8uUtltLMeKhxx0LdXsAlALro3UdhuvcnrqyW1pebSttQWhQ3SpJ3BHtiuRHVTahG9COqm29NqEb02oR1U2ptQjeuqVJZgxnZEl1uPHaSVuOuqCUISB1kk9QA9s1C+ERYlaLomtHniT79e5sV3bqeYduklbTifbSpCkqB8YIPjq6aUpSlKUpSlKUpSlKUpSlKUpSlV1rnqNM0+w5tuxMtzsxvklFpsEFzwOzHAdlqH8W0kLdWfEls+2K2+lOnMPSnA7ZjkR5c1xhKnZc9/57NlOKK35Dh8anHFKUfa328AFSW5WyHebfIgz4rE6FIbLT0aS2HG3UEbFKkncEEeEGqYvvCxohjtpn3afiUC02uEy5KkONSpEdhhpCSpauRDgSlIAJ2AFdsDhO0ZusCNNh4szKiSW0vMvtXGUpDiFAFKkkO9YIIIP2a7+4/0h2/Wej3/L87Wgc4ddDm88Yw84kr1aetjl2SgTJfJ0CHUNKPN0vh5nE9X9db/uP9If6Ho+EJfnadx/pD/Q9HwhL87TuP8ASHb9Z6PhCX52h4P9IT+89HwhL87TuP8ASH+h6PhCX52h4P8ASE/vPR8IS/O07j/SH+h6PhCX52ncf6Q/0PR8IS/O1oIHDrodc83vOKMYko3e0wos6SlUyWEBqQp5Leyul6zvHc3Hi6vbqRMcIejzL7bi8Ggy+jUFBqc69JaJHg3bcWpKvvg1bseM1DjtMR2kMsNJCG2m0hKUJA2AAHUAB1bV2UpSlKUpSlKUpSlKUpSlKUpSlFEJSSSAB4z4qo3SlJ1m1LuWqsn5pjttS9ZMPbUO9WyFcsu4Ae28tAbQf4trf/mGrypVCcb+C5HqHw7ZRbLDfI9jjsRnZ9zW60pa5MZhtTpYRsRy86kJ3J8QI2IJrZ8IGDZFp1oDi9jyG+sZB0UZDsCQ0yptTUVxCVoZXuTuUcykgjYcoSNuqroqnZvVxg2f7OCTviuEX/eripSlKUpVOYj1cWGpY9vFsfP/AKi51cdKUpSlKUpSlKUpSlKUpSlKUpSlU1r/AHqdlEmz6TY9KcjXrLErVcpjB2XbbOggSn9/2q3OYMNn+E4SPWGrYslkg43ZYFptkZuFboDCI0aM0NkNNISEpSB7QAArNri863HZW66tLbaElSlrOwSAOsk+IVRuda24ZqTppq9Y7Ff7dOn2uz3CN0bE1l0ykm3JeLrIQslTY6cIKthstCx4qsnSVXPpVhh9uywj/gIqV1Td0WGuL6wqJ6v1CXHfq9qfDqf4PqNYNSIUmVj8t+WxGcDTin4T8YhRG+wDqEk9XjAIqS0pSlKVTuL97xZ6hj+FiVhP90q5iripSlKUpSlKUpSlKUpSlKUpSlKVq8qye2YVjV0v95lohWq2RnJUqQ54G20JKlH7PUPB46rXh+xe5y4931HyiKuLleYqRI7DeHf2y3IBEOH9gpQorWOr5o6v2hVvUr4pIWkpICgRsQR4aqrUXTyyYrplq1cbdEQ1KvdsmzJCuVPerFvTH2RsByp5GEHb2yo+OpPo6rn0iwc+3Y4J/wDToqX146yvimw+zcb1osj8O+Ku0S0SsZXERAJWuXIlxHGSjvuttSEFXP4NiD469i0pSlKUrx1gfFRh1+41ckscSHfPVa52uBjyIjkApW1KiSJy5Bc77vUJQ6k83X1A+1XsWlKUpSlKUpSlKUpSlY9xfVGgPuo250IJG/t7V5Y4ceHvB9SdGrBkmRQbncb1cFynJMk364N85Ep1I71D4SNkpA2AHgqy+5E0s/mS5+UVz9IoeETSwj6SXPyiufpFDwiaWH9xLn5RXP0inciaWbfSS5+UVz9IoeETSw/uJc/KK5+kU7kTSzb6SXPyiufpFVrxKcOGBYJoHnmQ2KBdIF5ttpekxJSb/cFlpxI3CuVT5SfvgivVEJwvQ2Fq9cpAJ/r2qktQB27tXoGnrXzXEsXUxecoUPWSZO/PBgH2xukSHE/wUNA+vq86UqK6jan45pRZY10yWY9EiSZSITAjw3pTjrywSlCW2ULWSQk+AeKqszfiS09y7C7/AGNuVksddzt8iEl5WHXdQbLjakcxHY3WBzb7V8wfiS0+xLCsfsTkvJZLlrt8eEp5OHXdIcLbaUFQHY3Vvy77fZrdniy07/jMm8kLv6LVb3TNdD7trnaNVn4+THJ7bbXbc2r9R925FcxHK6R2L69KC6gH2nP/ACjayO6y07P/ADMm8kLv6LQ8WWnf8Zk3khd/Ra6pfF5prBivSZMrIo8dlCnHHncSuyUNpA3KlExtgABuSa7E8WmnS0hSXclUkjcEYhdtj/6avvdZadn/AJmTeSF39FoeLLTvb55k3khd/Rad1lp2f+Zk3khd/Rad1lp2R88ybyQu/otVvaM00OsmuN71UjR8mTk92tzUB1X6j7tyJ5T3zqR2L69aUtJJ9psfwlb2QeLLTv8AjMm8kLv6LVgaf6g2LVHF2MhxuW5NtTzrzKXHozsdYcacU24lTbqUrSUrQoEEDwVI6UpXnrX7H4Wea5aRYteeyXbHMN1dkRY0x6L0qkReZG6mlpV1EA7b1Ie5E0s/mS5+UVz9IoeETSw/uJc/KK5+kU7kTSz+ZLn5RXP0ih4RNLD+4lz8orn6RTuRNLP5kuflFc/SKHhE0sI+klz8orn6RTuRNLP5kuflFc/SK0OiGJ23TviG1SxqxCUxZWrRZJTcWTOflBtxZmhakqeWtQ35U7jfbqFegqUpSsO8/SqX/ZK/7V5k0xym+YXwL227Y07EZvzPStw3J7RcYS45c1tgrSCCU9/1gEGpdaOJF643J2Oq0q9Vw1AtisdLiUOxrw7IlNvMuPHqDaUsBwL2PM3stIVzpB3Fo1/l5JdnrNZ8OmT77bOlVeoAmtI7DS3IUxs0tWweUsoWtA7wFKe+KFEJPXhmus3LJYhWjHpd+MNzmukovsx1Rm3J0mM2UNk7OFIjOLUOZOyACOZZ5K0uNcW0LLLIm52/FLnJamtwX7UhpxP/ABLUqYxFa6VSglDK95LThRzLHISQolKkjCyXiXu8G/OQfUVFtjwLZdHLqUupkPMTIsqC0lLJPKlxtSJgVuQkkLTuEFCkmQZFxR2XHoL63LPNemxVuxpURK0bxpIuKIDDThG+wecUtaVbEdG0tWx70GN656hI1K4O9VLiLc/bH41tmQ3mXQooUtCQedpZSkuNqCkkK5Unr2IBBAtLVHUhvSvTNy8ojG5XVwNQrVa0Hv5850hDDCf+pZG58SQo+AV90V02XphgzMGdKTcsjnvOXO+XMDYzJ7x5nnP+kHZCB4kIQPFU9ri44hpsrWpKEJG5Uo7ACuVUTxNH/wCo9Gh/+bwf8rlXtVXagzr5cNXsRxi3ZTPxi2TMfvNzlOW9iItxx2PItjbW5kMugJCZT24SBuSNz1VDsH4ir3c8atU6dZ4NwiNSLZb51zamFhyS5MkdAzJjxwhYLKudh3rcHeuL236Pvu+x8Sd3cbxKTkOKwrFbr5brfcHJiLm9Kaidl8/RtOLRF5W1gpQN3S2hZWQhaikg8sS18u2c5njVp9Tm7Kh6fGW/0ReWmVFkW25PITtIjMrTs7CSrmSnZQCdlbFQq+agmvY30L1F+5y5fkrlS+zddng/2Df+UVodUX73BwO6zscW56sW9CJzMdpCVqlpZWl1cYBQIHTIQprfwjpNwQQCKLyDiiu1ru0u72OAjI8buUadNtClrLLBh24MIkO9IGyUqdfkOJ51nkQiOhW3fmuOfcTeWw8Yvsm12a2WwqF6j2yW9KU8427b5TbKlut9GEgLSpZAClcpSPXBR5drk3E1frBjl2uYsdoddjSb6mNCEuSp16PapK48h1fKwUtJKg336lBKAtO5USE1ZWjWW3DMrdk0qfIMgR7/ADIsYFKR0bCSnkR3oG+wPhPX9mrBqleEzr02vn3XZD+dZNXVSlcVuIQpCVKSkrPKkE7FR2J2H3gf7qo7U4f+KLRb/ovH5HV51QuGaw3y73fKZlznyE26yXW8R+wI2Lyg05HhuvITtPKuhKylsE9Y77dOwNd73FbaLfd7dbLpj9wtc5+PEmSIciRGVIjsynVtsENocJdVsgrWlvm5E+NSuqsnFuKOx5rc5sSzWuRNCI0mXEdRMigPtsOpbcLm7g6DcK6RPSbcyEqJ5VAprAPFC2mNarirFr4hu6R2VQbSW4wfldNcWITDyXS+EJQtT6FgK23QebcdQOXduJ1i1WO7zf1HXmVLsEKdcb5AYejdJBYivOMrUkqcAd51MuqQE+FKCTyq5Um7GnA80hY8CgCAapbBhvxY6rfaCwf5p1XXSlKVh3n6VS/7JX/aqS4Wcdt2V8LGK2y6xkzILqpC1sqUpIJRPdWk7gg9SkpP3qsW86NYbfr3eLxKsqBd7uiIiZPjPOx33DFWpcZYW2pJS42Vq5XE7K2O2+wAGtnaB4C9BYbkWhxDEZDwdWLjJQqWhxzpnky1BwGSha91KS+Vgkq3HfHfHtWlGnWSOw50C1PrZYQzIbU29LZiy0KeVMaKxzBuSlLrq3EhQWEKcVty7kViZDw1Ylc8eNqtiJdnbMu2upUibJdEaPEnsTBGjJU7/wAMhRYCfmPIE96QDyJA+zdLNNDdI9hftcuVcYTC+dCXpzzi2p7qlKXJdCiXUuOwioqdUrlUyg7p73eUXHSLELo9kjz9la7IyJ2M/cn23HG3XnY3L2O4FpUFNrbKEqSpBSUqAUDv11W3ExiVswvhJ1Pt9qaeQyq0S33HJMl2S864sbqW466pS1qPtqUeoAeAADaal6L5FqJleF5LZs3bxxWOMOqiwpFnROa7IdRyGRspxA5w2ShO4PKFK22Kq5nTbWEj2Z4Y/qxBjz9fDppq+fDrSx97EY3nah+sWjOsOR6TZna2dUxkD02zy46LQjG4sczVKZUkMhzpO859+Xm8W+9d/Bro1qtpBhnYepGdHIELaSmJY9uyPU7wdXZSu/V1dXIO8Tt3pNbbia/XJo1928H/ACuVe1aPJ8DxrNjFOQ47ab8YvN2P6pwWpPQ823Nyc6Ty78qd9vDyj2qpK0664bf2cey+56fiLcZOMKv1glLRDkzVxvmCUx0FKitlxapjKEpOySpwp5geoyI5ZiWQpwafYsCh3vIpVrdkW5h+PHYcs8VkobeQt5QPQ8i3UtdGjclXMAOVKlJmmneNYy3YLfcbVhULE3Vd/wBgiBHZdjOI6RsjdrdO4DjwCkkgpcVsdlneZ1BNe/YL1F+5y5fkrlS+zddng/2Df+UVmVTOruY49hj0WzScOsN4hWi3rvbbNxkxoqIyWlEDsdLqCnpBsdutAG474b7jhE1pwS9TMgiXGxJhWW32V+7vTrhEb6J1osMyJ7ZRsSFIblRi4D64uqHWUKrbY0mLl8q2Wu/6XRLTbiw5c4HZSYskRXCpJUh1oD5g+rpObvecEpc3XuBzWXBtsS2MlmHFZiNElRbYbCEk+3sPH1CsivPGgFqvt50XyGLjl+axy6qzG/lFwegpmJQkXaSVDoitIO46t9+qpKdN9YT9eaF97EGPP187WusB+vRF+9iEfz1Dpnq8fr1ND+rEo3nKpfio4ddcdRcJsNts2o36pbi1fGJLaU21i0iDytujsovtrKwE822yQSefqBIrdY/h+YYNrDoPac5zA5vf2W7x0lzMZLOw7C6kdXWvb+GrvleE16xqJWmz4q1+qLDoKE8zvS3C5wA44Vf8c4+tayoncBxaX+pJ6tjsANq0+ZafYRbnmMivJk21uOmFBcWxPkssyUofAitvttrAdCXXABzg+vIO6SRXRcuHTB7va7pbJcO5O2q4NuMuW9N5mNsNNuPpfcQ0hLoDSVuITzJTsCndHrCUnvuWkGFWyMLtcYtynm1oRJTImXKbOebSw+1LQEhTi1q5XY7awgb78u2xCiDFdZOGtnUqHMhWqcxYo90hz4NxeSqWJCky1BTqkqakNpWN+c9E6lbe6gQAApK7tbbDTaUJ8CQAN/sVS2DfssdVvtBYP806rrpSlKw7z9Kpf9kr/tVR8HH7G7EP/wCz8sfq6K885toper7rk7kMSywpUGaQ3Kl3NUeQz2KYK47iEAtiQyslXL0SVrYUlRWpIWpVRnDeGu7S5uKM5Fi9pYxyEmGmXYnHGn446G0PxVfMwORY6dbah1e0rYEVxwvh5yzT5vEblBtER2Ra7VjLt0gRZSG3LhcYzVyauDpUdkrdUJcdXSLI6To9iobCt3iOieZwLnZZlwZitOMXOHMkhuX0gCG7heH1gHYFXeTo+24G+56hsa9J1T3GF+xd1O+0cj/LVs276Xxv7NP/AGrIpSleeOLzILdib2lN6u8pEG1W/MYciVKcB5WW0pc3UdtzsK3vdoaJH641p/xPk0PGfol7o1p/xPk1UlnzXhesun+L4fHzexotlnTBTMW1GDTt3ERvZrspSGwVfNQ28SNu/QPETXSdQeH+0peOOat26zpU5OS1DkRezISYszolSoi2Fo79pbzRe6yFBTiwFciigzXS3iI4f9JcHhYvaNSLWuDFdkvpKm+iHO/Icfc5UNtJQhPO6rlQlICU7AeCpZ3aGiR+uNaf8T5NQ/WPi60dv+kOcWu359a5dwm2KdGjx0dJzOOLjrSlI73wkkD79Sa18ZeijNshtr1EtKVoZQlST0nUQkbj1tZR40NEvdGtP+J8mozceI/h6vGYt5FPzyxzpTUEQmmpLJcS2Ok6TnTu2SFE7D7wqGSc64YZsRhMnPLU/OMydKnXAtlL9xTMS8iUw+oNgqaWh8p5erl5G9tuQVnYprJoXj2bQsnnayx79coEBdtjPTIrKHi0oo3LzzcdLjqgG0gbqA8JKSo71ZZ40NEdvZGtP+J8mh40NEvdGtP+J8msbg4uMa86RT7jCdD8Kbk99kx3kggONLuchSFjfxFJBH9dXnSlKozU7r4otF/+i8fkdXnXnfUzSS+X/WK8X2BjQlvTodiYteRpmNtm0uRZkh2SspKw4N23UAdGlXSb8i9k7mq+Z4cs9uMW5QrpFlSZL+7dynrmRmmrwTdYsgO7tkvOFLLTpSp0tqZ5ujQFhRKZBqdoRkcn1TsePYvHdxd6RcnLc0w4wRb3HosFLSmmnnQ0yjpkzVFxLbjrZ2LaAHFbsn0P1HvGK5HaGnlqiycVlzoiV3DZ/wDVE/a+wSyVc2waT81eK99i7I5htyb1xzzRTOr0xlMe02osZJKeyJ9WWeqDaPVOHKhzG4EAd/0qeiW/DGy0pbR2HzpUSqvS+M45AxOww7VbIjcKHHSQhlrfYEkqUdz1klRJJPWSST4aqjBv2WOq32gsH+adV10pSlYd5+lUv+yV/wBq8q8MfEzpXg+h+OWK/wCd2a03iEqWiRClP8rjSjKeUARt1dRB+/Vp92Lon7pmP++v0U7sXRP3TMf99fop3YuifumY/wC+v0U7sXRP3TMf99fop3YuifumY/76/RTuxdE/dMx/31+iqv4n+KLSbL+HrUCy2XPrLcrrOtD7EaHHkczjrhT1JSNusmvVdu+l8b+zT/2rIpSlR3M8FtmeQmot0QpxhtRUEA9RP2ahp4bMKP1B8QqsuIvRPG8O0xNztTK4stN5s7HSNnlPRu3KM04nq8RQtQP2DVm9zZhW30B8Q/2oeGzCT9QfEP8Aah4bMJP1B8Qp3NmEkfQHxCh4bMKI+gPiFO5swoj6A+If7U7mzCT9QfEP9qHhswkj6A+If7UPDZhR+oPiH+1O5swr+QfEP9qdzZhW30B8Qp3NmFfyD4h/tU/xnG4mKWhq3QuYR29+ULO5H2K2tKUrzhxA5vYdO+ILR6+5LdY1ltDAuqHZktXK2gqi8qQT9kkD79Sw8YuifumY/wC+v0U7sXRP3TMf99fooeMXRP3TMf8AfX6Kd2LomfrmY/76/RTuxdEyPZMx/wB9fop3YuifumY/76/RTuxdEyPZMx/31+iovodn+Oal8Smqt6xa8Rb7ajZbGwJcNfM3zpVN5k7+2Nx/fXoulKUrrkx0yo7jK9+VaSk7VAHtDscfcU4tnnWo7lSkjcmuJ0Kxk/U4/AFDoTjJ+px+CKdorGT9Tj8EUOhWMn6nH4AodCsZI+hx+AKdorGf5OPwBXZH0Rx2K8h1poocQdwpKQCDU/aaSy0htPrUAJG/2K5UpSlKp3i0G+iMxX8C9WNX912iVcVKUpSlKUpSlKUpUbyzA7bmSmjPSVhsbBO24qPdorGdvocfgCh0Kxk/U4/AFDoVjJ+px+AKdorGf5OPwBTtFYz/ACcfgCh0Kxkj6HH4Ip2isZ2+hx+AKkmKYPbsOS6mAkpS54U+ACpDSlKUpSlKUpSlKUpSleWOPjhuyDXnA7RJw2S+1k9rmNoEVEhTTUqO44gK5wDtu2oJdBPgCV7dZFX7pXgEbS3TuwYrFkvTkWyKllcySsqckOeFx1RJJ3UsqVt4t9h1CpVSlKUpSlKUpSlKUpSlKUpSlKUpSlKUpSlKUrzvmmKJ1O4ppePXO+ZHAtEDD481qLZL7LtyC8qY8krUGHEcx5QB1+ICpKeFnFz++TUDy4uvpFO5Zxfb9cmoHlvdfSKHhZxcj9cmoHlxdfSKHhaxcj9cmoHlxdfSKdy1i+365NQPLi6+kU7lnF/6SageXF19IoeFnFyP1yageXF19Ip3LOL/ANJNQPLi6+kU7lnF/wCkmoHlxdfSKHhZxc/vk1A8t7r6RTuWcX/pJqB5cXX0ih4WcXP75NQPLi6+kUPCzi5H65NQPLi6+kU7lnFz++TUDy4uvpFDws4uf3yageXF19Ip3LOL/wBJNQPLi6+kUPCzi5/fJqB5cXX0incs4v8A0k1A8uLr6RWl0mxv9QHEdmuMQr1f7lZkYxarg3Hvl5lXHon3JM5C1IL7iyndLTYIG3rRV+0pSlKUpSlKUpSlKUpSlKUpSqTtw34zb39wsT8ueqZa2agdrPTa6XlqRFjXFZag29cwjoUy33EssqcG43QlawtfWO8So7jbeoHg/EVJyfFcdjW21ozDMXmriiaxbZLcdhZtzzTEp1ClkgBxT7CmkHwh9G6kjdQ1zHEpdLfer8q/WJ2LAtcu7cseKUKWYcNMNTjrhKjstCJC1lKdwdiB1gb7TLeKu049dJEGDYpt5dZcnEFhwJ6dmI4hl5TQ2JUsvl1pCTsklhZK0gp5pxlmpy7TKx+32Oyv5Bdr1DkXJiIHkxuWIwGulcJX+23kMISg7bqcG5SAVCqtL+LL9VVnxKJJs79zvMq3Whu6SIgKEouE2DHlJCGwk7M7SWuZZUOTnHUoJUU4mnfFLPatGPRMmtkq7X2bCgXW5LgpTyw25yyWG2W0J3cDaAFqCiFBBGxcXumpM1xQlUZ+S5hN2THdgT7hbSy62+5LbhzGIcgrbb5lNpSuS0sHZR6PnUUgp5TnWriLgXGLaLoqOhdvlhLb5t8xiZHjlU9MPpumQetAUtKiCEqSkkrShSVJGsncWtpZuseLEx64TmCymc860oFRhuS347D7KEhXS9ImO49y7pIbKdiVqCK3mp+rd+wDUOPBg2I36ys43Ovc5pl5tl5sR3mASgrPfq5FrCUdQJ23UmolmfFrFRaMxGN2uU+bdDurUG7FtS2xMiRHnlKda5RyM87Kmwsq3K07FKQpKlSGVxDyrLaFi9Yy3Zr0l6K2iPcrxHZjrRIZdcbWXQVHm+YOJLbaHFBW23MndY7MY4mrPldutDjFnnx5t3RbXIUJ0o53kylPBakkEg9CIspa/wDytbj1wrCwbiUayPHrBdPUaZIsrqrNbrheXFtNrZn3FiI4wnoASVJJnRgtSTsku9QUEqKce0cVAuFuwi4zMSfslvyyPFmwX7ncmGUuR5BbDXIsnolPbOBRYLiVcu3LzqIRV90qmsf/AGXubfcXZfyy41ctKUpSlKUpSlKUpSlKUpSlKUqk7d+zNvf3DRPy56rau+N22/TbRKnxUyX7TKM2EpSlAMvFpxrn2B2J5HXANwdubcdYBEYv2ieHZHcp9wlW19m4T3VPSZUC4SYbrqlMtsLBUy4k8qm2WUqT4FdEgkEpBHba9GsNst4fuUOyNtSHkvJU2XnVMJS62026lDJUW0BaGGgQlIB5N/CSTgSNAMEfsFnsrdnehW60wnLbFbgXCTFV2K4UlxhxbbiVOtrKUlSXCoKI3O5663uW6cY/m7MBu6RHgYHOIzsGY9DdaSpPKtAWytCuRSdgpG/Kdk7jqG2ns+hGEY/Ntki22dyELc3FbYiszpAjExmksx1uMdJ0brjbaEJS44lSwG0dfeJ27HtEsOeftTgtj7AtkaJDZZj3CSyy4zFVzR0PNocCXw2dykOhW259s12ytGcOmQYsVVn5G4keVFjqZkvNOMIkSGZL3ItKwpKi9HZcCwQpKmxylNcYmi2Gw7Rcram0F6Pc40iJOVJlPPPSm31czvSOrWVqUo7d8Vcw2ABAA24ztEsNuDtuWq1ORkwIkaA2xCmyIzLkaOoqYZebbcSl5tslXKhwKA51DbZSt8jNtI8W1EnNTL7AfkyG4jsAqYnyIwdjOqSp1hxLTiQ42soTzIWCk7dY2rCuGhWE3WbcpEm0uuouKZAkQ+z5KYilPsqYecTHDgaQ4tta0lxKQrZauvdRJzsi0lxfKJ8adNhSG58ZbS2ZkGfIiPtltt1tHK4y4lQHI+8kjfYhZ336tvlo0ew6wvY47AsbLDuOxn4lrXzuKMVp7bpEjdR33223O5AJAIBO+BbNBsGs0y2PwbMuMm3JiBiIibI7FKoraWozi2Ok6NxxtCG0pcWlSx0bffboTt9k6EYRKgWSCbS83Bs8GPbI0RifJaZVFjkFll5CXAH0IKRsHQrwq/hHef0qmsf/AGXubfcXZfyy41ctKUpSlKUpSlKUpSlKUpSlKUrz3rbohdsx1KbyqwZnd8QuXqY3bXXLVIDXSspcU4Aoch/bKJ8PtVDO0JqOfr25h7/Hm6HQPUf3bsw9/jzdDoJqPt7NuYe/x5uh0D1HP17cw9/jzdO0HqPt7NuYe/x5uh0E1G29m3MPf483TtCajkezbmHv8ebp2hNRj9e3MPf483TtCajH69uYe/x5uh0E1H923MPf483TtB6jkezbmHv8ebp2hNRj9e3MPf483Q6Caje7bmHv8ebodBNRtvZtzD3+PN07Qmox+vbmHv8AHm6doTUc/XtzD3+PN0Ogmo/u25h7/Hm6HQPUfb2bcw9/jzdWVoFo3NwHI7/kF6ym7ZbfLjFjwVTLrIDqkMNLdWhCe8TsAp1Z8fhq7aUpSlKUpSlKV//Z)
![[[@Pellman2016-xu]](https://doi.org/10.1016/j.tins.2016.04.001)](data:image/jpeg;base64,/9j/4AAQSkZJRgABAQEAcQBxAAD/2wBDAAMCAgICAgMCAgIDAwMDBAYEBAQEBAgGBgUGCQgKCgkICQkKDA8MCgsOCwkJDRENDg8QEBEQCgwSExIQEw8QEBD/2wBDAQMDAwQDBAgEBAgQCwkLEBAQEBAQEBAQEBAQEBAQEBAQEBAQEBAQEBAQEBAQEBAQEBAQEBAQEBAQEBAQEBAQEBD/wAARCAGBAjADASIAAhEBAxEB/8QAHQABAAIDAQEBAQAAAAAAAAAAAAYHBAUIAwECCf/EAGkQAAEDBAECAwUDBgUKDw0ECwECAwQABQYRBxIhCBMxFCJBUWEVMnEWI0JSgZEXM2JyoQkkJUNTY3OCkqIYJjQ1Njc4VXaFlLG1wdMnRHeDhpOVsrO00dLjdYSWtkVGVGVmeMLF1OHx/8QAGwEBAAIDAQEAAAAAAAAAAAAAAAMEAgUGAQf/xABDEQACAQIDBQQIBAIJAwUAAAAAAQIDEQQhMQUSQVFhE3GBkRQiMqGxwdHwI0Ji4QZSFiQzcoKSstLxFTSiQ1OTwsP/2gAMAwEAAhEDEQA/AP6p0pSgFKUoBSlKAUpSgFKUoBSlKAUpSgFKUoBSlKAUrXXvI8exmGbhkl+t1pip9X50pDDY/wAZZAqGnn/iV/tY8qVkhHr+TVvlXrX4+xNu6/bQFh0qvP4U8kuHfGuFM2nNq7IkzBCtrP8AjIkyESAP/En8KfaHP109xnF8Fx5CvR6ReJV0cA/lMIjx07/B4j60BYdKrz8iuXLn/r7zWII+WNY3Gif++qmf9VP4FbNN97Jc3z2+OH7ynsolwkK/nMwVMNEfQo19KAnc24QLawqVcZ0eKwnupx91KEj8STqobO534RtrxjTuYMKZfHbyVX6L5hPyCOvqJ+gFfIPBHC9vfTMa4sxd2Wnv7XKtbMiST9XnUqcP7VVMoNtt1sZEe2wI0RodghhpLaR+wCgIL/DtgcjtY4+UX0n7i7Ti1ylMrPy89DHkj9qwKfwlZxN7WfgfLyFfceuM21xGiPqBKW8n9rW6sOlAV79tc+Sv4jjvB4KFeipOWynXE/i2i3hP7nK+fZ3iBle8cv4+tgP9rGOzZxH+P7az/wCrVh0oCvfyS5mk/wCruYrYxv1+zcVQ1+7z5D3/AF0/g3zx3u/4gs1bPyjW2yIT/nwFn+mrCpQFe/wRzn/9c+YOQZvz/shGi7/5NHa/op/AdiTneVkfIEhR9Sc7vTYP+K3KSn+irCpQFe/wC8ZL/wBWW28T/n7fkVyl7/Hzn1bp/oe+Dlfx/FOMST85VubfJ/EuAk1YVKAr3/Q7+H/48G8fH0PfGYX/AGdfT4eeAVK6jwdx+TrWzjML0/8AN1YNKAr3/Q9cGo/1JxNi0FXwVBtjUVQ/BTQSR+yvh4Lwpn3rTdcztSh9wQsxuqG0H6MmQWv2FBH0qw6UBXZ495HtX5zGeb7w8U9kR8itMK4Rkj5fmERn1fiXia+flJzRjf8Asi49teUxU9jKxi4hiUr+UYU0oQkfzZTivkD8bFpQENxzl3A8luiMebu7lrvqwVJs15jOW6esD7ykMPpSp1A/XbCkfJRqZVqslxTGMyta7JluP268wHCFKjToyH2+oeiulQIBHqD6j4VCjx9nOE/n+LMzdlwm+/5O5RIdlxVD9ViaeqVGJ+azIbSBpLQoCyqVBce5ZtU27sYlmNrl4hk0glLFtuhR5c0gbJhyUEtSRoE9KFeakaK20b1U6oBSlKAUpSgFKUoBSlKAUpSgFKUoBSlKAUpSgFKUoBSlKAUpSgFKUoBSlKAUpSgFKUoBSlKAUpXjKlxYEZyZOktR47KStx11YQhCR6kqPYD6mgPalV2vnTDritUfAYd4zuQCUj8m4ftMUqHqkzllENKh8UqfB+lfPaed8m/1Nb8WweIruFzFuXqcpJ+BabLDLKx8w6+n6GgLFrSZPm+GYTFE3Mcts9ijqOku3Gc1GSo/IFahs/QVFf4GY929/PeQMxyk/wBweuf2fF+qSxASwh1H0eDn1JreYzxXxlhkoz8S4+xyzy1DS5MK2MsvL/nOJSFK/aaA0v8ADphckf2DteYXsq/ilW7E7m4w7/NkFgMEfXzNfWvn5dcqXf3cb4Ukwv77lF9iwW1D5pEP2xz9ikJP4VYlKArv7B5xvfa7chY7jsdfqzYrIqTKb+iZUtxTSv2xRT+Ba2z/AH8sz3PMhc+JfyF6A2r6KZt/s7Kh9CgirEpQEMsnDPEmOTBcrLxrjUWeO5mi2MqlKPzU8pJcUfqVGpkAANAaAr7SgFKUoCF5nzDx5x9mOIYHl1+FvvGdyJEWxNLYcUiS6yG+tBcSkobJLraU9ZHUpYSnZOq3GN5pjuWyr9Dsc1T7uM3RVmuQU0pAalpZaeKAVAdQ8t9o9Q2O+t7Bqm/ENw4/zFyjhFmlwZzdnGL5RGdu8Zs/2KnrdtTkF9K/RDqXY5db+amT8jVFRsI5+vnHdzm8iYNJhvSOXVXLL7cmyLuzFzgNWRmJ7W3BbWhU2Iqc008GkknpSD0r8spIHehUkDqJAHzoSB6muMInGrFjYwE53heT55xax+Urrlj/ACMkoRbrhJkRlwSbMouvCM22me2yFJUGvOT2QOkjCx/ju82dvCk89cR5Xl+KMY3do9ks7UBy7vWWW7dnXYjT6EKV5UhNvVEZRIUQGiw4nzEb2QO3NjetjfyrW5FktixO0u3zI7mzAgMraacfdOkpU64lpsHX6y1pSPqRXIV24+yCblt4ZXxhkiuVnuSYl0suZGA4qPEx1M6O4E/aOy22wiAl2OuJ17W4V/m1dfXX5yfw4W268C8k3m78RrvOYXDPbtdUpkwTJnP25vJXH20x0r2ry1ROtaG0aCvNUUglw9QHaJIBAJHf0psfOuH+TOLLnk+fXEsWK9WHGbhj9mj4DKg8eSrjKx8NoX1ojFDjf2RJQ70uEutoCgWwpRDZQJrj2EZpE8VDmZyMPv6eM5V9uCbfBUFFEPI/Y20O3tTHTtEV5KJTCCT0h1Sntf1wFJA6spSlAKUpQClKUApSlAKUpQClKUApSlAa3Isbx/LrQ/YMoskK7W2SAHYsxhLrS9HYJSoEbBAIPqCARUEOJcjcdfnuO725k9lR3VjmQTFKkNJ+UO4K6nPnpuT5gJ0A6ykVZtKAi2F8j47m6pUCH7Vbr1bgn7Rslya8ifCKt9Jca2epBIPS6gqaXo9C1AbqU1Fs346sOcoiypa5NuvNtKl2u9W9YanQFq1stuEEFJ0OppYU2sABaFDtWkxvPb7Yr5F4/wCV240e7y1Fu0XqMgtwL50gkpSCT7PKCQVKjqUeoAqbUsJWGwLEpSlAKUpQClKUApSlAKUpQClKUApSlAKUpQClKUApSlAKUpQClVLc4Ny4JJyKwvzJ/HqD1Xe0OuLkO2Jv4zIalbWY6PV2MSQhAK2enoLTlrR5DEthuVFfbeZeQHG3G1BSVpI2FAjsQR33QHpSlKAUpUbzPkLF8EZjC+THVzbgpTdvtkNlUmdPcA2UsMNgrXrYKiB0oHvKKUgmgJJUUy/k/DsKls2i53FyVepaPMiWW3MLmXGSneupEdoFfRvsXFANp/SUkd6jwtfLPIn5y/3BfH1hc9LbbXW37y+j5Pyx1MxdjsURw4sdimQk9hLcQwLEMDiPRMTsUeD7UvzZb46nJMt3WvMffWS6+5r9NxSlfWgIsJHNuZ94sO2ceWxz9OZ0XS8KT/g21eyxl/IlcofNIr2i8G4M9JauWZpn5vcWlBxEnJ5PtyG3B6LaikCKwr6ssoNWFSgPyhCG0JbbQEpSNJSBoAfIV+qUoBSlKAUpSgFKUoBSlVzzBzzgnDMWIzfnZdzv92V5Vnx21M+03O5u+gSyynvrfqtWkj570CBYqlJQkrWoJSkbJJ0AKo3MPFxxjCusvC+PWL/yNk7QLblvw2CZ3syz2Bdk9o7Wj69SyU67j4VH2uIOYfEQoXXxGXl3FcPeIWxx5YJpSXm/gLnNRpTxPxaaKUenfexV84jhmJYFZGMawrG7dY7XGH5uJAjpZbB+J0kDaj8Se5+JoDjbw98heP7kHkm72a9IslvxGxXN2JNuORWxl51PQsgxmlQlMokPp+6tSNNpUlWyDpJ7gccQy2p15xKEISVLUo6CQPUkn0FaXMczxDjfGJ2XZne4VlstuQXZEqQoIQnZ9APVSlE6CQCpROgCTXPbdp5H8YjiZmUR7tgvCqlBbFoKlR7vljfqFySPejRFfBsHrWDskApIAl2Pc/5Dy3yVHsHBuPQ7thFklqbyXMJylphOlIIMW3dP+qHQoglz+LSBrv1JNXnWvsNgsmLWaHjuN2mLbLZb2ksRYkVoNtMtj0SlI7AVmuuNstreecShtCSpSlHQSB6kn4CgKs8Q3NFz4Ww8X21Yi9eJMhxMZhxbqG47TquyQsdXmLJ9QhCSSAdlPrWX4dL9l2W8S2jMc4nmTdr8p6esBAQhppTig0hCR6JDaUH59ySSTuozg7C+dOQTy1dG1Kw/GnnYeIxXE+7MfB6XrkpJ9RsFDW/QAnQPrc9rtkCy22LaLVEbiwoTKI8dlsaS22kAJSB8gABWtw6q4jEPE734drRXl63jna/DPiVaW/Uqdrf1eC+ZlVyYfGqzgHivyzw98pQyi0CRFcsV6jsE+ypeiMulqSlI/i+pxWnQPd/T933k9Z1zh4n+IItntc3xG8cWVP8ACJh9xjZQp9P8bcIkVgMyYJPr5a4oc90dyodvWtkWjo1C0OIS42oKSobSoHYI+Yr9Vp8PyqzZzilnzPHpPn2y+QWbhEc+KmnUBad/I6PcfA7FbigFKUoBSlKAUpSgFKUoBSlKAUpSgFKUoBWqyjF7DmdilY3ktubnW+YkBxpZKSFAhSVoUkhSFpUEqStJCkqSFJIIBra0oCtsUyK/YVkcXjDkG4OTxNCxjWQPAA3NCElSosjWgJraElWwAHm0qcSAUupRZNQzmPG15TxnkFviaRcmITk60v8A6UW4sDzYr6T8FIeQ2r9mvQmt/i99YyjGbRk0VBSzd4Mec2k/BLraVgfuVQG0pSlAKUpQClfNj51ospz7B8HYRIzPMLLY23Tpo3Gc1H8w/JAWoFR+g2aA31Krsc02u4/7EMFzjJB6+ZFsTkNlSf1kPTjHacH1QtW/hXjE8QfHdzaQ3Y1Xu8XP30ybRbbPJlTYDqFqbW1LQ2giKsLQtP55SASg9JI70BZVKrxXNlghd8gxHObOj+6yMXmvtJHzU5GQ6hsfVZSKkuK57hGcsOScNy6z3ttg9L3sE1t8sq/VWEklCvmFaIoDfUpSgFKUoBSlKAUpSgFKUoD4pKVApUAQexBqs+OEnj7LLjw27tNqTHVesTJ9E24uBMiEn6RXnEBI7BLMmOgb6Cas2q45vQbLYLfyfGHTIwKei8vKH6VtILVwQf1gIrjzgT6Fxlo/oigLHpXwEEAg7B7giq0yTJb/AJ/f5nHXHFxct8a3r8nJMlZAUYBIBMOJsFK5hSQVKIKWEqBUFLKUUBlZNyBebrfZHH/FTEWdfYxSi63SUhS7dYgoAjzukgvSCkhSYyFBWilTimkqSpW2wnjax4W7Ju3nyrxkNxSE3K/XJSXJsvR2EFQAS20CSUstpQ2nZ6Ugkk7bFsVsGFWONjmM21uDAihXQ2klSlKUSpbi1qJU44tRKlLUSpSlFSiSSa21AKUpQClKUApSlAKUpQClKUApXm++xFYclSnm2WWUFxxxxQSlCQNlRJ7AAd91zRds/wA88VV2lYXwheJmNcaRHlRr7nrKemRdCk6ciWkkenqlUn0Hfp3odYEg5G59yS/5XL4Z8N1riZHmUfTd4vUgk2fGUnt1SXE/xr/Y9LCNnYPV90pMl4c8PWM8WSpWW3W5Sstz28J/svld20uXIJ9W2h3EdgeiWkaAASCToVLuOONMJ4lxSLheAWFi1WuJ3Dbfdbrh+866s+844rXdSiSf3VKKAVCuWuXsK4XxVeVZpPWhC3BGgwYyPNmXGUrsiPHaHdxxR0AB2HqSACa1XNXOWPcOW2FHVb5V/wAqvzhi49jVuHVMukj5JH6DafVbqvdSPmdAxTiXgvIpGVI5x8QE+NfeQnWym3QmdqtuLx1f97w0HsXNdlvn3lHsDrZUBqMH4dzPmbJ4PMviYt7bSYS/aMWwLr82HZB+jIl/oyJhHzHS38BvsnoylKAVTHMN2uPImTRfD/iUxxj29kTMsnsHvBte/wCJB+Dr/wB0D1CdkjR3U35W5Eh8Y4dJyJ2MqZOcWiHa4CO7k6c4elllAHckq9deiQo/Ctdwvx1MwTHZE7JZCZuWZG+bnkE0d/Mkq9Gkn+5tj3Ej00CQBvVUcS3iJ+ix01l3cu+Xwv0IKr7SXZLx7uXj8Ca2i026w2uJZbRDbiQYLKI8dhsaS22kAJSPoABWZSlXUlFWROlbJCvy42282pl1CVoWkpUlQ2FA+oIr9V8JABJOgO5Jr0HP3hFLmHM8hcBvrV08a5Q+xa0qOym0TR7XDBPx0HHE/ggV0FXP/GLjLvjD5ndtywthePYsZSkEFJeLcooOx/e9a+lXleL5Zcegrud/vEK2Q0EBUiZIQy2kn02pZAFAZ1Kh38JUS4dsRxm/5H8nosP2eMR8FpkSi006n6tKWfoaeTyteP4+Zj2MsnspEZDtzkFP6yXVhlttX0LTo/GgJjSoeONYMr85kOU5TeXj2K3ru7EQR8izD8lkj8UEn41+v4I+Lj7z3HuPyV60Vybe0+s/ipYJP76Al1KiJ4n4/b/1vsH2Sr9e0ynrer/KjrQaxX4ua4MDMt06Zllkb7vQJISq5R0frR3QAJASP7W6PMV3IdUQG1ATilYlpu1uvtsi3m0S0SoU1pL7DyPurQobB/8A9HvWXQClKUApSlAKUpQClfFKShJUpQAA2ST2FVzP5khXiQ7ZeJbSvOLqhZZckRHvLtENwHR9pn6U2Ck/ebZDrw/ueu4AyeYsgnxMdbwzGngMnzJS7PaQB1GOFp/PzVD+5x2ip070FKDbe+pxIMwsdngY9ZYFgtbRbhWyK1DjIJ30tNoCEjf0AFRnB+P5NjuUvMcuvCb7l1zaDEmelnymIscHqESI0SryWArudqUtagFLUrSQmaUArSZplltwbGJ+U3ZD7rEJCeliOnqekvLUEMsNJ2OpxxxSG0J2NqWkfGt3VecqIEzJuMbQ/wC9Fm5f1vtn0cMe1z5TW/5r7DLg+rYoDFhcfZxmbKLpyjnN3gGQA5+T2Mz12+LDB7htUxnplvuJ9FOB1ttRGw0msscA8Rr7zsRFzX/dLnOkznPx633Fq39d7qwqUBXv+h/4b10nAbeU730EuFP+T1a/oreYvxjxxhD7krDsCx6ySHRpx+BbWWHXP5y0JClftJqTUoBWNDttutypCrfAjxjLeVJkFlpKC86oAFxeh7yiAASe/YVk0oBUXyzjDj/N325uT4pAlz2B0x7glHkzo31Zkt9LzR+qFpNSilAVucH5LxH87x/yG5eIiPSy5b1SkFP6jU9A9pbJ+K3hK/m1+hzGmwfmeT8JvuIqT2VOLH2ha1fNftcYKDLf8qSlg/SrGpQEMtnNPDt7V5dn5Yw6cveumNfYrit/LSVk1Lo0qNMZTJhyGn2ljaXG1hSVD6EdjWJc8fsN6R5d5skCenWumTGQ6P3KBqu7/wAD2lu8Wy88UzI3HMovrbvcmwW9hly5QVNLBaKCgtFwO+SpLy0LUgJWE66zQFqUqujwfYGUB205nyBb7gnuicnMLjKUFfrFmU67HX/NW0pP0rxVeOWuPveyS3Iz2xI+9cLPGEe7x0D9J6Hvy5Wh3UqOULPoiOqgLLpWpxfK8dzSztX/ABa7sXGC8pSA60TtC0nS21pOlNuJUClSFAKSQQQCCK21AKUpQCse42+HdrfKtVxjokRJjK477SxtLja0lKkn6EEisilAUVasoyZrg3GcTt96ejZK9d2sBeuekqebciylxZctIUCPMLEV95vqBT1KQSCOxuDFsXseGWCHjOOQUxLfBR0NNhRUokkqUta1EqWtSipSlqJUpSlKUSSTVG5KBjnKky2rH9btZvjuUt/qtoucZ61dI/GTHUs/V7fxroegFKUoBSufLNZ0cqRZObZldrzI9tnS0QYEe6yYkaDGafW02hKGFoCllKApS1bJUogaAAr9v8VYUj7rd7H/AJQ3D/t6A6ApXOD3GmII9Bex/wCUFw/7avD+DrEd63e//wAQXD/tqA6WpXOLPGmHr9Re/wD8QXD/ALas9jirCl623ez/AOUNw/7egL/pVAXC1fwWOWrLcQu95ZZTdoEG426TdJEuNMjSZLcdXuPrX0OILoWladH3dHYNX/QCsC+36y4vZ5eQ5HdYtttkBovypcp0NtMtj1UpSuwFYuYZji+AY3Py/M75FtFmtjRelTJK+lDafQfUkkgBI2SSAASQK5isWLZh42L5FzjlC0zLBwrbpCZeN4u8S1JyRQ7omzwDsMfFDXbYIPce8sDYtt5Z41ZaX5IuONcEMu7bZ2qPcM06T95XopiDsdh2U4O/bY6OmbPZ7Tj1qiWOxW2Nb7fBZSxGixmg20y2kaSlKU9gAPgK940aPDjtRIjDbDDCEttNNpCUIQBoJSB2AAGgBUZ5A5T4+4tthu2e5XAtDJSVNoec289r1DbSdrcP0SDWUISqSUIK7fBHqTk7IldVVzrz3ZuHoEK0W+3qyLN8hUY+PY1FcAfmu6O3Fn+1MIAKluq91ISfjVG8reN/PUXCyYnw1w3dX7tlbpYsbl7a8p+aPi81DCgryQNnznFJQACSOxFb/GfA1Yslvq+S/EHlNzy7Mbq0k3FtiQY8JsEd46CgBwtp+6AlSEkD7g3V30KNL/uZqPRetLyWS8WmS9ko+27e9mNx3kPDvEF5ncn848w2HKeWshbDU563LM0W1ne0W6CwyFqaZTvW9BTh95X0sf8A0RWRXz3eP/D5yJeur+LfnxGrTGcHwIckLB19emrCw/i/jrAGUsYVhFlswSOnriQ0IcV/OcA6lH6qJNSise0wtP2IOX95/KNv9R5vU1or9/7fUpYZN4tL171t4vwHG0q9BeMgfmKT+IjNAE/ga+Lx/wAWctCnrhyhx5ZkgFS/YrBIfSgfHu88P6auqqS5+yC75Xc7V4e8KlrZu2WIL15mNdzbbMk6ecPyU5otpB9dkdiQar4naawtNzjTjfgrXu3ovWvqR1cR2Ud5RX333KtwnBOdudb8jkmTzrEbt2LT5EXGZ/5JsLamKAKHpSGC70gdQKULUVKOiR01aw438Tcb84x4lbXLV/c5OER0pP7UPA1a+OY9aMTsMDGrDDTFt9tjojRmk/ooSND8T8SfUnZNMgyTHsTtbt8ym+2+z25jXmy58lDDKN+m1rIA/fTD4qtTh66g5PV7kM3/AJfBdBTnOK9a1+OS+hVH2d4v7T70bJOLr+lPqiXAmwlqH0U2tYB/ZX5Vytz/AI37+Z+HVy4xEfxkvF76zMX/AIsZ0NuH9hNSY8qXrJCWeL+PbpfEE6F1uvVZ7WPqHHkF94H1C2WHEH9cdt6rI7FcGbYu+858zi1WlPZyBZJBsUH5hK5IcMxxY9PcfbSv4tfCrMcUqj3ZUoyfRNP/AMWvgSqe87OKf30IlfvHDw/jk22wb4xfrTLemJj3OBdLU/Fm25paFlMhTZSQ4gKSEkIUVe9sA60bMhZ1keZ21q4YJhqXbZOaDke53eeiKw+yoe64020HXldjvpcQ1v5iuas1xHw9ZlO8/BPD87eelJSifNmSrLauo/edDaVB15aiNqcDW19trPY1+8WxHJMYsf5Nw86uVksvmKeRZsdkPRYzJV94JedcdkgH1911AJJPTsmtlU2bQq0ouF4T4qVn5WzXikTyoQcU1k+uf34kR/qf3CvLMDGrvyTyZn93sWOZGmMiFb0PIYlXGPHb8qO87JUPOaYS0OhpDa0EpPV2T09XXloi8KWK4puMO4445dkbSJ8y4olz/qDIeWp4/gVVzkrj7BCtLkzG4lydSAA7cuqa52/lvlSv6a9hiOEpBSnD7IAfUC3s6/8AVqNbKpcZvyX1MVQhzfl+52Ew+xJZQ/GeQ604ApC0KCkqHzBHqK9K4dvHD+C3lry48edZyCoo+zJrjDaCfUhoEtD5/c9a3uIjDcDDNt5O49tN9sKAEjIokRXtUUfrTGAT1p+brXp8UAbNQ1dlTSvRlvdNH80/O/Qxlh/5Hf4nYtKrq38O8KXu3xrrZcWtLsWShL8aZb3lI60nulaHWlA6PzB717OcMWGOkrxzK82sMgb8p2Lk8yQhre/uxpa3o2u/oWiPpWqaayZWJ/SqzgZjlnHt4h4zyxMiT7Xc3kxbTlbDSYyFvq30RZzW+ll5etIdRpp1Z6OllZbQ5NpWV4zCss7I5N/t6bXbGXH5ksSEqaYbQkqWpagdAAAk0s7XFjS8fhES4ZnZYxAi23I3PJQP0DJixpjo/a9KdV/jVMKgfDUO4Lxmdlt3t8iDOzC6yr8uNIb8t5phwhuGhxBAKHBEZihaD3SsKB9KnleAUqIZ/wAsYNxqmKzk12WblciUW60QY7ky5XBYHdMeKyFOua+KgnpT6qKR3qJIuviMz5Pm2ex2DjK0u/cevY+2LypB+JisOIjR1emtvv6/SR8KAtyorlPK/GGDu+RmPImN2R49ksz7oww6o/AJQpQUo/QDdQ5fhysN9koncl59nGcOpbU2pi43gw4KgpSVEGHASwwtIKBoLQvt6lR71it+EHw9224OXrD8CThl3cSUm5YnPk2WSfj7yoriA53Po4FJPxBoCQ/w0225How3BM3ydfzi2NcJlQ+aZFwMdhY+qFqr4q586ZJ7lsxnG8MjL7iVd5a7rMSPkYkctsg/yhKUB+qfjpnE8v8AEK0zZF5mcl4a0CZYkRmm8htrYBJdQWEIauCE9ttBtt/pBKVPrIQbAm51iMDDDyE/fopx32JNwRcGleY26wpIUhTfTsr6gU9ISCVFQABJAoCJyOG8dnsOXHljKbtmjbKS681e5KGbU2gDZ6oLKW4y0jWwp5Dihr7/AMa+QuYLLNitReK8ByLLoDCA2zJs0RiLbQgDSfJky3GGXm/gDHLg/dWPbsNvfK8hnJ+WbcuLY0rS9asOdIU2gA7Q/cgCUvv+igx3ZaOv4xxIcTaQASAlIAA7AD4UBXoyTnaR+dh8UYkw0e4RcMzeaeA+qWbe6jf4LI+tfhzlXIMaBXyRxherPER/GXW1LTeIDf1V5IEpKfiVqjhCR3UoCrGpQGHaLxacgtka92G5xLjb5rYejS4jyXWXmz6KQtJIUD8warbky5zcgzrFMSwy1OXG+41d4OQ3CQXUtxbZBc82M55qj3U67HdmBptCSSUbUUJ0TqeSLpb/AA4SpvLECG8MOuanPyktURO0tT1pJjzWEeiXHnuiO6kdlrfacOihZXOuLsQuGK487KyN1p/Jr/JVd7/IaO0LmOJSPLQT3LTLaW2G99/LZRvvskCY0pSgFKUoBSlKAUpSgFKUoBSlKAUpSgK6yrjvJmswj5vxVd7Pj90n9UfIPb4bsmJcGQ3pp1cdp1rrkNqShKXOtJ8sqSSoBIB2z+ICGgyo2fYLdHWx1exPYxKhof1+gH0znSzv06y27r16VelWLSgItgmexc0YmxJNtfs9+s7qY93s8lQU9DdUNpIUn3XWlj3m3U+6sb9FJUlMpqs+W2Bh822c1W8eW5jhEW/9PYSbE6seeV/P2YkSknuQGnkp15qt2X696A+0pSgOf/EHEks5PMuFvYU9MRhEvIIzSBtT0ixXODNYR9SVvaH841fUOZGuENifCeS9HktpeacQdpWhQ2lQ+hBBqveUfKs+YcfZdJSkQ27s/j9wcWPcRGuDCkNhX0XMagt/i4K9eB3HIvHUXEpKyZGHSpWML6jtZahOqZjrV9XIyWHf/GCgLDrGuVxg2i3ybtdJTcaHCZXIkPOK0httCSpSifgAATWTVJcvPyeV84t/h/s7ribYlDd4zOS0SPLgBW2YfUPRb6x3HqEJJ7gmgNfwtNjXDi+03CE6HY8p2a80sAjqQqW8UnR79wRUllqPeo9w3HZj8aW2PHaQ200/OQ2hA0lKRMeAAHwAFSOUgndAU14nL7dcd4NzG9WW5TIE2LA62pMNxSH2z5iQShSfeCtE613+VU2/e+QbE3MlWu55tBxKTmWHw7OcjeeFwWXbi23cEAvH2j2ZaFNJCXfU+bodJrpzNcPsucY7PxXI4qpNtuLflSGkuKbKk7B+8kgjuB6GtTm/HuP8hWT8nslZlLiCVHmIVEmOxXmn2HUutOIdaUlaFJWhKgQR6UBKYqydVuYqj2qK4fisTE7S3ZoE26y2W1qWHbncn575KjsgvPrWsj5AnQ+FSyKj0oDQ8qHeHsf/AG9Y/wDpSNVrZlnOJcf4xdMxzC+xbZZ7M0XZsl5fZoaBCdDuVHqSEpAKlFSQASRVP813KHY+Onb1cnfKiQLtZ5T69b6G0XKMpR0PXQBri7i2dyTyvzfkXInPlpiwuNrVf05XNseVXY2yNGfkt+VCWqOW1uSlpjRQG2VJCVdAOwFnrzhTlUdoK/cepOWh07heCZb4v8jh8u84Wd+18ZW2R7XheEyAUmfr+LuVxTv3+od0NH3dK+KSS70PnvJvH3FVm+2M6yaBZogBDSHV/nHdfotNJ2tw/RINVEeYeXeblG3+HzFvsHHF7QrNcijFDa0frQoqved+iljp+CgmpVx/4acJxO7ozPLZk7Ocy7KXfr8vz3G1DuPIaJKGEj9EJ2R6dVXPRKeHzxUrP+Ve148I+N30JezjD+0fhx/b7yIr+XniB51T5XFmPL42xV06OSZDGC7jJbP6UWH6JBHcKWdEHYINRvPbDw74WYEPK7jbLlyTyzkTwhY6m7vGdc7pcDoIDSVdmWkKUnqWkApSQNklIN084c14twVhLmWZA29NlyHUwrPaIo6pd1nL7NxmUgElRPqdHQ2e/YGAcA8IZUnI5PiC59cYuXJd8a6YkUe/GxiArZTBjD0C9E+Y4O5JUNnalLxnjZ7rp0VuR5LV971fw6Hjqu1o5L74mb4euBr1iVwncx8yTmb7yvlDf9kZifeYtUb1Rb4Y9ENoGgoj7yge5Gib0pSqREKVEci5b42xa4myXjMbeLvrqTaYqzKuCx/IiMhby/8AFQa1Ks+5AyQBGA8YTGGHPu3TKXvs1gJ/WTGAXLUoevQ40zv9cetASbOsys3HuI3XM7+8G4Nqjqfc76KyOyUJ/lKUQkD5qFUHwbmFotyL1nNzj3DMOScxeEu4WzH4/ta7Wx/3vAdfJTHiFLeiUvutE+h30g1hTMTynmXnFGC5jnb+T41hyW7hk0RiC3EtC5yu7ENtgdbigAOpfnPPfpJHR6Hpy1Wm12K3MWiyW2Lb4MVAbYixWUtNNIHolKEgBI+gFa+n/WsQ6v5YXS6y4vw9lf4ivH8Wpv8ACOS7+L+XmQX2bmvLyDLuFqwC2qPdqElN0uyk/A+a6kRY6/gU+VJT8l1lWzizjnEJJzK7tG5XWChTy7/kU1U2THSBtakOvkpjI9SUtBtA/VAFTK5XO22a3yLteLhGgwojanZEmS6lpppAGypa1EBIHzJrni53F7nSd+UN2e68EjPk2S1JP5q6dCtCdJH9sQVDbTR90J6VqBJHTucNh+3bcnaK1fyXVlynT383kkb++835FmIXD4hgssW0npVk90YUWVj4mHH7Kf8Ao4spb+I8wVDGsMtouicjyCXMyO+p9LndnA+638wynQbYT/JaSkfjUwf0kdKQAANAD0ArVSVetbmju0lu0lZe997+WnQsJ7qtDI11xlezsOv635aFK189DdV4zycxPseGXlFuAOXuR0eSJAJi+bFcf2Tr39eX0+g9d/Spjdp8FbciD9oRkvFCkdCnUggkfEevxqn7Nw7bLDasDdt1ix+Nd8fcjrus6LHQhyQExHWnOlxKApfU4tKve1vWz3FbPDU6bi3U1/Z/OxNCMbesZ9u5dfkyID15x5u32u6yp0SJMRO85QXF84qLrfQnoBSw4QUlXoAdbrYYxmmRZGYdyXiSYdmuLfnRpK54VIDZT1IU6x0AIChrslaiOobA760GO8Y2m02aQzeIjTtzlG4NrkpeW55bUh5xWm+vs2ShaQekDZB9a/eIjOLRHt+P3iNaFwrawmMZ7MpwuyEoR0oPklsBCjpJV76h669e16dOi0+zWn76Z930JWo/lLMZk7+NbCO/vXftUbjPE671uIrhOqoThYhaMqwxMnw5+Q5xvl72Px5iy7Ity4qJcEuk7LiGl6LSjv3vLUkK9SCe9SRGW85vDpPJFiQkjXU3jOlj8CqQR/RWpiK9K28Y9hVOraTvKKb5tJvzsYyk3m7eSMBzAoWSyhcORrtPzOUD1JTd1JVFaP8Ae4iEpYT+PQVfyjWpzrA8MxaFF5Es+I2xo4tKYuc6ExEQiPPhsrCnEutJHQ4ptO3WlKBKHG0EarZ5ryViHGttbuOU3PylSFhmJEZQXZMx0kBLbLSfeWokgDXxI7jdbnHuNuTOVrAqTnUteDWO6trbVYo7CHbo5FWCCJD6iUMKWn1QhBWkK0VBQIEbquCUqsrQ5cGuKSX0sgpNZzdo/fAszNuaONeP1RIuQZK0u53FHmW+z29pc65zhre2IbAW84PT3koKRsbIFRZd0575QKWbHaE8VY64ffuNz8idkD7ev7TET1xYm/ULeW8oeimEn0muA8V8d8Xw3YeBYhbrP7SeqU+y31SZSv1n31bdeV6DqWpR0AN9qldc2+hQIZx/xFg/GypU6w296TebiB9o3y5yFzLnPI9POkukrUkHuGwQ2n0QlI7VM6UoBSlKAVQmE4TebnytkmJ3GF5WAYJkP21Z2Cdol3KZHZlBBH9zivPSXUpO09ciOU6LAq+6gMCLds9uV3uYyS52mxxZ71tiRrcptlctcdRakPuuFBcB85DjSUpUkBLQV36x0gT6lQS8cePWu3rumE3jIBe4KkyYrU7I58mPKKFBSo7qHnVoCXUhTfX0Eo6+tI2kVK8fvUPJbDbcit3X7JdYjM1jrGleW6gLTsfA6UKA2FKUoCuvEY20rgPkR95CVex4zcZqNjenGI63UKH1CkJI+oFWLVe+IQhzhLNLcPv3e0P2hofrOyx7M2n8Sp5I/bVhUApSlAKUpQClKUApSlAKUpQClKUApSlAKUpQEP5hulosvE+Y3O/Npdt7FinF9ojfnJLKh5YHxKthIHxKgPjW5xCDcbZidltt3e82fEt0ZiU5vfW8lpIWd/Hagaz7hb4N2gSbXdIbEuHMaWxIjvthbbzSwUqQpJ7KSQSCD2INQPiCRMsyb7xddJb0l7DJiGID76yt1+0PI8yEtSjsqKB5kYrUSpaoilHuqgLEpSlAajLcXtOa4zc8Tvja1wbrGXFe8tXQ4gKGgtCh3StJ0pKh3SoAjuKqDw6ZJdpmY8gY3kjiFX2K5bpd4KUhAdnpQ7bnXkp/RQ63a48hKfgiSjfrV7VzrfSeNfGdjdzWktWrlazTLWCPuKu0VlDqlOH0CjGhsIbHx/O/KgL7v17tuNWS4ZFeJAYg2yM5LkuH9BttJUo/uBqs/DfZLirEp/JeSRy1feQpyr9JSrupiModMRjf6qGQjQ+BUa8PEyty945jnF8ZSg5neQw7VI6TpQhIV58lX4dDWj9FVb7TTTDSGGW0tttpCEISNBKQNAAfAUBzDkU3kPGMrv2P8FWuNklpYuDz01m6ISzHtsx1XmvMMSPNSXR1LKijyyGysp6/0Rp3so8Vp+/xjiI/C4//AFasbi5ZVYb0tRJUvK8gKj8SftSR61v5Tnr3oCjnMm8Um/e41xP/ANIf/Vrx/KXxP7/2tsU/9If/AFauGS+Ruq0sPOOM5FPsECFBuaHMiuF6t0UutoAQ5bHnGnyvSzpKlNKKNbJBGwn0oDXt5N4pB93jTEz/AMYf/VrMZynxWj7nGOIn/jH/AOrVnRnydd620VwnVAU+9dOZbxLt0flCzWrHGBMZNnXHHn25d16x7IZ7iXVOJbS70KShKAlbiUJUtINY/G/gYv8AYuSG+S+R+QrNl0pyUqXNhz7KuQ3IdUdlzqU8kBY79JKCB290+lWTy6QrjS8gjeksKH0IfbIP7CKvSrmGx9fCQnTou29k8le3fw8CWFadNNR4n5QhLaQhCQlKRoADQA+VRvknkbEuJsKumf5xdEQLRaWS684e6lq9EtoT+ktStJSkepIre3G4wLPb5N2us1mHChMrkSZD6whtlpAKlLUo9gkAEkn0ArkzHnmvFTmJ5+5IeRaeD8Aedk4pBuH5lq8SWdhy7ywvQDKOkhtKvkd60sLpkRJODONcu5ZzZnxS88Wwxrk40U4Ri73vN47b1naXnEkDcxwaJV6pBHp7qW7dvXLVqZvEjFcLs87Mb/EX5UuJaijybevW9S5K1JZYOiD5ZUXSDtLaq07DOZcyJEuc5dMQwd4BTENtS4t4vLZ9FvLGnILCh3DSOmQRrrUz7zRsOwY/YsVtEawY1Z4dqtsNHRHiQ2EstNJ3shKUgAbJJPzJJoCGtWbmzIh1X3L7FiMdZO4uPQzPltj4amzEhpX4GGPxr9J4UxecfMy+9ZPla1dnUXi9PqivD+XCZLcQ/wDmasCtJlma4tg1uTdMqvTEBlxflMpVtb0l0js0y0gFx5w/BttKlH4A0BkY/jGNYnAFqxbHrZZoSTsRrfEbjtA/zEAD+io3zPyTD4m44u+aSAhyRGa8qCwr+3y1+60jXqR1HZ1+iFH4VrU5Hy3nPT+SONR8MtK9EXTJWi/OdT/erc0tPl7GtKfeQtJ+8wfSqnTx4xyD4jo+O3vJb9l1rwGGm5Xx28Sw5GfuT4/rZgRG0oithCNr2hoKPcLKqqY2rKnT3aftSdl3vj4K78CGvNxjaOryX301N3wbmGE4HgrVns8m453llzecul+exqEu4oeuTx6nEOTED2VtSfuaceR9wn13VgGfzhlSim3WKxYLb1E6fuj32pcun5ezMKTHaVruFe0PD5oPxsRtttltLTLaUIQAlKUjQSB6AD4CoTzRkV0x7j6cjHZfst9vb0ew2d7XUWZ015Edp7p/SDRd81Q/VaV6AbqejSjQpqnDRZEkIKnFRWiKTPG0/m3LpUK2Zbf5VhxmcuJcc0urjb0ybcGVadj2eP5aYcMNKCm3ZrbAc60qbbPUhTidlh1q/g+ut34ffdeUjH1+1WZb7qnHJFpfUpTJK1EqWW1hxlROz+bST96r7xHFbJg2L2rD8biCNbLNEahRWh6htCQASfio62T6kkk9zXOHiGzjGrveLZmPHP25csswia/DdRCsM56Jc4pV0S4KpDbJbKgpAKVdRSlxBB1skbbZqnUm6STafufBv4eNy1h05NwS1+0TKT8a1UketbCNLRcrfGuLbD7CJTKH0tvtFt1AUkHpWg90qG9EHuDWHIR61fhdOxmiAX/jbAL5Len3bDLNKlPHqcfchoLiz81K1sn8aisrhrjhslUTGkQl/BcOQ8wofgW1irUlN+taiSzWypYmrFWUn5k8as1o2VJcMOzDHFCbhuVzJ7TZ2u13l3z0Op/VQ8R1tn5bJG/XtWyx6+QsjYW/GDjMiOvyZcR0adjOj1Qsf8x9CO4qbPxiSe1VxyDbncXuUPP7BBlSbn5rcKTbobSnHLoyrfuJbSCVOoAK0kAnSSPT0vwqqst2evP6/UnjLtMpa8/qTaKgjXat1ESe1RrDcrxrNbai643dWZjKh76UqHmNK+KVp9UkH51KfNiw0ebLkNMIHqpxYSP3mqlW97MhldOzNrET6Vk3S8Qsdsk2+3JZTFt8dcl0j16UpJIHzPbQHzqCXnmvinFVNM3TOLYuS+4lliLFdEl951R0ltLbfUSokgAfEkVZmFcYZfyLcLde85szuO4tDeanNWeVoz7k6hQW17SgEpYaSoJV5RJWopAX0gFJpVd2l61bJe99333kbju5zyX3oZvA/h8TZrsrmnlNtF1z+7I64yHffZx6Koe7FjJPZKwk6cd11ElSQdbKr7pSuarVpV5upPV/du5FKc3OW8xSlKiMRSlKAUpSgFRHicAcf2kEfnQHhIPxU/5y/NJ+pc6yfqal1RHi7tjMtPwTkF+SB8gLrKAH7qAyc8yCbZ7Ui22FKXcgvSzBtLShtKXikkvOfJppILiz8QnpG1KSk7XH7LExuw23Hbf1+y2uIzCY6ztXltICE7PxOkio9ZEC9ck5FenfeTj7EewxUq9WnHG0S5K0/RxLsMH6sCpjQClK+KUlKSpRAAGyT8BQFc8jq/KnNsN42Z95n2wZTd9eiYkBaFx0k/BS5qoqgD95LD3yNWPVbcOpVk5vfMEoEqzJ5v7JKvVFjj9SYIH8l3rel9+49s6T90VZNAKUpQClKUApSlAKUpQClKUApSlAKUpQClKUAqvJY+yvEBa3GuwyXEJrUk/NVumR1MD91ykmrDqvLsr7T56xyGxopsOM3ObLI9UKlSIrUcH6KEeWfxaFAWHSlKAVWHiIxJ7IuPRkFqiqfvmDXGLl9nSgbW5Igr8xbCf8Ox58c/R41Z9fCARo+hoClbxcYOa+I7jlVvkok2+1YrcsljOIO0uCSWY7ax9OlaiPxNTzljkKLxhg1wyx6KqZJaCI8CEg+/MmOqCGWU/H3lkb13ABPwqjOBGXrZ4i8gwWU2tJ4/xQY7F2DoQftBb8IJPx1DdiJJ/WQofCrA5rSmZyhw1abh3trmRypbiT90ymITq42/r1k6oCL8CJvyeNynKVtKvQvd5FxU1roMr7Qf8ANKddtdfVrVTOUD3FRKwZLjnHk/IMLza9Q7FPRfrncY32i+mO3MiSpTkhp1layEuAB3oUASUqQoEDtvNkcm8ZK3rkXGD/AMbx/wD56A95aSd6rl+04FybhL+DX1rjy4XxyxZFmEuZCgToKHksXCZIXGcBffbbIUhxCiArqG9EA7FdDv8AI3GyvTkHGj/xsx/89Y38IXG+/wDbAxv/ANKsf/NQG3sUiVNt0WZMtr9vffZQ47EkKbU7HUQCW1ltSkFST2JSpSdjsSO9SKID2qHMcjcbJI3yDjQ/42Y/+atlH5N4zTrfImMD/jeP/wDPQH75bH/c1vX8xn/2yKvWudczy3GM6swwLCL9b79eb2/HYbYtslEnyGvOQpx94tkhttCApRUrWyAkbJAr28SfLGWSLzbfDhwlJT/CJmDKlyZ6e6MctXo7OdI+6vWw2OxKu479IUBFuTrtc/FryjK8PeGT34/GuJSEL5DvUZZT9oSEnaLOwsfUbdI9NEbGgHLSt9itmf5ajHIUGPH4+42eYhRreykCPPu7KUqSkoHYsQ0loJT6GR1bAVGST7WLEsT8LfBMi04VaVyWcfguPNtnvIu1xWNJK1fpOvvFCd/AqSBoAATHjrEU4JhNpxYyRKkQ2OqbK1oy5jii5JkK/lOvLccP1WaAklKVXGdXm+ZbkY4mwq5P25wsIl5Jeoyul21wlkhDDCtECU+ErCT/AGpCVOHSi0Fgfb3n+Q5NeJuF8RR4Umbb3PZ7rkE5JcttpcB0pkJQpKpUod9soUlKP7Y4glKF7XEeLcexa4rySU/Nv+TPtlp+/XdxL0xSD3LbegluO0SAfKZQ22SNlJOyZBjuO2PE7JDxzG7YxbrZAaDMeMwnpQhPr+0kkkk9ySSSSSa2NAa3JL7BxfHrnktzV0xLVDemvn4+W2grVr66FVl4XsfnQ+Nzm19bAveeTXsknK+On1bZSP5Ia6CB8Oo15eLie/G4NvNtiuKadvUmFaw4D91L0lsL39CgKH7atu3QItqt8W1wmg3HhsoYZQPRKEJCUgfgAKof2uNz0hH3yb+Cj7yD2q/91fH/AI95k1W2Y/6YOZ8Bxn7zFjj3LK5Ou4DqGkwoyVD5K9ukrH1j79QKsmq6wgKvHLXI2Sq30W9VqxZkH0IjxzNWtP4ruZQfqzo+gq+Tk+mzIluhv3CfIbjxorSnnnXFdKG20glSlE+gABJNfy98Mn9UMwPL+Qcuw3MyzYLbeMkuNxxyQvaWkMSJC3EtObJ6SerqJ9ApSj2Sfd6D/qqHPKeHPDBcsZtc3yr/AMiLVj0RKVaWmIpO5rmv1fK/NE/Avpr+RPh68HPiD8TD5k8W4W6u0sPeU/fJ7oi29lY9R5qu7ih22lsLUNjYG6sYfEdg3dXT1X3xJKc9zXNH9zvMjy2ESorzbzLqQttxtQUlaT6EEdiPrWBIbqjfDz/U/vEBxRam49/8W0xCNDdrtto9pjNK+JDshwdfx7loH/mq7Z/heuUppyVk3iPz9phsKW6uAbfAQlAT6k+zq1r1J/8A+1so4nDJX38u7P6e8m3qaz3vcYb7O99qj14u9itIJut5gQgO59okob1/lEVFpPE/h8lSFsw79y9y2pH5tZbyBbcFSh2O5iDGaV3Hfy3FEfL0FbWz8eYjZ1bsvhI458tJ6kqv9+XOf3o/FcR7R7/BZqB7dwMPZcpd0cvNNlR7VwcfZk5d0W15q6I9L5g4xTPTaYWVxbpcHd+VDtaFzX3D8koZCia+8f8ALGJ4pyM5yFzBhHI+LWWyQ1s2aVcsIuhiIdcB8+Y842wpDWmwG0lR7BbpOt9rnx7lLI8NY9lHh7t8KCNdacVusZzsPQhp1uNvXyBJ+VWHhfM/H+czvsS33R2BegkrVZ7rHXDmgD1KWnAC4kfrN9SfrUVTb9KvF0qS3XLLO9/C6XzPFtahV/Dg7N87p+F0vmReRw34Zud2WeSoGP4/f0XUeam+WKctgzPh1LfiOILih6HqJI1o+mqjWU+Evwh4ZZ7lneY4U1FtlniuTJ0u4X64usNMtjrUpSVyCk9h6aJPp33o+/JPDc7jG43DnTw621dvyGP1TshxWCkiBl0dI242Y4IQ3P6QS1IQkKK9IX1pV2/dg4jzzmSfb898Rs9ce3sSWrlaOOYLg+zYCkHrYVcnAOqfKQQhZSSGELT7qF9IWYI4qvGO5GbtyuyVVZpWTdiL+G7w4YW5k7fiEu3EllxB5xpTeF483bGmJFmt6/8AvqYQOpdwkJ0pRWVFlspaSQfMKupKUqFtt3ZgKUrzffYisOSZLzbLLSStxxxQSlCR3JJPYD614D0pVePc02G7OGHxpabjncr082ypSbc2d6JXPcKY3un7yELW6Pg2fSsdWB59nZS5yhlabba1p9/GsZfcaZXv1RJnkIkSE/RtMdJBKVpWKAzrxy7ahdZOMYLZp+Z32IotyI9q6RFhOD9GVMWQwyodiW+pT2jsNqrHGH8o5afNzjPk2CGok/ZGJp6D0nt0O3B5Jec+BCmERlA/P4zCHCxfBseTEgxrZYbHamD0ttpbjRYrSe5OhpKEjuT6CokOaLNeNDj7GMjzVJPuybRDS3BUneutE2UtmM6n/BOLP0NAfpXCWJpa6YuQ55HeTsofTnF5cUk73vpckqQrv8FJUPhrVfk45zFjg6sez+3ZTHR39kyeAliSvt90TYSUIbH1VFcP1+J9EZFzY4PPHFmMtskjTbuXOCTogeqUwVN7BOtBw+nqa8H+YHMa6XOTcEvWJwvRd4cWxMtbR+JdfYWpTCB8XH220fyqA9ovJWR255MPOOK8jta+4My1JTeIK1Ab0gx/651/Kcjtitzx1bbhbMTYF2iqizJ0qZdH4yiCqMuXKdkllRBIKked0EgkEpOu1SGLKizozUyFJakR30Bxp1pYWhaSNhSVDsQfmK9aAiOC6F5zdtQ/OIyL3z89wIak/uQpI/ZUuqI4h/W+YZ1DX9966RJ6R/e12+MyD/lRnB+ypdQCq55kmybzBgcTWV9xu6ZwpyI+40ohcO0o6fb5Ox3SQ0sMoUPR6Qz8N6n1wuEG0wJN1uctqLDhsrkSH3VBKGmkJKlLUT2AABJPyFQHie3zr69ceXshiOx7jlaWk26K+gpct9mbKjFZUk90OL61vug9wt7yzsNJoCwYsWNBiswobDbEeO2lppptISlCEjSUgDsAAAAK9aUoBSlKAUpSgFKUoBSlKAUpSgFKUoBSlKAUpSgFVrxTIbyXLeQc6WrTrt6GNsMrHS7HiW1JQEuIPdJVIemPI3rqafaUOygTZVVrkbCcG5YsWXwvzVvzVwY9fGx2QZaGluQJZ+AV+bcjE+q/OjgnTSdAWVSlKAUpSgKDlZLa8X8aTFkmNFpeYYSy1HeSn3VymZEhYQo/MtNOEE/3MD4ipb4h8VvF/wAATfcWaLmRYfOYyO0oHq69GJK2tfHraLiNfEkVBOYcPkZN4gbRDt0xuDdbjhclVmmrSSmNcoM9iVHcUB3UgL7LSPvIUtPoo1c/HuZMZ7icLIkQ1wZK+uPPgOKCnIM1pZbkRlkdipt1K0bHY6BGwQaA/OG5PjPKOGWjMrWy1Lt11jpkspfbSotqPZSFDuApKgpJ18Umtt9gWL/eWB/yZH/wqjMXkHw98xP8e3BwtYLyBKcn448r+Lt10UdvQifRKXD7yB6bIA2So10FQGB9gWL/AHlgf8mR/wDCn2BYv95YH/Jkf/Cs+lAYH2BYv95YH/Jkf/Cn2BYv95YH/Jkf/Cs+sS7XW22G1zL3eZzMKBb2HJMqS8sJbZaQkqWtRPoAAST9KArrnXlvGfD5x9KysWhqVdprqLfY7PFbCXrrcnOzEdASNnau5IBISDoE6B0vhm4UvHG9lueccjy27nyZnTwueTXAe8GlH+LhNHvpllJ6QAdEgkdukCBcH2i5+JXlNXipzWE8zillL1v40tElJHSzvpeuziD2DjpGkb9Ej49KFHqegK/5YQLhNwLGnf8AU94y+KXuxI/rOPJuLfp/fYLXr2qwKgWeFtPIXGinkHSrtPbbUUkgOm2SiBv4HoS5+wKqe0BGuQ8zbwTFZN9TBVcJyltw7bb0L6Vzpzyw3HjpPfp63FJBVrSU9Sj2Sa8uN8LcwnHPZLjOFxvdxfXcr3cekp9tnu681wAklKAAlttBJ6Gm20DskVHrV08jcqyMiKw7j+AOO222jXuv3paCiXIB9CGGl+zJIPZbsxJG0g1ZdAKUrxkS4sRBclSWmUD1U4sJA/aaArDxP4xc8o4XvrVkjl+42ssXeM2NkrVGdS6pIA9SUJWAPidVN8FzKzcg4jasysEhLsK6xkvo0dlCj95CvkpKgUkfMGs+4X2x2q3i7XW8QYcFXTqTIkIbaPV9331EDv8ADv3qiGcOumLZJccj8Lud41cI85wzLth8mel6EtxWup5hTZKo6yfh90nXwATVOrSrUKrxUIOUWkpW1Vr2a56u610sVpvs6m+s7rNcejOhqr/hBtxeHTrrIPU/dskv05Svj0KucgMpP1SylpH+JVeR/GLiuPznLBy1iN5xG7RXPJk+WEXGIhfxAeYJJPr26divXGedeDMNwS9s4FyTBvEkO3W82623Oa3EWuTIedkiIhTyWghHnOlKOs+6kgFWk7GSxlB2TlZvg8n5OzMo4qjLJSXwOTOYeHbn/VCfHXcsZmzJDHE3CrTVnu0pklPtU4nzJMVk/B1S/wA0tQ+4iPv1KN/0cxXFcbwfHbfiOIWSJaLNamExoUKI0G2mG0jslKR/z+pOydk1S3hlh8UcJ8SW3Fn+WMQul9nvP3nI7q1eIxFxu8pZclPb6+46z0JJ79CEA+lW81yFgMhsPMZxj7iFeikXNkg/tCqlVek9JLzJVOL0Z8zvOse46xuRlGSSFojMlLbbTSC49JfWdNsMoHdbi1aASP6ACRQ1wiZNyjITfOVgGrf1B2DijTvVDij1SqUR2lPem+r82k9kpJ94+9wvKeVuQpWaOPB/HMXfetmONg7bekJ2iVP+SiVBTLZ+CULI+/W1fe9dmtfUn6U7v2OC59XzXJaceVtNVqemNuX9mtFz6vmuS048renmtsoS0yhKEIASlKRoJA+AHwrzMrv96tc/K0fWsVUw7+9RzsHOxvEyd/pVhX7HsfyyEmBkFtaltoUHGlHaXGHB6ONOJ0ptY+CkkEfOsJuYSfvVnMSer415dTW7LNHjcai3ZK6NhhnJt/45uMTF+R7s7dscmupjW3I5BHnw3VHSI84jQUFEhKJGhs6C+5CjfFc/y4UC9W+RaLrEblQ5jSmH2XBtLiFDRSR9RUl4Fym4IauXFuRzXZVyxcNLgy3lbcnWpzYjuKJ+84goWys/EtpUfv1Nh6rpzVKTunp06fTy5FjC1pUZqjJ3i9HxT5eWnlyLcpSlbA2hCs7zu5We4wcLwq1M3fLbu0t+PHfcLcWDFSQlcyWtIJS0lSglKEjrdWelOgFrb18Xhi2XiQ3deVbxIzq4IUHEM3BsN2qOsEkFi3pJZBST7q3fNeH90NTpm02uPdJN7Yt7DdwmstR5ElLYDjrTRWW0KV6kJLrhA+HWr51H845GsuEmJb1xpl3v106xa7HbkJcmzinXUUpUQlttPUnqecUhpHUnqUNjYEnUpiIwVKLbLLKNknSUoSB+4ACq4Xypds1WqFwrZGb4ySULyacpTVkZI+LSx784j5MDytpUlTzahqvjPG19z5xF15qlx5cTYcYxGC4pVpjkdx7SohKrg4P74EsghJSyFJDhspCENIS20hKEIASlKRoAD0AFAQGFw9arlKjXrk25vZvdoy0vNG4tJTb4jo7hUaCNtNlJ30uL8x4A6Lqqn4Guwr7SgFfCARojYNfaUBB0cJ8Wx5bk224fGtTrq1OOfZTrsBDi1dytSGFISpROz1Eb2SfXvWQMFvFs74vyFfobae6YtwWi5sb+JUt8GSfwD4FTClAR7GcZuFquFxvt9vLdyutzSwy64xF9mYbZZ6/LbbbK1kaLjiiVLUSVnuAEpEhpUE5Gyy/NTIXHmA+X+VV+ZcdRKdQFsWiEhSUuznkn7xSVhLTXq44QOyEuLQBp8wkHlvI3OLbQhTmNWqS2vMZ39qe6QlxFpQf01ObQp/1CWSWz7zw6bTrS4diVnwbG4WL2Jt0RYaVbceWXHn3VqK3XnVnut1xalLWs91KUon1rdUApSlAKUpQClKUApSlAKUpQClKUApSlAKUpQClKUAqveek+XxpLugHezXK03gf/AHS4x5B/eGiPwJqwqr3xAac4gyKF+ncG2Le2Pm5IfbZQPxKnE0BYVKUoBSlKAp7lb+xnOPDuQHs05Lu9ndPzMiH1IH+UyK2GWok8T5VL5Qt0dx3F7x5f5XRWUFSojiEBDd2bSO6gltKW5CQNlpDbg7sqS5r/ABM/2Ox/DswPZvGc1s899fyYU6WF/s0/VwEAjRGwaAiHIeB4rzJgcjG7q6h+BcmkSIc6MsKUy5rqZksrHbY2CCDogkdwTUJ4Z5Pv8e8L4S5fUmPnNoZ6oko/xV/hJ2EymVH1XpJ60+uwT8FBOC/bLn4YpEi743AuN34pmSFSLhaIzZfkYmtatrkw20jqcgbKlOR07UydraBR1NolvI3HWJc5YnbbvZ762zOYCLljeS2t5Li4zhAUh1pxB0ttXu7SDpQ16EAgCx6VTnFfMd7XkB4h5mhsWfPIrZXHda7Q75HHpIjK9OrQPU36ggkAaUlFx0ArlrnK4y/Ejy3F8KuLynk4pZPIvHJNwjqIHkghce1JWPRbqgFL0dhI7fdUmrR8SXM44S41fvlsh/aWT3d9uzYza0jqXOuj56WUBPxSk7Wr091JG9kV+fDbwsnhPjlq03WZ9pZXe313nKLso9Tk+5vHqdUVepSknoT9BvW1GgLNt1ugWi3xrVa4bMSFCZRHjx2UBDbTSAEpQlI7AAAAAfAVk1qsmynHcNtD1+ym8xbZb2PvvyHAlO/gB8ST8ANk/CozyBfswlYbDv3F95xqKxMSmS/drytQjRoKmyv2hOtAnXT97t32ewNS06Mp24J5XenmYSqKN+LXBanzmCHc2rLas1ssJ6dNwm6ovyYTI6lymAw9HlNoT+k57LJkFtPxcSgdt1Ccu8RuL5FdbJxpxJlsOVkGWhKWruy37RGtLCk9ReUNaW/078to9gvXmdKeyv3wbyJjGVYNmF3xbm2FyvKtEt5mfLU43FgNSkx0rDDa0pKEMkKTtxJWjuojuFCuZeJeTvFNeOQIH5B4bx7brJAW57Xi+GWvz4rvU2pKUXC8u9KAtClJcBj9SV9AG+5rCtiMHg95VpOT/Lu8fBpyfdbyIalaUVF5K/B5vwSOmOQc+k+Hew2vGMZxmwRsdhQU/wBnckymPbozbnUrr81TxC3nVH84te9qU5skkmsPkjxBxcP8M55ZvGRMOzZ8Nl+I/hTTd0U8l99KGnIjMhaA+OlSSrZ7DrPwqucs8BEXn/kZPLfiPuVqeu3srURNtsTCiw0y2VFCet8qBV7x2fLJOh73YauXEPCZ4fsKZZat3HFullhAQlVy6pY0PTSHCWx+ASBVf02vUhFUaCjbjJ5vvXrLwy6mCVWbbV7Pm7eVlfzdzmDw2eMBi753crtyU/yNBtogFNvcvFxZkpmvqcQOgWuG2osqCdqC1Oa7ED1qDcuWbmbmLlHIMwxmAtm2T5W7a9A47uFxk+ypQlLXnJkyG2PM6Up6ilPSTs+h7/0ntVgsVia8iyWWBb2/Toix0NJ/ckCs+svSNpufaqsou1vVjbLl7XyPPRW47kmrd1/i2cfc2T+XuVuMrZxxxtxnnGGogyI6HZMrG7ZMbfjNsqQGfImKUhr3i2oLT3T0aCvWtZ4a8a8SvCrt8TkHFErNHbghhuHIMWw2ByK02V9aVLjJHndZU2SFqJBb7eprtSlQ9ni91x9IlZ6pKNv9LJPR3dS3s+6P0P5u5H4c/GhkfIErLZ1sRLgXC6qnyIEm24wFKZU71GOZAQXT7nueZvr+PrVk+JfizxOc9Wm0WvF+LLdgSoEhx+VJjXS1z3pm0BKElTrQKAn3j2PfqHyFds0rPdxd4yeJneOns5d3q5GPocc03rrlH/acHcFeHbmTi7HLvaOReALDyXPuMoOx7lc7rbo7kRkNhIaSgNrH3+tZUOknqAP3Qao29+FPxb4VAkZXmViw921xCHJa2Q005pSgnSEIfUnZKgAAnsSNDtX9Y6rHxH9+J5aT9xd4sSHP5hu0QK/zSagxEK0YVa8qrlJrNyjB3ssr3gU8bgaccNNr8qbWSXDoiBY5YIeI4xasXt6QI9riNRUEDXV0JAKj9SQSfqTXyY7rfetpK3utJM3s1XaUI7sdERuKhFRjoiGTeRccZupsi3Lj7aCr80LTLOwlQSVAhvRQFEDqB6e47969rbkMS8xTNhKWWg+9HPUNHradU2vt8upBrV3KNKPJMSaI7pjpsclpT3QegLMhkhPV6bIBOvoar63WKfaXLTeIluuAmu5DcxKHU4NxVLlqSCknpSgkNKB0Bsg+qtnWzrVIzs1df8d/MpTm0y2Lnf4ljtMy9T1LEaCwuQ8UjqIQkbOh8ToVIIj+9Hdc4sRb5OtWQCLZprCJ+LyQ5ERClJ1L2nTalOk+c6ApQ6kgb7+vw6Fg76RUmGrus3ll/wAmdKbZI4rmwKxFzDjnJ+CZY0elMmc7jk3XbrjzGyW9/PUhmPr+cfnXrD323Wrz7YtticR/Gt5Xjxa169X2rGH/ADE1bqu1Ny5Z+WZNWdqTnys/J3OnaViyrnbYICptwjRwo6BddSjZ/aayQd9xW5OgszQ5/laMGwa/5kuKZX2LbZE5McK6S+ttsqS2D8CpQCd/Dda3jzj1rD2JF4vE0XjLb0lDl8vS0aXJcTshppJJ8qM2VKDTIOkgknqWpa1brL8at+aYnesOuxcEG+26TbZJbV0rDTzam19J+B0o6NaPijLLhlGKJYyJKGslsLyrPf2E+iJ7KU9S0j18t1Km3299y28gnuTQ8JnSlKAUpSgFKUoBSlKAVXPFaRf8izjkZ0df2peV2OAs+qIFsKo3R+Htn2g4D8Q6Kntxcls2+U9AaS7JQytTLavRbgSekH8TqoZwM1Db4TwRcF9T7cjHYEpT6h7z7jrCHHHVfylrWpR+qjQE8pSlAKUpQClKUApSlAKUpQClKUApSlAKUpQClKUApSlAKrnlt1N5uWFcex/fk3nIYl0fSPVqFbHUTXHiP1POaisk/rSUD41Y1Vhf2l4RzNbs5nkybTl8SPi7jzvc2mWhbrkboPolmSpxTa/iXkxR36vdAs+lKUApSlAVx4jbA5k3Bma2tlJU8LS7LaA9S4xp5Gvr1NipZhGQN5ZhlhyhpQUm722NOBH98aSv/rrbSozE2M7Dkthxl9CmnEH0UlQ0R+41U3hYkvNcSs4nNcK5mH3S4Y7JJ9QY8hYQP/NqboC3qwrNZLNjtuas+P2mHbIDBWWosNhLLKCpRWopQkADalKUdDuST8azaUBDuT+KcR5ZsSbNk8VxLsZfnwLhGX5cuA+PuusuDukggdvQ6GxVZ4/yvmHC94jcf+ISSJFskOeRZM3QjpjSh+i1M/uL2v0j7p7knsVm/aovxn5mvFeCrnZrbbItyv8AmkqNilhhyWw4l2fMX0IV0kEdSEhbie33kJoCIcao/wBEh4krxzVMHn4Pxat/GsNQoBTUy6HQnXBPzCdJaQe4I0RopNXnnHKeK4DcbLZbs5Kk3S/y0RYUCEz50hYKtKdKAdhtPqVfTsCe1c/R7dmvgttmPQrZfjmeIXNxqE/YnelFxanrT1OuwelIC0rc6lFsgaKgPVRVWuzbxJ8U2PI4N+4DxxzkjmfklSYVpsbr6gu2tNJAdclFXeFGbAKla11nej0gqTYpQhBdpXT3Wna3F9/JcdeRFOUperTavx6Ikviiz7gzivMLPkmexrxnOcT2UR8SwCCsy3ZcjqIDzcUAhAJGlOr933DoKUNV98QD/GHJvFuE2HxCcfXd7I5rkS8t8d2u6refenBpQMZ8xlBLzCFqO1HSSUAgEgpMj5NzWOM8g2DjXA7FeOZHbYITt08hDqMfhOHrX50soC/L6j1Jb7dR0SnZCVTHirhSzcduycjulwfyPMLoOq53+cOp90n1Q2P7U0PQJT8AN70Nayvja+LaoUXfdy3nmo9EuL9y48iLWTjRWfF/er+HuK3wvw43TLLTCj8oQoGMYnEIXbuPMZSmJboqQdp9pU1rzl/EgaGxsHuRXQdnstox62sWew2yLb4MZPQzGjNJbbbT8glIAFZtKyoYWnQvJZyerebfj8llyRNTpRp5rXnxItnPJGM4FY7teLnK9qds8T26RAiLQuV5PUE9fllQ6UbUNrVpKRskgAmq/wCWL3cr9wze5F4uLVmutmXCvNxiWG8qXIiW9ElDp6nWwlQUqO273A6SQekqA3WnwHMONMCsOR4/ySbZEzGNOnovDMptKp18S+8tTLjKT78pDzam0pSneiPL0OnVag2SXw3gszFr2xbImFXGQ8zBtsWN51+vyZTZKbesdkIW2FqbU91LJQ2CS2ElRgqV3KLbeTTv07/vhxudtgdlwoV4whFupGcXFtf2iVm9zhbjxupJtxUXedcX8g5sh4WXObDeJFmc81VnyyTCEYS47aOrqmsA9UVet6WtKEua30oJCTlXPxJcbMvuRMZVdsvfaUUrGPQFymQofAyT0x9/TzKpye1eMrlxGeXVKFqabaTa7C0+py1RgkaCJCvWS8kBA63vdKlaQnts+/J05y04LJNpdXCLL0JDZjKLfQkyWgQnp1oaJGh8DqqjxtVQe5ouev7dL3fM38f4YwNTEReIV5SsrQdoJ31vZ736lDcimnuu1iyVeITIlnrj8HZP5Prt65Wxtev5okH/AJ6yIfiXxZsBWVYbl+ONH1kybamXHT9S5DW8Ej6q0PnquHPHzerfc1Ylx3m/Ms3jDCL1HuEu4XiNZpNzE2WwqOlmEtqOpKwkpdec7npJQNjYGq28EErFePObonHvC/iDm8o4tfLPPmXyM5jky0s2h5gteQ8lMhSupThWpBKdfDq37urMfSOz7VVPBpfKxo6y2QsasBLC2vZb0ZzTu/7zmvcf12xrK8ZzK2IvWJ3+Bd4LnZMiFIS6jfxBKSdEfEHuK2tcvy8QjpupyjELi/jGR9j9pW8BIf16JktfckI+iwT+qpJ71aPFnLz+TXBeD5zCj2nLorJeCGVH2W5sDsZEUq76Gx1tn3kEjewQoyUMZvyUKqs+D4P6Pp5NlXav8NPDU5YnBSc4RzkmrTiueWUo/qVusYq17PqEc3Y/NyfibKbTbGy5PNuckwkAfeks6eZH7XG0ipvXyrdSCqQcHxVjkKtNVqcqb0aa8zny13aHkVkt9/ty+uLcorUtlX8hxIUP6DWNMaJ32rEi2pXG2cXPi6Wny7dKW9eMYcPZLkNxfU9FSf1mHFkBPr5a2z8DW4kM77arVwbnH1tVk+/706Gmpyc4et7SyfetfqulmRmUxv4VrnIm1elSV+KdntWIqJ/JqOUCKVO5qWI3SewraxWSNdq/bcTv6VnsRta7V7GBlCFj3it6A7Vrr1ao+WZrg2CSIzcliZdVXWey4gLQYcNpTiutJ7FJeVGRo9vfrdJ8qO0uRIcS000krWtatJSkDZJJ9ABW04Csb9/n3TmO4sLbavDKbdjzbiSFJtTairz9HuDIc/Of4NDNSOPaSjSXHXuWv08SRxdScaK1bu+5O7+niVdZ/AvimE+Iy0chWa0MXjDJCpLz1rmq8w2qZ0FTTqeo/nmusEAK2UqUk+8B1JtPPeTudePsqmvt8IDMMK2gxpWPTwq5sp6E9fmRXAPNV19eg3oa1s7q5qVdjhYUk1S9W7vlb7sdlW2xWxlSM8alV3Y7vrX0ve901n148bmnfyyxW622y45DcI9j+1lNNR2bm+3HcL60FYY0VaLoCVe6kk+6rW9VFstx7IrDlrPJmAWz7TkyGW4GQWZL6GVXOKgksvMqcKW0yWStfT1qSlxtakKUClpSN5nnG+Ccn2X8ns/xeBe4AX5iGpTey0vRHW2oaU2rRI6kkHRPetRxhxDYuHbRcrPhlyvciHLd8+LCut0clMQtJ0GmOrZbb33P3js+p0BUt5qVrZe/yKW7h5Ud5SanfS2Vu+9/NeJ+BlvLtyUpdm4fhwGk70MiyZqK6r5aTCamJ7/VQ/6q+JyHnGOz7RcOLMTeCSetm2Zi888Rs66A/b2UE612UtI2fX41FMK5U5zhZXb8I5f4TW0q4OFlnI8YkiXa9hJUVOocIcjp0k91kknQA71cKbpbVXFdnTcIxntMpkLih1PnJaUSErKN7CSUqAOtEg/KkKkaiuvfkMRhamGluys7q901JW7039SM4zydZb/efyVudtumN5F5S302i8soaeeaQQFOMONqWzISnqT1FlxfR1J6ukkCphUQ5SxCXl2KrFjDDeR2d5F2sEh49KWbgz7zQUsAqS257zLuu5adcT+lXvgfImP5/blvW11UW5wulq62eSUpnWuRr3mZDW9pUCDpXdKxpSFKQpKjIViUUpSgFKUoBVccRH8nJ2VcWvHp/Jq5KmWxJ9TaZylvx9D4Ibd9qjJH6sUVY9VryUoYVl2Ocsp9yBHJx/IlegTAlOJ8mSr5+RJDWyeyGpElR9DQFlUpSgFKUoBSlKAUpSgFKUoBSlKAUpSgFKUoBSlKAUpSgFV74g2lHhLNZ7I/rq0WaReYR+KJcNPtUdY+qXWW1D8KsKoB4gXixwTyKtABcVit1Q2n9ZxUVxKE/iVED9tAT+lfCpIIBUAVeg3619oBSlKAVTeB/wClDxE5/hh9yLlMGHlsFH6IcH9bS9fUrS0o/jVyVTPPX+k7KcA5ka9xmw3cWe7r9ALdP00pa/mG3fKUPqTQFzUpSgFc2Zu0eT/GxhGHOJLtn4ux2Tl0xJ/i1XGUv2eKkj9dCUqdT8O5rpOuYOLnrk9zv4n85t0B6ZcYarTaLc0hIK1qi2xS/LQN/pOLSfrsfXXsY7zSR43ZXN54i8rwfgxxzxLZlJl5HMgw2rFiONsAdUq6SFK6W4xAUS676FXSSlCVnStACE8zYja7jyXiOR8Q4zJxLnXJYDb05+KplPsVvUAp4XMpC0OJSvaQRvqUj1VpAPlOx7h7DuNOP77nUgXXOuG0JvUXGGbgnz27tc0q8gS4o2rq61FSFKAKS2op3og2RiFqVwfht45f5P8ANvGd5Q6wZyI/SpxT7qwiNbYwJ0EpUpKfXXYknpSNV9qYmar+jQndQ1lwUE3ay5yzt4vviwtGrjKsMNho/iVGsuN3wz4vTp4EZ4OzGycHOnjLmDHXcVyi6ylvu5JLfMiFkL6iT5ntZ+6rv9xetb9epRFdNpUlaQtCgpKhsEHYIqqLXnGFcwJl8X8mYT9mXSQyp02W7eW81MaHq9FeQelzp33KelxB7kDsaqzkZeaeEWxuXTEM1avOESeqOxjt6kn263qUNAwX9ErQgkHoUNAD4k9VMO6XZpUfZRfxWz62zKnYV4brtdaNNPimrpp802nzOq6VTnhh55hc5YE3LluNNZJaQiPeI6dDa9e6+kfqOaJ+hCh8ATbkSdCnoW5BmMSEtrLa1NOBYSseqTr0I+VTlezaueU5m1M/2YuLMVJgtrWJLqE7ZRraj1n7o0O9ce3zMsxz7kS05tBRb2k3+LPRjybmy44m22xnyil4NJUnbsnq617IIQW079whV8+JO4vJ46RicV1Tb2Y3ONYCpJ0oR3SVydf/AHZp8ftqiecOQce4jsVuy+Tj/t86O4q3Wtltzyko8xIKwTohKelofok9gBrZNaTaleMXaTtGNm+/9l8VyPqH8CbLq1YxlRhv1q7lCCyyilna+VpSdm8rKEk8pNGLjOe5Nkj2PDJ4FnEHJYUiQ2xGS4XIz0coOytR0sK2VDSUlBA7q9az79HYZtq7HfXXHrGtTRS+XdOR/LUFpStR7lO20e8SVFStH4VRtx8WvDmD8XwOQMhx2Vb5dpku2m22iI557y3FoStflKUUjo6dFSl66da7kp6szgXxj8d+IWfMx+zwLhZr3EZMn2Gf0Hz2QQCttaSQrRI2CAe+xsbIo01KtR7eGcXx++p1eNnR2ZtN7MrpU6sXnC+ad21Zq6vu2s7565lpcw4vgGd4DdLbyRi7F/s8RhyeqKvYWFNIUoKbWkhSF62ApJB7kehNUD4PcgwLFsrm4Xh3F9qx5F3bW8JcR19+QoNgqCHnXlrUpOt60UpB/R71YGd5DPjSxxni1xQx9oNBUyQRs2mIohCiNADahoIB2oqWSSO1SjHMS444rhSb9brZFtqY8bUmc4oqX5aQN7Uon10CQnWzrt6VQniK9SqlTnaEfaXM6jD7I2Tgtn1J4zDdpiay/Ckvaiub728lm3bgmm7hjyd671i5PYHMigMuW6cq3Xq2PCbaLige/ElJHuq+qDspWn0UhSgfWqx4veyTLLxK5Lvj0uFBms+y2W0lwpS3E6t+e6n0LiyAR8k/PdW/Ec3qtjRqLEQu1k/u5x20MJLZWJ3IyTnHW2aT4x5O2j4PNZrW2OKM+TyPhcXIH4ghXJpbkG6wgrfsk5k9Dzf83qHUk/FCkn41MKoTiKace5lvePJV0w8ttCL02j0SJkRaGHyPqpp2Nv8Awe6vut7hKrq0k5arJ+H11PlH8Q4CngMfKNFWpySlFclLh13XeN+NrkR5N43tXJePptUyS7Anw3hMtdzjgefAlJB6XUb7EdyFIPZSSpJ9apSPfrvj93awrk+E1aL8slESUjYgXgD9OK4e3WR3Uyr30/JQ0o9M1DuYbdbblxblTd0tkOc0xZ5klDUuOh5sONsrUhXSoEbCgCD8NVlPCOtUTpO0nl0ff9fjocpisLdutTdpWz5O3P5P4rIrVyN9N14GKP1TVe8DSo3KlkMbA+Q5OPZDb2wZeP3lo3SEpI7ebHUtaJIbPYdJeV0dhrWiqyXMF5+irKE2fArkn4OpvEuIT9Sgxndf5RqLF4XEYGq6GJptSXj5Wzt1sa+nVdSCnuOz4rNe7PzSPJMX5Jr5Ol26ywXrpd50eFDjJ63n33A222kfEqPYVFeU/wCGnjXDHs3yI4lbbVEkx2pqramRc5LDLriWy4hLns6CQVJ7Hsd1OeIePuIs5gwuQmsuf5GkNqC2ZNzWkswXvXSISUpbYcHbupBcHY9XxqNUa8478IO2l3kvr7l3mce1qS3KcLP9WXu193ijS4xhl25tfak3WBKtnHjag4pEhCmpOQ6O0p6DpTUT0JKtKdHYAIJKrizTkPEON4MYXuSpD0j81AtsNkvSpRSPuMso7kAa2eyUjWyBW8vt3i49Y7hfpvV7NbYrst7p9ehtBUrX10DXPeKwJ09lXIGUafyXImUSJLhPUIjCvebhs/qttggaH3ldSjsmt5snZcJKVSq7rK/NvkuSXj5u5JPewb7OlnUlm5PRJdPHJX5ttu99zdeX+Wb5s4/jtmxSIr7rl2UqfMI+ZZZUhps/TzV1FJ165PmKKrlzHf0/3u3w4UVA/D8ypf71GtxcXCAe9ULZORrq6uJOm5C3co6mpr11ZTHQBbkNdRQrrQBrfSE9Ktk72PQ12uBwNKabp04q3S/vd37zXV96/wCJOTf95r3RsvcWq3LzRKtt8u5olXzMthX9CmSP6K3dty3mO3EKgcnouYT6NXuzsOpV9CqN5Cv296o61ch5BAsV5fvzbL10iNMzI7QAQny5AAbQrX6jnWgn5J38amWMXWdDyVmzXbNXJk11tSlwl20NNKISFEsuBI7DfoVLOvkat4jARUXvQi/8KfyIadr+rKS7pSXzLutPPt0s/Sjk7DlQYo+9ebM4qZDQP1nWikPMj5kJWkfFQFZ/IHA3FHNrkPOkPyYV88lCrblOO3Ax5iGwD0FDyCUrTonWwrse2qjEBZOq00LL7nwhlUZjH8Wul+x7K1vFVmtiQt6HPbQXVux2z2KXG0uKW2Ne831DupW+VxmyKOLTjSjaWtuDtrro1rrblbjusHtTFbMqKr2jtpf8yvlnbVPRq3fdaWtk93zLijjW3OWXG73yVcrYmPGm9D7Tc2S0EnzZOj2WvtsNpGyVAfM1H8QyPgnxOsSLonFkSrzjqhFlNXa1qiXazuL6iEBzQcaUelXdpfwPfdZ9g8THEF6li1z8jXjty7BUG/RlwHUE+gJcAR+5RqxWvYLnDdl2mY1qa3oTIhQsntoKCtFKiPhvYrQVsJiMNNdomlya+D/5N3DF4bEUmrJzvfeT8045rutu266EGc40y7GgX+M+S7tHCO4teSuuXqC4fjt11ftjZPoCJBQn18tWtGA8m868tcaiLf5/FlyfYhvsRLzbIscSYSmHCsquMe7daEMtN9PStEllKtlBKWkEvVuuPbP4jePcziYllF7g8gYRM83y7/IUmLdbZ0oKkIfQB0yASAkKT72ySopGhVxR7lb5cmVCiz470iEtLclpt1KlsKUkKSlaQdpJSQQD6gg1DTqb6u1bvM8Th+wlaMlNWvePLro0+jSfg0fLXdLZe7dHu9luMWfBlth2PKivJdaeQfRSFpJSoH5g6rKqAyeKIlovDuT8a3RWKXGS4p6ZFYa8y13FavVUiJtKesnuXWi26SB1LUn3TsrTnLrdzYxvNbT9g3eSSiKfO86FPUASRGf0nqXoE+UtKHNJUQlSR11IViWVi3O2W+9W2XZ7vCZmQZzC40mO8gLbeaWkpWhST2KSCQQfgayqUBVVhyC4cQTYuCZ/PekY2+6mLjmTSVlQSFHTcCe4fuvDshp9R08OlKj538ZatY1ytluvNvk2i8QI86DMaUxJjSWkuNPNqGlIWhQIUkgkEEaNVy3ifIHGA3xzI/KfG0f/AKtXWYUyoaP1YM1e9pH6LEjae4CXmkJCaAs+lR/C83sudWx6faRJYfhyFQ7hAmNeVLgSkgFTD7f6KgFJUCCUqSpK0KUhSVGQUApSlAKV5SZMeHHdly322GGUFx11xQShCQNlSiewAHxNfwr8e3jK5QzHxCZzjvF/Ot/PHbL7EKDHst1WzCeS3GbS/wBJZIDqFPB07JUFb2NjVAf1R8QPj18NXhzTJt+XZy1dshYBH2BYumZNCx+i4AQ2wf8ACrQfkDUG8D3jD5P8YuY5rk68GtGMcbY60zBt7RcckXGRcHFde1vbDZSlpJKkBsaLrelK7mv4MqUpaipSiSTsknuTX9z/AOpB2q02/wAHUKZb+gyLnkNzkz9evnJUhpIP/immv30B2xSlKAUpSgFKUoBSlKAUpSgFVxy05+UdzxXi2Kep2/XRq6XAD+1Wu3utyHlKHxS48IsYj4iST+ialWaZpYMCsTuQZFKU2ylSWWGWkFyRLkLOm47DY9511atJShOySa0PGuLXxiTcuQc5ZQ3lOSJaDsVDgcRaYLfUY8BCh2V0da1uLHZbrjhHuBCUgbvMsGx/OoLEO9tSEPQnxKgTYchceXBfCSkOsvIIUhXSpST30pKlJUFJUUmLqt3OeJNeZbshsedwo/pEuUP7MubyPn7Wyoxlua9E+zNJUfVSAeoWRSgIxhPIWP503KZt4lQbpbVJbudnuDXkzre4oHSXWtnsdHpcSVNrA2hak96k9RHOOObbmDsW9Q50ix5Na0qFsvsEJEiMCQVNLB915hRA62XAUK0D2UlK04WFcg3CVeFYByDBj2nL47SnkJZKvY7vHToGVCUruUjY8xkkuMqUArqSpt1wCd1oM9w62cg4ZecKvCf60vENyKtQGy2SPdWPqlXSofVIrf0oCrPDnnN1y3AfsXKlkZTiEpzH72hR99T7B6Uunfr5iOlXV6ElWvSrTqgeVGX+EOT4fPVpYcONXwNWnNo7SSQ2neo9w6R8UEhKj+qQANqJq+IcyLcIjM+DJbkRpLaXWXWlBSHEKG0qSR2IIIINAe1cn8f5Xc8A5G8Vt4hsxpsmx3O23hqMpWgoLtoX7xGjrpR+9J1uusK5S5chQeK/E/EynJmk/kDzjZk4Tf3yotpi3RpKxEcWv4BxpRaSfhpR2NVJScY1Iuaurq/dxMZpuLUdSAZxwZzVnHI/JHKeM4q2uJndrwy72aY3dI7JbnwY7iXEJSpXWQA926ukEkHZKdVYjeSZtl54wxLkywzrZfrCq4XS7NSm9JlOxGm47D6VD3VhRmde0nQWhXyry8VmO5fyTxI94dPDbyLjuN3+xyrXHnuXa6vRn48ZsoVGaZWGHVLWpwMfnErQpKkpHUSrVarB184Rcxtdi8QqsekZjYMNZjvz7JLceYnodlrAkLStpvy3V+z++lIKdp6hoHoTqNp4PsqzxCbW9ZWejV95Pvt10Zv/AOEKCntJ4hSu40535XcXFeKcif5jj8DLLeiLJfeiSorqZMCfGV0SIMlP3HmlfBQ/cQSCCCRU04f5Nk5ciXhObIjs5dZW0mWhCelm4xidImMJP6Cj2Unv0L2k9ikmGvytE96i2TxLg+/ByTGZqIOSWJ0yLZKVvoJI0th0DuplxPurH4KHdIqqqzoT7SPiua+q4eR3M9m09p4b0Ku7cYS/kl/tlpJcPaSbVn7PQYvI1+yG8zG7fi1ntlxnWaVAs0dMOfLQw6Uq9umJAd8tYSlwNNlKelSSpS91F7JzTwjw5n1qveH5ZjkfHLy2m1X232txCkNdIJjzQ21v3kHbaz6lDgPfoFbyfmVpvD0Pmyyw1QrbkD6Mcza1ukKVars3pth5z4aOwypXYKSuOr0Br3n45hpiSrf+TNqQxMaWw+lqI2jrQoEKBKQD3BNVak6kmp05LeWd3d35WzyTWTXejeYPDYSjSeFxlKaoyW66cHGKi1lNSvGW9OMrSjN3bW5K9mjWcx+JPjrMsiwZeCM5FlDdsuEyStu3WWQPNcMN1tHQXUoSsgOLJ0ew2aiF+wnMOcX455KswxvFIbhkR7Oh9Lk6U90qSlx5xPutABR0hPfud1KJGZzH8Ix62ZBLU9eOMMmhMyZLh96XZpjbsNiUfnrz0IcP67KifUVM7/drRZIqpt6ukSBHT952S+lpA/aogVHOLxTlKtK6dm1ay0tnm3k01rbIvYWrHYMKVDZlFxnDejGblvzXrOScbRhFOUZxknubyUlazVzkfxDcE5RhtoxfMPD5hNhv1xxJc9L9lvsBm5NzWpaGkre8qRtC3UeQjQ9dE9PcAGo+O7nzxdOQLHmPI/E+I4pcrRHnRsftVlxuLaJNxkSUJbW5IQwAosNJSVdTmu/ZPqop64u3K8rLHnLTxBZl31/Zbcu76FNWyKfiS4QC6R+qje/nX6xXjlrGXZN6u09y8ZDcAPbbm+nSlD+5tp9G2x8Ej6b+GvPTJ9j6Nhkt3S/BLpwb9y9x7/Rugtprbm2pSda6l2bfrSktHP8ANGPO7UpaJWbkqfRyHifE12kY9fjNu19nLS/fLi2hJHmrTsJGyCUpSRpIHYH5kipDYrbceRLjb05JFlQMRZUZFttshASqcUkFvztdghI7pbOyoJJOwKqnxSZp4feN87jP5s/kEu+zWm35VrsyWlbaHupW6pwpDfUE60FbIG9Deze3E/ImC854E1k3HUxZYjueSlp5vynoUhtIIbcSN9Pukd0kgpV2PetdTwOJV3VX4d8uvf8AebO0xv8AFOxJuFPAVL4tx/Eb/I7W9RaK2it7MbWs81bUD3UpAGgANAVIIROhUax6Qm4Qm5AGl/ddQdAoWOygQCdHfw3sVKYiCNVt6WeZ86x2UnF6n5tC1Nc28eqR951F3YV/gzFCz/nNo/oro2ud8CjKvviBgBA2ximNyZTyh3CX5rzbbST9fLjvmuiK2+z/AGJvnL5JfFHz3+MGliMPT4xpK/jOcl5xkn4isG+2eJkNkuFguBc9lucV2G/5atK8txBQrR+B0TWdStgm4u6OQaTVmVJg/EMvjDk2XKwmLCZwi8wkuSIK1nrgz2wEJWwNHYcR97Z9QST2ANt1+XEJcbU2relApOiQdH6juKqfEcCzziq9NRoPIUSfx435zj0S9pJl21GlKSlmRv30dWh+cPup3rZ71bnUeMvUrTSkktb+tbrz011531hhDsGoU43TfDhfpy+BL+VMFZ5L48v2CPSRHF4iKYQ8U9QacBCkL18dKSk/sqj+I7DYvDHylO4wvECI1Z8rCpWM5G8y2h5zp0XLfIeAG1JPvI367+oA3WaeMDGYM26WLjDFLrntyt6WwHrSA7B6l9veeb6ldj27IIJ7A1CMy4x5D5k4yXyBzper68yY4k2vDsVtPS5DkOfm21uId048tPXtSSRoBWla3UKqYiFONOnByjU0zSjfg975ZvobSls6jvyqY2qqW5qmm5vjZQyemjk4x/UemS+M52/5hM4cxniJGTzbi6u1JVFyBtceSlY0opcbaUOjpKiTsdIB3rR1urE/eMI9h4z5D8uLeIrCWbdMCiY12YQkBKmnCBt1I0FoICtjqA6VCtF4assx2y55bsIzzjiHiOaW/H2LVHlPFEYy0eYpxAajdIIcdSpSnFhRBU2EnRIFSXm7hfmTlGZkLV+z2KzhkCO5OtNthRh5776EKU2hzsDsKA94qP0AJJF/YuHxOArTobSqrdlmunJxsnd55p2uuTRU/iTFbNx8aVXYmHcHBWk223NvXeTdorLK2jvqj3uLRINVvMwOD+SycRMmQqGFDrUenqcR5vmKQe2tK7pPb0JqdcY8TX7KOL8ZyvEeTZzLs62suSId6jC4x/PCelwIX1NvIHWlXYuKA+A+FZkzizm2KSDY8RuqfguPd34yj/iOMKA/yzXT0cbQoTdNVVdPjlmu/LybOVmqsoqU6cs+XrL3XfmkVIvjDGnZa3WbazFZkRHYUliM0lpD7ayk7V0gHqSU+6R3GzW9x7BpEW7wbrccmuVyNsQ43EbkIZHSFgJUVKQgKWdADZP7z3qZN8dc0KUQOPLSj5FzIUhP+ayT/RW2gcPczzyEzLniGPNK9VM+0XN1I+gIYTv94/GrFbadLd9arG3en8Lsjp05X9WnJv8Autf6rL3nxuRFt0VybPlNRo7CSt155YQhCR6kqPYCohe3uc74YvK/C+C2u8RLAzIbs7N3fWw5PdeR0rmMtHp620o6kI6loKutah1Dp3cOP+HvE4kpm6ZncbhmM9hQca+1VJERlY9FNxWwloEfBSgtQ/Wq0QAAEgAD4CuXx+0KcqcqWHbvJW3tLJ62T5rK7txy4m5wNCpGtCtiILdi09x53a03rNZJ52Teds7ZPinw/cg898sX662vm3htrK4f2q1bZ/2g5Ehox5sp6t/Z7jfmr3sq8wklXSACOk7lvLNg8P3CWQRW/t3OON3bi0JH2rj5lKtqD1FIS/oOJBJT93Q7aNdRPuwIRVMkusMFSQlTrignYGyASfgNn95rA/KHFLgFwvty1SQ4ChbXtLa+oH1BTvvWjwlTGYOl2dGq/HNeK4+J0WPxGA2hiu3q4dRjbSDs9NU3e3gl55lL4re+ePsONk/HueYhy7jkjflOOpECWoA6KEutktdQI0evuD6iq2ziXiqs6Zz15vL+B+QHnG0TLquD7VaLqgEbbf6D5T/YdlkJ0dE70NdPZLhJuGCTsOwa8qwxchs+xzrRGaSYa1OeYpaG9dJ6ldXV6E9ajsE7qJ8Vp5tW5d8H50stgvMKIygxMiglKWrshRILbsQj3HEgbVoBHcAA+tTzxVHENU8XRT/VH1Wn4ZL/AC+JDh6dXCqeIwFfdtluT9a8XzurSXNX67tiQxeZuLZWT2rC4ueWiTeL1EVMt7LT4WJTaTo9C07QVdj7m+rQJ1oGpRebLachtj9nvdvZmwpICXWXk9SVaIIP0IIBBHcEAjRFUnnng44Sz1Sbnarc7js4K85mVZ3QhtLgO0qDZ2lOiAfzfQe3rWtv/L2e+GeFZrdy+wjK8ckOpt8TIrcHEzAoD3RKbc2grUPQ+YOrR9TupPRqdRt0aityl6r+O6/NN8itvbyjFQlv8besu9WzWWqadufKzvtO88ZnyskmSLtiaezd4dJclWtPwTMPq4yP/wBp+8kaL2wFvGdNOtvNoeZcS424kKStJ2FA+hB+IqLYLyjgvJEdxzE78zKeY7SYbgLUmOfQhxlYC09+2yNfImsB3Hrzx8tVwwSIudYyorlY4lQSWfiXIBUQEH5x1ENq9UFs9XmV6lOdKW5UTT5MxjOM1vRd0Tula+w3+0ZNa2rzY5qZMV7qSFBJSpC0kpWhaFAKQtKgUqQoBSVAggEEVsKwMiruWrZfsMXK5t49szt0vdogKTd7GwelWQW9rqWGkn4SmiVqZUQd9TjR0HepEI4b8f3hV5sjsJx/lK3WW6ugBVoyFabdLQs/oDzD5bp/wS110RX8qPGt/Uo86y7lOXyR4aolpct2SyFSbpY5MtET2CWo7cdZK/dLKztRRvaVEhIKSAkD+oM/LcVtVmdyO55LaolpYR5js56Y2iOhPzLhPSB3HxqCyuTM5zDcXh7AnH46+35RZMHbdbUj9Zlgp9qlfMaQ20sej1UB4Fv6nXiHhmtLWZ8jR7XkvJMhXme2JSXYtoT8G4vWB7/r1PdIV36U6G+rs+gKfPh2hZktM3njL7jyM4FhwWmSgQ8faUDsBNtaJQ8AfT2pchQ+ChUpyDg7hbK7amz5NxJht0hNthptiXY4zqG0gaASFI93Xw1rVTelAcich/1KzwbZ426uDgc/Epjuz7Tj9zda6Sfk095jI/AIFZHhr8PuTeBK037FrfdbvyBx1ebgLo0uJA3dLG+UBDqlx0EmU0tKGtlgeYlSOzSgoqT1nSgNTjOV41mdpbvuKX2FdoDqlID8R5LiQtJ0pCtfdWk9ik6IPYgGttUKyXiTFL/dnMntyp2N5I4lIVfLG/7LKc6RpIeGi1KSn4IkIdQPgmtX9r8x4N7t+scbPrSj/v6ypRCurafm5DdX5L+h3Utp1Cj6JY+FAWTSoziHJOFZ0uRGxy+Nuz4YBmW2Q2uLPh79PPivBLzW/h1oG/hupNQClKUApStPieXYznNiYyXEb1FutskqWhuTGX1IKkLKFp+ikqSpJB7ggg0BuKUqHcjZ6rEIca1WKE3dcsvilx7Hauvp890AdTzpGy3GaBC3XNe6nQHUtaEKAj2L2ay5LznmucPWqHKkY7GtmMwpjrKVuxnkNOypPkrI2gKRPjpX0kbLej93QtKo3x7hreCYpEx8zl3CWFOy7hPcQErmzXnFOyJCgOyet1a1BI7JBCR2SKklAKUpQCo/muD2HPbSm1Xxp5CmHUyoU2M4WpUCSkHofjuju24nZGx2IKkqCkqUkyClAVvjucX7FL1EwDlh1n22Y55FkyJpoNQ70dbDS0j3Y83QO2vuOaK2j99tqyK12Q47Y8sssvHcktce422c35ciM+jqQsb2PwIIBBHcEAgggGq7bvOW8Mj2PLnbhk+EN9o9+ShUi42hv4InoSCqQykdvakArA7vJ91b5Asy52233m3SbTdYbUuFMaWxIYdT1IdbUNKSofEEEiue8Wvt08L+TxuM81fek8bXeSW8Yv7yifspxZ37BKUfRAO+hZ7a+m/L6CtN3tV+tsa82O5xLjb5jYejS4jyXmXmz6KQtJKVA/MHVY2S4zYMxscvGsntUe42yc35ciM+naVj1H4EEAgjuCARo0Bs0qStIUlQII2CD2IqI8tcXYtzNx9eeOMxjFy3XhgtlxAHmR3R3bebJ9FoUAoH5jR2CRVStJ5M8L5UwzDuWd8Wtnqb8s+bd7A18U6/t7CR6fFIH6IHe6sJz3D+RrG1keFX+JdYDoHvsL2ptWt9DiT7yFfNKgD9KA4nj4zcOQMnRwRztm0HE+S8EglUHIjDURlVnbBW1LDqnUAqbS3taTtSVJWrfZYTZvsmS27lu8Qssv0e9XG3YrZYirgy0WhJT585SVqSSdKII6tHRPf46qX+Lrwwu+JfEbdbLTlUfHbzZXnH4U5cEPKPWjpU15iSlxtCh2V0kg9tpOhqp+ObJyHiWQzMQ5TXEfyOx4xY7e/LiPl1mY0y5ObZeSogK2ptKOrqAV1dWx8TBtXGVquGpYebvGMssllk+OvvOq/grD044zE1IrN0s/8A5KZOpcrp33rUSJ2j96vee4Ruo7NkKTs7rnKkz6lg8MpI1N0nwcPvM7JrhCVMxPI4v2TmtvQkq82GQUomoA7+axvewNlG9dwCNbeLLdLmF4Zds2vUWdjik9E+0zEtC7QXUBUWX1dKgpLjYBOvRYcHwrdIkJfCmXEhaFgpUlQ2CD6gioYtUjG32bIpK3HMajvTLKRtS5lhKuuZA+anIitSGh3PleYkVSm8rPT7v9V1uvzHS4Wm1NTj7aS5cFaLz4perL9G7JtKkiOZbxmI1vdv35XZndW4yQ3c467oS7It3WFPNI0BsjpDgSdpKmwNfGrB4j4v4pvllRc7rYIt4v8AbilqdJnyHZiXiUhbUlCXlKHlvNqQ6jt6L16g15XLO8etklu2WNt3IbtIQFswLaA6opI2FOKHutp7g7UfQ71UaxXiXML/AJAvDr1JbgSV25U6yY6zKLUa6w0SFuyLY5KR0q62/N6mwepKQs9wneoqdOEaqcIKX17+fv5LMtY3aeJxGBlTq15Ut3O6e7ePG6VrxWrbsrZuSjA3PK3MU3H0ew8TJt13bt3S1PDNvceZhlSglADyFhvZUpKQgAnfx+FTbB4OaLx5mZnkhlV1lfnVx2WghEZJHZvt3KvmSfXsPTZkmIjD5lpVYrFZWrWbU4hEqzPRgy/AfSQoB1r4K2AoK7hWgpKiNGt45E38K2UKMnLtJSunwWiOMrY+FKmsNGlutPOT9p8vrlqrZvV8DeJDGOVuNeVb/neK+GnBuXbRmJhvB++Yy9eJVrdYjoYLCEtOJUhtXQF76SCVa2CKtnwhYFnFtt2TcgZ5xxjvH8vL5ERTGMWGCYUWGzHZKA4WCtZbccKipSSd9gTreh017H3+7XoiJr4VflXnOkqTWRzFHZmHw+Olj4t7zu7cE3qR62xTbr69G0oMz0ee369KXU9lpHwGx0q0O5PWTUhnXC32G1yrzdpKI0OEyp991fohCRsmtVmD0Gx2f8p50lmK1Z3EyXHnD0gNb04nfQpXdJPYAEkAbA3X4VxpfeabWm+XrIpuDYdCdamQHHGUIlXFxtQUiQ6h4aaYBAKELHUo6UQAE7rxjK/Z0leXBffA29erR3Fi8ZPcpLKUuq4RVm3Jq2STtq8k2YV84P5Qyfge/ZnheR3vE+UL1K/KS3exzFxlpabb6I1tfCSAtKmB3SsFIeWSfSoZ4J/EjyjcrzFwjxOZzPjXjIIwfxRi6WViK1c2uwV5ctvXmPJUFJU0tIUDruVEpF5o8PWWXEedN8TXJL7D6UqSYc1mPsdiCFJQRoj5VgXXwW8a5KI/5a5jn2T+yOqfj/a1+LwZdJ2VoHQAhX1Gq3+ExVShhfRXQuuDbV0+Odnq834nzvaccDtHGTxlbF5yekYSaSWSit5xyikkuiRdl6yfG8bYMnIsgttrZA35k2U2wnXz2sgVWuQeK3g2yOCHCzJGQXBZ6WoNiYXPedPyT5YKP3qFU3kmPeGrhXmO34byZwuqFZMibaFpza9TnZ8GXcST1xX/ADVKSwoAApUvQV73oBs9P2S3YHjVulScVtNlgxYaVB8WqK2kI6U9RSUtD11o9Ot9x2rCVLHWTajFPTWX+0qKWxaP/uVH/hpr/wDR/Aqz+E3xGZ77nHXDMfFoTn3Lpmcotr6T8fZGfziTrv3JFIvhmfy99u68+ch3fOZCVhxNrQswbSyr4dMdojqI9OonuPUVJMT5x/LzIYdvxLjfK5FlfUfOv0yGIkRCekkKR5hCnAToaABG91l59xjlue3wJc5WvNjxgsJQ5a7Q0hh91zv1FUruoJI0OnXzqb/pKjUUMdPrnmu7djx6S8yP+kdSnBvZdONPheF97/PJuS67rS6G0vd3wLhPDfbG7Ki12iOtDLMO027ZW6s6QhLbSfvKPbZ0N+pry44zXLM6bn3C+8d3DFrZ7n2YZ76PapSD1dSnGU92SPd0CTvdSbH7FBxmxwcftpfMW3MIjMl95TrnQkaHUtRJJ1862NTqVGFN04xz58l0Sy7736czTydWrU7Scu/jd9W9fcUJk/hOx9dvVf8ACciu8LkCJLFyg5LcJi5L6307029vsWlA9JATvWt9WiDJ+GeZnM5cm4RnFsFgz+wAIutqWdJdSNASY5/TaVsHsTrqHcgpUbUqtuXeGIXI5g5JZLo5j2aWE+ZZr5GT77R7nynR/bGVbIKT8zrsVBUrxDxGWId+T5ft04cCNUlSzpLw5/uZX8LdotvJz/GGSQTZXnY7T9llyHQGbqCNOIbOgErQrQ6Sdnvr4bn9URima2Pkm4t8QeITDLfBzi0rRJYjyEBUW5BB2JMJw+oPTtSAd62DsBQTYvIOFZRlpgO4xyTdcTdheb1GGw08iR1dGvMQ4O/T0nX841lUpUnKMPYds27td6td592T6ZLyE5pN+1y4PufDImC1pbSVrUEpSNkk6AFfUqSoBSSCD3BFRPELBmeP4zOt+Z52rKZ6nHVsTjamoym2S2kJbLTZIcIUFK36nq18K5q48438S3FeM5BmsjNU2m0WmDMuTGPvtplmQGm1uJbDIUW4wVoD82skemu1Z0MFTrRqN1opxta9/Wvyyv7vIxqYicHFbjd9dMu/h7y5uTOeTj1/d4943x0ZVmDLaHZbSpHkW+0IWNoXNkAKKSoe8llCVOKHfSUnqqq7xaOUMv6n+R+ZL6UL7m14ss2SC2D6pDjRMtY+G1P6P6o9Kz+I7HGsvGtmkpme3zr5GRe7pcVd13CdKQHXpCj8epSu3ySEpHYCo14gb9dcZ4myi/WWe7CnQoKnGZDSQpbR2B1JBBBOifga3eDwdKi91K75v7yKVatOpx14Gok8KcPed7RcOP7Vdnvi9d0G4uH8VyStR/aa/KuIuGHx5TvEuFqT8jYYvb/MqpMnzHObZimWzLTkOVSbGzPsLFpuVwtwj3Ba35rbc1tlK2Wy4joUgJUpH3lrAJAGtPKzjkJ/Hb/j9ly27266NZVZ7bbX7xDaTPjRpS2E9TyAgIW2pwvdJ0SUggnY7dDCn6upr5RdzoC18P4FbVB3E2rvij4O0u47eZdt6T/MZcS2ofRSSPpUzteU89YCjrhZAzyZZUDTtsvSGoV2DfxDE1lKWXFa9EvNgn4uj1qneKOTLvm2fIhyVqiJYxpKrjazo+yXRuY4y+kn17FGh30U9KvjXQNvJOq1eMownlNJ9/11LNGU4PJ2NR4cccTCv5vnCWbvJ42kOyWr7hd/bc9vx24hPV5LIVtTR61AqQslJSepJWFJUL6vFtw7kWw3XF7u1br5a3i5b7jFK0uoCxrqbXo+6tOwfgpJ0ex1VC27/Sn4jcJu1kUGl5zGn2O+R0ekpuNFXKjSVD4raU2toK9emT0/KrCf4RcsXKrHJ/GeQDHDdH/9Ndq8jzIV4b0fzvlgjy5IOvzo9dkkHZCuLxmEjgpKFJXi87ck8vj7vI6mhjZbRk62Jnu1IrKVtWs1drO7X5s3e1+ZFc84z4P4wxDFomT3S+Wl2NMastpypuQ57fCW51FkPS0jQaTry0+ZtCR0jt61uVZZy1xCB/CBC/LfFW/W/wBqj9FwiN/rSYo7OJHxW36AEkb7VY99tuG8iWa94TeRb7xCcSYN1heYlZbK0BQQsA7bX0qSoehHuqHwNaTF0Yrw7YMU40vGcPyn5KlWyzuXiQgyphQFLDQIACihGkjtvQSNkkbypYydNKlVSlSWieq6J6r4X1TIa2HhiI9rByVdtt8VJNXvbpxvwzTWZixRb8nbHJ/Dd9t8l+cAZbPmEQrsEgAJe0Cpl9KQEh4JK0gBK0rSlKRJcYzO05Op+EhD8C7QgkzrVNSG5cXewCpIJCkEghLiCptWj0qVqqB5n4wzPiW+yOauCpzlvacV52Q2dtsuR3E+qnwyOyk+pWB7ye6kkd9TLBuSsE5qRb7HnNjZteVtMCXBT56m1PNrHeTbpaClfSoDv0KCxohXoTV2vs78FYrDPehx/mj0a+ay45Gup4r1+xrK0vc+76F10qIfkFdIfu2LknKYLKe6I7zsecjf8pclpx9Q+nmj9lSthLqGW0Puhx1KQFrCekKVrudd9b+W61hcPSlKUApSlAKUpQClKUBHMv47wvPEMHKLCxKkQyVQ5qFKYmQ1H1VHktFLzCv5Ta0n61GDYuXMEHm4vkLec2lv1tV/Wli5IR8mJ7aQhzQ9ESGypR+8+n1qyqUBXKOfOOoSfKzOZOwqWns7HyaGuAlCvilMhQ9ne/nMurSfgTXxzmq2X5XsfFVguGcyT29qg6YtTX8pye4A0ob9Us+c4PXy9VY1faAq+4cdcjcgwZELkjkFdntsxtTTtmw8qi7bUNKQ7cHAZDnr2WwmKfpUWh+HvMeK7jIyTgTkV5kux2mH8WyVtD1nlpZQG2iHGG0Px3ghKUeft33EISpCwhATfNKAofGuYua8luwwe9YLhOC5WtCymLdr9LlrcQke8/FaREaamtgdyGpIKQQHPLUdVZOD8dxcTlTshulzdv2UXjp+0b1KZQhxxCfuR2kJGmY6O/Q0nfcqUorWpa1bXLcOxvObOuxZTa25sUrS83tSkOsPJ7oeZcQQtp1J7pcQUqSe4INQyz5Fk3G16g4byJcXLvZrm+mHYsocQlLheV2bhXAJASl5XZLb6QEOnSCEOFAdAs2lKUApSlAKUpQClKUBU904Nfx+6y8p4Nyo4NdJrypU62GL7XYbm8o7U4/C6k+W4o/eejrZWonay5oCvWz8qZzZcis2Jcu8cN2V2+y1W6BfbRdW5tpkSg0t1LavMDUhhTiWl9IU0pPUAjzCop6rTqPZ9g9l5FxSbiV+89EeX5bjUiO55b8SQ0tLjEhlf6DrTqEOIV8FIB70BIap/MvDjZJt9XnXF1+l8f5aruudakAxZnffTJinTboJ7n0JPc9WqzeNuSMhi3pPEvMSIsHNWG1rt85nSIeTxG/WXFB+66kaL0b7zROx1NqSs2nQHLnIviM5p8P9tYY5XwzHrwuS+23Au1onhtuYlK0l1K4y/wA4hfldXvpBQFlPYg1rByFivJ/K9wzDD54kwbniVoWQezjDqJM5K2nE/orTsbH1BBIIJ60UhKvvJB/EVQHK8URud7c/0BIuWJOoT9THmJJ/d7SP31Q2ivw4vk1818zrv4Nnu42pD+anL3OMvhEjE9onfao9MjFRPappLi9W+1ah+Ds+laWpC59MwmKUUQe7y2setr11ehy5KWukeTEZLrq1EgAJSPiSR9KiV6xHkrkm3+1vOMYsmEFyLXHQormKf6FJSXXUkeUCFEEI76UQat9Nv7/drPiwta7VXeH7TJvI2a2s8PacEt7m8/C2mfErzgoY3CaVZLPaW7exc0OXGChSfzyFIX5cuE6s+8tyM/tIKu5acZV8TVkZph11vFqj3LGHkRcksMlF0skhXoiU3vSFf3txJU2sehSs1BLhid0tmepi470tvZG8btYus9LTWQR2j5jC1eiG5kRLjaj+u2lXqBVs2208t58oRLPjr2BW0AJlXO8ttvTirXvJjRkKUjt6ea6rp+IQsVYw1NuDpOLbWWS+0ua5JrianbWKhSxMcfTqxjFree++Gjus5TTzjJJNycZ5brRKI+MYF4g8Rs2fPQZdovS45bTPgPeRcLc+lRS9GU4B74Q6laVIWFIJST01GrhxbzbjyiLNdsbzGIn7on9dqm6+RW2l1lZ+vQ2PoKt3BMJs/H2OM41ZHJTzTbjsh2RLd8x+S+6suOvOK0NrUtSidADvoAAAVIK3focasVKplO2bXP4PxR8v/pHVwNedLBvfw6k9yNRXtG+ST9qPXdkrvU5tVB5rQry18Hylq9OpnIIBb/epaVf5tZcXCefL2oNt41iuLsnsp+4XJy4PJ+oZYQhJ/a6K6HpWK2fHjOT8vkkyxL+MK9vUw9KL52qP3SqSi/FNFVYp4frBbrhHyDOrzMzK8xVByOuehLcOI4PRTERH5tKh8Fr61j4KqrOaPCbmGU5DHyjHOQLxeI6JTbrtru0vznGW+oFXsy3D5Z0PRCwB27qNdUUrcbNxM9kz38LZN65Xv33z8dTltr4mttxqWOm5W04JdIpWSXRK3QiWA4fk2JtzU5HyHcspMktlkzYzLPswT1bCfLA31bG9/qioy5w3l8q9Kus7nfM/I9r9pRDilhhpKArYaOkHaddu/qPWrTpXscXVjJzVrv8ASvdll4FN0INKLvl1f1ILyrwrx9zVb4dn5GtT1zt0Na3BDEpxpp0q6fvhBBVrpGu/bvr1qB5qznXh3gWtfAnB1tyTDWQ89kFrt85TV369NhD0ZLm0vdKEKBQSVrPSBrWze1K8hiqkYqnJ3gvytu3kmZOlDe30s+fEiXHHJlh5LwSDyDboV0tECYHApi9RFQ5Edbbim1pcQv0IWlQ2CQddialtaXMcOxrkDGLjhmYWtFys11Z8iZEWtSEut7B0SghQ7gehFV7xR4X+MuFcicyDAJOTRULiKhJt0q/SZUFtsqSoFLTqlAKHSADvsCfmawtSabu0+Ctf33+RlmW5Sqaz3w3P53ls/K0eITmTGxPLZ+y7BkrcWBH6G0o/NNFhRT1dPUfeO1KUfjqpXlvGE3JeNInHUHkzMbK/EZiMjIYM9Kbs75ASCtx4o0pbnT+cPSArqV2G6OFPL1tdctBd8idUqtOKOH77xpPnzbvzTnObomspaQxkUpl1uOQd9bYbbQQo+h3vtUazPw3ZTl2RXS+RfE3ytYY9yeU6i3Wu4x2o8VJ/Qa2yVJSPh72/rXqp03Kznlzsxd20J7yfxPh/LFlRasniuIkRV+db7jFX5UuA8PRxlwd0kEA67g6GwdCqztXKWdcGXGPiPPrhuePPOCPa84jtENnfZDc9A35S/h1+h9STpShYfKfD9k5dxaDiWR5Hk0CLCkolKds10XCfklLS2+h1xHdSCHCSnttQSfhWu408OHEXFFpvllxXHX3Y2TeX9sC6T5Fw9t6AoJ6xIWsei1DsBvffehUtOpTVPdqNvpbTqnfLyaMJQblvRyZrrp4ufDXab/Dxd7mLH5V0ny24LEa3PKnKL7iwhKVGOlYR7xGyogD1JArYcz5/yjiAtlp4w4Qlcgy7wl9LqjdWIEOCEdA3IcdB7K6zpIHcIUKr1zhHJ/D7f5ud8A2WFeLPNc8664rLSkSNAAFUKSR1hWh/FrJHy6vdAtji/mPCeWYLzuNzHWLjCPRcLTNb8mdBc3opdaPcdwR1DY323sEVlOFOFqlJb0eN38UrNeefM8jNv1ZZM5lwi2czcAYk3A5kw6EnEvOecgS8aeeuSccjKWVIhy0FAcLLYPSh5AWlCQErIACjJcggYjy1hUqBGu7F0sd5Z8sybdKQtK0bB91xOx8BXVtVZlfhl4byq5P35OLuWC9SD1O3THZj1qkuq/WcVGUgPH/CBVbDD7XinetHPmvo/qVamDf/AKbKMz3CIGbWJ7Hro7KZYdeYkB2M4EOtuMvIebUkkEAhbaT6H0qLReFsbbLj0yTdJ81+4wbk/Olyut91yG4HI6SdBIbSoH3UpA95XxJNXZM8KuRRyRjXiGzJhr4NXWBbbgE/Tr9nbcP7VE/WsVHhg5RJ0/4inOn5s4lESv8AepxQ/orcQ21hVH2/c/oUngq97Je9EFxvi/GLLnV25Et0R1m73yM1GmkOHynA36L6PQLICQT8QkVJb9yDi2Gux4FymOSrvM7QbNb2lSrjMV8A1Hb2tX1VoJHqogd6lVv8J0F8hWacx8hX9J+/HZnMWphQ+X9YstO6/wDGVZmCcUcccZMvNYJhtss65PeTIZa3Jkn5vPq246fqtRNUMTtig/YvL3L6+4sUsFU/Nkc12rP7Pw9yrA5C8TkWbjFxv9rdhYowlr2uDZoZcSZDb7rRUTNcUlguFKS2hHlpSs++T1PimZYnnVpRfcNyO3Xq3rPSJEGSh5AVoEpJSTpQ2Ng9xUN584LxjnzD42LZGC2YVwjz48hOwpHQsB1Gx30torR+JSfVIr8cQYDeuFMeuuN3zN41yxC0nzLE7MaDUm3wgkqcbkvbCFpR+ivQISDs+gTy1bEYitiHKpbd4W4dNdPtnTrD7PWAi6Larp5p6Nc1lrnmm+DtyPHIeGZcfla28ucb3liw3OS63GymKtpSot6gj4rQkjUhH6Dn7Dsdq3nLvGlk5hwe4YdOlezykqS/b57J/PW2e3pbL6CO6VJJSexBKVEeiqmcWVGmxmpkKQ0/HfQlxp1pYUhxBGwpJHYgjuCKqg8aZbhvNyeQOPHI68ey8lvMLVIfKEtvobPlXCP2P5w9IbWkfe2kn4qTFOEYppRupPPx4/XzPaGIqVZqUqm7OmvVbX8uaTfde178I6FkY5EvETHbdb8nuLFyurMRpqdKaZ8puQ8EALWEbPSFHZ1v41z9knG+M4dlTHH2UxHE4NlM5T+MXFhZbfxu8LPUWGnR3bQ4odTfw6gUkEbNTfk7DsztnJ2Jcv8AHMZ+fKacbsGR2oPpSiXaXXN+ckLUEhxhZ6wexIJG9DRnXIeD2zkXDrliN2JQ3Oa028n78d5J6m3U/JSVhKh+Gq2WzsY8PUcJXUck38JLqvqtGazaGFjUpxqwak3d20s75xa5PK3DS2aygFhz/KeLr3FwTmaaiXAmrDFjy0I8tqUr4MSx6NP/ACV91f4gk3FVWcbXVjlXA7ngPJ1sjy71Y3DZcihvJ2l1xI9yQn49LielxKhrvvXpWkgXrIfD1PjY9mU+Rd+PJTqY9svrxKpFmUo6RHln9Jn0CXfh2B7a1cr4ZV5uEUlVWqWkuser1to/y8ilTq9nFSbvB8eK6P68OPMu+lfltxt5tLrS0rQsBSVJOwQfQg1+q1JdFKUoBSlKAUpSgFKUoBSlKAUpSgFarKMZs2ZY9PxfIYntNvuTKmH0BRSrR9FJUNKQtJAUlSSFJUAQQQDW1pQEB4yyW9JkT+Ns3l+fk+NIbPtikhP2vb1kiPPSBoBSulSHUgAJeQvQCFNlU+qD8nYleLmzBzPCw0nLsXLr9sDiuhucysJ9ogPK+DT4QgdXfocQy5pXl9JkOI5PbM1xa0ZfZi4YN6hMz44dT0uJQ6gLCVp/RUN6I+BBFAbelKUApSlAKUpQClKUBHM84/xXkmwqx3Lbb7THDqJEd5p1TMmHIR3bkR3kEOMvIPdLiCFD51Xacx5I4S/rPlJqZmWGtdmcvtsIruEFsen2pDZG1pA9ZUdPT8XGmgCs3PSgMCw3+x5TZ4mQ41eIV1tc9oPRZsJ9LzD7Z9FIWglKh9QaqPxBxfYco47ywjTSLlLsT6v1UzGOpG/oXozSfxUK2964IiQLvLy3h7JZHH1/muF+YmFHS/aLk6fVUy3qIbWo/F1pTLx+Luu1RDkq9chTsMn4hzLxtPZjlTUiNlmDtrvEeM+y4l1mQuDoTWylxCSUNtvp0FAuaO6r4qm6tFxjrqu9O6NxsHG09n7Rp1qztDOMnyjNOMn1sm3Y9X4u/hVGSc1yGPmEpld8uPkM5U3aDHetaRbm4ikt7KpXlDpd2s9ILhJUUjp71Y+N59frvaPtePjrWY2ttamVXrCpCLgz5ifvJdilQksOD9JsoWUnsTUOlq4xVcJ675mOV22Fcbki6y7NcrS7DYVISUEbLkVLvT1NIPT5mjr4jtWiru1k3Z9cvj+59Y2TFy3mo9rFrJwtPj+nes7cHuvnYkWH3Z+XbL/cr5LT5VrvFxY8xSQkNx2XCADoD0SPU9/nWr4lyq+X6fPt2SymXX5kZm+25CEoBZhPlQEdXT6qaKQFE9/zg3Wqfc4mmtXi3Ncx3Q2y+SJEiZbI6WFtqL6tupBEcuhJ2Rrr9DUlgIwh3IbfkWAYFlEqbAbfY6bHjL0diS26E7Q666020QFIQoe+NEeuid4Rkm1aSy66+RarUZwp1FOlJb2jcWlG2au5JJXeT4WN5yTAlpwi43i2EIuVhSm929f6smIoPI/YSjpPzCiPjVp3PLb/AJDmuLY9jl+ctMDJMVn3gPIjNOutuNvQfKUPMSofckOAjWve36gVDGOMOTOS2zbswiM4Zi8gdM2G3KTJus5o/eZUtvbUZCh2UUqcWRsAp3ureVhFr/K6z5cw44w5ZLVKtEeM2EhnyX1x1k61sFPsyANHWifpW0wtKo3KSTSdumjz66HBbcx2DjClRlKM5x7R5WklvRSim84y9ZXybS45tpVFa885Js/H94y6bmP21cnMmk4la4suBHajNv8A2uYDEhwspStWkgLUnqAPcDXbVuYlYswsrkgZLnS8iadQgtebbWYzjLg316LWgUH3dJKSoaPvK321K+IcckYPdsDmSpzsK7XSZd1PpcDb8eS/NVMStpQHuqbdUCgkH7g3vvvbYljF/sC5D1/z67ZI46hDbYmMRmUMpTvuEsNI2pW/eUd+g0B33bpQnFreu8lx48b5nP47FYavTqdjuxblLLcSbj6u7utRytZ39nXjwklKUqyaIUpSgFKUoBSlKAUpSgFKUoBSlKAUpWBfr/Y8Ws0zIslu8O1Wu3sqflzZjyWWWG0+qlrUQEgfMmgM+qy5P4Jx7P5zWWWW4ScWzOCNwshtnuPggdkPJGg838ClXw7AgEg+SvFL4b2ULM3nPCIC0AlTM+9x4jw76/i3VJX8Pl8vmKm2H5zhnINqVfMFyq05Bb0PLjLk22WiQ2l1OupsqQSAobGwe/cfOs4TlTe9F2ZjKKkrMqfHecsk4/vEbA/EbbmLPMfV5NuymMD9k3Q/DqVr+t3ddylWk+p90a3eSFocQlxtQUlQBSoHYI+YrX5Fjdhy2zycfya0RbnbpaOh6NJbC0KH4H0I9QR3B7jvVDybZnPhadNxx4XPLeKhtUu2LWX7hjyfi4wo93I47koPdIG9+qjNuwr+xlLlwfdy7vLkR3lT1zXvOiqVqcVyvHc2sMTJsVuzFytk5AcZkMq2CPkR6pUPQpIBB7EA1tqrtNOzJk75oUpSvAKx7hb4F2gyLXdIbMuHLaUy+w+gLbdbUNKSpJ7EEEgg1kUpqeptO6Kx4q4pncNzL3a7Zl638CcCZNotE1JW5Z1kqLzaH1K7x/QpSoEp2e/YlViW25269W+PdbRPjzYUttL0eTHdS406hQ2FJUkkKBHxFe7zLMllyPIaQ606koWhaQpKkkaIIPYgj4VWXF3ENw4jye+RMZyFA4/uSfbINgdbUpdqmqWS6I7m9JjqHveWQdKPbQ31QqLpWjBer8P2Ls6ixinWrz/Ey4e1wea/Nxu9c7u+uDxXL5CxTkbKuLMycut8tPfIMcyCQ2pwGI+6Q5Bed10hxpwnoBOy2ewASAMnhjkXJL/fc0455Bcj/lRiF1UOtlrykTLZIJchyEp/mbQdE6KO52alXJWQZJYeOb7lGB22Jd7tb7e5NgxXlKLUkoT1lIKDskpB6dHudDfeowOZeN4uJYny+q3H2fOnbbam7hHjIU4wZBPlIkubBS2hwqSfXpUT271HlSko72mefJ/R2LXrYunKoqSblaKtqpqzvb9SvdaN5rSxgZ3/ANzfmPHuSWvzdoysIxi+/BKHySqFIV/jdTZUewSoVa90tduvVvkWm7QmZcKW2pl9h5AUhxChopIPqKj3KmGN8hceX3EVaDs+IsRlnt5chPvsr38NOJQf2Vj8O5m5n3G1iyWVtM52N5E9ChpSJbRLbwI+HvoV2+RFbmo3Vw8Ky9qL3X8Yv4ruSOegtyrKm9JZ/X5PxZAbdcLl4cboxjuQy5E3jWe8GrXc3lFblgcUfdjSFepjk9kOH7v3T20au9C0OoS42sLQoBSVJOwQfQg1jXW1W2+W2TZ7xBZmQpjSmX2HkhSHEKGikg/Cqdxi6XHgbJInG+WTXZOFXV7ycXvD6uowXD6W+Qs/D+5rPqBr4e7lJLHxc1/arX9S5r9S489db3xX9Ve6/YenTp3cuWmhdtKUrXFsUpSgFKUoBSlKAUpSgFKUoBSlKAVr7DYbTjNqYsdjiCLBjdXlMhalBHUoqOiok+qj8a2FKAUpSgFKUoBSlKAUpSgFKUoBSlKAh2VcT4blVy/KJcORachSgNovlnkKhXAJH3Uqdb15yAe/lOhbZ+KTWoCubMK91xm38jWtHotCmrXekp+qTqHJWfmDFSPkasilAV0Od8Bt/uZp9rYW6ns4cktjsGMg/L2xQMRZ/mPKqYWTKsXyaIJ+OZJa7rGUNh6FMbfbI/nIJFbWojeuIOJcklmfkXF2I3SSo7L02yRn3Cf5y0E0BBuO7BkfJuMIzDIeVstYmyJs+K7FtUiNGhRXI8t2OtEfoZ6nGgpo9K3FrUpOlb76EgVYOZsQPnY5mMPN4Se6rfkjTcOZr5NzYjYb7D0S5GUVH1dT3NT+Bb4FqhMWy1wmIcOK2lpiPHbS220hI0EpSkAJAHYAdq1ec5ZCwPC77m1xjvPxbDbpFxeaZ0FrQ02VlIKiEgnp9VEAepIAJoDSY3y1j15uzWLX2HPxXJXgot2a9tpZekdI2oxnEqUzKAHclhxfSCOrpPapvVRX/DuXOTcfNvyiTxlFts5KXvs96yzLylo/eQpEkSou1pOilxDaCCAU6IBqx8Ss0/HcWs9gul9k3qZbYLESRcpI07McbQEqeX3PvKIKj3J2fU+tAbalKUApSlAKUpQClKUApSlAKUpQClKUAqvOeQE8eJmuJ2zbsgx64yN+nkR7xDedP7ENqP7KsOq65tX9p2Wx4Gx70rLr/AghHzisuiXMJ+Q9ljPpB/WWgepFAWLUIyji+NdL05mOJ32biuTuIQ29cYKUramoQNIRMjL23ISB2CiA6lOwhxGzW5y/OcUwSA3ccqvDUJEh0MRmghTsiW8RsNMMthTj7hAOkNpUo67Cou3lXMGU/ncW4/t+OW9f8XMymYr2pQ+ChBjBWkkfBx9pY9FIB7ADDunJHIPG9qmXblDCI9xs1sYcky7/AIzJSW2mG0lS3n4UlaXWQEgnpacknQ9asm2z2LrbotzjIeSzMZQ+2l9hbLgStIIC21gKQrR7pUAQexANUrnquWJd4wzA88s1luuOX/JIvt14sqXWEMpjIcloYkxXVLKW3XozLfWl1SVdRQpKeodV50BReY4fd+Dr3I5X4qtr8iwSHC9leLRvuOt/pTYiPRD6B3UkaC0j4Ebq4MZyax5jYYOT41cWp9suLQejvtHYUk/0gg7BB7ggg9xW0qhrrG/0OGdt5JbSWeNcumhm7xB/E2O5OkBuU3+ow4r3VjsEqIPyFWU/SFuv2lp16d/Ly5ETXZO60+BfNK+AggEHYPoRX2qxKKUpQCvCfBh3ODIttwjtyIstpbD7Tg2lxtQIUkj4ggkV70oeptO6Kp4T4+zTiqRfcBlTm7lgsNxt/FX3pBXMiMudRchOAj3kNEDoUTvpUB8NJxMowHiHIsXvHhYs7rdjel2lV4ZhRmV7iNrlFSZTZV7qumSOrpCux+QIqfciY3dsuwi9Y5YcjmWC5zoi24dyiOFDsZ7W0LBHfXUBsDuUkjt61UmD2LNOU5vHfLl8iO4vmWFP3CxZNGlRFpRcWSktvBo9gtsuoQ62obSCVDZIqpOKhako3XyvmvD4G6o1Z13LG1Km6075fzqLcZNcd5pptaN3eud626O9Et8aJJmLlvMMobckLSAp1SUgFZA7Ak99D51VvFI/JblLkjj1XuMPTmMnt6T26m5aNP8ASPgEvNkf41fnhOw5Nh2ccpYtcrdNFjfyFN/ss55J8p5E1lK32W1H4NuoUNfDrr95sPye8QPH+Rp9xrIIFxx2Uv0G0pTJYB/FSFgVtdnz7aFSk1a8X5xz+Ca8TSbRorD1YuMt5Jp36SS8tVdcGi2q1GV4rYs2x+bi+SwETLdPbLTzSv6FJPqlQOiCO4IBrb0qvGUoSUouzR40pKz0Kg46yi/YBk7fC/I9wXLWttS8Xvj3b7UjJ9WHT6e0NjQP6w0fXuq36i3I/Htn5Jxpyw3Rbkd5C0yYE5g6fgykd2321eoUk/XuNj41HeJ+QbxcpU3jjkNDcXNcfQPaOkabucXekTWPmlXbqA+6rsQN6F2tFYqDxFNWkvaX/wBl0fFcH0eVem3Rl2UtHo/k/lzRZdKUqgWRSlKAUpSgFKUoBSlKAUpSgFKUoBSlKAUpSgFKUoBSlKAUpSgFKUoBSlKAUpSgFflaEOIU24hK0LBSpKhsEH1BFfqlAVw5wnZrItUziy93DA5O+r2a1FK7W4fXS7e4CwAT95TSWnD/AHQUGZcpYifJzrj8X+In/wDTGJHzOw9Vu299Yeb+iWVySfpVj0oCHY5y/wAbZVcE2S15ZEavBGzaJ6VwbikfNUSQEPpH1KKmNarI8UxfMbcbRluOWu9wVHZjXGI3IaJ+fSsEbqH/AMCtrtPv4DmeW4eR6M2+5mVESP1UxJqX47Sfo0hB+ooCxaVXn2fz5Zvei5LhWTtI7JYm2yTaX1D+XIadkIJ+qY6R9Kfl9ybavdyLhC4ydfeexu9Q5zKR8/65VFdP4JbJ+lAWHSq8HPfG0QhOT3C54od6UvJbRLtTAV8hIkNpYWf5rihU5ttztt4ht3G0XCNOiPDqbfjOpdbWPmFJJB/ZQGVSlKAUpSgFKUoBSlUfyF4rMOsF4ODcaWyVyLmjvUhq02QlxllST0kvyEhSW0pV7q+kLKCR1hCT1AC2spyrHsLskjIsnujUCBG6QpxeyVLUQlDaEJBU44pRCUoSCpSiEpBJAqi8ezTNOSc7m5ViWOtPXaMw7Z7cbgSbbjEVa0qfXLW2f64uDqm2yqIyo+UlpttxxlSlqX8x3gjknk69x858ReTdKmepULG7Q6WmoaVDRQXUKPl7SelflKU4ra0mQtlZZHQFptFqsNsjWWx22Lb7fCaSzGixWUtMstpGglCEgBIHyAoCMYbxhaMYuDmTXWbJyLK5TZblX64hKpBQSCWWEgBEZjYGmmglJ0FK617WZlSlAfFJSoaUkEbB7j4ivtKUArW5Hj1oyyxT8av8JEu3XJhcaSysdloUNH8D8QR3BAIrZUr1Np3QavkU/wAG3y843c7vwRmM5cu6Yk229aJzp9+5WZZIZcPzW2R5Sz8wn17mrgqnfEFbJ+N/YfOmOxlu3LBnlLuLLQ96ZZ3dJlNH5lA06nfYFCj61bFsuUG822Jd7ZJRIhzmG5Md5B2lxtaQpKh9CCDU1ZbyVVcde/j56kdP1bwfD4GVSlKgJBSlKAVTviAzPIrG5YsGgdVugZ+ifj7V9jyVsyLZdVx1Kg9JTrQWtKhve9gVcVaHM8RxXMrQiDmFtRMhQpTNyQFLWgtPsLDjbgUghQKSnfY9xsHYJFR1YynBqLsy1gqtOjXjOqrpfTJ+Dzto7ZlO8f8AO2SuWDhFnIbKUDOWpNovEqWFofj3KNHV06SfXzXGXfX4Ea9d1J/Ed/WGNY1lvoMayy03Favk2XvJWPwIe1WlY5W4a5gvF2GQeyIs3G0m1ZTbb47cC1Ge8xpzy5SSOnpShXmNqSokE9iD6VJfEdGRd+BsvMZaXEptntja0nYIaUl0KH+RurGx5f1mneV05JeHsv5mW3aajTf4Tpys21wzbnHPpFxVsrWWVyzqVg2Oem6WWBc0naZcVp8fgtAV/wBdZ1YNbrsykndXFV9y3xzMy+JCyXEpSLdmWOLMqzTj2Cla9+M782nB7qh8N7+YNg0qSjWnQmqkNV92fR8TGpCNSLjIh/F/IsLkjHftERF2+6wXVQrvbHf42DMR2W2ofLfdJ+II+OwJhVQcoY7ecEyMc44FBckvsNJZye0sjvdICf7akfF9obKT6lIKd/A2bjmRWfLLFCySwTkS7dcWUvx3keikn/mI9CD3BBB7ip8TRjZV6PsS/wDF8U/k+K6pkdGpK7p1PaXvXP69fA2VKUqmTilKUApSlAKUpQClKUApSlAKUpQClKUApSlAKUpQClKUApSlAKUpQClKUApSlAKUpQClKUApSlAfFJSpJSpIII0QR2NQW5cG8TXGY5dG8HgWu4vHqcuNlK7XNWfrJiKbdP8AlVO6UBXf8FN+tn+xTmXNra2nuiLMejXVkn+UuYy5II/B4H6199i5/tfvN5DgeRIH3WnrXLtLhH8p1L8lJP1DQH0qw6UBXn5Wc0Qf9dOHrVMH/wC5cqS+f3So8Yf019/hMzVPZ3w956Neqm5thUn9n9kQo/uqwqUBXp5TyYa3wNyECRvXVaDr90+tHmPNGd49ZlXVvhq4WtKnW47P5QXWMlUh5w9LbTDNuVNeecUT2R0J332UgEi3qjOe4X+WluhIi3iRaLrZ5yLnargy2lwxpSULb2ptfuuIU2662tB1tLitFKulSQKRXxJzzzmevmfN38Uxd372OWVLbL8lH6j/AEreQkEbSpK3ZCFg7CGVDtdfH3GGBcWWf7CwLGYlpjK6fNU2Ct6QpI0FPOqJcdUB2BWokAAegrRJyjmXGx5WTcbwsoZR7vt2K3Btp5z+WqHNW2Gh/JTJeNfoc9cbw/dymdc8SUOy1ZLaZVsYSfkJL7aWF/ihxQ+tAWJStPj+Y4jljAlYrlVovLJ7hy3zmpCf3oURW4oBSlKAUpSgFK0WSZ3hGGtefl+Y2Oxtn9O5XBmMn97ihUIyPnmxybFOVxTFuOZXsMLNsbttnmSbfKkAbQ0uchv2ZpKj7vmKdATvq0QNECzpsONcIb8Cawh6PJbUy62sbStChpST9CCRVSeG2ZJs1ivvEN1eWufx7dXLY2pw++7bnPzsN0/i0rpH+DrdNct3WxpSzyXxlklhdAHVLtcZy+W9avj0OxEKeSkfrPsMiqwuHMfFWP8AiNtOXRs7tES1ZPj8i1Xl2XIERuPIjLDsd1/zenoKkqcbSVa36etT0vWjKn4+K/a5HPJqXh5nS1Kj+M8g4FmqnE4bm9gvxaT1OC2XJmUUD5ny1HQqQVASClKUAr4RsaPxr7SgOX8e8KDeLYnyNZ3rswq5ZRbLzj2PQ2pQEcW5116TEDiVICvPbceWCQopCAPrq2ZGP3lHh5dxbJkNi6ow5UCeG19aPaBCKHNK+I6t96pbmRfI0jxm8b220CRcrTAYYvLcZKW0iGwtb8Sc4FHRUOlyOojZ+gq0OGc1yLkjhW6TsufafvMWXerRMcaaDaFqYkPNo0kdh+bCB+IJqpgZQp4hRgmmn8LP5nQ7Yhiq+CjiK01NTSk+aclJW8o59eZLeF56rnxDhc5fdbtggdZ+agwgH+kGpnVc+HV3zuDcKXverQyn9w1/1VY1bTGR3cTUiuEn8TmcO70ovovgKUpVYlFUgd+HrOv1OOMvmfg3Yro4f3IjvH9iVfIet31rcjx6z5ZYp2N3+CiXb7iypiQysdlJP/MR6gjuCAR6VZw1dUm41FeEsmvmuq1Xlo2Q1qbmk45SWn3yZsfXvX2qh4qyG8YVkLnBudzVyJcJkv43dHj3utuT6IUfi+0NJUPUpAV9Tb1Y4ig8PPdbutU+a4P70eWplSqKrG+j4rkxSlKgJBSlYF1v1jsXs323eYNv9tfTFje1SENee8r7raOojqUdHSR37UBn0pSgFK0+UZjimE203fL8jt1mh9QQl6bJS0laz6IT1H3lH0CRsk9gDUGfvWf8rbgYlFueF4u52kX2fGLF1mN/FMKK4OqOD6efISFjv0NHaXUgTG0ZvZr9lF3xWzpkS3bEltNwltoBisSF9xFLm+7wRpakAHpStHUQVpBkFarGMXsOGWOLjeM21uDb4YIbaQSolSlFS1rUolS1qUVKUtRKlKUpSiSSa2tAKUpQClKUApSlAKUpQClKUApSlAKUpQClKUApSlAKUpQClKUApSlAKUpQClKUApSlAKUpQClKUBFL/wATcWZW/wC1ZRxrit4fPfzZ9njyF/5S0E1qP4AeKGv9bcaftI+AtF0mW8D8BHdQB+yrCpQFe/wF4YP4u+8goPw1yDfjr9hmEUPCGMHf+mrkIb//AI5u/wD/AJNWFSgK9/gNw9Xd3IOQVn/h/fU/+rLFP4BOMHf9c7Tdbyn4ovN/uFyQr8UyX3Af2irCpQEYxvi/jTDnfPxHjzGrI78XLdaWIyj+1CAak9KUAqnfFEk2vBLVyCz2fwnIrbekkeqmvPDLqN/ItvK3+FXFUE52sf5R8M5rZwjqW9Y5i2h83ENKWj/OSKmw8t2rFvS5hUV4M2mUca4DmyGjkuK2+Y8wSuNMDflyoyz+mxIRp1lf8pCkn61Hk4hyphqt4NmzOSW5P3bRlq1l1A+CWrk0lToA+JfakLP64qRcY3w5Nxvi2QlfUq5WaHKUf5S2UqP9JNSaopRcW0zJO6uV2OSs2tPuZZwrkjQR/GS7JKiXSKP5qQ43KV+yPX0c74AntJjZhEI9RKwu8s9P4lcUD9vpVh0rw9IAzz9wu48iNK5KsVtfcV0oZukoQHFn5JRI6FE/QCpxCnQbjGRMt8xiVHdG0OsuBaFD5hQ7Gv2+wxJaXHksodaWOlSFpCkqHyIPrUHmcDcKzpLk5zivFmpbp25JjWtlh5Z/lONpSo/tNARTxQ83wfD/AIXFzZFhZnXibNatcBx2P1Np6lBbqXFhQUkeUhwjW9qSNipZjd948m4tlTnHjUJqLap9yi3NMWEY6Rcm9mT1ApT1qKjsrGwonezVe8xeFbhPJsVYVcItvxq3Wa4MXe4THVueWuIzsvNLUXU+WlaCoFYIKfWtLY/DfxzhXGOfX2949jl6RMlXjIbRJbbLyWILjRcjoCljuUgb2NjvsE1BRVV4tRt6rt3/AGzbVfQ/+lKSbVVN35NZW8lfz6ln+HFryOC8KQfjaml9/wCUSf8ArqyK4H5v5H4V8L/BPHM9zgqJmuf5laGXYFtiMlp14ojtLkSX3G0qX0pLiN6BKirXYAqGP4Mee8B8SGQzePs44YyrjPKWork6CIWR3eLDuLCCkOBspcaKXEdaSUe9tOyCNEVsMbLexNSX6n8TR4dWowXRfA/oBSq+PBeD/C652n1+7n19H/NMr4eFLCz79szPkKC5+v8AllcpX+bJedR/m1WJiwqVXn8F2WNdoXPvIDCR6IUzZnh+0uwFKP8AlV9/InlmL/rfza5I+X2pjsR79/kFj/qoDN5W45b5Dx9tmFNNtv8AanhPslzQPfhy0d0n6oV91SfQg+mwK8uJeRnM8ssiJe4QtuUWF72G+20nuxIH6afm04B1IV3BB1s6rH+xef0dk8kYA6PgV4XMSf6Lpo/0VW3JHHPN9kmv8w2XN7O9fYEMRpsbHccVDenQAoFY/rmTKbcebAKm9t+o0ersmr+HksRD0Wo7fyvk+T6P3PPS5Wqp0pdtHxXTn3r3rwOi6gt35r47ttwesduvLmRXlhXQ7a8ejOXOU0v4B1McKDG/1nihPzIqJ2HDOJ85s9iv2TZhe87YyXaYLd/ualxZSw2txTaoDIbh9SUtubBY2Ckj6VbVps9osFvZtNitcO3QY6elmNEYSy02n5JQkAAfgKpThKm92asyxGSkrxZAzN5szP3IFpt3HttX6v3FTdzu6k/EJYaUYrCviFqdkD5t1mWnhTAYipMzIbarLbrOYVGmXPJOmfIeaV95oBafLZaUQCWWUNt7G+jdTylYnpXaeCcIg9sXnZPjKB/Fx7NkU2NEaHybieYY6B9Etiv0eG48jaLpyVyFNaPq3+UTsX9nXFDS/wDOqwqUBEca4l46xK5fblnxaMq8dJQbtNWubcVJPqlUuQpb6h9CsipdSlAKUpQClKUApSlAKUpQClKUApSlAKUpQClKUApSlAKUpQClKUApSlAKUpQClKUApSlAKUpQClKUApSlAKUpQClKUApSlAKUpQCtdkcX27HrpC1v2iG+1r+cgj/rqL8j81cc8S+Qc+u0+3NyAgoeas02WyOtwNoSpxhlaEKUtSUhKiCSoaB2KkeMZNZ8zsTGQWNUtUGX1hsy4L8R09KihW2n0IcT3SddSRsaI2CDXqdncEG8Mcr2zgHBXd76bQ01/kEo/wD6as+tZjeN2PEbJExzG7c1AtsFBRHjNb6W0kkkDez6kn9tbOsqslOcpLi2YwW7FJilKVgZClK1eT5Pj+GY/PyrKrtHtlptbCpEuXIV0oabHqT8/kAO5JAGyaAgPiitU29eHjkGBb1vJfNikvJDJIWoNp8xSe3chQQUkfEEj41zLd+V+SsW8GPH8ObIXPk5pCmwp1wkNA+TC2pLLKdAAFTCkgKOzpCvj3HVls5x44vwsbdvnXBZyK9uY5Eal2iVDdE5MJ2YW3GpDaHEDyGVqClJ0dp0TuphPx3H7rbPsS6WO3zLdoD2SRGQ4z29PcUCnt+FS4CVPC4+GMqx3ox1jz5PwLGIxMq2y5bOirNy3lLisrNePecCci8O88eKDhTj3l7hDKDh/JXHXt9lgOsSVwkXO3OIbbWEOjflrHlhPf3VHzASntqVeCfwz+LTHc7HK/i65TuN7lWeJIiY9Y37wbgYzr4Sl6StYJQk+WnoCUlW+ok66QD23Dhw7dFag2+IzGjMJCGmWWwhCEj0CUjsB9BWox/NLFk95yWw2l51cvE7i3a7mlbZSESFxGJSQkn7w8qS0dj4kj4VliqsK9edWEd1NtpcinRhKnTjCTu0tTe0pSoCQUpSgFYl2TdVW6QLG5ERP6D7OqUhSmev4dYSQrX4Ht9fSohYObOOMpy6Vg+P3afNusGZKt8jy7NN9makxyoPNmUWQx1JKFA/nPUaGzoVOq9Ts7h5n89MvkcrcRcz2TIc2x9Fks0fIftRpm2FxVrJdKEyVMFRPSVoSSUnRGz7oB1X9CkqStIUkgpI2CPQisK9WSz5FbX7NfrZFuEGSnoejyWg42sfUHtWREisQIjMKMgpZjtpabSVE6SkaA2e57D1NbXaW0o7RhTvDdlFNO2jXDrfXmUsJhHhZTtK6eefA9qVrMbyG15ZYoeR2VchUGe2HmDIiuxnCnf6TTqUuIPb0UkGtnWpLopSlAKUpQClKUApSlAKUpQClKUApSlAKUpQClKUApSlAKUpQClKUApSlAKUpQClKUApSlAKUpQClKUApSlAKUpQClKUApSlAKUpQClKUBUniqtVzvXCs+32e3Sp0pd7x5xLEZlTrhSi8wlrUEpBOkoSpRPwCST2FUZfIuTSMzvSDBzwcvjkyKuySW2rl9moxkT2CCHEj2IQ/s4PB1JOy8V7BdKa7OrQ5bn+CYDHjzM7zWw44xLc8mO7drkzDQ85+qguqSFK+g70BzBjGEZbaL7inIjLeYnIJnM+SQJ6n5k1xpGPuSrsG2zHUotIi6TGcQegJ6ihST3FYfhdsfISs7sF2znL7nDy2Mxck5hancbvSFXB9ZP8fLfeXC6EOBK2FtISC37rekqIrqaVyTx1Cv8AbsUmZ9jjF7vDaXrdbXbowmVMbV91bLRV1uJOjopBBqR0ApSlAKg/Ntqw++cWZBaM+g3mVYJbCG5qbNGkSJrSS6jpeZRHSp0qbX0ubQklPQVaIBqcUoDiti359y9c8GxfL5WW5Vhdk5OSm15PKtUmx3G4W38m7iXTKDaGVBtMhYjl8IaQ919OiSSpGtt4swxnG+UYGcPcV2XLM3t7rTSLnJWlKJqRZRJLHVIdiBgy0tqV1N9QY2eyK7SWtDSFOOLShCAVKUo6AA9STWuRk+NOQLZdW8htioV6U0m2yUy2y1NU6nqbDK96cK0glPSTsdxugONbHFvUdOGNc927kl/j823JTYWWmrw9OadN33akzxD3KEkWwthrzveCvMB07W9sVoze0+J/IL/frNkznHM/NIaYzcZqQhbd3Vj1pbiTZutqkxAUPs730NSNKc6tBTXUFp5BwG/ZFPxCx5vYLjfrUCZ9riXJl6XEAIB81lKitvuQPeA7kVIKAUpSgFKUoDmXijCOR7ZY+WL45lt+iQJOVZk9CxxVqYbS75kl8svtveX7QrqJStJSvR7a7VXt9wDKMT4u4UbDt+bsV2gfaOeP3dm8XhTl3ctzHkKmsxn0SEt9YfT07DSHAyCkaTrtyvytaG0KccWEoSCpSlHQAHqSaA4UyrGeRG2sMseWcgXq4YmnD5bVtvcrFsiWRc1TnSkqYjPolNSW4pipYcklalBtxST1KVufWWLDneI3C4OSy819vxSwQFTb0bVeI8fKbw9DDaStASY0dlpvqdcS4UkvPNpJ2ysHoCBzFxHdLVc77bOU8QmW2yKSm5zGL5FcYglSulIecSspaJPYdRHftW7xvKsXzK2JvWIZJa75blqKEy7bMbkslQ9QFtkp2PiN0Bxrh/HGXZ1jthh56xnBTA4gdeS17dPif2ZEt7y1L8tSSuSlOikK2obB1sJIhuWX7IcqvcNvlGbe4UO24xg0q95C8q9f6XHAhMq4pU1BYWw2uQk9DplOsKQlQUrqbCK/ohUDyXgvijMMgkZPkeIMzJ03yPbgZDyI8/ydBr2qOhYak9IAA81CtAAegAoCdp6QkBPprt+FfaUoBSlKAUpSgFKUoBSlKAUpSgFKUoBSlKAUpSgFKUoBSlKAUpSgFKUoBSlKAUpSgFKUoBSlKAUpSgFKUoBSlKAUpSgFKUoBSlKAUpSgFc13vJeN8J8VmW3fnS8WGzRp+H2ljEZ+QvtMQ1R0PSzcmGHXiGw75io6nEA9SkeUdFKe3SleEqFDnIS3NiMyEoUFpS62FhKh6Eb9CPnQHIN7zDjvjTlJjK+IMwwTJoma3fF4Nx4/Ww0m6x9tRo8OZa0oIW2hqKpl8sraLYQ24tK2z1b7FrwMGEqWmeqGwZKUeWl4tjzAn9UK9dfSvegFKUoBSlKAwr1Hdl2efFYT1OvRnW0DetqKCAP31xtgGb4jlfEnhf4ux3IYM/MMeutgTerCy8lU+1/ZtueRNMpge+wGlp6CXAkdSkAbKk77WrwagwmZLsxmGw3If0HXUtgLXr06lep/bQHKvAeU47hnK9p4i49zTDuRsUuse93SDPtgbVesYBkJedYnraUpLjTjrpQlaw051oCVhZBUOsa8GIMKK68/GhsNOSFdTy0NhKnD81Eep/GvegFKUoBSlKAVTfi/i3OZ4d8rYt0SZKZKreq6sQ0KW89aEz46rkhKUe8rqhCSCkdyCR8auSlAcneIHMeDM24LvzPCmRYHdLrHXjLaXbMGJSIsZV7hBht7yCClvqGw0VJJAVrWiRLvDPb7jZeSOZLVmv2Wzmy71bptwas0UxLY/b1wkogy47KlKWkrDT6HStayXWFgHpCavpq3W+O2tliBHbbcX5i0oaSApe99RAHc7AO69g02HFPBtIcUAlS9dyBvQJ+Q2f3mgP3SlKAUpSgFKUoBSlKAUpSgFKUoBSlKAUpSgFKUoBSlKAUpSgFKUoBSlKAUpSgFKUoBSlKAUpSgFKUoBSlKAUpSgFKUoBSlKAUpSgFKUoBSlKAUpSgFKUoBSlKAUpSgFKUoBSlKAUpSgFKVx74us85x4v5Tx2ThvNl1tmO3uwZNfH7GzZbW4Gvsa2IlBpuQ/GccHnq6gpSirp6vdHbVAdhUrimV/VCMzs93et9+8P5aTBYtzs8RLzIkqY+0IhlRD5ghBjpDZbDvU6lQUpzy0uholWysHjozy+ZLYcCc4ZsFvyTLItnu1o9qy9abcm3z7dMnAyJPsfU2+lEFxAbS2sKUsaVpJJA7EpXEvhj8b2dZs/x3hvI2MRZCslYjQ3cpVLKRNuL0V2UEIajRjGaUEt9HlOPNOLG3EJUn1/WB+OjNpTMhqVxyi8Wuw363W6+Xibf22piG7pkE62RizHZhIac8sxQSCpBKAAVKVtZA7YpX8/eUfF7zvgGfcm48/cAmwL5CsuJ4fdGbews2+QHbY7NgvAtkKD8OXIW24vagWHQCD0amuCf1RF7ML9jEKXw7JtlpzK7QIlquL899pLcWTKcjlb/nRW0eclSEK6GVutnrUnzQpBBA7NpXFts/qg+TZG1PnY1wrbX7db5tvtq50nK/Ka9ouF3mWyGSRFV0tKcieYtz0ShRAC1dIVlP+P6823IWLJeuJoMaLa7s5ZcmurF8elw4klFx9i2wpmIpRbUSFockJjtqVtrq60nQHZFK4jyPx8X6HiOJ8jXHDoVssl6eZvkWJZcoiTrnJtBiXF1TUyG7G8xlZMNP8UenrPR54KFpV72nx/cgX5+LYLXwHDTfZkSZc2vbb/KiQVRGbcJwKXHoCXVLKAtBHlBBUG1JWpC+pIHatK4zT/VEFOYRceWEcYW1GI29nyPJfytlm9PXD7JauHQ3CWyA4zp5KCtDinAlK3fL6EnWJcP6oXlVmjld54ORCFqkyRfZjtzmpixYraYikvIQYHtSUqTL0XH47TSFN6K9OIVQHa9KUoBSlKAUpSgFKUoBSlKAUpSgFKUoBSlKAUpSgFKUoBSlKAUpSgFKUoBSlKAUpSgFKUoBSlKAUpSgFKUoBSlKAUpSgFKUoBSlKAUpSgFKUoBSlKAUpSgFKUoBXI3jj/2WYZ/wI5G/6ETSlAcx8/f7rbiX/wADz/8A0ZKqPeKz/aUX/wAGeJf/AHe50pQHh4XP92/wp/wJT/7pJqf8df7FeSP+FOD/AP52u1KUBYXMH+xfl3/w+47/AOxtVchZp/te+MH/AIc2n/pJ6lKAtrjH/cgeIP8A4FW7/pm+VXVp/jPBJ/O//vBpSgJPwB/toci/+GuL/wC43qvngl/2HWP/AO1uQf8AoNmlKA+4R/umh/8Ay7Rv/wAutVTjP+5x8O3/AA+vP/todKUB/9k=)
- BLA, basolateral complex of the amygdala
- CEA, central nucleus of the amygdala
- ITC, intercalated cells of the amygdala
- PL, prelimbic cortex
- IL, infralimbic cortex
- HPC, hippocampus
- Thal, thalamus
- PAG, periaqueductal gray
- PBN, parabrachial nucleus
Stress types
- Acute stress
- Short duration
- Brain detects threat
- Mobilizes physiological, behavioral responses
- HPA (Cortisol), SAM (NE/Epi) axes
- vs. Chronic or stress
- Long duration, persistent
![[[@Kim2015-dd]](http://dx.doi.org/10.1101/lm.037291.114)](data:image/jpeg;base64,/9j/4AAQSkZJRgABAQEBLAEsAAD/7R+iUGhvdG9zaG9wIDMuMAA4QklNBCUAAAAAABCloxzzoeUqjatJhLeoNI9yOEJJTQQ6AAAAAADlAAAAEAAAAAEAAAAAAAtwcmludE91dHB1dAAAAAUAAAAAUHN0U2Jvb2wBAAAAAEludGVlbnVtAAAAAEludGUAAAAAQ2xybQAAAA9wcmludFNpeHRlZW5CaXRib29sAAAAAAtwcmludGVyTmFtZVRFWFQAAAABAAAAAAAPcHJpbnRQcm9vZlNldHVwT2JqYwAAAAwAUAByAG8AbwBmACAAUwBlAHQAdQBwAAAAAAAKcHJvb2ZTZXR1cAAAAAEAAAAAQmx0bmVudW0AAAAMYnVpbHRpblByb29mAAAACXByb29mQ01ZSwA4QklNBDsAAAAAAi0AAAAQAAAAAQAAAAAAEnByaW50T3V0cHV0T3B0aW9ucwAAABcAAAAAQ3B0bmJvb2wAAAAAAENsYnJib29sAAAAAABSZ3NNYm9vbAAAAAAAQ3JuQ2Jvb2wAAAAAAENudENib29sAAAAAABMYmxzYm9vbAAAAAAATmd0dmJvb2wAAAAAAEVtbERib29sAAAAAABJbnRyYm9vbAAAAAAAQmNrZ09iamMAAAABAAAAAAAAUkdCQwAAAAMAAAAAUmQgIGRvdWJAb+AAAAAAAAAAAABHcm4gZG91YkBv4AAAAAAAAAAAAEJsICBkb3ViQG/gAAAAAAAAAAAAQnJkVFVudEYjUmx0AAAAAAAAAAAAAAAAQmxkIFVudEYjUmx0AAAAAAAAAAAAAAAAUnNsdFVudEYjUHhsQHLAAAAAAAAAAAAKdmVjdG9yRGF0YWJvb2wBAAAAAFBnUHNlbnVtAAAAAFBnUHMAAAAAUGdQQwAAAABMZWZ0VW50RiNSbHQAAAAAAAAAAAAAAABUb3AgVW50RiNSbHQAAAAAAAAAAAAAAABTY2wgVW50RiNQcmNAWQAAAAAAAAAAABBjcm9wV2hlblByaW50aW5nYm9vbAAAAAAOY3JvcFJlY3RCb3R0b21sb25nAAAAAAAAAAxjcm9wUmVjdExlZnRsb25nAAAAAAAAAA1jcm9wUmVjdFJpZ2h0bG9uZwAAAAAAAAALY3JvcFJlY3RUb3Bsb25nAAAAAAA4QklNA+0AAAAAABABLAAAAAEAAgEsAAAAAQACOEJJTQQmAAAAAAAOAAAAAAAAAAAAAD+AAAA4QklNBA0AAAAAAAQAAAAeOEJJTQQZAAAAAAAEAAAAHjhCSU0D8wAAAAAACQAAAAAAAAAAAQA4QklNJxAAAAAAAAoAAQAAAAAAAAABOEJJTQP1AAAAAABIAC9mZgABAGxmZgAGAAAAAAABAC9mZgABAKGZmgAGAAAAAAABADIAAAABAFoAAAAGAAAAAAABADUAAAABAC0AAAAGAAAAAAABOEJJTQP4AAAAAABwAAD/////////////////////////////A+gAAAAA/////////////////////////////wPoAAAAAP////////////////////////////8D6AAAAAD/////////////////////////////A+gAADhCSU0ECAAAAAAAEAAAAAEAAAJAAAACQAAAAAA4QklNBB4AAAAAAAQAAAAAOEJJTQQaAAAAAANRAAAABgAAAAAAAAAAAAAB5gAAArwAAAAOAEsAaQBtAEwATQAwADMANwAyADkAMQBfAEYAMQAAAAEAAAAAAAAAAAAAAAAAAAAAAAAAAQAAAAAAAAAAAAACvAAAAeYAAAAAAAAAAAAAAAAAAAAAAQAAAAAAAAAAAAAAAAAAAAAAAAAQAAAAAQAAAAAAAG51bGwAAAACAAAABmJvdW5kc09iamMAAAABAAAAAAAAUmN0MQAAAAQAAAAAVG9wIGxvbmcAAAAAAAAAAExlZnRsb25nAAAAAAAAAABCdG9tbG9uZwAAAeYAAAAAUmdodGxvbmcAAAK8AAAABnNsaWNlc1ZsTHMAAAABT2JqYwAAAAEAAAAAAAVzbGljZQAAABIAAAAHc2xpY2VJRGxvbmcAAAAAAAAAB2dyb3VwSURsb25nAAAAAAAAAAZvcmlnaW5lbnVtAAAADEVTbGljZU9yaWdpbgAAAA1hdXRvR2VuZXJhdGVkAAAAAFR5cGVlbnVtAAAACkVTbGljZVR5cGUAAAAASW1nIAAAAAZib3VuZHNPYmpjAAAAAQAAAAAAAFJjdDEAAAAEAAAAAFRvcCBsb25nAAAAAAAAAABMZWZ0bG9uZwAAAAAAAAAAQnRvbWxvbmcAAAHmAAAAAFJnaHRsb25nAAACvAAAAAN1cmxURVhUAAAAAQAAAAAAAG51bGxURVhUAAAAAQAAAAAAAE1zZ2VURVhUAAAAAQAAAAAABmFsdFRhZ1RFWFQAAAABAAAAAAAOY2VsbFRleHRJc0hUTUxib29sAQAAAAhjZWxsVGV4dFRFWFQAAAABAAAAAAAJaG9yekFsaWduZW51bQAAAA9FU2xpY2VIb3J6QWxpZ24AAAAHZGVmYXVsdAAAAAl2ZXJ0QWxpZ25lbnVtAAAAD0VTbGljZVZlcnRBbGlnbgAAAAdkZWZhdWx0AAAAC2JnQ29sb3JUeXBlZW51bQAAABFFU2xpY2VCR0NvbG9yVHlwZQAAAABOb25lAAAACXRvcE91dHNldGxvbmcAAAAAAAAACmxlZnRPdXRzZXRsb25nAAAAAAAAAAxib3R0b21PdXRzZXRsb25nAAAAAAAAAAtyaWdodE91dHNldGxvbmcAAAAAADhCSU0EKAAAAAAADAAAAAI/8AAAAAAAADhCSU0EFAAAAAAABAAAAAU4QklNBAwAAAAAFtgAAAABAAAAoAAAAG8AAAHgAADQIAAAFrwAGAAB/9j/7QAMQWRvYmVfQ00AAf/uAA5BZG9iZQBkgAAAAAH/2wCEAAwICAgJCAwJCQwRCwoLERUPDAwPFRgTExUTExgRDAwMDAwMEQwMDAwMDAwMDAwMDAwMDAwMDAwMDAwMDAwMDAwBDQsLDQ4NEA4OEBQODg4UFA4ODg4UEQwMDAwMEREMDAwMDAwRDAwMDAwMDAwMDAwMDAwMDAwMDAwMDAwMDAwMDP/AABEIAG8AoAMBIgACEQEDEQH/3QAEAAr/xAE/AAABBQEBAQEBAQAAAAAAAAADAAECBAUGBwgJCgsBAAEFAQEBAQEBAAAAAAAAAAEAAgMEBQYHCAkKCxAAAQQBAwIEAgUHBggFAwwzAQACEQMEIRIxBUFRYRMicYEyBhSRobFCIyQVUsFiMzRygtFDByWSU/Dh8WNzNRaisoMmRJNUZEXCo3Q2F9JV4mXys4TD03Xj80YnlKSFtJXE1OT0pbXF1eX1VmZ2hpamtsbW5vY3R1dnd4eXp7fH1+f3EQACAgECBAQDBAUGBwcGBTUBAAIRAyExEgRBUWFxIhMFMoGRFKGxQiPBUtHwMyRi4XKCkkNTFWNzNPElBhaisoMHJjXC0kSTVKMXZEVVNnRl4vKzhMPTdePzRpSkhbSVxNTk9KW1xdXl9VZmdoaWprbG1ub2JzdHV2d3h5ent8f/2gAMAwEAAhEDEQA/APVVz20v+tkO9UemQ9ge5+2DTtsfjsbjOp9Cx/pMt9TO/pGP9D9N+sPi5Lv+ct7LLLH1ve6ukmx4aHtqqtfjDDbd6XsZuu+12Yf+E/nvU9P1wsY2z61C51b2ltxa4lpB3toe1r677KG7sX0P+01eR77/AFMj0/T9b1Ep0updXyMPqODh14VuQzMe5rrmGsNbDLLNPUtrdvZ6e9/s/mv5r1Lv0Su4l7sip1jgGkWW1wNdK7LKWn+16aOqnTP6M/8A8MZP/n+5JTbVK3rfRqc0dPtz8avNJa0YrrmC2XQa2+i53qbn7m7Pah/WLqVnS+iZmbSN2RXWW4zSJDr7CKcRm2W/TybKmKPT/q/03D6S3pdlTMup3uynXtFhyLne67KyvV9T1br7P0j96SnTSXPO+0fVWtrmmzL+r9elgcXWX4TO1rXu3XZfTa/8Jv35ODX7/wBNi/0bfrsrtrbbU4PreA5j2kFpaRLXNcPpNckpkkkkkpSSSSSlJJJJKUkkkkpSSSSSlJJJJKf/0O2xepOyPrHZXU3FcxpfjvewbryKxvPqWtdvrZTf+i2W1enZZbZ/3Hr+2Cmv/nearC1suFldQc073jH2faXV/bGvbkMY19P/ACfZ+rs/67jyxnWu+szw1lxx9xtY+CWH1KQx1pyfR9J+K70am14rcy39N6dvp/ofTw541Yu636l/UK8p1V1no0Bl7S3a19ewt+1/YPXp3Wb7vsXv/rpKd9VOmf0Z/wD4Yyf/AD/craqdM/oz/wDwxk/+f7klOf8AWg2Pb0rEZ/2q6njB/wDVoL+pu/8AbFbaw+pudkfWno2Ix4jFZk59zZ1ADB06iW/y3Z923/iVuJKUsDYPqxaX1iPq/c6Xs7YVjj/OV/u9Ltd/OM/7z7P0n9C/om+mc1r2ljwHNcIc06gg9ikpQM6jhOsBjrPqy8VWkv8Aq+4xVadXYJP+Bu/e6X/obv8AvP8A5q39S/SYm8CCAQZB1BCSl0kkklKSSSSUpJJJJSkkkklKSSSSU//R6zpGA3p/1hux6ca3ZvfZ676q2iLW732faa8Gljm+oxtWyvqFmQ/fvyK/570pZTq6vrXXfYy1oa8MNha9876dtfoxQ6uvB3b632faf6bbZ+jVinOyH/WZ+Mcx7qmOcwY4YAwTWLfSt/wnqv2+tTb9Cuqizf8A8o4yC0Mb9cbHO95eWhrmskM/Qfzdl78J/pPd9P06uqU1/pa/0HqWWfaUpu9Uw+o3dTxbccONTDUWvbZsbVttbZmG6vc31vteH+rVba7v+sst9VVsbH+tH6Z2FnYX2d+VkFleRi2OfW31rG7RbRl0Nu9wc/31MXQqp0z+jP8A/DGT/wCf7klNLp/SupYVluQ/Jry8zJ2/acqxjmlwYW+lVVTXZ6ePj01vyvSpb/hLPWtsttsv9S5t6vH08ef6j+Yf/wAJ+96X/giuJJKae3q37+P/AJj/APg/+E/4/wD8CS29W/fx/wDMf/wn/Cf8R/4KriSSmk5nVnNLS/GMgggseQZDef0n73q/9BZH2TrP1f32dPYMzpRl1nT6wTZjQSfU6Yxz91+P6X0+mb/Zb/yf7P1NdIkkpysDPyupYjcvAysTIqeNHta/R21nssZ6m+qxlm/1KbP0tf8ANv8A0itbeq7vp4+2f3HzG53/AAn0vS2f9cVTP+ruNkZLs/Dts6b1JwG7MxSAbIaWMGZRY2zGzWM/M+00vsr/AMDZUgHqH1l6aD+0MFvVKGz+tdOO22AGx6vTMp/9f+i5uU//AIBJTobesbfp426P3HxO0f8ACfR9bf8A9bUtvVd306Ns/uPnbu/4z6Xo/wDgqr4X1l6HnX/ZactrMudpxLw6jInn+iZTacj/AMDWmkppbesbf5zG3R+4+N23/jPo+t/4Epbeq7vp4+2f3HzG5v8Awn0vR3/9cVtJJTS29Y2/Txt0fuPidrv+E+j6vp/9bUtvVd308fbP7j5jc3/hP9H6n9tW0klNIt6xtMWY26NPY+Jiz/hP9J6H/gqupJJKf//S7DCyGW/Wm+Mzcze+tlLXtfudXWPUr9mW9rK6/U99H7Oov+0Y/wCkv/7kTt2u+tFZLWuay+Bk7WhzbHYuvTtv8471KP177d9D/tEj9P6V1ajOZkZF4IBeLdtj3B7NvsnGextLb8nJe/Lut/7RV043Tcb1sb9IqGXf6f1sbYy6ptoezHFlp2wx9XrO6ds9Xc71P6dVlfZv579U9X/RpT1SqdM/oz//AAxk/wDn+5SyLOpNyKW4tFNmO4/rFltzq3tE/wCBpZj3su9v799Cp4vUMfFZbRe25tjb7yQKLniH22Wsc19dT2O3Vva76SRIG6qJdVJUf2zg+F//ALDX/wDpFL9s4Phf/wCw1/8A6RQ4h3CaPYt5JZ56509vPrifHHv/APSKX7e6d43f+w9//pFDjj+8PtVwy7H7G2Mmg5FmMHj1qmMtsbroyw2MrfP0fc7Hu/zEhlYxrFouYaiNweHDaWwX7t07duxu9YPUHdMzr8hz8nKrozcYYmVS3Gt9zB9o2uZYcffXZ+uWKrZgdBsZ77ckWEPL3Mxrmtda605bMgt9Av30usyKWfpf6Nk3V2+p/OJccf3h9qeCXY/Y9K3qGE5l1guZ6ePHrWEwxu5jb2u9R3s2+lax+9M3qOE52SPWaBhvbXkOd7Wse5rLmsNj9te707qne13+EWDXX0erDdj13ZDXG6vJa/7LcWttq9MtPpehtdW59W9zHf8AW7GW+naq93/NrH6c7phyr6g91drHPx7N7TXXVisezbj17Hbcbfvr9PZZ/N/of0KRyQG8h9qhjmdBEnyDvZZ6B1Kh1ed9ly6K3PDmX+m9rXVO9C722ztdTd+if/LWf/ze6ThuDem9RyekmlwaaaMndU02fzdf2LqH2zEq3/4JldFazLn/AFXt9R5z7ja57XssOPZoWvyb9rxVRV6jX25+R/4H+4oPH1WIcyvqF1TCWGsNxngsLWvqNtZ+z+zIcy57fUZ+jt/7VU5P6b1h7uP9+P8AjBd7OX9yX+LJ2vsv1xxWAU5+H1EA6jKofj2Fvnk4dltO7/2nqX7X+sND3Ny+hPsYBIswcmm4H+xm/su3/wADU/8And0D/uQ//ti7/wBIqLvrh9XmNLn5LmtHJNN0f+eUvdx/vx/xgr2cv7kv8UsR9b+m11l+bj52BtMO+0YeRtH/AF+iq/G/8HR8f61fVnKLRR1XDe9/0WevWH/9tOcLP+iq3/Pn6rf9zf8AwK7/ANJIGV9avqTms9PMtqya/wBy7HseP82yhyPHH94faj25/uy+x6RrmuAc0gg8EahOuHtt/wAVtjg9lVWO8cPxab8Z3+fhV471tU/XT6r/AKOmvNJJ21sBruJJMMZL31f9J6XHH94faj25/uy+x//T9VWDTh5dnXnXX4lzMVji2p73MexzQ0n1Ld2bZZ7sgu9Fn2L9HX6H83/gt5JJSlm2EfaXTxv1+C0lg9VyM6jJnFxjlVku9UN0eDuqazZJazb6brvU/c/nf8HYos4sDzZcJ1l5OLRT9ZG147bPWcaQ9rnOsBNnrvqv327bGuc7p7PUxKf/AEYxXelftc5Dvt4saxlDWAODdm8P0eyxrnPvtsx9j8ix/wDhd9f/AAdcx1LOLQf2dY07dz9xdAnbt27KXvtd7t2xlfqV/wDGK/W5z6w59bqnESa3Fpc3+sanWM/6ShJNagMoAvQnRhd9Jk8d/vC5xrPrCw1n9K4UvufJc15e282OY19bn/pf2e1v6pU/2fp8X/Rrb6rbk1VMfjV+rZugt2uedvfa2st930fpO9NUT1LL2epXgWWMJIbtdJO07Hu+ht9P6fo2f4T+R6ii1BNAG+7LoQLJFdmrZb9YS5rmsfLNxY3YxrXVkFjLsiHud9u/P+xNZ6PqempPv+sTaPUbXvtJAZWK2AQ2v1N1u52/9YuPov8A5n0tn/adWsfNzbrWB2E5lNuzbb74AcNXOmpr/wDPrr/4b0v0fqQHUs50Or6fZYx2jSNw4L9znF1f/F/4P9/0n3Ja/uxVp+8WGE/rTctlWQC7FHqbrHtbuIm4tfvr+i7f6FdP7+L+k9L/AAlVfr39Mr/4of8AVPVs5/UtjyMB4fE1g7tfBrob9L/SfQ/4H1bP0ay+s5V1tldzaDIGw1zJLGWWMttr+h+c1/o7lDnBMdgNQGxypAnuToS5WU3NN7nUepqwhkGGA7H+9vv9Pd6mz9FfRv8Aoelek6zqYEtbuJ3FjSxoH0nNDbnbvZ+hbXaz/SXP9NTbk5Qa31MY7tA6CYLj+bX7Hf8AkP37P5xP9oygGg4xnTcQSRxy32/+Ts/4NQ+qgOGBrT9FtVGyRKY4tf0v+9R/aMtmVVRYQd7wBDWy9h3eo98O/R+nHs9P/rqNm/0S34D8rU1GW+2w1GpzOd0mY2xLXe36W57Pof8AfE+b/RLfgP8Aqmpp+eFxEdtvP+quHyTqRkNfm6abepxXb/VBh5bDduwwAZ9/qf2dv0kNv2sNAOsNBJgEk+zc3n8z9L/XU/UuBg1EjxHhLv8AvqRst7VH8eP81XRfYFpGu8h9rGco9u8RAGmh3uIP+e1WME2HIx/Und61UyA38+ufol3t3IPqWf6I/wCo+CNhPLsqjc3aRdUD4TvYT/1SRvsPokV3J83/1PVUkkklKXO9ZcPtIY7N+yNO57wCWuc1llDXPY5v7rrK6P8A0IXRLmuvOwhksGXivymzZJZuPptgM92z2fp7nVUN9R7P9N/gVFl/R82TF18kFdeW9zmu6w2x7Pc4MDA2D++G/m6s/P8A/Piga8ywAN6xWBWCC5pbq8NNm/d9Bu2v9J6XvZXWhPu6G3cbMS82B2oJc50gOY+zf6x91X0Lv8Kz9B/hPSRun4/Rcpz2U4zq20kBosc8b9rt3trL/fVXYyi3/jn/AOkUX8tgzfy3SdSdaMXHnKZTa87W3j6Lzss9NzB7tznbftHpf+fEKp2Q3ID7cquzGFQrc7e0E2y1u7bp+k9tn/bnp+mreVg4r6G41jN9ADoYSYAI9Pa3Xd7WP/R/6NVWdJ6awgsp2xJADngAkFu6N30mte5tf+jUEiLP8GaIlQ/i1zhsvrrNuYA4MZTZbVY73msOse5jrH7PVtqfXY922z6CJe6rIruc7LqFBqbXY4WOOx7HWObaLGOZ9Nv89/xSJ+yOnGllBp/RVOL2N3Pjc6Nz/pe76DVI9LwHOLnVbnGSXOc8ulxDne/fu+k3d/6sehxDufsTwnsPtR42LfXf6rLm2VOc4loc4ja7c9v0jZ+l2W0/R/R/q1X+mWb9ZQ85Ne2wVu9NvvP9azx/le7atvHx6sar0qp27nPJPJc9xsedP5TlhfWc0i+v1mF9ZraC0CfpOsZu/wCkospsd9R0Z+XFS+h6uYx1jLWF97HVNZD5cBLjrMf1fzkMV3gmc0aySBtMEaTr+bu/9JILn4JqcW1WtbtEPZqQHe/2O3PZ+k/PexTs+w1vNXoWQYcXN3TJb/X3/wA27ao+HwP+LDp/6M2OLxH+PPr/AOisyyw6HNG6S7kEARHj+9/Y/MU8ifsNkvFhAhzgZ1DggGzBa4uNFgIJdY4ky2A4us0ed22HN9v/AFtGvrqZg2+k3Y0tb7J0EFrY7+5u3Y7+ogd43e46RH/RTHadUdDfqkf+k4/p2gyLdOYInu53/fkxZfHtsk+HARUlZ4j4fY1uEeP2o/Tu/wBNrPgOEXDZaMzHLrNw9WuRH8tkJkTG/pWP/wAdV/1bEuI+H2K4R4/aX//V9VSSSSUpYfVKerPvnAcK2tLzZuB95cPSYGu2P2elvfkbvz7a6qvob1uLF6tT1OvqDcnp5tf+r2kVuc51HrB+PXjmzHZZVv8A0NmVZ6fqM9TZ/pNibOHFXguhLhvxaja/rMdrn2VNPuDmMYdv7zHe7c/6X6P6X817/wCdTNZ9Zi5psFDhLd0NcNN/6UNnf/gfoO/9G/zc7eo/Wn1MhlGM0tqc703vqPu9MZn6NrfXb/SH42Bst3/QzP8ASfo8V7c/6xiy0txpbXY9gsDHhrajZS31BVu9XJvox/Vf+jqfXa/+jWXfpKkz2fH8F/u+H4p8iuwlsMeRr+af7kL0rf8ARv8A8139yj9s+tD31G2oU1ufR6gqqLiGh+B9oG6yx3svZk5/+C9THrxv+D/SaNWTnHq763Mf9hfUQwlkBllThL3Wf4T7ay/9H/o/sX/DqM8qCb4vwZBzRArh/Foelb/o3/5rv7kvSt/0b/8ANd/ctm92c1w+zVVWMjU2WOYZ8gym5C9Tq3/cfH/7ff8A+8qH3Qfvfgn72f3fxcv0rf8ARv8A8139ywvrBjdRdk1nFosc702gk1vLY3P3tljT+aux9Tq3/cfH/wC33/8AvKl6vVv+42P/ANvv/wDeRCXJCQri/BdDnTE3w9K3fOfsfXo0ofoIDfRf4eOxOcTrsiMewNhsxVZ9KP02jme5u76C9F9Xq3/cbH/9iH/+8iqdQyPrQw437Pw8SzdcBkizIeAKtljnO3/Z2OY71W1s9lWT/wAR/hK2fcI/vD/EZP8ASUv3T/j/APoLxFOH1YgjIxrToNG1WbZl3Et/d2KObhZxxbQMW8kgaCmwnlv8hd/1unNvxqqsPcHOvq9XY91X6IOm3dbS5lzG7f8ARLNr/wCdWPZ9iDhdWz0625Lm7iWOdR6uRL3fzlHq5dXp3XW3Prxqsi31H/0hf6PjxXx9boRT/pOXCY+30qzLV89/Z/Uv+4WT/wBsW/8ApNN+z+pf9wsn/ti3/wBJr0b1/rQxnqWNDw5o3VsraHML22FzwfWs3/ZX11fmWPf61v6HJ/RUqDcn62/Zxa+poeYrfXsEtbFLrMwOa+1z7fdkenR6P/Wv9JL91H7xYvvp/cH2vnn7P6l/3Cyf+2Lf/SaLjdP6iMmgnDyQBbWSTRbEB7f+DXfet9arGsdY30276d4qrbu2NdhW3vHqWOd+lbZn02V7PoV/o/0n89dvs6gOtY9tTbXYwc7Gupj9H6b2DIZ1Dd9H1K8mv7Js/wBFdY9L7qP3ir76f3B9r//ZOEJJTQQhAAAAAABVAAAAAQEAAAAPAEEAZABvAGIAZQAgAFAAaABvAHQAbwBzAGgAbwBwAAAAEwBBAGQAbwBiAGUAIABQAGgAbwB0AG8AcwBoAG8AcAAgAEMAUwA2AAAAAQD/4gxYSUNDX1BST0ZJTEUAAQEAAAxITGlubwIQAABtbnRyUkdCIFhZWiAHzgACAAkABgAxAABhY3NwTVNGVAAAAABJRUMgc1JHQgAAAAAAAAAAAAAAAQAA9tYAAQAAAADTLUhQICAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABFjcHJ0AAABUAAAADNkZXNjAAABhAAAAGx3dHB0AAAB8AAAABRia3B0AAACBAAAABRyWFlaAAACGAAAABRnWFlaAAACLAAAABRiWFlaAAACQAAAABRkbW5kAAACVAAAAHBkbWRkAAACxAAAAIh2dWVkAAADTAAAAIZ2aWV3AAAD1AAAACRsdW1pAAAD+AAAABRtZWFzAAAEDAAAACR0ZWNoAAAEMAAAAAxyVFJDAAAEPAAACAxnVFJDAAAEPAAACAxiVFJDAAAEPAAACAx0ZXh0AAAAAENvcHlyaWdodCAoYykgMTk5OCBIZXdsZXR0LVBhY2thcmQgQ29tcGFueQAAZGVzYwAAAAAAAAASc1JHQiBJRUM2MTk2Ni0yLjEAAAAAAAAAAAAAABJzUkdCIElFQzYxOTY2LTIuMQAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAWFlaIAAAAAAAAPNRAAEAAAABFsxYWVogAAAAAAAAAAAAAAAAAAAAAFhZWiAAAAAAAABvogAAOPUAAAOQWFlaIAAAAAAAAGKZAAC3hQAAGNpYWVogAAAAAAAAJKAAAA+EAAC2z2Rlc2MAAAAAAAAAFklFQyBodHRwOi8vd3d3LmllYy5jaAAAAAAAAAAAAAAAFklFQyBodHRwOi8vd3d3LmllYy5jaAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABkZXNjAAAAAAAAAC5JRUMgNjE5NjYtMi4xIERlZmF1bHQgUkdCIGNvbG91ciBzcGFjZSAtIHNSR0IAAAAAAAAAAAAAAC5JRUMgNjE5NjYtMi4xIERlZmF1bHQgUkdCIGNvbG91ciBzcGFjZSAtIHNSR0IAAAAAAAAAAAAAAAAAAAAAAAAAAAAAZGVzYwAAAAAAAAAsUmVmZXJlbmNlIFZpZXdpbmcgQ29uZGl0aW9uIGluIElFQzYxOTY2LTIuMQAAAAAAAAAAAAAALFJlZmVyZW5jZSBWaWV3aW5nIENvbmRpdGlvbiBpbiBJRUM2MTk2Ni0yLjEAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHZpZXcAAAAAABOk/gAUXy4AEM8UAAPtzAAEEwsAA1yeAAAAAVhZWiAAAAAAAEwJVgBQAAAAVx/nbWVhcwAAAAAAAAABAAAAAAAAAAAAAAAAAAAAAAAAAo8AAAACc2lnIAAAAABDUlQgY3VydgAAAAAAAAQAAAAABQAKAA8AFAAZAB4AIwAoAC0AMgA3ADsAQABFAEoATwBUAFkAXgBjAGgAbQByAHcAfACBAIYAiwCQAJUAmgCfAKQAqQCuALIAtwC8AMEAxgDLANAA1QDbAOAA5QDrAPAA9gD7AQEBBwENARMBGQEfASUBKwEyATgBPgFFAUwBUgFZAWABZwFuAXUBfAGDAYsBkgGaAaEBqQGxAbkBwQHJAdEB2QHhAekB8gH6AgMCDAIUAh0CJgIvAjgCQQJLAlQCXQJnAnECegKEAo4CmAKiAqwCtgLBAssC1QLgAusC9QMAAwsDFgMhAy0DOANDA08DWgNmA3IDfgOKA5YDogOuA7oDxwPTA+AD7AP5BAYEEwQgBC0EOwRIBFUEYwRxBH4EjASaBKgEtgTEBNME4QTwBP4FDQUcBSsFOgVJBVgFZwV3BYYFlgWmBbUFxQXVBeUF9gYGBhYGJwY3BkgGWQZqBnsGjAadBq8GwAbRBuMG9QcHBxkHKwc9B08HYQd0B4YHmQesB78H0gflB/gICwgfCDIIRghaCG4IggiWCKoIvgjSCOcI+wkQCSUJOglPCWQJeQmPCaQJugnPCeUJ+woRCicKPQpUCmoKgQqYCq4KxQrcCvMLCwsiCzkLUQtpC4ALmAuwC8gL4Qv5DBIMKgxDDFwMdQyODKcMwAzZDPMNDQ0mDUANWg10DY4NqQ3DDd4N+A4TDi4OSQ5kDn8Omw62DtIO7g8JDyUPQQ9eD3oPlg+zD88P7BAJECYQQxBhEH4QmxC5ENcQ9RETETERTxFtEYwRqhHJEegSBxImEkUSZBKEEqMSwxLjEwMTIxNDE2MTgxOkE8UT5RQGFCcUSRRqFIsUrRTOFPAVEhU0FVYVeBWbFb0V4BYDFiYWSRZsFo8WshbWFvoXHRdBF2UXiReuF9IX9xgbGEAYZRiKGK8Y1Rj6GSAZRRlrGZEZtxndGgQaKhpRGncanhrFGuwbFBs7G2MbihuyG9ocAhwqHFIcexyjHMwc9R0eHUcdcB2ZHcMd7B4WHkAeah6UHr4e6R8THz4faR+UH78f6iAVIEEgbCCYIMQg8CEcIUghdSGhIc4h+yInIlUigiKvIt0jCiM4I2YjlCPCI/AkHyRNJHwkqyTaJQklOCVoJZclxyX3JicmVyaHJrcm6CcYJ0kneierJ9woDSg/KHEooijUKQYpOClrKZ0p0CoCKjUqaCqbKs8rAis2K2krnSvRLAUsOSxuLKIs1y0MLUEtdi2rLeEuFi5MLoIuty7uLyQvWi+RL8cv/jA1MGwwpDDbMRIxSjGCMbox8jIqMmMymzLUMw0zRjN/M7gz8TQrNGU0njTYNRM1TTWHNcI1/TY3NnI2rjbpNyQ3YDecN9c4FDhQOIw4yDkFOUI5fzm8Ofk6Njp0OrI67zstO2s7qjvoPCc8ZTykPOM9Ij1hPaE94D4gPmA+oD7gPyE/YT+iP+JAI0BkQKZA50EpQWpBrEHuQjBCckK1QvdDOkN9Q8BEA0RHRIpEzkUSRVVFmkXeRiJGZ0arRvBHNUd7R8BIBUhLSJFI10kdSWNJqUnwSjdKfUrESwxLU0uaS+JMKkxyTLpNAk1KTZNN3E4lTm5Ot08AT0lPk0/dUCdQcVC7UQZRUFGbUeZSMVJ8UsdTE1NfU6pT9lRCVI9U21UoVXVVwlYPVlxWqVb3V0RXklfgWC9YfVjLWRpZaVm4WgdaVlqmWvVbRVuVW+VcNVyGXNZdJ114XcleGl5sXr1fD19hX7NgBWBXYKpg/GFPYaJh9WJJYpxi8GNDY5dj62RAZJRk6WU9ZZJl52Y9ZpJm6Gc9Z5Nn6Wg/aJZo7GlDaZpp8WpIap9q92tPa6dr/2xXbK9tCG1gbbluEm5rbsRvHm94b9FwK3CGcOBxOnGVcfByS3KmcwFzXXO4dBR0cHTMdSh1hXXhdj52m3b4d1Z3s3gReG54zHkqeYl553pGeqV7BHtje8J8IXyBfOF9QX2hfgF+Yn7CfyN/hH/lgEeAqIEKgWuBzYIwgpKC9INXg7qEHYSAhOOFR4Wrhg6GcobXhzuHn4gEiGmIzokziZmJ/opkisqLMIuWi/yMY4zKjTGNmI3/jmaOzo82j56QBpBukNaRP5GokhGSepLjk02TtpQglIqU9JVflcmWNJaflwqXdZfgmEyYuJkkmZCZ/JpomtWbQpuvnByciZz3nWSd0p5Anq6fHZ+Ln/qgaaDYoUehtqImopajBqN2o+akVqTHpTilqaYapoum/adup+CoUqjEqTepqaocqo+rAqt1q+msXKzQrUStuK4trqGvFq+LsACwdbDqsWCx1rJLssKzOLOutCW0nLUTtYq2AbZ5tvC3aLfguFm40blKucK6O7q1uy67p7whvJu9Fb2Pvgq+hL7/v3q/9cBwwOzBZ8Hjwl/C28NYw9TEUcTOxUvFyMZGxsPHQce/yD3IvMk6ybnKOMq3yzbLtsw1zLXNNc21zjbOts83z7jQOdC60TzRvtI/0sHTRNPG1EnUy9VO1dHWVdbY11zX4Nhk2OjZbNnx2nba+9uA3AXcit0Q3ZbeHN6i3ynfr+A24L3hROHM4lPi2+Nj4+vkc+T85YTmDeaW5x/nqegy6LzpRunQ6lvq5etw6/vshu0R7ZzuKO6070DvzPBY8OXxcvH/8ozzGfOn9DT0wvVQ9d72bfb794r4Gfio+Tj5x/pX+uf7d/wH/Jj9Kf26/kv+3P9t////4Tt7aHR0cDovL25zLmFkb2JlLmNvbS94YXAvMS4wLwA8P3hwYWNrZXQgYmVnaW49Iu+7vyIgaWQ9Ilc1TTBNcENlaGlIenJlU3pOVGN6a2M5ZCI/Pgo8eDp4bXBtZXRhIHhtbG5zOng9ImFkb2JlOm5zOm1ldGEvIiB4OnhtcHRrPSJBZG9iZSBYTVAgQ29yZSA1LjMtYzAxMSA2Ni4xNDU2NjEsIDIwMTIvMDIvMDYtMTQ6NTY6MjcgICAgICAgICI+CiAgIDxyZGY6UkRGIHhtbG5zOnJkZj0iaHR0cDovL3d3dy53My5vcmcvMTk5OS8wMi8yMi1yZGYtc3ludGF4LW5zIyI+CiAgICAgIDxyZGY6RGVzY3JpcHRpb24gcmRmOmFib3V0PSIiCiAgICAgICAgICAgIHhtbG5zOnhtcD0iaHR0cDovL25zLmFkb2JlLmNvbS94YXAvMS4wLyI+CiAgICAgICAgIDx4bXA6Q3JlYXRvclRvb2w+QWRvYmUgUGhvdG9zaG9wIENTNiAoV2luZG93cyk8L3htcDpDcmVhdG9yVG9vbD4KICAgICAgICAgPHhtcDpDcmVhdGVEYXRlPjIwMTUtMDMtMTNUMDk6MTY6NDUtMDc6MDA8L3htcDpDcmVhdGVEYXRlPgogICAgICAgICA8eG1wOk1vZGlmeURhdGU+MjAxNS0wOC0xMVQxODo1NToyMCswNTozMDwveG1wOk1vZGlmeURhdGU+CiAgICAgICAgIDx4bXA6TWV0YWRhdGFEYXRlPjIwMTUtMDgtMTFUMTg6NTU6MjArMDU6MzA8L3htcDpNZXRhZGF0YURhdGU+CiAgICAgIDwvcmRmOkRlc2NyaXB0aW9uPgogICAgICA8cmRmOkRlc2NyaXB0aW9uIHJkZjphYm91dD0iIgogICAgICAgICAgICB4bWxuczpkYz0iaHR0cDovL3B1cmwub3JnL2RjL2VsZW1lbnRzLzEuMS8iPgogICAgICAgICA8ZGM6Zm9ybWF0PmltYWdlL3RpZmY8L2RjOmZvcm1hdD4KICAgICAgPC9yZGY6RGVzY3JpcHRpb24+CiAgICAgIDxyZGY6RGVzY3JpcHRpb24gcmRmOmFib3V0PSIiCiAgICAgICAgICAgIHhtbG5zOnBob3Rvc2hvcD0iaHR0cDovL25zLmFkb2JlLmNvbS9waG90b3Nob3AvMS4wLyI+CiAgICAgICAgIDxwaG90b3Nob3A6Q29sb3JNb2RlPjM8L3Bob3Rvc2hvcDpDb2xvck1vZGU+CiAgICAgICAgIDxwaG90b3Nob3A6SUNDUHJvZmlsZT5zUkdCIElFQzYxOTY2LTIuMTwvcGhvdG9zaG9wOklDQ1Byb2ZpbGU+CiAgICAgIDwvcmRmOkRlc2NyaXB0aW9uPgogICAgICA8cmRmOkRlc2NyaXB0aW9uIHJkZjphYm91dD0iIgogICAgICAgICAgICB4bWxuczp4bXBNTT0iaHR0cDovL25zLmFkb2JlLmNvbS94YXAvMS4wL21tLyIKICAgICAgICAgICAgeG1sbnM6c3RFdnQ9Imh0dHA6Ly9ucy5hZG9iZS5jb20veGFwLzEuMC9zVHlwZS9SZXNvdXJjZUV2ZW50IyI+CiAgICAgICAgIDx4bXBNTTpJbnN0YW5jZUlEPnhtcC5paWQ6NEQzQjY3NEIyQzQwRTUxMUExNTlBNjZDMzlFQTk3MkI8L3htcE1NOkluc3RhbmNlSUQ+CiAgICAgICAgIDx4bXBNTTpEb2N1bWVudElEPnhtcC5kaWQ6MDNGQ0UwODY5Q0M5RTQxMUIzNTBBNUQ0ODdGQThEMjM8L3htcE1NOkRvY3VtZW50SUQ+CiAgICAgICAgIDx4bXBNTTpPcmlnaW5hbERvY3VtZW50SUQ+eG1wLmRpZDowM0ZDRTA4NjlDQzlFNDExQjM1MEE1RDQ4N0ZBOEQyMzwveG1wTU06T3JpZ2luYWxEb2N1bWVudElEPgogICAgICAgICA8eG1wTU06SGlzdG9yeT4KICAgICAgICAgICAgPHJkZjpTZXE+CiAgICAgICAgICAgICAgIDxyZGY6bGkgcmRmOnBhcnNlVHlwZT0iUmVzb3VyY2UiPgogICAgICAgICAgICAgICAgICA8c3RFdnQ6YWN0aW9uPmNyZWF0ZWQ8L3N0RXZ0OmFjdGlvbj4KICAgICAgICAgICAgICAgICAgPHN0RXZ0Omluc3RhbmNlSUQ+eG1wLmlpZDowM0ZDRTA4NjlDQzlFNDExQjM1MEE1RDQ4N0ZBOEQyMzwvc3RFdnQ6aW5zdGFuY2VJRD4KICAgICAgICAgICAgICAgICAgPHN0RXZ0OndoZW4+MjAxNS0wMy0xM1QwOToxNjo0NS0wNzowMDwvc3RFdnQ6d2hlbj4KICAgICAgICAgICAgICAgICAgPHN0RXZ0OnNvZnR3YXJlQWdlbnQ+QWRvYmUgUGhvdG9zaG9wIENTNiAoV2luZG93cyk8L3N0RXZ0OnNvZnR3YXJlQWdlbnQ+CiAgICAgICAgICAgICAgIDwvcmRmOmxpPgogICAgICAgICAgICAgICA8cmRmOmxpIHJkZjpwYXJzZVR5cGU9IlJlc291cmNlIj4KICAgICAgICAgICAgICAgICAgPHN0RXZ0OmFjdGlvbj5jb252ZXJ0ZWQ8L3N0RXZ0OmFjdGlvbj4KICAgICAgICAgICAgICAgICAgPHN0RXZ0OnBhcmFtZXRlcnM+ZnJvbSBhcHBsaWNhdGlvbi92bmQuYWRvYmUucGhvdG9zaG9wIHRvIGltYWdlL3RpZmY8L3N0RXZ0OnBhcmFtZXRlcnM+CiAgICAgICAgICAgICAgIDwvcmRmOmxpPgogICAgICAgICAgICAgICA8cmRmOmxpIHJkZjpwYXJzZVR5cGU9IlJlc291cmNlIj4KICAgICAgICAgICAgICAgICAgPHN0RXZ0OmFjdGlvbj5zYXZlZDwvc3RFdnQ6YWN0aW9uPgogICAgICAgICAgICAgICAgICA8c3RFdnQ6aW5zdGFuY2VJRD54bXAuaWlkOjA0RkNFMDg2OUNDOUU0MTFCMzUwQTVENDg3RkE4RDIzPC9zdEV2dDppbnN0YW5jZUlEPgogICAgICAgICAgICAgICAgICA8c3RFdnQ6d2hlbj4yMDE1LTAzLTEzVDA5OjE4OjI3LTA3OjAwPC9zdEV2dDp3aGVuPgogICAgICAgICAgICAgICAgICA8c3RFdnQ6c29mdHdhcmVBZ2VudD5BZG9iZSBQaG90b3Nob3AgQ1M2IChXaW5kb3dzKTwvc3RFdnQ6c29mdHdhcmVBZ2VudD4KICAgICAgICAgICAgICAgICAgPHN0RXZ0OmNoYW5nZWQ+Lzwvc3RFdnQ6Y2hhbmdlZD4KICAgICAgICAgICAgICAgPC9yZGY6bGk+CiAgICAgICAgICAgICAgIDxyZGY6bGkgcmRmOnBhcnNlVHlwZT0iUmVzb3VyY2UiPgogICAgICAgICAgICAgICAgICA8c3RFdnQ6YWN0aW9uPnNhdmVkPC9zdEV2dDphY3Rpb24+CiAgICAgICAgICAgICAgICAgIDxzdEV2dDppbnN0YW5jZUlEPnhtcC5paWQ6NEQzQjY3NEIyQzQwRTUxMUExNTlBNjZDMzlFQTk3MkI8L3N0RXZ0Omluc3RhbmNlSUQ+CiAgICAgICAgICAgICAgICAgIDxzdEV2dDp3aGVuPjIwMTUtMDgtMTFUMTg6NTU6MjArMDU6MzA8L3N0RXZ0OndoZW4+CiAgICAgICAgICAgICAgICAgIDxzdEV2dDpzb2Z0d2FyZUFnZW50PkFkb2JlIFBob3Rvc2hvcCBDUzYgKFdpbmRvd3MpPC9zdEV2dDpzb2Z0d2FyZUFnZW50PgogICAgICAgICAgICAgICAgICA8c3RFdnQ6Y2hhbmdlZD4vPC9zdEV2dDpjaGFuZ2VkPgogICAgICAgICAgICAgICA8L3JkZjpsaT4KICAgICAgICAgICAgPC9yZGY6U2VxPgogICAgICAgICA8L3htcE1NOkhpc3Rvcnk+CiAgICAgIDwvcmRmOkRlc2NyaXB0aW9uPgogICA8L3JkZjpSREY+CjwveDp4bXBtZXRhPgogICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgCiAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAKICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgIAogICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgCiAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAKICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgIAogICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgCiAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAKICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgIAogICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgCiAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAKICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgIAogICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgCiAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAKICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgIAogICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgCiAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAKICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgIAogICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgCiAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAKICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgIAogICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgCiAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAKICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgIAogICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgCiAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAKICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgIAogICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgCiAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAKICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgIAogICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgCiAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAKICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgIAogICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgCiAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAKICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgIAogICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgCiAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAKICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgIAogICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgCiAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAKICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgIAogICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgCiAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAKICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgIAogICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgCiAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAKICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgIAogICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgCiAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAKICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgIAogICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgCiAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAKICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgIAogICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgCiAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAKICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgIAogICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgCiAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAKICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgIAogICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgCiAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAKICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgIAogICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgCiAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAKICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgIAogICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgCiAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAKICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgIAogICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgCiAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAKICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgIAogICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgCiAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAKICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgIAogICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgCiAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAKICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgIAogICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgCiAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAKICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgIAogICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgCiAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAKICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgIAogICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgCiAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAKICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgIAogICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgCiAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAKICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgIAogICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgCiAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAKICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgIAogICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgCiAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAKICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgIAogICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgCiAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAKICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgIAogICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgCiAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAKICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgIAogICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgCiAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAKICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgIAogICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgCiAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAKICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgIAogICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgCiAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAKICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgIAogICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgCiAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAKICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgIAogICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgCiAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAKICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgIAogICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgCiAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAKICAgICAgICAgICAgICAgICAgICAgICAgICAgIAo8P3hwYWNrZXQgZW5kPSJ3Ij8+/9sAQwADAgICAgIDAgICAwMDAwQGBAQEBAQIBgYFBgkICgoJCAkJCgwPDAoLDgsJCQ0RDQ4PEBAREAoMEhMSEBMPEBAQ/9sAQwEDAwMEAwQIBAQIEAsJCxAQEBAQEBAQEBAQEBAQEBAQEBAQEBAQEBAQEBAQEBAQEBAQEBAQEBAQEBAQEBAQEBAQ/8AAEQgB5gK8AwEiAAIRAQMRAf/EAB0AAQACAgMBAQAAAAAAAAAAAAAGBwUIAwQJAQL/xABpEAABAwMCAwMGAxANBwkGBAcBAgMEAAUGBxEIEiETMUEJFBUiUWEycdMWFxgjMzhCVVZzdoGRlrG0NTdScnWTlJWhsrPR0hk0NldYYoIkJjlDU3SDkqIlY3ejpMNEZNTwJ0VUZrXBwv/EABsBAQACAwEBAAAAAAAAAAAAAAABAwIEBQYH/8QAPREAAQMCAwQFCgYCAwEBAQAAAQACEQMhBBIxBUFRcRUiYYGhExQyM1KRkrHB0QYWNFNy8EJUI2LhwvGy/9oADAMBAAIRAxEAPwD1TpSlESlKURKpDi710yTQrTKHcMDsce8ZllN5iY3jsORuWlTZBOy1gEcwSlKjtuNyU7nbervqguMrR7OtVtPLHdNLPNXMzwPI4WVWWNKWENTHY5VzMKUSAOZKjsSQN0gEjfcEVT8Rms/EnguEaPaE2jIrKxrVqXKTCud1gtJVHiIQQHHWkqRskEuJ3VyHYNubDfY1IOHrPtfdNtfJfC9xD5lCzV24Y8clxrI2IwYdeaQ72bzDqQB1B3IJ3I5e8hQ5Y1oxpvxCa2cVEXia4gNM2cFgYbZF2rHrIuYiQ45JWFhTu6SdkjtXTzEDqUAA8pNZ/QLENc9WeJS58UGtmBfMJb7NY3cXxawvPdpKLa3udyQ50B2OytiQknnGw2TuSlbd0pSihKUrEzctxW3XqLjdwya1RbvNAVFgPTW0SXwd9ihonmUPVV3A9x9lEWWpSuGXLiQIrs2dKajR2EFx151YQhtI6lSlHoAPaaIualcbD7MlluTGeQ6y6kLbcQoKStJG4II6EEeNclESlKURKUpREpSlESlKURKUpREpSlESlKURKUpREpSlESlKURKUpREpSlESlKURKUpREpSlESlKURKUpREpSlESlKURKUpREpSlESlKURKUpREpSlESlKURKUpREpSlESlKURKUpREpSlESlKURKUpREpSlESlKURKUpREpSlESlKURKUpREqg+MzXrK9BtMLfL07s7FzzLLL1GxywsyElTSZL/ADHtFJ3HNsEkAEgcyk79AavytYvKDWG5XjRS2TcZxHKL5lFlyWDdMfVj0Lzt6DPZDi0Putd6mOhQvbr66SO7aiKFRdI/KfTY7cudxPYDBfeSFuRkWRtwNKI6p5hF2O3d0qzdBNPeMfFs2euWveuOM5djaoDrTUC22tMd1MorQUOFQZR6oSFjbf7IdKqWw8fPEAi0xm8n4E9R3bmlsCS5AZfSwte3VSErj8yQT4Enb2mrY0C4o8/1kzd7E8n4Ys40/iNW92aLrekqEda0rQkMjdpI5lBZI69yT0opWw9KUooWBvue4Ni8tEDJs0sVolONh1DM64sx3FIJICglagSNwRv7jXn/AMRmZYTc/KT6H5LAyuxyrXBtrKZFwZnsrYYIdm9FuhXKk+sO8jvHtrZviC4HNE+JfMYecakLyEXKDb0Wxr0fPQw32KXFuDdJbVurmcV139laE6xcGejmA8Z2mPD9YlX44tmMNuRci/OSuTzKXJSezcCAEjZlH2J8fbRSF6p27U/TW8TmbXaNQ8ZnTJKuRmPGu0d111XfslKVkqPQ9AKhPF2Arhb1XCgCDiF03B/7suqx0x8mxw5aS6gWLUrFHMrN4x6V53D86uaHGu05VJ9ZIaBI2UfEVZ3F19a5qv8AghdP1ZdEUl0MAGienwA2Axa0/qjVTeoRob+0pp/+C1q/VGqm9FCUpSiJSlKIlKUoiUpVdak8ROhukDojak6pY/YpRG/mj8sKk7e3sUbube/loisWlUFZ+PPhDvktMKFrnYmnVq5UmYh+Kgn9+82lP9NXbYshsOUW1q8Y1e4F2gPdW5UGSh9lfxLQSD+WiLIUpSiJSlKIlKUoiUpSiJSlKIlKUoiUpSiJSlKIlKUoiUpSiJSlKIlKUoiUpSiJSlKIlKUoiUpSiJSlKIlKUoiUpSiJSlKIlKUoiUpSiJSlKIlKUoiUpSiJSlKIlKUoiUpSiJSlKIlKUoiUpSiJSuvcJiLdAk3BxClojMreUlPeQlJJA9/SvPbRTTDiY4z8Xla9ZNxV5XglqvtxlIstix7mSzHjNOqbG4S4gDYpKRuFKPLzFXXYEXojStLxwCawAg/R4arHYg7do51/+orcuM0piO0wt1Tim0JQVq71EDbc/HRFy1VnEpr3ZeHLSyZqFdLY9dZan2rdabWwdnJ894kNMg7HYdConYnZJ2BOwNp1q/5QLE8tuWl2MajYZYnL7L0xy235dItTaSpUqLHKg6AkbklIUFdAfVCj4URQGFG8qzmcMZY1fNMcNRKSH2Mfkxu0daSeoQ4rs3SFbd+7m4Pft3VY/DBxJakZnnGQaBcQ+FxcY1MxmIi4gwlbw7tBUoJ84Z9ZXUKKd9lEHm+xKVJH7xfyinCHkmPMX2Rq1DszrjQW9b7lGeblMK8UFIQQog+KCoHwNV5ofnDXFTxnzOIHArPOY09wTFHcXiXiVHUz6XmPPdorkSrrypBUdj1ACSQCvailbr0pSihKqPNeGPTnPdcMV4gL3JvScmw9hEe3tx5aURFJSp1Q7RsoJUd3l9yh4VblKIlVHxdfWuar/ghdP1ZdW5VR8XP1rmq/4IXT9WXRFJdDf2lNP/wWtX6o1U3qEaG/tKaf/gtav1RqpvREpSlESlKURKUqjuMvXxPDtoPe80gvNi/zdrVYW1bHec8CEr28Q2kLcP7zbxoi1g4/ePq9YbeZmhGhF0Uxe2D5vfr7G9d2I4rp5pF23+ndRzLG5QSEp9bcpqzQzyWWpeqFvRnWuGYycRF0/wCUiD2Pnd2e5uvPIU4rlaUd99jzr/dBJ6Vk/Je8NDeoOUXDiR1EjLuESyzVtWMS/X87um/M9MXv8ItlWyT/ANopR70CvVCiy0sF565D5HjT1+EoYrrLksKYE+qq4QY8lpSvelAbUB8RrXPLeF7jR4Jrm7nenl2uEm0Rj2j12xZ1bzBQOv8AyuEob8vtKkLQP3Vey1KKJK8/uF/ypuM5i5Dw3iFjRMburpS0xkUUEW2SruHbpJJjKJ+y3LfXqUDpW/kaTHmR2pcR9t9h9CXGnW1BSFoI3CkkdCCDuCK034sPJv6e6zNTcz0qahYfmywp1aG2+S23NfiHm0j6Us/9qgd/wkq7xqBoFxX65cDecO6Oay49dZeMQneSVYpat5FuQo9JEBwnlW2eqggEtrG/KUnc0SJ0XsZSozpxqThOrWIQM70+yCNeLLcUczUhlXVKh8JtaT1QtJ6KQoAg94qTUUJSlKIlKUoiUpSiJSlKIlKUoiUpSiJSlKIlKUoiUpSiJSlKIlKUoiUpSiJSlKIlKUoiUpSiJSlKIlKUoiUpSiJSlKIlKUoiUpSiJSlKIlKUoiUpSiJSlKIlKUoiUpSiJSlKIlKUoi/LgJbUAkK3SfVPcfdXnXpNbfKH6ESb/ZtNuHPHhhtzuki5wbBcb/HfRbFuq5nEx3kvoWG1K9YIWFcu/TxJ9E3nWo7S33lhDbaStaj3JSBuTWhuO3Xi644Z12zzTjV0aQ6VRbi/b7EYcXtbjdEsrKFSFEFKtiR+7SkH1QlWxUSkKUDWbymW434UMGA3G/8A7fR3fymtyIynlx2lyEBDqkJLiQeiVbdR+WtDcqlcYnBE5A1LzfWI6waXpnMRciZnROyn29l1YQJCFEqVsCofZkb7ApG/MN8osqPOiszYjqXWJDaXWlp7loUNwR8YNFCxWaZBJxPD73lEOyS7y/aLfInNW6ICX5im2ysMtgA7rURyjoepFUldeKnL7boHZdaWuG7O5Vzu1zVb3cSaYX6SiICnR27iez3CPpQPwR9UT1rYR1TiWlqabC1hJKUlXKFHwG/h8dee+v8A5QHVO3Gy2awaF6k4NOt2Xw2LlIlwW3GbnHbcWl6AysoUla3ttkFHU7bg0QLG3vXPBMku67/ffJX5ROuLi+0ckO402VuK/dKPm3rH3netieGriUvOpeTjTn6FfMNMbRAtrkqPKuMHzaCkoWhIYQkNISFHnJAHgk9KgR8o1fQSPoONZOnttSv8Nd7h+1n4g9euJqXlUvT/ADPBdKrZjKoyrXf4vYokXIup5XUkoSVL5SroCQlKeuxUBRStw6UpRQlKUoiVUfF19a5qv+CF0/Vl1blVHxdfWuar/ghdP1ZdEUl0N/aU0/8AwWtX6o1U3qEaG/tKaf8A4LWr9Uaqb0RKUpREpSlESvJzyq2o9y1A13xnRHHSqQnG4zQ83R17W6T1J5E9PEN9iB7O0VXrC4tDSFOuLCUIBUpROwAHeTXjpw0ML4lfKKOZ3NbU/Aav1xypW45gmPGJTEHxBRjAfFRSOK9VND9MLZoxpLi2mNpSnsrBbmozq0/9dI25nnT71uKWr8dTmlKKEpSlESqf4kuF/TTibxE4/mkLza6REqNpvkZA87t7h/ck/DbJ25m1eqr3HZQuClEXi1Ysh4ivJpa1uWe6xTNslxXzvxAtQtmQxEnYPMqI+lvpB79uZB9VQUk9fWXRDXHT7iBwSJn2nl1EmI99LlRXNkyYMgDdTD6N/VWPyEbEEgg1+tbdENP9f8El4BqJaRKhv/TI0hvZMiC+Bsl9he3qLG/xEbgggkV5L3S1cQHk0Ndmp8F8z7NcVEMv7KTbsjgpO5acHXs30A93VSCdxzIV6xZar2mpVb6B6+YBxF4BFz3Ap/MhWzU+A6QJNuk7bqZeSO4jvCu5Q2I6GrIosUpSlESlKURKUpREpSlESlKURKUpREpSlESlKURKUpREpSlESlKURKUpREpSlESlKURKUpREpSlESlKURKUpREpSlESlKURKUpREpSlESlKURKUpREpSlESlKURKUpREpSlEXBOhs3CFIgSAS1JaWyvY7HlUCD/Qa8+tHOI9/gKtkvhw4jsGyNiz2K4SnMZyi1wfOIlwguvKcTv1HrArUfVJI35VJBTufQyuGVDiTmTHnRWZDSu9t1AWk/iPSiLz61p4oV8cuOnhr4Z8FyOdGyiTHayHJbnB83hWyAh5Li1b7nqeQfC5T05UhRUNt/7NbGLJaINmilRZgRmorZV3lKEhI3/EK5YkKHAZEeBEZjNDqG2WwhI/EOlc9ESqB409R8b0y0rtt6uWnkHN8hkZDBi4jZ5bfOhd7UVebu+0FvZat07HfYAjfcX9WvPG5pRnupGmNmyDSmM3LzLT7IoWWWiEsjaauOVc7A3IBUUqJA3G5Tt40RT3S9rXaRowyjVaRYI2pL8SWVrgDmhMPrUsxwQAdwhJbCtubcpOxV3mp+HDXXWtrVW68M/E/a7QnN7fahe7RfLQOWHe4IWELWEbDlWlR8AnuUClJT60Kg+VJ0ztcNNv1I0g1LxvKGE8ku0izh3leHQpQpa0KI37uZKT7q7/AA7o1K4iuJOTxYZhp9ccIxKy44vGsSt90TyTZqXXe0dkuJ2GyfhAeHrJCSrlJJStx6UpRQlKUoiVUfFz9a5qv+CF0/Vl1blVHxdfWuar/ghdP1ZdEUl0N/aU0/8AwWtX6o1U3qEaG/tKaf8A4LWr9Uaqb0RKUpREpSlEVX8UGanTzh31EzBDnI9Ax6YGFA7EPuNltrb38601or5HPCUrvGo2oTqOsSLBscdRH7sqed2P/AzV/eVMylWP8KM+1NO8i8jvdvtu2/VSAsvqH5GK6HkpMVFj4X135bPK7keQzpnNt1U23yR0/wBLKvy0U7luXSlKKEpSlESlKURKhOsOjuB66YJP091DtCZ1tmjmQseq9EeAPI+yvvQ4knofjBBBIM2pRF4sXi0a8eTQ19auFvfVcLLPJDD5CkQMit4VuWnANw2+gHu6qQohSeZKvW9Y9CddcC4htPoWoOAz+0jvfSpkN0gSIEkAc7DyR3KG/Q9ygQobgiuzrTovgmveAT9O9QbYJUCYOdl5GwfhyADyPsr+xWknv7iCQQQSD5JoVrr5NDiC5VBVxs0/vHVEHJLaFd47w2+jf3qbUfskK9YstV7T0qEaNax4LrtgNv1F0+ugl26cnlcbVsHoj4A52HkfYOJJ2I8ehBIIJm9FilKUoiUpSiJSlKIlKUoiUpSiJSlKIlKUoiUpSiJSlKIlKUoiUpSiJSlKIlKUoiUpSiJSlKIlKUoiUpSiJSlKIlKUoiUpSiJSlKIlKUoiUpSiJSlKIlKUoiUpSiJSlKIvytaG0KccUEpSCVKJ2AHtNax6O8YN71RjalaoTtOlWXSHDYslyz5A9IBfuzkUr7chskbJKUgp2BAJ5SoncDZW5QkXK3yrc4tSEymVsqUnvAUkjcflry1ulw4h8X4dZXk/LboNlLuUP3R22sZDHjKVa5NqcmF/tw8ByjmB5VEkAIJ3IPq0UhWHjPGrxfw8SgcT+daZ4wvRS6XJMZ2LCKk3KBCU/wBiJQUVHnAX03I2UR3IBCq9C4smPNjMzIjqXWH20utrT3KQobgj4wa0Y4oYeVWzSbEfJ86Rac3e83a/2G1xZd7QxtbYMBp5KXX1ufYqK2FKPMEgBXQqJArd2xWpuxWO3WRpwuIt8RmKlZ71BtASD/RRF36qTie4hrLw06YOZ9crNIvU2TMZtdptUdfIubMd35Ec2xKUgJUokAnpsASQKtutTPKSLsdl0bxnUG432Hb7nhmY2292ZmY04tm4ymu0V5oooSrkK0JWQtQ5QUgEjfeihRljiC8oxc2mrjH4L8fbbdSHGkyr0hDyEnuCkqdCkn2ggH3CrR0B1P4vcxzd60668P8AZsKx1NvdebuMO6okrVKC0BDXKHVdCkrO+32NQOw+VX4Trnao0y73fIbNMcbSX4T9oddUyvbqnna5kKAPiD19g7qs/RPjV0B4g8vdwbTHIrhOu7EFy4Lbftj0dIZQpCVHmWkDfdxPT30Uq9aUpRQlKUoiVUfF19a5qv8AghdP1ZdW5VR8XP1rmq/4IXT9WXRFJdDf2lNP/wAFrV+qNVN6hGhv7Smn/wCC1q/VGqm9ESlKURKUpRF50eWMyPscX00xJDv+dXGfc3Ub+DLSG0n/AOeqtqeCbG1Ypwo6Y2lbfItdhZnLG2x5pJU+d/42tB/K+XpV01vw7FmHOY27GS7yDwckyVgflDKa9R8AsacYwPG8aSnlFptEOCB7OyZSj/8A5opOiz9KUooSlKURKUpREpSlESq4170GwPiJ0+maf55C5mnN3YM5oDzi3yQCEPtKPcRvsR3KBIPQ1Y9KIvFvEsq1w8mnxBSbDkENy4WOcUmdDQoph3+3hWyJUcnoh5HXY96VboVuk169aYanYVrFhFs1C0/vLdys10b52nE9FtrHw2nE96HEnopJ6gj8dRLiS4csF4l9PH8IzBnsJbPM/aLs0gGRbZW2wcR7UnoFoPRSenQgEeW2k2qmsvk5Ndbhgme2uRJx+U8k3i2NKJYnxidm7hCUrYc4A92+xbXsQCkstV7QUrA4LnWKal4lbM5wi9R7rZbuwJEWUyrcKSe8Ed6VA7hSTsQQQQCKz1FilKUoiUpSiJSlKIlKUoiUpSiJSlKIlKUoiUpSiJSlKIlKUoiUpSiJSlKIlKUoiUpSiJSlKIlKUoiUpSiJSlKIlKUoiUpSiJSlKIlKUoiUpSiJSlKIlKUoiUpSiKheIPiE1W0byCDasQ4Zso1FtlxhqfFzsUonzZ1KiFNOthlRQduVQO+ygT4pNax8OXHFxDx9KLci4cMupOqT3nU3fJozpU3JHnC9mwUsKG7Q2bPXvRXolJeYjR3ZElYQy0hS3FK7gkDck/irU/gyzvVrVu9XfUW1Y3i+G6FqM2BiuPwISGZMh5MkbzlcidhzFLoUNwOZR2SduYlKrLWHiW4wdZ8MXp1pVwo6naeX66y4yEZC5JW0IjYcBX65ZQEBQ6FRUNkk95rfe1MS41rhxrhID8pqO2h90f8AWOBIClfjO5rVjie1J4k+HLNzrtb7laMk0YaVBh3zHDH5J1tQpQbcltu7bndax05iPggp23UNqoM2NcoUe4wnQ7HlNIeaWO5SFAFJ/GCKIueqo4k9WLLo5gUbKb7pfec8jvXNmELZaYCJjyFKQ4oPFCugSOQgn/eHtq16wOZZ7hOndqRfM9yy049bnHkxkSrnLRHaU6oEpQFLIHMQlRA9xooWliuPPTZCStfBNqMlKRuScVY2A/JVmcLHFhp5xA5nPs2CaD3/ABgW+K6qTeZFtjtx23EqQDFW40SUuHm5gg7bhBPhVp/RT8Nf+vnA/wCf43+OtdNL9RsN1B8overroRdGLnjj2C8mYzbed4Uue3IHYO8w9VbiUqSjnG+/rgE7Gilbu0pSihKUpREqo+Lr61zVf8ELp+rLq3KqPi5+tc1X/BC6fqy6IpLob+0pp/8Agtav1RqpvUI0N/aU0/8AwWtX6o1U3oiUpSiJSlKIvHvjSB1H8ovDxFs9qhN0xuxhPfsFFlax+V5VewlePWFA6m+VTcmNEvNMZ7OlJV3/AEuA25yn/wCQmvYWikpSlKKEpSlESlKURKUpREpSlESqU4qeFvC+KHAV47ewi33+3hb1jvSG+ZyE+R8FXitlewC0ePQjZQBF10oi8aeHvXvVbyf2slz0q1YtM1WMOygL1aUkrDXN0RcYRPRYKQCQNg4kbHZaRt7A4tlOO5vjluy7ErxFutnu0dMqHMjL5m3m1DoQf6CD1BBBAIIql+LvhJxDikwjzKQWbXl9pbWuxXrk3LSj1LD23VbCz3jvSfWT1BB87+F/iY1J4GNUbjozrPabg1ivnvZ3a2L3cctbyj0mxfBbahspQT0cT6yfWHUstV7G0rH4/kFkyuxwclxq6xrlarmwiTDlxnAtp5pQ3SpKh3gishRYpSlKIlKUoiUpSiJSlKIlKUoiUpSiJSlKIlKUoiUpSiJSlKIlKUoiUpSiJSlKIlKUoiUpSiJSlKIlKUoiUpSiJSlKIlKUoiUpSiJSlKIlKUoiUpSiJSlKIuGXFZnRHoUlPM1IbU04n2pUNiPyGtANOdWdX+AS3TtEtTNEMpzHBbZcJL2MZRjbHbgw3XVOBp5HwQoFROxUlQJI2UNlV6CKUlCStaglKRuSTsAK8+9VeMHinz/Fc61V4b7Bj+P6WYGZLaskvDQel3hTB5XFRmlbp5d+7dPxqB3SCkJqVrVq5x5WBOhGkmhuVYpiV9lxxkuVZLH83bYgtupcUhpPcpZKB0Cio7bbAEqG/dqt0az2yHaISSmPBYbjNAnchCEhKf6AK1E1I4odc9GbZpXq9kWKW+/6T5BjdnTls9hIRNt9zlJClyUpTsEt7LR6uxSTundBKd9wWH2ZTDcmO6lxp5AcbWk7hSSNwR7iKIoRrtnN70z0azTULG4DM26Y7ZZVyix30qU2txpsqHOEkEp6bnYg7A9a1a1G4oNANXeHXS+8atWTDM1kZVfbRHu9gRexGFllPBaHpS2youpQwSsEL2BCgebuNbtyY0eZHdhzI7b7D6FNutOICkOIUNilQPQggkEGtTNWPJscPmYN2l3T7CbLiUxi/RZ91dbTJcRNt6FEvw0th0JaDgIAUkDl26URcJ0E8mKDt5lpN0//ALmb/wD1FXJofB4YMQTIxDQCVgUdx9PnUmJj86O9IdSnYdo5yLU4sJ5gN1Egc3hvUM/yd/Bn/qOtv84TflqmOlfCdw96JZK5mGlum0Sw3h2IuCuU1KkOEsLUlSkbOOKT1KEnu36URW5SlKKEpSlESqj4ufrXNV/wQun6surcqo+Lr61zVf8ABC6fqy6IpLob+0pp/wDgtav1RqpvUI0N/aU0/wDwWtX6o1U3oiUpSiJXWuU+Pa7dKuctXKxDZW+4o+CEJKifyCuzVRcXOZfMDwzak5Mlzkdax6VGZO+306QnsG9vfzOpoi83vJiwH864wrxnb4KzCtN2vC1HwclPoQPyh5dev1eafkcsPCU6lZ440QAbfZY69u/YOPOAf+ZqvSyil2qUpSihKUpREpSlESlKURKUpREpSlEStc+Mfg8xbiixAPRjHtWc2dpXoa7lHRQ7/NpG3VTKj+NBPMn7IK2MpRF488KPFVqDwV6iztEdbrZcWMTbmlm4W95JW9ZJCj/nMfb4bKtwpSU7hQPOjruFevFjvlnyazwshx65xrjbLiwiTElxnA4080obpWlQ6EEGteuMvg0xfigxb0jbvN7TntoZULTdinZLyRufNZO3VTRPcepQTuOhUlWhvCnxX6h8FmoE3RHW+03JrE2ZpZn294Fb9jeUd/OI4G/OyrfmUlO4UDzo67hRZar2FpWPsGQWTKrJByTG7rFudruTCJMSXGcDjTzShulSVDoQRWQosUpSlESlKURKUpREpSlESlKURKUpREpSlESlKURKUpREpSlESlKURKUpREpSlESlKURKUpREpSlESlKURKUpREpSlESlKURKUpREpSlESlKURKUpRF1rjDRcbfKt7i1ITKZWypSe8BSSNx+WvLK4a023QnhO1K4ItZbVOx7NLTCuUPHpCoi1RL7HffU60424kEAnnPVWydtuoUCkXfd+Md7hy4qtVMU4jL/kMfE7i3AmYWEwDIitNdiO17PkTzHmUevUgKQoHY98W088olw15vplb4PFaxb73lTUmWX2FYn5zGbaLy+wKAUqSCWuz32O++9FIWF1J4k8W4geHfDeDnQKJNy7Msns1ktF2eahuIiWdphDCpDjjiwN+VTWxUN0gcx5t9gfRuw2pFisVusjbpcTb4jMVKz3qDaAnf8AHtWm2JeUM4BMEaXGweP6AbfIDiLZiSooc9nN2aBv+Ot1I77clhuS0SUOoC07jboRuKIVw3O526y26VeLvOYhQYTK5EmTIcDbTLSAVKWtR6JSACST0AFV59E7w5HqNdsC/OCL/jqe5BYLPlViuOM5DBRNtd2iuwpsZZIS8w4kpWg7EHYpJHQ+Na255wgcBGl2Jzs3z3SfF7LY7YgKky5D8nlRuQlKQA4SpRJACUgkkgAUUK1foneHL/Xrgf5wRf8AHWbxHWbSTProqx4PqZjF/uKGVSFRbbdWZLoaSQCsoQonlBUkb+8VoCrULyO6eYjBYpSk8vMLPdNv01cXCTmXAfe9XHrbww6fvwMl9CyHpFyRAksNJhhxoKbKnl9SpRbIAT9ieo8SmFudSlKKEpSovcNUtM7Tfk4tddRMZh3lSggW6Rdo7ckqPcOyUsK3Ps2oilFVHxc/Wuar/ghdP1ZdW5VR8XX1rmq/4IXT9WXRFJdDf2lNP/wWtX6o1U3qEaG/tKaf/gtav1RqpvREpSlEStFvK2akt49ohYdNo0gJmZfeUvPICuvmkQdorcewuqYH5a3eu11tlitcu93mexCgQGFyZUl9YQ2y0hJUpalHoAACSa8Z9Ycvv/lCeMC2Y3hKJCbA46mz2lakH/k1qaWVyZzg+xKgVr2PX6knvopC378mlp6vBOFLH58pns5eWypOQOgp2VyOqCGd/jZabI/fVtRWPx+xWzF7DbcasscR7faYjMGI0O5tlpAQhP4kpFZCihKUpREpSlESlKURKUpREpSlESlKURKUpRErXLjD4NsQ4osX86YMez5zamVC0Xnk6LHf5tJ26rZUfHvQTzJ+ySrY2lEXjlw0cUOqfAvqRO0Z1msdyOKtS+S52hz13rWtR/zyGe5bah6xSk8qx6ydlb7+u2J5ZjWdY5b8vw+9RbvZrqwmRDmxXOdt5s+IPgfAg9QQQQCCKpziy4RcF4pMREW5Bu05ZbG1ehb821u4wT17F4Dq4wo96e8H1k7Hffze0Y101y8nlqxO0y1IsUuRjjj4cudjLnM24hR2E+3uH1SSB4bJXtyq5VDdJZar2bpUW001NwjV/DYGe6e36PdrNcUbtvNHZSFj4TbiT1Q4k9FJVsQalNFilKUoiUpSiJSlKIlKUoiUpSiJSlKIlKUoiUpSiJSlKIlKUoiUpSiJSlKIlKUoiUpSiJSlKIlKUoiUpSiJSlKIlKUoiUpSiJSlKIlKUoiUpSiLQ/JeK+0aLcUuqeD8U1wnHEZTcGZhKJFnTLjMNlkB0tlLZX6yj1O5AUhYOxqveETjE4RdKtC7RhWqMplOQxZtxefCseclHs3ZbrjX0wNkH6WpPTfp3eFekN3xywZAlCL5ZIFwDe/Z+dRkO8m/fy8wO1aJ3ziG4SeD+2zNEcH07vep03EnJUi7PtW+NKFvU46pxxEiWpCUjkUsp2CSE7BJO4Iop1UxHlCOAMkAS4+5IA/5pO9/8VW5Ed1p+O0+x9TcQlaOm3qkbjpWskviN4arDm+n2DZpp9AsEjUjHoN8tU+TaI6oLbkrfkiOvBPqub7Dm25fWG5TuK2eACQEpAAA2AHhRQvtaheUkiW+fgul8LKXOzxCRqTZ2shUpXK2Iig6D2h8E9TufCtvaiWq+n+A6oafXnCtTrfGl43Ojkzg+72SWko9cOhzcdmpBSFBe4223oi1c1pxHT7TzjJ0BvELEsfi47l1uu+JyYzcBlMR09mlyMOQJ5CedaQk7e6szhlqxzQ/jylaaacQ4ttx7UjC15DdLNEQlDEK5RpBQmQ22no0HGyoFI2BI32qps+yryc0vRazcMl04gJi4+KS1ybLf4wfly7fJLrigtMltktrSA4UbD1SkJ7iARdXBnw6aPYOu461YZrDO1YvWQxxAVk02YmQWo6SklhIBUpCt0o5uclXqpGyR0JStpqUpRQqL42tX7vofw05hnOOSTGvfYN262PDvZkSHEtBwf7yEqUse9IqsNLfJ4cPdz0Ktls1Dw1N5y7IbW3Pu+SSHnFXFM59sLUtt0q9XkUrYDuPL6wVud+byqMGTL4R7pJYQpSIF7tkl/Ydzfa8m5/4lpraTC5ke4YdYrhEUlTEm2RXmlJ7ihTSSCPxGilaz+Ts1Cyu+acZXpJnV1fud60oySTjXnj6ipx6IgnsOYnqSnlcSN/sUpHhVrcXAKuF7VZIG5OI3QD+Trqh/J9EXDVPiayKL60GdqC40wsfBUUOSSrb8Tify1e/Fygr4X9UwHltbYnclFSdt9gwokdQeh22PuNE3rH8L2tekmfaUYZYsP1EsNzultsFvhS4DUxAlMvNxkIWlTKiFjZSSN9tunQ1dleHWk/ATxDapYVdM+s1hcsLtpQzJgRbwxIiPXVhxouJdhuBBC+gAHduVJ69akWGveU10/tiX8ZtusjFvjEIEeXCdmJR032DMhKzt7wNvCimF7R1gs1zrDdOMelZZneS2+xWiGkqelzn0tIHTuG/VSj4JG5PcAa8ml8R3lQ7yg2hi258l1Xq7x8GQ27/AObzbp8dRa48KnHJrnZbzqTqTasqmmxMPOtR8kfeVPlOISD2MOHsVbq3ABCUIPXr02oohTLi+418w4sL9H0L0Js91+ZWfLRHQwy0oTsifB9UKQOrbAI5gg9+3MvYDlG7nA7we27hiwpy65F5vNz7IWkG7y29lIiND1kw2VeKEnqpQ+Gsb9wTtSfCTpJl3D/wn5VrDZdCJR1raXKaiRLtbX3JkhlLqA22hncLQhSSokN8vNy7knarxtetXExJ4Q7hqzN0f7HVWO4tMfF/RUodokTUtJPm5X2p3ZJX0V4b91EPALZilazy9a+JlrhCj6ssaP8APqq44lLmLeipR5UmaWifN+ftfqOznwvf3V8yfWviatvCVYtVbLo953qlNcjpnYx6KlK7BKpC0LPYBfap2bCVdVeO/dRQtmaVrNqFrXxNWDhYxPU7EtHvSepd0chpu2N+ipTnmiXEuF49glYdTylKPhKO2/XvprFrXxNYjw44PqHp7o96c1AvXmXpyw+ipT3mPaRlre+koWHEcjqUp9YnbfY9aItmaVrNr/rXxNYHolgWZ6U6PfNJmN8bjKv1o9FSpHmClxC44OzbWFo5XvU9Ynbu76+cS+tnE1pxp1geQaM6PfNXf742FX6B6KlSfMFeboXtyNLCkfTCpPrE923fRFs1StauKXWniW0xsuCTdENIfmvmXwO+nmfRUqV5hyoZKejK0lG5W4PW3+D7jX74p9Z+JPTG84ND0P0j+a+Jey56de9FypXmAC2QnqytIRulbp9bf4HuNEWyVK1v4mNZeJHTjUfBce0Z0k+avH72tIvs/wBFyZPmAMhtBPO0sJR9LUtXrA/B37q/Wv2snEfgmtuBYZpVpL80mHX0xRf7v6MkyPR4XL7Nz6Y2sIRytev6wO3eelEWx1K1y1h1j4jcS4jsG080+0m9N6f3vzL07fvRcl7zDtJK0PfTkLDaOVtKFesDtvuelfnUbWTiQx7ikxTTLENJfSmmt0TEN2yP0XJc80LinQ6O3SsNJ5Qlv4STtzde+iLY+la4ZRrLxIWzi2s2lVk0k870umIYVNyf0XJV2ClMOLWO3C+yGziUJ6p6c23fX6l6x8RrXF+xpKxpNz6UraSteU+i5J5VmEXSPOOfsfq4CPg+O3fRFsbStcbVrHxHSuL2bpNO0l7HSplpxTGUejJI7RQhpdSPOCvsju8VI+D4bd9fnDdZeJG8cWF/0syLSTzHTCAiSbfk3ouSjzgobbU2O3UvslbqUsdE9eXpRFshStcNMdZOI/JeJ7MNNc10l9E6cWkTTZ8i9FyWvPC240ln6ctZaXzJUs+qkb8vTur9aKax8R2ZcQmd4DqRpN6BwSx+e+gb56LksekOzlIbZ+muLLa+ZoqX6oG+246URbG1UvEfwz6ccTOFKxbN4ZYnRgpdpvMdA86tzxHwkE/CQdhzNn1VD2EAiE8POsvEjn+sWeYjq1pJ8zOJ2NUgWG6+i5Mf0gEyy2j6Y6soXu0Av1QN99+6nDBrLxI6l51nNk1r0k+ZKy2RYFimei5MXz8du6jfmeWpK/UQ2r1QPhb9xoi86bXd+JDyaWsq7fOY87styXzOxypforIYqTt2jSv+qfSD37c6CdlBST19V9AOInTXiQwtvMNPrrzLa5W7jbHyEzLc8R9TeRv8fKsbpUOoJ67UTphJ1U4vcU1F0+4udA2bHZreGTYFG1SobzzpL47Zlx1aiHEcjRCkbfDIO4VtXm3jmO8VPCpdI+s0DCswwpEGSmGudPtrjUZ8K6hiQhQAW2vb7Ibb7bEK2ostV700rSPhJ49Mi18xrObblbGMQs/tUN6ZjVjgNPpVdEojqVslK3FF1XagAoQQrY77eNWHoTrXxM5xoNnec6oaPfM7m1jRLNhsvoqUx5+UREuNfSnFla+Z0lHqkb7bDrRYwtmaVrPpRrXxM5Twz5pqPnej3obUSzqnCy4/6KlNeehthtbP0lay4vncUtPqkb8uw6ivmC62cTV74VMn1QyfR70dqbbXJabXjXoqUjzpKCjsj2Cll1XMFL7lDfbp3URbM0rWew618TM7hFu+q900f821TiOOpiYv6KlJ7ZKZaW0HsCvtTu0VL6K8N+6g1r4mPoQfnsnR/wD/AIq9py/Mt6KlfB8+7Lfzfn7X6h9M+F7+7pRFsxStZr/rXxNQeEa0ar2rR7znVOW40mZi/oqUrsUqlKbWewC+1GzQSvqrx37q+ZzrZxN2ThTxjVHGdHvSGptyciJumNeipS/NUrLgdPYJWHU8oSjvUdt+vfRFs1StZtVta+JrFuGfC9R8F0e9M6iXhUIXrHvRUp3zIOMOLe+koWHEci0oT6yjtzbHvprrrZxNYRoNgec6YaPfNFm18REN+svoqU/5gVxFOO/Sm1haOV0BHrE7b7HrRFszStZuJDWzia080qwPJ9HtHvmpya+Ntqv1t9FSpHmCjFDihyNLCkbOko9Yn2d9frih1q4l9M8UwO6aJ6QfNdc74lw36N6KlSvMCGmlJ9VlaSjdS3B62/wfcaItl6VrZxUaz8SmmD2DI0O0i+bBF77X06fRcqV5hylnl+orTybhbvwt/ge41ycT+svEhppnWDWTRTST5rbLe1lN9m+i5MrzBPbtI35mVpSj1FuK9YH4O/cDRFsfStcOIbWTiPwDWHAsR0l0l+abE76pgX+6+i5Mj0eFSktrPaNLCEbNEr9YHu37q/WtesfEbhvEHgeA6b6TensEvnmXp6+ejJL/AKP7SUtt76a2sNo5WglfrA7b7npRFsbStcdTtZOI/GuJ7D9NcK0l9LacXcQjeci9FyXfMy446l76chYaRypSg+sk7c3Xvr85nrLxIWfiwx/SzHdJPPtMJ6IxuGTei5K/Nytt1Tg7dKw0nlUlsdU9Obr3iiLZCla43XWPiOi8XsLSaDpL22lTzTan8o9FyT2ajDU4oecBfZDZ4JR1T47d9IusfEc7xfvaSv6S8mlKG1KRlPoySOZQgh0Dzjn7L6vu38H3d9EWx1K1wxfWXiQufFtedKr3pJ5ppbDQ+qFk/ouSnt1JYbWgduV9kd3FLT0T15du+vmnWsvEjkHFLlWmWX6Sei9NbWmYbVkfouS352W1NBodupZaVzBTnwUjfl6dxoi2QpWuWj+sfEdlvEdnOnmoOk3oTT+yefegr96Lks+f9nJQhn6ctZbXzNqUr1QN9tx0r86A6ycR+d63Z7heqmkvzN4dYzK9AXf0XJj+kAiX2bf0xxZQvma9f1QN+8dKItj6Vrfw0ay8SOo+pGdY7rNpJ8ymP2RahYbh6LkxvPwJLiAed1akr+lpQr1QPhb91fOFjWbiS1OvWcw9cNI/mQh2RTYsT3ouTF8/BW8FHd5agvZKGj6u3w/eKItkaVrVws60cSup1mzqZrhpD8yEuyBr0E16KlRfP+ZDxV0eWor2KGx6u3wveK4+GjWviZ1H06zzINZtHvmUv9jQVWGB6KlRvPz5ute3I6sqX9MCU+qR37d9EWzNK1m4fta+JnPdE88zPVfR75msxsbclVhtHoqVH8/UiIHGx2biytfM96nqkb9w600d1r4mcu4cM31D1B0e9B6gWXz70HYPRUpnz7s4yFs/SVrLi+dxSk+qRvtsOtEWzNK1m081r4mr/wALGWanZbo96M1LtbkxNpxz0VKb87S2lssnsFLLquYqX8FQ35endTGNbOJq5cJV91VvWj3mmqMJx9MDGPRUpPbpTIQhB7Ar7VW7ZUrorw37qItmaVrPE1r4mXeEKRqy/o/yaqtuqS3i3oqUOZImhoHzfn7X6huv4Xv7ulLprXxMxuEOBqxC0f7bVR9xCZGL+ipR7NJmKaUfNwvtRsyAvqrx37qItmKVrLl2tnE3aeE7HdU7Bo757qhcHY6bjjPoqUvzdK3VpcPYJX2qeVKUnqrx99fdTNa+JnG+F/D9S8M0e9LakXZcJN4x30VKc8zDjTinj2KFh1HKpKB6yjtzde+iLZmlaza1a2cTWG8POC5/pvo96ezy9iF6dsXoqU95gXIi3HvpTaw4jldCUesTtvsetOInWvia0/0fwLLdI9Hvmnyu+NsKv1q9FSpHmClRO0WOzaWFo2d9T1ifZ30RbM0rWfic1q4mNNsGwS96LaP/ADW3q+IJv0P0VKk+YHsG1j1WlpUj11LT6xPdt4VycU+tHEpphFwV3Q7SH5sHL323p1PouVK8w5Qzy9GVp5N+dz4W/wAD3GiLZSla38UusvEjpllODWzRDST5r7be1rF9k+i5MrzAB1lIPMytIRuhbh9bf4HuNbEIdfM91hTezCWW1pXseqipYUN/cAn8tEXYpSlESlKURda5pmLtstFuUEy1MOBgnuDnKeX+navNXSm44UnyZ2sOPQlR2c7gRr182Ud0gTjN84Vs48D65TycqQT03Ch3g16Z1rPrp5PrQHXTJ5ecXCPeMayG4p2nzrDKSwJp7ip5paVIWo7DdQAJ8SaKQqj4wbtgSPJ9YVjV+VFfyy62LHG8SiAhUwz+zj/TGU/CACOcKUOnrBPeoCt4cZauTGN2pi8KKp7cFhEpRO5LwbSFn/zb1rrov5PTQXRvLIWeld/y7IbZy+j5eQzEyEQlJ+CpppKUoCk/Ykg8venY9a2dooStUPKMuyZeleG4lcL3JsuIZTnNps+WXFhZR2FscUsqC1dyUFaUbk9NwN+/atr6qfiZ0PufELpi9prA1ClYixMlNuTpDEFEsSoyUrCo621qSOUlSVb77goG1EXYxrhk4dcZx5jHbJoxhhtzbQQntrQxIW6nb4S3XEqW4T+6USTWvmluD43oZ5QG7ab6LN+Z4pk+EG+5JYoyyqLa5yHwlhxKdyGytJ6J9jh29XYDEY9wW6lYXZm8XxvyiGW2u2xk9k1DaSgIYSPsUBUolAHsTttWNxDyeOW2K73W44Nx1ZTEut7UH7pItzA85mlPcp5aZXOvbm+yJ23opW/lKqjh40czXRnHbpZc31ryDUqVPnCWzOvKVByK32aU9inmcX6u6SrvHVR6Va9FCiWrOmuP6xab5DpjlAX6MyKCuG8tvbnaJ6ocTv05kLCVj3pFan4ziflH9LcEb0NxSFp1f7dbmPRlmzaXcXGpESGBytlcc9VLbTsE+qrbYA8+253dpRFUXC3w/W7ht0nh4CzdTdrpIkO3O93NSSkzZ723aLAPUJACUp367JBPUmvvFz9a5qv+CF0/Vl1blVHxc/Wuar/ghdP1ZdEUl0N/aU0//Ba1fqjVTeoRob+0pp/+C1q/VGqm9ESlKURKUpREpSlESlKURKUpREpSlESlKURKUpREpSlESlKURKUpREpSlESulerLaMjtMuw3+2RbjbZ7SmJUSU0l1p5tQ2KVpVuCD7DXdpRF5ZcWfk3Mn03nvatcMXpGVbITvnzlijPL9I2paTzdpCWDzuoSeoRv2ifArHdJuEnyn6f+SadcTcjsnUERouWhrlBIOwTObA9RXgXkjb92kdVV6T1qVxY+T1054gvO8yw5UfEc8cBWqa01/wAjuS9u6U0n7I/9qn1/aF9BRTPFbWwLhAusJi52ubHmQ5TaXmJEdxLjbqFDdKkqSSFAjqCOldivGHANbOKnyeWajT/NbFJfx5ThcNhuLilwJTe/rPW+SNw2T/u7p3+Gjfu9NeHfi50a4lLYleEX0RL602FzMfuBS1Oj+0hO+zqB+7QVDu32PSiEQrppSlFCUpSiJSlKIlKUoiUpSiJSlKIlKUoiUpSiJSlKIlKUoiUpSiJSlKIlKUoiUpSiJSlKIlKUoiUpSiJSlKIlKUoiUpSiJSlKIlYeFmGKXLIZ+I27JLZKvdrabfnW5mUhciK2skIU62DzICtjtuBvXeur8iLa5kqG12j7MdxxpG2/MsJJA/Gdq81dI7xh+kfAFqDxOsTQ/qbn3pK23S6vSFKkmc/LW02wATskoCu27ubvJO22xFu1jvFjw45ZnytL8d1fx+bkwfVGRCbeUA68kkFtt0gNuL3BHKhRJ26VbVeZepvDlpho/wCTesmX3LHrdbNR4jFsvsC9IQG7iq6yJDbiWQ78NQS0op7PfYdnzbAjevSHGJU+bjVpm3VHJNkQWHZKdttnVNpKxt4dSaKVk61O8o/ccqRpBi2L2TI5WPWfLMztliyG7R1lBjQHysHmUCNkFfJv1AOwSeiiDtjWrXlEcwyO06K2vTvFbXa5c/U/IoeIBy5sJdjx0yOYlZCgUhW6EhKiDy7lQ6pFFAWBY8lRwmtsobegZZJcSkBTzl9cCln2kJSBufcAKsfQzgj0J4d8yezvTa33pi6vwXLctUy5rkI7Fa0KUOVXTfdtPWqbxrgS4m8esUKzQuPPM4TMRlLaI0eI6tpkAbciCuRzco7hvt8Qq29BOHbWvSrNnsl1C4p8m1FtjkB2Im0XKKW2kOqWhSXgS6v1khKh3fZnrRSthaUpRQlKUoiVUfFz9a5qv+CF0/Vl1blVHxc/Wuar/ghdP1ZdEUl0N/aU0/8AwWtX6o1U3qEaG/tKaf8A4LWr9Uaqb0RKUpREpSlESlKURKUpREpSlESlKURKUpREpSlESlKURKUpREpSlESlKURKUpREpSlEUW1I0uwDV7F5GG6kYrBvtpkdSzKb3La/BbaxsptY8FJII9teZ/EF5MXUzS65nULhovdxv0OC550zb0yOxvUBQ6gsOpKQ/t4bcrnhss9a9WaUUgwvKnQTyo+pGnExOC8SOOTcgiwV+bPXJtgR7zDKehD7KuVLxHjvyL8SVGvRjSTXfSXXOyi+aX5vbr22lIU/HbXySoxPg8wrZxs+HrJ2PgTUe114UND+IiGRqJiDSrohHIxeoJ83uDHs2dSPXA/crCk+6vPHVfyafENonePm50AyiVlDEBRejuW182+9xgOvRKVBLv8A4agT+4opsV620ryY0m8qBrvpPc/mM19xFzKG4KgzIMlj0beo23T1wUhDhHsUhJP7qt9NFeNTh212DMPEs7jwLy6B/wCxbxtDm83sSlZ5Xf8Aw1KooIhXnSlKKEpSlESlKURKUpREpSlESlKURKUpREpSlESlKURKUpREpSlESlKURKUpREpSlESlKURKUpREpSlESlKURKUpRErTFfBnwS6g675Aq2ZDJl3mxzEXzIMOgXj/ANmMSFrPrvMhJ7PmUFczYWPYQAdq3Eujkxq2S3bejnlIYcUwnbfmcCTyj8u1eYumOZ6caQ+TtzjVW2S7enVHM3LlZbxMcWlVycnvylt9kdzzgIaUXuXu71d53opVr5PnfAdxKcUOMKyHVaddLpjSW7bZ7E52rNgmzm3lKQsKUjkcXuoJACkpXypHrjYHe2vN3WrRzTPRjyalitF+s1qhZ0uNbbla5AZQi4OXp99t1YQrbtCUtrWggdAlA9gr0Pxddycxq0uXkKFwVBYMvm7+27NPPv8A8W9EKydUHxr6Xag6w6LKwnTTGbJd7y9dY0lC7nNXENv7ILUmXHdQQUvoWEcu+4IUoEEGr8rXrja1nz7SDTGzxNJ22Pm0zbIYeMWZ19tK0R3X+Yl3lV6pICdhzAgFW5B22JQqhsLnlZbLao9slW7S27rjoDfnc58du4ANt1ltSElXtISN6trQC4ccUrOHm+IuxYDDxYW90tOWJ1SpBmc6OQEFZ9Tl7Tfp37VrJfMP468f11xPQy9cYzzVwzKyTLtCuLVrbVGD8Y7uxeUoCj6vrBfTp9jVj8J0LiCxribynT3iR1vvN9vdlsAm2a17A226QH3UJM9pQ5Ppja2+zKFN7jtD12opW7tKUooSlKURKqPi6+tc1X/BC6fqy6tyqj4uvrXNV/wQun6suiKS6G/tKaf/AILWr9Uaqb1CNDf2lNP/AMFrV+qNVN6IlKUoiUpSiJSlKIlKUoiUpSiJSlKIlKUoiUpSiJSlKIlKUoiUpSiJSlKIlKUoiUpSiJSlKIlKUoigGq+gukGt9s9GaoYFa74EpKWZDrXJKY+9vo2cR+JQHtrRDWjyRDiS9d9BM+CgndaLNkXeCOoDcttP5OdHxqr0upRSCQvGiDrNx88E0tqz5izfRYo6g23FyJk3K1uJH2LMtKjyD2BDo/e1tDpB5W3SvI0x7drDiFyxCYvZK58HefAJ8VEJAebHu5F/HW982DCuUR2BcYbEqM+kodZfbC23EnvCknoR7jWsOrvk3uGTVMyJ9uxh7C7u9urzzHVhhsrPiqMQWSP3qUn30SQdVfOn2rOmeq1sF303zqy5FFKQpRgS0OLb9y0A8yD7lAGpZXknqB5LniN0uuZybRPMouS+aqK2HIUpVouqAOo23VyE/E6N/ZWEsfHJxw8Ndxax3Vu2Trmw0eXzTMbWtt5Y/wDdy0BKl+5RLg+OiRwXsPStFdMPK16K5IGYep2J3zDZa9kqkMp9Iwt/E8zYDqR/4Z+Ots9O9b9IdWYyZWnGo9gyAKG/ZQ5qFPpH+80SHE/8SRRREKb0pSiJSlKIlKUoiUpSiJSlKIlKUoiUpSiJSlKIlKUoiUpSiJSlKIlKUoiUpSiJSlKIlKUoiUpSiJXnxxEx/JoaQa3PZZqDYn7lmzcoXK4WOxh6THTJ35+1kMBSWErJ2UUFQ333Uk79d/rmuW1bZbtvbC5SGHFMJP2TgSeUfl2rzR0xx/BJ3k2NXdQ0x4k/UC8R70vLLjIQlc9E3zhWzS1H10J5ORQT0BKie80UhbBZm7wbam8SWmOUakyZBzm7YvBu2JW+6qU3AdZddW5H50DdBlBRVshatjsnYKIFba1oTxg2HT93yfuFZZfY8RnLrTYcb+ZOclITNE4tx/pTSx6xBR2iikdPV5u9INbyYy5cnsctT15SU3BcFhUsEbEPFsc//q3ooWSrX/jT0jznVHTG03XS1liRmeB5DCyuyxXlBKZbsYq5mNyQAVJUdtyASkDcb7jYCqB4zdZM50l03s9u0sajnNs7yCHitiekpCmor8jm3fIIIPKlJ23BG6gSCBsSKnNdrjrZlWJ6J8YNq0TvEPKdPbnNcyDDN1LnCC/uw9yDkCiNmuYDlJCXQrYgE1zaA5bqBxP8WjXEK9pbfcIwvD8SkY9DN5ZLT9wkvvBagAQAQnqTy7hPKnc7q2Gyel+neoOLaMM4Bnuqc3JcpXDltScm7IoeDr5WpK0AqO/Zc4CT03CE9E9wozh6yjWrRriDl8KWtGoD2fwLjj6sjxPJJbfLNWy272brEg7kqI6ndRURy95CgElK27pSlFCUpSiJVR8XP1rmq/4IXT9WXVuVUfF19a5qv+CF0/Vl0RSXQ39pTT/8FrV+qNVN6hGhv7Smn/4LWr9Uaqb0RKUpREpSlESlKURKUpREpSlESlKURKUpREpSlESlKURKUpREpSlESlKURKUpREpSlESlKURKUpREpSlESlKURK6F7sFiyW3O2jI7LBusF4bORZsdD7Sx70LBB/JXfpRFqrqf5NPhb1EL0u1YvLwy4Obnt8fk9i1zHxMdYU1t7kpTWpuofkkdYMXkqvGkOo1oyHsDzMNTAu1zk+zlcSVtk+/mRXq5SimSvG9OqflIeFUhnJRmZtMU8m17h+mbfyjwEkc/KPicTVs6c+WGuTYbi6r6RR5QBCXJuOzeQj2nsH9xv7u1FemxAUCCAQehBqp9RuFDhz1WK3s30hx6XKc33mx43mkrc+PbM8iyfjJokhQDT3yjHCdqB2bKtRDjUxwgCNkMVcPYn/3vVn/11sLYMnxvKoKbni+QW28Q1/BkQJbchs/8SCRWjeoPkh9Irz2kjTfULIsZdVuUx5qUXGMD4Ab8jgHxrNa937yZ3F3pTON50qyW23dbRK0P2G8uWyZ0/wB1woG/uDhokBevdK8c08SnlHuHhQZzhnLHITJ2PzT2Hz1hQH/5pKeYj3h2rPwTyxN+a5WNSNHIE0DYLk2G5KZUPaexeCh+LtBRIK9PaVqPhHlQ+FTKy2zer5fMTfWBum8WtZbB9naMdonb3natgML1y0a1FbbXg2qWL3tTnwWod0ZW7+Nvm5x+MUUKcUpSiJSlKIlKUoiUpSiJSlKIlKUoiUpSiJSlKIlKUoiUpSiJSlKIlKUoi4pUliFGemSXAhlhtTrij9ilI3J/IK8+7TwxaNcbNkyDXHQTN820uYyybKtuQ2/sEmDdnUEFxa46XQFJUVAn1uUq39UHff0GeQFtLQUJXzJI5VDcHp3H3V53aaaHeUX0XuF8b0ji6ZWTG7zcXLj8zyp65MCM8vopTAcSXWgoAbpDnL07ulFIVraVeTsxzGMtsOZ6watZPqjKxVLQsUC7Et2+B2QAa5WSte4Ryp5U8wT6o3SdhW31aVPTvKqRmy/KZ0XZZR1WtbriQB49Sdq3Qil5UZkyCkultJc5Pg823Xb3b0QrlqkuLnQi969aWtWfDb23Z8vxu6xsixyc6SENz45JSlZAJCVBShvsdjynYgEVdtUlxc68XjQPStF6xCyovGXZFc42PY5BcG6HJ8gkIUsAglKQlR23G52G433BQqSicWXG7i0VOO5rwO3i932OkNLuNmnnzGQsdO0HIhxKQe/ovbr4VLeG/SjXTLtZrpxUcSttgWC+PWcWDGsYhu9qLVBUsOOLcWCRzqPhuT6yyeX1UiIQOEHjQyiEMjznjlyKzZDJT2y7fZoqvMYyyN+zHI60lQHd0QB8ffUs4ZtVNc8R1ivPCrxK3WHf79EtIv8AjWTRmw36VgBYbWlYAAK0k9+wPqrBKtkqJStsKUpRQlKUoiVUfFz9a5qv+CF0/Vl1blVHxdfWuar/AIIXT9WXRFJdDf2lNP8A8FrV+qNVN6hGhv7Smn/4LWr9Uaqb0RKUpREpSlESlKURKUpREpSlESlKURKUpREpSlESlKURKUpREpUI1o1fxTQjTm6aoZs1cHLNaCyJCYDAefPaupbTyoKkg+ssb9e7es/h2U2zOMSsuaWVL6bffrfHuUUPoCHAy82lxHMkE7K5VDcbnrRFmKUpREpSlESlKURKUqtdE9f8F17YyWTg7F1bRil6esU/z+MlkmS18It7KVzI69D0+KiKyqUpREpStdNcuOTS3RDPk6XO4zl+X5S3FRMl27GrYJS4jShzJLm6k+sU7K2TvsCCdtxRFsXSsdjl5byPH7ZkLUKVDRdIbM1MeW3yPshxAWEOJ3PKsc2xG52INZGiJSlRiHqZgtw1Bm6WW/JYknKrZb03OdbWVFbkWOpSUoU7t0QVFQISTzEddtutEUnpSlESlKURKUpRF8UlK0lKkgpI2II6EVWOd8MPD3qXzqzXR7Fri85uVSRb0MyCfb2zXK5/6qs+lEWlmb+Sf4bsiDr2JXLKsTkL6oTGniWwn/gfSpW3xLFUBmnkfNR7e4uVp7qzj93DfrNN3SE7Be9w52y6nf39K9UqUUyV46/OA8pdoP1xN7N1w2j6osGQC4MED/8ALlZO3uLddiH5Q/jh0lkpg6kWZqYGfVU1k2MOQnDt7VtBrr79jXsFXBMhQ7hHXEnxGZLDg2W082FoUPeD0NElebWHeWNCkIbz3REqP2ciyXcKHxhp5A/r1d+I+VM4VMjLbd5umRYw6rbm9J2ha0JP7+OXBt7+lXBmPCPwzZ6pTmT6I4m86v4T0eAmI6ffzscit/x1SOXeSl4YL/2juPuZXjLq+qRCunbtJP7yQlw7f8QollfOIcUvDnngSMV1rxCY4v4LK7o0w6f/AA3Slf8ARVmRZcWcwmTCktSGVjdLjSwtKviI6GvM/L/I43EBS8G1uiyP3LN6sxT+IuNLP9SqzleTr44NL5Cpenl6jSuy6pcx3KHISzt7Eudl+TeiQF7BUrx4Gf8AlSdHd13JrUt6Ox0UqdaUXlnYe1zkd6f8VZKy+Vb4m8QdELOsNxW5rQdlpmW6Rbn/AP0r2B/4KJC9daV5xYt5YywPJbRmeh9xjn7N603duQn8SHUN/wBarexjyp/CnfShu7XDJ8eWr4XpGzLWhJ/fMFyiQVt/SqbxfjG4W8xUlFj11xErX3Ny56Ya/wDyv8hq0rNkuOZEyJGP3+23NojcOQ5TbySPjQSKKFkqUpREpSlESlKURKUpREpSlESlKURa/wDES/xnwMhgvcOETBLpYZENQnMX5K25EaQk96VhwBaFJI6bbgpPeCNtUeFmT5RW0aMWuBpTh2CKxxEyeqOciU61P7QynC7zpUtJCe0K+Xp8Har14h+JPXOdrQjhk4T8YtU/MIlvRc79ert1iWplexQkgnlB2Ugknm+qJSlJO+2V4yeI7UTRXD8KwPTiHDnaqaiS2rVbgUB1qO4AhLrwQdgr6Y4hKOYcvrFRBCSCUqoNVNJvKQcR2JnSnUyFphYsaucyO5cJkCQvtkIbcC+gClk7EA7AAnYDcAmt+bXBTa7ZEtiHVOpiMNsBa/hKCEhO5952rQy+51xp8GUrHdQtedSbPqXp7eLixbsgRHiJakWdbx6OtqDaCUjrt9iduUpSVJVW/LD7MlhuTHcS406kLQtJ3Ckkbgj8VEK5K1443tJ871K0utN+0rjol5lp9kMPLLRDXttMXG5uZgbkAqKVEgbjcp28a2HqqeJfX208OemT2dTbPIvVxlS2bVZbRHOzlwnvbhpoEAkDookgE7JOwJIBKFr/AGzyqOj8C3phakabahYzlDCezm2g2cOFDw70oUpaCRv3cyUn3V3OHJWo3EhxJTOLPK8DuOGYhZcdXjOIQLmjkmTUuu9o7JWnYer8Ib/B9ZISVcpNYpETypmZoay8I0mxXtEh5ixS4/bPNJPUIW5yuEK9v0wdfZVncNfEtn2bZzfNBuIDA4+Iam47DRcezhu88G6wSoJ84jklXcSncBSh17wQpKSlbI0pSihKUpREqo+Ln61zVf8ABC6fqy6tyqj4uvrXNV/wQun6suiKS6G/tKaf/gtav1RqpvUI0N/aU0//AAWtX6o1U3oiUpSiJSlKIlKUoiUpSiJSlKIlKUoiUpSiJSlKIlKUoiUpSiLWHylH1m2dfvrd+vMVbXDl9b9pp+CNo/VG6qXylH1m2dfvrd+vMVbXDn04ftNPwRtH6o3RFLstyqxYNi91zLJ56IVossN2dNkL7m2W0lSjt4nYdAOpOwFaj4lxGca+vFpd1I0H0Swi2YM4456IOW3F5E+7toUU86A0oJbBIIG/q777KUBvVleUBh3Sdwe6ls2hC1uotrLziUAklhElpb3d4dmlZPuBqlOHzRXiby/RDCMh0642F2zHZVki+Y29nEIjqYSEoCTH5yrdRbUFIJPUlJ3opVw6QcU+Ra06aZ4iz4K3YdWtP2341zxS5uKW2JyELLQStOylNOKbUkEdQfaNiqV8Jevv0SWilq1Kl26Pbbst+RAu0COpRRFltLIKBzetsUFtQ36+vVMcJel9qxDiX1OyC48S0DUnOXbe1b8ngMWTzFyK8hxvs3HChRbJAQUbJHfvudwarm26ixuBvW/iHw2aERrBfLG5qHiTKxs2uYrdCo6AenV1fLt+5Yoi2I024n7pqDrjq3i4tdphab6UsJZmZAtbhednJTzPJPXkDbYbf3IBPqp9tV1i3E3xc8Q6Z+ZcM+j2Hw8BiyXY9uueZTX25F4LZIUpptojkSSNuu4B3HNuCBHcD0byTA/JnZsVsvuZhnGO3LK7urb6c45Jb7TlPiSI4SCPaVVhOEzR/iPzfh5wu/6X8ZJx7H1wVNMWdnFIkgW5aXFhxguKVzKUF8x3VsTvv4iiLZDhm4nJes9wybTnUHC3ML1Lwh1LV9sanu2aU2r4EiO59k2Tt7duZJ3UFA1Uvky/2K1r/wDiVcf0CvvDxpO1i3GHk2R5XxTwtQ9Ro2OeYX20NWIQn245MctOOKbUW/UAbG22/rDfqK+eTL/YrWv/AOJVx/QKIt0H32IrDkmS8hpllBcccWoBKEgbkknuAFab2Xis4mOIi8XqZwl6U4qvCLHMXATkuXy3mkXN5HwhHaaIITsQevNsCnmKSeWtl9boVzuWjGe2+ypWq4ScZujMVKBuouqiuBIHvJI2qj/JnXSwXDg8w6NZFtdtbnZ8W4oTtzIledOLIWPaULbV18CKIu1oXxXZrnOY5PoPqxp1Fw3V3HoDk6NATKLluuzfL6rjDnUhO6kEjdXqkkHooJ1k0kv/ABXI44tUrladO9P388ftFtbyKC9dnxBhxNo+yozu3MtZSEEpOw3+Krb1MkRb15UrSuFjbiFz7HiE1d8LXe00tuUUJcI7ujiNt/8AtE+0V2dDv+kt17/Bu1/1ItEW6VaoZnxMa853rXlOiXC3p/i1wdwRLKMhv2UTHWozch0bpZaaa2WojZQ5uvVKugABO19aR5zw8YhrJrfm+ofCpxLXDBNS7S83Cy6Hb0rcjKkjdKQ+jdBBPZHm2507p35QSSShWFolxI6yXTV+58PnEBpXBx7Lo9rN2tl0sjzr9puLI6bBSgS2o9SN1deRYISQN9bdIL7xSJ49NS5sTAMFVl0q12tvJoSrq8IcS37xwHYzm3M47yBJ5VbDckVauluufFHpBxGYtw2cTUvHMtj5nFfdsuQ2hvsnwWkLVu6kJQCN2ylQKEkFQIUobiu1ox/0mOu34K2r+pFopX71U4ytaLLxEZdw2aTaS2vKMkjRYDmPKcfW2j6Yyh6S9MUVpQlptKtkgFO5Unr4GQ5RxY6m6JaHWXIdedMIY1UyW7vWWxYjYZJcE90L5W18+7nKkgpUeUr+G2B1VsIzpVHZc8qHrK+42lTjOFW7s1EdU8yYW+3x7Coh5QW1ZHK4n+Hl63Zn8x7UqTMhW+/uRESWrdcFONcqy24QhRO7Y9Y+O/hRFMsk1+479I8cOqurOhGCXDDoiRJu9ux66PG622N9k4StSm18oO6uXcdDuQNyLO1h4v8AC9O+Hi0a94pbnsnYytUSLjkFCwyZcqSDyNuKO/Z8vKvn6EgoKe81WWecP3FDbsIv8zUDj0DGMt22T6YclYZEDQhltQe5iF77FBUOnXr061H7XpjwvM8EGH6Qama6R7rid5vDqMXymNGciKM9Ul1TYYQoL2KSpxBC/VI599u8EWczHXvj40dxlWqWpuh+nl2xaIW3bnbMfuck3OEypQG+6ipCyCoA8oUPHoNyLL134tIemGk+GZvhuISshv8AqVIhwcXs0lRiqckSUBafOCRu2EcwCh38xA3A3UKMynS/jy4VsWumaYFxE27UbE8ahrmSLTlEQ+ciGynmUEqUVE8qEnol5Pd0HhUs1CyLQXjH4dNKpmseRrwG+Z7LS9irkd1RkMXhtZYWllRSQpBUdvX5d0qSdwQCCLizTiJ439BrZF1B1r0UwW94cqUyzcm8RnSXLhb0uKCUnlc5g51IHQEE7AlO+9bjWq5Rbza4d4glZjTo7clkrQUKKFpCk7pPUHYjoeorQnPLDx7cH2JXLUa267WXVLC8eSmRNg5HFUmYIwUATzklZ23G+zxPiEnurdPSTUCNqtphi2pUS3uQWsltUa5CK4rmUyXUBRRv47Ekb+IG9EUtpSlFCUpSiJSlKIlKUoiVjL1jON5IwY2RY/bboyRsW5sRt9O3xLBFZOlEVK5TwXcK2ZLW7etDMWS4vvcgxPMl7+3eOUGqiybyVHCze+0XZm8rx5xXwfMbuXUJ/wCGQlz9Nbj0okrzdyfyONrcSteH65zG/wBwzdbKh0fjW04j+rVW3jyUPEzirhl4TmmJ3JaeqDEuEm3vf0t7A/8AHXrpSimSvHk6N+VF0mATZZOojjDXcm2ZGi5NH4my6vf4uWvp4yvKMaXEM5lbr6ptn4Qv+GbA/G4htsn/AM1ewtKJK8oMf8sDrBbloj5dpbiNzKOizFkSILh/EouAH8VWrjnliNP5JQjLNGMjt+/w12+4MSwPiCw0a3kyHTHTbLgpOVafY3eOforz+1MPk/jWk1VWRcCHCPk5WufofYYzi/s7d2sIj3jsFpH9FEkKB455UfhLvhQi4ZBkNhUrv9I2R0hPxqZ7QVaeNcY/C1lqkt2XXbEStfciVPTEV/5X+Q1SuReSg4YLsFrssrMbC4r4Ii3YPIT/AML7az/TVWZJ5HG0uJWrEtdJrZ+waulkbeH41tuI/q0Sy9C7LluKZI2Hcdya03VB6hUKa2+D+NBNZavI+8+ST4iMfWqVh+d4dcVpO6OzkyYDx/8AlkA/8Vbv8B+jurWi2kFxxzWidJkX+TfX5LaXbsbgluJ2TSGwhZUeUEpWeXptv3UQhbI0pSihKUpRFpBpTk+PaUeUQ1ssOol2iWiTnNutdxsEuc6GW5TTbYCmkLVsN9+YAb9S0od4rEs3i0cSflMrVcMblsXnF9HMeWtyZGcDkcz1c49VY9UkOvpHTvMdXsraPXHhl0W4ioUSLqthzVzft4UIc5p1ceXHSTupKXWyFcpPXlO6d+u2/WshotoFpRw+449jGlWKM2iLKcD0t0uLekSnANgp11ZKlbDcAb7Dc7AbmimVrt5UXOLGxoCjSGNIal5Znd2gQ7VbGyFSFBEhK1O8g6hPMlKAfFSwB41ttilreseL2eyyV87tvgR4rit991NtpST+UVSOAcD+i2Dav3TXOUi75Jlk+4yLlFfvMoPM21bril7R2wkAcnMQlSuYpHdsetbB0UJWp/lHbhasV0kxPUx+8MQrvhOaW292ViTHcdYuEtoOK81c5ASjnQF7LI2BSAdt962wqvdeNWtO9EdNrjqLqaEuWm3qQluOGUuuypKtw2y0hXQrUd9uoAG5JABNEWvONeVY4W7tYWbjkEnJLDcS0FP2520uSFIc26pQ41uhY37junfxA7qoa0ce2hmS8XL/ABA5iq+WSxY5jK8Zx2G1bVPy5/avFx2TI5CUtpHUJRuT1B9tWoxxIcXWVMoyfBeAGIMcfSHo6bpLbamPMkbhQQoII3HUeoe/pvVy8NvEPp/r7LveH3fS5zB8+xflN5xm7QkB5lCjsHW1FCStvcgElKSOZPTZSSSlTfQLiP004lMfueTaYyLk7CtM0QJJnQ1R1B3s0r6JJ6jlUOtWjXXh2+BbkKbt8GPGQs8ykstJQCfaQBXYooUb1B1GwjSrGH8z1DyOLY7JGcbadmyebs0LcUEoB5QT1UQO7xqqfo8OEL/Xvjv/AM7/AAVOddtE8V4g9OJumOZzblEtc5+PIcdtzqG3wplwOJAUtKhtukb9O6tYv8kdw4fdZn384xv/ANPRSIW0mluuOk2tjFxlaV5xb8katLjbU1cTn2YU4CUBXMkd4Sr8lR3i6+tc1X/BC6fqy66XDVwo6ecLUG/wNP7rfZreRPx35Rusht0oUylSU8nI2jYbLO++/hXd4uvrXNV/wQun6suiKS6G/tKaf/gtav1RqpvUI0O/aU0//Ba1fqjVTeihKUpREpSlESlKURKUpREpSlESlKURKUpREpSlESlKURKUpRFXev8AoxaeIHSm8aUXu9TLVDvJjlyXEQhTqOyeQ6NgsFPUoAO47jWuUPybs63RGLfb+LrWKLFitpZYYZunI202kbJQlIOyUgAAAdAK3RpRFS2g3DWNGLPk1iv2qeWaixMnDSHm8pkedJZaShaFtoB3HKsOHmHjsKq5rgAnYXKuUPQfiZ1E02xq6PLfdsEFxEmMytfwuwUshTXsB6q6Dqdq26pRFVHDxw16d8NmMzLFhInTZ13fEu8Xi5PdrNuL/XZTi9gNhzK2SAAOYnqSSYzxM8G+nfFDesSvuX3S4W6Rizq9zCS2fPoy1oWqO6Vg7I3QdiNiOdXtq/aURcPmkURfMfNmvNuz7LseQcnJtty8vdtt02rVGRwCnE77drhw+cQudaU2q+PKkzbHa1IkQQ6e9TKHCC106DqSBsAQAANtKURU5w68Lmn/AA4wLq7j0u53zI8ieEi95FeHu2nT1gkgKVsAlPMpR5R3kkkqPWufh54dLFw8RsujWPI7jdxl+QP5A+ZjbaSw67tu2jkA3SNvHrVuUoiVqtf+A+JaszvGZ6A64ZjpGvI3S/drbZezegPukklaGV7Bs7knoSBueUJHStqaURUpw+cKWBcP0u8ZPCu15ynM8jO95ye+yO3myhvzFAPchBUASBuSQNydhtkMO4dbFhvEHm3EJFyO4SLlm1vjW+TbnG2xHjpZDYCkKA5iT2Q33PiatulEStbdVOCu1ZdqdK1o0s1WyrSzMro0lm6zLCpC2LikAAKeYXsCrZKdzvseUEjfrWyVKItfNGeD6yabahOax55qPk2pefebKhRr1f3EgQmFAhSI7KfVb3BUN9z0UoDbmO+N1T4KYGda3DXrDNYMvwHIZTMaPdRZnEhu4NM8gSlXcQFJbQlQ3KTyg7b7k7K0oiqbGeHmx4xxFZdxFx8iuD1zy60R7Q/bVttiOwhkMgLQoDnKj2I33O3rGstrpoPp3xD4M7gWo9scfidqJMSVHc7OVBkJBCXmV7HlUASOoIIJBBFWHSiLT2X5PzIMpt8XD9TeLPU7K8HirQfmffcbaD7aCCht18bqWBsO8bjbpsdiLw1H4atI9TdHGdC7zjiYeLQWWWra1AV2TluUyNmnGVddlpBI3O/MFK5t+Y72lSiLUB7gFy6/WhvBs74wNUMgwZIQ07YVraaMiOkjlZcf6qUnoBsR4eFW3qlwnaO6qaQ2nRi4WZ20WbHEsmwP2xzspNqcaTyocaWQdzsTzc2/NuSeuxFyUoi1DlcA+SZhGjYzrDxY6l5vhzDiFuWCQtuMiUEEFCX3UkrcAIHf18QQetbYWaz2vHrRCsFkgswrdbY7cSJGZTyoZZbSEoQkeACQAPiruUoiUpSiJSlKIlKUoiUpSiJSlKIlKUoiUpSiJSlKIlKUoiUpSiJSlKIlKUoiUpSiJSlKIlKUoiVql5RNiLbtMcL1EnuW6TDwTOLZf5NonSm2RdmWu0547XOQlbvKVKSjvUEq29lbK5jlVrwbEr1ml87b0dYYD9yl9g3zudiy2Vr5U/ZHZJ2HjWueqfElwq55w4WLWXVLEZ+Raf3e9CLBiTbMH3kzUF9AWpgq9Xbs3QFb9x99EUwxnjh4U8psLOQRtbcbgNuNBxcW5yhElMnbcoW05srmHd03HsJHWtarPxP8PeYcdEjWePqRYsexXGMOVji7lPdMZV/luv8AOC0hQClNNJG3OoDqBt0IqN/RG+Sq7/nAn8zk/wCOrG0Iunk5+IrNHsC070Ct3pRiA5cV+kMZTHa7FC0JVsrmPXdxPT46KdFtzpvq5ppq/bZd40xzS15JCgv+ayX4DvaIad5QrkJ9vKoH8dS6opp7pTpvpPbpVp01wu1Y3Dmv+cyGLdHDSHXeUJ51Ad55QBv7qldFCUpSiJWEzaxWfKMPvWOZBb2Z1suUB+LLjPDdDzS0EKSoewgms3XTvH7EzPvC/wCqawqmGEjgs6Ql7QeIWm7PBZo3HZbYj3fUFpppIQ223mc9KUJA2CUgObAAdAB3V+/oMdIft5qL+etw+Uq9h3Urx/nuI9s+9eu8yw/sD3KifoMdIft5qL+etw+Up9BjpD9vNRfz1uHylXtUa1H1CxvSrCrnn+XuyW7RaEIXKVHZLrgC1pQNkDqfWUKluLxLjlDzPNQ7CYZoksEclV30GOkP281F/PW4fKU+gx0h+3mov563D5Sp5o9rTgOuuMP5bp5PkyIEWYuC8JMcsOtupSlWxQeuxCgQfGpJluU2TB8XuuY5JK82tVliOTZboTzFLSE7nYeJ8APEkCpOJxbXZC4zwUDDYVzc4aIVP/QY6Q/bzUX89bh8pT6DHSH7eai/nrcPlKlml3ELptrBhd7z7CpVxctGPrdRNVKhKZcSW2Q8rlQT63qH8vSuvE4lNLZ2iDvELHmXJWHMhRW95irzj1Xwwdmd9/hn293WszWxoMZnaxv1OgWAo4MiQ0aT3BRv6DHSH7eai/nrcPlKfQY6Q/bzUX89bh8pVzY9fbflFgtmTWlTioN2hszoxcRyLLTqAtG6fA7KHSvzkuQW3EscumVXhTiYFnhvT5RaRzrDTSCtZSnxOwPSq/O8TOXOZ5qzzTDROQRyVN/QY6Q/bzUX89bh8pT6DHSH7eai/nrcPlKtPTrUDHNU8KtWoGIuyHLPeWlPRFSGS04UpWpB5kHqOqTUjo7F4lpgvMjtRuEwzgCGCOSon6DHSH7eai/nrcPlKfQY6Q/bzUX89bh8pV7UqPPcR7Z96nzLD+wPctc7hwh6Vxpa2mr9qGEgAjfM7gfD75XW+hK0v+3+oX553D5Sr0vH+fufvU/orp1yau0sYHkCq7XiV1qWzsIWAmm3TgqX+hK0v+3+oX553D5Svn0JWl/2/wBQvzzuHylXUkFSgkd56VU+nPE7pJqnm0zTzE7lcFXqEiQ4tqVCUyhQZWEOBCidlEE77ewE1LMbtCo1zmPcQNbmyh+D2fTcGvY0E6WF1jvoStL/ALf6hfnncPlKfQlaX/b/AFC/PO4fKVdA3JFVhjfEdpZluqk7RyxXKdIyO3LktvjzNQj8zH1UB3uOx6fGDUMxu0KoJZUcYubmwUvwWApkB7GibCwuVhvoStL/ALf6hfnncPlKfQlaX/b/AFC/PO4fKVdFKr6Txn7rveVn0bg/2m+4Kl/oStL/ALf6hfnncPlKfQlaX/b/AFC/PO4fKVdFRDVHVTENHcZby3NnpbVucmMwEqjRy8vtXN+Uco8PVPWs2bQx9RwYyo4k9pWL8BgabS99NoA7AoL9CVpf9v8AUL887h8pT6ErS/7f6hfnncPlKuhCgtKVjuUAofEa+1h0njP3Xe8rLo3B/tN9wVL/AEJWl/2/1C/PO4fKU+hK0v8At/qF+edw+Uq6KU6Txn7rveU6Nwf7TfcFS30JWl/2/wBQvzzuHylffoStLvt/qF+edw+Uq6KU6Txn7rveU6Nwf7TfcFS/0JWl/wBv9QvzzuHylPoStLvt/qF+edw+Uq0YGXYxdciuWJ2y+RJV4s7TT1wiMuc64qXCQjtNuiSeU+qeu3XasvWR2jjm2NR3vKgbPwTrim33BUv9CVpf9v8AUL887h8pT6ErS/7f6hfnncPlKuilY9J4z913vKno3B/tN9wVL/QlaX/b/UL887h8pWDzbhg07sWK3G727IdQEyYzaVNleYz1AErSOoLnsJrYSovqd/oFefvKP7RNVV9qY1tJxFV2h3ngtjCbLwT8RTa6k0guG4cVqD86+0fdTm350zflKfOvtH3U5t+dM35SphX2vJ9P7U/2H/EV9B/LmyP9Znwj7KHfOvtH3U5t+dM35Snzr7R91ObfnTN+UqYV97+6nT+1P9h/xFPy5sj/AFmfCPsod86+0fdTm350zflKfOvtH3U5t+dM35Ssbkeu+nuOXR2yGXNuc5hRS81bYpf7NQ7wVdBuPHYnashheruDZ5LVbLJcnWrghJUYUxksvEDvIB6K29xrovxH4kp0fOHOqhmsy7Tjy7VzGYb8L1K/mzGUS+YiG68OfZqv186+0fdTm350zflKfOvtH3U5t+dM75SpHcr1abOqKi6XFiMua+mNGQ4rZTzqjsEJHeT+iu7WgdubWa0ONd8HTrG66I/D+xnOLRh6cjXqttzUP+dfaPupzb86ZvylPnX2j7qc2/Omd8pUwpWPT+1P9h/xFZflzZH+sz4R9lD/AJ19o+6nNvzpm/KU+dfaPupzb86ZvylTGvlOn9qf7D/iKflzZH+sz4R9lD/nX2j7qc2/Omd8pX3519o+6nNvzpm/KVL6U6f2p/sP+Iqfy5sj/WZ8I+yh/wA6+0fdTm350zvlKfOvtH3U5t+dM35SpjSnT+1P9h/xFR+XNkf6zPhH2UO+dfaPupzb86Z3ylPnX2j7qc2/Omb8pUwpTp/an+w/4in5c2R/rM+EfZRAaX2jcD5qc2/Omb8pVXO22al1aRmGX7JUQP8AnHM8D98rYEd4+OqMf+ru/v1fprs7I2ztCsX+UrOMRqT2rjbX2FsyiGeTw7BM6NHYsf6Pnfdhl/5xzPlKej533YZf+ccz5Su7TvrtdJYz913vK4vROA/Zb8IXS9Hzvuwy/wDOOZ8pT0fO+7DL/wA45nylYq6Z/jVqlqgLkvSpLfRbURkulJ9hI6f01zWPNcfyCQYUKUtuUNz5vIbLbhHuB7/xVtGrtRtPypL8vG60xR2M6p5ENp5tIga8OfYu/wCj533YZf8AnHM+Up6Pnfdhl/5xzPlK+XC9W+2S4MGY4tL1xcLMcJQSCobd58O+u9VDsfjmgONR0HS5Wy3ZuznuLW0mSNbC29dL0fO+7DL/AM45nylPR877sMv/ADjmfKV3aVh0ljP3Xe8rPonAfst+ELpej533YZf+ccz5Sno+d92GX/nHM+Uru0p0ljP3Xe8p0TgP2W/CF0vR877sMv8AzjmfKU9Hzvuwy/8AOOZ8pXdpTpLGfuu95TonAfst+ELpej533YZf+ccz5Sno+d92GX/nHM+Ur8x75bpV3l2Jl1ZmQkJceSUEJCVbbbHx7679Zvx+OpmHVHDfqd+iwZs3Z1QEspMMEjQajVdL0fO+7DL/AM45nylPR877sMv/ADjmfKV3aVh0ljP3Xe8rPonAfst+ELpej533YZf+ccz5SvTfgJbca4c7Yh2fOmKFyuH02bKckOn6eehW4SogeHXpXmrXpdwIfW72z+Ep/wDbGu1sHGYiviiyq8kQdT2hed/E2BwuGwQfRphpzC4AG4rYWlKV7FeBSlKURflaEOIU24hKkKGykqG4I9hFVpr5rJpzw76Yy8/z2IV2yK8iPFgxY6Fuy5Tm/I00k7J5jsokkgABRPdU/vt8tOM2Wfkd/ntQbZa4zkyZJdOyGWW0lS1qPsCQSfirQ3je4iuHbVHT/Frxg2ruKZFcsEyy3ZQqxed7KusdlRDrCOZOxWUr3APfsR40QKQReOXUm5R259p8nzqLKhSEhxh7sCO0QeoVt5se8e81aPD5xGZ1qtnT2M5JwnZbptEbtzssXi6tlLK1pWhIYH0hHrKCirv+wPSu3YeP3hEv1qjXQa02a3l9tK1RbilyPIZJHVC0KT0UO47Ej2EjrU5064lNB9XL8vF9NdULJkN2ajLlriQnSpxLKVJSpexA6ArSPxiiKy6UpREpSlESuneP2JmfeF/1TXcrp3j9iZn3hf8AVNV1fVu5FWUfWN5hVgO6lB3Urwy9ulUVxyHbhTz8j/8ApI362zV61RPHL9ann/8A3SN+ts1fhfXs5j5qjE+pfyPyVKcG5OkOs3ztXB2Nn1Owi0ZdaE9yPO0Rk+cIT7ye2J/eCrV4zJ0vJ7Tg/D9ZnimfqhkceFK5T1btkdSXZKz7uiPyGqw1ihysL0P4eOI+0MqMrTpmzJuBQOq7bJYbQ6k+7cAf+IasXTWTG1n4xMt1NjOCVj2mdpj4xZHgd0LmyR2sl1PvCSU7+wiui+HVPOuAM/yFh9CuewltPzbiRHI3P1UI4C342N6M6vyYkBp6NaMlui2orvVC22YoKW1f7pCQk+6vzqLqS3q55OTJM9axK14ym4M8otltG0dns7k2jdPQd/Luenea4eCz9orXP+H75+qGohYf+iiuX7x7/wDyqate0GuX787fkq6biKIbuyOWw9210x/QHhjwLK7vbpF1nTrHabfaLVGOz0+WuKgpQDsdkgDcnY+AAJIFVxqprTxP2bSXJbjrHoDarZit9ssuCuTZrr5xMtC5DKkNLlMqJ3RzKSFEbcu/X2VHtYFogy+Dm8XxSU4/HdgIkLc+pIfLMQtlW/Qd2/X2GtrdeHrVG0Vz92/qbTBTjlxD/a/B6sLAHxlRSB7yK1oZRLDlkuJ48YstiX1Q4ZoDQPlN1B+CMEcKungPhb3h/wDUu1eFUdwRbjhU085u/wBHvb/H5y7V41pYr17+Z+a3cP6lnIfJKUpVCuUevH+fr+JP6K6dd28f5+v4k/orpVxq3rHc12aXq28l+mvqqPjFeYenfaaeXJjiGiJKW8Z1QlWa8qT3qt8pIBKvcPpg+NYr08a+qJ/fCtIuHXT5rVPQPXTA1oCnbpk1wTFJHwZKEBbJ/jEprr7Jqto0qrn+iS0HkSQfArk7VpOq1aTWel1iOYAI8VtjqlnkHTbTbItQZDqFM2a2uy2Tv0cc5dmUj28yygfjrSzhnwmbg/E/grF55jeL5gsi/wB0Ur4SpMsuunm94SUD8VZqLqBK4gtKtEtD3HVquV6uvY5Ujf124doP0zn/AH4CD18RVg3BLbflErIhCAhtODrCUgbBKQHNgPxVsUKRwVGrQd6Ra8nk2w95nwWvXqjGVaVdvogsA5uufcIUr1G4isoZ1Kd0X0O0/bzHLILIkXV6XK83gWxBAIDix1KtlJ3G42KgBudwOlj3Edn+K6hWfTXiK03hYrIyRfZWW9WuaZNvkvbgBpRO5Qokgb79CpO4AO9YDhMW1G1u1/tt1IF+OSJfUlf1RUTtHuUjxKfWT/5k+6vz5QJcZ3AcItcfY3uXmEQ2xCfqhISoLKfHvU2PjKapbQoecNwRYIIHWvMlszrEDhGiudXrebuxme4J6togGI0me2dVYGtuv1z0/wApsmlunOFqy/PshbL8a3F7sWIscEjtn1+A9VXTp0SSSOm+uPFzqTrDJ04g4FrTphCsMybeoc62XSzzfOoEgNFXaMrJ9ZtwBYIG/Ub+zerTx4+ZeUKylu+qAkT8NY9Elf2aUpZ7QI/8ju+3sVXa8oG7bEaExGpqmxKdyS3+ZAn1isFZXy/8HNv8dWYMUsPiaFMMBLgDN5k8LxA5cVXizVxGHr1C8gAkRaIHG0yefBWTrbrhZND8Ut1xl2qVerzenkQLLZon1adJIHTfY8qRuNzsTupIAJNVteteOJvTe1JzvVTQSyoxJspXcBZbuXp9uaUQOdxBJSrbfrtsPaRUS4tIOSy+IfRBNkyWPjrr7T7FtusqKJDEWcVp2Km1eqondsDfxIPhUszPS3ia+ZG9pzXilxxqwGA+i6LkYq0htMUoIc5j3gcpPUdfZ1qujh8PTpUzUykvuZzTrENyiPrPYs62Ir1KtQMzAMsIyxpMmTP/AIpvrNxBQtPNFYGs2IRIl/t1ykQUx+2cU2hUeQernqgkKSPsfaCKwumet2tOeZcxdbpo0zYNMJ8eRMi36ZL5ZKYzaCpt5xvc7doACElI2B33O3WldasYt2IcANnxy05pByy3xrzG81usJCksPtKlOqASFEnZJJT394NbgTr3aML04cyK5tEWyx2MS3m20gkstMAlIHd1A2AqurSo0KMMZmLnOaCZmBEWte+8dyspVa1etLn5QGtcQIiTM34KjbFxEa76wom3/QDR6yS8TiynIsa7ZFdDHVPUg7KU22nbYfGTt3E77gSXRfiDyjUS8ZdpzmOnyMb1AxGP27luErtI0oEbIKXPsQVFHiRyrBB79oNptkHFdrjjEfNcFu2CaZ4jcnXTbIqbX55KLQcUkuEbcm5UFdfV3IJ2rC8Oltvdm4ztSbTkucOZddomOstTLquOhguu88YlAbR6qQjcI2/3etX1MPRyVW5WgsEgAuLhBA6x9E9sdyppYitnpOzOIcYJIABkE2Go7PFYfh2v+tY4ldUHW9ObGZdyusAZY16YHLaUbq3Uwrb/AJQdio7dO4e2rlz/AIh8uc1LlaN6EafsZfktqaD15lzpfm9vtgO2yFrHVS+o3G46nYbkHaOcOO30VHEGT3ekYO//AK6q7Q7FtZ77q3rRD0+1bt2HXWPk7rlzjTLMia7JbLr3ZOBSuqUDcjYdPWB8RVtanSrVn1KgaMjGa5ouG6xJgbo7JVNGpVo0WU2EnM52kTYnSYEn7wrxwziEzm1akW3SXiA09iYpeb+lRsdztswybdcFjvaCj1QvwG57yAQNxvfVafam6RakTMnwFWtfFLjBeh31qZYI7liTEekSULb5m21NkEk+oNj03I8a3CWd1qO225PSuVj6dFoY+lFwZiYkHdmE/Yrq4GpVcXsqzYiJibjfH9hfKi+p3+gV5+8o/tE1KKi+p3+gV5+8o/tE1yMR6l/I/JdnBfqaf8m/MLXGlKV4tfTErguDUl63y2ITnZyHGHEMr325XCkhJ/Edq56x+RXc4/YLjfRBdmej4y5JYaIC3AkbkAnx2Bqyi1zqjWsEkkRz3Kus5rKbnPMAAzy3qjtDs/wjT20SMMzP/m/kbEt0zXZbJHnBKvVJc2O2w6deniN96sDIMJxjUi82LNcayOIzOsspL5mQOR4voBB7NZB6dx6nwJFd6yKwbWXE4OST8dgTmZSCFNyG0uOx1g7FsrHUEe7bvFVbqngNn0cXas/05mSbTNNxajLgB9S25SVbkpAPUjpsQdxsfA7V7Rj6WP2k7ybn0cU4kEGHMmCCOIB7QQO5eGdTq7P2WwVWsr4NoaQRLHhsgh3Aka2LSeZWQ10nZanUDCkxMfhOMR7qldqWqXymU+QjmbcG30tIOw5vGpZm2q130+t2MS8kxthMm8POtT48d8umOUjcBogeuTuB19tY7WglWZ6Xkp5Sb7uU+w/S+lfNc2m3ss00adSFJVkHUHx6t0wzaGLZgKFemC3LUJiZ6uffO8iT29llGKfiMHU2jiMPVIdmpATBAzZN0bgco7NZN1l8dz/NmIl3vmp2GsY1aLfEEtp5MjtHFAn6mU7ndW23s6kDbrWKt+o2sGUW/wCaTE9MYHoZzdcZE6eUSpLfgpKR0G/h/RvXc4k0SXNJLsWAohL8ZT3L/wBn2o3/ABb7VPcbdhyMdtL1tKFRXITBYKPg8nZjbaua6rhqeDGPGHaS9xbHWytDQD7Uy6dSd1guq2liqmNOzjiXhrGB2bq5nFznDXLGVsaAb7lR/BdSbdneOzbtEhOwp9r7Rqfb3z68d5KSeUnxB2Ox28D03FQzGNY9QtQbQ1KwXT2G8+0CJz8yYWorbm55WmyditXLyknuHNXFgC2pGqerMq3EKg9mltSkfBL4Qrm/HuF1leGZCUaSQClIHPLlKV7zz7b/AJAK3MThMHs6lXrtpBxBpFocT1c7C4gwQTHb2StHCY3HbSq0MOaxaCKocWhsu8m9rQRIIE6mO2N0ZjTPUqZmky7Y7kFgNlyCxrSmXFDnOhSVdApJ+P4+8EE71iX9U8wyXIrjY9K8Rh3SNZ3CxMuVwkllgujvQ3t39x6/j222J6WJpcVxB6goZ6OKtcYI/fcre39NcnDItoaeyoR2E2Ld5SZqD8MOEjbm/EP6DWOJwmEwjauNZSB6tIhpJyg1GyTrJAiBJ3rLC43GYx1HAPqkS6sC8AZnCm6ANCATMkgbua7GOaxXy6agQNO75hvoi4rZeXPC3ysIUhJUhTRA2WhQ8fD31aVVFkEiE7xMYuwwpJksWWQJO3eNw4UA+/br8Rq3a5O2qVGn5CpQp5M9MOIubku4yYtbsXY2HWr1BiKdepnLKhaDAFg1tjAAm5ntnklKUrirur6O8VRj/wBXd/fq/TV5jvFUa/8AV3f36v0mu7sTV/d9VwtuaU+/6LjqOag3l+x4nMlxFlL7nKw2od6Ss7Ej37b1I6i2plueuWHTEx0lTkcokgDvISfW/oJP4q9XgAw4qmKmkj5ryO0jUbg6ppellMe5d/EMfh45ZY0aM0kPuNpckO7es44Rudz7BvsK48nxKPkbkSY3KMKdDeS43KbRuvlH2J7t/D4vx13seuke9WSHcoywtLrKebY/BWBspJ94NdXK8ojYrAalux1SXX3UtNR0KCVuE95Hxf8A+xVrX4p2MJZPlCT/AG9o8FU9mDbgAKkeSAB7txteeV5WA1GnItt4xae6246GJbiyhtO6lnZPQD2k1zXHL8wsjAu14xFhu2hQ7QNSud5pJOwKh3b0zQdpkWG9q3ylU5Sig9djsk7fiNZTUD/Qu8f92P8AWFbzHU8mGpVGB2aQZnQuItBXOqNq+UxdanULcsERFyGA3kH3WX6ybKW7DYmL6wymQy86yn1lFOza+vN09g8K4LLkORXGaX52OJg2ZbS3mpTr30wIA3SVJ8Nx18NhUfzBIVphZ0q6gmED/wCWpdlrbqsVuzcYHn8ycCQPZy/3VWaNGnSazKCXOc2TNgCBOut1cK9erWfUzkNYxrsoi5IJg2JiywsXLslyDtZWKY5Hdt7aihEibILZeI7+VI8KyeM5Sm+vSbbNguW+5wSBIirVzbA9yknxH949tfMDWw5htoMYp5BGCTt+6BPN+PfesS0UuavL8269lagmRt+63GwPv6pqH06NR1ai2mG5ASDeeqd94M8lNOpXpNoV3VC7yhaCLR1hNoEiOZkar7Fzi83aVOtdjx1EiZDkuNKU49yMobSdgpStvhEg+qPZXdsOV3CVe3cayG0JgXBDXbtlpznbdR7Qf/33HurpacJAkZMoDqbu4Cfy1+5+3z1rV77W7+lVW1qVDytTDtpgZWyDeZAB4xHZCooVsT5GjinVCS54aRAiC4jhM75lZO33wS8tutiFvZbMJlpzzhJ9d3mA6Hp4b+2ulIy27XC7SbRiVmammCrkkypD3ZspX+5G3UmuGw/tnZH/AN0j/oTXHpUUiyXBpfSQ3c3g+D8Lm6bb/wBNYVKFKix1bLMNZYzEuFyb/wBlWU8RWrPbQz5cz6kkRMNNgLRv1ibLJWHKZM26v47fbYLfdGG+2CEOc7bzf7pB/wD3/QakVdFy52lu9NWlbjYuTrJdQns91Fsb7+tt0Hf0JrvVzMTlLg5rMoIBjdzE7veuvhcwaWOfnIJE2nkY3jkOSV6XcCH1vFt/hKf/AG5rzRr0u4EPreLb/CU/+3Ndf8OfrD/E/MLg/i39AP5D5FbC0pSvcr5slKUoi6d3tFrv9qmWO9wGJ1vuDDkWVGfQFtvMrSUrQpJ6FJBII9hqrRwhcLo6DQPB/wCZmf7qt6lEVQ/Qh8Lv+oPB/wCZmf7qkOD6C6LaaXleQ6faX43j1zcYVGVLt1vbYdLSiCpBUkb8pKUnb3Cp7SiJSlKIlKUoiV07x+xMz7wv+qa7ldO8fsTM+8L/AKpqur6t3Iqyj6xvMKsB3UoO6leGXt0qFa0aYQ9ZtMb7plPu79rj3xptpcthpLi2gh1Dm4SogHfk26+2prSsmuLCHN1Cxc0PaWnQqAXHRyx3jQ06F3Wc9Ithx9uwmYW0h3ZtpKEPhPcFBSUrA7txXV4e9DrFw9aft4JY7pIupVNenyp8lpLbsh1ZA3KUkgBKUpSOvcKsmlZ+WeWlk2JnvWAosDg+LgR3KntH+HC1aQ4VmmFwcrm3NrM50yc9IejIbVGVIa7MpSEkhQT3jesXB4UrLC4YZHDGnM7gu3yAsG7mK2HxzShI+p78veOXv7utXrSsjiapMk3kHvGixGGpAQBuI7jqq0y/QDB9QNGrfotmSX51ttkGJFjTW9mpLL0dsIbkNnqEr2B3HUEKIO4NVW7wZ5bklpTh+pvExmWV4lFaUiJZ3Y7bCSsJIZVIcSoqfDZ5VBJIBKR1FbP0qWYqrTs09u7w4dyPw1J5lw/v1UM0b00i6O6ZWHTSFd37oxYWFsImPtJbceCnFL3KUkgdV7dPZUzpSqXOL3Fx1Kta0MAaNAlKUrFZKP3j/P1/En9FdKu7eP8AP1/En9FdKuLW9Y7muzS9W3kvqTyqCtu471Wmh2iVv0PgZDBtuQyrsMhvLt5Wt9hLZZWsAcieUncDbvPWrKpRtV7WOpg2MT3aI6kxzw8i4mO9U1ppwwYdphq5k2rdpukmTJyHt+wguMpS1b+3dDjvZqHU8xG3XbYdKzknRO3yNfImvRyCUmZEsyrMLaGE9kpBCvphXvzb+t3bbdKsmlWuxld7i9zrkZe7gqm4Si1oa1tgZ7+KpnVHhqtub5ozqlhOb3jA82aZEd262sJWmW2BsEvtKIC+gA336gDcHYbdLCuF5qBnkLVDVfUm96i5LahtbF3BpDESCfBTbCNxzA9R12B67bgEXnSshjsQGeTDrRGgmOE6x2SsTgcOX+ULbzO+J4xpPbCq3WnQDH9YX7Rf0X25YxlePKK7Tf7WoCRHBO5QoH4aN+u24I3Ox6kGt8l4L7jqLC7XVbXfJcnvEdbXo6W5EaaYgNpUFLDcdJ5StzYArJ32Hd41s1SsqW0MRQaGsdEaWEjkYmOzRRVwGHrOLntmdbm/dMd6gmr2jGGa14gjEcwafSIziZEGfFXySYT6RsHG1bHw6EHcEfECKnuHCRnGWQWcV1I4mswyLE2VoK7UIrTDkhCSClDrwUSvuHUg+3v61snSsaOOr0G5WOtusDHKRbuWVXBUK7sz235kTzjXvVXar6AYxqZpJF0cgTXMas0F2IqKYTCXOxRH35UBKj1336knfx8asGZYbbc8eexe7MJmW+VCNvktL6B5lTfIoHbu3G9ZClVOr1HNDSbAk951PgrW0KbXFwFyAO4aLXHG+E/OsCiu4vp5xMZZYcScdW43bEwGHno6VndSWn1fA+MJHXrtvUk0u4WsU0h1Lf1DxHI7qpM20m2zYU4iQqU6Vha5S31HmLilpCj0279tquqlXv2hiagIc7XWwvztfnqqGbPw9Mgtbppc25XtyVGXThhfTrTK1lwrVe/Yw7d5MaTerXGaSti4dkU7oUeYEIWE9QQdt1bd+1dvU3hot+YZynVTAs6vOn+ZloMSbla0pcbmtgAAPNKICjsAN9+oA3B2Bq6KVAx+IBDs1wI0GnA2v3ypOBoEFuXUzqdeI4d0KjMG4XkWzUCJqpqpqVetRMmtieW2OXBlDEaCf3TbKSRzDckdwB67b7EXnSlU1sRUxDg6oZi3ADkBYK2jQp4cZaYie/3k3Sovqd/oFefvKP7RNSiovqd/oFefvKP7RNaeI9S/kfkt/Bfqaf8AJvzC1xpSvteLX0xfKEJUClSQUqBBBG4I9lfa+URVRL0Dat10kXXTzObxiZlq53Y0Y9oxv7k7jYe477eFdywaJRo9+jZPm2XXTLLjBPPF89PKyyodQoI3O5B2PftuO6rLpXYdt/aL2ZHVNREwM0cM0ZvFcVn4d2Yx4e2loZiXZZ45Jy+CiGpGnETUWHAbXeJdqm2qT51DmRgCpte3XodvYD3ggiupN0vdu7OJG95bOnTMVmGYJTjSeeYoqB2X16dwG4qdUrXpbVxdCmykx8BsxYWkEG8TBk2036rYrbIwdeq+tUZLnxmub5SCJAMSCBeJ3aLr3K3wbvAk2u5xUSIkttTTzSxulaFDYg1WcbRS/WWO5ZsU1bv1qsbhPLC7JDqmknvS24SCkfFVqUrHCbSxOCaWUXWN4IDhI0MOBE9uqzxmzMLj3B9dvWFgQS0wdRLSDHZoo1iGn2P4Pjj2OWFDqUSQsyJLqud59xSditZ8T7B3CvunmERtPMWYxaJcHZrTDjjgedQEqPOrfbYdOlSSlYVcfiazXtqPJzkOdO8iYPdJWdLZ+GoOY6kwDIC1sbgYkRpeB2qL2fAotpz69Z43c3nH72w1HXGUgBDQQEgEK7zvy+PtqvbbYMLz7I7zkeCZne8NvTUlca7Rmlttl1xJ2Ky0SR1O/rDpvv0Bq6qiOS6SadZdcFXW+4xHdmr+HIaWtlbn74oI5vjNdPAbW8m9zsQ9wJa1oLQ0wG7i0wHCI1uCJXL2jsfyjGtw1NpAc5xDi5sl15a8S5pmdBBBiyq7D8bsULiBixsYukm8C02t568XJ9/t1uynOZPrr7t9lJGw7ttvA1f9YjGcQxjDYSrfjFljW5hZ5lhpPrOH2qUdyr8ZrL1r7Z2iNpVmvbJDWhoJiTEmTFhc6DQLY2Jsw7LoOY6AXuLiGzAmBAm5gAXNyZKUpSuSuwvo7xVGP/V3fviv0mrzHeKox/6u7+/V+mu7sTV/d9VwtuaU+/6L8UIBBBG4PQg+IpSvQLz6iK9PUw5TsvGMhnWXtzzLZa2W0T7knurntWCRotzRe7zdZd5ntfUnJOwQ2falPtqT0redtHFOblLtbTAmOcT4rnt2VhGuDgzQzEmJ45ZjwWKvGPsXi42q4uSXG1Wp8voSlIIcJ26Hfu7q7F8tTd8tMu0PPLaRLb7NS0AEp677jf4q7tK1xXqDJf0dOy8/NbJw1I55Hp+l22j5cFgrriUa6Y9Ex1ya621ELJS6lIKldmNhuO7rWdIB3BAII2IPXcUpUPr1KgDXGwJPedVNPD06Ti5ggkAdw090qKJwR+3PPHGconWiPIWVrjIQl1tJPeUc3wayeOYvAxtLy47r0mVKVzyJT6uZx0+8+A91ZilXVMdXrMLHusdbCTzOp71TS2fhqDxUY240uYE6wCYHcAsTYMeYsC7gtmS48bjKVKVzpA5CfAbeFfX8fZfyaNkxkuB2LGVGS0EjlUFb9Se/frWVpVZxNUvNQm5EHlEfJWDCUWsbSDeq0yNdZn5rFQ8eYhZDPyJElxTtwabaU0UjlQE7dQe/wroT8MK7o9erDe5Vnlyf847FKVtun2lB6b++pJSsm4ysx2YO3AaCIGgI0KwfgcO9uQttJdqQZOpBBka7isHYMUj2WVIukmfIuNylAJdlyNublH2KQOiR3fkrOUpVdWs+u7PUMlXUaFPDsyUhA/vvSvS7gQ+t3tn8JT/7c15o16XcCH1u9s/hKf8A25rtfhz9Yf4n5hed/Fv6AfyHyK2FpSle5XzZKUpREpSlESlKURKUpREpSlESuneP2JmfeF/1TXcrp3j9iZn3hf8AVNV1fVu5FWUfWN5hVgO6lB3VqbxA6w6qaYcSdvumNSZtywzHcQaveT2Bs8wdhKmLYelNo26utBSF77/BQfDevGUKJruyt1Xsa1YUW5nLbKlUfrXqHOEnRG74FlTybRlub29h56E7s3cLe6w4vkV7UK2SdvaKkOZcQ2F4nlU3CYVjyvKr1amESbrGxqzOT/RraxugvqSQlKinqEAlRHXap83eQCBrPgYUeXYCZOn/AOqz6VXvz+dNHcWxPM7feHp9ozS7x7Ha3o0dSj548VJDbqTspopUhSVcw3SRsRWdvOoWOWLPMb04nqlC85W1MetwQzzNFMVAU7zr39XoobdOtYeSeDEcfDX3LPyrCJnh46KS0qn8T4qNL82yVGN45FyWRyuy2JtyNoWm325yP2naIkySeRo8rSljcn1SknbmFdOLxeaTyTEuTkHLYuLz5aYUTLJVheasrzqlciSJB7kKV0DikhJ9tZ+bVhbKVh5xSicwV2Up7wQR7RSqFelKUoiUpSiKPXj/AD9f71P6K6exNd28f5+v4k/orVSRfndTdaM4wvPNdr3p8MbntQ7Fj9ruLVscmxi0lXnqnXEkv86idkpOyQO6uc3DmvUeZgC53740C6LsQKFNgiSbDdunUrZ6lU7jd4z/AEWxnNbnq9fZeU4ljnZTrLfW223rlIgqT9NS+03tzFpX2f2Sdz4VOMs1LxTDsXt2X3OS+/Au8mFEt4hs9s7KdlqSGEtpHfvzA+4Anwqp2HeHZWdYGwI36H3303K1uIYWy/qkXIO7d9FKqVUl24nNPrddL5ZLfY8vvs/GZj8S8MWexuSjC7L4TrhSeUIPXl67q5VEDYVm7nrvppbcEsmoqby/PteTLbYsrUGK4/LuL699mWWAOdTg5VbggcvKd9qk4SuIlhvpZQMXQMw8WVgUqvcI1wxTNcokYOqzZJjuRMQzcBbb/a1Q3n4oUEl1rclK0gkA7HceyovH4uNK5toXksC2ZfKsEVfZ3K8MWJ1cO2L5ikpkLB6EdCrlCuUKBO29Bg8QSQGG0eOnv3KDjKAAJeN/hr7ldVKozUDXW4Y9rhp9h1mt96uGP3qBLmy1221GSmfzNI83Uy4PhNo5itwp+CCknpWfxzUnDcatmpOU5BqBdZVrxvJJMe4OXVkJRbXEob/5JFCdy41upPJ0KipZFZHB1QwOjUSPfCxGMpF5bOhg+6ValKrXEtfcNyvJYOIyLLlGN3O8MLk2lrILQuCLk2gcyiwpRIUoJ9YoOytuu1SvPM2sOnGI3LOMncfbtVpbS7JUy32i0pK0o3Cem/VQ/FVTqFVjxTLbnTtVza9N7C9rrDXsWfp1qrbLxHafXq+Wi0Kg5JbI2RPGPY7vc7Q5Gt10d23CGHld5UBujmCeYfB36VjMM1CetmouuMrNspWzjmITLa4yZbn0i3xzBDjvKPAFW5I7yas8zqgOzCCBPO4Fu8qvzukSMpmTHKxPyCuWlVHbuJzAplxssW5Y7mdjg5HIbiWi7XewuxYMt1z6kkOE7p5/seYDfesnlmvOJ4rkdxxWPj+WZHPsrTb13Fgs7kxFuStPOkPLBACin1uQbq267VHmdcOy5TKnzuhlzZhCsmlUBqzxHsWyz6ZZFpwbtdbNmF9il6Xb7SqT5xBBX2kZG/VMhSkgBvbn2Svbbapxj+W2aTqxlMdzLb62uHjttuUqzXFhLMG2MOBxQeSo9Q6oA9oD8HkFZHB1WszuEa+BjuusRjKTn5GmdPET3qx6VUNt4pNMrk/b3/M8nh2G7zEwLdksyzOs2iU+pXKhKX1dwUobJUpIST41GtQtXH8ExDWvIcUyPJLzesfnJitx5NuD0WxyVRUqQGwO+MBstS1dApXWpZga7nZC2DbXmB9Vi/HUWtzAyOzlP0WwdRfU7/QK8/eUf2iajuC60WG+rw7GrtCv1vvmT2h2bEF0tqo3nKoyUh89e4nfnSNvWQdx0pqJnuPzjlGm0RUl28WqzxrpM5WfpDDTz4S2lTm/w1cqiE7fBG9aeLw9SnSeHDRpPdpPvsuhs+vTfiKRB1c332Me5UnX2vlVBEy7JIGt9xhzrxIdx56c3Z0RFn6VHfcipeaWn2cykrT+OvKYHZ78eKnkyBkaXc4iw7br6Dj9o09nml5QE53Btt0g3PZa6t+lVPr1luRWaHEtmK3V63yWGzc5zzJ2UI4cQ0hG/hzrc/8AQamt/wA8suPXCPZFx7jcrrIZ84TBtsVUh5LXd2igOiU79NyRv4VZ0TiDQpVmCfKZoA1AbEk8BeeSqG2MMMRWoPOXyeWSdCXTAHE2jnZSOlRVOpuIqxeXl3nb6IVueEeahxhSH4rpUE8jjatikgqBPu610laxYezBTc5rV4hxHJaYbD0i3OID6lIUtKmweq0qCTsQOpKRt1qtuysa+ctJ1jGm/hzuFY/a+AZGas0SJ1GnHlYqb19qN2DPbNfrsuwGHc7Zc0s+coiXKIqO46zvt2iN9woA7b7HceIqCYJrHb4eEx5mTKvtxciuPpuNyagreZjfTlhIdcHTonl3232G29W09jYyo1xawy0tEbzmmCOy0cyAFVU23gqb2BzxlcHGZsMpbIPb1p7ACSrer7Udv+d4/jwt6HVyZ8q7J54ES3sF9+QjYErSkfYgEesdh1rE3HPrffMLymVYX5sG6WaBIL0eSyWJUR3slKQooV8W4UNwdqppbNxNUNfkIaSBMWuYnlNp421V9XaeFpFzM4LgCYm9hmjnF44X0U3r7ULYz222HF8ZXenZs+63eAwtmLEYL8qUvsklawkeA33KjsOtZOwZ3j2QM3BbTr8F60dbhFuDJjvRU7EhS0q+xIBIUNx0qKmzsTTaX5CWgxMW1ieU2njbVTS2lhajhTzgOImJvpMc4vHC+ikFKgTmtmGx4aLpKiXyPbpBCYk162OIYlkkABpR7999xvtuASN6lNtyS2XW9XTH4xeTOs5a85bcb5d0up5kLQfskkb9faNqVtnYvDtLqtMgcuBAPuJAPAmEobTweJcGUqgJPA8QSPeASOIEhZSlYyxZHbMjTOctSnXG7fMcgOuKRypU63tz8h+yAJ239oNZOtWpTfScWPEEblt06rKzQ+mZB3pX2vlKwWa+jvFUY/8AV3f36v0mrzHeKox76u7+/V+mu7sTV/d9VwtuaU+/6L8UqLZxNv8AFfszePP8sh6Q6S0T6r4Q3zdmfj2I+M1+r5kXnmBycgsshxham0FKgdltL50hST7COoNetZgaj203g2eY5Xi/OD7l4+ptGnTdVYQZpieYgExykTzCk9Kx1yvsG2PMRHUSJEqQkrbjxmi44pI71bDuHvO1cScntCrVLu5ceS1BJTJbW0UutKG3qqQeoPUVSMNWIDg0wdPl81sHFUGuLS4SNfdPyvyWWpXVn3OJbWmHpSlhMl9uO3yp39dZ2Tv7B766MjK7Wxc3LQG5j8ppxDbiGI6l8nMAQpRHcn1h1+P2VDMNVqeg0n/xTUxVGlZ7gP8A3T5LMUrCSMvtMdySlDU2S1DUUSZEeMpxppQ7wVDv28dt9q617yhUK5WNqCh+RFuClrWuOx2odb5CUhBHjvsSB4VazBV3uDcsTOvYJ+SqqbQw9NpdmBggW7THzUkpWJm5Nb4cxy3tsTZkhhIW8iJHU72IPUcxHQEjw76+ryeypt8W7CSpUOW6llLyUHlQsnYBfijr0O/ce+sPNa0A5TfT+9u7is/O6EkZxbW/cfdv4b1laVi516gJenWoSH0PxYapD7jKN/N0kHY793P4ge6uP5oLdb7ZbXHZEqWua0jzZKWiuRI9UHm5R47dSegG9BhargCAZO6OyZ933Q4uk1xBcIGpntiPfbnbVZilR+dklrl2ec6J063LhrbRIAZ5ZDBUocvqnwV3b9RsTXxWRvpzB+yKiSzHahpWOSMSO0Kjuvm/c7DYHu33qxuBrEExBE+EfdVO2hQa4CZBiI7Z+x+XFSGlY9u/2x2x/NGl5QghlT5WU7KCR3gj27jbb213WXQ+y2+lKkhxAWAobEAjfqPA1rOpvZ6Qi8d/BbTKrKkZTMie46FfuvS7gQ+t4tv8JT/7c15o16XcCH1vFt/hKf8A25rufhz9Yf4n5hec/Fv6AfyHyK2FpSle5XzZKUpREpSlESlKURKUpREpSlESuneP2JmfeF/1TXcrp3j9iZn3hf8AVNV1fVu5FWUfWN5hVgO6qFdZZk8cD0aSyh5l7SwtuNuJCkLQbiQUqB6EEEgir6HdX47Fnte37FHa8vL2nKObl9m/ft7q8VTqeTntEL2dSn5SOwytFcvxjJNFta9LNEPNn5OBSNRImSYdMUSrzBtQcTKtqifBDjqVo/3VH8WXs7sbTzV3Vqz59xG3vSeZcsmeyGEOwhJi3eA62ns32npDKy4tASWy2lW45RsnvrdNxll0oU6y2stq5kFSASlXtG/cfeK4J1rtd0S2m6WyHMDSuZsSWEOhB9o5gdj8Vbnn0iHDdftvM3B71qHBR6J3292i0jkRrBjOhOIalWx3L5GLtawxcruFyv8ACaafMVTikOz0ssJARHWvZY3SD6xOwBFWRctU8E1M4vtH1af5BHv8K12/IESLjB3ciB1yMhSWUu7cq3AlPMpKSeUKTvtvWzi223W1MutoW2tPIpCkgpKe7Yg9CPdXDFtttgttMwbdEjNs8xaQywlCW9+/lAA238du+sTi2uklt77+I5LIYVzYANrbuHetU9JsVvOW8FmpOLYm0fS97uOXMx0NnlU++qW6kJ39qgkIHxiupnGumkuW8IzmmeNzWZuW3jG2MXh4c0yr0m1cuzQ12S423M32biSorICQE7791beNMssJ5GGW2k7lXKhISNz3nYVwotlsbnrurdshomuDlXKSwgPKHsK9uY/lqPOmlxc5u/MLp5q4NDWndBsqfRpBratNncgcRt0sESHb7dHetDNhhyW0rZYbQ8O2c9dXOpCyT4c3SrpPU7gbUpWs+o6pr8gFssptp6fMpSlKrViUpSiKP3fb0gvfu2T+itXsl1F0TybJMh0+4r8Rxey3S0T3UWeRdYqgxcLWerL7EpQ3Cj15khQ2PcO+toLx/n6/iT+isVOtttujaWbnbYk1tJ3SiSwl0A+4KB2rnNrNo1XFwOuoMEX3FdF9F1Wk0NI5ESDzWuHDdacbuWUajY7p1Kulz0YlQ48SAJzjrsYznErTLbhre9ZTPIUg945iNt6jui0LIsm1HxbRbJWXnIegjs9+W84DyzHistWlW579o61L+NNbVXeNdPQMuFir8KBcBGWi3uPx+eOy7t6hW2kjdAO24BHSohpJpvdsFj3y9Zbf2L7lmV3D0jebhHjlhklKA2ywygklLTaEgDc77knxrZ8+Dm1HnUwBvMxBcTHAme2FrDBFrqbNwknhEzAE8QI7JUT4b2203vWd1KEhbmolwC1bdVAMtbb+3bc/lqk8Ozq54foTo7bodztGMw7zfrzHk5dc7ciWix8kh8o7ML9Rtx3coC1EAAGt1UNNNlRbaQgrVzK5Ugcx9p27zXE7AgvxVQX4MZyMv4TC2UqbPj1SRt3+6q249ocXOZIJB9zSOHbPC0FWOwLi0BroIB8XA8eyO+Vqhgt4t9x4ssYVD1YuuoSE4rd46rxJZZRD7fmbUqPFLKEtq5UgKXylWxI3PhXU0Y1x0mxrhYl4neLrHZvjMe7QzYi2TLub0h54N9g0Bu8HCtI5k7gEEHbatu24UNlptliHHaQwkpZShpKQ0COoSAPV/FUG0Z0sa0uwCzYjcX4N2nWdcpTdwTECFAPSHHdkc26k7c+3Q9dqudjqNSn12m2WACBMZr2Eb9IVLcFWpv6jhfNJidcttZ3ayqJt6fnN5Lw0I1OuTdpTa8XudpmSZSiG2ZjkdkNsKV3BW5CAPaKw+S224y8O1Tu0W2yLhExzWtm+XWIw0XFuQY5YU8QgdV8oIWQPBJ9lbkSYkSYgNzIjEhCVBaUutpWAodxAI6H31+0NttFRabQgrPMopSBzH2n2msBtOCH5b7729Iu7tSFmdmyC3Nblf0Q36SqoGuulucZ3iWN4Km353PmuvTDMglDqbBHDR3lOrUk9kpXRsI3Ssk7VwcXuw4a88Km+0AgNEo/dDzhrp+OrYh263W8uG326LF7ZXM72DKW+c+1XKBufjqJ6z6fSdVdLsh09iXRq2vXuOhhEp1ouIa2cQvcpBBPwdu/xrXo1aTcRSc0ENaRMmd88PkFfVpVXYeo1xlxB0Ebo/sqlNX9S8D1lwbDdONLruzc8lut9s8iPboyD5xZ24rqHHnZCNt44aQhSTzbdTsN66uRZTcsOuHFDk1nxuLfpMObZlIhS43nDCgYTaVOONbeuhAJWU+IRWztvtcK3pC2IcZElTSG3322UpW7ypA3UoDc93ia7KWmkKWtDSEqc+GQkAr6bdfb09tXNx1OmMjWS3tP/AGaTuHsx4qp2BfUOdzod2D/qQN/bPgtH9Wssg3bHsOdVxHydRJKslsct+32qDFYtcBkSUbuvhlBU0ASEIDiwdz3HY7T7UObgdr1lzKbbNbb3ovlqPNnpqrh2C7VkLYZAblNsu7pd5QOzVyqC9x8Gtmo9ptMNlyPEtUFhp1XO421GQhK1e1QA2J95r9TLZbLl2fpG2xJfZHdvzhhDnIfaOYHb8VZ9IssA0wJ9neQdMuXdw7VX0c+5LrmOO6Rrmnfx7FqTftQLvddF9GNStRocKzRrbqHGfmzY8NUWKqE2qQhub2W27SHE7K2I+y38azGYW2dqBqVrvbcKfEyTf9MLULYtlfSR2qJBb5D484IA/fCto348eUyqPKjtPMrGym3EBSCPYQeho3HjsnmZjtNnlCN0ICTyjuHTwHgPCsRtBrbtZBvF7XcHcOyFn0e50Bz5FptezS36rXq0cQukS9JcSxq3W2NlF/dZttnbwgNJ8+blt9mhaXWHEnsgyUqWVqAA5NweoqJZJGkzMP4u40Vhbzy5h5UNpKlK2trROwHf0Bra5Futzcxdyat8VEtwcq5CWEh1Q9hWBzH8tcqWmkFakNNpLp3WQkDnPd19v46xbjadIk02G5m5n/IHh2RxusnYKpUAFR4taw7COPatdNQLtZ8s0JwrW3Tq6MXeRppIg3hD0QlXOwy2lm4R/bv2Sl7p9qBXJpvFfvmkmday3JlaJupNzXdmA4PWbtjbiWYLfuHZI59v/eVYerenmaZxj7eFYTlNqxewXRDsS/8ALbiuS5Fc5QoRVJUlLa1J50kqB+Fv3isrnNrt9j0tmWS0xkx4VugsRYzKe5tptSEoSPiAFUY7EsOBexupzcw3WDYDW9ltbNw7htGm92gLeRdMSO6y17qmblY5V+m6rs28f+0Ic+3XGCR3iQxHS4jb4+Uj8dXPX5CUglQSkFXeQOp+OvFbP2g7Z5c5okmPBzXd85Y719L2js5u0WsY8wBmntljmd0ZpnsVDZHPOYaY5jqWtlbaL05BiQULTspEVh5sHp73VOn8QqXR71asL1XyKTlktq3MX+FBXbp0k8jK0soKXGe0PRKgo83KT133qy+zb5Oz7JHJ+55Rt+Svy8wxJb7KSw08jffkcQFJ3+I10H7ap1WOoOpkMMiAbhv/AB5QDGo8mJMXk6arm09h1KT2121QagymS2xcPKZiROjvKmBPVgXOioLK3Wr/AI9qbl9saWbJdZFqjxXigpTMWy4hLrqQe9JJA5vHap1rFcYFlXhFyuTalQ4eRMrcCUFZQgMuetyjqQn4XT2VYpbbKA2W0FAAATyjlA+KsBleLuZJMx6U3NRHFjuqLkpKkFXahKFp5B16H1u/3VZT2xSrV6XlG5WMzb7x5NrAJjXq6xqdBCrq7FrUaFU0nZnvy7oEiq6oTE6dc2nQaklRWXe7Tm+qOKO4lPZuTVhamSrhMjHnaabdbCENFY6FSldeXv2G9Y3DUITw53AJQBzW+7KI27yVvdTVsssR4yS3GjtMoJ5iltASCfbsK/QQ2E8gQkJ/chI2/JWsdrMbTZRpsOVhYRJk9UvJmw1LzyA3rabsd5qvr1HgueHgwIHWFNoi50FMTe5O7RU3YblCxPJ8TyXJXhFtdyw2Lbo097oyxISUrU2tXcjnSRsTtvttX5vU+JlV7z3KseX5xaI2IOWtya2D2UqSOdeyFfZ8iem46ddquVxll1osOstuNEbFCkApI+I9KIaabaDDbSEtpGwQlICQPZt3Vd03T8p5fyZzwG62jNm0jWBGsb4VPQVTyfm/lBkku9HrZi3LrMRJnSY6swqftVwhYnlOIZNkboi2mdh7FsYnOj6THkgpWULV3I509xOwO21dTKI7upV6zWfgxMyKMVFp87a37KZL7XtOyQruXsgFO46bq2q61tMutFh1ltbRGxQpIKSPiPSutcLcmZapNqjSXbeH2VstvRNkLYJGwUjboCD1FZU9tsbWGIDIqQGzMtAzZpyxJMW17dVjV2C91E4cvmmSXQBDicuWM0wBN9J3TCpzUbUjDb9pYizWp4vT3PMW1QAwoOQSh1vmLqSPpfKRyjfvJAHfWc1UuUvT/IIeo1tjrd8+t71jktoG/M+UlcNR2/8AeAp39iq78nAc3yGNEseYZba5VojvsvSVQ7epqXP7JQUhLqyopSCpIKuUddvCszf8TvWS5LAkXO8RPmct0huei3IjHtnpKAeXtHCdigKPMABvuBvW43F4DDvpsa4Fg8oXCSZDg0BoOVt7SLWsSVouwe0MQypUe0h5FJrCA1uUsLiXEZ3WGaDeTJAEXPdwTG/mSxC12Baud+MwFSV+LkhZ53VH41qVWdr7176+V5StWfiKjqtTVxJPMr11CizDUm0aYhrQAOQsEpSlVq1fR3j46ox/6u7+/V+mrzHeKox/6u7+/V+mu7sTV/d9VwtuaU+/6KOZD/pFi/8A3x/+xNRvP4cmwxLi/CaUu23zlEhtPcxKCkkOD3LAIPv2qxOh2JG+3d7qEAjYgEewivYYfHnDupnLIaII49YuHKDpyXjMVs0YplQZoLjIPDqhp5yAZ59kqF3ZPmOZOTJt8kWmPNgNNMSUpb7NSkE8zalLSQk9QR3b11JEZm4WLK5lrm3C6LejoaVIcbQEPlsb/SuQDm2HQnb8tT5aEOJ5HEJWk94UNx+SvqQEgBIAA7gOm1WN2lla2G3GUbo6pB4TuG+N6rfsrO50u6pzHfILgQd8bzundzhd/wAis93jWRm1zUSlquUN1Ya3PZJCvs/3J3O2x671lrAAMnydY6KMmMOb3diP76ziW20b8jaE7nc7JA3Ptr9bDffYde/31U/Fs8kaVNpAI3mf8geA4QrqeCqeVFao4Egg2EaNc3ieMqHYzfrRj2PqtV6lNxpsBx5L8dzo46StRCkJ718wI2I3roRkmwW7DHryfM22JEguF3oGQ4hZQlXs6ECp+W0KUFqbQpSfgqKQSPiNFJSsbLSFD2EbirfP2Zi7KesSXX4gi1rekTedyp6NfkazOOoAG24Oab3v6IFo3nlFbPdbbYLnfIl7mNQnn5y5rTjyuVL7Kkp5SlR79tiNh1FY9mWyxjN0cdtqnxkFxfFuhOJKS+HNgkkd6R0KyfAdanS223Nu0bQvlO45kg7H8dfSATuQCR3HbqKxGOZObKZMTf2dIta/2CzOz6kZM4gZotfrazeDAmLdpUEs7K7BaL5i1z2VcTEeliTuT562WyObc9d0Ecu3s2PjXJBkM2mZjN3uSw1CcsghpfX8Bp48igFH7HmA23PsqbkA9SkH4xXwpSpJQpKSk9CkjcfkqXbRDyS5vpa37Ita3Hw0WDdlmmGhjvR9G3B0ib34TY79VC8qvEC72C9m2xw60x5shU1AHI8rtU7oSe9XL7e7rWScmRoWoRRKfS0qZbG2o/N07RYdUSke07eFSIJSEhAQkJHQJAG35KFKVEFSQSnuJG5HxVh54wM8mGmL773Dez/r4xZW+Y1DU8qXjN1TpaxcePB0d0mVBHozyby5gYbV5pMnpuYO3QRPhuI/jUgf8VTRVxh+kRay+BLUyZAa2O/Z77c2/d39K6NqtE5i4ybxd5zMqW62I7XYtFtDTIJOwBJO5J3J9wrIGG2qcmeoqLiGiykb+qkEgk/Gdh+SpxdenWcGkzA1G9283HKeSxwWHqUWlwES7Q7mAmBY7pMa6wuavS7gQ+t3tv8ACU/+3NeaNel3Ah9bxbP4Sn/2xre/Dn6w/wAT8wuZ+LP0A/kPkVsLSlK9yvmyUpSiJSlKIlKUoiUpSiJSlKIldO8fsTM+8L/qmu5XTvH7EzPvC/6pqur6t3Iqyj6xvMKsB3VCtRNZtN9K3YETNci81nXTnMKBGivS5khKfhKQwylThSPFW23vqajurXq1XWy4vxp5jIzafEt0i94dakYvJnOpaQ7Hacc88ZZWsgcwcKVKSDuRse6vG0GNeTm3Cbb17CtULAI3mF2NWOIizMYTgWoem2eW8Y9dc4gWi8T1oSENwz2hktOh1IUypIQObcJUnb31O8A4gNI9T7/IxfDMr85uzDBliHJhPxHXo++3bNJeQkut9R6ydx1rX7Vi86RZ4myfMBjbJtzut9mjXq4JZT5lfJ4bX2rrSgoh4D1UKVsASNuvfVm6roP0WOib8fkRKctmUMpcI2UQIrZSkn9yFHfburadRp5Q2CDDju3Cb8fBaza1TMXSCJA95iykuS8TuhuI5FMxi+5001LtjqWLi6zDkPxbe6e5EmQ2hTTKuvULUNvHas5m2s+l+nKrcMzzGHbhd4rs23qKVuJlNN8nMWy2lQWT2iOVI9ZXMOUGtSND38js+guQWu88QWG4lGtMq7s5dYrxjDL8xmQp5ztu3UuQlbynEkFJ5fWCkpG+1SXBMdtMDOuFK0IuMy9woGO5BIt0q6W8xJCm0sJUwtTC1KLZShSQnck7AGsnYOkwkSbT3wCeEbu1YsxdVw0F48SBx7exXSjiz0Acsbl8bzsqDUpUJyCm2yjcEPJTzqBidn2wASQoqKOUDxrtZvqpjd70hhai4Lq1AsVpuNwgtxr/AOj1TW1BcpLao5ZI5kqcO7W5AKCdzttUa0xgwE8ZOstxREZEwWPG0dsEDn5VtuFQ37/W5Eb+3lHsqi5SWmeD/I2WglDTesTiEJT0Skenm+gHgPdUMw9IuGWdW8D6QnhuR2IqBpzR/lxGnets9RdcNL9LbnFsuY5Gpi6TW1SGLbDhvzZamUnYulphC1pQCCOYgDoa+ydctJImn0bVN7PLb8y01aWo09ClLD7xJSGUNpBcU7uCOzCeYEHcdKrTArzYcT4stZWs2ukK13m8RrNLsb895DPnFpbjlK0srWQClDwVzpB6Hqag2T6p6aSnsOjaK4HisB3INQLsm2ZRkkcptEe5MtAyLjHCFjtVOk8jZ3QFFKvZWDcK1xDYOgM7rif7rvssziXAF0jeI36x/dOav/TzXfSzVG8S8dxDI3HbvBZEl+3TYMiFKSyTsHQ0+hClI3IHMAR1FQTR3X+DH4fLLqdrTlrLUm4XKfAS+I30yU4iY820y0wykqcXyNjohJJ2JPiar7FrtepXG3iUDKdVrHm15iYdd25K7PbW4keBzLbUmPulxwuKIBWQpW6QR0G/WJ4NFxCZw+aOm76iT8DyNrLr2vFsibiIfhxp3nMkKaldp9LCXEEpHMRuegPfV3mtMCNxg9ujuyd3D3qrzqoTO8Ty1b29vFbaad6wad6qpnpwfIfPJNqUhE+E/Gdiy4pUN0dow8lK0hWx2O2x276mVa46NZzlTOuuS4PqLIwPKLxExhm5yM1xqN2DvmyHilEScN1BCuqlpAVtsCduvS/ceyOwZbZYuR4veYd2tU5JXGmRHQ4y8kEpJSodCNwR+KtGvS8k6BpbxW7Qq+UbfVY+8f5+v4k/oqrLPl16zDWq9Y9Zp6mMbwSG1HuaUJSTPu0pPOloqI3CGGQFEDbdbo36J2q1Lv8Asgv4k/orXvQ5q4k67W2Ioov3zbXUoJOyuZyK2Yp+Lbl2+KuS1gJqvOo+pifd811nPIFJo3/QTHvUruXEnolab8/js7OmEPxJQhSpKYr64UaQTt2TspKCyhW52IKxse/arLBCgFJUCFDcEHcEe0VpDps/LjcLTttveveJ2HHoVulwMhsMzF2np0aQVLD7LgU+lbr6lElJ5d1EpIraHS+649iWB6e4ddss7WfPtLEe1i5t+aTbilplJJ7BRKgsN8pUnckeNW43BMoD/iJJBI37t/oj3X5qrBYypXP/ACAAEA7t+7U++3JceVcQGkOF5I7ieRZg2xcowQqYhqK8+3BSv4BkuNoUhgHff1yOnU7CpTiOYY3nliZybErq3crXIcdaaktpUErU2soXtzAHopJG/cdunSqI0LzTAcAtWp+Pao3+02m/x8tu06+s3J5Dbs2K8rmjuJSvq82pkhKQnf2Ada7/AAnZRieMcNeMXK8XaJZbbKuk6JDMxwNDndnPBlkb/ZkbAJ79xTEYJtOmSwGQWidxkEyLdlrmymhjHVKgDyIIJjeIIF79vAXWwSACtIPcTtVU8NOc5RqHpvJyDLrimbPbyC7QUuBlDYDDElSG07JAHRIA37z41a6Bs6kHoQoVrPwn6kad41pbOtWRZ7jtrmoym+LVGm3Nll0JVMWQShSgQCO47VRRpeUw9QtEkFv/ANK6tU8niKYJgEO/+VaWpOJaqXeau8YRrK5ikCNCJXATYY00Ouo5lFztHDzDcbDbuG2/jVecOY1z1Jw7ENU8k1zU7BuhMmZZE45ESl1tDq0Frt07KTvy78wG43q8lXa1X3GJF3slzi3CDJhvKZkxXkutOp5VDdK0kgjcEdPZVYcGZB4acBIO480e7v8AvTtWtqubhHS0SCB6LZgh03id3NVuptdimw4wQT6RixbG+N/JQzSs8QGsFvyHJY2vq7BHgZNdLPHhIxiHJCWo7xSg86tiTsR3jw76nWhufZ1eckzzTHUedb7resFnxmRd4EfzdudHkNFxsraBIQ6kAhQHTr7tzUvD1o21nmP5bfVao6gWH/ntfGDDsd781i+rI+Fych9Y79Tv12FX5heA6c6D4pdXra+qBBU4u6Xm73aap599zb1n5D6+p2HxAflrYxzqLS+kACbAANAi43i5tbfqtfBNquDKpJAvJLiZtwNhe+5Tuqu03zfJcg1k1ZxK7XBL1rxeVaW7WyGkJLCXopccBUBurdXX1idvCrKhTYdzhR7lbpLcmJLaQ+w80rmQ62obpUkjvBBBBrX7T7NsMxHiK1xRleW2azKkzLIWRcJzUcuhML1innUN9txvt7RWjhqWdlURJDR//TVu4ipkfSMwCf8A5crb1CxnPMljQWcE1KXhzrDi1SXU2lmf5ykgBKdnSAjY7ncd+9UfpizxE6h3XN7c7xFrgjD8kesKVpxWE550ltCFdqR05Cebbbr3d9bFY/lGM5ZFcnYtkNsvMZpzsXHoEpEhCF7A8pUgkA7EHb31UPDUQcm1rA8NRJv9i1V1Co6nQqAtEtiJa0m57Qqa7GvrUyHGHTo4gWHYVi3J2tOfa6agYLjOsZxa1YlGtK2G0WGLMLqpMfmWSpzYj1kk95+Ft4VlsYzbVPTvVmzaUauX63ZPb8ujSHceyCLAEJ4SWE8zsWQyklG5R6yVJ+L4sLh+Z4fiPFLrMcryuz2YSomPBj0hNaj9qUxlc3Lzkc224329or83/KrBrZxHadW3Tu6R73bdO1Tr5fbpBWHYrDjrPYsRg6PVU4o7kgE7D4jW05hJyOYMmQGcoEHIDOaJu7tvMLWD8oztec+ciMxMjPERMej2WiVYmHZfeoWquVaU5TPVMcQw3kePynEpSty2vKKHGFcoAUWHgUg95QtG+5BJseqVyoKe4u9P0Qt+0i4be3ZxT4R1PMpb5v8AxAdviq2MeyTHsttib3i96h3W3rcW0mTEdDjZWhRStIUOm4UCD7xXOxNOA2oBqATzuPGJXQwz7upk6EgcrHwmFkqi+p3+gV5+8o/tE1KKi+p3+gV5+8o/tE1zsR6l/I/JdPBfqaf8m/MLXGov6bn3XUE47bZJag2OImVcykAl557cMsbnuASFLO3X4I9tSmoDgIWjN9RmFnaUbrGcST/2SoqezPxdDXm8DTaadaqRJa23e5rZ7gTHAwV7vHVHCrQpNMBz78mtc6O8tE8RI3rJ3PVLA7PcX7XPyBCHoqw3JWhlxxqMo9yXXUpKEH98Rt41KULQ6hLjS0rQsBSVJO4UD3EHxFUNgLtyhad3CJP1FsloahuzEXqBNtSHHm3StXadoVOArKgdwduoIAqy9PpFnx/BsWtT99U4mVHSxb1zW/N3pI2KkpDaiSFBG3T2CujtXZFHBtPkCXOa7KdTNiZHUaBpMAusZm0nmbI2zXxrgcQA1rm5hdoi4EHruJ1iSGXBESYHbvuo2GY1cvRF4vQblpbDrrbbDj3YNnuW6UJIbT71bVlbHf7PkkE3OxT25kUOuMds3vylaDsoAnv6+I6HwqA4LfbBjV+zuFlVzh2+5LvTs14zHEtl+EpCexUkq+GgJBGw326ivmjF4sFs04eujkpi3WtV7nhhT6g2lCVyCG09e4ncAD8VRitlU6WHc6m1xc0sAOrXZ2k9URPK5kKcJtepVxDW1XMDXCoSNHMyOA6xzRvvYQVZ9Q2/6pYba13G0pyBtNxiNuNlQaWplmRyEoQt0JLaVb7eqT7qmR5uoSdlbdCfbVP4Pd8Wtmjd2tGRy4jMqGbgxeYr6kh1UlS3O9B6qUrdPKdjv02rW2ZhKVdrqtRrnQ5ohuvWm5sbCIjeSLhbW1MZVoObRpOayWvdmdMdWLCCLmZmbAGx3SzHc6hQ9NsfyzNLu0y9cIbCnHCj1nn1p35UNoG5Uf3KRWYsWa4xkkeXIs9zDvmA3ltONLaeYG2+621gKAIB26dapuMJ0djSG4em4toiiyOsMTpkYPsNTFITyhQKkhKlJBCVE94NSeEwX9QbpLm5zGvV2i43IZlNwbYGmQyo7o7V1K1DnBBKUnrsTXXxWxsL13yQTncImBDy3LAZG7XMIJFo142D23i/+OnlBaMjTMSSabXZpLwZvpkMgG86Scaw6bqdjNjJ2imV2YQ92LvYpUsAoStzl5UKII6KII361lskzXGcSVHavty7J+Xv5vHaaW886B3lLbYKiB4nbaqx81itcJ5bbYbS2bAHyABsXCoKK/j3671nbPMh2rV55++yWo7lxxmCi0vPqCUqCCovoQo9ObcpUR3kVTU2ThAajqYcRTNQESJdky3By29KTYwAr6W2MaRTZULAajabgYMNz57EZr+iADLZLhZdzFtR4dyn5rd5l8Zex6yuRVxX0N9GmVMBS99hzE8++4PUHpXYnajY1eWS1jmatQ3IdziRpD5hLdQ52jnKGUkjY85BTzgnYiq/u02yz8f1ul2F1lyI4lklxnbkW52ADikkdCCoHqO871LdRo8eHgGJxozSGmmrxZEISkAAAOJ2AraqbOwgxDDlILi1oHVgRTpuOYFpkmTOl7rTp7Sxhw1TrtcGh7ietJmrUaA0hwgDKI1tbtHftupkW56hX/DfOEtNQI7YiKEV3mLwSsvFaiNtk8o27gduhNZG2ZlZLTg1syTIsujzWJDaQm4iOWvPFqJ25GQObc7fBA36VhIM2Kzq3nsN+Y02/JtEAstLcAW6EtO78oPVW3u7qh2JvxYFu0au16cbbtLUOWyHnSA01MWj6SVE9EkgKAJ8ards3D12jK0tGWmYES7/AInPMHLMkiN4mTBsBa3amJoOOZwcc1QSZAb/AMzGCRmghodO4xAkXJt7HMxxvLBIFiuPbOxCEyGHGlsvM793M2sBQB8DtsazNRmBecRuOezYlrjNyr1HtzfnlwYCVoQyVnkYW4D8LfdQTt3VJ683jKTaNQBjS0EAw6JuOIiRwsLbl6fBVnVqZL3NcQSJbMWPAkweIk33oO8VRj/1d39+r9NXmO8fHVGv/V3f36v0munsPV/d9VzduaU+/wCiiua3q+Wdy1eg20vLffc7VgpBLzaEc5SCe47A7bVy33Iv+Zb+SWKQN1NIcZWUg8u6wCCD4jcgj219yE7ZHi3X/wDGP/2JqMZwy7jEG5xWWybTeyFNpSOkaXzpKh7krAJ+MV7zCUKVcUGQM2v8hmIIPcLd44L59jcRWw/nNQOOXT+JyAgjmTB7YPFTm43q2WhLXpCV2a3+jTaUKW45t38qUgk/kr8t5BZnbY7eUT2zDY37VzY/SyDsQpO24PXu23qPTRNbzxxSLtHt637a0mKuRHDgcAUe0QglQ2O5BI8RWPuDKHbPmMlm7puLy2G0yCxF7NoOIG5KSCQpXL8Lb2VgzZ9FwbJMnKd+8gR6MCJ1zG4iFZV2lXY58NEDMN3+LSZ9KTMaZRYzPGdS58OChp2W+G0vuoZbJBPMtfwR09tdKXk9hgzPR8m4pTJ7VLJaCFKUFEAjfYd3rDr3daw2U3a2SolhaizmXlyLlDcQG1hR5Arqo7dw6gdfE7V3bA2381WTvBKe0L8ZHN47diDt+WqW4SmykatUHQ20/wAgOB4rYdjaj6wo0SLkCddWuduI4D3rtTMsx6BJciSrklLjBCXiltakMn2LUkFKfxmu1cLxbLVGbmTpiG2nSEtEbqLhPcEBO5V+Ko9h8+0wMVej3SSwy5FdkC5IeUArnK1ElQPU7gjb21i8fJtjmJy7yewi+ZSmo6n+gacWsFsEn4JLfQb/ABVacBSlwg9UkfygE2tbTtsR30DaVXK09XrgHf1Zc0da99f+twe6Y27ILPdXjGt85LryUFxTXKpK0JB2PMCAQd/A9a4E5dji5IipuiCoudiHORXZFzu5Q5tyb7+G9R+ZJj3LK72LE6h2T6AW0pxo7hTvMdgFDvIBA/orpJU29gTbMjLILVtVFTHUwi3guoV0HIE8+5cCvdvv1rNuzqJgukTltvEidzTPYLTxVbtqVxIblMZr2g5TG9wjtMmOCmlwyCz2t8xp05Lb4QlYaCFKWoEkDlABKuoPQeyv3AvVqucJdwhTW3I7ZUHFn1ezI7woHYpI99YaCwEZzu6S46zYmUhxadldXCCfcTt1rCX5l5yJm7cZJ287iOOhKeb1ORBcOw7+gJI8aqZgaNQimCQYaZ3XIGkds6q6ptCvSBqkAiXCLz1Wl2sxujTtUsg5TYLlJREh3AKcdBLXM2tAd27+RSgAv8W9J2U2G2yFxZc7lcZALoQ0tYaBG45ykEJ6e2o/dt5rFrRKzCG+29LYXCTEgAuFaTunl5VkpG3QnboK5XQtq53p/HclixVB8rnwbiyOyK+QbqB3CglQ269RWYwNCZkx38QNcn013rA7QxMRAmRw0gn0c/Z7WmgUgenNruNsQxdW0ty0OrQyG+fzoBIIKV/YhO+/v3rgXl2OIkGMu5pBS52Jc7NfZBzfbl7Tbk336d9YO3zk3Gfhk1EFEMOsTClhA2QgBsD1f907bj3EV0DINqxyTMst7gT7M0pxSrXcmQHEnnJU0CDvzc3cFA+FZNwFMkMfM6cL5nDUA3tadeMALF20qgBeyI13m2Rh0LgYveLi3VJJVhV6XcCH1vFt/hKf/bmvM5hwvMNOlstlxtKuQ96dxvsfir0x4EPrd7Z/CU/+3NW/h0RjSD7J+YWt+KzOzwR7Q+RWwtKUr3C+bpSlKIlKUoiUpSiJSlKIlKUoiV07x+xMz7wv+qa7ldO8fsTM+8L/AKpqur6t3Iqyj6xvMKsB3Vg8swbC89gt2zN8Ss9/iNL7Rtm5Q25CW1fukhYPKfeKzg7q1v1k4jM00x4jMUwNqHa14RNhQX77IdZUZMbzqWuKhxK+bYIDhZ33Se8+2vGUKb6rop6i69jWqMptl+ivhrDcQZtltsrOK2hu32d5Ei3REwmwzDdRvyONIA2QobnZSQCNzXck2WzzbnDvUy1Q37hbg4mHLcZSp6OHBs4G1kbo5gADseu3Wq24mdV75o9phIvuJRIsvJ50tq32eNJSVNLePM44paQQSlDLbqz1HwRXbwjWGyHQ7EtWNUckstgRerREmy5Mh5MaP27rYUUoCjv7dkgk1Ip1CwVOJhR5WmHmnwEqQXzSjS/J761lGSac41dLwzylE+Za2Xnxy/B9dSSTt4b91Z2RZLNLucO9SrTDeuFuS4iHLcYSp6MlY2WG1kboCgACARuB1qLK1n0yk6f3XUux5xYrpYbSy4t6YxPR2KXEj1Wlr/6tSlFKQFDfdQ6VA8K16k6oaY4BnONZZhNnn3u8QYd+gzZKnA0p0KUu3sEHcSynk5Qr3nbrUilWcJMwLclBqUmmBqbq5o9ms8S6S75FtURm4z0NtypjbKUvPpbBDaVrA5lBIJ2BPTfpXSOF4cq1uWNWJ2c256X5+5DMJvsVyefn7Yo25S5zgK5tt9+u+9c9hyXHspYkSsavUO5sxJbsGQuK8HAzJbOzjStvgrSehSeorCztWtLrZjBzW46hY/GsHnDkQXJye2mOp9CilbaV77KWFJIKRudwfZVYFSYEqwlkSYXdy7T7A9QGGI2dYZZMhbjKK2E3OC3I7InvKecHl38du+v1d8Ewi/463iF7w6yzrEyEpatkiA0uK2E/B5GynlTt4bAbVgWtcdKLjg961Bx7P8fu1nsTKnJciPcEdm0vb1G3Ff8AVlatkjcdSem9QjCte39T9KsHzzG8swqzXG8XiBDvsOdJU6hgu8yl29og7iWU8nIFe8kVY2lWjeAD7iqzUozuJI94VpWbT7AscFvTj+E2K2eie18wMS3tNGL2oAc7MpSCnnAAVt37DfeuR/BMIlY2vDpOHWR2wuFSlWtcBoxCVKKiey5eXcqJO+2+5JrG5xq9pXprJjw8/wBQrDj8iYOZhm4TUNOLTvtzBJO/Lv8AZbbe+srLzPEINogX+ZlFqatd0dZYgzVS0dhJcd+pJbc35VFXhsetYf8ALZ17rOad22suHGsCwrCbVJs2EYjZLDEkhRcZgwG2m3FEEbrSkDn7+4+HSuHTfBrfprhFqwi1vl9i2NrHalpLXaLW4pxxQbR6qEla1bIHRI2A7q6uH6waVagXSXZMG1Fx++3CCCqRGgT23XEJB2KuUHcpB6cw3HvqX1DzUEtfPG6lgYYcxR68fsgv4k/oqt4uC3SxaxTc+sC4/onKLY3FyCMtZStMyN/mspsbbKJbUppY3B2CD12NWTdxvPX8Sf0VrhgfEjd5+v8Alekeb2+LEtabw/asYubTZQl6SwhK3IjqiSC6ULSpJ6b7EbHcVy2U6r3VTSvAM8p+mvcum+pSY2kKlpIjnH1071bk/TDTW65CjLbnp/jsu9oUFpuD1tZXI5h3K5ynckeBPWs1LstmuFwhXa4WmHJnW1S1QpLzCVuxisbKLayN0EjodttxUEf1BvzfETE0rSiJ6GfxB2+qV2R7fzlMoNABW+3Jynu27/GsvN1i0ntuUJwm4akY5GvxcDPo924NpeDh7kEb9FH9yevuqt1OuY1NptJgfRWtqUBOgvG4SVXmTYJrf81FzmwbBppmTbstcjH7zkTJan2JtfcyUoZV2yW1blBCkqPjU1020gsGD6YWHTi9NRMhTZ1edOPzIqVJdmqdU6p9KFbhJDi1FPiBt1qXuX+yMX1nGHrtFRd5EZcxmCp0B9xhCglTgR3lIUQCfaa6svM8RgPXiPNyW2sO4/HRLuyHJCQYLC0lSXHh9gkgEgnwBrJ+IrVGBgECxsO4Hx3Rc8VgzD0aby8mdRc95Hhv3BZnc7779fbUQe0d0jkPLkSNLsSdddUVrWuzRypSidySSjcknxrmtuqumd3lXeHa8/sEl6wNB+6pbntnzJojot077JT7z8XfXDiGsOlGfXNdlwvUSwXqe2kuKiw5qFu8g71BPeoD2jcVW1lemCWgiNdfFWufQqEBxB4aKR22z2iy2xqy2e1Q4NuYQW2okZlLbKEHfdKUJAAB3PQDxNfbTabVYbezabHbYtugxgUsxorKWmmwSSQlKQAOpJ6e2oNoxn99z9rNFX1EVJx/L7lYonm7ZRvGYKOQr3J3X6x3PT4qleV5niOC2v01mmTW2xwOcNiROkpZQpZ7kp5j6x9w3NRUpVG1DSNz71LKlNzBUFgu7a7LZ7Gw7Gslqh29l55clxuKylpK3lndbhCQAVKPUnvNdbLLAjKcXu2NOPNNJusN2GXHYyZCEBaSnmLa/VXtvvsem+1dbDs+wjUKC5c8Fyy136KyvsnXIElLvZr8EqA6pPxgVjYOsWlFzyk4RbtR8dk3/tCz6PauDani4O9AAPVQ/cg7+6oDKweTBkXNtOaF9IsAkQbC+vJSGwWaHjlituO27n80tUNmExzndXZtICE7nxOyRvWIu+mOm2QXB2737T7G7lOfILsmXa2HnXNgAOZakknYADr4CoJjnEJj2X51qDgVpv8AjsaRjDCRaZDk0OCY4I63JDq0j/q2VgJWE7kcqtz4CaYHmcK82mxWu75djt0yabZmro+m0vfSZTRPKqTHSr1iwVdAqrX0a9AlxkG3HeJVbK1CsA0QRfwMLM4/i2M4nFcgYtjtss0Z1ztXGYEREdC17AcxSgAE7ADf3Cuxb7NZ7S5LdtVqiQ3Lg+ZUtUdlLZkPEAFxwgessgD1j16VFdRs4j2awZBBx7NcXtOTWqA3O3vbx82htLcCUvSUpPMltXrJB8TtUdncQGMWzWu36Qz7zY2u2shmyZapoCxPU4hLURKPBSkKLgB6kFOwqG0K9YFwk6k8hBnxUur0aRDTA0HeZEeCml50004yO4OXbIcAxy5zngkOSZlsZedWANhutSSTsAAPcKy1msVjx2Em249ZYFqhg7hiFGQw2D7eVAA3riteT45e7CjKrPfYM2zONLfTPYfStgto351846bJ5Vbnw2Ndi03e13+1xb3ZLgxOt85pL8aSwsLbebV3KSodCD4GqnGpGV0wLK1opzmbElQ7DMEucLUHLdTcqVHXdL2pq2W1llZWmHaY+5bQVED13HFLdXt0BKRueXesxp7g8DT3Gk45b3kvJVLlTnVoYQwhTr7ynV8jaPVbSCrYJHcAO+pJSj6z6libW8BAUMospwRrfxuUqL6nf6BXn7yj+0TUoqL6nf6BXn7yj+0TWpiPUv5H5LewX6mn/JvzC1xqPPY7Lj5wzllrU0GpkMwLq0pWxWlBKmHU+1SSVJI8Uq91SGsPLzPEbfMRbp2S21iU5J8zSy5ISFl/YHs9u/m2Unp/vD215TCmuC4UWl0ggiJseXDUcCAV9CxYoFrTXcGwQQSQLjnx0PEEhfufieK3Sei63PGrXLmo25ZD0RC3Bt3esRv0rvSIMGW7Hflw2HnIi+0jrcbCi0vbbmQT8E7dNxWLn5vhtruqbHcsqtcW4KIT5s7JSlYJ7gQT0J9hrv3i9WfH4SrjfbpFt8VJCS9IdCE7nuAJ7z7hWTm4s5GuDr+jrp/17OSxa7BjO5pbY9bSxHtdvNQ++WHO1XyVLjWrFciircD1uXd0dm9bTsN0ApbV2iOb1h1B8KyuIYPEx/EmcbvBj3Zan3JktbrILbkhxwuKUEHfYBR6eI2FZW25Njl4tjt6tN9gy4DAUXZDLyVIbCRurmI+DsOvWuKFl+KXGVJgwMktsiRDZEmQ23JSS00QCFq69E7EdffW5VxONqUvIZMoaRMNIMiwnhruiSZMladLC4GnW848pmLwYlwIh1zHGY3kwBAgWWYrGScYxubc03qZj9tfuCByplORUKdA22+ERv3V1LbnuEXhYbtWW2qWtTyY4S1JSolxW/KkDxJ2O3t2r9XvOcMxuUmFf8ptlvkKAUGn5CUr2PcSO8D3mtRmHxlKpkYxwcRoAQY321hbdTE4KrT8pUewsB1JBE7r6Su+9ZrPItossi1Q3beEhAiLYSWQkdwCCNgBX5tdhsVkiLgWazQoMZwkrajsJbSvcbHcAdento9frJGjRpki8Qm483/NnlPpCHvVKvVVvsfVBPxA117Fl2LZOp5OO5Db7kqP9VTGfSso9hIHXb391RlxXknGHZJk6xPE7plZZsJ5VolueIGkxrA3xHgu56KtZt3of0bF8w5Oy817JPY8n7nk222921cd0sNjvkVEG9WaFPjtkFDUlhLiUkdBsCOn4qwluzS1W6wQp+YZdj6Xpa3kokx3wiO9yLI2RzHclI2CvfvWSs+YYpkD64tiyS23B5pvtVojyErUlG+3MQD0G5HWrH4fGUCXgOhpPWGaJ0Jn66qpmJwWIaKZLZcB1SWzESBHfpouyiw2JqM/CassFEeShLbzSY6Ah1CRslKk7bEAdAD4VzyIEGWy3HlwmHmmVIW2hxsKShSfgqAPcR4HwrDw9QcFuFzTZYGX2iROUooSw3KSVKUPAddifcKx0XUi0S9QbhgiZkBK4cRtba/OAVuyVFXO0E+1CUgkd/XwrIYTHPLiWulozGZFrCb93dyWJxmAYGgObDjlEQb3MW7+/tKkkiyWaXcGrvKtMN6cwkoalOMJU6hJBBAWRuAQT+U0XZbO7bPQjlphrt3LyeaKYSWQn2cm2234qxlmyJlnEY+QZPkNlUnkUp+fFd5IavXIHIpR9wHxg13LJk+OZLGcl4/fYVxZZOzq47wX2Z/3gOo/HVdSliqc+kWsMSJgEHcd3Zobq2lVwlWIyhzxMGJIIvI39uosue02WzWGKYVktUS3xyrnLcZlLaSr2kJHU13KwCtQcERKjQl5jZ0vzUpXHQZaN3Eq+CR18fD21IKpr067HZq4IJ3mb+/VXYerh3tyYdzSG7mkW92iDvFUY/8AV3f36v01eY7xVGP/AFd374r9NdbYmr+76rk7c0p9/wBFxLZZcWhxxlC1tEqbUpIJQSNiQfDpXx+OxKbLMlht5s7EocSFJO3d0NfulehDiIvovPFoMgjVcEuDCuDXYT4bElsHcIdbCwD7djX7YixorCY0WM0yykbBttASkfiHSuSlTndlyzZRkbmzxfiuoxZ7RF5vNbVDZ51BauRhKd1A7g9B3g91dlLTLa1uttISt0guKCQCsgbAk+PSv1SpdUe70iSobTYyzQAuq/abVKkJmSrZEefRtyuuMpUobd3UjeutfIl2k9iq3eZPtJ5hIhzE/S30nuPNsSkg+7brWTpWbK72ODtY43/vcsH4dj2luk6xY/8AvfrobLB2SyzI1yfvFwRDYcWwiKxFhg9my0lRUepA3JJ37gKyPoi0+d+kPRcTzrfm7fsU8+/t5tt9/fXbpU1MRUqOLpjdbhwWNLC0qTAwCYM3vfj/APncvyGmQ8ZAaR2pTyFzlHMU777b9+3uohpltbjjbSErdILigkArIGw3Pj06V+qVTmPFX5RwXUjWi0wn1SodriMPL35nG2UpUd+/qBSXaLTPdS/OtcSQ6jolbrKVKH4yK7dKz8rUzZsxnmsPI08uTKI4QvwWGVLQ6plsraBDaikbo3Gx2Ph09ldddntC5fn67VDVKB5u2LCSvf277b7126VAqPboSpdTY70gClel3Ah9bvbP4Sn/ANua80a9LuBD63i2/wAJT/7c13Pw5+sP8T8wvN/iz9AP5D5FbC0pSvcr5slKUoiUpSiJSlKIlKUoiUpSiJXTvH7EzPvC/wCqa7ldO8fsTM+8L/qmq6vq3cirKPrG8wqwHdWrWr+AtapcR+W6fuEJcvWji2Izni1JFy5mHB7Cl1KD+KtpR3ViBiONJy456LOwMhNvFq9Ide1807TtOx79uXn9bu33rxlCr5FxcNYXsa1LyoDVqVbM5kcRFo+aaehXJpnpjcnLs2ofAyaZGejOIP8AvNtR3j7u3FcekEaz3TUHhzt2bssP2tnScysdYmAKjrvAU2HVJSr1VPJY25fEDqK2qtGmWAWCBkNrsmJwIUXLJD8u9NMoKROeeTyurc696kkg7bd52ro33RjSzJsMtmn19wi3S8fsiG0WyGoKHmIbTyoLLgIcbIHTdKgdq2/O6d2gED5SD9StXzSpZxIJ+dx9AqSyGBYofFLnNsw+HDTGmaWSZOWxozaew8+S6oRFuoHqh4t82xI5inrULt7cNPDJwsORW2EqczDGlOqaSkFS9ngSrbvV08etbV4JpZp5pla5VnwXEoNqjT1lyZyAuOS1kbbvOOFS3DsSPWJ6E1h7Dw+6MYxAatdg0+t0KHHvLWQssNqd7Nq4tAht9CSohKkgkAD1fdQYpgEXtHfAIv7+1DhXkza891wbKhdVMxufDnqDqpYrA0rtNWLWzecRaT3HIXHEQZDaf94l1l8+5JNZO94va9Osq0d0MwPTqy5RnWN43Ll26ZkFxcj2yC0OVEqSW0JV277jpURsnmSNzuK2LyLBMOy662O+5NjkO5XDGZZnWiQ+glcN8gAuI69/Qd+/cPZWN1C0h021UTB+b7FI11dtilLhSC44zIjFXwuR1pSVpB2G4B2NYtxTOrmB7fdA4bvmVk7Cu60Hl75PHetbMFZvJ4jtZbdly8NeuqtOGFXONjUZaITbwW4UJcDhJW8lJSSogHZSeg2rH21uGnhT4Zlxm2AXMwxdTymgkFS+d0Eq271eHXrW0OKaOaW4NPTdMQwa12mWICrYp6M2Qp2MpztFIc3J7TdfrFS91E+NdCxcP2jGMW9NpsGn1uhQkXlnIG47anezbuLW4afQkq2SU7nYDZPurM4umTv3eAI4rAYR4G7f4kFVzw/W3Hrzq1rvMyyDBm5Y3lyojwnNIcdbsojt+aJSFg7MFJWenqk99UNcLZZLtopfMWtbaZGBHX2Jb7E0hRMc29clAeaZP/Y9op0Dbp1O1bh59oNpDqfdWr9nODQrlcmmfN/PA46w8tn/ALJa2lJLiP8AdVuKy0jTDT2TjVpw5eH2xFjsUmPMttvaa7JiK+wrmaWhKNtilXX3nqd6huLY05r7u6OH9Cl2Ee4Zbb++eP8ASql1bx6w4/xIaAzLDZYFteVNvltUqHHQzzRRAJDJ5QN0AjcDuFX1HnwJjr7EOdHfdir7OQhp1K1Mr7+VYB3SfcdjWPu+JY3fr3ZckvFnYl3PHXXn7XJc354jjqOzcUjY7bqR0O+/SsDgWmsTCcoznKGTES9ml3buTjUVpSEoDbKWwpZUSVOqIUpShsOoAHTrrPe2owSbge+5+hWwxjqbzAsT7rD6hZW7/sgv4k/orVDHNLrfq61rzismWqBPb1CXOs9yb6O2+4NRmVMvoI6jZXQ+1JIra+7/AOfr+JP6KjtkxXHMbl3adYrQxCkX2abjcnG995MkpCS4rc9/KkDpt3Vx24g4d1TLqTbuMrsOw4xDaebQC/eIWqWmuo+WZ5r9JN5ti4Ge4vpvcrLeIoRsPSbMoFDjfgUOgtuJ2/dfFWG03xrK71wnOLlP6QtYnPtsp++XC6sTFT2ZRUrt3pK0/wD4lDnUbddwnYd1bfNYNh7GZvaiMY9DbyWTDFveuaUkPORwQQ2o77EDlHUjfoOvSordeHPQ+9ZA9k9002tT8+RIEuR9USw++Dv2jjAUGlq367qSd63RtKjoGlo6psJuJsLi17HdwK0js2tqXBx62pIsYvob2uN/FVfg634muuj7N4yJi7yn9K5TDdyCVtpuC0vMnnQHAFkqQObYgK8SKwGeXOBcck4q3bbMZkJjYbbozqmlhQS8iI9zoJHiD0I8D0rYvO9K9PNTYkOFnWKxLoi3LLkNaitp2MogA9m42UrQCAAQCAdhXXtmjGldmtV0sdpwW2RIF7gItdxjsoUlMqKjn5W19ev1Re5+ESokkmq2Y6iDnIMwBFos8O15WiFY7A1SMgIiSZvPo5f7dU3mlo0ywLhvwm2PaaQbyvIDYLbFgJeEFE+e4EutGZISN1NBzmWoL5gonuO9YjM288ga/wCiCc/i6fWq4LvkpMWBjaHjKRH81WF9q64E7tb8qQkIA38fCtj7/gOGZTiYwXIsbhXCwJZaYTBfQVNoQ0AG+U77pKdhsoEEbd9R+x6CaPY2/CmWfA4LUy3zG58aY4469KbfQkpQrtlqKyAFKASVcvU9KUsfTa0l8knN2+kI427bX4pVwFRxAZAAy9mhnhfsvbgorwxf5vqf/wDEi9/pbrqav3RE3W3DMUxTALPkWcxrRNukCVf7g4xbrdEK0tuLDSArtXlEADZPMkDvAq37BiuO4qm4Jx20sQBdZztzmhrf6fKd27R1W5PrK2G+3TpWIzvSjTzUwwl5vi8e5PW0qMOR2jjL7HN8IIdaUlYB2G432NUDFUziTWcDBB+UcRPKVsOw1TzYUmkSPvPAxzhawNyM1t2o+v6WJONnLGtOWnHW8VZcbjtSkh3lGyiVKfShQ3Udj1HQVZMGz8PVv4X8MuGTQoScUjxbRKjSIKT5ybiVN8q2ltfTS+XieblPN8IHpvVr4rpVpxg1wF1w/DrdaJghej1PRUFKlx+0LnKvr655yVcyt1EnvrD23h70Vs+TN5fbNObUxdGZCpjKkhZZZkHqXW2CrskL368yUg71sVMfSqEekIg2gEwI427DeJNitangatMH0TM6yYkzwv2i08VWdjjW9GovEuUxYyHkQYSm/paUrSFWlwq26bjc9/tPfUatjacJ0g4e9d2fpbeLQIFovrgH/wDJ57aWnFK9zbvZL93WthrhpLptdMwe1AnYjCdyGTCXbn5+60uPR1tltSFhKglW6CU7kE7dxrvqwLDV4T87hzHYi8YEIW30WoEs+bAbBvv32AA8d6x8/piLH/GeQblI137ll5i8zcf5RzLswPdvWrWWRncl4ZNa9b7g0RI1BfLsArHVFoiyEMQ0j3KSlbn/ABg1Zrce3ji3xxb0aL2itNFupK207qcExvZQ3HVQHj37VbM/BMPumGnTyfj0R7G/NW4PowghnzdG3I3sDvsOVPj4V1Mi0t09y+92PI8kxaHMumNrSu1TFFaXYvKoKASpJG4BSDyq3G47qg45jgWkEA5u4ENAGu6FIwL2kOBBIy95BJPvlaw5HIueFNZzwlWRS2JOZ5LF+Zop3HZWa6qU7NKf91nspKfdzitvbXa4FjtcOyWphLEK3x24kZtI2CGm0hKR+IAVUmMYXmuZa4/Pe1FwuDjrWMWt+yY/HRObmPyVOukuy1rQAEJ5ByoQeo5171ctV46sH5WDXV0GesQAbjsA7yVZgaJZmedNBIjqid3MnuASlKVzl0EqL6nf6BXn7yj+0TUoqL6nf6BXn7yj+0TVOI9S/kfktnBfqaf8m/MLXGq0wSzW2Tnmo11etsaROau7LbDjrYWUbR0KSE7/AAfW2PT2D2VZldODZ7ZbZM2bAhNsP3J4SJbid93nAkJCj79gB+KvM4TF+bUqrBMvAAjscDfuC97i8H51Wo1DEMcSZ7WubbtkqkcBtuQ3HS+c68MKXClqmKvTtzbfMlL3OrtS+oHopPh7ABtWRxaMw7l+n0HI7rHvMZjGHl2qUtKuxkyg4kdokOAErDO2243261YNz0xwC83Jy7XPFob0l9QW+r1kpeUO4uISQlZ96gayd9xbHcmgN2y+2eNLjMqCmUKTyllQGwKCnYoIHToRXfr7foVXOgOAeXEwAC3M0ixnrRP/AFkCI4eew/4exFFrQS0lgaBJcQ7K4OuIhsxwfDjM7jWuTtw4moWWM2NDTaHsKkO3hDIAR24KgypYHQLKeb37Vj7xZrTa9CMZWzAZYhSTafTLyGwFuRFuIU92ix1KSSObc91WracLxWx2yVZ7VY40eJOChKQASqRzDY86ySpXQkdTXDkNouLOIuWXDodtDjDLbMeHNb5ozjKdgplQ8AUApB67dKrpbZp56NKnMNcy7jEhoiTExrbWABeysq7EqeTrVakS5r7NEwXGYExIMX0zEmwlQ/UyNirWS6dqhNQG7h6eYTE83CATF5Fc23L3o35NvDfurgtMqdeLjldxwzHMbh28XJ9i6XK+POPOPutpCXFFsD1WwBsElQHuFcVo08kzMjskqNptbsPt9omi4yXEy0SH5TiEKS20jk35WwVE9SPiqbTNNsEuF2dvkzGYrkx9YceUSoIeWO5S2weRZ96gauq4zCYOmyg55eQ03s6OuTBDX5TxHWdGsSbU0cFjMbVfiGsawFwtds9QNkFzMw4HqtJFgYF6fx6HCu+nOmECa21Khu5W8go5fpa2wqQQkA/YdB09nSrEuUOJD1xxp2HFZjqkWG4IeLSAjtEoW3yhW3ftudqlLGH4vFZhx49kjttQJirhFQkEJZkK35nEjfoTzK93U13XbRbH7rHvj0Jtc+I04wxIO/M22vbnSPcdh+StXFbbp16rnNDspFW1tahcRv3SJ5LawewamHotY4tLgaN76Uw0HdvgxzuqpwvKLdi+kFmcetiLpc5kqZHtduCEqckvmQ5sBv8ABSO9Su4AVlFY7asBwHJcozRAulzuURbt5XDPZc6COURmSnbs208wSCNvFRqTy9M8BnxYcKZi0Nxi3hwRUHmAa7RXMvl2P2Supr7bNNcCszj7ltxaEyZLC4zwIUsONK+EghRIIOwrOrtXBvc57C8ZnlzhAuM0hubN1RvMA9a94Cwo7Ix1NjKbwwhjAxpk2OXKXFuSHHcJI6trSZqvUJnIYWB2VV0tGJWO3In230dFhqW9LB7VBSEObJSCE7lRAO439u9TmC1HGtuVbtNBfzPwVp9Ub787u5Hv9prKRtKNOYrLkdrE4im3EhOzqluciQoKAQVKPINwDsnburKzcRxq43yJk02zsO3WCjs2JZ3DiE9em4PUdT3799K22cLUpmk3NEPEwB6RYRbMT/jBJJN57EobExdOqKzi2QWGMxPoh7TfIBo8EANAtFtVTOGtxJMLSGHfUoVaVouS0IeA7Fc5KldiFA9CduYpB8aml+ZgxNZ7MLM201MkWK4el0MgDmYSE9ipwDx59wknrUvewrE38fbxR+wRHLQz1biKSShs7k7p8QdyTuDv1r7j+GYvizchFhszEUy+khzdS3HR4BS1EqI929MRtrD1qjqwDgYqANtlIe5xk31GbSDJaLphdhYihTZQJaQDTcXXzA02tbAEXBy6yIDiI41PabBZRwuSnfRcYuv2aRMccLQK1PBSilfN37jlG3sAq4LFJbVZrSh6Qjt3oLKwlSxzr+lp3IHeffXxvGbA1j5xRu1MJs5ZVGMMb9n2St909++x3NdSVikSRkdivraGGk2KO+wykIJc2cSEBAVvsEADfbbckJ69K08XtCltDOKhIl9R4m+o6o13kAHcFuYLZ1bZopmmAYZTpmLeiTmdpuBJG8rPDvFUY/8AV3f36v0mrzHePjqjH/q7v79X6TU7E1f3fVZbc0p9/wBF+KUpXoF59KUpREpSlESlKURKd1KURKUpREpSlESlKURK9LuBD63e2fwlP/tzXmjXpdwIfW72z+Ep/wDbmu/+HP1h/ifmF5j8WfoB/IfIrYWlKV7lfNkpSlESlKURKUpREpSlESlKURK6d4/YmZ94X/VNdyuneP2JmfeF/wBU1XV9W7kVZR9Y3mFWA7qg+tOpTekmm12zdMJE6bH7KLbYK1lImTn3EtMM7jr6y1jfbrsCanA7q1i4hr9lWYa8YFpthGFLzBGDgZzfbYi4tQ0qc3LUFK3XfVHKsqc5diT0rxuGpipUAOgue776L2GIqGmy2psFbuhmqEnVnA03+7Wlq0X63z5dmvlsbcK0wp8Z0ocbBPUpI5VAnvChUvRkuNuTI9ubyK1qly+cR46ZrRceKDsvkTzbq5SCDsOhB3rWnSLJszwbipvdlz3T9zC7frDGF3t0NV0ZnNqu8JCUyClxrZKS60QopI33SO/eojo5gOM2jQ3U/XG14tEnZ/ZLzlz9lubzfavwFMrfS2mPv0QAVuL2A6qUSetbL8K3MXTAMRF9e3sNlrsxLoDYkiZm2nZ2hbhjJ8ZVeTjicktRuwG5gCa15z/Fc3P/AEV3PPYfnno/zxjzvs+2837RPa9nvtz8m+/Lv03223rUe5aKaLw+Cs55FtNvRe2MSTlDWXJ29Jm7diHhI87+qFZePLtzbdeXapJpddLtfuJzGL7kKeW7XTRK3ypfMnlUp5cxCnOn74kkeG9VnDNLS5p0ndwWbcS6Q1w1jxWyjU6E+8/GZmx3HopAkNodSVMkjcc4B3TuOvXbpXStGUYxkDr8ewZJabm7FOz7cKa0+po93rBCiU/jrVO7WiDld84s8e+bq34om5y7HAN4lPhphh0wkJ7N1YIIStQ7JWx32WQOvSvuK22Pppqrpu3qpw32jBLquaqxWLJ8HuDfoydIeZKRHlspSl0oWEkgOc2x6+BNZeaCDe8adwPG+u7msfOzItb/ANI/sram65bililtQL5lNntsp/YtMTJ7TLjm/dypWoE/iqJaiamz8Lz3TXEIdrjS2M6u0q3SH3HFBUZDUVTwW2B0USU7demxrXSHY8Z1GiagZLgfDna82tF1ulz9IZrnl5YbLjjW6HRGSGlvIjslJDYHJty/jHV0ynSrjhvBxKmS3ZTnpK5t9q4sqUUohyEoG56nZKQB7gKsbhGtue21vZJ3G2m9YOxbnWHZ8wPruW69KbUrmropSlKIo/eP2QX+9T+iulXdu/7IL/ep/RXSrjVvWO5rs0vVt5JSlKqViUpSiJT8VKURKUpREpSlESlKURKUpREpSlESlKURKi+p3+gV5+8o/tE1KKi+p3+gV5+8o/tE1TiPUv5H5LZwX6mn/JvzC1xrrSbpa4b7cWZc4cd536m06+lC1/ECdzXaql7tis/0xlV0awyw5/a7nMcMhxMpKbjBKUAKjpKwQOTbdISQRvXndm4KljHOFV+WB2CTMRLiGjjc30C9ztPHVcExppMzEntMACZhoc48LAxMlXI7IjxyBIkNNEgqHOsJ3AG5PXwA764G7hGmwXZlplxZiUpVyLbeSpsrA6AqBIHXvqoZrWJ5rM0nbQJLtlktTUtMznCpx0NsgBl07+v6yNiO5XL41lp9tt+NanyLdjcRmFGu2LTZNyiR0hDXO2Qlp3kHRKjupO+3UCt7oem2GOcfKQ50Ftoa5zSDeZOU99t8jQ6aqPl7Wg08zWyHXl7GuBFogZgOUndBsG0XGQ9aIUq+GDFmvsdq80xJDjSVAbq5Fn4SR7a7cOdBuLXnFvmx5TW/LzsOpcTv7N0k1Rlms1uv1v0WtV0jh+G5bJZdZJ9V0JZQoIUB3pJA3HcdutZu4RWcJzzLvmOgNQAvDVXER4zYQ2ZLbi0pcCB0CtvYKtrbFpGo6mx/XOYgR1YFQsiZJ3TvjS+qpobdreTbVezqDKCZl0mk2pMZQN8aidbaK1hc7aZvo0XKIZgG5j9sntf/ACb7/wBFditfY+JXOVpbHmxsKxuK45CbuDeTu3zlkokEBXnS3C1uDzd6Srbwq+LU7LctUJ6Ypp2UqM2p1TSuZC3OQElJ8QT3H2VpbT2bTwQBpvzdYtPo6iL2JieBghb+ytqVMeSKlPL1Q4eloZt1mtmOIkHcuG63qDbWZKTNi+etRnJDcVbyQ4vlQVfB3326eyupimSC/YbasruCWIQnwm5boLmzbXMN9uZXgPaarXDMRxbJ9Kbjk2RwI793uPpCVOuTv+cMPoW4AEufCb5AlIABA2+OsK0Lld4WlOOJs0S7wnLCud6OmSvN48qQ2lITznlUF8gJUEEbePhXTbsXDua+g1xzMeQ5xAFmscTEuiCRviNTZcp23MS1zMQ5gyvYC1oJN3PY0ZobMgOvEzcC4V8xpsKbH87hzY8hg7/TWnUrR07/AFgdq/HpW1ecNRPSkPt30hbTXbo53EnuKU77kfFVWWqzXmzZbeXFWWxY9EuGPSFyrTAuQdLrqQeSSGQhPL0KklQHXpUeewvGYvDezkjdoYN4atUe4IuJTvJQ+FJ5SHPhAAdAkHYAbbVS3YmHLmh1Uw5zGiADd86kOixG4mexXO27iQxxbSEsa9zpLm2ZGgLZuDvAg8dTes24262oS5crhFiIWeVKn3kthR9g5iN66t5n3KMxCesrMGT28tlt0yJIbSGFH1loP2S9u5PjVeZRYLrcc1+aO12bHcwWi0R2JVmuT6Uvwwd1do1zApHabnfcDfbvrBOyrG7hlgt1ks8+0ei86iRpNumO9qqG/wBoVKbQoEjkHMOUDoBTD7GpuFN7XZiYkQIEg6jMHDdBiDuPGcTtuow1GOZlAnKZMmCNDlLDvkTLYuNYtC25xabjk19xwOMsmxhjnfckIAdLiSo8o36BOwB9/sqQOPMtNF915ttpI3LilgJA9u56VVtgw7GrtqLqTAlWSEG3mojHMlhIU2l+OrtSk7dConc+099YS3z5mVWPGNIrmormw7o7EvYPeYlvUFAn3ObsD37mlTZFCs8eQcQGhhdI0Bp5y4X7DbiQAopbZxFFh8u0FznVAyDq5tTI1ptbUXvYEnerv3BG4pX0+3bYV8rzS9Qvo7xVGv8A1d39+r9Jq8h3j46ox/6u7+/V+mu7sTV/d9VwtuaU+/6L8UpSvQLz6UpSiJSlKIlKUoiUpSiJSlKIlKUoiUpSiJXpdwIfW8W3+Ep/9ua80a9LuBD63i2fwlP/ALY13/w5+sP8T8wvMfi39AP5D5FbC0pSvcr5slKUoiUpSiJSlKIlKUoiUpSiJXTvH7EzPvC/6pruV07x+xMz7wv+qarq+rdyKso+sbzCrAd1RnHtOMUxfLsmzq1Q3/TeXOR13SS9IU6VhhHI0hAV0bQlJ+CnYb9akw7qiWq+ott0n09vOf3OIuYi1tJLMNtYQ5LkLWltlhBO+ylrWlIOx233rxLMxORu+y9o/KBmduuuTNNNMRz+fjd1yOG+ubiV0ReLRIjyFsuR5KRtvuk+skjopJ6HxrlwfTzFdOrJLx3FoLjMCdcJdzfafeU9zvyXCt47q39UqJ9XuA6VjtGdToWsOCw8wjWp60yVyZEC4Wx9wOO2+Yw6W3WFqAAJSU777DcEV+NHNT2tXsSfyuPZXLYli8XC09gt4OlRiyFMlzmAHRXLvtt0323NWOFVrS06A/36qtppOcHDUqIt8I2ijchLXoy9qsTcz0gjGV3ySqyJf5ubmEIq7Pbm9bk+Bv4VJtR9DMB1QutqyG+Iu9uvdlaXHg3WyXN63y2mF/DZ7Rojds/uT3eG1WACCOZJBG+24O/WuOS83EjuyXyQhltbqthueVI3Ow8elR5eqSDmMqfIUgCMohVxYOHHR3G7Hk+M2/Eg7asyZZZvcWXJdkJmdmgpC1FairtDuVKXvzFZ5t966+I8Nmm2IZBaslbkZNepePhQsqL7fpNwZtZKeXmjtuqKUK5fVCiCQO41LdMs/tOquB2bUPH4spi231hUmK3KSEuhsLUgFQBIBPLvtudt6k3fvsQeU7HY9x9lS6tWaS1zjO/5f+KG0qLgHACN3zVRK4V9IjcrjIbj5AzarxMXcJ+PMX2S3Z5MhZ3WpcRKgghR6qT8E+I26Vm7HoLprjsHC7da7bMRH0+lyZuPoXNcV5s4+FhwHc+unZxQCVbgeHdVheITuN1fBHifioegKidgO8nuFQa9U2Lj/bKRQpC4aFA8A07kYjnOoWXOuJQ1mNzjS2Y6ZTj/AChlgIU4efYNlaifpaPVSEp6kk1PKbHpt493voNjvsQeU7HY77H31W5xeZKza0MEBKUpWKzUeu/7IL+JP6K6dd28fsgv4k/orpVxq3rHc12aXq28kpSlVKxKUpREpSlESlKURKUpREpSlESlKURKUpREpSlESovqd/oFefvKP7RNSiovqd/oFefvKP7RNU4j1L+R+S2cF+pp/wAm/MLXGonc9L8Vulxm3PmukF25bGci33F2M3LO227iEEAkjoT0JqWV92PT393vryWHxNbCuLqDi0ngvouIwtDFtDa7A4DiJ/vBVxl+nkO7X7CLTEsa02C0CYh3zZZaEQdiAypKgQpKucDYjrvUmseCY9j4nuRW5UiVdEdlMmTJS35DqNiAkuKO4SATsBsKkOx9lPYfb3Vs1dqYqrRbQzkNA4m8uLpPbJ8AtWlsnCUq78QGAucQdBaGhsDgIHidxWAg4Njdu9AeaRHU/Myy5Htu7yj2aFpCVBX7roPGu78z1q+aBeUFhRuDkIW9SyslJYC+fl5e7vPfWS2Pdt79qbHr7u/3Vruxdd5lzybEa7iZI7yZ5rZZhKDAA1gABB03gAA9wAA7BChPznMF37HzWf6P7fzn0V5+75h2m++/Yb8u2/Xl7vdU1GyQAkAAdAANtq+7bbb+Pd76d3QimIxmIxUeXeXRpJn+9qjD4LD4SfN2Bs6wAOXu3KHz9KcMuE2XLdizW2ri5202EzNdbiSXPFbjIPKSe8+09+9dubp5ik/H7bjciE8ItnCPR7rchaJEYpGwUh0HmB26b79fGpLt3e/u99dCdOuMa5W6HFsrsqPLW4mVJS4lKYaUp3SpST1VzH1endWw3H42qWgVT1bjrREA8TrFuJ07Fru2fgaQc40m9axhszJGsDSb8Bqd5WLsmAY3YFT34rUuTLujXYS5s2Ut+Q43sRydoo7hI37hsK5nMLx53Dxga4rpswiph9j2yufsk7bDn79+g61neU+ym3efZ3+6qXY7EvdndUJMg67xoe7dwVzcBhWMyNpgCCNBobkd+/jvUaven2O3y4M3dw3CDcGGBFEu3zXIzy2R3IWpJ9YDw37qMad4lHtMKyM29xMaDPRdG/p6y4uUk7h1xZPMsk9+561nLk9NjW6TIt0NMuU0ypxlhS+QOqA3CebY7b9wO1dfHr5ByWyQ79bVKMea0HEhQ2Ug9ykKHgpKgUke0Grhi8b5AOFR2RpjXQwY+sd8b1ScJgfLlpptzuE6aiRPO8T3TuWOk4jZYuRyM9jRZyrt5uUutx5KkolBKCEhTe/IpW3QE+6sFgFhlSssyHUm6449ZZF5DMWLEk8vbpYbSOZxwJJCVLUB037kCp9X2obtGsKTqZMlwDZJMhoIOUXiJA7rKXbNourMqgQGuL4AEF5BGY2mYJ77lYC2445CzK85OV8qLlGix0th1SuZTfNuspPRHeE7Dv5dz31nqUrVrVn1yHP3ADuAAHgFt0aDMO0tZvJPeSSfEr6O8VRj31d39+r9NXmO8VRr/wBXc/fq/TXY2Jq/u+q4+3NKff8ARcdKUr0C8+lKUoiUpSiJSlKIlKUoiUpSiJSlKIlKUoiV6XcCH1vFs/hKf/bGvNGvS7gQ+t3tn8JT/wC2Nd/8OfrD/E/MLzH4t/QD+Q+RWwtKUr3K+bJSlKIlKUoiUpSiJSlKIlKUoiV07x+xMz7wv+qa7ldO8fsTM+8L/qmq6vq3cirKPrG8wqwHdWsXE9lsy86t6caW2rDb7lsSxyRnGRWyyNNuvqaYUUQkKDi0pCC+So7nuSNga2dHdUNxnS2wYxqDluprMydMveYCK1KXKWlSI0eOgpbYYASOVHUqIJJJ67143D1G0nF7twtz/t17Cux1VoY3jdUXoNnU+wcTeYYheMEyTDrVqcgZTZoN+ZbacNyZSlE8N9mtaSFp5XO/fdJ6VVcfI8mh6DYngmPR7u+xm+rN+tdzYtExESZMiIlvuqiNPrUlLRdKQkq5h0BAPWtxs+0ox/UK/YflFxmz4F1we6+lrXKhLSlRUU8rjLnMk8zS07BSehO3eKjZ4Z9OXdNH9LpT13dgLvcjIos5MoNToFwdfU+Ho7qEjkUhayE9D0Ox3rcbiqVnEXtPdIn5LTdhavog2vHfBj5qptPcUy7CtbMLuGnOhGQab45cFyIGUQbhkcSRDnMdnuy+hgSFrL7bgG6kjcpUd96xWgOhOAaj4HlWoOoEe5X26RsiyaFAD90kpZhxUSHh2SG0LCdipSlknc7keAAq8sQ4f7dY83t+o2Y5/lWeX+yMORrM/f3mS3bUuDZxTTTLaE9opPQuKBVtUl040vx/TLGJuJWWRMlwrhcp9zeMxaVLLkt1TjqQUgeruogeO3iaipiwAchva4nt3kzvWVPCGRmFr2Mdm4CFptYsNtkLhB0P+ZyTOsU3MM1snpSfAluIfWtbkhorSSohCuToOUAA7Hbered05xTQ7ie0wiaYRJlogZdb77FvcFM995mf5tHS6y4tLq1buhR+H3mphjnCtimO4taMKGb5ZcLJjmRw8jssSZJZWLeuMpakRmyGwexJcJIPrd2xFT/ItOLLkuf4lqNNlTG7lhonCC20tIZc87aDbnaAgk7ADbYjr371NTFNc4gGxzeIt7vBKeFcGgkXGXw1960xwC0XjWbC7nqRmOgudZpkmRzZ7kXKYOTxYgtQbeW2y1CbVIQY6WeQAgpBJBJ3Bqb5PjGtecYDpHeM+w1rUFdmt01OVYY1kTMV+5vghtieFNr7OSUpTuUcxTzOHx7rWufCzY1yL7GxHU3OMPsGTyHZV3sFlmtIhvOu/VlNc7alx+068wbUAdz3VlL/AMNmCzIOIt4ZcLvg1xwWIu32K52F5CH48RYAcYWHErQ82ogKIWDurc77k1m7F0y4EHwNrRGtu7mq24SoGkH5i9+V+9VDgjdtvelWr+m+jt2y7TzJ+ybdRjmXPmOMbceRsBFeJUUsPcqgFhZCVKBAHjitNo2Gadao4NHyfR7ONGsifkm1olR7j6RseSvuNFIjyX+dYUpSgVoJSFc32VXlbuGnA/mdy+yZlcL1mMzPWmmL/drzJSZchtofSENltKUspaPrICEjZXXrXBYuHCHEvtgvGZap5tmzGJvpl2O33uUyY8SQlJS28sNNoU+4hJISpwnbv76w85pw4TY8+Ed9+I7dVkMPU6pjT7/bh8lcFKUrmLpqP3j/AD9z4k/orpV3Lx/n6/3qf0V0641b1jua7NL1beSUpSqlYlKUoiUpSiJSlKIlKUoiUpSiJSlKIlPx0pREpSlESovqd/oFefvKP7RNSiovqd/oFefvKP7RNU4j1L+R+S2cF+pp/wAm/MLXKqSyjHpbOQZDc8swi55TBmSFLhXa0T+aRbGQgANJY5gUKQdzunv8auyobL0zjG4XGdY8rvtibvDhenxoDzYaddI2UtPOhRbUod5SR7a4mxsazBVHl7ssgD/LiDq0gjxB0Ihex25gX46mxrG5oMx1eBGjwWkX7CNQZUVdkwM6u2C4m3f58/G5lokXJ5xxwtPXJTJShKHinY9CSVJG25HWuw5bo+DZ65jWNreYtF7x2dMdgdqpbcd9nYJdbCiSjmCiCB0JFSidpljUiz2e025Uy1Lx/wDYuZCe5ZEYkbK9YghQV9kFAg1yWXT+3WuVcLrNutxu91ucYw3p85xJcQx1+ltpSAlCeu+wHU99dA7UwoYQx5yQ4ZCNSXEh5jqyAQZ1kQBC5w2Viy8F7Bnlpzg6ANAcwT1oJBAGkOzE5lUTuKxIWh9iz6POuCcmS3AebunnThdbC3Uo5Egnl5Ag7cu3hudzU1m4tasI1KwtzG/OmF3dybEuJXJcc88CWOcKc5id18w33qUPaeWV/BYmny5EwW6G2w2hwLT2xDKwtO52270jfpWTuuNwbve7NfpLryZFjddejJQoBClOI5Fcw269O7bbrWVbboqlwc8lpNa3/VzYpjkDMDRuqwo/h80Q0tY0OAoXtOZrpqHmWwCdXaFVXMgY1lN0v0hjEsizd9Ux5n0iqQmLFhFI27FhanEgBs96kpJJ9tTbRmfOuemGPzLlKckyFR1IU64vmWoJcUkbq8TsAN6/LWlkCK5Oj2/KL9CtFxkOSZNqjyEJZUtw7uBK+XtEJUe8JUO81nsTxi34bYI2N2px5cOGV9j2xBUlKllXLuANwCoge6qdp7Rw2IwfkKTiSHNImbANII9kGYs0Adp3X7K2bisNjfOKrQ0Frg6Iu4uaQbdYiAbvJN9Bvq7K8dmIybILrlGD3HLbbMdBhXC1T95FraCAOySxzApUk7q3T1O/Wsii6wrjdNJpVmvc24w3XJqBJlHlef5IxH00DbdYI2PTvFSebpswu6XG6WLKr5YfS6+1nsQHWw285tsXAFoVyLIHUp2rtRtPMdg/M4i3iRGbxftjBQhzfmLqChZcJBKidyd/aavdtXCupMDnEkNIsDAmm5twSQDJHoGCJJEwtdmyMU2s8taAC4EkkSYqtfYgAkZQ70wXAwASJVe5Tg+KZLkysFwu0dhLbUJF9uyJLyhb21Hfs0br5S+vwH2I6mv3k8dyZqE1hPzK3S/2LHrJHdYtkechpKnFqKe1eK1pLuwSEgbnruTUmtGkDFhZdj2bPsrhtvvLkOhuSzu46o7qWolvdRPtJrJ3LTuJcHLbcmsivMS9WyOYqLuw6jzh9oncoeBSUODfr1T0PdVvS9Cm9rfLF7Q0gE55DjEvJkEaZQGmWg2kzNXQ1eoxzvIBjnOBIbkgtEwwCCDrmJcIcReBAGJ0tiZDabvfbZIsc202L6RItsObObkuRVqBDqElK1FLZICkg9252rm0qUWJWa2hv/Nrfk0kMDwSHEocUkf8SlflrK2bG7Pp9Bu99L9wuc2SDLnzZB7aVKKE+qgBIA2HclKRt1r86bWCdY8bL14QEXS8SnrtPT39m88rm7P/AIE8qfxGufjMVTr069UaOyAWIkgXdck7jJJk5pMErpYLCVKFTD0TMs8o43BhrjAbYAbxAAgZSBIAKlVfK+18rzy9GvtfKV9oiDvHx1Rj/wBXd/fq/SavNPeKo1/6u798V+k13diav7vquFtzSn3/AEXHSlK9AvPpSlKIlKUoiUpSiJSlKIlKUoiUpSiJSlKIlel3Ah9bxbf4Sn/25rzRr0u4EPrd7Z/CU/8AtjXf/Dn6w/xPzC8x+Lf0A/kPkVsLSlK9yvmyUpSiJSlKIlKUoiUpSiJSlKIlcE2OZcN+KlQSXW1IBI323G29c9KggOEFS0lpkKGfMHJ+2TX8Wf76+/MHJ+2TX8Wf76i+rfFboBoVkEXFdVtRI1guk2IJ7Ed2JIdK2CtSAvdttQ+EhQ2336VCf8orwacvP8+yDy92/o2dt/Y1z+isL7PiV0OlMV7XgFb3zByftk1/Fn++nzByftk1/Fn++o7pBxRaEa9Xe4WLSXUCPkE61xky5bTUWQ0WmlK5Qol1tIO6unQ71alOisL7PiVHSmK9rwChvzByftk1/Fn++nzByftk1/Fn++plSnRWF9nxKdKYr2vAKG/MHJ+2TX8Wf76fMHJ+2TX8Wf76mVKdFYX2fEp0piva8Aob8wcn7ZNfxZ/vp8wcn7ZNfxZ/vqZUp0VhfZ8SnSmK9rwChvzByftk1/Fn++nzByftk1/Fn++vkTWHTidqnN0Vi5K25mlut6brJtfYOhTcVXLs5zlPZkfTEdArfr3VM6dFYX2fEp0piva8Aob8wcn7ZNfxZ/vp8wcn7ZNfxZ/vqZUp0VhfZ8SnSmK9rwCrubpbNlSVPpvDCQoAbFk+A+Ouv86Wd9umP4k/31ZlY68ZFYMfMJN9vMKAq5Sm4MNMl9LZkSHDshpsE7rWT3JG5qh2wcC4yWeJ+6vG3cc0QH+A+ygnzpZ326Y/iVf30+dLP+3TH8Sf76sylR0BgPY8T91PT2P9vwH2VZ/Olnfbpj+JV/fT50s77dMfxJ/vqye3Z7bzftkdqU8/Z8w5uXfbfbv238a/dOgMB7HifunT2P8Ab8B9lWnzpZ326Y/iVf318+dLO+3TH8Sr++rMpToDAex4n7p09j/b8B9lWfzpZ326Y/iVf30+dLO+3TH8Sf76sylOgMB7HifunT2P9vwH2VafOlnfbpj+JV/fT50s/wC3TH8Sr++rLpToDAex4n7p09j/AG/AfZVn86Wd9umP4k/319+dLO+3Uf8AiVf31ZdKdAYD2PE/dOnsf7fgPsq0+dLO+3TH8Sr++vnzpZ326Y/iVf31ZlKdAYD2PE/dOnsf7fgPsqz+dLO+3TH8Sr++nzpZ326Y/iVf31ZlKdAYD2PE/dOnsf7fgPsqz+dLO+3TH8Sr++nzpZ326Y/iVf31ZlKdAYD2PE/dOnsf7fgPsqz+dLO+3TH8Sr++sVlehdyyPHZ1kayKKwuWhKA4qOpQTsoHu5vdVw0rF34e2e9paWWPafus6f4h2hTeHtfcGdBu7lqh9BbkP3e27+QL/wAdPoLcg+723fyBf+Otr6Vo/k3Y37R+J33XV/Pe3f3R8LfstUPoLMg+723fyBf+On0FuQfd7bv5Av8Ax1tfSn5M2N+0fid90/Pe3f3R8LfstUPoLcg+723fyBf+On0FuQfd7bv5Av8Ax1tfSn5M2N+0fid90/Pe3f3R8LfstUPoLMg+723fyBf+On0FuQfd7bv5Av8Ax1tfSn5M2N+0fid90/Pe3f3R8LfstUPoLcg+723fyFf+On0FuQfd7b/5Cv8Ax1tfSn5M2N+0fid90/Pe3f3R8LfstUPoLcg+723fyBf+On0FuQfd7bv5Av8Ax1tfSn5M2N+0fid90/Pe3f3R8LfstUPoLchHUZ9b9/8AuK/8dPoLch+723fyBf8Ajra+lPyZsb9o/E77p+e9u/uj4W/ZaofQWZB93tu/kC/8dPoLcg+723fyBf8AjqV6+cbelnDvncLTvMMdy+53a4W5FzZTZbciSktKWtG3VxKuYFtRIA7tutV9/lSdA2Ppt2wbVC2xh8OTJxsJabHtUQ6Tt+Kn5M2N+0fid90/Pe3f3R8Lfsst9BZkH3e27+QL/wAdPoLcg+723fyBf+OthNMtUcD1jw+HnmnGRR7zZZpUlD7QKShxPRTbiFAKQtJ70qAI6eBFSun5N2N+0fid90/Pe3f3R8LfstUBwW5ADv8AN7bv5Av/AB1X7nk4ctW4tY1TtA5lE/sY74n75W99K2MP+Ftl4afJUyJ/7O+618R+MdsYmPK1AY/6t+y0O/yb+Xf607P/ADY78pT/ACb+Xf607R/Njvylb40rZ6BwPseJ+61vzNtL2x8I+y0O/wAm/l3+tOz/AM2O/KU/yb+Xf607R/Njvylb40p0DgfY8T90/M20vbHwj7LQ7/Jv5d/rTs/82O/KU/yb+Xf607R/Njvylb40p0DgfY8T90/M20vbHwj7LQ7/ACb+Xf607P8AzY78pT/Jv5d/rTtH82O/KVvjSnQOB9jxP3T8zbS9sfCPstDv8m/l3+tOz/zW78pT/Jv5d/rTs/8ANbvylb40p0DgfY8T90/M20vbHwj7LQ7/ACb+Xf607R/NjvylP8m/l3+tO0fzY78pW+NKdA4H2PE/dPzNtL2x8I+y0O/yb+Xf607R/NjvylP8m/l3+tOz/wA1u/KVuNjWqeBZjmGSYHi+RxrlesRLCL2xH3UITjvPyNLXty8/0tW6QSU9N9t6ldOgcD7Hifun5m2l7Y+EfZaHf5N/Lv8AWnaP5sd+Up/k38u/1p2f+bHflK3xqicJ4qMXzTijzThxhhjtcXtceQxLC+smWknzxkD2thxgdOu4d9lOgcD7Hifun5m2n7Y+EfZUJ/k38u/1p2j+bHflK2n4e9J52iumkXA7jeWLo9HlSJBksMlpJDrhUBykk9N/bVlUrYwuzMNg3+Uotg6alamM2zjMfT8lXdImdAPkEpSldBctKUpREpSlESlKURKUpREpSlESlKURR/INPcBy2Yi4ZVg+P3mU02GUP3C2MyHEt7k8oUtJIG5J27tya0etGn2BL8qVfMUXhGPqsjeCtvotptjJipc5GfXDXLyBXU9dt+tegNaO2X/pab/+ADf9RiiLYXVq96f8LulGV6yY1pbZe0skNDj8W1RWIDstBeQgILqW+gBXv1BHSqp0243s21dz3HLfgXDTlM7Ar3IZgv5gp5SIsd8o5n1DdrlW00oLRz8yeYp6AEgVL/KAnbg61OI+1jP60zUr4XI9sx7hg01DSW4sOPiNukOHuSneMlxxR/GVE/GaKVVd4418uy/N8hwjhl4f7vqenFJSoN2vRujNutzclJIU0244D2hBBG/TfYkAjYnPaBcYLuqOpl10P1M0qu+m+oNrh+fi2TpCZDUtgbcymnUhO+wUlQ6EFO5CjsQKh0n1r1+1eRf5vBZoLpth2nnph9oXq/FUc3GUAkOSBHjBPUpCN9wrwBVuCBGdOoGrNr8pjjcTW3MsfyLKDg8pbjljgmNFisqDpbYCVesoj1lcyupCx7BRFcuY8eTdg1ezHQrGtFsiyrMrBIjRrRAtclKjdS4yHXXVko5YrTSSnmWoq3KhsO/bl0z41cjnaxWvQvXzQy76Y5JkTanLI67PRNiTSATyBxKUgE8qgCnmHNsk8pI3h3DnEjOeUY4ipzjKVPsW63NtuEdUJWhkqA+PkT+Su3xrNoHFHwpvBIDhy2Sjm268vNF6fFRFZWtvFfPwLUuHofpNpRddSdQJED0pJt0SW3EjW+KTslx99YITv7Nu4p3O6gDidMuMe9XLV63aD68aL3TTHL77HckWUvXBqdCuQQCpSW3kADm2SrYDcbjYkEgGNa8cPuW5XxAv6wcL+vdjxbVSFZ2oV8sktxuQiTETt2anmwFqbBHZj12yk8qCCkjcxiwcR2v2lusGCYJxl6LYq6vJJ3onH8zsobcLchxSU9xKuUFSm+YJ7NQBCuVQFEVw2LUzTyXxtZJpZF0lgxsxg4ozcZOYpW35xKiqLO0Ujk5+Uc6e9ZHqDpWN1O4yZ1t1Sm6IaCaQ3bVPMrO2HL0mJMbhwLXvtsh6QsFPP1AI6AE7blW6RBMR/wClYzj/AOHUb+tFqkuDG2cV11uGr100TyjTe3THc2lpyFvKIkp2cqSFrKSC0Ng36y9gevMF0RbbaNcXa801Kc0P1g0vu2mOoRjGZBts+QiTGubKQSpUaQgBKyAlR227kq2JIIGJ1X45bZphrZedCWdKb/kuQx7bDlWVi0vBx67ypGxEdLfJ9KShHOtTqlEBKD0qrcw0V4nsr120hzTW/V3RSBccUvaZVrjW92RCnXFgutecMNJdT9OJSAAkdxX176zWOQosnyr2VyH2ELciaeMOMKI6tqPm6CR7DyqUPiJoikWMccGZ2XVPHNL+Irh4vGmbmZSBFsdzVc250V19RCUtrUhKQklSkpJBJSVJ3AB3qpOObP8AUGLxY6IsxNHLlMbxXIFyMfWm5MpGTPLTGUtpkEfSChXqFTnQ77jpU88pYhPNoO7yjnTqVAAV4gHbfr+Ifkrl41PrquFP8KpX9eLRFtlgd+v2T4dacgyjEZGL3afGS9Ls0iQh9yC4e9pTiPVUR7R0rPUqq+IzK71ZsEaxDDpRYyzPZzeMWNxPVUd18K7aXt37MR0vvb+1tI8aKFqdkPEDmOOcauE6zXeR2Wk2cuT9O7Q6F/S1IjyEoVLV4ALmeshXi0kmvQWtZOL/AIbLTmnCNL03wm3dlLwOAxcMaQ2PpiXITe3ICOpUtoOJ38VKB76mfB7rSjXrh7xTPH5AduqYwtt4G/VM9jZDpP7/AGS58TgopV0UpSihKVg84xqTmOJXXF4mTXbHnrlHUwi6Wl4NTIhP/WMrIISoeBINUN9B1l4+Dxla7A+031g/0djRFsrStafoPM1/2zdc/wCeY/yNPoPM1/2zdc/55j/I0RbLUrWn6D3Nx8HjO1yB994jn/7NPoP88/20Nb/51jfI0RbLUrWn6D/PP9tDW/8AnWN8jXz6EDPh8DjR1uHt3ucU/wD2aItl6VrR9CDqF/tp62fzhF+Rp9CDqF/tp62fzhF+Roi2XpWtB4QtRh8DjV1pA986If8A7NfPoQ9Sv9tfWj+WRPkaItmKVrP9CHqV/tr60fyuJ8jXw8Imp4+Bxtayge+RDP8AT2NEWzNK1m+hF1S/229ZP46H8jT6EXVL/bb1k/jofyNEWzNK1l+hI1ZHRPG9rAB4bqhE/l7Gn0JGrX+2/q/+WD8jRFs1StZfoSNWv9t/V/8ALB+Rr79CbrD4ccGrP8VA+Roi2ZpWs30JusX+3Dqz/FQPkafQm6xeHHDqzv8AeoHyNEWzNay8Z/Fvd+E+RgV1j4jGyG05HLnR7nHU+WJCENIaUlbK+qQr11bhSSD06p76+/Qoa1f7ceqf8kt/yVaW+Ur0mzfS+y4C9mOvOV6ht3CZcEsN31qO2mEUNslSm+yQnqrmAO+/wRRSBdegGg3GDoTxEMNsYPlrca9lHM7YrntGntnx5UE7Ogfumyoe3arrrwo4feC3iD16lQ71iOPPY/ZEuJcRkl1K4jCNj8NjYdq6oeBbG2/2Qr2V0L01y3SrAYmJZnqvfNQLgwdzc7s22laRsPpaOUc5QPAuLWr/AHvCiEQrCpSlFC0b1buEC1+VL0rnXOdHhxm8KlBbz7qW0J3RPA3UogDc9K2xyfVHSC12GdLy3PcUatCWFiX55cY6mlNcp5kqSVHmBG45djv3bGtKuJ/S7C9ZvKO6b6c6hWxdwsNzwt5UmOh9bJWWvPXEeughQ2UkHoauaB5NHg2gS25fzrHJJbO4bkXiYtB+Mdr1opVReTTu9nxXDdddSULdtOmDeSvz7O5ISpLTUVhDq3VJB/csmOkgdd0gd42qX4fxCcavETb5GougGl+CWDA+2dRZ3cvkyDNvCG1FJWlLJCW0kgjqNgdwFK2JqyuLXA4GO8FuoeE6aY7GtcK34675rb7cwGm22G1JcdCUpHihLhPidyT31Q/C9odrnn2gOE5Lp1xt32yWN+1tts2mLjcN1FtWjdLkbnKuZRQsKG6up7/GiLYXhg4mZetzuTYLnmHqw/UjBpKYuQWQvdq3srfkfYX3qbUQfbt6vVQUknXfB+NPjE14gXW36D6J4pcLti9zlt3uZMkLZhdilZTGjtdo8kqfWlC1LPNsBy9B3ma8MGl2M4nxWZ7e5vFC9qXqJHsibZklvcsQiLYSFsdmtx1slpSkhCEco69Tv1Sa63ksWWk6U6ivpbSHHNQrkFqA6qAaZ23+Lc0RdzUPjF1yw/XqHw94/pHZcjym74pAnQYzElxpDd2dBVILzylcqYjSUOq3A5jskbjfesBf+Mfid4dc/tWI8TelGOXiNlsZ4487hTrpcenJ5UtxdnlHfmcW2gkgFIcChzAEVk2mm3PKvvLWgKU3pqFIJ+xPaAbj8RI/HXY44EIc4keFNDiQpJzZ3cEf78Q0RYnUnig41OHxu06m646TYCNP7hcGYc+HZJ7z1wtaXT6oW4pXItYAPVKSgqG26dwaunib4m5OilkxO34FhzmY5nqDOTb8atgcLTLiiEkuvL29VA7RA26b83eACRX3lT/rQ7z/AAza/wC3Fcev/EJqXh900f4e9ELfZUZvqBb2XWrte2y5FtsdDYHOED4S/VcPUHYI25VFQ2IsLqRrZ5QbQvFZGq+oenelN9xa28j12gWSXLRLiMKUAVBbh2OxIBUAvbv22BNWHxC8W0vTbhgxziM0/ssGcxkL9qWiNdErIajS08y9w2pJ7RA6d+24qi+K3RziGxvh4zTM9ceMGfeIseAG0Y/arLHtsKfIcWlDbC1D13ElR+CACQPcawPEmUr8lXpftsQUY6P/AEKoite/a78fF2xKRrVguiWDwMJajKukSyXeY85fJVuCecPLCFJQham/WDY9Yb7bKPfcWnHFfgmZ8MaOJy9tOWWzRYD8m6RebtVx32VltbKDsO0KnAAju5udO+2/Sx7o2hGmUtpCEhCbC4kJA6AebnpXmRj9vu1w8j9ePRTS3Exb+5JloR4x0XJBXv7h0J9womq2RxPW3j81mxxGq2lukmnNixKakybNasjnSDcrlG39RZWgpbRzgeqVco6g9Rso2toBxQxtctM8nv7uMu45mWELkwcix6WorMKa0hRA5tgVNqKFbHod0qHhuaf0k0A4g8x0uxPJsK48shZsVws0R2AxHxiEpuO12SQGQeb/AKvbkO/XdJ361+eGPTnDMSyXXu+WPiNVqhks2IuLlSfQvmQizmxIHOpaSW3FKPaD1OnqmiLG6PcWvGLxP4LGu+iekuD296AXGb3e77Jfbt65fMVIjQ2kqLiilotla1EgKXtsPGyeHDig1T1VnajaQ6g4DaMe1a0/jlXmrchardNUtKgyvfdSkI5+z3IUrdDiVJPgML5K1CEcIVmUlASV3m5qUQO89ttufxAfkrHaJADyluvWw23xm1H/ANEWiKruC+48Rw4ptZ+xxvBSqVlUT5v95kjaEed/f0d0+md7n1T2Jr0frS/gq+up4rR4/NXF/ryq3QohUC101MGkelt8zZiL55co7SYtogj4U25PrDMRhI8St5aB08Nz4V588RWk134Ksx0Q4lbQ47OmQXhAzqW3vvcZ7qlvSnlnxLyHpSB7mmx4VuPkpOsPE9ZcMQS7jekDDeS3cDq29fpKFIt7KvaWWS9I9ynGTUq4ndHo2u+hmWabONoMyfCU9bHFAfSpzX0yOrc93rpSCf3KlUQKx7TdLffLVDvdplIkwbhHblRnkHdLrTiQpCh7ikg/jrt1p95MbWCTnugitOcgcWjIdNpZssll36oIhJMfm36+rs417uxrcGihKUpREpSlESlKURKUpREpSlESlKURKUpREqhoPDRPicYNw4oTl0dUSdjqbGLN5moOJUEtjtO259iPU7uXx76UoimXEVpPJ1z0VyrSeHe27O9kcVEZE5xgvJZ5XUOblAUkq+Bt3jvrNaa4InBtK8a01uMtu5psdji2Z94NlCJIaZS0pXLueUK2PTc9/fSlEWsOFcHnEjoNJu2NcOPEdaLPgt0nOT2bZfseE9+3qX3hpfNso7ADc8oPKCRvuTmdPeB29afcQ2O8Qi9ZLhk96aiS2cndvUTmdurz6FIC2S2oIjIQktpS2EqADYG/XopRJVg6ZcOU7T/iQ1N14eytibH1AYiMtW1MQoXD7FKRupwrIXvy+CRtvX71z4dp2sGqukeo8XKmLW1pleXbq9EciF1U4LLR5ErCx2ZHZHqQrv7qUoii2tfCXlGSauscQ2gmqatPdQDCTbrk47ATMg3VhIASH2ie8AJG+ygQhHQFINYvFuEnVfMdUMZ1W4pda42bv4S+ZdgsdotKYFujyehD7nXdxQKUnu70p3Ow5SpRJU6s3DnOtXFxfuJlWVsORbzjTVgTZxEIcaUgtHtS7z7EHsj6vKO/vqE6g8H2c2rVq7a5cMGr409yLJADf7bNt4m2u5Od5dU2T6iydyeh9YqIKSpW6lEX70w4Pcwc1hgcQHEtqwNRMwsjRasUWNb0w7bat9/Xbb8VDmJB2TsTzHmIBE0tPDnOtnFxe+JlWVsORbvjLePps4iEONqSWj2pd59iD2XweXx76Uoi+cUHDlO4hRgHmWVsWT5isoj5C52sQv8AnSWv+qGy08hP7rr8VY/ix4Yr5xAnC8kwjUJWG5fgNyXcbRcDE84b5l8m4UncEEFtBB6joQQQeilEVsaYWXN8dwGzWTUjLWcnyWJHKLjd2YojIlucxIWGh0T6pSNh7KgWpOlermQ6u2bU7CM2xWEzYLPItsCBerLImpYfkOJMiSktSGtlqQ222Nwdk8/7o0pRFy+geLQ9DqTpaR4g4lO6/wD19RPhM4X8w4bbrmxuGd2m7WPL54uzVqgWx2K1bpZUrn7Lndc+llBSnlPUdmnqaUoi2NpSlESlKURKUpREpSlESlKURKUpREpSlESlKURKUpREpSlESlKURKUpREqIZtpHptqReLDfM9w+3X6VjDjz1qE9vtmozjoSFrDavUUrZCdioHbbptSlEUtQhDaEttpCUpACUgbAD2Cv1SlESlKURVJkHDji+Q8RmN8Scm/XVq94zaXbRHgN9n5o62tLwKl7p5+b6eruUB0FW3SlEXG+wxKYcjSWUOsupKHG1pCkrSRsQQehBHhWqEnyflqxu8XSXoZr3qNpZab08qRNsljnBULnPeWkr6t9Og6kgdBsAAFKIrb4e+GjTbhusE61YO1OmXC8PCTd7zc3+3nXB4b7KcXsBsCpRCQAN1E9SST+uHfh4xnhwxq94xi98ul0Yvl7kX15y4dnzoeeShKkJ5EpHKOQbb7nqetKUREcPGMo4j3OJYXy6enHLAMfMD6X5p2PMFc/wefn6futvdX61a4fMa1fzzTjP71e7nCmaa3ZV3t7EXs+ykuKLZKXeZJPL9KHwSD1NKURc/EToPjvEhplK0uyi83K1wJcqPLVIt/Z9sFMr5kgc6VJ2J7+lRnXXhHwLXSz4si4Xy949keFJQLFkdnfS1Ni8oT0J22UCUJVt0IUN0kbndSiKFP8A1gzK3yYuuetmoepr3mj8e2+mJ6URrY64go85ajtjkU+nfdK18wB8KkmVcGmHZhw2WDhmu+aZAbLjrsdyNckBgTFpYWtTaFepybAL5eiR0SPHelKIr0k2diTYHcfW64GXYaoZWNuYJKOTf2b7VW2h3DfhOiGjitEIcmXkWPvLmGQLuhtapDckkuNrCEpSU7EjbbupSiKnU+T0j40mfZNJeJPVLA8SuTq3Hsdt1ySuM3z/DS0pQ5kAjpv1PtJq6dH+HDTPQzTSXpjp9AkRodyQ6Z859wOzJjziORTzrhGylbbAAAJAGwA60pRFy8Oug2O8N2mMTS3FrzcrpAiSpEpEi4dn2xU6vmUDyJSnYHu6V18T4esaxHXvMuIGFfLo9eM0t8a3S4TvZ+asoZDYSpvZIXueyG+6j3mlKIoJP4JsZa4gneILDtTcyxafcbjHud5tFulJEG6OtKSSl1JG5bXy+sgkjdSiNt62QpSiLXq38KWUWK85HecX4mNQbKvKbxIvlwaixbYpK5LuwOxcjKVypQhCEgnYJQAKyH0POqf+19qd/JLR/8Ao6UoixuhXBtYtCdU7/qzadTMovVzyll1F4YuDcVDEtxbgd7UpZaRyrC+Yjl2Hrq6da2HpSiL/9k=)
Glucocorticoids
- Released by
- Adrenal cortex
- Other areas in small amounts
- Cortisol (hydrocortisone)
- Increases blood glucose levels
- Aids in fat, protein, carbohydrate metabolism
- Suppresses immune system
- Reduces inflammation
- Receptors in body and brain
![[[@Kadmiel2013-zz]](10.1016/j.tips.2013.07.003)](data:image/jpeg; charset=UTF-8;base64,/9j/4AAQSkZJRgABAQEAcQBxAAD//gAuSGFuZG1hZGUgU29mdHdhcmUsIEluYy4gSW1hZ2UgQWxjaGVteSB2MS4xMwr/2wCEAAMCAgMCAgMDAwMEAwMEBQgFBQQEBQoHBwYIDAoMDAsKCwsNDhIQDQ4RDgsLEBYQERMUFRUVDA8XGBYUGBIUFRQBAwQEBQQFCQUFCRQNCw0UFBQUFBQUFBQUFBQUFBQUFBQUFBQUFBQUFBQUFBQUFBQUFBQUFBQUFBQUFBQUFBQUFP/AABEIAeEDLQMBIQACEQEDEQH/xADWAAEAAgMBAQEAAAAAAAAAAAAABQYEBwgDAgEBAQABBQEAAAAAAAAAAAAAAAABAgMEBQYHEAABAwMCAwMECREDBwsCAgsBAgMEAAURBhIHEyEUIjFBUWFxFRYXMlWBkpXTCAkjN0JSU1Z2hZGhsbTBxNJXlNEkJTNicnOCNDVDk6KywtTh4vBEVBhFRmPxJ0eEZIOjs8MRAAIBAgMFBAgFBAEDBAMBAAABAgMRBBIhBRMxQVFhcbHwFCIygZGhwdEVIzNS4QY0QvFiJHKSJYKy0lOi4sL/2gAMAwEAAhEDEQA/AO5eDXB7S2oOD+hbpcIs6XcJtigyZEhy6y9zrq46FLUfsviSSfjq4e4Por4Nl/Osz6WgY9wfRXwbL+dZn0tPcH0V8Gy/nWZ9LQD3B9FfBsv51mfS09wfRXwbL+dZn0tAPcH0V8Gy/nWZ9LT3B9FfBsv51mfS0A9wfRXwbL+dZn0tPcH0V8Gy/nWZ9LQD3B9FfBsv51mfS09wfRXwbL+dZn0tAPcH0V8Gy/nWZ9LT3B9FfBsv51mfS0A9wfRXwbL+dZn0tPcH0V8Gy/nWZ9LQD3B9FfBsv51mfS09wfRXwbL+dZn0tAPcH0V8Gy/nWZ9LT3B9FfBsv51mfS0A9wfRXwbL+dZn0tPcH0V8Gy/nWZ9LQD3B9FfBsv51mfS09wfRXwbL+dZn0tAPcH0V8Gy/nWZ9LT3B9FfBsv51mfS0A9wfRXwbL+dZn0tPcH0V8Gy/nWZ9LQD3B9FfBsv51mfS09wfRXwbL+dZn0tAPcH0V8Gy/nWZ9LT3B9FfBsv51mfS0A9wfRXwbL+dZn0tPcH0V8Gy/nWZ9LQD3B9FfBsv51mfS09wfRXwbL+dZn0tAPcH0V8Gy/nWZ9LT3B9FfBsv51mfS0A9wfRXwbL+dZn0tPcH0V8Gy/nWZ9LQD3B9FfBsv51mfS09wfRXwbL+dZn0tAPcH0V8Gy/nWZ9LT3B9FfBsv51mfS0A9wfRXwbL+dZn0tPcH0V8Gy/nWZ9LQD3B9FfBsv51mfS09wfRXwbL+dZn0tAPcH0V8Gy/nWZ9LT3B9FfBsv51mfS0APAfRWP+bZfzrM+lqCvPDfh5aVFsW6ZIfHTYi7S8A+k82rNWrGlHNIyaFCeInkgQD+ktGNnAsTwPmN6mZ/8A9lZls0jw8lOpal2mZDUroFm7y1I/TzelYUcdCUsrVjb1NkVI088Hd9C3NcDNDvICkW+UoEeIusv6Wvr3B9FfBsv51mfS1szn2PcH0V8Gy/nWZ9LT3B9FfBsv51mfS0A9wfRXwbL+dZn0tPcH0V8Gy/nWZ9LQD3B9FfBsv51mfS09wfRXwbL+dZn0tAPcH0V8Gy/nWZ9LT3B9FfBsv51mfS0A9wfRXwbL+dZn0tPcH0Uf/wAtl/Osz6WgHuD6K+DZfzrM+lp7g+ivg2X86zPpaAe4Por4Nl/Osz6WnuD6K+DZfzrM+loB7g+ivg2X86zPpae4Por4Nl/Osz6WgHuD6K+DZfzrM+lp7g+ivg2X86zPpaAe4Por4Nl/Osz6WnuD6K+DZfzrM+loB7g+ivg2X86zPpae4Por4Nl/Osz6WgHuD6K+DZfzrM+lp7g+ivg2X86zPpaAe4Por4Nl/Osz6WnuD6K+DZfzrM+loB7g+ivg2X86zPpae4Por4Nl/Osz6WgHuD6K+DZfzrM+lp7g+ivg2X86zPpaAe4Por4Nl/Osz6WnuD6K+DZfzrM+loB7g+ivg2X86zPpae4Por4Nl/Osz6WgHuD6K+DZfzrM+lp7g+ivg2X86zPpaAe4Por4Nl/Osz6WnuD6K+DZfzrM+loB7g+ivg2X86zPpae4Por4Nl/Osz6WgHuD6K+DZfzrM+lp7g+ivg2X86zPpaAe4Por4Nl/Osz6WnuD6K+DZfzrM+loB7g+ivg2X86zPpa4z+uJ2kcJvc+9qE+7WH2Q9kO1dlu0r7Ly+zbM5cPhvV8o0JR2hwC+0Tw4/Ju2/urdXyhDFKAUoBSgFKAUoBSgFKA1ld7lqJF1mJY1A4yyl5YQ0IjJ2JycDJTk9PPWJ7Kan/GV3+5sf01oJY+opNJI3scBTlFO7Hspqf8AGV3+5sf009lNT/jK7/c2P6ap/EKvRFX4fS6seymp/wAZXf7mx/TUbqXWd+0rp253mZqOSqJbozkp1LUFgrKEJKjtBSMnA84p+IVXyRVHZ1OTSTZW9M8cZGrhp42/U00i9MSX2ebAjp5PZygOod6d1QKwMdR0PXwzNx+Jy5cB2cxxFt70JpwMuSUdkLaHD4JKgMA+iqnjqy5Iuz2Sou2vl2+h7Q+IMu4xGZMXX0STGeeTGaeZTFUhbpGQ2kgYKiCCE+PWvJniW5J7fyeIkB32PBMvZ2Q9mA6EudO4PXio9PrdEUfhcejPW48QZVobkrna+iQ0RtgfVITFQGisZRuyO7uAOM+OOlely1xcLNa0XGfrmPCt69u2XJajNtK3DKcKIwcjw89PT6vREfhkXbjqfjeu5zzXMb13GW2H0Rt6W4pAeVja3nHvjuThPicjz1BXnjlEscdp6TxIhKaXMRAKmURXA28o4wvAO0DBJJwABk09PrPkiqOylN2imTaOIExy6+xaNexFXIN83sYRFL2zGd2zGcY65x4Vgni2yG1L90217EFAUrfDwCsZR8odR56en1uiC2UnwT8+4sXsrqYjpqV0/wD8mx/TT2U1P+Mrv9zY/pqPxCr0Ra/D6XVj2U1P+Mrv9zY/pp7Kan/GV3+5sf00/EKvRD8PpdWPZTU/4yu/3Nj+mpPTlwv717iIlX1yVHKjvZVFaRuGD5UpBFV08dUnNRaWrKKmBpxg5JvRGxaVvjRilAKUApQClAKUBTOIGsEWSMuO26G17cuLzgpHmHpNaOl3y43pbwaKorYOApGd2PX/AIZ9NcvtKveWVcjvNi4VQpb2S4kc2yp0EPyFL2AHClkk4Pf8vjipLe7b2yWVBbYzltSyoH9JrRpuLudLnU/VZsbhPrcPy2rY8tQbdBDAc8UKHUoPxZxW3a7fB1d7RTfHgeabUw/o+KlFcHr8f5FKzTUilAKUApQCtecS9PxLxcIa5BkbkNEDkynWRjPmQoA/HWFjJShRbi7MzMJCM6qjJXKd7SLX553zjJ+kp7SLX553zjJ+krQekVv3M3vo1L9qHtItfnnfOMn6SntItfnnfOMn6SnpFb9zHo1L9qMW6aMis2yW5BRLfmoZWphpy5yQlbgSdoJ5nQE4FapTr2BdNPafm22K8l+bZJt4ndouUtSIIjt7SCEublDnnZjocJV5aqVes/8ANmVSwNGoruK8p/YsNn1XpO4R5KVG7yHbdb0TbhJivzOzsksB8pBLmQSg5APnxnPSs9V80itRbjov06R7HRrmmPEemOuLZfUUtkJC/HKTnzDqSBTfV/3MPZ8E/ZREQ9SWbUGqdHRLOJyrdd1XFqWmXMltyGHYyU5RgujaQokHIPkxUhdtT6OtE2/x1ovz6bC0XLg/HelraYwgLKSoOeOxQV5vTnpUutX4ZmVPZ1O6ioq9tfjY+Lrq/Q1rluxQ9eJslEiPFQ1DlSnC8680XkIQQ5gnljcfNkeUio1vX2lmdQaqgXG33+2w7Aww85OekzNrnMAwnbuylRJASnqVdceFN9X/AHsLZsWvZV7fb7mU5rrQ7URpakai7WuYu3+xyVzVSkyEtc3YWwsnqjBBHTr4+OMnRmrtD69ubEG1OXvmyIqpcdyU7NZbfQlQS5sUpYCihSglWPL58GjrV7e0xLZsVFyyrQvPtItfnnfOMn6SntItfnnfOMn6SqPSK37mYfo1L9qHtItfnnfOMn6SntItfnnfOMn6SnpFb9zHo1L9qHtItfnnfOMn6SrxwxsUWzLuJjF8lwN7ufJce8N3hvUcePkrLwtarOtGMpXX8GLiqFOFGTjFX/kvlK6E0ApQClAKUApQClAKUArgH66v48L/AM6fylCUdhcAvtE8OPybtv7q3V8oQxSgFKAUoBSgFKAUoBSgNa3ZIN0l/wC9X+01ilIrlJxvJnWQl6qG0U2gireUrzDaKr/ELTsjVmgtRWSGtpuXcbfIiMrfJCErW2pIKiASBk9cA1KiVwnlkm+RqeZ9TrczqmfJgXWNDs8+yy4rjCd+9idIZQ048gYwUq5SFHJBznp1rxtvAG7PWdmPcI1sTJE20qklVwdlNyY8RwladimUhOUkhKevjgqAqu5s/TYtLroTr/BGavik9dGJcWLpFRM9uAzuS61cOzmMFpSBtCQg7gc53AdPLVYjfU86hm6eNrmOWS3GFpyRYosiApxSpq3FIIdfBQnaBsBKRvO5SjmlyI4yCSve6S971uZly4O6xvZvlynNWZN5muwXofYrk+z2JyOw61zA4WVbj3x3SnBClDpgE3HXvD++al0BYbc0uJK1HbuU4LgJS4QakJZUgvN7G1gjKj9jKcFKiOnSjsUSxNNyjbl2crW6lRZ4aXVXGuzIkrMq2It0a73RaIxbjPXJhtcZtSSBtGQoL2DqOWD5qx2eBep2tNqgpFoDMG6wbha7RIluSGmkMqUXWi+WQsIXu7qSlW3HicmmhdeKhG1+i8/AkrbwSvEbXMy4SxDlwXrm/dmH03F1tcdxxko2ckNYVjO3O8DbjpkYqKY+pslt6dbhqZshlo0a9Y+YEnAnKXuD2eXnb49/33Xwpcj02K4X5fXtN8WeEuBaIMV1QW6wwhtakkkFSUgEj9FZm0VbymqcrsbRTaKZSMw2ipHTqf8APUT/AGj+w1dpR/Mj3otVpXpy7mbDpXVHKilAKUApQClAK+XnA00tZPRIJPxVDCV9DmbXUqRqm/8AL3FLCHOcVAnO/qQPHwCf1msaans8B1yK0l5aUZSk4AWfPXBYiTnVbZ6vQgqdGMFyRQrjPnBxxT7ZUV5ALashOfEHwxmrJpWe5c7c+uWgMuNAp75HeT6PPiqZQurlEKsVKx5wpr1tccuURW5uLIbUFIBACgckY/8AnjXXUGSJkRp4eC0g10+y3ei12nK/1Ck68JLoe9K3JyopQClAKUAqn63TmZG/3Z/bWDjVeizOwWlZe8rW3I8abfTXO5Tosw2+mm300yjMNufLWtbTwJtNoc1opmdJUNSMPRghYGILTpcUtDQ8xW6pWD6KqSsX6dd000lx+h4p4C2928sTZN0febYta7UltqO0y4tpTAZIccSnc4AAVJCs7SfQKjB9Ti09AdZmapuE95USHCQt+OzsSzGcK221NhIC0HOFJVnd1JNVXMlY1r/Hp1JzSHBC36Pm2KSxcHnlWmRPkNoLLbaVqlbd42oACQnb0CQBWVN4Rw7lr53U82WqSVRnovYkRmmuY24gIUh1xICnUgA7Qo9CfHpUXLbxTcnK3Jr4u5TdH/U/vN8HGdL3aWmNNlPrkTS4y1M8U8ttIUtJwpDaWwFpxgp6dOlTl0+p/t91avUVy9Tzb7tb4cOTHcS24pTkXbyX96kklQ2jIPRRJzU3LssZ6zcVz+qt4GdY+DES0P2N9U1ov2q4OT0iHbmIaHFKYUztKGkgdAonJyc+XGAMjSXCCFpGXpx9ifIkKskOXDbS4lI5gfdS4pRx5QU4HrqG7lmWKcla3nX7l92+mm301RlMTMNvppt9NMozDb6atWhhhU31I/8AFWZhI/nR9/gYeLd6EvPMtdK6Q5sUoBSgFKAUoBSgFKAVwD9dX8eF/wCdP5ShKOwuAX2ieHH5N2391bq+UIYpQClAKUApQClAKUApQHOf1S8qTC4dX9yIqUl9U6I0EwXyw8tK5rKFIQ4FJKSpKinO4e+8RWrY+ntbWnWOlYrE42Vc25XN62xb6+5dlwIghsjluKDwLii6lxYHNIQHAATtxWjSWt+rOgu3a3RElA486gmW/SSFwLe3dNYWm3SrM2lCyjtC3EpnJV3sqQ024h4AEHbuBzjNej/Em9yLsi135EVq5tagt3IgsMuNITDeluNtvpfbfUl8KSgdFBGFJVuRggCndJFW8bK3q/i1qK/8LrlIlP2tyDqbS99kswYTC25drLEdZTzHC4oLxnYs7EbXCMeara/xX1BFucp2O7aharZqC26dXZ3WFmdJ7QmMC+lzmAJI7TvCeWoFDSiVd7u1OkuHnoQqjKkxxp1NrTQ+r7uv2OkWWJZlySw1DkRVsyS8QhoPh8KdTy05K0JTknxHvRe2uKd0Np1ZqGberLarTbZdwt8e3OW91+W05HfLKXF7X08zeU7+WEp7rrffHUmJUktEFUbK9pjXl61jr7TEW8oQzMs+p59sWWmgxzUi0c8FbaXXUpUC9tICz70HunIGRqXU+otI8T9aXa3SICrTGfsDEqDKjuOOPJfd5KuWsOJDRSHN2SleSADgVDgr27PqM7av2/Q92eMeo1SbfcSu0O2253a7WhuzojrEyGYaJRS845zSFZMUb07E45ycK6d6Cc4xcRINil3WRI0261E0vC1W4y1bn0qUh5TgXFSS+cEBokOkeJ95ip3UQ6kraFkPFrWcnW18TB08iXpyz3R23SWwllLuxEbmc0OGSFlalFOGwwcoUCFeWo+38WdcTIFvEM2C/wBwvenUX6G1AZLYhkvx0FpRW/teKkSFcslTW9bKgdoV3Y3USd5J8D2s3Em73TWtouJ1RCTpxOmp82Yw9aHWSXo8lLTpKC8SlSFZBxu96oDO4EYNr4ya3uzF+iw4kSbKgi2zESBbktOmHJS+pSkRTLVvUkMBQSXUK2rPdJSAqd3Hz3kbx8jKXxNvc1V5umlkxbo26xaJSpqGHVqcivRXXVvNRXJCBnCUkNpUlW0qJ3qSAd0aZuzGotN2q6xpLc2POiNSWpLbSmkupWgKCwhRJSCCDtJJGcE1anCyLkJ3JLb6BTb6BVqxcuNvoFfKGLw7cIYs0iFGkBZ3KnMLdTjB8AlaT+urtJLeRv1LVZt05W6Fg7FxA+FtN/Nj/wBPTsXED4W0382P/T10RzQ7FxA+FtN/Nj/09OxcQPhbTfzY/wDT0A7FxA+FtN/Nj/09OxcQPhbTfzY/9PQDsXED4W0382P/AE9OxcQPhbTfzY/9PQDsXED4W0382P8A09OxcQPhbTfzY/8AT0A7FxA+FtN/Nj/09YF/Y16xZJq3Ltp4oDKshFufCsY8h55qmbSiy5TTc0l1NC3mJfrbNU6/JgYfwrmIiO4SAkg/9J09R84qWalOx7S65cEtylJycRYygMY8ylevy1w9Z0nL1E796+x6ZTWIcVnlG1uSafiym2SLG1JMXGRBdXHSglZbSQU4IAzgn+Plq5W60uIcZc7I6IsTcrb13q6EYx4+BPm8BVp6tFdKMcrdynzeKU6dc73YJmi58KGxlli5IUHEvEpJC9uAcdACQVYJFdG6U4msnT8LNg1E6eWMqatbiknp5CK6bZ0FBSSknw4dxym2006d0+fFdv8ABLe6Yx+Lmpvmh2numMfi5qb5odrc2OXuPdMY/FzU3zQ7T3TGPxc1N80O0sLj3TGPxc1N80O090xj8XNTfNDtLC490xj8XNTfNDtPdMY/FzU3zQ7SwuPdMY/FzU3zQ7VT1vxKiN7ZL9mv0VhhlS3HH7a4gJSOpUc+QCsbExzU2jKw0stRM5s0FxXvNslakvExV2PtisczUVtjXqK+3HjPMFxSI7W8AFJjLjEhBxllxX3VSup+IuvPYS4Ql3i0QbiyNNXNudDt7iUNszp6mHGFJU+dwTyeq8jclShtSSFDVOlC+q88DcKcsuhddB6ovUS76vhzba7qKWxqFENyZZkBlpKVQIbgdW3IlK2AB3GGiQduduSSaEOLOrLbpm7vWX2IjR7LZ7vfXW5zMiSuQY8+SjkpUXwUBaWuqiVbSeicYApVOL49hU5tLTtMLW9skXXiHqK+T3YcpcC/aVYtx7KoSIaHZ0YqSh0uEBKgtYUAkbt3XoMVLXL6oK/QTql+GYN0gMaaul9tUlyAqMhSojzbQSR2ha3EEuYKilrJQSkEHu15FNJPzoinO4t2JeTxG15YNQ3KNcplgmwrTerRb3xGtzzTklue6y13SX1BstF7IJCt4GCE+JxrbxH4h6gulrYiztORGLw9emY/Ntj7ioogyS0hSsSAHC4B1Hc25yM+Bo3dO17ebXKs8+Gnl2PFn6oG73L2kT4QiORbqLGi5wBCP+SruGzAEhT6ckBwKCUtL6J7xGcp9NL691ixa4Fvk3+1SbleNYXaztXGVAWG4Tcd6aoJKOf3yoR0obTuTtB6le3vS6UUuBCqSb0ZmaY4ma31zdrZardIsMN4Rbo5JmuQnXmpS4k7siFMpDydiHPf5Kl7fAbvGrLwe4oTuKbs2R2VmJBt8WLFlthJ5jV1KVKlsZzja0C0nwzlSuvSqJU4pO3IrjOTaNm7RTaKx7F4bRTaKWA2im0UsBtFWfRAwqZ6kfxrLwq/Oj7/AAMXFfoy88y1UrfnPClAKUApQClAKUApQCuAfrq/jwv/ADp/KUJR2FwC+0Tw4/Ju2/urdXyhDFKAUoBSgFKAUoBSgFKA0Rx21Q1orRt8vLsKDPTHlMAM3JeyOFLlNoC1qwcBBUF5x02+SqjpfjJa75a/ZO7wI0ifHur1qtr9iC7i3cFBlLi3IiwgKKAnelZxgKaWCo4GdK6bd2upvVO1l2HtAvWhdTaw0a/aRLkvWy2rk2+NBgLEWGzJy0lx0Bv7ErEd5tKSU7fsgUnwxhab4l8KTb5Vyt9uYtUV+Mi/B9dicjduQFpSmQ0C2C8sOOISCkFW5xOM7hmlwm9Cc8TJRrXhtb2rtONhTDuFwmJtNxiGwLTOlPOtlxDbrQb3uBxGSCcpOT18cZcDWPD/AFDqKw3OHaBO1DOadbjy27MtUyI2092d0Or2b2UodJbUFEYIV5ASDhPjcnPHgUPScXhZoSbJlO3K5T4lutUyFHcuttT2Z2I062JKGltx0GUoLbaGVlxRwSknKib/AKwuWlNP6A1TxCf0cmS83bJDk1mRaQ3NltNpILTu5G4oO0dVZTt73vRmqpRm2QnFLQxGtfcO9GNRW2bKbMiM03PeEexONotiXQWUOvlLeGNyUFOVYOxOThIzVo1VfNN2K7wrfcLcqbcrz9lTHjW9Ulx1EdTf2VYSk91tTjfU+BUMVQ4SuVZ42KPA4m6EYuepLzJ0q7bLm1djYHJqbItcm4ulPdQlSW9zm7bjb1x3c9CKn3NdaENzNnehJbS4hu0uuP2paYqCGi8iG44UbAQhRPLJwCrb0UcGXTl1IU4mGnXGgJzto1gmwOyLrcl9nts32AcM+Sjkl3c0eXzC3ysnI6Y6ePSvx678MbbeLjpr2Agtt3Ca1BuK27H/AJC7McCVtsvuhvllw70YCj75aQcKUAWSfC4zRZjwuIvDi4MWR2FYnZSYrzsOzhjT7pKdqFB/so5ediUoUlRR06AdSQDdBwv0am2uW8aSsQgOKStcT2NZ5SlJKiklG3BIK1kdOm4+c1DU48wnGXBGZdNEadvkZUa42C1z46y2VMyobbiCUAhBIUCO6CQPMCcVMNsoZbS22lKEIASlCRgJA8ABVtpvQrvbgfW0U2ioyk5htFSFgSPZiL/tH9hq5Tj68e8t1JXg+5l8pW+NAKUApQClAKUAqI1arbpuf/uiP4VbqexLuZeoq9WHevE05MZZmRZSH0hbKklopPlHh+2qxO0o63Z3+zzZTigkhKAhKlHPTHhXCSVz0tSai0TumNOo05aG2koCXF990gdVK9PqqRKFJiL5aXc7cYT1FVcCwmmV+5uoEF0lICSk90+Q1ubhU8ZGgbOsncSwjqfL3RW92UtZ+40u3/Zprv8AoW2ldAceKUApQClAKpuvYbE9SY0lluRHeZU24y6kKQtJyClQPQgjoQaxcT+kzLwv6qNM2HiLpXXFz1PbpVsaYjaXKyJNwabLLjH2aO68144QFMyWVeHRJ8ihnG1BxS4XI0zcbi/Ls95tanIdpmJjtNvhaHXNrCFp8FNArWoeKcBZTk5FarJO9kbjPC12Wbh7f7DqJm8K08zAbgxJiYpdgKRh1SY7JBUlIBQoJKEBKuu1CT4EVgJ1bw4W3emROsKWocZ1NwStLaUJjqcId35GFNlxSt3inco56mqcsr2ROaNj9OvOHkyzv3j2Usr8FUxqO4+AhW6S0A40kjGStKQlxPTISAodOtRcO9cNkOXO4sQ9NM2aZbEz5F3QlhKZbUh1TZCxtypK1NJ6nIWcDqRVSjNEOUGSszXnD72E9nJN0s6oEuSEqkuBJLkhjvDcMbt7XL3dRlGzPTFesbWWgWp9njsXGzNypiOdASjYkkSVEhSTju85QOPDeQfE1GSZOaHUwLRceFuoNQoYt3tZm3q0x9rfKZZLsVqOsApQrGUpaXt6A9wlPhkV4O6g4UXmy3eU4vTc63S5DRnhcdpwSnlDLSlo2kulQBKVYOQCQSAanLUIvTJBnWekrA/IU4bVa4dvZjRokllbeVtvI5iGkISNyUkIyEjxCcgYFTWiNLWvS1mdRanVyWbhLkXRyU4pKlPuSHFOqXlIAI7+B06JCR5KpaaWpKcW9CwbRTaKt2K7jaKbRSwuNoptFLC42irPooYVL9SP41k4ZfmrzyMbEu9JlopW8NCKUApQClAKUApQClAK4B+ur+PC/wDOn8pQlHYXAL7RPDj8m7b+6t1fKEMUoBSgFKAUoBSgFKAUIyKA03xa07B1Np27xLlPTbLe1Ibmvy142toYfQ+SrJwE4awSfAEmtV+5DpLiOZc3T99tVyYj3pd1YhvQ2p8OI87HDb7S2SoZS4TzsZSoLUVA4JB16p1Msqii8qer6G2unZX1LI9wTQ5d9LTG5sGC3YUJ5aLbam4rqiNxU2hxChy2FlQ3NbSDgdQcmqFprgS7D0Foy2a31K81e2rNFsVpZjx2wYElJZlKG5O5LykrgtkKICdjODkqKjTBOTtFa+bkvvLzE4WMK1RHlXfUXshqhy4s3t1TUYMIW3HZVHQ2hvcragc4kkqUSpR64wBjTuB0ZJsUdV9Zjoi3uVdWXlQ0CbzX5zk1bcd/cFNghSm1gBW9tJ6Dqahpq2hOnUxLVwKs+hFymY0mxMNXFUiLBXK0+w5K3vqU4WnXs5fRt3p2kJJT75RIzVmtfB+ND4SXbQj9xddiXONNiuPMthtMdEneChhslQbbQF4QjJwABk1Er2TfMKy0Kjq/htHD1xg3rW4io1DaA3fo6LeEiZGjENrcaO48glMhDas78ggpAIJq68TuGB4kx7dHNwatzUV3m84Q0uSmlZThyO9uBYcABG4buijkGqpQlFRm1o+HgE73VzDTweIvsiWbytUFeoG9QsxOzAKafDRbWgr3d5KuhHQFOD1OekQ9wislg4lm8ok2pqXfrguUyiXZWpExMoRzu5Mk9UI2tFeFA4OQFAKAqIpzdorl4ah2Rhv/AFOPP0pdLL7MQWm7jMMtaGbOhEeIrlBsORGuZ/k72QV8xKvfqJ29cVMngeRdXUe2KSvTr92jXx+1usJU65LY5SkHn5yEFxhtxSdpJUDhQScVTmuLHlfOBCLtw5t2kGrs21GiKeV2x6Al19tS1KUHWFbgWXkbztcGceUGtpNMcppCNxVtAG5ZyTjymqZesipOx9cv0inL9IqjKVZhy/SKcv0imUZhy/SKzrGjF2jf7R/YarhH1kUTl6r7i8UrbmmFKAUoBSgFKAVEavaL2mrglPVXKURj0dat1FeDXYXaTtUi+1Gl3XBJtbxT75SioD0g5/hWTAWChJzkVxElyPR+MWSCXOelQB2kdAa8HmmgnCpTgV5wo4FQWY3TslcquqCEwUgqCyV9VjyjBP8ACt3cLmkNcPrCls5R2VGP0Vvtl/5Gm25rCn7/AKFppW+OSFKAUoBSgFUviK3MdiuJt62m56ozgjrfBKEuYO0qA64zjNY2IV6Zk4f9RHOzf1OTmnrXBbtmpJK1CwTLDOVdHCtDqH28hxIAG1QfCV9c9Fr8p6yl/wCClxnt3JcK4Q25DtrsEWMl1tWwPW2Y7JyvHUIXvSnp1GCcHoK17kmbPJbRFgsnC7bdr9cbxKeU/cbu1dm27ZOkxUNqTCixyhfLWnmJ3RlEBYIwoZHjWvpn1PWqL5Luz941GxPflWmZa0vvOyXS4XpTDyXC2tRbZASzsLbSQPA5PglGSQlFs8+MHDrUMbVCL3aJUphubfkXBUy3QnZT0JKLWuL1baIcVvUcZT0AV18le9r4M3m+2LT10ZYiaXn262WlmJZ5AW820uDKcdQlw7t21aFJHiVIV1O4p6zmSSIyttkHxF4Y6ph3OHcjIeRLu1wn3C5SbHBkSWoxXEjRW46ENKDpC22ernQbgokJJRif07wTuM+4W29TbZGgRbgxapE2wPXOYlNteittgNoQ04GnwnlIxvHdUknKgcCXJW0IytslZHAF6fp6BaJF0baZbi36I+9HbIXtuK1KCkelAV1z4kVhscDLmmwyEPxbW/fFdkbbnqu9yLoSwHtjjbynFOMKSp5RSlGQAtxJJCsinMirIZNx4KX2VBmrfucO93iQ3bv84SFvQnkSI8ZxpclC2DubcUpZVgdNpUk9FVtjT8CZbbDbYdwnG6T48ZtqROU2GzIcSgBbm0dE7iCcDwzVE7SVkVRTRIFJoU9KtZSu58oWl0EoUlYBKSUnOCOhFfW2mUXG2m2mUXG2rJo0YVL9SP41kYeNqi88jHxD/KZZqVuTTClAKUApQClAKUApQCuAfrq/jwv/ADp/KUJR2FwC+0Tw4/Ju2/urdXyhDFKAUoBSgFKAUoBSgFKA0jxzs8/UOjb1Zra2tyRdJDUBZS3vDbLr6EPOKGRlKWlLUeo6Ctea20LdbVPYlsy7rf75cpjTsxiJHfixZTSUhpLXNYWOzhtKlOJU4pQKs5ydu3KwlWEIqm7XbldvktOT0burrutzM6Sb17jBuTus7ncNesWJ6c6zp2VMfhPNPbzKkvxmyhlAJwoMc18lCum/lD7k4zblotm9yrAYMHVHsJAYn3R0TpUtEl+ShlDDTeVr5qC4l147Rt3bTkYUc5eanSy5MqdtbpcXG6+DbT5aWZTrLj51MBdq1lCg2hi5m/ybexEs1sua4anVSHQhl92U6jYd5K3VR2lrT3toV16E1+WPSFxenaHk3y26i5LM68XBLC5cl5yEreG4bLikrOPsBVgqJHQpyd+FVupRim6Tir35K98rvy4cF3si0nxv5Z6o0zrWxaSscm3Ku8zVC7LOu00ypTjiDcOz7WIpSpWwALkr2owAeSCeozV3tEN618N77L0vHvjl4MJ0xW9QOPl6RKQyditj6iU7l4B6JBI8MYNYmIlSqJOOVJuz0V7Xdre7i+65XG6NdWfSd4fv6plth6mjsOC0QhOurz/PdSJKpE93DiippK0stNkYSCT4YIzltx9Rpj3lzkajkWqTeGFSbilEtmeuGtbrjrKI5UVJ5R5TXMZSnchZ2jKc1mTnRm2pON1bpa+mnDhe92vqUK66khpnRl9u98tsecb7A000q43BmG9PeQvapxlEVh1wL3+CX3dhV3dyUnoCmqlFsesZ0VN7i2/ULGpomnJCpUq4FxSTdH1tJX2Zha9v2FAfCNgCVAoAJAqYVaDk03Gy0eiV7uV7ad3ckGpW088C02bQdzubtjh+y+oXLPLuDsyeoiZA2tNRi0lrDzq3kh1xxKlZX3igkAeNWXhZBuCNUanekt3R6AtaXIk66CSw4N61lUbkuqKVBkJbCXUABQVjqQonCxFSE6c0lFO3K175rclror6W+DK4ppo2bs9Jps9JrQZTJuNnpNNnpNMouNnpNNnpNMouNnpNZ1kRi6x+v3R/ZVUY6oom/VZdKVsDVilAKUApQClAK8pbIkRnGiMhSSMUauFoaAucJVuflQXMjY4QP9k+X9lflidKIwac9+33Ovo6VxdaOVyj0Z6VSkqtKMlzRJhxJyCcZr2IjcsAkE1jCSkvZKDqJLt4vsWxW37LJkr2pSOuwnpk+gDJNdN2K0t2Kywbez1bjMpaBPoGK6XZtNxpuT5nNbbrKpUhBcl4/wCjPpW4OaFKAUoBSgFVXWYy61jxDRqxXV4GRQdpo4X03oi+RvqeI2pENQ7HCnaWtbQZalOSfZCUt+O4Jr7ZCEpUBkYBKlcxWVDArZs7iPqSzX6Tpa56njQWGdSi1OapkRWWlNMqtbc1CCk/YkuKcWW0qUMbRjBUQaxZJSM5ScSKtfEXUly1Loq8SdQqLLtn1ClmDGShqNfXIs1huM4hJzlT7WFp25ICjs6KVnGHGrVA08ZFq1Va9SSZlhi3RyQzEbLVplOS47QZUlCs7FpedwhZ3jkK73mjJF8ic76k7qjiDrTT/EI6YYu8Rbluj291DtyVEipunOeWl1SwtSVYCU7EhkdFjKt2QmsaZxmvcbRGpdQNajtz97YlPRjpnlMhy0tIuKIpkLysKw20rmLLmEEkHKE+MZIuwzsL4pajjxtPRbjq61WeHctQSLd7OOOQ5Dgjpgl9IcLauQh7mgpHTBSUEpyrFQcHirqoPsanevZlSfaVPuMKxIQlEe8Px3XA24231UeYhLbvcJISsAHBqckegc31JmZxX1HGhXiLY9YW3VYS1Y3Gb41EaW1FemXBMdxpSW1BKkls70pJC0g9VHKTV+4tX69aD0JbH2L6yJDMhKZ8x5UWPKkspacUvs6XtrJdylKtqsDYHMYOMUuCulYlTbXEqJ47vKuEiEi7RUy5GpbHDt0SS0lqS9AlRoTji+Ue91U7I72O6QR4pryPFXUmktK+3/UN3al6Wa1BPt8+EzCSjscRqTKjMuJWMqUsuojhRPTCj0GMlu1wG8bMVGvNT6PnXG931MKO/boGn5moy3GCVJjvrmIkDd47WMpWD47WV/fGo7VHHjUNks1kmOXBUS7yY0a8KtbkeK0z2SXNWlhlZcWHVrDICCGQSlSdyjhSU1ORMjO0bZ4Ko/zJqX8qLx++u1sPZ66tSWpcUtBs9dNnrqnKTmGz11YtIJwqV6k/xq9RXros15XpsslK2hqRSgFKAUoBSgFKAUoBXAP11fx4X/nT+UoSjsLgF9onhx+Tdt/dW6vlCGKUApQClAKUApQClAKUBDv6aZfeW4XXAVqKsDHTNfB0qwf+mc/VVp00y/vXwPGHoe325jkxEiM0Vqc5bLaUJ3KUVKVgDxKiST5SSa9vaqx+Gc/VUuGZ3b1G9Y9qrH4Zz9VPaqx+Gc/VUbtDfMe1Vj8M5+qntVY/DOfqpu0N8wdKsH/pnP1U9qrH4Zz9VN2hvmPaqx+Gc/VT2qsfhnP1U3aG+Y9qrH4Zz9VPaqx+Gc/VTdob5j2qsfhnP1U9qrH4Zz9VRu0N8x7VWPwzn6qe1Vj8M5+qm7Q3zHtVY/DOfqp7VWPwzn6qbtDfMe1Vj8M5+qvaJp5mJIbeS6tSkHIBxUqmkQ6rasStKulkUoBSgFKAUoBSgNZ8VNNnli5x09UAhwDzeeqJanhJV5Nwxn9Fc1tGllnmXM7XZNbPQyc0S6I6HTg9DXhc4qI7Rws9R5PGtMbeMm5KJK8D7BEbuV3ua0hUzeGWirxSjAJI9fT9FbhAOT1HoFdrhtaMe44LH/3M10Z+0rJMAUoBSgFKAVWdWAmSxj7w/tqzV1gX6Pto1fH1uzI0zZpiLYnk3OeIMaMFjHLDiwHB3fDltlzbj0Z8tZcnV2mZNuS69JjvxZkVqcEqYUvnNOFKWlbdpKio7UpTjcSMAdK1qmiqOMptXnpon8r/AC0I+7cRbHGgt3BKW5USHHly3EKjupkMCOUoXtaLeUkKWEndtwDnqM4/IGu7IVyQYZgMhDDshh6A+1KU+6ohCQyWgXCdiu8kqOUnp0zUbyN7Ft7QpZsvv4Pha/Tjx0/0W1tiLdm4c0x0ulKQ6wuQwUuN5AOQFAKQcYyCAfPVNsfECDe51yXFtrCI8Z+S1dLitxKWmWmVLQFLVt7ylbdwR9yg7iRlO6qUkrIv1cRGm4RtfN7uFr/BamPdLloS4ptU6UOUizzCiNEVBdaPOfYWgJ5BQFL3trXtwkgnwyanF6502q6xohdUuahSmmMQXlbXOWVqaQrZjmBAJKAdwA6ijqRfMt+nYf8Ad8n3dOwr8HirYF2VE2NbnosAxH7q809b32lqZbKTzWxytjmSpskhXdCgSakU8QrNd0qTNirbjNMx3XI06DITJS86shpKWVNZXkoWApBPVJwPLVCqxZZhtGlOy668Hwtfpx42XM+OJ+uI2hdEnViYEZ/LsRjtFxzFbjNuvoRzX1lBW222V71ZTkY646kZvDbUrmu9GR58+1M2195bu+GCVoWgPLSh9IUhKtjqUh1O5IOFjIrIy3jc2ikpcCLTr6bfp8j2HsVumWjtK4CpdyuQjOTVNrLboYb5S+YlKtye8pAJBx0IJx/dY0403Nl38MxkRJk1iMtuK5JUiNHc5Lr69iFctvmJUCpWE9B1raegOXq023Lha1lfpe+vBrlqi3vFxfAtSNaWFlL22ahHLlR4riQ0sEPydhaSRt98rmoPo3ZOOtYiuKOmktznDcHORDVy1viG/wApxfMDexleza8rmEI2tlR3HGM1jLB1pcI/Nc7fcqc4rmY0jjDpOLAYlO3J1IfW+0iP2GQZJWzgupLAb5gUkEEgpBA6+HWvuFxd0lcYc6XHvCXI0NlqQt0sOhLjbqihpTRKfswUtJQOXuyobfHpVx7OxCWbL2cV1t16+dCN7DhcmdMastmsYsl+1PuPIivmK+l6O4w406EpUUKQ4lKgQFpyCOmceOavWkgQqVnzJ/jWOqcqVXJLiimq1Km2ixUrLNcKUApQClAKUApQClAK4B+ur+PC/wDOn8pQlHYXAL7RPDj8m7b+6t1fKEMUoBSgFKAUoBSgFKAUoDUV51XxHYu85qE9pcQ0PuJZD8OSXAgKITuIeAJxjOABmsP24cUPw+kf7lK+mrTy2gotrKbeOz8yTzD24cUPw+kf7lK+mp7cOKH4fSP9ylfTVT+Ir9pP4d/y+R8M634lyEktS9HuhJKSUQ5JwR4jo9X37cOKH4fSP9ylfTU/EV+0PZzX+XyMSTxH4gw3ltSLpolh1DYeU27HkJUlBO0LIL/ROemfDNZftw4ofh9I/wBylfTU/EV+0l7Na4y+Q9uHFD8PpH+5Svpqe3Dih+H0j/cpX01PxFftI/Dv+XyHtw4ofh9I/wBylfTV4ytfcR4RZEidoxgvOBloOxZKeYs5wlOX+pOD0Hmp+Ip/4krZzeil8j29uHFD8PpH+5Svpqe3Dih+H0j/AHKV9NT8RX7SPw7/AJfIe3Dih+H0j/cpX01PbhxQ/D6R/uUr6an4iv2j8O/5fIe3Dih+H0j/AHKV9NT24cUPw+kf7lK+mp+Ir9o/Dv8Al8h7cOKH4fSP9ylfTU9uHFD8PpH+5SvpqfiK/aPw7/l8h7cOKH4fSP8AcpX01TOkNS69n6ihsXh3Tircsq5ohRZCHsbSRtKnSB1x4g9M1XDHqclHLxKJ4DJFyzcDaFK25qhSgFKAUoBSgFKAxrlCbuMF6O4kKStJTg1o2RYzp66yGCrcAvA9ArUbSinSzHQbGm964crEjHSFKyDWFdlBxJ8uBiuYfA6uCvNH7ovUCtLTXnQ3zm38b05wR6RW3bHqi33tpIYfHOA6tL6KHxV02AxMZwVJ8Uc3tjASjUliIap8ewmfEUrcHMClAKUApQCqzqxO59kZIygjI8fGrdRXiXaftGrNMcO5lmFgj3C7M3CBp9vZbmGYZZVu5ZaDjqitW9QbUodAkZUTjwxEtcFER9Pswhc25UxiW0829LilTXJZStLMctpWDsSlajndneSry4rW7i6s2YbwLlFKU7tX5acElpfotet+RIXHhYJ1umx25cOEp+1pt6G4sHlsNkuqcdVy9+SlzKAU5z3SdxJ6fd94cTL4zd+ZcYKpFzWyXHHoK1BkNtBKS1h5KkKDm9xKtxxux5MmXR6eeJXLBtppSXw05rr0fyLHpyPdY7Mpu4OpW224Gom/vOlpCEp3uKBwVLUFK6YwFAeOaprPBlMXTs20sXbY1cLc3Hm7o2UvykEHtJTv6bhlK0Z7w2jcNvWZUnNK76lyrhpV4xzys0mvjp4fMmrZw/TEfszzqre0q3ynZim4EIsIedUyppKjlxRylK1dSTnp4YrAhcMZsee0ty+pXCiyJ8qKw3E2uJdlFwlxays7lIDqwMBPQ9ajc8LPzp9ih4R6ZZJa8l/28NdNY/M8nOFL8nThtr11Y5qLGmyNLahlLaEBWVK2FwkhSUtpKdw96TnrgZV84dTL21di7cILkm5cgOLegLUloNN4SWgHgpKg4VuJVuON2PJuqNy7WuUPBSy2zLh07GuvR/I+eIfDJzW+nLDb0XJkv2iW3LSbvE7bHmlLLjRTIa3o3ghwq98MLShXXGKwODXCGZwn9l0P39N8ZnlpSAuKttcUISUJZbUp1w8hCdoQhWVJ72VqyMZSVlY2q0tqY0PgrJZk2eI/d4MrT1ouirnCYctf+Wtnml5LXaObjYFkHIbBISkE9CTHyOAM82uXbo2qEMRrnam7ZdFqt+914B9951TR5oDYdMlwKBCumMEHrW8W0Yp6w0vfjzvfTTRX5dOepaydpOL4TzndYLuTl+bNnXe03xVuTCw646mKlhCFO8wgpSUNrThA97g56EY1u4S3y3aQ0/ZRqSC6vTsiO9bHV2pQSoNIW3iQnn5cUpK8lSSjCkhWPJVr02m0ouHC3Pmk03w6PTpZcSrK+p9I4LupRcX1X3fdptuuMZUzsmA3JmKQpx9KN/RKQ00hKM5CUdVEkmvW88HDJldqtlzZgvRxa0QWn4ZdZZbhLcUhCkhxJUCXVHoU4IT44p6er+zp38rp9O/4jJ2ln0HpF3R1nlR5VxN2ny50idImFkNcxbrhUAEAnASnagdfBIrYelBlUn1J/jWvnNVazmla/LoTN2p2LDSrphClAKUApQClAKUApQCuAfrq/jwv/On8pQlHYXAL7RPDj8m7b+6t1fKEMUoBSgFKAUoBSgFKAUoDWd0/5yl/75f/AHjWLXHVF6zOvgvVXcKofHZm7yOEWpm7GmQu4qjAJTFzzVN7080Ix1yW9+MdapildGTSUd5G/C6Na3HVuitP6Yjjh4p+yImSWI8qXbI4itpIadKEvOPMuAK6EHagrJ2gnqK/NG6z13qpVukruclDsXSabquA3DbAmzQ6+2ErJTlIUEpylOOuMY8t3LpqbJ0llcqyvLz2lBfv9xvUK+3j2Wkanmr0lED7k+2t7GZJmoU7HCC2EKCCo9CFEZ8egxsaRrvU7/EdcBu8zGbj7ZPY4aeTDRyRatnWXuLe7OO/v34z3cVLii5UpwejXC+l+Gi1IyBxP1guzX1V1lXKP7WYqLZPfiR2kLkTlzCjnpUttSUoDKEqKtpADpODgVgI4qarY03bpkm+SXmY94mRTHi8oTrgyktcktKVH2Pbd5ykJbKwQR4GoyIj0elwST1+l+vDUm7nr7Ujlt4jPM6guDGqbd24Q9PogoLceO2tPJfSS2SolHXJUQrccDpWPddbydfa0guRZD06wxdVWZUBRjFtISqM+XVJJSCRvHicjI6UyooVGCvJK1v4s/fqQ8bi/qyVH1c/b7ncXGm9PSJ8dE5tlb0WU2+hASUoZSEHarq0SsjoSetba1y9ftKcKEra1QPZZLjanbpPS2wXEqc3LbCg2pDRKcoSooIHTPXrUOKVkUVaVOEoxy8Xy/2UJXEbV7tx0rbIci5NtaxjRFQX57DPPgqaczMKtqAlRUzhQJGOuQBUDqvVGotQ6G19Dev1xmyra/GlCbbA2YoZMsjlpHJS40tCACpCio/Ywc4JBqUUi9GjTi00le67uNuvVFhGvdUv8TYltiajQLa29bkwu3lLYusVxtBedCExyVrUSrqlaAgpGUgZrAt9+1/eBZHFavuMZN3ZvRcbbgx/8m7I7hnYS3kFXlJzkeGPGoyx6FO6oxSbjy+jfXqrG8uG94mai4e6audxINwmW6O/IO3blxTaSo48nUnpVjqy0amcUpNIVKaZ/wCfIvrV/wB01eor82PevExqy/Kl3PwNhUrrjkxSgFKAUoBSgPGRMYipBefbaHnWoCsCRqq0xh35zRx5EHcf1VRKcY8WXoUalT2ItlU1BxNZbQpqAlRPhzFDr8QrXNzve5uTNkrQywyhTzrzywkISBlSifMAM1zWPxCrSUIvRHcbM2e8PDPP2meFk1rbLhPTDYlIedUx2gFs5SUbtpIPoOAfWKx75rK0wrsba7MQiSEpWoKBAQkpWQonGAO4R6yB5RWujTzaJm3nh6lKpaUXy072l4s+nVqbCSlIcSobkqQcgjz18N3OQy4FNtlJHgrdgj9FRC0Xe5kOCqKzLnYOJ90hBDclpExoffLwsfHj9tbCtGurXdtiOaYzyunLe6dfQfCumw+NhUShJ6nDbQ2VKk3Uor1enQsAUFJBBBFftbQ5sUoBSgFRd41TZNPuttXS8W+2uOJ3IRMkoaKh4ZAURkUBH+6TpD8arJ84s/1U90nSH41WT5xZ/qqLomw90nSH41WT5xZ/qp7pOkPxqsnziz/VS6Fh7pOkPxqsnziz/VT3SdIfjVZPnFn+ql0LD3SdIfjVZPnFn+qnuk6Q/GqyfOLP9VLoWHuk6Q/GqyfOLP8AVT3SdIfjVZPnFn+ql0LD3SdIfjVZPnFn+qnuk6Q/GqyfOLP9VLoWHuk6Q/GqyfOLP9VPdJ0h+NVk+cWf6qXQsPdJ0h+NVk+cWf6qe6TpD8arJ84s/wBVLoWHuk6Q/GqyfOLP9VPdJ0h+NVk+cWf6qXQsPdJ0h+NVk+cWf6qkrPqWz6iLotV1g3Ms45ghyEO7M5xnaTjOD4+Y0uRYkqVIFKAUoBSgFKAUoBSgFcA/XV/Hhf8AnT+UoSjsLgF9onhx+Tdt/dW6vlCGKUApQClAKUApQClAKUBq6/yWYD9xkyHUsx2VOOOOLOEoSCSSfQAKrGjdf2TXek4mo7ZLItkkuJSqWgsLQpClIWlSVYKVAoVkHzVysoXbfadXCaSS7DJm60sNvuMCBIu0VuXPjuSoranB9mabKAtST4Yy4jy9c9M4OPpWpmE6TRf+yTOzqipldl5X+UBJSDt2Z9916jNU5GV51yJBVxhpk9nMtgSMKVyi4N+BjJxnOBkZ9dUTR/Gm061v7Nui264Ro0mNMlxLnIUx2eS1GeaadUnY6pae88ggLQnIOalU8ybIdSzSLuLxb1QkzBPimIoEpkB5PLOAScKzjoAf0GvtFyhuKdSmYwosoC3AHE9xJGQo9egI8Capysqzny1doD620Nzozi3FLbQlLqSVqT75IGepHlHkrEvOq7NYLZdZ8+5xo8W1MqkTVlYUY7aUlRKkjJHQeGMnyUyt6BzSRlC8W9TcRYnRtss4jkupHOPmT16n0Cvs3GGmSIxmMCQf+hLid/m8M58o/TTKM6PdC0OlQQ4lZQdqgk52nzHzHwr62+mosTmI2Xpm3Tr7AvL8cO3KA261GeUpX2JLmN+E5xkhIGcZ9NSW300Jc20k+Q2+mhTny1FiMw29PGm300sMw2+mpTTScXyL61f901eor8yPeizWd6cu5+BsGldScqKUApQClAFEJTkkADynyVR9R685KlMwiAB05nlPq8wrCxdfcU7ribLA4X0qpZ8Ea/uV3dlulx1alLV5Scmop59bv3ah8dclOtKbu2ehUKUacUktEeAircJIVkDxJ8lUPWXEyPbocmLb4TV1bIUy+qUPsLgIwpIT4rHiD5PXUQWZ3M1LePKjUemNZxNEXeNqGLb1wdPdo7I5AaeL646HACEpATktqI3IGDtU0pGcKSRB3+7W3in9Utb4XbHGWHFturhrJSoR2W8rQ4EnAUsBw9fJgYBNdXhNmKnGviMRpCEWk+smrJL4nPbV/qGtVrYWjgfWxFS2Zccsacszk+9Rv8ew6FncT24bxU3ASqE2QgIwUqA8+R0HTyYqz2e9QdTQe125xLiB0W2rops+YiuUlE6F0nTipXujLbPfCfeq9PlqQSje3VCuixULBpPXMmxy0Q5rhdiKOEqX4o/9K27HkIlMpcQQUkV1uBr76lrxRwG1sMsPXzQ4S1+56UrYmkFKAVR9fwY8udFLzDTxDZALiAcdfTWBjv0H7jOwSvWXvKt7DwP/ALKP/wBUn/CnsPA/+yj/APVJ/wAK5rXqdHlQ9h4H/wBlH/6pP+FUZjibox/Uy7GYzrUhNwVahJdtyxGVLCdxZDu3bux1AJGalJvgXYUXUvl5FwQzZXFSEoTAUqN/pkgIJa/2vvfjrzUvT6YyJBVbQwtO9LpLe0pyBkHwxkgesilmUbt9DFYnWZ663KCuAiMYIa3SJDKEMu8xJUA2r7rGOvhistPsCt1DSTblOrb5qUDl7lIxncB5sdc+FLMl02uBgWa/aT1BY03iBItz1sUtTYkqSlCQpKikg7gMHIPjUi+3Y4oTzhb2twCk8zYnIOcEZ8nQ/oNLSEqUouzR6ogWpxwNojw1LKA4EhCSSk+Cseb015Nosbzqmmxb1upXylITsKgvGdpHnwD09FNSMjfIKRY0uttkW8OOKUhCDsypSffADykeUeSo60X7Sd/tj9wgyLc/CjvLYdfKUoShaVFKgdwGOoIz4HyZpaRO6k1exIvN2RgoDot7ZXt2BYQN27onHnzjp569I8K0zErMdmG+G1FtRbShW1Q8QceB9FLMhwdr2Pb2Hgf/AGUf/qk/4U9h4H/2Uf8A6pP+FRr1KcqHsPA/+yj/APVJ/wAKt/D6FHiKn8hhtncEZ5aAnPvvNWbg77+Pv8DDxi/Il55lypXTnNClAKUApQClAKUApQCuAfrq/jwv/On8pQlHYXAL7RPDj8m7b+6t1fKEMUoBSgFKAUoBSgFKAUoDQnG7TN11ppe76etLiI6rrITDlyFkfYoi3QJBA+6UWt6QPOoVrF3gnfV3t6BPMG/6ce1FC1AS+0hpIUGXG5DfJ6jG5DDg85dX5q57Mo3Xazo8rkk+w+bPwTlWq+aYflabtd0gQpd/juR18oiNFlzw/FWlKhgpQ2kjYOqd+APGswcHtRDgAxpj2Wkm6I0ozZzZitjsSZCY6WyoL5XM98k9d+OvhRzTsSoPh55Ho7wYkr1c/fPYy3mc7rBN1MwhPOMDsQYUgqxnBJWNnhhR89QUXgpqSwabsrFotdmEyHp+8QXY76W1MuPyZcNxvKSkpVlthYyoEAhIIIqVUWibIcHyPvSfA+7LnW4322RpFqb1e9fFQpxjOFMdVpMdBUhltDW/tGFFKU4B72VHvGLX9TldoGhG7Va7XbYUx3SN6s0vkqS3zn33o6owUoDvAJbdGT73OB41O8V+PnUjdtrt/wBFjuXANEbUN8uFlsVpgrcvtinW9xhtDSmWIzsftO3A7mW23Bge+BwfGq3d+Bur9VXXVSp1utsZNxsF+taSgRm4zj8l5pcVaUttBwgBvcpTylqC8kAZyYjUXFvX+BKm+CJLWHCS+6ij3ZDOj7Z/nrTjFnhpfkNJ9r0hC3yp1O1J7pLzbmWu9uaAPgkjP1PwrFi0ZxOv0hiIzfjNN8t925YU9/krMd1ncoDcE82OcoHiFHp3qZ1ok/OhLg3fQ2Lwk0/MsWhIJubQavdxU5dLkjxKJMhZdcRnyhBXyx/qoTVy21jTScnYvx0SQ2021TlKrjbTbTKLjbTbTKLjbUlpsYvcX1q/7pq7Sj+ZHvRaqv8ALl3M2BSumOYFKAUoBSgK5rq6Kt9oDaDhb6tmfMMdf8PjrVikl9ZJNcxtOd6qh0R2WyKajQc+rI2c3slLBPRIHT4qx+YlSghOMmtNa51MdYplf1DN9m3jZYjm1sY57gPvv9UHP/7apWuNNJZaLTScNtpwUkjaD5wB/wCtXYK0r8jKpyyWhzepzbrCTMjIszVnjNG8S1qYRICcqZQrxUnzE+f01k6fgWXS+ulwoyDIvTba5D84E4bePkHoG7H6q3+3NoVKlKlgocIK77Xd29yXiZH9G7BoYeWM2vO2ereK04RSinbo5S07kdNvacXN020+FANraC0lZ8pHh5/LVSs1xk6B1Cma2VusrIQ+3jAUgnw9flFaRO6sVwUasZQ83N3F1qbFZlxlhbLyQ42seY1OobS6w26nHfSFfqqy1Y0lVtJEfdY4cZyB1FbD4S6hVdLX2d1W5xklBz6K3WypevKPYc3tuOahTn0bXxX8GwqV0ZxwpQCqfrdOZcb/AHZ/bWFjFeizNwelZe8re0U2iudsjo8w2itERuC99g61XqNJTIV7bXrh7HvSiY5hutJQH0tnupfQckHxwMZ8KqVkZVCsqeZS5qxBaZ+p/u8Gy3OFebfInSlW56CqRGucdlE0LkIc3DEcL3d3dl5SsHI6hRIwL7oLUbly0haLjpq03+4ptF3T2RfLYZbC3W+WtRQ3y+aAoE7UpBO4g58a7pmxWJhOV81uL56aPt88ibZ4BX4xBb56Ylziqf08l8uO5D7URsok5B6kEHAB8RUhfOEF9uHE63zWLTao9gg3OO60qK3HaBhoj8pTakhrmqV9zjeEbABt80XRa9Lg29Xa3zskRNk4KX2wWbTTDmk7NdWbJOuCpFtXIbS3ckvZ5Mg5QU7mwdgCwSB4Y6CpvQnAN2BdrL7Z4cC7xIOm/Y3KzzA0+qQ4spbChkBLbmwK6HA8maNrkKmLi08snfX6/e3uMj6m/TFwiR73c7s8ZTsdwWC3PqSQVwohUhCxn75SlE+faKqN04b3vQ+hdUXIQo9tRaJMW72Vtam5MntDTq1LQp1tCVONqDikJC8q+yKyRTS5O+jvpR5NpefmTNo4M3y0XHQs2HBYduEdlk3ibOcacaWVvrfkDlKQVBwLWShxCh1IB6JFRi+BF/g2pqHDs0BpmBqCTPLcNyOk3CO4HQ0cONLQFNBxICXEkYzgpwDU3RPpcL8fHt7eV7/yet6+p3nXKzPxkW1h4s6U9jLeLhJQ87Hl9pccGFhCQNqVgBSUjA6Dp47L4a8Ol6E1PqtyPDjQLPPEIxGIuEpC22NjqikeBKsdfL41DaasWauKVSDhfzdfybD2im0VbsjW5htFWnQ4wqZ6kf8AirMwiW+j7/Aw8W70JeeZa6V0ZzYpQClAKUApQClAKUArgH66v48L/wA6fylCUdhcAvtE8OPybtv7q3V8oQxSgFKAUoBSgFKAUoBSgOfOP2oZ2m9KTXrdMNtelXaFbl3FKUkw2pE1phx4bgQClDiiCRgHBPQGq3qy5vcI7Y1Ds96k3O4TlOOsxb32q6vENt7l7A2C4EnpuUpW1ORgdQDoHG7t2s6JSsu5IgnePt6ds1y1DHtEBNjtekoepZDLry+0OLksvrQwggbUpSpkblkHoT3fNHjVWprZxJvMa/SY774m6bjIatMt9qOgPPSQpWwnIJ6BSSSFhKSfHAq3aV/PQh1G7GTJ4/3+BpVnUr9rtjltu1lul1tkVpbnPYMVkuoTIJ6KC0jCikJ2KIT3s5rKlcX9a2a53Fq42ywqi2t+0qlGM88VuMz3gyEN5AAW0dxKj0WABtRnpG6j188Cd6xefqhZ1lReoyrPHfuun03Jy6xw6pCGkNLbTDO7B2h9L7LmSDtSlzx21lz+LOqLNqY6OlxLO/qORLgsxrgzzUwkIktS3CXEElW9AhOjaFDdvb6pycRukN6+BmXXizfLDqdrSk2BbnNRznLeLb2dayxJQ4pYlr6977Chh5wjzKbGepNViycX9ZQdJ6fbniJdr5e7xdIjMmNb3nEMMxnnwdzLR3LP2NKQAU93KlElJ3SqUbB1Hc2BdNa6oa4XW+/R9NOM354Mdrtq2lyFQwpQDqw0nat3YMqCE4URjy9Krsrjq41ZL9PiIgXFm3Wm0XBiS0VobkqmOOoVgK7yUjljaD164PUVQqSZU6jR9ni3qCI/Ou06FamdKQtSmwvuJW52htsulkSVE91I5i2QRg4TuVnyCDa42XRCzqR22pRb1WiHdXWVSHdrVtduDyOfyydocTG2uqOM9CnwAqtUk+BS6rPrV/1QtysulJt/gxoLrUdu43FmAY0l96VAiuFtLpW2NjCXNqlBxfdwpHQ4URfeG9wkT9V8S0vPuvNMagZbYQ4sqDSDa4K9qQfejcpRwOmVE+U1TKnlj57CVUzMvtKsWL1xSlhcV7wrm3Z5TcxxmTIQ1klqIwt51WRjuoQCo+PkFXKS9ePei1Vf5cu5kv7qED4G1L8wS/o6e6hA+BtS/MEv6Oujsc1ce6hA+BtS/MEv6OnuoQPgbUvzBL+jpYXHuoQPgbUvzBL+jp7qED4G1L8wS/o6WFx7qED4G1L8wS/o6e6hA+BtS/MEv6OlhcpnETiJEm9iSi135oJC8h+zSW8+HhlAzVTY1lFSkf5uvOfRa3/6a5rHUXOs3de9pHX4DERhhYxyvnwTfMgtQa4ZVODbUG5IKkg4cgPJUB1BOCnNYrusWYcRbqYFzW5jDYVBdTuV6ynFY/ozUV6y+KN5SxsXC2WXT2ZfYi7FeGLWhxTsG5K53eyYLvj5x3fJ8XrqL1hqpufGJEKYygAkrejrAI9ZTRYeTt6y+K+5ceNg6mZRlb/tl9jTUS2OezbE5touCJB5aQlJOHcZGfR6apHB/Qd3W7edU3yE7IRK3NxkKcCSr7IncopJyOo8tbZbiji6lXEtWy2jz105d1zKVfGYrY1LC4BST3ic204rL6zVm+N3a9vedU8PHVTNIJLmSkvKUhKnSQ0kBI25J6EEHp6aitWWNExIc5qmySSdhyPV1HkrUVZwdZzgrroXcPCpGlunJqXXt96ZlcM0zJrEi0KvcqL2YFbDTKGikjPeGVIJzk5xnz1sa06fuEiIpI1Nc08tRSEhuPjyHytemqZVYf8A418/uazFYarFtOs+PSP2PqRpe4FBB1Lclegtx/oq+uFVluUm+TIzOqbnC2r98y3HJPTx7zRrO2dUjKq1GCWnb9zQbTpTjhk5VG9Vxt0fRG4vaLevx9vv/UQv/L09ot6/H2+/9RC/8vXRnJj2i3r8fb7/ANRC/wDL09ot6/H2+/8AUQv/AC9APaLevx9vv/UQv/L1E6jt8zTsJXPuM7UT6G1uoMlLSXVYHRtPLQhPUjpkeJ8aw8XrSZm4TSqjUXDrjIjUumI95vL9pZTKlxYKIlsdddfhynlBHZpKFISptxKlJBJA8SSlIAzNXPjFp23SkxEi4z5q5MqG3FgQHX3Vux9nOACQegC0nJwD5607pa2RulUVk2YUnj7o1hth1EqbKiOQItzXMjQHnGY8WQpSWnnVhOG0koVkKwRgkgBKiMx3jJptpy8J/wA4rTapYt7ziIDpQ5LLiWxHaO37I4VKT0TnorJIAOI3LJ3kWfUDjDp66TrdBiC4SLhMW82qEiE5zoxZcQ26Xk47gSpxAz5QoKGU9a8pHFi2zLY5MthdEZq7xbWZkyI6lh5Tk1MVYaUB3iF7k58AcE905pumuI3ifA/Uca9LLmPMl6ahlIlhma5CdTGlKjBRfQy4RhakhtZwPfBCinIScSOiuJVk19IlMWztrT8eOxL5U6G5GU5He38p5AWkFSFFtwA+dJzjpUOk0rkqom7ERJ43adQ5KajouEhSBLTFeTBdDE52MFF5phzbhak8tfQeOxRTkJOPHRXE5bnCCy6x1UsNPXFphaY8O3PNLLj6kpaZbaKlrWoqWEpUPfZCsJFVOlZdpTvLvQk3eLmno0W4OyzOt7sCG1NeizITrT/LcdcZQEoUMqUXG1JAGc5QR0WkmLs3HK03G23GVKtl1grjXh2yx4xiOLdmvpWsANJCcqOG1KUPuACVEAE1G6bJdRGFL492dnUdkQh1Qs8633Ba2nIronCdHlRI6YyWffbyp9wFG0klIIO3JMm7xhtdrkyIs0SZs1Vzk2+LEtMF591ZZbQtSSkJPeCVZJ976al0SN4j1e44aRRHgSGpkmVDlw41wMuPEcW1GjyFFLLjytv2MKIV0PUbSSAATU9pzW1t1ZcLlGtyJbiID7sVyWuMtDC3Wllt1CFkYJStJSfUcZHWqXSaV2VKom7IsO0U2irdkV3G0VZ9EDCpnqR/4qysKvzo+eRi4p/ky88y1UrfnPClAKUApQClAKUApQCuAfrq/jwv/On8pQlHYXAL7RPDj8m7b+6t1fKEMUoBSgFKAUoBSgFKAUoDTPFK7Wiy2S7PX6ELlaXpCIb8RTKXUuh95LISpCjgpy4Mg+TPj4VQnNJ8LLZOvFgl6as1ujWSPGu0qTMjtNx0pkF9pJLijnwjrSoK6bSkdR0GheZN2Z0KytK6Jxu48N7RpgXBM/TUbT06Ei3iUqSwIkiKyFISzuJ2KQgLWnb1A3EeWsbTVm4XwEXBuxN6ZR7H9nlzexuMkx+XuWw66Qcp2gqKVK8B4HFU2mVXg7HgzbuE0Gy3TUjftUZs91S9DmXUOsCM+l0nmtFzO3vqzuSD3j1OTUxcJegnJEdEyZYC/e22HWEvSGgqchg8xlSAT9kSg94EZA8aNTZN4cin6a1LwsuR1TrNWqLDLg6qdbt8pdwkMNtKSw0WhGUFHvZy4vCskh0fc4qZnac4Z2W2T9LC26fU4417JrsaVsJkP8tOUu7VqByAgBKyQE7R1AHSpqadvOhSnBrzzFosujLjrnSsqHLtsW7WOzPIgaeYdj82K1J5JU5sQonAS2lIKe7hajk5BrIuWneGaborTU1jTrV0uEpVzFrdcaRIdfUFFT6G8hW45XlSR1yvzmoefgifUK1b9eaM1zwq1TcI9kt9x0lpftQjxoslp3mCIHASlKD9izyyUZPVC0q8FYr1vS+Ft0sNsvcex6f1EmDJt1iQ1CSw6YPOktMtNEJyEBtT24J6Ed7GM1VlmnZMpzQa4Fxb9oupBftJNvWK5reLrl2syHWnVkrP2RTzQJIyT1Kh4mvFi78PL3arvPYuOnJ1tjQ/Y24yW5LC2WYw3fYXVA7Uo7yu6rA6mqLTK7wK/ZYHDXifYJq12KyS9P6aU5aWJDqWnIoipZacJQod0M7VI6eA2A+QGrbatT6MZZvVxtt5sgZTLSLnKjy2tqJG1DKQ8oKwF7UNoAV16JFTJTehCcOJlxteaZmzbdDj6itL8u4siRCjtzWlOSmiCQ42kKytJAJBGR0NeTvEfSTEWfJc1RZW49veEeW8u4NBEZ0kgIcO7CFEgjBwehqjJIqzR6kl7YbUW1r9k4e1tbTSldoRhK3doaSevQr3o2j7rcnGcisB3iDpZlFyWvUtoQi2FInKVOaAiFSlISHe93MqSpI3Y6pI8QaZJMOUSXt9wi3eCxNgyWZsN9AcZkR3A424k+CkqBIIPnFTWnR/nqN61f8AdNVU168e8oqtbuXcy+UroDnRSgFKApF24u2Kz67b0s+twyyyHHX0Jy2wo42oWfEEjr4dBjPjV2QtLqEqQoKSRkEeBqxTrQqylGL1i7GxxWArYOFKdThUipLuf8WfvKHxRTg25Xk74/ZVNjK3N59dcztFfnv3HTbN1wkff4lfusREu/gkZKWUj9KlVB6vnNp1DDtrag23HaC1ZGBuV5f0YrEleWnRG+w9k0u9mBcLgW47bScuoB2lY8+PNg9fCqdqpoP2x6CuSjbJUGlupHUAjOADnyZFXYyUGpdDLpU3Ulkvx0Nd6rntW23JjQjy1FWCU5wE5AKzjxPTy+Y162+OLJGU1uL7C2csvJdWG056lGwlQB6KOQR4Hxq825xqZ9bp279PobetlpOjCk3HLJXS4OLT5djS7veWvhrdH4jrcVlaG403LbilDO1QJIIPmIynPq9FXLUMJS4uUpxywckqyFkjrj/55asNZUra6I1id6rvzbKdZrguzaot8w5bKHgFp/1D0P6ia33YnlJdltqTgc3I9RAA/ZVDV4NljGq7VuhJShhs+isDhCd2r5+Pv/4Cs/ZetZ932OV2t/aL/uXgzoCldQcYKUAqm69ZVIUhpt5cdxbKkpebAKmyegUAoEZHj1BHnFYuK/SZl4X9VGg7rwaadlzLnetX41Pc5FubiXARGY6A5DdW+wnk5w6skubsnJT0TtArDd4KahtWpbDKs+p3kvNyLrOm3Z+Kys82UGcI5OACnKFYxgjanJPXOsU+q82Ns4dGYjX1OslNyudkYvMmDot/T1rsjzexp164IYclF4KURltRDqQVAYIdXgAgFNwv3BG2ag0tdbNIlFwTL0b6249HbdQ0/wA0OBKm1ApWjptKT4hR6g4Ih1OiCpq1myIf4M267NWuzN3e1RXrLJbnvNWm0x4zzC1OBaVs7DujlYbKCrvb07h6a9YnBSDdZ9zu4vqJAuM+BOJt0VpllxUSYJKVuBBIcdUUBtTvQ7QBjpTO1yJyJ8z9RwDaUy3Af1BKkWOEZ7lrgFhCTEclNvNrUpwdXAhEh1KAQMBXeKiARbdN8Po+m9QKurUt15w2aDZuWtIA2RlvqSv1q55yP9UVTKebSxKglzKPprhnY5919jomtG7zb9PSZj0WzR1Mrctz8lLyFc5SSVHYl95KEqCcBXUqIBFruPC5ibw7sel2bk/EesqYSoVxQ2lS0OxSgtrKD3VA7MKT5QogEdCJlJ3WgUVbRlW1xw3em6l0LfL5d0PQdPmVKvV1kFqK060lTb7La0DoEofbZcSc90MHJJWScxHClmfJuC7VqsolwNROXmGptht026Y404H2nRnvpW3JV3TtUkLSQfA1ObRWRGXXjqYkr6nC2zXrfOdvU32eg9ukM3VDaEutzZMhl9UlIAwnaWOWEeBbWpKsgnMpYNE2FnWUS4x9RMzbim43O4CK2tGVuKQ1HkJABJw0oJB+9UsA46Uc3JWsFBJ8SEt31M1ntMiyvMSIctcK2w7W+q6WliYp1Ecq2LbKxlpZC1AkZBwnpkZqRuPDa42HVFw1Ha9QRLfdLvIYiKWYrUdBZMhC1lwDIkP8tJabUQCAr1mmfNxQyJcGbV2im0VYsXRtFWfRScKmepH8aycMvzUY2J/SfnmWilbw0IpQClAKUApQClAKUArgH66v48L/AM6fylCUdhcAvtE8OPybtv7q3V8oQxSgFKAUoBSgFKAUoBSgNG8atFu8Q9K3awM8jMmZHWsSSQhTbcptxaTgE9UIUB08SPDxrWt5+p/ct2obtctKRLTEircssuPbH3HG2pD0J+U44l0hCtgUl9spUArCkAlPTrpVLLddv2N5lukyQ09wfuqNU2y/3ZFqQ77Ozr1Jt8danmoxehpjIQypTad6u5vUopR3nF4Hnq7n1Nl3Og7DYo8q2QnrfpdFpeMda0IekolxpAGQgHlKLLiVKxuHNJ2nJqVNIOF0TkPhPqG1TLXqGDbbcu+Rru9cZNvuF9kSWpXMhpih3tCo+UOpSlIADZG0EZBV089M8HNTaTuWmXrc5bob7Kyq7TWJrnLcYXMfkrhpiqZKVoRz1Jac3tqTknGBtUck1YZWYdx4TcQF6KY01DkWhmE0q6x3OVcHGVSWpDhWw6pfZlKTt3LC2U9Fd08zAxWazwh1VElNswXbfb4b9mEC6Oie48m4upgdnQosKZw0pLm37KheS2jCkkq7q8RlZLaD4bak0jrKHJbVCg2XsTTdybYnOSPZCQiKywh0MrZTyFJDQSVJcIUlKcpycpxb3wn1DcNR3phoWlVku+o7fqFd0dfcE6N2bsx5CWg2UqyYoSlfMTtS6runHepur3Jyu1iaj8NbkzwZ1VpEvxPZK6+zfIdClclPbJEl1rcduRhL6ArAOCDjPQmrN8GtS3N56XNbsdpfT7XYrMS2vuLZWxbZ/aVuqJaRtUpKlJQ3ghO0ArwcgmlcOLdjDb4DakuVjhadmybZbYNqg3eJFvMF9xyZLVMbcbC3Gy2kNkczmLwte5aEkYr0ufBzVuoXUXaTE09bLjb2LTHi2qHLdciTBDliQeassJLaTjahIQvZ4knOKrzIpysl1aGvdh4R8V250SD7JX72SuDMGzFbyUl2IlAbBKElaypByQkbic4BOBCR+C+odVWpx66wrFai9G07CTbYrjjjLkeBOEp1TgU0napSFKQlraQnGCshWRCaWpLi3oTU7g3cH+KFzvDjbNyslxusG7AKu78VcRyM2yhKeQhpSXQFMBYytOd6knp1MT7kGr2bTNiRY9vjR48pmVYobV/eCrS+EyEuvtSDEKtqg8lIjrQtISXOuCE0zKwyvkZNz4Ta2l3B1vtNkmRrjcLBdLhcHHXWHUuwXI6n0tsJbUkhfZwUkrGNxBHga9rjwRuo0TbYcF2Gi7QNVztRKQzKciolpekS1ISp5LZUlaW5KDu2KwpoDw71My0SGVs2Nw50t7TtIxbYYqIbodfkOstzXJiQ468t1ZDq0IUrKlk+9GCcAYFXfT3/ADzG9Z/7pqiCvUT7SZ6U2uwvVK3hohSgFV/XeqkaN0xMuRb7Q+gBEeOPF55RwhA9ZxnzDJ8lUTlki5dDJw1H0mvCiv8AJpfE45l8TLfoG+sdsegXa/3OQXZb8pw/Z3FK6ttYPTr3Qo5AOAARW2JfHV3SnsXJtbJudunHamC4dq0qBIWnPXapJBz4jp8dclSxG6k5Q4rj234nbQq09v7QqYaTy00nl7FFcfgja/ExPNtEB4DGHfDzZSa17AkboqlA+CiP11f2jG+IXakYuzH/ANHfo2YUlaUXxkHxcRgH0g/+tUmSkX27TZidylNvqQkDAKQDjHmI6eH/AO2sGK4M6CjbXsRE3OJMLjpQWXgkhRPMKN2PA7Qk/t8/pqna0jT4UNmQFh3a8jDKMdcnByo+OfUP21ehBVJqC56GZGqqC3j5a/A03rS4dsVPc5o2JIARggoO4dPHOf8A55aik6leTYjb475CkkrKgTlasDqB6h5KsSzQzKWlpLT4nRpwqxThqnGTv0VlZ/T3lu0nreVEYiwpQBLawtLycAhQPl/QK6QlsNzLUl1G8cxIWknPTzdP8KuTg4TaNJUacYT6+X8DWGp97CUpSBu3gdB4KJxn4v4V0FpuT2xhEoEHnMtudPSkGonG0DExLTXuJG5TEtxVlWBgGnAtky7tMlY6KcODWw2VG9SUuw5DbLy0IR6vwX8m+qV0pyApQCqRxKdlx4LzsBpMic3EcVHaV4LcAJSk+gnArGxCvTMnDfqI5WjmTrrS2hEtao1HPu673b3bo6/EDTlvfMWRzkoBYAbOcpUjrs7uNpIJwLzq/X7NvtMB69yLZGR7NxWrzNcVHU7Ij3FxiKXVIiuhauQhCwghAdyo97HTCyp6NGxbfJlgv72upl9ur6NS3eD2fUFmtjbFvjt9n5EiPFEpxIcaUT3nXFAqzsKfAd4HFVqi/QpMS1al1Jf7Rp2NIvcdN6jRwZMh5mU2mG24sNKyeSp0pG0cwoGdxBCqcsWtES2+pg6MvOuGuJdknajjzLXDudssib5IixsOdvLL3LZcGCG2VOKUFlPVKy2klKVKNYrt21yNHXudDul3tj1k0s9dYkOHEbQ3ImomTNqVpLZKgUNtgoGMhQJ64NVOMSlORaZGp9ZPcZrlCdu7dojxbq2iLBfW6pMm3dmQpSm2ExiHCVqcPM5wKVI2nABCrFwR1Rd52kr4me7M1HcbYsD2QDpVHuLnJCiWNzTZbyropoghtR2hRAqiUFbQrUnc09wGvkzSutdOxXYj1wRcW0RPZIQXm2LLFcS88ba6NiSmSJRbSHHSregddiuity6sv8qPxhjW653u7WOziDDdtbduj727hLVIdTIbcVy152oTHG3phLqleTKZnG8iIy9U1A7e9b6u4Z65bvVwD053TFxFxsG9119mZlIbCG+zNhlKRzEBIWveChQKsFR3Dp6TK07beMNy7HcHFs3iTLZagtBUl4Jt0XHJCgQpRKSE5BBUMHPWkorgvPARb4s1jadf6pft1xbk3e5R9Ni929D94jOLluRobsdxTpQ+qM2SnnoaQpQQeXzF4UMd3C0jpm73WZZHYl21BALbes5CLmmMliU/uuUVTJcC2sJK8BeAlO7b4YyKqcVHz2FN3Lz2kzbdb6/vmrLCmbc0WN1+NZX4zElTjTctLrba5gDKYyw4oqLrZy4jl7UKwkZKsq36p1BKvUm3uTJl+cZ1LbXDcUNlcVLDkxwFkNLZSY7qGwkLSCvACF7gVGocI8icz5kQrXWtF6dnSoN3vUnU4sN5fvdseiYatUltpRjBkcsbVBwJQgZVzEEqO7G6tucNfZ236y1LZ7ndbjeYLUC3TmJFxQgKDz3aEvpSUISNv2Fs7cd0qOMAgVTOCS0KoybZsjFWXRo70v1I/jTDxtUXnkU4h/lMs1K3JpRSgFKAUoBSgFKAUoBXAP11fx4X/nT+UoSjsLgF9onhx+Tdt/dW6vlCGKUApQClAKUApQClAKUBpfizqZWkrHPnNSm4styY1EjKXEVK3vOvpbQgNhaMqUVhIytIBIJOAa0xaOMeudURrbCgrs1uuQY1AuXImQFOhSrfJYZbCW25JCSoPd/DiwCDg1qcid2zdZrJJEPrXivfddaYW83MtVjtrEjTS3Le62tUqUuW7GfJbd5iQlI37ANit3LcyR5NmcYOKVz4ZXCIhqLGlR7tBfjWtLiVBTl3CmxGjqIPvXQtXgMjlK69ehwWkRm4sgXeMV+Y4kQrLHet90t8mRNtinFQjEbalxoi31AOqkKWoBbe1R5QSAsYUSnvY9n4uaouyrJp96dbLRqiTPfi3J64WtSWoC2oiJHIShMkpdUsOBSVpdxsz03JNMiGdkmrjNPYnS4Tki0PSW9W22wsloEB+O/EivLcQkrJJy86UkZG1I8cEmg27jDdtC8OLGLU+xNdhMCZPhPwis8l24OspUp9T7YRu2rSkJQ4rcnJGOhlQXAOZujh9qLUGqNQ6scnv29FmttzetkSJHirD5KAhXNcdLhBzuI2hA8Ac9cVrfR+v9ZmDarRJ1FaXbleNS3uCi5y7evbFbiyZWGg3zxvUoNp2AqG1CTneU5NKiuBLk9NTCmcd9ay9LXG7wPYJgWbTcu+SQ5EddRPVGlSGSGSHk8tt5MfelR3lIWPfeNWtrifrO7cT7lbLXY2XbBarvGtkouBhKuW5HadW/zFSUrBHOG1CWFBQQcKyruS6cSM7ZuXA9FMD0VYsXbjA9FMD0UsLjA9FMD0UsLjA9FMD0UsLjA9FSOnx/niN6z/AN01cpr113lFR+o+5l4pW5NGKUAPhXN/GfU7utOIqNMQ3yiDamVpecR1HanEEZ/4EqHxqUK1uPqZKNurS+v0Om2BhfSMRUk+EISfvayr5u/uNHz+BsxWoDd7245DjsBsKbefbdYdLSipCkgDeepJx0B6Zx1FWOwW5p60dsaQvsjZcbiKd98vJ+yOY8mSNoHkCfTXMU6WWVnxM7ZOz6mFoVMdU0fsR9/H5JnUvElwN6QjKz1CkkfINaqtUgFtbGckP4x6Op/hW1x6vWj3eCKdmf2k+9fN2PWWErntKX1CSR6jjP8ACqxp6ItifMQ7jkzFqU2PKojyevp09VayDukjoXZRb7j3uUVCm0jYC42QnYAAVjy7QPAftrW2qpSRHcbcdS0lsd5SjhPx+P8AjVxvVW4mVRtJa8CqaQ0VbOIWqo9rukRSrY9vBU2S24VBtRCwR4dUjocjzjqamtX/AFJ1h07Y7leLXfLmFQo7j/IlJbcCglJyMpCceWrc4RlUV+P1L8sfVw/5UbOLXDpcpGnOHPs5cGYzcguqwCo7An/tZrom4wBbbIw0VjKWg2Ac93GP/WsiUbNIivUzSS86mqNRhuStyW51ZbJCMZ994H9Hh8Z81bs4fOczTUFWQTyGxnHmGKisrQMWtrEk7+wZEBxIOMj9FXLgVbm4tlyMcwEg+us/ZT9tdxye243p05dr+htaldCckKUAqqazWGnWlq96hoqPqFY9dXgZOHdqiNCNfVN6Ydtnsh7FaiRC9jG73z3LYUp9j1ZzLzu6Npx1B75HVKVJ61Zrhxeslu1E7a1sXBxhiexa5N1aj5hx5bwRymVrzncrmtDISUguJClAnFYDpM2W8TKnavqh7ZbNKRJmog65O5UubMFvZBREiNSnWQ8sKUDjDZ7qdyjtWQnAOLPP4zWSBH1JM7Jc5Fo0+xJdmXViNujbo7XMdbQrdkqCQRnATuBTuyMUdJhVFY8dZ8Tk2q8w7RbMKnoudqjzA+2SkMS3FpBSc++w0r1dPGsAfVG6QMeXIUuYiK1FdmRpCmk7JzLbqGVLZwrIG91ofZAjIWFDu5UCp3QdRI2LZbiq82uPMVEkQFOgkxpYSHG8EjB2qUnyeIJBBBBrO2+qqHErzXBTkU21GUZht9VNvqplGYFOfNTb6qZRmG31U2+qmUZgU581NvSmUZht9VWLR4wqV6k/xq/RX5iLFd3psstK2hqBSgFKAUoBSgFKAUoBXAP11fx4X/nT+UoSjsLgF9onhx+Tdt/dW6vlCGKUApQClAKUApQClAKHwoDVXEAWZuyX57ULEWTY2Gnn5jUxpLrRaRlaipKgQQAnPXzVrzRXEDQ7vD+BqKXbIOiY6H5FrVAnstMuRH95DkfCRjK+UlW1PvwEHr0rVtN3sbdNJIxrrxB4TM3vTRfassqRMtzztsuCYbbiGmI7iEFsL25RhasBAHQoVnBFXWJe7LqLRFo1Rdm7eiB2Zm7IeecQ+1GUUbg4hzGCQFHC04znp41DjJcScyfArz2p+FjbZ1U69p1Ds2SqEq4uR2w+9IDSgWVEp3lzl7hsPXacYwaxrleuEFp0vbbfK9qqNPTwifEhiKyqK4FqCUPBASUjcohIUR1JwDmlpEXie8nUvD+2vLv18jWC2T2pj1tYlrS06+sRnD0SpKdwCNoUU/8AR+XGM193m8cK03OyxbidNPzXEN+xoXHadIQ+SUctQSQkOEKIwRuwSM4NLS5E3iTEPiHomO7ekRb3amVQEvzbgW1pSEBs7XnVEdFbCAlahnacA4NQrl54UX+wXmes6YuNqcmN+yK1x2nEPyVAcorTtPMWoKTsOCVAjbnNMsuQzRZJjVGgJbFrjol2WQ1eoxhRGUIQtMiOFcstYAP2MLVsKTgBStvicVg26/6Gv15sVzuESxo1VJStEJwoakSUJS4psbXgnKUlSSB1AzlIyQaWkHKJsTZ6qbPVVFiq42eqmz1UsLjZ6qbPVSwuNnqps9VLC42eqpCwpxd4/rP/AHTVcF6yKJv1WXWlbU1ApQEJrXUzGjdKXS9SMKbhMKdCD92rGEpHpKiB8dc48MNOTFx3LzcQXJk1apLzih1WtRKlH9JNaLaN5zjBck39Du9h5cPs/EV3xm1Fe5NvxRE8SJz+orxEsMNZS9MdDW4fcJ8VK+JIJ+Kpi925m2W1qHGQEMMNhttA8iQMD9lauirqUvcbvaMtzh8Nhl2yfv0XgzbPFOcBZ7NFB/0hQSPk/wAM1rG0EJu5UVe/deSE/wCz5f8AtGsraDe+Vuj8DQ7LUfRZX52/+TZlTpaGn2QpQClugDNQtoT7JWh5CVKTIafPhjKVJPQjPxfo8PGtWrxyPvN67NTXS31Nd8ReLbukk9mdQzOuK8JZaOUqPXvEgZ6dPP5Kodqt9y1NKF0vzm4lW9phKgEIz5k+TpWypxWXNzfAmGjyrguP2Nu8JbCV35NxaaxDjoXh3b0KyCnaD58E/wDw1ta525q82udb3lFDUyO5HUoeKQtJST+usCo8s1bkWK8s8nY5rumj7hotvsEvnNuoUSh8juOq8ikq8oxj1Z8lfg1zd2ozEe6pVKitKP2aOnc6oY6BQJx5uvj08PLW09Wrr8DOTUoqUdTEvurLXe4OID+0sY50dWQofEeuP/ma33w9Ty9MwE4Ayyg+rKQaxsRFxp6lirJTyqJY5bfNjqSfNUlwduvZrnJhE9N2QDVzZkrVmuqNDtaObB36NfY3TSupOHFKAVVdZNh5xts5wtpSSR5M9Ks1leBfo+2jR/uLack2F/SaLzIU41o5Gk3G0utl9uIUFCXinHRZwcEjaSD0rPn8G4MnUEiSq+XBq0TrrGvEmxjk8h+awGi2rcUcwJzHaWUBWCpGfAqBw8yZmrLJXTIONwF09PcjSrVdnRJgokW5992HFlcxJkuPFGHmlBKkOOuAFI+6IUFdMT0jgzFkx9VWxV8uadNajRM7XZEBkNoXKbKHlIc5fMAO5SgndgKUT4YAZriKjJXi9DHgcEGW7k5crnqO63q5OybdKW/JQw31hKcU0kJbbSAk8w7vKfHIzUNbuGOmuGLDtmY1SzaG5kdUWDHkQoHMaaW6k7SpTO51HTljeTkKwSVbVBKSS1KakoUo55ysu0ueh9AscNbPChxbzKVaILEgKivpZQxvcfLxc7qBywjKkJQkhCUHGOgImmdbaek2t65M322O25hWx2WiW2Wm1eZSwcA9R0PnqiTjfViVWnSeWcktL6vkuLDutNPsWhm6OXy2t2x5fLbmrlthlauvdCycE9D0z5DSdrXT9sTEVMvtsiCYjfGL8xtHPT07yMnvDqOo89UXh1KHisOld1FwT4rg+D9/ImAsKSCMEHwIr9z6KryoyRn0Uz6KZQM+imfRTKBn0Uz6KZQM+irDpHqqV6k/xq7SVpos1vYZY6VsDWilAKUApQClAKUApQCuAfrq/jwv/On8pQlHYXAL7RPDj8m7b+6t1fKEMUoBSgFKAUoBSgFKAUIyKA0vxe0W/wAQ7DO0+iZ2GJNmNic4nO9cVLwU62g+QrSkoyfAKNUSPwMl23VLkuLe3JVqcvUK/LZuBLr/AGlplxl07vDC0CNjzFtXnrAva6Nqo3SMiy8JLvp2/wBiuUSfBe7HPvrkhp5Cxuj3GaJPcI+7RsSnB6HJORgV+W3g5LsPDG2WK13+TA1XDsEe0t3BubIEbLTbaFLTH5mxO7bjelO9O7IOai6YymFovghdLBeYNynXJh5xnU71/W1zZEkhC7V2INB19SlrUFd7co9R4BPQDWmoOF2qtI6iixYkGRfIsa2REtw48Z7s11ebuEqSGXHm1gMBBcaO50FJycAjcFVJpkOJtBnhHqPTuo29Q2K42t65Jk3jMe5NOcnkzpLT4IKDuC0FlAI8FAkZHQ1rS18LdRaA15a7bEhTb5a7auyx2GFxXW480MNoQuYp5tfKaLJU4oNuAklsAZGzBNMOJaYv1N0qHpC+abcmw3jIstxs1rukiZMdeQmSPKwtwtIwEp3csd4oBASOlWHXvA6bqrUM68xLk3Hf7ZbpsWOHXo4UqMzIZWhbjKkrSFJkHCknoUjII6GHInKfKeDN1hq0y5aHLfYZkCSp+VPiSpbj5S5L58hrK1ntCXQSDzveqUVgZwB5WbgzerPM0s7brpCiuWt09qucZbyX32e1uvLjKbCuW62oL2DeMtqK1J6noumhlN0dadatWKx1p1pYDrTrSwHWnWlgOtZ9hz7LR/Wf+6argvWRRP2WXWlbE1QpQGpfqgL5Ei2eLBlkGIN0ySggnclsZQnHpXg/8Fc56W41X+93l8lpFutAbCilhJBbQFAJScnBBBxnoM+XCgau4bCwrSnUmuz6nVUpOODp0+WsrdrdvBIvuhLUL1qa56lUAWQwhmNgHAUtIW4Rnr0G0A+k1n6pRlpdcs6W6i49r8TbY/Eb/FK3CMYr5Jv5tlr4lzebLtRJwhjkBWfJkH/EVSdOuGey66OhKHFDPkUVjB/7NY+Mler7pfT7kYGnlwy74fV/Q8Y0d3UuomWmQcNNb9vmJGf8K8rjoiXKdnG1X520pmkqdQhkKUCfHCien6KipCMKFKUl1LtKrKWJxEIvhZfBGvTwHsNkuq5d31g0ZCzlapKQt0nznKs+brWwdK6P0K4pDcaVGv8AKT5JTyVn4mxgY9YNWKmKzO0dGZipVo0+Dt1L8EBlCG0NpbaQNqUITgJHmAHhX6VACsKWpbSVtDHmJblxlMSGm5DKhhTTqQpJ9YNVO5cNdO3IKKYrkNSvExnCB8k5A+ICr1OrKnwLkU46xZR75wC0+5LanOzpKeSdxDaEpUoeYq83xeWtgaPnNy4e5sBLaVFKUgYAHm/hWXVqutRu+RTly1NeMk/lYtKxuT082Kh9GSjC1/ywcBY8Kq2d/cL3+BrtpK+Cqe7xR0W2dzaT5xX1XXHBClAKqGv5/sVFcmch6UY8dbgYjNlbrhGTtSkdST4AemrVX2GXISyvN0NFWay6j09qVy63aGmLJutsmGXcLXzZy23ytksgthoYDaNyEpBVkI9efmzWmVc52ivZeLfEojSJrxkiRPUHHUJQ204oKO9oOJU4oJXgDChnCju1ChLg/PA1Kp1VanUT4pu17auLbukr31v07rswYlj1Na7GHoKryzcLraZkiQgl3ZHlzJLZaAb96hbXMeUSAFDBKjjwmhp2+Rb6+pi4X91hu/Q2YwfkvKQWUtNuSXVDOFIXhxGD3En3oSTUKEuGvn+RClXSSTlpbr0d/nb/AEfuhF3q6artUmYL8w4I0uRdBM5zcQPlaEtMNoVhGEJUvBQMK2g5UckQHEA3yTq+53Bq0TppaxCFpahP8mQw0C6xJW8lJS4EurWS0CCQcYUoAFJScOHP6EVHWeGWkm817NO/s6rsTd+xF/4kMPzNOWbmRpFyt6Z8Zy5sx4ylrcjpyonkgFShvDZUgAnbkYPWo+93Jh/V2nL2/arkuxsx5RQUWt9biJe5pLS1spQXE/Y+cElSRjcfDIq7Jau/YZ9aVpycot+y+Deif3vpxsQ1u0zqGdqaFLjNJsvaZtwvn+Ww1PIjlTbUZlCkpWgBxTanHCNxwSrIPWvuBb06KvF2tjNmud65FpjW2A2qCtTMtZLrjqlO7Q0gKU4gKyoAbfDpirag16z+HuMONKdNqrOLersrdY8+y6S5JGxdF2N3TOj7HaH3u0PwITMVbvXvqQgJJ9RIqarKULJI39NZIRg+SSFKqyl24pTKLilMouKn9KeMn1J/jVymrSRaqu8GWGlZhgClAKUApQClAKUApQCuAfrq/jwv/On8pQlHYXAL7RPDj8m7b+6t1fKEMUoBSgFKAUoBSgFKAUoDR3He9ytP6HvMqJKXb1KksRnp7fRUNh2S228+D5OW2ta8+TbnyVQrjfdJaMftcXR97tdjh3NbyLhfWX0yQBHaC9gKlKSt9RdT1UFKKd3icYy8NRnOlaMbpuV7c7JNK/eZspJPj0I6z8UdXzNLyNQPzmlxbNpdrUEuO1DSlcx57nutsdSeWkMtt7sZV3gc+OYxd4lX/U105t+g6vPLtVhRIRGb7Nvlyt85tG0lK0pZZQRnJHgokpNbCOFo0pSdOL0534ar6P5XKHNtK7JNPFPU19jLl2S8xJM+VBushVlbjtrFqSyhYjqcI7/MLqW0qSs4UVL2pG2sx7jDPvRts5F4jab01e319iukhpBUmOxHCnVo39Ctx5W1GQQUNFQB3CrT2fSXCDurq13q1dr4qL+OhO8fUx9McWb7Md0Y9c7q3Iaulv7WqLa2mTIU8pDslKH2FEuNtmMGtqkdQonccEZzLVxVkjT+lb1K1jaZLd8nRUTG2wymPa0qYdkuNczORlDezKznPUYzgU1MBTTahBtcuN1rLxtZcmu0Ko+b86FfvPE64XNuyakS5HPZWZ86I5JbKGo6JM9uDBecGQdqWVuuKyR0CuoqatOvdTX2aq1WjU8e5NydRi0xbuqE2SYzUIyJLoCcIWd5DaVAbchPQjJN2WDoxhecbqN1x6O9vemyM7b0fEsvFHXb2j5lmtbuoWbCyu3S58m7yGW1uOmPyUhptCu6VrU9uIAJwghIBII1tYNS3PSmjnpSZNvhXGBbbcmU86plqSp5/fOuCWC8dil7XmylK+7nJPjVvDYaEsOrxfrNPnr61mu5J68yZz9bjwLXaOIupNTa4RbLdco0aLb34bS2rjyWpE5tUduQ+tTP+kCyhwhCUBKQUKJJGUiI09xgv2og1Lhansy3JmnX7suHLS23FtklxTSIUZx4HIJLhCwo7tyOm0KAqfQaFvYbsle1+ab0666Lk+/UOpLqfiuLWor2zarZZbuiJKkwXpQn3pcNlTj4kLjoayAW1tJW0slTSVKUlbW0jdk9BMBwMNh0pU6EjeUAhJVjrgeatdj8NToZYxWut+NuPK/T/fZcpyctWelK1OUvXFKZRcVnWP8A51j+s/sNVxj6yKZP1WXKlZxrBSgOZdYXU8QeJGqLYJr8KIwtuAh9gZO1sHmJIyOhWpf6BWupnDNnhpHkTZN0aWxzVu9lh7krKR/o08zAIycFW3GNgx45GBQ2tDD0qkWtbu30PSHsyU6+Hw0f8owv2aK5unSNrVatF2xhxvlPKYS44jr3VKGSnr16Z2/FUBqdGW1+qtfO7jrxNI5KeJnKPBt2+J98TLzGTanXpCjsXywkJ98QnaSR8SSahtHzW5cW5PtEFlKyhJHh79xXT4lprU1bzlOfLXxj90dDRioUKcOd4/C0vqmXDgrahcLrOnKSCCSE9PAeA/VVT4n332suGCy4WpEh5xO9PRSG0nqR6TkD462O0lu8JC3L7Gp2M/SNo1Yvm2/gylW7TQvjZUnGVdcDy+k+f1moDU2gnIBK0goUnqFDoQa4pqdOKqHpMasHU3RIaL4i3yHCctjktciUh9tll2QSvCVBRwc+PvT+qtgXDVd906lJmsxJ6CMnZlCh+j/Cs6daSjmXBJGmng6W+cHo23bs4fcx4XFq1zH240uO/DkOEJSEjmJJJx4jr+qrgR1I81XadRVI5kYtfDSwzyyd+hDaoXttb+PIk146Ntxh2hpf3Lo3D1+WttRjmwtR9GjT4ieXFUF1UvoWttQW0POKhIiEwtURp56Dm7D6v/21awc8leL93xGLpb3DVY9nhqdFQHkyIbS0nIKRWRXannIpQCsSbao9wWlToUSkYGDioaurEp5dUY3tbhfer+VQ6bhEe9X8qqN3EubyQOm4RHvV/KodNwiPer+VTdxG8kPa3C+9X8o0Om4RHvV/Kpu4jeSHtbhfer+Uae1uF96v5Rpu4jeSHtbhfer+UaHTcI/cr+VTdxG8kPa3C+9X8o09rcL71fyjTdxG8kPa3C+9X8o09rcL71fyjTdxG8kPa3C+9X8o09rcL71fyjTdxG8kPa3C+9X8o09rcL71fyjTdxG8kPa3C+9X8o1lQrYxbyvkgjfjOTnwqVBLVFLm5KzMqlVlApQClAKUApQClAKUArgH66v48L/zp/KUJR2FwC+0Tw4/Ju2/urdXyhDFKAUoBSgFKAUoBSgFKAq0zT0uRIfUG0KQtSjhSh1BNYidGuJZaaESOGmjubQAnCD5wPJVjLJcDLVSNj7TpWS2khLDSQQAQCBkAYA/RXy1pB1hCENxWG0oxtSnaAnAwMfF0pll1J3sT9b0k+yt1TcZlCnTlwpwCs+c+ei9JPLbaQqMypDRBbSduEEeBA8lMs+pG8ifiNIPNvB1EZhLoQGwtO0KCR9znzeivkaNWlrlCHHDe/mbAE43Zzux589c0tPqTvYn0rSLy21IMZgoUnYUnbgp82PN1PT019I0pIb27I7KdvhjAx0x0plk9BvYn49pN+SUc2Oy6UElO/B2nGDj4unx18u6PdfThyKwsbgvCgk94DAV6wAOtFGa4Mb2J9HSTxkiR2ZntATs5vd3482fHHor49pi+U432OPsczvRhOFZOTkeXJpln1G8ifq9HuuFndEYUWP9FkJPL8nd83xV7+1yb94j5QqHCT4jexHtcm/eI+UKe1yb94j5QqN2xvYj2uTfvEfKFPa5N+8R8oU3bG9iPa5N+8R8oVlWuySos9l1xKQhJOcKHmNSoNMiVSLVix0rIMQVi3a4N2m1y5rpw1GZW8s+ZKUkn9QqG7K5XCLnJRXFnMPCuzuiGq8XN5LUq4OLlLCgSSpaionp6TWJxDtTt71dYLQrC2J0tAUU9QpsHcv/ALINcJCpTqy3UZXkrXXeez79R2lUqperBSt/7U/sbgnpy2cVQtTJw0v1Gt/V4Hm+G4oiZ4TdLjHQ6gOMx2ua4FDIAIIAPoKUrz/tJqI0FIbPD1TrYCVqcdK+n3WcfsArms0s6S4O/iv/AKnfuMd00+Ky+Ev/ALI3hwPt6Y2mw7jqvrmubvqiJ27W8Z7cUtcte30kuK/wFdHtj+3SOT/pj1sdJ9jIrS+uhZlo5mQkY6+PWrBqbWbF6gb0qSRjxFcHUrN0t1xuelvCXrqsigWx9aYz8xKTyhNQVLKfDaggEH/jq4c+8aqbKbZBfnhA6r6IQP8AiUQM+jNXp0p1JRprRNK76FmpUp01OtLVxb05vsR+WWzxNOyRctVW2W3KYXujtBlS2UdffrUnIPoHh6/JnyuMqy4osrZabGMIWjqoHy+eujoYBwpWXrd3m5zFbaNLFVm6t4Lknp8+HzLTHvsbVun3JDBTuAKHEA5wfR6DVp0u0mRoiM4nG9hYSfj6f4Vl4eFqNaD6Gq2g0q2HmnfVr42MltIA6forCuMQLjEDx6nPprSxdnc2i1dmbT4ZX32WsbaFqy42NpHpHSrnXfxlmipLmeY1Ibubg+TaFKqLYqh8RY9wfnxDDv1xtKQ2dzcItALOfE70KOaxcVUlSpOUeJlYanGrUUZcCh3ifK09E7VdOIl0tsbO3nTJMVpGfNlTQGa8o15enS4caNxGukl+YwuTHQy/GXzmkkBS0kNYIBUkZ9NaX06v5RvFs6m1dRdiT7DfPx0v3yo/0NOw3z8dL98qP9DUenVuvyKPQaPQGBfPx0v3yo/0NOw3z8dL98qP9DT06t1+Q9Bo9CGs+pkahuEqBa+J864zouefGizIjjjWDg7kpaJHXp66+7tqA2GQ4xcuJNzgOtx+1LTJkRW9rW8I3klroN6kpz5yKn02vw+hc/DYZsuV3JbsN8/HS/fKj/Q07DfD/wDppfvlR/oaj06t1+Rb9Bo9B2G+fjpfvlR/oadhvn46X75Uf6Gnp1br8h6DR6H4uJem05Vra+pGQMqXHHXw/A1+qhXxKSTrS/ADylcf6Gnp1br8h6DR6GJaZNwv1sj3C3a9vUyFIRvafacjlK0+cHk0ucqfZW2FztfXqKh99EZouuRxvdWdqED7D4k9AKn06vwv8ir8PpXy2dz9tkife4pkW/X93nRwtbRdjvRlpC0qKVJyGvEEEEeisrsN8/HS/fKj/Q1Hp1br8iHgKSdmmOw3z8dL98qP9DV04cR5zKp/bb3Pu+Q3sE4tnl++zt2IT49M5z4CsnDYurVqqEnp/Bi4jC0qdJyitUXalbw0opQClAKUApQClAKUArgH66v48L/zp/KUJR2FwC+0Tw4/Ju2/urdXyhDFKAUoBSgFKAUoBSgFKAps/jPw/tU6RCm6405EmR3FNPR37qwhxpaThSVJK8ggjBBrw93bht/aDpf54j/11TmXUnK+g93bht/aDpf54j/1093bht/aDpf54j/10zLqTlfQe7tw2/tB0v8APEf+unu7cNv7QdL/ADxH/rpmXUZX0Hu7cNv7QdL/ADxH/rp7u3Db+0HS/wA8R/66Zl1GV9B7u3Db+0HS/wA8R/66e7tw2/tB0v8APEf+umZdRlfQe7tw2/tB0v8APEf+unu7cNv7QdL/ADxH/rpmXUZX0Hu7cNv7QdL/ADxH/rp7u3Db+0HS/wA8R/66Zl1GV9B7u3Db+0HS/wA8R/66e7tw2/tB0v8APEf+umZdRlfQe7tw2/tB0v8APEf+unu7cNv7QdL/ADxH/rpmXUZX0Hu7cNv7QdL/ADxH/rp7u3Db+0HS/wA8R/66Zl1GV9B7u3Db+0HS/wA8R/66zLRxe0LqG5MW616zsFyuD5KWYsS5suuuHGcJSlRJ6Anp5qZk+ZGV9C2nwqsXbiVpqzyjEcurUqcP/ooKVSX8+blthSh8YqogwjqzU15GLNpRyK0fCVfpCYyfWGkb3D6iE1TeL8PUNt4e3SffNUENq5bBt9pipYZVzHEtlKlLK3FDCj4FOatVpKFOUuiZn7PpOvjKNJf5Sivi0ax1Hc70qRMtsG3yUW9UNDsefEbyUjHUJORk+CdowR1OfJU9w4jO3CdZDc2nUXCBEeeCZC97g3ENgqPkJBV0ryHZVOq9ryUuV7/E9Uxu6p7OnKEk52d+usWmu779hsScO4fVVD1QPsSvPXo9X2Tz3C6yRr/VN8ch6PdbaXy7heue9nPVEdtByfjQlCfW5X5pRRhcPYyB0MhKCB6V97PySmtLCl6tJc5NP3NyX/8AlnY1atpV2npG8f8AxUH4SR0toJr2F0HzVdNjBX19Vcp/VCxCtqwTPKVuMrPxAj+NbTbL/KS7zS/0sv8AqHL3fJmnnLuuS+3BjsvTJbmS21HbU4s+TwSCcZqxaf03cYkhL2om5NrYDgCYr7Skl7pk5Jx0Hh08fIR4nmcFhN5Fymd5tDaNTC1qWGpQzZ813eyjZad7bdku98jcFr4h22BDjttWuOymMtRYZZ7vinqT6eo8+TUpI4uxkDuxgoFOAsKykK2kgerPTNbZ0Ivmc+6Mm2278TW2r9bS5zr0h93uo281hCiApIyQU+bx6j1+atZXm8i7SGmI6nG+/wBwKG1JHTcAr0EDpjy+IxWXBW4ciatowUFzNzcMdIxJ+m5cybEQpanuW0onCgEpGSFDyEn9VXfRmnEyLNc2o91ukJTC8htmUVIxuH3Kwofqq9h60pupGTvZPiaHaOGpU1SnTWV50tPtw+RIN2q+x/8Ak9/Q/jwTNhJVn40FFfrr2pmWzvhWycnzsyFsq/QpKh+utRehPinF9mq+HH5mZlxNN6NS79H8Vp8jO4a64l2C9SY8mwXJaVHdthht/b8QVk/orbHuuaZZ6TpMu0LHim6QH4wH/EtAT+uuww9nRjZ30OIxt1iJ5lZ34d5N2rWFhvoT7G3q3zyfARpSHD+gE1L1kGHxFU7W6czI3+7P7awcar0WZ2Cdqy95zrxbt0iJxHs18nW9+fZGrVIiR32rau4ot01S0kPLjo7xCkApyP0jNUGXF1lcNO2m6QbJKtN7Rpm8hCLbb1xOU6Zbam9rYHcWtCd4T4kkny1oUlY7qlKGSLk9OFr999PPEmr1qvVV9fv06G3qeFpp242lK1Nw32pTcLlOCSthG3ePsgRuKRnBJ8uaxtWXDVXa5qLVK1kHkwoR0uns722S4Vq5xl9zGR0yHsYT18anKiqMaSaXq9vDjpf5X95l3u6a6iXXiIxZzeLtNMV92FKDclpqIQ6hPIbZcRy1rCN5Q40SVAZIJIq68JZF8b01qGQ5LmXkJWV2+LcGpTbiVBoZa5klCVqBWPHBA3EZ6VDimizU3W70tfTppw/n6mmuDw1Fo7WFhzZLjKYeS1DfmP2mW0mysurWt6GElPfy8W8PblYSO9gVf+OFsvqtXXSbZ4U51waXSw0/EYWv7J7IMkoBAPe2BRx44BNS0r3MipKnv1K6s11Xnt9553N3iKjWuoLDbjc3I9tNwvMKe6FcmUHI6RFiBZ6KCHnF90+RtPkqAsy9cTtJuJZvOoedImWll5Co0wSYilO7ZKwt9sDG05UlO5CcZ6A0yopToZU7Ll089jNqcSrReGLNou0We43lCFXePEmzYrqlSTG5LoWpxzB8SE5WfKQehxWtLPH4jWiNbpUSfqG5XGRHvsUx7kFLaQWNwhKIKQApRSkhauq93iRUKKtqWaU6ThaSWt+/n/BB3Cz6m1Nw0ua5lw1DPbjSrVKXCMeaJLLiXcSCC62FLwDuKG9yUqQCMdKtL951M9xStLdsZ1K3bETWozwmmUtp+IqJnmqQWw0gFWOpUXNwVnHgJsmZEnTldaWV+nRdpTtCI17YtHJh6aa1MmdG0y/2uHdIziGY8rmI5IjJWkAr2FZATnPTPWrGYt9vMhCII1JddMs6hsT0R2+MPdoS4l0mSocxIXy09wkkbQc46VLSKpulmctL/wA6W917/M+rdM1dbp+mXXfZ2UDdZbblnbZlR9yFXBYS+XUILakpbAJQ6Qko6g9c17wLBrO8321dqvurIca43O8xpaWnVtoYjNqWqMU5T3MlIAX4kHAOMCosi25Uo3dk+PTt/g29wklXe4cM9NP34Pi8LhN9qEtBQ7vAwSsEAhXTrmtr6GGFTfUj/wAVZGEj+fH3+Bz20Gt3NR4X+pa6V0ZywpQClAKUApQClAKUArgH66v48L/zp/KUJR2FwC+0Tw4/Ju2/urdXyhDFKAUoBSgFKAUoBSgFKA1deIbC7tMJZbJLyySUjzmsTsMf8A18gVxtT2mdfD2F3DsMf8A18gU7DH/ANfIFUFY7DH/ANfIFVziRPVpjh7qW7wWWEzYFtkSWS42FJC0NqUnI8oyB0qUrsrhHNJJmv5fHNm0JDTulZlz7I3b0zpcQspSlyW2kthKFKBJKlbceA8Say08aWnm4sZnR0x6+PXGXa1WtLrALb8dsOLy4VbSkoIOQfiNV5O0zXhOeZfPTS/2MCN9URaXrLKuj2lZ0SKmzKvUUvFnMllLiW1gBKjtIUoYz4jr0qwak4tW7T92udtRYZFwlw58G3JbY5aS87KbK29u4gADGDkjxo4PqQ8I07Zl5t9yDn/VC2O32mE69YyxdXn5sd23yZLDQZXFUEujmk7VEkpCQPfZ8mDVl1TxQtGndEWLUqLaX4l4UwlnnlEdDIdbKwp5au62MDGT5SB5c1DgyJYSSaV+LMWTxctUS8LtLtmc9lO1wIzEdCm189MpBWHUKGQUoCHdxGf8ARnHiKq87jy/P09brnYNLsyDIvUe2LjSJTPOQHFFOxaAoFp04wAvoMgnIzUqFyqGEvZyemnLqTbPHSwP60Xp8Wvvty3Lep0PMqWmQ22VqBaB3hvKSjmYxuHmOahkfVFMOW3tqdB3Ism0i+IVzo3WEDtcdPf6bT4J8VZBwKZO0lYJvjJLh1N0Q0Q58NiS0y2Wnm0uIJQAcEZH6q9uwx/wDXyBVs17VtB2GP+Aa+QKdhj/gGvkChA7DH/ANfIFfse1TJNwhotEqNbJRc6yXIoeKU7TnanckZ8OpyPRV6h+rG/VFmvfdSt0ZZ/cuh3Lvagut01Io++amSS3Hz/uWtiCPQoGrPabHbrDFEa2QI1vjjwaispbT+hIFdeclYza0x9VPKKNEWaGnr2u7soUP9VLbi/2pTWHjHbDz7jodgRzbUodkr/DX6ERa5b8DSiMKPcTlO4Z2nHiKg+Crrt0v2rrg+4p1e9iOlSjnGAtRH/aFc7QbdaC88Dr68YLAYyouLsv/AN0bLnDKD6q1zr5ambPMWgkLDZSnHjuUQlOPjIrNxTy0pNdDmdmQU8RTi+bRqvUcJyVb7nJUkc4lNot7X4NtCkII9ZUpzP8AseYVcLXZy5d7XZG+8iMQkgeZADaT8lAPx1YilLaEaUeEIxX/AIqXi22bWrJx2VOtL2pym/8AycforHQmrHU2TRCYyeintrQHo8T+ofrrnHjJbF3LRzC20hxUeYlZQSASkpUnpn0kH4qr2qnUkqceNij+nrUYb1/uNb6fajaLs6prBD06Wkc1bf3AAxsB9YJPpJqy6a1U1qNiVEltl1kYQ6053gQR0IPxfFilKmqdNRNnXrOtWlN82a9mxLpbL7NhRWnJaEOlDK2yCSnHdz8WM+SsiXadRQrSta7e6tCAFLCFoUQARnoDn9FHAuKurdp56YhQdROdsuq3XthHLjNuFOcHPfPievkBFbA1DeRqHTyrWiCmeVONtNNR8ZaUSAkg/cnyfH16E1U45YllVM8rmzYFsRYtOwoCG0tlhhKVIR1G/HeOfLk5OfTXrwmirlT7vHWOrm4EebIq3s1KVSd+aNbthuOGhJfu+5KpBbcKFe/QSkg+cV6ZycVo5KzaNvdSV0YOlpBia+ZAOAtGP110K2d7aT5xXaYB3w0PPM4PaitjJ+7wRDXXQ+nb7n2RsVtnKPiqREQs/pIzUR7kunmOtvTPs6h4G23GQwB/wpXt/VWeamw9o96hD/N2trugfg57UeUn9JbC/wDtVFXSHfIklKL1coVy7g5LkWGqOoDJzvBcWD5PDHlrExdnRZmYT9ZFJg8U9I3S7uWuHf4cqagup5bSioLU2CXEoUO6tSQlW5KSSNpyOhqesd3h6kstvu1ue7RAnx25Ud7aU721pCkKwQCMgg4IBrROm48Ub9TUuDM3bkeNNvpqnKVXG300Kc+WmUXG30029PGmUXI+332BdLndLfGf5su2OIalN7FDlqW2lxIyRg5SpJ6Z8a+514gW0PiRLabWxHVKca3ZcS0n3y9g7xA8MgeNTk5EXMmM83LjNPtK3tOoC0KwRkEZBx6q9NvTxqMpPEbfTTb6aZRcbfTTb6aZRcFOfLTb6aZRcbfTVo0QMKmepH/irKwqtWj55GJitaMvPMtVK6A54UoBSgFKAUoBSgFKAVwD9dX8eF/50/lKEo7C4BfaJ4cfk3bf3Vur5QhilAKUApQClAKUApQClAa0uwzdJf8Avl/tNYmK5GpH1mdfBrIu4YpiqMpXdDFYGoLFF1PYrhaJyVLhT47kZ5KFFJKFpKVYPkODSxKllaaK27wj0683MQph/EtcFbo5yupibeRjzY2jPnr3i8L7DDvjd2bZeE1ufJuSVF0kc99oNOHHmKQMDyVVqX3iJNWv5tYwBwS0r7FMW1UN5yIzanbMltbyv+TLWlahn77ckEK8RXlE4OaXsSFSnXJrq0zItxdmXCctxano4IaWtaz4AHGPDGPNU3b0KvSp2auez3BvTrym5EZc+DLRLlTm5sGYtp1K5BCngFA+8UQnunoMDFTWotEQ9Taeas0mXcmoraA2pTExaHHUbCgpcVnKwQeu7OT18etQ7lLxDbTdtOwrh4PwEcSNN6hZbYYg2G0KtsRhO4ubveoJJ+5Q2VgeXKzX27wP05IiT23nLm/KmPx5K7g7OcVKQ4xnklLmcjbk4Pj1Oc1N2VvFSVrdPrcybLoDTzd6m3C1XKaee6p2XDjXJao7jykbFOLbCiCogdc+JGcZFfjfBrTTdsTATHkdnTZF6fALys9kUQSnP32R76juin0mXFW/0XOHDbgQ2IzIIaZbS2gHqQAMD9Qr1xVGUxm0ximKZRdDFSWnR/nuJ/tH9hq7RX5ke9Fms1u5dz8DYlK6w5MVpX6olDUq56NjPZU0l6TKUgfdcttPT/tfrrFxUc9Jx62XzRvNi1NzjY1F/ipv4QkzRzXGi/OJalOMQzY3ZvYEwgRzR3tgOMZ8fjHnNba4OW1uDbdQrayUvXVZTnxCQ03gftrO2ts6jg6lKVHtT71bU57+mdvYvaeAxtPFWesJK3STenfoXOaMoNU7idCTZ7fZQ6O+472p4H71AKwP+wP01zmKhnpSXcvi0jrdmTyYmD6XfwTZr9Nu5EnTsV/GWCqY/n79KCsq+W8f0VfODNlVe7/IurqcpKiUk+QeT9VWdnre4yvW7fFIzNrvdbOw9Ds8G/4LpxXuG+dEhIIw02VkeknA/UP11pvie+0zo4suoUsyXkNpKD3k4ySR6cDHx1bxjzYpLoZuy4ZcJDtNFsKW5O9j2E9yQ4drSl9WunU9euMDJ9VWJc2BpS3lptwOO/dKHvnFfwrIi7oyasVF6GPGamxbki4Ow3w2tsKW4EnCT58eOPTU5Av6xPRudQ7FdGEkeTzg/FVVrlpOxGTtBy5N5cVHU2i1OEOb0nCz50j4/L5j5am7Q5bNN3O320nK3pLa1MtKwrAWCP8AtAevBq3UfqMrpwec3HPZ3BWfGvLh7LFr1sponCZCAcekdD/CsXZsrV0u8s7VjnwMuyz+ZY9YW/2Nv7hSMNvjmJ9fl/8AnpqMByoeqsTGQyV5rt8S7g573DU5dlvhoRMRYj62t6/OSmujIqt8Zo+dIrpdnO+Gj7/E5Ha6ti5dy8D1pWzNMKo3E2C7c4L0SPIMWQ/EdabfT4tqUCAoeonPxVi4n9NmXhf1Uc16Nb1lAZ0Nptiz3mzRbXblW29W9caMbbtahuoQ8w+BzFlbwZwEq96TuQk7qgYlj1vp3RkextWS+OypendOR4yoigGojsc4lpWd45agnbkD3wOBnBxrrRv56mz9a3noTEvT+pl2fXLTNp1GdbPSJ6k3NqapuPIgqlhTLUdRdCEuGLtQjCQULSokpJyqNvWlby7ab2/ZrZqOBYBPjSLXZZbT7/aHkRnUvJdaDyXW2FqW1g7gEuthwgg5UTXYGmySh6Y1lO4qIm3dVytQM2BJhCIw9LbaipjMh+MqQJCW0pLokJXvaKlbgoFR27bZrrRV9v2vr9cbcqW09G0uG7K6ZS24qLkVyQFqQFBKlJCm+qgcBXpqluKat0Kkm0akm2/UWmNE9VXhcm53C0QjZJTD0JL7yFurfTzHZbhcU4lIDi0uIQoIScr7wMhp8XuDpJlywxr/AH9t+yXu0LTFUSqNdDKBShW95WEowttLu9Qw377JGbrSaKFdaEtcNM6nhx7qHrBKmtXG4W3nvqS8+W0ItSELcLLTzSnhzkBsgrABVvOdlQrPDO/zLXYpV9s17lagd0O/bu1tyHeYzPbO5sOqS6cKIAwokhSvEk9ahNJaBxbJuZpjVr16cDtvvzmo1TbQ5Zrq1JWIcSGhuN2tDuHAkHemXvSpJLgcQBnptndBaHv9k1Zp+7vt3USJVyvybqqTMdcb7MqS6qGChSylKdobKNoGNx++OaW42tp5X3Kkm356/Y3ftFNorFsZA2im0UsBtFNopYDaKs+ihhUz1I/jWThl+avPIxsT+k/PMtFK3hoRSgFKAUoBSgFKAUoBXAP11fx4X/nT+UoSjsLgF9onhx+Tdt/dW6vlCGKUApQClAKUApQClAKUBz59UBbY130feYcq/QtOtuTGSJdzXtiuKTIQsMPd5OW3dvLUkKBIWR5cHTVi43sacsLTbKLRpSztWW9mK3HfS5CkT4slpCTEdUE8xtXMWUoAGc4x3a57I53Xazo8yik+w+3+NWoBdnEjU0BufGdsSY2nTHa51xExDHPxk7+nMWpJQBsIJVuT0GRA4x317UymWtTQblIN+vlqf0+1HbDsKLFTLUzJUQd+QWWUlSsIIdSMbupl0o24ebcRvHfj5uYd04na5tlj0Y2/qKK3LvticvZuUhESGx2jYwURhzjt5SeYpSgCXFA9FJCTU1euKeooOprxbnNQWaJPftSnLWyiTGcgRX0xG1r7Wo/ZmjzVKKVnLSkKbBwo9YdOD4LxG8l1IafxentwIN+toN0uULTmoCqXcozDjqHY8uCgnfG7i2k7lKPLICw2M4I6SK+JWo3bs9ZLJrCJqCEu52qK1qRuKw7sMkP89jDeG1LQlppaTjoHk7gryt2uaG8fJ+bFnY4iagt3A/Vd+cW1dL7YpF1hokLYwl0RZTrKHltox4IbClBOM4OMZ6a91pebhrhl7TiNYM6msUe+aeUm9Ro0VxDrj8pXMjLCUlpfL5bLoGM4dSFZHjEYJNu3MmUm0lfkbI4r6xe0lqTR9kb1VB0ZarjGnc6fJZZ7qmUsltKC53EnvK6EEEZAAOCKTbuLWu7npu7aiLrLLln0VBvirMiDkSpb7UvJUTlxLYLKFhCcK8mfHMRpxcU2vN7fImU2pWT82PaXr+bMkyLc3qCBru0xbrpl1m8JjsqS29JuIQ6zlruFSEIbcSQNyeanJPdNR07XWsbppKQqbqFK2r7YdQuobjwm2jCVDcShpTaupJUhSgrdnrgp2461qEea82KXNvg/NzF0/LicJnL32aSq3Lf0/piOJUCJFaUXXX5yASF7GUdMJ3r6AAdFHAPtB4va11NpZmTBv8eE7Es2obguSmIy/wBrVAnBhjJBKAlaOqyjocnYU5BEuCl60l5sQpOKyxfm5Ny+IutLbHuUA3uM/cHvazIjzVwEBMYXKcqO82Gwe8hIbJTuJV3uqjjNfdp4gaust9gqu+oUXS2o1JcNPusi2oStbLMKRKQ+dnUugspSQkJSQfeg9aoyRaenmxVnlda+bktwG4pz9f6i1BCeuS7tb49tt1xjPv8AZOcgyDICm1dmUpCQAy2oJUStO45JyDW6KsVIZZWRdhLNG4qR07/z1E/2j+w1NJfmR70RVf5cu5mwqV1By4rQf1ScpcXV2g1tnqO2Hr4H/Q9KwMc3HDya7PFHTf05BT2lCMuDU/8A4SKtcdAaagoN9atie3HLgSQnYF48fDP66sPAqQuZo6c84rc45cnySfP3axqm0a20K0N6/ZT+hXhthYTY2ysTLDKznUjf5mzLRbfZK4oSoZab768+UDyfHVO45MibdILAGd21r1BYUD+qpxDUKGZ9V4oxdnpzxKiv2y+aZq3UckrvMpCMbktCMnHnWrer9Qx8VdCcKdPpsummCUgLWnJqzsqnlpzl1Zk7eqZp0qfSPia/1ZcPZXUMx8HKS4Up9Q6D9lay4xPPRrdai2tTbZW4FKBwN2E4z+v9FaypLPimzpsJDd4enHs+hqFiHc7iVuWpkvKbV3lg7Up6eBJ6Dp5zV6RYF2nQTapzLfb0IU+42Qkndu3AFQPXpgVmw4EVrORjac1OLpcwhDnc2JJCvEk5FQ0jRrytTTXlS+wWsSUDlJT33NyColseAwQASene8uKrMZtySsZ2qZz+j3oSES1SIUnISpQ74x+o/oqP1FF7BcbZNZcQmJNaUpaVn/pU469BlRIUMZ8Np6gVD1RdhpNc7G6NFXpN708yhbhXKipDbufL06H1Y6fFX2477G363zAcbHQkn0Hp+3Fa7DPd4mPeXcZTz4WrDsf3Nv6zgC7abjz2hucZAUceUeWqC0vcU1f2rC1VS6rwNPsapnwzh+1+JDTCW9UWxQ8ruK6Qtp3QGP8AZFbfZv8AbL3mk2x/dPuRk0raGkFVPWhKX2SPENn9tY2IV6bMrDfqI0TpjiHcbhwy4SXq4SXfZDUYt4lrjNtpS6t2Gt1YUCk7UlSc93BGBg4yDAyPqhpFx0I9dXrHN04J+l39S2qQ3IYfdcZaDXMBSUlKFjntFIUFAhXXBBSNdurs2bqWRI3fixeLtrXS8O0212Jp9eqn7JIuSn2yZSmI0rmtlojclHNawFA5JaOQARn61pxVvGheJmpUqtr120zbbFa50kIfba7HzJMxt1xII3OKKW2ztyBho9QSAoqaelw5viTLfGMuX9pr2DdGnnb25p5F37QncZiCpB+w4zyuYhTe/dncM7dveqpx/qlrhIsDV3GhJaYr2nzqdvNyZyYKAC6SMdHBlJSjqFBQypByBCo35h1OwlL5xpsF61g5pGRY2b3bhOiW2VzylxXOfbbcbUI5Sd7aQ81uWSNuSQCEkjztnHFi22KFKi6NksWCdbpkywt21SFuTER0FfL5CQOUXEAqQAVdB12nAqd07WuN4uhl2/i9er5qfQ0a2WyzzLTfGZrkqTEunaA1yFtpJbWEAHAWcpUkHPd7u0k4bH1R0Z6LqIotLE2RaY8SUBbLkiYyWn31M7nHG0nl8spK3MBYSgEgqwRTdX4Pzcbzz7j1uHG42p26TwyLjHatNpmJajS23IDIlSJTRf7Shsq5QDIUpwggJSkhI71bTsdyF7skC4J5JEphDw7O8Hm+8kHuOJ6LT16KHQjrVEqdkVRlczsUxVvKVXGKYplFximKZRcYqy6NHel+pH8ayMPG1ReeRj4h3pMs1K3JphSgFKAUoBSgFKAUoBXAP11fx4X/AJ0/lKEo7C4BfaJ4cfk3bf3Vur5QhilAKUApQClAKUApQClAa1vbDcmdNadbQ62txYUhaQQRk9CK1oOLmlU3TU1unRpcFvS8hmO4/Itj/JWtxDexLJDeFLUXkoShOVLzlIIIJ5xxcpOx0ykoxVzDja+4eWR+VqREiQmZd7imAplVtlKliW1H/wBCI3L5qF8lAVt2DIweoIqY0Xc9GwxFj2OQhx29yrlKaCm1lxxxMlSpedycoCHnCCFYAJA81HGdtfNgnBvQXnihoyBLm2+fJJTbHEtyHTbn3Isdzu4QXg2WgtIWklO7KQckAAkY2oOK+mLJrSBYFRZdwudwmqtUhyFbXn0sOCMJAQ6tCCCOWpBxk4C8nASoiFTkyXOKPK6cYdLQYF3atDyX7jboU1yK2uE81GkLjJJdaZeKA25tKMKS2okbTnwOPS08S7LNgWJNvMDMie1BkthLiG2X1xlvqQ0pLakrcTt6jKQOuVBQCTO7dtSM8W7I9LLxu0Le7ZLnQbviExERcVOvQn2EusOKwl1re2OcCru/Y93eIT4kCvRripo2PbmSy5ITumLiC3tWiV2pD6UB1QVGDXNRhCkrKlIAwpJzhQJOnJaE54vmWO56Utl5vVuusyOH5cBp5lgL6oCXdhXlPgf9EjGfDFSwbQFlQSkKIAKsdSB4ftP6atN30K7I8mYMaO0Gmo7TbYVv2IQAN2c5wPLnrX0YrJSBym8AEAbR0B8R8dCT8dhx3m1ocYacQtIQpKkAhSR4Ajyjr4emv0RGAnAZbAwRjaPA9SPj8tBY/TGaUcltBPd8UjyHI/QfCnZ2gQQ2jIUVg7R0VjGfXigPliGxFBDLLbQPiG0BOepPk9JJ+OvXA9FQ9QMD0VI6eA9mYvh74/sNXKS9ePei1V/Tl3M2BSulOZFaH+qbYabuOkbjIc5USEJrj7mM7UBDaicepB6ViYqk61J048W0vmje7FxMMHjViKj9WMZt9yhI5yt/1Y2ntRXNmyv2R+DaZDhjs3MvFXXwCiNuCPOAfUVdM7/4FsKh6cvUNZG+PdnkEetDZ/jVzaGyFsqvSyyzKSfK2qtf3GJsX+o5be2Vi4VKeSUJwdr30k2lyWqaaa+epvOwQOxQgVD7K53lejzCtV8Tk+yGoXWwerRBSfMpKcj9ZrXbRVsLlM7Y7vi8y5I1tp22q1DrJpG3IU4XlDzZPT9X7a6anOJsmmpDicDkMKI9eOlXsAsuFi3zu/iW9rSVTHyguCtH4JI0UO+6c9SPKa8b9p6Lqizu26WCG19UOJHebWPBQ/8AnlrkpztUzdp3NssUlyNXPWj2uudiddBjwTtCMYDrhGSsjPx/GPJVX1Xq+RNS3GZdBaDqSsqHvzn3tb6NpJWMCcrXMa721/T02PeIjRKX29q2U5HUEEkDy9PJUjeNVvz9NuyUMOqSxhWcDPiOo6+HnqpopTtoe8OdbNZWVtiUEPcshxCc95pwDoRjyftqF1XIjXBi3xoziJCYDhJb5oSF93BwcjJHd6A56+B64jiXI8dC/wDB+RiVNbQkhCo6Vr3JOdwVgdfjP6Ktuq1cu3rWPFJBB9VarhXXejY1UnGa7Pobz0C6L3o9tC8LBbxg+qtc3OEq03R+IrwQo7c+UeStztanmpRl0Zxew6lqs6fVX+H+yEvH2K6W57714f4V0TZXA7a46h96Ku7Ld8PbtZY20rYldy8WZ1K25oRVW1ejmSGB5Cgj9dY9dXgZGHdqiOetM2fhtpyZZtPwbjeJDNvntR7YJj86RBjymW3GkMNPryyFBKlp5YV1KQCMpAHlwn4H6TTw4Ran3Jt8U1bHdLy5MmW+UbGl8qSiOlSzym1utE93HvE+RKQKKuHrUY5prp07fg9NVxXMzlKMnZF2b4NaZb1U1f0szBManrubTHbnuytyltqbceSxu2BSkrVk48ST0KlE/motM6Uv1zvjN2adRJvLEaySS9zWUyUo5rzTbSugURznSSgnygnu4GNGMpv1eRcbSR6s8IdOR9T+ziGJXPExVxTEVMdMREtSSlUgMbtgWQT1x4kqxuJNVSXZuHNomRtGrYmfY7UNJdxEhbLTD7aSmOt4ZSHFISkgk7gCOo3DNylSqV21Dkr9OBTKUY8Sw3HSGnNK6lgXVLtzgzLpMYjdnhTXkMy30NbUKdaSoJUQ0zgkjqlsBWQBX7bOCOmbO+t2Gm5M7Wn2YqE3OQEwEvKCnBGG/wCw5IHVOMAbRhPSrcoyjFN8/poTdPQ9IvBjTcQ21bbc3tMGU/MEpU50uvuPY53OVu+yJXtTlKumEgYAGKxrJwN03pyM61bXrxEWqGxb230XaSXGI7CiplptRWdqE7iMDxBwrOTVF3wJsjIg8GdOWiK4zbBcLWtcZiIZEK4PNvFDLjziMrCsklcl4qJzuK+ucDErw7asreirUxpxlbFjjNmNEQvcTsbUUZyoknO0ncSSc5PjVThKUHLkmvnf7BNJ2LHszTZkVayleYxrjNbtkXnupeWgLQjawyt1WVKCR3UAnGVDJxgDJOACaydmaqcLK5GY/eXTl1TlJzDl1YtIJwqV6k/xq9RVqiLFeV6bLJStoakUoBSgFKAUoBSgFKAVwD9dX8eF/wCdP5ShKOwuAX2ieHH5N2391bq+UIYpQClAKUApQClAKUApQGptcWS039q4RL1HYlW4vFTjcn3mQrIJ+OtaXfg09dZ16lWq/RoUOdLt90ht9hLxjTIgZS2SoOgOMlDCUlvCVd4kLHTGgcssmdEo5ooy7Nwffi6hg3+43puZeU3x29zFR4ZZZeWq3mChttBcUW0pb2HJUskpPhu6Y+g+HLumuKOvdSzN0e1vupTamnlI2MocQh6a6jB7qXX8EhWDlonGCCYc00+4nJax7ROGjjsTU0ZN4i3fRGo+1TF2xuGFSHDJb76USebsLZJKk9zPeHewKgdB8Jrza9H8PnblemY2tYM5y83Jc1kSO1vvsONyGyEOI6oQ6EpWkkDlJ6KFVZ1a1iMjvxMgcAJb1vZtErUjTtitqLj7EsIt5Q+wuU081l53mkOhtEhxICUIzkFRJFZmnuBCdNW+y2qJe1ewtnuqbrChri9WVFh5t5pKt/vFOPKcAIOzKkjIKdp1E9LBU+dyIv31P9sb0JbLfddRtRoNk0xHsypslhKGgY7rDyJDgUvARujJ3IJ6gkbh41+yeDTV/wBDx4Nvu+lYL1xkKnMXaz2dxtW5TCUokRVpl7kvJQkkOhagU4G3aOs7zTgQ6a6m3LTBnw1zBNuCJzS3QqKAxy1MthtCdijuO9RWFq3YT78DHTJkNorHdmXrDaKbRTQWG0U2imgseMqZGhcntEhpjnOBlvmrCd6z4JTnxJx4UhTI1yityYkhqVHcBKHmVhaFdcdCOh8KW7Ae20U2imgsNoqR0+nF5i/7R/YauU7Z495bq+xLuZfqV0JzgrTP1UMVh7R9qVJQpcV2cYcjb0Ulp1lxJI9OQmrFao6MHUjxjZ/B3NpszDxxmKjhp8KilH/yi19TkrTn1GCGLpGmytRNSdNRXee1Fa3c1WDkJ27cJ+Ucemulvqe3TdNTaviOYCUSmpZT6FJKf/8AmKvbU2vT2piKEaSaST49WvDQsf07/TlfYezNoyxUk5twXq3taM+OvNuTvyXU6EIx1zWi9XTUv3Ke/nruXg+jw/gK0+1NaSj2m22JG9aT7PqfPBCyiZdJVwWkY3kJ9A8n6q2fxJldm0pISD3nVIbH6QT+oVnpbvDpdF9DXN77Gt9ZfU0Ze72zp20vz3klYRhKGx4rWegFViyyNVa0D0mPd2bdGbVs2spScHxwBgk+smuTpwgoOrNXPR5KNryPa/6Zu7cRMq4vR7w2gbXTywlWMeJAAz5OprXFx0RZ1ylPiRLiNlYc5AAKQc5ODnw/ZWzoShVgsuhrpxTlZaH5rvUT89Fth2plsqbfDhSkgkICSP4jyeQ17Wdt+DdmIKGlOqnhbqGQB3SkZUn9RIHxVfnaEbjIuR8alsrLcGbNhQFmSsBDiY7RICj90ceBxmoCzaWvd/cDTFvkOAHdkoLaEKPj1OAB0HSsWVRRvqX6cVxN7aI0e1pG0rZ5iX5r+FPugYGQOiU+gZPryT6Bj60d5dpcHn6Vrabz1l3mTN/lzfYzefBvrpZn/ZrA4p2fkvsT209D3F4/V/8APTXV42G8oTXnQ862ZU3eLpvrp8dDW2okkxEODxbUFfoNb50VLEzT0ZQOe6P2VgbJd6Ul2mz25G1SEuzwf8k9St4cyKqWuWXn2y3HcDT6mFpbcI94o5wfiNWqlsupeo+2aA0zpTUc7SmhtLzNNPWWJpxUWTPlPyWHEy3oyQUJY5bilHe8ErK3AjpnIyTinW3hNq5rTAdg2CTp6/otTsa5yUS45lXR+TKZdkcpxLhG1DaHw2XCnCnRgJwo1vKeKw9OUk6iabu9Hrdu/LldN35rS+hecJStoTlz4UyWNLQY1n0zPelPXJVwVb7o1b/Y9Kw2ljZIjtuBAbKMrBaClhQ3HKuhk3eGcu46nsc696ZZukUahuU95DgYd7Mgt9nirIWr3haQhZCckKSju5HS28ZG2beet6y0zc07PVXu2ld8WrX5k5OzTToZfBOxzrhJfvMpSV2i3PXCBYlJXkPx3Ji3DI9RQlhtPobUQSFiqi/oXXitdK1Amyyl3KRMUm4ynHohZaYDyUtvQE80EO9mBbUXEhRST1ykJVMa2HhiKqm7K1ufO7a0v/lb/wBtw4ycVbz5XzNo8RtFu6z1BppMq2Iulktolz3o7i07HpAaDTLSkk94KS88eo293veIzrLTuj7lBvVs03JdSm1wNPWd7Ujod3Bh2Ip5wMDGRlwqbyM/6Jo598nNjDYiG43d/ZV7dqk2vi8qJnH1rkbpfQeqb5pJu+2eySbBf5NlnPu3J6SyJNwkTVpdQhtQWSlCElZTzNgSsowB3jVos3BdqUqyx3LLOjWd25GddYdw7I0hYairaaQWo6ilSVqdyrO5StnfOMCs2vjadNyjCabV1fXtcbaW0emvDloURg5WuvPMq2nYq7lxAYRbLPDm6jtDt5uL9xizI7pfCeYxCigoWS22EuthKHdmCycDuqNTOl9B3ZjUytLW5bS7RaLdaZM+QV5Sq5R2Vpbjqx5SpMZ9R8yUgjviqq1WEFJTd7RvwelpPLx71/oKLfDr9NQvhzeHtEWBhrSM6BO7Sx7ZCFwnZl22MOFS1lTpbeQZCkKIcUCRnu4GD7ai4HSbnYZdujWp96NF092O0tz32dzMyU+4uQ5tQeWhTIDRTtASnwb96KtenQpyTVRPW9/WvxtpdcFF2XPTsRO7uuHn/ZMscLblbNSyEWu3Ih2d2/xJRJeThxmPCKw+vvblLXK2JUTlR2bj0614cIOH13tmqbXd7tpw2aVFsao0+c++y4/c7i642p55fLWrcBy1bFKOcOkYSMCseeLpzpS9f1mldWers013LR/AqUGmtNDdm0+im0+iucsjKG0+irDpIEKlepP8au0ks6LNb2GWKlZ5rRSgFKAUoBSgFKAUoBXAP11fx4X/AJ0/lKEo7C4BfaJ4cfk3bf3Vur5QhilAKUApQClAKUApQChGRQHOf1TlvcunDa/RGGmn3XrhCQlt9outnM9j36ARuT5xkZGeorSjE/UHDeBOs8dyDpou6tkC8yYqhbrbFZVCSuMWFKZfDDTu1sqUUn7KXU7gVVp1Z3T6s3bbVmuh63viVq9u+6fsszV4hOC12qU/ebbyBbzzJjyZDrzjrGTvYaSEbUoAWScAEEWF/iZd7jPj6aduDztz9ndQxrjD7L3kQUNTVwgshGEgoTHKDkFYHirCqOEeSJU31K/auIN7tMnhzZLbPnw1Mx7DClW+QtpKHWX2W0uONM9nU4tKd2FOqdSErTtweoVi6Yn6i09aba/abvcNUX6C5qVpVquMdh1TEpoPrYRlLSXEKcPLONwCgsYG0ipcURmbJg671JKgymtMa0uOoYTybGh28uw2C5BlyLk0zIaQAylGSypSlNqSot4Gcbq2Jp++aot2heIqG3ZOor1YJc5i0qmNID0vbGQ6yhQbSkK77mzIAyAPLk1blGPQrUm+ZqbU13uutdK3S3W3WN11PZTHs0mTPMZhK481U9tLjA2shIyjvqaIJbKE5wFEHcvEd+bZ7zw/aiKMhQmS23ZDsdtxzCbZKUFZ29wlaUZKduc48CRUyS0Xf4EJt63NQLv/ABDhaOlXY6zvMiVH0PE1QGVwYoSueQ4VsYDGeUQgAoHe8u7NWi63DWlx1g6WNV3W3QntYGxJiR4kYttQjA5xWkraUrfzB0WSQPDFS1DoReXUh3+LmpNN6bfVeLvJS+bDe2ID7kNJcl3CLNcZYUEoRhTpbShWwDByohOAcS9z1LrIXadc2r7Pbj2646djN2xEVnkPolrjtyislsrPR1ZG1Q2kZ9FHCKJzSelz34Uaw1hqPiG81d73bmwh+4NzdPOSwqTHQh4pjqQwIyFNgJSnvqeWHAvcM5GPPU2vtQQ7rxF7JfJqdVWpEoWLSaIramZTKIKXGnscouOZcKzuCwNyQ345BjLHNZLz1GaVuJE2m7XzUbNkcl6hgant7ep7WqO5CmiY7HVtd5wdWiMwkDOwhO0lJKgSBtA2lwFYWxwi06h1Cm3EtuZQsEEfZV+SqZ2y2RMG2zYG0U2irBduNoqQsCR7MRf9o/sNXKftx7yio/Ufcy+UrfnPiq9rvREHiFp5dnuC3m46nW3d7BAWlSFBQwSCOuMHp4E1bqQVSDg+D0MrC4ieErwxFP2otNX7DztPDuyWeGmM3GU82kY+zrKif4fqqXtthttnU4qBAiw1OY3qYZSgrx5yB1+OrVPD0qXsLUu18diMRmzzdpcUtF8D0u0kQ7ZKez/o2lK/QK55nkvrWPEHoc+WtVtWWkUdB/T8VepLuLjwiubFslvW5Q2lffQT5R5RVh4uSNltgMg+/eUr9Cf/AFrKVXeYPN2GFLDqjtRQXC9179TRfES0yLtpRwRUF1+M6mQGwOqgAQcenCifirVE7i9p9iwtQIcWbbdRRmktmTCw22sDAJUoKBOfSM58taOgnOmlF8Hqd1u3U0XUs+k9QXFbTUlN8kOBYHR14uJUT5Nqs1bJ+h3b/AWX32LZIcGUlmPk/wDEndtHxCqnONCWaxj1KaekeJqfXOmjw5vDSIj/AG555kOKfeBHUkjGMnIGPPXhoyZLl6ss0hZDr6ZDSR1xgbxkAeTpmsipU3tO5TCPU6NcfW65tSemfCgYccON24jyZrSqLa0LzcYJI+XG1JGR5KrWt4i3bUSlPiQav4dWqx7yiq06Uu5+BvThC3y9LsA+O3wqd1vbxcNOS0AZUlBUPWOv8K7WpHNCS7GeYUJZKsJdGjR1wb7RbFdPFOa2dwduPa9OttE5U2Np9YrRbKdnOPcdTt2N4U5dr+hsGldCcgKreqUlUljAJ7h/bVuorxLtJ2kQmxXmP6KbFeY/orFymbmQ2K8x/RQtkj3p/RSwzI/EtbEhKUYAGAAMYr92K8x/RS1xmQ2K8x/RXzyOiu577x6ePTHWlhmR9BsgABJAHgMU2K8x/RSwzI+UsBBUUthJUckhOMnz1+hnbnCMZOTgeJo1cZkfuxXmP6KbFeY/oplGZDYrzH9FNivMf0UsMyGxXmP6KbFeY/opYZkNivMf0VP6VSQqTkEdE+Px1XBesi1UleDRYKVlmEKUApQClAKUApQClAK4B+ur+PC/86fylCUdhcAvtE8OPybtv7q3V8oQxSgFKAUoBSgFKAUoBQ+FAal4h6st2hYFwvN1W8mCzJbaUY7K3nCp15LSAEIBUolS0jABPWqbY+OGnb/e4tqQxd4ct6a5bVGbbXWm2ZaW1O9nWsjaFltBWMEggjrlQB0rhmbaN4pJJIo3ETVeiJDV8vs7S19l3eE1AMq1PdphdtjOPqQw46yFBDqAoO9HEkgjBAyK2YeKFjQ1KU8qVHciT4FsfZejqStuRL5HJQR//MNAnyZOfCpcG0E0jGt3Fq13uBcJdstd8uLEOYuDuj21zDzyFuNuBsnAUlKmlAqyE5wM5OKql5+qNtrQlG2Wq5S4Y08b6zdBDcW0hOVjDrYAWAkoO7yjBGMioVO5LmizOcZLBFuTUF9Utza9FhyrgxDcMONJkJbLLS3PuVK5rXnA5iNxG4V62DjBp/UmpE2WILgh9yRMiMyJEJxuO+/FcUh9ttwjCikpUenQgHBJSoCHTdicyehM6y1pbtCwYcm4plO9slIhR2Ycdb7rrywopQEpBPXaevgPKQOtVO3cd9PX9h5EAXCM+Ykx5l24255llTsXpIayQMrbV0UkeZQBJSrBQurhySdj2tfGyyTbhZrZsmypk92PDMqLDX2VEp2KJQaKj708khzrnAIBOelekHjVZ7vps3232rUE625VsdZtTv2RKQStac4ylJSRnykd3d0qXTIzple1L9UPCtz+oGoFtmvRLfZIV5ZvRiOOxFNSQ8UrWlOFBADWcjqSVDA29ba5xbsDephZiZpInC1m4CIvsaZhSFCOXcY34IH3u47c7ulS6YzpnxojjBp/iBcGIdsFwbckw1T4q5sJxhEphC0ocW2VAbgha0JP+2kjIOap0/jdcY3Gm4aUT7Ft221OIXPS8lYcYgmIHVT1yN4aaQl1Qa5SklR99kA9Cp6tMOeiaN07cjxoU58tWrFY2+mm300sBt9NSFhTi7xuv3R/Yarpr113lFT2H3F6pW7NEKUApQEXqlO/TlxA8rC/2VoRCklG5agARnJrntrL2fedhsF+rU9xjW64KiX+0yG3BlTmCEnyEVszio5zoVnV5ws/qFVUH/0Ml3leKX/qtN9i+pQmTgdD4VRNccDtPa558pLKrVdXB/yuKMBSvHK0eB9JGD6a5+nVdOV0dPmcHmR9p0ajQ3Dhm1Je7Y4w6zukqQElRL6CcDrgdfCru6FBHWrlRucVLtf0LEZKVeT6peLKBq/T104iOLtlptAl3CIcofUsJAScZBJ8h9PlqG0fo256V1EI8q2MrvjQPceeBbjnr1G3IJI8vkzWzVLdUVLjf6mEsWqtWeHjo43v3IvN11QxFgMuvpcjjtLbUlTSgCyCVZyfMVIKfUfTVWnXhdxvq3LAhbstpQcaYZURlORlSvQQMdfHpVlSjRpORqcVvK2KjFPhzNhRbxCvTqWoay6+UFS2QQFNkYyFAnof8DUbqUdosaltL3JUjKTWDTazJo6GM1UTh2G3uDEvtWmI5JydtXu5lKbdIK/epbUTn0Cu7eiPMEruxoKeRsU2kDbjAqw8G7h2WfKhqOO9uAPprmdmytXkup2+2Kd8Gn0a+xuWldOcMKUApQClAKUApQClAKUApQClAKUApigFKAUoBSgFKAUoBSgFKAVwD9dX8eF/50/lKEo7C4BfaJ4cfk3bf3Vur5QhilAKUApQClAKUApQClAak4jaSY1tCk2yQ+5HbFwjS97QBVuYlNvpHXyEtAH0E1VpHCCDInOSTcJSFOagOofseElLpidm2A+IG3vZ8c1qW7No3K1SKfYvqXbXY7NfISL08XrnbolvMpmCwysdmdU42+van7K6VKyta87seSpy58EHLtfFT3tUTUNSZ9su06I3GZCJUuEpkoXkgqQhQYbyhJ8QCD4g1OdyMtlY9LtwQg3bRltsKLmsx4d3k3dPaYzchiQp5x9xTbrKhtcQDJUUg+CkNq8RUYx9T5b7Fpxq3Mahkxbc1p2Tp+Wp1loB2MvmKDmQAG1ILij0ASQMYAqM3IlxMxngxAurqpELUkh7S90lwbzIt7KGnGpb7CGOU4h4AkNrEdhSkjIVt6EBRzmPcJV2i3Q37LOU5drVLu9zgiVhLa5E1T69rhCThCVP+IBOE+Bpm5CxYLholy+W3SzV0ubkqdZZTE1yWhpLfanm2lIUopHRIUVlWB4eAqBHBK3iNFZ9kZJEd+8vpO1OSbi46twepBdIT6hmqb24EtXNfac4Kak0pxWbusNlD8ZqbGDc2WY7jCYTcJqMshJRz0ySG1AFJCAF+OCoG33v6nq3XnSGmdPqujpi2Rt5lIlRGZLbyXE7StTTiSjmJ+4Xg7cnoQSKrcuDRSo6WEn6n+K7Z5NrZvstmDM0vG0xKb5Lai43HS6GngfFKhz3MjwPd8MdZE8GGjqFUj2clmxKvIv5spab29tBCgebjfy+YA5s+++6292ocrk2M7SHCaFo+VpZ9ic/IVYLNJszQcSkc1DzsdwrVjwIMZIAHTvGqdq36mGBq2dMUvUc+FDeuD90bajx2Oc0+8AHUF1SSVsLIBUyoEKwATtAAKVncNXVjc0dpxuO0h10POpQAtzbt3Kx1OPJnzV6bfTVqxczDb6abfTTKMw2+ms+xJ/ztG6/dH9hquC9ZFE36r7i8UrbmlFKAUoCPvyC7ZZyE9SWVD9Vc26lhOzIXIbJG0p3bR4gEVpMe0qlJy4XOq2PFyo1lHjb6Mstg4TSnZNsltvb46QFY81XPiswWIFoT94Vp/UP8KzK9ONPDzUO1muwdadbG05VHqrL4FAYTuI8wHWsNu9GS+hpkp2uLIRj7wDx/bXGKyZ6E4ZrvoYGvUuo0bLeT1Wh9kpSfL9lR/GvSBf0XOLEkI/0UjuqSrxQrwx+kYrMqJKEV3/Qw6Ec85T6WX/yLZwkfQ3q+5M4G5TCVfrqv6/KbDxRmuK/0bu1RVjwykH+NbSrf0SEuljS0l/6nVh1T+hWbQ5AvV01DGkKYkMylISqIpQ3EAElWPHxI6+cVg3bTbfD+Qi/WguGErDU1h1RVsGMJcSfHxwCPSD5DWmr3ksr6J/I27pQazR1eq+YsuoIVyhiGl9VsuDyO0pn43A944CvPlI8D5DU3GjvM6TQ1IfRJcJcVzm04SsFaiCB5sGsSirTKqC9extDgBK5tgQ3n3uRWz780X7LNbT75TKkj14r0JrND3HnGkavc/qaGe27huIGfPX7aZJsV9jTUn7GTtX6vPXGYWru8RFvqei4ynvsNOn2fNao39BlJmxG3UkEKArIrtzzMVrvixo6w6rdthvNmgXZTCXA0ZsZDpbztzt3A4zgZ9QrCxjcaMmn5uZmESlWSfb4Gv8A3IdDfifYvm5n+mnuQ6G/E+xfNzP9Nc3van7mdFuoftRrrjNadJcOYen1xNKaQii43EQ3Zd1tiCywjlLXuIRg+KAPjqEtWqOHMbR1muV/4fWlu4z40iYY1vtbKkiM0vaZI5m07FDapI6qO7oDVxSqNXUn8TOhgYVKcZRSu37uf2JmZe+EcS7qhDQ8V+O29EZeuTVkZMVkyUIUwpS8ZAVvSPDIPiAME/nszwrcEnk8O0PqbuCrUwluxMf5XKStaVNsk4CsBsqJJCQCMnOQGar+75lPoGl7Lz5uYl2v/C5FjQ7bdAxpd1eamHsDdiZU9EVH7rpfT9yEqKQcEk56Zr90/cuGo0O3c7xoy0qnQ7LbbncOzWhjYTKGE8sH/WByD4DHjU5qlvafxKngEo8Fe/1se2q7jw8sls1VItvDONePYBDyXZTVnYTEL7YBW0XMZG3PeO3AwcZIxUZbb7oFV0kuXbh9BtNubiQXDHk2FhLrapDxbDxcCyC1nA96CME9fCmapb2n8SY4CLjfS/T4fclbzf8AhDZ3JDR0TBlvMSZUctx7TGyoRtvOcSVFIKElQT45KgQAcVNajh8LdO2Kx3Q6NtU9m9rbRARFtkcKeK2y4nq5tSnujPUjzDJ6VTmq/ufxLTwNsvqrUxpSuFUKdIgv6Jt7U9i4RraqKq0McwuPt8xtYHlRtCiT4jYrp0qt3LVHDz2AsV4tHCyNcoV0ubUBtQtkUFaVg95ASVd44wEK2nOc7alSqP8AyZVHAKVrpJfwTSLpwiXqV2ze022hxt56KJJtUflKkMtlbjQ+63AJUMlISSkgKJqFOuuEQiGSeG7oZEJu5BZ08zgxFHBkf7CSMEnqfuQodaZqr/y+ZK2e3yRttrhLoR9pDiNIWFSFpCkqFuZ6gjx97X17kOhvxPsXzcz/AE1Rvan7mYTpQ/ah7kOhvxPsXzcz/TWzuFemLRpezy49ntcO1sOP71tQ2EtJUraBkhIGTgAfFWdgpzlWSbZg4ynGNFtJF2pXRHPilAKUApQClAKUApQCuAfrq/jwv/On8pQlHYXAL7RPDj8m7b+6t1fKEMUoBSgFKAUoBSgFKAUoDSPG+7ybNpO7PxIc+a6qUyypNuedZdbQt9CVuFbSFuJQlJKlFCSraFYx4jniHq+9j2rW7VF21hb7e8zqRSU2tqaqW8hmXFENSjy+eUhDhCVLAKspCidxBwFFGzvoiSVeeIK4bAvb2o42ukwrMbTEgMPdgedU00Zhf5aSyfspfS5zD3EJSU7cgnOsV8vOouIbkOLdtTyZidV3KBNYw+i3s2lLbw7rgSGwpLnK2qCuYF4SDt6BlQ1InQwkaf4e8OLRc39Z2vT0azSGLmYDM7tTV1bTGS2yShBdS2E9o2hP2JSkgdegM1p69a3ZvWnBfTeLnd5thbbmRGm5MdFskCE4tbriUo7M+FubUk7gpDhCU5GQDimTqZPDzUN+tGqbEq8m/XZt2wxnJTXImtC1qRAbW4lbXL5L+9wKwpJ5ocXswQk7d+wZrdwhR5TSXEtvtpcSl5tTawFAEBSVAFJ69QQCPLVuUehVHVHvuFNwqjKVDcKbhTKBuFNwplA3Cm4UygbhTcKZQNwpuFMoG4VnWMg3WN/tH9hquEfWRRP2WXalbM1QpQClAebrXNSpJ8FDFaH1PbXLReZbCUjCVFSQfKk9R/hWm2pBSpKXRnSbDqZa8odV4Enw714qLdU2tRWphQ6JWPeH0eirVxbSHbNAdHgH/L6UmqoZ5YJ5+NiJqnHacd3wv8+ZqG+3A2+zOKR/pHSGk+snFYmnG985CUD3h5KSfMDk/wDz0Vya1mkd9bLRkyxastKLpp6VC5imd2FBxIyUlKgodPWBWpITVyhOFpNz2IU4p1P+TpPeSrB8vTyVnTqwjZSjf32NdhcPVqRlKnUy8OSfC5sPh9ab47rmQ1G1CIz3ZSS72JC8jPhtJrI4habuz+o5SJt+RNdRs+yKt6En3ifMfDwrcznBYRPLp0v3nOwpVZbSnHPrbjbsXIqsnR0m7R0NvXNra1nlqRCQhTfpSoEFPxV+3bT2o0aZnQGrgi+MOsKbWzITyniMfcrT771HHrrVSrUqi3co26O97fx2G2qYWtSbqxnd81a1/wCe0gdMTrVfrJLUy40xIkpS3GYX3VIR70bc+IAwMjPhWw7y0liCWW0hDbbexKR5ABgVgRpypSyy89xlYOpGreS/0XD6npWbaoeZSv21ud5sOtKT5wRXfR9lHm1X9SXe/E5r4kRJMR19qM4WnW3sDHmJ/wDWvi3plR7QpFxIU6hOQojGRjoa5CpRiqEpr2oysd5SxE5YiFO/quCfvNq8INTi82ZLKl71N5Tn1eFbFrrabbgm+Njg6yjGpJR4XYqp65GVw/Uv+FY2MV6L88zIwf60ff4FWxk4yMjyU2+qucynSZiA1NouHqudYpUtx5C7PN7cyhsp2rXy1owvIOU4WfDHgOtUWT9T/YLZChusXibbE2tMlLT7qYzqGYzrnNU0Q80pIQhXVJIyB5SKqWmiMqGKdNKNtP8Af3MdfA1Vx1pqC5Xa7SWbA/JgSW7fHdQluSIzKAC+CjKcLRnCCAR4+TExcOFtgj6VbUm+ybY1Furt/jXhLzQMd51S1EhSklBbw4oYUCCCPXU8eBXPF8EkrK3hZmKeAVmciQlwr3d4ssIlCTcY7zRdnolKC3uYSgp7xAIKAnHkryuX1OdmuEJiGi9XmHEFriWqQzHdaAlNxurKnMtk7gfHbgHzUuVLGyWtl51+RLvcGoDi9TMi8XRuz6gElcu1IcbDCXX0bXHUnZvBPiAVFIPXFeQ4I2qZbbjGn3O4XQ3CzN2V194tBXKQta0LTtQAFgudDjHdT0JySKPSnyS8/wCjFd+p/sqbZYI8W4To0uztvttzlIYfcfDy97pdS62tCiV97O0EGrDrHhzG1ppRrTr1xkRYbbYad5TMdZdRs24KVtqSk+UFISQR0xTiQ8U5NSa4O/Pv6ldPBxtPFmyahaw1bbTZRCbJeKnXJCdzba1JIwShpbnfJJyvw6V+J+p+tgiTcXq5puUm4xrp7ItIjtrbfYBDag2loN5wo7iUEqJyaXK/S7Wsr6JfO5nwOCltteoLhco1xloanuvyXoZZjKSXnklLiw4WuakEkq2hYGfJjpXkvgRZHLObcZs/lHTidNbt6N3Z0qyF+99/6fD0UuU+lvjbp15GwIaWGEJhNPJWuM2hKkbgVpGMJJA8M4P6DXlCvVuucuVEh3CJKlRFBEhhl5K1sq8y0g5SfXVOS5hOaM3b6quWiRi3v/73+ArNwStWRg413ossdK6I50UoBSgFKAUoBSgFKAVwD9dX8eF/50/lKEo7C4BfaJ4cfk3bf3Vur5QhilAKUApQClAKUApQClAas4g39OlrdcriGDKeS8lliOFbea844G20Z8gK1pBPkzmqDdtRSNH6rjTNUNW+5vM2eVIZk2uEtD8ZPOjJcZAU4reHFKax73Ja8PNqaknGTfJFyviZUXdWyxSb6u7tprytfnfsJmfxPjW4LD1ju+9jlCUhKWCYqnV7Gkrw71Us7ThG7AUkq25ryga1sNlua4UK0S48OZPlg3JptsRnZLaVuSFHv7+hbWCopwVAgE1TvOVh+IQvlcHfTpzdr8eCej+xJ6R13E1i+ppm33CAvsbE9sTm0I5jLpWEKAStRHVtQwrB6eUVExOKG6PIelWC4oSu7OWqA2wWXFy3Eb0qwA5gYU05kqwAADnxw3midg8enCM1B2d+l1bTrzdvqZ8fiPCk3eJBTbLkESpa4CJhS1yRIbQpbrZw5u7gbWCoJKcoIBNR0bjLZpFrXcF2+5xYnYm5zDkhttHaGluBtJT9k7veUkZXtGDuztyaOouhS9owXGL59OSTvx53VjLvfE+HYG1GTZ7qt1mAq5yWmUsrMaOFKBWtQd2n3qiAkqJAOAcGvJrX6mr3d43ZZN0InGLAhQUo5q0Nx2lvuErUlISlbm3qodcAZJqHU1tYSx6zWUXo1fhrdPRa9bLoebvGG0JgxpTVuusll2Azc1qbab+wR3VqSha8rHU7SdoyojwBwcTWmdUSNQ3q/wARVqfiRLbLMVqW4tspfIQgqwErJHVXTp4YyQcpEqpmaSRVTxyrTjGEXr17n29VYsmwU2Cr1jZXGwU2ClhcbBTYKWFxsFNgpYXGwVnWRAF1j+hX8KqitUUzfqsulK2BqxSgFKAVQ+Jml3bnHTOiJBksg9B90PNWPXpKvTdN8zLwtd4atGquXlmp7fqJq03iKVo2upcAU2tPhk9etba4gqTcNDokI8ELbX+vH8a1mEjNYadOa4X8De42pTljqVWm+NvE0Frp1TbdqSCdinlZA84AI/Yal9LJDEhG73zYAP8AtHqa5eOlQ7urrh7LtJ+ZJEiK6emMkVq25xlIgRnE53F5Y+USP8KuVNWi1hPUjJeeDNrcJ2+dr25r8eXCSM+kqFZfE1Bi6olKP3bKHP8As4/8NdBON8Gl3HK0pW2rP3+CK1ETiOgeXAqQidHU1zsn6x081oa00lYo6+HLsWQyl1MW4vISFDqnDxHQ+I6DyVOXq2XOzMuiFMM6KAf8lnKJWkf6rvj8Ss+ur1ConeE+F/h3fVGrp0pZnUpe0vg+x/R8u4uf1PerLcypNvmPi3znVHlMyu5zT/qKPRfqBJroF11DLZW4tKEJHVSjgCu5VlFHnk7yqPTW7NNcUNPIutxbkwn2nC4RkJWD4eWqdrib2a3raCgXNgR08/hWgxSpr8qDu5SudTgXVd6tRWUINI2TwR08LbYW3j75Yz1rZ9dCzkUKpHFHtgtSzbgk3AR3uzBfvebtGzPozisTFK9J+eZmYT9Ve/wOO9B6l0/Y5ei1WGzWO9a1kW98XRTzhGomp6ILzr3ak437VOtpbO843OJ29Amsxri9qM6XuUuy6za1Q8dNN3KTK7JHUm0XBTzSAxtbSnAUlbv2J0qcTyequtax08z9b/X+zaqdl6pN6g4i3jSK77YbrqufzY9+Zgw72pECKoIXb0ySh5x1rkISFbwk8sqPdT1OVVr3iHxVvWr+Ddxbvmp29PuTNAtT2YSI7P8AnuU8mQiSgBaSrCEtt91opKOduVlOBSFKN00vHz51JlN2cb+f5LxqvixPkcUJOnoV3fegvvXC2TLVLMUcrl2559LjbaEc9KSptIC3F7VBRwnG01Iatszt74B8LYiJ8m3ocl6dQ52dtle8FxgDIdbWk4OFDp4pGcjINORQylWbPmPWx8XJz2pbHpo3eOu7J1Je4M63pabDrUJhExcTekDuAobjqSrA3gZ6jNVmFxR1xZ9FWy4zdQquL140rbb09KfgMITai9JYbkPNpQgZQ208pwhzdgtZztJTTdR4NDeNrR+dDN1Bc1T9aWExOK06fa5mnru1HmNG3KZnyG34pDWOzlp1e1Swdic4bOMfZN0dF4q32DYoCGL+xZbpDtNkVZtLsxI6EXvnMtl3ago3lO5S2gGCgN8vJyKlU1JJNePnQpcmuEvDp9S5WnWGqjrq2vSL6uRa5ur5+nvYkxGUtIjtxZLyFhYTzC4FsJGd20pOCnPer41rqK9W/iDfoNmlM2h+VOsEJU5qEyt4NvrfS5kqSdxCQNu7ISfJgkGjJG/u+pXmlbz0MmNxRvuntBJMl2NeLyrVEnTce4XVaYrJCZDqG3pBbQEg7EAYQkBSykAJ3dKRwi4dXHUHFJniEwm228QL1OtUyBY3VdhcZTHdSuQFHq+4uQpvxACAkpAyFE1KKgpNcyltzcV0LVq7ixPt3Gu22K3Xd9LabvFtsy0yjES2427H5hcab2GQoAqR9l3hAUCjBwarieKGvLNpdm8m7rvcq56bvVybhOQGg3EeiPMIaU2G0haxsdUVpUpW4pG3b4UVKNldEupK7s+BZOE1+jHWfEm5M6nka4hswbYpN0hx23XXQESSUoEZAQ4QSQNif9U5IJOufqfIotnEm2W9VhkW46VjOWRVzh6fmMv3hchDDoenKW0A1gYWQtSsrUVghJGasukuWi8Cm98vP/Z13tq4aKGID/8AvP4CowatVRTjHeiyxUrfmgFKAUoBSgFKAUoBSgFcA/XV/Hhf+dP5ShKOwuAX2ieHH5N2391bq+UIYpQClAKUApQClAKUApQGs9a6ejapjTbfKW60hT6XUOsK2uNONuBaFpOCAUqSkjII6dQRVeHDiG+XHZ9wnXWc67GWuZKU2FlLDodbaCUIShKN6ckJSCrPUnpjXzpqUrsu1MNCrLNJvha3Lnr82Q164d3C7637Yl4RrM5Pi3GQ2Je4PuMISEEtcoEHKED/AEpThIO3d4Za+ENscD6F3K5uMKamssMF1ARFEokulvCM7sqVgqKiM48Ks7m7dzEWBU5SlUbV27WfK9+nXW3zZZ7fp2JbLrMuDO/nSmWY5SojahtoK2JSMdB31H46jLToGHaXYKhMmSUwZkmbGbfKMNreC947qQSBzHMEknvnJPTF1007Ga8PB5dXpd/Fp+KMFrhXb2pin/ZK5KQhUxcaOXUBuKqSVF1SMIyTlaiCoqxnA6dKzHOH8VBJhz5lvX2KPb0lkNLCWGuZhO1xtSe9zDuyOu1PhiqFRS4MsxwUIrST+XS3TpYgYPB9huZLYcnSmrH2WJbm7ey4kpkRmEkhDxKNwytbmQhQyDg9DgSk3hbAkqC2bjcoLxM0OPRnEBbqJTgcdQSUHAylASU4UAkdfGoVFWsWYbPgouLk+zsV7q2nm7PdHDW0twnYqVyEsuIgtFIWno3FUFNtjp70kHcPLuVjGalbBpxrTy7jyZUh9ubKcllp/YQ0tZKlBJCQcEn7onHgMCrippO6MunhoU5KUW9P5f1JelV2My4pSwuKUsLilLC4rNsv/Okf1n9lVRWqKZP1WXOlZhrxSgFKAV+LQHElKgCD5DQFTu/Da1XaamSplIcSc5ArL1TaUnRs6I2M7GSUj0gZH7KpkrxaLlN5ZxfRnOWr2O022E4P+iltn4lZT/4hWXYXedGUseJfWB6cEgfqFcA1abPXm70kS01XZ7W5noapmoI5btsNAHeCmyR8YJqt6tIt0X6r9/gbP4Fsc/UGoHvEJLLX6iakOOMbkzYj4A+zR1NZ9IUP6q6pxvhUu44WM7bTb7X4FNYT0A8wrLZGFprjpcTuZlTt7Qj6TvwGBsuckgD/AHqjUtqdzZEfP+qaqo6y95h4fjIt3BTT8G+6YVFucNifEdHeZkNhaT08cHy+mrlO0LeLDEdRpq69pglODZb0pTzOPM291cb9R3j0CvQbJxys8xlJqeZcbmn9QXadZ7lyZsF6wSHFbUtyyC0s+Zt0d1XqyD6BUE1En6mvrMZTLm0OAqKh41qY7PVOvGcPZWv+jfT2s6uFlTqe29Pd2nUulrWm1WZhkDBCQMVL1uDnhVU1wdpinxwlZx+isXEq9JmXhf1V55HPvDjji1qbQ8fVV7VZYkeYxDWzCskxy4TEOSB3GHGktBQcyQAlIPUK8ACamWuNdilak05aIUS5SjezKQHkW98dmdYWltxDydmUKClEK3Y2gAnoQa1DpO7SNyqt0rmPqPjPb2LLcXLUxIN0hPW/fDu1vkRFKZky0MB1KXEoKh/pMEeBSM+myaw4h2PRMiOxclS3ZTrLslMeBBelupZb2hx1SWkKKUJ3JBJx1IAyajdvgTvERr3GjR7UqchNwdfRCgouUmXHhPOx2o62ua2tTqUFPfRkpGcqOQAT0r9j8YbBMgT5LEW+vGBK7FJjosUwvMu8sO4U2GsgbFJVnGO8nrlQBbqRO8R4PccdFtTIjKbi++JbMOSmVHgPuMIalHbGccdSgobSs9AVEenGDiX1txDs+hRHanqkuTJTTzrEaHCelOFtoJ5jhS0lRS2negFRwBuSPEio3UrpDeKzZWtJ8b7HcdM6UkXZaotyudst0ySmLDeciwnJaE8pC3QkpaClq2p3qBwRnoQTMv8AF7SLFui3B25hEd9mW+2ox3NyURnEtP5Tt3JKXFJRtIyVKCQCTipdKV9CFUTRHyuLsKTdNNR7YlQTMva7TcWrlGdjPw8W+RLBKFhJBIabIJBBSs49H2xx30Y9b5U5VwlRokeGm4B6Vb5DKX4pWlAfZ3IHNRuWjvIzjek+CgTO6b4DeLmeN34s6Hu9kdjXRmVMYkThaHLPNsshb7khTKpCG1RlNbyFNoKwSnaceOa+NLcZeHxtKWbTI9i7REtz09rdbHocYRmSkPcrc2lJLalpStCeqVKwRnIpuptWI3kbme7xn0rHhqkvu3CO8mY3bzDdtUlMvnONlxpAY5fMO9KSUnGCQRnIIHhB42WK7an0xaLfGuUxF9izJDUtEB8IYVHfbZcbdBRlCgtakq3Y2FA3Y3JzG6kTvUTHFDWC+HPD686iZityTbmQ6UOqKG0J3AKWogE7UAlZAGSEkDqah+Dl9b1RbbpeHYUOJcbjK5634ilhNxYSkNMTEtud9CFoawnIwQ3kEg5pk9S9w5esomw8Vb9GD/IH/wDefwFX8KvzUY+Ld6TLDSt2aIUoBSgFKAUoBSgFKAVwD9dX8eF/50/lKEo7C4BfaJ4cfk3bf3Vur5QhilAKUApQClAKUApQClAfBYbUcltBJ86RX52dr8Gj5IqLIm7BjNEf6NHyRTs7X4NHyRSyF2Oztfg0fJFOztfg0fJFLIXY7O1+DR8kU7O1+DR8kUshdjs7X4NHyRTs7X4NHyRSyF2Oztfg0fJFOztfg0fJFLIXY7O1+DR8kU7O1+DR8kUshdjs7X4NHyRTs7X4NHyRSyF2Oztfg0fJFOztfg0fJFLIXY7O1+DR8kU7O1+DR8kUshdjs7X4NHyRX6GG0kEISCPKBSyF2fdKkgUoBSgFKAV5yWg/HcbOCFJIwfVQHNd7hcqPNirH+gcx1/1Fg/wqL0md1uZJ8StR/STXBVllqtd565Slnw6fd4EteXdzJR5FqCQPWQKgtStlcyC19ytwD9RNUrWRNNWjbsfgbQ+p6Z5lvu0npl2YofJTj+NTXHCFzrDbngMluUlBPmCv/UCu0a/It2HnClfHX/5PxNbMDrWSyMrFcJLiejTK6y3y9J39YwQqZJWMf7xVfmtHy3bnvvinFXMOr1Ldpg0XZTfebe4HQuz6caPnTWzq9CZ5jxMS52mHeYbsWbGalR3RtW08gLSoeYg+NUIcJFaakGXpKemCodfY2egvxF+hPXe3/wAJwPvTQhok2OI6LQ63E1Vb3NNPkhCZTiubBcPk2vgYTnzOBBq5tuIfbS42tLiFDKVJOQR5xQk+qq2tkcwxU58UrH7KxsT+kzKw2lVGipXAK0yOFGnNGByMtdjRC5MyTAQ62+7GSEhTzJOFpUNwKdw98cKBwayIPB5dsd0vIt862WiVaHJXObtNnTFjPtyCnmhDSXPsa+4jCypfUEkKz01Tnc22XoVqwfUyt2mPNTJv6JMiVGt0Zclm3hpx4RJQkB15RcUXXnD0WskZ8QkeFXfWfDy4XzUTF9sd+TYrmmA7bHlOwxKQ4wtaV5SkrTtcSpOUqyR3juSrpiXNSfAKNlZMqOouBqrJww1fYdMrclqudqh2yNEdUlKkNR2EsABaiEqUUAnvbQT0JAORAaU4P3bVNgeiS5TNvgw7u662xdGWp7k0qYDTpmobkLQt1CwQhRcVt2J3IOABUpq1+ZS462RarX9T5FteknbEm8vONuWuzWznKYAITb1ZSvG7xX5R5PTVm1rw8mak1DbL3ar4bJcYkOVbXHDFTIS5GfU0pYCSoBLgUwgpUdwHXKVA1Q53d7FeXSyKZZfqabbYbhZpDTlnuQiQLdCkqvNiblvLMNtLaHGHCsFhSkoTkYWAQCMHJOXc/qcbRdpOuVSLi+5G1IkBiKplCkW1RcDzpQDkLDr4S6oKGCQB4VO9fEpyK1jLg8CoSIdrZk+w0XslzcuDrdgsybc1JC4T8TYpAcUc7XyreVE90DAHhWdVcBL4/od+Ii/Kvt0t9mTY7KERW4qmWC9HWtbhUpSXHSIzXewhPc6I7xoqivqHG6si2W3g28nUUTUF2v3sjem7yi6vOswww04EQXobbKUb1FICX1LKipRKs+AIAj7l9TrbbzpyHZZl0fcisW6628qSykKUJr7bxX1JGWy0AAQQc9fNUbyzVkS43VmZ1j4JsWtFrUpdkhyIN1auZVYbGi3If5bLjSUuJDiiT9lJ3Z9AAr3s/CBzT16sNxgXrDttlXh11D0TeH2bhNEtxsYWNikqShIX16A5T16HO4UbE/xE0P7fbCzBRO9j5EabHnx3lMh5vmsuBxAcbJAWglPVOQfAgggGqxwo4Kq4W3q7T270J7dzZQ25BTE5TETY44tCIo3qLTILzp5RKsFZIUB3ahStHKS1eWY2dt9VW7RwxBf/AN5/AVewy/MRYxTvSZP0rcmlFKAUoBSgFKAUoBSgFcA/XV/Hhf8AnT+UoSjsLgF9onhx+Tdt/dW6vlCGKUApQClAKUApQClAKEZFAaI1Jom4zNQXJ9vXGrIiHJLi0x41y2NNAqJ2oTt6JHgBUd7Qbp/aDrL51/8AbXNzxtZSaTOgjg6LimPaDdP7QdZfOv8A7aiG7ey9qZ3TqOKeqlXxpkPrgezH2UN9O9jb4dR+mo9OrvmXY4ClK9k9D9sVua1O9cGbTxT1VcHLe8Y8pLF43clweKT3fHof0VLe0G6f2g6y+df/AG0eOrrmJYClB2adx7Qbp/aDrL51/wDbUbd7I5YpNsjzeJGsmXrlJ7JFT7JKVzHdil7eiDjuoUcnA6UWOrvmRHAUpOyRJe0G6f2g6y+df/bUbIsbkXUEOyOcSNZJucthySyx7JKO5tspCznZgYK09CfLRY6u+YWApS0SJFeg7mhJJ4g6ywBk/wCdf/bUfp2xO6sscO72viRrKVb5bYcZe9klI3J8+FIBHxinp1e3EegUmr2MmNpKTOdkNR+JWrJDsZfKfQ1eApTS8A7VAJ7pwQcHyEVizbG5b75bbO/xI1ki43FDrkZn2SUeYloJKzkIwMBSfEjOelPTa/UlbPpN2sSR0DdMfbB1l86/+2ntBuh//iDrL51/9tR6dW6lHoVEx2dJSpUuTFZ4lasdkxtvPZReAVtbhlO5ITlOR1GfGsg6BumPtg6y+df/AG09Or9SXgaK4oe0G6f2g6y+df8A209oN0/tB1l86/8Atp6dW6kehUR7Qbp/aDrL51/9tbE4SWCVY/ZTtOoLzfebytvsvK5/Kxuzs6DGc9fUKysNi6tWqoSehj4jC0qdJyjxNiUreGlFKAUoBSgFRV/1TZ9Lxw9drlGgIV0QH3AFLPmSnxUfQAaA5213e5NwvF3VabVJEV9CnUyLghUVO0p6qCFDeckHHdHrqJ0c6pMFtDpTvSTu2jAz08K43HQhCs7PW56Zs2dSrhU2rRSXe/4MyZK7VdIrCTlPNyR6gT+0Vj3hXMvkVPiG0OOH4k4/aRWBB3+JtZRy2XYbc+p6jcrRfNx1efcX+sVaOJ7CX9JP7gDtcbIz5DvFdzJWovu+h5bB3xcf+76mm204UayYwy6muBl7R6ZPgVeMsvcO5riRuLrjqgPWtRrH1y7vW0wDkrcAx8dZOBjmrJdqNXKWShWl2M6H4Ywex6ZjDGCUCrhXennApQHw+w3JZW062h1pYKVIWnIUD4gjzVTXeHBsjipGkbk5p5wkqMAp50Bw+lkkbM+dsp+OgPlPEGTp5Qa1fal2hI6eykUl+Ar0lYG5r/8AuJA/1jXtrKY1Ot0aTCfbkNLacW06yoLSroMEEdDVjEK9NmRh/wBRHHaZHESPoE3X22alduSdAN6k5K2Get1Sgq5W0M52nokteXGTk5Jtl91lqCdxvgWeA9d4UJy6G3TWlvKKRHVbluc5toR9qG+bs2vKeKitKk429BgOMXyNjeXUh+F2qpehdF8M25VwvLltTaJ0K5NPxXXltXJCIvKjlKW9wUNr4SnHU5GScVIaLvur5lhfv95uOo3zadCWu4i2QGW0uS5zsWQZBCVNnc9uQjCPAKIyk+FHGOr88Qm9F54FSVd73qOIh+63m8M2K06qgOtT4ch2Ry47kVQcUHlRm1LQl04zsISVkZIAxYI15u9mcjW2+36+2PTQnX8G7wow7Q/IbmpTFQtaWj75ovLHd+yFIyTjCpcVwSCb43Plq9cSJdoVeLlcrvb7tbbXp2Qq1sR0IZdkvOlMwOI2EnKRhSAQEZJ6EAjpYpz5qtTjHkXItviNvqpt9VWrIrG31U2+qlkBt9VNvqpZAbfVTb6qWQG31U2+qlkBt9VWzSAxCe/3n8BWVh0t4Y2J/TZPUramoFKAUoBSgFKAUoBSgFcA/XV/Hhf+dP5ShKOwuAX2ieHH5N2391bq+UIYpQClAKUApQClAKUApQGtLv8A86TP98v9prErjqi9ZnXwXqLuFc98RdE6ti8QtTaz01a1ybvCXbzbgcASm1MPMyEDJ+53tqOfvBSFkZ+GlCMmpvRq3xa+hH23htqPh1IfVbrZPnRYN7tEp7sShzZzTcMokLSCob8uryQfT5q/Vac1oYdgmuWm8Sbk1dpr3sTKWTH5bssKSpbrbyS2tCOqVd9OMjHWq209TOdSlN5m1f8Ag9pml9UK07q9HsLf3dYGYuQbkmYezS46ZzTjbbCebjPJTgDaMYUCRuwc1uwat1DqRi6LtFwgtnVwnx2p5SVR43scpsLIClBKd56gHoT5800Kc9Kzd1zt3WViFsGi9WuQlNuRNUWeUbJKjX6b2rnLmTFLRy3I6S7hRGHDlO3unaCCRWHddEa9uUDT0iJZJEFUCHPafYYdWy5LjmSwrlpKnFqYW82heBuOOo6Z6TdFxVKKlq15T7OSsWW1aW1VI4qOz5Yu8OP7Ic+GtuOpbRglgJTHcWZAQhI6hSOUVbxuBOelTY4ea4Ysuk4L7V3gRo9iaYYNvYLzkKYHlqWraJDQQop5ffVuTgEdOuV0QqlFW4cF9ez795YZ2iLtp66cQhD0xcZ7tzuseQiWJDymFxVBrmKCEPoU4pKwslvKcjpnb0qR4Z6R1TEveg37vCmJbti722tck5LDTi2+Qk5WsgFIwBuVgDGTjNQ2mvPQolUpuF21f/8Am3Q9dQaIv194iapjMN3MW+6wn0C6yVrZ9jnA0gM8haXSlxsrTu2lAKTkk9etNhxtWcQOHbOsVs3KddrlOLzVqjFTkR1DLHIShwJdbUhKnA4sLScAkEg9KK1iqEqeVN25Luv7uz4sl9R8P9TRr7xGlWazXKNdrxCguxZceWpbRQOX2tkKLqcuHCgnwOAcFINZlm4d3q6HTcGUb4bC7eZTsyO4HYHZ45iYSgASHFhougdCvxUrpg9TasRvaeW+jf8A/OmlutyQ4M6K1Lpq+6YmXFN2zIsUlq6qnS3Hk89L7fIBSpRCSG9wGAOmfTW9atzSb0MHEyjOd4+dX9BVr0L4zf8Ag/8AFWXg1+fH3+BqMYvyJeeZa6V05zIpQClAY8+4RbXFXJmSWYkdsZU8+sIQkeknoKqSuJrN2Jb0xap2pV+AkMI5MQH0vuYSR/sbqWA9gNXaiGbtfWrFGPjCsSMuY8ypDgz8lCfXUrYdA2HTb5kw7ehU9Xv50lSn5K/W6slXxZxQgoXGiAUTWpCR/p4zjR9YGR+01pe0TBHccSD1IBA9YFcdtRZazZ6fsJ7zBqPS/iSFjWZN5UtX3AVjPn6f40uRxd5C/vYjmPlJrXUdbd5t6+k33G9OB8fkcP7Z06qQVfpUal+JatulXvS4gf8AaFd1U0oy7meT0NcVD/uXiaaAyTWQ2oN5V5Egn9FcBLWR6dPgVWzlQ4e29Kk958IBSfOoj/GsW6INz1dDjDqAvOK2OzI5q67zRYuWXB1n7vidT6aiiJaGGx5Ejp8VSldscCKUApQH4pIWkpIBBGCDVGuuk7Tp2Up21wWoJlkqebYyltSh90EA7QTk5IAz5c4qxW9hl+h+ojTdk4zGQpydLct8qziJLnOC3blPwmm3AloO94gqcSeg7pyDgEA4ssvijaYL8diRGnMvuFrmMuNJC44ccLbRWndnKiMhKcqA6lIrTRqK2pYo7UjKF6mvdw19/Fa37n3HjH4pRcXBUu2y4qWriu3QwSgqmrQDv25UAnaUOZ3EABIOeuBkRuKFnm9k7O1NeEiELipSWe6wxlSd7iicAZQrwznGRkdaqVRF+O0qb0ad/wCftZ9zMNrjFZnU57BdkJDMeSVORNoSw8opbdPXokqBGD3uhISR1qQ1nrB3TN6sENiK9NMwyHXmIzXMdU001129QE99bfUnGMjxIpnTV0PxGM6TnTT0a0trq7afMxnuJttaIlocclW5caE402xGUp5xyUpQaCevXKUglOMjOc+Soy+8VJMaDqLs9llw3Ld2WO2/JQlYMh8tgNltCiSoB5s4Gc5Ph0zDqK2hbq7SSi3BN8dbaLSVuevsl005qCPqaE8+wy/GUw+uM6xJSErbcQcEHBI83gT41K7KuqzVzaU6qqwU1zGymypsXMw2U2UsMw2U2UsMw2U2UsMw2VadJJxCe/3n8BV+gvXMfEO9MnaVsjWClAKUApQClAKUApQCuAfrq/jwv/On8pQlHYXAL7RPDj8m7b+6t1fKEMUoBSgFKAUoBSgFKAUoDWl4wm5TFEgAOrJJ8nU1DWG+23VNojXWzz410tslJUzLiOhxpwAkEpUOh6gj4q5Scbyb7TrYSWVLsMp2Swy+hlbzaHlpUtDalAKUlONxA8SBuTk+TI89YB1PaRptN/NwY9hVR0ykzSr7GWiAQvPmIINUZGyvOiU25qq2DidpvVF/VZrbPckTw2+6gKiPttPIZcS26pp1SAhwJWtKSUKOCalQck7EOaVkWrbmm3PmqmxVcFNecl9qHHdffdbZYaQVuOuKCUoSBkqJPQADy0y3GY9ANwBBBBpt9VMozDb6qbfVSxGY/FthaSlQCkkYII6EV5xYbMGM3HjMtx2G07UNNJCUpA8gA6AUsTm5Hrt9VNvqpYjMNvqpt9VLDMNvqq1aGGO2f8H/AIqzMIvzo+eRh4x3oS88y10rozmxX4pQSkkkAAdSfJQFSncULI1JXDtipGorgg4VFs7RkFB8y1juI/4lCvAjXGo/LB0jDPk6TZmP1NIPy6mxHE8ZPBjTd3aze0zdQyj/APV3KY4pxB/1NpSlv/gCa9xoe9WgD2D1dObQkdIt3bTOa9W47Xcf8ZqBYG/aysw/zjpuNeWh4v2SWAvHn5L239AWa9GOK2nQ8hi4yXrBKUcBm9R1xMn0KWAhX/Co0sDE4oss3PTDM2O4h9tl1Kw42oKBSe6cEeuuc7fGSu/KhH3wQcZ86TXM7UhepHtPQP6fquOHnblcsHY/Yu4IBHRaCQfTnB/UBUbdVntUxWMBERQz6Sof4VqKcbVLG/lPeQz9UdEcJmeToS0p/wD1Cf417cTE7tKvY64cR/3hXa1f0pdzPLcP/cw714mm0DJr4vDnIsdxcBwsMLSk/wCsRgfrIrgJe0em1PZMCXGEe22OGg4Adb6Y8ie9/wCGsXSLAn8R8K6hvAGfjrb7IV6t+853aH9jLvXidTREBEZtI8gr1rsDiBSgFKAVV9cMGSw2ylxTRcbcQHEe+TkAZHpqzVV4Mu0leVjUzx0dHgW7QXtgt0aXH7K2i3draTKdDKkLSkt5ydwbAPTJBOKmBodprV79/ZlONOyNheYLLSwooRsGFqSVpBAGQCPDyZOcKeHlTy5k1fVXXFFx4WnLLlfs26cr/dkW/wAKWHWWEi5SSuNOlTWFOsMuhHPUpTiClSCFDcokE94dOuPGRuui4kmFqEypr6BdbcmBIkHYktMoQ4MpwMA/ZFq8MZPQCrSorgvPIojgYxTWZ/Lpl6dPmeSdI2vUsZ24x5TrsS6KgS0KbwElphSXGkJyM7FEZIPXvnwrxuenIuvJvsrb79LjJaZk2lwwg2UlPN2voBWglKt7QTuGMFFVyoNJ9j1+ZVPCRnFxjJq/Tvb+ob0XZLZfoDYmciSqUmdFgFaRuSxFEcISnxKEBYX6FKHXHSsW7WixWW6z37lflNJXOZvjkJ1TfcUkJZSrATvKCsN4znvJAFTHDSm1GKeuq8CPQ6cVZS0u38uHcWLSUSBDt0pu2zEz2zOlOPPBaVkPqfWpxBKegKVEox4jaAfCptRKU5OAB5T5Klwyeq+Rm04KEFFcjERdobk5qGiWwuW6yZLbCXAVqaBAKwM5KcqSM+HUVl4PmqZQceJc4jB81MHzVTYDB81MHzUsBg+amD5qWAwfNVo0oP8AI3v95/AVepL1ixX9gnKVnGuFKAUoBSgFKAUoBSgFcA/XV/Hhf+dP5ShKOwuAX2ieHH5N2391bq+UIYpQClAKUApQClAKUAoRkUBoDj0zeZ+kL9atPsLeut3eFsbcSSkR0POBt15SgCUhDZWrOD1A6HNagt+idVWl5ekVW2TarGrUsa5Nr07LfQwiE+092hgPJCFAB5tS1DpgSEY8hrntFdPjdnR2btboedv4fzntT6Mdv8TU0iNEe1FaW5DM2bzGWVXBBgFxbbgWEFhs/ZFHBARuUe7U97GcRP8A8PDDZmuG7+1JlswxFe9lxP7OkKUZHO6q37sjl7s/dUk4u1/OrJSavbzoj8e0pqJetn7xzL+VK1f2cNidIEcWpUIJXhoL2BHMJO/GQoDBGBVSsmiL9onTmnHoWnrxOfiaavrT0Yy5CXUvOzIJQgOBRcQS22tYSggnYrb3jmpTTsvPBkOL4+eRIaS0xqm+exlsuLmoUaf9t76ipC50FSrd7EKUkHmOqfQ0ZRxha87jjCfeiOOktd2fQxMJ7VL94naOvDUpcibIedE1t6OmFtClEId5antpSAVdScnrU3je2hFpWvr5sWGbw/vth1Jd5Nqkaneag6gsirah66Sn2jGcdjpnnatZDiCgulW7ITgkbetVTU+ntYa1la6gotN9ixLlpy/xnbet2dy1TA612LDzrvLWVpDhSGUpQErUglQ6BFxert5/kST4K/n+CY1hAvMmLNRa4WtEhenWEaTEdya2Y9y3v8wytygUnd2Y5k9zYCB90DNXnTeoLdZOI2q3Z12avlsuIn29DtweRFcixm476mUNFQbDbqm3kFRGe+euB0jTTh5sTZvqbE4RG4zNDRLtdlyDPvTjl1UxJWSqM28sraYAJ7vLaLaCB0yknymrpWLNes7GRF6IUqizKrilLMXFKWYuKtOh+pmf8H/irLwi/Oj55GJi/wBGXnmWqq9ftf2HTkgRZdwQuer3kCKlT8lfqaQCr48YrojnSL9ntXai6Wmxs2GKrwm31e53HnTHbOflrT6q/U8MmLqQvU11namX49nkL5MQH0MN4SR/t7vXUkcS2wbfFtkVuNDjsxI7YwhlhAQhI9AHQV71BIpQCvORGalsqaebQ80oYUhxIUCPSDQGu9c8K9ON6fuUu3wTZ5SWyvfa3Vxgojr3kIISr4wa5wltXCw64iJZmplrdSVtmajGcpUCCpAHm83mrS7QlBOGdX8TrNiwqyp1XSlZ24Pg9Pl3lul3eatDTk+ySE8s5D0FYkII8vTov/s1D3K/WydGmpjzG+ctCUBlzLbnlz3VYP6q1+49bPB3Xz+BtKWK03VX1Xrbo+5+WdOcOkbNG2sYx9gR0+IV+cRmyvSUzHikoP6FDNdJUV6cl2M4ig7V4PtXiaXQMqrE1KsptbUcdVyn0NgDxxnJ/YK89no2enVXaJ835HLvNjQkEpQpeceQ8pVY+gk44hE+U4NbnZOlW3eaDHr/ANPl3rxOoGB9hR6q+6684cUoBSgFVfXIfMQCKUiVynOUV+9C8DGfRnFUTSa1LtL2kcz6Onaitdl0zabJZpyJkWC8/fYV1szjYcnpYcWXe1LAQ4tySW+qFKBGTkeTxjMarnWeSqyy9WJXNtsSBMl3JElDgnvyW0uvstOgcoMtl1Si2lLfVOM7TjeyjQU3Kbi7vXVPi3a3S6s3bglZ24F27tZEhLjX20THoElWq3tLv6heSVRnJb80sNwmwhCXUkuobckhxW4KSkYAylKqqdziaxumiPYm5tavXcZdhZVbm2FScoly33ucZTqMJPIbUwnlunG0Kwkk9K6Xo/qy9S+j/wAeOql7rapcL8NSHm4a+eBZ3E3z26xI7bGqI5hX0tFEUSW7exao8dS20ADDTpeU2kEjcrLhQSBhJnW4V90lwN01JjxLgu9xXoNxuECChRkOlyShyY1sHVRw49kHzdaxam5e7heOrjfhw1V3bvu78LlSb1feQ90tuvTGlRTLuzVxkwI6FvxVrW3HkT7h9l5SvAiIygAK+5Sc9Nxzl3vTt3XfHrS23eZliRdbTGQqW49Jy3HLk998rWSSFq5bJVnGUpQPAAVKeHXsZVbVcL6ZWl4q32FpczA0+5fbzc9GxLijVMC4w4SbpNdxORHlyXAp4QlKT9gCUE4WXOvRDafFWK+hnVN60re5KEawXNVo94zkzDMZQ9dninux2CQU8nY4NrQCSFgDd4m/FUIy/wALaft4Nvw0vfVW10ZS3Jrn8ycetFw005Kasntpj6QJtMFR3zn5LUYIdcfcZbWVOoyFR2lctIKMKwElJI+PYvVMyG8lbmqmbFFiXe5xoqZMlMx9GWkw463d3O3na84EFW8BSEnypqnNRfrzyXfZH2svTovg31sTZ8Fc3bpWLNhaXs8e5PqlXFmGy3Jfc9846EALUfSVAn46lK5iok5Nx4GSnoKVRlJuKUyi4qzaW/5G6P8AX/gKu01aRZrO8CapWWYIpQClAKUApQClAKUArgH66v48L/zp/KUJR2FwC+0Tw4/Ju2/urdXyhDFKAUoBSgFKAUoBSgFKA0Dx91Dd9K6LvtwsDsdi8iUxHjOymuY2hTsptrcpORkALPlrWN0473i5s8PRZWY0aVNlQ/bEy8grMMOTEQlxh17qy92gBXX/AJMvz1oN2pNvtZ0WdxSS6IuTvG+2tWaDcE2m4SEym7q4iPHQlx49gUsOJSkHvKXsO0DzjOK84fGj2Xi2JFosSr5c7szJltRbbcYzrQjsKbS45zt4Rnc82kJ6Hco5wATVvddpc3l+CK9J4+N6SvN9t13U3KmrvUpi3x5MqPDQ1GYjRVrBcWUpJ3yAAOpJX4hIJF+ncSITXDu36vjMJVBnR2JLSbhJahBtDoSUl1bqglGNwBHU56AGplS4PqFUvddCkufVKQpGn2btbdN3C6xxaZV5lGPIj7Y7EZ5TL+FFeHDuQrbtyFDrkZqxTeM1sgcQ7RpV1hvfdHUsRnkTmFPFRjqfClRwvmpbKUKTuIHex0wQqjo25+UQqiZStCfVDPw+H9gmavtFwTNmWL2Sjy0chRua21stLShCFANqUuQztCtoIXk7cECduPFa5StSWOyGA/p67NX2LFuEN1bT6XYr0WS4hSFpyMFTJB8FAtkeBBNTpK71KVUukeuguNDWoZOkrQth1+ZdrQzcjNmOMxlOhYXnltbvsik8vKwj3gWg9c9IGT9UTHvml1yJfD25y7TN0+u/KYkOxFoetyRh8rSXME4UnDZB3BYzt6gRuteJLqacLlh478RntC6JtVxtkqQwmXMbS77HRkPzjEDa3HVx2lgoKkpSFKKwUpQFk9QKn+FGq3tSaMjrulxh3C9QkNt3J6HgtJcWyh9OFJ7ijyXmlEoynKjjp0qlw9S5Vm9exEROMy5mikaqOnnIFlltsP2+Xc7lFitSGXclDi1Kc+xZG07SCfsifLuCcHTHG2VrPVOlmLTYi7YbzZJV0XJXIb5rK2X2mlJGF7VJBWRkZzuSQcA1O6WupGfgrEDePqhXLzCdZsEQKu8C7Wdt+NAmxJokMS5Sm+UHErKEOHlOJUCRtyCDgg1Z4nG1d0lNWiBpebK1SHpjUm0GQygRxGLPMWXSraQe0MbMdTzBkJwrEujZcSFUvyIFrjQ7etURJdvlyWbBcIGnJEaMqM2XELmzpbLiV56jIbbSrBO3YSnr4zMbj3GmWOTcY9ikyVourNoagNSmDKLzjpbSl1srHIUCASleO6oHr1AOlcKoZGkuNftlvtutsrTNws6Zsyba0SZDzK0dsi7y82AhZJThpzavAztPQZGdt224XyIpxmx2lm4PPAbpEqUGWWMZwVYClqznwSPJ1Iq9QhkrRVyxiJZqEn55Gf7R7xf+9qTUkh1o+NuswMKPjzFYJdUP+MD0VYbDpa0aXjlm022NAQrqrkNhKlnzqPio+k5rdmhsSlKEilAKUApQETq1AXpi6D/+nX+yucp9qFwMd9KdzrBI6eOD/wDsrRbTdnBnW7Evu6iXYSUOXuidVAdPDHWoPU1vjyrU6mQw29gKXhxAODWqU7SWV6m9pUlNSUldWNt6G4csJ0tbpFpvF3sMhTKVFMSUVsk48Sy6Fo/QBWbqO2a5j2ObGMm0ajjKaUMutrhSB08cp3oUfiTXYP1o2Z5wnlkmuRpqNqpLCQbjb51vAHedLJdaHp3N7sD14r6RdWL5PVc2lpXZ7a2oNvg9HXCO8R6B4euuCxFCVNOSd1fivOh6LLFRqrI1aXR/Tqe0F1V0ttknLO4rddys+coUAK+9ApzxC9QH8a2Oylasvf4Gv2g16BNdq8TpxgYZR1z0r7rrjhxSgFKAVh3C1M3IoLqljZnG0gePxVDV1YlPK7oxDpeIR7935Q/wp7V4mPfvfKH+FW92i7vZD2rxPv3vlD/Ch0vEI9+98of4U3aG9kDpeIR7935Q/wAKe1eJ9+98of4U3aG9kPavE+/e+UP8Ke1eJ9+78of4U3cRvZD2rxPv3vlD/CntXiffvfKH+FN3Eb2Q9q8THv3vlD/Ch0vEI9+98of4U3aG9kPavE+/e+UP8Ke1eJ9+98of4U3aG9kPavE+/e+UP8Ke1eJ9+98of4U3cRvZD2rxPv3vlD/CntXiffvfKH+FN3Eb2Q9q8T7975Q/wrNgW5u3NKQ0pSgo575qpQS1RTKbkrMyqVWWxSgFKAUoBSgFKAUoBXAP11fx4X/nT+UoSjsLgF9onhx+Tdt/dW6vlCGKUApQClAKUApQClAKUBpfirEsky1XBrUTxYtRnMFSwSCHBJbLI6Anq6Gx8dQDPBnS0addpbMJbUi6XeNepK0uq70lhaXG8eZO9JWUjoVLWfujWgk3Fu3U6KMVJLuMaHwPsFvuyrhGlXZl1BmKjNouDgbhmUoqfLKc4SVKJUD1IPQYHSq9cNA6Lst7at6rjfYuomt89y5Qnnu1vJmPNsL3qbTjatbTIOAAjlpVlOCqim2S4JFpmcHrJInruMZ+42y6qluyxPhSil5KnW2m3UZIIKFJYaykg9UAjBGaltSaDgaps1ut8uRNQq3PtSYsxmQRIaebBSlzec7jgqB3A53HNUOd7FWTiQkLghpm32eXbW25a48q2y7U6XZS1rUxJdW69lRySorcUdx69ax7Zwr0w9rB+5xXLomVbLmiauIt11EUTOxpZDoSQA4SwpKSQSkHPgoKqrO3cjItCPsfAnQwhTbO1ImXdiDDNk7LIubjxtraw08ptvB3NLVhhzOcjDe3CQBU/beEFkgS2Zrrs+5XNuc3cFT50kuPOOttLabCjgDYlDi8JAAyonxJJObejIVNcj4g8G7HA9rjTb9wXAsAaMG3uSiWEuNJUlt0pxkqAUfKAemQcCv1ngzptiyR7UhmR2RiwOabQkvHcIawkKTn77uJ71Q5snIiR1Lw7tupmLQFvTIEq0KKoM2A+WnmctltQz1CkqScEKBHgfEAjw4d8KrDwrt8yBp5uRHgSnee5FfkLeQHSMLWncSUlfQqAOMjoB1qM145ScmtyHj8CLDDtkKBGn3pmPbX237WgXBZFt5aHEJSwDkBOx1aMKCjtIGe6nGVD4J6ct7NnZj9ubatkWXBQntizz48hYW626SSVgqSlWc5BHj1OZdRsjdoj2fqe9MtQXYy5V4fUtuA0iQu4uB5lMJxa4obUnGwIU4rw8fE5JJOWOB1haYiKjTLvDujDkl03ePOUmY8ZBQXw4vwUF8tvpju8tG3btFS6jZG7RlNcGdLxnIRjw3IyIbFtjsNNvK2obgvOPRh1yThbqsk9VeWvmPwbsTdyduEl+4XG4OSIb/apsouOARXFuR284HdSpxZ65J3dSemIzsnIiRhcNLLAmwJTLboehXOZdmSp0kCRJDodJHlBDy8DydPNW0NFDBmf8H8avYd5qsSxiVaiy0UrdmiFKAUoBSgFKAidXLCNMXM/wD6hY/SMVoSMS2+4PSCP0Vz21npFHY7BjeFR9xlreOw+A6VXL+vNtfz1Ku6Pj6VoKHrSXejqHaNOT7GdIaPjGJpu3skYKGEJ6+faKl3kcxpSfHIIrv1wPJ2c2a2cOnmZxLRdZjzEBxCfHl8wdfVjFVLiXJuF/eYi259FvtCEhTriUglRPXAHlPo+M+IrhK1PNiJU+37nfTqKNKFZ66L42QsYnacgC0OvGbFlNhyI6UFK0OgjAwCcZJHXPrq1aFa/wD3ir9QrZbOhu8Q49DX4yaq4CVRc2vE6VZA5SMeavuuqOQFKAUoBVE1xedU2+8NtWWfZ40QshRRPtrshzfk5IUmQ2MYx02+fr16Y2IrbiGdK5kUKSrTyN2K97Z+IHwxpr5jkf8Anae2fiAf/wA4018xyP8Aztav8Rf7TZfh8f3fIgbHxf1PqC3PTo2pNLtxmZD8Za5FkkNYW0vYv300dAry+keeshPFLVK9RmwJ1PpU3gRhL7J7CSN3KJICs9sx4g9M58vhU/iEv2l57KabV3p2GWjXusnIrklOpdJKjNkpW8LQ8UJI8QT23Ar9b15rN9srb1JpNxAQXCpNoeICQcFX/LfAeen4jL9pb/De1/AK11rRDqGlaj0mHVHCUGzvBROM9B23zdfVX4vX2skQ0S1al0kIqzhL5tDwQTnGArtuKfiMv2j8N7X8D6d1xrVh1DTmotKIdWoJShdnfCiT4ADtvicH9FR1v4s6kurt0bi6r0k4u1vGPMBsshPIWACQSZg8ih1HTyZzmn4hJ/4lS2XdXTfwM9evNZNsNuq1JpJLTidyHFWh4JUMgZB7b1GSB8Yr1b1lrl4thu/aWWXEb0BNlfO5PTvD/Leo6jr6afiMv2lL2av3P4Hn7e9Zl9bHtj0nzkKShTfsO9uSo+AI7bkE+QUd13rRggOaj0m2S5yQF2d4ZX97/wAt8fRT8Rl+0fhq6v4GBa+LGpb2m5Khar0k8m2vqjS1ewshIZcTjcCTMHTqOvh6aznde6yZZQ65qTSSGlpC0rVaHglScgZB7b4ZIGfSKfiMv2lT2XZ2bfwPVnWWuZS3UM37Szq2lbHEosr5KFeY4m9D6K9vbPxA+GNNfMcj/wA7UfiMv2lD2dFf5fIe2fiB8Maa+Y5H/na2Fo+Xcp1hZduz8SROKlBTkKOphsjPTCFOLI6f6x+KsrDYt155GraGLiMIqMMydyapWzNcKUApQClAKUApQClAK4B+ur+PC/8AOn8pQlHYXAL7RPDj8m7b+6t1fKEMUoBSgFKAUoBSgFKAUoDRHHOwz9S6VuUC2xzKlquUN1LSVAEpbmsuLOSQOiUKPxVq6PwwvUHVEO+ot88TndT3hcyRHmAPG2ORZfJQjcvaEl4xilPQBZCiB3jWkulddrN9Zu3uMDSGmdV6RgaZfa0tNet1kvzqyzGQxHnzoy7ctkPvNc8tlYeUkKUFjdt37R5cK2cN9b+xlqcftclmemLa23gZKFLQW76JDqCoK64Yyo46EdBk9Km8epFpWsWfR3DO8WLWdkvpgSGpr2ob8u5yVSioqgOvSVxUkbyNn+gUlAHdJzgEqpxA4Y3++3TiVdbYw8m8SbfGj2OSZO3A5REhLQ3gNrWO4Vd3rt7wAyKcyzX88fsVZXlt54FYn8NbsjQkgQbLen3hdROiWKRb4qYJcTFLexyMJWAypRySFpKXQHMeJOTrfQ2rJ67/ADva/KuM5V2anW62EtSbe8oWqKyUPBbzakp5yXQl0dUKSV46jdWpK/EpadtDLumg7rZZ3E5cDQzc6TfbvCmiQpLbrL0VTUNEgBHObU4tDjT6y2opCvOd20w9g4O6ivVpt0LUVmlGFEtupEx4jkhLaWXXpzK4CdjbqgCGgsoG5Qb2jBBCaKSS4+bBxb0t5uWDTekdTN6gfkX2w3GfqGRAhi1agMhtTNsKYKEPIcy6FA9oDq1BKVBfNT47Ttz/AKnrQ150st1+7Rrpb5irdHjTGZLTKWJElJJW9vQ+4p5wknLpA3ApySRgUyccrSKkndNm7NtNtY1i9cbabaWFxtptpYXG2m2lhcbas2ixgy/+D+NZOGX5qMbEu9JlnpW7NGKUApQClAKUBXOIMoRtLyRnq4UoA+PP7BWkkj7ISK5jazvOK7DuNgxtQk+36Hq+cNE+eoV6Gq73i1WxsZVIkJ3DzJByTWrwcc1aC7TdYuWTC1H2M6ehNBqK2keQeFe9d0uB5czSnEWCmPqSYhxsLZkoBKSOhGMEfqqt26BGtqT2dshRyStxRWryeUk9Og6eiuJxyyYiR6Ngoxq4alN8l4aHy/DjOzES+Wec2CE5Udoz5dvhnpXhpR3smv46if8ASI/Yf/WsnZ874iPnkWdpUY08DUUex/NHSrKgtlBHlAr7rrjz4UoBSgFUnWgzdkf7kftNa/HK9H3mwwP63uIDbTbXO5Toro0DE4E3KbKtKLxboE6FGlX59xl9SXE/5UtKo52kYJ6Z/wBUgeWsKJwK1CwmBi328T3tGewTtzLqebCmJQtIdB2lSspKW9yTkJHmAFXrm39Mhwu7fy389Ctak4e3TQ+kIrr9sVCkybpbkOplPRnYYDDT3fUlmOlCEZKRucSoq7ucEA1nWbh1frroqM9pWwIbh3HTE2wONXKSlhxpxT6ldo6NgLSvqU4SnxT4DrUl514OKk5aX+neT954ATb3PucuXarfKfenWVTLrykqWmOw00iUkEjIBCVDH3Qx6KxLzwHvyS0iJCZcs8e83R9u0xXY6AI8hKA0pCXmltp27VjbtBAWcEVFyzHGQ0Tei+1j1vX1P1ynW68cmBHenItNniWmRMkJdfjuR3CXsO7Rg7QkbgBux4DwrNuXCS/W+brRNq03ZXm7reGbg1MWiOp1cfCOY0hDrakpWlSVKBWCnKicZ60vcj0uEtHJ/Ps+zGiuBdwjSdFt6itkGbAs67ut2O8pDyEF95C2MJCUpOACeiQAcYA6Yy+AmjZds1XqZ2UvnW7T7jmnrMopIIjh5Ty/HxI3tN5HT7FjyVD1TKamIjUhOz7ve39GQuoOFV2t1v4gzFRWLVbOS7doMh1bciSie2+ZCVocQgOcolI7iyogqwOlfDfCrUV40jpS8xbaw7qqS9Iust6atsxmnJTja1odYcQoqAbSlIKCFJ5Yx41OhcWIp2Tb5/TWx6XjgbfQzqSNbrPb2G3tTt3pp6O4yhc2Lj/kxSttSU7Fd8BYKM+Ty16ucAJU6BBjv2lp2MxZbsy3GuUhl8x5chxKminY2hCQO8RtSAjOB4Zpcj0uCs1LX334eUXDhRwtlaA1G9JTCiwociw2+M+mMQObNa5nOWoDxJ3Dvnqc1tXbVuSuzX16qqzzJgp6VfdJj/MbP+0r9tbDARtVfd9jTY9p0l3kxSugOfFKAUoBSgFKAUoBSgFcA/XV/Hhf+dP5ShKOwuAX2ieHH5N2391bq+UIYpQClAKUApQClAKUApQGieOOp7hpTTsp+0uNR7jMusO1synkb0RlSpjUfnFPgdgdKgD0JAB6GtfXrW2peE+qJ1senT9fxl2dNzZTLZabkRlpltR3CtUVgZZ2v8w4aUtIZcxu6AaXIm37zeKTSR7jjTdWdRmW9GssjR7GlnL/ACpdtuS5KwG1K3Fk8lIcHdwASjxJOCNtedp+qDn3RhDB0ktq7THobFubVJeREfXIDqtq33I6ChTaWFqWEocGCjaVFWA3XaTvOwx5fFXVF61dpmHabOwi7MzLrbblaHrmURFOstNLSvnhkqUjatKknlZyvBAwSPOx8bn7pcbzfmiwxb3dPWOVGtl2m9naZkPyZzbqeYlCyVEtIT3UKKihIA60dJW086kZ7vz0JSwfVALv69L5ske3R7tJkwn5k+c41HbfYlqiqZZXyMOOKUhSkIc5JUMAd7ITP8J9a6g1Ro66XbUkO2QXYtyuUVsw5iloKI8t9nvlbaNuA0Bu67gNxCM7RTKnlRUp5mVWD9UW9Ng3YNWGJOuMOVao7DdvuanI0lM98stLDzjDZ6KSonCFJxghRz0koPGm8Xe7DTcHTENesGpE1mVCeuykQ2kRkxlKcTIDBUrcJkbaOUDlas425MulbmRvL8iKe+qRffs8q82/SpkWi2WKJf7o7IuAaejsvLkIW22gNqDriOzOHG5KVY6KGRnMg8XtSxX9TNz7JbZTrWpkWCzMxrgtPNWtttY5yiz3EpSpSyobz4pCTgFU7q3Mby/I+3OPEyLJiQ5tgi26S3dH7VcpUu5LTAiPNhhSEpkBg7lOokJUgOJaBKVpJCgAflrjLJtUyPHcic6LKvtyt7l0vEwMRo3JlJaQ0HG45SFL3nloc25DZBcKsZjdDeG4qVZyl24pTKLilMouKsujR/yv/g/jWRh1+YjGxDvSZZqVuTTClAKUApQClAUTitL2W+JHB6qWpZHqGP41q9KMpHnrkdqu9a3RHf7GjlwifVsPpykCvTh5HTN4mMbuvZ46lJB856VZ2cs1de8v7UlbBT93idDgYSAPJSu0POTWXFqMkS4LoxuUhY/RtqhtjKcVx21ElXZ6Fsl3wcff4hSNqQPKahVr7Fqa1v8Ahh0oJ9Yz/CrOAdsRAytoLPg6q7PA6Ztboft7Cgc5SKyq7c8zFKAUoBVL1mM3Vv8A3I/aawcYr0jOwf6vuOftG8cZF0uGrHrxGjos0aC/ebIuGCXZUBl11l1StysFWWm3BjaNslsHqCa+r19UEY1jlSYGkrwq4x3rSrsEwxmluRZ8nktPpIeIGSlxISohQUE7kpSSoap0NeJuN7pwLDpPijDuUu+tX2QjTsmLd021i33ZUdh1KzGjuhsKQ+4l4nnBWUkHvhO3u5Nd/wDxBw7RZ3XpVru97VEizrjMk2+IyyiPEjynWXHFJW/k7eUeiSpSgMhPUpTG5uS6tiG1ZxN1YnX13agrk2yy2i62KAlCmIq2ZiJktlt0qUVKdBKHjtwEgbTu6kCrZcuPlkss7UkS4224xH7HbJN3cbC4zy3o7CkpcKUtvKUlWVowlwIJCh5jiXRTSylKqWvc+WOPEQ3hdvmaW1DbFsT4lvlvS243LirlFCYyllLyiUuKcSnuBRST3wmvBP1QMWTObiwtIalnLkLnNQ1stRgmWqG6W5AQVPjaEkEgr2hX3OT0qNx2one9hISOOVkYe0orskxVt1KmGYNw5kdKCZO0MgtF0PHJUkEpbITuGTgEjA07xwfudmD83Sd0Zuci+zrLb7aw5GW5LXHdkJUQedsRtRHUVlakjIISVZSTG504k73WyRkx+OsG5P2+LbNN366T5LMt56Gw0wlyH2Z/kPpdK3kp3Jd7uEFW7xTuHWrHpHiPaNcy0s2jnSG1WuJde0lIDYbk7y0gnOd+Gyopx0BT16iolRaV7kqopaFr2nFNtWspcuNtNtMouNtNtMouNtXvSgxZGv8AaV+2s/BK1T3Gvx2tJd5L0reGjFKAUoBSgFKAUoBSgFcA/XV/Hhf+dP5ShKOwuAX2ieHH5N2391bq+UIYpQClAKUApQClAKUAofCgNOcVzptOm9QHVqWlaewoSkvJUoEbxtCQnvFe7btCe9u27euK15om1aBvDt8tkJGobfeVMMPTF3qZcY9zMZKlFlSH3lh3lBQWMIXtBKgoZUQdO82tjdrK0rk5D0JoKLfbXa4zKE3GFa3UNREzHiH4byiHOcnftfSpZUr7LuO4lXic17NcJdI+1962f5fItyHGloL96luqhrZJLZZcU6VMFGT/AKMp6dD0qHKRXliz4lcHdFSLVbWjHkMtw3Hn405i6yWZPNfI5rhkJdDi1r8CpSiSDjODUdbNEcPtRxblFjWtdvatD8WyFbcl2KppUIlyOGVocBTsMheFAhR3EHIpmk0RliZ7vCrRFjgxW3G5DEG2OLuQiu3WUWVLD5kqedbLpDpDxLm5YVg48wqT09prSjNhmrtu12y6kW5KcadlOOR3zI3LXy0LUUoDhWpRSgJBKiSM1DcmicqRVr7wC0rKtC7RBdkQXZUq1vSXJNwkyX3mIUkOttJWt0rR0K0pUkgp3Z64FTB4OaMdt8eI2xKaXCkPPCbHu0puaHXsc4rkpdDqivCQoKUc7U/epxOeViMkSId4D6Rlajfu8lKPYNFqt9qjWuPIdjxm24rr6trgQ4EPIJeQNjgIBb8u41Yrnws0nc5N17TGeDt5kNTXkN3B9rMhoJCH2UpcHKdAQjK29qjtGSaOUmFBEZbeGWjL9pu2RxCuTMJ/fLMGXcJbTspS1JWsykKc3PklKch7d5vAkVLXDhJpi6N8p+HK7OqU7NditXCS2w+646HVl1tLgS4CsA7VggdQAASDDkxliy4bRim0VRYrG0U2ilgNoptFLAbRVk0cMGX/AMH8av0F+YjHxH6bLLStsacUoBSgFKAUoDVXFOVzb02yD0aZAx6SSf8ACqagdB+yuJ2jLNiJHo+zY5cHDuPiQe76q++Giy3xNR19/HI/RVzZv6y95TtRXwU/d4nQ9K7I86NWcVpYdvMaOD/oWCT61H/BIqlNeFcbtN3rs9D2XHLg4n04nI9VV3UiuWYro8UPIP68ViYR2rwfajOxKvhqi/4vwOjtIP8AabDGVnPdH7Kmq708uFKAUoBVC4jQBdO0wzIeiCRDUz2iOoJca3BQ3JJBAUM5BIPUVh4r9MzcJ+p7jnzS2h+DusUwoWjptqYkxLU/DIsamWpEmE8zyVFzCdziPeKC/DelJyeoNl1nw30vGt8l+5XKdbRcWrNZW5LKgVNOx5hVCUgbFDfz305KgUnCcgDOdY5SvZ8TbKMWroten9DwdPyLtIDjs+VdJaJ0h6YEKUXkx2Y+5ISlITlMdBIA8ScYGAK4OBmnxarxAD0/k3S2TbS+eancGZTzrzhT3eigp5eCcgADIPiaFOxU4pn1qzQ2mLXDud2u82VEiyp9qlPuBWQl6K+yYwACScKcQ2FefJ6p8REMfU2acaj3OMq43Z6LNtk60JZW4yBHjy3UOOhBS0FKVuQMLcKz5yaqU2kQ4KTLTdeFtovM26SX3ZQcuM+3XF4IWkAOwnG3GQnu9EktJ3DygnBFflp4VWizSbS8w7LUu2OXFxkOOJIJmul17d3euFHu+GB45qnPpbz0Jy8/PUrML6m/Ttufs6o9xuzbFtRa0pjlxkpeVbyjs6lqLRX9wNyUqSk9TtBOa9LlwysNvukC3J1Jd7bdJl3mXu0FgtFcSQsOrlBoqZUkoWJDpKXd473dxgYr3jkU5EuZJaG0Rpmw6jfFpmypV3szL0KaZCsqW5KdTMcccO0BS1qIXlPdG4jAwAPvg/wyHDS0XplfK7TdLvLuCkMrUtDLS3CGGUFQB2oZS2nGMA7sdKplK6aZUopWtyLbd79AsK4CJ8kR1T5IiRgUqPMdKFLCegOO62o5OB0r3tdxjXq2RLhCeTIhymkvsupBAWhQBSoZ69QRVvLpcrvyMrb6qbfVUWRNxt9VNvqpZC42+qrxpYYsrX+0r9tZuDX5nuMDG/prvJelbk0opQClAKUApQClAKUArgH66v48L/zp/KUJR2FwC+0Tw4/Ju2/urdXyhDFKAUoBSgFKAUoBSgFKA0bxv0pcNX6dkx7QGXLnCusO6R2JKyht9cWW1I5SlYO0L5W3ODgkHyVrTWWg77xPuNxvV50vOtkVm1t26Lao8yG5Ofc7YzJW6dylR9iDHawhxRCwXAoAHB1l0mbfVpIwI/CHUb06DOe0/abffZWk5dnF2trTDPsTLJXyXdoORlCwk8rcArcB3SDUVZ+Bt0t9gbcb05PcUzJtarjp+e/bERbm1G525LSYyEIUQp1Cwp/BXymwrbipzLqRlZnscIbvBdtU2ZoqHfLAmbdn06MU9HLdv7SWeQsBZ5JKQ2/uCSdval7N2OvzrPhJqi5267QG9OwZ0G5XW4SdqUxHnmA5GYbYKDJSUIbKkuBZCS4AE7R1NMy6hp2PZjg9fZN4tiZFiZkrlWFu3XqbeOyyWCpMBTQMdXV9DnOISpOC0pBWr3x64C+D929rVhjSOHUK5sNaWXZE2d16KlFunbu/K98U7HehLiMuDYO7kkBmXUZX0KnabMY3G1mBfHYt1ukG7xWY0ltDS7oZKbQ1G7SpLh5vZUrLjpKRjeN+fEGf079T3eY+gLjapFsuCb25bIdtlLkSLemFPU3JbccdQWEIdWe64oLkd/7IoHJUTVTaViEm/mS+qOBd1ZuM8Wm1BvTDepxdGbHaUwk8xhVrZYK22pKFMApkJcVsWE5ypYIVjMlZ+E1x05qnQsm02SW77Hw48KVKvrkKS2xFQ48vYNgDjT6OaQksDlkFKVdEAinMmirKzF4ecJtQ6T1FoOUbGgS4NtjwbrNmKivx22m2nh/k6s89t4Ld29z7GpKlZ610JtFW52k9CqN0htFNoqixXcbRTaKWFxtFNopYXG0VY9IDBlf8P8avUV66LFd3psslK2ZqhSgFKAUoBSgNPcSDnU0geZKP+6KraB3RXC4/9efeelYH+0p9yPGSelffDhHM4lM467I6iayNnL85e8p2p/ZT93ijoih8K7I83NKa8k9q1NPOchCg2PiSB+2oNodB6K4fHPNWl3npmCjlwtNdiPpYyk1V9Wq2xWRnqXUD/tCrOEX50O9F6v8A29TufgdE8P1FWm4v+wP2VZK748tFKAUoBVM1mnfPAHUlkDHxmsTEq9My8LpUOVdGae1RedI8Jrdb7BPsV40fZ1JlXG6sBhCX/Y1cZMdAJ3OAvLQskDZhkdc7awNK6J1U/p6F2p29SpiLjp1ydbptvktBDzNxZckP8x+Q6HVBAVvW1hBCUnrgYw9F57TP1fnsMi36Z1vqnVmoG2mbtpZubZ7mkrW5K5Ue4IlMKhLLzrykvd0LVlpCEbFKR1AwnCQ5rHiJpjTetHfZGLZdRy5NwlW1hmTNMRnkttQgGoz7SygpbccJQogLfSog4CkrR8+ewXkz51VojU82L7H3SNqrU9yTHsXsVcGy6yxy2ltqlqkNh3YlwqStS0r3KOUbdxT02XxzsGqblOso0y5cAzd23tPXFUF4pMJmQptRnDr3VNJadAUOuXR8VLtdFWtmUi3WrXU/T9sumroV9ciqu7MW8Wy3Ou89cOPBWyHUobUFqQ5Ny8QjvKQpGQQCK/ER9c6dsxWq26lnMTbLfIVsitOKfkxlOSwuAl8lfdWGMAOLOU7SlSt3jU1F6EXkSCdFaqXcHbyv2d9kot406mKntzwaEXbDRPPK37FApMjeSD70nxGa8dA6WvZ4n6Wn3O06h9l4Ui7+zNynPOKhLLhUGCyFLKNpRtCeWkBKcJVg9KjRrTzoNbknqTQF4PEDUmpLfHurc46ksBirjynUNORAqK3MUWwoIWnlF4KKgeifRUDpyx66tr+pLhO9mLlf2Id25tpbYlxmbipS1GOETDJU0O6EcssoQpAUQdpzTRol3T0PrR2mNVPzonPhXFdrjavjTonaI0hgNR/Y1xDi0tvuuOIRzjghSh3lEhI3ViRdC62maPbmyxqEX62aO092FCZz6P8AOSFvmXuQFhLjmA0F7wchWD0JqW0U2bRkcObrIunG9M66ybizHevF7g214yXnGbittxxKWigObEJZaYd25QMny56r6W2irNSOqsXYS0G0U2irWVldxtFXbTA/zM1/tK/bWZhVafuMPFu9P3ktStsakUoBSgFKAUoBSgFKAVwD9dX8eF/50/lKEo7C4BfaJ4cfk3bf3Vur5QhilAKUApQClAKUApQClAae4uXeVZdK6gkQFKTcVhUaIW8bg+6sNNkZ6Z3rTjPStZxL3qCU03EhSZpXJ1C1Djw7pJHaI4joU6/zXGwe4otjugqylWM9/A0VXNnsi1i6tVVlCm3wXdrdd972a7EyWj8WrxdY8Rm12JM65mImU822HS0vc6422lKwnajeGlL3LIABHRWTieTxE3zo7AYaCJF4kW5tSnDnksNLU66fUttSfN1T56hTb1sVU8fOaUnGydvs/g2v9lYXxb1ILM7cDY7e2hNkF85a5S9zbZ3bG1AI6rc29AOicHOcYM6viHcZF+hMRYMM2+TdV2lBdfUH1LaQpTzgSE42pLa04z5Ac9cVCnJ8iiGPrTsnBJ6c3wbt8b2RhaY1xqO5221NLRbpFyvBlzYzi1KQyxDbcGFKwMqOHGUgDGQrJPTr4W/ijKvC4MqNB/yyXDhNMR1SVdnL8lTiiT3eqUNs792MlKsADNM70uQsbVyxUkrvp1sn8HdL4mdp7V8uBpfVupbqUy+TOfQzHivKU0QylDGxoq8At1tZ9BWepxmvh7iLfokm4W9y222TdGJcKEyiPKXyluvZWtKiU5Ty2wFdAcjrgZxU5mki56XVpwjZJtpvjzeZq3ZpbusRWo9TarkRNVRg7E7j8KzRUQnFtrEp3lFxSF4yBtf8fudnTy1Mx+IF9nPRLbGt0By6vT5cRTin1iOlpgDe773ccLUlGPP5R5IUpXt54luOKxMajTSd9OLtfM14Wv7iOY1pfJl0hLtrsVuE8m53CQJziljkMPBlvBA7qVAhYA8x+OT0XrK5TZ1lts6KIQmW9ucFS1uOOPqWje4lte3b9jKkpKSc46gAAZKTbKqWKrOcb+y2uul1Hj79F3mw8jzimR5xWTlN6MjzimR5xTKBkecUyPOKZQMjzirDpLqZX/D/ABq7SVpos1vYZYqVsTWilAKUApQClAah4mtbNSqP37KD+0fwqsIGR0rh9oK2IkekbPd8JT7jHlGpPg3G7Zre5ycAhlpLYPpJzWTsxXre4s7YlbBtdWjfWOlD4V1556aCvzvaLrNcByFPuEfKNYrRBGK4LFO9WXeepUFahBdiPtYwmqdq9zfJgsDqVOjp6utVYFXxEF2lrFyy4Sq+x+B0rodrladip/1R0+KrBXdnmQpQClAKp+sMieCMZDI8fWaxsQrwMrDO0zk/hT9ULdXdNSr9qWc9dLZC0pFvs8PWsQHmZT3vWo4VsDzK8LAdxsykfZDk4t6vqlICbfk22OJ5n9hSVXRlMFR7MuRuEogJ942tO3Gd4xjb3qw5UrvQz1U01JGNx1hT9KXK6S7DJZRFsttu7sN1aSVNzVOpQ2cj3yQ33gfvseSoeRxiVw6tOqnZjMKRBtd5kw4cZ2bHgBqMxGYWGWUqxvV3lbUgetSRioVPkS6i4mVN4+vWa5asmXCztjS1qt9tmRJaH0pfeXLJS2hQUQlO5RSMkgJwSSQels0XxWt+q9M368PMphM2N1xuYtl4SGFBDKHitpxIAcTsWB4AhQUCOlQ6el0SqiehTrV9UzBu1qdlR7C/KfJt3Zo0OYy9ze2SAw2hSwQht1KlJK0E4AUCFHrj1vHGqeGkiPa3oV7gyrjBlWlx1pxlb7ELtKcu4zsKFNrBTtPewQOtTurcyN5dEQ/xr1VbnrFd5Wnlyojuj3r9cLZDktJS1sW0S6la+pJQpW1GfFWCRjNWU/VC2V3XKNPxYplNibGtzr6ZCA8l59ptxBSx79TYDrYUvptJPQhKiDpX4DedSD0JxgniyPyrtLE5+NZ2pyxMejw4+Vz5TAWp1QG3o0nOcjCRtBUSDLaT4uOcQNWaNVblCLbZrV6ZmRUONvtrfiPMNBSHUjvIypZSRjIWMjPQHT4hT4IyZXFiVbNQ3K0RbZL1BcHdSKssSOXGY6Gim2NTCd5/6MJKiScqyTgEACo+1ceZV0vlvkGyJiaVc02/fJkl94doillzY4nakkKCSCnp4+IOOlN1fmHUIKyccNIh6bfoukbbDvkkwkicy9GAdEtakoTIlJGG1JLffSd2Mo2leRVt0jxsk6znachwtLv8y7xnpy3e2tFqNGakBhTm7/pMkhSQkEKBznFHTfNhVFyRWdLcer69BkTH7E5d7JB0fA1IuWlxtE17miQXCW0jYThggJSB1SepCgBtjSmtImsp97at7Zdh2yQ3GE4KBbkLUy28dmPEJS6gE+fI8hqJU7cCYzuWPb6BVz0yMWhr/aV+2ruHVpljEu8CVpWyNWKUApQClAKUApQClAK4B+ur+PC/86fylCUdhcAvtE8OPybtv7q3V8oQxSgFKAUoBSgFKAUoBSgNYa3XaocW4Tr0tlu3QHDNcdf960Wlbwv1pKQR6QKpuntTaauepFtm0y7LdFsu3Vpy6RFRy+3htp15JJ6EDlBQVtUApOQM1ixwsq0ZVFbT4u3TrbizPcacmnJa6fLgZCGNDMX3T8Rly3puTkT/ADahhz/SMMqAABScKSlSxtCs9Scdc1mMaa0euTcrkyxbFPJLyZclK0nllY+yhRz3M5JUOniSfGrMsLKKV4eb28UW/RsM/wDFaGRIh6XNvMl5dtEKTHaa5y3UhtxlpRLYBzgpSpZx5Mq9NfidOaXteomZvZ4Ea9SXFusrUoJdWtScLUgE+JB64HXxNUug/wBvy6cfgVuhQbTcVpqvHxZgyNJ6KFuEN5i2ph20LUW1OgJjpWdqge93UEp2lJ7vdxjpWbcLFpabHablN28tXBxpbIK0p5y0JCWy2QRkhIAG3yeuno7/AG/Ls+xQsLhkmlFeVbw0PKBH027ZFWEQ4sG2vPSIbdufCEJfIWrmctAPUFW49OvoFSFu0VZbSG+x26OwW5Ha0lKevO5ZbLhPlUUEjJ85pOioNXXcV7ii3F5V6qsuy3A+xpC0i4PThBZ7U8+iStzB7zqE7Urx4bgOmfV5q9omnLdAktyI8Rpp9vn7XEp6jnOBx35SwFH0iqMqKlQpRd1Ht+d/Ew3NC2J1uMg21jbGjLhtAAjaysYUjoeoI89e8PSdqgXNVwjwmm5hSUB0A5SDjISPBOdozjGcDNRkj0KVhqKaajr9uBK8v0inL9Iqsybjl+kU5fpFBccv0inL9IoLjl+kVYtJJ2iV/wAP8au0vaRZrewyw0rONcKUApQClAKUBq/iwxtukNz79oj9B/8AWqW2O58VcXtNWxD88j0PZjvgoe/xMKarCVeqrhwBhb4Vzmkd56SQD6E9P41mbKj67fYY23HbDRj2/Rm4a/Fe9PqrqDhTnu55Tc5bShhTbq0kenca+WetcDiVapJPqeqUmnRi10R6OHCapUxPslrWDHHUIOcenNZWzVfERNftKWXBVH7vmdU2KP2e1R0YxhIrPrtDzsUoBSgFVPVYBuCUnytAfrNWKyvEyKHtmj7To/hvfG4Gm4zTzyIFkf06yy6H2kvwkKbQtAcISHShTSMLSSUkkgjJrx13wPfv8exph3aZcRbpTslbV4ucpK3dzJbTtfZUFoCfHaOij1V1ANYcZ5tUZFOpCvHNTfivPlnrL4I2d/Tsd/Vlxu15uLFsYi3OVDffb9kUsKU42XGW1EuFClrKc7lHd13VM33gfpbUEuVLeRPjSZa5SpDsOa6wp1ElDSH2yUkHYsR2egxgoGCMnM5nfQu2Tdj8u3CXSjrxRIYuHLuMKPaFsMPvctaWNy2HFbD3HG8Epdykg46521Z7RpeLarNItr0qbd2JG7nKushUhSwpISU97oE4HvQAOpPiSTS9SVa7tyNUjQmnNb6ZRpbT9+vZjNSbZPaE+TLwiHGlocHZXFYOCGylLiST707iAKsB0hobTVzh2l5q53C6NuPXBxZblTHXVykLYW6+tCVDvJSpA3EABHTAT0jeJxuvAxFiaMqaqpuz04O/wtf5cNSYg8HtOwrQi3ETpTKbO9YguVLW452Nwgqb3E56BKQD4gAVlQOGNntN+N0t79ygLcLS5EWNNcRHkuNtpaQtxsHBVsQhJ8NwQndnFVcTMsiMd4GaScg9mRGlM7Wo7bbzUpaXGSxKXKZWlWeikvOKVny9AcjpUlpvhZp/SsuBKhNylyoa5rjb0mU48tS5S0OPqUVElRUpCT18PJUttoZUZDfDmxNagF5THc7eLou77y6rHaVQxDKsZxjkgDHhnr41HReDumYTlsUzHkoRAiSYAZ7Qstvx31bnGnU5wtO7qM+FRqLIresOBDdy0tAtFqus95iHNYkJi3a5SVILLSHEJZQ6hQca/wBICVpJUQkJUSDVj4ecOV6VZgzLtcHbtfo8eRDTKU44pLcZx/mpYBWoqWEAIQFrJUQjJxnAlu6sRl1PawcKNP6Wl22Ta25LCoFpbsyGTJWWnozW/lIdSSQvaXHMEjPfPj0x+cHOHTHCfhvZdMNLQ85DaJfdbCtq3lqK1lO4lW3cohIJJCQkeSoeqJStwLoVAirjpzraWvWr9tXaKtIsYj2CTpWca8UoBSgFKAUoBSgFKAVwD9dX8eF/50/lKEo7C4BfaJ4cfk3bf3Vur5QhilAKUApQClAKUApQClAaa4x6an6t05Pg20MuTET40tLElZQ3IDEpt5TKlAHaFhspzg43VRdVaGvuvn5d0vdlQ2lMA2yHaolxTz20uOtuvPqcUjZu3MMAN4KcIVkndtrIw1anSim5Wkm7aXWttfgnp3GbJOWnKxgXDhhqycyHHGrV7OHST1sZusfayIlwcUpSnEpCAQCrlkLSBghZ2jIFecbhFLt9ptkiLp9LrrVwjKn2mZNZ/wAsiR2XksI+xtpaHLedDgG3rsyTnAGb6bSUcsJNLtTbXTnyevX5lOR9CAXw3vjmpX7ajT1quUhi2TZAS87thW6TcpLiu4C39kDaGAFYAJ3ZAG84sSODt2RcXbY5Dhy4RnWt5nUD7wMliJDRHIYQjblKy6y4chW37OpWc903Z42lGyzPhfh2qSXHnzZCg3yPSLwouNks+lpKLFbrvcG50m6XqEt5LfaZbwcUlwrUkhwNrdVgK8OhHVIFfth4U3uw3fSKmYkMOW7mGbKDqFxVNOvLkOMNMqRuQUOlAbWkp6JG7wCasSxtOad5PW+n/lbn0a7rWehUoNcF508+8/dH8Mb/AGa/6SlvQ4zT0SKEXiUt9D7UjdzX1pbbKN6HEyniQtKkgp6ncdqRumtdjqsK81KDv/t/SxcgnFWYpWuyly4pTKLilMouKUyi4pTKLirBpT/6r/h/jVymvWRaqu8GWClZZgClAKUApQClAa/4tRt0SA+PuXFI/SM/wrXbYyn1CuQ2rG1e/Yd7siV8Gl0bIu7ucuO6fMk1tXgfB7LoeIsjCndyz8Z/9KzNlL2n3GNt52p049/0Nh0IyK6M4s0lruAIOqZhSMBwJc+PGP4VDMjIriMerV5d56VgZZsJTfYJKtjaj5hVE0TZ7zfdbOKj3SO062rAdXD3jp/q7x+2szZKTqt25Gt2y5LCpJ6NnRjOndbNsoSnV1tAA6D2EPT/APz19+wGt/xvtvzGfp66s4YewGt/xvtvzGfp6ewGt/xvtvzGfp6Cw9gNb/jfbfmM/T09gNb/AI3235jP09BYewGt/wAb7b8xn6eoS62/UtuuC3rheIl4CI5KIzFv7Opauu0bi6oDr+2rVWziXaV1LQ1lE4XPwNBw27hIu90uybd2XlRVRwYandqny2O4FZWnBK1KJTkA9TmPuOjL5cLU3BmaYYdLlmEeEzDU01Et01bjpdeUjflJALKgUbyNqwk9euodFpWtfQ1c8JUhFRUXLRJ3tx5W4acL9iXFks5w9nSEXKW/GW/dJ13jN9pddBU1AaUwlZT3sJ5iGCSB1JWM+HSOtukry5qCPcJ1gmIlxjcX5dyZks86W4slthtr7J70NuEp37QnYB0Ocy6UrrTzcqlhqiknlb5vXnmvfj0uv4Puy6MvES3Qo71pkRocaPPyq3lliW6taWmWXFAObOdyy8oqzjKQehO2rDY7Tf7Dwvlx7fbYsK/JDvZ2WmmmisbsIWpIUWw6UAEjdt3eJAziY05LlyLlCjWp3ajqou3DjZW56Nta8u0hOFuk7zpbUi44tsyHp0NOcmRNWwuU57zltPFK1YQ3lzlhPTCuoTtyqbt1nuKeJ16uMmBdW2H3GWo0liQyIimG2QRzEczeTzVvDG3yg+U1MabUUrc/P8lVClUhSpwcXpLsvazV3rz/AMu25WUacup0R2d7RciRqOby2LpPdVFdU4okrdfSFO4WEqSOWFYxuT0wCKyY/Dp9zVkZh+DcmbJDTDFsU0phQjIaSFqClqWVoWXAoLKASoFI3Y8KN03bQx1hpzy3ptcE+GvG749bPs48TBTwtmvQmZsu0KlXFVnnyJTDshKu0Tn3EuNsL72FJQQvAPcBIIOetes7QV5l3JmFJtL1wLMm2tR7y5IRtjwmktF8JyvfvWtLwUAnvJWMk4ApuZJcPNil4OpFWUb3tfXstfjyevezc2B6KYHorMynTZhgeimB6KZRmGB6KYHoplGYYHoq36dH+am/Wr9tXqSsyxWd4klSsowhSgFKAUoBSgFKAUoBXAP11fx4X/nT+UoSjsLgF9onhx+Tdt/dW6vlCGKUApQClAKUApQClAKUBVplhmPS3lpbSUqWojKh4Zry9rs78Gn5QrHdNsy1UjYe12d+DT8oU9r07H+jT8oVG7ZO9iPa7O/Bp+UKe12d+DT8oU3bG8iPa7O/Bp+UKe12d+DT8oU3bG9iPa7O/Bp+UKe12d+DT8oU3bG9iPa7O/Bp+UKe12d+DT8oU3bG9iPa7O/Bp+UKe12d+DT8oU3bG9iPa7O/Bp+UKe12d+DT8oU3bG9iPa7O/Bp+UKe12d+DT8oU3bG9iPa7O/Bp+UKe12d+DT8oU3bG9iPa7O/Bp+UKl7Bbn4HP5yQnftxgg+GaqjBp3KJzUo2RL0q+YwpQClAKUApQFX4kRO1aWfUBlTKkuD0YIz+rNakaGQR6K5ba8fXi+w7XYkr4eUej+iIHUy9lvf8A9k1vvhzFEPR1sbAxhhPT1jP8aytlL1ZMs7ff6S7H9CzUrfnIGpuKKAnUQ9MdJ/7SqqjA7oritor8+R6Ls7+zgYV8kiNb3l58Emsn6n+1mTMdmKGdyiQT662GyI6ykazbsrUacOrbOiMdKV0pxopQClAKw5lpjTnQ46kqWBt6EjpUNKSsSm46o8Pa7B+8V8o0OnYJ+4V8o1Ru4lzeSHtdg494r5RodOwT9wr5Rpu4jeSB07BI94r5Rp7XYP3ivlGm7iN5Ie1yD+DV8o0OnYJ+4V8o03cRvJD2uwce8V8o09rsH7xXyjTdxG8kPa7B+8V8o09rsH7xXyjTdxG8kPa7B+8V8o09rsH7xXyjTdxG8kPa7B+8V8o09rsH7xXyjTdxG8kPa7B+8V8o09rsH7xXyjTdxG8kPa7B+8V8o1nRYrcNkNNAhAzgE5qVFR4FMpuSsz1pVZQKUApQClAKUApQClAK4B+ur+PC/wDOn8pQlHYXAL7RPDj8m7b+6t1fKEMUoBSgFKAUoBSgFKAUoBSgFKAUoBSgFKAUoBSgFKAUoBSgFKAUoBSgFKAUoBSgI/UMXttjnMeJWytI9eK0YwdyAfOK53a60g+863Yb9WpHu+pXNWnEYIHitaU/rrpPTkcRbJDb+8aSnHqAFXtlL8pst7ed6lNdn1JOlbw5Y1bxVZxeY6/Hcxj9Cj/jVRZSAj0+auM2kv8AqJeeR6Fs13wcPf4squvpfKthaSe84doFbb4H2XsGn23CnBUM1t9kxtSlLtNJt2V6lOHRP5mz6VvDmBSgFKAUoBSgFKAUoBSgFKAUoBSgFKAUoBSgFKAUoBSgFKAUoBSgFKAVwD9dX8eF/wCdP5ShKOwuAX2ieHH5N2391bq+UIYpQClAKUApQClAKUApQClAKUApQClAKUApQClAKUApQClAKUApQClAKUApQClAfLqQttQ8cgjFaFdY7PKea/BrUnB9BxWh2sr04vtOn2G/WqLsRWtQMmTdrVH8eZJQnFdNQkbIrafMKu7MX5PxLe3HevFdi+p70rcnOGteKaAq4Q/92R+uqWAAgVxu0f7iXnkegbM/tIe/xZRNTE3TUkGEnrhWSK6d0dbhbrFGbAxhI6Vv9nRy4aPbc5rbMs2La6JInKVszSClAKUApQClAKUApQClAKUApQClAKUApQClAKUApQClAKUApQClAKUArgH66v48L/zp/KUJR2FwC+0Tw4/Ju2/urdXyhDFKAUoBSgFKAUoBSgFKAUoBSgFKAUoBSgFKAUoBSgFKAUoBSgFKAUoBSgFKAUoARkVou6YN4mYxjnuf941o9q/pR7zpNifqT7iCSwJGsrCg9QJKTg10cwnaygeiruzF+R72W9t/3K7l9T7pW3OfNbcT1brpGT5Qzn9ZqkPHY0T5hXGbQ1xEj0HZq/6Wn55lX0RBN+1+pwjcltQArqOM0GY7aB9yMYrqcLHLQguw47aEs+LqPt8ND1pWUa8UoBSgFKAUoBSgFKAUoBSgFKAUoBSgFKAUoBSgFKAUoBSgFKAUoBSgFcA/XV/Hhf8AnT+UoSjsLgF9onhx+Tdt/dW6vlCGKUApQClAKUApQClAKUApQClAKUApQClAKUApQClAKUApQClAKUApQClAKUApQHw+4GWVuEjCElRz6K0I6/zpjiicqJJOfOa0O1X6sUdTsON3Ul3fUj3FiNqG0yfwclH6ziui2SFMoI81Xtlu9G3aWNuK1eL/AOK8WfdK3BzprDiO4HL6Eg52NJHx9TVHvDvJhOHyhJri8driJHomzlbD012HvwEtQkzZExQyVLPU+uugMdK7GCywiuxHA1pZ6spdW/EUqssilAKUApQClAKUApQClAKUApQClAKUApQClAKUApQClAKUApQClAKUArgH66v48L/zp/KUJR2FwC+0Tw4/Ju2/urdXyhDFKAUoBSgFKAUoBSgFKAUoBSgFKAUoBSgFKAUoBSgFKAUoBSgFKAUoBSgFKAUoCN1M+I2nri5kDbHXg+nFaFbfy6o5zXPbV/xOx2DG8KnuMOS8ZV3tcZHVb0ptIHxg/wAK6Tjf8nb9VZGzI2ot9v2MLbrXpEY9F9WetD4VuTmzTmqJon3uU6CCCsgeodB+yqnqI7oLyQepBFcNiJZ6zfaemYSGSnCPRIuvASIlqybsd7y1tqu4WqPNZKzaFKkpFKAUoBSgFKAUoBSgFKAUoBSgFKAUoBSgFKAUoBSgFKAUoBSgFKAUoBXAP11fx4X/AJ0/lKEo7C4BfaJ4cfk3bf3Vur5QhilAKUApQClAKUApQClAKUApQClAKUApQClAKUApQClAKUApQClAKUApQClAKUBr3ijqNKIotbCwVEhT2PR4J/ifUK1aSEJ9JrldpVM9VRXI9C2NQdLDZnxk7nlp5SZPEewx/fEKU6R5sCul2wA2kDyCtxs+NqCOa21LNi5Lol4H1Xy4Mtqx5jWyNEaEmulqY8hRyULUnr5wcVCXqThCyod3FcJKP5jXaer4dJxjLsLrwGuYkRHmgegWrH6a3HXb0/Yjfojy/EW307dX4ilXCwKUApQClAKUApQClAKUApQClAKUApQClAKUApQClAKUApQClAKUApQCuAfrq/jwv/On8pQlHYXAL7RPDj8m7b+6t1fKEMUoBSgFKAUoBSgFKAUoBSgFKAUoBSgFKAUoBSgFKAUoBSgFKAUoBSgFKAUoBVT1nrVqxMrjsLCpZGCfvP8A1rHr1VRpuRm4PDPFVlTXv7jTcqcu4yStaypSyTg+J9Px1iOuZQpYIIA/RXGTblJyZ6lTgoRUVyJnghpt6936VqR8ENpBjxkn9Z/R+2uggMADzV2VCGSmo9EeXY6rvsROfaxX4oZSfVWSYJz7dZLcqfLcbPdMhzHyzUZKQHkFJAPoNcNiFatLvZ6rhNaMH2LwJnhNNbsF+cjKIS0+dzefIfKK34lQWkEHINddham9oxZ5/tTDvD4qceT1XvP2lZZqhSgFKAUoBSgFKAUoBSgFKAUoBSgFKAUoBSgFKAUoBSgFKAUoBSgFKAVwD9dX8eF/50/lKEo7C4BfaJ4cfk3bf3Vur5QhilAKUApQClAKUApQClAKUApQClAKUApQClAKUApQClAKUApQClAKUApQClACMitH8RrVLt16kyXY7r8ValOBxCSQnPkOK1uPozrU1k4o3WycVTwtZupwatfoUk3OKHgjmpb2pzlQPUnyf+npr3tljuurpZj2yOpuGoBLkpxJShPXrjz9PJWkpYWpUmlNNLmddidqUaVJypyTb4HQWj9OMaZskaCwO4yjaCRgqPlPxmp2urirI85k7sV+KGUkDzVUUnN2poUjTGqJsV5JMWS8p2O55txyU/Ea8XiQAvr08Qa4/HU93Wfaem7LrRrYaFuWh5LBbcStBIUO8kjx+L01trQXEFue21BmrCZIGErPgv8A9ay9n18jyPgzD21hPSKW9j7UfDmbCSoLSCCCDX7XSnnwpQClAKUApQClAKUApQClAKUApQClAKUApQClAKUApQClAKUApQClAK4B+ur+PC/86fylCUdhcAvtE8OPybtv7q3V8oQxSgFKAUoBSgFKAUoBSgFKAUoBSgFKAUoBSgFKAUoBSgFKAUoBSgFKAUoBSgFfDzCH04UAahq44GEuyR3BgpSR5igGvdi3tRwAkDA8BjAFU5EVZjJx0pVZSKUBTeIulG71alvNtgyWSFoJ8461pRVxRIldkWC26kHclXQjHSuf2nRlJqquCR2GwsTCMXRk9b3R9KUEqabcTgDIQv01iSrkzb29wWErTnAB8FDr+utKm21lOulKKTvwNz8KdYK1JaW0LUVrbG3crxOPLV/rt4XyrNxseTVcu8lk4XdhSqy0KUApQClAKUApQClAKUApQClAKUApQClAKUApQClAKUApQClAKUArgH66v48L/wA6fylCUdhcAvtE8OPybtv7q3V8oQxSgFKAUoBSgFKAUoBSgFKAUoBSgFKAUoBSgFKAUoBSgFKAUoBSgFKAUoBSgFKAUoBSgFKA+XW0vNlCuoIxiteau4UQ7xvfjjlSeuFI6HNUyipJxfAqjKUJKUXZo1fL4O6kemBPOdLaDlJx+uvWLwJus2SO2vOLbzkhWasU8NQptOMdUZVXG4isnGc9Gbn0ToljSUNLTXjjrVprJMOwpQClAKUApQClAKUApQClAKUApQClAKUApQClAKUApQClAKUApQClAK4B+ur+PC/86fylCUdhcAvtE8OPybtv7q3V8oQxSgFKAUoBSgFKAUoBSgFKAUoBSgFKAUoBSgFKAUoBSgFKAUoBSgFKAUoBSgFKAUoBSgFKAUoBSgFKAUoBSgFQGqNWN6aXHSpth1TwUcOzGY5GMfhFDPj5PNQEWzxBekym4zVqadkuAKQyi6RitQIyCAF5II6+qvlPEVa3nWU21kvNBRcbF1i7kBPviRv6Ywc58MUFjJl60nQGyuTY+zISkKKnrjHQACcA5K/AnpXlK1/Ig21NwkWltiArATKducZLRz0GFFeOtAZZ1PdghtR046EuEBBM5jCiRkY73XpX2vUV6bQVK0w+lKRklU1gADz++oDCh67lXFsriWdEpAUGypm5RlgKPgMhfifNWd7P3z8V5H98Y/qoB7P3z8V5H98Y/qp7P3z8V5H98Y/qoB7P3z8V5H98Y/qr4TqS8OLWhOmnlLRjckTWMpz4ZG7pQB7Ul4jtLcd00822gFSlrmsAJA8STu6Cvv2fvn4ryP74x/VQHwNS3hTqmhpp4uIAUpAmsZAOcEjd4HB/QaxrfriZd1SEwbKmaqOstPJj3KM4Wl/eqws4PoNAZKdS3hbq206aeU4gAqQJrGU58Mjd5cV9+z98/FeR/fGP6qAez98/FeR/fGP6qkbRPmzg72y1uW0oxtDjyHN/jn3pOMdPHz0BI0oBSgFKAUoBSgFKAUoBXAP11fx4X/nT+UoSjsLgF9onhx+Tdt/dW6vlCGKUApQClAKUApQClAKUApQClAKUApQClAKUApQClAKUApQClAKUApQClAKUApQClAKUApQClAKUApQClAKUArXPFfQrOuL7oVh6yxrnDj3kSp777CF8phqO8tAyrrhT4jjA8euaA1I5wbv7dyhXx20xXo0nWL1zetTFraRLjxYpdVBKZIVuSkiHCGxIHRzYcAqqN0dwsu1w4Uo07cLJOduVwZgwbqJ1jjxMNvyWvZFQfSd7g5RkZyeoI89Co/IugdVW3Udua1Npi43+yWa5xLOlTLSJSp1sgx5rkR9SSrqVuy4wXn7uOvyYNZN14d3u2uGXG0xcrRIkS592tNth26LcoEdb6W2uyyI5IQ2tSWEOlxDiEhUh9PMwSVAXzXel9XRtB6H09piALRIhQS06u14U1b3ezpiIDRcycNmSt1JOTiN1z4HJvOmNWL4R8S7LHduU2bdJEqBZjOe5z7DDrTcfm7iclIWXngCeiVYGAAAIK7cfqeXLZqOySm3XHtzqHJ7mn4iLYhEeJHlchpIQoq5i3pQO8qJw2NpRjJrseRxjYdsriol+duVstymJSHF7mZjqLQ4UnIUlv7LMKOpStQKeriAQig4luf4fa3GoYMIal1O5bY8yDBXNE4JLzSWHn5klWB15ilojgADYUgoCSAoVO0O8YhP00exXwTIDSEPNTnith7/NzrwDigpKFbpCm46ioOKBb37m9wyGjJF2Nrp6NH5aNfPwZtve5ocfbZkOXZDbXKUMEKjMLU4/uGUt7o6egQr7LjTdOcTUTNRpUm7s3S7IcXb51pUlpo3JpliI09JVnuxwGC+EHurDzgKVKCU0JIuLqXVvETUq4tln3S6vqS45d2BIactimHbmyiOGsFSdnYm5feHvyoFW5QITLPvcRHNMQ3EQdau3O4MtG6CQ8ENpmNsuKcbZbbWlxptbjiUBSXWm/sCTuIKi4ILdomx6pg8OtYXO/W+53XUc2BEhhDLqI82U21b2UOKQo9EOF9ctSc46keFUjTultTaZdb7FZLurTClRbcu5W61N2y+PxGmJKgyttpSQEB4x0h4IaUOY4MBPfoD8vth4sqluwor98iMS0rajz0IQ7LL7UeKwyp91pbadoWmU9lR5aysgjolJn7tbeKEFZu8aVfHpEr2S7Ww2tC0xoxuUduPyGDlPORCMh5PQkrTtOQQkCdDDFr4kSdZW+Jb5Oqbfp4OxpMD2RIkOpSZThkdocDqcp5KGwlp4rUEvdAVjCLvwKTrCRc9RTNTpuyWXwyuObmS1hxSnVLbEcqWGy2ktIK218pzoUjIUSIZt6lCBSgFKAUoBSgFKAUoBXAP11fx4X/nT+UoSjsLgGR7hPDjr/wDo3bf3Vur5kecUIYyPOKZHnFAMjzimR5xQDI84pkecUAyPOKZHnFAMjzimR5xQDI84pkecUAyPOKZHnFAMjzimR5xQDI84pkecUAyPOKZHnFAMjzimR5xQDI84pkecUAyPOKZHnFAMjzimR5xQDI84pkecUAyPOKZHnFAMjzimR5xQDI84pkecUAyPOKZHnFAMjzimR5xQDI84pkecUAyPOKZHnFAMjzimR5xQDI84pkecUAyPOKZHnFAMjzimR5xQDI84pkecUAyPOKZHnFAMjzimR5xQDI84pkecUAyPOKZHnFAMjzimR5xQDI84pkecUAyPOKZHnFAMjzimR5xQDI84pkecUAyPOKZHnFAMjzimR5xQGNCtsO2pdEOKxFDqy44GWwjes+KjjxJ89ZOR5xQDI84pkecUAyPOKZHnFAMjzimR5xQDI84pkecUAyPOKZHnFAMjzimR5xQDI84pkecUAyPOKZHnFAMjzimR5xQDI84pkecUAyPOK4B+ur9fcv8Azp/KUJRwhZ/+Z4P+4R/3RWZQqYpQClAKUApQClAKUApQClAKUApQClAKUApQClAKUApQClAKUApQClAKUApQClAKUApQClAKUApQClAKUApQClAKUApQClAKUApQClAKUApQClAKUApQClAKUApQClAKUApQCq3rH/6P/j/8NAf/2Q==)
- Multiple feedback loops
- Diurnal pattern

Impacts of chronic stress
- Major depressive disorder (MDD) & Post-traumatic Stress Disorder (PTSD)
- Hippocampus and PFC volume reductions
- Synapse loss
- Reduced dendritic density
![[[@Schmaal2016-gb]](http://dx.doi.org/10.1038/mp.2015.69)](data:image/webp;base64,UklGRjoqAABXRUJQVlA4IC4qAADwkgCdASqHAfIAPjEWiUMiISET+v1sIAMEs7d8L93/3+hEqpN+Uawp2X7Q8Ce0E4N/wv9Q/JT50f4D/AexDzAP1V6TfmA/hX9L/ZD3zP6r6gP8B6gH/S9ID+vf//3MPQA/Xb0yv3R+E791v2W9oP//6z144/on4zd+P83/FX92vWP8M+P/n/4v/1j/mf534Gcw/RF/Efjp7jfxX6xfQ/x6/vP/y/2Xwh/bP7D+xvm/7zv378g/yv+wL8T/jH9L/p367f1j9m/qT+c7TPYPMF9Jvjv9d/vX7Q/2H9wvXs/Tf6B+vHub+R/0D+z/jT9AP8N/kP9W/rn67f1//3/Rn9m/0P8t8oT7R/Zv8r+YH+7+wH+L/zH+y/3D/Lf5r+0f//7Rv3P/Q/3b/O/9n/J///3rflv9p/zn97/yP/O/yH///7n6B/xr+Y/4X+1f4//d/4D///9T7gfXR+0//a9y/9Zf+3+f5k0qLmkk3nQCpHNMBzvIlqYzir0xe0B9Cla6Aoa3BuEF2KDpZ+iLg3LnaHQ8BxrAZkWC7r+LOMe8AV045qpWiUO6CInTtGpDiQwjf08xHVCVP31x7NkoHp5XjnsMWB2CBlo2B1IOPPOyQ+t1IVn0RT6m6yc508yo7J4nNTFvItUvSQg/JAmklcRiaoff0mjJUq/AFrplnuhJMTTTPxF+d9qzp2u2by6yQeMkZ7l0l1hjQPsnb7MWvpzUxbyOLhswYMqrvn22ROiWf+8yyY4XlHqg59LvLV1OMbxNe4qkorYFc3LJGjDwu2C1hhFSJMMGw02IX9qRZMYrRTURLaSjrAj45qYc+JHFT8OH2DCxxOlgdrlu3DtQr2pVO+9Bp11pgb7yEg5NtSgqBB+glS45fwlE03qox+lDX0pJEoVcYUUSzTUM0dUY+e67uG2zRjD6VkLmpCjDuqqMAzpfg6ZSWaTp5VNzYdmG/ZAh7Mq2hjofnFRzMUuEgEPoAqOwMVTHlJyiTFPWuIoRQJzIxE9/nvHOQId/6gmmtxujk4P4DsHQWvpzUxbyHO4y7f/MQMUxsBfoZdR8YIt23Iz4FhoJs3ULa02V/CSYp61jjYfJA9XzKL25r1wIP2j+gXT+RnhVe2vjzyDSHjinmHwjPYOI1P09a46S6qwn1D8/GJUWCydSN9CC0D0d7XEoo5fY8PLxP9d8O6DL0f3YSioow9a46TDBpLNe10WG5N4n9uYpmAp0BtOJSPrqFJZI5JjB9gQzTdOC2ZYmOLAly7DfuU1mMv9X8G9gi7nDKov3wosdb3v7L+PLX0Y7WoIgBEWWQ0v4yfdjlSjkBUBOcJ269FOqDIsnQaXFMjkTmLZkPmf/Cci4yaGzCCvwJ2PAroViz40VREYONHWb7EtQ7FhtAI1qn/awKuv0zFLVjrApJCu9eSzbtzYFEjrJ48a10jE7r1v6191LxKpU1PwalPp5l0BXeF6u/FcJk5sBGR2l6v86B4QtfTmpK8TWfGobSoqtKmS3Wh3O0eKAod7qaqxFXot3DVb5zNt/oAhQuHvcTbZoc8inrXHSYYNiIIKebeJv0yGorx1oMo6WWc3kmKeAAAD+/8C7aGY1hBW8LqikdrGE18yKrJxothnaGh+RreJxUNYThoTMq8GsXW2/70ei538k1s5aGraKD5zuRerQfJU3fAlUEVT7OwLA4mDxQIOfhSaSt4REKOexOw1UxzT+G2FEWO9fl1ATKAchg/DxHJSj1CpGPLrkdvx/BcsEb5x6bXyMAFAAyDDV3qpL85H0PJ+yQKOFsuc8iomOClc+fN5lZtJ6y/BrH36xEKhLH6s96KMyDLvYbEVHObbkle4bHW5BNCrDQh8p9U/JXkgrJBD/drVyxQQuGqFbjvdpoNR0PuUpPH0/ag/c1u09A6kLE+qJRY8r6XksDpegP/GXQA6pyZBlOUrrXT++Z84f3MsDpXILdDKDa1n2YYGk1+QvqKF6ZyZnoNcADf/FlRqHh2Kb7pd90RwA7gwgz5sIwWUSsGmqy3hu76mBI/Wi0OzvbhtteeyOEbuXUCdy5Yd0GphLagnGIU4mnHANshaGvE41V3rpbLuVECzCrOtwFTDN78vjMbpSPhW8SRC2OPtdyndo4OGqZ3nMWyWkEB/VVJIiys3cgdWsqDHtYeiZ+rBbGEv0ilPdQQobRkx7hiVTzILJKXuMp31YRLshX9STF2+Jim3A5L4kFSorXkgHC7DwUiCpAETPNIj2BWNqz8KurPSsfyQnTNrR5P5/YgD7tcyLDUZ+0rlzOCBykELXRt/s8Ozy7bUGHJszvbWsmgUhPZXd/xgA9MqaQyaJCNdPMkOlPtmxro40MQLc5cNOW4WC4duS26a/WaHJB+fzAcfWvUEHi/F1eINzQdaZtOHGJ1CAIVtlFsxVaniDL4QHsMOIgcGMKg8pzWegeEW8KYaxcyyykfKElDFNiHt+zWa3nS99zn7mfm4jRDXOG3zoQZgzIqB5a/ubrVYmYBF34ibGV2OmTnMjH/+IH2IPXAURIc/510F/44hjZNUnrFsrWNYutbBftaLvkTecAqhOc1BnLJIjx57HscfE3omqmONJHMY96bbM2lxTTdPSg8ZI+NVwC+mZJJhYQ1guSqe2NtjPW38r/Ts94JwjHgewGAlRcIeiXfJSEJ6iU1lNB27Z7hTDHxXoEOIx2JcvbhBd6Apx0DtDPZlPiyyJyPMifBLZLN9hZ+pvq6Vew9g/bi//jQpidt+WxIu2+atLd/cAdjlJ3L04RoWOuCVJTkdjSzXNquqlWXLpa+xAYxGPipQKvh03xVvRf5t6CqAkgBfBJJgGw9Hx3ltEDl1eDDjItYZ8xlBKHfDc/DV0oK8Ytx4GWB0AbxSYaYt1mbuUFwokKVaK0yyejiYjqLR4AxSc+bk8IuI9Zdgv6VpZPl+8buxCL5MrJ9xdBzM4Krt+MYEDutA1ZjFTva7q+2FdmYg68zt4zwk9mn8Oqbvxpt17QfIev0jlHBSlfKfZZ8aOistfntG+bF66+U40X4uZCQBMwLicpX/cl+KdsYeZkFbhfNY2FJ4IQJ+LQv3GC2tiXbUDKIinGhieN1KfBUg7Hch3rEqfhw0UFe205LJaVpVOo+d/q4TBas7GBFB0kbMsu/KHDFJjEHL85YS/PSVXIZQ6+osCi7Px0LQTZ+achb1QNMJ2seRBCebKfG1L7ospnZQdcY4NY2f1ExSNIF/EeHH/Atf+QeHZrbrbbk8ONfVeQIUv7cEJ2RRuiKUP7Yz2hf1oRZTSKN0RQbxr9JPe2AusZUKHURqs4E/kJG/MxQ2w/aeaUUfa0d7Ilqf5M6CTVBoJAOjzVjDA8RhHsTPIkLVRPp1T8Vk1W3LgDofj0DVejkSmW2PSiHitAHw4JUstHsO4FKBsRqfqgHSsKXfcJawee9omkSfPY326H4xWyQgw5g8Jrk8rznVsCx4Uc/4OK/aiwpkYPBBxWEaKltvgIhIUTdI8sOL6r6lX40A7ujdlcUr5+P8B7GiNOajwfzHXuhhKBh6DAwCF2+s9Zh3aQULbvM1Cxpxxmc9yEJvFmusBz8Q1WszKNnDbJKXUHG9LY8B/WTk8zRk91A4gzlkaClOPUD3txFc35Ia1nn9MOSXrGgP8kv/Zpe8dmJye+QuySHbMBQh35zsrgLLXq2lPRb8KbX6Sv6e8ID9+VYrfQKr7dgWUZvKg/spcOUdchcVCQtc9BcdMlp/p6IW3sdLtjcrLofgaxF8VvGRXZLSexVkZHD+xS0jX+MISJoEQIo1FeOztTd59SgkNFoCr+rmlHMdVGM28+AGcRxMxwBQWLn039f0Ov8IkVtr2+HnzhTtQCjuUEAR6F4YghCo9TePT8i5Cl1n0fbbgMF2xXwMHOJgUlrlQEP3gw9nsHqukC4xiWKeFZNdLco7Nu9SJ8hGNnuMiXRBuVtnu8uMRdDHIcIqECri4D3bFUp2qcOGgYksA3MdyOmL5HAOqgpvmXUtiOukTsrjxnmd1XCkNApnNULYqHmYx0wAwO0weX1gOTLsq/IOaYeBL3uzbl93ncJx4LLVFzKstU04BKoxB5uwdQsqtHckQoqVw+ub6C36Cpw57sVxf3ebjHw5fDvPJ4fBgBtzhKTLo9xkDDfvKWRoC1o6HzDojhmahPHDNM1ou9VdBHSjOnRt+GrGCxGWGFlEGGZwnLUoXllLYC8wC7SG49T/pn9BubfERRxrHZcJ1ykhfYBZCTQwe6INA473WSMGHLdPmulpuEDfLqswaXlIzBBnfeaBp9qZqodAlmizqD8Z+Gdvrj1UOK1edatoOMqQ/kjB2SHMVSUkMjL20kMumB+mN22X9WFKW4G4EnqjYZvzk8VEALxmxX2EG4/hugfzNbDBIqCQlrdhcYlOwVaX4+ZLU74SLOygsmvgFxf7jelIphvcHPNeVgU//13vGL6Ejzpbt7R7A5eBUds7H9S19gAth4/aHqpYx5+zuUHu0rLCPB3FIyvTl+G5lMyAISYAUHgB2+j85/TVU7KCsX4+PmLBauf2VbR2ptpo90zsHpn8nIy4FyfLvAPF6jyHIOtZ+LLa1LOfRzcniZbeOqVH1trhm3yXpHQVGXtjBdpUyeDBnF1jrDDyHiLYOp4XpbaGqj6Qz0B/odiYxBNI9riEAFYcWaEeYPhWe4VDkBqt6n7RTxFgLUgAa6H9iOHvZR8mSg1Fu5/gHh0h92LbD5t9xOq1FKiOUIn+4qdO+gpHTmTBBLVd8r2W9/lHY23QACAglHRUwZkcFTPdmfR0xN/kg9OvwPUTOkN3ZKV9qL82NRADdeoPPbQC24jIrYX2NJEjXwiwfUj4ZrQyboA3w6D2Tu+ghe1cExe3mOSoQaoPtir9BzdZB8srcgrABzJKxgF/zQtoyOYlHrTyXKfmrnB8Wf8nAm5Z6GL+gFvwgQouKnhvIKfn4rPW5Kp2Lzi7/pShB2BuoAUawyTZrQwCX0M6rI8nGSfOkdkGlUJff+FHbVZfLU3G5qbj1AcWIKAwsMcvSkgXXPBsB+pfT6/QYnmY2x7Wc4MoHODZAgWraN646hXCmEn5vzr0+l/l3Nogl4U5Ck9V0Zfa5jD3IU4nbMWtnFYA8qRY0lXYRQ+ctTaiLFpQQLTrHDiUNu1TjMpFSMOMCPlfUvUkoOkPGXx9xzZFntjZ1uJA/15kreqxxO84n1Sr285GzWt9+cSdKcrrN7yd1U/XPZ+dk0hFFNj/TvLwxWektABlo6MO1/vI9ADIjevRYH/i6QJZtPvGvIWTxDhYDAA+RSM4cTUJOH+ZujfXrMhdGg0rTGy7m85Vg9O416pP/4XFWLdb6BVCIIGW57JEdcM1Nq/SIHQRVwy749Hst4RkxiWCEkHTWa8nxE5S4IsjefrQXOcU2PnOvVWoI57tBtQJnecDcodz9sFlwqayJUhtQDkqCHebD9AHf/DTKFsreyFnyw1ux0G+MKs658gKHUh3l10WIpJCakq3JbBmTgSNhfo19Gqadfoxf6qB/nVlwEZ79i5Itj/elNjDVa7ITTVzQfIxw+y3CjaK7hz7MSIWVPa0aL2crHWn4/A6KoxSM/T+wWK3+bFFKPPNGThLViBmII9PiU2vvaicFaj1NI4oPbaj+8cEJGhXgqE06w97YdZHm1/Y3PomjoHUr/WcRT465dM0z1t2L03ZtdhPN8Q6Q7tIa0/Ku8SH6JlB/gY8PEZCbdks+dbv3GZGGQrsrrXxhGIb36bfZ+rrwE37ZSv5aM+bMBKwFdIkreZNbNpHfNvQ1NbH1w5WmwufTMembUN7o9gfmO34ArTyVCXACVsjrgmXGI1ExO11vHo5kBAS8mxM7/m0xNqrxf363QV+teSu65KezioQOXFze5pttlak+JHWlfQMk5Tgn72wOchGeQLSASh+KRfT6E5ZExmaru0NLZhn4WiyskjDyemjfDOazCVtjiyoEiQ7v3F/fBVXbSdiOuI2jxMfqswDUAfmrYDBJeNJBlYHkIVuP7W2d8U5t7TuN3DTsV0a6sj5iWRSXNlPmdnZDHrx3//MvSkwWZHQUyDbxcbpp4benOvR60jlhyJLH6KQi+6QlxjiOE9nH2yEEdbg5AZIDkM0SztDiD4T7RnoqpZcFa5hyX+auiiE6NJG90RZV3I/xhOkm0mWNj9z7HLztYuQr5tjFH4wjduenhbcyi7kh2gtZry3hcu+BZH6AGTJDHAdFaLs4jcpjIov3J5+3G+GMNakR6Og9WIWcRBr5XyhV3zspBjqOeAx7pi7cYufLgBWnhnNrBwNu5gEgExOMFXcp9AvMwXo823hmrfGdprySUdMKwHPAIqPF3Bo2ACpkE1E+7KhjbIJt0JauFaSrtB/TEjkmSkxjh7CmeYvCk8+4aIAe8dxG097jOicPkSuMwjF9Mu2a10DLxEeg5mJVxYnfeXUV3yEngBayGLm7dAjFtYllLJqRQsCLdgmukmMRCnadnUE2aCdogJOjcX3ugAA8gGlji7hecaSkAK9K4FqpExeX0nV3fPomN4sgia+g2NuI5AveynrJB/1qC2BBeQ39IXLgT9tdjkvNGsz95E7cTOHc1ZMDQomdIw5FLvawUlBWsWAHThXAPQrwUzIFBicKhgqAE+vCf+q8kdFcgAj4lligZwWBDWNU8KZu7mdTsGYXJINi50nnOtacxJhNOKiDEGQYCUab0PJHv8YWRnJxlL1pDoC0XuljOMYeVfPihnBGuHn5VlGRpHK8QnstnSPff7Pv/hQ+kWwq4+ervh4FIk+/xEhtMG3Rf2khPz5R8xBcNeJjCMGOupSDm0v3PytWZB7QKZCCBSVRARNIO/J7dKP9OkRxUHswCGLm/GquAJ0ZjMP41BhViV7ab6PMDcpNXSW+a83GHzfw5iuXs29FT5WgJy8qZzhHb3FXxNWhNma5VaVzwRYj8y/HFNdbJXrihfKfw3hzT/UPyblAj+1DmHUi99iYw+cmK6+l8XCm3tFBf4cCm/yNxkpIe9Vi/rsJ5i60UBocUWGoDupQ7PvgMDXcvLjV6i/haJNQeXMlul7/s1FEMm/lsSYIYYLzJ5Cqlw1bjfncLCpWJatUMK0HdGa7S9WrdmkjObktQKRYgAyp3ql1XCcC5RosS+WtV7WQnsBidGAwHiObLFeZSHLhOs1wDqy3qUEbPxJIzZY1zzFdN6myqNbaXI72tUXcbssBjOmzgcEcL9y3rMncx5cpoTugDHB8ywZeJA4W8w1W8N7ux1JIfwBNGfuRe3kn24RvdiL+mxKQwsAS3hyTObNWnfSF6KAKDQSJAJwYwWzBUogHPsjEDZbLFU6Z4QbL9a84Xz3mNszQqQdazvflIaLMXwtb2gB0vqrYg951j9MaVVMRlQNEbAQIxBcfDt3xcFBH6JMJfRoYuz1X9RbM8uRq/rDAxmNESkK5quiSwfwAlwYW2vtH3P2zJDwQ6G9chnw5tcPUgpo4wW4FS+NhJCauv8VJmrGbs00DXvrFnonVcrx5WIIFbfyGotiXG7g0nfb5gRgUwn/McltDbBIUzgRPcdNbLUqIF+eT0dVM3sNi5MKfwtg58H6n99loNYCzxrQz8cFAYOMYDfprpPSQ6o+nNjdxw0A4dwuTVNcgl3LFDsHutHDbepeyJaLrfJt4TAOKG5MFGKws6dUdal/k1COjYgojcJgvKcw6Eoy1ts4pCjvpoWPeNw/gSHRvMOAUjcoeNj+5zWL+gh2r/IdzdPJvf0rlyBMAriHzis07ekFipS3YyBuKyt8wpHWnVOB5yInjm8mG7kmzIHFGm7keweY3h9dfM3VUEvhL86i85TtAXaemPUXaiWAlTpD6X4LGf2PcyD/bmbct9+csbroK7iRKqknqocruhk6GNrl8kdaLjh88TWsFwwSiJktEJVWRnB3odjLDJMwWKPm67hJ911RjxwYQSDiWMXEpR8HovCV4m2JKh4/Jq8BAgkuTI3Mvwo7RC0TyEp5/RVo8LJGM+W9+RCEzwPnDjyspTQCXIjVU9m5o2qg2HwAi/StLSjAF+E6y04d/dotngZBy5KdnlBY9ngSD+DzR13O8s6JflYsXjH1fATrzC0j/hNEUdA2o4bfGskVui6JVe7baF9AISeNRSQBRb9iAMJLc/s+PuYGCvO5SrewVdJUvIGEkpYWd3Ske31DwQq4gLdiAKH0tQgH7Gy9uSnaV3t6P1Lsrd4gq1tMHbN679cUCfuTPmnDMF7s5MhgLQWKBfUlFl98N2G/xAkxbSqK18XQ/wLpkIEyFdBvwU3WKf68tihkcaVakwCFlHmEViCMouItpkjo9zhobiQqxCtjTqfZne82RxvzHpEWWth71dVdkI3c28ncLw/FzVG5n+HougVnlt2lHl6CAPXgfBZz76mAiobUYz2UuJLhgcvP4IxBnCrlCkHc/xBhYj2lkgqr5R6pwWiURlr2ZXNfsR1IE5hdbYU2ASQjnxw6S7XrdvFBTIxvBrGWy5c5epC/rLTCAZuzh+vxzQOl6hY/eDoO9FB8655oUv7o69ATvP/IeXAgvq0fLXr1zBGdsomSKY1QbxTSiCfiZNw/oaSMKJMhtM/rzryWYStTvB6UvqLrtULmRgMKh1syjuaUqdvD3R3QqcbPjkSs25N5bjIglC5vDCvHYAynKRRZu5fqfg9QGBaOWDAtwpKcRu0DNlsWe+lELyAv4ZqsIDbzST3/+YrIKu0Ys3wWYrG4NDwGQ45T/QuYhhrIlxuxf4jbO+zNyRNBCImg5k8MgZV4vSwsEZTDu/eNTGkQqoMtxOvtO9TJ74xmoSH3OALZu+ftk3lT3/GlunT8JQoYyxKTgftLxL5Ck6VzjBM1FooPuy+51CZCp+BQGWU+fOJvH460InQHyiVbSlPPr9poK7FOGxwV+9szUnnEyJ7BfckOE1wo6RA6OtGJ8RNHikqXQp37Dr0eOV/Egw89Jizf0VWOualhZ/MkYLvhQI+2QXbPwGxfoKzYgBsVePSN7sUqOSbSbZKy2Um6RSOUAid5KF0zxnKj3oZcMVbxEWZ3cubrlTnLM/eoroPVeXyUWruL73zxEucMmr4cURLjFLR+hZ+5aBZG3LAaUgpC9x0mp2XXf1XsPV6Lybm1iRptXIIouS2rD8Lonj4iebpcpyqlEuttFcLSj2qxP7457Nuia5z7uQPJdsUwTzdcv32yBYEO22N+C8tFvOXnFHgEbbgA1ClqWn2De7HFO4vwmjJd2psiEneFYdtYTayIXPEwqzUIWjAGkXdPtKmeI0Z7k1iji3G6A87qACKgpK+gU3SvnOgvJStVL08gc8+le7H9FDBRvlcXjBPTtcWIGDBdQgQPtBboGRqHhMTLEenye5N4geVp3B/kHNF5a0DF+E7Vm5mgl+zMRU5uWxc4q/zF9pWxpoWrzY62FFxKB1Zkdbetxt7X8yFS+XXHBDHjvW/Uw1mYgnW2m235jfePnldcmk3vxLbHQFdNSE6J2U3FVFeSU32GEl2P5I+yFRPHHwTaHf8zd2fkiY+V/ciXznNbjzAVs31RjvJu8fL2EtTjL35Xmp+iNk2MhRZchKimWggWwfg9rg/Wa/pwbCaADaIRjOWirnyrXJ785MbZPpKHoDwjZFkKhigeRqxBCUsdxEZE0iEz+47LYu3OXI1DTxj3ItPZBVvJGpkjipaOMIezGD35H+IJCCDNWFRqxC8ecA9AsQsiEYE5UO8Gz57VK4LqUIUqtKISFaYtN73TlcmkyQ94i1cWo06IP5ICDeGpxMXakVG9mY379nRx6UoJeHGLOeAKQ2teTdIWdIBXkv5JTzB/hwAVtbxO4t4peEvbnb0tx6htmYmzvXsjI/HnngJxKUsAnAMCr218gbRL3cuTlEwJvv2J61B88lS05EPh8KQlsPCBgBRwAL0ImDpECIVEaDT6z0LsDucYlWhbFKwhEENma/jQbatQ4u5jp1T5x0tEftvcwXS571XRsjtDesDCPwGr0MhyTbeYZ1SVSpA7bffghj6Cxez56ulNceXeEXoviLYZgm4giJSQGSvVKDKopdK9InRyDxY7q9bmJtNGQ+ScENkb4D4g5vjy6LpUqJaFzScywACegFP9zM/4UQ65VjPeKHd804rPtULxAvTdbnAPSLTm0T2mzdMA/uymBKDYiplsMkMInbUnjO/XrDAbI6Uc/qg3FUfSCWJ6AI4NVO0KfsfT71ijtqWUK1kvl35Pht38oD+fZyCHHCFeUoyiDxAZdYRSZOMBkwid0R7DsNzS23f2qn4DrBIny3CnprpKVJo0fLd53fFs7itEQCxSeV0LBd0P7Fl29Zo434dmXtZb5AplXyxEyTxwepmJp6aqimBogR63mSg61Hmn4ayB6pHq68ibk3TIHhVa3tSnfVOphaePzzEYIuMlnRXiJ++Vt5IWCS6a0Wyymttpdqtx+mjEfdcHgJ10AMuqznHLBru3xnq11D4fyyoOMewgLQY+HLcHSkUw9MjkrfpAl4gwwzVZkPXWKbP8pYGSm9KKnT5LRQ1HdTnPEHK/IN/m0e2PFNscklbE4GTXV11WqdTHQ5G3GctphV5PTwMnHen6Dv53tzglpHCoUd+5PqEdCt8Dv1yIOeO3a46HUwWB59GkTQG3Jd8+nXJ0D+KJQhB3G345ozySFeJmAYcxgErku0ylgtFiTXMsO2r7b2/9tuVTlReesp0s6gmRTrGdy/TToPQb67Ixm9B6KhdqICM9hb2Z4iL2MTrish2Whzeta9NOaXLhBePywOgpfWezyRh8PAvEJgfdTRPwfbgmxMKomw6GItS/3s596lbSGU4dkguOQqvgOHiGLWjV14PPPLAPk7HYhSc3nOefFaInf/u8rd9b/hjrBVdxOVuPUTmW9huVIeX/gr+WxERfyA5OviTYsXxDYedztaMdcwKiQ2nmh0TmcgVDRgvHJw7Vc9wx29gjCtvivjBxBeg1B/Lf8qZYHz2hqiQS1nmZIoWRlmFp7pXxt6/pyhkXL6fMoVq3ELxQclrMqtkHGmhy2rWma125pA/JEMEmUTtvBIn2nXwU6w7sqgLZ9mwuM79UtO2OWJEoTqWPhmpxUfpsxi0JoYBD+boLDvyEXeEHKdmu+ZHd0DNeI+NV54KQ9/aeFAPfu/Zs8Dk8YQEYC5lTU9aGU8aSdzUhDfZWvK1OCicHIsS5oWB6quDYF3GetnqY3Ao0dOgZBI1XGA3aZbJdh6D9GmbQFdIZ/96yMe3dFIExT1ZgwMXZBSrN9OFJT16eCTj+3itixZB9+i7zyQ1ztIX16eNqKuez1ae8BU4jtFd6oX5wI/7Jelap+KPqBt1Y9gHgvoIUxNClyOuCNEBRfhOryvz2VT/pf0KzcWi+O63pFnHAZysLzqp1+0llzafXHI5C090/Xo11ot0ySrj7vZfWiryEbsNqnSUymftAWu6pgXUKiYvb7jKC6CCi1XxIwGwXZJP6s5f+RrXUJeq+Bk7UIeqFs94Hj9+XoeNQUE3qmJ8BzE1g6u5Ahr2Ha5h/7hBledKOkH5SFXBLGHsfnMSZjW3XPL5u/ytCxQ3eC38edLCMOkI/A7bA+avhTYZx/VVH4G0n3iQN1jwa1ui1GLYwRZ2QILUOZzyV4YDWlM0WiAwDS2nL0p1c7NSFc8ZPKHiRvJ43nTW0v4KxUkUvgjL7o5gSa2GPTa0+8peRh7Gm9iXzBGFmiH0KfjJRe9Gde2+ELkRAyzrwmRG7BKGM6B46O43wZjSfJXAhyGElFqsTVwqcmA8IuTQCbfmk9f4fFo23ceNqXgtZUZSVaMXfae7A5hSsUWYmreXutBtPBTaj9qeuyIy9YTGVg7QFDQO3p59cvLnux0iLHIBI2QOmNr22IGbJH32MTBuzo9SYoCx7XQqLSGiEvLRjDGRniN4EcpPhvZ2lrbxdiAP7ebMnEMA/iTji8aXrlALEBr5/V8g4sKRUe0OxFh3GSJqscQSLbBEvnb7W6Uy2f4K5RVYTCqV/rWYAZztDZOq8/GmZjDGeZduJwEUVxE9q6YTyosLB7TIZyf8ZPgKKMyx5C8ydaPkbcnqsGcFDGPLUqe1+piuTjGM/PEvAu2+61IT8D011yARPG21Pb+J73vIoZGGSMOlljLQYK6RuCQSWi02ZZ4hx8Y6L/mCOCJofWRPB8C1PcKZgyqnPG9mXq4QLDlIrZNRzOivw/g6sQ0yUD7+BZXP3G2fSV0ycQuGd2ER2ETqwz52ST4azE/GKffVkXag6JGFkO6cUCYwqajybmkIaNTcnnOnGDL8DakJ+Ftz4GCoqe6/yLd7tJ0ZesKwfBxjirJZtrJ7NINCm1UE9ccJLEoJba36hmuIfWP+G9DmGh89PhNdSBT8ubi6rkgMOkSt8OQmXViyQXdXrcxNSB/q7m2jDF351Uf1rnQl96dNpUDY9nqkOrOOIvxvzCydnFe8qrvCHVdXLYexXvsE4F0K5+FnAfncsDPmJNJsxl2uN4x360IyPny3btrYUdmBt7v6292iQnp+L9z5YTVIkJxU1F8+mlE50lStfgM1Ajy8YUhdOMszrSkRDwtOaE/PSFc+Jg9YVS9KL9MTOIZG4xJx5qfGbG/IEnXf2xsg8j0vz3dR0I2Pa8b5kbMxNmiP4pizXBh2UJHxgBbMKSeq4Pav1DMYIlfqXYeSaspn8amRQfoPw5kRqluLsbEshWjELk9AGGPqXJGFqbY8zReyjutRpW+0ilKGbeM4yZY/n2PSX1zZ6H7P/6OF8htAeIL/3iiGb2wTGjLEtamiqeLycd8xZkopkiibtTLlstnns+tAaMleHEsKd5vLIPMwKcmfJBgqB6IjmjPfkDjVPvsVWYr3C4klxmi5jhoRmgJsFl3h4NSwdsY8kIVTKEzdy6RHmQe9LwfaMUdGLUu3hflASN/25vmslFviAl42D1u0mGK7xqCF+6qmvZnIt2jumKz8Yk3fjgMdS8buGqpSRje+yFxDwyeVnqlElCWLOocqWE7n/nVooROv5wMfpnYO/scVr3S6oZ/Yn+5Mncdf8Nc8e9GUtL8qP4qu+ZzlmrJv7e3+ZE6drdA/6rKlk0ZwGUeaD330+9zDCokQtyZDWA2mgxG7ebQhIFbnlxdrtHz/QrQW0ZagEQ6CeFwQ8aQjQtcmVtq3L577hpccjMvo4gECH1lCvCZzRA+MxnuD8L2LVMnHGrkXZ64LrfI5zPmu0v/jyRjoJeszENxXsH3Yn6i6zmlKUFYa5I3Z/MhZQ5uaYtCjPCrjFe1nFk6r4qNTz1RFCj9u8R5EnfkFlVO59l16I9hpXA185xXtKltnWJYL8rD2oFLAagsbmlKSMal+UmPQjty0Bw0cEmTwOyfCMnBd7Jsi3kAyQJY6P0vjO2ibsOFgbRt13Gkom/MNGHJ1Xw2gUY9B2hyR/zhHTXvSRwr/mm8WiMnZestkUXAjGLk492vZs8ul8eM02pKRi8ltM0cdTIzkeobEUi/vTGdRoRLke3Bi9FAOZHbFsAIP9oluUgW1gcKHGGmHgEGBbCWyeD4z8JJtuP9iuW1igx9vDpiPPDC86FzyFRFOx3wkAr/IqvJntv70bmuPGNaG+0FLqu7yB+LkZl1X/WHP9YscUmcMg3GNrGhnLJFxEDpvSPULKNL5tN2doWrC5Orz5a8A9dDh8fOA9tjdA6Y78BB0/YbkSoHeL9vn6h/Zp7Z2AFQ1MAWV4ei3L19Rb5f81zdp4gATNT7UECGhICtieyqRGdaSvA/pXBTcu1in+BZogsTJXBk/N98OT2Sl7O6E95pLTfWvGyam8UQJZ50OQKxEekXhR3VZ/osTf9XRVMHjb6yr+vjGgDMKpcd5Vp/aJ/MwZ04OhqhV0RPflE4iQ3GOvN2s/ic27i2ddNjbrMVf/BmyyTPA6nZ2AGNnUCGuLBoD5e2Zr12WDa2iTmUZH6r5s2pK0FffdMbl9adKpVIh1V6rUdOsuBsr9kZW4KUBrgx04P36uSDo38a6rjOrsYwvtE8PRlDASds89xRBZSBkJPOveIidA81SDKvn5GWFJU9uF0XXcbhSmXX+wrlTJU6yZPBxKDunTttHznIlem+skD05PY4GnyG06RLSyokrcIwmb4l3ZacyPsKFYg5qk3XmcawLvWh4RU829di9R0D2iDawaEn50lMR8OKncIZDP6fq8Qw6vMYOIsGdUisGBcXZi3qynwJf9W4jBdvPfYZHBOj3uwwUvqCxHUeKbTIDFl2g0Aj3/JeyUOu9GtNsohmoky6E/R9S970UqfXK7R+rbrHbv+l6oJ9goHyYR8+U5/BhTDRaDCcaUT694JHphKh1JYDqk7hT9WVwWJIZzZ1luQI5YIjQSsVRoTDVJgCO9rZL67ifrpT64K4EzW7+yiSq+Ng9Vj9djWRQHvpl2RkrnVS+4umty9ACuPomL/lIZXVTUATAAAALWnYZKMQQfL7LyKPr9xAJsa5D3MvufP2CSNAAAA)
Cohen’s d-effect sizes 95% CI and for differences in subcortical brain volumes between major depressive disorder (MDD) patients and healthy control subjects. Effect sizes were corrected for age, sex and intracranial volume (ICV). The effect size for ICV was corrected for age and sex. P<0.05 corrected. CI, confidence interval.
![[[@Schmaal2016-gb]](http://dx.doi.org/10.1038/mp.2015.69)](data:image/webp;base64,UklGRnpkAABXRUJQVlA4IG5kAACQOAGdASrbAjoBPjEWiUOiISESGOWMIAMEtLd8ITIA/ACQ5Oi+P9D/rCSHwABKE/23+W/tPsoHcv6n+3vE7av1qJ6pkr8536b+s+0//Ff5L+ze438a/633AP04/3P9i60f81+wD4Bfxz+gf9//Ae9r+GfuJ/VT9av9p8gH8t/1XWG+gL+yn//9bf9ufhK/cX9t/ab//+tO+OP55+P/mc/O/6F/fv1i/rv/N9Z/w/5D+g/2H/Df4P+vf9j/M/Cx/O+SXoH/Pf5D1H/jH1c+q/2D/Bf5v+1ftv9z/2T+x/kt/mf2q9o/fT+5/1T9nP6x+zn2C/iv8Y/qX9b/Yv+5/tL9Qn0f947gfOf8n/jvUF9PvkX9w/uH+I/3H9z/dD16f2v+yfsF+5/yV+R/0v+2/2f9jf7d/6vwB/iP8n/sn9k/wX+X/wv/1+if87/lP7N5bH13/Nf7b/P/s39AP8k/oP92/u/+f/zv9s///+t/En91/1/+L/03/T/y3////PxT/M/7b/qf8T/l/+t/i////2v0D/kH84/w39u/yX/M/vP///9X3S/8z3BftR/8PdF/XP/q/n//3STdFW6o0qcY5QbzhFgPT3NsEZkhPOxIz3O2GB8ljmihHy1Y4NETkToDI+8zxeG/WZYzy0f18wTLZf29b2ieVj9TobTtgLGfcKZ2rcODEWBTv3edtOuijGBgg+pIfR6ENzWRiE2Y7ltoJtgaoKT8jJjHm/BVYy0rYj1YIW4hhJOobbRYz38bm2/9O3pafHNBarg81ENeGX3evMfBQIMe0S/HeA8w+9e5QM/ck8hdcpQHgzCh6FSA6bzCKK/dcy5F9UpjDvs2kyuTMcREVWHrR4utKTtXpvdmSENNzgAZrwS1s3ZBgD7JuIdvb/Thn3zRxBq1Jm4IQDnpgQERqF7gU9I5ed2T0hOeBysNK7Ke9vppLSr+M0esSD3O/67OH0v7wTA/aUiSAHW6lkCroai3DBZc8OZZ0jMrR36GJRegIgzV/dDEbo5pCKLHdKUCnoeSmvYQKzyczCBhApmFAW2rFPcRoPTyE5Qho1GLQCZBgLDIx0+omaMfdhI3cj8N0VEdAoCFd1pmBj093BUBj1N93hLJiIJkKjTIN2PdZka89OenPTnAO6/+23y5QzMIGEDCBg/vDM5coZdr2wZYMsGWC7GCHwKU19y7J+wAxXet2F6XRuWXm5FMTIG2T5DcPac5z0z96/LWLdu44yVxq41YRxPpPabV31m3QUpx5vss9uXDqpZpgIiCZuUuDeDLBlgyTodru6OgCuSu+0X37R9nufpEkdYH4xAwfH79K5uTWbk9/2t9xD4h7sxETbdfiXUdmXwqwA0kVSj83TqXEnof2X8LtZfgMIGEDCBUmiAoViLCnw5Hpit92o2uyFugvaEf70fl5cUvo6bYn6aO+3CLceQAIsm1cHmdl5Re1ZVK9Z5IoX28/rv8AHwFLIxXMT4/c+Ipe6rh6YKyG0WtrbQeXdNMWrEPeAQV3h+T5KXMi7EanOPgvzVUBRRO8Ec1BNFgd53tHtVYfRMLiUBDiExDXN7F5YZUq5RaVjzqQWEen0cdyIwigIMRHr9PY1VrClnR20ixsaArmDAEG6ubidRk11SpOQn2Sf7WU7Q349sQyz7UY9xVlpzLyaugIVK61PhqChj59bewphhV3Jh5CBw0hokPPS215Bb5iQMyzyEdJbEPPTLrTX0Uz0lDnNh78Nt6oRfbhrQuWr7XoSnn1gY6sc70fZtJOXfNeC2ZLOA23dx/cTzkj3PgIeuL2mmGk5OhH57D4RU9iC67BCHrZxd3YR57eBgSbfdH8B9qRjLk4pOAVYvnmetyWzk/gnTZNIaHq/+A7HeTb53b6ddibKSHOK3kGFdJC8Nc8L9obIarfJtawWVYxCMKkcnGGNk1CkRCKZJif6GWDJcysBicUTM7R2diMPxJYW+v9RViDd51zgIaQkU6AEG405cK0UzMCZbiSh6IJK91Y07pz07MSPJmNhDAnDn+7Kzk196uJ2OjePifxQBMrLKx0K/eCovKb6xmGRCr6GWpNp0fHPwO6PRTYzJI3jfX3+fk7BsOhws5AMsb3zrdEQyxkLO81EpBydNYgqjSbJz055h02LVBsgdyUbRz7DiIBdJQkeoHCp9p1Clzm4fkuMzIaRJiiJaoN3R/cwvYdLf+4o9PMH6YqfcVeukNesEUw+2C3VbE50doWm8lrLH526NDYR45czY0oMLeXQGqbAg0yPLjwmPnUgaKlJuKLEubhhUWDK7b03OpMd3cswy7qOfpMQzUPMrp2oGzBhesDVcHm3Rzz0F2xvelzLflQebQFYNKvB9ERW8VvT2tMxEYESA7nixQNOenPLtM80Rg6b+tP2TFZHMKOALD5XeOdOEvTZCFGrjVxq41EdiRWK5Wzb1XT/Mgq1xM+VFnNbUCvm9VvvTmEG7KPSG3EkQdFxWEB/1wjxD4gTaSPW8jgi0b7JitXjk1xWqgGb3VaOxVeJsjuR3I6QgbzoaeSQoHp1u167uMUnJdvRjfb8jmxH3p7RVHPJXyNzlx5KLh6N4fHv7u7lMjE1NOemceH0afdHcrsH2zFLjjcis8gqJRCTQCPXI1S1wOdVXB56c9HEcSz/c9nVGlTEbmSAuHB0ucPV+Fk+AGnXOouSWghHxWFyW0LGZUncYsGVgdICQC1afxY9wSen95ljE04QQu9xxz055nx+GcDmsXDNFA5nEUr6fbA9i9VGXwRDzpgkrFiVnnwzSRokaLpWTp0mz4Qdm5lmGq3PiUazdL9X9qOfXXp+m6JsEE9/wTXbWDT/YrUlgYCamrGAfG6DYCBuQvDWSnCakA8El27y3dgyh8LWGHqkT/OY6MtvExIjqsgaxR/q3uH6zjDaFLwrWOWMZWlkdm85HMd+tkq+eBhLXnlEf2Eng4mYo2c3B7holopAfb1oTwUcJLad6tc9Zf8Rc0zT+UTYOANPn83YugTEjBbjoJsACxUKbAFVJiQErj+pmQgaNsulcYjg2Su5OSx6fD/ubi8Dm5Qt+/kKq+A0eu9M4KsbO/wr/pR74e27zQrzJanuC/DilQBKEbA2wxxWnxU1cJQcSD8YcPmhrz7Y1DDDlVnCMWuwWUtNCVcaiioaHjldnIlA6SgFsTEzkdkNbmnt6tp7SlGUbFibgu7mv50xQrYhAJXqZR4Rw4aU4EZhq41bQ0jDMrTj/wh45bp1orsUnWUgpapOHwd+9W/z7Ush4X6ukNA4wYyJoztEiX0MhZALg89OenPMZDzL51N+D93GoUz4srwY+KtwWUuB+OcvH5ibIhkw+xW4dkipzD75vM4PDB6OJ7VEQAAP7/iaOtcrr5N1tp9/t5d397KIUVikmjWOEPLv6B4lEExewcVWomPFijk/4AhE+/059q0Iwifbmh2Bp1VnpIx+d4sMvreZtk0eIBt+9La/d+9YmAdr6gCtqRhImjRlMvqesFxkUqNMxPcvec7Jfw+G10EJFkASdiY+/y/9iiSSmRlBpXkWRGNfN+knFDBiJ9LykM11a/nplAyk2gKbN532ALqWS6Z/Kg7gixGrq1CGFrHfaxq0mivawzPWiNbkWNlTraGeBL7wXsnYHcAeYQsgCpBra/lv8U0lrgtkZ2n/xN1pWeKdHetgf7Fq14ZOGyMYgIlSuM52PbLHUev9lrXCR3cekNh2qD5NREpfdNGaAuznBs0gkhko9oMEn9uAMvTiqaL0/cjjv7E8ySBnFq3l2928DRMRyhLmRcEouMK4Wll0m9XljX5xME5+HHwGfIz5XivNNmWEn9r0gRh2Ze+WDU1saptALfoTe5gG1155TfRVXqIQCn4IAAR/2n1aHz9S5GCsl8z654tSSvcKuC7jSdAycvi+VPmi704T0yq9KrvJMbjDUIR2lSSWke61kAuKVWXyb2TNWoZu9QAx/C+LAFmQwqWZjLJJiKLA+WvGJgtT8sDegn7AxVwRopSzsBOrvtBJ4zXhfjbJ7PB71gLCVjsP2tUH81lqqwj/Q2crLY8zyzyQ61AXqCNYanek4vZz7bPdBe/lfvrz4259c7R0hSCecbBQThluOX6fuSxI/32jffC+vTcKUigjag3Oqcoowojewqph9UajVltVAgO9yvP0z+ZsDQ3WUM2HVvGsa/jj9M8CVh6asPsGvN38fKOpq8OTtcPQDhjy+gVSCpG6qOtSpfGWmXsmNuxQFeYwaadicNYSyKd7WGTEZr0S52tiDGxbwGFLfwB7XpulRbIYqA1dah8NeggIAwxmfKqJPG3Wu01QqKVqWf0s+Ze4idJft0BfgsnWHNu/LQqanOT9UJu9U2DyGD5qDqjFk7bwq8WoTFUZwh862XoYAUeHKq8QPxlL6FPrBIivG2/GjGDnhblguk1epTZWNfNHDxdLRXhvemDE72OR6eogNnOygYcrFIt1VIsrE0WM0LN1hAYvli3A95qgjO3wLQBPc0aH0zpw+jtWOOrqFKPqBH7UonPOmPcOQfZShUmSreqansffQuw0T8NX9x8rEcYrszvgynrYOaNiM4eThCsOFAuxAYl7UBBhh6G+rrV8oHaKt7wQof5P5z2qZiAuR+83mInncCKKL7FOJooKbRuoxoQSY/uF8MLVt5gyfi2AMPSydkz4dig3gBAfPzbJasnCv5wy6gbQIEJEy2X+ss8BmjLlpYbatyuxEzaInEq+K1CChznRecRIIai4FYeoxDXHfyegIra6LaIPeCizu633gd6iAcTA1RayLhrNPwXj++Ol3zi0eBhMBEb0HPikNt2JW2YlAdOlnQNCplBna3YteOTvUhUdkA5jI2dBNmbdUzqNvQk+1H2jblS9gCP//OUZIE+UPqB8u6ckXy5NDPIo8FN255scziUv5Cjui9kYDp0gaecOPgX0vnHgMKP2MecBniXAxGwM43h4nfk18kaDzm/sR4xPan9Ku0qmDNZgkkcep0N+PWs8BLcWzgUJx0VwSt9YQMY01EGZwQYqkm2r55tl8Fe7oayJJFjn0ZSkkatXtaMaAA2S2nCx1hyd6Qll1TJayL8bZlN71rV70rWD9EvMcM/iH4Pq8PbRslKGXTtKjMx0aBmbxHd9lQqiqj5RbaGwUHQYrUY8o8kvL2VBB4BmarJyCvkkKcqIxuMWHJ4ahU/Hhohz0elCHiklKEF1Sy4/o+D34vUgOipBf3iHPDHxRtJfAniRHojlDS2ACNRufW16zaDBm6M9t1PFUqEa4yN/g+KVlodNz6Grh/5TPCN2LqSS/0to097SpxR14Xh8Q+uN0qOA1NdgfWiLXSEPtsmo2zNlL//DXBoikIrY405aG5awrHtUh3w59WY+WmcUt/e5Q5NK0FmT7xmd+8SfFUsjWVwlV/eebg5xtQQMXTA0i121LkUrU68UvJlatILCMXXDjRGwRJ4RPE6h1NNE4byBoiBV0qHQUhtHP5kGwp/qjKM22LhvLTEF4RiMYc0AkrMnhUHvJESdbhi0Pvx0mKnO5Fnu6EpZg/khMtSOWrQN7uRDWiTS//JvuoGRo7niQGcwCfqhM6epRfYPIML76nmJXfLtbvDGnl9cs/7vBMwo53bCz03gYV4imyMAMJAaR/bxT6ImI+9jmTU/OSiDxvEbVwa2P2hdGmdF4O9p+LVUHLrGVc9zhxD0yj/y0StGQpWPGN1dd2YpKODrwinJL/vTwhuuVRGgX3Cmd+mnKNXCyCch6eeBrZ+Ebgp7bIKdPAAlVRtm/f097rMS2wh3Q4PpAQfxkKObE/3DPD51FzPyTN0XFyTSM8I4ZVp6s70GF1MApO8kb5rVRGiDmN3qufIGxGCOUNcZW9HVNJpA4qZawGqwf+zTLzep970oQ4m3VVk6V52JuV24roiy21zLt089jQN8wdpLzk52THaGaKHbwfs3+aItZj+be3Oa0ZSdFnWWnfZfE/RNJpZ/tGNJS6wrEHnnc2NCdAlSciRtx+4mVIG/DNjigkwOp9QbGkstkS2jPNOSOKPWIXNdW1VkM4fVSKEKZeQb9BArZmGAsQzvlQpyD0Y71gsj3OQ74V0YfBLAnb4WnD/ClYQZ7r2b0Db3aGoVFYc/Ho1bfZ26o9oZkoLYKfYm5AYi9/MoDuB5kaqfD+jpCFmq6egomp22DRN9hu2Luie2PjCQ+doLgSYjSULKvBpDOHWQc8OGCufRo+PDegqlfKgi+qSqtyZGk/YGZovtKOxNNgWclEyuRUdzSlNUSWqXDCX9brevVR88AxZF+1A6eFtVPbgg3wI+P2WIL8CvRZc4b3NGrLWAdub1uO/GTK1guY8uPH7urlPXMv+EYMM/kUCCx8eMnXUIC4FrVI2uPWiU+KkX0k+bAROfYUtJ6jHeL2cIkpjBYCEnm/kvkFYr5c4R3oN5suRPO9vHDH8aLW2D1UgGUoUr3F8vpJ8+w5Px9vUkjRHF6swDqX8gyAkHU7hgaqRjyXuV70XLviGzLzdmI5Tuv6eba8DJskMWs/93q9X2jqDUUA6isLC/OlNnqAHZWyOuXLyP+Ay+WH9i+J8OfGTb0hqJH1P6oHrV02AYYNhqpz1nDXEGBhf+4Ww9d9HbXEv2D1UqXbibeByadE6BIzEE+K/me+0Q23+mBnhxZA2I0Vr4ThX0AJ3rPEOpaasXBbM0UxpGGzqSPgmTAHIKmoA3DDku5F6R+BiqxpSCM9Z3QX+u21VJbyEgBw3oDmXKgedF1jzPfNaVxofD1YZB720Yknwxe1I32V0hrVSQAkGow5rewuNHq7zmDPnqIAqaNI74pChlpqUR5eaWZNiyrm70QYdVydFjerFIrm90Q7gVwTwMZORfqQuhSGcHrVh4RyAGm+2FsuYYDEArVaz66s4VLb7qZHzOm+8XGidKu8j12zsIL6r10lZsCv0+HlsU13opBmHtbCDzMIBZRPpegKdQY6lwJ+1oSVjR3goKF6YSvJ/xJyN6nL5x9vvOoWU//trehgB3dneCK2fWFndjNHgInGNJxJ7wTOHS42MPXxP1qRVHmC3iIVcVxBqbhBruCN/Jb5NIi/3HbaBTiOMedTMCl4WEp6U1fxPhIstUnFZtxNUsfwd6+rdsMMpK32Y3TW4FROqMmagE/ajgW3pPhY1fCuz9jgRIISnJuI8VRVbly+7KfmdfpOD5eGyXkoRTOCOdLa0wuYMoh/PgZx9FtkIhOYepmNwMMXTgI0Fle+kjSQNUToBpIiZ3uw9x93rxlPhdqZOkGtBZy0v6FPMkd6D+C/cAt6kvS4oS1XnrgIfw4XIim05BZPOy/d7WQNm+cBo/8KrdCjfZdbKgXLiDmzOOJlTyMA1pSUTqxVVhYCDhGsXH1BOEBD6Xl1MqXvGOgJJYIqfmdXA9p2AudmmbuvDJQp/cfa0bV0ZoFNH0NsoKwB0M3A+0sMnGKgaF81FPEu4q6H8uC4wUXIvQTGhsJzMC37w+Puq/1+A1L5w4Ob+SyeOELD8vwBpauyjSLwwa6w3tQxkSCvcWzzu5NleMO+usvMqEs/K6F2gdesjgWP6I84OLyz/XNLfpmGPcAu9/jLJtJL5zUC+xEfQLpALolPXRcRsviNsVTx5dtms7kYPRmrmwGfn4ojfNQhjTulm4fUBCho5hE/7QWeZx4yr00AM6Sgym7RkhHsHgnLgVp3FGlnQdJYoxaCvoAo76i3S2/WBZ7WlidC4h15T5o9JoZLFotdyEdX3tLm3r31/sIz2YUx72ykgKLWNyJEtzvBN+Wl+ip7jOazFkneAjKLiSF1cFSVxQJRO7qbr8LUiJgYNwE6UXjSJu4qYt91LCCAHtH8BFa8clINAB5CXEpWLV53XoU5K3Kjqnf+w5QINBSAxrulmHII242UjZZ94CRGcgMbIcawX/OaMcVvPi/cNr2NcQDbhHBndDOoH7iGPQD8tPmnB5h6kPjqrqL2kCwJCG1TwKpZlunSevUfQXv8h6rzoAFKlSACM2Yf5a2hs+rEeV0ZfdHUJChw+3t8Fb8at/q4Zona5puqAsjbv4H4FHXoDIgoKXbZN3kH4sKOhe2zPwIGPkPLuxEEFiYAGdqeeg//F4LDYAG1mOp7aXbgkMs8KCQ0EAdeL8g4ZcVdxh2wRRYK8FJw/FrlhiNnmYo42Fi8xa9rD1SjGyWdVWhxvNGv5bU43W/CqlnAoMwyZ2tfHxR5gM6pFNgYKRo4PzX1QMKsf4CWWNuJg+LJmRLFBeSq7c3AkVnGRqRSC8L+kUC9KUGdhVlR6F0lHwIK2Nih/b5THR/90esUZvu1p9CUd3eohnk2DIVFSwx2acpbEJ7ev1H52Es6st76cui63w4C9RiNuzPX/r2iH32WYUu4EDAFWwuj3najXxk6quTjowAuAVK6iFnH9zqseksHyBHrCcN4fAgz74qv98OD2fEp8tAJaMwgT0IwPcj8cCudWE8r7O/az/JKwkhgctmuZyxCpAcmB63eX92Z0wak6JRWmj5XbbG49bgoxoX+pFdD84h8O3Xuc1EHm/1LEr13lpBboF0tu+jvj9sooR+7wTcbU+88RF6royAjlg+HKl8LeE0cg1E4R5QdqxK63L9bASzazc/I1qyE/VJBjCuSg4ShBSeD+/mOu3LlupQg/ebasiSJc7kLzfeF97/OOZu82byuL/9bZrYd4TABAr+g7iccyw5YuzRzWXkto1r+UNLcmKRCW+QiAffM/eSOqedxzt5qVd7oQxqELHF0BoMlpLAxNz38vhzu3+c60qKwyTP3tJibOaEJbepJoAQK0IRzfd+oz/x63NLzRc9WUq8R15vJEbiFltRMRqE1jT8vfYP9Q4N4rusl6bZFI6pLn95qh4AN1WpdJ/OeaszhQe8ORO0GEUtLzuf8W/ktHyRTr+jvTdattsT1NJcLzlqfLWpr0b+ijFiJDCrcnmqwXsViEQpfJvJbAcVDoA3AHN8hsvbKbtt275330kNtGzAVs4eFQQ7rcz8amR2ztd9GjmaTDajpTFL/vUbP/gmMuAypqglPcMtKnF9T5vJT512FTr05WIoWfP5WnILs9ZdazVFDasBGjzBvfcxGA1RiNF9ahN0k+k70O5cjaiW7koScsm2lu9qXkKX3f/YYBmiouYBIdGVWwIvg4w74v+Wg+/9wWt3X3/I6+BZ8f7lt6TGCHnIHv0ytqWwOWmSAoga1UCQQQXsbZHiTzs7M6YruutPXhbqrqIoAjcDi5fMI2XsCrwj35ENXEAriHI7uoDdQsD7dvo3skAyZegUTVnRjurZCBRYVSScknO6zSVxoHbhvF/tLgfaaysc0lKUFr6smD1PsHyTcNsbvwAOU8f+rLIB1JN+GgmggVW2EmrsHdGwePnXHTfFUuNtbdSn8DOeyxj7U1yfADI08lLvOV8YkfQ9M/LUdtN88o1szFeAVJhKCc0B3rG/urzWEw1dhqtUL9VGxgyE0xutJbrg1fDq0e3UUbGtfVQ/T2GG8SeEjzQ0FMhiinNdC0gCV3BYWj5o8ixUNsBbTPSheGAQdoPJKE3M8Wam8zFUwIO3UNibN8+Vj/zhChlf3ikcXwl6QBO444gc4PESh51yx5diOtzQLsaMGzSBu2SHoh0sb52Pez9HyxtV/DrEFSCUUcbzznUfNxp/+8C/TjgXF6XjgMCQV0v2qpJB0QRg7LVJfbeQp4Pwuk9n9P7qvkHkyN6lMnhfVmipx1i4AMs7QJukWuesUxuLpWZE2o3FKddCa9jZU/59huP3GArD+dBHA8eUPuxk0CA8tQASw6aTBgEP+XtO0GatC61cNj7YifGAndavbwchgH0jmj5IUrBlRwMt8lc2gtufTSPxAn9guyMj9qIEMTQzcgfDXZrN4uTfahYJpuNp5JxK69BnzUyLOJEfscqnwmMPNmrm8AA+4uVOBnyQKr2gunKJL+pVO3gGCzzsbsmmR1UkHII6WKQhkxBirnmHJQJ68wAymaRjriN/SG+sYjYL7JK4Ug64LxYC9hckT0s7+fFWB6cSyNEPj/gC/qOzVVtXRFUUh9rNQzct7YPUJgqgUxRD/J3jI5E+feWnWKvhPC0/q3K+a2UcTddhiR3hOHIGpl5HLrgL19vPhWH7Ty1gx5Pa+Y17hCIeeqRUw0aop4dKVuUWU0eBYY7IzIzz/c9biI63GgkFunnd5Q+W2bii1PyZMKTN8Kjk/DR3H5US3IeYM4gSQbYHXInJd80xXk7AAyO87qJU0VJAz3SE0JcNXEISGFsYhDVFzGs7GwQPmNW5De57nj3nWVIubp29ndmw5J+u2SISk2/wp4uspYjZ0Ns7zGlisoDQcHSTjB/t8IKHgpAlPOYjO+AwWV5tdpIlIn76caNpNwvg+/jQvmDSVnQY23R7aF3mJHHSd3fAqZ2ry6lhCZ5b5JTlmCvUOaKt4f0oqlPAZ2i6fh2cizntZAto7DPgTQ2jncJC1qG+AGKGcfWZFYdIzMtYv/MqSRx5rneT/n4ddScobB9MbosbnQ1AdqksqgbfcfY9BZFRwU/CZ0pG1Kzria8jnkxr9G0zhIS0gxytpRji7Zv+81fwmxQv33TGg0mv8G1uzn3xZuaaSAjVpYmmWx/B6o4tVz4ZTfRdW6bMQAIlY1apOJlQmCM64/fhRmA9j5eOzN7+jqHEH7bD0iHBzq3aS8ixsL3qrXq0BdFs0AY+TwYvXYguUSqCbpXr+VdpvstTOsd/A5Ech7jz/NcfAGGwWnxHTcwnn3lt2DXed8h58KnhgPvdLUUc6VmYm6GBFY2X4F7yHV2FxYvOi6kgEkc5x8D4VJChXu1tJDQKr5tKli14FV2sK/2jwYwsQ8f4IryTx5j3j5Kif3BuDS+keXnuzNsMyhRjoCAXRZifKi5LIasi/Bpf+uvn8nD4qGlSLIq12un9tWCaysZXBBPX9d1/HT/xIyh4xpoV5KoL+txrieV2A7/D9TbXf1HHe0sH4OgHNktW6JBg6NUkG/anwjWVb8W8wbDhLCaiwIZtYgSiHLxjSIs7fai47w7n5lXsWroQ/EKB1NJCvC3PuDB9T290REA0y/+isD2Q6MtU5IkAHMr8E7mVjSOeZs7KlkMX9B3llK+QavAsaqsvmt5oxhoQZQibKw2WpW6TPtnXQTyR+9J4tsAahajJYBAbNkQWPGhq4yQAqlQUla9611mYxWhHspk/MOY+A2UU1ic/+9N2bj7PlZljh1Fkw5z64Z1rxFG0AxmiInMB7KYyBX5ldUMMZ1DwFdGqP0lRRVVm5iuWuQvsA6j9GEdStGeRs7ZfW6Bl709wmPiXT0GQUuty9uvVdK+LUSNQ56CU5fav9CP9TAtP/G2x9CmG+sItejMELwuLYOjsMpRo6W/POXDVEI2UY8z57xi0KaKyqDE0jHNLjjuVUv6hpUoXI5aAYxz9dyChV7q+/p0H/wePPZ/Z28/0aPFLYW+mefnG+dg5GhCpd3zkmQwjx3l3oH8Jz+7H/PN8liUfTJ1BHOPQ7aq6q1aE03Y0+AwUHzWXRFop0EQeOChSKy6rOADYwHiqaFs0O2Zxb1x15XUmc9h675UQbwD2uQO11vW+9onxRmRN2CRkic/jM38Kfardw4WlJqlghvQVHo0XPUP8PL5HYejEal4AHCTOeOGO7M2taOcHIo3hU+WBm2MRfTjYBgL0kfUE/3a7jhP94tIS1J45SSwIaiH4+bT7bq5el9dEeCbqbwGHKSjxTgqCaN1eLWUMv8cYNRgas2zad1t6uPbgzjK6UcvgxCuOCj49a+5vYFfnXt2zJ+0mkCBIsc5KUvg2wFXOVV0Gn+ev9UtlkPxhjk2DiW4EMOUaiIcBBULOH99jfYNq2FLJilith6aoRc6KaPG/FmnpunPM7gwbAhpcYFaXEoXVzdvmGwIR1S6+/NDtJdN1CHASm7qRYTCOUS5XXkz7ElZk8CyWJsz3S8IP9Rwnr0dsLJ1PzyumdP8pFM3SDbVOw4HcxEE3z2vvYcpZbFERdXCTp7sSRAi4LzS9EUNi9vodf/4y6WAr7qs93791Wfhf5ZbMCOY1tMS/MVNuVjzWz4vvGjXckd3wai2uKUlE02F+LdJiH7ZbA4KJgEkuX/RaFiMNyHLdOXSzq4+KhsYUJ8Yqa9SLMQyVMbuHlBq6kVPmjyV8swKtEYvbkACKWklJxYG38XqdDvqCQqsJ7Ewwk2KToLSFMgkQZhn+WVcEdFoftxMSFXr7Eo6/Ev+Kr+NZbMASY9RdikUZu+o+gQDKiWqVsN6FOAc7AV4+gzXa5IKazfUzolDZput67B8xaoazAXIrUc8ZjgOcgVziyuk7DBoa8P8x/3YflQNyNbEl84z3vFv8sxKlDPqIsecZd3BPXwChCME8W/+7rxiusUZNCs0I7lUYbTemEKsvLybdx+8F0rowcIDnKZE8KmVLgXrH8XFc/Rf/09FrKrj7gvBBr/5SiXmHCXJEB83skNhAiklRjP67GAHBu9/XsJEGZRCa81MhyKWVDpYlxl7YOCAxaNaJ/f/m5v5D+SzNNhp5GaRaQpOYsbvMNEO6YOcMTWuzJKj+ARMSx6He/CP/nhwzL1/vYSPoy9UvQ/J6fD9eT3zZSqO9vOF/DJosde/lRUAQoXXNlT9z2S33XH5qUT46BVa4zOF3/2HRVzxJZus78Wu5R3m78BGyc7g0y596Qp8CmNWyqtdYpUR5PryRGvnxSghA7rE3Ea8d/hv8C35dsMkFf4nzO6pvybzo/wOt1bRQOjewWtYwS+yGAHRXQqZVlZDH6NUeK8MkRqcBhdMcRtmJlh5hI27O/PW4R8s1UAKz7q2Bh4y2wf51OU8eLVQ+LuPw99enaUa4mqOikrKKLh9mQXv0NLtjOsgGYG5y8jJCF1guXKQF6+9WkI8QTMquGGuey1PBCWMTTbvpzTEs+d3RXQ6QLJOHGKACQsvWqkDXq03fiCcpo82AlxqyzZmINK2tSvzpzR61UpT1TBl6rp3ULjrKQh+bj5pa2bjxgPhal8RFlAyyzJe0JeHJtmVI40gJzs5fN3qNYtwmQvFCK4moAr+qlSu/nJdg6geHXvIDWoJkEDzfrFJQspRcLM+LL4ayaCJ/pfmw0r8w6u52wS/twzwCFNP1MAA4UDhvA9dRJUuY3rQ8C4JK2tJ12UNfzi5EK3d6iubZikQkN0BYMHaEccE788IHkFv7plYJBt+ql7MyaV4Eu+YrbaFo9N5GrNjRPlTfFJacA0Oh322m/dpbSbmJGfNowkLvokoHr2EONhshgO0undWByag0xKAU1h9ANyqZFCWk1bG3HaUMLiAuyLRuRQRLjgrGSl2zUZjgqz4zzUQr8j2guFR3uJ4cI/uhGBcITuXOuK6gIBpYIkU3jfdDNy0FK1jwAGO3fw/3lQDAuyAvRN39jEHVohFRDFP4XoM5jAXuVfzjQKnb74V/gI/onTA7ElRs6rXJsxUNzAirCf8CypmF8kC7FDweK1U6+22fS+8fcUcOEeWm8e6yoqFieVGVDckbQugUW3AizaJbk73W8FMvDsTA5r0dNEkHK1jjaHlfnuZHZlqKPz95ysG7cR3mZDfo1fg2rYyq+d2enLduXaPGNY0wm0pELqj1DGVjrTFokqw9ACDSjZti8b72TUhkCYvZRY7pFKMmnivjOKabgsSUHdDy8oU3R/FtxcocSqxBSD5HQVkUFRQfCj8a3yjtuCMVzBd2GiZ1cS0TTzm3C8wXkDM4OtxMDBd6jrPhpsAfITSIozoqhD5V7rmZFtkP7eZuH8rqTFt4IZHgtCGUFJ0U86RbYk9dwg5gQWmESdDGYgUGihbHnNbp1S8wITrYQWqRDazPtOteWvHCW17y8xC1llEfwe1Oen9CLa8NZegxaF3eXjwhuSsUen/0L2aow7T5hEt+ZVan7LilCv3fgIWpJT8vIkz6P59FLrDLAbrvju7iVIhXsSjFnbyBkqHIrB8KcejW8jJYbjJHk79YBw8s4MBOrKe/0JPpOjjodA/v/vgifvl0Dl2C72bfb0WkWyo7V/VF+v6Mb3LX6MJsee6mTLCI/PQTSjmHt0F7Koh52fp4OS8JL9heE3KpB/LbSbCl7oyAFTfkqtED+qBUPwH5T1ovqAa7O4dmroHEz5XqiNHrU+DCI9+iWL44NLqqkRRGEcMTkyUAN745HiRIkutfZSA5TdMi8w246DDtbLm0+4/f4lOzpVQILJv1aa+rXrCV8/rXDgstLKB+21W+xCpR3fzT5bQcdvukv9T0NIgauk1srg8NmAN+4jz8Jg8i1M7fPmTqndvbS5aFCK6huzXRPRfUgiacFksavhAYPaKfq3sVZ8uXwVgAGnOuTj9vR2vE6/dviDovV2bWu0v+WKtBc+lMbQm8DZ1bzy1n1iGg1ylDrjwMhvMkCFeVCu7KcziTlY7RwVDNfNhAFKmiJLin3g/io6eRc5VEpI+nFbsFuTkPkEpQHAVganvhYK4cg5Now9s93WIqFg6iIesvNanaE2Lm6W1xmJdJpfYg6NTyACvkK3g9OXAPnSBzg3cW1R1Gzy+0kVwwT2QywVEAEE5NCpyuKqn0gLlLqFd3rsnuBA2o7N8NZwg1XRzRArQdZ6qjbT7/Yn/wjr/+GmPlmExXj0rso/EGb/V48+qM+koIjMecBku2kP7pY5S9Cp7xcFHPw2MFcToT46CmjPgaxutQk8U3h/Ppb/rNupU2Km0EZVaPpui+AQo5r7q0NY4TW2ExFsgUeKiiziFkQ7p/LucErof4x+2KezWvLHToAn1vvMfF5EU3rF/+GjBO4eutUshDhdrT8aOO4vTdHZ2N8LGoslZ44C0xBZa8L0nHxuGwcrcffmMUJXxSwfZVOSkxqJuMfeGiB/y/ZsyvSrQnfW+a9wBtfJKB0I6lAsr99BdyA8mY1zg9mVH8TDuD/taLLifPtoTckU8Y4+Ye6jkkCeSUsIpcIgwl57LJ8+ThotseGwlLqsTelXwXEob5PmvnbOWPf+29jAtD0FFQIL9xpHxhtFU+RHYGPdzgqpUqTMgyLqKH1CIYnpNFvOrsMrLSTN9Y28vweRzdtLs2A6XWXq3xhS1zWgphFtbFLAwByLYi6uGSlblMRVM1NsfYQD8LE5uAnqUriLa1vcXgO/64tpfaPURoBaK1MFUXFeor1Kv0y5Xv4hmJJFGZVHg0KdYW9+7rKkvnJwsSujP4D+LcNibpGzf77zx10DX+VVj+oRg3ibutVkboQRMr2Utq8RATcFCeZqqnj9K1KdpH+dthykHTobWOKCJuU6EjtVZwpFeKgN04Fl1tmlJGG5VYQSpRu1BwEf5ClAsnZJFS0/3xJv44g3bR71mD4YEI4Hg1MMHCeIZ3p9fG58jZdkHPlmGjplt2NkxUSVs/8pcncjssmqV4XV7z5iOluC6LAOCoR6W7BBungeHtmCizZ7aBZteXK70Usyb293arVYKPeiETz1Ol6k+JS/BiUF5giYLRfmKE/DPbJnOOb1wtEJ5E9IGL6tstkc0DN2Fr0Aw2CKRh7ydCoi0tQNPX/cBOx64LgFCLnWdRrnCMogzJOGsj1Bi6lgOhuzI0Zu8shJ4hJTGECj6HcTkmi+j6ryjhDyvkUhjV8ntBAygLtjqIWTNEbLbp5bMB8W1YHFw0ThQblrHiO+dClHUXPAdtGHwkHS0lgNpne56ALvPpXz27MTWaHbu/P0h4gHGBHuyDcF0k4ADGG2JD76Lg+N//xOmjU/m1O+k4kzJde798jkQUGVvwGylVFJo9H2KQfS0MWTd3ukiu5v+Ze2TGcbeuywJ3SbJAssYMJZUCdPPPsha1Iwjg32mdv3tp471f8dGkMKeiXUdEOi4CLYW19cNxRdwpe38bxGe1M7JVqy+z1fbXO0lpuHi77E26N171v7VC4TGmt7BB7AN12yN2jCqdmvVBZqC4u+nwDDoVZ0zjqgKC9uFpYZhu7vRFMoJIYcJYMZVumBR+aCC2fveZZiduGIU4e0TWYx5jEiCN9oPulDyuwm371l0w0iYskQZlkTkkd+EQ4A1uu/Pmm1BSUNtEl8T9ysOd+IUXN2iSwZoPto32Avyn64spLbfr682kjFK7/uyhGpmidvNkMxUvwy2jDTLbbbZCSU/1zAs8TserX2Smd+rg6PEvrjRGNaZk/nNOzcJY4gEaNvRbkg//bkLup4w7V0q7r0MwNQzGSxkq21GkPTOpSri1u39+RH0/U7aqw/zKJzxp34ZEAfg+43Gb6mq7Aihkm+9naLe+w/D4XWFlGAjBxzWInn35d+ZZIC06OEHedYCLljpIifGpDMh3GM13EvquCkoMCwKDhoZpFxeHDwPM2ZZAlPxAKyw1HeRTcAkK7f8a2zlTPG1pdkQt4W6DIIcVywu3QcazYbZnrJUzgQxwUm2S1vupavADMxkJzmkRIqxVF+z1dfFVeGwu+e+pi7krwna9BRK62UG2BhTW02tD9V0dWlfqmWBjCmiR+29NGlEobOMZGzPZhJYjBngArVPcGxE15dmixOJZJpBwbYCohYzJpz81Twblf631c5miriCxZiPlDuoX81WpRhddx3b8SKViR2tFv7mKguhEo5DyBC56PKoCvkrTAvaxw7HyZCD/1VLgYeqhIM6kxY9npWjlempwcxOfbcPifQva81+9VQufy70RdDSsvF1jVXMbsU+VUcvy62/mvOVDi3HQyGE2xE/5j0wUoS1MANeouoeNDfMErFTu5S+15Z5ygvO7LwIPltn+ZBou4kiJPq4nY4A2yUqTvnmA5CR0nyojQ18OyPETk0bi8fAJQtHcjFSJf6BMWKO//zWLci4qX/+d3pDvxHn0Sihf/4a6L+h/nfpxoUIgbllVH21VNGEetqynoGn4nHm4jzfD3I0PDFaEbUTpxvsh3AAucuDh5n+cG2zDTCfWdXldyYhpJQ/EgiSVcmF5vi0F7Jsu9VZ2fdwPMH/LFDi7kitcamb4s2OhhaW3CKZBk75lbWSgIHW98Jcd4lPrSfe5IR6fNgjDHL79rBaxdoLOIo645za806iS/cV3ie628IH9jqVRjwo4vdGhEPMocvegjGnTKHVXEF/tn8J37JvqlunWpDxemBz1qumxKgGAv5H1JBJh5QipQg14QVh0463EVxJIBu1D5AQUGzid+CJFMnav6olXMl4XVkODpiEP5qZo2BoF+YrovAScuUVMC45yjxVMKwE4oVvy2QvEW/035A7q1aoqOWrj+WQ3D9CM7CZsb0Hm7H2xaZ2zaVFGUpL6cpmP4V07UdzqVuuC6I4/wvIVqwu0KwYFZIj6bgZexzyO/BpFefIDCQnyNF8VLspt/HUG5eslvWaeSILITT42vBvLF8D6qfdR56xUNuBnEqzoxtC0ynURGHpPcOLgeHkf1kdnr332g1L3lVdQ4M8b6SPCvaBNckE2KYqVq4lRe/fHyVULjbeBVW1XfZ04wO7Rtt0GcXkxAmVkjTmcVdZYcnXz/Fap557Nc8iEhyNI52Ga08VhcFpmmRoVNwjokxy4ZqVO6csTNcPnXCTmNhBL3U28fPAKU6+9+3ij4NrDOQH7uyYnFVlklR9SH0+Q+ssMy3BQRCCki0orkHDKOTVEsAcrWhTp6mOB6zeJg90n501B0P05MjnW+LWrHAUUvOz5FSnYRo8OdApsFx5tte09lhGDYXr1rwyDxgFQwUy4YA6IPuTmfajR1dmTFmJvTy98pleZ8EcaMeoidykuHoBKYblgJFqeF2Ppv9CDpTQTZw7UP4w2+xaN5Hdap/X8eqKjFig2+Ux8IpxCtxDHtXCLnHDxZHaJL2gXRwwpvn1QJIYtfJAz1ipPJAPxM4Gs6eRD1AlhLExOQ4gxjei5psju6OPdQ+A98xM39tVoQUI4XlQflBj3QiMGljdmqHkOzwEHkT0elyJjxuPV0aMIoJnT0Jr3BUt7wzeMTQdGkxzunsGEUC+2O06EjM084X7rljbJlRuY5iLepJI3+itGC7lD0MpCSN/b7BmrewG8SU2P6FaQdFbAj5weXMTj6B9RDpmQ1cAf+G9S7zJ3UQCNGgzh695qIdTx4g2DVK1BFz9X04+Lq4c4imK3yftqV59M4KL2n+AO5Whljej5bl68yQnKTQfQ3ggjAB7aerb5YS/iBvW30SglnvKGbWIC0+AiNCoZIBLfEG3ovbN/xGPd43tMpekFulAUUcQJN4fmdVen/MJrO5qC5ZrG9DyuaNdpY166R3t6JR0BGugi8A+QrZTPI18E4RUDsui1YfbN2E7sxuk6FHJLyxnU70YSHaJGjla0BbsaZWK/+0CXsTzPRDng1lQkpuuzPxQgG/87scJ/sc0Zwufn/ySEt+r8PA1hYNlHqbowXY1mnYpUnUkjcpWIJ+Bzhkr2k9KSmGg3M81wAit9Iwc8h0pvl5fcRuMM/Xms7DCs58+vTLnJaPhDhYDCCji4ECCl5FakguhDnMAx7d8eiIZhSew5knqPaT9FLkPn5Kwzh3BandecZabrBw35628ZWEckDbzpNS8MYk41aRrMHM5aOvjv6Fxl9xYs+Iy/wNrhNZMbIL3Rfn2qLgqX5yTj0xGUgkFrO64IrnYixUJp+4y2gzE1eWBbhjxuNmcBL5keRngoJgbrlygRJkxrFtqtsqWOA9PToDxeqlAKavfz2ShdF6kqQfWpPfzVmP2XvDH5pOdmqRFFqnQs8eLet+LwJO9QylQatpV5uyHR6aqM3HmSmvDG+5gIkV/84Y5ZGwCpscy7Wg2APTj2ZTMicxpxg2wYjTYZYAk0lsZQ6IDDT8DfBxRehRIMY3CsdV6ol4YGsP/ZM4UMbtRAJveyaRYqnrFo7cS/qLdK20iOSjYRxD008ErQD5MWxg3Mj9hCZTZKWprgpLg3bkZWw/edliFh5bF4cZqgUxqgDqGjWJ7udDV8bp/4CyrQDOG4OYQhaV+U8KDZA9yptOXUy72pYEVv/lsG8STeRqqXTLRonKZoz3rn0vmauMODY7zrIF2UigV4Okxi19IYQueuFcXnIu1emWZ8bbFryJMpL2mgyIQLqtd4JxQc2SxJRpXKo3e5bavnyCmUH86ckzIV5WbMV06dpGLUZVKR2kDpUdyGMKO/ILSHFUID63Oc0IGKt5HL/UUej7mkPtd5wnJPO7VytWUkHM0btte8VWzy+nHWmdlp3g2sCoxrgoBn+ri3FeqbykbDgxdvbaGJzCVo66Bp8bcwdHlA/VN4Ar3z6VDSQWERx1CBfQj1agUYs+ZoxFZwxbnq4/fOHPjlpQNrfM6qOhb7fWmoxuoMc5cAeGryyzCZg91Rx2dlAbMfi4xHPrXmjODz27OANa8SR2pwoceOIqU20GchCuKgWwz1Ra6Ho8SJXpczoJ87s8P/GzPmOF7mhRbA8VnaVVZt1sE7+MdX91i/in0xI2QbMlXeToz8YCW0J7xShbtH7zTJ3xLysB6K3AyBcz32xyuToOhEO8y/kptwqVDOAHvEC9tfDkfkH1gqZ1+phvPgM0U/R12R/ba001zq7YG3SfLQM6PKsdScJM9pTFzb4Nx5UF+cqE41mQxIpHv1ozmmogzUPwDGyBdGZbNcIl5feEnOHPJkkAy5T1RNFAP2iGZEFQbJtzNOThwlY8U61u7lbIci4wkRyhrYm3EVq+EGHnEeCnQ7TxjZ2GqjGxc38mSKVUa979k+igTVT9fxmp1Z6IWLSdSFW8q0znheJbXJOXhWjSZmGgHQfV/NPnlpfO6R1yIi8N+3FhofrAtgA4LhKQeN0DYj8+y/ZRrcfzzb+maRtq7kxMsxjYdt+urgPgL2NqE5vCsTjC8+UZcCMItuX8WRnAVUfTkvYozJowBUg9QLhHpnQqFaX/a2yUrTs2oCosuJNBCDtRTPbq3H6QvckGVD8WjWLdbXkjfIbThUwBpIGv20WTPDvLcldo69F8xp7KhJ+e/6jQ8kr1fORpqqhu2AeDPiQZ/KHKN91BucAVxVLzREO/d8liOdnqQj6yOYqYdT4J2gpHQodc+RnEiQYEWteo8FXaPwi8y+vSt+vZ0qBTDGl27KZSjqJ+WfEGRRkFtb8Ok1WAPUVZJVBggGZPpaYD8lJ+dO0WaXxfBoLpEYTb4CTRkRbcPE6t3/hHwRA3LjDMbZ0WkTpOhPlZgnhD2MKDZY+ACv5W3vXmBgBFyJk67RK11zV08S/Nudhooa/6DPD2zTkao0XkZJCXgIo3wJDdlvjGJs6G5zMuh07EhoAVmmETwFw+4cyGMX8kLU5SXvURrpQ7/Sd5UKc+qw1uudr3HKZidfYxrCv65GYFnmSR3OmnHNPOfcXtz6DRRB1mmi4jbcxUiuvkVXqKU2vxuLN40kbHEQeBJhWihmAj3K2arkrbDHX9dXDyiqgMgk64fvo/Ny8k6/n17Nf6A+J/Hp90AnufmodpLt2UCAz6ov286a28tRXp7vtWPOLCGsZ57P240mwFOErFY8ZtGZb13O+h420jRzhBG7Ve3CVXebRUN4qnTU4QCNiPpUeGJuo5fUrGtQIjMvHRdEXGDdc/z1+gzghBtVJYmo0g8s8RB3PCOFFPnFpo9daHi0/fJDfwhP0J/Q10tp6l4JyJC1+m/Tk6MGkMDxziQ35b8Uj+Bk5LcGMI2keYNSQZ51t2k0JE+X7sZZ2p21CpV6I027GcZ4A2mm5S8hxAwsT3xT3YLnyrBV114howwMf7gBAVHOgEQjAtRR1COSzx5SY3pIIdzHkk0KsYov7PggSlsDoJfi7xX2T7fBW2FR4l2oxf14PIJ+g2S9tpZmuv5aD642Infg8IYBoqshRbl/9i0/2yrjdT+4MFzwt1Txtzhi+9hqjH2wRgATMlDdt8RVVD+RzwUDSWcEWNGeZEXBIx/aZOSQ8ocIxV2xBICIBL+S/WzBc2PH2YbJe3KuWu68oQE3Cmg2ulER/HMCrfvMDyaKoKt5Ode9N4lhyfN+XqRyiCm4kEXfdybHUyhKxu0T1uAaE2JoiHes78v6g1NxycMJ3LwLr7DaaF37N3HkzS7Xtnl1YMs5LhjPZtAw4qml1StPQ2kAYQRoVVypZZaQwW0IaHmhOeb1fIoP330WWv0sQHgfWYei5z/D9lzzvLyFQpDKcVyj01/wKB409ZGKMUjP8BCQ0+dDnjs5obk+iOyeT9p+fQJr16DF/O+t4AF21ckh7bHTt2ykFfr9dG2h7F3v9oH2sWO6zdKIAayv5UgbKf1c9fHxdvWN0McJZpg3mv30NMwLjIZOm4vdRyfdNpTZWv2l3JrSXnX7aPNO5SxCbRzwr6J8BWknAmQTmORDC+4F0WFw5ChYE2Qyrpt7MOfc3xJGKVQiz92Zt/7gk6Pt8Fa8biGZQWhXCrIOhCskKzmCwJl3zxE278MauIqUVw2GW6YmTAVckoEjmNTBf563OJiOdzBsjgbJEnoB2M1S9a01IhWsmve8M80hbsAMwjGhJsTtX8KNpJ+qjkynRcq/289acZ9wpbuqidZdPQSQzQAFEYp2QrSubFLSo7JKcbH5JtyVLatXZFOJp+m9bOf5Jnu1P5dzPikevLQ93rpIrKal5Ckis990q1OreYQD1S3lYeDxbsW3JphE9nyVICYQjnWf7Nm26pZG1lJBy5M1ppBM9gBv9xzM72WCQa/iizda0DwtnflPJaq2xcmZjopxEqyz8hil67nnl5PahhPq5uH6nHQyDue8DvpapkFd2puoNG0v7A6f2qnmDEa2FzGKI8esduPCUBW3x4p8L9R5+8WGmRCdZPlKFUUWrbR4Nah+x7I59F3zBJUPEJUrWYMJHN3aAuEVmAwhOE9V0WaIP0NiqU40F4E2BG7cxcHBhgGvguc1HQcPb4MmogGKdEzNUV07SKt7b87d5kAoQTHfWxtml05Vc+yhi072I2+jCwNxOwrqRwvzJ8jnZBtzkwY+bQQwbLbhQFqzUwnN77rqwFpcDWl2sLj7iuqZLP2rzr7Oq1oRSFe+2rnqTsUn9jUI5yo1UAdfZDoGaMGbONhZvbgpMSGsrGJqoGqxBiKDFgB76mNANl9nvIDZNMSSogYlMhHF/gNIJ8DXfFinsqP8p8oDcjcxCZhIvup0wczBDQ7sgY/uD/geveLY6JCqcwLGzqOo3NlOu+Mpp9og55kUfsV07t2woR5+dY9jqSQmLWFFI8KJvlM8eVv5bjpgEtVpQwBJPnrP0kqbGwlotpCY1RD1wfc+OjQNxNpcNBi5GS6p0VMnTplPSwzr85O+kMoHs7Nk/ax6NZPjwzD3/rWFeh4FQPDnZLVmycDZNkGdz3tp/vwBYkwIWvEenRP58Wb6lf1e+POZ3upFIPQc0RxpIiRj3+ip6qVHLEgoGEtfzjxzoW/Io3FVErdi1j8pv5XTjUloeAm/q9UZKRIWSICk/Djd2Pt0Px/F1+FzaVQ99Rz/odlHSWXZ7le8nYkY+J9NT5TtmcjhQFUCJ3ERuuTge/qmlx/rQijE001D47smYZxPtPbTXTBlKaeLsITrCcdjvW2jd8i5rZg1laLW8VJDtYl81kHM/ykSt9JvZc9I+dVo9SUIuLmU/xRc/CFAaX4qm7OOFRcBYsNCLPVaIZR1hM8Vp1xxGXtADnW6dCKJpR5vjvX/hB1o8nB7lu2ghRxM5RpK//vZrMzDTnziE32ZlWpAukiOi6bf8o/OjTxykJHiCtZUMJfeCOwW2P/EQ/5UhBNHH75RAPPMSEXfyGQKXfpKZRgE8tg7BBe8ueMlWeZ3h5+P4gqFaPHxHkUQJGVC6UwCeNf4ZzcJdNW1gzFVNbBi5yfY1I1pNjio7BJsyYkx3tleYhoui1IqjfaNtmZBKHNmegsewouNLcDVts07aWlDQvUPEOD9jOOepUrWOsPEz1Cj/skiYpis4uxs/z+8d0RUXxF2fEW8S+ia2or597nErQKGC+jjD7bNc5Wd0XY6N2iVSUHZZPbF0ltl2dyvqKlfBkBSdOItVBGG3KnXAnUy2y2KfUlkGGaAyrbgO3u9m/ZoeVePL2rrhey2HSl5Dg+12oIVCnWmD1J8m3We/7ON7tLXd/3N5dh/1zOArWpi6eMaMJX9+ho1S0Mg9VWab8t1crqQVVS6pd0LWdt7U1mbToeSkNRmckBa7p6Ia2j9nuKHPRqyoRGqtBzo+HUpFEw2UiBuJkanvNZlnPMm4KXkO9fBDgQ0qxdefaL/SdbxY4v5P2Ohl7/gpj1CwB6Av0gyP3kzPUfGkRHG1r5cMVz9a8S3cDui7Hka5JjUlDyeboleDhkw5l34zmH0/GLA2kYFEBSbtOGWlz+tiqPdGR7XlbQLaQXYeZXvYeeKfZEVqibEq6h5ygaJ68M2euD0fojVM6FLaVZgRGAF3xXG9wX4mi9bJMCVDDWRXjbm5yBCMzLkDAZsqM0ZRP9wYRfkajmcZ6acTGIF7c5nTPjBWziKwiV0d2JWnQu669h0n93ni4g22dK3c96EcuWov+GmuxoQG9CaqSeFPF/Uzoh+r/9DXvq1EUc2pcBkK7B1KElzjUpak7fzhRSZqGV4nuW9e4vsBdTYc5rw5JE19LQPw56d9YMmsHDO6wPcYz5ZwNxF1KY2YqpIgb2Pa7d8c1MX//jdJDDfNbi0kZBtLnVnKJLSJenoFT0xnn0E0bAqGgXtsHWtfdTczg8vwgcqHnSxCMrDRx5p7KiVpSGxlG8h3HqeHw0uy1MxsjgPt230rK9DzN2ELVVDf8oVhArksDGwkbqFiTYKZE3BjB7Rx1HZiAytpPwTTudIRLoZKhCPIP5L9GnDToygtLs3uc57ulMh7m0OZZ145y5o6IKu5yZOZ3t+f2ROzuhhVjp/WzK0Du8I2qugPx0JAG47y5wDLSorqz7TU5X8OByKdJRNRnkFAXf02Bjb20i5YAMCrAwTIMnQlFWv70C/P6/H/lttovtxUEvByp1sR+AHgtsjX+8waY1aCchwgxxWRct1AmSBkAe+InclgMi01gAhJ1pshbLTS5rs4u4FLpZVes3yMRrDpFGTXJ9VH7RIQWxzSxz8jutVcArPURnRoDo68g2Q/jlqsAr4WdUAHOmWLE2vZOoqreH5+DxZe5/hDVPsretYcCrpPpoAb1kxq5UFhLD5zxJQ9YSS/08EEQTyUdC20D70royU+6JPSM4puRNQQOurcq1Uj4hahuaIvVAl5VPr9o0do8kMdONHuUs19u2yJz+xTmQF5sQecbTknXAAlqSFwEil65guxmgaulmfyYdZTtHk31FbYLPwVvBfO9kJ8dsYUr/ldI4RKOy+UwUcu7NCWJPIYj/GPONdt22aqApbbwwIWQp5WxBcK30dTL/M9XnBUinpCmoEdBOpYgEqqlyVa2c+lfZWJJvpKpu73Xb1e0YUA51Jfd+enKn5zH+S4YUA1Y3JUB47rFiJh/BDilozYmAiFoTW8h6IVZxfwcIQuMH7kGXy7urMu8XtO1q1oWbmx+rlOOOzinxQcqNT1jXPzw9WaELBb3GviN2JzTi8Oh5V2CQLmIZW1V/9uxJgWSciwbZZnB+PPZSpsKk8zdOavR4Wmr6u96n3MwZtosR1e8C82QdBZSjS3+ReLehP/uaz/qfQY3EsOfrEEVmYNzkjGLaYApfMX0fTHuuEbeVG0s0FXPyqi5uEiaNsaLnQVapUStxVYZz3spzCOsonNaWyYSV/b/hGTIhwA806vDbrWGE/xchdUOA9BQ9qZKeI73xiG76Myz8/nt/q3FDrc/DDEBbbuRw9rBsCfwkmSD03WdyUPIy1EixXksPUOBlcmzDszkvTciJk4X9OJhQ9JUkEsI/89aSUdS0llnkjoNyIgzXHeFCLJBZtChvr79EOcHpqti4/oiZGVb3dhhWI7TecmKd/9gBXcax264TGq3Z2q5zumdRLPzt+qjUgF+BlIqftbTdXaRhdTyyZybwgya6moBd0wQ5bpy6VfnXbEwQCYjfN2DgLgJ3+gANDLqnvR2Rt3C3aw7zqAOCJYm4Anl4Vg/KJs21NwxneYOCDlZ1QyhF3EU95UlOWC5CDoTusY0OHC1i25zBlsVhvGW0S6z4kDfuMu32ER8lxlRsWG0jLLLMfT8LdjkgRsZFzY1SCzOT/Zgy6AThkr2cUKb5I0HkUnejsNia3szhHlZ+TvHX63xKLWZ7+kBqvroOcm5OGH3U9MKuPRdD9mUY66ueSTFwcYVdmd02F+MOSjv4frOzc3M1tCZC34phOUT9TkxwxXh/MyiJXCZsgRaI/mOpOvQcscRy/bqK3AUc+PheV7OLXau6KqjUMUDRcvmI4YSM218pIfAu7sO40gZU8o65pUckC6ZBwmZVM3UXxZp43AR2/8qb2MB/YY+JFYXz1RY+rQRD4Bp5y4cbljBL3rbW8tCQqRLLYvs+Med15DF8HH4I25cDAkPHK8KRE4v0hHWtHtrr/4ef/n6miY/n50Xy8D+0Wj/3KtJyn7eW/hb/1VmgVvblyK6GFhlWAgP0Dr125zmfErpCXCPLNsk6Z2qkQn81NIfoU2G7BY8E+Op/rY4fh997p2k6cHceZ2QfQRo/vBJ0ctLkgUlQExgmksYTD8PC6pKkZRVHwa508yYtMGlRG4t7MTuzF6fVuHRobKJG2Q5AYQKOmu7VQroNwv07NmZpVpJVSom48UNlUC6ODGB22VflkUcAk+tpshSMjOClGthAXJriszu8/8MhptJl4PaJkLTszhnYC9dQDDD5GamOgrwP8sXz0LL950AbFGUTisruo/SgGLlOfj+V65Bg3U3KSlsmnPyS0Kc3SN0Dji3Q0Y3xceiUHKh5FkD/B0SVAeRZ0m29PdjsaBBHOfBx7P3svwx5iqfi+4biWKqR+jOwX9xp4IewYM5aaMuz+q+5g4xJ572fSxaLNXeYxskw+lppYcfW5sAAUncAvaz5KeQMdgPps9lMZz8UJksMzP5LBqBy3JcMQhUbwzqG06orZqYPhqkigjKA2HTeH49odae4Ibb0RwfsxdrU1TtL1IAzRWR/wEytiB1VqHYh6BT+sJJ5FshN5DXUq3c7LbnJp7s/9+SqKmkXmOGL0DD8bFDI1Grf18OOaFH6mXP+1XfQ6nhuvk6sceLe3/Ww0LDhtmTKbMpeOWTxHRJJWkSjccGmx8PsoBSFBYUw5EOkEkMLrF96wdItlsSYxibSX5dLjVrMRipmOaszftwYNJv+os5fFlpMyZRy6/a3auVt+u7IIdA1GhyLTHBhLFeru/c2qbh7PCEXeclB+h4huuYnXsCMyX/11d+HNsy7pr3Kzy4XBYkIfy8lyWuKCJnMoP+BgAY/nYA6BxvyXZufTmZCJhmEO7OtPiOnbGwJ/IQK6Nvx3OosQm3vzmRuNS+t2/d+JFRb8/eNkow5S00m00SjMuF3ALyypZdLLnavJOjfdSDRRVOSgR51gX1YE4nYExXx9QY2J5hYVO5NOHY64/o1VDdSDkQCOWw8beHzTqkqXzK4j4g5QyA+1UbKlAcT9r6x5RiHi/B60xLWRiLJXAQ4olcWwlREzMYa69BxChYdNufu8Tx3mrS5Ti76BGIRl6kcS6hNEChmCcjoQ1fseFUG/LJQyndo3cAEic75pgcr8Ora2lnOb76lykmWJ7iNJP07VRHJ7xntUUOwB6SjFfSUUUM/b0sTCeFNZ8sz8BI0iLrVhUUeVo/FBiP3vPeBuNnkpcgehVsVXmXAcsIdx9DmuiJSGpCGoAxFNnDFxE0M4kPiEsIWxKFKW+g19m7R2J2lsKtIS8VJdJb/zoHc7Cb4cLxaEAImgiTKgjHagpKXhAO1x2fmS9bpsZfV1bbl9jzkyT00/7bNhiqybV5ZvO3SOoWEJfevDOIgZjFiZ4vAjF3T9LZupf9mdlTNjcxGqCdK3kKF5MgF4LOGSLv7fcYu6P+YnyJAGSFUWv3BguJFLB6DwFYv7VvfHNT88dmodHY7W8ZMPqSZLf1dXsqmhTDiOBA68/wiN7FYhcuj+6KnlCgWUk7hr9KtYpx97SK2VPf9IcPoz5gzMJQwSv7EbucB/VZ/A4TgKNg3W3MC9UfDRyzjgteZF0jEnKavOMVTZImgIjFcBkN38QBvxo1798ENG3522JWkmjCj0M0n5MpoRSum8Yybteop0vwrRG/R2eui75aBWnMuudYc3nrzsWTJA0yEey/dEezxgptKLgi4HhWDRGb0vqxmR4iCkO8xScG55u2tGWELzue3kOj9BCBwaoFh/QBFsg60+CH+7RlaJ7XS3j77A+5F4YtfG+/gB4MSV8m1K7J04GFWJva86o5kY0BeF11QXccX0+2eL2VxmtY4LKJai3cbWWDBzV3dh9HhPvZFbuKR1lyySfN5Laqlegfr+OrBFaD19QxK38nIuec3c0a3bENzSU09EW8b7ydyAUigeyeRQEO7MW61iSB9G52a6gJNckIVesJLw3krsz8KR48yGtW7pBXP1u/S/+UYgFIIQUAYB1jNT3SiCtBrXcVDFEbkrawy9kN+C8Rn9MKO16ym72Lbi0ZwoECF02aEXfVcII/Atb5xrxDVt6PKZiE/XI/CBnzKbYg6L8DVbuptM6afVUnBqnrhtChWB5sQvNOaG2sQubhGh/15nNfUlfIicT73FjdRNl8V76aEM0YmapTTxbqldtebsNMdUCiarX1noHhsFx+XJ1nqe/Zs9wgJNJDxg2vlK1PeZjGqV1GD8yiah2GlRBuCSug1VgwG0QYjhIAXLREideEFE+SwW5G+0W7rOOZBtuzegDnM73HG8cFiD2pEZoMDpQe/duq4aHHqcxH/vtWSHgDvRTV3wkpPSklC5ZN5DRPMQWhUWFJd/5xEUEbeHGBgtNFQKnzhDWx9jEVi15kmn6on1JSh5kNOiF4WSRYUaRe+Gt8qwUXK+5W/9VbnobkGL7gsdy5a67MSLgvLVaSOlFJ7UFigXTyOt8bYMQ0hu/f36PoZR+EwP5ak+KwRgPclyJG1yFEYpJzMKrtOLEMZ0qYKB4mX2WME4K9RYNecvvHZ+W+4glnLnUG/ZXltIEHdF0zWXUsk5Vih/KLTC4rl8VO5cU5c2aHxgt4l9l1UeqloVF58laUe1T76qvlq6DlZGxXbLWTTZLEYLy9xaBdhT3EDdhF17ceqo+5lNogspzIiBHKlWD0/ptyLjdc0384uZD4uQUw89jt5QFj/bzSv0PKNU/+bEqJlZW0LL78P491fQ/VaRrOK7+kiV4H+nzkAdmdOnbmV+NzyUpf/UND/zM734UqIGHS443kUjMRGGyH1uOTiizVGAf/VfJCFptaeUvVbP1MZxtGVjbUVDvmxoFl7/qpngIxxZXfHsfkuEv83jG75kpAX0nJZ+yN78dggX7deDdgQZ/oFY/B2kd8umPUuK55ww6kFsIpobpvdvwXfzJ1xtNaAQ94a7UsueiwN3ZcY7nMlFSc3J8pp9eumoi0On0cN3WbxG4y918gR5xfCOwgo64HU9gmHxhdXL5XvrMS+8UeXFywB9+t2U2aRv8tiJY4Po2W4fsuJdDbc9elO9dLZly5RsrzAhXgbUc/dRgrcF4rHz23ub0WrbnajI0Td5TasD71LcAwuAWeOsdT8HZG6vcdC9H/VzyulSkFveq3niLiBB+lTHapBNl9xZgPhKKLDydmdEmme/AsBhdxyfglKPifS99+BHdTqRGvvhN1Z7kM8f7WC9pQ2Nj/likRYzIFVjcENvcFjO1buRdSMCk29yM7kTca7ZDU3Oc6/ptwRmLk20k8qW0K+BcAlhrr/jw8IoiLILkwuyhVJKYhy92vP9EO9ukq2hDBKH+cMveWnpDDa5Xeiw0AjN4c4oUuhLM5jmyX0HRfv/VGs/8/ODPgE+hnuytzBpM3WJeL5RVrsgJdEbkOcIK6DWtyee8v26ANuPo2MyzO9nhIShS/YIj3lBCCjD+rFHtj/P5X8ExIHrv3Hxi9A4GtIZVxednV/VNIDVHa3g0ME5QCYCWLteK1U+n6DZehjNW1BCdi6LUhDo4oGQ+gOsgxmAz1e4vPL8wxFuqVc8arbBaHjD5EHcSg8odo5nELJmUoZulKvA/lfy819iLL1Lr6ertq1Tua5ZaCfwzZz/CuUv1V7Y7IdO3i4W0CWN4n3ve0AiGLf29XkpF6K/VeTQBts2a+VVFaaR4j00sjqgmCYUTeEiC/XCmHiqhWVPyPStnEjigzNaTm/vgPH7AkeymwojDLEzMVat1Onk20FIOH2x/LD6yTm5QxD0Q2H9/10TNzgk5BlPrFpkLvnuAVWWrgEEcVNlMUDn3QvARbCI4WL1avk91OOIQYFGaoL9DhDnBSLC3ABlnvfQ+BhQMHPEHTJPEFTheFFF1FucuaNdKcdoLGcHglTpFHBF1u+9c95lmYuIvtwfRArM8SyvmLCcEeacxMS7CGSQavbMXLInH3eib1qsmrrydKVqzxGFBlTOBui4FG2KuF7aWVst1JXW2BTtYyiN+E6H02RN3/+qKGXkRVJdHWR9MqyTlWw6ocp8YiQqfSeKTZqwL0opmXWlq5+iw+NO/FPhFe1zEPTW0au8Ioy4wOiA48Zj/3ThC4GdUJRtJsfsHYZNHWeGG4LK8nt0FjeRlgk33jpJpawBgWF/U7kvuARvl4WRk6uzOpRqlfxanVv5AQrI08jW/IVXUgqGHQD0vMEWJgExinPdXplehOT/JQ38TATheQhiuLrUx/oZFpjZXKo0lTiSySDENk1Tcy2Qmt9u+qHSspK8LFkWYPICKZjxJbyfDX2q4LkmeR86aDXbLZuuaDGq0dJb3ERK+2KbhIuh/6YGJZBlW8IA576lrl4ioIdvjWLveGRDIlhOm4RQpAVDG0eB2g/iZtDGb5yD1iivjYfuRvyqP6ySDWnCPj6MxKnx6vey4j3v2s16MzYMV+64NHMPMFzn3kvp5Y9/8lRnCFQrXPIPd4VptDZcdOW+l45HQUVIspeQYoSr6DISlX9a3k45mYgDybCvWSzxTsQEXCFeleJLl60cHKa8DTKoxsW1O78x9mwOLJAatYiTnWONDzvhTklcVxjW4FW2hJpYgKLluKYLMqKaoRJgV+gVF5UyKkBOGpyvwzud+33dGq05oIPJWsKMKLkdikhLN7B1pHg8ccF8nsQWMmWxXXbNZxtRHU47wNkPl9rd1zmynx/T3a9hV/Ns1u68UhndM/PEgsmv0cKGwTlMhMC+9BwKjcfPmqXquLbHAwYZxv7gO8q2wPYAmVqtfw3v4g+WoOB7i5u5THFwuU78yu3FAlJeiuLR51IzXsQyFBiQbUQkRsVPJYDZNVOGlNPX/qQQ44lfGRBfkBaG6n7wrIQ6s/NwJtITN7Immd1cbFMwyTz7hIEGU4fLhlxV63MMsV4w6a9AJCHNkmfF0s+ivpsB/K8i4XWxH9XIfxXLZtvstXEVWTIPZwZRVSwS4X30LoUCsWZBAqyvNeukCXZee+QunXs6mQUp9ogJzXzZIuRjVY8F8bt1zvdyIk8q5X6IuW+rVZr9CUCCEe5JI/xCaFJnrSaq6cdosWu1WXyUf9t3T9Y8MOREf2guQ3oZI2uIiCHnaFrktFc8JbfM4FX2c/l+tzS8iqszmcRt3yblO+AKpGv2MjCGJN+0FDSr1Ldv6x32usZDbr48IxaJEih+7YNmozZTVNOYmJdhDHYYIIzC4YN5rEULSAlAYgwivxDG3ZQYjCpMHtJePBszH5OoGZ3k2A1ncq4E5X7yWmXTsaMo06YwV/8FLejpfINvNaJ3D+0VrD3NA3FRjZojGDlcexQhs77nhQKQ3/dzFQs+SonI/cRHeYmSaTvgO9Nl+Hjjc3BmyzZt3ovDCEHCPA7vjKKJlB/MkJXd4Dksnf6o0v168qrYujMJ+d5Lz6Oxgl3up3SX8GNxUyfLTpPMuaCmtc8aaPsbt3PUkLghTv+GtCWSwdQwfNdDwYfkFZBKKlz4x54YXBhbSDxl5Zy9dhSMVevNSGNStj3jgpOBT7AeacZkWSFfxkoBXw5IU5kykK0n8XXJEaB/7d7TJFc4JXFByb7JfK3SN56KVAD8hGmu8CYHdIqxrVZ6QvglzKVWM3ncnrm0pA3buUhNnE0zNB+nu3y28HtI4cLbmXyGQXmy2WRJTlfCmlzhVpghOkQfuyUOGa1kqz/F1FQkeVuJEKdv1tMJGfYNSRX7rze8G7jXZyIOTaiM2F2MegNYmqpzOGySDlMQ13EHNADeQ9TEzCnkI4J5v6pi6guE4Pt4UjkDB6oOWtyhLd7N3xSn93KwaYdR9luXigWi5yniYzwHZ/OWli/V+YbF/05gVmeG/dZe3PUJa/qA9H0P5qzW0OPMo2ezEYbnYuuxctAqBGy9fqZQ3XBMK5wrm4WYj2IDYfZm8d5sMmE0tlB/xQcUa8RrOeBOF/xl7h6YB99G2qph/ZeW3H0r2ZwwFVXq+cNL91KEi9MtC6dKVTA1VZctSuThdZ64lVZjZ84jq1wrKnIhxzYGTS5UlgZSDe+S/dY/KW25x2iyNPwMgLzUiQilPfiOvBLtahnSv7ym4SHzRwsvjYug6PysX9oIUP3EDjhqwyViEppLcwAjyGap5cxJVyg8wSCdQHsC1t6EpSqqnbo/fx/j0EGSIXaOFgepwDWUYz2LSjHx+MWdcM3MAQ9kJm1YX97acRUvm6LpZcl0e8jB+6rwCzWhzjhBpretGsgUC+dmklEAqLEGr2W67SA04FufFbpRpbQnxrAX8SyoA06M4Y951UZDCPEW2POo3zPezk8/rX9v2fazBF9verJ9xEQZMf4tdxMBXSTDxLQMTSQ+VtlrKPg1SLfXTVPXNdr/wTAaIeqjqdsDIg31E0V0BEiZ3lWS4RQV1ZXWfWCVLyXv93rtKimn/G2/a5tyWMGXp6YlSlZtRJzGcyolzkfpg60fftiszxCOHfvFzuRdZiTQBCL8tbrowE2UWn6CRE9I/UjYUVba1XRrmD42xSdsKzQJPnD7o87gQkDYhD7B5JV8D9U4YI2+dEHEinerU4XeeCk3m+sytbPs6/egbNh58EnlGiuG3XbFSyiT8fnXd0sbKLT0G46A9GiFiL1Yu8cvgz2glOSke/CSJQ90o6xSt5Jnlq76nhPOo/9cBTj79r4jtsQDG+faEGyXLMdb5krcbtrRJT1o7Sr5RFDzky7eQHdhYuqdvuNVAvGEsnKWGAu1sianbslwNzWbTS6MEpR3eRnE+hW3CL4mybOTw0UwlV3ngoqh1GNQB2t+eaz5e+1rLWs9A2iOOX9S+viaaTJuVChDVcmDdoq4rIdseHnGC9Y5FrnsEpLwYTz0MJpD4HELRlYlA8/LGl6mOtVdtERGrsBdbNxDc0q9whXpusvAYrCVqGRJdJRBeML6mH8zwPN1I6ggtqMo+Pw0XGMPRd/U5Z/Yq9Gr9nHfxcxsO9CFbpnhhP9OqJwPoq5is+tJ8ih+wXhYwb9r1XqipBiXihwMeGHeYTZNIfDSGFinBfJTYeRC4OdxzRpBLYeVuO+2ZA1DVkuutZ+SnIytn5AtgGcOIKmAnr/uffynaWnom/c7jZlkMpE34uJvqmd7oPd/82qL1WcnutP6C9MjwDuG5zpKWZaCSIYpXdhAo6XzJ+oHL2rJtdFQ6dmZosKwjo20AOdlxIREfu1jcoWWEdybPIIfJc4ajUeDF/+xoes8AfZCO7VSvgAayeVEwKMnXrbp3xqKUqA4PoTfFfssO9ayRnwU+XVvVHzjnzrDjLQZ2xSHAx48O5kxIgz92TgDV8le9+f+ymCBiUpIMkFSRVUkrNdz7pN87Nf7SkfL5GdeFL6sn8b1NcArpkmpbKfsqVX3/SBAhfIKJMnCH2w80AoVpBxDbW657Dbka/g35MF1rzlAbZMYMGI0yc6alZw4pCEWHLhCME1Nhn4l7FwWeci5uTo+AdQVQwh5+4482p9DoK4EE+qLOp+v4BeZmhS+8+HI2D4PdmPGjmsZutENTN9YxVTjd/C7OJGLcAtbj9ed/5iPTJ8pcbCllZDb/+IFFidQYEOjQsZCKWmTK39hF1SlCdm3ad8/VKEBd9OiP/y3FaFWqTX6frVPvy4BRE83Q3eCrIlunYUwZvMRwaravGVCsyJApsAa/C+e/pD+TM/lBhspzYEtvhTPfHuJuHHSr1G5irkmzU/oR5b2KTrnHfae+VP5BFk7dm2ZXpmMdu/EdONbiNPq3petLH8d/dgvr+9k8mHhkI1oYJHaStEOR//gtBvYyoRt3d5ojm+oAtsJFRlJt7zTucTc3tD28SWN7/BiSotBX6/h2yP0ukFrQFTOmya7Q5AvJZLLKOQRfWuWxkGvpvHVG7uMcHj9bM/ai0A5TmFZ4FeYbXFYjXcQVSdN2wxgz8FRX5nc6RkeNeR05Un5vbqGI1myRi5HK05oMr7jVzf6oolJ1sZKL8eax1/E+xfcZM72TRvIvnMKOUebuN0V6vUbu3ISsCBQUTnijNmHE5TM/CmAloPE2C4W1O5ZJPOQqAaId1oT9gKNJpeHhki6VmYv3vizpewBvyZ6UMWVd8UabZpUDg1AdbKg00hdC2p7thMsUo3FUgqAvf2veaiUIP7TL5uaTgG/bmXpqWad87hNL0pqmUK+RSNobRF7jwSc7GtkJGDppWOSbN4n/NQyBrVZwlObkGAWf8XJ5cX7BCUQZB8iGzXL0vjhkHtZ9iA5TjXcdZaqyE5M58deCHIgAjTNtMs6ufiUPtp2kTnDP6ibKG013mvV3o36Tnfm5NgGqyao2GXql5atqXUTCGM2S8BoxcF/2nGZX9z9pucAK34Hcw+qdY2dL3bwUSa1E0EQNvA47QfnFXy/R/ASe2+J/PPYfqUZrzd5uiizTBKwiigKbvwTBq0KAIyqPEGGBpCZCb0bRb5zZjj/l/fyAAGLtnNBOQALqB2tePtDOKc175HaMprLCNsfcqmbFNHJx9Tsl4Haz0Sqd3mDTmlF6HW/aowd8L5kQV+ht5KRDAtZAATqa3ct8+9Foa5hQLuOP8AibkFewBO/dPxocMRICJiGcd92FnXnG8pZJ4FW9Q5kZqvjbwu79Q0BvSDRnOLHsmib1g1eB16Dy9mUMQ30+MB4SJyGk0LQu4pY6I+JpNM32EFKnk/gUojp56iIU0tQsGUV0W4DtNucHIce2kvmvgtFop2H7z7OGYEco3fI2HPwitePwdh381LkZdvMl60s46I4cbC+s+uZnoM9WA2+LOVKvXbni0nn97zw3gu50hq6GfB1PKWCL9n5nrOn9S4XSa7Ih0lU6maTtu5hTwYD9Dxnl9DPb1Wye7YIiAuUofu7XhYguV7w/i8GvSDqKtasjBnLVHv+J4EAilsA/3ERG6RWbTH0/fGss/KutFp6tBl8XcIiMQsVwlN9UrSdEGDaO572lWQcDiVkFC3uY4mORji+UK7H4gJdE99MH5V+oLdv37uMK0BfKVHCPD8vVIa4kL2Hht72Vu6mNz8DvJgAAAAA=)
(a) Cohen’s d-effect sizes 95% CI for differences in subcortical brain volumes between recurrent major depressive disorder (MDD) patients and healthy control subjects (striped pattern) and between first episode MDD patients and healthy controls (no pattern). (b) Cohen’s d-effect sizes 95% CI for differences in subcortical brain volumes between early onset (⩽21) MDD patients and healthy control subjects (no pattern) and between later onset (>21) MDD patients and healthy controls (striped pattern). Effect sizes were corrected for age, sex and intracranial volume (ICV). P<0.05 corrected, P<0.05. CI, confidence interval.
Impacts of acute stress
![[[@Musazzi2017-lr]](https://doi.org/10.1016/j.tins.2017.07.002)](data:image/jpeg;base64,/9j/4AAQSkZJRgABAQEB9AH0AAD/2wBDAAMCAgICAgMCAgIDAwMDBAYEBAQEBAgGBgUGCQgKCgkICQkKDA8MCgsOCwkJDRENDg8QEBEQCgwSExIQEw8QEBD/2wBDAQMDAwQDBAgEBAgQCwkLEBAQEBAQEBAQEBAQEBAQEBAQEBAQEBAQEBAQEBAQEBAQEBAQEBAQEBAQEBAQEBAQEBD/wAARCA7MBiYDASIAAhEBAxEB/8QAHgABAAEDBQEAAAAAAAAAAAAAAAMBAgQFBgcICQr/xAB6EAABAwMCBAMEAwgKCgkOAhsBAAIDBAURBiEHCBIxE0FRCRQiYTJxgRUjOFeRobG0FhgZQnaUtcHT1DM0UlRzdJOVltEXJDU3VnekpbIlKEdiZnJ1goWSs9Lh8CY2Q0ZTVWNkhMTxOURloqPCJ0VYg8NnhqbFKePk/8QAFwEBAQEBAAAAAAAAAAAAAAAAAAECA//EACURAQEBAQEBAAIDAQACAwEAAAABESExQQJhElFxIjJCA4Ghsf/aAAwDAQACEQMRAD8A9U0REBYFberNbpm09wu1HSyyDLGTTtY5wzjIBIJ39FkVdVT0NLNW1k7IKenjdLLLI4BrGNGXOJOwAAJJ+S+ffVF5vXPrzysbDLUe5651LFQUQweqjssR6Q4NPYspYnSOAwC7rO2Sg+g9FiW230dot9LardAIaSihZTwRAkiONjQ1rQTk4AAG5PZcd8yvGmh5euBuruL1ZTx1T7BQl9HTSuLWVNZI5sVPE4jfpdK9gONw3J8kHJ6LxZZ7armgEzXScPuFpiDgS1ttuIcW53APvpAOPPB+peufBziXaOMfCvSvFKxgMo9T2qnuIiDuowPewGSEnzMb+th+bSg3msCkvVnuFS+joLtR1M8YLnxQzte9oBAJIByMEgb+ayp4IqqCSnnYHxTNMb2ns5pGCPyFfP3U1N85AeeyqNrdU+5aI1GW+Fkl9bY6gBwjOThzn0kwwTkB+DuWhB9BKLFttxobxbqW72uqjqqOthZUU88ZyyWJ7Q5jmnzBBBB9CspAREQaffL1a9NWW4aivdWKW3WqllraydzS4RQRML3uIaCThrScAE7bArgF3tEeTRjix3G+gDmkgj7mV+xB3/8AkC5J5jo/F5euKEXUR16Mvbc+YzQzBfNBOzw5pI856HlufXBQfQt+6J8mP48aD/Ndf/QJ+6J8mP48aD/Ndf8A0C+eVEH0NfuifJj+PGg/zXX/ANAn7onyY/jxoP8ANdf/AEC+eVEH0NfuifJj+PGg/wA11/8AQJ+6J8mP48aD/Ndf/QL55UQfQ1+6J8mP48aD/Ndf/QJ+6J8mP48aD/Ndf/QL55UQfQ1+6J8mP48aD/Ndf/QJ+6J8mP48aD/Ndf8A0C+eVEH0NfuifJj+PGg/zXX/ANAn7onyY/jxoP8ANdf/AEC+eVEH0NfuifJj+PGg/wA11/8AQJ+6J8mP48aD/Ndf/QL55UQfQ1+6J8mP48aD/Ndf/QJ+6J8mP48aD/Ndf/QL55UQfQ3+6I8mf48KD/Ndf/QKn7olyZ/jwoP82V/9Avnn6DjOQqFhCD6Gf3RLky/HhQf5sr/6BP3RLky/HhQf5sr/AOgXzzdJTpKD6Gf3RPkx/HjQf5rr/wCgT90T5Mfx40H+a6/+gXzynZBug+hv90R5M/x4UH+bK/8AoFT90R5M84/2cKDJ/wDvXX/0C+ejwjjOQqCMlB9Do9oTycu3bxtoCP8AwbXf0Csd7Q7k2YcP43UAP/gyv/oF89IpiRnrCvFDIRkOBQfQl+6I8mf48KD/ADXX/wBAr2e0L5OHjLeN1AR/4Mrv6BfPI+IsOCQUbH1Z+IDCD6GT7Q/k1acO430A/wDJlf8A0CN9odybO+jxvoD/AOTK7+gXzy9BzjZVMZHchB9DR9obybDvxvoB/wCTK7+gVv7ojyZ/jwoP811/9Avnn8P/ALYKogcfMIPoX/dEeTP8eFB/muv/AKBP3RHkz/HfQf5sr/6BfPOYiO5Ctxsg+hr90R5M/wAeFB/muv8A6BVHtD+TU9uN1Af/ACZX/wBAvnjAyqlpHmEH0OfuhvJt+O+g/wA2V39AqH2h/JqO/G+g/wA2V/8AQL55A3PmFd4Lj2IKD6GP3RHkz/HhQf5rr/6BXN9obybO3HG+gI/8GV39Avnn93d/dBBA7zcEH0MfuhfJx+O6g/zZXf0CofaIcmjTg8b6Af8Akyv/AKBfPOYSPMIIHEZyEH0Ln2iXJmO/HCg/zZX/ANAqD2iXJke3HCg/zZX/ANAvnnMZb3IVWxF3YhB9DH7ojyZj/s4UH+bK/wDoFT90S5M/x4UH+bK/+gXz0GIjzCt6Sg+hr90R5M/x4UH+bK/+gT90R5M/x30H+bK/+gXzzxxOkIAOM+qvdSyN7kIPoWHtDeTY7jjfQf5srv6BVPtDOTYDJ430GP8AwZXf0C+eUxkdyFTw8/vgg+hke0Q5NHHA430BP/gyv/oFU+0P5NR3430H+bK/+gXzyBpPmpGU5fjDgM+qD6FT7RLkyHfjhQf5sr/6BP3RPkx/HjQf5rr/AOgXz2SUckbeokEKJsZccZAQfQx+6Jcmf48KD/Nlf/QJ+6Jcmf48KD/Ndf8A0C+ex9HJG3rJBCha0uOAg+hke0S5MnfR44UB/wDJlf8A0CvPtDeTYd+N9B/myv8A6BfPOaZ4APUN1b4Z/ugg+hj90R5M/wAeFB/muv8A6BP3RHkz/HhQf5rr/wCgXzylhHmmMnA80H0NfuiXJn+PCg/zZX/0Cp+6JcmX48KD/Nlf/QL563Uz2M6yQR3UQGSg+hke0T5Mj244UH+a6/8AoE/dEuTP8eFB/myv/oF89PgOAzkK3odnp2QfQwPaJcmZ7ccKD/Ndf/QK790P5NSf9++g/wA2V/8AQL56XUr2N6i4YUQac9x+VB9DjvaG8mzRk8bqAD/wZXf0CsHtEuTI9uOFB/myv/oF89TIC/s8KQ0Mg/fBB9CX7ojyZ/jwoP8ANlf/AECfuiPJn+PCg/zZX/0C+eh0DmnBcFb0Hq6RhB9DP7ojyZn/ALN9B/myv/oE/dEOTQf9m+g/zZX/ANAvns9xk6evqGFGYSPMIPoY/dD+TX8d9B/myv8A6BWn2iXJkO/HCg/zZX/0C+egQknAIVfAdjOQg+hb90T5Mfx40H+a6/8AoFX90S5M/wAeFB/myv8A6BfPQIXE4yAr/dH9+oIPoT/dE+TL8eFB/muv/oFUe0R5M3duN9Af/Jlf/QL56TTvHchRlhacFB9Dg9oZybntxuoP82V39ArXe0R5M2nDuN9AP/Jlf/QL55gzIzkKnTvgFB9DP7onyY/jxoP811/9An7onyZH/s40H+bK/wDoF89rKKV46sgKKSJ0Rw7CD6GR7Q/k1P0eN9Af/Jlf/QKv7obybD/s30H+bK7+gXzx4VWsLuxCD6GD7RLkzHfjhQf5sr/6BP3RPkx/HjQf5rr/AOgXzzdBzhXiBxHUCEH0LfuifJj+PGg/zXX/ANAtU0nzz8qWudTWzR+lOL1FX3i8VUVFRUrLfWMM08jg1jAXQhoyXAZJA33wvnSc0tOCuZOTWLxebDhCOst6daWd314q4zj7UH0hoiICIiDQNb620tw50vX611rdmWyyWxrX1dW+N7xEHPaxpIYC45c5o2B7rhD90T5Mj/2caD/Nlf8A0Cp7RWH3jkw4mRdZZ1UdFuO4xX05/mXzzgEnAQfQz+6J8mP48aD/ADXX/wBAn7onyY/jxoP811/9AvnlwVcIy7thB9DLfaI8mbjhvHCgJ/8ABlf/AECuPtDOTcbnjdQAf+DK7+gXzyGMjuVQjHmEH0M/uifJkP8As4UH+bK/+gT90T5Mfx40H+a6/wDoF88qIPoa/dE+TH8eNB/muv8A6BP3RPkx/HjQf5rr/wCgXzyog+hr90T5Mfx40H+a6/8AoE/dE+TH8eNB/muv/oF88qIPoa/dE+TH8eNB/muv/oE/dE+TH8eNB/muv/oF88qIPoa/dE+TH8eNB/muv/oE/dE+TH8eNB/muv8A6BfPKiD6Gv3RPkx/HjQf5rr/AOgT90T5Mfx40H+a6/8AoF88qIPoa/dE+TH8eNB/muv/AKBP3RPkx/HjQf5rr/6BfPKiD6Gv3RPkx/HjQf5rr/6BP3RPkx/HjQf5rr/6BfPKg3QfQ3+6I8mf48KD/Nlf/QKo9odyantxuoD/AOTK/wDoF883hHGchUEZ9QEH0OfuhXJz+O2g/wA2139ArXe0O5Nm9+N9AP8AyZXf0C+ejwD/AHYUbm9LunOUH0Nn2hvJsP8As30H+bK7+gVP3RDk0/HfQf5sr/6BfPW2le4dQI/OrHwuYdyCg+hf90R5M/x4UH+bK/8AoE/dEeTMnH+zhQf5sr/6BfPP4ZxnIVzKd7xkEBB9DH7oZyb/AI7qD/Nld/QIfaGcm43PG6g/zZX/ANAvno92dnHW1VFI47B4yg+hM+0T5MgcHjhQf5rr/wCgQe0T5Mj/ANnCg/zZX/0C+e9tvlduHDChkhdEcO9cIPoW/dEuTL8eFB/myv8A6BP3RPkyJwOOFBk//euv/oF883SVeIHu7YQfQ0PaE8nJGRxtoCP/AAbXf0Csd7Q/k1acO430AP8A4Mr/AOgXz0CE9i8BWuZg46gd8IPobHtC+Tg7jjdQY/8ABld/QKh9odyajvxuoB/5Mr/6BfPOISRnqCoY8eYQfQx+6JcmY/7OFB/myv8A6BU/dE+TL8eFB/muv/oF88qlZTPe3qDgg+hb90Q5ND/2b6D/ADZX/wBAqH2iXJkO/HCg/wA2V/8AQL56HROYcEhXMpnPGQ4IPoVHtEuTM9uOFB/myv8A6BD7RLkyHfjhQf5sr/6BfPS6Bzc5I2VnSUH0M/uifJj+PGg/zXX/ANAr2e0M5N5N2cbqA/8Akyu/oF88Suazq8wEH0Nv9oZybx7v43UA/wDJld/QKz90T5Mfx40H+a6/+gXz1spi/wCi8ZVksTondLkH0MN9ohyaPOGcb6An/wAGV/8AQKrvaHcmzPpcbqAf+TK/+gXzysZ1HuApm0T3Yw4boPoUHtD+TVxw3jfQE/8Agyv/AKBHe0Q5NG/S430A/wDJlf8A0C+emWmfFu5wP1KHHzQfQ1+6J8mP48aD/Ndf/QK5vtDuTV/0eN1Af/Jlf/QL541e2Eu8wEH0Mu9odybNGXcbqAf+TK/+gVv7onyY/jxoP811/wDQL56HROb5gqw7IPoaHtEuTI9uOFB/myv/AKBXD2hvJt+O+g/zZXf0C+eQNJ32+1OjfGQg+hs+0O5Nm/S43UA/8mV/9AqfuiPJn+PCg/zXX/0C+eYsI8wqBpJQfQz+6J8mQ/7OFB/muv8A6BB7RPkyPbjhQf5rr/6BfPQIXO7EKT3V+M5ag+hM+0S5Mx344UH+bK/+gVR7RHkzPbjhQf5sr/6BfPM9had8Kgb1HGcIPoaPtEOTUd+N9B/myv8A6BB7RHkzPbjfQf5sr/6BfPQ6EtGeoKzoPqEH0OH2hnJuBk8bqDH/AIMrv6BWH2iXJkO/HCg/zZX/ANAvnn6PmFQNJOEH0M/uifJl+PCg/wA11/8AQJ+6J8mQ/wCzhQf5sr/6BfPV7u8DOyjLCO5CD6Gf3RPkx/HjQf5rr/6BP3RPkx/HjQf5rr/6BfPKiD6Gv3RPkx/HjQf5rr/6BP3RPkx/HjQf5rr/AOgXzyog+hr90T5Mfx40H+a6/wDoE/dE+TH8eNB/muv/AKBfPKiD6Gv3RPkx/HjQf5rr/wCgT90T5Mfx40H+a6/+gXzyog+hr90T5Mfx40H+a6/+gT90T5Mfx40H+a6/+gXzyog+hr90T5Mfx40H+a6/+gT90T5Mfx40H+a6/wDoF88qIPoa/dE+TH8eNB/muv8A6BP3RPkx/HjQf5rr/wCgXzyog+hr90T5Mfx40H+a6/8AoE/dE+TH8eNB/muv/oF88qIPpN4T81nADjnqOp0lwp4i0uoLvSUT7jNTRUdTEWUzXxxukzLGxpAfLGMA5+LtsUXlj7FT8KbVP/F/XfyjbkQe1KIiDrd7RDiNNwy5POI15oal0Ndc7eyxUxacOJrZWU7+k+REUkrgRuOnIIOCvOH2NOhItRczd41lVRNdHpPTVRLA4gEsqamSOFpGe33o1AyPXHYlduvbO3OpoeVGy0tO7DLjregppxvuwUdbKMYOD8UTTvkbeuCOIPYb2v8A35LzJB/84aWKbP8AjzpG4z/gjuPTHmg9VF5f+2s4zGi0/ofgLbKvElzmk1JdmNcARDF1Q0rSO5Dnundg7Ziad/L1AXz2c1utrrzac6t8ZpKT3xt81BTaU060bsdDHI2lhc3Gfhe/qlOf/mhO3YBjcQuVa6aK5NeG/MnJBP4urL9cKWuY5pxFSPAbQEj96CaWqd1Y3E8YyNgfQb2MXGX9k/BvUnBe4VGavRNxFfQNLu9BWlzi1oP9xUMmJI2+/N2zueyPMdy4WjWvJrfuAGmqBrxZ9MQ0+n4+j4hU0EbX0oGMkFzoWsJGSQ92c5IPkD7NbjMzg1za6Tqa+r8C0asL9L3El2GhtUWiBxJ2AFSyAkns0OQfQGvGb202hIrLx70jr2mhDGan057vMRj75UUkzgXHG+fDmgG/9yPnj2ZXmD7cO2iXSvCW7mAE01wu9MJM/REsdM4txnfPhA5xt09xncOzvs1eJEvEnk40HVVlQZa3T8M2nagkk9IpJDHAMnv/ALX8D8uF2hXnn7FO5Vc/LrrG1yv6oKTWcskWc5aZKKl6gPID4AcDzJPmvQxAREQce8xH4P8AxN/gdev1KVfM/Wf21P8A4V36Svpg5iPwf+Jv8Dr1+pSr5n6z+2p/8K79JQQoiICIiAiIgIiICIiAiIgIiICqDhU7pjCCTqCocJgdKty5Be1wKqoxkEFSA5QRnugG+Srg3Ls+WVVw7YQXZGFYXhXeSoY98kFBJG4nCyo5g1oB7rEYMABXlpIJ3QRTnqdkdlECQpnjyKiIwQgkHZXBhcFcyPLckeSoXGNBcICSj/vWxKuinb5qOpd17tQRud1b5VqoM5wqoBOFLFCZAVb0Z7BZMDmxDB7oLBSH5hX+GIxgqWOYZVk5LyNkEL3dOSVZ4vdXPYSMeoUQjcPJBc14cfmr+oAbq1sXSckK4x53AQHM6xgKgHhg5UsYLTghWyhrigtYPEO3mVR8Bz57qWBgB2GVWZ3SR5II42iEglSvna4Y+SgkeHBWBu6C7HU7IQsIGUbkHsqueOkhBGxpccBTMaYyCVDEcOypS8uCCaapa5nT8lixtLXZUgjz2OVURnYYwgvnqQ6LoWGw4dkqeWE4zuohEcEoJ/G629KiMZyVbF9NZAGdkEcUJlPTnGVkfc5zSHnOO6ugaYSHuGyyn1jHRYyEGDJIHjwh3ChFMW7lXNGZy7yysqQ9TNkGIX4GP51Y1w6vtVryetG/SygzJGl8YA9FD7q4NJU0cjcDKudM0tIHmgw43mJ3fzWV72C0N81iStOS7yVjBlwQTynqBd6qFrsPDj5FZhgJj3Cw5G9LkGd700xdGR2wsZx6jkKOJocVkmItbnCCEO6DuqmXO+VGQ5ziEI6dkFwlAKmbU4bglQeHtkhWEYcgyC8E59VE8dRyFe2MkZAVrx0HCCgYVQNLXA/NSZ2yrDlxwgy46kNZ0qGb77lwUZidsd1JGwhu4KCBzS0DKuiGdklGEhKCcQlyteRF8JKnicsarOXoI3uDlzJyYgu5seEXy1naD/yuNcMLmnkt/Cx4S/wxtP63Gg+j9ERAREQdc/aHfgbcSv8AE6P9fp188jV9DvtCwTyb8SR60lH+vU6+eEeaC4Dr2Cvb8HdWwjLlLJGSgje8OBCjV5iOMqnQUFqJ2RAREQEREBERAREQEREBERAREHdBKw4CuIzuOyszsCrmEkIKdZ7K0n4gT6q90Z7+pV7IeppKC5lQA3HoEdH4oz5d1juwCd+yyYZWhu5CCIMJdhXiXwh05GVQvaHEhWPb1u6h27oLsOz1eWcq5vUD1eSs68bEqWPduPkgnZWsaME7qOWL3j4wsd8QydysiKTojwfTugxzH07dleyRrR0HurHSguPplBHlwcMoJBA5/wAQPcq19K4HKymytY0NPcK107CMEoMV56c/JRElxx81fIeo7eajA6Xb+SCdsHUPX6lI2UR/B5qkUo6d1DMMvyCgmdF42SPVUD/AAaVJE9rdj5qyePxT1DugjfJ1EqPufrVxj6cZVWtzv6IL2wZ3CskiLNvRZEbg3PzUczm5z5oLIJDE7JSdwld1BROIcRhTxxZb80ELR0kH5rLiqQ0b4WO9pb39VWKNzwMboL6moEgx6rGDSRlTS05YMkFWN3QWbjfCkbIAMKpj6uyie0tOCgvc8EKjIi87eatIwpoH9JAQVfAWMURjJ3WXUvBYFjNJOyCgHSmR6qshwFGCc90EjZMH/wBql94+HCxukqrWE7HIQXOPiHZXNhIKNaGbq8y4GwQWvaSO6hLSCVMHg5VjyM90FnSVexhBDj2CtaTlSk/BgeiCr5h04CgJycoRjKogIiICIiAiIgIiICIiAiIgIiICIiDv/wCxU/Cm1T/xf138o25E9ip+FNqn/i/rv5RtyIPalERB0N9s1aqi48ptprIcdFr1rb6ub/vDS1kI/wDwpmrhX2HN2hjr+MViknd4s8Niq4o87dLHVrZHAZ75kiBIHpnyXd/nv4azcVuUniVpOipTPWx2g3WjjYMvdNRPbVNa35u8EsA8+ojzXlz7H3iLDpDmudpOrn6INbWCstsTCcA1MJbUsPpnogmaM/3eBuQCHqfzs8ZDwJ5Ytda+pKoQXT7nm2WlwOHCtqiIInN9SwvMhHpGV4O8A+AHHXj1qGso+A+l6q7XawQsraiSnudPQupmOd0tcJZ5YwCTnAaS7YnGAcesftWOC3Mfx50pofRvBbQ0+obHQ11Vc702C4U8LxUhjY6YeHLIwuAa+oOQHYLhnHnuL2YHKlrfln4V6jq+KdijtGr9W3Rkk1I2phndDQ08fTTse+JzmdRfLO7AccBzc4OQA8+f2hftOf8Aglqj/T23/wBeXWjinwr4p8AeIEmjeJtjqdPapt4hrjF73FO5geBJHKyaB72OPnlrjggg4IIH03Lzl9qjyWcVOPmotH8TuCmjzqC7UFvns97p2VlPBIKdkni00jRM9gfh0tQ0hpLt2bYBIDuVyzcXIOO3AXRHFVksb6i+2mKSvEZbiOuZ97qWDGwAmZIAMDYDYdl0K9uJd4o7Fwjsfju8WervNWYgduljKVoc4Z75kIBx/dYPdc4ey14Ucf8Agpwf1JoDjdoufT9NHexX2Nk9ZBNI5ksYE7emJ7uhofG1w6sEmR5GV0m9srxCh1NzL2jQtHL1x6M07BFUN6s9FXVOdO4Y8swmmPrv9SDtN7FW0VNLy36tvE20dfrOdkIx3EdFSguz6EuI+tpXoQut3s8OGdRwq5QeHtjr6V8FwutC+/VjXsLXh9bI6dgcDuC2J8TSDvlu/ouyKAiIg495h/8AeA4m/wADrz+pTL5n6z+3J/8ACP8A0lfTBzEfg/8AE3+B16/UpV8z9Z/bU3+Ed+koIgRhVyFb80QXZCZCtRBdkJkK1EFSRhAdu6omMIB7qoOFRVAygrkJkKhbgdiqILshMhWoguDhnuqktViIJMgBUBBVO6qGjCC44wThR53GCqudnZWjugmaNs4RxAVGnZUz1HJQVB81dkKPONtk6vqQSq8TNbsoPEVjjvlBkuIdnHmrPBc49X5FRjsd8KZrx0790FzJmtHTjfso5YzIepqheD1E7rIhf0gbZOPRBD7tJ5AqgzGcOHkswPGMrFmHUc/NBaIevJCo5pYRlTwkNHT+lWTtB3BQXxN2yo6nIdscBXRyYGMK2oIccgoLoZCAApw9p7rEZthSg5QT/Cr2MaQoOsjYdlJHJjuUFZI87NUQe2IYd9am8QErEqRnCCYyNf22JVhY7PmooiR3WS17diUFGHwRkqKSQSuHThXTP6xgDuoYwWnuEB8bsIw9A3VxeDsfNWOHV29d0F/WH7BWvjIBPqFSMYcpiepvzCDHZ9LCmAzgKPpw4kKVnrg7IJQ3wwC5V8dn1qOSTbpKx3Eds/nQZfjxyfDkKh6cHGFiRu6XZUxeMbILQOl+fJSxvb1AntndY/Xl26kBGEGZU1DHwlrDvhacHvBGSVeSR29VbjJzugmaQAD5q4S9Q6fkoS44SP6YJygukgc34/VRZ8vNZ80jDF05GQMLTwcOQXujkY3JOyta4hwOfNTSyNdGAO+FjoMh0jHMx54ULBhwce2VRvdCcINQ98j8IN29FhSESOGPNRq6P6QOOyDJip3R/fHdlI6pjAAyqyzjwQ0YBwsA7k7oJTIzJIUZPU4YVqqO4KDIAy1QHZ6mDsDGyhdnqzhBnQ46QsWpI69vJSMkwMKKb4nZygAHAVzfhOSFRp2wUcgnbKwgD5J47AcLFa0/NUcCHeaDJdF4oy1Y72OidgrIhmDRuoaiTrf9SCSOcNxukrDKeoKNjQd89lIJAwdJQY7mlq5n5L39HNjwj+esrQPy1ca4bk33A7rmHk0BPNhwh+WtLOf+VxoPpDREQEREHXb2hLg3k54kk+VJR/r1OvnhacZX0Oe0O/A14lf4nR/r1OvniQSQvDHZKzA5sgyFp6lhl6MDKDIkYGAqAyNI2Us0ge3Y91i9KA4g9kBwqIguyEyFaiC7IVDhURBdkJkK1EAd1U4VEQAcKpwqIguyEyFaiCpIwqDcgIg2IQTiIloz2VPouGVM2Rpb9ix5Xb7IJjM0NAUkdSzo6PNYQ3O6HY7IJHRlziR2KNhkI2VzXjAB8leJg0bFBF4EhOCpGOEeGuAz2V4laM/NY8p6n9QQSugLj1DsrQ7w/hKmZKOjf0WNI7qdt5FBkBpduopM5wPVXNmHSAVY52XDt3QUFO874Kma8RMLXd+yuEwAwFiyv6znKA+RxccHurmxSPaXA7KId1kRyFrcA7ILOksOHeiubAZG9QCq7Du4V7H9LcIMU9TTjOMLJhpzI0uIVjmhzslTRyFjcDsggeDGftU0NQ3GCrJPiOT5qwMAQSylsm7f0KMODAepVBIVrx1boKGX5q5sLpzsVaIx5hZELizt9aCJ1M+L4nKWKZuFWV5kGD2ULWhpx5ZQXzPa4YCkppmRD4sK0tZ0588KAtO4GcIMqqnZM0ho3WGGlgOVI1mArcfFv2QXwytB381bM8OdkK3oA3wmG5wfRBYQTsFVrHAghSfCGbYyrY3fF8Xbsgq95IwQrWbbq+ToAzjfuomkg5PYoL5XA9vRRN7hXuIIwFa3Gcnsgma9uMbflQFvUrMNACo3uEEswLW7KABzuynmeHMwoWOIICCvhu8iqiF2d1MZG48lTrCCzo6Qqtkb9Eq5zm42WOPphBK6PO+FE4EEqcPGO+6heSTnH5EDITIVqILshWnurun61QscPJAJ2VFd0FOnHqgtRE6SfJBdkJkJ0/WnQfJAyEyFQMJ8iqEYOEFSdlREQXZCZCo1pcUc0tKDv/7FUj9tNqn+AFd/KNuRU9ip+FNqn/i/rv5RtyIPalERBY9jJWOjkYHNcC0tIyCPPI9F4L8ZeA3Ffk85ym3vh9oHUNws1i1HBqDTM9FbppoaqiMrZW0wexmHFoJgeB5g7YIz71ogw7VXsutso7rHT1NO2sgjqGw1MRjljD2h3S9h3a4ZwQdwQQsxEQEREGPWVLaKlmrJI5pGwRulLIYzJI4AEkNY0EuccYAGSdgN14O6C4H8WedTnRqbvrbQeo7PaNS6jmu9+nrbbNTx0FtY8uNOXuA6X+ExsDBknqIO+CV71Ighp4IaSCOmpoWQwwsEccbGhrWNAwGgDYAAYAHkpkRAREQce8xH4P8AxN/gdev1KVfM/Wf25P8A4R/6Svpg5iN+AHE0f9x15/UpV8z9btVzj/6I79JQRZ2wqIiAiIgIiIA7qpOVREBXsIAViZKC8uBCorfmq9RQUPdEO6IAGVUtIRn0lJjKCJVDseqoRgogr0k9vNOkpnZOooKg+R+pFb81XqKCh7oh3RARXBhPkU6CEFMhV6z5KnSe/wDMgaSgu69t+6uikxsowN8YVQ3HqgyTIMfPCjDt91aNgu1fJl7PziRzZyv1NPX/ALFdB0cxhmvc9OZX1cgPxxUkRIEhHZzyQ1pPdxBYg6rudgfDsVH8R7le0Vx5E/ZjcB6aCy8Y9Q2+S5PYMO1PrWSjq5Qf34hppYBj5hmBtk53WLJ7Or2eHMBaag8BNcRUFZC0kS6Z1ULqIXAAffoKiSV4GSCRlh3GCAQg8aAMJjPzXNPNdyuaw5TuKTuHGqrnS3WGqo2XK13OmjcyOspHvewOLHEmN4dG8OZk4IBBIIJ4aazugswR5K5XlmPJWEjyBQVBwq9X1qPq+SFyC8u/Ko35Pc+eyuxgZycq05d6oA2Cu6/rVoBHqq4HzQV6sqzqXYHjFyL8xXAvhlRcXOIWmbdS6fqpIInup7rBPLTumGYhIxrjnPbLC4A98Lr+I8+qChIwqZPplSBnoCrSz0KC1p3yVcX7KnT9aoO6C8PHmFcJAPkoyArUF0js+ay7NZbzqO509l0/aKy6XCqcWU9HRwOnnldgnDGNBc44BOAD2KwsZXan2XTS3nr4ZZ/+/P8AI9ag62al0jqvRde21ax0xdrHWujEzaa5UUlLKWHOHBkjQcHBwcY2K0jqJ2yV6Ke20/3+tCfwQH67ULzqQMHumT6q4jDR33VABhBc1wyq9TfVWEDCogu6t8nKv6xjbv6qJEFS9x7lUV3SPLKdI88oKbnZUwVe1oz5qvT9aCNFVwx29VRAVQ7CoqgBBcSSDk7KxVHUT5qvT9aC1VBGFXpHnlULd9kDrKdQTpVen60Dr9E6vrVOkJ0hBcX9/wDWjXjzVgGSq9IQSeI1WPcHHZWqoblBQEhM5VxYRvgqnT65QXNeAMZKtc7q9VcGZKtc0tKAHeuVzNyYn/rseEfz1naP1uNcMLmbky/Cx4Q/wztH63Gg+kJERAREQdc/aHfgbcS/8To/16nXzyN7r6G/aHfgbcS/8To/16nXzyN7oDuyp2VXdlRBc0nO6ucrG7FVc76kFqIiAiIgIiICIiAiIgIiICIiAiIgZI807oiAiIgZPqmT6oiCvU71QH1VEQMn1TOURAyfVMn1REDJTKIgdlID5KNXIJAcK4HzCsG67++zr9nXaeYm0O4zcZJauLQ8dVJS2y1UsroZbvLGemWR8rd2QNdlnwEOc9rwCwNy8OgqZK9mdQcwPsn+CF5rOH9Lw90Xcqig66Ssmt2jo7mzqBw6J1VIw+NuN8Pe3OcnOVpmsuX/ANnHzecMNT6q4DVNhsWpLNa6q5sOnmm3VFM5kZkAntsga0wlzQCRG0EZDJB3QePPdEXaflG9n9r7m70fftZ6X1zYLDSWSuNsEVwjmfJNUeE2TtG0hrMPaOrJOc/DsCQ6sItQ1DZKzTN/uenLg+J9VaqyahnMRJYZInlji0kAkZacEgHGNgtOJ8kFwAz8lUFWEgJkDzQSZyrSd1b1Z2VUF3Vn1VAPQfnVEQXYCoWhOop1ILSz5lWlhH1qXIKIIul2cE/mVS0+RUoaCeyqQEEHSfPKtLVkdI8v0KnT8kGMWEJgrJ6B6K0s9EELWlXYPqrunGSrScHGQgoQcf8AtViuLtvJWndBX4sef5VTceqr1FUO6Cp6vNOk9wqtOTur0Ebc/NVyFdgKxw3QURVLceqBu3mgua4Aq7rafRRDuhGCglMgKt6grekKoZnsgrjKuDmt22V3TgZwoX98IL+pvqrhK0DdQogyOtnyVvgmTcfoUTd9iVlRPDQAghdA4KIjCzXODh5dliyjDkF0ThGcuVszw92QrCSUQd//AGKn4U2qf+L+u/lG3InsVPwptU/8X9d/KNuRB7UoiICIiAiIgIiICIiAiIgIiIOPuYf/AHgOJn8Dr1+pSr5nq3+3aj/Cv/SV9MPMP/vAcTP4HXr9SlXzPVv9u1H+Ff8ApKCFERAREQEREBERAREQEREBERBVpwcq/qCjVW9kFD3RD3RAREQEREBERBK17RuqmRh8goUQS9bfknWxRIgvL25VQQSFGqsGSg5A4E8Kbpxx4waT4T2eR0U+pbnFRvmAyYIN3TTY8wyJr3keYavaLnc47WvkY5YrNpDhFRU9rvNfGzTmloWRtcKGGKPM1WQRh7mN6dyCTJKxzgQXLz+9j/YIrxzfR3GSmMjrHpm5V7HdOfCc4xU/Vny2qC3P/bY81vz21moqyr46aD0g9zjTWzSZuUYzsH1NZPG8gZ2OKRmTjfA9Ng89rxebxqO61d+1BdKu5XKvldPVVlXO6WaeRxyXPe4kuJO5JJJWTpzUuoNIXmk1Hpa+V9nutDIJaatoah0E8Lx5se0gg9xsexwtO6CqFuPNByTx05ieKXMbe7NqPiveobncbHaIrNBUR0zIDJEx739cgaADI50jiXAAHA2GFtXQWgta8T9T0mjeHul7hf73WuIgoqGEySOA7uONmsA3LiQANyQN1t/svbflG4c6D5E+Sys4760tTXagudlZqS+z9IFQ9soBorcxxHwgeJEzB28WR5O2AA6eaU9jjzQ3+3x1uo9RaG00+RuTSVNxnqamN22A7wIXRYwTu2Q7jtg5WwOMnsuOa/hBZKvUosFo1ja6FhlqZNM1b6mWKIDJcYJWRyuAA3DGOxuewJWgcUfaOc2/EnU9VfKfixdtK0MkrnUlqsEvukFLEScM6m4fIQDu57iSfQYA7Wezo9o3xS1fxStvArj1f/2RU2oy+CyXqaFjKumqw0ubBK6NoErJMFoc4F7XloLi0/CHlwQWkhwIIOCCMEH0XKfLhy38RuaTXVXw84YG1NudDa5bvO65VRgibTxyxRE9Qa4k9U8YAAPcnYArtN7XflwsfCni7ZuLGjbZHQ2riFHUPuEELcRR3WFzTLIANm+KyVjsAbvbK7zOLvYr5/bTapz/AMAK7+Ubcg65UPJrzA3njneuXvTujfu1qrTszI7o+inBoaNj2te2WWpd0sYwteCC7BJyAC7ZdjZ/Yycz0ViFxi1hw8muPSHm3NuFWHYxnoEhpgwvzgYJDe56sAZ7Qc+nO9pvlHuV54c8B7FbY+KWrpGXfUF1fTCRlAXxNZFLIHAiaoMbGBjHZYxgBIIIaekHBr2oHNJoTiPbtQa94i1mr9OS1Tfuxaa+nhLZKZzvjMJYxpikaCSzpIbkAEFuyDrlxV4UcQ+Ces6vh/xR0tV2C+UQD3U9RgtkjJIbJE9pLJYyQQHsJaSCM5BA5Q5eOR3j5zQaWuOs+FlqtMtptdwNsmnr7i2n6qkRskcxgIJOGSxknAHxDGd8ej/thOGunNa8tdn4w0UEL7npW6Upp65oAdJb6z4Hx5OCQZDA8emDgDJK8tuEXNbzCcB7BV6X4ScTa/Tlqrqw19RSwQQSsfUFjGGT77G4glsbAcEDDQg9rudvgBxD478rMfCPQNPQS39tTbJC2rqhDF0wY8T4yD6bbbryb41ezu5k+AXDm5cU9fWixCw2iSBlZJRXRs8kQmmbCx3RgEgySMacZI6gcYyR6mc/3GHiVwk5P4+IvDrVVRZdRmqtLDXxRRvf0y/2QYe1zd/Pb6sLx94kc6HM9xf0fWaA4j8W7letP3F0T6qhkpqaNsxikbIzJjia4gPY1wGcZaPRBp3Ljy4cROaTXVZw94Ym1NudDa5bvO65VJghbTxyRRE9Qa4k9c8YAA8ydgCt16Z5GOYXWHHS/wDADT+m6Sqv2lpGMvVe2pxbKFr2B7HyVHTsHAjDQ0vJBw09Jxzz7Fj8KPVP8AK7+Ubcu3PPrzg6d5NKCt0twbs9rHE/iDUOvVwqnsbIKNnQyEVc7DnxJHMiEcTD8IEbnEENDXh5/cyPs0uPHLVw4qeK2pr9pO+WC3vp4rg601VQZaV0sjYmOLJYWAsMj2NDgScuGWhdWbDYL7qu80enNMWetut0uEogpKKigdNPPIezGMaCXE+gC504j8+vNBxc4Y3nhJxL4gRX+w32aCep8W2U0E7TFM2drWvgYz4fEYw4II+EAYAwvQj2VvALR3BzgFceazX1LBHdr7SVlbDWzsybZY6bq6yzuWmQxSSOI3LBENt8h1V4fex/5rtY2yC7aiqNI6NZO0P90u1xklq2ggkZZTxSMBzgEOeCM7jIIWFxL9kbzZ6CtdTebFTaa1rBTNMhp7FXye9lg74hqI4up2N+lhcT2AJ2WkcwPtOOZXivrWvrNB67uehNKxzuba7ZaHtgmbAD8L552jxJJHAAuHUGAkgADvyHyU+0341ab4p2LQnHPWFRq7R2oKyG2vq7g1rqy1yyuDGVDZmtD5GBxb1teXfDktwchwdBayirLVW1FtudFPSVlLI6GeCeMxyRSNOHNc1wBaQQQQQCDsV2k9l49p56eGYHn92f5HrV2j9svy42KgpbBzKaYtbKatrK1ti1IYW4FQ50bnUtQ4DbqAifG52MkGIHsM9V/Zcfh18Mv/LP8j1qDmv22n+/1oT+CA/XahedS9Ffbaf7/WhP4ID9dqF51IL+sdICpkK1EFSRhUTBTBQETBTBQXh488qjnD5q3GEQXteB39VeZWkEKFEFXHJyqJjKYKADhXBwBVuCmCglD2YVC9qjxhEF5e09sqrXNxuo0QS9bfkqF7VGiCvUEJJPmqIgqTtsqZPqnzQAlAVzCARlW4KIJxKzHYKN7mk7KzGU7IJoyB3VkpBOyo0k9kfnbKC1czcmP4WPCL+Gdo/W41wyuZuTIH9tjwi+Ws7R+txoPpCREQEREHXP2h34G3Ev/E6P9ep188je6+hv2h34G3Ev/E6P9ep188ecIKu7KiEkogAZV+MYyFaz6SvcgjREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAVw7K1XDsguafJe9XLDaa/V3s0bJp3hnO2G8XXh/dbdbZIZAzoub2VMWQ7YNcKkkknsc5J7rwUb3XeL2entCG8sHj8MOJ9JW3Dh9c6o1UNRSt8Sos1S/AfIxhI8SF2AXsG4ILmgkua4Oldztdysdxq7NeaCpoLhQTPpqqlqIjHLBKwlr2PY4AtcCCCCAQQQVFT1FRSSeNS1EkMnS5nVG4tPS5pa4ZBzggkEeYJB2K95uJfLDybc/Wnv8AZE0/cLdV3OdnQzVOl6lkdW1+PhZVMwQ9wwMtmZ4gAwC0FeaHNh7NjjNy0UFVrS1zx620PT5fNdqCAxz0LM4BqqclxY3tmRjnNHmW5AQcEcvnBe9cw/GHTnBzT11o7ZX6ilnYyrqwTFCyGCSd7iG7k9ET8AdyQMjOV7g8h3KXqDlD4eag0XqHV1v1BNeb191I56OnfE2NvgRx9BDySTlhORtgrwHst8vWm7nBe9PXettdypCXU9XRVD4J4iQQSx7CHNJBIyCNiR5r2Z9jxrTWWt+COta/WerLzf6mn1UYYprpXy1T42e6QHoa6RxIGSTgHGST5oOGdZ+xm4k6p1hfNTQ8atNU8d3udTXtifbpyYxLK54aSDgkB2M+a83eImja/hxr/U3Dy61UFTW6XvFbZqmaAkxSS00zonuYTglpLCRkZwQuRuKvHjjhR8UNYUlHxl1zBBBfrhHFFHqKsYyNgqJAGgCQAAAAADYALhytrKq4VU9fX1UtTVVMjpp5ppC+SV7jlz3OJJcSSSSSSTnKDuZzH+zB4n8CKXRcWn9XUuu7xrW9NsNHa7dbnwSipML5eoOe8gsDY3kuPSGgFxwASObdLexF1HW6Sjq9YceaK1akmia80VFYnVdJTvxvGZnTxuf6FwYANyA7ZehXM5xr0Ty58Kbnxm1jb6etqLE10Vmp3geLUV8zSyOCJ2CWF+4c4DIjDycgEHwv1bzzc1mruIFTxFk426qtVbLM+SnorXc5qagpWE5EMdM13hlgAAw4OLukFxcd0HFHEHRtx4c6+1Lw9vEsUtfpe8VtmqnxbsdNTTOieWnzBcwkfLC7i8vPsmeOvGSwUWsNc3yh4d2W4xNnpWV1K+quMkThkPNKHMEYIwQJJGuxj4RnKwvZi8H4+Yjmuq9e8RYxeaTSsc2qbgasB4rbnLMBAZBtk+I98x2wTEAQQcHkL2rPOBxAufFyv5d9DajrbNpnTEUDLx7lM6F9zrZY2ylsj2kExRtexoZsC8PJ6sM6Q3dqv2IV0itjptDcwNLV3BjMNp7tYHU8Mj99zLFNI5g7beG89zk9l5+cb+AnFPl31nLoXitpiW014aZaaUOElNWw5wJoJR8MjCdjjdpyHBpBAxOEfGniXwM1fQ624Z6qrbRX0U7JjHFM73eqAO8c8QIbLG4ZBa4HY7YIBHrl7QLTmneZzkGtfHygtMcV1tNstur7aTvLTwVIiFXTF/oGSkkdi6Bh8gQHjHZbLdtR3ih0/YLbUXC53OojpKOkpoy+WonkcGsjY0AlznOIAA3JIC9E+E/sWuJeorPT3fi5xTtuj6iYCQ2u30Buc8YP72SXxY42u756DIO25yccQ+yhtenrnzmadN9ihlmpLXcqq2NlAINW2A4IBO7hGZXA4JBbkAEZHK3tlKzjNT8X7FBc6q8RcOKmzxMtLIpHigkrQ9xqA8NPSZhhhw7J6QwjbKCDjZ7G/iXoLTVy1Xwx4nWzWMNsp5ayW3VlA63Vbo2NLi2Ih8kcj8Ds4xg+W+AfPHBJDQCSTgDGSSuR+FnMhxx4Kw1lJw14l3qzUdfTS0tRQtn8WkkZI1zSTBIHRB4DiQ8N6mk5BG63LySWzT155teFVu1VFBLb5dS0pdHOAWPmaS6BpBIBBlEYwcg5xg5wQ7JcCvY+cZeI9gpdTcUtZUPDynro2TwW91A6uuIYQCPFiD42Qkgj4S8uachzQRg7x4jexP1rbLbNXcLONFsvtWxpc233i1voDIQB8LZ45JRk74yxoBwCQMkb69tBWcZqLT+h5NO1V4g4ev8Aeory6he5sDq9xYIW1JaRkFheGB3wk9YG+F5n8J+O3F/gbfItQcKuIF409UxvD3xU05NNPgg9M0DsxStOBlr2kbA4yAUG0rzabjp+711hvFK6lr7bUy0lXA4gmKaNxa9hIJBIcCMgkbbFYYdjzWVe71dNSXqv1De6t1XcrpVS1tZUOaAZZ5XF73EAAAlxJwABvsAsJBf1fMJnPfCsVWjzQJG7H6ljHusl30T9RWMe6AiIgIiIAODlXdX1q1O6C7q+tMj1VCNlRBf1tVQQRso1I0HCC1Wnur8K13dAJ9FcxwA+aswfRMH0QTCVo7qx5DnbKxB3QXiMlPCd6KSMjbspOofJBjOYWoHEeaklIIKhQSiX5q1/xHKsAJV4acdkFvSU6TjKqA7zCvAOOyDv17FX8KfVP/F/XfyjbkVfYrNI5ptUk/8AACu/lG3Ig9qEREBERAREQEREBERAREQEREHHvMR+D/xN/gdev1KVfM/Wf21P/hXfpK+mHmHGeAHEwf8Acfef1KVfM/WD/btQMdpH/pKCAHdXZCr0HpzhWlpQVyEyFQD1Tp+aCuQqHCp3V3QUFqq3snSc4VwbjzQUTIVS3burQ0lAJGFRXNYSVXwz6ILMFMYUojdtsjoSd8IIkVxY7dU6UFMFMFSNY4422VTGQgiRSeGT6K0xnKC1Ff4Z9FR0ZHogtwUxlSmM4/8AajYjvkIIsFMFSlmDjKp0/NBHgpjCl6EMRPogiwfRSw4zgq7w/qV8ce+EHdj2RWoaWy839HbZphG+/wCnbnb4W9ePEe1sdQW4/ffDTOOP+1z5Lkr21Okqqk4w8P8AXb4ZPdbrpqS0MkP0PEpaqSUgHyOKxp+Yx6FdFeDvEe88G+KOl+KVgYJK3TNzhr2ROdgTsY744iQMgPYXMJG4Djhe4XHnhZw59olyvW+v0PfoGSVjGXnTN0kb/ala1rmvgnAy5oOXxSNGS1wDgHFgBDwRy0eQUbwD2XLHE3lZ5geEF5qLJrvhRqKkdB1EVcFE+ppJWAkF7J4g6NwwM7HIBGQDss/hXyecyHGW7wWrRfCXUAileBJcrhRyUVBADjLnzyhrNgc9LSXEdmk4BDhXAxgr3J9q611o5J7vbLYDBSfdS0Uro2bt8Fs7S1h77AsZ+QLy35x+Uyt5RdVaX0bdNZQahrr3YW3arlgpDBFTzmeWN0UZc4mRgDBhxDSSSS0bAesdHT2b2gHIDHZrfdqRt21HYKemmlflwor7RmNxa8D4mDx4gT5mOQHBDsEPBJx/MuR+WyrrqPmJ4XVVre9tZHrOyOgLG9Tuv32LAAwc5O2MEHOMHK0DiVwu4h8IdTVOj+JekLnp67Uz3NdBXQFnWB+/jd9GRhyCHsJaQQQSCu4nsxOTniHxC4z6d446r0xWWrQ2j6lt2pauugdELpWsBNO2mDgDI1kgbI6QAtHh9OeojAdnvbY+7/7B2gXOEfjDVhDCcdQaaObqx54yG5xtnGfJdcfYsOa7ml1Tjv8AsArv5Rty1X2yvHm2a14o6b4JadrmVEGhYJqu7vieHMNwqQzphOCR1RRMBPYgzuB3G2j+xU/Cm1T/AMX9d/KNuQbE9q+8DnX1YCP/ANHWn9SiXT1xB7Lt/wC1h/DZ1b/4OtH6lEunyD3A9od/9rYbn+8tM/8ApadeH69wPaHf/a2B/iWmf/S068P0Ht/7UX8AiL/HbF+kLxCB/Kvdvn30XfOMHIHM/h9SS3manobNf4YKSMyyVVIzw3PdGG5LsRPMm2ctYcZOF4cwaV1RPZq3UkGmrpLabbI2Ksr2UchpqZ7nBrWySgdLCXEAAkEkgdyg72exYIPNHqn+AFb/ACjblsL2rzh+3W1aP/vdaf1KJb69iqf+um1T/wAX9d/KNuWwPawnHOxq3/wdaP1KJB1D7nq8l7paxDrP7Jan+5LTAZOD1sL+jfImoIPG75+kHvz9ZXhW13YYXudyGat0rzS8hreEV4roTV2yy1ehr3A34pKeF0b46aUNJBwad0Za4EDrjeAQWnAeGWRjfuqtkdG5skby1zSCCDggjsQfIrkjjxy88VOXPWtbo3iVpetofAqHx0dxMDvc7jECQ2aCXHS9rgM4B6m7hwBBA5Q5H+TjiFzK8UrFXS6ZrKfh/a6+Grvl5qYHMpZYInhz6aJ5GJZX46OluekO6nYA3D039qJLNVch13qbs1prZKmxyOMjA1wmNTEXkDAw7BfkAbDI7ZXmx7LnH7enhn6/9Wf5HrV2/wDbRcebXS6U0vy62auZJc66tZqK9NieCYKaJj2U8TwDsZHvdJgjIELD2cM9PfZcfh18M/8Ayz/I9ag5r9tp/v8AWhP4ID9dqF51L0V9tp/v9aE/ggP12oXnUgDuq53yhAwCCgAQXte0d8KvW35KMgY2Kogl62/JOtvyUSIL3lp7K0HCoiCpOyoiIL2kJ1NViIJetvyTrb8lEiC95aeytB27qiIB7oq/ChxnZBRVBGFREF2QqdzlUTJQX7Y+aqCAFbkKhdvsUF5x5foVhGVXOyp1FBcCB3VriD2VCSUQSsLR3Vsjg47eqoCD32VHY8igHthcz8mH4WPCPP8AwytH63GuF8lczcmRxzY8Iv4Z2gf8rjQfSEiIgIiIOuftDvwNuJf+J0f69Tr55G919DftDvwNuJf+J0f69Tr55G90AlUVXABUQVZ9JXuVjPpK9yCNERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBXDsrVVp8kFze6vAyVY3uvQTir7IbitbNJ2bWnAjU9Frumr7ZT1dTbqh0dFWNkfGHEwOc4RSxnJIy9jgCAA87oOkvD/AIla/wCFOoodV8N9YXbTl2gI6aq3VLonOHm14Bw9p7FrgWkEgggr269nhzfXLnA4Y6gs/Ee00TtU6UdBR3l8UAFLc6apbIIpjER0tc7wpWvYMty0EABwaPJWLkZ5vJa8W1nLzrQSuk8MOdbnNiznuZSQwD5k4+a9WPZ98rVZyVcINW604zXm3Wy96gEVxvY95a+mtNDRslMbHyglrnjxZnPc0lu7QC7pyQ8mecrhRaeCHM7xB4aWCDwLVa7oJ7fB1dXgUtTDHUxRg5JIYydrRkk4Azvlek3sUP8AeH13/C7/AOs4F5ic0XFuHjpzBa64rUTZG0V+ur30IlyH+5xNbDT9QIGD4MUeR5HI3xlelPsS7/bJ+FPEbS8dQ03Ci1DT3CaHO4hnphHG76i6nkH2fMIPK3i9/vr61/hFcf1mRbPcu2nMFyJcz1v5hdXWHTHBzVWobdc75W1lqu1Db3S0VRSyzOkic+oaPCif0uAc17mkOBHoT1f1Lpy9aP1FddJalt76G72Stnt1fSyEF0FTDIY5YyQSCWvaQcEjI2JCD2r9sf8Agj0v8L7d/wChqV4hN7r299sf+CPS/wAL7d/6GpXiG0HPZB6l+w59y9+4y+J4fvXg6f8ACzjr8PNw68eeM+Hn54W9+YnnM5HOHfG3V2i+JfJ9btS6mtVf4Nyu8ulrRUPrJSxrvEMk33x+Wkbu3wB5YXUb2WXMBY+CHMc2z6vuENBYNd0X3Enq53BkVNVh4kppHuOA1pcHRknYeKCSACR2J9p1yE8T9bcS6jmD4LaZqdSsvVNBHf7TQt66yKphjbEyeKIbytfGyMFrAXBzS4ghxIDSv3QD2cX/AOopaf8AQ2xf61i8wHtReCHEHly1JwM4YcGb9YGXa1ttFBE+OkpKChg6m7Mjgc7Aa0ENY1oGcDIG66MUXLZzEXKubbKHgNxDmqnuLRE3TFb1AjvkeHsBvknAGCTjC3jxV5I+YXgjwop+MPFPS1Jp+01Vwht0dFPXRvruuVj3sc6KMuDG4jcCHODwcAtHkHEGi9Zan4eartWuNG3ie13uyVTKyhq4cdUUrDkHBBBB3BBBBBIIIJC9WODvtaOB/FXTTdA82PD+C2Pq42w1lWy3i52WsxjL5ach0sWTuGhsoGCeobBeeHLRytcQuaq/6h0vw2rrPBc7BZ3XfwbjO+IVQErIxCx7WuAeS8YLyG7bkA5GHqzlP5mND3iWxaj4E63hqoTgup7NPVQv+bJoGuikHzY4j5oPSrjh7MTl349aDk4o8o1+oLTcauF1VQQ0Ff71ZLm4d4ty40zyR05YQ1hyHRjcjyLlbedMXsxuFVbLtaaog94p6Woif9ha5r2/Igj1C9ofZQcGeKvA3gzq+8cYbXWaXor9cYrjQW26nwJaaCKAiWpljcQYA/LQQ8NcBDkgAtJ81aPhfcucrnH1tpnhfeLXRHWOpNQ3y2VNyfIynMHjT1LATGxxHU3ABAIGc+SDuXy7e1+01ctOU+gOa/R8tSXU4o6jUNvpW1MFazBBdV0Z3BIA6jF1hxJxGwbDk3XfIHyX84WiJ+I/LRf7Xpy5VIc6Cu0+8m3GfGRFVUDsGA7jLWCJ7cgkOGx8zOInJjzR8L70+yan4Iarlc1xbHVWu3SXCkmAJALJ6cPYcgZwSHAEZAOy7/ex/wCX7jbw2vWteIXEDSl50vp6822noKKjukD6aWuqGzF/jCB+HhsbQ4B7mgO8b4CQHYDy64i6A1Nws11feHOsqIUl607XS2+sjDiW+IxxHUw4HUxww5rsYLXAjYrbq7Fe0N1lYNe85fEzUemK2Kqt5rqWgbNE8PZJJSUUFNKWkbEeJC/BGQRuuuqAqjzVEBwUFX/Qd9RWMsl27HfUsYjBQAcK7IVqILshMhWoguyEJBzurUQMZVQwlAr2uAwdkFWxHOVcXho6SglGDnCjkwTnKB1DOUzk7easVW/SCCXoCoQGhSYGMqxxaQd90EJ7qoYSMqh7qaMEtQWB3Sq+IqSNIO6sQVLiVURkjKtU0bvhx6oLRgAdSv8AEbjGFRwHdRO7oJvEYPJUMjT2UbGhyo5vSUHoD7Fh4PNLqn+AFd/KNuRWexU/Cm1T/wAX9d/KNuRB7UoiICIiAiIgIiICIiAiIgIiIOP+YTfgHxKHrpC8/qUq+aiWn8S5VjT+9kf/ANIr6WOYL/eE4lfwQvP6lKvmv3F2rv8ACP8A0lBCaPDBsVZ7n8itQzhu+FZnPbH2IME0h9CrPcz81qJGMkgKMyDOBhBhNpCD2Uopdu36VkNIJxgKZkMjx8Lc5KDCFESc4KOoyPIrUvd5292YCsMUzuzfzINN91J7qvufoCs009Q3dzcD1QBxztugw20m/YqZtED5FZDWuB3GFMyRo74QYnuQ/uSnuQ/uSs7xGdsjKqC0juEGmPt+RkA9lEKA5zgrVnAnZoBVghmJADe6DEbQ4GMFVND22K1HwJY29UjcBQmoiDuguGUGH7j/ANqrXW8ny/MtREczvia3LU6Jj8PRug04UBHkU9wJ7tK1ZtDWv3EZwrvudXYz4SDShRAfvSq+5Z8jutS9xq846Fc22V5IIjyB8kGkmzveOoA/YpI7E49wd1r0VHWgdPhfmWdBabnIMxwEjGxwg2yLC8+R3UjNOvcOx3K3MLZdGnDoCMfJSCKeDaWPCDa401JnsVczTcrSCAVuoOOx6Qr2mTyaO/bCDa/7H5gFzFy581XG7lWu09Tw4v0UtqrXiSusdyjM9BUuAA6ywOaY34AHXG5riAASQMLYoFSc4aCB5YWHU22Wpd8fwhB6aaS9sxZaqgazWPAq4QVzQA59rvDJYpD5kNkjaWfVl31rbfE72xmraignoeFPBuitNS9pEdxvlwdV9OdgRTxNjAcBuMyOGcZBAwfPu0WkUbg9z3PHoQsu5UYrG9DWlhPmAg0nirxH4j8bdZVWvOJ2q6y/XurAjM85AbFECS2KJjQGRRgkkMYA0FxOMkk7/wCWHmn4zcqOoKm5cPLhT1VpubmOudjuLXSUdX07B+GkGOQDIEjCDjAIcBhcfR6YrBL1xdT8dtlbU2+vhcA+Dp8sYQenFo9tLo/7nsfqbgPfKWuGz2UN4hniOw3DnsjIyc7EHG25XEfHj2w3FHWllrNN8EtCU+h2VTTE681lWK2vaw5BMLAxsULyMDJ8Qjcgg4I6Ny2iprGBrm9GfMKtNo+qIPR1Pz2OEGzLg+4XCsqLjcquaqq6qR88880hfJLI4kue9xJLnEkkkkknJK575IOaCLlD4tXHiVV6Jk1RBc7DPYpKWKvFI+ISVFPMJQ4xvDiDTdPSQM9ecjGDxpLw/ulQ4+EyQn06StHrrJV2UllW0tOcb7IOQebfj1+2b45XvjDFpR2norpDS08VA6sFU5jYIWRBxkDGAl3R1EBuBnGTjJ4abDI44DcrVpqOSraPDdgj080gs9bE7qO/ywg7o8w/tIWceOV+m5fBweks9b4FsgqLp92hPGPc3RuDmReA0nr8MbFw6eojLsZPRh0L2HDgtcdS1MYy5uAsKW3zSPLvFABPp2Qd7uUj2rereA+gLbwr4laEk1jYrJGKa1V9LXCnraSmA+CBwc1zJmsGzMlha3AyQABPzle1JoeZDg7deC2juFVXY6C+y0j7hcbjcWSy9EFRHO2OOKNmATJEzLi87Ajp3yOhsNvqOnw4/iOfRXjTtwd8RBGd+xQc4ckHNJDyicWrjxJqtEyaohudhnsUlJHXikfEJKinmEocY3hxBp+npIGevORjB29za8fG8znHO98YIdKu07FdIaWnjoH1gqnMbBC2IOMgYwEu6OogN2zjJxk8bUthrY39TgT5b5WU7SFzq5eqIOGcbAEoNCjoJHjqGT9i5P4A8e+LXLLrYa84WXttJVSM8Cto6lhlo6+HOfDniyOoA7gghzTktcCSVoUekr3RwDxKRxaB3I7q02WvmaWCH5ZA7IPS/SfttrA62MZrzgPcYbkwND32i8RywSnzcGysa6Pfs3L/AK1tLi97arVN0tNRaeCnCWCw1k7Cxl3vlYKt8JIALmUzGtZ1Dcgve9pOCWkZB88X6NrWSeLJ1YJ9FFPpWZ4OHEAfJBh6o1JqniDqS46x1hfKq8Xq71DqmtrauQvlmlPcknyAAAAwAAAAAAByHyt8YX8tnHjS3GqXTTr/AB6ddV9VvbV+6mZtRSTUxIk6HhpaJy4DpOS3GRnI2NSabqoX/C4vP1LPdY7g9pYYCM/I7IOYed/mvp+cXiTZNZ0GhZdLwWSzi1MgluAq3zHxpJTISI2Bu8nSGjOzc53wOvsVhnnGW5ytwW/Ql0fOJY2vec56cFbgZpPUMWOmicPLsg2N+xifHnlWu0zOPVb+dp2/xDL6Ygb+ShdZ7xj+1z+RBsGSw1DNyT+RWCyzEZyVvOqtN3Gc05/IsUWu7BhPu5wg2jJa5WHurRb5D6/kW4J6OtLy0xbjyx81QW6vaMmI4QaB9zpD6/kV4tUvqtXLZWHDm4IVzTISAG7oNIZaJXnAJ/IrzY5x5la9BSVo+MxHB+SyfCqCP7F5INsGyTDzKt+40vkT+RbkLZs46VY5k2MdG5+SDbf3KlB7p9ypfVa+6CpzkR91b4FV/wDM0Ghi0ynsT+RVbZpie5/ItfjpKw94zupmUtWCMsQbdNjqAMklRm0yg4JW7HsmLceHuFgyxzBxwzz+aDQPuVL6p9ypfVa54dR3LNlRzZht0dwg0E26UeZ/In3Ol+f5FrRZMDuPzJ4c57N/Mg0P3CVPcZFrJhnz9H8ytMUw/e7/AFINH9ykUrLXK8ZC1B1PUbEMWdSwzhn0EGgutczfNRmgkBxlbhlZKHHLFhSiQuwG90Gl+4Sp7hKtZZBPjIZ+ZX+51R3DEGii3yH1/IuXuTWB37bThIz+41naHH7KuNcd+71Le7Fydyaxyt5u+FeWnI1jaifkPe40H0aoiICIiDrn7Q78DbiX/idH+vU6+eMd19DntDvwNuJX+J0f6/Tr540Fx7K1XHsrcH0QVaCSriCFa04KuJz6ILEREBERAREQETBTCAirh3mFRARV6Hf3JTocPJBRFXod/clUII8kBFXpPoqYwgIiYQERMICKvS70VMYQERMICIiAiIgIqhrsZAKoQQgJ2RMH0QXBw7rtFy/e0a5m+X610elrHqik1HpuhaI6e0ahgNVFTxjYNila5szGgbBgf0NwMNxkHq50u9FTdB6eO9s3xdNu8RnBbSYnMe0hrqks68d+jIOM746s42z5rqXzH88vMLzMwmy8QNUwUenmTeMyw2eD3Wi6wSQXgl0kpGRgSPcARkAEkngD36qMIhNVL0jy6jhQtY6TcHJPdBMwdW4XJXAXmB4octeuWcQeFl6ZRVxhNPVU9RGZaWtgJyYp48jqbkAgghwIBBB3XG0UD2fFkn5DspnU8srPMZKD0NqvbYcbZrOaai4R6JguhZj3p0tW+APxu4Q9YOM7gF5+ZK8/tYalvWt9U3nWupKkVN1v9wqbpXzBoaJKmeR0krsDYZe8nA7ZWnChkHd2PsUvurzEWEk/Wg7QczntGOL/ADUcNKLhhrfSOkLXQwXCG5zVFqgqWzSzxMe1oHiTPa1h8R5IwST04IAIPVdrPPdUfSSMIJOfPYKpjkdnYoK7A/MLt9wG9qHzNcEbLR6Sqq62a3sNEwQ09NqCOSSpp4gAAyKpje1+AAABJ4gA2AAAx09dBLnJaVJFTPHxmQsI9M5QekNy9tbxiko5G2jgzo6lqiD4ctTVVU8bTg4JY10ZIzjYOH1rqxzE86HMFzRU9PaeJmpaZtjpagVVPZbZStpqOOcBwEhGS+RwDyAXvdgE4wSSeFYuurDYZKyUAHH0jut6abfTUEYaZS/bzQV4Kcb+KnLxrNmveE+pJLRc/BNNP96bLBVQFzXOhljeC17SWtO4BBAIIIBHdSye2s400lvjg1Fwi0bcK1mzqimnqqVjxgYywukwe5JDsb7ALqvQXGlnb0l4bv3BWpQzU8cjXtqZNj6oN0cx3tJeYzmM09VaHuVXa9K6WrR0VdssUL4zWMzkNnnkc6RzfVrS1jhs5pwuvHDbiRrjhFrS28QeHOoqiyX+0yF9JWQdLi3qaWuaWuBa5rmktLXAggkEELkLiBUxXGj8I1UgGMZyVxzbmR0YLRKXgnzQd7tJ+2k482u2No9W8NNGXyqYAG1cHvFE54AOS9ge9pJ23aGgei2Hxx9q1zKcX9P1mkbHHZtCWi4ROgqXWWOQ1ssTgQ5hqZHEsBBxmJrHdx1YJC6n3BrKpnT19I+R3WltoBEeoVbwQdsbIMBMZWXWRumGDUPOPIkkKOCDwiXFxOflsgh6SnQ4+Szc/IKmM+SDFEbnDpx3BCjNK/OyznE43Ct6253xlBh+6PG/8ye7H0/Ss3qz5BUz8ggw/dj6fpVDSv8A/cLNyPMBOpvoEGD7s/1VDA4HCyzIwBROeCcgoIRA4q9tHI5SMfv5LMhcNth2QYf3OlxnKC3Sk91qpkz5BXskaO+EGlfcqX1VzbPKdyThav47Pkpo6iPAzhBpItT8Y9FjT22SPf0W4/eIsHcdlg1tRG5pxjdBt8xkOwcqeIhrcHCo7HiE7K1x32PmgSjr7I2le4ZVWBxOcErKYSBghBjMpSTuPzKOYeG7GFqDHNBOQFh1bS5+WjKDHLyVNHSSSN6goCCO4WsUGfBPnsgra7DLWOPc49FDdLS+im6Nx9a3TpmVzXnDVp2qpQavcAFB3Y9i1A+Pmk1S4+egK0f8425Fkexgka7mf1PjH/xBre3/AIRt6IPZ1ERAREQEREBERAREQEREBERBsPj1H43AziLEP/kmk7u38tHKF8477C+K7XDrBx4rsfLdfR7xyOOCnEAny0tdT/ySVfPJXTiS715aM/fXfpKDb01nJj2B+xKPT0ku+D/rWtyRTzxhlOzLvPAWVbo6ukwKmMjPqEGinSc0/wALWnK0+o0xNRk9YP2rkKiroY35djBWl6gfNVHNMzqGfRBsuKyu6gcE7rcdqsrMDqbnZQxU1wGMwu/ItwWp/gge8NAwPRBE6wRTMIazdYE1kjpD8TVuyGrp3OxEQTjyWl3kyEHLMAoNtVtPC5nSxoz9SxqaxmQn4e6zJdnZcs6hrIWnDsINPk0y+RmGtKw5dJ1DQSAVvOG40oGARlVmrI3j4QCg2NHpioD9891mx6WmOBgrcDJpnSDDMrOjmlBB6PzINDodHyOcOphP2LWGaL6cP6Dha3QVjWY8QALWxdKMxEEjt6oOP7npnxIjFEzfC2z+wKs95D98Z7LkyqrabxSW4+Sg+6TM9m49UG2jp9lHRhr2fEB5haQKaOOcnAABW9LtWQPgOHDOFs6aRpkOCMZQbgoJaDwg10YJHqFqdLR0NU8RNjGXfJbYo5IW4y4ZW57JU0za2JxfsCEGux8P4jEKjwTg79lR+nqKlid1RDI+S5CgvVuFqDSW5DceS2nd7pb3RPb1jO6DY1UKKGbpDBj6luawV1ojhAljadvNbRuElM+oLg8YWZbZqQAdcmMoN0VE9rlkcWRtwtrXumgmf95bgZWp+80Iz98H5VgVNXRiTIcDugxaewmVoIYfRbqsvDt9eA8MO4yse3V1EGtPUM5XKOjrpC2JoaAdhug2tR8KXPJaYzkLRNRaLFol6XMI3x2XPFvuTTI7DBlcd8Ra2J1QS8AboNgUtmiDRlnf5KZ1ngJB8PstRpqunLBnClfU0wG2EFlupqCn/ssQ+eQtC1IbdJP0xMbgFavLWUTcl8obkLa93hnq580TuseoQbl0XouHVMzYIY8nbsFyc7hPS6fjBqafuFoHAakusFyaRESQRkLnHXl0jgijbXtDHdPnsg4enisFgJkqIGHb0XFeu9K02tKgy2uIBuc7Bb/19PR1cI8N4wR5FaJpCR1Kxwjb1emUHGh4cSWaIPnYdvULWbRpCnuDOpsWSPLC3lrCrfLAWvYB5dlj6OqY4onbD8iDa9bw6ZKC1kRBx5LSncM3x5zGcrl41zA84aCsKrujASOkDdBx9YuGzWzgyRErdh4d0oAaIFr1mukJm+JoAJW4n18JILADsg44n0BS0zevwcBZ1ltFrpZmvmhaQ075C3Zd61r4CA0efktsNOS4EYyg3fLRafvdGKSnpmCTGNhutLi4Y0sDup0JwTncKXSJjgqg/Od/NchT3Fj2hoAG3og2LHwtpLkPCigye3ZSP4BNAI8A5IXIemauSOqD+kYzst8yXSV5aGxgnZB1wqeBwtR8d8BA7rHZoCje7PgDbuuftXuuVTRHw6bAx6LjmGgvDS4mDb6kGg2PTFotcgdUQNI27hbiLdMyyNY2mjHbyWHVxzsBFQwt+xYMAp/eGlrxsRkoNU1BaLDHSeI2nbuPQLY/u9n6XHwm9/QLe2o3xzW3ohdl3ThcfCjrA1xIIGUGnXiSzRAgRMyAtFfW2j3Z48NucKuoI+nPUcFbdLIzC/EgzjtlBjRMoqqucAwYytYmttB4PQIxkjbZbfthZHXEucAMrdD6ikw09Q2CDY96sYpnmbpw0nbIWixGNkwlcBhpW9NT1dO6AgOHZbFdLEWO+MYznug1yfVFtgpvD6W9QHdaazVVE89G2SdlteujjleS1+SsNkbhKGnyPcIORqV8FXh7cHKzfcWBvX0D1Wk2B8TIW+IfTutxy1dG2nI6hnCDRZq2lp39D2jI9Vji80XijZqwLm6B87ix+crSPDaZdig33BcKGSMYaM+WFOZaQsOGb474W26CnlEYIBxhar4cgjyc4QUqaiFhO22Fpk1xgD+w/KrqtzOrBeAfPdaPO2MyZDh39UGtsrKd7QAB9aBsMrtu+VpsIYG91m0zx1hBluoGloIaPyLUKGyRyx9Rb+ZRsc4xg42Wu2zxvBw1hKDRXWaMOLcHZYU9sjbIG9I/9ytzSR1JefvZWm1VPVul2jPf0QYsVqicwFzN1ZLHT0w6ekA9uy1WGmrRHtGTstHuluucj+oRuxlBiTRRyklo7qsFlbM7PT+ZVZS1kQAew7LV7fL04Dhg/NBHDZomtA6T+RalTWaBzN2KRj8jIaPyLUKVkzhkMJ29EGmP05E9x6WbfUt88pNmZS83HDd/T9HVFrcPr96jWjxsqP8A5mfyLd/KvHUjmz4dF0RAGp7Z5dh71Gg97kREBERB129oSM8nPEketHR/r1OvnhZjfK+h72g/4HPEj/FKP9ep188IJGcIAx1b+qkADgcD8yi7lSsIGyCwsIVOkkqWR3w7fpVrCNifVBRzcDP8ytDcqSQ7YVrDgIDYy4q4wOAzuronjqU7ntIxtugwwwl2FcIzkbK9pb15OFIHNyDtsgtliDWggLHaRnJWVO9pZgFYiCV72FuB3wom7FVDSewTof8A3JQZTZog07eStdLHnIAWN2TCDJEsfmB2UMhaXZb2ViYKDIa5mMbKOQZ3H6EbgYKrkILcYCtOFe4EjZWEHzygoqtO6pg+iYPogl6mY8lG4g9lTB9ExhAVzSMY81aqtByEF3QcZ/mVRGSM4/MrwNsFVzgIMcjBQd1V3f6lRBOJGYxgbKOQg7j1VuHeQVMYQFIxzRuVGqhrj2CCUyMIxgK3bB7KPsqjOEFPqUsLw07/AKVEqsHxAHzQavFJEW5OFmsliAwMLSosAALIaRjugyXSw52AVvXGT2GCsd/T/dBWhw8igmc6N3kFJDTxu+LG3msbpWdTBxb2QDTREbDssWVrGHpwFqQjfjsVp1Y1wf2QQPkEO4wN/JZNLc5Wt+GTdadVdRaMJSNPTgkjdBuOivVTEfp7LVGaqlaQS8d1tWFvcElHsb26j9qDXL3qF1ewsa78hWgwyStBye52Vjow3cOJVzD2ygm8R57lUe8+apkeqsed8ILXOCp1BUeB5qgx5IJIzkqTA9FZHgFSBBY9oIysV8Ty7O+FmP7bKPBO6CxrSBuh27q5UcEER3CseSB3V7tio3nOcoID1uOxKqI3eeVe0AFXZ3QWtaR3UzZ+jGfJWYPorHNP2fUgyhWtOB/Oq+89XY+XqsHpA81LGMIMjxH+qr4r8Z8grQMo4gNwSgp7zL5lQvfI5w32Qub6q9nScdkDwiQPVXspHEZIPdXg4IWZG9gZvjsgwHFkHfCq14kGQoq9wcdvVXUZY1mHOCCszzGNlZFKx30sEhKtzSPhKhgYDuUE08XiAFg7Kamq200fQR8lVrmNbuRlYM27ycghBvjRb21kz2gdlo+t2OhuIbnGcrUuG0sMVU90rgAT2WFxBlilugMRBG6Dut7FYk80+qcnP/5v67+UbciexVBHNNqk+ugK7+UbciD2oREQEREBERAREQEREBERAREQbE49O6eBnER3mNJ3c/8AI5V87dPUB10uRdv99d+kr6JOPYLuBnEVre50ndwPr9zlXzqQ008F1uPiAjMrsbY80G59ITU3v594ALc9sLXNYGkfg0jQO3YLaummuNX9q3FeaOpeG9AJyEG0LhJLBGHtOFk2W/U0bcVQBI9VbfLfXMgy5mBj0W1Ymhpd4jwN+xQci09/tk8haI2rAvdUxwPu+2Vte1up2TZdMFq1TPA5oDJAdvVBSz1s1NUdcziQPVaheL/BK0NaB81ovU31AwsCsjBB6XZ/mQTVNzZKeluAViiolGelx3+awRE8v233WUI5AMYQZNBPUvk6eonf1W4YPEBBeTjzW37axwm7ea3GzsMoNRhnhiblwG6kbcYXOAAG/ZaZLNF0fE4D0WNHU07ZB8Xb5oNarKsiLMZxjzC0V1xrcnEhwsyoqqUQbPH5Vo8lVTjJDvzoM1twmcR1P3U1PVSS1DIc7nAWlwyRyu+9nJ+S1G1UdTJdoQxhJLht6oN2yaHrZ6EVXxEEZ7Lbs+kaiNjpMO2yudIbNfm6dY80bujo79PyWzLhS1rKWUvhIA77IOHp4ZqeYxdiFm0ktTB0zdRGN+6tus8TLiWvIBye5V7qqndTuaXDPrlBPNriqY00wlO23daXNqKtlJPiE5W36qMOrHOa7IJyp208xaCASPXCDPdcp3bucSUF0qm/QeQtPI6T8RwUJB3CDPN7rG5+M/aojeaonJkKwHNyDuoixxdhv2INy0N/qWYHiHuuU9HaudBCOtw7eq4UpaWcgHpPf0W99PxubCOokAIOYqLiNFTykue0em62TrvWjbhKXseDv6rQJn07HnqlI+1beu00Lieh+SEGqjVxhYB1jK1S13qW5uETXZJXHNRCXYIJO63Xo6aClqmOmdjHfJQcoWXhjdtV4bTOfvjtlcg2bgvPp9o+6MZJI/fea1fg7rvTVoLDWTRN3H0iFytqbW2n701ht8kbwP7kgoNo6Ehtmjqw1M0LQ3vutr8Z9Ru1NMH212GtP70rcF6hfd4TFQMJcBjZbBudqrrUx4rGEZPmg41vj6uOBrJXkkKGw6ohtzSyQjZZurJGhpJOBjuuPnTUxc4OlAweyDemoNTQXKItjxv6LRrZqH7nAtL8Z9StDbLBn4X5WJVuidnLsIN6fsyaCXdY3+aw6nVPjOy1y2c7wyBh3mr2MGNiUG7qbVToHAh/b5rXKXXXwDqeMj5rjcsHYuOFKw07AQX4OOyDkka1jqT0Fw7+ql+7cZG2NwuObfLStl3k2zvutfjraPI++D57oN92C6u8YFru5W9qW4PqKuKEuz1YHdcW2C52+KQeJIAM+q3zZb5ZxdqciYHcZ3Qcw0dpNutza5wxkZzj5KyLV8ELxI8ghn8y1iuuVFctMsionAu6ANvqXGVVa6uOCZ02QMEoOS6fiTZrk0UjmsJ7H1U1VdrJDTucImZIz2XBthko6a6/fpiCHLfVzudrdSbTjYfzINI1dfaSuldDSgA7rZ9PHO2U5cck7bqss9NNcCIH9RyswRyioZlh8kGpQU8pjDpiSFdU01MaYta0ZWcWvbSAvYRkeawXys8E43QcZ6rs7p5neGCB8ltVmn6joIy7BK5IvIlc8lrCQcrRRFMGn70fyIOOq+zTUTjKMg91pzZqmQ9PWVvTUsFU6MgREfYtp0tFVmQ/eyST2QadeaWqfAXdRIIW1nUU5jcclch3e31zaTJhOPq+S2pJT1AjeSw4B3OEG02UsonIf2ypxThrw4hZD8mctA3yr3QTebSAgujq5I2AMOMK59yqTGQXnCxXxvHcK0seWnYoMd9XKZMucSUhrcThztwD5qCaOQE5B7KFkbjIMjzQcg2q4QSwNaAMrWWGOSMRADJWzrG0dI33z6LdlGMzt9chBg1+mqlxMzc4K2vXUE1PIc52K5aq2T+5jEZIwuPL31CU9bcHJQaPEXtbjdZdNUFsgJOwWIjHfHjKDdLLrGIQD05W4LPqGlhjDXgLYbSzpAzus+lfDghz8fWg32dRURJO26khutFM8ZAK2SXQAbSLPoXxdQIeg33DcLe1gyxqzAbbUQF/htyB6LabHQujBDt89lqlLcrfTU5ZLKASPNBpt7loonuaxgGCttmsb4nwEYBWdfK6hke5zJAQVt1s0XWcOBGUG6aStbgZwtdpLxTwMHUAtm0VVTADqeB9qy5Kyk6dpN8IN3jVFHEfLK3ryp6noq3mv4dwMDep+p7YBg+ZqYwuDZ56Z7j98W/eTtkf7b7hq5js/wDwrtX63Gg+h9ERAREQdd/aDj/rOuJH+KUf69Tr54Wea+h/2gu/J5xIH/0nR/r1OvnfaCc47oKHdM4V3hu9E8N3ogtJJQA9wqljh5K5rXdsILTnzQNJVzmEBGdkBrHZ2z+RXOY8KSNuCkufJBj5KAuPYqh7qrdigvET3HfKr4BAJPkpo84yArnhzmkII4HMLsbLKkbGIyRjOFhRMd17LJkD+gjdBhPPx5VeoYR8b8kkK0tI8kFCd1kRM+DJ9FEI3kA42WTHgRkH9KDEcSHEIMnfyR/0j9aqOyCVsrOnCtc8HthRnclUwUF/WE6wrMFMFBIHjzVHOBKtDSU6cIKhwCva8YUfTlVAI2QXF+5wq+IMYP6VZ4bzvhPCf6IKHuSjfpBXCKT+4KCJ/UBhBOwDp+xQyNwfllZbYX9KgmieN8dygx1lQvjDMEbrFIwr2j4cIEuC4kdlcxnU3Pmo8E7Kdnws+pBA4YKuY0k/YrXHJUkWc7eiDJjBIV/Q4FIxgZA7qQAkdkETw4jCqxpHqr3IOyCRrwwDPqs6mrI2DB/OtKe4YwTj1UkTo8El31INabcIcLTq2pjkfkAHfyUQfFv8SxZnMDvhO2UEsrmvAVYo+7gsXr3AG/2rLgJLN/VBOxu2yjkjcScKSM4OFV7snt+dBjFhHfKpgjsVJIcBR9XyQU6iN0c8d89lQ9lE9xwUFrpsnv8AkVzZdtysfB8irg12BugymSbqUSdljRg7KcNxhBf1Z7q4OHZRHsqg5CCr3DGfmrC8YKqcHYqN+MbILS8AlWPdlWvPdWjsgtB6SFcCSQfJW9OSFcNvsQZAIKPaC1WRuG2VKXM6fsQYRa4O7lSRtII2VOrLj9ZUzAAEEnUA3Cx5ZOrYeimJYf8A7KxnnDsNQWFrgcq+PqAyrugkDZXlh6ED3kAY9Fa6qcdgVju7lACd/wA6CbpdL55UbuqMkA4WRA5rB8RA+tRVBY52RhBY1r5D5rIZC5rcDzVkTmjG47rKYHPA6RlBhzNewgZ7rfulOGNRqKk94Y0nIB7LZU1PO97C1hO/867IcILnBQ2lrKgAHpA3QcYVWhazTPW9vU3A3Wwr3NJLUkyEl2V2F4kXaikjlA6QCD2XXe7vY+pcWHIyUHfT2LDgeaTVAHcaArv5RtyKP2Kn4U2qf+L+u/lG3Ig9qUREBERAREQEREBERAREQEREGyuNnT/sM696uw0xdM/V7rIvAeWwMut0rzTswPFcdh8177ccXNbwV4gOccAaXuuf4pIvCrTN9sdPca7xZmD74cb/ADQabZdIvo5g57Tsc4wt0yUlOzHis7LMj1Pp50hxPH39Qo62vt1UM08gP1INE1BbqOspOiNgyQRsFty28GK2+uc6BrviK3XN1yACFpJz6ZXJ3Du/UVpjY2vDWEY+kg4NufL9d7ZF4xa8YGfNbWl0VW0bi2XPwnG5XcDWOutPy0LgJIgOnHkuBb9ebZUySOp3NILj2x6oONG6ankPSAQVbPpiWA/GCVvKmm8eXpp25OcKK7U9wA6nQkZ+SDYstrbTblv5ljPMbcj0K1240tc/bwj+RbdqbdciTiI758kFYKyKCTOB3WXLeWY2cAtFdabm13U+FwH1K11HUN+kCMIMyquL5MkOP1ZWC2pn6s9ZUZHRkPON/VVa6P8AuhlBLNV1RbgvJz+ZYXjVBd09ZKyXPYW4BysYf2QfWg1e0Okp3CSU5Hf61vKw6ko6G7U9VMB0scCfsW0IZITCAXAEDCpiBzg3r3PzQdvJOYDSZ0sy2tji8UM6T2z2XFt44lWmrpZoowzL84wAuIW2uMsDxM4hPcA0Za47IMW9CWsuLqiM4aSViubUAH4jhZ7z0bHyUEk8TdsjKDTQPDk6n+XdZbb9TwsEbsZOyhqR1Alq0Cpgl8Y/CTkoNYnnE7jIzseylhJ6Mu7rCpo3tiAIOwCzYmnw9wQgikqGh3TnzV9NUR+KAd8lYU7cSZyr6UN8YEnt80G86FkToAcd1uKhjDaYuaPJbctr4/AaC7fK3TRb0nw+iDaV/rZoZSGvI+S0COtlleS9xOD5rVtTNeZnLb9O0jq790GoitY0Yed1eLhKN4HEEbDC0uWN5I6QSsilY5uerIQZE971IwAUVU9mDnYrkLQnE+9WdgZdapziB5lbBifHHu8hJGRTSZD8D5FB2n4a8cqCmrw+4Oa5hcBuuSNV11v17TCptcbQCAfhXTHTFPRiVglnIHUF2x4X3ew0VlDDUMz0Y3I9EHEXEu1yW2NzNwQCD81xFTW6rrZHljiBnbBXO3Gi5W+rMhp5Gn6lxNpqspYC7xXAYJ7oNFq6OotrS+QnHqtEmujpnEBx2K3jq+spJ6c+E4ZOey47jjLXOJOx7INSjrngjJJWZHdmswCQtGY0Z3KOjJPdBrE15bjZywJa+WQ9THnB3WFJE7pOHJFFLg4OUGbDWz5HTIQfP5rLFdWAj74fyrTYYZi/bJOfRZfu1YezSgzfupXxgFkpH2qai1LdaSpFSZ3AMGe6080df0/2MkfUoZ6CuMLw5hGWlB2q5c9fVGs69loqJTJuG77rsnxB0FHbLGZmxYLmdX5l0z5KomUmtmyTk/2QbL0O4uXKhl001rMZ8L5eiDzs1peamwXx/Q8tAJwtEPE2snjcz3g7DCyeMcElVfZfByQSey4sp7PcA52zjnt3QcpaU1dJ90PFqJMguzuuQp9a0jXsk+HAA7rhLTmnL5PKPBgeRkeRW7qnR2pfAwaaXdvoUHJB4l0dbD7rG5uRtssObUXgQOlc7YZcuM7Npq80daX1MTw3OSSCt03One63OySMDdBhXDifRQ1BieW5GQsNvE+3udgFq4r1FSNFwf8AfCN1pbKcdf8AZD+VBzJX64t9XF2aStHg1LQRziQtaADnyWyYqV5jHS8nH+pPcpurzCDka9a0tU9AIWMZ1AfJbHmvVK+GRgAyc4K0ypo3uacOJOFgfc+qLSA0knyQZFpt5rrgXDcErd0+mQIQ/p2x3Wg6UobhR1YkqYnBme5XJFTXUbqPpDhkN9UHF9wpWQSlpb22WDmPdu3otWvkjHVDukjBJWi7ZOPVBJ7oybGADnZXtsfWdm7qSkeGkZWr09QwOGcIIKK0SUnxkHAWrwTtglbI7s1XmqhdFg4H2rGmfTmM/GEGvVGtKIUxh2JxhbFvNyjqpCWY9VjVjIfGPS/8hWDK3OcElAdNnYKnUVEGu2O/5FKGux2KC8SSDzV7J5QfpEKNVaW53IQZbZ5tj1FZ1NVSNAPUVp4cwNG4/KsmGSMM3cBsg1OK9OiPS560q61dfVPzBIQPkonPhLjl47qRk8LW/SHZBgn34gdchOBvurmyysGCSCppKiLf4gsWSSMk4cPNBd71O0npcR9qtM1ZI4ffDj61Z0SOx0qeKjrnjLGE4QVElQAAXHvvuuWeS+V37brhh1E5Oq7UD/GolxUKWsBw5hH2LlLkzjkbzecLw5pB/Zdaifq97iQfRgiIgIiIOvPtAt+T7iOP/pSj/XqdfPNRxiSR4Izgfzr6GvaBfgf8R/8AE6T9ep189NvOJZPmP50GQKZp7BV91b5hSg4V4JIzhBAKMEdlcyhDjjCyW5ONlNGS3ugwpLVluQCoG2xwPY/lWtPnbjBIUXjR9tkGCy3E4xuklrPSdsrUYpWA+XyUrpWluMhBt11rkyQqstbwRnJWrSyYO2N1Rkme+O/qgggthIwQVkOtPwglpWZBOwAdRHZZD6qHp8seuUGhe4eG/JaR9qv92acDH5VlzVVOSfiCtbU04IHUN0GLJQt7/wAywZKPD842WuSTRluW4WnSOJd5ILPAYI8BoyAtNqSWu6QVq5OWH6lpFUCZEEIaXbq8QPIz5Ixj/Q4U7Q/pIweyDGBAOD5K8Pb22VropMk9BQQyf3JQSgtPmPyq9sbXdsKHwZ/7kqWJtR26SgvMIHkqeB1bhSujqOnZhyrooqjH0CfsQQimPmE8DDgMLK8OoP7z8yt8KcPBe0480EjYGdOenzVwgYBjAUglhDekkZCqJYfMhBZ7vGRkNCuZSNcQMD8iu8eD+6Cmp5Y+oYwd0F7KAdOQFFVUDeg4butWjkZ0hY9TI3oOAEG2XUREhbjshpS3yWe933wkAK2R48wOyDBMQb3Ct8lNK5p7Y+vKhJGCgx39wronAEK1537qje6DUoXNLQpgW7f6ljQdgpur6kFHkeqt6hjCo45Ci6jnCCksb3fRCx3CVncnCz2nbyz6LFqSc/Ygg8R390fyq9kMko6snHqosErPpDiMAgII4KdwcC7y+SzQOnYK2M5OMBSYBQWCQNKeM0q2QBWhgIzlBWWUYwFD4o9Ve5uQVjPBB80Er5M9irPpZVGgnbfspGMI8kF0cB7nfKkFP8lc0u7YUgJx2QR+DhXNYR+X0V4ye4RBHICAocndZEnYqAg5QWPJxsqB2Rg91cRnYqgYAgtLM7p0fL8ykVRjz/SghMZ8lG5hGf8AUslRyd8IMf4lQOeXYye6kLTucFRtB8TsgnELukOwrXZaCsoNPhhY0rSAdkEAe4lSxxkuyQoWfSWTE52fl9SDKAZ0gbKjwHM6R5p1RYz1b/WqdTTgNOSgxXUr8k+qnp6cO+AjdSObLjIGyQw1ZmBDThBn0unJKwEsBWNVadnppfDOcreGm5XwNJkbjAWPe7nS+9jqc0INKtOh6i47tz28lPW2J1mcI5W4PzC3voq7Urf37DkLSNd1cVRVAtIxnyQbZZNTRYL2jY5WqUuu3W0CKGTDR6FbeqgCNvRaLNC4OLhuCUG59S6wqLxGWNkJzstriGR+7s5VGxuBBKzYzhu+PtQd8fYswOZzSapcfPQFcP8AnG3Ip/YvuJ5o9Uj/ALga3+UbciD2fREQEREBERAREQEREBERAREQbA5geo8BuJAacOOkbxj6/c5V841JZr/WXKtFPM8Ykfnf1JX0ecfTjgVxGP8A3JXf9TlXz66Xr4o7ncAQD99Pf60GgU2mNTNd8U79vmVrtJdq6xgNrJScbblbmFzjMhw0ea2Zq5zKhxLT39EG6LRxHo6eTMxaQfVSXnWVVcyHWyXpH/alcTtpGucAHkH61uqw6ev1QB7jE94PbAJQZt8uuoKim6DUPJA9VtSK83Ch6veZyd/MreV003qahgMlbSvYzGTkFceXqGaZzw0dj5IN16V19DQ1nXUuBGc7red04oWeoaGtazOy4Iho5PFAeC0A7lZ0tLHj4ZHHA9UHJtTxAtT/AN60j6lfRamtVUc9DTlcUOpG4GXFa7ZBTRkdb+yDkuSstdYzpijGe2QtMqbNDM0hjNz6BQ2uqtsWC+UAD1WvMutqwOiRp+pBsms0lK9xLQR8gtKqNN1EALiSuSZ6g1jSygb1nHYbrb9ztWoPDcXUrwDuNkGwZKWSJ/SSdsq0gjutQrrfc4JS6ojIHzWnudvuRkd0ASSg4BKkpnSvrGRknchRt77bqega/wC6UTyDjqG5Qb++474LaKh2cFufzLb81zjiBbkZG3Zb8rauA6fDDgEM/mXF1cGOe/Dt85CBUXIPccHAKw3mSR/UCcKB0b853wsmFrwMlpwglEwa3DlE50TjnA2SToOdxn0yoeh5yd0Eonjbt6KVtSxwwPNae/YnJOR81NCNu/mgrKwvJOMqxsbmPDhkKcnGdwrHEk9kGpUda+NoGexXIOnZRUUgzvkLjSDpIGTg5XJGk6q3xUYbK8ZxtkoND1JTB0rtvVbULRG4+S3rqJ7JZXmn3Bz2Wy6qkreonpIGUEjJIicnH5EkmYNm7LHbFKw9L8qkjHZ2ygvkEj8dJwroRIz6RKrCfIhXOe0EYwgq+rqKdodE8hcrcP6m91dF1RzuAxtuuJKktMYwfMLmbhZLHFbsHGcINI1ebgHPFRITj1K47mqamNzhE8jdcna7la57y3HmuLpnDrdk+ZQH1FTKMSvJHzVjYc7qplij3e4AHspYqStrBmiaXj/td0FkdK55GB3OFuyyaDmusYe1pIIytvUlpvjnhrYXkg7gBcz6BuNLZ6VrLrhjunHxbINnRcLJ5Hlnhkkeiw7lol1qPQ9hB7HZc00mq7EypJMsfSfM4Wztbahs1VUAwTMIz5EIOPY7GymxI5m3dajR01K9wHhjOcLKrLnbpaYNbM0kj1WPb6uibKC6QYz8kGqT0dHBB1mIDZaXK+hqInxNYMkHdarc7hbn0ha2UArb9NPRNeS54x33Qbx4PVh0ffBcWHpHV1b/AFrn/VnHI3W1inM4OG4/Muu1mmpa4iGB4Lu2Ad1qtbaH08DpJS4DG+UFlRLFqS8OfIAcuWqSaPpKVvjuh2Azutv6Wq6Fl0LGyguDu2VyLdaqM0BBIHw/zINN0pqvTtjqvCqYWEg+YW/K7ilo91MMQRfCN9guvda2kNzc6WXALvVZFV9yn0j2MqB1YPmg5Fr+IGnbzUGCiiYD22wtOr/DfSPjAGXdlxjpag8O7mUPJZ1dz9a5MrJIREPiGwGUHGF10ZPX1zpGNJBPkoP9jqoiHilpx3XIUF1o4JSJS0Aeq1CfUFofSloezOPkg40odPFknglpOCtSm00Aw/AfyLUaa50Iri4uaGk98rWJrxbCNpAg2FPZBTOLns2+pRRR0kMgmcwdLcFblvVxt0sZEcjc/JbWnkgdTva1wyQUGZdtT2t9N7tSsaJAPL1W2fulU9LsvOCSRutLfT4rC8OOAst3SY8A+SDBqZ3TOOTkkqJrDkZVXRSB5IBxnuqhsnUMBBPGwtHV2VRK9p7q7plDN2lRlrzn6kExq5MAB351G6adzS0uOSoQHg79sqVpbtjGUGI6kmLi4lXNgIGHLNMFXM0eCzOe2FE7Teppz1wU7y3vndBj+CMnYK4RAN7KT7jXuP4ZYXghXtt1xaz74woMMxnOArHQOc/pHmpnw1Ubj1tPyU1LpfVN1k67dSySN/7XKCSC0yyMacndajFpeomhLmk9lm0eg+IJZ0tt8pIO+GlbkoLRqC2UxZcaZzHYxuEHH0+mapspHUVWLTlSXBmTut31UNYZnHwj9eFFE+eKYGRmPsQaPS6ArKwZAcsg8NayDLnB2PRb+sN1iiGJA0YWrV2oLeY8Esz9iDiv9ixpiA9pyDutXoxQ0cBErBkDB2WrXC5UsjiWkflW3q2aBzSOrGSggq6+h8Q4aFvTk+qYn84PDFzAMO1bamj6/e41xlVBjnEh/wCdb+5NwRze8LBntrK1frcaD6NUREBERB169oAM8oHEYetHSfr1OvnpoAfFl+X+tfQxz/fgg8Rv8UpP16nXz1W4Zlmz/wC+5QZY7rIiYHYWPjB38lnUxi/fOHZBPT04ceylmojjLRhX0kkIds4LJmmjDe4wg0OpppGDPksRsMryd1q1ZKyTIZglYcbXDPwlBWkp5JDjPms/7myHG/l6qy2hwkJc3butaa4AjYBBpDrO7GcFRG0vaM/zrcMlTEGAEj6ljPrIMHdBt2emliBGTssaQzeGR5rVq2ogcTghac90fSSCEGkSMqC4/F5+qMhqeofF+dZgY6V+I25+pV92qmPDnRkDHogzKSmllaA491qAsT/DMhB237qO3VELCGkjI7rcQqI5KUxx4LvJBs+SndG8s9Fp9RTFr+ojIC1ipt12M7pBE7oJz2WDVZjBZMMO+fmgwWzRN2I3HyWQx8WPJYBjJeSCcZU7CA3CCcPiLsABTx+EcbBYg6Ac5UrJY9h1IM3rg9B+RBNTt/ejv6KDxIf7pWuDXbg7IM4VUB2LR+RXCrpmjdq00OiHd+6o/pcNnfkQamLjStO7QsaqutM5pa0DJGy04QPLiRlYs0ZjkwfNBI5srnl4JwSfNWukkYSCVlRujEYyRlYk5BcSPmgs8WQ93LOoJ3lwyStP6Hf3JWZQjDgCMINwMkf091HK4lpRj2dHfdWve0sODug09xxIVDUTBpxnzUrz8ePksKpHU7OSgjfKc91c1hkbkb7KFyzKYZjGyDFcwg/+xXMiJI2Ujxuduyo1/S4fXhBkRtLQFcqNORlXAHZAc0/nQRZ3wquB22UrOyCDGFa+EvOSFJIDk4z3VWdXYhBCKXJ7fmUrIugYwpQQ0kuwjnBxyOyC2MdJUzYy4ZUbQScNGSsuGGXoGGIIm0TpTupRanHAxusumgqXHDGEk9sBZrLRfZXAQwOJ8sBBoVVb3U8fUVg9AdvjK3FdrDqKOMunppGjv2I2Wg+DNEPvowc7/JBa2MZ7FSMj37K3raDuQFeyRpIwQftQTxwjA2V3hD0UkT29PdXP+QQY0jenyUXUFLMT6fmUB7IDiCqFoI8uypg+iu+LHnhBA7YlWl+Mq+UbHvlY7g7fugkD9/8A2oXb7KH4h6q5ucZIKCXrCo4Z3VGt6ir34azHmgh6h2V0YaX+Xksc+J1Z37qSHr8QZyg1IAeGNliz4xjbKymk+H2WFLnr80ETac9Q9Fl+B0xFyta4ADYLJ8RngEE7oNILnh3fcLJtsb6mrbHnv3WNJu4/WVnWMkXGMN7koN9Q6V6qRknTknvsrxaaelYOqMZC3RS09e63MLYjjHotLuMsMbC2fAd6INq192jt5LGEDOwU9u0Bc9WNNdA5xB37LQtSwulmY6HcLmnhFVT0NmDZI8jp8/qQbVs2gLnbMhxflndbR1jLNRVnhSuOQd12BN6omum966Wk59F1/wCJtRTVN3MlM4EZPZBtx9cHjuqskY/0OVp4a447rLh6WN+M4PzQSua3yA7q5sZO4H5FYC1xw05WTHsOyDvj7GBnTzRaoP8A3A138o25FJ7GT8KDU/8AAGt/lG3og9mkREBERAREQEREBERAREQEREGwOYE44DcST6aRvB/5FKvnBpL8+gudeevHVK7G/oV9HvMH/vCcSv4IXn9SlXzQXEH7o1QAP9mf/wBIoN60uqzI/d4OR6qC6XL3gF/UPVbTtwIm32C1iobmMhu/woNc0LaX6kuTaVgyeoZx9a7TaOt9j0TDAbnTtJwCeofJddeAcjKbUgkn7dQ7rn/iVebQ+OJpkDSAM4+pBrnEC/6V1LbDSW6nja8tI+EBdeK7QYhmle+M4eSRst9WK52hswImBOfNZd+raefAiAI23QcMX7SraeHqjYQQO+FtEUUkRPV5FcwaljlfTHwmb4K4xroZ43P624wfNBo1Q3bpCgjE7c9LiPULKlBzlytaRv2QRl9wfkRyEeuFNBLdIMufK4gd8lSUz2sfl3bKyamWJ0RDTug3DofW0Vlqw+4vDm9XZy5SqOKOmLoxkMcUWTscALrXPHI6QkNJGVqdhg8OrZJK4gDf5IOZNTNttzpTLSxgZHkFxtUaal+OQA4yThb6o6ujkoGxRODn4UE1JVvgc2OEkkeiDjl0Pujyx3l6qWmrYve2NHcFZF5sd4jnMr4Xhhyc4WjU1PKy4NByCD5oOQ6wVD7UHBx6ceq2NUNk8U9+65BlP/UQD1b/ADLYlQx/iE9O2UFImt6RkbrKaGdBYO57LGjLQB1HCljmha8dR+tBjuts3X15ODupg1jGkEDJWoGpiMIGQtNmcC7IIQYklK57sjOCnhOjGM7LNa7AChm3/Ogxi/CmiDS0k4WPIfIKWHqwUB7iHfD2yr4bjWQyBsUhACjeHA5AVjQfEBJQbvs1SZwDOcknzW/aDR8F0oDUNjzgZOy45s7mMYOo43XO2ipQ6xERgH4P5kHCGpaBluqnRYAxkdloTJGOz2OCt2cRIpTdJCG43K2VTvbGSJDgn1QSyyBv0VbGx8ozklRykP8AoHI8lmULHNG439UGNURua0ZPmuXeGUb5KENbncLiyuhlewGMZ3XLvCmCobSDqac9KDIvemprjI8bnvhcW6nsklomIORg7rsJHHOJ3ksGPmuIuJ1vuFRUuMEZI37INoWTSlVqt4p6fqJzvhcm2bTdPoiJsNzjBJH776licDo32i5tmujA1gIOXLd/GG52y5TsdQyNwMZ6SgwKS9WKkmM0kLMOHnhbN1rU1N4mLrO8sbns1Y9XDJUUwZASSB5LKstoujIzmBxz6hBodvsmoa4CFk7y7sd1mTcPNQdQM0khJPqVvfS9NV0dZ11EJAByFveqr6ZxYSG7IODpNAXmBvXI95H1rAmsdxp8kOIx6rnK6TMrIPDo2BziOwC2TctMakna4RUjz1Zx8KDjOcXFp6XSOxn1UMUdZI/wg85Oy3LcNBa3jf4klvlDO/0Sse3aQ1R74wmkfgHfYoNc0JQVdmqBWVhJYSDuuSa+8Ut9gFDTgB7xjbutt1NpulFam++wFgAGSVHpCaMXqLDicOHdBurQ/Ai9T3L7q/fDG53UPRcq3jhHXfcwvDXbDfZcnaCvtrt1iidX+GxoaNytx3bX+i5LNI2OqiJDTnBCDoNrzRdfbq17WFwIz6rZMFmuweZHvf0g7jyXPHFPU9hqrnKaaVhGT5hccMutu8CRpc3Jzjsg0u13NlAehxAcO59VqdTqcSRFofvjC2jWvjNW6QnDSdirDUUxaAJNygperrVyPLoZCMrQ47hdurBmJGfVajVAPBLe2FhMjPXuPNBkmrrRHlshB+tQvrrkWn78R9qmkw2MeShc4Fh3QY8FXcJJQJJcg7d1qQ8YjBzj61p9K9gm+LHdaoZY9gD3QYVREWfGQcrGY8+Jg9srUK1w6CRhaYxzfF7hBqZhYYgcb4UTY2NPUR2WX1MdAA3BKx3sf4Z2KCgqYZPgAGU8Bp3+a0+GGZs3Y4ytUbktx5oMZ8AecNzlWxWyV1QG74KyoWPZJ1OGBnus6kqIDVMaCM5GAg3hpDS0VQxhljyCRnZdjtKaBsIsJlno2lwZnt8lw7owSMhjke0BuQcrsXpy/wBjp9MvbUTMDgz1Hog6/wCprVaIbvLDHA0AOIxhbZuFnocEMiwtx6pv1kfqKYtmbgvON1pVTeLSc9Mjc5QbJueno5HNMbMYO+FzxwXq9KWOhb91qSNzgP3wHfC4nqbnQ9Wz2kZW4bPdbSKUtlnAOPIoOxVLxI4cUskjX0UHnj4QuDuMPETTNXUuFrhiY0nyAW06u42l07yKrbywVxxq8080xMEpdufNBuF2rbccHpbkj5LTKu709QS6IAbrZ3gnAw4rOpAGgAny80GsPuUrchj8fUtGrbnXvk2lOM+qnPST3WBU5DtiUE0dZVvGC85Pqo5ppnZHUdz6qkTXYG/kqytcDughDnd3Erkzk235veFv8MLV+txrjRwOFyVya5HN5ws/hhas/wAbjQfRoiIgIiIOvXtAT08n/Ed3pR0n69Tr567afvsp9R/OvoT9oIQOT3iQSf8A7jpP16nXz00Dg2R5JABH86DUX/EMN7q6OnlPYnHzUcUsYdu4LUIaiDAy8IK0dJN1d/zLJnppiMb5UlNV07T9MLIkqYHYIePyoNEdBJA/qkzhXxVcROBjZTXOWJ7AGHf0WkwN+I/Wg1+mqIgMjCy2ymVwY3uVokT2RgFzsD61qVBWUona5zxgY7oJbtRz08Pi5IHdbeM8z2uwTt6LeWoa2kmt/TG8EgLZMbgMgeaCB7pi7JJOD5o3xDnvupCx5OcbFXNYQeyDItzmUz+uXBWqVd1opYCxjRnt81o8o624Hf5LFFPI053IQTUVJPNVlzSeknYLe9ktMvjMlkJLNid1tqzvYxwMm2CFyJa7jQiEMLmgkYwg1SY2tlGWGJvWG9z3XD+r2A1pMIwMnYLkqre1/W5hyD2XHOpqOrdKZGtPSD3QaIzAjGcZUL3nsCruos+Fx3HdRlp6xncZQV8KVwzuq+BLjqzsspskYaBkZwry8GMgeaDTuqQbFxH2rJhqT0dGd1jPBLjsjNnjKCYwSvJcD3KniDoxl2cBSRyxhg37Y80kcHMPSUEjK2Juxx2WBWSNkcC3ChcHBx7p4byMhpIQUy75q5meoZKYPYjdUOQQgyeqMDdSwuAGW/YsQOyBusiFzW9zjyQZXjO7ZQzOwRlQlzfIqnWEF5d1O+tQyjO2B3VwcBvlWSOyduyCB0ZJ2WRTnw2bq1mDuVI4sDcjGUEc0jQRhY7Gue/YZSR3U4fJSwAAnHoEGXGwlowpmR4G/dWx7AAKYdkFmB6KoaqK4dkFR0eYCp0gu2G2VapGYxkIIJ4nOb8OyiYHMaQ45+pZjs42WM5khPY/kQXU8ojd1O7ZWqRXOEMwcLR3Rv7OGyt8M+pQbstV8o6eRrpQCM75XI1h19p2lc19RHGQMZ7Lg9kfxfSIWW2JhjILzkjzKDsNX6r01qqnNNSQxhxGNu64+veg/FeTDGQHHK0nh1PR0VYH1Eu2f3y5Yn1BZXPZiRnbfsg4cquHdS3docPNaVU6VnomF7gdiubqu92d7D0PYTj1WydR1MM9O9sGCT2wg4yEro5DGTuD5lTtqADucrGqqKsZVl7mENz3VAx59UGVLI2Ruw3wogCMqgY9u7hsrgCewQG4HkFd8OPLKt6XDuFRBY5gceyp4DfT9ClxlMFBEYWkYUT4w3YDyWVgqKVuRkIMQODDuniZcN9lR7SSRhR9Ls90GTmPp+xWxzRiTHzVnS7HdQBrusd+6DWA4FgO2FhzyMB8lkMz4YG602pz14z9iCRri47HzUjopC3bso4WO2J9dlm4IjOyDSnNIdhaxpynIuUMzuzTkhaVK1xfjC3DYaWqLw8MPSMboObqDU9titDYHMb1BuMn6lxPrO5SVVefAdhpd2WtMfTiAtc/cd/yLad4fGavZ2UGfb6ZtS1hlGcALe1u1fS2Ok92aQDjC2jY3NMePktMv7QZ/pEboNU1Tq6qq3PdSykdXfdbFlfU1LzJO8uJPms2cO6Rgk4WK1wGzsAoL4mtA3HkjqSSU5ZnCdbBj4gtQppoQ0ZIQaaIH0zup+cKZtYCMDCuuUsb2kNK05jXHsg9CPYvyh/NHqho/wCANaf+cbcig9iy0jmk1Tv/APKBW/yjbkQe0qIiAiIgIiICIiAiIgIiICIiDj/mE/3g+Jf8D7z+pSr5qHtD7lWA7/fH4/8AOK+lfmE/3g+JX8ELz+pSr5qC5jLnWFxwDI/H/nFBd0CIBw2WVT1AcC126x5ZY3swDvhR0wx3yg3Jp2/fseq/eWOwe6zdSa+uF+I8OckN9Ctn1zA9gIOCFZb2EElBrtHqO5ULg905wPmt42nW752N8aUHA81xxWRue3vhZtmbFGPjfj7UHMFmu0F+l92wCe2FpmsdNx0gL2sxkqLhrPQQ3LqfID8Q7lbt4gzxVUTfdwDgIOF6u1mR2GjGPJYT7W+HPVkZ7Lcc9NWNlLhGceSwqtlQdyAg0N9O7BAOCohTTN7uyFqLop87BWugq8f2M/kQYrY42fSaCqvna0ERDBxtgq2ohnjGXjAwsMbu2cEGs2G7VNFUB00hLAfPst+UPEK2MmjieGnsDnC4yLepmPVY0NNmrYS446gSg56u94s12tgdBEwOI7jHdcemysdV+KxuATlalbH0woWsjfl2OyyI4ap7x0Rkjy2QXVLOig8NbWqqdoDjhbrr6erZDl7CNltmqIDHdWBjug0CYu6yAexUT6Kod8YJx3Ur5IfGI6vNahG/qiPSMjCDSmySt+9ucSQpmtcd91DIPv5J23WQ2RgZud+yCx03Sen0Vd5GrGlBc8kHv6LIgDuk90ED4yO/6FNAQMA/kVr85xhI9nAnZBmCAPb2WO+nw/I8is6MgNGN1G879ggkgldGwBpxgrmHQeom01oLHuGQ31XDQ7D61v8A0tTvktzuk7dKDTNZ3annr5HdyT6rYFYx9RKTCSB3W4dTxllW7Ocg7LRacbn5FBdRQOGA/fHqs50rYXDHorYMb5wsWteDKB1DKDcVo8KtcGOaDuFzHoqSnoGMiDQMtXDOlLfX1sobRNL3ZHYLlyy2u5UEjHXCJzMgfS2Qcv6Q00zU05bFHkkei2HxgtVDo+qEdZEMk43C5i4D6gsVory+6zMa3/tiAuK+cWqo9SXQSaecJGdX7zdBxZcLjDV0bTasMeR+97rTqGyXasBdWSPfnffdY9ho6qip4/e2kYx3C3xbbpbxGA5zcjbcoNEoKBlrk6qloLRvuFqE2vrNb8R+GwEbHsq3u4W57COsYx5FcTajZTSTOMcmxJ80HJ9VxFtk7f8Aa3SHH0wtGn1hLJ1ES7H5rjughiBaA8n1WqhkQxlyDfumddsttUJa14LQc79lypa+Nuj2SRGeGIhuM5AXWqpZC9uC/A8isaGhp3f/ACY7/NB3BvHHPh5WW8QxUtOH9OOwytnxcWNGwOMnu8OQc9guu7LdG3cSvPpuraihjMZw89kHNWtuKtj1BSOpKBrAcY2wtjaZrH0l2ZK55wXAj5Lj+2URjrMhzjut621rjWRZyBsg56r9RXG76dZSW2Ytf042K2HU2LW8FvllkrJukgnufRbs0ixjaVnQclbkvT5xZpQY9uklB1D1BPfYLm9tTUPJDj3KwI6yvMozKcemVuHW/W69SADfJ/StAhZKH5xsg1WaZ8tMAD8XqtPhiqQ89Tzj0WYzIjHmcI0O6wS3YoMiNp8Ide5VPhClL2NjGe6h8SInugudEZtm+atdbZOg9wsqmLSQG4KzXg+GchBtZ9O+nlJJVzJ3dQPUcBZNf1eJkjAOfJYTdzhBqTWuq2dDTv2Q2KZg68H1V1pd0yA5zutxSyjwcfL+ZBtmEuhf0vPbyWUKhjnBmO6xaoOE5yMDKiYPvwOUGrOp42sMnSN/NY3ita/PkCs2Qg0w6e60uWGUgnBx6oM18rKiMMjGCoLTaqma9xMycFwUVBFPNKGw7nPZbg0/Zr+NQU8pp3dAcD2Qc/WnSbodOxzMYeoNzkLZeqLnerdRSww1D2jBHdczW+up4NLMiqAA8Mxv9S4c11cba6GVvW3OT5oOBrpUXqW5ulM78kqSOW6jd0zj9ay618bq0uZgjOxVS8EIIPe63BBkJx81BJdLpGS1k5A7d1P3Kwp2vdJlozuglFXc3jqMxJPnlTxU1RUAOmflIoZSwfCtRpmPbHgtxsgxPdcbY7K0wkHAWaQ4uOytcMOGQggZA4jP8yjlpQX5IytQZ27BWSHBOQO6CKClbj6P6FSopQPJZ0BH9yOyiqj8kGmup9uy5C5OY+nm/wCF4x21daj/AMriWxXLf/J1+GDwx/hbav1uJB9FSIiAiIg698/0BqeUHiNA3u+kpB/y6nXz7WmwzVj5cZ+AkH7CvoS57iG8pvEFx8qWk/XYF4K6TqomyVYODmU/pQaUNJVAGwcpotIVbv7rAW9TXUoaMgBZFLdaEDu1Bte16Arq1xa3q2U120JW22MucHbBcl6Vvdshly9zdlXWt5oKiAiItOx7IOAaunmglMbydvVQh4jBOey1i9Qz1VU73VpJPotHktF1wcxu3QY9RVvmHQx2FHG2qYQRIVUUNXTyB0sRaB3z5rJDhgHbAQVfU1Rj6XvJCiZKQc+iummieMNcMqEA9BQajT1Eco6MDI2WQYx6brSbc5rJS57sDK1h1TT5G4QQOb4Jy7cKhqoXNI6d1dVzwSMwHBaa0sBIDsnPqgyTOWuywkK6nudZDOHCQhoPZY4GULfnsg3nSaka6FrHEEnYrcNPa6e8UJcWBxIzlca0hga5vU7fO4XLekLlbY7eInSAvIwM+aDjqv0XKKt7mtIGey0uss/ujSHAgrlS4MqZpXvhi+E53wtm3uw32rcXQ0zzk7ABBsn3OUPxnZZjKMiIuJys86T1UHZ90f8AkKvksWoKeAunpntaO+Qg2+9oa8g+qGmdIOpoOyunBbKQ8YI7rMp3NERwg03w5M9OVlRgtZ8SjkkaHnsN1IJGFh33KCN5Zn0UjJ4ms6C3c+axHOPUd+5VvS5xyASgm6B1dXkqPAPb0V47D6lRwwMhBjkOBTqc3zVzs52VpBKC8SnzKr4hPmVFg+iqEExeSgPqrAUJwguMmAcKxz3O7E4VhJKvDXFoLQgsxhZVMCsbpefIrLpNsNP2oM6IbAKXoKtYGgf+1SZ2wgx5GkHZWtcQcFSPySosOBQSFpcFcxhASPq2CvPZBJTx9TtwpnwN6uwVlIQHblZTyC4FBiVELWsyAsMAYWdVu+BYbQMIKiP0Vjw4HzU0Z7hWzd0EIqKuHeneWn61V11u4zmoOUbnzVHgnJwgmpbvdeoB9Q4jPr2Wv2utlmna2ZxIJGVtqJrg4Z9fRa3aJI2VLTI7AyEG477S0rqIOawB2O+FtWCladunzW7ruY6yiDaLDnY3wttw0dbTnM7CBnzCDGrKdrGEgYwsKN7RklarXkGMg4z81ormSHZoOUE75GPGyjLCd/VWNgnZu4EBSg/DjHZBa0gbH0VepqsPdUyPVBISMKx2HKiIInQ5KoIB6FTIghMOUbTjIOFMgOHBBeIelvbyWBNSl0mfJat1NLR27LGkIzjCCJsTWtAHkrg9oHSVceyxi5viAZGUFzoB1tdjuQuS9O26n+4jpTGC4BcdPbJhjvJcp6dkibpx7ZDuWIOPqyumZXyxB2ACQAFpVSZJKjqccrLr+k3SUg7Fxx+VQux4n2oNas0rmMAPosW8uL5cqa3RyuaC3fbyWNc8skw/YoNPkIA3WC+ndI89JwsuoIbgKkDhuUELLdK8bE7LHmbJTv6CSMLXoJoowevC0e5Pa+bLe26DGAfIcEkjKzIYWgdljUxHWMkb+q1Bscrz8AyEHfb2MUQbzQ6oP/cDWj/nG3orvYzMlbzPanLm4/8AgFW/yhb0QezSIiAiIgIiICIiAiIgIiICIiDYHMGP/wAwnEof9yF5/UpV80NdG91xqw3uJXn/APCK+l7mCOOAnEont+xC8/qUq+aifDrpW+f3x+P/ADigxo4pQcuzhZjIsj4QVQAAAkABTRvYdmkIIZoXluD6KtH0xA9SyJC3py7GFjtLDnBQTyyNe0gY7eSxuqSMEMOM+ik+EdyrHhzjhgygyrPdqy2TibxCBkLe9PrMVjWid/Vj1XHUsMpb8Q2WVQNZGAHPIOEHKlqqqO7yeE1oJytSrdGNIH3s77gLa/Dusoaau6qiQAdQ3JXMNXf7EWxdMrNgM4wg4tuul22qIzyR4A33W3211G9xZ0jY4XJuvrjQ3G2GOgLXPLcAD1XDVNZbwKh5MTsEkhBuOnsDL997hb3UVw4bVFvaZHsPqt0cPaapoagOqmYAx3W9dVyispOilaC4jGyDr9W2t1K4tIxusDoDMkdwt2ai09f5JS+Ond0+RAW2Xad1AeomB4+woJ7PdJqSfMjz058yt923V9C2RjHBpJAXGhs13a/D43KemtNxbVMPS7GyDmquq6O5UP3poyR5LZ1bpuWWGR7QcHJWrWG23KWja2Nji7Gy186Z1JJRu6KVxyPRBwdW2uenrCDnYrIjrG00Ra70W4L/AKO1YKxzzSPxv5FbVuGn75TkvqKZ+B32wghfWNlkJae6uHWTndYUcUkcnS9pBB81qLQUF8eMAHusuOZjWkbZKxA+IH4nAK8OjI2d3QSPAc4kdlE5pDtlICD2VcZOcIJmPIbj0Ub5fi7q4D4cLHlyDgFBkGUNYDkblcycOre2ss7n9Ofg/mXCbyPCbv5rnzhNWUcdic2R4yW/zIOM9c2/wrhK0ZG5WzmuETiCVyFr+OSe5ymBuRk4IXHk7TFKRPscnugmM5jAdnGdsremleHNVqtonjYSCPJbEmjfJEzoGQSN12X4CQ1Mdub8AIxtt8kDhhoqm0hcPEucOWtO/V5rdvEC62uvLRbo2N6cDYLK1BPFDI81GGj5bLY1fWUMjyYZcgINGu0t98PptU72OA7tOFocWqJ7Zluo5DK8H99ufzreEFdRxuPW4dvNcScTZoJ6pxidt1b4OEGt37V9BXQllGGjI2AWz/u1WwFwExGStNoWsDBlxO3mVbXHJ+E5+pBlVt6rZ24ExWBHS1FUC55JJUZzgd1rFtLQwZwgwoKOWnIdkjByshwkfjBKy6l7N1BGcjZBjTQyyDAJz9arTUszXbk/astjRnJCkbLE0jJGyBI50cffGFi+9FwIJJWTWSwujwxwyfRaZGHdR6icINWs7GyTZxvndbxt9MDUxjG5wtoWJ8TJQHEblb4ttRT+9xYIzsg5AtFS6x0onlOG991n1vESir6F1GC0kjC0a/yR/cUF7sDp7rjy3y0z6rpjkJcXYAQYmtLRipdccfCclbPhqYXktwMhcna1oqx1nD2MJaW9/sXEVJR1nivcWHAKDV2uDdz2VxqIy0YG6gc4BnSTg/NRAZ9UEr2yTkhpO/ZU9xmaMnPZTUcUjnAtBOCtQfS1XRks2QafTPdTO+M7ZWc+6xkDBHZadVtLSQ7Y75WJkYODugzKuqjl7YysBo6ndI7lPAncctBKrFT1PjNJbtsg1GngNKBIe2MrKN3a4dJIwAqVEM3ug+HGy0VkMmSSCAg1VxZVfQ7qjbc9rg7dLeMH1K1Z2fDBx2QYbZOhoa7yV1LJFWVLaMAZfsFBM9hcQHbq2wkDUtNnt1jv9aDlfSnC59HEy6TxksJDlyJTssdDSeOadgewbHC3FS1lHFo2IS9I+AHP2LYl1u1oNDKwTNBOfNBoWpuIrg99HTygM7fUuINRXWuuFQQ2UkF2Ss2/PidcnvjflpJWjPOZs/NBYyhl6Q9xJOPNSsgf2OVqLHx+EASM4UfVH1dwgxm0Tj5IaIRkOcM4WosIDQRg7KGokZgl2Agtjnij+EgHCGvi6gwAb+SwnvjJ2csYlpqBg+aDcMcTZG9QA3Cxqlgjcdlm0G8AJ9FjVw+Pb1QRxOGPsWPUSAPyMBZEbds7rCqSOoZKDJhqA0bnKjqJus7KFm47q1+eoDJQXLkDk7/DB4Y/wttX63EuPx2XIPJ3+GDwx/hbav1uJB9FSIiAiIg6+c/kng8ofEWUfvaSkP8Ay6nXz5Wm6GlknPVjrOfzr6CPaDHHJ3xI3/8AuSj/AF6nXzxwt6iRnyQa/Pfp5PhbIfyrGF4q25xKVpjWfEcEqr2u7boNYpdS18D8tmIHyKypdV1c7cSzE7ea20IySOrP5UewgEAlBu6wX2mE/VUAHPdb0ZcLTWPaxkbfmuJKBuJvjOBtut6WaopI5m9bwO2d0G4r7Y6SspuqmjAJHotqt0bUSAxhpyduy5Diu1obSgOlZn61dT3W0mVvS9mcoOLa3Q9bbh4smcAZ7LBNuJaQB22XLesJ462h6aMAkjsFxzHaroQ4eCck7bINvVNFJEfhJCxvBqTuXHb5rXqyxX1uXGB2PtWD9xry7LWwvJPog07wqhx6clXiklZhxOwWezT19jcHvpXtHnlSyWi6OjIbEc/UgwI6hrfhzupDK17SwdyojZro129M/Kq22XFjg50LgAUERgn6uprjjOVqNsudfQ1bXiUhgI28laPhZ0vGCoZSwgtyAeyDk638QLfFSsim6S8Dc+q3JZeJmmmvDaiOMnsdguBBSyE5BOPrVIoHsqB1ZABzlB2th1to+ohyKaL4gPIKt8hsNy09LNTU7AS0kYHyXBllqqNsMbXTHqGM5XL9trLe/SskTJcvLDjfzwg623uPovFRE3bDyBhRxMcxhYc7+az7za7hLeKl8cDiDISD5YWNJTVUDgJ2kYQYppnudn1Q0kmM52CzmSR9IBdghXPmi8ItGCSMINHcMHBKyqdzBGAdzhY72OLzgKSJj8ZOQAgE9Ttuyu6Vafh8/NXRkk5+aC/wRjsqGAHfClzthMlBCYB6K10W+wWQd1a70QQCP0BVwjyNwVJjG+CgyR2QRmDJGPrWRDE1jMEbq0EjyUgcMfFsUFhDA45wojIA/wCHAUjukncjdY72PLx0ZIQZ0cp6RuspkmRv5lYMccoaDg5WQwOaMnP2oJtvkrhE0jKiD9x2WQ146Cgt+FvkFBLJvsR2Vzyc99lC9rnHZBLFKR2IWa2TIWnMY9p3ys2Nw7HCC2dxIIWPgrKeWd8qElu4BCC2Jrs/apjDnfBVsTmNPxHzWUJYs9wgxX0+BsregDZwWW57CPhwsV5O5CC0lrfRWOqHjPQd/krJerfurGDOUG9uHz3zVGKs9Tc9it1aohomtBhYGnC2rw/qKWCYGqcAM+a3PqesoahobSvBJHqg44ushbKd9liU1VHG7qfjA9VlXihrZZCYmEgrRhQ1kbT4jDgd0GuS3Cmlb0sAyRjZYb8b4WDAzoO5wVmAgjyQQvByVb0uUpGN91TB9EFojdnupWQOxlG91Ow7YQRuiwOyhkHSP9Sy5NhlYkwyDt6oIQ8gqUHbKxxnq81P+8QW+K4ef5lTrJO6idnJ790aSCMoMk7gj1WJDTSS1gbvgkLLHZSW+Cb35jujYEINzxaf66RjujcBZEta+3UDoAcDGMLW6R+KEN88Z/Mts6gkj6HAnB9EGzqidzqx8me5yroZ+qUAnzWO8ZlPfHqroGO8YfWg3zp9jHt+IA+i07UVGXVALBgeey1DTmejdWXx7BNvhBtG4sdEN+4CiozlmT3WVeT1D4QsSjp6l7D4bSfNBdUtkccNJUApJHAknJ7rL+510lOY4nEfJXClrIGls7CD80FbXZZKyQYycLcDLeygAZIMn5q7Scc7ZCQ38yzL5UQtqA2UgH0Qd3fY8CIczGpugAf/AAFrf5Qt6KnsdpIX8zOpvDOT+wStP2fdC3og9h0REBERAREQEREBERAREQEREHH3MNtwB4mH/uPvP6lKvmp73Gr+cj/+kvpX5hf94LiX/A+8/qUq+aeQltxq8eUj/wDpFBdVZbEC3uoqJ7nZyVI4mUBp3yqMZ4JyPyoJK15bGMZPqsWk6nE91kHM2GuUkcDIs4QQzBwHcrMtzWvGXblQVAaW7HdWU8ro/qyg1esjjbDtjJWkguBPSVlGpMo6Tn7VC4AZx6oKw1FZGQYJCD8lqMNfeiPiqXH6ytLY8s3HksplY4N7lBvHRU9XV1oZXS9TM9iuX4rPZzGwgMyQM9l14o79Lbn+JGSDlazDxMuAw0l2Bsg5quNDS08WaVzQfUJYaYVFQ1s8gIJ33XEcfESqnHTI449MqRvEWooiHxPdkeiDs0/T9gdQB0giLsZzgLb1VZNPtp3uEceR9S4Jfxoupb4XW/H1rFHFW4SvEb3Pwe+Sg37e6W1Q1BEcLTg+S0JjqH3oN92Pf0Wdpa9Wu8PBrpgCTvkrk+n09ol1vNT48XihuQMjKC7htbbfUFhmY0DI2IXYO06a04aJheyI7DvhdPq3WMmnbm6OhcTEDsQdlm0/MBdqeVsJfJ0DY7oOxurLJpWAvAp4iQD5Bde+IX3FggnbDQZIzjDVu6x8UrTfI2mvqB1EgblcvWbSPCrVGl5nVFXTmsewlgJGc4Qedd3kYbgSyItGexUZeC0+Wy5b4pcFdQUGpZ6i20bjQBxLXNGxC4uutorrc5zHsxg43CDSJXP6jgq+AybZUT/F7uaclT03USA4bIM+InAz3yp2jPmO/moWYA79kL5Adge6DJO3dY0x+L6lK2QFvxHBQxNcM+aDElJEYG/dcm6BfVe6ANlIHT2XG08Y6QFr+ntQS2/ELCR5bIN91zI3TvMoDiuK9ZOa2vxE3AyfJclW6o+6B8SQ7laPf9GVFwqBJCzIzk4CDZ9rlb7vGXxk7jK7c8Boqaa0NcWhpDex8tlwbYtIUMETGVvSCCDuuU9PX+m05TCG3vBGMHpKDVeLrXwCX3eTBGexXD9nmq5GyeLISc+q3HxA1k6sicZHHLgc7raem5pKtrnRbg7lBNcJqtjz0OIx6Lj3UU08kjvFJO65YipYpJCKkgfWuONex08FQRAQRlBt6nL9sH5FTSuyQDvssWKQtY0j61lwdMoBfjZBC85AHzWoUJPRscLCqmNY3qaVJQ1BDfi7oMmrLgDuTv5KShyW7g7qyN7Z3b9lksa2LcfmQJtgcbLTZnPAJB+pZ9Y8NjJB3+SwYCJHYd2KDHpZJXS4fnC1CXAb5KR9NDFH1tIyoGODz0uxhBbQPk8fbbdbptD5vuhDlxxkZWjxwwRNDmkZytUssjpLlE1uc5GEHJmrXPGmm9J36f5lxPpl1Q+/Rsc/4esZ9O65S1vFWQaYEnScdGfzLg+2XWenrHVLc5Y4kboOzWr6Gjh0ayUhr3eHnbv2XCNLPTmGbNKQQTgrf3DrWUGqui03qTEWQPiO2FytdtBcO4LO80s8BnezIAxnOEHUSpeXVh2IBPYqaP6QWv6w0jcKC7yS08RNPk4I7EZWhNp5x8Qb2QbgsMMTnjqx3W6ZqalFOdm5xsVx7RV09K8HcbrUpdQTGIjqPZBiXyNrZiWkYytDj6vE3zhZctRVVsx6QTuqU9trH1jIug/EUGp290fT0uYT9i1BjKczNHh/mW5KLSPulA2rqmBoIzurHQ2dkZkbI3IQYFxihFGCABsFtssjLCQtQu9yEhMMByAtF/2yGnY4+SC6hcfeO+2StfkaDAcEdltqke5s2XZBWsGt6mGMdzsg0SokeypIAJGVLYKlztSUzBGQS4ZP2rXrBYHV1UJKluGk9yuUKbQmnaK2m79UYljb1DtnIQbvvVNUR6JjmjqAMxjbPyXANxqroHyDxyRkqe+cW7gat9ka5xgjJZnOy0t93hnacuGTvhBivfI85kJJz3UZPxb+qu62vdlp2VzYwTnzygueXhmWlYviPMvcrNeB0LHZEDMMj8yDU4STEMgrT68uwcErWqaOIRbkflWl3OP4sMGyDRXOk6u/1pC53jjJKnMBBJwoi0xuD8dkG66AE0+OoZx6qOrb8WSQVoDLtPC0NbnYrVaGrbUsBlO/fdBNHv8ADhaZX5D1qwdC1xIcFpdwBe/73uEEcRyBuqu2dlWtD2gZGCq4c45wfyIJYxkLkLk7GOcLhl/Cy1frcS4+ZsFyFyeEHnC4ZfwstX63Eg+idERAREQdd/aD/gdcSf8AE6P9ep188cPc/Uvoc9oNg8nfEjP96Uf69Tr55qVvW4g+n86C+E5dhZBDcDsqNgDPiAVry4FAmaGjIULcE7qYZf8AS7J4QA2QHj4QWYz8laySqZ8THEEKrSc48lIgjmrri0Brp3AfWpLdX15qmhk7jk+Z2VHMMvwkKsUYpnh4wSEHLekIX1cLPe5Mg47lb3prNa/eY2kMwe+wXAdLrGttwAiJGPms6LiZc2PDyXbfNB2FvdlskVAHBjCceWFtq0260PrW9cbCOodwFxrS8UKyvaIKl78ZwclZsWso4JRI152IKDmTUNosDLYHRwsa7p7jC2XTW62b9TWn8i2rd+Ir56bwmPJ2xstvU+tKgS4c44yg5LraC0Rt2hb2Wg3eG3MonujgGR2WNQampapgE78Z2Wv0j7FW4hllZh2xyUHB9wqyKx4azABwsMyufMHHI37LsHUaC0nMw1DZIuojvsuPtS6Khhkc63NDtz2CDbcDIhACQMlQ1XhthLgMkLJ+4V7aQwRO6R8itQotNV0kgFQwhp7koNpU9VUslHQ9257Lk7TNZXe7APe4sLd25UlNoy1siD3dHXncLcGkbTTzX6G3bFjnBp+pBhxVFB1PElCS7vnHcrYesqmIPIhpi3J7kL0DsHLvpiq0/HcJhF1vYDnb0XF3FLlthZaprhb4WOazJBDUHR10jyc9lVjnucMeq3HeNLz0N7loTGR0Ejt2wtHraWSin6XNwAgkZG3p3G6Pa1sZxjKkj8MtGSMnyWPUyYJa3sgxTnqwewKyIvLsouh536VNEHDGQgkPZUHbsqnsqNc3tnfKCqo7urjHKRkAkK3pdnLhjCCoGQFVoGOyqHRbAkK8dOMgghBGWbnZY8zi12AVmZaNtlp9S7Mu3ogo95AAGe6z6INIy4ZOFimLLQcd8LXbRBTOAY8tB9EETOjGOlRTdIzjZbppbCajJibkeoWj3mhNHJ0vGDlBo5OPNTxZLe6gmBb2V8MhDfTCC9/f8qujxjcLHeZHE9IKyaduR8ex7ILj0+QUTiQdish7GtB3CwZpHA7IJC52O6ic92fMoyRxIB81L0NcQggLnH1VQ+QfWpnRtAyFb0BBWne8uwc4WUGgndYrPgKlbK7zwgpWABhWJD3H51lznrbufrUTIwHYHZBdLUVELR7s4tOVqVlqa2edoqHkjPmVhOYwt2wSp6WYU56weyDkakpqM0odKwE4ytFvsFG2BxjjA2Khs17ZLiOV+B23WqVjaSqb0NcCTsg4zcSKpwwQM+imBxha5drMyAmVg2O60EiRriMHYoJvJMD0UYe7bOU8Zg2JQTBoBypGjbyWKKlp81UVOOyDJcCAopBsTsqCfq2yqOd1DGUEG2SMfmV47I5gG6iMhDseSC4kZxhWObk52UoaCM7Kx5ABwgki7j61uK0NiJGWjK2xG53WAMrdlrpZG03vHTjG/wBSDccEZ8E/EMLZeqPEEpwdiSswaj6JHQl3bZbcvFwlqKgkE4zuUGN0jIzjKyaVrS7yyoQ6MtBLhlTUccr5/hb8KDdlkf0MOGErAvz3GYDcb+e3mt+6Ct9okiJrnsBx5lbR4ktpaa4hlvcCzOPhQbOubyA0HJW4dLtp5IgZGjt5rS47e+uaD05WtUXu9sb0PIBGyDfmn6G2PaTIxv5FtnV1LSsqy2EADKxo9URUYIY/8hW37tfZK2bxGklBvbSFFC9pyQDhbM1+18F2LWP+HfGCsen1VWW34YSclQVFf92XGapPxDvlB3p9i1JM/mh1OHkkfsArcZ/8I25FP7GJsbeaLVAYRtoGuGP/ACjbkQeziIiAiIgIiICIiAiIgIiICIiDj/mE/wB4LiX/AAPvP6lKvmmm/wB0av8Awj/+kV9LPMJ/vBcS/wCB95/UpV800u9xq/8ACP8A+kUF8YGB9SvVjNgFXqKC5VyfVWdRTqKCuCdj2VcBU6vkqdSCp7Km7vmmdlVqAeytLiNgr1TpCCzd2Ad1c1gGfhCYDTsrhuED4gMt2+pCZD9LJVzTnZX4CCMMady0ZVHtHSSAMj5KXAQNBQY1HVXOGcGCR7cHy2C3JSXbUjpmNFbIG+Y6jhaL4fhDxFay7TRSjB2GyDkKRsklIJJ3B0hHcrQKvLYnuDB1DtspbbeHVMYZIVqgp6WaP4nDdBx+66XiCoPgzyNwdgOy3XoLVWsf2X26nZdKkMdM0PZ1Hp6c7qaSxUpkL9vVZ+mKWnodU0lRkBrHg/Lug9F7tb9MScH4quqpWSVhgBc4gEk4XnlxNq6SOsqWw0ZwHkbN+a74af1RYL3oSG2TTt6vDDcE/Jcdak4E6evlrqKqmax0pyRjHdB0Mkc2TB6MZHZGADfGFyDrbgxrChvUsNvoHvhBOCAcYytm1ml77ZnFtygewg75CDEyR2WZTtYYySsMxvBw4d1DJUTxSBrQcZQZEziJC0ZU0RJGc91dE2F8Ye8/EVY8hrsN7ILZj5Ke3NzIO3fzUYYHjJG+VWJ/gygDuSg35ZMtjHlk+S5Y01TUM1v6p2gu6e5XDtkfPJG0tBP1Lf8Aapro2EMiaSCEGi6/fJSuf7mS3B8irdDPqqumLqiXJHbJWp1+m7xenkGFxz6hbYuNTWaPcYZQWnOMIIuIrZIoiWP2GVqPCoNlonGVuT6lbN1BepbvCDknqP1rffDGmMVAS4YyPRBPqPrZI/wXdPdcT6hdK6d3iuzvsuTNY1hppH/UVxTcah1VI53zQY8fYLIjzjDTsoGNIAGOwU7HdIwUCQEtwTsqRnA2VXvy3CozsgnpSeoeX1LPLjtutPgPS4YWcHdQCCypJwsduRnpWTI0SDZRCIjuEFjnSHOScK3J9VfMOkbeqhBcQUGRG+UYySVuPSMgff6djxgF4/StK09QS3KcRtaTvhbyh05PaJo60sILMHKDmTiZQUjNAMlaWFxi3x9S6q0QYXy7DZx8vmuUNXcRqivtP3Ic8kBvSuLabqY5/wD2xyg1ShlraaTqoZCx2e7dlqVsvOqpdQ00M9wkMReAQXbYytIppXxODtxusmO4Phq2VQO7DkEIOyGq7fa2aHjqXtY+cx5J7nsuA4Joj4oMRGHEBb101rpt+gFquMh8MYG/bC1+s05phtKWU0rDK/cDbOUHDU/S6UlrcfYo8D0W4NSaVutBIahkJEHcEdsLbAE8nVGwEvGfyoNUtBhjkBfF1b+S1qjraZ15gi90IBcB2Wp8HNJT3W6D7sR9MJeN3dsLsjeeEWiKK0fdamkhMsberbGc4Qca67pKeHRrJqZwa4x5279l1vFVcOqUeMcdXquQtf63qn3CSxQEmFhLRjt6LYLIpRIeobOOSSgktT5ZJsTE91uSSKnEWOkZWkNiiiYHw4L/AE77qySorfBJIO2d8eSCOdrWzEs3GVHTSu9+ZkEjq9FZZBU3OtMWCTnC3hQaNrPuvAZIiIiR1HHzQa9RSRNt7THEQ7HcBQXGruX3Oka2ocGkdsrky5WPTtt02x7ZGCYN3x3zhcQ3m7QtglhY4eY2QcU3Qu+6khcMu6ifrKjbUVAmAycLVKmjNRWmYN2JV7LXmQOIQZdE5zox1E5WoRty3OVFHA2OMDzCoZntPSOwQSknOMq5gwc+aia9zsE+qmYASgmD342ccIcOHx7qrBkKjzjKDGcGhxA7ZWNOGjP1rJd3KsdEHoNPcW57efosiBzg0BmQpzSNPkhiEecfYgtc6UD6R3WZTYc349yfVYbs7Z9Vk05+BBI9jc52VhDVa+QjOVH4pIygq7bst/8AJ0T+3A4YH11dav1uJcdSSH/3K5A5N3k83/C8nz1fah/yuJB9GKIiAiIg67e0Jz+054kgf3nR/r1Ovnpov7If+9X0Le0HI/adcSMn/wC5KP8AXqdfPTRf2R31fzoM1xGO6jIB7q53ZWoLsADYK3ICuPZWOA7oKN7q5RNc7qxupRnzQXo4En/WrA8lXDcIKOa0jcBWGNmMBoUwAPdOgIMRzXt3jGCreurPcrO8JvffCeE1BixvqM4eThSkkDYb4Kl8NqqYxg7D8iDGE9YD8DyPtU9NW3ZswDZ3flUEhLPoqEVMzHgjOyDfFHW3h0bQ6qfg481yFpWliqWtNZIDkDJJXDMF7qGsA3GFrFFriso2gMJGAg5/FpsnR9GPP1LTrpa7ZHSPdF0BwB7YXDbuJdxGficsefiTcZQYy52Dsg3RL4/vxY2b4c9itzaDpy7VNPmUAlw3z81xF+yerMnib7nJWbaNe1VtuLKtpcHMIPdB6fWi3VB0lTdFwAy0bB3yW6a+025nDermq3sleIzjsT2XnlFzT6hprfDRRSS9LSBjJXP3CXmJt2pbSLLfarpErcEOO26Dqvry408evq2JlG/pEhGcfMrYupXRyvLms6Su/tZwi4W3iWe9vnpy+QF25Gcrpzx30d9y7+W2OIyUwcfodsIOJi9wcACcD0VzcueCfzqZlFKGhr2kPH5Va9nhDc4cDsgyOhnSO3ZXtY3pzt2WnmSp8gcFUE9Q09JBQZZHxFYJcROBnsVmtyW9XmqmGDp8RxGQg1SlZG6nbkjOFhV7WsBLVg+/1DMtZnAUjKjxgGybH0KDAc95O5Ky6ZziwZz9ayTRxFvV327KrWQsHSSPtQYuHZOM7dljyAtlBKzi3JJA2ysWVodMAgyGEFgyFWkfL76A1xAyp4YB4YOAdlieIYq0Bvrsg5k0cyJ1MDLgnC2nxADW1pEfbK1nSlRP7uC3JGFtzW0dZNVdQYcZ3KDQH9JaM47KxuCeytk8VjQHAjbzUsABb1HuUE8TGYyQN91ZIcHDdvqTxOnb0UbnfFlBR5cB3P5VEQD3Uj3EhREnKCuQELneWUDQVe1gwgpknZV7DdXtjBV4i27IIGqqkMJx8ICjLZAdx+ZBXKoqlpHdARlBaHOHqjpHYI3WVC1jtnEBXuhhPYjdBi0Uk3WOlxH2rcdufUOnaHvJGfVaI1jYt2/oU8dwdD8YO47IN03ljG0mTucZWzpJW9Zb0Fa9abm25PENU7DTtuVuKewWV1IXROaZSNvrQcdTgdOQQPktPm6g0kFajeLdcKapdiM+GD3Wlhsxd0uBxndBFFI4uws1mSN1T3VrW9QG6oC4HGNkEjOoHG6lHZUZ4ZxkjKnb4JG2M+qDFc4kbgqzGTnCzvCYQrZI4RGcYzhBjHqx3UTuoZPfG6h/20ZCA12MnGyzLXba6qrYmvjd4Zd8RPbCCOnqCXtaYj37rky2QxSWFxIAPStUh0dp1lsZK97BKGgkbd1ti/XeC2U7qWlft22QbCq/EZc5B1H6Z/Ir5mhzCe5WJPUPkqHTb7lZUThIzpKDB6nB3crdVkDDT5c34sd8LbroB17AbkLddlhjbTfERnCC2SquELiKWZzR8itKqqiolkzVEuPfJWqVbhG7DTsT5LNobRHVxeI4D7UFNOCOQHqGNtlt/Vckkdf0seQN1rkz22kuDfsW3bjIK+XxSc7oNJ8aXzeVlUxL2/FurnQRjGcfJSRsa0bIMerAAwAPrUEZf+9JH1LNkYH5yschrDt2Qd+vYtdf7aTVOc//ABArv5RtyK72LZzzSap/gBW/yjbkQe0aIiAiIgIiICIiAiIgIiICIiDj/mE/3g+JX8ELz+pSr5ppf90qv/CP/wCkV9LPMJ/vB8S/4H3n9SlXzUSAfdOsHpI//pFBe36IVCcq4DYD5K0jCCio5VzsrSSUAbq4DCtzjdA4lBXqGcK8dlYGDKvOwwEFUIyrS4gZwqdZQVGx3Vyj6sndVLiNkEjfpBXKNricdld1FBcqt+kFZ1FOr6kGVLvF67LS3gCTdu61DrJbjyKiMDSeo4/Igzre5wa3Gy1qCSbb74fLzW32TiIdI7hSi4ytG2UG5epxG78/PKpTukbWMcx+HDzW2jeJgcElXNvMjHeJkjHmg7C6Iud693YwVxDMjs5djOHz5ajwWVdWCxwHUCe66DWviRWW9wjY52AVyRo/j5X0Vzp45JHiMYB3QeikOltFzUj5aqjikkLM5IBOcLpDzS09ot1bM22W44Dj9Buy5+0Rxv0zXWhhrqxge4AHLlv2Xh3wo4nWCWqmqqZ1U9pc0HBOcIPKyaUvLXeA4H6ihaxzCSzBXZriLy03uK/zNsFGX0ocektbthcJ6v4a6m03M9lZSvaAcZIIQbBkkkEnS0kAfNZkB6mjqU5s9S0AyRkElSMo3sbu0jCCEbHAOAotzVNCvncIskrGhkc+cOG4Qcp6RihMMYeB5d1zXpSloHRMD4Q7byC66Wm+R0UTOp+/UPNdn+DAt97o2OkeCenKDcmn6C1Ond10o/IuuvMhDTwXEiljDB1bbYXcfT+kW1NbI2mYDsey6mc2VontN2cx7CMO9Pmg4VoiDTx9e5yFy9oRjTQZbgfD2XFum7bLcIog1uchci26rNipxFMekkYQaRxAPSX437rjGP4nvJ9Vv7VVxirWkhwOVsuOOEZzjJQQ4Ctf32KyJGxj6OFiSvwdkFc5Usf0Vjh+TjZZMLcgfNBe3AKyo/iCxH5aftU8D/hKCaM/FupXBpChhIc7fCmcWgYBQQTDIwogxu6rUyYGyxWTvLiN0G/+GMUL6/EgGOpcp6rp6VltJYAD0+nyXEegZzS1Qkccbhcg6ivTJ6EtDv3uN0HDV6fJ903NP0clQMa3xBhTXhxdXucPM/zrFY93WEGoPDRGCMZUJGGlXSyAQ/NYDKlzn4JOEGTTz1NPJmneWnPkcFarp253ifU1LFPUuMReAclaZjw2iQ7eaip7i6mqxUxf2Rm4x8kHbXXVttMXDyOpaxj5vCycbnOF1nsMrZL0WSUjujrxuD6rlbg/eb3xCljslxa/wOoN+LJGFzzV8ttqtbI7m2NgacOcekfWg4wo6elprEyWhYIZC0bjYrFnud6ktErJbg4tAOxd5LO4vXay6atpoLZM0zsaRgeq69U3ES8y+LSydeCSBv5ILbuOu+SOlw5xcck753Uk8cXgktxnCwpYa6pkNUWHJ3Gyo6KvkaYWMJedgEEVpc99aWPJLc/zrdlVBAygc4R79P8AMtJ0rozUZqxU1VO4RE7HHkuQKfTMk1xggmZiAkB5Pkg2Fw6Mc2oCySnLW9fcjZdiqi222O3CVhYJAzPlkLStU6V0XpTTwuNuni98LeohpGcrhyXibc3skh6n4BIH1INS1lcLo2pfC2qJj326tsLjypqKh0pDnEjPqtRnv1Rc5i6XO5VopGPy5w790GNTtaRlw3+YWd0s8PsFF0Rty0EZVSXgYxsghcT1Y8lc1oxuFd97yMkZU7Ww9OdkEGAFJHjtlRPewEgHsrWy4IOUGa3tlUcovGOM5UjCJO5QQv7qg2wVkmNp7YyopIyM9KCmRjurHd1UB/bBVryWndBa5uR2U0IwxYzpN1LHMA1BSXz+tRqskrSe6s62+qCyTt9q5F5NfwvuF+3/AMt1q/W4lx27BH2rkXk2H/XfcL9v/lutX63Eg+i9ERAREQddfaE5/accScf3pR/r1Ovnpov7IfqX0Le0KP8A1nHEn/E6P9ep1889K7pkJKDOd2VrFSSQBmVGyXq7eqCZ24wFQN27KnUrmuaQgBjR5KpCvHQfNVAaUFnRhFIQCMD9Kt6QgsGQcq4H1VMKhJCCYHIQ9lEHuCeI7thBTrPXjKuyVFk5yniDZBe4AjdWGNmc4Cv2I7hXta0jyQRhjQOwVelvopOlvqnSweaCwhhH0QrOhnV9EKbDPVVLWYyMZQXGNoZ2B29Fprx9/AxtlZBlkzjfCkZAHfERugz4IgYgXMGQM7jdYdNebvQ3Ae5VT4nBwDQ04GFM2dzAGjssqKmo2s96cQHgZCDlWg1Lq77kRE3WQZaP35W9aOmpK/TctTdHNmnDCQXYJzhdcTrK4RDwI3HoacDfyW7dHcRJqiuitla4iGQgEk7YQbfu8rY79PHHSksBOMA4W3bk5zqrHhloz2I7ruZYeHnDa42z7pVlRAJXt6jkjOcLg7i5w8ijrC7TUYmYHfvB3H2IOOYIoXQAkA7LDqYmB+wACzI9F6yJDWW2ox+ZYldp3UNCSaullZjuSgjaAsKoe/xcZIHy81d1TMIjdnqWdFBBJCXyH4vmgihYzpBcBk4WJUZbKOkYCq98zXlrAcA4V8IDnAynBwgnie/wxuT9qw5pH+J9Ihag0wgYJWHUQl8nUwfUgyITloJ9FhzH7+MFZUQ6W4PcLFma4y5A2CDVIDmIBYAGLgOr1WTDN0sAPfzWPG5rq4OPbKDmHRkUZpGEgdvNU1TBTAkmME+uFp+m71S0lMxrngZ2O63o6go7vQOqGkHbKDhi7sjDj0t2+paZE4gYHmVuTVVLHT1L2MxtstsRuJyM9ignyVRMgogo7sqKriMdwgAwgqGkHClaw4UIdgjspWytwBlBLE0ealwB2UAkAOyr4yDLga1zj1Y+1VlYzq7AKCObByPMK/xOoglBj1IABwsEuIJWpTNa4YJCxfBGdsIIPEd5EqaJ78jJP5VHIwR7nYKscjSfhOQEGW7PTuoX+YV7ngtwoHPKCWJ8kbsxOwfVapZKyukuUcck5LCcHfyWgSTvZ9EnOVkWWrmjuLHu7AhBy1qWkoBZmvEYLy3JON8ri2oZGOrpYAc+i5Hmq47hbGxZyenC2pcLQIoy8jAKDbOSTgnZVfG3ozt2VJnMZMWAjbyVvjNceknvgBBhl0gcQM4+tXRPkMmMnHksx9O0tyPNWOiDW/CB1AeSCbLwwZPl/MsQSSePg5I81SKC4zPw1jiPqWtWjTtwmqGyTx4jyDkoL6eOF0bMx7nucLftpoaD7lGUNaHhuQfmofudZIaQB0jPEaNxstqXbUxoHGmpien6KCl0r7myqkijqHCME4wVti4T1MknTK8nf1WoSV75syk7u3Wn1AMrsjugh6GkDIVW/CduwQxzAHbsrKcufUNjPcuxgoMrPbIK1u3vcIsb/lWbT2DxKVspHcZUJg91f4fbOyCCd2SMgndbhtR/2r3xssakoIpxl5G26xbjcG253hMO3yQYOpHuycHK0CEuLCSVqtXUNrATsVpzmeGcDCDFnc4eqlpnFzQSrZWF3kpITGxuHHBQW1BIBwVjAEjdZUxa/wCicqDHTsg79exYP/XS6pyf/lArv5RtyKvsWPwpdU/wArv5RtyIPaVERAREQEREBERAREQEREBERBx/zCf7wfEr+CF5/UpV81EgP3TrP8I//pFfSvzCf7wfEr+CF5/UpV800riLlVn1kf8A9IoJQMAI5UDx0gq0vygY2VuQE6zntshAQVVW9lGCc4PZX5KC8HdCcq3qKA5QXFpwo8hSnsrOj1QUVQRhXFoA/wBSBg+aAD5qvUE6QnSPLKCqoRlV7Kmd8IJh9EBV7NKrGARurzH8Jwg0+NzvEPfGd1qLWjo7DssBrCJceq1FhZ0beiDFlA6jgKGXaM4CyZA0u7hGxNfscYKDSIz99G2clai0ys+OPAeO2ykdQxsy/bKtc7oOEE0F+1NCBHBWysaTtg7ZXZLl5uGtq690UM94kEDsdQ6zjGy60teWDI8t1uXSfFa86TuMU9IXBsbh2dvhB7icL9CaTrdMNnu3gyz+Hkl2Cc4XTrnEsemLTNP7hTMOHHHSBvuti8Ouc+uprIyCoqJASA3Bd8lkao1VDxVgdUSydQfucnKDqbeKmmZUAMgOPqW3rhcGAFrac+nZdg7joCxmoLHlgI79lFJww01NSPcHR9f2IOr1xqS7cMIz8lLacSNLj3XJeruGFQ+d0dsj6gO2AuPKuxXSwPcyriLMZHbCCKuM7AOh5G67O8uYq308ZMxx0+q6sVcs72NOCBlc78DNRV1HJFCwOwQM4KDvtwlEbbg/x29e3ouo/PjJAL4fBj6fj/nXcTlnjprzXk1xAz6rrj7Q7SMLb8Db2ggu8vLdB1f4YSwgRdbMjC1XiE4l492+ED7FrXCHSIf4QqGhowO4WHxrhitNSGQH8iDi+vfMYx1Oz6rSWh5J3P5VqE8z5omnc5Wmlz2Eg5QSYcB3P5VBKRuFeHuccFUfGXHKCOP6QK1GDsFhti6SCQsqN4a3dBdKkZICte/PZVjJAQZEJwftV8nUTkHZQMkDSrzP1DCCGY9Iwd1DC8eJ9HzWUxgmduMj5rIZRQ9Yx67oNcsDy0AsBB+S1q4zTmnIdnsrNKW1lS4NjGST5LcF+sMtPRueW7YPkg4prjmocSooyC7GN1dWPH3QdG44weylcyEbtI2QQzkhmDlYkRzKBhagWNk+pBSxxtL8jYZygtrneHSgtGThaLbp3G4sL2kjO4W6bFbJdQVIpYGdRzjGFvKi4QXKK5wOfTkNeR5FBzhyyUdtrHxBkLYn5Hxdl3R1fpqng4eT1LappkEJI388LpTBSVvDayMraBpEoaD8O2+Fpg5mtcXamfaKhs/gEdBJJxhB1/4o3m6jiDWU9RM98bJiAwnYglaXFXQGrYG0pGf+1XOkGj9L6irjeLpJGJ3nqOcZWtv4c6DaBM2WHIGdsIOLaJ9JJRNLocEjthVsnucmoqeF8XwF4BBHzW+rjadN0mYoJWYHbGFozaayUb/fmSN62fFnO6DsRJpnTg0dFUQRxNlLAc4Gey2HWWWlZZKiojkaJWglp8wuK7zxsr6eD7mQSPLGjpGCttP4t3SSnfTuL8O77oNu6juuoJNQyUk9Y90AcRjO2FQ0sYjLiBnzWm19bLXVbqtmS87qGOa5ulAc13QfNBmRNaJOkDGFnuyITghU8OBsAc7AeQsSSp26R5/NBZAXOnIJ2WrugHgk5GcLQjI+J3WB5qn3Vnd8GTg7IKTOk8cgE91kReIW7uKt8PqHieZ3yqeIWfCMoKEuyQcqrCQ7zVzfi3+akbEDuUEmfhU8Dcj0UONsK10/hDAQZoaQe6v6WnuQtMNc4Y7qaGsDsdRxlBnNDRnt+RYVZjJwPPyWYySMtzlYtU0O+jvug09xPqqhxx6q+SPCoxm2/wChBC8uz5qgLjnupJAD5K1owEFMkbnK5L5Njnm+4X/wutX63EuNTnthck8mv4X3DD+F1q/WokH0YIiICIiDrr7Qz8DfiV/idH+vU6+eSIZcvob9oZ+BvxK/xOj/AF6nXzyQ7OP1IJJc9PdUhd65SU5aqQ7j7UEpye2VVpcNsnumD6Kh6s7A/kQTtLiO6uBduoGOd23Uw3CCQH1Kr3VgOVdkoKkjCtVOruSULh8kFwGN9kdg47IXDBwrOooLZcYwoQSCfrU7su3UZjO+QgNkJwFK0u+e6gY34t/VThBcScd1G97uwypBg7HCu8Nrt0GI4u33P5VWORwdg5WV4TPkqGFgHWMZQStY3oB23CYxsFhipId0jI3U+JjGXAHCCYNyd8bLTqqWZsha17g3yVrqyRpI32Ky6eH3hnilBZFHG5gcWjPnt5rHZJLFVh0B6Xg7YWUXsjd0A9laGRg+MMZG6Dc771qyKkhbBcpA0kDHUey7H8CLPT3uCN99lbKS0E9Rzk/aupb75UFgj3w3tut3aX4v3rTQDacvAHo5B6L2Th9oeSmf1w0+QD3A9F1w5hrPp6zMlFDTMJBOOnC4nHNBqinjDYHS5P0viwt1WPWts4h0JffpQHuGT1Hz+1B1zraoGve9seADgDzWM+ole/YkDOwXYo6B0LJM+V8sW/Y7LZGsuG7WNdV2ZgfEN8gbYQceQMYYw5wGT3WFVktf8JwPkpp/Fpag0rwQ5uxWLIHveMg47ILPEd6lanQ4dFl2/wBajipI3MycZwruttP8AP1IEgAdgdlbt0knCo54dvlVPSR80ERJBIB2WOSRLsTnKncHZ2yrWwlzuo+qDN8WraxnRIRuFzFoyWc2Q+K8k9Hr8lwtNO9jWD5hc8cOaP3vTxcRk9Hp8kHFur5XCsl7n4itvUpDwfrW9dZ2lxrpQBsCVswNbSktccYQXkYVpd3G6tdJ6Kgd1boLhk7ZVc9IwfzKwOIVc5QXE52CYcrQcK7rKC5p3PdXZPqow45V3UUE0HfdTk75CipSwn4iFJM+Nv0XbYQWS5x3UTTvufNWyzE7NVIoqiR4DWkkoIrnkMGPRY1E84OSThahc7dWthD3MOPqWDRwStyXNIQZiteMb4V5+EfFthWl8ZGxGEFsYa47gfasiBrWvDm4BCxi5gHwkFTWounr2ROz0k4KDd9inlkaGOJAHyWfeo2e6EggnBWRVUdNbLc2oYR1YytqXG/CaFzAd+wwg2rWBzK15B2yVDG9xlHfZVkdJLOXkHBKymwMDeoY6sIMsZ8IEnKx4JCKoA5I9EgjrJX9PSenOPsWr2ayTSXBjpm4ZkZyNkG57KyhkhZ1QDO2+FyI22239j75mMYJOjI9ey0dlHYKK3NIkZ4gGSMrbN71q2lpnUcDyWnbZBsW7VVwbeJo2zu6ATjfbC0eokkdP98JJB81nTVXj1Tpz3cVa6Bkh6hjKC5mPCCMDS/B7ZVo62jBzhUc8NGfNBqkcMZZuPzrSGEC6tAbsHgKwVtSPI4+tboslopqqEVUmOsDI9coN60VNEbYx2QD05P5FtO7gMqAGnzWQbnWxA07WnpGw2Wl1cz3P6pc57lBrFuY57M9eMhbW1KJG1P0s4WSL1JS/CCcLTqypNc4SHf60FKTqeACfJUqG4fjKx5J3U+Ok7/JXRSvnHU7coLvLcLDn6uo7lZoGQcrDqHYdgILqbPmqS/SVKZxJwkx+NB389i0f+uk1Tv/APKBXfyjbkVPYr/hS6p/4v63+UbciD2mREQEREBERAREQEREBERAREQcf8wv+8FxL/gfef1KVfNHUOxcar5yP/SV9LnML/vA8S/4H3n9SlXzRVIH3Qqh/wDRH/pKAHbK9nZWNGwB7KQADsgZA8kBBVCwFXNaAgIrsBMBBTORgKob33TAzlVyUF5GO6t+l2VeonY9lQnHbCCgyDuVXIVDuqgbdkFVUHburSd8KoORlAPdFTqb5lC9udkE8e4wCspjPhOSFhCUAZBVBVvxgA/lQXStAfgKwl2D3VOsuIKrkoIup3VjfusqHJwM+ahc0d0EhaDhBlyAhuSfzrBlcOrsVVk8kjsOzhZTIY3NycEoIMjpAx5KEsZ1glgJU8gAcQOyNgc74/0oMqGpuMbGNgf0t6vIrtVwPpair04ZqicBwYcAn5LqWat0WG52BW/tC8WrjY6iO2sc8QuOCQdsIOTNV3Otpr7LDG5xAcRkEqK33C4SShjpnAFb1tDdKXy3/dKsmj8d4zuRnK2/fbLM13jWZvU0HO3og5D4eWq218xNw6Scb9S4Y5l4Ldaa/ooKcEF2MgbKWq1jqLTDGmNjw7ttlby0lSaY4jUvvGq5YxL05+PHf7UHVKarD6dh92IOR5LmXg3cAyaJvujjsP3pW+dScN9AU7+illhLQ7bGF2K5dOCWhbnQtqHvg6g3IzhBunl3rp31RMLXRn8i445xZuq6N97aZTn612UtOi6DS1ZILMGnH9wuHeNenaK/VDpbuWgh3mEHW/QT4ndIiYWfD57LjPjw2T33LnZHUudbhQWfTUZkp3tGGHGMLrRxdv5ula8scSA4oNrU3SYm5xsFBUtYSAlucXsbn8ihujjE8dOUFHMAbsQqsxjcZUMLy9oBWZExv75BCfPCsJIJCyZmxgHpwFinugujOT3UpOFFH3UhGSgpkq9uSO6jOR5K+MnsUEzAQfTsp4i7qxlYznOaMt7qxk8mSc9h2QcncMv7dHiDI6u2VyTrJ0H3KdiPB6T5fJcdcEw2ruYZU4A6huVzhr2021lmJjc3Jbv+RB09vLiby9rQQOpULpARus7VkEUV4f4GO57fWtIZ7yZPizjKDVIiSAcpUOIid9RVrHNawZ7qN8wl6oWbuIIAQb84DSRDUA95i6x4g2x8124mdaHz0zW0OCQN8LgflF0VHW6lZPfGhsBkBy7tjK9ANb6I4d2zTrbjRzQGWOPPcZyAg4A1paKGeyguc3p6PonC4nOnrTHbJ5WMiDwCc7ZWg8YOME9Dd5bTQyuMTCWjpOy4rZxWubopIHF56s+aCmpa27Ut2fHS1T2sBOwdstNbedRHvXPIz26itOrLzPW1BnfnJzlQtr5OoDff5oNYluV3eD11LifrWNJW3IsOZnEeYJUbKlz2gHO6q6UdBHmUGAXvdKTIAT9Ske1nhk9I7eiiLh1k/NVbJ1PEZ7H9CC+xNbLVhsjdgfMLfEtBQtoy8Nb1BpOdltJ8cdHF40JHV32WJ+yStcfCJOO3fyQUrJJG1ha3PSD9io0kuGfVV/sn3x53KuPS1hcMZHoEE0zG+F5LAY1viDYKx1VK5xac4VQ49WR3Qao1o8Mduyhe0Z7KBk78BqmBDm5zugqwb7HCnDHEZ6h+VYD3yA7Zwqe8zA4wcINQwTnf86gn227qIVDxvvlTRkTAF5GUGM879ikb3dQAysvwWYxhRvjaw5HcIJo3PDe4U8eXDf8AOVgeK5owCVNFMenfP2FBLIB8lGdsgKySR7s43VrS8jcFBSTvuqN81bIXZV0YJCCq5I5N/wAL7hf/AAutX63EuOOy5H5N/wAL7hf/AAutX63Eg+i5ERAREQddfaF7cm/Eo/8A0nR/r1Ovnkh+mvob9oXg8m/Eof8A0nR/r1Ovnmp2hz90F02OnZXU/ZXSR5bsroYunugkHT2wq4b5hXNYMo4AHZBZhvlhVVDgbhAfVBIrXd0LiqZzugtecDY7qLqPmrzklOgIAfnYq9p8la1gyq4DSgmAwM7YVHDJ2wjHZGD2Q4QWkBu6sPdSuA6dlH0oAJ8le1xA3JVmCCqgk5QXdRzsSrZHkMI3V+B6K14DtvJBhwHqk7ea3JTxR+5EnGek+a0NtO1h6tsrMirHBvhYOCMbINIqABM8eWVqtvH+1D5bZQUcEhLnYye6jkk93HhR9ig06Z7jI7PqVfG5zm4U/gtd8Rxk7lQkeG/oHbOEEUmAdgs6hijdH1OaCfmqNpmOaDtlQue+B/SztkBBkmONziDGCo23SvpJCylnfGPINOFI2VmOoncqKSJrz1AbhBlT3q+NZGXVsoydsOO5XanhTaaS5cN56u5SNfKIc/Ed84XUeR0khA3+Hdb+0XxVu9oENkLiKaVwY7fyOyDaerHNZqat6GYDJiACO4C09tQJHAFgBXbTT/DDh5f7cbxcpYBPO3qOcZzhcJcWdB09ouBfYGB8Acfoen2INgRucOxOCsOpeTKs5kb4gI5dnAbj0WDVN6X5PdBe0nHdVy7I74ULXOwO/wCVSxODnAFBP8JAyFcxrSQBjupY4o3Nz8lhVEpjfhp+aCaqi/sf1hdi+FMLBp05cB8OfzLrS6eZ4Gc7HK5h4farfSWk07nEHpx3+SDK1hFCKybbO5XEd7HTVHpzjJXJd1lqrnPI+MOIOVxxf4ZaaqLJW4OeyDFaCGDOVeOyowgsH1K8AAb/AJ0FAcK4HKNaHHZXdIHkgsd2Qdlc4BU6mN7lBaO6uG/ZA3xP7H5p4EzTuCEBxe0DGyp4jsbndUkiqHDDMoy33B/0WO3QSU7sv+Lfdbgs5gNSzqYD28lpNutVW2QGoaQM+a3hYaGkbWx+MQACMoM2+MozQAiLB6e62YI49yG4XKerKa0m1gUr2lwb2GFxfJE6MOyNxlBptxbgEN81pQc/pKz7hK45DcrDha/Pxds+aCtMXl3xZWs2lrffmEYzlYccIeB4eMrJoqWriqmy4OAe6DeGpZJWWsEOOC3tlcbtmkfIQexJyt9XWsE9CIXnfGCtpPp42kuGO6B4TfDDsb4UcQcJQSdh5K/rcR0j7EtkE09zZE4HoLsfYg3LajTva0GLBW8hQ0rbWZWYDwNvyLHq7PQ2+0sqIy3r6crZFw1dVwg0sZONx9iDBudfdW10kQnd0A7DOy0ioqKh7/vryT9ay3VD53GZ3dyx3x9b9wNz5oImOdt3WbTEk4J2+atEADR2ULpHMdgE90GrPjb0E43wtGmJFQRuQsttQ8jf9Kvp6ZlROARuTsgqBGYm5ZuQMrWLNVyMe2MRnB2xhZH3CPhMcBtsuSNOaNtYsrq2fpDw3O/fsg0CKGmdB1uaMkLad9YGzfBgfILXam60sFdLShw6WkgLad/ruuYeHuPkg0+uJ8u6tph8G6s6uv6XooJJ3RnpYTgILq0YPkp6EN8I7DzWIXmYfEsiNwjYQMoJy3v2WnVH01JLUPByPNXRNbIMvwgsoxl5zj7VWpbh/krpAISSz1UJ8SQZIyg7++xYOOaXVP8AACt/lG3InsWQ4c0mqf4AV38o25EHtMiIgIiICIiAiIgIiICIiAiIg4+5hv8AeB4mfwPvP6lKvmiqf90Kj/Cv/SV9LvMN/vA8TP4H3n9SlXzSTtzcKrPlI/8ASUFzPohXg7d1E122B2UiCpB7qirnZUQUD8nCvGwVmG5zkK8HIQVb9IKrlQeW6qcbYQWE+iA5TpCqBhARVHdVwEFp7K3dSFowoz3QQuLsk7/lVhkcNt/yqdzQQrPADt0FsT3E5OVkjdWsgDQO2PkpA0AoL2swAfkrgFQHAAVQceSCN5KtAyrnnKo1AGNthkKVriBjJ/Ko2jBV47oLC7LskHutUpomOgJOMgLHZDEW5OAfRYdVWy07vDizgoIbh8MpAGVJQO6PvgaA8ealhh94aHydzv2WU2GCJhBIz80GoWvUV9jLIo6t4Z1AY6iu4/Aqz0t6sAmuUrHv6M/EQfJdH/Eljky3OA7OVyHo7jjedLBlBC54Zs3YoOdeJ9nt1LXzMjha9ozsBsutGq9S3m2XF8FvnlpmAkfDkLsNoPV1o1lN418maOvv1lYPFThXYq9hqrN0v8/hwg68x6o1DPBG+atkOSO5O67p8rF6vElFEHV7gCz+6XTPUel7pbXCCKJ2G+gXNXAXVt3sAZDIHjAx5oPS7ho11fXSCrnD/rOVwZzXxvoKwikl6Bny2Wq8GNeXCpq3OLnHIXEnNxqi5vrx0hxy7yQcJa0uVcKIAzlxLfVcCX2WSed/inJ6id/rXJtyutRU0rfesjLe5XHN6ZE+VxjIznugxbcBtjAworqB1dwVa174QOlXHFTvJsQgxqfY4PkpnvwRhXOY1gJbjKjO/dAcSQcqxXndAwYQUZ9JTtxjdRhoHZSNdt2QXYHorT3Ver5JjO6CrDkjP51I1gBOMHZQuPQNlk2cGruUVMRnrIHqg39wmkeLiAwFhz37LlvW00/3IJ8fOGds/JbPi0y7SlkbeAwNyzqzj5LatdxGdc4JKd0hIGQg4+r3vmvz2yEkdR7rUpaWBkeW4yAtOr3MdVmpi3Oc7LH99q35BBwghrJHxPPSCR9SxrVUPdeacuBIdIB0n0W9dCaWl1PWCHoJy7HZcmt4C1FDc6atfABGHB5OEHJug6WO26YirLf94lLQertvhZt91VqWosssc11e5oaRgvWka21LadJ6RZR00zRO1mMA+YC4D/2WK2phnge9+5IG6DRNZ1M9Te5XVDw8knOfrWiRtaD9EKtXXPr6x1Q8kknuqOBbkoJXAYzsohu/ZQy1BAxvsomVB6kGsNGGgjCseTjusX3l3SNyqGoOD3QVyQ7urgTkEHdRxO639/yqZ0Zbugk8Rz24e4kK3woxv0jPqrOpw7hOsoLy4hqjLndvJHOJVMZQUDQT2G/mpGs/SqFhAyEDnBBJ0gbhVa7GysY8k4KmY0eePLuguDWnGQEMbDnYfkVx6QFbkILDH3VM9GwV5f6K3AcUFQ8n1Vc5VwY0AKvQAgjwCqjuN8KhIUUkoafNBnRNB3Pmr3Mb22WNDMMeakdN2wCgo9jTvgfkV8TBjsPyKDxc+qnid8PZBHIzBOCuQ+TYf9d9ww+WrbV+txLjyR5yuROTYn9t/wAMR/3W2r9biQfRaiIgIiIOuntDPwN+JP8AidH+vU6+eilHxn6l9DHtChnk44k4/vOj/XqdfPRSfTcPkgyQMq7YIBjdUIz5IL2knzPZHeStYd8K5yCxvdHNJOQUxjcJ1ILm9sFCcq3JVRuEAjvhU6ShOCqjcILcplEAyguPZW5KvLds/wAyt6QgvaSfPyVwGVEHbjZSB4wgO2yo3AqQ791aW+iCPxDnCuaSRlPCyc7fUpWxtAPkgjRVcMFWkkILutw7Eq1w6jlOoqo3CCmSNgo3MBOR3Uju2QrRuEFzXFowqSBpaTjJVA7J3Q7tIQYbnO6jgkDPZZtIS4blYroXE7BTxPEUZ9UGV0MydgsA5iqw6PAIII+tUNRLk4zhVY0veJHeSDdZ1XqOmo4o6eve1oIGOryXMekKGO8aRlrLm8SyhmfiOT2XXmesJa1g7Arkbh1rt0crLJO8iKT4Tk7INiagnMV8qY44cNY8gNPmtInkdI8ktI+S7taZ4KaEv1C67Vr4fEe3q3xnOF1v43aQotOag93tIDoeoj4eyDjxjG9OThQPJbJkZCridm2D9Smjja5vU8jPzQZERJjG/wCdYc7SZfrKGSRpIGcdgrCZXHJB2QZrImFoJAOSty2GZ0ZEbDgYWzvGmAA32Wu2Ksd4gDkHL+lKWGdjjK3Jx3XGfE+GOG79MbcDJXJuhpjJGc+i4+4rxsF0DvPKDZtNnwxn0Sd+AQFdHhsQDfRRSNc45x5IJaEOc7c7LJmbhw3CxqUvj8lHV1L+obIMvY53HZYFU9wO2ceqR1Ly4DHfZbrsmmxdmj4Qc7oNBsL+qpEb2kha/WQxNxiPyW5KXRcNpHjztAHdYtfHb3u+BwONs5QaHboYnyAOYPyLetptNFLjqaPqW2x7nAcgjbfuo5NVGiP3s9vRBu6+2mkp6fqjwCPMLZ88ssTHOjkIIysSp1nNVgse447blaXUXYuY4AnJyg1e33CvqJTHPKXN7BLu4MYSGY+pW6KdDPUk1RABPmt03+1Uc46aYgkhBxrE7xJyHDz9FNUtYyMloAWZcrJV0DzKYyB3C00mWZpbg5QTaed4lSGyDIyuQG0dOKbq6ADjvhbL09bpGTiRwxvlb8lBioSdsgINmXcvbIWh2wOO60Mk4KzbnWdVY9pJwMrDx1npj3JQXU27hsT9i121FgqWEQb5747LHsVqkkkY6cYaT5jyXL1HpzTsGn3Vj3sEwbnG3fCDZt3fMaHeXbp9VxtcSPeD57rVr3qGoNbUU7CTG1xaN/JaCZDPKC/zQZoAMYIHkrA4BwBWWGMEQwR2WK5mHdSCYfR+xY7h1Pycd1KJWAYJ8liyueZMR5QZgaMDGFLasm4Mb5ZWJDHVOxhpWoWugrY6ts7mnoByg5NipYfcY3EDOAVqNRLNFY3iKbAx2ytFpr1Qe5+FI8dQGMZ81s+/6wqGyOo6dxLPr2wg21X1dQLhK8yEkPKvZI6ZvU8gn5q8wxTgzOO79ysSV7ondLOwQXyfDkZ9VivOTlVc+R++6tId5hBfF5fWp85UMbXBSjPmgo8AhWbt7ZH2qRUI74HkgoD1EZ/OsuJo6R2WGwbnIVXzvacD9KD0D9jC0Dmi1RsP/iDXfyjbkWP7FqV8nNJqkHy0BXH/AJxtyIPaVERAREQEREBERAREQEREBERBx9zDf7wPEz+B95/UpV80VTn3+px/80f+kr6XeYb/AHgeJn8D7z+pSr5oqn/dCp/wkn6SgrHvhSqGNx27KUnCCoOCqPBJ2VW74yquAHmgiIcEDiD9aud2Vg2QSg9lUO9VF1K4OyCT5IJMb5RRNkdnB7KTqQXDdV7dyrOogoXEnKC8juVbkBOsqnfcoHc7JggoNih8kEo+iDjyRqsDzgZVzXDBO35UA90VhccqocSEFcEb/NVByqA52KqBhATsqA5QnHogqXP8nH8qdDXuBeAT6lFjyyPa7b7EGoMAa0hp28gsCpklEwAccZUsdQQ0DcIYhI7qPf5oM6BjTAHOwThYsrWiXLWAn6lcydzfgB2GymYInbvIyg1S2193po2e5VDoz1DscLstwmgnudn67rVeIenPxHJGy6o1FfNAB4G4B8lvXRHFi42h7aORzg0/Cg5e1rR2yCsePAD8HbAWNpSSnFW0Q02B8gszTlytGpQJ7hIMvO/UVyxpvh1bKkMqaANcMZyN0HJnASJks4Lmgbei2JzYingrADEHn5Bb105eKDQ7yZnhhG3dY+s7RQ8ToDVwkPxv6oOk2q3h1EPCYWHpXGvx9TvEdnJK7C8bNHfsdpnMiYAWg+S65Ucr5ZJfE8iQgyCAfJRSktdgH8iyW9AznHZYdW/pcS35IKZJ80xso2vJ7q8POMIJGlXKBrjlSB/6UF6IiCre6vBHZWN7qvZAl+is/R4D9T0jXDYvH6Vhxs8Ygd91q+n6V1PeoJwACHgjCDszxOpIIuGcb4sB/g/zLp7SyVPiT/Ee5z+VdlNeaimm0WykeTgR4/Mut8Jb1yluNyST9qCegL5JcSnI9MrV5Y4YmFwYCQ0rSKKGaWXEAycrXbRaq+pvEFNUxkRPcAc5xgoOUuWc09x1GyCeItHiAZdsO67waw0rZodKmeKSPxfDzsRkbLqkLTZuG2mmXy3PZ7z0de3fOFtaXmfvlzt8tFK+QgZaN/Lsg2RxhqLozUM9MalzoQSAM7YyuNIXO6nY2yfJa7qPUVTfbi+pnJPUSclaQxjQ7t3QTUg6nAYwcrNkaAzHko42NjYHDAVXzNLDv2QYM4wVEzd/n3U/9keR6q4QNa7OAgu/eKM9lleG3o7KIxg5AQKP6YB9VqLi0gZx2WDSRFsgLu31rUHdGB2QYzwPJQnusmTpOyi6RnKCxoyVe1oBycKjcA9wrg5udyEEgwQqPA9AqgtAzlMtPmEEeAq9TvVVfgA4x2WK+ctPmgyj1HYlWkYUbZifVSNJd3QM5V7OyBoV2wCC4OPqjnH1VheMFQyTFowCgo8nPf8AOoiHZ7+atfK7yOfsVWOLu/mgyIc4GFIXEbKyEYO4VzwcoLSTjusun3asTusuD6KCyT6S5G5Nvwv+GHz1bav1uJcdyDfK5D5NgP23/DD5attX63Eg+i5ERAREQddvaED/AKzniR/ilH+vU6+eij/sh+r+dfQx7Qf8DriT/idH+vU6+eiiH3x3yH86DMVMhVVOkILcgnZEwAUQU6gqHuq9IRwA3QURWBzs48lVzsILlVvZRtcc/wDtV+Sgq36QVzfpBUwFVv0ggld9BQqZ30FEBtlBYqgkHCodkGyCUdlUEAK1pPZVQXq7yVqF+Gnf86CB30ijeyic89Xy+tXsdlBXGThXDYKmAN1UbhAVHbYwqqjkEeM7Ju1wyUzjdW5JcB80GXG1haMgErAqMiUAHZZ8TmhuD9iwahpM3ZBlRtj8MfCM49FSRgDCWjCkiMYjAJGVSQBzCW9kGmOJJOfXss6xvlZdKfwnkEvAysMtHWfrV8MklNM2eP8AeHIQdrNNOvLbCwx3FzQWDI6vkuMOIvUZTJVuMsme53W0qTipfKSmFLGT0gY7rcNjudPqmHquDgH+h9UHH1VLG2Q4iP5Fpr5HPfkZHkFzGzQlHWueWBpAzjZcb6utLLPX+EwYGThBgxtZ4YJaM/V3V3RH6BInN8IbjOFizTuDsN7IMroYdsBZFAOmpAaQB8lp8criAclXU9U9tSMdkHN/D3eM/UuPuLLib2G743W9uHdypoox47wMjzWhcSbNJc673qlHUPkg4/pBmJueymMbO+31K6allom9DhjAWA+okycdkGY1jPLAWHVMxt3VDUvYAXA7qZgbO0k5QYLcB7Nj3XMXDWKF3heIB5fauK/doy9uO65H0TWsoiwOOMYQbz4jsZDaiachvw/zLhSGWqIcXSOO/quR+JN/a+29Ebt+nC4soKmSTrB9UGRUT1PbrPZafK+U7uOftWdNkDJCxXNDkGE+R2cZIVYy9zhkkhahDRxynfCklo442ZaRnGyCW3GSMAwv6T8tlvLT7pZqyJtRJ1ZIyFx42rlp348h2WtWm/up5hIXEdOMboOTNc0FHFaA9jQXYXFdEWmQksOxwt5y3436mFMHZ2x3WJDpsxvHw9yCgms8cJAw3BPqFrNxiAoCQ8DY9lpNWw2yPqGxAWj1WpXyQujJOeyDat3Lm1rxnCvsruqsY14JBWqWu0uu9V1OGcn0W6GaRjoAKgtAxg7IMtlPAyha+LDXYzkfUtKuNZc207mMqXdB8gdlbX3qOBpga7JG2FtasvdRI8xj6OUGBWHNQ4uGSf0qBhw8LPLGSjrd9I91Q00bW9Q7oL2ud0DcpKQIyfNQteQ7p8sqY4e3pJQacXPLsZO5Wo0cTc9TxnChFOMkjCzbHRVc1zgY5h8Jz/i9MINRp5Yh0t8A/kW94KKkfZHyBoDg3O/fst2t0lpaKyMqDJH43Tk7jOVxfqm/fc57qKldlh22OyDZ9XV1ba2VrZXfSIAzssWUvdJmTc+pWY4RveZjjqccqN0bXO6jjCCwPeAACQPJVO/dSYjx5ZwonOaHEIJWsZj6IVeiP0Cia/JOFXr/APfKC8hjcYACicMnZWySHG2VF4rvUoJ1cMAbrG8R/kcqWPrd3BQXFu+yqImkZONvkhDvIFROkkbtvsg9APYuNA5pNU4x/wDEGu8v/vjbkUfsWS880uqc/wDACu/lG3Ig9pkREBERAREQEREBERAREQEREHH/ADC/7wXEv+B95/UpV80FX/b9T/hH/pK+l/mG/wB4HiX/AAPvP6lKvmgqhmvqf8I/9JQWs7BTtOBuomDABUjd9ygvVrgSq7KjnY2QHHbGVGqnJ3KqAMILVXfyVQPiwrg0BAGAPLKtJydsq7pCBoCCrSOyZAQN3V3SEFFUEYVQ0ZG6qWt8igt6gm53wqhoKuDQBhBYSMYVMoe6IA7q5Ub9IKqC8eSub2QAYCdkFhBBPZWOUjvNWYBQXNOVHKQDjGSVeAR2ysiOBkjcuxnKDDBzusmIZH1Kj4WNPw9s+axZZpGShoyAgyHnpOQ1Yc07/EAB2C1OJkb4g52MnfCwKqnIdlo7boNQt0bJgPEwdvRYtw/2vVt8EAH6ljsq5qfAAxspoT768Pk7hBvOxXK5x00fhTluXBd4eXN76zTodWPLz4exP1LoD77LSRMbHnAcO31rvfyqVranSodMd/D7Z+SDQOOVbW0k8vukxYA4jYrfnLhUzV1iea15eQDhcScx9bUx1Eop846iuV+T/wD23p55rNj0+aDjrmdjia2fDDjBXTODHjz4GPiK788yWn2VhqGwtB79guid8oX2irna8Y+M/pQYcxcNxnZYpJOepZAcZWAgd1C9nSd9s+qACFVVY0HfKOABQW9lc07IW7I3sgkar1C15BClachBdjZWnYI5xAUb5CGk4QZdudiUEjO/ZbltT2m6QDwyMuHcfNaLoel+6VwEcmMdX865TqtMU9AxtWwAFoBQZvEKLo0g17XDPh9vsXXilkd1vGTvv+dcnaw1TU1NMba8noHw4WwKejY5zgzBcQUG4NEBnvYMkfUA7cYXJ3+1BUROjpC122DhaVwO0j7/AHVpuTQIi8bkbYyuzWp+HmlaGwmup3ReIxnVsBnOEHXvXlTWusoY6oJZ0/QJXDFK49Umwx1Hstz8QtVVBvUtriJMTXFowdu62fDK9sh+ZQZk3TjIAB+pYzCS8b+aypunws5GcLAjeS/I9UGfM8+FtnssUPeAckrJcCYxn09FD4fqEFKY5eMrJcooogzf19FM7HTkdwgE+aMPxYP51AJiXFvopY93jKDLIHR8IwVH8XqfyqeUNbFlp3wsMSP/AD7IJCT5lCT6qwOd5qpdgdkFjiQrHOJ3yUe45VvUUEjC4Y3KmBJA3WMHkKokdkAFBO/PSsOX6ay3OJbusZzQX7oJ4xlo+pZDG/D5LHYcNCmEmG/NBKBjfKskPzULah3Vv2Vkkzi7bsUEmfmrHndR+IVcz4+6ChIPkr2NyQRhVMbfJVb8J2QZEY2wFUjHfCpFuAkmyC1/dZNOMs3wsN7yNlJFUdDd9kEshwTv5rkXk2Oeb/hhv/8ALbav1uJcYyz5zgrknk0ef23nC0nz1hagf43Gg+jNERAREQdd/aD/AIHXEn/E6P8AXqdfPRRH7475j+dfQt7Qg45OeJJH96Uf69Tr56KP+yH6v50GadwrTkbZVysc/wDQgtJJRWBxJwr0DJ9UyqZHqrXE5wEFS4b+qt3Jz+lV6Ad1Q/D2QVaDlXqxpOdwr0FyqO6onZBL5Kwg/kQv8lTOyCxBursBUOx2QXtB7quQFYHlVJygqXZHcqx5OEByoZpXA4CA7urmuA2VgORn1V4aCMoL8n1VW9lGHb4KkBIQVyCqP7KgPmrXuOUFjnbnureoKpGVY4YORuglBcBkE4+tX9IczJ3OFieM/BCyad/U3pJCDFc92SMkBZ9KS6A5/OozTNJOCO/oqGXwWhgzvsgxZTiR31rKp2NdCcjJwsd8ZcerHdZVPhrCD9uUGC8YeR81rWmqipZWNbFIWt2zjZYXgREnOPyqlNUPpKtvh9gQg7D6QiE1IXPkBOM91xFxVY2O99De261+x6umpKZoBIyMd1De7UdQNNe8ZI3yg4za5+AASrXH4t91nXCk92qHRAdisRjA4/EgkYPhACviYBMNuxU8ccYA7K4RsB6hjP1oNz2OadjWCOTG481yXQU0MttL6hoe7p7rg43ielcGxE7LnfhrLFd9OGWpwXdHmUHFuq442VUjY2YAJ8ltIloJ281yBralhbXTBu4BK2O6Bm+UGBUyBwAaMKWmcfDIzgqR9PERt9qxnvMPwtQZtG0mQZ3GVuu3Pewt6DjdbXsvVPP0nGxW4pJDSuGD+RBbquWV9MGvdkehW37TGCNwPPutSutS6ojLXdgPNaPBK6BpxlBqVZE3wydhv3WnYwCFZUV8rxgZ3UcUkjh8Q/MgyoCc91M4OzuchYL3yMHw5/Ir4ZpS4dQ2+aCtY1oGcd1hB7g048/ktRrWdUfwYJwsGOH4D1BBujQTnPrQJMlocFyhPDES0gdguNNBiJlSC4gbrketq4WMBa4ZHbdBtrV7RHAej+5XGhnk8cjG2fRciXYy3AlgBI+a28NNytn6y0BufRBuPQFPDO5vWA3fdb41PQwxWpz43jIb6/JceNrmWKIGA4cNzhYFdryurITTvLsHbcoNq180puEoLicOIUnQ1zeogZVs0fiSOmI3JJUD5JQcYOPqQUa5/V05OFkMJLfpLHHqO6oXzDyOAgyS0eRGVG9zmAkZKhEsjjgd1qVhttTcrrBSuY4te4A57IMFlU8OGY8jK5I0+2nNrM3gEPDcg481yjbeCdoks0NY9rOsgE5Cnr9NadstpfG50YeBjGyDgmvvl295fE2oeGDPw5W2rlUzTyffck5zkredfTULrpIGkFpcd8rPg0/ZqloMhZk+qDjEved89lkRPL4znuOy5Lj0bZnNcSWeeOy2dqK3RUVR4VIARnGyDbrnODjnuqdTjuclagbZK4B/T3+St9wczuMFBjMcNs5+1XFwKl93x6BRuaGnZBY/srWjI7KZrGuwCjmMbsOyCkTATkhZIY0DbAWN1hg2PkrXVLs7E7IMzpb6q0saT2Cw/eH+SyYpOpgLu6Dv37F4NHNFqjGP/iDW/wAo25Fb7F12eaTVI/7gK3+UbciD2hREQEREBERAREQEREBERAREQcfcw3+8DxM/gfef1KVfNFU/7oVP+Ek/SV9LvMN/vA8TP4H3n9SlXzRVRAr6n/CP/SUFjHdgpAD5KxjW42I+pTMxjGUFA1wOSjvpBXFWHqPkgq4gt27q1rSfyIQQrmIAaQVKOyqACeyEADZBQ7hUAwgOVUnCAioXbKgeguOfJBnzVOsJk+iCqrgq3LlcCQNwgtIOSqYKvO6ogdu6qN+xVpOdkBLfJBMCMYyrVZ1En/2p1kILiRhUHdUzlEGWyJpbk+ixqiQxP6WlXtncMAdvrVHsEm7u6CrCXtDj+cqyVjc5IBKuaSxpAPZQyveT5/YgqJHNAw4jCyYWNkaS4rHYwOxk+WVFLPLC8Nb2KCte1rR8I2yqW4nrIWZBCypaDKqT07KbDod/qQam2Jj4WZIPxBd0eWZ7odPBjJAAY/X5Lok+4zsAaD23XaLl84gwW62NgqpMEtx3Qb340U8dTLJ4jevc/Nck8tcXu1nc2N4YOnt2XHt6li1RO8wuBBdstQ09crzpRpgpWvDT5AIN/cVKcVL5w5pl79t10Q4v04prjK1sJj+M7EY816DcKbVU6+ubobm0kOH75dW+eHQFPo6+sZRtaGufvgIOttuw9gzjt5qle0A/CVjQvlp6ZjiCM5Ukb3VWduyCkefNXqjmGPyRpc7sgqeyoCMJgj6Ww+aAA9kFVI0jG6jVwOAgq7f8qjfs07FSMLSfi7KZ0AkYenGcINa4bjNzBA/feQXMN6Y40gznHT5ri7hjR+Hcg+bZvVncfNcwaqfRRWd0sbgS1nr8kHCerw1jiRgn0A3W2tOSOlvEUTmkhzgCPtWpRSVWotQvoowXAuIA7+a5EsPDSS33imqqiIBmQSSg5N01aI7ZYmVsDfDeWg5GykuuorhNZpY31xIAIxlZXE2/WqwaIbDQSN8ZsZ2b3zhdZIuIl2l6oJXu6X5GyDD1O8zX173gE9R39d1h4aNxj8iy6wNqj7yT8ZGd1hMbIT8WUFDI5x6SThVjHS8KR8TWjI7qJnUXg5QakWjwvIKENBzuPyq185EYwDnCxveH52QZ2PPIKtd9EqBszyPn808VxCCrMde4WS0HI2WPAMuHV6rMcGBuRjYIJHOyzCiwFZHJ1OLd1MWjG3ogsy0dv0KhwrTkdlTqKC0jIVOn5K8d1XAQR9PyCBuPRXkDCoguyMKE/S+1XPcRlRh5Lt0GQ36IVcq1pGApo2B3dBAdlG879is33YfJUdTtAGwQYW/opG5G6mETAOyskAaPhHmgoHH1VwcCRkqHL8+f5Fc0nIygy4PVXSgq2F7WjBKukcCNkGOT5KnYHCo5wBTIIyEEbjvuuUOTQ/8AXd8K8f8AC+1frca4td1ZGxXKXJn+F3ws/hhav1uNB9GqIiAiIg66+0KOOTjiSf8A6To/16nXz0Un0z9X86+hb2hn4G3Er/FKL9ep1889OcSZ+SDNPZRP7qTq237KJ5BOyC5qOBPZUYd8K5xwEEZyD3VzSMYKj6iXYVUF4GDknZXH4twqAgjHy9VXYeaC7oCqNgqdSdYQO25Q74wqFwI8k6w3I2ygq3uriMKNjndWcbKTOTugpnCocK5wGFYgoTjdVHxbqNz85GVfEWgblBXBUUoy7ssguaVaWNKCHpw0bK9pyMKu2MFBgDZBa5uNwgdscoXdwrEFzTg5KOIJ2VqIKO7I36O6qjtgceiDGOxOFJA49QCjI3OxUsTQ0dWd/mg1ONh6MkjfdabU5EuFX3uVo6fJU3kPU5BlMYCzP8yx539BIb5qQSEDCj6A+Tf1QQiV4Ox+xZVI1r3hxAWRHRQFpJ7rFmcaZ4bH2CDcMUnSxgDh5DuuTbBQsqLA+R2CehcI/dCYADOw7LlvRWoIXWF0EjviLcY+xBx1f2NZdJWAdiVoUp6X+i3ZeLbPUXKaaNpIcSdgtv1FqrhOWNhJycbBBiidwAG/2KaOVxG/5FqtLobUNYzrgpHEfUVg19mrrPL4VdGWHtj5oMGoOXLmvhlO+PT+GyBo6fX5LiBtPG4AuI3W4LPqOot0fukBPTjHdBq2q5HGrlOSSSdwVsud7hnII39VvKnpp7s4veCSd91t2/W80UhYRj1QaRE8uJ7qGo7jbzKy4Imh2ceXmoayJxf8I2QZ+mW5qTnbcLcFxaA8bhbaschp6gudsFqlwri946N0EVYNjuOy014AB7FXVtTKRtnsoqZxkBDggpCwOfgj8oWQ6NrdgAAqtiazfG6gqJH747IMqGNjzuMqtREyNnW3AIWHSTydZBB7rKne5w6e+fmgx6aYSyFjtwFJVxtY09O2AoYWNhf15+eFLNI2Zp+aDL0vUObUhodg5W9J6mUDJJdjyXHdJK+jlD2d8rkPQj4LtdYaetI6HOAOfIIM2yN97lDXxEb98LclwsNPHQGUEZ6djlb91dpzTOndPMrqKSMylnUenGVwfedcSupnwsce5A3QbWvs7o618ZdkAnzWiiUF2OkfYFWerkrKsySA4JVz442sy07oLwMjOQrZGM6DsMrGZPIHYwcZWQXF0ZJQQwDqkA9VqLqeMwk4GSFo4e5jj057rJiqamdwhZkk7BBLQwNdOAW5GcdlyfoGzQzXWJxhIwRutqWLTNcXR1E8ZDSQTkLm6zRWWy6dNc57GzsbkDbOcIOT3UAhsQ6aoABnbI22XWbine7hR1boY6olpcc4Kxbrxr1AaiekgefBDy0b+XZbFu99qr7N4lUTknPdBCbvOXl7nHqKngvdZ4jQJSMkDuoBSQlmfNY8kPhuBYc4KDeMNxqjEHCY7rTpCamqHiuzkrQ23KcNDQeykiuD2nxCNxug35Fa4DT9WR2z+ZaDdImxPLWszn0WANUVLQGDJHY7rdFpZRXGn8WocOrGd0G0ZRgE9J7ei0yd56tshbzuFJRNLgwjC25U0Ic8mPsg00SOHb85QyuPdZ7bTK/Ba0nPZSNsk4G7T39EGldRyd1QnJytXNpLfpNCnZbKfHxYyg0Nh3+jlXGR42DcD6luWhtVJI7BwQVqElhoDgnpQdyPYrvLuaXVI6f/AJQK4/8AONuRa97Ha30tHzNanlhx1HQla0/V90KD/UiD2JREQEREBERAREQEREBERAREQcf8wv8AvBcS/wCB95/UpV8z1ccV1R/hX/pK+mHmF/3guJf8D7z+pSr5oKwdVfU/4V5/OUELXEY3U7HHGVE1mw791MwABBc2QHZXYHoqBgByqOcQdkFxAOw2QNwqde3fdWOe7tsglzhM5UbX9RHqryenCC5v0gjlE15J7qbY90EbuyjLiNlM8fCcKHpz5IKx5Ls+SnUcbcYUh7ICZAVoJz32VSQUFwGVXp+aoDsE6kFekeqtTqJOPJUJx5oKqhOVXyTAQUxjdUJyr8b7hV6W+iCNr9wpA/bZQP2xhVY49soJierb1TpH1qg2TqQVAwchC1rsZG6p1eioXEFBI1zmYDTgK8OLx8W6jG4VzCRlBDPGzA2W59I3WspKtkcEpY3GFt4t6tlJT1LqOYSNJ2KDuJwapjduj3moG5G5IXOtbpq3wBmAx+R3GCvP7T/GG66VYw0pfkEdiu0XB7ijcdbUjJatziQ0HcoO0/BK100N0cYwGnGy6o+0Gja28HLS4h537rshwkvc8F3eBnbZcQ832n2aiqvGmAJzlB5/XPAooi1hb8PoobNhwJccLfWtNNxW+mDQAMNXHNPO6mc5rc7FBq1YzYkEK2kaC3BHdYZqpJT8Xb5qaOUsCC+uwxuAN1iQOJBBU07jI3f1SniDQSfNAb3UUry04HZTvAadlfDDHLkuH2oMPxHEAYxhanbiXPHVuCFh18LYmdUfdZum2GaoYJNxlBvjTjWx4cwdJWu6gqpzaJcyk4Yds/JYdTTNo7YJacfHgLQ6SquFwm9zlB6X7HKCLgbSxXDXDYqhmQXZPy3Xa/Xem6GgtLaiDpDwzIPbyXCek9LU2kCL2wAPPxHAWtam4oz3G3PhLiQAWoOJdfXm4y1z6OaoL4hkYJyFsdkMbCSGAH1wtTvtc6tujpHZ3PmsbobhBZE4vd0nsp3xNDcjyWDJIYzlqMqnvPQfPugFxdJ0581L0ANzhSiBgYH5GTuqHB2KCAkqmB6KRzQUDR5hBY3urkaBlXYCCvZV63YxlV2VMNQX0+7srKPksSI9JU4eSQgpLsCowcrIe0Obuo+gAHZBC0klSDYK0ANOyr1bHt2QH7BRFxJ+tHyE7KwOIIGUEh2BURyXZCnABbv5qhjA3A7IEZI7rJiPplQAdvkr43EHugy8jpVkjsDzQPyB2Ucju+6Cx7iDsVYSXd0JJOSrckO7oJQwY7eShk2dgKUP7bqx+5ygsDnNwcqRsjiO6jO4VWDAQUe4k91fEMjurHfzq+InCC5wHouS+TX8LzhX/DG1frcS4zJJK5M5NfwvOFf8MbV+txIPo2REQEREHXX2hn4G/Er/ABOj/XqdfPNT/SK+hn2hf4G/En/E6P8AXqdfPLBs/dBkTkhhPmseN7icEqef6H1qOBre6CZowA4+iteS491cScYVuCgsB3VwOSrQN8FXAYOQguLcDIKq0F25Kp1EhUDiEEjuyjyfVV6nHZUIIQMlWnqLs+SqTsjSe6CdhHTjzVVFG/JwqukLTj1QXl2fVRv88K4HACEBwQYpeSUL3DsVI6MA5AVhZv2QXMeTjdSh5AKjDQPJSMAIyUET3EnOSqtJxuVUgAnZAPRATBJCvLcDOFaNkFzgA3ZWK4O8ihxjyQUaMpIOkFWteQ7GUc7OEEJ7qrScK4tBz6qgGPJBdkHyV47bKwD1Qvx5oL1Qj4s9lQuOMqgOe6CVszgMZP5VFI7qO5yT6qhJ33VCfUoI3AA7LcmmKqdsjYmyENO2Mrb5ZnBx5rNoqk0nxtJyEHMNttNNUwCSRzcnfOVLR6doJLsxjmtIJHouL49c3CnaI4s4HzwtfsWtKh8raiQnI33KDuhwo4Zaer6AvnZESG5GQPRdXeaSy0dh1OKShjAYXkEjtstesfMHdtPQiGB7wD8PfyU11gpOKDDdLkQZB8W/qg63yTPMbMAj5rWtOxidwL+63zd9A0zHujiAw3bZbCr6ettFw8KHOAcBByhpmjhcDkDttstl8QwIKkhrfNbq0dJVGFrnZycfoWPqqwSV84c4ZyUHFsEjnOG2MrObG13fBWo3+wSW2PraMY+S24KuRvdBm1ThTt6o+6Usrp93eSxo5DVO6HFZsUTYdmoKzRtLT8gsQuEY22CmrZS1vwrDhLpCQUGTFOXnCvIB2Pmo4ogzfKrISe3kgngjYT5BSSsHcYWnPqZYuxUkNW+QgOz80FtYSzttlQwyOJ33ws2RjJW4OCrBBG3YbILmYdjI81rlhqJaWsZJC4ggjBGy0CR3hAdJwtZ00TUVzGk7ZAQby1PqO7T2wQyzvLenG5K4wfWyPkLXbglcwajs0P3ID/Mtz9a4sq7a2Mue0AYKDHljAjDmjBxlYYlc44yftKyGyukcY3H4Rt3VXQRtGcboLWdPfG6leR4ZI9CoWHcg4wr2vD3+G3zQY0Jy8fDn1C1bTQE1+gidFlr3hpGOy1vTul21b2OcBucLmW1cKrZR2c3trWCVjertvkINWl0tQQabZUxlgcWZ8s5XCOsbxcaWR1NFUER9WCAdiFnXziXeoK2Wzt6/CYS0b+S2fcqyS4OMs2c90GmujY9xke0EnfPzWJL8Mmwws3J7LDnbvlBIJXkDBKkbvjKxYyScLIDseaCvgt9ArJAG7K/xM+asecn7EEGcOGd8LVKOtqGMwyQgDyWnFmdyFQSOj2ag1KWsncRl5JPzWp2+JszcuIJW2TM84yeyyYbnPD9ElByDZKCnlID8YytVq7TStblvT6rjyi1JU0+4ysmbWFTI3BJ7eqDV7nSxx5Ax9i25UPeCQ1/b5qKqvs9RkklYD6uRxz6oNXo5JA76Z8/NZE1TMNusrRqSrkJIJU8lS7IOUHf/ANjVNJJzPana52caDrT/AM4W9Fi+xgkc/mi1QDnH7Aa0/wDONuRB7PIiICIiAiIgIiICIiAiIgIiIOPuYb/eB4mfwPvP6lKvmiqf90Kn/CSfpK+l7mF/3geJf8D7z+pSr5oKvavqf8I/9JQAcABXNHV2VjB8IJClYQAguGytegdl3krZDv3QURUBJ7qqC1oLXKRzuoYVAzfsqvwO3dBYz6SyR2WNHnKmJ7ILiO5VFd3GSrUFW/SCqrR3VyCjuyjDsKR+Ok77rGOQe6CYSAFXdQd2WOqtcR2QZDfpBVVjTsCrxuEF3Tt3Tp+aoCe2Uy5Bcq42yqJ1eSDHk2OEZ2KmcwO8t1QMx5ICo7upAB6K1wGUFvVsqg5VQweYV7WgdkFFVp8lcAM9ldgIKsZvlWzDBz3VwcW+eyka1kgPV6+iDS6yXqa1mPPK7acrcjZrewkgfD/MuqlfTNDQWnfsuwPAG/Ms1KIuojDUHd3hg0Nu8hGDutscxlAKj4jkHKz+X++U13vhbI/IJ8ysvmdg8DHu4BB9EHRfifRCKAj0acrgsgiZ+Bndc+cVevwCSNy0rg2OMPkkJHmgtY4Y7b+anYdu3oqsiYPRHAN7bIK46tlcBgYVjXYO6u6vRBbMekdsqyOYgdyFMG+JkEZUE8XRjpBQXhxnw12+VrVohEL2vH1hbeY90eHY/MtzWBr6qRrANz6BByBpxjrqBTuYXjYYW7KfRTIaqJ/uxGcHstV4J6XgNa2a4gBnUDuFzzrG2aXobIaumdF4jGZ2IznCDgHXlK2z2EPbKAenOCuCjqCScSsc8dytw8Vdf1lbdJLXG4mIEj5YXGlMXskcST8RzhBnTnxJi/G5Ksc4taSFeMdG3dRv3yPVBAD1OOd90Dfvo7LIpYQ6TJ9VnOpo2kHCCMtxCMlQdJOSsmYgN6R5LHBIQWHZFRxx+VUyUADJwq5A2UfUUyScoJRuq9PzVgcewKr1uQXNOCqiTcD5qwjZUaCHoM9o6m5+StLdjurmH4B9SEjBQYrzg9laXbdlc4ZO+Va5u3mgiL8nCA/ECqFpB7KrQeoIMhv0QrsFXRsy0YUgaAMIIelVAwrnAKmEF4d32Vr90AIPZXhmQCggVru6nMfdRvZhBQdla7umSE7oLepXtII2VhGyq0kDZBRxyVJGPhULjupYjkIKnbOy5N5NDnm74V/wwtX63GuMjjdcm8muP23fCv8Ahhav1uJB9GyIiAiIg66+0MOOTfiT/idH+vU6+eanGZMZX0M+0LweTfiUP/pOj/XqdfPNTEh5x6IJ5xhuPT/UoIyQVPPuzdQRtJOUE52ajHDBVHfRwO6tAwgO2OUacqpAPdXsa0DHqgsRXdO+42Qj0QGnCOdlWogK15wqucBsCFGSd0FWPwfRXE5OVC0HqVT1B2AO6DKz1DAVzQQMKOLYKTIQWub5qgbkKRU2CC3oV7WYblWlwCt6yMZKCh7oNlcektVvdBUvyMYVqlDRgbKx4AQUaMqrmhvmqA4UcjznCADvhUcdwFTIVHHdBI1mwJ80PdW+JtgKjXEnCC49lYQpukYzhOjPZBGDhOr5K/oA8lQtHogK0tycqnV81a55ygyABhRvf0bZUYlcPMoGukcNkFHP6vLdZ9HWmBoxthG0kZaDjyVjoWDYbZQZLry4EbA43XK2gb06elEYl6QR2yuIBRtdvnuFmUV+q7OQymccD54QdjLdYo7k4ufMMk9yVpV54Z00tUJSWE9/JcR0XFK+0O0bj+VZTuMF/kcDISR/3yDmm1aOp6KMND2jCyptKxVEo+EHdcOQcWbq4M6uoZIHdcuaG1pS1tIJax+HBud0GzeKmn22uhMkdMX/AA+QyuCZCS4lzMEn0wvQzhlYdFcRaiWkvskXQAQOvH864H5lOD9m03fxDpcMdE5/7zGPzIOt9IcSDb7VqRIzha1cdH1VupmylhBIytsyx1bc7HY4QX1Q+H1VKVgI7fJS26GSWQtqM4+YU9dHHAPvOPsQQzjobkEK2ACTc+vmsZsr5XFjs4R8zoSOnsgyqmFvSe2fmsRo6eyo6qlk+EZKvga5xw4FBa+Ut75KvilLgO+M9lkOpo3NGf0K1sUbD8OEENUfhH1rWtJPxXsOcbhaLVDqADVqemmzNq2loPdBytfnF9obuT8K42rIsMflpxkrkItmmoGtlzjpWg3O3wtpnEYyg44mf4cpwrfedsb/ADWRVxsdVOYOw2UbqQkHpAz9SDHMhycHupKIu96j6WF5LgMdyVrulbCLlVsbO0Bpdg59Mrs7w14KaRrDFV1phBABOQEHGmj7QXUkUzoHNJ3OQdlzBTU/VYnU5mGC3GCVujV9h0bp22eHQviLmDbBC4B1XxFlt0MkVI87ZAwUGjak0jTtuM0/wE5JOy48vcIpZCxoxusmt4gXOsnc5xOHHHdaTU1j649cnfOUGOThY8p6uyneQoS3fO6CxrSw5zlXH4t1eRnYq13wgYQWjumcuyoi7dSRboJCCQMKgbvgqdob0qGU4dkIBh6hsAgiDcZClheMd1eek90GOZAzOyhdL1HIWVLGwjKhEbc47/WgjdJnspGRAjcBSsgjxnCvw1uwQWRkRnso5ZviV87g36PdYpy7coPQP2LMvXzSapGf/lArT/zjbkUXsVR/10+qcj/sf138o25EHtSiIgIiICIiAiIgIiICIiAiIg4/5hf94LiX/A+8/qUq+Z+t/t6p/wAI/wDSV9MHML/vBcS/4H3n9SlXzP139vVP+Ff+lBa0jpCvZuFjjOQN1kM7IKomTnfsrXPxsCguJwMo12SrA7qOD2KvAwglHZWOYSr2EFVcQB5II2bHCvJwowd8gqTGUFOoJ1hUcO6twgvDgSrwRhRsBzlXoB3Ubo8+avJwgcCEEbW74Rw6VcM9WQri0HugM3CvAwgAHZXDGPJAAxvlOoKvkVYguyDsgGFZk5yCnWR5oJEVocShdjuguO6p0/NU6/mqggoLxgjCAYyrRnOVIAMIAHmqoThW9XzQVccI2RwzhWnfzRoOEFZR1AZyt76LvElAQ1kgG3qtkE5Cvp66WmkPQSNkHeXlX1BNU34B1QAC4Dv81znx8h8ekY4jxD0g7DK6GcDuIFVpy4snD3DLh5/Nd89O1DeImn2VE+HEMB338kHSHjHD4UTvgI+EhdeWPIlkGcfEV3A5kNNR2908bAB0grp+Y3MnlDvJxQZEZJ3JP5Uf3VgdgAjCvb8QyQgtJwr4zkK9kTXuxthUlb4ZAadigvj7q50QfjJWOZegZPf5IZZjEXMzsCUCvj93jaQMnK3lwxhNfdYIXxkdTgOy0XQ9qk1JdBSTDLcgbhc8WnRFHpeeCqb0gtwSg5IutmbpfSjLjTP8N3Rn08lxDcOI9zraGeCSuLgMjBctd4u8R3HTf3Op3nZhAx9S6zw32s6y0ucQ7ORlBqd4n9+uj5XkEk91C6lABI+xQsk6j4h7k7qb3gu2QQsJDyCpnNCjkYWZeFH4rz67IMqEdG/2qYyE9iViQy9WxyFkOIAGECR2WqFC4kogsd/OrVI4D5K3AQRIr3NxvjzViCrfpBXI3GfJXhoJxhBVn8yuDB1AqvTgAgKrTvugnGzVaeyFxDc58lH4mRjKB3KFgKsa85zlX9X1ILHMG/ZWgYcAFc5x+StzuDkIMphAaEcRlQh5wPqVer5hBcBlV7bKnVg52VQcnKCRXDsrR2VQR5oKkZUcoxkKbLfko5cZ+xBj43yqOVzsAKIk57oKK4dlar2DIQRu/nUsXZWOG+4UkQGEA7Z2XJvJr+F3wr/hhav1uJcZEZXJvJrj9t3wrx/wwtX63Eg+jZERAREQddfaGHHJvxJ/xOj/AF6nXzzU/wBIn5L6GfaF4/ab8Sc/3nR/r1Ovnmgz1HA8kEszgW4CsieG7eqrJsN0ha0n5j0QX4zuqK54IGytbnzQFc1pO6uABCrsNkFAR9FVxgEIRjcDzTOxyUEZ7q1wyFce6oTgEZQRYIcqgZ80IPdVafJADcFMjO439Vf0Z3CFmDuEBXNaTurScKrHH1QSA+XorH91Uux2KtcSd0FFR3dW9RzhUcXZQSEkAKoOfLzVBuBlNggkDgVR6s6sDYqhe49ygqrHDOT8lU9RHmqDtuUEZ2VFeQCTsrSPRBYNlex3xA4VOhXNZ54QT9QI7K9nZRjsFew4CAWZUcg6R9eylLgFFIc+e6CAggqh3OEPVk91TfPzQSthPfOxWVHE0M6iQMBYYfJ5FXe8SgdJJQTe9EHAPbZRGoLn5UTh5+qzqamjczqOMoKtf1NB88qdlMx4y/B+xYMznRkhp7KsdZI0HugnlhhYcdI/IkTIvpdIysUyvkO+Qqskc13Sc49UGcJ2Nx8IwO2y1Gl1VWUMZZDKWjGFgwRRyN3xv6rGraeNrvhPZBvzSXFW92aqZ7tWuYXuAJBIXZnQWmncTIY6y6VQleQD8TsrpHFCA8ODjkHK5G0rx01Ho2JsFtc/DRjc4QdkOKXB+jtdA90cjDgbAYXXyfSUcbnAU5dhx7BZ0nMPqTUkraS5l2HnB+LIwuXOHcGn7w1jri5gLhkg47oOt+qqB9njDo6Zzc7ZwcfWtpirleMOZ1fYu+GouG2gLxREPdB2x5Li+5cHtKQPIpBEcuxthB1iYSd/CLfsUU+O+FzPrfhpHRwF1taCQD9FcVVGnblDlszDkE+SDTqMBz8OC1B8LW7t2WHLR1FCepwIVGVj3kd0F9VKWDYlYvvLlmuh8ZucDKiNC7sB+ZApMTvDXLe2lrbGa2NpA3IWyXQS0p62g7ei13Tl1q2VbC3OQQg5mu9rjpbSHNcAenK46vEzhTPw4efmtavOobg+2hsgOMYWwK+4yyxPYM5JKDQpJHGrc7c/EtQogZpwwRncq+wWee4VYLm93ea5Y0joSl+6cXvLWhhI7oNGsFoMEbJhC4bg7rk2mv1ba7SXw1hYQ3tnHktz6isOmrVZA6ndGJAzOxGey69aw1fNAH0lO49JcRt6IJtRcSbzVV0sEta5wBI+kto3C7y1TSZHBxOfNaFJJJUTGZ2cuOcq9h36XH8qCzxG+L1dKyWnLc+ip7vGPixnzyqOPSCAgofP61RROlwfVGSZPZBKrJVerXjIQY57q5mzlQg57KrMdW6DKaHFuxOFZJscK4OIAGdla/B3PdBYHlu6qZ8f/YUbz3VgBKCQzHGMq0vJOclXsjDvJTCCP5ZQRtlIHdXeLkZJVr4xvgK3YbZQHuLtyUY0OGCrx0nY4VpcG7NQd/vYtMA5pNU/wArv5RtyK32LD880mqc/8AK7+UbciD2mREQEREBERAREQEREBERAREQcf8wv+8FxL/gfef1KVfNBW/29Vf4R/wCkr6XuYb/eB4mfwPvP6lKvmiqhm4VX+Ef+koIo25wSpQMK4MaGAgeSq0DG6CNxBUTgQd1M4DfChf3QVb5KRvZR9hsjXuzhBkMG+cpIDgH0VI3eRVJnEdvRBbGVOsJjzlZbHdQQVPZWEZUxAwdlZgHyQWtdnbCk6UDGjyRxwEFpHcKJzunbKuc9zTsfrUDnFzt0GQx3UFerIgA3KvQVB7bK7BVABscKoJCCpIGytVcFUQWnuqEZ3V+2/ZRuJzgIJOw+pWu3QOJHkmEFQ04Cq0HKo07gKTHoguAwrgceSADHZVwEFrvPCsV5Gcqx4x2BQFe1QknCq2QhBN0hVYxvV9ii8XO4O6zaRodu4INf01WOoXxvaQD1juvR7lhmdc9Jhz3ZPhDt9S8zJ5hSxMLdsuHn816QclNQ6r0iMnI8L+ZBxfzR0obU1A6CdiujtbGBUT7Y+I/pXoPzNW5ktVV5GcZXQPVUfuddM1vbrP6UGmDsFNH9FRQgOjBO+VcXdJwAMIMqAAuVlXgOGPJQCZzTkBSAmUb5KDFmeQ0ZWqWxjn0zj7s5/wAJ7AlaVXtDWN6dslduOWHg9aNa2tklcxhL2Dv80HDXAyjFdqLw5Kd0eHjcjHmudOJtC21UbZI5wcDyPyW5OJ/Cu1cJoZbra42h4BPw4C606g4p3HUImgmc/DCQMlBt/VV7mral1NJJ1DcYW120jI8uAAJU1TIZqkyknJ3Vr5CW/Ugx3u8POOyRSlzgqAGR+HeanEDWEEYQTTEeFkd1hh+AchZDJOv4D2VzqRgbnbdBj0py7ss57QG7FYJHguPT5J7y9xxv80Ewd8XbzV6saPh6vNC4oKoqHYKmSgo5wIwrERBc3+dTMUTFIHYIQSn6KsHdXt+IYVQwZGyA4kswollGNvT5/lUZjbg5ygxs74Vw2Cu6AHdlUgAeSCBzsnsqA5VHdyg7oJQcYVwOVRoz3CdiMIJFe04CsQuICCQuACB433/IoC8nyVA8hBltJ7qjzgKISI55IO6C1x7hWO7oScq1zkAnCkYdlBlxx8lPH9FBR/dXxA4Vj+6lhOwQUc35+a5M5Nfwu+Fef+GFq/W4lxs/Bz281yTybfhd8K/4Y2r9bjQfRsiIgIiIOuntDATybcSv8To/16nXzz0+A459F9DPtC/wN+JP+J0f69Tr55YPpH6kEkx2yqQjq3+aSDLVSJ3Tsgne4AYVoHUMhXuAI8lYNuyC4fDuUx19lbuVewYCCgf5YQt6u3qrukd02AQWFmB3UbhkqQuJyFagjLhhUacnCucB6eataN0GUCAzdRuIOT8lf+8UbjgbFBYHAnCuBxsoMkPVS49QKCdFRhy3JVUFOn5qx+xwVIrJG53wgq1wIQuycKJriHYCu+aCQDG6qRndWB5yFXqKCoPlhUeCd1cBjBwiCJBursBVa0ZCB0/UmMbKYNGELW4OUEZOBugOdwqO3J9FGT0oJcj1Ubxk7Huo+s/JVDye/kgodlRo6nD5q/APkqtYBuEGZDA3pyfNYdS3D9uyk96ez4R6q8NbM3qxuggDctGe6zYAejAOPrKwXvc1xA2CvZUOaMb/AJUEsoDisYuwcYVzpjnzWTSUzZW9ZxnugpTsBGeyslaGu+xZYZ4ZLR2z6LBnkJfgkY+pBNHUluQCQFFPO52Mkn0VzWNwPmpRTNcPix39EENM8l2FPLDG456RlVZTtj3Cjklc3ugmpQ2meJG4BBytzUGvbja8NgqnNx81s90xcPNQvBJ6iTn5oOToOLN7mLYzXPxtncrfemNQVdzIMtUTnfcrrzTsPiBwJ232W5KDVlRaWjwy7PbYoO5GgNE02rQWVMzXYGMEhbS4rcLKKxVgZDCHAn96uGtHcwN901M3wC/BIHdc46Z1zLxFkjmuQyTg/Eg69a/sjLdCSynI23OFxsw9Ls4z8l3L4qaGtlTazJExmen0+S661+jYaSN7wBkE+SDa9uYJQAfyLWIrc1zuwK0OpkdQTljTsCtZ09XuqqtkZ3zgboLrpbWRQ56PrWRpOhjluMbekYJC3Fqe3tjtrZQACRlbb07O+CrbIO4cNkHI2sbVDSWUPY0Z6c5x8lw7G4yVRjLc/FjGFypfLlPcLaInnbGFtHT1iZPfWRvwQXD9KDeHD7TUNQGPfH09tyuQrraY7XS+8xSgFrfJQ36mg0np9lTSNAd0dWy4hvHE641kD6d5dgHHdBkal1rcJJ30hqiWgFuOpcc3OtNTMQ7BJPdKmudU1DpnOOTuoBE2R4Pf60FWxgb4H5FR8fSOoFZLmgM7dlhvkcXdHllBYJiSRgq/ORlTikZ09eBnuoZR0ZDUGO4ZKoO4TJyUB33QThwAVC7PkqNORgq8NBCCNU6dwcq/ZWuODgIJmj4cfUoZX4OArmuOO6skw52T+ZBYTlXxtzthVYzO2FkNY0emUFjQAr8ZGc7YUcrunKjE7gCEErtgoHndXeIT281cI+sZP50EPW5UJJO6ndEG+X1bK0MbjzQd+vYqfhTap/4v67+Ubciv9iywDml1T3/+IFd/KNuRB7TIiICIiAiIgIiICIiAiIgIiIOPuYb/AHgeJn8D7z+pSr5oak/7fqfnI/8ASV9L3MN/vA8TP4H3n9SlXzQVZ/29UY/+aP8A0lBUOwAnV6K1p2CuAygqRkKwMyPVSN7q52PJBA5hxsqM7FTNwTur+hqCEHCtkcTssjoCsexoBQYud8qaKQnZRuDfJXMGEGUHZVwIwoQT5KvxIJer5KhOTlUHz7qvbKCx7h04WMQerspyMnBVwa3zQXRt+EI52FeMdgrXNHogt8THdVEoJxhRuG5CoBg7IMlrg7bKr0/NY7XEdlXregkLcZKBuRurS49OVYHkIBcGkKjpwDgD86o7fdWdLSd0EwlHdSxv6j2UTWt6d1LCADsgy2s2Hnsq+EVRjzjuFeH7b4QWOZhQv+HCmcXEqGUBBEd1E6UA4wpVaY2k5QUjDjhalTS4wPksMNxsAr2Et3QTXafogj8/jyvSbkQe2p0eD1//ACId/qXmZcnl0bM+q788lt+qrbpoRRdWPDGfyIN2cyFJ11dWM52K8+te0vhV82dvjPl81394sVj7lV1BnyMg5JXRXi4xsF1eyPG8hQbPhw2MBUe7dQ0ry5oz5bKV/dBUN6h6J4ohaQR2UM8hjaCPNSU3TURuLyAQDhBqFhtcmoqptLGwuIcNgu3vBfUreG9JFTzS+FsBhxwuCuXGy0901KY34IDvNbk5jrjUaWu8FNRZaCfI48kHLXHvidBqCyyNZUteXNPnldOmVg8Sclo+Jxx+VZ82rq260Qinc4kDB3Wj9AGfU7oKmTqflXF2RhWBoCqgtY3od1KR8hIwPNWpgIKRP6Xg4281kGpBGFj9ITpCCrx1E/NWCP4sq9EF7X9gr1Cq9RQSOIKsJxsqZPdM5QURFQk52QXsOCql2CoxnO6qgy4XA4CnAwsEOLdwnvDgg1B3bCjWMJnnzKr4jkEkgxv6KB8mB9Ske4kKCXsUERnGexVzXguGyxj3VQSDkIM/96DlAcnKxGyu2Cla4nCDJDxhM5UZJwjScIKomEwgr1bdlTxf/fKoeyjd3QXdatJycqiILy8YUsRyMrHV7HEDAQSv7q+L6P2rHc453CmgJLUFXPIyuTOTU55u+FXz1lav1uNcZubnsFybyaD/AK7rhV8tY2r9bjQfRuiIgIiIOuvtDDjk34k/4nR/r1Ovnlg+kfqX0Ne0M/A34lf4nR/r1Ovnlg+nhBIWn86NYO6nAHdWO+XZBTJ9VRVZguxlVeMIKMdur3ODdseShBwrh8RyUFzDk5KSOx2V2OluR6KF5JO/qgqTsrQ7Y7Kr3fDsVGD6lAzl2Vc0eaoG+YCrh3oUE5fhqjIJB8kHz9FUjGcIIXNwVQDJ+pXkb7hVAAQMbYVW7DcK4AYBwqEeiB1fJWueM9PmqqxwHUD80F4g/fZ+aoW/NSdZ6MKNzvmgtOyvYcqw791VuQRhBIDvhWvd04VT227lROBJQVzjBU0YLm5Hoo+nYZCyIgBH9iCJsgB7I+XI9PkonD4+yuA2IQW+ICceaO+JWhuHdlUkoLehVDcZVRnzQnCChdnZVa7AwrfmiA7cqWObob0H0USo7ugPd1OJVzWkqxSs7BBa6NZlLN4bMEeXqoG4JUrWtKCR0/UsWRnW5TFgxsrcY+1AaOkb7qdj9iQoQc7FN8/JBktdnKx5Yuo7KRhwNihJ6kEIjLMk/oUbo89lkykkLHPVnO+EF8WIzufzK2YGTYAd1a4nCq0khBWBpila8gHByuTtH8SRp6NrvEDekDbOFxbI5wx81JFE2ZuOrB80HZrT/EVmvj7i6cbnpUmpOH4ZCel4JO66+6cvdRpaq97p3EuyD3wuSbHxTut9roqaUOOcDdBsTXmnZLTN1Fhx6rG0HE6qujIukn4huuaNcaehrbS2sq2hvU3JyFs/SdnoKKsbLTvYSD5YQa9rmz+52Bk3f4Qd1x7pSndV1oYGE5djAC5G4h3QS2pkDnADGO6x+B1hpbtqinpz0kPeB+dBu63cPJrnb2PfCWAjuQkvCr7h/wDVXrA6PiXZnivpFuhtAx3KjhAPgh2QPkul+puMN6npJ6NwcG5Lc5QW694gMlhdafFDiwFvfK4imrBLkADc+SxrlWyVtbJUvJJccnKjpvieBlBL4RyXZ2Vvi+G7fKy5QAzYeSw5Ggg57oJm1AkGFY+HpPiFQREB2yy3EmM5QWGqBb0gkFWE9QyfRY7gOvCmZ9FBE5mSVaW481Mfl6+Sjd3KCrVd1EKwH0Kubugua0+qFu6qCQqFxygHdWlmfNXHsqZKCrSAq+J81Yduyt3yNkF0rsj7FGBlXuGQrNwcD1QXtPyUnXjt5BWAYTPzQXdfVt/OqEYOEYAju6Dv57Ff8KXVP8AK7+UbciexXP8A10uqf4AV38o25EHtMiIgIiICIiAiIgIiICIiAiIg4+5hv94HiZ/A+8/qUq+Z+s2raj/CP/SV9MHMN/vA8TP4H3n9SlXzQVn9u1Of/mj/ANJQWNd8I2VzThWN+iFc0jdBIioDlSBoPYILQMqVrMdyo2d1JkoK9PzUcsfV5+SlVHAkIMXwiCrun5q8j4t0ONsIKNPThXh4KoAPRAAdx5IK43yrla0lXILCxBGSpCBjsg7d0FAPJV6fmrS4g7FA47oLXNOSVQNJUxGU+H1CCAbFX9YVXNBzt5q0NGd8oLtyArOgqYNHkFe1uRugx+j1Vhhydip3d1aThBaIzsFJG3pO6N/OqoMlmwH1KuQoevsE6iglLwFC92cKuSqYBQWIqnurSSEErR5o5/SqRknuqSAEoIK1/VGwfNd2OU7UkNtsbYX4z0Ab/UuktZsxv1rtRy2Hqtzepx+gNgUHOmq3fshq5xCM5B7BdLeOtnktV5LJAQTIfJd5tJUDKi5Sg4PfuupfN1RtpdSMaBjMh7fag6/08vRsSsrPibhaf2wQs6iy5qCOsBDB81qFntstXE4sYTsc4WDcAWho9VzTwS0rTXqh6pQ3ceYQZPLbRyW/UpdI1zAD5qvNdUsqL1B0uB+LuPqW/wCCxQaSq5KqmwCM5wMLgvjHeZrzcg+TJ6HHug2FS1LWxCMjBBWSJQViQwAMEh7lTgNx/wC1BL1hOsKAOyVIO25QSg57IrGE57q9AOwTuqO2CsEmEF5cAqdasLspkeqC8vwPJA/I8lHt2VQMIJQcorWq5AJwFQOBR2MKiCocCrh3Ubf51IO6CTyVhBJ2UjACPVVLCEEbXY8vJVDxsrXABWoJnHPZWOZsjHeSuyPVBhvjIJVpaQsxzAVaYsg4CDHG2FMw4H5EEYzuriMIKg5V47K0AbHCuHZBeBjdCM7oq4PogsG6o5qvwArXFBEdlRX4HomAfJBYrmuDQchCBhWoKPk38lk07gW5WG9oypoTgIMkkZXJnJr+F3wr/hjav1uNcWl4wuUOTM/9dzwp+esbV+txoPo5REQEREHXX2hn4G/Er/E6P9ep188sA+M/JfQz7Q38DbiUf/pOj/XqdfPPTgF5z6IJy7IwrVcQD2Vj8tCC6NuHZyqyfzKJjz1KQnPdBY05OFeNlGfhOydRJQSh4IwfRWO77IMDfKo47oLSMBU6coXbeSqw5CC9owcK8EbK1gyd1L0t9EFpbtlWKR3bCjPZBY4gqgOPJWknPdVyPVBcDlXjso2/SCuyUFSO+6sI3ypFa4YGyC0SfvVa45KoAOpXEZQB2V7SAFaGq9oGOyB2ySrXHJV/fZWPGCguIy3CljPTHuo2bgZUn7woMbqy85V3UoT9JXg7IKndEVHEjZBQvCZzuqYBKIB2VOr5Kp7K1BcqO7qo7Kju6CivacqxXMQSgeaua7BA9SqNUrQ3AOUFxONyrD8Tkc4lU7IKubhXM7KjT1FXtAGyADg5QnfKqQAFRBR7tuysJBCvcMhRkYKCxwwMoHDCuc3ZW4wMeqC9sJqMBqnjg932JUMc3u++ypNWF/2IJZagDAI2zutb0ze4rbWMq3tADCCVtyBvjuwTup56fwoXDcBB2V0xPHxdo2WijkAcMN2K12Tl8uGliHyPc7J6sd1wfwQ1lUaSvAnpyclwOM7LtZb+JtZqurigqgACAMkoOHNZaEq6+IUzI3ZHfZZvCPS8ukb/AAXCocWhjwST9a531FY6OO2++xlheW5OBuuHNT3SW30kkjRgtz2QcxcfuNluuGhm2ls7HObFjAIzsF0Lu+ooKjxWNaCS8rWtXavqLrUyU0kjyO2CVsk0gc4uzuTlBjuHjSlzRgHsp4oDGQSFMynEW+Ar35xkoKPcOlYUzwCWqWSQbjIWM7d2580F0DcuyszHw9Kx4sDH1qYO33/SggMPxEkK8Mw1SPdtj0VnUcYKCLGCVZJvspCfMqN2M7ILWjBUjfpBWN7qRoBwgvHZWOG+cq8nCtJygoDnZCM7pgBVQRkZTONsKqoQPNAPZUHdVPZU+pBcqEZ3QEk7oSQgq3Yo53yRWnug7/exXb/10uqf4AV38o25FX2K/wCFLqn+AFd/KNuRB7TIiICIiAiIgIiICIiAiIgIiIOPuYb/AHgeJn8D7z+pSr5oKz+3an/CP/Svpf5hv94HiZ/A+8/qUq+aGrGa6p/wj/0lBGGnpCdCuaMAZVwxvlBRo7KVrgBhWbfJWSEjzQSg/FlV61EwnG6uyPVBIHgHdV8Uf+4UWR6q1xwgq546iqg5UY3Pqp2NGEDvt6q5o6VVoGRsrsD0QWjdV6UOxyqjcIKEjGFRHd9vzKgO25QRnYqRpyrCMn5K8ADsguznbCoQQrtlbnKAisLiqdbkGQCFVQh/YKQOJQUf5q1XHfumAgoB2VURBUbKoOVXY7bKrQEFFXBVdlc0DHZBEW7qCSXoccjssrbqWFUNHWgyYXh/YKR0ecH1UVO0ABZDj2QYVeA1rRnuV2f5aJw+3hvUPo75XWC4nIZn1K7B8udY+npw1oJ2Qdv+HtL7xc5QM9vJdSOdGlNNqZgII++H9C7bcHql012k6vmurHPOANVQ4/8Amh/QUHV2KLxRt6rNph4DcHuoKA/EWlXVshY7AQUuD+pzQPLK7bcpGiX6ltYcSWAjHfGV1e0zam3qoEL8bHByu13CHUFTw5pYoKQHBA7IN28aNHjR9HNVOkBABO66T6svjLpVy9ODh5G31rtPx84jV1/szxP1AOac/kXT1rGzSyHO/USPyoJqaYSgRjYhZEkXQFDBC2J3UDup5Hl2xQQNG+FeGlUYBnKk2QXNbgDdVVMp1IDxkYyoun5qr3k9j6qPqcEFQN+6uDSjCPPCv8wgtLNsoB5KQ7tVvTvtlBe1pKvDPmrWq8E9kFjmnCt6SpX46fmo0FjdirwcpgeiIJIyAd/VSl4x9ixwfROooD3D0VvV8lU7qnSEFQ7fsr27o1oyrg0DCC7pLghZgKQDAGEPphBj9JzlCzJUuB6KhHogsDVUbBXEbIBt2QVVeoDCsyUzlALsnACKmAFVBYrm9lQjZXMGyC0tyqCMlSgeiuA27IMN7d1fG7pbhXSNA3wrCAM4QULsZ9FyryZHPNvwoPrrG1frca4nJzlcrcmP4W3Cj+GVq/W40H0eIiICIiDrr7QwZ5N+JP8AidH+vU6+ean+kfqX0M+0L/A34k/4nR/r1Ovnlg+kfqQThwb3VHuDkIyonEgoLmfSV6sjV6CwnJx81Vrdu6ADKuQWnZWuBIP1KTYnGQqHCCEDfClAxtnzVMeY7oCe5QTNbgBXqxjj2KuJ2280FrnDsrCQqPOFZkn1QWkZVCN8K8Ku2d0FYx+VVIyje6qgIT+dFR/ZBCWZOcqZkecfUoWkl2D2WVH5IKeH8lQjpU+B6KyRo6SUGP1BWlwyrXu3OCrQd8oJuoNwpGP6mlQdxur2/CEFnSQ7OVXICZJOCrXEeqCvUFa52+MImAUFAMblOoKmSiAiIgu7K091ceytwgK9jTsfmrFI3sEEoGRjPZVGQO6tZt3/ADq4YQCcKrd1f0BWHAPwoJGs88q7pKozOMFXIKEeaoqPJ8lZ1nOCgkVvTuCq5Hqhdg7IKP7KNS7FRO2OAgq2A1GzVSSidGDnOVJDL4JyrKisLzsSgUrfCfk+ayqmVskRaPNYUMnU5TvIDfsQZWmribbXMkOMFw7rtPwHik1nqijtkXeVwAwPXC6qWijbVVEbTj6Q/Su4nKdSttmu7bUMxkPafzhB214vcAazQ/D6O/TdQa+Lq/NldD9c3RjqGfDgekkFen3ODrmpk4NR0waAPdwCR5/CvIPUV3lnpKlpzu8/pQcaXGr8S5yOGCM4TqGftWDPk1T89y7KyBJkd0EzpQBhWOeHNwo35xlWNdg4QPBJKkFLncK5vZSF5a3ZBivj8P8AIozOBsFSaZznEYUTB1OwUEzZie4V+erdXGEBmcfmVoGEEbxnOFYRhS59So3kZQUHdSNPZR/UpGAZGUF6o71VTsFGScnKB1fJCcnKrhp7fpQjbZBadlYTk4VwG++VQgZ2QVPZW9WCq5I7qhwSgu6kI6twqDY7q7IQGjBRxGVUK1wJKDv97Ff8KXVP8AK7+UbciexX/Cl1T/ACu/lG3Ig9pkREBERAREQEREBERAREQEREHH3MN/vA8TP4H3n9SlXzRVA/2/UD/wCiP/SV9LvMP/vAcTP4H3n9SlXzRT719Tj/AOaP/SUFP3quYzKuY0dIQ5HZBZjB3Vj1OACrHtCC0N2GE6SrgD2AVQMdwgjOytc3KlIG+ypgII2bd1LlW4A8kyPMoJGv+xXB+25Cx5HYBIKjEhzuUGYZPLZVY4EKBpyrgSOyCU7q1zcq5vzV23oEAMGELcdiqgkbK4b90ESYKuIGUQRmMqnhlTAHKu6fkgg8MhXg4UnT8lToA8kFMFOkqYMGBsjmgHsgi6U6VVxx+VGb90Fo2Kkb2VvRvnyV4B8ggpjGSnXjZV7KF5OUF4IzlQyw+Icq/qIAUkeXDsgtiZ0gDyCve8A4yFVYNVM4O27oK1xDg1cxcEtRR2toje4N2wuGOoygdS3bpGd1NUtDCfsQegvLvcm3u8PbGQc57Lr1z60TqPVcAcCMvP6FzdyUB1ZfdwSVxj7R+m921dSDGMvP6Cg6a0Z6XFx9VlTUctSS9jHEAZzjusLq6YGkHck5XKvC/TlPe6TrmIyR5hBLy/6Ll1Lf/d3sIAcO65V4uXGLhlVwUxeMnATQVKzRF1fVUQBcPRcV8w+qarUt5jmqsgsJxlBja24mRX+2CKN7XEjGAuOLbTunLnHzKxaWETOwSdvJZxk9xwGIJKqE0reon8ixopxKcfnV76l1W3pdnCtjpxGdvNBMW4GVQHHkrz9FW4CAXAK1zvRUd/OrUFOpUPdV6Qq9KA0YKvGyNaMqQMCA0+SrkBU7BUJJBygvHdV6gog4quT6oJRv2ToykRJKkIOchBEWkBWqZ428lCdsoKN7q4bqME9SlZglBVjCSr/CwpGtwBsqnsghxgqo7qMvOVQSEOxlBmNZkd1c2Iu7KOJziFmQjJAQY5gI3Ub2Y+xak9nwjZYE22UEPZU6ghJyqICKh7INwguxsgbkZVVezAHkgs6FXGNlJgeiseBnZBRv0gpB2Uf1K8dWAghkbnZWdBUrwfNXRgFpygxHM/SuVOTIf9dvwoHprK1frca4ycBkrk/k0A/bc8KvlrG1H/lcaD6OUREBERB119oXtyb8Sj/9J0f69Tr55IThx+YX0N+0M/A34k/4nR/r1OvnlhGXFBMrHnJV6ied9iguYQDurjuMhRtO+6kyUFBurgPJUb9IK76kFBERurXg5UoLvMbK1w7II+ydQR2wVuR6oJeoBVD1j9ZPmrgTjcoKuOSgIwo+og4Vevtv+ZBI0fNCw5yqsIP5FK0DG6C1rDgK7oUjRvjCPGB2wggO3dWvIV7t8rHkcQe6A0/EsqJYkf0srKjzsAgyA3KimOGkKZp8j6KGdBhEEHKo7uqvOCqsaHHJQVbuFedgrmtDUc0HcD8iCEEDJVHbqp2VEBEVCSEFEREBERBXIxhUPdVAyqHugKRmwCjUjeyC8b9lKxpAyo2D5KQOOMBBVzs7BRB3xgK/DvMKnQAQSgnB2BwhIwVQO2ACscT2ygF4Bwo3HLshUkJxnKiLiAcIJwcjGSq9WO6x2yEHdXBxIz6oJ2uwd1a8jJUfUVaX74QTBnibBU9zeT3O6sjlLD3V5rHDtugoYjBuqeKXA7KSOU1B6XqSWlbG0kAZQatpWn8WpiycfEP0ruVyzW1p1lbx4gHxt7/WF0htdykop2dB7FdiOBPEert2qaKVryOlwwg9FOdENtvCON5maf8Aa+cZ+S8irhe2TQ1DQQT1n9K75c1XFau1Bw2jpJXuI8HG5+S85IZC/wAVpycvJQQF3i1RcOxKn8LB3KoYRF8QChkqXZ27/WgnfgAhQ4wc5UYlc47+fzVS8+ZQTeNjb+dUdUjGCo8AqPpBfj1KC0kuOcKSGMucD81O2nb0g7K8s8NuR6IJXNHh91jOHdWCpcTjyCvJ6tz5oIyMlWOGCpFFIfQoDe6kAOQVE0nKlb5IK52wqEeavwFY4Hy7IKDuq9QQ9uytQFQjKHYICgO7KwNOVee3dUHdBVwJGFQNI7q5EBUzhVwhHqg7++xX/Cl1T/ACu/lG3InsV/wpdU/wArv5RtyIPaZERAREQEREBERAREQEREBERBx9zD78AOJn8D7z+pSr5pJsNr6n/CP/AElfS3zDf7wPEz+B95/UpV80VScV9T/hH/pKCVpBCqoWP2Cka4EZQWg/Ej9yqHYp3QSDsrXbq0P8lUEeZQUAyVd0K3ODsr2uyN0FOhWuiyc5UqIIvCwPVWGHfbCyFY/O5AQRhvSqqnxZ+1XYygkbvsrwMK1m3fZXIL1aTlC44VmSgrnfCqqb91TJQStOFcDlRgnATqKCVWuI7hUJ7oASNwgePjbPZC/qGVAR8ROFe3sgkVQPNUVzeyBkDZVDxhWHuiC5zgeyt6coqgkIInnBxhTQnIwo3NB3UkQAGyCj3AZWJJCZCSsmVWsJyghbT4wABstbsNUKeraD2WmqSjP+2gUHfTkgv8UF+Bc4ALY/tHKltdqmmqGYIEnf7FHygXFtLeg5xPf1WNzvEXS6ROaf3w7oOoE8Do6WJ57PJK7Pcsug5NTWsSsB3C6/Xy3+FaKdwI+FdtuUK5/cyytZ0g/Cgt15ph+hXS1M2QBk7ldVteXxmorg6WMghjiNl275lbwa23zgjGx7fUukOQ2on3z8R/SgrSxGF3UVfU5mOQq9YcAnb5II4Yy3usgEDCjBycoScoJPEzthU6vkqNAKqcIAGThWubvspAPRVA8ygtEeMHKuDfmqq8NyOyC3pxuqEZ7K9wwFYgq4YGVGDlXuJIUfbYIKgYVUwfREEkZ6e/opQ8FQ5IVzTvugvcDjPqrHNy3KkfgtUeSAgiDfi7qUDBByox9L7VIgvEwAAVfFyFF0/Wq4xsgt7lVEed8q5rRnfKuQXxO6dispk+HAjZYY7q7sgzzUgjBKwpnhzvtVHEAHdRdXxdwgkEefNPCxvurmvAABQuyNkEDzg4wqB2B280eCSrQN0EzNypQ3Az9qjaNtleCcboKGTfCsfLv5KrgMkqF386C8S+ZVwqG4WM/sjQCEE75weyvieC0rEI32HmsmAbbhBRztyuUOTQ/9dzwpP/djav1uNcXSD5LlDkzB/bb8Kdv/AJcrV+txoPo6REQEREHXT2hpxybcSj/9J0f69Tr55YTh31r6GfaH/gacSv8AE6P9fp1880X00E6ie057qVRyHBQGj8yv6gMZUQcfMquc+aCofgjspGuB3UQb5q4ZAQTB4wo3PwjUcBjOEFj3A7hWAZVe537ZQ7H9KCrIj5q4sx5qrcnshB80EZZk4ygix5q5oKuQWtPSrw/zVhG3ZVG3dBM2UNCo6fIxnKicMhUaBhBeXb9lG9uT3Vx2CpuUFjW9JWQx+BlWtb6hV2CC8zgK10nXlRkDKbBBaW5PdOnpOVUbnKOQVLsjCNdgEFWogtcdycK3q+Sq7+dUAGN0FTsqfS3VT2VG9kDp+adPzQk5KqDsgp0/NOn5qqIKZHZUJycoe6IK9PzUjdsKxXAjHdBIHhOsKMnOwVzRsgmE2TgBUJ6jsrGgK8YyMIKtJaUJ6iFV5bjZWID2ZCj8L5qUklUQR+EPRXtj27qqr1Y22QWlufNWmIndX5BVclBGIS5V91cVPDudt1P0n0QY0EJjcSVNM4FhSQho3WNLK0sIBQRU7DJOMeq5k4PUzxqGlJ83Dy+pcQ25uZx9YXNnCUFl/pT6OH8yDm/mGbINBxg4/sXp8l0ihf0l5+a7ucwsviaFYP8A6Fj8y6RxsP3zPkSglkl6m4wsZ7chSOIVmSgtYzdX+Hkq5mPqTqx2QOjA7q3w/izlSEkqmR6oL2zdLcYSSYEdIUZAwrcb5QWFmD1K8OwMIe3dWoKuf8u6s6cquxVewQUAxurgRjsrQSTuhOPNBeH4RzgVYCSd1VARUPZUySgqDnZCMquAFQlA6UAwqogHcINgioSR2QXDurHu37K5WO/nQd//AGKzs802qf4AV38o25Fb7FT8KbVP/F/XfyjbkQe1KIiAiIgIiICIiAiIgIiICIiDj/mF/wB4HiX/AAPvP6lKvmgq/wC3qn/CP/SV9L3MP/vAcTP4H3n9SlXzQVeffaj/AAj/ANJQWNBwNlIHEK1v0QqoKh4QuAVgO6qcHscoKg+ao8791YCdu6rlBeHjYK4HPZRDupW/zoJR2VcFUaRhXNOxQUO3dUByrnAHyVuMIBGQqNGPNVyFVAREQWCQk9OFereloOfNUcT5IJT2VqqSenI7qwFxzn0QV8QDb+ZPFCid9NMgeaDIa7Jyrw8YUTfrVyC0kE4VQcKNzsHZVac+aCbqCuaRjsVEM581e3sgqd0Jwio5BVU6gqEnGFZuPVBkMj6leI+n6lbG44wFeXO80Fj4+obq0RYypMlUQWFuPNQtqfCqQcKdY0kQdICcoOd+A+uG6brmzmQNyQNyuV+Ithk4qSRVTGmQDB2XVvTNQ2mcz4jjqG67u8A7bJdrZE6NoflmTkZQdf8AU/BeojoWwGB2GDzBWtcOLv8A7H9OaaR4YRsc7LsdrbTc8fit8EZAJ7LqZxOmNtrntfkfH5beaDP4wa9ZeKJ4EgJcD5rrZ19Ukh/uiSt8aorWz0QwScj8q2HGfiKDKpyXEZUsmxUdMBn55Uso3QGAjdXKkePUK/AQWsBypOlUb32CuQAMborclXDcIL2N3V5GNgrGOIPdX5ygtf2Uakf2UZ7IKdWdggGFYPpKQZ80A9lariNlQDbsgN7q4d1a3urh3QSbkbeipgqVrdtwrugEdkGOxuHZJ7qTp+aFvScgKmSgr0/NOn5oCSd0JIQCMKiuPZWoKFwCp1n1Vrv51aguMpIVvXvlU6QqHugm8TZVEvzUQ7Iglcc9vVUG/wBior2Db60F7dhgqvUFaqE480AuznZRP7qbAUbwM9kEZGUAwrjjG2Ea3KC1ZEIyFjrIhJA2QWSDBXKHJn+Fzwp/hjav1uNcXv7rlDkz/C54U/wxtX63Gg+jlERAREQdc/aHD/rNeJQ/+k6L9fp1880O7j9S+hr2h34GvEr/ABOj/X6dfPNAAXlBKBjcqjviKucCB2VqCzpLld4ZAKM+krz5oLWkZVyjH0lINwgqBk4Vrtiq59O6sdknfKCnTt3VCzPZXF2B5K0OOEEzAGlXOHVuFjh7ipGEgIKHZUJxsrndtlaMHcoHUFb09W49VUDKrkj6KAdgjeyfWmwQVRWOJHZU63IJshRuOSjSqOyTsguBCo71VASEzlAa7fsjndtlQj0QDzKCqd1aSd8qnURsEA7d1QjO4VT2QDZBQ7BG9lXYogHcIBhEQUByqqhHoqjPmgtPdBsh7p3QXHcKnSVU9lQElBVu3dSB2Rt9Sswrm7ILgMK4vAVnUqd0F4BPmrhsFaDj0VC8+SC9D2VuSqFxJwgq05Kq/fZW/R7KnUUFzRgq5WZcN1UOJCCWOUQ7lXurWu3HksOYk4UsEDXtJQJqgvGAsfcrNFM12yqaZrQfqQLY3pmaT5kLnDhHD41/pmjclw7LhCmIjlZnbcZXOnA0mXU9GGb5cP5kHNfMNbH0uhWPIIHhZ3+pdHmuBbKcfvivQfmboXs4dRvczAMIOfsXnu1vwzHfAcf0oMcPHVv6q4jO6iOzzhXtJ7Z80F4GVQnpKrkgbKg37hBd3BVOkg5V2MKhdhAJBCtPZDv2/Mqb/NBRUIyqqhKCnSVcNgrclMlBcit6vmqFzs7IKgZVewVjSQVcXE/UgrsdlUDCtBwmSguJwitzlU6nZwEF6pkBUyVac5zugk7J3VnUTsU6iDgIL1Y7+dA4kqju6Dv97FT8KbVP/F/XfyjbkT2Kn4U2qf8Ai/rv5RtyIPalERAREQEREBERAREQEREBERBx9zDf7wPEz+B95/UpV80NSM19SP8A6I/9JX0vcw3+8DxM/gfef1KVfNFUf7oVP+Ef+koLGs2Crjp2VSSBsqA/3SCpbkd1aG9I3VxcFTOfRBQHJwrun5pgDsN1UbhBb0b5yrgMKhJyqjcIKg4UjTlRtIyrgQPRBISFaeyta4+auJ8s90FmcOVetUPZWoJA/Pkq91aANjhVzjbZBQsKuaMK7Ax2VpOEFC8J1jyUZPqVVhBPdBd4efiVroyTlTD6OyAILcbYTq6QQriDvsrHAHGUEbgSchXsTpHor2NA7oLlc3srVc0IKqjlf0/WqOb27oI1Rw81dgqjgcdkFoqQwj5qaOdsiwpY+rGFLTNLR3QZXWqGQBW4VrvP6kEgcCqeEXvyPko2vII3U8btwQgzIZHUscbs4+IL0d5LqWK52KN0oBPhj9C82quT7yzP90CvQLky1Iy3WGNhJ/sY/Qg5319ZaYyzta0ZAK86+ZGb7m3iSFm2ZCPzr0B1BqRtwrqhrTsWnuV5781BH7Ij6mQ/zoOH6mudWUjW9WTjHdaaync3OSVJSH72B6EqdzSCgigj6HZ+amlHUjsgZ7I3ceqCkTfiwSpizAyrI9jnCkJygtb9IKqo36QVT2QWZCqHjCsPmrSd+6Cdr1IHhY7CFIHAIJH9lGqlxKogoG4OVVEQOyp1BVPZWoDTkq8EA4UTTgq8HJ7hBlNdkK8EYUAdhuxQSH1QSSOAz9Sh60e/OVYgv6061YiC/rVC/KtRBedwqBpV7W5xsVd0jOcIIyzZU6VM5mQdlGWkIIgMKqu6PrTAQXA4Kva4YURPoVcw5G6CUDKFmSqtVUFn0VY7fKuJySFG5xHZBadlezsrDur2dkFHN81LF9FWO7K+P6KCyQ74XJ/Jof8AruOFP8MrUP8Alca4ukO/dcocmf4XHCj+GVq/W40H0doiICIiDrp7Q78DXiV/idH+v06+eWE4fn5L6GvaHfga8Sv8To/1+nXzzQ/Tx6oMrIeMBWOZjKvaMbqj+6CMNwUd2yqqjzgIIwdwpR2UI+l9qmHZBbnp7oSCNkf27KxpIyMoKO7YVAcK47gq360F7Xb7+qv6grAABt3VRlBcHdW2E6UDQNwqoLXDbuqN7K89lGdjsgrnfCo7uq/PG6scSD3QVRUGc+aEkILwMKqt6icJkoKHumQFac790388oLsgoThWg4RxGyCjt0AwqA77qqAiE7IDsgtzg/UrgcpsmwQDuEAwiICE42VuT6qowdygod0GyHuiCvfYKo2CYAVCUFw7oXb4wrRnPmqkDugr1YVw3UWSVIw4CC7BVR2VC71wrS4hBIqHbdU6imSdkDq6vyquMb5Vh27eqr1EjCB1Z2Vw7KzACuyUFC3r7eSy4GdDcFYrXdJWQyXOMIJi4MGVG+YHKtkeCN1A9w6TuglicZpWho7Y/SuZ+C10badSUk8pwGuC4isFKampDQM7hcmadh+59bFLISACN0HZzmK17R3rQDKKN7SRD07H5Loo1nwStP75x/SucOImo4KuyiBkhJDe2fkuDy8hrifUlBhvh6HE5VvnlSyS9ZwoHOPVjOyCTqCqN+ysHZXN7Z+aC9Uc3KqCCcZRBYBjcq3r3xhXSKMY8+6C4nCtO6u7q07HCChOFTq+SqmAgp0lM42VUwEA7hBsMIdgm+PmgoBhVJwqDPmqkZycIBOFTqCDc75VcBARD2TO2UFOlV7BUByqnsgo3ujvpFG90d9IoO/3sVPwptU/8X9d/KNuRPYqfhTap/4v67+UbciD2pREQEREBERAREQEREBERAREQcfcw3+8DxM/gfef1KVfNDUn/b1Sf/oj/wBJX0vcw/8AvAcTP4H3n9SlXzP1f9u1H+Ef+koKtcMBUO/bZUaBgHCuQW7nzVQMJkeXdAfVBUHBVS4BW5HqrHZJQSdWRhUJwrWEeZVXEY2QUDzlTNPXusZpyd+yma4N2CCXucK13w7Ko7ZHdWuOe6B5ZVC7CtzvhNj3QXCQnZVByVYMZCvBA7IJRsFY52Va2QF2Fc7HcIIiCThXsaR3QD0CkYNtwgubt3VyhL/JXMeQgkzvhRvZ5qTbuFXGfJBGxpHmr1bkAp1j1CCQDtur2jIUYccDdXsJ9UEiEZREFp2zhRnyVxcMnf8AMonvAKC5sfVspGRhowFSI5wVJkILMAbkq126q5wx3VWAPGyCGT4RnKuhk3z38lWRvfbZRMADsBBkznxY2gDsQu0PL9rZ1gtgj8UNwzG5XWCJhcAPqXLvD6QQ0nSSRlqDtRpLWrr9d5Y/EBJyF1g5sITFqJh9Xkrl/gi9s+oXhpJ38yuLecGIxaghyMZef0IOvtECSPTKznYOFh0I+HOPNZmdwgtkGGq1jtlWXsrGdigvDulV8VRvcAMnyVoc09igl6vknV8laTtkIDsgq7fsgZsqtGSrsH0QWAEFMn1Vzlagva7fceSr1jKj6seauBB8/NBcTtlWeIRsrnnDcAqIEHz3QSh+QitarkFoGUGxVQN/NMfEgvG4G6K5jSR2VfDwgsRXFpCtQEREBVGyYPomCgkYcbK7q+Ssaq5CCQvAUZkBOEJGO6iJw/GfNBkhuW/YrXRnvupG/QCoeyDGIwroxkqj+6ui7oMljdke34e6kjblo2SRuGnIQYLnbnZWOdv2SQgPOCqAgjdBVSM7KPZSM7IKu7K5n0VYSScK9gPSfqQRyDK5Q5M9ubfhR/DK1frca4vk7n61yhyafhccKf4Z2r9bjQfR2iIgIiIOuntDd+TbiV/ilH+vU6+eWEZfn0X0Ne0MOOTbiUT/AHnR/r1Ovnmg+kfqQZbHDpwrXowY3Ko/ugNb1FHt6ThXMOD88KriD9aCAMwVePJCDlMFBVzB07qIt9MKZ2cKzp+SCxzNlaGZUrmnCjwQgvDceauDR3Ctb1E4KkAOEFju2FQdiriO+VTsgsLt0Byri0DyVEFo2Ko74j3Q90QU6h6KhOTlUTIQXqhONkJ2yFTOUFyt68eSuVrgMoKHdU6VVUJI7IKncKn0e4VchUKCqE42Q9lbnKCoHZCMqudkG4QUznbCqBhMBEBETICChGVQjBwqk+hVdiEFOr5Kh3REFepOrZURAHdVDvkqIgkJGEBACsBz5qqCvVnZAceSpgBEFSc+StJwqoQNzhBd1BOoKwHPmhJyguIL9gpI2Fo3VIhk+qkJA7oKPBI2UXhE5Oe26lLgfyq0uAadxug3jw1tgra74twSAuSb5YjSdJj27eS2Hwmq2QVoDjjf+dcvXmVlYWNZgkjZBw7qqWpia5sjiQPmtjvn6g4fWuTeINrmhhMjm7Eei4uYAQR69kEYJJ7q4d1cYi3fCtGxCC5VG4wqITj60AAt3KqZCNkaS44VekIKFwwrHNzuFV2+Aqb4wgoHY2KoTk5QhUQVJyqDZEQVJygOFREFepOoKiIK9SdXyVEQV6lQ7oiCpOUzthURABwcqvVsqIgA4OUO6Ig7/wDsVPwptU/8X9d/KNuRPYqfhTap/wCL+u/lG3Ig9qUREBERAREQEREBERAREQEREHHvMT+D/wATf4HXn9SlXzQVe1ZP/hH/AKSvpg5h/wDeA4mfwPvP6lKvmerf7cn/AMK79JQWB23ZV6vkrBuQFdhAGyuByrQPUK5Bb2VQco3c7q4sx2QULDuqBpKkVWt27IIOg5VwGFIW7nZVEZxuPzIDXdtlVW9ODk+quQWFh3Kp4ZcpD2VucIKdBCdBVxPw5Rjsnf1QWiBwPVlTNjJHdXB3w4Vc47ILQzpKqRkYVgeeshXl3ogsMZJVRGSruofL8iqD5hBUAjCvBCDGPJWoKOiLsKwwkKft2VpP5kEY+HOUE2PkVa9xzhQv+kEGfHIHDJUob1jusSLPQN1kwvICAYDlQzUzuoHKyesqyR5JQIY8NAKtny0qWN3fZRz4PkggeC4YCkpz4Y+JWK4dkFZZA7Ix8lGzuri0nyQNPUAPNBmUhAwCPNck6WlLYMNOPhXG9M3G+PNb803MI4Tn+5Qc4cBK3wNQvc8gjK2BzhVbarUUJb/dfzLd3ATqrNQPZG0k58lszm5pX01/hDwQS7YfYg4Eongjw/QrMcC3Gd8rCpHRxsDyRnPZZXieL23QUduoz8AKkOQCseZ+AcnyQXxD3p3hgkk7LJFtkhGSDv6qzT0TpqsNaMnK3NdKSSCIF7SEG2ZB0bK1pyFWokAeRnvlRtO3dBMw4KvyFC12/dXZKC8tyrehXdRTqKCIsx5qhG+Mq5zsDyVvdAcS5WgYKB2+CqoL2uV3UowcKpdvsUEjSFdjfKiDiMbqviIMlsgaqmUHyUAPqq5CCWSUFuFB1hHKxBKCCisaSq5KCTIQuAUfUVRxJOyCTqB2CDdRgnPdV6sd0F+NlGWEu6lIXghUQTMkGOlXhwIwsdvdSjOAUFXRZ80ZH075V2dkzsgmjnDdiVSepaW4z5LHwc9jlWPac9kETmlzsqnQVeiC07KWP6Ks6fkpGDAQVIypGfRUZPoQrmuJCCGR+CVyjyaH/rt+FHz1naf1uNcXyNBJ2XKHJmP+u34T/LWVp/W40H0eIiICIiDrn7Q4f9ZrxKH/ANJ0X6/Tr556f6a+hn2h34GvEr/E6P8AX6dfPNAcP+xBkul6VaHeIdklaOnZWwkAoJWtwcq1/dXyOw3IVrCCMkoKNOFUyAK048lQjKC4SjzVfFaoy3AyrHIJTMCMKzxBnCiHdXYyUE7MEq8nyUcRI7q8nKCw7qmQFeQMd1C8nKA5+RhUBxsqZCDugu6D3QMJV47Ko2QWGDAyozGclZBeSMeijzk5KCgjOE6FOcdPko0FnT81Y8Y3V2TlUfuO6CPq+SoTkoiCvT806fmqqo7hBXp+adPzV+Bjt5KgxgoIgMKqqe6tOQUFUQ7BG7ndBd0/NU8PIzlSDGEccDZBAdlUHCoe5+tEBBuiDugr0/NUOyuPZUwCM+aB0/NVDFQEk7q7OyCiHsh7KgJz3QG91VMAIgA52RUO3ZUyUAHByqnfdVwE2wgkp3YJVJ3ZOyupwCTt5KlQAHD60FIWl5wpZoC2MnA23VlIQH7rJqZWujI+SDVtDzyQXD4TgZC5foriTOwyOzjC4Z0h/bn2hctWuEzVEbR3IQQ8Ta2Ca2YY0A9Pf7Fwcx3S5c38Sra6G29bgcEfzLhJkfX1bdkEr6hrgAFERndVMRbuAVQE9igqqdOVVXN7IHZUJyjkAGEFuMbkpkeqO/nVhJygE5VERAREQEREBERAREQEREBERAREQEREHf8A9ip+FNqn/i/rv5RtyJ7FT8KbVP8Axf138o25EHtSiIgIiICIiAiIgIiICIiAiIg4+5h/94DiZ/A69fqUq+Z6s/tyf/CP/SV9MPMP/vAcTP4HXr9SlXzP1YzWzj/6I/8ASUETfLKv7qgbsO6ua0DyQUBypA3HmrGgZ7KTI9UFA3BzlVTIKICvYcBWtwThSfCPRBZ3KkVAG+WFcMb5QRkdWw806FIWgIO3ZBEWYCtUr+xwoXEg4QUPdFaCSd1Vxwgu6j9iubIB3UXUCPmqZ+aCUuCdX1qEP3xlDJv3QS5OcglVbLgbqLrBG/omR6oMlsuVV78YUDHYI3VxfnugkM+AjX9XdYjnbkZVzZMfYglkcBsoHuw7ZHPJyrCc7lBmxSjpAU0cmRt+hYLX4x8ip4pWgHJQZPWrJJMH61Z4rVFLICcgoMtkux/1qOao3AyFjCQgfZ5KKRznFBmeKMZ2VzZRjcj8qwOp/ZVEjhsgz/Fb6hXMlaO+PyrTRI7OclV8Z/r+dBrMdS1mNx3W4bbeBFEcOAOMbLZIqCMd1lQVYDcFxHog514LcQ6fSl697qJGgE/viFqnHIO4qV0VdbQXlpBywZXXaS5SR48JxBBznK7Zcs9sptQ2lj6zBOP326DrbfdBVtlpeuWJ4IzkkLbNNIYy4OPbZduuYDTlFb6KUwtYCAcYHyXUJ1NNJLII2k4cc/lQZPjA77brIZbZKxuWNJysSG110xDYoySt66coTSRhta3BPqg0jTtvdbq7xpmnAK1vUVxhnhwzGcYUl48CBhczAB3ytp1Na2RjgHZQaTUSdVQfr9VK1Yzw7xC4jzUzJO26CUHBV4eAFC54xsjXjzQT9QTqCi6z1dwhkG6CR5yFbkdKiLzuVTxQgub9JSAhQh2+cp1n1QSndVBGFD4hynWfVBPkFFB4uPVUMuD2KDJ8TCCXO+VjGTHyQSjHf86DKDs7ZVHOHoscTAnYq8PB7lBK12+48lXqCiD2jdPFB7IJHvDQrPEyrHvBCj6wAUGQ1+D3COk37rFMu+2VTxUGb1keauEuBvusJswJV3iAjIQZonCuFSAMZAWneKcnOVa6Yg43Qan703+6/MnvTf7r8y0rxz808c/NBq3vQ9QqOqGntgrTPG+aqJchBn+N8h+VPG+Q/KsIPPclXB2e2EGZ4w88BVFQ0D6QWnSyEdla2Z2PNBqXjg+YCvbUADuFpLpnE7KWOU9ODnZBnSVDT5hcq8mRzzbcJz66ytP63GuGJJTnbP5VzLyXHPNlwlP/AHY2j9ajQfR8iIgIiIOuntDvwNeJX+J0f6/Tr55Ysly+hr2h34GvEr/E6P8AX6dfPPTjLigvlPw7qyLf8qum7FWReX1oJy7bBVvV6I8gDurA4EHdBVrvi3Ug2UAJDtlks6SMnCChcCMYVuNzlVGMqjjgoLHM2VrdipDuFbgILwcgfUitB9EyUFxJx3Vh7K53ZWZI7oIy34s/NVHdURBKOyuBCi6nDZVBJG6C7OT2VD3VFUbkIJGEuGFUjH2q0bDZV6sjcoLCMlWPGNypD3UUmcoLEV3T9atIwUFyAfEMIeyDKCXsFHguV432PZUOAMhBaG9O+UJygcCcHCEenZBb1bKrWkhUOPLurmbD0QVPZVadsK3z3zhSDGPJBC4bkqikcAVb0/WgtRVDT6KvTjbCCje6rkBMAKh7oK9kVpJKZKC4nCoRncKuMp2BQMgpnfCt+pXADue6B2REQCMpjbCoCSd1VBLTDDyqVP0kicGnPr81SXMhyEFsZIVZCXN38lRrHZ7IWuwe6DV9Mz+DWA+rguZdIPbV3GBuxyQNlwfaXiKpaXbbhczcNp45b1TsB7kIN3cZLU2HTwkDcfBnP2LrLBgF4x5ruBx3pBHo8P6QMx5/Munsbg1zvmUGQ9oxnHksZ2M7LJfgtB+SxnIDe6O7qn1J9aC9FTI9Udny7II5HZOFYrnA7bK1AREQEREBERAREQEREBERAREQEREBERB3/wDYqfhTap/4v67+UbciexU/Cm1T/wAX9d/KNuRB7UoiICIiAiIgIiICIiAiIgIiIOPuYf8A3gOJn8D7z+pSr5oan+3aj/CP/SV9L3MP/vAcTP4H3n9SlXzRVABrqjP/AM0f+koLWt+EbodjhGu7DIVH7lBRv0gqkZQAAfNWufjZBcPh3R0norOsqmc+aC9rjkFXeJ81F1EeadRPmgyAfM5UnUFj9Xw/NVa87oJ+oYQHIyo+r5oH9OyC8OGcK2VpVSW42IyjSHfSQRkYGcqMguUsgzsFYAR3QR9Badz2VFeeokqgb6oLCPPKplXuz+dWYQMn1VQcqmD6KoGEFw7q4nCtB9E3KCw7oASq9JRoOeyC4s2QN2UnkjW58kEPScplzftUj24yQoiSUFetyuGXBWK9gcM4QOoZxhXgAjJUR9fNSROHYnsgk8MO7BRyRFpx6rIY5uM5CpI4ZGcBBjeGVUQk7qXqaFe1zMIMfwj6KhY5p7lZmW74woXYz5IMYgjuu3XK3UltpYGnGG+S6kS4yF2o5Y6uKC1jrOMNQb84q2ObUccsLQXHBBxuus130s3Tk8jJ4iMuPddyqCeirrhKx+MHPddcuYV1HQ3IshIyXbYQbCtM1FQvE8rBg77+SwNRX+GeUGlIaAR2WnVVxgdSABwz0rQJC6dxLckINYrri+pgEZdk+e60QxPBJJO5UjOqPBeDhVfOwjyKCF2Hbeasx0qmcu2KEkn5ILjlvcqgJx3VXDIVmD5ILy/G6tLiTlXFpxuNlb0nuEFwd1DCtdGSVVowVcgsDS0boT5YVzuys3O6Bu3dVacoQCqduyABvhUOyvDT3x3VCDlBa9pxsrRkdyVIM9iFXw8nOyCJmc7qpfvgK8s6fLcq0s3zhALiR3QEgd1aCcq7I9UBxJ81TB9fzquFXpKCHBzhV6SrywjdULSc7ILcEdipWZxurGA53UuWtGPkgsc3zVnT1FX9QPnsr29AG2EERjx6qgjJCyAWZ7hXgx47hBiGM4yqtjJwpyW57hWl7R2wgFu2Mq5rcBWiUZV/iMLdwOyCGRudlRrMjzVznAlSxvaGHJQY7oy3zVzGnCSPBPl3UsZYGHsggcwg91zNyW/hZcJR/wB2Np/W41wy92XnHbK5m5LT/wBdlwl/hjaf1uNB9H6IiAiIg65+0MH/AFm3Eof/AEnRfr1Ovnop/pFfQz7Qz8DfiV/idH+vU6+ean+kUF03YqyLy+tSTNdg7KyLYIKyk+vmomnupX7gjzUeMILgM+alZnGMqFmSVO0YQXFuBlAzIV2RjdVBA7IIX7DCjWQ8NIyocBAAwmd8KqdPnhAI2yVT6R2V5GUDMbgIIxGRunT81e7sqAOx2KCzHVsrmxkZ7q4NwVVBTp+auEeRn86oeyvYT0H6kELndOytL/RUccvIJTAKC4kKioM581XCCmQNkwCgaSd1XGNkEeCCqqvdWnIKCQpg47qjSVVBb0HP2q4bIqEoKBuDkqrvIBCfQqm5QVPZUGTsEznYnZVBwdkF2MDc9lRXOyBnso8uQXKhHdVTyQWKhGVdhWl2DsgEYVEJJRBch7Ih7IKN7qqoBjyVUAHOyKmMdlUFAAxuioCSd1VBQnCyaduRkhY5BdjAWVTnpBz6IL3dIyAAonAEnZXSvHkVEJNkFwy1wLdt1ytwhe92oKYOJPxDzXFJOS3G+65U4Rnp1BTHH74fpQc+cf2Buh2Eb5i/mXSxkTnlxA2BXdnjnG6p0OxrR/8AI/5l039yliZLkbFx3QYjnYaGnyUD3bqryestJ3CtON/VBc1VIz5q1h8lcSewQWgZV2cbKgyN8KuxGSgsPZRnuryTuFYe6AiYTB9EBEwfRMH0QERMH0QERMH0QETB9EwgIiICIiAiIgIiIO//ALFT8KbVP/F/XfyjbkT2Kn4U2qf+L+u/lG3Ig9qUREBERAREQEREBERAREQEREHH3MP/ALwHEz+B95/UpV80FWcVtQcf/JH/AKSvpf5h/wDeA4mfwPvP6lKvmfrDitqP8I/9JQWDcBXA4VjD2GVcgFwKt8Mk5yq49FVvVgoBZkYVvQW7KZqqW53QQdJOyBhG3qpwwhXtjyM4QRCMkBV8Pp7nup+nG2FZIMIIlY47jClLQBk/pUbsE5CC8EgAoDlW5+HA8lVgwgkAwq9IIGFbkq4OIG/50ETtlTurpMHOFaBhBadgVYRndXO/nVqAiKhJCBj5q9pVrd8KpPT2OMoJOkKN5wcK9pJGSrJPpH60DrCvjeOyhUkfTt6oL3nKhIypiMqJxwdigp0/NSs7KMFXMJyB6oK+GS7ZBCRssqINx23VXFgIyQgijiPmVSSEkhXidoTxWnfKCB0JAVgBHn5rKJ6hsoXndBaXEBG77q1+cK6PJbugtf3/ACrmHhLq51koegSdOGnG64ek8vrK1/T9SIYXDJHwnsg7ZcIr1PrK+vpopC4nbuuK+a6xVViv0UcxID3HZb45LZ46nWjg7cE+f1qnPzA2PUtIWAYz5D5IOpbXSP8Ah6iQs2lmbTgB/l6qClZnLilSxznDA2CDKqahk7elnmsMQPHfKQNLHAu23WX4jD2wghbCRuVRwwpnvGFAXZ7oCvazOCqNGVeBhAIyMKgaMYV2EwfRBY5uNwrFe8nsSqAbdkFjhkK36O2FdIfRW49UADHmn0iFa4kq5hI7+SC8nCd1aXeWVTrA2ygvV7XADBUXV8068eYQXPx3CtVQ4E4yFUgDOyCEjGVaquJPmqIJAcHKkZ8WFG0AjKqD0nZBK6MAbK0D1VeokKiC0+eFYYXHdXZHqr2u22wggMRCubCSFf5qVrg1BB4R+aqYnDzU3iD1VrnAlBD0H1VpYSVk/CqEDOyCEwOAyrehw2ysjJ7FQyEA7II3NI3UkcTnt77K0uB7kbKeF7WsxsgidEc4VWROwcFXvcAfJSMcOjYoMR7ekkLmbkuz+2x4SY/4ZWj9bjXDUvdczclv4WPCX+GNp/W40H0foiICIiDrr7QkZ5OOJI/+lKP9ep1889KMyH6l9DHtCc/tOOJOP70o/wBep189FJ9M/V/OgyJQOhQNb6KeTOMFWsbt2QROYe6sLPRZMgPSoh5oLY+6vVA3BBwqoKkYVCceSqTkYUbzugq5+R3VnVjZFa7ugkbsVXrx5IwAjZVMe6Co3VwcA3CtVe6C0j4sqRrgNlXAA+xWHGdkAkE7KisLun7ED853QXqVo+DKiUgJ6Dugw5Bh5VWnAR+C/wDSqILi4d8K5jshWAZ2Vw27BBeqEZVHdkachBaBhD5Kp7q13dBRXN7K1XN7IGd8KqdPnhMH0QGtJO+yqRhVwArX58kA7hWkYVytcgdWdlUEK3ARBeRlBsERAIyrCzcq9UJOUDp+adPzQZ81XBxlBYib+aIKdXyVRumAiAqEZVUQFQjKZKr9Z3QXx98HyV7nEdlYzAO/mqnc7ILZHEhR5UjmOd5K10Tm9wgkhcS5oXJvDStbSXmCQkDBHdcYwnpcCfIremkayOKujeTgDBQdmtcXdl40syn6gcMx+Zddr1Z2QQSO6CNyQuW47rT1tsEUb8kDG5WyNVQdNG/6ig4YqB01Lx6HCsIypK7asfj1UY3CCrRgq9WZwq5cQUFe+2VTp+aow+pV4PmEET9wrenbur5DhWDcFAb3VchUb3Qg5QVREQWgZOFcNk2G6IKdPzVRsqAkndMnKCqo7uqnsrc5QEREBERAREQEREHf/wBip+FNqn/i/rv5RtyJ7FT8KbVP/F/XfyjbkQe1KIiAiIgIiICIiAiIgIiICIiDj7mH/wB4DiZ/A68/qUq+Z6t/tyf/AArv0lfTBzE/g/8AE3+B15/UpV8z9Z/bU/8AhXfpKCJv0grwRhR9kyUEoIyFeHALHyQcqvUUE4OT9qlaQO6ww8g7KviFBm9TfVU6wsPxHJ4jkGohzcd1FJhYvjvAwqeK4oJ3/RUQGFYZCVTrcgv8ypA4BQdR9U6igyQ4d1UkEZCxetyqJDjCCTq+LdCcqLqKdR8kF7jsQFGRhMnumcoCImEFw7KhGVUeSIJGvAVkm5yrSSCqEkoCqDgoB6oR6ILxIDsrHDzVQBsVdse+EFnSVJGNslWgb9tlcNkE0buklRzkkghWF5yVcwh+CfIoItwrm5x3UxY0+StLQCMBAYfIq2YZKvOGjIxlWEgndBGc4CkjyG7qvwE+WVUYHogjk/nWdQyFsLxnGGlYkvT0nGMqxsrmggeYwg7K8l1cYdZnLsHqH6VubnmhfXXemlYC45ByFxvyp36isOsDUVzg1hcBkrm3mJtbdfVdPNZsPbt23QdOxbXQULHvYQSM5Kw2FriQfJcra+0PcNPWUSVEfThuTthcOwveXEAnKCeqaA09PcLFYX/nU4LnHEnZHsaQejv8kFjJC7zVXNLjsrGRuY7LuwKymPiyNx+VBYB0gEqvWFJUM6oyW98bLD6JRknKDJa4ZV+QsWHqDviWTluRhBbI3bKi77KeXHT5KBn0s/NAczpGSoicrIqJWmPAO4WH1EIJGuCvwSCo42Fx81kOic1nZBA1vU5XOid33SDZ5ztv5rNkaAwoNPaDnCo9pJ2CubjxDk7DspukdQQRsjLRlX+WFLI0tbsogTugie1WFh7rILPVU+FBFH3UoGVb0Y3wq5xtkIDnAK3qBBVsjifNWgnHdBXqJOFI3ON1Cz6SyGjbcIKAZVr8jIB8lJt5IQCggaTn/wBquUhYAMqw4zsgoHFXB2B5q3AVCceaC/qCifnzVcn1VpBJQA0lXtBaO6kjj2VkuWlBY7JORlXMc5ox8/VUaD5hS9LcboIngk5XM3Jb+Fjwk/hjaP1uNcPHpI7rmHkvIHNjwkH/AHZWgD+NxoPo/REQEREHXX2hX4HHEn/FKP8AXqdfPTR7SHPp/OvoV9oaccm3Eo//AEpR/r1OvnlhJDtkGZM4YKQuH5FC52QcqPrLSgypHDBCiUbXuLt1KTgAoKZBVC4A7qIuOVUZIyguyChGd1Y0nPmrsuHqgoN0LSVUd1e3B9ECMdJypHOyFYc42VAdt0AblVJwUyD2QgEHKCpkBGBsrc7HdRPPT2VA4+aCrhuSqDureok9/NXt9SgkZ5KX94o24wOyu6sAjZBjPHxH5lVa04wr8DqVdggpjZAceSvPSfROlqC3uCqEEK5oGVV2PkgiBBR3qqDuju/dBRA4Z7IeyoAglyAFUOBGVHlMoLyRhAdu6jBOeyrkoBOSqKgznfKqgHcINgqDOd8qqAThXN7KgGVd2CArgOysBJO6uyUFSMhU6gNiqg+qtcN0FHgHJVnbYq5x22Kt6cjJygqdwmMd1QZz5puSgdSqqFuB5qo7IBOFQjO4VcZTGBgIKOUsW6ja1z1M0dGxG6CWPHmpJOjo2HdY5kDR3VRMDsUGO/ZxK1e0TviPUHYIWnyMa7t3UsRdFG52/ZBzDw9lqLkREXF4zjC1zXNpdTUL3FhGW+i0jl0idcrn4ThnfH51zHxf00YLS5zGYPQf0IOmNe0iulb6OwrA0gZWXc6WWO6TMeMESFWyBgb+lBiZBVzTv04URJDjhTQNDnglBd4BxkKhHSFnuDQzyWDORugizk7eSoQcFWtODklTtwQggAIO6qpCW9jhRvIB+FAIyg2CtyUyUFxGUGwVuSmSguRW5KZKC4jKtIwcJkpnKAiIgIiICIiAiIg7/wDsVPwptU/8X9d/KNuRPYqfhTap/wCL+u/lG3Ig9qUREBERAREQEREBERAREQEREHHvMR+D/wATf4HXr9SlXzP1n9tT/wCFd+kr6YOYj8H/AIm/wOvX6lKvmfrP7an/AMK79JQQoiICIiAiIgIiICIiAiIgIiICIiAiIgIiICqCMKiqBlAxvlVTZEFp7oh7ogu8kBynkgGEBVAJVFUH0QXIiIKEZCqxuEVzSMIKgZQjAyVRh+aud6II3ZOysc3B2KvcHDsCjWl2MoIxnKkAOFd4R/8AcK4RnCCJ7Tjv5q0RkhTyRkt+HukUbh3CDOtV1qrO5ktJIY3lwyQcLvRy0U37K7dBJdW+OQ0bnddCKkFvSM+pXoZyLinq7LEZcbNH6EGl82GmaGj07N4FMGEMPljyXQmkw2Z+fLb869MOcmjhdp2dtOBnoP19l5mFroah7XjByR+dBNVEFvw7KOmeWn4ldK9rmLGb1+WUGfKGys6WgZ+SxxSPDgclSW+nqayTohBO61WWwXSFvW+MgIMNhDWBp3x6o5zCOw/IrJmvhPTKMEeqsEgPZBRwBPw7KPJBAyVMRtkBVa0dzsgFmRhWGA74UzHxjY42V/iRDbIyg02SFzTkk4Vgbv281nTdLgMFY/SEEkAGw/mWQ97WsOwWLks3VxeHMIyghj+KUgLNkb8JCwaf+y/aVnvIQYDWESfapg05V5aATlNvRBdI7LQPkoB3Ur3ZGPRR4KCpPUMYUfQc/apS0AZ88KxAP0cKN3dXO7bFWgd8hBb0/NULdlIe3ZWHPkgo3Z2/qpx2GFCWkBXscdgguGxIVr3EOGFft3VrgHIKeJnbCtV3SEwEFo3Ven5qpHT2ygyQgp0/NUOxV6jcfi7oJ4XAqGo+mpoQAMqGo3ego05VS7AVrOyPz88ILety5k5Mzjmx4RfPWdo/W41w0uZuTP8ACx4RfwztH63Gg+kJERAREQdc/aG7cmvEr/E6L9ep188sZw7K+hr2h/4GnEr/ABOj/X6dfPLGMuQSkZVjj0nBV4z81HJ3QSM3KlDMhQRu+ayWkY7oIHMwUAwMKR3dUI88IKdIVCMHCNcScFVd3QWYKq3tlVVhyDgA4KCQkK0jKeWVTqKCmenzVQ7I7Kzdx3KuGyC1zcpjGyuwfROnKCzpxurh2V/TtuFacDYIKglC7CoqOQOoKhPzVDv2/Mg+v8qC7qKuDvVWYPoqgbIBd3TqOCCqHurTnPmgpnfKE5REFx7KjeyqiBnfCKh+XdAdt0FchEwFQnHmgqdwqZLdiFU7BU77lBUjKDYJkIgK7IxhR5PqrgdslBVvdXK0HCuG4QFXO2FRUOSdsoAbv8lf0+WdkAwqoKdITpGPmqq0nugpjOyoWkFXN7qu3yQUIx5oBlVTOUF0Ja0klWTuDjt+ZUfsMgKM5PrugNaXHCvdE4DOSroGHqBU8rT09kEFNu/B3+tZcxDYXjHcLFpceJv8lkVTh4ZAI7IOZOWasZS3olxAHWuxPFOvp6u1luQfg/LsuqvAqpdDeyBnHUFz/rF9RW0gjjySW7fkQdY9W00bbtK9rcAkraU7yCR81yDrKw3KmqpZp2ENOTk5XHdRnrIPfO6CPBU0GR3CpG0OwpOjp3QTFxIUEjCe6u6z6oD1EBBjiPKru34VO9nS3I9FF05ByEERyN8/nVFc4HOAFbjCAiIgIiICIiAiIgIiICIiAiIgIiIO/wD7FT8KbVP/ABf138o25E9ip+FNqn/i/rv5RtyIPalERAREQEREBERAREQEREBERBx7zEfg/wDE3+B16/UpV8z9Z/bU/wDhXfpK+mDmI/B/4m/wOvX6lKvmfrP7an/wrv0lBCiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgKrVRVGN8oGN8qqHsrclAPdERBUDsqoPJEA9lRqqqdjsUEioThVGfNMZQWndVZ3Veken51cwfLzQXhUd9Iqp7KgPmfVBU480BH5FQkY7qgcO2e6CVrw7ZXYHorWdIOykbjYlAZ0juArsB30Qoqn6PwK+jfgYk7oIa4YLPqK74cjM72WZg68YC6IXFzSWdJ9V3R5NLkaS0N3xhqDljmekM9qqGvd1jpO2fkvN2/MEdwqA1vSOs7favRbi/BV6np56akaXOLSF0K4k6brNP3WSKrYWlzz3+tBszLj6rKp48tOW5zurqeFr4gdsrcFntPvErWADJwEGZoCni9+HjQ5GfRco3ynt/uOWxAHp9PktDtFgjs8QqpgAO6yrjfrfPSOY2QZGR3QcUanDG1b/DGBlaFC4h/c/lWt6kkZNUvcw7eS0OMEPGyDUAfgCo7ZpOUZ/Ywqu+ggwetxkIye6rh5dnJ/KrTE/r7eakDH5CC8hwGSfzq0EdeypIXBuCFFG89YQZEv0M/JQZcFkyEGMdlYAwtPqgip/pjdZhOMb/nWDhwcejyV7fGJwUGQ/YZVgm6UfHKG9juFCGOzugmb8SvJDRgo3GAAquAKCPOUwVVjSScjZVdt2QR46TuhGdwh3Gcq3J+xABycK7p33QADfzV3dBa7thWt7KRzcjYKPpcDj5oK4zsqfR2PqrtsfPCt3Jz80FSMIrhuFQj0QVO6Y2ITIRBb0nJUUgw5TZco3tc53bzQXx/RUc30vtU8TNv9ajnjPVgII2dkefJSMjI3KpIwZ2CCFcy8mX4WPCL+Gdo/W41w5gLmTkz/Cx4RfwztH63Gg+kJERAREQdc/aH/gacSv8AE6P9fp188sZw5fQ17Q/8DTiV/idH+v06+eWPHVugk6tlG/upHbDKiduUFY/pKcO/98qCP6SmQXIT5KhcPIqzJQVafiyjnb9lRpHqq7HdBVUJx3VvV80zlBQHKqreyZKA36QV2NwrW/SCuJwfkgqD5KVrMhWNb6q8HG3kgo8Y2URae6nkALRjurG5wcoIs74VHuxsriBnZWuCC3cboNzn0VSMqoaB5ILlQnGyrg+iFvyQWdWShOFd0hUc0bYBQWImMIgA4OVXq+SEbbKmD6IKgdkPdVHZMBARUBVUFM52wqjYKgByhz5IKHurh2Vv1pkoLkVuSrgUFAMK8dlaq9WNkAnKq048lQDKuw30QXA5TIUfUVaScoJuyodzt5qzqKZKCQDG6Y3yrOo+adRQXuBIwqD4dlb1HyVjnElBK4jt3RrQcHCsbucE7LKZ0BuMoEQ37KaVo6MZG6hMjQNlaJHOIx2QQR5Dz5K6YktySr52dA6h3WOXEjCDkbgxIIr3kuA+Id/sXZps1NJLF4jmkYGey6iaHuotVf4uSNwVzHbuIELqiPrk2GM7oNwcYoaAW17oYmglp7BdXKrPjPGP3xXP/ELUtHdLYWwuBJGO/wAlwXVUT/jkx55/OgxKf6WVlvAwCP0LDicWOIPkpHykt2QUcQCqM3conFxOVVjj1AfNBmPA6VjuxuppHEx5WIXElBIQPPdRO7qQ9WFG7ugoiIgIiICIiAiIgIiICIiAiIgIiIO//sVPwptU/wDF/XfyjbkT2Kn4U2qf+L+u/lG3Ig9qUREBERAREQEREBERAREQEREHHvMR+D/xN/gdev1KVfM/Wf21P/hXfpK+mDmI/B/4m/wOvX6lKvmfrP7an/wrv0lBCiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIBlEBKC5EyFQn0QUPdERBVv0gqk42TA2KbHdA7hVa3uVRVBIQVyFUHKsVzUF+x2VzRgK04b9YRr8jCC9WvBKu+eVa4HOyC3BVOkudlSBpdgEKRkZxlBYGEKVo2wrhGdlI2B2NggsDBjfdRSxPLvvbDhZQikcQGg98bLclltNPNGDOBnHmEGyp2SDBc0jK7fcqs3ulnHW4M+DO+y4VtenrLNMG1ZaGg53wuQbZqy36SpjDbZBgNxsUHbjh+2lvl+mgnxKN9hvldSudexQ2jVDPd6UxsLzk4wFzjyg69pL3rItr5AQ52Nzt3W7uczhvZ9R3KGamDHZwdkHmxa6aeRgLad7hnIIBwd1v/AEtRPZUxGWBzQcbkLsHovgfaHMYyWNmw32UevtDWjS9O6aAMBZug4x19PHT6fHgODXBnl9S4Op7nWPMjXTEjdbv19qp1Q91AxxLQCO64+icWvyPNBnSl0pJecn5qIRDPbsVkRgdIJVHPjaM5CC1vbCuDsbeqiEzCSGuCvGNslBJ0s9FUBqtccDKhe92DgboFSGlpwsRjcPGVIC8u+IbJhrngN9UF0jSG9lAC7OPmtUdRSyw5Zvt6KKO01PVu0/kQQ0rATkjO6zmRt6x8IU0NufCMvbgeamYxnUDtgILZo2CL6IWA6MYccLV6t0LYvLIWl+LGWkZ3KDCYcyY8lMdgogwiQnyU+M+SCMvAVHfEDhHtAyVRvZBZ0/NBtsqqn75Bf04Gcqg7qQDITpHogDsrH/UpOypgeiCAghV6h6KR7dtlY1u+CEFACDuqqTAwrDhBZgtOU6wq9yrXDBQXB4KuAB3IUIxlSMe0bZQTx77KyVvxZKkic0enZQzvAdgIJMAjZQyNLfyKRjsjdWy7jZBjO/nXMXJp+Fjwh/hrZ/1uNcPuA9FzDyafhYcIv4a2f9bjQfSGiIgIiIOuftDvwNuJf+J0f69Tr55GfSX0N+0O/A24l/4nR/r1OvnkZ9JBIeyjd3Uh7KN3dAb3UncKNgyVedsYQUaCT3Vyj6inW5BeBhVUfWU63IKorclMlBcioCqoKtOSq5AVjTgq7Od0EzewKqojJhoQSbIJOvfdCRhR9Q9U60FDsc/NUJyhOVaSQgub3VytB806vmgkyEJ9FEXfPKq05CCuN8qqt6vmFQvPkgEZ8/NWkYOEyUzlBcmQETAKAqE4VUwEA7hAMIiChIQEYVcBMBBad0Q90QEG6IO6CuOncpud1VEBVAyqKoJygohOFcR6J4ZIygtRVAyq4CC1Ff058k6fLCCxFUghVDcjsgtb8XZVPU3ufJTU7W5y7sqVHTk9KDHySpoHgOAKiDS7srvDkbv0lBlVQHh91hK90j3DDldG0FAhlfCephwVnQ3SsDh0yEELELWK+IMG+eyDcFJV1tUA2WQvHosyqoc0pd4RBxnKyNFULbjKGAZJOFybcdDvjs7pwwYDcn8iDrxOwsnc3sQVVjfmsq9w+73OWIDHS7CxotzugOaAPJRN+mFmuYC37FiEASDCDJecMWJnLht5rNe0GPKxS0B32oJfJY8n0lMXgDv5KB5yUFEREBERAREQEREBERAREQEREBERB3/9ip+FNqn/AIv67+UbciexU/Cm1T/xf138o25EHtSiIgIiICIiAiIgIiICIiAiIg495iPwf+Jv8Dr1+pSr5n6z+2p/8K79JX0wcxH4P/E3+B16/UpV8z9Z/bU/+Fd+koIUREBERAREQEREBERAREQEREBERAREQEVQPVVwEFqq1CNuyuYO+yC0jJVRsFdjf5KuB6IIj3RXYGUwPRA8vsQDCoSR5qmSgqd9gqt2TbumR6oLj2RqoSSNldGNiSgteVSMZKqWOLt1UNIwBlBIB2UsY9fkomjOFkRt2zgoK9IV7eytGcnujnFoQTswO+FIJGN9Fp7pXAnB7fNWmZ+DufyoNZo5Yy/fBC1b31sQAY8D6ltOlncx5Pz9VfV1k/UOjOEG4K+4VDWZhmIP1rFguVcYn+LOT8JG60eCpmkd0yZwp5puiJwb5g+aDn3lGr679lw8GXB8QbZ+a7f8V5ZJpKcVjy4kDufkunHJpVQRa6DagjHWO67g8bKuF9dRikILTjKDaXv0Vnp/G6+gdOd1wjxa1q24U07GVId37Fb0423WezaTFRBkO8Inbv2XTeu1Zca7rEziQ7P77KDT7tO6or5XvcTk7LHbs/dCHTuLu5KOp5gckFBmNd1sDW9/ko/dJZT0AEknZTWegqayoDGZIOy37pzSUhvNM2rA8MuHVt5INkRWCsaBK+neB64OFM62y4yI3LtxfdF6Op9IMliMXj9APYZzhcQ/cO1mnlPw5GcIOHZYnMyCCMeRUTRjutf1BbJIapxYPgyfyLR/dyGOPmEEM0bXNw0DPyWPS07veGgtPf0UtGyWWcsPZa7Q2wm5RNI2Lgg1e2UAbTh74SR6kLLdTwDDvD7Le09ooqaxskGOrp3W2T7uaR7sjIyNkG2bvLC2MtZgHG60Fk3c9QVl8rZPe3xtOwPqtMFRINgUGdVSveDh2ftWA3qD+5V7JXPOHKfoCC+PHQNt1VUYPLyUjsdOR3QQydz9qiVs0hycKLxCglBwVUblWs37qQAZGyCQdlcBndGduyuHcIKHcK0jCkPZWoLVa5oPZXKjicZCCM5G2VRWOcclOtyCp2VO6pnJ3V4AwgheCPyq0Zypi3OVe2IYygtjDh3P51SQHqByriSDtsqE574QVac4VSdjn0VoPoVa9xQUA37rmDk025suEWf+GloH/K41w+CQBhcwcmm/Nlwi/hpaP1uNB9IaIiAiIg65+0O/A24l/wCJ0f69Tr55GfSX0N+0O/A24l/4nR/r1Ovnkb3QXu7KNXOJwrUFW/SCqqN7oc580FEREBERAREQVb9IKqt7JkoCIiAiIgZPqiIgIiICIiAmURAREQEREF2QUVucJkoLshU6gqIguO4QDCtyUyUFyK3JTJQD3RVIHdBjHkgoiqB6quB6ILVcOyoB6qqCnV8lUbphp7fpRBI1XKzOFXJQUI6e6tO5281V7sj/AFKPJQSg9I3Vc5OVF1EqoJxsgveRhVZ2VjT1d1cDjbZAkJaNlESSd1fISQN1aBt2QTUgBfghZ0zWBucBafE7oOfmpHTl2B5fWgjdgvONlTONgquAG47q3ugtcT6qRpxGdz22UWcqvUQMIOSeEjh78zrPd/Zdl7v7mdKP+EZ8P+ZdU+G1U+G4Bo7B4XYSsukj7AYzndmPzIOsGq2dOoKkN7F60lrul2Vr+qYXG8TSHuSVt97S0/aUGV1/D3HZYxI8UEK3rdhVa13UCgzC4eH5rDfu7ZT9fw4UYA6u3mgsLHYzlWEYWaQ3p8liyDdBYiIgIiICIiAiIgIiICIiAiIgIiIO/wD7FT8KbVP/ABf138o25E9ip+FNqn/i/rv5RtyIPalERAREQEREBERAREQEREBERBx7zEfg/wDE3+B16/UpV8z9Z/bU/wDhXfpK+mDmI/B/4m/wOvX6lKvmfrP7an/wrv0lBCiIgIiICIiAiIgIiICIiAiIgIiICIiC4dkQdkQFUHCorgBhAJCN9VQ7JkoDvpFWu7q4gnOxVhz5oCK7AVCPRBXyVoBKuxtgqrcAdggo0jIyFKzuo+nfz7q9vqgnDAd8J4YVofgYyhlwgvDAFKzsoBMD3UzXDG2EF6jeM/lV3UVHI4g/lQWubjfKswPRUkeQO6pG4kHKCr3dAyB3WVSlszTkLHDGvIDllRM8P6KC58bWgkbLDnds4ZysiqMhacZ7ZWnvL9w7zQctcutwfb9UtmZJ0HrG+V3Bvlxmus1K45lO2MbroRoa+Gx3JlRkj4wvR7lv0m7iTSUtQ1vUOkE53QcM8wAkGlC18RaBGe4+S6TNGXyYGcZ2+1eoXOtwudpzRUroWAObEfLzwvNGxWeorKt8ZYR07H68oMK3NkfMAIi4Z8h2WvOo3lv9rHt6LkrQWibW2USXMNAyDuuRrvpvRMdJmJ0WcY8kHD/C6wQ3C5Bk7A0dXmMLmHUWlqG0UQqoZGh7W5zt6Li2+Xqk0rUmW0kZGT8JW3avi3errG6mqHP6Tt3QatdNb3F9Y63mrJjBwBnZQ09xkdUCLxxhxHmtjV1SHymqBPWd/tWPHe543iUZy3cIOUtSWqEW0VLcOcW52+pcfxxOex4MR2zstwaRvtTqCUUFSSWjAwTst7v0hSU9RHlo8M4J2QcUWymf7zvTkDPmFuelpMVkbvDOxHkuS6zTOnKegD4CzxceWNlt9tBBG10pAy0lAv1QY7SG9eMN9VsaOrPu0o8Rahq69dERhadgFsVl2eGvGTugxrl98q3b5WMI8Dsq+IZZy4+ZWQ5oIGyDGjbv5brIAJ7KxrMO+1TNxgIKO+HcqwvODuqy53x6KLDseaCOY9yo2jJUhaSTlWdJDhsgyWNyBhX+GRv/ADowAN27q7J7FBZ1Y2Vwkx3KskwBt3UOXeiDK8X5pnq3WO0OzuFIOoDZBc/sqBwxgqnUTsrXZCCw7nKqmD6KoG3ZBQbq4DyTAV7GklA6Fd2bhXYwFaQD3QQHclWkEqdzQBnCswPRBEB091d3+xX4HogYO4CCwDC5e5NvwseEX8NbP+txriQjywuXOTf8LLhF/DWz/rcaD6QkREBERB1z9od+BtxL/wATo/16nXzyN7r6G/aHfgbcS/8AE6P9ep188gQHdlRXHsrUFWjJVTscKje6ucCdzlBYiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgu7hU6Sq+SICIc+Sb4+aAioCSd1VBQDCrnfCKn75BImd8KhOEx5lBQ7d1YTkqQjqGFb0ILVUEKvQqFpB7IL2pjfKozKuQUd2Vo2CvO4UZJzhBVOrdDkBW+aCQ7jZVa04V8QacZ9Fe4AeiDGc0j7FappAMbKHBQbv4eY+6Q3x8QXP0gabKcOBPT2z8l100dUGnrQ4f3QXMtFeJKmBtOCdxhBxbqmH/qlKeg4yd8La04AB2XJ2s7I+BjqktG4JyuLql+7m9vkggYOpyy2xjGVjQAFyzwB0oMOQkEqMPwVLKBkqENJcgvdIQFETk5WV4QLclY8jQ04CC1ERAREQEREBERAREQEREBERAREQd/8A2Kn4U2qf+L+u/lG3InsVPwptU/8AF/XfyjbkQe1KIiAiIgIiICIiAiIgIiICIiDj3mI/B/4m/wADr1+pSr5n6z+2p/8ACu/SV9MHMR+D/wATf4HXr9SlXzP1n9tT/wCFd+koIUREBERAREQEREBERAREQEREBERAREQXdggOVTJ7INhugZ3wrgceSpgeiILsfNAMKvkEyPVBQHKo9XYCoQD3QWjsiHHqiAqgEquAq4+XdAA8le1vzVikadgCgtJwo3uOVO1gJ37FUkjBIwgxx1BZUJPScq3oGM5UkYAGyC5r98K2R2fsV3SFFKcHZBa7cYCN2G4QHtlStAcMhBa1ZsGD+lY2AFPE7A2QZccTHkgrS7nEGSfCNsrJlqXRAEKsDW1ozIg0kOc3dpIwvZH2XNBSXbScM9Xh7mRA4J7kBePdygZB0Bnnlel3s5uJEmmtMCLqIAjx3Qdm+fvT9DLpOcRxhxMRJaPLZeTmm9OxQ11U58HRiQ9x816X8ZuIH7PDUUFS7LcEb7+S6PcT7LFpqKpnpAASS7YINhaweLXby6knDHYzscFcV/sqvErXsfWPIGcbrHvmrrjcKt9PM49Ad04ytN6WsJ37ndAq7nU1EpE8hdv5lQ+K0dgMn0VKuNoaXN7rGiDycOygyHuc75hRFhwThZ1GxkmzlqLqKF56B3IwgzeGRP3TAA/fDdc33iIi3iVoIIbt+Rca8M7CYa4TSD4eoHsua7rBSSUAYMY6d8fUg4ThvFc66uhkld0B2MEla7WTNFKS2TuFpmqqSnttU6ohxnutsO1BNJG5pJ2yEGl6pnc6Rw6sra4cRnfutUutUah7j5laa2IuQXQfTWWfJQtjLNwpWnI3QVG6uAwreyZKCuztlUgAbKje6rkeqCAgAlRvdh2MLI6QngtO5HZBZH81eeyuEeFUtHoggxk7K4M+SkDMHKuDST2QWhox2VCMdlM5owoSd0Fobg5yjm53UgHbIVHDA2QQnZWk4V+MlWPGCgqpGds/UoS4gKWIkt3QSZzsqEYOFVvdHd0Fj+ysAz5qTAKtdkdkFiqH9OyvDR6Kjo8koLM5XLfJsP8ArsuEX8NbP+uRrilrW4GSuV+Tf8LPhHj/AIa2j9cjQfSEiIgIiIOuftDvwNuJf+J0f69Tr54wMr6HPaHfgbcS/wDE6P8AXqdfPI3ugqeytVXdlRBVpw5XE5VrBkq4jCCxERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERBd3CDYIOyoSQgqThPJMZTsCgtAyruwVAqoAOUBwUAwmN8oLj2VM7YVQSSqkAZ2QWt7qpOFRvdXEDc4QEVASTuqoKE4VQcj60xlW7jbKC9gwTuqPG+VRrj6qUDIQRZyDhWlhKnDBnZC3CCyMH1Uh3UbndIVpl9EEgZg91RzR0n6ldTu6jush0Y3CDK06C2YHzyuUtMzGS4xRE5zgLjG1Dw5A4bYK5A0BUMm1RTQvPwlwH50G9uJluENi8UREfBn8y641WTK8+QcV3Y412Ojh0NHNEB1GLJ/IumM0A635/uigxacEnssw4a3CoI2sGVbLIOk4QQyDqJOVRowe3dRteS7ClO2+N+6CVw+HY+Sw5fpKrpnZ2OyjJygIiICIiAiIgIiICIiAiIgIiICIiDv8A+xU/Cm1T/wAX9d/KNuRPYqfhTap/4v67+UbciD2pREQEREBERAREQEREBERAREQce8xH4P8AxN/gdev1KVfM/Wf21P8A4V36Svpg5iPwf+Jv8Dr1+pSr5n6z+2p/8K79JQQoiICIiAiIgIiICIiAiIgIiICIiAiIgu7Kh+LcKvlumMILSCFe05Coqj5ILjuFb0lXHYKgOUADCE42VwG+6ODRhBGW758lUDPmqg/UmcfRQWg4KlHZR43yQpARjugoPpEK4sz2VAB5BXjPmgkDcgb+SoY8DKMec7lJJD2QRn4TurmuBUbnEjcq5nbZBKRjzUErt+ylJcdlE9uSgtOSBhSREgDI/OrA0+imjZkZI3QUc7fZZFMTjso/ALuwWfR0T+ny3+SDArCQzGFZRzvjBAaVrbLWZ3YPr6LWqHSIljyMbj0QbIrZXTFuQds4Xa/la1pcNP2UxRwvADTg4O64l0rw8guNayKpAI68bjyXoBy88F9GRWeP3xsQJjGdh3wg41tmurje71LHJC/BJ3wVx/xrZIKKXMZ3BXdUcINGUlfJLQeEXbnYDuuvXMNo+mpaGoLQNicIPPC8R9F2cenGTk4VhccfUtV1ZSCC9PbjYE/pWluaMHZBHE7xX9BWQ6maGny2WLEOh/UFNLUP8Mn0CC2F7opPhyd/JZttqZ5rrFAYyA5wCydD25t4uggk3BIXLFRw5pqF0dZGB1AZ7INxWiwRW6xsrmbOLQ44VjrrJLTODnnDeywY73UxQCgmz4Y2+xSVE1Gy2yPYRnBPdBx9rSry9xMm/wBa2GKh2HYI3JWXqe6TTXJ8YJwCfNaNlwYe+6CoJllIO+SpxEG42+axaTPi5+a1GTsPqQY0nwhUj3U3QHlXCIDGEETgQ3KiLhnZZFQOlpwFhNcSTugme7pGVY2QuUkjQWZUQAHZBKwHOVIO6sapG4zvjuglDdhkqhZ6K4dkQRluFTIHfyVS76lY7JcMfmQXuIwVGr/JRnOfNBcBk4QjCoNlXcoLAMnCslHzUuwGR3UUhz3QQeakjPTj61QtHcBSRtGUEjd1VXBoAyB5KxxIOEB52VjdxnCuO+xVWgAeSCiK7AVDhBYGkrljk3OObPhGP+7W0D/lka4pHfK5W5N/wtOEf8NbR+txoPpCREQEREHXP2h34G3Ev/E6P9ep188je6+hv2h34G3Ev/E6P9ep188je6ARjdUVXdlRBVn0le5WN7q7fzQWIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiIKt+kFVABscIgp1KqocKo7IB3CYx3VBnPmqoKZzsgGE/71VHz7oLwfJWu7qoyqnBGcILW91f07fWrWDfspgBgbeSCINIVexV5GydI8wgsd2VnSVI7A2KtQCcKRpBACicr2IJWjzVHIM+SvAGxJQYkgO6sWXM1p7KAtAH2IJaY5Kyw35rAjd0nZZUchyMnugz6Y+GQc+i3PoCs6dWUx6sfGP0rZkkzmjOfzrL0vXyU98hnbnII80HcDi3U+PoWNrZA8+F5H5LqBVxOa57ukj4juuwtffJr3p1lLKSR0Ad/kuL79YY6emfIAM5J7IOO5HbkZ7LFe856VdVOLZ3tB7HCsjBdIAfVBcxpByQpHnZagaVohBWFMzGwQYR7or3NCsIwUBERAREQEREBERAREQEREBERAREQd//AGKn4U2qf+L+u/lG3InsVPwptU/8X9d/KNuRB7UoiICIiAiIgIiICIiAiIgIiIOPeYj8H/ib/A69fqUq+Z+s/tqf/Cu/SV9MHMR+D/xN/gdev1KVfM/Wf21P/hXfpKCFERAREQEREBERAREQEREBERAREQEREF3cIBhUBOQqoCqDhW5OftVUF2c7BVAwgA2wrhjzCCqseMqwvcNsq5juo7oKdJVR2UrA3O+FJiPBO2UGOdwgGFQl3UR5ZQnCCRvkricKNhKyGNa5u/fKCMbqyR+CqzdTT8AOFJBCJG5f3CCD6QCmhjwM581SaMsJDR2UTDLkAZP5UGVgeijkJBwG5VzGSOAx3yty2W0wVDAZ8Zx6INsN6+/QfyKviyNziFx+wrfrdOUriA3Hdb201oC01cOZ2syfPCDiPTdI+61IgdEcZAzhb+/YeKUNwzOQFum4aTtmnQZ6UMBG+QqUF1hqwRJjbZBots0kJnt+9nd2F2S4UculJqmiZLLI1hLc4JXD9or6SKc9RGBus26cyeoNCObT2Z7wwbDBQdgLny3UGlS6pbUMHRuCCFhR8QjoiB9NFcwBECB8QXWvUvN1rW8UnTJJLlwwclcTXfilqC8Pe6oldl5JPxIPSLgZxPfrTUJpZbkHhzsYLlqnM3p1kFrkcxwcXAnbzXRHlo4lXOya0YfEfgkEYPzXcXiFrWfVlHDBMSepoG/1IOgPEK3vp708mIgZP6VtFoBBK7B8cdHNoKJ1wDACWkrrjTyvdIW+qDIDcO8gqT7QuI3PbZS1FPIYsxjdSWK11NdcY6aRp6XkA5Qby4E2p921ZHS+GXAkb47LuDe+GDYaOJxjOC3+Zce8GdE2rTEcd6lDA/IduADsubtTa/tc2n3uY5vWxpxj6kHWXijY4tNQOlZIAQCVw7T6zmmhkhMpwCQFqnFvXtxvN7mpHOcYskd9iFxtCxzX5Gd0GpVwFRUmY75OVjOi+A9lMTiPPnhQ9RO3qUGNTnEv1FZxdlRthaCHDuVegvjGT9qlDBkbqBri07FXeK4oK1jB0EgrSWu6XYWoySueMFQinaXZQXn4mBWBmPNTys6I8jusYOcSUEre6uAzurATsVcHYHkgl68BC8EKPKpn5oBOThM4TG+UIBQXEghW5ARCAgK4dlB1kHugkII/1oJXeahf3UuSftVOkHughUkY3CuLGjyVWAZ7ILwc7KjxncKUMb3VS0IMZVAyrnNAJ2+pY8j3NdgdkE+QVR3qo2PJPdSgZxlBaO+FytycfhacI/4a2j9bjXFvSFynycD/AK7ThJ8tbWj9cjQfSCiIgIiIOuftDvwNuJf+J0f69Tr55G919DftDvwNuJf+J0f69Tr5484QD3RN0QVYQDuricq1gyVcRhBYiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgu7KnUFTJRBXpKqNgiICHsiuABCCxp8lUAkqvSB2TceqCp7KzO/ZC53YoAScoJW+qkyFB1EJ1kg4QT5BRY7XnKv8RyA/4uyo0EDCp1FOr6kAnKvaMKM9kD3AIMqNw7fJVJydisQPf5K4OcNygnc3YEqJ/ZVD3E4PZCMn5IImfCd1K2YNIPorXNGNggbkHIQXST9YAAWVYnBtyjJ9f51iwxNLt1ksDafMzO4GdkHYPTLBW21jGkOPTjZYur7C+K2SSGEgAE9lpnAu8OuFaymqT8IdjcrnHiDY6J+nZHQ9OSwkfkQdIa4dFdK3HZxCtp2vdMD4ZOT6LW6yyzu1DNE5vweITnHkt323TFGIhJIG9XzCDaroyYBkEHC0qduHY+a3tdbcyJpbGBgBbNr6Kpa/LQ4jugwzCAO6x5AAdlefHzg5z9qtLHnvlBYiqGEnsVKI/h37/UghRSRxFz8Hss1tI3w87INORZYgHUc9so+GMN2wgxEV5j3KtcMHCCiIiAiIgIiICIpY2tcMnZB369ip+FNqn/i/rv5RtyK/2LDQOabVP8AK7+UbciD2mREQEREBERAREQEREBERAREQce8xH4P/ABN/gdev1KVfM/Wf21P/AIV36Svpg5iPwf8Aib/A69fqUq+Z+s/tqf8Awrv0lBCiIgIiICIiAiIgIiICIiAiIgIiICIiADgq7urR3VyAqgEqiuHmgux07+ir3GyHsg2yEEbvNW5LSpMDKuDW53QRmRxHfCt6nf3RKy2QM9Ff7uz0QY47Z8ygaT3U/htydla8Bu3zQRgdOE8TBxkqjnZGxULu6DUI4w9uT+ZWyS+Celo+xYrKiRg2KzKKAVRBed0EfWXD6JK1CipWvG7PzLUqC1QvwHALPlo46YYZ6boMGgtbJXbgYWoyRe5ODWOxn8yxDVPpSXM9FpVbdKiaXzQb7swE+A5xK33bXtpmMb43T9uFxTpSvndKASdlk6p1TW0D2iInA22Qb+1dWNFKXeODt6rj2nvkkTnASYWhVmq66tjDZHEgj1WlsqnkknO/llBvZ+ppoQXCXG3qtq3u+yVshMhDjnz3WHNO5zT39FCymZKOooL/AH4TARlox9SslijETiGgHGRhSMpY4zkY2UVU7YtB280G6+Eld7hqJk3UG7jf7V3EsVzbd6ikiM7XZwNjldELVXy26oE0Wc5HZdhOEOsK2ovFI2QkgOHz9EHMfMrpn3XRJq4oS8+GTtv5LonS+KyVxbCXHB7A7L0813b6XVmhWw1IG8fTvv5LrWzgvZYGyyZYSSf3qDgnR1tN4qBDLCRkjuuWrNw4pYrlTvLQMkbqN+nKTTdWXUwGQc9lZXayrqWEzMO8fbb0Qcoa8jj0npVstNVNBDM46guAWcVbjVQTUz6skAkd1o+rOKd8v5NtqXPMQ+Hc7YWzI6dsRc4E5dug1C8Ti4Vbp3YJJ7rBbC1vorPEdnCq+RwYSgTHpbjOVjiTft2VkUrny9LsYWaYGZGEEfXgKhecdlkeEzGMBDC3BIGyDEZIS/fKl6gqmJrTkKmAgdPqqhoyrJ3Fo2UAmeBk/kQZsrQWYz5KDoHkoYKhz3hruyyiBtsgjAwhGVJgeitI9EFDuEGwVcKoAwgtO4QDCrhUz80DIKOOB2VucKvcboIT3VzG5IKvcxvoqtAAQVGwVCMqqoSQgormd1G7ONldGTtlBktKqeyjDiPNV6yRsgsk7/Ysd4GVO477qxwGUFjI/NTN2xlWA42GFRzigmBBXKXJv+Fpwk/hraP1yNcSeI4DuuWuTXfmz4R/w0tH63Gg+kJERAREQdc/aHfgbcS/8To/16nXzxgZX0Oe0O/A24l/4nR/r1Ovnkb3QVPZWqpz59lRBVv0gqqjBkq52AgsREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEGyIguO4QbDCIgoBhSeSsV47ILR3V2dsq0DKu+SCM/EQFUbBMBEA7hBsERARD2QbhAVHfSKqmxQBv2V3hZOcqsTR1dlKgswB3QY8lf05HZVbH9XdBYB6K4DAUga1qHpwggJTOQryzHcJgeiCwEs3VHVBLS312UzIw87hVnp2siLh3G6DkTg1Vmlrmua4A9a7IT1El3pG0riSHNwV1Y4WzFt0DPIPC7Q6ee0zwjHog2Hf+G8VNK+t8HBO+VsG6ZoHljXHbyXYrXbzHbHObgfCf0LrHU1MtZqtlJKfgdIGn8qDLpoZa1wDoHYPmQVq9RoqF9udUOYAcZ7fJctHRVqotNxVrGNDywEkY9Fxvfb3LStdSx/R7bIOIbjb2w1skTWbD5LT6iEMGendb1rKaOaUzuAydytKqqOJxLTsg2i05fjpU8cZJxha062wsJI7rCfEGSYAQR+7AAEBXkBjDk/WsksPQVpNXUPa4tCC/IcSM9ljSPPWBkqMTPGcHuqgF/xeaDKZE1zc+axZx0u2KqJ3tGAo3vLzkoKIiICIiAiIgK5ryB3VqIPQH2Kzs802qf4AV38o25FT2KoH7abVP8AACu/lG3Ig9qEREBERAREQEREBERAREQEREHHvMR+D/xN/gdev1KVfM/Wf21P/hXfpK+mDmI/B/4m/wADr1+pSr5n6z+2p/8ACu/SUEKIiAiIgIiICIiAiIgIiICIiAiIgIiIA2VwOVaO6u2CCmVUHKoRsqsAKCUEIThUb3VTkkBBF1EOyr2kuI2x9qyW0rSMkjdVELWDZBGwlqq6foVHbFRvHUQAgGp381a+YuOf51OymYW7kKaKjjJCDCaSW4x8lLHTGRwBad1qsNDEQO3dalT0EQZ1bbINNptPCdoOcZ+a3NbdJMEYf1Yx55Whz3CWkPTH5Dur4tV10bC1ucZ9UGszUQoXkdfZaPX3MM2Ds4K0uuv9ZM7JcclYUcjqknrzlBnVN2y0D1Wp2K1m8YeIyc+gW3Z6cBoIOPJcucMbbD7j1kDOEEGndN+DOQ4Yx6rbXEONlJUhmQd1vq/1z7VLIYRjv2XEmprpNcakumzkOKCOINliHbsseZ/gnsoqSZ7T052CnmjEuCdtkFni+I3A7kZWZTRZaMnyWLHG0HbyWQ2UsxhBfNGWAnKx/C8UEd8qtTUPc3AGFfQ5I375QY74PdsOx2OVvzQmsGWapjqSQ3w/iz27LaFU0PaWlYj8QwvDSdwQg9C+WzVEPG2pZpg1Q2cI8Z9dl2W4g8m8emNNS3RsoJAJwPqyvMvlC1/ctDa8bVUBdkuaQAcea9HdUcyuqNSU1NZ6nq8KUBrsn1CDz5401dRo2/z0JhcGsJGd1xFLrl1UHRFux9V3Z5iuGlqu1mffpunxXsJO2+V0rk0rRRCd4xljiB+VBodQ9lQ8ygYJ3UKileI6swNeCAcKbBQWGLuVTw+odKysDp7K17elhcO/kgwjRiI+J81eJM9/JQNqZHzGNx2+tZJiGM/LKC5ruod1eCMYyscEg7FXdRzlBcWbqrY9xurev1VBIesAIJX05Ld91A+lPScDyWpwkOaMqR0bent3QbfZB4b8n1WQcbKasb0ZI8lhB7t9ignHdXd1E0k4ypB2QVAyq9JVzQMq7AQQu2CsI7qVwH51a4bFBFnfCuGwUTiQ7HzV7OrGEDOThXdJVcD0VcZQUVrj5q49lE4nyQXBwJwrseYVjW43V2SgkJBCN7KwHPmrgSEFCM/lVC0hSYQjKCFgwVc/sqkAElQyPIO6Cxx7hcucmv4WPCL+Gtn/AFuNcRLl3k1/Cx4Rfw1s/wCtxoPpDREQEREHXP2h34G3Ev8AxOj/AF6nXzyN7r6G/aHfgbcS/wDE6P8AXqdfPI3ugOIKoqkKiCrPpK54J3VrPpK9yCNERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQXA5RUamTlBVXDsrT2VzTsgOQdkO4yqZKCiITsrclBUDCqqAkndCSEFUVASTunnhA3dsFXGNlcG4VSBucILoW5OVMWYUMTiDgKYvJCCjSAVeSMKP5qvUe386C3qJKvDcb5VnSQcq4uICCrnAjAUTu6uGTklWSEjsgvif0nZXzy5iI+Ssp2hzhnzKzBTscQD27boMrR11FqrxMTj4gcrnXR2v2VV0gphLnJAxldfamBtO0OYVuLhlWOdqumY7OC4efzQdxdR251wsInDSQ5n8y69XTS5or22uLMdL85x812qlii/YbA8AZMYzn6lwTrwMiopZWgZGTsgw7zr9tPaBQ+Pu1uO/yXD961p48xbnOT3Wg3a81E9xfG4uwDjusOWjZIHSEnOMoNZk1A1zc9XceSwX3vrkxlaKXFpLd8DburOx2QbmNUHMDg4bjKxYy6eoDQ0nJC0uGpkJDPJbysFtil6ZXd++EFwtBMAeQckbrbt1tj48vDTsVyQWRiLowMALQrrTROjcNkHHRBBII7FZMTA6MuI8lnuoIzKe2MrBqj4LjG3sgxX/SPyVEJyiAiIgIiICIiAiIg7/8AsVPwptU/8X9d/KNuRPYqfhTap/4v67+UbciD2pREQEREBERAREQEREBERAREQce8xH4P/E3+B16/UpV8z9Z/bU/+Fd+kr6YOYj8H/ib/AAOvX6lKvmfrP7an/wAK79JQQoiICIiAiIgIiICIiAiIgIiICIiAiIgq36QQjKoO6uQUOwVzOxVFVoAQSAY3KqDvnHYqh7I3PUAfNBf72RgfzKhrCRj+ZZbKKNzA443T3GL0QYD5y7sEjc4uG2crU22+I+QWXFbYQRjHdBp7GOLQcEbKaJhG5BWrNooRgDGPqUcsLWZ6QNggxWzdO26v+6YjaW9X2FQytAcdvJaVVucH7EhBnz1YmJPfKsbI0DcBQ07QWAnurJ3dBACCSVjZSdgoGv8AAdjHdTQuzgjsoKhpLgUGQXeIwEeq5N0Ncfc6PDndOwG64/slOyc9L+2fNbo6/cYwI84wg1LU10EhceoHPmFsd9vbXyF+cZOdlnXSqdJnqJ3Wgm4zwOLYyQgyqygFDH1DchRUY95BJ8lG2slrXeHKSQsyFgp24b5oLfdehyilb0uwr62d7WktH51hxSvkBLs5CDIEXXjbKkZ952Ax9itondcmCs2eBpIwgw5ZS4b7KCQdcZHqsmvjEUQc3usGle55IJQbx4VX4aavja15Aw4H8i7Q6X4yRah1VbrX4zCXua3y+S6f0cYa4HON1vjhfM6n4g22VrjkStx+VB6a8ZOFgruEsV6Dz98hB/MujWn+FLb3cpbcX7SydJ37ZK7w8V9bXJnAmCBuQBTgDf5LoHauIV5tNVU18DyXxvJGCe4KDd3FzlNGhNN/snikDyWF+M5XWWnNTMJgaV+IyR9E7Ls5p7jjqbihINMXzqNMCGfEcjHZbou3BvTVBbXzxNi6pG9R28yg6YNrZPHMTmY3xgrL8TqcIj5re3EDSdDaK55pw3bJ7LYsIBeX+YQZM1qEMQqB3O6w2yEtOWn0zhanFVPqGiF/bturpLfGxmQRv8kGjNOXKQgnsFKyACQjIU4haHAFBgvBDc4VrHY3wtQqYGtjyMLADQGlBksnIx8lI6pIZnPZYbO+5Urh8GEGPJU+LIW998KwjHkq08IdKSVn+7MIQYQx5KoByFMYgPJOkA5QGjAVVQEoc580FrvNRv8AoqQ9jlRv7IIBs7dXiQKM90HdBNgndXNIGytachVQWudvsO6oDlMZVUBVxsqYVw22JQMhO6tPdVHbuguG6uGwVo2TJ9UFFDM3J2V73EKBzyThBXuMei5c5NNubLhF8taWj9bjXEkTTnfK5c5NtubThH/DW0frcaD6QkREBERB1z9od+BtxL/xOj/XqdfPI3uvob9od+BtxL/xOj/XqdfPIEB3ZUVSdzuqIKs+kr3Kxn0le5BGiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiILkPZMj1RBRvdSeSswArx5IKAYVHOwcK5UIGCSgsd6qiqTuVRAHdXA5Vqu8kDIKIwDO6uwEFVUDIRuCcKvbZAYcFXdQVjvhwQqBxIQSB4VzTuCoBnPmpGnYBBK52RhWKhO2xVRuN0Fpf8la49Sv6QrHDHZBWOXwiCp2VpLwAoI2B/0lOynjyCAgkq5eqME/JaroOpFLqCGfYYcD2Wj1mGxhaho4B13jB7ZCDttU8RyNMR04JPSwDH2LhnV2tKmrhfB4DiCTvhcqaf07RXC0RCUA5AC3Bd+Edg/Y3LXYZ1hpI2HfCDppVReJWPnc3BJzhJDiMgla3qekjor9PSxAdDHEDC0CocerpztlBgO+J5281c6n+Au9ApTG0bqxkj3Shh7FBHTnplaMZyQuWNN2Vr7UavOCG5/MuP4bfFhsm2Tut5Ud4qKS0PgZnpxhBp9ZfzBVvp+vZpwtPmu3vT/CD9itt19S+StllOclyto5n+8sO5yUG622svaZhkg7rbd2p3RT4wTlbyhncKRm3cLTLhRxzMMhxkINrimJbkg5UUjek4WpEBshaPLZYNSMPQQoiICIiAiIgKoaSFRXtQd/vYrNxzTap/gBXfyjbkVfYr/hS6p/gBXfyjbkQe0yIiAiIgIiICIiAiIgIiICIiDj3mI/B/4m/wOvX6lKvmfrP7an/wrv0lfTBzEfg/8Tf4HXr9SlXzP1n9tT/4V36SghREQEREBERAREQEREBERAREQEREBEHdXYCC3urhsFe1rc5wrulvogiVzVf0t9EwAgZCHYgq1+W7hUaSRkoNQjmwwNzuApGvz5/nWleK4bBXtnf6lBqolx5qYVXSM5Wi+8v9fzJ7w8jzQas+7dJx1fnUcl4zt5H5rSXEu3JKdGfMoM19w6jn1WPIRM4uG/2KwRN2ySp4mABBfC/pHSoKp3x5Cn6MHKilYHHdBWnd8IyFP7v4rh+VQsAaBhZsBOAUGdZoDG/vjfzWdc69sGGdQK0kVUkOenPn2WLU1Bmd8ZIKCarqTI3q75CwvdxKS7A/Iq5BHfZZFNjG32oLKemDHE/zLIk2wPkrXPLRlWF5cgSgSDpPphUipekHHmquJbgrIppWlp6yAcoMdjDTv6iCshk8kxBEZOPNXyuhOOtwAK33pGwW2upS9xadshBx7XCWSPpbC4nfyS1WupmeA6meM/Jc36Q0Pa7rczBI1pGcYK3hqThrbLREJKaNoIGdgg4k0poAXR4D/hyQuTNLcJoLVqSkuJmaBG8E7jbByti329VumXZpBgjfZaCzjJqQh7gX5YD6oPR+thpNaaDj07DUNleIw3pBBPbC66ay4AzaZtNXXOpHAYLuyweTjibqLVerBbqsvezrAO+R3Xd3jdppg0qfGiYGviy7b1CDyk0rqn9j+s30T4/DxJgHGPNc2VvEVldHHQiqBL8DuuH+L1ptNk1RNVUkgEvUTgeuVsexamrJb3CJHHHWANz6oOWOJ2nJPuebo0Fwc0u9VwhSvkeZPvR2J8l2cvMsNw0gxtU4AeHjf6lxTQWWyGsMYlYep2Mfag27pCzSXqrEToi0EgZxjzXIdfwxbBTCQPBJGVm1lno9PW8V1vA6i3Ow81tp+vrrURPjc49IyEG3Llpr3KoIaMkHdYYtUx38M7LcNorjda4ioIO/ot4T2WiZACMZIyg4iuUToI8EELRhJ8JJW9tY0sEIPSQMZWxmlhY8E7BBSOYOfj1KyyDhadTub4+52zstWHh9YHUB6oI6SLqk7d/ktQMBwFc00zIw5jwSPRUknYIi7PZBjTRBuSCFB07d1hCve+cgk9JOAsxriZA3yKCPPS4DCGTuMLUnUbPD6tsrDmha1hd5hBjkg7hWSHAUEUpdN0+SzpI2+GThBp/UCVQyYIGPNWPOJMBSsja5pce4CCePcZyFdjbKwTO5pwDsFkQzF4AJ8kF52VC4A4V+Ad1E/Ad3QSNHmri3O+Va0uxsnU4bIKHZFeADvhRyEtdgILuoIXDBVGjIVr+rqx5ILHP7jCiccu7LLjhB7o+BuCUEcRHTjzXLPJv+Fpwj/htaP1uNcRk9B28ly5yanPNlwi+etbQf+VxoPpCREQEREHXP2h34G3Ev/E6P9ep188YGV9DntDvwNuJf+J0f69Tr55G90FT2Vqq7sqIKt7q5zgrW91c5oQWIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiABlXDYKgGPJSMAPcILFUA91N4Q74VRHtjCCEHOyoWZOcqRzA0bDdROcRsgo5UVRv3VcBBarh9HCqGg+SrhoQUYCN0c7BwrtvJOgHdAYCN1djfKHYbK0uKBI7AwqNOyviaH9+6vdG0DAQRtIJVenJVjssJwqeI4b/zoJwMKqjY4u7qdobndBZ0dO6tIOeyyC0OHZULGjsEELWHIUoJYBugAHZXSfQygxaibxNvQrUtLze73FkvotLhYHvx81qlNGID1DuN9kHYzSmrmtoIovFzjGwK5NrLxJPpKU5JBYT+ZdYtEVskszI3uOA4LtTb7dSv0HLK8/8AyIn8yDpjquq69RVJPcuK0KpJwXgZxvha5q51I3VVUxrxgSEdlp1MyCerEJdkEoNKZLI9wb0Eg+mVqlrtL6upDHRkA+q35RaQoHUrJgASd+y1B9hioqU1ETR1AINIGkGspRJ1gYGSFty63QW8GjBHoVm1mpriyd9MM9IyPsW0LpM+eoMj85OUCSATPM3k45UDfvEwPocqgqXtaGjsFY5xkOT3Qa63UHTEIs7BalTyitpi/qC2csuC4TQN6GnA7INSfTgSOWlVremRZhqpHN6vMjdYk2XnqcEGMiq4AFUQEV8bcuwQsgU7SMoMRFfKzp7LPo6VksfUcdkGmq5rgAsutp2wg4HmsJB6A+xWdnmm1T/ACu/lG3IrfYqfhTap/wCL+u/lG3Ig9qUREBERAREQEREBERAREQEREHHvMR+D/wATf4HXr9SlXzP1n9tT/wCFd+kr6YOYj8H/AIm/wOvX6lKvmfrP7an/AMK79JQQoiICIiAiIgIiICIiAiIgIiICIiCrfpBVVG/SCqguZ9JXk4VrQFVyAHZOFUjKtaMFXoLEQncjKILXbKrCDsqHcnKq3bsgu6R/7hUVx7K1BQjvujVVVaAgq1pKyGDGCo24AUmcoDpAMhRvdk9lL0N8whjBQQ9fyWTFUgbZwoXRgdljyHpJwUGpNmBJPqopY/EcC3ssRkjhjPyWfTnIBQWe7kNCmgi6RurslWueW7YQSiESZGVBNGYiMNJz6KGpqpIgOg4J7rXtNU0VxYTOQSAg0ConkYBhnfzwVuzRelZtQwmR7C0AHy7reehdEWu+V/g1IaQDtlbo1PaodFPZBbGANO2yDa9g4OG+VhpHP6cHzWt3XR40ERSicHPw91pddrq66dh98pMh5Gdu62Vc+It41PIZa4uy0oORrZem6dkFwMw3Oe6XvjPHXxlnig9Oy4sqL3U1kQgkJ6QMDdYEdFE7OcnKDXr7rBl2cRgEElbdFQxjJPgG4KyPubCz4m5z9aClY54bjY7FBy9yq8Q6bRGqG1knQwCQHJAHmu9Gp+OtBxBgp7BBVsLp2hmA4eYwvMmnjbagJ4Mg5zsuQeFOtLi7Xdub1SbSNxn60HOfMLysV9ssztatie9j2GTzIPmuoul7BVS6gwaN4EcmM9PzXsLrLr1RwZghuDW9JgxuPkun54XWC20FZcYBF4jcnbGcoMbRfCn9ndijoGzAOIDcefZbik5Hqi1UL7w6Y7fF/OuNeH3Ey96e1Y+30PUWMkwAPrXYG98ctWTWoUb2noczBQdXeIdkdYJn2Z7iejI3+S40jt7JHOj232XIvF27T1lRLXSbSHJK4YbfJ42Syh24QcgaZ0cx84lEoG/qt71OlGmAffx2HmuCLDr67x1fhhxIyt5VOv7sIQPi7IGtdLANdiTP2rYMOmHOLmHOCcLcE2qa64Sls2cHuquqJImGRo375QaZScPnOcJC/YLUqTQfvFeyl6x8ZAysB2qa+GToH6VfTasr4ahtQwnraQQg5KuPAT7n2dtxMwORnutmO0EHxPb1jYkLLruMWo6ugFBIXmMDC23BrO5uqPDycOKDTLto+W1ymQNJHcei0jxJWuJ8InB+a5Wp2G704dUAEkKlJpCgkrmROaMOdug40ZcKh4DDA4D7Uka+RhBaRlc/XXhnZqa0ipjYwOLMhcY3WzQU0Ej2gDpz5INgNp/DkL1PJJiMgLHqJi2rMZOwOFcXNdt1eSDCcC6U/WsqJmR0+qqYmgdWFC+Z0bgAUF7qE5zk4KNhLTj0VwqHFg+pWmXJQZbIfh791a6lycrHdUvAGFmU8rnsJPogxJHiHI2V8BEwyseryXnJU9H8MeUEjm4UT2lxVsszg8qsZL8FBK1myse4A4KlB2x9ijkaC7KC5hyqu2ByrW7AKjyT5oIJBk59Vy3ya7c2PCIf92tn/W41xJJt3XLfJpvzZcIv4aWj9bjQfSGiIgIiIOuftDvwNuJf+J0f69Tr55G919DftDvwNuJf+J0f69Tr55G90AjG6oqu7KiCrPpK9ysZ9JXPONggsREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERARE7oK42VMFSADAUjWByCDq+Sq1+FN4IVHRtBxhBTx1eKhoGFA5oAyFRreooLzJ1FV6Sd/JXMi9O4UwjwM4QYuCrXfSKnkaBkEKLAz280Bru+yqRndU6cKoO25QVa3fcq9WZwrhuEFqr07KmMK4dkFIxgqQnfsrRt2V2xCCOQEj61H0FZAaCcK/obhBFCwqUMKtGWnAUjckboLunbuqgbYVASTupGgHGyC0Mx3R7eodP2JUuLBkLDZUODs/NBJ4PgO6z2ysykmErunPksSqe4xjPy3VKBxbJlBy1w7swqpoTnu4fpXdbRnDx180mbe1xy+PHf5LpPw0uE0c8PTsA4LvRwt1DWwWyNzCchg80HXLX3KRU0d1qbk4uAc4uXEF24XfcGpMjpB8LsfkXcjjFr27xU07AMjpO4XSnXmu7rNK9riRlxCDNNybQRCAyglq0q5a7ZFEacOznbAK2RLfKuoeXPcd+6i8FtQ7rcTk/NBmTXpsk75ekbkrTauoFRJ14wsj3KNWyUzWNJQYbW9RWQyAEYWPnpccK8TuCCX3Qq0wdG58lb7y9Wumc4IJ21AAxlRvm6tseahWZT07XtyUEEcRk77YVkjOh2FPNmLIaMbpGGy/E9wBQY/VhZEcxDdwTupo6eI7ZC1KkoIHuDTgoNCkc5+/SQM+imgrJYG9IZkfUuRtP6Voa3BkA39VqNboi2skwGjug4nmnlqdvDO/wBamp7TLM3qLSPkuWLZoe3HJ6W7DzC0a/W2C3VHhxAAZQdvfYw0D6Pmh1RK7ODoGtb/AM42/wD1Itc9j5G1vMrqRw89C1o/5wt6IPYFERAREQEREBERAREQEREBERBx7zEfg/8AE3+B16/UpV8z9Z/bU/8AhXfpK+mDmI/B/wCJv8Dr1+pSr5n6z+2p/wDCu/SUEKIiAiIgIiICIiAiIgIiICIiAiIgq36QVSMqjfpBVQSM/mVytZ/MquznzQOk5+1CcKqpjKCzG+VUnCO2yrd0A7qrVar2DPkgvG6Oae4KqAQQr2gEHzQY+d8K4beSvLG5KENP/wBlADSRlTMacKFriOykEhQSoqA57lRyydJACC9zcgqF0RO6u8RxHdSx/EN0ELaffKyoPgABVWqjs529UEpcO+VFI8DJHkjT1d1ksgjc0k+iC212yS9SiGOMuOcYC5V0zw2dBC0zfeuoeeypy/6fprvfjFI0EA7ea3FzA3iu0XcYaS3ANBwMg4H5kETKWPRUhq2zjbfutk6t4nC5zZLuroPc7ratZre53iDwqpxOcjKxaS1QVDS5/nug1Ko1S27RCFwHbG4WAImsJIAGfRVktsFIOqMjIOysEjn7eiCXoG5CtMvhlWve4DdUjAkcOooKyV5AIAUcdaTINsYK1KO3xPbkgKsttiZGXgbj0QXtmbJEMkFbm4dVMdBq+kqiB8Dw7t81sRtSWzGPPY4W6dFuL9SUrCO7gg9Nrbr5uo+HUFoY4ZEYG31Ljm9aVlo9NVlSSSC0nutU4f0UdNpSGfcHpBP5FqOoa1s2m6iFzxjoI7fJB0Jrtf8A7FdbzBzMdMn863e/mHir5Y6EADOAey2jxK05a36pmmmcAS4nt81tuOwWSFpqo5gXs3CDlTU0EWorcKxsjSXjPdccT6SEVLLIME7nyWmN1tWU7/cY3nw27BZdo1HNXXmGge4lkhAOUG1KdlVR3MsFG4gHGcLcM9ZVinyKRxIHoV2t0bwU07d7LHcpo4y9wznAytYh4G6dlr4oHxM6C4A5AQdFn3utgnJfSFoBPdpBUsmqJegt8Ild6eMPLlo2x6XFwpWRCUs6tgO+F1etvD62Tyyte0YDiEHGNDWy11SWuiOCtedbzGwPLdu631X6Ktdqj8WIAEDOy0eqZAaJ56hkAoNm19eyEdAAyO60uO7Bk/UANj6KCvl8S4uZnbqxlXGlhwCO/wBaDeNDrBkEIb1AYHqpqbXTG1zH9YGCFsV7GgYGfyqzoDQXjOQg5+fryO42wU/jg7Y7rbNRC24Ewh4+M77+q4woLxUQyBgcSCtzUl7mY5rwdwAUGsVHC4S/7bDhg7rEqOHhhp3TA5LfILUINY18kYh3IG3dTUl9rKusZSSfQecFBxpWQVUFSacU7jg47HdPubVuYZPdH4HyK7O2DhjZ7pSMrJmML3DJyFvlnBix/cJ8wjZkNJ7BB0dzIH+H4RBBxj0U4ppy3r8I47rl+96HoKa+ywsYMB2Pzqd+jqJtMT0DYeiDhF73gkdBGD6KeCs6fhIwt/S6YoDVPYcbZWzdR0MVDVBkRGAUERpfefiG/mnh+7DpPksihLvCGPNYlwkd1b7boMeQ9btvVSw/DgH0UTS3AOd1exw2wUEytcfiwr2kEb4RzRnOEADKEYCq0klVIzsUGHKMk4XLvJn+Fjwi/hnaP1uNcSSgZOy5a5NPwsuEX8NLP+txoPpDREQEREHXP2h34G3Ev/E6P9ep188je6+hv2h34G3Ev/E6P9ep188f1IKu7Kib/NEFWfSVX91Rn0lc8Dz/AEoLEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEGyIgvDxhXNlA2CiRBP43zVDICM/JQq5m6CvVnYKo7K5rG+iv6QPJAbJ0+XdXGpGMKN7RhQ43wgkMnWURjW4yq437bIKdlTBO4VxDT2VEFCcK9vZW4yrhsED6XZD8OyMxndSdLSBkILAMqRrDhW4AUrOwQWtYcqTpOFcB6BU39EEX0CcqvWFbITk5URdugyGuBKmZtjKxYjnzUzXH1QXSgPCxTAWu6llnsqHPYoMcu8UdCua33cF/oFIyIMOQlRhzHZOBhBvDh9ffBroos9nj9K7xcOL2IdPNqT2bHn8y6G8P4YH3GIk79YXevh1b45NMYI+Ex/wAyDjPivxCZVzT0uB5jC6uanqW1s7xgZLj5LsBxVtNNDW1L299yuul8AjncWns4/pQaYaDp+LHzVpn8E9OFktnLgAfRRvgjeOo/pQRe+5VTKJtvVRPjiaTuB9qh6yx3wlBlGhyOrPdRvpQ0Z/nVnvcuME7KhqZCNygiOyKrsHcZ+azKGGKQ4ce6DCwT5KeOpliGA3Za06hgDQdtz6rd2ndJUNfEHvYCSPRBxu90suB4R7+hW6bFo99xhEjmluy5HotAW5wJLW7b9lp938Swu8GmADe2wQaXa+GgqSep4GFh3TTLbNUBniA/Uo7jrW7W7JhBGe+62tX6ruVe/rnfkoN+2uubQMGZB29VDcdWMbJ0+ICuPnXqrdsT+dVjnNQC55OQg30NeiiGxzn0VPfBfv8AbDnY81tSgoIq5xEmdlmzVElqIigBx22QegHsgohHzK6lw7P/AMBq0f8AOFAi0r2NldLU8zup2SdhoOtP2/dC3/60QeyKIiAiIgIiICIiAiIgIiICIiDj3mI/B/4m/wADr1+pSr5n6z+2p/8ACu/SV9MHMR+D/wATf4HXr9SlXzP1n9tT/wCFd+koIUREBERAREQEREBERAREQEREBERBVv0gqq0d1dkeqC9oxgq7uqN37qp27IKq1yAocbYQURFQnCCwtOVLHtuo1c0nb0QS5zsqgYVGjsfNXdJQMZ2VHMwrzhoBHdWdROcoKAYVwGUAB7qQADsgvbHkBRSR5dhZUQKuLAdyEGH4eBt5KSMdI3UpaMKPGCgu6gFaXZKtkO2xV0IB3cUF7AQQcLVqKhdMw/NYcTGOwB5FbltEQDB9SDX+GWoRoe5mrkPSCc91Xi9qhnECqZPG8EsI7LQLrE1zD3WgsqPdyQ1xz80EElv91haCMkDdZFvmIBGCMKemf72el5yMrUm26JmOlo3xlBpc7zIDnKwnP8LO3ZazXU7YmF2AFhxQRzA5wfrQYcUnvDukA+izG0joSCWkfYsi30kUc2XAAeqz6uSkDcl42QacavwW4BUbq4zAxjcntjzVk9RRvd0tkBKz9M0tJVX6CnkIIe4BBo8tDXRnxm0byDvnpK1rSFXW09+gqHUTwGOByWldyNH8HdKXDTkVVVtiBLQdwPRZU3CHQ1HA+eIwhzWk7AeiDeHBG9Saus8NpYwl2zcea5d1RwmrKTTMsrqdwBjJO3yWx+U+12Rurm0cEjSBIB8u6748WNO2+m4dVk7GMBjgyDjv8KDx8q+Ch1lrOWi8TBMhb3+a3veeSGS2aflufX2YXfS+S2S7iBe7VxmqaS3FxDZyNvrXPF84ra3qrYKCWN/hvbj7MIOjldwumptUy2ksOGOIz9q3DTcIPuXWxXDIwwg91yHrWlqKOsfeHRhszsk47rZMusbrUQvjeTjsg5RsXEWKxW9ttdUhvQMd1mW3i3TT3+npffB8TwPpfNdY9UX6she6UF2T6LaNr1fco7xFVB7+oPB7790HoVx4vDI+Hra+Op8QmEnAOfJdHbPxAmdWyweG4fGR+dczWvWN11pp1lquPWYukAdR+S0VnCq109Y2VkbMvOSgitNul1VAAWkA+a1W48HDDZ5akPyeknGVuJ9rbp23tdRgAhudgtFGtL1VO9yeXeGTg/Ug653jTVRS3ySn8J2A702WVNpx8dOZsHYdlzbqTTVD7sbi7p8UjJ9VsVjGSSGBwGDsUHEkj52zmNsZJBxjGVcTVn4fdHb7fRK7I6B4X2O+VDZJ2MJcfQLmOr5e9Mx2s1QhiyG57D0QdHLRZJamQOkiLcnzW9aLSbHRh5cAt2a/09SabrJYqQABucY8lxnPq2vp3uYwnAOBug3M+yMpf3wOFDSFkNcyQkAAhbdbqOuqN3dRyrnVswj8TfI3Qc7WjWkNtt7QagDA9Vm1fHiCktj6T3kZLS3uustTqa4OcYy9wAOMZwtOkrZahxMrnZPzQck3LX7Ky6vqg/PUSVkRa7ZUD3frG+y4xYwHByVlUTMVLXAknyCDkyCj96JqQ84duFs/VNlldKZWsLgPMLXaWrrIaRnQDgretmstNc7O+eqaOoNzug4SjkNMBG4YI8lDMz3k5C1nUtJDT3SWJmAASBj61pHV0EhvZBD7m4jAVpgMZCnMxbuqF5fuUFIxtkqriOyuAxsFDIT17IJWHdVk3aSFY3PSO6PLg3CDHfIOoj0XLfJof+ux4Rfw0s/63GuIJPpE/Jcv8mn4WPCL+Gtn/W40H0hoiICIiDrn7Q78DbiX/idH+vU6+eRvdfQ37Q78DbiX/idH+vU6+eRvdAd2VFV3ZUQVZ9JXPBO6tZ9JXu8soI0REBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEG6INkEnglDEQM5VPFcrmSdRGUERGPJVBwswRxdOSRkqvgwkZ27IMQSkeSkEmRlSGKLP0lG8xj6JQULtlEd3K8Fp2KuxHjKA1uQFdjCozvhX4BQR4KtyFc8nfdRoL0VASTurwBhBRvdSt8lY0bqVoHfZA6Fc3YDKqiC8O3KHurRsd9kJIOyCObuoiCW5UrxlGNGOyCyFpz9qyAN+6sDendSs3OUFsr+ho+ShZVBxxjzWTIxrxuohTMac7flQSHdod6q6mp3Vkvu7cknYBXhox0+S1zR1Iye+RNdg5I7/AFoN26E0DUQSRVcjHD4g7867Z6IvMNBZxQGUBxaG4z8lx7S2UUlhimiaCekHZaHZLzcxqaKlIf0dYH50Go8WbPUP8eqZC5weCcgFdVtRNq2Vj2GleBk9wV6V1OjbXd9MxTVYZ1PYCc9+y4mvnB/SE0hJ8IvJ9B3QdHo2VTgM0zht36SsmOmmkIYWEErtZeeD1lpWOfDGzAGRgBcTX/S1Pb6zpjYAAcbBBxp+xySQB/Sd1kwaOfI3JB/Kt/G1uEQ6WqCqinp4C5oxgINnt0Pn9/8AZlSM0F1kDr7/ADUk97ropnMwRhYj9TXKN22Qg1FvDckZ8TbHqjtDikHX1AY+awv2Y3UAgdS0+s1ddJAWkkA+pQau60NB6C8fDusyl1M2x4i6+2y2b9363JJduUy6tb4rycoOR6TiNGxpzIB9q0G+a3ZVy5BB3+tbLqG+ESAT381ZHEHjJJQapXXplXkYG/yWCylEwLm7Dvssd8Yb2yr4ql8QAbkBBe+kIPms6koyW7LFZUh2OojK1uhZ1sBbvsgjp5DQEuKw667CaTKy7iw9LtitHMDD3Qd/fYv1Ym5o9UMx20DXH/nG3IqexchbHzR6pc3z0BWj/nG3Ig9oUREBERAREQEREBERAREQEREHHvMR+D/xN/gdev1KVfM/Wf21P/hXfpK+mDmI/B/4m/wOvX6lKvmfrP7an/wrv0lBCiIgIiICIiAiIgIiICIiAiIgIiIA3VcFUHdSNwdkFR2VzeyFhAyFaHkdygrjJwhBCs68O7qQEOCCNv0griMqpZjdWZKC0nywpI+31K4RZ3VrgWnO6CVmxypQQeyx/EbjdXRv9D3QSFhPmrS3CkG4TGfJBEr2u6SEc0AIwA5BQStqA3y/IpBN1kb4WOWbEhUDxGQCgySRhWOV0J8Qqkw8M77IIyD+VTwU5kO2wUBcCAcrU7eC5oIGdkElNRO7E/aFuW3U5YwBaIJPD7havbqvLRkoK3r7zEXHfIWkUVtFxJcFm3+oDoelx2WFZbiynBAcgyTazbvi9N+6NuLz5Zwr6u5NqRguG+y1Ox6e9/Z1x75GeyDRamd9SwjpJyoI4pImlxBx3W8H6Wkicfh2VkmnpDG74TgAoNG0xbpL1ViBjSTnC5MpOBtXdHMjLHDxMYysPg/paervYZEzJ6wuy9Xb6/T7YZnxgYAKDrvqbloqLDQfdF2f7rutq6Z0E+K8MqS4gRHff0K584ncSJpLYaN5AAaRj7FwjZ9WFlY8Z2JPmg7b8JbRJqW3xWdk7h2b3+xcvnlprKhghE7j4re2fULrhwV4hRWOobUF+CCDuu0ls5g4paqBrZwSANsoN88A+Vmr0Zexd3l5HV1Zyux/FyhfJw5raYEkshLT89sLZ3BniZPqYMhGHA4AXJfEKlfW6UrYGDJLO32FB4XcR6o6B4s1VzqI8ASl2Xdu61ip5l7dWU4Y3oywYOy1rnE0S6j1JU1cwwMuPb5ldRrZSUjjM0OJIJGEHNd54s0+pZDCwjB2xhaLJVsjj3APUthWijjpqjrbnBOQtxyVIkw0eWyDS9SBtSDgYGPRbRt9B03ePbOXD9K3jdYi2MvOwW3rU9kt8hj6u7x+lBzFT3pumrGycsAw3JPZaHR8aI6m4Mg6gcuxlbg1vY//AIGsnd2Mf8y4DsFEyS+xxEnAeMflQdoq3UwqbM2c9i3OStmw6lgM/cZ6lq94oW0OkmSE4HRn8y4ko7xTvuBjD9w7GAg5XutxNdRBoJxgY3W0mW4seZMnvkLXaaJ8lvEjRkELFMcpaRjug1LTOrDpiUPdIQAc7lb9m5gopqE0XjjJGO/yXC15iIaesY29VtgCASEgnOUG8ta3v7uTPqQ4u6s+a4zqqLLnSY7lbmqZx4PSM4WjSEvPSD3KDSxIIti0bKQ1zTGW47hZFXb3NZ4h2BWmPY1pJygjfE17y7AUL4QD2CyBKzI+JXP6SwuygxmgNUsMohlEp7BYj5wHdPzVzXh7S0lBuMawijiEOBt8lytoWq+7Nhe8Ox8OQPsXAEtN0gOb5rlbh/eRa7QYScZbjug27qm2E3iXcnB7rbFSw07y0+uFu681rZ6+SUkbklbZrojK/PfdBi+H1NBVRHhStGAB6I7ugsAJUUrg07hTbDsoZG9R3CCjJB2Vs0uNsK9rAPJQzgH8qCCR+QuYeTPfmx4RfwztH63GuHHei5j5Mz/12HCH+Gln/W40H0hoiICIiDrn7Q78DbiX/idH+vU6+eMd19DntDvwNuJf+J0f69Tr54wMoLj2VquPZWoKs+kqv7qjO+6ucAcFBYiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAg32ROyC8s2ysinoXTnuVAHZGPRajRT9Hn9aCZloc5ucrKbYXdOcqsdf0jv8AkUzbs7p75QYbtPuJO/51iyWR7TgfpWq/dN/fOMqnvRk32QaR9xnq77jOAyfJar4pQyEgjA3QaFJTmAlqjDs+RWtPoZJzkN7lY8lufGMEeqDS3DAUZ7kYPdajLRuaMlQmn7nG31IMMnfsrmuIH0Vqdvtj6t4axuT2Wq/sWqQMFhG3mg2yx5J+ipmg+hwtbk05NFv0n61jutsrCQWlBpxPSBsjX79lnvojg5HZYssTIu57ILSQQqKzxWE4ypWFhA38/RBQjAQNyqTvDNyVSGVjuxQShmVe1hxn5pHue2VJ0n0QW+GXeaqYS3clStbjKP3aEELPjcB9i17S7jQ3WOpJwAQtBp/7J9q1Wml8OQOPYIOfxxHhZZ2UxkBIbghbcsOvYptWQsDQT1jyWwoK2OeMMyQey1rh/Y212sqdo3JeNvtQd6bVUy3XS8BbkBzBjv6Lb2pNCVdLaZL2XPAYC7K5L0tpCpptIU0giOAwY/IsfiF7xBoWpiczA6CPzIOk2p+NjKO5TWd0gJiy0/YuML9xFZVVXWHZBOThaVrikgdrKsd1HJkJP1ra9fTNY8uYT9SDereIcfQG57DCgqteRzRlgwcrYgGVK2AuGcoNWnvjZZC/A3+Sx33RriD0j8iwTAQM91GWgHGUGoi5tH70KySoZO3sM/UsPo80aQw4yguMOTnyWRHVCBvRhWdQx37/ACVrous5QJ5hIo2ShgwqOZ0jdWfUgue/qU1NSOn3wQEgpzJha3S0jo2ggIMelsLp/Pspn1YtThD8sLPgqXUwOPRbeu8xmqOpBPVXUTA9jlYQmyc5WPgY2O6vYNkHoF7FuTr5o9UD00BW/wAo25FZ7Fof9dJqnb/5QK7+UbciD2lREQEREBERAREQEREBERAREQce8xH4P/E3+B16/UpV8z9Z/bU/+Fd+kr6YOYj8H/ib/A69fqUq+Z+s/tqf/Cu/SUEKIiAiIgIiICIiAiIgIiICIiAiIgq0ZKkDCCFYzupxjHkgvc74MLFf3U7yAMrHdklADSpGMOcq0A7bKSPJIyguOwUeD1ZWQ5gx9qgLd8YQXNna3v5Kx8gfgeqo2nkd2CoY3xn4hjHmgvFO5w6t91LFCW7+ivjmjEYGeyr48fScnZBTqA29FaZAFa6RhccH5qx+HHY/kQTdYcMK5kJO6xxIGYBKnZVxgblBe/EXdQuiMx6gdlWWojfkZV0ErACGuQSxOEAGUkPjkFvko5JGOGOrupqWF79mboIpGFrQPQBS0t092+AhTGhmfkHP1BQ/cOrlf8LSfXZBmMrfePi9Vlw1roAPtVbVpK6z4EUTjvjstaPDbUcwBbTvwe2Ag02khffZPBA77Bag/RktGOxGRkLXNKcPtRUFUHyQOABz2W6rrYLsQAYjsPRBtDTGgZb3UiHBPxdlzrprhTHa4GNkj7j0W0eH1uuNurg+SIgB2c+i5YreIlstQYyte1pGBuQg0C66Ahja6Twx2JWyrla4KMuhLBucdvVb8u3FOwzREMnYcj1C45vGrbZWVBeyQEdWRug1jR16pdCVn3TmaMZzutS1vzH0d2pjHCQC0Yzlceaj1BbKihLJHgDGO64ruM9od4gik7koNwap4iTXuYtbNkHZbeobhNHP1h59dytFghgqKj7y8nJWtUlnqJp2QsBJdsEHMPC2pq75M2nilIOQNiucJLDdbFG2vdUHDG9Xf5LiHg/pm6acmbcKuMiPIduFzZqXWFBWWl0THA4jIPy2QcvcrHHiai1Ey1zT5LZADv8ANd5LnxWpq9kdr6W5qcD5nK8uuVqyMvmvgaQkudKAMfWvRUcO7m250FQ2F2IuknKDr3zxcEY6nSsmo4qfd8ReDj5ZXmRYuHkzhVzlhIY8+XzXtjzcadvmoeGYtdujJf4JbgD5LzasvL9xCp6GullpZekuJHwnsg6t3RhttQYAMEHCw23MxgyOdnHdb34j8PL5YblIbgx7SMk5BXHnuvih8TH5O4wg1+y1A1PN7kBkk9K3DbeFstNqOmeWHBeD+daVws07WtuoljBwXZ7LnWGzXRt6pJ3NPQCCg1nibo5lHw8jxHv4XfHyXVLSlkLtUsZ0H+ygfnXePiPS1V90SygomdUojxgd84XWjTXCTV9LqVlRLSvDfFB3B9UHLOrNEe86Cje1pBMQ3x8l1zHDeoopn3E5wx2cruLq2mrrZodlPPGWuEe/5F16rK6WajngwMkkINgz8QWWqMWxxGWZCw/9kiMfD81t/U2mK0V76p8bgxxznC0L7lDBccgAoN5V+r23Fha07laMKpwcTnvv3WkM8GmIBdgqZtdTE46x6INbiqnTjpJz5LMgosvDz9a0u2tY9wc1wK3HTMe7DQPJBI+ibVxeEGjJ2WE7RrpDjHf5LWoWPpvjd27rIjvMbHAkjZBt79gDwOoBUn0SYqVzyOy3aNS05Z0B4zjCgrLu2opnRRkEkbAIOIqu2uhqzHnYFVFIWNyFrtwtNaat05YQz1IWmVUzIWmN+xQYD6hjT0nyWs2i6yY8Jr8A7LbkgLnlwGc7rULW3oeHOyg3VFbn1ZMhOT5rTrjTikcWHfK1WhvNNTM6XkDy3UVfbqq7gz0rSWjc49EGgFpcc+RVrm4BWW6inid4TgeoLEqJGQEiQ4QQF2/ZUPdRuqYM5zlU95h9UEijkjLuye8w+quFRD5OGUEBpnHcBcu8mzSObPhEPTWto/W41xT7xFnHUuV+TZwPNpwkPrrW0frcaD6QUREBERB1z9od+BtxL/xOj/XqdfPI3uvob9od+BtxL/xOj/XqdfPHnCCruyohJRBVoJKuIIVre6uzlBYiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgKWKUtGM+aiVRnyQZBmcexVBUOae5/KogD5K13dBl+9u7ZU8NWcYJWnbZysmMMHn6INQFX8wqGvLCNx332WH1s9QqExHu5Bq8V6jjAB2+vzWWyqjrAH47LbTxF64WsW57Gs2I7INUioG1gIAWnXGi90eY/sWtWqpaxxAwtPv0rXy9RIygxrbcorO8SyNBzuFrD9d0hxhgGFtK4DraMHOFgiJ57BBvV2rqeqPSAPqVpqopd8d1tSjgcJgXDGFrTZ4Yhufkg12129te/px32WTWaEdM4dI7jOcLF07e6SllBc4Df1W7f2U0r3AtcMAeWEGya7QL6VvUVgw6dcNsHbZb9u16bWRdMRBWjU8VTI/AbuSg27W6Ze5mR5LTvuI+nyXZz81yDPbK90RIjP5FpU2n7rUEtbEck+iDZ7yackEdlWOrDnAYK3BV6Dv0oLhC7H1LAdo260nxzRkAeoQQNOQCqvGWbeitkDab4JHYIUbq+mwWh4ygjY0sdn0KknqjHG4g4ICpE3xyPDdkEqeWy1M8ZawHcIJtNVclTVtiJzkgLsdwr0eKe6095fHkMw4n8i4F0Roy8z18cjIyMuA2+tdxeH+htQ0tmFVLC4RBmST6YQcyf7NtFbrBHacAGJvT+Zccay4yxXq0yWhjxl4IwCtl6gr7VFUSU0rwJWkg7+a2XNJQGsBjf8QO3yQbA1Vw5fUXSe8dBIfly4t1HbfcZDGRvnC7J32eZ1EWt3HTsuAdbUtQ+qc7BxklBsp0Bzsr2TNj2KkdkHpPdY0kbuonGyCbx2v8lQU7nnqULI39Q2I3WoRZazJ7IIRBgELGkaRJj1Wd7xGCd1hyHqmBb6oJRES0H5KgmDD0n6llsYfDAUf3Jqp3dcTSQd0EXu/jYwFmQ2F0gDt8LIgtNYwDLewWqwOdDH0yDyQabBQCH4SOyumuTaYeHj5LKfMATnzK0OvdHI/IKCSouROcHutMmk8R2c5VXsLvoglRkEHcIL4Y+sqVzOg4VlPI1jtzhSyOD3ZbvlB339iy3/rpdU5/4AV38o25Fd7Fn8KTVP8AACu/lG3Ig9pEREBERAREQEREBERAREQEREHHvMR+D/xN/gdev1KVfM/Wf21P/hXfpK+mDmI/B/4m/wADr1+pSr5n6z+2p/8ACu/SUEKIiAiIgIiICIiAiIgIiICIiAiIgq0gYV3iEdlYiCpcXHCq0YPdW9lc077oJmtGFexnmrW9ld19OyC4uGMKzIacq0yDJVpkBGEE7KyNmxHYqKWVsxwBjKgwVdH0h4ygmZSPcAQdsqRtvef3xWRC8dI3HdZcIc8gNAOfNBpv3Nd81NHb87ELWG2+ocMhhwpWWmsI6gw479kGiGzuk3bn0WXT6UnnGQTutXjoalnwlu/1LV6KR8ADHgA7INrO0dM3OSdljSWF9OS0grkCKGrqDljcg/JaLeKeaCT4xj12Qbfp9OunI74PzW4LfYG07cuGfrUVNVeEBv27LNF6ia0dTgEFH26KLJ6Rsr4Kimp3dT2DZYlZfabBy4b+a011yglJw4EH5oN+WbWdttbgZImnG+63jS8aLFCwNdBHt3yFwbNPSuGC7f61AIqeTJaSR9aDnqXjfZG58OJg+wLTKrjHbqg/Rbj6lwu+npwCXEqHFI0n4j+VBzOzjHR05LowATt2WwNY6ruWpZzLS1TmgnYBbVeykfsHlapbre97AYQSD2QYZnvLY2tfVvONu6yqWSv8MvdM4kb91JWUksDeqQYB9Vqdns1TX0j5IckAEnZBBY7TcNVVQo2vc7JxjJXIlr5a7lVuYHxPIeM/lUnL9oG96m1SKK1xF8nigds75XfC2cAuINCaZ0lG8AAA/Ce6DqDaeVCooYxUyQOPn5rWYuB0drmbcH0/wxEHt6LuNqXQuqdP2nqrabpAbnt8lxVco66pt88TYxncIODtba3oLNahaqWIMkYMbbHOFxlYtUV13rzQGVxEuW4+tbo4l6Srpax8j43DJO+Ft7QGj5zqSEtJJDhtgoO83JFw5+4d5hv1XFkGQP3C9HJb/bInMaYgS8AjYbLqFyu6aukNrg62Et28l2lqrFO+SF3ScNAyg3BVUFv1BRiOrgbJGdsEZ2WlycOtJmgko47RCA8Hfp3ytYowaKmAI8vNXsuUb3hgAyTjug6S80PKjbbtSSXKht7W9YJ+Fq6Vx8qElHBU1jqc4YSd167caY66bTrmUrMjB8s74XU2ts97ktFZCI9znGyDpfoTRlPab+aF8ABa/HZc3V+k6OKkbUiEAtbnOPktvM0be7VqmStqYSGdZOSPmt93SqlNkdJ0/Cxu+PqQcOX3iLTaQqHRVhBY396d1orOZLTBPiR08Qcw98DuuMuNeo7ZJcJYHyAPGQQuFaettmXt6z8R23Qdkte8xFHqKi9yge0DpxgLhxurg2R0jnDBOVtGVlICXxuJHfKhdLGWkZOfJBv196otQsFM2MdRGCoDopnhkdIy7cLbWnamOiqfFPbOVvb9lcJLQCNtu6Dadx4dSPcXtGMrS6bh1Uy1rIS44LlyNNfWviBBGCsGC7+FVNqNsA5QalR8Jfcbe2oPfGd1j/cJtIMkD4fmtwVPFC3e4CkdM3Ibj7VtSq1nRTBwEjST80GPeZmMYY2d8LafVNLN4fWdzjZarW3OCpcS05z2Wmx5bUh47AgoNVh0zVOgE3WcFUFJLRPD3uPSD5rW6W6NNG2IYJUVTRVFXETG0nKDS66+U0sPgiMdWMLZdyt0lTKZG5wTlbkqbBWNlLiwrHkoKloLOjdBtkUvR8OOyhfUeB8DRhbjNgr3uLww4+paXWadrQ/JYRug06QTygOa44znut6aV1VT22mFFUtDnEYyVt+G0VYAb0K+Cw1Lqpry05BQbufSxVshqQ0YecrGqdCyXMl7Gkea1i30NS2kY3oOy1ulqpaSLpcADhBx5Nw4nZkZIIWDJoeoj7krkqouT3OOAFpNVWPJOcINiO0hMPMqCXTUkXfOy3nJVu9AtLrKpxBGyDar7PI0kErlbk2iLebThIw/vNaWj81XGuPZJyXHZcjcnLv+u54Tn11paf1uNB9HiIiAiIg65+0O/A24l/4nR/r1OvnjAycL6HPaHfgbcS/8To/16nXzxgkdkFSR2VFUgd1RBVhAO6uJyrW91djCCxERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAV7D5KxXN+fqgkBwrXbnZMhUyMoLSwhXNBx5/lVSRjurm4x5IIw1x9VT4gcb58lPj0VhY5ztvJBVlO5wzkqdk5pgR1b9lfFlrdwsap79/mg1G33J7X4ytVfSOryHE5W16Z7WSBzvVbwtUwcwFmCMIMKptIhaS4dhlaVM9sJORlbpqxLUgtYMlaFV2mcndp3Qab74B9EAH1UkcT6kbHGUltjoT8YOFJTTMgI+LZBj1FJPSjqDyAqQXGph/+SnHllZNfWR1DOlrgfIrTQ0D6Rx8kGsUeoJYHZkeSPLK1ml1tFDIHEdvktp09M2od0tcck7BZZs0nzQb6HE+m8MRlg2+SzrfxMt7HF7o2nAychcaG0vbu4kBSwWd7wQ0EkjCDnvT3ESz3YiEwMyduwW622K3agc2ligaC/wCS4J0bapKCZssjSACDuuY7Bq6ktFVHPI8AMwclBk3vlmfPCa6Nhw4ZWy6vl0qYAZegrsFRcZrVdKFtFDOwuGG4yFJJc5blH4cGCT2QdXKvh46wu6Xxn4Vk2WjglqxEYgQTjsuXNZaNvVSHTeGek5PZbSseiLpHXh5YTh2cYQckaA07b6RkM76YZyD2XZxuorTQaAmhjpmB4iIzgei4S0rp+uZSxNcw7Y8lyddLFUt0dM5zT/Yz+hB0/v8AV1dy1lUmOQhhlO32rclJo2odTe/knbfK2/JRyN1dOBnaQ9/rXLdvp6mWymFrckswEHEl3vYjc+jdv0jC471EIqlrvhGSdlyLqTSteyvmmdG7BJOVxvd8RVPuzvp9WEGzJrOfGL+k4zssCoiEJxjC37Lp+uMRmDDgtyNlsu+R+7S4k8ig0/pGc4VzjlvQBuoxUROGAd/qU0ERnkDWHJPZBiSUcgOerudlqNDauoB7hnG61AWOrkY13TsVmwUs1NHhzfJBpzqMDAxsCtdtk1PTQ/fGjOPMLAd1ZOyo6ZrG4Pog1Ga70rM4YPyLQa+5CR+WbBQ1U8YyCVpss8RPclBkVNY53Zy0t73OdkklTv6Xj4fRGUcjxlBbHUBo3GVVzRMeoeqOopAVJCwswCgsbRl24JCGN0JAO6zI5OkbqKcteSUHfX2Lf4Umqf4AVv8AKNuRPYtgjmk1T/ACt/lG3Ig9okREBERAREQEREBERAREQEREHHvMR+D/AMTf4HXr9SlXzP1n9tT/AOFd+kr6YOYj8H/ib/A69fqUq+Z+s/tqf/Cu/SUEKIiAiIgIiICIiAiIgIiICIiAiIgu6NlaRhXjJHZULfUIDWnvlXdPzVGo4kdkF3iADGVa+TPZWfWiBurwxxIBV0cJO6kETxIMDzQSso+poOPJXi3dRz/Os6GN3hjbyV4cWDsgx2URAG52WXT4gIcRnHyVpq4wN1DLWxHIyg1qPUFPC0RkDKkGsqWJvT0jOFtZzonOLgfzrEmjaSS0oN2u1jTlxcMb/JQHUgmkD2OwM+S2oWADOVk0wY0jJ80HIVs1lBTMAeB6FLlXR3geJGBgraMVOx7AQO63VZLXUSU4MYJCDRawugyN9tlpwjmqScOIwVrt4ttSyQhwI3Kw6OjmZnAQac6y1NRhoeeylhsM8Gzs9lrtJT1HXsxZ5oKuRw+BBolFpaercO/1KSrsEtvPSQRlb2sFmuD3jpjOfqVdQaduL3gujI39EHH77ZJKBjO4xjK1O26EmrWdWCc7rWRp2vjY0lnbzwtw2V09HGGvaMjZBtIcOZWnBB2WoUlqZagI3t7bdluaruxhBc7ABW3a66R1Ly4EYz3QYGoaeOogHQ0brk/hJpWCrtErZY8lzCO3yXGc5NVG1sQyc9lzNwtuLLZQeHOQCWoOYeVFtq4b69N0uFO1zBMHHqG2Mrv3f+afRcVPGI6SEEEHsPReb2mtT2+juhlq3gMLsg5W+rhq7TtyYxkEuTgDuEHOvG/misN5tjqakjjaekgEALq0/jNTQUtTK8tI3KxNdU1JPTGSLJGDuFxQ+1e9wT00TSXPBCDmDhDWUfHTVBsbIQ/MnTsPmu3Ft5IINNVtPc/dNiA4/Irq1yLaEu+mdfMuzonCMzB24OO69Xr7qZzqKBrWtGWBzvPfCDbPDey0WkKGOiEABZgHZcjxX2CSVsRaPi2ythWipbcarpbuSVuOsphbYDXSghsW53Qalq24MtlsdUdYbjJ+tcY2/iGHVQd44w138623xP476ajjfaZJ29YBGOoLh+k4r6bi65HSgAHPcIOxPEDihROsphJaSQc/M4XBdbxCoqa2VNUYmgNySsWn1tZuITza7NIHyn4cAg7q3V3CO/W/SlXPUU7w3pJJwdtsoOtfEfmItL7nJQwMYx4JGwWy3ccoqmgktXijMu35VxLxRpaO1axnincQ8POxPzW1qK42910jAdv1DzQTcU9CV9ylffGvcWyAu7riaHSdYSX5OGHddldX1kk+mmsYB09C4ip6eZsEr9sb+SDZs0UlMzwSdwFhCR7cuPZahdZ2vqXRg7jOVgOhcWlvqgtbcX7Ma/B7K5tdUB4cZT6rFbb52v6iDjusj3d7sN88YQaxS3+WRojL1lC5vm/2qHnLzgLQY6U02HvyB3U1NW07KxkhPYoNbdoK51LffBM7pdvjKjm0TXQxGXxHHHzW7qTVVO23tiDh2wFBLfoqmI07HAudsAg48mdNQSmJzjsgv/S7G+Vq9605cJHmr8M9B3yFtmS0zMJc4HA3Qa7TagfEA4u2G+MrXKTiJBSx9DsHC2G58LR4fVuNlA+Fu7m5Qb+n4iUspPwjf5LEOrIJZOvAxlbIZEXOwVnR0vS3IzhBviPW9FEwNc0ZWPLqikqjs1q2a9sI2Oc/Wr4TG07HzQbyZdqbAPSFG/UlNTvB6Rse60SGRhb37fNafcHxdRGfNBviLiBTxt6cjssGt122Vw6HAA+i2T949T+VXNp45PiblBul+rzjPWN1YdTeMD8e5+a2w6DyV8cQaPMINe+7ZJ3cVjz3EyZOe605rBnfdVcAMIJHTk5K5Q5Nz/12vCQ+utbR+txricuA7lcr8mu/NnwjP/dpaP1uNB9ISIiAiIg65+0O/A24l/4nR/r1OvnjHdfQ57Q78DbiX/idH+vU6+ePCC49larj5q1BVn0lc4+Stb3VTv3QWoiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiIKhpKEEK9rSqSAjyQWZPqmUTBQMqocRsgaT5IWnOEErHY3wsqNre5CxGgflWXGSWoLsK00hl3x2VynjmYzIcQMoMQ21wGxK1GgrDQx9BPlhRvqmYwCCSrG0z6j4hkhBqtPfWROzIAQfValFcIa3DgAtqvoJj3ytYtjfAZh22O6DVpLYy4N6Wjf5LBqNHStaekEZWq2+4RxPwCN/Nasah9Rs3Bz2QbJp9HTeL8eSFr1Jw2fXDAYd1r1NQVkpBYzO63zp62XBrR97OfXHdBx3R8K30J8R7SQN91kVWlWQNL/DOy5Xq7fcXRnqZstu3K11xhdlmO6DjM2ZlTIYmswRt2VwtrLeQ97OxW46a11MdUcjH1hUutqnnYd9/kg27LqGOAeFG0AjbZX0za2/u93ieQXjGR5KEaRrKufDAd9zsVuzS+lK+3VjJHA4BHkg1zQXCi80krK6eoeWlwdgnbC5zslrFtcyeZ2Q1o7lbVpNURWW3sZVEDpwe6iquLNkFO5rpRsMdwg5moKi136IUfhsJ2BWRVaNt1siNU2nAxv2XEvDrifZX3DEcoJ6txkLli9a6t9Xby1jwct/mQY9n1JRsqhShoHScDZcp1lZS12kJY2gHMZ8vkuuFsu1ILoZeru7P51zFSanoHafdCHjJZj8yDrje7O2h1NUVIbgdZOftWrU+tILdEGOxgbH8ik1KwVN0mka4YLj5LaV3tjWwOkcTjugm1NrugqWSNa1uSPL6lwJeq73nUIka74S/OPtW77xWUrZnxAkEDdbArSG3ESZ26soOWYqykfZmN6AXFv8y4r1VZXV9STE3HxZGFuumv1IKJkXUM4AUbaGa4SiSDcZQbCj0bO0h7g4j5rUaOxe5yCR7e3qt7SWeuZFks2AWgXGoNKC2XA9UGTHcKZkAi6RkLQrreImEhuB5dlH90KbqOTv3WhXWphlkwHY3QTOuuXF2e6x5qx0h2cVg9Ufm7ClZEX4LTlBSaN9R2J3Rtpe4ZJJWRG3oxkrJZVsYMFBiR23w9zk7+qlMzIG4wO6lfXRAHJWm1M8cj9j3QSy1zN8ALGdVhx2GFR0PVuFEad2UE7JTJsFV2Q7cqOIeCfjUhc2T4h2wg79exdcDzR6oH/cBW/yjbkT2LgA5pNU/wAAK3+UbciD2hREQEREBERAREQEREBERAREQce8xH4P/E3+B16/UpV8z9Z/bU/+Fd+kr6YOYj8H/ib/AAOvX6lKvmfrP7an/wAK79JQQoiICIiAiIgIiICIiAiIgIiICIiCVrgFR7hsgAIx5qngyHsMoKBwBVesKvu8v9yqeDIDjpQWE5Kkij63gZ2yngSeilhp5+sEA4yg1CCmb0DI3x6LNZQtLfEwNlFB1NYA4bhZzapjYS3bOEGmTV7aZ3R6dliSXLrGAorgA+YuCxOk+iCZ0znknq8vJUEUjt+pWMBCyonBox6oIhE8DcqgBGxWQ9wOMKB2c7IKEZV0bHZ6h5KgBx2UsewOUGTHX+AwArkTSF/gbSdD2A7d1xa/oOC71W8tMVFJFTgPcM4Qbhu1bBUyOIYPyLatZeY6F5ZgblaxJNFLI7o7H0W1L5b6iScSRsJH1INQg1fFFv07rWaDW0D9ywbBbBdQVDN3tIWZRU8bdnE5PdBypaOJtJQyElrQc4WoV/E+hrSCGt29FxQKCOQgjOVkxWzpHn9qDkCXiFQvGA1u2y0up1jG92YyBlbVdbQO2d1jysiiJD3EINeu2pX1EXSH+WFplFXyOaep/V9q06oEUjMRvyVPQU0oaTug1qjvHucge85HzXK+kKqS6UwkhdgAZ2XC7bZUVxEcAJOcdlzVw2tlTbqARzsIJb5oNWFtrLlMaeGUhwOMrMZa7pYnCWaoeQN+6yrbP9yq91RUjDc53UuotYWiqhID27DGMhBpF6104we6vfkjZaTpvUsb73Ex+C0uGR9q29dqy3185FO8E+gVdO0ng3mGWQkAOCDvlwi1hbNPWuKrhgayTAPUBhcywccX3FzKczA7dPddXNHXyyRWOKKeQA4Axn5LemnbvYJ7hHDBKC9xAAHqg7mcHrsbrWCd7uoFwO65O4kuMejq90ezgw4I+orh3gfTyWuNlbO0iHY5+S5R4iX+21Oj66OGYOc+MgD0OCg8iePWqb/Qa5qWtuDwwPOwd81xkeIN7kp5Ym178kEA5W/OYyhlj1lU1UxIjLie3zXC0FXb5nyRRuHWcjYoOw/KBru42HWgrbrVGWMyB2HOyO69D9f8drHetLSWqKBgdOwN8vTC8yeX61SQ3kVFS0iIvBH1ZXbm7T2YUkTo5AXBo+xB085kOHcpvc+oYQ5rXkuC6626gqW1bqsyHETs913F5hrl71bHwwYx0nt3XU2hgqBHODkg5/Sg3nY9Q/sgpxaHHJHwrcFZoBtHaJJejGWk5WwdCugtd18efYdWVzJddaWqrtBpI5GmQtwBkeiDqvqKhdQ3mRpyAHFYbalo3ONluTW9nuAub69zT4RJOfktnmWI5aDv2KDUTcI3jpDRnt2VjalrXB2BscrAawB2fJSZaRjKDMrKxtTEI2DBHmFpDaKUP6w45z6rMYA05UniD5IKROqY2hviHAU9JVTU9U2d7yWt3KiL2nzVHuYWkeqDfDtbUVZQij6Gl2MH5LSAyKrl93DBl+2ceq27RMijlDvM/Na9b6uCKujlJ2BHmg1CfhjIYffQDgjK2tcrSaBxjI7HC5jk1RSSWsQMeCenHdcaaho6use6aNpLckoNrNpww5wrnVrY/vfmr5XhjvCd9IbLAkhe+Q4GxKCR0TpHF+ThSR0zgOrfZSsgl8MDH5lNH1NYWu2KDEkqXQ/DnsjKV9b8YJ2UdXE5zupqz7UTFGQdsIMM2x4JGTsVNFAYW4Pks50o6j2UEsjTkIMdxGTn1VMhHbnZROyHIJWuGeyq4dX2q1uNjlXCRoGCgifCSuWOTUY5s+EYPlrS0frca4u8Znn2XKfJwc82vCQjsda2j9bjQfSAiIgIiIOuftDvwNuJf+J0f69Tr55G919DftDvwNuJf+J0f69Tr5484QVIxuqK49lagqz6SvcrGd91ecbYQRoiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgINkRBK2YAK17w7J9VaGuPkhaQgopomghQqaJwA3IQSsYDlRygMKvbI1ud1DK4yHI3CCocAQVlROBYCsPpJ8lNGcNQXPqA091WNrpviaTgLGkOT+VZVHKyNh6jugsljdG3OVmUd1ZA3pd5DCxquRsjMN81jtgc4E7oNaju0Up6cdysyJ3ijDdshbep4S14JBWt0k8ceM+QQTSeJT/ABdWMbrOoNQiADrdsFg1VRFJHjzPbdYLGxnclBvmj4gQUoHUBgFbkoOM1HSgH4RgeS4glpopBhuSfPBSOip2tIeCPtQc3jjfR1IEYx6LPoNd0t3kFOGgl3ouCKSmo45ARnvut76VrbfSVzJJHDA9UHPmnNExXlrZxET1fkW5peEEZaeqnPr2WLw+4maXttJFHUTRgjHmFyMeLOlatvRBNGSR2yEGl8OeAVNeavofSg4+S3Lr7gVSaXoXVTaUN6RnOMLfHBrWtG+tMkPQQXLdHGa6zXWzvhhYCXNOMfUg83eLd0qKaploqeQs6QR3wuCam5XMucySqfgncArnfjTpm6U13qKiaMhpyc/Jdf6tjmyOJPY4Qa5pLUVwtNwE0dS8NyCcu81zBaeJlbWyMp3VRO2MdS4HtzomzDxTgZW9dPVttpq5kkjxgd0HYvTbaitDKgOO5HZct6PsNbdauOg8RxDsDC4c0VrXT7aaKMSMyMDcrnzhhqGhqb5AKYguyMYKDkKfltdJbG15iJLxnJC2LxF4HfcbSVRcfCILGk5x2wF23kudbHpiNzxhgYFw9xm1TBLoKsow9vW6MjH2IPJ/U15kp9UVNF1nDJCzuoZ4zLCZPPyUWsbRWHWlbIGktfOXAqVpxCaYP+M7Yxug0mGepFUyPxDgux3+a7LcONHMn02bpNHnpZ1Zx8l12ZpG8vqI6lsbujqDuxXa3h1dordoaShqyA8xkbnzwg4sv2rKSnrpreGAFhIC4u1HcjW1RjjdgEnst0aot757/UzxA/E4kLZ1baquOsEhacAoIvuLM5gkDjurqbSstXMAcnK1iOcCFsZxkYWRbr7SU1WyOR+HZ7ILouF75IRJ0n1UMmkPcPvZaSVyJDqKEUYLXDGP5ltu6X+kc/LnAlBtWXThGSG+S0mpscjHZ3W7Zb5RnPxDt6rSau6UzyCCEG3Ki0SkYGVgPoXROGSditwy1rHk4IWlzydTidvVBAAGDfyCrlp3wkj2jvhWeLGgpND4o+HZRCMxjpKyWSs3AKhme0u7oO/PsW/wpNU/wArf5RtyKnsW3A80mqf4AVv8o25EHtGiIgIiICIiAiIgIiICIiAiIg495iPwf+Jv8Dr1+pSr5n6z+2p/8K79JX0wcxH4P/E3+B16/UpV8z9Z/bU/+Fd+koIUREBERAREQEREBERAREQEREBERBfG7pcM9lmMlYSMrBZjO6na5oPfsg1ASMLceqicY89lY17cDJVHPZnugnAjx2WTFIwAbLDbI3bJV4kaBjOUGRJUg7DZYU1Q4ZwTv81c+Ru+6w5nDJQVfJnJJyrA4eaAZGyt6XeiC/rBTrx2VA0jcp0FyC4POVcHAjKjLSNgrmgjY57ILi8BPEA7K3pyfNC052CC18ZduFkU9RNARh5CMhkcM42R0JBHUEGrU1zezDnOzn5rWaetgqWgOaCfqW12Na1ozt9q1W3zxRjc9kGpzUkNT8LWDJ7LVbRoU1o62g77rTKavgY7cjv6rfumtSUVPGA5w29Sggt/DkucepmSFLW6KbSEN6Fuii1hbo3kl7dz5kLEvGr7bI8Ykb+ZBteXSzWMDunut1aX4Cv1ZCJmRk5Gey0yp1RbnxNDXt7+q5+4J3xxom+AAQWoOMYuV2WKQtdETjdbE1toAaSmNOY8YOMLuVLqGSGolMjQBgrrVxvu0VXcC7I+kUHF1CyCyYq5mZHfda/TcTadmBFgBvpstt3mqjkt4DiMYW0o57fF1db8FBybe+IYulOYYHgOxjI2Wz3T3GVjy6cnOT3Wi01XQvkHRLnJ23W5Lfa57jiOlcSXDbGUGp8ObBU3e5hszi4FwGMrmit4dmijbVNZggZW1uFGg9RW64Crmjd4fVkHpPZc4X6nrTbSSzYN3/Ig2jw/slw1DcBa45nYD8YXPmnuDNysV2pbhLI8tBBwT3XEPA2rEGriMAuEg2+1dv7nfDDDA+YANDRk9kG5rhxJi0xpRlDC4MlazBI+pcdUnG6su0htMlWXCV2MZWj631LZrlSGnieDJjdccadgp6S+xVL8gB4P50GTzO8NGVGnH39kWHPjLur7F0bsOnKplxlqHPJaxx8/LK9HePV7i1Bw+bbbcQZBF04G57LpLa+H+paUVNRNE/oJJzg9soOV+BhiuczKCJoDwQPtXYy+aEq6S2ioc84Dc/mXWnl7rYLVqQQzkB4kA3+tdxNTalpp7MyFjgSWAd/kg6bcXw9rn0shJwCFws20xx0U04bjGT2XOPGWknbVSVLmkMOTlcOe9RyWyeFvcghBxbU3pxuTqWF3SQcd91kQVdwjuMTn1DiwOGQStFraKWivb6mbIaXZ3WZFdqOaujja4E5AQb61jX0tXp4RCMdfTu7G5OFwm2geJXEuOMn9K5fv9FKbKJmg4LM9lxh4cgc/PkTlBAYiGADurBEc53WQ4+XmrUEMp6G4ysUzO3+I9/VZNT2KwS057n8qDIimc7z8lMQfXusaFpad1kgkkAIKsY9pyCpAZWnqDiD65VW/MKqDOt1VVZAdKSM9iVv22sgnt5Y9gJI81x5RPDXDJ81vmy3GmbCGEjqI23QbVu2nT76+VrcNz/OtJlhbTvLSOy33do5T1SAbHPktjXOQCU9RAQSRyRhvbyWNUPGfhRkjC0Yd3UcmHEn5oLSWuG47K11QIvhacJuoZGgu890EhqCfM7qN0jnEb7K9sGW5Csc0tIHzQSxnIwop3AOx81NHjGVBOB1IKA/WqEnyVYwCN1V+AMZQQOc71XMHJnn9tjwi/hpaP1uNcPP7rmLkz/Cy4Rfw0tH63Gg+kJERAREQdc/aHfgbcS/8To/16nXzxjuvoc9od+BtxL/xOj/XqdfPGBlBceytVT2VEFWgkq4ghWs77q842wgjREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAQbIiCVsjQMEI5wd2CiVwxsMoKdJVQ0+qrgq9oAHbdBaWOOO6q0hmxV6ikPxbFBc54wCqeJgYCsJGFRAJyqguxsqKRkT3/RB2QUa4tILvVTiqY0YA/MoZIZIxlysa0uQZsc7X7D1U3x52KwoGlrt1nNmYCCe/1oLX9bBkuKrG8v2CrI9sowO4VsXTFu5BKZzTDqf9mVC+s8fJaceqtrpGSNwO4WPTxuyThBlRve3fqKvfV1LBlkpB8t1YGuA3Cte3qCCn3buzDhtdK3Ho7C1aza2vlBN1vuEhHzctAdA4u2VRT/AN3kIOw/DbmMqtMFniVZyDncrlKq5vI7iGsmnDgO+Tsul1NFSZHiOPzWrUsdtDx0vJ8u6DsVqrXlv4isMcUbSXgjPquPbnwgPuT6oM75d2WHo66261va9zgADnuuSazX9qqLV7vHIwuLcd0HW282R9tqnwtyOk4WnNFQ36LyMfNbx1TBNVV0lQ36JJK2s+CSPJd2Qalp2uudLUNc2peGgjYkrsXwJ4h1VPrSkp5qgkdQBGfmF10tTo3yAE75C5W4LUjJuIFGxhOS8Y3+aD14priy7aCpntAy6MfoXDHETRFXXWOZ/USzBPdcyaS07Xnh/RyMYS0xjG3yC0bV1hurtOTtDCMtKDzyunCqCv1DKwU4JLyM47rVf2q7/dTfPAPQ0dWMFb/uNHXWnUcs1RsGvLjkdlu2p436aotOPs01TEJi0txkZzhB1rq6W3WyZ1qfTtDoj09vRaFc73LStMED+lhOMBa/fqeS43ue4QHLJXEtIG260G66FvlREa9kbzENycHGEG2qytjBdI8ZJGSStl3zVFOxxgDBncLcN0qIYXvpJHYkaCDuuO7xQSz1ZdECeooL33gxuD+rZ61Wi0pVXWL7qRvIAGditFGnq6RrCGEgd9lv+y3mntNkNFUEB5bjH2INsOvtRTE0JlOWbbrQLhcamSXaV3f1UtY+Oa4SzNOQ5xIWnVQxIcFANZUnvKVT3qc93kqLpPorxE4oJWVTwd3FXOq+rzWMWuHcKmMoJJJS7bKs6neqqInHyV3gP9EFrXlvmUc8uOVXwX+bSPrVrhgoO/3sVT/10+qcn/sf138o25E9ip+FNqn/AIv67+UbciD2pREQEREBERAREQEREBERAREQce8xH4P/ABN/gdev1KVfM/Wf21P/AIV36Svpg5iPwf8Aib/A69fqUq+Z+s/tqf8Awrv0lBCiIgIiICIiAiIgIiICIiAiIgIiIKkY3VMn1VcO+aogva8nAV2HHdRDuMKdgONwUDJA7oZS3z80PVkqOQEYQC8n7VTpLtyqNByr8OHkgkb6d8q7A9Fa0gd+6r1N9UFSPQBGgDujXNzuVf1RoLDgnsiuLmYVGuZjfBQXNb5o5oG/orvFYqEscCUFfe2xtDcDKkZM2bcALT5AerYFZVIOkZQTOORgKN0r4zgOwryQO5UEpBcgk95mGD1Hv5rOpa6qABbK4LTWgOxv6LLgw1oQaiLlXZ2nKuaa6pwTO4/WsOMs7krUqSpgjZgkZQWxU9Y4hvik4Oe65L0dxnfoeAUr5dwMYK2BBXU7XEuwFot5p4qubxIic/LzQc5VfMea0lzJ8F2y0eoq5dak1HWXZ328lwvHbmggv6ic7rljQl8ttrpw2Z7RgYQQ33TcrKcR4PotZ0XwAqdW0rqhgJGCQcql/wBRW6vZ000gyXYC5+4C2zUFZaT7iwuDmbY+pBxnovlRqbndfdSwnDsHddj9H8oIs9XTPmpCRkZBC1bQbb7pK/urLzGWxB/Vlwxtlc/UvHzRkE1OaqeECPAdkhBxLxb0da+FWkRWtoRG8Rk5x6BdUWcdae8MnoQQSCWrt5ziccuHWrNDGgttTC6bwi3AcM5x8l5p6bmtMFwnke4fFISPyoOYLFrw6PuRvfX0gnqyt71PNc6+MFEypGcdI3XCOpqu31Ns8NrhjGNitmafp7bBcQ9xOA4efzQdq7Dqivu0/vklQXMJzuVu1t5PisDHYOy400LV0dXRMgpSOrYLe0Voqveo2jJLseSDkjT0U90aGVbzIw42JyFu6+6UtMGk6mWOhYHeGd+nfstA0haq6zU7auuYREcHJ9Fvi/aotNRo+pijewuEZ8x6IOj9uvMun+IM3hOLGiU4H2rmC5cV6htO15nJDGjP5FwZdIxVcQJ3s3aZDjH1rd14tzWW4vcw4DT+hBp3EHitDqCN1E0guxhcY01TJEXBxyHFRXKe3fdN0cbvjBVT0Nw53ZBpOq6FlXCTEwBxWyKDTtRTVTatzzhjs4z81yFcHivjMNIAXDbC0Z+nr3DE+R8bgzHfyAQbjt94jvlC204BIGFfW8MxBbX1gjH0SVominRRXIR5++h2/wBeVzBd3VTtPPbjuz+ZB1YukppLjJT9gxxao/efmr9SU07L1UdbTu/KwQ05GPtQagAJcBX+4t7kFR0mdsLUhnpzhBp7qXoO32KwfCd1lTl2TssRxySSgkMm3dWmTuMqxWu7oJBI4HKzrbXTCtjYHnGQFp47KS3ENuDC7sHboOVJWNmtbT05JauOb3bz4rng7Z7rkhtVAbQ0NIJ6Vse7wzyFzmt2yg274TmAD+dACNipXRyh3S4bqxzSDugoFiTP6ZMfNZzQO60+rY4yZ8kGVHUNDRsNwoppA92VZGzLQCVZLkHbKCeJ+RjKin+kr4G7Z3Vk/wBJBRpyFbK4hSR4IxsrZmgg4/MggB37LmTkz/Cy4Rfw0tH63GuGgCD9S5l5M/wsuEX8NLR+txoPpCREQEREHXP2h34G3Ev/ABOj/XqdfPGO6+hz2h34G3Ev/E6P9ep188Y7oKkHuqK49lagq36QVVRvdHZGyCiIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiArmtVqvagkB2Az2Vjn4OxVHE+Ss7oL3PJ81aAXHKorowc9kEkcJJ37bLIFHlXRj4dgslhcW4wg02WnLRkK+nnbCNxlZM1FUyk9LSVgTQSwHEjcIJ5qhs46QPqV9NS5HrlYcYJcMZzlanTzsiAbJsggqGeEMrEJc7ss64TxyN+EjOFixUk8ozG0lBNQsd4m/n81NWNIGyUlvq2OyWkD6llS2WuqWgRMJKDRA4h25WQyqazyWU/Td1Z3pzhSwaSvVQMx0pIQYrKlsp6e26yAzOyy4tCajlI8OjcT9RWWOH+qWHL6d7fsKDEpKJsp7d1qLNPeOenp7qSm05d6Eg1EZGD5rXKKOZhBwNkGkR8P5J8OYCr/wBgU9NhxJwDvut7W+omIDW7lZFXFWSRnDc4KDj2ew1UY6GPIx81HbbPc3VzGvncW5GxK3TU264yOJYw/YordbLnFXMe5h6QRlBrlRpVslva57MktBytqXPSrI4Xu6MY+S5TbR11fQNipWkvxjb1WiXPQWq56d4jhfk+gKDj3S2k3Vlc2MHOXAd/mu0nArgv4WrKS7PjJDCHDb6lwvofhnrWmubJHwyYDgex9V3M4MWm/W+tg97jIaAM5+xB3b03eaSj0ZTW50XxRtAP5FgXC6UVwpjQeED17YwsG2zUz7NGzPx4Gd1DDRv95Ewbt1ZKDiziHwVjuYnrYIMF7Sc9vJdLdecuuohqgVEdTIIhJnGT6r0r1PeRBQOjIHwsOfyLq1rfV0H3a8HLc9fbPzQceWvhMKe1QRSw5kaBkla/frRbLPoueB1K3r6CM437LesNe99C2RrBgt2K23xBcH6Vnkc3HwnP5Cg8/wDVdPIdV1LmHDOsnGVpMpjp3GR7Qcea13VFdTDU9SwkZDytvXSrpjGQCN0GXHqqihYYiwbDbZbSvVykq6ovjkIZ6A9lYTCXkkrDqOjrIYchBcyYhoyclRyO635T4cbOVoGSgmjAwFkNxjcKGIsAHUpvEjxhBjz9zhQtODlTTEH8igwUE7ZmgDICk97ZjGPzLEwVXpd6IMh1Swjssd7uoqoje7sFRzS04KDv97FT8KbVP/F/XfyjbkT2Kn4U2qf+L+u/lG3Ig9qUREBERAREQEREBERAREQEREHHvMR+D/xN/gdev1KVfM/Wf21P/hXfpK+mDmI/B/4m/wADr1+pSr5n6z+2p/8ACu/SUEKIiAiIgIiICIiAiIgIiICIiAg2REEvU3HzUROSrj2VqCrSAd1OHjCgb33V+Qgv8UZVHkOOU+HuhxnZBRjcKbIxjG6jb5ZUrenBygxpNzkdlZk+qvkaerYK3pPogYcmXDzVwacBULSUFA4kjJKuGfIqgYRuQr2g4QWdLhvlXNLt91Tc7K4Ndjsgr1DzUjJhjp7KB7HDfuCVWMEd0GScvGxRsDnearHuFM3IQWMg6VJ0uBVfEDfpJ7xD5oBDlafEH74/lV3isPYqx7snLSgoDNn6ZVPfDEQHnKdYZ3PdXw2O43F3iUsJeD6IKx17HkeW+Fua06brLrCZIXkDGVt4aUvAcM05B+orkjRrprVSmOqGCGkboNT4Y8K6y/3ZtLM4vDXg4zldtdM6wt/BeOChqImnYA5GVwpwOuE0uoT7tuS7yW+uMElD79AbrgHIKDXuNPH6jrrE6a3NbE90ZwWjG66e3Hilqe4mfwbrICXHpw8+q5R4myWOXT4bTkE9GPzLrzSwwiWXpPcnCDIrb1qu4yE111lkafIvJGFjRGphkD2yEHOe6ynxODfhUQhlJQar91Z6iAQGQ+ndQAy0rXzhxBAJUEMJgw5+QpqqaKalljY7J6CPzIOe+UF8+sdWMtMzy8eIBgr0SvvAqCyw0t1MGAGhy87+QqkqqbiMyrwQ0yt8vRetevrxJJp+BrnDpEYzn6kHVLi5r6mtFqNopmBj2NIyNjlcBRcRK+S31NO+pJD8jGVyFxzrLVJVy5cOs5XBVNLQGKXDhnJ80GhG4mlvj65/cuJytcuWt21tC6kB3c3C2vcqikZUuMhw3Oywvf7W7aN4LsoNu11jqGXF1eXEgnPdTzyOdSOwewwtRuLpnMJH0Fpkhb7o93lhBt6y3/7m3QumOQHZ3W+K3iJQVVIaNsLQXN6ey4uM9IbiRIQN8LUuu3Nma4EZygzqN7rTcTdQcNLs4W6BxajrsW3qBzhq2lc62mfR9BIIwtuWv3CO7Ry526wd0G/tT6OjqqU3ZseOsdS2H9xMMe70XL13u8dVptkFOQT042+pccOpK0U8jnA43yg2o2XwJzHjsVmCtGB3Wnz4bWEPO+VN1R4G5QSSVIeFjkjKOLR2KjaSXbIJ+kgKw7nKm7sUDtjsguBCRnolDxthW98KvSSg3BS354iERfsNsZWqQ1TJ4PiAJPyW0qeNodnK3JQyQiHBwSgw5qdj5S4BaTX4ifjC1stkdK4t7LRLo1zZD1eqCMPBAKr4LZdz6KFssbQOoqeKZnTt5IIvd8bAqGSLBOQsl1THnuopHtkOW9kFrNsqGoaXFZABx2UMv0kEcbD2BVz2n1QZ7hCSchBjkEFcxcmf4WXCL+Glo/W41w+8HK5h5M/wsuEX8NLR+txoPpCREQEREHXP2h34G3Ev/E6P9ep188je6+hv2h34G3Ev/E6P9ep188je6A4gqiqQqIKs+kqv7qjPpK5wG2UFiIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAr2uHZWKrfpBBL05xnCtMe6vHYISR2QRFmAqtcGn6k3Ktd3QTsqenA32WTFWguGx7LBDRuVVnS12UG5aO4RMALmjfbcKyso2XB2YwBnbZYFPLAGgOI/StYo45p8GmyR6/JBotTazbx4jt8LDcHVGXNOMfNbgvlBXPi+JpwFpVPp27VDS6GncR5oLrNaHXKUNJzgrd0FkioMBzAtP0tpm+xVZcIHNA8lue5UldC8e8MIIQaZMIAMCMLU7TU00YHXGD9YWlT4A379ypaOspYSDKRgd8lBuC4XagbEMQjb5K+16wttM3pfAw+WcLQbhc7VLF0scCcYwtKjfRk56h38kHIzOKFmtvxvpmEfUoqnjZY5WkNp2A/UuMrvFSSxYBzlaAaGENO57oOUanXtDe3FkUbR5DZYprmR5Ab3O2y2bYGU9PL1OPnvlbmlrqLIPU3CDLOpRbz1E4xur28RIcbgHy7Lbd2q6CZvSCPktDjip35a3uUHI8PECldgOaPyKaTXtH0HDGg+vouPae2OcQWg7rPdZpWxOc5hxhBzFw111BLWtErQ8dQ2Iz5rsdpe+2a5VUVO+iYQ4Adl054bVdstlc01jwAHAkE+S7MaF17oylukD5J48DG2yDsRBZbDRULattuYDjIPSoLTrehhvDKGCJrSSAMbeanm4m6Kq7GyOOeMnp8iPRce2u92GfVkUkT24LwfzoO2tojm+5UdbvhzQ75LIfqP3aMNLd+3ZQ2nV1gptNQNmezAaM5I9FpNXxG0JLilEkfik4G47oNta+1XKIJy15A6TsukPFfW9Tarq+4eMQGPJ/Ou4PECSjmppqiMDwnMJBB8l0T461lrkbNB1jJcRhBWPm2NLTtozMT0YGVbeOaBt9s0lu8YEvGF1iqLfmrcyMkMJyPqWp0VvooW5cSX/WgzLo+a43aW4hxxISfyqBtslqpQwknPzU7fhOB2CyKKvhgqA5xGAfNBINEPMYkwdwtNl0k8T9GCt+x6otraUMc5oOFpUlaKmqE8WC0HP50Gjs4evfF4mD2ytFuNg9wcWOGPLdcnw36nigEbiMgeq2bqGkrrlMZKZpIJzkeSDaElEQCQcLFe10Zxnutcks1zGeph2C06a11gecsJKDDDidiq9QA3UzrfVDvGVGaWYd2oLesK4StGxH5lQ08o7tVhY4HBCDIiqGtzt+ZRTvD39QGFa1u+6o4DuEHf72Kn4U2qf+L+u/lG3InsVPwptU/8X9d/KNuRB7UoiICIiAiIgIiICIiAiIgIiIOPeYj8H/ib/A69fqUq+Z+s/tqf/Cu/SV9MHMR+D/xN/gdev1KVfM/Wf21P/hXfpKCFERAREQEREBERAREQEREBERAREQAN1f4RxlWA7qdrhjyQWOZgFR5UshONiokAZ7BSMafMqNSxnKCpGN1b4hG2VfIfRQYJ3QS9YIQOHmrMHGACgBxugv8AEGcK5r24+1QHz+tXsPllBKXgoHAKIO3wSqknbGUF47q/I7KIE53UjfpBBOGNc3ssabDD0rJZNGwblY1Q7rflv5kF0U/T3xnOyyWyhwysBrC7GFPG9kY6XHBQXVMuxaEo6F9VuD23wopfvpHRuMrV7JSVIGQCMoMu36fdPhpBPYLctJoEzMBLSdvRWWcSxOHWMb53W9KPUtDRMDJ3tBxug2fJw5dJsGEY3WvWWOj01GIqiEEgY3C1n9l9s3IkaAVty9XihrpMtcDv6oMq6alt1MDKIWjPyW16vU7awPdAQ0b9tlj3+qpH02Bv9q0O3Ujpo3+FnfOPRBzxyyXzp1KRKc4ctR5sNVTQ3SBtPIWbgbbLaHLmWWrUplrdm9WVLzU3KhuN3hfSOBw7y38kHG8up6u70DaeSckAYIJWnw0TondR8zlafaHxMaMnzWsuqo8DsguyA0ZRsrATkdlE6eIhWAh5Iae4Qa1areL28QsHc4W5oeGUkJ6ntJBbvlaVoUOo6wSybDOd1yybxDVMEMLx1luPtQcs8lWi44dUsayLDhKO31r0k4gaSkbpRr+kjEWc/YuhfJ7M7TepWXG7/DD4gdk7DC9BtdcXtGV2l3UtNURukdHgDI2OMYCDzF462qqiv0zPEOASMZ+a4gZb54qCeo6z8IJXPHHtslRe566L+xEkhcNum67HVD1B2CDiQ3CW9Xl1tY85B6cBa8NC1dHIyZ73EHfdbSsFbTUGspJZSB8ZP51zBWaqts7I4mPaScDZBsvVNU2127w8YIauPW6rEkEkHUMnbC3zxGHi0Rl/ekZBXDVOIzK7B8/50GQ+lmnqjM15ALs91kGmqG/GXk43UkDukD0U0kzTGRsg0+eqmlaYuo7bd1ix008cjXh5BBz3U7o/vpcB3Kvy7JyPJBvPTNylqI208ryQNsLeNbRQi1vd4YyQT+Zcf6QDTUDHfK5Lr4ZRaXEg46f5kHBV4b0XR4Gwyoep6yLxg3V+/ZyhLSCNkFCHDuVWNwDgCrnEOGB5BWNY8Pz5FBlPeGsWI6pw7Hl5qec/e8LTiMvx6oM6OUEgqUzBoA9VjMjd0jCuMUhHURsEEoqN8jZTU1zeJhH1YHZYLnNbtndRwY95YfLqQcj26lbLTiUjORnOFtvUdOGyHAwtz2i4U8dA1rzvhaFqCGSsd1wbj5INqywuG++VLA1wjP1K+WCYHpPl8lRj2saWv74QYkjX9Z+ZWRTMIG5Vr43OcXN7ZUkJLfpBBOBnKhlZ8WcqZrhvurJBk7ILAB2Vkgwpg3zwopWuJxjughe7C5e5M/wsuEX8NLR+txriJ0LiFy7yaZHNlwiHprS0D/lcaD6QkREBERB1z9od+BtxL/xOj/XqdfPGDg5X0Oe0O/A24l/4nR/r1OvnjAyguPZWq49lagqz6SvcrG91cT6oLEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQE7og2IKC8RnCBpA38lKxwIxkKjy3GxQRl4Cr4oVmHeYVRG4+SC4SAHKrjr+IdlaYnBXR7DB2QW9BPZULHDsrw4BXBpdg4QR9Mmdsrc1ivMdui++YOAtuOcG91mU1BNWxkxAnKDkGx1lNqaTwAwHG2y3MZ7fpxwp5KduSMdlszh1G6yVpmrBhvfda1rC/Wusq2vY4YHzQarVa+tlnaZRCwE/JbSvHEGC7uLmDB9Oy0LUtTSVkAELgSN9vVbZhHS4l23kg3NLevFzv5YWl1lVLKD0PI/MoY+lww07o9hHbKCGGOqc8OLzgeru6yXTSxfviEpw4HJGFHWAkHZBa+vectLiQrWVJccE/nWKI3lXxwyFwx3QZTp3wjqacZVpqqiUbSn8qpPTTBgLmnA+Siia5u57IJoqaeodu8n7VqVLbXxvHV2Kst0b5XARjfK1oUlTHhz84G/ZBJFVR0UY6mg4U0epIKpwpQ0ZOyxnWWtu4EVG0ud22CkoOFmrY6hlQaN4Y0gk4PZBr1v0LV3XE9M4t6txhazTcNb/RPE7KqQEb9ytx6XlkskDIazYtxsVuifV1tMJaXs2GO6Db1lpdQxdNPJXyEAgY6iuQ9K2+409wiq5JnnBByVs6g1LbBUB5e3c/zrftk1dZnPZF1tz9aDeGteJ9ytNk93jrHN6WY+l8l1jquOOoKfVbJjdX9DZNx1/NcscTamiqbc+RmCC1dULxBAL34rs9Ifk/lQd0hx+kvenIqV1Rl5YATnfsuKta6KqtTUcl1LiRu7utr6YrrUaKCNjh17Df1XMkNJXS6TfJA0lhZ3x8kHU2uoRSV8lKW/EzI7ei00wv97DWuIBPqtw6hjeNQztd9IPP5Voxa6Ota5wwAUGtfc/NK1w8wtuXKnkYT0uIOe+Vu736D3RrQR2W27m1z8vA2zlBoTm1eceI4/atVob26hhML3ZOO5WD4rASD5LEmAMmAUGpyXqofJ1NkIHfutwWrWdPSQiOYBxPmVs3wSW5HZW+G0fSJQb3l1fSSk4Y0ZWHJeaaY56QtssjZlZLWtaEGqyXCB2waFQeFIMhgWmhzGkZWZFVwtGMhBKaZj8gNH2BYNRQgPOAO61BlZCPMbqKeUPdlp+aDSamAMacDCw1qlZG+UHpGVp7qeVpwWoO/fsVPwptU/wDF/XfyjbkVfYrxvbzS6pyP/lArv5RtyIPahERAREQEREBERAREQEREBERBx7zEfg/8Tf4HXr9SlXzP1n9tT/4V36Svpg5iPwf+Jv8AA69fqUq+Z+s/tqf/AArv0lBCiIgIiICIiAiIgIiICIiAiIgIiICZPqiIK9R8wqYKqBvuFVAVzVadgqsPfJQVI7nKAIeyNBwgoXgK5pDjsrTG8nPTsjcsOHbIJPAyM75VRT4GckKZs8eDkhUfO0jDSN0GGdiQqtcAMFVMcjiT0908GT+5QUzn7VkxN6tsrHLHNCyaWZjR8ZQUdSPecgqJzTE7pdus4VlOP3wUD6aWtk6qcdQKC6CMEA+q1qh0nJcwJIwRkb4WDTWS4vaA1hyCt96dqRaYw2rAacY3QaZbtBkP6HMyQVS6mLTxEZZjBx2W86LU1timL5HjBWxeIFzorlMTA4EZ8igxJNTsjb1tOPJaNWXKruDy6KUgZ8jhYcsXVG3oBJCvpOmEHr2JQTMkrsAGodt81kwz1DPpykrHbMwu2O+VKDuN0EFzmlLBl5wTut3aFpWVNOS8Z29Fs+5DLG433W+uHcMvupx5hBvbSNGYrg4046CPsWzeMNNUmvY6Z5d8Wd1v7SnVT3F7pRtkrZfGCdk1YOgjAKDjmChfGwSbjO6vkc/yceyy+rFM0fILDd8WfNAY92d3Ej0WXTTdMoJ7BYbWPByQVPGcPDj2HdByJpSP7p9MUYwRtkLkK1aZqKKuhnkJLRg4XHvDWsgFUGNIzkLmo1ccfhPlwAAN8oN2V2vxpuxtjoneDI1vcbbrath476hr71FQzXV7ml4GC7yz2Wj63vdnqLaYWuBfjyXG+kH0UeqoHk7GUEb/ADQdsOKtc12iWXGTBeY+rq+xdZaPV3i26pg6hvkd12J4qXCik4aRxsIz4P8AMuo9vdC2GpJce5QbVuZlbeX1MbyCXZ/Osyhu1aa+IundgEeawrhMw1rhnbPqraOSNtbGT2yEHIOrLh73YQzuelcUU1OWvc7OVyPfZYpbO1sZBPThbChpp29RIOM/NBZLJ0Nx2UDatzjguyM9ldUtOSDnIWKyPDsn1QaiHtIBPdHOHSfVRMJ6VVxPSUGu6OqjFWDO46lyhWXlj7aYMA5bj8y4l0vI1lUC87ZXIU1XSPphG0jqIQca3uhLa90+NicrBc4ObjzWv6goapshmcD0HfK24HDJGUFsLT1nKyzjAUUYGchSHsghk32WN0/GPrWTJv8AkUIA6xkIMpjg1gVHVTcFhAz2VAR0b+ixJHDxdj5oL/dnP+PO3fCNg6Hh2+xzlZUYcGZ8tla4OJJIQZLayVjAxr8AeWVq9DcWeCWSYJI7rbxkYNid1nUksRbnKDLmLJJS4NGCtHrabqk627AeS1UEE7FYlTkHtsggje1sfSe6imdvgKj3AbKzOQd0APLSN1MyQEbrFcTn5ZU0RGO6CdrxlUdI0dwretucZCte5vYlBd4zf7n865W5Nz/12fCT561tH65GuIepvr+Zcu8m34WXCL+Gtn/W40H0hIiICIiDrn7Q78DbiX/idH+vU6+eMHByvoc9od+BtxL/AMTo/wBep188Y7oLj2VqqSeyogqz6SueCd1aw4KucRsgsREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAQboqt7oJGQudnBIwrXtLDg5WZDJGBv3WNUva5/wAKC3rGAr2TtaOwKgwUIIQZBnaT2CiccnIKsVQRhAAwsiGRoHSoRvt6q6OF5eMAoMplA6fDm+a3FZZora0NkYD27rAoIJukYWRMOhw8QYwgy9Q3tpgIpfgPqFs99RUSkufK9x9crU7q6Pw9jnK0pjHuB6coJIDI54DnEj0ValuDkequpmOa/wCPsq1TmkDB80EdLJ0PyTssmWqHfHZYUW7sKSSJ2M4P5EGTBVB7sAYUkmHLCgb4b8v2CzAQ45G4QW9IxjClg6WvBwrX4jGXBUZNH3yBhBlV9XGIsBozhaeyRrmHsNlPPTyVxAp/iOfLzWTS6Nv1W4MgpiSfrQa5oO3MrakNcMguXKT9GRzgRNiGSMLafD7QGqLZUipqqZ7GZBGRsuW2VX3PeyWqADWBBty02CDSDxV1MIIB6t1uSp4w6fioDTilYHgY7BaVrDWun66jNM2VnWBjGRnK4lr57c8O6XbnOMINU1PrcV1VJJTHoBzjC2s++VsgI8c4J9VhVYh6z0HKxmYzjJQa5R3Ot6siY9/VbjstzuMVWyX3l2Ac91tKjZnGAtzWyMtexx7IN26n1RNNbPBdKSQ3z+pcI3qtdJO/HcuJz6Lkq/T0/uxbnfpI7riq648ckepQa1pa71FPPH1SktDh3K7WaU15TnSD6KQAuMeMn6l1Csj42TMc/wDulzzpi72ptm8LqAf0AYQbH1BRtmvtRVtbsXEraN3d4c2QcHK3zdIpJKyWRo+A5IWwr9tOQfXsgjjmldgdZwtRdGHUZJwTjv8AYtKgJ2WseG91ESO2Cg2i+Mmoc30KGItdnOVJK17al226q5ri0kjZBTxw0dJGfrUMmXnqGVa/HV37K9qC5mR5qdpOO6jYGk+SlA9Agika78yxyXg9yst5wMZCxJM5wgCSQY+I/lWoU8x6N9ytOGDjKy4XsaBkoM2Koa0nqGc7Kc08dQQQwBYHiRZH1+i1KlqImsHZB3u9jTTsj5n9TkNxnQVaP+cLeim9jdOyTmd1O1pGRoOtJ/zhb0QeyCIiAiIgIiICIiAiIgIiICIiDj3mI/B/4m/wOvX6lKvmfrP7an/wrv0lfTBzEfg/8Tf4HXr9SlXzP1n9tT/4V36SghREQEREBERAREQEREBERAREQEREBERBXOdlUDCADY4RAyM4VcFW4OftV7eyCvZAd+3Yqh7IBhBM2drRjGUdF42SPJQkeayaeRo74QQ+6vHqo3xuj3Wd4rC47hY1Q9ru2EFW1LWtxjsnvQJx091jhpKubE5x2GUGYYw9mQrobW6oeGjbPokEM5YBg4+patbqWqbIH42HyQTW7QtRXtBaT9i1L7kM04A2ZmfmVubTt7pKAdNQQMLStY3eir3F0LgcHyKDBj1NSURLjGPtW3b7qN1fITTu6QCoqpsUgwfRaVLTlrj05IQSmrrJgGiYnHfdQSGUEdbifRX07HNdkhXVDC8ghAiqQ3OeysmlEhyzbdR+E/0VfDePIoLoCQ4rIfVdGBn8ixmAtOXbJIOs5acoMky+PgfNcpaCcyClAx3C4op6aaVwEYOcrljRcL4KcCUb47oN10codUvcwd1sPiBA6ap+Lfdb5oSwTuO2Pl6LZuuZme9Dy3QbQqaUxUwO+AFpsbsZz6rWq+QPpRj0WhsGAcoJxI12wCvDC7LAdz5rHjc1rtysqJ7ZJhHGcuPZBuXRcT7dUioLiBnK5GuGpHzUvQx5BAwti2S2VlOwTztIYd91rE8sErOiI74wg0atqKyrqTmYkE+qksdNLHfaeUOOQ4FXGinpXmaYEMO+Vk2avpZLzC1pGeoZ7IOf77FPd9Gx0peSPDx3+S4sj4bvgtlRUAHzOVy6wF+mozEM/Dt+RaPNHWiw1Hw7dJQdTNQtdRXeWnB+icb/AFrBhncJmuDu261DWTXDUM/VsS7+daQxrg/JzhBvKgq31cAie7IxhZr7fG2Enox/OtGsVRC3APdbmnqoxAcY3CDZtxpg2Y4GFhimGCcb+q1O5PDpSQAsMEhp9EGPjBwqkZYVaHtLyPPzUji3p8kEVFO6CUkOx8wtYt92lnu0EBecFwBC0SJrQ/LuxWTaC1t+p3A5HWB9qDmPVun4o9LNrBGASzOcfJcFuaQ9+fIldjtYukfolgAyPDHb6l10dE/qlJB7lBdA89ieyme7DchYUD/j3WW/dndBB4mXEEq47DKgJw5S7lu6C0vz6JFAZHdR7KwD4uyyost8kGT1BsfSoy8dJCOeDsVG4jyQYskbi7usilL2AbkK17T3wjS5pwdkGcyfp3JVk84cCofEb5lUc9jtsoIHZLs+quawhqr053Hqr25xugic09kaC0YypHgAZwrcH0QWdJyrHhwOFKc+SjcTndBZ8S5i5NPwseEX8NbP+txriDpJHYrl/k1OObDhF/DWz/rcaD6Q0REBERB1z9od+BtxL/xOj/XqdfPI3uvob9od+BtxL/xOj/XqdfPIEB3ZUVx7K1AVcEowZKuIwgsREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQVyfUqaGAzepUABKzaKRsZAJQZEdv6mj4VKLVn96sllRGGjdZEdQHAhuMoNJfZzvgFRm1OBwcrWXGR3Zvn6KJzZSc4KDDjtWAMjzWoQ2sNZ1dKua94wMLUoahjYS12xQYLatlJkEYwtKuNxErssON1PXzRdbsn1WjTuY7PSQgq6R1ScZJwqskEB6SoYCQ44SfJfkhBLJUtdnpGFGGGQZUYaXHDd1kRU83kDj0wgtjaI3A91lCRrhlQyRvYPiGyrG9uAOoIJTF456W/WnSabOVdT1EcLsvcBkK6pjfV58Hcd8oMSao8X4QpqGgdUSeEM7qxlorpDhkRP2LWrLZ7hT1DZJWEAHKDeOjNIiMtnmj6gCDuuQ4LrarBOyaWmbhmNsLQtOXimoYAyqIGB5qPUl9s9VGelzSe3dBu67ccLJHS+7QQMY4DGekLjbUfEs3amfDBJguyAtm3h9vlmJjOc/NaTG2EH4TvlBFO+4vqHTPmecnOepSsmnOA55KulcC3AIKhYSHfagyDI7G5yjJMnOVbkbZKBzR5hBq1vqgCBud1u2CUNo+sYGB3WyKF7Mg58wt3RPaLccnOyDbN0vEkk74i/YLbNY8vlOVqFdJGax+D5labUbv23QVp5HMcMHGFuOw3iqZVsi8c4JAxlbaiBJ81qlkAFewk9iEHLha2SgEhAJLe6441HB9+Lh6rkeGWP7mAEfvQuP9SMke8lowM+SDRIpekgBbppHNfbjkeX8y2jHgvAPfK3VRSsFAWkjOEG2Khg96fgYwVZMWhh2V9Tn3l5B2JUM72lpb5oNPcfiJV7HANwhhkJJwqdDmjcILhIWn/wBqk96HllQd1TpCCV03UdlTurMBXDsgo5vzVuSD3Kvd2VhBJJQMn1KmZUva0AO7BQEEKoaT2CD0B9izM9/NLqkF237AK4/8425Fb7FYEc02qc/i/rv5RtyIPahERAREQEREBERAREQEREBERBx7zEfg/wDE3+B16/UpV8z9Z/bU/wDhXfpK+mDmI/B/4m/wOvX6lKvmfrP7an/wrv0lBCiIgIiICIiAiIgIiICIiAiIgIiICIrgMBBQDCEZWRV0dXQy+71tLNTygBxjlYWOAIyDggHcEEKBAPZGnCIguBycYQnClfR1cVNFWPppWwTEiOVzCGPI7gEjBI88dlHgFBTpzv6qhyNsq8DdCwHzQR5d/dfmUsdOZn4VoiHllZlKAxwKCaO2/BnpWZT2wdOS38yyKeQdIGVmRzta09kGG1jItulZTLvDTRlhAB7brHlmaXEjC0ypDXPyR5oL66eoqJC+GQgfWsPNQw/fXk7eayYy0AD0VkzWv80GO6QgHJKj94GfLKpLHucOI+WVjOb0lBkGcDsVNA8SN33WAsyiifJszdBI8tadhhG9JaTj61K63VUmzQTn5KaHTV2lYSyMkYQYsNOKt3Q3crUY7EYhlzScrIsmmbrHUZfERuPJbhq6aSmwyRuCAg0ezW1kcvxsyMrdQuDKMBrCG7eS0qnnhpT1ykAfNRXC7UUx+9uG3zQbjoLz98J6xuFs3W1zMlTs7zWbbq+GSQhp7Lb+qmufUBw3CDHbWGWINJzt2UJcMKCCN7WAkKbORugRwOmcMLOttG6G4RSP3AIKstsTqiUMiGTla59yauB4ke0gAoN5XG904sbYY2APDQOy2nari8VoDnZBcq3Cvp/dxBkFwGO60q3zMjqmyO7ZQciX+aKezjwwAenuFsSwsmivTJC846h5rddVVxVttbDAcu6cbLRbbaa2O4Mc5hALgg7AUWpm0un2Ne8HDR+hYMuuad9qlpw4fECFtusmbTWVrZMghq2rFc6WSN7ARk9t0GydY0onuslY1owSStuuj+E477rd+pIJCDIOx+S2kd2O+1BFQVL4p9nYGVrb7i5zMdS27EQyYl3lus11RGQB8kEz5/EdnOVR8nTGQAsdjwXZadipJfoFBgiQ+Kd1kNJd5rEG8x+1ZILgUEjmHGyjt7jHdYHekgKleT0du4UFJltcyR3YOBKDni8XltXpNlNkEhmPzLiKahHS89Pckre/3RpaizMhiILwMLb81BP4D34OMEoNlGMtnLcYA8lOfo4Sob01Ra4bqvfKCBkeH5O6mLdu/bKtGQ7JV3U0bEjdBCRg/arxLgYx5KUsbuoXhoKChy931q9sRByrI/pLI8ggeX2LHlfg4UwIBWJUfTQVc0uwQCfqV7YHY6sFXR/RCyAPgOyDFa/pOD6q/wAUehUUuA8hW5zugmc/P1K9h2UGR6qRjsDZBf075Vj2gHYq7rOMK1zslAGy5a5N/wALLhF/DWz/AK5GuJMjC5c5NfwseEX8NbP+txoPpCREQEREHXP2h34G3Ev/ABOj/XqdfPGBlfQ57Q78DbiX/idH+vU6+eRvdBU9laqu7KiCrPpK9ysZ33V7kEaIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiC8HHdA85y04VmSrgxzvo75QZHiuIA6jhZNPUOYQeo4AWn+FL6FSMEjcZyg1plwaBuQrjcGHO4WkNe0dyqGVoORug1R1e3yPmo31r3A4cQFgue3HdUDHO+iEFanrkyepYwhPms1lLO7Yg4+pH0czdgD+RBBTsw4k7qyq+msuGklJOxUU9BUl+zSUGPTvDHZK1GKsZsFgOoahgyWK6Onk7kIMiunbJGsanhL8nOwV76aeTAAJWZRWqsLThp3HbCBS2eWvIazOVr1JavufhsjMn5rI07F9zpOqrGB33Wddrvb5JB4ZGPPCCJlfSUI6nRD1Vs2q6NzSGMAK0e619LKzDSO2DutvPcHNPQTklBuCvvc1ST4MxGfRaU99dK09Uzj9qxKVspfvkj5rO6+j6Xkgx2UswcS8k575KkEDgfNTCriA+JwH2p71E47OB+1BZ4JwASrfCw4BTl4I23Vo3OcILJIz09/JQdBOVlyn4Vjjz+tBlUMR6m7g7jst6Np3fclxH9z/Mto27+yNW+mkfcd3/elBxTWdTayQZ3DlQMyFfWke/yE/3W6B7O2yCjWhpU1FUiGsa4eoUJkb9SjicBOHE7Z7oOR4b0TSNb1dwsaqDamnJLck5K0OCti8FjQRkDcZWv0r430ZxjPzQbTfTOFQcZ7+i1aIOZTkA+SxZWn3p2O2VmRub4WD3KDTPA8SV2fyrS6tpimG+y14RvDiQFolxik8XqO43QXNqGBoGPzKKUiQ5Cxeojb0WVBgsyd0EJjI3UZacrKkHdY5BygswVVp7BHdlQd0EhIwgO3dW5Hqnlsgu2PmrhgDCiaTlXEn1QegHsWSP20uqcf8AK7+Ubcit9isc802qf4AV38o25EHtOiIgIiICIiAiIgIiICIiAiIg495iPwf8Aib/A69fqUq+Z+s/tqf8Awrv0lfTBzEfg/wDE3+B16/UpV8z9Z/bU/wDhXfpKCFERAREQEREBERAREQEREHIXBLgRxP5h9axaC4Uaaku90dGaidxeI4KSnDmtdNNI44YwFzRk7kkAAkgH1p5buQLlp5ULtpm4cbdT2XV3E3UVU2js9PXAGkbUnJAoqNwL5HNA3nkBDcAgRZOfLjlw5qeKPKzcdSXnhZ9yY7hqW1i2S1FfSe8GmxI17ZogSB1jBADw5h6t2nAxyByd8QdccUOfDhlrLiHqq5ahvdbf2matr53SyEeHIQwZOGsGThrQGtGwAGyDtx7bOwWK1aa4U1VrstBRzSV12jfJBTMjc5ojpiGktAJAJJwdt15Rr1q9uL/8U+Ev/hG7/wDoqZeSoGSg5w5PuWO/c1vGa38OKCqlt9ohjdcL7c44+v3OiYQHFoOxke4tjYDkdTgSCGleqnEfjXyVezQobboHSvDttTqqalE3ulop4prk6M7CetrJXBzQ/DsDJJ8mBpyNkexU4fU1q4Ma24mSwMFZqHUDLWx/SC73akga4YPkDJUyAjbJYCc7LzC5meI9x4t8wGv+INyqXTOu1+qzTlxJ6KVkhjp4xnBwyFkbRkDZvYdkHrXwd9o7yu82+oqfgxxG4cyWqqvr/dqCk1LTU1dbq2VwwIQ/cNkdkhocwBxwA7qIB6Ye0y5GbJy2Xq3cUuFlNLDoPU9W6jlt7nF4tFf0F7YmPcS50UjGyOYDktMbwTgtC6O0FdWWuup7nbqmSnqqSVk8E0Ti18cjCHNc0jcEEAg+RC93eaZ1NzCezXuutK1kc1RctEW7WDXtaCYqiKOGrfjp2aQGyMIHYFwQeGejtJX/AF9qq0aJ0pbZK+83ytioKGmjGTLNK4NaPkMnJJ2ABJ2BXs9prgfyf+zR4O0XEDi3b6HUOrZemCW6zULKuvr60jqdDb4JSGxMaM7gtPSAZH5IC6Vex/4e02r+a92p62COSLRmn6y6Ql4Dh7zK6OmZgHzDJ5XA+RaOxwVqntkOI9y1NzLW3h8al33N0ZYYGsgyekVVWTNLJg7ZMfu7ds7RjfOwDs3p72zXL7qi9t09rXhdqi02OueIJK6YU9bGxhIHVNA0g9AychvWcDYE7LQvaEchfCzWfCat5meXS0Wy3VttoPu5cKOysY2gvNtLRI+piYzDGSMjJlywASNDsguIJ8i17leyc1w/iVyeN0pqKRtezS13r9OuinIcXUb2Rzsa4dyzFS9gz5MwNhgB4cdle0F3dbg4jaX/AGD8RNUaKy4iwXmttYLsg/eJ3x5OfP4FoAIwgqNicdgp4iVunhDw6rOLvE/S3DC3XKC31OqLpT2uKqmYXshdK4NDnAbkAnJA3XY3mz9nhq/lI4cW3iNqDiPZ7/TXK9w2RtNR0csL2PkgnmEhLyQQBTkY75cPmg6rwykYCmMxDcZWEx24Xdvl19mDrnmK4Oae4x2finY7RR6g978OiqaKaSSLwKuWnOXNIByYS4Y7Agdwg6Uve/J7qLDn4LlmNazp+tdi+RTlh0tzW8X7rw71ZqG6Waht2nai9Ca2tjMr3sqaaEMzI1wAIqCScE/CBtnKDrMQWnuoJpCB/wC1egOl/ZT6x1zx/wBZ6Ktmpau0cN9I3COibqS4UgfU3BxhjkdFTxNLWyPb4ha6TIY0jsSCxdhYvZD8pWq7JNQ6L4vauqLrSACSthu1BWsY89vFhjhGG7HADmnbcnByHjg58hOQFb0PcexXa3iB7PLjzo3mDo+X2yWduoqu8QmutV5hYYqKagDul9TMST4AjOA9pJIJaG9fWwv7o6E9kRy76co6Sy8YeMl3uGq6+JoFNba6lt8Qe84HgxSsklkIIIDiQHb5YCcAPImGhklIHSdytz2ezNjj6yCQfVd3Ocj2alw5dNMScUOGeoa7U2kaORrblBXRMFdbWOIayVzowGTRlxAc4MYWkgkEElvTulqYYocnHb1QY58Ck3cwbeSzKTWVBRAMdE3IGMrSblcqV4LeoZx+RbaqJIHuJz9iDf7tf0MZL2xtGfNaJc9Tx3R/iMIAW0Krw+kdPdSUJbg52JQahd7m6SEMY8gn0WJbaeaoBd1HHlurKxniABgyQVqNkBhZ8Ywgy7VSywzlxcdvmsbUNQ0TAHvnyWqwVETXlziFt/UMscs4czcAoMQ1jSAMKenZ4+APNaa1rnnA3K1e0wTdbQQfqQazZKUW6UVDxkZytfr7/TzUrmtaAcYytKm+8U+ZthjutLlraUxkNIyRsg0mvqpTVl/WcHKpHWHIw7cfNQVX3yY9HbdWRwvPluUG69N3N8UwMjstz2K3vR3mnnuMMAjGSRvhcbWcCneHzbBbntFwo33ymDCCS4Y3Qcra0tLodNCrZkAs6vzLhm3TympcS84BwuwGuGSSaIYWt28LuPqXX62wvdUPYBuXoNR1JWsFCBtkD+ZbEbVAkj+6W89T2urZQ+I5pDSFsSNhc8beaCaaLAL/ALVjgnI3WZKHGMtA8liiGTI+EhBl00bjuT5rJkblhGVHT/C3cflUrnAhBgNjw/PqfRTkBoyhbvnCOB6SEFGzB56cgq5zQ1peNj3ysZgcyTLu2VPJKx0RAO6DV9LVM01UInPJGcYyuSKiiY22OJj3LfRcYaScILgHybDbuuVau9251sMYcOvpwEHD1zAbc3j5nZUV1yilkub5GtOC7YqpjcB2KCCU9IysbxT1Zz5rIqDgYWF5/agy/Ey0DKqYiWl2+FFGQe6yw7MRHyQYXWWOwO6nZNkY2yVjvY4vO3mr4o3BwODsgyGxk5OVjTjDt1mtcAMHCw6kFzhjdAZKGgYIGPVZbJA6P7O4Wn9LsYWXC8CIg98IMWUkvOFLDH1DJUT93nHZTRuIGEFCw5O6q0EDCFxyeyubgjdBQHOyseTlX4AJVHDPZBQAkLl7k225suEX8NbP+txriJrXA+a5c5N/wsuEX8NbP+uRoPpDREQEREHXP2h34G3Ev/E6P9ep188je6+hv2h34G3Ev/E6P9ep188je6ATnZUVXABUQVaCSqnY4IVGfSVzhnCCxERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERBUjAU0MzY24IUWxVDhBle9MPcKpnaRt6LDUjeyCrz1HI+ajPUNiVIc42UZBJJQXZPmVqNI5obk/UtOA33WVE8NYB39d0Gotr44ttsqKS7RlaZM8uA+pQ4KDXaK5Rl+4+S1qJ0MuCWZJ+S2jRPY2QF3qty0dfSsaOo7/AFoJ6xkPQcMAWCymY4YDQPqWTVXGkeDgjP1qGG4U+N3j8qCWmpo2HLmg49VmfdanpCPhGB3CxWXKlHdwWmXGop5Aehw3QZV5vgrIy2B3SVtsyTjvI7f5qR5GfhKjeHHJQUzJJ++JU0Y6MdSjiDg7GFM4DHZBI2rYwbBHVHjYAWE9pz2U9O0AoKvppHnIyqsp5GOyc7FZbJABgn8yo5wJ7oKRkjYqUPGyhJAGyeIBtkd0EznZG6hPdVMrW/SPdWhzZCejc5Qajb/ptW9A4/clw/7X+ZbNoIZGFrnA4yt2Grgba3NJGS0+aDjav2rJf++UABJwp649dXIR5uUTGOyDjAQSNpHOblVfSOazqWTG74QNldKHGM+eyDBgke14HUcZ9VuShryynwXeQW2QCJN1q0DwIMZ3QSzXBgkJ2zuomXUmUN6tlhSse55OTjKx2AMmHVsAUG6mytcwPwdwr5rcyemMob+ZYbKymELW5GRha5TVMUtvdGzBdjCDY0lOfHeztgqmTAekrOfbK01T5Aw4yTlYNUx8chY8EEZQDLnurSQT9asGPJSCJx3AKCwjCtIypMOGzvRWktB2QW9JVwGw3VpPbCka0kIDW+aseN8fNXjIJ6lY875CDv77FT8KbVP/ABf138o25E9ip+FNqn/i/rv5RtyIPalERAREQEREBERAREQEREBERBx7zEfg/wDE3+B16/UpV8z9Z/bU/wDhXfpK+mDmI/B/4m/wOvX6lKvmfrP7an/wrv0lBCiIgIiICIiAiIgIiICIiAuwXs/vwy+FH/h5v/opF19XYL2f34ZfCj/w83/0UiDvT7cT/wCKfCT/AMI3f/0VMvJYDC9afbif/FPhL/4Ru/8A6OmXksg93vZM0kdPyWabljc4uqrrdZX57AiqezA+WGA+e5P1Dwje98jjJI4ue4lznE5JJ7kk9yvbr2Omoqa78plTaInN8exaqr6WVgO+HxwTNcR6HxSAf+1PovGjiTpiXRPEXVOjaiAwS2G9V1skic0jw3QTvjLTkkgjoIwSe3coNtL3Y0DUTVXsmKqWd/U5vBu8xg4Aw1luqGtG3oGgfZuvCde7PELx+D3sp57fd5BTVVPwspLRUB2B0T1tNFTuj7EE9dSWA9ycHIJyA6uexBpYn654qVxLuuK02yIAYwQ+aYknbOcsGN/Xv5ddPajvc/np4lBziQwWYNydgPuRRnA9NyT9ZK5p9ijqOlouO+uNMTOa2W6aVFVDnYuMFVEC0fPE5OPRpPquOPa26YlsPObe7rJAYxqSy2u5xuIIEjWQClLhucjNKRkY3aRjIJIdMl7CexJqJn8H+IlK5+Yo9SwSNbgbOdStBOe5yGN/J9a8e17Uexh0zWWnlm1DqGrjLG33VtS+m2+nDDTU8fWD/hBK3Hl0/NB5X81/4U3GL/jA1D/KM64uAyt38Z9URa24xa61pDO2aO/6lud0ZI3GJBPVSSBwIAGCH52AG/YLZ4PmEHNnJVtza8I/4XW3/wBM1enftoNuV3S/8PqL+TrivL7k1rKei5sOEM1Q8tY7WdphBAJ+N9SxjR9rnAZ8s5XqB7aI45XNLfw+ov5OuKDxlGwyCvfD2ZEUsXI9w0ZNE+Nxbd3hrgQS112rCDg+RBBB8wc+a8De4xlfRTyWaSm0Nyq8MNMVbeirp9O0tTVRZy6GWpb7y5jgScOBnwR+TbCD52TLMOwJC76+xhke/mj1Q1wOP2A1p/5xty6MRsiMYJwu+Xsa2sHM9qcs/wCAdbv/AOULeg3d7Wbm94jUet6rlg0VUVNjsdJR09Rfqunl6Z7oZ42ysg6hgsha0jqAOXkkH4Rg9GeWPinqrgtxz0dr3TdwqqY0t2po66GJ5aKuifK1s9O8DZzXsLhg5wcEYIBHP/tTI4nc52qy/ubfav1OJdZ9Hxwfsps3wj/dCn7f4RqD3e55+YG+8tPAWv4i6TtVNWagqauKy22aob1MpJZw5xmc39+GiIkM2BcGZyAQfBbUep9U6y1HW6v1bfq+63u4zmoqq+rndJPLITnqLic7bAAYAAAAAAC9mfa39P7VamLsf/Gug/8AQ1C8XpwzdwxjCD2/5NNVX7mU5EYbZryaW8XKttd20tWT1Lg59Yxokiic9zju7wnxtLiclzS4nfK8Kqi7PYC0E/Yvcf2T2DygWvH/AM/Ln/6QLwjqz99IQWzTvleXFx3VGhzt9yFTGWE4UsJAGCQgtc0kblGSGPscFXuIxthQvQZUdSBjrwcrKbW7fAcfMFaSp4XNDcFBnPq3kbO8vJafUPc93xHJUnUDnBUMndBdA8MeCVrFHXRtcDkDBWhtAJwVlQPY0jJ2Qa/drmKum8Nh3xjZbc8GbPc4CznTRdOFG6aMsO+Sgx4pAHdLhnCmErWuDjjb5rDDgJCQdldI/b1QZlVXh8fRG7BVlprJKW4wVHX9BwJ+pYQBcdlJHG8PBIwEHNl64pR1mmGW0vBIZjGe+y4zoL+yCqMpOxOfzrTamWJ0HQCc/WtMwQg3/f8AWlPcbcKZoGQMbLY0MoaXE43UOUQZXvDfUJ7w35flWKiDJFSD54Vfec93DbssVEGX7y0eYKe8tPmAsREE8sweNtlCDgg5KoAUwQg1Gln6AOlwB9fRZBrarIzMcLTYHgbFZDpWbb4wgynVkbm5OM47qCWsbgjIUDnM6TgrHeQfNBdLKXknPdRJ3TBQAcFZMUvSADhY2MquSEGSJADlVMwwcfoWNn5pn5oJTNud8K0y5OSPzqJEEviBWuec7KxEFepSNkA2USuaB3JQSOcDgq9h2ULjgYV0TwBgoJcgqhcAd1QO+Y3VrnZPcIJvEGBsuWuTX8LLhGf+7W0frca4gJwM57Ll7k0/Cy4RfPWlo/W40H0hoiICIiDrn7Q78DbiX/idH+vU6+eRvdfQ37Q78DbiX/idH+vU6+eQIDiCqKpByqIKs+kr3KxvdXE+qCxERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQFcOytVw7IK5x3Vdu6twCqgHGB2QXDfsrxkDurWjGVcgsO25QAO3VxAPdMEdhsgjLT5FAJcbE/lV+CqjbYoI8yHzP5UzIBs4gKTA9EwEEXVJ/dFXBryNyVcW+iqDjYoKNaR3V+R2Vp2CDOcnKC8AA5wit6vqVev6kFzRkq8RkHIVGeqlHZBRrSdgVd4ZPmVfGN1IB6BBCYiN8qzwiX+fdZLmux2VmCHdvNBBUQuc3Y4VtIwxyjqPmst+7VBGMSjI2Qa6xzfCGAraqWX3c4cQEY5vgjcZVlU9hpyPP60G2n7znPqspuOj5rFI+/n61lDZuUEbZOknPZSPqm9PTkdljPdud1YWEguwUF3iAvz6rUKRhkIGe60xg3H1rVqB4a4IMp9E1rD8JK0arh6HZGwW5nhzoytDroX7nyzuUGm9b/AO6Oy16y1D2EdbjjHqtBIPVj5rVLfI1jh1dkG7m1tMIiOgZ9VoVytJrJfHYMArINTB0DfdahTnrhy0bYQbQnt7qfPUoTO1m3dazd4ZXk9I33WguglacFvdAkkDuwV0dOZBncqMxPHcLNpHtY3fvugx5IvCI/KqCYDGylq3h/ZYmD6IJS7ryAMfUo3DpV0QyVSTPVug7++xU/Cm1T/wAX9d/KNuRPYqfhTap/4v67+UbciD2pREQEREBERAREQEREBERAREQce8xH4P8AxN/gdev1KVfM/Wf21P8A4V36Svpg5iPwf+Jv8Dr1+pSr5n6z+2p/8K79JQQoiICIiAiIgIiICIiAiIg5S5e+W7inzPawq9EcKbbR1Vfb6B9yqn1lU2nhiga9jMl7u5LpGANAJO5xgEju9ypezN5oeD/MVoTiZrG26bZZdP3RtXWuprw2WUR9Dmktb0jqOSNsrojwW4+cW+XjU1Tq/g5rGXT11raN1vqJm0kFS2WAva8tdHOx7D8TGkHpyMHBGTnmke1G56z/ANnP/wDpmz/1RB6R+0z5T+LvNPYtBW7hPSWueXT1XcJ6019cKYNbKyEM6cg9RJY7Ppj5ryv5iuSXjvyuWG06n4qWu0x228VjqCnmoLiyo6ZwwvDXgAEZaHEEAj4Tkg4zu791F56vx5//ANM2f+qLjjjbzecxHMZZbfp3jJxEdqC3WqqNbSU/3LoqQMnLCwvJp4Yy49JIAcSBk4Ayg7MeyP5mrDwi4p3jhJri6MobLxB93bb6qeQNhp7pF1CNjicBomY8s6s/TZEMYORzV7QH2ZXEXihxPuHGzl8prdcajUPTNerBNVR0kvvYaGmeB8pbE4SAZe172EPyQXBxDfJpdneFftIObzhNZodO2fic682umYGQU9+pI68wgDADZnjxsAAANLy0AbAIOd+Vz2SXGa6cRLXf+YyzW7Tek7TVMqqq1fdCnraq6BhDhAPd3vjZG4gB7i8O6cgNycje3tgeauw3WloOV7Ql2iq5KOsZcdVy07g6OCSMEQUJIyC8EmR4GOktiGSS4Dq/xE9pxzj8RbTNY5+JrbBR1EZjmbYKCKimkBzkicAzMOD+8e1dWZppaiZ9RUSukllcXve92XOcTkkk7kknJJ7oOYeULjt+1w5hNJcValk8troKh9Ld4YfpyUE7DFNgfviwO8RrcjLo2jI7r1q57OUK288nDfS/Evg1qWzS6itlKZ7PWvlPud3t8+H+CZWg9JBHUxxBALntdgOLm+GC5n4Hc4fMVy6wm3cK+JNbb7S9xe+01UcdXQkk5JbDM1wjJO5dH0uPmSg5m0R7JfnF1JqaGz6o0hatJ2p0gbPd629UdTGyMHBcyKmlklccZIaQ0E4BLe47481vFLh3yAcnlDwI4e3Rn7KLhZ5LJYacFoqfvvV73dJWjIaOp8rwcYdK4NAwHFvQm9+1l5zrxbn0NLrKxWh8mxqaGxQeK0YIIBlD2jOe4GRjYhdVdZa21hxD1FV6u11qa5X+81zuqorrhUunmfjsC5xJAA2DRgAAAAAAINEbknsrw3G2/wCRGkfbj0V+R5hBylyoAjmk4ObH/wCP+nv5RgXq77Ruz3Dmc5IrLrnhPbqy+Q0V5otSupaKIzTeAyGpp52ljRkuidOQ8AEjw3HsCR5S8qODzR8He3/x/wBPfyjAuyHF/j/zI8hHMtxK4b8N9XGn01cL5PfqK1XGlbV0ZhrT48ckQkHU1wD+hxYQC6Mh2SEHEXLHyv37iNqQ674n2av07wm0eDdtV32vgdTwupIfidSwOcB4k8pAjDWZILwSM4B9aPZ3cw0PMRwu1XqKo6Ke50msLk6WhBy6lpJ3iakZtt0Nid4TSMA+CRjbJ8feP3ObzDcydJDaOJ2uHz2Wne2aO0UEDKSj8UA4kexgBkcMnBeXEZOMZW1uBXMJxa5cNWO1jwn1TLaaueMQVcD42zU1bEDkRzROBa4A5wcBzSSWkEkoOWOGHAi66VtvHW78XdET01t0BpSvt0dVcqV8UcV9lqYoKIRF2AZCS9zcZ+EE75GeZ/YyytfzP6nAOT+wKtP/ADjb11q5hOdTmD5m7fRWLijq2CWzW+YVMNrt9IylpjOAWiV4bvI8BxALiQ0E4Ayc7P4G8f8Aipy5awm13wk1DHabtU0Mlsnklo4alktK98cjoyyVrgAXxRuyACC0AHBIIdi/apTBnOjqtpP/AOj7V+pxLq9pOsa3VNmAdv8AdCmH/wCMapeLfF/iFxz13X8SeJl6bdL9chEyeoZTRU7eiNgjjYGRNawBrGgA4ycZJJJJ2jTVVVR1EVVTymOWB4kje07tcCCCPmCAUHt17YWYwcplLIO/7LbeP/xNSvEmS5yOOxJC5n4286/MjzD6Oo9B8WNfMu1loqplcII7XSUplqGMc1sj3QxtLiA9+2Q3JyRkAjgwAeeEHut7I+Qy8m9qe7ub7dP/AEoXhSSZXF2N1zzwd54eZbgLoSfhtwu4hNtNhlmkqY4XWukqH08smPEdHJLE5wJwDgkgEZABJJ4KaB042QRtyMg/Wrc4KncwAZHkoHNIJQGnBVXblW5VQdu6AAScBC0gqVvSNx+lUcQgsblpyjhk5VXkEK0HbcoBGFTJ9VUlA0lBTJ9Sq5d6lOkqrWEuAAygq5uADlWgFxx3U8jCIwPJWQEdSCaGn6D1EKZ5HQSB2TxGhgBwrTKwtI9UGI0kybk91K9gI2Hko27P6j2zlSeIz1CCFzDnZW9PzUznNO43VnUEFnT806fmr+oKjjkoLen5qh2UnUFQ4z5ILen5p0/NX9QVCcoLWHBVziDsrMpk+qBlMn1KIgrknz/OmCe5VBsVICMILS3p3QHKuLgRurdvUILRsUJycoiAiIgIiICqBlOkpgjsUDp+adjsrsH0Vrgc9kFM5TKJg+iCvW5MklURBcTt3XMfJn+Fhwh/hpZ/1uNcO4PouYuTT8LHhF/DWz/rcaD6Q0REBERB1z9od+BtxL/xOj/XqdfPGBk4X0Oe0O/A24l/4nR/r1OvnjBwguPZWq49lagqz6SucMq1n0lc/PzQWIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAg3RB3QV+jurg7ZU29Qm3qEF3V8lUbqxXDsguLseSB3yVhOVUEYQXdXyVDuqdQTq+tBcDg5VC85VocPUlVyPRBd1fJUO6p1BOoILwcoTjZWhw3VSQT8kFDsFQb75V5wR2CtQSxyAbFTtcDssMOAUjZPPsgzGvDT6/UrxJ54WJHM3O/51IJ2A7oMl0zceaj691E6eMjYK1s7eoHyz6oMh0mBlWMILgc+aimmZjAUDZ3B3fI80Gqic4x1DH1qOaZxYd/yLGFSzz7o+oYWYCDFG8pwO5WST8B2WPER4m+6yJHDp2QYoZ1PysrwwIzt5LHY8B2fJZJnYWYQYB+kR81mUTyJASR3WG76RO6lpw8SA749UG52zt8IZ74WLVua6MjGfsULKqMRgE7q2WrjLcZCDTwzqkPdVcCw7EhXiWMEkAZUM8oJJCCXxnZHx/nWu264hkXSSDstq9bvVZdLUdJw47INymVkpJxnPZYNRTNLshpx9SUtdAAMkfasl1fSuGeoINIqYWsH0cLTnk52JC1iunjkz0EfYtGeHdXdBcCTgE5UzIQ8b9/JRtwMeqyo5o24z3QQOiEAzv8AUsd7uo5CyaqVrwcLEQd//YqfhTap/wCL+u/lG3InsVPwptU/8X9d/KNuRB7UoiICIiAiIgIiICIiAiIgIiIOPeYj8H/ib/A69fqUq+Z+s/tqf/Cu/SV9MHMR+D/xN/gdev1KVfM/Wf21P/hXfpKCFERAREQEREBERAREQEREBXAYCtG6uQEREBERAREQEREFW/SCuVrfpBXE4QXM+krz2VrVUjKDk7ld1LYNI8yHDDVGqZ4ae0WrVtqqqyonIEdPEyqjJmcT2DMdZPkG7Lu97aCLhnd9R8NdX6b1Jaq3U1TQ1tDXQ0dRHM91Ax0clNI8sJLQHyzhpP0g44+iV5pDAPkFdkeqCh2PcqmVQvGSqdSCjvNVYfI+qtLjugOUEpI3wo85Vx7Kwk9uyCjsDPqrR5qqA9wguALt1JgDH51GH9IV4cDugtLyCe6tJJ7qQ481G84QWkDCAZRrS7bdHNLcILmHOyo/uqDI3wqE5QEQDKva0AboKMHUcK8/Dsg+HdWuOTnKC9mHKVgaCM/YsUPcPNV8R/qgyqjHQMHKxWD4gr2vc84d2VzgG7hBdK3EeQVjb/NXukcQR5K1mOoZQXOdluArN1mvEXhg7ZKxwG+mUEW/mikf042CjwfRARMJhARMH0TB9EBETB9EBEAJVekoKIruj5KuMbYQWIiICJjKYQEVekqiAiIgKrQCqJlBI0jsqqIEhV6igkVryqdfzVzcH57oLMbZVR2R+3bsqsxhBa36QQ9wrwAFR2M+SC4dlzByafhY8Iv4a2f9bjXD47LmDk0/Cx4Rfw1s/wCtxoPpDREQEREHXP2h34G3Ev8AxOj/AF6nXzxgZOF9DntDvwNuJf8AidH+vU6+eRvdBU9laqu7KiCrPpK9ysZ9JXuQRoiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiIKtVVblMn1QEREBERAREQEREFWjJUiiBwq9RQSKmd8KjXZ7q7IQWkYVpOFe4geat2KC3KZPqmCmD6IGT6qoySN1TCqAQQUFzgQ0b/AJ1YrnOJGN9lbg+iBk+qZPqiIL4vpLILR0HdYrTgq8zOxgFBYdnEAqmT6od1UNJQXYysqIAN+fqoGjZVdJ0bBBe5pOfixjssd5cDguJVTM8537qjWueRsSgtyfVPrWQIDjsrHsDfJBEgz2yh7q9pbjPmgAP8ipGxSu7yYP1qPJ2weyqJZBtkoJSxzB8Ts+isd9XZWOkfjByp4iwsOcZKCEHfsrX5z3UsnSOwUJySUFMoq9JVMYQd/wD2Kn4U2qf+L+u/lG3InsVPwptU/wDF/XfyjbkQe1KIiAiIgIiICIiAiIgIiICIiDj3mI/B/wCJv8Dr1+pSr5n6z+2p/wDCu/SV9MHMR+D/AMTf4HXr9SlXzP1n9tT/AOFd+koIUREBERAREQEREBERAREQB3Vyo36QX0T8l+l9M1PKdwlqanTtslll0lbnPkkpI3OcTC3JJIySg+dlF9R/7ENJf8F7T/EY/wD1U/YhpL/gvaf4jH/6qD5cEX1H/sQ0l/wXtP8AEY//AFU/YhpL/gvaf4jH/wCqg+XBF9R/7ENJf8F7T/EY/wD1U/YhpL/gvaf4jH/6qD5cEX1H/sQ0l/wXtP8AEY//AFU/YhpL/gvaf4jH/wCqg+XFv0gquwvqN/YhpL/gvaf4jH/6qfsQ0l/wXtP8Rj/9VB8urSVUn1K+on9iGkv+C9p/iMf/AKqfsQ0l/wAF7T/EY/8A1UHy643yhOF9RX7ENJf8F7T/ABGP/wBVP2IaS/4L2n+Ix/8AqoPlywM5Qn0X1G/sQ0l/wXtP8Rj/APVXA3PbpnTdJygcVqmk0/bIJo9PSuZJHSRtc09TexAyPsQfPSqg4VvUrmb7ILxuRlC0HdUOWqrXDBygiJJJCHbzV5Ldz3VjnZQXAA91JgKDOFUE4O5QSk+SsfjHdUDvrQHPcoL4MdW6rPguCj6sdsKhcSgqRthWq5mCcHssjEZQY7O6uIyr5OhoIafmoSTnugOdnZURSxdH75BEBkoe6yZfD6fhIysfud0F0X0sFSSAY2VjiA34cIx+SM/mQRnuiypBF0HpIJWKgdRPmiIMZ3QVAOVVXP6enbuo8lBdgBMAq3JTJQXIrclMn1QXYCK3JTJ9UF7fpBSjGBnKx8lVD3DzQSHurXd1b1FAfVA6VRStLSN1a/pz5ILR3VQBlUOMeSpkoJiRhRO+kVTJKICK7b5Jt8kFqKpxjyVEBERATJRVA27IBOfJGjzQ9uyuZG542BQXK13f6lcY3DyKjcSCQgk7D6lzByafhY8Iv4a2f9bjXDnV6lcx8mjermx4RY8taWc/8rjQfSGiIgIiIOuftDvwNuJf+J0f69Tr55G919DftDvwNuJf+J0f69Tr55GjJQHdlRXuY70VhBCCrPpK5xwrWd1V5GdkFqIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiCre6qrcpk+qC4jKdgrclMlBcDnZD2VoO6kBGO6CxvdX+SoSMK0n0QB3Kqrcn1TJ9UBEGM7q7b5ILUVTjG2FT60BSN7K3b5KmcHv5oJWqN/dXh+EJaW5PdBG0ZKyoWNzk/WsbIBVfFcDsUGpNa0NI/nWHUYzt2yohO8A791Rry546jsgswfRFnsbT9IyRlQyiME9OO6CJpwVJ0h25CgJ3BVfEf2ygvkOyj32xlXAgn7FcOnHln60EbuykjAIVryPJWhxHYoJSBhRPO6F7j5qmcoO//sVPwptU/wDF/XfyjbkT2Kn4U2qf+L+u/lG3Ig9qUREBERAREQEREBERAREQEREHHvMR+D/xN/gdev1KVfM/Wf21P/hXfpK+mDmH/wB4Dib/AAOvP6lMvmfrP7cn/wAI/wDSUEKKoxjyVdvkgtRXbfJUd3QURXbfJUd3QURXbfJNvkgtRVOMeSogIrtvkqO7oKDZb3tfG3jLY7dTWiy8XNaUFDRxiGmpaW/VcUUMYGAxjGyANAGwAAAWytvkmR6oN/8A7YHjz+O3X3+ktb/SJ+2B48/jt19/pLW/0i2Ae2xVBjfKDkD9sDx5/Hbr7/SWt/pE/bA8efx26+/0lrf6RbAyPVMj1Qb/AP2wPHn8duvv9Ja3+kT9sDx5/Hbr7/SWt/pFsDI9UyPVBv8A/bA8efx26+/0lrf6RUPMDx6x/v3a+/0lrP6RbByPVUPog38OYHj1n/fu19/pJWf0iu/bA8efx26+/wBJa3+kXH+w3VQQfNByB+2B48AZPG3X31fskrf6RVHMDx5IyeNuvfs1JW/0i4/BGfsVHHJGEHIP7YDjx+OzX3+ktZ/SKh5gePIxjjbr7/SWs/pFx8HEKpfnGyDf/wC2B48/jt19/pLW/wBIsS78aeMl/ttRZr7xZ1lcqCrYYqikq79VTQzMPdr2OeQ4fIghbK6gmQcIKHZXsVNvVNsoLi7PfsVb2Qn6lbnKC8b7YKOa0dla0/PCrsfRBacIrtvkm3yQWoqnGNsIMY8kFEV23yTb5ILUyfVXbfJNvkgtzlFdt8k2+SC1FU4xthBjHkgplERARVOMbYQYx5IKZRVOMbYQYx5IKIqnGNsIMY8kFEV23yTb5ILUV23yTb5ILUQYzurtvkgtRXbfJWnGdkBFdt8k2+SC1EGM7q7b5ILUREBEVRjHkgoiu2+SbfJBaiqcY2wgxjyQURVOMbYQYx5IKIrtvkm3yQWqreyp9anj6Mb9yghcsyjAwc4UMkZP0fJWNErPogoMyUN8sBYMn0irj4zvIrJhbGG/fcAoMJc08luP22PCTP8AwwtP63GuHZxGPo4XMHJiHHmw4R9PlrO0E/xuNB9IKIiAiIg65+0P/A04lf4nR/r9Ovnmix1jK+hr2hrS/k24lNHc0dH+vU6+eNmc5HkgytsHuoJRvsqGR31FT07PFG/c+aDGb3R3dZdRA2MEgfUsPKAiKoxjyQURXbfJNvkgtRXbfJNvkgtRVOMeSogIqnGPJBjHkgoiqcY2wgxjyQURVOMeSogIgxndXbfJBaiu2+So7ugoiu2+SbfJBaiu2+SbfJBaiqcY8kGMeSCiIMZ3V23yQWoh7qoxjyQURXbfJNvkgtRBjO6u2+SC1FU4xthBjHkgoiu2+SbfJBaiu2+SbfJBaiqceSDGPJBRFU48kGMeSCiIMZ3V23yQWoqnGNsKn1oCKpxjbCDGPJBRFU48kGMeSCiIMZ3Q4zsgIrtvkm3yQWortvkm3yQWoqnGNsIMY8kFEV23yTb5ILUV23yTb5ILUV23yTbB7ILUQYzurtvkgtRVOPJBjHkgoiqceSDGPJBRFU4xthBjHkgpk+pTOUGM7q7b5ILUV23yTbB7ILUQYzurtvkgtRXbfJNvkgtRXbfJWnGdkHf/ANip+FNqn/i/rv5RtyKvsVsftptU4/4AV38o25EHtQiIgIiICIiAiIgIiICIiAiIg465jTK3l64oGCMvkGjL2WNHdx9xmwPtK+aeejr3TSOfRThxcSR4Z2Od/JfUhdrXQXy11llu1MKiiuEElLUwuJAkie0tewkEEAgkbEHdcQO5MOWFzi53CS2kkkk+81O5P/7VB85PuNb/AHnP/kynuNb/AHnP/kyvo2/aXcsP4o7b/Gan+lT9pdyw/ijtv8Zqf6VB85PuNb/ec/8AkynuNb/ec/8Akyvo2/aXcsP4o7b/ABmp/pU/aXcsP4o7b/Gan+lQfOT7jW/3nP8A5Mp7jW/3nP8A5Mr6Nv2l3LD+KO2/xmp/pU/aXcsP4o7b/Gan+lQfOT7jW/3nP/kynuNb/ec/+TK+jb9pdyw/ijtv8Zqf6VP2l3LD+KO2/wAZqf6VB85PuNb/AHnP/kynuNb/AHnP/kyvo2/aXcsP4o7b/Gan+lT9pdyw/ijtv8Zqf6VB85PuNb/ec/8AkynuNb/ec/8Akyvo2/aXcsP4o7b/ABmp/pU/aXcsP4o7b/Gan+lQfOT7jW/3nP8A5Mp7jW/3nP8A5Mr6Nv2l3LD+KO2/xmp/pU/aXcsP4o7b/Gan+lQfOT7jW/3nP/kynuNb/ec/+TK+jb9pdyw/ijtv8Zqf6VP2l3LD+KO2/wAZqf6VB85PuNb/AHnP/kynuNb/AHnP/kyvo2/aXcsP4o7b/Gan+lT9pdyw/ijtv8Zqf6VB85PuNb/ec/8AkynuNb/ec/8Akyvo2/aXcsP4o7b/ABmp/pU/aXcsP4o7b/Gan+lQfOT7jW/3nP8A5Mp7jW/3nP8A5Mr6Nv2l3LD+KO2/xmp/pU/aXcsP4o7b/Gan+lQfOT7jW/3nP/kynuNb/ec/+TK+jb9pdyw/ijtv8Zqf6VP2l3LD+KO2/wAZqf6VB85PuNb/AHnP/kynuNb/AHnP/kyvo2/aXcsP4o7b/Gan+lT9pdyw/ijtv8Zqf6VB85PuNb/ec/8AkynuNb/ec/8Akyvo2/aXcsP4o7b/ABmp/pU/aXcsP4o7b/Gan+lQfOT7jW/3nP8A5Mp7jW/3nP8A5Mr6Nv2l3LD+KO2/xmp/pU/aXcsP4o7b/Gan+lQfOT7jW/3nP/kynuNb/ec/+TK+jb9pdyw/ijtv8Zqf6VP2l3LD+KO2/wAZqf6VB85PuNb/AHnP/kynuNb/AHnP/kyvo2/aXcsP4o7b/Gan+lT9pdyw/ijtv8Zqf6VB85PuNb/ec/8AkynuNb/ec/8Akyvo2/aXcsP4o7b/ABmp/pU/aXcsP4o7b/Gan+lQfOT7jW/3nP8A5Mp7jW/3nP8A5Mr6Nv2l3LD+KO2/xmp/pU/aXcsP4o7b/Gan+lQfOT7jW/3nP/kynuNb/ec/+TK+jb9pdyw/ijtv8Zqf6VP2l3LD+KO2/wAZqf6VB85PuNb/AHnP/kynuNb/AHnP/kyvo2/aXcsP4o7b/Gan+lT9pdyw/ijtv8Zqf6VB85PuNb/ec/8AkynuNb/ec/8Akyvo2/aXcsP4o7b/ABmp/pU/aXcsP4o7b/Gan+lQfOT7jW/3nP8A5Mp7jW/3nP8A5Mr6Nv2l3LD+KO2/xmp/pU/aXcsP4o7b/Gan+lQfOT7jW/3nP/kynuNb/ec/+TK+jb9pdyw/ijtv8Zqf6VP2l3LD+KO2/wAZqf6VB85PuNb/AHnP/kynuNb/AHnP/kyvo2/aXcsP4o7b/Gan+lT9pdyw/ijtv8Zqf6VB85PuNb/ec/8AkynuNb/ec/8Akyvo2/aXcsP4o7b/ABmp/pU/aXcsP4o7b/Gan+lQfOT7jW/3nP8A5Mp7jW/3nP8A5Mr6Nv2l3LD+KO2/xmp/pU/aXcsP4o7b/Gan+lQfOT7jW/3nP/kynuNb/ec/+TK+jb9pdyw/ijtv8Zqf6VP2l3LD+KO2/wAZqf6VB85PuNb/AHnP/kynuNb/AHnP/kyvo2/aXcsP4o7b/Gan+lT9pdyw/ijtv8Zqf6VB85PuNb/ec/8AkynuNb/ec/8Akyvo2/aXcsP4o7b/ABmp/pU/aXcsP4o7b/Gan+lQfOT7jW/3nP8A5Mp7jW/3nP8A5Mr6Nv2l3LD+KO2/xmp/pU/aXcsP4o7b/Gan+lQfOT7jW/3nP/kynuNb/ec/+TK+jb9pdyw/ijtv8Zqf6VP2l3LD+KO2/wAZqf6VB85PuNb/AHnP/kynuNb/AHnP/kyvo2/aXcsP4o7b/Gan+lT9pdyw/ijtv8Zqf6VB85PuNb/ec/8AkynuNb/ec/8Akyvo2/aXcsP4o7b/ABmp/pU/aXcsP4o7b/Gan+lQfOT7jW/3nP8A5Mp7jW/3nP8A5Mr6Nv2l3LD+KO2/xmp/pU/aXcsP4o7b/Gan+lQfOT7jW/3nP/kynuNb/ec/+TK+jb9pdyw/ijtv8Zqf6VP2l3LD+KO2/wAZqf6VB85PuNb/AHnP/kynuNb/AHnP/kyvo2/aXcsP4o7b/Gan+lT9pdyw/ijtv8Zqf6VB85PuNb/ec/8AkynuNb/ec/8Akyvo2/aXcsP4o7b/ABmp/pU/aXcsP4o7b/Gan+lQfOT7jW/3nP8A5Mp7jW/3nP8A5Mr6Nv2l3LD+KO2/xmp/pU/aXcsP4o7b/Gan+lQfOT7jW/3nP/kynuNb/ec/+TK+jb9pdyw/ijtv8Zqf6VP2l3LD+KO2/wAZqf6VB85PuNb/AHnP/kyq+5Vo/wDuOf8AyZX0a/tLuWH8Udt/jNT/AEqftLuWH8Udt/jNT/SoPnNbBcB2pZv8mf8AUq+Fcv71m/yR/wBS+jH9pbywfijtv8aqf6VP2l3LD+KO2/xmp/pUHzniK5Z2pJv8kf8AUrHUte45dST/AOTP+pfRn+0t5YPxR23+M1P9Kn7S7lgzn/Yjtv8AGqn+lQfOV7lWn/7jn/yZXMvJrBcKfmu4RvbRSkHWdoa7MZADTVx5OfLA3Xuh+0u5YfxR23+M1P8ASrP09yn8vOlL5Q6k0/wyoKK52yoZVUlQyonJilY4Oa4BzyCQQDuD2QcuoiICIiDrj7RB9THyacSn0kDppRR0XSwDJP8At+nzt9WT9i+extHXDOKSff8A+hn/AFL6idV6U0/rjT9ZpbVNsjuFqr2tbUUz3Oa2QNcHAEtIOzmg7EdlxX+0u5YPxR23+NVP9Kg+cr3KtP8A9xz/AOTKkjguUf0KSf8AyZ/1L6Mf2l3LB+KO2/xqp/pUHJdywjtwktv8Zqf6VB850kNzlGH0s5H+CP8AqUXuNb/ec/8Akyvo3/aX8sJ78JLb/Gqn+kVP2l3LD+KO2/xmp/pUHzk+41v95z/5Mp7jW/3nP/kyvo2/aXcsP4o7b/Gan+lT9pdyw/ijtv8AGan+lQfOT7jW/wB5z/5Mp7jW/wB5z/5Mr6Nv2l3LD+KO2/xmp/pU/aXcsP4o7b/Gan+lQfOT7jW/3nP/AJMp7jW/3nP/AJMr6Nv2l3LD+KO2/wAZqf6VP2l3LD+KO2/xmp/pUHzk+41v95z/AOTKe41v95z/AOTK+jb9pdyw/ijtv8Zqf6VP2l3LD+KO2/xmp/pUHzk+41v95z/5Mp7jW/3nP/kyvo2/aXcsP4o7b/Gan+lT9pdyw/ijtv8AGan+lQfOT7jW/wB5z/5Mp7jW/wB5z/5Mr6Nv2l3LD+KO2/xmp/pU/aXcsP4o7b/Gan+lQfOT7jW/3nP/AJMp7jW/3nP/AJMr6Nv2l3LD+KO2/wAZqf6VP2l3LD+KO2/xmp/pUHzk+41v95z/AOTKe41v95z/AOTK+jb9pdyw/ijtv8Zqf6VP2l3LD+KO2/xmp/pUHzk+41v95z/5Mp7jW/3nP/kyvo2/aXcsP4o7b/Gan+lT9pdyw/ijtv8AGan+lQfOT7jW/wB5z/5Mp7jW/wB5z/5Mr6Nv2l3LD+KO2/xmp/pU/aXcsP4o7b/Gan+lQfOT7jW/3nP/AJMp7jW/3nP/AJMr6Nv2l3LD+KO2/wAZqf6VP2l3LD+KO2/xmp/pUHzk+41v95z/AOTKe41v95z/AOTK+jb9pdyw/ijtv8Zqf6VP2l3LD+KO2/xmp/pUHzk+41v95z/5Mp7jW/3nP/kyvo2/aXcsP4o7b/Gan+lT9pdyw/ijtv8AGan+lQfOT7jW/wB5z/5Mp7jW/wB5z/5Mr6Nv2l3LD+KO2/xmp/pU/aXcsP4o7b/Gan+lQfOT7jW/3nP/AJMp7jW/3nP/AJMr6Nv2l3LD+KO2/wAZqf6VP2l3LD+KO2/xmp/pUHzk+41v95z/AOTKe41v95z/AOTK+jb9pdyw/ijtv8Zqf6VP2l3LD+KO2/xmp/pUHzk+41v95z/5Mp7jW/3nP/kyvo2/aXcsP4o7b/Gan+lT9pdyw/ijtv8AGan+lQfOT7jW/wB5z/5Mp7jW/wB5z/5Mr6Nv2l3LD+KO2/xmp/pU/aXcsP4o7b/Gan+lQfOT7jW/3nP/AJMp7jW/3nP/AJMr6Nv2l3LD+KO2/wAZqf6VP2l3LD+KO2/xmp/pUHzk+41v95z/AOTKe41v95z/AOTK+jb9pdyw/ijtv8Zqf6VP2l3LD+KO2/xmp/pUHzk+41v95z/5Mp7jW/3nP/kyvo2/aXcsP4o7b/Gan+lT9pdyw/ijtv8AGan+lQfOT7jW/wB5z/5Mp7jW/wB5z/5Mr6Nv2l3LD+KO2/xmp/pU/aXcsP4o7b/Gan+lQfOT7jW/3nP/AJMp7jW/3nP/AJMr6Nv2l3LD+KO2/wAZqf6VP2l3LD+KO2/xmp/pUHzk+41v95z/AOTKe41v95z/AOTK+jb9pdyw/ijtv8Zqf6VP2l3LD+KO2/xmp/pUHzk+41v95z/5Mp7jW/3nP/kyvo2/aXcsP4o7b/Gan+lT9pdyw/ijtv8AGan+lQfOT7jW/wB5z/5Mp7jW/wB5z/5Mr6Nv2l3LD+KO2/xmp/pU/aXcsP4o7b/Gan+lQfOT7jW/3nP/AJMp7jW/3nP/AJMr6Nv2l3LD+KO2/wAZqf6VP2l3LD+KO2/xmp/pUHzk+41v95z/AOTKe41v95z/AOTK+jb9pdyw/ijtv8Zqf6VP2l3LD+KO2/xmp/pUHzk+41v95z/5Mp7jW/3nP/kyvo2/aXcsP4o7b/Gan+lT9pdyw/ijtv8AGan+lQfOT7jW/wB5z/5Mp7jW/wB5z/5Mr6Nv2l3LD+KO2/xmp/pU/aXcsP4o7b/Gan+lQfOT7jW/3nP/AJMp7jW/3nP/AJMr6Nv2l3LD+KO2/wAZqf6VP2l3LD+KO2/xmp/pUHzk+41v95z/AOTKe41v95z/AOTK+jb9pdyw/ijtv8Zqf6VP2l3LD+KO2/xmp/pUHzk+41v95z/5Mp7jW/3nP/kyvo2/aXcsP4o7b/Gan+lT9pdyw/ijtv8AGan+lQfOT7jW/wB5z/5Mp7jW/wB5z/5Mr6Nv2l3LD+KO2/xmp/pU/aXcsP4o7b/Gan+lQfOT7jW/3nP/AJMp7jW/3nP/AJMr6Nv2l3LD+KO2/wAZqf6VP2l3LD+KO2/xmp/pUHzk+41v95z/AOTKe41v95z/AOTK+jb9pdyw/ijtv8Zqf6VP2l3LD+KO2/xmp/pUHzk+41v95z/5Mp7jW/3nP/kyvo2/aXcsP4o7b/Gan+lT9pdyw/ijtv8AGan+lQfOT7jW/wB5z/5Mp7jW/wB5z/5Mr6Nv2l3LD+KO2/xmp/pU/aXcsP4o7b/Gan+lQfOT7jW/3nP/AJMp7jW/3nP/AJMr6Nv2l3LD+KO2/wAZqf6VP2l3LD+KO2/xmp/pUHzk+41v95z/AOTKe41v95z/AOTK+jb9pdyw/ijtv8Zqf6VP2l3LD+KO2/xmp/pUHzk+41v95z/5Mp7jW/3nP/kyvo2/aXcsP4o7b/Gan+lT9pdyw/ijtv8AGan+lQeY3sWqeoh5pNUumgkYDoCuGXMI3+6Nu27IvV/h7y+8HOFF6n1Fw+0PSWW41NK6ilqIZpXF0DnseWYe8jBdGw5xn4QiDkZERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERARF1x53ecO08nnDy1andp2PUN8v9w9xttqfWe7B7GML5p3PDXHoYCwEAZ6pWDIBJQdjkXXTki5o9T82vDG58SL9w2h0lTUt2fbKLwriaptYGRsc94zGwtDS8NzuCQcEEELlLWPHHgpw7uYsvEHjBonTFwcwSCkvOoKSimLSAQ4Mlka7BBBzjHZBvlFxjQ8zvLXdJXQ2zmF4aVkjR1ObT6soJCBnGSGykgbjf5rf1lvtk1JQMuunrxQ3SikJaypoqlk8TiDggPYSCR2O6DUEUNQ6VkEj4IhLK1pLGF3T1OxsM74z2yvPXgL7V+p4kcwFs4IcTeDlPok3W5S2QVX3ZdUPpLiHFkUErHRNB6pW+FkEYe9pxjKD0QREQEREGl6j1DZtJaeumq9RVoo7VZaKe419QWOeIaeGMvkeWtBccNaTgAk42BOy62n2n3IwCR/s6R5H/c9dv6quXuZVjZOXPipG7s7RV8afqNDMvmhc3Di0b4JCD6AP3T/kX/HrH/o7dv6qn7p9yL/j0i/0eu39VXgNHRPcOp5LR3zhWywRNHwPJP1IPfv90/5F/wAekX+j12/qqq32nnIy4bcdYvt09dh/9ar5/iwoI3ncBB9AJ9p7yMDY8dYv9Hrt/VVT90/5F/x6x/6O3b+qr5/uh2cYRzHN7hB9AI9p9yMHYcdIvt09dv6qqn2nvIwO/HSL/R67f1VfP8xjpDhoyVV8T4z8Qwg9/wD90/5F/wAesf8Ao7dv6qn7p/yL/j1j/wBHbt/VV8/zWl5w0Kro3M7oPoA/dPuRj8esX+j12/qqH2n3Ix+PWL/R67f1VfP6pGQSyNLmtyAg9/v3T/kX/HrH/o7dv6qqj2nvIwRtx0i/0eu39VXz/CNxf0Abq8wFoy44Qe/v7p9yMD/s6Rf6PXb+qp+6f8i/49Y/9Hbt/VV8/wAGknDd0cxze4wg+gH90/5F/wAesf8Ao7dv6qn7p/yL/j1j/wBHbt/VV8/uMqWOJrh8TiD6IPf390/5F/x6x/6O3b+qoPafcjB2/wBnSL/R67f1VfP84YJGc481NDDC9pL5CD5IPf0+075GgMnjrD9mn7qf/rVW/un/ACL/AI9Y/wDR27f1VfP84AOIaSQELHDuEH0A/un/ACL/AI9Yv9Hrt/VU/dP+Rf8AHrF/o9dv6qvn9wScIQR3QfQH+6fcjH49Yv8AR67f1VU/dP8AkX/HrH/o7dv6qvn92+auDHEZHZB9AH7p/wAi/wCPWP8A0du39VVR7TzkZP8A2dIv9Hrt/VV8/vSSceaOaW90H0A/un3Iv2/2dIv9Hrt/VUPtPuRj8esX+j12/qq+f7BAzhUQfQF+6f8AIv8Aj1j/ANHbt/VU/dP+Rf8AHpH/AKO3b+qr5/VNBCyVwD3kZ+SD38HtPuRg9uOsX26eu39VVT7T3kZbseOsX2aeux/+tV4F1Nvjhj62SOdtnBAWAg+gL90+5F/x6Rf6PXb+qp+6fci/49Iv9Hrt/VV8/vdSNhyMk4Qe/wB+6f8AIv8Aj1j/ANHbt/VVUe075GX9uOsP26fuo/8ArVfP64YOFRB9AR9p9yMD/s6Rf6PXb+qp+6f8i/49Y/8AR27f1VfP8Gk9kLSO6D6Af3T/AJF/x6x/6O3b+qqo9p7yMH/s6Rf6PXb+qr5/E2+aD6Av3T7kYH/Z1j/0eu39VT90/wCRf8esf+jt2/qq+f1AMoPoC/dP+Rf8esf+jt2/qqqPaecjLhtx0i+3T92H/wBar5/zDI1vUWkK+OONzSXvIPlhB7+fun3Ix+PSL/R67f1VP3T/AJF/x6x/6O3b+qrwBjja6Toc4gKs0TGODWOJz8kHv7+6f8i/49Y/9Hbt/VU/dP8AkX/HrH/o7dv6qvAAxPDc42ViD6Av3T/kX/HrH/o7dv6qn7p/yL/j1i/0eu39VXz+og+gL90/5F/x6x/6O3b+qp+6f8i/49Y/9Hbt/VV8/qIPoC/dP+Rf8esf+jt2/qqqPae8jB/7OkX+j12/qq+fwDKqBvgnCD6Af3T7kX/HrF/o9dv6qn7p/wAi/wCPWP8A0du39VXgE6JmMteSfIKIgoPoC/dP+Rf8esf+jt2/qqfun/Iv+PWP/R27f1VeAETGvOHEhXPh6T8JyEHv7+6f8i/49Y/9Hbt/VU/dP+Rf8esf+jt2/qq+f3siD6Av3T/kX/HrH/o7dv6qn7p/yL/j1j/0du39VXz+4yhGEH0Bfun/ACL/AI9Y/wDR27f1VP3T/kX/AB6x/wCjt2/qq+f4scO4VAMoPoC/dP8AkX/HrH/o7dv6qn7p/wAi/wCPWP8A0du39VXz+og+gL90/wCRf8esf+jt2/qqfun/ACL/AI9Y/wDR27f1VfP6qtY53YIPoCHtPeRg7DjrF/o9dh/9aofae8jA2PHWLPy09dv6qvn/AHwvYMkHCs+tB9Ah9p1yNNHV/s6w/wCYLqf/AK2VG+095GCNuOkX26euw/8ArVeAAjDvok5VjmlpwUH0A/un/Iv+PSL/AEeu39VQe0+5GDsOOkW//c9dv6qvn9VQwkZCD6Bn+015HWN63cdafB7YsV0J/IKbKi/dP+Rf8esf+jt2/qq+f1EH0Bfun/Iv+PWL/R67f1VP3T/kX/HrH/o7dv6qvn9TBQfQF+6f8i/49Y/9Hbt/VVUe085GXDI46Rfbp+7D/wCtV8/ibfNB9AX7p/yL/j0j/wBHbt/VU/dP+Rf8esX+j12/qq+f1EH0Bfun/Iv+PWP/AEdu39VT90/5F/x6R/6O3b+qr5/QMqvSQRnbKD6AR7T7kYP/AGdIv9Hrt/VU/dPuRf8AHpF/o9dv6qvAJ0MbWhweST5YUSD6Av3T/kX/AB6x/wCjt2/qqfun/Iv+PWP/AEdu39VXz+og+gP9085GMZ/2dIv9Hrt/VVT90/5F/wAesf8Ao7dv6qvn+IwFRB9KvA/md4G8x7Ly/gvrpuoxp807bl02+rpfAM/ieF/bEUfV1eDJ9HOOnfGRnlNeWPsNY2ik4zyb9TpNPNP1AXAj9JXqcgIiINtcROIejuFGjbnxB1/em2nT9nYyStrHQySiJr5GxtPRE1zzl72jYHvk7Arr9+6gci/49I/9Hbt/VVqvtF4WVHJfxMhkGWmjoifsr6c/zL576WGOYvEjy3DcjHmUHv3+6f8AIv8Aj1j/ANHbt/VVUe075GTuOOsP26fuo/8ArVfP+6IhxDdwFNFSsc3MjiCg9+f3T7kYH/Z0i/0eu39VT90/5F/x6x/6O3b+qrwCmijj+i8n7FFjKD6Av3T/AJF/x6x/6O3b+qp+6f8AIv8Aj1j/ANHbt/VV8/zWl3ZVdE5vcIPoA/dP+Rf8esf+jt2/qqfun/Iv+PWP/R27f1VfP6qhjj2CD6Af3T/kX/HrH/o7dv6qn7p/yL/j1j/0du39VXz/ADWOecNG6q5jmHDtkH0Afun/ACL/AI9Y/wDR27f1VP3T/kX/AB6x/wCjt2/qq+f4gAZBVEH0Bfun/Iv+PWP/AEdu39VT90/5F/x6x/6O3b+qr5/VVjepwBOMoPoB/dP+Rf8AHrH/AKO3b+qp+6f8i/49Y/8AR27f1VeAs1KI2dYeSoGAOOCUH0A/un/Iv+PWP/R27f1VP3T/AJF/x6x/6O3b+qrwGZT07gPvpz5jZRyQEH4MkIPf390/5F/x6x/6O3b+qp+6f8i/49Y/9Hbt/VV8/uDnClbSyvHU1uQg9/f3T/kX/HrH/o7dv6qn7p/yL/j1j/0eu39VXgVR211U/pL8E+gytctegLzc6yOCOI+G5wBeAcgeqD3Z/dQORf8AHpH/AKO3b+qqo9p7yMntx0i+3T12/qq8eqjlufXWeGazOlNUQM9zk/MLXtNcmWpbrbnyVEU5nIy3DSMfYg9Z3e0+5GG9+OsX+j12P/1qg9p7yMOGRx1i+3T12H/1qvI1vIxxZlqXMio5DFvh3hHJC2zf+VfX1hrRQTwPMzj0gGM90Hsv+6fcjA/7OkX+j12/qqfuoHIv+PSP/R27f1VeJWreXziFo2hFxu1uLICOoO6SNlsWjtTJiRPIWlpxgIPe/wDdP+Rf8ekf+jt2/qqfun/Iv+PWP/R27f1VeCFbaoqZvVFK53yOFppY4eSD6Af3T/kX/HrH/o7dv6qn7p/yL/j1j/0du39VXz/FjhuVRB9AX7p/yL/j1j/0du39VT90/wCRf8esf+jt2/qq+f1EH0Bfun/Iv+PWP/R27f1VP3T/AJF/x6x/6O3b+qrwBhja84c4gKkjGtf0tJIQe/8A+6f8i/49Y/8AR27f1VP3T/kX/HrH/o7dv6qvAaGmY76bz9iimYxj+lriR6oPf790/wCRf8esf+jt2/qqfun/ACL/AI9Y/wDR27f1VeAckUbWBzXknz2UKD6Av3T/AJF/x6x/6O3b+qp+6f8AIv8Aj1j/ANHbt/VV4ARsa4/E4hHsAI6TlB7/AP7p/wAi/wCPWP8A0du39VT90/5F/wAesf8Ao7dv6qvn9RB9AX7p/wAi/wCPWP8A0du39VT90/5F/wAesf8Ao7dv6qvn9xlSsiYWFznEFB7+/un/ACL/AI9Y/wDR27f1VP3T/kX/AB6x/wCjt2/qq+f47EqiD6Av3T/kX/HrH/o7dv6qn7p/yL/j1j/0du39VXz/AAAPyVCADsg+gL90/wCRf8esf+jt2/qqfun/ACL/AI9Y/wDR27f1VfP6iD6Av3T/AJF/x6x/6O3b+qp+6f8AIv8Aj1j/ANHbt/VV8/qIPoC/dP8AkX/HrH/o7dv6qn7p/wAi/wCPWP8A0du39VXz/NaD3OEcADscoPoB/dP+Rf8AHrH/AKO3b+qp+6f8i/49Y/8AR27f1VeAMLGvPxuICpKxrHYYSUHv/wDun/Iv+PWP/R27f1VP3T/kX/HrH/o7dv6qvABsL3DICk93AaS95B+QQe/f7p/yL/j1j/0du39VT90/5F/x6x/6O3b+qr5/iMnZXCFzgg9//wB0/wCRf8esf+jt2/qqfun/ACL/AI9Y/wDR67f1VeADo3NGT2WXBSU0kJkfM5rvIDCD30/dP+Rf8esf+jt2/qqfun/Iv+PWP/R27f1VfP8Aub0uIG4BxlWoPoC/dP8AkX/HrH/o7dv6qn7p/wAi/wCPWP8A0du39VXz+og+gL90/wCRf8esf+jt2/qqfun/ACL/AI9Y/wDR27f1VfP6iD6Av3T/AJF/x6xf6PXb+qp+6f8AIv8Aj1j/ANHbt/VV8/qkbBI8ZDSUHv8Afun/ACL/AI9Y/wDR27f1VP3T/kX/AB6x/wCjt2/qq8BoaXrcRIS0D0CjqImRP6WOJCD39/dP+Rf8esf+jt2/qqfun/Iv+PWP/R27f1VfP6iD6Av3T/kX/HrH/o7dv6qn7p/yL/j1j/0du39VXz+og+gL90/5F/x6xf6PXb+qp+6gci/49I/9Hbt/VV8/q1ujsdPU0ZqHTPDsZHbA2Qe8/wC6f8i/49Y/9Hbt/VU/dP8AkX/HrH/o7dv6qvAB8REjo2nODj608GTIBGEH0ecGOcblw5hdUVWi+D/ElmoLzRUD7pPTC011N0UrJI43SdU8LGnD5oxgEn4s4wDgvML2LMD4+aTVLnNODoCuA/zjbkQe0yIiAvCv2svGKbiTzVV+kKSqc+0cPaGKywMB+A1TwJqqQA9ndb2xH18AY9T7qL5uqOQ8cub2nkq3sqhr7iPGHkkFsja25gY74IIl8jjHnjdB708p3CyDgvy48PuHUdOIai22SCSuHSRmtnHj1JwQDvNLJjIyBgeS8SfaRSSS87XFJ8sjnuFfSNBcckAUNOAPqAAAHkAAvoSXz1e0e/DZ4p/+EaX9Sp0HOmkPY48WNbaDset7Rxd0lEL/AGilutNTVNLUt6BPC2RrHva12CA8AkA9tgV1j0trnjzyP8c6+htF2qLFqXS1wNJdrcJzJRVzWkExysB6ZYntIIPcBwc0tIBHpnw29q/ypaF4QaT01cJdX1d10/py30FRTUtnBMk8FNGx7WPfI1hy5pAJIGPNeeVwtHEP2hnNtfbzouxwW6p1hdGzvFRUN8K2ULGNiY6V56etzIYwS1o6nlp6WkkBB77cPtYUXELQWm9f2yN0dFqa0Ud4pmF3UWxVMLJWAnAyQHgZwPqXh17T/h3Nwg5zb3f7B4lAzVEVJq6hkhy0x1DyWzPa4fvjUwSyZG4LgvcjQmkrfw/0Rp7QdoLnUGm7VSWilJABMNPC2JmQNh8LBsvL724Wl4ortwm1rFGPEqae7Wuof2PTE6nkiHfcZlm7D6zuMB6NcvHFOLjZwP0TxViDGyajs1PV1TGfRjq+npqIx8mzNkaPkPJcirox7HbV8uoeUqWwTzlx0tqivt0UZIyyKVkVUCAPIvqZe/mCu86AiIg455jxnl44oj10Xex/yGZfNA49Mzj6OP6V9L3Mj+DvxS+Wi73+ozL5oJN5H/8AfH9KDMkrg+HwmjBx6LCGSQPVOkqseA7dBqUVO1zAdtxuoJHsgHT3ypPePvYbGd/qWFKJCSXoL43h0mT2/wDalTg4x6qAHCqXF3dBkUDwyX4lJcZGyOGMLCBIOQqucXd0F8MnhP6sZ2V08olII+rsoUQXlnwjBWTS1jYWFjhuRhYzH7jq7K97A7dqCQFrJPHG4PcKskrZ2nGyxy92Oh3YKRuw+FBG13hvz6bKssniHYKx30z9ali6OnfGcIIhsd1I05GQo346jjsjXYQHfSKkZCXt6s4Cp8KlZS1D2l0QJAQRMb986TvlSyNbjI2UOCx+HbEI6Qkj0CC5g+LtsqSgZ28leS0s+DurRFK7cg49UESy6YNMZDgoadodJ0uP1KSozAegeaC3AZI4+Stld1uACsDnOP1qvQ4kEIJpIumPqaN+6xsYWbEfhDZN1BUhgOGdkEKq0lrgeykp4/EfhX1MXhYHYlBlmUTQhhIwdgtPkj6Djv8AUjHSZw1ShvnIgupqcOPU4qWoiDG7brG8dwPSz1U8Uhe3755oMN/0iqKWSJ7nktGQSo3NLThwwUFWODTlVe/r8uxVivZ04JKCxSxwh4zlRHGdlc2VzeyC6SLoVjThwPop3EzNw3vlWeBI3BLcBBPLUtdD09O/qsPJ9Vk+G2Roa3GVA+J8Zw4IKdR8tkaT1DJKdJ9Fc1hxlBkyvYYceaw1eOt/wjuqOY5pAIwSgtRXGJ4b1EbK1AREQXM7o/uqxEB26mIa4ZQQNJB81R25V78Z+Hur44S9h23QQZWZTAGM9XdQOhcwkuGAFRsj8hrPPZBbKMPPorVnOtk5j8UtODvusRzeiTDkFoyN8IN3D5rKlEPgggDKxW/SGEGTNGGx5woWAFpUsof4YJzhWxMPQThBAdiUV3SXPIHdUc1zDg7FBRZ1HG3p63HssNjOryWRG57MNBQTSysl+94+sqF1H0sL8qpiezLzkb7K7Mr2EN3+pBHSENlw71U9VB1/G0HCw3MkicXdsFanRzNfATJ6INMEQzglZjImNiz3OFhyuPiO6e2VJDUu6gHHb1QQvB6yAPNAzK1GQUxj6ts4ysFzXO3aNkEZACAhHAjuqIB3QboMZ3Q90F3RtnKtVS4nbKuZjG6C+mYHvwVWqYGOGComyFhy3ZXOc6Td26CPJ9UQ90QEG6qcY8lRBeQMKxVLidsqiD1c9hr/ALncZf8ADWD/AKNevUxeWfsNf9zuMv8AhrB/0a9epiAiIg65+0O/A04l/wCJ0f69Tr55ASOxwvoc9oa3q5NuJQ/+k6P9ep188be4QZAd0N6sZKsdO522MZUzHROaOrfCskEWD0jdBdFS+Lv1BRVEYhPSEiqHMOM7Kk0viv6iUGTa4WzSlr+3qp7jA1jg1gyOywopnU+HsWU2qE7cyHdBiSwdA6gFYyTpBBWV1ZPxdljT9HV8CBBKInlxGQlRKJX5AUZaRvhU7oCKpY4dwg7FBTCuZkOB9FWHp6j1Kd5iA2AygpNIHRgd/koooy/tlWvd1bDtlZED2MYcnBQRPgcxvV1KragtaQRnbGVkRUs9e/ohGSCsz7kmkH+2Bg/NBpULHPlAxnJW/dL6d+6WIjCTlvotAoqamdI0NxnI+a5n4fWqMgOaB1BuRsg5E4H8r7NbVUbjATlw8l2+sPJta7MY4JLe3rLRv0j0WzeVqtqrVWsLnANBC7aXPWk4uMBEoIDQg0Dh3yt2y2ETVFuEjB8WOnIW7X2jTmmtRQ0P3HYGhwbjoC5A0fxGt0VEGV8rR23BGVsDXWv9LR6njuMr2eEx+XHIQc50Fp0+2giqIbTSxxlgdvCMjbz2XF/EHg7pO+XSLUDLRFiNwLiGDBPmpavmM4fy2r3e3V7Hzlgb0Bww3ZbTvHM3pWhsclBPPGJXAgfEMoOtnPRe9JwaTfZLdbYmSxRFp6WjOcLyemjcx1Q7pxh5/Su/HM9rui1JVVNQXgxPBIyfJdG70aVz5RBjBeSg2zJVOmd4bvLuseWNoO3n6LIrIWRElndaeZXu2ccoKyPDgQPJRorxE8jIGyCxEIIOCqgHICCmcKoODkq4wuAyrCMIL3SkjAVWx9TS4nfuo1UPIGEFDnsUwfRO6mZ0dG+MoIcqvV67qvSXuIaELSwjI7oLVUAK5+MbKzJQAcJklXR9Od1cYi4/D5+SCNMK50bm9wpGeH0kYGUEKkEQLeoq+Onc92w2KSwzMOMEBBAdlVo6nYVOyua7G/mEEr6cMb1Z8sqBXule4YJVre4QU3+anjpw9nVlXNDCO2+FY6VzPhB2QRuYWkgFA1436SQEc7qxv3W4LcyhdRnxiOrGd0Gl0s3cObv6KGsdl4xsFlRxxmocG9t1jVbfv3Q0b7oMdpwVnU3S5uT3UAopukOLdirWvfC8NJwPMIJ6ohwxjHzWH1OAx1HH1rOqZYzFluOohYGcoCnbThzesuwoFK0zBuG9kEZGCQqDdVPc57qndBUgAd1RXhru57K1+AUFFlwVYiaGlufsWIsiMRGM57+SBLVdZ+HZQFxd3V4j6TuNvJWvxnZBaiIgJg+ivj6c/Epz4RG3dBHTw+M4gnGFqIuPukJpgfktN8R0Tss2Ub3l7up3dBIyXEpkIzuSslj2vw/YfWsFXCQhvT5IPQj2MFU2bmi1RG1uMaBrT/zjbv8AWixPYq/hT6p/4v67+UbciD2pREQF82nKG+kj5q+Drq2F0kZ11YwA04IkNdEGH6g8tJHoCvpLXzeahpZuX/m7rIqnMB0DxBMzXNaRiOkuAex7QDnBYxrhg5wRug+kNfPV7R78Nnin/wCEaX9Sp19CTHslY2SN4c1wDg4HII8sH0Xz2+0e/DZ4p/8AhGl/UqdB6BcK/ZKcrOtOF2kNX3au1yyvvtgt9yqvAu8LWCaemZI/oaYDgdTjgEnbG66E883KNVcmPFOz27Teq626WK+U5udjr5QIq2nfFIA+J5jwC9hMbhI0NBDxgAghdueG/tkuHGhuHeltFVPBfUlVNp+y0VrknjuMDWyuggZGXAEZAJZkA7gFdMubnmm1xzt8WrVeINIS0FNSQttOntPUTnVs+Xvy45awOlmkeWjDWgYaxoBIJIeuns2OP+qOYXljoL/reslrtQabuc+m6+vlOX1roY4pY5nHu55hniDnHdzmucdyuuPtwH0o0NwqjfG73k3W5mN47Bghh6wd+5JZjY9juPPs57O/l5v/AC3ctNq0lq+EU+o79Wzahu9KDn3SedkbGQk9i9kMMIdjYPDgCQAT0u9t/q1lTq/hZoOOTDrdbbld5mA/SFRLFFGSM+XusuDjzPfyDlf2Jn+8Lrv+F5/Uqdeiq6R+yC0XNpblAp73URFp1bqK43eMluCY2eHSD7M0jiPLcnzXdxAREQcccyX4O3FP+BV7/UZl80En9kd/3x/Svpf5kvwduKf8Cr3+ozL5oJP7I/6z+lAL8q1EQXwu6XKaR/iODcdyo6ZnXJhSVLDC8Ed0E01v8OLxO+ywDss6WsL4OgrBO6Ag3TBTsgl8IYyo3DBUjpMNwFESSgKeFwDSDsrIenq+JXyt6t2DZBZKMklWteR2VDnsVNF0dPxd0EBOTlFdJjqOOyozHWM9soKYPogGVqEwgEALcZxusAd0FSzA7rU6G6x00Jjc0EkYyVpgDpDgI+NzDhwQS1BE0jpG+ZyoMYVzCM7qVxYQgji2OSsmOWSSUQxxlxcQAsdkMkh+AErf+g7TbffYJLkGgAgnKDTabR1W2nFa6ncAd9wVot2oZGElsZ2OF2kvVTo/9jgp6V0XidOMDC4lm03DXGSRgBBOcoOIMOac9J2+SyYyOnJ2W8NQWWkoWHYAjutkyvw8hvbKCr5TkgeShJJOSq5yclDjyQSU83gv6sK6pnE7gRthWwR9b8eSrUxiNwA80FkcnQcq58plwAFYY3BvUVn0TacwkyYyEGOIRG0PJ3woxKS4KtRKS9zQdsqEbFBnucI4wfyqANE+XfNWgvmb0jyVuHwnBGMoLCCCR6KiypDGY8juVioCAZICIO6DNYwQND+/yVstYHt6QDlXUr/G+B/ZWV0TI3AMGyCKnm8KTrV1VUCd+R2+agVQPMoKg5GMdlUOd9HCmjDMA4Uh8M4AaMnzQQRkRnqP1q2WXxHAgdlmy0BMZkb9iwTGWOw7ugvfUdUYYArWxtc3JOCqugc0dXljKtDiEFpYQSBuhBHcELJpXMa8l52VagMkkDY8blBiKWPJ2zjKnfb5WR+IRt3WOA7q6RsUEkkDWDqBz67qSCTpxsrHRSgZd2VrT+ZBPK/xG4Pmo4ow14cQDg5VS7ZUB8wg1Q3Fxi8PAwBhaY9gnmAO2ShecbqwuIIIOCEGfW2eOnpxM2XJIzjK0poOR6rJdVTyDpe8lv6FaIS74h5oEkhcwNVGSFrC0hVjAyWlUl2dhuwygpTkiTqI7lZ0tC2dokBWGzsrzPM1uGuOPrQY/wDYpC0+SzKfpfI3tjKwHEk5PmrmTPYct7oNbuEcbabIO60qGr8PYq2Wsmmb0OO31qxkEj29TRsgyHTNnPT6qwuMWWA91BvG4+qua/qcC5BPDSePvndRVNP4DunOVM6oMTB0LGklfKcvOSgt6iRjJWRC8CPdYyvY17thlBSQ5eSgaMZyrnQSNGS0gKzJ7IKY8kwpadoc7DlWpa0PAb2QQoN1Ieno8sqxuM7oLxFkKgwHdPqrnFzR22UROTlBO+ANZ1Z7qBXmVxb0q1uOoZ7IKYPomFkyeH4W3dRw9Hn6oIsIsmXo6du6xkHq57DX/c7jL/hrB+ivXqYvLf2HDQLRxid5mosQ/I2u/wBa9SEBERB109oa7p5NuJTvSjo/16nXzxYX0Pe0Lx+034lZ7e50f69Tr55oOn4urthBFlFV+Oo4VEF8UfiHGVV8YY8DPdWs6s/D3R/XnLu6CeSLpjyFjgkHIWRTQz1R6GjICkno3U4xIEEcTvF+E4CvfS9J/wBSxmlwcfD7/JZbJyGnr7/NBFOwMYPVVgga9nWTjG6hlkc9xHllUbK+MFo7ILpnjJbhRKoaXnZXBha8BwQWYPomFlSwfACB3WbSUDX05kcBkDvhBp9PCJFbURGN2PIKRjvCnLT2U73wv+nhBkadr20M5c9uQey1m5VLLkR0NAz3Wk25tI525CypaqmZKIYnfEdgEEtJRSU7mvAzkrnPhTDV1VVGxtK4ggDt3XGelrHPVyQvqASxzx3Hlld8uWjg/RXiWB7Ymklo7hByny66Cnro45HU5YTg5xhcwcStOO01bjXiTdjc5z6LkjRXDWXSlrEtPABgZ2C2TxrpbtV6dnkew+GAQfqwg6u/7NNVNeZrX90TEGZGOrC4q4vcXaunglpILuTI7IB698rjvipfJLDqWpNFkSgnOD57rrre9e19fqBouLn9Afg5PYZQc2aI17qmnvD6urvUpiduAXnC0XiTxSvsl8Y6C8v8MPGcPK0a76gtjLCx9tIMxYOx37Limuq62vlJlB687IOUNXa5qL3aGxOqy9/RjPV32XEUtQ+Fxa9xOTndZcMNxib1ThwZj8yxq5jZGF7RugxKqXrBLTlYBDj5Kene3xemQ7ZU8oi6h0YA88IMDGFn00nTF04UEkXUMs7hTwuZHD0v2KDDlOZCVbnfKq/d5ICtwc4QTeOOnG/bCiJ6ihY4DKogqRhUQklEBEVQ0kZCDIow3q381SswHjp7AYULXuYdlRzi45KCmFUNyd1dEWg/F2Vz3sxhvdBY5oAyFfHN0YBCiJJRBLLIHjsrGtzuN8K1XMd0nfsUGRDViLYg7KstYHggA5UT4gGeJ5ndQoLmgOJJIH1qjmhvmqxtL3ABZLKTp+lugxFUDKyqmBkbMtxn5LFYcHHqgNcWoT1FZDaQu3yrfB8JwBwSgR05cAcFTU0Er5xEHFoJwrhIA0YHdZVHKyN4kc3JCDVvuC2npxMNyRnK0ialHvAe/bBWui7e8xiAHCwKul2LnPyTugsmqIY4ekAZA8loM7zI/OFmSA5LSSQsd4Y0gEd/VBEyIv3OVbI3odhTu6g3LBthY7i4nJQUAycKds4awt9VAAT2UohJbn5IIicklBsQUIwSPRVb3QXmYkYwoycrI+9dKgdjOyCinjgyOvPZQK8SvA6QgulkDst9FEN07p2QXOaAMhWoSSiAnZEQVLiQqK+Lpyco7HiDCCzCKeXo6Nu+AoEHoB7FYY5p9U/8X9d/KNuRU9iqf+un1Tk/9j+u/lG3Ig9qUREBeIXtfODE+gOZWPiVRUbmWjiHb46zxGtIYK+ma2GoYD2yWCCQ75JlK9vVwpzWcq2g+bXh3BoPW1bW2yW31rbhbbnQtjM9LMGuaRh4Icx7XEOZtkhpyC0EBg8jXFuLjRyr8PtYPn8WvgtbLRc8n4hV0f8AteRzsdi/wxIB6SBR8VuRLlR42a1rOInEvhLBddQ3BkUdVWx3avozOI2BjC9lPPGxzg0BvURkhoBJAGL+ULlPsvKLoi7aG0/rq9ajpLtcjcz90GRxsp5DGyN3hMYNuoMZ1Ek56R2xvz0g6qt9l3yKtcCOBoyPXUt4I/W1y3wt5ZuAPBSpNfwt4Tac0/XOaYzXU9IH1fQRgt8eTql6T5jqwfNcnogLwC5/9f3DmI51tR2zSLTc20lwptF2OGFwf474XCEtYexD6p8xaRsQ8bnuffWsp31VLPSsqpqZ00boxNDgSRkgjqb1AjIzkZBGQMgjZdL+X72WfCngTxhoOMk2vtQ6suVollqaCnucULY2VD2uaJ5C0EyPaHkg7Ydh2MgYDtFwW4a2/g5wl0jwttbxJBpi0U1uMo/+TSsYBJKdhu9/W87Ddx2HZb2REBERBxxzI78u/FL56Lvf6jMvmgk/sj/rP6V9MfMX0/tfOJ3V2/Ybes/V7jMvmfnGamUN7dbsfVkoIlUDJA9VUxuAyVRuBugy2RCBvi5zlRVE3juz2wjZy4dLuysl6AfhQIYvEd052CrPD4WMKxrnMPU04KvDZJ9zk480EedsK9kQcCSeysc0tOCrow8j4eyCe30nvk3hk4ClulvFA8NBzlQxVD6V3XGcOVtRVy1RBmOcdkEClim6AQclRHGdkQVccuyqZVzBlwCyPcJ3jrY3IQYqYVSxwcWEbg4WTE1rGkPGCUGM3qdsCqujLcfNTxRlsnU7spZnROGBjOPkgxYJRE7Kunl8dwwFYYnd8bIB0vH1oJPAIbnBHzUJbgkZWrTmI0w6QM4WkknzQZdFUshILluqiJqIDJDJ0kDIIK2TkhbgtNybDCYidyMBBvzhlaavVF6+51TWnoDsbnZcm6600zRdEGwt8QkZ2+pcJ6TuV1sleK+gJBLgcjK5+slXJrG3D7tHJ6ezvqQdbtTXaprKksMZaCtOo7EaqMvJx3Pdcta/0VQwyvNI0ZGcYXGomfauuKZ2MHGEGg1dC6nkLAM4WKWuHcLWn1EE8hccEFY1SIA74cAIMOmd0P3HdXVbupwOOylzE0Z2yseX74ct3QHTB0fQo2k5ABIyqxxPld0sGSrpIZISOsYQXOh6W9WQoVcXvI3Oyt7oJYJRE/JH5FWpnExBAxhWCF5GcK1zS04KCmUVek4yqICK7b5KWOkmlb1Mbt3QRxSmJ3UFWaYzHJV0TWsl6ZRsq1AiLh4YGB6IIRgDJGVIxgdjyRoGNxn5IQR9EbIJOgAZyrQ4tcD5Aq0F2O63fo7hFxZ4iQOqOH3C7V2p4mOLXPs1kqa1oIG4JiY4Aj0Qba+6Dg0Nx2URHinxFu/VvA/jRoCiNx13wh1rpukADjPd7BV0ceCcAl0sbRjO3futoxzsbH0nugo6braGHZR9ADwO4VMdTj0qh6mnfZBLNA1jeoOUcbuh4d6KreuU4yhYQQw9ygy5LkXxBnT22ysIO+PqUjqaRreo9kFM9w6h2QSSVJkYGkbKAHCqWluAVWPAcCe2UFd++CPrRZNQ+J0YDQM4WGglwMq9sQfj0UAa49ld1Ob5+aCcUzCDuNla5z4/gaMhRGVyzaWWEx/fMEj1QYPxB2cYyr+gOGSr6l7CT0D6lACe2UFxwO2+EBJHbsqmFwb1OPfdR90FpH5Fb0hbr4aaFrOJfEfSnDigroqKp1Xe6GyQ1MzSY4X1M7IWvcBuQC8EgbkA4XPvOhyI37k4otLXK6cQ7fqiDU8tVAz3egfSvgfAIychz3BwIkGCCCCDt5oOrLmgD5rNpp2MgwcZWCQqtJyG52QXtZ48p8spLB4bw0HO6n92lY3xGA9tiFjuLxIDIT3QTvp3eFnBWJjDsFao6oiMGMDOFpshHXkIL3QBrOrKrTzCPcqIyPIwSjQcIMmasbI3pwVC2HqaXKzpIOSsmORgjxnfCDHDuhxx5FUe/rKucwucS0bFWFpacFBRXNb++VqZIGEEvWHfD81Y9vSreyqXE90FFcG5GVbhVyRsgoc+aIgGUDOUVWsLjgKr43M7oPVz2Gz82vjGz+5nsJ/K2u/1L1KXln7DX/c/jKfWawfor16mICIiDrn7Q78DTiX/AInR/r1OvnjX0Oe0O/A04l/4nR/r1OvnjQEVTjHkqAEoL4n9ByQkj/EIVpY4dwqINStdYykdlwCvuVcyseAwjdaW0OdsFcYpG74xhBksgbEA87qOQGT4u2FY2Z5OHHZJH4OAgjOx+pUJycp3V0eOsZ7ZQZVuZ1y9JGyXFgjmAHkskSQwR9UYAJWHK2Sof17kZQXQz9fwkHbzWYa4RwljdzhYLuiMYGAR6KahjE0zfEHwoMXEkshcGkkn0WsU1jdUQGVxwcdluGmpbPDTh0haHHfsFj1EM8zvDt4JafT0QY2mdNirqjHI/AzjK3Qzhuz7qxPDwWkg91oNNYdSwkTUbH57nGVrdHe71ayHXEuDh6oOTZ7ZQ2K1wFrmhzMErsxwA43W7R1pFdLUtDomZwT6LovetdVVbF4JedvmrbFxBrKQe5SOd4b9j8Xkg9X9M8+tFfKh1n94DQD0hZ/EjmKs9w0nPSGsYXvYcbj0XmVZdV2m0NFdESJXDJPV5rF1Nxjr6yIxRSP6ew+I9kGbxX1Syo1PU1gmy17iRvnzXF1XYmX3qq2SBrskrTr5fp7pMXvcST55Wn0l2udOeiMnoKDUi6otTjHLOXMG3fOywn3sCoErWbA+Sju1aaiMFx+Iqlqgp3jMwGEGp1OqYq2AQeGGkDC0x0zSOkuByVNcaajDM0uC75LSPvjN35z5IL6iAMy8KBr3D5q4vkl+HJKz4I6cQESY6kGHFUdBJIJVz2Gb4wceeFZH4ZmPV2yrnShswDfojCCAgtJyEzvlarUU8U0OYQOpaZJC+IgPGCUFXSdTcY8lGrnRuaMkbK1ATB9FVrS7ssmOIeHuN0EEUYecHZSvZ4bcfarYYnuf8Pqsh+GNLZO+EGJG3xCc/WrXDBwFeyToccAHKvbA6RpkxjdBC0OPZZEFMHDLwomv8MkY+Sk96wNggsljDHEfkUSvkk68ZViAirjz7pudsIKmRxb0nsre6zLZabre6+G12a21VfW1LuiGmpoXSyyuwThrGglxwDsAey5N/am80LLV93HcuvEgUQGTIdL1oIbjq6+nw+rpxv1Yx80HFMQcHggLKLpXNyO+FdcKG5WSvmtl3t1TQ1lO7pmp6mJ0UsZwDhzHAEHGDgjzVjKxgABb+lBCW1Ez+nBP6FQ00oeGlu5WoU9RG09RA7qlRWMEwe0ZAKC59FVQ0wkAO4ysRmXA+J+QrVpr7HLTCHp3x6LTGtMmXtHz2QW9D25O+B5rKpmBzCSoZJ9ugtwQr6dwDPrQSMkdHKejOQUqKyY/CeygZKGSF2PNUqJhKQcdkF1PIDITJ2yqVcHXIDENlCxhecA4WT4wp4+hwySEFJI2+AGs3cFitiLARLtn1VYpyybrO4KrVTtneC0YCCPwXZy0ZGVORIyM/ASMKUPayIZG+MqsVUyTEbu52Qaa7OST3RoyQFqNbQtZEZge+600Z7jyQSuhwM5UYblwHqrvvjh54VfBkA6vRBWSHob1ZzlWhmQTnsj5HOGCeytDiNsoDW5OFMab4c52UAOFf4r8YygsOxIVwZkZyqFjhuQgccYCCmPJMeSy6JsZcfEGVZVhgePD7BBB0lUU9KWZPXuElLDOOnsgg3+aDdZdR4PhDoxkhYiD0B9iwwjmk1SfXQFd/KNuRU9isSeabVIP4v67+UbciD2oREQEREBERAREQEREBERAREQcccyef2uvFPG3/wACr3+ozL5ogcTEnc5K+l3mS/B24pfwLvn6jMvmhccSu/74/pQXPlJGOlRLJf4XR8PdYyAqg5KoiCRzfgyp6WUMYQQoYD1Ow7thX1ADNmoKT4cS4FIHYafhzuoS9x7lZVMYy3pI3OyDFecuJVFlzW+ojBkLPhPmseEN8Vod2ygswfQotbqhRCjBjx14WiIKtd0nK1Wlu0cUXhubknbstJAyr2hoOHZz8kGYYmPkM2Rvusepdl4LewQCfGG5woiHdWD37IJTUExhgByoviDgXZWp2ekYJxJVNwzCkvsNM6UOomjp80GJEA5u48vRY07C15wDssuCSNkYDtiFV74ZGntlBhidwHSSVkQ0YmYX9QHmVjvhdkkDZXRvqWNLWE4QRys8N5b6K6JkgIe3O2/dWODy4l3crUKBrXMId5oN06Pu8Uz200jRkHG67D6LoDPBEyJuQ4D7F1coJG26ZszdiXDddyOAtnqNR2AVELcvDMj68INscQ9Px2uhdVhoe4tJx3XCsuj5dRU89WYizoJPbC7CaztN1ornLFfGn3ZucZ7YWybrerBQ0EsNuc0E5BwRlB1qu1FJbKx9KSR0ZCwS4k7krcmsJYJ7g+ZhBJJyVt5kMkjctGyCkbPEd05wPPdSSNEIAG+VEOpr8N79lc7qOPEO6C+knFPKHEZCkq6ltVI3AwFB4DzuBsqdLo3AkYIKDOmpGspw/bt5LAjAL2j5qeWre+IRnsFA0OyC3v3Qa1JTRw0olBGSPVYDYBUAvBwR5eqo6aqmj8PfAH5VFE+SneGnYEoLJOppLCMBUZH1NzlavWCkNJ1tx143WjB7gMAoKxs63dK1ajrIqSMxOwSey0yDJdkd0nz1glBqDqH315lacA7lYz6bwSW98FTUtaWN6QdyMKkj+sk+qDFLQFc0fD80ct88CeHD+L/GfRPDEOeyPUt8pLfUSM7xwPkaJXj5tj63fYg9APZw+zp0vq7TNDzEcwlohuFprWGp07p6rH+15YAdqyrBwHsOCWRn4XN+JwcHALlnjZ7XDg7wkukug+CPDz9mUNnPufvkVWy3WqPo26aYNjeZWDBAIaxh2LS4YJ3n7UTirNwM5TodBaB/6kz6tng0xStpR0e62yOIunazyAMcbIcf3MpxggEeHD5Jc4cT2QetPDv21mibvc22ni7wVrrLbqhwjfX2m5NuDWMdsTJBJHGS0dz0ucSCcNJAB4R9pJoflau0Wg+NfLrWaeZHreOvdcaeylscEpiMJbM6nGPd5cyPa9pY0uIBIDg4noASSpo6mpYPCjkIB2wgpKBDM9rd8HGyjc7qIXLPLvy08SOZniNScO9A00TZZIzVV9wqcimt9KCA6aUgE4yQ0NAJc5wA7kj0tdyH+zo5VbDb38y+t4rzeapgkbJervPSeM4Hd8FFRvbIYwRghxkG5BO4ADx8Y/w91R0nU/qwvZXT/KT7LHmfbU2Tgxd6CmvbYXOxYNQVUVcxg28RtJVucHNacZd4RG4ydwvPHnL5L9dcoOsqa33WtF80te+t1lvkUJjbL0n4oJm7iOZoIJaCQ4EFpOCGh19fVl8YZjsjKotZ04XN/I3wl0Xxz5pdEcLeIdFUVen7064msggqHQPkEFvqZ2APaQWjriZnBBIyM75Xd3W/sjtOai5kai36Rmr9FcHrXZqWsr7hNUmpqairc6XxYKV0xOAGMYXSP6ms6hs8ktAeVxf1E580AJGV7M6M5bfZGXa5xcKLJd9Iag1BVSCmY9+sqt9VVT5GGxSxztic8kgBsWA7BGDuurftAfZ20PLRaYeK3Cq4XC4aGqKltJXUlc4SVNpneT4Z8QAeJC85aCQHNd0Al3VkB0JXpNdvZ/8AAWl9nvHzEwP1INZnSNPqQzm4NMBneGudF4Xh48P4iAPpdviK4G5EeGPKLxGuWszzV69g07FboaH7hxT3cW9lS6R03ju6zu4sDIQAD2kJIOBj2JrdEcv03KezQdVqWBvB/wDY3DRi6fdMCM2sBvRJ7z2wQB8fnlB85jHFuxCo7JOcHC9M+NfLz7LWx8IdZXnhzxjt9RqugsdbU2Sng1aKp89cyFzoIhFv1h8ga0jHZx3HddSeRrhTojjlzO6K4WcQqKersN6NwNZBBUPgfIILfUTsAewgtHXEzOCCRkZGUHBltpWVcha/GEuNK2kmEbDsV6t3j2TmiLtzO1UFibdNL8HbLZqOtrJHVbpp6+ue6XxKaCWXJYxrGMc95z0hwABLsty+P/LV7NSs4F8QavhRqDR7dYaL0/X11HLatZSVtWaqGFz2RviNQ8Sh8gbGSWnBcQCCCEHkuaYeGHZ7rOp7ZG+LxfMb91v/AJcOXriJzPcSaThtw9pYxK5hqq+vqMimt1I0gOmlIBOMkNDQCXOcAO5I9ORyP+z15VbDbzzI66ivF4qWCRr71d5qQzuB3dT0VI4SGMEYIcZBuQScgIPHuqy3LC04G2Vitac7L2csnKf7LvmijqrJweu9BT3sQucBYb/UxV0bBt4jaSrc4Oa04y4REbjJ3C89OcTkz1tyhavprdd6wXzS968R1lvkUJjbL0n4oZm5IjmaCCQCQ4EEE4IaGxuVI/8AXR8HNv8A5f8AT38owL0P9t+M6U4Tf+ELv/6OmXndypYPNNwdI7fs/wBPfyjAvRX23MZk0rwnA8rhd8/5OmQeTVPCJHEHZJaUMcHDfClja5p2PmpcgEB/YoIW3IMb4bmHZQPDajLxt5rKqqeB0fVF3K04+JFt2QWuy0kZVFXDnH5oWEHBQUU8QHTuomxOf2VSHx7HZBWR+5CjBPllV3KyI6clnUgtikGMFWTHJyFQfC/dJCCgsREQEwfRAMlTNDA3fGUETBko4HPZCcOyFqdEKZ0BMuMgbINL7KVgy0k+itmLTIensrQ4jZBdG/w3Eqs0viqkXSXZckvTn4UHqz7DX/c/jKPSawfor16mLyz9hr/udxl/w1g/6NevUxAREQdc/aHfgacS/wDE6P8AXqdfPGvob9of+BpxK/xOj/XqdfPK1hcTjyQUwqg9JUkTcP8Ai7Ks7R1At7fJBYX5GFYnZEEkLul3ZTyTAt2GFBC4NO6unLT9FBYzdxRzSXhvqrASOylha50jXHsDuUE/uJDOoggeqxXsLXEDJwtWrKmP3cNYRnHksKF0RiJkwDugijY6UYyVP7wIAYzufkFHBK0SYGQCj2ddTvnHfKCwxSPd1lp+1ajRRjIGwOPVbmsWha7V0QgszS+Q7YHqs6v4E6+soFRV0TmRjfqweyDDsumn3g9Jlw0H1Woyti0tOIngP+vdQUbrhbj7pBnxW7EAbrGudRJKcXH6fofVBuWPiRb7XFl0bSSPzrZWptYtvs3VA3oBOwxhaNd4XE9eDgditKiP3wfWg1GaR8bfEc4nKxhUyOf1tJBHmpqp4dAAMbBYURAdv2QZzrlWPZ0CYgAK6Oql6Ol8uVBmINyMZKgfIQ7bsgyJD0nqJ81LHc42x9BbusN8vW3HmrWQSP8AohBOWiZxcTsT2yqPmMHwMOPqUZZNCMkEBROc55ydygyYZZGODnEkFSSOFQ7t052VlD0Pf0vGwV1YwseDENkFvhNp/jO6gllMjsgkD0VHSvcMOKsQPmmT6ogBKDKoqz3Z+XZIHkrayoFRJ1tGAFj4wpoaeSTBA2QT07BUt6XDBxjKhmp/DlEYOSSsmV7KeMCPY9isaGXrqGvkOyDIfSmGMP6fJVYOuMu7bLV6ioo30obkE4WjSSsGWRnugUcvTIdsqlbmRwI2wpqKEMcXv8/VQ1sjfGHR2QTUVCwYmm7DdZNVROnjJoxkAb4URe6emEbNjhbi09E2loiJm9Rx57oNkzQyQu6ZBgq0NJ7LVL25j6olgwPqWBE4MygjLenv3Vqvkf1HsrEE1MA9/Se2FqVLbZbjX01soKd89VVysggiYMukkeQ1rQPMkkAfWtKhd0vBWv6L1FDpfXGndU1MLpobNdaSvfGACXtimbIQAdiSGkYO3qg9vNIaD4CezA5a5OIGpLSyv1GIIIbpcoYWvrrvc5W5FJA9wHhwhwf0tyA1jC53U7JPVxntwdZC8eK/l/sxtXi58AX2UVHheni+EW9Xz6MfJdwOfPghcOcLlWgpOE1zpLnWwVVHq6wdEgEV0YIJWiNjzgAvhqXlhOAXBoJaCXDwa1NpfUmi79WaX1dYa+y3e3yeFV0NfTugngePJ7HAEbEEZG4IPZB6Hc7/ADg8qvNty2Q3iyWI2rilbbrRinpLjb8V8FOesTiOqjBZLARnLS8b9BLGu6ceb8rQHAN8x5KWNoeekldqeRThdyicRLlrL9tbr2DTsVuhofuFFPeBb2VLpHTeO7rO7iwMhAAPaQkg4GA57u/IDwGpPZ7R8xFO/Ug1mdI0+pDObi0wGd7WOdF4XRjw/iIA+kNviPn5w0vhP2lx9q+iuv0Ty+S8prdCVep4GcHhpqGjF1+6YEZtYDeiT3ntggD4/PK8/eNHLx7LOxcItZ3rhvxkoKjVlBYq2qscEGrRVPnr2QudBGIt+sPkDWkejjuO6DzYqoqdrcx4ys+zupRA4TkZwcLl3kd4TaN4580miOFvEOjqKzT96NxNZBBUOgfIILfUzsAe0gtHXEzOCCRkZGcrvZfPZLaFvnM1U01lF00vwcslno62se6sdLPX1z3y+JTQSy5LGBjGOe856Q4ADLssDyrrGwmc+H2z5LGLyw4HZetfHnlk9mlJwL4gVPCW+aQi1dovT9dXUctq1lJW1ZqoYXPjjfE6oeJQ+QNYSWnBcQCCCF5zctvLhxB5oeJNLw60DTMYen3m5XGcH3e3UgcA+aQjc7kBrRu5xAGBkgOLnFhbnzwoicr13u3J/wCzL5RqKht/MfrN991BPE2Yi51taZXAnHiMobd8bYyQQPEDxsfiOCRdRck3s4ubuxXN3LBrV1ivlHD4pdbK2rlMDnEhsk9BXnxPCJ2+DwxkABwPcPIqMgbjurJnFzhlciceuBeuOXPihdOFev6eIXC3dEsNTT9RgraZ4JjqIi4AlrgCNwCHNc0gFpA5L5JuTa/84HEOsswu0lj0rp+FlTe7syESPZ1kiKCJpIBlkLXkE7NbG9xyQGuDrxFFE8fNQTUxa7LRsvXbUfCP2PnL1c5+HPEi5U9fqGjBpa91XW3e4VMb8DIl9xHhQyDO4DWEEkYGMDhHm/5UeUe38CKrmL5VOI0dZb7fcaSirbVFdRXQNE7y0NAkPvEEgJDumQklgJAHch58SbRhp7jyWO13ScjuubOWHlp1VzTcXbfwy0xVMoIHxvrrrc3xGRlvomEB8paCC5xLmMY3IBe9oJAy4eid+5ZvZT8qElNpHjnfRd9TiBkszrpWXGrqsPBIc6lt46ImnBLQ5mcY3OckPIZ9ZLI3w3nbCjbTveOpu4XpXx65R+QviLwJ1lxp5S+I8VJcdG2192ntEF1kqY3xMLctlpqs+9QlwJDHEhvUcYPYebVPUiFhaRnOyC2ncOvoeMYV007WfAAsfqDpS7yJzuqSHLsoLScoiujx1jPZBLTQ+KcHsppqPo+MK50kcTAWndSQvdMN+yDAkeT8OFa1jsdQGQsuqpwceGN1l01BikL3jfyQaV1OzgHBQ5IJPdXsaDOQe2VSf4X4HZBSGLxD3wqSs6DjOVa17mfRKqXOkO5yUFCCBnKopBDIR22VpiePJB399ip+FNqn/i/rv5RtyKvsVWlvNNqkn8X9d/KNuRB7UIiICIiAiIgIiICIiAiIgIiIOOOZL8Hbin/Aq9/qMy+aFw6pCMd3H9K+l7mS/B14p/wKvn6jMvmljIE7y7tv+lAdTdLOolQKSSUuJAOysDSeyCiYTss6j8AxnrxkBBhscWOyj5C9VmIMh6eypGQJAXeqC3B9Cro3Frw70OVrNQ2iFGOjHVjfC0djHOIwEGtVN1jfRCLAyBjOFofSXErKfD1MAHdWxMETvvm2UGPl3Y5wqLPmbFIzDMZKhbb6hzS4MOyDGBwVlwUonb4nVjG6xiwtf0nuFKyV8b/Dae+AgmdVeHmPHbZRtaJHeIRjByst1AWR+PINisQxuecxZxnyQTVFf1w+CzIxtlY0VQ6MEOyc+qlxCBh3cd91GTH5II3El2RnBVuTnJys2IQ79WMH1UNV4fUPDxhA95+DpwFdHVhrS0t9Vi/WqnGR6ILnydTi7HdSQ1DmOGDjKscW9OwVrY3OGQgzauYmBnSd85Xbnlf4n0OnbUyGqnaMNAOSunLnOI6T5LemjKPU08YfaWPLB5tyg7bcZ9eWzVVLLFRysa4tO7e66p18VZSSysNQ5weT3OVn3eq1HQxn3rxA4d8krSqSu96jLqndw9UGg3ihcGeO525GStMp6oQsLcd9lq18quoFo7BaAgkL8SGT1KSPMruoBWkjpABU9N4fQerugqyrDWhpA2UEsniHKpJjrOOyuZA+QdTRsgjV7JOnHyVrmlpwVVkbn9kGRHWdB7KKonEzsgYVHxdDclRoLwXOwwu2HkrjAfJRZwtftmnK6vpTUMaS0DO3kg0ON/hOORuslkQqsHOCoauF1PUPhcMFpwrI5XRkY7IMo0ZiJIJITfqAORlTQ1TXD4iFZUOaXZZsEFJomsaHNP512O9m7DRz87XC6Otk6IxW1rwejrzI2gqHMGPm8NGfLOfJday9xGCVylyq8RaThPzH8OeIFyl8KgtOoKQ10nV0+HSSPEU787do3vOCQDjBIBKD0F9uDW1EFLwapWOd4Er9Qve3Gxe0W8NJ+YD34+srykkcXOyvbH2wXCS56+5cbbxBsVO6pm0DdRXVbGN6iLfUM8KWQY3PS8QOONg0PJ2GV4mD6QKBg+iqx/Q8O9FPKY3M+HGVj43wg9sfZK6Gsmg+Va48YbjC2Kp1ZcKysqKtzQCKChL4WMBP71r2VLu+MuPovIjjfxf1Vx34pah4paxrZp6691j5o43vLm0tPkiKnYD2ZGzDQB5DJySSfZ32dMJ1T7OnT+mbS9slaaLUVsLSQA2eWurHNBPYZEzDvjYrwtmgmpKh9NUwvhlheY5I3tLXMcDgtcDuCCCCDvkINW0bq3U2gNU2rW2j7tUWy9WWqZWUNXTuLXxSsOQRjuDuCDsQSCCCQvb7nOt9n5l/Z2VfESWmgFQ7Tdt1zQuaARSztjZLK0ZJIIiknjIzkdRBydj4cUs1O0fGfJe4nEuL/Yy9lNUW7UjvdpqfhhQ2+Vj/AIeipqoIoWRnvuJZmt+Z9MoPNT2XLAeejhqf7n7s4/zPWrtv7ZPmE4i6Vk05wD0xcjbNP6ktTrrepad5bPXN8Z8TaZzgRiEeGXOaPpkgHYEHqT7Lgn9vPw2+f3Z/ketXNPts/wDf40J/BH/68nQedsMksMrJoHvZIxwcxzSQWkHIII3BHfK909c6kq+MXsra/Vmsi6rr7jw3ZXVkr9zNV00bXCY5zuZYQ8/M7YXhjRuY156+y9ubOR+5Dzlvb/YwrMf5ORB4kzQsazqDskr21vb+n2QsL/ThhR/+jjXiD4riOknIXt5f/wD7UFF/xX0f/o40HiTGwSuJce67UezAjDOerhrgjb7s/wAj1q6qxdWdiQu1Hsv+r9vXw0z5/dn+R61B3X9r/wAxurdEaXs/AbSNTUWyPVdG+43uthk6Hz0Qe6NtICNw17muMnbLWtbuHPC8k7ZbIaqnMkhGQvQH210zm8ctDQg7O0kCf45OvPKCtkhjLGOIBQe0fsluG1g4a8rt14uVkLI6vVtxq6upq3AAigoS+FjATjLWvZUu74y8+i8kOOvFnVPHHipqDifq+ulqK691j5mMe8ubSwAkRQMz2ZGzDQB5DPckn2Y9nYDq72dlg07aZGyVzqLUVtc0kANnlrqxzASe20rDvjuvDaahqoppKerhkimhcY5I5GFrmPBIIIIyCCCCDuCEGoaK1lqTh9qi1600jdqi2Xmy1LKyiqoHlr4pWHIIPmDuCDsQSCCCQvbjnPobPzKezqq+Iz6aD3h2m7brmgLQCKWZsbJZWjJJB8KSeMjJI6iNyMHwvkhdGTnyXubxCpn8M/ZSyWrU7hT1EHDCgt87HjpMdTUwRRNjPfcSzNZ8z6Z2DyH5TYyeaLg8SDga908f+cYF6Ne2uIGleFOcb3C7d/8AB0687OVN7RzP8HWjGf2faeH/ADjAvQv23Li3SvCfHncLt/6OmQeV0VMx7snz+a1SLT7KqmfKCB0t6u60GOoc056lknUFRTx+E0nDtj9SDCgaW17qV5+EHG6jusbInhrMb+itnqQH+O36Z3ysaWd85y/ugpE4NKuld1OyAos4U7Cwswe6C2Kbw+wyVSaUyOzjCvp/D8Q9eMJV+H1AR9kETBkrUY5GiLGd1gDBG3dVAmx8kFkp++HCtQgg4KIGEwpafoz8STlhd8CCIHByhJPdFVvcZQUwrmgucGgnfZSyGMx7d1CDggjyQZctD4cXidQPnhYalfUyPaGE7BRICIiD1c9hr/udxl/w1g/6NevUxeWfsNf9zuMv+GsH/Rr16mICIiDrl7Q/8DXiX/idH+vU6+eeKTwySd8jC+hn2h34GnEv/E6P9ep188fdBe+TqO2yz6alZJAXuOTjK0/oI3I2U0FU6NwaDsTugikaQ87Hb5KxahVeC6MObjqPmsAAk480F0UZkdhJY/DOFkRxuiHUdgoZnB7soIlLHN0N6cKjunp2xlWNx1DPZAJc7vkrJoqM1c7YgcZwrZujwx091m2aOTxDM3OwzlBuOHQdQYGTwxGQnBGBlb50vwZuV4jD3W5+4/uSt6cv9HS6grIqW59PT1gb/Wu/WjOENi92gdSxxkFoPl6IOkugbIzhDP79cqHDW/F8TVuLXXMZpu/2mShgpIg/pLdmjK5b5sdFUNkskromtBDD2GPJefEjf7OQ07OIQZF31IyhvMtxYwFrySAAtNr6oX8GtyGdO+FG1tNVvLKjGB2WDcHupMxUezDscIMOvrw8GDGS34crTAcFSvwSSe5yowMuA+aC4yuIweykZTh7OrKuliayMO27KJglI+HsgsOxx6LIp6XxgDnusdwIO61OhttZPF40LSQEERoGxjLj2VoqvAcB05APcKlS6oY8xHORscq6FsZjIf8AS8soLamtEzelo8vNQsgBYXZ3Vr4XNJwNlaJHAYzsgviJY/GSpX1I+id1i5OcoTlBe5oPxA9yrMH0TJWfSwsc0Od288oMDB9FczcgepWTViIDEeMjuoIxt23QTGmaQCCVqdIyOOmOSM49Vo7nSjbyVW1MrW9OcjthAqiXTOGSQocFZsMQeOp3n3VtQxjMgY7IMXrd/dFajQUImAe4jHcrTfqWfTzSxQkt8ggyrhiCPEZyQtOijMx63k5HZT00/vEpFQQQVbWgRyBsXb5IMulZ4Tep2w7rc9vudAyjLJC3ONt/NbOdWuMXht74WMXy9upw+1Bk3adstW8sOQsRpHmd1I6AhniOOTjPdWRReI3Oe3kgsPfZAN9wrmtw/pKukAbsgqekM2O6iG7hlBk7eSmig6m5Qd0uTL2k+u+WO103D3WFsk1hoCJ7jBSiborrWHHLhTSO+F0ZJJMT8DJy1zMnPpBa9XciftE7Ay1VUen9TXSOAkUFwjNDfqAYJPhkFs3S05JMT3R5xkleBTgWnBWZZr1d9N3akv2n7rV225UErZ6Wso5nRTQSNOQ9j2kFpB3BBBCDuzzzezVv/LTbKjipwwutXqPQDJQKyOpaDXWcPcGsMpaAJYi4hviANIJAc3HxHo055d3X0I8rmuK7mm5IrHqDiyaeWfVlhudnvc8kbfDqGRTVFHJO5oAbl7YutwAADiQMABfPjJD0PDSe6D25v2/sgYv+K+j/APRxrxE7L29vzceyFjaPLhhRj/8AFxrxDwSeyDtT7Ls/9fTw0Hn/ANWf5HrV3Q9sLzNav0HZbFy+6PnqbazVtC+53yvik6HTUQkdE2kaRuGvcx5k7ZaGt3DnhdMPZexlvPPwzcR/8+f5HrVzT7bH/f50L/BEfrtQg89aV7Xv++EYyvaP2THD+g0Dys3ritBa5qq56tuVXVEQxkyz01EHQxQsABLj4jagjAJJkIwSN/FRuR8l7tezRvlVU8gWmo9NMbNd7My/U8Ubx1NdV+/VU8TSAckESx5HfdB5RcTOC3OfxZ13fOIus+X3itW3i/1klbUyO0hciGlx2jbmI4YxuGtaNg1oA2AW/OUHhLzY8GuY7QOuabgLxRtlLDeqWjussulbhFC63TyNiqRK50QaGCN7nZccNLQ7YtBG9T7YbmxyY/ufoQPGxzZptj/l1FN7Yfm4iIHuGgt//vNN/ToOdPbaaKthtvDDiLFG1lwbUV9lmcBvLAWxzRgn0Y4Skf4Uqb2KuuNLwWTiHw2mrIodQTVlNeYIXHD6ilEZieWb4IjeG5GAR4oO47dGeZLnY42c1lDZLTxPlscVBYZpammp7XQGBrpXtDS95c5ziQ0EAAgbnIJwRxJpLXWquH2pLfq/RGoK2yXu1yiajraKYxyxOwQcEHcEEgtOQ4EgggkEOy3OnyPcf+EPEzVutv2KXLU2jbrdKu609/tsLqlscU0jpCKpjMvhe3rw5zwGEjLXHOB1MjFQ5jmxl3QSHFudsjIBI9Rk/lK9KOAHtl9b2iSisHMHomm1FRNLY5b7ZQKauazYGSSnP3mV3fIYYRjsCRvzXzucsPAjmB5cbpzVcFrZbqK9UNmfqZlxt8AporxQRNMlQ2piwAZWxtkIcQJA6MMcSNgHXH2Muv8ATOneO2qtGX6tipbnqixsZaTLt48sEviPhac46ywl4BG4iOCCMF7SPkj4+xcbtVcctF6XuGsdK6mlZXyyWyI1FVbniJrHxSwNzIWDoJEjQWhuAS0jB8/6G63Kz3SmvNmuFTQV9HMyopaqmmdFLBKwhzHse0gtcCAQQQQQCCu/XAT2xPGbQkNJYeNWnKPX1rhLYzcYnCiurWdiXOaDFMQMYBYxxIPU8k5Aefj2zwSSwSB8TwTHKw5BBB3BHyIGx7EfJREYXuTxc4F8t/tHuAU/F/hbaqan1bPSTutF7ZTNpK1tfED/ALSrwMiRpeA09Rd0h3VG4g5d4cFoCC0DKlFOHNzlWYPoqgTOBDScIITsSFJHF1DI7p4D/RTU0MjZWk9kFjqeQn6JGPNSUspbIIXDYnC158NPLTjwwOohabNTCHLv3w7IJ6yJlLEJc991ji5l8JY1pxjCx3Nra0lgBIHktQpoKampiyox1ntn1QaMHFshee5KtkPW7OVkSUcssjjG3LTvlQSQSQnDhgoLOkoD0nJClgxn4/VWy9Pi/D2QSirIbgNypGkSDqIwVLTR0/SPExn5qyVuXgxfRCDvv7Fv8KTVP8Aa7+Ubcik9i+G/todUev7Aa3P+cbeiD2eREQEREBERAREQEREBERAREQcccyX4O/FIf9xd7/UZl80Lx98dv3J/Svpe5kvwduKX8C75+ozL5oJDiR3/AHx/SglNPiMPUTXYGEMryOnOytQDugz2CIgvLMNyr4qcygkeSjc/IxlZFJO2I4cdigge17D0knHkqxy9Aws6qfTuiy0jKwIw398gljquk5IVlRN4pBGyPYzuovqQXxPLHgknC1aK8xxw9BbuR6LRkQSSy9cxk9TlU68SCQeRBVifWg1SpuolpBABuEthY2I9Q3K03Axkd1fDM+Nw32ygnkpXyzOLQcEq2WjMQy7Za3b6mj6MvLcrT7xOx7w2HHzQablxPS1Wua5p+IbrKo4eiUPlGB81dcjE54MePmghp6fxjjKVNOITgHKjZI9n0SrnGaX4nb4QIouvfKl6xA3p9Vjh72dtlUdcrw0bknZAaPEefmuXOFutYdNhlBLSh/VsSQCuM47HcOls/hHpJ74XN3DTh0y70TauRmXtAPZBqGvTSXC1uuEULQXjOMBcJteQX74OTsuadWWmuog+je0iIZH2LiK72OsNQ91O0lud9kGgVbnyOIOSFp72hpwFrlwpxTU4a4YfhaE5xcclBk01GJ/32FbVQe7ODQe6jjnkjPwlVc987up26CNreo4WZDVCmZ0YzlHeGI8N743WG7qzugkx40hPbJUkbxA8NOMErHa5w7I5zi7qPdBqNSxhh6x3IWnNGXAeqv8AFkeAzOysIcw77IMmSlDIxJlbpsOtYrXb3UbmAkt6QcLaD6h7h052U1PFC5mX7lBfVSi418k3YPOVBND4bwweatc7wpD4Z2VD4kh6zuQgzvc/DiEoOQQsd3oqGrl6DH9ijDifPuguJ8gqfNCcK4OaW790HsX7PPn20BxX4dUHLrx5u9vpdT0VH9xqOa7FvumoKDoEbInvkPSajoPhuY/HijBHUS4DQeNnsXNLah1DPfeB3Es6YoquV8j7Ld6R1VBT53DYZ2uDwwHYNeHkA/SOMHyO+pcsaE5seZbhpQR2jQ/HHWFst8TBHFRi6SS08TQAAGRSFzGbAD4QOyDvnw79iRXi509RxX430/3PYQZ6PT9ucZZR5tbPOQI/LcxOz6DuuI/aS27lL0LbtAcGeW+m0+65aVfXyX6ptfTUSEytgaxtTWDJnmJjeS0ucWYwQwENPWbX3NHzJcS6GW268426xu1vnaWzUMl2lZSyAjBD4GFsbtsjdp7n1OeKg942Dig9HfZRc4Ok+ElzunAbibd4bVZNS1wuFluVTKGQU1wLGxyQyvJwxkjWR9LiQA5hB+mCOWebL2R83FXiBcuJ3ADWVhsUmoah1bX2W8NliomTvIL5aeaBkha15JcYywgEnDgCGt8iup43JK5Z4e81vMbwttTLHoPjTqy022Fnhw0LLi+SmhbntHFIXMj7d2gIO/3Ln7G+46W1pR6u5jtb6du1ptE7apljsTp5Ya4s+IConnjiLIwQOpjWEuGR1NGc7M9qtzr6S4nw0nLrwjvcV0s1qrm1uorrSSB9LVVEYIipYnjaRjCS57hlpcGAE9JXS/iFzVcx3FW2vsmv+NWrLvbJGFktBJcXx00wOdpIoy1kncjLgTg47Linsg7Yey9Y1nPNw0x5/dn+R61cye2yP/5+tCD/ALkf/r2ddA9Kaw1XoXUNHqzRepbnYb1b3OdS3C3VT6eogLmFruiRhDgC1zmkA4LXEHIJB1DiBxQ4jcV7zFqDiZrm96ouMEIpoam610lS+KIEkRsLyeluST0jAy4nGSSQ2vuvb7T4/wD8QUmfxX1n/o5F4iOLS3but80/HrjbSaBdwspuLerotHvidA6xsvE4ovBdu6LwurpEZJJLAOkkkkZJyGwzsvdTkyuOhuar2fdDwjmukUc0GnZ9GXqGIh89vlax8cMxZkZJZ4UzM7EjGSWleFS3Tw/4ocReFV5/ZDw21xe9MXAtDHz2qukpnSMBz0v6CA9uf3rgQfRB3U1v7Ivixwy0LrjiLq3ivo42nSVmrrzTMoo6mSetZTwvlEb2yMjbC54YGjD5BlwG/nxn7MM/9fXw0A8vuz/I9auKuIfNjzH8VbG/TWvuNGqrvaZhiegkrnMp5gDkCSOPpbIAQCA4EZAPkFx/pHWOqdCaio9W6M1HcrFe7e5z6S4W6qfT1EJc0td0SMIIBa5zSAcEOIOQSCHfz217M8d9Cu/7kQP+WzrzqW7eInFTiHxXu0V+4la3vmp7hBCKaGoutbJUviiBJEbC8npbkk4GBlxOMkk7RyEHot7JznE0pwiud14C8TrxDarJqaubcbLc6mQMgpbgWNjkhleThjZWsi6XEgB7CD9MEc0c1nsm6nihr65cTeAOsbFZH6gqHVtdZbw2WKjZO8gvlgmgZIWteSXGMsIBJw4AhrfIJcq8P+avmQ4WWuOx6C416ttNshZ4cNAy4vkpoW+kcUhcyPt+9AKD0K5evY83HTmtKPVvMdrbT11tNpmbVNsljdPLDXFnxAVE88cRbGCPiY1hLhkdTRnOyfao87Wk+JMNLy78JL1FdLPa60VmorpRvD6WpqIwRFSxPGRIxhJc9wJaXBgBPSV0s4gc1XMfxUtj7Hr/AI1asu9skYWS0ElxfHTTA52kijLWSdzu4E427LisO9UHLHKkD+2l4OnP/wAv+nv5RgXoj7bwkaU4TY/+eF3/APR0y8obXdbpY7nSXuyXKqt9xt88dVSVdJM6KennY4OZJG9pDmua4AhwIIIBBBC3XxC408XOLhoBxR4l6l1Uy2B4o2Xa5S1LKcvx1ljXuIaXYaCQMkNAJIAwGzxn8yx53ZcB6LKkLSz4e+Fjup5CerG3qgjjYZDgpJH0OA9VL1sY3AO6s6HyHqwSEB0Qa3q+SsEb3DLQcK+QPDQ0hZ9EadtORJjOPNBpe4V4Z1DJKTFpkcWnbKNLsbFBaD0n6lO2pAb048ljnckpjCCrj1OJ9VREQFLHD1jJKiVzZHgYBQUcMEhUG6E5RBd0nzKtOyvYcpIN8oLWt6ij29JwjXdKOd1HKCiIiD1c9hr/ALncZf8ADWD/AKNevUxeWfsNf9zuMv8AhrB/0a9epiAiIg65+0P/AANOJfmfc6P9ep188Y2X0P8AtB+n9p1xJ6u3ulH+vU6+eBBL1h4wo3DCoNkJygqCRvuqh2HA+itVQC4gDuUGRLUiSLpHcLGUj4JGDqLcAq+GMP3QQK9rMjKlkjDPy7KyBrpZWxNGcnGEFjWOc4DBWu2lzovg6O4V0lqNDTiombgHdataBQzwl7cEjzCDk3hDd/uPWRzCTpPWD6Lvpw24w22mt0QqbgwODR3cPRecOln1AqSIwcA7KfUPEbU1irWw0rpAztsSAg7v8etW23W9FLTsrWOBaQPiHoui+rLXFZ5ZYY3A9TyrrnxZvj6IEuf1kb5JW0arVE11YZKtxLjknKDBrQY3dbHd1qVJRx1VC6R53AytAdUGplLRuApnV1TSxGJmcFBptS3pne0eTtkijyA5X08ZqJCT3O6rUtMOGjZBbLL1t6M9ldDOI29J/KsYnzzundBVx6nE+pW6LPfIaSgMEjRktxuFtyma1zj1LKkaxrdvzII6qrEtS+QdidljOkJd1AkK/oEh+Dcq18RjzkeSCcSiVob5gKF8eCqQuDXbnCmkBeMjdBjvYGgH1V7IHPaXY7JEx0j+nBO61eMQxU5DiM4QaIRgkeiyIJ+lpYdsqsLGy1BGxBKtq2eHIAPrQWSswS4+aja7pxhXOkLmgKWKNpjLj5IKwnxT0eakkpQz4sYI3UdFKyOXLuxWXM4yPDm7tCC2my4lpGBhY9YPvwYBkLJe9hbiHd2PJWtp3u+N43+aDHmpBHGH57qkM5aBH67K6rdIB0O7K+mpHSxeIB5ILPCEZ8Qbeaj8bLxndJnva8sdthVp6V8xBGcIJoI4zKC/sErmxAgxd8pUwvhb5ghYrXHO+/yQVcZSzB7K1sjmbBXunLh09ICjaMkBBXpePiIKoXFyyXkGLGygaNwe+6CvhuA7bLlXlYk4XN5g9DR8a2UTtEyXRsV3Fa4imEbmOawykdoxIWFxJAABJIGVxe6UdGMYKhAz9SD114y+xy4ba9ndrHl04nM09SXEe8wWytabhbnNfuDBUsd4jI8HI6hKT5EDAXHmivYk6/lv1P8A7IvGfT1NZWuDqj7i0s89VI0d2s8ZsbGE9g49WM56TjB6G8OOPvGvhA0xcMeKmp9NQOd1vprfcpY6Z7s5y6HPhuOfMtPc+pW8tQc73Nvqa2zWi7cwWsjSzAtkbT15pi9pBBaXRBriCCQRnBHcIPUTnR5kuEXJjy6O5buE9XTDVc9j+4VqtUEviy2ukljLX1tS7fDy0vc0O+KSRwdgt6iPE1z3OOT3V9TU1NbUy1dXPJPPO8ySyyOL3veSSXOJySSSSSdySo8FB7pcmdz0NzV+z9ouEk10ijmg07Poy9QxEPnt8rWPjhmLMjJLBFMzOxORklpx0n1t7Ivirwy0JrfiJq/iro42rSVmrrxTMoo6mWetZTwvlEb2yMjbC54YGjDpBlwG/n0u4ecSuI3C29fd/hrrm96YuJaGPqLXXSUzpGA5DH9JAe3P71wIPot5cQ+armR4qWd+l+IHGnVV4tM+BNQSVzo6ecAggSRx9LZACAQHA4IB7gIOV/ZivjPPPw0aw/8Az5/ketXMXtrmdfHrQnoNIj9cqF0S0xfNVcP71R6x0VqS5WK929zn0twt1U+nqYC5pa7okYQ4Asc5pAOC1xByCQZ9ecUuIvFW8R3/AIma4veqLjDCKaKputdJUviiBJEbC8npZkk9IwCXE4ySSG1yAG9xlehHspOcjS/By73XgZxTvUVq05qesZXWe5VMgZTUVxLRG+OZ5OI45WMiAccNa6PfZ5I89HHJyqIPWHmg9kbc9c69unErl41bYqGkv077hUWK7OkihhmkPU8000THgxuJLgxzQG5wHEYA4j037GXmLvV0jZq3XWh7Dbg4CaeCoqa2cA9yyIRMa4geTpG+W/cjqbw65pOYnhPbm2bh/wAZtWWe2MaGx0EdxkfSxD/tIXlzGHYAloBIx6BazqjnS5p9YURt994+a1fTuBD4qa6SUrZAQQQ8QlvUDnscjt6BBuLnl5Y9E8qPEixcOtIa/qNU1E9jjrrtJOImPp6oyyN6fDjJMYLWNcGuJcA7OSCCuW+T/wBn1oDm65dbnq60cS6mwcQrXf56N0T/AA6qkFKIonRCWnHTLGXlz8SB5Gx+F2MDo1V1c1ZPLU1Mz5ppnmSSSRxc57iSSSTuSSSSTuSVqukdY6x0Hdmah0Nqu8aeukQIZW2qukpJ2gkEgPjcHAEgbZwcIO7dL7Gjml+7LKKfVHD6KgMwa+uZc6l4Eed3hnu4cTjs04ydiQNx2+5ptX8OeSHkP/a5nVDbrqK86XqdL2unLQJ601LXsq6wxdRMUI8aVwySASxgLjuvMSTnw5w5KI293MLq8RmMR9TasNlwBjPiAB+f+2zk985XDGoNTai1dd6i/wCq7/cb1das9VRXXCrkqaiU4xl8khLnHG2SSg7achnJjw25wNK8Q7XfdfVunNYWI0L7IYHxSs8N4l8SSWmdh8rA5rASx7Okkb74O8rp7GfmgpLtJR2nVOgK+h8TpirH3GphJYTs58ZpyWkDcgF2PIldGLJeb5pm509905ea61XGld1wVlFUPgnidjGWPYQ5pwe4IXNFFz284NDQMt0HMLrB0UbOgOmrBNKQc7mV4Lyd+5JPz2Qer2laXQ3svuTmW2ax1nTXW+mWsrKdsbOg3S8TtHRBTxOJcY2BkQc49mtc8huekeFJa5pHUtw6v4ga34hXp2o+IGsL1qS5vb0mrutdLVyhuSQ0OkcSGgk4AIA8gFoE84cRgILnlmzRuSsmINYzB7lYrjGGh/mr2CSTcDIQJZww7KaKdr2Y81jzRebhuoInODwAdkGpwVL4HElxIyqyzid/V2VgaAwFywZZSJPh7AoNxW2eCEfG0En1WFdIBUVHiRuwPPCwfenNYOk7/WohVTvcGgndBq1PVR0zOkgZ7LT7jO2Z+WhR1DJmMD3Z3UTT1sJPdBbG3xDjsqSM8NyqyKR7j4apKyRhxJnKAXPdsCSB6KWOoMbS1wOVNa2wuefFxhWXBsYm+94x8kHff2LUpk5ptUjf/wCIFcf+cbciu9iyWfto9U9Pf9gFb/KNuRB7SIiICIiAiIgIiICIiAiIgIiIOOOZL8Hbin/Aq9/qMy+aCT+yP+s/pX0v8yX4O3FP+BV7/UZl80En9kf9Z/SgRt63YWR7o3pyHb/WsZvUDspBLI0gOJwgjc3pcR6K6OMPCnkMbmZ2ysYOI7FBe6INU9PRCZvVnGFAC6TIG6uEk9PsCRlBWamMRPxZULdyFc+Z8h+I5Vn1INQjpmyRgFw3+axaiAQu6QcqjZ5mjYnCqHmXJeclBCg3ICqdnY+aHYghBIYQGdWVErjI4jGVagqTkKrG9bsKjWF3ZXBro3AnbdBPJSvjYJA4gLHDnFwcd8LOfOJIgzIyArIIGlpDu57IIZKnrZ07gqEk+ZU81HLHlxGyx0AHBU7ZhsCFDthBnbHqgyJYGtZ4g891bRENqWPPYHKue2Xo+LOFDGHNdkDsg33Pf6aO2sjawF22Thc2cHdXUdLY3ySuAIautUFQyVoif6redjvM1DQOggzgtxhBvnW/EamuVzkpYyCASMhbQkvlNFE9rwCTk5IW1nxTz3J9Q4HJOcqy5eKCM5wEGPfKjxpC9p2PotC3O+FqVW8GMZO6x4RE5hJ7oMRXxydAIx3RzS6QtaPNUdG5n0kAPId1Ekqrj148lYmT6oAODlV7lGNLjgKr2FhQTU8Yz1HCpUEF4a1WtEmPgRoIkHiILnQFsXWVE15AxnC1CYB0ADcLTuk9QB80DGfNSiYNHThXGmk6OoA47rHIwd0AnJJUsTcjOVErm9XYeaCV7G42KjOB5qha8bFA1xCC+BviEgpIPDePNWElh2OCqOcXHJKCaScvbhRjZAM4HyUwYC0+qCIuyEDQRnKodimSgHZUREBZkFLHJGXO2IWGrxK9o6Q7ZBWSPoOAcqzG26yaSMzyEHdUrYvBeG+u6C2nibK4tKrLC1jsA/kULXOb9E4yrupx7nJQVkiDRkKPGFPHJueo7Kyfpz8PYoIkRMgICuBz9atyPVVbs4ZQXYwimfIwtw3AKgyCgrnCuByrXAgZVseS4Z7IJeox74UzKvxAIgzOdlZO6MxgDGVlW73VsZdJjI3GUENVQeHGZc4zvhY8U7mN6Q3I+pT3CrMp8Np+FajZ20Bpne8AdWNuyDRpJSTuNyqxRGXJDsBXV/heO7wcdI7KBsjmfRKCWSnEYznP2qOI5cGHsSjpXuGCVYCQQR3CDPnoBFCJeruFgE5OVPJVyyM8MnZQICIiAiIgIgBPZVLSO6A13SjndRVEQFe1gLSSVa1jn/RCqWvZ8J2ygkpoRM/pJwr6qlbB2OVBHK6I9TDurpJ5JNnHKD1Y9hr/ALncZf8ADWD/AKNevUxeWfsNf9zuMv8AhrB/0a9epiAiIg65e0POOTTiXjv7nR/r1Ovnnhj8RxBOMDK+hj2h/wCBpxK/xOj/AF6nXzyteW56fNBdJH0easVS5x7lXMxjdBLR0wqXlpOFWppnUsgxk4KiimfC4uYVqUNRBMzM5BPzQYU1WJIwwtwQrYJCwbBX1QhyehWxeHgg9z2QUmm6xt5K2lmFPO2UjIBR8JZ8ThsVVlNJMC6MEhBuWS4fd6mFHG3ceixIYqi0PMTsgFahoOnijry6p2B2Wt6vpKOWYOgx5dggxLdqRtpb4u2TusK9aliupMpAJBWnVop44+mQkgBaO5hkJMP0coJai4SVDvDHbt2UsND1xHLsZCsgFOPpYyo56yRjixh2QWge6SE5BVJqvxtgPzKJznz/ADKnpKR3iB7x8IKCKnndA/qLSqVE/juBx2WrXRlC2mHhY6seS0aPpwerugsRVO5JCoguY8sKkdMXDpz3UKIJo3eFl2e6vkkE4yO4WP1E7KrXFuw80F8MQkdjKzI4m+IIz2IWFG50ZznushrJ5R1x5JCDNnpY6SPxWkEndaXLO+U4yQFmMFW/4ajOAPNYlS1rX/B2QVid4J6vVWTSmV3VurMl2wU7Gsa34hv/ADoLKeHxndOcLPjpWhwiJAzssSlJEvwrIMdV47ZG5wPNBlT2ExRiUOwO/dVhgb4JZkEnbur66ulfTCIHcLT6KsdHKBMdiUF0VK+CUvcDgKsldmUNDdlqlfU0ppgWuGT3WktZE6MyDv5IL61rJIg4EZUdHXeC0xEdz3WK+WV58POwU8LYmEGTuguq4erDwO+6yaCRsUWSMqCoqGPHS3sFfA4GIs8yEEVXUid2AdwsNwAfg9vkp5KSZji4ggZ7qz3eQjrxsgy3ik92y0gnG608B2ctyqdlk0mHO6SggJeNiVkU/hlvx7lZs1vY5nW3GStOkj8M4z8kF72BziGK10bm/SGFWJ/Q4EjKzGhk7CT6eiDB6Sr2hvmArntIcR5BW9KCh7p3UkUXiHp3VZoTERgoKDLRlvdWFzs9WexVwcR3GytccoJ31sro/DPb61j5PqirjZBRVDHEZAQNJUsT+huDvughIIOPRSsbGWnq7qx+SS7HdWk4BwgoW/EQ3tnZXN6gFa14HcK7xgfLugozd3xJI3pdgKuAN1aXZdnKCTD3AeiBnyQVDWgA+Smj6HNLyfJBG2Euz5LHMeJg3vkqR8z3OLYlJEAN5MdSBU0wZEHZ7fNW01YIm9J/Qq1LnubjyWMInkZAQTT1BkBx+hQMJa8EqSAN6sPCpN09WGIJZakuYGtUTInOOXZ3U9CyMOJmAwpauSFv9ixsgw5B0KSCMAiUnssdzi7uqiRwGM7IMysrhMwRtHYYWNHtGfmrWROf2CudFI31wgvpqnwHE9OVbUzeO/qxhS0MTHv++K2tYxk3SzGEEcOQe+Mqkpycd0EEuMgKoheXDqGyDv17FZrhzS6p/gBXfyjbkWT7F2Po5odUH/uBrR/zjbkQe0CIiAiIgIiICIiAiIgIiICIiDjjmS/B24p/wKvf6jMvmgk/sj/rP6V9L/Ml+DtxT/gVe/1GZfNA7+yO+soDHdJyrnP684CsLCN1kU5Z0/F3QY5JzuSqYUxYOs+ipKGkgsQUhk8J3URsVdUTCXGBhXPbH4fbdY6Ag2RVa0ndBIxwd8OEePD8kwMYHdWmOQ/S8kFh3RCMK9jchBYmMK9uA49Suk6MjG2/ogvo8GTpd2V9cB1Do3HyUbWOcPvQ3+SyI2Fo+/8Af5oMJri053WZTyg4cSBhRzQg5cwd1j9TmbIM+quLZYzEBvjGcLTsK9jMnJ7KsgBIDAgjVW5yCpDTyNb1uGAr4vDx8XdBJLUtMQbgZVaVoew7d1iy99uyyaV4I8Nvc9tkF8NKfGz1fkW4qJ74IgCMj5rS7fYrlJIJek9Pqt1Q2WVtP1PB2CDRJa8RPLgN1ptbdDLnY7LOroGxTObIFpk9K2UExjAQYgeapwYVO6ibCw4dnKx5GGA5bsR6K33uUjDiSgsDvClJ74KrNMJO22yjdkkn1QAYygomCr4sdW6ym+D1AYH5EGLG50bsgH8iq95lcNu61NvunTuArJIIXsJjA27IIB96jy0ZPmsaR7nuBcMeiz7X0e8dFTu35q69Mp/EHuuMfJBDTHraGE5PooatnhyBwG2VdQxSvfhgx6rJq4hG3EoyfJBbBUiYeFgfasSri8N+B5lUgcGTZ8lWrlEjhhBa6IBvVny9VRkgbjPkrep7tsqhaW90F75A/bCkiADMlQsxndSFkjhlnZBG/PURuQCrcH0U8fSDhw3zupHNjI2AQQ07et3Sp5j4W3fPooXNfEetuyvZIJB987oIC4EkpkFVe0hxI7IxjnEY7Z9UBwLVYsyUxCPY7rEa0nPyQXNHUhBBwVa0uYcjurgHSOBPZBNTzmnd1AblUqKgzuBPcK6WNojGO6xSCEF7e6yo4WubkkZx5rHpmdb8FX1HVE4FrigldA8ZIGwWO/Y4V5rpy3oyMfUoC4uOSgdRVERA2+aZwiuYwkg+SCmSBuEB3ypJAwN2xlRIJHyBzcBWtdgYVY4nSHZUkjdGcOCChJPqUy4eqlgjy74uynlEIbgYygws5VQ5w2BICq5hGT5KjWlxwEFM5RS+A9vxOGFYMeIPTKCmD6H8iotVbFBNEAwDOFgVMJidgjdBCiIgIiqGOIyAgoidkQXMd0nJVZHhysRAwmFfH053wkvTnZBWGXwzkpNL4pBCjV7cYQWIhxnZEHq57DX/AHO4y/4awfor16mLyz9hr/udxl/w1g/6NevUxAREQdc/aHfgacS/8To/16nXzxr6HPaHfgacS/8AE6P9ep188aAiubG5xwFV0fR3CCxVGc4yUAJ7Jgt3QZEdPkdTj+UqN7fCeHA7Aq0SSn4WnurjDM7uCUGS1wq2iLHb0WTQPbBVNpXAfEcLHo4JInl7hhXSQzSVgfGDkEEIOUrHpCOanZUwuGXHPdcgaP4RP1PdYaKVnV4hAW1uFFj1DqExU0DXkZAxgruBws4Ea3gvNJcI4nhow7PSUHEnGblC/YtpT7tRU+CY+vI+pdOaykmtPiwviILXEdl7L8ZtAapv+ivuU4OLmxdOA0+i80uJ3BO7WSqmNVG8ZeT9H5oOAHeI13Wc77qg++PA9VuC/aflt4LXAggLQWRu3djcIMjwW07A875Vwrx4ZYAAfVYj5nuHQ45AVGR9QJQVaHzPwSTlVfD0EDPdXUzmseeoqtQ7rdlnZBG6MAA5UeFe7rA+Iq9nT0b4z6oIcZVek5xgqekDPE+PGFk1BpwMsxlBispw4d1HJGWOwAUMjgT0lZFO6J39kxlBiuJI7YwsykuHu7QOnOPkrKrwsfAraaNjsF4yMoMma6CXOG4z8lB4fiN6vUqWoiiLMRAA/JQxNka0g5zn1QWBvhuyrJHFxyOwSRj2nLuxKmho3zNL2jZBHTSCN+StT+6UbWdOBkrS3wPY4gjsrCD6IMh9W5zydzlSQ03vLg4/CVZQNiMh8Xt5KWsm8KUCE7BBPJbfgA8TOFEYzC0t+xWQ1U+fjzghZEjfFZ1fL0QQUMDJZT1EKOvi8OXpb2URlkhkPSVXxnSyAv7oBgexvVudspFOY3dXos97oxH8WCMLT3NaX5b2QZvv7Z2eGW4wO+FG+pbGCwNG4KjBhjbkAZWNI/rdlBRx6iXepU1NI2MnPfdRMie8/CMqroXsGXDCCd1dMSQDsFZ1de7u6siYXFXSDodjCBjdSxyOaMDsrIWiR4GcLLNKGjuggPxHJ2Ven5KTowrhHn1QWwENfnsrpXh8gHkrHAqxxOEGbMyn8DYjON1ppG+FL8R2JKjfsN0Fxx0/NUYfiAPZUiHiODcd1JJD4YyThBqUMMEkY2GVj1NOGdwBlR0L3F+xysirc5xAwUGnkHOwVrmHz2Cy+mKNuTufRRPkicR5DKDHk6Q3BG6h6lkVDGdPwjdY4GXAIJGOc7v2VuD148sqbw+huRlW5afr+pBWWIdIKlpmF48MnAKhBcTv2WUxwjj6h3CBLStpB4mQVhyTlzw4bKs1TLLlrjsomN6nAeqCd1R1jGOyMnGzCO+3ZXmmDGFxHdYzh8XwoJ54+loc091ZHEHNLirg4lvx9vmrPELX4B29EFJJHduyQRmaQRk7E7q+RrXNBb3V8OIWh/ZwQahLYwyLxA4H5LSJYjG/pWoRXWR/wPJwsOqcJH5agQShhxhSS1QIwAM48liKanj6pA5w+EHdBRk7mEnGMq18hfJ1ndahWNpnRARYzstPEbgfiG3mgyG1gDcEBU95DjkNUMgYAOnup6WFpb1O+tB389i5MZOaPVLSMY0DWn/nG3Ir/YwCMc0WqOjv+wGt/lG3Ig9nkREBERAREQEREBERAREQEREHHHMl+DtxT/gVe/1GZfNC/wDsrv8Avj+lfS9zJfg7cU/4FXv9RmXzQSf2R3/fH9KDJe1vg5A3wsVucgequErsBp7I0hsoPkCEE0jHiPPScY7qOEd8rWppKR1EAMF/yWig9GcILZCVYquOTlUQXRtDnYKmc0MBxuoG9WfhVznOz8SC6DeTJWc6Nhj2wSsQ9LWBw7qxk7sjJ+tBZID1HIUkTSW5CzZjTGmyPpEbqOjkiaCHYQYT2uDiCOytAJWpzGmeD04yfRYjw1ufrQW0s4gk6jur6yq94cHDIAWO7uqIMqKcOAYdlHUNDXAjz3UIODlZMdLNUjraNh5oLesdP1qJpw8fI5V5p5WuLCDlSNt9Q5vUBsEGVPOx9P0DvhaYr3FzCWHOQUjifI7DRuUFOrIDSsmhilFQx7WEgFDbKoNDi3Y/JclaD4c1V6g8UNJ+HP0Sgw4r5LT0IZT0Ze/5NytYtddU1lE91VSOiOMjLcLmXgZwft9yv5pb5GDEDj4gtb5g+H9g0nKyOytAZtnpAQdSb+ZXVb2+GQBnyWjNrPBcYzvuuSNUUNvbR9ceDKRvsuL5aaQTkkbZygyqqHqiMwHdaeyPrySexWqVEzW0oZ8lpHUR2KCR46dgolO1zT9L1VH9CCFMn1Q4zsgBKBk+pWZTS9DD1HssZmx+Lsrnu6iGM80EniGWUhpxn0UwhHSS92TjbdQuoqiKMTFu3dRePIfMoNasbY/Hw8j7VZqQNbM3oOy0+iqJY5epp3+tSVszpnAy90GCAco7up5DGWDp7rH+tBJAMvV1QBsomuLTkIXF3dBRZdPKGxkY8ljRxukOG91kiIwsPX3QYzzl5KyaaPrwScKFhb4pJ7ZV0k3S7EZ2QTVJHR0jyWG36QVXSOd3VvZBkyj7381C2TpGFaXkjBVEEkcfiu3PdXyNEO3qo43EHbukpc5wz3QGDqOVnQwNczJI2WPHEWN6nDb1VX1LmjDEEpjHUQDlY07MPwOyrFOQ/Luyvkljf2QSUsYbh2d1FWnLwArWOkj3PZRyyCQ5QCwBuVYq5cRjyTpKCiYPopIsA/Esv/a/T5ZQYCva/AwrX4LjjsgyTgeaCh3JTCkfC9reryUsPh+GervhBHBMIjkhJ5RK7qUb/pHHZUQZEUgd8KyYqLxZAfIlaeCQchaxQF7oC8dwEFa+gZDTkg7gLSIndLwVkVdXPI90b3HY4Chj6enJQZE84fF0jvhYSuc7cgdlJH0dG/dApqgwP6jkj0Vauo8d/UoXd1ezp6fnhBGiq7uqICkZIGjCsxsqIB3JKIiAiIgIrmMc84b3R8bmHDkFqIpWdHT8WMoIsKURfey87EK6lY18hBV1X8B6G9ig9VPYa/7m8ZP8NYP0V69S15aew1/3O4yj0msH6K9epaAiIg65+0O/A04l/wCJ0f69Tr548L6HfaGRul5N+JUbe5o6PH8ep1889P09R6vRBWOXw91bNL4hyrH7uOPsVEEtOAXEFJmO68AEhWxP6HA581rdE+ikAMuM/NBp1JAWkPe3APqtxUVup5mhxcPyrAuDGGPNOB28lpsdfW0x6ASg1u9QxUUeY8ZPop9F00dzubIpeziAtAqqmonYPHJWqaZrjb6oVEf73fKDuhwot1No2kpa+npBO4vBOG5XqTy5VNu1JpOOuntLGStY3dzPUfNeYnKNxB0pcq2Cj1fJGYmyDZ5Hr816faP4tcNbRBBbNPzQiIsAPQR6fJBuuvtNDcK+ekkoWlhJH0V1945cslu1Bbpa6mtTS85OAzddprReLNeQKqicwud6YzlTVtE+tkEEuDDncY7oPBbmZ4U12iqubNsdHG0E9XSQusNHE6WKR3TnB/IvbT2h3CjT3+x++808TWzGN4OwzsvHSlsjYIavOdpCBt80GyjGTKW/NZjYgyInCmqaFwqSGZzlWvPhRlryMoNPDC+QgeqzGQCOPJ7qGjwZSSr6mVzZQM7ZQQTPcSW4wFEs+pdAYR0/SWC36QQBnyR2fM5U0gYGDChacuHUdsoLooy92/YJIzof0hZJ6OgeHjON1iv6+r4+6Cd0A6Ac/NR9ZZsPqVpmeRjKs7oM2lHx9TjkHyV1RI0PGOwWGxz+zCjg/ILu6DImIkaMKSmrxTM6MfmWI5xAGPtVGgOG6DK8cTvJ7BRzN6dgoSegjpWRHIxzD1d0GKHFrstVesk5Ko76R9MqrQOsDyygnjkdJhob2/OtQjHRCcqGJsTWgjGfNSSVUTYi0ndBpsruuUj5qPBa7Poqvdl5I8ysmBrHNy5BC6V8oDe2FI1vTFv37qsvhnPh91EZCG9JQWsZ4j8E+ay3UDRGXZ3HzWJGXdRLdypBUyhwa8nGe3yQVpnmKQtxkeanfIJXBmMb4U4dTBnUcZI9FjOe10mWoJnU7YGeIPNYkjvEy5T1b5REOo7KGmiL29WNggpA09WRsVkBzw7DnZCic5uelhGRsoDI8O+JBqbGtcM5GyvLRnHqtN96LRt3VpqpM5Bwg1CVoaMkhY56ck57LGM0sm2Vl09O/p63BBawgnCrI0dGcHdJJYmHB7hZbY2SwF48kGBRvDZd2g4V9VP1ShgGyip3MjqD1dlLNLAZOr0QXMe6lAe1hOd1K2uM7g1zcZPotUtDKKuHhyYJ+ahvFHBSyYhAG/kgxHU3W3q6lpk7PDkxnzWZ7w6IYedljS/f35Z3ygpJ8Ddt0gi63B3oVV1PLEOqXICyIZI+npb3QVqWARYHcLTx8J+pakxrjkydvJY81G8uLmj4fNBCJC7yVzJs/AfPZZlJRCf4QNwoKyhlp5QPXsgsnpwxvWDlR04zID6JK2ZrR4mcKrHtY3I7oNQq3AwAN3OMLSuzsqeOoycP7KOoLS/4UCR+QAFSOPq3yrFe3qIw1BIcRhRl5e4Dturw1w/snZV6Bjqb5II5I+jBWVBTNki6id/msdpMrseiqZpIyWA4CCk0QY47qrJsN6PVUBMpV/Sxjd+6Cz4mEOLsgrLYGywl22cLBLuo7nZXxzOaQPLKCN4PUfrU0VQWgNUsxiLAW4yVZExj2n1Qd/PYtj/rpNUnP/ygVv8AKNuRPYuU8sXNLql7ux0BXAf5xtyIPaNERAREQEREBERAREQEREBERBxxzJfg7cU/4FXv9RmXzQP3kd/3x/Svpf5kvwduKf8AAq9/qMy+aCT+yO+s/pQWnZZNPS+KwuJwsYd1I2ZzBhh2QUd1MeW5JwrXOyhcXEk+ap3QEwpXU0rW9RGylh8Lo+LGUGOx3SclZccLahvWSBhYsgy8gKSLxmghucILH/A7p9FY4eau8N73HbdDGWvDT5oDGve4NbkpIwxkDstYpKONkYldjCw66MPlBYNggwh1t+LdUc4u7rU2+7+CAQMgbrCm8PJwggRPqVRuMIKLU7bcG07fDLe/mtPMbhueytZkOBQaxUFmPGx33Ucd3a2Pwy37cKA1HXH4edvJYb2hqC+YiSQyA7Eq6nqBA8OxnCgyeyvh6fFZ1ds7oN7aWbLfJWUzaYuBeG9l6S8rnLfHfNKCultgJMPV9H5LpJwPm0tTTQvrizq6wd8eq9k+VHU+mXaXipKJ8QDogNseiDrjYeDtRa9YzUsFrcwB5AIb5ZWVxd5bxeLNJX1MBBaCcn6l3Sq59DWC6z3a4eA3OTk47rgnmE4z6cqrFU09gliOAW/CQg8nOK+h49OXeSj6wA3IXEdZRwYkII29FypzAahra++TTRuJJJ7H5rgk19W5xa9x3O6C6dhlmMWcAeqxp4xC7AOc/NZvuk87A+IHqWZSaNvNc3xGRkg+ZBKDQPmq4J37rVq3S90oM+PEQB8itMAbGSHZz2wgtY3rKq9vRsN1QEl/weZ2R4cD8XdBfBGJn9J2V74hTyNcTkA5UUXX1fB3V0jJj8UmftQapVXGKSkEQAyVpMYzkq0k9kBxt5FBNFJ4bt/mqTyeK8Y7KwxuDerdXRtz8SC99N0M6w7yWOTk5V7pXkFpKtb3GUFEWTIYvDAGMrGQTU0/guyRlXVNV4/YYWOr2wvcOoBAhj8R3TlXTw+EQM5yrA50bjjYoXue4F26C1FOSzGFaIHv3agiVWjJAVz4Xs7hWsOHA+iDJNP4bRIscvy7KzJZ2uhDfRYOD6IMmSq6o+gBY7Q7yGcoxjnnAWqUMELGZm749UGI2lLm57Eq/wBx6W9YOVfWzsaS2LH1hX0crnREOP5UEDHCd3g9vJSvtga3qznbKxZg6KXraqioqZR0gkoLHkNcW+mysLxhXup5R8RHdWxgCRod2ygtw7uQVJHCXtJ6lqlV7n7oOgDrwtIa942B7oKPb0uIRp6SHehUsbCD1SdldJGHDLOyCslSHxhgByFCGEgnOFYRgkeiqHEbBBJCzxSWq+WmEZwraZ4jdkqSp65D1N7ILBTAjOVkx1nusZiB3WG2VzTglWyO6zlBWaTxXl/qrERBd0bZyqduxQhw3VEBERAQbIiC4uGMYQMyM5VoBPZVPU3ZBQ7IiICJ3V4heR1Y2QVhl8J3VjKrPMZt8YASCMOf0uCzXsgYzcDOEGmp8ldLjr27K1BLE7wj1ZVJpDIQSo8lMbZQernsNf8Ac7jL/hrB/wBGvXqYvLP2Gv8Audxl/wANYP8Ao169TEBERB149oLMKfk74kSns2jo/wBep187+5OfVfQ37Q/B5NOJYP8AedH+vU6+eNBUtwMqrWZBOVQknb0TqLdkF0UfiO6cq6Rj4XDDj8t1ZGHk/B3UrYpXSDxBtndBkUta4fA8EhXTyRueHjCvqo4WQfewMrTg92wPZBkVVT1sDcdllW4EQHBwfVYE3QWgt7+a1OgkYykd1d8INa0verlbKyJ1JXviJePokjzXofy3U8t2tUdzr9RHqZEDh0m/ZeZtFO/3vLewOQuRNNcd9YaQmbS0M72wDALeojZB7X8GdcwQV3uEl0Dw14G7l2YjutDLC2RlQwkjOM7rxi4C8x1RLLDLUSkyl4J+Nd19BcytHW3WmtlRMMvAGOpBunnHgqtW6XktFPTOlDI3NwBnJK8pNRcKdQWySeFtilDXyHJ6D6r3VptOWHWFngrauAPEzc+RW0NV8v3DqotNXPLQMY8MLmuLRsUHgjqzRdVY4zUS0ZY7GcELjWqjlqXOd0EYK9DOZXg/DHeKqC3NHgtyG9LfrXT3UPDK4W2CaXoIAJ8kHGMEPhuJJ7eqhqzl6krWzUtS+FxOQscP6j8SC+ng8d2CUqIRC/pG6CXwvoHdWEvmPUfyoMiKm8Roy7uoZoTG/pwSFQPlj81qtA+lkgJnxlBpMb3sdsCtQhofe2eI7YrMoBQeOfGxj54V1e5hd4dFjHkg0aqpTA4gbgFRMZ1nB2WqmjkaDJONiM7rTqhzQ/72dkE0ELWHqO6VPSdwOwWKJpPIqhe52xOUFHHJQHAVPrVWgHYdz2QCO+6plS+7yYzjZXCmd09SCDupGR5I38wrCMEj0QOcMYQZ8cOR3VHUfiPGHbd1i+JMzfJAU8Mz/pZ7IJZ7c2NoOe/zWo0VojlpS8uAOFpktVJO3oadwqMqq+CMtBOEEU8Pu85aDkZQwiQdecK6mjlrJT5lVq4JqZ3QQQghpxiQg9knwZQAMKkRLCSdklJc7qCC+WEtZ1dWVDG8tcHZR0jnDpKowZcPRBqDZBWMEZyMeZVBK2m/2v67ZUT3iFmWbFQYkmPUe6DKkgELfHz3UbYveB1dsBRSSTFoY8nAU0D8RHGyCrKEOz8SilpnRnYEhUFRI15wdsrVI5qeSAmT6Xkg06ma0O3Wp+KzwC0YGy0kv6Jic7ZVHzvecA7INwaZ03HfKkiR4AzlSalomWGYUkTgQdlo9vrrnbj4lISCfMKyvr6m4P8AErSS8eaCyGnFS7qyBk9lrUWmYpIDIZBkDPdaDGyobvGDjyWSLpcIm+H1HH2oJ4o3WyoPS4kZxstcbSsuEHvD34I8itv0Mjp5i6ffKlqq6ogmEUBIZ8kF8tEKiUxZwBsoX0jaF3XscfNX1E7xGHxH4yMlY8L5ZyRNndBIKgXD7z04UU1D7qerOwWUKYUwMjAAVgVM8sx6STj0QTsqhNhg7jsr3VzYh4RaN/NYlKwsd1FKqNzn9YQZ1LWtoXmU753wo6q6Nqpg4jAHyWmue52AfJVa04yg1GZ7J4uloGeyw/d/iwCoxK9qkgZK9wduQgv9yJGcqPwMHpJ81qTmER7AZC02dzmv77oKzwCNocCqwAAdXoopDIRl3ZXte0MIB7oLnyCY9PbCoHBg6M91CctOyFxPmgyqdga7J81lSW5kkfi9WMDstPjL3HA3wpHVk7QY8kDsgiB8F5HfGyE+K/HYFWuY4fE7zVoOEE0sXQ0HKvipfEjL8qB0jnDBVzJntb0DsUFjgQSPTZXwv8NwceytcxwGXeaoAT2QehXsYK1tTzP6ni6e2ga13/ONvRYnsVmkc02qT/8Aw/rv5RtyIPahERAREQEREBERAREQEREBERBxxzJfg7cU/wCBV7/UZl80En9kf9Z/Svpf5kvwduKf8Cr3+ozL5oJP7I/6z+lBaqtblU7q9uwwUFh2V8Y3DlYe6kjYSMhBkS1ofF4Ybv64WIc/MKeNjA74gqTBpIDMboIRtuVkR1AaOnHl6K19FMyPxHDZWwBmfi7oJPF6HF5GxVC4Su6x5bq6Yx9GG4yFjtf0gj1QanHXiRgg7YWPPIIstzklYjHFrg4HdVlkMmEFCXOccHurvDJI/wBatjBc7AG6n8N4GSPNBX3TbJKgewMd3U75yGluVjOcXd0E3ih7QwhRuAZ9qs7KpcT3QASCHKVrBNjJwo+42V7Y5gMtBwgvfAGN2OT8lAAc4AOVMyRxd0vPn5rKijjLm9kG4tDUdTJUMxUOYA4HY9l3B4a8xdfwrtrGNujsMaB3PkF02gubrVGJIjgkLdVruLL9bHtnBLunHdB2mvHOlc+INS+1wXV4J+H6S451jxautogkiqbm94kyfidnuuBtL242rUJqekhgJx8hlZ/Ea4i4AFuT0/NBh6j1Yb7VukfJ1k53WzqtzWSF2MbqlK8CUNO31pX7uGEGq6frmGZkb2g/EAu53BXhWzU1gFYLZ4oDM56c+S6RWkiCWOV3/wA0C9duRS66Sr9CC3VpiNTJD0jOM5wg6tX3g7BdrxLbfcAwgluMYwuOdccsFxt8T6mmtzyNzlrSvQW7cA9R1mtJ7tbYne7SSlzcN2xlcsUnAaGo0zKLvE0ydG3U3zwg8K9S6PuGm6gsmpnNLfVaA1rqhxLhghehvHnlZuV4vlQy0QEgZIw3610q4i8NLrw4rZaa5Mc0h3TuMINkxsEMhc7sFWoqWyfAGqaWMuhEjR3GVp+/V9qC+RnSA7KyqOiE7DIfLfCxpGPDAT2U9LVOhaG52PdBbUuDCYgNh5qBsnSDhTVL2yZIxlYyCSJnivI7ZVZ4vCOAqQv6HZVZ5PEcEFrGmRwaN0ljMZAIxlT0bQx5fINsearXOjkeOjzQYuMb5U8VSI2dKsdBI1nUQcKwMc4ZAQUe7qcSq9G2cq0jBwqhxG2UFY29bsFZImEHw4ysZhIOyPJJ3QTS1AlyAO6hLMdireyu69vmgpkqrfiOD5qseM/EhIDwW9gUGSyMU4EhGQrJal0g2KlmmY6ANzusNh3QUJIOTusinmcDgKN4GMqejMQI6sIKztL29u6thcISCQs+UQtZ1bYWnTSMcdigkkrWuGAPLyWG49TtkcMKojcR1AbIKua8Dc7KwbEFVJPYqaGnMg6h9qC185cwNAwkc3S0g75V0zWNGB3UI7jKAdzn1VFkxMY8YA3wopoyx2EEeVOypLW9JGVC1pccBTxxNxuN0EDjkkqinfFjsoXDBQUQbIiCQvDh0qM91XpcN1TOUBERARFUYx5IKscGnJSR4eqFhVOyAiIAT2QAcFZDKnoYW47/ACUBY4eSlZTPe0uAOAgtbL0vLgqSSF5yrS0hxGNwVQgjugJg+iKeKPqZnCCBVBwCPVHDDiFeGZjLvRB6sew1/wBzuMv+GsH/AEa9epi8tfYbY+5vGT18awZ/JXr1KQEREHXP2h34GnEv/E6P9ep188a+hz2h34GnEv8AxOj/AF6nXzxoKt7q4tJGVYsmIs6PixnCC2kIDyXfJTz1DeoBvmsMuLXHCp1FzsuQZkryY89wsMnJypjKCzpB8lB2KBv55WXA8iI5OFWAQ4+LH2qOpeGvxHjCC2KYwSEhST1fjA9gfVY43PxIW75ag3Jo/VVbpysZLFUOaC4bA9l244B3Wv1JqujuE1wcGNIO7tvJdL7ayJ8rfH3Adtkrn/hLrqOy1cVPQvAlwAMHzQe3XDrXFLRaNpaeKrbLIxob3yQcLeb459T2shxcOods4yuk/K7qe+XVsD7xk05cCMnbC7v0WqLJStho4HMBIA2I74Qdb+K/COGrlqZX0YeQ0knC84eYSuptMV0tpbTBnVIW9vmvaHWFhdeqKaWnaMSxkDH1Lye5x+Cd+jvU1z8J3htlJyB80HSXVGnBITcGN+mM7LZUkRYSCMYOFyjf62OgP3OqjhzG9O/yXHN0kifKTF2yUGKI+od1IwNjaoWvdnGe6q95QVlkDtgqRscQSHEBViAc45Gd1JJFI0fB29UGOSQTuc/WtatY6IvFeclu+Fop7nK1ChlIHhk7HCDWqOQXiR1N0gAHC029WYW95wVrun6PocZYgMndYGqpZBKWyd+yDbbY8uxlXPjEYHqVQE9WR3VJHklBYe6uj+mPrVquax2xCDPlnayIAYVrKhr4znuFiPkJHST2VGskIJGcILXn4ifXdXMZnDsq0g5we6ujd0kA9kF8rgW9OMfYqMl6W9KvmLCzY7qAAEIL4JTG/q7rLkr2uYWBo3WAqgAjY7oMq31nucxlIzlS19xFY8EDHqsSOMA5eNlI5kRAazugikIIGO4VGuPTjCk8CRh6ng4UhdCGY2zhBiE5VWnBCrguJI9VVrSHYPdBdKctCvp5A3AxlDGSBtsqDpZ37oMl0bZxsAonRmFnT3z5q1tR0HAKm8WORu5GUGC4HJQOcNgSsiPw+o5Ix5KKbp6/h7fJBLTRtqHFpwPrVJohBMGg5AKiZI+I5bsUfI6QhzzlBqza6KniGQCVp1VUieTrAwM9lE+QvABVgGUGbBWCMAdOVJLMyRpdsNlFBSOPxOGx3UVQ1zH9I80EtNLmT4VmvY0jJ7rDpIS09TxtlZfu88r+pgPSgsgDnPIIyN1ZUuNO/qAwtVgijp25lxladcHwzyYZj8iCjK33gdG4WO9obKB3CkFP4I6h6KGQlx6vMIMiRjQwFvdTRQh8GXAZ77rBhn6iA8nAWf1ExERnbCDSJmhshAVGvI8sq+aGRriXDue6lggDgHkbIIWsMh3HZZcErYT07KyV0bAejGQsbrd1Bx8ig1Yytc0labUH76CpmylwwDlRStxufrQVk6XMAHcLH3acYKkiZJO7pZlZDo2xNPiYBQQRs8XZXGmw7Cjik6HegJU+XucHeQKC+OAQ5c7zWNM4dYI8lkTzgtwO+FiNPU4ZQSh5mb0YxhWuhwpneGxoI7qM9bwXDsEET2dIypIowR1fJGsc84IVXOMYLeyCk0oI6R5LWaCzMmoTUOxkDK0A7kla1R3R0dKYAe4x3Qd6vYu4HNVqtg/e6Brh/wA425E9i2x45qNVyO7O0DXfyjbkQe0iIiAiIgIiICIiAiIgIiICIiDjjmS/B24p/wACr3+ozL5oJP7I/wCs/pX0wcyP4O3FL+Bd7/UZl80Lh99cPmUFjTgo52VUNLnYAR8bmdwgoRhSRTeGOkqxoc89I3VXsMZHUMIL5Pi+IKJjulwd6HKuLxjAKsQajPc/EgETW79lj09MXtL3ZCrQ+D4n33H2rJrZ44x0wkINPljLHkZyrMYUsTw6T41fUeHn4MYQQFu2VQbqZzm9GFCgliIiPV3Uj6oOGFjgnO5Q4QHHqOVnUdEyeIvJ3+awFLFUywtww4BQWys6JSweRwrXNwhJe4k9zujmuH0kBruk9srJZWdLOkhQw9OT1KswYD8KCxz8vLgpI6hzSDnsVCrmRuf2CDNqp3Twhoct5aMYYaMue49u2Vsmnp5HENx5rdNor20bRC7bOyDPqrsfe3MYzBz3C0q5zOljd4ric+pWsBtJGTUzYwd91ty+1sVQ/wAOkIJJ2wg0F7vDnJHkUkn8TGQp5LZVxsE0jMNIzlYh6c7ds7oNYskDa6RsR2w4brujyu6kOmdQ2+nddDHGS0FvXgYXSaiqXU2HQ7OG/wA1vDSnEK72W8wTse4FhGDkhB9F/Dq/acuGk6SqhqadxDB1k4JzhZGpLlQzUT2UrA4EfvR3Xlbwa5wLhYbTDT3WqcIgQMOd5Lu9wk5itKaupIaieWIsDQXEuCC++1MFvuEtRNaPEHSTkx5/mXmbztQwahv0r6aiEJ8Q/RbjzXrnWcTOE+o2y2+GSmdUBhA3AIOOy8vOd2rsds1I/wADoDHS58vVB0uvFilt9rjf4OMM9Fs3fqyQe/oucdQ3rT1XYI2Ncwu6MbY7riKpNJ1u6D5lBBLIDAB54WDg+WcLIqXbDp7Kaj8DwvvmM4QYQbnuULAB3V0xHiHp7Kxp3G6CmCPIqoODlZI8Pp+fmoH9PVt2QXuqCWdAGPqUbCS8Z8lJ0B7fhG6jcxzDug1GokYaYAd1DSMaYySFiGRzgAeynhlEbCghl/shVoBJGyPd1OJ9VkQ9HRv38kEfQGAOUbjlXyhxPyUaAqtHU7CNYXHZSCCRo6kFz6cNb1AntlQgZIHqrnSPOxVrTj68oMiWm6Y+sO+xY3ZTsnLvhd2VkvT1DpQGZeensr2wEOBB2yqB7Q0dPdXeN8J33QZXhiWPo6lC+kEeSHLH8d7SSCpG1LnHDkEUgIwCs6hiY+E9WPtUZayUbYyoTJLT5aDjKClRGGykA7ZUlPUiMdB89lAA+d3qUkjfEQHDBQZ0lG1zTL1eWVimn3OOyGacMAdnCkilaI/ixlBbCPBd1ZykzhO8eW6jZIOsl3ZJXjqBYUE4pvDb1DJJWZTUTZmF7tiFiUUpe/oedlJV1T4XhkRwD3QUrGNiyG74WASTupmukqSQdyrpIvCGHd0GMg2VXd1e3pwc4ygo6TLcYViq7uVeynke3qaMhBGqgZVCMEg+SZKAdkGyIgk8Tb7FY45KoiCSKMPO5V7mCM4HYqFry07KaN/UR1ILpBhgPosmnqAIiCN1YXx9OCsWR+DhvZBRzgZS7yyqSHJVmTnKZygKWOYsb0huVEs2mEPhEvxnHmgwycnKvbLiMsQsL3kM3Hkj4HsGXDAQerPsNm/9TeMjvWawD8ja/wD1r1LXln7DU5t/GVvpNYP0V69TEBERB1z9od+BpxL/AMTo/wBep188a+hz2h34GnEv/E6P9ep188YxndARS/DjyUbsZ2QUVWjqcB6qidkE8kAiYHZOT5KsUAkYXE4KidK5zek9lWMShpLBsgsPwkgEjCEknfdUOcnPdSNALckILCcqrD5Kju5VzcEfNBMwEYcDgrUrDeaiyXRlxEpwwg4z3WkgPaST2VHdUjsDsg7q8F+b82uGC0Ol8ItIGTsu7PCrjBTarr6aaS8tJIBAL142aepzDUMqI8l+RjHkux3Byr4k0V+pblTNmFDGQ5xBOMIPc/Reo6OvtA8SpYRGM9RPcYXRnnh4l2mmlntlNAyQvcQCBndbk0dzAQHS7LVTS/7dawNd8W+cLrLx3vsd/uRdc8GRzyRkoOmnEeyy1dyfcWtLQ8E7bLjOSnMbnAuzuQuwfFOnhoaIytADS3Y/YuAC41MznNO3VlBC+Exjr8lC52VqFWAIQMbgLTkFWPLDkLKbV9TRGRudlBD0dXx9lK2LMoc36OcoLZoS0dYBwTlZVG0FviZxgKaokgdAGgjK03xXNyxmwO2yDW6DURoCR6eiwbxdDc5evyzkrCkgkY3rd2O6vi8IRnPdBADg5QkuO+yki6Os5xhUl6esdPZBb0bZVzZA1pbj7Vc9w6Rg/Woe6Cp3KmjqOhvSWq2Dp6j1Kkwb1fCgsccuJVenIyrVVhIcN9soBBB3VMH0U8xZ0DHdXRGPw9+6DGQHdX9Be8hqmFM5rSXjGyCJ8uRgKxjulwd6Kh2KqGk9kGTNViSMMDVSKk8SMvzg+ijg6cnrHZXmdwf0sO3ZBG0+E8g+RVTJ1vBA7KadgEfV5rEb3CDUYg3o+L081h1IAfsdlUzENAb5qwO6nguQSUtP7w8tJI+apUwCB/SHEhSvmbE0GM4JWNJK+V3U45KC1BsqhpPZCOk7oBOVRVJBCqInEZQWqrSWuBxnCuYADg4UrOjqGe2fJBk09V1NDCMYCx6mT76CBkK6odExo8PGSrqd0UjSJMZQZVuxV/ARjBWTUV4t7hAG58lDRtNM4vbsCpJ3007svwXIMSurXzMy3IysGBzvFAJO53WvU9ubUtw0ZA7bLTa+kNLPgbYQagYBLTjA3wtNniMZLcYWdBUiOPDj3UdUY5GlzT9WEGpaX03DeJCJHgYUN/pPuHcBSx7tJwtKorzXW1592kwcrVzWMuUXvNY4GUbhBfU2+GakEmQCRlYbaSOOAgEZWBPcalzzHG49OcAKEVs4PS87ILnUZMh3OCSoJI3RnGDj6ll+9N6Qc7k7rPiZTzUxe4DKDRIpCwqTqM7ukDvso5cCVwHbKzaKIEhyCsY9xHidOSfNRZNdMM7BajN4TmYesE9MLuqPG3ogvrbX7tF4gOViRTHHhkd/NZj67x2hkh2Gyjd4Ab1NO49EGNLB0DqyTlRDIPzWTG/xCQ45Hko+lrZ9+yCSmgdOcEHCumcKclnfKz/FgiiHh98LSamTxHknugq2cNVpd4rwO2So1UBw3HkgmlpxGzqyVExxB2OFV0r3DBViD0H9i5OJOaHU7dttAVp/5xtyLH9ip+FNqn/i/rv5RtyIPalERAREQEREBERAREQEREBERBxxzI/g78Uvlou9/qMy+aJ5xK8/9sf0r6X+Y5vXy9cUGjz0Zex/yGZfNJ7u+WeVrR9AnP5UFtKQJN1JWkEhIGBshae6VowQgghk8J/UrpphKRjZRJ2QSvg6W9SugpnS74V1M8Pd0SHZZ5dDGwluMoNPnpvCGx7KDJPqVJNO97zk7ZSnDXPAKC3w3YyAVac9itWNM3wg5YcjWNzuMhBjRs63YWSaQBufkscuLXZaVXx5PM7ILHDBIVEJyiAqgZIHqqKrdiEGR4Qjb1/JQyyB+MeSmkf1x4UDR5lBamUOM7KrcfagopqeR7XYa3OVfTW6pqiBEwnK3lpzRjp5mRzAguGeyDbkXvg+KKlc8n0GVWOnvMtUwmjkbv26V2u4L8ujtV1LQ+NzmnG/Tlcxaj5QY7ZPCWU5GcfvEHRO62+4G1gOgcw9PnstmRB9NN4jzuxd4eNnBCLSGmfeHR4IYT9H5LpBciPe5mNOzXkdvmg1Gtv7Kik93azBGw2WhhpPdXwNa+QB3ZSzgMcANslBFFIYX5xsshtY7x2PaexCtlYwxAjc98rG3BQbrud7rnW1kVPWPZjfAdgrf/C7j5fdGWx9A67TAlpa3LyuHGSSEYkz0lXmmjc0uYDsg7A6T5kNQ2bUEl2mvszmPOcGQra3GzijUcT6ttQ6qLyHZ3PdcSR+GXmMtP5VfK2WLBYCAgyHU9UYQw1BIHllY5oy1pPVk/MqMVco7lWuqZD5oLZHH6PojY3OGxwFYcnc+akYyRwy30QRnYlFe4YODjKtGOrfsgpusmOmD25ykgYIw4d1E2WUD4eyAx3gvPnhSvf4rer0UUUbpX4AJz3V0kboXhh80FkUfW/pV0sPhyBgOcqZ0To4xINs/JYzpHOIc7chBlvoA2ESAkk+Sw92nHosmGrefgeR0qssPivBjGR5oMd0uQBhSw0wkZnKndSgsAA3+pYpdJCehAafDlI+azGzNe0AhY7Y8Drf57qN8mD8KCSeANy4eahDMgo6WR+xOVb1OCCh2OPRSRxdfcqPur2vIGAdygo5hacKgG4BWRTRkuzJ9HKVYjDh4fr5ILZIA1nVnyyoWtLlc50hGHZwjTgfNBLAfBPUSraiXxSCOwKjcXHPorUGTQvEcmT2V1fI2SQOCxQSOyr1EuGfVBK6YPj6PsUB281O7oDNjuoM5QTU0Anf0k4Vaqm8B4YDnKijldEctO6rJK+Ugu3IQTNb4LRIPNQyymQ5KlgeZD0O7Kk0WHAMHdAp5fAcXHdUqZ/HdkDsrX08sbepwwFk0jIi34u6DCwizKtsYaejCxmAY3QWE5WXTVYij8MjOQsV30ijMdQygmmizmT1UAGVlzSNMYA9FjMOxQWnZAMkBVAy7YZQgtI2QSvhDWhyhV5kcRjOwViAqg4VEQVLyfNURSwxOe4EDbKCLCLUKiOJsAxjqwtPQEBIGEV8LeuVjR5kBBWGQxO6sEhTT1Imjxjf/wBq1d9pbHTCV4GFodQGtcQzsg9V/Ya/2hxl/wANYP0V69TF5aew2H/U3jIfWawf9GvXqWgIiIOuftDvwNeJf+JUf69Tr5419D3tCsftOOJOe3udH+vU6+eHCAiIgIid0BTxVPRGWH6layF7SHOBASQM7hBG45cSgdgK5kT5CQwZwkkT4jhwwgsO6q04IKoiCV8wc0N9FbHJ0eSsV7I3OwSNs7oNSt13NC9jsbB2V2G0FzI0NmsTrM6BolczpBwM5wut5ZERjzU1AyBk7XEEkboOzmiuJ1fQXuW7vr3+FI4kNzsFZrviLJqO7RziYkBwJXFdiqQ6FrMbLX36ar6mH3yFh6QM5QTcSKwXazNja/J6MbLiGkofdY3l43+a3xXV56zR1DiC3buto3ZxbIfD3B9EGjVriSR5LBWo1fT0/PG60490F8UfiHCnMpgb0evoooHljsnYLNJhlaQcdWNkGnElzs5O6q0ljg7vhXyQPjy7Gymg8IsAd3OyCstWJY/DxjAwoRBlvVlHxeG7q8j2V7aeokb1Rg4QY7hgkeikjh62lyjcC12HdwpmShjSMoICMEj0UzIepvUrOnrcQ0ZV33yIEHYYQRkYOyomcogImMpjCBlMn1RVY3JGeyC+CQRklTS1bpAQ1uR9Sim6MDHdZ9G2lNMevHVjzQacxvW45VziGfCqPcBKensrHO6jlAJyTjzRmzwqIDjcIM9zRKzGe6xZIQw4BVGySdmqj3PJw7ugsOyK9mCfiVHjfAQU3PmqHZVw5u6vBb074QWsd0qpIe4Kz6kBwgnkg6G5ykcwDOnCjdK9wwTsrEFX7nPqpWRZjLgdwoc5UkchBA8igpHGZHYOVlmhewCQZwN1Izw8As7rLbPH4BidjJBQae6sLW+H2woep7ndfUfqSWnkDi4Dbuo4yQ8NKDWrZdhT4a/thR3GpbVyB7VizUxMYdGN1JTQvEfx7HyQXxwe8DAOCFjVJdTO8PvnyWVBKKZ5LjgK33OWrqRMBlgOUGA4vaQ4sOPmtTpYBUQdRdj6lqVXHQGmEbMeIAtvvqJoHGNhwAguniFK8433WP8A2YnyU0XXIS6bsfMqOdpY8mPsgkjouodWVV0rovvWdilHM9zulW1JDpOj98giljx8We6vgqTGMdlWSjqIWCSRpDe6x3YzsgzTIZezle6MiInvssAPLeyz6Rxfjq7FBhxxOkk6TlZrreGxdQdv6ZV1b4UTA6PGcLFhq5TI1r3bdkEA6onHY7KQjrHUtWnZSPgy0DqK0eU9Di1vZBZ1vG3UVeyPxDkpCGk/Er3skacsGyC2aARjKngga+LJxlY3W52zlUyPYOkfoQWSDDyPRW4PoqklxyVm08LHR790HfH2Kn4U2qf+L+u/lG3IpvYuUzo+aXVLndjoGuA/zjbkQe0aIiAiIgIiICIiAiIgIiICIiDjvmLd0cvnE939zo29n/kMy+a2lqGsrKkuxiQux+VfSfzJfg7cUv4F3z9RmXzTw001RM/whu0kn8qCd0YEhlA2ysaplMh+pZXiCMGKTuFiTmM56fNBCBkqpbhV8N4HVhXxFhHxdwgjBLTncK50rj5lXydHkoe6B3VWu6HAjyVTG4DJCoGk9kGp09WJW9Du3ZYtY3pdt2KxmudGcjYqr5XSHLt0FqqDhURAO6DdEQSuhAZ1ZUSu63kdOVbjfCCaD43dJ9FWpYGEBvYqPokjHXjCOe6UgHcoLFUB3cA7KQUspGenZZlM2BlO8TYDuk4+tBr2hT7zXNg8LqPUBhdyeC/A6r1bdaI/colj8ZPTsuq/AOzsvOroqdwyDI3y+a9yuV/hXZbdpimuUkTTKI242Gc47oMjgty52rR9sp6me3xiRwBIIGVyffeG1nuoZ/tGPLMeQW9WMEbAxvZowrkHULmX4KW6/WKWidSsiYIiM9vJePXHDhTR6HudWymeD99PbfzXt3zn3ip09w1mudEcShjxnz2C8KeIWtLvqq61/wB0HlxExAz9aDj18XgAPB7qJzjKcnyWTUxSgBz84CxWH4xjsgZe3uDj5q+OLxAXenksqf3c046cdX86gpntawgnugtlmy3wwMYV9LUeG0sd5rHkx1n61QNd3CCY/A8yj1UhqvEYW9KhjILul/bKzWin6NsZQac7JJJCoNldJjrOOytQV3Pkpoqjw29JCtpojLJ0gZV1VAYXAeqCF7upxKmhp/EZ1eagU1NKWSsBPw5GUFrmyZ6XAgZ8wrm4a3H5FuW5OtZtg8Igy4+3K2qHbjKCWCYwvLiPzKtTUCeQHsAqv8Lw/nhY/ZBmSVAdCGDusPB9FXJOyz6NkRjy8DKDALHNGSCFkQVIjGDulW6LJDFjIMwV2Dv2+pQTy+K7qwokQSvnywNwokVwjcRkDZBRpwjjlU7Jg+iAqjYgnyV0YGd+yS9OdkGoW9rax3hHAPkrLlTCkkAG+fmsejqDTydbVfWTPqnCQ74QRSSBzdvRQjZDkbZRBISCFToyM5VmT6qoLjsEFW4BVHnJV7oJGgOI2KjygZPqgBOyfWsmMxBnxYyghdGA3KsVz3ZJA7K0AlBdG/oOVK2ceIHHsPVQIgz6mrZJF0tAzhYbPEH0QSFZnK122GiFKfGx1YKDRXvc4YOfqVmSpqsNEzjH9EnZQ90DumFLHHggu7K2XpzsgsyfVERBVhwUduVTKlYARkoIkG6ufhWoJREMZKjc3BwnW5Xt+I5KCPCniqPDb04SRjWtBx3UCC+WV0hJzsrWjJA9VRSwloIygq6DpZ1KyJ/hytf6HKmmma5nSFjINckvTainFOdtlo0wxIRnvurASDkeSvLHuBeRsg9Y/YeQCOwcW5vOWosmfsFcF6hLy19hvM99r4xwE5bHNYC37RX5/QvUpAREQdc/aHnHJrxKI/vOj/XqdfPI3uvob9od+BpxL/xOj/XqdfPHlAVelx3wVRZkDoREQ7GUGH2VWnpcD6KsmOs4VqCeSo64w0d1BlEwUE1NUCEkkZyq1Mwnd1AYULWlxwFIG+GMHuUER2RVd9IqiANlPHMAzp81AiCpcc5ydlcyUskD8nZWIg3Ha9SGk6Wl+MLkOw8WY/DbanMB6/hzhcM4ytzadt0Yc2qcN27hByVc9Mx1kT7ox27/AIsBca6gJopCzB7rfdLqg+CaRzvhAxjK2fqWOOqlL24OSg2nLK6Rx77q1sbiCcHZZxihiOH4Vzn03hnpI/Ig07uelXAGN4Oc4VOlznno9Va4OacHugzpJWzxhoGSsVrHMkAO26ntz42vPidldWM65g+LcBAqgTCCB23Vaa4iGLw3N39cKYPifGGHBOFG+nibGXYGUGKQJ5C4bZSSme0bAlIM+KWt7LVWvpxH0y4zhBo8bzE7JH5le+bxCB5q+q8MuPQsbcEFBkOph09Q7rHc0t7qeCY5w47AKyoLS7LeyCIHByhOUVWDLwEFMK9uOnCyJ2MEWR3KhZBK5vWB8IQWObg7lUDiOxKOJGx8kDSd8IKIqjG+cKRtNI/dgyEFsbA84PkkjOggAq4xSx+SjcXH6WcoLo39Byqud4pz2UaqOruPJBeIwrXfC9U63KRpb05OM4QUkOQjIuoZyoycqoe4digq9oarRsqkk91RBUnKogBPZMHsgq1uUI6fNXeG9u6tJJ7oL4pTGc5Ko6Vzn9QJ+SsxlVAw4ZQalHUtkjDSMH6lhTANly3sVdJI1rcMO6iaHPcCUGa2sEbQHDOyyIahsrcjbCwJ2gtGO6tjm8NpaO6DKrGh+Oh261e3TiOiMZbv6rQI/Ge4Ek4W4aWppIaQseQH4QaJMZY6hz+skZzhYr5HOd1dKzXSxPncXbgq1/u5PS3zQUgeJ8MO2NlkTUzGRHcE4/mWI8+B8TNsqnvj5G4cUFtK4RzH61q8VqjlxWdQwN8ZWhOfg5b3U7bjUsZ4bXbINTuVfHPGaVoG2y0R7Ok4G6F7i4vPcq8Oa8fF3QRLUYAGwB2dwsHw3nOArmTPZhhO3mEB73ySdJJx2WXFRNMfUCMqGYxhgczuVbDVva4AnY90FwD2OLXOOM+qnpqBtXMGl3dWVUkZZlhGVBFVywu6mOwUGs3CxsooRKxwJIzsVpjZQIyCN1NFdZpwWVD8hYs2HzhsfY+iCIEuefJXOzjAGVkVdI6GMPxjKU/hvjwfpYQY3RjJKubO5jtspURSMOd8KSPwjFl/fCDv97F6rE/NDqeP00DWn/nG3f60WN7FfH7ajVWO3+x/XfyjbkQe1CIiAiIgIiICIiAiIgIiICIiDjnmPGeXjikP+4u9/qMy+bSzVbKWqqOoAh7SN/rX0l8yO3LvxSI8tF3v9RmXzRFzhO4juXEfnQXVbzNUveNgSpo6Jr2F7ndlNJbpRB4wbv3WGJ54vhJOAgvB6neFj5bqOeDwSAD3Uznxhge3HVjusV8rpMdRzhBblAcIiCczBzelRg9GVZ2V4Id37oLDuSUUpp5MZA2URBHdARXRkBw6uyrKWk/CgsRE+tABwVXOHdXornY6BhZNJEx7cux5ILRL4zPDIwr6elHWDlW1BjZnoG6tgqnMkBcdh80Gq1IZDBs0bLR+maeTEbHEnsBlav48NVEIgdycLkPhxo+33Ovhp5wCXYxkZ3Qb/wCUPQ1dU6kjrpKMuzI05I+a9reEVTNabNSUZgLAYwCMfJdNOTzgfT09TFVPjHh9QOS1eiVDp+30McTYogCwAZ+pBqUb/EYHeqiqa6npceNKG5ON1M0NYA0bYXDnGbVk9oppJKZ2Cz0Qccc92oLceHUtC2sYXeE8kA+oXhTqeqEV3rulwIMx7fWV3+5v+L96uVvkpJHuI8Mt+kvOiSR9bUzvlOS4k/blBMKkVg8I4BxhYdTAKdwAOcqhjkgd1N2A81RxkmILjnCCxuXHGTj61dIOjsrmNz8LRk4VsjXN2egj7q5j8DGFQtPfGymh8Pp+LGSghJycoHO8iVIYHlx6RkK3oLHgP9d0ElHTmomDD2Kyqu2tgHU13l6qQFkUIfFs7HksGWrneS15P1IFNN7vIXYzhXVdV7wQQo+jxAOnurmU0ge0ubtkIIek4zgqi1usFEKMeHjrA3wtPoqfxMud2QYxc/G5P5VRrS4geqzqxsLG9IAyraKnEjC7AyN0FklMGxded/rWKs18NSXdG/TlY0sZjcMoIxnyUscjgQ0ZGSrYi0O+IbKZ/hd2jdBc+kAj8Qu3x2WL0/NTPdOR3OFYyN5IPl9aCPB9EWQ9sZZgDcKAAhwygphSsmDW4V0nheH81BjKCWFokfgpPGI3gAqxj3RnZHPdIQT3QSvjAYCDhQKRr3HZx2V3wYOMIIcrUaKJksZDz81gOAJ+FXRzyRbNOEEtXTiMlzTsfmrIoPEaTnsqte+d2HbpIXwnpGwKCF7elxCN2IPkry4OGfMqWPwiw9QGUFZKsPj6A3ClpaBk8ReTjG6wXAdR6RspY6uaJvSw4CClRD4Ly3OQroabxGFwconvdI4ucckqrZnsHS0kBBa9vS4hARhUJJOSr2wyOGWjZBYiEYOFUEAIKKuXeRP5VIzo7HCmAi22CCGMGTY/nVr2eG4Y3WQ5gcSIu6hkhmG7/wBKC57/AIBhQZV7MDYqri3GEEYGThCMHCqCAfkhwgoqhxCoiAd0REBXsd0hWJk+qCR0uRhRoqt7jKCmFVoJICmk8Pw/h74UA2KCZ8HQwPJ7q6GmEjC/0+aidI5zQ09ldFK9vwA7FBG4dLyPRS+MPCMeNyr5adwb4h7HdYyD1d9hs3Fu4yO9ZrAPyCv/ANa9S15Z+w2JNv4yjyE1g/RXr1MQEREHXP2h34GnEv8AxOj/AF6nXzyxs6ifkMr6GvaH7cmnEs//AElR/r1Ovnka4tOQgodiQqtaSNinSTuVVvU4hoO52wgt+tMLIkop4mB7wACkJiDD1d8IMdVB2VXgF56VQtI7oLonhjuopLJ4hyNla1hccBXlnQPi7oI0wfRSQdPV8SyiISMDGUGDhFNLEBuOyhQEwgGVPGIw34u6COPuNuy1mhvDoQKdg77BaREwufsNlrOm7W273qG2xtJllcA0fNBmmOvA8eOMkHfZXEPkhJqB0keq7I2bl4ulosLLzd4SIHsBGRtjC4g19oGvqK0iyscYwcbD/Ug2BTWxtfMWOcAAtPu1v9wm8NjsgrctVoXUdriEga8HGTsVodVR1bWk1mSQg02lcA74vVZslC2VpkGPVaaT0uPT6rMhrXCIsJKDDeCx5b6KeKo6GFpGVBI4ueT6q6NvifAO5QUErg7qBKldVPkHTjv5KdtoqgA9zR0q9tGGP6iNh3CDEj6oSXkED5hWTSukdnJws+t8Is6WYytNx0/SQOooTlD3yjfpDPqgpv8ANMrNPg9G2MrFkxk4wgsQHBBREEr5S8BvfCnirWxRlmM5WGiCr3dTicd1Xr2xhWq9sTnb42QWkE+SzKSrbHhjh3VobE4Y8wOyx3sLSSB2QalP0OYXN7kZWmSA9RwDhSwyve4NJ2WS8QhhyBnCDBjb1FVdhnwqrHdMhPkqyNLj1AbIIjupmQdcfVlQ9leJXAYzsgtIwSPRUVx3+vKt7IJIYvFcR2wqSx+GcKjHlhyFRzy85KA12FUElwwrVKOgNJxugkkkb0DHfCxid8hVJz27Iz6QygbhB8Tseuyy+iNzcsbn7FivHSe6CSWDob1ZVrZQ0YVrpXOGCVfHBI/s3OEFplJysyGgbLEZiQPPCxpWdAwWkFUZVSsb0NOAgyYpmtcYyBtsoXdU1QGBxwThRYe0h+dyr4WyF4kA7boNRrbX4FMJQckrT44nY6yTstTNf4sIhkOQB2WmTy4eWM2CC7qM5LSeyhlj8N2Mq5vVjMffCsf15+NBQNLuyrgN7jdTUjWuccqtRAfE+EbIMdreoqbwAG9WVZ4MvcD8ioXyD4ST9SDKoi2R/QR8lqE1mjdEZg4A/WtKo5RE8uKnnuFS74GOPT9aDCe0tcW5JxsrVkRdB3kG6jn6ev4eyCPJ9U+tEQV6j5bK6KTokDzvhWd1Kynkee2yDMmrhVRiIgDbCxYPgqA3O2d1c+mcwfB3xukcMhOQPiQajc44xTAtwTjyWjta9xwAd1muFQxrfeCek4/Is1kNM+AvZjICDvV7FyAM5otUOzudAVw/5xtyK32LrpDzUaqaew0DXY/zjbkQe0iIiAiIgIiICIiAiIgIiICIiDjjmS/B24p/wKvf6jMvmiJ6ZifRxX0u8yX4O3FP+BV7/UZl80En9kf9Z/Sg1k3dhpxEANgtJml6yTjurGYzur5OnfCCIZPmqkYVOymi6cfEghwinf0Y2UCAgOCD6IqgZP1oMoVg6C3HkoCQ/dXOpy1nWoQSEFzGdbsKskfQRv3VocQcjujnl3dBRERAUscxY0tHmozjHkr4mhxAPclBY4l5J3JKqGOc4NA3KzfcHRtEruxIW+NJ8IL3rKIy2uN5JaSMAlBpehtJVd6rGe7QmZxcB0gZXeLgDy33O5XSiq6m0vaMBxywrbXJfyxahl1o39kMDhAJm92HGM/Newul+F2mdM01PHRUoD4mAZAAycboNA4PcNqXSNkiYaVrHjG+MHsuUlZHG2JgYwYAVziGgk+SChaNyV195inWah0/WVM1xjY9rScFwXKWqtf2+zRyRBzesMJz1DYryr54OOWsX1FXSWuV/gl7h8Lj2yfRB1x5j+ITK/UMtuiqxIzJaCHZ2yuA6OnYfFlJ7kn86sv10r7pVirryTKdyT3WLT1rmNLc90ElXI1riwYUDZg0EYVkjzK85Vj2kHsgvik8OUuPbzVamUSuBHZQogu68twkQ6nhvqrQCTgKWOJ/WMINWfEyCmD+5xlaRK/xHZx5rU5mTtg++dvqWmu6QCG/nQSwPyME7KWWlEnxhYcfV1Yb6rUo4pjET8kGnsd4Mpz2CyH1zXNIDViy5Ehz3VB0477oHxPO5O/qVlRVIgZ0eZCxAcHZVDsuHV280CRznuL3Z3WTSVXgjB/Qpn+7+AMYzhYTQ3Jz67IM83BoySPtwsOWQTyjHYlWyFnTt3VI8AZPcboMx1tHhCQO3I7LFMBac57FVNZNjpzspIX+I3pzudkFpmHT0eamj6PDPqk9nrYI/Gcz4O4KxGSuBwTsUF8bCZifJKloaQAVe+VoblvdYznFx3QVjb1uwrnsDAqBwaMg7q1zy7ugod0GyIguLs7K0jCqzAKuy3qyUFoyD2RxJ7rNZ7v0DOMqCbo36UEcT3MOWgkqsr3vcOsELLs/gCoBnxhSXt1KZh7tj54QaeY8NzlWtHUQ3PdC44wUG3mEEr4vDYHeeFfDTCZpOcYUDpHOGCrmTvYMNQSMpg55aqTwCLzUYmeHdQKo+Vz+6C1ZMNUImdGFjtaXHAR7HMOCgPPU4n1RoyqKoa47hBc5nSM+it6j6q8B0nwjdGs6Jmh+2+6C6nlMMgeQSFPPWCUYaFnV0dKKIFmOvHktNpRGQS5BjnJOcd1LHTlzS47BZ9BFC6QhwGMqS4Uxa3MPZBo7hgkK5seRlU6XF3T5q4h0ex2QWOHSVQboTkqrfpD0yguZGT3VHs6SsmR7BH8PdYpcT3QUREQEREDJ9UG6K9vT0/NAczAzlViYSQ7HYqwuyd+yyonM6C098ILp6lrovDHcLCU3u8jiSBsrHRmMEO7oPVn2Gv8Audxl/wANYP8Ao169TF5Z+w1/3O4y/wCGsH/Rr16mICIiDrr7QpvXyccSW+tHR/r1OvnmgiEjjnyGV9DHtDXdPJtxKcPKjo/16nXzzwSFhPzGEF8jQwYUULumVrj2ByqvbI4kkKPBHcINbuFfHLSCNgHpstKip/EaXZxhIcyHpPZVkc6I9DfNBER0O23VC7O2FkMhcB1yDYqOYM/eILYpPDOVdI/xnjHmVEqt2cD6IJpIehnWFE0EknPZZb5WOiwViggPHplBcZC4BuPtURGDhZkph8MFuM+axQxztwNkFoyN8K/pLgSRhSUwYHnxAqzub4oaztlApHlsjWkdzhcocMLAykv9NqGRoLIHB5BG3dccObE1jHNIyO65G0lqqloLW+KRwBDD37oO91l4oWniTpaLSdPCwSRMEflnstm3Dh7JY68UzrWZGPd36M91xvypy1epNTBtACcyjsdsZXf670GlLVaWQ3oRitLBjOM5wg6d660RbILSZvdWNeWZI6R6LqXr63MhlkbHEAA4j6l3T4tUVyfUzyUwIpiCW+mF1M1lHE6ofTyYLy8hBw2aFzpCCCFjzw+C7AK3leLBPQsNUBhhGey2hM/rlJd2B3QRxQmTc7BXj/a0zH9wDlZD5IxEBH381BHSz1DgCDv8kGqu1B1QCJsYyPksN9YXtIIwVqFutUcXxTtHbO6xrjSNfOGQdsoNOy7ryeytkGdx9a1Kqtc1PCJCDgj0WNHTl0RPogxY29YLT5KkrAx2AqtJjefkqyOa/f5ILCwhvVnZWqpcTtlUQEREBEVWjJ3QUUzJuhnT8lKKfrGyx5YzG7BQXQ/E85PzV0rw04ChDi3sqjqee+6CWkGXnKVQ+MYOfJWjMZGD5KpcSQe6CItcO4KvZL0tI9UyZCchXCMAeSCI7nPqp46brj6jsoizpd8vkpnSENAGyCBw6HEd1ad1kNjY8ZKoYQSA3dBABlMFTOY2M/ErmFjwQ1v5UGOqtHU7Cq9paSrQcbhBPJCGAH18lSmgM0nSNlY6VzhglX0rnNf8PdBqrqWOnh3IzjyWjOy5/wBZWRUVUrnhrjsPJbm0rw213xDkdR8PNCah1NWNx1w2e1z1sjcgndsTXEbAnt5INqMgcSCNwO61a1TU0UuJyMDyK3pqflw5h+H1AbvrHgdrqz0AjErqursFVHAwYJ+OQs6WkAEkEggDJAC4yc9znF52yg1W/wA1LJKPdsYPcBaQMeavZG6TO/ZUDCX9B7oKl2cBSMcWjGFZLH4RClDQGB47oLoy0gk7LHePiOPNZtotF21Fd6KwWG21NwuVyqI6SjpKWMyTVE73BrI2MaCXOc4gAAEkkALkLily08deCNBQXXixwzvGm6K6SmClqKxjCySUN6izLXEB2MnBwSASOxwHHNI2MA9ZwVFUjqkOOwV0sRj+r61GgrEHNOWqYSOJ3womOxgLIdBhnidkFWyd87rGnaHOz2WVDEJBnz9VFIwtJA3QYpbg91Ox/RGCTugiLiVE5pHfyQWvd1O9FeYmdHX17qMgk5V0cZkcG52QGRPkz0gnHdZUNHHJtI4gqSBzKb4CM9SlLTgzDZBjS0fgqWGVrWYIV007HQ9J8ljNBLC4IMrxGny+ah96bFMHY2ChbLgkFQynLkGse8MujfCxghQTN9yHhh2Vp0U0kJLmHBKufO+ZwMhyg7/+xbl6+abVP8AK4/8AONuRXexbDRzRaoI89AVv8o25EHtEiIgIiICIiAiIgIiICIiAiIg455kvwduKX8C75+ozL5oSOuVw+ZK+l7mP/B44o57fsMvf6jMvmgkJEryP7o/pQWuGCQqITlO6Aqgeau8F4HUQrW9wgoizHxMERPmsM90BVGxz6Kiq3BcB5IJ31ZfH0dPlhY61FrIPD+awpg0H4cII0REAboRhVa1zjhvdXOic3uEFoBOwyVdGC1wcdsFZtIacDL8du6irnR5xERhBkT3R09MynY3fI8l6D8i2nfujSQe82gShzBklufJdA9HWpt1u0UDhkF4C9tOQbh3Y6PScU8kTHSiAEDAznAQc2cKtE2y11HvMdoZC4OByGALmnssWlttLSDELMLLQadern9y6U1GM4ytr3LiHZqfTdZXVVeyJ7GHAJAOcLVNe3GltdjlqqvHQ1p7/AFLzH5neN2ooW1tHpyR4gDiCGOPbPyQY3MZzZyWLUtRS0t2JYHEDD9u66ba940/s594dUTdZJPffO62RxG1FW3utM9y6jKc9WTvlbAZI6PPQcZQajXyNranDfJY0tH0DOfJWUsobJ1PPdX1lSXkBuMY3wgx2noccqjnF57FXwQSVMnQwblao2iZSRET4BPqg0ZXMb1uwqzdPiu6e3kjI3n4m+SDJbTBg68g4V7JWNeM+SxDLKR0FXCmnc3rAKDWampjmp/DHdaHLH0HCkhlcx3S/sqTkPd1BBZC/odlZ4unQzoAyFp7WOedt1kwxxNjPijdBD/Z5ie2TlTOowGl2ewWO5wZISw7K9rp5Bt2+pBE4dJI9FLFB4jSScFROa4HdXNlcwYbsgo7LT05zhWk5OVLHG6d2G91SaB8Jw8II85VWDLgqIDjcIMw04dGDkA4UDCYZWu7hpBVBLK74QU8GQjJCDXKzUbKmgFKG42x2W3zuhBBwUxlAyg3ITuq7hBI+HpblRK4yOI3VBj5IKIr3ROaM42UkXR0fFjsggWVT0gmaXZ7LGOMnHbKvZPIwYaUFZoTETvsos5Vz5HPOSUEb3DIGyC3JHms+jt4qYzI53YeqwOyljqZY29DTsgtlZ4chZ3wVYqucXkuJ3VzInvOGjKCxFlx0joz1S7DGVHP0GUdA2QQYPoi1CaKIU4IwDhaeBkgDzQT0xDXZPmlS7qcCBsFaYZY29RGApYXRmM9WMoI6aETP6ScK+dgp/hG+VEJHRyFzNt1QyGV4MhQXQy+E/qIUz45Kn741pwPkrZRGWDHda/aX0TaBwkx1EfnQbcfLIR4W58lYOtnkQs+lEJubuvHRlZlxFG14DMY88YQaPHUPZg5Kz47g1zPDcck+qhqhAY/gxnCwQS0oMt7BE4y+RUE0niOyrXSucOknZVZG5x2CBFGZDgKssRiIB81ltMcbc7AqN72POSgxMn1RTylhHw91AgIiki6cjOEEeMIsufwug9IGVihjj2CCiDdOyIL3R4bnKtDiCChcSMKiDMjrgxuCM/YoZ5BN8Q8lCmUHq57DX/c7jL/hrB/0a9epi8s/Ya/2hxl/w1g/RXr1MQEREHXT2hozybcSh/8ASdH+vU6+eNvfPovoa9oftya8SyO/udH+vU6+euk6Ot3Xjtt9aCpnHTghQOd1FTyxYJI7KFw32CCsUnhuyqyS9T+oD5qNVbjIz9qDIlqzJGGYwsYHdZcwhEQ6AMrEQVIwqYPomT6rJh8Pw/ixlBDGzxD05wFV0fTIGfNW9Ra49JVC8l3V5oMiWk8OMPz3UlMAYySsZ08j29JOyzqRhdTkj0QYEhy442xsrRnv6KsoxIVagzrdF7zIWPOAErYpaacxxyHB8srEimfCephwSs6mcZniSbfCDsXyma9bw+ubayoI3fnfyXY3iNxVl1dXxXSnrXMYwB2A7ZdINH6gpKB4aSPh+a5Ji4lUDWCmLgCQBjKDstYNYUetITaql4Ja3pLj+RddeN+lKaxXkzU7wQHdX51daeITbFKaumfgv7brbuqLvd9cXATFxcwn1yg2Xqa/mooBSiPGG4zhccujle44Y4jPkFy3qzT0dLQNc5uHgbrTLXQ2dtokfOG+IGnuPNBx5HTluHOO/mFqNPOyFodjssC4TAVsoiPwB2ApoHBzM+SCervJcPDaMD5Lcml7XTXCEVMzh2zuey2bO2J5Jb3HzU9LeqygjMULiB27oN0arkp4ITHG4HGy25RTNfTu6husOavqKx3392eoq0zGIFjexQQz7Su+tWtGXAeqvja6V2+5KSs8NwGPmgnNHhgcN8hYvSQ7BCniqXdnnIWb7kZoTMwbAZ7IMGWEMZkH5qENJGVfI9+SwntspqUx5Ad5jCDFwfQrJhpw47nCz5hTCMEYz9i018rg8hh2ygyzKKcErDnm8Z3UBhXS9RYCcqHpPdBPDRvlb1D8iq+mkjz/AKlJR1Zj6WeXZZlSWOj6sAEhBpZaezhuqkYUojdI8kDZSOpy1ucbj1QYvS7BcAdlGXOOxJWTJHI1uQ3ZYpzndBK146cHvhWOJzhVbGSVc6IuwQUFPj6cg7K+me7qO+VQPaG9J7q+Bm/UMd0FkjZZJMuacZVzmtgALTue6zZaiFsfRgZx3WluJcSUE+GyM6iQCrGRNdnJVgDvLOEIc3bJQTso8tLidh6KFjzC843WVDMHQlmdzsoWUz3ShpGxQdoeQDlGh5t+LlRQ6lqKmm0bpaCKvvslOcSTh7iIaRr/AN4ZSyQlw3DI34wcEeh3M5z58GeRKWm4CcGuFduuN7t1PHLPa6FzaC32sSMDmeK5jHOlmc3oeWgAkODnPycHjX2JV2skOn+KWlx0MvLa221787OlpjHKxuPUMeHZ9PEHquontKuDHEPhzzT6w1Pqm2Vcll1pcX3ay3XocaeoieATAHkYEkR+AszkANOOlzSQ7U8NvbaR1V+jpeLPBcUVnlIDqyw3AzTU+4yTDKGiQYO+HtIxsDnC6b8+HEbgtxZ5hbxrXgbR0kOnayjpOqemt7qJtXV+GDPMYnNaQ4ud0OJaOosLt89R69CikZH1OBG2eyxHdQcQSQgNLx9HzVA5zXZ816Scn/I3wW4icluo+PXEzS15vOoZ2Xiexsoq2aIuZTRmOGOKKMjxZH1EcjQCHFxLWgeR3Fwo9mty+cEdFW7iTz4cSKG3VdxIEenzdhRUkRJB8F8sZE1RKBguEJaG5O7gA5B5dkvl+eE63N+Er2si5CPZ68znD6sufL5PRUE8LDSw3nT14qak0lQB1MFTTVEjge4Ja9rHuadnDYjyB4ucMdS8G+Jeo+F2r4WR3bTdfJRTmPJZKBgslYSASx7Cx7SQCWuBIB2Qbr5VuL1l4CcfdH8X9Raalv1u05VTTTUMMjWSP66eWJr2Fwx1MdIJADgEsAJGcjsDz/e0EtPNtYdNaI0Toy6WGxWSufdamS5zxumqqrwzHHhkZLWtY18u5cS7r7DG8XsxeVDh7zP661j/ALKdsuFdYNL2yncIaWpfTB1XUSuEfW9hDsdEM2GgjJySdsHhbnL0Fw64XczGueHnCulqafTdgrIqKniqKp072Stp4jO3rduQJjKACSQABlBwu55f3VMeq9B/Zm8lXBzmK0NrniHxrtVdW26018Nrt3g3CSkjieyEy1L3GMguIbLBjJwBnY5ytw8DvZpcMbRpOTj1zeayOiNHVjzV2vT1RcG0ksVJLl0Da2peARKWH+wRgSZAy4OywB5sAYUviuc3pPkvajS/JV7M/mU0rc7dwSpqCSa0f7XmuOn7/WPq6F7w4RueyokeHglji0yMcHdJ6TjK8muYjgre+XnjNqfhDfaltXPYKoNhqmtLW1VNJG2WCUDyLo3sJAJwSRk4Qceslczsroz1v+Jbm4W8L9b8ZtdWnhxw8skt0vt5m8KngZgNaACXyPcdmRsaC5zjsACV6bWH2dHJtyxaNodWc5PFBlyuVYMOpTXzUVE6QDLo6aKnxV1BaCAXAjOx6G5wg8rHmFjMDGSFgOaXvJb2J2XsTozl39lBzUT1WjuDckdPf46dz43W2tulDWNY0bvjirh4c2M5J8N5AAzgArzy5yeVS/co/Ff9hNdc/uvZbpTm4WO5mMMdPTF5aWSNBIbKwjDgDg5a4YDgAHAbYwwHJ7+qt6ulxLQF3A5FOQa883FdW6t1Lep7Bw/slSKOqq6VrTV11SGtcaen6gWt6WPYXyOBA6mgNcSentfeNA+xg4TXOo4d6uuVtr7xC4UtZUPr73XubICQQ6ekJgicCSHdBZgjfBGweSPUXuBcs9xPgdLfzLuvz78sHKlwx4Y6T41ctGvff6DU93NsZbIboLhTOiEMkr5o5DmVjo3CKN7HuJBlbs0g56RMqQ1uCPsQYszXgbpHP0sLT3KkmlEuwG/1LHMLwOojZBa52TlGjrcAqtjc44AVehzHhvmgyoqBrxnPkseeIQuABypS+eJoO+FAXOlcOooO/wB7Fd/VzSapBztoCu/lG3IrvYsQlnNJqhx89AVo/wCcbciD2lREQEREBERAREQEREBERAREQcc8yO/LvxRHrou9/qMy+aEt6pXt+Z/Svpe5kfwd+KR/7i73+ozL5oHnEryP7o/pQWuGCQqDYgodyiDIfUNLOjHljKxxsiIJnT9TOkqFACeyq5jm9wgonZMH0TGEF/iu9SqHLvNWqrSB3QUIKqAMK8luM+ajyewQXxv8N5OFJJOJMDGFZE1ucv7KsrWdQEe5PogvdDhnU13llQbuONysiSCojhEjvokKa022a4VcccbcguGfqQb24QWqtqL1FLFSOe0yNGcfNe4PJXbKin0vDLNCYumAbEY7gLzo5UOHlkrJIBXNZ1h48h3yF668ILBb7HY6eKgADTEMgDHkg5EUVRKIIXzO7MaSpVpmoTILTOWZyGkn6kHWTmj4vmjsktohn6CGuzjbyXmLrfXbJI61tTVdZc44yc+a7c832oIqKScSgAhrl5ha31VUVdbUxwE9PiHfPbdBo2r7kKy4Pe0jBJ7Lbud9ldJK+Z3U7cqrQGg9WyCPf5qaCnMzgCcAlSQRtlf0gLIkb7uAdgglgYy3Hx+59Fj3G5GtII2+ShbUukf0yOJBVsjI8joCCDf5qeKYNb0Y77JJ0Bm2MqFrScIMgwhuZCPNTx17WxlmBlYr5iWdOUhY1zST3QRyO6nl3bKpv6H8iukYWE7bLUqCGJ9OS7GfVBp0MojOcJNKZHeYCrLH1TlsfZWvjdEQHjCCxTxVAjb0gZUJI9FcxgcDsgrJIHuJ9VGq46nYarjC/vhBJSTiB/VjKurKoVLsgYWMdiiCpAVMH0TJU0fQWfFhBWlAMg6uwKzZ3xsb2HZad1ljyWKrpnO7nyQABLJ37o9nhnBVIndJykrutwQWjYZTd3YIQ4DfssiFjCwnAygjEQIyrCOlwVXvcHEA7A7K5sbpAT3QVfP1M6Qocn1VSMEj0VEBEAyhGEBTRzBjen1UKILn7kn1KvZD1NJ3VISM/F2WR4kbdgQgxC0gkeimp5xFgEK4tY4beahkb0lBl1FwE0ZY1uD6rDZnqHfurVPEWBuTjOdkE05+89/JYbT0uB9Fc+RxJGdlaGkjKDKkqhJH0AeXosXcbKrQA7fySTBOyC1Va3JCuj6c/EryB15b2QHxkNBVrZ5GN6Q44UssgcwAd1bFEDGSe6CNkjmO6h3yj5HSHclWuHxEKiDIgi8Q4LtvrV81I1o6mrGbK9nYqX3lzhhxQWRRdby3PZSkinw3vlQB5a7LSq9fXu47oDnFzu6tOR3Q7HZXNjc7c9kFmUAypHgAfNRjOcgIJXsAbn1UWVUvce5QYx5IKZPqpGSBrcFRogHckoiICImEBEwmEHq97DZuLZxkf/dT2AfkbXf616lLy09hsSbdxkaewmsBH2iv/wBS9S0BERB1z9od+BpxL/xOj/XqdfPGCQvoc9od+BpxL/xOj/XqdfPNGMk/IIJmS+IOk91ZKwM+1WRuDH5V8uZMEdkEKyIqbxGdZOPNQswCQ5SGV7Bhh2QRPHS4tzlVDcjOVQnJyUAcRsgEAKmT6qpz2Koglp2dbsJURiN2FWmeGOJKrVPEj85QQYWVDWPii8MDbGFCenpGO6z6b3X3ch+Ov5oNNe7qcSqK+VuHkjtlWt7oAB9CpxUFjekbHGOylhZE5o7ZWPUNDX4CCkU8sTg5jyDn1WZTSVM87ZjOdj6rT1LDM6N43OAUG65rnPJTNibMct+a1ixa8+4sJif8TgO62e2tiEYz381fQNp6utYx4BBOEHLGmrl+zyZ0E7cN8s7LQde2CPT83usEuzu+CtbscDLFRCooQA4jORstsajpr3fq4VMgcWg580Gz6uzOYwzd87rT+qSEFoG3Zb4rbTUPpREM9QGCtFltbYInCUHqPqg25lzXFx33QHreAdsrLZQvbKesfD8wrKqkc09bOw7oKS07Y4+sEbrFcS4+ZV753ub0OOQFe0M6N+6C2GURlUll8Q5VGROleWsGSqSRPiPS8YKC1alR3PwYfBI77brTj04GM5VCMbFBkVEQ3kac53x6KAZzhSxlx/shOPJVNM9w62DIHmgteXYx1bKMHBB9FNK4FobtkKFm7gD2ygyBKZcMI2JWRLE2OMOyO3qrWxxgtx3JV1ZC8sbgnGyDEjYfEyMYz3WbI1j2ANkGcdgq01vfURhjThysns9ZSfE/YILm9cbcNBx8lHJM5vcqeGZgwxw3xjdd0OR/2dOp+ZKvpOIvEqGrsPDWCUPa7Bjqr30kExwZwWwkEh0+/YhmTksDgThLy08Q+LPD/WXFeCAWjROh7VV3CvvVXGTFPUQxdTKKADHiTPcWN2Ia0OBcQS0O4XeBnYbr3G9ppUaa4IciNZw70PaKSx2q619s01b6OkYGMhiEhqXNA7kllK8EnJPU4kkkleIFNTlzsuKCrIT4ZcR9ii8UNyN+6zXPAb4Y7rfnC3lq468b61lNwq4XX6/xyP6DWQ0xjooznGH1MnTCzfP0njsfQ4DjPYnOylhk6dsr0n4N+xW15dpKe48c+Jdu0/Sn4pLZYGGsqyP7l08gbFGe5y1so7eu3eng57PXlP4JugrdP8L6S93aDBF11E77o1HUOzmtkHhRu+ccbCg8R+EPKdzD8ep4Dwy4VXu50UxDRc5Yfdbe0eZNTMWxHA3IDi4+QJIB7ycG/Yo3qqMFy498VYKGIgOktOmIvFl3OcGqnaGNIGxAieMnYkDf1bnqKO2Uj6iqngpaWmjLnySODI4mAdyTgNAA79tl1j4x+0o5TODslRbp+IA1Zd6ckOt+l4xXkOHcGcObTtIOxBlyDkEZGEGxOMXs1eXaw8tOuNN8KOGsDdWMtElbbbvVyyVdwkqqciZrGSPJEfieH4REYaCHnI3JXh5MRkgeS9HOMHtnOLOo3VFt4M6GtGkKR2Wx19xP3RrseTw0hsLD/wBqWSD5leddxlnrKuevqSwyVMjpXlkbWNLnEk4a3AaMnYAADsAAgsp2taOolZTZmNw8DcBYDHEDAU0bx59kHJXArjzxB5fuI1v4n8OLiKa40WYqinlBdT11M4gyU87AR1MdgdiCCA5pDmgj2P4G85XK3z06THDDX9mtVHqCvYG1WkdQtY9lRKAR10UrsCUjJLSzpmbgnpAAcfC4TwxxlrcZKW+qmo62OtpaiSGeF4kikjcWvjeCCHAgggggEEHIIQegHPz7N+XgTYKvi/wVlrrpommd1Xa11B8Wqs7HOAErHjBmpwSASR1xjBcXguc3zxkGQXFfQjwN1Pc+LvIfar/xclNbLfdEVsN5nqm5dVQCOaIzSAghxfE0PJIOS4nByvBzhRoWq4jcUdI8PYYnPfqO+UFq6WnG087Iyc+QAcST2ABPYIPd7hBdtOcpfIXpTUusIhFR6R0ZTXWtgDg0yVs7RMYGk7db6mfw2n+6cCvDbj5xy19zD8SrnxI4g3eWrrK2RwpqYPcaegp8nop4GEkMjaDjA3Jy4kuJJ9UvbOa4bpvl60jw5t720x1NqFsr4WYa11JRwuJYB2AEktMfl0heNDNnA/mQejPsTLrd4eOGvrJE94tlXpRlVUN68AzxVkTYiW43IbPPg5GMkYOduLfawwWuHnW1JJRNxNParTJWnpxmcUjGg5wM/e2xDO/bGdsDtr7F3hHXWXQ2tuNFyp5IotSVUFmtfUMCSGm6nTyD1BkkawHyMLwvPHnQ4i03F3mg4jcQLfUiooa68vpqGZrgRLS0zG00LwRth0cLHD5FB6X+xq0ZT6c5edWcQax0cB1FqN8YkdhoFLSQMAcXHGAJJZwc7ANzncryP4pasqeIvEzVvEGfqL9S3yuuzuruPHnfJj5Y68Y8sYXsvoIO5dfZOtufgCmrf9j+quTC4gOFXdeuSFzz3JD6uIYO4DQ3bAx4lUzjO5sDGFznEANaCSSTgADzJQe53s09P2fhPyJ2bVWppIqGjuQuuqbrLMPgjpxI9okdtuPd6eNx+S8neb/mq1jzVcVq/Vd2rauDTNFPJBpyyulPg0NIDgPLAceNIAHSO3JJAB6WtA9ReeWf9r77NqDhtRy+7VclpsejIy0gFzgIzUA+pfFBOD69RK8Ren1QejHsUKG5y8bNf3SCSUW2n0syCojaB0Onkq4nREnvkNimA+RdnyXC3tRb3Q3rnY12KCTxG0EVsoZHZ2MrKGAvA+oktPzafLBXdX2KehfuXwd17xDlh6JNQ6ghtrHEbvio4A4EfLrq5B9bSvL/AI/awdxN42a94h+8ieO/6iuFdTuDstED53GJoPmBH0AH0AQeqPsluANo4a8C67mLvdrnqNQaxZUCk8OmMk8Npp5HNEcTAC9z5pYnPIA+MNgAG2T0f4+cPeeDml4tXriLqXl/4nAVM5ittFV6eq4IrdRdZ8Gnj8RjWgNBy4ju4ucd3Er0+4ka+vnKb7PK26z0BTUM100dpTTtLRtq4y+B8kslJTvke0FpdnxnvIBGSV521HthebeaB0UUGh4Hu7SR2aQubgg7B0xG/bcHv67oOZPZ6ez249aA4zWHjbxdtkWk7Zp4VE9JbZKqOWurJZYJImhzIy4RRgSEnrIecAdOCSOJPa3cfdEcX+Ndi0hoO6wXak0HQ1FFXV9O4PhdXTSgyxMeMh4jEUYLgcdReP3uTwhxZ57uazjTQ1Fm1nxducdoqWmOW22pkdup5GHux4p2tMrT3IkLs/UABwFkIPdH2c7LTrD2fdp0voC8x268upb5a6uqYfvtFdJZ5y2R5G+QyWCQdz0Fg8sLx4438uXGXl61JLp7ivoqvtbzKWU9w6DJQ1vfDoKgDokyATjIcB9IA5AzuXnmh4y8sGpZdRcKNSCljregXG2Vcfj0Ne1mekTREg5GThzC14BIDgCQfXvlN51+FfPlY7twk4icPKOj1FHbnVVzsdY1tZbrhSh7WPlhc8Aghz2EscOpvUC1zsFwDwt6XN7hXRhpd8XZdh+fXl+sXLZzG3jh/pOSU6fq6aC8WmKV5e+np5+rMJcclwY9kjQScloaSSSSeu2+UF8vQx2WKx1SS0tAztjsq9OT6rJpqRheHHBAKDCikMTuotODujpC+QPx5+i1eqp6d0eIwMhYIZDF9PGUEc8/XH043UEYwQ7PZZMsbJtoRuoH00sfcIPQL2Lk4k5otUMz20BWn/nG3IofYrxvbzS6pJGP/gBXD/nG3Ig9p0REBERAREQEREBERAREQEREHHHMl+DtxT/gVe/1GZfNBJ/ZH/Wf0r6X+ZL8Hbin/Aq9/qMy+aB4zI7/AL4/pQWoqhpPkpW0srt2jKCFFI+B0eeryVrGFxA+aAxxYQ7CrJJ4nYLIlYwQ7d1jR9O/UgoDhUJyrnNPcdlVmMboLFkwUolZ1ZWMcZ2WTDMY2EDsggkb0OLfRWjZXSHLiVaASguL8jCowlrg7GcbqizabwPCIfjJCC6e4+NTiHpwR8llaevDbZVMc5oI8ytKliLXEgbJCwuePTOUHohyKzUeqL5BBUzBjfFHc4HcL1909ZqW0UMMNK/qa1gAP2BeBXKzr6t0zeITbnlrxIOx+a9qOAfECo1JYIDdJMyviadzvnAQcyLStS1UFLYq6eWRrQyFxzntsob/AKpt9gpnVNU8YAJ7rrvxc48Ukmmq9lDKMFpacO8kHnTzscUpKvVdVbYagluXDY/NdGZal8k0znkkvcSfyrnbmL1TR3zVtRKSC4uP6VwHJgyuLe2UF30fiAV7YzO0uOyla1nR8XdVjI+ixBjwTGnlJ9FfUVT5+w2CVMBaOs+ayqAU3hHrx1IMCNvW7B2KyxStDSSc/aoZm9UxEH5lRzp4gWvzuggeOlxHoUa7CuDHPJKoY3A7oKbndXMkLNsYWREI8fF381DO0AjpCCkkvWMYV8VU+JvS1QLNp4WPaSQggik8OQyHz8lWeYVD27YSfpBIHko2xvI6ggnkpAyMPBOSscPwOyudPI4dBOyjQSQ/SCnlqGgdOO6xo85VZAc7oLTuSfVAMnHqpKdoc/BUk1O5jgR2QWvpulnWCoFlyTAxdBP1rEQMJhS0/T1fF2Sbo6x09soIt0B3WTJ4fh/NYyC5z8gDCqyUtGArFc2NzhkIKE5JPqpYp/Db04VjB0vw5XP6EEb3dTi71VE+pSx9HTv3QRsd0nKPd1HKudjPkrRjrHogpg+iljh62lxypJRH4e3fCgEjgMBBQjBI9EDSRkFNymSNkF0bywqkj+s5VG4zujgAdkFEREBXh/SMEKSl6Ov48YUlR4J/sfmgxDuSUVxjI3xsqNx1D60FMH0V7ZOkYwsmQw9AAxnG6ib4ZbjzQQZ3yVKJsNwFa6Nw8tlYgHckoiICIrgwkZCC1E7IgKVkoY3px5KJEFXHJJQHCormxOcMhBad0Q7FACUBEIIVQ0kZQUVWjqcG+qdJVBsUGXJSNjj61iE5OVKaiRw6SdlG7ugoDg5QnJyiIPVz2Gv+53GX/DWD/o169TF5Z+w1/tDjL/hrB+ivXqYgIiIOuftDvwNOJf8AidH+vU6+eRjsH69l9DftDvwNOJf+J0f69Tr55GjLggq5pxnCvZL0jBCyRG3w+3ksSQAHb1QWE5JKuD8DGFaiC9jPEdj1UpHhDCtpnBjsnCuqT17goIHO6iqIQVc2JzhkBBamUIwcJgoL42dRVekiQNBO5VgLmdtlI2KWTLwOyDKqKdscIcDk+awWtyr3zyOHQ45AUYJCC9jywqcRCcg9vqWM3c5KmZL0Yx5IL30gZ5lYzm9JUslS5+yhJygZPqVmUD3U8gqN8jssNZdIMkdf0fNBuSHXEzY2wOJAGB2W7LDqumqIhC/pJPzXHFTFRysAhA6/ktUsFkuEkodEDgeiDkOp8IDxmNBB8lPRaSivLBM9oGfLsrrLp+uqI2tmBwcALfVt4aamqouq3Nf04zsCg4k1fpR1uhPukPWR6BbSoKZ0pMNXGGEnGCF250Fy/aj1NUyRXGN7g3OcgrhLmC4Z13D+9iGnY5oa4jthBxhe9PRUMJnY8HIyBlbeDHkbNJH1LWbg65yU7GzkkYCzLZT0bqF7X48QggeuUGh0LnMkBazqJPZby01o6bU1xipZYC1jyMk7YWl6ZtHTcg+rbiIu7nt3XZbRHD997pWiwt/2yW/Dgb5+xBt6u5Z7db7M24wyte8tzjqBK2PJwcqXTdfuh6AcZxsu02ieXHjZcqsNuLak0bnZblpxhc0VXKLqf9jz5GQP8UM6iek5zhB57VnCulgg+LpDgOxIWwb5QR2kup2YOMjK7Pa25a+K0F6qY4Y5vCbk4wRsuuvEHR170rWmlvTXCQOI+LYoNlik6n9R891jyM6JekeuFqhop44xJjY9tlps4d43zQSv6m9PTk+akxWTgBkZIASORrD8e/butbt14oIGYlYMj5INIoKqppKlrZWkDPmtduMxrKYCJuThYM2b3cYKC0UktRVVUrYYIYYy98j3EBrGtAJcSSAAASSQAvWfkI9mlDoaG38XuYq1RVeoPgqrTpqdofFbjs5k1UOz5h5REFsfc5dgMDifkM9mJV62dbeMvMZapqTTuWVdo01MCyW5D4XMmqR3bTkE4jOHSdzhmA/1uoqKkttHT263UkNLSUsbIIIIYwyOKNoAaxjQAGtAAAAGABhZCIPO72suiOKnGiXhpwg4UaCvupqoTV16rhQ0r3w0+BHDA6WXaOMHqnGXloG2DuVwNwb9jNxi1Aae5cZtfWbR9G4h0lvtw+6VdjzY5wLYGE9g5r5APQ9j7FLgbjLzxcsHArxqXXHFW2S3WHY2m0k19b1Z+i5kPUIj3/shYPn2QbU4PezZ5TeD0lPcYOH/AOyu705DhcNTSiud1DcEQYbTtIO4IiyNt12cggo7bSMpqWGGlpaeMNYxjQyONgGwAGAAAO3ovKTjJ7ay81bprbwD4Vw0UZy1l11PJ4suCcBwpYHBjSBuCZXjJGQQMHo/xp5seYjjs944mcVL1caGVxP3MglFLQNB7D3aENjOBgAuBdjuSSSQ9uOL/tAeVTgxJLQ37ifR3q7Q5BtengLjP1Du1zoz4Ubh6SSNK6S8a/bJa3rY5qDgdw9t1jheOllyvrjV1Xn8TYWFsUZ7DDjIO/qMeYbagtZgE9XqrHTTSfSeSg5W4t8wnHLjrLJV8VeKN9v8bndYopagx0UZzkFlNGGwsOcbtYDsPQLijL2EjHZTx1Tmt6HHbzVDLG52wQRBr89WNwr5JHyABw7LOija5nVgKCo6QTgIMUscG9S7J+z54fcE+J/MfbdJcepbe7TtTbaswUldXPpIqyuLWthh8Vr2EOPW9zQHAlzABkkA9dDNGIunG6pThucnsUHqlxf9ijS1t5kuXAviuy30E7i4WrUcD5fd8ncMqYQS5uCQA6PIAGXOJJEPCH2KtVR6ip7lxt4sUVZaKaVsj7Zp+nkD6toOeh1RKGmJpxg9LCSCcFpwR0W4fc13Mdw3t4tOiuNurrdb42COKj+6UktPE0YwGRSFzGbAD4QDgY7KziFzV8yHEu3us+s+Nur7jb5GGOWi+6UkVPK05yHxRlrH7Ej4gdjjsg9FPaGc8vDLh/wuquVzgNcqGvuNbQNsdzqLXIHUdltrGCN1Ix7SWukfGPCLQSGMLwcOwB1B9mPolmtucnRs0rC6nsEdbfJgBuDFA5sRz5ATSRE57gY2JBXVSCGPALgMr0v9ipoh1RrXiVxIfCAy3WujscEhG7veZnTSAH5CliJ/75vzwHcbm04AcE+c6kquCd41jT2viFo2Fl2ts8IElVbo6oY63Qkt8Wnk8Joe0EEGNhy09OejmjfYn8Wn6vgj1/xZ0jT6XZMDNNZ/eZ6+WIHcNilhZHG4jbJe8NJzh2MHr3z0cXNT1vPDxE1xo/Ulys1xsF5Fnoqy31clPPTGhiZSu6JGEOaS+J5OCN3H1W07tzvc397tUlor+YLWXuszAx/gV5gkLcYx4kYa/fsd9/PKD0l55+afhlyicBY+VrgRW0rNUzWv7iR09HKHusdA5pbLPM8DapkBfgEh/VIZTjA6vH3TFmr9Y6ntGlqD+27zXwW+DYn75LI2Nuw3O7hsFptVNU1lTJV1k8k887zJLJI4ue95yS4k7kkkkk7klZNruF0sVxo75ZLlVW+42+eOro6ulmdFPTzscHMkje0hzHtcAQ4EEEAgghB7Qe1p1Db+G3JtaeGVnlfFDfLxbLLDT9W7qOkjdPk9gQ11PAD83NOO+PKzlN0N/skcyvDPRj4xJBX6loXVTSCeqmikEs4/ycb/AJeuy2zxD40cXOLZo/8AZP4l6l1S23gikZdrnNUsgJADixr3ENJAAJABOBknAWmaQ1LqjQt/otXaK1JcrFere5zqW4W6qfT1EBc0td0PYQ4Asc5pAOCHEHIJBD1I9trrhkGleGXDaGoLn11wrr5URA7NEEbIYnEfM1EwH/euz5Lyd6ftW69fcSeIvFO7svvEnXN91RcIYhBFUXavkqXxRAk9DC8npbkk4GBkk4ySVtktzsEHtzy9Y5ePZas1W5ng1zdF3XUjXZLTJUVniy0uSNxkS07cjyAK8SJJfvRGVve6cduNl40JBwvu3FjVtZpGmhip47JPd6h9E2GLHhReEXdJjZ0t6WEFreluAMDHH0jsE7nCD3X5bNY8OOerkii4VXy9NFzZp+DTWoqdkjXVdHVU7GthrAxxJIc6KOdhOWkgtJy1wHSDUHsaOZykvc1NYNV6CuVu68Q1ktfU073MJIBfEYHFpxuQC8DOxK6NaV1pq7Qd5i1HofVN309dYAWxV1qrZKWdgOMgSRuDgDgZGcHC5kHPtzitpRSDmF1Z0CPw8moaX4xjPWW9Wcfvs5zvnO6DsNxF9lVPwK4G6v4vcZ+OFop6mw2qSahttno3SQ1NcRiCE1E5jJD5C1gAiyQ7IxjB6+cjvArh/wAxvH+38MOJGpqmy2mst9ZOw0lTFBU1VQyPEUMLpGuaX5d146SS2NwA3yOKNdcVeJvFCsbcOJPELUeqKiIkxvu9zmqzFk5IYJHEMG5wG4A7AYW3YDLA+OqgmfFLG4PY9hw5rgdiCNwQQCCOyD0O1r7GXjtZ7/NFw/11pHUFke5xp56+aahqmtzsJYhHIzOCN2vOTkkN2B7M8mnJbYuRGl1Hx749cRNPw3M2p1C6SGVwoLZSukbJJiWVrXSzPMcbQGsBGC1vWXrzB07zvc22l7dFZ7RzBay91gw2NtTcHVRY0bBodMHODQNgM4AAAGFtTiLxr4t8XJWV3EziJqDU0kbuqJtxr5Jooic/2OMnojG52aANz6oN4c5vHqn5kuYfU3FC1U8sFmnfFQ2iOZvTIKKBgjjc8eTnkOkIJODIRkgBcJBp2OFJG1jndT8hXGSL6OOw7oI2gN3KvbUdGQNgn3osJG5WM/YnB28kGQ6qLCXZyCoej3l3VnCs8MvbkK375FuEGbTR+7kknO6TVgc7HThW0E7S4+MdlFXeG6bMeN0Hf/2Lzg7mi1Rj/gDW/wAo25FD7FmKVnNHqlzhsdAVoH+cbciD2lREQEREBERAREQEREBERAREQcccyX4OvFP+BV8/UZl80bd5nA+pX0ucyX4O3FMf9xV7/UZl80LiRK7H90f0oBIa/q+a1Cnqm9P0c4WmE5Wp2yJjmfEM5QQ9ZqpSzCTRCl28ytQZSeHKXtHnufVYNyPVMAeyDDe9zhk9ipIKYyjqWXNAw0w6AMgKKmlEDC12xOcIKeED979FjzReEcK9kpM2c7KtU7qKDHVW9lT5oCQgq76RUkYGFF3VQ4hBV4wVQEjceSZLirxA8jqxsgq6YFnTjdIZSwEAZyCo8HOAN8rVLfbTOekjJIQchcBqqanvzHNJ2kBXplws4x3DT0FJCyRww0DGe+y8/eXTh9X3e+tELCfjHl816Bac4VzWmooai4QkRANJ28sIOVL9xLuus7cYJXvjaR09W4XDWvLFS0ulq6Sa7DrLScF++VzJrK46UtuljTWoRirER7HfK89OOmvtbUs1TTQyP8AvI2JxjKDrJxbD26xq2+IXsDj0nOQd1spv0lu7VFUKx/vFVgzvBJytpNBc/A9coJ5TiPY4VaNwGSSqytHh5IWK15b2QZtVUB7egeiwsub5kKWAh0mH+arUMb1DoQW08wik6yMrKmlbUAEDBCw3QvY3qcFRjiCMIJmP8NxCnIbIwuwrXRtEfWfRY7piNm9kFGNLpC0K6dj2nBbt9SvonNEuX9lqE8tLJHjbKDRy3AzlSxz+G0tCskOHEDsrO6CrndTic91NDMGN6VY6FzG9Thso0Ekjf32e6jwrgHuGB2CngawY8Tudu6CBjug7hHv6llVELQwuasNBfFJ4bsqd9WHN6cLHjYXnCrJGYzgoLSckn1VERAyiKoY4jONkFMn1RVaxzjgBHMc04cMIKKWOYMaRjuokQVe7qcSrmRF4yFYs2mfE2Ih+MoMI7beivbH1NykxBkJb2V0bJHNJb2HzQRY3wp2QgtzlR9JJIHdXNe9jg09soLXtc3OeysWdO6Ew+XUsFBfGN0kABVu7UJJ7oKIid0BFe6J7QHEbFWIGcK5mXPAPmVQgYQZB6h5INTqYGNpuod8ZWlqV1RI5vS5xIxhRIG6DYqWn6er4uyVHT1fD2QHTBzelRImCgIiqMY8kFFIyUNb04UZ7ogHckoiyIqV8repo2QY6K6SMsOCrUBTRzhjS3ChAJ7IQR3QXP3cT6qjeypkoCQguPZVa/pGFZkoguc8HsFaiICIg3KAivdE9oDiNirEHq97DZv8A1M4yP9ZrAPyNr/8AWvUpeWnsNnH7ncZR6TWD84r16loCIiDrn7Q78DTiX/idH+vU6+eeFpL9gSvoX9ogf+sz4ln/AOk6L9ep18+lpYx3j9XcMQWnBGMrElGDlXzvIkcB2BURJPdBRERAV7TturEygqTlTR1AYzpP1KBXRgF4DuyCjsuJcQd1Vm5wsyoMHggRkZHosSNjnH4UF0jR0gqSGqEbOjHyyo5A5owQokFXHqcSPNURVDHHsEFFUDJA9VfF0hx6klIDh0+SC+SnDG9WVCwdTgPVXOlc4YKvipZpB1MCC+alEcYcMnKrTOkcfC6e6yKWCXqDJs4JA3W66PRdRV0hroGktaM7BBt2C2OgLZvpEnOFyVof3qaVkUNCXnYbDK2rZqP3itZRSk5D8Ywu5XLPwkt921FRmriBiJaT1AfJBxw60akoqWCeOxS9JcNxGV3Y5cNBOvulffa+zDrbEDhzPku1du5b+HFw0zS077fGXmMHxA0HB9FvfR/DSxaQoTQ0MTegjGwxsg4B4WaUp4b3V081na1oyM9GF0p59dJQMv730tGA4SHYAZ7r1rpNLWmhlknpYA1785PzK6Bc2XDO4XPWgqKlhNMJsnPbGUHldqOhuFN0tnoHRsxjqLcBaRR0kYidUF+OnchdwOZrTWkLPpJrbe2MVTYt8AA5wukcFdUNikj6iQUG+dKuivFXHSjbDwF3X5Y2UNm1XQw1LWyAluQdx3XQrRt6Zaa1s0hA+LPddqOA3ESF2taKUvBY1zSTn5hB7daWitVTYKKemoYGRmIbeGBg43W39b6/s+monUxpWSZ2PwjCwuGOsaXUmh6Zltc0ysiDcB3nhcbcXri20wPdcw0Pye5QcX8X+NlgsYqagWyIl7D+8HovL/mE1SzXF5lrqemDGiQnAbgd12n46a9s8kk0MpaRgjGdl1E1LqLT1RHLHH0B5ccdkHHNRVtMTadzcENwtEmjJf1huQFm32QeOZId2lVoHwupnCQguI2QY0MLJx8ZGVq+mtC3/XOo7dpHRlnq7vervO2moqGljL5ZpD2AA8gMkk4AAJJABI0aQkOxGVzJy581/ETlYuFyvfDiwaUnudziEEtfdra6pnjiyCY43iRpjYSASBjqOM5w3AeqnIl7OPSvLfTUnEjibDR3/iXNGHxnpEtLYg4DLKfIw6Ybh03kMtZgEuf3dXi5afbS8zUEkcVz0Dw1rY2t6XFlBXRSvPkSfey0fMBoH1Ld1F7bDihTxxC78E9LVLwT4hprhUQhwycABwf0nGBkk7g7eQD11ReWFn9ttXSYZdeW6GUmTeSm1WYw1m37x1I7JG5+kAdht3W6ovbVaAiY9114F6hhIx0Cnu8EuR556mNx+f7EHolqHT1k1ZY6/TWpLXT3G1XOB9LV0lQwOjmieMFpB8iP/fK8VOfL2bup+XuWt4ocJoq2/wDDdzjJUxkGWrsQONpsbyQZziYD4RgPwQHP7ZW/21nLvK8i6cMOI9Ozp2MFPQzEu9CHVLMDvvn7FuSn9r9ye3mB1HdqLWtLT1UJbNHW2OKRnS5uDG8RzPByCQQAQd9yEHiPHAWjr74UwaahpGM4XYzm9byf36+HX3KxqyspI7tOXXDSVXZ56ZlE8lxMtLIW+G2I4H3on4c/AcfAzrtSvEWcjugwZYzG8tKtYd8eqy52tkeXYGVCYcZP5EFjm57Hcqemg6jl42UUQa5+CcLKnc2JoHUMnzQZjDEwBoWHVhg7KKJxDi57tlfKYJSC12SPLKCngZi68LH6izICy+mQM6dyAsOQFriEE0U7oxj181NCS52Xb4Cx2u+AE+iRyOD/AIdwgynvIf0jsu+HIdz68M+UzhRf9H6g4d3+83y73l9097opoWxSR+BFHHC8vILA0secgO/shPkAuhcgcGh6sNVKzZpx9SDVtZX2t1Nqq76orzmqvFdPcJ/iJ++SyOkdudzu47nutOM48MZG5V1NNFK3MxGVbVSwkdLOw7IL4YGyt8QnJG6glkAJYNlVksgjJYdvksVuXvHqUF+CTjyKyAJGtBD8D0VpAiGHdz2UcrpBgjsfkgzYi1wAcRnzUro2gZBWmMqC1XmplI22QZr2DpJyNvmsGUjKjdPKcguVrWOedt/mgo4jOB2VSchVkb0nBG6tbsceR2QUGyyXzjwQ1rcEjGVjuGHEKpeCwNxuEBucl3fCnbXSBob0jASikjZIGyDYrVq6gomUomY0ZIzlBp8YE4BzjKhqYzHgg5yoWyOYfhccDsqyTOkAB2QVjPwnfcKwkvd9qq2GRw6mtJCkY3oaXYJKCSPMTRkq/DZWkkK2FwmJa5WTuMbyxqCKVhY7bskG7wT2BVZQ7py4KMEjsg9DfYxvY7md1OBjbQVb/KNvRYfsWPFPNJqnq7fsArcf5xtyIPaZERAREQEREBERAREQEREBERBxxzJfg7cU/wCBV7/UZl80En9kf9Z/Svpg5kvwduKX8Cr5+ozL5o48eO/q7b/pQRNGThZEdQ6DGMqyWF7cvA2UsAY8dJxk7BBMLq7pwVJBEKxhkd5bqCa01UTDKW/B3WNHUSRNLWk4KDIdL0SmPOcHCjliLztsrYHtM3VIsmchwzH+ZBFDA0O3O6uqIBjIO/1rHMkrHeYVPFkeQ3OclBYfh+FUwsl9FIGeIeykp/ADC1/fyygwkUkrCHEjsowCeyCrXdJyshlQHYYR32WMMeaqDhwIQZ0tG2NgmAz54WTbLqYp2/CFBHMZ4/CO+B2S3xxx3eGOT6JcAUHZLl74lyaVuMdQ+lOOsbkL0J03xO/ZrY4jFBh3QNwPkuinDbS9mqrPDLHG0vJBPqu5/A+t0/ZaSClrQwEgAZIQcdcUtQXXTdTLVTyPbGATgk4wupnEjiRBfo52AhxBOThd4uba32mo0vLV21rcmIn4cei8u7nWmlnq4pwR8ZHf5oNtXirdWVx+I4BwFiFnhHI3VtQ4Onc9vYnIV8T2Obh53QUdO57ekDP2KSlozMCXZAU9vEDZfvuMLKrKmCNwEGMDyQafNS+ASR5LHEpyCd8eqzqupZLHt3WEyEuBQSvqPEZ0Y8lSOnJOcdkYxrCQ5ZHjxtYQCMoKlolb4YO6w54DCcFS0sxE2SfNX172vILftwgxC/bAyFbk+qIgKoOCD6K5sEjtwEcwx5DggnE3jN8PG6gkjfG7dp/IpaZ0TXjPmtVa+kdAS7BJGyDSaZ3xdLkqdngtKtlIbM7o7ZVhL3bnfCCV1R1R9BUCkhgknd0xjJVZ6aSnIEgwUFaUgSZPZXVjgSMKBrnA/Cdyruh79ygswfRFI8AN+ajQFe2QtGAFYsqnMIZ8fchBBG8Nf1EeaSyeI7OFSTHWensrUBV6SewKotVoRTeAfExnHmg0pMn1UlR0+M7p7KNAU8VQImluM5UCIL2yYkLj5lJH9ZyArFkRNjLST3QQAFyEdJ7fUqv2ccdsqeIxlh6sZQY5PkRuqKr8dZx2yqIKsb1HCuwGOCsBwrxG94yBsgmkma5mPP5LGVXAt2KDJOB5oBOUBGFe+nkY3qI2UeD6ICK5mM7o/GcBBblV3JwSqIguLMDOVVh7D1VpcTtlU7IJXxBreofWolcZHEY8lagqxvUcKr2hpwrQcISSclBUgAd1lU9b4LOnCxmfEcFVcwjsEF0jxK8kbZUbm9KNPSd0cclBVjg05KkwJRlQq6NxBx5IKOaWkqobkZypZSwt2xlQgu8iUFCMFTxU4ezqJUCubK9owDgIKOGHFvzVFc34ic+ao/AKCiq04IKdJG6NBJAQSvn6m9Kh+ayJYQyPqx81Y0s8Ig90Hqx7DZv/AFN4yP8AWawD8ja//WvUteWfsNTm38ZR5CawY/JXr1MQEREHXP2h4zyacS/8To/16nXz0QTuhLsH6QwvoX9od+BpxL/xOj/XqdfPIzGTlBe0eNIc+akdAGt2O6gDiw7FSiUuHxIITsSsmKlEjOv5Z7rHd3KubM9o6QcBBa9vS4hUxlVJJOVkQhgjPVjPkgg6Nt1aRvgK57tzgq3ugq0FxxlTMlEQxjdQA4V3Q5w6kEksokbsocH0UkXT1Yf2ytRbTxvj6mjIwg0rCkZIGtwVWYNDiBhRgEnA80AnfIVM5UjoJGNDiNirWkYx5oLcLULfWiJwYW9/VYTA0Oy7spGuj8QFvfKDc7Io5mNd23yt96a1PSUtH9ypAD1jH1Li8VckUYwTjyWTSe+Pd7zGDsM5yg550JoC23KvFwkkYMu6sH61z5pLiDT8O7vCynOzMDI9F0201xDvFqe2FshABwd1yzQaup7hbDWVhBlDc7ndB6icHuaCkrqCnppq8OyQC0nOF2isWqrPe6GKpgr4S5zQXNLwCD9S8KuG3G5llvLoXvwxj9vi2XNdj5qdWx6opobfO/3MOAd8RxjKD1V1Drmns8jw2QFrATnuF1N478TbTfHyxvczrLyM7ZC1H/ZwtV/0g3qew1Ji+L4t84XTTi5rKvuF++5lC4+LLJgDPzQbC5iqaKqifO2rDmEHbqyupD6RsbZceTjhdk+M+i9bW2xi7XJsggfH1DIOMYXWmOpMkMgJ3DigxWU7ppMNfjC39oC/zacukU/vJaQRvnsuPopXiY4PmtTbHVkeK0HYZQeiXBjnHk0HbWRTXQuAA2LlovGrnRk1oXCGucN+2V0Utl7qmP8ACmDiB8yrrjdoy8AtIJPqg37rniJV6llkc+sccgj6S4jrfGZO5zpi7JznqKyqqtDwTEMfatOdJI9w8Qk7+aDJZUunYInnJHmVE6V8TukH4fkpWOiY0FuMnurZOh/xbZQZFI6N4y4jKtr+nG2FgsJY7LTjHdSlzqnDR3CCWmjYRluCVqENPG/+2NgPVabSMfFUDq2C1K5NdNE3wTgj86CSWSCkYXRkbZWk1NwlqDg7D61QwTlha4krHfG6M9LhgoM2GMPh6gMlYchIcWuHY9lmUUzhEWtBJ7LGqGu6i4txugsEuBgNClgc957faoo2dbw38q1WngjZFkYyAgxPDxuXbqZtP1x7KKOlq6+pMNMwuOcYC1F9BU2mHFa0gkbAoNGY3E/TnCmqoMuBBz9qgky6YuZk75BWQygr529YYSAgyqS1itjDGkZ9VHV2aahIL3ZxulumqaGpAkyAD5rWLgJLjGHRgnHfAQaNHUtA6CB9qxazBfkKeopHsPQ1p6xnZQsppWvHijA+aCHrLmhuO2wWRTwgDqP6VkTRU5hxGQX/ACWFieLHUCAUE8sviDw2jHksVzXg/EssOjMeQR1qjjEYyXkA+SDDBI7Km/zVzMdRypD4fScYzhBY2WRo6QdlRjix4d5qSm6Os9XZXSQ9cgEY280E0IbVuAdnI81SqLYz4ZGQVY+N9MOoZB88LHe90h6nHJQU22ONllRywNjwWEu+tYmSiCRjPFeT2CkLhF8I7qKMP/eKjw4H4u6CpzK4uOxKo5obsCVQOI7KhOUAnJ9ERB3QACVqDKh0kXgPdtjusHICNeQcoJ6imETcs3BCxsYWbDJ4ww7yWPO0MkwOyC5tU5kfhtGPrUfinoLScqrunp274UaCSGUxOLgqvk8SUPUSDZBkzSNdGAo42bB6iJJ7q9ryBgIPQH2LUodzSapaB20BWn/nG3Io/YrAjmm1ST+L+u/lG3Ig9qEREBERAREQEREBERAREQEREHHHMl+DtxT/AIFXv9RmXzQv2kcQf3x/Svpf5kRnl34pAdzou9j/AJDMvmgkGJXjz6iPzoJnVIdF0YCgjeWPDh5HKvdTSNZ1kbKL60GuTX8VFIKUsxgY3WjOb0nPfKYYBkd0B6s9XkgqYsNDh3VY5yzZ2SkUgLsO7Ks4jJAYN0FR/th+AEkjNO8E52UtEwxSdcgwFdXubMQ6PcBBV1xD4/D6QPmsaOJ8rupoOFCditSt00DAGv7lBFOwsiAIwVhtdhatcGsMfW3sRlaS3BeM+qCrWOeex/Ij2FhWptkpWw+Rd9SwpHMMoz2QWQvfA4PwQD2WbRRmqq2zjIwQfyKlS6ndTANxn1WZp6mfUyCGP6RQc08NtdyW2NlL4hJaQMZXaHRl6fcaWKvfUmMMAPfC6scLeF94ra0TvjcWE9XbZc1XyordKW4UEJIcW9I3xvhBqPHLjGx1sfaffPELWFuM5XRrUFxdX100nkXk/XuuVtd22/S1D7jXNf4LgXb9iFxFcmslqSYgNjugw44zI4DBwVdLD4fYrPe6AQBrCOv5LFFJUyjrIOCgx+t3ZUy7zJ/KrzGY3YfsVd8GNxugthb1uwT2WS+RsOAFidRY7LdkfI55yUF739ZzlWNaSe6tyq9RCDI8Dob1A749VD1OLgD6qWCb48PO3mqzxgyB0Y2z3CA+lIj6w0nbOygDDn4gQtYo6iAM6Zcdt1hXF8XWPB2CBSztz0EBW1QD5Q1vmVjRte52Gd1LEDHO0y+XzQZk9qMNMJ9xtnK04PdsMrX664wyUIiGM4wtvtx1D0QHZO5Cua4Bp2WTKYfCwO+FisIwQUGfaJmRT9TgPqKvvUzZntc0AfUtOje6MkjZVmlMpyUFIwCd1M6VsYwMHPyWO3q8kf1Z+LugqAXu9E6MP6SrQSOyqHfF1FBkSUoZEHjufmoWRl4O/ZTPnc9gZnsoAXs8+6CjhhxHoqKpJJyfNUQFcJHAY8laiBuSmFJH0+aq4MxsgiRD3RAVwcRtlWogqTn8qAHCoqgjCCn1or2Rukd0t75SSJ8Rw4YKCxTxThjOkjKgV7YnPBcOwQHu63beayIKXLes74KxN2n5hatQNc+Ikjb5oMSeqJa6LpAxt2VkMHWwuxlX1rY2yHpABV1K7ERx6IMN4w4hZEVMHxlx7qGT+yKolePhaSgrFEJJejyCVEIiOAVcIZmffMfao5XuecnKCxERAREAyQPVBfHH1pJH0KZsMkI63dio5nh3ZBG13SVnQQNljLzjssFjHPOAsuGZ0DCwjugxZAGvICtV5a6WQ43JVHsdGcOGCgtAycKo2OFVgJOyOBaQSgq6M46s7KSGMO2Kt6i8dI7qsbJWSAEEILpqfw29WfrWOtQqY3iEdXmsKOMuPbYd0FBlu5CoTk5WVKYhFgfSHyWJ9aCRzstxhUjBDxsVPCYv32PJZBdTbYwgt6fGYGn6lFNR+FGZMnZTePG3GAlRMJKZ2PQfpQepfsNf9z+Mv+GsH6K9epi8s/Ybf2hxm/w1g/RXr1MQEREHXP2h34GnEv8AxOj/AF6nXzx/Uvob9of+BrxL/wATo/16nXz1UxZlwf6bfWghAycIQR5q8MLpCGpIwswCgsVQMkD1VWYzup20csvxxjbugtdB0s6s9/mowSexU0/WxnQ7uFGynkc0ub2CCalpBM7Dj5KyrpxA/A3CjZNJETg4KlEvjMPiYJCDGUscwY3pISIM6z14wlR0gjp7IIyd8hZ1NU9MPQT5LAUsbgBj5ILZclxJ81SPZwd6FXSRPb8R7FWAjCDIlqg9hZ04x6BRshy3q9FF9SuEjgMA7IMu30wq5DGdh81mVVnZTDrDwcb91pUFQ+nd1M7qaa41Eww5xwgvdP4v3rOwWfBehRw+CG5J81ogcQ7IOCp4YxUEM/fnbuglNwf43igkHOVrEWsKuKkNMJXAEY81pNTZqyli8aRmGdwsEY6hnsg1Ckr6s1fjioc3fPfC31p/iJV22pZEagkjA6s5XHcksYAEQwfPbC3BpazfdWUHpJee2yDs7oLibXugDn1zyCPo9RWddbv03uLUUkgcInBxz+VccaV01X2yNr5mu6DuFu+uslxr6EuhB8PG4+SDVuKnH6i4g6dbpYQtBiZ4eQAPLC6q3Systrnsa4fGSdj5Lfl+tjbXVSOawh++Vx7daqoravwMkuJwEGlTMFO4OHmVr1sndLSkdGR6rFrtK3OKnbVOYS0jO61nTk9vpaM01R/ZcY3CDQ5HeDM49sZWBVh8zg8O2Ws3uz1D5TNAPgduFiQWCuNM6VwOBug0iJ3S7DjsqzOBOwVsjHRyOYe4OE2xv3QWZPqVXqd6qh7ogdldHIY3dQ7q1VDSd8IMmOqBOX91L7+8kNYsOIDJDlV5DXDp7hBqTJ2t+OXb61iV08Uzh4Q7LHdK9wwSfyqrBhpJ29EF9PU+Ce2y1FlPHWR9YOMbrR/qU8NZLC0tadigOAinLPTbKmNUYx0g+WMKAh0z8juVZJG9jgH+aDWNP3xlqrBUSxhwBydsrK1XqSK/lphYGBuMjtlYtttgr5IaWCCSeedzY4o42Fz5HkgBoABJJJAAG5JXK3MByuaw5cdJaGuvEaVtBqHW0dZWiwdOZ7dSRGJsbqh2cCSQyP8AvYB6RHhx6iWtDhOCbwT2ytdo9RxU8XQ6MZ7dlpEDIHD48ZV8tNAd2Z+xBPNWNuFQQwYyfRahDcxaW+C/cuWiUxbFICO6y6mL3nD3d/kgyJqlhd72RkHfCxpa9tUC1rcLLsdjvep7lBp7Tlmr7tcKl3TDSUNO+eaQ+jWMBc4/IArtfwd9lVzVcTZIK6+ado9BWmTDjU6imLKgt2yG0sYdKHD0kEYPqg6fQQOik8RxyApKysY9vS1oz27L2m4PeyE5ftEmCv4oXq8cQLhFgugkcbfby4bg+FE4yuwfJ0paR3buQus/tduWDRvCibh9xJ4X6LtmndPVVPLpuupbZSMp4I6mMungeWtA6pJGPqMuOSfBBJJ3QecceTIPr3V1SCHj5hSiHob1gK9kHvDC7HZBh5G2BuqZPqrpG9Dy0+StQVaCSsmKcR9wD9axQcId0GdLO2oZ07LCe3pOEDi3shJcdygkihEnmrZY/DOEbIWfRKBxc4dZygrFKY8npyqPd4rxjbKzI4Y5WYA3WLLGYpQ38iC4U2W5UL29JwssFwbv2WNKcuQWK8N+HKsV7XE4Z6oEUfiHCndSBoz/ADqIxywgO7ZVDNI4YLjugrEfDcVbK7qdlXPjcwZcoiclAyfVMbZQHCqT5BBfBCZXYWTJQtY3Izn61FTSNjOVWoqnvcQHbIMcjBI9FVrfPKtO5V4I6cZQd/8A2K7weaXVI9NAV38o25FZ7FX8KfVX/F/XfyjbkQe1KIiAiIgIiICIiAiIgIiICIiDjnmPPTy88UHemi72f+QzL5oZSTM93q4n86+l3mS/B14p/wACr5+ozL5ox0+M/q7ZP6UEz63qhEfSsRSRxGaToZ5q6opnU7gHeaCH5pjKyQ1jmYCkpxEz+yYzlBhbqeGHq+I+SyKhkYZluPsWPE8gEAoLp58gxjbCrTY6Dnf0WM8kuOVJTv6X79kFwpp5ZCGRl31LVKbTtfK0SNgIOMjZbu4eWmjvVSIXNBPUAF2q0Byo3jV1XSzUMDjC/BJDcjCDpLPatQOzF9zZy0bfDGTlYMlqucR++UM7frYQvZDTXJbpe0WgTXylaJA3LstHouD+MHAvRNHVOobZCwPLukANHdB5usoKnqHXE5g9SMK6alEY88hdnuLHLrcNNWb7tRQuERYXZ6cLq/MJRNJG4k9JIOyDHO2y3RoqURXOF7gCARkFbadC9reojY/Javp6rZDVMbg5yMIO4WitdUdltDXRRNLwBuBvlbX1vr0XerE8jugMOcdloGiKyGKAPry4xEeYW2+JrhWvJsuenP71Bl694g0lys33Oi6eoN6c+a4RiZgylxySThalU09c04nDsj1ytHqJHRvLQgvgiPj7nbKzZbg2nb4YGVp1O5/VnClmj8QjqG580Ec7vFPidsjssbCypoHsjDh2VaYxCM9WMlBiJjKukx1nCnp/DDfiIQY2CsiKAOaSe+FLI2LuFjulLT0tPZAhi65ehal7syOI75P1rS2uLR1DuVcaqU7E7ILZSRIcEqzOVM4ZaSRvhQoJIZRE/qxlVnn8Yg4xhRAE9lUtIOCEFMn1VQMuAVxicG9XkrASCD6IJ3w4ZnPllQDZXmRzhhWIKk5VERBcxwacpI/rKtRAVwbkZVvdV6iBhBJTNDn4V9U0MO3moGuLDsjnueckoKIBuilZGS3IwgOYOnZRHZX5cXdKtcCDuEFMZU0cJczKU3R1gPWU+SNo6WlBg9PxFqo9vScKSQtzkd1GST3QURE7oCqBkgIWOHkqbtPzCDUII2U7RI4ZzvuoKqYTuyNsKEzPIxnZWdkE9NA2Z2CVkSBlK0tHmsJkr2fRKu8Rz3DrOR80FjiXuLvVahR1QhhLScZVI/ALQCRkLGnIa7DeyC1w8aUk9iVlNY2OPGd8LE6sDIO6p4r+26Cj93E/NUaelwd6K4kEeWVZjKDKNblnR0/asZzuo5VExlAVWjLgFTsnZBNJH0tzlRMPS4H0VS8kYVAxx7BBlSVYmj8PGFikYOEwWlUJygvik8M5VZZfEOVGqggIJad4Y7JSpkEhyFDnB2Vd3HJQTUu78HsppoQ5wA8ysfq6Bkd0ZM4vGSe6DNZSNhAfscrUKejimhMwxtutPqpXCAK2lrpWxOjB27IFTUukkMDG5wcIxnhxnqGCsi0xxGpMk4BGVS9uYZA2DtnyQaYG9chaT5qj4y12ACpDTzRgPPZXRlriOs7hBC6MtaHFWLLnb8O3ZYiCoye2VkBp91d1d9v0qlIYw775hS1cbnAuj+gNyg9T/Ybf2hxm/wANYP0V69TF5Z+w2H/U/jKfWawfor16mICIiDrn7Q78DTiX/idH+vU6+ePK+hz2h34GnEv/ABOj/XqdfPGgvhk8N2VWaTxSCrDjHkqIC1q31UUVMWuxnC0VXiR4GAgkqJvElJHbKyqWVpiLD3wsExPA6iO6uicWnYoNVt9nbXykE4H1rFu1vFumEbT3U9uuRpXZz3UdxmfcJfE7oNMTKyp4PDjBxusVAVQelwPoqIgnkqOtnTjsoERAREQERPrQFJTy+DMyX+5OVUhpHw91YWEH5INduepW11EKUR4IGM4WnWihFwrWUx/fEBY8cBlwGHdajaKeopa1kgBBG42Qbj1Jw8mtltiraSMyF4GQN1y5wR4bVFVafurV0ZaWNzuFrfBnTbddthoq34mhwG4Xby28FnWTTrqegjADo8YAx5IOvun7ZNfqt9qpqIvMZ6dm5U2oJavTDxaJaItc/wCHcLsBwG0Hb9P6qnqL+xoYXE/Eq8ddB2a56piutuY000bgXFo2wEHVG7cOKy8NfXvoiGvaSPhXXrUmnqyzaxipzTER+KBnyxlepFl01p7WNoZZbI1hqmsDSAATnGFw1xL5M9RyVZrG08nik9TfgPqg67ans9LFpKB8MDTI6MZAxnstu2LhNBc7Q+8yvEb2AnpJwueI+XHW9KDTXVkvgMGwLTjC2JrPRWrNMxujpfFbSjIcMEbIOv1+vcltrpLb4RLIjjPkcLFGsT7qafwsZ2yte1SbSJnsmA95P0vXK2PUR0pDujAI7FBBn3modIfM5VtRGGO2OypES1/w/nSQue8N7k7ILY29RKSDpOApnUNRG3xCNu6x3FxOXIKKeJuWb+Sth6cnq9VdLIAMN7FBCdnHCmbAHMLsqA7qeOGZ7OpucBBY0NLiD2CyPDD2bBYwikc7pAycq4iaDZwIB9UAQlzy3OB6qk0XhnCoJXg9WVR8jn90F9PKIpA5axYNO6i19qO3aU0hY6y73m6ztpqOho4jJLPKezWtAz2ySewAJOACVq3CbhJr7jfrm38OeGenprxe7k49EUezIYwR1zSvO0cbQQS8kAZA3JAPuhyV8h3D3lL0+27T+BqDiDcIAy5358WWwAgdVPRhwzHDkDLj8chGXYHSxgbB5D/Zyad5d6Wh4l8WIaS+cSXs8SnjGJaWw5BBZCe0k2CA6XGGnIZgZe/o17YDXX7KebiTTUU2YtH6eoLY5gOQJZQ6rcfrLamMH5NC9yF4k8TeSbmt5tuZfiFxEsPDqexadvWpKs0V51LIaCF1EyUxU8gjcDO9phjY4GONwx2JBGQ6HsY9xAGVmUVBcbjWQ2y10VRWVdS8RwwU8TpJJHHsGtaCST6AEr174M+xj4ZaedT3LjVxCueqqlnxPttpj9wo8/3LpSXTSDG+WmI5I9N+7vC7l/4KcFKUU3CzhlYNOuDPDfU0tI01UrfSSodmWT/xnlB4l8GPZk82nFyWmrKnQf7CrPMQXXDVDzRuDe5IpgHVBOO2YwCSAXAZI738HfY5cF9KGmuPGDWN41vVxgOkoaUG20BOclp6HOncBsARKzO5IGQB3g11xK4e8MbUb3xE1tZNN0IBImuddHTh+MbM6iC85IADckkgAEkLpvxh9rzy+6GfPbuG1lvGvK+IlrZ2NNvt5I2P32VplOD6REEDIdggkO3fDfg1wp4QW/7mcMeHdg0zA5vTIbdRRxSzDIP3yQDrkOw3eSdh6BOJHGXhRwgtv3W4n8Q7DpmAt6oxca1kUkwyRiOMnrkOQdmAnY+i8YeNHtT+aTiSJ6PTupqTQtqlyBT6fh6J+nyzVSF0od84ywH0XT6/alv2qbrPfNSXmtulwqndU9XW1D55pT2y97yXOOMbklB7DcYfbK8FtLOqLbwe0Xetb1bOprK6r/6mUBOcBzetrp3DG+DEzIwMjJx58czXPtx85oba7S+t6+02/S4q2VkdktlAxkTZWZDHmWTqmc4AkfTAOT8IzgcAiAupw7G+PJYEjC0kHZBIybxG+GsuEiGM5Cw6Ggqq6QR07CTlZlTST2twjrWkE9soNMnd1yud81YtTmtpfD48QJB32CwDE5rSXNxhBGromdbww+ZVqq1xachBnVVvZBAJGuycZWApZKqaRvQ52QokBFLBH1k/JWzN6X9KC+CoMJJ7qksviyB+PsUSq3uEEr5j0hoUsNIJW9RKie1oakZmwS3OAgl9zGcKppRF8ee3zUL53g48wsmPrlhI9UFniCp+94xjZVfRmI9QGwVaWFzZc/NZ880TYel5GfNBpb5BKOgeSge3pOFUn74S3sga5z/rQI2dRwqPYWnZT1DPCaC3zUlM6JzT4gGUGF2Vehx3DSQsmojD3Dwxt5rXbebe2gLJgPEwg2yRjuqLJnp3CZzgPhyoHgB32oO/vsVPwptU/wDF/XfyjbkV3sWB/wBdJqnb/sf1v8o25EHtOiIgIiICIiAiIgIiICIiAiIg445kvwduKX8Cr5+ozL5ouh0kpa0ZJK+l/mPx+144o5/4GXv9RmXzV0To21kviEY3x9eUE8VM2jb4zsZxusOtq/eXZxjCya5s4b1/vDuFHRUJqGl7UGHG4g4SQk4Pks5rI2zGM9wcbqCuYGvAb2QQGUlvSkZwDlTU1MXnJGQqVcYjPSBhBjnclSxxk5x6FRtb1KWmJ94YD2LgCg5M4KQyx3hj3sPT4oOftXrvyu61p7dBQUf3KbMCxoLunONl5d8LdP8AvNPFJSNBeXA7L1O5RrlpWyW6Cm1AGCpMYDerGc4QdsblZKLU9qDm0zYzK30xhddtd8uPj31lc2m62+IHeoxldpLfNTVFJHLSY8Jw+H6lJPTxVAAkbnCDo1zaaANHwy9wt9lD5W05GWs3zheSNTwz1JHV10tVZpYwJCd2Eea+jrUGjLFqanNLdqUSxkYxgLrVxw5WNGuoZZrLRsjdKCS0NGR+RB4fy6Vu8k3usVvcSDjsVuHSPCHVldfKZossxY9w/eHGF6I6J5Q4arUw95pyYy/f4NsZXcfRPKloGzQwVVTRsfKwA4DB3wg83YOXG+P0iyVlpeJOgHZpznC4c1Bw/uumvEjuFA9pBwOpp/nXueeHmjrfRGOaljZA0b9WAMLoFzm0ug6O6SRWfwwevB6QPVB5vaqtb4gXug6AR6LjCvAE7gPXC7JcWqe3Q2UTUwGSw7rrbD9+neX7jJIz9aBTkgDIISaV4kHS04Wow07ZGhrQMlalHZ2+AXSN3HbZBock7ZIQCN8LTjkEj1WTXAxTuY3tlY7ficAe2UE1JTmeQN6SR81LXUvu5GPzLUqbwYIhJtnC0+vc+od1jcBBhtc47ZVHDHmqY6Hb+SrnLgPLKC796o1lSdAiBB3UUMDpSPTzQR9RxjCzKWiZNEXk7+im9zE8YZGPiUL6arowQcgHugx3YhlI9FIyLxx1jyWOSXO33OVlQPEUZa7YoLJph0+HjssdSxlpmy47fNKgsLssQRIN0RBUjComSr2wveMjsgsRVILTgqiCv0d8qhOTlXHsrcFBIxviHHqksXh90he1jgXeSrPKJCOlBEpWTBjOnCiRBcHYcXeqE9bh5K1OyCZ0PSzrB3UJJ8ypY5M7OOySdHkgiWVT0rZWF5PZYp7q9sz2t6QcBA6MSFnoVmw0TC0En61gdRznzUgqJW7ZQT1DBH2WGd1c6V7vpHKNjc4EtGwQWonZEF8UfiOwTsqzRCM4ByrGucw/CcI57n7uKA1xac5Rzuo5VMFEFWnpKuyHYHqrFUEAfNBI+Lpb1KNp2Kq6RzhjyVuT6oJIIhK7BKvlYIT05ULXuZ9Eo57nnLjlBQ7oN0U0XSWnPcIInNwAfVZlKwOj3WI85JHkFk0zXlnwjZBDOzDiouyyw3reQUloZAS5o2QYinhpvEGfJQlpaSD5LKgnDGYzughkh6Cd1GDjZSHrmeQPVWPjdGcOCC0klVZs8H5q+Hp6viVZenqBagzHjxYekKNlOImF2VJSOy3dUq45D9EbIKUknXIWZwqVjvClG+ViffIXZ7FUfI6Q5cUGYa1sjBGW4P1LEdlryftVpGMELIiczoPX3IQWOmywNPooRuqvADjhGglwCCrmFoB8is2GcChlYdyWkAqKUN8MduygDiIyB5j+dB6uew2GLVxjPrPYf+jXL1JXlt7DbP3L4x/4ew/9GuXqSgIiIOuftDvwNOJf+J0f69Tr55GN6s/JfQ37Q78DTiX/AInR/r1OvnjBwgHZMEeSmpgHP3WTMyNrc7D7UGAN1fEzLwCVafpK8RvOHNQZdU0eEAzusRmWgggjKyKOZvWWznZSVroD/Y+2EGA7IPdZFPUCNuDuQsdxyUbsRnsgyJqp0o6cLG7LNjNOGjKgmDXSZbuMoIQMqpBCyh4Ybgd8K13Sd9t0GMiq7uqICmjpy9vUPTKiDSey1Klmijpix3dBprhgkKivlIMjiOxKsQXRnByrnydQxhWN2O6mjLC4ZQKWc08gdjIPqt0aU/6rX2KkezDXkDOFosdNFO0dIBW59KiKkr2CLAmyMb+aDuLw2slFoGzwXaEte/ZxAXYTTPG+2VlAI6yRrMNx8Rwuo+i7xdaagBvhJpgMjPbC2rxI4lPoGvbY3EAZHwn/AFIO3usOJNjid41HcGRuef3rgCuQdCmx6v0rMKuuY97oz8ROSNl5M1fFbVdZVATTvwHdiTlczaI5ib9YNPS0zJ3B/h7fEfRB6S8uGg7VYdavq21bKhplJDc5HddyrtYrRXweLPb4pCwZb8A7Lyp5BOLWpdZ65DbhM50bphs4kjGV6yy1kELmRSuALwMIONKrRFm1LNLSG0xMABH0QF1U5m+E4stiqYqC0h5OdwzOO67z3ClNNGamiHS47nC2pq6zWC66elmv3QdifiCD53eJ2j77b9U1DpqGVjSSRkEY3Wz20LhGXS5BGdl6Q81Wn+H0dfVi3NhEgaewHfdeeOrgIKySOn2b1kbINvdXhyEY7bKjZB4wee2VV9PK1vW4bHdRdkGtOuLZ4BEANvzLSpxh35VfA5rdyrJHh0mfLPYIIle2PqGSVfJJG5ga0HKsYSR0+qCw7LUaWcMhLT5jCwXxPZueyvZIAzHmgzYg2MiY7j61BW1DZ3Dp8lCZnFvQDt9aswgpgei5Y5b+WbifzQ69h0Pw5tZMcfTJc7rO1worZATjxJngHc4IawZc4ggDAJG9uTjkp4jc22rPCtcclm0ZbZgy9ailizHDsCYIASPFnIIIaDhoILiAQHe7HBPgdw25fNCUfDvhfp2K2WymHXNKQHVFbPgB09RIADJIcDJOAAA1oDQAA2pyt8pvC7lR0ONL6EoverrWNY6836pYBV3OYDu7c+HGMnoiaeloJPxOLnu5rREBaFq7W2jdAWiS/wCudWWfT1siz11l0rY6WEHvjrkcBn5ZytdXCfNHym8L+a7RR01rmiNJdqNr3Wa/UsY97t0pHcHbxIicdcTj0uwCC1wa4BwXxl9rfyz8OGz0WhGXfiFco8gfc2I0tCHjydUzAEjt8Uccg+a6McX/AGs/NDxOdPQaHrLbw7tUg6RHaYRPWObvs6pmBIPbeNsZwPmc9feZPlq4qcsGvJdDcR7U4QzF77XdqdrjRXSAEffIXkbkZAew/E0kAjBBPErPGiOQCMlBuO86s1ZrK/y6j1tqa63+6T/2WtudZJVTvGScF8hLiMknGe5PqsS5wmr6TG7GPLKxI52Njy7GVBJXS9WGk4CCYUEj2BuSSFhz076Z+HBa1aKuFzh4pGfmo78IXuBjIQW0UzXRYdhQVkTTlzcbLGDniMCInYK0zyAFrwclBqWnr5HZ6kTSR9Q/KtR1BXx6mc2op2hvR8sLap3WRTVE0LXCHO6DUBdG0sIpMZLdisGSYTA4GAsfPW8l53JyVVzgNh2QWHug2V7GdbvtWV4TWxnI3+tBh9WyopoekPPV2yqTlvXlqBDL4ZJ9VNJGJsyD0WIs2KVjIDn0QYRGCQr4W9UrQfNXRwvqHkR7lJYpKZ4Dtig1OqoI/AD2kdsrA8fwmmPCtNZO5vSX7KrCxzMu7lBATkk+q1GmkayDfyC09zCMnGyCRwGM7IMuOrDZCMdyo6p7pXAjJCxy1zdysmGRn74+SCymi63HOyrIRE8AK+eVjW/ezvlYrnOccuOSgyHS+OOk+She1zNvJWhxb2U7S18Z6u+EFkU/hjcZUga6RxlDiAN1ju7lVEjgMA7IMmary3oxv5lYpznOCpqVjXv+LfdZE4ha0t2Bwg74+xWdnml1T/ACu/lG3InsVvwqNVY/4AV38o25EHtQiIgIiICIiAiIgIiICIiAiIg445kvwduKX8C75+ozL5oXEtmJHcOP6V9L/MiM8u/FFo7nRd7H/IZl8z8oIlcP+2IQZs9x8amEJG48yq0FyFLEWEd9srT0QTvlJmM4Pc5VJZTKcnuFGc4wrdwgy4qzw+wGyiqJvGOSoUQVa7CmpW9VQ05xg5UZheG9R2CRvMfxDug5k0VxBh0jTxSmTfqAwvSjk7ttt4sGjub7oI3MaHdPVjO3ZeOz6qaVoY5xIBzhdq+UHjrqfQepaCgp5nsgLmtPxEDHZB702a3stVvioWP6xEMdXqs5cf8ACriLQ6w0rRVjpWmdzAHjqHfC3+DkA+qCq065WimuIInjDwR5rLfUxMk8IuGcZ7rTLvqSitlNK+V4Dg04GfPCDEtWnbJQVRfFTRh4ORsNitxgADAGy4Ps3E9sWpHCplBhLiNz81y9a7/QXdodSyZyMhBZfbUb1SOpQ4tyCM5x3XUDj/ywW+upam7VdW3LSXbu+1d0n5DSWd8HC6Uc3mpNa0dHVU9BNKGEkYGUHndxv0vb7bUPs8dS0hgLcdS6xXS1R2yWUxuDsOXKvFmLWVVd31VS+Yk5JJBXD9VPUxufHWEknbdBDT3A08hducLVGah8SIsOR5Lb7ulzic7K49LNm9ygkqpBPMXeqhAAkAzgZRzXNw71U0NFPUNMrGkgbkoNQqY+mkBZuSPVYlI/Mbmyd/LIWRZmyVVWKWTJA2+pS6gtpoHjwwQD6BBpLmdcpDe2VbJGYzur6VwbL8Xn6qeric8hzBsgxC8uAC1Kg6GRHqIytNETySANwry6WL4TkZ7IMuGv92qC7uAdlLVXQVpDC0AHZYMRje7MgG/qqTRgOBjCDNntvRF48WHHvgLT39ZPxAj7MLX7HOwHprfofNY2o30bph7pjAO6DSSzDcj0yrM5VcuI+SogIBkgIgz3wgyH04awOyVSKoEbOkgFRmZ5GCVYgq93U4n1VEwmEBVBCNaXHARzS04KCakgFRJ0k4+tXVtKKZ+GuyoI5XxHqYcFVlmkmOXnKCxBuQETsgyo6VrxnJUU8XhEYVomeBgFUe9z+6C1Mq5mOxUhDPXyQWxRh/dZbaFhZkkZWF1lp2KvFTIBjJQWSN6HlvogaCrSeo59VUOI2ygvgjEjukqeQtpvhG+Vise5hyFV8jn/AEjlBPSwNqZSHbD61fW0TabHS4H6isWOV8TuppwVdJPJL9M5QRoiIKkhUREBVAJIHqqYPosiMxiM9XfCCj6V3SCwEqEsc04cCFuDSAgnr+irwW4U2toaKCpYKPAGfL6kG3Hxhoyo1Uvce5QNJ3CCWmhEzsOOFWphEDgGuyFG0vYctSR73nLiThBdBGJXEFZBlFM0saMlYbXuZ9E4R73POXFBKyfok68eayzcsxlmButOwfREFz3dby71Vu6K+NuXD86CWlJjf1Fu3llZz6OOoYZcgEKCd8Ihw3usVtVM0FodgFBbIzokLR5K07H1VXdTviKpuUEsdQY+yn996mkOwSFhK9jc522QXSSl5OG/mWZR20Ts6nbKOnbEdjglZUlWYGdMZwUGLWUrYNgewWG3JIClnqHyuJJ81SKF7z1Abd0GbTUAnaOrAWPVU5pn4YMj1U81S6KMNjOCp6SaCSA+8EF2EGmGQyDpVmenLVfIW+OS3tlWyN/fAbIPVz2G/wDuTxi9fHsP/Rrl6kryz9hsT9zuMo9JrB+ivXqYgIiIOuntDRnk14lZ/vOj/XqdfPLFH4hIPkMr6GfaHE/tNeJRH950f69Tr55A4tOW7ILmnw3EAq90pcO6hJycplA81OyoDW9OPtUCILietxI2yhafVWg4VeooKKobkZVFVrsfUgkp4XTv6d8K+eIwHowSsikniiOT3Usk8EjwThBpjQ7O+ceaq5xHZZ9SIZGdMOMrAfG9n0gUFoY4/RGULS07ghalZHUzZz70AW/Wl8NM6oBpsdPyQQ22BtRIWuIGOxV9yoxA4FjtlhRTPhd1MOCrpKmWU5e7KBBF4riCUqIfCfgbhRse5hy0rOghNREXkEkBBiQxeK7C1JlsYI+skA9wsOlxHMQ5ST1sgl6AfhB7IL6WofTzluMgLKoLu6kvMVV1EBrgVYRCYw4Y6iFhCklfJ4jRkA5Qcyag4s+8WCOihkLCG9ORsVs+nq33GgfNLOXE5O5W0ayZj4RGBuB3V1DWzxwGKMnB27oLZanprHN2wHYyp2VkxeYxMQCMYBWlSFxkLjscqgke1wOdwg7XcqXFCPhfdYK11UGEyAnB+a9gODXFvT3Eynpqua9M8URjDC8ZJwvnotOoqqgmYckgOBAyuw3L9x04hWHXlE6mmlbbmvHUOogYyEH0BReHLB0sPUzGAfULjLjTR1EelKn3XqDsHGPqWHwj476W1jpq3MbVN98EIbK3qHcBazrjW1gdS/c2tLD4m27gg8i+M1tutXqmsbXue1m/fI9V1R11aGU1e7wcOHXvheknOFo+impZ7hpxjQ9zC7Le/b5Lztudqr4IJ/unkyhxx1Dfug2TdBG2naBgHG+FpMMPiAlZldSVfjO6s9Odsq2KB8cZ2OceiDEkb0HAK1CzWoXOrZTAglxA7qlssVdeJiylZkk+i3no3hzqBmp6RskRa0PBJAO4ygg1Dw1nsttFe5hAc3q3K2SyLuW4JBwN13V4hcFbzqHREUduY8yGMD4Wk74XB9Hyua9hxWVtM9kDDknoIyEHDgjqJyGGEgfVhXSUXhNySVzVdOG0dHTmjZGROwYO2+VxXqOz1Fqe5kucA+iDbv0HFSBnUOrKjwXOx6qTDm4aD3Qdt+G3tP8AmZ4T6StGhNHwaJprHY6f3eko/wBj7GMDNzl3hOaS4klxdsXOJLiSSTyDbfbQ80lGImV+jOGte0PBkc+2VrJHNzuAW1YaDjIB6TjzB7LoP0nOSmB6IPSGh9ttxcijcLlwZ0fUSdWzoKqqhAGOxDnOJOc75H1LcdB7ca7xF5unLbR1DTjo931W6Ej1z1Uj8+XbGPmvLstAGcqxB64W724GhZX4u/AG/UzOjOae+Qznr22w6Jm3ffOdhtvtuO2e2w4ATPjF34V8QKVpB8Q0zKKctPkAHTsyPmcfUV41Yyq7jfCD2G4qe0Z5BeY7Qk2gOLOiNdSWuudlrqqz04qKCUZDaiKWKoeY3juCzJIJaQQS0+Wuu7Joi1aquVBw+1TPqHT0cubfcKiidSTyREZAkicT0vByDgkHGQd8DZcFS7qDXdldLWPY4NjOB5oIJwWyuGexVgd5LUGNinj6iAXEbrCmj8N2B2KC7okiaJGuI+pXNe+cEPJJCmjfG6ANfjGPNXRtiaPvePrQQ07wyXDhsDhR1bw+UluMBVqWFpyPNQtAJwQSfJBkGkPu4m8QZO+MqBrnN2G2VkNie0Zl7Dy8gopunqBYgtLCMHcq0g9ysile0O++YwrqmIPf1RjbzQR0o+I9XZSVEoaekHYhROcGD4e6iJc877lAyfLKp9ayIIsEl6sqOnr+FBEpIQZHtiJwDsrDsAjXFjg5uxB7oNehpYbczx8gkhaXcKptVJ1NGAo5KyeRnQ52QoEBXMyXABW4V8P9kCCSUYZhRRDMgB9VmVTAIgR3IWKyOT6bew3QZVXCGxtLRn6liMjcQTuMLLhqWOHRL5Kkzc/FH2+SDDIKqxvUcFXgsBPUrXOwctKCV9OGtyP0qwAhpVpke4YysuKnc+InzwgxooxITlJogzsnhyh5Dc90aXNkAk7ZQWxvdGcjI+xJHl5zlZ1Q+mdCA3GVpxGCg7/+xU/Cm1T/AMX9d/KNuRPYqfhTap/4v67+UbciD2pREQEREBERAREQEREBERAREQcdcx7unl54oP8ATRl7P/IZl8z8x6pXn1cT+dfS9zJfg7cUv4F3v9RmXzSNj8SdzR5En86CHBKr0/NZMoY0dIG6xslA6vIq4N6lYpoeyCtLAJ5OhxwFmz22OJvX1dsFae2V0byWHG6vdV1EowXEgILpZwWmPHbzWMhzndBuguY4McD6Fbi07q+psNwiroHlhiIO3yWhNhLm7d1aYenOSg7icMOfjUGhqNlGaqoLWEY74wF2m0H7T9two46errW9fTj4sZBXktTsjdIBMT0/JanTVgo6thomEAH1Qew9o52WXqtNSbngPd/dLsFonVlm4k2kVE14YHPbn6a8SNN6zq6alY1mQ7IPfdc68POYHWFgjbFSzyNjDf7ooO/fFGlp9PyPmtlaHuDv3rsreHAXWV3rHMiqA8jtvldSuF/FWt4j14o7nIXlzwDk5XfrhHpix2e3RP8ADaJHsBH14QcpUkhmgbI7uVsHiHwooNcxvbPEw9XfqxuuQowwMAZ2VyDz85h+Uy3Wugkr4LfGR0EhwAIyF5YcY9FTWW8TxxwdIZIRsPmvoR4y6YqtT6afRUmesscNhk7ryK5m+ENVperrZrkwh3WSMj5oOklTaPdqQTEbkZWnQM8RxJ8luXUtQ2GX3fq+EArbDHESfDkAlBkzxksDWgbFbr0raa2ppjFBSOlLhgYGVplHaHV0LOnOcgbBd0uVLgNDqWemNXHlrwO7UHVqz8PNUx13vMdllI6s56CuRKXgrqTUULXPscgyP7gr100Tyh6LbbWyVdNEXHB3YFyLbuW/QdvpxFHSMyPPoGEHgrrzgdqDS8BqRaJBgE/QK4rNNdIupklI8Y2OW4wV748ZOV/TV0oX+DTsILD+8C6JcUuUyG00FbXUlMAGEnZvkg89afrbMfFaQfyKlYet46QcfILc+u7GLLfH29jcOa4jGPmtJp7e4xlzx29Qg02SlDIusHfGe6ijkI774U80M7Zi056QdvqUbw1uR5oJTMZm9LDg9lYaYjdzsq2GJ3V1Y2V08jhgZKCF5xsFq1ts0VZAZXuwQM91pYicPid2Pmpm3CogBZE/AQR1UAgnMTTkA91CDjYq6SR0p63nJJ3KNje9pcNwEFnnsmFLA0eJ0lSzU+2W9ggxR3Q91UNJOFcI3ZBxsgRuMburCSSdZGBjClf4fh48womQveCR2CCxEIxsRurmxlwyEFqmiha9vUcqHsrhI4DAQUc3pcR6KSKLxPNWuaT8SMe5pGD+RBJLThg6gVBk+qnkc8sAJ2KgQEREBFdH09XxI/GdkFqIiAiKSMsBGUEeMIp5PD6fh77KBAAycIRvhM4TJzlBeR8OVaAfVDnzQNcUE9HVvpJOtpIPyV9ZWSVsgL3E79ysQjCDYoMmWmDIw8Zye6hY8dj5o6V7h0knCMY4nOEGXFG0j4sLGqGhrsDsVcZH/RYTn5KJ/XnD+4QWoro43SnDRus1tvkZGXuB9d0GCTlVEZIzgq8hrZCHDzWQCwx/CgwwN8KQHpGFZn4iqOOfNBc9rs5zlWDZVGTsqEYQSdQcOlW46dijCAd0kIJ2QWndXh+BjCsUscXW3KCxry12clVe57ztkhUcxzSVPTGPADsZKDHII7grIhqRGzpxvjCknhDm/Dj7FjimkG5GyC0vcXl2CrXOce+QsmHwmkh+CrJywu+AABBB2V/ifAW43V3wlvbdHQ4YX+iD1X9hr/udxl/w1g/6NevUxeWfsNf7Q4y/4awfor16mICIiDrn7Q78DTiX/idH+vU6+ePGV9DftD/wNeJf+J0f69Tr566RgcX58hlBAq4J9VWTHWfrWVTRAxlxA7ZQYaqBlXFpc/AVrg5hwUFDsqtblUQEhAOyvbH1NyrACSpY43gg+SCPDh6qgyfNZkgjazJ7rGLeo5aNkE1O3oPU4jdKqRrnbKMRyu2BVro3h2Hd0FnZMk+amki6GZUKCpGEwe+D+RAC44WeyKNkJLsZA9EEFHSsqHYc7H2rOe4ULDHHg5HksGlp6iqn8KlBy442C5G07wT1LeaYVxjeWYB7FBsWgo31Mpc5uMq6utzYpB2zlb6rdEVVmc6AtIe3YraN2oapryHZ2KDTKhvhsHxdsZ9VSK4+HEY+nOfkraqGaNuX5x5LGYGdO4QWyOL3l2+6ljqDGzpUB2JARBfI7qOcd1YiAZ2CCaCMyvAbtg5W99O6+fp4e7xjD+3UPJbMpCYX9Tu2FRxZLV9R7E5KDtrwO5g7zo6rZVS3WToe8YHUcALudZ+JFTxCtrLya8kMYHHf5LzB03Gyuigip2glrhndd0eDF/bS6d+4AJE8kfSB55wg3RrrXM10M9ujBqSxpGO66Q8Zq27UtxkaKB0YLz+9wO69AuGXBu4RXiW96ijcaWZxILhsQfrWk8ceV626vca2yU4LSST0t2QebdmZNd3hs8GCduy3JXaRp4aJ0gAz07Bcma54KVXD+d5ex7ejf6K47mmramJ8bXOLRkIND0XXutF2DRD1jrx2z5rtLwy9xvV0hlkpGB4AwC3dcCaKtFHFWiataPpZ3C5m4cahpKTXdJSwYERcGkfag9FOB3Dyk1Jb4Yq62h0ewy5u2Fy3rngppyPR1cyis8DpYoi5vS0ZzjyWHwsv9ptOiaSqpDGJS0E7jPZb8o+J2natgpK6VgfIOlwyMHPqg8qLrwyuk+vK6nks7xC15G7NgMlcM8buGNvoS90jWxPGfh2BXrNxWj4baXhqr29lOHysLgds5IXk5zR1t51dfJqnTbyaYSn6PbGfkg6xXm1NttWWs3aNwsNlP4gMvVjC3BfbXW08TffN5A3fK2yJ3MaWDt2QHODnFnoriwjySlDXSEu81lT+GBsQgwXDGQrCcKQub1HKseR1bdkFrTgo52VMAwx+ShcMHZBTKZQHCqfJBMwGIB2ThRyyGR2VaXEjBOVRBeQegenokUhjJKsySqtGXYQXPlc/udlawlrg4DOFO+mcG5CijaBKA7sglmqnyN6enA7LHWpVMMJhDmAZwsJtO8jPkgj3JUzJyxnR3/mUbwWYHmrmNBYSRugjccnKyoKZr2GQ+W6xD3OFeJntGAdkEkk2MsbthQd1XI3ynSTuOyBudgFc2Jx8ipIywEZWaHQmLpHfCDTHNwqKSZhDyfJWN7oA2Rp6XB3oVJJ0luw3VscLpDgIL5agygNV0M4a3wiO+yubTmL4pVFIwFxcwbZQTGka5vXnGVaZfDb4ZURlkb8O+FYSXHJ7oMqhpRVyEE9lbXUwppOgFXUz30x6gcZG6jq53TvDnElBbFGDuVkmtdEOgDIWPTtfI7DVnFsDY+l+Os5wgippQ52XDc/JQVXxTdIHmrmxuik6gPhKvDQ53V80FPcelvWSsdsfVN0eWcLJqZ3BnSD8lixE+ID5oPQv2MlC2l5ndTzg/S0DWj8twt5/mRV9jIZzzO6n6+37Aq3H+cLeiD2ZREQEREBERAREQEREBERAREQcccyX4O3FMf8AcVe/1GZfNIyXwZnuxnJI/Ovpb5kvwduKf8Cr3+ozL5oJP7I/6z+lAe7reXeqtJyikZ043wgsa3qKvHwkN9UijdLJ0sV08L4XAuyCeyCWSlHhh7dz8lA1zo8tcCFmW2dgnDZyCz5lXXg0zngwYQac45OVdE3JVivYyR2S3OAgyw8RtysaWbxFa55x0nurEFSdgPRTU9S+A5aCcKBa5aLY2qpzI4dgSgydP3Z7qoRyPIGR5rmGx19MIQ33kAluMLgeokFHUnwdnA91mUGo7lHVRYmOOoDHyQd2eXOvhtWo2VVZVhsZlGCXYGMr1h4UVVov1spp6Wua8tiBwHDfZeGdDqK7UtgintLiJ9icHdd4OS3j9e7fUUtqvsxy/DcOd/rQen0bAxvSDkK5adYK8XO1wVzSCJW5H1LUUFrg0j4gCPmvOT2htGK8VclFRjAJy5rfyr0UrhKYiIs5wV0w5o4bNW0FZb7hg1EhLW+Zyg8UtYUszbr4Tm4IyMfasO12eaoqms6CT6LsNrLlj1VeNRPudBFKaZ7iQQ3bGVNYOWnUdLd4nzRShgIzsUGqcuvBep1pWRU8tAXNLxuRnzXrXwD5f6DQ9vpah9ExhEYO4xvhdYuXzT9k4X0kFbdsMIIPxBd9NA67tOsaCJ1qka5jGDJHoAg3XFDHDGI42BrQMABSIuPuIXGCxcO5mxXh7GhxHd2DgoN3X4UXuDzWAYDTgkLohzP8VLBp2019tgdGS4uHkuVuN/NvpC16Xe611LC58ZOS8ZzheTnHbj7JrWsq/DlJDpDjfPmg4n4qXKKu1NLc4jkOcXbfMrQLdcGTBzXbfWsC63F9e4Oed/rWnxyvi3YcINauUsQBLcZWhuOXZU7G1Fa/w2nJUr6N1KMVDcE9soKUswPwO2VlVG4vy1pIUBd0PPT2BWuWr3eaHolwXeSDTHyB0PTjBHcLFa0lwb67LVau2SwSGYj4MrTpS0ODmIMqegayHxA4A4WNHOY2FoGcqr6p7mBh7YVYTEGfGBlApWiSbJOAsirkEWGt3ysLrMbz0bbqj5HPwXHJQA7DifVZsLGyxknvhYCvbK9gw04CCkg6XkKWKoEbC3HdQklxyVRBV7upxKkZKGt6cKJEFXHqcT6lZNPTtkYXnuoIy0fSVzp3DZhwECV2CWDfBUe4OSpIC0yZf2V1SYy4Fg+tA8TxB0/nULm9KoDhVLie6A0ZTpJOBkqgOFlQOjDCX4ygo+mDYg8d1jKeeZz3FoO2FGzGN0FiqBlUOM7JkoB2U8NOJGl2cKBSNmcwdIyEFjmlri0qiucerfzKq2JzgXAbBBYg2Q7FEFznAgAKrX4GMKxEA7klERAxlZEBJ+AjCsp+nq+PGFkyPiABbsQgulgZC3xB3VaekZWAuc7BHzWG6WSXLSchGTzQbMJCDWrbb445TnfHqq3irbA4QtbjywFhW6vex+XuUV1mE8ocEFjacTHrzud1eYBEw7/nUEE5YQM7AK6Z0ku7c4AQY7h8RCoqkEEg91RBfF9JTGDrb1DusdhAOSs6KaPw8HzQYbYy5/SkkTmOxg7rJa+Jry4KV0sDmHtlBHDQiVgOdyPVWyH3Z3Tjuq01UWSnfZWVsrZXgtQJcFufVY7Thw3VetxGFb9SDVIw0RjJB27KpkYWFo7rT2yyN+lnHzTx3FwPkgpMxwcT5FRrKlfG6PbGVjDHmgAkHIUpqMxGP1VsMZkdhJ4zG7CD1Y9hr/aHGX/DWD9FevUxeWnsNsfc3jJ/hrB+ivXqWgIiIOuXtD/wNeJf+J0f69Tr554pPDJPqvoZ9od+BpxL/wATo/16nXzyxgFwz2QUPUSTgqSOcsZ0rILoC3G2ViyBvUcIDH9Ly5JZA85VYYHzEtYNwk0EkBDZBgoI0RVDCTgIDTgqdk+4GO6idE9vcKzsg1SGjZUjJOPRY1SxtK/oByFCyqlZ9F2Fd4c9SS87oKsquk5VH1HU7OFG+J0ZwQrR3GeyDJa81Dejp7KGWLwyB5rJppIWD5qOpe2ScdPZBAwlhBIOFrel6L7t3yltjx8Ezg0n0WHUwNdAzwwM+a1XSLX0FwZWjZ8Zy0+hQdjbny/0mlNMU1/oIWzSPaHEDBOVzJwVmfWWQUNVaAHY6d27laZy6Vlw4oRwWO5P64g4NAcc7LurZeW0aZghucMDRAGhxPljCDodxt0DVWpk1yhthAcC7Zi6tV5ke6Q1EJYQ49wvU3jnc9Dx26a1Vfg+MxhBzjOcLzX4sz2unrZhbOnpMh7fWg41vTmlpaxvYeS0HBHfIWsy1kMxw/B23Wm1Bj6j0YQQKrRlwb81RAcEEeSDJmgaxnUMZWO04OVf1yS/DkkKpp5ACSEB8pc0Ad1Y3qbkgFUaMOGVqscUL6UkAdQGxQbs4W1/hXGJriSC8DC9AuX7QNtu93pLrUVDWhoBLSdl53cOmPF0a7GwcD3XZvh5xvrrLqGn07RTYlfhgAPn2Qer12t1ortKQW61wxmRjQCWYyVujh5oGiFmMVwo2vcRsXNC435atN6qu9oprxfg800uHDq8xsV2PNN7qGNpRgZwUHWzjlys2zWdDUT0lvjLugn4Wj0Xlvxw4U3XhtdJaGC2P+mQMN+a91rte6CzwNluMgYx2266v8WOAFp4qX1t+pImvphJ1OwBjGUHjZdDfLVGJamgkhYRnJaQsfTGsH268MuZqSDG4HOfQrvPzmcK9H6U0w6ktcMbamKMg4AzkBeZr3VVM6ZmXbPIH5UHoBoLm4LLZDapLq4AYGOr7Fy7/stUM1gfqFuoAHxt68CT5ZXlDT3avonB0Ty0jfzW4GcUtXMoTbhcHeAR0luT2QdrtXc0FVxIu8+mXXd4ZESzPWcEDZcY6q1/S6VgfROkE5eTgk53XXylulXR1hroXkSuJOfme6rcbtWXV4krJC9w8yg1vUWrvuzO+QMIDicLbDjkkqrAD3V5gc4FzewQUgHxZV8zzkDP51CCWE+qkbBLI0yAZCCI7oh2KIAz5ZVekrIp3RN3fhXyUj5T1xD4fkgw1fE0PkDT2Kklh6PhxuEZSzFviNHZBkVVCyGHrackjKwFLJPM4eG93bZRICqx3S4FUUzKWWRnW0bIJDWZb04WO5/U/qVCCCQe4VEE/vBI6SThXCrw3pwsYDKEYQXSP6ygc4DGDhWrUqWGN1OS4DONkGm4JOwVek7k7KaHobMQ/sCrpHRmQdJGMoMqz2xtwlLZHYAP5Uu1GyglEMZyDnOFGax9OAaY4J8wrRUmcl1Scu8kGGASfRZUcfS3q6lHM6M56FGJXAYygvllLyWgZ8lF0nz2U1N0Bxc/t81SdzOrLNkEOT6rJpJwyQZWMqg4OUGqXF7HQ5bjPyWmskcwbDKOle4YJ2WRCYRGOvGUGO9+diFPDTtLOs9xuopgzOWqjTN0/DnCC6WXqBb6bKFD81I2CRzeoDZBJRy+GSVbUyF0vUPLsrqVoLyCN0na1su42QVdOXNDcdgrBK9u3TtlT9cHQNgpgad0eBjqKDBP3wgOPdajBanGIVDWkgDPyUENoq6ol0LNvJbmo6ukt1vNHVkeKRj7UHdf2MVUJuaLVNOR9HQVafyXG3/60VPYyU/RzUarnA+F+gq7H23G3FEHs6iIgIiICIiAiIgIiICIiAiIg445kvwduKf8Cr3+ozL5oX7yOHzP6V9L/Mfj9rxxRz2/YZe/1GZfNFloneXduo/pQSvpWth6877LFypXzucC0HZRIJqWfwH9WMq+qqfeiCBuFjKSItGepBZuPkqE5Ur+g9lG3dwQUwfRZtKWCIhw3Vr4miEnG6xQ9w7FBdLjxDjsrEJyiAtXtl5NHCYfUYWkKo+kMoM2ri8Vzqj+6KxqZxZUMcPIrJkqGugDW4z5LGpWh1QwHzKDka0aufbaJniZwCNly7wZ4hGXWtueysMTGvBODjzXAt3gjZaQ4DfHl9Sh0Lf5rTeoHMLuouAaQexQfRfwL1tZ7zoWhYLlG6WNgB6nDJ2C5ObKyRvVG8OB7Y3yvKzlZ1JxCraSAQvn93JGO+MbL0n4em5G1QuuJJeWDv8AUg3d3XAfGrgYdXzSXiNgd0Hq6Vz6rJI2ysMbxlrtig6daV01Ttrhp+ssjSIz09bmBU1hpK22e7xU8FsjAeQMhoXZu86LtccT62jgDJ9znbdcf3TQVZc2PrJw4vYSWnHog2VcuB9r1VpuN7JmQP6eo4OMbLR9KaktvA2b3Ke7NIz0kF2QoNcak1TpYSUtPLI1jGkDuF0K5hOKurXVc7/EeS15x8R9UHpBq7m505p61CubdISenOMhdCOY7mRdxUrZqmjvRaGvJAa7GBn5LqBqbinrq/xe4h8rm9OPpFbEder9bXubV9Y6u+coOU+IetbtcqAUX3alcA3p+mSuFZIpyXOkl6jvkkk5WdW3SoqI+t5O/qtLMrznfugtcMHGVRXNILsuVZMbYQT2+sFHN4uMq+5XD354cBgDyWEqhpd2CCiz6L4CJertvhYPSScKWMyMcM5xkZ+pBqlVeH1Ufuxbgds4WkyxeGe+crVah9GKUFgHiY3Wjue553QCMIM+RVMlASEFRud1Q91cxjnnDe6q+N0ZDXILCMLLp6Rk0ZeXdt1int3UjKiSNvS04CCyRvQ8t9FaqucXHJVEBERBLBEJXdJKrUQiEgA5BUTXuYcgqr3uefiOUFqZPqiICIqjGPJBREPdVb3QUQbqZ8YLcjuosFqC5zMDKsVS4kKojc4EgdkFqqBlC0jyVASEA7KZk/QzpxlQ91XAwgOOSSqIASdkIIO6AO6r07gDuqZwp6MA1MfX2yM59EEZhkaOoscB64Vi3xefuV9xh4XT4uN/VbHQEyfVFVrS44AQXRv6CSkj+vCPhewZIVrWOd2CA1xb2VXFzzkglW9lqdCyEwkvAyg0xZdO8iI5Gfmo6trQ89PbKyqKMPhI2JIQae/dxKosmoo5YnFxG3ksZBVreoqYRHpyHdgoASFd4j8YyguZGXuLc4VJWeGcZyqNc7yO6OLjuUFuT6pk+qIgINiCqtaXHA7o5had0F75A4Ywo0RAyfVEVWsc7sgvhl8I5U0rXTsMwacDcqAQvPktYgkpo7XLE/HWWED60HqD7DX/AHP4yn1msH6K9epi8s/Ya/2hxl/w1g/RXr1MQEREHXP2h/4GnEvH95Uf69Tr54/qX0Qe0H6RydcSOrt7pR5/j1Ovnga0uOAgdJAzlU3V5a8bEYUkfRjB7oK0dT7s/qxnKrV1IqndQGFEYXHJA2VOno2KCz+Yq9smCCB2Vh7qWPw+g9XcoKvn6xjGFDg+iq7usiF0Qj+LGUGMsumqhHhhAx2WK/6RwqdkGoVbWub1jzWCxnW7GVUySdIBOyMY93xNQTTUojjDw7fzWODgg+iudLIRglWAFBOyqLABuVqVHchE0nIBIwtGxhSxwyv3bnCDsPy9cdBw4uLKiSbAY8HcrtVqP2nLXWj7hQynJZ0ZHltheaQdJS4yDk91lMFJURGWSMh477oOxmruMVRr25TXOS5uxLk46j5rhbWEviSuHjdZJzuVtaO51NM4tgeWgbKOe4VFQ7qleSgskhLTkHuqsgywuJVHy9Q33VgkcNs7ILo4HSOIAJCTRGJ2CCFnW+WGMl0hH2qC5SMlkBjOQgxopPDOcKZ1XkEY7rGVQ0kZCAficSpmVDmN6PI91BuCqtaXnA3JQazZr660SeIzOTuuTeFdA65atp9VOqDmIh/ST6FcNvidHgv8yuSuD9wqhfoKKMnwnYGM+WUHsby38y09dp6h0yykP3jEYd0/Yu3tmuD7jSMmkaQS0O7YXRLlji0lpy0UlfcWMEjiCScd9l3msF6tl6twntTgWBoxj6kGHrTSo1VRNpmzdBaT54XHt+1LPwu09La20plJyOrGfzrlGmfUwzuMpPR55C0nV9rsF9tc7KxsbpC3AzjOUHlTzT6rm1JXVMlQ4tD8nB8l1BqtD22a3T1wewOyXY813P51tCVdlnq6qijAjDSQR2xhed1Rq69U3jUb5HdPWRglBpdeGG4PpMYaxxAWHUwCI4BUtVKXP94z8btysV8j3/SOUFqIqhpPZBRTx1HQ0txnPyUBBHdO6CrjlxI81lU9V0gRFvc4UMQDSS7srpHMOCzGyCWqpQ1viNOc7rDWVFM5+WvO3zUc7B15b2QXUdN7w8tJwAs81TaH/a43+a08TOhH3s7qNznykvccn1QZVV0kCQee+EjrwyIx9PdYhc94Dc5wqFpHdBVxL3FwB3VCCDg5CzbWITIfGxhVuMLPE6oRsgwFm09cIoTHjywsJVDHHyQTMYJ5CTtk5Uj6RrQTnssZr3M7bKpmkIwSgsIwiq5rhuQqICmbUFjOgZURaRvhGDLgCgkZE+ZxIB7+ipJCYytRpxHE3JwDhYdbKHSfD2CCGN3SVUgyybZwo8ZWbA+NkfxYygglg6G5yoRsp55g/YdlCGk7hBfGzxDjOFSSPoQB7dwrSXHYoKDdZMVM14ySomQPf2VXOkiPTnCC6aERbqHc7qrpHu+kcoBtnyQWgZKmbP0M6AO6sjidIcNCyPAEbSH9ygxDuSVkNqelhZjyWO7Yq6MAuGeyCrHvY7raDv8AJVc50zhkd1qMRpQwdQGfrUM4iA6o8IIH0/S0ODlC3rYQQDsVUyvJxnbK1yjoopaQvdjPSgtteoBQANczJ+pawbJBfIzcfFDcDOMraE0RbM5o7ArIp7tX00fgxPIb2wg9CvY3hkfM/qiFpHwaErR/zhb0WD7F+SSTmj1S6Q7nQNcT/nG3Ig9nkREBERAREQEREBERAREQEREHHXMe3q5eeKDfXRl7/UZl80gg8SolYT9En9K+lrmRz+134o47/sLvf6jMvmhe97ZXuBwS4/pQWPb0uIVFe5jyOsjYqxAUkcReqwAOdgq+f70QG+aCx8PSogcK50jnKgGTjzQTGcuZ0nsrIow4ElXuo5mR+I4bYUTXuGwQUcMOwqYV7mHp6yp4I2OaSUGKiuePjICz6ax11XGZYmZA3Qadk+qvif4Tw/zCulgfTTGOUYIWXR2eprh4kQyzug5Q4I6Xj4kX1tjrAOgkN37bruzpr2c9K+tor1R0olGA/YZHqukfCvV9PoGvZVscGTskBO+Oy9m+SXitBxN05E6SZkhZDsMg+SDV+BPByHRNLFbX25rfDwDsF2Up4I6eJkcbA0NAGAo4qCnhf4kbcFZKAiIgtexsgw4ZCt8GLo8PoHSdsYUip2QdfuYPTDo6CWrpKYEmMnYfJeTnH+evp6yrZNTuaRIR2+ZXuVfbFRakgfS1IBAYW4PqV5Rc9dhsGhr7LHKxgDpSPL1QdD23Wot9R4tRAQ3vkhbd1Neo7rL1RgDB8lyLru86ZqbM0UHh+L0eWM5XDpySSgyo3icCIjsFHUQiB224Kjik8N2VfNL4hBd3QQ53yEVwjcfJULHDyQUU8IHSSfNQK5r3DYIK56JCe+FcZsjturYnN8QF/ZXSFnigt7ILRHK49iphSjGScfIrUPFphT7Y6sLBf4k7wyLt8kEEkYYdiqRxh5xlZE9tqoWeLINlBHHK8FzM7IJ2tbAfE9FDPMJiCBjCtc927XK1u7ggpg+iLKljaI8gb+axUBBuqnGPJUQVIwqJkogIN1VrS47IWuad0EghyMqx7ek90Ejh5qrXdR+IoKNaXHG+Fc6LHYq9zmNb8PfChL3HzQCMKg2VdyqEEIJWSHZuMq2RxOxGFJRlgky/GPmrqwxkjwwAEGMpY5QxvTjKiVW9wgue7JzjYqSGnEjepXSOiMeBjIULZXsOAdkFHNLHEYVpOVnt8KRm2CVizNDDhBG13SUc7qKoiApYjj4gcbKJVDiBhBkeM+f7255I9MqGVgYcBWtcW9kc4u7oKK+J/S4KxXx9OQT5IMiomD4gAMLKoaVkkBcRusNz4y3Awpoqzwoiwefogw529Mrm/NXRSEYbnGdlZK7reXeqlippZcOaNu+UElRTksD27k7q2mqHQOAO261S1UrqpxhduQsW9UBo5QMADsgkrquOanwMZIWkK8l+Bnseys7oCqBlXvhcxvUeyjyUFWnpKv6utwHqVGpGQSPHU0IMk0jPD6tsrEe3pPdXmWVvwEnZRlxPdBJT469+yrUkFwwoQSOyqXE90FFVo6nAeqoqsOHA+iDLmo2xwh+fJYzHluwGfsWTNU+JD0egVlIYw4F+NkFS9zW9XQoXudJl2du61aaSjMYwAT6LSpnDJDexQerHsNh/1N4yH1msA/NXr1LXln7Db/c/jKPITWD9FevUxAREQdc/aH5/aacS8d/c6P8AXqdfPNG/oJJGV9DPtD8Dk14l/wCJUf69Tr54+6CV0wf5YRsZPxZVhjcBnCdbhsgnjmGekj5ZUdQQXZHZRgkHI7hCSTugomUV8UTpHhvqUFpBHkQpI4epvVnb0ysmpphFECe6w2yOagq1nU8t9FV0JbuAThWBxyT5lahTyw+Dh+MkblBgufkYV0cwa3pIyrJMdZ6e2Vagvc3qOfVXNaAO6j6inWUBxyVk09Z4LOnpWKiCeeo8UKNkrmAgFWKrMdQz2QCHHchSsh6m5PoppjE6LEffCxhI9ux8vJBYRgkeiKpyTk+aogqXEqWODraXk9lCrmyOaMBA6fi6VM1vwqEk9wd8rMgkj8Ih2M4QYbm/Ftv5LU6KjZ0eI8gLAaWeISe2VO+re1nTGdu2yCarj8chkYyQVu3hZNJQapgbK3pHUNzt5ra1mnZDUh1X9Ekd1zPoThrXasqW1FljJkLQR0jdB6E8GLE3VmnKOKiqcvyNmu3XenhVpyo05Y20tQ4klo7rpPyO8KtbWSsgkvwl92a8H4gcY+1ehscTImhrBgDZBSWISMLNt9l0K5reYbV3C3VIt9vpp3Uwl6S5rSQBld+CQAT6brrVzQVnBWTSlzdqWGlfc+k9BcBnO/qg83+YrmYi1np8RzSDx3xYLT3zhdGK6rNbPJLjHU4lcgcZZxPrqsbQNLaAPPhgdsZK44d0MeQPVBfKQIwPNWxwhzSc+Stc7qCCRzRgeSCjGdTiD5K5x6D0qwOIyfMqhJPdBNGwS7KssIjIP291C1xb2U7aaomYZDkgb7oELROejsqyUojcADlQNc6Nx6Tg9lcXyuO+coJpIS1o6BkqLxSAWuGD9SzreWtOajGFHcY43zdUGMfJBiwx+K7BKmfC2MFo81jhz4z6IZXnzQP7G/KPk69gFaS532K6HpEo6u2UBhcw53HzUwnLh046vms6rigngaKcDqCU9vLYC543IQadLD0jIH5FRkn70hZEVLLJKWnOPRJqcQSDqxjKCCaDw2h3qroacPYX+ivqJmPj6R3wsdszmjAQXSPH0QOyuigDm9R8lE1jpDt3Vxc+P4CgpI4fR9FaDg5VWMdKcDdVDfDkHX2BQZEbnSABxOFBM0tdjOVm1MkLYQY8B2PJaeSXHKCrfVBlzsBW5VR1D4ggmfTdLcjJOMqxr3MGC0qekmaXYlwR81kVDqbB6cZQYjJA7vsopMB6EkPJZ2zsqFrnblBNFVdHYbq2QiY5Gyi6cd06iDsUFzmdIzlGHPw+qNa+U4Co5ro3YOxCDMhDab4jjJCsmm8Zwwdvksd0jzsc4+aNcR8Q7hBkupGlvVnf9Kxnt8M7FXtfNJ2JUbw4H4+6CmSfMq4OIZ0klUY0kqQ00hGcIIhscrPhujooxGM4xhYToXt8lMygqHs8QDb5oJAGzOL8ZypmwgEE4/IsaB5gd0yDsVm4c+MyM7IO+/sZGtbzQaox/wAAq3+Ubeix/Yuyl/NNqpp8tA1p/wCcbciD2iREQEREBERAREQEREBERAREQcc8x5xy8cUT6aLvZ/5DMvmgk/sr/wDvj+lfS9zJfg7cUz/3FXv9RmXzQu/sp9Oo/pQSxOMmGEbYVs8Ij7FZJMXhDw9nLDk684egox3Q7KrLJ4hB81YqgAoLoY/EfjyUksYheHA5wQVHhzB1DZUc9z8dRygzZbl4kHg47DCwBsrwGY37q3bPyQZVMPHd4Z7eS1yG0QNpi8ObnGVokcZLQYtipBJco4z1F3SBvugxalojqHNHYFa3bdUGhg8ARggjB2WhNbJUzBjQS9xx6reVq4Y3a403vPQ/GM7NQaDUxtuUpnGxK1Ciu0dmpjBgEkYUFbZayyzmGXIDT6LR62XxJMZJIQK2pdUVL5g44ccrvp7PTja3QFdSWutuhYJHABpd2z5LoXSQeK7ftlbh0nqi56U1JSVVJI5oila4jcZGyD6YdHakpNUWSG6UtQ2QSDJIK11dEOTDmgtt103SWe4TgyABuCd84AXeO3XOmucDJ6Z3U1zQ4IMxERAVrt2lXKhGUGwdb6yk0YyarxkdBO/yXjbz+a/qNeX2onDjhkxP14K9juKulZNRUEsUTCT4Z8vkvGnnR0qNL3mqZUsLcyHy+aDpkTJ++LiPnlZNLC2QfFhahcG03u7HxYyRlaOyZ7OxQZFTTtZu1RRwBwJJ7Kx00j+7kY9wIGfPdBeZXN2A7bKx8nVjZZnTH4XU4b4WG/GRhBYr4o+sgeqoY3AZ8lfE/pwR3GUGX9z2dHV1ALDlj8N2BurveZnfC07FXtIDCJPpY2QY4DnbNySfILIpah9LKHPb29VqGn44GVYdVY6D2WbqKmpJ8PoWjA74QYFbfPe6fwRHg4x2WnwVBhaWkd/kkDWNkxINwlUYy4dCCP8AsryT5o5vhuCCN+Opqtd1Z+LOUEzpepgaoCMHCdkzlAVWjLgPVUQHCDLlpGti6wd/RYivM0jh0k7KzGEFzHhhzhZTI2zs6jhYaubK9ow0oEg6XFvorUJySfVSRdHUOrCCwg+hVFmzGAx/CBnCwkFWnBRxyVREDKZREEkMYkdgqs8QjIx5qNr3M3aUc9zzlxygoiqcYQYx5IJqUZf8kq2gOBHmo2h5J6PzK52cYf8AnQRIq7Z+SO7oKKrR1OA9UbjzV+QN/RBLLTNZEHg7rGV7pnuGCdvRWYKAqgZOFRVacFBc6ItGe6sysphDxjChlb0nA9UEazKau8CMswsMghahR2aprYvFjBwBk7IL6G8GjmdIAdzthW3K5SXJwf0nZWMoxFL0SjcHfK1JhoI2AOwD9SDQnPcQGkHZXRsBw7PZZVa6nOfDwsEPcOxQTSz9Q8MD5KDspqbo6z14+1X1EYLh4YG6DGAycLJiqRE3pUYgfjbuoyC04KC+STqJKjO6q1hd2UhppAM4QRsb1HCq9nSqDIPzUkQBI6/zoIsH0KYwtRa6m6dwMrEqCwn4MYQQ7lNx6qeljD379lkVEbGY2HbugwCT55UgjzGXnur5ejG2FD1uDenOxQerfsNj/wBT+Mo9JrB+ivXqYvLP2Gv+5/GX/DWD9FevUxAREQddvaDxGfk64kxNGS6jo9v/AKup188Iy1247HdfRVz4yxQ8pfEKSbHQ2kpM/wAdgXzuTmMyzuYNi8lv1ZQWGbLcKI7oiCrTgrJjgbIzrP1rGAJ7K4SSRjAOAgte3pdj0UkM3hEH0UeHPPqSqujc3uEGTPWeO3owViEYV7W43KY6yANyUFib9lPLSujZ1kHCg7IJXRAMBUSuLyRjJVG4Dt+yC6Nhecb4WQaZgZ1HurZHsawFgGfNQ+NIRjKC12ziqKpB7nzVEBFVuM7qTwXOHU3sUFIBl26rO0NIx2UYJYUc4u7oKKaKEObkjKhVzZHN2BQVMYBKtcMFCXdyqE5QFXB9CqLPpnQeF8WM4QYCqM5zvsrpsGQ47LJpnQBhDwM+SCsMRq3RN7HIH513g5NJ3UmoqKjloxLE4tDiW5Hluukccb5Zofd8/THb616K8mwsNLTQSVbW+9BgwTjOUHq3oyjtNu0zSzUVNFH1RBzukDOcLX6So94j6yMLgXS3ER9io43XSQ+7EgNB7YXM2mNQ0Oo6MVtvIMeB2Qa07dpGcZBC8ifaGjVrNZSQ2185pjMQQ3OMZXrTcJXyRPigPxtBzhdWuYfh1pW/WapqrtEx1XuRkAnO6Dxk1tb3Q0DJqgffiwFxPfOFxcKZ8gc8DsSuznGjhddam/VEFua7wGZ6QBthcIVtgfp5j4K4Yfv3QbL3YSCNwmHHfpU9YzExc3sdwtRofdDSHxMdWEGjIr58eK7p7ZVndAGy1GmuIZGInDvsSsFsTn9lXwJAcY3QZE8AaPFbg532WOJMHJCyYmyM2mz0481DO1r5PvfYoLZJ3OaB2x6JHOWtIOSkkJY0HbPmqNp3ub1BBR7+s7BWYwpoOhrj1jsr5og8/ewEGMDhCd1c6NzO6p0nGUE1LUmB2SSQsx936hgDC0+KF0zsNSSIxO6XbFBmx3IMJcRuoJ6k1T8YwCo2xHAceykZF1kNZgE7ILZacRt6urOQoFlz0NVC3rlBwFjBnW7DcIKxSGM5AySksniOzjCngg6Tl+MKOpa0OywbIKQS+E7qWosoo6yEz9WCBnC0ojACkZVTRs6GvICCyRpY4sznBwqA4T4nnPclTCLDCXDceSCBXB2AQQr4WguII7K2YAO2QWZ80yfVEQVa4jsMrIaMtyRg+irQMa95D+yrWkRu6Wn8iDHe7qPSN8Kzssuiaxzj1AKysa1smGgYQWQTeESfVZZiZUMMud+6wDjAx3UsMsjSBv0oLXB7nFoB9OyoWuZ3aVq0Puz252yVDUsj3AAzlBiU0nScFWVDg56yGRhpJOCsafHVsgtjeGLKjrNunH2rEa0u7KdjWsaesYKDNjjZKOo4V77k2nb4AGfmsAVJbswqGV5e7qPdBWZ/iyF+O6ngrXRx+Fj5LHjLQd1mxxRuZ1DfZB329i7GBzSapf66Brv5RtyK72Lzs80mqW+mgK3+UbciD2gREQEREBERAREQEREBERAREQcccyX4O3FL+BV8/UZl80Ls+I7bfJX0wcxxA5euKBPYaMvf6jMvmppohNVytHYdRH5UGNASHqSpD3HZhx9SmbGyCb4sYC1eKOmngLhjIQba7JupapobM8DsFLTtYWfF3QQeJtjCsUrITLJ0t7Ks1O6N4bjcoIVPDTmQdSklopI4g/HzV1HMyNhB7lAZKKY4+a3dp+y3O/gQ0dC6UOHcDK2XUtOz/InZeg3Ifwis2taCH36JjpXsBGQO+EHBvCLlru18ubJ57Y4/ENi0nG67zWDlW9w0W+odZm+IIc46PPC7M8NuWel05cDWSRMEJdkbDsud/wBjNrbQmh8IeEW9JGPJB4DcZOGeo6XVtRRU1jkDATuGHHdcPXrQN7oS99Tb3s6TvluML331Hy88P9QXWWeSCIykEn4R3XSDmv4U6R0XBVthja0gnGGgIPNG2UbmVPgzM6cHHbC1KWzia5RxxAEnbbyWrX21irvBjtY7uxsPmt26T4ZXuoqWVMzXEAZCDnrlX0tcLXUwVfvjwOsHAcfVeuvB+qdUWGAPlLiIh558l5g8AbNVQVEdIO4eBhekfCf3u3UVNFJ0gFgG5+SDl5Fi++xtZ1yEDfHdTxvEjA8bgjKC9FhsuDDVGky3rB9Vln5IMWtq6Wla51Q4ABhzn0XkX7Raz2+93CqqKVzRiQnbHqvT7ifDdfcnvoSd4z2+peT/ADZ2bUlfW1bKkyEGQ+p80HQm6wyU8raduSGjGy09zHN7tIXId10u2xPFbdQ8MxnJC2vdK611bwKNhH1jCDQwd84TO+cLMqYAyMPHp5rDGM7oJHzFzejyVjPpD61fDH4jy0foV9RC6IgkIMueNgp9sZK00HCvM0jh0k5CviYMfEO/ZBSmID8laiaRkzfEBwQtLcelxwp46qQMLQdkCecj70zIwcLVLUWmmJkdn5LQ3HLic7kqSOoljHS04BQVqh/th+M9/RIoeodTjjCyYvDA6pfPdQ1MzeoCI7BBZ43R8IGcKxz/ABHAdkZ9LLle6PqIMe4HkgPgDWdWd1CpGyHPS7sqSYJGCgsVWjJwqKWOJxId5BBe+mLGdY3WOSfNaiyZgaWPxttusSpLC4dHZBCqsGXAHzV0eAclHkZy075QTSUwZH1g7+igYwOGc4VTK9w6SdlaHEbIKHZEWRDSulYXAZ+xBA1vUqvaGnCO6mOLeytJJ7oCJg+iAEoA3WbBRskj6yRkLCwQpG1ErB0tdsgpKzokLQc4ViqXFxJPcqiCeBwYdwSrah4e7IGFbEQHZd2SQgnZBYqt3ICrGzrOMKrmGN4z6oJH0xYzr+We6gWXNOx0XSO5CxEBXxnfCsVQ13cBBlClD25HcqCWIR9iqxzOad3FXuY+dwDTlBCx5YchVy6R4Jb3Kym0bom9cvbY7qZr6cMLhjIQQVMDWwtcDuFqtmvjKKldE5oJx+RaLJI+d/Q3OFGWOicA4IMqsrXT1BkDcBxUBY6QdfUpJGx+F1YGcKCNzgQM7BBaWu8wdlTGFqEboA34sZ81BNGJXZjxhBjDPksqBwJAcopGFrR6hRhzgdkGo1BbCwEb5UlLb462B07jg91htf1tHiHIUgrXwDw4zgFBFG3wqgx+QKnqakN+ABWeNGT17Fx7qB7XTO23QVp4/GlWVUUha0uGBhRwQupyJHBZD6lssZGUGlnYkKivdjxCT2VrsdXyQZFER1rIqmBzcg9lih7Wj4e6Cdx2JJCCEjcqinaWZyVbMG74QerHsNjm3cZB6TWD9FevUteWnsNv9zuMv+GsH6K9epaAiIg65e0Qz+0z4l9Jwfc6PB/+rqdfPIvoe9oW3q5N+JLR/edH+vU6+eJrS7sOyA1vUVcYwOyozY7+Sq95JQVgGX7qtQMEKIEjsq9RcR1boL4XBrslSSvBKoHMA+eFE92TsUFXSZGFSM9MgPoVaBkqo+F31INSqp+uAABaaBvhZfjxlmDj7VjPI6stKC4xBrerKiJycqUyZGMqI4zsgINldtjyyrUGfb6dlW4sccD5qO40jKSUNYcgqCGeSA5jdgpLPJOcvOSgjWVFMBGRj5LFVzN3AepQHZLicH8ioc+a1KNkLG5eBlYtWY3HMeMIMZTxQB7OslQK9kj24aDsfJAkOD0jyViy2sDR1vx2UE3T1ZadkEaZREF8cfiHCnjoy6Tp8vrUVOQHbrWKWmdM17mZyG52Qb/4XaDkv07GwQ+KeobDddseEmmdRaa1XSQGhkjhyAT0kDGVtnkD4dT6x1FFBO3LTKBv9a9T7lwA01aaJlYyJnjsYBkNHcBBtis0Y3Umh6UQS4mawEgd1rHDvVEWgaU2atqx1HAAcVoltv8ANpGslprm4imaCGg9sLiDiLqO43jUrayzOJga7JIPllB25h1cZM1cDusSb7b91tPiBpal1NZJa2peGOIJwThaTwa1PS11AyluHQXsaM5d54W0eNfFptturdO26QDxHdIAPqg6ecdnU+lrjVMipWvABGQAV0d4i1Ul8nkqWxdADjtj5r0V4z8PLhcrY691rSWSs6skd8hdLtc6BdHRz1EEeGgk9kHXGpc7r8Mt7bKxjntBGSMrV6uCKO5Pp3DcHC0+vYI5OlowEGIfP60GyuAxufNTinc9nWBkAILI5+gnbKmZUBxDsdliOGCR6KgJQZlTUGVnS0fkWNG4scCQe6zbXC2oeQ4dlk11HHD2G4QafNMC0DGMrUqJkTqU9WMrSZW5OB5IyoliaWNOAgpUNDZnAdlWObwx0q0MfKc5ySqGMg4QVkk6/JXMJ6ekDuFY1vxBp81qMFLCWZLhlBDRAxOyCMqWpp3yyB58ionh8c2WAkD0HzWRJO7ABGAfNBC9nU0M7KaipxDKJHEkDy9FPHFGYvE2yN85WP44JLR2CDU7pVR1VN4QAHzHdaPHTNiy4OB2V+XO+kSR5ZVJGAAfGUFpc52QQAMDdY8w6D6jCzYyzsfJUlijd5glBppOVMync9hd6BTGmYBnIWTTlrgIhuTsEGmNJY7GFlhrXxlyrXWuamaZndjusLxXgY6tkFcljiPsVrndRVDuiAASdt1I2F5BJBAUtF0B+ZMY+a1CeamEZDcINMp5PCcc7FUqHdbupWOPxkjzJVCSe6C6J/hk5SV/W7KsV7QMb+iCxXh3wYwq9HWR0qcRBsZ6hughhkLHZBKukmL35O31KyPpDvi9Ul6SdkEz5R0gDIysZxyVQnKuBGN+6BG/oOVVzvFcBnurPqVQHDceSCWSEMGc+ShVxe53cq1BLDF4pIzhSumMA8MHKxwXsORsp2M8SPrO5Qd9/YsOL+abVOc//ECu/lG3InsVhjmo1UP/AOH9d/KNuRB7UIiICIiAiIgIiICIiAiIgIiIOO+Ytok5fOJzD2do29D/AJDKvmtpCIK+obnIaXAfPBX0o8xxLeXrig5vcaMvZ/5DMvmmZKWVUjndyTn8qDJmjEzidgDuoxUSUoMbckFRz1I6cMO6kpHxOaTNglBhSPMjy49ypIy4AjCvzGJj6ZIU0j4eghuAgxopnRSFzRkrK8TxyHvGCN+yioBHJUYeMgrIuRjiAbFgZCCOor+uLwgAcbZWC0EOA7KaCMF4e8ZCrVFnWOgIM73dktM1pwCCD9i7b8rnH6DhX4LjUdAiaOx9AunjKl5Z0tzlajS3aqpYy0ZOQUHufy486lm4pXOOxzVTA7rDcnGfRdt/HppW9InYQ4eThuCvnT5bOLlboHVDK7rLQJA7PUR5r1A0DziU10ipBLUMJLWg5f8AIIO7JtVroah1bUTNaD/dHC4N5hOAGjeJmm6+5+/RdbIy4DqG5XGvGHmtgo9OOlop2B4Yezvkuml5509a1UFXb6eueInkjHUeyDhriDoml0Hr6WhpwHxxyEDG+wK3jpjXFujlipJmMYDgE4xhccar1828XF90ucgdK/Lsk+eVsK5axfLUf9TXfHnbBQd04te2jQdCL3TVTQ/6WAftWsaU9oA5tzioX15Y1rg3vjZdG5LxrCvpg2udIYMbZJxhbJu1S+gqxLTtLXg5Jz5oPaKLm4tNXp5tYb0A8t6sdfyWx672jMGmuu3MuIfnIBOCR9S8pWcTtVw0wpRVOEYGMEnssAXu4XVxlqAXOByTk7oPYfhVzlUOqrz77V3jAec/E/5re+tOdK22e+0tuproCyRwBwQvFSz8R9QaZm/2k9zMYx8RC3XRcUb3qCdlXVF5lYMg9RO6D3ssnFvSuq9LRXCW6ROkkZ9EuHchcR6v4H23ifW+LTUjJ2PdnOARgleavC7jFxHq5IrZTSzCFrw3Zxxheq3K3qWc6cjF8lAndED8XfOEHC/Ej2eFt1PZXQw0UQkEZ2bjOcLzj488qlz4R180cdK9zY3ludyBuvf2S9UskLhDIOog4yV0y5p+H1t1BbqyuqsOIJd2CDxhuNpniibHJCWkhbbqIHQOLSMYXOnFKittovslIwABuQPyrhu8mKWd3g7nq8kGBC8Q4k9dlWebxyB81Y+nkY3qI2UfZBlNpwG9Rwfko3v6cAA7K0zOxgfpVGMdJkjyQVhZ4r8bblZbqZkYysWnyJNtipKmSQEDKC0QgkjON+ylFO1oJ2z3CxfEcPPdXB8rvUoJQ3xT0Zwo5ovCIGcqhL2bjbdVD/EBLjuEEfV6K+OXoGFY7AJwrmdPmgtOS4kZ3VPrWXD4ZOHKOoDMgNQQLLgcBF33WKWkDOFUdeNs4QVlOZDg90azqBOUY0yOx3Kuex0QwRjKCI7HHoid0QERVb9IIKYWVBWeCwtwSrJCws+Huo2BuDnugpI7reXYxlW4PopaeIyPwM7LKkiZCMOxlBhHsqA4UoLeoq18fVuzcILCc7AKmFLG3od8ewVJywu+FBGiJ3QEUvu8mOrGyjIIO4QXRyeGc4VXyeI4bY3UaAEnZBM+HpZ1dWdlZFGHuwSrnMmDep2cKNri07FBkS0zWN6gfJQB+BjCu65JT05yrHNLDhwQUJ3yFNBOYiCRlQ4V8YHV8XZBmTV5qWeHjG2Oyw8ODunJGVkSGEN+DGVjtI6uo7lBktYKZvi9yVE94neDjCva2SqIjYo5IZKaQB+yCSZvTEN/JWQxBzckq6R4ljDWndWNLomnJwgr7u55w0kqpLqfDXBTWydjJiZTkLNuEUVS0vhAP1INLBM7i3B3VJIDGMnZS0TmwznxNsK64zNkeOg7eaDFL9sA4VmSVVrS84AWQ2nc1pcRj5IMZSw1BjO4yo3DDiFJGwOG6DIdV+OAzGFNDSNMZOe4WCYXA/BkrIiqXxsLXdwgiqIBG44Kx8H0V8krnk5WVTmHw8OxkoMQtICt+pSSscHE+SvgLBjqQRN6wcgFUfnO+VntNOBusSoLS/LeyD1X9hrn7n8Zf8NYP0V69TF5b+w4A+5HGI+fvFi/6NcvUhAREQddfaFv8Pk34kv9KOj/AF6nXzyRuDSc+YX0M+0O35NeJef7yo/16nXzx4QVO5JVERBLTxCVxDjhKiEROAHmo2vcw5COe5+7iguiZ1kDyV0sQZ2KjaXA/Cr/AIi4dfZBqOn7fHX1JZLgAeqrqG3xUFQI4iCCoG1jqRvVTnDvl5LGqaueqd1TOJKCyKPxHYJVXxhsoZ5FWs6s/CN0eXh2Xd0GRNTNjj6gdyrIoA9pJKjMr3DpLtlRsjmjAQVkZ0HCswfQq9rskdR2ystroOnfGUGE1uSjhhXyOAcS0qzBdugoro/pj61aqtaSchBqT4WyR4DuwWnyR+GcZyrvFmZtkhRueXHJQUVRsQfRVjx1bq+Tp8igPm629O4USK9oBCCxVaMuAVXNxvjzVoJByO6DKNN0hrmnc481vfSVuZUghwBJbgBbIpZ8PAeRgdlvnR8kjK+KoB+9AjPphB3O5ONbR8MLtBVzxhg8QHJ+teomkeJdu4gRQkSDoIyfTBXi7/snWOyW6COBzPGaRnB3XbHgZzU2WzaVMk0rBKyPAy7zwg7z8TuG9o1DbZZKGoaJGNJwDv2Xn7xe4nQ8J7nLaJ6jDustGfrXIWi+eSgmvdZTXKrYYTloy/bC6Lc6fE+n1trP3u0SAxmQuy05Qdh9Oc0LrMRVR3ItEm+Mq/8A2TKPWup4L5U3cEseDgu+eV0Tg1RUx0EYlBy1g8/NYlt17qWlujJKOV4iDh8IzjHrlB6ecUOL0N10gy1UDRKWR9OQM52XU7VV4vD7LURyW14Ds7lpW7eCfEiy3Slhg1D0FxwD1H/WuwGqNOaB1NpOWntMcBnfGcdOM5IQeV94iqPu5LKyP4i4khaXW+MZSZmlpz5hdlq7l5u9BqqruNXG8UpcSMjbC4a4pWamtFz8GAYw4goNjOdkALIjqzHH0YUL4nMaHO7FSwMZIwg9wggcepxPqpY4A9nVnBVronRu+IbeqoZHDZpwEGRRVLqaQgAndZFXWGcb7ZUdubE9x6wCc7qlxYGyfe+3yKDF8T4jkKRsPiMLwFA7H2rMpziHtnPZBjsk8MkHyVQXSPVHtcXZIxlZEbGsZ1ZBICCGWJzCHHO6jMsgJHUVM9759u2FA6NzScoNQpauEMaZRkhWTVEbiMYA+SwBnOyvwUGaXvbCSHHHmoqdpkkAafrVsbZ5WSdDHvbG3qeWgkNGQMn0GSBk+ZA81ax5jf1DIIQZj3CPEZzkrHnEjTlxOCsqmDJ8ueACPNQ1UgDjHgEIIBO9ux33VSZnDqycK3IBBIz8lkGqb4fQGAeWUEDS493FahQtIcH+nqFiU7GuJLnYAUxn8P4W+XmgzrlXe8RCAgYG2VoMrAx2AswuDwSTgrCkB6jkoKdO3dU81UE+SogvcMDZWZPqq5JV3hPxnCCwZG+FMyMO7q6ljD3EY3VJ8xvw3YfUgqKcYO6iI6XBoTqkbuSd0Zl0gJ9UEmDAASO6zomRzQdZOCFj1TR4QP2/mWOyokYOkO2QWyNw9wHkSrWtJcAspjonD4u/mo3lgkBb2BQJ6cRNDldFStfH1k7hJ5hIzA8lHDM5jgCds9kEbmlpIPksqKJpiLjjKVDo3M+HvhYwlcG9OdkF8MQlkIJwFkvomMHUCDjdYbXuacg4yrjUSuGC5BSR2ct9FfHP0NDMKE7q8QvIzjZB3+9iyz/rptUuz30BXfyjbkT2K7j+2l1SD5aArv5RtyIPadERAREQEREBERAREQEREBERBx3zG/g9cUP4GXv9RmXzPzD7/IB36j+lfTBzGEN5e+J5Pb9ht7P/ACGVfNG4B9XL6dTiPyoDKUuYHE4yontLDsdvrWXLOGs6WncLD+KQ47oLcq5jepwBOyuMD2jqI2Vjc5wPNBnRU7Ym+IHb/WoZ+qV3fOCqdNQGdTielT07BI3qGMnKDHdIBH0+YVYaYzMLsknyCsnjLXdjhTUU/hPDT2J3ygg6ZIXZLDt8le6pcduy12qdQOpsgDqI3W3pA3rOOxKDUbSZXS/BKWfPst9WviJdNMuYGXST4e3xFcfCpEMf3vYqHonqyXncDzQcuXnjPddQ0Ro5bnKQR04yVtiKsmjifI6qJJye62hE1jD0sb8X1qd1TURs6HE4KC673ieqe6LxSQD3B7rDoKx1HVMnySAclY793k+qoMZ3QchScQmy28UobvjGcLbNRK2uc6Rx3zkLT6c047gZSpnIcBFsEF1XIHjwgO3nhUpa73RpYN1Y+RrmfP6ljxjqlAdvkoMqd0lWerGPPss601pt0wy/86mqKaNlAHxgA48losYkdKHnJwRklBzlw64hu07Kypxn4gR+VegHLzxt+7klJA6rMYwAd8LzDs12t8NOxkoBIIzldiuC3EOntdbBHS4BGBsUHsPYrjHV2sVFPUeI8jtnOdltrXGk2aksNRDVRgOeCBnzytjcruqqvU7mQVTsxkgDJyudOIGnK6o8FtqBaw4zhB5L8y/L0+2XSe4sgw3dwK6i1ukGwSTFwGWE+a9kOZzhFc7npUzRNPi+EcnBznC81tR8AdYQw1td0S+GxxPYoOuV0Y2PMIb22WlNps5yey1/UdMbbc32+oz4rT0nK0aRj2OwPP5oIGwEkgkK8ubT/CBnPmrnQygdQysaTq6vi7oJqUgzZPYqtb09Q6Ssdri3cHdXta+bfOcILACTsMqQSdAwW7qWJrWH48Z+ajqC1xBYgse8O8la0OPYKrY3OGQsmIMjaQ/vhBiHuivd0+IfrVrsZ2QUVcnO+6omD3QZD2jw1E14aCMK0vJGCqIL4pPDd1Y7q6WXx3AYxkqJOyDLlpGsiEgIysRXmWV46S4kKwgg4KAiIgIr48B2+ElLSfhwgugm8F3Vj8yuqJzPg9sKBAcFAUjJukYIyqEA7DurmdIHxAZQVb9/OMYVs0XhkDOcq/rawktwCrHl0hH6EFrG9RwrzGGkHKtAczfsqF5O2cj5oMj3v4enp27LHe/rOcYVqlbA4gkeSCJVacEFUIIOCq4KCd9SHx9HThQEE+SpjCyIjH4Z6sZwgiikEZyQslsLahviHZYpb1OOMYUrJHxNIzhBE4dDyPQ4VO5AOyuYQXku80kxkFqC50PSOrKiV5e5zcE9lZjCDIo6oU0nWRlZE7xXuD8Yx6rT1k07iG/CcFBNDShjs5Bwoaz6XbGCpYnyNcS7OCoKqQPfsggysynrfBiLDk+iwxjO6u6CdwEFXHxHlw2zurS0g4yqDIO3dDk90EsZ8PcjKkdVlzSMLH+I+uEA3GRsgOPUSfUrJpGNe4Nd2OyjeY+gAYyrIpHMcCCg1apgZBF1jc+i0mSTrOQMLImqpJI+lx2CxmMc87IKNb1FXhhDh6Z9UMT4vicNlfHTyTHqaMj1QZLIhKzpKxqinMJ2O31rKY/wB8XdRTzNk322QYeT6opXAPwGYVcBsTgcdX/tQeq/sNj/ANTeMgz2msH/AEa9epa8s/YbA/c7jK4+c1gA+wV/+tepiAiIg65+0O/A04l/4nR/r1OvnjHdfQ57Q78DTiX/AInR/r1OvnjxlAREwUBExhEFWnBVxkBGwwrEQM5U8dOHs6sqDB9FLHI7HQPs3QUY8RPPmryBMerGMqx8L2guIVjXOb2OEF74g0d1nU9ujlgMpO4WAZC7uSpY5akMwwnpQQyN6JC0eRVWRlwznAVrslx6u/mrg5zANyEFpbg4yqg9Oyqw5d8RSQZ3CC3IysmJgazPmsXspYnPJwScFAe8SHpxhWPb0nvlZErOluRjKxiSe6CiIiCSGPxCQfIKsjBE7AOVG17mnLThSCKWUdeMhBYX5HZWoRgkeiqGOPYILoovEOM4Wu2+/Ot0YpQTvsT6LQo3ujcqPdl3V5+aDVrq6eUCcVRIO+MlT2zVl2oKV1NFcJWA7AAlaN4k0zRHnICiewsOCg1FmoLzFOZ47jKHE5J6ik91qq+QSVc7pXDzO5Wmqrc52QZ1TXOfGI2uI+Sz7VWNbHhwyfVaNMwgA4WVSzMEJa36WEG6LfqOqtMrXwVjm4OwBXNnCjjjcqe8U9LU1L3xggbk4XXO2Uc9RUDxAS0uwuXOHul4qq6xRQR/fTjpI9UHdy+6no9QaWa6kpGvlfHjIAznC6m8TOGctx8WulgDXhxcAdiu0nD+xHSNqjqtTNIgcBjq7YWoas4HXLiXSG86XY80QHU7obkY+xB50jSE9TVGkkg6WsyAcYWkXmyNs0nQ0jv6rsxxO0RDoqaSmkaROwEHbByuBbjpy5X575YuogHOUGzZYzMMNwVgyRmN3TkFb3j0JcG9LXtcC44GAt1Wzlp1teqE3WiheYAOonoJ2QcPxyPiPUMgFSOndKQCNlvW7cNLhb5TQOaRLGcHbzWkT6Sq6CIyTtOR54QaBJB0M6sqSnkDR8ezQqTNkfKYRnY4xlWtjcSYnbEbIMuofBVNDIQOpYpilhcA7OB32U0ETqZ3ikElRVVWZj04wEEhljcwBmMg74Vs0kXhlp+kuauX7kv5ieY6aKr4daCqRZJHlj79cyaS3MwQCRK4ZlIJGWxB7h6L0p4A+x34O6HdBfeON/qdfXVmH/c6n6qK1xO9CGnxZsHGCXMadwWEFB5PcJeB3FvjlfP2PcKNAXfUlWCBKaSH7xTgkAOmmcRHEMkbvcBv3XoxwB9jG54pb7zIa5DRtI7T+nX5Pr0S1bxgeQLY2HzxJ2K9OtLaS0toeyU+mtGactlitNI3pgobdSR00EY7fCyMADsPJbQ5huNWneXng7qXi1qUh8NkpC6mpi7DqurfhkEDfPL5HNBI7N6nHYEoPK/2l2p+D/BintPKDy/aRtOnLdRCK8askoG5mqpyM0tNUTOJllLW4mcJHOBLoCMFgx5+Ng8aTbYFanrTWOouImsLzrzVtxdXXm/101wrqh23iTSvLnEDs0AnAaNgAAAAAFpbKsxDAQXzxvgA6SQCsYkuOTusmSbxxglYxxnAQUzlBssiGFjxk/nUUjA12G7oL443uGWnAVHZaSDuVq0GmNWsaC3TF2LSMgiikOR6/RWrt4ScVpiHf7GWqyDuD9xanBH/AJiDaPUVaIw87rUq+01lqrKi3XOjnpKyklfBUU88ZjlilYS1zHtcAWuBBBBAIIIIyFp7hhxAQROb4ZON1Zu45OyyGxlpzJ5qNwD5A1h7nCCMHBypfH+EjpUtRbZ6eMSvBwRlWQ+F0/EBlBZFMYnF2O6Of40oKTFpPwgd1axjiR090GVLE10YIxkBYo+9u+orJ6ZWD4jssaU9TtjlBdJOZG4OVF3VS0gAnzU9PEC4FwGPNBAeod0AycArNqzD4eGYBWNHA92HY2QUMbmjq75UZJ81miSPp6XdxtusWUjq+Hsgs7qvTtlSQ9Gfix3Usgb0/CBjzQRU8IlJBKTxCJ/SN0gL+s9Bwk/UXfEclBEp2VAa3oxlQIBlB6A+xYb/ANdLql2e+gK7+UbciexXbIOaXVJPnoCu/lG3Ig9p0REBERAREQEREBERAREQEREHHPMeM8vHFIeui72P+QzL5os9M7/rI/Ovpd5kATy8cUQO50Xex/yGZfNI2CSaeRre7SSfyoIXnLir4nBpBKueGtHScdQUIJCDLmmBjxhRU7MnqJVgDndky+PYdj6IMies8SPwwFZTTPjeG4OCooQ0vAK1VnuzYsux1D9KBVQRug68jJWkEEE48lNNUyvcWNcS3PZTw0hMfW4bkZQYXW8jHUcKivlADyB5KSNrek5HkggWXT1vgxlgb32yscsLnYaMqjmOYcHYoJI5cTmTyO5UtTU+I0AD5LH8N4HVjZVa5u/V3wgjJyq4J9Vd0OeT0bhZULGRtPiDfGyDD6SBlTwNDwerPdWSMd1E42Vpf07M2QZwpGkZ6u6xJWCKQFvYFBPM74Qd8KyQvP00GW+5PfF4QBwFA2oLQdu6tgYHO+LssiSBpb8IHZBiF7z++PquSeG1zno6qOpdOWhm+crjprWNeA7H2rc1oukVJCQwjOMYQel3Jtxtt1Fc4qSuuQjAkA+J2F6bWvU9gu1LFPS3OnkD2Bw+MZ7L5utJa/1DYK/x7U94cHA5BK7WcMub3W9ndBFcqqQMYAPpFB66cU5bVVaefG4xyuIOACDthdUtV262nS1xhdbWAvDgPgA9VtPQvNhBqmIUtfO1xOBhzluLWGs7bX2SVtK5n3xudig8tePukPufrCprYouhpeTjt5riB1QXvcQOy7TcxlA2WtmqdsHJ2XWihtrap8rGtJJccIMOKsL/AIC1W1EIf8Y2wtSl0zV0mZng9I3GVplVLh3SNsd0GMGFzukDdZDXGAYLSpaGSEHL8faldJC7ZmPsQYcknWTjYeisGyqGknATpLSA4IJI5+gYwrZHmRwIH1BXyRdLOpVpDEJAZMYQQ9DvMEKhGDhaxWPpDTkRgdWPJaOST3QFkQRtczJWOpIxK76OcILXtPWQN1b2WXF0A5eBnG+VG8NM4Ix0jB+xBJQUclRKB4ZI9VqH7HLxPVshpKCSXrIA6W5C3tw/03Fq2RlBQDM2QD0913b4FcvNmoq6km1LC0g4PxD/AFoOit04X6tt9pFcbHMBjqJ6Cth1FNVQOIqIXsI2PU3GF71XvgdwtvulTRUVPAZRERgAZzheXHNbwWg0Zdpxbog1niHGBjbKDqthFqtdbHUkLS4b4WlYygIgBKEEICIrxgt+eEF0DQ526VDQHAjsrPiZnGQq9Yd9IoLFe1/R5IyN0rulgypfBMQIeN0FhcHjthRuGFcY3t3A2VhJPdBk0MLZpelxwFkVpZTHoacrEhbKXfedj8ldPBUjeXJ+tBcyFrwH777q50LPqUEUpZseyrJM5xHSSgvkiaG5BWMRg4UvhzP33wfmrHMeNyEBj+go9/UUjjdI7pb3VXwvj+kEFiDdFUAgjZBMGYG6ifklSPlHQAO6vhaHRlxxkIMctIx81n0dMXR9Z/OFj0zQ+bpcNgtUfJHDGWgjceSDTppSXmJo+pYr2lpw7utQpDE6cueB3Udy8PxB0AYz5IMJXsk6RjCsRBXO+VWMdUgHqVaBlXiOQfEB23Qat7hCIQ/IzhaXUNDXYHZVNXP09HVsonOc4/EgoRhEyVVrHO7BAALtlJG/wiMhXQNDHZd2VtQWlwLUGTJI2aMAdwqRVYpW9IGSoaZ+HYPZKnpc7bugrUTeKOoeagHp6q9sEjhnCtLC1wae6C9oEfxEFWyO6zkBSSwSNYHnsoQR0n1QervsNnZtnGRn9zPYD+Vtd/qXqUvLP2Go/wCp/GU//RrB+ivXqYgIiIOuftDvwNOJf+J0f69Tr54wcHK+hz2h34GnEv8AxOj/AF6nXzx4ygK5h3DfmrVIzpAyfJBLLC0M6gd8LGV7pHnIJ2VY4XvwQNkEeD6IN1kyiMNxgZWN9SC8j4cq1julwKbqiCeSp62BmFArg3JwFVrC05cNkFmD6KeOoLG9OPkkvhhmGgAqsXhdHxDdBGwCR+eyTNAKpgl5EarvnD+/kgsaMlXF3Tt3QxubuOytyD3QUO6va8jsFYe6nYI/DJONkB82RgZKhdnO4UkLow89fYqtT0F3wDZBCirj4fmqIK9PzWXDVCKPoPmsPJTOUFXnqeT6qeIEx+p8ljrMp3sY3pfsgxcO6ux7qhB6sFagyOJx2wdljTROEufLOyCrJDE3JHceiikf4hJU03R4QAxkKOmpZKh4Y0dyghwqt7g/NazWafqIIBJ0481p4t8xYX42CCF0pcOnHyWp26hY6MyPcAfTKxrbbpK2boAOxws6qsdzpN2ZDO6CVlwdSSRsiZnL8HC7a8q+nLVfNRUlRcZWNPU0kOPzXVGxR0/ihtY0Et33XIGjuJF10nqKJ9rlLY4yD8Jxsg9guJXB6x6s0DSQWuSMPa0fRI3/ACLlbly0XTaV0SdPVVK17i3BLmg5GMY3XVjlZ4oag4pW+ktYmLyCAcld3LbcKPR7I7bcntEr2jzGcoOoXNpyvUNY2p1BT0rA2UF46QO/oug1z0fV6ZiqIae3mQtJAw3OV69cxV3hq9Jtjp5NntJP2rqbo3hZbdRV7pbhGHRl+Tkd90HWvgJwtufES8sp7jZiyMSDdzMDuu/Fdwst+geHUtHTWSOSQwkAhgJzhb10Bwk0pYqWKSwxMZOMdgFzFT6YprlbfdbszxARjBHlhB4ccTLbf6PW1bL9wntiLyR8G2N1wvrO53PpfG+gLBkg/D2XuNxV5V9I3yOeuoqRole0n6Izlef3HXl5t1hinhZBiXqIGyDz2aZBUGQs374UL53Ccue3G+4wuTbxwyrbRdZHzMIiBJG3ktk6ho4qV58No2JGEHaPl69m3zHcwVDQ6imssGitKVzGTxXe+gsfUQuwQ6CmbmWQEEOa5wYxwIIeV6R8v3suuWbgmKa76hsjuIepIcOdX6hja+ljf5mKi3iaMgEeJ4jwezgvGvg5zOceeAdU2fhTxOvVkpw/rfb2zeNQSnOSX00odE499+nIycEZXfTgn7au6UvgWrmB4YMrYwA1940y8RyjGd3UkzuhxORktlYBg4acgAPV2GGKniZT08TY4o2hjGMbhrGjYAAbAADGB2Uq4Z4Lc3/LpzAMhi4Z8ULTWXKYA/cireaS4A75Ap5Q178YOSwOb23wQTzMgLZnE3hBw04zWan07xS0hQ6ktdLUirho60OdE2YNLRJ0ggFwDnAE9g447lbzXE3M/eeNWmODV+1bwCjtlTqyxRG4R0FwpDUR11PGCZomNa9pEnRlzME5LenBLgQGmN5KeUhpBHLnoHI33skJH/RWr/tT+Vn/APVq4V/6G27+hXkNL7XznEfI97bhpCNriXBjbGMNB8hl5OB8yT8ytOb7V7nSqqpzv2f2aFj3EhjNP0fSwE5ABcwnA7DJJ9STug9of2v3Aby4JaB/0bo/6Nat/sWcMfxc6X/zRT/+ovD27e0954GSOmpuNjIWvcSI49N2ktaCew6qYnA+ZJ+a21P7SPnaqZpJ5OPVzDpCXEMtlAxoz6NbAAB8gAEHvx+xHSn/AAXtH8Sj/wDVWqtDWgNaAAOwxgBfPBL7QLnLlkfK7mB1Nl5LiGmFoBPoAwAD5AADyWl1HO/zd1U755eYnXQc89REd2kjaD8mtIAHyAAQfRqi+biTm75p55Xzy8yHE/rkPUQzVlexoPya2UAD5AALR6jmL5gK2pfV1XHPiDNLIcvkk1NWuc4+pJlyT9aDc3N9SSWzmn4vU8kgeX63vVQC0YAEtZLIBv5gPAPzC4ZLgXZCy7teLpfbhUXa9XGqr66rkMtRVVUzpZZnk5L3vcSXEnckkkqxtPG6IuyMoIZ5A9gDRvhY8TjFI2THYqaDpbIeoDAKlndAWnpwgnrbt7zAIQ3cDC05sZ6SSqxdIcSewKSvGS1vbKCIg5PyUsEnQ8ZCkp4w/uFZUNDHY8wgzjIydmM4WnzM8N+26tErx2Ko5znn4jlBVz+oAeivZMWtIAUX1rJb4Ij3xugxic91ksq+iMsx9Sx3YySOyADCChOST6oid0BXB/w4yq+G4DLgrThBfFL4Z7KksniHOFYqsx1DKCgBVzQQc4yFLN0dI6MZ2V8To/Cw7vhB359ixN180eqG4xjQFcf+cbcis9itj9tRqrHb/Y/rv5RtyIPahERAREQEREBERAREQEREBERBx1zHEN5euKDj5aMvf6jMvmoiqhBUzPxkPJH5SvpV5kvwduKX8Cr3+ozL5oiWiVxPqUEs0YcTLnvusZXkue4hvb0Qxua4A+ZQXQk9QyNlfUgFw6f/ALKmfT9EQeO/dY8bx2f3QDGGNDs7qzre4gFxGVe/qePh3V0cDhuRugyYKVhZ1kjJUvjsDhF5HZYL/Hi7ZAwomyOMgcTkhBrE1nYIvHHnutJlBaSBtg42Wre/k04YT5LC6oyTnG6DEY90burCrLL4jg7GFPOYi09OMrEQSmfqZ0YUsFL4zS5YqzKaoDIy1BAHeBIRjOFSWQyOyAQFbI7qeTnup4XRhhDsZwgiEmW9BUrKYObnO/1qENL5DjClc90RAPYoImkxv7dikr+vBwsiaSIsGME4WId0FzXluyy4Jg4YJ7n0WEqhxachBPUMH0m+vZZFDCXRlxeQsaCQF+JDt81kSPc0jwu3mgyKO8Pt05wM481uCn1DNcQCKgsI/Mtq1BhMe+C/1WPE6dgJiyAg5U0rrau09UiVtyeADn6RXLls5g6yVjKWSte4YA+kV1TZVzl46iT8luW2VsUUPW4AOAQdib3W0OuYPvswJfscn1U2n+BlrbQPrx0E4Jx6rr/ZdY3OCsDIXENBGN9u6570drW61FCKd0pIcMHdBxlxKpRapX0EMOwBGQuIHU0sj3ue0jf0XZPWVoirC+qqAC45K4Sv7aSlqXMj/ukG0pIHxkktIHkkMJlI7lazcKdklKHsxkjusKgmihBY/wCkeyCF8DYR1ZyfrUQjlmflrCQFlTU8xcJDno7rXbL7hMwU7gPEfsM+qDSIqZs0Ya/Y4wsapoDFks3wtx3rSdxtcP3RIcIjuCO2Fj2aiN1aXgEgd8oNtBknUBg5VzoXbEDJ9At1VVjiP3uMO6x6Bac6jbQnE+dz5oNGigkc7BYcKV7vdx047rVfHo2gdOFiVIZUE9OM/Ug0556z1IzrPwAZzsrnU72nLgceoWpUNvEzesA7DOcIOfuUC3W+DVLa24vb0F4OCu+WrtaWKhdTMoJ2MIaBkH5Ly30vq65aSqA+ikLSDnAOFyRSccbtXOjdWyuPTgdyg9B7Pxco7JCZprhlpbnBd8l095quK9v1PWyine1/xHfutiak4y1tZbzDRvPX042K4Vvt6uV0ke6sJJJJGSgx73cG1LB047LQQMLId1g/HnGFE9zTnCCwHByhOTlUUzIS9uRhBCAT2V3SQcnZS0wDZSHeiuq+nI6PVBBnq2VMZdj1VWHpduqvcOoFuyDKj6aXEnqM4UFTUeO8OxgBRmR7hgnZWoJXTZZ048lEd0RBlW+cQydRGQsmurBKMNbstNBIOQspkjDH8XdBiE7qoODlHYLtuyogyRVkNx0qN8viHtjKiRBkxkQEOG6tnqfF8lCerzQNJQUGyy4Y2ubv3WO1mO6uEvQdkEjKXrf8lbK10B6G+fmpWVDW4PdSiaB8Z68ZwUGJTkh+fNSTyE91ExwExd5EpM8Of8PZBWJuHdWcJMM5OcquC5nw91QU8hGSEFKeISuwVfUQNh7bqFrnxk4OCEfI9/0jlBRrulTNqNg3p2KgTsgyZIGhniD61jK8yPcOknIVBsEF0LA84KvfiIgAZUIcWk4Tqycu3QXOkL9lfFB4gzlUc5vTtjKyKdpLcjsEGMWlhVnxZzvssioe3OB3CvhdCIiHgEoLGVfS3px2UZf4kodghRvILiR2VzXADPnhBqRcJ4vDA8sLTpojE4AjGVk0FQ1knxnZLlIySQFmMIPU/wBhr/ufxlHpNYP0V69TF5Z+w1/3O4y/4awf9GvXqYgIiIOuftD9+TXiWB39yo/16nXzxg4OV9EPtBozLyd8SIx3NHR/r1OvneIwUBERAU0dQWM6cfJQqeIsLQ04ygtjZ4rzkqWSm6SOnGFG+F8R6xsFPBOwxkOO/mgp4THjB8ljSsDDgKrpXhx6XbZVjnF3coDXFpyAry9zgfh/MrqfoDyH9lkOdC0DBG6DFij6z8WcKksYYcBSySNGejCgLi47oLopBGckKZwEnx9sHKxlc15btnZBe+Xbpx2VjW5GVK9zentuoRk7DzQSQRiR2CpoadslWynJwCcFY/TJFh3ZGyyNeJAfiHmg3JdNMU9HQipZICSM91tj5FZk12rZ4vCllJaBjCw0EpaOnKiPdVyTsqAEnCAp4IBKQHHGSqwxdJJeNvVUll6XjwzjCDMNvYxvUDkrAnaWv7YWXRVL3ydMhyFFXlpk+HCCKKcx9srKZIJYyd8kLBbgHdSRy9Dx6ILXMeXHDSfTZbr03QxNg95mABAzurLObe8ZmxnG+Vi3mvdTv8Ojdhh74KDVa++Qzn3XAwDgFZttoKOqi6SW5ctkEyTYfHnq/nWZTVVypGkhxGAg3my10tpcZ2lp89lot31W12YGMB7jstOivNbW5ikcT5LS66Ppn++DGUEhq3+J442B8lk22ve6tGZMZ27rBmki8INYBn5LHieY3h7c5HZB234Ccx9XwMfBWwTveS8HAOV3l4V8x9Xx01DR1lZUviZsfiyAvKXQ1Tb6uSJl5aHMDh9L/wBq7b8OOI9i0fQtfZehkjGbYO/ZB6ZcY7fZpdExSU1cyWQMGW9WT2XFfCp9tM/udRI1jnOxuuu2keYa9almdQ3KoJgBwMk4wobvxpOmdSRS08oEYeCd9kHpfpCw2m1UjKwTMJfuMuGAtyfdK3Db32Af+OF5+XLnGDtPR01HUt8UMxs7fOFxjDzZarqrw2mjqnYc8DHWfVB6mVd2tMERdU1kAYQc/EDkLrBx90rom+UU1z98gBDicEhcD6q4+asp7D7y6oO8efpn0XUjinzE8RKynmjgqpfCyR9I9kEvHq42Sgrp6SjdG7oBaCMLqbeqttRJI4Ozhxws/UGu73f657q6RxeSQdytvSQTyS5OdygxXuLj2wrVqEsLYowXYz5rBduS5o2yg1Ky2etuUwdRuLXMIIcCQQQfIjsV2Y4R+0H5rOX6pgtFNxBn1NaIcD7l6lDq+INB2ayRzhPGANgGSADPbYLrZYNRzWN5dHH1A91kXOdt+f7+44IG4KD2G4Oe2A4Paojp6LjHpG56JrH9LXV1KTcaAknBcelomYN84DH4Gdzhd1uH3FPhvxXswv8Aw11xZdS0G3XLbKxk/hEjIbIGkmN3q1wBHmF8ysl3k6PAaMtA6crM0xrTVuibzBqTRmprrYLrTYMNbbauSmnZgg4D4yCBkA4zjYIO33tSeUz/AGB+Lx4naQt/haK1/US1UbIxhlBcz8c9Pjs1ryTKwbDBe0DEa6RscWHLSQV2kvftDONHEjhFeOCnHaktPEKw3OBograuIUl0oKmMh0E8VREA1xY4AnxI3l4y0uw456t4cN8FBI+SR4+JxP1qxVBc7YDKdkE9PF1gqKRvS8t9FdDMYtsd1NJCHtMnrugxc4UkfxOCjOymo2g1DQ44CC6UAYGNyjYpuguHY91qVfT0/hh0ZGRusETlo6MbIMcN6yQVFI3oOM5Urn9OcbKFzi45QUQbK5kbn9kexzDuguinMZypSxszeskrGVwL8HGcIKOGCjRlwB81VjHSE9IyjmOjcAe6DLbRsxnPdY/hAziPOxKp48mMdSsD3B3VndBqFXb44IRIHb4z37rTlLJUyyN6XuJCtZC9/ZBYrg1wOcKXwTEOp4UjHw53IQQvlcRghXtia6MuI3wr6gxY+HCxQ9wGAUBwwVRO6ICDcgLKobbU3BxbTtyQra2hnoZPDmGCg79exYja3mj1S710BXfyjbkVPYrte3ml1ST2OgK7H+cbciD2nREQEREBERAREQEREBERAREQcccyX4O3FL+BV8/UZl80Rb1yvA9T+lfS9zIjPLvxSHrou9j/AJDMvmkYD7w8fM/pQRtPhv3HZZLGtmw/HZX+DGcl+FE4PaC2DsdjhBfPVN6DEMkhYrGF+6lghLpPvowFfOWMna2PGNs4QW07vjDSDjPdZhDBg7LLnhpGUQlZjrIz2Wi+PK/YZ3QT1U4LOgdwsNvcLOoaGSWbMrcAd0ucDIJAGYGEFnSRHk+ixnOIPmslzneAPqWGSSd0DOUREBVAcRsgY49hlZlKyIAeIcFBhfWizKtkIALMbq+ipmSxlxAygx6bAccqtZ9IYVpIjmI8gqVDupwI7IKQxGZ4blXVEPhEABWRvfGepqSyvlPxeSC0DJwsqOj6mdRx+VYgONwpm1crW9OdkEb29Dy30Usc3S0gkklQucXOJPcqiC52SSfVTQztY3pKgAce2Sha5p32QXZ+MuWZHK4MI6j9eVheG/GenZVBk7gHCDMoqt0U4dk91yJpfXDaSqigM7wTgYBH+pcb26Fss/S7yWrUsFNBeoC84HUPNB2PMrL1aw8HYjzXFerdHPHXUMYwgEnIz/rW/K68U1DpJht7sy9A7fUuP6XU9XWRSU9bkF2RglBslozUe6EdtsHsr59Pv8QShrQO5O/+tanX2aenqDWhuxOey0mrvkgaYx3Bwg1FlIyaIQgAkeS06GhkobtFKRgAg+axrVd3xVXXUPw0rcVtibf9QU9JSyAl5AGN+6Dl602+TWtijtlJROmk6QMFufL5LTafh1dtNXNluqLQ5hmcB9AgbrvXyUcs5uUMNxukDTEcE9Q2wuyPFvlX0vO6C6W+CLrZ2HSAchB5w0vLzcX2YXkWiJwc3qz0HPb61wfxI4eXGh8Qm2lga4j6JC9huH/DMyvFnuELPdmjpGRthcNc0PAS001LLJbIYzuTsAg8gqu0VUM3Q6IjHrlYThPTv6fDcQPzrn7iNoP7j1uOgDvlZuiuGFrvlplqJ2tLw0nt5oOGtMWZ18cIxECTtuFv6bQstroTI6kYB074af8AWr4rP+w6/P6mhsQf57bZXLFPqPTN105KJpIzKGEdx3wg6sXWN0de6IDByey0qpqKmAloe4fb2WvanGdTymH+xdRxjthYNdBTvYXnGR80Gm0VxmilBkmcR8ypa24NncCHH5rAezMhazcKjo3MOHDCCd0gmAaFb4GCrWNd3YDlOuUPHVkIKSQlrQ71Wo29jXwHtsPNQzYfAMbnCx4p5YmmMZGQgo8YnONhlWyEvf0gkq7wKiUlwaSSs2no+mPqmGD5ZQQy0PRCJdgMZWEsuoqJ3ExbkDZYhBBwe6AsumojNGXbfasRTR1M0bOlp2QRyNLHlvoVdFH17eqsc4vcSdyVc17ozkFBkS0hazqxhYrc9QHzwpXVUj29JOysbHIR1NBOPMIJpKQsiD9u2VjKZ1S9zeh3bso2Y6xntlBfDEZDjCrND4Weyz5TBDAHMIz8lp73yzZO5AQRkjCB2AqEEHdVDSd0FS9XMiL98KPss2nAERPYhBjyQmMZKRwue3OdkkkdI4t8lRsj4sjPdBY4YJCN3IQ5PxFU7IMtg6QCVnQFskeDgnHmtOp39Zw87BXzTmJwDOyBV03QS4YH1LEUslTJIMEqMMJ7boDW9RUgi7bfnVgDmn0U7C8jOOyCx0QYM4UbclwCrI9xy0qaBjC3r8wPylBZJD0N6z+lWsiLgSB5Ksszn5bnZZlvMWAH4QYUUTnuwQszIgjLc4/mUtUIo2l0RGVpr5XyHBQUfu8n1KngpXys62jIUBjeB1EbFbhsslKKbolI6iEG3pGFjiCFbjK1G6wtbIXs7FQU8THxl3mEEbIT3CpIxwGXeSkgl6Zel3ZX1Tg5h6cIPVD2Gv8Audxl/wANYP8Ao169TF5Z+w1/tDjL/hrB+ivXqYgIiIOvHtBJBDye8SJD5UlHn+PU6+d/6TvrX0Ne0Px+004l/wCJ0f69Tr55WguO3kgnbTdTQdlFKzoOPzK5s72jGeyse9zzugo0ZKrjocDnsrQcITlBkzVLZIw0HdY31K+INLviWV4cHTtjKDHgi8V2PRTS0nQBsO3qoGyGJ5LDsrnVUjxgoInN6SqZJ81X4nlULSO4QFMyDrb1faoVe2Z7R0g7IL6eDxnlgI2SpgMDwNvsVkcz4nFzTuVkdXjt639wEGIqg4IPojx8RAVEGSHNmHSB29VFI3wzgBWMe5hyFe13iSDr7IEEZkfjGR5qs0XhkDYLUo/do2dTSAQtPqpA92WnZBH0YblW9WHdSEuwAe3kqDGd0E8k4cwNGRhWxwmRvUpYIY5PRVneYctb5+aCOmOJME4wpqiny3xBuQsUOLTkHdZFNUOdMxkm7SQCgxcHOMHusuOhc6Ev2z3yVun7n2p9GJGFvXj8627XySwPMcf0T5hBjUheJejqIxsd1lVDMjc5JC05sj2PLgVeamV22fzoJoJhTPyfVZklwbNEWtytKcyUjqcCtat8FL7mXSEdWEGm0c/gTlzs7lUr5hPKCO3kopi3xnBvbPkrDjqAQXiB2MkjCMjw8Z9VMSQ0E9sKHpkz174G+UGqRTPgYCx5BGCty6S1LcH3SOibLMeogYDtv0LZj5vFDI2vOScLsRwW4c2u4wx3CcNMoaHb+qDdMVVV6ZsrLhJI9hc3qyDg/nWzr3qupvkL6hlTISP7p2/6Fy3qbR9VdKB9ExpMTGkfYuBdVUTdMSOo3bZdjHZBjWzUVzNW5j66XpBxjrVtRr+Sy3iOofWygNOdnD/UttyVDKZxnYTkjOFtG71fvdSX5JIz3Qdh7vx/p7tZ20LLlOSGgEF49PqXHN31rFVUckRqHEu33IP8y4vBwQfRXOe5+ASThBM6XrrHS5zlxKynVzWEbZwtO3G6zoKaOWEuI3x3QUklFUOkElY7m+Eek7K6Fro5jgbBKlj3O6sHCCMDqO3orxJJHGWsJA88KEEt7J1OP2oKsAJ38lRzsnY7IQRv6q5sZIyBlBWFnUQfmsmRrRH2GcKGAhpPUe3ZUmlySGnZBWncxueoq2Vwc847KLJ9VVpJ2QXDus0ysFP0k+SxA30GVJBGJpRE92AUFIaaWoJEYzhWvZJBIWuyCFrbBDaoy4EElaPV1AqJTIBgHyQW+PIdnOJCnD2GLAIyfJYeQpWBhGS4DHkgieD3Vqq7ucbjKp3QSQyCMklVleJX7KMtI7hUBwgyo6Xqbnb7VHIPD+An8iNqpGjAKsd4kp6tygugkEbslUnkEjshWmN43IVqAiqSCFRAWTTTNYQHHusctI3wqdkGozBsrBjcrFfTuaCRjZWNll7NJVfFkLgHk90ERz2OUWTNGwsyzusfpOcY3QUWSym6o+vA7eqh8GTv0nCvbPLG3oOceiDVdOXSG2VD3SuwCVBqC4RXCr8WM5C0xxzv5lXCCVw6g0kIPQH2LModzRaob6aArf5RtyKL2Ku3NPqkf/w/rv5RtyIPalERAREQEREBERAREQEREBERBxzzHnHLxxRPpoy9n/kMy+aPxA2oe4j98f0r6XOZEZ5d+KQHnou9j/kMy+aB4PiPGNwTn8qC6acvwATgKSmqGxfSGfrUMTC9/SFWWIx9/NBLPUtefh2+pUhp3SjqJ7fNQMaXnA3KmEkkBx5IL5xMxnSXkhUpA0O6ndgpJZGSR5zk/wDsWKC9uQM4KDWBdoWN6GtAOMZWHNGaomTO3cLBIOMrJppXBmAdgguiHju8DtjZR1dKaZwaTnKGTwXmRndWTVD5zl/dBV1P0x9ZKhUjpZC3pPYqwNJ7BBfHKG+SlDTMCWnGyxiMd1ex72j4c4QWnqBIJ7LIgqfCYWdsq2nDHPPiED7UqWxhw6DthBC93U8u9Sr42dQJUjhEI/hxnCgDnDYFBNE0OcQlS3pI9FE2RzXZBWW10cjCXHf86DCRSxhhlwcYV1S1gcAzH2IKQxeN54VJofCeG57qjfFiGQCAqSSukOT3QZLYmxsEh3ysaV4e7IG3oqmd7m9JOyjxhBP44LOnG/ZGzNAIPooFVo+IAoMmgm8Kfr7ArLe51TcIy13mOx7LHkETIstxk/NW2+YtroXuO3WM/Ug5SpZailtTH1Li5mMbq2htAuzve6cgBhyQFm3W4Ws6ZY1jx19I/QtE4fXtz75Fbuv4JHhpH2oNWvEsk1N7hHQvc4NxkNXHtwsNZDK5z6Z7fPcFejPDfl+09qSxMuczYzI9gd2HouM+LfLzPT3T3e1xAh5xgBB1U0Lw4uGuallFS0cjnF4blrTldzOBfI3c4tQUFzrbfKWAhx6mnGNlytyacq1xprtBX3alAj6wSXN2xnK9I7bo+2WhkbaOFgMYwPhA8kGjcJNDUGiNK01vp6URyBoDtsHYLdN1tcFxj+/HZvkeynq62mttP49VIGMbsT5BbIves/eqtjLQ8viBAcR5oNN1LUUlg63QRhhaD8S6/cWNV0lbZauSeUPc0HbOSua+KeorJS2Iy1crWy+GSe2c4XQXW/FW2zal+5Lp8wSy9J32wSg4V1DTfs81ZJbRRuYwPI6iDjus2t07Hw7pTGZdiMlc5cRqPh/o/RTdU2iSH318XWcYznGV0/v3Ftmqaeq97k+JhIbug2/xTvcFc13ujgHnJ2K4korveaNkrXVbwwk5BK1Grv4rbu+OVx6Oogei0280kssgNI0lh3JCBTV/v1QWybuJ7lYt4a6B3SHbFXNibTMBh/svmB3WDVGqkw+cHb1QR08jY5Otw2V1VO2ZwLRjChAHmcK4AdWB+lBfTTNifl3ZVqZmyuBYMK2Rga0eqiGM7oJ2zlrcHdU8YF2cKx/T0jHdWINVpq+CIfEAVHV3Bsp+AYA8gtPaxzjhoyjmuadxhBkuqWEdlaIfG+LsoYwC7dSulMezSgsliMeclT0rGuYQR3KgcZH4684V0bpGbt8kF81P4ZLh2WO53UVe+d7xglXxhhaerGcIIAMnCyoahkTOgjJWMfpHHbKkaGluT3QWOPU4ntndUA3Ayn75CcFBK9khaDkkAK6OURs6XN3U1uex0vTMdvmp6+Gl6x4ZCDTQDI4/NXE+HthSytbG0EdyqRMbKwlx3CDGJys2mHVGR22VtMyIydMmNvVKqQROAiIwgt6BE8uJyCj4vF+Jo7KEyud9JZlM9gb05GSgwXZBIwrmR9TcrKqqYMaX477rFZ4uPhzhALiw4CkjZ45HUe6RMEh6Xd1V5dTvGEEk9D4TOrKggeOoN9dlNLWuljDT9Sx4WOfIA3ug1R9EDF14G4WH4gjd4eFqng1EVOHSA4xtlaROxz5C5gyEElRCOjrHnusZjiPh9VJJM/wwwlWx08kjetvYboKyQFrev1UQcW/RJCkbI4/A7yUnQzG3z3QRCZx2JVA7MgPzR4aNgqMB6gfLKDLqHt8EY7/UqUdLNJh7XEAHdWTuaYwMjIUtJUmKIgIJ52CVvhk7gKFsXgxkZ7rHdUPEhcFkR1Albh5H2oMFxPWfJOt2CCc5WVVMjAJbjKjYxhgc4kZCD1Y9hvF02rjFL/dz2EfkFd/rXqSvLb2G8hdauMUR7MnsJ/KK7/UvUlAREQddPaGt6uTbiU3/AOk6P9ep188sb+gk+owvoa9oa7p5NuJRHlR0f69Tr55YgHEgoLScnKopPBeD2ICo7AQWIr48ZwUkDcgBBYq9Tv7oqRzA1gPqrGxPeMtGQgtTCqQRse6zKVkTmkuwgxGHpPURsqyPDzlZVSyLpxGRnKxPDf3wUFqJ2RBUHBypBNgY9e6iVwYXDI7ILSclXtjyMqjWOkcQ0ZKv8KWMgOBAKCI7EqoBcQB3KzGsiwc4yjhC1pcCMhBA+F8berqOFDnfJ3UxnLyGnt2VYI2unAwOnKCN3UAAQcKxbinp6E0rS0jqC0KWItccDIygQyFhJBKTyeI7JGFJRtjLyJMJWtia4eHhBjKoyDkZ+Sos2BsJhPURlBDFWTxH+yOI9MqaWtbKw9Q3VtJFHJMQ8jCtq4o45w1vZBjk5JPqjT0uDvRZUrImxAtxlYowgypaljow0D61AJpGghriAoz3UsbWlmSgiJzuUGxV7OnqIPZXmLqOIxk+aC+MSVBEccZeSQOy3/pvQUt2pg11O4OLc4IWXwS0ay/XvFawCPIAyO67L2rhPdjqWmo7PTkxPIGwQdZ6nhHdbdIyobbpZG9WRhpK7DcEtM1zGxiSjkiAAByCF344R8qVqr7HBVasp4mAgH42j+dZmv8AlzoLJG6TSETHNb2MY/1IOvhsVPTW95dT9ZLT5b9l07462GaovLnRQFg6yMYx5r0V0bw7u9NWyjUsJZTAHHUNsfautHMvYrDR3/NCWFgfvjG26DpDqOjntdMzqYRkAZVlo0JJeba+5tkIABcQuTOJ1LY5LcGxOYXhucfPC4lpNZXC10j7fSnDCSPsQaFVwmkqZKcnJY4jKh6vkr6iZ1RM+Z5+J5yfrUaC9revZZDZ/CaGA4wsVri3shD3bnJQTe8AO6sK81bXNLS3fyUDAM4d6rI8OEt2IzhBBFGZnnfCn9zw7BULS6F5d2CkdUSu3CCtRCGN2VsNQ1kZaRkqN875BhxVoic4ZAQXsYJXnyVroul4afNGGRpPSDlUe55ILu/kgyo6Avb1Z7qCWLwXgFXMrJmjAKPf4nxPO6CQSsa0ZCxzI4P62kgq1xJ28ggaT2CCSWofKMOcSrGDJwsqkgbnMuw+avqWQswWYz8kGG9nTjfusyG2OmgMwdjG6wnvLjlZcE1Y1nTGD0lBikdDy354VucOyppmhu7hhx7qDOUGQwCcFvbCslg8PuVYyQxnIVxmc8jJ80EZ2Usc3R3bkKkgaMYPfutXoaWlmpiXY68bINLkmEmwHdWRxdZAz3Uk1HNG53wHAOypGyRg6jkYQUlp/DGcqxjPP0V0kr5sNGThWglmxHmgFwdtjsrCMHCv6c/RGSrS0tOCEF0UgYckZSV4e7IGFdA1rnHqVswaHYb2QGSdJ3JwqdR6+oeqRsL3YWsw0lN7qXOwHYyMoMEVTGx4LclYsknW7OMD0WVRwRTVBY47A91LWwU8Owx2Qac0ZKz46yKOIxkAnssENLnHpVxglG/SUHfz2K7geajVR/7gK7+UbciexXje3ml1Tkf/ACgV38o25EHtQiIgIiICIiAiIgIiICIiAiIg465jndPLzxQcew0Zez/yGVfNJgSVEpwMEuI/Kvpa5j2GTl54oRju7Rl7b+WhlXzTNYWVMrCN2kj6sFBZC8QS5IzhXVU7ZyCBjCtEL3vy5pAKrLCA4BmSfRBZBII39RV1RIJXDCubRzAB72kN9VHMGgjpQUALd/L0VfFCt6sjCtQSPcC3HzWTSRdcZd57rGiAJ+LsstkzIh0twgw5MtkI9FQ/F8lmCKOR2dt1G+jl6stGRnv6IIHP26cIyQNBHqs6WniEPS3HWsB0bmHDhhBQnJyrmPDRurmhh7kdlbKGg7ILoonTydLPrVZqd8MgY7uVPansZUZfjClvEkb5Q6MjI9EEUludHCJc5GMrCWbHXPeBFKcNxhSuhp3sywhBp7GdSOaWbZ7q/IY/HkrZHBzgPJBaMk7d1XdrgTk4U5Y3w8txlRjpIIcd8oJjNHMwRgYKx5YvDwM5JVAel3UzyVesveOvsgsGyqTlZ80cApw5uM48lp6AiDGd1M0MOCSEEQ6nEDcrIijwes7EbhZ1HDTPdvhY9wIhkDWYwe+EGtadpKu/yto3znpLg3crtRwc5VTdZIL4JASwB+MrqFb7xNaCyWlOHg5yu2/LTxt1TXXGms8Rd4bwGHBKDvDwR03LC9mnnRvPQQzP5lz/AD8vduqaiK4VNGH9ADjnfPmtK4F6Jmdbo9QTMHiuIdnG+V2LpnOfC3rbg4AQbB0xVWHSsYttJbxG9mBnGFuSp1D0QirAIY3c+inm0tb56g1JbhxOTssbUdohZp2ubAD1iEluPXCDjXX3EKm1DSPslDWsilJIOHAHK0DSdUNO0UjK6tbK924JOV054paz1fo/Xk8uZmwCQ774xlajozjNdtRXunppZXuaXAO3KDnbiVbbrrN8sNNK8RlpGx2wuifGXhld9M3V9YZ3dQf1A5+a9LKGlg/YuK6BgMpjz674XRPmSvU/3UkZVsDWCQg/VlB1d4kaz1E+xi2S1sjmBhGMnC4Cira5rpW+KQHE53+a7G8RJ9MVFoHhSsM5ZvgjuuvlU2CN0oaRkk4QYcNJLUyFwfh2crWYrrFbKc087Q5xBAK0OGpmp5urfpWe80tW3xJCMhBhw1XTVmqf9AnsVm1M0NezqYAMeixvudLcJRT0EfW4eQ81FVUNxs7+iqiMZO2Cgx/d8yFg2wrZIzCRk5yrmznqLgdyo5Hukdv3QSxN8c9CsqKcwuxuQqN8WEh4BCz4C2pjLpMZA9UGC2nLhnOFa+MtUpkdHIWncZVeprnDJGCgtpT0vGW5U1VEHkOAAWRSNpwSXkBQ3CZvWGR4wfRBiQjL8KszelwP6FI6J0cfiAeXdRtJld8SCQztLMdO/wBSyKaJjoyT5qCWFjI8tG4CpBUOa0tHdBHO0NecbKLKklEhJc8HB81Y1jndggomVcY3jyVBsd0FFUDKleGdO2MhQ5KCocW9jhOtxIJJOFWMAndXyBgG2EF8YNQejKvePdcsOd1BDIYndSrPN4rgfRBGXHqJBVQ8/vt1RmM7o7GdkAAuOyuj6myAdt1dBjO6rKAXAMOSdhhBqNaR7oCNzhYEE7WN6C3JKyGU1WG9dQ0iP1PZXOjpenqBGQEEDGGJ5kJ27qGaQTSDH1KZzzIegdgcbBTRUrAOo9wgxZaV0bA4qe2hrZBI7sD5qWUgt6XYA+axSyUf2FpI9UG4rjdYJ6QQRgZAxstLo2NILXDJK0+MS+IA7OVmxsmY4PDTgDKC6528RR+IwZ+pWUFQxsDmvG42Wp00grAYZcbbLCrqVlO/oZ2J7oNJmcHSOPbfsroiXfCVlVMEYj6gNyAseGlqHjxGNOB5oE0HSOrKia/pBHqpJHvd97IyQVfFEzo++jB3KDH3d5qocW4HkqO2cQ3tnZVbu4A+qC5xy3t5KPJ9Vn1DYBCOkg7eihpaOSdpc0bIMcuce5ymSBt2U1RTuh2cD3UjGxGle446gNkHqj7DX/c7jL/hrB+ivXqYvLT2GzSLdxkcexmsGPsFevUtAREQdc/aHfga8S/8So/16nXzyNPSQV9D3tCQ08nPEkO7e50f69Tr54AMoMt9Sx0Ybjf1WIdySr/BkxnGyuYxuD1Yz80ESqDg574R3dZlMyAxHqI6sIIARIOlZkRZDD0uwThYB+9vJA2VzpnPwEFkhy8kdlVrnAYBIVCx30iO6oDvv2QXxlznbkqd8rGDpxvhQuLWty3G6taHSOyfyoLXZO+NlRZUjYxHtjOFjNA6sHsgvhiMrukLNbStZGRtnCjbEXNHu/0h6I+OvZvI0hBSLFI/qcAcrUzHHWQmVoAIC05nTJtN3CuE8sThHFksJ3wgwpg5kjmknup4KR84z1HH1qetijEXXgBx+axqerdCCMoLqmiNOMk5x3ULJS1uMHPqsp1WJ9nlXeFAYycjKDGhMsjiOokeeSpJT0jGdyoYniOQnO26TyB7sgoIydyQSN1QknucqrcZ3VenreGsG57ILVexrnkNDlNLbqyBniyREN75WO17mHIQZBidAA/OPtUZzL8WckKkk75GhpOwVjXEZA80Al3Yk7KikEErtwMq4RhrT1gA790EKkiOXBnkSo3blZlNE1zA8H4h2QKigMMQkByqW6QCoAfuCrzLUPyyUEMzjdVi8COUOGBjdBzjwqudNbXwytw1xcDkbea72cCdVWE3Wmqq1sb3MAPxYK85NFvqq0xsowXFpH0V2H4Va6/Y5qGmpLq5zGnA3OEHsHbb/btaadgt9pIgIAGWnC3bp7TcFqoRTVhZO93fr3/SuquiuNGmbHpqnqKOpb4pAOxHdb6uHGSvrNJz3qkecxMLgQfkg3px6oLdT6QmmoY4oqkMdgMABxjvsvIfjtcrka6eKo6+oykAnPqu7PC3i9f+MGsavTlZI+RjHGPBJIxuFwtzi8MKTS1Q+eRoYSS7thB5+a0jrmAyPncQR2ythxwulBfvgfnW+Nb3KGetfRxSA4yPqW0ozHBEWP7lBgBu5HorvD+LGVRzvjJHmVQPIOUEslOWN6icq1sgDelV8Yvw16tdG7OWjZBb9I7JlzSMk/lUkHT1dLlNK2MA7jKCOWRr24HcK2OQBnSo3YJ2VRG4jONkB7cbqWOdrG9Pn5KINe/Yb4VHNLDhwQZFO4Oec9iqVIH70AKBri05CvaXPcCc4QWtcAe2yo52ST6rJkiY9g8MAlY7mOYR1DCBjqGyrGel4yNsqSIsxvhJOgbtIQTVDh4Q6Tv8lhlxPcqrpHOGM7K+MMI+LGyC2NnWcLcdG6nhoiHgZI2WgFpGTH9aGqmA6XfnQUqniSdxB2yoeyrkk5JWSwQlvxYJIQYqr0nGcFZQbDnfGFd4TDu3GPkgwlPTTyQvDg8gD5pO2No+HGVC1pd9SDX21cNXGGYGQFjvjaXeCAN9lprJPCx4ZOcrUqaRpHiyYyNwgjntxoW+M85Cw+kVD/hWbXVctaPCYMgFRQ22tY0yCMhBG6n91HWTlY0j/EdkbKeSR5d0TjABxur/AAoXM+DGQgw9x5rJpaJ1R3yEpqVz3/GNgs0yiD4IvpIIaimbSNBysc1MjmloccYwr6j3t2DOCGnsqU9I6Zw6BsgigmdA7qxuSp3RPqn5zsVkTWxzGgkHspaePw4vhG6DD8IUfxOGVM2pjkZ1dOFV4EjiJtgOyse2FowwjHqg76+xeqGyc0uqWgYxoGuP/ONuRW+xcbG3ml1SWnc6Arcj/wAo25EHtGiIgIiICIiAiIgIiICIiAiIg495h3iPgBxMkPZujry77BRSr5qeoS3GpcBs573AD6yvpR5ktuXbikf+4q+fqMy+amkcG1Ti47HP6UGYOlw6AN8YWNIBDMHO7Z7LNhlga8lxCxLifGeDFuEGdNXQzUnhsABWjmnIdudvVXxQyxnLwQFdP1vcBH9uEFsdIXnYlXPoSwZyrqdlQx+XA4KpVTyB2M4HmgxXZaSFJBA6Ujv3UvTE5oIIz5qWAeEM9kFZIjBGDlZtDVRupj1tBIWBLUtk+F/b5qalpZJCPBBLfVBhOmLakuPYH8ypUyiZ4DR5+S1K4UsMUJJwH+axrfFA5hdJjI7Z9UEDqNzYvEJxtlY+CTglZFRUv6nRj6IOyhGNzn8iC50LmMDwVGXOd3cT9ZWVSl08gid2Va6lMJGBsUCjtxq3Ya7CzZbc6jYeok7LToK2aDeMgFavTV7aqA+8Yz5INDl+krRnvlSVPSZndPbyVjASgvEhARsRlGQrXRuGTjZGSuYCATugqY+g7lWE9R2CkIfLjAJU0VPhhLxgoIMvLQCchRkYV73YccdgVPTRseB1Yygx2MMjsBVkjMZAz3Wa+ONo+9dysR8czsucCgviL4PvmcD0VlRMZnZKt6nvAj3OEbG4vDOxJwgrFDLM4NjYXEnAwF3p5KOGE9TcqOsmoHEbOz0rrdwl0B926iJ8wBBkHcZ817F8nPCK2W+ywVj448MiB+judgg7GcL6SCh09HSCEMLcAjHyW9AABgDCxaK209C3phGAstAWNcGNfRTMcMgsIKyVa9ge0tPYjCDqjxO4JUvEa6y09LQfHv8AEGrrRqjhlcODV+b10TyPE79PbdenFJaaWjndURNw9wIJwtk8UuE9k13bqmWriaakRksPT5gIOl2ouYx2jdKNeXdRMf0O57Lp9xO4q/7KENTO370/JIyMFdsZeXG76l1tPZ7rC8UDCQMg4xldTua/hY7hXqRlt09kMfJhwaPmg6van+7EFe4SyvMQOPPGFtaWV8s30juVz1rDh9WjSUV1kjJkfH1HbfsuC4IWMqXQyHDw7pH1oNQNCJaUEDdaRJFNC/w8n0W7YLJc4YBUSRnwiMgnsQtODaOW6MhkeAOrdBkaKuDrFcffqmldI042xlazrWsOq5GTUdudGG75DSMLnvghwPoeJEUVPSBsjyQNguedTcllVYNNyvhpAJ3RnoHTvnCDzVqrdNSHdpJHcY7LFDXl4+A/UuwlJyycQ4dZSNu1vkbRCQnqLTgjK0viboS0aUrWQ4Y0ggO8t0HD0jC+EAsIPqsZhMBIzjPdb21DR26mtbZaYtLi3bH1LYmXyEk5OOyCfoExyNyrJKd7B2P2qtF1eIGrcMdHG6LqkA+0INsN8Rx6WkqcUjxh7z27rUXMpI5SARsViVcj3SgRbjO6CyeoaY/CA7LFjf0HK1CopWmAFuOrvhad4b+2EF75y9vSSrG7OH1hVdDI0ZcMBWD1QZ9Q9pgGACSsankDXAO81l2owyz9FQQGq+7U0LZQaQAjzwgsmazo6hjdYpg6m9QKkeJvCAOVE2RwBYO6CI5zgpjfCkMEv0sbHzVh2PzQXmItHVlRnJOCVeDI8YOcKnTh2D3QUcDt81qlvsb62HxMkbZUtnoo5Xg1GzfLKyblUz0REdDuzzwg0WroTSyFhPZRspy5pcD2V0888zyZMgqxkrm5b6/mQS0UPjS9BOPJTy0fu0wlJ2ByVjB74MSMzlS+NNUsPUM7INamulNcKUUcbQH4wtEnoJIXAdec/NQMMkEnWO4KmZVSyzAydsoL2wOiaHBhKt97LSMtx9a3fYo7ZUsLagtB8srQtU0VPBVD3XdvbZBpE9UZQAB2WRR1scERZIASrZI4W04II6gsIDKDN94Y+Tqa3zysltxYxvQWjONli0UB6svGAVfVW6YuL4mEhBNDUGJ5lbuCUqaoy/EQdlNYIGPqfCqxgZxus2/UdNHtTY+xBt6WcuGCc4WqUFxhjpzE5oyQtHfG9meoYVgJG4QZU3wTGYYIJJCjc507tgQPRZtqYypm6J/orKucFNB8FPjJ9EGlMphncq91GcFwJSSlrIW+I9hDfUqsVXkdDiQTsgihgfM/w8nAWsU8sVvh6XjJwsWGN0RMg7d1Wdj6z4WDLjsAgslkbcJC1u3msOpp3Uzugk4WWy219C7x5Yi1o8/IrGrZ/HkDsoPVv2HRH3D4vDz94sefyVy9RF5bew3JNp4xZ8p7Dj/za5epKAiIg65+0P8AwNOJeP7yo/16nXzyNOHA+i+hv2h34GnEv/E6P9ep188gBdsEGWauPo6elYr39RyOyt7J3QFLHC57cg4AGVaIZCMgKrZXxgt+zCC4DxD0eaq6nMeDkqlOCZMjusucgRkOAyAgxHzAt6QFPS2508JlzsFdaI6eWcio7LKudUymf4NKR079kGkSNLHFh8jhXslDW9OMq3q63Fzj37qRrIzjsghcXE+aNb1ED1WoMjh88ALEnDWvHRjYoM23vbQP8WUAj0WZWXumnbhjBn6lpc8rXxAeixQxxGcbIJJZC+QuacZU1NUNj+FwGfVQBoxhvcd1RjCJAHBBmSMdO3PkofcndslZUrhFCCNtlbHUt6O/xIIXUD2t6upWQxSPlbDkgHZTNNVM/AzjspTBJCfE8wgyavT/AINOJw7fGSMrRHMc0kEHbZag+61r2iKTPT2WrUUVrkpS6ZzevGd/VBt2CAzOwDhZcFMKapZI85DSCVbUjwahxpzlp9PNQPqZTkOyg3JfL1SVFvFPEwB4GMhbdhozKzrzj0Csg6ZH/Hn8qllmfFhsfb5IMZ7C1xbnOFNFB1Dr3wPLKsjw+QmTfO6kdMGENadj3QTMrI4x0lo2WNPIJX/D2KlliY5oMfc7rGLXMIJGEEppsM6iVdTy+C4Z7BR+LI8BoJwgjcW9jkjZBlVdbHKzpYAPJRxUL5WdQJyscwyDfpOyzIKswxEHvjbKDe3DrUsOlqgyVIDhlarqXW013u7K6gk8PoIO2y2RpttPWVBFYQAfmp7o6lpbiIaV2Wnug7U8EtQ6h1a2CgNe8tY4DHUu5+lbzU09vGjJx1vqWdHruV0d5brtQWGaCeZwBc4fpXf7hjaKLUuoaS7QgEsw4fmQci8AeXZ/Dq8TayqoMCoBmAPz3XEnOZYZNf1bqejpXkFxGQCu91vAqbNHD5+EGH7Bhce6/wBDaYZZ5ayr8ITjJ+LGcoPDLjBwGrdEyvu8sTsEF2N1wWaB9WHSgEBpIXo5zTilrKqponFnhMBaMBdCdRQw0Hix02MdR7INkvj6JCz0VpbjzUrg4zFz/M53Vkh+LAOyCxZMMo6PDIyVjYV8RAcMhBI5hiPWPNWdD5BkKeoYTGFnUQphB8ZAI+SDSfDcPIrLiDejpO5wrphG+QiPBChiL2yjPbKCWmiDHlxGAoKwh0vw9hss2tmhbD8BHUdlpoPW8EoLFl046oyCMErJpoqUsBkIUU72RvxEdvPCCyAGB5c4ZGfNRVMrZpR0jA+Symujkb8WFhzhrXjpQVfTlg6gowx7jjBU0T3ynpKlLGsJB7jdBjSRdAyo91kSB0v0RlROiez6Qwgvhl8J2TuqyAS/GNlbAGPOHqszgx2I+yCE7KdlM97C8Ht81B3U8MkmQwA4KCSkoXVTyzJBCzJqP3OMsJyVaXSUrRJENyMqNz6uob1yBBjspHSuJzspjT+G3p81HDU+G4g9+3ZSSVDX9juUEtBbfHfue6nq6IwAsDsfUoKOqmhJLcqKrrppJN8oJqQNpnB0hzutZGoaTwhEGDPZbXc+d+MgqRlPgdZJGN0Govom3KQvZtn0WHPSOoCeo5CkpbiaU4YVJ4NVcKgPeCWZ8+2EChmbUnww3BVk1O6CqEh3AO61WoZQ0UAdER1+nzWM6eCWEvkIBAQR3WvhqKZsUTBkADsrbHK1jgx7N8/zrJs8NDVTFrnA7+az322GCsa5gHTnyQXXNrBT9QbjbK0BtZiQMDcjK3VevAfSNZFjON1gW+jt/h9VQQHd90Gj1sJliBaCCsOOB/RhxW4K+SkaC1hGAtHcS5x6OyDvj7FyndFzS6pcT30BWj/nG3Isj2MQeOaHVBP/AABrf5Rt6IPZtERAREQEREBERAREQEREBERBx1zGs8Tl64oR/wB3oy9t/LQzL5oZmmOolYDgsc4fkJX0u8xzizl54oOb3GjL2f8AkMy+aGQl873eZcT+dAjEkpwHFZtOzwZGtk3yViUruiTdZE7J3yiRgJxug1e4U0fufWwAHC0SknEL8SDOTjdayK2M0ohk7gea0qSKMv6mEd8oNVdJEYQRgKalsQuELpQdxuFpUswjhAB7eSzbbeZ6enc1hOMINMqKM0lSWF2w2WbHRidgIPktOrKqWpnLz3KrFW1MA6d8fNBdNRO6+kHss+33NlthMT2gk7LT5KxxHV5lZNLTw1URe/uEGJXVjqqUuz8J8lJS0sj2FzCcY8lizsEcpa3sFqlrlDYSHeiDTZ/hcWkb+arDAZGlwaSp6hrJZz04JWbSRtji6X4BI9EGlwTGmmLunOPJTVFYKrALcFapbaOklqianABPmsfUMNFTzhtGWn1wg010PSOs9vJRmR3lkBXCYuHS7ssmJlP0EkjKCClh94l6XFSVNP7o4ADOVEyYwSlzOyyTO2q3kO4QY75g9uAMHzVohLh1ZwsoR07TkkBWkh72sYdigjppfCfhzdgVmva2aPLdgFebb97EhG+FCHGNhZnftsg06UYkIWTTxmQEjzUUkUnUXuG3qsykLekdJ8kEZa6A9TgT9aq6uY9haWgLUIYoqk9DiCVj1lne12Yxt5lBp9L8U4ONite03ZHXrVVHbWxktleG9vUrFobcXuDWbvzjZcz8MtA3D3yG9NgJfFhw+xB2v0dy7U+jtBU2pwwFzgH7d+2V3w5RbiK/TTm9BaY4wMLpHw64pXK8UMek73kQMIZg9sdl3z4C3HSdgsbaallYx0kYJIwg5wRYVFd6CvOKWcPWagIiICoQHAtcMg/nVUQaZNYbQ4vmFDEJS0jqAwV1Y4+cpkXECpmvppPEMJMoB3O267dK17GyMcxwyHAg/ag8TeY8jRFI7TTLS/7y0s2afqXVSx8PK3UMst6FM9kbHl5GCBsV7g8xHLLovVFJLfKqOIPIJOWgbrpBq3gkbBZK9lgiBjGRlrfLdB0q1HrehpaB1ibEBJCOknG+Vyny6cqcvGauiqmyYEuHbH1XB3EjSVwtWopw5hLnuOR6HK7F8p/FLWOgJYPc2v8ADjwg9I+Wnk7oeCoZdrrKC1hBw4rs9X6e01qqKJ/hxSMiPZoG49CujNy5xr1U2BtHVyEP7EA9lvrhXzKMFlfPUyn6OSSeyDk7mEsGhLDpmV8Ftpoarwzu0DPbbPzXjhzAWw3i7VU8EuA2UkeWBld5OPfM9adQPnt4qQSGkYDgvOvizrKWeqqPdXkh7yQc/NBsmrt0skDad0xOBjutFfbxSAh3ftuqR3WtMollyAfM+ayJZnVYyNygxqWINlDg3JK1R885j8NkJwdksMMbajNZgN6ly3bLbpGe0mV74/EDM74znCDheGwz1kpdkjJycpVUrba8MfjPb5rXbnXMortJHTkeGDsfJbevNQ6sqAQfzoLu8Yf3BCxyGh4JA2Wo0kPVAA4bYWHXNDX4Ygjq5GeAWtAJ+pac3Oe3dai2lknAa0HJVwtj4wC5pBG+6DBEDyA6M4JWdRNcMCbdWgGM4OwUjD1EBp3QZk1LG5hIHdaPLD01IaG4C1bFQxuXAhqhkpHTt64xkj0CDImhiZRAtAJwtJjpPHfkggfUtQt8FYZeiqYRGPMqy7Sx0sgbTlBFNCymiOwOAsaGLxn9R2A+SloqgVE3ROR0+SyK10EBHgkYPoUF0tQI4uiN2CPPKrT17GwkSjqPqd1pspfMMMyT5qWBgawtm2JQR1FSx8hIaAFBG0yPGdlke65cXYwCrXlsQ27oMkwtEeTg/UqwyNb8GN/qWF71I7AzssqmjOQ5yCd1KwjrOAFjyQNGQ3v5Kapnd0dMe5CwGzSmQB3clBKX1VIQ9khA+RWXE81seZCScJUU7nU4c4eSjpA5jD09ggxZKWQOPcjPksqntwc0yO2xusyghNVKWgeapcXS0jxCBjqQYE9UIz4bG7hajQXqGOAslaCSPRRMtZnAlc3YrBrII6clmd/IIJJazExki2JO6zYnvlj65XE/WtKghLjkjZZTqgRMLB57IMeokbJIWNHn3UBHh7FVljcCZB5nKjPU4dR8kGRBM6M5YcFUM8hqRJIScEH7Eogxz+h3msyopYw3bYoNUqrpTVNAIGtHV22UFFpd9RT+89ZAG/2LSaWnm8UDBwFqH3drKP8A2s3IZ2O/kghqqsRF1Pjdm2VHQXMUtS2R7OpoV1ayKePxo93u32WRbKOjkgIqSA89kGq3vVNHcbeKeGNrX4xlbS8M9Jd6LLrbeYXkxgliwiSAWkoPV32GzwbXxjZ5iewn8ra7/UvUpeWfsNR/1P4yn/6NYP0V69TEBERB1z9od+BpxL/xOj/XqdfPNG4A7+YwvoZ9od+BpxL/AMTo/wBep188Y7oLnN7lWtOCCrzuMDurS0hBmCrja3GFn01iNfAapjsbZwtFYwvOFqFJdqujAgbszOMIMZhNHUOY4djhW1Eviu6mnZahcW07ofGBHURn7VpccbnkdKC0Oc3dpIQuc76RJWRJD0szjfCxwxx8kFFKyIuAcCogCVeJHsHT5IHU4npDla4EHcpkg581d0vfuASgoxpeenKz44GiPpdjPqsGM9DtzhSOmlcfgz+RBd0eC8ucBjKym0wmYJRtjdYTvEcB1g7KenqnMHhg7HugjqZycxY7eayaG2GoaJQ7YblRzsiLTJtkqlPXzQjw2du3dBqsb4afLHAZGyw6q4NMwaG7HYqz77Keog5K1CG1xyQl7/pYyEGM6nZNGC0bn5LNotHVNZH4rZCB37rGp2vjmLXDDR6rWBf5qNnhxE9OPJBpVxtgtQPWckLQ5pBI7LRgd1rFxugryRK7utJlEYJawdygtp2FztjhTSYjAB3UHRJCQ5SszMzffCDHLt8jYhU3Pqr3xuZ38+y3VZLdb6i3OklI6wM7oNu0AzJ8XZUrwBLhqvrI3QVT2wjbO2yxnF73BzwguiAbhxCnbUsLg3t5FXRiMN+IhYkpAky1BqVQ6OOIPA7rFjpjU/EDgKaloK26t8OmYXYxsrJ6ettEng1DC0nyQZVNA+HAjODnfBWtW/TUtZMKl7snv3Wi08z3APB+a3BYdQ+71bYJXYbkAoNxR6iuGlm07aVrwQ8Z2PqvSrki1tNf6WAVeS/oGMrpbofRVi1tFT9YY55cO4Hqu+XLTw+i0pWU0VIwNaQOyDvfp5rhbmOd++3C2bxA0dW6kkFPBK4NecYyt82lpZbIG7ZDFoldV3GO4MdC0kB2+3kg85eenhPV6EsUt36HOJiLie+divK+svlRUySeIz6Tie/bde7nPDYn6m0e+KtjAjEJ8vkvHPWWibFZxPgsBa8gbfNBxFLUCUYDcFY+4O6zbnHBFO4QkELEEMjh1BpI9UFpOVkU9I6RviYOBur6GBsjul43WrSeDBAWN7kYQaNPOC0sxuNlbHTzSN6mk4CNhMk2wyCVqTYTGzCDTR1wPy4E5Wa2EVERkGxCuayGQkSYGPVQSzuhl6IvodigUtumq5S3fA7KGto3UcwjOd/Va9bqqmgb4hIye6027zNqZ+tnfO2EGLLTSRxhzSd/JY2XYOcrUYWTStDXjYdtlHUU3SDgIMEPc3sVeWOe0u9EZCXOx5LJPQxvScA4/KghpZAxx6kqn9b8g7KOQdJOPNUbG94yBlBNTVAiPxbrLcIqpoc3bK08QyH96VI1tRCMgHCC2aF0Tts49VHgk+a1qwiCqqSyrIwT5latd7bb4Y+uAt+xBtN0PS3KrDOIyMhKmTMha05A2WXTUImgLsboElc3AGM4U0FxjfH4PQAT8lZR0QMhEw2zss19BTxnrZjI+aDT6i2uwZW9juscUr24e44wtYil7slwAPVYNfPG13SwjCCkD2u+HbKuFOySUA47rBbJ0HqCq2rka/qCDVq2mjpYRIAFhRSe8fABhWVFbLVxhnlsFfSGODd2xQWzW5zB1577rV6O5QU1CYXAF2NjhaZV1b5WlrOw9Fi0x6pgyTOD3QTRRzVtQ4kkAkncrJpLZPWVzLe0Eh5A27KatDaaFj6bBccdlujThpI6L3+XAnbuPXKCer4by6foG3J0xyQDjK27Pcwxh3yRsp7/AK8vNwLqFziYmnpAWjxQvnhORuRlBkQV7p3EPdkLBuJnbJmNxA9AqRU89PIS4EBXOkEknxHbKDTnTTHZzzssimrPCOHDK1aK2RTtBYASraq1wQREu2Pog73exjqmTc0Gp2jy0DWn/nG3osP2LGBzU6rA7fsArv5RtyIPadERAREQEREBERAREQEREBERBx1zHfg88UP4GXv9RmXzR5BqJDjuTj8q+lrmR25d+KXy0Xe/1GZfNGP7YOexcf0oKRtc6QYB/ItXiqo4YSHgZPyWbQ0VLJEHDHVgLSrxA6N4ABwgwZpXOlcWuOCfVS02SMk9ljEYb23VzHPGzexQTVBJ2zsqQVJjYWeoKvaMj41ZLGz975d8II2yBsnXjIWSZWSt2AB79lBS0z6mToYMlZ8lA6kbh4IJ9UGntjdI8tWUwvpmEA4z6K2OCRsnWG7LMkiDmfH5INIkPU8u9VfFO6NpaD3V5a0y9KpJTSE5Y3IQWxTdEnW7JWQ6tdI4BuytpIGud0yfkKyqinghYHNxnugjqS9kXXG4gnzWqWTSdTeaZ1U95GBkZK0MTeIQx30fmtzWrUbrbTGCI7OGEG27lQuoKl1Od8KOnp5JzgEgLUquRldUmom8ySsSXqiePA7IKutjhk5ysSWJ0R3OFqDqmVsQLx5LAe8yyDPmgkoqV9ZKIgTus2ahNulHUcq2EGjZ40Y37qyWpmrJQX+qDUjcGGAN2Gy0eebM46TtlT1AbHDgd8LEjYZD1eiDUpwySlAa0ZwrbdRSFp+HcqOk68hju2VvvTlgkuQbFAzL3jbbzQaDY7NNPVEFhOD6LPvTZLc4Qe6uOds4XKFh4YXy04uFdTubCTnJG2Fu6ey6Gq7c51ZJEKho2zjOUHX22wNoiKuRpAznC5a0NxttVqmitEjWgvwxbA1NbzDcHtpx/tYE4IG2FsOugZS3JtREThrw4j5goO6V1v0dntA1JRztb14eMY2W9uCvMdca+5RULroWAkN3euo1DxIbdbKyzVch6GtxgrDteq6bS90ZUUMhDgc7FB7g8K9ZMhtsdzqrsx4ODguC50sV/pr5B41M9rhjOxyvF7QXMpfjbYaJsshZkDAcV6T8q/EWmvGlxUXGfp+9B2Sg7IItjVvGTQlFK6nfd2GUHHQCM5/KrIuMujXM8SWtaxn911AhBvskAb7BA5p7ELgziXzS8ONN2mRlFeY5ql2wAcBj862Vw05l6TUtSS+oywnb4vJB2oLgO5QOB7ELZ1r4mafulOBTzAy4x0gjurXXa4S1TRTtPQ47b+SDM19pZ+qrQbfGSCc+eO64pqOBNHbdNVsVVSiVz2k+vdc3VVRVQ24TRDMmB33UVNJUXK1ytqWAPcC3sg8lOMPLfFWaqnkjtxIDiRhvzXEd3npeFkclM6mETgSBkYXqNrjQswu89ZUMaIyCQSF56c12gn1VZPU0jepjXknpHzQdd9QcYK2WUujqMMJzjK3VpfjXcGWOWnir+klpAHV8lwvqq3UVK0Uod99AwR5rRrTVGijLHkgH5oNy3fU1/uuoJaiaveWlx7u8lpN8ifMPFkf143O+Vp9VcWNcZITkrCdd5p3eHJkA7HdBq2n7ZFf6gUTQBjbKk1NY26WlDC/qz6LCori+y/7bpPpndafetQVl+eHVZOQgtq7h7xEBCcE98bKS31VziicPenhh8srTw1rQOnchDUysPTvhBkS1Ukkx63Hq9VqdHbhNH4riDjfdaM/pcwOYfi9FLT3KqiYYxnB2QZtZXCmJjbjY7LHik95Bed8LHdC+Z3iT5337o6dsPwsIwgzYLhHSSfGAcK+pvkUo6Wt3PyWjzOa/DhnKzbdTQSDrk8vzIJ6eMVru2MlXSxe4zBxOQrppGw592OSAsGSplkJ8fOUGrVd2ikphG1oyFZbK9kLgZO2fNaZEwSOACyPDZER1HCDWLread0GIWgH1wtssbLWSEuJIJWc/3eQEOKjzFCCWEfJBZUUToY+phxjvusAvee7iVl1FW+XLTnBUEcJc8bbZQSUVQ2CXqeMjCnqP9tnxY9gPRWy00YYD2PooGNqGgiPJCDLhqGO+9E4I2+tWTUbnyAZ2J8lhgvifkjB+azIqs9PUTuMIJo7UY8PJUU1R4J8MDHl2V0Vxke/ocTjKlmp4pWl+2UGI2Xo+NxzlQSStfMHgYAQsd1lruwVXsYG/NBrdNPHXQiAbEDChqmChYWeq0umqX0ry5qy5pJK1vW4Hsgnste2nnL3disq5yNq6hsjQMDcrRqfpjkIdtgrMfFUS/wBgBIPog1umqYWwBhIzjCx6uwmsaalrthuVoZFbTOzKCAFmR6gnZEYRnpIwghyIZDTgbg4UU1K53x57b4V0b2yTGV5wScqyoneZQxv0eyCPxevEfTuFNFTB7hH2ysqCOla0PJHUVDPO2N/XHjIQS1FndQRioLshVomiqHW47ArFqrjVVUYjdkgKGmmmpwcAgFBqk1VFTAgDcLSqicVEgwO6nbJHOcykKOdsUZBYex8kFY43U4Ejzkeio1z55wYyQAfVUHj1mIomk4WRT08tG8mdvTnyQalUvYKQZAJAWhPiLup47DJWoVk4fHhpzlYga4U7zjyQeq/sOo+mycXpf7uosf5hXf616iry39hxI51p4xRnsyewkfaK7P6F6kICIiDrn7Q/P7TTiXj+86P9ep188a+iD2gxjHJ3xI8U4b7nR5/j1OvngwSdkFzfhKq94xjCszvgqux3QSUo+NZsrGiMuAGVi0Za1/xbLLfNGXhuRglBhRRyVMgZk4/MFmGNtI0NOCVkzGlgj64CC4hYJqROfj2wghmqOo4A2RmHNKkkpTIMxDKi8CaL6TSAUFkf9kKTd8Y7KWENLvmVmOtFTK3xmsPSN0GnwQOnOAp3BtMOh3dT0DoqeQiQgHtuoLnKySbLCMfJBZS0zqyQtaceinlhFCQ14BVKSZsGHMIyppGSV7+nBJPZBiSztk2a1QdRb5LVHWSopG+LUMIb8/RRNpo5XhwGWg9wg090jyMeSrHL0HOMrVKyKiZEPDILvRWUkdK5nxkZQRxXBrAMhZAvmBgNJCgqIqbqHSR3Ws2u22yemLnkdSDDpq1tc/oAwfqVa5ngxnLtyrqimjopC6DGPJaZNWSTy9D890GMxr5pCATuqPY+GQDfY7LVW2qokYH0zSdlq0FphZRF1aAHgfnQbZnqWyxhoGDjBUlIOiMvP2KelslVcat8NIwuAPkr6211VqeIappagx2tFc/w2DGFeZpraPCDyAVl0lBLN8VE3Lj6BW1FrqY5eqtaW49UGC2q8Q5c3JKuijFTUtixjJ8luC3ad+6oMdC0vdjsBlYVw01dbNVCSWMgg57IJbnp91HRicO7jK2zgk4GSc4W56p93ulO2mjjccDsFiR2v3FmK1oa/wCfkg1HSV8pdPv8WpYCPmqapr6fUUprKVoaButDkpH1MmGE4HbAWo2/T12D8sjJiPfbbCCCzsMsngubgDbdR19FLFXgRkjK3BUU1PbYg5rgJfP61p8XVVyGQnJCDk3hxrip0T7pLI50gLx55XrRyd3aHXtvguDosERg/mC8vOBHCir4lV1NSmEvDZAO3zXq/wAvOlRwopKe1uZ0OMYABHyQdq4I2xQiJvZowngQtPWWAn1WBaKiaoiEj+z91qZGRhB185qI4rvpyS3BuCIiMryd4t8Gqo0lVWRF2Osu/OvW3mi07drhpoy2RjnTmM5wvOvX1u1TbtMVrbvA5rxnGRhB56363vtlxkpHkktKhgqmRxFjmgkrVNXxVU2o6oPBJ6yB9S01lve0ZkB+pBDHUdEheNlkh0lQQerI9FjVEbG4wsuhYXMLW98IMmKNjADgZSomaBjqAQU9TGSZGkDyWBVNlc/DASEEU8juokOO6sEp6SDufVWva9v0wVRgy4IHW/8Auismmhc8h7twFZNG1jAQO6lpRKW5aCQEGd71HG3pDRssKesDsjzU8JhdkSYGNlZPTRk9TBsPRAtzRUPLf0qG4RmKfp8sK+mf4TvvX0sqefolaXSn4seaCOK3mqY0t7lbhp9PRQUJmeQMNJ3Wm2CUNmIkxgHupb3dqxrzDDvGR5IMe3NjqqswEAAHzWddaeKkiLWgEkbLRI5n0rhLH9M7ndalFUGtizUdkGjUwkfUfeyQSfJbqs9rlutUy3veSX7ZWiObBG8+DjIK1i2XOWgPvMX9lbu1BuDVfCN1gtoufjZBbnAPyWwoLg2mBjx22W5bvxA1BdYTRV4IhG262s+mjlcXMPc74QXS3Ed2DGVay5OB3JI+ZVvurGHDu3zVzIacncjCCr6s1BwzYqM0LyC4uyp5hTxN6oyMqSIyyRdTRkINOkiLdlEQRndZT3AuLXbEFROgleS4AkILYZfDdupHNdN8Tdt1QMZ9Fx3zhZbWBsRLRlBFTxY+kcqypaGP6m7bqwzPEh6Va/xH5Ls4QZEVaW4bIcgeqzKetle4MZIQ30Wj4GcFXxSOY74PPsg1mrdFG0OLQSrqSrY1niHsPJYT455IwZQcd91iSSkZjb2PzQazJcYKrMbQAe2cK91ozAZw4bbrQG+JC4PwQsxl4qQwRE/CdigyrdchTTGN52GyvulaKgZY7YhabUBgHiA/EVj+M47Hsg7++xW/Co1V/ACu/lK3IrvYsdP7aXVP8AK3+UbciD2mREQEREBERAREQEREBERAREQcccyX4O3FP+BV7/UZl80D/wCyu/74/pX0v8yX4OvFP+BV8/UZl80sYY6Z4f233+1BNQ3B9O7d2yyKqvbVt7ZK0yTHWentnZZtC2HwyX4zv3QYUn0ismma0syR6rHmI8UnyypmSNaMAoL2HxnlgOFlG2kRkl26wg4gh8fdXGvqGjpcUGTbJG0NWHO3GVlXm5MqSCwYx3wtPoJGS1P3/GCR3WRdI6cEeBvn0QXUFUyXER79t1dcXdBDG9iFpLXmFwcw7rOgnjnA8ZwyEEEsZY0SY3WZQTMkjLXDfturiIp/vbSCMLBqWvpHgN2BQZZiHiks2CxqvxM4zkKaGqYWbuyVL7nLUsMjGkgDJOEGmPgLW9eVdC2R56urYeqTF4eYvMbJEyZjgCCASgy44nzfB5/NZLKQQgdZyfmkskcFOHtPxHv6rTXV8z9iSgyq9zejpbj6sLTmfTB+azaMmqkLH4+1W3GBtO8NZ5oMoyDwMDcqGJ/S0npwVBSSdUga/stTMdO34cjdBpNRUOkPTjYLJtzerIe3A9cLc2nbNQVM3+2A3HzVurKKlt5/2kBgf3KDTGxMZjB3XIXDu/QWu708k4BY1wzkLiukrZHSgPPmt+WU0rYhK44cBkBB3Au+uNPag0gKGlbGybw8Z2B7LqbrWhu9BcnzRVrxEX5wHHGMq5mtKqgkMUTn9I7b7Lb+oNVy3AESk5+aDMumqoW2xsD3AygYz55W142CuifMTk7laXUF1TICScLLoZzAREdg7ZBFbIKmeuEDHkYJzus240FXS3GNnUXE+XdZoNNRtFTER1kZ2WXpCaO+azoqWs3bJIG7/Wg5f4UWO4VkEPXSuA6geotOF3h4faxq9HaNfTRVRjd4RGxx5LB09wq0/YeGkF6ijjDzEHZwPRce112FXQT0cDz17taAUGybtxK1dVa3nf8AdmQRF5IBecYys3VHGfUFDbnULL24Pc3GQ/zXFOsbTfNO3OS61AkZDu4EgrhXWOr7hdKpzqSeQhh8soNy8RtW6+bUm5G+zSxudnAeSPyLnXlq4l3ieiDamrLXdONzhdddA1LNQVLaO/S/ewd+pcstvWmdGzRUtpnZ1uAHw7boO4uleLN009WCpmuRczrB+l5Lt9we4tUerWQCeVuekbleXumr+6qhZU3BxEJIOSfJdh9AcZNNaYtJdR1A8VrNviGc4QemFPVUdVEBFPG8Y8iNlO1rGDpY0ALodwL5j7rqXVxopZHmnMoG7tsZXeqgq4qymjmjOQ5od9WwQaTqvTUF+oHU7Y2h5BGe3defHNtpgaMoa2CShdI5xJB6cr0l33GFwjx94XWnW9lqam5NZ1tBxkAlB4EaypKt2on1D4XsYSSBuPNbZuUhDiIxjfH1LtvzIcLKOx3aeK39I6c9mrqpVUBgklgnPx5wMjdBobRK09TnEk+WVHJIXSjGy1NtiuMDveJ4iISc58iFg3ERNlBi3QTiUthAJz9axXnrBLR+ZHygwgA7pTuaWkO7oIQ9zSc5V4kDzjHfucK6ojAy4eqpAGkfPyQZZpTBH4rjnzWM6pb1ghuwUo96l+FzT0K808DI/iwCgrJWNni6GAArH9zIHUXZyoCC156OwKyGyyubg52QYz2FpwT5q+Od7AWtJGVbLnOT5qSKHrb1AdkF9NUmJ/VIchUrKgTvBaMBY7hhxHocK9keSPNBPTzCnwXd1LNKahpc0/MbqKWncWDA3VtMTG/peMAoICZMkEnKyqSB8wJJP2rUY4qF0fUSMlYlT4kcgbTAkILKin8IH5LHjnazvnZZVUycw9UjSD5rTxjfPdBr1johdpfCJGyz7xb4bOBGQCStBtFylt03XGTup7vd5LjvKd/JBgVLmucXjbKg+Lturvjk2G61+10lF7oXVJAfjbPqg0FoMeHeayYqslwYTscBQ1nSah7Y/og7KJuQ8DzBQarWUrWQeOD3WDTx+Nk/NSVEtS6ANePhUMAlaC5gJCCaWjAaCCpIZ2wxFp79ljvqJAC05BUBLn7eqCaCI1M+AcZK3JRzQUDQJQDt5rbUPjQu62A5V0lVNK4BxOUG4LnV01ZGWxMAK0B1K6POSp3NkhjEhB9VH7yJGHJ3+aCMd8A4+1ZAgzH1dzjusQRTPJMYLt1nU/XHHiYY8kGnPdIHFuTseyyqSB8reo5IUnhQyvIbglxWsUkUEEBD8AoNKk8OLLSMYWM6pYQWgb/Up7jiSQ+GchQUdNmQGUYGUEbYJH/EARnfKjkY9h6XFa9VCmFOGwEF2PJaX4YIPi9/mgytP1EVLUeJMAQVmXuoirZA6ABoWhSNfEcjICzaN3WwuOdh5oLTA5rAXFXSEe6Px/cqsk7XnoBCsqHAUzm+uP0oPU32G3+5nGP/AA9h/wCjXL1KXln7DX/c/jKP/o1g/RXr1MQEREHAXPlazeuUriHbAcePSUoz9VbAf5l88VwojbKuWnceoxkt/OvoZ5+bhLa+UXiLXQ/TipKQj7a2Afzr56Y5n3OqklqTkuy4/WSgxm0rntD8YB3VroSwdlrTzBFGBsCAsX73IDuEGlAOJwM5JUjYpHEAnfy3Wc1lODsRlQOjldUgRjOT5ILoqKZ27nHHzKslo3NcMFapV09TBTB4aRtnssGke6V/3w/JBlW5zYQC8E/Yo7nUskdhjMfMLO8OJkYcquioXU7nFw6sbINBiYWPDz69luWjv9LFSGne0AkYW36bDqoh/wBHOypXtiY/71+byQSy0TqmZ8rHYDiSFBPRuhBLiSVWCukj2yrZqp8r9ycHugspmufIG52C1eme2jmbM8fCN1jRiGNoeCMnCyiY5acg43QandNQUt2pBSRABwGPrWiUzDC73Y7lxxlac0ugmDm52Oy12mfTGDx5MB4GQggrrBLBEagvJB3wtKEEnSS07BZtVe6mfMRPwZx9iU9PLKOtrctPcoMKGCSZ3Tk4CmE89E8MDzj5KaYeB/Y9jnJUUfRK8unIBQTzVr/DDiMk+q0/rLpQ7tutbggpqhvS1wJx2ysSrs1Q1/3phLfqQa9p27wUjR4zQR81jahuUtdITTREM+QWLQ0ZhDRUbAkeS5ZsOl7DV2J9S8s6wzPl3QbA0XeaayzGSrYCT6rUtWeBqNpqqVgAG+Qtu3iCCK6PhjdlodgYW4bfJCLW+GMgvIwg0TTFc20VhbLH148sZWfe66O9VrYIYejqIGMYWu8LdNU9denvvTAIi7PxDbCzeJFustnvcclrLS0OH0cILdM0kWi4xcasBzSM7rE1TrC337MtPANttgob7c/uvbI6VpyQ0DZY+l6O0U7hBXuALj++KDT7Pqant1QTPTjBON2q28R/skqxJSRkNO+y3tftCW+6UrZLQ0OcRn4VkaV4fXCjiLJWkP8ALI3QbfsmiJZI2ObTOeR6DOVr09RNaIfueLJI5xGM9BK7IcunByu1Fcooq2PLHSAHLfLK762Pkc0BX00NZdIYetzQcCME9kHixPpC9XWY1AtM4a7cDoKxjpivtsjjLQSNAG+WkL3Dn5KdDwRvFJFAWgHGWALrhxl5T6GibKaCFmQ4gYag63cnevLZou7ROuULWjrH0hjzXoLaNe0+sdR0kttIEW2SOy87L1wO1hYbnGbTBKQH/vWkefyXeDlY4e6mhtbK26wyCRjAR1A90HdmzNa22U2N/vY3WctmaRqbkHikqQ4Bhxv2wt5oNHv0lAylqDX04lY2MkNIz5Lyq52eL9ttVZUWiioREHvLdm488L1hr4KOWJ4rCA14IOfReanPHwUsd7nqblb+lzw4kYA75QeZd5kirLi64dI+MZWhV9SzBDWY39FuvUWmLjZ7s+mqGFsTMjsts3COjBLRKAcoNvz9RdkhZVunMDvEI7HPZW1IhjcPjz5jAWdR00NRSu8MgnHbG6DKnvUFVH4LGjIWOCyOMueBnuse10zIqs+8YABS8zsM3hwHLfPCCyKFtxlLG7H5LHraM0UgYXZKvpDNTnxIgckKyrNTO8ySg7IIxmYAE9llRVTaeMxFoJIwsNgeDiPup2Rsc8GU7hBDIHhxduAd1LDUnoMZ3JGN1PUQmVgEQyrIKCRjg+RuAECnYYnGR/YqGqmMkp6ScLLr54GxCOMjq88LTmMfI7DRklBrVohM+zTg+ZytTrqRkUJc7GQFpNtZV0nxlpA7qlyuksp8LJydkE9ot7blUmM4wFNd6b7nu8BmMkYWmUNTWWt4qMEA75WdJVm64cTl6DCpKR7ZDI95wfmsxjzFUNf3aPJRy09XTAGRpDfJZMNM+opjIAgybrV01ZSiKNrQ7GFoEJNO4NdvnzVk0s0MrmjOFPEY5Iy6TAI7IKVH35mGnfGywfjDunJznCy2v6XnqO3kpXtgLS8EZQYZp5OkOLtis2nuLKenMRAJOywTJKcgE7FXiOPpy47hBBI/rkLxtk5WbBXRshLC0ErBI32UzYmFmScFBE93U8uGd91NHU9MZYc5UUYb1HPZUkADsNQVY7D847rVY6dksBcBvhaZGzBBctUpJ2tAZnZBpb4iJS3sMrMp7eMCcnIG+FdcPBa0lhGT2WPHVziIt3xhBl1dyjfEYmgAjZaY0nrDjvvlW93fEs6FkDmZJGcIEpZNG1owDhYr4Swbb4UkzXsdlmwVzXh0Z6u6COngNQSOo7K2eAwP6e6RzuheSw9ysnpNSOt26Dvj7FXP7abVOfxf138o25Fkexdja3mk1SR/wBrR/wA425EHtCiIgIiICIiAiIgIiICIiAiIg455jx1cvHFFvroy9j/kMy+aPwy+ofGNjk/pX0ucyJxy78USPLRd7P8AyGZfNEZCyZ7x3JP6UE7qAtAcTlY5a5pwCQsmlq3PkDZCMJXlrXDw8euUEIpXEdRJUTmlhxlX+8PAwP0qNzi4oJIJRG/LhkKs0jZXANAG6j8NwGSNkY1x3HkgyZKN8UQlDt/kscSu/fEkKWSrkcwxHt27qBrHO7BBQ7klZMFDNNGZIwSB+dIac7F+wK33pPTslaWlrMxkfFjthBsOGaenkyGnPYghb4s+j5tQ0BqvdyCBnsuUdOcPNLXBwjmMfikjbA7rlS08FroaQNssWYnDyaeyDpXdrRUWytdTujcADjOFq1tuEdPSmN7QTghc3cYOElbpqmfXV8IY7BO4XANNSSVhl8AkhhPZBgVEo97dKBtnssps4kjLgzBHkpqGijdVGOo23wtbbp7x6ljKZuWkjOEG0ZJJXnBBAz6LLgoHPiMnScgLkm4aBjbaxPEweIRutJtum6qOB4lYcD1CDZNu6o6rp6T6brWKuzurAJGnB7rUo7NGyrPwjKtrKS5MqmNgYfDyMkDyQbeFqlim6ek7fJWTwSxyZ6jgLkCqo6WC3CSUgS9O4PdbFrnl0zunsCgkp7rJTNAa4gjZSTVrqxmJXZPzWFKyIxg+eN1hid0ZwEE8sIp3eIFnUd2mGGh+AtJfUPlHS/sjHtYQUGuz3Itb1EjK0SpqHzyE52Kkle2RgDd1ilrmkZ2QZ7C1kYzuceix3znq227KeAggBxyFSWKFpyO4QX/fOgFzzj8y1TTLHUV7prqH4EDw/v6LQ5J3Nb0jsqx11Q1pijzh22B3Qd79JcybdR6Vh0jHUguaBHgHf0XPHCXglPqShF3kaXNLfEIPn5rzr4DwMo79HWV4IZ4oxkr105db4+5U1HQWxofE+MA4+pBxTrrgUziBG/TVJaiJWZZ1Nbv6LZtl9mbXQvPvFukd4pyMtzjK9MbDw+tNsmbcfBb7y74nfCO63bjsg8jOJns9rloW2OudDQSDDS74f/Yur924A6upqmS7yUNSWUrydwSNivoDvlko7/QuoK1gdE/uCMrju4cu+iK6inpDTMAnBBPQPNB4R3vibcrdTjTQopIZGDpyWkdloNDri/0PW99e8NwTjqXo/wA1/JtpHSsE1+t0EQeWF4LW4XmLrGh+5l2qKUghrHkfZlB2g5S+Kc1VqWKCSUBxlALj57r2V4Y3E3Cx08jpw8mIHvnyC8H+X51HZ66Cspj98MgO3rlexXLJfbrd7VTdQcWCIZJ7Ywg7Frb+rNPG+26SkGfjytfGcbqqDozx25U5Kxkt490dLlpO268xONfCq46Zv0tQy1ytjjlO/ScbFfQvcKCnuVK+kqWhzHjBGMrqjzNcuelKrTdbdfChDyC7doz2QeMmpL7RPsTLe2EMlDMdsHOFxj4DjkuySuVeNGnoLNq6Wgo3DpY4jA+tcaPjfF1h+xQYLoi1XwxE7gqhLnP37IZXMOGoJJPiBb5qNrTG8OPYblXRMe7cg7+ayPDBZhwQZEdyi8MR9IyFhzB8z/h2BUHhlr9xsFqUAj8MuwMgd0GM6EQtBIyVEJwTgBVknL5C09gVR0HYtCCYQidm3fKoJBTDoLe6sglfC7DsgLP6aadhe4jKDTZotjID33WVb4Wvb1HyWNNKOsszloRtU6IFsfYoMySqZE4tOD9awpX+8zAM2zsoiXSuJ7koC6NwI2I3CDWRY5hAJvFz591CypbSuAkwT6qEXiq8MRknAGFhzSmV/Ue6DU6m4Mqm9DGgZWG2iLsHJ3VlP0B3Vn8qmkqiwdLe6COSI0wyN90jgdUML+2FkUDm1MobPjHzUlwHgHophsUGHSkRzFrvqKrVTPL8McQ30yoXMkaetwOe5WbSMjliLndwEENLSmT4nHurahngzA/NXslkZKWtBxkqKdxfKOrsUGTU1rJacRtbg47q2knZHGQcElUliiEILe+Fhhru6CWZ+XktHdUieesAjutc0vSUlVP01eMZ81qF+tFEydkdFgk+QQaS1jGxhxaNwoxQtlImHYbrNuFor6OlEsjCGYyNlgUlU97C0dgguqqlksXu7W7jbOFjw217iCSRv2WfSR07nkuxn+dR107opAI+2UFWujoG9TgCoJqhtYMR7fUrKiT3hmD3xusJrnwOyEGTG11I8Pc7I+tZDpXVQ+F+B9a06Sd8uziqxVD4hhoygynfeT1OOVUVLZWlrNisXrkmO/ZWnMJ+FBkRl9O/xHuJHlurpZBO7racAb4CxJJnPGCjDI0EDODsgmqJ2uYGAbhUp5+hpYT37KMBo3crSQXZagyTTmP78XbFRSzl7S1XOfO9gbg4UfguDC5w7IPVj2Gv+53GU+s1g/RXr1MXlp7DYEW7jIT28awY/JXr1LQEREHXX2hRxyccST3xR0f69Tr57bacPkzt8K+hT2gzQ/k74kNPY0dH+vU6+eWnkMbnEeYwUGRO2SVxaHHCjaxzCGEnJ2WVFLGd9sqKpxkvb37oJYbW8/fC/wCeFm0cccNSzrAONsrS4q2qkPhx5OVQz1EMwMmRvlBuu9VELKLIaDstptqgHDpAC1SSpbWwCLOTjCwRQdDwHDAygvdVukbjssZ7Jn7h5x9ay6yBkcOWAZ+SwYZndYa/OCUFr3OZ2UTnOdu4krWnQ074sgjOOywZYGNyR+ZBj08XiuwlRGI3YCuY4sdlvfKpKyaQl5BKCLrfjHUcKeOpeB05+SgDd8EbqSBrTO0PG2UEjuofGWndQmWQ9nEZ8gt1GG3upM7ZwtuVsTRIfDIIQWUtMap5aCcrXKeojt8Bp3gEkYGVo9ukMUpI8lLLN4tezr+jkIJpI3dZlIOHb7qF9KZyDGcb7rXbu6ibbWmFw6wPzrblLUyNd0eqDKpWSUcvUSSMrdlrq46rDXQZ8uy0SjiZIGunAAzlckacpLEKMbs8THn6oNGl022vYHRjpJ3W69O2CaOn9xdOQH7YytHr7gbZIS0YZnbdY8VdqOoIraBjyxm+RnGEGdqXhaKUPr/FBJHUFsCeSW0VPm5gK3HfNYaoucYoI2vLh8JAyrLdaBLRudeG4eRnfuEGEzXIjhDKX72/sfI5Wm1VZV3WTqmlLj3BJ7JU2ugbUuERBx6KNskUEnhg79kGsWSlcx3312x7ZWPd7HVz1nj0zyADnAUkcs8bGuaDgrcFkiqKpxc8fBj0Qbn4XV9RRSwxVdO6cF4b2zsu7/CbgVS8QBFXCh6AWA46cZXEHK7pzQdwuEQ1K+IESDHURuV340pb22q700Oi2NfSEAfB2x9iDXuEXAig0p0O9xLSxwOcei7DU0DKaFsTBgAAKCzxyR26DxgBIWAu+tZqCjmhzS0+a2jqHQlBenEywB+SVu9UJDRknYIOGr5wr05aWCrltTJCN8luVuTRl4s1BAKOmoWxdm7Nwt61QoK6M007gR5grRZoNMWtpf8AB4g7YI7oNZgZTNHvMLAOrfYLLjf1tytuWvUVHNL4IA6PLdbkYWuYHMxg7oNOvlvluFKYoXEEghdfONXCJ8ml6y6VMZlMYLj5+S7K5GcLRdZW0XbTVfQEZ8WIjGMoPDniLp+K/wCrqizwUBaQ8tyB81i23lErrwz3gwPAO5XcrVHLlcWayqLnR02S55d9H5rXafQWsrZQmGCmOQMDYoOkJ5Iq65h4ga8OjBJIOFwhxL4a1nCipfRTRPc7q6TkZyvT+zaU4mU9XKfdnhr8/vT2XAXHfgDqq/15ut3pCIA7qcS04QdAIGT3F/S2FzMjudljVttkpJPvjicbrsBrDR1htMbqO1hhqo24IAGcrgnUVNcKepc2oaQMkIIqSaNgw4A4VtZWxEFjWjJWFF1gkklVeGOf80CJ4if1OGxOVe9pqHdbThWSRHGfyKyCR7HhoPc4Qa5aYW7CQbg+a1CoiikPgMxk7LTYp2wRB+TkhR+PUzuDqfJdnZBrdv4ezXIl4lIB32UtboptjBlkdkhY9sv9+s3x1AIZ5Z7KK76vqLmwtcSR8igwZrxCMwNYAQcbLDZSeLOJyfhzlQNjikkLycnOVIaiSMhreyDVa4xVdMII2gEeawKNptswkecjOTlIanpGR3UdTIZgRjc+SDUrtfoKqnELAMjbZX2muY2lMb9yR5rbT4nsGXBahRvaIekfSIQTVUUcshcMDJWlyB7JCG5G+2FlzwV8B8R7SGnzWvW2ntk9L1zFviAefqg27Mw+H1disXrcD3K1G5xlsjmxA9K04xvAyQgzaEMmcWuH1K2up/DeOjsVixTOhJLO61FjvHiLpO6DCji6iB6LJNPhhId9ixXeLG8kZwqiqk7O7foQUZDlxGVeafDgfLKsMpzlndX+K/p+LuUFzgCMKwvLBsTlReIclW5cd0Avc47klZDKhjY+nAzhQNYXZ9VRzek4QVx1OOPVZcNJIG+KCcDyWLF9JatTskfH0DsUGDJUjBYR22WM5znZ74WfW290PxYxncqSnipvd8ucOrHog0sjzWZSTDHSsWUYefTKyKeIlnW3ug9A/YyRBvM/qd/91oGt/lG3oofYvSyv5pNUsf2GgK0j/ONuRB7QIiICIiAiIgIiICIiAiIgIiIOOOZH8Hfil/Au9/qMy+aCT+yP+s/pX0v8yW/LtxS/gVfP1GZfNXRRxyVMjZcY3x9eUGGHEHIRz3OOXHKyaqEMeSOyxT3QVBwrmt6jlWKeFzQ3dBWSYFnRjfCshfj4fVWP3ccbqjctcCR2QZEtMWgyZwO6sglDO4U80wdB0gqykYxwPX9iCUyh7RgY81vzRGr6eiljtcmOqXDcnyJWxXyU7BhpGVjU1R7vWx1DScscHA/Ug7eae4dzQ0bNRiq+F7g7GV6LcreiNP3vh9Ldq5sb3QRF2XYPYLye05xqrnWmK0SSHoaQF3J4E80NPpPRE1qfUYEsJafix3CDROYyeza91lV6MomsZ0OLNsDzwur+qeE0fDeOdrx19ZJHn3W4uJvFemi1vU6itUmZXvJ2PzW27lxLl1lSvN3d8Q7AoOH73TOpqk1EYIBOVnWHVsFEfv4BLfVVu0kM1a5riPDB+xbbuNLH4x92AIJQcrWfX9LdJBSvx09luGrloRRkx9GSM7LhWga2gaJo8l53W4qLVD5GiGZxz2AKCW4VbYK0v7Ala3bLtRzQ4ewF3YHC2ve8vi8YdjumnK+mjB8Z2CEGbqQyPaXsJDSDgZWzS4gPyMnJ3W8L7X0s9OWxEE4W14RD0uEuBk+aDSnuk68b4UngGQdXoswQwmUk9iserf4bw2PtnfCDEe3oOPRXxQOkGw7qR7mOjz5rIoXsa34iNuyCAQGA5duoSfFkHz2WdJI2V5AWMad4cHNCDJfCWQhwwFgOfIT3Kz3Pd4QaVqNDR0MlK98mOrGQg0KGM1B6PPyWbDRe7ytkeMgbqGICKuIZ2BWsTNMkJICDVIdWttlNGylIY8OB27rvxyXcw9NZDTQ3GtZlrQPiK81HDqnLXkgZW7dLa5qtMV0ZpXvAG3wlB9FfDri/aNbRMEFXE4nAyHBck5B814+covM1S2ariN6qXNb1D6TvJeh1DzWcP6w0zY6yMhzR1fGO+PrQc7KGqnZTU8k8jg1rGkkk9sLY1v43cPrhEJGXhjSfIkH+dcecZOZTRthsk8FHXMe4gjOQD2+tBw1ze8WrdVwz2gVDSWsLe68leJsUNTcKueJ7Tl5OftXYTmO40UGobpNJbpy5zgdg7K6jXW91tVPOZy7DnEjKDc3DjVkun62LLzgSA9/mvWTk/wCYelbS0tpkDSXsDd/qC8g9KR0tRUxmbGesfpXoTyn6h0HaLpQRV8zGzbAAkd9kHrHaq9tyoIqxgwJBlZi0LRtzt9ysNNPbXAwhgAwtdQFx9xa0hJqfTtTSNDndYIwPqXIKjmjEsZZgHPqg8Tua7l6qtJXee+PpH4OXZIPzK6V1bfeZpmBvT0OIxj5r3F52OGVfqbT8zqOIH70ezd+xXjDr/Q920jc6xlQ0tHikbgjzQbHqIBCM+fmsIbnqWoVQcYx1HJIWPBSveeoA4CCsVU1rcEI6qBO3ZWTsjYOnzz2UI7d0EskodkBXQyODenJ3WOe6qCR2QZLaYucXZCq9/g7HcKKOdwO/ZSOLH9ygp1ePsBjKkjpZBt1HB+axuvwz8Cy4KgujJPcILm2tz9wd/rWLUUr4DggrJguL2S9LthlSzyNqRkbkhBhwxhnxE9/JUlb1O6gNlSSCoZnIOFkUzMxEyDdBivkaWgY380ZB1jKsk2e4DtlGyOYMBBd0mNyyYqE1A684woYmOLuqTssj3p8Y6IuyCyZjqbZpwR6KkVbj+yfF9aVMc5b4jwcH1SnjikZhx38kFKmpZIAGhQxSuYcBxAKmZRv69x8OdlFUMDHDCDI6wxvV3KhkDpj1AHCsY/JDXHZa1RtoxCS4jOPNBpUbHOw1x/KsptM0MJOFHVYEhEH5lAKmZnwu28kGTDK+CQ9DyFkR3J8VWyomcXBpzvusGSRvhhwUkUbJoTk90G8rhqqjvluFvjaA8Nx2W0Xw/c/Mbjnq7LCa91LN1Rncdisk1Yqd5jgjtlBa0vjcJerYrMjZ48JeRk+qwXSNcenPZa1RSU7KYg4zhBorAROW91HUYLw0eqzGuhFSScYCx6wNfKDHgj5II5KYsYHnsomDJz6LMmLvBDT6KGOBxaSAglgcHfCfJQ1Yw8K3pkY49Oc5U4Z1M6pO/wA0GGpWShrcfJWSNw44GytQSsYZ3YGyldSmIgkrHjkMbshXyVL5BugzKeVuwIBwo62QDLAMZUEDul+XKSpcx7cjug9VPYbPza+MbPMT2E/lbXf6l6lLyz9hr/aHGb18Wwfor16mICIiDrt7Ql3Ryc8SXelJR/r1OvnjjO7vmF9DXtDTjk24lf4nR/r1OvnmgaXEgeiC0Eh3c/lUwn26HZJ7ZVnhkO3VTESOoeSDOtzGUr/GkwQeyrc3sqcvjaBjfZaeZpHDoH1d1nUboxGGy4Bwghtshjm6XdlqFXMDgt8h5LAqRGw5ixk+YWTRlkkf33v80GJNVOOxyQFjOy457Y7LWqe3MmkJHZYdxoJKeTDR8PyQYwlfGB8RWQ1hli6s7kZWNBE6R5a7OyymkQyBhO23dBggmKQ5B2K1WB0clOSe+FHWQQPj6oiC4+iwopZIT0E4BO4QRyZEp+tN3fEDghZVSIfDD24ye+FhtOMhBNHJPJ8IkOAkjiz4TuSoWuLT8KyIg2QZk/OgtpvpHyWQ+mMj9tj5FWO8JnYoyqAeCDsEEs9NPHH8b8j5qtJRdZEpxgK+R8la0MhGSB2V0LKmkaWytIBQTVE7ugRx7EbbLLtM9ZA4SmYho8iVhwy0zjmRwznsqz10TAWRn6sINcul6+6MLYGP+IY81vjSmt7dZbE+iqY2OeW4BI3yuJ4A/q62jcrVBJTGEtlJ6sINzWzU9BFeJaqWNhY4kgY7LB1PqaK51ggo3BjScbLab2vjlLxkNz5KF5LZBMzPUEG6X2V0MAqXSbkZ3K0U0L5KwPDtg5RfshrXsEMuQwBUZdGs3Dt+6DdMsscdKxuRkYW5dK18MrPdwB1OGFxiLnJOQ3JIC3VoS6U8N+hiqTgEgblBzPp2z3+ilgqrfPLEOsH4SR5r0u5SdbRUNrhgvDxLMGAZecnOPmuk1tv+kLZYKaarljDjjHbuuwHA+5OqHx3i3tLqGMAucO2EHoha79Bcow+PAB7fUtVBz2XCek+OWgaqjjtFHUxmtZhrgHDIK5SteoqOqphI54bkZB+SDWiQO5UVS5op3kvAGO+Vol11daqOIjxQ5x+YWzNS64nhtEtZCD4YBOfkgyrneWU9RP1VzWhoLhkhdMeP/M9WaHvD4IK4uYHkZDtu6s4r8b7vV189Fp+R75QCCGlda9f1tlu1onn1g/oqzkjrO+ftQc6aH51m3CSKN9waHEjPxb5yu1HD/mdt9zp44J6qN5IGCSDheIsF8pKDUhZbJHGNsmBh3lldnOH/ABZt1ps/XPMRMGbZO+UHrxpjXduvUnjfdCMhx7dQW7qyopvcpZHzM6OkknqGOy8jdCc2NTarmY6iocIg7bLtsZXL8HOJWXSdtHDO8wOAB+LbCDuRab5YK+9vpX07HkOIzgFbqNJp/wB7Y00UeD5dK638L+K2lp3MrauZgleRnJHdc2Ra4sNSWVkUoMYGTuOyDkGOz2ctDo7fT4PowLrRzV6r0zbNOVdohoI2zPBAcG43wVzVScYtFPzA+4sY+MbjIXVLmv1ro28UVRLTTsMuSRuEHmVrOKaz6wq7rPKXQyOJDSTjC4y1bdqW8TF8LAME9lvTindZK2+TsYT4QyBv5LiyQtjkIztkoMZ0IJJxhYMzOiYn5rVZ5o+j4DutIlEj5CSPP0QapTUoq4wB3GyhmohTzBp232U9qmdTZ8QkD1KguVZ4tUCw5AQZUtMZoWhv5lk26VlreJpWjDdyMKOOti8EN6hlSudT1UJiJHUfyoNckutJqmIUUDGteBjOMLQa/TjraC1zsk53VbRb6y31JngBI8io7tW3GrqRG9pyTjsg0+Kie1+fLKtnaGuDStWbbKyKISzNIBHfCxZtPXaozVRQuLMZz6IIaemLt87d1DUPFPOARthTx1ApwY37OGxHzWPUxPqT1sGcIKVFU2aMtY3J+QUFK90NQx5BIDtwsigga2fNRsFuSC10FSAY8Z+SCC43OCqoWxMiAcB3xv2W2g+eN+z3AZW66i0MhaS4YHdbarmAThrTsglfXDww13f1WPJUte0gN7qdlF1MDiO4VstI2NhOUGNTsEjzkKUymNwHke6ip39MmAdisySFr2dXnhBFLMC3YZ9VjsPiuDO2TupqdoDy2XYeWVILZVPf4tOwlg3z6IE1CaSMSk5GMqxsfjM6x5KWslmfGIXA5A3CshkEUJY7Y4QYRHxY+ayY4mlu6gBHiEntlXl0n73OEGZS07XEk4WLVx4mDRuqRVEzThgJWbSxCSQPm9coLILc57Q7tlZkNWylkEZGVNW1UMUIbDgnHktPpmsmf1Sd8+aDVqhzKqMYAGR6LCFmkJ6g44V1XL4cYEG5VtLdXsHRIdygxrjQ+7N2GSFJQQtMAc44OFlOmjqATNgBYU1SIneFFggoO/HsYnNPNJqkAbjQNdv/AOUbciexhaBzQ6od5nQNb/KNvRB7OIiICIiAiIgIiICIiAiIgIiIOOeY/wDB44pfwLvf6jMvmhL3RzPc3Y5P6V9L3MeP+t44pfwLvf6jMvmi2M7g7t1FBc+cvaQcknurI4esE57KeSFrWdQGx+axxI5pw07ILXjpcQqZVd3O+aubC9xAwcFBa0YOUc7KypYg2P54WJ9SBk+pUsBJe2MfvjhRKeCN+0je4OUGtv0o9tIKoybHfC0KaHwXEZ3C1B2oa4weBn4cYWmvkdI4k+ZygkpqqankDmPIwtei1teoIRTU9Y5rTtgOW2lPDHGW9bjuD2yg1Gapr5JDWVM5fnc5OVWK51EhAZKQB3wtNlqXvb4efh7K2OTwslp7oNRuNS8xAdRye5VbUA6NzpN/rWGJWy/C4/lKlbO2Bhaw4QJqwMlc0ZwCo2zvfIJWkgA5IUDB40p6vPcrIkDImdIxkoNSlvjZ6YU5OTjCggopZIzLHJj5ArScEnLfJZDK+eJnh52QXGqmilLHyEgbbqGeodI/LSQFYSZHl7+5OSVacAjCCYVLg0DJz6q10nWN9yrSGFox3ViCpO+EyR2JH2q4Ru7kbBUfjyQXwyFrjkndZRrWtHmVgtwCsyGOB7R1dz80FktYHjsQrY6iUERtcQDtsrqqKKP6Csp4HyHqA2QZpp/BjE/mVJFcgYXMI+Sj6pZmiHHZVhpWxPDZNslBpsri6QvwRkq+BjjI12cYwtwT26lbT9YxnC0Gc+FIfDIwEG5aG8XehgH3PrHROHocLdlk4p60tbA+W9TEsG3xlcYw10jXAHsVkuqmSDoccA7boOa7LzNaxo5jG+9ygZx9MqTVPFzVOqaF7nXp7gQdi/K4HqaeGNvWxxOVJR3aemZ4QJIIx3QZFVeLk64ufWVT3kEg5ORhVrJG1LepmMkb4WFUmOQGXO7t1bTzBrS0nGEElurpKGoDmuIAK5Z4ZXG/y6oo7jS3B7GRuDsdWBsuIYqOepeRG3K5G0FdDZa6KCc4JwMFB7RcoPFyS82OmsVXWB8jcNyTn0XbheVfKRq6Oz1UNZI4hpeDnOy9M9Laqob9QwvhkBc5gPfPkg3Cip3VUGi6m03Qajt76Srp2ydQI3+YXk7zxcF6Ky3ioFJQBgdITsPmvXxdFednRl0v0s9VBHkB2cgIPI3VmiTbKds3Rj4e2FsyimYwuhIydxsucONVsuNnZ4NQwgNaQchcCUjw4yOz8WcoMyrsrsGcO2O60iWMxu6T5LP+6dT1mN/0QcKr2RTN6tiSg0xVDSRkbqWeLoIwNlSMt6DnCCJMlVd3KyIYmyAtAyT2QUo6Y1UnQtSmtvusZOdvPf5LDpoqiklEpb8PmVqrpn1kZ6RnyQaPT0hrJywHBU1RTm2yta453RgmoJjLgjBPkoK6sdVvDneSDKlucb4ujpGcKyAidpAWI2AEZ3U0UrYGYB3QWVVIYsuzndVo6Q1BHfAKMqPGeGyYxlatDJTQQnoIz9aDHrqdkdOWtxkLGo4QPjecgHKjq618shx2GyhE8gBaPNBnVtwjfGYWAZ7bLTmucxwIJGFm22nY+bNQMN27q67x08cgEGMHugq64s8IR4BI81gPd4hyrEBIQFUPcP3x/Krouku+I7LKcyHw87ZQQU83hydTjkJUStlcC0KIjLjjsgGBlBcGOd57LKjBjjwCsPqI7LKglaQA5BjSElxysyltclTEZRkAK8RwbE439Vr1uqaOCjLCW5wUG1ZITFIWnyVfHlaMBxx2WRcCJahxi3yfJY4pZtnFh7oEET6iTAO5PfK1BlGIGdTznAWJnwAOj6QVzameRhDsoI6icFxaM7FXQ1bWNIKpSwtdJmYbFK5kTXjwjn6kFW1LA7JCtmqOs/D2WNg+iyYBEWfF3QQueHDCtaMuAV7onEktGQrC1zDvsgnfS9EfWSoGjLgFIZpZR0bYQROaOojCCR8GGZHosbJ7ZU8Uri7pf2Vk2A8huMfJB6wew8ozDZOLtWf/AJPPYh/5orv9a9RF5e+w9qTJYOLdOf8A5DUWQ/lFd/qXqEgIiIOuvtC2GTk34ksHnR0f69Tr55qd/hvJI7jC+h32gbg3k94juOwFJR5/j1Ovnih6eo9XbH86CaY5b1Ad1qVFQskpDISOyxI3QOADyMBUFxdE/wAJhPRnHdBiOJhndtsCRgqj5HSOAaSAdsBaxLHROhLyR1FaS9sbX/Aex9UE8NO4NDnb77qURnYt2weyrTTMDMSEKskhc8dGMZHZBl0dW2DZxx81bV3OGdwZsSdliVkjGxjBwT5Bae0PcepoJIOcoNcpqBo++jG6xq2AyTdLcDfGVFSXJ7CGSnYKWaXxHeIzy3QJaM0cQle4kFYEhExJb5Karr5J2+Ec4CjpnMaC12Mn1QRxQvld052HzUslH4YySrpXiI5ZhRGqe/AcBhApYvEl6Srqpvgu6QftVWSNiAeDuVE90lVINiSUF9JTuqpOjqKmnt5hPSCDlWx09ZSHxukgDzVXVj5ZAHH8yDMs8jaCYyygEE+azrhWRXAgRAD0AWNFE18fx9j6KNturBOJImksGM4QY8tqmYC8EgHdRMoZA4OLs4K1q61TI6UMaR1gbhaHFWPwQ7v5FBnCpbSMwd1jufJUO8ZriAN1iv8AEmJOCrRJLA0sGwPdBqQrmyt8I9x3WPJMIiN1hN6y74c5VZOskdaDKfM2cdLcZOyqLW8t6+r54WE1xY7qC1CmrndHS7HogupI/CcWnyUjDPT17KqJ5b0nOVjvlLj97Jz8vRZkMjPBIkOHYQa5ftVXi6UMFJBXPAjxkBxC7QcGeaSDQPDCo05VVDXVb4C0OJ3zjC6ZyySMl6os4Pos+zy089U1la8gZ3ycIOe+EfHbVdBxKlvNZd5BSy1PU1pccYJXploTj+28WOOdtzacRjOHj0XjzfaenpKCKaykmRuD8Pdcx8KeM77Dp00Nwmc2YMwATv2Qd5Ne8x81rqZcXQENJOOpbSg5zI7xRu0u6paXz/B1Z9dl0tvWt6u/XGSWR7zG9xI3OMK2E2y3x/dVjyJ4/ibv5oOyWptRQcPZJtYXCrZKydpkAcQRvuuofF/ifWcTLuZbXOY4eo/C1xAKj1rxYueqIzZLjI/3do6RvthbKobd4LvFoyXA75JQZltt1RaSKuZ5c47nO612K4XGrIkhqyxoPYHAWjur3GPw6s4A9ViSXYwbUxJb+hBqt/v9xo4x4FSQ7O5BW7tCa+uUFIXz1ZLw3zK4ymqo6/aU5PplT2mcQ1ggaT0FB2q4XcQtRXSubDBWSYD/ACJ9V29suuLpSae91mqnCR0eAS7fsusXKxoFmo5o30o63lw2AyuxmuNEXfTssYlY5rABnuAg4g1lrnVFhuctULpK1jyT9IrZV1uF91lRPqZbi+QDJI6s5WscXLRVXiAw25xdKB2b6rjvR4vVjm9xuJc1jnYIdlBx5rOyTMqJQ+IkjOSQuKLtSPje5oBByu2XEC32iO1uqzIzrc3Pb5LrVqSGN7pHwkEAoNnRwvDsuJ7qTDGHqICte53iFgKx5Xyh+N8IJKuq629DCR9SxGkk43Kuk6TuCtToYqYw5cR1fNBp/hzMwcnB+azqGF4cJ3OOG77pD0OkLX46fVRVNZ0S+HFjpQbnpNS00LPBc0ZAxlYE94gfWiXpBAIPZaV7sx7PEYSThWtYxrwH7b+aDdly1LTVVGyKKMAgBblseorYywyUksbC9zSNxvlceM9ya0dTt/JVdUFjSYiSAgxLtRudXTSs2a9xIUlvf4QEb2EhWvqTK8AnfzWt0TLe2m65SA/HmggdaBXgOiwD8lk0NE+3PAe8keix6e6dE5ZCds4G6yZKO6VT/eOg+GN8oMu7VbXQhjdydlbb9HCvpjWOcNt1i1NVTNaIpCOsbd+xT7vXGnpXR0ocY/ke6DBuIZRyGBpBxtssmmtIrqUynH2rRW1ZrKomfZxPmtbhqZ6aDEQJZ6oNEmtZp5zvsPUKoIjGVfXXASOO++VHCRI3B7oMeoHjOAYMEei3JaLvS0dF7vKwFxGAtAkDYTnP5VUOjc0uzuB6oJK+VnjPmxgHOy0t7y9xLc4UtVP4h6QcgKWmZE6I5xlBhKZkwEZaQMlJGBrj6KI4LtuyCaCdsb+pzcqZ9X4gDW7Hso4mRObvhGw4mBA+EboDY5XOBcSQs+KmJb1NIGykkfTNhABGQFiurC2MtZhBkwNDXEOOd0fbXTTCRowPksCnrHGQdZ7lbkjnjFF1tIJ6coNGukZgjDWu7Adlh0cBmJc4/UpDLLW1DmHJGSo5fFpH9IGB9SD0E9jI0N5n9TgH/wCUKt/lG3osP2LVRJLzSapa7y0BXH/nG3Ig9pUREBERAREQEREBERAREQEREHHXMeenl54oH00Xe/1GZfM/Kcyvd/2xP519L3Ml+DtxT/gVe/1GZfNA/wDsjs+pQSmoJjDPRWxxhwyVFjfAV7RJ2b5oKH4HEYWbTyNLd1E+BxYXAb4UccUrc7EBBLUvBBwdvkseOMOVZXE7FWBzhsCgq9nR5rUKJzPBIK07qJIyVe2Z0ezTt9aCrmCScgbAlUli8MoH4+Id1SSQyHdBYmcIqhpO+EFFUDJA9UYOpwCuewtI9UFzoC1vVnZRbn1UrvF6BnONlSLpOAUFI3Fjursr5HeMAfRSGJob1Y2UXUB27YQVpsdeCMq+pYM5G2AracfHkKtQ49QGdiggyR5IBkgeqyHRsMfw91AAQ4Z9UEhhLWhxUQ3cPmsyUkwjdYbTgoMybp8A47rDDc91LFJ1ODH9kmw0gM80EJ2VQXDsSgO+SsiNsZ7777IIHPc4AEnZalbXtbGQcZWJOxjG5ChZK5mzSfyoNUbUMglLzjHmoKutE7w6M4xvgLDfKX5ByrGnBGeyDVWVrpY/DcT2wVg1bMOznOVeXRxxdTD8RUTQ+d2cbIIVLDC6QjBU0lPhuw3VIXGIEH8iCtRGWMxnKx2RF7SQeylLnyyHqGyp1CN4YPNBCcjb0WTT0jpgHdt91mGjZJEHsGSe6x2mSlBBGPrQa1Z54KJ/3xoOFWsry+7x1MDi1rSDsVo0NT4r++5Pkr6gPY0vaDsEHcjl41w6phgoIqoB4cBsV6fcCbrKynpmT1gOWDbq+S8Q+AOo6q36kZGCcB4Py7r0s4UcYRSXi3UEkhBeGtxn6kHo1TvEkIdnOVKtH0nVmusFHVn/AOSxgrWEBcQ8cbPb6yx1M88AeQCcrl5cQ8bbpDTWmeKVwAIPmg8gubttOyumhp4A0DqAOPrXT+hZh7ydgNl3k5qxaJ6qaUOaXEE4P2ro/O9kdRM2EgguO6CKpw5xa0AHKvp4nNaT1eSjnIDOoHdWxT4aWk4QJJQ5xaoHt6T3VHHLy4KhJPdA7rOomBjg89gsSIAu3UxkczYA4QarWVsM1OIWDBCpbZ204w8ggrR+pxPw5Qunbucj6kGrXSeN8Z6MAn0Wis3cFc+Z7hgk/lVzAACfRBnMjb4fcZUUtIZN2kYHzWO2peNvJXNq3g/JBX3Q5wDgqOQSRHp6jhZMk2YwWfSKlpoBKwvm7hBpn1qrThwKmqWsY8hqiEb3DIGQgyZasGMNZscYKhjjdMcuJP2qNzenY91kU5IYelBHJAWBRtbkqciQuPXnAUTiA/4UB0ZABCtyfU/lUxDntw0b/JRFhb9IYQWg7qvchUPdTxR5b1eiC3wvhzlRn4SrnPdkhUG4QW5PqVMwTOaS15woVJFI4EN8iguhkMcmX5Kz3XGHwyzAyoaiBjYevzKwWjLgEGTSt8acl3YlT1RZDs3HzwoT94YHM7lRmbxc+J3QJp+r6Gyh6nZznOfVD3OOy1KhghkhLn98INNJ8lNFTF7eoFVmjZ4ha3sCrPGfFlg7IJGzNiJa4ZwrzCKhviDbCwyckn1WTDUeHGW/JApQPGLSpKyQNcGjssUSOa/rb6qQPEu7u6CsrQGAt7lQhuQSfJZEUUsjsEHHkrKiF8WRjbzQeq/sNmEWvjHJ5OnsI/IK7/WvUpeW3sN3E2vjG0+U9hP5W13+pepKAiIg66+0MBdyb8Smg4Jo6Pf/AOrqdfPRSwmaQt7YC+hf2hmRyb8SiO/udH+vU6+eaCV0Ti5vfsg1SCxvqBs/Cwa+idRv6Tk791L90qyAAg7FZ8AFfTGSbuBnsg0emgkqX9AcQAlTTmmf0uOd/VXxTe7VLuntnCtrJ/Hf1ILzEXMy126pHJ4Pwu3PqoRO8NxlVa7r3cd0GoU9uNwOevHoo6qFtvJiO+c7qGC4T0xxGdgsnomuEZneCSEGFDA6pkIb5lZzofdovDJycLFpXmCoI7EKR1Q6WsaJMdJcB9iDELXukz0lT+5PcA4ZAG5W7zb7bPQgwFpkwtJqIvdYXNeMFBoUjSPhzlXCjcW9ZyAArBIPHy7tlZ3jtLCGEHKDAZGZXlgPZZNvaIrjHG8ZBcAVjvf4b+pmM5WbSFm1U/ALd0G9dR2+mp7OyaNoyW+XdaFZdLC4wOqiQMbqJuoH3Boo5XHpGwB81A7UFZbXGmpzhnmEGRU04pXmEH6Pn9SyItSU1JTmmc0F3bcLCkrWzReNKcFwycrR6gRSEyMcgvrZXTzOl6sgrDz8WQrmvI+E9soQHuAZuSUGfTPj6NwFBORNOIgNicKnuNaxvX4bgD5rUKKgd4fvEgOW7oL3WYUkAqXHJIzgrT5YvHOWjGD5LMluclQfdndhsowWxDZBjG3HGepQSNdEegeazXVD9iBsrXU7pWGQDJAygtpYjGesjJ7q2oL5JdsgA+qybTC+ebw3jYLVK6iihZkDBwg02mgYGDqwdlWotb3NNRGSMbqOllPjFrvohT1tbKxhZH9EhBk6avHutQYax3W0bAOOVk1sdXcrwx1FIWxEjYHAxlbWif0TB7sjfOy3XaLhTw4f1DqCDkyhsoZa2FzgX9I3+a0G6sro8sjDi3PkraDV+OmKV+G5wuY9J2WwX+1eJI9hl6QcHc5wg65XOw1FYepseHdzsseCuFkjMEwyRsuZtY6disk0snSBGM42XD98gprhI4QkF+dsINIrrgbi8ti2z2PZKandGwse7JKR6cusZ8RsR6R5gdwseplqKaTolByEF8lG+F4eHHc5C3Fo6zSXW7Mh6SScBaRQMmrg0HcA+i3/AMNqiii1TBbRIBO9wAGN85QegvJVpVukxT1lTTF4c4bd13f1jw9oOINA0U9EA4s7huMbLhDlF4a3OW0UdXdIiIH4c0lvku5lDQQW+IRQDAGyDqAzlSNHcX1NXRF7HE9xkLRNVcmcd2Z7/Q28sxvsMLvC9jXt6XAEH7VUNAaGjGAMIPJDjPy13m2wS07KOQiNp7A+i6bat0JcbHLJTT0Mg+IjJB9V9BGquG9p1KyUzxs6ntI3aO+F0a5iOXuzUDpZmRxghxOzUHk/c9PyULjM+ItyM7rb1R2wGrsPxo0abTNJDTgYAOMDC4Vp7FK+B75AdiT2QbUdkOyQpY2v6eoOOFlXCiMTyGjOFhtjnDSADhBK15mPS12D65T3RwI6jk/WoWCSN2QN1l0z5JJQHefyQZNO7wW/ENsLGqGvqJw1gIyVqk9NiEP9VdTwQiF0px1AZQYM1onEIkY4kjySDqhiMcg3I81uPTTH3GoMMm7RtuotU2oU0+Im9zhBolHRGWQuJwDlRXGGoif0sccFTRyS0ZBkGAfVTSPFUwyDsB3QYtrjfHIHSHfPmt+UmqaGGgNGY2l5GM/NcempDZOlh3B8j2WrU1OHw+M4fEN0EN0tss9S6pbIQHEnAWdT1ENNbnxvAJ6dsrSKy5TscYgTgdlhxVUs0rY3HIccILqOF1VXEjYdRK12WpZTx+7HBJ2BWLW0brZTipYNz5rSfe3ykvkJ6gdt0EldQuhJn6gQd8KtM/731+mFE6qlqPgeTgIZAxpjaUFZpPeHFoKtEbo24zuoMuiPUPNXdcsm4GyC+KlMvmpW0xiOCVZTPkaT9avmqD1bgoKywBw27rDewsdjCymTnbq81IYWSN6kGACQfNZ0JD4+kndQug32UfW+N+B3BQTOopck9exKgeHMPR3Uz6mUNHphWCN0kZkPdBWKmdJu361luq3QQmBx+xY9HKWPIPkVHWODpSQgkpZ208viHz3UtVUtqRloCwC4kAHyWVCxpiyO6Dvn7Ff8KfVP/F/XfyjbkV/sV29PNRqr+AFd/KNuRB7ToiICIiAiIgIiICIiAiIgIiIOOOZL8Hbil/Au+fqMy+aCT+yP+s/pX0w8xwzy88UB66Mvf6jMvmgkY4zPaBkhx/SgjGy1CjY17Oo42WC1vS4dXZXiZzPhYdvNBkvqmMcW+hVffIizGN/LZYJOST6qiC6R3U4lVjj63AZ7qxXxOLXjHqgmlo3Rx9Z7FRRReJ5rPl65acgeQWn5fCUF8tP0DOVE1vUVeXyS7K0tczuMILTssqmjEjceZWMAXHA3Kngc6GQA7Z8ign9xdGS8jZRvaHHPoVqpkY6DLsY7rSqiVuSGY3QWy1DCzwwBkeeFj7g+akihc8jIOFrVFTUfQPFIBPqg0cSl7OgjceaiezpwFqtwpYosviwtJcXH6SCelI6sEpVj4hvlQNcWHLdlc57pDg90BshaMHcK9jfFIOMYRlNIdyCB9SvA8Nw6eyCSfLYgPsUMVOZWFwPbyWVO+J0I7EnyWPSOeJg0btJ3QQODmE5B29VTdzgPmty3Kjo2UAlbjrxk+q24yF8n0BndBK+lLY+sHKgBI3BKmkFSxvTJkD5qADKDKpozVO8NUq6T3UgZzlWUsxhlDgcKare+p+I74QYamipzKMqIDfCyIpOgdI80GO7LSQSdlNBUNi2wrJI3jLiDvvur4xGWfH3QSe9tz6+igfJ1OGOyscB1YCp3QZpLWRZwMrEc4uf1fNVc6QtAd2VGxucMjsgzIri6NoaG9RVktU+pIa5nST2Wt6FpbbU3Xw7pjw9hutX4hWyz0r2utHSQNzhBtRtF7swTl2RjKtfXMe0sPnsseStmezwndsYUA2KDdGkL/wDsVrG12cEuBXa3gtrWbUmpaC4+/BjIcHHVhdOKdjZ2htQ7AB2K3ZpXXlbpO4xNtz3BgI7FB9BvAniXa7/pyktb6+HxYGBoy4An5Ll5rmuHwuB+o5XiHw75jtUWe2wy2eSYzOIzhxXpRyl8QtW66tcdXfhLgxh3xEnGyDssuJOOOkKi9acqqqAEloJ2+pcsyZ6DhaNqAwzWGpgqHABzC3dB4fcytmu8t/noWwSu6cjsfUrqVcLNd7dUStfbZwOo7+GSP0L2V4h8ELRq/UsskDWPe95yOnPmtbh5LuGsWg6y5X6OmZUiMlvU0Zzj5oPEdtJPMAJIyw/PZTNtDsbu7/Ncycw2iKLR+uauitJApmPIbgbYyuGXV8+XhucAkd0FH2gjPxLEnp/CyCc4UjrjUHOSseSZ0hJd5oKMd0nKvMocMYUWMqrcdQz2ygljPhnrIyFe6drx2WTIyJ8IDQMkLTnMLDgoKO3KuDj9H1VoBPZZtNGzpy8DZBjuhIbnfcqaGgfLGXjIU7DG49Pop3ziGItb5oNLyYJCDvg+q1amcJoC4bYWBQtZPUnxCMH1U9fKKZ4jh7HugwzGZagtOdzhZhDKePBGfnhRSPjZEJGEdZ3+eVG2cytPiHJQQSHrecKSKXwiAVbGR4pz2VJ+ku2QTVFS2RuG91Axpd5qxSsdgfNBLG8Qkk9lFPKJHZaMKNzi47qiAsun3YQQo6WB07+lo3WouiZTRdLgAfJBpczOlxx6qwHyUzfvkx9N1bUMDCAEFjG9RVSPDcCCrQ4t7IXF3dBkSVXXGGbq2KDqb157bqBXtleB0Dz2QSOk6/vfocLOprIahnX142z3WnOhkYOstwO6yYbrUwgNadsYQUqKAwOLSeyiZO6IFoJwVNU1TpW9bu5WLGzxDgoL4SXvOT3SpGHDClbCYh1FHM8ZuQgxmM6yhYQ4Nz3VxikjPp81UQSuHWGkgdygvkpfDj6y7OVbDCXDrB2HdUMk0uIjvjZSMbLFGcgjKDKpKoA9DWElWV8+csczBPZaroplvfWn38gD5q7W8FCysDreR0b5wg9O/Ycx4s3F+T+6qLF+YVy9Rl5b+w4lLrTxii/uJ7CR9ra7/UvUhAREQddPaGOLeTbiU7GcUdH+vU6+eWIEvGB23X0Pe0Fa13J5xID+xo6PP8ep1881M6NjnlxA22yg1LojnhDMDICoJhSQmMEBYsVT0u2O31qOoc+V+c5HmgxpCXPLj5lW5WXKIhHt3+axO6CpGFVvb7UMbwMkbK3f5oKu7LU6S5sp6cw4GSMLTWsc87bq+OP74A/sgPkJmMgGMnKtfIXOyBg/JZ9VTxeCDH3KipqTqaXu8igmtNxko5uqR56PQq+73X3t33vHzWBO1vV0tG+VfDDH0/HjKC+3W91wkLerCya63Otw6erOywoKqahkLojgkqSarqrgSXnOO6DDdkuK1awUBulYygLseIQO6xaVkRdiXuFM2pnt1Wyro+7DnKDfOpuFp0raW3czdRLerGc4XG8sxlf1kbrdNx4hXm+Ugt1e/qix0jdbanpwHZiHwoMilifXjwgcAbKOrpjRO8MnOcq621Hushce5VLjUmqqQT2QY0UfjOIJUkYEFUzq3AIypZAIog5gwcLHHVI4k98oN01l2pGULWNaOrHooqepjmo3luBkbLbUhkcAHZwO2Vk01UY4jGCd0EbD/ttxH90q1T3B/wAKia7EpcdslZjImyjqcNggxDUOLcFu/qtTopw6Etc1Y8kFOQA3BPms6njhipy44z5ILIqllE8ydslTms9+OAditDqHvlkI8gdln0hEMORscIL6ylFO3xGEZWAaw9Ja8brJFW+aUsmyG7+ajnhgA+HBJ7IMNrPFfttkrUqa3yNHX14A+a08RyMcHN9VMLhUMBbn5FBvLTVhkvkrIWyYIcB3XYzROhJLJbxVSVeABnHUusmkNQT22cStJBBz3XI7eKGopsU8Rf4RGPPsg3dxFrG3Zj6CnHUQCMjdcRN01NbpDUzNJw7JBXKOna+31DxUXBw63d8rAv8Aa5a25NdStzTF25HbCDYlVq+lpYTSupgCNuohbRnibfK8eFsCdyFybxBsOnIrUDSFhqQ3fGM5wuMLfFUUUT52g5buEG7obHHZqWGUAOLyBsuxXK7yqV3ELW1FqjpeIGOEhb5dwV110LJX6oroaOcEta8Abdt16acpUF+05WUtNTNIiLW74QegnD/TVHpfSlttNNTtYaeENO2+fNblWDZJnT2unkf9Is3+tZyAiKjnBoLjsAgquFONHDxmo4nubGSCVy4680Ic5omBLe4Wxtd8T9MWagcK2dgcCdiQg8xeabhfHp0SzmInAJXU2C1U8tsml6QCCV3m5q+JWnNTGohgmYctIGMLoRe76KLxaeA/A5xH50HH+oHtp6x7A3YH0WPSSRSQE4GcLIvToZ3GZ+5O60Nsz2OLY/okoJppmxyEBmQtSt1CJ/v2MAd1iU0cLwDNgFanAJoxmAZYPRBPU4EfQfJYhY4RnDsjG6VdSHtDIyOsd1HTyFrSJvNBqlguYoJicb+qzblcI6+cPdvvvlafbY6edxJxlYN6m91lxF64CCe+QtmhAgbkgeSwKDqZCYXjBI2yta0tVUk7y24EADtlaZqmanjuGKBwLRnsggitBEhlcds5ClfcnU7xThmx2U9IysqIWhgPbfZa5FY6JtI6aqwJQMhBtmajM4MuM5WJDSGnnbK7YNOd1rdJS1FVUvggBLBsNlHctM3oydMcRIPkPNBPU1UV2pRTN7jZaJUWl1KC4nIHfKyW2i7Wo+JNE5o7qyoqJ52nq7INOiIe7oA+3CjmYY37lZUVM8OLmjdWT0dSXFxaSPX5IMeSUPAAUsEjQ0tIGSE6I2bOGCFC0gS58soJzJ0ZONlBJIXu6lnVLInxAMxnvssExlpGfVAdITjA7KaCcghh81G5jTgN7qrGhm5G4QZkvTEwPznKwyPEd1+WVR5llAHcDsqwtc2QNI2JQVll6mhvTjCujqA1hZ5/Usmqp4hD1N74WnAbgYQSA4JI81ZIclXGN4GcKjY3OztugughMxwFqVNRAEMccZUdtiMLvEmGB80ra5zJ8xEYHzQegfsa7c2k5m9TTA56tB1o/LcLef5kWF7F64T1XNFqiKU5A0DWuH1/dG3f60Qez6IiAiIgIiICIiAiIgIiICIiDjrmOcG8vPFBx8tGXs/8hmXzSCQCpleexLj+dfSzzJfg7cUx/wBxV7/UZl80D9pHfWf0oJP7M8t7bqk0HheasY8td1KR8hlG/fCCEDKq5papIG4eOrsr6vpyOlBZTwGd/QFJUUxpXNB7lVt7xHNkqW5TMlcMHOEFsdUA3BJWPPK2R2QrDjpHqqxsLnDbZAjk6PJVc7xXgDz2Wf4NOIcnGfNYrhG3cEZCCdtH7uzxnHOViTyiR4cPJXy1kkjPD8goWxvduBlBlMmdMwReiikpyzcnfKpF96dv3V8glmOW5IQGVIjA2Ub55HOyHEBZEcUIGJe4UM7WD6HoglNb1xBjichQD48lRYypovh2KCxkZe/pCufH4LgfmsmGLDurCirAS4fJBlRVkb2eGO/qfNWvjyQMdytPBIOR3WpUMckhD39ggiqaRzIuo9zvhWUJGST5LVqmaF0ZjJGRt3WjSB0bj0digza+R7oenrJb5DKttkrImkv7DzWK8TSsGxJUkI8KMtfsTnZBlTVMVa4wtAB7dlgTwe7O6T59lbHJ4U3UPVVqJTM8Hug1K0Wf39wVbzRC2OEePpFT2SuNFgv2CtvtbFXPDs9ig0EnJyVPBH1YckrWNj2xlSULh2d2QWz1HU0xgbjbso4qd0jcrUzBSObtjJ+aw5n+FljfsCDFbGS/oJR8ZY4BTMhf1dZCtlY9zwcZQXygGIDG4CjhmDB047q+Z2I8eapAI+nLu4QW9UsbjJG8tJ32OFNDXTPcGTSOeDtvuomuDpC3yVJQ0OHQRnKDKqaAMZ7wDsd1jQReKer0UshrJIg0glhHZKORkL+iQ4JKC9zHOb0N2PyV9spnfdCONxBJ2U1Z0Mg8RmCT2WNbahza1krv3u+yDsxwutpttLBVSMEo6genGV6s8omr6N9mhoxA2IujDRtjOy8wOXDVOnaqeGk1BK1sfWB8R/1r0x4SW22vNJU6TIfCGhzujtjb0QdsJJmtgMoORjyWg3GkfeKWSna4jryMLUrQySS2sZU5Dj3WnXSOrpagGkBIyg2ZZOF7bXeHXKrYXtJzusriJw/n1JY5ILc97Glp+Fpxlbynr6iOjDqjAyNytPbra10UI94kAa04Jz2CDyQ5xOBM+naie4TUzwQCS4groaKHpNTtnpcRn7V7Je0G1RpS+6TlitE8T6rwiDjGScLyFtlkr2yVr6oFrS8nf6yg2dI0mUjHnhZcdrfJGX5IxutVktEdRUGOCQF/op/2K3yIdQDjH57Hsg25DATOYyCB2V9TSljgANytUuUENFGQHjxvP61i0UUlVGXvGSPNBjNYYG9bvtWPNIJHZCnrC/qMatghHd4QY7XdJUhmOMBTTtiDT04ysRBfFKWOzkqSSZ0n1KIRvIyAp2Ma2P4xughjkdG7qad1cXuqJACdycJC0Pk6fVVmHhSDp7hBmVFqdDTiXq7jstNyst1xmkjETjsFCIHudlrSRnugiTKyJWNazvuoG9xlBRFkvja5vwYJUHQ4EAjugtAyrg09QaVkxxhoy5WzBn0m9wgyqMsovvr8eqguFb71IC3YBY8kznjpJ2CjQXxydCPcZHD5qxZFPGDhyCklOWN6ie6jZH1tLvRZdTK0s6R3UFM8NyCgshhMr+nspXU/gkOJ7b4V0jmRDqZ3Ki8Z8rgHdiUGRPWski8MNwQMZwoYabxGdfyU9RTRsi6x3IWIyd7G9LeyCkrjkt9CsmjYPpHyVkUYJLnjYnzVJZjG7EewQTVso6Q1p7Ka3xAwl5K0xzy45K1CkbMYCWA4QXUzW1c5h+fdahPDFQxdBAJIPktEgmkp6gub3WovqXVDQZkGnMkEM5kPYlSz1bZiGtB+eypVNjd9DGVWmpHGMvc3sgrFRTO+OF/SfkcKyodNE0xTPLie2+VQVksTixvYFUkd4kZe8/F5IPVX2GzCLbxkf5GewD8ja/8A1r1KXlp7DZxNu4yN8hNYCPtFf/qXqWgIiIOuftDnFnJrxKcDuKOjx/HqdfPnbLW64slcHY8LB+vK+g/2hDWu5OeJLX9jSUef49Tr56qKunow9sPZ+xQROjdFMY87g4KldJ4Y6SFRrg55kk7k5+1Umcx475+aCJjTK4jKuczw5AD6hVpXtZJl3ZSTRulkD2DIQSzkeEMDyWD1fJZ5czo6XdwoDExwJaeyCynlEZ3CTy9bshWFhJwNyrSxze4QZNPI55Ae4kBaiXtbAQPRabTQyF2QDj1U0spjcI3H5II6ZhfUHI2yqyNzWNhBwMrJYWRjrBG4WE+Qip8b0OUHIen+GQ1JA1zZAw4zkbLRdU6eZo+U0jnB7jtnuUtmvLxbIhHbiSe3mtLvF3uN+n8e556gc7oNEc8l5eMjJysykqmtAjf3Pr5pJFEGfBglRimOOoDcb5QZdTSs6PGjA3+Sij2Zh4UttqDI7wpuw2VLm0NcPB7IMSdvSepqRUz5B1nyVsT+p/TItZpzTiHAIyQg0l0hyYz5KMydBxhZkzGCUuWFLgyDHYoJYyJ8jCjmZ4b+kdisnwHMYHRA5PorGwyPJfINwgilh6WBw+tSU1QQ0ReZ2SeQOaG5+Stp4iHh57A5QTPpH048dxOD+ZZEUZnhLw7t3VtdUtkhDBucLCimqGN6G5wUFC/wpCDvgqUTFzcjsopYnY6vMlZFO2IQHr7oMaSYu2Gx9VGHODhk5wfVVwDKfTKlNHO4F7WkhBNHUsc3pxusaZpzt9ataHB+PMbLOpRC+UMl7bBBDQTPjeAOwOVv+w3enDRFIRnGMralxpKaGAPg+l3Wl09dPC8EE5CDnCzWCS9fHDUBgBzsVqFdqSn06BZZXB8j9urvhcU2fXd1tTR4ZIB27rNrLi++D7pTZMjfiH1oNU1VZ6inYbq+oL2P+Lp6srakVxilgezpAJyMeqtn1Pca8+4VJPhN2H1LD8ARSdTR8GMn5IOQ+EVXBRXaCSRgAMo7/WvYzlKsdrvNrgqx0F4jB8vReNOi7TXXqenhsjS6USD6O/mvU/k3puIGn4qaCtglERYAc57IO/VNAKOn8NvZvb6lMyTqGVi2+fxaKJ85w8ty761dUXOjpGF80oa0IJZqhsIJeQB81o9zv9JHTSM8djCRgElaJqbXtkip5Iqeoa94BzuF1q4occ6Sy08rBNh4JxgoOUtQambaKmac3NmCDgF4XWXje+4ajoZqukvOACSAHrhjW3MFerpWSxU0ryzB7FcQam4936mhkpJ3vDTsckoNj8Vaq70Fymjnr3PDcj6WVwfcrw95d1Oyeo9yt6a41kL5K+Z78ud3yuLap4dIcdicoL6mrfUHGThRRP6DjGVH2WbTNhczLgMhAlDngFjsEd1rVHcIqe3uieQXYwCe62/LMQ4hh2VrZnEgOOyDMt5Mla6R5yCSsq4nLg2M428lglxYA6Hv5rJppuo5nOD80ElBLJRu6y44Pqoq6o97m6s5U1XLD0bFaQ6Qhx6UGbLDMGgwvIz5A4WuW3Sr56I187y4gZGTlbdhquwcey1mi1PNEBSE4jOxQahp65xxVjqWRgw04/IptQVznSFkLtj6eSx3e4MaKmBw8R25381WCopJHj3gjKDIsFyZbniSVmcbk47rcLuIlsilaHwtJGO60Z4tpjx1Df5rRKzT7qp5lpgSM+SDcGq9Y2+403TBG0EjC2pTdMsDnlY9bap6YASggD1Ucb3sgLWdkGRBUsbIQc7KWpu8LWGIRjPrhabSxSOmLnA481kyQ05fl/dBpsjnPeXHO5UkdM6RuQMrKqII3ACEZ+pZFIzw2jrGEGmgvpyOrO63JSaeZX0BrMgYGSMrSK9rJRhmCR6KSmvlZR0xpGZwdu6DTZB7vUPYTnpJCtz4kgxsMq6Rk0r3SFpJJyVZ4cjd8YwgzJOmJgON1jmoBOQFb4pk+F52Toa4hrNyUE7ZXTDpznyVj4eg9ayKWjkgPiTNw0+qjrS57vve4QRmpBHThWxz+HKHFuwKthYA8da1D3OOSHrbgoIau4CaMNj2KwtycnO6yYIovFIfjA9VNPHA1vwY+xB3v9iu/PNLqn+AFd/KNuRPYrDHNPqr+AFd/KNuRB7UIiICIiAiIgIiICIiAiIgIiIOOeZL8Hbil/Au9/qMy+aB4zK4f9sf0r6XuZL8Hbil/Au+fqMy+aJzHOleG98k/nQStpOsZzhRvZ4TgBurwJ2tDRkK9oHSTN3QVcwNi6xsViF5PdSPlc49LTsgp5Tv07IIw4tOQVkR075m9X2rHDSXdPmtRgDo4SDtsg0+RvQ4t9FdHL0AjCqekz5d2J3WcyGmdGXEINPdI95xkgK1zXDz7q+UASfB2ysykbA+ImTuEGnqaKfw29JaSFZNjxT09lnUNE2eEuIzjdBp7ndTi4eazaKqZGBG8bnbsseVoZOWeQKtfgOBb3QajW0REXvLScEZWlZJ9VrNJO58QjmGG9lDVMpQQIyDlBpoPSdwshjfEBd2wpJaT731NCxep8RI9UE4qQD0/nUvh+I0O7ghYLQ5x27rWraYRAWS46sflQabFCBJjuc9lqbJmQxlmME+ixDTTR1JlIwwnOUqp49seXogtZTyTTF2SR3Sq+8OALe/ZZNonjEv3zGPJXXmnM7w6BuQEGHDWMadwVBVTeK7IGAqwQhsoE2wHdXVgiBHhhBFTxGZ/TlTTwCHzBKx4XPY8FndSTGeQZdnCC4TOlHhtdg+qsdDICA5yjj6ur4e6vke8OHV3QSPhwzPV5K2HIaQFaHSyDHcK5gMQw7YlBTrkjd1dRIVHzdTw474VJI37uOcJEwuIPkgzKeobIBHjdUqXtj2xuVSIxR7jvhQVUge4EdkFkjs7+qsGfIoScfJUBPYIKglp7qaGPrd4hOw3wrHMwMlGPf9Bvc7INUddIWxeEGDI+SmoLCbpCatr8Ab4WlignDRLIwhh3WRBeaiiiMFOR0lBFXPdC91LnPQcKKlB3O/1qhcZpDPL3ccrVIKeL3Z8g2AaTsg3Vw2tl3vNbDDbqh7HeKOxI817Z8jNlqrPpOOnuuZJXQDpL9/T1XjtywRtrdXxUfVv4oOD6Er3L5d7K+3WWkcRgGAHt8kHNoDWjDRgKhYxxyWglXIg0+7W/36n8Jm3cLqnx3vF40rLJbKYvBkcQCM7Lt4uO+JnDK16tpZa6oYDLG0uG3og8xuL2mdSVlL91at0tQyQZ6SSdvqWwtC8B6jXIfCyzvYX5/eELvxaeGNLqW/Gw1jGmFj+kdTdgMrnbSPArSukg11HEzqGM/AMZQecGjPZ3VlTWe/TW5+M5+iVqWu+U6PTVKaJlnJcfhB6CvVCjoYaKPwoWNA+rC0i96Ks9+k8StiBOc/RBQeDPGTlrvGmGvujLVL0kF2A0rgyioquBskU1C9haSNxhfQ5xC5fNL6xtD6N8MeQwjdg32Xnjxw5J7rbbhPJYaQmLxCfhZ5ZQebt1o5oHGQwEDv2WBE50rCOkjyXZriXwRq9L0PRXxFr2tOQRjC4LmpKCjZM1xHUDhBtOaJzCTlIYPEGfRZbXxy1hDz8BKsrnsieGwHZBCJhGS3HZXdBmYXBRdI6Q5/cqeKWNseBhBjMd4byfRavBZ/fYDODjAytIIdLIekZJ9Fl09yqqMeDk422z5IMSaIwyuj9DhZVJUsY3ocNzsr6lrHxmYjc7rAa0uIAG5QahV0g8PxgdjvhYsVN4jS4FTSU1x8LL2Hox+ZY7ZHxAt+aBG/wn4JyAshzWSjrGNlh4c87bkqRj3t+DdBIXGQ9DfJRPa6LYnOVkRMEZMjvNQ1L2vfkIITuiIgKZk3Q3pGVDhEFS4kkoAfJVYx0hwwZPoqmN8bg1wxlBbvhGnpcD6LOZBG1nU8DBCx5Yep2YhkfJBdJV9cYZ6LHbnY+iv8PIAHdTRsaxpDwMoInTZGBso9z6lXSRlpJA2UkQYWHOMlBZBD4zunK1eGoio6cxHBJGFpAe6N5LVd1PnkGcoKOkPjmQDzVz6lz9gMLUoaOB0Y68A4UNXDTRg9PdBp7HFruo7rNbcGNi6PNYDjknHZXshfJ9EZQU8T74X47pI8vOR2UklO6OPJCjHT0HPdB6s+w1/3O4y/4awf9GvXqYvLP2Gv+53GX/DWD/o169TEBERB119oYwv5N+JLR3NHR/r1OvnnpnASgOGc7L6GvaDHHJ3xIJ/vSj/XqdfPCzqLh09wcoNadaOuLxQe+4WkTwuheWlaxS1VT4Qa7t2WJUwmaUADJJQY0dI54BBCzYiyCEtcMkD61ZUU1TSxB+CBjuoqaoY/4ZXbkoMSRxc899ytWoLf41IZM+W6ifSxuGWYUlPJUwRmJp2QadnwahwI2BwqTyF0hIGB9S1GG3PneXFuSSppqOmgPTKQHZQVtYjkAaRgqC50QM4LSrg18Xx042WG6pqHTgSdupBdUsdHGBuMhahQ2NtXSOnLhkDI3S5Ni9yDx3Wn094qqeIwsOxGEGRZ5mUVa6KbBAPmMqS+VsM8obCAB22C0p5k6jOSQScqzpkly/coMqOA4Dicq91UIx04PbCxoppCegnKyi2Dwy5/cBBEw9TupuxJWVC3xHCJ2+Vp8cmJBjYErUaR7RO1x8igtuNrdTRiYDGfzLcFh0s6vtrqsuAwMhQ3aWOppWsbjI8lDb9R1VuonUbCcEY7oNDuYMNXJCCfgOFiAPyHAHb5LLmhnqql0xaT1uyVr1FQ0TKMmbAOPNBo9JWDAa5pKyp+l0J2wfqS3U9PPWOjb2BVt7Y6nf4URxnYoNJawyS9I3GVq0VOBF23wsShopeoSPaQPmtUM0UbS3IGEGiTRSRyknOMrXLfbI56cyuABxnsoIIGVLznHfZWVtwmovvEWQD33QY9a1sMpY0ZCg6HPZ1DYBX003vMxM++VfVyRRDw2dkGJSxCWXpJ+1bjZ4EFIWkAnC2yyV0T+tqldXTvb0nt2QW7PqSfIlTGnJcHtdj5rDDiHZ81mQTEN+PGEEkkzw0McSQsNrz4oIaO/os+ERzuxkZWbFTUbcB+Mn86DFkcDCAABhZNLc/AhMbjsfLKx7kGxNBjOyw4ZI3Rnq7oMn3hs8pLAM+qnqarppJI/PGMrGtkIdMS7ssuqNJ1lj3ADCDnXk0dENWtfV0pnjEoIBGfML1O0jxxtNjvVJYqa1CIvAbkNx6BeffIbUaFt19hk1FPE1r5B9LHr816j2/hLoLVl6pL/YHRyBjQ7LcH0KDmywyi5WiCtPwidgdj0yto8Q454KF7YJ98HsVruoWVdlsMVNbM/eW4GPkuLLhermYnTXbqDGnfPog4F13qS86erZ3yVDy1wON11q19XV2rKlzjXloLjtnC7ScVdSaKroJ4HyxmZrD5jOcLzr43cQqzT10kbZiegSHGD5ZQbovdtZYI3zy1LXkDPfK6+6+1Ea+rdFC8DLiEuHFq53SMR1z3YI88rY90qzVzmaIk5OUE1TQzFvjPkJyPVaJUM6HEZzgrOlr6lzBHvjGFpzhI4lzsklBYnUR2JVwjcewVHNLTuMIKbkqZsBLCT39VbD0g/FjCyjIzpLWFBBTzeC89W4Som8V4c3IH5FRkfxEvG3zVZYj3YDhBCXvPdxKNHU4A+ap2QHBygyH0pY3qBUbI3Odhp38kM8hGMq1sjmnqHdBn07Z6cl75CR9eVFI+WapAa4jJ9cKkVSXHD+ypJ1Nf4kXlug1WognigY/xCe3mt6aaqqaK2GScgkDzK4/hrppvglzgK6W71MQNPESG9ig129XKmuM74o8DfC0SSIUrSDvstPEs8TxNvk7qc1XjMJk7/Wguiq25Iao6hr5D1glRU3T4mT2V0zndfwn4UCmmMUnx5wtQbK2od4bTjK02Toc34O6sjlfC8OHcINUqaM0cfilwIIysBswklGe3zV8lZUVrfCdkhYxBikx5hBrBEbIsgZysVz2ynp6cA/JXU0he3Enb5qSSFvT1x7lBg1NJ4Q6hvlZFtojJIyV2cAqJ3iyOAk7BarRyRwxY2AQS3moifTCKMAEADZaLC/oYQ7crMqXtncWs7qANjYMO7oMObd22yy4Kl7KctIJ2VsjG5BxsVuWhoKF9tMjgOrCDZ5cfEJBIJKuY5z3hpJIOyuqWg1L2xds4VjY3scCRjBQeg/sYKZsXM/qd4PfQNaP+cbeit9i7OZeaDU7D5aBrT/zjbkQez6IiAiIgIiICIiAiIgIiICIiDjjmS/B34pH/ALi73+ozL5poXiOpc5wGCSPzr6WuZEZ5d+KQHnou9j/kMy+aEtPjOHm1xJ+woNZlMUUPi4BJWmnNY/DdhlUjqHyARO3AWQ1ghb1N7oLH0Pu7fEc7Ko2sYW9HTudlFNVSzHwj27YUfguZjPfKCbwmxnxc7I+rLvhb57Kx/iuaAMq+njiDT4n0vIIIpIfg8Qb5VjZHtGATgrKpj1TYk+gqXBsLXDwiEEEURmOSk0ToTgE4+SsZI5hyCtRoo2VTcydx6oNMOfms+juAp4yw+eypXQsiJDQMBQQMiLSXndBbO7reZAe6uggLx157bqNzgXY8srJjf0sLWnYoJY5BUfeAcEfYoKmmkpHgFxO+26ibI6GUvad1c+pfO8GXyQZzZmiEZ9FhSNEpy1XyPZ4YAI2VtMCWk4QIAGPwVdMHCQPadgfVWwjMp6vVXVTXjHRnCDNqbtFLSCFoHVjGVgQUjp2l3V2WOGOc7pG5WdTSmBpa/bIQYbuqJxa0kEHyWqWy4t6mwzb9RAysVkInlJG+UmpTTkSAfEN8BBue86cjioPf4zgkZ7rZmC4gEn0Wqy6hr6inFLK4lg2xlad0/H1em6DJFOIo/EVhqGv+DCmklzCGKKGOPBcTugtbEIiJFFK7xXbA4Bwrw9z5Ok5wp2wNG4wfPsgtiIhaHEdgoZpBM8ObthZFQG9GFgglp2QTyTtMfRjcDCvpm5HSd87LEWZTgmMuHcILaildGC8H7AoIwX7EbK+aeR2WE7BXwdAYScZKCJ8JaMqwZBGyyqdpkkPX9HPolZExrh0YwEFjR4rcK3p8CVrjuAQVVrgxox3UjWiRhL/Lsg1Squ0E1EIGNAdjG3qtCLSDv3ypIA0zYPZXThrZg0dsoLZHHpDQMLkPhfo6XV9Qy3CIuMw6QMd8rbtus8VdEwtxkkA7Lsly36UFLrG3N2wXN8vmg5h4C8ody0TPFqyahlLS8OBIJ2XqJwVmEliijMXQY4wO2FJw80bbKrQlugqo2HrjBJDR6LedmsNDY4jFRNwCMeiDU0RUQVUFYyOWmkikIAe0gqYHK2NxK15Q6Ptkr6ohrg0uHxYzsgx7Hoyior864NkBJdlchLqzw1421Gq9Xmgp3EsMmO+fNdo4+rw29QwcBBeiIgoQCMEbFYNdZ7VWwvZV0MMjXgh2WDJWeqEAjBQdEObngrbbi2oqKakbEwsJBxjyXlZxH4ZGy1VU6OXYPPY/Ne0vPNVVdr4ePq7Yz7+Y39u/YrxD1VqnU1fUV4uDXACR2Cc+qDYVXQGnf4bXZI7kLE8F3V8We/mp3Vkk1VmTzOFHVSYflvmgzaS3NrR0AgELFuNAaGUR9Wc/NW01wmpndTCraqrkrJA6TuglgY2naJnbghQ1U7ZnhzR2V0hkMXRg4CxwMkD1QTPnL4+j5YVkLhFMx7hkAgqV0XS3qI2PZY7sE7INyT6hpZKIU4jGcYW3ZHdbyQO5VHMIAPqp6eFrxlyC2mHxbqtQQ2QEeSvm6W/2Lv8AJUZGZGFz++EFPEMzQxux+Sl+5rujrLvsUVKBHLl+w+anq6mQ/DEfhPfCDAe3pcWqiq4knJ7qsWDI3PbKCSOIyeRVX0xa4AAkeq1KVsUNOHNxk91HCY3wlzsZAQR0sDYCZHEfaoK6UPmDmDsqGeR0hYDtnYKb3cFuXILGvNSwRA4wsiMNp4ix2CSsJ7hA/LO6ujmdK7DjugqB0SmU9vRRTyOe/qGQAsp0Dy3JGyyYqaDwSXY6sIMAyCVnTjdQFrmOxuFkxxCOc57Z2VlSWeKMdggsLD0jYhSQlsYLsdlJKWGEdGMrGAdg5zghBnU85nd0AkJU0LyQS7y9VDQYbJlS11VICOnIQY8lIWD1KQVHgbOYdj6KWknD5MTdsrPqYaJ0fwEE/JBptRVtlGBsscRkxmTyVz4HBx6RtlZAj6aR+e+P50Hqf7DX+0OMv+GsH6K9epi8s/Ya/wC53GX/AA1g/RXr1MQEREHAnPZb33blO4g2+Nhe6akpAAO5xWQH+ZfPx+xmagdO6oYWmJxGHfIr6NuZKa103BHVU16c0UTaeLxS7tjx48Z+3C8EOONdZnXapj06WGN8pd8GO2fkg4oqa9sP3pjdx2wsmhb1MNS8dt1lUNtp5wHTkdRG+ynrYY6eIxw/RIwSgut80F/f7jgNI27JcNCupJOpkhIzkBbehrn2up8amPx+uVuCg1LW3B7ROSQDgoKRWgxAMkGAFlMtsIb5FX3itEdP1RkdWPJaHSXeoLsSds+qDVOqKiccjZaVcKI1xNQx4AG/dXXWq8aL4CST6LS4q6pjjMZzjHqgyqStbG8wvwcbbrFr5W+MCzG26xmMlqJT4YJJ3VJo5Yn9EoIKCeesdNGGb4G2FJT28Pi8VxxjdR0lK5zuqRuymqHVEe0QJZ5oI2tbNIYcD4VIGNikEOBvsooyWnLT8R7rJgafGbNKNhugnltXgR+N04B33WnwU8ldWNpmk/EQFuCprIqumEEJBIGFp1ocygu0clRthwO6DUL1omez29tc/qwRlaLRRmWF8gJy1ck631Jb7hp9lNC4F4bg4I9Fx1ZJWNeYnuwHFBbQzyy1AifnGcLcn7G2ywGpDxtv3UFVS0dPEZ4AOrvt6rSJNRXBgNO0kN7YyglfWRU0xg2y3bKw56588hhY4gOOFQwukJmeNzuViSgxy9TTuCg1eGlNpaKsvySM91O1v3U/2z05A3WjTVdVVRtieTgbLcWnRFE0Mk+ifVBjmqY0GER4xt2WFPTukBeHHC3Jd6ClmiL6PBf8ltpwnpgWzggD1QRw1hpHEbnCxKupdVSdRB/IsyD3aZ33wjupZYKQkNjAyf0oNJBczcEgqQROmb1F3YZWXNa52Drc34e/ZYbnuZ8LfqQWOAB6UYd+jGclCx4HUdgsm3CI1DfF7Z80EUlO+MCQtOFJEzxoz5FbkuTKCWjDYiOoflW143Pjn8MeuEGRSsdBIS4kD5qGqmkM3UCQAdlnVtPLHTiTcZxkrV7DaYK6A9eC4oNvmd1UwRuG+FjSROiOx2WvXyyPtgMjW4b+hbfdI5/0kGrWUiaURdsnC3tZOF1x1JXMjgpXvD8btBK4/tUc/vUL4ASS8Zx6ZXoZypV3DaipYpdVTU7JQwfTxnOPmg494dcruq6eOlq7dFPCQ8EkZHmF6r8rGkbrpjTDYLu575REBlxz6LSeFNz4ZaoiZSWOemcQ4AdOD+hdgLbbaa1QiGAADYdsILbvWUlNTv8Ae3ta3Gd1wTxVv1uqLPNSUdQwPcSPhIWvcympK3T2mTNQh3X0E7bLobduMV4fSzOrS8Frj3cUHGXH6svGl7jU14uTiw5OA7ZdUb/qN2omSTzPDyCe/muYeM2sKrWZlp43FxIIwuvtTSy2iB7JgQST3CDRpKE19QY2uAx81jzx/c5/hOOSe6sFXUsqHOp85yoak1Ur+ucHKC51XnsM/Yql3W0npxjvslFFG6T76s+oFIyLDCMkeSDTI5hG45CslkEjttgrZPpn61eGs6TvugQQeMcZwVWSN1PIOo5VjJHRu+D1Vzy6UdT8k4QXSz9bQAkU4a0tIzlQY3wFLHC9xBxtnugtkYQS713VmD6LLqCwRho74UUYYWHqxnyQQoAfIK5+M7FalRU0M0O+OpBppaWjIUsc/S0NO6T000ZJc04Hmo2xlw2QSCfBJaMKPqc54cQTv6KvgvBGRstXpqemdTHrxnyyg02eobKwADBWNv8ANSzQuje7bYErJgZAYSXgZAQYsLC4nfGFK0jq8M+eyhLi1x6TtlSxPb9I9wglko3U7TId8rEcS93bC1SGdsrQ2fAH1qyppGvGYACPkgw4XiA5Izn1WV7oKgeM3G2+FhPglZu9pH1rJonVPW2MZ6CcFAY4k+GGkY+SyWyeEzB3ws+rpoIoQ9uOo9/rWDGwPGX9kEAl8Z2OjH2LUBRH3cuDt8ZwO6iijp/3gGVNBNN7wI3ZLD3yg0eOZ9NMS8E48ir5HGc+I3sCtz3CktXu/VloeVtiSN/XiL6OfJBYZCcNPksxlwkZD4QfgYWPUxiOPI7+awy4koJmnw5DI7fJSSoDhgNUb2uAHVnCzKenjezKDvh7FcO/bTapzn/4gV38o25Fk+xfYxvNJqnGP/iDXD/nG3Ig9oEREBERAREQEREBERAREQEREHHXMc7p5eeKDvTRl7P/ACGZfNE5+J5HY7k/nK+lzmR/B34pfwLvf6jMvmfk/sj/AKz+lBUHpd1KYVOWkEqANO2eykbSySDqYMhBRjwyTrUk1SJCMBY7gWktV8bQDlyDU6ANqj0EK6vtLmHxM4HdXWeeCGXLiAMq++V/iuDITkY/Ig0pzjtE3uFG+F43cc/ap4m4HiP2VJZmOGxCClPR+OcZU780GG53KxI6mSN2WJPLLK/qlygrNOZck7qAZzj1WTJ4Xg/D38ljdkE/u3wZ81a09AIP1IycgYPZVkwdx5oIXHJJROyd0FzGl5xlTg+Bhuc5UDXFjsqsr+sgoMmJgJLsq6aYsBHTnAUETywb9lqUFOyogLz3CDSo5QyTrIVZ5vFdkbBHsDZywjYFZQZCGZ2ygrbXtY4dRU9wqWk4G59FpjpC156TtnZVZIZJWeIdsoL5J8jHhgZ77KWkYJcAjutUrYKAUAMZBeAtKo2SNy9o2BQZVVRGOMvDtlhsY8gkDZTzVb5fvR8lm0rKdtOfGIzjbdBplPh8pae4WVOPBZ6rAleI6hzoztlZLZmyR4ed8IMWaUvcVEr5Bhxx2ykbQSMjZBRrCe/ZZEc4hb0Y77KV/hMZ8PcDyWC85cSgukHU4uz3VoJBB9FTJxhVaMuAQalQNFSQztup6yhEWQSCSsNj3UrBIzuPmrfuhLK8eKdkFHwFpzuonSY2CzaieMw7d1pww4nPmUEtIzrkU1RTEPLgc4GVFB1QOD37ArU7faq27VzBTsJaT3AQcmcBdIy6wukdG+EkGQDt816X8FOVR1JcaC6tpHEANdsF1E5Y9DVNLUwS0jAZg8ZwN+69cuCtaLdYIYbq5scvhgD17IORdLW37kWOmt2MeC3pK1ZaZDXtmqSYzlnqPNakCD2QVVFVEGyOJPEKl4dWt1yq3NDOkuBdjGy83eZnmyrNXyVbLTVZjjJaOg5AA+pdwOeu33Wt4YuFpa8zBrx8PfGF5j6G0/ZYdPXUawf0VZ6+kSd87oN3csPMFU2rWLJqqq6iJRsT816l6M420GpWU7OqMOkYPtOF4j8IrZBT8TZHtBNIJst3wMZXoDpLippmwXm30DJQHnpbjqHyQehVNO2pgZMwghwypVtvQN1beNOU9dH9B4GD67LPrNSWmhmEFTUhjycYQaqtp6p11R6bk6KiRjTnHxLVLnqqzWilFbXVIjiIz1Ejsun/ADO8wekXSPhtNW1748AlrhuR5oNP51OL9AdLvDaiORnhnDQQcbLyG1Dq+jvNRWCONrSZD2HzXZ/jVxRj1TRGnqZHGIsI7rpzfqKNtXK6gBILydvrQabV04bKZWnbOcLEe4v3z2WRJFVFnxg/NQtZ0uHUEF1NAZ3dH51dUUrqVwPdZBljgZ1sxlXRyxVLT4ux790GG6p6mdOFC04IKzfcjI49AUMlHKx4bjcoD6gSRhmOwwoG/SA+ayn26oYwPLTj5KNjABv3QXTtxGDlZlBT+JTlx7hYD2SvxkbLVbfIIYS1x7hBg0lM6SrMZBwCsiujFK4AY+pTQSwsmL24JJysa6OfO4P7gd0EYhNSPh74Vrs0g6HblSW2dkbsOO6tucjJJAW+R3QYTjlxPqpIoz9PyCU8Zld0hZbw2JnQTuUGNLUOc3oydlkUsD5YsjOPVYpgkO+NluG1NhZRHrIzjZBoZZ7vIT3KvdVhzdhjCSsfJVuB3blUljjjcAfNBFT07qqTGcZUk9OaN4djOCp45ooMOad1KZI6oEuO6BS1bagCPG/1LGrHOhkDQ44zupGxiElzFi1T+t+XdwgpJP1Nx5+qRUxlYX5VjIJJPohZsUMsMR6hj5IMFuWvx3wcKR0mRjCkpmNfOetW1rWslAb2QUpyWODlWqe5x7LIhbG9gA+ktco7fSPpi6TGcINrxsMuw2KkMUsZBc8kfWsiZscFSQ3sCqveyUYG6CKOoDgIyN1kTtHuryPILEkp3NPUwbqcS9NLIx/cgjdB6m+w2b/1L4xv9Z7CPyNrv9a9Sl5bew3d/wBSuMbPMT2E/lbXf6l6koCIiDrx7QRtU/k94jtopTHMaSj6XDuP9vU+fzZXz5UFZP40pqZXPwMfEc75+a+gz2g0zqfk64kTM7to6P8AXqdfPPSnqe8k9xk/lQa7TVPjvIaSB5KO5VphaYznJWHDUshOxx6q6WWGp3kO6DT443Ty9R7HdanBOKTDQcHCsY2OLdvYdlh1LnPf1DtlBuS2QC6uLXu29MrHvNvbbnFrSO/ktNo7pLRD7ySDhKy5zVpzPnKCAzub3yR6FWGo8Q+EGYJOFNE6A/TICm6KUOEjSDhBJQRttp94mAIPqFZXujuEgqGDACrU1UUsfhk9lgCbwwWg7HKCaSua2IRN7jbZX01exzPCc3JOyxBAag5j3KmjiZTY8U4cgnkp/AzMexVYXiscIBsScKKarEreg9go4XuhlEkfcHIQaybS6xxiskfkHyKgNG68g1ER6S3O6grrpU1kIhnz0j1Kso7l7jCY2HugxayCqp3GOSQuA27rNtdvM7BI0gFYwqTVzEydvJTNrn0p8OE/LZBk15fTMI6yQB9a0oVZ6g7ozusqaWSb4pwcYU0dPROgJ26kEUda2QFuwWNUYD9t/NRvp5A89AOM+StPWHjxNkGXBI0gDA2WSat8LD0E5WP1wsjGO6mtcTa6tZT9y4gYQahpuvlfUEVH0fLKu1I6OeoEUQABPkte1Bpl9htYrmtxkZytm09Uakl79nBBHU2p8EYlDsZGVi0znNnY4kkNOe6zqyeoeOg9hssBsMob1AEDPog3DX3eB1EI2gZxhaHE0SAvPrlY8j+oY88qscpY0tHmglfKJAYwOyoaVzW9QOCFZTkB+XLJlqG9BaD3QUt7pZJulzyR5qaopjHVCUN27qG3Atm6jsD2K3VS6Oud5AkpgSCP7koNMqq1s1K2IM+IAKWy3J1FIM7DtuVnV2lKnT7fEueWN+YWHHDRVcZfSvBx3QZ2pbtDcKPobjPYraVBRe9VQgPnssitilLixhOyxKed1HL1hxDxuPrQb1ntEOmaWKrLQ8vxgLsRwT4CX/ilb/ulbro+nHRkAPIXWq3XGe9iKnrjmMEYyu0PBLWGt9NtZQaaa8wloG2eyDulyn8EdR8P7q19wucszY3g/E8nsV2/1Hr19qqIY25w3AJx3XV7l41tqGlfHU6qBDXuHV1bfpXZW61WlL0Ia/xGFgwXHIPkg4G5meK4dZ5GyuAaYz3HyXntqXWwujZoIZgA5x7fWu4fOjf9KSWeWls72mVsZB6SO+F5tVF5NLHO6RxD+s4z9aDVrrO20SPrKiQPyMri3Vt5iu0hMQDQCey1eqvk9ylfFU5LOw3WzL5TuimzTg9JJygjoPDp5A5wB+azKsQ1HxMaBt5BaKyo6fpHcKbx5CMs7II6uPwj8OfsWKXPPclZjJWSH76s0Q0boiQRn60GjEYVO+ylmic1xONsqNo6iGgb5QVb8O5VTIT2Gymlopg0PDdisqnhgFOTLgPA2yg0+JnW7A9eyy/eGws8Mjcqyj6feDntlUr2gS9TOyCCTJJd6qwAk7Ko6nbAKdga1pzsUEToy1uSVNSVZgeCTsoZHFxx6K5rG9BJ74QahXXKKePpaBn1Cw4JAxu/qoWQvkPwAlVcHRnocMFBO+racgBRx1EnWPiOConMLQDjYq9hjA+aDKq5WvjwBvhYLck4BO/zVXv6tvJSwtDsDzQTQUPiDOeyhqIDA4b7FZbvGpm9WDhRPeJ2ZPdBbTwGsPQ04IWSJzbyI3NJ2791iUsj6WQP7DO61aD3OreHTEZQT2elZfpjCWYx59lPdKWKyv8AAABJ22Ck6fuUzxrf3O+AsB9TLXzB9aTlBVkLqndxIHoVj1bRC3w2nJ+Syq2rgp4wIiMjthabTyuqpgX9soI6SOQS9Ts4z5rUp52shLm4yFJUwRxw5j743WjyOlc4tIOMoMq2Us94nMZk+EeWVPcYRaCITuSrLXUm3u64+6yK2Vtxd1SnfyQaNJI6c7DYq6Gjy4Z/OtQZSMYcuAwoJqmKJ3S07oIq+IsYOlv5Asenncz735Lc1kFDUf26RjG2Vh3uCgjqA6lIx8kHen2MMeOaHVDj56Brf5Rt6K/2McjHcz2p2t8tBVv8o29EHs0iIgIiICIiAiIgIiICIiAiIg465jWeJy9cUI/7rRl7b+WhmXzS+BmqkiJ+g4j8hwvpa5ji5vLzxQc0ZI0ZeyPr9xmXzRPfIZ5HD6RcSfyoMt1M3w9isukkjigLXYz81pPi1DQCc4VQ6eQfACR5oLahwM7iO2VNHB4zOrPYZURppidxup6aKeMEYIBQYmHMd0gkH5LOgp3uZ1OycbqGoiLPiI3zlara5YTSlshHUAg00Hx3mFvfspDaXtwXZGfVX0nhw3AyPwG52WXdKgzYNNg4Qac6lFMS4nOAsaaQSH4Ve41U7vDwSVU0ksQLpWkY8kGRRWx1T3cMKK4UXujw3vlUpK2aF4DCsqd5qCHy+XqgwnUpEXib9sqFrsd1qFRKx0XhNIzhYTYXdQ6u2UE9soTcKjws4B81n3SzNtrc5B2+tRsqY6GMSwEB6x6isrrn9IFwHogxYITUS9I81qBszmgOLvnhQ0cEtNN4krcALIr7hK84h3A2QYlTGIm4BVaWuMMZYc7rHdM97vvndWvA26SgSyF8hf6nKtLyfPZV6ds4VqDPtltNwk6AcbrIudjdbxkuz6brBo66ajf1xd1JWXaprABKeyDF6pMhvUTj5rMirGxR9G2SFgEnOfNO6CUS4mL/ACJUj5XTkNa4jO3dYyqxxa4Ed0Gey3fD1k5WLUtLHdOdlmCokZFk7jHosGaTxHZQWl+RjG6Nd0hXu6OjY74UY9SgpknzWbTUHvEfX1YxusL6lPFVyRMLGHYoI5WeHIWeisBwQVPGxxcZJOx3ypHMjcQG4QRvqOpnSroaQyxl+cYVJoA1uRlWR1EkY6QdkFvQ4ydGc4VXs8N4/KniY+L98VOyhqapnisYSAEEz4hPC1o7hcgcK6uNl/prVLT9fiuAJI7ZXGgklid4Z2I2XO3B1+maaWK43B7BUR4cM4zlB334SaHtmg9OwarlLAH4d0ny812y4bXCn11bBcKSsbEIWZ6Q4DsF5wxcfWyW4WSWX/ajMNAzthcpcL+YSWytFttU56JBg4cg796X1UJbubL4weWO6cg5XKMbS0ALq1wFvYvt1bcpiS97w4rtONwEFURUByg4345R08mlntqKUTtLXDpxnyXnZxE5cbprChrb5aqOWCJhLi1oIGO69S71ZKK+U3uta3LN/LK2xetBWyk0pX0FviaC+MkZaPRB4Mamu9Vwsvr6EUzxLE4gnBySFp+n+K9/vWtKS4uq3sZE8HBJHYrt1xd5ajede1VXcIfvTnEj4du5XAOseWTUluvrajTdO90DX5PS09kHfzhbzeNsGg6a3yzh8jIwM53Bwr7xxvr9Swm/RVhayI9WzsDZdZ9AcOqqltMVLdw8SgDIIIwt83+stGnNJVNE0gOLC3v8kG2ePPPDXTUL9M0VzIljaWZDt89vVdIb3xd1Pdq2Wesucjw55OC89sqTXlFSVmtJ6uZzuhzydzt3Wy9RUtDHKfdnZAQbguetaq80opzOSQMZytBgqm0riJnB3UVpLYstHu5Jf/OrxHO1pfUZBQajXVkLoyWALRHyeID0hPFMrujyKnjpHA5xsUGGeo7HyVzOoEOyQAtTjp4c4djP1rHrKfpyYzsgup7g2HyyqyXMPkD+g4CxKaNrpQJNgtYbTUHR3GUFTd4qiIRYAOMLSan4Jct9VLU0wjPVAD3U9LA2SI+P3QYfvY6QA3JUjHmSI9wpaW3OdOeofCCrLi0U7wyLbKDHpusy7k7LUCWuIiO+VprYph8TQVXxpGODndx6hBn1toMEXjtcN91pRJPcrMmuNRNH4bycYVIhEYzuMoIqaYQu6iFWWbxpQ7sFGA0yYPZH9LXfB5INbhjjfCAcZVzojHEXNOAtIbWSsaA1ZdPXF0fTKe4QQGrEbztuCo5AZx1g9lfPHG8ks7qDplZloBwUF1JTuqZOjOMeqlqI3UTw3Oc91HTtnhf1sByPRSVDzIcy9wgt972xv2VW0/jtMmdljmIuJ6Nwr2umiBb2BQTwVbaYlpGcFZEleyduAMfJYtPT+K8l47lR1TGxPAYguc7w3dY81UsNS3rzuFHEC8/H2VXyeE/pYdkFsUroJPqWd91nhnQ04+1WRtgczL8ZWLUNYH4j3yUCaUyEuz5q2KUsduSqeBIB1Y2VhBB3QZ7atrRk+axZpTKcjOFaI3u3wsqOKNtO8v8ApdJ/Kg9U/YcRFtp4xS+T57CB9grv9a9SF5c+w5eHWXi/GO7aixH8orv9S9RkBERB1z9oeccmvEr/ABOj/XqdfPPA/ocfmML6GPaH7cmvEv8AxKj/AF6nXzxjYgoJ3wOHxh2xVaanfPKG5VHzEtABVaOfwpg49kGrVtCaamD/AJenyWnxYfE4nvutRulwZNSBgIJ7fmWl01LUSRufGDjugjh/sp6uyvn+JwaB3UWHMlLT3VxyJRlBqFusb68gdRCguNC+2T+B1Zycd1mx1NXDHmkBJI8gon09XUHx6xpBG+4QYVTSPhjbKezlrVn04LhQmoc7GBlYT6iGVohkOzeyoy+1NGw09M74eyCMdNqqyx+4CznWwXaH3uN2AAThaQ8z3CQvAy5ZlNcai3RmmdkA+qDEpqQyVRp89jgqWrYKOQNzlXeM1rzNEfiJyoagvn+OTOUFk1T4jQ1oSGjdKzrJIU1sigkkLZj27K+4zNgkEcBGEGCMseW57HGyqZSHh3fBVrCC7LsKZkPXJn97lBK+o96Z4YGMDChbHPE8AZIPp2WTUmnhYDEfi81uSxw2+ehL6gtDwEGg+9NgjGW7n5LGcwVh6m7LPubKcTuAI6QVplNMIqprWn4OoILJqaSIbg4WXYKgU11gmcdg4ErWryykfbmuhwXkb/Wtvw26t6fHbGQG75Qcp611HTXXTzKSMgkMx+ZcbUdEY4ny5zso5bm8tELyctOCsq3SPk2Oeg99kEEE3vEphczAG3ZXzvEbxA1merzWdWMo6RniR46yoIZKWePxHkdY7ZQYNdbfd4/GB77q2ltpqIDN1YACVz6t2WyA9A7bKGCulgb4YPwlBA9hY8t9DhT09JJK0v6SfTZalR0TK8ARgFx7rclrt9NSuEVSAMj0QYOktPTXiqggEBPU8N/OvULlv5NLZqDQn3fq4GF7IQ/BAz2yum3CKwWqeqikjawuDwR275XohwYu/Eu00cVHbqeX7mOYA4tzjGEHAuseT/8A2R71V2Ght5YISQHNb6LrFxF5bK3hNqAWGeF7jJJ05IPqvYCg1rpLSsUtXJLCLg8HrBxnOF075jjDrW8fdyFrHFjy74R80HRriPwvj0fZobq9oBlAdj61uzhZyt0vEbScupzO1pjYXkZHplYfMBdbncLc22AHpibgBbC0FzEav0DY5dP0r3CF46cZxgIF/wBFjT99faIQD4EmNvPBXZPl71Nb7LVwUtbTMd2HxNXW7T2tYL9eTcbrIOuV+T1epK7I6IsVNURNulC0FrR1Zag7I8TuJEdl05Tz2RgD3gfQ7/mXNvCKkuep+ClbqWsqnsligLx1Eg9srrvwxfpjVk7bZqKRnTE4DDlzZrvi/pXh7w+qdI2CdgZLEWYa4emEHnFxV4p3a5cS7lYKuufJHFI5gBdkYBIXEmr4yxzpY3YGerZbh4h0sTNdVuoXZxO8vz9ZWw7/AHhtYCGEEbhBoct36pPCacEbK19ZHJGWPIJPZafLC10pewbn5rFd4rJATkDKDKNtMhc8HAKhe91MPC6ScrKhrwxoa47+ayC2knHW7GUGjNY55J7bquJI3Dc4WXVMDP7FuOyhDmOjPWd0FJqlr4+kDfGMqGAgStJ9VdAIzJh2MfNTTeE3dhGQg1SepibTAYGStLfG6cF7Tgeix3zPeME7LVaK21c1P4rGnpxlBgUcfVKR2wq1o6HdPclUlJppXNbsRkKEyOkeC7dAjeGqjnFxJGcLLbRSVhApGdR8wFqtNaWUtOffQA/0KDQ4Y+o7qkjSJOkHYqSqPhznw+yuj6JIyf3/AJIMiFzaNof0k58lmfch1whNZ9EAbjtlWWKKOSbFfswdsrPu10hpG+7UZHSdkG3Z3DqMAHY4WpU+nXz0xqGk4AzstJnDw8yOH0jnPzW57XVTm2Ox2wg2tLGY5HR98HCQvMcgd2ws2jbFJWvFQRuUuVM1sgMABHnhBdWXBk8AjaN8brAjeWHfsrDsUQTyziQANGCsikoXvxJ1kDusFndZDa2WLZp2Qa77+ykjDJHg4GN91gvqW1soDDgeS0yWodL9MlXRudF8bM7INVmtTnNDusHPzWp0dhY2lM/WMgLbwuNZKQxm+FkxXyshb4EhIadkFJ610czoTuAVA2pbLKI8d/kppDTvBkO7itPkLWPD4+6DVpabwYhI3fPktPFW8S7kjBWpWWsZO8w1ZHSB5lYtzhhdXiOmwQ4oNy6etTb00NLsZ+akuugfdpQ4SAjv3WnOmuNhpmzQAgEZWHJq28VTw6V5IHfugivdJLbulrHkDtstWsmln3K2Gtkec4yFpU1Q+64DtyO61Sh1DLbKc0HVgEYQd3fYy0jqXmp1W09hoKuH/ONuRZXsbS6Tmh1TM79/oOtP/OFvRB7JIiICIiAiIgIiICIiAiIgIiIOOuY53Ty9cUHHsNGXs/8AIZl80rHA1jieznn85X0scyX4O3FMf9xV7/UZl80LiWykg7hxx+VBuG4WuJtAJ2OGSM4ytNtkkbWuDwFFJdaqWEQOPwgYWI17m56fNBnyVrBIdhstSozFOzJwFt3IJy491NHWSw7RnZBqF0ia3PTjbzWmwyPY8Na4gEqaWrMzB1d1jNzkH7UGrT0niUwe12+Fl2qla2ncZXZOD5rTG1Mz2eGzOQr4qiqp2Fr8gFAFVHSVriRsDlX1tfFVjDMD7FgzDxXl3mfNUbC9u+6DIoqYPl6idgr7g4QnpYe/oomzOYMRndWuLnyAzILqWne8h7uykqS2IdA7rKZNC2MBuNliPgkqJQ4AkZQYskUnT1nOD6rKt9dHSjDxlZNU2JlMGbZC0ZBqtfco6hpbGACVh0zwGnq3ysfpKAuYgrJu8kDAyrc75Kd0QSxjxCGhUmi8I4yrGlzTlvdXOc55HV6oLPmiyHhnhjHfCx0BAMkD1RVaR380GSyj62dWcfaonx+G/A8kE8oGB2CdRdu/ugynkOiAB3woIqXrOCcbqxj3uPSFV0kkTh5FBJPRGJnWN1bTU5lznYLULOfeZ+mp+gfMq++MippAKTBHmQg099F0k4OygEZEoaBncLLD5ZogG5JU0FL4cZfMBkIJp6VhpQQQCtOjj8PJJ7FJayZ7jGw7dgoXOkZs/wA0F8s/VsrWRF7SR5BRtaXHA7rPhi6IyCgwmR9T+j5rcVDcqeipjC8DJGN1oMbgyYnZVnf4sga3OeyCWoYJKgzNGxOcBTUdTcGVTBT1D2DbYHAwpqW2VLGCaVhDD2ytSodJXa5SCegjLmDc4CDWbrebpR2iJ8MznPJG4O67acpPD6p1lQNuldV9Ba0Ow52D2XXPS1jpG9NNqDDQ0j6Q/wBa5d0zrm/aTniotGOeYCACWZxj7EHf3hpeWaHuHugqARG8DOV2XpOLFC60+/PqWfe2ZI232XnVoLWNxnibWXt7xKcF2T5rdl6400tthFIJyIjs7fCDtZDzU2mpvJtDpoosP6eo4C5r0vqC23u3x1dLcYpy8Bxw8bZ+S8ceNfF6y2qhNw09UZrfpHpdvlbY4N882urPIaWunnDBloy49kHugHxn6L2/YVHMYJGOilkYA/Ygkbroly582tTrWuEFdMXZIHxOyuW9ecbJbdqOko6ZxDJCMgO23Qcg6+4UWy8h1RBRgvO/UB3W1LJwysFuhNJXWlkj3nGSzK5o0jd2X2wUtaCCXsHUO6y5LRSSSCUt3Bz2QdUOLfCeHTtJLdqC3lrSwuADfkvPTjRqvUUE9RSwUExaHkD4SvbS/adt+oaM0VezqYRjtnZcO6i5Q+HF/lfLNA0F5ycxgoPCW9Ut/u8viNs03W7z6CtU01wZv99b1S2iYl3n0le10fJDwyjxiCLI/wDoQW5bPyrcP7QAIqdhx/8AQwg8PLvwPvOmYjVPsspA3+gVx7d7NcJusOtkkQbt9EhfQZeuWHQN6hdDPTMwQRvGF015luT22aatdXcLNAwMYCchqDyVqbc+hd1yDBG+CVa24fD0hpK3PxQofuVqB9sGzmEggfWtPsdohlpXST4yMoMe20nvr8l2N/MqW6UrKJpGQStPnrZaCreynIABUctZU12XSZOAgxovjmONs+iunEkbs9Zx9ahDjG447qYMknYXY2QSwVYAAdushknVgjb5LBpg0S4f2CyHTMbIA07IMqS4+C3DYyT6gKDoNY0zPGCNwtftNNba1gEpBP8AOtNvrYqGcRQH4exwg0+CoaHmNzdhsoKvp8QBv5llzin8APjx1keSgpaSWd4c7OEBsXiMAIwseRjoj0g/nWr1ZhpoAAcOWl9Dp2l43wgg3JUsdO6QE4KkooPEm6HDzW4mUsEUBztsg2v8TTgj5dlaXHPos+obD4pDcLEkaM/D2QX0py/DjstREDHEbBaSzrYQ4ZC1KGY+EXeY9UE1SWUzMgAn5LS3uNQ4uxhZFNIaqfw5dwra6JsEgbH2KCFknhEAtytWoqEVsXihuMb9lhwxwStBJGVuKhMdPRFrcdkG36h7aWQtHkeytbR+9RmbPZY9c8y1T/PcqWlqXwx9BOAUFKZgkmMPbBxlW18AgeADnJR8nhuMkZ+InOyjdK6V4Mp80EPU7GMnCqw5eM+qnlEQZluMrG80Gpl0ZixtlafLjq2+xBK8jARjeqUB3mUEkc/R3b+ZVmlL2kjYbLPrKSGOlEgG5HqtJAdgnyQerfsNf9z+Mvp41g/RXr1MXln7DX/c/jL/AIawfor16mICIiDrr7Qpgk5OOJLPWjo/16nXzw4X0O+0NeWcm3Ep47ijo/16nXzzQtDic+iCzp2yqKSXbICoyJz9wNkFYmOld09RwPmtfpKyGipDE4ZOFomC0fB3UkbnOx4v1IK07BV1bnYwCcqSvp2wEOGCqiSOn+KM4Kxp6l8+xJJ8kGpWO4iGQNfH17rc1WyOspy4RBhx6YWncP6W3ury644DNsZK3PrKstNMwx25zTtjZBxvU0ThUPa0nv5FRPo+gEk7hTulkNQZHZwfNRzulmk6Is4KBbaptJMXObkE4WoVlKLgDUMIGBnCxaa2uaPFnacd1bNXOil8OE/B2QLbSGSpMb+wK1C4UHgxEsGSAqMcxkQkj+mfRbj07SisqWGtx4ZIyT6INgtEzX5a1wJ+RCndTukBe87hc5XPTGlZ7bmh8MzhvljuuHNRUs1vqTEAQMkYQaK4EOx2WZRO8RwgxnO2VDM3pY1x7kLVbTa5JIDVRjOBnOOyClw0/LTQipblwO/qsejFT0Z6i0D7FufTVypqyoNHciA1u26rqoW2md0UJaQfRBs6rlf1FpJyrqaiMrPEyQRup2QNfJ1P7FRyTmB/Qw/D54QSROm6vDc4kA9luGiulO2n90dE3qcMbhaZbHUzyC/GSswxU3vDSzGUGLVabkke6duQHHKthiFGzwj3HmtdrKt0VMBnGy0CWqZK0kHJQQ1VI6XLuskHtkrHioJI3CTr2B33UU1TUtcc5A8vqUfvs5HTnug1C410UsHgsaOoDC0fCy6aLqd1SDY9/mrqlkQBDMZQZenK5tFVF8h2K1K9XMzyePC8gD0K2x8UZyM4WVHO2Rha4nfZBybwr4iSWWvgEsxAZICcnyyvU3gNzX6VZo9un53Qe8SRhoccZBxheNVK5kEgkjzlpyuTdG8Q5rK9j2Pf1tAxh2EHf3i5ca0V09/pbxiGUlwaH7YO64guXFSGCzy+9Voe8ZAJOVwjqPjjfbnb/dZZH9GMdz2XFVx1hdKtxhy/ocTnfZBvbVevae+3SaF5BaSQCcLjCut/3Qu7aWFoaHuxkD5q6pkY0CaPPWe61C31EMcXvbiPFYMhBq154eTWS1wXGCow7ZxAK5g4CcWfDli0tVu3lwzqP5FwZ+y+8XaQUNQT4LTj7FvfTb7JYIhemuAqYh1DB3yg7yy8NX2Oy/spo7kGmVvXhr9+2V174g3zUU1WXOrJHxtdg5JIwtL0Dxn19xGnGm6DxX07D0bEkY7LkTUWlWWm1OivTQ2oe3PxDfOEHXrV8hucJYN5CN/VcbVdqlpIHl+Sd1yPqCjqKG6SyPBEOSR6YW17s6OphcWEEIOPGVDopz1AkA47Kd2Khwy3Hks1tJTNqD4mFZVOp4ngx42QYk1tIb1NP51h9EjH4Djt81mVVbI5vSwqCInBLxugknPTCM7nCwhlzw07ZKz4+iYlpKx54DHJ1NGwQJqR0TA8ZKx8O9T9q1u0ywTnwqjHyysO8MgiqOmnxjfsg09bntmpaejtxpXsBcRjOFtk9gqiNxGQEF9TL4075fJxJUeD6H8iOGDhbnttNbn24vkx148/VBp2n7tHaqgyTM6gfIhahd7ky7/foT0gDcDZaLBTx1Na9mfgyVfVNdSSiKPIaTgoIWxGZ5YfJXRweDUta45Gd1LLlkfXH9LvssN00nX1O7hBrdwiApQYTg47grSGxSHEj3E49d1c2qnnHh5OArJGzs+HBwSgrI8T4Z6LUaS6tpac0p7laQSY9x3PdBHI8GQDIygmqGODzMwkZOe6yYZQ6Ah5BPzWA6RxHSfLZBI4DA7fWgPaDIceZ2VpbjzVQ7fJQnbKC1FVrS44RzS04KCiyo2tMZyViq8SuaMBBlUcjaV5c7G6irJmzS9TQAAPIKAlzyqEEd0Fep3bJVY29bw31QDq7BMljgR3CDKfRPhYJGOIJHkVHTzOgqWTSAnpOfVVFW9+GvOyyI6dlU3oYMuI/Og3RHdaa+0/unSAWjC0e508dvaYWtBJ9Fhx0lfaXe8lhA+ruFN78yud1T9/mgwKSrdSSlzgSCq1EjqiYTtOB6ZWZNDSEdgsMM+MMYNkHf72MFb7xzP6ngI+joGtP/ONv/1oq+xipmRcz+p5ANzoGtB/zjb/APUiD2cREQEREBERAREQEREBERAREQcccyX4O3FP+BV7/UZl80En9kf9Z/Svpf5kvwduKZ/7ir3+ozL5oJP7I76z+lBaqhwAOyyIIf3z+xVsjWOma1v0SUFkUTpXYwqzQ+C4Nxla1LTwwUQkYB1Y9FpccjZSTLuQgt91Phh2Vaxgzgq6aoO7WnZQCR2e6DUKciA9ZwQrK2sZKfhA29AoHzdTMHOVHHGXb42z5oKNlwckLMgmbL8OO/yVrI4nnpxkqX3bw2ktG6C73YQgy9wVhVMpkfkNwB8lmUTaipm8N4OPrWVXUkEAAIGSg0ukDpJOguIH1rVy9lNFnAK0yZjoMSx7Z7lZVHOyeIibGQEGn1NQ6SQkE4PkoBspqpjWynp7KFBNE4OIarpINsqOnc1r8lS1MuSAwnsgxiMHCYPopY4z1dTxsshogLhntlBhty05wrnPBWqzxUvg/BgkjstM6GjcoEMRmdjJwk8JidgZPzVrZHRnLdlm04ZPGXSbkINPQbK+fp8QhvbKtaxzvojKC7rGFTOR2wqtaGn49keW/vOyCsMnhP6iO3yWW2JtVh+e25WApYp5Ih0t7FBmCUQ/C0gEKOScuPxHq+Z3UEgfjxHDvurYjl46kGq25jM9TsYHkVWuqGunFPH2JwsaaZ0ceWHv6LNttqqKyI1XSSWjKCeXTQpKYVpcCCM4JWiS/wC2HHAxhaqLnXzTe4TZ6B8OFj1tMIDiLGXd8IMSmhDHguWbI1pjJBH2LAmiqIQHuzg9lPS+NUREAk4CDAkBDyFmUNIZCJyNmnKiZEyOb787AB3WrUlTRvmZSRu3fgZxsSUGqQXdlzhFuZF8TdicLkLSWpqTS1Kaappg8ub3LVpth4aVlvhF5nYRE7DskbYWustdnuU7IsgvPw4+aDWdK6ePFK5+70v3kOd9S7CaN4Ts0JUQUlTAKgyAbkZXDFmo7hw6gZdqFhDcgjyXargRryy63bFLfHMMzABvjOUG6WcJpK+1CrpYCwvbnA2XDnEHhXdY4JYw5weSQF3Mor/aaSmMYLPBaNu3ZcG8UtcWGG5gdbAzr+LcY7oOiWsuFeoLdUvqK0yvi3OHZIXFV1pJaCocadwjLTj03XfXipqfRF206WUEkJqDGe2M5wuiuuOuOrn6execYPllBvngvxbuGgLi2qfXFoDge65guXNhJfdU0RNwBDCBu4LpnVGdrex7euFgU9VLTVLalpPUw53O6D3m5cuYOguWnaaGe6Rk7NwSF2ltF+pbtEySCVrw8A7LwD5feMmoKSsht0L5T0vA+kfVewHLXq6uu9vpPfycujB+I/JB2SRWxuDm9XqrkBERAWyOK+maG/aLuFLLSiR7mHp2yclb3WJdWMkt87JMdJYcoPA3ms4Mz6c1tV3T3V7Ii4kHBx3K6609yMccsLcjpJbsvUrnu0tS3WOoZbQwzFhGAN8rzPj0FdKBlVLWsLcOPkfmg2VVyGWdztySVNSShjegt3PyV8tM1lcYD5FTPpo43jYIMWelc0GTyKQVbI2FhGcrMrHM8DAK0ljepwHqgvkB6i8bAqwBziAMklZ1RQyxwCQg4WqWWK3vg6piOseqDHt1HUU+Jg477rIrqF1XGZHHBA8ytStToXVRZN/Y8+foq32GTrDaIZYg2dh8chjOdjg7rVqOribiPHfZKygAjDyMPO5+tabE7wieruEGt1FoNUzxGuz9q0yX/aGYXDJOyy6C4zjZx+H61jXOWOWUF3cIII5PAf42O+61qB3vlMXdeMBaJO+N0QaDuArqOudTxmMEgFBa+BxqC3qzj5/NWuAZKGHzUgm+IyHud1BI4yShwQZ9TStipxLjusFk5HwD1U9RUSOgEZzgKOj8IOBkwgvjaYAZcY8+ysdL71M1vqcZWXWSQuh6WEZx5LBhaWnxB3ByEGrVFqNHSCp6h2z3VKOtdLAWdR+1ROuM1WwU7ySBthIKUskAYNigwahrmTmQtOCVGXdYyB2W4qyhjfADgZxutDexsOQdkEdMwvfvuPrUtTT4I6dvqVInfFlndTHrJBd5IMN8D2DJyqNhLxkFZpcx46c5KjdGWbDzQY7QY3dsqZsRfiTpxjdXUrWF/wB89fNZ8slPGwtYR6bIMGSqdO3wBnYLGfiMGPG5UzPCEhd81HVOY5xIO6D1X9hs3Fs4yP8A7qewD8ja7/WvUpeW3sN3f9SuMbPSewn8ra7/AFL1JQEREHXP2hwzya8Sh/8ASdH+v06+eqkHU9wPovoV9ocT+014lEf3nR/r1OvnlY9zDlvcoMmWmzuFRknhN6A3P2KIyzYyScLUqOlZLTmR3fCDGpo2mQud5+StrXhrgG7KJ8z45S0HYHCslf4jhjugtMjiMZKRkB4PllXupJ2t63MwFYMAEHug1ATSNYBA8tP1qIGrklHjSudk+ZVtM14OXdlmHAcC3uEE01MPBGMZWJTO8CYOczIB81nxufKA0Z2Ugipw7pkx1FAr6+OamDGDBx6Lbjg7qyck5Wu11N4bepo2WmRmBxw/CCShmdlrXEkLeVHVtjoC1jsOLe47ra0LYSPveCVM2eeJwac4CDNoL9WWquc+onJYTsCdlq9dQR6kiNbHg4HUtp3d8T42kfSW5tLsr22p7o2ks6cn6kGNZdItvlV7oXBvQcDyW4bnaqbS9I6iDmkkY75WznX+vtle/wBxJ68nKhqdQXS51TDcHE5KDHnpagVbp4iWB2+RsrHxTySAyykgepWuVr4Pcw5hHVhbedUloPfOSg1IwNMfwuGVp1RFh2PNKepmc45JwhjnkqAd+nKCEGWn+IEhahSVJc0TOf2+akr46f3YDbqwtHZO5mWg/CUGq1d5NQ3wQSMbLCYyYHqwSFDM1jQHMO57rdVkioZbYXzgdYHmg23UStkb0YwVHFBgdffCV5aKt/hHbOyuimHhkHuUFJKkYLQMEeeFAHOc7JOd8o5vU8hoUzKZ3SXfL0QUmx4Y9VZHE5zS7P51bJ1BxafJGvdjoHnthBkUI65OkqaWrnpakFmcA5woaeCaJweRgLXrdHbqiRvvOO/mguprpJcIRG8EY7krErK5lKTFsSfQLV76610NL1UBHV8itnPkfUuLnnLvJBqLZg774XDB7LHlrpGyYYT0+gWK4zRtw7IBWo0VNFVRHb4seaC9l1EQBawEk4Jwt36esFbqMeG2QgPb2ytN0rw8umoqkR00RILtsLm/RPLxxKZcY30NPL4WAc9B/wBSDfHLbZo+Gld90K2h8XJzktyt98ZbhcdZVbbjb7ZI2Jm5DWkAhcwcD+AdzqaaCPUcGCCOrqBG3mu0R4Q8KaK1Nt1QacTubjBAznCDx71wZ6lhpH0To3hvSSW4Wxm2V0dG8v777Fd9OaTgPb7F49xssTPDLS5paF0vn0xfDDK0RuwHEDYoOJbhRSCqeACMZWh1LXxy9LiVyDeLHV26V8lWwj1yti3OJ8s5fEMjtsgi8LoYHHclQmTqd0gYyVNA57T0zbAKGcxicFmMZQSgOhAefPdVNW2QdJ7n5KSoLZoQ2PchaeWuY7DhghBO6N8XxseRtthQOe55y4k/Wqukc4YPZWoCnZO1rC3GT9SgRBU/ESQCfsV7ZpmNLA9wafJarpyOklqCKsjp+amvUVBHVgQEdPmg0emnNO/qOd1NLUidwJOSFkVcNNJCPBILgPRab0vieA7YoMkzdI3WO4CQ9Q2V8rmluB3UIcQCB5oJYJhC7JGcfJTy1rXjAblYSy6VsBHx4ygxXuLjkjCyYqprGFnT388KtYIWtHh4yVHE2PwyT3CCnhdbi7132Vwptx/rVadxLj6eSrUGVrsg7ZQRTQeEO6vpqfx8Anuonyvf9JVinfF9FBqLre2nHWTlafN8TyAOyvfWVEgwScKSmaw4LsZ8ygjZSFwydlE6PpeGZ7lZtVKI24adysAvcXdR7oMrwPDaHd1Z4XiZ8lJSTBxLZTsPVWVMgbJ977ILejwdyoyfEeMeaO8R+5VGZDwgvkgMYB3IKy7TL7vMJnnYFXxdD2YkwfRY0z2iQMb2Qbju99pq+lFPG0AgYOB3W2Cx8ZwCST6KaSneWh0YOSN1lWqISVDY5+5Qac98wA6gQFPTygDJ3K3DqG2U1LRiRgAyM7LbNK+NpLX+fZB6CexhqPE5odUN9NA1p/5xt6K/2MbIhzP6nczGToKt/J90beiD2aREQEREBERAREQEREBERAREQcc8x46uXjii310Xex/yGZfNBIMTP+Tj+lfS/wAyP4O/FHHf9hd7/UZl80LgXTOz36ifzoJ2zeIwR9ioXMMTg4nOFKDEwfC74lA97nHBQZbq10sXhEnCxHtLdv0K73eXHV07LKphEWffe/zQYLWlxwrnR9OMqZ4aH/CdlHK4eSC3w9s5VWzFrcdKug+N+CdlnGKmbHvjJCDTo5SyQPWYK7qA22UJpZJXkQjKyI6F0DCZxglBfTXRlO8uxv8AUoLhXGskDhkBYsg++ED1U0EGRkhBI6oD4BGe4GFjMLmyYBIBKukjIccBR7tPxdx2QZ/uofHknc+ZUQoMjuofepAMBXsq3DY5QJ6UwjPn9aU1KZgX57K2aZ05wN8Kele+FpaRjKAAJCYh3Gyx6iAwOAz3KnB8OQvHnurKgvqDkDYIIoC9zukuOPrWVJS/D1BYQLoz8ws2CpDmEOQYRaS8tHfKzaaNwHQRjKsgdD4pc/GPmsuSaAY6CMhA+43Xl/XgH5rFlYKQ9Hf5q+S4Ts+jnBWFLM+Z2X90GVT0gqyT2yo6yk91d05ykdS6ADoO6jmqHznL9ygja3qOFXHQ4I3LDn0UjWGTDgNggmle0wjbBVKKl8Vwz5q2Z7OjoaQCFkUE7YmFzvLdBmS0AjiGfJZ1t1FDb4DTloyRgLRKq5Pm+CPPfZRwRxBwNR3JQan7wyaoNR0gZKx62pBmDhuAr3mJ8YZT7n0WII3slHjDb5oNQ8RlygELW4Prhbu0Ro0V9bFQub1GUgfXlbct/uzWgxYyuR+HFTPDqWjkcPhDgd0HLbeTuS6WVl0jpXfGAfo7LdOifZ7XCspP2TSxyGKm++dJzjbddktM66tdNoWCKd8YcGjPb0XIeluONno9GVNtZJF8bCO49EHTG72e2mR+hBE2OSEeHnt22XFGqOH50RXe/tm62tOTg5W6uOOsRZdX1N9tzsOc4nI+vK4uqOKz9V2+ZlwkBcAe5QZ2suMNHVWVtnjc0PaMZWq8G9X1tteKltxLGjf6WF101D8VyfMzPSTkH7Vm2bVtZbYzFGXYO2yDvpHx8kmpzQC5jqAxnrXCHEziDX1tW6JtwJL3d+tcI0et54pjM5z8n5rEvGppLhOJ2k5G43Qb7rq6826AV09ye9hGenqzsuPr5qkXKYk+XmpW6olrohSVJPQNt1t+6RU8cmYOxQZL7iyVgGM4+S02cFzi8NwFWm6RJh/ZZ0hgMZAIzhBvDg3e/uLf45yAR4gXsByq63fdoaJkQ26G9vqXj1wqsNRebtFHTtJJlA/OvZXkz0E620FJJVsIIjGMj5IO5VueZKRjj3KylFBGIoxG3sFKgIiICxLrGZbdURju6MhS1FVDSs65nhrfUrQK3WdmY7wPGBJOD2QdLuPWj55LvPPPTOlYcntkYXU7XHDyiqrTVyx0bYiCdsYXp5xVrNE0tvfX3l0TWFhI6sLodxgmpb7TVf7DyJIuo7tG2EHnhqvTJtl/kAbsHFbfrWDrx6LlbXtguVPcnyVkZBGc5XEd6kdHUFoO4KCCrBDNjlYUTuh4cfLdTRymQ4k3CjnawPHT280Go1V3ZPTCAN3xjssCBkpcOl5Az5JK2JsYLT8StglLHDKDWnVDoIRhxBA3OVJS6jbFGWSnJxjOMrS55fEjAHfCxRTyu8kG5KSRlylOexWLdrQ2BwcwrDtrp6KXqIIGfVTV90dNKMnYd0GOJOgBg2UrbaapviF26pVOhfDmI5djyWPT1s8QMeTv2QWVFEYCRnOFi7g7FZdUZyC9+cFQMhkkBc1uUFnWcYyVJC398o+h2enG6mijka8E7AndBWaT4cEKOKF8h+H8yzq0QeAOgjqUts8HwT1EZ8gg099PIxvUTlXxuIYdsbeazo+h0xEhHTlT1FJG+MuiAx32QaPHN4cmfms+K5ta0HG+FgCLomIkGwR3hGQY7eaDOkvBeC3yWLIDU/EPVSSRU/hDGM4SJ8cUZBKCGMeA/JWUZGyw7d8LT3vMj9u2VcXPjHSOyAHmOUuys6NzZ4+ohYlIwTS4dupKkmnIYzYHKCCZ5DyATtsrWtkkOMn8qtIccux3U8GcZ/8AfsgpLCY25UbYy5hf6KefqkGAEbDKyne4jbCD1T9hsz/qbxkf6z2Af/g1/wDrXqWvLX2G2fuXxjz28ew/9GuXqUgIiIOuftD8/tNOJeO/udH+vU6+eMbEFfRF7QPo/ae8R/E+j7pR5/j1Ovnjhp5Jy7wxkN3JQXNeJfhwp21ZpmeF5HZHiGJo6cBwG/1qWnp2ztL34QaY9xe4u9VlUtP1gSHsN1dVQMBwxRxQ1eCyMHBQZlVXxOjELW7jbYK2ntZnYZjsBvhWRWese4EsIHfstbgpKmGAsaDjHog02JjXPMRIBCjnAimEYPdXTW6udKXQNJPyWPLSVkJ66gEHPmg1iGNkEfWXD1WjVdRI6tDmuOARjdWSV0zh4QJx9anpmxFodKfiQarNK2oowzzwtMFo6wXB2PtSWpxszssd1bUMyATj60EjWGgfkuyApTcxMQzpO+ywTK+pcGO3ysuGlYxvUe43CDOZZfeGiUuyM5xla7T6pgs1ufb8DJGFtuO5VEJLNw3sFiVZbUu8TO6DJttSyS5vqZQCHHO6zLt0TSh0TAMHyWm2y111ZKPdmE+uFuJ1A6hhxWNw7Hmg0CqnkjYGuJIKzaCyiupjUdW4GcLHmo3VUhEYJAV9HPXW6pbE8ERZ328kFrKZ0cxicwgDzwrqmqZTDpDQT2W5a8W+oow6jIM7hvhaHT6Ru1W7xXxHoznceSDQZqmaYkuJA9FBg+hW7qmxwQR+GWgPChZRUUcJ8TGQEG36WEzv6XLJkllgeKZkhASZ7KeYmLGD8ljuinmJmAP1oJ6q3eDCJi7JO60/KyJaqdzfCf2Gyg6SgvhkEbskLKFa3pxjyWCpI6eSQdTG5CBLJ1uJCsY7ocHeiq+N8Zw4YU8TGGIlBkuuDHxhmCCsKSWTOQ4gfWo3bOOEG5G6C4ySPGHPcR9aRnpkB9Fc8MDcjurIxl4J7ZQamY2VUYbjBWfR0ElHCahrC4BpOwWkzzeE1nhFcjaKrbNUWp0VyLA4jHxFBv3lv1lbDeqekr4Wj76G7geq9n+AVh0BdtMQ1TaWjlndGCQ7GcY7rwkZU0ena5tXZvpB/UOn6/ku0HAvmX4i2uohoYHz+CAB9I9kHrBqmDTlie/3ERRZyMNIAC4/m4X12pZhqCC5uEbHdXSH7Lg4a71prS0RyU7ZTK7BduScrdWk9aa903TikuglbARvnPZBpnMFWMNpks5pjM+JhGcZ7BdB9WaumsU0lObK/BeRnoK9LITpXWbi2ukYZ35DurGVoGvuW/hlXaelqnsg94LS4bDug8i+JWrHXCPMVKWF3/a9ltSx25tTbn1M4w7GRkLs7xc4S2WyXmpY+NggZnp+xdbdS1kVsqnUtBjwuotPog2tUA1NW+na3AGywqinMEnRuVr9S6kZGJoyPEO5WGHwStLpSOodkGJCw04D3bgqdtGK0hwOMrHqpQ8hrdwOyQ1r6dnwHdBdWW73VpdnP2rAWRPWy1AId2KxxjO6Aiv6OoDp7jyVrmOb3CAHuYctcR9RWRTU8lVIGuecntusdrHO7BSRzSQOBGxCDcVDYRAPEkfnI23Wj3cNbPhnYKv3arOno6tvrWJJK6d/U/ugjOcbqindEXNHRvhRtZ0vDXIEUZkOApnUpbuDj7VO10LG5BGVjS1Li4hp2QTUlvdUkguOyiq6Y0snRn86rBWPgyWndWzSvq5B5k7BBZHL0HIWX4gkZlQzW+pgYJJIyGnzVrZAIyMoIXfSKDYgqemDS4l6sqQwP+Dsgp4pIw1qrG8l4G4ys+1Mpifv+MHtlWXOGNsrXUwH2IM6mtDa9merBCwK+2e5PA6s7qynuNbTjpjJUdRU1NQ7qlySgjdG4fRCj3B3Wdb3ROcfFxhVq2QmbEeMIMaN/VkEKzB8UbLOdAxrQ4YU0MUErPDaR4p2H1oMWqBZEOk7qCGAub4h8itQdaa2mJlq2ER+WfRUZH4jg2Ldud0EENeIvhc3OFYK7pqBMzbByp66h6GdTAMhYMLW+KGSbBBq9Vd3XSNtMc77brXbbw7NZQmtM2MDOMhaPT21joxJAPiG6yhqi825nugyGHZB3s9jfQ+5c0WqYyc9Og61v/OFvRU9jbVOqeZ7U7n/AEjoOtJ/zhb0QeySIiAiIgIiICIiAiIgIiICIiDj7mHYJeAHEyI/v9H3lv5aKVfNM+HouNRB/cve386+lfmJf4XL9xNk/udHXp35KKUr5pjN1XCaY/vnuP5Sgyai0Ogg94znIytPb8b8ny3Wq+/umiET+w2Wm1HSx+GeaDJZWMLfDI+SxJyQ4dJOCpvAAiD8bq6lpXz/ABOBwEFtLTOnA2P5FJU0HhDJWbTBkbuhuMhW1koHwv8AP1QadQxF8pbgqtdHJHMG74WZSvhY/qGM7LOmdRys6nEFwCClsjipYhNJgn5rCu9wFQ7pibgA74WPLWSOeY2ZLfJZ1PHR+ATMR1YQYVnohXVQjKzb1TNtrwxv1LDoZ309wLoM4z5LNu03vADpjuEGmNeHDJxkrOhtDaqEyg42WkueQSGnZZdLdJoB4YOx2QYs0Rhlcw52OOyjIwtXjEE5Lz3PdYtbSkP+9jIQY0EgjdkjKlmqPEcCNlE+mljGXNwqRxPk+iNggyJc+FsrIakMaWuGcqWEAnokVlTAQ7LBsgx5HdbyR5q3KkiDQ/Dlkupw9nUwdkEdNRmoIAdglKqlfSOALs/aqU8z4JcNzsVfVOlndlyCN8wfH09yoFc6J7e4V8UXWNkESq04O6uML84wqOY5ndBlx0omaMd1QDwHCIjJJ8lDFPK0hrSsuNofIDIN/JBbUW50cRnOSDuobfCaqripM48Vwbn61q1fIDS9AxgBaNTTOp6hk7O8bg4fYg5KvXCX7haeZf3z9QLerpz8lsajoPusJHNOOhbgu/Eq6XazNs8rj0AdOFtm11slF1dOR1HdBLDTm3VJEhyAcbq+uc2qbmPYhQ1tT4+XA5JWHDM9jul2dygkpKuSklGScAreml9Y+BdIOnYhw3Wz6in64xIzc91kac8CC5RSVJGGuCDtk7XlwfptjWVZaA3IGcLarOK10oIXxi5uA7Y61odx1NaRpwQ07wXhnr8lwtdrvUyVD2tccElBv/XGsZdRlzXTB7jkE91xmZKmhe4NlIBJ2HmsmCWZjRJJnf1WFWSePKMeaDU7W0XafwJO58yr7xZ22yQNY7OVhwPfbmiphJBx5KGqulVWvDpnZwgvqaV7YRIDjbJCxI53MBGc5WVNO+WERjyUMVOekl2N0EBkd1Eg4Qlz3DJJKmjpJJpOlgOCdlkmiMABe3dBiPgLWB/ZRZPqVmVBPT0jssPB3Qc78rz426gi64fF++jbGfNe2vLxEyW10bmUxixED2x5Lxy5GqO3V+u2w3HHR4zcZx8l7raAtdmttrom2wM3hGcfUg3iBgYVURAVFVUQaJqKP3ymMHiBnffOFwjrOzS2mlmuTbkHGIl2zvRck8XvuzFYzJZGuM3Sfo910r1bqLiRC2qhuombASQc5xhBh67v994uVb9JW6pkLm5Z8JP8y4/u1il4KWOohvpL3vBx17nP2rlvltvejqHVr6q7zME5d+/I7rYvPhS1GqKn3iwAvpQcks7Y+xB0O4q8Tqe83qWOCEBriewXDlyzUyOk6e5yNlyXrKj05bgGTygVQHxDzyuNqmopX9TYZM+myDSnbHHoqZysiOimmf8AA3IKyfuFXgdXhHpAzlBFbaB1fP4QJwFk3O0ut3bJJWVbXx0z+mPHiZwVqlXLDLAXVRGQPNBtBkpY7LsrIZXNaQensrJGwy1BEbj0k7HClfTxRNw47lBmQ1bKw+E0AFQVdBIxwbjOfl2WPRB8UweOwW5o308tMXSY6x2QbdfSOpW9bnZCxTNmQPDcAYWVUunqZzE0EtBwFR1E6NmXDGyClTWNlhEYAypLfMxrCxze6wHsLXHKyqZmW5AOR2QanSWuOaQv2wd1S5QMpWkNAysSCtnp3kHOAsg1PvMo8Xt80GkxskmkAwSCd1qbKBzBkEgLVOm3RQhzSOpYks/iMLYdyghjphM4sB3Hmk1T7h96cck7KCljrI5y8g4KsuMUsj/EdlBC9/vUh6RhTNt3TGXk7qKnLGnDfpeivqKyRuYxndBj08bnzdGfNao2zeIBvjPzWBRRSCQSEbd1qpuJiw3PYIMaW1NpvjJzhadO8E9I+pZ1dXySNOCd1poa5x6kE0J93PiZUdRP4zsq5xL29A7hVZTO6SUEReOkDCqx+NwPNUMburGFNDASNxugtjqek7jKyH17XQvi6cZGFJTU8TXffcYUFdFEHdcPYd0Hqt7DlgFk4vu83VFi/MK5eoy8tfYbSF1t4yRns2awEfaK/wD1L1KQEREHXP2h4LuTXiUG9/c6PH8ep18+tnqGQmVjxnraAvoM9oWWt5OOJJd29zo/16nXz42mgnqnvkjaSIxkoLayje0uk3wTspaaVzIScYwFlzSxSfeM/GDhXwQRNYY34yUGDa2Gsqel/bK1moay3YcIsgBQUlsnjl8SnbtnyWvR0sE0PRWAdR2QQ6duEF0lEBjAycLelXp6kgoTOCM9JPdbFjpBaJjPAMDuMJWaruVTEYGE4O3dAbfKahrnwvaCAdsrS77doa0kRNAyfktLuFPO2Q1EwOXb5VLXB7xWMDvo53ygrR0skkhcYjjPfCuqqWQSdLWkBchmitcNva5vSH4ycLQqh9vcC0Edfkg2w+AwMDnLIpqRlRCXKS50NQ5pcxpLe4WNS1LqaMsdkH5oEdI2GUkn86vnkDBkEbfNYctY5zjg9z3UL53vyCgy4ZBUPDO2VuC06aZVSta9wHVjuts0T+h/V5haiL9WUkwdESAEHIhoqLR1L71IGu2ytj37UQvlY3wR0gn0V9Zfqy9U4gqSensVprKenpj1AjI+e6DclBSxUtOJXkE43Wm3Ksiqn+BG0AnbIHZYE13mc3wwTgbKkE0X03H4gEGfQ0MlpcKyaQlp3wSt10/EegiozTeEOrHT2+S2HV3OWVojeT05wPTCkordBNGZj2QalU3U1tSZm9iewCxaymfJGZA8gYzjKmtxo3yGJu5BWHeZpYZPDjJDUGjyOcHlp8tlmQ17Io+gjdY8nQ4dXmVjfWgyC4TSEgBWPJZluO6loYHzyYaCd/JZs9CIsGUY+tBpcbDI7C1ikdFBF0uAysYMiH0MZWLNM8PLQTsgurntc/LexKpTsc5pB2UbnNeBnusqOSNsR6cZQY89OYhkb+agOSs6mDppD4nb5qKsYyOUBuMIMbf5qWNhLc4Ky4GU/QOvGVe4whwazGCg01xdn4vJSwGpJDYJHD6iQAp6uFrWdQSm2i6mjcINUpquSjEZlkLiHAnddjuEeqKKloxUuDA5rc7gLrBSiWeUCQEgHZch6W1ALU4QuJDCMFB6RctXF+3T3BlHWBj2dYGDjGMrt7f7ZpzVlq6aSWGKV7OwwDnC8suB/ETTNnlE8kzRIDnuO65DPN5Vx8RaLT9rqCY3yBgAdnbKDt9p/hFXW28S1b6kiIEuznZbC43avn0hSPxWksYSCA5cq3nWN0oeHcF/LSDNCHflC6f8WK69a1tNTK1jnZz2KDrRxr4v/shrJaSnOXkEEjuuCpLa+ppZKiYnqJJGe65GuGi5aO+zzXSMgAkjqWwdZV7KOpMNGfgzg47INnOEnvLo3OJAJCjnDmPIycLKqpI+kSM+kVbCYnxEyYygjpIfEcclKqDw34B2UfiujefDOytfM+Q5cUFHR4Co1uTjspI3dR+IqkmGuy1BK1rYB1HzSQ+IMgKBz3HYqencwfS7IIopOg4I2VJX9ZBHkFkzxxub96G6xCxzThwwgnpKX3l3TnCrV0vuzgM5UcU7oDlndXGV88gc/t6oEU/h9wrZ5BI7LRjCzKhkHhZGOrC076kF8cbpDjOMKkjOg4yjXOZuFRzi7ugoBkrJp+mGVsrtw05WO3YqrpHEYzsg12vvENfTCna0AgYytHNP0nOdlC1xachXGZ57oDzjtkYVGfE4ZVpJPdSNikI6w3tugklicwAtOyyKN4c3pecnON1hOleRg+SNe8H4e6DUZjHAC7AKxnzNkBIarTBVSgZBIVzYhGOl/coMdpcThpwgLmPyTlSysDRlqiEUjt8ZQZ1NKJh0LLoaEwVzKhzvha7OFgUkT2O6sbqeavcz4SfJBubVV9pa22tpYWgODQNgtrUFSKeF3V3Jysd05lO+d/VW+DK5wABwUGXJchIS13Y7KE0/UfFbt5qKaERAE91JFI4xFoQZ9su4pj0POQpa64x1JJa0fWtCe0h2Ss6mdD4I6u6Dv37GF/VzR6o/gDW/yjbkVnsXiDzS6px/wBrf5RtyIPaBERAREQEREBERAREQEREBERBxxzJfg7cUsd/2F3v9RmXzQvJErjnfqP6V9L/MeM8vHFEeui72P+QzL5oH/wBlfnt1H9KC7xjjucq+KAzN6ySsc4zssmldJ5dggePjMe5xss6mqmxM6CACVCBT5JcRlQzjqdmPt8kGo0MfiVBfnIUF7aWyAtO2VbRzSU56nZA9VJVzw1LDkjKDTqcPe/paSVdUMljPxEgFZ1hpfFrPvg2HqsnU1M2KRvhjbzwg0WOYsOcZKy46aSsYZA4tAHZXWuljmf8Afcefkpbm73Jwjh2B74QYdK4U9Vh25HmVJcqjxngDtsseNruvxJAcH1V8sEk3xRjPqgjdT4Z1qINJUwkIHhv7hZUFH4reqMZ80GJG50R6uo4WQ2uAIJHZRyUdQXkBufqUEsL4Th4wgzamtjnj8MAA47qGCdsDS0gHKxRsq/E7f0QSOmPX1DZStqQcAjJKxiAMjzRpw4FBkT0xazxR2O6mopw2Itdv5bpNUMdBgdwMLDjbId2g4QZHwmYu7b+iyB0kgbFYL3Fg+ayKWZuMv7oJq2JoiyBusejc0A5wpKupD2FrViwxTPz0A4+SDIfIxr/JY80ofnCPhlzhwKjc0tOD3QXRP6HZUpqndQIzsoGtyQT2WoQR0zmdJxkj5IIZawyR9JJWO0HfAP5Fli1zj4y09HfKmEcLRg4yEGm5IO+e/ZXiUkgD1WVJBG4/CQVi9IbMG/MIMh0TmM8RwJH1LFc8PkDgMLWpoeulBx5ei0Ys6JQ1BnmbwoASM7LGax87xIw9JG6yxB40IGxVgIpR0nZANZWwsLHzkgbYysY1AfIHOHcqSZ7ZQceaxWtw8A9soNUq5waUBoAOO6wbdAKqsihd2cQFmTGJ1OGjGVi0svujhOPpM3CDft/0hBQWAVrXAktzj7Fx7TweL1b9lq1dq66V9KKSV/3sDHdaMyV8eek90E1O7pl6DuApqiURkALEjkAf1n7VWaQSHCDU7RVxtm+PH5FLeqqNx+9/LstHa2WHEgBAVfvtQcnOAgnpyal/QRlW1NOInlvbKy6AQscNx1KK6hxkEg7fJBzxymu+5+pYqv3jwyZRvnHmF7gcCrhT3Gy0kr7i172wjDeoZJwF89/DDVlTYLzG2OQtBcCMfWu/3BjmYvttrKO2w1MvxAN7n0Qet4cw7hwP2qvdcFcM9Y6gv1pirpush2DnK5otTqh9KyScYLgDv3QZyIiCySKKYdMsbXj/ALYZXFnFbh5a9QW6anpbewSPBGWt81ysopadk2790HQuk5WdQUeonXKkEkbOrq2yNk4g6TqaG0y2WuoHVErmloc4E79l3w9xg3+Ab7dlt67cOdPXmbx6uAF2c/RCDxz1LyS37WN9ku0NBKInkuDcEBbE1ryY3zSNtmrxa5XdAJ7Fe5lJoKxUbBHFA0ADH0QtNv3CXS+oad1NXU7HRv7joG4QfNpV092sd4fQVNqkj6HluHMIJGe+Vqs97e2A0zKEkkYOG9l6/c13Jbw/g0vJqLT9IyOpAd1DoAJIGc7LoLaOH+jrSyppL6+NtQ1xawOaM5+1B1bZRVcVUat0LgHHOFvDTmjavV1bHRRQu++EDGPVcqXnhfFNOZ6NgNMTkYG2FyJwX0dQ0GqKUTNbgOA7INl/tQLxQ2gXgWyV46c7NJ8lwtrfQFysszxLQPi6DjdpC939EjhfbNFRHVppWxlgHx4G2PmulPNfpHh1qupnfoMQPBef7Hg/oQeYfRJG7ofGQVe58jWnDjhcsa/4Y1dk+/mLAxnIC4zjtlQ/rJacNJB2QYdFUBkpLm757kKtwrWOIaAFNMynp/hdjqAWk1Dmvl27HCCOSTxDsFNTTmMhuO+ykbDGGBxx2WRE2l6QSRkILKkt8PrwASoYC58ZIzn5JWzNcOlp2CyLfG10JBxkoNOc+TJBcfzrIpKx0DvjyQtVttFA2cuq8Bqxb5BTCcGjwW/JBlQXaGX4OkA+pVlRURyOEe260Vgex2QDsr2vlMoznOdkGoy2z3Ue8E7YysbwPeH+J5BZVZLUClAlzhW0EMs0J8HdBQTMjAj2yAsSXqfIACcZVKinqYpCXg90jd0ty7ugmni6Yg7OSrYQHRkEKNtQXu6Xnb61OJImAAEIMSMnxt+2VqrWMMXVjyWBMIwcs7rIp5sRlr0GI6UNedlUVXYAK6Rkcjz0nJKkZSs8PLu6A9xqGgMOD8ioJC6Jhjdkk7bq+l6mTEHspK4xOYSMZCD1N9hr/aHGX/DWD9FevUxeWfsNf7Q4y/4awfor16mICIiDgPnvtou/KZxCtxOBNSUg/JWQH+ZeA1tbFYhW0b8OcXloJ+Wy99efaoqKXlH4iT0ufFZSUnTj51sAP5iV891HXSVFa99Yc5OT9eUGpUNjfPWGqecNJyrbxT+6Sh0ZyB6LOrbvHFSBtLjqxjutJhqn1pIqO58ig1KyX2Nh6JGDPqsa93vFWDE/ABzgKKeibSNMrdvqWiT9U0hduQg12tvvj0oaD8WMErGtda3xQ14ByfNYTWRPZ0t7qSGIw/H5hBuO70cNXSAsxn5LS6SmbRROecAjdRQ3d4+9vJwFDUVb5ZPDYdjsgyqa51FZMafrOO2FSrtksMoqDISBur6GljpgJSBk75UdbPUyv2B6AUF0uoGeEacs3G3ZaLNKZ5B07AlZdVDTOj+9kdawfClZh2MYQZotR8ISFx7ZKxZaYx+qkir5mHpcduylkkZM3uOyDBa4sK1uit7aqnM7sEgZ3WkthPUS/stQtc87axkQz4RIB9MIM600r6ipdC6MgZxkhYOoKc0lSI2k4+S5HNtpJLeJLcB45G+MZytm19mrJKrFW09RO2yDQKZjpQOr8qvlpHtcHNccei1ytss1BTiTpwPVadFHNKwvI2CDDqI+tobncKajrjC00pO52yrZiAceYWI2KTxw7G2coNZpbZJSu97LtnbrMqaSOqgMhcCQsSSrlkphGO4C0w3GrhcYySB6FBi1EZjmczfY7IyEyN6lqJjhkZ4j8ZI3WC+QNk6WnbKCW3Vgopep7chTXG6Nq/ojChqGQuiBYR1LCQT08pD9yVqDra2oiM2QMD1Wl9D2YfjHzU8VfUMHQ0kj0QY8jTG4s9Eb1DfKmqOgguHcpBSzTjLAcfJBLBUD6IGCFDUkk5PcqT3Z0BJdscKCV3W7O5+SCzLj5n8quiJMjRk9/VHNLWjHmpaboyC7vkIM6ujApw4dwFp0U5iGN1lVlT1Rhqio4WyfE7sEF8df4ZBDcfYt12yhNxpDIJMHC28YqMDfAwpWXyWiYYafOEGpwSXey1BMFa8DP90tT0rPcqDVlNquqqiRTyCTJPfC29a659XMTVHb5rLuFyMR93iP3s7HCDvjS858GrdHQaObVN6oIwzuPIYW1bjx4o7FQvpJyHlwPououmKqhtk4qISetxyVuyqZ+yPEbQ4vPZBuXV3EGl1XUyGmAaXZ3C4sv1lcWPlkJJOSt82fhVqKN5q2wP8AD79itq64dUWeQ0tSSHH4cIOPxC585iGTgqR1K6M4yQsigkhNQZHeZ81fcZo+sdB/Ig0+SHpA9VcKclvUsmFok2crah3hg9OyDCcOkkeioDvkqu73bdyrjDINy1Bcxgl2HkrXsdGVRriw7d1qMLYZYT1Y6kGBFKWHJ3CpLJ4jgQMYWRFSlshMow31UdQGCUBm4QQYV7XgNxjdZLo4wwZxnAWM6M92jZBaXE+ZVPmhBCnY1jmYPdBCTlUUoj6D8fZUfGXHLdwgjRXho7FC0eiCxO6ef2qZgYG52ygiLcDKyY6voj6OjOVBId8DstUpI6R1KTJjq/nQaS49TiR5nK3fYdKR11Ca17gCBndbYp/C94PVjp3WqN1LU0TTTUzvvfY7oK11ZHRTGn6AcbdlqFLYY7jRGsBAIGVoc74qo+M/ue6yaa91NJEaePPQdu/kgw/CAqXQns04yrpCyAYABVtTK3qMrD8R74UcDZJpB4gOEFwrQ0kdOFa+EzDxMrKrKaBkWRgHCxofE6CG9igxQSx31LUoqhgiLukZHqsN8DnnYbqeOIdBZnchBGf9tydPZarHaWx0xk6gT6LSTDJTO6xnHdStulQR4R7HZBizn745o8jhWxteSMZxlZjoGgeI/G+6uhEReMAIO+vsW2lvNHqnP/ACt/lG3Isz2NDGN5ndTkY/+IVb/KFvRB7LIiICIiAiIgIiICIiAiIgIiIOOuY/bl54oH/uMvf6jMvmicwyTyBu+HE/nX0t8yRxy7cUj6aLvn6jMvmut3huqpvExuDjP1oMEMPX0nusynkEOI+5Kx6kdM7i3yKpFJ98DneSDKqKBzWmYE4Kx4JXR5+HK1YSiogDArYqaCNp8fAJQYPje9YjAwcKCeF8LgM5zusySKMO+8YJ+Sp4Ly0mYdvVBWguQpseRVa65e9uAzkrT3tzIQ0K5sbmuHUNgUGXFJ4ADxsop6l1TIHbnGPsUkw8SEMZ5KaigijicZ8A+SCKbD4OhvdKOsbTMMT27nbdWQytNUQT8Pln0VLiIusGIg/UgrNT+K4yg7HdZluqGUzCHHJC0sVEgHT5JHLmQdXYndBqrrpGx5JYME98LFqpG1z+puBhX1LKYwAtIzhYdOyQnLQcFBT3c9XSc/Wr2xhh6c91PI5oZt9LCw/EIeCSdkGTLQdLfEzsVbDQmUE5Oyumq3SRBozkLKt8Uz4z0g5PZBiNt7i4jq2V7umjb0HclVngrYZC7Bx9SwpZHyO+PuEFZMyO6h5qgjceyyadjXDCmLGMG6DAYwvd05/Os6nqWUbcEAk+vmoZelgLmndYznOkOTvhBmyVscj8gbeitNOJ3AjbKw+hyljklZ28kGdLbhHDkd1ZS29xcJC7Zu+FD75Mch5OAhrpGgtjOxQa3W3aBlMIABkDC2+1zppg3OzjhWPEjyXuV8LHDDx3bug3DV2ZlLQCpLxnGe622XOc8OA81mTXirnh8B7vg7YytVtUVv8AdSZsdWEEVNMJIAwn7CsGrpfjD2t8/RJZwyqLIj8I2C1Omlgc3plIBI80GnMqTTt3Cw6if3hw8lrVYyj6C4uAGNlokvhl4EJzugvbD0N6ifmrugOBIG4VrqWrYzqcCGq2J5DC090FrHPD+nuqSdRfjtlZNujY6ozL2Ul1ZEJR4BB+pBB7kSzrGSovAOcHZZ1B4mSJvoj1VlwLDMGw+Z3QQMonOwQdir/ci0gk+a1GOB7acPI+tYoqQJQ13bO+UCZ2Y+jpxhRRzMiYW+fktTrTSvpR4OC/HktFhpKmpmEEbCXnsEFYRM6YOhYXEnt3XK2iuGVfq2IMNveS5pxsd1qHBHgTqbVd9ilqKMtpgR3BIO69I+EPKxJQVdDI2nAjLR1fDt2QdSOBPJHedVXlr57bMG9YAJae2fLK7hW/kUrtJVlNdmUzsRYJx5YXdbQXDSzaNo4TSwsE4aC4taBut5TRMnjdFIAWuGDlB1ks2sH6CtTbO+mLXRAdx6LbepueG16KgkZWSQgxAjDgNsLmXizYtJWyhlra1jGvMZO+AV5Cc2eobZNe6mntjiWGUj4T80HfDSntI7BqO5igHu4y8Nzgeq7VcPOJ9DrmCKankYRI0O2x6LwK0Q6jtMlPXN6g8vBzn5r1V5NNVyXCipRuW+GBufkEHd4HKqoKSobNCCO4U6AiIgIiINta60rBqy0vt08fWHNIwe266J8WuQi4Xi5yXy3QPbGJC8tbkDGc+S9EVjV8scNHLJKQGhpznsg8otdcOI9CWptrqaYtfE3pyRvstnaUoaKB5uoqmsMLurvjsuxvObd7PTx1D6ZzPFDCfhxnK80Ltxav9tkqqSne8Rl5Gx8soOw3HHjZerzZv2O2a8PYY2lo6HkLhnQ3FS56Qikbf7g6cnJ+N2f0rY1g1K+8V5lrHOLnH98VtviFUt99EcWwJPZBy7q/jDbNYRupWdAJBC4ouN1it5fAxgPiHbC2sxs1LC2eIHJwStdtstHWUjpa4jxADjJ80Gk19O6oPvIdjIysOOlPWMnzU8lWfezE0/ADgKGpmLXAsKCarZ0QjpOSsFjJH7jOPrWVTeLUO6XdlnC2vY3qxsg0+jpXTvId9SyJnmgkAA2UzZI6Z2Ad/ko52ioPW7yQUfWGsHhx7FVZTuiYfEOSfMqynbHE/busqRsk27ewQYGAJNmjCzqaja9wlLRtuqU8LZHdJ7rNY4QvDDsCgjrR73F4DWYIHfCso6plpjMbwMn1WsxCkjYZHdyMrbt3gkqJ+uFuWoLpbiyskIDQM/JabVMLJMDsVlsbBAwb/GFdiKZhJO+EGmEEAFXshc9vWD2V72fERjZTxOYG9A80FbVSe9z+GfXCzLnbxSkMae6x4vGpHGaLuVFPXzVEoEmSc4QIaYsPU7ssvoBGQdsKOfrEAc3vhYbKpzWkO74QS1X3sfD3WKGue0vLjsMqemPjSYf2KurYvD2YNvNB6oew2bi3cZHes1gH5BX/AOtepa8tPYbZ+53GTP8A82sH6K9epaAiIg4F563MZyn8QXyfRFJS5/jkC+d2V/XUVPTt1SEjHpkr6E/aDukbydcSDCPjFHR4/j1OvnkjLi/O+UGbDFI1oL3Ej61cZSx4kaNgdwrhBUSxgMBTw+iMsl2cdvtQX1l4E8HhA4IGFiU5zGc9yCon0U7B1FuxWbA1ghI8wOyCGjYRMXHfdZMsvVIIgMA43WPQu6ag+J2z5rJqmtMokjI2QW3CgMMAmBxnfZLbT9UJmcMkbqOurHvhbG7t2UtDUsZSlod8jsgOuHW/wWnsp2VrHAU5AJdtlaKHhs5f5ZKnhikfUtkAOA4HKDMqbY+jHvRzg7hRRTNqPgx2Wr3qqjkt7YgckDBC0OgfFE0ucQCguqqERt624GVFBC76WeyVFY55x5AoyYCIhBSpl2DW9/NZ9tkaIT1DBx3WnU5Y+X75jGfNZMzwyPEO6DeGjb17jUkzv6mA9vJV1ZqqnfcWvjYABvhbVslUIZyZzgZWTdmU1XMHRnOT5INYrdRw3OjbEAAQPNaWypjggcw4JIWDPAKaMeGMbLD94Jb8WcoLJpz45cN91LHWta3duCraCmNTKdsjKlrqQQHoxugloKrqkIduPqWPci3xgRgeqpTQSROEhBCtq8vPUOyC6F7ph4fkoqiExuDc5VaSRsbuonzWRIwzODxu1BjMgfsXHYrNhtXiNMwOw3UVQ4dAazuFLS1z44TE/byQRS4kJhaO2yhLRBlpxupWva6QuGMkqs8ZePmgxmQmV/fAK1Clr2ULfCcATjutOc2WE9W4ClhjbN8b/JBk1M4qclu2Vp7mFj8d8LMc6NgwFdEYXuBeRn5oIvAMjMEfb6LHex0RA7LWm+AG/DhadVwOe/I7fUggijdUHp9FM9wph4Y7lVpj4JydjhR1Z8R3UEEIEjt+o7n1WdDQExmUntusWlcA7DuynnrXtb0RnY7FBNRO8WUxNOCFWbJrG0xcfiIG6waSV8chc3uqzTu8cTA/EN0G7/2Nvo6VlW2QHIzjK1XSWovubd43zfRa4bYW2bRf6qpcKaodlgwButZp4qT39kmRgEFB2u05xJs9VZxT+6s6yzp7fJcIcXtLxX2d1xh+ABxdgbLc+iJrVNTCOMgvAC2zxHluviGnow7pJwMIOCq6lfQVBh6u3mr6KlfVyDqd5rW7pp24McamqYRnfstMhJp3Yj7goM6qtraODrBWlOZ4w2Kz6l9bJEPFB6cea0tspZKGkbZQDGac9Thsp/GbJEThX15a6AEd1FSNBhPrhBhP+mVdG9zHh2TgHKudG4PJd2T4MYQZdVXtmh6GjBWBnfJVMFAN8FBc6QkAZV7Jg0dOFfGI/NSuZD05BCDEc4O7BXxxOH3zyCjdjJA7LLhe1sJBO5QB01OGA47LLjpWRMIJ3K0xsjmPJj9VJ75K47oL6mHoyQsXPyWXK5xZ1O7LDOx2QVIwgacZUkJaXYedllGJpjLm4wgwmsycKrevqDA4gKRuziAjQDMB55QZDbY5zA4O77qz7nvDwCSQStREVVFE1zmnp9cKgqo2j4yMoNOqqd0DQcqkbsxHPdZb2urCWs3WHLG6nd4btkEAcQ/JWUysDNgN1jyFuBjupaZsWxeUCWZ0x3zhXR1QiYWEBTSeAGnpIWDJhx2KDIgqAZNx3WrxUjHR+OSBjdaLAwdyFK+4TMBjacBBlTzCYmJjQcbbLDbTFsgcdt+yzrD4Jmc+pIAKjvc0QqsU5GPl2QKpoliDWncLTgXQvxlT00xcSHKypjJkyBsUHf72LdSZeaLVDDnA0BWn/nG3Io/YsMA5pNU47/sArv5RtyIPaZERAREQEREBERAREQEREBERBxzzI78u/FEeui73+ozL5ousxzPLcjcj86+lzmS/B24pfwKvf6jMvmjazrnIPmSfzoL5AS3rPcqyKAyDIU88ErYs42V9Dswg990ClkML+k7qSse+cjpJChpone8nxBtnzWVW+G0DwyEGPAfdvjec/WlVXiYYbsoRBU1J6WsJwrjSOpxmoGCfVBBG7pf1n61fLUB/YeStjYJJegdlkSUzI9j3KCOmqBG74tx81kzu94HVH2CxDTuHxY2U0UrWMLUGG4FriD3CoST3KnEZMhc8bK2cM6gGbhBHnbCBud8qZzYww4IyoQT2CAXO8yVlU9WIY+kjdRxQOBDnjAVtQ1gd8CC17+qQv8iVI2PxiMeagWTT9Qb1BBIyl6PiJysynubaQgdPYrT21Tw/DthlSSCCQZyCfrQZ9Xe4Z4+no3PnhaaIfFdkDusZwAOAsiCoawAO23QSBhp90a8zkA7K5srJDlxGFDO9sTx4eEGTNR4j6s5UMMIAPZRiskIDXnZZEdREAR1DdAp4myP6TgJVhtOQAPzKx8vT8UJBPyUD5JJHgzZxnzQXM/2w7oDcfNWzQ+A8A7rNkMMUIdEfiAWC7xpviIJwgmfI3wdh81SCRvQR6rGJcPhPkrmuIGyAB98/Kqvc9p+kQPrVoJJw38iOBx8QwUElMSX581Wpe9sgwSNtlfQAGXfstRqaaFwDtsoJrDaXX0iB0mM+pU160t9w5WvMnUAQe60+G5zWlwNK7B9R6q2svlwuh/2w/OPtQZlRcIZKYQADOPILToqXYuxsogAw5dkFZkVTGIyMjJQIKYSu6QcFTS2tsI8Rzs4WmGpnhkL25A9VcbhVzkNzn5IMls4leYGjHkCsWeM08wJOVmU9I4YkaNysKtL/ABx4mQEGpTV7vdQxkZJ+paZ4Ms2XOYWnyW9NGQWirc2OsLe3mrtY2qmpagOoMBnnhBsljJoXguBIB9VuLTboY7rFVyNHSzBOVbI63up+jqHiY7fNaeJXU7j05weyD0G5XOJ2lZBBb5ooWPDwC44+S9T+GdXpyrsVO+ilp3SlgJGRnGPJfO5oHW1x0zVMnpHPBDwe/wA13S4N85l4tFRSUNXUPAwAcuQexHiRAf2RoH1haffbnS2+0VdbLVMY2GMu6uobEBdJrhzex0mnxXGpIJbnPV8lwDrXnevFzgqKGnqXmKXLXYd5IN4cx/M5UXXUFTpuluJDWgs2dt5hdAOKVxmqrjLLPVeIXPJyTlbw15rq03mtkuzHn3l+Sd98rg7VN1rbhI95BIyT3QZsupHUFPEGSAEOHmu93Jfxwnt0lNRF5OQB+heb7ZXTyMjlzgOGV3T5UdSaK0/XUkl2kYCwDPUR8kHsbw21O+/29tQST1Y7rf64T4HcVdB6htkdBY5ojLkNwCDuua0FUREBERAWj6rY1+n63qk6OmIuznHZas5wYCTsBuuufMjx2t2jLNVUjJgHlpZ3QdBubfUkx1JVUZrepoJb9L5ldL7jQw1UdRKXgnJOVyTzCcSTqS/zVUDyS8kjfv3XX199uQ62ZdgkoMqjrHW+4ENf2OMqC+zGunExdnHmtLkne55kJPUe6uEkkjDjOEGQ65N8ERYBwMdljNfNK7pa4tB+arRtj8bE2wHqsmrdTxbwuGfkgsdRmNgf3JWVBbfGhMru49VBb5zPKIpDsSsm4yT04DIc4O2yDDjqW08pYR2OFyBZrVT3G1Omc4ZAzgrjR0UriXFpJO+Vrtpvdwo4PAjLuk7FBpt3idTXGVjc4B2SGYvHS4YWfII6mYyzkdR3WnVjo43kQkZQJmmP4gfNZlNcmCAtIGcYWl9Urx8WcK5vSG9PmUGVS1eKku8sqSprS+cALBgYWu6nA4UkhjIy07+SDWmNdUQjDx9QU7PDggLH4J8iVodHUzsdg5xlUrK2UuwCgirY3CZzmk4JUUcj27b4WbbcTzffztnzWTW09KyUBmMfJBiUsLqk9PSVJJQvhkBAOFqNPHEGjwsE/JZR8IMIlIDvmgx4IWSxhrsA4WjXCEQVQ6RtlZVbJURPLoQceSipopao+JMDt6oMsYNOAR3C0+ek6gXjbC1OaSGKLoyM4WkS1RLy0HbKClMcSdPbCyKuQNiLO5IxlQu6GN6291R0jZIHE9/JB6p+w2I+5vGQek1g/wCjXr1LXln7DX+0OMv+GsH6K9epiAiIg4A59en9qPxE6gCPdKTv/jsC+eSlibJNKfIE/pX0K+0CkEXJ9xHkPZtHSfr1Ovnigle2T4TsTkoN026SNoDXNB29FZXW1k8gnY4ADfCxo6qBsI6T8XnusWe6yMaWAkhBJcK2FkQgAGRssOmZ1sLurA7rDd4lRISBklXNM8B6d8FBkY6nlrfyqY/eYi4nJAyo4vhHW7uVc9kk4w0EgoNY0dp5urK40jnAAELWNWaCZpppayTK27aLrVael8ehJDzjspLlrK5XicCvcSOx3QaIKR7pi0ggd1mwzsgeIDjJ2WRXPhEAkiI6yFo7WTyO8Yg7boNWudKYoRN15B37rS4aYzMLwcY+akNZPUtEDzkBRtdJFIIxsDsghbGXP6FMaYjACyJ6YwRCbzKpSwzz/fcEgIMZ9K9oyAVRkj2H4gStTMsHT0uIyNkMVO9hI7oNLkmL/ojCvgqHxnLiTj1WsWWlo5JiJsYyse/UsEU4FOBj5IIHVxmb0YyfkFiSMOcYwtUtNLCDmbG6juDYRVtbHjpJ7IIKKqNEeotO/wAlbVVvvMwk8h5LVn0tKacOOM4WkSsha7DCMZ8kGT742aIR4wQsd/wNI7q1wYxuWnf61A6QvOCgo5u5IKmiq/DjLDnKhd1gbg4U8MUb2dR/SgibKQ8vJ2O6tkf1uyFdM0NOw2VjGlx2CC9mYzkkrIiqQ+UA+eyiMMj2gAdlGGuhkDnjGDlBq9xhY2nDwBn1wtLimDWFvYlZU9S6phDGb4CwXQyN3c3CC0kk59VNFA95BBKpTQumd0tG61PwHQMOe+EFIqUhu7jsVkmJhZlwWBDLOZCDuFO+V4+FBhVoLHHp7FY7X7EHyWoRx+I4mXzUFSyJknwkAZ9UGFnzV7Dtup6hsIjHSRlY4jeRkA4QZNKzLs5U01F4h6s+SwWyvZsPJSsrJcgE7IM2liEZAbt55WtULTLKGeJudgtBke/w+phOfkFdba2eOtjLsjDkHKFprajTUYqHSnHdZkmuG3KZr3wdePPGVjU9vk1DQRxxDJ2Gy3PZNOWOz04bc+lsh/uh5oNq6ku1NXUnSIQwkYzjC42naIZjKNwDlch8Q32+GJ33PcPPGFxhHJLI14lzug1GW7RVEYhAGRsoxaxLGZ87gZ3WDTQDxSSCp5q+SEiIdsoMGokcXGPyacKsE/hjCzMUzml5xk7lYc8QLi6PsgrNUNkGFRtN1MLs9lB2UsczxhvkUFhyCRjsrTusx0cfR1eZWI7HVt2QUyfVMn1Kud04GO6q2Nz8YGyCxMn1Uz6d4AwMqRtDK6IvI+aDHjeGHcIX/EDjspIWtDy1+2FV0PVIOgZBQXeIZm9AGFcKAluclSzwsiiBaN/NUp6rbpeUGLJCYypIqjpiLCVJVDxforE8NwPSQgB+HE+qzqCl65BMT2OcLFMfSB1dllwVPRH0t80GtVdxhdTiFmMgYW3Z45C4uBWoU7DKSXZ3VKpgaekBBBaqkQvPX+dR3OZs84LeyinjMJyNsowscwl2MoApi5odlWOiczOD2UkEx6i04wraiQl2AdkEJLvMlAPMqSAMLj1kBXSxFzh4YyEFBL0hWN++SjPmhgkb3ajYpM5AO3ogzJ2uZGCxxBWC4uccuJJUj3T4w8HCjG5GUFWPLDstQp+mVmPNYUgZ0gg7q6nn8Lug9BfYxU3g80WqZPXQNaP+cbf/AKkVvsYKsTc0OqG+mga0/wDONvRB7PIiICIiAiIgIiICIiAiIgIiIOPuYaIVHAHiXAe0mj7yz8tFKF801RAaW5VMPfwZHD86+lbmLkMXL5xPlb3j0benD6xQylfNOZnT1k8km5kLifrygzZJ45YQ3Ayo4IA1hd6LAMxa84zgFZEVXtg/nQVFS10nQBj5qyqkLXfSzlVfCZMui7n0Vnu05kHitOMoMu23NlM/L2flUlyljuLg6PAAVTS0xgHT9LC08iWGQdIOAgvbAaY+Iew81HPUmRwLc7LMqJY30/TtlYdOIcnrI+0oMqnkbUt6MYWLWQOgkGDlSiWKE5jP5FRkraiZviHbKCNkhmb4Rbj5hRyRGI7nK1usZQxUnVFjr+S0F8jnnLkFDnPmjSWuBx2KmpYfEfg9llTUzA0NbjJ2QQyVbXQhjRgrGY0vcGnO6yJbbVQs8VzD0+qpThnT338kEs1u8KDxerO2cK2kwYznyVZ/feg9eQz5qGDr6T09kFvT4kvSNlSSN0Rxkn1VOpzJM5IIWQJGyD4+6CtsoTcKgQ5IJxutxy6GDIvEEp7ZW3YKv3OXxYTgrLl1PcpQGdZwEEFwofciWB3ZadknzU89XNUOLpTklQAZQTUtOamToBwtX/Y4RD4vUdt1ptE50Uoedt1qU10qXAMYThBpBDqaYtcM4O+QqTSeIQQFl1UbXtLz9IjdY8bYw057oLadpkeG52+tZMsjIAGgDdYfWWPLmFVL/Ex1ncILHnLiR5oM+Suc3bICyaUQY+PGc7IMdgLPiI8sqksnWR8lnS04OcDbCwZGdJAwgmoQPEyThasyMSStiJyCQtDY57XfDv8AUs2GrkjkDnEgg7INYu2nmwUnvIOSd1oFI4NJa71WqS3ueoj8GUnoxjdac6illPiQNJH6EF9ZHhvUwZz3VlDSPnfkkgfpWdYQyoq/d6rGB6rJ1DGKCVraTYH0QY9TRs8LpbgkKlHRMjjc92MjPdRUxqpPpg4UdVVSxyCFmf8AWglFzEDywAYClbRi6t8QbEKSk0vWVsYmDTgjKsn8eyu8J22dkGI9tRaZQ6ORwGcbLf8ApemZqKjDahwcSMbrYFRWRVQHWRlatZ77Na4emlcc+WEGrXnQptk7qhs2RknAK25UsLScDOFm1eo7xcX9Ly4tJ3V1GYAC2pIye+Sgw6GvdTvGRjK3BaayulucMsFUWgEHvjC0G7xQRx9cOFjWu7PpPiLtx80HOWuNYXSHSjaeO5nqDMfS+S4ctup7o9ssctW9xO+cqG5alqLjEad5JHbdafTGGFpc44JQZUlbXNqDK+dxGfVJ7w2RhY5oJKx45o5JCH9vrVtRQSSuMtO3LfPHkgxXS5lMg23yFrtlrrzG4zUVc+LAJ2OFokkTQ0RgfHlaraKhlLG5kmxII7oO/fs1dWXqv1vDSXW7udGJ2jDn7dwvY9r2kbOB+1fPhyrcQZdB6mjuMDyPvoOxx5heuPCbmGh1G6kinlBc9oz8Q74QdoEWJba1lwo2VTOzh9ay0BEVr3tjaXu2AGSg0fVNzjtlulnkmEYDDvnHkvJTnb1tVVlfVR0t1JHiEYDvmu//ADUcVbZprR80FLUZn6HnAOCDg7LxK418R7pqa8VZkc8t8U+edslBse+1E8swknlL89yVpbWRywuIaMjKVU0lRTtJyThR0pcyNzXeaDSXxF87m9leXGn+DpP14WS6FzZTIBtnKkkdSGI+KQHAdkGnxxuqJPgyCVqlPp98uBISCVBYZII63qmIDfmtculY4uD6I5A9EGFLYxbB7wX5xuN1gPuLJngEZ3x2V9xutTLH4UhIytLgx4rertlBuCGlZNGHFoCTGOljIAGVZLcI4acNjIzhac6WWoaXHcIIqioPWS091kUtsfVxGcknAWAd5MOPYrWrfXsgiEZIwe6DS55OgmLpxjZY+d8rU7iKd4649yVpjcDIKCYzdTekDCtijc6Qd+6o1od2WdBTu6esDcIMsQMbDnYbLSak4fjup6iqkblu/osdrHTPz+VBnWaH3mXw848srcc2mGmPrMm5Ge627Tl9CRIzYq+XUle4dAcQB80GX4X3PlILwQCsWqqTPJ1NdgA+qwxVz1DyZTnKinkLDhpO6DW4Jo52eG7BKx56v3aUQMZ3OMrAopJGPDt+616JtDLH1y46wM7+qCGS0meHx+vGd+60wW0+KGk9ysituU8LixmfDGwUcNY6Rpfn4h2QTXC1imphIHZJ3xlaKGuIJ3wFqJrKqZxZPno+apU+7tgcGEdR/wBaD1L9hr/udxl/w1g/6NevUxeWfsNf7Q4y/wCGsH6K9epiAiIg66e0MaXcm/EpoOCaOj/XqdfPXRgEyH0avoU9oa4s5NuJTh3FHR/r1Ovnopi4FwHmEGVaaZ1VU9DnYB2Ut5oTTP23CxI6l9JJ1RHBzutQFbHVwHx/pYQadRStilJcBj5qWrqGPcHNA2+SxSR4xx2yrx0OcBnugkikM2GdgtYiEcdMRtnC02SNkEYeMZKUM8k0oa7JblAhnPvDutuB81ZVx+LL1RjG/daxd6elgpBJHjrO60ykka6BznfSCC6KBz2AOfn1GVLI6OCEsIGeywoatwnLSdj2UlTTVU/3xrSWhBjU7wJy7bBK1qO2sli96GO2VoLIpHSdDR8QK1mnrpKaHwJOx27oLqVrbnIaTGOk4WoTRxWyL3XYk7ZWl0Mvu85miO5PcKa4GpqszHJwMoNMraR7CZg7IO+FSkLnsILirDUySOMch27YKtDzHIGs7H0QSukkp3FzSRt6qx1Y+U5eC4rUG0Lqhmcd1R1LTwsLHgByDCNa5oy3ZRGZ0kgeSSQrpKR3USOykhgAB6ggnhmfUtEfUQsSrpzC7HUpJHCn3j7n0WPJNJO74u5KCWGEydzsop4/CfgKVokhb1E4CticJqgF/wBHKCN8pc3BCo2R7RgdlrFxhpPdR4OOvG+FfbKKnkpnGUDqQaI9xcASMbq+IhoJI/KtSFLCyc+JjAysOsMLZOmI7eaDJtbhM8tLchR3aMNlwBhajZGQRYfJgZUF6ayafqj3CDTYcwkPcMrIllbKwux+ZXxwdcYBUU5ZC0szuUENJUCkmLiMrPkrmVADhstMiDZJD1dirpHeE7DOyDIdWtjOOn8yljlE4LuwwooooZGguxlXvfHCA1vmgVFQGDDTjIWE7MpLid1fUZIyVC15aCEFDknBJKmbUBrOkt3VsDOtyy308TWEnuEGJDF47z5I+Ixyhu57KjJDE/LewKzWuhlb1ux1YQZlNA0xgu3ysVwDKwYbjfZVjqnMIBOBlXv6XtMg3PcIOaOEtXSsA96e0g+RKzOJdtuFVKai1wvLO+2cLhmw3y50lTC2m6sdYB+rK71cGINGXbSYm1M6IS+Hkl+O+Pmg6a0tsr7lOYK2J7QNjlaDqO2NtchZGARnyXZDi3FpOkrqkaakjc4E7Nx/MuvN4kMpea3Z2TjIQbZbU+GfrUMsnjSD5qWpax78RHOFA1jmHJG4QSy07o2B/Vt9aRTBrC07nyyq9NRO0NAJAUMkT4j0vGEFrjkkqgOCD6KrBupCGEY80Fr5S4YyrE+pPrQSwQ+KcZUxIpz0rHbK6M/Cd1R73SOy7coNSo5GVD+l2PtU9VWRU7TEBnPotIjkfAQW7Eqkj3zEvdugo9/U4u7ZWTBIGsy7GVBC0OJB7qR8EmPhzhBf43jEtJVRRkuBBwsYNkYdhgrUIWTuh6gDnCCKRogG5yVjmcF3VhVnbMXnxAcZUXSgvlm6wMDCsY8tOclGsLvojKeGeoNPmg1a3ztcN9tlDXVAbLscrGLnU7QW7ZUJk8R4dIUGR4nvPwrHkjLH9Ge6me9jG5jO6x3Pc93Ue6CQsLAHZUZJJyUL3EYJTbHzQXRsL8gKZsghOCM/WrKZwa4kqlQ4OfkIJpatrxsB+RWxVLW9wPyKOCPrKtmaGvwEE1RO2VpwAsbCLVaazzVNMZWNJQaVuVVrS4hozusuKmMUpbUNwAVmmGmx1MxkBB3t9i3TOh5otUPPnoCtH/ONuRS+xhkL+aPVDfIaBrsf5xtyIPZ1ERAREQEREBERAREQEREBERBxzzHnHLxxSP8A3F3v9RmXzRDJnfj1P6V9LvMeM8vPFAeui73+ozL5pG4FTJnyJ/SgyaW2e9E74UVdRmikDc/nV0NTPC8mMnH1qyrqJJ3h0ncIM+la2CMSPwQd8lR1NxjkOGNGQPJRSTgwBvljZY8EQeC4oMuF2dydj81meAySEuwM422Wiulc1xaOwKzKWv6Y+gk57IMOcuEhaSdlfFSOkxh2MqksT3yF4GQUbNLA8Z8kEz7e9jerOw+ahawsB9Qs+OqbKzB7rFmikzlo2KDG6nuPSXE/aqFmH9KnfGI25PfCga74slBrdtoWuaHHAyo65ogqA/uAcrHjuD4mANPZHVPvDT4p3KDUK29wTUQp2tAIGM4WgxOLXtO+xyjmnrIA2yqMwHAu7ZQa3UVsc9EIgwA49FpcTzC0jpys6WSlFKOgjKut0dPKwmXGUGk4Mj+2MlXugcMbrU5mUzXERYyFiSODe/qgx3wFjeo7qekiDoySBso/F8U9IKuMpgAbnugjlAc8twAQqsiLDnurcOB8T1WbSdMuM+aC2nZ4zg3GFkyNFO3BAJwpKmNtNH1x4BWJHVMmafF7hBE9xkcR5f8AtUT4DkYP51Jlrn4YfPbCo4uZIC47eaCOWkdGzqwSoGg9QGCtwU8tLLGGyEZWPVMpGZMZGUGBJD0x9XmpaK3OqB4mcAK4RyTHpGSFY+omoyI2kgFBkzSCFvhYyRstMld1v7YytRp4Hz/fZMnIWLWwtjcC3b6kFkbBH8ZKq8+K4uG2FAZHOGCkbi04zgHugklcSAMHYrU7Vco6eAwyNBJVaenhkiDngZKxqima2YFnbO5QUY99PVOqW5A7qWpr/e8OduR6rLeaX3cBxGcLSZGDJEY2QZtPcWEhgAH2LGqyW1IlG4Byoo4+ncd/VZ8NL7wPiGQg1+2a2p6KmELoxkDC0i71jL7VNdGAMnK06uo44BkHBHfCst0hjflvcFBqkumDHB4xedxnC0xhNLJ0ncA4WoVt0rXQdG+PLBWn072zu++dydkGpMrIoYustGT8lpVXWOmk6muICzp6STw874WB7m5xy0HGUF4qXzxiNxJwO5ULoHNOx7qaWNkEYcO6pDHJMwuwdvNBB0OZuVPHSOnb1BxGFHHkzdD+2cLNMohZhpA2QYDoXNcWgrU7fdW0kRp3jJO3ZaYZHSSbHv2WUylBw93fugmkpMPNVtgnOFH4JqZAWnA+SkDpXN8N2ekKF4mjJ8L8yDc9ju09hDH07yHAg5C7IcIeYC5WSphqprgWeEAcF2Oy6waafAanF0OGZ81qVxuNJFdY4La8lhOHdKD2R5ZecGn1lNHZK2uaB1huS5d16Wrp6yFk1PM2RrwCC05yF4S8GrbqEUtPU6RbKaovB+DOc/YvWnlYl1m/TTW6vbKJWwgDrzny9UHPCiqojPTyRN2Lm4UqIOk3Orw0r49OTXsVBLHMecdXbGV40a4ndT3Kqjk3IlI7fNe5HPFqRlu0hJR1DgI/Bcdz6grwy4h1VNcLzVyUmCBMe31oNAZcPhADcj6le2d8hHSw9+2Fl2a3QSsaJCMlbht9lpKi5RUceMvIAA+tBtSrfUsjy2nOD54Wp2bRVzvlK6sbSvDQM5APZdgqLgjFHZ4rlXsHhEAjLdsLmnR/DXTTeH9TUU7IzIyM+Qz2QeflbaJqOpdT5Ic04IzupIKt1DGY5dyfVa5rWknj11WUcQPSJiBj0ytIvNC6Fw6++UENHTi71JZnGSrbtaDbiAHE/arKQz0b/EhzlSVNa+qOKlxJ2QaXlxOMndaxQxAUpy3OR3WDJC12DH6+i3DavAjoyJsZwg2tMMTOHbdXMje8Eg9lPcPCNS7w+xKhZIWAjKCMuc07lapRWf3yHxckbLT4C1zvvnZaiytlgb0QE49Agw5qU00pb3wshlaGM6cI9xlPU/uVfGynDD1kA49EGBM4yuyB3WqUFGwQGR2MhY8cLJHkM7Aq2eaop8xtzg7IJi4SymILFrYBA7yV8Ac0+L5lQVcj5XDKC2OUdgFR46379layN2c4VHlzHehCDUGNaIx2yoS2UyjDyBn1UUMrv3xOEMx8YYPyQanPTeLCAAM47rHhpXQAvI2CzhMxtOCT3H8yCRksBxjKDS6iqbJmNowfqWKWPwSXbBXTRPZK78qtLn4wc480Hq37DZuLZxkf/dT2AfkbXf616lLy09hsf+p3GVvpNYCPtFf/AKl6loCIiDrl7Q/8DXiX/idH+vU6+eqlcGvIPnsvoW9od+BrxL/xKj/XqdfPNCx73gMG4QZr6HqHX6qF9HI0F7TsPmp5zURRDOcY2SjkMrSx+STsgxqSn94lLT+dTVVD7u4EH5q6ZjqP75Htn0UHvUk8jRIUFks7nN6CNh2UlLUeC0+v1LIrI6dsAdHjJCxKZgeSXdkE8lTJV/A4nCoyMx/ACcH5q+YRRs+9fSWIXzE5JOUE8tN4IEmd+6zqa9xxU5hczJOx2WnZnlHQ4ZCtayJrwJD9aCanqGxVJnc3YnO6jrqn3iTqbsFNUsi8JpZhX0tkqqiIztaekDKC+zgyPwStQuFeymaYQBvstNgkbRylvmDhUqZop3dTu6DClJMhkHnusiCn8Vvjk9t0cYSMd8K4SFsZbH2O2EGpWy4Rh5hdjbZY90iL5hIw7ZzgFaX98hcXAYWVHW9YAegvkqmtjDMbgYOyjbICNlFUua7dqkpmtMRHmghmfnOfVWQAOmaD2V7qSZzj0tyMo2lnYesjAHmg1arpmvpR0bnHkFpTIXMz5FZ9FVGTEbz223KguDS12WHA88IMZnWD8TiR9azoaggDpJC0+Lqe/p7rNbRykfCDuEFlbUlw+EnJUdNRunHWT2+apNRVDd3MOAsihdJE4Nd2KC49bQGNJGFPE0OYS9386juDmMj6mjcrCp55nO6QSQg1KnIc8txsq1VpdMfEAPZS0fhMAe/GfXCzPeWucA3GD6INIpbS5zyN9ljXClNPIG4ytxy1UNPGXjAJWk1VRBUuLiQSgwII3YznuoJi4PyT28srUIS0khvYeSw60dMuEEb5utuMK1rOoZyrFUOcNgcIL4ZPDcSVfLU9eME4Chdj7VRBUAuPburwx7SDnYb91LT+GPp4VKj4zmPsEFWOMuB2wpWyOj+AglQ0jHOecd1qvuzPD6nAdQQZ1ifHBIx7mgkuC514eMvGpKmGzUs0jGS4bsSBhcEWNoNTE1/bqH6V294LQWelpo6uLo95a0EY75QYHE/l9uOhLCdSPe+oMjC/GSfLK6vXOkqLxK8ywmIBxByML0io75FqOmlt2rnD3QNLW9fYjHzXUrj/AKfsdFc3M0mWFheT8H1/JB1+rrELe3xM5+taUQH5ctc1BQXmFgdUA9GB6rb9OyTJz2zugyoKxlNnqb+ZYtZUNqH9bRhX1EbXAdI3WOIJD5II0yfVSGCRoyQrW4DviCC1AMqV7Qdmqgw0YPdBGBk4VW7OTfJwVPCI8guQSmESMCgdiIdOMkqaedrW4YdyqRuZJGXP3IQY8b+h3UsxlYwjpx+ZYRGXEDtlUaQHAnsCgzX4ODhavSOjZSlxA7ZWmQwuqgGxbkeSve+Wl+8S7Z2QWyTR1MxZ0gb4WLUxeHkBXTDwT4je53SmlEs48U7EoIYZDEclu3datBb46qH3gYGBlSVtNRywAQkFw9AtOiqamnHgDIBQRVUg6jEB2OMqyCmMxwFdUxkfG4bndSUs7YwSTugpNROhHUTkLH6fhOAs+WpbM3pLlYGM6fkUGJDF4pxlSyUhaNiqOIG0PdUbLIXdDyUEJBaVQbkBagLVV1Q64WZCxJqeWlk6JW4KCrD4QB9VJ4HjMMvbCgfuAAVVssrGdI7IKNZ1PLc9lr9Df46Cm93LRnGFt4PIJPmVcxviPGexKDMr60Vj3OY3GSoI5HRtwSVlPp4oog4Yz6rGfGXjZB359i1KXc0uqR/3AVx/5xtyKnsVmObzTapyP/lArh/zjbkQe06IiAiIgIiICIiAiIgIiICIiDjzmKGeXzic310beh/yGVfNG9nVWTD0e79K+lvmMOOXzie700Zez/yGVfNNHl1dLjzc4/nQZcMTekZA7eihqYAckK+eobHH0tO4UNPK6TJec5QY2XE+GewWbTxYIZ6qGfobks7pTz9PxPO4QZ1VagyLxQdytMbA9xJGcAqWS4VE2Y8kg9gFn0EQEBM2ASCd0GE2oELekjdUMXvLTIBjzUcgjNSWk4blarC2NseGkHPog0eMuhkI9FltrB1CMj8ytmiDXkkYCoGREF2RkDKDIrKUe7+M3fzWmsi6wfLCzKarMj/DlPwK2tMbHhsGMH0QR0cBmm6PJZNXRmFwA2Ulsp3xuErhsd1PcX+INu4QafPEIYg7zKyLbZ/ug0v6sLCnbP0/GD0jtlZ9suTqSEsacEjyQRVVpdTOLevIHzWJ4j4cta4jyU1TV1NRK525BWOY35+MblBfTB739RJP1pVZMgZnZXU8ckTupw2VtXI1zwW9wgy4rf4bBNnPyVXUQqCHg9litmqhHgk9CvhqpQ4AZx5oLqmHoj6PRQUrzG8brOqh40Xw/SKxIqd0bXF4wUGpNeKpvh57rAqqJ0Uga04yVHBUOim27K6qqXvkDsnZBkR0PgMEpOfNQylsn1hJK5zoQwFYrJD1j0JQV6JWH4SRjzyrS+TPxOJK1hsUb4QcbkLTqinf1ZaNkEtLcRCMOGVWV7at4eB2WHHTyyO6WjJWQyKSlB8QYJQaj7yyGERjuAsGcmZpdjPmsdzZpHEtBws+jjDYD4o3QaYG9bw3z81ksonY6t9t1EcCpPT2ytYhc3wTnvhBiUz3vd4LWknONkq3z07uh0RGfMrV9MCiFeTVY6Sdluy+UViqKbqgLC/HlhBxpJDI9vidRGd1dTx9MZz3UtbQVjJndLT0A7K2A4eIT3O2EEcRzKW481ltuBhmbC1uQSAdlPW6croqYVUbDg77eamssFI6M+94Eo7Z9UGRX2pk1EKjO53W34o/AcXemy1a4xXaUlsAcYvLHotNFNUQMInG59UFJK5rx0Y/MsaMHxg8ds5UUgw4+izqTw/CJcdwg1KorYxSCMHcBabDXtYektzn5LFlkdK8tb2ysuihhA+/fSPbKC+oh8dnUPTKshqm0sRjIyTssz3eYAuA+ErFMcL5xG/GSQEGLTtMs/VggLLqYPhABWp19sFNRCeJuNsrRYKhzyRIe2wygthh8N/UVkGcFwHbCqHxYy5wUFQ3r+KHdBq1Oxk7QNsrX7Xp+OZvW8A+a2ZQzTxSjqLgAQt3Wy/Nhc2Pr7jCDeWkuFbNXVXurHdHlnstcv8AwIi0hJ7w+YPIbkHOVotn1tVaWaK6meQTvssC9cY7tqqsFGZXOLwWj60HdD2fb7cdYQUFfTsnYJhs4AjuvXCnpqanYG00EcQx2a0Db7F5I+zy0Zfo9TQXeqid4ZlDs79l65N+iPqCC5bfumpDQXBlGI89bg3K3AtPqrRRVE7audoLmfFn6kHVH2hGnX1/DyW4RydJ8FwOPkCvEigsT6i+VNLMS4GYj6t17ac/N/8AfdDPs1vky8ROzj1IK8Y301ZZa2tqKvIf4hIz9aDlLTHBClr6FtU14JIyQtb05wXp6fW1EJXAN8QZH2rYXDzitd6esFLNIRENhnstxVfFeuh17RGCQ9HiDOD80Hojc+BduunDGFtNAOoQgkj1wuq+sL7JwxtdXZZJC0Oyzcrtrw/4y0p4dQtrpG5MIBJPyXSnmgqmakqp6u3tDh1lxx9aDrvcKaC4ailuzsEykuJW0dZytbPiMZAKya67VNFOICSCNiFhVXTXxmSTc/NBo0dY1jASPJXCkNcetuyhfE10pjHYFalR1EVLhriBlBZTUPhkh/l6rHr53ROMTHHB22WbXVTZYyaY5djy9VoksNST1StOUFHxuAEhOSVVkBlaXHyUtI5pd0zHYequmkY2cMiO2d8IMWKJ0j+gZCzmYgcGHfKmqYo6enbKwjqO605vjSuEgBwD3Qau2la9nX5ELCqKRweel2AhrnmPw4ycjbCRCpJDn5xnKCagjMJDnb5U1S1kjhgZKhfUxsb0g7jyV1IXSzB+PhHcoKmFwb9ArDki+LsFuipq7Y2mDQ4dY2+1aBLJE8ktP1IMR8jYg3butZtmn47pTGpJxjfutFqGF4yPJahbrlV0cBhjJAIwgxKunFPOYAexwrGUfUQ/PnlUqnyvlMrs5O+6jD6g/QzsgnrHkRBgKmtzCYS4k4WnubNIdwVkQvliiLeyC2qqAXuaBkqAuLmHIwsmjib4pdOMA+qlr20/hF0G/bKD1K9hr/udxl/w1g/6NevUxeWfsNf7Q4y/4awfor16mICIiDrn7Q445NeJRP8AedH+vU6+e+3ODJHuO+G/zr6D/aH/AIGnEv8AxOj/AF6nXz10n0n/AFIM6atjqB4QHZYrHiCcN8iVjuPRISPJXiOaQh+Ox7oNVuBa+mGBv3/MtHa4Nysx9SHM8M91imB7jloyCgtc97zjJPoMqWJ3Qwt8yrI4nB2TthSOADwPJBJT05Li552z2WY+nYGdYA2WJNP4cQ6O5U0EkstM4lBD703r6QBucdlq8GmxW05q+rBxkhaPb4myVOHgbFalVXSqox4MJIYdsINInLoJXRdwxy3Db9RwU1AadzAXEYWlMbFUEvdjqKxJoC2oA8iUEdTKZZ3y7gOOUZC9zevfC1CujpmUw6MByipJIxAQ87oMDDs4z2WZS4GGu81FA1rpzntnZVqXdD/h+xBkVjAGZaAfksRlM5zC87YWXbpGzSlkx2+auueIXdMI2QaexpL+klajDThjfEz2WmguLst7qcGrLekZwg1CKuiDvDIGRssiWeB0BAA3C0J8b4z1O2OVc2d5w09vrQXR9QqCWg4JWriidUQF7muPzUFAyE4dJjuO63LFW2yOkMXUA7GMfNBs9kfhTkHyK1Btyjhb2yAsS4003vBkiB6Sdiom0sj4SSN8IM/7ptqT0MiyT6KKTrALzGW/WMLUtE0VMbgXVwAYMd1rmr6e2NbijLe3lhBsYPNU4sJWRFAIMdlhPbJDKSz7FK2oe5uX9wNkGfK0vYOklRslfC3fO3mobdM6WXpO61yWGl93OcdWEGiuL6w9PURlR/c57ZAzqznG6gc+SCZxZkAFZ9HJLI8SO7Ad0F01N7lH1n0WA5vvOZM9lqV1rI5oTGCMjbutIhe8Hpb2PdBGQQSPRUwtShhiA6pMKk8ERBdGRgDyQYMUfiHCrJF0Z+So1zmOPT5Kj3ud3QXwRGV3SDhahHSCJhaTklabHI6I5apW1Mz3ZycfWgnJ90d1BuQs62PNxqRE4YBwCpbYynrMNmwT81LVMioZA+mwCPRBvSn0nTQ07KgSAHIOy5A4e32a1XCJjZyQCBjK4Mdq27RNbFIXdHzW+uGVzdcr9DC8klxAQdtrrUVOo7GyOlcY3FoGRtlccVvDuohBqa0GXBzl265ot2k6tumoJ6WM7sByB8lfVaHvVXZXyOYc4yNvkg6fcT46WKB1PHCGloxt9S4ZjaAXMx5rnzixpG60tfOJmkDfyXC8lhq4ZXSlp6Q7vhBiRUII63bBYdTM2CXpA2HyWfcqxsMPhRH4gPIrTYWCdpfL3+aCdz2SxZDdyFp07MHIWoMaMlo7BQ1MfSCUGHG/oOSsplH7w3xAcfasPGTspo5poh0gnB+aC2aAxZ+SjGfJTSmSTfCtbC8HJGyCxzHAZKtysqUeI3DBnCi6Gsb8ZwUEbe+PVZjKHxIzID81hhpcfhClFTKxpYDhBqVhqGUtQ7xN/RW32Rs9QHxgY7rBhp6p5LowTnutSigIgPvOA4IMHBmYG+YCgfC5h2WRTtPjkA7Zws2OldPMGY3KDFtpe2Q9ecHHdapVUUbmeO3Gwzsl1tM1FTiWMEZGdlhUctXIz75np7FBDEPfZTCRjyWY3T4LwwOOD81HK6Kn+KD6fmp6atqXNzkl31oLptOiBnWH9t8LANO7xPBwcZxlZVXVV4wZAen1WZQyUr4syYDygxBaG0rBK7cnc79lEaASP8UDAUtfWSg4cT0DOFSmqDNH0R7koM+gu8FtBbI0E4wtJvM7bhN48bAAobhQVcX3yRpwVLSmJ1MWD6RQaXnBxjspoCJHCPCTUk0eXFuxKkpmhuHjuOyCSqtpgjEjSTnfCuoaQyNBc0j5rPppmztDZuwWoSMp4qcuiIyg0Kvb4bMA9ljwTgMIO+6sqpZJJXNcdgdlCw7geRQegXsXJg/mm1SPTQFcf+cbciv9i3GxvNFqhze/7AK0f8425EHtCiIgIiICIiAiIgIiICIiAiIg455kTjl34pH00Xez/wAhmXzSsm8KeRx88j86+lnmS/B24p/wKvf6jMvmgk/sjvrP6UCR5e8uJ7lSRTY+HtnzV1MI84eqVIY12WYQVdARh5dsr46UzjqacBHSfecE+SrSySNBDckIK+7imd1k5VZK/LelnbsoqiZzjhxVKemMxyATug1OxWcXWo6XbZWffLU2zAYcDt6rEo5qm2/HDkELHudfWXI/fSSgwZZ/HOANyonNdGTk91kU9BO456SrauJ0TgH7b7oMdrS922yuc10TwXHOFPhhj+97lY72SZ+MFBqTbo1sIjaACB3WOa0l2XHIyomxx4yVC4Dqx5INQfUtqI+gD7VC2hPUPi281QmJkQLcZwr4Ko9OXIMx8cdNCCcErCjkbUTbDAyraiofKMEnCxonuY4FvdBr1xgZFRB7cZwobPY/uhGZnOO2+FiyS1E0QbJnCvpLtLQxmKM4BygmuBjp80wxkbLT2SiPIO+eypUTuqZC8nJKlgoZZB1luw+aBFV+Gep24U8lQ2pjONliVDWNHT2IVkDi1pHqgloYPeKjpzjKnudKKYDB+xQ0Mwp5uvspLhO6rkGNwgwWMLz5qrmdDw1ZsjY44Bj6WN1htzIS4+SDUWZZD1Fw9e6o2pY49GM7LBfO/HTk4CpC/wCMEoNftdLGyTxHAYPzVl6jjeepmFF750QgR7n5LT56qaVxac4JQXsrY4x0FvZXmfxmnp2WE+nka3rcNldCT0HCCkYIl381qQb97yD5eS0vLmv6iPmpW1MjvhGcIKieVsxDHEYPqtSgqapjg4vLgN/MrSpm4AeM5WvadmostFWRv5FBmQ3BlU3wXxgEDutDr4TS1zZm7gOytzXt9sp6UyUhHX3W045zVFxl3wg3rFqmlqrY2jcwdXTjdbVrqd0dR48b8NBzhQNAYcxA5+tSe8uLemQ90GtW7VMDYxTSRgntnCxrtLHVkujaB57LRTTsDvEZnOc91nQzNEZD+6DTqmIN+sLH8RzctBWbO10sh6exULqV2CcdgghgkEb+pwyslpdLM17M488FYb2lriCtUoIQ+LLdwg1SCtZJD4DsZ7ZysGW3dVSJg47EFRCOQydMWcq2eargeGvBQa3W3GP3AUxAOBgrQYKfx5cDbJWUYpKiIEZJIVaWlmEnQwEPPZBrNBpL36P4Tk91o91tlwtNX7uyke4Ht8BOflsuUeFXDDWesKuOO2slcHPAwGkhd3OF3I3W3SWmn1LSHJAJLmoOgWktB3a+xskmtkjASNywhcmUnLnc5aX7oMonkNHVnpK9Jb5ybW+y2Rgs9O0SAfvWbrVtJ8FobNpSphvTGiQMIHU35IPJ+9aHrHzOtZgc1zPhxgrZA0hVabvsU8jSRG4uIwu6/FnRVHprU9TWFrREXEg4wMZXANaKDUGr4qSLBY6QNOPrQd3/AGf/ABDjr56a0ig6CHhpd049F6ZDcBdJeTLhFZ7Fbaa707AJNjsF3aGwCCqiqIzLC+MHBcMKVEHUnm40TJS6fmvU2ZGOY75jbK8kNf26G41dW6JoYGyEHy817bc1lmqr3oCWkpgSSx/YfJeMHE3QmpbRXVh6HhpkPcH1QcTCGS1ygw5z8goG1dQ25suUjyDG4HJ+RWu0lN7tKPupgDHmtJ1FTtmeTbhmPzwg5Cl5jrnRWZtogrXgMGAAStBPGZ9ypn09ZKXuftly4vuNJAxuW5Eo7/NYdPG0Zc/II7BBuK6SC4VZqmEAHfCwpJ/AjLC5YcNRM1xG/SoqqRz3fJBaybEpcVWZr5nAsKhkIA2O6yKWOaduGZyUE1Az3d4MjsjPZa2aSKsi6wBsFoRt9eCCWnutVpX1EFKWnOceaDSqqixIQw4OVSlt5dO0PJ7+atdNKak9fbKldVlrh0/S8sFBqV3tvg0Qc0k7ZWlW57S0wObudlqNJNUztDaskM+agrGwRTB8OPsQWS233T/bBJI74Vn3TY4eGG4ztlTVlcJacMJ+S0mNjTIAO6CaopXgGYEkHdS0NZ0AQEYJ2WqW+hlnaA8fCPVYdzpYqScFmAQgyZNOySRmpMpwd+/ZaNPE+lf09WcbLUHXKvEIY3PSoWUstV98kyfPsgggl6z0uCmdVNjkDOnupI4Yo342yo6iOPrz5hBPMxskYeNs7qCKobFlhbny7KR0mWANO2FaGR9JJ7oJGeG89WAPtR7GdxjZYTWVMzyyHJx6KT3SujIMjSB5oJXQ+P8AC0kfNWTUboKZxLs//ZWTEDj4O6xa2eUNMT/NB6m+w2/tDjN/hrB+ivXqYvLX2GzcWzjI/wDup7APyNrv9a9SkBERB1y9ohtyacS/8To/1+nXz3UMfiOk37Nz+dfQj7Q/8DXiXnt7nR/r1OvntonOa5/T5t3/ACoIZB0yH61qFNUMdCWFu+MZWDKMvJPcrIhfCyMnzQU91LpC7OyvMzISGYBPZQxvmmk6YlaYZGVAZMMElBkTPw3rAxlYnWXHYEn0Wty0QbTh7hgY2WDaGQm7wifHhdYz9SDDe2ctBfE7HlsVn0c7WUzmnvhcv6hpNGt04w07ozOY84GM5wuGJZGNrC1v0Or8yClK97akuwcE5Oyzqt4ezYglahVNt4o2mAjrK0NsuHhrz+VBbSh7ZSTkD0UlVUNB6QNwsnELW9TcZwsSUMcSSghiZJUuxk4Vs0Lon9G+6yaV7GOy37VlTMjezxDjI80GF4BgYJM7kLHe4yuAAySpJKhzndB7DZZNvbA6oaXkdOUETYJqdvi9Dhnzx2VpqXSkRuGc7LeFZ9zPcxgt6sLb5gpvptI2OUGIaT3domcfmsuCpidHnAJWPPN4wMQOQFisJgd0E7FBfV1DZHEAY3WMDg5WbJC2Zo8HBcVPS2Oqc8SSM+AblBhQzS56QCsyCgnml8TxDjOVn1cdEyIMix1jY4UUDpooS8eSDNqfBhpww4JA3UdpEdXVsgIwHHdaNNUz1Mhjbk4PksmljqqFwqACCNwg3tf9Nx2m2isgkAcRnZbJZV1FQT4ji75ndZVTqm4XGMUlQ4lnbGfJZFLDSspd8dR/Og0uoDGDOMnCx44jINhsFfUCQzEfvVJG5rAgswKP4wr2TyVI6gTlRVsrZAATslBPDE4NecAndAlBOAW/NXMrWxs8PHdZ1wmozT5icMgLQXHJJQSzRv3eTkE5VIpQzuM7qSmka49Mp2AVk4j8UBnZAmqDIA0ZAWfQQ+LF0ud3CwJRGGDHdXQVboRgZ27boJqmlFM4u7gqBrBKC4AbK2aqkn2cVWJ4b8OdiUEbznbHZSxStaOnHdXOi8XHhgEqx9JPGMubgfWgzKQOhPWH4BVJ65xnGTkLGhdK53RvspjRPx1lp2QZU8jKqJrW4BC3XwxqTbdRwzPdgNcD+dbNp8lwa3yWt6frRR3dj5DgAjKD0y4X6yoLrp6mpJSw4Ab5LnWlttunsBEcLXkt7Yz5LoFw64i0tFRU0cT/AIi4Duu/HAm2XfWFthqA1xhLQT6Ywg6kcwNh6amocy3EAZ3DV1UuTI+h9I6ENJJHbC9VuYDhzZhbZ8sYZgw52HfC86eIXDW6UVVLcYIyIGPLiQNsIOAtSadfb3GrIODutEgAmhLwcALkHVtVT3eP7l0ZBnYMEfNbP/YreaGBzpYy1p+SDR4Z8SlvopJCZXdI81DHF01BY76QJytSjpRs8II6S0BxDn5x3UtVQRxMLh3CummmibhhOAFpctdMXdLyfmgvo2tlmLCNldWPZB96A3PoreroaJIu5CsbDJO8SPz9qCyjdiXLhnKzprd7xh7dhjyUL6UuAbCMn5LVqKzXQwGRzSGgeiDQHA0ri1zc5Wp0dnNbCanoOAj4YJJQx+OoHC3nabSfuc4xgY6c9kGzoaoUMgiEecHfZatS0Au7genpB+xa5p7Rrr3WuY1mSD2V99stVpysEDQQfkEGnVWkjRxeLG3J7jZaXSB0Fa0SREAHzC5G05BLWxhtaMgjzWFqWxU0EniwMGAc5QaBqSqg+5zfhGcLatLWxvhMTWjJPktVvcU9Y3wIsnGywLRY6iCpDp2kN88hBps9HVwvMghc4H5KtNcnRPHWzGFy3a49NGlDK4s6sefqtjat0wX1ZqrVGDBk5x2AQaZLcGVkRaGAZWjudJFUDDyBlazR00RaIW/2QDB+taZcaWWGfLge6DOqWtqacAEZA3WHTO9wlD3HIBzhIJJJG9EeSVZLBMJPvwICDUblfo6unELWDPrhaPSPcKhpGcErLMdM1oIxkd1kMp4RCZW4yNwgza4RPpRjGcLRujoBcOytFbJ19DicfWpWRPqSGsGcoMd9U4EBu2FrVvaZ6b4n+XbK0urt0lO3reMAqyirpIJWNz8OcIJLjTmJ5PSVhRbyAeq3JVUZu1OPcxlw7rQ5qSagkLKhvS5B6B+xih8Pme1O/Oc6CrR/zjb0UPsXZJHc0WqA/t+wGtI/zjbkQe0CIiAiIgIiICIiAiIgIiICIiDjjmS/B24pn/uKvf6jMvmkjb1zPB+Z/Ovpd5jx1cvPFBvroy9/qMy+aJwe2d4b36iPzoLCCJC0FH5Dhk5V76edjfEc3AKi6iXAlBM4ZjwqxTCEYPmrnPYIhjGVA1jpCSEF0r/EOR6rOtzxG3JasWijBmAftha1JHA2EluM/JBG+qjOxwozLE0F2y0uR8j5SxmSc7K51PVDcg4KDXbFWRVVV4TgAFLqW2wtcDERvvstAhfLQOErMgq+e71VSR4jiSgvp6YRHLiCEqmMcdsfYoHsrGN8RwOPXKibJK8jbv5oKyAtbgKDJ9VqXurfC63uOcdliyMa0HpwggyfUo3JIABKE5OcK+POeoA7IMuWENgDj3WLAPi6j2CyHzOezocVihjwfgBwgypq0OZ0Nbv2WGck75+1ajQU0QeHzH7EujYsgwgDHogxKRoMm/ktUdXx00XhhoyR5LRmPcw5GcqdrhICZO6CzBqJSSe/zU/u3Q3I/SsUu8N3wKaOaR/cnA7oIH5DiOyvjmcwY6cqWcR9PlnKzbVT08jOqTGUGmyOe74iDuro3DpWo3CKAAiMjtsFpAPSSEF2Ot5HzRzTE4DfOEhcGvyVmF1O9uSRnyQWRZDR1FRvlAkGw8lZIXtccdj2VscUkxJaMlBnVFSySn6WgZ+SxIXYaVQRyMcGvBAUwMQ2yghe4P2G2yMcIzjGcqsga3Lge42VjGOe4HuM7oMvoDm5PmkdG97w6N5HoqyysZEGjuB2UENXJHI3fsUGfWU87KcdbyfrJWlse9mela5cahj6IdOOr5KC0No3RkTkdR9fVBi0cpLviGc+qsrZD4gDey1N1JHDKZMAMWFVup3kFpBIQZ9qpBVDDioLzT+5yAMO3yUdLcDSj4T2WNcKx9W8dRzhBm2RrauURvAxlZV7hjovhbjdaTDVCkaHxH4z6KKqrZqx3VKcoIXffHEgd1n0tWaaItI8vNQ0zIyd8ZCnkjix5Zwgmt1awVPiOxjvusu5OiqSJGYWhhkjXnwxn0UkNRKJmsfsCfNBq1FM9hDRGSM47Le2hNPPvuqKWkNMSJHAHb1VNJWKkr4mOe0ZJAyuxvBfhLUVuqqGqpIyQC09soO9nJzy/Waw2imvNdaGPyQcluV3OZbLcwNDKKFob26WgYWxOCdlqbNoymppwAQ0DGMeS5FGwQRmngc3pMLCPQtC2vrXSVDdrZKWhsPQ0k42BW7Vw5xq1XebHZ6mChB8R4LW4+aDofzkW2htrZoaadgeAex3810N0DXSxa7j8SQvImGB3812E5pK7XctxmrbmyUU7sncnGF1l4XXSldxJo31RHhmYdRPbYoPbflIrZJtK0oMJaC0AHB9Au0Y7LhDlkuOkJ+HltZbXRmfoHVjGc4C5wQEREG2tc2yK52mSGWISDpO2M+S83OZfTFJbhVuZbWtPWf3vzXqJLCyZhY8ZB8l055sdLWsvlYcAuJ8kHkbrajlq7g6FkRiAJ3xhaNDTRW6gkbK4PdjzOVztxo0F7ox9TbR8RBOQF1tqZ66j8WGuJByQEG26gme6OBOGklQ1jRC8dJ2zupa8eG8zRk5PmsdkFRVtMm5A80FRVAjp7H1Q5e3IGfsWK4FrsHuCtYtDIpG/Hj55QaTJ1ZwW4ws6214ppGgtBA9VmXKnhAJYB9ixaWgD2l+Nwg3FLdab3YODRkrJoYaevpnPyBt2ytnVc0jD4e+3zV9Hd6qmIYx3w53CDULhQiOd3S3b1WIygy7xtyRutcbC+5QB0YJeQsFtPLSVQZVOIadiMIDZZqhnu8UByB3wVpNaypp5OiRhBPrlcu6RpNOCMS1b2AkZWPfdOWm7XiKKhcCHOAwAg4jeyo6QXMdg+arAwteH9wO65y13wlktemmV0LDksz2+S4rtWlLlURvmMZDW9+6CS23drR4RaB5ZVlwonVj/GAyBv2WNJSMop+h2Q4HfZbit4jloyNs/NBtupmZBG2LpGQtzWC2U9bbnyvIBAytq3SmkNQ7vjyV1LWXaniMVMXBh9EEV4Z7pXPYw5AJ81jR9VQcnKyJIJ3PM1YDknJyr42xhwMRyPNBY2MtGDlVMXWCc4WRPNA2MgEZwtMfVP6iGdkGsWkxU8mXgFa1Vvp6iElrQDj0W04H1EX3yQEDust1e50eGZQXAshnJJGFp1xkEsvU3GFZJNL4hL8qKQl2/og9XfYbn/qRxiH/ANMWL/o1y9SF5Z+w1Obfxlb6TWD9FevUxAREQddvaERiXk54kRnzpKP9ep188sMvgl22SRhfQz7Qpsj+TjiS2L6Zo6PH8ep189lBGx0j/G7NH58oIZA53xYO/wAlk01tfOzqyQVmxyUIPS5wwPLZTQMfJUh1PvHnJ+pBDaqY0tQTK3b1Kx7vJmsEkbcgHIwFua7mkioR4ePEA39VptCaCaAipx174z6oMGS7e8Uwh7EBaaQ4P+A4J7FahUUTGyl8X0c7bqKGlcagOcPhB7oIZjcWxjxZpCz6yoI4TI0uz27brddU63GlDSR1ALbMsgZL0xnYlBdRskMvSScfWsyW3NeQ8Hf5KJ8sbIuuMguWKK6fOOo4z6oMuSBzB09R7KNlN4h791bPJOIw453V1PVhkRJO/wBaC99v8AdbXFRGY9PTlUZXSSPLZHfCVFM8CUdJGEE9Pb/eHEk4yVc+iNPIA1xUZrXRMaIzg+auZWOkYXSHdvZBkTwSujBEhPyV1LSPLOk5OcrFo6qSWYMcditdgj6cPxtjKDS5KI0zi89u/ZYk8Yly5u2CtZuVVA9nQCM9u60joeWEsBwgsopjTy5duAtd/ZLCaYwBoBO3ZbfgAkkLT3Vs0QjlAGUEs3iRzePkkE5W4rfFHV0Rc7YkdlpVPG2WMeIBgDzVs9wdSfeoTt8kEL8UNW52Adys43OOenOGDIHotNe/3ogu3JWdRQRRkB+APmgsstH7/Vlr24BPmFq1fa/dJA1sm3yUc1VTUTeqkIDj6eq0qa411W8lxOQgyK+NkcZcMZG/dafBmU4z3WVHaLrc8eDGXY8kdYrlQv8Av0ZYfLYoLZLflvVlZ9NpYz0xnBwQM91jOZVQNzLnHzWfb7zJFiMn4B3QaDPbqiGYxYJAPdYr43sdgtIK34ZLdVMDgWlxWj3Kjp2MLgBkINBZSlzeoHCikjLHYJzlZEZkmlMcJJUckT4ZwJgQfmgsdE9rQ7BIUeFrrH0hgAcQCButMeIfehjHTnyQY3Q4AEggH5KvScZytauPufug8Ejq81ozHb4d2QTUU/gyfE3Pmsua4RzkM6QM7KDpiDc7ZWK/AflpQai2JkOJdjnyUza6ORnh4G6wIXy1LhEMkeiyG2SvDg9sZ6RvlBXBp3GXpyD2U1IPeqjqDukrLjpmvZ4co3AwtHnmko6lzYjgAoOVtKC6wPpBS9coEg2GT5he0XJXdKSHhqx1wY2KYRA/GMHt815V8pV+0J90IG6zliDQ8ACTHr816SWe9we5RnQhzQhgJ8PtjHyQbS5oNdVFHXVYpWuezqOzckLqxdtZw3vTFTRVFF0SPaQCRvldyJLHYNaPmgvHSZRnqBx3XXPj1pvS+jaCdtD0AjJGMIOhV6t8uldSzXR+XxlxcB3wpqviHTXWlMIpw04x2WfdLlQXu+y09S8FgJAytt3+gtFvcW05AOcoNMpLU2uq3T7AE7BZktMKaYRY2yobRNI+UNgJO61eopJDIJJG7hBivtgMfiBu5GVpNRYDKesNwtysqGlvQ87ALGrKqJsZDCM/JBtqa3mnADB1Y+S1OkomS0hc4BrvqWTaRDJMTVkdPllRXWoEVUGUv9j88BBoklRJb6neMkA53C5N0zcaK5WgxSNYHFuMea2/RWeiu8YJwX4WmVtuvNlrG+7BwiB+zCCe76cFPXGoDsAuyMfWuQ9GUtNPQ+A9zcub05K40vF5qJ6ZkeSXjuPmtY09cbrSW10zOoEDIQckMq6LQ0rq3LXdW60muutJrCoFaS1vSe2y49bqmoutW+muZPS043WLVXmot9QGW4uDM74PzQcnyVcFDCWMeAQMZW2bpqHxgY89Wdlonvtyu8QbAHFx74WdT2Semp/ErWkHvugutLI5JTLLgjusXUN6gpsxwtGfkFgT3T3aQxQntlafUAVR8WbsN90EcRrLlNlk5aPQHC1/9lbbRb3W6aMvcR05xnC061UE9yk8G2Alw74C12o0s2CAi5MAmI2z6oNq2p5dXvqj2cc4V18mEmQGeu+FlRUX3OqC6YER9xnthSVr6KohJYRlBoNreIZSXjIz2Ky66pZKMBoyViRhomPoFfO+MDAO6DAlje1x7kfJTRVj2RGM5WdaWwyvJnxjtupLjFRNkxHjCDR2xPmcelpyStet0RpmCSRnb1CstvugI6yMZPda1MaV0PwkbBBoV1r21bDE1gBAwtIFLL3a0/WtZlpGyynwxndZ0YoIacxzYD+26DA09evuM9zpW9QJ7FQ3uvbeKsTxt6QsqkoqaaVzpcdOc5Kx65lNHMIqfGc42Qd9fYxtaOZvUwwMjQVb5f8A3wt6K/2M9HUU/M5qeaYHpdoKtA+v7oW9EHssiIgIiICIiAiIgIiICIiAiIg485i/wfOJ+f8Agbev1GZfNGXiKsleRt1O/SvpZ5kT08u/FJ3pou9n/kMy+Z+Q9Ujz6k/pQZ8tc2ojEQZg/UsUUrj3BCntTI3TZkxgeqy7mY2ECIjt5INPbSZ7uKZMBDAM5RjKskENOFmUkcZnYZ8AA+aDHfHI1ni9BGfPClp5ZJGEHOOy3caOgrKMRwAF+FpT7a2gid4rRk57oNFpoxFUdbtwd91kVdwjbhvSNvksVsmasjOytr4wXAs3QSSFtQzI9FbDQZw8u7b4WOxkzewICuE8zSWg4Hmg1SerjfB4HSNh3WnBzGdWw/Ioo5C+TD/MrUG0kMjC4u+aDBfNIThpOFA8OHcnBU07Qx5DT2K3rS8HuJ88Ie7hpqvtkEWapwR/5iDYSyYJWMbhw3W6ZeEHFYSEN4YatIH/AN5Kn/1FjXPh/q7T1PFUan0peLSyclsTq6hlpxIQASGl7RkgEZA7ZQbbe/qcSD3WbQywgBjxknZYkkYZKWHyR4DHgjy3QahV0ssbPHaTg77LT3SOdu45WsUtT7xCIZRhp23WLVUtPGcRkHKDTW4zupWs8TJGwCllpSGdTQscPfGCN8FBcXt6sBTRsw3YbY8lh7uO3da1bI4n058U/EAg0owuc87d1n0rGxNILiDjyUQhlZVkuaQzPdS1Do2kYIz5oLTBJI49yM+SlNmDozISQQMrJsz43y4lIwsi8OexwEGSPPCDbZhxOYvQ4WWLYRh3UfVQ+C90xdu091nxSuZGWvdnbZBC2nbUEReY81kx00dC3cAk/nWnRzSioPQD3WVI2pleHOBwgjrnhwJa37Vj01MZnDJwtXmZTinw4jqI8+60d0j4n/evIoM2qtoihDg7JwsKKURgx+pWQ+slmi8MnKwjHIDkhBnfc7rb4ue6iNDvt6+qiFVOwBpyAFlwVBdGSTv6IIqnqZF05yEpKJ8g8TqwBv3VKY+PUdLtwp62V1KPDjOAUF01b4rPdvMbZWO6gIb1dZKxuiQESYye6laaqRuQCQggflpxlUaOpwaT32UjmFp+MbqwOAeCPIoM6S2dEPi9XlnCwSzufRZclVNLEI25KpCzoYRJ3KCCnBc/CkqOqM91GCY5ersMqYSRyuHUUFsFV4bskAq5wE8olbt57KSekBjBjwSo6Y+ECx4wT2Qb00lqB9I6KJmTh4GPtXqFyV26z3hlJVXDoa8NB+LY9l5hcMm2yK4tluxaIw/Iyu2HDvjozRmoKGOyS4pmlod0uwMbIPaGxPpGUDIacjojGM+Sz/GhHeVg/wDGC6pWDm20cdFRNbVM98MYz8QznC2HqLmtmZQTSUs5yMkYcg7oan1ZaNNW2StrK6Fha0loLhvhdAOY7m5t1FVzeHcY3siedg4eRXX/AIvc2usr1PLQU88zmEFuA4rp3xA1RqC/zze/CUh7iTklBzZx15oLfxDt33LpI2PkLenIG64T4XaTqLhqinnkzGerIJOO5Wl6EsVLPcGy3IENztkbd1zLT6NubXtr9OhxDG5y0eiD075NLTLQ2mmY+5CQAj4erOOy7pjsvLfkQ1VrKbVENouPiljZQCHE+q9SG9ggqiIgLiLi3wg/ZyJZw0uIyQM91y6iDzP41cH5bIZYXW1zwxhG4JXRLiXw8qXTzTR0Low15/e4817u8RuGtu1VQSPMTTLg5JA3XRPjty61VFZa2tp4sAEkfCg8uavT0bZPAkwC3uFhSxxW2IxtaDnZarxOprnp/VE1IXPGCQRg+q2xT1zJ2n3p24QYNTEXSl/YFZ1FCWxF2cYUM8sLz8BysiGpZFCWE7lBbFmplMTnHYqSoq/cD4TW5BWlmd8UznxnGStYoXUlVTl1SR1gIMN8Taz4+xKup7YPEAJz6ZWLUeLDK4x5DM7FZVrtt1u1SyGl6y9xAGEG4KOV9oDJAwvDnAdsrtRwY5XJuMlk+6zaA5DOsYGPLK4j0lwluNPS09ZqEEQl4PxA/L1XqvyUaapY7BEy1lpgEQa4geWEHUnQfIPddQX19rNFIxjHdOcEbLmym9m83TzmXJ0AlMRBIDsld/7JpWgslQ+qga3xZM5IGO61Z5Lz4fSOkggoPLfjXwcba7M60OtpAiZjPT6BdQbxbqHTNNUUbomh3URuvYfmk0ra4dHzXVjGiUteOwyTheN3Fj3ue6VIiyQ2Ujbt3QcT322NnrXztGGkkrSxWvoT4YJwDhbhubJqaFrpMg47laJ4cE7HPOCQgmhjjr8EgZ81K+SmtwDCAtENY+lkLYjsCsGtuEszsuJyg1a4yx1zemPAJHksOOm91icHHJPqobZUjxOl5VbpO4vDWdkGnzOcZCC4ndXQxEkPOQBusm3QtfKTMNlfc3RxnphIx2QR1Fa2SLwmjB7ZVlG4tOD5rE+az6WFz4+sDOAgrWsaG9Q81jMjBp3vJ3CTeK55a4HGVHmSNhb5FB6tew2GLdxlPrNYP0V69S15Z+w1/wBzuMo9JrB+ivXqYgIiIOvfP7MYOUHiNM1hcW0lJ8Pr/t6nXz10EElVJMOkjPxHbtkr6L+cukpa7ln1xS1mPBkpqYOz22q4SPzgLwH1BDaKGaqjoS0FshaMY9UG26TSxrHH75j7VuWkskdrpjG4ZJC25DJdopRJBkNP19lr1NV1NVF4MpPiHbBQaHcKWQTOJdkHcDK0eallDi9riACt5O0Tf6lxnbG7oO+cHC0i46cuVHJiRrg3z2QaM2vIAjcM4HmpxWARkAblSS2+MDGAHLEfSva75IMOV8vUS5xwSkcT3t6unIC1A0oe0YxlarTUMTaIlwAOCg21Ax0j+jJwsp1EGOD/ACHksaVxiqH9G2CVX3iZ+2SQgyauoa6Pwh9Sx4aQyjOcKNx6/rV8RmY4YzjKBNTGIZyoWtL3BudycLOqnZiGe4WLTECZhd2BQZL7fI2IP3I7rDLXAlgBzlbpmq6N1IGgjOFp9DDTvqw9+OnIJ+pBJZLM6VwllaR2Wdc3No2mNh7DGy1e5XK20lvDaVw6wNwDutqPrhVg+KTnO26DTpnPdKXF2xKzIahohMZ7lYssMj3ZYMgfJQHrjdh2xQZkEXRMX52Khqnjxg4KITv8irHOLjkoM9tUHMDRt9SxJ/pZznKjBIQkk5KC+GUxOyc49Fnsm8WMgHfC0xSxxzlvUwHCDOt1I+pnIe7I+ZWdPHFRuztstLiqnw7Md8RUc1VNNJ98OxKDkHQ99ooZi2drQM+in1beqWeva2CMYJ8gtgMdPC0Ops588LXrcPeYQ6p/sg9e6DLu8EU1K1zAMkKy36YZPQOm6wDjI9VYaatmJa0O6fLzWHU3S5UGYGkhpONigxaWhmpq1zDISASrrgHyOEQ8xhTUc7pHiSQ5JWZJReL99b3G6DTaO3ihPjv3yFh3Rral5lZthaw1kkxMThsFpNyttXG8uY0lnyQaW1skjuhpJKvfTvaMuzlZNrEYqCJNiPVSXNnXIGQjOSg03qcdi4q7wyVLLRzwND5G4B81lU0LZGdQ3CDDZG6TIJwrJY/DdgnKyZ3NhJDDvhYpLpHfNBl2ydkE3U/styHVVNFB4TWAktwts01sq6vAgjJJWu0uhLnJH400ZAxnsUGni5mWcu3AcVr+mdHDVV4ipc4EhAO+O60WstApHCPqIcDgd1vrhvZL225xVtIX4YQ7OCg5D1Hy23Kw2Slutrq3ROGHZY7HzXbPlQ4pTadszNLXabxZXsEfU85PbHdcaWyw6513ZobZSeI/GBsCVyBw25ctc2a7QVVTTyh3UDnpIQdjqrTf3Ngk1EKjpbPlwGcd1164t6POuOuMVGQSRjqXN+ubLrS3aaZSytkAa3A2PouCa3TuuhTSV8LZehmTkgoOturOXV9infXxAuJ32XF124cTTTEPJBBwAT3XaqqvdxrKiWguh+IAtwQuKdXaVvrLmK+mjPuof1EjthBxTBo02NplkjIAGdwts3fUAbWClY3zwuVtdaktBtwoYHtNSG9LgCM5XE1DZPfavx5Acl2RkfNBPUxSNgZPuOpbi0poqO/Oa6SQZJGxKhudnkkoQyHJ6RtstNseobnp+uZGSQ0FBuTXHD82Kh8amBJAzkLYFFTvljc2UZcFzdFdmartwhlIcS3G+64+vOl6q3XISMaREXZOBthBotqmks0niSghvff0WsVOqKO4ReCGNLiMZWHrGooPucIaYjxenBA75W17Haa+SMzhpwNwgyrjbpmy+8RRl474G6zKTUIp6N1LNH0EjG4W8NCvtBldFe+gNG2HYK2zxJjs7bgDaC0sB3IQbZfSmqqXTMOAd1LGwMk8F257BQNkk6AIgc43wlA2cV7JJwegHJyg3rpqSOz/AH+dgI77hS3/AFTBXnwYcDIworlcKB9ubHC4dYG+FtQwSuJe0HOThBdWUDw/3hmXZ3IUcrZHUj2lhBxjOFuLTbGSvEdcMDO2Vq15sMLoTNTNBZjJIQaLwrr4bRc3TVjRjPZy1nXWooK+7NkpnAMHcBbEra0W6YshPxDbbyUJrJKthfkl/kg3BqGeKtog2JwDsDOFtuihkAMb3ZyrGmu6iZAen5qcS+GCR9LCDMFpaG9YO5WlVdI9kmcnusuG5SsJDycK+oqYZmHpcMlBpxLoh8JwVjPkkeclxKukbLkkg4PZS22IPq2Nk+jndBB1ysHmAtVpqiQwEdRK1nUNDQR25r4Onr6fJbdtbwJA1+wB3QSQXJ9LIetpwfVZzbW66N98EmAN8ZUd5hpDAHREdQ32WmU9yqoIjDE44Iwg1uCNrSYQ8AgYWmVcD6erEwJIByscVFY0mQZyVqFvkbNKPesYQd/fY13z7oczGpaPw+nw9BVrs+uLhbx/OizfY8wWuPmU1I6jA8U6ErAcenv9vz+fCIPYNERAREQEREBERAREQEREBERBxzzHND+Xnig310Zex+WhmXzRPi+/ys/uS78xX0vcx2Ry9cUCO/7DL3j+IzL5o+o+8S58y7P5UEluYXz9IOFl3GN0TwXHKxqBr4ZxI5pA9Vn1tTDNjJ3CDH99ayMANGR8lfFGaoZbkZ9FZHSCtPhQbn0CymRPtTg2cYz2ygko6mW0yCSR5IG4S53sXV2GbYS4zU1TB0sIJwtCYHxSeYPZBqlJbxO7J8llVNtETOrGfmsGCWojHWM4WqQ1Qlpz4h3x5/Ug0kzM6ywAbKCcta4gDc7Kyp2qiWHbyU0TGSNL3uwR2QYxhIb4m+FdHUysHT1HHop6YiaUxP2b2VK+CKFwEZB+pBvXgHBHW8c+HcNREyWKTVloa9j2gtc01kQIIOxBBIIPfK97ecXmot3KJwytnEq5aNqNSxXK/Q2MUkFa2lcx0lPUTeJ1uY4ED3cjGB9LOdsHwS5e5C3jxw3Hrq6zj/lsS9d/bF2euvXLLpmkt9DUVUjdd0UjmQROeQBb7gMkNGcZIH2hBxh+7h6T/wD1eLt/pFF/V11751faNWnm14eWXQVs4XTaZba7yLvJVVNybVPeWwSxNjYGxt6QfGcTknJa3bZdNLzpq82eIT3Cz1tJEXhjXzU72NLiCQASAM4BOPkfRaZDC17SXOwR2QUqT1yGQdiqxQuk+M9gVG45PT5LKif4bC0eaCZj/Fb4MRw4eixp4p6d48Uk77LKsNM2uv1DQOkLG1dTFAXAAloe8NJGfTK9FOb32X+geXzl+1Vxms/FHUN2rdPe4eFR1VJAyKXx66CnPUW7jAmLhjzAHZB53CVhh39FgysEhy1SSkCIAFd5+Q32eOiubXhDd+JGpOIV7sVTbtSVFjZTUNNDJG6OOmpZg8l+4JNQRgbYaPUoOilO1rZPiwpJHSxyh0ZIaPILdnF3RFJw24w644b0NdLWU2lNSXOxw1EzQHzspqqSFr3AbAkMBIGwJOFyTyS8JtG8ceaXRHCziFRVFXp69G4msggqHwPeILfUzsAe0hzR1xMzggkZGRlBw1VV9M6jDGgeJjdaZFTzTtLtyMeq9TNceyP0zqDmQqaHSs9forg5arLS1lwuE1UaioqKtzpfFgpnzZAAYxjnyPy1nUMBxJA5S0Jy0+yf1TXDhNpG46Xv2oXl1O1zNXVb62pmblpMT2zCOR2QSGxgtOCQ0jKDxgZJJAelriCFrNur4psRT4LnHAXezmc9lFrzSPFjTdq5foq3UOl9Y1T6Zkle4dVilaC9/vcrWgGDoDnMkx1HoLCHP6DJzjp7kp9nVyww2+xcyvEaz37Wk0DZ5xd7zLSRxkjHVHSU72lkZycGYuJIJBGMAPLG76afBSe/M+jjIW1w95IaXEnsvX7mm9mvwx1RwjquInKnUSR1dJRG5UtrguTrhQXilDC4imle57xI5u7CHuY4gNwMhw8gg0NkBzsN0GTHB4I8UjKnFwj6ekt+Sslma6EMBGcLBdC97CWndBLO90rtjssZ58MdJ3V0TJmu+IFJ29urugjik6D1FSmoa4jYKEML9h5q9rY4zh5IKC6dwMew7lZFBTGWEu9FHmEgFx2CyYa2GGIsZhBhMJp6krIlb7zh58gseNzZJy+Q7ZV9ROGYbEdsoD6hjcNwDjur469rGkdI3WLC0SSfEqzxNYQG9ygSO8eQkeajewsUkcT2YkcMBXSOY5pwQUCll6CM77qacmUBwGMDsrLdE2SYB42ytQrabp2jHfZBp8EYqXiIj7VkS2oQ4d1H6lDFBPSSCV4wPVKmtmnOx2CDIbOIm9Lt8LBqJOuXrb27q2WQvAB7lZtrtc1wmFLEMveMBBufQVjqNV1IoafIcDjYrlCPQd60/cIKFjHudLgA4Jxlaxy5cA9a3G6x18FO/oc4YIacYXd6zcqt/r7tRVdfTHDcE5afkg622vhJrOhtDbs6ecRub1BpJwFsHU2qNUWuqNu6JnNLuk7Er1ereX+Z+kWW+OAFwZjAb8l1p1/wc0bpZs7tQxxNqQ44DgAcoOktwt9VFSfdWracub1YK45utSLhUua2nBAO+GrsbxM0w65tNLYmAw4PT09sLiy1aTp7HO77sgNJJ2cgg4e8PJtSyRsjpyD1Ds0rudoHhlbNMaXe+viYZPCP0u52XFnBao07BWNEcjRkjy81y1rF2oqjodbC80mNyO2EGncH+IUOgdeumipwxgmznGPNeg2ieMdJqwQtZIwF7QT5HsvNau1Ho+wMa+5SMFWDl2cA5XMPBzmK0ZbJox47MMwPpBB6MQStmibI1wIPmpFwRp3mn0DPSMbLUsBGAPjAXLWltX2nVtIKy1SB8eAc5BQa6iIgoWhwwQCFtfW2irZqixz219GxxlaR29VulEHklzdcnctmrp77R2kuDgXDDMrqba+XK73G1VNwfbJGCPJ3aQvfTXuhLfri3GjrmsIDSPiGVwhqbltt1DpirpLZEwPkacAN890Hg1cdJVdsv0tulhLQxxGCsG6Wp1PKGN233XdvivyXcQp9WVNfbqOUsJLgWsPbK6w8ROF2odEXIw3xr2EOwcg+qDjqrtzoYBJ0/MrTmvew7OI+oreV7ipoLcwuk3I9Fs5oaSc9gg3Bp2i+7dSKXpycgLnHhToylptZUFHPA1we8ZGFxLw3dTtuDQ0jrLgAF264QcBuIGrNXUN+tNNI6mYWvJAOMIO7tLyrW3XPDeklore1shaCCAATsFzvy8cJHcK7C+3SxlrnNA3C3Twgtdws2iKK23FpEsLQN/qC3ugKnzVUQcD82cwGiXw9WMxuP6V5Ba1ggfcqlshBzKc5+tet3N3S1dRpR5gB/sJH6V488R7Vfqepq6oNdhshOftQbX1vaIGUAfC4E9OdlxM2WWF7o3ZGc7Fbyi1DVVtT7jVuO22CtEvbaOnqgBjcoNHmieGiQtIypaGzC4PDcdytQrZKUUjSCM4VLPcYojlhGR2QXVmkvudD4zTkrbcsmJMO3wVvmesnuEZjzkEYW2rjp2uaTMyMkZyg090vTHlvfHksOR7nn4jlTxNLJPClGCDhSOts8juqNpLT5oIYqfxMb9lmQztpGiPHfZWT5pow3s5Y9O11RKC89kGTI8OPX091izyh2WgbrWXQRCDBAyAtEqmdMm3ZB6r+w2bi3cZHes1gH5BX/wCtepa8ufYc9P3G4wY7+8WLP/m1y9RkBERBwJz21TqPlN4hVLTgspKTB9M1kA/nXz42qOe7XKd73kgyZOTnuSvoH5+2GTlE4iRju6kpP16nXgLpeExx1cg2e15A29EG666Kltdva4AF+Fi2RjKmYVzsDp3Wjtrp5agxVxPhjtlTvu9PSPEVO7DD6IOWbNrm3Og9wfCzLRjK4/19qgeOYaWkL8nGQFiQ1kAaySH6T8ZOVzLw74Qw6spWXKthBYACSQg4O01Z5rxN4lTAWA+owrNWWyG1P6YyCMrmvihaLPpGkfBaulsrAQcBcATXKa4yPNY4kh22UGlsqnMeMMJBK3VQ0cdXb3SOdg4zhYMFHRytBBGfrVKiK5QMIpgfD+Xog29VUbvfHsGSOoqaKlbGw574/OsyN8RkPi7SeaxK6RweQw7eWEGE5pY/IGVNHO36JABWRTNicMyHdYlb0Cb7z2KCSZuW9Q81iCMkEg75WoBhEQMgwCsWV7W56TlBB1P7FxU8VQ5jCMkH1WN3RBe6WRxw57j9qtD3DsVREGZFWeG3BAJVJGtqXZG26so2xF+ZcLLktdXLmanaSzvlBhSweGPsyocLIBc15jmO4OFnw+5loyRn5oNIxhFqNdHE0dTMfWFhRwSSkBo7lBGtXoHMMBY4DOO6x2UJhAdNsAd9lSWdsX9i3ygSxeDIX9x3UkMDaghxCxWzmQ9L9wVmxTxwsLWHfCDPpoo6cAvIIHbKSVYbKHsOAN8LRZqufrOTseyyaOVszel538wg3tp+6Q1X3uQAHssO+wwPrGxtcMErQoqh1vJkjBCxqqurKyTxmEkgoNZulCy307Jo3ZJ9FrVgYyrtr5HYLsea2dFV1lQRHWE9IPmtwWe5NpZmRh2Ij3wgxqiofR1bgYiAD5hapTzw10BjcwEkbbLN1DNZqijBgLTKe60KirYKOnc5zsEdt0ENVpsQyGoa4AHJwtMkYaeQPcMhpWvWqgvetqs0NmDnvBxtusq8aIuul2mO9xljyPMINq1dwbXR+C1gyNuygjkfSxGMNy5ajBS0/jHoxlahbrVHU3OOKQAtJHdBtmCjmqpeqSMgHvthZDre2F4AOVyXrPT1FabIyopgA8tySFxjRVjpHlspyT2Qbt0k4U80YFP1guwdsrtJwx0DBrMRUUtvDOsAE9PquGuC9stU1XC66dBaXjGfrXeTh5ara1sbdPtYZugdPT3yg4u1/wAmIoaCK60VGZS85IaMlcz8uPKEy5W8SVlpLD04+JmF2N4N6A1Pf3tGqoyaQHI6hkYC7D2rTlHpyNsdpiDGdnNx3/Ig4q4Vcutm0NMJaigifh2QCAVzA6x2WMiX7mwAs8wwLUGnIBOxKEBwIPmg2Xq7TVu1PTupWUTT0jH0QMrr7xQFHo2wz29lqaXOyB8O/wChdso4I4i5zBglcacR9KWK8MJuIYcu3QeSPEm+VNBf5qp9OYWFxPbAWl0vFe13K0usjgx0j8tDtiQVzbzw6W07p2glltPQH9BPwronoyluczpbjhxDHkgn60Gua14dVVJWOvjZXPZK4uDQdsLVNBaZpLo9jKkhhJwckBZ8er6eugNBcXA+GMb+q421PrWssVy/6jSFrQe47IOcNVaMttgtpqGysflue4XCZo23m+NpmgNDn4/Osmg19qPV8Io5HvcAMHdRyyGxVAkk2nG49coOTKzS0WibIy5CcPJYHYBWx7vrqnuVFIzpAeMjturJdY3S7U3ut0cfAAwM+i2TdZ6SOpcyn+iTvug3Pw50TLri7yCZxcwO2C1viDR03D/NA1gBxjspuEesLfph5le8AkZ3K0PjFqmj1TcvGY8EZ8ig2vaIZ9R1R8GUxh3zwrdRWGW1EtllMh9c5WNaqiWiIdQk5AGcLLr7ualpZWu+M9soNJtID5el24CkuNU2OoELO52ylPG6KQyM7HOFDKwSVQleNwUFKhlRSxtm6yQdxutXtk7JqQyvABCPfTT04jkI2WmzTinHREcN+SDOiuPXOWMd0hp9cLPm1r7rSmid8RO3qtsNnYH9UW5KyRTU0sfiTY68bZKDTqiN9TUGozs85WsWu2t2kdnbfdYDcdfSO3kst1e+CHAOAAgyLvUQQxlrGjI27YWjwAzHqztlG1Tap5Evl6qsk0UAIZhBHXjobhqwmve1wcMrU6ICskw8bfWsqaCkiPScAoNJfVAtA6dwtSt1IySPxgcHusSemiGXDGFDBXSwPDWn4RthBqFwmfgxucSBt3WHBGSMg4WRVzRTRdTTkrFEvhRHPfyQZIp3zHBJIV3uTYskjBVLRWRiUibzO2VPXQVM9QHwjLM+SDGmkZG3AaNliuqC4HpJGVqktK0R/H3A3WmTMjz0sIz6IO+vsWXyu5pdU9byR+wCuxk//fG3IpPYtU00PNJqh72kNOgK0A/P7o25EHtGiIgIiICIiAiIgIiICIiAiIg455jz08vHFF3poy9n/kMy+aeCPxqqQnyJP519K/MkM8u3FIeuir2P+QzL5pYy9kz+jORn9KCeapDAYm4281hFzneZVZCes575WVT03iR9XljKCWx17aGrEr8EehWbqG4i5Oa+IAAei0Rw6ZOn0WdEYg0B53KDDY58bsnOAkkoe8EBSVLQB1N7FW04iLvjQZ1JVeN96c0DbCtqJnQyiJrtj6Khja9v+1QC/wBQqMp5GDrqRuNxlBqQtUToBUFwyd1p1XQyxnqYSB3UbrnUZEfVhgI2UlbXucxsbT5IMYPwAxuzx5hUe2V2DISVWCMk+I5TSOZjZBrPDy/zaM13pzWkNI2rdp+7Ul0EDnFglMEzZAwnBxnoxnBxnK9Sr97bfS1CG/cTl7utY446hWaijpgDvnBbTyZxtjtnJ7Y38mIat0L8tGd1SqqH1DwXggfNB7JcN/bC8vvEe5w6S4rcPLtpGjujm076qpkiuduZ1ED7+Q1r2szsSI3AdzgAkbX9oz7PvhlV8MLrzE8A7BQ2K4WOnN0vFttTWtoLjQYDpKiKNvwRPjb98JYA1zA8kF2CfJZ8cYiyHDK97+AFfUH2a9krtdyB8MXDOt94fL1Ee4MpZhFnqwSPdwwemBtkYKDwN93d09SNPSDn6lVlQcdJ7KkoGMjz32QalpM51bZT63Gm/wDSNXvJ7Ub8BXiZ/wCRv5Yol4JWSujtd6oLnMxz46SqiqHNbjJDHBxAztkgL3t9qN+ArxM/8jfyxRIPAZvU89IK9q/YxUVTScq2oXzx9LanXVfNCcg9TBQ0DM7Hb4mOGDg7Z7EE+Kkbuh2ey97vZeaSGjeULSlJVFrLle31GoamEDBZFUzPFM4+vXBFE4Z3wR5YJDxt5qWH9tLxjcP+H+of5RnXK/sxJG/t5uGjMb/9Wf5HrVxNzVSFvNJxj9P2f6h/lGdcvezAha7ne4bVHp92Pz2itCDtT7Y3mH4haTqtN8BNNXF1s0/qK0uu95lp3ubPXN8Z8TaZ5BGIR0Fzm/viQDs3B8raS711ruVLdrRWS0dZRTMqKaeF5a+KVjg5j2kbgggEEbggL0E9te3q4+aEB/4Ij9dqF58Np4mt6s7oPoW4x8d9V6J5Lq3j7YoKRuoXaTt91gEzOuKKpq2wDq6ezgwzlwB2JaAQRkLwH1pqq9avv1fqnVF3qbrd7pUPqq2sqpC+WeVxyXOJ3JJ+wDAGAAF7V8zbiz2WVU4eWgdO/pol4YiXx5Whx2Jwd0Hsv7GbWt5vvADVGk7jVSTUWmtRkW8PdnwIp4WPfE0eTRIHvx6yOK8qOOmnINL8duI+lKaONkVn1bd7fG2P6DWw1ksYDdhsA0Y2G3kF6kexfpoafhRxB8Eg9V/pi76/dgvNDmic9vNLxkeGkga/1D/KM6DjGekkawuPZY8RcMnDtllT1vit8Mbb9llUdNC6AulwCAg0mSsAcR0jI81uSz6diulE6pcQCBlbfmhgEjzsQOyyqe711LCYaQno7FBg1bW0lY+EbhjsbLUYbI2ugM/VghucZWkv8SScyVGcu3JWU2uq2tMVNnpOx9EEFPRmarNL1bA4WbW2J1Kwua4lYLnT08njA4edyVqFPdTNEWVB+QQaZDGXyeH2wlVCYnAE5KklPhyGRnbKhL3zSAuyRkILY3ujdnBwVWSTxHh3otQqRTe7ANI6seSwqdjXZ6hsEEwnbOzww3BWPKwwnGc5UspZHuwAHChLzI4dR7FBdBOYTnGFqMde1+HO8liyQx+CD5rDDiAQCg1OvubJ4/DaMEbEhYtKB0OLhnI2WP4TwM42WbStHT0oMenj8SoDD2JW/NG0sVLfIJy0ODcHC2lHSPYQ+Mb91ujR1V4N+gNWcRgjOe2EHqlyQaosVYKeiqrawdLgOosC9DPCtsMAn8CAMa3OekbBeW/L1xN0Vo60wVDJIhOMH6QG67WUfNLYKi2db5mFnRh3x+QCDmW5cW7RbK99JKGhjD5rqBzRW23a8mlutFcWxAEuwH4W3+LfNFolssvudRGZ+k9njOV0t4kcy1/uFbNR2yZ5icT2d5ZQbyu+tItHVApCBUFu3rlarpzh7VcZZ2y09E6Pq32GFx3wybFrarimvW5e8Zyc+fzXo7y9cIoKaOlqbXE3wukOJA+SDjHg3yXXKOdkskL2hpBJOey5k1poCg0VYn2qeFplLekE98rsxR1Vu03QGJzmCQDcbDdcBcYLRqTU87rxTMcaSJ3U4jtgIPObmB4UXh1dNc6aWSOJ2XAAkALqtLqHUOlri6njucoAfj6R9V6D8x/E7SdDZH2MyxCtZGWkbZzhedmpnR3KqqKlozl5I/Kg5Gt/ErUlNQxzC/SAgg4612k4Ec/s/DahFHcbn4vS3pIe4HP5V531FyqmfeA4ho8ljsljdkyhxJ88oPbHhb7RC1a3uDaeWtiAe8NG42XcTSnEDT2pLfHVQ3Wm63tBLesDyXzW6O1rU6Uqm1FF1ghwcN1z9pPnR1zZpooIZ5hEwAfTOMIPe+C92qqlMUFdE9w8g4LKknazAByT6bryl4M861VUvidc6k9ZIz1PXc3SHOVw2np4aa73COKUtHxdYzn7UHZAHqG4VHRxuGHMBHzGVxZBzLcLJ4xKy9N6T23H+tbn0/xS0lqVzWWqvEpd2wR/rQbjltNrnyJrdTPzscxA5C6pcxHJRZeK9ZJXUFvijJd1BrQAu3DSHAOHmqoPJ3Xfs6JYYnU4oSBGw74OF0p4zcALtw1qJ4IbdI8NcRnBOF9B2vrbV19rPueOvBHbddZdc8slNrOz1dVd4mmXBOS0IPFrhtZK5t6ifPTvYA8HBGF6kcrnMdb9EPpdMS2Vsz5YxH19GcH61wXqflaudJqQ09ihdtJj4WfNdyeWHlctVBTxXbVNMTUxMBaHDcn7UHbrT11jvVnprpFEI21DA4NxjC1JY1voILbSR0VMMRRDDR6BZKAiLSL1qO32aB8lTIAWg4HzQbQ412mivGl5Yahzchrts+WF5KcxUlt04a2kjawkvIBH1rvJzD8wEFljnijmwwNOPiXlrxv4n0+sKyoDHhzi8+efNBxBVyNF0dUtcACcjBWkXnrqZesOzg5zlZMluuNTIJGNJaVDPTSwMLZhglBgvL5IvD6uw7KCJ01NJtn8iHxYpcu2Ge61SlkopGZeQT80EtJfXUjB17+e4WqP1pTvpTC5gycjstt3E0xBER7LCgja879gguqZ/GqXTtGMnK1SkvkcMXQ5u5Cx2xQSDpZgkd1izUpY8j17IL6uX3l/W3sT5LdFj07DNb3VDyA4AlbdpY2RtBk7BZR1BU048CnJ6O3dBjVtU6GpkhHZriPrWBLJ4hWbWGKZnin6Z3WnIPV32GxJt3GUek1g/RXr1LXln7DX/c7jL/hrB/0a9epiAiIg4M53adlXyta8p34AfS0o/JWQH+ZeEFrskdO6qLCCPEPb617sc9bKuXlQ4gMof7OaWk6P45Bn82V4SaZFzpqWtdcQeoSHv65QabqCijaMRnpJ2yCoLfpXx6J9U6YFzRnBK0++3SZ9UYwdsrWNP6V1ldKcz0TJDT4ycZxhBoEE89JWiFxJDHfzrn7Q/Hym0zZDa3AB5Z0/bhcSy2EUzyyqGJh9IY3ytEqoaSOqDXA5zjug5aNYeIFzlnmn+CRxOM+q0DUPDqKhqOiEg9W+y0e21ddRQNdaw7Jx2K3Laqq9V0zHXAE4Pmg2jedOyWSATAHAGVi26+RTwGmcwEnbcLf+umRutvS4DPT2XGNvjp4JDI7AAKClfZix5q8kB3ZYPujZASSCR2Wt3mtFbT+BSOyQPJbXhlqqKoaypJDfmgVTHw5ABA9Vj08Je8PccgFa9c30dRRjwSC9aVS0NU8FrAST2CCtbMPBDG4yFpu5O+VrIsNY0+JMwgFYNVQyRuJaNggiipjJ5+Stmh8M4U1NT1UpLYwVZVwywvDJcg/NBj4ymMLLt1vluEvhxAkhSXK1VNvfiZpCDAyQt12m9wU1vMErQT0rbELWudgqd8UgblmcDzQVmaKure9uACcqJ8D4ngAnurWuka7DMqZkm4Mx3QJw4xAnurrdO2OdjXAYJwpHyQuZ0kjssOTpY8Oj7hBua8RxPoQ6MgEjdbVGScb91kuuE72iNztvrUbOkOB+aCr4PDYH5UbXOLxudysiolD48fJY8JaJGl3YFBr1FZ23GMNdsR2Kw7hQfcqUBjiVmS3L3emApDlxG6xYXT1p6qrf60GJPWvlZ047K+luAp8te3Pzws2enpWNw0DPplY3u8MgLG4Lj2QKq5MmaBGME+i3NZLL75bXTF+HBuRutpm3TwESTNwwH8q1u13uancI43HwhsfqQYho6uGtcyV5LQcblahJZjVQFrX4yPVZlyEV0g/6n4M3njvlaPHUXChLqWYkPOwGEG8OGGs6fhbeX19TEJQ4ggEZWs8SuJlPxLl95pKcR48gMYWzrNou4X1/i1bXFhOfqW6DoZlmg6o2HpxvlBs61Wx75yXg7K+quP3JuDCBuCtXbXUdHK6MkBx2+1aBdKGW41gqWg+GDnI7YQbmq76/UNAKVxJAbjdbMjsbortHATsXb/lW5bdPQUsQjjI6xgbrFrIKk1Yro27NOQUHI9FaHWm1w1FPKQQA7YrmLgTxsn07qmmirZS5gIBydu64BsmtaepjbRVTxtgFbltl807QVzarLA9pB7hB7UcP+P2m63TFE5kkUby0A7gZXMOn77SX6jbVU8zX5AJxjZeMujuLlyuMVNQ2V7yA4AdLj6r0E5aNfXI0cVDdSepzB9IoO1iKKml8aBko7OGVKgoey2nqzTD7rSyFrjkZPdbtUFaHupntZ3IwEHmhzl8O6usjnZ0ueGtO3ddG6YnTlvnoH0paS4jJH1r1e5k7XTUsVTPc8BhYTv6Lzh4m1+kWif3d0fWHnthB121CKmGrdURylocTsFh0Nm+64Jmdknz7rP1HPHPVOEZHRvhYcNRVUlI6Sn8vRBqdFJT6HzUEtd8gtrak1W+8XBtVHs0HOPVaZX3mquMpjq3HpyRj0UMNG2WUBuS38qDfWmKpupx7i8hnQMZ7LQdY2wWSt8BknXk985UIqptPsE1ESHu7n0KxZWXa/wAnvk4Lh6/JBhzTVcbQ5kjmg+hUTZp5CS+RzvrOVm10bRGIG/TA7KGloZH7Ebk7INSsdSKckv3B9VDc2OrK4OYcNyFa2lmpfidnAVonD34afiQa4II4qUHqGQPVbfr6lzJSBnf8yyp31kUYMhPQoP8Aa87SDguI2+tBitrJHHHUfyrNjPjNDCckhYMlG+Bxe4HpG4KR1Xhu2PmgyJYDRHxM5BOcJFJ7ych2FWeV1ZH0Dc4wsPw6inJABx8kGrxU7SMl2/1rGrGANLQ4HPoViuq52Nxk5UXvUjnjrORlBPSUjnPJ3UlVby0F/UVLT1UMbe+4WPV1skri1pOD80FtDP7s85VlbO6WbqDjhWCN7CXv2yrCQ6QE9soJRUlwDTlV91dI0vA2CsmEQA6MZHdZ1LPEabw8jqOwQaaS6N2CchXsJnkEfYEqskL2yFzxgEoAAeqPuEGuUeng5gmEmD37q+a5x24eAfiIBGRutJZdq6FvTkgKJ1PVVhM5aTlBPPczM842BU9Pb2vjNQ52cb91pMkMkJw4YIU0ddMxvhg/D6IPQb2M1ayo5odUU4b9DQVafyXG3j+dFi+xbDP20mqXjudAVv8AKNuRB7RoiICIiAiIgIiICIiAiIgIiIOOeZE45d+KR9NF3s/8hmXzUU5HvEhPY5xn619K/Mj+DvxRz/wLvf6jMvmiw8zvDO+T+lBbN/ZTj1WTTVohjLCPkrG07mnqkHfurJmNJyzCChIklLwD6rUIqDx2eITjzWFRgeKerZZNVVPhwyI4CCQ0wkPhZysWqojTnYkJSVZZL1POVkVlUybB2QYtFVOpZhIGlwzuFn1dYa0ZDC3AU9gjoJJg2pIx9S1C/C1wACmLc/LCDa8jTnAGT8lNBRyPb1yNJGM7rIgfTufl5H1rW6KKmkpjnGQEG3+kSEws79tlQ2yfAzndZlHCxlxPXjpz3WZdpy0jwBnHog5u5FOA/DHj7x3bwy4tXmpt1uuFnrHULqarZTzSV46BExheCHOAL3BuDksxgjIPZjiD7E/inBcpDwv4v6UudvdIegX6GpoJo2EdiYGThxBwMgNB74HZecJq6l7w6MuY9pBBaSCCDkEHyK5e0jzf81WhbeLbpzj1rWmo4wGxwT3WSpjiA8mNmLgwfJoA3Pqg7ucGvYq6gg1FR3Hj1xPs01nppBJUWrTQnkfWAOyIzUzMiMTSMZIYXYJAIOHDeXtMudHQGieFlVyr8Fq6hq7lc6SO1XeW2va6ks9tYA00bXNJb4r2tEZYMhkZeDhxbjzu1Xzg8zGv6N1p1lxt1dcaGVhjlpPulJDBK05yHxxlrX9z9IH8wXFtU/3x4fIMYQae6me2PxD2xlWB+x6lqNS5gh6GEZxhae2NznjI2KCaho310/hR7Er3/wCYOhPNzy1cYuDei/AOrLTXzWmS2yTCJ4qqKtZUUuS76LZ2QROa44aS8jOGkjwQppYraBUR4Lsdl379pp/srctXNmzixwo1dedMR8QLXFWtqrZUugElTThsFRC8A4kGGwykOBBMwOMhBwvwp9nvxquuo5bxx20zW8MeHWnXe96m1BfemmEVIwgvZTtJLpZHgFrCwObkjJJIDu/ns5eZ+w8WOKnFvSUFPFZaGoqaKt0daSQwQ2akp20TIGNJyDHDDTOc0ZGZHkdiT5V6/wCYrjxxjbBRcVeKWo9RUVMeuKkrKx3uzX/3XhNwwu/7YgnHmtrW3WuodIXui1FpK9V1outukEtJXUNQ6CeB4GA5j2kFpwSMgjYkeaDvzqblg1/wu51eKHMDxL0mKXhfYavUesp7zXtjNHWMq4ah1NTRZyJJnT1MUYYAXAgnGcA8H+y5qh+3e4bQZ3d92fzWitK4d4n8zvH3jTbILFxQ4rahv9sgkEzKKpqiKcyDs8xNAa5wycEgkZOCMlbG0tqzVOgtQ0WrNE6kuVhvVvc51JcLdVPpqiEuaWu6JGEOALHOaQDghxByCQQ79+2xOOPehCDuNIj9cqF52uleduo4W5+IHE/iNxXu8eoOJmub5qm4wQimhqbrXSVL4ogSRGwvJ6W5JOBgZcTjJJO1UHuzzJU7qv2Ws1Oz6UmgdOgfloivDius1RQfE879xst7T8xPG646Lh4cXrixq24aUp4ooY7NU3ieSkbFEQYo/Cc4tLGFrS1pBDeluAMDGy7je5a8AOb29UHrd7EqSSThPxH8RxONRUw38v8AawXnHzT1EUfM/wAY2Oxk6+1Dn/OM62hw942cX+ErK6HhhxO1NpWK5498itNzmpmTkAhrntY4AuAJAcRkAkAjJztK5XO43m41V3u9fUVtfWzPqaqqqJXSyzyvcXPke9xJc5ziSXEkkkkkkoLBKBP1HtlSy1Uj8MieRnY4Kwuoox5YQR5IJ5YZ2sLy4keazbZKxhDHgb+ah8bqiw8bFQ+KGPDmnt2QazdLc0weKzGSM7LAoJGRRnrG6miufiR+HIdu3dYdQ4OlHhdigtqqnrkIDRjPooImufIGjO53WrMpoXQgkDqx5qF7YoB5Zx6IFTSNZTh2d1p8b8beZ2Uk1ZJJ8OfhVkdPJIOpoyAgudTvx1dWx+asDzHsFkSyhrOnzCxASXAlAJJO5KN3cFkAR9AVzBH5Yz5IJpoCKfqz5ZWnDY59FqQZM0ffAeg/oWNJEHTZYAR5oLTUfB09GB5bJFO9rxhu2fRZtTFTupg2PHWprZFSsgd7xgPQBcGRRAkbqtPdC5/Ux3S4dj2Wm1bT4p6foZ2WRS2yomb4sIJaEG8rPrO9UYa2K4vYAe3UuTLZxMvRsj4Dd3AlmAepcBCdsLvDfnqaVltvckERjaT29UGRqW732e6yzzXKV+XEg9fZYtmulTJc4WVEhf1vDclaeKp085dLuD5KRssEFUydpwWu6kHZq1VkOjtPwXaKcBxId3HyXoZyT8w1qu1tht9bVxhxYGguPbZeRp1XcL/bmWlhJDcDY+S5U4W8VanhfAMOcx47bkboPc640Niv0Tq37uxMB+LHWFxlxT4xaU0HoG42kXCCWZzHNDuoE5wV5Uam559fU0LobZUy9J2GJD2XFV95l9a6vglhu00ruvJyXkjdBpvH3Ulxv/EWsuEVwe+CSQlo69gMrYDK57GEOJJOck75WPebnU3CsdUyl2ST3V1DJTmE+L3QYc9S2R/UWhY5yewOFLL0eOentlZ9OKXwsvI27ZQafA1z3ho2Wex00J6Wv3PbChcGmUiDzOy1CGB3gl8g3wgybfertZ3ePDXOb54B7Kyp11quprRNHe6lhBHT0yEALRxL11BjlPwZIV8tMzrDoRt3Qcjs4wazt1tjYL/MXAD9+V2L5bua682S600dzvTyBgHqeun1DFTS4ZVAYCkluUFonEtuJ6wdsIPd3S3OBYaiw0001yge/ABJIyVzbw94n2fW9KJoK2IvIyBkDK+ejTXGDU9GI4ZHS+EHDGXHC7ZcEeb6bTUkFNNUEENAILkHs51Qy7BzH/nUVRR089PJA6Jpa9pBGF1L4Oc3Vgr42PvdS0RyYGS8ZGfrXaLTGrbPq2j99s1QJYsA5BB7/Ug27a+GVkpbw+4z26N+T1btzut4+5U9OwGmhbH0/wBztsstUIzsgxIqsl/hnfHmsvIwMnCjFPG09QG623qu6TUMZEJOewQbn62f/NB+ULj3ilY6KutMtS65MhLcnBeAth3jiXcNNSyT3B5EQGck4H510u5nObi/wU1TTWSZ/QCR8Lv9SDj/AJ0K/wBx8eCiuweQ0jZ+SvPuCWqnqJKiaZzsOJOTnJyt8a24wah1tdZPutI8tcS0guJK2bPNBBlrMDrO6DWKfV0NNF4LmAkbdlot0vT66UujZgA+ixW0MlY//a4y4+S3HbrbQUdIWVoAlIOAfVBhWmgbeB4bgAVh3yzPtMga07LOoXzW6udLF/Yy7Y/JL3UPuXx5yQg0OGkdLu5351SUe7ktHdXMM0Enxk4V73xyvBcQgghkfA4SYOCtxUVDHXweMcAgZWnOFNJGGtxkJHcJKKMxx9jsgtrYvvhiaQMHGyxxD4TTk5UU9RI5/Xnc/NSQRVE46xnAQYz3u6iD2z2VpGQSsp9P4pDGD4h3UMsMkILZBgnCD1X9hr/udxl/w1g/6NevUxeWfsNf9zuMv+GsH/Rr16mICIiDgznducdm5Wde3OUAsgpaYn7ayAfzrwIq9ctuHvLIGdIe4uxjHmveD2hQzyccSR60lH+vU6+frT8MMoqDJjIaAMoMqmpvuhVCRxzggn8q5w0nxStOlLC+1yQMc8s6RkAnsuDKaCrhqiYs4JWr09vlqqhvjg790G9La9mrr5LUNaGse4kD0Wj6y0h9zqoPYcgnyW4bFQvt0IfRjDvkotRuq6mme+pJ6gD5INJ0vUw0zmxSlpI23W47nqSlo4w+MNBA8lsLTWnr9froYbe17sHAwCtZ1Jw41ZbAH1sUgb3PdBo1+1m65OMPUcAELQmdUmY2n6XzUNzom0j+gg+IO6yba3qhLjjrHbKDIoqc21xnkd1h2+5ytOur23GoBYA3dZkUdVLI4TZ6M7fUsO40s0LvEhzgfLsgs9wfSxh5dkei3PYYoHRiaQDbyW1qKpkqn+DI7O+FqzG1seIIM77bINUv19p44zDG0Ejbt3W3WVEdTG4uGD9SzKux1rGmeqaQDutHma5m0QwAPIoMikr4qCQkj5JX9FzkEzcAALSXNe92DnurmzyQnpBOAg1vTtXHaKp0soBGfrTU96iuUoMTRgfJaUWy1bQIck53V7bdMxp8UEO8hhBhta4ODmgkla7SwF9GXPaRt5qOz0jI5eqrx0gjutZrauga3wacjfZBtKPDKohw2yVmy28TsMrTuBlWV9vlDvGjGx3UdLVyMxFITgnCDCcHteWnORsqOaW9ytRq4AWCRgGSsEwyndyCNMn1VTjGPNUQVwfVZcFD4zcl26xot3YKmM8kZAYTjCC9zH0p2+ILLpKt0/wdGCdhsprNA2rcBPghahJQQUlaxzQOgdyAgxZrc8M8R+QMLDghkjqRIAXNB3K3Bqa40Zt7Y6UjrA3VliZTPtT5KnBeGkjPdBWobDdKUQMADgPLutAdTupJjRnYu2Cy7Gamquj44MludsLKvNvlgrmvlBDgg1bTtrfZ8VkoLw4Zx3WbUWoXevZWtiwBv2U9jmZJAxlWR0j1W5I3UsceKbGMboJI9R01jomRMgBcBjss5+oaa8Wp4dhriOxWHbNPi+yOYW+Xotp6rp63TlwFLHkNLsHGUG07/SVDbi+VpPTklanZ6+N1MaWQAuIwp7pJS+6iSUgFwyVolFR1VTJ4lKCQN8hBkXG1y0bjVMc4gnqwCrqXUYkgNG9uCds4WpOZJ4AZWAgAY3WiV0NIxpkpwOofpQWmyzwyGtimJ6t8A9liyi5vlJEjx9qztPVk8lR4VYT4YO2VuqSC2Fwc3pxj5IOUuV6YQ3BhuUow14Pxf+1dvtO8a5bLr+itNA8mIuDSW9sZC8/6LVZ066N1rcQ4uA2PzXbnltgtWsLjBX3LDqsYLcnfKD1q0heW3HSNDcusEvhBO/mtcpp/HZ1EYXGXCikucNogpKkPEAA6c5xhcoRxMiGGDAQXqhIA3VVo+qp6mmss89LnrYM7IOsPPz4dNoY1lJIBO+IghuM7LyTk0dc73R1NxqJnjDyfiz6r0H5heJM1zq6mz6gf94jBaOo7YXTXXOpLRRWqogtD2dLiexCDrrfG+51j6XqyWEjutPfemxUroT3PZY9yuDqq9SyP7ElaXcXM68N80EDh7xM5w2yVm087aXHVuVp7Q9m4yqPe4ncoNakqG1jQMjZZ0Go4bfSuoxGCSMZAW2WzvYPhKyY4PeIzI7OceqCyWpL6gznJBOVm090ZGQ/AGPktJeOlxb6IMk4HmeyDWqm9Nqm9AG/1LDpmFtQJXO2zkrFdTyxjrcNgqCd4GMn8qDX7tcYp6URNxkLQqbqErXZ2ByrH+JjLgcFWh5GyDW6udlTTCNoGcYWlvo3sHVuprfM1snx9shZ9XUUzoz0kZQaXTTe7vy4rLkqY5G5xhabIepxI7ZVOt2MIMjqErukD5Kx1Oc47KWjDGO6n9lfLI0zjp7HzQYskT4wM5wfmp6eDLfFdvhTVjWeFkd8LCbO9regE4KCaeobICwDGFjs3OPNUw4b+qoCQcoJ3U/QOouykcb2kSDcNOVGZnkYJOFm0ssIb0vwgskqPeW+HjBCsYzwjlxz9akqDCz4o8ZWKXSPGfIIJp5mHAA7fJZdLdY4IvDLVprI3yn4RlPDLHhr9kE9XUidx6W4yVY2mLmF+cYWVG2nDN8ZVHyNblreyDvh7FYn9tPqkHy4f138o25FL7FvwzzSap6cZ/YBW5/zjbkQe0aIiAiIgIiICIiAiIgIiICIiDjjmS35duKX8Cr5+ozL5p6eQRVLnH5j86+ljmS/B24pfwLvf6jMvmhcCZXADu4/pQZ1VO18fw91hQu3I9VJ7rO1oe4bKrQwAHzQSMhBy/GEkhDsZ3R8/SzDSscPlduM4HogkFKXnDRv8k90ka7DgQCsy2VUDZA2U4381n1r6edoFPjJ7YQYNPbXNHU2TB77KSa2STML3SZx81B7vcYj1uB6f5lksrXiMtz9iDTKemLqnws4we61LqkpXiJrzg7FadMZWS+IzvnyVfGkeQ52c/NBqlVTPdCJWHDu6ybbSGSBxqNz5ZWA2tkMfSzv6K+GvniYWvGMoLY5IaetIeBgHzUtfPBOMQ4+xadOfEkL98lUYHM/+wgvpaUyTZxsFLXn3f4RjJCsZOYQS3uopZDUSDxNsoKU7JJXBzuynmDIxgAZU0TY2MGHdgoZBJK8ANyPVBbQVk1uuNJchEJRSzsnDH9nFrgcH5HGF6k+0z5muWDjnwB0g7Qeu7fftTtvMNwoqamyamipn08gnbUNIxFkmIFpIcXNaRkAleX1TCxlP5ZWjuOSg1m5V9O9hEGMkeSwKeQOaes5+tYmD6K5ryzZBdI4dZx2VvVkjZUJJOSqIJ2ffT0juqSRGPuVHG8sd1NV8kviuBKCzbPdCR6KZ0bGsDtsrHO6ASSg3RVBH0sIJmUjnt6gdlG6MxuAKkFW9owFaXGQ9Z7+qDIl3h+xYZBAyVIJHv+EDKNyX4OeyBBA6c4acLIMb6Y4c0nKutgxU9J7LWqw0YjBfgnHdBogqSzJJ8s4WNPO6V2fIKaoaJZOmLcZV0ULYmHxsAkbZQYS1q2Fgpj14BHqsCjYx1QQ7GFPXvMRDITsUGFUn7+4eWVERg4V7mvPxHuVNHQVErS9jcgIMbJ9VfHkODidhuqvZ4eWOG47qjWPf2QZ0twEsIgaMEbKKF3hsIO5KxnRvjwXDCNEjxlvkgkhe5suSTjPqppY3SvDmuwB81FTRudJghTykxODfU+SCOokHQGY3C1Wy3yOlgNK9o+LbJWnOh629XqsN7DG8YQazX2xshNW14w7daS9j999gsiK4zOxHIctV8ga5pczcFBp2SD6LW7bYDXwmYvx5jdaXBD4svSNt/Naw24zW+Dw4z8kE1rlbp2tL5HAhX326zXyQSU8pa1v2LQJ6iatkydz3V0T5KVpbuCUGoU8U0gEckvUQlVGabIBGPPbyWn09VJ4uQTusqpkdK3fzQYrpBOekrHkaY3YBKqWSROzhWkueQPM7IKFpG6mjp5XsLgCMLKprfKG+I/t5bLUY/AYADjHnsg0SF7qeTLmn7Vqor2yQkDuFHXOpHMIZjJC02B+JWg9icFBZIXGQuwe/dZENT0M33OFrNfTUgoA9mOshbdj3cB5ZQZbJpKh3S3O/5lDKx8EoLiTg5WqUYpwwdOOrCwLl/ZAgzReB4IiYAD9SrQm5e8Cqhq3N6TnZy0VanQ1oib0u7IOX9F65vk8cFCy8PiLJAD8ePNes/JZxDbR2CK1190E7pImjLnZwcBeHzrtPQSMlo3lrgc5yu5HKbzBuszIqa4zHxQ0BvxeaD3Bpqunq4hJBMx4PoQd1OurvL1xVuOr6lge5xhL/AIcuXaJAWh3yy+/Ay4zjfC1xUI6hjyKDohzs6nuelrJMLfC8EREZa35FeRmqeKl9u89VDW9ZzIR8X1n1X0A8e+C9p4j6flilhDpS0t3Gc5C8peNPItfqK5TvtFI8B8hx0sPmUHSItlq5jUuGM7pPQOLS/qycbBcxa45cdY6ApPerlA9rA3qyQQuKYIpnVRa7PSx2HflQY9ouTrXP1yRktzvkJeLq641ImjJDR2wcLX7pBaXUI6OnxMeS2x0RMDuw9EGTJdQ6mEO+QMZwrqKqyMPd3WlOGDt2UkLi0bd0GpVzWvYXMxkrScHOPP61nML8fGCAfUqGSme9xdGMoJqamLcvLs58lFWbO2KoZJWN6N8hQlznuAcgq374A3C1GGvbSwmItySFHBC1rQ491HVQdWXtHZBfRSvbUGVrC7J7BUucrpn9To+n7MLWdFS2xlSRcS0Nz5rI1pHbpJXS20joAJQelvsNgRbuMpI2M1g3+yvXqWvMP2ITYRpzis6PHWZ7J1/krcL08QEREHXn2gmBye8R8/3pR/r1Ovnwsok8aXw2l2wzj619BntCGudydcSGt7mjo8fx6nXgXoee3QOqPfiN8BqDLpp3RYzCSfXC16lnjLOstwQM4WZTTWFzi5xbg/UsSrEMlUBRgeGTvj0QZlBql9HL0PYensFvOjoafUVKXuwOpu62x9xqSppW+GAZBvt6q6G+zWSI0jCQ/sN0G8NI3G2cPLgamZjH4Od8Fbq1HxRs2sqc08dPG0uGMgBcJ1EF6u0hlma4xk57q91LU2mldKxpBAQZuodBUs8z66NwORnGVxxdi+z1BY1mRnstbh15XMq3U1W8hgOMH0WPcpKG6PExwQd8oMGlu5qWhrYjkjHZTSO8Rhiczc/JZlnFpgfiTAIS6PpnTh9PjpByg0c2z3B/vGNjusmkvsMUwkeB8O+Cq3O5QPp/DJGcY2W2QJJZcMBxnvnyQbwvus466l93ijGe2cLQqctdA5z8ZO6yaShpGRB8wGfmVplxnAm8OnPw5xhBdSRtqKgscNgr6i3NNSI29io301RBEJ2A5O6vpa1wIknJDgg12jtUNrgFQ/ByM+q0+tuEc1QOjAAUNbepZovC6tgtJ8bDuo57oNbuD2upwYjg4WhtdK2QOLicHOcqY1bnjBJVW9BYer6RQarFcI54BEQCfPZY09C0jxW7EbrAgL4XlzgceSvlrpHS4Dvhz2QXeOWHoIzjZSgtkZjCpmAs3IJ81jSzdGzcoL2UYc85UL4gKgReWcbK0VMoz8RVPvjj4oBJBQanU2llNTiYOyS3KxKePryTjbZWSV9RJGI3u2CyKWeOOM9XfugzKZ7aUdWcEK6qrnyxkDJOO60meqc93w5wFq1A6mNIXSYLsINJhbI+YeI4nHffK1qITGLwo2kAhaO6ZsdQSO2fzLfFifbn0fiSkdQCDQ7VV/sfnFQ+PdxytyB8OosVewI3WkXKSgq5DG3BAOBhQU0tTROEVPnoPzQaldZzSM6IzjCwaHU88JLHPOfrWQ9pmbmfv9a0p1NA6pEbAMuOEHLvC/U8Hjn3lwAPqsbioYq6o95pmBwBzkLjyunq9PU8c9PlucLeWmdTWy9WZ8Ve4GfpIwe+UGzaS3yX+YUpJHT5ZWvRQU+mWeFIASAsajY613OWpYMRkkj6lpGqdQMqpjh3Y+SDVKmqiu2WtcGjGy2ldA+3z9Gc7qtJc5GuzETuj546iraaog7oMq29VRjbpJWtPo5GxEiQ7BYU01JBTA02zsKkF1xTvdK7AAQbi0fpt14rYo3vzl4A/Ku/fK5wtrKK+UlVHE4xgA/LyXRzhDbLnqu8U8FjLi/xQCAD6r2V5SOF9bZ7NBU36PEoiBGRvnCDsnpqCOGyUcbYw3oiDTtvkLPfN0uDVdDCyGMRMGABhHmJvxOwgGVrWF7iAAMrhjivxmptO0s1v8RoL8tyt56711btLU0r6l4A6Cd9vJdKON/EqyXaCWtjeCWOJz1BB145stTe/wAk9bTVQaXgnY4XSp9/qpKeWKWqL8uIxlcx8Z9ZSarq5aGgJOAW4BXXqaKe1yugq8glxQYdaXsndNjuSsTrdLICStWmfBOzAxn0WlSxmJ/w9kE7y2Nu3osV7g45R0jnDBVG4yM9kFBsQVmRVfS3oA2Oyx5AwNGO6jAPcIJZh1O6h5qtOwiRrnDsVLROZ1/fOykrHxDHh4+pBJW1DHwiNo37ZUlFZ2zQ+O5w23xlaU55ccnyKyYa6rjjMcRIBQSVbWBxjHlsNlgluHdKudJI5xc7OT3Vud8oLzG5oyFTpkJ3BU1PI3bxO2Vmu92MJ6SMoNMdEWgEd1aDgjPksiCRrpD4hGFSpY3r6mYIKCJ0mRgHCtaSCHZ7FU7K9rQW5QSGUyDpJURHQ4KgPSdlIzDu6Cxz8gDCtUsjQAcKJAG5AWYyh6ojIHY2WNHC+Q/AFP4VYB0AHBQYzsglpPbZZsIb4R2WNLTyxfE8YVGyP2HkglilFO8kjKsqJRO/qGyO6XDfurWwPf8ARGQgs6iPNSRlzx0/NUkgkj3eMLKp2N8IuPfCDvx7Fyn8Pmi1Q/PfQFaP+cbcip7F2Vz+aXVIPloGu/lG3Ig9o0REBERAREQEREBERAREQEREHHPMf+DxxS/gXe/1GZfNF1BlQXEZw4/pX0u8yO/LvxSA89F3v9RmXzQlpdM5o75P6UGfJcmzQeCG4OO6097XNO5WRHQzAhzm7KOp2dhBASSMLPt0TZnCIgbnuseCjmnx4bScrIYySgIc/Y+iDVZ9LhsJnbJjzxlYNPCaWcdTiQCopL5WvZ4fX8PosVtS8yAv7FBuysr4ZKIMawZx5La7pnCXp7ZK1innhcwBxWLV0Qlk8WLsDlBdNTNZS+MG7ndadATIT5LWBUxOh92djIGFhSRsiOWY9UElMGxOLngELlTl04Aai5o+KMPCrR17tdpuEtFUVwqLj4ng9EQBIPhtccnIxthcSTS/eyM9/RdgOQfj5oflu5gqbidxEiuklnitVZROFup2zTeJK1oaQ1zmjGxyc7eiDsW32KnHkHJ4r6CP21v9CpR7Fnjt04PFTQX2Gs/oV2iHth+Up2wodef5nh/rCvPtg+U1o6jb9e4/8Dw/06Dquz2KfHVry48VtBHPzrP6FWVHsU+PMrss4raBb9Zrf6FdqYPbC8ptQ/w47dr7Pzs0I/8A36nn9r3yoU/9loddj/yPD/ToOp0fsU+PjdncWtBEfI1n9Cs2H2L/AByiZ0/7KehCfXNZ/Qrs5J7YjlLiAL7fr7B9LND/AFhXQe2E5S6gEx0OvAB3zZ4R/wDv0HVep9ivx8ncSOLGggD5E1n9Csf9xL49fjY0D+Wt/oV2sk9sZykRu6XUGvsj/wC80P8ATqz92Q5Rv7w19/maH+nQdWR7E/jyBvxX0D+Ws/oVa72JnHl3bixoEfbWf0K7Ut9sdyjuOBQa++2zQ/1hVf7Y3lIYcGh16fqs0P8AToOqf7iXx6/GzoH8tb/Qp+4l8evxs6B/LW/0K7Wt9sZyku7W/Xw+uzQ/1hXD2w/KUSAKLXm//wB54f6dB1Sb7Ezj0Dk8WNAn7a3+hVXexO49HtxY0D+Wt/oV2xf7X7lOY3rNFrsj/wADw/06g/djOUgDeg17/maH+sIOqn7ibx7xg8WtA4+us/oVT9xL48+XFnQP5az+hXar92Q5Rh/9wa+/zND/AFhSxe2G5S5mlzKDXuB62eEf/v0HU/8AcS+PX42dA/lrf6FVHsS+PIP++xoH8tb/AEK7Vu9sbykNJaaDXxI9LND/AFhB7Y3lIP8A+j9f/wCZof6wg6r/ALidx4xj/ZX0D+Ws/oVafYm8eT/2WNAj7az+hXase2M5SCce4a+H/kaH+sIfbGcpDf8A7h16fqs0P9YQeSPMTwG1FyzcW7lwk1ZebbdLjbIKaokqbeZPAcJoWytA8RrXZAeAcjuDjK43kb4mJGDbzwud+ebjdovmK5jr9xU0BFcmWW5UlBBCLhA2Gfqhpo435Y1zgPiacbnIx2XA8T+hvQexQXRvMLfFAxndQz1ElQ7JccemVneJAYek42C0+TpbL8PZBdTyGB3U5uQppXe+fG04wkxhMIx381ixyujBx5oDXGJ5wdx5rOizOzqIzssHHiH5larQuihZ0yHBQYQeHv8ADLcAHutQjubKKIxdGcjuoql1Mz42YBPorI2QzMc54GQgwKiXxpXSYxlXwTCPAwCrJQGyEDACsGzvtQZtU4OjyBvhWU0rWNId3wksrDFjzWOzIzjKCcVIieSG+ajnqHTO6iMYUlIxkjyHrJlihY8AAb+SCOkndIfDI2CjrmdLw3tlapTx08DRKcDZYFfieTrjxgFBAaQiMPBKkif0Rlp3UjZBJH4TdyFEYnsPxDYoIoajwZi7Gd1dVVbpz6D6ldMyPp+axR9JBLBOInAkdkqJ/FOwwpX0/VGHAbqKKPpdiQYQWxSGMjZTmrzv3wpKlsPhEtxlYbI3OIwNsoJzL7x8HSBnzwrm0wjIeT233V7DFG35hQTVBc/DScIM993a6EQNbgjZYBqHk9OTucJ7segP3yVfEI2gF3dBSaleyMShxOVHDEXfF2xgrJNQH5Y47DsFGZGsOB2QJq2RzTEScDbCxg4DOFkSticwlm5/nWMWOHkgvimdE7qBKk6zVzNa491ZT+H1/Gr5i2N4fF5IM2rs4p6cTde+MrSx1H6OdlkS3ColZ4b3bYwsqgFP4REmM+SDTw8uw12+Cty6Sra223aOrgqjE1m+M4WgVQY2TLfXZVhNU1hljyWjv9SD1f8AZ/cUGXuvp7VLWBzxIB3Xp8Oy8HvZ2aqqbfxLZB1EgzNPfbyXuzb6gVNDBP8A3cbXflCDJREQWuY1ww5oI+Yytv6m0/Yp6CasqrVDK+JvV9AZOFuJRzwsqIXwSD4XjB+pB5n85V8pr3FU2W26eDehhaC2P5FeY2prZXWSSpZLROYS8nJGMbr6D+IvAPSmp6aerNO0Tlp7gblebXM1yrz0UdXXUMH3trydm7Yyg86KNk1ZUgPccHc7pdKb3aQNadity6nsTtO3V9GARI35eeVtatfNLKfEzkeqDGzthXRPDHDKr7vL09XSrOgg4Qah4rZmBowCp4pWQREO3OFp8ZMe57Kkkr3nDc90EwlbJKdgoahoZICPXsr6SB7n5A3VauCRrx1A4ygrDOX/AAK58/SfD8z3V9IYW/XjCnbY62snEkLMsJBz6BBfbtPTXA9UbizPmtSrrIbdb5Wyv6z0HzWowudb6YMbgPAWh3e8yVEL6d7iXHbug9QPYcg/cbjAc7Gexbem1cvUZeXnsP6CqpbTxfmmbhk8thLPngV2f0heoaAiIg69+0AcGcoHEZx7CkpP16nXzyUtNLUueYSR07r6FfaFO6eTjiS70o6P9ep18+lmnZD43XjdoQOupjAZ4xyDjut86fLG0JlllBd09yey2HM/xZT0euyyGXOspYDE0kAjCDdUOsRb6x7C/I8t1ot31VPVXJtRHnoBBW3PFMk4fJkkndbrtsFpmpel4b4p2Hbug3RScRKeCgbC4DqDcFQV2s466jcGgbjZbKuFlqKeQzOafC7j5qtNNT+EYwdyg0+re6srHFpxuVdHUS0uIerOdlfPTPjeZGdiqxCJw65cZCCyoEzGeK2QjPkqw3GXwi1ziT6q2rf4oDIzkBWQMjjaRLgFBjySySvJLicntlSwVPgYBaCrZGgu+BTRNi8Ih/c+vqgrUXGSZoawkLEa4teHO3wc7rVbJDTNnLqoDp8iVZfhSCYe64wguF3Y6MRdPyWJPH4jS9u2d8JFTNkA6BvhVcTCeh/ZBDSwmaTo3wsmqoBGOoHdX00kTMkEZUNZUSPcQOyDFiaS/CvkY5jwcHA81PSw+JuBv5qapYxjSH9ygxJajrZ0gY8u6x8q9vT1nP2KsnRjZBZ1u/uiqEk90UrGtLCSgiU7Jw1vT0j7VC76RVO6Cp+Ikt8ypo6Yvd09WEijLXdTu3dJJi1wLCgkqaIQM6g7KvpGufHjqOFiyVEkg6XHZXRSuYMDsgz6O0irkILytTno30FKWskP2FaG24TQnMbiCVP90pp4yJCSfrQRUHiPq8uedtyVrrq8QOA6cjtlbahe5svU3PfyWq+PC6Il/wBLCDUKi4h0Y6SMkeS0cVzoasTZ7EFRQ1Q8Uh3bySQxPnAbuD3Qbwhlj1RTCkOxAxnC0yqsU1gqOtk5DfTKxoq42aMSQHBKxqm4XG7NMjiSAg1av1D4tOIGOHV2JCw4dPyV8RqXuOe60ujdGJSJfpA4Wutub4YhHEdsboNGZTe5zlrjkA43Vs9OZ5OtjsDKkmhqq+QmEEklXxQTUg6KgYJ9UGNM+SBoy4nHkVrGiLVLqq/wWMDIqSG/nWmzeE8t6yNyuS+BVgfPrqjqKckljwRt80Hdngfy8RcHLVRatqaMSiRwk+IZ+a9IuCWpqfU1jbUQ0wi6IwMAYXBmgNHXPV2g6CjqwXMa0Dt8guxfDHSEWk7QKSNuDgDsg3qtAuVwjjrWQPlDQXY3WvriXirHeaSJ9zog7piOThBxnzi5odOmpoa7DnRnIa7ft8l0bqrNJdtD1tbVXD74ASMu81ynx241Oq3TWm8SkhjSMErp7rHiTeGUc9LanO91cSD0nbCDheqv8tk1nVQ1Dy9geQCdwtH1U+K+VHvUOBjPZR3+ppq+ufUEgzO3PrlaUKt0A6HbZQQUtM4SkF3Yqapp2Bp2yVC+fqJMfdWNnf1YlPdBiOGHFAMkD1WVUeD4eW4yfRY7Y3kdYHZBdJD0NzlWBwAwjnucMHy2QRvPYIKZ32KplCMIgd1k08wB6HADOyut3hGQ+L2VK4RiUeCgvqIGsYXj61hZ3yp3uke0A5wsmCGPw8uxnCCOhpffH+H28u6z57T7uz6WVhjxInE0/f1Cjnrqp2WSE/agx5R0PLc9lVsuG4O6sc4uOSroQDIM9kFpyTnB/IrmvwMFa3FBSGHqdjK0mtDBL8HZBjndASDsiILnP6grVczGd1V/T5IJqaqbBgkZwssXZmfofmWlgE9lmxRwiI9YGfrQXVNUys+FowVCIgxv1qIFrJMg7KRs3VIGncIIHZyc5CyqerEIGWglXVTIxHlvfCxIw3rAd280E9VV+OMdICsimLQGDzUtS2ARjoxlQU/T4rS7sN0HoB7Fpjm80mqXlvfQFb/KNuRaj7Gl9M7mZ1KIcdQ0FW5x6fdC3og9k0REBERAREQEREBERAREQEREHHPMeccvHFE+mjL2f+QzL5o+tzKhz2jJ6jt9q+lzmPaXcvHFFje7tF3sD6zQzL5s7PFD7/MypxhoI+3OEFhuxMfQ6MjZQMhFSesnC1a6R21sZ8HGceS0Fsz2ZDDgINXpqplAfiAIGyw7jWiukHSMDKxhK6TAecq13SxwLfIoMl9GIohITnssdrBK7Y4wsiWqL4OgnyWJG5zTkIMx3UxuzuylgungxmJwySO6wDK4k9RV7Yg/Ligq9z/G8bOxOVkscZo8nbGVFD0ud0kjZVqpPCIZHgZ74QQSv3IyqNm6W4xuj2OI6z5qrIWvaXDKCkUzmPDycj0WYK5jgAcLT3DBwsuipDO1zyNggy6O4U9PIXkBY9zuBq5AWOwAsOZnhyFvoqMZ1oMp9Q2SnDTuRsoIpHxu6WuwCoz8JIVASDlBnOpXSN6u5KiFC8+qNrXhobkqRlYQPiKCGWmfCM7pBTPn+LuArqioM2wUlHMYmlpGAUFSA8+E0bjZY0scsJ+IkZKyYz0TF/rurKrqmcHNGQEEcUsr3BpcSFNJSuDeoZWI1zo3dt1nw1LXxkO+rdBp5BLiMbrNpWux0bjKtgZE6YlxGMrNd4MeCxwOCgiNomeS8HAKiMLaf4JMZWQ+6yR/AAcFYcs7qh+XbIJoqE1JyzG/ZWVFI6nd0u3Knpqk07QRglW1M76kgu7oMdjC5wCnNMCwknHoomNcHbDdWzzysPSgxn5a8jKBhcOpAQXZd5lSB7c9IG527ILI4zI7p3VZo/DOFmNpXQDxyDgqN8kcmSSMoMaN3S4EhZkUJqIy/JGB5LDeB1Yb22WZTTmKIt9dkGGWu8ToyT9qkc90WysL8zF6yY3RPcOrG6DDcS45OVUMJb1LOrI4WxhzcZWNA5vSQ5BCcg4JVzX9LcfJJCCThWswXDPZBdFIY3Z9VkNBmIf1Ks3g+F8OMqlLQVdS3rhaSB6IJ3h0rOjONlFkU46HHOfVWS+LTfA7II2OVAfEmPbKDIgd4b/Ed2KlqKkPAw0D6ljeFK0DOcLIhg64ySOyDCe8kqjBlwVZG9LyFfFE5wLgNwg1SmY0xjqI+1YVd8L/AIfVRR1EjHFuSs0UxqB1oNNLyRhZNIG4ION1HUQ+ESMYUTZHN7FBk1NORlzSSc9ljxgmRoOdyO6yqKYPkDZz8KrWtjMoMGMAoM98LW0wPyWiyn4sLVPGe6EMJ8lpcoPVugsyfVVbkkDPdS01JNVv6IW5KrU0dRROAmbglBeYPCb4hJOfIqIPa9wb6lWume5vSTsrQS0gjuEGZPQiKLxQ75rFa7Iwd1JJPO5ga4nHZWRhuQSgjO6yIYnObkFJfDDfhIyrI3SAEtBwgmZT+M7pJ7LMpKlsTjQ9GRJ8OfTK00TvYdjutXo5KTwveJCPEAyPrQc08Fb1HwjrodTCpDHF4fnPovV3lp5s6XX8NLRT17XERhuCc+S8NKzUlwq4xTSSExNOwyey7UcqOqrnp6+0lZG14pmYLjk4wg94aGrZW0sdSwgh4yshcRcHeNWltZ2Gkt9FUtNbG0RlmRuVyzE17W4eQSgkREQQ1DWOiIe3IIxhcT8YdE2y9aPrG/c1r5HNPlv2K5dc0OGCsO426Ovo30j8dLhjsg8D+ZLhhUWHV1TVmgMUfUT22xldepLYJ+uUNB6SV65c+XAiSa0z3S2x7mMuyGryokoXad96pbo5weHkAEfNBs6pnbE4w9I223WIZOo5AWZdWQmYyscSHbrDhfE146wceaCr3ktx0kfYrY5OjOcFahV+6imEkZySAtMa1zs4QZcFaIjnAVZ6wVBx09wsXo6iA3crKpo42tLX7OPkgh8MxkSZPfK3Ja9XxUNN4TosnGFoFRTTt+NwPR5KKnhbLMGk/D5oNdZd5a+oLmtJaXLU4dJNquqtkeAGguIKyLFQ25rB1AAhafqC9VdNKaWjcQwjGPkg9WfY0VEEmmeJdPCBmGa0BxHnkVmP0FekC8xfYjwVcdg4szVQIEs9kLc/IVuf0henSAiIg65e0Q25NOJeO/udH+vU6+eqlaXuIBxsvod9oBTmq5QOI0DRu6kpPzV1Of5l88kB8KR7T3Hw/nQZNGzpkPVusir6S3pHmsWBspcX42PYqlXLgjOcoLJqQNHUFFTzPgnY8OI6SD3VWSSzO6BurXxOY/pd3QbruGoYa63NpmgdQGDstApaU5MnWfqUMdLMz4iDhTBlQW5ZnAQTVNaxrDHgHyzhaa+R0mcHzV9QQ7PqFSGPqaSB5ILYZfCcSRn61JKDOQ8bBY7vpEYWRBI1rME7oLYcNeQd1bO775nyCteSHZCp0uec4QZbOqoaGtcQR6LHmjdG7pc4ndWhz4TsVVxdN8Tu4QZ1E9sQy45+Sirj4j+tmwysXxHDb0UjZA5uHd8II2FzX9yry7qwDuowCXfCpmREYJCCaCb3UdZ81ZUSmrd1DIGVbK5rxgHcbK2IiMfNAFLtnKjewt7lSSSvA27KFz3O7oKKSMEt2KjV7HYGPVBadiQjThwKOx5IMeaCaScPZgABQZyn1KrR8QCCikY4BuFJNG1seQoGoKd1kxx/eyVBt8lIJiGdI9EF1J/ZOk4wq1LHdeAdlAHlriW+albKXEE90FvgHpBzuVdHE5pD++O6q+XYAd/rU0UjDEWv74wgzaGnF2IgLsYO261iSmgtFKYDgkhbbt76mCUvgzlVuFwq5pC2YnIQYtTnx3ludznZXxVT2NLXE77KShYHyF02wPqlbTjxOqIbIM+2XVlA4SSNyD6q6vrfum8yRNwB6LR58AAZ3WtWKBksbhgZI/Og3Xw80INW1McLx3cAu3XBPgcyy6npZRTdXY9l134SUVfT18LqXIy8b/avRfl70pcbjc6WpqwSA0E7fUg7ocHLRBbtLU8L6YAgDBI+S5DADRsMLTNO0MVDaaeFg7MC1RAWx+J11EGnamlbB1ve0gbZ8lvhafcbLRXRhZVM6gUHjXzWaevpvVVWRwSRsOTsCAur8V4qYLbLRTsLnElu+69hOcHhRYKuxyTUMTGymIkjAznC8vdT6AitBmlnbjDz5Y80HXivttRTVr6stJDiTgj1Wj1UrppckEeS5SvT7X1up3EZA81xxeWwRzkQHzQQMZ4PxE5yFDLKHnYY+pHyOe0A9lZGOp4HqgrHG6Q4AJU+Xwt6OjPzwtQgpGxMD3bZWpmGhkpSTjrP1ZQbTdkO+3Ky4eksOyhqYXRyO22yrI5CCEEphEjs9lDIzocW+inlmAaOjuqsDZIi53dBjZI7Ej7UafiBO6OaeohUx5IMl0zQ0ABXseXM9FBGwu3cNlM6RjGkA7oLo6kwOJDSfkseomM7+vox9izLU2GSU+PjCyq6GkH9iwPVBogBJwFe1pBBCmIZ1YaUIACC33h7RjJVzYPFaXuO6sZF4pwFcRNEQ3fCCyODqcQfJWyx+G7GVNI50Q6vMo2nlnZ4nogxgCeyqGkkZ81e1ha7D9le4tAyCEEsFOCN1BUNLHkDOFc2oc3ABxupXPikaS47lBisYZDgI5pjdjzU1KPjPor5oDJJ8KDHdI5wx8lLFTdcfXur20bmAFwUrmvbEekbIMBwIJafJUAKnpul0h68KWcRNHwkZQd9PYrh37abVOT/APKBXfyjbkT2K7j+2m1T/ACu/lG3Ig9qEREBERAREQEREBERAREQEREHHnMW8M5fOJ8h7N0bej+ShlK+aKad5qZ5GEjre47ehOV9LPMl+DrxT/gVfP1GZfNhaoopqmcTY2aSPryg08ue7u4k/WsmCjMjS7cqGqAZO4N7ArUKCojbAWvO+6DTZAGuLVZ9ayXxl05ONirahjQ4BqCEZJWQyIFp7KR9N0Q9ZWKJXDIBQUkGHkBSQkjIx37Kaho5KqTICz56NtKwB+xwg0wtMbi/J3VuTNIM+qpOX9Rz2Kvp4i5pe3yygyJ2NEAAGSsWGXw8tI77KakeZZxG45BPmsmttE3UHxNyCgxJKfLTJn7FkUdc2ljMTm9/PCsbBPGMS5wPVWTQgtLgN0FJYTO8y5774UZhfHk5wVkWuKWrnELcnZTXmjloXhjwRlBprGGR/T2V08XhuGNxhRte5pyCs2nfE+M+JjI7ZQYkLOt2FdOzwzhUe7okJZ28la575Dg7lBWN4a7JGVc+cuI6R2Vpie0ZcMBS0oY54DsIKtJc3p3B9VM2RkILZBkkKepjZHCHMAzjK0zr6z8SBLh7y5vmrQ4jsU7nA81f4Eg3IwgrFC+V2Gnc/NXSRzQuAcSfNUjcYTnOCFkZM7ck5I9UEb3tdHjbOFE14HdWyEtcW4wrEGdT4mcGqeoj8AZOFiUDHulyzususEjQBJlBBDUAvwQoq0guBBUJJjeS3ZVDJJtwMoKRR+I7C1OC0jpEpd23x6rTB1xntup2XCoYA3q+H0QZ0lUJ80uMY2ytOqac07ukEnKymR+8AGnH3wo+jnjBNTn5ZQYXR0s6laXk9tle54Li3yyrHtA3CBGzrOFV7OhwCozqJw3uVV4cHDr7oNSpaP3qPpJWJWUnu0ojBzn5qWOeRkYEIOVjze8SO8SQHPzQZzbN1QCUE5PkFhy0bogScjCzrPcSyYRzu+D5lSXtomcHUwBb54QaRBEZpA3K1yiu0dqhMDm5JGFpVuglmn6IwSVfcqSamqGicYygpWymskMoGATsrIcRuA9VlyGD3UNiI6vRaaS9p37oNWk6DFnbKugZmEkNytJMs3TvnC161SwClLZSMkINDEJlnLey1BtOKeEg75ChcAKsuZ2JV1RP8Qa490GEAXzEgE7+izWVzoCIjGd8eSyLVBA5/U/C1SsjtjWlx6eodig0qqpGzQibzIzgrGjt4cwu7qSpqy93REds4GFl0cUzoHHHcINClb4chb6KsUjmuHnkq+rjeyYlwUkAiLMEjJQSeKA3Oyxnt8T4lIIJevselUdFIH7DZBNaLgLdUeI5uVLebmy5yB7G4x8lA6mbIwdI+JYz4JI93DCCwAuOwJ+pVDXNcCWn7VrelKWnqK7pqcdO3ktY1PbaGAj3YDsg221jZ4+kjCw54fBdgFZM7nU4GMjKxHyOkO/dBaAT2U8Ti34SO6rTxljuuTACnldCRluM4QY80AY0yA91AC7HnhTNkdI7pPZZIEDGkOxn6kGANiFzBwz4pCyxC1h/Q97ekO+fZcT4jLidsLNtrqamrGT+bTn5IO8/A/jDceF1wZqOqvDjE94eGl+RjK9C+EvOPp3X8kEM1fAwvAB7DdeI911rJVWllHE4npGBg/JX6S4zal0aA+3PlD2/R+IhB9GjtdaVZQiuN8pugjI++DOVlWvVFhu0Ilo7rTyeuJBkL57Z+criy6EU3v0ojB2BkK3Tpfnf4nW2ieG1U+QDjEhQe/Da+iccMrIXH5SAqQTw/wDzZh+0Lxa4Cc53E/VuoRSVdRO5hkAHxk7ZXeSy8W9VVD6eOQvy9gPc98IOy3EPQ9s15ZJbbUwxyktLR2OxXnFx25BZa25y1VDbSGvkLtm7Yyu/3DS93mvDZKzqLX987rkGvtdLcQBUMBA9QCg8OeIPJldrDRsEVnkccY+iVHozkGv2o7FLeJbfO3oBON17U3bhjpe8t6a2ka//AMULKtWgbBZ6F1vo6ZrYnDBHSEHzzcReBGpNLXGa1sss3TCSAQ0riets92tUj46mgkZg4JLSvot13yx6D1XDPUmiY2pc0nJYNyuhXMVyTPprdV1dppcOBJGGfWg8vWs8H74e/mPRROmJkDxtjfZbw1vww1Fou5zU10gc1jHn4i0jIytvU8dPM7AIICC2ougnpxD04IGMqlvgaWl5O6yKu3xmPMI3wsaliqIpMEEDKDNZXyUpIBIAWFLcTLUGZ2+Fqzrf73CAwZPmtHrLXUUmXPBwEHrZ7FS8RXLTXFGnYADSz2YO+0VmP0L0vXln7DV2bfxlb6TWA/lFf/qXqYgIiIOAOfSZsHKPxEmd2bSUmf47Avnamf1zPkHZzifylfQt7RKobS8mXEudxwG0dF+evpx/OvnlQZ0NaGsDekbDCo6L3g9WceawlPHUeGMYOfVBNEwUri7vsoZ5hJMHgYwVRtRl33zJHorZHMe7LWkBBnSXRr4xGG9gkNzDYvDLe6xmS0wbhzHZTxKXv0nPqgPh63F+e5yglETOhWvqQRhoICgJJOSgmhjE0hzsEqGeE/AOFE17mHIKq95ecuKAXZGPNXxyhgxhRfWh37BBlRsFU7HYZWS6jbBGd8ladHI6I9TTup/fXub0vyUEPT1PIHqqOb0uwCqZ3LgqEk90EzGiPDie6ym4kZkd1gF5IwpYp+jY5wgscHMeUz1vHkpjNTk9RySe+yhe5pdlgIA7IJ5Yw2IHY9liq90pc3pOVYgr07d1Mym6mdRdhQZPqrxK9owDsgtIwSPRTx0wezqyVAXA5OO6ubM9o6QdkFr29JwqA4KqSSclUQSOl6hgklRg4REA7qrR1OA9VRVBIOR5IMv3FvR1Z/OsWRvQ7Cu94l7Z2VhcXHJQAT3Ru7gPVStkiaN2klRucOoOaMYQa1QyRwNy7GSsO44mk62YWJ7xJjGVe2pHR0uBJ9UFWTdQ6RsQr3VIYOnvlY8cjWOLiDuqSvDzkAhBlUlN79LjOPJatRt+5spYDnA7LRaOr91d1brVrDM2536CCUkNncGDbO5Qd0uSHhy7iReKenlo+oeIAdvLK9ZtC8HaDSIj8OBgLGhuQun3s9eGkmnqeku7G4DiHZxheiSCKniEETYx2ClREENRP4DOpYVVcqYUhe+oDCPnhZlVH4kT/wDvThcA8WNUXKwskdE54YCdwSgm4m0lJqHxoZJhK0gjGcrprxw4QWZlgqqqORjJBk4yAVvi6cxdutNVURXGpAc0HuV06478z1VdL0aKhq3Gkc8h2DthB1n1tbaq3alnp9xG0kA+RC29LazODL1ZXIXELVGnLjRtqad4dVPb5AZJK46prwyOneyTPUe2Agx/dw1xj9Faabw3CQeRyojVOdKZDnBOcKYVkeMPBIQTfdDxW+D2xt2UDqqSNwHWcFU8enGSxjgVjyPD3ZAQZFTO2Rgx3PdYoOFTJRBJDH4hwSQrn5hPSOyjY9zOyuMgcPiBJ9UFOv5K3PxZVWkA5IJCdTQ8HBx5oLjKcYxhZMNB40fi58u2VFLJC5gaxhBVI6ySNhY3sUCXqgd0sOMLKijMsHW5xyAtPe8vPUVeyeRjS0HYoKZ6Hn5EqXHiM7+ixyckn1VQ9w7FBLDMYHZxnZTGoEzgSMFW00tM0nx2kqyeSIyZgBDQgvqsFox5K+mr2wReGW7/AFLHZMM/GCQkj4HD4WkH1QJH+K4uAxnKjcCO6p2OyqCPMZQUAyp46fq3yVACQr2yub2QZD2+CNvJX09QA8ZCg94aW4e0qMvHV1NyEGsTSskjAb3wsM1TwfCDDjOM4WPFUuY4F2S3zWtQ1tl90JkJEoGwxvlBpE8HhN8QE5Kx8FwySpJ5zK89+nO31KLqxsOyDv8A+xWP/XTap/gBXfylbkVPYq/hT6p/4v67+UbciD2pREQEREBERAREQEREBERAREQcccyX4O3FL+BV8/UZl80sMxhne7OM5H519L3MVSS1/L7xOoYc+JU6OvUTMerqGUD85XzlN4XarfI9jaIksJB2Pkg214DZiX5GTv3UEjDC/GfyLd7OFGricGjcPsKuk4TasbuKVx/8UoNs5YIerIJwoo2CVwcSNlulnCjVzjg0RA+oq/8A2JNWBwb7qd/+1KDbFTUO6PCaMrDawnOdseq5MtfBW/v+OpjeAf8AtCsO8cHtR08/+1YHvB/7U7INm0FxFG/doIHZSV1eKwgggDK3TTcGdV1IBFO4E/8AaFKrgxq6mODSuOf+1KDZ4jZMAzIz9a1Kgoo2t6CRutXbwj1jneicB69JWQ3hRq5mCGygj0BQaBU2ltvzVdY23xlWs1AwN6XNBx8lrlTwv1lJs+KR4HYEFQt4T6td3oyP/FKDblTcxOdhjPyWOKkklp7HZbvbwh1Ue9MR/wCKVQ8ItV9YaKYnP/alBoNrlbbJBVEAjHqor3d/urIH9OMLdw4NapMYLo348h0nAULuDuqWgkQk/LoKDZdFTiom6CRj5qS4UraVwDHA59Fuc8KdXsd8NE768EKh4VawcfioXH7CUGzs7YWTSRMfgvIC3bDwh1fK7BoyB69JVKjhHq6B2BRud9hQbcqWMLMNwfXBWn4exwdg7HZb0p+Emq5SOqmcwHvsVqP+wpqN0XUGSZG+7Cg2FLUB8QYTkrHij8R4aTjJW9XcIdXeKWNpCcefSVR/CHV0Z3pHf+aUG3WWtrG+K4j1wSoJ52N+jg4W6n8KtXhgxBI4emDhRM4T6veQPcXDJx2KDa9FTmumEewC1CqohbxgDIPfdbnbwd1XTxiZsbw/5NKlHCbVNTCTM2QnH9yUHHs0gc7ICsjb1vAW83cJdWCUsFI4gHv0lTjg/qjb7y7/AM0oNvUkbKVolON/mrauaOoaT1AELczuEWqy3p8OQj5tOFjS8JdWxn+1HH/xSg2ZK7Llm0Jj6MHGVuVvCTVzsZo3D/xSpYuEGrnTsYaRwBxl2Dsg2jPG/qJLMBWNp2PHcD55XKF14OXtttb4UbzKBkgtO5W1/wDYq1eO1AfyH/Ug2/Qztts/i5DhtssmvujLkMjDSPJa5Bwj1bM7D6QsHrglVqOEeq4XYZSl/wD4pCDZLu53VzXeS3f/ALFWrv7xP/mlU/2KtXf3ifyH/Ug22yFkAEpcD8sqOYibLhgEfNbqbwp1c49LqNw/8U/6lf8A7EmrNv8Aah/80oNo0swjeA47KepqmEYAC3M/hJq1o6hRk/8AilWDhTq496E/kP8AqQbPAJdscZWZHWeFF0OPUey3KeE+rgM+5O/IVb/sVau86E/kP+pBt213AUNUagtyD5KS83Vt0kDmsxhbii4S6skODRuA/wC9KS8JdWRuw2lLv/FKDZ0bnMOc7fWqmQSOG2N1vAcJdWkf2oR/4pVruE+rgTiiJA+RQaEyCN8OcgkoKdzIyWuxhbhh4S6vkPSKV7T6dJWQeEOrGjDopB/4pQbZtEAqqnw3Y22KkvltFK4ODh+VbmouEmqoJRII5G48w0hVuPCvVtQ4Hw5Hn0IKDadoxNII+oD7VLd6BzXBzX7Y9Vr9Nwi1i2QPNM5gHmAVPWcK9WS4JjlOPVpQbFBMB3IJC1Cku4biIjY7LX28ItVudg0zh/4pVXcINVNGRTk/+IUGi3WmiNMJmuGTutFgYXO6vILd3+xbrEnoNI8geoOFIOEeq8Z92cD/AN6UG1zXjp6AFkU5bM3tknZa6OE2rerHuhA9cFZ9Bwf1P4rXPieBncBhQbTmY6jHikbZVY/DrmdRwCB5rkC68IL9NTARMeXAZA6StAbwl1cwke7Pb9QO6DbLJzbpepjsH61dPeZalwDiSPmtxv4Sar7imcST5tOVk0/BrUkjC6WJ7T5DoKDbMFJHcMNc4Ageqx622Mo3D4wd991uSbhVq6mkIhgeQOxAIUR4X6xlOX0b3fM5KDb0hZJGGhwyPmsN7PDyOoH6it2jhRq4k/7RIx8iq/7FGrc49yP/AJpQbRgx1YJwsiWna9vUHjZbndwm1aAMUhOfkVZ/sV6v/vA/kP8AqQbPcOk4znCdTvIlbx/2KtXf3if/ADSqt4UaucQ33IgE9+koNs0VTJG743kj0ysuSsa5wLXDA7rck3B/VEUfWyBzj6dJWJ/sVavx/aJ/IUGkyVkc7AzDcj1wrmVRghcwSAZBGNlrNPwk1dO/pFE5v2FVn4R6vil6HUjjv36Sg1rg7r92i7w2sc4AdYOV3J0BzRQ3XUFHSuq242bjP1LpdS8HNQtjD3RyB3fZpW4dG8PtTW3UsEzGzDocD2KD3e4F6mor/p+CpilY9zgO2PRcwLpDyY3C9w0FNTXBz+gED4s/Jd3RvugqiIgoQDsVo2pLBbLxbpKWroY5esYHwjK1pUIyg8yOefgHQx0tRXUFoDMxk5a3C8vKzTbrLNO2bLXNeR39Cvfvmz03HetJyEgdRhI7LxU4v8JdTPvdQbfE97DKdsHBGUHDwnDHYLgR9alMzHM6gBstxu4JayjhEslO8ZGSOk7JBwg1M6MtLZBn0aUG3KG7tpZviwQEvN5irYXsa0AkLWn8HdWtdvTEj16So5eEmqo4zIKYnAz9EoPSn2HEXTaeMU3m+ewj8grv9a9SF5kexT0zdtOUHF43OIsbUy2LwgfPpFdn9IXpugIiIOMeZHhPU8cuCWqOFVJU01PNf4YImSVLnCJvRURy7lrXEf2PyB3wvOeD2MGtYyfF1ppR4IwP9t1X9AvWZEHkwfYva0LsjW2lgP8AG6r+rqv7i9rPH/x20tn/ABuq/q69ZkQeTB9i7rU9tb6WH/1XVf1dUHsXda431vpb+N1X9XXrQiDyZj9i/rRpy7W2lXD/ABuq/q6vf7GHWTvo6z0oP/qqq/q69Y0QeTB9i9rQ/wDy7aVH1VdV/V0/cXtaf8N9Lfxuq/q69Z0QeTcPsYNZRuy/WmlXj0NXVf1dJPYwaxe7qbrTSrR6e91X9XXrIiDydf7GPWDmBo1lpUH196qv6BQj2LutfPXGlv43Vf1detCIPJk+xe1mRj9m2ls/43Vf1dB7F/Wg7610qf8A6rqv6uvWZEHkyfYvazPbW2lv43Vf1dUPsXta+Wt9Lfxuq/q69Z0QeTH7i9rX/hvpb+N1X9XUsPsY9YR7v1lpV/11VUf/AK3XrEiDyin9jPq2VuI9XaUaflU1Q/8A3CxR7F7WmRnW+lv43Vf1des6IPJ0exj1e0f/ABx0ofn71Vf0Csd7F/WTjtrXSwHoKuq/q69ZUQeTY9jDrEd9Z6VP/wBVVX9ArX+xf1m45GttLD/6rqv6uvWZEHkx+4va0/4b6W/jdV/V0/cXtaf8N9Lfxuq/q69Z0QeTH7i7rT/hvpb+N1X9XT9xd1p/w30t/G6r+rr1nRB5MfuLutP+G+lv43Vf1dP3F7Wv/DfS38bqv6uvWdEHk7D7GLWEf09ZaVf9dXVf1dXP9jDqtzg4az0u3HkKqqx/6BesCIPJ6X2MWrXNxHrLSwPr71Vf0Ch/cXtaeet9Lfxuq/q69Z0QeTTvYv6zLcN1tpUH197qv6urB7F3WvnrfS38bqv6uvWhEHkx+4u60/4b6W/jdV/V1Q+xd1rnbW+lv43Vf1detCIPJf8AcXdb/wDDjSv8aqv6ujfYu61By7XGlj/9V1X9XXrQiDyf/cY9XYA/ZhpXPn/tqq/oFq2kPY86i0/qGmu9fq3TU0UDw7oZUVJPfyBhA/OvUpEHG/CfhDRcM7BTWmLwHSQtALoiSDj0yAfzLkhEQEREFCMgj1XHuv8Ahc7WVJJTsmp4y/O7yR+gFchog6K8QvZ23/VtVJU27U1mg687STTD9ERXFNd7IvVlc1xm1bpd7ySeo1FTt/8AiV6gIg8nqn2M+tKiTr/ZppQN/wAaqgf/AECo32MWsA3pOstKn5mrqs/q69YkQeTTPYwaya7Lta6VI9Pe6r+rqV3sZNXEYbrHSgP+NVX9AvWBEHky32L+tA4l2tdKkenvdV/V0d7F/WbjtrbSw/8Aquq/q69ZkQeTJ9i9rPbGttLfxuq/q6p+4u60/wCG+lv43Vf1des6IPJcexe1r5630t/G6r+rqv7i7rT/AIb6W/jdV/V16zog8mW+xe1mO+ttLH/6rqv6uqO9i9rU9tb6VH/1XVf1des6IPJf9xd1t/w40t/G6r+rp+4u62z/APHjS38bqv6uvWhEHk0z2L+smnLta6Wd/wDVdV/V1R/sX9ZudlutdLAenvdV/V16zIg8mD7F7WmNtb6Wz/jdV/V1Kz2MWsA3DtZaVJ9feqr+gXrEiDyaPsX9ZZ21tpbH+NVX9XVjvYva1JyNb6VA/wAbqv6uvWhEHkwfYu61xtrfS38bqv6ur2+xg1iG4OtNKk+vvdV/V16yIg8mo/Yv6yacu1ppYj097qv6uknsX9ZOdlutdLAenvdV/V16yog8l/3F7Wv/AA30t/G6r+rqv7i9rX/hvpb+N1X9XXrOiDyXPsXdantrjSw/+q6r+rq5vsX9Zgb610qT/jdV/V16zIg8mP3F7Wn/AA30t/G6r+rqQexi1iG4/ZlpQn196qv6uvWNEHkufYva1Of/AIb6VH/1XVf1dV/cXdZ/8N9Lfxuq/q69Z0QdHeR72feoeVHixduIl11FZq+G46ensrYqKeZ7w6Spppg4iSNgwBTkZBzkjbui7xIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiIC48498btHcu3Cu9cWtde9PtdmbGPd6RrXVFTLJI1kcUTXOaC4ucO5AABJ2BXIa8nPbW8Z5Km9aI4A2ys+8UUD9T3eNpyHTPL4KRpx2LWtqXEHylYdtsh3Q5R+eHQnODW6jo9D6G1VZm6ZippKmpukcHgOMxeGMD45HYf97ecEbgE5XZNdNfZR8JYOG/KVZtQz07WXTXlZPf6p2AXCEu8GmZkd2+FE2QDyMrvMlcq84nNNpvlN4Q1OvrlSsuV5rZfcLDajJ0e+VZBOXkbiJgBe8jfADRguCDnCoqIKWF9RUzMhijaXPfI4Na0DzJOwC0u16v0nfayW32TVFpuFVTkiaClrY5ZIyO4c1riQdj39F89epOJHNdz1cSo9PT3K/wCtLxcHGSksNA7wqCkjb3c2EEQwsbkAyvwexe8k5PIV59l5zuaMtrNUUHD6GrnpWifwrPfKeSsgcMfRaHgucCe0ZcdjjO2Q95aiUwQSTiKSQxtLuiMZc7AzgDPc9gunHAr2o/BLjnxdtvByk0fq3TN1vEk1NRz3mKnZE6pY0uEDwyVxY9wa4AEH4sN7kLbHsnOMPGfiFobXuh+M95u9yr9C3Oko6Z16a/7o07ZY5euCd0mJHdDohjry4dRBOAAPP72gmh7hy787t+v+j3utrq2vpNb2WWMAGGeV4lc9oGwDauObAGwDQg99UWzuD3Ee28X+Fek+KFpYI6bU9opbmIg7PgvkjBfET6seXMPzaVvFAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQF89XtGNYVOtOc3ibWzSl0duuUdnhbviNlJBHAQAewL2PcfLLifNfQqvm05oWUcnNnxbjuEr4qV3ES/id7BlzIzcp+ogb7gZIGCg+hvhDpKPQXCjRmhooDC3T+n7fa+jB+HwadjCDkA5y05JGc9915A+2U4jV+pOZS06AFTJ9ztH6fg6YC74RV1TnSyyAZ2JiFM05AP3v0IXtcvCH2tNvqqLnS1DU1MTmR19otVRA4g4ewUzYyRnuOuN4yMjIPnkIO9XseuEFm0hy1ScVnUEDr1ry6VTjWFoMgoaWV1PHCD5NEsU7yB3LhnOBjvoupvst9RUl+5JNB08Do/GtEtzt1S1jcBsja+eRox5kxyRuJ8y4ldskGLDQUNLUVNXTUUEM9Y5r6iVkYa+ZzWhrS8gZcQ0AAnOAAOy8nvbf6Rjp9XcLNdxxEvuNuuVomeBsBTywyxg7bZ96lI38j2xv61rzI9uAylOhuFUj5He8i63MRsHYsMMPWTt3BDMbjudj5By/7IfWs+quT2is1RMZDpLUFxszM5yGOMdWBv3A97IHkBgeS7sLzq9iZ/vC67/hef1KnXoqgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIo5JI4Y3SyvaxjAXOc4gBoG5JPkF181Rz98qmkdYfsGu3FOhddBMKeRsDTJHFISB0ueNgclB2HRYtuuNDd6CnultqY6ikq42zQysOWyMcMgg+hBWUgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAvnY5+9IzaK5xuK1pmh8L3q/y3doHZza1rasOz558ff0OR5L6J148+2m4Qz2PinpDjTQ059y1PbHWatc0bNrKRxcwk+r4ZQAPSByD1j4daoj1tw+0xrOGRskd+s1Fc2vaRhwngZICNhsQ/wBB9S89fbGcs971lp2x8xmkbc+rm0rSOtOoo4xl7bcZDJDUAebY5JJQ/GSBKDgBriOePZe8VouJ/KBpWjlqfEuOjJJtM1rc7tEBDqfAO+PdpYBntlpA7YHbCaGKoifT1ETZIpGlj2Pblr2nYgg7EEHGD3QeDHITz73PlCud001qayVeodB3+ZlVVUdI9raqhqgA01FOHkMcXMAa9ji0O6GEOb0nq9AL57YzlPt9nkr7RQ63utaGgx0EdqjieXHyc98oY0A9yCT6A9lq/GL2TPKzxQu1TqDT1NetBXCqc6SSPT80YoXSHu73aVjmsH/axGNvyXHmm/YocDqC5x1Wp+Kus7vRxkONLTx01IZCD2c/okOD2OADjsQdwHKvIDzqaz5xbnxNrdQaOtun7Rpiotgs0NK+SWTw6kVXW2aV2BI8eAw5a1o+PGBsT1i9uDqmOW8cJ9ExPAkpqa63Wdu24ldTxxHtkY8Kbz3z8l6P8FuA/Cnl70n+wrhJpCmsVudIJ6gtc6SeqmxgyTSvJfI7AwMnAGwAGAvFX2nXEqbi/wA5WoLPZPFrYNKMptI0MUY63STxEmdjWjOXe8zTNAG56R57APQH2OWkZdP8ptTqCeHpdqfVNfXxPxu6GKOGmA+oPgl/KV3qXGvLhwpZwQ4E6H4VDoM+nrPBT1bmYDX1bh11LxgkYdM+Rw3PfuVyUgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgLhTmp5j6Tlo4cVOuqnT811fE09ETSQwu8gT8zjzH1rmtce8ftCaY4j8HNXaU1dRxz26ptFU55eB96c2JxDwT2IIzn0z6oOk3KF7U68cd+NVBwr17oy3WaO/vfDa6iklc778AS2N2fMgH17HfsvRtfNJy2vvdPzA6AdpnrdcW6hoxTlmQSTIASPMfDkr6WGdfQ3xMdWBnp7Z8+6VJzi9ERFEREBERAREQEREBERAREQEREBERAVrnNY0vcQANyTsFxzxk5huD/AK0C88VNbUVlZICYYHkvnnI8mRtBJPp2HzXkpzue0z1Pxvnl0NwVr7ppzRrcsnqQTBV3Aepwcsb8s52+02T6OxXtOOfKk0lYn8DeDWpYZ73cgW3u40Uwd7nDjHgtc0463efoPyHoByk8ovEPm41zNbLFKKKy297JbzeKkOLIg45LGnfqlcMkDyzk+QO6eTvka4l8y+vKCq1FYLxaNDslE90vNRA6ITs7lkLnAdbn9sjbvvnce5HCXg5w64HaRg0Rwy01TWe1w/E5kTfjmf5vkf3c47kk+ZKl74fprGhNJ0mhdHWbR1DPJNBZ6KKjZJJ9J4Y0DJ+vC19EQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAXDfNjy52Xmm4LXXhRdri22VM0sVda7kYTN7lWxE9EnQHN6gWukjIyPhkdjfC5kRB075AuSniJyeSavp9TcTbXqC16lbSvjoKGjkYIaiHrHjdch2Ja8tIA3w0k/CAu4iIgIiIMesjqpaSeKiqGQVD4nNilfH4jY3kHDi3I6gDg4yM+o7rzp5d/ZQXzhzx/tvGni5xRtOrorRcZbzFR09vkY+ruBLnRTSl5w3okIlwAcuaBnGc+jyICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiIIppoYInTTytjjYMuc5wAaPmT2UdJXUVfCKmgq4amInAfDIHtJ+sEhdGPa/wDFLU/D3l5sti0vc6i3y6qvzaKqnp5THJ7vHDJI5oI3wSGA79lw17FbXmtb7euI+lbve62us1DRUVZDHUSvkEVQ+R7TguJxlrTt8iid16qrqd7RTmXtPAjgjd7FFUY1BqqiloKBgOCGvBaXfPbIPoMldsV4re2W1u+88xtn0TG8+Fp6wQSPAOxknc5+49Q0D7HKXd4rjL2YGgp9a832ka73Z0lNpwzXaV2Pha5kbgwH55JI/wC9XvsvOX2Q/LpNpLh2/jleWllTqXrbQsI38AEtDs+hGSPmT6L0aVBERAREQEREBERAREQEREBERAREQFxnzBcd9Fcu/DS68Q9Z3SmgFJA/3KlkeBJWVGPgiY3OXEkjOOwPqQtc4q8WNBcFNE1/ELiVqCGz2S3ACSeTJL3n6EbGjd73HYNHcrwd5xOY7U3Ofx7NfpO3XWazRlls01Z2sc+UsGcyGNpOHvJLjjsMdsFDWx9a624x84/G2GerNTfNTamrRS22gjJLIGuPwxsHZrWgbnA2bnsAB6NcrnshafQWq7Tr3jtqi3X2W3uZUssVFC404mGCBK92PEAPoAPtwRzfyE8jejOXXQVm1nqjTsU/Ey5UgnuFZUfE+3+IM+7xA7NwCATgEkb4wu4GcKWauoaalpqKnjpKOnjgghaGRxxNDGMaOwAAwAPQKdEVQREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAUU80VNDJUTO6Y4ml7z6ADJP5FKtt8SK9tq4eaoubndIpLNWz59OmB5/mQeG/PXzY6n5tuKdNomx2/psVjuUlFZqWPd087neH1n5k7Z8xjsAF6Yeze5Ub1yx8Jq+TWLYhqfVk8VbWxsGfd42MIjiz326jkeu+BnA8muQLR0nEHnE4a2ian95ijuzrnVB248OmikncT8iYwPrIXvRxj4ycP8AgToau4g8R75BbbXRtIb1kddRKR8MUbe7nk7AD1UyH1pXMXx40py58KrvxN1XURdFFH0UdK5+H1dS7ZkTB3JJ3OPIFeDk9Lxe57eYm43S226Ssveo6oSzdDSYqKmGGjONgGtGwGMnOMDOK80XNBxN5ueJkl1u01Y+1tqnRWCwwAuZTxk4YAxueqQjGTuckgE5JPqD7LTlI1FwG0HdOIXEaze4ao1X4YhppWjxaSjAyGu22c4nPfbfyOTeTg7gcItAUHCvhfpfh1bIwyn09aqegaAdupjAHEfW7J+1bvRFJMBERUEREBERAREQEREBERAREQFtfiRxD0twq0Td9f6zuUVDabNSyVM8kjwOrpaSGN9XEjAHqtA47ceOHfLtoKq4gcRrsKWihyyCBnxTVUvlHG3uT2yfLPqQD4PczXNTxa5u+JE89XU3T7jz1PhWTTVM9z4omZ+AGNuz5DgEnBOc4PclMG7uarnJ4u882sLPoa06fNDY2XHw7Hp+iJfLUzyHoZJO8nDpMEbDDW5d9a9HfZ5cgZ5ZLfV6/wCJn3PuGu7xCyONkLRIy0wdzGx5G8jjjqcMDAA38sTkA9nvozghp2w8XOIdrfcOI9XTiqYyoI8Kz9Y2ZGwbGQNO7zkgk4xgY70K78SQREUUREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEXHXHzjDZeBXC+7cR745vgW9gDA799IQcD8x/IvNjh37UDiJxH4zW2w00M9Nba2r6I4+oAObntjfGR5H6knbhePWxFj0M7qqjp6l7el0sTJCPQkAn9KyEBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAXVL2ifHej4ScBr7Y6eYtu2o6GWkgAOCI3AtJHnk7j6srtavK/2w1/isurtEwXCF01HPEJHMB+kwPcHDH2H7SpuLP24X9kTpr3XmVfre/D3Kht9krIaaepHhsdPKGt2c7HZvUPtwsv2v/HSDiHxus3DXTt6ZWWLRttD5RTzh8L7hO4mQ4G2WxtibnfufVbJ5k+YvhzXcK9JaU4Izm2174gbtLSHw5GNDfolzcFpJI884BXLnspOUzQ/GmbUXGfi1Y33yCy10VLaoK0udFLUAdb5nA/2TpOBudj9uUlt2+JXPfszuRXT3D7RFo468S7I2q1ffIRV22lqosi20zvoHpcNpHDDjttkd9sehiihiip4mQQRsjijaGMY0ANa0DAAA7AAYwFKqCIiAiIgIiICIiAiIgIiICIiAthcaeMmieBXD+68QdcXanpKO3U75I4nyASVMgHwxxtO7iSQMAHuuDOZ72jfAjlxqLjpP7pS6l1lSsc37k29vU2CXGzZ5T8LCMgluc4z2K8aeInErjxzgcTJq+4NvmprpXzk0dpoxJPFSMJw1rIxs0AHBcQPmQMABrvM1zScV+cbiJTS3aKU0bak09hsVICWxdZw0YBPVIc4J8s49SfTL2c3IAOA9udxT4u2qiqdaXOJvuVK9gkFqiO5wSMeIdskdseoBWkez69nBQ8JI6DjDxrtgn1oPvlBbJcGO257OcPOX6+3l8/Q0HKZ8FUREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQef3tm79Ja+XDT9pZOWC76kiic0bdYZE+Tf6ukLqf7JrlvpOLfEy8cS7+0m0aOEMbGjvJUyZcG+gwGg59MrkL2yPHrRGs5dH8HtK3eG41tgrai53OSCQPjhe6MRsjJGxOC87Fcy+xe0hcbJwB1VqatpzHDqHUXVSuLSC+OGFrCc+Y6i7H/tV/8UnXoQ1jWNDGtAa0AAegHZXIiiiIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAvJT23NbS/st4c28SD3g2+plLPPoEhGfyleta8JfazcQItZ83F1s1HVial0pbaS1dI7Mn6PElH15eB9ig4M5WuA935jeNWnuGduhm91rJxLc6iNpxT0jN5HkjtsMD5nZfQzwj4S6J4JaGt/D/AEFaY6G2UDcANGHSPPd7j5klcL8gXL9w74Ocvukb7pqy0xv2qLLSXS7XVzAZ55JoxIWBx3axvUAGg+W/YY5B5geZ/g/y26efe+JWqKalq5IXyUNsY7qqqxwBw1jBk4J2yRgb98LW74jloDCqvAax+0a4/wBr49v4vVeqa+rtU9xdLLYnPAg9zLtoQOwIZgZzgn0GMepGgPae8n2tbVFWV/EpumqpzGmWlu9LLE6N+N2hwaQ4A7ZB3Wartii484U8wHBzjhFVy8KtfWvUXuOPeG0ryXRgnAJBAI3I/KPULkNAREVBERAREQEREBEWHdLpbrJbqm73etho6KjjdNPPM8NZGwDJcSdgAEGYvP72hXtG6vl9uj+DnB+GhrtYTUnXcrlK4vjtIfnpY1oIDpiATgnDQQSM4C485/fab6Tq9Gy8LOWTWtVU3aunMV11DQsfFFT0wG8cEpALnPJwXNGAAcHJXVPko5H9c82+uJNR63N3t2iqd3vFxvMwd41wkJBEcT5Aesu3Jfk7djnJFzBsfl15U+N3OXrK41Wm3MEHvJnvOoLm8iGOR56nEkAmR5JJ6QPyL2m5Q+T3QfKbop1lsr23W/158S53mWECSZ3kxnm1g8h57Z3XJvCTg/w84G6LpNBcNNPQWm0Ug2Yzd8rz3kkcd3vJ3JPmt6qAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAtL1OJjpq7NpnlkpoZ/DcO7XeG7B+w4WqLDu0RntVZCO8lPI38rSFKPmS1hTVtx4q3ehu88k1RPfZoJ5Huy5xM5aTk/mX0N8qGl7NpDgFpKzWKlZTUzKMP8NgwAT/7AF8+nE+GW0cb9QNlGHQaglk//AB3UPzEL6CuU66G78B9MVZzn3YN3+QH+tS3sHL6Ii0CIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiDrN7QLmSu/LLwEqNVaXdE3UF4rGWq2OkGRG97XOdIB6ta3P8AqOCvJblI5Xtac9PFy+XPUOpnwUNDIy4aguco6pZTK44jYP7o9Jx6AD6x2n9tNxZ0/eItF8JLbWMmr7VXTXStaw56CYzG1p9COo/lPot5+xIs7qbhbxGvckRBrr5SQscR9JkUDu32vP5E79T9x6H6Q0xa9F6WtOkLLEY6CzUUNDTtPcRxsDRn54GSvDj2plw1RqbnCvNFVU1ZLFBTU1Da4AxxDmNGD0NHclxOcbnbPkvd/stlah4N8L9V6ip9Wah0VbK+7Ujg+Kqliy8Edj6HcKTivNnh17G6g1ZwStF71Nr+tseu7nSsrXRCAS0tP1gObC8ZBJAIyRvn07DirWPsceZHT1JUVdg1BpnUPgglscEskL5APQPGcn0XtgAAMDyVVR85eh9Z8feRbi+26wW6aw36nZ4dTRVsZdBWQE7tcAcOGxwRuD+f2a5H+c60c32ibhcJ7JFY9TWGVkVzt8cxkYWvBLJYyQD0EgjB3BCxOe/k0sHNTw7fLbYYqPW1ja6a0VwaAZdt4JD3LTjb0IHyI8heXHjfxE5JOPNS+sozTvgqBatQW+UHD4g7c/YCXA+hz3V+J/j6H0Wn2C70+oLFbr9Sf2C5UkNZFvn4JGBw3+ohagooiIgIiICimmhp4nz1ErIo42lz3vcA1oG5JJ2AXDXNpzJ6d5XeEdfxBvIbUV0jvdbTRA/FU1LhsMeg7n0HfbdeHHE3mu5meYnUMtNcta6kq21krnU9ltMkoja3chjY4t3YHfIIPfAQepnNB7UzhnwI1LNozR1jbrO7U3w1DoKoNp4n47F4B7H0z8hjdefnMv7TXjlzG6Xq9Am3WnSmmrgAyrpLd1vmqGA/RfK45LT5gAArSOCvs5OZni7dKaW56GuWmbPPIDLX3SEwuLTuS1jsEn68fUV6ccEvZa8snCmS3Xq+2Go1bfqMNkNRc53OpxKMEObCMNyCMgn5bJn9m/06U8knsvtb8SLpYOK3Gulis+jGyMrobTKSau5MBy1r2do4zgZyckHt3C9krZa7dZbfBarRQwUdFSsEcFPAwMZG0dg0DYBTQxRU8LIKeJscUbQxjGN6WsaBgAAbAAADAUg3Vt0xVERQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQFpWqbmyyaYu96ecMoKCoqnH0DI3OJ/MtVWyuNj5GcG9eSQ/wBkbpm6Fv1ilkwp+Xg+dO1U1w4z8eIaeijNRUan1BmNoByWPlyBjvswfmX0WcHtCt4bcOLLpAY66Gna2TpGB1kbgfVsPsXhr7MaxUV+50dBsr2BzaI1ldGP/okVNIWn7Duvf4YIwrn1PKqiIiiIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiIC03Ud2ZYtP3O+SY6bdRz1Zz2xGwuP6FqS455i7lJaOA2v7hE4h8enq5rTnzdC5v86D5+4ItVc1/MjT22S4Sz3PW9+93hke4kxxPeSAAe2GZONhnJxuvfblx5fdG8tnDej4eaOjc6OM+LVVD/AKU8x7uPyznGfU+u3h37OimFTzpcLw7GI7nLLg+ZFNKR+fB+xfQwgIiICIiAvC72t2iJ9Nc2Vwv8FrdT0GobZSVTJg3DJpwzplOfM5AJ+v5L3RWx+JvBThRxlt0dq4o6CtOpKaI5jbWw9RZvn4XAhw332KDy/wCQL2m130vNprgNxyNPUafZ0Wy16ie8tmom7CKOozkSMH0Q/ZwGAc4yvXGnqaetp46ukqI5oJmiSOWNwcx7SMhwI2II3BC8m/aYcgXC7hNwzpuM3AvSc9nioK5lPfLfDPLPAKeQENmY15cW9LwAdwMOyey032WfPFQaDqazgZxo1hJFZ657H6crq+Vz2Us5OHUxefosdkFuc4IwMDAVySepv16+oooZYqiJk8EjJIpGh7HtILXNIyCCO4IOchSqKLTdQX20aXslfqO/VsVHbrZTvqqqeR2GxxMBLiSfkFFqnVWntE6fr9V6svNLabPa4XVFXWVUgZFDGO7nH0/nXiT7Qfn7u3MVqiXQXC6+XCi4bW8eE5jS6E3ibJ6ppR3MYzhrTgYySN0iVsLmF4/8W+fTjtbdM2ljpaOruht2lbNGS2ONj39LZH7ZLy0dTiRkDIGdyfUrkX9n/pzlSo6nVOpbjTai1vc4mxyVjYAIqGMbmODqyQSe7tifqwB1u9lfyN3q03O2czvEyidSFkcjtN26ZmJPjaW+8vB3bsT0jvvv3IHqii+CIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAisc9rGlznANAySfILrJrn2jHK1w/wCIjeGl51nPLdBOKeeWlpjJTwPOMB8gOGnfzwg7PIoKOrprhSQV1HM2anqY2yxSNOQ9jgCCPkQQVOgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgLj3mGubbNwH4h3JzciDTFzOPrppAP0rkJcMc5F/odN8rnE253GZscf7HKyFvUR8TnxlgAB7k57J6PG/2W5cOdTRIaQMxV4P1e6SH+Ze+68C/ZZgft0tGPIz0U9wP1E00g/nXvnkIKoiIKYCqtkWPjRwx1JrGs0DZNYW+rvlCS2akikBcCO4B7Ej/7GVvdSWUzBERUEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBcL85N2pbNyxcRKqqlEYfZZo2knGXEbD9K5oXnX7YHjfNo/hza+E9A8CbU7HyzgOwfCBLTkemx+0oa6Dezagkm5z+HLo258Osne75AU8gJ/OvoNXhl7JDRkuoOaij1E9jzT2K31MvV05Akewhu/1A/lXuaoKZyqOc1uA4gE7DJXXDn45j6zln4A3DVun54majulQy12cP3xM/JdIB5hjQSfr2XiVYeIHNBxx4jU8WmdXa21HqqunE0MNHWTuLXAj4uhp6GMBxkkAbjPdUfSMi2PwTt+tLVwm0rbuIsxl1LT2yJlye53UTMBuCfMgYB+YW+FJQREVGFd7Ra7/bKmy3qggraGsjMNRTzsD2SMI3BB2IXz48/wDwPs3L/wAzuo9F6Yp209jrIoLvbYGg4hhnaSYx8g9rwMbAADyX0NryM9tLwmudNrfSXGSmiD7fXUH3GqXDuyWN5czO+4w/8pUwehHJdfLnqXlS4W3u81clTWVOm6QzTSHLnkN6ck/UAt+cUuJ+j+D2hrtxA1vdYKG12mnfO8yPDXSkDaNgO7nE4AAz3XljoT2r1j4S8t2m+GOh9CVMup7FbGW6Gect92i6QcPIP0iCe2CPULpffNZcxXNjrkUtxuWptb3mvmHhUMTpJYoS4/CGxj4I2jOATjYDJKTIWOYOcL2ivEzmqtx0LSWen0xotlUJhQU7nPqK0tJ8MzvJwcZB6GjAI7lc78gXsy7nqSqtPGnj9bvdbKwsrLTYJB99qyCCyScfvWeYadztnvt2Q5S/ZgcGuFem7Fq3itp4am10I2VVQ2rlLqSimOCGRxDAcWn987Jz5bZPeKKKOGNkMUbWMYA1rWjAaBsAAPIK2aRbT08FJTxUtNCyGGFgjjjjaGtY0AANAGwAAAAG2ApkRAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQQVdNFW0s1HNnw543RuwcHBBBx88FfOrzs8GL9wO5jdX6Xu4mkpqyufdLbVSA4qKac9bSDvuCS0jO2PqX0Yrgrmt5TeHPNHoWpsuprXFHfqWB5tF2YAJqaXBLR1ebCe4O35wRrZHs6uYbTXGPly0lZJtS01RqvTVEy0XKkknb7y4wDpZJ0HBILA05A27nGQu1i+cfVfKzzR8HdYVdvh4Z62irLdI9sVxs1BUva9gO0jJIQSARg9wcLXOFfO7zN8HdYUMtbxL1LW0lvqWx11ru1RJLlgcA9jhJlwcBkYJ2PcJkzhuvocRaPo+/xar0nZdUQBoju9vp65oacgCWNr8fZnC1hAREQEREBERAREQEREBERAREQEREBERAREQEREFPmvGL2rfNhqvVvFe6cv2m7q6n0tpvw4blHES11VWFoc9rz5tbkDbv223B9UOY7iLd+FvCO96xsdOJaylj6Y8jPSSD8X1jC8L+DfBzWvNVzEsmv8Uvut5vXvN1q5tssMmSzJ7kgAfUp+N/6yDv17LnkdqeH1FaOZfXVS9t3vNvc+z27owaemlbgSPzv1OBBA9MfWfSRbN1frzhnwP0ZBdNbamtmmNP2yGOlhkq5QxjWMaGtY0bl2AAMAErhF/tLeSxjJHHjPSHwyRgW+qJdj0xHurm+I5A5peYmwcsPCWv4nX2kNa6GVlLR0bXYNRO/PS36tt+2PMjuvMLiD7ZvjjqfT9dZNH8P9N6Ynq2OijuLZZqmeFhyCWBxDQ/B2ODg7hYntMeejhtzLWfTvDvhFLX1dns9a+4VtwqaZ0Ank6CxrGMd8RaMk5IG+FyT7MLkW4K8V+E9Rxp4waaGpqiuuM9BbrfVSSMpqeKLAdIWtI63OcTgk4AGw3ynnV9cJeyxtOtdZ83FNq509ZVQ00FRVXWreXESPduA89iTuR6eWF7mLZPDfgtwp4RUj6HhnoS06dgk+myih6c/acn863snPgIsG8Xi16ftdVer3cIKKhoozNUVE7wxkbB3JJXQ3iP7X3gnp3X1PpTRdrrb5a46gQ1d6DQ2nG+CWAkEtHfqAOR6IPQBFp9hvNHqGyUF+t7w+muNNHVQnOcse0OH5itQQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQUyF4ne2Tv8A79zO2uwiVzhatOUzi3OQ10r3nb0JDWn8i9ob/e7dpuzVt+u07YaOghdPM8ns0D9J7favnW5x+L8nHfmO1brqA+JTT1YoqJrcnEMQ6GAeu4OPrCg9WvZV8v1n4ecv9o4nVdM1171jE6uEpaMsp3HDB65IGPqHzXdevrqW10FTcq2URU9JE+eZ57NYwFzifqAK2Hy7acZpHgNw/wBNsaGih05QR4HkTC0n85K6/wDtEOcrSPL7w5uvDile+r1rq20T09FTxEYpYpQYzNIfIYLsDYnuOytvU248wOcjmu1zzl8WaSw2qnLdPUNxdQ6btkRyZXyPEbZXHsXv2wfIH7B6nezx5On8rPDSprdWQUkut9TuZPcpo2gupoAPvdMHkZwCSSPMnOARgeaPs2uWLXvFvjzpXiN+x6dmi9J3JlxrrjURHwJZISXMhYTs5xeADjIGDnsce8ecIqqIiAiLrNzP8/fAvlopq6zXS+NvWsooXGCw0A8SRspHweO4YETSc5yc7EYBTRyBzKcx2heWLhpWcRNbTGUsPg0Fuie1s9dOe0bAT9pPYDcrxR5u+fLiZzcCj0/dbDQWLT1BOZqS3UhdLI952DnvduSRjYADIGPnx5xE4mccOcDiuyqvFRdNR3y7VJjt1rpy58VMxzto4mDZrQDu7A2G5wMD1i5KfZrcPOCun6LWXFuyUepNdVUbZXR1LfEprbkA+Gxh2c4di4g9yPTAeffK57NrjRzEMg1BdGN0lpdzx1VtcwieVvc+HGRnOPM+fcL2X5fuXDhry4aKpdIaCs0LHxRgVVwdEPeKp/m57tzgnfGT9q5OpaSloYGUlFTRU8EY6WRxMDGMHoAMAD6lOoCIioIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgpnfC8Ffaq2e02bnP1Yy1UUNMKyit1XUNjGA+d9MwvefmdiV7d8VOK2g+C2iq7iDxIv8NnslvAEk8mSXPOemNjRu57sYAHcr5++arizPzWczWodd6OstZLHf6qCis9H4eZ5IYomRR5aCcEhnUfQHfGCifXvJyvzvqeXHhlPJIHufpO1ku9T7szK5QXG/Lho678PuAnD7RN/yLjZNO0NFUh3cSMhaCD8wdj8wuSFN1fBERUEREBERAREQEVjZGPJDHtd0nBwQcH0KvQEREBERAREQEREBERAREQdYvaN6xZoblS1NeeoCV8sNNDk95XlwaPyheKfCzme4k8JJZ6jTc9MZ5D1NlezBY7vnbuvRj2x3MJpSDQFs5erTXxVl9rq+G63OKM590giyYw4+TnOJ222GfI46UciPJveOa/iG9txbLSaLsTmvvNcw4LicEQMP92R6bgHPbJFlv4+J762dqfiTzG85esrZpy6XC46nuJd0UlDCCIYiTu4gkgfWT8hud+fNM+x45przHFNea7SllZK0OIlrnyubnyLWtyD6het/CTlt4J8D6KOl4a8PrXapWNDXVbYQ6oef7oyHfJ8yMZXJyyryV0j7EnVXvUc2ueM1p92yDJDbKKQv77jqft2+S9LuCfB/S/Arhza+GukGvFutjSA9ww6R57uO53OB5lb8UFTUwUcElVVTxwwxNLnySODWtA8yTsAqJ1aXNY0uc4ADuScbLrjxH9oVyl8L6ya2ah4qU1VW05xLTW2mlqpGn0IY3GftXTLm89rNpTW/Dq6cPuX+ivlLV3hhppr1VxCAxQH6YY3JcHEZAP19vNOoj9rBzn2i/0cXLpws1LHVwtlMupqqjflhLfoUweDgjuXAHBGxGCCuvPJH7PPWnNU064vdyGntC0VUIX1b4i6a4Pb9OOAbDA2BcTgbgbrSeRvkw1hzO8T7dcNUWK50+gaKX3y8XSWN7GVTRuIY3kfE552JGcDOSO4949I6Q01oLTlBpHR9mprVZ7ZC2CkpKaMMjiYBsAP/cp5PTe8SaW09RaT03a9MW3qFJa6SKkhz3LGNDQT89lqyIiiIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIixLnWG322rrwzrNNBJN0+vS0nH24QdRvapcUajhvypXOktd1FJdNT3CntMAD8PfESXTFuPRre/kSF5rezZ5W6bmP40vrtTU8rtLaOjjuFeQMCaYu+8wg4xkkEkdwBn5Hj/jtxq4wc2vGhlq1LfKmtNReHW6zW5ufApQ+Usb0sG2cHJP14wvbvlH5ZdJ8qvCOm0ZaOl1yrA2uvtweQHVFV0YJJ7BjBkDyAz64T5o3fxq4waF5c+Flx4jaxlFLZ7LCyKGniwHTSH4YoIwfMkAD0AJ8sL5+eOfFfXfNtx5uWt32uepumoqllLa7ZTtLzBTt+GGFoGTgDcknuSc43XYj2l3OMOZDX9Lwm4diefSmlq+SKN8TSXXSvJ8Pra0buaN2sBGSTnAPftl7LfkluHCWxVnGbi7pZtNqq8Bkdmo6tgMtBSABxkIIPS95II3yA0bZwVP2fp3B5aOGsXCDgNobh42lbBNaLNTR1bQ0AuqSwOlLsAZcXk5PyXJyIqC2ZxW4s6G4L6Nrtca/vtLbbfRROkHiyhrpnAfQYDu4kkDA9VxlzQ86nBrlYtnh6yujq7UVRCZKKyUWH1EpxsX+UbTkbn5eoK8R+LnFbjfzocXqm5Nobzfq2vqD9y7HQtfMyjhzhjWMbsMAgF5A3O5AwAy3wdn+Mntj+Neo6+ttvCLTll0xaOp8UFZUwuqq2Rh2D/iIZG7zGAcfIrhvlr5LuOHPJeL5xAdqOnt1Aasm4X+7NfIaipduWxsbvIQO+CAMAbeXdzlX9knwuoNB23UnMdarhdNV1uKmW1x1roaaiadxE8RkF7sYzk4Bz9Q7/6G0Bozhnpik0boHTdFY7LQgiCjpGdDGZ7n1JO25JPb0CT+x1+5OuQzh5ym0lRdYK/9keq65vRPd56drDEzP0IRkloPmc5Pn557RoiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgLbuvtdaY4Z6Pu2vNY3OKgtFlpX1dVPI4ABrQTgZ7k4wB5khbiXh17Ujmb1jxH463vhJbr/PFo7SUjKRtDDIWxVNSGhz5X4OH4JAGe2PPAwGxOcznc4gc42qKLTtDbTbNJUFY5tns9Pl8tTK89LJJj3fIQQA0AAZOBkrvx7NbkFrOCtG/jFxjtNK7VtziaLXQSMEhtcJ3LnEjHiu2zjtgY3AJ2L7NH2fNspLXp/mT4tQGeuqmit0/Z5I/ggYfoVEoPdxG4aRsDuPX1CVvE9ERFFEREBERAREQFs7jBqq4aI4War1daKV1RW2m01NVTRtGS6RsZLcDzwcHHyW8VDPBDVQSU1RCyWGVhjkje0Oa9pGCCDsQQSCPmg+fzh57Rnmg4ea2qdYU2sxdYa6YyVVqr4g+lewnJaAMFpwSAckjvuvY3k45r9O82nDD9mdvoWWu82+f3S72wSdfu82AQ5p7ljgcgnfHfB2XWrnp9mfwx1LoG88TOBOlWaf1da2vr56Cic73a5RjLpAIiSGSblwLcA43XnTyf8yeuOVvi/BX2t8kdDcp2W682+YHpe3r6cuafNpJ+Yx6gK88H0TosGyXOK9WagvMIxHX00VUwegewOH5is5QEREBERAREQERQVdTT0NLNXVk7IIKeN0ssjzhrGNBJcT5AAEkoJ10H5+PaR0PAGaThfwfNvvGs54T73XGQSwWnOQAQ04dL59JO2+cHC4F9oX7Sq267tbuD/LhqW5Q0PjvF71BTF0AqmDIENO4Yf0E7l+2RgD1HAfJ7yA8UuZLWtPc9c2S96d0SxxqK+7VkDopavO/RD4gBc5xO7sEYJ37kJNHH3AfgxxU53uOzrPLeaqsrrjI64X++VWXimgz8T3HtknZrfM9gcYXvJy+8A9C8uHDeg4baDpnCkpcy1FVKB41XO7d0jyPMnsOwGwVnBngBwZ5bNKTWPhlpeisNE1ni11bI/qmqOhu8k87jl2AMkkgDHYLqlzYe1R4ScO7HdtIcGLs7U2ri19Oysp2H3OjfuC7xDs9wORgDYj8ijv2uP+LvHHhvwU01V6k11qWho2U0Ze2mdUNE0rvIBpOftIx9uAfnzrOK3M9xXu9Reo9YcQb5U1Dz4n3OqKt7AST8IZEcDv6Ka2cunNPxLuUcLeFev7nUSOAEtwt9SAM+ZfMBtt6pm+DvPxE9thfKe61FHwx4SWuooWktjq7rVS9TvQhjMbd+5H1LrRxn9ppzPcaLJU6YuF3tNhtNUcPgs9I6J5HoZHOcSF275U/ZHaOOmItR8y1JXVV2qD1R2imq3wMgHo8tIJ9Mdz32GAe2en/Z9cnWmZ4qu3cDLI+eEhzJamSedzSOxHW8jP2KZLdp48eeWvkE4780cL9RWGKjs1jc89d2u73gSOJ3LWgFz8nJ9fPcbrt3wz9ipX2rVtvunEzi1a7rZaWVktRQ26gkY+fBB6C6Q46T2O3b8i9QbFp+yaZtkNm09a6W3UMAxHT00YjY0Yx2Hn8/ktRyFRp1gsFm0vZ6TT+n7bBQW+hibDT08EYYxjAMAADz9T3JyfNakiongqiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiII5ZY4Y3SzSNjY0Zc5zsAD5k7K2nqKepjEtPNHKw/vo3hwP2hdBva2WDj5fOHmnTwppL7U2GlnkkvcVnEhlO3wl4j+IsGBsPn5ZXl7wH5m+L3LpxHt2pbRqW9CnoKkC42ioqZPCqYs4fG+N5wDjODgEEDO2Ql/GzqS6+kFFxDy680XCPmc0yb/wANdQNmqaZjDcLZN8FVRPI7PYdy3OQHDY7dsgLl5FEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAWnaiPTp+5uxnFHMcf8AiFaitK1VM2n0veKh+OmKgqHuz6CNx/mQfP3yO0MeoueHh1FPAJ2S6llqXMIyCGMlkJI+XTn7F66+0S5h6LgJy83llNcBT6i1XTzWm0NB+Lre3D3gZzs135z6Ly09mPX2ig55tHy3V0bWzG6U9MXf/N30k7WAepOSB9efJcxe2s1lFcuNGiNE09UXCy6ffVzxhxw2WomdjI7Z6IgfqcnlHB3sz+HNTr7m50bUvthq7dp+WW6VZfH1RN6In9AdnbPWQQD36V7+nsupfs2eAen+EnLfpfUxtTWaj1ZQsulfUPYBKxsvxMjz3wBj8oHku2qAuo/PTz5ae5ULPDp2w0tNetd3WAyUtE6T4KOMjAmmAye+4ae+PRZfPvznWflZ4dPorDV09Try+sMVpoyQ73duN6iQdwBkYHmSO22fGbh/w94385vGKSitzq/UWorxMJ7lcqkl0dLETvJK47NaN8DbONvMh7wV0xpjjhzsccn0lLJPf9WaindPU1U7iIaWIHJc47iONgJwP0kkr2v5KuS3SnKPoyopG1sN71Zdy191u4g6AQBtDEDktjBye+53O/fcvKvym8NuVjQ8Gn9KW6Gpvc8TfutepGAz1kvnuRlrAc4aMDGNuwHOKeAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiDavFDU8ejeH991LJUCA0VG97ZC7HS4jAIPrkr5vdWXWTX3GqvuVY41JvGoMOyc+I10waB8wRgL239qDrufQfKZfqqjkcypudZT26ItOCDIXAn7MAryV5EOFDeLnMRYLRUhroKKRtZJ1eZDgB3+sn6wENfQJoqngpNG2GlpoWxRQ2yljjjaMBjRE0AAegAwtbUUEEdNBHTQtDY4mhjR6ADAH5FKgIiICIiAiIgIiICIiDgnnN5iLXy08D7rrqtpRV1tW4Wy302ceLPKDjPyABJ/8Asrww4Q6D1nzN8fKWjs1s66y63IV9Z7vGeiCIPBJ27eQH5fIru17ZjjvpXVFZpTghp24MrK+wVk10u5heHMgkMfhsidj99gvJH5fJcg+xb4SR2nh3q/i/c7aG1l6uDLZb5ntOfdoW9TyMjze/GR5D5obr0Y05a/uHp612XqDvufRQUufXoYG5/MtSREBERAREQERRVFRBSwvqKmeOGKMdTnyODWtHqSdggkAwvMn2m/P3VaPqL/yx8Moo3V1TRCmv93bN8VMJW5NPGBuH9BHUTgjI+a5x50vaFcOOX/R9ZZdBagt+odeVrDFR0tJKJYqPIIM0rm5G3kAck/Vg+Hl4ueruJOqrhqCvFder5d6iSsqnxxullke45c7ABOMkD5DARLNerXIt7L3S2nDpTj5xf1BTakq5qSC72qwwQA0UD5GNfFJO9xPjloOQwBrQcE5xhelrWta0NYAANgAMBfO9pXjRzyWXR1Nw80ZqnilSWChjMFPQ0FPUgQR7noa9rOtrdzgA4HkttXbixzaWiGS337iBxUomSjDo6u43CIkf+M4FCft6X+165mptD8P7VwR0RqYQXnU0r5r02lmHiw0LAMRuLTlhe8jY4JAyF0k5GOQrUXNzW1+o7nffuBoyyVDKerrGxdc9VMQHGGEEgZAIJcdhnHfC444M8s3HbmX1zBbbTYb1UGqkaa283KOUxxszu90j93EDyyfngL3m5Z+AmnuW3hFaeF2nZDOKPqnq6hwwaipfgvf9uB+T7E/1WvcIuD+heCWirfoXQNlgoaGghbF1hgEszh3e9wGXEn1W91QHKqgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAupXO/wAiGjOZrR9RddMW6gs+vaBrpaG4RxBgqyASYZiMZB7AnscZ7AjtqqHPkg+cThlxM41ckXG+a52+kfa9QWaR9Fc7ZWMJgrIM4dG8DGWnGWuG4OCPML3B5P8Amz0lzacNhq20UzLXeqCU013tBlD300gAIeD3Mbgcg/YcFaHzc8ivCnmptT7hcqX7jawpoXNor1SgB7iB8LJh2ezO3qM53wMeI1n1Xxi5ROM90o9O32tsWo9MXJ9DXRxvLY6kRSfQkZnDmOAyAd8O2IJVH0oItrcL9YR8QeG+l9cx9GL9aKS4ODOwdLE1zgPqJI+xbozvhQVREQEREBERAREQEREBERARFFNLFTxPqJ5WxxRtL3ve7paxoGSSTsAME5KCVec3tBvaH6t4A8TIeFXDFlPLX0lLFUXGV52jMgyG533xjby7nyzzVxW9ppyn8LqmttL9by6gutH1M91tNLJM0yt/eOlx0t32J3x814j8YuI194+8ZdQcQaqmebhqy6ulhpgcloe4NijG+MgdI22z2VnE9e1nIBzQaz5kdI1N31VQvDoch0uMtD2npIB8xkLt2uH+VHg3Z+BvAjSWiLZSsjqY7bBPcJQ0B81S9gc8uPqCSFzAsyYvvoiIqCIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiIC6686PMPpjgnw0rbXX3GOG7ahpZaWlYXYIa8FpOPU5IC7FLxO9sfqeureZu3aYdM8Utp0/Szxt6sDrlc8k49R09++5VmfR1bobTxF4H6wsvFRtqqqWGlrhV0NY0Ya4nOwPkSCQt+26j1/zz8xNJcaiCernrZKaGse92XiFpORsSdxkfLOV3y5++H+maX2c+ktR0Ntp2VtHHYp2zhgDyKhjS8EgZO7gPqC479ihoJtwv8Ar/iFVQdUVsZS0EDiNhM/qcT9fTn6sgqTPad8equmLJDpvTVp07SsayK2UUFGwNGwEcYaMfkXG/MzzGaK5bOGN01vqi5U7a1lPILXQOeBLWVOCGMa09xnGT2wFv8A1rrbSvDnS9w1nra+UtostrhM9XWVL+lkbB6+ZPoBuV4Hc8vMhVc2vMFPdtINr6nTtGI7TpujLT1ysBwZBH3D5HknG5xgKzN6NjUreMHOdx8pqKWrqLzqnV1f0NfK4mKliLiST5Mijbk+Xb1K9zeUDlK0bymcPDpexzC5Xq4uE94uz4w19TKBs1vmI274H2ndba5GOUHQnLhwrsN0fpumOvrtboqi93WUdc7ZJWhxgYSPvbGZDcADJBJJyu0CXJ4giIooiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiKhGRhBwDq/nW4K6C1c/SWq73HQSMk8MzSytAzn0P+tcxaT1tpLXVqjvWjtR2+8UUjQ4S0dQ2UAHsCAcg/I4K8g+fvkl5hb3xar9daJ0RddS2mqafhoYzK6LBJJAHcHO4G+w7+XULQnEDjfyva9pLraZdRaTutFMJZaCqZNTNqGg7tkicAHAgYzj6irZJxmb6+lZF0h0F7W/lQvek6C4a11BdtP3wwM99oHWionDJsfF0Pia5rm57bg/X3O76L2onJZXR+I3ipLD8prTVMP5DGs2z8eNO1wGEOy6rn2nfJaHFv+y2Djz+5lTj/AKCwq/2pvJXQs6v9k6rqDjtBZat5/wChhX08dtkXTN/tbeS9rukau1C4eosFRj84BVf3W3ku/wCF2of8wVH+pB3LRdNGe1r5MHvaz9l2oG9RAy6wVGBnzOy7O8L+Kmg+Mmj6TXfDnUNPeLNWEtZUQn6Lx9Jjgd2uGdwU0cMe0H4K3Pjry033S1jGblQTRXSmaO7jFkkY9Dnf5ArxH5eOMOoOW3jTbdYRwOikttUaW5U7xuGB2HjbzaRnzzgjzX0kyxRzRPhlYHseC1zT2IPcFfOtz1aEj4c81WvtOU7Q2mNwFVTgDH3uRjXA48iTk/aoPoc09eINRaftuoKVuIbnRw1kYJzhsjA4fmIWorZfBV4k4O6GkAID9OW12D33poyt6JAREVBERAREQEREBdPefrnosHLPo2q0ppGtgreIV3gfFRQNcHC3hwx48oByCM5DdiTjcLc/P7zQ1fK9wVff7B4R1JfZzbrSJNwx5aS6THn0jf8AL54XjPwd4N8bOdjjBLBQGsu1bWVDai93qpJMVFA52C97jsMDIa0dzgJmoxOWrhbfuaLmP09o+7z1dyffbia291jiXvFO0l80jz3AIGNu3UMYX0O6F0NpbhrpW36L0baYbbabbEIoIImgAAADJ9Scblcc8uvKfwb5ZtPw2zh5pmFtzfA2OuvE466useB8Rc8/RBOcNGABt5LmdX/D9iIiiiIiAiLhLjxzkcvvLhPHb+KGt20tzmAey20lO+pqi0jIcWMGw3G5I7hByZr3W+n+HGkrnrXVFbHS221QOnme9waDjsAT5k7LxN5tfaT8TuN1VXaU0RXzWHSwkcxrqdxZLOwEgHI3AI8ySd/Jb29oD7RjS3Mbo6n4U8HaG8wWaWpbNX1dXF4L6oD6MYYCTjOO/cEjzwut/Bbki5jeONzo4NOcO7pRWupc3xLrcIHQU0TCd3ZdgnbJGBg47hJpjinRGiNZcVtYUWkdKW2pu97u84jjY3L3OJIBe87kAZySfzkgL3E5KuQDRPLbppt11dSUl+1jcI2urJ5Yg+ODbeNgOQQMn1H1k5W/OVjkx4Tcr+m6WHT1nirtTPhaLheqhodNLIR8QYT9Fuc4A/NnC7Bq6MWjtlutrSy3W+mpWu3IhhawH6wAFjXXTenb4WuvVgt1wLNmmqpY5SB8uoHC1NFBi0Ntt9rhFNbKCnpIhuI4ImxtH2NACykRAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQUIXjR7W7lp1RYeL9Xx7sFhdJpvUFNT/dKeBmRDVsYIy9/wD3wa3J8sA9jt7LriTmx0/ZdS8tvEm2X+FklINN19R8TQel8cLntIz2OWgZG+CUneJbk11I9lJzew8QtIUfLrqaMsvWlqA/cybqz7xRsx8Jz3LQfs+ojHomvAL2YOpjprnR0KzOG3f322OOf/mlNIR+UsA+1e/qtIIiKKIiICIiAiIgIiICLHrK2kt1LNXV1RHT08DC+SWRwaxjRuSSdgFwfqfno5TNHzupb9xv0/DOwlroY3SSvBHfZjSU09c8Lyc9pV7QvUceor1y68GblFSUFG0Ut+vVO8mWaUjL6eJw2a0ZALhnJyM+mu873tU9G3LRE3Drlh1BXVdzumYrhqAUz6dlLTkbtg8QBznuzjIAAGdz2PUjki5KNX83WtpL1e5qqi0XbqgSXi6yZL6p5OTDG4nLnu3y7ORv55IfsWcp3s9uMfNPSfsqoJ6TTekmymN13r2ucZ3Du2GMbv8AME5ABx6r0V4Ceya4PcH9V2/WmoNU3HVdwt0gmhiqIGRQtkGMEAE5wR5jPzXc3Q+idNcOtJ2zRGj7XDbrPZ6dlLSU8TQA1jRjJwNye5PmSVryXota1rWhrWgADYAYVyIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiIC8RPbG2qSl5qqW7OaQyu03RtafUxueD/wBIL27XmB7ZXgfe9Q0eluMVjonVEdnp5aCvDNyIy7qDseeNvsB9FYODtJ8xWpubLltsnKBU0xF5txpYIagDLqiOAgQu8u2wI9R6YXpfyh8ummeUXgbHpiqr4BVyF921BcpiGNdMWDOXHADGNAAz6FeIvJ7xvsnL5x1svErUlvlrLfRNfHMyIZc3qwQ4D1BHl6rlfnK53uJvNprU6Q4dVF9pdFBjYqWxULZOuufg9T5WMHVJ3wGkYG+3bE+nGqe0M5+LnzK3ufhloR5o+HVlri5j2uPXd5oyQ2d/kIwcljd+4JOcLnv2QvKqx7LrzBcQdMte1wZS6bbWQZx5yVDA4eewB9Nx3UfIX7L+krqWl4r8ydhqA4PbLbdN1ALGkDcPqB3I7Yb2+0ZXqdbLZb7PQQWu00UNJR0zBHDBCwMZGwdg0DYBS9GWiIqCIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgouEuablS4ec0+hZNK6ugbR3KA+JbrvBE01FK8ehPdp8wfn6kHm5EHkJW+xF4mCtkFu426YdSdR8MzUFQJMeQIGR288/YtMuPsTuNdOM2/ixpCsHoYKiM/nBH517HIg8ZIfYscfZCRNxE0fEB2J8Y5/I1TR+xT47vmDHcTtGsjPd5bUbfYGkr2VRB5CRexC4mujBn46aYZJ/cttdQ4flLgfzKrPYhcSuoB/HTTIb6i1VBP5Or+devSIPGPiL7GLjlpfT8940TxB05qypp2F7reIJaOaQAEkRlxc1x22BIJJAC4O5b+czjryT3S+aQtNooqmklqT90LHeopOmGobsXNLXAsJHfBIOx88n6DVwrxR5NuWrjLdn3/iHwntNxukhy+tYZIJn/AFujc0n7fMkpz6PNuX22fG57Hsj4R6IjLmkNd4lWS0+u8mDj0wumurdean5l+O8eqtfV0Dbjq2700FS+NvhxxMe9rA1o8gAcD5+i9t6r2aXJZU0j6VnBekp3Pb0+LFcKsPbt3BMhAP2LyG51+WO68p/G2rsdmjrf2NVMjK7T9wfknoOHCMv83sII9dvPBKmTR9AmlLRDp/S9nsFMQYbbQU9JGR2LY42tBH2ALVl1k5IecLQHMvw5tVspbtHT61stuhivNqlOJetjQ0zMz9NjiM5HYkgjZdm1bP48SXRERFEREBERAREQeWvtu6eoFp4aVXWfANRWRludusNBB/ISt5exPbQHgZrp8NOG1f7KWtnl83sFLEWDPoMu2+Z9Vtv23czTo/hrTgjqbcqt5HngxAD9BW+fYv6XqbTy36j1JMR4d+1RM6EefTDBFGT+XP5E2j0DRcbcT+YrgnwZmZScTeItosNRIwSNhqZT4hafPABI9d/LdcYye0e5M4nmM8a7Y7Hm2GYg/UQ1TR2XRdaI/aPcmL85422tmP7qCff6sMW3tQe1R5MLD8LOItdc3f3NvtFRL+ctA/OreDtvgLafE7iZpHhHoy6a51rd6egt1rpn1DzLKGGQtGehgO5JOBsD3z2XkRzs+07v3GIU+juAVzvmm9PNDjV1ocaeqqiezR0nLR9uRtjzXVCwcMuZHjVcaa02vTOutTSVb2lj6iKqmi3Iw4vflo7jfKel47g8RfbQ8bLlcqyn4a6F0vZbcHubSz10UtVUFmTh7h1tYCRjbBA+a68cM+CfMX7QLijfdTUNTDcLi94nut3uMhipoM9mAgHGB2aB2+Z39DOXT2R3BSzaLtd147224X/VE7G1FVSMr5IKWncd/D6YyC/Hnk/Ihd3uGvCfhxwe0+NK8MdH27TtrD+s09HH0hzvVxJJce+5J7n1Q/boLy2ex/oeHOtbdrfjHryh1G61ytqILXbqZ7ad0gOR4j5MFwB9AB8s4I9I4IIaWGOnpoWRRRNDWRsaGtaBsAANgB8lMiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiKKeaKmhkqJ5AyKJpe9x7NaBkk/UAglWk6r05bdYaZu2k7zGZKC80U9BUtHnFKwsdj54ccLp1xe9q3wB4YajrdK0lFdL5XUEhjmdTswxrhkEZ7fnz8ltPSftluAl0uDKbVGlr9Z6Z7g01DY/G6BnuWt3I+0JO9hZnK6Lcz/ACvcTPZ+cWdNa80tqIVlufXurNO3drC18csRB8GUf3QaRnB3BPbz9ZORvmrp+bDg9+yyto4aHUVnqfufeqWI5aJukObK0eTXgkgeoPbsOi3tMec3l25g+EVl0Vw3u016u9Nc2XGOd1K5jaYBha4ZPYlriCF1M5NebLU/KjxMZqShElZp65lkF7twdtNCDs9o7B7ckg+fZN4j6KEXAen+evlP1Dp+j1FFxr09RxVcYd4FVP0TxOIyWPZgkOHmFyrojiXoLiTQG56E1Xbr3TAAl9LL1YB8yO4H2Irc6IiAiIgIi4o5h+ZThfy06Ln1bxEvccMjo3+4W+Mg1NbKBsxjO+57nsBk74wh65XW0eJ/FDRnB/Rdx11rq809utlthdK90rw10mBsxgO7iTjYZXgNxe5wOYvj/wAQau502ttTUjK+qcLbZrPWTRiJhJ6I2MiILjj68nsFHNy586/ECmibc+GXE67wSESRtucNSW5PmBORv80n7HMXN17T3iXx8pq3Q2gKU6R0dKXRv8N5NbWszsXv26AR5Aeh2IXD3LNyX8Y+aisqX6Io6eitVIcT3W4FzIA4ncNIBLyPPH5znHaHk09lXq3VOoWat5lLBV2WxUThJHZnuDZqx4wQHkHZue4GQQCCd8L1r0dojSPD6xwac0Vp6hs1tp2hsdPSRBjQB2zjcn5nJV1LP6eWGm/YjayFbDJq7jdZRShwMsdut8xeRkZAfIQBtnuF6acGuEWkOBfDu08NdEUQgttqhDA7pAfNIQOqR583Ejckk9sk91vlFFEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQF5w+175kZ9B6ZsvBWwRxuuOoYX11XM7B8CAEsGB/dHfGdsHO+MH0XqqqmoaaWsrKiOCCFpfJLI4NYxoGSSTsAPmvAP2kPG+xcc+aC93rStX7zZbHTQ2Skma7LJjFkySN+Re5w/8UfWg4Y4KcFtdcfeINt4ccPraaq43B4DpHbRU8QPxSyO7BoG69wuTPkM0FyqWd9xq5oNRawrQ01V0kpwGwkfvIQcloHrse/qSYvZ38tui+CvAHTGpaO0RO1Tqu2Q3O63B7QZSZWh7YmkjIY1pAA898rtaroIiKAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiIC4Y5seX6w8xnBe/6FuFvhkuj6R8toqXMBfT1TRlhae4yRjHb1XM6JR82nDPXfE3k+49UWqGW6e36h0vVugrKGcOY2ogOWyxO7Ese3OD9R8l728sXMho3mi4YU3EbSEc1LiQ01dRTEGSlqGgEsOO432PmPXueoftgeXO06k4X0HHXTdlY2/wCnaxtPdJIIvjqKGQOy5+O5Y8A9RycEhcC+yM5ntFcJtQam4Ta9usVrptWVFNV22qmJDPe2AxujJ7DLS3B7kj0BSo9lEUbHskY2SN4cx46g4HII8sH0UiKIiICIiAiKKeZsEEkzu0bC8/UBlB5Ge2i13brvrDSOi6Wpa+a1RySzMB+iSDnPz+ID7FwNyo+0b4o8qvD+v4b2XSll1DaJqmStohXPkifRzPA6yCz6bSQDg4wQcHfC4n5puI924q8dtUXu4VT5xHcpqOnzvhrJC049ckH6xhesPKD7O/lptvCGwaq1roGi1dfb3Ssq56i6h0rIiScNjZnpHbc43/ObuciSc68oLtW8ceczi/WXuSmqb/qO8zdTxEwtp6ZpOA0DcMYNgAN9uxXYG3eyD5qLhTxzv/Y9TF7Q4xzVvS5vyII7r2R0JwY4UcMWkaA4fWSxHGOqjpWscB6A9x9hW9U3ivD+o9j3zUQkeFLpqX6q8D+Za9YPYv8AMPcgDfNcaQs/r1SSz/8AQavaJFmfseeHK77JTTvCDWdLrnivq+h1fU0Di+loaejLKYP8i4PySR/r7ZyvQmmpqejgZTUlPHBDE3pZHG0Na0eQAGwH1KZFTBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQFi3KhiuduqrbM4iOrhfA8juGuaQcfPBWUiDy+4n+xln1brK6ak0pxeprdTXGpfUeBWULpXMLiScFpHme2T9a29SexBvfSffuO1Dny8K0vA/O5esSIPLmy+xDsgfnUPHGtc3zFHbWA//hErR+LnsW47Fo6vvfCfipX3e8UUL52W+6UccbajpGehj4zs442yPy9l6vIg+WWstFwt91msldTOp62nqHU00UmAY5A7pLT5DB29FyxwU5iONvKxqRt30Nc30bXn75SVUZkpahuQTgZAIPqD57hdjfalcplTwg4if7LmmKV79NapmJqC1u1NVHGxwMDPr9W25XPfJhfOUnjPyaT8KOJ7tMN1JS0tVS3KK4eG2vDt/AqYHnD8j4S0sOQ5pz5peEm+uVORP2j45pNWTcMNa6Rp7JqdlG+sppaKUvp6tke8mzt2uAIOO3pnfHeUnC+frlG1FJyvc22ndUa5hqKCzW+qqaGorZGFkclPKx0fW0nAIIIOMr3u0tqrT2tbFS6k0td6a52ytb1wVNO/qY4fzEeY7plTdawise9kTHSSODGNBJcTgAeZJ9F5mc/vtNqfS7a/g7y+3MyXlrnU90v0ZBZTDsY4T5v7gnsPswSu1nNjzscLuVewGS9VMd31JUNd7lZaaZplcQNjJv8AA3Prv9WQvEPjVxo4tc4PF9l+v0ctwu11nbRWi0UmXRUrHHDIYm/ky44zgk7dsLhzwe46cz2syzTFjveprhWTf7buUzXyRxZO7nynIAGScA7DyAXtHyf+z+4V8tFst+pLjbIb5r3wg6outQA8UryBlkAOzQD5jfYEbjJWfYNrchXs9LFy10cXETiA6nvHECtp24Phgw2oOGTHETuX9gX7bg48sd2URAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEXXfnF5vNP8p2k7ffLnbX3CtucxipaVveQgZIByB5E7nyWw+V32mHBbmIukOjrqybSOqZj0wUle5vg1R9IpASM/InOxS86ku8juIiIiiIiAiIgIiICIiAiIg6t+0t1heNFcoWr7nY62Wkqal9NQmWJxa4MlkDTgj7F8/a94vao3yy0/KhqDT1XURmur5YJKeHIL8Ru6i7HfAx3+teDqD6c+CkUcPB7Q8UI6WN07bgB6f7XYt6rZ/B0Y4TaMAII+4FB27f2uxbwU/GZMBERUEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQafe7LatSWmrsV8t8Ffb66J0FRTztDmSMOxBB/wDsggEdl87vOrwjpOAnM7rPQdgidSW2jrI661hriHR080bZWYI7YLiBjtgYX0ZLx99r3y668HFQ8wNptU1bp2uttNSV0sTer3R8LekOdjyI7ny2PbJDfiPUDl61C3VfAjh9qJtUKk1+mbdM6Yfv3mnZ1H8uVyEvMX2SvN3X6lo6Xlj1U0PltFHLNZKgvGTTs+IxY9Ggn8oPmQvTpSc4tERFQREQFsLjrru3cM+DusdcXSpbBDarNVTBxIH3zw3BgHzLiFv1eS3tduay7yalPLTpebw6Clp4qq+TNd/ZHvHU2LA9ARnPby77B0v5SOFk3MLzNaU0Zc3OdBc7i+4XJ47iKMGWQ4+ZGMfNfRRYLHbdNWaisFopxBRUELYIWZzhoGBk+ZPc/NeRvsdeBN8uHEms49VdO5lqtdLUW2leRtJJIOl5B+WMfYV7BoCIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIg2fxV4WaO4y6IuOgddWxlbarjGWva4bsdggPafIjK8EucflX1Nyh8VxaIK6SpslY73uyXBoLXGPOzXejgQR3OcH5r6HV0V9rBwUOvOCE/EaD4pNIQOmkbjcRgk9Q+WTg/YrB5j803G608TbTovT9nooI22e3h9ZM1vxSzEADJ+QDs/WOy9NvY73etr+VmqoqysfMKDUNUyBjiT4cRZGRj5ZDgB5YXRr2YfLdwt5k+ImqbTxQt0ldSaftsVdFTtkLPFL5OjBI8hsTt5geeRuP2hdRqHlL4m0fB3gXqC56S0vc7HFc3U9vqDE54kkkjLS5uCADG7cEHceiTs7Un/ADyOyXtTudeq4b2aDgZwp1HCzUF2jc++VVLIHPoqY7CIEfRe45z6AeoIXSrkj5CdX83lZXatu9+dYtH2yrENZcHxGSetmPxPjhBIBIByXkkAnHdafyN8pl75uOLPj6vNzdpKhcam9XMyPL6hwxiISnJLndic5GQvdrh3w60Zwp0lQaG0BYaaz2W3M6IKaBuBnzcT3c49yT+gJ4rB4ScJNE8E9DW3QGg7TFQ223RNjBa0dczgN3vPdzicnJW9URS3UkwRERRERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERB1x5z+TjT/N5o632Wu1HLYbtZpXz2+tbD4rA9wwWvZkZac74OfTfC8SuYnl94kcoPFtmjdS3GF1fTMjuVrutAXCOoiLiGSM6gC0gtILTnBGDlfSGvLT20fCi83h2j+LFvp3yUtoopLbVlrchrXSl4cfqJH2E+iu/Ekzx3i5PeMMPHHl50frp9U2WvloGU1xb1AllTGOl4PzOAcnuSVzSvG32QXMBd9P8UJeBlfUOdab7FLU0rD2ZM0ZIH1k5+0r2SUUREQEREBERAREQFj19XHQUNTXzHEdNE+V/wBTQSfzBZC0HXjZH6G1EyHPW601Yb9ZhdhB41cI9c6m51+dKXQ+ur1LUacuM1e6KmLi5rYIAXZGT5taSB27D6+IOf3lotvLBx3k0jp18rrBeKCO72wyDdjHvex8Y9elzD8t8Dso/Z869peGPOJoS93uZlPSy1lTbKuWU4EYngkiJP1OcFzh7Y2/UuquYnSlvsszKz3LTsdG0wODw+R9RI/AIOCfjA2Uk7U2716xcvFyN34EcPrkSD7xpq3OOO39rsH8y5EXHfLzY59NcC9A2KqBE1Fp6hicD3BELVyIqoiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgLYHHzTVNrDglrvTlTRNqxW6er2shLc9UggeWYHr1AYW/wBWua17S1wBBGCDuEHzt8iGtqbhtzc8O75erk220Aupoa6WX4GsZNG6PD89h1ubn0wvohhmimiZNDI18cjQ5rmnIcDuCCO4K8gva2cqXDnhM3T3GnhvZhaHajuk1Dd6SnBEBnLDK2Zo7NJ6XggYGceuBzB7G/jlrHXWkdY8MNX3yqukemXUtXa5KqUyPhgl6mvjBO/SHNBGTtnCmXd+Hx6RIiKgiIg6/c6nNPQ8p3CJ2t/uay43m5VIt9oo3v6WvnLS4ucfMNAyQPz4wfHDg7wN4z+0H43ai1GyupYqmsqRcL9dajIipmyHDQ0DJJAbgD0A+pdofbO8ZNPakuejOE1kr46qosU9Rca7w3AiOR7AwNPzxn865Q9ilpKe3cIdc6uqoA0Xm9ww07z3dFDEQcfLrcR9ifjZ9S78d2+AfBfTXADhXY+F2lx101pgDZagjD6mY/Tkd8yfzLkRERRERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQFsfjfoyHiJwf1noiaLxfuzZKylYwdy8xO6P8A8IBb4RB88nJxzL3Pky451up7vYZ7lQSU1RY75bopAyUtEjSCwuGOtkkQwDgYJUfM1xp1Rzycx7NSaY0tU077jHTWSx2sEyyxQMJx4hbkZL5HucQMAH5Le3tPuDVs4S8yFbVWWJsVHqeI3IRN7MeSM/lBH2gru97H/gpw9ouA3+zK+x0tVqq8XWrpjXTRh76WGF4a1kZP0SdySN9/mcz7p/juHy+cIbBwO4Raa4d2G2Q0n3Nt8DassaOqWp6AZXvd3cS8ncrkhEVBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBcTc1HD2g4mcANcaZq6KOpkkstXLTB4yWysjLgR8zjH2rllY9bSw19HNQ1LOqGpidFIPVrgQR+QpR8+Ps6brDZ+czhpJV1AginuMtM9ziAMvgkDQSe2X9I+1fQsvnr5xOW7XXKRxrfUwTzQ22vrZLnp+5wZGAJC4NyOzmnGB6D5Feovs1ecK98zegLrp7Xs8curtJmJtRMCA6rpnghkxHfOQQT5lB3PREQEREBERAREQFxrx+4j2Xhzw3vFbc5mCerpJqamiccFzntLSfqGc/M4H1cklefPtS9eSael0nZGvLWVmA7cjJLyPL5AfkTxLceQ+t7ddtL62ragslppPfn1dJK3I28Qua5p9QcfaF3p5CuFh5stbU+stegV79Lvb48so6unpwRjPmcgY+a4i519FUFk0poTUdNGGyV8T2uIGMgtDj+cLuD7Ealidw84k15H31l5pYQf+1MGSPygJu9iTsj0tggipoI6aBgZHCwRsaOzWgYAH2BSoiNCIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIg6pe0x4V3XivysXm2WG2vrLjaqynukDGDLgI+oOx9YIH2ryu9nzzBnlz47xtvkj6W3317LbXCQFoY4OIAeD23JG4yCB2Xv1JFHNG6KWNr2PBa5rgCHA7EEdsY8l8+3tHOHFBwu5vNZWez0zaWirzTXamijGGxtnia4huOwDg77cpB9A9PUQ1cEVVTyCSKZgkje3cOaQCCPkRuplwPyRcXqLjPy0aI1MyvinuNNa4bdc2Mdl0dTC0RkEd9w0HJ77/Nc8KQFpeprh9ydN3W6h3SaKhnqAfTojc7P5lqi4I5yuPOjeBfArVV01DdIY7jcbTU0VsovEAlqJ5Y3Rt6RnOAXZz22SzeFuPCGwW+88xvMNQ2msrZaqr1hfhEHucc+G+QkNHphgwP8AWvoZ4KcItMcEOHdr4e6UpIoaSgjHWWNx4khA6nfbheGvs0tJV2qucnQEtJEXxWWomutQQ3IayOF+CT/3zgvoGTICIioIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIg8bvbRVFM/jFpSnYczMtshd8h972/OF2t9j1VPm5SRTO+jBqKvDftLSV57+064mU3ETmQrIqOQPgs0Jpm4IOCXbg488NH5l6J+yLsdTaeUilqqhhabheq2oaPVhLcH7RhSdm0d2URFQREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQcHc3HK1pXmw4Xu0Hfqx1ur6Kf3203JsYe6lqA0jcHuxwOHAb47eh8WKO88avZ1cytfaqSuibd7JIyKsY3JprnRPAe04Pk5jsgncHPfz+hheU/tX+TziZq7W9RzH6GtTrvaoLRDBeKeDBmp2wAgShndw6TvjPYY32V94mPSvhPxDtHFnhrpriVYng0Wo7ZBcIgM/CXtBczffLXZafmFuzOF5A+yo5y9QWLVVl5X9WEVVku00rbJO4/HTTEF/hZPdpIOB2Hb0z6/YylVVERQEREBERAXkN7ZrUZh4raHtDJz94ofenMHlh/c/lXryvEf2x1ZJNzS0VEfoU2nKQt+XU55P6EiWa3n7RXh19yuVfg/rCFu0ssccvyD6dxH5Tj/ANwuWPYhOB4ZcTWeYv1Efy07v9St9oTQe9ezh4a3GQtMlM6yO33OX0xBx9vdR+xBDhw94ou8jebeB9fgSf6wpLxXpmiIqCIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgLoR7TnkspuNGk6zjfpSUw6o0tbXvqIenIraWIFxH1gZx6AZ9Qe+60XWVr+7mkL5ZOkO+6FtqaXGM564nNx+dJc6PFv2SnGq7aM5iqPhlWXiRlj1bTzwspXPPR721hczpHqcHP/ej5r2+XzH6bvmoOB/F+ivlEZYLpo+9B4/euLoZCCPlkAj7V9GHAnixauNnC6x8RLQ5vh3OBrpGtOQ2XALgPyg/LOEz6b8cgLwo9rLxAqNV829503Ddn1Fu0xQ0VDHAHZZDOYWyS7eRy8A/V8yvWXnK5gX8tfAu78R6SnZPcBIyioI39jPKCGnHngheHXCrhJxf51eNdc63QzXC53mtNderk7ZlOx7t3EnYYGwB9BlB65ezg5UdI8EuC+nuIlVbevWesLXDca2qlALoIpm9bIWegDHDJ+Z+ZPcVaRpKxR6W0pZtMQvDo7Rb6egaRsCIo2sB/I1auiQRERRERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQFpmpKttv07dK97wwU1FPMXH970sJz+Zamth8ea6e2cEOIFxpTiWm0xc5WH0IppCEHzs6jgufF/jnX0duD6iq1BfJIYcHJLDIQCP8AxRlfQbyx8NBwk4J6a0QaZsElHSh0kYGOlxA2x9QC8RPZw6Yg1Vzk8PKGpibLFTVNRWvY4ZBEVPI7B/IF9CCH6EREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAWJc6Cmu1tq7XWRNkgrIHwSscMhzHtLSCPQglZaIPmn0ze7jwC5jbffYmvpqjRWqmyEEEERwVGCN/IsB+whfRdw24iac4qaPt+tNLVQmorhEHjfJjcRktPzGV4ue1K5fRwo4yv1tQR9FBquV8oAGAZME5z5nAIPzC7weyC1tJqnlxrrTPIHy6fuhpHfESQCzLc/+KApuzieY72oiKqIiICIiAvNP2qvKPqriberZxn0lF4ot9CKO4NDckMaSQT6Y8j23K9LFousrdT3fSV6tlWxr4aqgnje0jOxYR+XzQeD3M3zj3nixwf0nwFp4THbtLFgqpSB99fE0ta31ODvnywB5ld6PYr2CS38CtZXx42u+oW9B/7WKENx+UleP+qoxFqe7xNOQyvqGj6hI5e63ss9MN03ykWB3R0uuNVNWOGMfT6Tv88kp+kjt6iIiiIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiIPGj2u/Ldp7hjrCw8XNJ0DaSk1bNLTV0bRhoq2t6sj62gn82+F2X9jTrI3/l2v2mZqx0k2nb++NrHPyWRSxte3APYZ6gPLbHkuVfaRcBbrx55b663acpvGvWnqpl3o2AZLwxpD2gYzktOPkMnfC8pOQnmQ1Jyy8frfQ1TXix6irobLfqKTI6MydAkx5OY47/nzjCSDvz7aq4VNLy+6Po4iRFW6qayXHYhtNK4A/aB+Rbd9iLQUf+xrxJunu7PezfKWDxekdXhiDq6c98ZOcfJche2QsDLpyoUV4Lcvs2qqGYH0bJHNEfzvati+xEz/ALE3Ejbb9kVN+rBIPSdERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAXVb2ifH/SXCLl01dp2qvUMeo9UWqa2W2kDsyu8YFhfjvgAu/T2BXajv2XgB7THXV31lzha4oa+pc+m07PFaqNhcS1jGRMJIHYEuJzj0Cnd4Nz+yWstxuHOLZLnSUr5Ke2Wu4yVMgaemMPgcxuT2BJdt64K92VwFya8vPD7gVwY0zHpaxU0V3u9opKy63ExDx6mWSNryHO74BOMdtvqXPqtzeG6IiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIg87/bTabgrOAuk9TeEzxrbqNsHXjfplhk2z6ZaStt+xEuVE7QHE6zCpZ73HeKGqMOR1iN0D2h2O+MsIz8l2P9o1wB1nzD8ulRpPQFKKu+2y5092p6UvDTUCNr2vYCduoh5wPVeQ3Lfx24ncivG+puN20xXU8nh/c+/wBiq2mKSaHqDhsf3zTktOcHffzUkzT19DyLiPl25n+FHM5pMam4bXwSywBor7dN8FVRvIGWvYdyMnZw2OQfMLlxUEREBERAWNcoRPbqqDykhez8rSFkrEudXHQW2rrpSAymgklcfQNaSfzBB8weu6R8XEDUNBGCXMvFXC0epE7gAvoe5OrB+xvl20hazF0FlE12MYzkD/Vj7F4I6F0zVcYeZG2acsrfEk1Pqs+EAO7H1Bef/wAHK+j3R+nKbSGl7Zpmkd1RW6mZAHduogbn7TlBrSIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiC1zWuaWuaCCNwRleEHtN+Aly4L8yNy1dZLVLSad1U9l0oZ4WFscVQQPEYCBhpDhkb5JyV7wrifme4SaN4ycFdU6X1jaoKmNlsqqmlmcwGSnmZG5zXsPcHIGcfpAIeeDyG5pPaL3LmP5dNO8G6jS76G5Qy0s98rXSBzKl9O0hpZvkdTviOfP07LvB7Hrh9XaS5bLjqO4Uz4XapvT6yMO7ujZG1jHD5EAEfWvH7gpw+quK3FrSfDqkZ1SX+6wUZGCcsLh19t/ogr6TuH2iLLw30ba9F2CARUVrp2wMAGOogYLj9Z/NgeSfltxJzjciIiKIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIg0HXGr7VoLStx1XeZRHS2+B0riTjqIBwPtK+cPivqWfjTzCX/UWXSSap1EQ0jcuD5QwEfWMFe2ftK71XWDlP1LX0Epjk8RkeQcEZa/8A1Lxd5TLRSXvmD0bS1zeuJlwZK4EZBI7Z/Ln6wFfln086+jHSFH9z9J2Sg6en3a3U0OPTpjaMfmWsK1rWtaGsGABgD0VyzOTAREVBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQF0z59/Z/WvmqpIdcaOuNPZte2qlMMckrPvFyiG7YZSN2uByGu3AzvtldzEQfN/wZ4s8UuTbjoLzBTVduutiq3UN7tMuWioiBw+J4Ox23afmCDgnPvpy/ce9DcxnDqi4h6FrRJBOBHU07j98pph9KN48iDn8/oupHtWeUyx8QeF0/HDRemh+zLTb2PuD6SH466gOQ/rDRlzmEgg4zuRldJPZsc3tLy18TKnS+sHSO0hrCSGCpc07UVTkNZPjzBBAPyA+ZEvJo93kUVPUQ1UEdTTSNkimYJI3tOQ5pGQR8iCFKqCIiAuOeYjV9BoXgfrfUtwqWwMp7HWMjcXdJMr4XNYB6nJBx8iuRTsvLf2xvHu+WN9k4I2aqMVNc6I1teGu7sJIAI+YOPqBSDpf7Ouilrec/hh4cBkEF0knf2+FraeX4t/QkL6GV5Bex85fn3bXFVx2ukJNPaYZaSgBBx1u+Eu+ZyNvQA+q9fU9NEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREFCMrYPH7UVLpTghrzUNZKI4qPT1e4u9CYHNb+chb+JwvOz2tfM9BofQh4CWZ2bnqmm661w/+R05O4PpkdvUkehIDoZ7NC1sufOjw8EkYeKSeqqhnyLKaTB+wkL6B14keyK4Uahv3MbTcSjb5W2WwUNU01DmEMfM9haAD54zv8zjyK9t03eAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIg6G+2M1mdO8stu05HKWyakv0NOMdy2JjpHfZjb7V1K9kjy80vE7iNeeJt6YTbdJGGKPb6U7/jDfrwAfqB7rlb23Or4ZKbhpoVj2mSOWtujwHDIy1sYyPLbJ+1cm+xeskNDy46kvLR98umqJer6ooImj9JS9nB6CIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiCCppqetp5aSqgZNDMwxyRvaHNe0jBBB2IIPYrwp9pjyrVHAjjRV6v0tY3QaM1URWUzomfeqapP9kiOBhuSCRnvk/LPu6tscQeHGieKmmKvR2v9OUd6tNawslp6mMOG/mD3afmCEHm57Ob2kFwvVfYeXfjdPTDpp20Vi1FLKWOeWNAjp6jOxOBhr8jPY4wM+pAIIyNwV4k8+/s6rvy+VzeI3Bqhul10VUPLp4mgzT2p+cjJAyWeju4+oHGj8kntGtZ8v8Afv2P8WLvfdU6HqmeGIpKkzz0D87Pj6zkt9W57DA+TOnx7oIup1i9qHyZX6nbOzifLQudsYq+2zwPafnluPtBIXYLh3xY4d8V7UL1w91ZQXukLQ7rppMkA9iR3x8+yDd68MPa9XOSt5wq2iMpcygsFtiDc7NLmOecemeoL3PXgZ7UyaafnP1iZiT0U9CxvyaKdoCg9buQjQNv0Byn8OqOmpWxVF0stPdatw7ySzsEmSfTBH5/UrsKuKeVJ0juWjhf4v0xpS2A/WKdg/mXKySYkERFVEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQF4I+1P1DUX7nJ1VDO8ltrpaKhjGdmhsIOw+fVuve5eA/tQ7a+3c5+tS9rgKuOiqW5HcOgaMj5bIPYTkr4W2bhVy26EtFuoYoqqtslJXVszWAOmlljEmSR32cPtyfNc6LjLlm1NQ6v5e+HV/t8gfBPpugYCDndkLYz+dhXJqegiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAoauphoqWasqHBkUDHSvd6NAyT+QFTLiLmx4oWzhFy+a41hcKmOOSKzVMFIxzsGWeSMsY1vqcuz9ilHhtzucd67j/AMeLxfDO6ahts0lvoG7kdLXYPSPQkAD5AL1x9l/w6u/DjlOslDfKKSmrbpXVNzcx7cODZS3pz9QGPsXkNyQcJmcb+Z3Rujrgx01D72643DO5MEIL3HfvuG919Elvt9FaaCC2W2mZT0tNGI4omDDWMAwAAqn6ZSIiKIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiCCrpKWup5KOupoqiCZpZJFKwPY9p7gg5BB9CvPrmK9kLw/wCJ+pKzV/CvWDdF1dc8yy0MlGZqTrPcgBwI3zsPXfJ3XoYiDw14v+yV5jeGVtnvVjuNk1dQ0zS+R1A6SKUNAP8A8jcCfz4+a67cK+OfG7ll1bLU6I1BX2G40cpZU0FQwmJzhkFskTtsHzxjO25C+lIjOx7LibiJyp8u3FZ0suvOEWnblUSgj3n3QRzNJGOoPZgg+efX1QdWeTj2pml+M1UzRHGmlt+ltRBn3iujkIpKwjuCDuxx9BkfZkjoZ7T2+6a1Zzc3u76QucFypaq30LHS07utrpmsLSAR3Ow/Kt1c/fs+bxy4XU8QOGFDXXHQFU4dTiTLLbJCThjyBkswNnfLOwyB1m5f9K/s640aUsVWHSxT3KKSck79DD1bk+RIA+1XkiZdfRBy92uey8C9AWqpZ0SUunaCNzfTEDcfmwuQlh2mhitlqorbC0COkp44GAdgGNDR+YLMWZxRERUEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQF5E+2L4CX5mvbfx4tdC+W21VBDQV8jWk9Do/hYSfkDv8iPReupK4Y5x9GW7XfLJxFs9wpmSmOwVdVC5w3Y+KMvBB8jhpH2qxK6HeyH5pL3XXY8t2o6kzUsdLLV2hzjvGGDLmD6hgY9Mei9WV4C+y8mki50tDRxuI8dlcw/MCmkcR/8Agr36WfuKIiKgiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiIC8mPbMcU7ozUFg4V01fIykdAysqKcOPS4DcEjtnJH2Bes68Mfa81s0/N7W0j3feqexW8sHoXMOf0BByH7GLQFTV8XNR8RJ6LMFBan0cExHZz3APwfn8I+wr2LXSL2TulLTZ+XSmvdLE0VVxcHSuAx3JJB9cnBXd1SXQREVBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERBwtzjakpdK8tut7rVW8VoFuMbYS0EOcSMEg+mM/WAvEDkuvVmpeYayz3gRwNq6oGLsGtcXZ6RkjA32+pfQLxC0VauImjrro29Rh9Lc4HQuyM4JGx+wr5z+N+kK7gLzBah0zauunm01d+ul3x0gEPaNsbb4x6J+qPpQjMbo2uiILCAW47Y8sfYr1s/g/fXan4U6P1C85dcLHRVDj6l0LSTv88reCkBERUEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQFtnidZ36h4b6rsEcfW+5WSupGt9TJA9oH5SFuZEHzkcpevoOAvNRozVmqSaSmsF4fS3MEbxsc18MgOe2C4/kX0Rad1JY9XWWm1Bpu5wXC31jBJDPA/qa4H+f5Lx29rny3aX4Va4sHFLR1C2jpdYyTx10LRhvvTAHZH1tP6B5b9k/Yw6wvuoeBerLFdq2SoprBfY6eh8RxcWRvhDywZ3wD28gMAJan7ehaIiKIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAvC72vIxzhV59bDbv8AoOXuivDH2vZ/676rG21ht3/QKDv97Kd5PLNQsOdnMO/bs7/3+xd0F0p9lExzeWykc47EswM/98u6yn4ngiIqCIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAvAb2oVsjtnOdrVsQ2qWUdS75l8DSfzr35Xgr7VppZzo6qPrQW4/wDJ2oPZblWldPy2cMZnnJfpW2uJ+uBi5UXE3KX+DFwt/gnbP1di5ZT0EREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERB5re26cP9irhtH05J1BVEH0xTdvtz+ZZPsR/95riH/CeH9UYtC9txqa2P0fw50gydjrgy6VNwcwOyWxeD0AkeWSf0LXvYlQvZwZ4hSOaQyTU0XSfI4pWZwpJkHo+iIqCIiAiIgIiICIiAiIgIiICIuq3MB7RTgVy8cS4uF2rPulWXNrY310lGwOZRh4Bb1+pwckdwg7UotO09fbZqixW/UdlqW1FBc6aOrppR2fG9oc0/kIWoqS6CIioIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAvCz2u34YVePSxW7/oOXumvCz2u34Ydw/8BW7/AKDkHoH7KWTq5ZqEYxhzB/0l3QXSz2Upb+1qomjvlh/6S7pqRJdERFVEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBeCvtWnF/Ohqn5UFub/ydq96l4Ie1RnbPzoat6CT4dJQMPyIp25QezHKjGIeWfhdH/c6Utg/5Oxcrd1xXyqtDeWvhg3vjSls/V2LlTslSb9VRERRERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBYtyrobXbqu51H9ipIHzvx/csaXH8wWUtj8cbp9xeC+u7qO9Npy5SN+sUz8fnwg8BeYTihrnmt5g6qZkstwmrrk62Wan8mxmTpBAHYHAJ9AB6L2p5GuW6Xlm4IUejbk5r7xXzGvuLm+UrgMNPzA/JsPJeOvs4rNS3vnQ4a0tYxro4a6oqg1wyC6OlleBg/MA/YvoTV+J9ERFFEREBERAREQEREBERARFoettW2nQekLzrS+ztit9kopq6oc5wA6I2FxGT2JxgfMhBxpzWcy+kOVrhVV6+1K9s9dKTS2e3h2JK2qIyGgdy0Ddx8h5jOR4OUNg4u843HO511mtU921Lqu4PratzGkx07XuABc7ya1oAHmQ3YLW+YvmE4rc5nF2KsqYKyv8WoNFp2x0jC8Qsc7DWsYM5e7Yk/n7k+rXsy+UK/ct/De56k4kWiKj1nqmdsksGQ6SipGD4IS4dnEklw8jtkpP2l347S8IdEnhrwu0roF0xlfYLTTUD5Cc9T2RgOOfPfK3giKSYoiIqCIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgLws9rsSOcK4N9LFbj+Vjl7prws9rt+GLct//wBA23/0bkHoD7KVmOWqhfn6XR/+Uu6S6U+yic48tlGCcgFm3pu5d1lICIioIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiIKAYXz+e0ylMvOZrwlxPTLTtHyAhbsvoEXz4+0ic885PELr/AL8YB9QjACsg9vuWCLweXThrEf3ul7cP+TtXKC405aWlvL5w6a4YI01b/wD0DVyWsfjtnSiIi0CIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiIKY2wuHOcK9Udi5ZOI9bWVTadrrDVQsc4gZe9hAA+ZyfyLmMbry19sZx1vdpNo4LWqqfFS3GnFVWBpx1tyCQfXILR9RKS5U9dUPZfUrqnnQ0Q9rc+7MrZT8h7u9uf/wAJe/S8bPY8cHbrceMNVxdq6Z7bfaqGelp3kEBznjpJz2O+B9hXsmiiIiAiIgIiICIiAiIgIiIKDZeTftXudK6zXu4cr3D24Rst0ETG6nqonAullOHe6gjsAMdQPmcb+XYHn19ohpfgHZK/hxwxucF04h1THQOdEeqK0gjBe89useTRvn0IJHlHwI5fuMHNzxLlotO0lXcZaur95vd5qCfDpw9+XyPedi45OAPMjsElwd8vZM8ml5tFwpeaHX1LDHBU0L2aYpJG9TyJNnVRBGB8OQ3G4yTnOQPU5bf0Do+3cP8ARNh0RaWNbR2K3wUEXSMAiNgbnHqSCT8ytwJQREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQF4We13/AAxbj/4Ctv8A6Mr3TXhb7XgY5xLgfWw23/oFIO/vsoR/1tdGfmwfncu6q6V+yjc08tdGwDcFhP8A+F/rXdRZ/G7AREWgREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQF8+vtKN+cbXn+MM/wCivoKXz6+0njkj5yNf9ZyH1Mbm/IFg2/Sg9w+XD/eB4dY7fsZt3/oGrkdca8tbi7l94cuduf2M24H7IGhclIkEREUREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAXid7Y+kuLOZW31U0Lm0klkgbA/GziAOoZ9c4XtiuoPtHuVqh498F7pqaz0XVqvStK+toSwfFMxgLnR/aM/ZnzAQcf8Asitc6MvHBqo0rbamAXy2ECrh7SdOSCcfMkH6t/Vd/c4Xzz8hXMHTcuPMZY9U6gqnwaduPXar13xHDICBKR5lj8HfGBklfQPYb/ZdUWimvunrpT3G31jBJBU07w5kjSNiCP0d0zPE/TUkREUREQEREBERAREQF0p5/wDn+snLZZ5+HehZY7hxCuVM7pAOWWtjwQJX4/fb5A77fWRy7zp8xx5XuBly4jUdHHV3WWeO3WyCRwAdUyh2HEEHIaGknY7Dsey8ONB8OeOXOxxnrvuPS1F8v94qPfLrcJARBRxOdjrkcdmtHYDOTj5EhBDwK4HcUebbi0zTlkdUVldc6g1V2u9QC9lOwnL5ZHds+g8zgDA7e9/Lby7aJ5Z+GlFw80bD1FgEtfWvaBJWTnu9x/LgeQKg5ZuW3QnLLw2t2h9JW+B1a2Bhulz6AJq+oxl73OwDjOcDYAY2C5eTfgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAvCz2u3iHnEuJc3DfuDbg0/Ux2V7prx19svw7raXirZeJEULvdqygjo5XgbZaMNJP1gj7Qg7deynfQP5Z6AU0rXTNLfGb1ZLTg+Xku6C8mfYy8Wa+TUeouE1XKXwCjNbTZJ+EB2Tj6iD9hC9ZlJJDMERFQREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQFTO+EPyW0eKPFTRHB3SVZrbX15jt1romFz5HDLnYGcNHmUiW43ei80NZ+2t4f0VzdSaI4V3e4Ukbi33urmZH4g8i1mQR9RW/dIe2L5YLpZWVWr6HU9juXSPEpY7calpON8Pa4D8v5UWea75K0va0hpcAT2C8W+bL2qut+J9Y+w8DH3HSthYDG6qe4Nqaj/tsAkDPlnsNsHcnqQ3UHMXqbr1RFduIVwZnxHV0MlY9jfmHt2A+ohOD6XVxhxx5heGvAfQ921hq/Utujlt0D3w0AqWGoqJgD0sawHq3OMnGw+eAvn7quZDmQv1ti0fPxZ1jU04PhMpI66USEj97luHk+ozv5rWNAcp3M9xov8AS09q4XatndWysbJc7nRTRQxsJwZHyygZaM5JBP51c0drKD20fGVmsvfrjoDT8mmjNg0MYcKhsWe4kJwXY8jtnz816kcB+O+gOYfh9Q8QuH91jqaWoaBU0/UPFo5sbxyN7gg5xkb4PmCB5+cdvZAWa3cFKOv4K1NXW8QLTTskrqaoqfvV0Ib99bH1bMeDkt7A4xtkBdMeWLmW4ocj3Fyvjr7JXNphN7nqLTtXmJxLTuQ07NkHcHsRjfGCpmJLr6GEW0eFPEzTHGLh9Y+JOjqrx7VfaRtVAf3zM7Fjh5OByCDg7LdyKinnhp4X1E8jY44ml73uOA0AZJJXzvc+erKLW/NZrq92x/iwyVojYRv2aNh+Ve5HNjqa5aS4Basvdp6/eYqMhpacEA99/JfPDpepl1XxPtVXd5et9yvMEk7n756pgSD8sbJLtwr6PuAdtltHBHQdtmBD6fTtAx2fXwGrfy0jSUEdLpWy00OBHFb6eNoHYARtA/MtXSAiIgIiICIiAiIgIiICIiAiIgIiICIiAseeuoqaRkNTWQRSS7Ma+QNLvqBO6yF4je1W19xRsXNpU0UOprxbbdRW+jntTKapfFGMtyXDpIBOQM5zj7TmXfg9uUXVD2e/NhZ+Yng3abTe71G/XFgpxSXWnkcBJOGHDZmjO+W4J8/P1x2vVIIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAo5Io5o3RSxtex4LXNcAQ4HYgjtjHkpEQeLPtT+TnSHAi9Wri9w7BpLPrC4TU1ZbMZZTVfSZOph8muAdt6j7T2g9jNrO7ah4A6m0/cauaaHTl9bT0gkeXCOOSEP6RnsAcgD5Bbe9tzUTN4UcN6VufCk1DUyO9OptNgfmcVP7EiMDgzxCl83aniafqFJGR+kpUekCIiKIiICIiAiIgLCvF0obFaa2+XOcQ0dvp5auokP7yKNpc4/YASs1dAvadc7Wm+Feir5y9aYEtZrLU9r8Ctljd0stlJOMEk9zI5mcAdgQT32f4OinPrzw3Tm81JbNIaYsj6LSOn66V1thGXz19Q/wC9iZwHmWkhrR5POwJK9F/Za8uN/wCA/Aqru+trT7hqLWVcLhLDIB4kNMxgbCwkbjILnEepPcYK6E+zX5ONb8V+LunOLmqdLz0/D/TtT90Pe6uPEdwqIxmKKIHBcA/pJcNhgYz5e43byTzhuqoiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgLqR7TnhhT685WtT3uOibNW6bg+6DHAZcI2EE49cEA4+tdt1tniZpaHXHDvU2jp4hIy9Wmqoek+skTmg/lIP2IPFj2Qd0ioeb2moppwxtdp+4RsaTgOkAY4D5nAcvc1fNLoXVGr+WXjxb9RRskp7zo27ESMILfEa0lrxg9w5pJAO24X0PcEeK9l42cNLLxHsIaKa6wB7mB2Qx+BkA/aD/rQb8REQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBcNc0fLlaOZjhxVaCut4mtpkBMU8YyGv2IJHyIHkVzIDlVTzxLN9eR1L7EPXJumK3jnYmW4PzmK1zOnLM9sFwaDjzz9i1S5+w/uJnabPx9pmw7dQqbI4v+eC2UD8y9XUUxXnVwq9jVwr0lcKe76/4gXLU00D2v91ZStgpyQc9skn6jkbArvrpzQekdJ6dh0rYbBRU1rhiEPgNhb0vaBj4hj4ifPK3AiWS+jr3pLkg4G6Q4r1XFi36ZpH19Q90jIX07CyN5Oc/PHkMfWuwQAaA0DYdlciSZw0XQn2iPs9YuYE1XGnhtWx0OtLbbyKygez71d44mktGQfgmA2BwQQACCcFd9kVHhH7P3na1py68QrPwy1TdXP4eXi5ClrqOpac22WV3SZ487tw4guacDuTjBz7qUdXTV9NFWUdRHPTzsbJHLG8OY9pGQQR3BHmF49e1V5MNFcHfcuOvDWjloqHUd2fT3igaSYYKmRrpBKzP0Q8hwI9TnzK5a9jdx+1VrC1at4Natvc9wi0/DBcbQ6plL3xwucWPiaSckAhpx5Z9EtI9Fta6TteutLXLSd5j66S5wOhftkjPYj6jgr51eZLhzLwD5itRaQoyP+oV0bUU++wGQ8fZnt8sL6Rs+a+f/ANptSz0fOZrpkzSPEfTSs+bXQtIP505um/HurwnuzL9wv0jemO6hW2Ohnz6l0DCfz5W7FxjyxTCp5deGs+c9el7c7P1wMXJykBERUEREBERAREQEREBERAREQEREBERAXV7ns5RtGcyHC67XSWijp9X2G3T1NpuLW/FljS/wn43IOMfb9WO0K2Dx14gWHhfwh1ZrfUddFTUlutVS4GRwHiSeG4MYAe5JwMDfGfRIleHPs0dT3LTvOToKkoaiRkV5qJ7fUta4gPjMD3YI7HBaPylfQGOy+e72cbY3c6vDKaoI6IrhUPJJxgmkma3/APCc1e2fHbmv4F8uFPBJxW1pFb6mqHVBQQRuqKuRvm4RMycfM47qb3FcvouhuqfbG8r1rp3O0zbdVXucA4YaAUzSfrcSfzBdddbe2a1ldKl37D9D/c6nB+Fsr2F2PmQSc/b9ivo9e91VeJl+9r1x4rqZkFot9NSvGeqQvwT+QfzrudyDc/T+YuSfRWt4RTahpmhzXdQIkB7YOxIPbJGchM+RPm13lRERRERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREHm77bhgPB3h3L++bqWdo+o0rv9QVnsRZi7hHxHpz2ZqOmf+WmA/wDyVL7bc/8A5muHg9dTy/qj1B7EVrRwm4kO8zqKmB+ymH+tPB6ToiICIiAiLgnjjzr8unLzcPuNxH1wIrnjqNBQ076mob27saNu/mdvPCZo52ReV/NB7X3TV40rUaW5dbZeI66saWPvFfCIPCaRjDGZJz3+sY3G4PQKr5puZO9eLTO4vaqeJyXOZT1jmHc746MEd/JJ0e5vOLzbaR5UOG0mpa/3e46irneBZ7OZQJJ5CDl7gNxG3uT9gyV4g2DS/GTnb4911VR0kt01FqivNXcalkZ8GjicQOo+TWMaAAO+G/IkYGh+DPMRzH6ogorLYNT6lrpiGOr7gZpY4mk5JdNJnAHc4JPyXuByT8omnuVPhrFaD4Nbqm6NE15uAaPikIz4bDv8I2GxwcDvjJXZyEcz8N9FWvhvoDTug7NTQwUdgttPb4mRN6W4jjDSQB6kEn5krcyIiSYIiIoiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiD58/aIz2ufmWvslso2QE7yFjcdRLjgn5916l+ypp56fk+sDqiUvMtfVPaCc9LD04A9F5S8/TS3mOvrj3P8z3L1h9lh1nk8031nP+3KnGfT4Um503eu3SIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiDiHmr4EUHMdwRv/C2rlEM9a1lRQzHtFVRnMbt/nkfavBinunHLkr401kFpuNZprVNkldTvf0fe6mEnG7XbPY4D9Iz3X0irjDjHy1cEePdEaPinw/tl4lDC2KsMfRVQ5GAWStw4EDtkkfJB1u5AvaG0nMzFUaB4nR26z66oYxJE+FwigukfYujaT8Lwe7QSMEY+fRb2vtNZWc2zq6018M89Vp2hNcyNwcYp2GRgB+ZY1m3yWXzRezQ40cGNZ1GouBdmvOo9MNLp6WegeTWUTfMOwQ7YEjI3+sLpTfpb/wDdqcasNfJc4ZOiqbcHPM4cO7X9fxAjtg9kH0dcpbfD5ZeGERzmLS9BGcnzbC1p/OCuWl0x9mlzJUfGbhWzR7ohDWaXhbD0dQy1gwOnHpvkfIn7O5ykv8upBERVRERAREQEREBFsXirxr4Y8FbL93+JeraOzUhBLPGf8cmO/S0bldebf7Vjk6ueoYdOw6vvDHzyiFtVLaZGU3UTgEvJ2HzxhTdHcBFjUFdSXOip7lb6hk9LVxMnhlYctkjcAWuB8wQQVkqgiIgIiICKh2XCvNVzO6G5YuG1fqnUdzh+7E1PI2z24OBmqqjB6MNP70Hck7bEepD1NRcy/Ntwn5YtLT3fWN6gnvDmH3GzQSg1NQ/GRluctHzPl22yR4ocw3Nxxz5wtaQWq7180Nqq6pkFq05QuLadhc7DA4DeRxyMk7bD0yuPaSi4uc03F/wKOKv1Nq/VFYXYJc/p6nZJJOeiNue52A9Sd/c7lb5IeC/Ldou1j9iNoumsI6Vr7pqCtpmSzmfALxE5+fDY05A6cbDJ74F35Fn7dR+O3J1wu5IuV88ZtH1dwbxHtwoqY3GoqOoPq53Br2xsGA0NJJ2O4YT9XSXgjwR46c9/FKtpIdRyXC4U9OKi5Xm7SufHTRZw0ED1OcNAA7+Z31bjTxn4186PMCzh/Lqy4XG2XbUz7fp61MefdaaJ83hxyCIEAkMAcScnvjHZe0PKpyqaA5U9BDSmkWOqrjXFk12ucwHi1cwGPsYMnA/+wJJJ2+joPZPYhakO+o+O9taPMUVpkP53vH6Fvm1exH4bRMDrzxp1DPJ++bDb4WN+zJyvS1EHmldvYkcNpWZs3GfUMDx5TUEL2/mOVzZys+zj4f8ALTenamg1VX326OwHSywtjBAOw2Ow37Y+3zPcDGVVJsL3giIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIg83/bcZ/wBhnh5/CeX9UeovYjf70XEf+EdP+rBZ/tsaZ0vAnQtS0ZEGqj1H0BpJR+kBaf7EeZjuEfEenB+Nmo6Z5+o0wA/6JU9HpMiIqCIti8ZeMmheBGgLlxH4hXVtFa7dGTgbyTyH6Mcbe7nE7ABBw57QLmequWDgZLf9PSRfsnv9SLZZ2v36XlpdJLj/ALRoz9ZHY4I8YeEHBjjbzqcV6+hsFSLpe5wa26XO4TERQMJx1PIBwM5AAHlgLdfNVzN8Q+efi9aYLRpupjpYnm36csVOTLKTI4ZLsbGR2Bk9gAd8L1P9m5yh3rle4YXKu13DAzWGrJ46isjjw40lOxv3uDqHc5LnEep+wOxI67cOPYl26DwqvirxikqXNIc+ks1D0MI82mSR2ftAC7g8KuQ3l04TQRR2XR7KyWMAOkrMPLz6nbOftXYgZ80O/ZJbFvWn2awWTT1IKGxWikt8Ax97p4Wxg49cDc/MrUURAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQdOPadce+JPAbgpbrpw1mko6273EUs9cxmfdmAA5z5ZJx9ZC6BcFPa1cxnD+4UtJxEqqPWtjDwJ21UAZVtYTuWStIyQOwIwvanV2jNJ6+sVRprWunqC9Wuqb0zUlbC2WNw+o9j8xuvMTn29mFo3S2jLrxj5ebfPb/uS11ZdNPB7pYjAPpPpwcuaWjfoyRgH1JFk03Ho7wc4vaN446AtfETQ9wZU2+4xNeWdQL4HkbxvHkQdv/chb4wF4HchXPJX8p+rJ7VqmlrrtoW8kCupKd4MtHJnaeJriAcebcjIzjc7+vvBLnX5ceYO5NsnDfX0dRdXs622+sp301QR5gNeBkj5HfyyoOdUREBERAREQEREBERAREQEREBUPyVVY97Y2l73ANaCSTsAB3QfPPz8NnHMZfDMwtJGQMY/fOyvWL2WUol5PdN4/eVdQ0/WAxeTHPdr+zcQOYjUNdYWN92opXUxeP3zw45/Jt9uV60ey4tdTaeUHTjKljmmoqqicNcMEB3SQr7NSedduERFFEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBeR3tcuVG70WqWcxeirF1Wmrp2Q3/3dn9hmaSBM4DsCO5/1FeuK2pxUsFo1Tw01Tp6/wBNHPb6+0VcNQyRoLS0xO3wfQ4I+YCDxY9kzxJm0dzT2/S0tSW0OrKSejdGXHDp2sc6Pb12P5AvdJfPx7NTS1dqfnT4eR0DwGWqpqrnO/tiKGmlJ/KS1v8A4y9sOYfmX4U8sOk6bV3FS8T00FfOaWipqWAzVFVKBkhjBjIAIJJIABCYfpyui83bx7avhLHcfBsnDLUU1GD/AGed0bXO+podt+dZMXtp+DRaPG4c6hJ8+l7AP51cS16MovOj92n4LZ/3udRY/wAIxVHtp+Cnnw61H/57P9SYr0WRedbPbS8EyW9fDrUeCdz4jNh+RZ1w9szy/RWSertuktSzXMMcYqV8QDXPxsC/YYz37fWmJa793K6W2zUclxvFxpqGkhGZKipmbFGwepc4gD7SvLPm89rZqzTuu67QvLaLHLbrW8wT3+qgNT7xKNnCFpIZ0g9nEHO+3p0r5ieazjVzY63f90a66vt1RIYrbp2gfI+MDOQPCZ/ZHHv2OPLzJ7g8hnsvv2Qwz8RuajRlXT0Z6BaNPVMroXy+ZmnDCCGkEAMJHfJ9A5FdKNUcQ+YfnG13SUF/u9x1TeJ39MEDGdMMIPn0tGGgdgT27Bcua09llzQ6K0HUa7qbXaqyOjgNRPQ01SXVLWAZOGYySPQ4+eF7TcOeAnBnhJEIeG3DSwafGc9dJRtD8+vWcuz88rfckbJWOilYHseCHNIyCMbgj0U+DwQ4Ye025ruD+kqHQFnvFkraCzsFNTtu9sM08TG7dBeHtOBjGD2Xd7kv9qhTcYtTjh7x3prFpq51DM0N0hkMNLUPH7xweSGE+W+PPOM47Pat5GeUrXF2qr9qPgZp2e4VrzJPPEySBz3nu4iJzRn7F5rc6fswte6H1yy98tWhLrqLSlyj6jQUpM89BKM5aQTlzD5dyMjvvhnE349mKKuorlTtrLdWQVUEgyyWGQPY4fIjIKyF87tTy288PDWzuuzuHXEmw26lb1GSndNEyMD1Ebtu3ot/8vPtLOPXBC6R0OsLjV6ussJMctHXzETR4OCAT3I7YOCPVMsWZePeNF0S0z7YjlYuVohq9SU2qrNXOaDJSNtvvAacbgPa4A7rWpPa6cmzKZ1Q2/6oe9u4hFhlDz9RJDfylB3Pe9kTHSSODWNBcXE4AHmSV4Ue1b4oWviNzUVdDp7UUN2tWm7XTW9hp5RJDHU4Lpw0jYnLmg4OMtWRzse0N1lzPXal0fwujvmmtIQnw20jJy2quczjgGQRE7HsGAnOcEZzndfIp7NjXfE3VdDxC466Wr7Boug6KqCirWGKour+7R0H4mxg7kuAzjGMbG8zpnXdT2XfLZZuEnAG0cRLtZomau1pCbhPUSR/fYaRxPgwjIy34ACcd8j5rQfaYc8beA2nZ+C+hWiXWWpra41FV1bWyklyzqx5yPAcB5Ab/V3poaKktlFT22gp2QUtLEyCGJgw2ONoAa0D0AAH2L5/vaS61Ot+cbXtUyVskFqmgtMDmuy0shhaNiPmXKRHJ/snOA2ptYcwds4uVlgn/Y3paOolirnxkRuqzG5jQw+ZGSD8z6g49uh2XEXKToqz6A5bOHOnbJTNiiGnaGplIYGuknlhbJI92O7i5xyT6ALl5FEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQdMvav8Pq7XPKnV1Nrp3T1VgulPcQ1oJPhgODyAO5wV5wezo5uJ+WvidJYbrA6o01q+WCmrGB2DFMDhkg+e+PyeRK9y9faVodbaOu2l7jCJYLhSviLSAcnGRsfmAvm64p2Km4dcbr5ZaZvTBZL2ehvo1rw7H1bp7yj6X6Krgr6KCupndUNTG2WN3q1wBB/IVOtvcPZxVaC01VN7TWeikH2wtP8AOuIuaznJ4V8qemzU6srvfNR1sD32qyU/xT1DhsHO3HTGDjLiR5DbOUG8OP3MFw75ctA1mu+IF4hgZCwikog8CetmwcRxt7knG5xgfXgHwo5nubfi9zf62iZeXTRWkVXh2TT1ECWRlxwzIGTJKcgZOe+B6nbHEfiPxs5veLEt3uMV21LfbpM4UNrpGvmbTRE4EcTBs1oyATgDtnAwB6y+z89n1p7gPpel4icWdOUdw4iXDE7G1DRK20REfDGwHYSb5c7BI7ZGMBidbd9nj7OWbgZcKDjjxbqIanV8tITbbSxgdHahK0Aue456pukkbYDckd8r0JREUREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAWPW0dJcqKe3V9OyemqonwzRPGWyMcCHNI9CCQfrWQiDzK157FPS981TcbtovjDPY7XVzPngoJ7UJ/d+ok+GHCRvwjOB54XRPmD5duM3ItxZtokvzmysIrbFqO2NdEyfB3ADslrwcgsOQR5kZx9EK4+41cDuHHMBoir0HxL0/DcaGdjvBkIxNSykYEsTxu1wODsd8DPZJ/Q6b+z59pFUcdrsOEXG2e2UGrBD12u5MIhiunSPijIJwJsbgAjqwcAnYehK8B+M3s4OaHhVre42/SHD296sslJO6S33e0wmUviByxzg3Ba8DGcDuMjHYcUUnE7mU4J6np6qr1NrjT1zo5ARTXOaqjDsEEgxykAg4xkDt2KfOD6T0XnTwU9sPwUqdG0FJxmob/atQU0LY6makoveoZ3AYLstIIJ+Y377Lf1f7Xnk7pYHSUd41RWygZbGyySMyfTLyAEHdhF58Wj2zfACour6e8aR1LSUGcMqIomyvx6lmRn8oW9af2ufJpMAZdQ6np89xJYZTj/zSUSXXdJF1HovancmVZEZTxFrYA3fE1qnaT9QwStCvXtdOTy2Ryfc+96lusrPosp7LIwOP/fSFoA+e6Tqu6iHdecVT7aXhG25FlJw8vr6EHZzi0SEevfH2Y2+ay3+2l4I7dHDzUhHnmRnf8izbnweiaLzs/dpeCPlw+1F/wCez/Uran20vBNtMXU3D3UTp8bNc9mM/kH6VJ+X6o9FUXnfpb20XAmuqWwas0Jqm1xl2DUQRxzgD16A4H8hK7qcG+NfDjj5oyn17wx1Ay62md5jJ6SyWF47skYd2uHof9eNjfi2/wAQa42vQWpbkHdJpLRWT59OiFzs/mWvk4XGXM3dH2Xl54j3KNxDodM3DBHzgc3+dDx89PCTRtZxs442DSABdNqi9ASDGSQ95e8faAR9q+jzhpoO08M9D2jRFliaymtdO2LLRgOdjc/aV4V+zB0+dQc5+hxgn7nCsuG3/wBDp39/yr38QEREBERAREQEREBERAREQEREBERAREQEXGfHrmE4act+i/2c8Trs+koZJxTwRQsMk1RKRnpYwbkgDJx2UfADmL4ZcymkJNZcMbpLU0lPN7vURTxGOWCTuA5p7ZAOETXKCIiKIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgLSdV0zazS15pHnDZ7fURn6jG4H9K1VaTq2Geo0reaelIE8tvqGR/9+Y3AfnITNS3Hzk8FONWqOVnjrBxI0VT0tXVWCqqqN1LVA+FVU7uqN8biNxkbgjcEArmHitxS4/e064w2Czae0VS0jrVTOpqKgppnOpaMPIdLNLK4A5d0tycZw3YYG3DHBThBceOHMHYeErql0M19vbqasqGtJ8KNrnOmf8ALDWuwfUjuvoE4IcuPCHl509Fp/hlpOltxEYZUVhHXU1JHdz5DuSfPGB29Eswn9ugGjvYjUElihk17xvq4bw8ZkitdsY6CPIz09UjsuI7E4AK1M+xC0fnDeOt5HzNqiO31dS9O0RXmH+4g6Px/v73n/NMX/rIfYhaRAOOO94J8v8AqREN/wDz16d4wqqYPL/9xB0ye/Ha5/5qj/8AWUMnsQrHk+Fx1ryP+2tTAf8ApFeoyKjqByj+zi4acrmo59by3qXVWoi0xUlXV0zWCkae5Y0EgOPr3GAc7DHb9EQEREFFVcO8wfNZwY5Z7RHc+J2onQT1G1NQUkRmqZzg9mAjbbuSMDfsugXH32x1s1Toi86U4NaEvNouVfE6miu1wljHhMdsXsawkg47Z9fLuk7wrnv2gvP7pDgVpOt4ZcPq6hvmur1Tvp3sjkEsNsheCHSSkHBcQSAzOc5zjBx5xcnvJNrnnSvt+vh1JHYbLbZw6vuc1MZTNUSkuMcbQQOrG5ycAFcbcAeBPEXml4rUmlbHDW1clfUiW63R4L208ZOXyPedurGcZ+vsCvoB4BcCtD8u/Dmg4c6EoRDSUw8SeYgeJUzkYdI89yTjz9fUlO5iZN2Og1w9iBpz3LFq4+XIVfT3qLLGY8/U2QHH2rZU3sQeIjXn3fjxp17fLrtE7T+Z5XruiK6E8oHsstNcvmuoeJfEXVtLrG9W8E2yCGiMVLTSEY8Uh5Jc8DOOw3/L3wqamno4H1VXURwQRNLpJJXhrGgdySdh9ZWyeNvGbRPALhxdeJ+v6409qtjQOlgzJPK44ZEwebnHYD614Yc0/O9xg5qdXvprfXXWz6ZdIYbfYKGd+ZATgeIGH748jGRuM+uyDur7QX2l1Touvl4Rcu2oKSe4CMtu99pyJW05O3hQuG3WPNw7fMHfqbyJ8od45yuI131Xre61X7HbNUsnu9W74n1lQ89XhdR8yBkj0I7BZXLb7Mbj3xlrqK7azsNTo3S8hbI+pr2eHUTRnzjjO+D6kZ+XmvZbgPwH4f8ALroKl4e8O7Z7vRQnxJ5nby1MxG8jz3JP17Z+1L5iT1vqy2mhsFoobFbIRDR2+njpaeMfvY2NDWj7AAs5ERRERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBfPJ7Q3TEGlOcTiPbaZnTDLcIqtjP7kSwRvx+fP2r6G14F+1AgdR87Os5qqMuZMy3zBv8AdM92jGPt6SrB7Xfs0sfCLgHSa21lP7rbtMaZpqmtcdiBHTtBaPmTgD5kLwD5g+Lus+bfj9ctZQ22pqay+1TKKzWyIF7ooG/DFE0fVuSe2Tk4GV6te054xWWn5GgbFdIf/wA4clrpaERvBMlMXNqZMY2wGRdJ/wC+wuvnsXeDWltQ3LWvGG926GsuNhmprXbPGYHCne9pkfI3PZ2A0Z+ojBCfB3F5EOTXTvK/w1o6m7UEFTrq8wtnu9a5gLoHOGfd4ydwGg4JGMkH5k9pkRQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERARcaceOYPhry5aPbrTibdn0dDLMIIGRRmSWZ+MkNaNzjbON9+yzeC3G3h7x/0RT8QOGt6Ffap5HQu6h0yRSjGWPb3aRnsUG/kREBERAREQFxNzB8tHDLmR0m/TGv7SxzxvBXRMAnhPydscfLI37ELllFLNHlPrP2I08lY6Th/xwgipiSRFdrW5zgPIdUbvL5grb9s9iJxFdVR/djjjpyOn6h1+7W2d7y3zx1EDP1r14RUeXd19iDYDbMWTjvcG3Hp71dpYYOrHo14djPzXHV09ibxipRm18XdK1w8g6lnid+Q5H517Eog8V5fY0cw8cgZHqrTkgPdwfgD8rs/mWvWH2KHGKtka6/8WdL22LI6hHTTTvxnfABA7fNexSLPf7HmTT+xD0IKFoquOl+NZgdRjtcIiz8gXE/nWKPYg6Yyc8drpjy/6lR9vn8S9QEWh5gfuIOl/wAe10/zTH/6yli9iFoz4fG46Xr/ALbptcX5suXp0iDyb4g+xKvVJQuq+GPGiC4VMbCfdbxbvA8V2Ng2SNxDc/MfaupfDjjzzNcg2utR6Fsk0FqrhUBlztdypTPSyvbsJGty07gYDgQcbbHIX0MLini7yucBOOrxU8UeGlqvNa1gYytc0x1LB8pGEH8pKbiXrzJ0j7azjLQvij1nwv0tdI8/fZaN81O/HqGlzhn6yt0cxHtUNE8YuXrUehtO2CqtN5v9C6imjmPUWh2MgEDGPXc7efry3xR9jvwOvUU9VwxuFzsVQWOLYJ6t00fVjbBdnAz6j7SugnMR7PHjxy+Was1fdrXHc9OURJlraV4eYmZ2LwMgDHc7eeyZs6uuS/Y5WF1fzSVd98MubatP1Qz5NMpDQfyAj7SvbleLXsZNQ0du5h9QafmIFRd7C8wZ7kRP6nY+whe0qAiIgIiICIiAiIgIiICIiAiIgIiIC2Rxj4uaN4H8O7xxK1zco6W22imdN0OeGvqJAPghjHdz3HAAAJ37bK3jDxm4ecCNFVev+Jl/itdqpBgEjqlmee0cbBu957ADzwvBTm+5q9bc2vFipu5fXxacjqBTaesTXFwhjz0sJY3Z0zyckgZ3wPnc+jI5kuaLjHzv8SbVQVlB/tVlUaXTun6BpLYjK7ALtz1yEAZcdgAewBK9a/Z18qeoOVvhFW2/WVVFJqHU1Uy410ERJZS4YGtjBPc4xnbuPsHGfs6/Z+Wfg1ZbXxo4n0JqNc10AnpKSUDotbHgEbecuO58vJd/BjyTZid1VERRRERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREHlPzve0a5geFPFCfQPD2npLJS0oL/eKmjEplGSMNzjOMDJ3HbbdcASe1p5tqnS9w01cLnpypdXU7qZtf9yhHURNcC0kFrg0nB2JGQd917M8QOA3BrioSeIvDSw6gLu7qyka8n5k98/PuuGuIXs5OUnVGkLpZ7Pwds9juE9PJ7tXW8vilhl6T0kHqIxnGRjstb3jOfa8wvZS3S0Rc51kq9R1UYmqrbcWUr5ju+rewBoB83EF4HzK93/LZfL1Wm78O9dVbLLdKikuWnbnLDBVwu6JGSQyFocCOxy36tyF76ez5426t4+8s9j11ridtReYquptlTUDvOYHACQ/Mggn/ANypWnZHbCsfJHEOp72tHqTheXnPP7Ua4aa1PfeDPAeICe0zS2+5agMgLfeGEtkZAG5yGuBBdkAkHuNl53u1ZzJcSnT3uC9a/vrXEmSWkkqpIgcnO0fwjfOwCniPpShnhnb1wyskGcZY4EZ9NlKvn95PucjXvKxxairtbVuoLlpmpBprxaamaR0jGkbSMZIdntODjbPz7L0Z1N7YTlTt2n6mv01+ym73RsRdTULrSYA6TGwe9zg0DPcglXF13jmq6alGaqpiiHq94b+lQC8Wh/0brRn6p2n+dfOlrfitzD81fFK4XS3T6pvdyu1Q59NabU6aRlPEXHoYI4zgAA4yQMn0GANXm5Oed+NvXJwT4iuB32p5XH8gcSsZR7f8w/NHwv5dtCV+rNTait81bFG4UdsjqWmeplxs0NBJAz548sBeV2sfbG8z12uU0mj7dpawUJe7woXW/wB5kDc7Zc92M477HdcI2bkP5xtV3KKB/A3VkLpXiM1NxpzFGwernPOcD5Ar1R5afZkcAeG3Du3w8VtAWnWOrqqNstyqbkwzRQyEbxxMJ6QB2zjcj7Tc70ed0vtYecqc/FrCyNz/AHFmib+jCrU+1i5yqmhfQ/svskQcOnxo7PGJR8w7J3Xrc3kZ5QQeocu+iQf/AAa1Wyci3KBJku5d9FZPpbwP0FXzweIugdKceee/jVS2Ct1HWXu+VodJU3GuJdDRQA5c8hoAaPkME488Lvjo32I+mqWWnn13xurq8MIdLT261NhY/fdvW6QuAIzuACvQXhtwN4Q8H4pouGPDuyabFR/ZTQUoY5/yJ74+WVvpNHGnA3l84Z8vmlmaW4c2GKjjIHj1BaDLOe5Lj3xnfHr6rktEUkwFhXi409mtVbd6vPgUNPJUyY79DGlxx9gWYThbd4j1tJbuH2pq6ue1tPBZ6ySQuOB0iFxKo8I+cbnc4kc4OpabStJRst+k6OvItFopcl9TKT0MklJOXPIIw3sCdvLHev2ens4JOEFVT8ZONkFJV6llga612npEjLe1wB65CRgyYxtjbttg586OQ3Slt1pzfcMLJd6RlTRfdptXLE8Za7wI3ytBHmOtjV9FaAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiIC8LPa708EPOJcZIj8c1htr3j59Dhn8gXul3C8EvapXKS485+rWvfkUlJb6ZvyAp2HH5SVO6a2Py18MuJnN9r/TnAifV1yNhtcT6gGVxfFbqdgwSwEEA4OADnbIAXuZy28tugOWDh+3QGgmTyRSTGprKuoI8WpmIA6nY7bDYZOMnywBxH7OXlk0hwQ4E6e1hHbAdW6xtcFxulZIAXtbK0PZCw+TACPrP5+2y1Yk0REUUREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREHQX2wnCvVnEDgXpzUGl7dPXN0pd5KuthgYXv8GSLpLgBucYyfllee/IjzqX/lO1xJRXCN1fofUU8TbzRg/FA4fCKmP/tmg4I8wNu2D7+VFNT1cD6aqgjmilaWvjkaHNcD3BB2I+teSvtIPZ01tiuFbx24CaadNaJg6e/WOjYS6mePpVELBuWEbuaOxBIG+4erWmNTWLWWnrfqrTNzguFrulOyppKmF4cySNwyCCPPfceRBB3C1ZeEHINz53nlh1O3SWvaqvuPDu4u6Z6dpMj7ZIT/AGaJp7j+6aO47br3A0PrjSnEjS1u1rom9012s10iE9LV07w5kjT+gjsR3Qa+iIgIiICIiAiIgIiICIiAiIgIiICIiAuK+aaKln5cuJEVZCyWI6ariWOGQSInEEj5EA/YuVFsPjzZn6i4J68skf06vTtwjb8z4DyB9pGFKPFb2T9c2i5z9NNdL0Cptlyg/wC+zASB+UL3lXzl8kWqn6I5reHN/Li1kF3EUwO2WPjc0g/lX0YseyRokY4Oa4AgjcEHsVcF6IiAiIgIiICIiAiIgIiICIiAtv691nZOHWir5rvUdQIbZYaGavqXk4+CNpJAPqSAB8yFuBeY3ta+b52nrfVcr2lGg1V1pIp77Uh39jgeepsOPMuABI/1AEOkPNdzdcTuc7XtDSTUL6azU1UYbDYabLsPeelr3/3UrgQM+WSAcbr0G5BPZqUPCZ9Dxe44UlNcNVljZrdaXND4bZkZ6n52dJg+mB29c9e/ZPco961jxHpuYLWljdHpnTge+ze8MwKytILRI0Hu1gJIOCM4O2xPsql6giIiiIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIujXtE+avUfLvV6fNgbI502HdLXYDnEk7+Q2Hf5IO8qxrjK+C31M8bS50cL3geZIaSAvN3hn7aHhjNQUtv4naA1HSVTQGTVtC2KaM9vi6C4Env2/Iu+fBzjLw949aEo+I3DO9i52Wtc6MPLCySKVuOqORh3a8ZGQfUJg+dKfTNRxF46XDTTauOgmveoquIyyj4Yi6d5JPbtvttuuzFBzh8VOSO1Xfly4X1NsrqSle+V9xqGl7oqiZuXviAIAOC05z3A22XA/NhpCXhfzO8QdOUL3wfc7UFRNTSRuIID3eI1zSNwQXbHyIW4uT/hHa+Yrjtb7Rr+51M9G+eOWsLndTqj4gOl5O+MDf12Cs/LJqfx2uy/LN7KDUnG3Ttm4w8XuIrrVbdSMF0+51LSl9bOyR3V1SSOIazryScAnfO2y9V+F/CHQXB/SNFozQ2n6WhoKOIRDpjb1yYABLjjcn07LdNqtlDZbZSWe2QNgo6CCOmp4m9o42NDWtHyAACzFL3qzkdSubz2ePDLmgbFfKCqZpLVFM0tbX0tK10c422lYCM/WN/qJJXUW3+xB1r7437q8ebIKUO+L3ezzeIR8up+Afyr1vVMZUPHAfKfyecOeU/S89o0q+S6XavcHVt2qomiaXA+iMfRbnyHoFz6iJJgIiKgiIgIiICIiAugHtWeaWu4UaDi4Q6bm8O6aupXiolacOjgJwf/AH9SF3/XiT7ZC8Gq5orfZgTig05SyfLMrnnb/wAwKWaNI9kzw3umqOZ636zbSuNv03TTyvlI+ESuYQAD6gE/lC9zV069l7wes+gOWbTusY6Roueraf7oSSuHxeG4kgZ+sfkA+pdxMhUVREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREFjnNY0uc4NA3JPkF86fPTrBuuebDiNqCF2YH3UwQ/JkcbWAH/zSvoc1M58enbm+P6TaOYj6+gr5t+KNjuGouYO96eBJrLrqI0rSe/XLKGjP/nBZl/6xc5r6K+DzQ3hHohrW9IGnLaMen+1o9lvBaPo+zv07pKyaflcC+2W6mo3EdiY4msJH2hawtIIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgKOSOOaN0UrGvY8FrmuAIcDsQR5hSIg8nPaE+zSqLfUXbjpwCt5mpJHPq7zp6JvxRE5L5oAO47ks7+Y3znrlyW+0E4g8qUg0VcaGO+6Fqqzxqi3zOLZaF7iBJJA4diRuWkEE79172vYyRrmSNDmuBBBGQQfLC8J/accrs3A/jpcNY6Wsb4NG6vcLhTuij+9UtU/wDs0Www0F+XDJ/fEemRmPcuw3ij1DY7dqC3kmkudLFWQE7ExyMD2k/YQtQXUP2d3Nvorj5wms2gWVLqXWOj7TT0dwope8sUTRG2aM/vmkAAjuPygduxulNVREQEREBERAREQEREBERAREQEREBaFrpzW6J1C54y0WqrJ+Y8Fy11aBr/AP8AiHqT/wAEVn/oXoPnB4Nt8bj9psUvwh1/YWeWB4hx+ZfSZp4Obp+2tefiFHCD9fQF82fAxrncfdJhgJJvse3/AI5X0n2RpZZqBju4pYgfrDApndGciIqCIiAiIgIiICIiAiIgIi0LW+sLFw+0hd9balq201rslJJWVMrjgBjBnH1nYD5kINu8auNWhOAega/iDxAu0VHQ0bD4UZdiSplwemNg7kk7dvz4B+eHinrfVnMpx0vWsBRzVN31ldyaSlYCXNa9wbFGAM4DWBoONhglcj85nN9rHm44gxSiGel03bpTBZbVHklxccCRzRuXu2wNyM488D0V9mHyTUPCrQ8XGHijpaE61vh8W3xVcYc+20mPhwD9F7+5OMjbBOxAdz+DuiqXh1wq0loeko2UrbNZ6SlfEwYDZWxt69vUv6ifrW8kRAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQFwTzP8AKLw65prHBadY1FVQz0pzBV0zQXsPkcEjOPr37LnZFM0eS/EL2JWo6WN9Vww4yUVd0NJFLeKEwvefICSMkD7R9q4Z4Tcx3Gj2a961ZwbvmnqS4VtZK2rfSvlLoIpeksbIwkA7gZ2GDgZBXuaNl5H+2J4A39+vrPxtsFrkqaGtt7aK4uiYSWPiOGkgd9jv57j0K1vxHntxX4mah4wcQLzxG1Q9puV6qDUTBpyGkjGATvj616Rex44KWW+W278VLjEJJaCr8CId/vgyAD6AAE/XheWknheFG1jHB4z4hPmc7AfYvYb2J1RNLwZ15BIHdEGoYQzPbBgycfLJWbNV6OoiKgiIgIiICIiAiIgIiICIiAvCr2u0/jc5V0j/APmFgtcf5Yy7/wDKXuqvCL2tvV+3Rv8Akbfce1Y+r3cfz5Qet/JH0/tR+Epa3AdpWhdj5mME/nK5uXC/JeAOU7hKB2/Ylbf/AELVzON0iaqiIiiIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIg4S5uuY2k5YOD9ZxJnt3v87Z46Smp/J8r8gZ3HnhcScj3P5S81ddcdM3mwxWi9ULPFDI3ZD2EZB79tiM7bhbY9seWDlYo8t+I6jpMH0HS/P8y6oexls8lfxz1LcOg+FRWljnHyyXOA/P+lS8i8ezyIiqCIiAiIgIiINC1xVOodH3qqZ3jopnD/zSvm34t6groON2oNR26Y09bS3l1RBI3vHJG8Frh8wWgr6S9aQsqNJXmGT6LqGYH6ugr5v+IFlo7rzC3KwVEhipa7UjaWV47tZJM1riPqBJWZn8tXeY7Q1ftieZ+XR1Np2itWlaa6RRCOa9e6SSTy4GCehz+hrj6481ufgD7YDi5ZNQ0lp420Ftv9jqJWxy1sEPgVFO0kZccEhwHc58vIr0l0Zyd8semdDUmj6TgxpCtofdmMlmq7VDPNUHpGZHSvaX5J3JBG5Xj17Srlv0vy7ce/dNAWz7n6Y1HRNuNHSAkspZMkSxsJ7MBwQMkgE+i3us2PdzS+pLTq/Ttu1RYqptTb7pTsqaeVp2cxwyPtHY/MFasvP/ANknx2rdb8IhwzvMxlqNNl0dM9xyfCGMAnv9HH5D9vf8brP3i+KoiKgiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiIC4i5q+DtHxy4C6x0A+giqa+rtk77Y57Op0VW1pdGWejiQAPrXLqIPmw4F8XNecqPHC3a6obdNTXawVD6S5W2oaWGeB3wywPHcAjBBHmAQcL6AeXnmA0LzJcN6LiPoSq6qecmKqpXn77STgfFE8dwR5HGCNwuqftMuRg8bNKM4r8ItL07td2VxNdTUsYZJdqQgkjAwHStOCCdyCRkZyvPLkm5wNU8nPEisprvbKio0zdpm0t/tcgcyWCRjukytaRlsjNwQRkgY8sF74PoHRaXpzUFq1ZYLdqex1Tai3XWlirKWZpBD4pGhzTt54I28itUQEREBERAREQEREBERAREQEREBbf4hf/EHUp74s9Z/6F63AtF1nG2XR99jcMh9tqWkeuYnIPnb5VLPHeeaTRdvqHARC+F8hPkG9Rz+UBfReLtZoJYqAXKjZKWgRwmdoeRjbAzk7L5o7DrSq4Z8XpNXWuImS03WocxmcHHW5uB6HBWbrnmB4qa31pNrOq1reKap8XrpmQVkjGQNB+EAA4+vOc5KJr6XkXlNyq+15sWnNJ2/RXMTarvUVNCBCy/UMYnL4gMAyx5DiR6jO3rsB6ZcOeI+jeLGkLfrvQF9p7vZblH1wVMJ2z5tcO7XDzB/QQU8VudERAREQEREBERARaNqvVunNEWGr1Pqu701stdBGZKipqH9LGN/nPyG681eZr2wFhgjqtLcvlrqayUExvvVSPCYcHBMQ3OPMHG/kQg9Q14++1k5v79fdc1nLZoe8ti07Z44vu++ndvVVZAcYCR+9YCAR2JJHkc9TNQc7XNHqGunrZeMN/pBOCDFS1JYwA+XmfzrdHKLyc8TebrW33YqGVUemIasSXm9VTnEzEuy9rXk5c875Oc98ZIOE52jtZ7Nj2eM1dPp7mS4uCB9A6MV9gsro8mQn6E8wIxjzDcfM5yCPWAAAYGwC03Tdit2l7BbdN2mIRUdrpIqOnYABiONoaO3ngBamgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgoThcC87+oNG6a5a9Y3LWT6fwm0LxSMlALnT/AL0MHcnv29cea56AwvHb2zfFC/13FrT/AArir5Y7Ra7Uyulp2OIZLNK4kFw8yAB3+R9FZy7Es3jp9ytcA7nzNcbrHwpobh9z47iZKmurA0EwUsTS6V7Qdi7GwB8yO/Y+/fL5y88POWrQcWgeHdLOyl6/GqKiok65aiYjd7j29cDyzjJXkT7Hw045t2eJjxjp6vEee+MNJx+QL3GUUREQEREBERAREQEREBERAREQF4S+1va4c517LhgGy2sj5jwAP05Xu0vAz2o+ubDrvnD1PUaeqWVFPZ6OitEsrCC188UeX4IOCAXkZHog9i+TDo/an8Juk5/+Cdu/L4Lc/nXNC4U5KoZYOUrhLDM0tkZpO3hzTsQfCGy5rQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQdKva4acrL7ykVlTRQGV1rvFJVvx3bGOvrP2DK6E+ym40WThXx1q7PqCpbBTampmUrHnYdYJ8z9YOPkV7TcQ9B2HiZo656I1JAJaC5wmKQFoPSfJwB22K+dnj5w9unLdzEag0db5XxTaYurZqKQEjMe0kZyMHGCB8x9aZsNfSSHNcMtIIO+e+Vctn8INSt1lwq0fqprur7q2Oiq3E+bnwtJ/PlbwSAiIgIiICIiDbPEutfb9AX+siHxx0MpH1kY/nXze6tuD6rjlW3F2zjqEP+oiYf6l9I3EKnbV6HvlO/s+hlB/80r5uddxNp+N1ziiGAy/7D0+/BZn/AJD6TtGSmbR9ilduX22lcfrMTSvK/wBtbdrfLqbRVpaxhq4KVzy7s4NJeftG4/KF6maCDhobToeRkWmkBPz8Fq82fbB8u+rtRtoOOdjhdVWyzUop69jckwM2BeR2xsDn5Fak/leJuettexgttQ7UGpLi1x8FsTuoY2zgAb/WR+ZetK8VfZFcebRw/wCM1Twv1BIyCDWURioqh7gA2paA4RnPbq6dvt7eftUpFERFQREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAXl57TH2fIvJ1PzQ8MKuGCWnpTX6hsxiI8YRt++VMRGfiIGXNI3IyMbr1DWi6y03S6y0hfNI1pAp73bqm3ykjOGyxujJx8g5NzweSvsvOeLXlr15pjlk1vVsuOmLmH0Njle379QzBrnMiDh3jOCAD2Prnb2EXzocQeG/E/kb5hbZLd6IC46er2XK1VDto6yFjtnAjsSNiRnBIIXuNymcylh5peEdJxJs9L7lUsnfRXGiL8mnqWBpI9cEOBB8wln1P8c0oiIoiIgIiICIiAiIgIiICIiAoKylirqOeinGY6iN0T8eYcMH8xU6IPnd53eXq7cv3Gm7W2dmbbdqmWsonegc7qI38t8j5HHkvQjkG5JeAPETlYtGptfaSpr5XanbLLLO92HwjJbhpHZwIO5zjAwuHPbWtij4jaFDMB0ttne8epDwAfyLsp7Hi+XG58qc9srJHOhtWoayGmJJJDHBjy0E+QJOB5ZRJ11p5svZG3XQVjuXELl/v1Te7dQRuqKiwVrc1ccQySYZBtIABnDgD33wF1i5WOd3jLymXKS2acqI7jpuoqRJcLDcGEx9Q2cYzkGJ/rjYnORkkr6GXMa9rmPaC1wwQRkEHyXSLjV7K3gnxP4oR8RrNJLYWVlSKm622n2pqh+clwbg9PUc5AwDn5qX8sX13A0Fqun11oqxaypIHQxXu3wVzYnd4/EYHFp+okj7FuBYNltFDp+z0NitkIipLdTx0sEY7NjY0NaPyALOVBERAREQFa5wY0uccADJVy4v5j+OGluXvhDf+JOqaprG0VM9lHThwElVVOBEUTASMkuIz6DKDyf9ptzq3PjBrCbg1oqrlpNKaencyu6SQa6pB/fHzaBjbtk48jmfkd9mM3mM0HHxa4n6qrrFYK2ZzLXRUUIM9Yxhw6R73bNYTkDAJOM53261cvGiqPmO5o9N6U1ZI5lNrC+Sy13hk5w7rlc0HvjbH1L6JNHaSsGhNL2zR2l6BlFarRTtpaWBnZjB/OTkn5kptMx0/q/ZKcrs1so7fTUtzhfTY6qgy9UkuP7ogjOfnldrOFvDDSHB3RNu0Doe1x0NrtzOljGNAL3YGXnHcnA+wAeS3ciGCIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiCOSSOGN0ssjWMYC5znEANA3JJ8gF4c+1iutr1RzGDUVhq21lGygiopJonBzBJGMYyO2d8fUvRj2nHE3UXC3lVvF20tVy0lfca6ltwnjODGyQkOI+YwCPmF5oezn1RpHV/MLbuH/ABkibeqTUZdHRmucZOqrAJbGSTk9XYD6/knfhWzvZ38S7Zwo5qNMaqvlYKW3PjnoaqZ3ZkcrOkk/mX0DWu6W69UENztFdBWUdQwPingkD2PB8wRkFedHtVOWbhjpXlwHEXQGjqCx1tju9HHUmiiDGyQTPMfxfMPczGPXfsFX2LnEHVWpuHOvdJ3261NdRafuFE+gM8heYWzMk6owSc4zHkemcDYYSk69IkREBERAREQEREBERAREQERcL8y3Ndwp5XdJSag19dw64zxuNttEB6qmsk3ADR+9bnALjsBk74QcW+0z5h67gNy7VMOl74KDVOrKplqt5YfvrIiC6eVvp0sGMnbLgPNecHs+OSek5wNS6i1jxC1DWQad07UwisETg6ouFVL1PLC924GASXbklw+3hfi1xW4yc53Ghtzr46y8Xm71JpbNaKYF0dJE53wxRN7AAYLnHvjJOAAPZX2dnKxqLla4N1dl1nUQu1BqOtbc6+GF3UynIjDWxg+ZAGD88nbOAHZnTmn7VpSwW7TVjphT2+100dJTRA56ImNDWj8gC1NEQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQF4O+1jpzT86GpCY+hstqtcg22dmmaCfyg/aF7wk4Xmr7Urkh1zxYuw4+8MaT7qV1vtrKe7W2Mff3wRAkSMH74gE7d8Z9Bm6n7dyuUG72C98sXDKr05cI6ykZpm3wGRrskSsha2Rrh5EPBBB3XMK+fnkj50NXcpvEKOkvFRcKzQ1fN4N6sxcSICTgzxMP0ZGHJIGCRkb9j7ucN+JWiuLmkKDXfD6/093stxZ1RTwnOHebHDu1wJwQf0YUP9bqRERRERAREQbb4jPlZoW+Ogz4gopenYnfC+bzW5J443I1A6T+yAdXyHjD+ZfS/W0kFdSTUVQzqinY6N49QRgr5yOcHR83Djmf19plo6TQXgyRO8i1zWvBHy3We/wAl+Poj0Q4HRdgLTlptdLg/LwWqPXmkLZr7Rd70VeIGS0V7oZqKVrxkYe0gHHyJB+xYfCavjunCzRtyjcHNq9P2+YH1Dqdh/nW7Fpn181fG3hFxB5YuL1ZpW7R1ltr7TWGe117CW+LG12Y5WPGxI2zjz+RGfV/2aHPJeuYCy13DXixdKaTV1kax1HVkBjrhTnYdQ7GQY3xudz5gDs1zAcsPCHmT099w+JWmoqqaEH3S4RjoqaZxGAWvGDjftn8mSvMOq9m1zE8FOPNtunCmSprbLT1okguMTiHthyCWvwMHI8jg9vMZUvPxN69l0Wn2GO4Q2O3xXZ/XXMpYm1Ls5zKGgPOfPfK1BVRERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERB069pNyiXHmZ4YUN50XDEdX6RfJPRsIGauncPjgz3G4DgB5+vY+TnAXmt5geTC+32w6NdT0fvU7W3azXijMsXjR7B3Tlpa/G3UDuMdwvooXDPGjlC5euPdJUs4g8N7VPcJ43MZdaaLwK2In982VmCTn1zlOjgrkb9o5p/mXlqdGcR4LTpfWNOA+mijnLKe4M7ExmQ56wcfDnzx6Z7tr55ubfk54mcp+uZ3S0VdV6VkqDJZ7/A13R05y1r3tA6JBkemTjG+w7mcgHtMdGWHQkHCjmQ1XVUtdbJfDtd+qw+WOamPaOd4yWlhyA45yMZOcp74PU1Fpem9TWDV9lpdRaYu1Pc7bWsElPU07+pkjfUFaogIiICIiAiIgIi2/rbXujuHFim1NrnUVHZrZB9Opqn9LR+kn7Ag3Ai85uav2svCqzaQr9LcAK+p1BqOsYYG3MQuipaQEfTa44Lz6YG325HmjUc3fMjUVMtT/ALL1/iMzy5zY5gAMnOBt2QfSNsUGy+b6DnG5l6fHTxevrsf3UoP8y31wi9oZzHcONbUOorvrq4X+2xyt98oKl4LZosgloxgZ+vP2d1cSft2b9ttYaiLVvDvUTY3e7z0VVTF/l4geDj68ALn72N/4K9dtv+yasz/5ka6Ve0M56NA82GldG6d0Xp6to32Spkr6yaqbgh74+gxt8yN85+S71eyHs77byg0ldK0h1yv9wnGf7kOa0H7ekqEd2kREUREQEREBFj1lbS2+llrq6ojp6eBhlllkcGtY0DJJJ2AAXnzzXe1i4baFt130RwO8fUWqB1033UDemipH7gua47yEb4wBg98HcB2j5qeaTQXKxw5qdY6pqoqm6TNMVptDZQJ62c5wAO4YMEl3YAHtuR4d8deZrjxzk65oaLUdVLV+8VQgs2n7ezEEL3nDQ1vdzsEAuPoTtutt0WneYbmn1nJcqG06o11eqyUh9R0STMjJIy3rPwRgbbZAXsRyF8g2kuXHStDrPXVjo7lxJrWeLPVyASi2tIBEMGdmkebhuTjtjCYY2lyJezTo+Xu9W3jDxKvv3U1mymPu9vp2Yprc6RoBy47vkAJBOAB5eYPfhEQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQFBU1NPRU8tVVTMhhhaXySPIDWtAySSewCnXF/Mzo7V/EDgTrLSGgqx1NfrlbJYaJ7XdJMmMhoPkThBJovmP4I8RdWVehtGcRbRdL5RZElJDOC8474HnhcmL5rNJXfilyn8cLLqi8aduNl1Jpa4sqpKKujdEZ2Nd8bCTs5rxkZGRvnfC9s+X32iPLjx/pWU1DqN+mr62MOntV5xE9pPfokBLJAM9wR9SZfp+naBFwjrnnP5aOHjzHqbiraYXg9PTG8vOfQEbH7CtD057QHlO1TVihtXFi3mZ2MMkBaTupLvg7FItsad4l6C1XCyfT+q7dWNkx09EwBJ9MHG/yW51QREQEREBERAREQEREBERAREQbA438F9G8feHVz4Z66hlfbLi0HriOHxSD6L2n1BP8A9juvCvmf5Y+JPJDxgt1XRVtTUW+CpjuWn79FGWtc5jw5rXkbB7SBkZGR2819Ca2nxJ4Y6F4t6VrNG8QNO0l3ttZE+N0dRGHGMkY6mEg9J7HI9BkEbIPJTmi9pbpbmQ5T38KqjTNZbNXXV9E664w6mLoJmSF0ZwNnOYCATkDbHprHsfuNGgeG9w1Vo7VV/goqzVNRTupmSPA3iBDcjvv1u3HyXEPHn2YnMXw+1rd4+HuiarU+l2SyS0FVSSNe9sG5DXgkEEDbJ749crqax2odCaoa98M9tvNkqwTHICx8U0btwRse43HmPkr/APSY+o3IcA4HI7gqo7LZPBLU41pwf0Xqsu6jdLHRVLj6uMLcn8uVvdRRERAREQEREBERARFQkAZOwCDgjm+5q9LcqXDSTV91ijr7xWONPabb14dUS4O5xuGDzP8AqK8LOIXEPjJzh8ZW3a7NqL5qS/TimoKCmaTHAwn4Y429mtA7n5ZO2ANd5vePevOYjjbeam+V81TSW+5T260ULN2RsbIWDpA7lxAx8iB659XfZx8mNi4BcN6PX+rLNDNr3UUDaiaeZgc+ggcAWwx5HwkjckbnbtuFJPtTc8ZPIp7P7S/LBbYtbaskivPECvpw2aoLcxW9rgCYoQfMdi7uTnyxjuQiK+rBFtHiTxV0Bwj0/LqfiFqSls9viaXGSd2CcDcAea6p6k9rxyi2QzRW6t1ReZoiWgUtqwxxHo97gCPmMoO7SLzqpPbW8C5q8U9Xwu1lTUxOPeOunfgevSHZXdbgrxu4e8f9CUnEPhrdzXWqqcYyHt6JYZG92PYd2uHp/wC1Bv8AREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBU7qqIPJX2qfI/btMRz8yXCy1Np6KeYN1Jb4GYbE92cVTAOwJGHDGBnOR2XX32f3OxeuV/WTtL3wSV+idR1EbaymLzmknJwJ4/Tvgjtjf1XuzqPT1n1ZYq7TeoKCKst1xhdT1MErctkYdiCD+Y+RwV84fNVw+sHCnmG1zoHSr/APqXZ7o+KlAP9jYQHdIPoOrA+WFZ4l7x9IttuFNdrdS3Whf4lPWwsqIXYx1Me0OafyELKW0+Epc7hVo1z/pHT9uJ+v3aPK3YszsUREVBERBTtgLwG9qG2Ac6mufAb0gsoC75u90iyV78nzK8A/ae/ho67P8AiX6rGp5U+vbrl2Y6PgJw6Y85cNLWvP8AFY1yGtgcvpzwJ4dH10rav1WNb/SbnVERFQREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERBoWs9GaY4g6artI6vtFPc7VcYjFUU87A4EEEZGexGdiN14k87Ps6ddcvl0q9a6DpJ7/oaomc5joGF09ADuGyNGSWjfBH5wCR7pAYUFZRUlxppKK4UcNTTyt6ZIZow9jx6EEEEfWkp/j5+OWDn9438r0Isen6qmv2mS7qNmuZcY2HbPhvG7M48s7bDC7RVXtutYED3LglZ8+fi18v5sLt1zH+zn4AcarJXVth0XQab1Q4GSGvtzBAJX+j2j4TnyOMevqOpWkPY0XqZ0rtU6sMLeohgEjdwD3+EZ3HqnBp49tzxE/Erp4/XXTKOo9txxLdD003BjTbZP7p1bPgfpW9X+xetYP3vXMn2OP8AOFmUnsYNMl4971vKGjuQ52/5Ag0zg97Z2a8amprXxd4fW+02yplDHV1vme8RA7ZcHdgM5Ox2HkvSrQ3ETRHEmywag0Nqi3XmhqI2yNkpKhkhaD26gCS0/I4XkHzA+yK4uaNj+6/Bot1ZSNH3yiErW1A2/eB2C7cdj69/Xq9Ucv8AzZcMah8X+xrxCsMrnYf7nTVDOoj1MWx7eqGY+kFdJec72kumeWfVNDoTSVoo9U3voE9xY2o+90rSdmEtP0jg5Gc+W2CvN3hvyv8AtBeKn33TVm4gQUrgf9tXS9S0EORjIzPK0k79gCuEeNPCHinwX1vU6T4vWOrt99LRO91RMJvHae0jZQSHg+uf5kTXpZD7bLTDLWJKng/Xvry36EVU0Ma755JOM/NdIObLna4oc112hbqER2jTtG8uo7PSvd4YOfpvJ+kdh37HzOBjZXBblg428wNW6DhhoqquULD0yVbvvdOw9jl52wDsSM7rtPQext5ip7I6sr9Q6eprl0AtoxP1jOOxkBA+3CX9mY0f2efIRofmmpbjrTiLraaC02mcQmzWyVrKuc75MjyCY2beQzuN99vSbT/s4eU3TNOKe18PCD0hpklnMkjsZ3LnAkncrx+1twH5veSq+RatqrfqDShilDYr7ZqpzqV7hnAMkZwRv2eMbjzK7ocp/tfqQ01HormepZveQWww6nooW9Dh2BqYhjB7Ze3bYkjfZfFdr6j2c/LHPM+YaRewvJJAc04z/wCKtra19lZyt6ttb6SitNfZ6xwHTWU0jeoEeZAAz9RK7e2u52+9W2lvFqq4qqjrYWT088ZyySNwBa4H0IIKy1Pxn8eQvfXz887vJDfeUO+22VmoW3zTl8c9tDVlnRKx7RkskA2zjJyPT7V389kHxx0bd+BL+D1ZfKOm1Fp64zzspJZgySenmd1NcwH6WDkHH8xxsT221ZN9wuH1AD95NVLKP++6Xg/mAXmroHh/xfvNLV604c6b1BPBZh11Fyt0b2iADc5e3H2gE/NWS3kP9fTei82PZQ81uu+J9RduEvEK9TXSotdOKijmnOZegdwSfTBBH1L0nQEREBEWm6ibcpLBcmWd3TXuo5hSHOMTdB6P/wALCDoB7XbmVl0Dw5tfBrR2ovAvmqJXS3RtNLiWChYBsSN2l7jjBwcbjsF0j5GOQG/c3Lrhqu8aifpzRtnqW0k1THB11FXNgOMcIOGgAEZccgE4xlcCcd5tfzcWNSniXU1k9+bXStmfVOLj0dR6Okn97jGMfPO+V7hezRi0jDyh6QZpBgbF9+fW7DqdVkgyF2PM5CZzRzTwX4LaD4DaEt/D/h9aGUdBQxhrpOkGWof5ySO7uJJJ3/8Aad+E4VVTAQVREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREHAPNpyc8OObTSkFn1RLJabzb3+JQXmlia6eHbBY4HHWw+YJ27heMPNxybcRuTvVlBT3S7su9nuTTJbb3RRvhBIJBjeMkseB5Z33I2xn6HFoGrtC6P19QMtWtNN2+80kb/EZDWQCRrXeoyNvL8gV/wBHzncL+XTjTxzk970pp6trojt73UdZa8DfYkEkfPstS4g8oXHzho4SXrRNXIzORJStc/GPlgHP1L6KNP6W01pWjbb9NWGgtdPG0NEdJTtiGB5HpAz9qzK23W65ReDcaCnqo/7ieJrx+QghKPmmouJfG/hx0UdNqTUNk8N2GMkDmbj06xuuyHBH2rHMnw0uNvodZ3Wl1lp6F7GVMFdABVeFkAmOVuCCBnAIwSBnHde1N24VcMb7EYL1w803XMd3E9rgfn8rV0r5zvZf8OOI+kqrVPL9pG36Z1pRkzCjpD4NJcW5Jcwsz0sec7EAdvPsZh47mcLOLGhOMmjrdrfQGoqO6UFxp2VAEE7XvgJG8cjQcscDkEEDt9S3kvm8u2heZ7lI1LRX+86b1boOujmbLT1RbJFBO5p2HiMJZINjsSds7L0v5f8A2wnB/Udptdg412m66avrY44J7lFGJ6GeTYGQkEPjB7kkEDfdP8HooiwLLebXqK1Ud9slfDXW+vhZUU1TC7qZLG4ZDgfQgrPQEREBERAREQEREBERAREQF4T+1j4YwaA5ra++W2gbTUGrrdT3RpbgB9QB4cxAHbdoP2r3XJwupPtDuUu28xnC+q1FRyui1LpOhnqqB2NpGMBe5h+vB288+uFO7xLcZXs1OK1i4g8q2jrDSXKKW6aWoW2ushL8yNEZIa4g74xgfYPULtavBv2WnEy+6N5rtNaTpK6Vtt1UZ6Gopur4HP8ABc5r8dsgMIz37ei94x2VWcVREQEREBERARbS4ncUtDcHdIVeuuId+htNnogPEml7ucezWjuScHAXVOT2vfJ0yRzG3PV0gacBzbE7Dvqy8H8oCDuyulHP3z8aM4CaMufD/Qd6o7txBu1M+mjhp5RI21te0tM0padnAE4bkHI8sLrTzte1S03xJ4dO4dcuU+obdLdHgXO81EXuksdOM5iiAcSC7bLtiB2we/Vrlv5EOYPmbvlJc6XTNfZ9L1c3i12pbuwxRFhcC90Qf8c7yHEjpBGdyQmjP9nhwG1Bxz5ldNXGaxzV+nNN1zbvfKuVpMQEfxMY557vc/pwO/mvf9744Iy+R7Y2MGSSQA0D1PkuIdN6f4F8lfBKOnfV27S2lNPwMFVXT4a+qmO3W8jLpJXuJwNyc4GwXkxz6e0HvfMrfqXRPCKrvVn0Nb3kNa1zoKi7Tk4D3sachoGA1hJO5yATvB6d83vOZoPli4c1N+iuVrvWp6g+BbLOyra58kh/fPDTlrB3J2PpnBXmLdfbAc3VbNMbdUaRt0TyfDZHZvEdGPIdT3nJ+ZH2LanBT2anNRxnnoL1dNGzaXsFa+OSS432ZsEzoXbl7IHHxXHG+7R38+y9TeFvs2uUzh1p2mtVy4WWvVVxYwe83G9NdUvlfgZIa4lrBnsANk+4TzXjDxO4/cwnNPf6W36y1DcNQ1Ujw2noKaLpiBJ2wxowACfPYLtpwb9jXxH1rYKLUXEbiTQ6VbWxiQUUFA6qqGNIyCSXMAyPI7r1P0hy/wDBHQLmP0Zwp0vaHswWupbZE0tI8wcZB+a5BT/B5P649iNcaa1SVHDrjlFX3CNjnNprtaPAZKQNmh8cjukk4GSCN/tXUjhRzD8ynITxBvmiba4UMlPWCO82C5wmSmnewgdYGxaS0YD2kbHcHsvcnWXMnwO4f6jZpXWPEi02u5vd0eBLIctd6HAOD+jzW1OYnlO4G82Wj53aksdvku9VROFo1RQtb71TOLR4bxK3+yxghuWOJBGQMdxrRJyr82/DXmi0FQag09daKi1D4YZdLC+ob7zSTgfFhhw5zDjIcARjz9edV4P8OeRnnO4V8xVkttk0HeaOa03eJ37IKJ3+0JKVsgLpBMDgscwfROCCcEAgke7VKJ200LapwdMGNEhb2LsDJHyzlQTIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIusHGH2jfK7wVv9RpbU2qq+uutK4smp7VRGpMbh3BOQB9Z2PllDXZ9F145eOezl95mb9NpTh1e7jFeoYnVHuFypDTyyRjcuZuQ7A3ODsuw6aCIiAiK1zmsaXvIAGSSewQWy+J4T/Cx19J6c9s42z9q+b/m9tGqbLzI68ptZW6ejuUt1kmc2ZpBfGQOhzSe4IAwRtsR5L2d1n7TDlC0Fqyr0df8AiDVGsoZXQzy0lrnqYGPGxHXG07jt2Xn17S7mw5Z+ZGksDeE1uqrnqG2SOE19lt7qX7wf/kX3wB7hkZGRjf5BMu6ePYHhTXW+5cMtJ1tpqWT0ctlozDIxwILRC0dx5jGCPIghbrXhbyAc+Gq+BGvbXojiLqWsrOHVxPuksU7+sW17tmTMJ7NBwCO2PMAFe4lou9svtsprzZq6GtoayMTQTwv6mSMPYgojORERRERBT5LwF9qC0t50tdfNtAf+SRL36K8AvafSGTnU17nPwe4tH1CkiWfqevb7l96f9gnh30HI/YrasfxWNcgLjnlwDm8v/Ddr85GlbXnP+LRrkZWTFgiIqCIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIvOvn19pjbOFkt34N8FJn1Wr4gaetvLHDwbc892sIOXSAefYfkJDs/zW83XDblP0T+yLV0v3RvNaTHarFTzNbU1j/NxznojHm8gjsACTheKfE/ifxe5/eYO2TVFki+6d0kjt1ut1ujc6Kipurzccl2Aclxxk42GcLTNG8u/NbzRXP9mFo0ZqvVX3QmDJL7cOswbnOTPIcdIJJw3IG+AvZbkk5KNHcqmh4Zaujpblrq5xh92uxYHOYSP7DCT9FjckbdyScnOSvL0zeuWOAfBzTnAnhdY+HunaGCAUFLG2qljaAZ58fG4nz3yB8h6lcjIiDb+udEaZ4j6Tuei9YWyG4Wm7QPp6iGVoIwQRkZ7EZ2P8y8gOaL2Smu+FlmvPEDhJqSLU1hoOupda5YnMroIBknB3bJgYHkcDJJXs+o5Yo5onwyxtex4LXNc0EOB2IIOxB7KUfPxwn9otzW8FNJ0mgtK61pZrRbG+DS090t0dS6nYCfvbXOHUGgk7ZXYjgl7ZDixHq62W3jXp7T1fYqqoZDVVtvp3Us1MwkAyY6i0gZye22V6N3bky5VL5eKi/3jgFoyrr6t5knmltjD1uO5JHbP2Lqvza+ya4f6/t79TcuVBQaQ1BCPjtPW5lBWADs3JIidnzGxz5YC1qZG1vbBas4Ya24KaFvlh1PbbjcX3My0Qpp2ve6B8eSSASWgDOxwclcqezhtsVm9n7JcvuY0SVcN7qnDoGagDxAMnzB6SB9q8eeMPB/irwT1N+wnivpy4WivpmnwGVBL4nx5+lE8EtLTsdj2IJAyF7r8hdopZ+R3htasMbFW6elEnoTLLKXE/8AnHKk/ssleXnso9R1FPzo2lrnuY29W+5slYzZpcYXPGR6Ag4+te668E/Za0MzedzSUA3NNT3MvI7YbSyAn6sr3r3wpn1VURFQWl6mvLNOabu2oZWdbLXQz1rm/wB0Io3PI+3pWqLqrz284vDHl64aag0hcLk24a11BZ6iktlmg+J7TPG6MTTHtHGA4u33OAADkJR423uu1VzicykVLboI47lrC5mlo2MZ9CPLnAkbZPTkn8i93eVjgFQ8t/B61cNKOsNXLTF01VOezpXYzj5bflz5YXjh7LHSEuqecrSdWI3GGxU9bc5HdJIaWQOa0E4wCXPGF73IecEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQbC428G9G8e+G934Y66pPGtt1iwJGAeJTyjdkrCezmncfb2Xk/xv9jvxT4fadvGr+Huvrbquktsb6gW59K+nrXQNBJIOSxzgAdhjK9nAMIRnY9kP8fP5wN9opzN8u+nqbQFgu1vuFjtj/Djt15ovFdTtB3iY4Fr2AHO2+D5L1J5MvaDaF5mbNJbdVSWzS2r6PHi0ElT0xztx9KIvIyMg7Zz8lrvHf2d3LPxzbX3Ws0RDYdR1fVILtanGne6UjZ0jGnoePsXknxw9nxzLcFL5XS0egrtf7DSyl1NdrVEZssz8PU1mSHY9ARtnbsL/pn9PoFhnhqI2y08zJWO7OY4EH6iNlKvm74Zcx/H7l91VDWWjVV/o5aSRrprZcZpgx7Qd2ljzluRkZAHzB7L1U4Ce1t4CcQqWgsvE733Rt/kDY5JZ4uuikf5uEjc9AJxgEevbsp9HfBFp1jv9m1Laqe96fudPcKCqb1w1FO8PY8eoIWooCIiAiIgIiICIiAsW5Uja631VC5ocKiF8RB7EOaRj6t1lLanEDijoHhXZZdQa+1TQWejjYX9VTKGueB5Nb3PbyGPqQfPVwf1gOX7muserLxA6FmkNVPbVseCOljJXRvBGxADSc/JejnM37XS16EmpbHwO0tSXusnj8SauuT3CKEEbBrGHJOfU4xnttnzi5wNd6I4mcxOstd8PmAWe81zqiMtGGvcdi4Y2wcA7bZyu0nss7Dy98Rq+8aG4xacs12umCbbDc2hzJGnBwASM+YA8zt6K5vqbnW3qn2x3NlI/qgt+hoWZ7fcmVx/KZv5lqNF7ZzmZgh6a3SuiKp/937nMz8wlXpVWez/AOTauJdLy+6XYXf/ADGOSLH1BrwFtit9mPybVk/jt4XCmA/+RwVkgZ+ck/nUV5tat9rrzcakpJKK3Vml7AyQYMlvth8UfU+R7sfYAVpej/avc4Gko/AqNU2a/RF2SLtbRK8fIPa5px9q9XtMcgPJ/pRzZaDgZp6pmYciWuidUuB+QkJH5lNrLkM5RtcUstPdeBunKWSRvSKm20/ukzR5EOjI3+sFTJujqRyse1xq+IGtKTRPHHT1oszK94jhulEXsia89g8OJx+ft3XpM6+Wdtnff/unTG2RwGpdVtlBi8IDJf1jbAAJz8l5O8zfsftQWR82o+W+uN0oWtLnWWun/wBsMA8mPP0z6Dufl59GNR685heG9LcuEWp9XatstOzMNbZqmtlawDsW9JJHSd+xwd1an+Oc+c7m41xzj8WIeHGlJfC0hBdxQWOjjccVTy8MbPIRnOTuO+2DtsB2F4e+xJuNfa4q7iXxsFuqpo2PNFarQJTESMlrpJJACR22aui3B223XhdqrTnGfVdlnhsdqrGVUDpG71Dx9ENHcjJz9YXo3qn20Wg4tMv/AGI6ArZb4YiG+9OIhD8d8YB777k/ar01yNwW9kVy88L9SQaq1beb3rqqo5Gy0tLcRHBRseM4c6KMZkIOCAXYBA2K5t5oebLhdyj6E+6V5iirLn0CC1WCieyN8hwOkEDaKMDzx2Gwxkjx9vnNpz18x1/qf2Oay1pJDVvIZbdN+JTU0Q7Bo8IDJA7knPcqCh5UOYGsulPrHmFs+oaOwwOMtXPeKt8s72DcgEucQDvvnPyU55qruMnM3x+5/wDiPp7QNw90t9DVV4itVkoA4U1O5+xklcTmRwGfiOPQAbL0p5XPZg8FeXqak17ryvdrPVlAwTiprWiK30DwAXOihJIOMH45Cds7BdPrfzR8lHA+yPfwc4bzz6vgj6G3IQHDnjsQ92MjJJzk/wAy68cRecbmw48++acqeIWqKi1VxLXWa1OkDHsJ2Y8RDqkG4BBJB9N1cSXXqjD7VrloPGCq4X1U9ygt8FW6hZqQtaaCSVp6SRg9QZkbOIwRv8lzFxQ5xeAXC/SU2qq/iFZ7hiLxIKWjq2PklJGQMDOPt3+S8H7TyocyN9pWVtp4L6pnhfjpcKIjOe2xwfzLe2lPZ5c4OqqiOGHgtebfE/8A+6bh0wRAZ75JJP2BRa7Ma69tNxSqLpPFw94c6epbe1zmwy1/iySPHkS0OAH5fPsuMdQe125vL1Rz0VJW6UtLJmlvi0VpIlYD/cufI7BHkQMrtHy4+yC0Ta7ZHdeYGaW6XFx6jQ0lSWRNHoS09vryfqXaKy+z35ObG6KWn4F2GoliILZKrxZjkdiQ52Cc/JEeCF7l4m8U7pWayu1JfNQ1dU8vnrW08kwzncAtBAAJ7D5rsLy/e0p5j+XWwxaJo32nUlkoi5sNFfoZXyU++7GSMe17QDnY5GV7n2fhtw909QMtdk0PYqKkY3pEMNvia3HoQG7/AGrjribya8s3FyKb9mnCGwyVUzCz32kpxS1LM9y18eCD9eU2q628mPtSLVzBazk4c8WNPWfR95qmF9qqaaqcKSqcO8R8U5a/G43OcH7O/TXNe0OaQQRkEbheIfOb7M/iDwPu8msOC9ou2p9GEGVzYB41XbiPJwb8Tm9sEAnPrvjirlg54OMXLRraKrq71dr9YOvwrjZK+qe8FoOD0dZPQ4Y2Gwz6HdIPoTRdQ+GHtR+U/iPFSwVeq6vTFyn6WupLvSuYGvP70SNy131j8nbPafT2ptP6rt0d203eKW5UcoBZLTyB7SD2zjt5908NaqiIgIiICIiAiIgIiICIiAiIgItua14haL4dWt951rqOitFGwF3iVD8ZwMnAG5/IuqOvfaz8pmkI6mGz3m86mroQeiG30JEUhHl4riGgfPdB3RXWnm554OG3K7pgzGsor7qedxZS2eCoa54I7mXpOWj5HB8/TPkpzPc//GfmE1XNNY79dtK6cx4dLa6CrdG4gnu9zCCSdhgH1yTnbK4Iezr5n+YCODWVdp2qs9jrCJTcrzOI56ph7ujY89buxGTgZ9UG/uIHtgeY3V9BVWqxWPTmnKeqY6NzqeOSWUNOQQHuIwceYC1Dlv8AZ0Tc4HDyq4zXDi0+03G61Mhcw0gqR43n1jLSBnbv5LiHml0Jy68KaD/Y70jDWT60tbxFWzCVzg14xkPySN9zjvvsB3WVyP8APFqrlZ1THa7m6S4aIuEo+6FCMkx5O8jPQjJOcE9+4JBTvSd6k4z8sHMd7PfX1l4j226snpqWfqtupLXG8wdfYxzMcPgJBx0uJB8jnC5LsvtmeZyip2wXnTGiblIBjxvcZoXOPqQ2XH5AAvVHRPF7l45pdGzUNiv2ndW2m5xGKrtNZ4cj8EHLXwOycjfcA4PY5XHNy9mjyaXCpfUs4SU9GZHdRZTVUoYPkA5xAHyCbR59ye2e5kXSF0ejtGMYTkN92mOPlnrVJPbO8yLo3Ni0fotr/Jxppjj7PEC7/wAfsyOTmMdP+xeHfN1U4lP3Mrk4D+v/AGLWnBzg1T8LEg87G+2Q5sWyl/3M0MWHsw2mXb7RMFt7XftYebXXOna/Tf3T03ZKe4wup5ZrXbCyoDHDB6Xve/pJGRsOxK9SIfZx8mMTBGeB9pkx++fPOSfrw8LWLPyF8n1hqoq63cAtMCeEhzHyxySkEeeHuI/MtSmR49cr3s/eNfNXBNqigq6XT9gc8l13ujXvM7ycksYN35Od8jPfscruFZPYg6XbE39kXHe6vkx8XuVpjYM/LreSvTK0Wa0WC3xWqx22moKOBvTHBTRNjYwD0DQAFnKjwt56PZ1VvKfp22cQNI6qrdTaXqqkUNZLU0zY5qKZwJYX9Hw9DiCAe+frXYf2QHNF4tBeOAGvNSNb7r0VunX1k4HwHIkp2lx8iAQPmAPRel2s9F6W4g6ardIa0sdJd7PcYzFU0lSwPY8H5eRHkRuvnw5yuHmlOBfMrqXR/C2snpLbbJmyUroqg9dO92SWNeDkY7bHb5dkkn0fRYi+Z2n5i+PNLRRW6Di/q1lNEA1kYus2wHYZ6sn7VyLwJ53OOfCviFadQXDiHd7pbI6lja2mrKgysdCSA47nyHrkYyMKj6HUWyuEnFfSfGXRNBrbSNxiqaarjaZWMcC6GQjJYcfmPp88gblvd9s+m7XPer9coKChpW9c087g1jB8yVBqC8Ffasfcs86Oq3WueOQmhtvvXQ7IbOKZoc0+hwBkeWV6G8yXtSOBnC+xXC18NL9FqzVIY+KnbSsLqaGXGAXuIwSDvgjH19l4s6p1je+JWvq7WutbhJW3K/XA1VfUPO7nPcM7+gGw+QCJO9fSLwDhmp+B3D6CcdMjNL2trh6H3WNb+W2uG1VQVnDzTNVbHxvpH2ik8EsIIDRE0ADHp2x5YW5U8WCIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICItOul/stldA27XSmpDVP8OESyBpe70Ge/dBqKKgIIyPNVQEREBERAREQEREBERAREQERdc+bjnT4Xcrelq6G73eOs1lUUb3WmywEOlfKRhj5O4YwEgnO5GNtwUG8OZDmS4e8tmgLhrHWF4pBWRQuNBbPGAqKyYj4WtZnJGe5xj5heVep/bJ8zNfcZZdLWLSFpoy8mOGahfUvDM7AuLxvjvsurlttPHbm54qyR0VPeNY6qvNQZZXEvkZTteSS5xORFGBn0GBtk9/WLgD7JrgLo/Q1NFxlsp1ZqmqiDq2U1MkcEDjv0RBhHbtnJz8+5ZBxHyve17vWp9Z0WkeYO02W3Ude8RMvFCx0TInnt1sJIaO2+T6bHGfTqxaisOp7fHdNO3iiudHKA5s9LO2VhBGRu0n8i8kucz2Ud+0hINacsdorb3aCD75YnTeLVU53PVEXHMjfUZyPIbDPUKzcEObmwRy0lg0LxEtUcZPiR0rKmnbnudmkA+quW+EvH0eq0vaDguAJ7L5w3cPeb6Wo8N2n+J75XHG5rCSfryt1Wnli5+dQxCptvD3ibMzydJUzRgf8AnyBSSj6FEXz23jlf5+7BTuq7pw94mxQsBLnx1UsoA8/oSFca1t85jeH9Z7zcrtxCsU9O7PXPPWQAEH1cQD+cIPf/AJouO2l+Xvg9fNc6hu0VJUCmkp7ZGXYknqnNwwMA3JBOTjtgeoXgLwx4X8QuZ/i8bFpihmrLlfK99VWzk5FOySQl8jifTJwPM+gyRp2o9c8bOPFyoqTUd/1NrOspsRUsMjpKl0edhhoyATkDOF60eyr5StXcEtLXniPxNsbrZfdQ+HHRUczcSwUwGcu32JJ/ISrIl47u8O9F2vh1oSwaFs1PFDRWK3QUETImhrcRsDSQB6kEn5lbjVMKqn+r4IiICIiAujXOh7TK1cr+vf8AYv0xoNup79BTx1NZJUVhgp4A/drMtaSXYGT/ADbE95CcL5/faXmd3OHrV8/d0kfR/wB6AQP0JKNC5u+crVvNzeLVc9R6Yt1jitDC2CCkkdJ3z3c4AnuVydwY9ppr7g5y3zcDLdpmnq6ympp6O03WSUj3OKXP73O5aXEjbuTuPLpnSUjZ2vc5+On86pNR+H9B3UVcyM7PHeX2OlqtNXzTVd3uFU1lVb9O1fuUbiAZJZHMa7HqQwP/ACr26Xy7aK1vqzhzqWj1bou+VVou9veHwVNO8se05BwcdxtuD6fJe3HINz7ae5jNN0Whte3amouIdHH4b43EMFxa0f2Rg7FxHcDv9fdWndFca8cuP3DXl+0XX6y19qGjpBSwl0FG6ZonqpMfCxjM5JJwM4/KcA7M54OPtdy4cvN94h2drTdpJYrbbur97UTEgO+wAn5Yz5Lwptdj5gubbiFP9y6HUGutRVLjNMWl0radjjuSSemJu3qM4PcqDtdrX2zPMTcrrVP0NpfSdmtrnOFOyro5KmZrM/CSfEADsbkYIz6hdfeHvDPj9z78a66vFW+53m5y+PdLvVgtpqOPyGwwAB2YP0kk+jHLB7JXhJpvSVuvnMFaZdQ6omaJp6BtY9lJT538MhhHWR2OT/qHeTh/wr4c8KrS2x8OdF2nT1CAG+FQ07Y8gdskbn7SUvPBwvyf8kPDrlKs00tnqJLzqi5RNjuN3naAXAb9Ebf3rAc48yO+5JPZNETwEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERB1n5nuQfgvzPmO432KewXyFpay52yJgec/3TSAHeXfbbffdeW/Nn7NLiry2W9+sLHdI9Y6V6y11TTU7mVNMPISxjIO2TkHG2w749385UFbRUVxpZKK40cNVTSjpkhmjD2PHoWkEEfWEo+ePgNz18x3LtQiy6G1gyps4IItt1g96gYM79AJBb6bHbyXpVyhe1L0hxoqnaW4x01s0hfGMzFUsmIpajHnlxyCfTyPyOV2C4mcj3K7xWpKmPUfCGxU9ZUNIFfQU4pamMnJyHR48z5heWnML7KrjvoLU9XJwj07Uav0+95fTCncDUMaTs0gkZwPUg/X3TZPR7P27iNw+vIa60650/WdX0RBcoXk/YHZW4Ipop42ywytkY4ZDmEEH6iF859y5KubzTERuVZwF1pTMi+Iyw0Tn9OPPMZJC0OzcY+ZDgxXmlptY6w0/NG8A01bJM0ZGduiXsO/bCSX34PpQReBujfaa8zOmZmuuGpn3SLqBc2SRzCQDn5ruBoH2z+hP2LlvEDQd1F7hjIaaQt8OV2PP03+rbvvuk6Xj0x2AyFjPuVvZnxK6nbjv1StGPzrxG4h83HNHzzcTIeHvBltxs8FSJH09toarwneEB8TnvB+EAEZIJPnnGVwXxu4X80/L7c4bdxak1XaTWAup6n7rSzQVAB36ZGvIO/kcH5Iks8vr6LqavoazIpK2Cct7iOQOx9eCslfMlp7ivxj0JX0moLNrXUtvkDhJDI6rm8OTG+wccOC9XOXL2tnB658OaOi47V1faNV22BsdRLFSumjrcDaRpGMOOBkHfO/zVxXbTml4+Wflu4MX3ibcwyWopIxDb6ZxGaiqfsxuMjbPf8A9q8BOOfMbxU5htTTak4i6imqQ55dDRscRBA3OwDexwMDJ9NgOy5k58eeC7c1OqY7Rp9tRQaItLy6gpZMtfO7/wCaPHqdu+/btgLqjT00lQ8NbsM7u8goLY4JZgXMYSB3Woae1He9KXinv2nrjNQ11K8PimicWlpG/l5fJWeMaZxhiwAMgk759VFLS9bHTMw053b/AKlbMR7DezP59NR8a7nPwY4sVkMt9paT3i1VxIDqtjCA6M5OS4Agjv2AHfA9DK640FspzV3Ktp6WBv0pZ5WsaPrLiAvl70lq/U2hL/S6n0leam1XSid1wVNO7D2H5eX5Vzba9Pc7HOOKm8W2DXPECmosQTSuqT7qwjcM+NzYyfkMn1U7R7/N4i8P3lzW650+S3JIFzgJA+fxLq3zN+0y4L8Ap/uFYHM1lfsfFTUFQDFF/wB+8ZH5/I4yvKaj5B+dFtWI6bgJqqCTOOvEbAP/AB+sDH2rd9n9ltzm3tpmk4eUtDt1H326Qsd+QEklNnn1crsXB7brUhrnCp4I21tHv0llfIZflkHA9PNdL+bHmIi5ktffs5+4jLdK8OD2Ab7nO581t3jhwD1dy/3mLTGveiK7ygvNOzOGtGMnJ79x6d1rvJvwrtvGbmE0voK8RNfRVs/VOHbjoBGcjz7q2Z6mNN0w/jVzDS2jhPYIai7tge0QQxU+GxgDALy0bD9JWRxq5TOOnANjKviHoqrpaCUAtrY2OdCM7YJwMb7b7L6EeHfB7hlwot0Vs4f6LtVnZEwMMlPTNEr9sEufjqOfMZx6BZfEvh9p7inoW9aB1TQQ1duvVHLSyNlb1dBewgPHoQTkH5Y7EqEfPhy+82/FDlwe92hRbpGucXhtXB1lrj6HPbf0W9OK/PxzO8yFENA3isoHU9ycKeOjtlCWPkLsDAIJ3PmcDbOVyFws9lrxxuXH6DRnEPS9VRaIt9xLq29Ne3w6mjY4kCMj989oA+WTg9ivXDQ3LDy/cN3Usui+EWmbdPRta2KoZQMdKMDuXuBJPz7rMn47/KerucedHA/2NFTqOwW3U3F3iRUWt9ZEyoNrt9IDI1pAIY97jscd8b/UV6CcE+UzgpwHtMNu0dpOlfPE3Bq6mJskrj5ncdz3PnnzXMqKmKAADA2AVURUEREBERBQjOx7LpNzR+y44S8f9Q1WudL3qXReoasdU5paVslLUSf3T48jp+ZG/l2wB3aRB4D8xvs3+O/L3C66uZTarsoyRW2yN4IA7kxnJGPMZz2xnK4u4W81fMFwRrYnaF4h3SgbSEsNJPiWLAO7HMeCcZ7jb0X0hVVJS10DqatpoqiF+zo5WB7XD5g7FcYaw5VuXLXsUserOC+k64zZLnm3RxyZPmHsAIPzBQdDOVD2ul31Pqui0TzGUFnoYK5wjjv9GwwMY8nAErMkNHbcfPtsD6X2TWekdSRRzae1RabkyYB0ZpayOXqB9A0kryc5v/ZParsd9fqflkskt2ss7TJLZn1IM8D8kkRF5HUPQEgY8xjB6FXWwcW+COshabra9RaQ1JSSt6IXskpp+sEYwNusZ22yD23ROvpyRcd8vNw1LduB2iLnrFz3XqpstNJWuf8ASMhaNz88YXIiKIiICIiAiIgIuD+OnOXy+8u07bfxK1n4Fwc0uFDRwmoqMbfvG/WPq88LpVx79stpWr09cLBwH0fe23KdhjhvF0EcLIge7mRtc52R5Zx9ndTdHpjedTac03TuqtQ3+3WyFrS4yVlUyFuB3OXEei8xOb32teo9L6zrNCctzLNUUtteYqi/1MPvLJpB3ETDhpA9Tnf7QPPMR8f+ZTU001NSas1zdKiUuc2COapYxxGQMDLIwB2Gy738lPsprld6ubWnNRpuooKGMAUOnzUdM0r+5dOWH4QO3Tnf7dmmfXRjitzGcdeYK7B+v9ZXO9yzO6YqKEFsf/etjYN/q3XYzl59lJxp40WGDVmqL7Q6LtdSA6JtXTvmqJW/JgIAPzJI+edl676I5YuXzhzGyPRfB7S1t6Nw5lvY9+fXqeCSdu+VyXHHFBG2GJjWMYA1rWjAaBsAB6LW2pn9Oj3L/wCyd4LcHtQUmrdVX2t1pcqN/iRR1lOyKmjeMYIYCc/bvuRld16+sorDZ6mvkYIqS3Uz5nNYAA2ONpJAHYAAbBZ62txSbI/hlq5sWzzYq8N+s078fnUV86Qseq+ZfmHrbXpeJ1ZeNbagqZKYEZAbJI54Jx5NZ8+w7+at5h+AGsuW/iHLw91rEBVCnZVRSNHwyRuzgjy7hc8eyXdQN50NPMrGtMj7TdRTZ3xKKZxyPn0h67z+0+5JtZ8xNJZOKHDN1JNfNL0M9LW0Ezuh1XS9XiBzHY3ew9ex3IIA+Vp+nlF+xLi1wZsWneLNiutXa4Lu0S01XRSEGN2MtDz2OR27j8oz6dcgftJv9kWhl4c8fLpBFfqFjTRXbGPfI9hiQf3YPmPt7jGyPZy6h4T8ftD3Dla4uabpq2v0/RvMVPUAZljY8Nd0H1bnJ9PyZ67+0Q5VLZyj8UbHeuGNRU02nr/G+pow6Ql9JOx+8ZJ7jGCPLA7DspLsMx7GXfmT4MadibUai1tRWuneQGz1bgyM57YO63vpjVemtZ2eHUGk75R3a21H9iqaSUSMdjyyOx7bd9x6rxj1Ny28wXN3wj0lrjhpbYryyhp/9tUjquOHreWjDgZCAXAAgAkbOP1LcvJTzN3L2f8AqDUfBDmj0vqGxUtwkhuNPEIhO+ieWkFwY1xBY8YJLScnffzn4y+pLr2PRceaL4/8Hdf6WptZ6W4g2istdS0ObKZwxwyM4LTggjzGFva1Xi13ukbXWi4QVlO7tJC8PaftCarNRFoGq9daR0NTxVertQUlrind0RunfjrPywMq+jqT7UbmT4lcvPCKyR8Mp3W64arr5aCW6tYC+kjZH1kMJyA92cAnyBI7ELoJyKcmNVztaj1TxA4q6mvEVktU8UdRWxPBnr6yQFxj6nA4DW9JJHbqAXpfzWcwXJ3S8N57NxwvFm1Ha6wh8NrjZ7zLJKAS1zAN2uG+4OQuHuDHtL+Q7h3pWh0FpS23jRdpo3ERwR2Rzoepx+KRzo8uLidySM9t8dp/LuSH8bja3M17L7l64acCtSa34ewXmO6afonVrnV9eZxK1uAcAAAHfOO23l5+RVPFTyRzOmqPDcxuY29Oes+mfJeyPOj7RrlzvnL/AKm0Vwu1i3U181TQOoIWU9PIxlO1/wBJ0heGkEAdsefrgHyT4bcLNfcXtSQaT4daZrb1c5yAIqeMkMB83HsB3PrscAq+o5s5POd/XHKheaptLSuvenq8AVNtlkIAI7FmTsc+hH2gkLe3N97SbXXMxp2HRNish0tYA7rqY45sy1B9HEHt5d+2dtyth8YfZ+cyHA3QT+IuvdOUENqhaHVDqasEzoAe3WGj4T9eOxXW1rS4gBXsVcyN8juljS4+gCrLFLAQJGlpO4ysyAe6xF7sAvxv6hWTOfWvYwPzjZM/pj+V3HcLlx9qDxo4FWWi0hdLfRansFH0sZFUuLZY4wMYa4ef2gL2F5cOYHSHMnwyoeIukj4bZvvdXSudl9PMO7T+fH2+i+bKqpJKR4ZJvkZBC7b+zw5ur1y+cS6TSNwf4+ltS1TIKmNzsCKRxADhnYZOBnyJ9CVPG3vUiigmiqYY6mCQSRStD2ObuHNIyCD6EYUqAiIgIiICIiAiIgIiICIiAiIgIuNuN3MLwp5edON1NxS1My2UsriyGNrDJPM4eTGDcldX7x7YrlOoaOaW1Ums7nUMGY4RamxCQ+nU6Tb7Qmmu9Kgqayko2ddXVQwN/upHho/KV4fcyPtTOMPFuulo+HXj6NseOmONk2ZyMEZcQenO+c7/AFALrKy7cf8AiXcBDDcNcahqapwIjifVTBxPYhrcgD7MJ8H0mnVOmOroOo7X1envkefyZVBqrS7pBE3UlrL+wYKyMnP1Zyvnvh5IedCshbXxcCdbPbKA4OdCQ4g75wXA/mWkX/lU5tNA0Umo77wb11aqeAdT6sUcvwgb5JZkjtlJ+x9HTHNe0PY4OBGQQcgq7GV4PcoPtE+JHLjenWfXVRddVaWlIZNRz1BdPTnOCWF5zkfPcYI3Bwu5vFH2xnBMcOLi7hdZtRVOrqqlfFRw1lIIYKaVwIbI6TqPV05zgDy8+xSW3KluOz3NBzf8KOV/SdTdNVXqnrL86MigsdPKHVNRJjbLRu1ucZJxt9pHiPxm5yuPHHHiO3XV11XW0T6eoDrXbKGQsp6VgPwRho+me2SQcnJwMrYNrsvFjmK4jOprTQ3fV+q77OZHBgdNI4uJJJJyGMBJ7kAdgvZXkr9nJw14H6PtupeK2lLZqLiHUEVM81W0TwW4/vYoWnLcgd3EEk+gCvPhjs9wNvF81Bwe0betS074LpV2alkqmPBDg8xjOfme/wBq30o442RMbFEwMYwANaBgAY2AHopFmftRERUEREBERAREQERcU8euZvg5y12WlvXFjU4tza55bS00MRmqJ8dyyNu5A9f9RwHKy25rHiFofh7ap73rbVlrslHTRmSSWsqmxYaNyQCck/IAlecvMd7YrQ9RpGv05y9WG9zXmuhMTLzcoG00VIDsXMj6i9zgOxOB37HBHnrprhNzP8zVRU6k09pHWOui2Vxmrel80TXknID3kMG+dgQB2wplvIO8vML7Y/VNLqO4ae5f9N2n7mUznRQ3m5RumdMRt1sjBDS3OcZ/ON157XTU+qePnFqK9cSdZddz1LcY46y6Vz8Mga9wGQOzWtB2AwPzlemXs8PZuHTAuXEfmg4c0ktyeWQWax3NrZ2U7e75pWAlvWTgAHIAyQthe0S9nTqyPiNRa/5ZeFktbZL1T9FytFmhAFHWNJzIyMHZjwQTjYOzgYO1nD249EOWrgLwf5cOF9BZ+HbKA00tOyauvrntL7g8gEyvl7dJPYA4AAHkt5X3jPwl01SPrb3xJ03SxRjLi65QuI/8VriT9gXhDHyY8+brWy0x8IeIAt7G9LaQzlsTR6CMyAD6sLS2cgXOVNKI/wBr7qkOcfpOjjA+0l+Ev9mPYl/tF+Vht5fZo9dNlew9JkYGFhPyPVuFujT/ADp8vmpLlHbbfrWEPlIaHydIbk+uCSF5B0HstecatijkOgqOmMn7yevY0t+vAIH5Vve/+yU5j9E8N7rxAk1JZDc7RSyVslnop3md7I2lzw2QYaSADgZ3x81n+Ns2Ue0dFqzS9ypPfqDUVtng6eoyMqmEAepOdvtXRnnO9qLauAWq4tAcJbHadW3aKPruFXPUk0tOSdmNMZy53qc47+gJ8hdKy8VeIF+oNE6Sr9SXq63WZtPR2+lqppHzPPkGh2O2ST2ABJwAu9fBf2OnFHVltlu3GjUsOlpqhmYaSnlbUztJGcvIy3PkRnbfutTnqa7Hcm3tTLFx51d/sdcWrHbNI3urGbZVwVJ90qnjvGTJu15G43xt9ZHdzVugdHa6oJbbqzTlDc4JW9J8eFrnAfJ3cflXiDzXezY4u8sNmPEOy3aLVelaV4dUV1Ex0dTbjn4XysO/TkfTbkDbOO65o5Gfak27hxpT/Y05krhd7hRUOBab3HEamaOLsIZQD1OA2w7vjvk7meK9LOHHLpwh4U1stx0Zo+jpaqV3V4zo2uew5z8JwMH591yYthcION/DHjtpv9lfC7VVNereHdEpiyHwv/uXtO7Tst+qgiIgIiICIiDjLmL4zWvgFwkvfEu6MD226PpiYQSHSuz0jA79jt8l873GnireuNHEe78Q76CKi5SlzWk5LWZJAJ8zuc/WvZD2vl4bb+Uee3NeGy3C/ULGjOCWtc4uwPPuAfrXhvG3rka3OMnCTqMyjPRG4jJz5ZVA9wlLnjIJ9VcIwwGJrfQnJ/OrHRkOOM4GCVrnlYlkvVKqne9/XHvkZxndalonWF/4fastWs9M10tJdLRUsqqeVji0hzTnGfQjY/IrDe7oZnqGw7ZCitduqr1daS00TOuor6iOniHq97g0D8pCl73Gvxu3k47Nc2nPrr/mv0np/R+obVT2yis0/vkwgOPeajoLQ5wG2ACSB2GdgF6jezA4WaM4ecrtku+n6mirrvqd77jeayEhzvFJwyEkeTGhuB88rpHQexm4z3bSNJfaTiBp6luNRTtmdbqxkrXAloIb1NBDTv2PbscLr7a9fc0Xs/eLVXov7tVdluNskjkq7UZzNQVsLviDgOzmOGSHADJ79sKf2v19DSLqbyd+0H4Zc0kcGlauN2ntcRwdc1rmIMdRgbvgf++Hng7jb1AXbJFEREBEW09fcUtB8MaFlw1xqSktcUgJZ4z8OcB3IHf7fr9FNG7EWyuH3GLhxxRifLofVNJc/D3cyN/xY9ceY+pb1VBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQFxHxp5XeEHHilMWuNNU8lQdjUxxtEjt87nG5+Z3XLiIOhWoPY78ul4eZKC93y2OPbwCCPyE4/MtnXn2J/CuSz1LLDxb1JFcywmmkqaeF0PXg4DwBnBOMkHPovSZUJwng+fHTdRxt9m/zJw3u/aTa64WsT0wZUNcKS6Ucgw4xSAYwQAcjJBH1Falzf8AP7rrmrltlvl0zQabstocZIqWJ5nklkOCXPe4DbYbAeQ+a70e2E4qcG/9hmDhdXVtvuOv5LjTVlup4umSe3RNcfFkeRkxh7QW9JwXZBxgAri/2UXJ7w64lcP9Q8ZeKOmYrwKm4PtFminOWRsjY0yygDfqLnhoPb4T8wry+pn11l1BrrTXGLg3adL11PRW69U3QGva0Ah4wMgd8H9BK7Z8AfZF8J9c8J6PVmtOI95qrveqYy077Y6NsFITkDIIJeR3IJC6H85eiLdw05l9c6JsnwUVouJipg0AdLC0OAwO3dbe0Lxq4/2iSm0poDiVq2D3yQQQ0FHcZi2RzjgNDM4P5Oyfl2SEaNxg4dVnCXiXqHh1XVTaqSxV0lK2oaMNmYD8Lx8iMfnW1Karlpj8PbuQvcrT3s7+GvETg1azxZtr5tdXG3MmrLg7BdDUPYDgjGSRkZ3BBz9vQniV7Jzmc01qCop9F2Sk1DazKfd5o6trH9Gdsg/LzOPqUlV0uM0D3FzuoZJPrjKymyRtiModsDn/ANi5f4yclfMVwGsTNTcRtCSUlrLg11VBK2eOM/8AbFvZcGgkLV36kyzF0rw95cBhe7fspdQ6MuHKVYrHp24Ub7nbqqrN1pY3gSxzOlPxvZ3wR04JGDheS/AXkt49cx0ctXw703CaKEZdV104gh+WHEHvg49fLKg1jw+5kuSLibRS3mmu2kb3TPZUUVfRzu91rWtP7yVhDZW5BBafnkealvyn6j6OVh3S6W2yUE91vFwpqGipWGSeoqZWxxRMHcuc4gAfMldStB+0/wCVa58LbRq3W3EeC032SijdcLP7rLLUsqQ0CQNbG0gguzjcfk3Xm/zn81HELnU4pO0/wlodT12jLexkNus9HTSvfUOzvPLFGCSSewcNsd9hifcVFzna/pecnnANm4V1MVbQveyzWuqGTFUljcOlae5aejIPn9W62xrvgXzIch+srRxBEElJUx70t0hgLomOODh2dhuB3PfbuucvZ+chHHqm46aa4qcSND1+ldP6bqPfh902+FNUvDSGtbGfiwcnJIHl5ZXsRfNO2HU1vfatS2SgutG/6dPW07J4j5ZLXAjsT5eabL/iSY8c+FHtj+Plo1BQ0/FSzacv9kfIyOqkp6J1LUxsJAc8Fji1xA3wW+vfsvXjh/xE0hxN03Rap0be6W4UddAyceFK1zo+oZ6XgHLSO2D9my6Ee015Z+WHh1y83PiBp7hrYdNam9+ggoZrXAKZ08j3YILGYBAG5ONs7915ccG+OvErgZrG16w0LqSupZLbUMldSe8O93qGAgujezOCCBg7eac+K+mVYtfcrdaqc1dzr6ekgbu6WolbGwfWXEBdGaj2wfLM3hs3UlOy+v1W6kBGnzbn5bU9O7TP/YywH98HZI8ge3nK7S/Odz5ahvmttMaf1DqqhNW5ssTLhHBQ0ucubEPGkYwkDb1OPLsGD3Odx14NNuAtTOKGmZavOPBhuUUrs+h6ScLe1LVU1bAyppJ2TRSDqa9hyCPkV4VcNPZZc6F11NSSXXRlNo6CmnY59wrLzSOdGM7uY2CSRziBnyH1r2t4WaJqeHmhbTpKtu0lzqKCAMlqXZ+N+BnGd8Z7Z/8AYlsvidbuRERRERAREQEREBERBQdlsDifwR4ecW3W6fWNgpKuqtcokpqh8LXSMGc9Icd8Z8u3f1XICpkKWaS4ho6SnoKSChpYwyCnjbFG0dmtaAAPsACnRFQREQEREGk6pvbtOaduF9ZSPqnUUDphCwbvI7ALxX5hPal80N/1Pe9L6RulNou2UlVLSsZS0bfewGuIyZH5IyBsMeec+nt29jZGlj2hzXAggjIIPcYXDms+Tnlf4g3l+odYcEdMXG4ykF9Q+mLHOPz6CAftCYPDHg5wU45c6vEyWkt1fUXOvkw+4Xi4vcYoG99zjvuT0jHrtnfvNoz2ItLFNFNr/jm+djSDJBabT0AjzaHySE4PrgFekmgeGHDrhbam2Ph1oy0aeoWjHg0FK2IEeWSBk/aVunICXdWZHGfATl94dcumiYdE8PbWIIGfFNUPaDNUO9Xu7nzO5O5K5NRFMxBFwLx153OXjl0uzbDxJ1fLFcywPNFQ0zqmZoPbLW9v5vPC4fZ7X3k6fL4brlq5je3iOsZ6fzPJ/MrOju0uLOZribpPhRwQ1hqfVt1gooPuPVwQNe8B88z4nMYxg7uJLhsFxLbPaf8AJhcqM1f+yo6lw3q8KpttQyQ/IDpO/wBq8uPaDc345reKcFPoiauboixRCmtcErSx1VMSS+d0ee5Jw0EZAHnlLCMr2VMtgpuc3S1RfrjT0hZQXFtF48gYJap9O5jIwTjLiHPIHmQvVbnZ5wNHcrfDWsnlqIa/V13gkp7PamSjrMjm4E0g3LY25zkjfGBleO1h5BOcO66aoNd2Dg1en01QGVFL0TRRVXSd2SCNzw8eRBxnsVv3hj7Ofm24t63om8UNIX+y2/xGMq7jfJi6UQgjLWFziexOPTdLeQn9uufCvjTxB4NcS6Li3oO8e6aiop5ZhK9nXHMJARIyRmQHMcCchbv5lebni7zU3a3XLibV29kNpYWUVFb6YwwRdXd2CSS4+pPbZe3mi+RTlf0npOi0vUcINOXc08TWTVNdRtlkmfjdxJ9TkreOn+V/l00tj7gcEtGUnScjptELsH/xmlW3+j3rxO5QfaA8UOU8zWGktlLqfSdY8PmtVXI6N8LgfpwyjPScE/CQQcjthdwOJvDjg/7UDRg4+aX1zDofUem6B9FVW+tdHKWBgLw2bdpxknDhgYI8iF2P5w+QbhXzAcPa39iGkLPp7W1BGZbVcKClZTCR4H9hlDAA5ju24yDjcdx5HXTkl5yNHVtZamcHNXNbJmGWSgY50NQwZ82n4mnfAI+xTh64jZqjVuiKyv07Y9V1TKalq5Iz7tMfBlLHFvW0dsHGQR3BC5/4Ge0O428Ea6E0NRFdbcMNnpKlzi2Rvn6798fpHdYekPZuc4usAXwcI6u1sAyHXSeOmDh8sk5XCvFfhJr7gnrGp0HxHsMlqvFK0PdE8ghzD2e0jYg74KX8f5TKT+3oxrD22Vxfp6OHQ3CGnjvUkZEk1wqSYInEbFrWnLiDvgnB9V0h40c0nMBzMXxty1pqGrrDACYqG2QuighbnyYzJxv5laPw14eWy23Gxa24uW2op9FVM7XOduPeWZzgY3wR6YJ28u/vby96H5aK3hlaL/wR0ZpZ2nbhA10U9PQwve44HU2RxBPWD3BOM7jYhJv4ziceE3BDlv4y8y+tabSlhoq4vPwyVtx8TwoGDucu8h3IBGwK780XsQ7P+xh33R48Vn7IvBJaYbOwUQlwcAgv8QtzjfIOPJemNo0rpnT75JLDp62W50v0zS0kcRd9fSAtWRXy9cRdEXXhrru/6Avjo3V+n7hPbqh0Zyxz43FpI+Rxn7V7L+yF0Doyzctrda26npZdQXm41La+oAaZWMY7DGerRgH68D0Xj3xdrLnqLjJrCrrJHT11dqOu6iTkueah4G/5FrVp1xx95fK33bTGsdUaR8b42uoauWCOUHfIwQD8x+VD9Pbf2kuqLLp3k+17R3SaJs94o20NHG5w6nSmRpyB3OADnHbIz3Xz9Rg9Y+tb54gcdOL/ABVY2PiPxGvmoWsIIFfVGQZHbv8AzrY8Ujonh4wSPIp+Oz0ahVB0rersAMfLb/3Khgd4B6wNxv8AYr/efHhOMNcD2+v0VjIXPYdu6352eOf4y/8Auknnjqo8Sdw34T6beuFhQzS08zJ4JHRyxOD2PacFrgcggjzHdSTtdCQzYE+nktxcKtOU2r+JOmNMVzC6mud1pqecAZJidIA8f+blT8sxr8Y7O6Q9q3za6N0vQaUortpuup7dC2CGor7UZZ+hoAaC4PAOAPMdtlzpy7e2I11V63t2n+YKzWL7hV8ogmu9tgdTvpCTgPcwuILd9zkY77DJHojDymctTdLU+jqnghoyst1PE2JvvVngllPSMBxkLevPnkEYPbC88+eX2WdxorpFrvlW0fJUUFRn7oacp5S50L/J0AeSek9unOB8sb5+NZr1TsGpdP6qtcF701e6K50FSwSRVFLO2SN7SMgggkdvJamHtPYhfO/aOTnndo3SUdl4LcQ6MRElzYoZIGA+eD1AE/UStvaO42cwXLfxSguFTfdTW692CrArbTdKqYCQAjqjkjeSMEDY4OxBGR3SXNOePpDRcJcr/Njwv5pdHRX/AETdGxXamiabrZ5fhqKOQ4ByD9JuezhkdvUZ5tQEREBERAREQEW1eJXEzRPCLR9drziFfoLRZbe3qlqJifpeTGgbucSMABePXPR7S+8cc3N0HwQqLxp3SVNI41FeZDT1VyPYbNOY2AeROc+nnKPSjnG5vNJcrPDGs1IKy3XPVNQRT2izOqB1zSn9+9oPUGNG5P5M4IXl/cfbB83dU6UUcmjqJjiejosxe5g8hlzyCR8wuK+CHJNzL8yN2t91otG3uKxV8jRNqG7B0cIiyMuY6UgyHByMZB9V63cLfZo8pnDzTlNbbxwwt+qrm2NvvNfeHPqDJJj4i1hPS0ZzgALX+jx7uGrOY3nr4sWnT97vlXqW+VsvhU0ZYI6akYTu7pYA1jR64+Q7ruVpj2Imop6eObWHHigpZXNBfBQWV8vQSNx1vlaD9YGNvNekvDjgDwX4RSST8M+Gen9OSzAh8tBSNY8g9/i7jOT2PmuQcYUHn9wp9jtwU0Nd6e96x1reNWSwOD208tNHTw5B9AST5d8ruto3hZoDQEMcWk9LUFA6JgY2VkQ8THb6WM7+eMLdioThSyeiqjlijmifDLG17Hgtc1zQQ4HYgg7EHspEVHRbjt7Jrgvxg1fXazsWqLnpGruEhlngo6dksLnnucOIx9m/qSuJ6P2IWmRVF1fx3ujqbyEVqja/GfMlxHb5L1BRDMcFcr/J/wAK+VSxVNu0NTSVlxryDV3WraPeJgOzc74HyBwudURAREQEREBERAREQUHZUc5rWlznAAdyfRbe19r7SPDDSdw1xrq+09pstsjMtTVTuw1o8gB3JPkBuvFjnv8AaJai5hr83SPCa5XfT+hLeSA5srqeoub8/wBkkDTlrMYw0nPr55Dv9z98/Fv5YdMUdh4b1Vlveurw94jhkl8eO3QtG80rGkZJJADSQe5IxuvJOsvfMFz18a7fb7tc59S6ourvAp2loipqOHOSQ1o6Y4xkZOPQZOy5c5IuQDiBzF60tur+JmmrtbOG7Saqrr6lzoJbngnEcBPxkE93gYAOxOdvZPhRy58EeB0bm8LOG1msEsjAySop4eqokAGPildl5JHffdXidefGivYjQCnpajiBxwkM5IdUU1rtQDAO5YJJHkk+WcD5L0h4UcL9J8G9A2jhzomgbS2mzwCGIdIDpD5vefNxO5J+S3eiiiIiAiLzY9p5z3X/AIX17+BHCa5Po73NTiS7XKF2H0rHjaNhH74j8hznsAUmj0kD2OcWhwJacEA5x9a87faK+0Yh4UNu/AThHBTV2pq+hdTXa8OkD4rYyVpa6ONo2dMWk7k4bkHBOF5t2Lh1zg6ssn+yDp+ycSa+21YMgucM1SWytPmHdQJBHYjYjtstX5a4+FejeN1LfOba010tqpn+NLS10b5Hzyg46pWH4ngbbHv+cP8ADz12j9jNwXvlTxQ1DxovOnquO1WyzPt9rrZoi2OWpne3r8MnAdiNrgSMj4iM916zaq1bprQ1hrdU6vvlHaLVb4nz1NXVSiONjWgk7nucA4A3PYAldA+Yf2p/ATS/CKv0zy3XaWq1JPS+6Wz3a1yUlLbg4Y6x1NaMtB2a0d+x2Xl9R2fmO5ib5/tWh1vrmuuEpf1FtRUsc8nOcnLGjPbcAeWE7anza7Y83ftV9XcY7ZfuF/CzS1FZNHXDrpJK+tBmrq6DP0unZkQJAIGHHHc57deNG8jvMlrvhzPxP0/w+qpLPFEZmNkyyaZgGSWMI32wdyPsXopyJ+zH03oK103EnmG01DdNVPeJaKz1JElPQAbgvZ2fJkA75A9PX0Rpqamo4GUlJBHBBE0NjjjYGtaB2AA2A+QSK+bLgrzB8aeWHVtReeGuoamyV2fBr6Cph64J8H6E0LxvggehBA3XqDybe1XtXFy/DQXHyCx6UuksY9xusMjoqSqkzgseHkiNxztvj8+O3utOVflx4i1sty1pwV0ldayb6dRNbYxKfn1tAOfmDleWXPl7NfVfDDUk+vuX7Rlfd9E1MZmqbfRdVRPbHjJdhm73R9sYBIyBvvhmxJXszb7lbbvSMr7VcKatppBlk1PK2Rjh8nNJB+xZJGV8/nJBzl6m5SuJnTqU3av0XXB1HeLOJCXU5BPTNFG84a9ru42JGR3wF6/8D+e/lq5g78zS3D7W0gvUreqKguNK+kmlx3DA/Zx3xsTvt3IyXrsKiLj3mCv2pNLcDtd6j0e8svVssFbVUTx3bKyJzgR6EYJB9Qg4h57Oci18qHDdtXZHW+4a1u8ogtdtnf1Bgxl88jQchrRjGcAkgZXmj+6/c4eHf7e0hudv+oY2+r4/05XE3LNwS17ztcwNHpW96juVS2YPuWob1UvdPJS0THDrILsjqc54YxuwDnjbAK9mrBwB5L+UDRYvNw0tozTlBT9Ect91EIZqqZ+TgGecF5cSThrMfIIY8T+P/N/x25lm0sHFTU8VTRUZ64KKkpm09O13910t7nbvlcO0BiE48UgDB7nG69TfaOcz3JjxW4Iy6S4c3m0ai1cKuGW31FttkkXurWu+MGV0bdiNukEg79vPyskjkid0SMcxw8iMEfYVczxPeVnTAxODukjO2Qc5UbXOc4Z2zt2UIqpmt6CQ4DyKlgmY89Bw3JyMnYJIn5SSciSV3RAS7O4IHb/381vflytk954+8PLZT03jyVGpbe0R4z1Dx2k7fUCthV4DZGta7Ixn5ZXL/JhqOz6S5quGGor8/ooKPUNO6Z+M9OSWh2PkSD9ifk1v172cyfMBpXll4TXPixq6jqq6loZIqaCipiBLVVEpwyME7NzuSTnABK8er/Q8XPaocebjqXSOmLdYZLXQxQPa+YvigpgXdBe8gZOc5+ZwPILvN7ZDUtrt3KrbrFNO33u96noxSs7lzYo5ZHuHyA6R/wCMFsj2JWnIKfhdxC1W+nAqKy+QUTJcbmKOAOIB9Op5/Is3ZODodx35ZuPnIvrGwajuF0FJPM/xbVfLTIehszdywkjZ2MnBzkA7YXobyA+0kj4vQ1HDvj/fbTbNS0bWm33R5FPFcI9gWvycCQbEnYHue+B3d4ocJ+HvGbSdRoriVpejvlpqRkw1DMmN/k9ju7XA4IIPcBeOXNF7LjjRoHX1SeBmjLlq7SdUQ+jFO9r56cHux/UQSAdsk5+sbrWTE2+PWnihzK8G+FGmKrU2odcWiWOniMjIaetjkfJjyHSSB9ZXnZxK9tTqYVNTQ8LOGVqMTXlsVbc5JCSPIhjSM/bhcDaY9llzo6yoI6u56VorRE1vwQ3a7xtlacbAMBdj8oC6/ScKLloTjlRcJuLNDLbpaS+Utuu8THbtjfK0OLXDGxa7II7D5jCzJftXeOyNw9r1zh1jn+7XPSdE05AENkacA9t3uduFwDxH42cfeZe/+Pq6/wB21HVkfDS00Z8Nje30GDAHlvsvde2cjfKHQWimtcPL9oqpghY1rZqi2RyzSYH0nSuBc4nuTnf6lv8A0fwQ4PcP6cU2iuGWm7PG3sKW2xNI+o4z+dXk8Nr51tMcQON/L/fo6jTl9v8ApK4xkPY0gsBx5hrgQR5EgfJd8OU/2uOuf2YW3RvMhJbaux1rxT/d6GAQS0bjsHTAHpczPd2ARt5ZK9P9X8HOE+vqGS3a14baavVPK0tc2ttkMpIPcAluR9hXlTz/APs0btoW7S8UOXDSFTW6XmjdLcbHRh0stueMkuiZu50RHkMkYx9cHr1Z71aNQ22C8WK501woKpgkhqaaUSRyNO4IcCQVnLwO5Eudy/8AKbryaz62dd7hoS4h0FxtbHEyUM4O00THkAOaQQ5uxIJGx7+s/BPn55Z+PuootJaC1rM28zgmGiuVI+kklIxkMDtnHfyJVHYpERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERARF5u+0p9oDeOFNxl4H8HLkINQOiDrvc4yCaRrhtE3/tyM59PPbAId9uIXFHh/wrsVVqPX+q7bZqKliMrnVVQxjngeTGkguPlt+jdeV/Hn2yPEW4XC6ae4HaXtFqtoc+GmvNax09S5uSC9jMhg27Eg774XS7TXDPmV5mbjPddOaW1hrudryZavpknjY4nceI89Dd+4BGF3i5TfZG3m4V8WquZyB1HQxYfFYqapHiSnOwlew5A23AI9N85Etwn7dReW3l44pc7HGaWkmudZNHUzmu1HqGqzIYWE5ccnYvOQGt7DI2xgH3j4c8P9B8t/B+i0bpyH3LTuk7fJI+QgF8gaDJLK8/vnuOSSTnJAWqcN+FHDrhDYWaZ4a6Ptun7azBMNHCGdZ9XHu4/MkrrL7TbmTo+CPA6r0nQv6r9rSGShpWjvHERhzz8u/wCQjzCvvB4x8wvEp3GHjdrTiW5jWMv94nqomtOQIurpZj/xQF6c+yn5PrVZtFt46cRtLMdd7jL1WZlZCC6KEf8AyQA7jOAR8yfTC6K+z84E23j5zKaf03qGlM9itnVdbjGfoysi+IRk+jiPzFfQZQ0FHa6KG3W6mjpqWmjEUMUbQ1rGDYADyACsuJYyURFFbZ4h6C05xQ0ZddCasoY6q2XendTzMewHGRs4Z8wf9Xmvng5q+Xu/8tXGK8cPLtBI6iZIZ7XVFp6KilcctIJG5AIBGSRtnuvpE2wuEOavla0FzP8ADuu05qK2xMvcMDzaLm1o8WlnwS3B/uSe4P6Mgk8de/ZJcXtF6l4Dt4dxXGki1RZ6l5qaUvAmmhx8DwO7gACT9Z9Cu0XMbwG0lzE8Lbxw81TbaeZ9RA91uqZGAvpKoD4JGO7t3wDjy+YC+f2ut/GLlI4xsMsdw03qnTdZ1wyEOY2ZjXdx2643ADI/QRt7tcmfM7aeajg/Ta2hZHTXmgkFDeaRh/sVSGg5A8g4bj7fLCcs2G5Xz5a90NqPhtrC66H1ZbpqK6WeqkpZ4pWFpy1xHUAfI4yD6L2q9lNwP0NoDl7o+IdsqqO6aj1c81Fwq4yHGlYNmUwO5bgbnfckHAU3tNeU+y8Z+DVx4h6V0xC/XWl2irinp4gJqymH9lifjd5DdwSdsb5AGPH/AIQ8x3HDl8raocM9dXWxNmcW1VCHkwPcCQeqJ2wcD5gA7DPZXVfStnA3K6H89ftJaTl4r4+HvCSC1X7VksRkqaiZ/i09AD2yGnDn7dicdxsQvOrX3MBztcYtKRakuep9U1Fiewhxtb5Gt74JIYeoA+gOMd1wZw8rtMjiZY67ilFVV1kFyiN4a57jK+Lqw8uJOTjufPAI+ShK57tOs+Zr2i/F+xcPtXamq7lTTVHiSspaZsVJbYsEul6WjDRgHuT642JXdiP2JPCZ1LEJ+Muq21HSPEMdJTlvVjyBGcZXajTfEvk+4CcJYtY6Ku2itN6S91bNF9yzCyWcEZDS1p63vJ2+LfOMldYL77ZjhdHPUN0/pCrlijcRG6oa/MgHY4AGMq58S864M5l/ZB6q4X6Nq9c8Hdb1WsobZGZay1VVCIqwxju6IsJD8AbjAO+ewJXDHKLz88TOTeiu2i49F0V/stbVComttfJJST004GHFrw0luQN2uad913g0P7aHg1cas0uvND320RdWBVUjBO3H/eZz+cLXdS+0B9mjq+ilfqizUl1fOCZGVGiXvmcT3+Mx9z69X2qeK4Sr/bg6ldGfuXy/WyOTb+2L9I8D12bCCVlcNfbR6ivOurZbOInDCyWqwVc7IaipoaqV74A4gdZ69sDO5/N3K2Jq3j97Me6X6pmtPAeWGleD0PZbfDBcR36QSRv5bro/xVuWgbvre4V/DSzz2ywSuzT08x3b3zgZOB2wE8H002a8W2/2qkvdmrY6uiromz080TgWvY7cEFZ68NuQ72h2pOXy9UuguJlxq7toCqeI2lzi+S2E4+NmdyztkeQ+WMe1+kdXab13p2h1ZpK709ztNyibPTVUDw5j2EZ7+R9Qg1pERAREQEREBERAREQEREBERAREQEREBEVCQBk+SDRtX6u09oXTdfqzVVzht9rtsLp6iolcA1rQM4HqT2AXkzzC+2K4kXG9XDT3ASyWq0WiJ74YrvWQmepnbuC5jSQxg8wcE/LsVH7UnnksHE9p5feFVykqbTa65z79cYnER1U8eWiBhH0mNJJJ7EjG++NP9mRyGaY41U1w4wcb9Mz1umKaRtPZbfM58UVfL3fK/pILowCABkZJzuMFIOsPCbgzx253eKtW+iq6i63OtkM10vVwJMMGfUjbYHIaMYHpkZ7m03sQNQfc4Oq+Ptu9/Ld2R2R/gg77BxlyfLfA+penuhOGXD3hhaWWPh7o206foGNDRDQUrYgQOwJAyftK3Oc42S9Hzc8zfLZqblg1+7QWp73QXSUxmWOekBaHN23LCSW5BBG57rul7Knkp0vxBpBzF8SKVlwpLfXOp7HbntzG6ePHVNID3wSMDt5rhLnE5euY3WXMPqO91umbneBcKwiCcDLImA7NOTsBuds916jezw4Sam4Mct9r0pqpjoqyWqlrDE4Y6S/BOPlkY+xSWWe9K7NABoAAAA7K5EVBERAREQF5He2e4Y6mquIGlOJNs0/NNao7QaKtq4Yi4MkEhLS8gbADbJ7beq9cVpGqNK2DWdkqtO6ltkFdQVbDHJFK0EYIxkZ7EZ2KlHz0f7LFNxS4TWXg7faiC2T2PHulU8AMkDQQAfnjuNvPC3/yu8+/EDk8pLloWy2m3arsUtU6YwTzuYxkh7ujLQSM753wT3zgLkjmd9lfxo07rq8Xzg7Y479pipkfVQRRyATQgnPR0bk/mx64XQ+/WG86Yu9VYNQ26eguFFIY6innZ0vjeO4IK1+NkT+L1NovbcWs0hdcOC9Q2p8hDWAsP2kgrj7iX7Z/ijqPT9fYtA8NbPp+ethdCy5zVMk80AcMFzGbNDsE4JJwcHGy84vkvR/2aXIrwm5gtAXjiZxYoa24Qw3B1BQ00c5ij+EDrccdyCR3HmO2N0/GG46ncouj7bxQ5ktK23V9a33GW5CsrppzkPIdnJJ7kvIP2Fe9WuuWrghxJ0ydK6r4fWqpoTEImFsIbIwYwCHjfPzOV5W+0X5TdNcoselddcGqmrtdHeK6SkLmyHxYpWsc8Dq7kYHfAz6bLvB7MDjrrDjhy6mq1zczcbrpy5vtLqpxBkkiDGuZ1/MB3/uMLM29XI4Z1L7FThpcL9NW6e4p3q2WyV3UykdTskMeT2DjkkfX+ZcP8wPsh73w30LcNY8OtZVGoZLdGZpKSWJrXuYBkkAAfmJ+pew+D6qOWKOaN8MrGvjeC1zHNBDgdiCD3BWtTHyw1EM1NPJTzxujmieWPY4YLXA4II9RghZdPWwtYGyEg+Z9V6xc/Hs0oNTyVvFjgZbmQXF+Zrja2bNkOCS5oH6dyAN8jceWOqeHettFOe3VGnKu3iOQxOMrQB1A9shNniWT8uVoFTI2SZzmZ6SfNel3smeUnS+vYqvj3rSmFUy1VxpbVTnPSZGD4nH6jsfkQPMrobwN4Law4/cRrbw10TAx9xuLiS9/0Ioxjqe75AHJ3C+gTlR5fqLlo4N2vhhTXEV01M99TVVAbgSTPx1Y2GRt6d8pZ9X9OY0RFFF5L+2J5etT1mtLTx001p19TbnW1tFeKiniJLHxk9D348g3bPp8gcetC2zxJtFnv2gNRWm/UkVRQT2ypE7JGhw6RG45wfMYyPmEHzi8BOPOvuXPiHRcRuHtwbFW04LJ6eXJgqoT9KOQZ3B3we4+rIPohpr22LJ6djdVcJIqaoDPjdS1Lnxl3yycgfWF0P5XuFWmuM3M1pThleJSyyXe7SRS4JBdC0PcGjfuQ0DGd+2d16R8dfY76A1U2Ku4L6jbpqpjZ0vpqxhfDIQO+W+ZO/YfWm58M1uHhH7XPhBri9NtWs7cdONkcAyWRxAGTjckkE/LIXaCn5seXKqphVR8XdPBjh1YNSMgY9AvILXvsnObDSEjn2Ox23U1OM4fQVbQ84/7RxyM/WuP4/Z2c50ryxvAe9twcdTpacA/P+yJydqfrXufp3mQ4EapnFLZOK2m55iSAx1cyMk/LqIyuQqWtpK+FtTQ1UNRC/6MkTw9p+ojYr55Lr7P3nIs8Tqip4B6jkjYC4up2xzEADfZjyfzLZ9FxB5j+B1c6gj1FrPSs8R6DTVTpomjHkGSDA7eQ8kV9Ka2nxQ4k6U4R6Fu/ELWlziorVZqZ9RK+R2DIQCRG31c4jAAB7+mV48csntSeJfC6SqoOK9bU6moHt6ovFBLwSMjGOx7b7DGPt4g5i+ZfjPzucUILNbIbjUUFRO2Cy6cpD8GfJxaDguJ8ydvXO6ufU1NzYc6vFrnF1JTWCaA2/TUNb02ew0YOZHuPSx0p7ySEEbdgTtnAK7acofskK1lXZuJPMRdImRxmOsg0zTN6i4ghzRUSHbHYloHcEHIK5O5FPZj23gzXW/izxtNNddXxNEtDamYfTW15GznEj45AD9Q/KD6FKaIKWlp6GmioqSCOCCBjYooo2hrGMAwGgDYAAAADsp0RFEREBERAREQEREBERAREQEREBERAXHPMHxZi4GcHdT8VJbW64/sfoX1LKYHAkcNgCR2GTv/ADd1yMtv680XY+Imj7tonUdM2e3XimfSzsIzlp8/sOD9iFeAHMnzt8eObKamsGrq6GlsjKkPo7DaoS2J8pOGde5dK8HYZ8zsM4XcDk49kxV1L7JxR5iK5kdPmOtp9LwNy6QbOZ7zITgAgglgGdiCSCux3AP2XfA7gvxBHESuM2oKuilMtrpqwl8FK7Ozi0/SI8s5+vGQe6KaIKWmp6Gmio6SCOCCCNsUUUbQ1kbAAA0AbAAAAAeinREBERARFsXjBxp4c8CNG1Gu+JmoYrVa6choLvilmeezI2d3OPoPl6hBsXnI5i2csPBW4cRYqJtZXmZlFQwvHwmZ+cE/IED5eq+fXiJrzUXFjW984harr2zXS8VDquoc9/ck4DW574GB9mV3E5+/aG6b5o9M0/DfQmka2hsdFWCqNdXua2Wd7TsQxpPSMDsdxk99lwZwV5IuYzjpJTT6R0DWQWqoLc3OtHgwNaT3ydztkgbA47puTpnXNXDT2rvGbh1w3tnDK1aGsFZDbqQUVPUOLxJ07gfCAQTuutV4dxV5luLbQLJNV6hvtU2GKmigcyOEOccA7fC0ZJJPofkF7Xcrns/uC/AXS9DJetKWzUOrHMa6suNbCJ+l+N2sDgRgZPlj0279iKDh5oK1XJl5tei7HR18Ywypp6CKORv1Oa0EKTzkN2686eDvsXNLW2G33njJxIqrlXNLJai1WymDKYHYmMyuJc7B2JAAOD3BXoxovQ2lOHen6XTOjrHS2y30kbY2RQRhuQBgFxAGT8ytwItaeiIigIiIOlPNp7MXhjzGXqfXOlb1+wvVM0eJZIKQSUlU/wAnSxggg+pacleT3GrgBxz5MuJlI3UEdRba2inFVZ79by73efB2fG/GM47sO/cHO6+jdcJcxvG3lg4dW6LT3MHftO+HXtJittwp21Mj2nYuEeCWjtvt5IPOvhT7aHiLY7dSWnirw6t1/dAwMkuVDK6CeXHYujJLM47kHf0V/Mh7XR/Evhpd+H/DfQtRaJr7Svo6muqpQ8xxPGHhgHmQT3BGDjPrwXzx1/KBdbrDcuWyhpaYzuBkZRAsiHcuPRkgfbuutfD3SVTrvXFj0dS9Qku9dFSgt3IDiMkfMDJRJ/bkvlQ5q9ccpPEGp1zo22W66x3Kidb7hb64OEdREXBzcPaQ5jmuaCCMjuCCCtwcxPM7xv56+I9kt9faGNZC4Uli05aQ90MUkmA+Q9RJfI4gZecAAAAAA59btD+zY5ULHpG2WnUPCq3Xa4RQR+81VT1eI+XA6t2kHGdtycrlvhvyv8v3CKvF04ccJtPWOuA2qqemBlB23D3EkHYbhF/bo9ys+yDtOl6i1674/wCpG3O4xNbO3T1CzFPA8gECWV28hB9ABv6gEdjOLvs3eWTi1FG6q0xNZKuJoayotsnQQB6g7n8uPku06Jhjx15oPZI3jhrpur1nwc1LW6ipKQGSa31UQE7W4zsW9x5ZyfnhedVwt1faayS33OinpamFxbJDMwse0jyIOCF9Tj2MkaWPaHNcCCCMgg99lxvqflt4D6yr/upqXhVp+uqg7q8V9KGnPqenGUHzRfavUrkN9m9oPiJwq0/x11veq51wuhdV2yniAEUQY8tDiM7nLT88gnbYK/2tnKxw50HoLTvFzh1peksckNeLZcY6VvTHJG9pMZxnY9QO/wDrUnJD7RLhVwo5VH8Ode3WSi1PpCKrjtMJiyysY9zpIQ09shzyCDvt27ZW/B1Z5tOK2reYDi5RcIbZc5bhT6duEtqt4fISx0rSWHG57YI+zC9duRTgBLy6cALTo64Ydc62V9yrngYJkkA2P1AbDyBwvJ72dPArVfHvmXt2t6umk+4en62S8XetcD0PkJJEYJzlznO+eMjPde83SA0NA7K+TGZMv6XIi8s/aGe0g1ZpPVV24G8Fat1tqrc4090vUZ++NeRuyL0ODnPbBHfOBGncrm45yOHHKhox91vk0d11LVgstVihlAlmeRkPk7lkY2JJGTsB3yPD6o1NxI5ruZOPVFRQOq9Q6nvMM74aOM9FPEHt2A8msaBue5+ZAWRwO4HcZucvim2w2m4VdzrJD41zvNznfJHRwl2S97nEk9zhg7nYBe3fK5yYcIuV3T0FNpm1R3HUT42iuvlVGDUTP8+nb4G57AfPtnCWDnKyUs1DZaCiqDmWnpYopDnPxNaAd/rBWciICIiDo3zcey64f8wmoajiBoS/R6L1NUtHvLW0gfR1bx2e9rSC047lvfz7DHlpxz5aeO3Jzrahq9RxSUUkEwmtd9tj3GF7h2LX4HSfkfmN919F62xxB4b6J4paaqtIa/03RXu1VjHMkp6mMOGCMZae7T8xhE/15ncmntYtQ3bUts4a8x77e6lrC2mptSsaYnskOA33gbgg+bhg5P1A+p1BcKC60kdfbK2CrpZmh0c0Egex4PYggkELxn5tvZT8S9A6grdT8v8AZqjU2k5QZxQNlaayi8y0BxBkaBnfuAPPsONOT/nd4kcpGuJNOa5F3uOl/EFPcLNVPd4tIRt1MDt2kA7Dt2wCCQSve1Fw/wACOazgjzGW1tXw11jT1NWG5mt0/wB6qoTgEgsO/n5b48guYEBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERARFtbiZxB09ws0Jetf6orIqa3WSjlqpXSPDQ4taSGgnzJAA+tBwfz9c0DeWTgdWXuyV0DNWXqQW6ywuOXh7geqbp9GAE59e24XirwV4M8VecPjE+yWmea4XW6TGtu90qSS2BhPxSPP1AgD5YGwVvFHihxY5wuNrK+51VXdbvqG4iis1u6iY6WOR+I4mNGQ0AY6iB5E9gAPbLkh5PbBym8OnW90rK/Vd6DJrzXhox1AbQsP9w0/lPr3L/COVOBfCDTnAnhbYeGOmIWNpbPSsikla3BqJsfHK49ySfM74AW/wBEQbe13rnTfDbSdy1rq64MorVa4TPUTOI2A8hnuT5BeAvPTzPy80nGifVVAJItPWmEW+zwPcTiIHLn/W87n6vLOF6fe17vVfauUaWnopHxtuOoaGlmLTjMZEji0+oPSNl1w9kxy4cJeMfC3iTe+I+k6S8zS3BlkhdO3JhiNOHuLPR2XjB+W+Uzulcf+xpvNDRcx15s9QWipuNjkNPnufDJLsfYQvaxfPryk6yoOXvniszqmqMFqt+o6vTtVK87infI+AE588hhX0DRyRzRslika9jwHNc05DgdwQfQhTBIiIqCIiDzr9srwooL7wcsXFOjt7BctPXIU087GDrfTyjAaT3IBGfsXnryW83mqeUziMbzSRur9M3kxwXy2lxxJEDtKweUjQSQfMbHI2XeT2kHOfwu1hpu78uumqt1fWulMdXNGCWNmAIaMjtgnO+5yDheS5pqgVHuhhf43X4fhhp6urOMY75zthPx/wCfEt3lfT3w/wCIOjOLGj6DWuh7zS3ey3WESRSxPDxggEseBnDgCAWnt9RBXjT7Wnl+tXCXjbbdc6UtEVDZdbUr53xwR9MUdZEQJBgAAFwIdgbYws/k9ZzG8i99j4ycWtFahsfCi7U4hukUzmmOYygGCVsQcS2QEAgkAkHG4K7Mc3Ny4e+0T5dH3rgFdn3W46Gq33LwaiAwyx5jw+NzDuCWj9B3S/jfVl3xx17HXilbdRVGo+C+p6Kkqn0dH79b/Hia/rj6x1sw7PbJPbthch+0E9m/pzXlkr+L/BGyx23U1FG6ouFrpmARV7AMlzWj6LwB5bYHZeeXInxDufCfmt0ReoHOhbPXG11zHfCXQSgtew57HOPyL6Ixgjy3QzHy03SnvNsnfYrwyrp5aGRzHUk/U0wv8x0HsfsW4eEmj7VrziRYNIX68RWq33KsZDU1cjwwRMPc5O2T2+1fSNU8HeE1bdTe6zhppievd3qJbVA95PrktO+/fuuk3tHOQSp4t2G3a44B6OoItR2yQtrbZRxsgFZCRsWAYAeCRt2wBsMkoM0ex25U7xpuE2vVesBUzRBzLlR3KF8Uno5rHRuBB+s/Wtj1nsQdBOk6rfx4v7IydmzWiF5A+sPAP5AujVr1Bzr8kV8t1zqWa50RAZAY6O4NlNsqwHAmN0biYXg4xgb4zghejfAj2wHA/XNPQWbizarhoy+yhkU1QGePbnykbuDweqNuf7ofak2HrZlN7ELh23BrOOuo3+ojtUDc/aXFb1s3saOW+hiDLtqbVNyeO7zO2En7G7Lvja7nb71bqa72mriqqKsibPBNE7LJGOGQ4H0IKzFIPILna9ljZOEXDeo4pcCa+83OGzuD7paKsiaUU52M0TgAT0ncg747Z8uEuSD2gWteVuvj0fqBkl80BW1IfUUT3Ey0BJw6WA+QxuW9j3wSMH3lmhiqIn09RE2SKRpY9j25a9p2IIOxBBxg9143e0o5Ab/oDV1y438GdJun0TcGGqvFFRMybTUZPiSBg38FxwcgfCSc7HIpuPYLTWobVq3T9u1RYqptTbrrSx1lLM0gh8T2hzTt54IyPI5WqLzS9ljzvWfUen7Dyta2idTXq1Uz4bBW5Hh1cDCXCA+Ye1uceoG3nj0tSAiIgIiICIiAiIgIiICIiAiIgK1zmsaXPIAG5J8grlBWwGqo56ZrukyxuYD6EgjP50HnzzT+1VpuC+r6vQ+gtIUl6r6QkPlqZXBjTuNy09s+gJ2XT7iX7Wzmg4gacr9L22LTumaa4wvp5qm3UjzVNjcMEMke8hpIJGQ3O+2Fy7xJ9lnxY4jcabnf6i4CC03GqMsk2Wk9JPkSSBtj1XbXhH7MDlV4dUEDr/oSHVl1DQZai6yvliDxjJZGSAN/UfYFP+fyHl5yFcod85n+LFHW36imboqzTirvFXIwltT0nIgaTs4uOAe+xOe5I97rFYrTpmz0lgsNvhobfQxNgp6eFgayNg2AAH5Se5JJO5WPpfSGltFWuOyaQ09b7PQRgBtPRQNiZtsMho3OPMrVw4HsQcKi5ERBjyUNHM/xZqSF7+3U6ME/lIUwaAAAMAdlciAiIgIiICIiAiIgLqlzSezv4J8yc9bquemqLBq+WEiO5ULg1szwD0+KwjDgTjJ8sk4JzntaurXPZzlaS5YuHldaYKttTre/UM0VnomO3ic5paJ3kbtDSQR8wPkCo8NdNcKb5rDi9Bwe025tTdKy8yWenfjIe5kjml+B3GGkr6EOVPgLQctvBOycLaSo94mo+uprJx2kqZDl7vzAem22y8dPZc6GuOu+cPTd9kjmmh062qu1VUOyQJPCeGdR33cXEjPcgr3qSxHnb7aqgulVwG0XVUdFJLSUepjLVytbkQg00jWdR8gXOA+vC63+yc5q9GcE9R6o4a8S9RUtmsWpzBXUVXUnpiirWAscHO7APYWAfNq9jtRaa0/q60VFg1RZaK622qaWTUtZA2WJ4O27XAjzOD5eS8gufX2a2sNL6/GsuWvh1VXPSl1j66i1W1viOt04PxBrCc+GdsAdu2+5C2Z0evOm9XaW1hQtuelNR2270rgHCaiqWTNwe2S0nH2rWF5K+zH5b+arhpxdGo9WaZvml9I+A9tVTXAmNlQ44ILWE4B7g7ZOd+wI9aQmz4uWesC93SnslprLrV7w0sLpHD1AHb7V4Wc+nG6z671BV6etVvhhIrC+QsYBgNdny9TgL274mkM0BfnOBIFFIcfUF889ZZBxE5pafS1UQI7nqOGiIPbpMgBH27/lWJd/+T+K/wDrrkP2cnFixcDuYy26z1n/ALTsdZQz2+WrlBayPxcAOB7HG5P1L3p07qOxastFPftN3amuNvqm9cNRTSB7HA/MfoO684/ab8o+htG8slBrLh5ZW0c+j62mbXeE3+z08n3oyO32w9zSceR+W/WLkB9oJ+1cFy0TxFo7leNHXBzZqcUpD5qCYbEsDiMsI/ejG/5tX8b/AC2XiT/9e5gz5qq4E5dOdjgNzQVtZZ+Gt/q23ahj8aW3XGm93nMecF7ASQ8DIzg/oOOe1QWFeLdHd7TW2mU4ZW08lO7z2e0tP5is1EHzkcUdLcReUHmPnntfi2y76duj6+z1JjPSWdR6SO2Rglpwc49MhdyeEHto9asvdutXGTh1Zam1yvbHU3O1SSQTx5wPEMbi5hA3JAxst5+2o03pGm0ho3VMlKxl/qq59LHI1uHPjazJ6j5jGRv6D5Lh3kJ5DeF3M9wH1Rq6/XSoh1Eyslt1HICTHSyBmWEtHkcgk4J3OM4ASd9Hrbovi1w34g2Sk1BpLWlor6StjEkYZWR9bcjPS5mctI8wR+Zbsimhnb1wyskb6scCPzL569Wez85wtFalqrJBwW1LdRSSlkVwtMPj087QdnsewnYjyOCFtS88M+bTh/P7jddK8S7PJ28MMrGf9E4TvqPpC7rZHEvhNwv4mWieDiJpe110DInF1VURta+Boafi8Q4IAGTuceq+fGCw82TQ2pp6biczOMPbLWg/pysrVNJzh2zSc9w1hU8VYtNgf7YfW1dcaUNxuXhzi0DHmRhTfx8tPY2zxutdql416pseg2OuNBDdJaWgFKwvMrGbfCG5J3B7KHhXxG1xy9cUbRr2z0D6S92KobN7rXQFnW3zY9rgCAQO+O4B3Gy7b+x0tuhbrzFXyDVNDS1V1isD57MyojDwJGysEpAIO4YQfljK9Q+YblG4Ncxun5bfrHSVAy6sa40l1ghbHUxPI83tALgcDvvtt5g0+OO+Wr2jPAvj1ZaZl6vdJpDUuGtqbdcJw2PrPcskOAQTnGewxuV2aptXaUrGCSk1RaZmOGQ6OtjcD9RDl4c8Z/ZdczGhNR1kehNE1OrbL15ppaGRjpQ0nYFpI7euc+o9eNYORnnOid4dPwC1rH5bU3SPy9QCf4r6D36x0jG4Mk1VZ2uds0GuiBJ9AC5arFJHMxssT2vY8Atc0ggg9sEeS+bPiNyzcyHCO3M1HxG4T6qsVCHbV09M4xRkb5MjCQ0/MkLuNyJe0/snBbRL+GHHyO/3S3UkpktV2pGipmgjIGYZGvcC4A7gg7DOQTuaPYpFwjy884vArmeNdT8LNSVE1dbWiSooK6nNPUtYTgPDCTlufME/mOOblAREQEREBERAREQEREBERAREQEREBEXF/HbmS4QcuFgg1FxY1Q22Q1bzHTQRxmWoqCBkhkbd3Y/1+hQcoLDqbvaqLPvlzpIOk4PizNZj68kLzK5n/a+aCuug7jpLl6td+kvNzhMP3Zr4G0sdI07EsZ1F7nY7ZAHcehXnnpLh1zNcwctVW6S05rfWnxOfPNCJp4sknPxE9HfOwO3og+i9+vNDRHEms7Gzy+K4wj/8pYl04o8NrLQy3K569sFPTRNLnvNwiOw74AcST8gCV8/8XITzlSH4OX3VwI/uoGt/S4LcFh9nBzsamq2ULuDdyoWOP9luVfTwRNPzLpM/mQd/ON3tfOHXD7UFXYNAaY/ZS6mJY2oE3TEXA7EkYGD8iSP0+bPHjmQ44c53EWgGoPGrZp6gU1k09bWOMMDnnAaxndzzndx37nYErs/oP2LfG+7tgqde8QNNWBpcPGpqcyVUob5gPaAzP5R9a9C+CnKxy38nWjzqGlttno623U5dcdU3YMFQ4AEuIkefvYO+GtIO+N+yQdT+X/2Num7ZS2TVfHPW1VXXRvhVVVYbfE1tNG7Z3hPlJLnkHYkAA77EL0osdjtem7RSWKyUUdHQ0MTYYIYhhrGAYA/9vcrqFqn2tfJ5pq61Frpb/qO++7vLDU2u0F8Dz6sdI5nUPmAtBHtj+U8k/wDU/W4A7ZtUf80pWf5ymY72Iui8ftjOUh2Q+l1wz/yPGc/klUcvtkOU5hPh27XD/wDyTGP0ypo72IuonDX2pHKZxKvdLp2m1HebHXVjxHE28W7wWFxOAOtjnDJ+tds6OspLjSxV1DURz087Q+OSNwLXNPYgjuFr9jIRFwJx253+Xfl0vUOnOJWrporpKwPNJQ0rqmWNp7F7W7hE3HPaLorxK9rny12PStTV8Pq65329PicKWB9C+JjXkHBcXeh8jgfNeVXFLm/5gOK2qanU144k32j8WUyQUtDXSQRwNzs0dJBOBjc/ZgbJZVe2HPDzaW3lQ4UP1FSQ09dqi7Se6WagkfsZCCTM8d+hgBPzI88ELwo1fqXi1zHa8ues7425amv1afFqHwQueImZOGgDZjRuAPr7ndbb1TrvWmuJ4anWOq7te5YG9MT6+rfOWD5dROF375H+dflO5eeG8li1XoOviv8AOQ6rq20QqTUOAGT1YOxOwzjAAGAEtTrzyutoudjrX2670M9HUx/ThmYWuH2Fds/ZncGtQcROYS0app7Q+e0aemEtRO6PLGPyDnPbIAP5Qttc2PFLQnM9xaoK3g/pWWhmrJDBHEYvD6+o7AgDyO+cbDK9e+Qbl+i5feX+z2StpWNvd2BuFxl6MPc5+7WnPkBuB6EIk/J2TRERoREQEREHSj2u1da6bk/r6Wuc3x6q+W9tK3zLw5xJH1AEn5LxL05oXWmroqmo0tpO73eKiHVUSUVHJMyEd/jLQQ3b1K9Sfbc6tli07w20NG9zWVFZWXSUAnDuhjY2g+RwXOIH+pa77MTmd5atA8rx0ZrLXGn9K6gs9bXVN4iuUrYX1sb5C5kzMjMoEbms6RkgtIxgjIdeeQnnv0Dyxabm0Trmx1I94qXColigPUz4jgkgE5GSCCPXPqvXDhJxp4d8b9MQ6s4dahp7nRytBe1jx4kJPk4A7LwPs/AriPzb8dNcO4A6NfX26ovVdXxyueyCmpaWSd7oi+RxAbkEYA33Gy1nhfxP46+z5481Fj1BbqmgqLdPGy9WWV/VDVQOAIkjIOHAsILXA4I2JHcL7lT/AB9CS88ubb2UVl4yaovvFLhnraWz6iujnVUturIRJSzygdg8EOaTgAE5A2yu9nD7W1k4kaJsmvNN1LZ7ZfaKKtp3tdkdL2g4z6g5B+YK3CivnQ4S8e+PvJHxCvls0qaa13WGV1HdLfcaPxYZSw43GWuIOAQQRtghdzNE+2z1B7uYeIfCC2+OGgNntVRIGuONyWSOJG/kCfsXo5rvl04EcT651z4g8JdL3+rf9KorrdHJIT6lxGc/POV5784/spTc9R0upuWSwwW+kq8ittPiHwoH57x5JIGPLfzHplv9jnPls9qBwp42X6XTmrGU2kqp/wDaxqZeljznsSSR9oO22e+3buLiFoGoYHwa3sEjXAEFlyhIIPbBDl4cat9ljzbacomVls0bDfOpnU6GjqGeK0+gBOD9pC49n5CuciAffOXvVrh/2lOx/wD0XFE6+h6gulrusZmtdypayMbF0EzZAD8y0kLMXzv6A1ZzNcj/ABGs2o77p7VWmI4ph4ttuUMkMFZF+/Z0u+AkjOMeXyyvS/Sfti+V66WmGfVdDqqy3AsHiwMtoqGB2N8Pa718iPtKkurmeu+Kp1LoVqf2ynLBbKSR+mrFrK9VIB6IzQR0zCcbAudISPyLzl5jedLjdzOa5lqbTdL3abWXFtDZrXPKC1mdsiM5cd8Y7euScqzvB7W8y3M9w35ZtA12rNYXqkdcWxFtutImaamsnIPQ0MBz05GScAAA7r55OJOvLxxT17fNf3mCOOsvVW+qkihHwRgnZo+QGBnbPdckaS5TOa7jJXQVNs4V6vuPjkMNfcYJI42NJ7ufLg9IznbPngL2i5QuTTh/y88I7Zpq96Wst21RVRiovdwnpI5jJO4bsaXA4Y3sMencjCtll6m/08CdGa51dw71DS6p0TqCss90onh8NTSydDgR5EdnD5EEfJekPLZ7YnUsNwoNL8w1jo6ujkLITfaFpikZ5B0rNwR2JcPPOcDdekmouXbgNqylmotRcHNHVkM7SyQOs1OC4Hvu1oI+sEFeLXtC+S65ctXEqp1DoyxVR4dXxwmt87QXsoZT9One7fAB+iSdwQNtst5ivdHSurtO62sNLqbTF2p6+21kYlinieHDBGcHB2I8wVmTXi0wO6JrpSRu/uXztB/ISvmbsXGvi1pqxN0zp/iHe6C1sBaymgqnNa0HyBG4HyBAWmy8TOI08hll19qN7zvk3SfOf/OUvvCPpqq9ZaRoG9Vdqm0Uw9Zq6Jg/O4LGZxE4fSDqj11p9w9W3OAj/pL5/eDfLHzXc1FuqLnoS33W92uB/hvrbld/Cpy8Zy0Olfgkb7D7Fq+o/Zyc6+las0b+C11rge0trraepjd9schI+0BLB7sXfjVwgsQH3X4oaWpS44AfdoMk/V1ZW5rPerPqCgjuthulLcKOYZjnppWyRu+pzSQvnduHIxzf28CWu5fdY4dj4mUXi/lLCcfavVf2WfB3jbwd4RXi1cX7dXWplbXtntlsrT99p4ugA7ZPSCcnHzA8tm/jfDK7tIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiIC8c/bD8ddYVfGCn4GUN2nptP2u101bV08by1tRNMC4dQB3AAGx9R6L2MXhX7XelNPzk3OUg4qbBa5R9Qjcz/APIKDtD7LTkZtNhsun+afXc7ay63OnfU6doA0FlJE7LRO4+byASNhgH7T6ZrgbkTuFNc+T/hPNSuaWxaapad2DnD4x0OB+eQVzwPmgqiIg6I+2Pn6OVKhiyB4uqaIb9ziOYrZnsSKqJ3BriHRB33yLU0Mrh6B9KwD/oFc1+0y4G6y468ts1n0Hb5bheLHdILtHRR/SnYxr2vAz3IDyR+kDK6Bezs5r9Kcmt111o/jdb7va6e8vpZ2xsonOmimhEjTlhwSCH47+WRlIRsb2lHAWt4G8yt41LaYjBZ9XVT75b5IgWiGZ7uuRoPqHkkHOc5+S9OPZq8cbpxt5eqeovtR49xsE4t8z3HJOBtn8hI+ted3PxzV6Q5yr/pyz8LLVXSutchjp2zQuZLN1dWdiB3Ls48gPNd+/Ze8BtW8DuBdYzWVMaav1HXNr/APeNgaQB+Qj8iflPMqfj+Wyu5SIiKLaHF69XHTvC3Vt8tMT5K2is9XLTtaN/EETukj6jg/Yt3qKeGGphkpqiNskUrSx7HDIc0jBBHoQUHz88hXCOo5g+bWw23VMMtXQU01TfL66QH42xtc7pcQNi6UsG/qfRcn+1J4EWLl+496b1voC1MorVqSkbXNp2sJiiraaQBwye4I6Dj0yvYTQvBbhXwzutzvmg9DWuyV94Oa2opYul82SDgnPbIBwPQei4N9oly3Wzj7wJude1pF70hS1N0tzwMk9LOp7T8sNP2ZQY3E42Pn15HbpQcNqyjqLpfrVR1UFMXhppa+GSOUxOGct+Jjmj5H1yF5TcO+JnHbkG1Lq7SF90pUW2v1HQGjmpKs9LSB1ASt75BDu+MkYXNXsbuJ94s3MHcuGU1bI+1agsVTNHA55LWVMDmSAtHkSwPz9XzK5d9t1pe0MsPDPWbKZjbnJW1ttfKGjLoRGyQAnucHP5Umynx5rcJ70WcYtO3+4ShjjeoquV42AcZOokegyV9KeirvTX/AElaLxRyiSKpo4nh3qekA/nBXy7QTS0s8dRA8skicHscO4cDkEfaF9BHs6eJFRxN5XdPXmtlDqmkkkopcHOCwNyPykoOziIiDYPG/gvonj/w5unDLX9B7xbLkwdMjceJTyjdk0ZPZ7TuD9YXg7zncnuqOUPX1Np+vuzb3YLzG+os9zbH0OkjDsFkrASGyDbODg9xsvogXlp7bC4UD6HQtseM1Mb5JWEDtnqBBP1D8/5B47B+y/41W7iNy3WHSFTcjNeNLwmhkbI7L3RNJ6TnucfPfBHcDK7j4C8cvYyG8ycWdQRxzO+50VE58jOrbrLSM4/838i9jcH1UkwVWm6js8GodP3OwVcbXw3KjmpJGu3BbIwtIPfbB9FqSKj5p6mn4gcrHHmCpkoKq16h0RexUUwnaWGVsUnwnJG7HtBBIHZx7Fe+XKvzM6N5puGMGv8ASrXU1TC8Ut0oHn46SpAyWn5Hcg+nrsTwJ7UnlWfxs4PN4gaH042q1npKYVB93jHj1lCQRLGT3cW/C4Dv8OMgLoB7O3nItnKVr692DiFQVx0vqUxRVxiYfFt9TESBKYzgnYlpHcYHzIvqPeBFo+k9U2HW+m7dq3S9xir7VdYG1NJUxHLZGHsQfyj68rWFFEREBERAREQEREBERAREQEREBERBQgEEEbFUjYyNoaxuAPJXIgIiICIiAiIgIiICIiAiIgoNgvnz9o1xIm4k822tK0VL5KOzzR2mjycgRRMG4HYZJOV7gcxvGC08CuEGoOI93diOggLIh5ulcMNAxvnuRjzC+eVsV748cZnupKOR9bqq7mUxsGSxj35JOM4w386vxPr3U5EuWjSXL9wP086itsDtR363QXC71/T8ckkrBJ0AncNAIGPl6ALsmtG0fbZbNpKyWedpEtDbqameD3DmRNac/aFrKiiIiAiIg0XWVonv2lrpZqUtE1ZTPiYXHAyRtkr58eYHTGr+XPmXku1ZRup7hQ3Jl1p2P2yWuBxn0OO/zX0J6l1Np/R1jrdS6pu9La7Vb4nTVVXVSBkcTANySfzAbk7DdeCPtEuY7R/Mnx8l1HoGJ77BZ6Nltpax7C11YWkl8oB3DScAZ3IGexCSSXfqdclc1XtP9R8xvB4cIrLocWQXUwi8VLpOt1R0ODuiNozgFwBPmt58sPsjZuLXCSm4g8VNcXbStyvcZntdvpaSN5igP0HzB5zl2M9IwcH6idd9llyO2jVNG3mE4sWOCtohJ06foKgdTHkYJnc3sRvtn5eRIXrLFFHDGyGKNrGMAa1rRgNA2AAHkEl/pXzwcTuG3HP2fvHymZS311FerYfe7ReqNp93r6YnGel2QQRs+M5IyM74K9MORP2lNq4++PoXjVUWPTerqZjXUlUJBT01zb++wHnDJAf3oOCMY32XbnirwR4U8bLN9wuKWh7ZqGlaCI/eosvhJ82PGHNI77HuvGTnf9ndrzgBqe4at4aacuV94cS/foqiEGomtoJ3jmAy7pG2H4xjOcYyU/aV7oU9RBVQsqKaeOaKQdTXxuDmuHqCNipD2Xg/yF8+t/5a9ZfcHiJdrrd+H90AjqYDI6d9veO00TSScDsWjuO2DuvcHQ2udKcStKW7W+iL1T3ay3WET0tVA7LXtP6CNwR6hFdBvbXWCnq+COiNQl7Wz2/UjoAPNzJad+R9hYD9pW2fYiX+pfpXiZpV0TjBHcKO4Mk8g90ZjcPrIa38i0X22PEJ80uguGMMg8OB893maO/WW9Az8sO2+ZK5H9izpunoOBer9SBg94u2oAwu6d+iKENAB9MlxQ3HokoKmkpa2PwqulinZnPTIwOH5Cp0QYwt9CGtYKKANb2HhjA+rbZbW4wW2iunCbWNtrqSKenlsNc10T2gtOIHkbHbYgEehAW8ltniYf8A83Oqt8f9RK7f/wDYPUye0eDXs2L5VWPnP4de7TGP7oVU9DJj98x9PJkH5HAX0Fr52/Z+hx5y+FAZ3+7o/J4UmfzZX0SKgiIgwL3ZbXqO01divdvgrrfXROgqKeZvUyRh2II/9/Jeb/GP2Luj9QV1fe+D/EubT8tRK6WG13GjE1LHkk9Aewh7Rk4GxAGMDbC9L0V0fOPdrVx+5EuOQYJ5dO6rspJhqYfvlPWQO74J2kjcMAg/6ivX3kO56rLzSaNqKDWk9psuurQ9sdVRMnEbK2MgdM8TXHOCcgtGcHOMgZXM/HPln4NcxdlNn4paOpbjJHG5lLXNHh1dKT5xyjcfVuF4mc1nI7xh5YdZV9xtdiu9y0dHOZLbf6KN7xHGRkCVzBmNwGQScDbvvhN51M7r6Bu6qvE/kN9pLW8BHVehuOFfe7/pOqLZKOra41FTbn+YAccvYR5ZyD54AC7ofuwHKPJdYLfDLq4wTPDX1j7QGRRgnuQX9WB57KK7v9Sd1pWmdSWbWOn7fqjT1cyrttzgZU0s7Oz2OGQVqrUSXVUREUREQEREBERAREQU8wrJZY4Y3SzSNYxoy5zjgADuSfJYV+v1n0xaKq/3+4wUNvoozLUVEzw1kbAMkkleJfP/AM+2p+NGuKjR3C3VFdbtFWsmEOpJ3ROrX5w5xLcHG3kd84zgbk3uO5nPb7SlnL9XU2g+DMNlv2paiN0lVWzvM9NQjyAawgPdnyJx39N/LvU+tuYnng4t2+lu9XXas1NcHimoqWCIMp6SMnJ6WNHTFGM5JP5TsFuvkIl4GV3MPQs5jKagr7FU00oiddvvlN73t0GUOOCPrz2XuHpC58tWj4BPoKu4bWSOVg+K0PoabqYd8fesEj5KeRfrz+4Z+xMp/dqS4cWeMUnvJ6JJ7fZqAdDexMZmkdk+YJDR8vVeknDLhppDhFou26C0Ra4qC1WuIRRMY0AuIABc7GMk4GT8goG8ZOFbmSPbr6ydMQLnn3tmAB9v6F0V5xvasaS0TT1/D7gFL92r9h0E96aemmpDuD4eRl7h6jt8jgq+pndejT3xxt6pJGsHqSAuAeZjnV4MctGlKm7XzUNFer8QW0NhoKtjqmok9HYz4bRtkkeY27keFNTqTmM4z3aovMVZrnU9RVykyOoxVTxhx7gCPLW/V8lyHwl5AuaLirqy222u4V6islrrKhnvl2ulMaeOGIkF7syYJdjOBg5OMornuu9tbzByXKSa2cMOH9PQnIjgnirZZGj1MgnaCfqYB8lwLxs5wuZfnKuVBom+1MLqSecClsdkpnRQOeTsXguc5+PVxIAC9fNJezq5P9MacobFVcFrJeJqWBsc1dXiSWeoeBu9zi7Yk52GNtlyVoHlo4A8LqgVmgOEembJUDcTU9C3rB9Q52SD9qX09eX3Dj2L3FDVFgor5rTirZ9OS1cbZXUMdukqpYwRkAnrYM4Pb8uFvs+w8ZjI5hDn+D4x/wCmXqmiDyqHsPD58wg/0f8A/wDcsmk9h3QGT/b/ADD1Aj3/ALDp1pPy71C9Qbnc7fZ6Ce6XWsipKSlYZZp5XBrI2Dckk9guovFD2qfKfw699oaDUNz1TdaQvY2ltNGXMe9pILTM4hjdwd8k/JN3g6T81fsubLy3cLaripaeM9Tc47Y9okhq7e2ne55yR4Za92Tt9ecLS+DPtT9b8KeGg0pLbpbnXUkfhUhqMPYNtjk9gPMfmK4x5lebTjZzz63otK2q0VMFmZMfuTpyh+PLs7SSkfTdjG52Bx6DGmWD2c/OLqG5wW2Dg1cqNlRjpq62SOGnAPm55Ow+sLUtkxOWxuW7e1M5vrhW1VTR64pKCKoyGwxUbSIge3STnBHquE9SWHjDxdmu/FfWTqurkk+/VFfcCYzMNyAzI3A8uw+ZOV638kfs0NIcAqZ+tuMtDZdWa1mA93jkgE9Ha2YIPQJAQ6Q5IL8bDt326qe1c5luHOs9SW/g5wlnoaqKxF4vtwoGtEDpgdqZjm7PDcfERsDsM74z+orzvjjfM9scbC97yA1oGSSewHzXpXyh+yZoOKXDik4i8cr/AHmySXgeLb7TRNayVsB7SSl4JBPcADsd/Ind3szfZ/0gpKLmA402GOodUxiawWqrjyGA9p3sI747A/oyD6kRxxwxtiiY1jGANa1owABsAB5BPD686qj2KPBOSYvpuKmrIYs58MwQOIHpnA/QsLiT7HHgnaeGN6r9I661LBqO20M1ZBV1skbqaR0cZd0vjDQQ04wSDkL0mXm17U3nW1ZwvFTy+cP+miq77bf+qlxafvsdNK3djD+9LmnGRvjO+Ni0dDfZ3abh1TzgcPrbU0oqKeOslqJ4y3ILGROcc/LYL6FQA0BoAAHb0Xjn7HvgNqK58XKjjhcqB8NmstvnpaJ724E00o6S4ZHkM4Od9/QZ9jk6CIiAiIgIiIOF+ZTlM4Sc09ipbPxMoa1s9u63UNfQziKenLhvgkEEZ3IIwV4a85XLdRcrnGSr4b2rVzNQUHgMqqaocGtnja4bsma3YOB9MZ9B5+l3tNeeW4cGdPU/Cvg1quOm1jdJHC51tK8Ga204G4Yd+mQnAz3A7b5C8kdD6yof9lax604oS1l/oo7pDVXV1TI6aWeMOBcSXElxA3xnO2AhHqd7FqwaqsXDjXVTeNNVdHbrzcaWqoa2aEsE4ZGWkNJG43yD22PqtD9sHyvaj1ZLZeYfRdp98Fqt/wByr/FBGTL4THufDOQO4HW9p27Ab9ge/wBwY4r8HuKekKK4cHNTWe42iGFrY6ahexrqVoA+B8Q3YQSAcjGc7lb7q6Slr6aWjrqWKop5m9EkMzA9j2nuCDkEfIqYPBTlC9ofxZ5ZnUOkK6duodBtmzJbKneWlY47mB/doGSensduwAC9wOE3FbRnGrQls4h6DujK21XKMOY4H4o34+Jjh5OBOCF5xe0l9ndbY7a3jTy76IdDPE951BZbez4HsO/vMUY7EHPUBsQcjyC69+zn51Krlm1tLw61xHUy6O1JWMina5xDrbVE9PiBp7AkgOG2MHPckX0x7poooJoamGOpp5GyRStD2PachzSMgg+hBClQERbN4r8WdDcFdFV+v+IV6jttot7ep73bvkd5MY3u5x8gEHTP2zDLF+1ssj64xi5/sjhZQdWOogxvMoHn2AJ+pdQ+Sv2ZX7aPha/ivqXie/Tttqqyait9LRUIqZnGJ3S98hc5oaM5AAyTjJIyFsfnv5y67nJ1zZLRpCwXCk03YXSRWuhd98nrKiUgGYsaD8RAaGgb4OCMr059mJwi4jcG+WamsPEihlt1Zc7pPdaagmH3ymhlawAOHkT05x5En1yWDrZL7D6gbU/eOYKofTZ/f2FrX4+yYjK7R8rvs8uDPLPNJeKWWo1RfZQB90LlAweHjP0GDIHfbf54zuu1KoThNTFA1rRhoAA2x2wrkRFFtviBoHSvFDSNz0NrW0wXGz3WB0FRDKwEYII6hnsRnIP82VuREHm9J7FDhFJf5q4cWdSR2t0hfFQspIi5jT+8MhJJA9cZUfEr2NnCOk0Jd6zh7qvUbtQ0tJJPRx1UjXxzSNaSGYAyCcYHfc488j0lRPB4D8q3P1xi5Q6ubRU1mo73pmKrLK+y1zTFPA9pLXiKVu7Hgg5Dg4E7Edse0fL3zJ8LeZbRNLrLhzfYpXSMHvlsme0VlDKMdUcrAcjBOzhkEEEHfC4V5uvZz8IOYW0XXUul7JTab1++N0sFypAI4quUDZs7Bhrs4A6tiPUZyPGFkvGTlU4uOjD7npXVum6wF7GudGJOh2RnGBJG7H1EehGzd6c+PpdRcPcs/MloTmS4d23VelrzSyXL3SJ11t7XjxaSfpAe0t746s4P1LmFAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQF4r+2btHuvMfYryI8e/6dii6vXwpHfo617ULyY9thYP/AIR6F1G1mT7o+nJ9AXuP6QFL6OwnsldfR37lvpNJS1HXNZp5egeYYXkEfZsu8a8hPY28RTT64vHDyR3wzwvqGD5YJ/SD+Zeva1bvSCIigLxq9svwxodNcYNM6/tdJFCzUdtfHV9DQ3M8byASANyRkk+q9lCcLyY9tveozqfhzp4Oy80NRWEegEhb/Og6Ycll10XZOZTRdz4gVcNNZIa4GeSYgMGRgAk7DOSvortdfbbpb4LhZ6ynqqKdgdBNTvDo3N8i0jYj6l8znCfgpxO44X2XTfC7SdVfbhAwSyRQYHQw53JJAA2K7tcpvNfxE5EdcVHArmXs13p7PU+FKyKaTxH20PyBIMk/e9jsM7Z7AAhibj2VRYVoutvvtqo73aqhtRRXCCOqppWbiSN7Q5rh8iCCs1FEREBbd4iNa7h/qZkjQWmz1ocPUeA/K3EtP1BbvuxYblaM49+pJqbJ8uthb/Og8IPZTucOd7RAacA0l3DvmPudUH9IC7d+2+mA0Fwtp8fSu9xf+SGIfzrqvyi2C6crvPJbJeIlvqbZQWCor6SSqnjIYYZYpIWvBOActfnHcLkb2t3Mbw84wXXRujuH98hu0OnzUVM9RDno65QBgZ37NHcA91NHnavav2NNzbU8uF4tocSaK+ynHkA8Ej9BXiovWr2MOpPd9Oaj0y+T4aqZ0zG5/fNOc4+on86tuD0+REQF0o9p7y1T8Y+DlXrexQmW86Tp3VTYm/SkibucfPGQR6HPlt3XWNX0NLdKGpttdC2amq4nwTRu7PY8FrgfkQSEo8B/Zx8e4uBnMnYpbzWCCwajf9ybg5zsMjMm0ch+pxA+35L39iljmiZNFI18bwHNc12Q4HcEEeR7rwH59OTfUXKvxEN6t7nVOjdR1kstnrWDp8CTPWad2OzmgkjHkPkCfRz2TnHDVXF/gNdLNq+5SV9Zo65Mt0c8rsvdA+IPYCc+QBH1gojvAiIiqEZXjb7VHk0v2jdd1/MFoGwtk0peQ2S7x0rP7RqgAHSFoGAx2xyMAY89yPZLPktucRNM2nWeg9QaVvtLFUUF0ttRTTxyNy0tcw7kH0OCPmAg8o/ZPc32orHrG3ctGqqk1Vku7pTZpJHEvpp8F3hN+RwSB2AB9AvX/wAyvnF5V9QU3DHmy0HeKl5dBZ9UR073HAJaXuiyfLfqX0bxyMljbLG4OY8AtcNwQdwQVbEnqRERRRERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQdCvbI3t1u5YLbaGz9P3V1HTMLf7oRse/82F1S9jvwkptacYtQa9utvZPR6YoohE97QQJ5HHDdxjyBI9AuevbcXAM4U8Ora1+HS6gqJnN9Q2nIB/KVJ7Eelpm8IOIla1o8eTUkETj59DaVpA/K5yko9JERFQREQFpepdQ2rSen7jqe+VTae32qmkq6mVx2ZGxpcTv54G3zwtUXm97WvmwqtD6aPLvpeTor9SUYlu0wJzHSuOQz/xgPz58lZNpXT3nV9oHrvmprX6C0tRyWfQ8NYfdqKEuM9xIOGOlwATnYhmO5GwJIW7+FvspeJ174TVnFriNdoLF4NukudLY8E1MkTGF+ZDghuWjZvfzyQRm72XPKDeOKHFOg4z63068aL0y51TSe8R4jr60bRgAjDmsJ6jtuQMdl7L63pff9F3+iAz7xa6qL/zonD+dTepnHmD7M7nvvt413ZuW3Vtpo47VcWSx2Soh2fFKyMuERHmC1h+rBP1+rK+cLlL1dScIuarQOptRu8GCx6gbDWZOOkEOhfv8usr6M7bcqC8UEFztlXHU0tSwSRSxuBa9p3BBCkknispQVNPBWU8tJVQsmgnY6OSN7ctewggtIOxBBwR6KdFR8+HP7y63Pl+483eGO2+Bp/UFRLcLU+NmIwxxy5gxsME5x8yPIrv37HjinaBy56l0nervHAdL3mer6ZX4LaeWNryRnyBBOB5kqT20WmrXWcCtLaolgHv1uvwp4pAN+iSN2QT6bE/WV5W8KeNuq+ElDqCg07UObFf6T3eVvXgA+RI89j+YJtwcoc6PFu58z/M3dJ9OwvqadlUbVaourJLGnGc+mwzsOxXsByAcFa7gby6WfTF1hMdfWSvrqgEYOXgHceW+V5U+y+4c0PE3mloRe2tmp7XRz3CVr9+sjBHzySMZ+a95WMZG0MjaGtaAAAMAD0wqm3V6Iiii2txUyOGGryO4sNwP/J3rdK25xIj8Xh3qmLBPXZa1uB33geEHgB7PiRkXObwpdIQB92i3f1MEoH5yF9ES+bHlTu8umeZPh7d2vbE+jv1OSXbBpyWnP5V9JbJGSsbJG4OY4AgjsR6qfReiIqCIiAsavoKO6Uc1vuNLFU0tQwxywytDmSNPcEHYhZKIPOPj37HXRWvdSV2qeE+vDpSS4SGZ9uq6XxqVjycksLSHAEnt+kklcH2v2KnGR9+jprxxQ0tHaRKPFqYI5jM6MHfpYW4BIyBkkA4yCvYxUyAplONrcLtA23hZw9sHDy0zSTUlgoY6KKST6Tg0dz9ZJW6kRVJMEREUREQFpV01RpmxvEd71DbLe52MCqq44ic+gcQtUJXhFz2cC+aO58wGoblftH6o1Fbq6sL7VNR08tVCIiNmtawHpPfJxvnv5CfcHuENbaMIa4atsuH/AET7/Fg/V8S1eGeGqiZPTzMljeMtexwc1w+RHdfN1c+Vvmf0/aW3u5cGNcUtBE0vE33NmwxoGScAZG3yXYTko9pBqflobU6F4pUF41PpaSX720TA1dukBwQ0SEZaN8tJB2x5DGslZle4yLpvR+1l5M6qhbWSaxvlM8jeCayTeI0+hDQQfrBIWBqb2uPKJadO1d2sN8v16uMcbjTW6K0SxPmkAyGl8gDGgnbJKjTjD2yXHcae0DYOClgu7o7hfah1bdYopCHCkY3DWux5OJ3B7grrp7OnkI0lzOaV1BxE4j1tVBaqGs+51BDASDLKGBz3nBGw6gME9x88jrRrzWPE3nE4+S3d1G6s1BqyuENDRRkllPGT8LB/2rRkk49cDsF7p8mXL1Jyz8CbTw2raplTcfFkr7hIz6PvEuC4D5DAHn6bgBVM+PMbmH9mDrXRmoamPhvTTVdvbl0LiHPD2ncb749Pr2XUDU3DDi3w7uElrvGnL5QyRkjqhjkLDjzBbt+XC+mzAK0e4aP0ndnOddNMWmsce7p6ON5P2kErH48mXq3r5ln1fEekYY5J9SQtlHSWudO0OB8t+6535X+QbjJzI3eGWO3v09pxrw6qudbGWkMzv0MIySRnGfyFe8NTwr4Z1nT7zw+04/oOR/1Mh2P2NWv261Wyz0zaK026moqdv0YqeJsbB9jQAtT9n+Ni8DeB+ieAPD+2aA0Tb4ooKGERy1JjAlqH43e93c5Izgk/l3XIqImAiIgIi0HXWoo9IaJv+q5XMa2z2yprsu7Ziic8Z+0BB55e115qXaW0xQcAdDXwx3S7uNRfJKeQdUVMBgREjcFxO49M9iF0p5LuQvWnN991r1TalptN6bsszKeorpoDM+aYjq8OJgIBIBBJJwO3dcV08HEDmu49Q0EEslZqHWl1McPiuLhC1xJA/wC9Y3JwMZx6le+PKly7WTli4P23hraag1U7Huq6+qcN56l+Os/Vtgf6tkzib8cd8qPs9uEfKzXSakt1fWal1HIzpFyr4mM8EeYjY3IHyOc+ffBHZe+X2z6YtFXf9QXKnt9toInT1FVUPDY4mDuST/7k4AWbJIyJhkkeGNaCXOJwAPMnPkvDr2lPOZqXjPxTu/C7Ruo54dA6bmNEYaaXEdxqWZEkryPpNDstAPoc52w/S43vzye1HvnE+O6cJ+Ack9m0s8upq6+h/TV3JuSHNix/YoiMDIPU7cggELj3kH5A9Tcx19puIWtoJLZoO21LJHOlYeu5uBz4bAe7Djc+h+Yzyr7Ob2ckPEGC38dOO1n6tOStE9jss2R7/wCk0w7+HtkDz/6PrlabPa7DboLTZLdTUFFTMEcNPTxiOONo7ANGwTwS0NFSW2ip7bQQMhpqSJkEMbRhrGNADWj5AABZCIgLwf8Aayvkdzl34Od1Blptwb8h4ROPykr3gXg/7WhuOcy/bd7RbT/+JQer3IZYLZp/lV0DFboWM95tkc8zmtALnnYkkd9gF2CXAXIg6R/Klw+Mmci1RgZPlgLn1Sd9STBERVRERBQDC67c6PNXpXls4S3q5MvNM7VlTTOp7PQCT76Z3ghriBu0Dvn5Z7Arl3inxFsHCbQF74hanqo4KCzUr6iQveG9RA2aCT3J/Nn0XzocwvG3UfH7iheeIN/qZXNrKh5pKdziWwQ5+EAdgSME/PbyCf6n1yZyX8EJecHmXhsWv7hW1dBJFU3q+1Qc4ySMYR8HX+9LnPaB6AYGwXfzmM9k3y/0/CzUepeFP3WsOobNbp7hT+PXOnp5zCwvMbmuHw9QaRkdiR2C4Z9iXU2BvEXiFSzNaLy+z07oS7GXQCYdQb8wcE48iF6scQ7HVam0DqbTVEemou1nrKGE5xh8sLmN3+twSX4PC32YGvbpo/m60jZ6Wumio9SPmt1RC1x6JCY3OaSOxIwcH5r30XzdcN75qblL5krFqPVunKhlx0ReBNUUcjSx0rB1MJbnGQWkkHsfyr6A+BvG3Q3MHw5t/Erh/Wunttd1McyQYkgmbjriePIgn/7HZFcg4BHbuvnn9oZw7h4ac3WvrNQ0UdJQ19Wy7UkMbelrYqiMPOB6dZevoZXjV7Z3Q5t3GrTuu4oMMu1qZSTSY+k6LPQM+eAHKD0v5euNnDjV/A/RWoIdcWdvVY6KGf3ivjje2ZkLWvDg5wIPUD3/AErlanvtjq6c1lJeaGaDuZY6hjmAeuQcL5m+F/DziZxg1PS8OOGNmud7utZ1SR0FK8hoY3d8jskNa0ZyXEgD1XLWseTXnY4cxmlvPC/WIpDgddunNXA70H3h7h9hGVrNvE3PXq9zj+0M4d8tNnFr0rUW3VmsakERW+GqD4qcY2dM5hOB8s5XkDzA82nHXmmu8DeIN/fUUjJR7jZrfD4dNG4nA6Y25LnHYZOSSM91uThX7P3ml4qXylpP9i29WS3zyhs9yusBgjjBIycOwScHPbG25Xrry3ez44A8vtBRXFmlqa/6qiY0zXe4tEzmyYGfCa7ZoyPTyzgdlLLOUl3x1U9mz7PO+6cv9s5geM9vigdBH41js0zOp7XuGBPKDsCAdh9ffO3qYrWtaxoYwAAYAA7BXIoiIgIiICIiAiIgLp97Sflk01xo4Dag1rRWOJ2sdJUT7jQ1UUY8aaKIdT4SRuQWg4z2x9WO4Kxblb6S7W+qtdfCJaathfTzMPZ7HtLXA/WCQhXgv7MPjDa+E3NPZRqW+C22PUVNPaqh0rwyHx3sPgF5Oww8YBP91jzXvdHJHNG2WKRr2PAc17SCHA7gg9iMLwZ59eRnVHLRq+o1XpqgnuGgrtM6Smq44yRQvJz4MmPojfY9vL0zy/yE+05h4SWaHhJx/nuVwsLJQ213xpM8tCw7GOYE9TowcEOBJAJyNiTaPYtFptgv9m1VZaLUenbnBcLZcYW1FLUwODmSxuGQQf0g7g5BwdlqSgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgLyb9tZq63OvuitGwzMdWtpHVUrM5LGdbsE+mSRj7V6yLx99qty58ada8xkOudI6Oul+s9wtFJRwGkiLxDJGCHAjO2Sc5+vPkpltmJsnav9jZw4kqtf3jiK933ump307RntsQPty4/YvX4dyuk/sxuWfW/AXhhV3HiDQPoLre5DMKN5+KFpIxkeRwBn5krux2VvOHL1VERFY1wuNDaqKa43KripaWnYZJZpnhrGNHcknYBeFvtSuM+mOM3MFTVOi7tFdLTYra23snhOWOk6iX4+RPbsu+HteOL9Xw/5cqTRlluMtLcdZXWOkkdDJ0vFJG1z5AfPDiGjK61+x04H6P4hV/EDW+tNP0t2p7SaKhpYqpgfGJXiR5OD5gAfLcJLJ6OOvZU8adLcE+LuoZ9d1Lbfar9amUnvcrdopGyFwI+vsceXkVoXtNdW0/GPmUn1toajqrnYaex0dBHW08DnxvdEZC8kgEDd+2fLC7L+135ddKaS4Z6e4n6C05TWqKC7NorpHSx9EZEjHeG7A2B6hjHbYY81vb2RR4f3/le1NBqC326rqrTe6qK5Oq4WyFlGYY5GZJBIbgy9vT5BEn91y17NnjzpPiVy66S0Ky/xS6n0rbWUFdSSS5mLYyQ15B3O2AfPbPrjt0vnx5SeNdFwk50bNrShrXUenLrqOehq2tyG+41UrmNyO2G9bDv2APqvoJiljnjZNFI2SORoc17XAhwO4II7g+qYu6lREQEREHQ32uWgrF+1uquI1FQw016oLnR0r6mJga+WKaQNIcR3x6nffvsF478M+Hl54l6oi09ZqSWc9Jlm8MfRYNjv2Gcj8696PaEcKL5xk5W9TaR05CZa+OWnuEUQ7vML+rH1ZwT8gV5ocjmqdBctdVqy/wDGOjio7nUR+6wRVGAWhoPbPzJO38wxP5Ty3prqnxa0O7QN6p7I+ExvbGS9p75BGcrt57JzilYdKcXH6Xv9yZTG4H/aokd0hxcMYz8iQftXXXjjqek47cV664aSjaIXyPMRAIaWnz+fYbriy2XC96E1VT3KjkfS3Oz1bZGFriC17Dnv3we3zBVkl9S/9ePqORbA4Da9g4mcH9J62inZK652uB8xDs/fQ0B+fnkE/at/DdCXiqIiK6he1J4Z3viZyo3Sm01ZJLlc7Nc6S6RxxML5GxMLhKWgDOS13Yd9l5c8h3OHc+UniRVi7UUlXpXURiprxS5LXwOY4hszQezmgkEYyR9WD7/yRslY6ORge14LXNIyCD3BXhf7VrgjaOE/Mg6/6Xs7KCzaxoGXERws6YmVYJbN0gbAEgHA8yfmg9w9P3y3amsdBqGzziehuVNHV00gx8Ub2hzT574Iz81qK6h+zI46Wnizyz2DTTq9j79oyAWquhc/MhjYfvchGc7tIHp29V28SAoK6D3qjnpdvv0T4/ygj+dTog+aDU1DFobmFr6a9SGngtGqnSyvAJLYm1PWDjv9HBX0O8FOKfDvijoW0XTQGsLbe4GUMLZPdqpkkkZawAh7QctOR5gLy354PZt8crhxZvfEfhDpd2p7Ve5feZKallaKiOQ7HDHEZGAM7jcbZyunU+kOZHlf1JTanr9L6u0NcaSZjmVUtNNBG5wOQ1z/AKDgSexJ7pun7fSei89uWD2tXC7X0Nn0Txno6vTGpJvCpDc8CSgqZTsHOcCDGSdzkYycBegdNUQVlPHVU07JoZmiSORjg5r2kZBBGxBBByEvOJOpkREUREQEREBERAREQEREBERAREQEREBERAREQEREBERARFBV1MdHSzVcv0II3SO+oDJ/Qg8bfbCcY6XWnEuwcPrbL10+nI5XyYOQZScZ+3JH1BdkfYvWF9t5e9T3lwIF31G6RpPbEcTY9vtC8tOYjWNZxU486muzACaq7SUVM07ABshYPsJye3Yhe5XIVwrdwi5btPael3lqeqte7YZLwBnA9SCftU5LkI7EoiKgiIgjkkjhjfLLI1jGAuc5xwGgbkk+gC+eX2gfEmPinzZa71BR1fvNBSVUdsopM5Bhgja0Y9Pi6tl7S872sb5oTlu1Xf8AT8j2VbIfD6mEghpDs4x9QXgZwis7Nf8AGLTlsvkzpW3W7xvqnHcvy7qdn1yRg/Wk6Poa5X9KQaK5eOHem4KNlN7tp2hL42tA+N8LXOJA8yXHK5PexkjHRvaC1wLSPIg9wsWzUUNttFDbqcNEVLTRQMA7BrWgD8wCzUSPnw5/OBNz4Hce7oXUjoKC+zSV9G9rSGkl2SQe2+QdvMlen/so+JV14hcskcF6rX1VVYrlLQue92T0gAtB+wA/auKfbZ6aop+FGgdViFgq6W/yULpMfEYn073BufQOZn7SrPYi1xk4V8SbeXEiC/0kwHp105H/AOQlz1delKIiDjrjtwM0HzD8PazhrxDopZ7ZVObMySF/RLBM3PRIw42IyvBPnI5c7Xyx8X6vh1ZtVfd2jbGJopnholjBOzHhuwIBAPzBXsZ7R7jXrfgVyzXLVnD64GgvFbcKa1x1bMdcDJerqcz0cOnY/wCteB1+1BfNUXWovuo7tV3K4Vby+apqpTJI8k9ySSfPt29FZfiZ1yNy0cwmq+WjifRcStKQx1MkTHU9TTSEhk8LsdTTj5BewnL57T7gXxjfFbNT1kWj7o9o+CtnAje/bIBI7b98n+debfs6uWDSvMxxIu9o1g1z7baaMTyNG+ASRnGdznAGfVb/AOdX2Y+vODTbjxK4TU37INFU0ZnrKeH+27e0d3GPu9gG5Lc4xk98Bknpu3I9n7LqCxajo23HT95oblSuxiakqGzMOe27SRlakvDL2UHFjVWmOaCy6CjvdSbDqSmqqeahfM4wiVsTnse1vYHLT2xnbOcBe5qhBYl0oI7pbKu2ynDKuCSBx+T2lp/MVloivnB5nODOteXHjjdbNdKSegc2vkuNpqg0gPjMnWxzCRg9JI7Z2x6r2T9nJzFVPMJy+0lXfa1s2otNzm1XMZ32aDG/1w5pJHpjGVtf2qfAmm4scus+rLZaxPqLR1Q2tpJY48yOgORLHkbkEEEAnAO+MrzF5CeZrUXLrxnoY6ep/wCoepJ4rfc6aQnoJLgGPx2BBOM99wc7IPoNRRQTR1MEdRE7qZK0PafUEZClUl3oIiKgiIgIiICIiAiIgIiICIiC1zWvaWuAIIwQdwvO3mi9khZeMHEG48ReFmvKXSdReJfHrbdV0RlpfFI+J0ZYQW5O5GCPluSvRREHjdV+xU41QvLKXifpqpGcdQp5GAj13dn8yyLV7FLi/UzBt04saboou5c2klkPzAAcvYlFJo6b8nvs3NBcrepXcQLjqSXVOqBC6GCeSmEUNIHbExtyTkjzO4/JjuQiKgiilmigjfNNK2OONpc97iAGgbkknYADzXVTiN7TrlM4a3yp07c9W3G5VtJIY5Ra6A1DGuB3BdkAfag7YIuCeX/nT4Bcy9dPaeGWpqiS50zDI+gr6Y085YO7g05yBg7g+RXOyAiIgIiIKYC89farc3UPDfRNZy/aabIb7quh6a6YEjwKR/cA+RIx9ecdgV6FAY814Qe1cq6q7c6d/o2ZkfBbrZSRMbucmEEAD1Jf+dQbp9j5w4uOouZV+vn24y2vTFqqg6Z0eWtqJmhjMHsCAT/5wXtq5waC5zgANyTthcE8mnLnY+W/gjYtKUtPG69VdLFV3qqDcGWpe0Oc0eYDSSAPXJ9AOBed32k/Crhbp3VvCPh1X1F+15PSzWt01NH/ALTtkkjCxz3ykgPewE4azO+MkBX9I4N9pJ7RGsr62o4GcAtUOho4euHUF6on4dK8HBponjcAYPUQfPHftxZ7PL2fLOZIScXOKVVV0mjLfXCKno2M++XmVuHPHWe0QyATuSdu2Vtb2dHJxXcyPFWn1br/AE5VVHDqxPfU3CebqZFcqkZ6KYO2LwXbvwewIPc49zrDYLJpez0lg05aaS2W2hjEVNSUkQiiiYOwa1oAH+vfugyLfQUdqoKa126mZT0lHCynghjGGxxsAa1oHoAAB9SykRFEREBeEHtafwzL7/4Itv8A6Fe768JPa3Qui5y70SQeuy2x4+own/Ug9XuRE55UuHp/+9Uf6AufVwHyJjHKpw+G4ItUY3+oLnxSfsERFQWj6u1BT6S0vdtUVULpYrVRzVjo2d3hjS7A+Zxj7VrCw7vbaS82urtFdGH09bA+nlafNj2kEfkKDwB5tufTi9zRVk1iuNUyyaOgnc6ms9GC1sgB+F07ju92PI7DJ7rtZyp+yl0LxQ5eKXXHFq43q2ap1RTvqrYKaTwxboTtEXxuGHuIHUQQMZG5XZbh97Lvlv0nrqfXl7sDrzP7yammop5HGlid1dQJZnBwQCBgY9e4XcKnp4KSCKlpoWRQwsEccbAGtY0DAAA2AAAAHom6PnroK3jP7OnmclLqaP7sWFz4XNe1zaa7UEnYjOT0PABBGSCO5xv7bcqXMbY+aLhFRcT7LbpLc+SeSiraJ7uowVEeOoA+YOQR/P3XSz22mhbRLobh9xGhoGi6091ntM1Q1u7qZ8JkAcfQPZt/3xW3vY6cyGh9OWDUHAPV16gttzrbr917K6od0R1QfGyOWEOOweDGHAHGQ4+YTnqedrcXtjuXaW9aZsHHrSdhdLVWmV9vvzqaLLjTvAMczwBk9LgQT2AOSVx37H3mT0hoOt1HwV1tfo7d+yGsirrM+pk6YnT9Ba+IEnDSQAfmftI9b7na7febfUWq60UNXR1UZimgmYHskYRuCDsQvnq55uEsfL/zUaq05pqmfbbaKuK72YREtMMUoEjek+rX5xjtgKfVfQ+Oy6g+0/4MW3iXyw6i1OKdv3W0fB91aeUDLjGw5e37Rn8pWrcgPN1p7mZ4UUdtqqtsWtdMUcNLe6Nxw6TpAa2pZ6sfgZPkcjvsOauPlhGp+CWvLAR/bmnq+PGM5PgPIGPmQrMt6l548ifY26ptdk5o7nYrhFF49/0xVU9HI/GWyxyxSkAnzLGP7d8L21XzAcM+IepuEmvrJxE0lUmmu9gq21VO45AyNi0/IgkH5FfRTyw8a6fmD4K6d4pQ07KeS6QkVELTtHMw4ePlv5IrlZERAREQEREBERAREQEREBERBomsdHab19pqv0jq21QXK1XOF0FRTzMDmuaRjz7EeRXl7zJex1obFpvUGvOBmt62omoIZa9mna+na4yMYC4xQzNIPVgHAcDnGM7herqIPBDk69oDxT5ZL3btHahr57xoBlQIqy1VQLpKFhOHugJ3YR3LdxtgDyPuRw84i6M4q6UodbaCv9Nd7RcIxJFPA8OAyAelwHZwzuD+cbrz79qPyPaYuuhZOOvBzQsdLqS21XiX+mtcGBX0rwQZzE3A8RjsEkDJBIwThdTPZx84d05deJceitSVM02jdVVMVLURPd8NFUF2GzNHlknB/RuSrm9S17vIrGPZKxskbw5rgHBwOQR5YPor1FEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBEUc0giifK7GGNLvyDKDw69qxxgPEPjnHpmlrjNR6cZJC2MH4WkkAEeWSAfyn1XdP2OujJdN8uN1vc8HhyagvJqs/3TWxhrD/5uD9q8muJs1y4n8wV/hYXS1N51JLRxdO+xn8NuPsAK+gTlh4VM4OcGrBowtDZooGyzAbdLnNG31gAZ+eVPys/lhP24u9prb6O5cnGtmVkbXiBsFRH1eUjH5aR8wQvNX2X94vVz4p3XhRT3KentmqqUMqmxuIBaAQ4keZwR+Reh3tXby+zcmuoRG8B1dcqCi+sPkOR+QE/YuinsdNLtvXMRdby9hIstq8XqA+iXlwGfrICW3OGa1H2gHs4bPy8aKHGfhVerhX2hleyG7UFQ0E0QkJ6Jo3N36OvDSD26gQQFzb7Jfm+r9X2y5cBOKOrjUXS2iOfTU1dOPEngORJThzjl7mnBGfJ2Nz39GtUaW09rfTtw0nqyz011s11p3UtZRVLA+KeJ2xa4H/7IIBG6+f3nb4FO5U+ZW56a0TPX0NoPhXewVLZHCSGJ4z0CUYJcxwIyNwMZ9Td/sfQxlVXVfkF5uNPcyHCCzW67XyE68slIykvVHI8CWZ0Y6RUNGckPADj3Oc588dqFJdBERUWuaHDBG35l4g+140pa9Jcx1Ayz00VNT3KzsrXRRNDWiQvIccDbcjP2r3AXj77ZrQOoJOLGmddwUMktuks4onyMYSGEPJGcDtkn83qpk3aOpeieHerNEcLncaam1SCgqHFtPI5pAc0bA57EE7j6wuGrvdKm83OputY4GaqkMj8bDJ8h+hexFup+FnMPyP2vgdw9npW3yltkVOyFgaHslY0gnI8ycbY8l5T3Ph7X8NOLdNofiTQGD3G508ddG7s6EyDLt+4Izn7QkupJ9emfseuJHE66WO5aGvNNVT6Xoml1FPKw9EeBkBpOxGfh2+XovThbO4UaQ0fovQVlteh7PSUFsdQwSRinjDRIHMBDye5JznftnAwFvFMkvF9ERFQXVz2hXLzQcdOAN8mo7Syp1Jp6lfW2x4bl5LRktB7+p+rPqu0asexsjSx7Q5rgQQdwQe4wk5dHzScDuO/Ezlu1/Brfh3dpLfcKZxhq6SZpMNVGD8UU0Z7jv8wc4813utvtt9beHE278GLKHhoEj6eslIJ9QHEd/rXePiZ7PzlU4pXKqvV84X0VHdKxxfNWW9xgfI8j6TgDgn6gOy6pcUvY3abqoamfhhqJ9PKSTDHUSnA9AQ44Hz3Tnujl/ls9qFwh4z14setJKbSFykOIW1MvRG4+mSSPtBwNs/Lt/Sa30XcGCSh1fZalrsEOir4ng57dnLwy1j7LDm501USttWi6e/QMyWPoquPreP8AvSQM/LK4e1Nytcz/AA/nxf8AgxrW3yNAw+K3SyDB9HRBw/IVN3sH0iQVVPVx+JS1EUzP7qN4cPyhbW4q8LdH8ZtC3Th7rq2srbTdYTFI0gdbCRs9h/euB3BC8uPZbaN5m7VxZFdqOn1LQ6QbATPFdXSNa53o1khyNhgjA3x8166nZSdg8IOev2e155TKah1xpvUkuotF3SrNH4s1P4dRQTEFzGS4Ja5rgHAOyNwARkrtv7HfmC4hcQbLqvhJrG4VN1oNLU1PWWurqHF74o3vLDAXncgYBGf/ALPoJxB4d6M4p6VrNFa/0/S3qy1zQJ6Spblpx2PqCD2IW3eD3L5wg4B2+qtvCnRVHYoq54fUuhyXzEdupxJO3+r0C1pHI6IiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAoK2ljrqKoopfoVETonfU4EH9KnVrnBrS5xwBuSg+a7mK0bV8KOPep7LE/46K7SVlO474BkLxn6jkfUAvcPkD4tzcXuXGwXuqhMdRQg0Tx8mgY389yfzLxK5ttTO1nzF6yuMXx/9UpKaPpOc9LiMfXkkfWvan2dehhoTlh09b5GETVLnTyZGMkhu/wCXKg7OIiKgiIg4F54mU7uWbWLqoAxx0vW7Pyz/AK14Kcv0og416Om6ukC6xbnyzkL179rFzA6f4fcBqjhXS10UupNZPbA2ma/44KUbvkcO4zsB88eoXnT7Pnln1NzAcZaW40TTT2TS0kdZX1bgekOz8LB6k9yAQe3qiePfa2vElvpZP7uFjvytBWUoKOnbSUsFI0kiGNsYJ8wAB/Mp0V5+e2iiMnLfpl7R/YtVwuP1GnmH6SFxl7EG5OFNxPtGThz6Cp+WQHt/LuudPa7aXrNQ8qMlfRwmV1mvVNWSAfvYwHBzvsBK6Seyb5gtMcIuKt30dqqoZTQavjiggne7AErCSB6Z7fMjPokm8Tx7borWua5vU0ggjIOcgrRNdXuTTOiNQ6jhH3y1WqrrmfXFC54/O1FefvtnuItrpeEGmuHFHcIZK+uvbKuqp2vBfHFHE4sLgO2Sex+R9F109n7yhcOOLvB7iFxP4pysijpIZKW2+I5oDGtjLnS7+rh0/YMd1x1ykWS5c5/N9R27i5daq6UdfHW3SsimlL2tjjHUI2g7DcgA4+ZXKftOrBcOW++6e4X8MLrU2fSOpLXJVS0lPIWdXQ/oMbiMAjfJwNwQPUEl8Xex/wBS0en+Z3VOjKOqbLS3W11DKd+QfEZBIXBw9cgBext4o47jaq23TQNmjqaeSF0bhkPDmkFpHoc4Xhr7I6llqOcS2SRt+GGyXFzj6Doavdbtup9V82lDetV8sHMe3UNFb3UV00dfpJ6eB7cB0Qe4ADyIMZIHlv8AYvfvlx48ab5jeFds4maajMUdXmKogJyYZ2gdbfqyfPtuN8ZPnN7aThbpqw3LRPEu1UEVPcL3LU0VY5gx4pY0P6j89x+f1K5e9izeJq/l91ZbJHEstmpjHGPIB8Ebzj7SVR6GIiIMW5W6iu9DPbLlTMqKWpjMcsTxlr2nuCF85XN3oB/CDmc1zpK3wmkit14dUULmt6R0PxIx7flknH1L6P15oe1n5Qjqiy1PMxpMYuFmpGR3uAN2kpmYAk2Hdo/n9dg73cAda23iLwU0TrG1VbamG5WOikdI12fvgiaHg/MOBXIK8T/Zc83+o+G3FG08D9WXgy6P1NMaakE8m1DVuBLCCdgwkYPpnHmMe2CkmAiIqCIiAiIgIiICIiAiIgIiICIiAiIgLzo51fadap4A8TKrhfw80fbq2st4zU1NeXlgzkDABGdwdttvPsvRdec3OR7N7UXH7jj/ALIOnrkKOkuQYyse3pJGDknfsRk99t04SbXSnjF7UDml4wafqdKzXy16atdWOiojstIYpZW+bTK5znYPmBj9GJOVj2cPFnmm0lU8QItRUemLTJK5lLU3KmkkNY7uXNAIJGT33znPmM+oPCH2bPKtwxs9FFceHNDqe8QBr5rhduqcmUdyxpOGgHsMf6l2ettrttloIbXZ7fT0NHTtDIqeniEccbR5Na0AAfUg+e/inwZ5g+Qbitbrs25y2+vgPi22+W8EwTtzu05BG+N2nPb1Bxzfw39sVzG2S9UY4iW/TuorQ1zG1AioPdqjo83BzHYJxvggZPmvYzWegdE8RLPLp/XelLXfbfK0tdT19MyZmD3x1A4PzGCujHNZ7KPhPq7SddfuX7T7NM6qgBlioI6h/udSAN2BjiQwnyx/7CMdyuEPG7hzxs0jbtX6E1NQV0FfC2Q07KhpmhcRux7O4IORnG+Fv5fNfqfgDzJcHrvLS3jh1rKx1UTi0zUtLP0kA7ESRZBG+Qc+a0putuP9K/3caq18x428N1bWZH2EpwfTKi+czR+lOcniE8u0qziXXhm5d77VRtA9cvcAR9SmvXFbnD4D3SO3ag1vrPT1XuWR1VWX5xjOCS4fnynqPoU1Xq3TehrDV6m1beqW12uijMk1TUyBjGgDPfzOxwBuvne5wOMFu4zcyWreJ2mKl5oKuuZ7hLnfoiaGscDgdunY/LPZbb4l8w3HHjYIKHiNxEveoImODYaWaYlmSdgGNADjntkE57Lm3lz9m3zA8bbjb7le9MVOltKzubJLcLgBG+SLIz4bDuSRkb7j0Kvw/bdepfa6czF54bU+hLVQads1eKQUdTfqeCR9ZK0M6S9ge4sjeR++wTncYO60bk09nrxA5thNxG1VqF+n9Ie+uE1fLGZau5SZ6pfCBIGcnd7idyduxPp9of2bvKJpGwUlpuPCS1agqoGNElbcg+SSVwG5PxAYJ8jldjNN6a09o+y02nNL2eltVsomeHT0tLEGRxt9AApMnfptrTeG/DzS/CnRFn4f6Mt0dFaLLSspaeJrQCQ0AFziO7iQST5krc6IiiIiAiIgLwK9qPq61aw5ytXTWerZUwWumoLY6Rjg5viRU7esAjY4c4jbzyvRj2r/ABt1/wAGeBFmbw/vdRaavU15NsqKund0ytiEL5CAe4z0kfkXnp7P3k6qebniBddS6zuNS3TGnJoprnM8mSSuqZCXCIuJJJIBc4nuD37gh66cj9vr7bym8MKe6xOirHWCCSZrhggkEj82FzosO02qhsdrpLLbIGwUdDAyngib2ZGxoa0D6gAsxAREQEREBERBx1x14E8PuYjQNVw74jW19Vbqhwljkid0S08oz0yMd5ELwE5k+Dd55UOYW8cPaG9zTy6eqYK213DAZJJC9jZYXkDYOAOCBtkFfSAvBX2rNY6t50NUlxz4NDboR9TadoQe2PA/Vc2t+Dui9W1WfHutjo6mUuOSXmJvUc/M5K4c51uSfRnNdpB87I2W7Wtsp3/cm5taPiOMiGX1YSANztt6AjlXlzhgg4C6AhphiJmnqEN+rwmrkbKJ6+cfhtxA4w8j3Hx1ybb3UF/sEz6O5W2oyIayAnDmH1a4DIcBkH0OQve3gnxV09zIcELLxIs9O6Ch1VbpBNSvcHOp5PijliJGxIcHD5jBXnh7bHhxpq3O4fcUKGjigvF0mq7XWyMaAZ442Mexzj5kZIHyK7Aey3uM1LyG0VS15DqGrvhjPpiV7xj7Sl54v+vFe6afuN71/cNOaats9wq6i6VFPSU1LGXySnxHABrRknYZ+QBJ7Lvd7P8A53ZeVypk5d+Mmn62itst06opZB0vt80pGQ9pOwJIJ+zceei+xvsFp1JzUX293mFlTW2fSlXcKMvaD0TvqqaJ0gyNiGyvAI7dS297Xa0W+z84VZUW6COB9x09bq2o8Nob1TEyMLjjuSI2knuTur//AFL17lU1RDV08VXTSNkhnYJI3tOQ5pGQR8iDlTLZHA+rqq/gtoGurSTUVGmLVLKT5vdSRkn8pK3uooiIgIiICIiAiIgIiICIiAiIgilijnjfFNG2SORpa5jmghwOxBB2IPovnt9oJwiPBrmn1haaC1ihtN0qhdrY2NhawRTAOIae2zyc47ZC+hdcXcX+Wrgxx1dTScTtFUd4lpABFLIwdYAzgZIOe5Vl+I6yezO5xdT8d9GwcO9c218t201TNpW3YZAqo42DpLx2L+kAEjG47brvYtg8MOBnC3g5Rmi4d6Ro7SxwwTE34iPTPl9i38sxYIiKgiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAtN1HI6HT10mZ9JlHO4fWGEhaksG9xNnstfA7GJKWVh+osIT08fPJyc2f9nnOXw9oJ4DO2v1QKqZnn4bC+Z5+xrCfsX0MX2+2bS9nrNQahuVPbrZb4nT1NVUSBkcTB3JJ/8AfJAGSvnz5SuLml+WTm6tmvdb000tmsVdcrfWOgZ4kkDJI5YDI1o3cW9WcDcjK5q9pDz4WzmRrLNwx4MXW5u0XQ/7YrpDE+A3SsccMaWHBLGDIAI3c4nyCZ3Rt72ivPRJzN6lj0BoYuh0Dp6rdLBJuHXKoALfGdvgsAJ6R23z8z2t9jLwcvmltEaw4qX+0T0Q1HNBRW4zRlpmpowXF4BA2LiMHzGD2K4n9nZ7OO7arvVHxk4/aVlpdPUwE9os1a0sfWy5BbJKw4IYO4B7n0I29dqCgorXRxW+20cFJSwNDIoYIwxkbR2AaMAD5If4yMb5XAfN7ynaC5ouHdda75bms1JQUkrrJcoxiWCcAljCfNhdsR8z8wefUQfOhyc8RajgXzV6Mvt6rprbSUd6FsvLWuwPBe4xSMfjYgOIJ8vh9F9FNNUQVlPFVUs7JoJmiSORjg5r2kZBBGxBBByF4ge0y5M6ngJrd3FnTMpqdKayuUznMDMGirH5kLHYGAHfEQdhnt3wO2nsg+Y++8R9AX3hBrK9SV9x0iYqi2yTv6pXUUhILSTu7peMDucHJ7pn0eiKIiAuPuN3CHTXGXQVz0nf7bDUSzU0gpJXNHVFL0npIJ7DOAVyCtOv18tumrJX6hvFSynobbTyVVRK4gBrGNLicn5BKPm90TxR1zy2cXa+5aQrnxVNkudTSS07nnw5RHI5pDgP+97haHxn4sX/AI18QrlxD1HHHHW3DpBZH2Y1owBnG/mc/Nbn0q3T/F/mlpnXdrRZ9Vatkke1xwDFLUOc0H6wQD65K788+Xs5dIaY4d1fFHhdTGCaz03iVkLQAHho747eXfy/St7g7e8hfHWycZeX7S0UNcH3iyW2CiroXHLh4bQ0H6sAD7F2SXid7H/iLdLPzGu4fuqHm3Xy21U3h52bLGzIwPnt+Re2KEEREBERAREQEREFrWNb9FoGfQYVyIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAtO1FKafT9znacGOjmeD8wwlaitF1o/w9HX2T+5tlU78kTkHzVMZLqXjX0byuuWpzn1IfVZP5ivpA4S2JumuG2nrM2IRGChiy0DGCRn9BC+djl8t37IeYvRVA5pPvWo4C71P33qP6F9KMMUdPDHTxjDYmhjR6ADA/QrcEqIigLBvdwFps1fdSMiippagj16GF38yzlpWqYhPpi7wu7SUFQ0/UY3IPm+4va81pzD8bbhdrlXy11ferq6joGSv+GNjpS2No9BuCfrK9zORvlkt/LDwWpdM+IyovV5e243apDceJK5oDW/U0bD8nlleEmhsW7jvaRFsKfUrWtPpiowF9J+k6h1Vpe0VLjl0lDA5x9SYxn86cnDWrIiINu690RYeI2krlozUtIyot9zgdDI1wB6cggOHzBK+czj7w8uHL9zAam0PSyOim0xd+qjkHk0ESREH0AIH2L6VV4g+184eVunOZ12to6R7bdqa10zmy9B6TPEzoeCe2SACB8j6Ij2L4NakptY8JdG6opZxNHcrHRVHXtuTC3q/Pla/qeyx6j03dtPSuDWXShnonE7gCSNzCfyFdA/ZOc11Lr7QdLy938Fl40lRn7nPJ/s1I07NHr0gn5jB8sL0QUxXhjoHTeuvZucz8evdf6cllsVOyrooJW5PvNPLgAAgH4sAHO42ytg89/NxS823EW06jtVkltlqsNE+io45SDI5r39RJI7nP1fUF6i+1M4SScSuWC43K10Qnu+nKqGuhIaC4xA/fGg99xjG+BuvCOSnniyZIZGdJ6TlpGD6H5/JX8eTKj0I9jFpaor+OuotU+7F0Fss7ovF8muecEfkI/KvZteJ3sneZDRHBviTdtEa6qY7fTatYyOlr5CAxk4LcMefIHAx9Z+S9rIZoqiJlRTytkikaHsex2WvadwQRsQQc5HdFebHtvYgeGPDObG4v1Y3PyNOP8AUs72JH+8pxBGf/lpj/VI1ontvLhGdFcMrSHDrF0rahwz5GFrQfzFa77EyCWPglr6ZzXBkuqGdB8jikjBI+1SXR6OIiKgtI1Zpm0600zdNJX6mbUW+70klHUxuAIcx7S09/PfI+YWrog+e/nI5PNd8ouvYp4Jaip0zWz+PZbxTlwMRa7LWOcN2yNOMHIO2fme7Hs4/aMHU8b+EPMVrWkguEDGNsl6uEjYhUtGxhleSB1jbBOMj6iR6O6t0XpLXtmn07rPTluvdsqAWyUtdTtmjORjs4HB37jdeOHtBPZ16o4VatruJfAzRVVXcP6uL3mpo6BpmfaJBs8Fg+LwicEEA4zgnbZo9oqKtorjSx1tvrIKqmmb1RzQSB7Hj1DhkEfMLIXhF7PHnavHLtxFptJa81BW1HDy9ltNUQzSl7LbKcBk7Ac9LR2IGBg523K9zrPebXqC2Ut6slwgrqGtjE1PUQO6mSMO4II/9/Lugz0REBERAREQEREBERAREQEREBERAREQEREBERBDUU1PVxOp6qCOaN3dkjA5p+sHZbePDLh4ag1btD2Myk5LjQxnf6sYWj8Z+OXDTgBoybXfFDUUdqtsTvDjb0l81RJjPRFGN3u+Q+XqM+O3Oh7SnXnMLcW6R4ST3jSWjKd5aGxTmKtuT84D5DGctafJgPmM75zL+M/LlP8AHqlzBczvBnle0RX3W5XWyxXRkbhR2akfG2aeUDYGNm4H1gZ8vMjwl44ccOJHNBxJfqHUDHVNXX1HhW+20rMiPrdhrGgbuccgZ/JgLfGhuQrnA4sGivkHCq+RUd2DJW3O7vEIcx3Z5EjvEIxvuF6/8qnIvwd5dtJ2mabSFsuutGQtdX3qrhbNL4x3IjLshgB7Y8xkHsr5yeGT1wxyXezA0Dwut1j4mcXon33WL4Y6ttvkx7pb3uaCG4/fvHYk+eR2yF39YxkbBHG0Na0ABoGAB5YHor0QEREBERAREQFHLLHDG6WV4axgLnOPYAbklSLhrm44zWXgXy/6y1vcqyKKrba56W2Quf0unrJWFkTW+uHOBPyHzCDyu9qZzeac4+avtPDHQb3T2LRdVUPqKs9qmtIDCWfJoBH1k+eQO4/sgOHN/wBD8uV3uuoLRPQSalvz66Bs7Cx74WwxxtJB3A+EkfWV55ezT4c6f4s83unLdrOibcqKhpq28zQzjrbPLFGSzrB7jrcCc+YXvtT08FJBHTU0McMMTQxkcbQ1rWgbAAbAAeQS9EyIiAiIgIiICIiCmOy8B/ahPe7nO1t1jGGUYH1CnZhe/GV4E+1EmZNzn6z6BjojomH5kQNz+lTfhn17YcubBHwH0BGDkN09Qj/8U1cjY3ytgcAaCS18D9B0Erg58WnqDJHnmBh/nW/yqks/LseZntv344f8Lo/M3i4O/JBH/rXKPswGNdyBxsJx11F+BP1yPC4s9uAD+wPhafIXe4j/APExLlj2U9ObtyNU1sZ9KW53mmH1vecf9JJL9Xjpd7F0O/bVag6c4/YLXdXpj36h/nWje2FDm84DnHsdL20j6uqYfpBWs+yHr6fRXN/f9OXqWOCpqtM3G1DxHBo8aOrpnloJ8/vJAHmth+1R4g2TiBzgX4WGobUQadt1FY5JmEFr5o2ukkwRsQHSlufVpTPqa9uOC4xwd0IO2NM2sf8AJY1vNbG4GVbK/gpoGsjBDZdMWtwGMd6WNb5QnRERFEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQUcuOuYTiJQcKuDOrdc3GdsTLdbJ3MJOMvLCG4+e+fsXIy8xfbI8wVXYbTYuAdkmAkvVObnc3NduyEPLWMI9XYONuxO/ZP8T9PL/TGltW8aeJdHpfS1uNbqDVlzcyngBwHTSuLiSfIDJJPoCvYblM9ldoPgjdqDXvE68R6s1NSgSRUwgAoqWXAOQHZLyNxkj1wcEhebns/OL3DzgbzL2TiDxNm8CzU1HV0/vHheJ4EssfS2TGCdgT2332XsQ/2iPJm2jNXHx2scuG9XhNZMJD8sOYN/klmq7Gta1jQxgAAwAB2CuXTS6+1p5NrZUPp2ao1DWlhx10tlkcx31EkArqVzie1fZxM0xNoHl/ob1Y6WpOKu8VfTDPIz+5YxpJaPrPofLCYPXGS+WWGUwTXeiZINix1QwO/ITlXC8Wh7/DbdaMuO3SJ2k/kyvnH01wO5quKdKNSaa4fcQNQwS7itZBUSB49Q93cfMFay3lL50myjp4NcRRJnY+7TA5+vKk36c+PdnmW4KaX5g+C+o+Geq3iGnr6czU1XkZpKmP4opgTsCD3z5EheCXDLiXrvlC48TXPTt1a+rsdY6hrRC7MVXAHDqGOxyACAc4JI9Vr104bc8XC2ifWXqxcSrHRtYXOFTNOIunG+WucR2+WVwa28VT9RMvl68SsnFW2oqRKfilIeC4HPbOCPkrPTOPp90jen6k0pZdRSQ+E+6W+mrXR/wBwZI2vx9mcLV11/wCBHOFy+8UtCWOusGvrZR1HukME1uqX+FNTyhgBYWnyBGB8sdlz3DNDURMnglbJHIA5r2OBa4HcEEbEH1TUiQ7rx99q7zg6hvOv38AeG2sOjTlrpGi/OoJRiqq37mFzwdwxuAR6nHrnmf2nPPZYdI6GqeB3BvWIm1Xd5vAvVZb5jm3UgyXxiRvaR5wPhIIbnffC6q+zI5Q7VzH8RrnxA4mW+ordJ6WdHK6KQnouNc9xcI3u7uaAC5w884z3Sr42DfuRnjnwz5fbFzSx5jicILn7pDGfeaGneQ6Kd47gYwTkYAOD8+X9C+0w1bqrgRq3hTxfngq66S1SU9DX9ADpmdJwwjO7gQPzY9B7MT2m1VVsfZKm3U0tukh93fSviaYjFjHQWEY6cbYxjC8CfaQ8M+HXCjmfvGluGltht1sdSQVklJAMRQzydRe1o8gCCMfIJ/qNq8ivEmz8KOafQesNRXBlFaoq59NWzvIDY4pY3MySfLJavohtd0t16t9PdbRXQ1lHVMEkM8Lw9kjT2II2IXzT6itukK/hnp66aYoybzBI6C7BgJdnBw4j0JAIPzXpR7I3mbtsGjbjwQ4hanZT1dLXCSwCtmwTE5o6oQXHOxzgbAAYUuSrOvTxERUEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQFomtg06Mv/WcN+5lVn6vCdla2tF1nSS3DR99oIATLU22phZgZPU6JwH5yEHzrcprj+2l4eOiYZP8A4RwFoG/UOor6Q18z3CrVcvAvjxp7Vd1o3yO0hfmS1UJBDi2KTpfgeuMkDz2X0bcMeJOk+Luh7VxA0Vco621XaFs0T2OyWkgEsOOxBOCEG60REBaZqV3Rpy6v74oZz+SMrUh2Wlar20veDvtQVH1/2NylI+arTbpJeN9A6Bpe9+pmFoHzqV9JuhRI3RtkEzel3uEGRjH7wY/Mvmrs98j0jxip7/UxjwrXqAzStcOzGTnq2+QzsvpK4Z6ntusuH+n9TWiZktJX2+GWNzSCPogH84KmdT63QiItKLgznC5ftLcf+COpdPXa2xPu1LbZ6m1VfQDLBUMYXsAPfBIxj5/XnnNRzRR1EL6eVuWStLHD1BGD+lB87nIzxL/2Gea3RGo7rVuoaI3B1ruRdsRFMDG5p/8AGLfyL6Iaaogq6eOqpZWSwzMEkb2Oy1zSMggjyPdfP9z88sd15a+M9RJTvc6zaknmuVsnbkdJL+pzQfIgnIx23HkvT72WHGDUfFvlsDdUVr6yt03cn2vx3uy5zAxrmZ9NiPtJU+6N2+0Z4lT8LOVXU98poxJNXSwW2MOGR1SuOCfkCAvC7RctRr6+w6Muc7Ou9TiOCXpALJicgem523+rzXsZ7YKvip+UR9I+RofV6jtzWNJ3cW+I44HngDdeR/Kzw+v3E3mC0HpXT1PJLM++UdRM5gz4UEUzXyPJ8gAO52yQtT/ntTNbj5juTvi7yw0llv8ArOjDrVendNJXQbBsoHUGOwT0uwMjfy8ts+rfsp+Nt44ucuL7RqOtkrLno+4G2PnkJLnxOaHxZPmQCc/Wsb2vlTZ4OT+rhuPh+91OoLay3h2OrxQ5znkef9jEmceq2b7FfS1TbOAGrtUz9Ph3zU5ih9cQU8YP53n8nyS3+l/11R9rXxgqde8cqbRkeBQ6VifBGPVxOCftIJ+0LvL7I/ScGneU+mubSDNe7vU1cpHphoaPsGAvMHnNv/8Asz83OqLbo6jbK916ltVOGEffZWyFrjkeQII+wr2W5H+DV+4F8vdh0PqWbruAzUyt8mF4G3y7dlmf88J5rn9ERUEREBRyxRTRPhlja+OQFrmOaCHA7EEHYgqREHjn7TnkKtvCrxeP3CK2ys09cKwi+WuGMllvlfkidmNxGTkEdgSCcbk6B7Nzn3uHBq+03Brind5ajRNzmDLdUzvLja5nHGASf7ETjbyPbyA9oLlbbfeKGe2Xahp62jqWGOanqIxJHKw92ua4EEfIheAvtI+E2hODXNTfdKcOrTHa7RUUNHchRRbR08szCXtYPJuRkDyyg9/aSrpq+lhraOdk9PUMbLFIw5a9hGQQfMEHIWQur/s4LjrG78pejbjrGvlqpZYXCjfK4ud7s04YSTuc4P5/VdoEBERAREQEREBERAREQEREBERAREQERcMcwXNzwQ5ZIqL/AGVdSTUtVcAXU1FSU5qKh7RsXdAOwz5koOZsYXAfNTzgcMeWTQlxvV2vturtStYY7bY46hr6iec9g5jTlrRnJJxt8skdO+aT2vGgbxw9r9J8u1JfnX26xGB14r6YUzKJh2cWNLi5zyOxwAN156cKeX/j/wA0Gpnu0RpW86iqKmYGsu9T1e7xEnBfLO/bY9wCSPRPBq3GXmA4886HEa10uqKyS6VtRUils1loYyyngLzgBjB3OO7jvgE7Aleh3KB7Jii4e36ycUOO98p7tdaAsrKbT1NFmmgmAyPGe7eQtPYAAeu427Fco3Inwn5ZdO225OsVJd9deA0198nZ1vZKQC5sIORG0HAGN9s5XZ5PRaAGgAAADsrkRAREQEREBERARFxnzJ8S/wDYe4E634jxyhlRZbNUzUvxYJnLCIwPn1EEfUg688+ftB7Zyv0lPo7h/T26+a5rgS6Kd5fBbox+/lDSCXE9m5Hbfzx5A8eOaDjVzIXaO58VtYS3GOncTTUMMYhpKfP9xE3bPfc5O533W5eVbhhWc2HNLp7SGuK6tuMN9rZrhfakzESvp4mOllw7uM4DRjGAQBjZe2+r+Wvl20fwJ1To+g4V6Ytmn4bHWvnMdvj8RrWQucZTKQXl4x1dRJOQrZPiT9vLD2PBpP22rhNjxzpuv8H8sfV+bC9w14K+ymr6ih50tKMgziroLlTyD1YaZ5/S0Fe9SiiIiAiIgIiICIiAvCL2sWka3TvNzebxNCRTX6hpKqGTycRGGuA+rA/Kvd1dN/aLcmlx5m9CQXvQkcR1hYAZKWKQ9IqoxkmPPkSCcZzvjb1aNS9nxzZ6H488KbLomK5Mh1fpe2Q0tfQSHD3sjaGCRmfpDAAz8s+oHbZfOjwY15xM5HeYm16q1Vo65W25WV74LnaKphhfUUrx0yMBOx9QQSMjGRuvcflr5sOEnNLpua+cN7pKKqh6RcLZVsEdTSk+oyepudg4bHbtlSTJo6Ye3AcP2CcLW5//AEtcTj/9jGuVfY+XClquUNlJDKHS0WprjHM0HdpcInDI+YcCofawcvGuON3Byx6i0FbpLjX6KrZ6yooYx1Sy00kYD3MHmWlgJHcj8o6k+yb5o7Pwc17dOBWunGgoNa18Roqib4BTXIDw+iTOOkPAa3fGCBk7AG7vg4i58+HGqeW3my1FctMz1Vmp79M+9WippXln3qYnraCO+HZyPUrkjkM9n1Tc2NqrONHE7WNZDYmXaSnNLTsDp7hOwtdL1yE/C09e5Azk7eeO0ftm+E0WpeCOnOK1FRsdX6Ru4pKmUMJf7lVNIIJ8miVkZ32+IrY3shOZ/h1p7Qdz4Aayv9LaLy+9SXKzmpd0Mq2TRxtfGHnYODoycHuCT5JqPTqy2e36es9DYbVTiCit1NHS08Y7MiY0NaPsAAWciIoiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIg4t5k+O1h5cOEV44ragpJKuG3dEUNOwgGaeQ9MbMntk/+47jw4uL+MntFuZx9Vb7dH91746OLYE09roY9gXHv0tBJ8sknsMkd+vbKccdOWrhRaOBFFUw1F9v1yhudZE1+XUtJACWlwGcF7yzAODgEraHsQ9N2d9HxQ1c+nabpDNbreyUgEtgc2V5APzc0Z+xBy3rn2TnBat4Bt0DoxklLrSgphNBf5jl1XVgZPijYBjjkYBGAR6ZHjPrTRt/0Bqu6aL1PRmluloqX0tTETkB7dsg+YOxH1+XZfUWvn29pPFboucPXQtrGtaZ43SdIGPELfiO32JvwdhuHfsZ9Va04b2jVt14xUNout4o4q1lGLY6eKJkjQ5oc8PaScEdh+TsuUuEPsYNL6Z1BR3zilxQfqGGjlZMaChofAimLSDh5c4nHy8/MEL0B4Pl54SaJMjsuOnbaXH1Pu0eVvBL0YFls9s0/aqWx2aiipKKiibDBDE0Naxg2AAH6fXJWeiINPvdkteorVU2W8UjKmjq4zFLE8Agg/X5+hXz688PLxfuB/MBfrJTWaoNouU5q7bLFAeh7X79Ix55OcfP619DJI81oeoND6P1ZNT1OpdNW65yUpzC+qp2yFm+diR2z5dlM7sWX4+dGbl4486P0TTcUG6Vu9Bb5SHNkha9szW9w8gbgdjjvuMjvjddn5+eaewaEm4dW/ibVstkkLoPEdGDURsOQQ1/lsSO35919Cc9ntNVQfcmptdJNRdPT7s+BrosenQRjH2LYdfy3cArpLJPcOD2kp5Jv7I51riy768BXm7U2x4E8rHL5qfml402zQ1LJUmlqZzV3q5O6nmGAHqkc55zl7twMnJJJ3wvoK4R8ItDcEdD0HD/h7ZYrda6FgGGN+KV/76R57lxPclZOiOFfDfhtHLHoHRFmsDZ9pfcaRkRcM5wSBkjPl2W68hXURVT5WU0r6dnXI1jixvq4DYflwvn59oTT3E8w13uN5LxXVORIx/cAOJG3l3IX0Fg5Xih7Yjh5X6c5iqHWsdKWWzUdpiLHgHHjsLg8E+p74+RUGm+zy4AT3PUsutuJFlfT6WmiaY5KmMhjwM/Fv5b/AJlwzzlUto0nzIXmp0BKaS3wPimt89MegNI3ywjsQfTtsu1PMhzt8Mafk20Xwh4P3CMaorqSmZcX0zA00MTGYkJcOznk4AO4x2IC3/7LnhNwq5guXPWNBxT0rTagq4r1JbJKisb1yxxOgZI1zHnJDgXnB8i0fayew765t9npzx6b4/6GtvDzWt7gp+IVnp2wSxTODTcY2AASszsXEYyBv9ZyT3SJwvne4/cCeLvI9xxilj97pIqGvdW6Yv0efDrIGuyw9Q26g0hr2HHmOy9iORLm9t3NlwymuVbSsodU6ffHS3mlDhh7i34ZmDv0uwfLAIxt2FzFdmkRFAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERB5L+1n5O9P6UoafmL4d2r3RlVWil1DSxD4A+QExzgDtuCD5AEb4AA2t7KDm+tfDC+VvAriBcnQ2W/1AqLPPI7LKaqOz2Y8g/Y7eeT5YPrVxH4f6Y4p6LumgtYW9lZarvAYJ43DOx7EZ8wV86vMXwurOXfj/AKm4eUFVM06auYfQVH74x4bJE8HzxkYJ323TNP0+k9rmuaHMIIO4IPkrlsbgdqQ6v4N6I1M6QPfcrBQVD3A5BeYG9Rz9eVvgbpUl1VRyxxyxvilYHMeC1zT2IPcKREV40+1B5HbFwbLOO3DdksdkvNf4N2oXbilqJMlsjT5Ncdsep+07i9lbzss0tVM5duJVyJt1ZKHafrJXZMEhwDAc92kDI8wBsNjn0+4x8KNL8beHV34cauphLb7tCWE+cbx9F4+YP6SvnX4qaNvPLzx3vmkIal4r9HXnpglyWkhjg9hJB82kA+uSno+l0HIyCqranCrUkesOGelNUxyiT7rWajqy4ebnwtLvzkrdWd8IKoiIPMX23lNTnRnDSsETfGFzrIjJjfoMQIGfTIJXJ/scrRSUHKbVXOE5nueqq98xz26I4WNH5Bn/AMZbV9tJpOW6cFNKapjJ6bLeS14+UrQwfnK6ScqHPhqPlr4Raz4a0lL7yLx4lRa3EH/a88kYY4gjt2Byf5gnqMn2jPOFfOY7ipWaLtMgp9DaMuE9LbYW96uoYTG+qefU4IaBsG77k7dz/Y68BaHTPCq6cb7xbWG8amq30tvmkYOuKii2JaT5PcScjuNl5icunBi/cyHGmy8N7fM4TXeodPW1BGTHAD1SyH7D+fK+ibhZw5sXCXQFl4e6cj6bfZaZtPFtjqx3OB2yfJLd5R0U9tjSVMnAvQtXGT4MGqnNkx2y6llwT+Q/lW6PY26iobnyq19hhkHvVl1RWsnZ5hsscUjD9uXD7FqHtfNOS3vlNdcYWF77RfqOpwPJpD2uP1AErqz7GHihNY+JGqeGNROBR36njrI2F2Pv8YduB5nAA+1SVa6d1WoLhwp5pazVGoaN7Kix6xnrKuN7T1FvvLnOOO5y0kgeeQvoe4XcSdM8W9C2nX+kq6Oqtt1gbKx8bgQ1xG7TjzBP5MFebvtnuCmkLPpzSXGixWGGkvFdd32m6VELOkTsdC+RjngbF2YyMnvn1Jzv32LWrLneOBOsNNVtS6SnsWom+6sJP3tk0DXED63NJVHoiiIgIiICIiAvGz2xPBnVFFxnpOMtLa55rJdbZTUdRUMaXMhliBaOogbZHmfl6r2TWjaq0lprW9kqNO6sstLdLdVtLJaepjD2uB+vsfmN0HidyZe0r1ry72+28N9aW6K+aJp5AxhAIqaKMnfoI+kAPLBwPIle0XDfiNpLixo22680TdIrhabpEJYZoyDjIBLT6EZ3C8e/aDezvuXBW6P4mcGbFVV2iqrLqykgaZH2x+fQDPhkeY7eQxnD2afPhYuXx1Xwj4pvqRpa81jZ6CvZ8Qt07tnBw/8AmbiQSfI5JIAwWbeH7r2sRY1DXUdzoae5W+oZPS1cTJoJWHLZGOALXA+hBBCyUBERAREQEREBERAREQEREBYF8usNis1feqlrjFQU8lS8DuQxpcQPrws9adqC2C82K42dx2rqSWmyfLrYW5/Og8guLfti+OI1Rc7Zwz0zpu2W6lqJIIZq2mfUyvDXY6sdQAzjtv3+xdZrZZ+ZH2gPGOWqkml1Bf6loE1TI3wqSihHZuGjDGgZwPQbnA270cOvZEWO6cULjqDihca0abNS+oZQU8oYZi5+enqG4Gc/LB8+y9BuFnBPhZwTsg0/wv0TbbBSEASe7RYklO2S95y5xJGTk4z5Kb8hkebnDb2J1f4tLW8VeL9OI8sfNQ2iicTjzYZXuwfTIA+RXplw14b6R4S6Mtug9D2mG3Wm1wiKKONgBfgAF7iBu443J+Q7ALdSKgiIgIiICIiAiIgIiIC87vbA8Z6XTvCqk4UUlwDK6+vbNLA127og7fI9MA/lXogPVeEftZdWzai5v73ahVeJTWC3UVExgORG8wte/wCokuGfqUw1yL7FzQlRdeO2p9fS0rvdrDYH0scxG3izyMBaD69LSfqPzXcL2pfMbb+EHAat4eUU7v2Qa+p5bfC1hwYqY7SyH0yMgH5Eea3L7NfgLBwU5arJX1lIYr5rGKO9V5e3D2te3MTD57MOftHovNj2snFJnEDmwuOm6OpdLRaJoILO0EYDZyPFnA9QHPA+tpVJ3rX/AGPnD2537maOuxSONt05aasPlLfh8aVnQ0A+oBJ+0L25XV32eHAW08GOXXTNcKUNvepbfDcq+QjceKOsNHywQfyei7Q5CUVREQEREBERAREQEREHWPnX5ItJ83WmKMm5ssOrbKH/AHNuvgeI1zXYzDMBglhIB2OQfsx406f1Txq5DeYatp6KpNDqHTVX7pcaYPJprhT7HpO2HMe0gh2Dgn6wfoyXRD2jnIXFx/tc/Fzh3CI9c2mkDZqdowLlCwEhp/7cDIB/9oOfP8MdquA3G3RnMJwxs/EvRddHPS3KBpqacPBfSVGMSQvHkWuyN+4wV4T89FgZwy5zuINPp6nFvjo73FcqFsQwIy+OOYFvph5JC3hyA8yN+5UuN9ZZtYPqqLTl1ifS3q3VGWCOZm7JOk7NeMEZxuCBnC455p+Klv5jOaS/63skfTbr5c6ajo/V0TQyJpJ8847+mFvPsTX0AXDTGnuLnCtmmtc2yK5WrUtohbX00oyJBJG1x+ogkEHyIC+e3mc4UyctfMfqjh5Ya+YxabuUc9rqScSeC9jJoSSD3AeAd85C+jHTFH9z9NWmgO/u1DBD/wCbG0fzLwP9prLPLzs8RvHaQWS0DW582ihgwfyKTs6vde73DO4SXjhvpS7TSGSStslDUPf36i+BjiftJytzLZfBXH+w3oPp7fsZtePq91jW9EBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQERcCc1nODwv5VdLOrtV3AVGoa6nkfabND8U1S8AgOI26WB2AXH6tu4Ucq6x4l8P+HtFJcNc60s1igjYZHGurI4iWjuQ0nLvsBK8peaf2ufEW4anuukeXV9DarBTudTsvssAmqanGxkiDstYO2Dgn8xPSi81HG7ms4m3C+01n1BrDUN4q3SeDSQyVPgdbiWxjAwxg2AzgbZO+SvRzkh9lbZ7XbJNcc1Gk4q641GPcNPSzZjp2Y3dOGHdx/uc7fZuieujHL/wK4w89XGWajrNQ1lVM8e9XvUFf1SiniB/J1EnAaMDJ+eD7ZcqPKjoPlL0LVaP0bXVlxnudQ2quVwqgBJUytb0g9LdmgAkAD1XIugOFnDnhXavuLw50VaNOURxmG30rYg7HqQMn7SVutFF88/tELHdrFzf8QobrG5oqa5lTSk5HVA6Noa4fIkFfQwulftE+Ryk5kNIycQNFQth15YKVzoW+VwhaM+CT/dYG3/uCHLXJRxk0xxk5cdE3azXmlqrhbrNS266UzJAZaaphjbG5r29xnpBB7HPfIOOel85/KZx41RytcfLRqKpnraK3Q1wodQ25ziwPgLumQPYSB1MO+SMjBx3X0J6N1npvX+nKLVekbtBcbZXxiSGeF4IIIBwcdiM7hPODXUREBERAREQEREBeaHtpLxp0aC0nZnvgdeDWlzQCC9seM4PmBsT9vzXoZxDvtbpnRV4v1uYHVNHTGSIEZHVkDJHnjOV4P8ANazjRxe1ZcNZX+KqrqShe8huS7oGdyM/LP50zUtk9bK5POW+Tmm410HC51+Nmon00tdXVjI+uRkEWOoMadi85AGdvVe8PLpy28OOWHQn7AuG9NVe7TTmqq6qrl65qqcgAveRgdhgADYbLwp5IuKdz4Pcx+m9V27Ac5z6KdjsgPikx1tPyIC+h2x3Wnvloo7xS/2KthZM3fOARnGfkpvcazmuEOdbljpeangpWaDhmp6S+0U7bjZayVoIiqWAgtJ7hr2ktOPl27jxY4RcXuMnIZx4ukEVKKa6Wyb7mX+0zHqiqomkO6T88Hqae46uwPb6Jl48e155adV27imOYDTdjkqdP3ighiu0tPHkUtTEOjreB5OYGknyxk+ZDcR6vcLtfWnipw707xGsRPuGordDcIATktD25Iz8jkfYt0rze9lZznaf1PpGycsOqI3Ul9slPLHZ5yR4dXTtJf4fycAT9ewHy9IVcwgiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAvFT2xfDe42PmHo+IrKNwt2orTTwmVrfhE0ILTk+pH/R+e/tUdl1d9orwisHErlt1PdbjSsdWacon11PKRlzA3c4+YOCPtRNcGeyh5thr/AEbScvWpInfdXTFIfudUZJ8WkaCQ05PcAH/3Ix6KErwf9lBeza+cOw0ZPwXK31tOR6uERcD9gBXvDjKKqiIgLxV9r5wTuOlOPLOLFBQEWjVVFB48jG7Nqox0EuI83ADv6D1C9qScLrF7RjQFm1pyqa0r7jSsfUWGgkr6aQtyWEYzg49cH7FZ24lcCeyk5vv2e6WouXXU0R+62mqI/c2oJP36lZuGnPcgZ+f2EY9GV4Keyqu/3K5y9LR5wLhRV9J9eYS7/wDIXvWsyZxREUc0sVPC+omeGxxtL3uPYADJP5AqOj3tfLtQ0nK0bbO9nvFXdqZ0LT3Ia4A4H/jBeHi7k823M3rTm84xv4Z2qMCzsujrfboo8kuLHkAgdsZBOfPvlcHcYuWjilwScyXV9jljpHgOE4YQAD2yD2+sZCkuenzXaX2NNDDPzIXmvfE1z6WwStY4jdvW7fH1gY+1e1XmV41exfonHjXqe49WzbSIgPXdxP6QvZUdyqjgrnY4eXbify4as0pZKR1VVy0/iRxNGS7AIOB5nf8A9xleGXLJxLruXjmO0tq68tloorNeI6a7xO+Etg8QNlDs+Q7n6jhfSGvHn2qHJVqDTGsrlzHcO7J7xpe6NbLf4aZu9BUABpmLR2jdtkgYae+2cWTeK9GeangbZuazl7u2gaO4Ql9zghulirmnqjbUsHXBJkYyxwJB3GzzuvGzlk5lOJ3ITxkvulb5auug9+FBqS1TNOQ6JxHiM7EOAJI7ZGPqPaf2S3OVequ8Rcr3ES7Gppn0z5dKVU8g6onRgufR5JyQWBzmDfdpAG++m+1S5Idc1Ou7nzNcNLK252Svo2S6lpKZv+2KKeJgaano/fxuY1pcRu0tJIwQVD/XqtpjUNr1dp216psdS2e33ekirqWRpyHRSNDmnbzwR9RWqrzP9lTztQaotVm5WNbU4iudoo5Gaerw7DamniBd4Dgez2tzjfcDAz2HpggIiICIiAiIgx62jpLjSTUNdTRVFPOwxyxSNDmvaRggg7EYXhp7RfkqvfAXiLcNfaLs002hL9M6rjdDGS23yE5fG7HZuSSCe2fIEL3TWm3zT9k1NbJrNqG1UtxoagYkp6mISMcMeYP1lB5Y+ys5zOKF91VbuXDWDZL3ZIKOR1tr3tLpqJjcYjc7zbvgZ7AH5lesC2BoHgJwa4XXKovHD3h1ZbDW1WfFmpIOl7s99yds/LC3+gIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIsG9XSCyWevvNUcQ0FNLVSb4HSxpcd/qBQcL84HNLpjlU4V1Gs7qI6u81rjS2W3F2HVNQQTk/9q0bkrxN4TaH4kc7/ADQxuuEMldW6lu7bjf6sMPhUtIHDxHOI7AMHSB32HfdadzBceOKXN7xebcLg+ruUlVVmh0/aIASImPfhjGMG3U74cn+YL2M9nlyjO5WuEbv2UU8Dtbane2svEjACadgH3qla7vhgyTvguJOBhUbx5wOOVJyqct921rZKOP3yjhhs1ipv3jaiQdEXzwxrS7z+iO68Q+XzhVqnmy5gqOyXO4GoqL1Xm43irmcSXsc/qk39Tkgeg+oBdxPbE8yOm9YXGxcA9KV4q36drHXK9SRPyxtR0FjIT6loc4n0J+orgH2Yd9lsfNPZHNf0sqIjE4eoLmjH5yppXvLZ7XS2S0UVloW9NNQU8VLCPRjGhrR+QBZqIgIiICIiAiIgIiICIiAiKx72RtL5HBrWgkknAAHc5QePfthuGWl9I6xseq7Lb6ekqb84umETQ0uOHZ2HkS0n6yF0i0XwZ4rX/SFw4u6V0tV1Fi0zIJ6iva09Eboz1EjHfGMnt6Ddc+e0u5jqbjtxvktdjl6rNpTroYiDlskoOHHPnjB+0keS9PPZ88M7DR8j2kNMXS1tdT6nt1XU3SCVmDIaiWQPBB/7XA+StsnE21tL2cvPPFzLafqOHGtYqeh1tpqkZI3oeA240YwzxGA79bD0hwGdnA+pPQX2u9qoLdzj3Cqoy3xbnp+2VVUB5Shjohn/AMSJh+1bf478tvMH7PjipS8RdL3GogtEdfK2wajoZMtfGc4hnb3a4sJBaRg9wdxjr/rriRrHjVxFdrXiPeZbpdrpUQx1E7/7gENDQPIAZwFFfRly/VElTwK4fySjDxpq3Md9badjf5lyAey2jwkpYaLhbpKlgGI47NSBo+XhNwt3p71J4IiIoiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIg0LWmrLdojS9x1Tder3a3wume1vd2OwH1lfOrzV8cdScxPHLUXEC9Pf0S1Jo7dSg5bTUsRLY42D54ye5yTuV9Euv9JU+uNI3HTFSQGVsRZk9gV1x4dezd5ctIX9msL1pGnu94jnFSzxd4Y3g5B6fM53zsm5w+a1f2e3Ami4H8tGlaWqszKTUN+pBdrvI9g8UyTHrbGXDchrC0Adl2ZUccUcMbYoY2sYwBrWtGA0DYADsApEBQS1DYnMa5hw84z5BTq3pQXIiIPF32vHL5R8OuKFr4t2C2x01r1iXx1JiYGt98YOojAGMkZP1YHkuxnsWdXVl54L6z0vW1rpRYr5CaeNzyfDiliLth6dTXfkHyXJPtYtCt1jylXO4RUokrNO3KmuULunJYwFzZMfW1xXnz7LjmStnArjHXad1JKWWbWUMVK92QBHMxxLHfX8R+wEbZTDsj3TRRQTxVMMdTTyB8UzRIxzdw5pGQR8iCCpUBERAREQEREG0OLbms4b6ge8gNFG4k/aF5q3fUFtZw81KPdIpDh+/SCRsf51355s9Wx6K4Caovkj2saynDCT5ZOf5l51cBqR/GLgzqu8UQEngsmzg58j2/KPyLNt+Fks66H8IYjdeM1m8MdPjXMvAHkCScfnX0c8K6SWh4eWGlmdl7KJmflkZx+dfOhwY/6kceLJTzbeBd3Qu+WHOH8y+kDRkjJdJ2h8eC11HEQR/3oVv+r3P01tbY4l6QodfcPtRaLuNJHUwXm21FH4cgBBc+Mhp32yHEEfMLc6JZqPmn4Z6mvvLfzC2PUdyp5ae46H1Aw1kRYQ7pjk6ZQAdyCwux6gjyK+iThPxb0Hxq0bRa54eX+multq2BxdE8F0TiN2PHdrgcgg+i6L+055EKviZRDjdwb02J9TUYcb1QUrMProcZ8Vo83twc+oJ9duh3Jdzhar5O+IlV90LXVV2m7k8U97tDyY5Y3tOPEYHfRkbuCCAT2PbBs86Y+g1Fs3hNxY0Xxr0JbOIegrqyttVziEjD2fE7G7Hj964HYhbyQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAWwOP8AZhqHgfr2zlgeajTtwDWnsXCB7h+cBb/Wn3+jjuFiuNvmALKmkmhcMZyHMIIx9RQfP37OS6MtXOhwykkf0tqLjNTEk+clNK0D7TgL6FF8zLL3duCfHGW+WFhhrtJ6glkpmn4dopnAD5Zb+TK9/uVDmLs3M1wmoeINsp3U1TtT10Bx97nAw7tsNwdvl6Jb8TPrmhERFFxhzOaYq9Z8vfELTFDAZqi4afrI4owMlzhGSBjz3C5PVj2MkaWPaHNcCCCMgg9xhB4Cezb07qR/Ojof3W3VDTZ6uofcQ6MjwI/BkYQ/bY5IGDjsfRe/y2rp7hfw80neKjUGm9G2q3XKryJqqnp2tkfnvv33/Qt1IC6u+0J48aj4JcAL/Po+3Ty3e7Uj6OOqawltJHJ8D5fTIBJ+Xddolpd/01p/VVuktGpLNSXKjlBa+GpiD2nPfv2+xSjwr9lhwzqOIvN1YbvPQy1Fv0lS1N6rJOjqYx4jMcQee28kgwD3wfRd2fbNXils/BfTELYoxV3a5y0bZCB1eGGNcRnvjGfyld39A8I+GXC1lVFw80PadPiud1VPuNOIzKR6nuR8uy85fbcVk5tfDug6j4LaiaYDy6y14J/IAm+DhD2Ouo5qDmjqNOAuMV1sVVId9gYgCNvXDj+Re3K8I/ZKVcVJzl2QTPDRPZrnC3Pm4wggD57L3cV8BabqCxWrVFjr9OXykZVW+500lJUwvGWyRvaWuBH1ErUkQeCvODyp6x5DuK+n9c6G1PLJaayvfW6duDAWT0ksLg4QvOdyARvncbHO5PrRydcxFo5uuXyk1hdaKm+6ZEtk1LbxgsFS1gD8NySGSRva8A42eR5Lhb2u3CC/8SeXu16k05RTVdRo26mvmhhYXOdBJGWPIA3OMNO2/YLoF7OjnIk5YuIk+mtRsfPo7WE8ENcAd6ScEtZOB9TsH5AemQvewn7cZ6vseuuSjmplFLTPp7loe/mrtckmeirpBIfDdkjdr4jg7HuRuvdXle5iNPczfCqh4k2CjfRGRxgrKZxz4M7QOtoPmM5/1nuumvtl9GaOuvCPSHFOkhpjdvukykhrIwA6oppIi4Anu4DuPs9At++x0tz6blTqLk54IrNR1rWt9AwM/SXFP8HexERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBeUntJPaIa2sWr9U8tPCcU1BRU1ObZf7uWl1TJJLGDLBCc4YA1waXAE5Lht5d1+eLmLqeWfgVcdd2qKOW71MzLfb2v7CZ4OHfZjPY+Z8l4r8v3A7iRzo8cZacySzG4V33R1Dc3AlsMb35eQfUjIAzsB54AKbuF867peyq5H7pT3Ozc1PENsTKUQSS6YtzmZe8vBYKtx8gAT0Y9cn5enuua+rtWitQXSgc5tTR2uqqIS3uHshc5pHzyAptKabtejtMWrSdkgbBb7PRw0NLG0ABscbQ1u3rgbriHm35neGnLXw0uN01tcmPudzpJqe1WqIh09ZK5haMN8mAndx2wD6HDNHgFp60XrjJxYgtU9W83HU90eZZXkl3U9xc45PmBnGfku1fCbhEeXrmh09bBUvkkdNFgudk4JH+tda+Al1fFx30xeWsER+6pn6W7hoIcSB8hldttWaldf+bvTFV47Xjx4g4NwfMJ+ksty69o2kOaHDsRlXKOAh0MbhsC0H8ykRRERAREQEREBERAREQF1t58OYu2cv3AXUNdT1bRqG7Ub6C1w5yRJKOjrI74AJ3+3yXZJeTPtmdL6vdebHqgRTusDfDjLhnobIW9O/l3/OVP8AB1E5G+BUfMnzKWPSGoHSTWqN0t3vD3EkyQxfEWl3q9xaN++SvoGp6XTvD7SIp6Onjt9lsFC5wZG3DYYImlxwPkASvJ32JdHaZ+JnEGrkjBuFLZoDGT5RumwcfbnP1heiPObxIoeFfLDxF1VVzxRymxVVBRse7p8WpqIzFG0fPL8/UCn0ednNvzpQ880TeWzgpoavlZNXipp6ypcC6skhOWuY1uS1uCe4PrkjC8/tRaM1Lwz1zJpLXFontd0tVWxtVBK3DmYcDkZ2II3BXcr2NWn4bvzV3S61EPULJpCtqYnEZ6ZX1FNED/5sj1D7ZC32ui5saGehY1tRW6SoKiswMZkE9RGCfU9DGfYAtbzEeyPCqaGo4ZaUlgdmN1louk+oELR/Mt1ri/linmq+Xrh5VVBJkl09RvdnvvGFygpFgiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiDRNZaSsmutM3HSWoqRlTb7nCYJ43DIIPY/WDgr51+avhbHwB5idT6EsdR0wWeubPRPZsY2nDm49MHsPIYHkvo4ulxprRbqm6Vjw2CljdLI70AGV88fNxf5+MnNlqWpt0rHy3i7RUMBGS0Fxa0fYCd8IPe3gZd5b7wa0ReJ3F0lXYKGVzj3JMLclb6W2OGOnBo/h1pjSwABtNopKNw/7ZkTWn84K3OhRERAREQEREHUH2ql0ntfJ7qF9O8tfUV9JTnBIPS8uB/MV1Z9ku24XbhLxSoHR9dNSxvEeRsCYg4gfPJJXfHnI4E13MTwQufDm2ziKqqJ4543EgYczOMZ2yCQVtXkq5UByy8F7ho+6TsqrzfXSz1zxg4ywta3IHodwM+XopL3pZx4j6SYI+Y6BjtsammHpj78/1X0baEiEOjbLGNwKKL/ogr5xYHik5kZSx+AzV8zATtsatw/nX0ccPj1aIsZL+r/aEO/r8IUs/71f/AFxuFERaQXnD7Qj2bVPxKlvfHjg0+Gj1FHTuq7rZfDAjuJY3LpIiPoykDJB2OPmMejyoRkYIQfPlyVc4OtOVjiTRU9dcKyTRlZViG92p5OI2l2HSMafovbuTt5HbK989Hay0zr/TdDq7R93p7nabjE2amqYHhzXtIB+w+oXlz7Vrkp0zpezx8w/CjTfuXXVGPU1JSNPhAPGW1IYM9J6sh2MADc+QGxfZbc7WnuC9wrOCfFW6vpdO36qZNaK+QkxUFUch0b/7lj8g5x3HzAIezaKKGWKoiZPTyMkikaHsew5a5pGQQR3BG+QpUBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQeffMb7JzSHGDW1brnR+rjp+quMhmqYjFlj3H5YIz89icb58uyXKPyzW3lZ4ZfsBt91NyklnNRPUHI6nnJz2HmT5Bc5IpgIiKgiIgIiICIiAvKD22N2ikuOhLOCPEiY+YjzwfEH869X14+e2kpqz/ZM0nWOY73Y0Jja7G3VucZ9djt8kHSPl34tXDgfxi01xKtx+O01YdIMAh0Z2cDnbGDk58gvoq4QcTbRxf0Ba9eWVhZT3CMFzDv0PAGQD5jcY+tfM42ppqmnpaH3NrZGyAOlB3cCfNfQvyK2x1r5a9MUzmdJewydsdw3/Un5f+UkJmOwKIiDGrqGjudHNb7hTR1FNUMMcsUjepr2nYgg915Pc7fswp7VqK4cWuD1RDS6fqHmpuVu6TmlcTlz2Y7AnPkRkjse/rUtmcZYoJeEetBUAFjLBXyHPkWwPcD9hAKl3ODwh5quaO88YdK6U4Vmrlnt2jx0uke4kPlazoAHrgZyfXH2ZnKr7QTi/wAqGmLlovStqtF9slfOauKkufiYpZ3ABz2FjgdwBkHYkfVjafJHw1sPGPmk0NoXVtOKu13GsllrIX7iVkcMkhB+ssBXr1rn2YPKjrO4wXGDRbrM6MjxY6CQxslAx3AIOdtySfqVqWuCeRn2knGPmC41w8NOIWmrO+juEMksdRbad0Xu3T2Dsk5G43J8ivS1cScGeVjgZwCa6ThloSjttW8dMlY4dc7xjG7z/qXLaKIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiDzY9tvc3Q8J+HNqbMWiq1BUzOYDs4R0+Nx54LwrvYl6bEHCPiBqqWkAdW6hioYpyzdzIqZjnNB8xmQbeq2t7bx9Q6j4Zx7+AyWtd8ushv8zVz77JykobLyT2a65a0Vd1u1bUuHqycx5Pz6Im/mVG7Oe3nUtfKVoalNnpqO660vjnR2y3zyfDEwD4p5QDnpBIwOxO2fXxybXceefXj7b7fd7nLe9S32YQtf0dNLb6YbucGDaONg37+QBPZYl2u3EjnR5mIo6u5z1171vezT0YkJLKSnc8lrGN2DWRxgnAwPhOe5K9xeVvk94T8rGmGUGj7Syqv9RE0XK91DQ6pqXYGQCfoMznDRgbn6hPODq3q72bvB3l04I3biBa7nW3HU+n6D3mpr6l3wyP2Duho+gMnbHcdxleenLrq+7az5m9NXK5VBd7xXBoaewaDkBex/tFL9V2HlN1lJSS9BrIm0z/m05cR+VoXixyeMDuYXSXUM9NUD+hP3fB9HkbemNrPQAK9WtOWg+oCuQEREBERAREQEREBERAXGfMRwYsPHvhJqHhxfaZjzcKORtJLgdcM4GWOaT23ABP2+QXJigrKqGhpJq2od0RU8bpXu9GtBJP5AUS9fPLy08wmseR/jtdrxDZ2XT3dtTY7vbpJDE2oY2QYcCOxDmBwztvutZ5pOdLjPzq6ktulvuX9zbE2pa216btrnSeNOcAPldsZH+mQGtycDbK4f5gL/AEeqONetb9b2gU9XealzMdiA8tJHyJBP2r1p9k7wA4T27gLaeNB01QXDWN2q6lr7nURiSWkbHJ0tjiznw8AAkjBOx9c2K5E9nxyUW/lW0VNqPUEjqrXWqaSH7qPOzKKIfGKZg+TiC4nckDsBheavtYb82986GpqaNxLbRbbZbx8iKdshH5ZSveVfPr7S0552eJRz/wDdNEP+RQLP0j3K4AUgouBfD6naAA3TFsOB23poz/OuQFsrgo0N4NaCZ6aZtY/5LGt6q26k5BERFEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERBxBzZVWpaPgBq6bSUEktxFE4RtjBJx8sfPC+cmqrr7R3+S51k1VBd4anx3ySAtlZMHdWTncHO6+pCaGGoifBPE2WOQFrmPaCHA9wQe4XHWr+W/gLryjnodV8I9K18dQD4jnWyJryT5hzQCDv3yrM+p10q5DfabWnX9NScK+YG60Fov1NGyC33qVwigrwBgNlzs2TAAz2PfzOPRilq6atp46uiqYqiCVodHLE8PY9vqCNiPmvDn2hPIRceXLULtf8NrVV1fDy5OJJaDI61y9yyQ7kMPkTgbfYN++zM5+bZwkll4J8ar7VjTlyqGOslzneZWW6U4aYXnOWxHYg7hpBGwwFNl8V7IIoYJ4aqCOpp5mSwysEkcjHBzXtIyCCNiCCCD81MgIiICIiAqYB7qqIPmq4/2G78OeYDVVPUU76eppb9NXwdTSMh0pka4eeMn8y9zeRLjHHxp5e7JqAvzUUQ9zmBIJaWgbHBO4OR9i6Je2Y4V2HTd/0nxFtdGyCpvkktPUua3HU4NLj+gH7VzV7FyukqeX/VVM52W0eojG0egMLXn87lfy5mH4249C0RFAREQYF6s9r1FaqqyXu3wV1vronQ1FPO3qZIw7EEH/AN+xHZeB/tBuU2t5aOMdbLYLVONEahca20VAYTHAXH46cu7AtOcA4yOwwF7/AK2pxK4Z6M4uaNuWhNe2OmulqucD4JI5ow4xkjAewn6LgcEEeY+xB0C9mbz/ANu1jZ7Ry7cXbq2n1Db4RS2G51EmG3CFg+CB7j/8laBgZOXADGdwPShfO7zK8nnGvlS1hUVd0stwk07TVxdaNSUYLoXhruqNxe3eOQANJyBg4x6LuHyw+2DoNO6ct+jOYjTt1rpaNjYGX+2hsr3xgbGWIkEnyyCfn8rR6vouOOCfMHwo5hdNnU/CzU8d0pmHpmiLTHPA4+T2HcFcjEZUoqiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAuAOczlm05zI8ILzZKigYdQ0VFLPZ6oN+Nk7AXNZt3BIwB6n5lc/qmESvlqfRVWntQG33yklp6i31YiqoHtw9jmOw9pHqMFfSJyxXXTV64DaMuWkqqGots1sjMb4nAgnz3Hnn+ZefXtheWTS1ks1n5hNF2CKhq5682/UJp2Bsc3iAmKZwG3X1AtJ2zkea0D2WfPLozhnZZOXri3dnW6kq7iZ9PXOUZgiklwH00p7tBcAWk7AuI22S/pZNeuyKxj2SsbJG8Oa4BwcDkEeWD6K9BQdlsLj5HcZOB3EKK0QGatk0vdI6eMDJc80sgA/KVv5RyxRzRPhlja+OQFrmOAIcDsQQdsHsg+en2eUtzo+c7hfLb4JXPF3dFOGg/DE+GRjy7HkA49/PC+hpcaaM5buBvDzVtVrvRfDOyWm/VhJmrqenAkJOcnPlnJ/KuS1biTRERRRERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQedPtqdNxV3BPR2pAweJbL86MuHfpliIwftC6XcqfP7qrgHwj1BwQntbK213fx3W2qzh9DLO3pkGNsgnfPkSe3dd+/bFU7ZOVSnnJGYdRUYH2h3+peL+irJUaj1hZLBSwulkuFwgp2ta0knqkAOwycAEk/IFIZr059ljyXay0jr2PmB1zSww0UFDNBZoiMukdKOky7jYhpOMdsnfdeqa0TRdjg0zpCy6epoRFHbqCCmDA3GC1gB29cgk/PK1tQdT/adTwwco2p3SSdLnSRtbjuSWu2XjtyY0dTW8xWlIqaMvIqQXYB2GQM7fWvbLnq4KX3jxy+3nR2m5D90IiKuGPf74WA7YHc/L614lctXECXgBzDWe5aloA1tuuYt9wjeMGHEga52+NgRk9ts/Ub/wCUyl84+jpo6WgegwrlDTVMNZTRVlO/rinY2SN47OaQCD9oKmUgIiKgiIgIiICIiAiIgLhLnB4tWnhBwE1Vfq+sEFTU2+alpW+b3PaQcD6iR9oXNq8lPbNcVLh+yjT/AAqp5y2nFK2snYM/EM5GfrJH2BWDoZwP4HcQeY7iANE6EozVXGaOSrnkePhjYNy5xHqT8vP0XYvgTzhcdOQjXlXwP1bbqet05Z7qY7paqmJzZYg8gulhdnIJYeoDGDsCVz57FDh9NBWa84k1VOQ2op4LZSucBuA8ue5v2jB+r5rkX2qfJi3ibpqbmG0HR/8Awm05RBl4gYN6+iZ2fjuXsBIGO4AHlkTTld/dMaks+sNO23VOn6yOrtt2pY6ylmY4EPie0OB2JGcHBHkcr5/PaJVD67nU4ouALi26xQtGN/hpoW4/KFrvKj7QrjDy1+66XkuDr7oxmWfcyq+M0wON4nHcAf3PpgAgDB3FaOFOqOejmgk4jaXoy60Xq4RVlxlLSPDDSOsHbYYGDnsAfrSXPR7YcLKc0fDDSFIW4MNht8ePTFOwfzLdKxLVQR2u2Udsh/sdHBHTs+pjQ0fmCy0BERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERBp19sVo1PaKuwagttPcLbXxOgqKaoYHMkYdiCD/wDZBwQvCv2hPJHdeWfX02qtGWuoqOHd9kMtFM1pcLdKTl1PIR2AJ+E7AggbbBe8q0TV+kNNa905X6S1dZ6a6Wm5ROgqaaojD2PaQQdj2PoRuEHmx7MXn8r9RT2Tlo4r1DJKmCnNLp28Sv8AjlawZZTSk7ucAMNcdyBgnZeoS+bnmB0LWctvMpqjSOkrnUU7tKXpz7VVNfiRsYIfEc99gQDnvg5zleyHs7+bqp5oeF9RBqt0TNXaZeynuHQf7YjIHTMB332BPqcZJBJHjtsiIgIiICIiDz49s5oi9ah4CaY1Ra6CSop9N34y1z42dRjhlhcwOONwA7GT2AySuqHsrebfTHAXXF54bcQ68UOnNaSQPp6xzcspa9mWtLyNw1zSB9YHc4B9qrrabXfbdUWi80FPXUNUwxz09RGHxyNPcFp2IXhd7SHk9ruX/i5W6u0PpWph4d6ixV0clPGXQUE5/stOSM9DQ7duT2cB6Jmj3YhmiqImTQyskjkaHse0ghwO4II7gjzUq6R+zw549A8aOH+meD2pLy+k4h2O2sopIakYbcWQt6WyxP7FxYBlvcEfMBd3EIIiICIiDb2vdC6Y4maQumhtZWyK4Wi7076apgkAILSMZGexHcHyIXmxxU9irZI7Ldbtwk4rV5ucbXz0drulE10MmMkQiVjuoE4DQSDud16kIg+czlp5gdf8oXGiK9RsqYoKOsNDqC0SEgSxtd0yNIBx1twcEemM+Y+gDhJxZ0Txs0Nb+IGgbvFX2uvYCHMOTE/A6o3jycCcELz29qFyHi/QTcwfBnTMsl3Dy7Uluo2Z94Z/fLWDu4fvsdx67Adc/Zv871s5X9RXTh9xMZVDSGoqmN5naMm2VbctL3NJ2YQQHAYwRk+ZF94PchFjUFdR3Ohprlb6hlRS1cTJ4JWHLZI3gOa4HzBBBCyVAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERBtjiJw70hxW0dctCa5tEVzs11iMVRBIB28nA+RHcFeEXPDyTaq5UNbOqbcypuehbrIX2m6dJJgJP8Aa8xHZ48ie4x59/oEW0eKnDLSfGLQV44da1tsdZarzTPgkD2gmJxBDZGHuHtJyCCPTsSg6E+zV9oO3iFBauXri/WNZqOlpxTWO7Svx90WMGGwyE//ACUNG2+4G3oPSRfOdzGctPF3k94lxU95gq6eniq/eLBf6bPh1AY7qY4PGzZAACWnByDtsQPS3kl9p/ojija7Vw34217bFrRjW0sVxkBFLcnAYBLt+iQ7ZB7k9zgk3B6CIrGvbI0PY4Oa4ZBByCD5q9QEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREHnj7afUEtv4AaRsUf0LvqUdf1RQPcPzkLhP2PHLhozXlw1Jxv1dRR3CfS9dFbbTTyjLI5zGJHzEdiQHNAB/RkHlj229OTwd4d1Wf7HqaePH/fUrz/+T+dY/sRaiV3C7iXSud96jv8ARyNHo51MQ78zWpDXpUiIgtJK+fj2jvCy5cMubHWr5LU6lteoKsXe2vEeInxysa5waRsSH9WR3G2V9BGQuvnPNwb0dxZ5b9c/d+yUtTcrHYqy52urdGDLTTwROkBY7uM9JGO26HjReQnme0jx+4J6ctsN5iOq9PWuChvFE44kD4mCPxAO5BABz88rs8vBX2VeoLvaOcvSlvt072wXelr6WrjBPQ+MU0kgJGdyCwYPln5le9SJOCIiKIiICIiAiIgIiIC8IPayXuS685F/pC8+HbbZb6ZrfIHwQ4n6zkfkC93XOa1pc5wAA3JOF873PrqyHiBzfcRLxa5PeIH3VlFSlu/W2KKOMY+sg4QeufszNFDSvKlpW5yUohmvtMysPw4L2EHBPrkkrtJdbdTXi2VlorGB9PXQSU8zT5se0tIx9RK2lwO0zTaM4N6I0vSx9EdtsNDD04xhwhaXbfWSt8qTs6Pm+5seX3UvLlxhu+jL7b3QUVRNJV2mYDEc1K556ek9vhyAQPLB816eexmt9oHLzfrtT+G64O1BLTVB26mNaxrmjPcAh4OPkFp3to+H9Hd+CuleIsVOwVun717nJN0/E6nnYR059A9rSPrPquMPYpcQH0N213w4qKhoguHgXKBhIBMzWlpI8/ojf6gs/lNxdx6yoiLaCIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAurPOxzy6V5SbVSWz7luu+qrzA+Wgow4NYxoPT4jz6A/zd84XZPUGoLLpSyVmo9R3Snttrt0JnqquoeGxxMHckn8nzJAG5Xz/APtA+YeycynMXc9X6QfNJp22UsNotb5M5nZHkvmA8g97nED0A7pmjZDtP8cObzipedWWPSlw1Be75WOqKp9NETFETjAL+wwMfPzxutZ4ccQuNnI1xoNQaOe0Xek6GXG3TD4KmAnOD6g74J+eQvb3kr4SWPg9y2aEsFutMFJX1Vmpq+5ytjDZJ6qZgke557nBeQB2AHzJPC3tF+RWv5nLVb9bcN4aSLW1nYYXNmeI210HcMLsdxjYn5AeYKkv1zzy4c0fC/mR0TQak0jqOhFzkiaK60umaKilnx8TSwnJGdwRnbGSuZF83eueCvMryoagotQaq0rf9H1bJv8Aad0hd96kcCD8MsZLSM42J39F345ePbJWiOgtmleP+kK1tQwR08l/tr2yNcAMGSWJ2Dk7ZIJ9Tnup5B6mItN07qC06rsVBqSw1jKu3XKBlTSzM7SRvGQR9hWpKgiIgLYXHfQ1u4kcHNZaKudBFWR3SyVkMccjQR4phd4ZGexDsHK36rXNa4YcMg+Xqg+cTk91LScO+a3htftQVJt9PbNSQR1j37eE1zjG8H/ziCvo4hmiqImVFPK2SKVoex7HZa5pGQQRsQQc5C8Tvaecmtv4AanpOLuiZX/se1dcZWTU+N6SsIMhwRsA7BOdskbb5XbX2PvGrV/EvhJqnR2sb3Pc5NH19NHQyTv65BTTMeQ0k7kAsIGf05U+6PQFERUEREBERBY5jXtLXNBaRgg+YXin7T/kzvnC7iNcuNmh7J16K1DIKisbTMwLdVu2eCBsGOO4I7ZJPnj2vWga70Vp/iLo+76I1VQx1drvNJJSVEUjQ4Fr2kZAPmM5HzCDzL9mJz+V9XXWHlj4qSRPi8E0mnLu95DyRuymlz3OMhrvlgntn1TXzxc0XKjxZ5MuIdLXVpk+5Ete6bTl/pXZbIY3BzQT+9laMEgjcDOMHA9UPZx86tbzQ6Nr9La58CPWml44veHxnArqcjpE4B3ByMHvucn1I8dzUREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREHHnHPgbw/5hOH1dw64iWsVVBVjqimbgTUso+jLG7yIP/vnBXz88z/Aa9crfHG8cMKi8e+utb4qy3V8Y6HyU8gD4XkD6LwNjjzBIx2H0krwv9rvbqyn5vq+4TRFsFZZLf4Lj2d0R4OPqyPypvwevnK5ryLiXy+aB1lHU+O+usdKJXZyfEYwNcD88tyfPdcqLpX7KaTUlPy1UNqvbJWwRSOlpRICOhjiSAAe2RgruopAREVBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQFQnG57Kq4O5zON1FwE5etWa1dVRMuT6J9Fa4nvwZamUdDQPPbOcgHBwg85vbAcyekeJGo9NcGdGXKG4x6TqJ6+61ERyxtW9oYyIHscM6ifTI7ggrmr2JVBLBwc4hXCRuGVWo4GM27hlMMn8riPsXlNoXQuveN3EGj0hpG3VF81JqCqPS0blz3El8j3dmtGSSTsPyL3y5HuWer5WuCNNoK8V8NZequqfcLlLAcxiV4A6AdshoGM/V3xlXhjsOiIoC2lxatjr1wr1lZ2N6nV2n7jTNA8y+mkaB+UrdqjlijmjfDIwOZIC1wPYgjBCD5x+UTiNDwN5ntE61v/VT01nurqe4NOAWMex8LwT5YL9z5YX0VWG/WrU9mpL/Y62Oroa6Js0E0bshzTuPt9QvFP2pfKpZuBPEC18QdIscyy6yln8SIjAiqmYc4DAA3Bzt8vmu7fsiuJNx11yy1FlutY+pn0teZbeHvcS4ROjY9gyd9gUHeNERAREQEREBERAREQcYcyWuBw+4M6i1GanwHMpjEx4O4Lgdh9YBH2r57+H9sruJvHi2QxNfUvuuoG1UhOXExmcOJJ+rbPzC9jfay6sqNL8qlVBTSFr7rc4KTIODg5yR9QJXSH2TnBCk4jcTbhrOvjLoNPOYT8IIBAB7+RJICnnSf1XtLaqcUdso6QDHgQRx49MNA/mWWiKjqL7Uu2PufKDqIMp/FdT1UE/bJaGh5J+S8VOBXGbVHAjiPatf6WrJYpKOZhqImOwJ4sjqYfnjOPn8iV9GfFrhrZuLmgLroK/EilucJjLgM9LsHBx9q+drmW4M1nALjLqHhlVvc9tsmBge799E4Zac+Y7gHzASc8H0GcB+O+heP+hKDWWjLzTVJmgY6qpmP++U8hAyC07gZ/wDfK5KAwvAj2anEvU2heZ3TtttdynZaru90FwpRIRHIw4+IjOMjtn0PngL34Sn6EREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQFDPNFTQyVNRKyKKJpe+R7g1rGgZJJOwAA3KmXlB7VvnS1FR6lqOWzhteJKKkpoGO1HWU0uHyveMimBG4AGOofPzzsGi+1R53dO8Raan4A8IdSMr7VS1Jn1FX0j8xTysJDaZruzmg5LiDjIxuFtf2XXJJauNN3q+NHFOzTy6XsU7IrRSyNLY7hVjdzzkfEyP4ex3J9QCNoez35DNU8fNcWniLxG0tNBwwt0hqZZKppYLw9n0IIhkF0ZcB1uG2NhnJx7gWOxWXTNrp7Hp610ttt9IwRwUtLE2KKNo7ANaAAmGsuCCKlgjpqeNscUTRHGxowGtAwAB6AAKVEQbD42cGtGceuHV14aa7ovHttzj6Q9uPEgkH0ZWHycD2K88v3ES1t1C2Ucc6l1jEgLoDagKksz26w/pzjz6V6komjb+gdGWnh1ouzaGsQf7hZKOOjg6zlxawYyfmdz9q3AiICIiAiIg6oe014V6k4tcqd5sukrTJcbpa7hS3eKCNuXuZEXB/SPXoefyryv5B+bL9qLxXuMuqrbUv0/qCNlvvEPSRLTOjeemToO4LSXAjGfL1Xv8AbOG47rxf9pPyX8SqbmLrdc8KOG1yuuntZMhqf+plP4jYK7p6JWFrRhgJaHDy+I5xsSzU17FaV1NZdaabtuq9N18dba7tTMq6SoYciSN4BB2+vceRyFqx7Lg3km4eay4V8sOhNDa+Y+K9W6gInp3nLqcOe5zYj82ggfmXOakuqIiKgiIgIiIOK+ZLl40VzOcL63hhrd89PTzyMqKSupgDPR1LM9MrM7HGSCDsQSCvD/Xtk4r+zr5nqyzaU1c2W5WdkU8FXC0sjrqKYBzWysycZAwRk7j0JC+hReGXtfKOeDnBq6qSJzY6nT9tMbiNndLXA4+oj86ej2V4J8RIuLfCLSHEyKHwW6ls9NcXRnbodIwFw+w5W9wMLrF7OXiLpzXPKZoK3WmvhkrdO2tlqrqdrh1xviJbkjvgjG/rldnQcqS6KoiKgiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiIC4Z41cqPCTjxeaG/a6srJ6yhAa2QNBLgO2cjy7fUAuZkTxLNaDozRWndAWKDTmmKBlJRU4wxjQM/bha8iIoiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAvEf2ufGa86v5iJuFtPcpDZNJUsDTTh/wAJqnxh7yR6gOGPr+QXtwvnd9oNJJLzm8VnSk5F76R9QhjA/MAg9EPZRcn0vDHSreYTWcURvuq7eGWiAty+jonkEuJPZzwBkem3kCfRJbW4XUlNQcM9JUVI0NggsdBHGBsA0U7APzBbpQEREBERB5ve23az/Yb4dv6fjGppgD6A0r8/oH5FB7EUk8J+JLD2GoaUj7ab/wBim9twCeDnDs/900w/5K9R+xFaBwl4kOzudRUw/JTD/WrnB6TIiKAiIgIiICIiAiIg87fbV3IUnAbRlv36q7Uxb9jKd7ifygflW1vYi0ch0jxJr/DHhtuVLD1/MxZx+YLi/wBsPzA6b4g6s0vwl0tXRVsWlX1FZXyxuBaKmQBgaCO+AD9uV2Q9jZoyo09y4XvU9TDJGdS6hkmj6gR1MiiZGCB6ZyQfPKeI79qnf7FVFL1VPJdM+fzkQ0TzF6eunFG21Ulo1zYrVK+KoYMw10cLC4RSjvnAwHDtsNskruauL+Z3XdHw15feIGs62VrBQafrfDycF0r4nMYB6kucMBXcHgTyiXSotfMboV9PJ0e8XSOnefVrsjH5cL6RACGgE5K+aTlruEdq4+aCuErepkV9pcj63gD85X0tRyCWNkrezwHD6iE34md1eiIiiIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiLiHmT5mOHPLHoOo1hrm5sFS+N7bbbmOzPWzgbNa3uBnGT2AyiW44c9o5zgycs3DCKzaKucDddamcYKFvUC+kgA+OoI7jGwB9T5d15nck/KbqPnd4p3rUOt9SVTLHapo62/wBwcS+eslkcSIWHsC4NOTthvbyxxJq7U/FrnB44yXB8NTedS6prTHR0jCXNgjJ+GNo/esYO5A8icZ2Xt5yK8qbOVHhE/TFwrGVmoL3Ui4XiZn0PFDQ1rG/JoyP9fcpMW75HPumtO2fSGn7dpfT1BHRWy000dHSU8bQ1scTGhrQAPkNz591qiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiIC6p8+PJxonmR4e3HVD4TR6w05bp57bXR95AxpcYXjzBxt88fIjtYtK1T0fsYu/i4DPcKjq+rw3Z/MrOJZrwr9l9xc1BoHmn0vpOmuE4tOrpn2yqpuv72XuY4sdgnAORjbfOPQL3pXzkcmIlHN5wv9xz1jVVL0dPfHWe32L6N1FEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAXzue0FOec3it/4c//AHMa+iNeAvtOdD3DR3OHrSuq4ZBBqN8N2p3kbOa+JrSAexwW7+mQg90+GEkc3DXScsJBjfY6BzT3yDTswtzrql7O7mS07xx4DaesPv7f2SaVt8FsuNO4/E4RNDGvHnggDY74wd8rtapmAiIqCIiDzt9tRYbrc+BOjbrQ0j5aW06jdLVva0kRtfTuY0n0GSB9q4g9jNxr0hpK46z4T6iu9PQ1uoKimuFtEzw0TSMa6NzAT54I/N6r1T1povTHELTdbpLWFmp7parhGY56edgc0gjGRnsR5FfOzzLaF/a88zmsNGaOrJ6Rulr11W2ZhLXxsLWyxkfUHgY7eW4Tfg+kMdlVcbcuHEM8WOBGheIbi4y3qx0tRNk5Pi9Aa/Pz6gT9q5JUl0ERFQREQEREBbY4nXWSxcN9V3uF/RJb7JXVTHf3JZA9wP2EBbmJwtj8dMf7CfEAOGR+xe6/qkiRLcfPLwV0TX8xPMJp7R90qZJKnV15/wBtSucSS09T37nsMAj5BfRRwz4c6Z4T6Jteg9JUTKW22uERRta3p6iBu448zj8mB5LwV9nKcc6HDMZH+6Mo/wCTyL6FVd+GCIiii8Xvauc0+t9UcXbzy92m5SUeldOGBlbTsODVVJjbIes/3IDxj68ds59oV893tIo2x86/E4tB+OupXb/4lAg7y8kPsvtMaeodI8ceKeoJ7jd56eG7Udogj6IKfxGB0Ze4nLnAOHljfOx7elQw34QAANgtocGyDwi0OR56bth/5LGt4Jct1J51VERFEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQERdaOYj2gfL1y4z1dl1HfKm86jp4y5tntUfiSl+Dhr3khseSO5JA+vZE1y3xm408PuA2iK3XvEe/QW63UrD4bXvHi1EnlHG3u5xJA29QvAfmr5jtac2vGSp1ZUwVLqMye5WG0xNLzBBnDWhgzl7jgkgZJwN8ZOXzG8x3FrnV4tU881BVyxzzik0/pyi6pWwAnAAA+k87kuxtv5ZJ9JfZyez4k4J0EvFPjhp2hm1pW9P3MoZg2b7kw4ySe48Uk74+j27gFFbg9m9yO27gHo2k4pa9t7Zde36mEgZK0H7mU7xkRj/tyMEn59+wHeREQEREBERAREQEREBERAREQEREBERAREQEREBERAWia4PTou/uzjFrqzn/8AYuWtrQdeN69DaiZ/dWmrH5YXIPn45Aqf3rnM4Uxub1Yv7Zf/ADI5HZ+zGV9Eq+dv2f1XHQc5fCmaY9LTfPCyTjd8MjB+dwX0SICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgLzi9sJwbotQaAtnFGmpWtrbM0xyTNG7mA5IPywcfYPRejq83Pa+cfrXp3R1HwVpHMmuV4iM87Ad4oycZP2AEepP5LLg68exrrqqPmKu1vbUvbBLZZZHxdWziAcHHyz+j0XtUvFv2NFoq5+YC93psLjT01nfE5+Ng52ds+vb8q9pFAREQEREBeRHtJOTfilr/mYl13oCw+90GpKWlbO8DAjlYwMc84G4wBnzyPnt67qN8UUnSZI2vLDkZAOD6j0U+6OK+VnhpVcIuAukOH1a5zprTQMif1dwe/n+Vcr43yqoqCIiAiIgIiIC2jxco3XLhVrO3MaXOqtPXGENHcl1NIAB+VbuWNX0kddQ1NFMMsqInxO+pwIP5ig+dPkr1PSaA5sOHeoL24QQW689FR17dGWPYQfQ5OF9F9LU09bTxVdLK2WGZgfG9pyHNIyCPrXzT8fbNDorjjqmgs05jbSXaSaB0ZwY3F3Vt6YJJH2L3J9nnrm/cQeV3TF91E98lUOuDrdn4mgNIx8skq8z9p3eeOyqIiii+fT2lrCznY4kg+dTRn8tHAvoLXz5+0qnE/OxxLIGOiqpGH6xRwDKD3b4NgDhDocDsNN2wf8ljW8Vszgtn/AGHNB57/ALGbX+qxreaJBERFEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAXHPMFxMuXB3g9qfiPaLM661lkoX1ENKAT1vA2yBuRn0XIyxq6gornRzW+4UsVTTVDDHLDK0OY9p7gg7EKwrwP117TznF1tNOIuJjdP0k3U0U9ooIIQ1hzt1lpeTg4znK0Hlt5SONPOnq+vudHdDTUhkMtx1DdA+VheTg4A3eQdsAgDsOxA9npeRPlBqLm+8TcANKPq5ZDK9zqdxa55OSSzq6c5+S5h0zpLS+jLVHZNJaet9noIgAynoqdsTBjts0DJ+ZUvvCOp3J/7Njh3yu6kHEC46kn1ZqpsJigqJqZsUFH1DDjEzJJJH74nI8l3JREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBafqClNbYblRgZNRSTRfa5hH861BUIyMEKWD5odC3qThJzBWi9zudTnS+qGOkOS0sbFUYcc+gAJ+pfSDonV9l17pe36ssNUyoorhA2VjmkHBI3B+YXjP7U/lSouCmuqPinptx+4+s6uZsrOk/eqkDqI9M4B389j3yu3XsdOJl01nwEv2krtWPnfpO7MggL3ZPgyx9TfsHTj7PtNz6O/iIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgpnOxC+en2gWodU6y5wNffduCpM9PchbrfTlhLhTsaGxNYBuc5JGM7nC+hbOO62DqHgLwZ1Zq6m15qPhtYrjqCkcHw3CelDpWuHY+hO3mFPo4v5G+WfT3LtwT0/SttsQ1NdrfDWXmq3LzNK0PdGCdwASBj5Y7ALscNkwAMDbHoqpAREVBERAREQEREBERAREQEREBaTquuktel7zc4vp0lBUTt+tkbnD84WrLbXEmuhtvDrVNyqHBsVLZa6d5PYNbA8n8wQfOK2hrOMnHuS3Vc3TLqK+yRud26WGQ/kw0YX0OcBOGts4ScKNP6JtTWiKjpWOd0jAL3AEnsPLA+xfPry7un1DzKaSfRwlktyvwcyNu5BkLjj7Mr6Q7bSOordTUTn9ToIWRl3qQ0DP5kvbtTvjKRERRfPj7SiDwOdjiYM/TrKST8tFAV9By+e32kNca7nV4nO6ceDX00H19FJCMoPeHg2McIdD7Y/8Ag3bP1WNbxWzOC8rZ+DuhJmnIfpq2OB+uljW808T0RERRERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQERYV3u9ssNrqrzea2GjoaKJ01RPK7pZGwbkknyQdLfa/2GmunKLNdnwB89n1Db54nYyWh5dG7HpkPGfqXU32NHFi0aS4n6t4fXq4Npxqajp5aJr34Dp4nOB2+pwGfmFle0Z9onoDjroip4F8I7dV1lrfXxVFffKgBkc3guJDIGbkjqGS44BGMLhL2d3L5xH4rcaLXqzTlJPSWaxSmWruBy1noWA+fff54AyeyTfE8e+iKOJhjiZG55eWNALj3JAxkqRFEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBeX3tO+fTUOkbpqHlg0DbYoHVVAynvN1kcesMmbl0UQHY9JAJ+fmCQvUFfPJ7Q3U1JqvnG4lXKgmjmpoblHRRyMcC13gwRxkgjY/E0qd3YeuznsteSbU151fYeZnW0DKWw20vqbJTu3fVS4LWy47BoySPX69l6/rYPATTlBpLgloPTttp2wU9Dp23xtjHYEwMLvyuJP2rfyTfpREXWPjb7Q7lw4B61foLW96ucl0gx7yygozP4G/77B+R2+RVHZxeHHtduHtp0jzRv1Napow/VlthrauFvdk7GtjLj/3wDT9i72a19rryn6atBrLBWag1LXPj64qKkoDFk/3L5JCGsP5V5P8AMtxg1xzV8T9Q8Z5tO1zLdCxkQjjY6WKgpmjDGucBgbHc/USfNB6tezC5qIeMnCui4Y3WB7b1o2hjpXTeUsTAA0n7MY+rHku8B3XjZ7F3U1LRcbNVaWne0TXGzePTtPcmN3x4+oEflXsnnfCW23qTiqIiKIiICLye9otzw80XAjmXuHD7hTxP+4dggtNBVR0f3Ft1TiSSMl7uueB7zkjOC7A8gF3w5KeI2tOLnK7oLiLxCvP3V1De6KeWurPd4oPFe2qmjB6ImtY3DWNHwtA2z3OUHOKIiAiIgIiICIiAiIgIiICLoB7Vnmj468tn+xd/sK65/Y5+yL7t/dP/AKmUdX7x7v7j4X9sxSdPT48v0cZ6t84GN3ey55hOMHMXwk1ZqfjJq/8AZDc7bqM0FLP9z6Wk8OD3aJ/R008UbT8T3HJBO+M4ACDuiiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiIC8jva5c1Wv6TiHJy3aXuT7bYaa3U9VdzCcPrJJ29YjJG4aGEZHnny3z64rwT9qpVNrOdDVc7WkBtFb4tx36IGsOPtaUHLfJ37KiDjdw5sPGDibr6otlovsfvdHardAHSywB5AMkrjhodg7AZH5CvV7hTwj0LwW0fSaH4fWWK3WykaAA0DrkIGOp5AGT/rPqVtHlAscumuV3hdYqhpEtJpihbIHDBDjECdvrJXMGMqT8v5dTFURFVEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREHTT2oPMdrPl94I21mgqx1Bd9W3B9sZWtGXwRNjLnlvoSNgfXHlleX3JByyaj5seM5kuVRK+zWidlzvtbLlxkJfkMye5cQSfPG3nkerntFuVvUfNDwltNm0fJGLzp+5e/UzH5w8Pb0PH5P058lP7PjlRruVrhPW2zUr4pNSahrBWV7mDZjWtDWMB8gBnI9d/ND/AF2hoaOnt1FT2+kZ0Q0sTIY298NaAAPsACyERAXjf7Rjkp4sXTjpd+Jeh7JLd7bfyJjHHkvjeBg48sbgY2xgnz29kFZJHHK3pkja8ejhlB4ecBvZQcb+K8Ud11fdqLRtp6wHvqIXTTvbncMYCN+/fb1Xoje+XvgByi8nOt9KMp6Q0LbBXGsul06PebjXPgcGEu/ui8NDWN2AAG4BK7Nat1dpjQWn6rU2rrzS2q1ULC+aoqHhjGgAnA9TgbAei8CudrmY1RzXceLgbJcqyr0pSVgt2mLXG53hlgwwS9GSDJK8k9Xo4AYCe8J+13s371qOyc4/D2XT0U0nvdXJR1zY2k/7Ukjc2Qux2A+E5O2QAvoOXWvkv5QuHvLhwzsE8emaN2uKu3RS3q7SR5nM72hz42k7ta0ktAHod8HC7KJeXEl0RERRERB4Ue12/DLuv/gG1/8AoisjgV7VTi/wF4T6d4R6d4daPuNt05BJBT1Nb7148gfM+Ul3RKG5y8jYDYBY/tdvwy7r/wCAbX/6Irs1yd+zX5ZONfLXofijre36jkvl+pJ5qx1NdnRRFzamWMdLA04HSweffJQcU/u2nHr8U2gfyVv9Mn7tpx6/FNoH8lb/AEy7efuQfJx/86tW/wCfXf8Aqp+5B8nH/wA6tW/59d/6qDqH+7acevxTaB/JW/0yfu2nHr8U2gfyVv8ATLt5+5B8nH/zq1b/AJ9d/wCqn7kHycf/ADq1b/n13/qoOof7tpx6/FNoH8lb/TJ+7acevxTaB/JW/wBMu3n7kHycf/OrVv8An13/AKqfuQfJx/8AOrVv+fXf+qg6h/u2nHr8U2gfyVv9Mn7tpx6/FNoH8lb/AEy7efuQfJx/86tW/wCfXf8Aqp+5B8nH/wA6tW/59d/6qDqH+7acevxTaB/JW/0yfu2nHr8U2gfyVv8ATLt5+5B8nH/zq1b/AJ9d/wCqn7kHycf/ADq1b/n13/qoOof7tpx6/FNoH8lb/TJ+7acevxTaB/JW/wBMu3n7kHycf/OrVv8An13/AKqfuQfJx/8AOrVv+fXf+qg8uebznZ17zi/sT/ZvpKwWT9iPv/u33KE3333r3fr6/Fe7t7szGMfSOc7Y9CPYmf7wuu/4Xn9Sp11U9p1yhcHOVP8A2Nf9ialu0P7KPuz90Pf641Ofdvc/C6cgdP8AZ5M+uR6LtX7Ez/eF13/C8/qVOg9FUREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAXW/jRyHcEOOXEOm4laut8v3UhLDL0fRm6TkdW4z9uV2QRBiWu20lnttJaaCIR01FCynhYP3rGNDQPyALLREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQefntl36mHALTkdlE/3PdenG5+FnHhiP4OrHl1Hz27rzO5LLDdbzzI6KnttkNxjoLiyoqG+EXMY0AgE+QOSMZ+fovoa1Vo/TGuLNLp/V1jpbrbp8eJT1MYe0/Ytt6H4EcH+G1QarQ3D2z2ecknxKeD4sk+ROcfYn+J1v5ERFEREBERB0j5rvZkWHml4v1XFq4cXa/T01TQ01F7jDZ2VDWiFpaHdZlaSTnOMbfNdleXng9S8AODemeD9HfJbzDpunlgbXywCB04fNJLksDnBuPEI7nt81yOiAiIgIiICIiAiIgIiICIiDq/zrcj1p5zP2G/dTiJV6X/AGIfdHo8C3Nq/efe/d856pGdPT7sMYznqPbC13k05R7byf6HvmibZreq1NHert91XVE9C2lMR8GOPoDWvcCPvec5HfGNl2DRAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQRyyxwxPmlkaxjAXOc5wAaBuSSdgB3XQjjn7YTgZw0v1Vpnhppa5cRqqie6Katp6xtDbnPBwRFO5kj5QCD8TY+g7FrnA5Wre1u49XThNy80uhNN1r6S7cSKyS2SSsf0vZbYmh1X0+fxdcMR/wC1ld54W1/ZZ8m/D7T3Be18d9faQt151bq8vq7a+4UzZxbLeHFkXhNeCGSSBrpDINyx7GgjDgQ2hpD23+kq26R0+veAN1tNuc/46q1X6OvlY0k4PgyQwA4GM/GMnJAHZehnCrixoDjZoi38ROGeo6e9WK5NPhTxAtcx4OHxyMcA6ORp2LXAEbbYIJ435nOULhRzGcMrxpW4aPstHqE0kpsd6ipGRVFBWBpMTvEYA4xl4aHsJIc3O2QCPOD2PvFu/aA5hL7wAvs8sNBqukqXsoXu/sN2ogXkgZwCYGVAdjv4bM5DRgPZdERARYF6vll01aqq+6jvFFarZRRmaqra2oZBBAwd3PkeQ1o37khcGu5++Tdl1+4zuYTSvvGcdYmkMGerp/s4b4WM+fVjG/bdB2BWnXq/2LTdEblqK9UNrpOsR+PW1LII+o5w3qeQMnfAznZWae1LpzV1np9Q6Tv9tvVrqx1U9dbqqOpp5h6skjJa4Z8wSvOL2vHGjg7xB5c9P6a0DxZ0bqW70euaSoqLfZ79S1tTDEyhr2PkfFE9zmtDnsaSQAC4AnJAQek9uuVuvFFDcrTX09bR1DeuGoppWyxyN9WuaSCPmCspdFPZx8wnAPQ/Jnw80vrbjhoDT96ofut7zbrpqWipKqDrutW9nXFJK17epj2OGQMhwI2IK7rab1PpvWNkpdS6Q1Dbb5Z65pfS3C21cdTTTtDi0lksZLHAEEEgncEeSDVUWxtMccODuttY3Lh9pDifpm8altHi+/WmiucU1VT+E8Ml6o2uJHQ8hrtvhJAODstn6v5y+VnQWrZdDat45aWt97ppDDUUrqvrFPIDgslewFkTge7XkEYOQMFBzSix6OrpbjSQXCgqoamlqY2zQTQvD45Y3AFr2uBIcCCCCDgjdbF4ncf+CfBgRDinxR05pqacB0VNXVzG1EjScdTIQTI5ue7g0geZCDkJFwvoDnL5W+KF3p7BonjhpeuudY7opqKWqNLPO7JAZGycML3HBw0AkjfGFzQgIoKmpp6Knlq6ueOCngY6WWWV4axjACS5xOwAGSSdsBcE3nnz5PLDdnWW4cwOk3VTXOY40tQ6piBAyQZYmujH/nbnYb7IOfUW3tFa/wBDcSbIzUfD7WFl1Ja3npFZaq6Oqi6sAlpdGSA4AjLTgjzAWtVNTT0VPLV1c8cFPAx0sssrw1jGAElzidgAMkk7YCCdFwFeefPk8sN2dZbhzA6TdVNc5jjS1DqmIEDJBlia6Mf+dudhvsuXNFa/0NxJsjNR8PtYWXUlreekVlqro6qLqwCWl0ZIDgCMtOCPMBBuFERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQFt3XWvtF8MdMVms+IGqLdYLJQN6qitr5xFGzPZoJ3c49g0Zc4kAAkgLcS8sZOSHm552uKtw1tzZaxrdG6Lt1yqI7VZ4nsfOKYSEBlJTtzFACxoBnlBe74SWyA5Acl272wvCfUnGO0cNtIcMNQXKz3m9Ulnp77LVx0xd40zYvGFM5pd0AuyA57XEdw07D0DXgBxJ4ZaR4N+0RtvDHQtHNS2LT+t9O01HHNO6WTBNI5znPduSXuc4nYZOBgYA9/0BFsbiPxu4QcIHUDOKXErTulX3MSe5Mutwjp3VAZjrLGuILg3qaCQMDqGe4W59PaisOrLJRak0veqK72m5QtqKOuop2zQVERGQ9j2khwPqCUGpItH1Xq3S+hdPVurdaaht9jsttYJayvr6hsEELSQ0Fz3EAZcWtAzkkgDJIC0rh3xX4Z8W7ZUXnhjr2xapoqSb3eomtVdHUthkxkMf0E9JIIIBxkb9kG7UREBFsPidx14O8GKeCo4qcStPaY96GaeK4VrI5pxnBMcWet4B7loIHnhbV4fc4/K7xTusNi0Pxw0tX3KpIbBRS1fus8ziSA2OOcMc9xx9FoJxvjG6DmZaNS6x0lXXqTTdHqq0VF3g6hLQRV0T6lhb9LqiDuoY88jbzWna44pcMeGMVJUcSuIumNJxXBz2Uj75d6egbUOYAXiMzPaHkBzScZxkZ7rxh5feIegbL7V66cRbxrjT9BpSTW+s6tl9qbnDFbnQTw3EQyipc4RFkhkYGO6sOL24JyMh7kItgaT4/cCNe3uLTWheNeg9RXioa98NvtOpKOrqZGsaXPLYopHPIABJwNgCStU4h8U+HHCW0Q3/AIm65sul7dUTilhqbrWMpmSzFpcI2FxHU7pa44GTgE9gg3Wi2TqXjRwl0foOm4oan4j6et+k66KOakvE1fGKaqbI0vj8F4OJS5oJaGZJAOAcLTeEvMZwP47e+M4ScS7NqWa3sbLVU9LKWzwsdsHuieGvDc7ZxjO2coOSEWLcLjQWiinud1r6ejo6VhlnqKiVscUTBuXPe4gNAG5JIC4Ku3PtydWW6us9bzB6UfUNe5jnU076mEFoyfv0TXRkb7HqwTkAkoOf0W3NC8RNB8TrEzU3DvWNm1LapHFgq7XWx1MQcO7SWE9Lh5tOCPMLcaAi2HxO468HeDFPBUcVOJWntMe9DNPFcK1kc04zgmOLPW8A9y0EDzwtq8Pucfld4p3WGxaH44aWr7lUkNgopav3WeZxJAbHHOGOe44+i0E43xjdBzMiLafEPipw14S2ZuoOJ2urJpi3vcWRzXStjgErwMlsYccyOxv0tBOPJBuxFwVpbnk5R9Z3ZtjsPH3Sbq2SQxRsq6s0YkeNsMfOGNcSSAME9R7ZXOTHslY2SN4c1wDg4HII8sH0QXoiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIg0HWOutF8O7LJqTXurbPpy1RHpfWXWtjpYQ7GQ3rkIBJwcDOTjYFddrj7Tvketdwdbp+N8Ur2EtklprDc54mkDyeymLXA52LSQuLeZv2fXEvm05n6nV2vuJj7Pwsttvo4LXR0spnqy/w/v7YYnDwoSZAXOkd1EgtAa4DDdxyeyI5OHafNmZZtUsrPBMQuv3dkNV1Y/snSR4HV8vC6fkg7N8K+OfCHjfa5bvwn4h2XU1PTkCcUVSDLAT2EsRxJHny62jPkt9r59tS2zXfs5Ocz3Sx6gqK0aVuFNUsnjBjbd7RO1rzDKzIB643OjcM4EjCWnZrl9AlPPDVQR1NPI2SKVokje05DmkZBB8wQUEqIiAiIgIiICIiAi86/a7cfuMnA9nCmPhLxBuml23x18dcTQOa01Bg9x8LqJBOG+NLgAgHq3zgY7n8uupL3rLl94Y6v1LcH114vmjrLcrhVPa1rqipmoYpJZCGgAFz3OJAAG+wCDkVERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERB4++24vNXUcW+HGnpHE01DpyprY2kjAknqix5AxncU7PM9h2xv6e8ulrisfL9wys8FMYGUWj7NAIz3b00UQIPnnI3J88kry69trQ1MfGnh7dHtxBUaXlp2Owd3x1cjnDtjYSs7HO+4G2fUrl+uTbzwF4bXlkolbX6Rs9SJAMB3XRROBA8gc5x80HIC8NOFNNNov2tslDb2xwtj4oXqniaHEhtPPLUtDck5z4cuNz3758/cteHnC4zaz9rtPNbuh//AOc69ztMeXtfDTSVLi8YB7xwl2ew7k43Qe4aIiDx89qBr3WfGTm/0jyrx6lNl0xT1FnoWskkLaY11wezNZOAQHCNksbR1fRDXkY63Z7T0XsguUGn00yy1dDqyquAiDH3g3pzKkvxgvDA3wQc5IBjIGfNaB7Rv2fOpuYm70fGngzWU7NbWyibR1tqqJhA26RREuhfDMcNjnYSW4eQ1zen4mFnx9bdC+0m5weVm7UnDrmf4bVt+paTEIN6ppKC6mJhALo6kNMdSACD1Oa4u2zJvkh6A8mvKHDygWbV+lbVr6r1JZr/AHaO429lVTCKWjYIgwteWuLHvJAy5rWAho2HYecvtBvZ6cK+VDhRbeJmgdXamuE911TDZhRXOSB8UFPLTVc2Q5kbXFwNOxoJO4JJGcY9TOXHmb4U80eiTrXhddppBSvbBcrZWRiKtt0zgSGTMBI3AJa9pcx2DhxLXAdWfbV/gs6W/wCMCh/k64oOBeT/ANl1wj5i+XXSXGXU/ELV1sueoPf/AB6WgNN4EfgV1RTN6OuJzt2wtJyTuTjbZenHATgvpjl54Saf4PaOrbhV2nT0czYaive188r5p5J5HOLWtG8kryAAABgb4yuFvZcfgKcM/wDyz/LFau1aD54J9f8AFTQ3NTxVt3BhtV+yvWl8vmlKJ1Fk1bfe7lgiAjBbK4M6A7Yt6y4EEAjvDoH2KOkavh9Sy8TeLF/pdb1ULZakWmOB9vo5CCTH0yML5yMgF4ewEg4GCCuD+Sikpar2qV4NTTRSmHUWrpYi9gd0PAq8OGexGTgjcL2uQdVubXjlJyP8pVrGnKuGv1JS0dBpLT0lTEMSVLKfpNS+PJyGRQvkxkjq6GnIdv0v5J/Z6R81lhqeZfmd1XqG4Q6orZ5qGjiqDHU3LpeWvqqidwLgwvD2sYwNJDA4ODSAdye3Gu9cDwfsLWSso3i91b3Y+CWUe5taAcd2Auzg9pBkdltvgjx29qFpLg/o3TvC/l6t9fpGjstI2yVZsL5DU0ZjDopS8VADi9pDicDJcdh2Qco80PsjOD8XDO96s5f3XqzalsdHLXQWuprnVdJcWRs6nw5kBkZIQ09Dg8t6jgjBBbrnsj+arVPF3Rd84LcQrvNdLxomCCptNdUOL5prY89BikeSS4wvDAHHctkYP3q2R+2Z9sB/+rXbv9HZP6ytI9lTys8xfB3j7ftbcSeG1y0vYZNLVFsfNcQxhnmlqaaRkcbA4uJ+8OcXYwAME5cAQyfbS8bNZWmp0ZwJslyqaCx3Wgkvt4ZDIWe/kTGKCJ5G5YwxyPLD8Jc5hIJY0jmDhv7IblYoNCW2m1qL3qe+1FHE+ru0N3fBEZnNy51OyLDBHk/D1h5IAJJJOd5e0L5HpubfSdpvWjbrS2zXWlWytoDWEimuFNJgvppHNBMbg5ocx+CAS5pGH9bOjei+crnq5EPufw2438PKu76at4bSUVNqCFzSIWjaOkuUXUyRoDSBkzBoBAAAAAd+OUvkPsnKHxG1hqPRPEO6XbTmpqCnpYbVcYW+PTSxyOd1vljLWS4BwD4bSA4j5nqz7aXjZrK01OjOBNkuVTQWO60El9vDIZCz38iYxQRPI3LGGOR5YfhLnMJBLGkd0eVHnT4Q829lqZtDzVVr1DbI2yXSwXENFVTsJDRKwtJbLCXHHW05BI6msLgDsD2hfI9Nzb6TtN60bdaW2a60q2VtAawkU1wppMF9NI5oJjcHNDmPwQCXNIw/rYGzeG/shuVig0JbabWovep77UUcT6u7Q3d8ERmc3LnU7IsMEeT8PWHkgAkkk55G5S+Q+ycofEbWGo9E8Q7pdtOamoKelhtVxhb49NLHI53W+WMtZLgHAPhtIDiPmeg+i+crnq5EPufw2438PKu76at4bSUVNqCFzSIWjaOkuUXUyRoDSBkzBoBAAAAHpHyo86fCHm3stTNoeaqteobZG2S6WC4hoqqdhIaJWFpLZYS4462nIJHU1hcAQ7AIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIg8KeZ3/wC2p1X/ABgac/RRL3WXhTzO/wD21Oq/4wNOfool7rIOtfPryr0nNNwNrrFbKWIax0713PTFQ44JqA0eJTEkgBkzW9BycBwjcc9GF019kVzT1enr3XcpHEaqlp/FmqKvS/vRLTTVTSXVVAQfo9RD5WjbDmzA5L2hery8TvataL0nwY5qrLxG4U6lFr1VfqdmornR0RLJbdcI5gIq0PGzTMWl3SNw6J7ztIEG/wD2n/MDqPj7xisXJnwabLdI7fdIoLpHTH+3r289DKcnsGU4cS4k4D3P6seECvRblb5fNOcsnBix8K7F4U1TSx+83euY3Hv9wkAM0xzvgkBrQdwxjB5Lzq9jJw+0DqzX+uOLupL1Hctd2QMioKKoyZYIakO8euyfpvecxZG7QX5/sjV64oC2/wAQdU/sG0FqXWvuvvX7H7RWXTwM48XwIXydGfLPRj7VuBYlyt1Fd7dVWm5UzKikrYZKeoheMtkie0te0/IgkH60HidyO8v1F7QjjTxA4icxGtrrcXWltPXVlPT1QiqK6aqfKGNaSCY6eIQkdDAOnqiaC0bHuFxT9jny5anszxwtvOodEXqJpNPM6rdcaRz/AC8WKU+IRnzZI3HfB7LrLxK5K+b3ka4oVvFzlTq7tqDTQdIYZrVEKqshoy4O91rqIgmdowB1sa5pDA8iJ2AOWuXv2x1BW3eDRnNBohunqkSilkv9nikMEMgPS41NI8mSMAg9RjLyCceGANg7QcyHInoPmj0doDTfEXWeoqWu0DSyU1PcLa6MOqvFigZO6RsrZMlzqaNwOcj4gSc5HkTwn5XNJcQOe2u5VLlqG702n6XUuo7I24weF74YrdHWOiectLOpxpmdXw4+I4A2x9AttuNvvNvpbvaq2Ctoa6FlTS1NPIHxTxPaHMexwOHNIIIIOCCCF4w8tP8A9uSvH/GBrv8A9BdEHd3l09l9wc5ceLdo4w6e1zq+8Xaxx1LaOnr5KYQB88D4HOeI4ml2I5ZMDI3IO+MHjb22f+8LoT+F4/Uqheiq86vbZ/7wuhP4Xj9SqEHVrle5auL/ALQ11gn4hanrNP8ACrhjaKbTNvlpmAlz4om9UNKx3wGV3wySzOBwDG3BAaG+h/Kn7PzQXKPxIv2utCa7v93pb3afuV7jdo4XSQjxmS9fjRNYHH4AMdA7k58lqPs2qSlpeSfhgKWmjh8Wiq5ZPDYG9TzXVBc447k+ZO67IXGploqCprIKWSpkghfK2Bgy6UhpIaNs5JGBse6Dx/5u+LXFLnu5toOULhfevctG2e8yW13h5ME89MHe919SWnEjIuiURtyAQ0EfFJldqNN+x/5SbTpqK0X+HVN8unhtE12kuzqeQyebmRRgRtBOcNIcQMAknc+XvJ1rfmO0/wAbbhrPl30XBrHW77VVy1UE9EanpppZohNOGNfHg9bmNyDsJCMb7d4f2zPtgP8A9Wu3f6Oyf1lBwNxS0fxC9lLzT2O/cOdU1930dfoW1TI6otBuVvbIG1NDVNaAx0rMgtkAABfG8Bpy0exmoOItst/CS5cWrMz3+30unJtRUrc9PvELaY1DB8upoH1ZXjzzOaQ9pFzbHTI4p8udbGdLe+e4fcu1GnB968HxOsvmdn+148YxjfOc7es/AXQl607y4aC4a8R7fAbjbdH26yXek6xIwFlIyGSFzmkh2AC0kEgkEgkHKDyS5HeX6i9oRxp4gcROYjW11uLrS2nrqynp6oRVFdNVPlDGtJBMdPEISOhgHT1RNBaNj3C4p+xz5ctT2Z44W3nUOiL1E0mnmdVuuNI5/l4sUp8QjPmyRuO+D2XWXiVyV83vI1xQreLnKnV3bUGmg6QwzWqIVVZDRlwd7rXURBM7RgDrY1zSGB5ETsActcvftjqCtu8GjOaDRDdPVIlFLJf7PFIYIZAelxqaR5MkYBB6jGXkE48MAbB6SaWtFTYNM2iw1dxfXz22gp6SWreCHVD442tMhBJOXEEnJJ37nuvFPhfpWs9pJz16kp+MGsLhR2SjZX3GKigna2WO3U07IoaCm6gWxn76xzyGkkNlefiJcvbe23G33m30t3tVbBW0NdCyppamnkD4p4ntDmPY4HDmkEEEHBBBC8oeaX2fvMPwU40V/MlyeVNfWwT1813bQWpwF0tM8pc6aNkTsiqp3FzgGNDnFr/DdG4NL3B2B1v7HrlS1BYH0GkXao0pdGMxDcILk6rBeBsZYp8hwz3DDGT5ELtLwA4Y3DgzwX0hwrumojfanTFtjt77h4ToxP0E4wxznFoAIaASdmj6l5z8F/bD620pfGaI5ruGMjJKV4p6u6WqkdSV1K/beoopSA44OT0GMgDZjicL090TrbSnEjSVr1zoe+U14sN6p21VDXUzj0TRnbODgtIIILSA5pBBAIIAa8iIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiDGra6it1Oaqvq4aaFpAdJNIGMBJ2yTsN1sbXXMFwP4Z2qqvOuuLOlbRT0beuRs1ziMp2yAyJpMkjiNw1jS4+QK2tzbctFs5ruEx4WXXVdZp2Ntzp7oytpqZs5D4g8dDmOc3IIkPmCCAd8YPRm/+w8g9we/S3MU81rWksiuGmwIpDtgF7KjLB3yel31IOvGpqHUntLueeuuOhdPVjNKz1dLBPVOi6PcLFT9MZqJ3dmySND3NaSSXSBgzjK9u9Y6XZq3Rt50dHdbhZ23a3T29tbbJ3U9TR+JGWCWGRhBY9mQWkEYIC8b9G8e+an2X/Ean4QcTNOWa7aOmLar3KmpIY4q+mz0uqaOrZGx5k2wRMHHIPU0FwcvY7QmttOcSNGWTX2ka4Vll1DQw3Ghnx0l8MrA5uQd2uAOC07ggg7hB4+8AOZHjTyR84Nx4Q8x+vtQXnS81Z9xrrNd7jPVQ08T3B1LdYBK93Qwtcxzsb+HI8EFzQB6rcwnHTSvLxwev/F3VEjZqW1U2aSma/Dq6rf8ADBTsIzu95AJAIa3qcdmldTvazcqdNxT4Uu486XpGN1Tw/pHyXADY11nBLpWkk46oSXyg9y0yjcloHnDBxU4785beDnKnU3eKaCxSttFsdI4jxS4kCpqSTl5gph0DG4ZG7ALnnIdx/Zn0nMRzJ8a9Q8zHE/ihrB2lrRWTiO3tu1THQXC4zNJ93bAHiMU8Eb2kxgdIJhGCAQvVNbI4M8JdKcC+GOnuFWiaYRWqwUjadry0B9RKT1Szvx3fI8ve75uONsBb3QF5T853NdzE8auaH9pryy6hqdOxQVrbPU1dDUupqmurRGZKl76lo8SGnhaHgtZgkRSOJcC1rfVheNPOtwu43cnnOLLzf8P7JJX6duF3N8pbmIXy01PUTsMdVRVfSQYxJ1ygEkBzJQGu6muDQ39L7Kfm001SHVWiubLOrI8TdDa240YdI3IGKpr3PJAyASwDfBwMldwuAmnuay9cpN90Pxovcln4ry0d5s1svUtRGZInmN8dFWPlpy7Ja8hweB1kMBIJOTxhwF9rby6cUn0lk4jtq+G98nwwuuTxNbHPIGwq2geGM53lZG0DHxEld3aOrpbjSQXCgqoamlqY2zQTQvD45Y3AFr2uBIcCCCCDgjdB8/3OvwQ5r+DX7Df2z/FKr1l92Puj9w/eNTVl3908L3f3jHvIHhdfiwfR+l0DP0Qud+FvJ57THU3DHSOpdBcx90tmmbtYaCustEziFdKdtLQS07H08QiYOiMNjcxvQ3ZuMDYBb+9ud/2E/wD+ZP8A+2Lv/wAp34LHBv8A4v8AT38nQINb03a9W6P4J2qy6s1C67ansmloKW53Zsj3GrroaQNmqA92HEvka5+SAd8kZXiLy9c/HHXhNpXXlmturNUat1prH7nUGnH3WtmubLfMXzeNURxSuf1zEPiaxoBDnEFwcGdDvd3WH/xSvf8A4Oqf/RuXjD7HPh9pXWHMtddRajtjK2q0jp6W5WkSYLIat88UIm6SN3NZI8NPkXZG4BAbo01yHe0kmu9l4z1/EKoF/qaynramln1jO26xMDmuIlLgIhgFwMYlOBlpA7H0S5y+azT3KTwlk1xcKFt0vlymNvsNrL+kVNUWl3VIQciFgHU8jfdrRguBHPa8avbWa0q7nx40VoMVLn0Ni0t90BH1HpZU1dTK2TYjGTHTQbgnOw8kGVwv4Bc+ntBLWeLXEHjlXaX0hcZJfua2eomignaCWONLQU5awRjBYZHlpdju/crE4s8C+er2d9FBxd0TxyrNSaRpZ4oK4wVE8tPD1ODWCroKguj8N3wsEjS4tLsAsJaTzJoX2w/LpoPRNg0RaOEOuoqKwWymtlOyNlE1rY4YmxtAAmGBho8loPHf2tfAfi/wV1xwvi4VazZU6nsFdbKSWqFJ4MNTLC5sEr+mYnDJCxxwCfh2B7IO8HJ9zNWXmt4MW/iXQULLddIpn22+W5knW2krow0vDCdzG5r2SMzv0vAOSCuo3tKOc7jDpjijZeVTl4uFTa7/AHeKlbc7jQkCulqax4bTUdM87wkgscZGkOJlYA5oa7q2v7Dm63IwcX7I+Od1Ax9lqo34+9xzEVbXjP8AdOa1m3pGtO9qby8cXtIccLRzicLrbWV9BRsoKmvqaKIyy2WvoC0w1EjBkiEtZEQ8Dpa5jg4t6mdQT0vsoubCqoG6quHNeINYSRiQxmtuMgbKSHdLqwP69n5JcIzuMgFdveRTT/Nlo3R+qNGc1lzmu9bZ7qyKw3WetjrH1lIYgXkTg+JI0OxgygPBJB7YHCPAH2xXBvWsNDY+OVirdD3t4ZFLcqdjqu1SSdi4luZoATg4LHhoJy/Aye/Gm9S6f1hZKPU2lL5QXiz3GIT0dfQ1DZ4J4ycBzHsJa4ZB3BO4Pog6R+1C51Nact9hsHDfhPVx0GrdWQzVdRdHRtkfbqBh6AYmOBb4kjy4B5B6RG/A6i1zeB9LezB5vOJtgo+IXErmdrbRqe4wCsbR1tXXV1TTmVueiaYyN6H4IDgwOA3GThcje145VeInFSh01xu4bWOrv0ul6KW2Xq20cZlqWUhkMsdTHGMmRrHOkDw0FwDmuwWteW6Ly1+2N0e6yWvSHMnpy5266UcLKWXUttjNVBUloDfGqIBiWN5AJd4YkyckNaDgBzlyK8OedPhBqzVvDzmS1VLqvSVLQ082nrxJc/f2un8RzXRxyy4qQ3oAyyRoDcN6cA79yltrQHEXQnFPTNPrLhxqy2ajstWSIq231DZo+oYyx2DljxkZY4BwOxAK3KgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIg87fbOcHLjq/g1pbi1Zbc+pk0Ncpqe4uiYS6OhrGsBldj962aGFvnjxSdhlbu9ldzOaQ4lcArJweud9o6bWmh4nW77nzTtbLW0Ac50E8LCcvaxhbG/GeksBOA9oXdS92Sz6ms1bp7UFsprjbLnTyUlZSVMQkiqIXtLXxvadnNIJBB8ivNfi77FjT1yv89/4HcWptN00kolhtF3o3VLKYk5PhVTHh4aP3ocxzthl57oO63NBzL6C5YOF911zqu7Uf3VbTSCyWh8w8e51hBEUbGAhxZ1Y63gYa3JO4APm77Hzg/qLXnG7U/MnqSnmkoLDDU0lNWPjAFTd6zBlLT5lkL5OoAbeOzcZwd76F9inW1d/juvGbj0+40bJWmals9A73ipjGNveZ3nwztj+xP27EL0h4Y8L9CcGtEWzh3w309T2Ww2mMsp6aLJJJOXPe9xLnvcSS5ziSSckoN2IiIOPdP8feDWquId84T2PiLZ6jWGnKgU1wsr5TFUsk6A8hjZA3xgA4ZdH1BpyCQQQMfmI07we1Hwd1PS8d6W1yaNhoZZ66WvDAKYNacSxOdu2YHHQWkO6sAbnB6s81XsqdGcfOIdz4t6H4j1+jdTXiVtVXRS0vvlHPUAAeKwB7JIXEAEkOcMjIaMlcMU/sc+Mep6yno+JfNIyqtETxI5kdLVVsmfRjJpWsaSMjqycZzg9kGx/YmN1GeOmvXUrphYBpMCuAP3v3w1kHu2fn0Crx8i5dlfbR0dVU8qunpqeB72Umu6GaZzRtGw0NewOPoOp7B9bgu0HLdyzcMOVvQY0Nw0t04FTI2ouVxq3h9XcKgNDfElcAAAB2Y0BrQTgZJJ3Jxk4RaK47cN71ws4g0L6qy3yERS+E7olhka4Ojmidg9L2Pa1wJBGRgggkEOtnsnta6d1ByZaY05bblBJcNJVtzobpTiQdcD5a6eqjLm9wHRztIPYkOAOQQO3tsulsvVEy5Wa5UtfSSlwZPTTNljcWuLXAOaSDhwIODsQQey8pbr7EvW1HfJ49JcwdtFmqGmMyVVomiqfCccmN7I5Cx4GB++AcRnA7L0X5auCFBy4cEdL8F7df571DpyKoabhNAIXVEk9TLUSO8MOcGDrmcA3qJAABJOSQ8peR//AO2p3z/w9q/9FUvaldJOBvs46vg5za3XmWk4tw3WlrK67V0FpbZTDIDXeJljpfGcMR+KQCGnq6QcNzgd20Hnv7ZfhBdta8DdNcT7NRvqXaBukormsaSYqGtbGx8u3kJYacH0Ds7AFbv9l3zOaE4ocv8ApzhHPfaWl1toikNtntU0rWTVVHGSYaiBpOZGCPpa/GS1zT1ABzSe5t0tdtvltqrLebfTV9vroX01VS1MbZIp4ngtex7HAhzSCQQRgg4K85eMfsZNEXy/VGpeBPFCs0YZJDPHabhTOrKeB57CGdr2yxtB7BwkI9cYAD0gqJ4aSCSpqZmRQwsMkkkjg1rGgElxJ2AAGSTstp6F4v8AC7ifcL3a+HOv7Fqap05LFDdW2qtZUilfIHFjXOYS3J6HjY7FjgdwQPNuD2OvGfU0zKHiNzUtntYcHOaykq69xI3wGTTMaDkDfJx3wcYPdjlJ5NeGnJ/p+82vQt0vF2uGpH00t2uNzkZ1TGASCJrGRta2NjfFlIHxOy85ccAAN+1vH7gza+KJ4K3XiLZ6DWvusNay0VkpgkljlLgwRueAyR56SfDY4vAwS0AgncmtbNo7UOlLraeINutVdpyeleLlDdWRupDABl5l8T4Q0AEknGMZyMLqxzkezd0DzYalj4iU2s7hpLV7KNlFJVMpxWUlXHHnw/EhLmkOGcdTHgY7tJGV1nPseOOt3DLFqPmnp5LCCGFnu1ZU9MbThpED5WsJAxhvWAOwPmg4f9ndS0NN7SOWm4P1E8uiKer1KyORryfEsYjnFIXk7kGT3M5O5OD3Xr3W8fuDNr4ongrdeItnoNa+6w1rLRWSmCSWOUuDBG54DJHnpJ8Nji8DBLQCCeP+U3kt4Uco1iq6bRvvV21BdmNZdb9Xge8VDGklsbGt+GKME56W5JIBcXEAjjznI9m7oHmw1LHxEptZ3DSWr2UbKKSqZTispKuOPPh+JCXNIcM46mPAx3aSMoO0+tbNo7UOlLraeINutVdpyeleLlDdWRupDABl5l8T4Q0AEknGMZyMLxg9ndS0NN7SOWm4P1E8uiKer1KyORryfEsYjnFIXk7kGT3M5O5OD3XMB9jxx1u4ZYtR809PJYQQws92rKnpjacNIgfK1hIGMN6wB2B813b5TeS3hRyjWKrptG+9XbUF2Y1l1v1eB7xUMaSWxsa34YowTnpbkkgFxcQCA7AoiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAtKn1RpqlmfT1WobZFLE7pfHJVxtc0juCCcg/JaqvMbXPsVo9Ya1v+rYuZaopmXu6VVxENRpIVMsYmldJ0vl99Z4jh1YLuhuSCcDOEHVzmXr6Gf2o9VcIa2nkpTr3Tz/HbIDH0gUeT1A4wMHJz5Fe5FPqbTlZOympNQWyeaQ4ZHHVxuc4+gAOSfqXl/8AuGI//Wi//on/AP71r2gPYtt0RrvTmszzL1FSLBdqS5+DT6S92lk8CZsgayX313huJbgO6XYODg4wg768d+NOk+X3hVqDixrOUe42SmL4qYSBktbUHaKmjz+/e8ho2OAS47AkeV/ITwU1HzucyupuafjrRx3aw2m4GqmhqIi6mrrmWj3eka0nBhp4gwlpJwGwNIIccd4Oebkw1xzht0vZaDjNDpTTdikkqKi2PszqsVFU/wCEVBcJmZLYy5rWkADqeer4jjm3gDwS0ly8cJ7Bwm0ZH1UVmp8TVTmBktdUuPVNUyYJ+J7yTjJDRho2aAA8d+J2ndX+zD53KLVWj4aqfSNTIa63RGQ4uNjmfiooHvIwZIyC0E5IcyGQjcL2t0LrbTXEnR1n17o25suFkv1HFXUNSzYPieARkd2uG4LTuCCDgghcR843KVpXm94aQaJvV2Niu9rqxXWe9MpPeH0ch+GVpj62dccjNi3rG4Y7u0BYPJPyx6z5UeGldw11JxVZrG3Pr3VtsiZbDSNtofkyxtJkeXNe/D8bBri4jJcSg7Erb+vNc6V4Z6Pu+vtc3dlrsNipn1lfWOjfIIYW9z0Rtc9x7ANa0kkgAEkLcC2xxK4eaY4saCvvDfWdLLU2XUNFJQ1kcUpjf0P82OG7XA4IO+4GQRsgu0JxF0JxQ09Bqzh3q61ajtFQAWVduqmzMBIz0u6Tlrh5tcA4HYgHZebntqtO8HaPTui9QwUtsp+JtbdDFI6ANbVVNobBJ1unDd3BswgDHPBx1SBpxkKO/exm1jpq8T1vBXmUqbZR1PwGOvo5YKhseSQ181M8CXG37xozk4C3Lwj9jfYaPVcOr+Ybi3V64McolltVHTyQR1JHbxqqSR0r2HbLWtYdsdRBwg7LezmbqJvJTwsGqXTOrTbKgxGU5PuZrJzSAfL3fwQPlheaHCnVNo4Ze12u2oNaVUdrof8AZJ1TTOmqXCNjPfjXQQOc47BpdURHJ2w7JIG69tqChpLXQ09tttJDS0lJEynp4IWBkcUbB0ta1o2AAAAA2AAwuk3OZ7MLSPMzrKfipozWI0dq2shYy5NloveaK5PY0MZI8Nc10UgYA0ub1Bwa3Lc5cQ7rVF1tdJXUlrq7nSw1teXilppJmtlqOhpc/oYTl2Ggk4BwNzsvPf22f+8LoT+F4/UqhZ/J/wCy81Dy98aLHxq11xip79WaZgqYLdbaGhkEZE1NLT5fLK/Ia1kz8Mazvg5ABB5155OUaq5weHNl0VQ67i0vUWS8C6snltxq2TjwZIjGQJGFu0nUHDP0cY3yAs9nB+BNws/8HVX67ULsouM+W/g67l/4JaU4Pu1AL47TVLJA64Cl92E5fNJKSI+p/SAZCAOo5Az54HJiDwv4M3+m9n37Qm5WriFTzUWm6SurbLUVIiJ6bTVkPpatrRkuYAKeRwGSAHgAkYPt/Yb/AGLVNnpNQaavFFdrXcIhPSVtFOyeCeM9nskYS1wPqCVwfzVclHB7m1tVKNcU9Va9Q2yMxW7UFt6W1cLCc+E8OBbNFnJ6HDIJcWlhcSekf7jlxr0vVVFJw+5oqaltkjusf7Vq6F7z5dccUrmkgeed99gg9KOI/GbhNwhp4KvifxG0/phlU5rKdtyr44ZJyTgdDCep4znJAOACTgAlarrzXOleGej7vr7XN3Za7DYqZ9ZX1jo3yCGFvc9EbXPcewDWtJJIABJC8+eGfsX9E0N6g1Dxs4yXnV8glbPNQ26jFHHMWkfBLPI+WSRpAwcCN2DgEYyu/wDxK4eaY4saCvvDfWdLLU2XUNFJQ1kcUpjf0P8ANjhu1wOCDvuBkEbILtCcRdCcUNPQas4d6utWo7RUAFlXbqpszASM9Luk5a4ebXAOB2IB2Xm57arTvB2j07ovUMFLbKfibW3QxSOgDW1VTaGwSdbpw3dwbMIAxzwcdUgacZCjv3sZtY6avE9bwV5lKm2UdT8Bjr6OWCobHkkNfNTPAlxt+8aM5OAty8I/Y32Gj1XDq/mG4t1euDHKJZbVR08kEdSR28aqkkdK9h2y1rWHbHUQcIOxvs+a67WXkZ4bXXXlW+B1HZquqfNUH+x29tVUPpnH/tBSiIj/ALUBcw8KONvCfjjp9mp+E+u7VqSgIBkNJKRNAT2bNC/EsLv+1ka0/Jbmm07ZJtPP0k62QMs8lEbcaKNvhxCmMfh+E0Nx0t6PhAGMDthea+uvYyS27UU2oOAnHuv07G97/BpLnTPMtOxx3Y2qge1zmgEjBZnAGSTkoOSPa/6d4OzctsmpdV0tsi15T19JT6VqCGNrZSZ2e8xAj43QiAyuIOWhwYdiRnN9jo3UY5RZjfHTGjdqy4mzh52FH4VOHBny94FSfrLlxNof2NFyuupae+8fuYCs1FSU72B9FbqaUzVEYOeg1U7yY29wQIycE4LTuvSLRWi9McOdJ2rQ2irPBarFZKZlHQ0cAPRFE0bDJySSckuJJJJJJJJQa6iIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiDAt96s13krIbRdqOtfb6h1LVtp6hspp5hjMcgaSWvGRlpwRkbLPXl9zH8hPN7pbjzqvmD5T+Ikr5dVXGW51Fvo7t9zK6N7z1Oid1ubBURdRcQHvGxALTjJ2X/sk+2pbR/scOm9QunJ93Fb+x20mQb4/soj8PHl1nyOc53Qbr9t7qTS8lr4XaQFRBLqKCouNx8IAGWCie2JmXHu0PkYAB++MR/uV269nTZLzp/kr4V0F+8T3mW1z1sfiDB92qKueenx8vBlix8sFdKOCHst+OnF/iQOLPOrqiZkUtQyqrLY+5iuudzLcYilmjc6OCHAAwx7nBoLQI9nD004j1urNF8LL3VcJdFRXzUFqtb47DZI5YqaOWZrQ2KPL3NY1jdiRkEtaQNyAg87Pa1c0tfdayg5POFk81ZcbnNTS6nbRZfLJI9zTSW0AZJc5xZK4DfeEAnLwut3NNyOcQ+S3R3C/jVpy+1b7jG2n+71XTEE2e/h7p4jG5uR4WMRtccgvgJJ++NaOzfIJyJ8Z6TjxduZXmvsE9LeaOeSutNPXVcFTPW3OdzjJWSiN7w0RgnoBwet7XDAYM+hnFrhfpTjRw41Bwv1tRips+oaN9LOMfFG7YslZns+N4Y9p8nNBQcf8nfMnZ+aXgfZuI1KYIL1EPufqGhjP9qXGMDxAAdwx4LZGbnDZACSQcc4Lyu5GOWbnQ5ROZertFXw7fdOHF+kfbbxcoLrRtpZaZj3e73BjDKZA9hyfDLesske3GSCPVFAUFVSU1dTy0dbTR1EE7DHLFKwPY9pG7SDkEEbEHZTryl1Ly/e0w5WOI2pr/wAu2pqrV+j7zd6u6spIamCpjcJ5XSu8WhqjlkpJw58OS4gHq3wA5Y58fZ38vdz4Paz4wcPtNUehdT6VtNXfHfcseBQVzKeN0r4X0w+9sc5ocGujDD1FueobLb3sVOJ2q9TcM9fcNL3WT1Vp0bX0FTaTK8u8Bta2oMsDCezA+mDw0bAyvPmuJtc6d9rdzY2V3C3WukpdP6arHs+6QmipLRTStDgR4z+ozSsBAJjjDgSAS04BHfzku5TbFyi8KjoulurbxfrrU/dC+XRsZjZUT9PSxkbSSWxxtGADuSXOIBdgB0r9ud/2E/8A+ZP/AO2LvzyjVVPWcqvB2WmmbIxug7DEXNOQHsoIWuH1hzSD8wVxf7QzlCuXNnwmoaHSFXS02sdKVUlfZxVO6YqpkjA2emc//wCRl4bG4OOR1RtBwCXDofwy0t7Xvg/pgcDOH2mdR22z08r4aUPhtc8VGHuLn+BWylzWMJcTtJhpJLekkkh7Caw/+KV7/wDB1T/6Ny8g/Yl/7/Wu/wCCB/XaderGjLFrem4IWLTOuK4XHWEWlKWgvFSZQ8VNzFG1k8nXsD1TB56sDOcroN7LDlK5hOX/AIuay1Lxg4dTadt1ZYDbKeaSvpJ/FqPeonlrRDK8kdLHHqx0nAwTkIPTReOvtsNC11v4z6E4jtpS2gvem3WjxWsGDU0lTJI7qP8AdFlXGBnybt2OPYpcQ80PLVojmn4V1nDTWTn0kgkFZarnCwOlt1a1rgyZoJAc3Di17CQHNcRkHDgGNwRtXA7i9wj0jxKsegNHVtNf7TT1bpGWalPTMWATRuwzAeyQPY4eTmkeS3z/ALEPCb8V2kv8yU3/AKi8pNK8uvtQeSa5V9r4IF2pNL1U5kMdrlpq+hqHHAEho6jEsMhAaHOYwdgC9wAW7n6n9tFxajfp6k09UaRppR0VFW2kttsLWnzEspMo7jeH4tttsoPUix6c09pikdQabsNutNNI/wAR0NDSxwMc8gAuLWAAnAAz32C1FzWuaWuaCCNwRlcAck/A/ixwF4P1OluNXER+sdU3W+VN6qas11RWCnEscLBCJpwHvwYnPJIA6pXYz3PVHmJ5buf7hdzCav478qOs6q62nV1Yy41Vqp66EOjeI2RmOajqz4M4aGAMe3Lg3YBuNw5+5m/ZzcufHWw3e627SVv0XrF8Ms9NfbQwUrDPjINTC3EUrCQOtxaH4yQ8Eknpz7FDidqxnEfXHBySsnm03UWE6jige4mOmrIqiCAlg7NMjKj4vXwWeiyNU3/2xXHOyVfDav0HW2K33mJ1NXTxUVFa+uneMPY6okeC1pBOQwhxGQM5we2/s/8Akbh5RNK3O7aou1Ld9danZE24z0gd7tRQM+JtLEXYL/jJc6QgdRDQAA0Eh24XX3j9yL8t3MVS1tRrDQFHbtQ1bXFmorOwUleyUggSPcwdM5Ge0zXjHpsR1+51eWbnJqOOzOZLlM1xUMrJbRTW6stNPcWUs4FOXlo8Oc+71MR6y7oednE4ac5XD9fxO9s7qyhk0fDw+r7bU1bfdXXCntNvpJBt0ucKh7hFGTuesFuM5aRsUHHfs2L3q7gbz73bl8ob6642W4Vl805dfCJFPUSW5lRJFVhmSA7qpi0HOzZnDO69p10P9np7Pa98uV6rOMnGO60tw17cqaSlpqOmlM8dsjkdmV75icSzPAAJAw0F4Bd1EjvggIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICLrZzI8/HAzlY1zQ8PuJ1NqeS6XC0xXmE2y3xzxCnkmmiaC50rSHdUD8jHbG++3FP7shyjf3hr7/ADND/ToO9KLot+7Ico394a+/zND/AE6575ZebrhVzY0F/uPC6C+xw6cmp4az7q0bICXTNeWdAbI/IxG7OceSDm5ERAREQEREBERARcE8zvOPwj5Sv2Nf7KkF/l/ZV757h9yqNlRj3bwfF6+qRnT/AGxHjGc79sb7y4D8cNGcxXDWg4q8P47kyy3GaeCFtwgbDOHQyOjflrXOAHU0437IOREREBEXU3XPtLuXTh/xer+Cd+o9XnUNvuzLNK6ntkT6Y1D3NaCHmYEty8b4B77IO2SIiAiIgIupHGH2nHLhwO4lX3hXrSj1i69afmZBVmitcUsBc+Nsg6HmZpI6XjuBvldj+GfEXSnFvQNi4k6HuHvtj1DSMrKOUt6XdJyCx436XscHMc3OzmkeSDdCLh7mV5ouGnKrpa16w4nQ3mSgu9w+5tOLXStnkE3hvky4Oe0AdLDvk74Tlq5ouGnNVpa6aw4Yw3mOgtFw+5tQLpStgkM3hsky0Ne4EdLxvkb5QcwoiICIiAiIgIiICIiAiIgIiICItscSdf2LhVoDUHEjU7ap1p03b5rlWiljD5jDE0ud0NJAJwDgEj60G50XXTlr56+CXNXqm6aP4YU2pYq+0W/7pVBulBHBGYfEbH8JbK8k9TxtgbZ3XYtAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREHVDmt9nfwz5teIlu4kaz1vqez11tssVkjgthp/CdFHPPMHnxI3HqJqHA4OMAbd8+YftC+TXQ3J/etFWzRWqb7emampa2eoddfBzEYHxNaGeGxowfEOc57DC9615Ke3F/+NnCX/wdd/8A0tMg2XyJ+zh4X81XBKbidq/XWqbRXx3yqtYp7b7v4JjijicHffI3HJMhB3xsF2S5PdZ8lXKXxP1Py66C4t6tvWrdR6lisNVTXa0SBkVwpJJacxsljhazp63PBcSQcAg4W5PY3/gjVX8L7j/6GmXlTzFU19rucfilb9LmoF3reJF+pKIU7yyR00tzmja1pBGCS4DOfNB7J1ntLOV88YrLwS03er1qa9Xu+0uno620ULJLfFVzzCFpdPJIzrjD3AF8QkGDkdQW5OP3Pry1ct94dpfXmtJKzULAHS2ey05rKqnBAI8bBDIiQ4ENe9riCCARuul+uvZ1cMeTPl/ruZS96+1PduJGgRb7zbpaN0ENsiu4qoW0rfAdDI+SEVD4w8ueC9gOPDJyOoPKFZeWziFxXveq+c3iTNSWaGM1vu9RLVma+V80hLjLPA10gaB1ueQ5j3OczDsBwIeqHC32rfKPxNv1Jp2e+X3R1VWuEcMmpqGOnpy8kgNfPFLLHHnGep7mt3GTnZdheM3HThtwC0G7iVxLvMtFYG1ENN7zT0slSS+XPRhsYJIODuBheNvPRpHkB/Yta9W8omtaKK/RV4p7pYYH3B8NRTPY4+8MNU09D2PawFoeGlrycZbvrFVxwv3FT2V960dqq4zV9w4f60tdrp55pOuR1uka6Sna4nf4CJYxnYMjYAdsAPRum9pHyiVPD258S4+JMrLVbKxtv8KW21EdXVVLmdYjggc0Pk+HBLgOhuR1OaCCti6Q9rzyh6pv0VkrqnV+mo5pvBbX3i0sFLuQA5zoJZXtaSe7mgDcnAGV0O9mPycaB5ptU6uvfFdtbVaZ0dDSMbbqaofTitqqoy9PVKzDgxjIHEhha4l8e+AQYPaecpHDnld1/pOt4UUVZb9Oaxoqp7bdUVMlS2lqaZ8YkEcshLywtniOHOcQc74IAD3EN5s7bOdQm7UYtQpvfTXGdvu4p+nr8bxM9PR0/F1ZxjfON1021r7XTlC0hqB9hoK/VeqGQy+FJcLLamOpQc4JD55YnPaMd2NcDkEZG66a8buYHV1r9lnwJ0BFdZo6nXUt1t9ZMyUhz7Vaa6WFlP3z0nNIDvjEZbjDsLensy+QTg5xl4WVPG3jbYptRNrrjPQ2e1uqpqemjigw187/AAnNdI8ydbQCSwBmSCT8IbQ9qvzDcJeZDSXBbWXCXVUd2o4JdRwVkD43Q1NFKRbSI5onAOaSM4OC12CWkgEruD7NHWelOHvIDYtZ641DQ2Sx2qsu81XXVswiihZ79KBknuSSAGjJcSAASQF0L9pxycaH5Xda6a1DwrgqKPSesoalrLdNUPn9wq6cx9bGSSEvMb2ysIDi4gtfvjAGbr+l1tU+ya4XSWGmqZdP0+t7lLfXQBx8P79VNpzL07CLrcQS4Y6zEMgluQ7tan9sXyj2C4uorXRa81HExzm+92uzQshcAQAQKqeF+D3Hw523xsuZeX3ns5b+ZW6HTfDzWMtNqHpfIyy3enNJVysaMl0QJLJcDJIY9zgASQAMryk5HuL3ItojTN30vzS8IfuteK+udJTagmoDcII6R0bGiExB3XC5jg9wfGxxcH7kFoB5o4Sck/BLiRx4i4p8ofN9pqjgtF6ivlo08KGd1xoYWObIYiyaVkz4xhzcuYQWnDid8h67LzK4xWv2YsvNLeZ+Ieo9ZxcSjqaF1bDTx1hphcfEj6AC2Is6erpzg477r01Xz980n/2xTVf/ABgUv/pokHr7xp58+Wnl91zNw64n6vr7ffYKaGqkghtFTUNEcoJYeuNhbkgds5C2nxk9pzyrcGa+GzXC/wB51JdX08VTLRafoGzvpmysa9jZXyyRxtf0uBMfWXN7OAOAfNv2u34Zd1/8A2v/ANEV295X/ZacuOreXPS+qeKtpvN21XrOyU95luEV0npnW73qNssTIYmkMLmMe0Eysky/rOMYADsry489fLzzQ3SfTnDrUNbSagghNQbNeKT3WqkhH0nx4c6OQDO4Y8uAGSAN12FXzs8DILtwD56dJafZcJBU6T4kRadq5o24dJC2v9zqQAD+/iMgxnHxYX0TIPnq9o9+GzxT/wDCNL+pU67E+yL5s/2Ca0l5bNbXLosWraj3jTssh2pbqcB0APk2doGMnAkY0AZkJXXb2j34bPFP/wAI0v6lTrkn2jPKzWcA9cad45cPqeSh0vrVkFYH0mYxar02Nsj2NLcdAkIMzCDkESgABoyHbf22f+8LoT+F4/UqhPYmf7wuu/4Xn9Sp11v5v+aik5puRThvqC4VUI1hp/V8dr1NTNAB95FBUdFSG+TJmgvGNg4SNGegrX+STjPdOAXs6uOfEnT84hvVJqJtFapfD6/Cq6mnpYI5cHbLDIZB1bEswQc4Id9ePntA+WTl1vculNZ6ynueoqfHvFnsVN75UUxPlKctiidgg9D3h2CDjBBO0eE3tUOUjitfqXTTtR3nSFdXOEdP+yahZTQvkJwGGeKSWKM/N72t8s5IC8sOS/TvKvrPiDqDVXOVxFdR2uiZHPTUFTLWeJeqyd7zJJLPA0ydLAzLh1Nc50rDkgOB3dz2aR5D4bRZdXcoGtaT7ourTS3iwQPrpInQuY5zKqM1TcsLXN6HND8HxGENHS4kPZzjhx64acu+io+IXFO8T2+yS1sVvbPBSSVJM0jXOYOiME4IY45xjZaZwB5m+EHMzabte+EN+qrpS2WoZS1jqigmpSyR7S5oAlaC4EA7jK8oNZ8cb5xh9lZRWfVVxlrrvoTiDRWI1E8xkmmo/dZpaZzyTn4WvdEM+UI3JyuxXsQf97Hib/4eov1dyDuJzAc3fAzljrLLb+MGpKy1z3+KeagEFtnqvEZEWB5JiaQ3Be3Y4znbstN1Vzt8vGi+D+l+OuoNVV0Gj9Y1D6S0VbbVUPkmkb4mQYmtL2D7y/cgDYeoXQ724v8A8bOEv/g67/8ApaZbA5lf/tVnLl/4eqf/AK/Qeit29ofyp2bhPRcY6vX85st0q6ihttM23TCurp4MeK2Knc0O6W9TcyO6WAkAuBIB3Hwl5xeCnFrgvcuPtPd6rS+j7TXz2+qq9SNhpHMkiDDnDJHgh3iMDACXOJA6c7Lyf5AeRB3OJT3TU/EHWV3tOh9KTC1U7LcWmpqKl48Z8UL5WvjiYwSh7j0OJMowNyRt/wBoDp2ycA9aUvKVw0vl/l0RpQt1BNTXSqjlfLda6FjnyExxxhzWwthawOBLS6XBw7CD0NvftiOUS03X7n0MOurzB1lvv1DZY2wYH77E80cuD/3mfUBdgOAHNxwF5mqep/2J9bR1txoYxNV2mrhfTV0DCQOsxPALmAkAvYXNBIBOSAet3Lh7LPlpp+CFgn4uaMn1Nq2/2uCuuVbJcqqn90kmYHiGBkT2BojyG9TgXOIJJwQwecnG3SOpOQTnIqKXhzf6wu0nX012slVI4NknoZo2yeBMQMPBa98Em2HAOOADgB7hcduY/g3y3ach1Lxe1hBZ4q1zo6Glax09XWvaB1NhhYC9wHU3LsBreodRGRnq432zHKc66e4O0/xHbBnHvptFJ4GMZzgVRk+X0M5+W66Se1mu2pdT8w2n9ZVsFaNM33RtsrNNySjMLqZ7XSStYRsXCSRxcM5HUwnYtW/uF3En2WvEbgVQ8MeIugW8ONYutDaKe/y22oqZGXERhvvbKuDre4GQeJ0yhrAMtIx3D1K4Kcf+EnMNph+ruEWsaW+0MD2xVTGtfFPSSluQyaKQB8ZIzgkYODgkDK29zA83XAXllgp28V9aMo7jWx+NSWmkhdU108eSOsRMB6WZBHW8taS0gEkYXUXkN5W7Ny23LUXMVovmf0zxD0YzTtZFd6SzUuGubGBO1z3eM4RyMMR2e0OAc8bZK6G8GNJ6p5/ecmlpuIV8qQ/Vtxnut8qY35fT2+BhkMEJIIbiNjII9iG5YSCAQg9LtO+2H5RL3ePuXcItcWGn8RsYuFxs0bqfBOC/FPNLKAO5yzOOwJ2XYjifq/gnxH5a9Uau1Bqhl14Y3fT1VLcbnaJTKX0BYRKYzGC4PADgWgdTXAggEELq9zIeyc4Fag4T1NPy+6Ofp3XNsjY63TPvFRJDcCHfHHU+8SPaMguIe0NIIaMlo6VxNwo5bOZvlh5SOY3SHF6G2R6Su+jquvtsVLc21Jp69sTmy9LW46RJGW9WxBMTMY3yHMfs+7fyFUfErUb+U6+6ordQusZbcmXZlSI20XvEW7fFjaOrxOjsScZXYXmF5x+AHLG2Gn4payEV3qohNTWWghNVXzRkkB/ht2Y0lrgHSFjSQQCSMLy09k5rej4Z6x4y8Rq+HxqfS3DauvMsWcGRtPNFKWA/PowPmQuDeDVdw14/8yVXrXm84nPstiuctTer1WnxjJXTdTeikiMbXujaS4AYADYoy1pBDSA9VNGe115QNW3tlnuNfqzS7JZjCytvVoaKZ24AcXU8spYwk93gAAEu6Ruu0+teK2hNAcNK/i9fr7E/SduoW3OS40QNWySmd09MkfhdXiNIcCC3IIII2XkFzkaM9mfW8JJ79yva4orXrazywOhtlPLdJ47tAXtjfG73oODHtYTIHgjJa4O6i4EWcrfGy96j5AeYjgffq6apg0rZYrtZTI7q8GmqJg2eEHyY2RrHgesz+2yD0W0z7RzlE1VpnUer6Licaa16XZA6vkrrbU07nOmMgijhY5gdNI7wn4YwE4aSQACRxtavbD8odyv/ANxqpmuLZSmVsYutXZYzS4Pd+IpnzdI88xZ9AV5x+zr5YtM803G+q0nr2rrGaWsNqfe7jS0lQYZK1zJGRRRdQBLQXTElwwekOALS4Ecx+1C5JODvLdp7SHEPgzbKuy0V3uEtouFsmrpauLxPCMsUsbpnOkaSGShwLyDlmAMHIew2mNT6f1pp+3ar0peKW62e607KqiraWQSRTxPGWua4dwR+fIOMFdceN/tJOVbgVf6nSd91ZXahvlC50dXQ6dpBVupnjux8rnMhD87FoeSD9IBdIeBHMprDhR7J/WNxsVwqKe702s6rRliqmPPVRMq4aeplkYRuxzWz1RYQch5aQc7LYvsweTDQPM3f9U634u0s9x0zpV1PSxWuKqfAK6smDnEyvjIkDGNYDgFvUZBvhrgQ7/cKfan8ovFO80tgfqi76Prq14ip26noW00T3kgBpnifLDHnPd72jbGckA9tayuobfQz3SvrYKajponVE9RNIGRRRNBc57nk4a0AEkk4AGSvL/nE9kjHdrvaL/yi6fpLbHUeNHebNX3d4giI6THNA+YucMkvDmFxAw3pAGQuLOdDiBzCcD+UjhXyk8UK4U19qRWyXmppK/x/e7PTStbQUzpAcOaOshzdsClhznJQdy9f+1s5QdD3may2+7al1a6nl8KSpsFrbJTZBwS2SeWIPaDn4mZBxkEjBPLfLzzq8vXM3NLbeGWsnfdyCMzS2S5wGlrmxA7vax2WytG2TG5wbkdWMheZvJ9on2Zdu4TUl85nNeUV31vefGdVW2okucMVqi63NZEz3VrQ6QsAeXlxILgG46ST14401vDbgTzJQ6x5Q+JUt0sFrnprzYa9olElDMD8dM8yNa6Roc1wPUMOjeGu6skkPePjnzGcHuXHTkWp+Lusaey09U8x0VOI3zVVY8Yy2GFgL34yMnHS3I6iAQurLfbM8pxugt507xHEHVj342ik8AbZzj3rxcZ2+hnPljddFfagaw1RxL4raB4o1tPVR6a1boC0XXT4cSYGsmYZaiNh7dbZZCHg/EMszt0rk3g9xM9ltr3gXbuGHE7h7FoHWJs8dDV3+otlRUPdchCGGsjq6cuf0mT750SBjBu0jGCQ9SOB/MHwj5i9LSav4R6ugvVHTyCCri8N8NRSSkZDJYngPYTvg46XYJaSBlF0d5GuSi88Lrxc+K3Anm50nq23Xe2G1VBtdu8SIOMscrTIBM/pe3oIDXNDgHntkgkHpSiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgLyU9uL/8AGzhL/wCDrv8A+lpl61rz99qBwV0txY1Bw/n1FX3WndbaOvZF7lLGzqD5IM9XWx2fojthBrPsb/wRqr+F9x/9DTLzomhiqPaiyU88TJY5ePpY9j2hzXNOosEEHYgjYherPs5OHll4Z8vdRp2w1VbPTHUNZU9VW9j39TooMjLWtGPhHln5rp/+1o0J+3p/2Qvutfvuj/stfdnwvHh8Hxvux4vTjwuro6tsdWceed0Hevnu4a3zi3ykcSdDabppam6VFtirqWnibmSd9HUxVfhNHm5/u5aANyXABeNfInYOU/WPEu46P5rXiit1zo2fcS4zXSWgpaera/4o5pWPaGh7Ds556QWYJBIz9CC8n/alcm/BPh9Zp+Omh7XXWO9XqqldW0FHMxtukmyC6YQlhLHuLiSGOa3O/SCSSFeY3hX7KHl901T3SmtDtc3itmYynsuntbTVExjJPVNI9krmRMABALsFxwADgkbJ4oWXgrP7NzU3EfghwlvmhLNqfW1sg8K7Xh9e+uFIZGCdhcSGsD5JY8DcmN2ewXD3s9+WHhtzMcTm2DiVNefudSCad0FvqmwCcRsa4Me4sc7pJJB6S047EHdepXPHwa0NeOVOLhbQ0DrFp2119vZR0tqEcIgZEXBjGhzXAD12yTvnKDrb7Dr/AOKfFr/wjaP/AEVStJ9ud/2E/wD+ZP8A+2LnH2YHCTTnCfT3ECDTtdcqhtyrKB8vvssby0sZPjp6GNx9I5zlYPtRuDmmOLR4ZDUdddKf7l/dnwfcpY2dXie5dXV1sdn+xjGMdz32wHSbjTw5ut/9l5y7cTKCldNS6Ru+pKGtczJMMVddajpkI7dPiUrGEnzkYB3K7Neya5sOE9l4KVHBDiDray6YvWn7lU1du+61bHSRV1HOfFPhySFrXSMkMuWZJ6S0jIz09oOUfhBoePkz05wgvFs+7umamnutLU0tzDZPeIpbhUvcH9LWju84IAIwCDkZXjhz1cAdEcuPHWv4fcP6m6y2lsbaiMXGdk0kfXg9Ac1jctGcDIJwBkk7oOw3tfOZLhxxe1dorhzw01NQahp9IR1tXc7lbp2z0pqanwmsgZK0lryxsJLi0kAyAZy1wG/uAnNlZ+Vb2fXDOq1Twjdrmxauu+obZVxOq44oInMqnObHKySKRsgkaZNjjaM7HO2y7lyNcE6blT4ca0gdfWX3VlZLXXGv95hMhDWSNbCzMRDIhguwBkk5cThuO6HLTyxcJtZ8lNt4F6ztVTe9NVFXXzg1UoFTBP73I5s0UkbW9D2lxwQO2QcgkEOCuEfLz7O3no4eTa507o9nC/U3jzxXK02fUIjqKAte7okEEgMHhuZ0vDmQtbu5uctJXnRxW00zl05j7pp3gxxKdqGTSF3hNl1Da3APknDGP6WmMkF8cjnRO6SQXMdgYOFkc2HBzS/AvjHeeH2kK251VtoJZGRSXGWOSYgSOaMuYxgOwHku9PstuTjgvra3UfHnWFDcbxfbFUQTUFFVTRut8U5Bc2YxBgc97C0FvU8tB3xkAgPU60y1s9qo5rlF4VXJTxunYBgMkLQXDGT2OR3XgPzSf/bFNV/8YFL/AOmiX0CLyV46cs2g7/zjXzWdZdr8ytqtWU1a+OKeERB/iRnABiJxkeZz80HCXtdvwy7r/wCAbX/6Ir2Y5ff94Xht/BCz/qUS88vaEcumieI/MbcNU3y6XyCrktFDGWUk8LY8MYQNnROP516O8IbbBZ+FOjLPTPkdDQaet1NG6QguLWU0bQTgAZwPIAfJB4M6v/8AtkN7/wCO+p/l5y+hVeQ2oOWTQdVzm3PW8l3v/v8AJxMmuxjFRD4PjG6ul6QPCz09XlnOPPzXryg+er2j34bPFP8A8I0v6lTr294mcFtJ8wXL5Lwn1lADRXuy07IqgMBko6lsbXQ1Eeez2PAcB2IBachxB83+czlj0Frfma11qm63e/xVdwrKd8rKeohbGCKWJo6Q6Ins0dyV6zadhZT6ftdOwnpjo4WDJ3wGDH6EHzM8U+HWseDWvdQ8KNa08lHc7FXmnq4Q4+HK5gPhTN8nNcx/Ux2Poybdyu6PLBw3vfFH2YnHuwacgkqLlR6lgvMMEYBMraSKknlaB3JMUcmANyQAM5we0XtMeV/hjxI1JpDiHdI7hb75UQ1Ftq6i3SRR+9wxdDovFDo3dTmdbwHbHpODkBuOQfZm8LdP8KeFGp7Rp2suFRDXahNTIa2Rj3B3u0TcDoY0Yw0dwUHmByD6Z5RNc67vmjOa6RlDFcKWCTT9yqbvLb6SKdj3CWGWVj2tBka9haXkNHhEZBcAeznMlw29lFy+WSiqqKwTa9vNfM1kdo07rWaeSOHDi6aWRsrmRtBAABPU4uGAQCRge1J5OuCvCuhPGPh9bK6yXG+VTzV22mmYLcJPhLpGRFnVGXFxJDXhuezRvnhL2c/Ktwv5mNfz0fE595lobX4k/ulDVNgjqejwz0SnoL+k9Zz0Oae2CEHLHGbRXC2q9mhX8SeCPCO+aFsF81zQ18tNdrrJXy1MEDZaVtSHOJ6GmWUsA2B6c5ORnVPY+cxPCLhdQa/4f8TNc2bSlVdaqjuduqbxWx0lNUtYx8csYmkIY17SYyGkguDjgHpOPUPVfB7hzrPhVW8Frxpikbo6utrbUbZTMEMUFO0ARiINx4ZYWtcwj6LmtI7L56eZLhJpvg9x9v3CzTFbcqi02y4mkhlrpWPqCzxC3LnMY1pOAOzQg7C+1h5huHXHLjRp608MtSwX+1aPtL6We40kolo5auaXreIHjIkAY2IGRpIJyBnpydb5lf8A7VZy5f8Ah6p/+v1vjjnyFcCdG8I+E0FgF+grLnT3K53G4mqhdVVsskdGWskcYunw4wCGNa1oHU8nLnFx5E41cA9HX3kR4OcPKu5Xhlusd3nkp5Y5ohO4k1eesmMtI+M9mjyQcg+xrp4IOUu4SRRhrqjWVwklI7ucKekaCfn0taPsC6Ve1/4aXbSnNVJr2Wge2164tFHU09UG/A+opom00sef7prY4SR6SNXo37OTh5ZeGfL3UadsNVWz0x1DWVPVVvY9/U6KDIy1rRj4R5Z+a5f478v/AAv5j9Ey6A4q2D7oW8vE9NPE/wAOpop8ECaCQDLHgEjzBBIcCNkHF/LXzpcAuI/AzTeqbzxY0tZLpQWiCG+0N1utPRz0dVFGGzEskc0lhc1zmOAwWkYwcgeRfOdxJp+bfnFutdwlo5LtTXarodOae8ONzX3Asa2FsgBwQJJC4tyAQwtyAQVxlzIcMrDwf4v3zh/pqrr6m3WyXohlrpGPmcMkfEWMa09vJoXrh7OPk44J8PtF2fjtR2qtuusrjFM2GuukzJRb25LHCmY1jWsLm5BeQ5+CQHAEghx1zTc5PB7gfdKflF408r0ev7Vo202qmpq+qrYxFUR+4xYnp2yQl8bhl8fU14OWOHV3VnFv2fPIHrfhRcOLXCTihTaKoxbZLjR18OoBW2wERlwbNHUOfKNxgsa9rwcjGR0rspzvco/BvmG0LV6r1zbK2m1Dpm3yyUF3tk7YakRty7wHlzXskjLiThzSQSektyc+C0mnqJmvBpcSze6/dNtF15HidBk6c5xjOPl9iDsN7N+PWd/5jm8MdN3Oop7ZrvTV+sl8Y1zvANK+2VBZJKADjomEJacA5IbkBxzhck3Eyl5VecKy3HirTPs9Lbqqt05f/HYeu3GRroXPeACcRyhhdgH4Q7Gdl6x8ifKFwa4B6Gotf6Nt1fV6m1RbYzW3W5zsmnZE4hxgi6WMbHH1AEgDqOB1E4GOOPaTcnnBTW+ibzx6qLTWWrWVvgYJqy2TMibcA0Yb7yxzHNeQMDrHS/AALiAAA5L5oufbg3wV4O3HWOgeJGjNV6qq4QzT1ro7hHcGVM7sYklZTyhwha0lzndTcgAA5IC6s6B5y+PnN5yu8xtXxG0jpG2ab03oipjhqrLQ1UL5a6WN5EZdNUyhwEbHEhrcguYSQCAfP/lV4R6b448bLDw31ZW3Kktl0kDJpbdJHHO0dbW/C6Rj2jZx7tK9yNacvXC/hdyf604J6Asj7NpyTT9fDN4MnVUzSSRESTySvBL5XYGXEEAAAANa1oDy39mFoSfihX8d+HNH0+9al4W3G10xcQA2eaSNkTjkgbPLTuQNtzhcL8qdk4Dxce4dG82VtqqPS87Kq2VUklTNR/cy4tI6HzujIe1ocx8bgchpkBdgNJHov7M7gTpHhTxW1ReNO3G71E1bp400grZYntDfeYnZHRG05y0dyQsn2nfJzwTuOjb1zE0VrrrPq9p/24+3Tsjpri8RPIknicxwL/vYBcwsLt+rqOCA2Xx04IeyX4I6Fm1dI+l1TXPAFvs1g1vNV1Va8kdvDmcI2AHJe/DcDYkkA7S4f0fLzqbkt5i+JXAXghqLQ1IyyRWOprbrfpK9tdIJI5nRxtcSGmMGIuO2fFaBnfHUjkv4FaO5heNNq4fa4rLrTWyrnjbK62zRxTEFr3EBz2PA+gB27E/Ij2j4wcBOGui+TnVXBDRdmNi0xFZHU4jo3ATEl7XOlc94d1yOcMue4EkkoOg3sRP99niT/B2m/WVzb7bP/eF0J/C8fqVQpPZjcDtJ8Kdd6yumnbhdqiWutFPBIK2aN7Q0T526I2nOfUlcm+0y4W6f4rcKNMWjUVZcKeGh1CKmM0UjGOLvdpW4PWxwxhx7AIOg3Crh5fdf+yY4kS6fpn1M+l+J7tQTQxty99NFb6GOZwHoyOZ0h/7WM/Ut++yA5muHHDCfWXCLiTqug042/wBRT3az1lxqWwUsk7GGOaF0jyGMeWiItyR1dLhnIAPcf2bPC/TvDTgNqDS1mmrKyirdVVNVMK90cjnOfRUjHN+FjQW4jGxB7n6l0V9p/wAoHBrgFXUWtuFtBcLQdQ1HXNam1DXUFOXF3V4DCzrjGR9HrLR2AAwEHaHng9p/a+BtfY9Lcu120Rri+zulnvUs5kr6KigAHhRiSmnjBmc4kkdTukN3ALgunPPTceNXHngTwc5teKGlbPbH3wXKzPZZ6OeCGCmE3XRPeJpJH5mDap7T1YLQ0gfFvT2Z/Kjwm5jNbVlfxSp7nX0tgaKxluhqWxUtUWvaAycdHW5m+4a9ufXGQfZ/WfDLQPEHQFZwv1fpW33DStfStoZbU6IMgELMeG1gZjw+gtYWFuCwtaW4ICDyg5H+C3s5eNHByhdxguNDaOIdrfNDeoLnquW2iob4rjFPC10rGOYYyxp6c9LgcgZBOhcZaj2XHDfiIOHmg+DGqOJT2iON9wsGrag0rqp7y0QRPMhE5Hw/EzLSXAAkg44J56OX3Q/LhxtrtA6Aq7vNamsM0YuVQyaSPLWuDQ5rGZaOogZBOAMknJPer2W/JxwVrdMW7mLvtur7zqmiqzHQRV8sclFRSNjjeJ44gwEygvOHPc7pw0tAcA5BTmS5weCXL2y38ovEjlEdqPTemrJazQU9zvENQ2KKSkjcAxz4nuD4nmSHxA/JMZIIBwr+Jvs/PZ/8R+FNXxe4TcU4NC26S3OuVPcYr4Ku2xAs6gJ4KhzpWkHYxtex7XAtxkdK7L873KXwe5iNAVWptdW2sp7/AKWoZpbdeLZKyGqbEMv93eXMe2SIuyelzSWkuLS0udnwOq7FSU+sH6fZJMadtyFGHEjr6OvpznGM4+X2INz8EuKHGzhbqGtunA6+3+3XasojBWfchj5HyU4ex3xsaDsHBu5G2cZ3OS9puQrk/wCCnBjh1RcQ9O2eruepNV0A99uN3lZO+OHrz4ETWsaxkfVG1x+EuJxlxAaAQf/Z)
“Figure 1. Neuroarchitectural Changes Induced by Repeated or Acute Stress in Rodents. (A) Repeated restraint stress (7 days) induces a reduction in the number and length of apical dendrites of pyramidal neurons (layer V) in the medial prefrontal cortex (PFC) of rats. (B) Magnified segment of dendrite from the same stressed rats, showing that repeated stress significantly decreases the number of spine synapses in medial PFC. (C) Reconstructions of representative infralimbic pyramidal neurons in mice exposed to zero (0), one (1×), or three (3×) unpredictable sessions of 10 min of forced swim stress. Apical dendritic branch length was significantly reduced after one or three stress episodes relative to controls. Adapted, with permission, from [24] (B) and [23] (C).”
![[[@Musazzi2017-lr]](https://doi.org/10.1016/j.tins.2017.07.002)](data:image/jpeg;base64,/9j/4AAQSkZJRgABAQEAcQBxAAD/2wBDAAMCAgICAgMCAgIDAwMDBAYEBAQEBAgGBgUGCQgKCgkICQkKDA8MCgsOCwkJDRENDg8QEBEQCgwSExIQEw8QEBD/2wBDAQMDAwQDBAgEBAgQCwkLEBAQEBAQEBAQEBAQEBAQEBAQEBAQEBAQEBAQEBAQEBAQEBAQEBAQEBAQEBAQEBAQEBD/wAARCAG3AjADASIAAhEBAxEB/8QAHgABAAICAwEBAQAAAAAAAAAAAAcIBQYBAwQJAgr/xABiEAAABgEDAgIEBgsJDAYGCwAAAQIDBAUGBxESCCETMRQiQVEVGSMyV2EJFhg4cXWBlZa11BckQlJUYpGToTlVcnZ3orGys8LS0zNDY4KS0SUmNFN0wSc1NjdERVZYZYSj/8QAGwEBAAIDAQEAAAAAAAAAAAAAAAMEAQIFBgf/xAA/EQACAQIDAwoCBwgCAwEAAAAAAQIDEQQSIRMxQQUUUWFxgZGhsdEiMhUjM1JTcsEGQmKSotLh8IKyJDTC8f/aAAwDAQACEQMRAD8A+qYAAADWNStM8G1fw2dp9qRj7V3j1mbRyoLrjjaXTbcS4jdTakqLZaEn2P2DZwAFQeizTvDNJ9fepXT7T2iapsfqLrHEwoLTi1pZJdWTi9lLUpR7rWo+5n5i3wrJ04ffX9U/48xj9TIFmwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAGqZhqvpjp9KYg53qFjuPSZTZusM2dkzGW6gj2NSUrURmW/bf3jaxrmU6b6eZw+xJzXBMev3oqTQw5aVjEpTSTPc0pNxJmkjP2ENZ5rfDvJaOyz/XXy9W/zNY+6X6d/pxwX8/xf+MRFV/ZF9B7LVFeny1zolZ6QqI1kshTSa9bidy5GfLklozLYnDLbuRmRF3E0fc/aDfQpgf6OQ/8AliJqz7Hz0+VuqDmpBVcyTGN9UpvHpBtLq23VGZ/9Fw3NsjPcmzUaS8tjLYhUqc7uslt+p2cI+RbT5wql7abt/d+uhJ33S/Tv9OOC/n+L/wAY2XDdStPNRClqwLOKLIigmgpXwXYNSfA578OfhqPjvxVtv58T9wwv3P2g30KYH+jkP/ljYcVwHBcFRIbwnDKLH0zDSchNXXMxSeNO/E1+GkuW257b+W5ieO2v8Vrd5zq3Msj2ObN12t5Fb9MdY+pzqFybI820rk6f0enWM5Y/jkWvuYMp+fdMxXEolSDfbcJMfcjV4RE2ruWyiMi3PTk9SnVVn+k+a9Uul56ewdP8UkWr1ZjtnXyXp9xW1y1lIfclIeSlhxRNOmhCWzIuJEZn5nIeHdLusekmZ5A1o3rxX0en2U5I5k0ukn4yibMgvPLSuSzEkG6lJIc47FzQrgR7kRnuatYc6JtW6bEsq0OwHqCh0mkWXTpsiRWu42T9tAiTFmqVCjSvGJHhr5LIlLbNSSWfmfnMUjQ9W+vXUCDlbTuD5npviFFJ0rrdQ6yJlsJ5+TbS5XNRVzKmpDRm4pJJIiQhR7kZ7beWz59rz1gwpWjNlTP4DjUfWV+BAaprjHpj8yjlOQEvvE+spLfiETiXCJPBCiI0kruRjd8l6EcEyfKcgkTpkVWMWemETTiuq3IJOPVvozilMzEPKV89BcOOySPknfcZ2V00ZjfU+g7WXalsWdxo9aM2E6w+DlEd0TcZbCexuGbazSaTUozVuZKPbuAJxxlnIo+O1rGXToM27bitJsZMCOpiM9IJJeItptSlKQg1bmSTUoyL2mMmAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAKydOH31/VP+PMY/UyBZsVk6cPvr+qf8eYx+pkCzYAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA1LNdUcO0+fixsnmyWHJiFLaJqG68RkkyI9zQkyLz8hFXr0sNB1K0lGK4t2RpOpGnHNN2XWbaAi37pXSX+/E/81Sf+AaC11iVis1OscxpSceOQbBTSWo5BJ32J02uPlv34/O2+vsOTX/aTkrD5c9ePxO2jv42vZdbKlTlLCU7XqLXo19CyACLvulNJf78WH5qk/8AANnwnUzEdQjlpxeZJfOCSDe8WI6zty347GtJEfzT8hco8qYHETVOjWjKT4KSbJ4YqhUllhNN9qNqAVpw/VHXEuuK90Xzu4x9WJLwN7Kaevq4iiWyn4UKKyp99wua3jaSpSyTs2Rr2Ij481R90tdQGo2Z6rxsZ1i19chXlg/cJi6fWGnq6tT7LDzqWnI9mokpkcWUIdNKCM9lGR/NMxfJy64Cv+ufVzB0Zz6RprB0oyzMrqNiS80eTTejk0zWtvONvOOLdcTx4eEZ7ERmo1oSkjM+2rUPXPU561PraPTjLcZetsEsczxC1vI8cmLZiM1upRNtuLUjZRpMiWRckly7EaeQFqQHz30e65tWj6cM4nalP19pqnDhUkzEiZhoYRaFfMNlXETKdkuG1JU6lzjtuTffbzOw/RrqhqRqdoNIybVG5iWmS1t5dVL8yNERHbd9EkuNJUTaCJJdke78IAsCA+dXT71yawas4RGyyddQyhaU4bZZhqQ96Cw1JvHiVKOFAiNGnZtvwmEqcfQnbcyTuR9zlXT3JOrWw0NX1NZbrNjrkG4wuwyhrEo2LNIarUrgOSIXgy/ENxxaFeCaydJSTI1p8yIzAuAAhDQ3qDxm70P08yXVHUXHIuWZBiMG+nRnpjEeQ+a4/iOOojEZK4+q4fqp29U9vIRtS/ZF8Eu4cmwY01yVEeVjlnlOPEmZAeeuIcBPiSE+C0+pcV3wvlUtvEk1JI9u/YwLcAK3ZN136RYzGetnK66n1MbA67O3pkJptwktWEhDEGCSeZGcl5bhbJ7JIiMzURCMOpDrKy+PoZqtSQcRyjSbVDEaqmvIzFg7EkLXXSrOOx6Qy6wtxs/nLbWk9jSatvMj2AvAArLY9dmBRsXPK6jELu1rrPJ/tSxSUT8SMxkstCV+kPx3XnUobiNKbWk33DSkzL1dx0RuvTELrH8aPDtNMoyLMslyG0xlnE4L0M5LUyuQTk01SDe9HU0htbaycS4ZKJxO23fYC0ICsXSXrlkuU9PGbauakSraWqiyPJnvAmsoblxoMR5a0RVIT2JbaE8Ntz2MttzEWYR1Dat3EfTvUrU/qmxXBHNSX4llS4EWGHLhu1r8gm2o6rAj8Un1o/hmpJJUfzdiAF8AFQNI+qG3x6yzKFqPJtcg+GOoiy0yx/w/CIq1hTKXI6Vb8d2keG571et7RP8ApTrHS6s2Od1tRVTYS8CymTikxUk0bSH2WmnFOt8TP1DJ4iLfY+x9gBIACp9T9kJxGy0+lamvaSZnBoXLRnHqSTLOI0m7uHJK2PRWFKeJKUp4GpTzhpQRJc77oMj1zMur+x1Ac0sTgTllidk1rXVYXl1WqTFlE5HdivuqbS+wpxp1lxPhqJaFb7pMu23cC6YCsuD9eWnGd6jU2HV2M2rNLk9xNoKHIVzIS2p02Nz5EcVDxyWWlm04TbjjZEsyLsW476jrZoU5pZYXnulmT4jJi45ZZREKRLr5z0iJBTzkNrZiPuKjvkg9yac2M+5bkfYAWTAVih9dOMK0dh6x2+nVvBhZBaQKjGIiLirkruJMxKlNN+K1INqKpBJUbqX1INoiPfc+wkzQHXzH9faC6saqnl09ljVu7R3FdIkMSDjykIQvdD0da2nm1IcSaVoUZH39wAlEAAAAAAAAAAAAAAVk6cPvr+qf8eYx+pkCzYrJ04ffX9U/48xj9TIFmwBr2b1+aWVQlnBcogUU9LxLXJm1hzkKaIj3QTZOt7GZ8T5bn5GW3caH0432qObYTX59n+W1FixdRfEjwYVOcQ4y0uqSZm4bq+ZGSfLiQ2LW/Vqj0Q0zuNSMghyJcatS2hMaPt4jzri0toQRn2L1lFuZ+REZ9/IQ/wBA+tdBqfpCjFK+BJh2WGcYs1DpkaHEvLcW24gy9h7KIyPuRp9u4rSqQVdQvrZ6f73nUp4atLk+eIUPgUks1lpvvrv35fEs2AALJywAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA4HIADjYvcNNb0e03ay886bxeOVybpv+NzXxJ0+/iE3vw579+W2+/fz7jcwENbDUcRl2sFKzurpOz6V1mk6UKls6TscbEA5ATG5XmP0w6gNdSyuo57qCmuvqhqo/gf7WoaWvgP0w5KYHikfLsoyLxtvEMi8x+sc6XMuf1bxXVjWLXm21AfwFE0sYhLo4dY3FclNE066+qOW8hfhkRF2QRGRHt572EAAQ7m/TjW5tq3darv5RJiv3OnEvTpcJEZKkNsvyVPnKJZq3NRGrbhtt7dxqlb0aU1dGwKMnOpqywXTuw0+bM4SC9KalMttHJP1/UUkm9yQW5Hv59u9jQAFW6noFwKBmeiuazMpnS5WjtJHpiaOKlDdycVKjhvOkSj4KZdWtxJFy7mRblt3lnQ7ROFongdjgsK/ftWrC5tLhUh1gmlIVNkLeNskkZ7knntvv3232LyElgAKoYd9j7wzC6XT+srs2mqfxOltMVvXvQUpRlFFPdecdgykEv1SSp9RoWRmaT77H22ymDdIGeYXiUjSxzqdye107ZpbCjqsfk00InIzEmM6whL0tKSdkJZJ3khHqFuhHsIiFmwAES4T0y6W4zpzjGC3uL0uRTsZxtjGWr6XVMpnrjNsGyfF3Y1tbkpfZKu3M9vMaRoL0cM6ITI8BWcVeQ43XwJFZChPYZXRp6mHC4kmTYNp8Z/igzT2JHIj9bfyFkQAFRMJ+xxafYpo9qDpHYZ9fWqc4egGzbqbQ3Kqo8BRLr2Wu6iUTKyM++3IlGWxeY9eUdCcvUjHs+LVfW+zybLs6qKzH1XxUkeG3X10KaiWllqK0riZrdRutRr7mfYi22O2AACpmRfY+8ZsI1hW45qDJqKxjLU5ni9a/TR58OimuNrRMZJh4zQ/FfNfPwTJJIUXqn3PfMWfRtdSa/Brii1iRjmd4FaWNhXX9PiFfFim1OZSzJYOvQRNGRttoIlqUpRGW5mfYis2AAifQjp8qdFNNbbTOVkczLId1a2dnMk2bKEOyPTVmp1DhI9VW/IyMyJJHufql5CHGOga4iV+OaesdRmSOaWYnkMXIanFZVPEefjLjvm82wmwPZ02SUZkSTSexHtv2FuwAFYcg6IotlQXsSi1UsqW9nasO6uVNs3WtPfBlkpBIJk2Vq4vtkk1/ONO5mW5bFscidPOgi9B4GXtSs9ssusM0yR7J7CwnxWmHTlPMMtulxa2RxNTJqIiIiIlEnb1dzloABWR7oaxaR03UvT69msxa8ayNzKai7XXsuGzPOY/JQbkVzk282RSFtqQrstJnvtuPZE6P3H6rC4uRagV78zDs+hZ027T4lCp2JCozC2kxTYjmRER81KNxSlr3Pby2IrHgAKv6XdD8HSbJzcxvP4h4m1NnzItY5iVedpG9K8QzaTbGk5BIQt01IMiJZcUp5bdhhNIfsfzWkOR0uT0+rq1Tcbobagqno2LQYzqETC9WTIWW/pUhtXrGt0lJXsRcU+tyt2AAp4f2ObG51DkqbnURKskvskqcqjza7GYcOshTq9t1tpRVhcmHCcS+74xKP5Q1EZ7bd7AaI6WT9JsXl0drkFVcy5k5UxcitxuJSMJI20IJtMeMWx7cDPmtSlHyMt9iSRSGAAAAAAAAAAAAAAAACsnTh99f1T/jzGP1MgWbFZOnD76/qn/HmMfqZAs2ANC13xbGMx0ey6lzCtROrPgqRKcaUo0mS2UG6haVF3JSVISZGXu9wr/wDYx6nGmNA5l1V17bVtLupEe0kEozU+bRJNojIz7ElDpbEXbuZ+ZmJn6o3MnjaD5nNxi7gVqo1NOdmKlw1SPGjejOEtpGziPDWrctlnyIv4pivv2NyqyLD2M4wGxtI7kOEmnukx/RzS6TlhCQ8SiXy24khJJ249zLff2ChUdsXDTg/98meiwyb5ErrP+9F214aPq/eXguguwAAL550AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAANYyvUjEMJksQ8ksHozshBuNkiG88RpI9vNtJkXf2GNnHGxCGvGrKFqMkpdLV14Jr1JKTpqV6qbXU7Pxs/Qjz937Sz+/sr81yv+WNFidV1W9l3wbJoFNUanjZTNJalPEnfYnDaJO+x/wAUu5EftPsJ92L3DU42lGn0TKFZlHxphFspw3vG5rNJOH5rJvfgSvbuRb79/McLHYTlypKnzbEQST+L4GrrvlK/Z8PadbCYjkmCnt6MndafEnZ9yjbt17DEfu/aWf39lfmuV/yxseJZ9i+cFKPGpzskoZoJ7nFdZ48t9tvESnf5p+Q2DYvcA6lGnjozTrVIuPQoNPxzv0KFWeFcWqcJJ9ck14ZF6lPrvUHqSwTrK0+0xXq7U5tT5sq2sbnFo2NsxSxunaL97SDkpUp1RmZ8CU4oiWtBkSfWIi1bQPqX1DdzSW3rjrpZRbltjIJsbT2x09+CvS48Nb5tqjWakJJ8yYbQ6ZJ3PZRke+xmJU076Rs7031ev9VqrqQuZS8suk2d7EmY3XPOzo6F+pC9LWlTzTKUeolLZpJJd0kRj21fSXeXWpVBqHrfrjdakJw+NYRserpNPCrmo3prPgPuPqjJI5Cza9UjPiRee24vlQiLQ3qC6hpObaCZFqPntbfY71Axbp46FilZipx5cdg5MYo76PlHiNBEhfimZ+Zl9Uta49ZUPRvOr/AIOkOV5fOxnFm8ys36t2M2xGq/EdS86tTziT3b8LfiRGpXLsXYzLyaN9FMXSzMMPvLjVq+yym0zjWMTBaabCjsJp25pcXjcebIlylk38mlS9iSnyIbXn3TBT57nef51JyuZEez7TtzT1+O3HSpMVla3lHJSZnupfy3zT2L1fPuAI4kdcRZXjWdw8f06yfD7eNpfZajYfYXjUZTdpAaZUTcjwm3F+H8obaibc7qR3Mi3IjjLTbrS1jk9N2R12W2FfP1qVPoKzGVFCbabnfbDHZfrHzaSRIPw0OSDURFttGPfuZid5nRrQy269s83npKBpBI0iTtER68Z1CEHNPv2cLh8zy7+Y8tN0Naf1Wq2lWrLt/Okz9L8Wh423FUylLFm5EjrYjTHiI+zraXXDLbfuae/q9wOzph18yW86NmdetYrF25saiLkE22kQILbbkhmvmy0fJsN8U8zajpIiLYjPzPzMcY31oVlxg+EZ5Z6bWMCvz/ADGoxGnVHuq+wQpVglSm31rjurJvw+BpcaVstJ9tjG9aQ6EHo1oSWiuL5tNJ1hNqcS8VDZN6O7MkvyCcJlZKbUban+xKI0q4FuXcyEONfY+a1vGrtDOrEyvy60zCpzeLdVVBDhQ6+yrkKRHW1XI+R7k44bm5+uoyUflsYG4ZP1pYhjjOWRmcJv7S6oM7jad1tVEUz411bvspdbS0paiS23xUozUsy2JBn7SIdVl1e3tXm2J6XSunTOPt1yisftV06JMFZ17DE047zjzyXTbNskpN1K0qPmSm0kXJexYI+g2M5jFzGk61ZI/l0/OomosHKXIMb0iFdMMJa5+AlJMuNK+UPwuKUkSkp/g7nIGF9OdrR6rY7rLmWq9pl2SUuMz8ckPyq6PFTLTJmlJ8QkMkSWyRsTaUER+qRbmZ7mYGn0nXRjd1kFDJTpdlLGnuVZWrCqTN3HYpw5lqTi2kF6OThvoZW40tCXVJ2My7kXfb9Y712YRc4vqBqNOwu2rsKwBE30i0VYQXZLz8eSUf0dUFD3pEdx1Z/JE6lKVJLczTuW/kxjoWg49b49TvavX07TXEMsXmlBhi4EZCIll4rjzZLmEXiustuuuLS2e3c+5nsPPddAeO51keb5JqjqNNvZOY46/jRuV9JCqXUMLktyEvyFx0bS5Da2WuK3EkWye6T37AbPpP1p6cZ7Y5BSZs1D0/n4/DhWTqrbIK9+G5ElKUlpRSmHlNJcJaTQppRkpJmnz37YbVzr50y0yyWxpKeley+Fj1RDvb2yrLevbZjwpO6mTjJeeSqa4bZG7wZI/UNJ77nsNt0k6YouEV1nU6i3mO57BnMxmG4r2D1dc0kmTUZLdJlvd9wzNJ7rPYjTulJGZmNaz/AKKKa81Lm6ladZfAw164rIVVZ17uI1tvEU3ESaI64zclBlGWls+HqkaDJKd09gB3W3XDgNZW5s6jFrmVaYlkFJQxattTRP3J25Nqr34+5kRIdQ4peytjIm1e3sPJM66MciZA/ITpblLunsPLywaTm6XYvobdt4pMntHNzx1RydMkG8SeO/kR9t8vlvRphuW9RmKdQcnJZ8deOMwjk0TTDZRLOZCQ+mFKd224rZ9IVxIk7eqki2Lca+roWhLvJNUrV6+/cyl5l9vbuE+gRuCrLxifNHpm3jejeKRK8L6vnb9wBqHTv1gvu5jn2muYS7bKbmt1By9+U+lTZM4xjEB5KW3pKj22bJXJttCSUtRkfuGy4T9kT0tyZ52TfYva49UScasssqJrs+FLXNgQUeI+TjEd5bkV42zJaWnSI1Fv3Iy2HorOgXDKfMJOdVubWTFvZ5LkFpbuIioJNnUXB7yqh4iV6zST9ZtzfkhRmoi38udPehWrw3F7TTm51FTeYbNx+bjUeGWK1kOxbiyGzbJTti234zziEGZEfqkZ7GolbbADX9LOpnUfVzqtwenmYTlWC4reaeT79iptnIzjdmlUiOcaYk2VK4qJtaiNtXFSd+5dyMe7WrW/Uu41vyzS3DNUIml+I6ZY7CvcpyUqBNxPffmKMmI7EdZKSTZILkpZIUrftsRdxtWjXSJcaZ6iY1qPlWuN7m03E8YexCsjzKyNFZZrTUybSS8EiNS0+EZKcWalL5FuZcdjzWqfTLY5Vqg1rbpRq3c6a5uusTS2M2HXxrCJZwkr5ITIiyCNCloM/VcIyMi7d9i2Ahio6gctm5503nU65t5/j+Rys4RdWddT/BTdy3AgG4wh6Mr5jjSyMj48UmotyLYxMWC9XOKZ2eiiYWK20Y9bYdtMrPFW2foCYDJOuE9sfc1EeyeO/wBY17T3ocxnA1aXOrz62tnNN5mSz3FyIrTfwq9dNKbkciRsTRJ5GaSTv7tx4tH+hl3SvMNO8hma45FklXpX8KR8XppldFZZiRJzK23G1ONkS3FkayPxFGe5ISkkpIAb1qD1Mpw3Xin6fqTS7JMpvbSmi5C9Kr3GERYFc5McjOyH1OrTxS0bfIyLc1c0pLuYjrFuv/A9SLqLjlBjtlXwctYuGsUvF2EJ30x+C04tRuxG3VPxUrJpamjdQXMiLsW4mleidY51EK6hHLp9UxeFFhaqw2U+CbPpxy/G5778t1ceO223cRVpF0SRNHJ6K3H9Rm38RhpsEV9W/ilac9hEpDpcF2fD0hxLZvGadjSoySlJqNO6TAjTSrruPDtB9L4mYV9nnWdXGGPZZcOLtIMA0QGpLjXiqdlONpdeWpBpQy3utXBR9vM5U1J62qTBsBodWKbTG8yDDLnHo+TOWqbSugmzGd3M2W2JD6XZEhCS5LbbSZESk+sZn21tn7HxT0ddgjuHalnBvcLxlWJOzrPGYNtHsYHpK5CeUWQRobdS44s0uJVvsZke5dh36tdAdVqpY/CL+qD8FyXhUfC7I1Y3Xvm40yalJkRSNBIgrWtZmtLCUpMj2TwPZRAbrRdXePZbrVJ0hxTDZ85qrhwJ1pbP2cKJ6OxLiFKaebiPOpkSGktrRzW2g+ClbbHtuMDpD14afavahY5hlbi1lXwM2+ESxe1esIT3p5wkqU740Vp1T8TkhC1t+MkuZF7D7D03HRnGyfUXB8vynUd2fU4AqE7U1qKGGxLSqPEJjwVWCC8dUZZ8nFsHuRqWZctiIi/egnR03oFkFcqg1DZsMXpDlFWVkjFK1M9tD3PZDtmlv0hwkeIexlxMyIkmZp9UwLHgAAAAAAKydOH31/VP+PMY/UyBZsVk6cPvr+qf8eYx+pkCzYAhvrEnHXdMeoj6TMjXTrY7f9otLf8AvCP9FYR4f1SWGPEkkt5HpdRWe5mW61w1Jif6pl/QNn67HjR0vZdFQrZc1ytioP61z2C/0bjH540WL9XOi9g2k0ovMfusfcVv22ZaQ+gv6dxRq/bZujL5to7+C1wLp/e2n9MYS/QsaAALxwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAANC1L1frtMX4LE/DMvu/T0OLSujqVS0NcTItnD5ESTPfsX1GJKVGdeap01ds1nOMFmluN9AQb91ljv0Rarfowr/mB91ljv0Rarfowr/mC79E4z8N+XuQ86o/eJyAQb91ljv0Rarfowr/mDeNM9Wq/U9VgmBh2W0ZV5NmpV7VHDJ3ny7Nmaj5GXHv7ty94iq8n4mhB1KkLJdhtHEU5vLF6m9APl/8AdW66afWOvH2851YSMYyaTnlLgtms0kqgvagn1sxUKIvVS4yps29/NbREkvnGdjcX6vbqmwfK2LbFfhefpzpDSZ4/OdsDQu2kSIK3VtLImz8LdTXzy5fOP1ewpkxbMBTjPeurUOietG8K0KhXrdBp1Wak278nJSiIjwX21uPspI2FKccSSSJGxbK9Yz47ElWn6udYGomZ6Uan4hb6cyMAm2Okq9RMTs4OQFIkqrnHUsp8Y2kI8B/daVbIUoiIzLfctzAvuApfivW/k+m1HOpNetK1U79JpixntS/CvU2LtrEQpuObT5m2kmpC3VoM+60lzPdR7bqxsP7JHIYwXP8AJbfTyhnz8Nx6uyZhGP5IubCfjyZjcZcV2QqM34Ulk3UmpJJUlRH2Mi7gC8QCH5+uGS6f9Pd7rjrZp+jG5VFBk2TtHAs0zlmyX/QN+MSUp8VfJCVduKVGfcy7iFsk6t9cIlLlGF5No9UYxmFhpvNzrFnoWTHLY9HaIiebfWcZJtyGUr8QiJKkLNPHknfcAXJAVI6Zc/m6K9BsbW3U6NbzFtUX21SlP5DIt5Nl4kdtSXCckERsKeVt8iW6GzUfc9zMYTD+v3LMtx7OXqrS3HLq9xPEms2jxaPKFyoj8EnSTJiOyFRUeFMZR63AkLSvsRKLzAF0gFOabr8t9RYNFI0h0kbtnM/yZ/HsIO1uDgontxIhP2E2Tsys2G2lGbaUp5qWaVH6vYjwGtuvWsOF5rnsupx6wps7pNCVZS5WrywpVHXOpsnW3H2opxjbekNpQbiXVGXiJJLSko3NRAXlAU2yTrG1Y0z0PwjOM4xXTZm8ucbO8ksWudJhvWKEMoWkojCIhqcedSfI08UttqUSOSt9x+cS6sNY8z6l8bg1mMVcfTC+0ngZ87Fk2KEyYsV51BuzTUmOa1OtmpbBRyXwUlJOciM+JAXLAVIwLrbzTIrHTu/yvRiNSYJq3ImRMVsmr/0mehxltxxn0yN4KUtk8lozLg4vjv624xuEddeo15plh2ueYaDwsf09y7IauiTYpyUpD8RqS48w7NcaJgiSy3IbaQXJRGonCV28gBckBEGkevcvVbC871Ag4U+VRjN9b1NH6JI9Ievo8D1DkNJ4pJJuOpdQhO6vm+fcQjhnX9cXeDU+pd5g2JIoZeU1FBdIqspdkzsZanqcb52Md2I0ppxp1LSFFvxX4hmhR8T3AuaAqNY9ceWynKtODaMR71GUaiW+B4245kBRUWCYTJ/v81GyZIaU+hxJkXIyQ2oy5Hsk/wBRetnJ5ukx5fKw3BcYyOuy20w66iZRmqYECPOgmXMo7xMLclGvdOyUNlt6xmexbmBbcBT2l67Mwz2g0Zd0w0Ui2t/rDBv3Y8KbkRRY9c/VOoQ7ze8FXiNGXiqJRJJRklJcd1HxxHxiVrNxzAJ8fTSjx+blL9rBtJGVZGuBUQLCBIJhcFE1uM6lTzh+ug3CbQSdtzMwBdkBTXPfsiVbiOrF5hcDEKedS4rkMLG7RS71TdzJffNtLr8KElhSXmWDdLkanUGrio0lsW4kLR3qmtdVNc8y0nlYzj1A1is6dAKHMvHEX7yY60pRM9AXHShUV0lckuNvL2I07/OIAWIAAAAAAAAAAAAAAFZOnD76/qn/AB5jH6mQLNisnTh99f1T/jzGP1MgWbAFfutrjK0npaJfzbrMqKAoveSpaVbf5g6+rFZ0eQaLZ03slVTqHBguubfMjzG3GnO/5EkPz1fEc2VozREfebqfTube8midUY9XXLEcPpyvL2Og1Scdn1lwxsW+ympjW5/+FShSra7R9CXlqd/Bac1h96Ul/NaJPoDzwJbdhBjT2T3bktIeQZe0lERl/pHoF04G4AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAACA8s6KNFc20syvSLIkXUmny3K5eZvyPSmylw7OQ74i1xl+HshJbqQRKSo+C1EZnvuMXnHQfpHnEpDzuVZ5SsO4nGwyyiU92UZm2royDRHKUXhmbikEfsNKVbbKSpJmk7FKmRESkQlymUyFpNaWTWRLUkvMyT5mX1juAEISOkPS6S3kjbk/INspwCNpxN2lterVstrbQtv5Ps/xWe6j3TuReqQ8+TdGulGVsHHsrHI0JPThOl/yMtoj+CEuocJzu0f745Np9f5u2/qCaoV1TWU2bXV9tDlS6xaW5rDL6VuRlqTySlxJHugzSZGRK23LuPaAK7470L6NU5ZMi/tMvzBGSY19pxFkdyqWqtpeXMocRZJSptBLJKiUo1LI0JMlee/pl9HONXel+R6Q5lq/qflGPZDAjVnh21xHcXBjsOodQTBpjpTy3bQRrcJajSW24m1OSY6rIVYim+rjvUQysVVhSkelpims0E+bO/MmzWRp57ceRGW+4/V9kFDitRJyDJ7uBT1cNJLkzZ8lEeOykzIiNbizJKS3Mi3M/MyAGOzjAsX1Gwa305y+v8ATqK9gOV01g1mk1srTxPZRd0qLzJRdyMiMvIRFh3Rjp3ix3Mm2zTOsusLbFnsKYsMhtm5L9ZTOpMlx4vFpCUb9jNakqUZpLczLcjn0jJREpJkZGW5GXtHIAj+JodgLWh0fp5nxJNliDGPN4ypuW98u7DQyTRGpxBJ2c4kR8kkWyi3LYaGjpCpZGnd1pbkGtmq97j93AYqVsWF2wpUeC2tKjjtqRHT2cSkm1qVyWbZqTyLczE13uRY/i1edtk15X1EEnW2TkzpKI7ROOLJDaOazIuSlKSki33MzIi7mPRY2VdTwJFrbT48KFEbU9IkyHUttMtpLdSlrUZElJF3MzPYAV2n9BGjCnpkjFbrL8QdPIk5TUqx+xai/ANgbHgPqgkbSibQ+gk+KhfNJmhOxJ22GyK6RtNZdhZ2t9d5Zey7rT93Tixfs7Qn3ZVY4+4846tw0Es5ClvL9ffiRbESS2EzFYQDgfCpTo5wjZ9I9J8RPheFx5c+e+3Hj333227jiusq64gR7Spnx5sKU2TrEmO6lxp1BluSkrSZkojLyMj2AFcHOgvTt2BSxXNUtTjkVGNyMOcnfDTHpM+jdXy9AeX6PsTSfmpNskL49jUY2as6QdOaTJ8ByulyLLIMvAcVYwpptme2TNvTsmSkRp6PC2dTyLkfDhufn22Ipgtckx2ilV0G7vq6vkXEn0OuZlSkNLmP8TV4TKVGRuL4pUfFO57EZ7dhkgBXnT/of0q09yChtomT5vb1uIKmLxagt7gpFZQqkkpLiozZNkszJK1pSbq3OJH277GPzn/S/W1PRde9L2mNa5dNJo366lRdTUoV6Qt43W3nHkoIiNt1ROFskvmEX1ixAACN9GdHoWlWguNaLxpr7BVFEitkzILptOqkrQZyJDay7pWp1bjhK8yNW4iHUHoxqmtIdYqzHLC8zzOdTqVmvcsMrsmSWpyMRlC9dlltCSZWonN+BqUaC3Ufbayk7JMdrLisx6yvq6Ja3XjfBsF+UhuRN8FJKd8Fsz5OcEmSlcSPiRkZ7EMkAIH0/wCkzC8a0/0Wxm1lTfT9HjRYRFwnkpZlWLjC0SXHSUgzUlbjrq9iNJ7mXf2DwzuiLTJ2exeUmX5tQXkPLbvMYttWWLCJMeXbISiayjmwpHgqShJERpNaduy9xYYABTbJug44mV6HY5prl2T0OF6bx8r9JuId2TF3FeslIdaNlzwzJwvF8RKuSTLh2Vy33G92HQtpfK0uqtGIObZ9WYfDYdYsq2LbNGi8N2Qb7rk03GVGp1ThmZuNm2rY+JGREkiscAAg+P0mYhS55PzjBdQs+w5m5lxZ1vR0Vs2zW2D7CEIStxC2luJNSG0JX4biOZF394yNF0z4tVaxs622+aZjkt5XNz2aaPc2DT0WoamK5PIjkhpLhp29VJOrc4p7Ft2EvgAAAAAAAAAAAAAAAArJ04ffX9U/48xj9TIFmxWTpw++v6p/x5jH6mQLNgCvXUl+/NZOn+n8/EyyTN4+/wACKat9vq3Eh9Q1F9smhOoFITfNcnHLDw0/9olhakf5ySEd6yrOZ1a6A1iS3KOjJJy/5u0JKSP+kzITXmtvjtDiVta5bPZhU7MRz0x50/VS2aeJlt5mZ77ERbmZmRERmYgpwdWVSC4u3kjp1qmwhhqnRG/9cvY1bp2v1ZPoRgF44s1uScdg+Io/a4llKVf5yTEiCkvSvF171B0bpcOxDMqvEMSx1cmuXcx2SlWE1ZPLXwQhR7NJSlxKdz4q3Lct99imMukfHZuz+Sar6l3EtXdx1+/NJGft2SSPVL6tz/COlTwkKUEsTUSlZXSWZrt4ebObjKubEVNhG8czs911fTr8idwEEl0k49CPxaDVvU+peSfJC4+QnsR+zcjR3L8pDj9yDqKxr5TDeo92zbRsSYeRU7b5KL+c+Rmv8pEN+bYeXyVl3pr0uV9pUW+Hg1/gncBAv7oXVPhZb5no1SZbFR8+VitkptZEXtJh7daz29hEX5BlMf6s9JLOcVLk8m0wu23IjhZLCVDUR+W/Pu2Rb+01EMPk+vbNBZl/C0/TVd6CxFO9paduhMwDzwLCBaRG59ZNYlxni5NvMOJcbWXvJSTMjHoFLdvJwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAjbVZ3qJamwP3Eoun70Q2l+nfbMuYThOblx8PwNi47b7799xJIDScc6te3YbRlld7XK9+ldfH969Df6y1/8w9K6+P72aG/1lr/5iwgCHmz+/Lx/wS7f+FeBXv0rr4/vZob/AFlr/wCYkHSd3X107L92+LgrJF4Xwd9rK5ZmfzvE8Xx/+5x4/wA7f2CQwG0KGSV8zfazWVXMrZUj5PatazkrqXuOryFS5nJZ06z2BjMGwjUct2mLForb0S13mJT4BKdkSXFJIz7cSLzUW8p66dU2u9BlXUBb4brnjNHU6PyaGbS0EimiSHLtiZGjrWwp1Rk54ZqUvipBGs1OkXMiIiF5I2lemsPBX9MImCUbWJSm3mnqREFsobiHVqW6SmtuJ8lqUo+3czMxFOO9FOi9dqpl2p+S4pj+SuX02qmU8OfSsqTQegw0RkIYWrlukybQrbZJEaU9j4kYnIiteQ9RGoGI5vqpGxBGPYbZZXqfheJqvnKWNzp2bCtJx6VJPikpLjfE0pN8zIuXsIb1rprJqjp3SVmC4x1H2mTZnAoby+lSscw+ndOTHiu7JemuSX0xmG2T+ScSyXiLPcyJJlxO1F1o1pPkcXI4N9pxjtgxl7rL983JrmnCsnWkklpb+5eupBJSSVH3TsW2wwcvpi6dp1RRUM3RHCn67GTcOoiuUrCmofiK5OcEmnYuSvWV/GV3Pc+4ArV0z59dap9XGL6j5GhhFrknThS2U0o6eLfjuWy1LNKe/EjUZmRb9t9hFnXDqxqfqBifUtjkjValxvFNPHqejYxB+tYVJuSkeA8qQchSieQs1nu2SN0mlsyNJ91D6HY7pfpziNjEuMXwilqp0Cnax+LIiQ0NOM1jazW3ESoi3JlKzNRI8iPuMLl/T1oVn+RvZfm+kOI3t3IiHBen2FSy++4xtx4KWpJmfq+qR+ZF28uwAqTqV1HdU0/WfUTEtGK+a41pm7SMRKdESoKDYofYbffXYyJkhuS0laVLS2qOnYuJGZmZmPDqd1HdTlDP1h1Do9S6uJjek2odVRNY45j8d1VlDlriNuMuyvnISjx+SVILmZqXuoy4km42YdPuhuoOSQ8xzjSXFL27gJQhifPqmXnkpQe6EmpSd1Ek/Ij3IvYMjP0g0stYV7XWWn1BJi5POas7pl2A2pFhLaNBtvPEZfKLSbTZko9zI0kAPnh1Oasan6qUOpsi81VparG8R1erMLhYIqtYKVKaizYq0zkyDUT/ADWozXtspHBtzt23TIupmpeuuXVnVVPsM9pHMI0sas62FjMrGYstNhzrUuIJ91wjI0NLMlkk0q5mpZLMyJJFba86d9CMnyqXnGRaQYjZZBPJopNnKqGXJDvhqSps1LNO5qSbaNlefqpLfYhm3dL9OX4eTV72D0i42ZqUvIWlQmzTaqU2TajkFt8qZoIk+tv2IAfPto9XrXV/LmafXGNj8Q9Dqa8bpV45CkwX4px3DVAbiuH4ZI3SvdaU78V8TLikhsulGtOquYN6V6TYvqtjGj9XE0cgZzKn/a/CWxPkOPGybCGXTQ0zGbSg1qJriruZbkWxldCz0Q0eubypya10yxqXa0VcqorZjta0p2LCU2ps46Fbbk3wcWnj5ES1EXmY8N/06aC5VW4/T5Lo9iFpBxVpMeljy6hl1uAynbZpolJ2SjsXqfN7F2AER9XZmrVXpkVzSvfUpB8k+R/vF/uQivp36keqnV7Nsbzf4NnP4de5FbVdzVvxKePX1MRk3kMnGeKR8IOSW1NINxLrWyiUrikiJKju7d4dimSzKewyDHa6xk4/L9PqnpMdLi4UngaPFaMy9RXFSi3LvsY1ev6ftDqnPnNUqvSXFImXPOOPLuWaplEo3XCNLjnMk7ktRKUSll6xko9zPcwBSzRLqf6m5EDQjU3ULU6kvKPVmJkkeZSKoWITcBdbGfcak+kt+upS1x91lsSCSoySnfuMJhGsnVVrxpVnGTq1ZuIGOt6WT72xdexeuioat1oUoocFSPlnI/o5L2fM+STMjM+R8SvZZaC6WScRg4fT4fWUMejiT42Pv1cRph6iVLacbeehGSdmXDJ1Z8iLzPvuIZ0G6DMZ0azOozCflFbcHQY8/jUOLX4tEqEzIzxIS45YqaNSp7ppQRcnNi3NStjM9wBXSoqdVcmndEEan1vbVkttQZI/FyF6nYlLqmF0kFSo5tcuL7qEc0+I6ZmalclEe2x3B6MdUc31d0FrMo1GnRp+RRbO0p502PHTHRLVEmusJe8NPqoNSUJMyLtvvsReQ3fFdCNGMGXTOYfpfjVMvHn5kmpVDrmmlQXZSEokraMi3QbiEJSoy80pSXkRDZMWxDFsIqvgLD8fgUtd470r0WCwllrxnVm46vikiLkpalKM/aZmAMuAAAAAAAAAAAAAAAAAAAAAAAAAKydOH31/VP8AjzGP1MgWbFZOnD76/qn/AB5jH6mQLNgCveWNfCXXDgbG+/wPg1pP293iPpZ3/D3HRkzKuo3WxzBFqNzT/Tp5D9ykj9S0tT34R1e9Dfrci95KI/nJMtN1j1HRp11QZXmZcXJNFpY1Fr2+xmqdJstmk7eZ9zJRkX8FJmJ26ftOHNMtMKulsOS7mbys7h5Z7rdmves5yP2mnsjf+YLOAfNqVTFfvOTjHtsrvuWi63fgT8p/WSo0OCgm+9tpd9yP+kxLVHkGtGCNbIRTagTJbLRFsTbEptDiCIvYXZWwsOK86aLLH+sjV/HiTwRkVJSX7ZeRGbSFR1mX4TMvy7iww5+H+S3Q2vNljlLWvn+9GL8Yq/ncAACcoAYrIsUxnLoJ1mU4/X20U9/kZsZDyS39pEoj2P6yGVAZjJxd4uzMNJqzIJsOlGjpJblvoxnGQ6eWCz5m3BkKkQHFf9pGcV63s7ctu3kPP+6B1KaWHx1G07i57TNfOuMWPjLQgv4TkRRFzV7dkElJe8T8AurHznpiEprr3/zLXxbXUQ83jHWm8vZu8NxHenmv2lWpqyh43lDLdnvxcq5xHGmNrLzT4S9jUZe3hyL6xIg0PUPQ7S/VFBqy7FIr00i+TsI5eBLbMvIyeRso9vYR7l9Qjs9PeorSPZ3THOm89ome5UWTr4zEIL+AzLLbc9vLmaUlt5GM7HDV/sZ5X0S3d0lp4pdpjPUp/OrrpXt7XLAAIWxbqlwuXaIxbUqrsdPMjPscO9b8Nhw/e1J2JCk7+Slcd/ZuJmadafaQ+w4lxtxJKQtB7pUR+RkZeZCtWw9XDu1WNvR9j3PuJYVIVFeLP2AAITcAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAKydOH31/VP+PMY/UyBZsVk6cPvr+qf8eYx+pkCzYApVkeDr1K+yDPRpSVuVdBX10+Y3v6qijs+K3uXkZeM+2W3uNQuqK8aCSouUdQmtuXcN3Ik+FSMq9yGEuNuF+VTKT/ILDi5i4qjkoL91ecvifm7dxhVZV26supd0Uor0v3lesmJOPdcWGWR7JTluD2NR/hORZBSP9UxYUV66klfAOsGgmcEjb0fLH6Ja9/4M+KpvY//AAiwo5lLSU49fqkdDGfFSoT/AIbeEpL0sAABOUAAAAAAAAAAADDZVhuKZxVLpMvx+Dbwl77tSmSWST96TPulX1kZH9Yg2RoplOlclyd096rtVsdCzWvFsgkelVyz37oQsz8Rn8m6jPzURCxYiTJdOMCkasY6y/h9Qtuyg2smWlURBk+6lUcyWvt3URqV3/nH7xrV5RxWBprYWkm0mpbtWlus+nt6yriYaKUVrdK97PV23owlL1PNY9YM4zr1hs/AbVw+Dc5wjfqpR+9uQnck7+ex7kReahN1fY19tDZsaudHmRJCebT8d1LjbifelSTMjL6yHmsseobmoVQXFPDnVq0E0qJJYS60aSLYiNKiMuwhWw6bbrBJjt/0653KxOQtRuO0U5SpVRKV7jQrdTRn/GLkZF2LiOjbC4nd9XLvcfdefcbfW09/xLz9n5E9gIGq+paww6ezjfUPhErC5rivCZuGCVJqJavel1O5tmf8U+WxfOMhN9ZaVt1AZtKewjToclPNmRHdS424n3pUkzIy/AIK+Gq4ezmtHue9Psa0ZJCrGp8r9z1AACuSAAAAcKUlPzlEX4THCVJV81RH+AxpmrOK49f4fbWNxVtSpNZWTHoji992V+EZ7lsfvSk/yD1abYtj+OYzBfpKtqIuwhxnpKkb7ur8Mj3Pc/rP+kU1Xq86dDKstr3u7+FunrLOyp7Da5ne9rW08b/obWAALhWAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA0nWzVGm0U0myvVW+NJxMarHpvhqVt47pFs0yR+9xw0IL61EAIRx/rPpLrrsvelRL0b4PhUDZRJJbcnLtojfkMEr2l6O4RbexcdZe0WkH84dA71F4fq3F6kJGFZIu8r5bOoEuU7AdT4kZ6WveS4W26Y7riHUczLiZGZb7GW/8AQ7gOa0epGEUOf4zI8eqyKuj2cRft8J5slpI/cot9jL2GRkAID6cPvr+qf8eYx+pkCy8h5uMw5IdUSUNINajP2ERbmK0dOH31/VP+PMY/UyBN2rtmdLpTmVun50OgsHk9/NSY6zIv6dhvThtJqC4uxiTypsrv9j4mP3dLqPk8nu7Y5KRrV71eAlw/7XjFtRVH7HDG8PRbIJh+crLZp7/4DEdH+6LXCTFVNriKk1xlLwu7eRNVp7Jxh0Riu/Kr+ZXvrgSmv0frM42PlhmWUl6ky9nCUls/7HTFgkKStCVpPdKiIyP6hE/VlQHkvTbqJWpTyUiikTEltv60cvHLb692xt+lF8WU6X4jkvI1HaUcGWZn/GWwhR/2mYox0rS60v1LVT4sDTfRKS8VFr9TawABOUAAAAAAAAAA6pEmPDYclS322WWkmpbjiiSlJF5mZn2IgbtqwcvtE+ytk1rQS0mk1IVsoty8yP2H9YrTYvY5R9RddjtrnV0mFCgkhpb9q6tSJbikGTSl7+qSyJO5HsXlv7Bv9vrVPyWc9jei9ArI5zZ8HrNzduuin71LPbmZe4vP2b+QhOVohb5DrkjF8tylL02fF+GrGVGY4bq5es22W/b2ESjLt58fYPGcv454nY8xp7RqpHXdHsvxu99rpW11KVXHYhU5RwUcyvG74K0laz4u9lpeybLjF5DkRKeg9nXq5YzrFmteRFsTb80pLZf90yIhyjBte6ojKr1jg2SU/NRZU6E7/hUgzUPQc9xMPnw8u5wfrJPyN9vVXzU33NP9USdaVNXeQHqq6ro0+FJTwejyWkuNuJ9ykqIyP8og+z6bbjB5z2SdOebP4jLcV4r1HMUqRTy1e40Hupoz/jFy2LskkjYDn9TlWoydosKukJ9sd95hav8AxmRF/QP3+6lqvXFte6D2Z7fOXXWTMn+hKS3/ALRYocvc1unGcU96cJNPtsnEjnWpT+0jJPpyv1RuOnNrnFvirEnUXHY1LfIccZkx4r5Osq4qMkutq3P1VlsoiMzMt9j7jZxhcRyN3KqRu4eobKnWta21RLBnw3kGk9tzT7j8yEcdQV1mOM/AGQ4XqRT49MjKkpOruGjOHcEfhnwWsu7akcT2Pt/0h+sXcW41qNb62Uowg9bu6ir7t+q6Ne8sOrGnS2jd1/vYTCAgTTLq5wzKZqMYz2MWI5CSib2efS9AkK7Fu1KT6nc/Yoy9xGoTXJyGghtePMu69hvbfm5JQlO34TMZqWpfM1bpTTXc1ozajVhiFek7mraxR8ldwW6kUN3DgsR6qauY2/BN9T6PBM+KFE4nwz2JRb7K8y7du/p0sYyVvD6x/IbqHPKRBiuRkx4Rx/BbNoj4qM1q5n3Lv6vl5DSta9a8JrMEt6qptodxNtIr8BLUSQlRNE42pJuKUW5bFv5eZmZfhLYNBs7h51p3XPx4qo7tY01XvtmrkXJttOyiP3GWx/Ue5fWPP0sbgqvKzpU6l55dyba3+G7W3eWZYLE0/rJxaho9d2t1e3dvsSKAAO+RAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAeC8vqPGKiVf5Lcwamrgtm9KmzpCGGGEF5qW4sySkvrMyIa6zbaS6wVsKPGtsYzCA4hi7istyWJrTjaXVEzKJBGolJJ1tfFexkS2z2PkntpHVvhDWoGkaKL4fxqtmN5BTTK5nJkmups5rU1pTECUgvWW2+vi3xSRnupOxHtscC9BuM2sXUi/ayvUTTWdc6e0TuHJx7DprslUNt2zdmOuSlOJQW6XTNpCWyNKSSojMlbkYEqVSEOfZBspbcQlSFaQVCTSZbkZHbze2wnzHsboMSqWqHF6aHVVrC3Fsw4jRNMtG4tTi+KE9kka1KPYi23MxAtP/dCco/yQ1H63mixgArJ04ffX9U/48xj9TIEldT8467QDOJBJ351a2NvqcUlv/eEa9OH31/VP+PMY/UyBtvWfMXD6cMrNB7eKcJo/wABy2d/7BbwEc2LpL+KPqiKu7UpdjNa+x7xfA6a62X23n29pIM/f++Vo/3BZMVi6Tctq9NelDA5VrU3MiPJizpzr0GAuQhlJynVmpxSeyCMlbkZ+4/cLA4plkTL4K7CHV20JtKiSkrGEuMpwjIjJSSV85PfzIculiqU5KnmWe17cddS9yhUgsdVpJ6qT07zvyqlayPGLfHpCSNq0gSISyMt90uNqQfb8ChBfShdZXedK2DO45YVkebVRn66V6fGW8njGedaJJEhaDI9kJ79xYgVa6csupdMdONUKO9cUhnFtQbmsYZaTutaXVJcaSkvr5q77+w/cIsZKnT+sqyyxs7u9rWs9/DcXMHGdXDTp01eWaDStf7y3drROOmFzmeTUMTJcllU6o1jFQ8wxCiuNrbUZ/wlKcURlt7iIbqIZ0g1h0+i6eVdba5DHrpVWymK81KUSFKURb8kF35J+sbFL6gtJIZHvliHjL2Mxnl/6EbChyfyvgY4OnOpiI3cU3eave2u99JnGcnYt4mcYUZaN7ovdw3LoJEARZ90Vhbxb1dLktjv5HGrTMj/AKTIclrPfTC3p9HMvfI/mqfjkwk/ynuLP01gH8lTN+VOXomQfReLXzQt22Xq0SkON9hAmX9SOV4o81Bm6YFHnSVEliI5aockKM/L5JCeRb+z+zceC/q+pzVmuS09HrsQqni9eIUhTb7yD9jii5LLt229Xz7pMVZftBQnmhhYTqTj+6oSXi5JJFKvCVGoqKacnwUou35mm1Hv1fBMkPL9baOmsVYziVfIyvIzPiVfXFyS0fvdcIjSgi9vmZe3bzGGY0ozHUV9NprTf8ofInGccq3Dbit+4nll6zhl9R9j8lbdhj8N0o1dxGuKrosixSgZMi8RUKvN910y9q1uJ3Ufn5n/AEDYC001Ulnvaa3ztttjTEq2mf8AOJX/AMhTzYvGfFi6E5L7qyRh33mnLvVv4UbfRcKuuKxEPyrO132jr36dRItVUVVHBarKavjwojJbNssNkhCS+oiEHR8qxyd1TIkRbqKtlFI7CNw3CJPjpMjUgjPsZkRH5e4xtZ6HSJZ73GquaS9/Mk2HhpP8mxiIqfpbr5uptpQWeUzHa2uaamGbatn3EPGvgjcyMkmXE9z779u3ftFyrW5SnLDwo4ZJKaeslwTdtNytfXXduLyw2GVCooVMzstysklKL4791ktO0sfYag4LVmZWGYUzCk+aVTW+X9G+41qw6gdKIG6U5OUte+xIixnXDM/qMk7f2jI1GjWmFK2hETDK1w0ERc5LXjqPYvMzc37jZoNHS1hca6ohRS222YYQjt+Qh2svKlT96nDulL9YGl8BHhOXeo/pIjo9e4c0yRjmnuX2yj9rdcaUfhM9z2L8g/Rahau2fq0+jTkcj/6yws229vwp2IxKIBzLGT+0xLX5YxXqpPzHOsNH5KC/5OT9HExWLvZFIo4z2WQokS0Vz8dmKs1tI9Y+OxmZ7+rx3+vca9qTp5O1BTAitZJ8GRI3im+2UJt83jVx47Gv5vHZXl58vqG7ALlXCwr0Ob1W2tE9Wm7dLVt/GxVjVcKm0il2WTXg7+ZBrHSHputZu2ljcyln3Pg8hhP9CEEf9ojHUjpByTHbA8l0vnKyCtQZrfx+e8hiSafaTEkk7GfuJZdvaa/IXAAVcPyJyXhrpYaEk96lFO/e9V2pplbE0oYqWeaWbpSSfkigc6kwS8xyZX41bXNJlVQZybLF8nNpiSltCFG4phfBBPGRdyItjNJGfEu29oumTDLnDdNW0XfhpctZJ2TLaFEo0MuNtkjkZe0yTvt7NyHu1501wjN8EuLTJcdiy59NXSJkCZx4vsOtNqWni4nZXHkRbpM+J+0jEYYzc6+aIY3U2U+G5qVgrsFh8zit8LeqaU2lXHj38dtBHt5mexdzQRDTB/s5ybT5VeI5PWzlk+RvTV/ut9m5272c6NF0MU61Rt/Da929L8Vv8NOos2A1TTnVDCNVqT4ewi7anMIMkPtGRoejLMvmOtn3Qfn59j27GZDax2JwlTk4TVmuDOpGSkrx3AAAamQAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAACCOtJ3FmdElOZGeXFNTfVH2u/akltVv8N+lt+hejE78mavF23Jfbjy9uwhDoSm6dT9Qo0XFNMs+xJ+jwNcCK9kq4R/CzKrZxUuWpbKjcffVKSolq24I47bJ5p3n/AKs7yjodKosi1wpeVT38mo49BXJtDrSVcqntHCcVLLuwlDxJUau/YjTsfLYQt0cP6dq1ceua3T6TSZXm2Hy8lmF9sj1hEqy+G32ZUKKw4giYackJJ8jI9lKUsiSkkdwJEp/7oTlH+SGo/W80WMFc6f8AuhOUf5Iaj9bzRYwAVk6cPvr+qf8AHmMfqZAzXXU+bPTrclv2cmQkn9ezxK/3RhenD76/qn/HmMfqZA7vsgko4/T1IbI9vSLWM3/QhxX+6LvJzti6b6Gn4EdVZoOPSZPSuH6B0PUTB9uWCk95f+8jGv8A3hOOO/8A2frP/g2P9QhCvpN3jXStjtRAxhc6OeCx2Jj5S22iifvJstzSruvzPsn3fWN3o9SG6PAWMizenOhhsRY5ReUtuQuZu2RlwSjvuexdj9+57ERjzUcVSoYlqo7Wgrtp2Vrvfa3n1by5XpTr46pKGrk9Emr6yfDf5G9WVnX00B6ztJjUWJHSa3XnVcUoT7zMUjhRm831Q1NiYiqzm1FxYw8kmV0WM2mQjdhTKHSN1ZJ4qU24oz33Lkn1T9lg63Gcl1mnsZJnrDtZi7LhPV1EZ7Kkl/Bdf+oy9n+gvnaLkCp+DdZiW8ZoWppZbp2UZqEmQmKjxYss9zIzI0+q0ZbJ+sxz8bGfK9PNUTjh+izzTvpe1rqPVa8uNlo+3gVzJVKdGf12W99MsbNPjo2rXvuXXwzfTfiGMz8bTOs8BkvrmtJklY2jEd1hexmngwW5qSXmZ7kW/wDQQnSLRUkEyOFTwY5l3Lwo6Ebf0EIw0Gv59RpRUKyiti1FRAhGaLF6wQaXPlFF6ydi4flMx1y9XMqz+S5T6J0PpLKVm29kNig24TXsM20n3cP8n/dMhvyXUwXJuAowivicVZKPxPpsrJ7+L04t8TzeJ5SrVrVMVUcpys9Nb36EtNPLiSLleZ4tg1Ydrk9vHgRy3JHM/XcP+KhJd1H9REI0+2fVjV75LCILmGY04fe4sGt5shHvZa8kkfv3+slF5DN4nofTV9iWUZxYv5dkSjJRy7AuTTB+5lr5qSI/Lz29mwkwX9ji8d9u9nD7sX8T/NJbuyP8xUyVq/2jyx6Fv73w7vE0rBdIsOwJS5tfFcnWr/rSLSerxpTqj33Pmfzd9/JO2/t38xuoAOhQoUsNBU6MVFLgizTpwpRywVkAABMbgaPSH/8AS9k/fyqa7/WeG8DRqVSf3Xsm9b/8prv9Z4UcX9pR/N/8yLWH+Sr+X/6ibyAALxVAAAAAAAAAAAI11vhXFpQfBVQ1lTi5kWUyaKYmTZWakEkkyPEI1cT3/g99uX1D0aMxLSvx86+0TlKVxm2GUovCZIk8W9jJjw+/Atv4Xfy+sSEAoLA/+ZzvO91rcDf6rL8nxfeu7+F7eRianFMborOyuaWliQZlwptc92O2SDkrRy4qWRdjV6yvW8z9pnsQywAOg5OTu2RpJbgAAMGQAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAACCutVu+f6f7eLQ6Z/ug+POr25uNpq3JzlhC9JbN9DaWzJTbhII1IdLfgpJK7bblo/RdUaL41b5BUaUdJOp2kzsmK3Im2WYVLrJTSSvZLDb70h5ZmRrNXAtk7bn5iROr6gtLzRKbKp8yx/G5FBaVl8T+RTjh1Mooktp70WY8RkaGXeBIP6zIhEvSfqDkOtevmUarZHnGl7UhGKRqNGL4XlSblbiW5SnfT5Sk7JSaTc8JBkW+yzI9ti3A3en/ALoTlH+SGo/W80WMFc6f+6E5R/khqP1vNFjABWTpw++v6p/x5jH6mQOj7Isrj0/tF7TvGdi9/wC9pA7+nD76/qn/AB5jH6mQPN9kKaOdpRjtOgz5TslZbLYt/OM+X+lRC5gP/Yi11+jMO2l+leqPV1SRrGn0qwyqiOPIgMG1GkIQZkhRoYImyV79uKtiP/5DUtFtKc/z2ih5PFzJMBmllLYrY0+H6WygtiWa0JWfEvWV5bdjL3l22rrHuXX28dwmDyUskvWjyE+xKEGlB+fuJ3+gTZpZZlNxKvhJxqxp0wYcdsky46WUun4ZbqbJJnuW+579vMfNXydQ5T/aGs6snlhGKsrq7snvT0to+0rUqKniauJjmUrKN1Jrf82is3e6W+2jujVvtU6jmS+S1ToH9vInahKS/wA1IgDqMq9Xsez/AE8yK/t2Zt5OOxoqqRQtqaWnxWSUttZGRGZqSk+PHuR7/ULsivXWYSarHdO86MjIsV1BpZ7pl7GVOKaWX5eZD0ON5Ipzw8oqpNaffk/Vst4HAxrVtjnksylHST4prp6zWenTSqZnNSdpqQi1mUMBxB00CTIWUVxZmrxF+GfmkjJJFt2MzVvuLSxIcSBGbhwYzUdhpJIbaaQSEISXkREXYiGl6H//AHU47/8ADq/2ixvQ35B5PpYDBU1DWTim297vrq+hbkuCKHJ+Hhh6Ecu9pXfH/egAADtF4AAAAAAAOt9luSy5HeTybdSaFlvtuRlsZDTI2i2mcOU3NjYylt9paVpWUp8zJST3SfdffYxu4CGrhqNdp1YKTW66Tt2EkK1SkmoSavvs7HA5ABMRgAAAAAAAAAAAAAAAAAAAAAAAAAAGsXepOFY3ldbheQX8autLhlT0BuSrgmRxVxNKVn6vPcy2SZ7nv23GzjaUJRScla+7rMKSeiAAA1MgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAFaOu1OH5bpMrTWXqNphQ5Q7YVt5WQ86v2oEN9MWWlwzdaUSlPNK8NaDRwNKtzLcj7liej+6tLTL7xqenpaJCK1Ki/cjn+PP38VP/tJcS2Z+v8AjcRPOcaKaO6m2DFtqNpXiWUTYrPo7Em4po8x1prkauCVOoUZJ3Mz2LtuZjnB9FtH9Mp0iz040txPF5ktrwJEinp48Nx1rclcFKaQkzTuRHsfbciAETU/90Jyj/JDUfreaLGCudP/AHQnKP8AJDUfreaLGACsnTh99f1T/jzGP1MgevrOiFYsaX1nElHLzeC1xP2krcj/ANI8nTh99f1T/jzGP1Mge3q7sotTfaSWcw/kK/LEWDhe9LBJcP8A0f2i5gZKNbNLclJ+EWQ4iWSnmfC3qjVM7u6PI9XdQra1uIkdFDj8uqrW33koU9I8I21IQRn6x8lO9iL2kLM4Nf0d9jkJykuIdgmNHZZeVGfS4TbhNp3SrifY/qMRD0uYfBtMMs8wyOqhzZN/ZuvkqQwlw+KTMty5Ee26+f8AYJ2h18CuQpqvgx4qFHyUllpKCM/eZEQ8b+z1Cs4PHSatVvK1tdW2tb9FuBrSpzjllLo3cbu8n4N27j0CEes+mdtunHLpLDZOuU8dFsllZbtu+A4lZksuxmRERqLYyPklJ+zY5uGia7Loy0ZzZjI7OLX18uimQ3ZMpwkNoN5pTSN1H5bqWki+syHoMRBVKUoS3NMv4Wyrwurq69To0axSvpcTqbinyS8nQLOsjyGIs2SlxllLiSc3bSSC4n63v9okIV96QNatPsu0ewLDIuZVb2Tw6FmM/VFJScpHo6PDUZo8/JBK/AYsENMJGnCjGNLdYxiMI8DVlQy2s2l2XAAAskIAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAavqDpphOqNIqgzehj2MbubSlFxdYV/GbcL1kK+sj7+R7l2EOpg669Ohb1apuqGAs//hnFb3Va1/MP/r0kXs8+3YkEW4sUAtUcXOlHZyWaHQ93dxT60RToqTzLR9P+7zTdNdXcB1ZqjtMKvWpSmyL0iI58nJjK/iuNH6ye/bfuR+wzG5CJ9SenXEs3tCzHHZsvD8yZM1s31OrwnVK/7ZBGROkft32UZduW3YanH1p1O0ZfRUdQ+NHOp+RNs5lRsG5GUW/Y5TKS3aV7zSRFv2JJ+YmeFp4jXCO7+6/m7uEu6z6jTayp6VV3rd39Hp1lhAGMxzJsey+pZvcXuYdpXyC3bkRXScQf1bl5GXtI+5e0ZMUGnF2e8nTTV0AABgyAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAFc6f8AuhOUf5Iaj9bzRYwaPF0mo4utVhriixnHb2ONRsXciGaPRkx2JLshLhFx5czU8oj9bbYi7b9xvAArJ04mSeq7qoUoyIiu8YMzP2f+hkDDdckhE+fp5EiuodQh6ykrNtXLY0oYSkj295uDIaCJr19TvVki2Jo4KrXHCkk7twNr4FTz5b+zbfcR5lzenEvqLpK3H5lVHxh+dFSa2HEpjElDRLd3VvsR7oMtz9p/WOHy7j62EobHDpOVVShvs1mi03a3BXe9WKmMi6ihSa+Gcoxb7X2dCZcLTXHSxPAaHHvDJC4cFpLiS9jhp5L/AM41DZRwQ5HYo0o0KcaUN0Uku4uzk5ycnxA1/UDKqnBsHvsyvmFvV9HXSLCS0hJKU420g1mlJH2Mz22Lf3jYBr2oNnitThdvLzhhT9CqMqPYNFEcleIy78mpBtNpUtZHz2MiI+xn7BtLSLNqSTqRTV9Vot77Ci/QV1UaSYhjVjplm0iJi5xpsmyr7KfISTT7bzhfIGrYuC0lt7dlFufbbY7u0GrOluUpJWN6jYzZ77dotqw4rv5diVuKvdJ+AdMabbKsfg4D6ZZ2l1YHAbu6CY838ENr3j7LlNG2j1T9pktR7b7nsJuvekPpoyLmdjo1jiDXvucOOcQ+/wBbJo2FHCbeNFJNOx6Hlp8n1cbOeWcL67k+12bXHrJcbdbdQTjTiVoV3JST3I/yj9ivLvQvonEIzxCfmuJL9i6TJpTRp9xl4ill2/AOv7lnUekIvtH6tNS4PEvURbuM2qCP2dlpTuX1CztKq3w8H72OVzfBy+Stb80Wv+rkWKAV0Rp51tY+slVHUBhmTIT/ANXe4z6KR/hVG9Y/6Rx9tfXXQL4WWlGmuUoT/Cqbp6EpRf8A9g9iMNvb5otd1/S5n6PUvs60H32/7KJYwBXUupLW6lT/AOuXR/mrXH5y6Owj2hH9ZJRxP8g5V1w6aVSSPOMD1JxAyMiWdxi76EJ/7yOW5Bzmlxdu1Nepj6Lxb+SOb8rUv+rZYkcbl7xClF1pdMOQKJuLq7UxVn5psEOw9j9xm8hJf2iveuOp3SGvXbDHnsiTaw7KVJk5RMq7SS/ENLsdSI3JbTvFPF0kqUlHzSLcy9g1qYqnGOaMk+8kw/JGJrVHTnTmtG/lb3K/G3/6XwARyzrd0/4xXxqhGsOExY8JlEdlpeRRlKQ2hJJSXdw1HsRF3Pcxi53Vp01VxbyNasVVse3yE5L3+z3Erq01vkvEqRwWIn8lOT7mS0AgWZ109LENXA9VY7yvYTFdLd3/ACpa2GP+726eX1m3U2OS2qy/gwsdlrM/6UENOc0fvrxJlyVjnrsZfyv2LFANX031CqNT8VYy+jr7aFDkOONoatISor5GhRpMzbV3IjMux+0hjdWNR7zTirgzqHTHI81fmSDYOLStpUtgiSZ83DUZElO5bfhMSOcVHPwKyoVHV2NrS3a6epvQCuyuojqCmFvSdG+TuFuZbzr+JFMvyGRjj91frKnf+w9LFLCSouypuZRlcT29pILcR84hwT8H7Fr6MrcXBf8AOH9xvPUZiyLrTDIL9rJMlp5uOUtlYQ3Ka6k1/J5EdS0m74K0+IRGgtiVuXn7zHv0PxCPjOCVk9vIcjtn7qBDnSXbq6k2CkuKZSZ+Gb6leGkzUZ7J2IVT6rcj64bPRu0+F8Co8fouZfCyscsXJU44nFROEvY+zHcuZp77F39XkJi6C3dVXtAIKtVisvHKY4VOdilRSDreDfhGrl6xp5eJxNX8Hjt22EEK0Z4jLle7e0X6+BqUOS1VdWLWe1lK/Dq8SxgAAvHAAAAAAAAAAAAA632GJTDkaSyh1l1JocbWklJWky2MjI+xkZewdgACDMi6Z26S2ezHQTKpGAXrp83ojJeJVTT9zsY90p9xGkti8yTv3Hlr+o3JNPpbNB1HYM/jTi1k0zkNchUmokqPyM1J3U0Z9ux7n5mfEhPo886BBtIb1fZwmJcWQg23WH2ycbcSfmSkq3Iy+oxeWN2iy4mOddO6S7+PY7kDoZXek7enh7HRSXtLklazcY/bRLKDILk1IivJdbWX1KSZkPeILu+l+JR2D+TaE5lZae3Dp+IuNGV41ZJV7nIy90l7i29VPsSPGnW/V7S397a7aXPS65rseS4sRyY3Eu3N5k/XaL2mZ7fUkZ5nGtrhZ5up6S8Nz7nfqMbZw+1Vuvev8d5YABqOCatab6mRik4RmFdaK481R23eMhsv57KtnE/lSQ24U5050pZZqz6GTxkpK8XcAADQyAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAEV4zqzd3fUdm+jUiugoq8Yx2nuI0pBL9IddmOSUrSszVx4kTCdtiI+57mYlQAVg6eEG71T9VjaVKSarnGiI0+Zb0yPL6xUbMMh+0d2Bd2FI1Zxay4iOTkLQy+34SHiUtJtu9nTUSVI47e0+5bC2mgSJ7nU91Yt1bzbMxVrjaY7jpboQ6dKniai9pEe242jHNGdXIObu5Xd5Lij6bJ+OuyJFalSnENH/AJTRElRkZ7mWxmZ7mY8p+0PJ1TG4rC1KcZPJJt2SaV7b7yjxXXpfsaFPCuvTrYiLbhK6s2uD36Pjl8OhskHSjW3TLWmm+GNPMojWHhkRyYij8OVFP3Osq9ZHfct9tj27GY3sQrqt0qYDqHcFnONy52DZ0yZuMZJQL8B9Sz/APfoTsl8j8j32UZduW3YaOzrvrX0+OoqupvEzvsaSom2s8xuOa2kpM9iObGIt2j7lupJEW/ZJL8x6LbSpaVlp0rd39Hp1lrYRq60Hr0Pf3dPr1FogGEw/NcSz+jYyXCsigXVZIL5OTDeJxG/tSe3dKi9qT2MvaQzYsJpq6KzTTszgcgAyYAAAAAAAAODIj7GQ5AAYC8wDBMmIyyPC6K13/ltcy//AK6TFHOoTph6bLHqBw+PCzOlxSvu5EiPkNXBkxY7UM40ZTqVcdyJhTppJB7l5nuXc+/0CGt2GmmnFtNesrXT/G5suQrm9IkVTDjjivepSkmZn9ZiviMPGvGzSOnybynV5PqOcZyWjWj6e3o3moU/TZ03IiR5dTo5hD7DjSVMvlVR3icQZbkolmk+RGR777nuNkhaO6SVvev0vxKMfvapYyT/ALEDa40aPDjtRIjDbDDCEttNNpJKEIItiSki7ERF2IiHYJVTgtyRUniq8/mm32tmOh47j9ckkV9FXxUl5EzGQgv7CGQSlKS4pSREXsLsOQG9iBtveAAAMAAAARX1I5nVY1pZkVJMhXEqZkdLZV8BuuqZM3k+qMpKUrNlCibIzWkiNWxefuMe/QvOKbMMCrItVEuY7tLAhwZabKpkwjJ1LKSMk+OhPMi27mncv6RIoCPI8+a+hY2sNhssrve97/pb9QAAJCuAAAAAAAAAAAAAAAAAAAcDkABGGddNmj+fyTtLHFm6235eIm0qFnDlJc/jmpvYlq+tZKGpp0u6kMAPfTnWeNk8BvuitzCMbi9i9npTe7iu3+CXYT2AuQx9eEcknmj0SV13X3d1iGVCDd1o+rQgYtdtZMS+S1M6dL1baO652MSEWTak+1XhF6yCL+coZKm6vdCbOR6BY5TIoZpbEuNcQHoykH7lKNJoIy/whM4x9xjuP5FH9DyCjr7Ng9y8KZGQ8jv59lkZDbb4Wp89K35ZW8pZvVGMlWPyyv2r2sY+g1CwPKiI8ZzSjtd/Iodg08f9CVGYz+5CKb/pX6f8jM1TdMquOo+/KAbkPb8jKkl/YMB9yJiNYX/qVqPqJi3HYkN1t+smiIvZxUkzMvyhs8HP5akl2xXqn+gzVlvin2P/AATsAgpOhmt1PseL9Ud+RIL1W7epYn8j+tS1b7fkMZPFsW6n6nIIDmS6o4tfUqXkemtqqDjSFNfwuBtlxJXu37DEsLSs3GtF/wAyfnG3mZVWV7OD8vcmIAEM5DqH1I0tzYx6rQCDe1jUp1EKUxkrEdbrBKPgtTayMyM07GZewzEFGhKu7RaXa0vVo3nNQ1d+5N+hMwCCl6662wkF8KdK+QpX7Sh3MeSX+akcK6j88Y2KV0x6iEZ/+5jpd2/oFj6OxHCz/wCUX+pHzmn1+D9idgEG/dPzWC3senfV1n3+Fj5OEX5fEIdKurelQfFzRPV1Hv5Y0ktv/wDUPo7FcI+a9xzml0k8AIKT1b4yZevpLqoj6lY0f/ycH6+61xf6KtUf0aP/AJgx9G4r7jHOaX3icwEYae6+U+ouRfa5BwDOahzwFv8ApVxTejRtk7erz5n6x79i29hiTxXq0Z0JZaisyWE41FeIAAERsAAAAAAAAAAAAEEdZV9nuO6UwZuFW2T1EBd9DZye0xiCUu1r6VSXPGejN8VnyJwmSUpKFGlClqIj2EedCWrep2raHLDIX8pl49VYTjtdJmX0Bcf0jJEek+nrYU4lK3S4HHJxfcuZFt57mBt2Aff26u/4jYt/tp4sYK54B9/bq7/iNi3+2nixgArJ04ffX9U/48xj9TIFmxWTpw++v6p/x5jH6mQLNgAPw8y1IaWw+0hxtxJoWhaSNKkmWxkZH5kY/YACu2YdI8aovH9QOnDLpGmWUuHzejxE86ewMv4L8Q90EX1pLYtzPgZ9x4qTqtyLTi1j4b1W4K5hk55ZMxclgpVIop6u+xk4W5smfnxVvt3NXAhZYeG7oqXJauRR5FUw7OuloNt+LLZS606k/YpCiMjFd0MjzUXZ9HDw9iysRnWWssy6eK7/AHOyrtay7r49tTWMafBlIJxiTGeS606g/JSVpMyUX1kY9QrPadLOZ6V2EjKelDUBzGVOrN6RiVwtcqkmK8zJJHuthR+8tz8iI0kPdivV7BpLpjBeo7Dpul+SuHwakTT8SnnGXmpmWXqEXv5HxLfbmZjCxGV5ayyvy8feweHzrNReZdHHw9rligHVGlRpsdqZDkNPsPoJxp1pZKQtJluSkmXYyMvaQ7RZKwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAEKdWUnP6bTyqzLTzK6GlmYtkES2lt314dRX2EZKHWzjPydyJKDcdac4q9VfhEk/MRl0SUdjSZPk5J6iKzU+PNoaiXdORcubuSTkLjstUx5ltKjOLHNKmW0EZJ5+FvsXEb31l0um87T/GMi1ayPH6vFcTzGsu7Fi9jqkRLJlBOtKiGylKjdWpLylIQSFbrbTuW25lFnQDppXYJe5FLqcwwGfXTsVx/wCCI+MVsiG/PqecxcO2sEPoT++nkLUhXDcvkfW7kQAkTAPv7dXf8RsW/wBtPFjBXPAPv7dXf8RsW/208WMAFZOnD76/qn/HmMfqZAs2KydOH31/VP8AjzGP1MgWbAAAAAAAAAGIynEcXzemfx3MKCBc1kktnYs1hLravceyi7GXsMu5ewZcBhpNWZlNp3RWSV0x6j6OSHbvpU1IdrIfM3XMMyJxcupePzMmlmZuMGfv3Mz37rIhkMb6wq2ht2ML6jcKstLshcM225E75aomGXmpmYkuPH2+t6pb7czMWLGLyTF8bzCofoMroYFxWyS2diTo6Hml/hSojLcvYfmQr7B09aLt1b1/juLPOFU0rK/Xuf8Anv8AE9kCfBtYbNjWTWJcSQgnGX2HCcbcQfkpKk7kZH7yHoFaJ3SXkmm0x6/6WdUrDCnXFm65jlktU6kkKPzLw18lNGf8YuRkXYthxG6sMw0vkN0/VLpPY4mRrJtOTUyFT6V4z8lGaN1s7/xT5K95EHOMmlZW6+Hj72HNtprRebq3Pw491yzADB4hnGH5/UIv8Jyatu69zsUiDIS6kj/iq4n6qveR7GXuGcFhNNXRWacXZgAAZMAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAV363XqaDp7iFvY5ZJxSfAziqcpcg+D2ZkGqsFk602/YNuuNpKJxdcQtfLdBrQotzIYHor0Ya03Ows7fXjGtSLqJj9TisVNAlluPV1MI3jYbUSHFqWta3nVGte3lsRbENh62zhtaY49PyDTeNmeKwsvrJGURnaP4Wch1JeIT8lqNsZmtO6UmoiM0ocWoiPYaX0Z2emN5qpqTbaBaUQcd00XX07UG3bxVVMuZYJOR6Qwk1oQt9tBG0rdSSNClqLyNIA23APv7dXf8RsW/wBtPFjBXPAPv7dXf8RsW/208WMAFZOnD76/qn/HmMfqZAs2KydOH31/VP8AjzGP1MgWbAAAAAAAAAAAAAAAAB1SI0eYw5FlsNvsvINtxtxJKStJlsZGR9jIy9g7QAFfss6MNOJNwvL9J7e30uyc9zKdjT5sx3D332ci7k2pH81PAj9u4wxZx1g6LbIz/A6/VzHmvnW+Ll6NaoQX8JyGZbOK9vFotveoWaAV3hop3pvK+r23FlYmTVqizLr3+O8h7Tjqz0N1LlfA8DL0Ul6lXhu0t8j4Pmtub7eHxc2StX1IUoTB59yGkaj6I6T6txTjah4JVXCuPBEl1nhJbL+Y+jZxP5FEIf8AuXtVdMflunTX65q4jfdGO5Sn4Trtv4iFmXNlPl81Jq8+/cYzVqfzLMurR+D9xkoVPleV9eq8V7FmAFafuidfdMi8LXfp1sZcJrsu/wAIdKwjGRea1RzPxGk+3dai/AJA086ptA9UFNx8V1KqvTlnw9AnrOFK5+1JNPElSzL+byL6xtHEU5O17Poej8zWWGqxWa110rVeRKwAAnIAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAgjrKk6gxdKYLuDSczjQfh6GWUP4W2Tl41S8XPGVDLY1cyc8DkaCNRN+IZF2EedCWd6v6gocssyhZzHoarCcdp3XcrjOsuTcgZ9J9OkR/G9daTQphK3D+ctJeZkZjRer3P9GrLqbZ0y6l9a73CtP6XDoltAqquZLipt7OTKkIW4+uMhS1JaaYQSUntsazMj7qI9o6DtRMKtNQtUNMdINW7nUDTeii01rQybaQ/IfrXZPpKJMMnX0pcU2SmG1pIy2TyPzMzMwJAwD7+3V3/EbFv9tPFjBXPAPv7dXf8RsW/wBtPFjABWTpw++v6p/x5jH6mQLNisnTh99f1T/jzGP1MgWbAAAAAAAAAAAAAAAAAAAAAAAAAAABH+oegOjWqqVqz3TqmtJCy4nLNjwpRF9T7fFwv/EJAAayhGatJXRtGcoO8XZlavuQslwX5bQLqEzPD0N+s1VWLibatSf8UmXduJH7z5H/AKByWXdb+nPqZJpnh+p0BrucrH7E66YpPvU28XE1fUhP/mLKAIebRj9m3Hs3eDuifnUpfaJS7Vr4qz8zU9MM4n6h4jHyazwm8xOU666y7V3LHhSWjQo07mXtSe26T9pHuNglXFTBmRq6baRI8qaSzjMOvpQ49w25cEme6tuSd9vLcvePYNA1Z0I0r1viQompmLItvgzxThOlIdYdjG5w5mhTakn38NG++5eqQledQ01fgRLJKfxaLq1N/AVtLo0fxg+WknULqXh6E924arP0+Cgy8vkHNtyLbyNRgWIdduIOF8C6r6e51GbLYk31Q5XvLItv5MW3I+/c1f0iLbVI/PB91n/nyJdhTl8lRd917rzLJAK2o1r6usaUacx6T2LdhPdUvHcmYV2278WF8lqP8pflH7+7Ur6lRNZx0/6vY6stvFcdxw3o6PefiJXuoi95JDnVPjddqaHNKr+Wz7Gn+pY8BHukGvGnOuUOxl4BYTXzqVttzWZcF2M4ypwlGjcnEkR78FfNM/LuJCE0Zxms0XdEE4Sg8slZgAAbGoAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB5pNZWzVk7Mr4z6yLiSnWkqMi925kNM1S1MwLQPDndQMxjO12PMSo8axnQoXiJhJdX4aHnko9fwycUhJmklGXMj223Mt8Gt6kYHRao4BkOnOTM+JV5JWyK2UW25pQ6g080+5STMlEfsMiMAVaoNe9F8X6p9WdXLzU7HY+HvYFizjFuU5DjEj5afslk0GZurPY9kII1GZGW24tRp/mtZqPhVNnlJEnR62+iInwkzWfCeVHc7tOKRuZp5oNKyI+5EotyI9yL+fbRTpZyfNur6u6aMjiOIercheh36kEZEiFEWpUlxJ+wlNtnwV7TWj3kP6JIkSNAiswYTDbEeO2lppptJJShCS2SkiLyIiIiIgBWrpw++v6p/x5jH6mQLNisnTh99f1T/jzGP1MgWa8gByA43L3kG5H5GAOQAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHBJSRmokkRn5nt5jkAAAAAAAAABF2r+mWqWeWFdN0812scBahsrQ+xGqWpqJSzURktXiKTtsXbbv5jQntG+saIkkVnV9DkpL+V4RCSZ/hMuQscAhlQjN3bfi/cnhiJwWVJd6T/QrkeBdcsNvjF15wOxV75mNmz/syP8A0D8JoOv9hB8c70dlKLy8aDNRv+VKBZABrzZcJS8Wbc5fGMfBFcWmvshEQjW7I0Jm/wA0/hVJ/wBiSIHcg6+45cSwHSCUftUzYTUl/QtRCxwBzfonLxHOemEfD/JXJrMuu1kt5Ojmm0j6mbx1v/WUY5d1K62WC2+5rxWSZeZs5a0kj/IrYWMANhLhN+XsOcR404+fuVtTq11sIP5XpMpnS/7PNIaf9Kh2/uydYyC9fo8hq/wc7g/+RixoBsJ/iPy9ht4fhx/q/uK2q1x6v2z2V0ZJV/g5xCP/AHATrz1a/wALosf/ACZtC/4BZIA2M/xH5ew29P8ADX9X9xXFOvPVX359FswvdtmsE/8AcHYnXnqh29fowsSP6szg/wDALFAGxqfiPy9ht6f4a/q9yu37vPU//wDswsv0yg/8IzGI6ydQl3k1bUZJ0pz6CrlyEtS7NeVQ5CYjZ+bhtoTyXt7i7icQGVSmn878vYw61Nq2zXjL3AgTKNaepSpyKzq8e6SZd1XRJjrEOxLLobBTGUrMkPE2pBqRyIiVxPuW+wnsBvUhKfyya7LfqmR05xg7yin23/RorarXnqz/AIHRZI/Lm0L/AIB+S116vV/M6LzL/CzeEX+4LKAI9jP8R+XsS7en+Gv6v7iuKdaOsNwt09HEZP8AhZ3C/wCEcOav9Z6v+h6Q61H+HnMI/wDyFjwDYT/Efl7Dbw/Dj/V/cVtRqr1trPv0rUDZfzsxin/oUPR+6H1tvl8l084bGP8A7fKkr/1RYoA2EvxH5ew28fw4+fuQrp5kvVlZZjAjalacYPUYysnfTZFfaOPym9m1G3wSZ7Hu4SCPt5GYmoAEsIZFZtvtIak1N3SS7AAANzQAAAAAAAAAAAAAACJca6b8IxjqPy3qTgtEV9llHCp3m/DIiaU0o/GdI/e6huIky27eAZ7nzPbb9UbfUSjwiws9KcRgZPk7Rs+h1c6wKEw+RupJzk8ZHx4tmtRdu5pIvaNrAAVs6UMB1tptRdYtUtbMKqsXn6hWVRLi19dbInttpiwvR1fKJIj78Un3IvnH7hsXWdpjm2r+gN1gunsBEy7mSoTrLS5KGCNLchC1+usySXqkftE4jRtX9btLdBcbjZfq3ljOPVEuaiuZkux3niXIUha0t8WULVuaW1nvtt6vn5DWcFOLi+JPhsRLCVoV4b4tNX3aHys+Lt6vf/0XF/P8T/mi7P2PrQPVbQbGswrtU6dqBItp0V+GlE5qTyQhtZKPdtSuPcy7GM38ZL0S/TrB/NFj+zh8ZN0Sn2LXaD2//ibH9nFalgqdGeeLdzu8o/tRjOU8O8NWjFRdtyaejv0sswArP8ZL0S/TrB/NFj+zh8ZL0S/TrB/NFj+zi2ebLMAKz/GTdEm+37u0Hf8AFNj+zh8ZL0S/TrB/NFj+zgCzACs/xkvRL9OsH80WP7OHxkvRL9OsH80WP7OALMAKz/GS9Ev06wfzRY/s4fGS9Ev06wfzRY/s4AswArP8ZL0S/TrB/NFj+zh8ZL0S/TrB/NFj+zgCzACs/wAZL0S/TrB/NFj+zgf2SbokLz12glv2/wDqmx/ZwBZgBWf4yXol+nWD+aLH9nD4yXol+nWD+aLH9nAFmAFZ/jJeiX6dYP5osf2cPjJeiX6dYP5osf2cAWYAQJol1l6UdQ2q19pzpRKVd19DSR7dy8QlxplxbjxtmwTTraFkpOxGavI+XYbH1KdQ2O9M+C1uoOV1j0urmX0Glkrbc4eiIkKVykK9VRqShKTUaSLc/YAJYAVn+Ml6Jfp1g/mix/Zw+Ml6Jfp1g/mix/ZwBZgBWf4yXol+naD+aLH9nAvsk3RKZblrtB2/FFj+zgCzACDOmfqxwzqhs8+bwSCfwThlqzXxrInlqTaNOtqWl9La20KaLdKi4nufYTmAAAAAAAAAAgbqc6s8X6XLvT5rNqlS6HMrGXCnWiFuqVWIZaSsnCYaacW/yUok8S228+41H4znow+lCx/Re1/ZgBacBVj4znow+lCx/Re1/Zg+M56MPpQsf0Xtf2YAWnAVY+M56MPpQsf0Xtf2YPjOejD6ULH9F7X9mAFpwFWPjOejD6ULH9F7X9mD4znow+lCx/Re1/ZgBacBU2N9lI6MZE+XBPUO3bKJ4ezysasTQ7yLf1SSyai28j5JT9W5dx6/jOejD6ULH9F7X9mAFpwFWPjOejD6ULH9F7X9mD4znow+lCx/Re1/ZgBacBVj4znow+lCx/Re1/Zg+M56MPpQsf0Xtf2YAWnAVY+M56MPpQsf0Xtf2Yanql9le6ZsQwqbfaeWM3Nb5hxhEalXXz6spBLdSlZ+kvRTQjig1L2MvW47F3MAXTAfhlzxmW3dtuaSVt7tyFQaP7Jz0/wshzfE9WJknD7fEcts8cZjswZ1mU2NFcJtMs1sR+LZrUTheEZmaeHmZGQAuCAqx8Zz0YfShY/ova/swfGc9GH0oWP6L2v7MALTgKsfGc9GH0oWP6L2v7MHxnPRh9KFj+i9r+zAC04CrHxnPRh9KFj+i9r+zB8Zz0YfShY/ova/swAtOAqbK+ykdGMWXEilqHbu+lKUnxEYzYklrYt91EpklHv5eqSj9+w9fxnPRh9KFj+i9r+zAC04CrHxnPRh9KFj+i9r+zB8Zz0YfShY/ova/swAtOAqx8Zz0YfShY/ova/swfGc9GH0oWP6L2v7MALTgKsfGc9GH0oWP6L2v7MPy79k76M0NLW1qZYLWlJmlP2sWpcj92/o3YAWpARf0y62l1G6HYxrOnGTx8skRKX8GnM9K8DwZTrG3i8Ect/C5fMLblt323OUAAAAAAAAABGGuWiTetLmny3cj+CU4Jm9ZmXH0P0gpvohOl6Mfrp8Pn4vz/W22+ae4k8AB0egwv5Gx/VkIz0p0Lh6ZZtqXmJ3bdn+6JfN3foxwSaKBwjoZ8Ilc1eJvw5ctk+e23tESdQXVRkldpneRsCpbTHcnqbJmPlBOFGl22J0S5Ztqu/QEG56Q2phta0EXzScQtRcS7750y5LrHPhT6DUe0r84oWI7Fhi2o1YthtnIYD/ACNCHo7Z/JyWyT6xoLgojSfY/nATX6DC/kbH9WQegwv5Gx/VkOx55mMyuRIdQ000k1rWtRJSlJFuZmZ9iIhXHq1y3qTwbF1ap9P8yuv6BmmkR7SoRBRIlRjc7t3ENRbnIUyXdUc90rSRmRGfcgN8xnQeHjuv+aa5/Djcgsvpqyp+CTgEkononP5QneZ8+fPy4J228zEnegwv5Gx/VkK99K+Z6uW6jr8hy2Jqpp9a1xXOLakRUx4rryTWSV186KjjxkIM1bLSnbZCiWSVkZFYsAdHoML+Rsf1ZB6DC/kbH9WQ7wAHR6DC/kbH9WQegwv5Gx/VkO8AB0egwv5Gx/VkHoML+Rsf1ZDvAAdHoML+Rsf1ZCNdcdDYes0LDIZXTdL9qOaVGX8kwSf9K9BdNfoxlzRwJe+3P1uP8Uxms3myc5x/LcC0t1Qr6LN66M2g5UcmJr1O+6XNhUiMrlslwkH2URGaTUae5EZU+wnVLrBsNUcgXa5FDc1IxSWym00geSzFrLnHvDaR8JVExwiUpxTviuc1rMk8iaWSdiJQF8fQYX8jY/qyD0GF/I2P6sh2oNSkJUpJpMyIzSfsP3D9ADo9BhfyNj+rIPQYX8jY/qyHeAA1iDppg1Zn8/VGBj7LGUWdazUS7BLjm7sRpZrQ1w5cC2Uoz3JJH37mOc901wjU+vrqrO6Fu2iVVpGuobS3XGyamx1cmXfUUkzNJnvse6T9pGNmAAdHoML+Rsf1ZB6DC/kbH9WQ7xirDKsYqbusxq0yGtiW92Tx1sB+UhEiZ4SSU74LZnyc4JMjVxI9iPuAPPl2JwsqxS6xjZmL8MV0mB45MEvwvFbUjnx7b7ct9ty328yGu6H6Sw9GtIsT0rOxauftXq2a34QVDJg5PAtuZt8l8d/dyP8ACKl6tudTeF6qU+nGoPUdcYtiVpJmvYLncGvYS1KuXnW1xKq9Tw8PghJPoQSSQh4lJ78kGkri6T2WpNtgNVL1dxuuo8tJC2bOJXS/SYqnELUgnml+ZIcSknCSe6kkskqPcjAHVgmkOCabZBl2T4jVuxJ+cWSba6WqQtxL0lKOBKSlRmSC4+xOxDcwAAAAAAAAAGp5bpZg2c5RieZZRTHMt8IlvTqN/wBIdb9Ffdb8NxXFCiSvdPbZZKIvZ3G17F7hyAA42L3BsXuHIADjYvcGxe4cgAONi9w/LjjTKSW64hCTUSSNRkRbmexF+EzMiL6zHEiQxEYclSnm2WWUG4444okpQki3NRmfYiIu+4rr11XdG/0t209yqh3dNaSqttVoTrqo9Qw9Ja43BLjmTikx90vEbai34l347gDftP8AQuBgWteqWs7GQvzJGp50xvwFx0oRB+D4px08FkZmvmSuR7kW3l3En7F7hWbo/wBXc+yKdkejWcX8HUL7RW4qIepFItC6+7ZdZbcQ0/xWrhMSh1BqJJqSot1Ge5bqs0AONi9wbF7hyAA42L3BsXuHIADjYvcNQ1Z0lwPXDBZ+m2pVMu0x+yWw5JiokuxzWpl1LrZ82lJWWy0JPsffbY+w3AAB+UIS2hLaC2SkiSX4CGqae6V4LpY3kLWDUp1yMpvpeTWpHIde8eylcfHe+UUrjy4J9VOyS27EQ20ABxsXuDYvcOQAHGxe4Ni9w5AAcbF7gPiRGZ7EReY5GHqMrx7KGLJWJX9XcOVUp6ullElodTHmN/PYdNBnwWkzLkk+5b+QAjXKdNcN19zLSjW3G8+jS4Gn9jYWMBdYbcuNZG+ycdafFSrZJINJ9y37kZHsJh2L3D5q9Kmb5/husVZQ4dMdlZbml7YOaq6WyKz4Mh4utCu9pBUSeDbfHw/4Sik809+e3H6VgDjYvcGxe4cgAONi9wbF7hyAA42L3D8vMtvtLZcTuhxJpUXvIy2MfsABqmlml2E6L4FVaZ6dVK63HaVLqYUVclx82ycdW6v5R1Slq3W4s+5n57eRENrAAAAAAAAHnsLCBUwJNraTWIcKGyuRJkPuEhtlpCTUpa1H2SkiIzMz7ERAD0Cs/Vh1E5liapuiWgmLycn1PsscnXK2Yctph+lrkElv01tDqTTJe5ufJMJ7qNCt/IiVpesut2WZ9qphNHpN1DxMEw/L8bXPwzJY0CNOrL/IkSVJXXTHXUn4aSZSji0ngpSnFluakpQNtw2jresPBKrJ9RKqXhOqWl2USaeRbY9JSl6ttYbiSklFeUSicivoNPJtZKSaVmkyM08gBGOiuj95rJkeOak02rVtlOM1KCn4vqI++03lEEifSmfjVwwpHGY0sjd2UtPyaiV2MjSR3bx3G8exGli45itHAp6qCk0RoMGOhhhlJmZmSEIIkpLczPsXmZmPXEgw4DSmYMRmO2txbyktNkglOLUalrMi81KUZqM/MzMzMd4Aj/WvRyk10xSJguVWc5nHztI062gxnDbTbRmjUr0N5STJRNLXwUriZGfhkXkZit/Rtkt5p/m+a9LOEYTZz8ZwfO7nxbOdJWiJQ0jrTb0GKytZKOQ8t5xzZsjLi2SlqV3TvdAfhDTTalrbbQlTquSzSREalbEW5+89iIvwEQAx+PYxjeI16qnFaCvp4S33pSo0GMhho3nVm465xQRFyUtSlGfmZmZmMmAAAAAAAAAAAAAD5xan431NaLZvmWKYrY57EiW9tZ5dgdrhdCie3kF3Nk+KUG+WpKjQhojJlPLg2bfrmr5MkD6B1lMxKVVZNkNFVlk0evKM5LbZSpyP4hIU+y06ZcybNaEntvsfBJn5EM0AAAAAAAAAAAAA0XWfVWJo7gr+Xu4/Y3812VGrKqnrkkcixnyXUtR46DV6qeS1FupXZKSM++2x0i1qO+6vKnGtVcY0MyiFqDhNxb4Db0sWybZscbuFJZkQbBEwiJJMsvtsmp0k7eHJd9VRD6KrbQ4RJcQlREZKIjLfuR7kf5DFeMU6VrSu6i8u1iyTPJErG7G8ayOixqGa2GmrM4DMR2VLUW3jqSllRNoMzQnxFKMuR9gNx0DxjWNjS1jHOpybSZRlMG0fWqxjpbcYmMof8WK+TZMtpaUj1SJPEzI20q5bnsUsAAAAAAAAAAAAAAAAAAAAAAAAAjjWzR1Ot1PWYddZbY1uK+mk/kNVBTwVexUpPjDcfIyW0ya+JuEjutJGjciMzEX9GzUvGaDP+lvLIrkxOlN47U16pbZuIm49NSciv5GouKzJpa2lJ77E2kj8xZcflLbaFKWlCSUs91GRd1Htt39/YgBhMOwTCdO6f7XsAxClxur8Vb/oVTAaiMeIr5y/DbSSeR7Fue257EM6AAAAAAAAAAAAAAAAAAAAAAAACGNYeqvTnQ7UbF8Cz1iyixsjYU9IvUs8q+oJTqWo5zHP+qQ64a0Es/VI0+sZEZmVVMc6eOo/TnX3JJ3TdQuYzWMZE5a3FnLsWmqjLqWXNalsRGUKae2kspdmsqkEkjQ2hCdzNZC/OW4nQ5nj1pjmQUldZw7WG5Ckx50cnWXmll3QtPmad++25fVsfcRp0udO8fpz07axiRlNlkVzJS0c+bKlvustk2nizGiturWbMdlB8EJ3MzIt1GZn2AmEmWUuqfJpBOqSSVLJJcjIvIjP3dzH7AAAAAAAAAAAAAAAAAAAAABjMnx2sy7GrbE7tgnq66gv18ts/wCGy82pC0/lSoxkwAHzr0J6JdXXLeHU6i4pj9Xpa26isyjCbCacljIZkeK6weQx0NpNMR11wmFJQlSDVspwzSfFJ30wPAMM0wxaHhWn+OQ6OkryUUeHERxQk1KNSlGZ7mpSlGZmpRmZmZmZmY2AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAQJZddXSzUZRIwuw1Meau4r7kd2H9r9opZLQ4batuMYyUklltyIzT9YnsVkvKuyV9kcxi4TXSTgI0jnx1SiZV4KXTtWzJBr24krbvtvvsAJ1wLULHtSKubb40mxKPX2Uqpe9OgPRF+PHXwc4pdSk1I38lkWx+w/MbMPmfi2MZ7qDkmmGF5dYZ0zSXGruoca59GsJsRblcls1stOPNqStLKjQaS2URGlSiSZbjz6NYLnOJlopqC1eaiv3idYbHDZJWNtOkMt40g5aEMKYcUaCaLw0nzNO+6t+XZJEB9OAHzO6YUan2mvWNzc+1Am0eo8bKbn7aauVDv3ZdpDM3+MdzlvXIipbJpbK08Up4oIjNR7Hvf2RG9lSMt+06PidwU5ODTrKiuysbsmV2ZOLS3Fgxa75NU4jJDniPnxJHHcuO+4FqM06idOMD1NgaT3blqu6lVDmQS3Y1et2JU1iFKQcya+XqR2eba081HsRl32IyM5BpbmpyOog5BQWUaxrLKO3LhzIzpOMyGHEkpDiFp7KSpJkZGXYyMfMyx0Ikas6g2mQ5VR5hZ3tx0xV12t9FnYsrlZEbRt+Grg4klKVwTyjH6ilGpRoNSjM8K84UHRHp2rsceu4mmqIE5nOfhhOQ+hM5IiDGSTcr0YykpaSvxSaS0ZRyWR7di7AfVoB87cA0wynP9SNBcEz3UjNMqxpnBbyVNs4T9pVNWRsWaVQkSPF4PnwSTO3ibKX4KFesk/WkD7Io69XOYRkTV3JsPgduxfThDjdy1GyVaiaSlLUqt2NqY3/1JOGaTNxR9iJRgC3cDLcXtMitMRrshr5N3SNx3bKuakJVIhofJRsqdbI+SCWSVGkzLuRHsMuPm5IraDTrqM1uzS3041G+2PIdO4N5i9NEtLZL8x5dZJ+EmClNGpCXGzVxSpW5trI/BSS9iOKnLzUVnTLXwtNjvoNRNwvGrarj0r948w1YnaNpfOO7YfLreJHJDqm/VUaD3L1TIgPrFPy3F6rIarErLIa+Ld3qJDlZXvSEIkTEsJJTxtNmfJZISpJq2I9iMtxlh8/tY9ANLtIuovpxtLRnO5ONJK7g2Ns5b28934RWTTsPxXGlqWhTsh101bElKy3SojQjZMldbMptnUXSFjUx7J2dFnX7j7bl0hzkoOaUdHwemWqF8uTRueJsRdjURcuxEAJz1i11wrRKPQ/bRFvLGfk9h8GVFXR1jk+dMeJtTi+DLfcyShJqUfsL3mGsmumFaH11LMyuLdz5eR2Sampq6WtcnTpsk0KWaW2W+57IQpRn9XvMiHz+xPSXIc9tunaZntbncumc1CyiLRvWU+xYnM4uhpx2uN9XNLjSjMj2WrZam+KdzRsQsT1p6T1WoetfTs/Y1d1KYXlcmDPcgTpbCWYvojjhGZsLT4R80l8oWytvV5bHsALdIWTiErIlESiI9lFsZfhL2D9D5f4/IzKZnePmUnUj7o5Or6ivWlqsygN4uUpfiEaT/eXwf6L4fHj6xmfq9gw+RmU7OMPUUnUj7o391tZZa06q0KA3jBSnfFJRK/eZQPRfB4ce+5+r7AB9Kcfy3F8s+EvtYyGvtfgaweqbH0OQl30Sa1t4sd3iZ8HE8k8kH3LctxhMg1YxHGtTMT0ltHZRX+aRrCXVoQwamlNwkIW/zXv6p7OJ29/cfO7G9O8Qw/DepbAGswy/T3KW9QpLjE2UzezIjmPqlxlxfGUyZmpqQaVtuPoV45oVupRo7HkcPwZOv2a9NlTlenmTYrjqYWdR5jVbcWrbT7STZNt1qU+pMtuK+ouSG1qSfH1e6exgfTIB8tcMtrt/BtCofVPZahK0pjVOUw5rzKrQluWzFm61BTOVD+XMkxUkTPLsZl23Lceqrk5rHw/QWP1Zv6lNaVuQskVIWlVmU1Uj0k/ghNkcL988vRuJtEfffz8lAD6HYrqziGY6hZvplSuylXenzlc3dJcYNDaTmx/SGPDXv6/yfn7j7Dch8t7uLl9L1QZll7ldmytFzyHT9WUJQiWzayGPgBCIT0le3iusNPmXpTfzzUaeXcjIWP8Ash/NnEsQuEZWuOipnS5543Jj23wfkxpY2TEefrdnWXiNRKZ5HxUvfcjJJ7AWiay3F3spfwdnIa9eQxoSLJ6rTIScpuItZoS8prfkSDWlSSVttuRkMuPmqrG6Ov6kE5zkGF53i1xl+h0abj0R6dbSlM3iIz6XYjrxKMlLaYSndL2xcy5mknF7nhbnB84wjpl0IyVdnlr9RmUmLP1NlXc+4loJSYSkwkSkRFekMxEqPgaWySndDPPcy3MD6iAPlrn8a8x3RDT66l6rTc9i09hkMmpxR+DlEONfRFutkxEYltkmSUmOfJEdT5mlSV7kfFJmcnrubWt62MZuDg3uVSMldp4ruOynLqNYYKgq1JuOpcTtBlw91Gbxr3UbijLupKjAF+wAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABxsXuAcgAA4HIADjYByAA4HIAAA42L3DkAAAAAAAAAAAABxsA5AAcbDkAAHGwbDkABxsXuHIAAONi9w5AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAcKSlaTQotyUWxl7yHzcVpZF066ockp9FG42BSZmpFHjMazr4UeQ9Bq5OMPSZTTCZKXEINbzaVcuJmSu+xlukwADHaY9SXVRneR1uIWOsBw5UnLanG8sKFDgvSaOa+/KQ9HjG5BSzwNprcj3lbG0hXinyWgazlnUHqxmOPVOD6s6oW/wpMssecxqsh1kEmMhai5StmXKmuJbJTTqPRWNkoWylRl2Solr2AANyxLqG1S1J6e9cGMgzVOZyMbi0xpnSq+GUdia7OUl+McRUVskKSlCDNPKS0WxGh4z3ISDplqbrbk8aQnOtRp19XZhp5nNg5XPwYLDUCRXWpQ4xsKYaQ53YcPlzUvdXft2IgACu0C86hNDazTwqG5RLZpdH1ZHR3UmQ2t+FWWMioXPZWhaFkpcThMNpRocI21tJJCzRwVurvWzrbjeKLetNTinpy3GnXtP7BitjuqnSGLpSHXnjXFZ2WmHukzcZZSokEZNk4ZkYABksh6iuoLEcEx7Oc316vYCMyr8j+15MGjqloeyOHYKj11Y4k45mmM40k1OOKNO6j7uNkREfTZ9WXUXV5vqHjs7UR1TmPmuTkaIUOEX2oVyLSG084yw5GNL7qIrsjwlJkyfE23caQvikAAGGyrqa1Oq7V/UXDdSpr+ISqmnamZrbU9dAtzplWd02098hFkmZKW1GQR+imvipSvR2lKWottoNeeryZDRlK9R0T7SJkOJ0kOiRXQGq2zbs6cn1OPL8EnkrW4bThcXEEg1rLjx2SkAA1q26xdV0abV+U45rxkEmjRNiQMsyC1pq6Gqou1wHXFQIpsxH1us+kpSSt4aiIkoSl9SVrWiVenzqJ1tyLqiqtL9Q86K2kTaj0u1pIkWPFYrElVx3ie4+jqNxC5Cl8VomGvdzZbKUJJQAAP/Z)
Figure 3. Graphic Summary of Short- and Long-Term Functional and Neuroarchitectural Effects in Prefrontal Cortex (PFC) Synapses after Acute Footshock (FS) Stress [44]. The fast and transient increase in corticosterone (CORT) release induced by acute (40 min) FS stress was accompanied by the rapid increase in both depolarization-evoked and hypertonic sucrose-evoked (readily releasable pool) glutamate release in PFC, and the number of small excitatory synapses. The enhancement of glutamate release was sustained for up to 24 h, as well as the increased number of excitatory synapses, which normalized between 24 h and 7 days after FS. Before 24 h had elapsed from the start of FS stress, retraction of apical dendrites began and was sustained for up to 14 days. The timing of actual FS stress (40 min) is indicated by the red marker. Number of excitatory synapses and apical dendrite length are indicative and not in scale with other readouts. CORT and glutamate release data adapted from [44].
Changes in neural architecture
- Hippocampus (rich in CORT receptors)
- Prefrontal cortex
Neurochemical factors
- Cortisol enhances glutamate release
- Corticosteroid antagonists block this
- Ketamine (NMDA receptor antagonist) may act via similar mechanisms

Sapolsky, Why Zebras Don’t Get Ulcers
Pleasure/reward
From conceptual category to brain circuitry
![[[@kringelbach2009towards]](http://dx.doi.org/10.1016/j.tics.2009.08.006)](data:image/jpeg;base64,/9j/4AAQSkZJRgABAQEAcQBxAAD//gAuSGFuZG1hZGUgU29mdHdhcmUsIEluYy4gSW1hZ2UgQWxjaGVteSB2MS4xMwr/7QAsUGhvdG9zaG9wIDMuMAA4QklNA+0AAAAAABAAcQAAAAEAAQBxAAAAAQAB/9sAhAADAgIDAgIDAwMDBAMDBAUIBQUEBAUKBwcGCAwKDAwLCgsLDQ4SEA0OEQ4LCxAWEBETFBUVFQwPFxgWFBgSFBUUAQMEBAUEBQkFBQkUDQsNFBQUFBQUFBQUFBQUFBQUFBQUFBQUFBQUFBQUFBQUFBQUFBQUFBQUFBQUFBQUFBQUFBT/wAARCAFfAywDASEAAhEBAxEB/8QA3QABAAIDAQEBAQAAAAAAAAAAAAYHBAUIAwIJAQEBAAIDAQEBAAAAAAAAAAAAAAEDAgQFBgcIEAABAwIEAAUMCwsJBwEHBAMCAQMEAAUGBxESCBMYIVcUFRYXIjFRlZbS09Q3QVVWdXaRkpS0tQkjMjhTYXGTpbPRNDU5QlR0gbGyJDNSYnJzdzYlQ2OCocHDJkRm4icpxBEAAgECAQUMCAUDAgYCAwAAAAECAxEEEiExUZEFExQVFjIzQVJhcbEGNFNygaGywSJC0eHwNbPCc8MjQ2KCotIkkmOD8f/aAAwDAQACEQMRAD8A7myRyQy5n5LYAkycAYXkSXsP29xx52zRiMzWM2qkSqGqqq86qtTXtDZZ9HeE/EcbzKAdobLPo7wn4jjeZTtDZZ9HeE/EcbzKAdobLPo7wn4jjeZTtDZZ9HeE/EcbzKAdobLPo7wn4jjeZTtDZZ9HeE/EcbzKAdobLPo7wn4jjeZTtDZZ9HeE/EcbzKAdobLPo7wn4jjeZTtDZZ9HeE/EcbzKAdobLPo7wn4jjeZTtDZZ9HeE/EcbzKAdobLPo7wn4jjeZTtDZZ9HeE/EcbzKAdobLPo7wn4jjeZTtDZZ9HeE/EcbzKAdobLPo7wn4jjeZTtDZZ9HeE/EcbzKAdobLPo7wn4jjeZTtDZZ9HeE/EcbzKAdobLPo7wn4jjeZTtDZZ9HeE/EcbzKAdobLPo7wn4jjeZTtDZZ9HeE/EcbzKAdobLPo7wn4jjeZTtDZZ9HeE/EcbzKAdobLPo7wn4jjeZTtDZZ9HeE/EcbzKAdobLPo7wn4jjeZTtDZZ9HeE/EcbzKAdobLPo7wn4jjeZTtDZZ9HeE/EcbzKAdobLPo7wn4jjeZTtDZZ9HeE/EcbzKAdobLPo7wn4jjeZTtDZZ9HeE/EcbzKAdobLPo7wn4jjeZTtDZZ9HeE/EcbzKAdobLPo7wn4jjeZTtDZZ9HeE/EcbzKAdobLPo7wn4jjeZTtDZZ9HeE/EcbzKAdobLPo7wn4jjeZTtDZZ9HeE/EcbzKAdobLPo7wn4jjeZTtDZZ9HeE/EcbzKAdobLPo7wn4jjeZTtDZZ9HeE/EcbzKAdobLPo7wn4jjeZTtDZZ9HeE/EcbzKAdobLPo7wn4jjeZTtDZZ9HeE/EcbzKAdobLPo7wn4jjeZTtDZZ9HeE/EcbzKAdobLPo7wn4jjeZTtDZZ9HeE/EcbzKAdobLPo7wn4jjeZTtDZZ9HeE/EcbzKAdobLPo7wn4jjeZTtDZZ9HeE/EcbzKAdobLPo7wn4jjeZTtDZZ9HeE/EcbzKAdobLPo7wn4jjeZTtDZZ9HeE/EcbzKAdobLPo7wn4jjeZTtDZZ9HeE/EcbzKAdobLPo7wn4jjeZTtDZZ9HeE/EcbzKAdobLPo7wn4jjeZTtDZZ9HeE/EcbzKAdobLPo7wn4jjeZTtDZZ9HeE/EcbzKAdobLPo7wn4jjeZTtDZZ9HeE/EcbzKAdobLPo7wn4jjeZTtDZZ9HeE/EcbzKAdobLPo7wn4jjeZTtDZZ9HeE/EcbzKAdobLPo7wn4jjeZTtDZZ9HeE/EcbzKAdobLPo7wn4jjeZTtDZZ9HeE/EcbzKAdobLPo7wn4jjeZTtDZZ9HeE/EcbzKAdobLPo7wn4jjeZTtDZZ9HeE/EcbzKAdobLPo7wn4jjeZTtDZZ9HeE/EcbzKAdobLPo7wn4jjeZTtDZZ9HeE/EcbzKAdobLPo7wn4jjeZTtDZZ9HeE/EcbzKAdobLPo7wn4jjeZTtDZZ9HeE/EcbzKAdobLPo7wn4jjeZTtDZZ9HeE/EcbzKAdobLPo7wn4jjeZTtDZZ9HeE/EcbzKAdobLPo7wn4jjeZX56/dCsucJ4dzossa04Ys1sjlh9lwmYdvaZBTWTJRSVBFE10RE1/MlAfoVkP7BuXXxct31Zup1QClAK/MrAnC0zMlzMvbbc8WvuudnUhy6vm2wCyLQTsdphgyUUEBUzlohaoukZVVUoC1I/D0dv8AYYd5veGpljG0YiiK83YbqZtSob1onTQ3K7FBXxQYxCotogGSAYOkKaFL7Vwxsa3DFWHcGO5cWpjGd8lw+p2ExEZQmoUqDKmNPG8kXVXBGE8BtoOmqIokqElAWRi/Pm+WjOZ7B9kwrDvFms0e3SMQ3SVeW4L0MZrr7bKstuigPbEjkZorgroSIKEXMvPkPh94vxXd8MpDwpbbPb28SQmrq43cHZKSbZJt86UKMqcUN5oMNwt7aq2RNgIGQuKYASPB3DvxVjeBaosLK1I2I7y83JtrVynyYcB6AUJ6aR9Uuwx3PCEcgUQAgUjEkNQ1WthYeHZccQ26w35nATLeF32bIzdHyu6rJizrpHB6O2y3xGjzQcY0JuKQL98VUFdhUBHZ/DZxBa42DcV3LBrzE/FeE2blZrBCvjkqGXVc+AxHWQIQ+N41OqVJSbQlQUUEAyNFGWYf4bl0uWMMurNdsv3sMR8TcXGkzLy9Kji3MKU/GVhhSioJKisC4gvkwZg+3tFVXSgOsKUApQClAK544amNMQYNwbgQcPXXEVqdu2LotslnhOOy/c3Y5xpJk2wDwGCkpNgvOPtUBAMM8KHMLLJnLLCePcLuv3S/Jtev2Iniglo9PdYhsu8RHcYblcQjDjgGTSKp7Q3EqomwsvDhvGO4doiYTwDEk4jucmHaW4dzvasMMXQokqXOiuOiwWiR2oopuRNTJ8e5FEVVAjdu4ZWL8E48xhccWYcF/AxXmJCRtu6tPzLO8eHGriUdoGWybkN72ntXEe51dTaiolMb8M3FFoHDEi/2McJP267Q7tdIVnnJcQn2aTZrrMba3k0Co6hQdCFE/CAVElEqAl0Lhg4xk3KBhNzLy1tZh3KbCGFb+yAltyxJMGVOFxyX1PqLgtw3gVtG11PaqEorqkPwPw277jm9ROxnBxTcUYrg2iRb7Fd8Ro3bownCny31FxIuoKgRNqqu7eRgugIK6gM5eHDiCAzjXDVtw61Ypg4SucqDeYt0Ga/b7kxZ+r1Fzi2ijLt3bUQXyJVBCUEAxVblzCz5xJg2Tguw4awvExhf7jYX8Qzwn3cLYLcOMsYHVbcMFBXSOUO0TUATaSkQpz0BXeLOHVdcKRbjd3Mv2JOHpT1yi4deS8qEia9AuDUGSklviFSOJG4RAqK5qjehIKklaweHviSzQsXliDLi3sS7FCurrLMC/k6EmRbrozb5QKZRh2ApyAID0VVQS1EdUoDbXThq4rt0jENhHLu2yMZYSC6T8RQgv5DDagQWYbxnFkLG1edMJzKC2QAiEJ7lRNFXCn/dAZFocvN4nYDAcGRlvrECaxdSObKftsdJK746sCLQONqiIu8lEtdU0TVQMOfw8cawrJK1ynZavdrtt6vFyZuF0lwGOo7c1EeNyNx0EXHVcCXtRCAERxokVVHuqwneHHjXCVkxXe7vhCFc7Qxi+Zbok3qx8GLfAahx3xGSTMRxQI+OFEcJOLQjLeYiCKYHa8CY1coEaWyYOMvti6BASEKiSIqKipqipovfTmr3oBSgFKAV+aN14VGaEW+3/DoYskpOjZqo+24jDSkGHOvDlrWIncfgo8Id1+F3Xf50oC2rR90GvV/wZNvMbLdYrkxyGeHlnyZjTM9iQr/MS9RqZvgMfcrUYH0VHUVC0AyTYQfugTs6baHXMDJbLTc8KjiCI9cbg60Ux5bac5WI5pGVk0BQ4pdXBd1VTRpRHnA3cjhn3m446w/hGw4EiS7peQtZtuzb0TLTSS7ZJuBqW2OSrxYRSHROc1JF7nvLG8KcPfEmNbNCkW3K4m5V+uNui4fO4zpUSJIZm9Uo2Tr5w9EcDqcVMWkcHR4dprotAYFl+6HXu8YOjzO1wy1f7t1mcssFm5SJTTrVwZmOiTysxTdEgS3yNRbaPXc3zp3SpIrBw38S4oxHbIsXLJu32yROtVnfK7XhyPMYuFwgdUtNEwsVdGwcRWzNS100MQXVRQDU5L8NfFd1hZL2zGeG4QSMZW6G7IxEUp1mM/IkPPNttsbYyso6gtNkTLjjZKrwi3xmmq9oUApQClAKUBRfDOxfiLBWSXVuFZ1zgXiRfbRbxcsqMdWm2/PZZcbY49FaRwgMhFT7lFVNdEqBtZ/YqyStuW2HrxhvEt+uuN7m7bILuObzb49xjyuPaFAfSI2rXEcSTjgm3uLcAtqOrgqgGmf4fF3PHF1wzb8B2+fKcukW3WR9L26EeSj1zW373nUiqCKjiCaowrwohKKkhiqVj4b4fGJ79bre2WW1uavl5WzdaIvZCXEGk6ZKgqTzvU2rex+G4qIInubMV1QkVKA1F54cWIrZi+23ELGTr0yzOWrsVbeceit3pu+Hb3HePaYN02kRlwkUGyUh2ogbl5p9hXhr3G/Y8y9sN0wC/hWLidlltyVe3pMdRmG5IbJhjdFRslAo6dy8bLjgvtqAL7YGyzT4VmIMI4xxNabLhCHOs1pnt4bW7SroTL3Xh+3pMjpxCMl/s2hsgbm7cimqoBCC6w+PmpmbauDJwZL3b5oXnGeLbtbBnjcp6A1cQlW+XI2PPIwqtgqo0SoDaqGzYinpuIDJsHDrvOK8OYnvkDAlti23DlphSZrlxxArZFOlPORwjNokdd4I6y4m9NSNNmxsiJBqO23hzXm8XvDk6FgyQ7frxHbs/WKRfTjW5uYV8ctvGaHD40V3Bu3EIkgrsVtCTWgLbj8J+73HI3DeKoeFYa4xvuIjwnHsr9zIIQ3AJj0Vwiko0pcSnUzriLxe5U0TTVagznCsxdCzRCwRcEq7iq+dYrelouOKWittvfkR7u84bZsR3NUTreqESEquIQdy2QKJAb/CHC+xBfwwjcp2AYkXDeMb1Dg2K4R7+DrpRZDkgEcfYVpDbcFI4ntRCBUdROMQgJFrXEnDGxhfsUYKmYRs6QJN7fs8aTAuF4Q4QMPYglW80AEiqSOucQKK7r3IOcwqrfdgS9jhyX+JhCyYku+XUKJBxQ1Paw0zGv5Ouy5zElmOzFe1jCjZPm44QkO/aLfOKquiddDqoJuRELTnRF1RFoD+0oBSgFKA5s4ZeLsXWCRlZacJTsTxnb7f34ctjB6wkuUloLfJeQGlmIrKaE0JLu07kS059KxnOEhesrcYZb5cXbDlwuVwxDaVuI3XEt7hsTmWmG3XZvVgMto0jrYg3tRtVBxTLugRsioCtof3RXEdxtMpuLltbXr40r8gWivr7UVYjdrcuSGLhw0Iz4tow7kOLItpC4QFqkss3DTxfiZ222O2Zc2pcY3K+La48SViIm4YNHZhuzLpvJGVd3EkgECBohouhKlAV/C4f1/K+3W42TDUzE6YgjWmbZsPPE8gwG1tRS5Yb48d1wzI0QB7jaqluUgFFq7stOFo9mDng3gWVhNzDMSXA6st713dfYmy0SMy+RA0TCNEKccQKIPE4CskpgKLzAV3n3wyMX2LCOYo4dwxFt0RtnEFpsGIDuarJGfbGTKU65HVhRAEFt4ml3FuJkULYjiKkvzp4TOI+DhgLAbPYwzi66P4WmXq6vzb2rRNhb2oayNppGXj3DSQWiqLSKo6qiIWiARjEnDxxFh20MxUyzW5YxYul2hTrRbZsmUyjVvbjOOEw41FMyMxmMCKG2AIW7cYoiKvvdeHpcIuMMZWi3ZeuXxi2W+XLtSwZjzj8s40iKw828CR1EFTqpDIGSecBGiEgQ1QaAyrXw8hn4WuN0DC0Ka7brLBujyQrs5xZOP3h62k0nGRwcBR4njVRwBNN2wgFU3LqcRcPHFdogRJkPK0boM+bexhRINxkypD8S1SljSnFFmGex0zUEbAu4513uCm3cBjYg+6A4jtV6vRR8toT2Hba9dCWU7fTblORbdcWoclxWVjaC4XVDJA2pf8aESaJrssUcPi6WC/43Yi5aSrlaLLPftVvnrIkMBKlsXCPBdB50oqstIRPmQKDjiqjKoSApJoBrT4ZeZA5gvxCwHBks2Kz4k662G1XM5Dsu4W+RGAOpnFjCRAqSGkRNqEvGuLtVWgFzong85xpnnlyGIyjwokluZIgyI8F91wAcaPauqPMsutkqaKrbjYEOvOipoqgWZSgFKAV+bv3SL2crH8XGPrMqgO6Mh/YNy6+Llu+rN1OqAUoBVXtcGHKtlxoxwRa9zfFbdwEv8Au3Xng1RS50RyQ8Wi9/fouqIiIBGHuB1gKDPwmuHoLdhtlnurN0mQUE5S3PiYciLHacN4yVAAJRppouooI8yImkswVwbMs8unoD2HcIw7a/AmlcIzwm4bjT6sHH1QjJV2oy4bYh+CIkqCiUBn4syKwDjvG1rxdf8AC8G64itjaNRpkhCXQEUlFDDXY5tUzUd6FsUlUdFVVrR2DgqZUYXZit23BkNpIsqHNZVx550geiC4MYkIzVfvYPOAKa6bC2abdEoCFZhcCfBt2wGOG8EWvD2FmiuQ3F8LrZiurDpCybQCgk8242gCa7BBwQRNRUVAyFZPg/gj5aYWg4MV/D0e8XnC9oi2eNdpW5HHgYZVoHHAEkAjRCPaSiqhvXaqUBv7rwdMt75YoNnnYShSLdBs4WGKwSmnEQQcacBoFQtR2mwyQki7kVsVRUVKxYPBiyutlzw9PiYOhR5VgbZbgK2bqCCNOE60Tgbtrxg4ZuCbiEQmZEioqqtAWjSgFKAUoBWoxDhGz4rdtDl2gNTnLROC5QSc1+8SQEwF0dF76C4ac/N3VAR7FmSWCMdYvtmKL7h+Pcb7bUaGPKcNwdUadV5lHAEkF1G3VVwEcQkAlUh0Vdag+GuCVg1rLa6YUxhFj4wW74hl4ouEvqdYPGT33CVXWxaPc1o2SN9yeqiioq6EqUBMWcg8vIzbbbWEra2y3MYuAMC2qNi+zE6iaJA126DGTikHTTb7VazDnBeyrwnbmoFrwVb2IjU5LiDTiuO6Pow5HFdTJVURZedbFte4ETVEFEoCG444GuD5mXPYrgi14fw0yV0aujw3i0ld2JBttk2AkJvA4KCJIgbHRQUTZpsIhXNw3wLcrrXljZ8G3jD7OIghQoMR64yCcZfknFacbbc1bNFBdr76aCqJtdIV1RaA30rgpZSy7lNnHgiAj8xh2M6DZugzxbsTqR0RaE0AN8dEbLaKbkEddVFFTe4/yJwDmmzYmsV4Zh3puxlrAF9TRGU7nUF2km9stgbmy1Etg6ouiUBqpHBhysl33EF5dwXb3LlfteuDxK5o6qug8SiO7a2puNNmagg7yBFLVa0+anBRwVmLgrElmhQI1guN7amNOXVtknyBJcxqXL7hTRC411kSJNU5+9onNQGTI4IeT0vDlvsT2BYD1sgy35zTbjjxGTr23jlccU97omgAhA4pCSACKioIokjk5DZfTFc4/ClvfF2RPlG26CkBuTWuKlqoqui8a2qiSaaad5EoDV2bgw5XYfsrlqgYPhtQnYU+3OgTjpm7HmC2MoDMjUi4wWWhVSVVRGxRFRERK8b5wVsqcR8f1dg2GayH1kOqy68yrhqyLJoSgaaibYABgvcmgpvQtKAtOPGahx2mGGwZYaBAbabFBEBRNERETvIic2lelAKUApQCq1d4N2Wb10euR4Ot5T3nSeckqh7yMpw3BV13e3LEXtO9uTwc1AasOCPlA0xMaawNAZGXPbuTisuPAQvN8YgbFQ0VsER99OKDQNHnE26GWuVauC3lVZLrBuELBkFqRCihDYBTdNkWwYWOCq0RqBGLBK0jhCpoC7ULTmoD+4F4L2VuWlxg3DDWEIttnwnhkMSeOedcBwWHY4luMyVdGX3W0110EtE7yae2G+DPlhhCQb9mwbAgOlcmbshNqfcSWeM4ogRSXYIK66otjoCK4SoKarQGPJ4LOVUvDp2M8GQktpRoURGm3HWzbbiK4sZAcE0MFb453QhVF0cJFVUWtvbchcvrMsbqDCsCGkaVCmsgwJALb0NlGIpiKLonFtIgIne0Tn1oDUWjgtZVWG6WK4W7BkKHIsmxYAsuOi00oOm80qt79h8W464Te9F4tSXZtq1aAUoBSgFKAjeYWXOG81sLv4cxZamb1ZX3G3XIjxEIkbZoYFqKoqKhCipz+1UJg8FDKa2wWIjGDIiMR9VYQ3njJgleZeUmyI1VsuMjsFuFUXVoefRKA9bVwWsq7Hexu8LB0Rm4BKGa27xrxI06MlJQKAqaiAi+nGIAogoqroibl1j2YfA5y+xvZbJa4drj2ONb5lvN1GAM1kQ4sp2SkTXjEUEJyQ8u9F3Cpqqa82gEie4LmVT+G41hLBcDrXGt42xloScEmmBkdUogmhb0Pj/vvGa796qW7VVr2t3BnywtF7sF2g4PgxZtiaZZgKyTgttozu4kjbQtjhgpmouOIRCpKqKiqq0Bn3nIPL7EGYKY3uGF4UrFKNcT1wPfqqcWTSEoIWxXEbMgRxR3oJKKEic1f3E2Q+AsYYCsOC7thyPKwzYUY61wEddbSGrLJMtE2YEhiQNkQiWuqa666oi0Bgt8GvLJjCtww2xhCDGss+LChyIscnGtzcNVKLoQkhCTZKpCYqhbl11156+bVwZssLJItz8LB8Jh23vDIimhuKrLgylliSakvekKrn/UtAbS7ZG4FvmAn8FzMORncNPTHLgUETcDbJOQUgngMSQwNXjI9wkioqrppWNZeD5l1h25WmfbMJQIMu0jEGE4whDxCRmn2WNqa6dw3Lkjz9/jSVdV56A0pcErKJYF8hjgiC0zepbU6ZxTrzZq824TjatkJoTKCbjhILaiKKZ83dLrmscGLKyLa4luZwXb2ocRhmNHaDenEttSjltICoWo7X3DcRU50Ul59OagMPEnBxsN5lZZRIKsWjCuB7yd+YsjURHOPloDiMkjpFq2gOPOOKiIu5VTnTSrcoBSgFKAUoCGZnZOYMzmtsCBjOxMX2LAkLKjNvG4HFO7FDeKgQrrtIk7/t1H2uC9lY21Bb7DILqQQZbjK+brpMg0rygAkRKqD/tL+o94kdJCRUWgPCx8FDKbDjXFwMGxQ1ZOOpOyH3TVsoxxSFSNxVX7w4bXOvMKoiaaJposw+Bxl/mFdsNOPW1qFabdcCn3C2so6PXJUthW9kVdFwSaVtvilQh5/vSJ7arQEhvvBWynxLbFt8/A9uOJpFQW2eMZ2DHYWOyIqBCoiLJK3tRUQhVUJFRa3NjyHwDhnHB4vtWGIkC/kJCkhlTFttSbFsjbZ3cW2ZAAARiKEoiiKqpQGvuXBnywu+IsRX2Zg6BIumIYr8O5vGrmj7bwID3cbtoE4IihmCIRIiaqulb3GmUGDsxGWGsR2GNdm2LfKtbYvKSbYskWxfaTRU5jRptF9vuU00oDQ4p4M+WGNYpx71g6BcGnLk7djFxXEU5LoA26RKhIqiYNgJtr3BIKIQrWJM4KOU0+4Xia/guE5JuyvrLPjXkQledbedUEQ9G1V1pt3UEHRwUNNC1WgPCZwRcoLiNpSTgiE+trExjk48+pKpPrIInV36vFxyq5uc3FuJV11Va2mIuDTljiyw22zXbB8CbbLbMkz4sc1cRAekuk7I1VCRSB0zJTbJVAtU1FURNAPWfwdct7mk5JOEoLyTkmjIRVNOMSW+3Ikouhf+8dabNfzimmiV4zeDPlhcMR32+yMHQHbreyAp8glP76QvNPbkHdtBSdYZMlFEU1bFS3aUB6Xrg35Z4jkypFywdb5j8pyc68biHq4UxASUq91z7+LbVU9pQFU0VNak2A8vMPZY2HrPhq3DboJPHJcRXXHnHnjXU3HHXCI3DXRNSIlXmRNdESgJHSgFKAV+bv3SL2crH8XGPrMqgO6Mh/YNy6+Llu+rN1OqAUoBSgFKAUoBSgFKAUoBSgFebz4MCikq8/NzJVdScaUXOWhEpXzHl1wZ8K/JTrgz4V+StDjHD63sMshjrgz4V+SnXBnwr8lOMcPrewZDHXBnwr8lOuDPhX5KcY4fW9gyGOuDPhX5KdcGfCvyU4xw+t7BkMdcGfCvyU64M+Ffkpxjh9b2DIY64M+Ffkp1wZ8K/JTjHD63sGQx1wZ8K/JTrgz4V+SnGOH1vYMhjrgz4V+SnXBnwr8lOMcPrewZDHXBnwr8lOuDPhX5KcY4fW9gyGOuDPhX5KdcGfCvyU4xw+t7BkMdcGfCvyU64M+Ffkpxjh9b2DIY64M+Ffkp1wZ8K/JTjHD63sGQx1wZ8K/JTrgz4V+SnGOH1vYMhjrgz4V+SnXBnwr8lOMcPrewZDHXBnwr8lOuDPhX5KcY4fW9gyGOuDPhX5KdcGfCvyU4xw+t7BkMdcGfCvyU64M+Ffkpxjh9b2DIY64M+Ffkp1wZ8K/JTjHD63sGQx1wZ8K/JTrgz4V+SnGOH1vYMhjrgz4V+SnXBnwr8lOMcPrewZDHXBnwr8lOuDPhX5KcY4fW9gyGOuDPhX5KdcGfCvyU4xw+t7BkMdcGfCvyU64M+Ffkpxjh9b2DIY64M+Ffkp1wZ8K/JTjHD63sGQx1wZ8K/JTrgz4V+SnGOH1vYMhjrgz4V+SnXBnwr8lOMcPrewZDHXBnwr8lOuDPhX5KcY4fW9gyGOuDPhX5KdcGfCvyU4xw+t7BkMdcGfCvyU64M+Ffkpxjh9b2DIY64M+Ffkp1wZ8K/JTjHD63sGQx1wZ8K/JTrgz4V+SnGOH1vYMhjrgz4V+SnXBnwr8lOMcPrewZDHXBnwr8lOuDPhX5KcY4fW9gyGOuDPhX5KdcGfCvyU4xw+t7BkMdcGfCvyU64M+Ffkpxjh9b2DIY64M+Ffkp1wZ8K/JTjHD63sGQx1wZ8K/JTrgz4V+SnGOH1vYMhjrgz4V+SnXBnwr8lOMcPrewZDHXBnwr8lOuDPhX5KcY4fW9gyGOuDPhX5K+2pjbxoAquq+FKzhjqFSSjF533BwaPavzd+6RezlY/i4x9ZlV0DA7oyH9g3Lr4uW76s3U6oBSgFKAUoBSgFKAUoBSgFKAVh3P8A3QfprSxvq8zKOkqPFWeluwzjWdhaPhnFOJLrBiR5kpLDallNsNvk6LW4kJNFJWXOb/lrC7fxdGOY/k6Xn156GBrVIqUbWZc5JZh2/i6Mcx/J0vPp2/i6Mcx/J0vPqzi6v3bSMtDt/F0Y5j+TpefTt/F0Y5j+TpefTi6v3bRlodv4ujHMfydLz6dv4ujHMfydLz6cXV+7aMtDt/F0Y5j+TpefTt/F0Y5j+TpefTi6v3bRlodv4ujHMfydLz6dv4ujHMfydLz6cXV+7aMtDt/F0Y5j+TpefTt/F0Y5j+TpefTi6v3bRlodv4ujHMfydLz6+HuEJxDJuOZZ5jA2AqREuHS0RETnX8OnF1fu2jLRYWDMVwMe4OsWJrUThWu9QGLjEJ4Nhqy82LgKQ+0u0k1T2q3NcxqzsZ6SP5gY3tuWuC7vii8cetstccpD6Rm+McUU9oR9tfzVB+38XRjmP5Ol59bVHC1MQm4dRDkkO38XRjmP5Ol59O38XRjmP5Ol59bPF1fu2mOWh2/i6Mcx/J0vPp2/i6Mcx/J0vPpxdX7toy0O38XRjmP5Ol59O38XRjmP5Ol59OLq/dtGWh2/i6Mcx/J0vPp2/i6Mcx/J0vPpxdX7toy0O38XRjmP5Ol59O38XRjmP5Ol59OLq/dtGWh2/i6Mcx/J0vPp2/i6Mcx/J0vPpxdX7toy0O38XRjmP5Ol59O38XRjmP5Ol59OLq/dtGWjZYLzut2M8ZrhY8O4lw5eFt7lybav9sWIjrAONtmQKpLqqE6Cafnqxq0a1KVGWRPSZp3WYVV9+z8t1nxhesNw8KYuxHOs6shNdsdoKSy0TrQugKmhJz7CFdPz1NGjKvLJhpIbSMft/F0Y5j+TpefTt/F0Y5j+TpefW7xdX7tpjlodv4ujHMfydLz6dv4ujHMfydLz6cXV+7aMtDt/F0Y5j+TpefTt/F0Y5j+TpefTi6v3bRlodv4ujHMfydLz6dv4ujHMfydLz6cXV+7aMtDt/F0Y5j+TpefTt/F0Y5j+TpefTi6v3bRlodv4ujHMfydLz6dv4ujHMfydLz6cXV+7aMtDt/F0Y5j+TpefTt/F0Y5j+TpefTi6v3bRloxbrwk41jtcy4z8ucxIkGGyciRIdw8SC02AqREq7+8iIqr+irWs90Yvtog3KKpLGmMBIaU00XYYoSap7S6KlatfDVMOk59ZkpKWg8MTT5Fqw5dJsRo35UeM46003HKQRmIqqIjYqhGqqn4KKir3tU79Vtbcw8aq3c0uFobA1jsHANm0zEUCIBJ3jQLTeQpxpI20SqvFqCkhEG6qEYyWdksxLfmLmJIukBmRh4GmHXIKPL1olfe+MFFdDfv059SLjFTazs2GhESKkih5nXy63K9RIGBLjKG1TSguvLPiAJmgA4hCiua6KLgLz6d/81WSpw6pfz4EXM3s0xX0eXDxlD9LTs0xX0eXDxlD9LVeRDteZI7NMV9Hlw8ZQ/S07NMV9Hlw8ZQ/S0yIdrzA7NMV9Hlw8ZQ/S07NMV9Hlw8ZQ/S0yIdrzA7NMV9Hlw8ZQ/S07NMV9Hlw8ZQ/S0yIdrzA7NMV9Hlw8ZQ/S07NMV9Hlw8ZQ/S0yIdrzA7NMV9Hlw8ZQ/S07NMV9Hlw8ZQ/S0yIdrzA7NMV9Hlw8ZQ/S07NMV9Hlw8ZQ/S0yIdrzA7NMV9Hlw8ZQ/S07NMV9Hlw8ZQ/S0yIdrzA7NMV9Hlw8ZQ/S07NMV9Hlw8ZQ/S0yIdrzA7NMV9Hlw8ZQ/S1IcI4kYxlhOyX+K06xGusFic00+icYAOtiYoWiqmqISIuiqmvt1EopK6dwbesiB/Kg/Qv+VW4bpoeKIloNrX5u/dIvZysfxcY+syq9qax3RkP7BuXXxct31Zup1QClAKUApQClAKUApQClAKUArDuf+6D9NaWN9XmZR0lQZbfjPZs/F/Dn7y6VMszcbzsDXDBLzSRus9xvoWy6OSBXcy07HfRowJFRBXqhI4qqoqaGveXRUywvQQ8BLSUngPhW4gxDZ8LybpaWBlyJ9wk3KFbYrrz62sYbUqEjIbtePIbjaxJV1HcbnMKKijOrlwq8PWqOovYaxQV1YC4uTrQzDZdkQAgpGKUTqi8reghLYNNhnuQtB3FoK7ZiaiTwq4duvOKnrpClW3D2HZ0xtXxhq+dxjM2tmcrjSi5qJaOKuiiu4VbTmIl2ypvhF2l5lWAwziUsQpdFtPY51Iyk7jEjDKVzRXeLRviDFzcriL3SBpxi7KAnWA8aW3MfBlmxPZ+P613aMEqN1S0rTmwk1TcC84r+Ze9W+oBSgFKAUoBWBiD+Ybl/dnf9C0BUfBe/Fmyj+KFo+pNVZ1eFqc9+JtLQVHwtPxb8f8AwaX+oaujEd+h4Vw7dL1cDIIFtiuzJBgKkQttgpmqInfXQV5q9BuX0cvEqqGgw5mdbL1bn5VxZdwyTIuPcVd3mB42OANGUhs23DbcZRH2kUxJUEl2roqaVgYezxwfiWyQrpGvUBuK82pyePnxwKBo0rii+PGaiSCnOia6d9dERVrtFRk4gzqwJhq0LcpuLLOkbqUZw8VNaMnI5GgI8IoWpBuXTcnN3+fmrMYzRwqVqKfKv9qgMtyCiOLIuDGjbyNq5xZEJqKFxacZt112rqqJQGRc8x8J2W02+6XDE9ng2y4JrDmSZ7TbMlNu7Vs1LQ+bn5lXm560Tue+BRubUNrE1qkIcNLiUoJ7CMBF1fEnt5GiEIlGdQtuqjt500RVQDas5r4JkBbSaxhYXRubitQVC5Mr1UaOI2otd13aoaoGia90unfrJDMXCjmGHcSBiazlh5ktjl2Sc0sUC3IOiu7tqLuVE0176ont0BrbNm5hi9v4pVq5xWbfhtxhuZc3pTKRvvsduQJiaGugcW6HdEgoqqumui154dzowZiSIjzOIrZGIlmE2zJmsibjMZ91lyQKIa6sqrDhIfe2pqumi6AerecOCzdDfiiytR3kFYsk7pH4uVqiqvF6OarptXXVE/Nroukjsd/tmJ7Y1cbPcYl1gOqSBKhPC80SiqiSIQqqLoqKi+BUoDPpQFJ4l/G3w38R7n9fgVZ9eU3R6d+CNiGgVWeTHs154fCdq+zGKz3M6Z+H6ET0Fn4uxUxg+0JNdjSJ7rr7UWNChoKvSXnDQAANxCOuq6qpEiCiESqiIqpD7fnT15xANjgYRvki7MoS3CKTkNsrcomI6OqT6IWokJpxamiiSL7eleoKDc4KzIDHEgOIw/eYFukMFLgXWYy31NOZQhFDBQcIg3IQkIuoBEK6oi7S24GJc7cPYUxlc8NTRmFPt9iK/GTDSGDjQuI2rIaLqTyqoKgac6GPPz0B5uZvPtKxDLA2Jkv7zrghZVCIjxNA2Bm+jqyOIJtONbBVR1V3kg6a1JpeMoEDDtuvUtDt8OYjRaXFRiGwjg7vvoukKiQp+EH4SKippqlAYTGbOB5ceI+zjGwPMy3XGY7gXNlRecBEUxBULulFCFVRO9uTwpXhd83cJ2O7BDnXuDFY6kdmO3B6U0EaOIFFFBcNSTaR9WMKKe2hIvtjqBurTjGwYguEiBa73brjNjtNvvRokoHXG23BQ2zIRVVQSEhIVXmVFRU79bigFKAUoCC58+wbmJ8XLj9WcrHyy9jbCfwTE/chXC3V5sPiW0yS0rzxcKg+XH89ZgfGEvqUSs1oZBOKVgSKUBHMwMYhgLDB3g4pTRCVEjcSJ7FVX5LbCFroveV3dp7emlVlibhXYbwbmNiPDF5iSGI1kAEduDCG7xjxtsOAyLaAiqex4zXRVTQE0VSJRG+nRdVZv5o/Uhuxn4s4R9rwRj16yXi3OxLQDCG1eFI1GQ6rCvo02iN8XuURLuCdFzmUuL2d3Ufh8NPAkopkkwuEeysNA83OOM4qvtnGhSBdEEFdrfFzm1VTIVRR7y61YsNKSTRFzNsHC/wVcwZYuAzLbdpCvLHtoNFIcdEbh1C0KKKaIbjhNKgrzIjnOWiKtf2Hwu8GuT7w1OZn22NbzMBN6M4rrwgDzpvI2Iro2LMd5xVVULRsk267UI8LPOLokUXhC4ZvFsusi0jKlv2+4w7WrD7JMI47JmrCZISVF1bV8XBUk10Rsl0Xm1g1+4ZVqw5FhPTsJ3iOUoZ4iyZARi/Gdmtq1qCkJLrb3N2wiVFdZQUc38yOGlLM3/LC5vMs+FfhbNC9SLVb477E5m2QZvUrrrfHlIkC4XUuzcm0w2ChKaiiEW1dqotRLEnDks2GrVZ5j2EL06d2s8S6xYzWhuGT6h96RAQtyih86ju5xVNNO6rJYWTlk3GVmLDwrnr2R4pj212yBHt024ja4VzYnC+L0goDk9BUUBNNI7aKSoRChGgopaFpbFa1SGQ7XJ0ilVkioPkZ7CWXvxet/wBWbrNc1/zWQTisiB/Kg/Qv+VXYbpoeKIloNrX5u/dIvZysfxcY+syq9qax3RkP7BuXXxct31Zup1QClAKUApQClAKUApQClAKUArDuf+6D9NaWN9XmZR0lNZXzY8vhR5xNMvtuuRrHhtp4AJFVo1K5HtJPaXaYFp4CRfbqbZ45WjnTldecIddXbE9O4lxi6MNI45EeaeB5t0RVU1VCbH20rPCr/gQ8BLSQHGnBUh4ixTiG82y+LaRuVgiWRi3HEVyOzxT7TjpGguAptvtRorDjaKOrbSpu7rm1+DeCYeF3r6pX20x49ytt5gNQLHYEgRYnXFq3Catto8SILa27UR76o7zlqKqW0Yn9v3BOkX9jEUNzFbTMG5i/xKBbFV1k3rOzbXFJeO0JESODiIiD3yFVXmKvfNnAlywXiZzHdhkz3r1MvqSgKJYHLs3EbK2BDMHozToOuASsASGCpsNQ3JsQ1oCweD9hm64NyPwNZb4BNXqHaI7cxo9u4HtiKYrtVR1RVVF2qqapzc1WBQClAKUApQCsDEH8w3L+7O/6FoDn3g0Zk4Yg8HDKmM/eorT7OE7S242SrqJJDaRU73hSrJ7aeEvd6J85f4V4mpSm5vN1mynmKr4U2YuGrpwescxIt5jPyXreQNtiq6ku4eZOaujcQW1282G42+PNdtr8qM4w3MZESOORCoo4KEiiqiq6oioqc3PXe3Ni405X1lVTOUgXBdktQpwW/EFosb1yZmxZke04f6ngIzJGKjnER0kLxTilDAiNSPcpn3KKqKOju3BXuk+Rhy3yL4spmEyw0F0gxigpFbhuG9FExCRxrjhuvFuJsm+4BdFbJUIuwVkjhcGeXbLBPtMXFTCMT3nLi+cm1lJc64EhpxouOvk4jWhc4blcVd2r2hmi+V34J8O7PC92RyGHWmGlaRlpxkRltzhlDJVW3QPdtAWV2mJbNdDFVTQCRT8hVmYJtFjav7kWVAjXBvq8WTMjeliaOOJvdJxO6cJedxTVF/3m7uqhz3BJkS7HMtkjGCOty2Jjbrh29xw9z/Vq67nJJkSCVwe/DIiJBDUtUIiA+sdZHYgm5gTys4g/acS3G2zbrMkRmXG47cOVx4tIhvISL+Hzo06hq4iEjSCpFMLBkjcMO4HYtEfErbl2iXVu6wpciG89FjkDYto1xLkgnCa2CXcq/wBypdwoCIAIH9sGQwYdwLdbBHvXGPyrnAvDMt6IigzJiBD2bm0NNwK5CElFCDQTUUVNEKoZE4InFyLg7LxSM7rk/OmzGHITyRjlSRkBuGOkriUbQZGiioEa7V0cFC0QDNjcFIHYVxYueLJc9ydbZtvN9GnONbWRv7sHHHjcXYh6JvMiVE7o1VVVbQyywGeX1gkQn7gt0mSpbkuRKXjl3mSCKaK8884ugACd04S83tJoiAS6lAUFmBiG24b4V+F37nMbhsngm5tibq6IpdXQV0+RFqa9tPCXu9E+cv8ACvL7oQlKu2l1IvhoHbTwl7vRPnL/AAqJZBXeFfc3c7ZlvkBKjHc7Ygut95VS2sIv/wBaz3OhKNZtrq/QiegtXG+FXMWWmO1FmpbblClNToUwmeOFp5stU3N7h3AQqQEKEKqJloQroSQyw5C2tjG54vxH1uxNiB1VfKQ/awEWZKE3sdjoRGrW1tlkERCUvvaKpKtelKTPwll/iPBLNshRcRsTrPamGrdAtzkImkbho4G5XjRwuNeBpsQA0EETutwrvVUjOLuDLBxdiO4YhcxBPi3yZcXJZPtG6rLbKw0jNsjHV1WkUFBh3jNuqm0mqbVUaAkD2W2Knbkxf+zKKuKWVfZbfO0EsFuK6DKGwEbqjcmpx2nd6uqu5F/qqgp9YoytuGNsvBwtdL4u6O6As3I2VdfebANqG8m5EVwl1UlTROfvJ3qAjGIODON8vfVreIygtOy233kjxSB5WhjQWVZExdQVA+oUUhcBwe7TaImAuVrR4MV5cd6sfx0PXPcmjkO2uxW9iNW5tB+9ykdTmtyEqi6KqTvf2ooEBYWTOVA5Q4adtPXNLsR9SD1QsbiV2x4EWGKKm4tVVIqGq6981TTmqf0ApQClAQXPn2DcxPi5cfqzlY+WXsbYT+CYn7kK4W6vNh8S2mSWleeLhUHy4/nrMD4wl9SiVmtDIJxSsCRSgMK8WW34itj9uusGNc7e+iC7EmMi604iKiohASKi86IvOntVp0yzwekN2ImFLGkV0QFxjrczsNAQUBFHboqCjYaeDYOneSslKSVkyLGTdcC4bvs12ZccPWq4THmOpXJEqE244bPf4tSIVVQ/5V5qxCywwacdWFwlYlZJsGlbW2s7VAEBAHTb3hRptETvJsHT8FNJy5LMmLH1Iy0whLfceewrZHnnWSjuOOW5kiNsjUyBVUdVFSVSVO9qqr369XMvcLOm8R4as5k+bbjqlAaVXDb04si7nnUdqbVXvaJpppTLnrYseknA+H5UKTFWzw2mZJNm71O0jJEQOq8Bbg0XcLiqaLrqhKqpzqq0kYGw3LLc9h+1PErb7OrkJsl2PEpPDzj+C4SqpJ3iVVVddaZctYsZ0CxW21K2sG3xIatsBFBY7At7WQ1UG00TmAdxaD3k1XTv1rJOXmFZjXFP4Zsz7axghbHIDRJxAEhg1oo/gCSISD3kVEVE1qFKSzpixl9iloSdCljb2G34brj7CtjsEHTDYbm1OZTUFUdypqiESIuirrtqhtvSBSoJFQfIz2Esvfi9b/qzdZrmv+ayCcVkQP5UH6F/yq7DdNDxREtBta/N37pF7OVj+LjH1mVXtTWO6Mh/YNy6+Llu+rN1OqAUoBSgFKAUoBSgFKAUoBSgFQ3N7H1uyty9vOK7ohnCtUcn1ZaTVx8+8DQJ7ZmaiAp7ZEiVqYuOVRlFdf6mUdJQHA8wPdcE5u5qFiN1H8XXu0WC931wSUhSa+7cyNoP+RoUBkf+Voau7N++XiM7g3DtjuZ2OZie8rbTurLLbrsRluJJluE0LgkHGEMXi0UxJB4zdtXTSrKDUqUWiJaTRO4mxdlebOGjdLMq+XSc8Vk6qfZgvhBbYbN1ZjgNIGoGqgJA3qSOsoSIu861Fp4U7OJDt86z4Tny8NP3G1WeTcnpTTTsObOCO4LZsc6qLYSmUMkLXeaCiKiESXkGptPCfnYutGHZC4auGGTvYWW6WsRlx5BTIkyQjSA7qKo1qumumq7HBVCEtRHGyi4TFyvbmD1xhEchPX+x2mTIVo2eooDshu6Oi9uQd+jowNqoRKgqrSJzqaqBscE8MfD+M8aWCwdQx7ad1OPFVqXd46T2JL0NJYCsPXjFb2GAK4necLbt0FSTeYnzrueHMyp+F7TZZeKbrKmsQocFySxEjxyWA7KIld2KW1RZVF13LuVNERKA8rZwmmr/AGy33y1YVnTMMja4F0vNwWQAuWxuWzxwJxKIqu8WG03VQh2CaKO/ukTQ4a4Y0DGEJW7NYot3vbsu3MRrfbb9FlAjc1H+JJ91vUWTFY7nGt6EoaIoq5qmoGvc4S2Ir3iBppmyu2a07LO24rUhl2Q1PdxCtsfjqpCQk0RNmCmiIqAikOhKO3bXnhj2G22vD7yW5mLKvMGDIAb1dGbfGiSJAyjWNIfPUQJsYUjdohLuQBRF3otAZuF+FjaMaYrwnZrPa231vdvizyR66x25Ai8brZLHa1XqsWTZNHTaLQUUVHfrV80ArAxB/MNy/uzv+haAqPgvfizZR/FC0fUmqs6vC1Oe/E2loKj4Wn4t+P8A4NL/AFDVqZmx7rLy3xWxYuN69uWmWEHiC2OcerJI3tXVNC3aaLqmi16Dcvo5eJVUKPu97XCNuk9rOw4gwvZzaMleZsD5jKuLbOrEZIjrakAuKu11/YG4gQOMQ1VUwYGYmZGJsw4eHAvlxtTsh10rm0xY21C2II3FABh5xlRdbVY0ZN679VJV1RHBEe0VEbsN9zSul1j3dqHPg3eZF6pj2x2zokJqSLN7cI0QmtW3CPqNsj3IRC8iLrqmk/y+zBzRv+ZlqC7QThYauLzshmNJhPNmlvcZdfZI9sVRbfbJWWSQ5AaKJIrakYFQGouOa2Nlx87aHbzeIVuJ+Z1zO34cV87Sy3JfbYOMvEH1Qh8XHEl0d28cZLsRRQNBKzEzVxLJjRrrZrnGMht01yEVod4uHNYmW0iEdjSbgJHJJqnVDqkLfea0VFAvvIvEGIMSYG4/EqSjuTUpxnqiVGVhXwRBVDFFZYJR1JRRSZbXuVTRdN5WHQClAKUApQFJ4l/G3w38R7n9fgVZ9eU3R6d+CNiGgVWeTHs154fCdq+zGKz3M6Z+H6ET0Egz7hwZuDraF1t/XS0jerec2KsMpYkyj4qSk0IkpCnfXmWqLvOPMcZd4Mv7uGY92tFqgxp83D9slWRxWTh8ZMMnXnHG1VjiQBo2mSIO4VsFBeMEQ9QUG6umamZMG/wWbcl9uVobuDDbM2Xh5yOU+M65HbeN1tIqqJMkchNFWMiCDZKT2qpX8gYgzVWNYzu8ubfWpN6tMSRDk2JkAbbNmBKekJtbRR2OuPtiqroCBzqphvoDBuGYeddps1kBlJNwfmWKJd350u1m3xM91pV6gVuPDeXi94kpIoiafg8cCkOvWIKqgO5EQtOdE9paA/tKAUoBSgFKAgufPsG5ifFy4/VnKxct3AZyywsZkIAFoikREuiCiMjqqrXC3V5sPiW0zNvOJI0PCdxvUOTFksRorsgXkNTaXYKrqqtoSqnNz7UVfAmtV7b84rpIancfbojZBHA2HWzIwRVBo+NNRVUUNrjpbQItEiPJuLvpxIU8pZy1mJb867/NucCM5h4GAkOQRNCF3c2rwopN/g6b1RVMPaQQLftXSt/lJc3rjfcxxetcy3I1iMkEpStaO/7JGTUdhlzaCi8+nMSe3qiZygop2ZGksilaxkKUApQClAKUBT2YfCTtGXWc2FcupdluMybfxjkFwjk1xEfjnnGQQ0IkJe6bXXai6IqVcNZNJdf8/jM505QUW/zK62teaYpWJgKUApQCoPkZ7CWXvxet/wBWbrNc1/zWQTisiB/Kg/Qv+VXYbpoeKIloNrX5u/dIvZysfxcY+syq9qax3RkP7BuXXxct31Zup1QClAKUApQClAKUApQClAKUAqgcxj7cGfViwY39+w1gbicR37/geuJ69boq+HZocok9pRjr/WrVxUsmhJ9xlFXYwhimy4b4T+aaXa7wLWr2H8O8Uk2SDO/Ry567dyprpqmv6UqX5hXLAuYNqhMLj63Wa426WFwt11t9yi9UQ5AiQbwRzeCooOONkJCqKLhJ7etRhegh4CWkhi4IwMyyE+Jm2kTGa3B24vYrGfbzlvuOMBHMCbJtWeK4ptoRBG0QeKBeckVSri4YMs9mx5h+w4UxA3a8FRLrZrpKkriO2uwZiwAaTjHxP/auqCSO03tDVs9rbhGJIQrtmJZdvy9ypt0TB8dvHkcgwvbrXbIand4mrjcB0HWSc5uciIEQtNqKirog9+tJ2l8pZOArhhSfmMM2HLtdus/VKXaG0+zGgyZD8ZAIBREJEkk2Rad0Ip3lUlUCWxLLgG25gHiOBmWxb4LstLi/h+LdIbcN6UkZIyGSonG7Nggqtb9imCEo6665btty1ezIbxquN4HXQJQy0ZS6xuJ3jEciomn4Wmx0l/C/CRPa5lAiUDKrK+1wbXaouZitYdYt0G3XCzjeYgtXYIjStMFIMRQ0VR2oaNkAuIAISbUVF2WHMH4Dsw27q/NlzEB22ZDkw+rrtCEWRig6DTexoAFdUeJTc04w1QdS0EUQDETLXK9u6MymsxmW2BktTHYvXSGouutXpbw0qrt1RBfJwNEVNWz0XukQq+ly2ytgtXF6zZhRrRdpOI38Ts3ILlCeciSXmjacbbBwSBWFB15OLIV0VxVRUVEVAPuVgXAM6dhopebj9xt1lcgyVt9xvMOSMmVEdR1qQrhirjRkaIriMkAmgomiJqi252z8G++2xeMmfOoB2z8G++2xeMmfOrBv2Z2DjsdxEcWWMiWM4iIlyZ1Vdq/81AQjgvfizZR/FC0fUmqs6vC1Oe/E2loKi4W5CHBszAIiQRG2EqqvMiJuHnq1O2fg3322Lxkz51eg3L6OXiVVB2z8G++2xeMmfOrUWTEGWOGptxl2m7YVt0q4ucdMeiy47ZyD1ItTVC1XujMuf2zJe+SqvaKjb9s/BvvtsXjJnzqds/BvvtsXjJnzqA09mxBljh243Gfa7thW3zbk5xsyRGlx2zkHuItTVC1XujMv+oyXvkqruO2fg3322Lxkz51AO2fg3322Lxkz51O2fg3322Lxkz51AO2fg3322Lxkz51O2fg3322Lxkz51AO2fg3322Lxkz51O2fg3322Lxkz51AO2fg3322Lxkz51O2fg3322Lxkz51AVPMxLaMScLTD52m6wroLWCLkjhQpAPICrPg6Iu1V010q4a8puj078EbENAqncscXWLDueOdjV1vVutjrlxtZA3NlNskSdbGE1RCVNUrPczpn4foRPQWz2z8G++2xeMmfOrGuePcB3m2yrfOxLh6XCltGw/HeuDBA62QqJCSKXOioqoqfnr1BQZPbPwb77bF4yZ86nbPwb77bF4yZ86gHbPwb77bF4yZ86nbPwb77bF4yZ86gHbPwb77bF4yZ86nbPwb77bF4yZ86gHbPwb77bF4yZ86nbPwb77bF4yZ86gHbPwb77bF4yZ86nbPwb77bF4yZ86gHbPwb77bF4yZ86nbPwb77bF4yZ86gIRnlmThGVknmCyziqyOvOYeuAA2FxZUiJYziIiIhc6r4KrHOkGR4LuEpElnD7sSO3aifPEwNuwWQIBbVw2HHWgf0U00bIlXn1ASMQFePuhzqfiWQ6zY8EKzWvsNut7gRcNb5FxeiBc8LRY8OPcWWtqI64xHMmxND40R3aOcWgb0AlIUvxRQlTVEVU5019pa8/WbdR3+ZctB/ag+XH89ZgfGEvqUSsFoYJxSsCRSgFKAUoBXy4YtgpESCKd9SXREoDh/hOOtjw5sp21MUcNuzqIKvOulxk66J/iny10vwhsaTcH5H5h3fD9yCHf7PZ5Ehl5tAcOM6jakBKBISa+2iEiovgWrVG7z615RN6taUKC/6f85Gg4H+N8QZh5FWy8Ynurl7vCz7jGOc8000bgNTHmw1FoADVAAU5hTvVUOPsYYhifdD8E2VjEV5YsbsdgHbO1cnxgu7oVyMlOOh8WSqTTa6qKrqA8/NURlpaX8zGaoQWIqUtKip2+CdvI7IpVZzhSgFQfIz2Esvfi9b/qzdZrmv+ayCcVkQP5UH6F/yq7DdNDxREtBta/N37pF7OVj+LjH1mVXtTWO6Mh/YNy6+Llu+rN1OqAUoBSgFKAUoBSgFKAUoBSgIxmfmDbcqsv79iy67zhWqKT6stJq4+febZbT2zcNRAU9sjRPbqG5H5fXHAeXTb+IlBzGmIZbl9xC8C6os59EUmxX/AIGQRtgP+Rka0sb6vMyjpNviHLLB+L56Tr7hSx3qagI0km425l9xATVUHcYquiarzfnrV9orLbo9wr4kjeZXklVmlZSe02LJjtFZbdHuFfEkbzKdorLbo9wr4kjeZU79U7T2iyHaKy26PcK+JI3mU7RWW3R7hXxJG8ym/VO09osh2istuj3CviSN5lO0Vlt0e4V8SRvMpv1TtPaLIdorLbo9wr4kjeZTtFZbdHuFfEkbzKb9U7T2iyHaKy26PcK+JI3mU7RWW3R7hXxJG8ym/VO09osh2istuj3CviSN5lO0Vlt0e4V8SRvMpv1TtPaLIdorLbo9wr4kjeZTtFZbdHuFfEkbzKb9U7T2iyJlBgxrZBjw4cdqJEjtiyzHYBAbaAURBERTmQURERETmTSveqnnJMW6WqFe7e/AuMRifBkArb0WU0Ljbgr3xISRUVPzLUQ7RWW3R7hXxJG8ys4zlHNF2IaTHaKy26PcK+JI3mU7RWW3R7hXxJG8yst+qdp7RZDtFZbdHuFfEkbzKdorLbo9wr4kjeZTfqnae0WQ7RWW3R7hXxJG8ynaKy26PcK+JI3mU36p2ntFkO0Vlt0e4V8SRvMp2istuj3CviSN5lN+qdp7RZDtFZbdHuFfEkbzKdorLbo9wr4kjeZTfqnae0WQ7RWW3R7hXxJG8ynaKy26PcK+JI3mU36p2ntFkO0Vlt0e4V8SRvMp2istuj3CviSN5lN+qdp7RZG3w5lzhPBstyVYML2WxynW+KN+229qOZhqi7VIBRVTVEXT8yVIqwlJyd27iwqLX3KvBWKbk5cLzhCw3e4OIgnLn2xh50kRNERTIVVdERETnpGTjni7Bq5r+0Vlt0e4V8SRvMp2istuj3CviSN5lZ79U7T2iyHaKy26PcK+JI3mU7RWW3R7hXxJG8ym/VO09osh2istuj3CviSN5lO0Vlt0e4V8SRvMpv1TtPaLIdorLbo9wr4kjeZTtFZbdHuFfEkbzKb9U7T2iyHaKy26PcK+JI3mU7RWW3R7hXxJG8ym/VO09osh2istuj3CviSN5lO0Vlt0e4V8SRvMpv1TtPaLIdorLbo9wr4kjeZTtFZbdHuFfEkbzKb9U7T2iyHaKy26PcK+JI3mVDeFkPWvJN9+K2EILfMiOBcWjcbK0iLiCkpsGnWTNQRdEbFwdyEqc6aiucJynOOU75yGlY/nBMZmsZcXELmDzl1W6uFKuEzjgl3AlZZUZD7LzzrrBkCgKNGa6ADaogiQol2VhW57sStAqD5cfz1mB8YS+pRKxWhgnFKwJFKAUoChs5uG7k9kphq4XG44ytl6uMU3GBsVkltSpzrwEoE3xYl3GhIqKRqKJovPrzVV194XGdWZa4Yk5K5GXQ7BcpIA5fcdNpDZNowUgcBpt3eLWibuOXVNEREFVMa6FPC2ip1nkx+fwMHLqRmW7JLhV4uxzdrjirPGz4NskyGyI27BtqSSDTgqWoNdVBub5udXdykSkiaIgppQ/Ce4H98yt4PcRi8Z2Y/xXKmX0Y70aVcyG2k24TzqF1MqkqnqKKpKapuVV0Tm0z4VSpL/AIVNZs933I2MPQ4RWhSk+c0trsV/iTg34eumc/B/S43nE11S/WKwzZxz7y+86rkt00dRt1S3tDzaogKmiqtauw8H20Sx4Tr7eIsUw+sUGccUI15eFHRYmTI4NyFVVV8OLZAdD175c/PVzxs08myte2juubUcHB0lUu75OV8cvJ8vmQrMnIi92/giYBkR8d3O4W5b5eG27FcBDqKMsdyRvdZ0FTEjEFVUVVTUl72vNb97mZ54J4beXj8uHZczb7HtcZyLFt5jaQlisKcjYGbikgOI2r666bV2inMq0eIoYiNqiyevuzsynhq2Hk3Td+eu+0dPyZaWc3D7k3XKDHVixfgzGGTGJxnMQrbKMJPFyECQysgQmNAKC4gI7rtXRRUdpLu0qeZX8L2Jlrwf8m3rkxc8ayMThNFbis7jHmm2Zjbe5wnVUzLSS1oirroPOta1XDumtKcW9Pw2mvShGtTUI8+72JHaVK55rCoPkZ7CWXvxet/1Zus1zX/NZBOKyIH8qD9C/wCVXYbpoeKIloNrX5u/dIvZysfxcY+syq9qax3RkP7BuXXxct31ZusrNHM635T4fi3W4QLldeq5zFtjQrSwL0h594trYiJEKc6/noCHcoiV0SZk+KGPWKcoiV0SZk+KGPWKAcoiV0SZk+KGPWKcoiV0SZk+KGPWKAcoiV0SZk+KGPWKcoiV0SZk+KGPWKAcoiV0SZk+KGPWKcoiV0SZk+KGPWKAcoiV0SZk+KGPWKcoiV0SZk+KGPWKAcoiV0SZk+KGPWKcoiV0SZk+KGPWKAcoiV0SZk+KGPWKcoiV0SZk+KGPWKAcoiV0SZk+KGPWKkOWWckDM673+0t2G/YcutjGMcuFfogMObX0NWyFBM0VF4svbTvUBC8bJ24+EBY8Fh99wzgXiMSX3T8F+4lr1tir/wBGhyiT2lCMv9arouf+6D9NaWN9XmZR0lQYsz3h4axxPwpDwlirFF0gQ482UVigtvNsNvk6LW4idHnVWHObT2qweUBL6KMxvFLHrFcGnufXqwU42syx1Ip2Y5QEvoozG8UsesU5QEvoozG8UsesVZxXiO4jfYDlAS+ijMbxSx6xTlAS+ijMbxSx6xTivEdw32A5QEvoozG8UsesU5QEvoozG8UsesU4rxHcN9gOUBL6KMxvFLHrFOUBL6KMxvFLHrFOK8R3DfYDlAS+ijMbxSx6xTlAS+ijMbxSx6xTivEdw32A5QEvoozG8UsesU5QEvoozG8UsesU4rxHcN9gOUBL6KMxvFLHrFfD3CFkR2TdcyqzFFtsVIiW0scyImqr/KKcV4juG+xLFwTi2Bj/AAXYMT2vjetl7t8e5RUfHa5xTzYuBuTVdF2kmqa9+t1XJksl2LdJHcw8c27LPBF5xTdgkO261RykvhEbQ3SFPaEVVNV5+9qlQXlAS+ijMbxSx6xW5QwdXExcqfUYSmo6RygJfRRmN4pY9YpygJfRRmN4pY9Yra4rxHcY77AcoCX0UZjeKWPWKcoCX0UZjeKWPWKcV4juG+wHKAl9FGY3ilj1inKAl9FGY3ilj1inFeI7hvsBygJfRRmN4pY9YpygJfRRmN4pY9YpxXiO4b7AcoCX0UZjeKWPWKcoCX0UZjeKWPWKcV4juG+wHKAl9FGY3ilj1inKAl9FGY3ilj1inFeI7hvsBygJfRRmN4pY9YpygJfRRmN4pY9YpxXiO4b7A2WCM8IeM8bLhR/C+JsMXdbc5c2wv0JtgXmAcbbNQUXD1VCdDm5u/VlVz61GVCeRPSWJqSuhVV3/AD/iWjGd7wzAwZi7E02ykyE1+y29p1hsnWhdAdxOiuuwkXvVNChPESyIaRKSirsx+UBL6KMxvFLHrFOUBL6KMxvFLHrFb/FeI7ivfYDlAS+ijMbxSx6xTlAS+ijMbxSx6xTivEdw32A5QEvoozG8UsesU5QEvoozG8UsesU4rxHcN9gOUBL6KMxvFLHrFOUBL6KMxvFLHrFOK8R3DfYDlAS+ijMbxSx6xTlAS+ijMbxSx6xTivEdw32A5QEvoozG8UsesU5QEvoozG8UsesU4rxHcN9gOUBL6KMxvFLHrFOUBL6KMxvFLHrFOK8R3DfYGJd+EqlhtM25T8r8w4sCEwciQ+5aWNrbYCpES/f+8iIq/wCFbbNxzBmLMsLXeMUXN60WPqmFc4r4sg66TqqhNN8STbqOqe7bxewl5+50VEVNephauGlFy0vQZKUZrMaPgo2jD9pykKfhs33HLo+M6faXH1PrXMSKw0UBveIkDbQstgAnzoKDz6aVtrdnuzMjzyft0aE6xAhzWW3biK7lfZbcJHVQF4kG+N1Ui51ADNBVBLSqcHUnK+ky6jAt/CMZuFzgQ0tDDfVRwQRSuKbw6oEV2qPF/wC8XdubDX76AkaKO3St/lLeo93v2ZAMNywVjEZAXVUN6OiqkSMPcq4IoSaivOOqaKi95UVYlTyE84vcseonmRmphPKGxsXjGF7j2K2vyEityJCEqE6oG5tRBRV/AbMv0Cta6i5OyMkm3ZEoYfblMNvNEhtuChgSe2ipqi1zBwPc9cYZt4tzOhYsuUeZDspRXIKNxgY4gXJE9shVR/CTbFb5y5+/z89ZK1nmzl9Okp05zvzbfN2PTP7h54SykvVnw1hG2vZr4zuLzYdZMNyRc6nA1FBJxwRNBIlIUENNV11XamirEAyK4RmfWPCu+ZePlyywDMh8U5g3BFzNZYIhczTkji9u4tVU3BVdU0FERF7nfhCGHgqlVXk9C+7NV/izI5IyQy7w/aOBtmXPYtrK3KXcsNsuzzFFk8VIei8cyjv4SNnvJFBF0VCVF1r9LeCjKfncGzLd6S+7JfOyx9zr5qZl3Omqkqqq/wCNa9arOtdzeryOtisPTowagtEmvhZGuxFwnsLvFmTYMNzHJOMcIWa4XFyPKgvBH3xgRCTjFQRNEM20VBL2+/7dco5kZy4mzt4Kr10xWtuK427HjFvbO2Rjjtk11uB9FUCcNd258k117yJzVRKKjFp6bSeyy+5dgMO1Vp1ZZmp01b3ru+xLaeF49mbgv/FbCf792tJhX+RcL34Nu/2rcaulz373+JbD1eP+mv7rI/mb+JlgD4fxP/qmVc18/pAsrPgy3/Zl0qhaPgvM262mXjW8kVhZZMjEeRPCVh3aZKukVy5WhFZmyDdFEO7viSChKu1FERTRNO8lU7mfwdMP2jLHIu+YONzCeIL2lzdnzGHHHQeOPcIzTJcUR7UUUcRV0RN2wdddNa36WJlSlnzq+g0J4WE1aGZ30/8AZc7Tb4UOdPB/xy5Zs8MCje8BQooq7mLg63SHWUUlXY880mqAiIKi4KIiiuioiiQ69O5V5y4Jzvw+9e8DYjh4jtjD5RnXoqkituj3xISRCHm0VNU50VFTVF1qutQioqpSzx8nqZw075mTSoPkZ7CWXvxet/1ZutVc1/zWSTisiB/Kg/Qv+VXYbpoeKIloNrX5u/dIvZysfxcY+syq9qax3RkP7BuXXxct31Zuoxwk/wCb8ufj1Zf39AW9Ic4phw00VQFV0X8yVy7g7hlvXrDOS12vFph2prF7Fzk4gc3mQWlqHCfkbx9tUcVjcO5OcEJU50oCU4j4WVqt10hWuPZbrAlvNSzkuXmA811vJkIzgo62KKqo43KBUJC5lVBXutyDLp3CGwrHgMvxGrrd33I7cjqK3wDcfATKQOhouiAorEk7kJU04kk76iigRO0cLXDt5v8Ad2xZkRrJbyJGJRxHzcuoFCgSWjjigJ3RLPAEbXUi3Nqn4eibwuFFg1qazDej3xiYKik5h21uItrQpHU2spe82nGaJzKuokhpqGpIBb1KAUoBSgFKAVztNzBtuVOa+f2LbshnCtNksT6stJq4+eyYjbLae2bhqICntkSJ7dAT3g+ZfXHAWXwvYiVt3GmIJTl9xC82uqLOf0UmxX2wZBG2A/5GRqwLn/ug/TWljfV5mUdJSmBvxn81vi/h395dKlWZGJ7hhu44HZgui2F1xA1b5SECFuZKPINUTXvLubDnTwfnrawXq0PA16nPZW2cHCDZyozHtzFyuxxLA1PiNXBk4raoTb1tu7rbbJa7yccfhxxQdE50ER13rWW/wgp9sxFcLQGFb3er49K4tqygURlYGy2xpbjZvK6glzPabtV7slFO5RCrcuY5NzLa4TdtuDltlWvDd3ueHpUizQnbw2rIBFkXPqco4G2RoeghKjkaoi6caCJu59t0VOkxasKVJApQClAKUArAv/8AMVx/uzn+laArvgvfizZR/FC0fUmqs6vnVTnvxOqtBUXC2/FtzA+DS/1DVoYlvbWGcOXW8PMvSWbdEdlmzGDe64LYKSiA+2SomiJ4Vr025HRS8fsalfSiG4Qzps+ILC7crhJtsJoGn5Tci3XAbhEkxWRZVx9p4BTcIrIbAkUUVCQkRFREJcTDvCEwrf7PbJyzmIZSGxORFeI+OZUgJREBQPvqkQkCbe+QkKakKjXeua9jKxDn9grDtoOedyenbWkdSLBiuuvr3airezbqjqKJqrS6GiNuLt0AtMuPnRhPrb1TKvEZkxmrb1ZZ4x0lkbOMFsUQEIiINuiIPORIKakqIq5FjMvGbWEbBZrbdZ97ZjwLih9SukBrxmwVJxNqCqioIJKSKibdpa6aLpHpvCIwfEuHEFMdYitQ1uEifMjPMMMsiskXBVSDVHQKG7uaNBJERV9pUpcWubVc8MDj1Ih39pkpDosID7LrZNGTvFCLqEKKzuPUR4zbu0VU1RFVNg1mhhl7CzmIQuRFa23RYUupneNVwlFABGdnGKRbwURQdSQxVEVFRaXFjTWjO7D1zaxRPN9I1gscmHFS6kLhNyiksMOtk3oGiivVLQptUl1VdUFNFX+QM9sJzLhLYO4Aww0YA0+Qmu/URVwjFB1ZFsiEDVzagEqISiq6UuLGaWdOCxZddW9ggNPowSlHeTvoZcYibOdna24vHJ970Al3aCunrNzgwbb1gpIv0dtZpMhHRRP74rvH8XpoPeVIshdV5kRolXRE1oLM3WF8XWjGcByXZ5aS2GnOJc1bNsgLaJIiiaISagYGiqmhCYkmqEiruKENFP3X8bbD3xHuX1+DVt143dT1l+COhS5iFVLlP7N+d3wja/sxirNyfWH4P7EVuYWDjPFC4UtTLzMQrjPlSWoUOELiN8c84WiIpLrtEU3GRaKqCBKiEqIi1+Gesp/GMfCvWuy26+NmrM6Pd791Nsd3NoIRtrBrIUwdbcFFQF0cFFQVVUT1zNFK5KsB48uGNuIlnh12BY7hF6utly6pB1H2dRQFdBERWjMSQxFFNNuupCSKKafFeedswlje64cft8qS5AshXZH2CDR91DEUhghKn34kNskRVRNDTnRNVoTa+Y9nsyMSMSYtnPBiJieQTrjcProPUpxmgaJx8ZHF6qiE821tVtF4wvaD75Uiu+N4thwxb73dGltDEjiVeZnlscjbx1IS2ISKY86KiLpqi8/t0IsalnO/BD7DDw30EbecJsVOO8O1URtdTRQ7gSR5naRaCfHN7VLeOvldM68NWq+NQTlK8wsRyQ5IYbcdIDRYfFtC0AqZkYzmSRBRV/N4FxY32G8f4exhNlRLLdGrk9FaZfd4gSURB5oXWl3abV3NmBJoveKpDTSGhSpIFKAg+evsI5hfF24/VnKqbOt5InBMsks244sRYdreeuDzxNHbW0FtClNKEmMXGBrzILza866KX4BcHdPnUvH9Dao6GbrgdSBn5OhNZ4iVFlz3XY93bfN166t7QFJT6nKlGjiqKgqE8a6NivcaoA3ebDbhApABEC6ipDqoroqap4OZVT/Fa81WzVJG2tB9bU8CfJUIy4/nrMD4wl9SiVgtDBOK48+6f+wXh34fd+yLlULr8H5G7gvWaXvLzReuNM9sA5H4Gt1wxtiq22EUt4PNRZD6dVSRERFeJYTVx1dSFO5Fe+mulfnDwRMB464UeKc2LDHxDJwBlxPWL14WLF23S5QykTiYYAz1RpC3yEcJEX8ER0VFLXfowjCLrVFo0LW/0KIzmoTjFZna/dnLwzgwsnAjiZS4MyjuD2G4E9Zjt3lpHjvSru43It4CUhw21Ve5kOpoG1E3IiaIiInWMPhCYNn5yy8r2ZMwsWRNUdbWG4jCEkcJCijum1V4twF019utacnWeU9Oculh7UoTgr3Tb+DsfnTlH+JPj/4Vwl++h1emFs+MWZXYO4LeGbCVuS04khwWLkMuMTjpAc6HGVGyQ0QF2yTXVUXnRKiKTdn3HXxVPfcqN/zTeyCf2InaE/8A8+cJb4rYr/exqhUH8UC9/wDkqP8AY0aqpc1+E/qRtUefD38P9DJhePZm4L/xWwn+/drSYV/kXC9+Dbv9q3Gr5c9+9/iaMPV4/wCmv7rI/mb+JlgD4fxP/qmVc18/pAsrPgy3/Zl0qhaPgvM262mXjW8kVZgz2F+Ej8J2X7ZkV5Zjewzwb/8As3z7Vg1Y9PxMY85eP+2dbYZz9xPdOGZjbLW4u2sMGWi3OPs6x1B8CCLbniI3VPao6zHebamiCPP36rbE/Bvw/OtF9xzwPMdWvDWN2yIZkTDl4ZmWi4qSqStPNKrrTTiISq2ugiOumiIqEOxRnvee34Hp/nkecrUt7aSfUntSZOsqOGUxa79ZMsc74L+X+bTxNRGwksKtuvJlqIvxXwRW0Fwh02ko6Eu1NV5kufJeS3CyIwJIdVUaZw3AcNUTXREitqv/ANErCtS3vRnT0eGcpTuQbDPDKy/xZdrTAhxcTtFczgDHky8PSmY+k1VSIZOECCIOqi7SVdF0XTvVf0D+VB+hf8qzpUpUq8FLWiG7pm1r83fukXs5WP4uMfWZVewNc7oyH9g3Lr4uW76s3UY4Sf8AN+XPx6sv7+gLfebR5o21XRCFR1T2tUrn2HwK8Hx7a1Aeul1kxUiRYTjRk2iE2zZ5Nq7nue5U25ZuEqd8xFe9zUBmXbgsBiaYVyvmMJ1yvcpXgnzkhstcewbTDYNCApoCCsVs9efUjd7yEiD64j4KFjvYXHibq+z1df3b2bUuGxMYBHG3UJkWXBUFFHJEh4CJCUHXVLnTuaA1dn4HVrtkNpk8UXB4ozEZIjoxmhKPJYiW+O3ITVFRVHrXEcQVTTcha6oWib6LwZbeYYkkXK/zrheMRstBc54stNca43JV4TEETQURNjSJz6A2OqquqqBdNKAUoBSgFKAVyRJwZccyOHXia2yADsLsMGx4iuIKXPLnNDKCA0o/8Amrr6/87DVAdb1h3P8A3QfprSxvq8zKOkpHAjoOcKDNkQMSJuwYcEkRdVFd9zXRfAuiov8AilTPMnADuP4FoCLepFgn2q4hcos2My26ouC243ooOIoqii6XteCtrBL/AONDwNepz2Qe78GW1YzuATcZXiTit0yDqpuVGZabkgMS4RUEgAUQfvdyc500XVoF8OufhPg/RMNX2NeZOIrnero0hcZJmI3vfJYEeEpntFNSUIoGqppqRF7WiJu2MLlYXTIzElixPhnCmGGb01haNNw5NnTjOGUGYlsWOiuu6qj7bqtw2W1BsVElFkkIUFwV6nokG7ilSYilAKUApQCsC/8A8xXH+7Of6VoCluDRmLYYXBwypjuyJKOs4TtLZoMCQSISQ2kXRUBUXvd9OarJ7Z2Hf7TK8XSfR18+qU5Ob8daOonmKp4U+YFjuXB5x1FjyJJPu28gATgvgiruH21BET/Favi+WtL5ZZ9u6rlQeq47kfqqE6rT7O4VHe2afgmmuqL7Soi16TcmLjTknrNWvpRVFw4N0W59XyHcT3Fq53MJLNwlxosZpJLTwRgMOLRvaKr1Gzqad0vdproo7NBd+CrGur9it826ybxa4jLbRyJrcfcw3GInIjQtI1tcRHXTUlNF1EERdddV7djXuSCBwa7bZ7FNtFtv8+226YpyH4kOLEZYOYSKnVGwGkRObb3CaCuwVXUtVXzu/BZwxfHOMmSpT5pDCMIOttONbwmpLF0myBRItwiC6ouoJpza61NhlG9k5GWt3B9rw/HuMu2tQI0xhuRb2mYx7pIkjjiC2AgK6kRaCKJz8+vPUUHgm2QLM/bAvk5uG8zKaJpqNHaEeO6rVdggAiKCs6RoiJ3tiLrtVVWFz+4yyLu1zx3IlWiacW03ufBn3h5Xm9VSJJR8GkbJkl01RdqiY87i7uYUQpPY8jomHMFtWO33qZGkRriNzhTWmGAGG8goG1pgQRoG1FCRQQUTVwyTaqoqLC5gysjuseAJ2H8N3OU3Im3u1XMpshW1daWO7CRxwdQUFNQiK5oo7VMlTRB5k1E/glYYus/q+dNenzXHJDkh2ZCiSBdWQ4DklUBxohBTMEVFFE2oqon5lhc8y4JlnNqYJYjuZvSiYI5ZR4qyRJpJCI8L3Fb0kKkglV/XjNwouu1VGttG4NloSa6c693K5wibSM3CeFoBajozcWuKQgBCXubo93Sqq/ew5+/rFhlEtyvyutuVlpmwreYO9VyOqHHAhR4uqo2DYoosNgKrtBFUlRVVSXnQdojM6kh5yj8X3+Dh7hXYbfnOONtngq5AKtsOOrr1dBXvAKqne79WD2zsO/2mV4uk+jryG6UHLENrUjfpP8CHbOw7/aZXi6T6OoHkldot8zizrlwzM2DuVsRFcaNtdUtrKL3JIi//AEqzcuDjXbep/Yxrcws3GuHI+JrJxL8521ORXm5rFxZUEOK60SEhpvRR00RRJFTRRIkXv1HsIZZWmx3l7FBXM75cJrKlIuElGlR8lISR1NgoIoIA2A6cyC2POq6qvqzSvmPjD2XTuD0ty2/Fc123wwag26FMVrqePDV0CVkUAR4wlAAbAzVSERTRVVTU9LiPg54WxffbjeJEqSN9fuLkt+4RjFHm90PqYWk5lQUBvijHVNdzYEutLXJubhzKaWckLn2Z3pMRgbqJddkdVFhxtsDYFlW1bEFVlpzmFF4wEJVVFUV9b3lQGJ8CDha5XaW7DYcBI0hS4yQjQAgAjhlqrh99VNedVXn176rC5or9wcLNiG8BPeuk5kuqwlGjLbKOaDHiMK2DqgrjYmMMENBJEMTMSRe5UdOPBptckBOTjK5T5D5q2jrrcZQeQW4QcUTaN7HBQbcCqKouqqar3kRFhlE+yvy0t+UOGXbXEnvS4opHXjpaACgDEKPEDXagj/u4oKq6Imqr3k0RJmj7ROI2jgK4o70DVNVHw6eCliHnPulSQKUBB89fYRzC+Ltx+rOVVmbVpnXrg1YLi2qI+9dS60FFmMA64VucQQ0lK2024RoHtpsJE3aloKKqcHdN2lS8X9jao6GbngrRbjbsLYniXix3Sx3Ru+mj63kkKROdSJF46QhiANuN8aptgbQABA2C7UVSq7a81Ws5uxtrQKoqw554JwVm7iXA94vDkXE96v4FBhjAkui4jkWO2G50G1bb3E2aJvIe9WMIuSdjJRcnaKv+yu/kXBivE9swThe74hvUnqOz2mI7OmSeLNzimWgU3D2gikWgiq6CiqunMi1+dX3R7hX5f5hZK4YtuCLymIr4d9dfK2LFfjyGWAgymHXHGnQEwRFkCoqQohaFpqglpdQo79K17LP5Z2XUZTpzjWjG+S1t0pfGxlZUYHhQeGBgOVnBGmY2zXucBl9iW28y3Z7IhRH3mI7UbapOK0EY0RxS03ubtFVN1f3g2Zhz8qLFwiMV2uPGl3C2sWw2WZiFxREVyugd1tVF07rXmVO9VleoqkkloVkblChKNGaf5lF/+VvsY/CKzKuOb+G8gsV3aNFh3Gat7aeZhISMorN2tzGooSqqIqNIuiqvfqfYX/pL8R/3x37Ch1qu3V3+RvKG90nDVCS/8yh8o/xJ8f8AwrhL99DqY3r8Dga/9i2fbFsrKOleMS6rpn41P7Zt7R7PnCW+K2K/3saoVB/FAvf/AJKj/Y0aq5c1+E/qRbR58Pfw/wBDJhePZm4L/wAVsJ/v3a0mFf5Fwvfg27/atxq+XPfvf4mjD1eP+mv7rI/mb+JlgD4fxP8A6plXNfP6QLKz4Mt/2ZdKoWj4LzNutpl41vJFWYM9hfhI/Cdl+2ZFeWY3sM8G/wD7N8+1YNWPT8TGPOXj/tlsNfj8Z1fFuf8AZdlrdfc1H2o2EMauPOA02J2zU3CQUT/ZUTvrWD/L4PzRq1PVan/6vpZ1hiLLXCGOb/h3EV5sNuvF3sLpSLTcJDIuORTJNFJsvkXwIqCqc6IqQ7B2LLBgrgx4PumJrzBsNnDDNvacm3GS3HaFTjNiIqbhCKKqqiIiqmqqlZpynFR06LfM4JQXBIubGZWI2LPe3surrDtFmt7kJjB11aly21t8hEhFNRuS4oG0JptFve0qmeri9yNdvwP5UH6F/wAq3bWxUFn0rTpMdMTa1+bv3SL2crH8XGPrMqvVmud0ZD+wbl18XLd9WbqMcJP+b8ufj1Zf39AXDSgFKAUoBSgFKAUoBSgFciW2+S8M/dKb2Bu/+yMSYXiWhW+9pNaB6Uyv59WWpnN/y0B13WDdjFuOhESCI6qpEuiImnfrSxvq8jKOk464F90m4vzlzpxtMdM28XsWe8wWz/8AdwFduTEPT/qjx2XP0uLV5ZzSZkyXgXDMe4zLTExJfVgzpdvfViRxDcKVKVtt0e6bUyjCCkKoSCRbVFdFTfw6tRil1Kxrz57I2t0u+Xd5DA2EL6xiu7XKe+9GZxLPdkLY4zcVlxxl57cbzqkbgECOLuQZOupCCIuhwfwmsQYyuECexhq3xcMHc7daJAHNMpwyJcJp9UFEBG1Rpx0QVVXu0VSTbtRCvMLXPDBnCXxPjcYlnYs9mgYuucxiKzbpzkply0kUaVIdbmsONg7qAxCETBNjpOJtVBAlrOtObuMp2MLjheDY4UTGkqWBPRrrdHH4EIGrbDef4tQHXRXJLYCgomu8nFTXUVXJsjOw3wl3cUYcC7xrAjQncrHBSKcjVwVngwRoq6aKTZPEKacxbddURa2+Q+dFyzWgXkrtBg2u4wGo7ztrbV9qZBJ0TVY8lh9sDEgVtURxE2Oaqo/grS9yGiuWuEBirEuGcHSbzCiYbdvfY1iCMVomOOqkOVNaB+O8qiOqoKjrpzEjqjp3Ope2G+Fxc8R4VmXdLTboTcmLBm2x+WMxphkJL6Nqy+psjxz4AbZoEffxpKojoiIark5KMix8Kq9XZ/CSybHAtMK5S3YL865JMjx5T7d0fgONMOGyiMOojCPI1I2kXHA2ndIRJtb9wmbrbsKWW5wcNxZs652mbcG4jswmg41m5QoQN79i6IXVikpaLpsTmXWlxknrhnhD3q55rR8IzbTDdbandZpz1vYmmrUoYnHOPi4rPFIxxn3lEI0PnQl0RdtX7U6TFqwrAv8A/MVx/uzn+lakgrvgvfizZR/FC0fUmqs6vnVTnvxOqtBUXC2/FtzA+DS/1DVxV6bcjopeP2NSvpQpXfNUUoBSgFKAUoBSgFKAp+6/jbYe+I9y+vwatuvGbqesvwR0KXMQqpcp/Zvzu+EbX9mMVZuT6w/B/YitzCU5uWl67YahoNvO8QYtyiSp1sabQylRwdQiRBX8Lau13Z3y4raiKpaLRBuYxhY2xJb8MR73Z7dM6ludosDMJoYzDLr5tyikAQqrSuizJeAVUedxEVEPQa9azSWgzHlzfcjRWYLNxO7tjq+xOitJbmSF5jqJWV2oq/euNV7nXaSFrt0brxw8uPbFig58QMXO2mXiQH5QXSG0y9KZSFa23XXDEFEQBG5iIKgKHtNAMSFpSZzLMWfmJe8UvXOyTsPM3x62vQxkRmLSyyivSFcbVQlK8n3sOJ3aKuiIqn3zRtFrXCrmbk4ROXMxE0NvNx+ODscRSWirbFEHd7QkY6lcE00BdNeYdoaGRmsYV7kZzWPDVrahu4luM9yxRLm5INltwgufEOcbFIAa5x3CC7VURRUVFUiMRXzbtuZsGzPxbC3iKOLMy5uKkyEyhRnzauisFF1DnBd0VVVdyam2n4SujTOTmNvi9vNaGzfbbb3r/cHGUcZsxOR2nGZbSvv9ULKLZzkkdWUa103Eg/hLvrUXyJmcF8l3FhcThdGY8iO863BbWOgrKMmGoxAKkIk2MZVNAd2rt3jzv7WcZjqm0uSHrVCOY0TEsmQV5oyElA1FNwqo8y6Lqmqc3grKrIrFKAg+evsI5hfF24/VnKiOIYWHHuDdap2KLVDu1ttdlizkblzAhcUYMCm8JJkCMGgkSIe8dNypuRFWvP7q3vTydN39jboaGabg1Yny2vcK6SMD4avlou6sC5JS/sPnMltCRCGya8bgSW0JCQVbeMEVe+muqze3ZzQLlGccC1zgLqRiQy24rWrputxzRvVDVBVOq2EUiVB1Ul10FVrz9SnJzak85spqxgQc/rVcJ0KM3aLknVSxlBwlaREB3ahEqb9e4IxFR75aqoIYoqpxfjq8RLx90AglEcJxGr/aWz3NkGhI4SKndImvOi89HTcIv4fUjfwbvVfuz+iRZHDO4XNuu+E8zMoMtreeM8YsWK4jiJ9vuIdjgiwqSnXHVVEN0RPaIDroa6FzpsLkUMprRgTgVtXUmGZ2KZWOZkSZezb+/vNN2acoN7lVVQE1VduvfXX9Gw/+BR3t6ZJvwVlm+N7luBpuU4Vb5lOCt3u+f4W+Z1Jev6QzLT+4Q/sq51U2XfsT8J3+72j7WuVaj07PI34dHH3Y/WYmNfYj4Ov94xL9vQ6uvC/9JfiP++O/YUOsP38i2pzZ+7P6yh8o/wASfH/wrhL99DqY3r8Dga/9i2fbFsrOOleMSaumfjU/tm3tHs+cJb4rYr/exqhUH8UC9/8AkqP9jRqrlzX4T+pFtHnw9/D/AEMmF49mbgv/ABWwn+/drSYV/kXC9+Dbv9q3Gr5c9+9/iaMPV4/6a/usj+Zv4mWAPh/E/wDqmVc18/pAsrPgy3/Zl0qhaPgvM262mXjW8kVZgz2F+Ej8J2X7ZkV5Zjewzwb/APs3z7Vg1Y9PxMY85eP+2Ww1+PxnV8W5/wBl2WqOwVGZm8EzHjD7QPsuX3DQm06KEJJ975lReZUrCTaSa1PzROHipKzzq9HyZ+h/BTFB4MmVKIiIiYXtqIie1/swVtMnm0dyCwOKhxmuGYPcaKuq9Sh4Of5OfwVnLTLx/U8u9JyfwVSxXYceNFEWZdrfAZi4eJu52a5N9aoKPCnU7KrHbRVFVRVJ9TMRFVUkTdr3zA/lQfoX/Kt92eJg1rRh+U2tfm790i9nKx/Fxj6zKr1JrndGQ/sG5dfFy3fVm6hvCqvEDD+HMB3K6To1tt0XG1mdflzHRaaZBH+cjMlRBRPCq0BJuUjlH0p4K8oYnpKcpHKPpTwV5QxPSUA5SOUfSngryhiekpykco+lPBXlDE9JQDlI5R9KeCvKGJ6SnKRyj6U8FeUMT0lAOUjlH0p4K8oYnpKcpHKPpTwV5QxPSUA5SOUfSngryhiekpykco+lPBXlDE9JQDlI5R9KeCvKGJ6SnKRyj6U8FeUMT0lAOUjlH0p4K8oYnpKcpHKPpTwV5QxPSUA5SOUfSngryhiekrmfMPFtgxDjvM/MbDV6tt/iYJnYSvvV1rlNyWhZZOU3NTjAVURUiPSdU19vnoDt7vpzc9U5wsLvMiZOT7JaZCx77iqQzhm3OB+EDsw0ZJxP+20Trq+BGlX2q1sQr0n/ADrMo6SucK4swLkzwg8dWW7YjseE4DOFsNRbdHutwZiITTJXENocYSbkFNiLp3tU8NS7G+amSWYNlG23XM7Cgg08EqPJh4ojx5EZ8F1B1p0HUICTnTVF50VUXVFVFuwbvh4X1FFRPLdiLOTuD8VkZgtZqWSNNanOXLr2xjRsbkclxvinHCkcdvLc3o2or3KCIIKDsDbm22+cHS0W4LfCxxguPBCXDmhGDEsfYLkVhqOxoiu/gi0y2OneXTVdVVa3LowzmkixODtDioDWatoSYwsdINyLHIFMt7bAPAy3HdV9SABCTIDTn3I6SFu5tPfZwcY1sZh2zMbDdiJh1p1iXasYtMSWVCKEXQXEe3aEy2Ikiqu5U3L3WhIzDOfLzHBscudukt5gYXjRoAweJtcfFzTcLdDFAjGTKPbTIARA7rVFFE1RdEVN3gXGGReXITitOaeHZEiVGZhI9dMXsyzajM8ZxDAE48qo2HGuaJ313Kqqq0zEO7IdlDDyKwBl7hq0XXNHCt2vNvhWluRJdxiEhsHYOjjYMb3dQZF5TNA0RFQtFTRERNnHj8HSHBKNHzQsjXF9TpAdTGwK5awYIiZbiqry8UAqZJtTmUVQV1FBRGYl3PG3Wzg4Wo2kj5o2dI6SFkPxXMci41MXqo5aI+hPrxqJIddc7rVVV00VVElSveHF4N0OfIldsfDr5OA4y2zIxk2bUZo5bUsmmQV7a2HHsgWiJzabdduiIzC7N+OL8hgx32VjmdhoZ/VCzepExcz1H1UrHELJ4jjtnGcV3Gun59N3PU05RmU/ShgzygiekpdENNjlGZT9KGDPKCJ6SsK+cInKl2yzwDM7BpmUdxEEb/EVVXavMn3ypuiLM+eC9+LNlH8ULR9Saqzq+d1Oe/E6i0FQcLpwGeDTmE44YgAWsyIiXRBRCHVVXwVIOUZlP0oYM8oInpK9LuQ1vUvH7GpXV2hyjMp+lDBnlBE9JTlGZT9KGDPKCJ6Su/dGtZjlGZT9KGDPKCJ6SnKMyn6UMGeUET0lLoWY5RmU/ShgzygiekpyjMp+lDBnlBE9JS6FmOUZlP0oYM8oInpKcozKfpQwZ5QRPSUuhZjlGZT9KGDPKCJ6SnKMyn6UMGeUET0lLoWY5RmU/ShgzygiekpyjMp+lDBnlBE9JS6FmOUZlP0oYM8oInpKcozKfpQwZ5QRPSUuhZkEs+YGFsf8LGzu4YxLZ8RtRsE3EXztM9qUjSrPgqiErZLtVdF018FX1XjN08+JfgjfpcxCufsJ5o4MwFnznPFxNi6xYdkvzrW40zdrkzFNwOtrKbhRwkVU15tUqzcrp34P7EVleBYXKMyn6UMGeUET0leacIPKEZJSEzLwSkgwRsnUv0PeooqqgqvGa6IpEqJ+dfDXr7o0bM9OUZlP0oYM8oInpKcozKfpQwZ5QRPSUuhZjlGZT9KGDPKCJ6SnKMyn6UMGeUET0lLoWY5RmU/ShgzygiekpyjMp+lDBnlBE9JS6FmOUZlP0oYM8oInpKcozKfpQwZ5QRPSUuhZjlGZT9KGDPKCJ6SnKMyn6UMGeUET0lLoWY5RmU/ShgzygiekpyjMp+lDBnlBE9JS6FmQ3OnhAZXXHJzHcSJmThCVKfsM9pphm+xTNwyjmgiIo5qqqqoiInhr3x4IlwRpKktzFAwuye6zmASxUWAXc2RAaAqaa7lEtqIq6LpXA3Ud3Str/Q2qKzM+eCXjm/ZgZZzJ+IplznXFm5uRlcuzkVx4ERpolb1jRYzfcERCSbFUTE0UlVNBtV7CNjkEZOWeAZGCNGqxg7sEAm0EubnFAMxRF5kQlT2685VWRNqJtLOj+jhOxg8Lo2a3o6BtOCaRQQhNodrRIunfAVVBX2kXRNK/MDhWhjG6cOa22HBj/Wq5Xa626Ol94njVthqqILyCqoKqiERIi99RRE51SrsOozk98eZW+TVl8dBfRlOEm6au7SXwcWm/grs6xxbwbsGcHjgj5uxMOQlfvMzCNzW64gmrxk+6PdSvETrzi6qqqREuneTWuSMwfxJYv/kK4fYs2qp1HVk5y0tS/wATq7nq1NL/APJT/wAy671/SGZaf3CH9lXOqmy79ifhO/3e0fa1yqHp2eRdDo4+7H6zExr7EfB1/vGJft6HV14X/pL8R/3x37Ch1h+/kW1ObP3Z/WUPlH+JPj/4Vwl++h1Mb1+BwNf+xbPti2VnHSvGJNXTPxqf2zb2j2fOEt8VsV/vY1QqD+KBe/8AyVH+xo1Vy5r8J/Ui2jz4e/h/oZMLx7M3Bf8AithP9+7Wkwr/ACLhe/Bt3+1bjV8ue/e/xNGHq8f9Nf3WR/M38TLAHw/if/VMq5r5/SBZWfBlv+zLpVC0fBeZt1tMvGt5IqzBnsL8JH4Tsv2zIryzG9hng3/9m+fasGrHp+JjHnLx/wBsthr8fjOr4tz/ALLstUngH8VPHPw9hn/8dVz5q8H5meG6vGj5M6pyu4Slqyjyx4OuDJlkuFxk4nsFmjtzIpto1GV4o8YSNCJCVN7oqu1F5kWr3yaVEyFwLrt0TDUHXc4rafyUO+Sc4/p9rv1dOKSbT6zzNSnKFpP82dbWvNM5P4KGKsOWDM6x2mxxsMBc5Fhas8Nq131x5ZkVqRxjsl1RtEcXXhaJNCM13bObarjhr3vA/lQfoX/KtxprFQb1rz8Wa/5Ta1+bv3SL2crH8XGPrMqvVFB3RkP7BuXXxct31ZuplPt0S6x1jzIzMxhVRVafbQxVU7y6LqlAa3sIw57gWv6E35tOwjDnuBa/oTfm0A7CMOe4Fr+hN+bTsIw57gWv6E35tAOwjDnuBa/oTfm07CMOe4Fr+hN+bQDsIw57gWv6E35tOwjDnuBa/oTfm0A7CMOe4Fr+hN+bTsIw57gWv6E35tAOwjDnuBa/oTfm07CMOe4Fr+hN+bQDsIw57gWv6E35tOwjDnuBa/oTfm0A7CMOe4Fr+hN+bWJiLLyx4hwbiDDS2+NDt17gvQJYRmBDe242TZaoiJrzEvfoCJ8GHFM3GGQeC5l0NTvUWF1ruaqqqvVsQyiydfb141lyo9i8+z3hN4ctA/fLbgS0uXyUic4pcJu+LET9IsNz1VP/AIoLza8+pi3k0JvuMoq7RYlww5abs+j021wpjyDtRyRHAy0T2tVRV0rG7CsPe4Ns+ht/wrx2U11mxYdhWHvcG2fQ2/4U7CsPe4Ns+ht/wplS1iw7CsPe4Ns+ht/wp2FYe9wbZ9Db/hTKlrFh2FYe9wbZ9Db/AIU7CsPe4Ns+ht/wplS1iw7CsPe4Ns+ht/wp2FYe9wbZ9Db/AIUypaxYdhWHvcG2fQ2/4U7CsPe4Ns+ht/wplS1iw7CsPe4Ns+ht/wAKdhWHvcG2fQ2/4UypaxYdhWHvcG2fQ2/4U7CsPe4Ns+ht/wAKZUtYsbdlhuMy2yyANNNigA2A6CKImiIiJ3k09qvusSTykxWZsc2H2m32XE2m26KEJJ4FRe+larsKw97g2z6G3/CpTa0MgdhWHvcG2fQ2/wCFOwrD3uDbPobf8KnKlrFh2FYe9wbZ9Db/AIU7CsPe4Ns+ht/wplS1iw7CsPe4Ns+ht/wp2FYe9wbZ9Db/AIUypaxYdhWHvcG2fQ2/4U7CsPe4Ns+ht/wplS1iw7CsPe4Ns+ht/wAKdhWHvcG2fQ2/4UypaxYdhWHvcG2fQ2/4U7CsPe4Ns+ht/wAKZUtYsOwrD3uDbPobf8KdhWHvcG2fQ2/4UypaxYyrfh+12h0nYNthwnSHapx2BbJU8GqJ3uZK2FQ3fSBWsnYZs9zkE/LtUGW+SIiuvxgMl0TROdU1om1oB4dhWHvcG2fQ2/4U7CsPe4Ns+ht/wqcqWsWHYVh73Btn0Nv+FOwrD3uDbPobf8KZUtYsOwrD3uDbPobf8KdhWHvcG2fQ2/4UypaxYdhWHvcG2fQ2/wCFOwrD3uDbPobf8KZUtYsOwrD3uDbPobf8KdhWHvcG2fQ2/wCFMqWsWHYVh73Btn0Nv+FOwrD3uDbPobf8KZUtYsOwrD3uDbPobf8ACnYVh73Btn0Nv+FMqWsWHYVh73Btn0Nv+FQThJWGDd8pLi0cSHLuMfQ7PDlzjhtuzVEgabRReZ3ku9UFtXBQl0TUfwkzhJuazh6CIcCCU9PyakSHXbdIbO7Po3ItT/HR3drbQOKBdVytUR0XBReMTVBQtg7tK6ErKuv+JILQK/OnMz+kDtnw5Z/9a1hHmv4fUjewfSv3Z/RI1NkfdO68MVCecNHLBipSEjVUVRnTQHm19oUQU8CJpUQzB/Eli/8AkK4fYs2spNybb1T80dqlCMMlRVvx0PnBt7WXXev6QzLT+4Q/sq51U2XfsT8J3+72j7WuVHp2eRTDo4+7H6zExr7EfB1/vGJft6HV14X/AKS/Ef8AfHfsKHWH7+RbU5s/dn9ZQ+Uf4k+P/hXCX76HUxvX4HA1/wCxbPti2VnHSvGJNXTPxqf2zb2j2fOEt8VsV/vY1QqD+KBe/wDyVH+xo1Vy5r8J/Ui2jz4e/h/oZMLx7M3Bf+K2E/37taTCv8i4Xvwbd/tW41fLnv3v8TRh6vH/AE1/dZH8zfxMsAfD+J/9Uyrmvn9IFlZ8GW/7MulULR8F5m3W0y8a3kirMGewvwkfhOy/bMivLMb2GeDf/wBm+fasGrHp+JjHnLx/2y2Gvx+M6vi3P+y7LVJ4B/FTxz8PYZ//AB1XPmrwfmZ4bq8aPkyW4h/lvA6+CMMfadvrvnI4UPJHL5FRFRcO29FRf7s3V09Eve+yOHiujo+6/rkTRuIw0ragy2CthxYKIImwebuU8Ccyc35krNgfyoP0L/lU4bPWh4o58tBta/N37pF7OVj+LjH1mVXtTWO6Mh/YNy6+Llu+rN1OqAUoBSgFKAUoBSgFKAUoBSgKYyTEMJZm5xYNVUbYYvLOJoYkumke4s7nF/R1WxNXX89a7g278U4XxJmS/uJzHd9kXWIp66jbmkSLARPAhR2Ad/S8S+3WljfV5mUdJ9Y24Q0LB+O52EouDsVYoucGHHmyXLJHjG0yD5Oi2iq6+2uqqw5zIi97v1q+Uy/0SZh/RLf65XIo7kYqvTVSCVmRPEUoPJk845TL/RJmH9Et/rlOUy/0SZh/RLf65V3EeM1LaY8Lo9ocpl/okzD+iW/1ynKZf6JMw/olv9cpxHjNS2jhdHtDlMv9EmYf0S3+uU5TL/RJmH9Et/rlOI8ZqW0cLo9ocpl/okzD+iW/1ynKZf6JMw/olv8AXKcR4zUto4XR7Q5TL/RJmH9Et/rlOUy/0SZh/RLf65TiPGaltHC6PaHKZf6JMw/olv8AXKcpl/okzD+iW/1ynEeM1LaOF0e0OUy/0SZh/RLf65Xw9wn3GGTccymzDEAFSIliQOZETnX+WU4jxmpbRwuj2i1MDYvg5hYJw9im2C8Ftvluj3OKEkUF0WnmhcBDRFVELaSaoirz+2tbyuBJZLsbekjeY+PLflfgW9YrurUmRbrTGKS81DATeMU9oEIhRV5/bVP01W3KZf6JMw/olv8AXK6OF3Pr42LlSWgpqVoUrZbHKZf6JMw/olv9cpymX+iTMP6Jb/XK3uI8ZqW0q4XR7Q5TL/RJmH9Et/rlOUy/0SZh/RLf65TiPGaltHC6PaHKZf6JMw/olv8AXKcpl/okzD+iW/1ynEeM1LaOF0e0OUy/0SZh/RLf65TlMv8ARJmH9Et/rlOI8ZqW0cLo9ocpl/okzD+iW/1ynKZf6JMw/olv9cpxHjNS2jhdHtDlMv8ARJmH9Et/rlOUy/0SZh/RLf65TiPGaltHC6PaHKZf6JMw/olv9cpymX+iTMP6Jb/XKcR4zUto4XR7Rusvs/oWPscFhR3CeJsL3brc5dG0vseOAOsg620W1Wn3OdCdDmVEq065GIw88LUdKppRswnGpHKjoFU/inhIQ8PY4vmF4WCMXYmm2VWRmSbPGikwBOtC6AorshslXaSa9zWWGwtTFz3ulpInONNZUtBhcpl/okzD+iW/1ynKZf6JMw/olv8AXK6vEeM1Laa/C6PaHKZf6JMw/olv9cpymX+iTMP6Jb/XKcR4zUto4XR7Q5TL/RJmH9Et/rlOUy/0SZh/RLf65TiPGaltHC6PaHKZf6JMw/olv9cpymX+iTMP6Jb/AFynEeM1LaOF0e0OUy/0SZh/RLf65TlMv9EmYf0S3+uU4jxmpbRwuj2hymX+iTMP6Jb/AFynKZf6JMw/olv9cpxHjNS2jhdHtDlMv9EmYf0S3+uU5TL/AESZh/RLf65TiPGaltHC6PaMW7cK4LHaplynZVZhR4UNk5D7xRICo22AqREukzXmRFWpRmjllbeEVl/ZA673KwtkqXGJMhC1x7YvRHWDTRwTFFVmU6iKiaiSiSLqKVpYjB1tz5RlVSzlsKsKyeQzf5a5Z27K+23OFbXnnmp085qi6ICjI8W2y0yAiiIgNssstprqq7NxKpEq19zsx4EC7Xq3OQp6SbaLW1FbAUmk5xaCLCkab9CdbAlXaIkaaqmutaLvVk2XaDRws97FPmRI7UC6ayuIJtwm2kHY5tQjX75rtbIwA+bXUk2oSaqnE2Or1b7390Agnb58acLN/tLThRnRcQDFwkIV0VdFRUVFT81Wb24Rfw+pG9g3eq/dn9EjCsX868ML4v4s+0J1RPMH8SWL/wCQrh9izaqel+E/NHdhpj71D6GXXev6QzLT+4Q/sq51U2XfsT8J3+72j7WuVZPTs8jWh0cfdj9ZiY19iPg6/wB4xL9vQ6uvC/8ASX4j/vjv2FDrD9/Itqc2fuz+sofKP8SfH/wrhL99DqY3r8Dga/8AYtn2xbKzjpXjEmrpn41P7Zt7R7PnCW+K2K/3saoVB/FAvf8A5Kj/AGNGquXNfhP6kW0efD38P9DJhePZm4L/AMVsJ/v3a0mFf5Fwvfg27/atxq+XPfvf4mjD1eP+mv7rI/mb+JlgD4fxP/qmVc18/pAsrPgy3/Zl0qhaPgvM262mXjW8kVZgz2F+Ej8J2X7ZkV5Zjewzwb/+zfPtWDVj0/Exjzl4/wC2Ww1+PxnV8W5/2XZapPAP4qeOfh7DP/46rnzV4PzM8N1eNHyZLcQ/y3gdfBGGPtO3131kZ7CWXvxet/1Zurp6Je99kcPFdHR91/XInFZED+VB+hf8qnDdNDxRz5aDa1+bv3SL2crH8XGPrMqvamsd0ZD+wbl18XLd9WbqdUApQClAKUApQClAKUApQClAcjcMqXfMG48w/Jw0LyXHMizyMuheaFV6nlvPtHEkKqd5GmnLifP4K6btuHYOEcL2exWtlI9ttcZqFFZHvNtNggAP+AiiVpY31eZlHSc+B+NDmV8XsP8A7y5V/MzbjLt9ywGEWU9HGTiJph8WnFFHW1jSFUC0XnHURXRebVE8Fej3M9Rp+H3OJiVevL+dRFcb533HD+aAYatlpcuEWIUEZyNW2bIdc6pdUVUHGWyaaRoERxeMXu0VUTbpuWLXHhL3yHcr8xFttsuqNQpsy3I0EplndHnRovFlKIFafU0lCSkzqjSioqhaoq78qrV7FSpJpZzcS828Z26/T4Vzg2Qrbb7sxYZbkJx8H3H3ra3K41rVdBATdEdF1UkVV1FR7rSYY4QGIIlog3GVbYR4WhuQra8pyHnri4btkYuCuby5i2k4gLu1U9VLVFTQodSSZKpxazFg5YZgYoxHiN604kg2phSskK9sO2s3FQUkG8Kslv76hxSJvTRC112j3ks6r4NyV2UzSTshSszAUoBSgFYd7/maf/d3P9K1DJMrgvfizZR/FC0fUmqs6vjFTnvxPXrQVBwu/wAWnMP4LP8A1DWFmHiCRhPAGJb5EFpyXbLZKmsg+KkBG20RihIioqpqKaoiov50r2/o67Uar715HHx6vKCK0a4QsfC8O7PYlcGfHhyW2UmRLeVqVEVlXT3MTHkMtojqmxSI92gAqgVZ1vz+bObdo8uxXI5UN2QI2+G0ybvEMSpbJyd6vIhDpFRVDRCRSRB4zcmnqVV6jnOlqPaXwjsPQ4MyYdtuYxGnXWo77pxGW5iNvuMOG2bj4iIobSonGqCluDahKSJX27ndGhYNfxRKEjtbc+UDYxIqm69HajOvpohODtNRa766przbU11Gd9TI3tmquvCQiWPFKxbla5Nrt7cdWTZmuRGn+rlcPRnjSko0KcUy45zqqEiom9CTYWS/wosHx5LQutzmojwg4xNfKOy2+2rTDpm2JvI4ewJLGoiCkqnoAmoltjfktJO9N6DZ2zPm13dWWo9gvxzH4rdwZhCwyTzsM0P/AGkRF1dQRWyTb/vFXREBdya7jL3N3D2Z0u5R7I+rxQRbcIuNaNHGzU0A04syUdVbLuHEA05tRTVNc1UUmkYum0rk1pVpUKUApQELw9+NhaPiTcfr8GugK+Ybt+uz+HkelwnQxFc74b9nvOb+/Wv7NZq7cD1z4MwxvQs3WOL9MsdthhbRYW5XCazBjnKEiZaI17ozEVRSQQElQUVNyoI6jrqldx80cTP5grhJ1+AxJgucTLlRsPzJrcg1VogIVbe0jDxbwaq6pIhIXdKgrX0SUmnmOFGKaJfl3iLEOJ4ttu9xcs/Wi8QEuEONFBxuVGEthABqRkjvcOJuNEb2kiJtJC1SHYz4QCYbzOu+FY3WySsW0qbLTr6i+ty7gxaMUVVRpWnWzUkHm0LnXTSolNximyVBSk0iQP33G7V7h4Z6vw2V4kC/MS4jCfVlIzQsIQlG4/cLiuPiiLxqptFS017mpBd8ZnhrCtquU6A5cZ0wo0fqSzGDqG+7omjZuE2ihuX8IlHm510rJNq9zFxTtYrW6cIORe0tHY9GkWbqyM5L23ayPXF11tWmnGCBqI9uETRxecl1RR0UU1TX7g5uYsmrdVcbhQJTM2Nbm7c/Y5itsPPuR2UU5vGIy5sORqrYIhKgqmo85JVvjejyLN7SWcs2BcbtbsPXR29yLdInQONVX7a0YgQICGKkyRqoFovOHGLrzLuTdokEHhIWKLYhuMy2XVlghd4mQ6EdhqUrMgY7xCRP7WhFwh53iBNCREUl5qslPJ0lahlaDOv+eMCHEglAjSCdflo2SSGeZGglRGX1RBLXdpLFRTwouqc2i5WDc77Pjy52uBarfcHnZvVauOCcZ1mILAxyInHG3iAkLqphBRpTXU11QdpaRvivYne3a5YtKuKiHZzew/jr4Bn/AFc6tjLL2NsJ/BMT9yFeJ9JP+V8fsdrc/RIktaWbguwXKXJlSrLb5EmUm2S65GBSkJxat6OLp3abCUdC1TRa8Um1oOseTeAMLtSG3ww5aAfbcYdBwYLSEJsjsZJF26orYqoiv9VF0TRK4IzMT/8A2B2z4cs/+tatUnKLu9X1I3sH0r92f0SNNYv514YXxfxZ9oTqieYP4ksX/wAhXD7Fm1i9L8J+aO7DTH3qH0Muu9f0hmWn9wh/ZVzqpsu/Yn4Tv93tH2tcqyenZ5GtDo4+7H6zExr7EfB1/vGJft6HV14X/pL8R/3x37Ch1h+/kW1ObP3Z/WUPlH+JPj/4Vwl++h1Mb1+BwNf+xbPti2VnHSvGJNXTPxqf2zb2j2fOEt8VsV/vY1QqD+KBe/8AyVH+xo1Vy5r8J/Ui2jz4e/h/oZMLx7M3Bf8AithP9+7Wkwr/ACLhe/Bt3+1bjV8ue/e/xNGHq8f9Nf3WR/M38TLAHw/if/VMq5r5/SBZWfBlv+zLpVC0fBeZt1tMvGt5IqzBnsL8JH4Tsv2zIryzG9hng3/9m+fasGrHp+JjHnLx/wBsthr8fjOr4tz/ALLstUngH8VPHPw9hn/8dVz5q8H5meG6vGj5MluIf5bwOvgjDH2nb676yM9hLL34vW/6s3V09Eve+yOHiujo+6/rkTisiB/Kg/Qv+VThumh4o58tBta/N37pF7OVj+LjH1mVXtTWO6Mh/YNy6+Llu+rN1OqAUoBSgFKAUoBSgFKAUoBSgOPeF7fLndMzLNOtUt1q2ZPxouOruDPOLxuSxZRovBpBauhL7fdh7S8/WlxMXI7ZCSEJLqipzoqad+tLG+rzMo6TmppwHOFHmYgkJKGH8PiSIvOK77kui+DmVF/xrd4zwPaceW6NEu4y+LiyBlsOwZz8N5p0RIUMXWTA05jJOZdOevSblq+Bpp6vucPFO1eTX8zEfDJDChzbfNMbrMdiKyaFLvUyQklWnSeYV/e6XH8W4SkHGbtvMicyIiYocHnBITUkrCnuKLbzDbDl1lEy0y46DxtNtq5tAOMabLaKIiKA6cyIidLeoso3yRJJ2XGHrk/NekQFccmXFu6vrxzib5IMBHBzmLm0abAdqc3Nqqaqq1gwsocLWq3BDg2sGmmpTM1sHXXXBR9mI3DaJUU+6RGGmwUVXRduq86qtS4RZjlvQRLI/Jy5Ze3y73e7JEYflW+Ha2IcG5S5zLTUcni1ApCITQLxyILA6i2g/hGpKtXBSEcmNhOWU7oUqwwFKAUoBWHe/wCZp/8Ad3P9K1DJNJwaMw7LC4OGVMd1bhxrOE7S2Wy1SjHVIbSLoQtqipzd9F0WrJ7Zli8Ny8TzPRV8cqU5Ob8daPXJ5ip+FZj+z3Lg6Y9isLP4522kI8Za5TY67h75E2iJ+lVSt9jNLQuEL2mICEbCUF4bgRKQokdQVHVVR7pE27tVTnTv17X0ejk0qieteRyMfnlCxG79l5grMS+3jq5h2XcmmRgT1jzJDCo2balxJK2YoqEDiKo+3qCr3hrHHKbAt8l3lpuK+5KblK3OJq4SmzFw0N8m1JHEXYaTTIgRdpcbzouiaeqcIvOcvLklY9pOWGB7zckgdREk61q5KQI0t9g4/VTjxmqEBpohqb6aIumhKOiJzVmrlFhRcKt4cW2EVnbU1Fg5TyrqbJMEqmp7l1bMk519vXv89TkRegOcjwk5e4OvOIbmnEOje0VqXIdjTJDD7SnxggYGBooKqI6PcKmqKSLzLz5J5TYXImlGC+yTZKSExNfbUkUGQUCUTTeCjHZRQLUV2Jqi8+rIixlSRrxyzwNeHWorMRXjgRGrehRpj47GGVdBGCMDTVEV11DAlXdqm9F0HSR4bwXacJOSStbLzKSEEVbclOug2AqSiDYGSi0Cby0AEEU173MlFCKzohyk1Zm8pVhgKUApQFds36Hh7hT2V+Yr/Fngy4AnU8Z19deroS95sSVE5u+qaVc3bMsXhuXieZ6Kvmm7UHLGya7vI9JhOhiO2ZYvDcvE8z0VUrga7R73nbnHLi8bxJz7YicewbJc1uZRe5NEJPkq3cKDjjLvUzDG9CyVY8GxJhC5v4lcRiyRGurJMhXDbVgWlRxHRMFQxIVBCRRXciomnPWDarFhbBUIb1F+8sm0gLPdkOvk8LriEhEZESmpGad0uq86c+iV9Daje/WcFN2sj+wMsrHZ5qS7aEqFIQxUSGY8aNNo6jhMtCRqLTRkibgBEFURE07kdPB/A2EMWBcojsMJ/FXN1+WJOuapKONxR7l11XVh7Zt/BQSRETm5oyI6BlN5z+llLhoois9TzkdV8pKzUukpJhOK2jSqshHeNVNgiO1S00EebmTTNTCdns9mjxnW3Ct0GS1IiRxU1GKobRaBsR50AdE0HvJz82nenISDk2fyx5b4dw5OhTLdAWPJhQwgMGrzhbGAAQEdFJUXQQFNV5+bv861lycGWeXbLjb3Ym6LcH1lSBR00JXdRXeJIu4CRQFUUVTaooqaLTJSViHJvOY1pwDa7KxJjRVldRymnG32H5TryvEaqpuGZkpEapoO5V1RERNdERE1TuVuDmWrZaRZchux25ZQm41yfYkCDrwOyFAgcQ9Fc2aqi8yFt5kLRYcI2Jyn1GvueReX7caQ9KshlDGMsdYgyZJMA1uYIhBgTUR1WMwq7RRS2c+uq6yHDmW9gwpciuFvjyeriR1CkSpz8ky4wWBPVXTJV1SKwn5tnNpqusKnFPMS5yasSelWlZDs5vYfx18Az/q51bGWXsbYT+CYn7kK8T6Sf8r4/Y7W5+iRJaV4k64r86czP6QO2fDln/1rVkea/h9SN3B9K/dn9EjTWL+deGF8X8WfaE6onmD+JLF/8hXD7Fm0el+E/NHdhpj71D6GXXev6QzLT+4Q/sq51U2XfsT8J3+72j7WuVZPTs8jWh0cfdj9ZiY19iPg6/3jEv29Dq68L/0l+I/7479hQ6w/fyLanNn7s/rKHyj/ABJ8f/CuEv30OpjevwOBr/2LZ9sWys46V4xJq6Z+NT+2be0ez5wlvitiv97GqFQfxQL3/wCSo/2NGquXNfhP6kW0efD38P8AQyYXj2ZuC/8AFbCf792tJhX+RcL34Nu/2rcavlz373+Jow9Xj/pr+6yP5m/iZYA+H8T/AOqZVzXz+kCys+DLf9mXSqFo+C8zbraZeNbyRVmDPYX4SPwnZftmRXlmN7DPBv8A+zfPtWDVj0/Exjzl4/7ZbDX4/GdXxbn/AGXZapPAP4qeOfh7DP8A+Oq581eD8zPDdXjR8mS3EP8ALeB18EYY+07fXfWRnsJZe/F63/Vm6unol732Rw8V0dH3X9cicVkQP5UH6F/yqcN00PFHPloNrX5u/dIvZysfxcY+syq9qax3RkP7BuXXxct31ZutpmDmRh3KywDesTTyt9uKQ3FBwIzsgzdcXQAEGhI1VV5uZKAgXK4yw91bx5MXT1anK4yw91bx5MXT1apsyLocrjLD3VvHkxdPVqcrjLD3VvHkxdPVqWYuhyuMsPdW8eTF09WpyuMsPdW8eTF09WpZi6HK4yw91bx5MXT1anK4yw91bx5MXT1almLocrjLD3VvHkxdPVqcrjLD3VvHkxdPVqWYuhyuMsPdW8eTF09WpyuMsPdW8eTF09WpZi6HK4yw91bx5MXT1anK4yw91bx5MXT1almLocrjLD3VvHkxdPVqluXWc2EM2HLq1hi5vTXrVxXVjMmBIhuM8YhK2qi+2CqhIBaKiKnNRoaSucjMNQs1svcwcVXRtHomZ0+Y4Oqc5WlG+ooaJ7W047Iu6eF8vDW84OGJZmJ8gMGncz33q2xyslyVe+syE4cR9V8GrjBrp+dK0cb6vMzjpOauD1fJOMeEVn1ih13jYV5ft52/wdSxnrhBbJPzF1Ipp/16+3Vh5zRI94l4DslzFHcP3XECRrlHcVUakAMOU6004ntgT7TKKK8xcwqioWi+mwCSwdOPdb52OJiOnl/OoiwdQ4axOmBcuLq3h0J11knNVIaOxrY61DYdWNFaJBbFXEcbdIUUkTV5dBJe5jeF87cZYvt9uxENwtlvhDerFZZFoGJvGSMtqI49IacU9yIqylRvvptbJVRVXUdtycc0dH6FaipZ5aTAsmZmML7ZsELf59tujmIoeHr2zst/FDAdkShAkFENd+iEBIpd4hJe8SCOsy6zUvGAbLhy+X6ezcLXHw1Yn79dHmF6pNiQl1RpVLcuqg8EcdV7/GGq86pphlyumzNwTTSJXgXNvMS5Y3tcK9wSbjtT49nuTIRo7UZHjgBIcMXSfR3jBdcREBAVFaDXTVdRkeKcw8U2vMS8DFuULrJaLpY7etqWKhOyBnvAy4aubtR2cYhjon9QkXVO9mpycbsrcI3siKWjMzMK7YYiTHcSWmDMcsM3FCuFbURlW2SbBuLzmq7E7onXEVC1cDbtSvabnfitMG3DHLE23x2HnpdthYUlx06pbdacRoHULeJOGhIThgqiPFkPOKipFG+StpMsiLNdds4sxbXa5DbqPxxtL80pk5yBEfngy2xEdackQmpCqjKLIdFwmdXEFGCQR4zWvrGObWIXDxZHupRnrQ+1LS1Ryhg9Blgy+2Ik1JbdUuMRN/GtPICoSaCmgKpQ5yzpkqEc1j2n5y45s7L0926W2QzdHL3FhsFA2hbli3ZqEy+ZIeriIDqm4i7UXYmm3RVW2Mqb7fLhccb2i+3OPeHbBeW7fHmMx0YJxooESR98EVVN+580VU0TvaIneqyEpN2f8zFcoxSzFgVh3v8Amaf/AHdz/StXspMrgvfizZR/FC0fUmqs6vjFTnvxPXrQVBwu/wAWnMP4LP8A1DWVdbZGvVrl2+Y0L8OWycd5ou8YEKiQr+ZUVUr3Ho4r0qnivI426GmPxOdrVkLjdi6QLnOu4vXFxGCmOxpYNiLzRkBOJuYM146MEZokAg12KhLpoqfLGQeJbTEmRbVBgR1cVk3Jp3N5TltLb2Y8iMQbefc8JPIZEqEQDvFU1SvS71I0t8iSHDGUmL7fgfFNrdnNQbjPjBGiPMykLa2EqS4LW9GkQE4l1ttFFvQefQNBRKheIsnMZWa1DMFtJkJmP1N1qK4Eh73JLhNuITDQCCNNmjKIDfcjJc0UkZb1OnKyCnG5nYgyKxvLWQ5bW7fGA2jOHHGcqLAdR6Q5HACJohQGweQUURQhXaLZAKKtZGKchMVzbPKiWrqIDnpxkk3ZhCSyEOcoPakBiqij8dNVAlEQTZoTbao3qQ3yJkhkriVq4SpR2q2zIb812ZLtZTiZC4G6jy7nCEF7oTdbLVUXVWEVOdBqX5UZWXrBF0t0y6TAuE5YlwZuk/jSU5jpyGDjGqL4AF/m/qq4Wn4SrWUabTv/AD+fqYymmrItmlbJrilAKUBC8PfjYWj4k3H6/BroCvmG7frs/h5HpcJ0MRXO+G/Z7zm/v1r+zWau3A9c+DMMb0LM7M7C9xxpYYdngSQhNPXCO7LkmKGrbLJ8dogLzHuNtttRXm2mS+1VSN5G4qdsUOy3AYd0ahtMwI9yO4OsvNwmCNsW+LAdFVwEadJVL8NV9ttta+hTg5O5xITUVYsK34RvEXL7EmEIoJbXTZnpbJrL6g20MiTKVhsVFNzfFNqx+D+ChIg/g1WjmTOKIDt5mx7ZAtbD8tuXBYi3Fw1tANkBPI0iAm9ZDYcUopogoKCmorUSg2kIySuYlryextMtGFJkOJDtjUYo01IPXIzcZdAYiuHucaXuneKf3aIJ6kiqaca9rsGeD3frdCsJwVYYuMdu3pMdSa5q4QRHwlLqvf3ulHVf+Lig1/AFEr3qTM3UjoPG7cH7Fa2yDHt8oWWUtsUZkVmcjSv3ARkiclXHGXdFHexoW3ciCijorQIthZo4Qv8Aj/L+Jb4LShc400+5lykaF1GxeabdPRstwkqtu6DsIdUICQwHXNQkk0YucW0yHW3IvFT+ITfu90KRHcuRypDqSxUJGvVKtucULIFqCuMigm4fMih+AA64C5D4mOTbphW21ikRviSgLPNzjF3RSkucaTeqdVDHcbNNO8aEWqmaJG9SsZb5EyUyJxQkCQ8LkZq6OzXlZUZjhJFgnEkAEQSVOcW3DYHvJuRkS/qiiWFlTl5OwNPuDsnikCZBjI7sdI1clC9KJxwtfbVt1gUXwAg8yAlZRptO7MJzTVkWNStgoIdnN7D+OvgGf9XOrYyy9jbCfwTE/chXifST/lfH7Ha3P0SJLSvEnXNPiDFduwy5ACe44JTXxYaRtonNFVUTcW1F2gikKKS6Im5NVTWvz1zWvVvhcPqC+/NjtMtXq1GZk4mgoJKpL/h7dXwi3F27vqRuYRpVHfsz+iRGrFjCxrcuFaSXWIo3Sx4qbhKjqffyKZPdFB/S2JEn5kqMY9xPaHuBuxbm7lFOaGO50kmEdTcjR2icAn+hSFUT86VLpyu83VPzR24TjeOf81D5QZcV9xxh4OHjgG7leYSWyDAa6plK8OxribZdQe1X/lISRfzpVYYFxLaYWWXCQjP3GM3ImN2pqO2TiauGNyuThIn6ARSX81S6cr6NXka8JR3uOf8ALH6zDxdiW0yssOD/AA2rlFclRn8Qq80Lqbm999iEGvP/AFkFVT8yVcOGMc4eT7oliG89eoXWpZLjiS1eHYorZIYouuve3c36ax3uerX5FlSccmef8s/rKSyrxLaYvA3xvBeuUVuZIueFyaZJ1EI0behcYqJr/V1TXwVMbhiW0zneCIwxcorrsBLYxJAXU1Zc66Wp3aXPzLsMS/QtZqErrN2TKrON5Z+up/bM6yY2sD2cvCHugXiGVvn4XxGkWQjybXlfeiIzt8O9SHTw61EYmJ7Q3wUbpBK5xEmP5htyWmVdHcbTdohgZpz95CcBFXwklVypzs83VP6kWUZxy4Z/z4f5QdyV3jGViXNvg4Seu0Tqe3YZww3KcV1NGSbdcJxC/wClFRV8FanDWKrMxD4VqHdIgrcIN2ZiorqffjW43JxBHn51UBUv0JV7pyy3m/N/iaUJR4PFX/J/usj+ZGJ7Q/wRcD25u5RTnMXnEUhxhHU3C265PFsv0ErZon/TVu37HOHh4dGXd3W8wktsG2x+qZSvIgNcRb7qL2q/8pCqL+iqFTnbR1LzNqrOLcs/XW8kVvhbEtphZQcIWO/corT8y7WhuO2TqauEN2lOEic/tAKkv5kryzAxLaZOVHB4hNXKK5LisXhXmUdTc2h3SEoa8/toJKn6FrN05X0dZjGcbrP1/wCBZ0bHOH3eG9m9dxvMJbZNw5IGPK45NjqvW6yNtIi/8xEgp+eqcwNia0M8GTGUA7lFGY/ecPPNsq6O4gbVlHCTn7wqYIvg3JWEqc8lZup+Znh5xVs/XR8mS28YltM268Elhi5RXXoNtwxHkgLqasudX21zaXPzLsMS/QtfobkZz5JZe6c//wCnrf8AVm6zqRaUr6/sji4lpwo27L+uROKyIH8qD9C/5Uw3TQ8UaEtBta/N37pF7OVj+LjH1mVXtTWO6Mh/YNy6+Llu+rN1FOE3/NOXnx5sv7+pWkh6CypDnFMOGmiqAqui/mSuVMvOGFccTM4NfkycI3uJfY8STOk4dlm4NiV1W90eXqZILqo4Sjzp/J3dRTRK2WzXSuWq5wlcLN2F+6Lb8QfeNrpw1tbiSBjE0boy1Be8yoNmu5VRdRUNN/c1nM8ITCMi7SYbR3B1hmVEgpcm4RlEckSHozLbQuImiFvmR9ULTmIlTVAPauMk8Lpwj8HWy4uwdbpLktFIbMYludcQTZlnDUFJE0QjkhxQJrqREOnNqqTbBuMIGOrC3dLeMhltXXWHI8xpWn2HWzVtxswXvKhCqe2ipoqKqKiq0kNWN5SpIFKAUoBSgFcsZm4im2XGGetts76x8Q4miYZw1anB/CbkzVlMI6n/AGgM3V/M0tVz0FkNJ1bhrD0HCWHLVYrWyke22yI1Cisp3m2mwQAH/ARRK5muuNXMh2+EhFbFXHLaY4xs8dPwnVnxlBGxT21KdFkf4up4a52KjlUXH+aTZjpK3wdcsG8HLNC5YTxNiux4cOFgrDMNs7vcWYqynGurxecHjCTcqmu4lT2z5+/UzxFnbkfi6zv2q9ZhYCuluf04yLLvsNwCVFQhXRXO+ioiovfRURU50rv7nVabwkMprR9zjYmE9+bimaSRjjg4y8Ls4cdxblwdkZdWQ3C68wkAXV13OJo5ruLcWpd9dxaquq1EsR3bJrEeY1nv7ubGWrVttb0SRGihJgjLZWNztNNyRfRBaQtC0VsiTUxQ0E1FN6UqL0SXyKUqq0pk9azWyBYbt4N43y7AbeyxHiCN5gokdtkkJkA7vuUAkRRRO8qc1YcXMDg6wrTItbOLsuBt0gAadileIJNmAOm8AKKuKm0XHXCFO8KkumlZ5dHWvkY5NXUzKczUyAdxYGJyxvl4WIWx2DclvULj0Taoa7uM112Ko69/aqprpzVpY2M8he2Tc8azcxcv7heJHEdSvPXiErkPim1DuTV1VXXVVRdE03Lp31qHOi/zLX1EqNVdT1Gxu+YPB2v9utkC4Yvy4mQrYqrDju3iEoMa6aoKcZogromo95dE1RdK+n8xeDvJvFzur2L8t3rjdI5RJ0hy7wSKS0QiJgep90hCACWvfQBRdUFNJy6L618iMmrqZr3sTcGeRaIdsdxLlo7BiOuPMsHd4SoJuacYq6uarv0Hci67tqa66JWWuPODmt2uNzXFuW6z7iJBLfW7wVJ5CUVLX75/WUAUv+JRHXXRKjKoa18icmtqYxJmbkjd8OzbfAzIy7t0p2PLZYkuXK3yAaWSW9/c0Tmhi4fOY82/n50XnTWZPY8ycyost0jN5rZfHJuk9bjJS3XWLEjC4rLTWjbavmSJtZFVUjJSJSVV59EjLpZSakvkMmpk2yWT3lI5SdKeCvKGJ6SsS8cI7KZ20zQDNHBhmTBoIjiCIqqu1eZPvlWOtT7S2mG9T7L2Fn8F78WbKP4oWj6k1VnV8dqc9+J6taCnuF+6DPBkzFccMQbC1OERmuiCiKOqqvtJUO5SOUnSngryhiekr2vo9OMKVTKds68jkY+EpOOSr6RykcpOlPBXlDE9JTlI5SdKeCvKGJ6SvW79T7S2nL3qfZewcpHKTpTwV5QxPSU5SOUnSngryhiekpv1PtLaN6n2XsHKRyk6U8FeUMT0lOUjlJ0p4K8oYnpKb9T7S2jep9l7BykcpOlPBXlDE9JTlI5SdKeCvKGJ6Sm/U+0to3qfZewcpHKTpTwV5QxPSU5SOUnSngryhiekpv1PtLaN6n2XsHKRyk6U8FeUMT0lOUjlJ0p4K8oYnpKb9T7S2jep9l7BykcpOlPBXlDE9JTlI5SdKeCvKGJ6Sm/U+0to3qfZewwcsMw8K5g8KqC7hbE1nxK1FwXPF87RPalI0qzoSohK2S7VXRdNfBXUFfNd2mpY2TT1eR6HCpqjFMVydKzTwVgDhC5vxcT4wsOHJL8u2OMs3e5sRTcDrcym4RcJFVNebVKu3Cko4u7fU/sYYyLlSsjf8pHKTpTwV5QxPSU5SOUnSngryhiekr6Jv1PtLacHep9l7BykcpOlPBXlDE9JTlI5SdKeCvKGJ6Sm/U+0to3qfZewcpHKTpTwV5QxPSU5SOUnSngryhiekpv1PtLaN6n2XsHKRyk6U8FeUMT0lOUjlJ0p4K8oYnpKb9T7S2jep9l7BykcpOlPBXlDE9JTlI5SdKeCvKGJ6Sm/U+0to3qfZewcpHKTpTwV5QxPSU5SOUnSngryhiekpv1PtLaN6n2XsHKRyk6U8FeUMT0lOUjlJ0p4K8oYnpKb9T7S2jep9l7CKZtcIXKu5ZVYziRMzMHypT9lmtMsM36KZuGTBoIiKOaqqqqIiJ4a6Yyy9jbCfwTE/chXjPSKcZ73ku+n7HXwMXFSuiS0rxh1TWXnDluxAcIp8ZJBQnxksLuIdrgrqi8ypqmqIu1dUVUTVOZKp7FOHcN27NqNYGsF4Zkwr4CTLi8/a0N9TcU2yNT9rXY2iIKEv4ZFtRFKr6cnoTIaNZBwtaXrNiObJwFg5piwI45Zm0szoKKto4yhOkYCLiKOuu1do685f1k1F3G3wMC2zFUbAGDCu8xqU3KNy1I4002wZkCojSmJc25CUDJO6U0IgbKthNt2yntMSRJgzCT+ZVxwP2C4XPDj6OJIVLWiPbn2TeJN+uupFxq9yO0UTTcJKIrro9mgTsCTMTXDAWDBuEFuPJgNBZjaECdIgLcrwtoSK2QqhKogJEW4kEN645Us34n1devSTY8LrFtdqs+HL1FwDg3qi6xZMi6PHad4gLCijaNiCKi7lc2qSESLqhCrgoir7u4dwuGIcUYXTAmFFsNrtkiTHRbT984xrYSAZfhLzqpLoKdyTe0l1VEm8u09uoix93ixwbdgxzEMzAWDUvcaesaM11p4ltpg2wIhJZHEjvQkVEMyACRE290SCvpOtFms13wz1LgLCCMX4bfJuT52glVHXjbYJAFQTamzboq/gqAoaIroqq7f5ntJsYbdiw20/imzN4Awm5asNwZD1tbW1KJK9DJtWgMucyRNW1Jdqbtw7Vc7rT7xRZ4NgwXa8RPYFwUd/WUcLQ7R1O02ybaGabX1a2rvaHnNUVRRNqbiQFm7ullPaQZ87CuHmMwIOHRwDhQbBdmQdnOnaSR5x19txFAdRRG/92q8/fRCRERQJU1ceDZZ0jEjQ4AwjJLD81pq1CloIUMyeWPtJREyVUBxBVdid3u0QkFVSE5P8z2kntii0W7C1is14HA+DHb1cSdiy3Ds3Etq006fFbW3VAkLRxVUE3OqqrsBxB5tg/g3DQ5hT8Hu4AwoOGZbrrW5LWqvvq7FckP6lt0DQ1RE05iRxUFUVtUqMqVrqT6+vUDR2di24mwW7eVwFg5+8w7i2xAULKfEKjgKBOKDYmZJ3RqiaISjoqiO4Vra4islowzLwc6zgbBxzb6LY3MytQimok2P3oDUS7zh9wIm5qo9yqISpLbvk5T2ix8s4Vw2OL7rhh7L/CjeHrRHJ2AIWolfccj8S6CqW3nQVUEUE1Ve5XnE0RdfbIVuvVhh3gcB4PcvI3g7SDrdjNxsYhxwfN0WmkMjJeLEh101HahK2qlou9OU9pBt5GH7JY8d4etjeBsHbL4xHl3IwtoEXHkPFkrSEqEo7Wg02Nlt0UneLRUJb0tFoh2C0wrZbo4RIEJgI0eO2nctNgKCIp+ZERE/wqio20s9zJIzKyIH8qD9C/5Vlhumh4oiWg2tfm790i9nKx/Fxj6zKr2prHdGQ/sG5dfFy3fVm6inCb/mnLz482X9/UrSQ9BZTzaPNG2q6IQqOqe1qlV1OyLst0yswngOXJkyLRh9mHHEi2ocluOzxKIfNom4e/pp3+atm1zWTsVivAew6li62hc4scT+8upGw9AaZcYRomhImRaQCkihmSSCQlFTLQURdqbR3gjxJFxC5O4pk9dI0aGESazbo7bovxnor7RvEKbpDYOwwUGnFVARxxEVdRUYsZZRmu8E6y3HD12t13vMq9v3NDcffnRI7jZPlczuSGTKhsIUfNE4tU2qCaL31WrHyuy5g5WYQZsUDqdQF52Q4USCzCaVxwlItjLIiACmqCiaKuiJqpLqqzYhu5LqVJiKUApQClAK5YsuFZ+M/uhl+N1pCw3hayW+9uKvOh3FxqVFioqf8rTs0kXw6eCq56CyGk6/rkjhY4UdufCMyOYYmR40DFMty13mO+SIslmA61dY6Int6ORnQVP/AI+nNrWjX6KT1K+zObC0nSEi3Qpjm9+Mw8emm5xsSXT/ABry6yWz+wRP1I/wrxGV3m1YdZLZ/YIn6kf4U6yWz+wRP1I/wpld4sOsls/sET9SP8KdZLZ/YIn6kf4Uyu8WHWS2f2CJ+pH+FOsls/sET9SP8KZXeLDrJbP7BE/Uj/CnWS2f2CJ+pH+FMrvFh1ktn9gifqR/hTrJbP7BE/Uj/CmV3iw6yWz+wRP1I/wp1ktn9gifqR/hTK7xYdZLZ/YIn6kf4U6yWz+wRP1I/wAKZXeLGYAg02IAgiApoIimiIie0lfWqeFKi6Fj5dbbfbJtwRcAk0UTTVFT9FYfWS2f2CJ+pH+FTld4sOsls/sET9SP8KdZLZ/YIn6kf4Uyu8WHWS2f2CJ+pH+FOsls/sET9SP8KZXeLDrJbP7BE/Uj/CnWS2f2CJ+pH+FMrvFh1ktn9gifqR/hTrJbP7BE/Uj/AApld4sOsls/sET9SP8ACnWS2f2CJ+pH+FMrvFh1ktn9gifqR/hTrJbP7BE/Uj/CmV3iw6yWz+wRP1I/wp1ktn9gifqR/hTK7xY9o9vhwzUo8Zhg1TRSbbQVVP8ACsjVPClQ2n1iw1TwpWK/bIMpxXHosd5xe+bjYqq/4qlLrWLHn1ktn9gifqR/hTrJbP7BE/Uj/Cpyu8WHWS2f2CJ+pH+FOsls/sET9SP8KZXeLDrJbP7BE/Uj/CnWS2f2CJ+pH+FMrvFh1ktn9gifqR/hTrJbP7BE/Uj/AApld4sOsls/sET9SP8ACnWS2f2CJ+pH+FMrvFh1ktn9gifqR/hTrJbP7BE/Uj/CmV3iw6yWz+wRP1I/wp1ktn9gifqR/hTK7xYdZLZ/YIn6kf4VmigiKCOiIiaIie1Ryv1ix/dU8KU1TwpUXQsNU8KVHLrhibc8UQ7kN+kMQI4t/wDsoGxVozEjUjVe+qqigKIuqIgqqJuUSDKMkhYxoGDblGfvTz2Kpzz1wAwYdFppDg6majxSEJD3IkCaEK6qKquqKIhgy8AXuVg1izjjeexcUeddkXNuHG1ki4ZErZNkCiiIJaIo6L3KKqqmqLnvkdSIsbg8Nz3MbNXksRSjtTcc2xsZsM8SLy7UR0TQOMRUFDTRVX8NdFRNUXWNYIv0e2YgZHG01+fcHwOHPkw46nBaHb96QQEENNd/Oun4fh1VYU46hY/lxwHc5VutsGPid+JHjLIV8UZ3rJQy1bQiU9V2f8ylv/r7tSRcpcI3Q8RXC4niZ9WHoz0eNGRgU6l37VQxVVVFIVQudR50UUX8HnnfI6hYwiwLfUw7NhN4udG4yZiS1ndTKibeLETaUUcQkFVFSTaYqOqIiog8/vNwRc5EjDaM4leZhWsWEkxljppLVtUXd3JCgqWmioqGiJ+Cgqm6m+R1Cx/W8GXdLxfJjmKH3G5sd5iG3xGhQd+ioqLu2rsUe50EVXXulPQdvzbcE3eJZYUV7FkopzUlZEmYxHBOqdWlDYouK53KroSoqrppoO0UFBOpHULG3bw/JC9jN67urEEGxG28S3xAqIqikmqKQqqqi8xInNpotYYYTuIxJTS4mmG88+jgSVZa4yOHH8YrTaoP4Kj3Hd7vD+aoy46ibGHinBF4vtohQYOK3rWrRuFIdSI2SykJdU3oKgiKnPrp3JKq7hVOaslzCd1dx0l7LEjpWsB0as6x02tFxagqoe721XdqoqqaaIqCpIU75G1rayLGnhZeYli4SudtPHcl28zHRMLz1EKONCip3KBvVO9qiaKiJzcyruUs2fgS6y4WGozWKHowWsUSUgMc04kQU1JN+iJzFzHvTuufVU1WXUhqFjLawjcwxdcrwWJHyiyY6sR4PEoqRS0TuxUiUdUXXvCmvNu3aarq2MvL2zheTa1xjJWW/NKSs4WjExBQQeKReN36ISbhVTXvCK7hRRWN8jqFj5nWG/BirDjDcibJtUOKwMiQAtAwTgEu5VbR4ectB11aNBRE4tQLVasHVPClROUXaxNhqnhSsiAqLKDnTvL/AJVbhmt+h4oiSzG2r83fukXs5WP4uMfWZVe2NU7oyH9g3Lr4uW76s3UU4Tf805efHmy/v6laSHoLNpW0aopQClAKUApQClAKUArla2Yin4K+6D3uS6+g4axJaLbh2QBLogT+KlSoZKve7oGJbaa/1jFE51quegshpOwa4g4XTb2Jc0bzjCLudHJq2Wy5soneR9+YMqeKJ4eoIbafofrkY92w0u+y+DaT+Rt01eSLRu2bF4PHNzwxhXL6942kWyDEny5Vtm2+Oy2ElXkaROqZDSkq9TuL3KKic3Pz199nGZXQTirxzZPXq+d4f0dxGJoxrRnFKSvnv+h054mMG42HZxmV0E4q8c2T16nZxmV0E4q8c2T16tjkviu3H5/oY8LhqY7OMyugnFXjmyevU7OMyugnFXjmyevU5L4rtx+f6DhcNTHZxmV0E4q8c2T16nZxmV0E4q8c2T16nJfFduPz/QcLhqY7OMyugnFXjmyevU7OMyugnFXjmyevU5L4rtx+f6DhcNTHZxmV0E4q8c2T16nZxmV0E4q8c2T16nJfFduPz/QcLhqY7OMyugnFXjmyevU7OMyugnFXjmyevU5L4rtx+f6DhcNTHZxmV0E4q8c2T16vOTj/ADHix3HncisVC20KmRdebIuiImqr/Lqcl8V24/P9BwuGpkswFi6LmDgXDmKYLL0eHe7bGuTDMhERxtt5oXBEtFVNyISIuiqmvt1va8hODhJwfVmN1O6uRrMnHMbLTAl6xRMiyJ0a1xykHGi7eNdRNE2juVB1XX21RPz1H+zjMroJxV45snr1drc/cetulTdSnJKztnv+hr1a8aTs0OzjMroJxV45snr1OzjMroJxV45snr1dXkviu3H5/oVcLhqY7OMyugnFXjmyevU7OMyugnFXjmyevU5L4rtx+f6DhcNTHZxmV0E4q8c2T16nZxmV0E4q8c2T16nJfFduPz/QcLhqY7OMyugnFXjmyevU7OMyugnFXjmyevU5L4rtx+f6DhcNTHZxmV0E4q8c2T16nZxmV0E4q8c2T16nJfFduPz/AEHC4amOzjMroJxV45snr1OzjMroJxV45snr1OS+K7cfn+g4XDUx2cZldBOKvHNk9ep2cZldBOKvHNk9epyXxXbj8/0HC4amfeGM0bvc8fBhHEWAr1gy5vWx26xyuUuDIbeabdaaNEWNId0VCeDmLT26sKvOY7BzwFZ0JtNrUbVOaqRykKrWbmvf5WNMRYdwxlpf8XlYTYamTYM63R2hcdZF4RRJElsl7k01VB019us8BgJ7o1XRptJpXz/D9SKlRUo3Z69nGZXQTirxzZPXqdnGZXQTirxzZPXq9ByXxXbj8/0NbhcNTHZxmV0E4q8c2T16nZxmV0E4q8c2T16nJfFduPz/AEHC4amOzjMroJxV45snr1OzjMroJxV45snr1OS+K7cfn+g4XDUx2cZldBOKvHNk9ep2cZldBOKvHNk9epyXxXbj8/0HC4amOzjMroJxV45snr1OzjMroJxV45snr1OS+K7cfn+g4XDUx2cZldBOKvHNk9ep2cZldBOKvHNk9epyXxXbj8/0HC4amOzjMroJxV45snr1OzjMroJxV45snr1OS+K7cfn+g4XDUzAxBmpj3DFhuV5uWR+Ko9ut0Z2XJe672UuLabBTMtozlVdBRV0RFWrGsV2av9kt9zYE22ZsduSAOabhExQkRdPb0WuPujuVV3NUXUknlX0X6i6lWVW9uozqVxTYFKAUoBSgFfxCElVEJFUV0VE9paA/tKAUoBSgFKAUoBSgFKAVtMN/zu1+gv8AJa6O5vrtH3o+aKqvRy8CZV+bv3SL2crH8XGPrMqvtpwTujIf2Dcuvi5bvqzdRThN/wA05efHmy/v6laSHoLNpW0aopQClAKUApQClAKUArl/MPBc/G2L8/G7IO7E1pt+G75ZVROfq+GsqSwKeBDJpG1/5XC8NVz0FkNJ03gTGVuzFwTYMU2d3jrXeoLNwjGvf4t0EMdfAui6KntLVEZR2aJmHldjLE1wbR6PmHiC5yyQudXICawYqa+AosZok9ru68/uw3HAVWtX3RvUFeoiG8Au8yrvifGbFwd4272fDlgsVwJfwlkwpF2iukqe0pEypf8AzV0tmiGLncOK3gx1mPczGSJSHdiqz/scjiTECEkNUk9TdyunNu1XRFRdvBRUcPBIwqZ5M5puPCrxZdo98u1lfit2qfgtkbECsCvF4iOPDkAiqqakJDdYybF1T/Zz5tVWrFn8LS34dvF2sc3Dtxl3S3sE6xGjy4hTJghOjQVJxjjEWMRuS2TbRzahAqr3KiopvFZpcwOFBfrZHJtjDkuyJFh38by6hx5L9ufgssuNmzqaA6JC8JIiouu8UVB0JKlT/Cmt9tkXp+6YXu0CxW9L2DFzVxhzq121vE3IBtoT3pu2FxalpuUSRdvcqQEgwXmHiDHmMX7W/ZJWEZGHjbK8QJjrEgZIPxzJninW1X8EkFVXuecSTnTRVtGgFKAUoBSgFYGIP5huX92d/wBC0Bz/AMGf8W/Kj4p2n6m1Vk18IxHTT8X5noY81FV8Kb8XrHXweX+oa6GxViKLhDC93vs1DKHa4b014WkRTUGwUyQUVUTXQV0r6H6L+rVPe+yOZi+cvAifbX7GYJvY8to4ScUx4lWpCz2nQUFMi3NghCjaCXGkQoAaa71FUKv4/n1gZkbhpe97kJ8oxtjEf1cdEnhNGu4++oKx30Ig3CHEmpKiCuntDRI1hLhQYbxL1AMkUtSvQmrg+++TixmWjbmHsR7ikEnQ6hd3Au1NNVEi2qlShjPfAsiWxG6+oy84COEMmK+z1Oim62iPqYIjCqbDo7Xdq7h001VNQMG+8IPCNqt8R2NNOdLkzGYYQljvMutk48w1udQm9Wh/2hohVxBQ+MBBVd467vEWbuEcJ3GZBu14GC9Da418nI7vFBzCW3jEFQU9piXFou/aSFpt56AwFz7wGLytHfxacCMcp0XYr4cQAq8hC7qCcW5rGkDxZ6GqsuIg6iqJ/Y+e+Bpbb5MXo3uJbFza3BkkTu42wQWhRvV09zzKKDaESK63qibx1AwsTcIXBtgtcaYxchuXHuQEAI7bi7glPx2hUVQF3OIMltziU1cUSHuU3Itb3EmZEC1ZW3XG9tQLpBiW524NAZlHR1ABS2kpCpNrqmi6jqKouqappQEKh59XC5lc4MKwWuXebcxOec4i9kcB1IowHCRuQMdSLUJ499pNDaIV5u6qyME3q4YkwpbLrc4Ma2y5rAyOpYkopIAJJqP3wm21Vdqpqm1NF1TVe+oG8pQFB4+/Gvwx8Sbn9eg1Nq+TekXr8vBeR2MN0aFRHIP2Xs7fhO1/ZrFbHoz67L3X5oxxfR/EsDNvMyPlJhEcQy4L1whhMjR5Ax1RDZacdEXHtNO6RsFI1FOdUFUTnVKjzGf1ufxbhexpbnF6+XC6QerAeEmovUjzjQGfN3nzbVARPbXTn0r6ickm+HMb2jGlvlysPzG7kkddmmhtoRKCGCopDqoGJCQmiKJCqKKqlVk7whLjc7RAn4ewtGnA7Cs0mSNyuhROp3bi+TAR+4ju6m0SCriLt0Qk0RV5qAluWeareYd2xNazt3W6bYpIxz2v8c3ITU2jdbXaK7Ekx5bKbhRV6n3aJu0T4h534ZPGF6w1NlLb7lbri3bgR1txW3zcCKobXEDYhKUxkdm7cm4VXRCSgP5Cz+wDcQklGxAD4xwE1UIr68YpcVtBruPvri8exo2G4vvzfN3Y66E+FHgscYpZOMmJEGKUo7sUN7qdE2MmiIqAuqILpK4q6cVxZb9vtAbq6Z74VgTWGG5hPtDNdiTHiadbSMIRpb/HChBq8CrCdbQm9UUkJEVVFUr+yuEHgKI7GZcvT3VEhtHAjjbpRPIquPNIBNo0pA4rkaQCNkiGpNGiDqmlAZ5504L60wbk1fW5cScLxRCiMuvlJRp5tlzigAVI1Rx1sdBRVXdqmqIqpL4M6PdIMeZEeCRFkNi6y82uomBIiiSL4FRUWgPelAQXPn2DcxPi5cfqzlRHLf2O8LfBUX9yNeD9KuZR8X9jo4P8xIqV89OkKUApQClAYzVyjPSDYRza8BbOLcFQVV074oqJuT86ap+eq0wAtiwFfsxnH5hQWbhionFcnTHXG+NK3Rnj0VwlRsdENdE2imiIiJzJWzTUnGcEtK+6MHbMyZrmHhjqwovX6AjwNg8ur4oKNkCmJ7u9tURVdddK2tsvMG8i+UGS3KBkhAzaXUdSbBwdF7xIoOAuqap3VVSpTiryRldMzK116xFbcPttlPlgwTqqLTSIpOvEia7W2x1I109oUVamlRnXmqdNXb/nwS63oSzsiUlBXZpZ2OJLANEzh6eoPOC0w5MNqOjpl3hQVNXEVfAoJomqrzJUgtEqTOtcZ+ZDW3SnAQnIhOI4rSr/AFVIeZVT83NW5iMJToUlJVVKTehJ2787STs82bNqbK4VHOVsmy/nUZdK5xcKUApQClAK2mG/53a/QX+S10dzfXaPvR80VVejl4Eyr83fukXs5WP4uMfWZVfbTgndGQ/sG5dfFy3fVm63OO8vcOZnWFbLim0x71a+NB/qWSi7UcBdRJNFRUVF71AV/wAkHJ33h2/57vn05IOTvvDt/wA93z6EWHJByd94dv8Anu+fTkg5O+8O3/Pd8+gsOSDk77w7f893z6ckHJ33h2/57vn0FhyQcnfeHb/nu+fTkg5O+8O3/Pd8+gsOSDk77w7f893z6ckHJ33h2/57vn0FhyQcnfeHb/nu+fTkg5O+8O3/AD3fPoLDkg5O+8O3/Pd8+nJByd94dv8Anu+fQWHJByd94dv+e759TDLzJ7BmU6XDsRw/Esa3FW1lrH3KryghIG5SVV5kItP0rQk59bxlLyQydz0wRblIbvhCQ8uGm9dFJm6qp20R/MEp9yOn5mEq9bPg2Ll1lphPCsHTqOxwI9tZVE01FlkW0X/FB1ri7tf0+r4fdF9DpEUrwYMEJgrhR8Ism1/2W8JYLsy3rrs4xuajn6NXQdL/AOb89dUVs7nT3zB0pf8ASvIxqq02ikoHBMwlb8M4Zsjcy5FGsGLOy2O4bgq44+jhm2wa7edkBJsEH/hYbTXmrVxOCFbYt0gv9lVyKDBJ9I8EYkUERt2fFnnvcFtDccV6GyiuEqqoov8AWVSXolRusbcGS14zO6qt9uEFLq7cyli022W5udFaYdAdR7nbxDRiXPzoqLqi833jDg9Wqfg25Q2xm3lwRxFJZt6yQjLJduhvOuNcdsXitDdUQPRVHRFXVUoDy4PGEsXW2741xHi87r1Ven4bcZu9jDCWjTDCBuIIhE0CKREmiEqqoqS6ISCN00ApQClAKUArAxB/MNy/uzv+haA5c4OOOIsTg8ZXMFbb04TWFrWCm1anzAlSI0mokg6KngVKsXs/ie5V98USPMr4hiKDdaf4lpfWtZ34y/CsxWPCZxpGn5C40jhbrw0TkFRQ37Y+2A90POpKOiJ+da6qxHYIeK8PXOy3ACcgXKK7DkAJKKq24CgSIqd5dFXnr33ozBww802ud1O/Ujm4t3kit5+SDOKozkyRjq/XCfJYdgHdBOKqlBcBAdjiAtcUO5RQ1cEUPeKLu2ogIe4O1qBy3vW6+XS0TLcUkokmKjJEzx7kkj0RxsxXuZboJqi6aCvOqc/sTSNDa+ChhiJEjQGb9dHWoLYsmAkzvFwWpiNHrsXaYpcXSRFTRdG9UVEXXZ3Tg3YeuNlxU3f7zPuCYhCId5mSSaaV5Iz7ryL3IiICou8WuiJoDY/1tSUDHtvBYsVka22y8zLUT0tiTN63W+BFCaLLrDrYOA1HEU0WOiKQohKjjia6bNnnmzwbQx1fZ+JbffJsW/vG2rCGEchYRGwaIQcNkjEEQONFvVQ4xTJRXeqIB4SODIl3veJWrliK4vWK7wmkfaFI6HKmcbNcN49GkUdhSgUBFUT72KKioi7ttifg7Wa/yTCTiK5R4j0hx6HbVSOTDbzjjb0hNptqrwuKwSk25uHQ3dERFTaAw5wcbXhG1RLexia8dZ4b0CZ1G71ODRvQyi8U6e1oV/3cFltURUDRTJBRVRRkbGVzMrAeIMJv3NyVYbs0+20ogHGNA+pm4u7TQtScXTVF5kTv68wG1umXdsuNwjSWkWCMe1TrSEeKAg2jco2CMtNOYkWOOntd0WutbyyWpux2WBbWjJxqHHbjgZ/hEgCgoq6e3olAZtKA5zzXvLVk4U+FXXY8ySh4MuYIMOMb5IvV0JdVQEVUTm79b/s/ie5V98USPMr5b6QUnPHSaa0LS0uo6+GdqaHZ/E9yr74okeZWt4N9yC7ZpZ1SW2ZDAldLYiNymCZcTS2spziSIqVd6N0nDGNtrmvQ0+tGOKd6fxLbx5bbRPtMR293Bq2QIM6PNV990G21NtxCESU+bQl0T/GqrwvwSsEWq2MlDmyp/EvsSrROcMHCt7bc92c02ySJoQb3yDUtVIEFFVe/X0w5RYmAsvpWX7EK3x8QzrrZo0ZYzca4C0pNCAstxwBWwHQQBtzXXVSJ1VVeZETWwcmrFh+y3yE3OkssXfEbWIXXHjHUJCSWXQZDVNEb3sgKD3+6XRdVoDDwdkLbMubjBuOGZbkG5LEbh3WQ42JLdUR8XieeTT/eqqyNFHREWSa6d5Kws0MikxXZsRO2me+xfrg6/JhvE4IDDlOMxmm5ArtVdWVjNPCmi6kOntpoBrbTwa2W5Vw468zYMGHcm5mHokRGFbtyttRRbdTc0pESdS7dpqQ7TPwptzY/BkskYr4vXq6PLebROtksneK3G5M0WTJ1QE0MyFC2omxF10FEXRANcfBLsZS7lKTEl6GRPNDdfBI4ukSNTWxU3BaQnS23B1CJxSUkEUVdNUXeYJ4OdkwK/FOHcpjoxpLcltpWWGQFRkTpG1AabARFTuL3MKJoggid5dQI/A4I9lt9gt9lTEd0lWu3QnYcOHOiw5DTSOvR33SUTYXepORhLutdu7udqoipdWH7O3h6w221NSJEtuDGaihImOcY86gAgoTh/wBYl01VfbVVWgM+lAQXPn2DcxPi5cfqzlRHLf2O8LfBUX9yNeD9KuZR8X9jo4P8xIqV89OkKUApQClAU1ivDcPFOckZiY2iRSZbVCFqOZOGAvIqi4TiPNKKKqaACpz7kVCVSH7lO4djzccysRySYjwcTDNZRtV3Ebdqi68yd9NpF+bvc9dRSnLJjHTk/dFVkf1zCuXdxi9VsXt0W33AdEmnULjCBoGxVBIF3psfDudFEkdTVCRalOFnMI5eYeZjwrs0MB1lh9o3nEXc11PsZUdqJqitRl00Tn2L31qqpKvOORKJKUUZdwzDt0qMMewy41zu8kwZisbl27zQl3F/yiIGRac6bFT8LRK87Vb2rVIfS3qN9xG5uamXWSSaNmmwlA1HVWx0NFFoU08OmqlW1SpujRyZZsrnPVBdS96WbP1pLMmyqTypZurR4/svMkMCytQ5JynHDmTiEm1lPIO9G1NSRtNqIiCm7RObXmTVVVNa2FcurUdWWU82palqRfGOSrClVGQpQClAKUAraYb/AJ3a/QX+S10dzfXaPvR80VVejl4Eyr83fukXs5WP4uMfWZVfbTgndGQ/sG5dfFy3fVm6nVAKUApQClAKUApQClAKUApQHI3CltFqZ4VXB+OVdBgNYjuBQrlEIFJJowHAmwFXTvbJhCKKvtya6cxb/I2P+v8A+1cXdr+n1fD7ovodIio8n/xk82vgHDn7y6VOc171Os90y8bhS3YwTsTMxZItlojrSxZJKBeFNwCun/KlWbk+o0fdRFbpJFVYT4UWKb9ZcPnNwjaoV3xZarRc8Pxmbq460iTTIVGSasio8WI8b3CFuRVDmVEIsm98KC+YWxjbsNXnDMCLdoyROvEVmW++4SSJbjAHD4tkhMUba6oVHVBdrghohIW3rFJHsVcJfMGVhDqi22CzWWbdYxzrO91yN5Uaj3aJCfF9Cjqgq4Mpsh2oWiKSLoqIqyLD3CHuFuO7Wp6ynKnuXaTGtPVFxJ0pjy4hk258NeKRQbYTqdxNEXa24g/1NygbnInhIrnRiI4rdtjs22Zbiu1ukRDfM22EcAEbk72QAHSF1o0QCNOdwf8A3e4tGXCouISsZx5FpsVrk2Z5WWoV2uzkKUwPV4xBdkNusipNmJo8BR+NQuZsVIjBSAjQcIa7Yvu2E7kaSIMd58bbJg22W6yy6+GJ7bb0fRXGhPYoOGqgQIqg4YaprvrNsPDLuOKbSk2zYZt9yW5rbXLM2k2Q0AtzLhHhiMxwo+1txBlNO6N8Yipxgp+BuICWWzPfGmJBbtFkwlap+LoPXM7pCO5m1HMYUzqVAjOE3zm+qKQ8YgiG1UJe8q++UnCSXNLMOTZG7ZHatbzc5yFJYN83mkjSBZ0koTQtirqErgbDLRAUV56AvOlAKwMQfzDcv7s7/oWgOf8Agz/i35UfFO0/U2qsmvhGI6afi/M9DHmoqvhTfi9Y6+Dy/wBQ1fmPp92tmCL9KsMVZ19bhPLb4yIqo5J2KjSL/wAu/bqvtJqtfQ/Rf1ap732RzMXzl4HN+EcPZlZP2UMGQbROctsJZLNvesy9UMkrz0B4XDcNAVFFSuWqqneTTnUgRfDDNozUw9hOHZ4LuKIkiHG6mUnmAkstQhhaGY7iEnJKTt21EPcrOijqm3X2hon3bbRmrarJdr+wxiJq73K6LHfgo80SlES0bBl6qzu38ayxpqCmm3ZtU1JS1Ei85t3SFiKyNOYldxNEsKrFhuxmupDbOXdgQ3kLcSuHGZiIGqqqmga66u0BMMSXDHWFLuxKu1+v0bD9yu106tQDjo+yy3KmLCbib0TTVpI5knOqtNkRcyEtTKy4oxFiXILDMuOV2dvQRrSN6djsoM4xJqO5KJlFTRT2uLrtTX8NA7tESgILebtnEUVu3wIuIAfa0dlTpkdlV6mJbdqiK0u0nkBZ2otc6KJ7edW92bgm25h3LHuB38WJf5iwbq5JbcdjMtQ2oK2mQ0BvIKqqSePdMV59VQk1EUUUQDWYmtOcKzXmG515l26fIlELJR23dBKdIBxktNog0kIY+xT1XUzIdx6Iuii3rMzD13tcC8Diu2WCVMahE1CbBx4RbVwUBktqaIbPFntAddqaIRugegG2kv54hIJt07wbbkKIVyfFgB6mRG4yOAyDW7jXCPjyI2CEhTjR2KvEFXR2Xq3ksCYfXEJuOX1YLPVpPNA04r2xNymIEQiWvfQVUdddOagJBSgKDx9+Nfhj4k3P69BqbV8m9IvX5eC8jsYbo0KiOQfsvZ2/Cdr+zWK2PRn12XuvzRji+j+JNs54hyMN2p1LdJurMS9QJUiNEjFJcVoHxUyRsUVS0Tn0RFrn/FZ5i4GwVimfYIl4wzCai3G6WaAwwyEOFGI5rr6yU0VQe28S42GvckbYomiPIn1E5JvLpPzZZv8ABW1N4pcsrVwYKI5cGmiedim5HCUMgAFNFD/aSEiVe5IOLFdqkn9tWH82LezY3ZN0xFdjO82pmZGniwbYxeIgPyXtEbRUVJHVQ669yiKIoiIlAa64Rc8LXZrIxDuN9fF6xRJkuQ7HbkSm7wbSorKCmxOKQh1ISVAQlHeSCRLXWIIuwd2m7Tn08NAf2lAKUApQClAQXPn2DcxPi5cfqzlRHLf2O8LfBUX9yNeD9KuZR8X9jo4P8xIqV89OkKUApQClAU5dH7bbOEPbWEgRDl3BtHOPbYYFxsxZcRTMyYVxVUUEUUXR5ubRe6RclMOhiGZmC11qW7OdkTbSsjPchLxTlvgi7q4HOo7V1UO8W3w6V0pSlBKV/wAq7+vvv/OoxgoylaTsrkjuWUdkfbbct/VFrnMsgwxIakvKjQijQ8w70RCUWGhU00PQdN2irr9zss8Khb4LdxFxWosJq1g49OdaQ2xBxkBJBMRUlR9wddNfvi/mqmOIqytGOd+HyIcUs7IbhfLKx2nMKB2P2orBbktLxPALrqSHBMgABMD52NdHFQhVDXaWu1auKNGaiR22GQRtpsUAQT2kRNE/+lbu6NVyVOm3dqOd63dvq6rPNrWfrKaMedLv/Q9KVxjZFKAUoBSgFKAVtMN/zu1+gv8AJa6O5vrtH3o+aKqvRy8CZV+bv3SL2crH8XGPrMqvtpwTujIf2Dcuvi5bvqzdTqgFKAUoBSgFKAUoBSgFKAUoDlLNjA0rPbMvN1bVol4wVhyBa8PyFTXibyrqXXci+13TNr19vRF8NXThzHcTNDK3CeLYIq3GvUJmcLRa7mlNtCJstedCFVUVRe8qLXF3a/p9Xw+6L6HSIpnD2cmA8p+EtmYONMZ2HCZTrDYFijeri1EV9AcuW9Q4wk3bdw66d7cnhqaX/hM8HrEz9nenZv4IcctM4bhFUcSRh2vC2baKujnOm1wuZfDVm5PqNH3URW6SRCXcd8Fo7FaLWzm7hSINntUG0W6VGxYwD8RqGaHHMD4z8MSFF1XXcmqEioqovjExpwXrfJtr0XOnD8fqbiilNt40aQbobck5QHL++ffS4911xV5tyuEhbh7musUmRdsfcFu72KJaizdwnHahwJVvjPMYrji6y2/JYlGQrvXukejMkiqi6bdNFRVSsyJmjwXYl3w5dEzWwY7PsFwuVzgvu4ojqovz3DclKScZoSEbikgqmgqgqmm1KA98CZxcG7Ly5vTLXnVhl0eIWJEiTcXMPR7fHU96sx21c0AN2nhVEEBRUEBFI5OxRwXrhNelu532JZIGTltcXGbBLZyKS1KPqTU12ITzDRKJbk0BAREDuaA9bTivgr2gmiTOLDU02pvV4OTsYsvGj3V0aeSqROKRbpURtxdyr+Eacwronvacc8F607QHObDcqLHehuQIcrGLLjNuCNKCUyzHBXNAbR1sF0XVVEBDXYIigDEGN+C3iMTR3OHDEVH3pxS0h4vYZ6sZmPI9KjPaOd0y4YpqPMqJqiKiEWu2wjnDwbcE4lkXi3Z04YIiGQEaBIxcw5Egi+6jz6MNK5oCG4KFz67dNo7R7mgJzywsiumPA3lDF9JTlhZFdMeBvKGL6SgHLCyK6Y8DeUMX0lYV74X2Rr1lntt5wYHNw47giI4giqqqoroid3QEc4M/4t+VHxTtP1NqrJr4RiOmn4vzPQx5qKo4Vjzcfg6Y9ddMW2m7aZEZrogohCqqq+Cp9ywsiumPA3lDF9JX0P0X9Wqe99kczF85eA5YWRXTHgbyhi+kpywsiumPA3lDF9JXtDRHLCyK6Y8DeUMX0leY8LnIUJBvjm9gJH3BEDdS/wATcQiqqKKu/VURSLRPa3L4aA8LpwrOD5fIZRLjmtl9cIpKhKxKvkNwFVFRUVRI1TmVEVPzpWXywsitPZjwL5QxfSUA5YWRXTHgbyhi+kpywsiumPA3lDF9JQDlhZFdMeBvKGL6SvN7hc5CyFbV3N7ATqtFvBTv8RdhaKm5O75l0VU1/PQHpywsiumPA3lDF9JTlhZFdMeBvKGL6SgHLCyK6Y8DeUMX0lOWFkV0x4G8oYvpKArZM2cE5r8KiyP4LxdZMWMwsGXEJLlmntShZIp0JRQ1bJdqqiLpr4Fq4q+T+kXr8vBeR2MN0aFU1gDPDLvKvOzOSFjLHOHsKzJU+2PMR7zc2YpugluZRSETJFVNU01Sr/Rn12XuvzRji+j+JZnLCyK6Y8DeUMX0lecnhc5CzIzsd/N7AT7DoK2407f4hCYqmioqKeioqc2lfUTknpywsiumPA3lDF9JTlhZFdMeBvKGL6SgHLCyK6Y8DeUMX0lOWFkV0x4G8oYvpKAcsLIrpjwN5QxfSU5YWRXTHgbyhi+koBywsiumPA3lDF9JTlhZFdMeBvKGL6SgHLCyK6Y8DeUMX0lOWFkV0x4G8oYvpKAcsLIrpjwN5QxfSU5YWRXTHgbyhi+koCG50cLHJS75O47gQc2sFTJsqwz2GI7F+jG464UcxEBFD1VVVURET21qU5b+x3hb4Ki/uRrwfpVzKPi/sdHB/mJFSvnp0hSgFKAUoCnbXg2+PZ5yr6EZ9uyMOmhib4w2kNWVRHBabVzqol3aKTvFbddU3aaLLsAf+o8w/h8Ps6FW/VnGa/D1RXmtv8XVcrSsSO53fqQkjxmSmzzElbYHVB1RNe7NEVG0Xm5y7/tItaC43WNarmrch07/AIhLcse1w9U4ttS1BSBSUQ00RFePTXRdNNUGrcNQlKyjzmr36ox65PTb5PRa+Uiuc0vh83q/n2NrhmxyLcMqbcnm5F4nEhyTaReLbRE0BpvXn2Airpr31Ii0RSVE3daeJqRq1XKCzaF4JWV++yVy2EcmKT0ila5mKUApQClAKUAraYb/AJ3a/QX+S10dzfXaPvR80VVejl4Eyr83fukXs5WP4uMfWZVfbTgndGQ/sG5dfFy3fVm6nVAKUApQClAKUApQClAKUAr+GQgCkRIIimqqvMiJ4aApngni5d8rZGMnxUZGN7zOxLzp3477yjD+SI3GT/Co9lchYKxRmNls73DFovXX60AvMnW65cY+iCngCUM5tETvII1xd2v6fV8Pui+h0iLBUULvoi/pSmwfAnyV8bO4Ng+BPkpsHwJ8lANg+BPkpsHwJ8lANg+BPkpsHwJ8lANg+BPkpsHwJ8lANg+BPkpsHwJ8lANg+BPkpsHwJ8lANg+BPkpsHwJ8lAf3TmpQBU1TTv1/Ng+BPkoBsHwJ8lNg+BPkoBsHwJ8lNg+BPkoBsHwJ8lNg+BPkoBsHwJ8lNg+BPkoBsHwJ8lNg+BPkoBsHwJ8lNg+BPkoBsHwJ8lNg+BPkoAgoneREr+0Ar+KCL7SKv50oBsHwJ8lNg+BPkoBsHwJ8lNg+BPkoBsHwJ8lNg+BPkoBsHwJ8lNg+BPkoBsHwJ8lNg+BPkoBsHwJ8lNg+BPkoBsHwJ8lNg+BPkoBsHwJ8lf3TmoBSgFKAUoBSgFVnEl3/AAxiPGrY4Nvlzi3O5jKiz7ZIgCKgsKM0qoj0kCQkNo05x9pF50Wtijku6lJLN13tpWpNmMu41sSPPjtkDuCcfS0PTep3a2tE4qCgoRk1NBTXRETUlXVE563loxDLsEVY9tyqxBBZVVJQjuWkEIl75LpN51Xwrz11MTiZYmLhvkIx1RUks2i/4bu3Vdu3VY14U1B3s2+//wDpndnt86N8T/SLX67Ts9vnRvif6Ra/Xa5u8w9rH/y/9S/Kery/Udnt86N8T/SLX67WDe8zMR2y1vSY2VmKp7ze3bHbk2tCPUkRdNJi+0uve9rnVE50mNCm2k6sf/L/ANSHJ6vL9SfsuK6y2ZNkyRCiq2em4F07y6Kqap+ZVT89fdajLBSgFKAUoBW0w3/O7X6C/wAlro7m+u0fej5oqq9HLwJlX5u/dIvZysfxcY+syq+2nBO6Mh/YNy6+Llu+rN1rOEBmXesrsH2ubh6DAuF2uV5h2hlu5uG2wCvns3koIpc35kqYrKaREnZXIB2ys8/cnL36ZO9HTtlZ5+5OXv0yd6OulwCetGhwyOodsrPP3Jy9+mTvR07ZWefuTl79MnejpwCetDhkdQ7ZWefuTl79Mnejp2ys8/cnL36ZO9HTgE9aHDI6h2ys8/cnL36ZO9HTtlZ5+5OXv0yd6OnAJ60OGR1DtlZ5+5OXv0yd6OnbKzz9ycvfpk70dOAT1ocMjqHbKzz9ycvfpk70dO2Vnn7k5e/TJ3o6cAnrQ4ZHUO2Vnn7k5e/TJ3o6dsrPP3Jy9+mTvR04BPWhwyOodsrPP3Jy9+mTvR1FMW58Y6vGWecWHbzBsttxVDh2602iRZHnnG3JV2M4rC6OCi6g4QLoiVTWwsqMcpstpYiNWWSkdR4Ww5Dwfhez2G3hxcC1w2YMcP8AhbaBAFPkFKpHhAR0wXnBlhjwFRuJNdewhdj10RW5WjkMy/6ZLItp/ei8Nee3UgqmCqxep7er5nQou1RGDfc1b81mBdsJ4ZwBcsXSrVCiTZkiNcIcVtsZBPC2Oj7gKS/7O5roi6c3hp2dZmdCl58fWv1ivBYX0bxGKowrxmkpK/WdCeLhCTi1oHZ1mZ0KXnx9a/WKdnWZnQpefH1r9Yra5KYr2kfmV8Op6mOzrMzoUvPj61+sU7OszOhS8+PrX6xTkpivaR+Y4dT1MdnWZnQpefH1r9Yp2dZmdCl58fWv1inJTFe0j8xw6nqY7OszOhS8+PrX6xTs6zM6FLz4+tfrFOSmK9pH5jh1PUx2dZmdCl58fWv1inZ1mZ0KXnx9a/WKclMV7SPzHDqepjs6zM6FLz4+tfrFOzrMzoUvPj61+sU5KYr2kfmOHU9THZ1mZ0KXnx9a/WK85OYOZMWO685kreUbaBTJev1r5kRNV/8A3FOSmK9pH5jh1PUyYYAxfHzCwHhvFMRh2NFvltjXNlh/Te2DzQuCJac2qISIulb6vF1IZEnB9WY6Kd1cjOZmOWMs8A3zFMmI9cGLVGKQUWOoo47pom0VJURFXX210qN9nWZnQpefH1r9Yru7m7jVt06cqlOSVnbPc1q2IjRaUh2dZmdCl58fWv1inZ1mZ0KXnx9a/WK6/JTFe0j8zX4dT1MdnWZnQpefH1r9Yp2dZmdCl58fWv1inJTFe0j8xw6nqY7OszOhS8+PrX6xTs6zM6FLz4+tfrFOSmK9pH5jh1PUx2dZmdCl58fWv1inZ1mZ0KXnx9a/WKclMV7SPzHDqepjs6zM6FLz4+tfrFOzrMzoUvPj61+sU5KYr2kfmOHU9THZ1mZ0KXnx9a/WKdnWZnQpefH1r9YpyUxXtI/McOp6mOzrMzoUvPj61+sU7OszOhS8+PrX6xTkpivaR+Y4dT1M9MK5pXq6ZghhDEeBrjhC4PWt26x3JU6LJB1pt1po0TiHD0XV4e/p7dWNXmMfgp7n13Qm02raO83aVRVY5SFVhPzZxHKxviPDuGMubpirrAcdqXNYuUKM2husi8IoLzokugmmq6aa1nudufPdKs6NNpNK+f4fqRVqqlHKZ69nWZnQpefH1r9Yp2dZmdCl58fWv1ivSclMV7SPzNPh1PUx2dZmdCl58fWv1inZ1mZ0KXnx9a/WKclMV7SPzHDqepjs6zM6FLz4+tfrFOzrMzoUvPj61+sU5KYr2kfmOHU9THZ1mZ0KXnx9a/WKdnWZnQpefH1r9YpyUxXtI/McOp6mOzrMzoUvPj61+sU7OszOhS8+PrX6xTkpivaR+Y4dT1MdnWZnQpefH1r9Yp2dZmdCl58fWv1inJTFe0j8xw6nqY7OszOhS8+PrX6xTs6zM6FLz4+tfrFOSmK9pH5jh1PUzAxDmvmBhewXK83HJi9M2+3RnZchwb5bCUGmwUzVER/VdEReZKsuw3ZvEFit1zaAm2psZuSAH+EKGKEiLp7ei1xd09yKu5ig6kk8q+juNmjXjWvk9RnUrhGyKUApQClAKieBsV3TEtzxfEuVsZtyWa8Fb4pMyON6oZ6nZdB0uZNpLx3OPPpzc689WRipRk286/UhvQSylVkilAKUApQClAKUApQCtphv+d2v0F/ktdHc312j70fNFVXo5eBMq/N37pF7OVj+LjH1mVX204J3RkP7BuXXxct31ZuoPwtf/SWCPjrZ/wB/VlPnx8TCfNfgesk1bjukK6EIqqL+fSqULPiRhXJ3LS8XBqNdsRYltUWQfVUhYbRn1ILzzik205tVVVEQRDTcaIu0dVT0s5ZOf+dRwIxysxhXHhXRIL05wLAsiE3Z3bpGAJydWGQRW5CtPM7NjCqLm1NzinqPOAiqEvzjjhEXnDduxK3OwytpC2o/b3J8K5tvPNTxsvXRUbbNhQIBDcHGH3yEfvaiq6VOtmzItVLvNi7wipzd0KH2Jt6zZk+32Uiun8sfiXNq3HxycUvEArjwGhIri7UPudUFCsbLjGL2PMLJdJNt60y25063vw0fR9AdiynYxqhog7hUmVJF0TmVOarIzynaxXKCir3JPSrioUoBSgFKAVz7ZhHFvDeHBuqmyydsxZNbH22YMaW2zr4E6qmRiTw8Uune5ufjei+Ju4TpPgd11VvCUwGeZuUN+w3He6mnzGCKDJ/IS29HI7v/AMjwNl/8teM3UnveDnPVZ7Gjv0VeaRUHBXx03mbmXjPFDbfEHc8KYZeejr32H99zR5pfzg4hgqeEVq68zTvcbD3V1mltRBgjJkzFNVQjaSG+gIGiKikjysFz6JoJfmRdzcyG94OnDUrbDWru9VsrrGOYVztGUmW1x6tmpdrxEc3SY7oAhPJYZ0rc6KgW8dzCFtFQXegLuURUCg9s4WJTsA2TGEK2zLxbY9vnqwTdxbZ67vRI7XHk42rSq2nHKYgqkKrpvUdpDr0rlVrk+i564om3lzCzWDrauNWLu9b3oa3s+oRYbhR5hPJI6m3qqhLYbQOKTuyXn2puqY5M4knY+wZDxnKR6JHxLGiXOLaXnhd63gcVrcyhII692hqvf51VdURdEnSQ1YnlKkxFKAUoBSgFYF//AJiuP92c/wBK0BS/Bn/Fvyo+Kdp+ptVZNfn3EdNPxfmesjzUVVwqPxecd/Bxf6hq1s08ZuYAwVIvTKRVcCVDjIc0yFltH5TTCuGo8+0UdUl0/wCGvpHon6tU977HHx+ecSNW7PvD0IY8S+XNkrg6ybwSLbEfOJI050bYJRVXHFFR7kdVUl0TVeavQc/8MsS0bmOTI7bs9u3Bvt74kw6SNCov7gRG14x4R/x150ElT3Nzm5JkRc6rRCwPgu/34HreuI4sZ4RYYceaYJ0W+YzRNBFCdFNV0VdddNEJUg2HuFjbCuUtrFFvdsEYIXVzKoy664bfVcxoiUUHmAG4oERf8TmiJ+DuXGSSbFHCKsNnAOoWZc1sykgs44jwxQFqPJd45D2/fG1WI6KKGuumqaooqWTI4SOA4rsJp66PNnJiNztFiuLxLRLoRO6J3GxVHehaKO8VXm1VFxkm1xVnVhTB1+ds1wmPrcGWVecajRXHtmjRuoCqKKm9W2zJA76oic2pghaguEngYYkuYsyctvjRnJazht7xR3GhYckCTbiColvZZccDRe6FEVPwx3LjJufwuErgUEeEps1uQ0uxIrkB4HXHEdBk2xFRRVMHTACHviS6e0untJ4QWExnK0E51uPHfcalyJEJ8ABAiPSTEV2fhiLK6gui99NNVTVdDJPtrhBYQO4rBcO5xZLRstywkWx8OoTdcVtoZCqP3pTLagoXfQxVNRXWpVgfHFqzEw+zebMT7kB1fvZyI5sqaKKEhChIm4VEhVFTVOfTvoqIvchokFKkgo/F3412G/iVcvr0GrCr496S/wBRl4LyPQ4PoUKgWSfsxZ1/CVs+zWa2PRb15+6/NGGN6L4kzzhzEfytweF9j2s7zsnRmHorRKjnEm6Ium2iIu4xDcQh/WVETVNdaiULhDNTsVYVtrMGO/brvcLjEkXFqTqEVtqQ6xEc73ddUG0opzomuvOulfWGzh2uiXN5xYYdwfNxN1RKbtEUmk41+G60rqO7OJJtDFFMXOMDaSc3dc6poukbh8JLDIXG+M3UZtriwpDTMSS/AfEZPGRIr4t84Jo+qytqNfhcyc3f0XGTc98HZ6RMd5gN2S1RxetbkZ50ZpqQOIbStibZAqcyiZGK/nCtfmFntOwUWJIrdoYlXG1PyDBk3lFHITdqOb1Suiaoiugsf/q56XFj2i51Siy1zBxJ/wCxLqmF4b8li42qYTlvmK3F45R4zTuVFe5LRSROZdddRGO2rhQJ2WQ7TOjWudahkSGpmIbPMJ2IIJFR6O63qPdIZhIYXQl0NhU1XVUGLjJuT3LrMmdiq8yLVd2bLDuIwWLikK33YJEmKDqa8VIZVBMSQSbXeiKBb+bTRN3piTPHC2E8QzLJcHZ43KMLRcS3AePjuMdjtIjWg/fF3ymEXbrzmqc6iSDNxa+g1sLhIYJnNPnx9yiq1FOXxcy2PsEYCjq6AhiikSrHkCgpzqrJ6apoq63E/CgwnacP3uXbhuNyuEK1ldY0UbbIRJbHFvGD4Kgc7C8QWrqdymo+2Qoq4yTYzeEBYm8RMWeOEpySxMNi4tuRHhNpoYk17jWg27nUU4LgIgpqvfTVFFSmuCsa2vMCwN3e0OOORSddYIXm1bNtxs1AwIV7yoQqngXvoqoutNJDVjSZ6+wjmF8Xbj9WcrT5b+x3hb4Ki/uRr596Xcyj4y+x1sB+YkVK+cHXFKAUoBSgFQfBEhuJe8yH3SUWmr6hkqIq6IlthKq6JzrV1OLkpJavujGWYmkaS1NjNPsOC8w6CG24C6iQqmqKi+BUr0qqUXF5L0onToFKgkUoBSgFKAUoBSgONMu0vGIc/o2Hbrd8ZNQnwnTwIp+IbeAqw41tFEmbGngXjdFRrciIid4VRa7fw3/O7X6C/wAlr00clY/DKKVrx0ZOvXHM9Bqvo5fHX9yZV+bv3SL2crH8XGPrMqvrBxjujIf2Dcuvi5bvqzdQfha/+ksEfHWz/v6sp8+PiYT5r8DIMEcBRLnRUVFSq4g8H3B9sssa1w+v0eJE4sYidktyM4gg2TaAwZyFJoFAyFRBRRU26ou0dPTygpaTz6k46D4f4OmA5D5EtrkjHIHm0gt3GSEVsHWVZdEGRcQAQx0VdqIqkKFrqmtbvEWUWEsWMXNm62lJTdyluTpQ9UPBxjxwVgGfcmmmsZVb0TRP62m7uqx3qNrEupJ9ZqMaZLWi9YeNi0RWIV1ackvxZD70ja2cic1Nkqig4JiRvMgSGK6tkiKKaJtXcZTYDLLXAkOwuSkmvhIlS3n0VxUJ2RJdkHorhEaohOqiKRKS6aquqrRQSllLUHO8clkvpVpWKUApQClAKqPJW9WK2cPHMOBOaNvEF0wpbBtkk00bNto3iksov/Gu9g9vti2S/wBWufjei+Ju4TpPgdi1osW/yNj/AK//ALV4fdr+n1fD7o9BQ6RHMXBiwGeXvCRz/hA9vtk7rPdYLP5AJB3B10ET2k6oKQSJ4CSumLtbWbzaplvkbuIlMmw5sXRdpCorovh0WuluZPfMFSnritvWaldWqyRWMLg6Wpq22q33DE+Jb5AtKEkGNcJLCjHQoUiEqDsZFV+8ynE51XnQV9rn8ZnBdwXNdnKq3JpmZGWO7GakILe5YzcY3UTbzOE0yyhL3lVtF01VdelYquYua+Us5yUd9wjFnO3+ZeeuUmTCujcKSwpQG4aqyTjTjagrbDKGBiq98w7sQSpxk9gp/LfKfB2FJT4SpdltEWA882REJuNtCBEKlzqiqi6KvPp36WDd0TClSYilAKUApQCsC/8A8xXH+7Of6VoDnTg441ai8HjK5lbRenVbwtaw3tW9wgLSI0moqnfTwLVi9njPuJffFrn8K+DYig3Wn+JaX1956mMvwrMVhwmsZNTshMax0tN4ZVyAoo4/AMAHuh51Je8ldFYut1rutpYZu8gY0QLhBfAydRvWQ3KacYHVf+J4Wx298t2ic619D9FoZGHqK6f4urwOTjneUSDX3I7B2OcZXG/OTJR3MJIrICJIBOIkjGFsCVUFTExBWXBFS0EhEkRN57/C58GjC93ntT5c+8v3EXSfOY9JBx1xxVZLdqQLt7uO0WgbU5lDTi1UK9rY51zPxDkBhnEljwxa33Z7LGHYSwITjLgcYjWjOmqkBd0ix2l3Doq7VFdQMxLW4g4PeBps6CcuRJiXB9pbZFd6oBHXG90iQ4y3uFeckcdJVFENEaQhUVDWlhc2Fw4P2HbjAeguTbukEjdVqKkvVqM241IaNpoVFUEFSW8v/F+Am7aACnlL4OGDpU92aLUtiW9cJVxeebdFSMpBNE83qQrsBVYa0UNpjt0EkQi1WFzPxbkbhzGOIZd4kuTokqZHJiSkF0WuO1ZJnepbVNCRsyHuSRPwVVFIAUYY9kBhLC7d9bvd8dgYSmNnHg212YDMeKHUbzSoO4U04plx4W0RdogIqqKrYqJoX6jf3rI7Bzc+6OOXGRanb0b5qyDzIgpOOcfIVAMFR1DJFIhdRxB1XagotY8ng14DhudcJqzFiwoRxQSVM0GPF6ldYMFd0RwhRt01RXDJQVE2KCaoqwuzb27JDDit3CUc243OXeHIMqZcnZScdKOK+j7DikAiiaKgD3CImwBRNNNakOAMvbZlxapcK2m+/wBWSimSH5KhvddUABSVAERRdrYIugpqqKRakREqxDdzfFc4YTQhrKZSWaEosK4m8tERV0Hv8yKi/wCKeGsmpIKAzHvA2XhTYYdKLMmIWDLkGyGwTpJ/t0JdVRO8nN36lXZ4z7iX3xa5/CvkvpFSy90JPKWhaX3HfwkrUkOzxn3Evvi1z+FR7g/XEbtmtnRJFiRGQrnbU4qU0rTiaW1lOcV50q/0ZpZGNbun+F6H3owxrvS+JbOM4tndtLUy+zG7fb7ZJauBSn3hZbbJo0IVMi5kHXTXWqvwpwd8qRw+lxs4RrlbuOGZAuovtSOoBamuzBGK9ovFtg8453KKqbUQV5k0r6k1c4qbRLYOXdgs+AzsLN8uA2ierDUSQ9OFxWgRACOyzuRQUEEAFAISRzVd/Gby10UXg8YBl2N3D8fjX41vntyDZCQJrGlhEjttqo6KgqjbbDiAqbdVRdu1UGlhc2liyLs2FrhEuNon3CNcmFRFkOGB8YCvNuPCoIKCiOI2o6CiIKGu1ERERNrijKTD+Lr3dbtPbf6uuVhew4+405t/2NwlIkRNF0LUl0X89LEXNtfsJhii2YhtdymPvWq8wyhHGDQFZAmyBxQLTXUkL2+9olRLEmVuAMf3fr3cwhXO5XCRGZhzzJo3WzhuE8LMc1RVREJt4jBNdfvmvNqiLC562zKa3YLeZu7OJLsB26KEQJFzlC6EaA2626THOiJt0a2q4ermi90ZbR0wZ2SWDrpmKeICnSRvkl1q59StSG0Q0adinu027yBXIkbnVV27dBUUMkJYm58XvICzzrjh52LxiNwLjDmPG8+W5BiuS3mxBBTRVJyY4JISomwlTnVOeN2bgxdXs3scR3iVH6qt7diiDZJZD1PbgF8SZAjDUAMZCpxa7yDiwVHSJNyRYXN25wWsGOy7i8R3JRnLIV1lHgEF45uUBd4EVVRJslUIlUl3ohKSAKDPMBYDt+XVkctdtekPR3JDkpVkqGqOOKinogCIiKlqW1ERB3KgogoIpNiG7mrz19hHML4u3H6s5Wny39jvC3wVF/cjXz70u5lHxl9jrYD8xIqV84OuKUApQClAVhiF7EZZhFb7DepDDai2+cdxk3owKouaiZLHPbuVBXRHg5hVEQVJCLd4A/8AUeYfw+H2dCrdnk5H4Vntn2rvf2K+s97WS4Kuo2d7XrJMcVba+qdzHcJdVil4E11Vte9ouzmUR3S2pxiy5quvzq/x0S+d3bU0RTzLI1fxfIUrRLRSgFKAUoBSgFa7EVum3ayS4luurtknOhtauDDLbpsLqndIDgkC+DQkVOepi0mm1ch5zg3g34qw5grP3dfMc4cWcAXGFOkz7rZdrr5yASMENWXVfVTFFVwSBrukRFROYa/RfDf87tfoL/Ja9XUjJbp4eTi0m42umuvqv3Wv1XNRW3qST1kyr83fukXs5WP4uMfWZVfUjkHdGQ/sG5dfFy3fVm6g/C1/9JYI+Otn/f1ZT58fEwnzX4GTSvVHnBSgFKAUoBSgFKAUoBVI3DBV1xLmBmxe8MMC/jXCKYexBY215lfeZbmo5F176DIYJ5hf+6i+1XPxvRfE3cJ0nwOucA42tWZWCbHimxv9UWm8Q25kYyTQkAxRdpJ7RJ3lT2lRUXvV/cW/yNj/AK//ALV4fdr+n1fD7o9BQ6RFL5X/AIyma3wDh395c62+f10vFpHL92xRRn3AsUMAMJyWsUH0WLJ1EnEEtE5tecV50Stzcj+n0fdRr4jppFQlnHjmwYqxna27YlsxDdMWmnFsw5GIGoTDFitZ7QaZVki3m6CquooO5xeddNfW75tY2zDtdnu5JAw/ZoOJ8LQp1nRlxZxvSBgTHSB5HEHYiygZ4tQXVAcVS50ROvcosjLTP7MVLbgkxcw05Jx7DgXS1qsB9W7Sy/LiMm08iP6yCRua2QmitIRNOaiiKm3ztWdmK5uJ2reTdnHFMt1cPJd0jyOphNm8z4hPrHV/TRW4imgbt280FXFHTRcmyLNxJiS7xcv8Kv3pyzXi7liiBbpMi1OPDFIkuIs8YCCaEBoiIpNERiBoQFxiDqtTYXz+zUuFmsN9uMrCZQZNhw3iOXCi2qQBqF0lLHKM2ZSS27BAzRxRXulRNqii6myLJm0Z4QmOIFlj3K7SsNNhfIVxdtgsW2WfUTsa5xoTYmAOm5KJ1JYaNgjaq6gtouh7h9MNZ5Zg4yGXZIUiw2a92gLpKlXLEVseiMSGopx0aA2ePUo29JGrjikexARUDu9oriyLL4P90xBeMP4ofxBd2Lu4GKLsxENhkwRmOEowBvUnD3Im1dqptRB2ppzaraNSYsUqSBWBf/5iuP8AdnP9K0BS/Bn/ABb8qPinafqbVWTX59xHTT8X5nrI81FVcKj8XnHfwcX+oatLNjDdwxbg8Lda3HWJi3W1yEfYVtHGAanx3XHR4xFBSAGyJEJCRVHTaWui/SPRP1ap732OPj+fEpyfgDM7DuIsWtWty9XyNerrbX491K4RIxNgy9FWQ46DatIm9oXG0RtvUka0JNFRSwiyqzMsdkI7dLxFcLg46xHdal4lfeRYqwofHkIlKBN/VIy1RRNol/BFxsCQk9xZnOujf5WYFzGgXzD9zxG/eEcjT1jyGJF5J1lYKRJYIRM8e4JqrvUi7i3ud5VLmNah3YhmRjPqi54cdusd2LeMQIEu73VJDRv/APtWNFdjAryqwDfGNNKKI3+GC6KjalQZjPi4MzkYvNneFLt1GzIAhRbntNlonTF4HBO4PCugKLiK51WuhaCrSggFmYRwvm1Z2GokuJepTMmXaxbkPXgFcissT23ZRvict5UVxlXB0accQ0DRUbQkbFnDaPTLHLjNrDN/tAS71MSM1AZU5V4lPXNsTWA2jzbyFPRXD6tV5xNrSaDtQXkBEbrT4wyzzivF0v7oypUoxeub0F1iXxDakYSRgmzvmHxZNg42CqLTGhFqqu7d6LMXRs8Y5a5g4ltl8gOxL8+L6SuqP/1CIsTy1kE0sdtHUVgdFYFEVQTVERRFBUz00/LXN6dYb7Hu63q8tPx3GigW+7dSFKlEzJEX23DnGiMoZs7m/vIqm1eJ7hUI0xdE8zOy/wAdXYbQ9h6VcIyW+1xGTixbi40264j48ePFtyWEM+KRURVcFO+iEi6KkNjYLzpZvcZx7rs/D6mZYk8VdthOx0CObjY7ppCD6oD7KGIb0MhNZBiSkBoXRkFllj+HfrlPt8DEkeDPcjPy4xYlQ5L0YOpwWMLnHjtfRpp0OM1TnMfvxLq4nsxlrmclomylm4jauD0+Q1HjjiDe5HtZW51GW1QnuKJ8H+pxJzXepipI4gkrlLC6PnDEK927PDA8bEYG3d28FXdHAdeJ40HrpE4vcROuru4vZqiuubV1TjD03Ld1fIPST+oy8F5HewnRIVAsk/Zizr+ErZ9ms1sei3rz91+aMMb0XxJ7mZZp92stvet0Rbm9bbnFuBW1DAFlg04hKCKaoO8eZwNyom9sEVRTukoWfhzHVwzNxR1riXyzzLkMK7xYEa8A1GtYE+bT3VDYPcW44+1GcXuEcQTd76ab0+rs4asZruA83nQabhS7vBvCASS7pKvKOw31V5hYytMcaWziWhcF3uA4wtV++79U8LHgLMKx4kC6QLRiOBbixEM56Fcr+EonY6RLY24484MvcS6x5SAJK8OiqitghiYLE3RZWYkPFt8udkudit92ejrDFxiKxdRtqRJKuNmSyxVV4wFbRQ2oDmioSaDu3jWuFsuM1SETuT1/YW3m4/AbO+mCEarbFFHR6tfVxNWp/cuuODoZJoiEIoYVrGFe8t84LZhm1w7M5fZcrrFEkSHnsQuPOheRYcF1NTmN9wpcXrqrjOqJ95PcRB6Lljme3aJsSzMX+xici5K82d+DR8nW7lxDsfa+qMghPRO50Bd+wlHVtTpZk3RssU5d5nMJeoVjcxC5t3hYZvZEStxG+qHzeSRxj+94nWTabaI0cVshRdWkFCXVX7LLM5+6zZkeNiIriLT7TlwC+AQS90lw2habSY0bTYt8ShCLjPOCf7xBVHGci6OpLSMobVDSagDNRkEfEDUxRzam7QlRFVNdefRNfAlZVZGApQEHz19hHML4u3H6s5Wny39jvC3wVF/cjXz30u5lHxl9jr4D8xIqV84OuKUApQClAU/crNMPPiz3N23wWYAibQTnVmFJI9jugCu1GEFdSXaqqSacyqioiSzAH/qPMP4fD7OhVvVGpRzP8q1a+4rS8yWz4Ee6QnoktkJEZ4VA2jTVCSow3KueCRRmakm9WME0bnNirsuMngeBOd0U/KDqXe3CvOa54ZxrReGm7Xzxb6panqUtD1NRbdkzGpeLy18fD9v1JLbrlEu8NuXBlMzIriag8waGBfoVOasmtCcJU5OE1ZrM09KZampK6FKxJFKAUoBSgFKA46yl67XHNiQ3fr/NG1zeuJORAxHL3WdWJgAwJmck23+OTnRW229qLpzotds4b/ndr9Bf5LXpY5K3QoKKsrx6kuvuS06e65qvopeDJlX5u/dIvZysfxcY+syq+snGO6Mh/YNy6+Llu+rN1DOFpAuUvAeG5Nss1yvp23FFsnvxLTFOTI4lt3UyFsUVV0TwVlBpSTZjJXi0iBdt53o7zF8lJfm07bzvR3mL5KS/NrvcMo6zjcFq6h23nejvMXyUl+bTtvO9HeYvkpL82nDKOscFq6h23nejvMXyUl+bTtvO9HeYvkpL82nDKOscFq6h23nejvMXyUl+bTtvO9HeYvkpL82nDKOscFq6h23nejvMXyUl+bTtvO9HeYvkpL82nDKOscFq6h23nejvMXyUl+bTtvO9HeYvkpL82nDKOscFq6h23nejvMXyUl+bTtvO9HeYvkpL82nDKOscFq6h23nejvMXyUl+bUh4NHXO75n5oYhlYbvuH7dcGLQxFW+25yE48TQSUc2iaIqom8ef89amKxFOrTyYvObOHoTpzvI2+VZllNnLinLN9eLsd7V/FmF1XmERcdTrjDH2vvchwXkFP6svTvAulq4t/kbH/X/9q8hu1/T6vh90dmh0iOaoOa2FMqeEhmK5iy7hZW7lYrEkQ3mXCF5W3LjxiCoiqdzxga/9SVLLpwm8kr07Acm4tgSTgSUlxlNh/wC9PIJChp3Hf2mSf41duTWpRwFFOSvZdZTiISdWTSI7iXNrg54vbmDdL7AeOZNS4vPMpLYeKQkcI3GI42gmKqwAtKiKiEKaKioq640rMjg0TL3a7qdztQTLX1MsRGWpTTLRR9Opz4oRRtSbRNoEoqojqKKgqqV1t/o9tbSjInqP7ZsyeDTh+U/IgXW1suuyGZWqtyjRo2nVdbRpCFUaAXFU0bBBBCVV26rXrPzT4N1zjTmX7vbFGaROPG21Kbc3rKOWpiYihAaSHDdQhVFQi1RU5qb/AEe2toyJ6jaQ8+cgIGHbXYY+Iba3abW6y/EioxI0adaNHAc12aqSGm5SVVVV1VdVVaw4+cfB1i25mAzebUERmDCtrbKR5GgxoZq5FaTue82RKqfp59ab/R7a2jInqIbgW/8ABxwthaZbJ+JLdc5c83Sly0bmAq7pSyRVpP8A3Co5sNVb2qpghr3SIqbNcT8FkoiRlnW4mVkHKMCWavGuOC2LvGe2Yu8U2rgFqLhDuNCLVab/AEe2tocJ6iaYf4RmRuFnLodrxZEi9c5jlwlCgSiE3zRN5oJCqBu01VBREUlUlTUlVdvyuMofftD/AFD/AJlTwij21tI3ub6hyuMofftD/UP+ZTlcZQ+/aH+of8ynCKPbW0b3LUOVxlD79of6h/zKw7zwsspJFnnNN40iG4bDgiKMP6qqiqIn4FOEUe2to3uWoxuDcw5G4O2VjLzZNPN4VtQG24KiQkkRpFRUXvL+arGr4LiM9afi/M9RHmoqvhTAR8HnHaABuEltNdoCpEuhCq6InOtSPlcZQ+/aH+of8yvonotUhDD1FKSX4vscnHRcpRshyuMofftD/UP+ZTlcZQ+/aH+of8yvbcIo9tbTm73LUOVxlD79of6h/wAynK4yh9+0P9Q/5lOEUe2to3uWocrjKH37Q/1D/mU5XGUPv2h/qH/Mpwij21tG9y1DlcZQ+/aH+of8ynK4yh9+0P8AUP8AmU4RR7a2je5ahyuMofftD/UP+ZTlcZQ+/aH+of8AMpwij21tG9y1DlcZQ+/aH+of8ynK4yh9+0P9Q/5lOEUe2to3uWocrjKH37Q/1D/mU5XGUPv2h/qH/Mpwij21tG9y1EHtuZuGc0+E/aJuFbqF5iQsHT2ZD7TTgi2ZzYaiKqQpzqgkun5lq6a+Sekcoz3Qk4u6svI72ETVJJiqQwnnFg7KrOzN2Piu9t2V2dNtr0YX2XVR0Et7IqQqIqiohIqf4VsejE4wxrcnb8L80YYxOVKy1k+5XGUPv2h/qH/MpyuMofftD/UP+ZX1PhFHtracTe5ahyuMofftD/UP+ZTlcZQ+/aH+of8AMpwij21tG9y1DlcZQ+/aH+of8ynK4yh9+0P9Q/5lOEUe2to3uWocrjKH37Q/1D/mU5XGUPv2h/qH/Mpwij21tG9y1DlcZQ+/aH+of8ynK4yh9+0P9Q/5lOEUe2to3uWocrjKH37Q/wBQ/wCZTlcZQ+/aH+of8ynCKPbW0b3LUOVxlD79of6h/wAynK4yh9+0P9Q/5lOEUe2to3uWoiOb/CjysvuUuNrbAxfGlT5ljnR47Dcd/c44bBiIp3HfVVRP8an2XbZN5fYYAxUTG1xUUSTRUVGh5q8D6V1IVIUchp6fsdXAxccq5IaV88OqKUApQClAaHsMhOXxLq+/OkyG3+PYbdnPKyyWxQ5mt+zvKX9Xm1/xrU4ATXEeYfw+H2dCrYU3OMr9S+6MWrWJpStcyNBPwVb5Mx2dDN+z3J1dxy7cfFk4XhcBUUHF9ruxLSvIWcV20doP2y+AnMnVCHCcRPCRCjgkv6AFK6scTSxEVDGJ3WZSXOSXU07KSXVnTWuySNd05Qd6ex6P2/mY+0xFd2dRewrPcNP60STGMF/QpugvyilbuJJKTEbedYciGac7L6jvBe9ou1VT5FWtavQpU4qVKqp36kpJrxuktjZnCUpZnG2z9fse1K0y0UoBSgFKAoezcF122Y1hYgdxq4+ASXH5lsawzZ2GJQEaGjO4IqOoCKic5GR8yKhIXPXRmG/53a/QX+S13cNiOEY7DtRtaS62+vvbNeccmnLP1Eyr83fukXs5WP4uMfWZVfYTiHdGQ/sG5dfFy3fVm6nVAKUApQClAKUApQClAKUApQFScJHCd1uGEbfi/C8cpWMsETEvtsjt8xTQECGVD/Q/HJ1tE729Wy/qpUnTFNtx1gmwYis0hJdpu0dqdEfRNN7TjaGC6e1zKnN7VcXdr+n1fD7ovodIjW0r42dwUoBSgFKAUoBSgFKAUoBSgFKAUoBSgFKAUoBSgFKAUoBSgFKAUoBSgFKAUoBSgFKAUoBSgFKAUoBSgFKAVWtqsuWmaF+xRJh26BdbrbbkVuuzvFEJpLbabRRLvblQFbHdzp3OmvNV9J1IKU6btbSYySeZm27TGCPe3C+av8adpjBHvbhfNX+NZ8Kr9tkZEdQ7TGCPe3C+av8AGnaYwR724XzV/jThVftsZEdQ7TGCPe3C+av8awr1wf8AL3EFset87C8R2K8iIYApgqoiov4QkipzoneWpWMrxaam8wcIvqJ8yyEdltptNoNigiPgRE0SvutR5zMUoBSgFKAVtMN/zu1+gv8AJa6O5vrtH3o+aKqvRy8CZV+bv3SL2crH8XGPrMqvtpwTujIf2Dcuvi5bvqzdTqgFKAUoBSgFKAUoBSgFKAUoBXN+AgXKnMDE2UzugWdTPE+FkVeYYL7i9VRRTwR5JKqJ7TclpPari7sq+59Xw+6L6HSRNfivOLE0HMi7YSwxgdnEZ2qBDnSZci8jCROqCfEBEVaPXTqctV19tKx+2fml0UW/yrD1evLYL0Tr47DQxMKiSkr2t+5bW3Rp0ZunJZ0O2fml0UW/yrD1enbPzS6KLf5Vh6vW9yIxPtVs/cq42pamO2fml0UW/wAqw9Xp2z80uii3+VYer05EYn2q2fuONqWpjtn5pdFFv8qw9Xp2z80uii3+VYer05EYn2q2fuONqWpjtn5pdFFv8qw9Xp2z80uii3+VYer05EYn2q2fuONqWpjtn5pdFFv8qw9Xp2z80uii3+VYer05EYn2q2fuONqWpjtn5pdFFv8AKsPV6ds/NLoot/lWHq9ORGJ9qtn7jjalqY7Z+aXRRb/KsPV685Oa+aEWM68eVEDY0CmWmK29dETVf/29ORGJ9qtn7jjalqZYmXOMG8xMvML4qajFCbvtri3MIpnvJlHmRcQFLRNVTdproneqRV85qQ3ubhqdjtp3VyKZrY8HLDLm/wCKiglc0tMUpHUYuo0rypom3cqLt117+i1CO2fml0UW/wAqw9Xr0+4+4FXdinKpTmo5LtnRoYnGQwrSktI7Z+aXRRb/ACrD1enbPzS6KLf5Vh6vXf5EYn2q2fuafG1LUx2z80uii3+VYer07Z+aXRRb/KsPV6ciMT7VbP3HG1LUx2z80uii3+VYer07Z+aXRRb/ACrD1enIjE+1Wz9xxtS1Mds/NLoot/lWHq9O2fml0UW/yrD1enIjE+1Wz9xxtS1Mds/NLoot/lWHq9O2fml0UW/yrD1enIjE+1Wz9xxtS1Mds/NLoot/lWHq9O2fml0UW/yrD1enIjE+1Wz9xxtS1Mds/NLoot/lWHq9O2fml0UW/wAqw9XpyIxPtVs/ccbUtTM3A2b2Ir7mOOD8S4Maw1KdtL12YkMXcZouA2800QqiNBtXV4V15+9Vq14vdPc+W5mJeGnK7VvmdShWWIpqpHrFU9e858Vrj/E2GsM4CYv7VhOO1ImyL4EPcbrAPIggrJLoiGia6+1Vm5O5k91sRweErOzeyxjiK8cPDLkfztn5pdFFv8qw9Xp2z80uii3+VYer17DkRifarZ+5zeNqWpjtn5pdFFv8qw9Xp2z80uii3+VYer05EYn2q2fuONqWpjtn5pdFFv8AKsPV6ds/NLoot/lWHq9ORGJ9qtn7jjalqY7Z+aXRRb/KsPV6ds/NLoot/lWHq9ORGJ9qtn7jjalqY7Z+aXRRb/KsPV6ds/NLoot/lWHq9ORGJ9qtn7jjalqY7Z+aXRRb/KsPV6ds/NLoot/lWHq9ORGJ9qtn7jjalqY7Z+aXRRb/ACrD1enbPzS6KLf5Vh6vTkRifarZ+442pama/EOdeZOGMP3O8TcqYSQ7dFdlvK3ilsi4tsFItE6n510Reariw9dxxBh+2XQW1ZGdFako0S6qCGCFpr7emuleb3Z3DqbjqDqTUsq+harG9hsXDFXyVoNhSvNG8KUApQClAahzF9kYuqWx66xI1yUkAIcl1GnXFXvbBLRSRefRR1RdF8FaLAH/AKjzD+Hw+zoVXqEoxlddX3Ri2nYmlKoMhSgFKAUoBSgFKAUoBW0w3/O7X6C/yWujub67R96Pmiqr0cvAmVfm790i9nKx/Fxj6zKr7acE7oyH9g3Lr4uW76s3U6oBSgFKAUoBSgFKAUoBSgFKAVRHC1sUyNhWy4/sccpGJMDyzurLDad3LhqG2dETw8YxuUU/KNtL7Vc3dFpYSaloeZ+DdmW0l+NWK7wVeoOJM/Md3a2SW5lun4Zw5KjSWl1F1oyuJAafmVFRf8akuZtpduOGnZTU1yIttbky1BsUXj06kfa2Kq86Iiuoeqc+oJ4VrvbhRa3KoRelL7nFxr/+VL+dRAL9j29YTyqywj2VuQ9cr63FhrJZgrOcZEYLj5kLW8N5LxO3nJERCIufbosYuGfeLwbw3JBqODgtWrrvAZthuiJyZqxnCceV1AYFRTc2Aq4eqrvTRE3diU5LMjXUIvSZcbMXMm+Jb0j3mwQFur99JlVtLjvUzEF5WmhX7+m8zVEUl5kRFVETXuq01jzcxfia4z0w+dnsD9xB26yHpEN2XqTdks8gRQeOFPwpagqpomwE5kLUlxc5kqEDe2jN/FuLHrZdG3rTDsc29QLG9aHIpm+IvwWpBuo6jiaFue0EdumxELXVavaw2dnD1it1qjmbjEGM3GbJ1dTUQFBRVX210Tnq6Dcs7KppRzIzqVcVClAKUArDvf8AM0/+7uf6VqGSaXgz/i35UfFO0/U2qsmvy3iOmn4vzPo0eaiqOFb+Lpj34NP/AFDXrmvfZuHMFOzLe+/HklNgxuNisi88IPTGWTVsCEkI9jhbUVF59OZa+sehLthK9tf2PN7rK9SmiuRz6uWDHBtt/s865OCkl4JZoDUlWAbkPgrzLYqLZqxFMtFUFJVDQOc9mWGf7o3fECt2pyfDhsPzmmhdFtUhRiMX5CEqaHv+9EAe2jic6Iiqn0TfeqxxHTT0M+i4RMeRJ3x7Y+gx4Zo+w68AAM5H2WjjEqipbm1eb/ARVLjNBAi2iu3wZngOMbta47dhkRIk15IfVLzyITclYrklQ4tRQtqCy4ikui67O55y25Krd2sYunZXufM3PA2W3Tj4cflBxL85guq2g46EyriOu6LzierabW17/GhqQ6OcXiw8+3LnNZhw8Mvuvz5Uhi2aywFJIsHJFxw9U+9c0Q1FF113gi7e62t86rDe82khmFuEfdFt9gm3iOr9tcthzZchjYj5vAxJcNtA2oPFqkdVRdULXRFXRVqZS+EB1pudyh3PDrsVbK7GavDrcsXRirIcAGuL0FFd1R1ol0QdEUvbFELGNXNdmUqSvZM1M/hETrrh6UVpwzPg3JuMUtTnCrTbTGrCI4PGAPGFq+KKIpomxzQi7jfJce52FgnFS2QMPS7magyjb7BKSG64DziBsADNNAjOKpbdNSBO8pEM77mvYh0+q5r43CAO63iLbLdhaYc2c+LUKPPeSG6YKw88Thg4OqCiMKmo701JEVRJFFPiLwiWrxFtEm1YdkSo19JQtBvyQZV8hfaYd40dFVpBN5OfulVAJUT8FCb7fqG9d54O8JeG0jaHh6a0ciLGdiA4pKUh13iEURQQJTAFf0I296orTiKKKrfGWnhPEIYsw3AuwRJMHqpvcsWW2TbrRIqoQkKoi8yovtc/Mvt1nGeW7GEoZKvciLH409l+Jlw+vQquevhHpZ/VanhHyPZbm+rR+PmKpTA3s3Zx/wB/tn2czW16G/1N+6/NFW6vq/xRtc4MWv4MwBPmwpTUO5Pm1BhyHmldFl99wWhdUE5yQN/GKPtoC1BLdwi5dxtNkkQ8LuXFybCjuOmMwGEbknFfkONbSRVTYkV8SVf62xERdSUftkp5MrHlIwyo3MtjhGM3OCbtrw5LmvtsuvEwcgGtvFFJ4wdyppu2RUIU9vj2u8KkQ4EzhBzxu0CQxYS61zI8sIkd2SPGyn2ZLbJKggJOJt2vKggJqQqJaJoWzF1brMid61s98K8IYLxfIkU4DsiJc5jCMOiqI5FZkA3xCGAoWndkqKRqCL3hU1EkSUY/zCuGC8Z2SM3B6us78GQ/NVHBAmdsmI0Lg6oqkqceS7U0RU150VERZVS8bkOnZ2Irb+EQ6lmtr5YclzAlvW22xpBSmhOVMlRor6CQoiIA7JP4Xe3NkmiIoquc9wh40ViRIfsEoYsOa9Z5Rg+CkFybFwlYAV03AvF6I6qin3wNURN6hG+9xLpd584j4QvYrLfjTsNvk/Hhy3nW4ssHtH2GJL3FbhTaKGEQ1EnFAu7DuPwtv9HPe5Ff7XY1wc4l0ukyZCiAlyb4s3IqFx5KW3uRTQdF01XUuZNE3N96rDe1rPsOEPDnWCFc7fYp0pJsd6QwwuqmgtuxG9TFsTJERJgkSiJbRbJdFqzcO3lrEmH7ZdWEQWZ0ZuSAo4DmiGCEibgVRLv98VVF9pVSrIzysxhKGSjYUqwrIdnN7D+OvgGf9XOpllv7HeFvgqL+5GvlXpzow/8A3fY9LuPon8CRUr5QekFKAUoBSgKoxHa3MQZrswZduuLlkfYDe43BdWObgg8Kqb/GCja7SUURAJFQ/CoqMjwB/wCo8w/h8Ps6FW9N3hZP8q80V9ZNKVolgpUAUoBSpApQClAKUAraYb/ndr9Bf5LXR3N9do+9HzRVV6OXgTKvzd+6RezlY/i4x9ZlV9tOCd0ZD+wbl18XLd9WbqdUApQClAKUApQClAKUApQClAK0WLkQobKLovdrzL+iuLu1/T6vh90X0OkRxnkLl/Iywz4zew6j4vWOPEs7tkaTvxoTrlwdFhfzNuG8A+ABBPaq97mkRbbKSeTQweKPj1fVBBG9q7lJV5kTTXXWvWbizVXc6jPWr/Hr+ZwMarYiS/mgg1tyQy7OyCzAw/BK2yRadaOO4ajoKKrZtGhajzEuhAqcxLz6LWVc8j8B3gYYysLwDCHFahx2wBWxaaaVVZQRFURFbVVUC74bl2qmq1197hbQau+S1m/YwZY4vU3E2uM31N1QjO0NOL48tz+n/WXOvhWtfZMDYQYQn7Va7doCOxFcjIiomjbUZxvVF76BFZaVPa4lEXvVlkxIynrIDOyMt8vNyyXdZlljw7c43Og25mEoTkGPHSOgIaObSbAjBVNW1NEJG9yAqJVz1EI5NyZyyrHiU2OMwIivtJLNtXRYU04xQRURSQe/oikKKv508Ne1ZlYrxbmx3Zb0UH2jksiJuMiaKYCWu1SHvoi7S0Ve/tXwUB7UqQKw73/M0/8Au7n+lahkkC4OOMupeDxlcz1jvT3F4WtYcY1D1AtIjSaiuvOi+1Vi9nP/APH779C//tX5jxFG9Wf4lpfX3n0SL/CsxV3Cfxh1dwf8cMdZLwxxlvIeNfibQHuh51XXmSp/iiTa4lsZO7ti7FWdDbASBS0fKS0LC6J4HVbXX2tNfar6r6FwUMNWTaf4lo8Dze6zvOFu81s/CmEL9eGL1KgWybchJGWphoJHu2EO1F8OwzHwqJqneVUrXWvCGX2LbHbFgWy0XG2IwyUMW2xUUZbDi20FP+FAXbp3tF0WvojjFnCypIz38EYOuivtHbLY8Ul12Q4IiO43SJo3HObn3bm2S3d9FEF1RUSvuwYSwjZUhtWaBbYqNurNjNxUFEElbNvjARO8mwzHm5tCVKnJje5GVK1jDkZe4DuZGjtms0gp8jqkk4sFV9375qv59d72qd5d7mqLuLXJnYHwde2Abk2q2TGnXHUATESQjJw3HEH86kTikif8Ra8yrUZEdQypazxby7wRb3IjY2a1NFDA2GWiAdAFxNihtXvoSGo6L/xqifhLrgna8BYTw5cZjlug26BBbckyQNru9kR4neMUeci2mCmi8+vN+aoyYIm8mIuXWXk60xxTD9oCMhi+LLjQCrZkDZIip7SoLbOg95Eab0TQB03l8wrhjEqH10gwJyySQVJ5BVTJtDFERfCIm6i6e0RIvMq1OTHUHKWsjt2ylwHiObCkpbrWiRZRTnW2ADa7oy8yqLp+CKK8pLpp3SIq8+tZ86xYCw2UzEEiHaIRMMJNeliIooNMoJ8YiJ3kRAbXmTn2h39EqMmCzjKk8xkjlZg11DcHD1uLjmQbUxaTnEdigqL7SpxTWhJz/e2+fuB0kdstcSy26NAgRmocKM2jTMdkEEGwRNEFETvJpWailoMXJvM2Vrcrv1m4T9id6imTt2DrgOyG1xhJ/tsNdVTVObmqz+zn/wDj99+hf/2r4X6VUsvdSbyksy0vuPZbnO2Gjm1+Y7Of/wCP336F/wD2qsMsrh11zhzgkdTSIm6fbU4qU3scTS3MpzprW16IU8jdK+Un+F6PgV7qO+H+KJrjW42bD1idxDfA3RLGh3ATFtXDbIWzDUATnI1EyFBRFVVLROdUrSvwMD2izDe0hwuo5R9Xi60GquE+pgrqJ3+66rc1X/4q+GvtLyb5zySyrZjSXrIiwYnwxcbczKBmJdrgVxfdjxmXBcEoyRREUIVRNGBAUJO+o6luQiFcy3YGy1vk2YbFktL0mO+sVx02h3GaNg6SCq/hIomikqcy7j113FrhkR6zLLk1mM+PlngFooD0ey2ls2TbbiOtiOomyv3tAXXnINq6e2PdImiEWufJtmGRtkVt1pi5BDipDaRSR53itzSqnf1XnFklX8wrWWTFaCHKTIq9kjgaW1djBiCjc6bAMyEG1RtuOMMQip/yGMRpFRedULTwVK+1dhFXGzXDltMm4xQ03xxJFaJCQhVF7+qOOIqrz6OHz90usKnFdQc5PrMY8ncEubNcMW1VBs2kXiE10MTE+fwkLriEXfVDJFVda+cUZR4ZxbMs7862sH1tmvzRBW0VHCebMXEX201Ihc1Tn3NitS6cbWsRly1ns/lLg2RKfklhq3JIfaRk3AYQF2IrRIiaabdFYZXVNOdoF9pKk0CBGtcGPChR2osSO2LTMdkEAGwFNBERTmRERERESslFLQiHJtWbPelZGJDs5vYfx18Az/q51Mst/Y7wt8FRf3I18q9OdGH/AO77Hpdx9E/gSKlfKD0gpQClAKUBUF3uUSDwhrXGOO2/KmRhFt5Eb3MoDbqqi72VJEXXm2OjruVFRdF1leAVQcRZiKqoiJfx519r/wBnQq36ieSvdXn4/oVr7mUxebziptHrKka3Wo/91cZjaunIH/jaaQhRBXvoZFz9/YqKir7pg5yRp1fiG8zu/wByMgYqJ+jiBbXT9Kr+mtmVSjgXvcIKdRaXLOk9UY6HbQ3JO/UktNSjKrnbstS+707LH32CWpfwzuTyL3xeuspxF/ShOqip+anYFYv7Gf0h3zqr4zxP5Wl4RivJIneKf8b/AFP4uAbIum2O+0Sd42pjwEn6FE0VK3cOG3Aitx2lcVttNBV1wnC/xIlVV/xWtevjK2Jio1WnbuSe1K9u7QZxpxhzfue1K0y0UoBSgFKAVtMN/wA7tfoL/Ja6O5vrtH3o+aKqvRy8CZV+bv3SL2crH8XGPrMqvtpwTujIf2Dcuvi5bvqzdTqgFKAUoBSgFKAUoBSgFKAUoBWixb/I2P8Ar/8AtXF3a/p9Xw+6L6HSI5usv4yuZXwBYP3lyrE4RE6UeAmcO2+GdyuGJ57NnCE24LZPsFq7LFCJURP9lakc6r39K9JuF/SKHunExqvi5eP2KBbzGxBgfCNosT1zew92GdXwptliT44TxaYdZOG4AuATcsRiOsiTAuCSq6Om5VHTfzc2MR2y+4rjnil13jnpIjNhSWJEW3wwu0eM68TJNi5Eeix3z1Q0Ns1BSUl00Xq5bWa+j+fzaVOCfUZEnMOSN5vNjm5lTbPhO09cnbZigTaeemuMswDRtS2KkjiXJEkFbFNznFoK6qBax/GucF9slkubsLEDlum2mVeZqCr8e3xy0vUtlpSAwI5JbWFFWUFO/uM9xiSS5tXz/wA/cKCdsxk26+3XCU+dJs9/fkhb5uKr1MBxGXS4iNercbzKKgaiJxieXTXnV1C9odPW85q44culvlhekhR5cKPfrU3JuDUQHxl3B8Wo5tKyZyEGOMRtQDQkJ9V/CIFFlSWZMZMXnaJpwicS3XCmNYM2zudTyjsixTlccDPU7L11t7TrnGGJCCoBloZIqAq7lRUFarXHGc2KbPl5LdYxSbdxs8G8ToskbvGRl7iXkBhHHyZ/2twDQmeJbDaf9c0JRWpqTcXKzIhBSSuWG5cMSYhxMwIY3u0eFd8cyLKLULiBBiCzb5T6A0vFqupm2Kq4qqSIg7VTTWozhXF+KcX4jwpZnsTzoS3J+BCnXCG2y3KkNhCvhlqfFroRlDZVVRE0VFUdutTeTazkWjbQYsvNXEB4VZuFzx3Jst1hWe0Hb2dWG0urr0x1mQ4YKH3wyRsAVA02Ku5ETeldeVZSk5aWVVIpaEKw73/M0/8Au7n+lavZUaXgz/i35UfFO0/U2qsmvy3iOmn4vzPo0eaiqOFb+Lpj34NP/UNbjGWH3cTWePDZdBo2rjAmKTmuiixLZfJOb21FpUT86pX1n0JV8LWX/UvI81uu7Th8SmWeDXNt0XD7NudtEUrfEgMmYNKgi+yCC7IRtQUHFLYPMQoS6CqONqPPNcqsqXsu5SPHHtROrCj25X4wqjgtMoemncJzGpCSjroiqvOWiKv0SNLJaZxJVFJWIKXBdeCWCtP27iXoZNSNUcb2uq3IEtqNoJkLiv8AdKjgKicZ+FvRR10fg8X64XO7WmSttgMvWqODd8YZ1kxDWRcXFaYMW20c5pAI4ejakhLqiqeqYby0Z76mSWFwfZJ3WddX2rHAlSJcCTHiQWSVm3IxJYcdBhdoqiOCwpLog6m4WvNzrp7DwZrta5tpedn29WIkpkgiNkW2G22TBITZ8WhumXEd81Ek+9IrjnFd3LpNkb6je5mcH17G99u13YdgLKmyPwJIqiEysMGO7JAJVUCEyQU01Rwk3Apbk1l34M0m5Wy/N9VW1yfPOWjMx5oicBt7rj3KkqKXenDqmvPtPv688uldtkKrZJGRJ4NztyuAi+7bIkBl95RciMrx0pDSUoPvIooPHNHIDZ30TaZapuQRx5fBzuc65wpmtjt4h1OKwrcLjTEXiTAuMbTaqkbmxEJEVtE2t68ZtXedK5O+o85PBaQLMMa3Ha7bKR+M4T0VjiyeZZiRGljkSCq7DcjGqoqEmjmu0l1Rf4XBhfdw7Nik5ahnyuqU45WycVsHYU1lGt6iiqIuSxXREFNAVUEeYUjeRvuY6CigQRWRJttkhBEVtpdQBdO8K6Jqie1zJ+hK9K2TWK7Y/GnsvxMuH16FVz18E9LP6rU8I+R7bc31aPx8xVKYG9m7OP8Av9s+zma2vQ3+pv3X5oq3V9X+KN/mHhKXje1QLZHuJ2uOlwjypUhhBV5AZLjgRveJDrxzbOu5FTbu/RVVM8HK5BaY9nlSLPdoTAtRmplwYI5TURhXG2Y47UQdqskG7m038Z3K70IPtk6eU7nlIzUVYnUXL2aOBMT4N/2aFAntXDqSZH12trLkyj2cWmmiNg4z3lTXUkTTTWq8lcH28I7ep3U9gjFcJLUwY0Bs1W2oyYu7YvcJqT+3i3F7hFRBXRdNtYyptpExmlcwrbwd8QzLVhiUKWS0PREivnbmG1bBlxkYaoSFxSnvPqVUJEUUHcGqucX3WxDgv9SwrCkPrPFnQW7eMiS1H2m8TEWQ06u5B1Xebwlz9/auvPpWCotmbqrQjwvHBelyYEBmFItjYs2uNCfibFbYkPiMoXZJ6AupaSB05kIkQxUg3ISdCxI/UsRhjeTitAIbzVVUtE01XVVWrYQyCqc8s9aVcVClAKUBDs5vYfx18Az/AKudTLLf2O8LfBUX9yNfKvTnRh/+77Hpdx9E/gSKlfKD0gpQClAKUBWN9iFIzjsFxYvE422N8R21DDfVpFVo141XNyNIic6c4KSqem7vImywhBaud0zMhvbuJkXriT2EoltK2wkXRU7y6L3638twSklZpL6itq+b+aCdNNAw0DbYiDYCgiIpoiInMiJX1Wg3fOywUoBSgFKAUoBSgFKAVtMN/wA7tfoL/Ja6O5vrtH3o+aKqvRy8CZV+bv3SL2crH8XGPrMqvtpwTujIf2Dcuvi5bvqzdTqgFKAUoBSgFKAUoBSgFKAUoBWixb/I2P8Ar/8AtXF3a/p9Xw+6L6HSI5Kv+YViy24R2OX8RSJMBi4WKyjFdGC+8DqtuT+MRFbAk1HjA1Rf+JK258JHLZw2zK8PkbaqoEVnmKorpoqp955uZdP8a7O4WOwtLcyhCdRJpaLnKxuHqzxE5RjmMWVn7lVNebdkTePdadR4DdscsiBxE0Q0VWeYkTm179f1vhAZVtSJT4T1B+WiJIdGyS0J5ETRENeJ1Lm5ueu7xhgvax2o0uDV+yz4HPjKcIcWIMoUixDFyOwlilIDJD3iAeI0FU9pU71f2Vn1lROIVkTEkECOIKu2KUSoh670TVn+tquvh156jjDBe1jtQ4NX7LPtjhAZWRTdNmerRuqquE3ZJYqaqgouujPPqgCnP7Qp4Er+v8IPK2S5FcduBOuRV3RzOyS1VldNNQXie55ubmqeMMF7WO1Dg1fss9H+EXlnKEkeurryGCtkjlmmLuBe+K6s95fbSsUs98pjZislKAmooqMdtbFK2soqbVQE4juUVObm9qj3Qwb/AObHahwav2WZDPCIyxjttg1c3GwbXcAhZZaIK6aapozzc3N+iv6HCIyxbISC6OCQ84qNmmIqLoqc33nwEXzl8NOMcH7WO0cFr9lkIxnjfKrHF8t8yZi+8sQIvE77QxaHljPK07xo/hRSNvcvcnxZBvFBEtURKnvKXy693JPimb6GsVj8Gm3vq2ol4au0lkDlL5de7knxTN9DWLdeEjl5Itcxpu9SScNkxEUtMznVRVE/9zWT3RwftY7SOC1uyyc8HGK9C4PGV0aQ0bEhnC1rbcadFRMCSI0iiqLzoqKmiotWJX5sxDvWnbW/M95HmoqvhTsOyeDxjxthpx91bae1tkFMy0VF5hTVV/QlaDlL5de7knxTN9DX1H0NxNDD4aqqs0ryWl9x57dWlOrKGQr5mOUvl17uSfFM30NOUvl17uSfFM30NfQuMcH7WO04fBa3ZY5S+XXu5J8UzfQ05S+XXu5J8UzfQ04xwftY7RwWt2WOUvl17uSfFM30NOUvl17uSfFM30NOMcH7WO0cFrdljlL5de7knxTN9DTlL5de7knxTN9DTjHB+1jtHBa3ZY5S+XXu5J8UzfQ05S+XXu5J8UzfQ04xwftY7RwWt2WOUvl17uSfFM30NOUvl17uSfFM30NOMcH7WO0cFrdljlL5de7knxTN9DTlL5de7knxTN9DTjHB+1jtHBa3ZZrMBY6s2YvCWgz8PvSJsKHhGaw++5CfYAHCmRCEdXAHVVQSXRPAtdDV8R9KKsK26c503dWWjwPXYCEoYeMZKzziubO2Xh7LbPPNVvEUqTb1nSrc9GJIEh4XQGAyKqhNtknMSKnf9qtn0SrU6G6LnUkksl6fFFe6UJVKGTBXdyQcpfLr3ck+KZvoacpfLr3ck+KZvoa+y8Y4P2sdp5XgtbsscpfLr3ck+KZvoacpfLr3ck+KZvoacY4P2sdo4LW7LHKXy693JPimb6GnKXy693JPimb6GnGOD9rHaOC1uyxyl8uvdyT4pm+hpyl8uvdyT4pm+hpxjg/ax2jgtbsscpfLr3ck+KZvoacpfLr3ck+KZvoacY4P2sdo4LW7LHKXy693JPimb6GnKXy693JPimb6GnGOD9rHaOC1uyxyl8uvdyT4pm+hpyl8uvdyT4pm+hpxjg/ax2jgtbssjGaPCDwJfMssXW6DdZcmbMs8yOwyNpmauOGyYiKate2qon+NX1l40bOX+GW3BIHAtcUSA00UVRodUVPar5p6Z4ijiI0N6mpWytD8D0G5VKdNTy1bQSClfMDvilAKUApQHkzEZjm6bTLbZulvcIBRFMtETVdO+uiImq+CoUuDcVWu/wB/m2PEdoixLtMGaUe4WV2S40aR2WVRDCU2ioqMov4PNqvOtXU5xjfLTaa12+zMWr6D360Zhe+rDXk3I9fp1ozC99WGvJuR6/VmVh+w/wD7L/1ItLX8v3HWjML31Ya8m5Hr9OtGYXvqw15NyPX6ZWH7D/8Asv8A1Fpa/l+460Zhe+rDXk3I9frR4gax+81JtcPMLCltu6NA9vXDbpGw2pqiGolOVNF2GiapoqoqIqaapnB4dvo5P/uX/qGpa/l+5YsSay80yHVbEh5UUVJpURDIUTdomq6fo1XTWsqtNqxmKVAFKAUoBW0w3/O7X6C/yWujub67R96Pmiqr0cvAmVfm790i9nKx/Fxj6zKr7acE7oyH9g3Lr4uW76s3U6oBSgFKAUoBSgFKAUoBSgFKAVosW/yNj/r/APtXF3a/p9Xw+6L6HSIi1K+NncFKAUoBSgFKAUoBSgFKAUoBSgFKAUoBSgFKAUoBSgFKAUoBSgFKAUoBSgFKAUoBSgFKAUoBSgFKAUoBSgFKAUoBSgFRHEWWVrxPiRq8zHHleBuM2rCICtmjL6vBqiiq85KqLz97wLz1ZTqOm7xIauaWwZDYfw3iW23qFInNvwQaBthHBFkuLj8QJKIinOo6qq+2q+DmqyKzrVpV2nLqViFFRzIUqgyFKAUoBW0w3/O7X6C/yWujub67R96Pmiqr0cvAmVfm790i9nKx/Fxj6zKr7acE7oyH9g3Lr4uW76s3U6oBSgFKAUoBSgFKAUoBSgFKAVjzrezcGxB5FJBXVERdOetfEUIYmk6NTQzKMnB3Rh9jUD8mXz1p2NQPyZfPWuFyd3P7L2s2OE1NY7GoH5MvnrTsagfky+etOTu5/Ze1jhNTWOxqB+TL5607GoH5MvnrTk7uf2XtY4TU1jsagfky+etOxqB+TL5605O7n9l7WOE1NY7GoH5MvnrTsagfky+etOTu5/Ze1jhNTWOxqB+TL5607GoH5MvnrTk7uf2XtY4TU1jsagfky+etOxqB+TL5605O7n9l7WOE1NY7GoH5MvnrTsagfky+etOTu5/Ze1jhNTWOxqB+TL5607GoH5MvnrTk7uf2XtY4TU1jsagfky+etOxqB+TL5605O7n9l7WOE1NY7GoH5MvnrTsagfky+etOTu5/Ze1jhNTWOxqB+TL5607GoH5MvnrTk7uf2XtY4TU1jsagfky+etOxqB+TL5605O7n9l7WOE1NY7GoH5MvnrTsagfky+etOTu5/Ze1jhNTWOxqB+TL5607GoH5MvnrTk7uf2XtY4TU1jsagfky+etOxqB+TL5605O7n9l7WOE1NY7GoH5MvnrTsagfky+etOTu5/Ze1jhNTWOxqB+TL5607GoH5MvnrTk7uf2XtY4TU1jsagfky+etOxqB+TL5605O7n9l7WOE1NY7GoH5MvnrTsagfky+etOTu5/Ze1jhNTWOxqB+TL5607GoH5MvnrTk7uf2XtY4TU1jsagfky+etOxqB+TL5605O7n9l7WOE1NY7GoH5MvnrTsagfky+etOTu5/Ze1jhNTWOxqB+TL5607GoH5MvnrTk7uf2XtY4TU1jsagfky+etOxqB+TL5605O7n9l7WOE1NY7GoH5MvnrTsagfky+etOTu5/Ze1jhNTWOxqB+TL5607GoH5MvnrTk7uf2XtY4TU1jsagfky+etOxqB+TL5605O7n9l7WOE1NY7GoH5MvnrTsagfky+etOTu5/Ze1jhNTWOxqB+TL5607GoH5MvnrTk7uf2XtY4TU1jsagfky+etOxqB+TL5605O7n9l7WOE1NY7GoH5MvnrTsagfky+etOTu5/Ze1jhNTWOxqB+TL5607GoH5MvnrTk7uf2XtY4TU1jsagfky+etOxqB+TL5605O7n9l7WOE1NY7GoH5MvnrTsagfky+etOTu5/Ze1jhNTWOxqB+TL5607GoH5MvnrTk7uf2XtY4TU1jsagfky+etOxqB+TL5605O7n9l7WOE1NY7GoH5MvnrTsagfky+etOTu5/Ze1jhNTWOxqB+TL5607GoH5MvnrTk7uf2XtY4TU1jsagfky+etesWyRIT4utASGOqIqkq1bS3CwNGpGrCLvF3Wd9REsRUkrNmfX5u/dIvZysfxcY+syq9Aax3RkP7BuXXxct31Zup1QClAKUApQClAKUApQClAKUApQClAKUApQClAKUApQClAKUApQClAKUApQClAKUApQClAKUApQClAKUApQClAKUApQClAKUApQClAKUApQClAKrXM/M13A+IbLbRlux3LsXERGgw1MuSvvoDrqgLjLgiio0y4W1U1RAVddFSgI/AzpeuFziwxxFFZGTcXrSzMkYOntxSltK4jjPHLI2IQky6POqJuBU110StfI4QjcDA7uMp2ImbbhVuKxMS7TcFXFpg2njbBpQUpHdKaut6InPouveRaA3WIc15eG72lnfxNbpl0J5mOEW3YVmSjccdYefARRuSuv3qM6a+BEHXTcOuEeeILZI9zZxXBmNOtynSjRMG3F6UwMZ1WpKvMC+rjXFuIoEhii7kVE1WgJqFwxK+1a3WcR2SRHuSIUd9mwSDbUVBTQyJJegiqImiroiqop31Sv7e7hiDDljl3i54sw3CtcRon35btmeQGwRNVJV6soDU4XxteMXSp0WHiG3xZcJpp96NdMJzoLotOqaNuID8gFUSVs01ROZRVF0VFSpI5Exe2qIeIrAKqiqiFZXk//AOygNViC93/DFv6uuWKLExBDjFdlJYZBNsC20bpm4SS1QBQWy7otE1UU75Ii5Ful4nuoyCYxDZUBg0BTdsEhtC1ESQhUpabh0LTcnNqhJ30VEAz0g4xLXTEFhXTmXSyPcy/TKhuFszZeNLwzbbViq1vPSIhz4jz2FZzMebGAgE3o7xyEB0EV1vugVUVHBJNRJFUCTi5is57kRL9Zt7bIPq4tgkcVtJSRND6r2qvcrza6omi+2lZKRMXk5xaYisCmmvc9ZXtfa9rqz86fKlAEh4vUdUxDYFTXbr1le7+umn8s8PNWysjN5afeG63O2z0RE2hCgnHIF8Jbn3NUVPzJQG3r83fukXs5WP4uMfWZVAd0ZD+wbl18XLd9WbqdUApQClAKUApQClAKUApQClAKUApQClAKUApQClAKUApQClAKUApQClAKUApQClAKUApQClAKUApQClAKUApQClAKUApQClAKUApQClAKUApQCohiHB0q/Zj4PvZPMparEzOdVglXjSmOg00yYppptFopaLqqLqY6Iqa6AUlcuCviBvA1jZtV7BMWMW+7OzXZt2mOwOu02K+CyGGS3g2gvyXS3g2B7VX/AIiFZRa8jbhItVhss23RbdYIVziSJcBzFNwvYPxo7LytNAEloRa0f6mVRFERRBdecRSgIxgHg+4/y0xDbLmxIsGKeoJE/iAuFwkRCBrqaFCt5KQx3d5hFiOo4nMinINUXnVV9rxwZ8SSY6RY9xty3J1uRIbxhEnS7Zc7XPlSHZEp1ttnVJLKuOAQR3XRBEbQC3oqrQE3zgyXm5q4qw67IeguYfgNg1KiytynIFZ0SU6igg7VQhgg3oqp3Lzn+PguSM6HlGmGLeVsjTmcTdkTUYEJuEQjeOrwjLtHUA2ILWqAqIqbtpd5QPjG2W2N8eOW6Zd28L3Zll94+xa5K49bGNzIg08R8Tukm2SOHtMGxXjdEUFBDKi14PeI8VTkwW1bklDZxlsJjW/w32ZvFjbFtkdsXFb2GP3xySPEuGKkCKaNGqoQFhYq4J8i5TMVMWmLh6Hh65hJbjWZENlkRdYtsfukFtUEuKizk3ohLrJDv7Vr7Tgr3xyXLRm9260RbhCkHMSE0ZEk0Z0yXb0EVQU4hhydv05lVYzQ/gqVAS3J3JO95X4UxOEedCgYgucVuPEINsmPHcaaMWnnNjMfjTUzVTVQQiEQFTLaipFIPByxNb2RW3tWizWZHYiyMCNXubKs00WjMj53WtIwqqt6NNM7FRrQ9yF3IGsvnBQxFPe4yNKsDEZXNnWeMCtwBb6nHaSNvsyEVGpD9wIWdNFGQOhtqOiZVy4JVz61AdtuFuZxEEuW8t6cJ1JbzKQEt0Vgn0FXBEowpxiovcuLvFCJEKgP5ijgp3XFFxZIAw/h60yxdSVaLNq3Ht7hk2JSGENg+MeVllod7fUqiQbhVFItZ7k9k3dcC45xNiO7FbRcue9AbhFx5kbj5vOuK6TIOoJKQ6MuG9s0VBcQeagLir83fukXs5WP4uMfWZVAX9k3wy8nbLlBga3TMYcTMiWKDHea62TC2OBHASTVGVRdFRU1TmqYct7JT36/sqb6GgHLeyU9+v7Km+hpy3slPfr+ypvoaAct7JT36/sqb6GnLeyU9+v7Km+hoBy3slPfr+ypvoact7JT36/sqb6GgHLeyU9+v7Km+hpy3slPfr+ypvoaAct7JT36/sqb6GnLeyU9+v7Km+hoBy3slPfr+ypvoact7JT36/sqb6GgHLeyU9+v7Km+hpy3slPfr+ypvoaAct7JT36/sqb6GnLeyU9+v7Km+hoBy3slPfr+ypvoact7JT36/sqb6GgHLeyU9+v7Km+hpy3slPfr+ypvoaAct7JT36/sqb6GnLeyU9+v7Km+hoBy3slPfr+ypvoact7JT36/sqb6GgHLeyU9+v7Km+hpy3slPfr+ypvoaAct7JT36/sqb6GnLeyU9+v7Km+hoBy3slPfr+ypvoact7JT36/sqb6GgHLeyU9+v7Km+hpy3slPfr+ypvoaAct7JT36/sqb6GnLeyU9+v7Km+hoBy3slPfr+ypvoact7JT36/sqb6GgHLeyU9+v7Km+hpy3slPfr+ypvoaAct7JT36/sqb6GnLeyU9+v7Km+hoBy3slPfr+ypvoact7JT36/sqb6GgHLeyU9+v7Km+hpy3slPfr+ypvoaAct7JT36/sqb6GnLeyU9+v7Km+hoBy3slPfr+ypvoact7JT36/sqb6GgHLeyU9+v7Km+hpy3slPfr+ypvoaAct7JT36/sqb6GnLeyU9+v7Km+hoBy3slPfr+ypvoact7JT36/sqb6GgHLeyU9+v7Km+hpy3slPfr+ypvoaAct7JT36/sqb6GnLeyU9+v7Km+hoBy3slPfr+ypvoact7JT36/sqb6GgHLeyU9+v7Km+hpy3slPfr+ypvoaAct7JT36/sqb6GnLeyU9+v7Km+hoBy3slPfr+ypvoact7JT36/sqb6GgHLeyU9+v7Km+hpy3slPfr+ypvoaAct7JT36/sqb6GnLeyU9+v7Km+hoBy3slPfr+ypvoact7JT36/sqb6GgHLeyU9+v7Km+hpy3slPfr+ypvoaAct7JT36/sqb6GnLeyU9+v7Km+hoBy3slPfr+ypvoact7JT36/sqb6GgHLeyU9+v7Km+hpy3slPfr+ypvoaAct7JT36/sqb6GnLeyU9+v7Km+hoBy3slPfr+ypvoact7JT36/sqb6GgHLeyU9+v7Km+hpy3slPfr+ypvoaAct7JT36/sqb6GnLeyU9+v7Km+hoBy3slPfr+ypvoact7JT36/sqb6GgHLeyU9+v7Km+hpy3slPfr+ypvoaAct7JT36/sqb6GnLeyU9+v7Km+hoBy3slPfr+ypvoact7JT36/sqb6GgHLeyU9+v7Km+hpy3slPfr+ypvoaAct7JT36/sqb6GuDeHjwgMA5gZv2i4WC/8AV8NqxMxzc6jkN6OJIkEqaG2i94h59NOegP/Z)
Figure 1. Measuring reward and hedonia. Reward and pleasure are multifaceted psychological concepts. Major processes within reward (first column) consist of motivation or wanting (white), learning (blue), and – most relevant to happiness – pleasure, liking or affect (light blue). Each of these contains explicit (top rows, light yellow) and implicit (bottom rows, yellow) psychological components (second column) that constantly interact and require careful scientific experimentation to tease apart. Explicit processes are consciously experienced (e.g. explicit pleasure and happiness, desire, or expectation), whereas implicit psychological processes are potentially unconscious in the sense that they can operate at a level not always directly accessible to conscious experience (implicit incentive salience, habits and ‘liking’ reactions), and must be further translated by other mechanisms into subjective feelings. Measurements or behavioral procedures that are especially sensitive markers of each of the processes are listed (third column). Examples of some of the brain regions and neurotransmitters are listed (fourth column), as well as specific examples of measurements (fifth column), such as an example of how highest subjective life satisfaction does not lead to the highest salaries (top) [93]. Another example shows the incentive-sensitization model of addiction and how ‘wanting’ to take drugs may grow over time independently of ‘liking’ and ‘learning’ drug pleasure as an individual becomes an addict (bottom) [94].
Neuroanatomy of ‘pleasure’
![[[@kringelbach2009towards]](http://dx.doi.org/10.1016/j.tics.2009.08.006)](data:image/jpeg;base64,/9j/4AAQSkZJRgABAQEAcQBxAAD//gAuSGFuZG1hZGUgU29mdHdhcmUsIEluYy4gSW1hZ2UgQWxjaGVteSB2MS4xMwr/7TR6UGhvdG9zaG9wIDMuMAA4QklNBCUAAAAAABAAAAAAAAAAAAAAAAAAAAAAOEJJTQPqAAAAAB2mPD94bWwgdmVyc2lvbj0iMS4wIiBlbmNvZGluZz0iVVRGLTgiPz4KPCFET0NUWVBFIHBsaXN0IFBVQkxJQyAiLS8vQXBwbGUgQ29tcHV0ZXIvL0RURCBQTElTVCAxLjAvL0VOIiAiaHR0cDovL3d3dy5hcHBsZS5jb20vRFREcy9Qcm9wZXJ0eUxpc3QtMS4wLmR0ZCI+CjxwbGlzdCB2ZXJzaW9uPSIxLjAiPgo8ZGljdD4KCTxrZXk+Y29tLmFwcGxlLnByaW50LlBhZ2VGb3JtYXQuUE1Ib3Jpem9udGFsUmVzPC9rZXk+Cgk8ZGljdD4KCQk8a2V5PmNvbS5hcHBsZS5wcmludC50aWNrZXQuY3JlYXRvcjwva2V5PgoJCTxzdHJpbmc+Y29tLmFwcGxlLnByaW50aW5nbWFuYWdlcjwvc3RyaW5nPgoJCTxrZXk+Y29tLmFwcGxlLnByaW50LnRpY2tldC5pdGVtQXJyYXk8L2tleT4KCQk8YXJyYXk+CgkJCTxkaWN0PgoJCQkJPGtleT5jb20uYXBwbGUucHJpbnQuUGFnZUZvcm1hdC5QTUhvcml6b250YWxSZXM8L2tleT4KCQkJCTxyZWFsPjcyPC9yZWFsPgoJCQkJPGtleT5jb20uYXBwbGUucHJpbnQudGlja2V0LmNsaWVudDwva2V5PgoJCQkJPHN0cmluZz5jb20uYXBwbGUucHJpbnRpbmdtYW5hZ2VyPC9zdHJpbmc+CgkJCQk8a2V5PmNvbS5hcHBsZS5wcmludC50aWNrZXQubW9kRGF0ZTwva2V5PgoJCQkJPGRhdGU+MjAwOS0wOC0zMVQxMjo0Mzo1NVo8L2RhdGU+CgkJCQk8a2V5PmNvbS5hcHBsZS5wcmludC50aWNrZXQuc3RhdGVGbGFnPC9rZXk+CgkJCQk8aW50ZWdlcj4wPC9pbnRlZ2VyPgoJCQk8L2RpY3Q+CgkJPC9hcnJheT4KCTwvZGljdD4KCTxrZXk+Y29tLmFwcGxlLnByaW50LlBhZ2VGb3JtYXQuUE1PcmllbnRhdGlvbjwva2V5PgoJPGRpY3Q+CgkJPGtleT5jb20uYXBwbGUucHJpbnQudGlja2V0LmNyZWF0b3I8L2tleT4KCQk8c3RyaW5nPmNvbS5hcHBsZS5wcmludGluZ21hbmFnZXI8L3N0cmluZz4KCQk8a2V5PmNvbS5hcHBsZS5wcmludC50aWNrZXQuaXRlbUFycmF5PC9rZXk+CgkJPGFycmF5PgoJCQk8ZGljdD4KCQkJCTxrZXk+Y29tLmFwcGxlLnByaW50LlBhZ2VGb3JtYXQuUE1PcmllbnRhdGlvbjwva2V5PgoJCQkJPGludGVnZXI+MTwvaW50ZWdlcj4KCQkJCTxrZXk+Y29tLmFwcGxlLnByaW50LnRpY2tldC5jbGllbnQ8L2tleT4KCQkJCTxzdHJpbmc+Y29tLmFwcGxlLnByaW50aW5nbWFuYWdlcjwvc3RyaW5nPgoJCQkJPGtleT5jb20uYXBwbGUucHJpbnQudGlja2V0Lm1vZERhdGU8L2tleT4KCQkJCTxkYXRlPjIwMDktMDgtMzFUMTI6NDM6NTVaPC9kYXRlPgoJCQkJPGtleT5jb20uYXBwbGUucHJpbnQudGlja2V0LnN0YXRlRmxhZzwva2V5PgoJCQkJPGludGVnZXI+MDwvaW50ZWdlcj4KCQkJPC9kaWN0PgoJCTwvYXJyYXk+Cgk8L2RpY3Q+Cgk8a2V5PmNvbS5hcHBsZS5wcmludC5QYWdlRm9ybWF0LlBNU2NhbGluZzwva2V5PgoJPGRpY3Q+CgkJPGtleT5jb20uYXBwbGUucHJpbnQudGlja2V0LmNyZWF0b3I8L2tleT4KCQk8c3RyaW5nPmNvbS5hcHBsZS5wcmludGluZ21hbmFnZXI8L3N0cmluZz4KCQk8a2V5PmNvbS5hcHBsZS5wcmludC50aWNrZXQuaXRlbUFycmF5PC9rZXk+CgkJPGFycmF5PgoJCQk8ZGljdD4KCQkJCTxrZXk+Y29tLmFwcGxlLnByaW50LlBhZ2VGb3JtYXQuUE1TY2FsaW5nPC9rZXk+CgkJCQk8cmVhbD4xPC9yZWFsPgoJCQkJPGtleT5jb20uYXBwbGUucHJpbnQudGlja2V0LmNsaWVudDwva2V5PgoJCQkJPHN0cmluZz5jb20uYXBwbGUucHJpbnRpbmdtYW5hZ2VyPC9zdHJpbmc+CgkJCQk8a2V5PmNvbS5hcHBsZS5wcmludC50aWNrZXQubW9kRGF0ZTwva2V5PgoJCQkJPGRhdGU+MjAwOS0wOC0zMVQxMjo0Mzo1NVo8L2RhdGU+CgkJCQk8a2V5PmNvbS5hcHBsZS5wcmludC50aWNrZXQuc3RhdGVGbGFnPC9rZXk+CgkJCQk8aW50ZWdlcj4wPC9pbnRlZ2VyPgoJCQk8L2RpY3Q+CgkJPC9hcnJheT4KCTwvZGljdD4KCTxrZXk+Y29tLmFwcGxlLnByaW50LlBhZ2VGb3JtYXQuUE1WZXJ0aWNhbFJlczwva2V5PgoJPGRpY3Q+CgkJPGtleT5jb20uYXBwbGUucHJpbnQudGlja2V0LmNyZWF0b3I8L2tleT4KCQk8c3RyaW5nPmNvbS5hcHBsZS5wcmludGluZ21hbmFnZXI8L3N0cmluZz4KCQk8a2V5PmNvbS5hcHBsZS5wcmludC50aWNrZXQuaXRlbUFycmF5PC9rZXk+CgkJPGFycmF5PgoJCQk8ZGljdD4KCQkJCTxrZXk+Y29tLmFwcGxlLnByaW50LlBhZ2VGb3JtYXQuUE1WZXJ0aWNhbFJlczwva2V5PgoJCQkJPHJlYWw+NzI8L3JlYWw+CgkJCQk8a2V5PmNvbS5hcHBsZS5wcmludC50aWNrZXQuY2xpZW50PC9rZXk+CgkJCQk8c3RyaW5nPmNvbS5hcHBsZS5wcmludGluZ21hbmFnZXI8L3N0cmluZz4KCQkJCTxrZXk+Y29tLmFwcGxlLnByaW50LnRpY2tldC5tb2REYXRlPC9rZXk+CgkJCQk8ZGF0ZT4yMDA5LTA4LTMxVDEyOjQzOjU1WjwvZGF0ZT4KCQkJCTxrZXk+Y29tLmFwcGxlLnByaW50LnRpY2tldC5zdGF0ZUZsYWc8L2tleT4KCQkJCTxpbnRlZ2VyPjA8L2ludGVnZXI+CgkJCTwvZGljdD4KCQk8L2FycmF5PgoJPC9kaWN0PgoJPGtleT5jb20uYXBwbGUucHJpbnQuUGFnZUZvcm1hdC5QTVZlcnRpY2FsU2NhbGluZzwva2V5PgoJPGRpY3Q+CgkJPGtleT5jb20uYXBwbGUucHJpbnQudGlja2V0LmNyZWF0b3I8L2tleT4KCQk8c3RyaW5nPmNvbS5hcHBsZS5wcmludGluZ21hbmFnZXI8L3N0cmluZz4KCQk8a2V5PmNvbS5hcHBsZS5wcmludC50aWNrZXQuaXRlbUFycmF5PC9rZXk+CgkJPGFycmF5PgoJCQk8ZGljdD4KCQkJCTxrZXk+Y29tLmFwcGxlLnByaW50LlBhZ2VGb3JtYXQuUE1WZXJ0aWNhbFNjYWxpbmc8L2tleT4KCQkJCTxyZWFsPjE8L3JlYWw+CgkJCQk8a2V5PmNvbS5hcHBsZS5wcmludC50aWNrZXQuY2xpZW50PC9rZXk+CgkJCQk8c3RyaW5nPmNvbS5hcHBsZS5wcmludGluZ21hbmFnZXI8L3N0cmluZz4KCQkJCTxrZXk+Y29tLmFwcGxlLnByaW50LnRpY2tldC5tb2REYXRlPC9rZXk+CgkJCQk8ZGF0ZT4yMDA5LTA4LTMxVDEyOjQzOjU1WjwvZGF0ZT4KCQkJCTxrZXk+Y29tLmFwcGxlLnByaW50LnRpY2tldC5zdGF0ZUZsYWc8L2tleT4KCQkJCTxpbnRlZ2VyPjA8L2ludGVnZXI+CgkJCTwvZGljdD4KCQk8L2FycmF5PgoJPC9kaWN0PgoJPGtleT5jb20uYXBwbGUucHJpbnQuc3ViVGlja2V0LnBhcGVyX2luZm9fdGlja2V0PC9rZXk+Cgk8ZGljdD4KCQk8a2V5PmNvbS5hcHBsZS5wcmludC5QYWdlRm9ybWF0LlBNQWRqdXN0ZWRQYWdlUmVjdDwva2V5PgoJCTxkaWN0PgoJCQk8a2V5PmNvbS5hcHBsZS5wcmludC50aWNrZXQuY3JlYXRvcjwva2V5PgoJCQk8c3RyaW5nPmNvbS5hcHBsZS5wcmludGluZ21hbmFnZXI8L3N0cmluZz4KCQkJPGtleT5jb20uYXBwbGUucHJpbnQudGlja2V0Lml0ZW1BcnJheTwva2V5PgoJCQk8YXJyYXk+CgkJCQk8ZGljdD4KCQkJCQk8a2V5PmNvbS5hcHBsZS5wcmludC5QYWdlRm9ybWF0LlBNQWRqdXN0ZWRQYWdlUmVjdDwva2V5PgoJCQkJCTxhcnJheT4KCQkJCQkJPHJlYWw+MC4wPC9yZWFsPgoJCQkJCQk8cmVhbD4wLjA8L3JlYWw+CgkJCQkJCTxyZWFsPjc4MzwvcmVhbD4KCQkJCQkJPHJlYWw+NTU5PC9yZWFsPgoJCQkJCTwvYXJyYXk+CgkJCQkJPGtleT5jb20uYXBwbGUucHJpbnQudGlja2V0LmNsaWVudDwva2V5PgoJCQkJCTxzdHJpbmc+Y29tLmFwcGxlLnByaW50aW5nbWFuYWdlcjwvc3RyaW5nPgoJCQkJCTxrZXk+Y29tLmFwcGxlLnByaW50LnRpY2tldC5tb2REYXRlPC9rZXk+CgkJCQkJPGRhdGU+MjAwOS0wOC0zMVQxMjo0Mzo1NVo8L2RhdGU+CgkJCQkJPGtleT5jb20uYXBwbGUucHJpbnQudGlja2V0LnN0YXRlRmxhZzwva2V5PgoJCQkJCTxpbnRlZ2VyPjA8L2ludGVnZXI+CgkJCQk8L2RpY3Q+CgkJCTwvYXJyYXk+CgkJPC9kaWN0PgoJCTxrZXk+Y29tLmFwcGxlLnByaW50LlBhZ2VGb3JtYXQuUE1BZGp1c3RlZFBhcGVyUmVjdDwva2V5PgoJCTxkaWN0PgoJCQk8a2V5PmNvbS5hcHBsZS5wcmludC50aWNrZXQuY3JlYXRvcjwva2V5PgoJCQk8c3RyaW5nPmNvbS5hcHBsZS5wcmludGluZ21hbmFnZXI8L3N0cmluZz4KCQkJPGtleT5jb20uYXBwbGUucHJpbnQudGlja2V0Lml0ZW1BcnJheTwva2V5PgoJCQk8YXJyYXk+CgkJCQk8ZGljdD4KCQkJCQk8a2V5PmNvbS5hcHBsZS5wcmludC5QYWdlRm9ybWF0LlBNQWRqdXN0ZWRQYXBlclJlY3Q8L2tleT4KCQkJCQk8YXJyYXk+CgkJCQkJCTxyZWFsPi0xODwvcmVhbD4KCQkJCQkJPHJlYWw+LTE4PC9yZWFsPgoJCQkJCQk8cmVhbD44MjQ8L3JlYWw+CgkJCQkJCTxyZWFsPjU3NzwvcmVhbD4KCQkJCQk8L2FycmF5PgoJCQkJCTxrZXk+Y29tLmFwcGxlLnByaW50LnRpY2tldC5jbGllbnQ8L2tleT4KCQkJCQk8c3RyaW5nPmNvbS5hcHBsZS5wcmludGluZ21hbmFnZXI8L3N0cmluZz4KCQkJCQk8a2V5PmNvbS5hcHBsZS5wcmludC50aWNrZXQubW9kRGF0ZTwva2V5PgoJCQkJCTxkYXRlPjIwMDktMDgtMzFUMTI6NDM6NTVaPC9kYXRlPgoJCQkJCTxrZXk+Y29tLmFwcGxlLnByaW50LnRpY2tldC5zdGF0ZUZsYWc8L2tleT4KCQkJCQk8aW50ZWdlcj4wPC9pbnRlZ2VyPgoJCQkJPC9kaWN0PgoJCQk8L2FycmF5PgoJCTwvZGljdD4KCQk8a2V5PmNvbS5hcHBsZS5wcmludC5QYXBlckluZm8uUE1QYXBlck5hbWU8L2tleT4KCQk8ZGljdD4KCQkJPGtleT5jb20uYXBwbGUucHJpbnQudGlja2V0LmNyZWF0b3I8L2tleT4KCQkJPHN0cmluZz5jb20uYXBwbGUucHJpbnQucG0uUG9zdFNjcmlwdDwvc3RyaW5nPgoJCQk8a2V5PmNvbS5hcHBsZS5wcmludC50aWNrZXQuaXRlbUFycmF5PC9rZXk+CgkJCTxhcnJheT4KCQkJCTxkaWN0PgoJCQkJCTxrZXk+Y29tLmFwcGxlLnByaW50LlBhcGVySW5mby5QTVBhcGVyTmFtZTwva2V5PgoJCQkJCTxzdHJpbmc+aXNvLWE0PC9zdHJpbmc+CgkJCQkJPGtleT5jb20uYXBwbGUucHJpbnQudGlja2V0LmNsaWVudDwva2V5PgoJCQkJCTxzdHJpbmc+Y29tLmFwcGxlLnByaW50LnBtLlBvc3RTY3JpcHQ8L3N0cmluZz4KCQkJCQk8a2V5PmNvbS5hcHBsZS5wcmludC50aWNrZXQubW9kRGF0ZTwva2V5PgoJCQkJCTxkYXRlPjIwMDMtMDctMDFUMTc6NDk6MzZaPC9kYXRlPgoJCQkJCTxrZXk+Y29tLmFwcGxlLnByaW50LnRpY2tldC5zdGF0ZUZsYWc8L2tleT4KCQkJCQk8aW50ZWdlcj4xPC9pbnRlZ2VyPgoJCQkJPC9kaWN0PgoJCQk8L2FycmF5PgoJCTwvZGljdD4KCQk8a2V5PmNvbS5hcHBsZS5wcmludC5QYXBlckluZm8uUE1VbmFkanVzdGVkUGFnZVJlY3Q8L2tleT4KCQk8ZGljdD4KCQkJPGtleT5jb20uYXBwbGUucHJpbnQudGlja2V0LmNyZWF0b3I8L2tleT4KCQkJPHN0cmluZz5jb20uYXBwbGUucHJpbnQucG0uUG9zdFNjcmlwdDwvc3RyaW5nPgoJCQk8a2V5PmNvbS5hcHBsZS5wcmludC50aWNrZXQuaXRlbUFycmF5PC9rZXk+CgkJCTxhcnJheT4KCQkJCTxkaWN0PgoJCQkJCTxrZXk+Y29tLmFwcGxlLnByaW50LlBhcGVySW5mby5QTVVuYWRqdXN0ZWRQYWdlUmVjdDwva2V5PgoJCQkJCTxhcnJheT4KCQkJCQkJPHJlYWw+MC4wPC9yZWFsPgoJCQkJCQk8cmVhbD4wLjA8L3JlYWw+CgkJCQkJCTxyZWFsPjc4MzwvcmVhbD4KCQkJCQkJPHJlYWw+NTU5PC9yZWFsPgoJCQkJCTwvYXJyYXk+CgkJCQkJPGtleT5jb20uYXBwbGUucHJpbnQudGlja2V0LmNsaWVudDwva2V5PgoJCQkJCTxzdHJpbmc+Y29tLmFwcGxlLnByaW50aW5nbWFuYWdlcjwvc3RyaW5nPgoJCQkJCTxrZXk+Y29tLmFwcGxlLnByaW50LnRpY2tldC5tb2REYXRlPC9rZXk+CgkJCQkJPGRhdGU+MjAwOS0wOC0zMVQxMjo0Mzo1NVo8L2RhdGU+CgkJCQkJPGtleT5jb20uYXBwbGUucHJpbnQudGlja2V0LnN0YXRlRmxhZzwva2V5PgoJCQkJCTxpbnRlZ2VyPjA8L2ludGVnZXI+CgkJCQk8L2RpY3Q+CgkJCTwvYXJyYXk+CgkJPC9kaWN0PgoJCTxrZXk+Y29tLmFwcGxlLnByaW50LlBhcGVySW5mby5QTVVuYWRqdXN0ZWRQYXBlclJlY3Q8L2tleT4KCQk8ZGljdD4KCQkJPGtleT5jb20uYXBwbGUucHJpbnQudGlja2V0LmNyZWF0b3I8L2tleT4KCQkJPHN0cmluZz5jb20uYXBwbGUucHJpbnQucG0uUG9zdFNjcmlwdDwvc3RyaW5nPgoJCQk8a2V5PmNvbS5hcHBsZS5wcmludC50aWNrZXQuaXRlbUFycmF5PC9rZXk+CgkJCTxhcnJheT4KCQkJCTxkaWN0PgoJCQkJCTxrZXk+Y29tLmFwcGxlLnByaW50LlBhcGVySW5mby5QTVVuYWRqdXN0ZWRQYXBlclJlY3Q8L2tleT4KCQkJCQk8YXJyYXk+CgkJCQkJCTxyZWFsPi0xODwvcmVhbD4KCQkJCQkJPHJlYWw+LTE4PC9yZWFsPgoJCQkJCQk8cmVhbD44MjQ8L3JlYWw+CgkJCQkJCTxyZWFsPjU3NzwvcmVhbD4KCQkJCQk8L2FycmF5PgoJCQkJCTxrZXk+Y29tLmFwcGxlLnByaW50LnRpY2tldC5jbGllbnQ8L2tleT4KCQkJCQk8c3RyaW5nPmNvbS5hcHBsZS5wcmludGluZ21hbmFnZXI8L3N0cmluZz4KCQkJCQk8a2V5PmNvbS5hcHBsZS5wcmludC50aWNrZXQubW9kRGF0ZTwva2V5PgoJCQkJCTxkYXRlPjIwMDktMDgtMzFUMTI6NDM6NTVaPC9kYXRlPgoJCQkJCTxrZXk+Y29tLmFwcGxlLnByaW50LnRpY2tldC5zdGF0ZUZsYWc8L2tleT4KCQkJCQk8aW50ZWdlcj4wPC9pbnRlZ2VyPgoJCQkJPC9kaWN0PgoJCQk8L2FycmF5PgoJCTwvZGljdD4KCQk8a2V5PmNvbS5hcHBsZS5wcmludC5QYXBlckluZm8ucHBkLlBNUGFwZXJOYW1lPC9rZXk+CgkJPGRpY3Q+CgkJCTxrZXk+Y29tLmFwcGxlLnByaW50LnRpY2tldC5jcmVhdG9yPC9rZXk+CgkJCTxzdHJpbmc+Y29tLmFwcGxlLnByaW50LnBtLlBvc3RTY3JpcHQ8L3N0cmluZz4KCQkJPGtleT5jb20uYXBwbGUucHJpbnQudGlja2V0Lml0ZW1BcnJheTwva2V5PgoJCQk8YXJyYXk+CgkJCQk8ZGljdD4KCQkJCQk8a2V5PmNvbS5hcHBsZS5wcmludC5QYXBlckluZm8ucHBkLlBNUGFwZXJOYW1lPC9rZXk+CgkJCQkJPHN0cmluZz5BNDwvc3RyaW5nPgoJCQkJCTxrZXk+Y29tLmFwcGxlLnByaW50LnRpY2tldC5jbGllbnQ8L2tleT4KCQkJCQk8c3RyaW5nPmNvbS5hcHBsZS5wcmludC5wbS5Qb3N0U2NyaXB0PC9zdHJpbmc+CgkJCQkJPGtleT5jb20uYXBwbGUucHJpbnQudGlja2V0Lm1vZERhdGU8L2tleT4KCQkJCQk8ZGF0ZT4yMDAzLTA3LTAxVDE3OjQ5OjM2WjwvZGF0ZT4KCQkJCQk8a2V5PmNvbS5hcHBsZS5wcmludC50aWNrZXQuc3RhdGVGbGFnPC9rZXk+CgkJCQkJPGludGVnZXI+MTwvaW50ZWdlcj4KCQkJCTwvZGljdD4KCQkJPC9hcnJheT4KCQk8L2RpY3Q+CgkJPGtleT5jb20uYXBwbGUucHJpbnQudGlja2V0LkFQSVZlcnNpb248L2tleT4KCQk8c3RyaW5nPjAwLjIwPC9zdHJpbmc+CgkJPGtleT5jb20uYXBwbGUucHJpbnQudGlja2V0LnByaXZhdGVMb2NrPC9rZXk+CgkJPGZhbHNlLz4KCQk8a2V5PmNvbS5hcHBsZS5wcmludC50aWNrZXQudHlwZTwva2V5PgoJCTxzdHJpbmc+Y29tLmFwcGxlLnByaW50LlBhcGVySW5mb1RpY2tldDwvc3RyaW5nPgoJPC9kaWN0PgoJPGtleT5jb20uYXBwbGUucHJpbnQudGlja2V0LkFQSVZlcnNpb248L2tleT4KCTxzdHJpbmc+MDAuMjA8L3N0cmluZz4KCTxrZXk+Y29tLmFwcGxlLnByaW50LnRpY2tldC5wcml2YXRlTG9jazwva2V5PgoJPGZhbHNlLz4KCTxrZXk+Y29tLmFwcGxlLnByaW50LnRpY2tldC50eXBlPC9rZXk+Cgk8c3RyaW5nPmNvbS5hcHBsZS5wcmludC5QYWdlRm9ybWF0VGlja2V0PC9zdHJpbmc+CjwvZGljdD4KPC9wbGlzdD4KOEJJTQPpAAAAAAB4AAMAAABIAEgAAAAAAw8CL//u/+4DOAJBA2cFewPgAAIAAABIAEgAAAAAAtgCKAABAAAAZAAAAAEAAwMDAAAAAX//AAEAAQAAAAAAAAAAAAAAAGgIABkBkAAAAAAAIAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAOEJJTQPtAAAAAAAQAHEAAAABAAEAcQAAAAEAAThCSU0EJgAAAAAADgAAAAAAAAAAAAA/gAAAOEJJTQQNAAAAAAAEAAAAeDhCSU0EGQAAAAAABAAAAB44QklNA/MAAAAAAAkAAAAAAAAAAAEAOEJJTQQKAAAAAAABAAA4QklNJxAAAAAAAAoAAQAAAAAAAAABOEJJTQP1AAAAAABIAC9mZgABAGxmZgAGAAAAAAABAC9mZgABAKGZmgAGAAAAAAABADIAAAABAFoAAAAGAAAAAAABADUAAAABAC0AAAAGAAAAAAABOEJJTQP4AAAAAABwAAD/////////////////////////////A+gAAAAA/////////////////////////////wPoAAAAAP////////////////////////////8D6AAAAAD/////////////////////////////A+gAADhCSU0ECAAAAAAAEAAAAAEAAAJAAAACQAAAAAA4QklNBB4AAAAAAAQAAAAAOEJJTQQaAAAAAAM7AAAABgAAAAAAAAAAAAAF2QAADg0AAAADAGcAcgAyAAAAAQAAAAAAAAAAAAAAAAAAAAAAAAABAAAAAAAAAAAAAA4NAAAF2QAAAAAAAAAAAAAAAAAAAAABAAAAAAAAAAAAAAAAAAAAAAAAABAAAAABAAAAAAAAbnVsbAAAAAIAAAAGYm91bmRzT2JqYwAAAAEAAAAAAABSY3QxAAAABAAAAABUb3AgbG9uZwAAAAAAAAAATGVmdGxvbmcAAAAAAAAAAEJ0b21sb25nAAAF2QAAAABSZ2h0bG9uZwAADg0AAAAGc2xpY2VzVmxMcwAAAAFPYmpjAAAAAQAAAAAABXNsaWNlAAAAEgAAAAdzbGljZUlEbG9uZwAAAAAAAAAHZ3JvdXBJRGxvbmcAAAAAAAAABm9yaWdpbmVudW0AAAAMRVNsaWNlT3JpZ2luAAAADWF1dG9HZW5lcmF0ZWQAAAAAVHlwZWVudW0AAAAKRVNsaWNlVHlwZQAAAABJbWcgAAAABmJvdW5kc09iamMAAAABAAAAAAAAUmN0MQAAAAQAAAAAVG9wIGxvbmcAAAAAAAAAAExlZnRsb25nAAAAAAAAAABCdG9tbG9uZwAABdkAAAAAUmdodGxvbmcAAA4NAAAAA3VybFRFWFQAAAABAAAAAAAAbnVsbFRFWFQAAAABAAAAAAAATXNnZVRFWFQAAAABAAAAAAAGYWx0VGFnVEVYVAAAAAEAAAAAAA5jZWxsVGV4dElzSFRNTGJvb2wBAAAACGNlbGxUZXh0VEVYVAAAAAEAAAAAAAlob3J6QWxpZ25lbnVtAAAAD0VTbGljZUhvcnpBbGlnbgAAAAdkZWZhdWx0AAAACXZlcnRBbGlnbmVudW0AAAAPRVNsaWNlVmVydEFsaWduAAAAB2RlZmF1bHQAAAALYmdDb2xvclR5cGVlbnVtAAAAEUVTbGljZUJHQ29sb3JUeXBlAAAAAE5vbmUAAAAJdG9wT3V0c2V0bG9uZwAAAAAAAAAKbGVmdE91dHNldGxvbmcAAAAAAAAADGJvdHRvbU91dHNldGxvbmcAAAAAAAAAC3JpZ2h0T3V0c2V0bG9uZwAAAAAAOEJJTQQoAAAAAAAMAAAAAT/wAAAAAAAAOEJJTQQUAAAAAAAEAAAAAjhCSU0EDAAAAAAQmgAAAAEAAACgAAAAQwAAAeAAAH2gAAAQfgAYAAH/2P/gABBKRklGAAECAABIAEgAAP/tAAxBZG9iZV9DTQAB/+4ADkFkb2JlAGSAAAAAAf/bAIQADAgICAkIDAkJDBELCgsRFQ8MDA8VGBMTFRMTGBEMDAwMDAwRDAwMDAwMDAwMDAwMDAwMDAwMDAwMDAwMDAwMDAENCwsNDg0QDg4QFA4ODhQUDg4ODhQRDAwMDAwREQwMDAwMDBEMDAwMDAwMDAwMDAwMDAwMDAwMDAwMDAwMDAwM/8AAEQgAQwCgAwEiAAIRAQMRAf/dAAQACv/EAT8AAAEFAQEBAQEBAAAAAAAAAAMAAQIEBQYHCAkKCwEAAQUBAQEBAQEAAAAAAAAAAQACAwQFBgcICQoLEAABBAEDAgQCBQcGCAUDDDMBAAIRAwQhEjEFQVFhEyJxgTIGFJGhsUIjJBVSwWIzNHKC0UMHJZJT8OHxY3M1FqKygyZEk1RkRcKjdDYX0lXiZfKzhMPTdePzRieUpIW0lcTU5PSltcXV5fVWZnaGlqa2xtbm9jdHV2d3h5ent8fX5/cRAAICAQIEBAMEBQYHBwYFNQEAAhEDITESBEFRYXEiEwUygZEUobFCI8FS0fAzJGLhcoKSQ1MVY3M08SUGFqKygwcmNcLSRJNUoxdkRVU2dGXi8rOEw9N14/NGlKSFtJXE1OT0pbXF1eX1VmZ2hpamtsbW5vYnN0dXZ3eHl6e3x//aAAwDAQACEQMRAD8A9IpqyTe3cyj7H6Y2kA+rugc6ensVr0av3G/cFzmT17q9PVLsHHrx/suPTVYbrdzdu5sv9V+9tf8AUWfV9c+u2Z2HQcfGYy7Krxslrg/1K5e2q1ulm31WtduYkp7P0av3G/cEvRq/cb9wXL/WT6zdc6d1l2D07HxrKK8NuXddkGxuwb7q7HvdTv8A0LG01/Qr+nYqrvrb9ZMa3Hoz8bBrvufUH0ssc5wrsc2v1WbDZu59n5iSnqhTlfbDLKPsW0bYB9Xd+dP+D2o5rpHLWidBoOVC/Lro3F7Xltbd9jmjdtGse0e930fzGKtflken6ljK9zmkMYDYef8ASS1n/RSU3fRq/cb9wS9Gr9xv3BZf7Xx3PbXTfbdc9hspqY2susaNu57Ki1tvpfpGfpbPTr/4RWq8vKeNWMBGhDiWyQ3cXf4T02bhY39/2f5iU2vRq/cb9wS9Gr9xv3BNRb61TbI2kyHNOsFp2PbI/lNRElIrKR6bvSYz1IOzcPbuj27o/NUMeqz0h9qZV6uu70x7fKN/uVhUh1XHLtsEaTLi0CP85JTaNVIEljQByYCCX0/m1A+ZAH8E9tge1scGSQfEdkCy1rNC0uJ7Tt0/rfSSUwozmW9Zt6aaGAVYtWSLBrJssyKCzbt/N+zbv7adlz8Zh/abag5xJrOPVY5oYA3+cdsftduVHp8n61ZLpL2/s3HAeYnTIzva/wD4Rv5y3La2WVuY8EtcNQCRI8JakpqNzunusNTWkvGpBqcBA0+m9jWf9JJuXjOJ21NcAYJABg+BXLfXDOyOn4+JTjTUL7iyxwc4uDfpMrZr+ffYz+p/glgYua9jKsiq/Y4GbG7iHPZG+K3H1P09nv8A+22KH3Sc0cW3GDwGr1H6Mv8AFZoctOfLz5niEceOXAf3uLT/ABfnfTce3CyAfRNbyOQ0tcR8dqfLqqGLcQxv827sP3SszBc7GxhkUVNe1xcBa8vabK5/V7WVtbZS31v+ss/wn6Pf6VVv18u/FuseyttDq37XNc4vkCIdW6trW/nf4RTDZhf/0Oq650LIzPtVuNcKrLqGs97C+sj0/Tcy1kO31vauaw6OpVdYoOVivJvz8S31amWvYTNbLrHWPZ+jax1X5/5n86vTaf5pn9UfkU0lPJfWbCys7M6rjVuuY23plOOwsbvZuvuyfWds9m+yuqhv+H/R+qsfo/T7a+k43TM85GTTj3MdTddU8WUvFjHN9CWud9k9v81a5/8Axv8ANr0VJJSK2kucLazttbpPZw/csH/Uf6NYfVa+oYb6b8ShpYS5tzXOaBqD6TKb3bPRqduey+y6r6H8z++uhVbMsoaGNss2OLhtDSd+sj2NZL3IEX9OqYyrx8Ds5uE5mJVh0sx3bryGmvG2tqpDWF+1jKnel9mrcz0f3Pf/AMIrHTOnZGHhVYrbGllO5jQQS7ZuPpsdf7H2OZ+fds9S/wD8ERQ3GnfYy14fDQ9zSTp+b7W+s1v/ABiNXjYTmfo62ObJ4AImZd/0khGkylxdAPJFbUWU11uf9O0yWF9eji50N9FzXKuAxzi02kgHWLcgGJ/r/SVvJ3UtpZRTvabGtcA4MDGmd1n8rZ+4mrbSWDfa5jpnb6pPH0dd30XJWCSOyKNA918WgAesXOLn6j9JY5sEafo7Xbf+iqh6LYbA8X7QIgAfL87+S1aqSKGu6gV0tDfzOfhx/wBFUs57aWeq76IEdhrMNbud7W7nPWm9zW6O79oJ/IqltGHkssqcdzK3gWMIdDXNDbG8bX/nssSU0OhV1252TnuEXuYymWztNbXWW1+0/n7rbP0n59XprUyr76QH11C1gDi8l+wiI28tdu3e5Rw6KMcenQ47YktdqSSdzn73/pHfS/fVkgEQeEACBqbKhdavE/W/HPU+nvvupaKaS5zXNs986bhWyGObkVbW2e79H/58XPdF6Ozr2VjtyLrLa6y9mTvqDAysA2H03sY1jPWc307L2v8AtHv/AOuV+m9RwcPNxH4mUAKbvaQIEk/u/wAtZXTel4/TWW14eR61bnh125zdzBH0Ru9jN7f5zf8ATUZgRkiQBRv1fpxv5uH++sHuCREZkY5D1QB9Mpf1ouxjNIc6SZIb7S3a2OG+m2XbP6ill/0S7/i3fkKVLb5DrthIaAHMnWefafo/56WX/RLv+Ld+QqVe/wD/0eq+sHXur4/UcLpHSX41dt1Xq22XA2vaIhv6rW6tzK3f9yLH+mtHoXUer32WYvVBjuuYA+q/F3tY9s7bGvpvL31WVy3/AAlnqf8ABqtnYeUM85tFLrZprbta0SSwP+hZ9J385t22JugU54zrMrKodSywFtDNjmlrC5rnNuk7N+/dZv8A9bEpudT60auofsnGkZIo+1W2RIZWXmmvbu9u+yxln9T0/wCWql/1ls6e6t+UDbjOfUyx4bBYLntxmXbg1rH7L7K23Vfz35//ABl7LwzX1mrqFdDrvXqGLkPaR7GVufkY7nMc4bq/Vttrf6TPU/SV/wCCWY3oOc3Ed07Jvv6k3OzBbbdcGs9GkObk2N9p/m/UobVjsr+h63+hrSU9Dk3FkMa7YXAlzoktaPaXNbr7tzlmOznNDG41Je/eJJ3GHRsL7JDbXbnbve//AAasdV9u8zt9VjWtf2Ba4n/P2v8Aobvf7/0lax8zqP2DGr2el6bHBvva/QE7N3qP9P8ASN/Ns2/men/wqBlEWZERAF6qAkSBEE26DM6+twGVT6bXDT0w4lon83d/ObdzP5tWxY9r23Ve5zm/pKeHPn6LgCfbY3bbs/wfp/4VYtlmLjlrXVux8ZzgMr1HB53H31Psqd/M22f4K/d9D/raOepZdLGvbhbqJDKxvLTHt275Ftbtn/gf6Sz6aUZA3v8AXT/vUmJG9fTV17n+vXRZRZDfUBMAOkCQ+t276Hu+ko1NqFcWY7nuE6mtgJn3fme1LHrF+Jj21wQ8tvJeCD7hv+jLtr9Uq8AbgLWUmsdmtcDPZ2ry1KhZ8UU2arfUBOxzI7PEf3qq/rGCx5aXPgHaXhji2Y3Da7b7938hXA1rWhjRDQIAHgudq6D1Ghm6t4YcbeKaqyRvDtdzXTW2j6b/AORvRU7vrUvFd7Heo1zd1ZbqCHQdzVQ6xdbjN+24ZG9jQMrTcfRBLxb6f71Ltzv+J+0f4T0kSjHOHiYuK4t31t1DNGzI37P5O96lYfs9/wBoP8yBJfMDbth2530W7Y3JKYdLz/tdr2Pr2WsbLnNMsMnlv9b81aaw+g1OZmZLqGbenvA+ykiC0A60Nb/oq3O/Rf8ABv8As36P7Kr3XM23p/ReoZ9Aa67Exrr62vBLS6ut9rA8NLXbNzf3kAb1U89U131gzX9RN7qGM3tw2Nlr/SqtfWzIduH6GvJ9GzbZbs9RUemX9M6hn5GMOpVTnWC2ptVrS57K93qbIP6Sv2Wb/wDi1yVHWMuyr1LsDHrFzje6um3Mqa59gDX33tGaG5D7mj9LZ/hv+EUXfW3PDbmt6bhY+RTayhluP9oxnsreyyx/pW05PrY7f1Wv9Hj2elZ/orUJxjI8Uugr/BC4SkDCj8hMof3pPpn1de/Ey8jpL7DZSB6+I5wcDtnbkVtD2t9lXqYzv3P0/wCjW1l/0S7/AIt35CvJul/XHrWN1XAa3Cxa2WXNxjt+0gNZkWt+0PZRbkuqfbba71H32epd6385/OL1nL/ol3/Fu/IU7jE/UDdoII0L/9L1Kn+aZ/VH5FNVrX5DMVn2es2PIA0LdNPpxY5jXe7+Wo4VmcS5uVWR3bZDWjt7S1lt3u3bnJKbaSHeLTS4Uu22R7SRu1H8klv0kDBPUHS7Lho1AZtAMmDu3122t2N+h9FJTac1rmlrgHNOhB1BCrvw2gbaIpa4g2Ma1u1wADfouafzG7VZQ7n2MrLqqza8EewENkEgOO53t9rfckppnp4obf6dQy2ZE+tXaQHEEBljK9zfT2Prb7KHejT/AMIsJnSrbc00VW6P/SUX2S2a2na6l1U7nX4znbL6Xel+Z/NLdrs6swtNlfqiNWBrGGTp9P7Q/wCj9L6KvpKJJ3YUVCiiukEuFbQ3ceTAjc5ESSSUpJJJJTWy63vLHMBLmg6diCW7mO/rbfpIDsQZLdl9IIGo9VrXQf8AptRmv6nw6mjvqLX+Gnt+z/vfyk7H9RL2h9NIYT7yLXEgfyW/Z27v85JS9DHstc0sLWgAB0jafJmu5uzb+4gdeotyeh9Rxqa/Vtuxbq66pjc51b2Mrn+W4q+kkp8byOhfWPMtNn7JtFXoNYLara5LmbW+3bk+2rbX6f0vU9P06v6PVXXWm/Vj6xlxN/T7K797LKg406itpYNwNzfV2er73f6T016obLaHNbbTiVtJO0+qWmB+6x1H/f0hkX2EF1WI4NEAi8ugn83+j/8AFppjceHpVfRMTRt80xfq/wDWO6zGdkdJfjDFyqbg6atYvZY97tr9/wDM+r67N3066fT/AOD9Vy/6Jd/xbvyFVhde1zf0WKBIFsXGRJjT9X939r6aPk2Vuxbg1wJ9N/B8koQERQ2VKRO7/9P0Cv7f6bf6ZwOPsccdlL9f/wC7v/smvmhJJT9L/r//AHd/9k0v1/8A7u/+ya+aEklP0v8Ar/8A3d/9k0v1/wD7u/8AsmvmhJJT9L/r/wD3d/8AZNL9f/7u/wDsmvmhJJT9L/r/AP3d/wDZNL9f/wC7v/smvmhJJT9L/r//AHd/9k0v1/8A7u/+ya+aEklP0v8Ar/8A3d/9k0v1/wD7u/8AsmvmhJJT9L/r/wD3d/8AZNL9f/7u/wDsmvmhJJT9HZn2+Gfz/I/nvsf/AEP5SrM+3+l3+ifo/Y9v5n/Q/wDUS+ekklP0BR9v2/mfQ/wf2OOa/wDwL/1Cnp+3w/j+af8AzP2Pdx/57/fXz8kkp//ZOEJJTQQhAAAAAABVAAAAAQEAAAAPAEEAZABvAGIAZQAgAFAAaABvAHQAbwBzAGgAbwBwAAAAEwBBAGQAbwBiAGUAIABQAGgAbwB0AG8AcwBoAG8AcAAgAEMAUwAyAAAAAQA4QklNBAYAAAAAAAcACAAAAAEBAP/bAIQAAwICAwICAwMDAwQDAwQFCAUFBAQFCgcHBggMCgwMCwoLCw0OEhANDhEOCwsQFhARExQVFRUMDxcYFhQYEhQVFAEDBAQFBAUJBQUJFA0LDRQUFBQUFBQUFBQUFBQUFBQUFBQUFBQUFBQUFBQUFBQUFBQUFBQUFBQUFBQUFBQUFBQU/8AAEQgBUgMsAwEhAAIRAQMRAf/EANEAAQACAgMBAQEAAAAAAAAAAAAGBwUIAwQJAgEKAQEAAgMBAQEAAAAAAAAAAAAABAUBAwYCBwgQAAEDBAECAgUGCQUKCAsFCQECAwQABQYREgchEzEIFCJBURcyV2GV0gkVIzdxdoGRtBYkQlKhMzZUYnJ0sbLB0yU0Q1WCotHwGSYnOUZTY5KTlOEYOERz1DVFR1ZkZYOz8REBAAIBAgQCCAUEAQQDAQAAAAECAwQRBRIhMUFREyJhcYGhsfAGFDKRwTNC0eEjFVJy8TRigrL/2gAMAwEAAhEDEQA/AN7ujPRjp9O6PYJIkYLjUiQ9YYDjjrtnjqWtRjoJUSUbJJ7kmpj8h3Tj6P8AFvsWN9ygfId04+j/ABb7Fjfcp8h3Tj6P8W+xY33KB8h3Tj6P8W+xY33KfId04+j/ABb7FjfcoHyHdOPo/wAW+xY33KfId04+j/FvsWN9ygfId04+j/FvsWN9ynyHdOPo/wAW+xY33KB8h3Tj6P8AFvsWN9ynyHdOPo/xb7FjfcoHyHdOPo/xb7Fjfcp8h3Tj6P8AFvsWN9ygfId04+j/ABb7Fjfcp8h3Tj6P8W+xY33KB8h3Tj6P8W+xY33KfId04+j/ABb7FjfcoHyHdOPo/wAW+xY33KfId04+j/FvsWN9ygfId04+j/FvsWN9ynyHdOPo/wAW+xY33KB8h3Tj6P8AFvsWN9ynyHdOPo/xb7FjfcoHyHdOPo/xb7Fjfcp8h3Tj6P8AFvsWN9ygfId04+j/ABb7Fjfcp8h3Tj6P8W+xY33KB8h3Tj6P8W+xY33KfId04+j/ABb7FjfcoHyHdOPo/wAW+xY33KfId04+j/FvsWN9ygfId04+j/FvsWN9ynyHdOPo/wAW+xY33KB8h3Tj6P8AFvsWN9ynyHdOPo/xb7FjfcoHyHdOPo/xb7Fjfcp8h3Tj6P8AFvsWN9ygfId04+j/ABb7Fjfcp8h3Tj6P8W+xY33KB8h3Tj6P8W+xY33KfId04+j/ABb7FjfcoHyHdOPo/wAW+xY33KfId04+j/FvsWN9ygfId04+j/FvsWN9ynyHdOPo/wAW+xY33KB8h3Tj6P8AFvsWN9ynyHdOPo/xb7FjfcoHyHdOPo/xb7Fjfcp8h3Tj6P8AFvsWN9ygfId04+j/ABb7Fjfcp8h3Tj6P8W+xY33KB8h3Tj6P8W+xY33KfId04+j/ABb7FjfcoHyHdOPo/wAW+xY33KfId04+j/FvsWN9ygfId04+j/FvsWN9ynyHdOPo/wAW+xY33KB8h3Tj6P8AFvsWN9ynyHdOPo/xb7FjfcoHyHdOPo/xb7Fjfcp8h3Tj6P8AFvsWN9ygfId04+j/ABb7Fjfcp8h3Tj6P8W+xY33KB8h3Tj6P8W+xY33KfId04+j/ABb7FjfcoHyH9ONfm/xb7Fjfcp8h3Tj6P8W+xY33KB8h3Tj6P8W+xY33KfId04+j/FvsWN9ygfId04+j/FvsWN9ynyHdOPo/xb7FjfcoHyHdOPo/xb7Fjfcp8h3Tj6P8W+xY33KB8h3Tj6P8W+xY33KfId04+j/FvsWN9ygfId04+j/FvsWN9ynyHdOPo/xb7FjfcoHyHdOPo/xb7Fjfcp8h3Tj6P8W+xY33KB8h3Tj6P8W+xY33KfId04+j/FvsWN9ygfId04+j/FvsWN9ynyHdOPo/xb7FjfcoHyHdOPo/xb7Fjfcp8h3Tj6P8W+xY33KB8h3Tj6P8W+xY33KfId04+j/FvsWN9ygfId04+j/FvsWN9ynyHdOPo/xb7FjfcoHyHdOPo/xb7Fjfcp8h3Tj6P8W+xY33KB8h3Tj6P8W+xY33KfId04+j/FvsWN9ygfId04+j/FvsWN9ynyHdOPo/xb7FjfcoHyHdOPo/xb7Fjfcp8h3Tj6P8W+xY33KB8h3Tj6P8W+xY33KfId04+j/FvsWN9ygfId04+j/FvsWN9ytCPT36fYtj/WGzxrXjdotsdVhZcUzDgNNIKjIkAqISkDegBv6hQb79D/zK9P8A9Xrf/DN1IMnyu0YZbEXC9T2bfEW+zFQ46T7bzriW2m0gbKlKWpKQAN7NB3585i2QJEySvw48dtTriwkq4pSNk6Gyew8hXXsN9t2U2OBeLRMZuNrnsIkxZcdXJt5paQpK0n3ggg0HfpQKUClApQKUClApQKUClBEsV6tYbnFzat1hyOBdJ7sNdwbjx3NrXGS+qOX0j3t+KhaAodiR2JrLxsrtE3Jp2Osz2V3yDHalyIJJDiGXStLbmj5pJbWNjY2kg96DLUoFKBSgUoFKBSgUoFKBSgUoFKBSgUoFKBSgVib7ldoxmRaWLnPZhv3WWmBBacJ5SHylSghAHcnihaj7gEknQFB95Nktsw3H598vUxFvtNvZVIlS3QeDLaRtS1aB0AO5PuHesgy83JZbdacS604kKQtB2lQI2CCPMEUH3SgUoFKBSgUoFKBSgUoFKDEy8rtELJYGPPT2UXufHelRoOyXHGWigOOaHkkFxA2dDagPOsRlfVnDsHnyYN+yKDbJkW3m7Px33NLahh1LJkKA7hsOKSkq8hvvoUEsSoLSCkggjYI99ftApqgUPlQPMUoFfilhPYmgBQUOxFfvmKD85AnWxv4V+0AjYpQKUClApQK88fwh356rL+rzP8TJoN2Oh/5len/6vW/+Gbqr/SJcekdffRwt8z+9p7Ibg/JC+yDNatzyoW/r58ykfEUGs3Rnq1mF+9LTEJDeT5I5bLvf75b7zZbtdZD4jtttPGO2/E8FEeGeSEltKFrWoeZ897M+g448Ojd2io72ODld7iWQjun1BE50NhJ/qg80j4BIHuoNhqUClApQKUClApQKUClAqtfSYuF2tXo7dTJlhU4m8MY5cHIy2d+IhYjrPJGu/IeY+sCg0TztWJYp1KwqVcMsyTBcWb6H21q3XbFXH2FPSRJcWw0XmUk+184JJAWoJ2TvRufptd8vmdWvRXuORB5OZ3bCru3lHipKH1xUtxnGFPp0NK8YpJBA0pSh270G59KBSgUoFKBSgUoFKBSgUoFKBSgUoFKBSgUoFaz9XZd1HpgYIhplMgQsJv06wsO9kO3XkwhQHxV4KgPqC1fE0FWeiPldqzZxES9Z5n2U3ybjEo53ZsiS47abdIKglxCvFCfVnBtaUttghSASR23V7eg1Pu109EfpdIvanFzzaEISp3fIspWpLB7/ABaDdBetKBSgUoFKBSg4ZcyPAjqfkvtR2UfOceWEJT+kntWFdzm0kaiuPXEn5phsqW2v9DmvD/61B1XMzlOn+bWdaOJ0fXZCG9/Wnw/E3+3VcX8qru539ShMdvm+Otz+3in/AEUH0jMbi0Ap+0NOIHmIsvk4f0JWhCf3qHb4+VaL2X00Op12/CGN9LPxow1g67wuJ+L3beyHktiMXOJcAKt8h5gn9OqD0VpQaZdSLtl8Tq/6Udwx4OrzK0YNaUYsGk83kxlIlOSFMp0eSvGTsAA7UlI71VPRs4plfW68yIOWZLneKv8ARmfHul0ytx9/w31SmTJYDryAdAEkoBISSoD3gBuJ6IM+73X0Xel0q+KcXcnLBEK1u75rR4YDalE9ySgIJJ891b9AofKgeYoRsUHy4sITs1xGSFfN1+2vMzsxLpypq2t+1rR/2Vin7u+SocxofpqPe8x2eLS6qcncbUArir6/KsoxkLLqRpej8DutdM+/SXmL7uVF0Q4rzG/jXOm5hO+RBFbou97u0xMQ8eIUCa7HmK3xO8PXcpWWSlApQK88fwh356rL+rzP8TJoN2Oh/wCZXp/+r1v/AIZusnnWAWfqLbIMK8MuLECfHucR5hwtux5LDgW24hQ7gggg+4pUoHYJoM5cIYuECTFLrrAfaU0XWF8HEbBHJKvcRvYPuNYrCMLs/TrELRjFgiCFZ7VGRFisAlRShI1sk91KPmVHuSST3NBnKUClApQKUClApQKUClAr5daQ80ptxKXELBSpChsEHzBHwoMBgGCWnpniFvxmxIdZs9vSpuIw86XCy2VlQbSo9+KeXFI9yQB7q+Y2AWeN1AnZp4LjuQSre1a/WHXCpLMZC1OeG2nySFLWVK13UQnfzRoJHSgUoFKBSgUoFKBSgUoFKBSgUoFKBSgUoFKBUcynALPmF4xu6z2XBcsdmmdbpTDhbW0tTamlpJHmhaFqSpJ7Ht7wCA7WaYnBzzErvjlzVITbbrFchyfVXS04ppaeK0hY7p2kkbHfvWQtdriWS2RLdAjtxIMNlEePHaTxQ02hISlKR7gAAAPqoOzSgUoFKBSgxt6yGFYUN+suFT7u/BjNJK3XSNb4pHu7jZOgNjZAqMSr/erqfYWiyx/MJbCXpB/SpQKE/ApCVeXZdB0WrRFRITIWhUmUn5siUtTzqfqC1kkD6gdV3KBSgV5SIdlx/wAK647AUkTG8gWtoL+apQhk8VfAK1onzAPbvQex1unNXW3RZjO/BktJeRyGjxUARsfoNdigjkrALPLz+BmZZcayCHb3bWJDThSHYzi0uFtxPkoBaApO+6SVa+cd8ud4Vbeo+H3bGLx46rTdWDFltx3lNLcaV89HJPcBQ2k68woj30GVbTDs8SNHR4EOMjhHYaGkJHklCEjy+AAFfqbhFXA9dTJZMLwy76yHB4fDW+XLeta778qDnWtLaCpSglKQSVKOgB8a4VT4qYaJRksiKsJKXiscFBRASQreu+xr47FB+szI8hgPNPtOMlRQHELBTyB4kbHv2Na+NdWfeWYiSAoKUN14vaKxuxM7QjMzLkvKUELBKfMb8q6KsuUhBAI37vOqy+p69Eebug9kzroPJalf9I11FXt07AJ19ZNRLZ5lrm27qOzXHD596/WZzjY1yKh9daeed93ndlYV5OwOXta1omu6LvxPFR9r4b8qmVy7w2RZ3Lfdyl1J5dt+QNTCDLTIbT3BOqn4L83RupO7tUqY2lKBSgV55fhDfz1WX9X2f4mTQbr9D/zK9P8A9Xrf/DN1NqBSgUoFKBSgUoFKBSgUoFKBSgUoFKBSgVEemOUXbL8enzbzbhbJbF6ukBtkNrb5x4859hh3Su/5Rptte/I89jsRQS6lApQKUClApQKUClApQKUClApQKUClApQKUClApQKjuS5Ou2vC329CJF1cQF/lAS1HQSR4jmiCR2Okg7UR5gBSkhHIdvTGW4844uVMe140t/RccI8t6AAA2dJAAG+wFdqgUoFKBXlTD/8AOyK/WBz+DNB684aAMSswGgBDa0P+gKzFAofKggvV/pj8q9gtVr/GsizphXiHdFSIpUl4hh0LKEKSoFCjrQUPLz0a1kc9ALMzhsrGU9bbkuzu2+PbkW12LJEUNogriqBbblo2OSg8lO+PJI5hwE7DNX/0ZLyzcpcS79epcadMkvXGFEfkvtHwxIafdWptUs+J+QaeaWAEshKgQ2nirlksC9GlcnpZesRT1SRkcO5ptiYc2Mp9S47dvlJK+CTKW2nSm/BBaS3x4J5cymgi0/0MbrGkG0NdXJTrESQ3dWrY8JKlI1c0zEeIhMwJ4FTTg8QIS4XFrUHO3ERBj0UepMN5iFK6z3Z2HDt3gJuHOWHpLqlzeaFoEoAI0+wSolS1eGAlTZTyMPPkinSYa72iFkdGuk0/pUb767kTmRu3RbDqpD6HQ4lbbQbI2464SjilATs8gB7SlnvVkE7+uqTJbntvCJM7yx93uQgNp0tKD5lSvcKirvUWCDr18n/JaP8A2VSavXU09uW1tmq1tknst1RcGteIHD5pUPeKyeqscN4yUiz3HWH6Do9j3rBZfnjOJJgofhXC4PTHPCZRBY5kq2kaKiQlJPLsCdnR1vR1Jr16Q9R1R1j0msFhw7TKNyddRcXEoZQhrS0BTbziVuJUQUAiO4nR9rY1r4TjFPSc6e3S1W64/wApYkKHNiJmNOy3AhPh9uWyCQngSEqJOgogbNWmCtq92+kTC1MIzy09QrfJnWV1yVBjyFxfW1NKQ264js4EcgCrgraFHXZSVJ80nUjqz7pBSshSgV55fhDU7602X9X2f4mTQbr9D/zK9P8A9Xrf/DN1NqBSgUoFKBSgUoFKBSgUoFKBSgUoFKBSgVW3QMsKwu7+r3J+6o/lTkAL0hotqQv8bS+TQBUraW1bbB3ohAIAB0AsmlApQKUClApQKUClApQKUClApQKUClApQKUClApQYXKb+qyw224yEvXKSS3GaXspBA9pxWu/BI7ny2SEg7UKisGCmE0v21vvuqLj8h3ut5wgArUf2AADsAAAAAAA7NKBSgUoFeVMP/zsiv1gc/gzQevOHf3p2b/M2v8AUFZigUI2KBSgpHqqrAuol5XCez224xf7CZUeZ4rjLcjwlxFocBDuiUJbkKUFp2lJJ79lCqyx/oJ0gskR9gdVoM5iPLk3T+cXKGpUd51LjPiFQOwG3HAUFXdC0pAI0AAlnQzCMf6aWXIMdayy0Xu53WSqUE26U36ymO0w1FHJSNLWoKZWtbmgfFdc2SolR611lZPgQInesZjjze9TWGR+M4yd/wDKNIAEhIH9JsBfkPDWdqqs1O1uiPk69HZTl1tutgRc7TPYnxXdht9hYUnYOlA/Ag9iD3B7HVQC49U2ojjiFypKyjey2AB2/aK4jiet/KWiszMe5Cvbl7qwzD0jbFGW7E9bmT5ziT/MorSpD2v8lO+Pw7686qu4dcrVIbQ5dbpLtUZZGrbbWeMtYPuW85xCAR5hA2N9l1I4PwzFr88ariFvUjtWf7p9s+EfOfc6DR8F1OetdRenqz1iPGfbt32+rJxOqVrUW/xJaZ9laT7KXo90ksrJHvUUO7WfiVEk+/dTrDert5lvpifyyuMV8k8GXkMSUL7e9TjZX7vLmK+y5eHYMmPe2OJ90bfONlrk0VLU3msTP7fRb9s6oPY5MYbyi7QZcGZCU9EkRIK2ni+24htbBQFr8RavGZKAgAq2oaOu3U6g5bIi4a1My7HR6u887ORb49wVGkQo7DKndreSoBTxCFeylSU7Xw5KAK1cNrNNXT5+XH2ntv19/wA3M58UY77V7MBAu3Q65NMykYrLksW9UdiOhMB9LTKX3vAYLKCQni44+UgoHx5a49sza8p6CO48m0MWF6BBmWoqS2u3KCWYi5LKeSkubQhHiLZc9scT59/aFbMVb7dZYrEth/RyVjyulNsGLu3aTaFOyH0Sb20pEqQt55b7jqtpTvmp1SgQNaV7tECz6sY7NxSsslKBXnj+EO/PVZf1eZ/iZNBux0P/ADK9P/1et/8ADN1NqBSgUoFKBSgUoFKBSgUoFKBSgUoFKBSgVU/SvIoGFYRdHMpmWfGeeTX5bfjSGYza2zdZRbcJ5a5LQUqUfPko70d0Cb6V3R6G8plvqNYLlIR5sWmWJ7o7f1GOav7Pj8DXEr0msamJ1ZceznIXO+kwsPuTaFa+Dr7LbfuI3y91B9fLVlk0H8W9Es4fSPJc2TaIgPwOlzuWvj7Ox8KK6t9QIySZHQ/JXQBsGDebQ5+nYXLQd/UAaDjV6S9ms3I5biWZ4M0ne5N4sa34yfrU/DL7SBrvta0irExDN8d6g2du64xfbdkFsX2TLtkpEhvevLkgkA/V50GbpQKUClApQKUClApQKUClApQKUClApQKUFdIl/j25ybuTybc2zE+CWEnsof5Z2vfvBQD82u1QKUCvhnxpz6mITJlPJ7KO+LbZ/wAZXkPd2G1d96oPu9dLBk0NLU+/XSIdHk3a3UsIP1HaVKUPdonR+Aqrr5j976HXiFM/Gj12xSW8GXvHJ2xv3kbIBABVyToK0QQDx2Fo15Uw/wDzsiv1gc/gzQevOHf3p2b/ADNr/UFZigUoFKCs776OeB5DfbtepVrfTd7nLjT3prE15txMiOh1DDqNL0hSA85rQA2QSDoVGc19Ffppkipblxs77r0uO5GlPCY6lx5DkhyQ5yWDyHJx50nRG+f+Knj4tO0bsT0hjbB0RwvCsukZNabWW7u62tpUpxwr0lbilnjv5ndavm63vvs96zs6V4zhSk+yO36apNRfmRLzu189IUWjB4svKRc/5KzlABcxhJUiYrWkpeYHZ49tA9lgA8VpG61YhdSZWcxoUnJJL2MY9PbUqMm3EF64lK1NlJdJ0wCpBHE+0e+iR3rmcugjV5fzGePVpO0f/bbb6bx7+3g6DgfDI12eM2aN6Vnx7TPhv7I8fhHbd07Y9bYokItbLduGyrwEBQPw7kklWviSTU36b5QiEubarmBPt0tBJQvaghWtH9hHb9gqm1+bek9Ovd+gNRpY9BOLbrG23wRO52NrFLn6m274ltkLWYKlAgpHn4SviQN6PvA+o1hMsvQscBp5kIVJLqUNhTnh7Pn2V7joHXxOh76+58C4nHEeEY9TP6ojaY9sdPn3+L5dxHH+WvfePalFqMnqbj9luc7J7jj17gyI0iLMtylMm1tKV4bikpOuSuDhUsq80oUkcQTu65vUjqH0OXFa6yY7E6iYRHcS4zm9khBT0LXk5Kja9kjsStGgPcVk6qp4pjra1JnpMxO0/wAfNwWtpHNXfvLaLpqxgXUGyW7JsZj2K8QH1mVHuMSO2rTpUCpW+O0uckp5b0oFI33FTy1dMMTaW2tGL2ZC0ABChbmgU6X4g0ePuX7X6e/nUDDWY6Sh1hL7RZYFhgpiW2FGt8ZJUoMRWktoCiSVHikAbJJJ+s13antxSgUoFeeP4Q789Vl/V5n+Jk0G7HQ/8yvT/wDV63/wzdac/hSr/kFjufSz8R3u52dLiLqXhb5bjCXCDDCSsIUOWtq1vy2aDVXGc4zQJSl3Lb+6CNEm5vk+74q/RVhW3McqATrJr04nXkq4Pa/1qDvScvyhnxCMnvKm9eyRPe3/AK1YqT1IyGNPbgO5ZeWZbmyhtyc8Cr9HtfVQdS45Zl6QpaMovfb3C4va/wBaofe81zJTR4ZffkHy2m5vgp7f5VBAbj1BzmI9w/llkJH9b8bP7P1/Prdv8FRkWQX+69VPx1frpemW0Wsx03Ka5IDJJl8wjmo63pO9fAfCg9Ca45JKY7pB0Qg6I93ag/nhPpadbdA/K5mnf/8Av0n79WtN9JjqjZsCvcdvrDkjst2S0iPLcv0pxxQBQXEtrU0lQ1oE64dleagSKDqZH6UHVC3YJi7kbqpmcy8zUvGSDfJIDSgsa0UhJV5ka5LAPw1qsVavSE65veC691UzFpO9qacvkjkE/HRX7+2qCb2X0iurkK3+s3HqNlXNY20BeZC0qI7HzWO3kf2H41L8I6v9YsxacnPdQsntdmig+PcXru8lJ1o9gV9yd+etDyO/Ihjn/SR6nXKc/Gi9QcnWlKjxcRdJDaUpAPtE8u2yd6PuFIvpM9SoVxjMxs8yW5LW8hHiO3eQpCtqA7J59/8ARQey9KBSgV+OLS2hS1qCUpBJUo6AHxoKinekjZ7tcJFr6f2W69TrmwtTTrmPoQLdHcHml2e6pDAI96ULWsa+buuM2nrbmveZfMa6awVEn1ezRlXmeB7vy74bZQfq8BwfWaDkR6NNnuSd5Pl+c5c6fnGdksmI0T8fAhqYa/Zw7VFfRy6E9KY+NXe5WzCLXIlpyS+RVT7vBZky9s3SS2Uh1SSrw0lGkAnfEJ3tWzQbBRIMe3x0sRWGozKfJtlAQkfoArmoFKBVH9bunvSjHWHsyvlxb6b34kJbyexSfULg85v2UaQP52SToNLQ4DvXE7oJl0RyDI770fx27ZswuBf3IqlyxKj+rOFIUoIdca/5Ja2wham/6ClFPbVROydfMozmzxLzh/SS/wB1sM9sP2+6XG4wIDcplQ2h0NqeU6lChop5NgkEHQoOFrq91XkZPNsjfSa0CVChsTXEOZilK1tureQktgRClWiwrltSdbT57rJfLHnbKFesdDcrKmd+MqNdbQtCh8WiqYkr7e5SUUH5hvpQYllr9galQMgxT+UAbFofyK1ORY09S08kIakDk0VqHdKCsKV/RBq36BSgUoFKBSgUoFKBSgUoFKBSgVgs1mLjY8+0ytSJEspiNqQSFp5nipSde9KeS/8Ao0EfbaQy0httKUIQAlKEjQSB5AD4V9UCvxRCU7JAA9591B27fZxPievT3/VLaE+IEcuClI/rLV/RTrvoaOtbI7prhk9RrXbG0xbRBXKbb7IU2AywP0E9yPrSkg786Dhg9Ugp5In2wxmT2LkZ4vFP1lPBJ1+jZ+qpBkljt3UDEpducdbkQZ7OkPtkLAPmhafcdKAPw7UFdYNdJKobljuw8O/WjUaUhR7uADSHkn3pWkA77d9/VXmhD/8AOyK/WBz+DNB684d/enZv8za/1BWYoFKBSg+Hl+G0pXwFQXI7opCDxX3V51E1NuWrVknaEYeuCnGuI9499Y2S+mOypalBISCSpR0B8TVFa3MiTO7yZ9Jzqfl/V+DieYXu5paxu9y7kbHYWGeKY0eOpLQdcV5rcWVKHfyAOtBWhlcFfbk9OLal9oGGUON6dG0LHI7Hfse58vrrZxuvotHh9H7f35pjd9m/BkY5w2xzHff/AG69lsk13J2fUpKfxekj8i5KJCEEHmAniSTvhx9rtogaCjqVKZRHUtpvXIKI8RJPeuF1eWmS9a0jaYiN/bO8voelw59POTHmtvG+9fZXaIiJ393zWRi+KMdT8bkW65POxVNABqU2PbbeT8xxPxKfePf3HvqlbjksjHrvcbLePAZuVvdXHcdbc2hakHRUneux8x9Rr6B+Ccs4q5NNHWLTMx7Nun0cFx2IrkmY8P5drpHkbOQX2Rjry1Ji3Nt6MnifmqWkhI/Rsjv7q9Guh+RKzHE7WqQPF9Ygsv8Atp2FJWgEg/o3rVdHxuu/o/ZM/Pb/AA+X8TjeaT7/AL+Spup2PRvQi6kY/wBS8QcRBwTKbw1asmw5pWmlOOg8JkNr3OJ4qJQnzA0AATx2+6p5zN6bWixy7dZH8genXVmAqDFKA8pK0rUSjxFoRyHAfOUBrf1VExxvG890GvZrPN/CPxm8ElSY+GzE5NGhwlOCY4hiCJD8NUjjtS+eiUKDadcnAUlPv1Lk+nhbbjHkG34PfFKZedZdedWx4cbTsRlBcAc3z8WcwlbQ9tvS+XkN7nplIfpf/wAoumOTZBarG3Bn2O4QrdIVd3wmPG8aX6s5JkJbKnGWm+K3iFgEtcFbAVsRO7+n2iy+pQv5LO3ic0hh+bLs/N2NIZLbLq1xW3A29xWh9AaW4hKVrQ6kE+HyIdfGvwi9pmGcxdsYW1JaDrzDsS4MGKto3MQWSX1K4BPtIWp4kI0SRsVJ8T9OS35xmeMWC3YbdIX43nworsi6PNNeqIkRXH0lxCVKKVlSUpQlWvECipJOgCG0NeeP4Q4b61WX9Xmf4mTQbsdD/wAyvT/9Xrf/AAzdahfhO0c7v0tIKuQbuugnfnuHQas49HaeCUKUska8+9WZYsXlXJtJjRwpQ7JKgUhX1fA/vBoLYX08RiuNSJaI5cmerOJI+clSyg90n3EE+8Vp5mUu83y32dCLPKXc7WkKmTi2lKEKQNlZVsAE+ZHcdzQWBZZrOTY/DnsaS3IaCij4E+f9tYC/2lTJ5pBIHnugr6+2hLoPs8tHtyFbtfgsoYhzOpieAQootvl7+8rvQb/VxyhuM6P8Q+f6KD+dZ/pK9dICp2K3CNla4rQVcbSw0WZ7AAJUvweSg6gaJ8RlS0gAFXHfe18ntV6yaDc7NMxORFuMp6FFbDMJmM1G4Adl8GE/lVeGrikEdk69oqIIXjA9AGJFwnGk5r1HtWKZNEQt4WmJaw4+PEIUEvuNKSoLA0DsL1sgEjQqtsr9HzM42URYWRJmuqfmKUq6lh5TDkZlCVeI04pseIClaeIAJUQEgb7UE7sHRuBd7N/KW/LctuExNtFpplQfmlJ4pYYA1tRUdcgO6gQjekqru5709tt6RYBmzbPSnHYqnFw7MmW2/eLglR0nhFCB4ex7IW5vXlxoMHk/TJ/LbcmJZMdT0/xeEriZ15/m70oAa5uN8eS1HtoqUE62B2A1CWbThOIXSJGk3Cfkt08ZCNxWfBjIPIDfZW/2hRHl2oPbilBXfUrq7/Iy823GbFY5OX5rc2lyItliOpZS0wkhKpEl5XssMhRCeRBKldkJUQQI/Kyfrlj8f8YzMJxHIoqAVPWuw3x9ualPnppT7CW3Va7aUWgT7xQTjpv1PsHVaxLuVikuFUd1UabAltKYlwJCfnsSGVe024n3g+fmCQQTUtmgo9LO8XS7XOY850gt8pcC22WO9wayF1pRS9KlFPdcbmODbJPFfArUFBSRQX5bLVCslujwLdEYgQYyA2zFitJbaaSPJKUpAAH1Cu1QYa85lY8fSo3C6xYyk/8AJqcBc+HZA2o/uqj+jnXCFDssiDIam3lyVkd9cbmQGEqZSyblKW0jY1taUcEqGtgg7Owd02u4rg0G1bRNp3iNqxvtv5rDS6HJqv0zERtPeduy3LZ1Txy5KKFzhAcSO6Zw8LX6VH2f7ajGT+kvgeNwhJTc13RlauDUiIkIiOq7bCZjpRG2NjsXR51L0msw66nPhnt3jxj3o+fT5NPblyQgUz0qmL3IS3EvFlxiCvumSqPKvktaffpuKgMoOvJQedHcEpI885YupZH4vu9lzq45hB/GsK13G1Xy2NQX0CW4ltp1hIjsLGlOJXtQWlTbbgB5DYnI7r9bOvmSxLO5C6SWBnLLn4yo8vIZUhli02rR0sl15xtEh0dwGm19iPbUnyNdYbl+J9Gp0XLOq2MX1eS3EKbi59dH2L83Ke0VCJFVF5eqlXE8GUNNpWRoFavMJ11A6u9Z2OneRZLYukkGNBjwH3Y0K831IuxHhqKXVRW2ltAJOlFov8yNjsrsc503xfJ+gmBY/Zba9I6kYdb7dHYbUgtoucZKUAEsp0lt9nWilBUlxAGkl3aUgJneMetnVG02fKMfuztuuqI5dtN+hp9pLbgSotuNqGnGlFKebSx5pBHBaUqTirvfrn1A9HK83CLFcayCdj0tpcKEFKWzODK23GUAe0VIeCkD37TQdjq7guM5DhUS2ZRLt9twC3uNP3GBLaQliQ2yUqYaUtR022FpQSANq4pTsAqCoX6N+YItN1v3TuZ+OIjcR9y5Ymzfmlofk2JQbSkpUv2yGnlON8XNOJbLHIe0DQX1SgUoFKBSgUoFKBWGzB5yPj0lbTq2XApsBbailQ24kHuO/lQVfesrXY46Fv3C4OOuq4Mx2ZLinXl63xSOXw7knQA7kgDdYVzJcpuBJFwVamTsBCZDr72u/flySlJ7+WlD6z2oOMzMiA2nLryVeYDjjZST37EBAOu/uIP1jQrOYVkmV5lYxLkXhyxtMSH4jbcBDTrjxadU2txxbyXNArQopSNaTrZJOkhIEu5XBG4mTpmqJ7pvFvadTr4J8DwSP0nl+isrZOoylXGNa8ht4s06SoIjyGnvGhyV/wBRLnFJSv8AxVpTvySV6Ogm1RbNFF2ZZ2AQAhxyT5dzxQUa+r+6/wBlBjaUCuMxRcpkSARtEhz8qP8A2SRyXsfA6CPq5igj2U5E5k89ftf8HsuER2h81XE68Q/EnWwfcNa0SScTQK7uHZUrB7z4Epz/AMXri77RUe0GQo65/U24T7XuSo8v6SiAn+Z9P4uVvRp7EhdrvkQaj3FlPJQT70LTsBaD/VJHv0Rs15Evqulq/ClzFKitzLuxenimPEC1odcEJWgka5aJ120SNnz1sh7CYO8mRh9nUhDqEJjIQnxkFK1BI4hRBAI5Ab0QD3rOUClAPlSg6tzBMNYT5kf7ary+RFKV3HlVfqo3hpyRuj7zXhK1sGo1nKHnsWvLTP8AdlQnkt99e0WyB393eqbbaUS3R5b9Voce4ei50LmRm0h2KuVFUop1/dXHOQ/eyo1w9OsoS90ybtHgJ4Q5i3lqI2VrWAB+gaQO3v4/VUnjVJy6HDMT2tb6/wC3138ETGS/Lv2mfrLINXgR3EqbaDRBHtJPcf2VLI0bxWEOL5Nkp2QR2FfP9TX0e0777vsmevJtO+6z+mWRNxrhHaI5R2zwCd637+9fWFTRafS2z5pTTSGbtZo85oaBGm/DSrW/rcV5V2f4IzT/ANUrgmO8TPyfGPxvi5dBmv7I/wD6h+dcIuJ2S52rLfCdtl8iK4tNQmUAz0pBJRxT3K0pBPL3DYUQNFN2eiN1awxnEPBm5XZoQS694aZk9phbbYdVwSUrUD8wpP76+l/iLFWJrFZ3neN4+E/f/t8l0We2o0dYt/bPT3dfoxV1yTAvSh9Ku6t5NlVib6Z9O7auBBEm5stN3K6ymyl19olQCwyj2Qob4rSkg+1W0PTX0gsKu2BWN6855jLV6RHEeely7R0bkt7beIBX80rQog+8EH31z9Y2jZYRCWQ+smAXGWxFi5zjcmU+tLTTDN3jrW4tR0EpSF7JJIAA+NR/q5meZYVcYr9htJvVsdt8w+rRbc/JfEtthxbIUpvYShaw0gbT5lXcdq9Mq7xDrz1alwGzc+lE1yU3NdjvIcEiKSyA6sPJ/IKSSUpbQlAOiVjktJJSJ56P/V66da7VeL3LtcayQYr7UFqC26484HvAQ86pTi22+w8dDfEJ0FNLIUoKToLPuNrh3eOhidFYmsodafS1IbC0pdbcS42sA79pC0IWk+YUkEaIFdmgV54/hDvz1WXv/wCjzP8AEyaDdjof+ZXp/wDq9b/4ZutT/wAJWlDl26ZJUAVeHdCn47Bidv2/6dUGvXTfD1XNbC3UhDHIJ35HX/ZW3PTyxWxllLaUNENpGgR5ke/fvoIJmfpD4Hbc9jYkuYr1mRJEQLKNMeICAU89+7Y3UN6uYYYLUlEVaI6JLavFAQSgq15LH9VXlv3EUGtGBZS1jl3RjJSGW0lQ8BRJLC9j2QRsFJ5DXkfdqrMusYSoh0o71+z99BW12hpCXEK0CN6Jrb/8F6lSXuo4VraU24dv0yaDfSuOSNxnf8g/6KDwKsuKPWt6M20Ex3isPR5GPXONKeJA2lSIS3kvJWf6zbiD568tVu16OnRZ/q5kEN/JrS9CuEB2PPkXtq2KhOlLfAtM/lGWVIUFbKVNBaQOXJa/ZNBV/pmYVe4HVSXdI0xh6yKmKaW4qMFrDbquJStJHtcCogKVtXcgEAADZP0Zut4teHwMTvL8e72+IwmKgvJDaQkdtFJQfZ1oaJNBaXUq2wbbY5GX49Du9weKEsiPaE83IzQQElDBAX6sghPdbbfL4qAAA0jvXpBQsSuc13CsftNgu3i/zqdcH7eue4FK0ecyTMW8VHffbI137DWwEMvWSN5RcXpVzexm+OqUXC3fcubcUj2RtCVMeGSOW9aUD7u42awVvtd3m3iN6njuISmS+kcbbe5jw7q3oAzyew+P1+7zD22pQU/03QhXpEdY1SuKrklFmTHJ+cmB6qstgb/o+seuHt23v37q4KCletnSG/XC5yM16cqhRcyctz1ruUGWssR73DUghDbjiQSh5pR5NOkHjyUk+yo6gnQR2+Yf0vxSK9BVjd8t1sYt861uhK0IdYbLRQsJPdO9qHFQOiO9Be9k6jQZ6kszkfi6QTxSVq5NK7kDSvcSADoga3oE1AOpF99Vud3U8+5Ma8RDbMYvFTR5Mp0OAOtELUT27g+/tXuM1NPS+a8b8tZnb2+H7tV6WyzXHSdt5iEIVGjWVcF24xXrvfLk94MO1RGwt11zRUUoQSE+ykFSlKISkAkkAbrC9Gs8yeBjFzxyzxolqnJyG9L/ABNbY67tdmSbjIDil8lNRooW4HVocecU2oKASFa768ekppMMVv1y29a8+2fD3R4f7Z9NbPkm1elI6Vj2R4/FDeub+UQZBiRzbo2QS57FnRcr5cBcJLlweR4rERC3S1GYWpBC3G2G0oQlQKXFLKW120jpN076ZZY8/mVgx68PzGecfIsnabkPJ4JA4KddCuCdDXsgAEg676FNqqfls1NVjjvMVt7YnpE/Cdlpht6bHbDbwjePfHh8YYW1elv0isuGXi83XB5eMLtaXHJdvRZmXHVpQoI8VtLftFpRI4uOJQNEb47rXnD/AEisd9KrqpNmWnAodkxy0j1e0xHoAU9cpDqV+K5JbjDxJISjsIbauCvF5OrCRyTdK9ttiPQq6XtMadeimyKabS2y/JjxZl2DY7BCdoVFgta7eBGbV2APict1LrH6N2GWW+G6KYl3F31pielia/tlMpnZbkFCQkLcSVHS18iNDWtDQZXq3dXJVoZw+2vKTfsn5QmvC+fGi9hKlf4oaaUeJPYuLaT5rFYyQLv0WisPeuPX/AoyUtvollJnWdoaAcS4APHYQNcgvbqQCoKc+aAm2OYpBxJNzTby8iNOmOz1RlL22y65pTvhj+iFr5OEeXNxZ7bqP9D23XOlliuD5R6xeUO3x1DY9ltc11ctTafiEl8pBPchOz50GMv2bdOrzkcOW8XMpullcWGPxTb5V1RCe8lK4x0OIQ6NFPIgLSCobAJBhHUDK3751i6KTnMYvOP29m+y2E3u5tsth7xrZKCYoaDpdQHFpQrbiEgFpA8zqg2EpQKUClApQKUClArBZt/e1J8uy2vP/wDNRQULal/jd5y9ukrclj+bhX/JR97QkD3FQ0tXv2rROkp11r1lTFskCI14bsw9tOLKEIJGwCQConXfihKlaIJAB3QY+05HNRkLMOdMgXC3zkLMKbDYXHKHm9FcdxC1qIXxIUO4JCV+yNVnrdlj+HdJ3JECMi4XyVdp8G1W9ZI9amOz30tI7d+O/aUR81CVqPZJNB3bd1Gk2F2PDvd4tGTuqX4TkzHob0ZCXtkFtptxbgkcdEHwnFObGvC89Tp9i3ZXY+Ci1cLZPZCkrbXtDragClSVA/DRCgfgQaDOdNbzKn2iVbbi+uVcrPIMJ2S5rlIRwStp068yW1oCjoDmlehoCubMU+HcLS6SQFeMwPgVKCV6/c2f3UGNpQKQpCoSr3ck6Krfb1Fo+7krkpQP/wAJv9/1igr+NHTFjMspJKW0BAJ89AarlNA8xXHIjtS47jDyEusuJKFtrGwpJGiCPhqgzPRrqV4kp3DbzIJuUBamYcl5XtSGh3QlR96wgpP1j6wd+c8H/wA8Sr9Y3P4FVB6z413xy1fExWj/ANQVkqBSgeYpQdea2VsEJAJ+uo3doAW2SvvseR891GzV3hrvG6H3eP4TafqqM3wpTDPLRBBTxNUGb1JmUO8PMPI7al/0K1Q1DcjEr/Ijp9rTgUJitg/9F0nyHlUJ6NMN35nJIbfsNsMGehB8ylK0p/eAv926m6yfTcOm0f23t9Il9L/BWf0We3/l9dmTUy4l/SUKB32SoeVTeyTBIt6WVkBTI0tRHu+P/f4VwOsjmpEx4PveqiLUiY8H5Duj1vuCJcXkEpVwQ2PNQPmKz17zCBZ/SKwtxdyjonuWJ+BcW5K/D9Xc5OupQsnQBIS0Nb7Ejeq6f8I1tXjGK8eUxP3+75n+OMNb8KyR47T8v97QkHV/JbfkeNs2lxEWYqU6ktthQUBr+l5/Xofp+qpf0o65z+gvRLIr1DZVc7xeZKbVi8JI5rnSPBbjx1ITrakhbalgHupA15kV9m/EvJXT03j1ptER8/43fnrh9Ix4I6+P39Jbw+ir0TPQLonZMZluiXkDvO43uby5GRPePN5RV/S0dIB94QmpJ00T6jeM9taTpmHkC3GUHsQmRGjylkD4F197v8Qfrri12nNayQbh6RuPSCIlnYvrLwSHU396GfDfDjylFkx3UFLCmiwkFfJaVpJ4aOyGZu+W+kHElqdtuJWS5W5bbimmn2kRpIUIzJSHAJy0J5PqfHslXstgHWwo9jp5kvXWTkWOw8jwmw2WxOJ53R+IpBLSyw0tSUJTJVx08p5PIBwK4j5u9kL6pQK88fwhp/8ALVZff/4vs/xMmg3Y6H/mV6f/AKvW/wDhm61Y/CPxUyLx0xJAPFNz7H9MTv8A2UFF41dV2SO2dHwvLZHlVq4VlIlSFJLwDDg4kJPuJ/7/ANlBQnpkdEQfWMosD6IT7bfrKkJHmpACtj4K7diO/wCwmrkyTO4d/wCldguTboducm3suuNjfIKU2OQO/rJ7Gg0k6gQP+HoD0p9MO6Nv84s1GwFg79hZ94IASCe/fv8ACr+sMxy4YzDecAU84yConzJ1rzoInlUNJ5OpAOwR+g+VbTfgvCpU3qaVABWrcP4mg32r4f34K9a3xOt/ooPLbBsGhIkvB/Iup8ELI8FS8obZiS1AFSix40bxFgHQ4oQrsfM6Irbz0d/xPaenN/kWCVMlR0SU8/XCjxUBLaRw34LThOh5up5d/PWtBpP6cWVvRcavjKVR0uXBaWlJA13JGxs+ZHl293vrWfpN6Qc7ErrDjXmWpBYI/nKwV79nWlAg99HXlqg9MfRn67WjImGgxNEmDJQEPR1oKRsjv7J7fu7HR+NWF1Q9FnEc3sD72NWeyQJPhqXHis2yM1GcdIPtKU22lwHat75efuoNFuoHSmJYZ622btAtF8tylrlm42FDrbS0bSrwn/V1kDfYALUToGqck2R6xX+K1dsfT3kIV6zDcITskeY2tCD5expB7eQoPdmlBTvV61XPBs3s3VWxQH7omDFVasktkNsrflWsq8RLzSB3W5Gc5LCANqQ68BtRSDZuLZVZ83x6DfLBc4t4s85sOxpsNwONup+II+vYI8wQQe4oMrVUdXrPLg3CNeIz/gw3uEeQrgV8HeQS2VAnXBW+J1xO+Pfv21ZbTWkzXvD1SItMRKGqRcXWyFSmGiQRttgn3Dv3Uffv3eRHw3UTv0JSMiZ/nvq7bLRkrWQQlCEk6ASDoaSNA6+B76rm9Xr72rWkR0m1frE/wtcGmrvMzPaJ+jvWzIpGP5bkU1Qbayt6y2624/62NttyJc1xl7gNkOBta4KnNe4J7gHtZvoyWCJjnTq5RI14XkDgyW+B+6PhfjvuJuchBDylpBW4gIDZVrR8P2Tx411l7zknmt3UtaRSOWvZpL6QnSbIPSH6xybFJQ2np905fuN+yJSnX21yHZVykqQ0ksMvOlfqjLGtIPFAOiOSSbPwX0W0YF0zazWAzFuSbtAYmuW125TnHGG3EoUloOLkONPn2tFRbRvZ0dGq3X1i2kyxv/bPyjdM007Zqe+Fp3vA7H6UHSVrDWsTl4fjj6HUovVr/F/gRykll1plCHFLSpQLqCFNJ4kHkARo3L0u6VYv0Zwq3YriVqYtVohI4pbbSObiyByccV5qWrQJUfP9gqZSeasS0WjaZhLapb0s/SUgeiz0nfyyVapl5lvPepQI0dsloyVIUpBeX5Nt+z3PmfIAny9vKgfQOsWX9S+lFy6zpzGW71Dya4yzIRdCZNucZadWG4ngkhTLaSVlJaUkp59wpKQk7Rpu1u66dObnboc02e7LaS1LhuhLkm0TEkKCH2idEocSDxPsuJGwSlQJDN4vebpleKSm7jb3rBf2PEhSm1IV4SXwkflWHCB4rKuSVJWPcdKCVpUlPw3ilotPTKFjV6eaTaIttZgSFF9UdtTbbaUkFQUCEkJ7jfcEg7BNBgonUeHNjsWfpnZY2SsRUpZEqI8mLZoSQNBHrCUqCiANcGEOFPYK4Ag1BOt8DMbZbsKyDIL7ZZcG15jZnF2u3Wt1grD8tEQDxVyFlZR6zzHFCeRbA0ASKDYSsXkWSW/FYUeVcniyy/MjQG1JQVbefeQy0nQB81uJG/Ib2e1BH7p1jxCy3iVa596jw5sZt5a0PqCEnwvC5pCj2K/yzek72d9h2NdsdVMM9UlyVZbY22IaktyluXFlIjLUSkJc2r2FEpUNHR2kj3Gg7txzvGrQ4hudkVphOLimclEia02VRx5vAFQ22P63l9ddFfVTETd8dtbOQ2+ZPyBxSLaxDkJeU+EsvOlY4E+xxjPe2fZ2njvZAoPp/qlhcdkuu5fYWm0p5la7myAE6bO98vLTzXf/ANoj+sN9a8dYsHsU1MKZldoROVI9U9URLQt0PcFrDakpJKSUtOfO180jzoOdnqrhj8KRKGWWRDMYNmQpy4sp9XKzpAc2r2CTsAHWyCK7Fz6j4lZQ0bhlFlgB2Mma361cGW+bCiEpeHJQ2gkgBXkSfOg453UzFLfdkWt3IrX+MysIVDTMbLze0hQUpHLaU8Sk7I17SfiKRup+GzVREx8tsT6pba3Y4buTKi8hPLkpGle0B4a9keXBXwNBmrPerfkVtZuFqnxbnAeBLUqG8l1pzRIPFSSQe4I7e8ViOobKpGG3FpCy2twIQlQ7EErSN/8Af+2gpCwPJk2G2uoAShyM2pKR5AFANQuxtofu99lrCVyjNcaLh+dwSRpP7Pm/9AfAUHBfJDVkvVredhpuEC6Sm4M2Fx5FZ0S08j+q42UkhQ7kdvMJKfqc9YrNbces+L2K72a45Hdp0N++XWY5Icjt+K4ua3EcLy/Acd4OjaOB0lSu5CVALHdsdudshtC4Uc2rwPVvU/DAaDQTx4BPkE67artdC5Tsvpnai4+ZXFJ1IJ3zKtLWP+gtS2//APHQT/AR6xlmUyEE+E0mJCUPd4qELdV/1H2qy+dN6gQJJ0URprZUPf7YUyNftdH7N0GIpQKRGVSbFlsBtJXIeYU6gJGyQtnw0j9O2lfvFBAm3EvNoWhQUhQBSoeRB99fVA12pQVncmJDXV23qhpSqWqdDkoQ4SAoJUjZOu+tNq7/AFH4VpjBnzV/hZzKIjsTjkDh0ApxtJ9TP+SSNfooPXjDVuOYjZC8pCnfUmeakJKUk8BsgEnQ+rZ/TWYoB8qUClB+KTyTqsXcYpU2dnyrXeN4YmN4Qe9t8wtvvpO9Gq+yzm3HQQSQdpI+H1/7K5fXxMUtMIN3nBe7LIlW3rZiEdBUUZFLlNJUQAC+gLbHwAJSP/pWtFkudxsM8rZLkK4wj4brPdKk6GjsfDXY/wDZU/h9I1Wk1OCs+tW0X29k1jd1P4c1MafUXjfrvE/KGwuBwpnVu1uybHFVLuUUJEiEz3d0f6SU+ZHx15bFWJjXQ7KZq24sizXCGXTtx56I4lKEjvrZGv8A61831uqx6e9sMz60T28Z8Y29779/1bTTh35o3jwmdpYDqvLtXR2QpqLd2p+WNoAZtyWj/M1EdnXD80ED2gk998SRo1rPNsQyq4oXLcdkS3VkqeUslS1KOypROye/evpf4O0WbDW2sz05bT0iO/SPHp57vnPH9d+e5cXaNv8A1+/+Eyj9DbFbm4T8tcp91xQWI5cHEoBG96Tvue3n7yfca3G9BzHIvWjr6xl9yaK8QwttcTG4yxplc4pSHJAB7HiOyfr4kaKa6Xiuo9LqcdJ7Vjf426fSPm+Zauta54pTwjf4z/p6dA7FVt05ySBeeqHVGDGdK5MCdCS6kpIBBiITySfJSeaHEEjyU0sHuKiNSyaEboFKBSgV54/hDhvrVZfL+95n+Jk0G7HQ/wDMr0//AFet/wDDN1rT+EIhCZd+nAPLQRchtPu/4r/2UFEWqI4izNt9ir3pcR84fX7vLVYK4Q7njalyLS6HVqPMxlez38zx+A1QV/lPXGLmVy/ElyU/HcZTpcWSdEggdv3e6rKs5tyMfY8JaVIQ2AEA/NGvIUGu/V+zycons2+Ijkpa0eCrtoEq0T5eWh/bV7WuMLZaY0dSgSy0En9IFBH8jQlbCh/SA936a2Y/BkNhu7dT0+agm27Px/4zqg3wr4fIDK+XkEnf7qDwYyvPc59IK8G14FY58SKgqRKTZluojzUghLa3kKOuwCAkOKURvzPbXqD6HeHScY9GqExcFvOTXEBctx9zxFmQn2XQDyI4pI4ADQ2hXmTsh5ten9mX4yz5mxtKBaZJkLIGu/dIH+mtVfVHvDSvw1BCklSVEaBA8yKCd9HerF76X5NHftkg+A6sIcYWSUdzrYG/OvX3pX1duTtkhOSHVaWgLIQ8OJ7d9EAnv+yglvUfC8Y6/W8bhGJk8ZvTEoJDTywkK4teKpBUU7Uo8fI/VXntmeDX3p1kXql5hqciJkpj8nG3FFgFWwlJUlKkq1vse2qD2cpQKpvL+i1xxe8zsx6Ty2cfyV9wyLhYZBIs98V5q8Zof3F9XukNgK388ODtQTnpf1ChdU8Ht2Rwo78H1jxGZECVoPwpLTimn47gBI5tuIWg+72djsakF4tMW/WqXbprSX4kppTLrah5pUNGgoaZY7zjMr8XSi1IWjkGXXtt+MlI3yBCSFfObB0BrZ89d4xkLExi9W2Y+liOyVpaWtD6yAkKCjs8Ukf0vq7d/MiuZ1+jjFSM0T0ras/DeN/kt9Nn57Tj26zEx8ksYvPTrHbnDumaZHbbRdmD61bYUi4hh8oKU/lg0lfJe9aAI7DY17RqCdF/SDxSzQHLV69dbM5Kv95ksNtQlPxH45uclaX1LcKnObqFJWohXzl7AAIFXmXiOHD6l+u3Tt/P/tjScM1OtiZxRHaZ6zt277LowEWXF1ZFk0dMeTHySe3PuF5gOKU14wZbY2ptW1MIShlskFSgkrWSUjdVBmWXwWpqLP0s/GE6HNUWpFrU8t23uhSgpBjIWo+AQsADhwbKVKBSTxUiDxbUUx6O04+vPG0fHp/PxbeH6G+TUev6sUned/Z4JHgHTDqddYVxk3G9OYYxcZhkG3slJeRpttrlppWk8g0FaCz3USdEms9/9l9qZdk3G5ZvkU15LRb8IP8AFrZ17XE8u/b41qroMuWI9Pkn3QnRxXT6XeNLhie/W3f7+Pwd4+jmzFHK3Zbeob3uWXAdH/o8T8ffVaekH0azvI+kGT4pc7s1lONz4ikrdkNeI/EUnSkPgKPLaFJSrso+Xes20mbSxz4LzO3hPixXX6bX/wDFqscVme1o8FC/g4+rivR/TN6R9Rwqxw7rOVNxu+yiUQZbikpDkdLitJQogIWlKtElagdEpB3zwvM5ObznLlbLI01jDvJDF6kSeD84IOkuNMhB2ySVcVqWkke0ElKgTcUvGSsXr2lzmXFbDecdu8O5lXUix4bk+JWC5yFtXLKJb0K3ISjYW42wt5XI+4cUa/SpI9/bpZrlVigTE2/J7BOkW1PF9q4OWpU6H4g35lsLLRT/AFnEoHfsT3rY1MnDy6FkmNSJ+GS7TkimQW2m2J6UsFwa22pxCV8Dr3cTrtsCqh6uZ7b87xHFYzaXbdc4+c48zcrNM4plwnUXFlxKVpBI0VNBSVpJStI2kkHdBsBUV6l47bMoxhMO63V+ysNTYk1qdGU2lxp9h9D7RHiIWg+22nYUkgjdBUeQdB+l+U3pF5uWTT5F7SZR/GKZjLbhVJYajyDpLYR+Uaa4kBIHtq0B7PH6yj0U7HkmOMqxHIHbTO5yls3OQ0JiUpkKmF7igKbGz68+jaidJOgAoBaQkmXejDi+czLRJu0+7OKtlrRa22WXm0tFCWH2efEoJCiiS5vRAJCNg8RXfsno7Yxj2aRMlhSbmiXHukq8GMp5BZdlPmbyWocOWgLjISkJUBrhvZTshGLj6GWAz3748mReort48QSSxLRoIW8p1TaUqQU8SC23og6QwyBooBrKveivhr0qFJD91RIiLkrbWl9GyX08Xdjho7TtP1A9u+iAw2S+iRZ37NFaxi8yLLd4i3jHnzWhLDYdK/F9gFsElLikgkkBKj23pSZJefRtxfI7BDt1ykT3HY1pt9pRKYcS2tKYa1LacSCkjkVLVsHaSNdu26CGL9DGwXLJZTl1ujj2JpiyYkDH4bJY9XRIQ0l7k8VqUrkGvcE6KjrQAFSqR6LWHXB64v3B65XOXclNrnPynGiZKkKcIKkpbCACHCkhKQCEp7b2SE76dYBA6Z4pHsFtlTZsZlx13xrg94rqlOOKWruAABtRASkAAa7V280Osbk99e00PPX/ACifrH/f4+VBRDEc2a7SrWoEMqKpURfuLalbWj/oKVrXkEqR9eolkEZixZw3NCiyzdITgf38wusFJQR/jFC3Nj3hv/FoImzeU5imJeYcxX8xW9MZjpBSlnwmVe04f6ZKloSdeyAogbIKjmX2kZLb7W9+MnokPGrzJlh2G4App+TOuMPxVAgg+GfZ0oFOnHOQOgQHwvqQx1DwS+2e33IG8+us485MhtLbJckLS14rSFDklSUqcUUHfAtL2SlPI3+hi2dP8SbYhRFNW22R0tR4kcclqCQEobQCfaWo6SNnZUod9mgmeA49Ix7HUJnlCrvLcVMnKbVyT4y9bQk+9KEhLaSf6Lad1k8htirzY5sNtSUPOtKDS1eSHANoV+xQB/ZQQy3zE3GBHkpSUB1AXwV85BI7pP1jyP6K7FAr8akrt01mc0krU0Chxsea2zrkB9fYEfWNdtmggzsRu3yX4jSgthlWmVp8lNEBTZB9/sKTv691+UCnnQRqxtRZfXRoLdQp6NZvFQgEckr8RaP9V1VacW7pblw/CAHJHcUnpnDJnJRlhhz1YQCVNh7n8zj4Gjvfz+3zvZqLnnJHJ6Pzjf3Og4Ri0GWNR+etttjtNOu3r9NvfPsepWHf3p2b/M2v9QVmKlOfKUAjdKAfKuhPd0ggkdxrfwrxfpDEoPfQQgn+kNioXeGEyGVMrHZWzs+6ue1NebeJQsjSdrFvG9JzqzZlEoN0tlvuTTZHzvDQWj5/42u9UJ1n6PS3ppuVuR4N5ikgqI0Hx7kq+B92z+g/ERtDlnh2urqJnpMViY9nLET9N/fCPXU/lc9c09o23923X/KtMUv0jFZpulvyCbgt6bSW32zHke38SgtoOh8Uq1o+XnoZi4ek91HEYRflRuz7K1e0qNIfbWAD58uKVfs3V/qvwnp9bqfzWOtLbTExPjG3WO3k+p4eIYctYmYi37SxMq/Y9LdMl64PTH3z4j0mU4C66s91KV7W9k7/APr51ksezPELZPbWsPyXB2Q2w1yUpR8gO/vrvMWkrp8UY69oj7lDy5IyWm9u6V5Gu7ZbOhY/ZWFPZdexr1Nja026NrQWsjy0k738VKV8AfUD0VekFt6V4LZ7U1wag2qOH5EhzSApXdRWo+Q2rks7+uuA9J+Zzzl/7p3j3R0r8o+biZt6XJbJ5z8vD5LRfzm8dR21RMCR6vbHNoXmExrcZKfeYbR7yVf1XCAz3BCnNFBhUy52nph1FsSbAy4/Y7DFXbMrurj3JMf1p1DjC31a24/4ylOuKJ/JtyHHF6DiTVi2r+pQKeYoFKBXnj+EO/PVZf1eZ/iZNBux0P8AzK9P/wBXrf8AwzdVD6YWHHKpmGOgkGH62rtrvvwO3/VoKjt2BOLilKU7KkHZPuPYf7f7KhOUdOJrbCZIK3igLPIpIAG+J7D3a3o/t3Qan9ZenT0hh2WhEj15rbjQT59hv6id9u/6KxfRvObm4pVouYe9n2ELUlQPl7/roOp1hzN3GrvbfUHihSFg+Inz1ruP09/7KnHQDO7pmuJT1XiT6zJhyPCS4UgFSSkEb122O9BJshngAp37Pmd/CtpfwY0kS7p1NWND2LaPL65VBvlXHJHKO6P8Q/6KDxXwC/WW02m02JLcmO9MQFNqYbStYSAUvPOEduAJKST58FpJAUrXqjHkQ8X6ZWOzRHC+01bmWkOrSlJWkIAB0kaBPY9vLdB5W+ln6OuU5v1SN0saGpgkNhBbUopCdE6IPlr939tUpN9HLKsUXHTev5ulathuIrmoHyI3rW/20GzXRToHhEKwsyLlZTOuRaJDk4BZSQf6I9w7/D41M03q42ptMW2sqLbYKG21qUlAHu127ACgtHDcpuFkioflP6WgAqKHCfIfVrR7H/trNdUcYtvX6HGXbJKo14ZWhHiu8wHtEEKXxQQtQJA7+739u4bwUoFKClbti+V9Hs4vGS4TZnMuxfIZHrl5xaPIZYlRZpSlKpkNTqkNq8QJT4jK1o2oc0q2pQPYf9Jyy2B9kZhimX4FDccS3+NL9awYDXI6CnZMdx1plO9DbikjZHu70FnXGzWvKobC5CEzGCgqZdadUNpUnXJKkEeY8iD9YqIZxgGNwMOyCZIaUhDEJ9/xX5awlninnyBKtJ0Uj2vMDffua1Zsdc2O2O3aYmHul5x2i0d4eWsfFZuW9ZId1bU2xAnRo11Lk51Th5rCfyjiilRIB3sciNb1r5o2m9FGNieFdJsjvl3zhEC2XDI7q65ZprUdxQbbkvtBPhkK2tS+RKwNqCGk9gnvzHCNXpLc3P1vMRvFum8bTtt+8+PWIjpGzpuJX1OSuGaRtSsdOXfpO/WZ9vn7feg0vqkzcepabBiEu5zMBuLrPipkxlMPPOAe0EtkJPnsJJTruDpWga216a9PLhh15F2mWBtEN1KAxDgvc1xFn2VOKQsjsEgE8SpWyTxJrVoox6/U8mOJilJ3mZ6x0n1a/v1nrKz4vedNpse8+tesb9NusxG8z8OkdIW7ar1CvbBchvcynXNpxCm3WifILbUApB+pQBrpZJnGO4c9bWb7fLfZ3bk8I0NE6ShoyHTocUBRHI7IHb4j4iuytE1naXB7bsrDmx7jGRIiSGpTCxtDrKwtKh9RHY1is0ySyYji9wumQymYtnYaV6wt/ulSSNcdf0iregkdyTqvFpisby9VrN5isd5aI9DJV8uV8j4VKwZvqD09utkJlWqS0he3meCQ+XZHFohJcQgpQ4VDkVEbCAdkOmuAdSehuI47j9iYs2V4+xGQh21Trg5FftLh0VNxpHhLD8dJJCUOJStIHz1DSExtJXlwVj769U7iFufU3n4ftEQ+OpHouK60ZPi2aZHld2sGZY0/6zZxj76FQYCuQVx8N5s+OTxSFrUEcwNcUp0kTPLs2zHpnYWJ0zGH8/YQ621JcxZrhNSlR14ghrUeaU+/g6T32EAA6lq9hMxj326ZFaskwfDZ8bIgWfWLhcpDNvhzYnL8pGlI5LeUoIKi2SyShZGlBJWFR/qHaL11B6t9KGp3Tl63zrPeXLq9k3JmVEYjNRnUlgPJ/KBTjjrXFK0IB4lQPJA0GwlRLqb/AHutf5y3/toK2qXm8TbPguPmK6qHFkTVMTrihoOGCwfFPi8SCPnpbRyUClIcKlDSTQQiH1qzRm8oZkY+JEGO9OjtqLbrT10DZZVGWhPhkIW42vYSSElROygJ1WMc6+58xgl3lDE/Fu6LeH4IWh0OKWqBHeALYb04ptx1anB+THEAJBIIASe7dbcoskG4k4Yqc9ERcPBSmQ4Fy/VlzEJUAGde0YjalfASmuPIkJV0m+veUQL/ACYlyxIKji7C3o8AupKWEyRGck8lJ0pO+ToA0eCF+YStaQzETq5kNvxPG3pdgVNucrH4lxkFfisl2Q4ytTqEIQ0viG1Ngub1xDqdBSuKVYud1yy9DSZ0bCw5CYubsBxlLjinJSEw1vJfaUUJCWlKCSFqB2jfshXaglHS/qrdc+vtzgXDHHLG1DYbdakOLUtMorA2G1cQk8OxUQSNOt+8qCbLoFYPNRvGpOvPk1//ALE0FSX+xm7qjyI8r1ObG5Bt0p5oUlWuSFp2NpPFJ7EHaR38wcA/KfiAJulreSps7DsVtUponR7pKElQ7b2VJT7/ADoI3ExBE+S9CskWNabU8srkratpjIVoqJCiSC8SoK7JCU91ciR7KpanoXIxOzN3nFXpMvxlSFXCBJSJLj4dcKnTxUpKXUEgEsniRyUW1oJUhwMphsPHsVtjcKx4q4xKU4qSuPaMZfhIU+oEFSkqQEtLIJG3Fg9z31U0tuG3jJZcSVfkotNsjvtymrSysOPuuNqC2y+4NpSEqSlXht72UjbhSVIIWLSggM+GbNkUqJ/+Hmc5sbt5EqHjJ/YtQXs/+t0Oya+6BSgh+TNCFdouvmyApCAPPXtLP6AlRUfrLvwA11KASAPMdvjVUXnO7hnjzkPF5Zt+PJPF2/tAF2WQdKRE2NBPbRfIO/6APZaYerzxp8U28fD3tWW/o67oLAwiDAzq9N2Xdouca3W6XFuje1yG5PizR4q1qJU4VAcV8yeaSUq2Ca2n6c5eM8wy3XgtJjynUrZlxkq5BiS2stvNg+8JcQsA+8AH31A4dmtkm1LTvPSfl1aMF5tvErIw7+9Ozf5m1/qCsxV2mFKDqXK8QLM2yu4To0FDziWW1SXUthbh8kJJI2o6Oh518Wy+2y+JUbdcYk8IAUoxX0ucQSQCeJPYlKh/0T8KDlVc4aJyYKpbAmqTzTGLgDhT37hO967H91Y/JJLUKOjk6yl99RajtOuBsvO8SoIST5khJPbfYE+414vG8MT2UyrM8svbSlW+x4zKQG0uKUMlWscVjaFDhFV2UNkHtseW6wEu4ZtcYqVt2bGmUqSFodbvT8gLB+H81R2I773+yqbNFd+qJeIno1v6w4F1NxzqUjqni4xy6XtFmVaZGOLDiDJjpdLpU2sqHJwEp/q9h9eqriHljvW+JJkw7zb4N6ZWUXOxyLO6xKhrHY8kKkHad9uQGvjo7Aj5q0yYuaI6x3/z2VuppS+OZ8u/+UKzLCL3FbcZTcELcOuKEtmOFaI2OQUpSfLWxXew3GnfHji5PJmuNbWp4tgb79k6HuHl37nVU9r09HyUrtO/f2OLvOPeMdK7W5t99/Dy+/5TfMMGgoxpTj8BkvKHNlwoG9HyA7eRqquoCEWjEok+LamVXt91uDBbDaUhT7hKUqOh8NnX7PfWznyRyYueYiek9Z7ePydvh5uSuPeesNuvRd6HW3oFiZvd7VIuGY5E54HFmOXptwdIKihhBIOgNnvpKQFKWUpBIvyB03z+5zVP5Tj1ivNj5pdi44L642y2rQIVLR6qpMhYI8istpOilBUPEPS6GZyU9Lbpv2jy+4W9OsbrEesue5Ygx7lcbbhtsKeK2cfcXMmuD/FkuttpaGuxAZUrv7K0kA1JbNhVjx/GTj8K2sptC0OIdjPAuh/xNlwulZJcUsqUVKWSVFRKiSTVm2GH4s3htjbtLE+ZPhsLUI3rzniuMNE+yyF65KSgeykrJVoDajrdZugHypQKUCvPH8Id+eqy/q8z/EyaDdjof+ZXp/8Aq9b/AOGbrF9YYJnyLK2Ndg+SSPIfk6CFRcfjsNlKx4nuSfLQr4k4zDk8QU6SlHHjrz7jv/ZQV1n/AEJtmSRXFpSlK9HsB27jWyK16vXQa1wZCWIclLUlXJxTjbJSGwNbJX3+IAHv/foKOz7oaqzrj5Df4L+RWdExTa2Yi+Ckp3oLX9X6PLfevrpTMiLtl5eiWhmxxnZQ8GIyoK0gNpAUVDsSfj//ANoP3LJo8NXFQHIgA/7a2x/BTPl+59VO+wEWsD98rvQehNccntHd158Dr91B4seiZjmX5Pn6IzcWDEiPtxoRbROhGUWkKSnTaFE8jyaW4VKQr2k+yUk7Ppn1WyKPjrYZShxbbaOPBsbICewAG6DV3NsslSOEi1l1pxIKuKmwCe3kQdjvv6u3v99Vrcs5EyAhctr1OWtRUqM+CQCD5hXkBrv/AN90HbtOYNMS4rEXwlKe7gIdSoD47I8tj41PrbIblMLjpQhnQ3xS1pP10GUlXm2PRY0SVFbhyxptm4NxzxWo+QXrRG/ifj5/CQ4XEuOEXBuZKuTKZLiAFtR1pKORGgSdq7aIPbz0KdxvNSgUoFcUqKzOiuxpDTciO6gtuMupCkLSRopIPYgjto0FQRvR5lYip5jp71Bv+B2R1ZWLBGYiTYEZRPteroksrUwk7J8NCg2D3CQNg4+15P1A6RXC8WbLrRkvVSyPOiTa8ktMOGt9LJbQHI0qO34OlJcDikqQhQUhaQfaTo4mN+g1GyfDMBs/U643C2XjMcTsZZZlNsZlj82Nara4ZTjqo6Vvsj1Zkq3o/wBy5bG+/eDY76OXWLPbvc7xa2Is+yXG7TVsS7BNgrtxT624CWy0riBsHY898uXtbqunTUrlm04ovExt18Okxuu8ObFlwxTLlmk137ePWNo+rL4Z6NXWbGfSLTP/ABK8m344iPdk/wA9bfUELK20ANhw7Kiw/rinfc77cAN3MS60WdyLHh3nJb7aJ7CEtOonR2FhSwjiSVJYJ7n2jyA7/V2qDos1eH0rjzY4iJiPPpMR17ea21eP/q+OMuC3NeszvE99pnpt4dPvskrWR4rKnsXFHUQJktlvalOxGy6hKVAtrHhAqSoq5EHulQ9kp2QdYPTL6gYzFTimRoyu1ZQ14jmPvRFpbAC3iHAsr2EJTtnisHyBB0dEVaajVabU45xxMbz22md/5VWl0uq02WM18c8te8zHTr0Z30dr9fOnSMgatFoReJF4Uw4m1WtlwRIbqPEC3vE0C4pxKmgo+G3/AHJPc+dXLA6U5N1FuLNy6g3AIiNK5tWaIrSU/USDofsJUR7xVTi9JrKVxRv6OO8z3laZ/Q8OvfUW2nLbtWO1d/H7/wBpg5DYgdYMdjRmkMR2ccnobabTxShIkQgAB7hU7roYiIjaHITM2neSlZYKUCo/nCLeuyAXJx9tjxUcVRhtfP3aBBHxrMVm0xEMTMVjeUE8DGCP+NXr/wCEj7lWLiqYacfhiApxcUBXBTw0snkd77DvvdbcmG+KN7xs1Uy0ydKTuy1K0txSgUoFKBSgUoFKBSgUoMPlVjVfLXxYUlE+OsPxXFnQS4ARo/4qgVJP1KOu+jUUgTE3CIh5KFtk7SppwaW2sEhSFAeRSQQR8QaDsUoItnzfGLbJCezrc1CQR5lKgUqH6PI/sHwrG0FQ55flZ3dpePtSfVcWhOFi5vpXwNwf8zFSr3NJH90I+cT4e9JcByMdDSI7aWAhLKUhKEtgBISOwA121XJcSyzly8sdq9Pj4qvUW5rbeEI1iChcb5lF1BCm3JwhMODyU2wgIUP2PGQn9lWr0DWptWbxgdtIvKXUJ9yOcOMVAfUVBSv0qNbeHTtqJj2f4etP+vb2L7w0axCybUFEwmTsDQPsCsxXULIpQQjrF0otXWXBZ+PXIiM+ttaoFzS2FvW6UW1IRJa35LRzOu48yN961xgfg64uONer451KvllhamISwhnkUIfjBkJSpLiT7Ci64N7HJ09hrdBk7x6HFixJy5Xy6dSZFmsjklpyQ9NWWQhoXFT7cdUhb40jg4mKkdjx18eNd3od6OF26X9VJF1yjLYGU2H8XxomPKnTHHprj6GG2y+UueylXBLoTwUo8XVd/OsTG4hyvQDYx2NaIlvzy+sxYbEOPITzdV44YQ2gqSS8fDJDZCUjaG0uLSlOj2gdw9Ba+2RiJBt/Uecm3FbLb6S2634DTUF+OlTSQ8fynN1tY9pKUlO9K0E1X5r8u+8NFp28EisPovnFcxTlDuVz75KZRMQhmYgqAS+G/a2XD+UBbBUoDShocU6BqgOvmDWWVc2r0qecZy2I54US7QNiQ6QPLgnu8Nf0db1sdhuqDPqb1zVtSPDbb2eKp1Nr1tF6R7NvNWEnrHmtkgNryvGfXQlPFF0tnElaR25rY80kjufm6+ArIY11ot2QPtGNcWpCAR4kcI8NwD39iAf9lMunraJy4Z3r84QbaOlv+XFO9fou2beo+SW+JCLni6R+R15LB8v9tQ7rlaIWLYJjOUzlrdh2zLLYoMEbXKCVKUtCQO5VxSo9vgaxixxlzU3WeH1rxD0f6OYbKU4/muRRfCyW5tBpmMvv+K4ewpMVHwPZJcI+c4D34pQlNhZDkNtxWzSbrd5rNvt8ZILkh9XFKdkAD6ySQAkdySAASRXV4q8tIhbx2RONmWZX9oS7JhUdi3qG2lZJdF2+S4PcoMNx3igEe5woWPegGu/jOfm63xzH7zaZGO5EhlUhuI+tLrMtlJSlTsd5PZxKVKSFAhK08klSEhSSdzKXe6hoBG6UClArzx/CGgHrVZf1fZ/iZNBux0P/ADK9P/1et/8ADN1x9SYtwkzLT6lAemtgPB0sp34eyjW/3H91BHUY9dF6H4vkA/4yDX07jd1bQVfi99WvclOzQV/mzucCK/HsmHXN1Winxltgb/QN7/0VTFvxLqtb5vgzMEu8mA+shTjbQKm9+R1vyB+HuoOg90s6kW61Sl/yHvFyZXNdjuQEt6UtlYGnEd+wBPffmAaqKF6KPUjH23YKumkych55x9mew0A62CsqCFHewQDr4HXuNBF7/wCi/wBaXmQ2308vkhSARzDA2fr862p/BldJc96Xz+pJzXE5+MNTU24QjOQEl8oMnxNaJ8uSP/eFBvZXxIBLDgA5HidD49qDxr9DH0dOqjXVVuZcsKyjD3G5CZQuN7tc1phptJ2UobW2ltxzYSE+IogaCuKuNejeTYXd7sVKfizpaHNDh6sdj4ny7fuoKyuPSC6NSHFM2C6Pe7SYrih/anvVW9RuguaZDDcRbsbvDUjmlSSqG9xIB9ry+ryoI3E9HXLUPuRn8QvS2SoAKbtDqeKknYUDw89cdn4g6qyLd0wyaI2j/wAV7+pYT35257X+rQF9MMrU7IWcavTAOwl4Wx5wp7e5HAk1AZ2L9Tl3pU264Df7hFhlLbaodllJeWNj2uHBQPl7vIE+VB6a0oFKBSgUoIQ0hLnWielQCknHo4KSOxHrL1apeijfcx6Gs5fCuOPZBc+mKMkvMa0Q8asyLgzbVNXF9pxLXhPGUG+Ta/YUwvzBCh3BC58S9IzppeOs+Q/+ONutb8myWphmJe1KtklxxEi4FaAzJDayQHG9jj/SHxq5rnjliyplDk63QLohSfYddZQ52+pWj/ZXi9K5I5bRvDZjyXxW5sc7T7Eef6JYPIVyVj0cH4IW4gfuChWs3pBQemOQWyd0w6YYTb856jKmx5KoNpbQuPaltvoWt2bJVttgFKVIKFbUrnxKCFGo1dHp6W5q0jdPvxPWZMc4rZJ5Z7wtn0ZOkWWdLZuVJvqI1nsL646bJjdqvcm5QLe2G9u+EZDaFo2s64j2QEgJAHnfFS9tlZM7oTP/AD1WL9Xrh/Ew6m1ZClApQKinUn/9gM/5yj/Qa3Yf6lffDVl/p290q891SZ5q5u4Pj/qKZjkJM9Rubduc8OSuL+W/uagQoac8Iq4kKKAsJ2SAbXiH6aqzQ/qshE/IurONR8lmQbOpyDBbZehR7w4h518GMyF+K4lwJTwV4hPDQJSTtWypXWVnHVLKLS2bTbXZdukTClm5MMtsuOR2356Sv+6aBUlq3kEDRDqyNA6TSLh9x8q63POIlrsDLfBlxpcVaUhB2mKsO9tqKwpb7YSO3FJJ5EclTzJb51AjTGEW+1NFtUSKp0sspeShalL9bVyLqTttIbLaQFcyojuCS2EUlXnrKpxqeLbGSW3QybchtCW1tqYhKU6o8lq5JUqbpKdgKbSn29gql3Su955d7hdUZla2rbHbYjKhKZbADq1JPrAUeRI4rHFIIHs6O1EnQWNSgUoFKBSgUoFKBUFusdEPKrkhvQS+0zKUNeS1c2z+zTSf27+NB+UoOje7Ozfbc5EeUtvkQpDrR0ttYO0qT5jYPxBB8iCCRUKS87GkuwpwQzNZHJXHYQ4j3OI3/RPv+B7H4kKA6G9S2I+Nw2JLUWDkMlv1l1EsqLiEulTxfQ2O7qHCoKKkqA5KKVKBSddu550u6l+2Y0tq75A4pQfkN+3FgOKJK1vLT7O0kkhoHkrsNBO1J5bVXjJjrSO0TMz/AJ+O6tyTFqxVIsdsUfGrHCtcZS3GorYR4jp244r+ktR96lHaifeSTU46GL8DEcpyXiVRbjcn5MXz2tlhluPsfUpbDikn3pUk70RWeFxN81r+z6yaaN7zLY2zwTbLRBhlQUY7CGir48Ugb/srt11CyKUAjYoRugr7rK7iV0x4Y5leQx8dYmFq4IkSHkMJ1ElR3P7ov2APEUykgkEhZ15Eikk9A+ksW83lyd1NiPT7i/PEsOzoKHgXg00toEDaUpI7p963NkDsKDJdMsBwfpb1Ku2QQ+oVik2SRCcYjxX7jGU8gOOMp1zSE8WmzFLbbe1BPJeuJ2Dc13ghJWAkL89A1Ez13hrvG8KiyLJV227vWu4wV2uJLX6tAvLbgeiuOqGg04QAplZOwOQ4k6AWVEJrUz0gseVNnNzmpT0CbbvEjpUyU7KVa5jSgQRtI8x5AaIrlNfHo+W3f76qrUxEUa93d29uRirxGLkln5yEt+E7x+o8ikny7aT+moc3gkDIVyVPW96MoPlbMlKFMOoJAKu/v0oq89jv28qYM0Yq82OdvZ994R8VvRxvRbnSLp11FvVxas9o6lJtEuN7cWHc7U0+t1nzC0uK7rTo+1ruk+fmN7IYz6M/US8dUekLfULNLVlOOW6+u3Rq3RLZ6s54zcZbqXF6PFSAtpoaP/rPrq90s4slualNp96yxTSbbxXaXofrtVaYvbz1PyRWXXZHOz2uY/Hx+3L3wCmlqacnuDyU4tSVhrz4N6UCFOKAuU9ZdQLq8ymDbLDkSNJk2G8xJAdI+Yw64I0nf1eA+6rXxSk+YFBPfMUoFKBSgV54/hDvz1WX9Xmf4mTQbsdD/wAyvT/9Xrf/AAzdTagUoFKBSgUoFKBSgUoFKBSgUoFKBSgUoISx+eqd+r0f+JerpdDXZL2H3Yyr2i/uDJr8lMlDrrgaQLrKCGNuJBBZSA0QBxHh6SSkAkON/HLTlHVzLYV5tcK7w1Y9ZwqNOjoebP8AOLn5pUCKxjnoo9KUvKct+JN48tRKj/JybJtI2fMgRXGwP+/xoIh1Q9D2Nl+LCBjueZrZ5aH0Olq6ZXd7hBmNJ3yjPsqlpV4a9jZQpKhoaPmDz4CMz6E46xj0Tojal2KL3DnTy6saWfe4qPM9XVzPv/KOKP8AWNBJh6RTCAlL3TrqKy+d/kjjTrmiPP20FSP+t39264H+qXU3KQEYj0nkWtBUP+EM5ujEBrQPfTMYyHiSBocko7kHuARQRSZ1azWwdW7M9lXSe8IcRY5zfLFZzF3bUC/EKlgEsu8QQBrw9+0NA99TM+kdbXUgRsF6iyXdbLX8kJrOj8OTqEJPw7E/u70H6PSFS2SmR026iR1jyQcfLux8dtrUP2b39VcT3XDJ7mkN490bzOdJUOy7sqDbI6frWtyQVgf5Laj9VBws4P1czd9dwyDqAjp8kI1GsmGRY0xLZ8+UiVNjrLx92m2mgPid7rvdHOo10mXK54Jmjry86sz0g+tG2rjM3SAlwBiY0QC0eSHGwtKFey5yGgNCgtesLlzUZ60cZUYS2/FR+TLymiDvzCk99/UK82v6OJv5df2ZivPPL5ocuDZQHuFnWop14QVcXhy+PLv7P7N/sqcY+uEiyxRDT4EbuhCFKJIVyII2e577rTi106ueXr083q2mjBG8RDJ1+JWlZUEqCik6IB8jUh4ftKBSgUoFKBSgUoFKBSgVHMuscmYqPcbegPTYqVIVGKgn1hpWiUgnsFgpBST28wSnkVAI/Dnsz0r8JRC21cHGnElDjStb0pJ0UnWjojyIPka7FAqr+v2Rw7FiaWUtKev81LrNs4L4FpXDS3FK0dNp2nkNHZKBrZBHulJyWiseL1Ws3tFYa+Y7leNXWzWawZFaRGnxy3BTFnwHHowdACEeG+pvwyF6HE7BPIAgK2kWKfUbFbSf5vb4EZBUT7LTTSB3J9wSBXD6vTZtLknFlj3e32wp82K+K3JdzYzg1/6qw2n3eWM4rJAUXfEP4zlskAgJQBqMFpIPNRLgSfmNqIUm9sfsrF0ZgWq0RWmMfgFtpa20hLAbaI1HbHkoeyEKA9kDkCd9q6XRaX8tj2nvPdPxY/R19qzaVYN5SgUoIrmnTDHOoUq3yL7BVMdgJcRHIeWgJC1NqUCEkb2WW/P4fWaih9GPp+uCzEctciRHjvh+MiRMddEf+67Qjko6QfWH9p7j8ofgNBhmPQ36WxVR1N2R9amElKS9Ndc57JKi4FKPiFQUoEr32WvWitRNtSrclZO+4PuNa715oeZjdTPUfovHuxukyxPN2ubcG1Nz4j8YSbfcgUhP84jqIClaAHNKkL7AFRSONaIdbsavuIrRJfmORYiFlhmazLdfjpcHzfFafK1tIPl2WoADzR2rn9dWJjlmN/v6qjXTtTZE5Lsy+Y6iXb2QJrSkouEYtF12P29oeGCkqI2DoH2gdp3sb61oi25EVD1zb4RSSkXOOpSoilA6IUfNlQ8ilegD25E1ws3nHWaY59aJ29/l+8dvPt5bwcVOantbL9JMes92ZSxdrWxcEsKCm0PJ0odvnoUO6FAdgpJBHuNbItYwx0sybFsw/GF0m4wGn7VJauUxUoW0SVMluTzXyc4hbCGlFSyEh7kdJSo11f4c1Pp8dqz4LnTdar2qD9E1celtgjkachNLguAgjTjLimV9vd7SD2rs05ONVD+skFy59Is2isjb7tkmoa7dwvwF8SPPuDo0Eot01u526LMaILUhpLqCDsaUAR3Hn512KBSgUoFeeP4Q389Vl9//AIvs/wATJoN2Oh/5len/AOr1v/hm6m1ApQKUClApQKUClApQKUClApQKUClApQR6TjEg5e5f4k1pl1yAiCpl6OXE6S4tYUCFp0fbI/ZVU4Tac8k9N5zeJC0YfdE5ffHHGbjCkLafZ/GE3bunFrX+XWWngUlKQFnj7PEUHShYv6SJTJel5H04ZuD7SGlSYltk8khIGgnls6BU6ocuQBUTrSuKcrfLD6QMqHEXbMowqJNMFTUppyC+ppMjkdOtHRVrjxGlbAPLse2gwjlm9KSXbpDaci6bwpiFgMuiDLWlwBLfcgk6BPPt5j4+0Cm65EPKHfCLF1tEceGkLQ7bHXjz0ORChIR2J3ocew+NBxeoZf8A8+2T7Fe//V09Qy//AJ9sn2K9/wDq6Dht2KXT+WEe/wB2usOY5HgPQWmIcBccAOONLUpRU85s/kUgAa8z51KqBSgUoFYHNChNmC1oSpCHUErWviG/dy/t1r6605+uK3un6PeP9cIcZLIJHio2E8yCodk/H9Ffud4dd+onS1mBYruzBuocL0a4FakBpaeYQpKkAkFJKe4/q7qn4dHr29ydqv0wwr3S7P42RGVAyxxdualoeZjS7pJXthEsPhgjie60AtqdWXCEniE6841E6HdUIYPhZmwyuRGbEx5i4SkLef8Axb6q64r2e58ZLLqVjiseGdn2tJvlcsjMMJye7XdTtpvKo8ANxkpaVcJDKuDbhLzXsb7up0kvklaNdgfOoe/0WzlxyPO/li67dmUeEHlT5CR4ZRb/ABEJHcJ5mJIBUE7HrHPuoEUEx6VYXlmJy7qvJchVfWZDUZEVC5LjpjlCVB35wAV4ivynLQIKygDihFWJQKUClApQKUClApQKUGIveLwr6pDrgXHmtji3MjHg6geet6IUnffioFO++t1FpkG62JP88YNxjJH/AByA0Sof5bOyoe4bRz95PEUH5FmMT2Q7GfbkNEkBbSgpOx2I2KwmWdP7DnLkJd6hKlrhc/AUl91op58eQPBSdg8U9jsdhWYmazvE9WYmYneGHmdC+nk+Ohl/CrE4hCkrH8wbCipKgoEkDZIKQe+/Kq2vfovNv3WbcGbrjVggIcU5HZhYnEQqM2k7STIKvE5gAEuBSTvZHHehneJne0bs77zvbqneE9OrZ0k6b5Jepjd0utjityL5KYVxEq6uJZBWspUpI4lDSQlC1ArOy4TvZk49JjBYmSKxpBujc9id+LC0i0v+G24P8YJ48OOlch213rFp5p3Ymd53W3SsMFKARsUoFKAfKvxaQpOvOgr7LcEvjt1cvWNXtcWYtKUv2q5lT9vlADXzd8mF67c2yB32pDmhWs3V62x5rb1ryuzOYlLeOmnbgtLkB1fkC3KGm1A908V+G4R34DdVWtwTkp6vdX6zB6bHMQ19u3Q7JsPCZ8OBIERtICDBkN+txkeYDRJKXWveGnNgb9k+SaxsOJOvEqRLgMOypzZDcmfZo6vET27JmwV+35aHbkvXlwBr5nrMNpybzHXtP+J9k+E/226T02mK/FW81jmjr9/fsWt6NcaNZ83bj3O6WawWhxHhJtkqS7HUqUVoCPCZfabLIV7QLQUob1oAhRVv/fVWix4fcnL14JscaE65O9Yb8RssBBLvJOjyTx5bGjsdtV3PA9P6LFbJMdZ89uvt6TMfHxnquMNdoUp0z6nXjAcikYtl1nk2fERFjTbLc58tLz9tjvuONtRLgQB4agpohCyVgBSELWV+0qxMEeGLZnk2IP7bQ9Idv9qKz/dmJC+UlIPvU3KW4VD+imQz8a6hIWCfKule4X4yss+IF+H48dxrnrfHkkjev20GD6UzPxj0tw6VxCC/ZobvDe+PJhB1/bUpoFKBSgV54/hDjrrVZf1eZ/iZNBux0P8AzK9P/wBXrf8AwzdTagUoFKBSgUoFKBSgUoFKBSgUoFKBSgUoFVn6PzUNnCbwmFOduLRyvIVKdeZ8IpcN3llxsDkrYQvkgK37QSDpO9ALMpQKUClApQKUClArAZxl0DB8edutyZefiNrQhTbCEqUSo6HZRA/trxe0UrNp7Q2Y6TlvGOveZ2/dXB9J3Dj/APu267I1/wAXZ8vh/dKtPGb5FyawQbpCQ41FlNhxtDqQlSR9YBIH76i4NXi1MzGPwTdVoM+jrFssd/aydKmq4pQKUClApQKUClApQKUClApQYq44taro+p9+E360oaMlrbTxHw8RBCtfVusXIwdSEn1C6yWCPmNSkpkNj9O9OH/36DgTil4c0DOgscRrYjrd5n465p4/o9r9NdyBhEVt9uRcXl3SQ2oLbQ6kJZaUDsFLY7bB1oqKiNdiO9BmZ1u9ccacTJfjONhSQplQGwdb2CCP6I/7muD8Tvf86zv3t/coOzBg+oodHjvSFOL5qW8oE74gaGgABpI7a/012aBSgUoFNUClAI2Kx96sMO/wlRZjQcaV5ggeXkR39xHavNqxaNpYmItGyp7/AOi3jTsdZxiVLxCUre029SfVF/5UUgtdzrakpSs/1hUBu3Q6PaXmzm2Lqusdj+5ZPijb632QfMqZQv1pnfbsyp4diVECqfPwzHmyRlidp++7V6Ku+6VYlh7ltsr8rErtbOq2IvpWzKtNwWyZXlpTaHx+TWQPZ8KQkHZ9p0Vgr05kE/DrpjGBlORssCOp/BsnkG33i3x0uoUtlt5wKDrS0JU2kubSORIeWkBFWWDBTT0jHTs2xERG0JlhF5hruE57N4ty/HuUpZt0hufZXmbY02kOBqClSkqQU7edHNxX5VbpA0FIbEcz+xyMGvuO2GxXC63cx1GVaYsSKqbcbB7JQF8yQHIS/aaU0+tKtE8HNoSESGWdt3pD3awsqRnnTjKrElnXiXi121dygrT/AOs4sc32x22UqbIT/XUO57d26/YnnuAXQ4JdP5UXadGeh2+LBjPKK5SklCUuewPDSFEc1L0EAEq0BQWpYrUixWO3WxtRW3CjNx0qV5kISEg/2V3qBSgUoFeeP4Q789Vl/V5n+Jk0G7HQ/wDMr0//AFet/wDDN1NqBSgUoFKBSgUoFKBSgUoFKBSgUoFKBSgVBOjVvuttxW5tXm2sWqUvIb082wwwhpK467lJWw6QjsVONKbcKj3UVkq7k0E7ro3u+W3GrTJud3uEW122KguPzJryWWWkj+kpaiAB9ZNBAXfSP6fMxvWDd5i4vEKEluzzVsqSfJSXEslJSe2iDo7qV4R1CxrqVZBdsXvcO+W8OKZU9EdCvDcHzm1p80LHvSoAjfcUEhpQKUClApQKq/0jxvpZN/zhn/XFRtT/AEL+6fomaL/5OL/yj6w1FI3W2FlgXqX0Kx2RjaEvX6AhmZEjuPllEgoX7bS1eWltlae/YFQPurn+D/rv7nW/iH+lj98/RF8n6M5iwp2Di09TLi7fBZ/Hcuc4lfjsIkKU6QklRUt/1NSxrS0oWlR0avq3vPybfFdlR/VJTjSVOx+YX4SyAVJ5DsdHY2PhXUuGdilApQKUClApQKUClApQKUClApQKUClApQKUClApqgeYpQKUClBFMg6YY5klzN1dgrg3opCDdrW+5CmKSPJKnmlJUtI/qKJT9VRDLej1+vseK2cki5AmIouxncgt4TOjL1rlHmQ1MKYVrsVhC1a8999hGzjnVdtVugZOyMnxyDOYnpas8mO7cX1MOpdZQ6+8IrfFLiEKIS2FEIAK1bO57jzF6vfUtWRP47MxmAm0G3yG7jJjremOB5K2SEMOOpCWgZHcqBJfPbQ3QWDUamdOcel5KzkCIAhXxtaVLnwHVxnZAT5IfLZHjI/xHOSfeADo0ElpQKUClArzy/CG6+Wmy/q+z/EyaDdfof8AmV6f/q9b/wCGbqbUClApQKUClApQKUClApQKUClApQKUChIA2SAB8aCmcp9LHA7JcUWeyu3HN8iff9Uh2zG4DslMqRoktJlaEZKkhKirk6OIQonWjVanrf1wh9FMxvIwBmfeYs+7oiXCNeYM1MFluY6lAdYZUkLMZoeGUJUpxSmCFAKJoJJfvRru8HBJc/FOpeZS+pDbHNV8fvi1ouKuO1MKjLKozKFgkIKGwWitKgTo7zOF+jz05yqTi2eLcyHKX2o/rMUZPfJlwbDqwkh1cd9xTaHm9KSAlKQglXs8kpKQvB59pgI8RxDfNQQnmoDko+QH1/VVddRvRx6Z9Wn3JGVYbbLnNcAC5wbLElWvm7ebKVnXu2e1BAMG6B2bG71fsStl2vuOS7WGJ9suljuC4yjEf8RKEOsd4zy23GHk7cZVtHh8tqJUqYWjIupuC2ptvLrBGzVlnaVXXFXQmWpI8lOQ3QgE68yytRJ+a0PKgzuJ9cMIzW8Is1vvzTF+UgrFlujLkC4cR5n1Z9KHSBruQnQqdUClApQKwmXqjm1Iak22Ldm3nko9Wma8MnudnaVeWvhUfUZYwYb5ZjeKxM/tG71WJm0RCJ/iW0kf3j45+5H+4qYWCbFTZoQSxGtiFLUw1FaUAgKSVDijsN9kk6AHvqi4VxfFxO9qY8fLtHsSM1L1iOa27Jx5DUtlLrDqHmlfNW2oKSfd2Ir8kymIbXiPvNsNjtzcUEj95rpUV+vyGoyUqedQ0lSkoSXFBO1E6AG/eSQAPrrkoFKBSgUoFKBSgUoFYC6RJ12uz8WNeZdpSww06DFbYVyUpToPLxG19vYT5a/TQY2RZcsiI5oyFdxA82m4zDCwPq2lQUfqJT+muCPFyua8lozLjASfnSH0wlBI9+glJJPw7a+PwISb8Uyv+ep3/uMf7qtbMUyvJujHpJXLpzmmV3Wfi2bPvXbDb1NW24W5BPKRbFrWg6KSQppI0OJCR3IADZP8Uyv+ep3/ALjH+6p+KZX/AD1O/wDcY/3VBEnJd4gzVwJV+lCUjZTtqOPFRvssDwvh5j3Hfu0SfuNxjNFx3IJLTafNa244A/b4dB1k3LIFTo/K6TWITzTikF+OyhxaklvvxLe0p9sjuNkg+Q0T3fWrt/z5L/8AhR/91QSjF5T0/GrVIkOF6Q7FbW44QAVKKQSdAADZ+AArJ0ClApQKUClBjchyK34ta3J9ykJjx0dgT85avclI95PwqJ23qW9kiUG2QA224oBLry+Skj3kpGgO2/6R7/Gt9MM3rN/AYvILxBst2aZnKRIfCApLzyQt0q357I7D6hoCsDcusa8Nnx1OurmQXHAHWnFlagCe5ST32B5DeqqNXqZw3rENV7cskDqk5ms9xxEl2Iyh1bbbLLq29AKI0riQSe3ffvqcRHmXIvhpmSm1KAHaY6VdvhtR1+ysaTUTntaLFLczpSc1k4urbri5sUeaHlgrA+pWt7/Tv9lSzGMuteYQfWLbJS6UaDrJIDjKvgpPu/0H3Eirm+Ga1i8dm1maVHClApQKUCvPH8Id+eqy/q8z/EyaDdjof+ZXp/8Aq9b/AOGbqbUClApQKUClApQKUClApQKUClApQKUFU5/14hWq8s4jhTUfNeoMvYbtESQCzAQDpUmc6nl6uyk/Ec1n2UJUfLihejtaL+j13qNNf6iXt1XN03JS0W5k/wDq48IK8JtseXtBa1D5610Ek6kQUWLEoFyt0RDTeNSmJ6I8ZrQbjN+xIDaEjzEZbwSkDudAedYT0dZVouXT+6v2Zt4W93KchO5EhD/iOG7y/FWlSUJHhqXyUlOiQlSQVKIKiGaviv5GZ1AvIPCz3oNWqcB2SzJCiIrx9wCipTCj5krjjyT2+bQ5/IrPZNjWAi039TtwtqydJblgcpUfXuK+8hI7lRMk9ggbDn6nvrt0bGriOKm4t/gocQob5B9z1UfuVISrtr5vw3WTuk6SvNLFbWnFNRjHlTnik/3Qt+G0lsj+qfHK/wBLafjQYuyhMjq7lj4IPhWm2Rdd9hQcmOH9hDqP3VI79kdrxiEJd2uEe3x1LDaFyHAnms+SEjzUo+5I2T7hQQ/qvYbNmKMbsd9tLNzt12mrjof9puVAfTHcfakMLHtNrSWSOSSFJKgQRo108Xv+Q4NlVsw7MLk1fWbmhxNkyENBl6StpHNcaUgex4/hhTiVtgJWltzaEFHtBZtKBSgVH8zdbYt8Rx1aW20ykFS1nQHZXmagcQrNtHmrXrPLb6S90/VDAG923/nCL/8AGT/21+ycQGaWHHCma5FagXVyeVsOKQtxID6AELSQRvxAd/AH41w34WwZcWbLOSkx0jvEx4pmpmJiNpVcOhGawrhb4EG/SRZ4cnTa3JjjSUByDIQ87xbcClpS6ISkJJB8RLyuwUFVmcl9HW75PODs7K3pzbctbyBKU6pXFRkkqACtIWEvtNAIAHFhJ8yAn6QgOvcOgGZS7lJcRnKvUvEWqJHe8ZaWAJLD0c65glTQZWgEKHZ464kKK/1Po+5Y5GdDudzRJQ041FdTKfHggouBQCEqSlXByXEIPEb9SQdD2QkMxY+jmW26+WiZJzN6SzAlNqLXiPadZQ2yg80lelKX4Sz2I0XjsrHJK7loFKBSgUoFKBSgVCskY55Sp1DjjL7cRng80ripPtu/vHl2OwdDYNBl8avUie9IhSuLj8ZCFh9I14iVFQGx5BXsHeux32A8hnqBVZ+kR0Qt/X/pnNxuTIVbbo04idZ7wz2ets9o8mX0Edxo9jrRKVKGwTsBgfRb61z+q+HTrRlbCbb1KxKT+KMmtvYcZCR7MhAHm08kc0kdvnAbA2bqoOCdb4tzY8GXGZlMk78N9sLTv46NYiXaLHjkc3Bmzwm3WiAhTEZCF8lEJAB122SO9BgrzeXb09bk/i5+M4w+pTjxW2pvgW1jQIVyO1FHbiO4+qlBI8N/vRsv+Ztf6grMUClApQKUCtX+uvpNXGwXa5WbHFCO1CWph6alAW4twdlhO9gAHY8t7HuqfosEajLFbdhr9J6oXW+lEm4XKVcVoVyHrb63eP6OROvM+VWl016veoKbQt1ITvXE1njGux6KYwUiFxpeHX1OObws+85dbboszY8ZCpCm+BcV3UBWsPVjK5U69IaYeU2mOS8VJOvaHzR/t/ZXC67LGTJW0eTndVWcV5rPg+OnuaSIctK3JKxzkKWok9yVKJJP7TWywzD8Xxo06QQlsD9Hu86zo88YrWyS96Sk57xSPFVuf9Y1znFtNOpKAryFQBfUiXbHPWo0hbD3lzaUUq/eK7Xg2vrrJnDfq6TXcNtpccXXT0f9JuXAucK25HKcuFrkgo9Zc249HX2IJVvaka3sHZHbXlo7WQZse5wmJcV5EiM+gONuoO0qSRsEVt12m/L5OkdJUXdz0quClApQK88fwh356rL+rzP8TJoN2Oh/5len/wCr1v8A4Zupm682yBzWE77DdB1ZV6gQkFT8pttI96jUYPWfBxKcjHKLcH2uy2y73T+mg6ny/dNzKbijNrIqS4SEsiWkr7eZI3sAe8nsKk8TMLHcCRGusR8jWwh0HVB32rnEkJBbkNrB8iDX23MYe3wdSrj5691B9l9sD5w7Vxsz48mQ6w26FutJStaQD7IVsA/t4n91Bz0oFfilBCSSQABsk+6grnM/SFwPA5Pq91vrfjje246S6R/7u6x1p9JvBr20lyLIuCkK8iqA4n/ZQZM9esTIOnZp0N69UXuszjnVHG8qUtEO4JbcRxBblJLKtq3oDlrZ7e6gllKBSgUoPh55uOyt11aW20JKlLWdBIHmSfcKoa8X5n0m8wm4lj17Q701snFOSXO0S/auspQCk21p5s7S2lBC3lJIJC0NgjbmgtDBbPiOJu3LGsVstvx8W0tF+Db4aIyOK0bbcCUABSTpSeXxQse411GMyya/SHfxJh6moDayhMzIpht5fAOipplLbruvP+6paJ12GiDQZnHo+SiRIdv0y0uMLSAzDt8RxKmTvvyeW4fE7a8m0ftqE+jXGTB6bTILUpEmLByS/QoqUFZTHYau0ptphPIAgNIQlsAdgEAAkAGgsW/2OJk1knWmeguQ5rK2HUpUUq4qGiQR3BHmCO4OiKr9pm457g9zsL01tnOcckpbTNUniBLaCXI0kpH/ACbyCgrSntpx1vfZVB381lv5FZMIgyYLlsmXm72992M6tKlxFR/5+pCikkEj1QtkjttWxXdymSqzdQ8PuDhV6nLRLsytb0h11Lb7S1e4D+aLQN/0nUD30Hdy242rALbf8ylNPLcYgJDzbBKlyEtFxTTbaN6LilOqSnXdRUke4axmCYl6pIj3vJlR5ueS43iSFlfMQkK1yjxQfmMpPFJKQCspClkqPYP21vpz7MxdWlBdix55+NEWDtMmdotPuj/FaSXGR8VLe2PZST0+r2KWPqKbBil0Tc0Tn5K7lBn2eWqJJti2EEGWh1JBHEvIb1pQJfAKSnloI70Z6jXBjMsj6Y5XfE3rI7CpK4N2fjiK9dIikJUStsAJU81zQHC2OJS6yvSeZSLmoFKBUW6k5LMxPGFz4AZVIDqGwJCCpOidHsCD/bWrLbkx2vHhEvNp5azKqD1xygj5lr/+Wc/3lWlas9jsYfZrrenC2/cFhhDcOM66XHfbPFDaAtR9lCj7/I1XaLWW1Vpi0bbI+HLOSZiX1D6qYrc2HHIV3bnFvwB4UVpx11Rea8ZoJbSkqUVN7XpIOglROuJ1I7Zc4t6tsafBfRJhyW0usvI8lpI2CKtkp2aUClApQKUClApQKUCoXkZcdysxoyA9Ldhs8GydAAOO7Uo+5I2Nn6wBskAhIbJYm7Oh1ZcMiU9rxXlJCdgb0kD3JGzod/M9zusnQKUGrvpQYvd+kGa230hcJhOTJdnYEHMbPGHe62bYKnQPe9H1zSf6o7ninRvuP1LxubgMPM4t0alY7Mity40xn2g8hYBRxHns7A15g+etGgjlg9ILE79cG4anJVsW52Q5PbShsq+BUlSgn9KtD66saRHamR1svNoeaWNKQsbChQRuThrzJ/4PnlDYH9xmIL2vqC+QUP0q5V0VWe9NdlwGXSfIxpIUB+nklJ/soM9hyVIxKzJWngsQ2gUk70eA2KzFApQKUClBW3W7qW3gmKSGoUlsXuSUsMoSocmQre3SPdoA6+J19decvUu+3BiStKXPWAtR2oklRJPcn411XBsPe7zbs69rakKtSVdyop77r9tEudGuKEgq0D7q+Y/ie9p1to9r67wDHjtouvk2IwidIftDpc2eLZHf9FUd1FmKamSFg91KJqji02iN3yjjtYpqrRD66duiW+lCtdydcvjV4dRbjIcx5hLRPHwxrX6K8ZJmImIbfw/EW1dd2uziZ0ueoHkRsispNtMhy0LWQQQPfV/+GLT+drD6f+IoxxpOnkjdivkyHcw2slKm1cm1HyreT0N+pDtyx+bjd6eSzPElci3NqXyDjJQCtCfrSpK1Ef45I8jr6xxnDzYuar5LXq2WpXFvRSgUoFeeP4Q4b61WX9Xmf4mTQbsdD/zK9P8A9Xrf/DN12s/uqrZHjFJA2lxaifclITs/2ig079KL0l5eGYxNTYYLt4nNNvLUEKCW0BsAuFSj7gCDrzO+1ebMzOs66i5IJrPr0dcsOuKahl1tpCWxtxSiCCriPPZAHbehQdWbiGSXG5x1WlLkx2Y6ttx3W08E6BLhHc99/wDu+/tXDZ+oOc9M78pq15BcoC0uFtKoHNDCzsaPEjSgdeWu9BtD0y9PXJsSulri5yWjDeJb9ZjubUjie6nEj2dHf9Hy0d16L9DerNq6mWdu52qa3LiyO6VtrCh2/R/37UFpynFEAb0PhXLYk6ck9v6vf99BlqUHDMlswIrkiQ4lpltJUpajoAVq/wBVOr0vqHclWKwevNW1lTqXX4w7OrSCEga7n2tD6qCHR+n1ng3CKtLS5T8hCFrVIa24lSvMd+6qlq7dGtzKAyNp8VbZ5I4EBOtdvie/b3UHVmNMqALCfyx/o8hsnddK4ssvs/zhnw1dwfd3HnQZ7C+vk3ppNTGzWUtzF3nG20XeQQPUVL0EBxX9RR0AfiRW0DD7cpht5lxLrTiQpC0HaVAjYIPvBFB90oFR/P8ANoHTrELjkFxS66xEQAiNHTzekvLUENMNJ/pOOOKQhKfepYFBXeNdJLt1DbZv3V9TF1mOqD8bDmVFVotI80oWjepbw37TrgKeQ/JpQO5zyIjGEdYYpZbbiWvKLamEhttIQ2ibEC1tpSB25OR1u+75sMfVsMplVjeu9zavmMTYicms5VFdacc/Iym1JStUSTx2UdlIWhWipskKAUlS0L6Nm684JcrS3LmZPaLFJDz0WRAutwYYfjSGXC28ytJX85C0qSSCQdAgkEEhH4vpPYm7lNzsq27hLbjpS7DulkhPXWFNbKQTp2MhYbcB2Chzj7ikqB2K/wCiXU242HAZVqxrpvIgMMXy8eCb3c1Rm3kKuMhQdKleO/zWCFq5IA5KPH2eNSMWnvm/TDRlzUxR60uSyelDccbvsqH1Pm2HEbm9z/F1rDhTFfaSQfEZlOEGQvRAUkJbKd929FKlYbIvSCymFlv8ssbwdE23PQ2bMo3OUbc5PddmNIhrDakrcS0hT0j2loST4pISod6lzpaVjabet5dvv2Iv5i9p3iOjKZ31iz6z37Db3OwOzPwYMxbcx6DkTikxw+0WkrWFxEkNhak7UN6B2QACRJs06xIvUTHcfvGPXLH7rcb/AGpMeSoofgFTUxp9X85QdIPFpQSl5LalqISlKt1Hzaa+LrPZIxaimXpHdbOWYj/K6XZEyJhbtdvmonvwUtA+tutEKYClE+ylDgS5oDZU2juAFBXQKfA6v7UXD61YtIBHsDwpHtaPxPjo/cPqqIku904tj1m6eYxBkxfUpUe2RmnoxOy24GkhaSR5kK3s+81hrTcY+U9XblIhOolRsetv4uckNkKQiU+6lx5nkP6aEMR1KHu8VG+/kFS9eMUyC59Y7ZKwuwMXPLGIkG7wZkmSIrEVUSQ8l9LrmirUiPKXH0kK3ySSAlvkm+8Ly6DnWMw7zA5oafCkOR3gA7GeQoodZcAJ0ttaVIUPcpJoM3SgVCur1tl3XDHGIUZ2W/47avDZQVK0D3Oh3rTmrN8Vq17zE/R4vG9ZiFHfyLyD/mO4/wDyq/8Asq5bTgDGSYRisS8NvsKtkgTfVztClLCXUBKveBpzfbR7Cqjh2ny4bWnJG3RFwY7UmeaGDgejXj9jvEe6WafPtUxtTqloZWFMOhXjBtHgnaUIabkvtoS2E6SsdyUg1Y2JY4MSx2HaEz5dzTGCkiVPWFvLBUVe0QAO29dgBoDtV6msvSgUoFKBSgUoFKBSgVimGUKyma4UJLiYTCQsjuAVvbG/gdD9woMrSgUoPiQ228w4h5KXGlJKVoWNpKSO4I+Gq847VmHyN5Dc+kqIS09O8rmqu2CvSJHEWuRy5u28niRxXyJQk+XMgFSjsBJ/nFSVJKVJPFSFjRSfgRU1wTq1f8BW2zGf9etadbt0pRKAPg2ruW/2bT32Umgv/Euu2K5Qhtt6X+JZqgAY9xIQkn4Jc3wPfyGwT8BViAhQBBBB8iKDG4z/AHu2z/N2/wDVFZKgUoFKBXHKktQ4zsh5YbZaQVrWrySkDZP7qDzWyzq3P6nRUZG3yXHuF3fcW2o7U00dpZT+hKABUUuVhkXd/k034yx3CSdV2XDrcmNiY3XBgHSKRkkJDQDbS0p7hXfv8KzbHQl6PNebkNpS+z3I15/AiuC4/op1Go56Oo4bxOdLinHMsn+L0WSG9b0gJe4EVrv1CtbiZ7qFnkQfOuY1OH0For7HIcQyTnyzfzMGgKEpngCdKB7VsH6j+NrJEYLfiqLY0AO/lWdNg9PM1Z4flnT5IvHg+LV0ZjIjKku8UKKieKvM1z5V0cujGJh1qE37ST7CT7ZTvz1+iuk4Po/ymf0lnT8Q4lOrpFIlQmR4o3AnKb4lt1I7pUNEV+4vmi+n8iyTWJJE+3zhKQFE6VpRPE6/okHRHwNfRs+WMuLaZ8HPbPTu13KPebZDnxHA7FlMofaWP6SFJCkn9oIrtVw0wyUoFKBXnj+EO/PVZe//AKPM/wATJoN2Oh/5len/AOr1v/hm6wnXqRKZx9hEMK8daXdqSCQlA4cioD+iPM+7tQeRnX7otldvynKXk51crvAXHlzNRI0kxfDCkFbTiweCfnDkCPcPPtXR6HdM3JWMIclZNMLCYtwectjLLpZUltwNuJS4SE6cQConWtDSvdQbTYv0vt8qwwIUSP4bIHt8eIJBH1fV2/bUuY6DWFn8qYaFOgAAKSkkDX1jdBBMt9Hqwznm3PUGFSGOQYc8NPJvZ2fPzrr+j5Eu/o5dRYRTLJwq4r8OVGWnSYj6yNODXkknST+kHt3oPSNpaZcZLg9oa8xXcs6eJe+PYf6aDJUoNKfTv9JpWJTbd0zsbjQvV4Qh595TxSphgOa2Ep77UUEA79yqhfSiROt1ijpXMVsIUUAeY5EqJ3572o96CeyZN9vBiuwGiFR1IUiQ4kJIKR2+sjvWVtcWc8XUXMguOul5St62o+ZoOa/RYtpjpkso5vJII8zoj31CnLFe7yjx4c/whtShHcB1s9z3oKw9IJU679O7xjd6ZUETGmxoaKXFtgFsg/AFIP7O9ff4Mf01myzE6L53PKJTKgzjdxfVsOJ3r1NZJ7EdvD+I2ntpOw9M6UCqsydKs06841YVKCrXi1vVkcxk9wuU8pcaDsfBIROX/lIbPmkGg7t2ijC+q1kujcmcm15GJFvmMvTnnY6JvBtyMtLS1ltnaGJCCUBPJTiN7JFZTOOmcbqHdLW5drpcm7VblCQzbrdKchFUoH2XlPtKS77I2AlKkp9pXLlscQru55E0qBJwzpolNitDbzrVyyxI5hlwrPjpi8tmVLKuYU6raEL2VqWtJbrhscvFej+Ps2XFbbFsUVpBBcA8WbIUSVKcdcO1LWpRKipRJJO91v8A+HT4p1OqttSPn7ka1smW8YMEb2n5K+u3XFMZ+abtOt8JaZCkNOzpIbPh6BRvkrz0dk71+6oJ0WvF+ybH56LEqbPt7l8uzpub49UjhS5761ISSStYSVFIUlJHs/0fIc7PEOJcbtOPQV9Hj8+3Tx6/t27eay/K6Ph0c+qnnt5d+v35pZaMMnP5vOh3p1pU9UREmPOaC3QprkUqaBUBxKVBJ7E8ue+2tV+4r02Rf4K7n+MHWUvurCG1AOEeE+oAk7134A692yNnW60ZPwZfUZKZMupnmifLf+erbT8RVw47Ux4Y2mPP/Ts5DZ37dcU2WbLAtl0YLDb76ihh9xRKVR1+YClJI4g/P9oDuO+UVPu9qtn4uv0NF0hOM+rv+OgFLySNKBPcEH4H417th4p+H/8Aki3pcMd469I9077e+N483iMmi4rtTbkyeE+3+fj8E16YdYE2G4W/Gb7Mel2uWrwLVepa9uNuaJESSSdlfFJ8N0/PCSlR5gKckHU3PLPgnWfpV+Mp7Md7IVXHH4zTigCpbqGX0rH1c4rbf+U+n41fVyY89K5sM71t1j/HvjtKHtakzjyfqjv9+1YeT4xBzC0qtlyDrkBxaVPMNOqbDyUnfBZSQSg6HJPkodjsEg9my2O241ao1stECLa7bGTwYhwmUsstJ+CUJAAH1AVllDMSW7lObZfkTCwmNHCbBbnFp5IKo6nFSHQNjYL7haI7d4v11W9x6bN5R03PUTFBPwHqDNiouF0XiiuBmyEaMlpxhaFtSHDxcQhbjalAlJB7nYd3Fet9wwPIrNjHUG5R79b77HMzH86tsXw4sxjm2jjMQjaY7gU+wkOD8kvxU64KPGr7oFYTL5ztvtKXWJJjOl1KQQnfLe/Z+r/6V4yTtSZjyYnshgvt1CQPxk+dL5bITv8AR83yrMXHqLAwvF7NcL4++83Pmpg+sNNAhpSis83Na4toShRUvyASSewJETT5bZJmLNVLTbu+7X1fxO6wUyEXVthRK0mM8PywUgsgp4DZKv5wx2GyfFT8a+rP1cxS9o2zd2W1GY5AShxQ9t1MoxQAQSDyc48RvZStJ0AanNzvWnqJjd/hSpluvEeZEixky3X2iSgNFHPmDrRHHz1vXkdGuNPU3FnEFSL1HWUjRQnkVBfiBvwinWw7zIT4euezrjug6k3rFhNviJkv5LARHJUA6HCpOk+PyVsA+ykRZBKvIBlZJABNcVq6zYjeJF1YZuqUrtskQ3A4hSS46UMkoQNbUsKkNtlGuYWeJTvWw5z1fww8uORQ18ewKFFQc0nkoo0PbCU+0op2Ep9pWh3qYUClAqOXTqTiNju5tVxyqyQLokpBgyriy0+CobSOClBXcdx270H5bOo+NXiaqJFuzKng2t1PiBTaXEIOlrbUoBLiR71JJA+NYnH+t+E5Pc1QIN9bD/fwlSmnI7cgDh3ZccSlLo042RwJ2FpUNhQJCdUoFY2P/fJO/wA0j/671Bz3O6xrSwHH16KjxbbSNrcVrySPef8AQO50ATUeGVXIuBfqkYNb/uPiKK9f5Wtb/Zr6/fQSWBOauUNqSwSW3B2BGiCOxBHuIIII+IrnoIbmMXM7uiVBtCLNDgOoLfrL8l0yFJI0SEhvig+Y81/HtWq/W7o8zn2N3jDrupuJcYriVxZsRRUYcpKQtl5tWknY5J32HYqFBAek2WzepWPTbZfG2oPUbGHRbrxGV7KXlAew8NDu26kckq12JVrt5yFaVNOrZdQpl5Hzm1jRH1/WPrHagEbFSzp51IuHTu8R5KXZMq0oSpt62+OfDKDrZbSTwSsEAg9t60SASQG1mAXH8cYLj0/wVxjKt7D/AIDvz2+TaVcT9Y3o1n6BSgUoFVl6S2bsdPeh2WXd6SiJuL6mh1agOCn1JZChvzI58tf4tIHmJ07vDN0yL+SWLvqyJlmW94KIRDiltpWUhZAPYAa2fIbrbvp50an5Iw28mGY7QUUqcePDZB0SAe5/dV/g1NcWPrIvnHOljWMxwI7+3R32D7/3Vw5fZ5T7QfYX/O2ho6Hzx8KqcmT0luZiVNZhbTcY/rsZzwJzAIWhzsFa8wapDJUfjkLWpgtubKVa7gH9Nc1xHHvaLIuWu7J9OrexaJzZd04Eq9kq99XdaZzUZtwMoBdIIbGuw+qscOx+vNjFGyycQxZmI01KmJVcLg57Z5/Mb+oCpTPtkm7K0W0BAHm4vikfsFdHvslKe6mdGoV3cVIEQyJ3Ehsw+5J+H1/trSHNID8fLxjAQVX6VITHjW9fsPqcUQlKeB7991ZRqN6bSy9M/R3uz916N401MS03cLYwbTKZaWFeG7GUWCDryJ8MK1/jCrHqot3kKVgKUCvPH8IcddarL+rzP8TJoN2Oh/5len/6vW/+GbqKekVdTa7fafyiGmnhIbdWs69gpQFDfwIoPPPrN6S+I4zFuWH40U3KZcWnY8tUYlQCXB+UBI7d9Dfeq+6J2m5ItrVtgtulEmQpuQkuklweEVLBHwUQAonz3QbidNrbKslubMtpW+KdJKe6joE+X1nWvqqbSLusMb9UCBrsVa2P3UEcfmJmPLLSQpX9Lt3FYSdbWLwy7GfQlaXElBCh270G0PQ2+LyHpjYpL5JkGMlCyruSU9t/tA3Vi29ISXdefb/bQdylB4AdQOs0zrN6RTeQyWPCdEgsgciVKCV9v2duwr0J6L2M3y0w5DgVwSkK0fIaAH+ygnt5yy5W6QY1nxqfduCCRIa8NDIPYa2pQPx8gfL9FY+33jIL84RcLSq1eGTsKdSvt8dp8h+mgljVtt92xd19NwilDqVNtSkuhTYc7pHtA99KGiPqNa6Xqw9b7a5KTb/xAtbKXFMspcWovqHHgOSuIAO1bPu0PPfYM3Y4uV9RcQft2b40myXpttRSEuJdbKh5aWnY76ry9u8qb086oOTYgSzcbRckymQ4nYS624FjY+HIeVB/SYyorZQo+ZSD/ZX3QdG+3qDjVkuF3uUhES3W+O5KkyHDpLTSElS1H6gkE1W/Qm2uz0XrNbxJJyvKhGmSrYtwFVphhCjDhlAJ4lDbilKJ+c446R2IADBekbKynDbZCyC1Rncts/49tCnsdSpDclDomsIQqMskJWFK47aWQAohYWAlSVdLqbacoyywy4+Q5zc8PefbO4GHSEMJhII0AuUtsuuu/Wjwk9wOB0FHbjrW0zN52rEbzPlENWS01j1Y3mekR7VSdLmb5imLwMCiTvxqzZ2EsQ7n4PhPOxRtKA4ORAWkDiVDXLsdAkip1ZenEC2Oyps1fjvvq8V3Z0gEADufMjtv9pqk02nyfiLP6fP009OlY7c2331/aPFKz5qcJxeixf1bdZny+/D92Et2R45JyG4y8Gx2fnl8dCGXF49FQ5HbKBxDZmOFEdsg+aS7y2SdVx9HsR6y5Bj09TdkwzEIqr3df+NynJb6SZ75UVsR0pbKt9iQ8SpQKjoqIHX31OLTxGPBWNo/aPcpKae+be2We/7rRj9CMznIH426nGMs+Yx+wsRgPqHrK5P6N1jo3ow5Dj7LyMf6s3ltLjz0nwbxaIEprxXXFOOEhpplWitROgoa2de7USdblmd90qNJi22RvJmcvw+BKh9RcSi33GloKH79jqFSovh+8yYawXmk68ynxkAd1KSPLHYt02sN1YNyN/nZhZXnA7a2pFzckRI7HFIDYAXwfAUFELcClAKA37NWeLJTVV2t38Y8/wDSuy47aafV7eE+SP8AVzAY0PHbhHtwXDRcmzHYUwooMeRsFpxGj7JQsJcCh3BRv3VPOmXS7GW8aRBzrfUKfcIzbcu/31Idf7aUAz/g7YV7SUtceJAVsq2o8tw/Szg1Gp0MT0rMWrHsn/HZdanURkxYdTt1neLfD73TTpzY85Ee+W1PUBcti2XR6G0q621uVIQxpLrADyVN8j4LrQ5OBZJBJJ3qpTmV0u+BdNbi6Lqu9ZE8Uw7e/IjtoCpkhxLMdPBsAcA44je9kJCiT2qRMPfdJMPxeJhWK2qwwStUW3xkR0OOna3OI0VrPvUo7UonzJJPnUIxawPThkONOz7rY5tnu8mXFmW9wNeMxLWuQ24EqSptxKS441paVDkwokdwSC/WSPjmI3BrNIlpv9kKDb1Tm4qIrojTH0NyPGR8wJ9pLji0KSFcSeCSkbx3SnqZOt93tvTvMihWRJhKctV9Ye8aJkMdkJSp5DmhxkAFKnWT3HIqSVI7gLgrEZTb5VytJZiaLgWlRQdDkB7hv3+VeLxzVmIYmN42RFGKXVZaBiFHPeypxHsa+Ole/wCrdSBjEYt5sVqj3uE28uE6p9DCjtIUUuN+0AdKBQ4oEHY71FwYbYpmbNdKTXujdq9HnB7GuKuBb5cd2K2W2HE3GRzaJDALiSV7CymKwkq8yEaJ9pW/uP6PeCRJjstmzrbluyDLXITLe8Qv+OHw6V8+RWHEpIJPkkDy7VNbXatPQ/D7JbJ8CJbnkRp9sbtD4XMeUpcVDYbSjkVk9kgDe9/XXHauhWH2KIuNb4UqGwqSZvhsz30pEguBzxgAvXMEABXmEgJ+b2oON3oFhT3rYXb5LiZTbjTiXJ8hfsrblNr1yWSCoTpRJ8yXN72Brjf9HnBJTl2U/aHJH4znu3R5D0t1aEy3FtrW8hBUUoUVNN+Q8k68lKBDkm+j/g8+1NW120uGC224ylhMt4Dg54fNJ9vuPySNb3opBGiAasNKAhCUjyA0N0H7SgobMrtm/WDKcsw/GZcbG7FaAxHk3JbqkSZKlklZRpCtIT4biCn2Sr+sAe1Ax42b9L+oaVsQbPLxwofs01qPb1kLcKQeUhKVcUlTgQpB9hJalJ4oJ2EhmLn0gTKhWqKZLV1vkSErIvVLVb0Rbb6vHYCW4jLeyp1O0tMhBWoht5wniXNq6ap1nsdzv0bG4do6j3K4eEm0zh6vKkD1jw0SA7tQSshDLjqiriPyY5El3QD96ez2ZGNQrpil7cxXIZS1NNvR32I0B11lmOHi60lOn1redcUgBBToq9gcEir69H30jbZ1Sjs2e4zU/wApSkuMEw3IwnRwkEOhJ2G178RJaKgs+EpfFKeyQvCoxd7wu2X+W2w2HJT0RjgVg8EALe2pXx1seyO537hsgMQQ6++qRJd9YlLGi5rQSN/NSP6KR8P2kk96+6Du49cDbrg3DUR6rKUooB/oO65aH1K0o/5X+VWVyrJW8ZtyXi36xIdV4bDHLjzVreyfckAbJ1+8kCgjNl6nPzH7bb5FndcuUhQaU5GcbSwVBJUpSeawrWkqVx0ToeZrp9SumX46XFlY/borNzdkKMt3YZS4goUStzQJUrkEAHRPtH3b0GlXXzFbr0/yeL1ZxyG4q82FComQ2lA0q4W4K/KJI97jWipJ+A94AFWbFkWrqNjFvvFplIfizGEyIc1seaVAHRHwPkUnuPqIBARtSFtOONOp8N5tXFaD/RP/AGa0R9RFZDHbei7ZBa4bg5NPymm3E/FBWAoftGxQbn4b/ejZf8za/wBQVmKBSgUoFeev4XbIMlvGO9Nem2IsPTbvf50u4LjsFI03GaSnktStJSgB9ZKlEAcdny2ArX0LcDs/QeBBjGc7fM3ylj12Ui3t8/BaHfi3ruB3G1dtn3DQq9mvTXwK25uvDYtyej5Ow6YxtcmO40sOD+h3TrdbttujHdN8bzDqbfc0gOKtEluGoPqdbLXFtpO0JRsn3nSj+01c8l96XFDcxlUSWoaBUn2FH4bHlSYGkHpEelbgmKXpdjZurLmQCQY0i3oVtSHEnRSsD5p38dVWjfVq35Hd4dmgzG27lcz4ceGvssnW1KA/paSCe3wqLnxRlh4tXeHTyTrvjvSa8pgOTDc4ElCn7dLQsL8ZAUUKPMDj2WFD93apP059O3BMbuTEXNXnLfIcT4jS/DU42E77AkDt5e+sYMcY6s1ptDc/oH1pgdbMUey62sqiYp4y2Ycx4aMsoPFSwNdkhQKfiSk+7W/y8ZanMZ9xxyLkCIEptRJdQRzAPcHRqX3epUp1WtvV3pJj82+4pcD1CZiIU6u2ttakkDz4pG+R+od6xcwZPkWMYH1mzLCmrfmeOB6RHiS/7uy2tpaCF9tjsrmAfIgHWxToMv8AgvOsV26rMdYk3WM5CVGyBuaiK6RyaMhLhI7f/lA799b01qnqyUrAUoFeeP4Q789Vl1//AC8z/EyaDdjof+ZXp/8Aq9b/AOGbrX/8JJZbhfuhy41qkORp2nloW0SFEJ4KKQR5bA1+2g0f6RejxikKRFjzbPLYubLLYevciXyQ7IW2lS2kt648dq0CNnsO53V09EsNlYvfLkLg2EIS+4hAA7EA9iP2UGxDHhvteEwAghPsrUN6/tqL5lJvFohlUe3tXJOvNC+Cv3Ht/bQYrFpj71ucckx1MyVjugjsk/DfvqKQ88izsgkWkvJE1l4pcYJ7p1r+3RBoNofRmhm3dMrfBW8JDsNbsdTgGt8VEDt+gD99XHDGlOfsoOzSg/nAwCMZXVuMhOyr19WgPrc1/tr2d6V4wiyYjCjaCVhpIX29+qCXLitqUUIWn4AbrE5Laob1olRZLTcmNIaUy9HdTtLiFDipJHwINBBUWnHHLVGsDdvgG2RnEvtwClJaQtKuSSlHu0e9T2HBantBbikOH5wUBriPqoPp+A0pPzQsjeifPdeRPpy4U1hnpAXYRmFMRLiyiYhJB1yVsL0T57Ukn6uWvdQe+8f/AIu1/kD/AEVyUFC+kjdLFPzjpbhuWXCNFxG9T5cm5RZjgQzOMdlPgMPEnXhF51pRSrspSEJO+WjMc1wfJ7n1CtF7xK9Q8ejyLe7BvctyKJLrzSFpXFDSSQkLQpcjS1ckhLqwUKJSUhGslNwt2dWm33S/yr7Bx2Kq/SHpjTLJ9YWFxoiFBpCUrSdTHdcRpTKCPdqns5z+XeMns9uSy665eJgjQwQripfdbqlqAIBSylxzR8w2QO/nV8Qm+auPh+KfWyz19lY7/v8AwkaeK0tbU37Y46e+fv5pbc5H4lvUGyYnjv8AKHMJDCvCjJcDKI8cqHJ+U+QfCa5JA3pSlFOkJVo6nFm9HOFdG2rh1LuQzGW3p02spLFljEd9CNs+MB/WkFzuNgI8h0uaaaXHXSYI2rWNvv8AlT4aWz2nUZesy5YXpGY9enHbR0yx+59RFwT6sXMeYbZtcZSe3AzHlNsEDXdLSlqH9WoT6PUHqVk2IXgxbnZMJsy8oyAuMtx1XS4pfN2leOEvKLbKUh3xAklpzaQnffdVywdmcza7zGuCoGR5/nyoJcRKyFOSqslrjLQSF8n45jsqCSFBXgtu8SkhQBBqAvZtmlueYewLNI1zcb2VsOZLKyK3rSn3rX+KSojX9WS17xyoLB6OemJasuyS3YXmyLTj2aS9tsKtV2ZnW+Y6P+TQtKvEZdI7hp5KSf6JXUyzf0eoVxuki/4XclYRkryi5IVGYDlvuC/jKi7CVqP/AK1BQ75e2QNV7pe2O0Wr3eL0rkry2UP1DyHIDlmP4lkNhcsF2iOP3J98OeLBnNIaLSTFe0CvapAUUqSlaA2eSQFJKpbgd3yWdd58GXZ2ImOwUtoiXFx1QelFTLayEt6I4JKlgr5DZHEJ7FVNNl9Nxm9qR09HG/7tGfFGLh9a2n+6dv2WZ6OUtV1sF+u5GkXi5rns/As8EsMrH1LajtL/AOnXWzawXPq51bt1rhX2RZsZw7+c3NcAJ8eXcHmiGo6VkHw/DYWXFKA5fzlopKVAKTt1EbZbffd7wTvjj77M/cOnvTu2T4Fnm47CuNyvRdQhycyZkt1KEc1uLfdKnOKfZHMq7KWgA7UN8qWL90x0GBNyvEkDXgkqkXS3j/FJ2qU0B/RO3ho6LuwlMdvS2z3yz5laDJt0uLdbe7ybWW1BadjspCx7lDyKVAEHsQK109KrEovS+12TqPi9siwH7Te4Um4R4jQaS9+VCQ8AkD8qUqXGKv6TcpYUTwRxDZ+on1OWpOKOBKlAKebCgn3jfv8Aq8q90je0Q9VjeYVMp5xW9uKJPLe1H3+f7/fVls5c3h2A2a5zk84HjtR5UhxwJ9XQ454aXPI7AWpsEdtJJO/Z0ZmprEVjaG7LERHRG7d6TWLybE1OlMzI8lKV+tw2kh1URTbT7j4X3BHhCJICgQFEoHFKgtBV2LR6RuNT7TGmy4021+LJahqbkeES264224ANLPPSXmieAJHPy7K1AR3bh9eLEMetF0uTEm3omWWFen1ABbURElKy2hS+xJKm1p2E67AnQNRiV6V2PR8hTF9TX+KRCdluXFTvIoU1KTGdbDaEqCiFlelJUUq49iQQaDNR/SMsLcRarlb7jb5ybg5bxB8MOucw/NZRsg8dq9QePYkDae5B3UxwXqHa+ocOQ/bUSWTHUgLalthCwFJ5JPYkaPcefuPu0SEnpQKhvWa+OY10lzC4sTFQJbNqk+rSUK4qQ+pspa4n3KKygD6yKDz9n5dmuFu3OO3Lu9pszcjwW40Vkt3KQ2ma+opkeAoKfUEKcClIKSFcdqHJXLZ/DG7Rc8YteSXGM2iYiOA5PuLKGVko9jxSNkJ5AbB3viQN0FfR7vDx29WaX4vr1tXNfahtxXGHmW0qSptDbBSS538VBKFn3aSkaCayfSCdY8wyh3N75MjXGz3FT9snqyLwDEijl6ylpl1xaS5pwsFICAgclqSlBUaC7+pvSTH3+mVwhW+wqlIipelNwIoQtUsqRpTS/EQ7zQoBIKOKuzaAkeykDTfp5d5WLNZFc4lpvyZuPCLMedlX+E3Atah4pSthSneTiSlT/JtagAl9bRJOzQehthvMfJLFbrtE8QRZ8ZuUyHUFCwhaQpPJPuOiNj3VHcg/vle/zNn/AF3aDCXm4uW/1FLSUKVIlIYJXv2UnZJ0PfoaH6d99arI0Hy0nneLQgd1KlDQ/Qhaj/YDWG6gzhPyxbQVyRBYSzon5ri/bX2+tPg/uoI44HUradjuliSysONOjYKVD9BB0QSCARsEj31bGD36VkdmXKmKj+Ol9bRbjoUjgE60FAqPcj2vPyUmgjvUrpj/ACoUq42wNIuZRwdadPFuQkDQ2dHStdt+8aB7AEaJ4s1J9GjrA70/uTLkLCclkuPWLxxoWyefadgk70Er3zb9x5DXLZNBamYQPAksz0D2FgMu/Uf6Cv7Sn9qa7PTJnxuolgT30HnVEj3aYcP+kCg26w3+9Gy/5m1/qCsxQKUClArUD0ycEf6j9UsWh47IjNZha8cujsduTvg746mUshY8ikLYWdd+47/X6r3Hndh/pnzfR3yRuJPx9Bza2Oqtt3Wgg+CG3FJWwlWyNBQG9diR5ntXoJjmbYT1OhWXqs9iNgm5A7HSWL8YTapTaNeXiEb2O4+rvqvcTuxKN3P8IXgmL9Q28Wuk96PJUkKW8v8AuaPgCfdWRzX8ID0+Rb5sa23YSHGG9rfa2tHcdtEeZPbsKz0Hm71i9Hzqj1t6vXfJrfja0C6FEjm66lC0eyAgODzS4UBCinzHIA99gWP0z9AXr701kp6gly1xn4MdxLaZExS1hKkFJ3sDQ0fKvO077ssH129EfrhkWMW9cLDkXjH8fektMSrdIQt5IWsFaVI5eQUPNO6rz0f/AEfOoPULO37XdLZGg2uMhJnzMjhF9tpPcJQ2kEFTh0rSQpPYEkgDdYmJmR619GJ2KQ+mOPwWLhFtWMxoLbUJtoeGgp4j2gn3bJJP1k7rp53K6SdOrfdcyauDD12is7kzG5JUQ2B22kHX9le2FN+jH6Wyutmc5E/BD0ew2vg0z4iO7ylb9r6hoeX11f8A1Qu8h7HEuvykhmQopTFdHn28v0edO7Eqf9F7P7J0z6t4niqZaWk3ky7K3rX5ZKU+sRQ4rzUUcXm0fAOaHvrf+tdu70UryFKBXnj+EOG+tVl8v73mf4mTQbsdD/zK9P8A9Xrf/DN1X/pX3aHasbtRmlosOqeQUu60r2U0Gn/T+Vit2tNoSi7MSLnaJfqkZllZW40tt3Q5J37RLaUDZ8gd1abyHHListEDmdgD3j40E1s8kR4/FzfL4VwX2Up+MQjvrsBQV7KvV7x51j/g6PIh+2Xny8Q4n+oEI46O++9ke6ola7da88vP44cguW+4vjbi2yEOpI7cSpJ0ewHxoNrPR2juWrGH21FSkrkrXyUdkknZJ/sq7Le4HC7og61QdylB4q2H0N8x6V9bJ7F+aaL0CWl9L8dXJp9pS+aVI3o6IHmQNEEa3uvRzE33J9kjIR58ACE/GglluswbQlT2yoe6tZfTt66vdC8Aa/FgaVers8qPES4T7ACdqXrzIAI/aR8aDzgxj0lsrtWU264yrk/IZDqVS0LWSFDl3KBv2dDR0PfXrr0nvqMtxK33GO6lxqQylxLifIgiglcpJikcu6T7685PTU6fzupvXvG40RbTbT0mJaNr7kLfeAB0Ds65jtQeyjaODaE+fEAV9UFH4NdbXlnpN9UGbs63+PrJBh2i32p9B3+K3GkSHZSQRpSXpDpaURv/AIogHXvlULB4dvX+OunL8SzqS44xItSEFu2TVNrU2tK2kj8i6laVJ8VtO9j2kuABICnOqWR3ZzqTMgzbJLsovMOC2n1ktrDjzDkoFKHG1KCkKDqVJCuKhxVtIJIGDn4Vk8PN8Ourk6I9jtpmuhVphQ1qe5vMOMplOPFXtcSsApShISlxZKlBNRuF4pz8RzZ57UitY+s/ftY1+SMWlx4v+7eZ+kLd9GZyDOg5zN5BeQKySVGuQX/dGktcUxUa8wj1fwXEjyJdUrzUayvpJ2iNful7kC6SHY2NPXKCm+raWUbt3rLfrCVqBBS0U9nFAjTZWd6Bqdl3nJbfzl4xxEUrt5LJtVtg2e2RYNsix4VvjtpbYjRW0ttNoA7JQlOgBryA7VRLuRXa2+jhn8s3ZD10Xer/AAIkyCoFTBdu0iOwlPED8o2FoTrz5I0ST3rU2Il1uskC+ZRa8ATFabxO0i22e3Y6tbTFsdkSGpKg7L8Rp1K22WYgS03w7urH9IoUjBdOeit9seU5hjODZjcsJv2NGG+uFEuJnWiQiQh1SE+E614bKtIIUhLO0p8MgnkkpDsdSznDDHqHWXpRjfU7H2wCzkls1Eehkd1K5LKvCIHcO84/ce7e6kPQn0kYVsy6J0yyydeWJMlKTjdyymCuJLmNkbTFfWfybzyU64vNqIeHnpY9sJl1OSjqzksPF4aiqNGcKjKb823B85xJ/wAUdh8SSO4NRvEm8kX0ZN5nRE3G9QDMtb0a3oUlUmXGkuw/yadeyHHGtjvoBYJOhuq3geqi+p1Opt+mZisf/mPpKRxTTz6HDir3iN5+Kw7NFPQ/pdbIAaRdr4GWLbAt8c+H65JDfFtpJ0eKfZUpStaQhK1HskmpZ0/xRzEMaaiS5CJ12kOOTLjNQniJEp1RW6sDzCNnihJ3xQlCd+zVvlv6S82hCx05KRVW72OX/LOpN36g47c1Jm2J1dgg2qS4UwLlEQEKlpWdEtuGTyQHUg8TFTsKSVJM7w3qlacvucizOMy7Dk0VvxZFhu7YZlIRvXiI0Sh5vZA8RpS0bOtg7Fam1hetWI2pOE5VkzDb1ryKFapD7N2tbq40sqbbKkJUttSS4naQOCiQd699Yn0gbDHvfT2w4GiVJem3292yLGUtwuPqRHlNS33VKOyeLEZ1RJ8zob2oUFw1gc1tH47snqwkMRiXUqDkjsnt7v016rPLMT5Mx0mFeIwcKSlQvtqIKw3sSN+370jt5/Ae/wCqrCt+MQpWHpsdzYh3eA60pp9h1oOMPoJOwpJ2FA777rdlzRliIiG29uaOz6k4HjUx2U6/j9sedl8vWFuRGyXeSFoVyOu+0uupO/MOLH9I74Xum2JyGCy5jNocaLqnyhUFsjxClKSvXHzIQgb/AMRPwFR2lwOdLcUfuImPWGA+pFubtLTD0dCmmYqPEAbQgjSUkPLSQOxGh5CvpHSzDG1KKcTsgKkrQr/g9rulS+awfZ8lL9oj3nue9B+t9L8OacYWjFbMhyOoraWmA0C2orWslJ49iVuuqJ+Liz5qNZm12G22NLgt0CLBDgSFiMylvlxGhvQ9woO9SgVWPpKW9d06LX9htCHCFxHClZPdKJbK1aA7k6SdJGyToAHeqDVzqei/yLbDtgt1qtuQSJE2KLzKBW9JC1pf8JT4T3afcfSNAALUjkWkt820fNmi362Y08w7dDc4rUxuS81PSCHNoQ74ylfOAKVJICt8ArZHJO6CIZpcV4PasiZirdg3GGwUxkONgucVJJbbdQdpcSg72DyBSArvyBrtYa7/AC1dv9+swfub8e0sxWbnMfc/ILm69nxCSQ6oFhLaEkBJWsninvQS/qG9nlguTBRny4Nqt9sjI4vvCLG8RRdbWxtCmwwlwJ024SAlfBB2CkoxUm1WLJLrdLnKYiT74/IdslvRfIqHIrbq22gXyH2nH1LivlaVd1Hi4CTwCEthvjY7NGx2yW+1QgtMOBHbisJcWVqCEJCU7USSToDue5qN5B/fK9/mbP8Aru0EXzI+HboTyfnt3KEAP8qQ22f7Fms9Qc1naMrI4SB5R0OSFK+Hs+GB9W/EJ/6BqEXpan8gu7y/7ouW4D+hB8NP/VQmg6uqyOLSTBym3r9YeYYfdS08GlkJWdKDYUneiOSgPLfceQ2QFy1S/pT+j7bOvnTK525xCmLwy0XoUxgadQ6jakFJ/rBXdJ7eZGwFK2Gp3Sfqt/KfBr1Y81WIuX4zqFd22xtUgnQYksjW1eN7JTod1HQHdO7T6AtS7nnEhM+Cu3zLVCS+8ytSVDb3JDakqSSCDwfB77BR3GiCdc5K1vGOe8/w8zaImKtrsN/vRsv+Ztf6grMVseilApQK0v8ASxxLqjN9J/p7dsAxeddLY/bUW66XdniGYSPWHFbUSQdgLKtD3Gsx0kQSV6NkbB4mZO3jo9O6pSLvd5M5xa4DReeDyySUqJHHRJIOwe9VLiWBdeeiByPE8b6T3+94BJKpdladbSp+F4nf1dZK+4STrv8ADe+9e5mI7HdrXnfoSde8paVdpPSq/wAnJLjJVKkuNtp4MtkaS2Pb8wNVnOhfoP8AXa05Mw9e+nd6t9sgq9aQ06wg+M8Pmf0u+tb+o6NefFlvP0LxPrBiXTq5LmYRKRlUia96i1PaHBoEhDS1qBOhxSCSa7sTod6Q2X4VmVpzi+RJF0vdukRonqh4xYyltqSgEgbABI2dGve8SwxuC9N/SL9HvopiVnsNni5TfLc2oz7eZIU28VuKWsB1WgVe15+RNZC+QepGS2pu5w+k92x24LltPS7elpslZKOKzySog9yTsHyH6aRMMbbtS+vfQb0irrfZsPFMAyh/HpaA+0wnSRDeUT4iU+180kcte7mfdqu56I3ox9aMdvF6tWf9Nb05YbqgJfVcmw4lY0QUk8idfVXnfqy3Lb6OM9CcL/GPTrpQ9+MkNkuWuAzoyHQNA6J7bAGzVE55Z+vfVy1evzemeS2e6NLCmovBAQjv7tK8tV63hh8+j/6LfV+zdWT1RyayPRI9omRXIWOuNBTjyhx8d5IB9khKnOHf5yj7h39QPMV4t1ZKV5ClArzx/CGjfWqy/q+z/EyaDdjof+ZXp/8Aq9b/AOGbqAelTjcHKLNZIk9hL7BU/tKvd2R5UGl1l6SI6R3qZLtQdftsmQZCkr9pcdRSByBHmOw+urWtc0yWGJLTofASDzT7/jQSRF2S6kDmEke9VYfKsqTaLTLlp9pMdpTgTv55A3oUGonUDr3ns29xnbY7CMAl1TkFKx4jAQARvet8knYq98VyBh2022etJQ7IbQ74QT3CikEjX6TQbadI5ipVnCY8Z/wBo+I42UJUo+et9zVwWNfNL3lsa7fDzoMpSg1A9M+13jA+oOH9ToxbOJeEbHkRCVFUbksmK+QAdp5rU2T7itHmCdSnppd2GX3IhVoqG0b94OqCy/MVp/6Z3ojO+kJktrurWQSbU7CYLKGiz47JBIJITyTxV27kefb4UGsb/wCDVujZ/J5kyvie/KBx7fV+UNb2+jH09V006dwMYW+5LTCTwS+6faX3JJ/efKgnGYXGNa7W/IlOpabZSpRJOtdvOtaekmNr6zeldiMhmGzJsdj8bJLk9JZJSpSEqYhpQruOYdWV6Ojpve/dQejFKCpYNsau/XPqMxGuEu0Xg2KyJ/GERLSnm2PFnlIQHW1pA5h3e0nz9xFdjCrF8hizZJd2uF3xy73J2REudzWHHos2S6pxxh1aUpHB11alNqIHtrKCdlsEKv8ASdsNxzPMo+KMreiP3dmOIFyiuFlUQpeHNwudykpPcEDvy1+nhsGaSseu0fEc3cbg5KPyMWctIajXjj/TZPkl3XdTPzgdlPJGlVq4BaKZdVWZ/v8ArHT79zxxevPjwzEf2u9fMNnt345JiWQv4hk6m0svy2WUyI05tPzUSo6iEuhOzxUClxOyErAJB57d1K6s3Nt+HIkdOr1H4lDjrEeWjxEka0pkuuAAjexzNdFn0cZb81Z23VOHV+jpy2jfZVGS4p1Gw22/jWx5XCxBUTfgwLLd57FvIHzW0RZPrbLY/wAVtoD6h51FumnpHzbp0IveKzcXl/y0g5DIvSH4zaXIkmUi7KnBbwUR4KXH0FJCeYCVbRvXEUGtvTRTFbTvPs8HQaDT5eIRzUjaO3XxlfnTvqK/1pvt0zdjEEqxSRaYtpu9gnOsyp7ykLedS+GEKWgtI8VYCTpbocUQkFtKF33h+I43h1oEbFrHbLBbX1eser2uE3FbWpQHtlCEpHIjXcjdYraL1i1ezXkx2x2mlo6wzbi0ttqUtQShIJKlHQA+Nad9Ws5wvJlTbZY4kez2j1j+fy7wUM2h1YUClxLSyW0L5cVh4eEs+yeRFVXE9VOnw8mP9dukR4/f8pejwxlyb2/THWXL0hwrqrmtl/G+I9QLRjuLznH47d3Tb/xlLlMIWpCXo3ioQlKVEEpWpToICVe0CKvGx3jF+juOW/BsfdmZPeLe3wTa4zol3B1xaitT0lewlrmtSlqcdKEkqOvcKl6LTV0mCuGvh9fFp1GWc+SbyrG+dcI/T/IcsybJ8Yv+TXvHWHYzsi1iI3bbYAwzIciRVSH2nHXCh1lTjgQVOEeyAlHBP5kfpg3G2xrhancPdtWRJfYhsPRnVXyKl15JLaT6khag8oFtbbLvheKlYPNI2RNR2b6b+k70px+FExKRebtj02C02X3sus0u2F5bq18pDrrzSW0l15L21EgKc5hO6tS6Jw7qe2i3fjKBdJcdKZrDlump9aib2EvtLbVzb94CgQD3HcEigg/VS4Kw7E7Na8wydqTZHrmhyZdpLbbMp+OypLrURLLfeQ+84lDZDTaeSC5pIVrlk+mmP3rK8rldR8st71onPR1QLFYZKgpy1QSoKWp3RIEh9SUKWATwS20jZKVEhalRXqW2V4uvTYc4vNkknXEb8/8AZ+2tWb+nb3S3Yf6tffCqCNipfkeZ3DBekkGTZYCZ1/lupiW6Ctp14POqWpSiUt+2QlpLrhA9yDVXof1StuIfor70cV6UbTzCpkPGJMy3KaS6ypEkCQ7ySxpPhcOyucltOirtpfvACuzL9JqA27a/AtSFsXR9tqE7InpaLrToStmXx4kiMpC21F3vxPMFJKe9yo0VtHpgNTYNgL8KGJL8Mzpy2VrU3wRAXIdCBradL8IBQ8QFKiex0KvPp7msbqHika9xWvAbcekR1N8+YC2XlsrIOhtJU2SCQCQRsA9gEjpQKUCsRl2Mxcyxm5WSYpbbE1hTRda1zaUfmuI2CApKgFAkHRSKDzzbsWP2MvSrG9bMglLsqYN7t6ylC7M6pSUSU7CkrQlJc4JS8pxQUtQ2viQJaepDr9zecuL0JiS80hRjOrSDNa5Kb5jfFKnBwIXriAlPzTrsEfv8L8bTETb0xuJDacTHiuhE5byvCcQltsucQoJ5p01sKJcHffEp7GCKulgx6cjHpkGXZbCt16RcXrNcPCgIbLjaXn2W1lCnUsp5hRBVw0la9jmQz1rOT3q1tyLxaGJDsHw7pKti40tuYWnGV+B64pC5PhaLagUnwk8HZKfDHPasR09jTMoyrFba6xbWY0y5man1GVGUuA0p1Dag6205yQVgIQlsNLS2kRUF32HNh6D1Dsg/vle/zNn/AF3aCL5mPEtcRtPdxdyg8UjzPGU0tX7kpUf2Vnj5UGaw+GRGfuCh7Uwp8Pfn4KR7H7yVK/QsVXGRo8LLL4jXFKZIKU+4cmm1Ej9JJP6TQdE+VcUmSYDaZiQVqiLRKCB/SLagsJ/bx1+2gvpKgtIKSCCNgj31+0HmZ6SWGXDG+q936j41bm3ZuDTVQ5drSj8pcYBAeUN+XJtp5stnufZUST7IE2uOeQo+HMdUMUu4jSkQQ9DkI2pu4IPdER5r/lApZ4BPz0rUeBSonfOajLadRXJj7xPL9+/qgZLT6SLV8OjeTDDyxCyke+G15j/EFZmujTylApQat/hF/SHy70ZOgkHLcKchN3d6+x7eoz4/jt+Etp9SvZ2O+209/wBNeaP/AIX/ANIT/C8b+yB9+gD8L96Qh/8AxeN/ZA+/Xetf4WX0kb1IQxDXjz7it61ZxrsPjz1Q2WDA9PL0vJ0dp8xccjsuDaVv2ngCPiCVAH99ZB704vSxTED7EnFZI3ri3akqIOt+SXDWdpNkByb8KD6UOIrUm5t4+wEnRUbP2H6fa2P21Hv/AAv/AKQn+F439kD79YJhKOlv4V7rzl3U7ELFPlY8YFzvEOFIDdqCVeG48hCtHn2OlHvUg9Ib8JN116d9duoOMWS929u0We/TYENtdrYWpDTby0IBUU7J0B3NYlmI3V3/AOFa9Iged+to/TZ4/wB2ifwrnpCqT2yC2A/XZ4/3a87vXL7X1/4Vr0hd/wB8Fs18fxPH+7X6fwrPpCD/ANIbZ9jx/u1ndiavw/hW/SD3/fFbPseP92v1H4Vr0g1rCRkds2fd+Jo/3axzHK9pumV8l5P02xO8T1pcnXC0xJchaUhIU44yhSiAOwGye1SWvbyUoFeeP4Q4b61WXy/veZ/iZNBux0P/ADK9P/1et/8ADN1FfSEUkNWFKla2t/8A0IoKWfhIkI0sBW/7PrqLycRVbnHn7S4mG84SpTagVNrP1p7a39WqCH3/ADZ/HwW7zFVBIP8AxlJ5sH/peY/aBULuXUSFmsQ2eZaL89DUlYMuzQnH+XHzCVIBHke+u42O1B+YjaumF0ltWqT0dyK2LeCiZl2t0kdh5HxSSRseQ2P2VbuB4XasAt3qdtW9Mtr0pLzPray4tkbBDYUe5SCNjfeg2tst1hxLJGKNMtKbBAA+r6qlWIXFq4Jl+FshBT3UNb8/+ygkVKDA57iEPP8AC73jk4bi3OI5GWfIp5JICh8CDoj6xXmF0n9Iq73TqvOxa7WeTjt6sO48qJKcCl+IhZQseQ8u3f37+FBvpil/ZvlsbcQscgO6feK790ipkM+0kq121/toMKuzQ1p7x0cv62qzMGExCjJU2kJ0nz1Qadenl1uTgOFuQYTqVT5hLaE7/Rs/oGxWw34PjGZlv9HGxZBdrb+LrvkIM1aFlRWpgEpZUdga5IHMfUsH30GzFKCpOpklnpt1NxnqDIWmPY5TCsbvslQ0iO244HIchxX9FCHubRJ7D1vkdAEhNbifjfKRirC8gt0Z3wMjxpxBXEkLdQVuiMV+wJHFaVLbB4L8QBXFaisBV+U4pDwqSvKIFyuV4w67MsvQbvPnyJn4o4pCVR1rdUpTTCtBxK1aCXPESsjk2BN3nsb6yY6LVkMS3S5jiEpcYuLSHI04DyJCgU8vLz/Z9XPY9THC+Lc2T+nmiI9m8dPv3rLJg/PaHlp+vHPylhovo0Yta20tN4BZlIb3x8WE06ED6ioHQrsfJDjl+cdiN45YpL7Pzm2W44daJB7jieSTreiNGvofptNEd4/Zx84tRM+KoepcFzCOoVhxy+vXF6wJjqltPXF3x1JJUUpYCgC46SUgDkVLO9bPuqHJpE3F7Pbb/b7tLkWyZk9xsGStW6S5Dek2f8cPPLZQ4ClSSzwWyngQOLzgBCVVzc5sPNnyWnrvt8IiNnTZcOaI0uDH2mu/umZtv+3RZ8O43GR1Vhx8As6sVt8iGW2HRIUz6u2EHioHYBT7CVcVc078wfKs/ds16r5dKvFrczdpmwMq9TL9taTHfmKABWtmQ0gKSgH2OaOBJSrj20o/OtPxnJTT2tzxG09vHr4R7H0DUaLQxkpXNWb2msbzHSImOm/y7OD5NMoYs7l3Nja9Tjtku3QPKbkLSdbUp1Sg4580eZUP3nddY1ncLEMsdt+Z2b8bYVcHQqFkNsKo92sq9EEtvM6cOtnaEkFQCikHfgVO4ZbNl1MajXx61o9X2fD6fv4qrW6bDOlt+QnpWfWj7+a5rC3CwvqNY8Tyr8XZliOTgT7XdnI7aos4uLShElKEjw0Seb7SHfCAQ8mQh4BCkuhUnznr5FwV9zFum2NNwrbbZjsG45D+JH12i3SEJClR22o4SZEg/NCUqQjnxQV81BNdy4xSEm6yYOE5hkF8TbZU9qVccsjvxUllpTJbiLkBHJRcTJYlx2FqaU6QGXh4Tiwlvjlsds4iZi/EsrJXNjMEQkbIcU7FlOKt7KiRyAMWxp48ykflFgnbhCgieSY6i/TzOm9QrvCdmWN2xxbo/JEy3SW4MZiSlUhtHsyWV+sTAhQSV8nG1IJUQowNMLHrdmy8NkYdjzD7Dq7pdMixdvxLTBYeakeqyo76VtrQtSZCEpb5pO4Tagpak8iEvs8id6JzFuziVdZ/Ul1DrYcyC/4882/MirWPFQ29JZD7TgbUpaC0++hfFXNCE+0PR+LKYnxWZMZ5uRHeQHGnmlBSFpI2FJI7EEaINBy1jMisDGSW0w33HGk8gsLb1yBH6QRXm1YtWaz4vVbctotHgjSelEEcN3GYrjvYPh+1/wBWpVZbUiyWqPBbdW8hkEBbuuR2Se+u3vrRiwUwzM1SM2ovniIt4OxMiNXCG9FfTzYeQptaQSNpI0Rsdx2roRsVs8O6IuLFujMzW2Qwh1tsJKEAAAADsOwA7e4AeQqSisrSgUoFKBSg146qdE8wg5Pecl6d3EtG6R3PWrYXg2tLxBJMcniEhx0Mrc043vgvZVyT4dOHCupGSSn7XMsFyh5cw80UoLCF211amtqlB0ttobU08ORWkr5cxxBKlIAbfyOmtnlraW8HHnGSS2twIUUEgglO09vP+wfXvt9QcaczLBMgsLLrbTtygPxEqd5cNrQU6VxIVxO9HiQdE6INBpXHumbp6k3mw3C05CvG5c52RKtloiSp8eLq4surWotx0cVLPjKRpJcKEgq1y2b66H9AGcectWQXtmZ69CBdt7FxkNvywpxoBb8xxLSSqQdqSUhSkDST7SxyAX1UOyD++V7/ADNn/XdoItcCpWX25p87irjuKaT/AO2SpJIP7CFd/ega178tFfgz5rrFykiDDbX4ZTIQpsSToEgLUAnj7tb2rR8gO4WJrtVfdScfUh5F6YSVICQ1LSP6KRvi5+zfFR+HE9gk0EK8xQjdBlMeyq5YyUMsqE23j2RDfXwDI/8AZqCSQP8AFOx37a1WfyrrhjmK4e7eXXFSJ/stMWNtSfXX5K+yGEo35k79r5oSCsniCqvNrRWJmfBiZ2jdrtb0XCS9cbneXWn71dpS5s0sAhpK1AJS2jffghtDbYJ7kIBPcmqI6UY/D6ZdYbQ1e4Zt/S/KJcpGNSHXSWbfLWHFK0CChovJ/uZ8+DvbRUvXNaK05c9snhvv9+7eTh8VzXy0tEetE7TO+8THWNtvGduX4vSrDGizh9jQpZcUmCwCpQAJPhp2e1ZmunZKUClBob+Ga/8Aul2v9aof8PKrxHYjuSnkMsoU46s8UoSNkmguvGumePYNbY11zd11+fIQHYljh6L7qfcok+y2g/11b3/RSfOphaMhyW8shFit8PEIA2nnEbC5K0+78u5sg/WkJ/RWfc9bMY3i8p+7OolxEXCWEBZemyVOqdG/MFW9/XXTdt9vkJSfxFzUrlpUQFCkhJ0fm/A02GUkXu4WqAGJrknIsdSnTtvuB5yIqSNFTSld+39U9qp7qFicK0usXOyvetWOZssuaIKVDzSR7iPh/wDSkkw7HQb8+XTr9Y7d/Et1Znpc9/Sl6t/rVc/4ldeZeVUMtl5SUISVrUdBIHcmrDgdObXZLaxcswuzdoafSFtQ0JLkl1J96W099f4xIFV+qz2xxFMfW0pOLHFus9n0qZ0vSCn1PJFM/ND5jM+fx489/wBtcczAbBkENx/EbqJ7rKQpyFIaUzITsb+YokqA95STqq+uXU4Ji+SYtXx28Em2Oto222QGTCMZ5TTzXBxPYpUK4w0gEEJGx5VfVmLRFoV8xMTtL+kvol+ZjAf1ft/8M3U0r28lKBXnj+EOG+tVl9//AIvM/wATJoN2Oh/5len/AOr1v/hm66XVrpfJ6krs3gXJNvTBU6pe0FRXzCNa/Rx/toIo96PEhmMgRLs2t8D2jISdH9Gh2rGP+j1kDiNIuFtCvcSpz7lBgLz6K2Q3VhSTcbSVEEArW79yqyf9CnrDj6Fow7McYtDPNTqY7yXi34iiCpRAZ95HfXxoJ/ifo9dY1Qg3k+TYmXhobt7MhzkNd+60p1+6pLP9HbJXIgRGuNq8YdwpxTgBP7EHtQZ7FOmGbQLcym6LsXraAARGlvON/s5NJ/0VZOIWa42huULj6pycKeHqqlKHbe97SPiKCRUoFVP1K9GLAup2RtZLMtKLdlLYCDe7clLUl1AGuDp0Q4NaA5AkaGiBQdOw9BZ2PPH1fIkLZ2eKFxDvj7gdL86mTOByW2VNquKFgjQJZOx/1qDqq6aySskXRAB93gH71fM/pxcJUQstXhto60FGOTr/AK1BVMj0FMHy7MIeSZ64/mEiEoKj218BuCCFbBW33Ln1pUrifek+7ZFlluOyhppCW220hKEIGgkAaAAHkKD7pQdW62qHfLZKt1wiszoEtpTD8aQgLbdbUNKSpJ7EEEgg/GtculEdzppb7pgFvdetsjHJslQt5cUVKhvSHHI8pPIkuIWhQBWNgOJcQdFNBzRp+X4ncrsqzPRbjZrjJVNXb33Cy6w6tI8Ytq4qQoLUCspIR7biyVHl2jV0btrze37DdsZcVvSYULm2rz9opiFaR+k6NQNbo8euwziyfCfKUnT57ae/PVHGbLKvd6L9sctGRx2EJS4qW23KcZ77CFJdBUhXbeu3kPhXayW2Tb1FNvew6FK4p/JyVxfDEdWuy21NkLSse4o4ka7KBrkcE8a4fNqWjnpE9Om/T4Tv+/0X2WOHaqK2rPLbbr4dfoig6WXJFgiJflXy9X6IhP8Aw3c33n5KyB30CeLYPnpCR389nZNFW2UxJkXGyZNNtVjU1OuSlpuMlMVjiJTqisqWrjzXzDhA7nkSAAO2rPm4nqKXtXHy7zHTbvtE+fujsttHOix15L2ieWJ6+MbzHl8V0Yhb7jm+T2yJYksZlMvduNwizH5ao8NTAUWy4VqQVKSghHIJQSA43oHmKt75G7o7kmF471Ds9gTibipDD0izTZKk3SZ4ChHadZUhHqzXDxlhIW5txtoch7PLbwLgUYafmNX1vv0r4Rt5+c/KFRxPiU5bejwxtXb47fwylluYtYvHo+5RfkSr+iIZGK3KfICnrrASQptt072ZDBSG17G1oCXBva+MN6k4SMa6PRIWS2NDFy9YeKXi6krLPJA4+yTsbWSO4II2PM1fcTpNN8+28RX9p36NOg1sYcMYfGbx084n1ZUrIyydhfT9OCZG8zDuuMXS15Ri10mKCWxCkSG239KOgEBiQ8vR7BbcjQCGUVaN/wAruGR5VensYtc9jHcluKbpYDb7qqz3+WstsNypDDLyENORH/DSUtSHG1uFtxaOQ4cLvHbnpFvOFHlrFL2rHhMs1bLjjWRS7CV2vx4sVSLfJt/NEN91yOhUdlp5uS82WZfhOeC4y8VJkMOkoWriEiU9MOgGZ2y0Ya/PFosl8xWAIseQ+FTU3BSmPViuS0haAFJjpZSQHFAuJUQePdWxrYu/dM5PSrJre5GZjSEWuPIkWy4Tx45bQ3GQyHlNoQVHw1PkFCQpbihEQjSQtSaYyGwzcfDWRIt34nkWh5V3fu/JYnMQYrkKFLjrcYWw4ttmK8nmhDx8STHd4+XYJTm3Q1TmNXm/WK7ScvxabGdiXRnEpyVtrR84uCPrkh9JJVpxckLVrxfD0Hm9ivQ5yuHmfov9OJ0A7ix7S3bUqJUeXqpMYq9ruORZ5aPcctHyoLlpQKUClApQKUClApQKUClApQKq7qVkz2OZSyW4iZbbkRvxE+JwWAFud09iCe57HXu7ig6mUPINpi3hg80w1olBSQQVNH5437gUnZ/R3rPgpdbGiFpUP0gigzGMXdLCUWySshSO0Zxfk4j3I3/WT5aPmAD30rUkWhLiChQCkqBBSobBFBWmRdPZduWp60NmbEOz6qVgOtfUnegpP1EgjX9LfaJKdS3JMZzkzKSORjvJLbgHx4nR19eqDqX2+QMas8u6XOSiJAiNl155e9JSPqHcn3ADZJIABJqpISJuX39OVXuOYrqG1NWm2ufOgML1yUv/ANu5xSVf1AAgeSlLqOJZvR4uSO9voi6i/LTaPF3I6Z/UC7PYtiDi5F4UoMTLhHQHGbOhWgp55R9kLSklSGj7Szrtx5KTblz6aYpeencWy3K0R51igIbfiRpO1JaSyjix79+y0lKfrAIO9mvHDdPy4pteP1fRs0VsmnmMtJ2tHWJXZaI3qdphR+JT4TCG+J8xpIGq7dXTcUoFKDQ38M1/90u1/rVD/h5VeTOCW5jCrUzepcQTb5NBFthuJ2lA8i6sf1QfIf0j28gaMxCyMW6eXa531qVcpPrdzmrTJV4oSS6rzKVKJAGu3wFSGY+VPPJDv5FIC0IWNKTskEdviPKsRaJtNPGOv77/AOHqY6buolYAQ0+0l3wQCwRvmDvts+73VwpQWpKHg0GijlptCt8io7V+jvrVe2H2/jq7upDqX2mHh3Vse0TojW99x38qqp+0pYn3PHX+CIU4lUcp2UtSU72E/UrWv06rEjB9EIy4fXvp+w4NONZLb0KB+IlN1ZHpdf8A3pOrf61XP+JcrzLEMRZIUDALCzd7iwJeQTU7t1vcHsoTv+7OD3p9wT/SP1VyQsTuuV39Mq9+I9NlI8Van3NKcHlrt5a8tdgBoAVzeXNETbNM9Z3iPdC0pXliIZ+6YBGdv7VuYYQ20hjmGeBKFL132f8AbWKu+BLhT479vUbTNYQXQhDpBbWNaUlQ7p3/ALPKoePVTExE9ejZMOnOHyhQZTcphEPMbc2XXG0p4JnNgbUsDyCwO5A7HzHvquyeWtVf6O21Zx+EdvdPZAz16xL+kvol+ZjAf1ft/wDDN1NKskUpQK88fwhqd9arL+r7P8TJoN2Oh/5len/6vW/+GbqbUClApQKUClApQKUClApQKUClAqM5p0yxPqMmIMmx23XtcNRVGdmR0rcjkjRLa/nI2PPiRugpnqZ0PyDCEwbr0tnS2oSXwi62CcF3RvwCgjxorbr7aw4hXAlpLyUqTy0krACoPOyrNUOqh41Ffzu8pHe3HC7rYUpOuwXJlKUy3v8AxlD9B1QSS54v1btltXfn8Cwpc+PBcU4W8okKfbSlHiKbB/F3fakgcQojeu/vr76fYZ1P6qYPj2Trv+OYbDvVvj3FuA1aX7jKZbebS4EKdW8ykLAUAfyZG90EyHovW+9xVMZdmOT5Qw4CFw25ibXHP1ahpacKfqW4qqDxD0S7dK6czH+n82UWLffr7bk2u4sNDg21cpTJQ0rZJ48SAtaypQAV7B9kQ9VinNi2r3jrHvhY6DPXT5t7/pmJifdLJej9ZrN0cuT6c7kXR7JY0NVugXmQw627CiniC0hpKy0gANtaU2yknieRVvvL+tUvDsk6Z3ppjqBfbpdpBYj28uXSRFEN9yQ2lqQG2PC5+Espd9sK/udRsOvwzXlv6sx4Sm6nhWpi3Pj9es9pj7/0pvEcG6EXi4ypV2izrvMts7xrfDjvOt3SQttwhNxnT0qQ8+88seIlPicG0KbAQFbq8svw/NfSHkQZMi3JsGPw1Esx5Kylx0K1s7I2T2HfQGvLZqHq8s8QrOm0/Xznw6N0aC3Dcf5nVdLberXzme0+5ifTJ9Gn5Q+gUZcFhMrJMWhrCFR0K5vwyjUhhIGyfYAUkAcjwKU68Q1QvTufmTPTKwRHbRkHUu22hgwTEtzUeY9HYdRyDTRRtQSpJac8GY06wrwmlMyOBbNXuOvJSK+UOajfbqvF2fn1u6fyrrfWsUiRpVrkRXlZS16tkCre2pSjtDhCFrQlWg248pG+ClOklRV+2z0hH7U7jGQWm+zsrtN6uki0N21T0dMBthE5UCO4HA2p3mXnoAcWVq2lbiwjyA2MoFk3UHJuoLGOXHKrcLlKagtvMQ3IIMJxQfUzOacjKJbkrS+gJDS1t8hHZW0oOOBCs3kuQ43mWG5fIs1siWpp7EbxYbwliKpVogNx47q2ZAWhoLYDS2CgxnfBWkv8gnY5LDk6MY3Z1RotwMa9XTLBBQiep65SIORrCUJU4pDyFtpmBBWklh1DbiEqSVBXiNhdo+hRHa+TfLJ0WXKuMC4ZleJMWfMhiIqU36x4fieEEoCCVNqChxT7YWeI3oBsHSgUoFKBSgUoFKBSgUoFKBSgVTPWZtT+V29hC/DU+wywHNb4Fbyk8te/W96+qgkjMJliC3ECQphDYaCF99pA1o/HtWHxtxVr52WSo84xIiuLO/GY80d/epI9kj/F35GgzjzKZDSm170fgSCPrBHcH66yMHNlJgMIdgy5kxALb6mkJQnkklPIcykEK1yHHYG6DhvvVS041Y5NxubUmEWkbbjOpSVvqPkhBSopKt+7Y0Nk9gTVEZh6S9yyNlUWLj9rYib7C4J9cUdeSgCEpSfqIVU/TaX0/rTPRLwYPS9ZnopvNXXuoMFMO8SXjHbeRIaTBX6mWnEHaFJUzwVsHuNk6IFWH0cwjH83auEC62v15yAllZlTp0uYXQsr0C086toEeGdnhohWtDWzu1mg08Y/SRXrHxbtRpsVac0R1hfNqWjDrZBxe1MxrbbVqCIrcGO3HDKEpWpxKUtpSkeSACAD7ajvYFcl7YRGxiey0kJbbhuISn4AIIAqqVyzqUClApQaVfhYbEMm9HnFrYSAmRmMFCifID1eVuvM7o3b7BmfWOLcsglPRcTRMTDQttQStpkey2TtJAHkVfXyPnUXV2zU0+S2nje8VnlifPbp82zHETaIt2bEelRabb07fx7Dsffeds7cVd2Ly+ClvrfWpIIWkDkAlsgfAHz71QTqW7bCEu5ymbbEPdKnT3V/kpHdRqj/AAzlyajhWLU5v15Oa0++1pn4bdtvDsk6raMs1r2jaP2h1rbktgmuDwV3ARk75TnWkNsgD3gqO/7N1ksgYgwI0GQzJDkeajm24vsD2BB/QQRXUd0R9xH4q2mmyhCXlFRJCRz4p9yfrPlWI6nWVu/2APW5gxJkH8sySOPZJ3y38ewGvjSeorzBmRJ9ILpjdkJCUXC/W1xQA0EuplNpWP3irD6/WZq++mb1OjyN+qoyu5vPa8yhMlwkft8v21F1F+TDa0eT3SN7xCIY6o59nNyuEgpRHac1FI9kJ4Di2lO/IADevrq0LZan3mo8mUlTEqKsjlr56PPX6K5DVzFLRTyiI+XVaR1hHjPel5GHoyFxXHdhBUN8iD7/AKjUpm2ASHG57raQ4kBx8dyTxG+KR+mo155Jjbvsz3VVn878V3y3ZFAIQ/DkDmhRGyknyIHmNbH6DUPzW2N2vJpaWE8Iz2pDKf6qVjev2HYrotBO00ifKY/bb/KLnjer+i7ol+ZjAf1ft/8ADN1NKvleUoFeeP4Q789Vl/V5n+Jk0G7HQ/8AMr0//V63/wAM3U2oFKBSgUoFKBSgUoFKBSgUoFKBSgUoPl1pD7S23EhaFpKVJUOxB7EVUuIWXMui1sj47Ht72fYfCQGLW9GkNNXWEwkaQw8l5aG30oACQ6FpXoJBQogrIZvAettj6h5XfMZiwbxa77ZEpVNhXeCqMttKtcVDZIUlW/ZUkkHirROjUW6RdTrZa7FfLfeZ0dy7MZNfE+rWeGH9NfjST4W0xEKSV+GUc9+3y5eJ7fOgld06j4rc2gxPsuQTWyAQh/Drm6nuN++MRWqvpGrvWfZdbMS6XYLBt9wgvN3cJn+HEkXEsKD3Bcb+6MRVJbU3zeS3zdcaSkceZOjJgxZZ3vWJlKxarPhjlx3mI8t1zehnkuJ5v0pYdtVnYt1+sMl2y3RmREDM5l1lRCPG2OYWpsoUd+RUoeYNbB1srStI2pGzTky3zTzZLTM+0rWPqJ0dw3p9fG514trsTEJLqvUsgtct6BNxuU4sqDHrMdSHUwnXFkpTvi04sgjgtPh+2trDiMGBluHxMout1hZhNatNpil6fJFwfkXR+Kh5cJa3vFC5fiNvKPiBxMZvw0tNKdcVw76LJlL2J3567vqnT7yYN5YmvhLG3Lm81IAYTyOmi5AcCd8TsbI5diGa6kWK5WzoreZ0tyQm3W+6W7LrXeF8ksfi+7PIMqKh1SXGlBEhXNTZTxKCNpKVFJh+fYFJsmEIu97tM27z89mluC/aJLAcXDakrecjJaUsOI3b2tIjvrlt+KhTZbQpQ8QLDy69t47g98zbCJkKJPs6PGcW4lUZiJNjNrWww/FU44iKtaFLSh6Ov1eSlxbJCQ42sbb+jxiTuGdHMdhyriu7XCYh67zZrjSWvFlTHly3yEJ7JT4j6wE+4AdzQWPSgUoFKBSgUoFKBSg/CgFaVbOwCANnXfXu/ZX7QKUClAqqOq+NysgvhMVbZUzCRthYKS5tbnzVg+yRrt289dx50GIs3UJhLIYu6XWJLZLangyopUQdHkEglCu3fYA+B76EgeTByKKDGlMvqRpTbzKwvgSOx7H/ALPiCCAQHXi30w5iLddR6vJV2ZkEfkZA+AVrQX8UnR94BHlm6DqTLSzkD8SA5EZnASGX1tPNpWlKEuAqUQe3kFD696+NY7OMQwCM7BT+JsdacRK4vpRGZStI8JeuQA2BvXn5HVe63tX9M7PUWtXtLF/Jbh89CXUWiO4hQJSppxYSQfeOKtVIbLYbfjsEQ7ZEahxwSrg0nW1HzJPmT9Zr1fLe8bWtu9WyXvG1p3dG4kIy60KcKQFMvtNAnuVEJUe36Ef213ci/vfuf+au/wCoa1Nay6UClApQai/hN0o+QKzOuHSWMhbe3r+rClmvPXpbZx0+xG0W6yWFjIc7u0Nq7yXpsQTG7YzIbS7HaRHUlTanFMradUtwEJ8VKUhJSpR8XyUxUm+SYiI7zPSIeoiZ6QmmTdQL9jOVxcc6w4yxdo8dhtpbTNvjx50FvvxcjOtpTyQNnTaips9xxB7iiOuXSK6xeqdzjfjli6wUvI9Xf7oAjLSFtLSk+SVNqSof5XfvutWmvhzYa5NP+i3WPDv7PD2+16tExO1u7Js4swbZ6k1DbebaTwJ0Ph3Peqwvlpvsm5tWd2Q4YzDaS0V90oT8E6+vt+wVJmN3lK7DPXY5zcO6nm2oqCJR7LGgPa/Z5b+upmk+IXEKlKWy8j2UvEcFFXIbJ+GkjQrIrjp9bmk9Qum7rS/G9XzmK0HPikyGSD+07qb9bnEs+ln1vWRtQv1zSlWvmbmkb/tqBrY3wWj77tuL+pDCdG4yRY1w5EIPcH1pWpaU6QsE7JO9/wBnuq02wyWw0242eI0EoUDoVxesnfNbafFZV/TEPzA8csFzzm4zryZf4qs0FU+ethwJIQlaG0NI3vSnHXWkciCEhRVo8dVPbjndwRaTKi4bjbeOFZaTHkWhLjaxoewZStv70R3DoPf3Vi2SMNaTbbefP6e+f/Ryzedo8FOekbitsRilovOPQHYNnvqSGoLiw4YcxDoQ6xzI2sDkhaVefB1O/aBqluprPhXC0E72YOtkeenXB/sq+0e0XpEecz+8Qj5Y3pMv6GeiX5mMB/V+3/wzdTSukVxSgV54/hDvz1WXy/veZ/iZNBux0P8AzK9P/wBXrf8AwzdTagUoFKBSgUoFKBSgUoFKBSgUoFKBSgUoInm/S7HuoD8KXc4rrN2gcvUrvb5DkSdF3rYbebIWEnQ5IJKFaHJJFU90BwbIpGHXVFpz+82q1N5Pf2FtPw4UmYt1F1lJde8ZTARtxxKnNFohPPj31ugsiV0Vj3hSTesxzK7pSQS2m9uQEq/SIYY2D8D2+qpZi2GWLCLeqFYLVEtUdxZdcTGaCS64fNxavNaz71KJJ95oKjz/AAK6dK+pUvq3g9vfuabk2zHzDGoiOTlxjtghubGSO5lMpJHD/lUbSNLCSbgxbKrRm+Pwr3YbgxdLVMRzYlR1ckqG9EfEEEEFJ0QQQQCCKDliZDap93nWqLc4cm6QEoXLgtPoU/HSsHgXEA8kBWjrYG9HXlXZuFvi3aBIhTY7UyHJbUy9HfQFtuoUNKSpJ7EEEgg/Gg82/SB6N576PWYZuMJvROA5PCkXqTAkuNCSlmPHImIivvNuFEpMYvgKBC1tr3+UU04akvTeNeEw1ZNivXS3XnIJ8Rq3z8flxzkDzwZCkMtJgtsxHoa2PEXr2UjkolzsSaDu2nGci6dY7AsvTXJpdveTJZtS7PILd1tQkJZQ1GiHxOYVNK2Q9ILCvDjtJd2CUIUvEYJh1vxdz+UFyeZvd/m5Hxk3eXxQEl+ZZ5ocZSSUssJffWpKQdaknfLZUQ6lmxuH6RdxxHF8JizL3js2Hjy8uySG0kW0RmY4E23OvLBC3SYtuUltAUUqU5so2vfo0lCUISlICUpGgkDQAoP2lApQKUClApQKUClApQKUClAqHZB/fK9/mbP+u7QV51AsYZP44YACQAmWAPd5Jc/Z5H6tEkBNd/p3BZZsapaQDIkvOeIr6kLUhKf0ADevion30EkmQ2J8ZbEhpLzKxooWNg1h0wrlYgBCWLhCHYR5CiHGx/iqAOx9Wj2AAG6DsRrq3LcU5FlP2yW4PV1KGgVe/gfNPMBRIHzk77gbIPcetcR+B6mthBjceIbA0B8CNeRHxHcUEaT04iPErlPpcdJ+c1EZTy+tXJKiVH3nYH1Cs/YrKxj1sbgxipTSFOLBWEg7WsrPZIA81HyFB3ykKI2ASPLfuroZF/e/c/8ANXf9Q0Fl0oFKBSg0/wDwobvH0drZHSQHZl+biNFR0Oa4UtI/tNedGMdVLx0Y6h4x1TssVd4sFwt0G23FllXaFcYsNuLIiPf1FLS14zfLzS4kjkUK1D1ulrrdLk01p2i9Zr7t423+DZS3LMT5J31r642j0rOqVk/E9knxpyoYjFMsNqJSlSlqddUlRS002CpSlqUAkBROgKpXqn1Xx/N+pM+Pizjs6HaosW2RJLiwwxLahx2oyXvaOwlYZCwOxIVrtUXhOlzaHQ49PqLRa8R1mO0zMzP8vWW0XvM17P3pdNuVzm3MT5gcSylLYQ3riO5IKSPd/wBor5zqezYMkgqdivPtymlNbZ7kEKB7JPv8jVx4NTtXiBGnvl/2FJcaCG+adJG9EhR8+9V/lc+baUt2mO4uS5KBcLbYPspB+aAfr/21iWUjwGzLsl+6TOvkJkXHM4Tnhg7CUofaHb9qtfsqycxx9jJPTN63QnyfDev1xY0B71zikH9h0ar9fPLprTH31bsPXJCqTZb3En3BdqQly5Nh2PLgubHJaRpS0aPZRSPKszjjd8tdtYvjlgkIhKW4kPtuBzkpPZZ/Z8D9dc5f0d699p7f4WMQzvTPq/iz93vFrv08QoeR21VtcnPsrUzEfD7TzDy0juG/EYQF63pKlEAkAVJb/wBHOsN9hK/FmJTptsjth5NyblIVa5CfMPJfSoxynyPLnoDzIrdbQ29Sckdunn8f9vFcsRM7Sw2fqSqDhWItXOJfpllbfdlP2xfiRnLhIUnxA0ryWltpplsKHYrDhT7JSaivpVYojEcgxyCgjkxALCx7+SVbUT+1RrbhtWNXStO0bx8njJH/ABy92+iX5mMB/V+3/wAM3U0rp1YUoFeeP4Q789Vl/V5n+Jk0G7HQ/wDMr0//AFet/wDDN1NqBSgUoFKBSgUoFKBSgUoFKBSgUoFKBSgVEemNhv2O49Pi5HcDc57t6uktl4vre4RHpz7sVragCODC2kcfJPHiNgCgl1KBVVZN6PFpvOQTbvZcmynBn7ivxblHxe4pisTnPIurbUhaUuEdi43wWrttRIBAVXk3ozK6PPs5D00U9b5MKQ9NbnhDkyahbvEvtTe5duER0oQVJUVSGlJ5NKWOKEW70M642rrTjzq20t27Jbbwau9m8UOKiukbC0KHZ1hwe026n2Vp+BCgA7/XLpZG6x9NbtjjhbamuNl2BJcHZiSlJCCdd+CgVNrA+c24tPko1o83lrULC7rl8i3zn5VmULX1IxJuGZTzc1CS23ckRS+0wQ8UhLylJWjkkLAIQtSgzHT7NLverw6LVYsxuGXeAu3QG3oBYasEYjS24bIbaZjyHBpKpL7cVCQRxS4AUu3DgPoTw57UOR1RuDOSxY4cXDxC3trj2mAHFtrLLntlUwN+DHbQpwJTxjtnwwoboNnLdbIlngMQYEViFCjoDbMaM2G220jySlI0APqFdmgUoFKBSgUoFKBSgUoFKBSgUoFQ7IP75Xv8zZ/13aDqLQlxBSoBSSNFKh2Iqvrxi9zx6Y5KsbkhEVZ5FuMrkpo+R22QUrGuwOiRvWu26DgY6hXS0rQ1cY6HuR4pEptcR1X1jY0s+7QCR9dSe255abg60yt1cGQ4oIS1LTw2onQSFjaCSfIBW/qoMvOtUa4pV4qNLUngXEeyrj8N+8b78TsfEVlLDYrdeYThS25AmRnCw56m6pDYVxCuSW9lACkqSdFJ1v4jdB3V4SCdpvE8EeQKWCN/X+T3/aK+f5GPf87vf/BR/wBlBX3WDNIPRi1NXC7OXm4xnG3HN22Kwso4raRohak9yXQR7tJVsg6Bjdg664Pld4tlqEzInE3F5hpKZFlU20sPOeG0Fq49kOKCk7+AO9edBsZSgUoFKDTL8KvBXP8AR2x9tvYUjK4bnJPmnjHkq3/Zr9teZ+KPZd0zduN5sinIS7r7UxtCeTchPLlweaUChxIOjxUCKzEbsw62U9U8qyi3y7RKu0W2WickJlW+0WyNbWZPckB5MZpsOjZ3pYVo9/OqJk9N7rIuhZY9VKlnklCXQND46PfX7KxszKwLFkdw6eSW0upEl0tJakJc9kK15FJ/R76n2RTWcjxaHeo3EeAoPnkNlAHZY/Z/srMMSg9+y6ZcJottmdakIeSnTo2Cg+/v5V22bAu3odukyWZcx1PgtuEeeuxIHw9wH6T76d2UjjhS+o/RBpB/JxL3AaUNeajNQon96qsO6KcPp2dXUNq1/wCM05xR+CUz9mq7iEb6a7dh/qQieQT2sk6hT7xZ0uwok115bSHBxUlYdKCSAexCkH99QLqB1Cu2Ku3DDYL8RbUWW6uTMabJU48oaUEknsE7I2NbP1VQaXBGfJ6O3hHX4dE3Jfkruri0WB2e29zU3sp9lal74kfWN6PY9v8AtrNWLP7nbrWu0SJCX7TIZEdxLjYUppO+ykHz2nZ0D2q5vSup5qz4dYapn0e2y5LNYkdObzBlraXc2rO2J20DXithIcJHw2PfXN6aUcycnst6QypmPc/HebQsdxy4K1+zeqpdJeb6ql59v0bcsepL296JfmYwH9X7f/DN1NK7BVFKBXnj+EO/PVZf1eZ/iZNBux0P/Mr0/wD1et/8M3U2oFKBSgUoFKBSgUoFKBSgUoFKBSgUoFKBSgUoFKBWuXpIdGLvCmR+q3TCEodQsfBd/F0QhsXdjxUuPR1AD2itIWOJOiV8teIlBAWz0p6vYv1oxdu94vckS20nwpcNwcJUB8fOYkNH2m3EkEFKh7tjY0aqvr/6PNwuGVsdVOm7cWP1BgsLYn2uQkCJksMpAXEkAnjyKUhKVqHuSCRxQpsI56OnpB2KPaPURNdiYhFcTCXb7wvhccNl8uH4unBXf1fkCll9XYa8NR+aa2r8xQKUClApQKUClApQKUClApQKUClAqP3vGpVyufrcWczG2ylpSHoxd3xUogghadfO/soOj/JC6/8AO0P/AOQX/vqfyQuv/O0P/wCQX/vqD5ewq4yWlNO3OA62sFKkLtyiFD4EeNWBufRUXFTSkT4UBaFAqVDt3h+IAd8VDxSCPr8x7iNmgkH8kLr/AM7Q/wD5Bf8AvqzGPWJdlZfL0hMmS+5zWtDfhp7ABICdkjsBvZPcn3aADLUoOhKskaZLVJWqSl5TaWz4Mp1sFKSoj2UqA37R763+4Vwv4zCksOMuOTlIcSUKHr742CNH+nQZWlApQKUFU+kh0UkdeMGg2CNdY1oci3Nq4F6VEMlCwhDieHALQe/ib3v3eXetSMp/BxXV1thif1esdrQre0qsqmy6PgCZfl5eVJtFY6kzs6Mb8FjIusNK4nUuxyY42kOM2Faxsf4wlGo/cvwN92mXSNPjdWYMF9hRVtrHVnlvXY/zry7U3i0dGYndm5H4JC4TifWupNue5J4qIsDiT5a7fzrtXFbPwR95tlouFtR1VhLiy0kBCsfX+T2NHX86rO5u61k/A9zbN4R+U6A+tJ9tZx9aSpO9kD+c9v01mb9+CkvF8meMOpdsitAjhHZx5YSgDWgP5z9XnTc3fcX8FTdmctxa8vdUIPq9iuUa4+rN2FaS6GXUucORknjvjrejr4VlXPwfcef16zjPoHVazPO5VKnPItarfzXH9ZWpekrTIBUpPLzCRvXuqLqaVzY5xWttu3YovvzVrM7PjEfwXlyxqSwuT1Eg3RlqauWGXLItGwtXJbZIkHsSN7+s/Gq9vn4F+bervPnfKzHZMt9x7wxj6jw5KJ1v1nv56qNp9HGnva8W33Zvm54iNnct34HGfa0uoZ6qxPDcSE8Tj69g9xy36z567fsrFPfgUpji1lHVuM2lRJCBjyiEg+7/AIzUjHg9Habb77sXy88bbLmy/wDBxXTJcFt2Mw84tNoQxFbhyZrdjcW/KaQB7BJkjikkbI777DyrC9W/wX926sWLFra/1IhwDY2XG/HTZVuKkFZ2VHcga/RUHDw6MV635t9pme3m92z80TGzeTB8cOHYTj9gMgS1Wq3x4JfCOHilptKOXHZ1vjvWzrfnWbq4RSlArzx/CGjfWqy/q+z/ABMmg3Y6H/mV6f8A6vW/+GbqbUClApQKUClApQKUClApQKUClApQKUClApQKUClApQVfnXQKzZJkisvx6W/hGfpQEDI7QlIXISPJuWyfYlNdh7Lg2APZUg6IsKxN3JqzQ0Xh6LJuqWwmS9CaU0y4seakIUpRSD58So63rZ86CreonoyYvnXUixZ5EU5juUwXkouEy3tp1eoJGnYUxB9l1taQBtQJAA1VusMNxWG2WW0NMtpCENoSEpSkDQAA8gB7qD7pQKUClApQKUClApQKUClApQKUClApQKUClApQKUClApQKUCtXfTI6VZX1HuuLu43Znrq3EZkJeU0tCeBUpGvnKHno1C1mO2XBNaRvPT6o+es3xzFX50+6S5/ZejuHRYMVFqyWx5E9djFmSOCHG/Vn2whRQSCFl1KdHy3vzAqS4h086r4hBFpRkTT9liqKWWgEKkrj+O2ogPL3p4oD/dSeIC0jYPtI24KzTFWtu8Q2Y4mKREsDZumPWO3YZZsfXcYhjItyYs5IfQVOL9WdQVcwhKuQdEZXLuSEr+b3Dky6tdMMny+3ZnFtHgt3S6jdsvSpRQqEwIyG1xeOt6cWlzsPZ/LlZ9pCQZDYzA6aTLPksVyHAhXG3paiJYWHBCEB5uS44+8G2UgbcbW2n2fn+CEr9hRNWlQde5NKft0ptA5LW0tKU/EkGtF+i3QDqBjXVbGLpc8ZkRIESYlx59bjZCEgHvoKJqj4hp8mbNhtjjeInr+8Ov4JrNPpdNqaZr7TaOnt6W/yt+T0b6jC8yHY1wjJisXu63GCHZBcSG5jz6yhxCtghGmFJHu8VYGuINSax4h1QtuMZMFXZn8dXK5Rn0OuKQ44kJjQ2nloUAlAbK2ZGm+AVxWDySfZq8cgxIwrrTNjWOU9krTN1aTLekoU6j1duQqLIaj+whAC20uOtrUlWx7AOiUgV3pOIdWpSbbIcvrTrke4tPKbCGELTD4yw6lAKVhL5Q5HQFFRAIJ5a5FQZ+JiWdTsTuke/XRu5z3ZtvdbjJcS007FabjGSyVIQnRdcTJB2OKuQGkoUQI+x0XyCbecsuHrMa0pl2oR7DGPF42p0mX8xziVNcS605togAniNpbTQWbglnds8KaldnYsTTr4W1DYlKf1ptCVKO/ZSSpKuyfMaWr21qAk9ApQK88fwho31qsv6vs+7/8AqZNBux0P/Mr0/wD1et/8M3U2oFKBSgUoFKBSgUoFKBSgUoFKBSgUoFKBSgUoFKBSgUoFKBSgUoFKBSgUoFKBSgUoFKBSgUoFKBSgUoFKBSgUoFKBSgUoFKBSgUoFKBSgUoFKBSgUoFKBXnj+EOG+tVl/V5n+Jk0G7HQ/8yvT/wDV63/wzdTagV8uoLjS0JWW1KBAWnzSfiKDV3LLlmWFW7rFjcNnMsjurlkjt41NVCmPtyXmrYErUJDKQhtxbqSTxUglau3cio7Av3W235Rf7fJs16dtbGWy5dmeDL7oehKnT9qdc2ClKEiKG2T+TLa0qHLybDCdQ/SG609LsFdlZE9Etj5tSpzNxfsgDjkz8VtviGGC4PYEguoW4NlHFIV86tx8QtZtdjb5yJch+U45MdMx1TikOOqLikJ381CSopSgdkpAHuoM1SgUoFdS8wF3W0ToTct+A5JYcZTLikB1gqSQFoJBAUN7Gwe4FBp1n1r6iQsjz0Y2xnM3JmrtJXDkOPTxa3rQmxoTwSOYYK1ywQkNAOhw7BSATWYmZV1svWfM2STZ7pHxyLeLI83JgwXmuLCZsPxkuPkjmFNKklxKS6OKDzLWuCwg+E3rr9CsN+iXOwZCmJOssGREC0S3XxcEWu1B5gqUvxGWwv1tW0q2t1LoUoEacnF16w9Y8Pta52S+DbbTKdYLtxfs3BdsbVcZLCkNtlY8dwsIjrCT304VBJGk0FzejbeLlmnSbH80vct+Re8kgR5stCipMdCg2EgstbIbSoAKIBOyd7NWlQKUClApQKUClBrz6SUDqbLyKScCdktRv5CX5tzimQptUsrierpa8JaeMogOeEo7I9vQ7mojauo3pAsO36NNx5Me1xJMeO1Katj8mXGietIbMhtBATKWqMVOKSlS1IWO6e3AhgcTzv0g8dwiHETYrnNubNufcaauNpddXJBZluKedfUslt5DyY7aI52VJUNBQVyRnZeZddrffn1OWeTc126NKaRcWbU622tpw2FxTgjpc4OuNJkXQNp7qUYpSDvnyDZDpvOu10we0Sr545ujrRLqpUEQXVe0eKlsBxzw1FPElPLzPkn5oktApQKUClApQKUClApQKUClApQKUClAqAnr309TeZtpVltubuMJ8xXoziylSXQ+iOpA2PaIddbQeO9Fad63QT6lApQY/H8gtuV2WHd7RMauFtlo8RiSydocT8R9Vcou8BV2VaxNjG5pYElUIOp8YNFRSHCjfLiVAjlrWwRQdpSglJJIAA2SfdXRx+/W7K7DbrzaJbVwtVxjtyoktk7Q80tIUhaT8Ckgj9NB36UClApQKUClArA5nndg6e2xi4ZDc2rZEffTFacdCj4jqgSlCQkEkkJUda91B84n1AxvPG3F49eod5Q2yzIUuG6HEht1JU0rY7aUASP0GpBQKUClB1Ltd4Fht7s65zY1uhNa8STLdS02jZCRtSiANkgD6yK/bbc4t4jKfiO+M0l1xkq4lOltrU2saI9ykqH7O3ag7VKDHxMgts+9XC0R5jTtzt7bTkqKk+2yl3l4ZUPgrgvX+SayFBhXMzsjWTfydNxa/HYQ04YQBK0pdS+ptR0OwUIr+if/AFZ+qs1QKUClArzy/CG/npsvf/0fZ/iZNBuv0P8AzK9P/wBXrf8AwzdTagVqPK9KbJMO6n5nEvL8K9Wu1quLyLdALHBiJFkNBbi1pWZDDjcda1rS8yW3VABtxIIKg4Ven64i1wJzvT5cZp+Y5BdEm7hvw32kIcdZSSzxU74T0dbaSUhfNe1I4bPNevTduL11mxLVjVvht2vJWrVKVcrmeao/iy2VhaA2Cw8pcUFAPNJCx7XIFIDgv3pv+o4zb5t+6dQjMcciym2F3oLjNtO29M5sh5yOkCRxUUJQUhPJJPigbI7mR+nkzYXMyaGLQnH7Bckwm0O3wpDrZdkILrhRHWEq/mxIaa8ZY56UEFK+IbQ41e2snxu1XhhJSzcIjUttJ3sJcQFAdwD5H3gfoFZKgUoFKDF5TPetWMXebHITIjQ3nmyobAUlBI7fpFaWdN/S6zi52iIJkpi8zZLNvfZWswwkOv2q6SnEOKi+KkN+JAa02fDfSFEq7EBQZS2enle7JaX5WQY/Y7o4t+ClgWy6lhtLblniznNrW2oFxa3nA0k8UngtJWPCK1bIdKM6kdZMTudyuuOxLbbBdZ1tjR1yvWzJbiynY6nHEltIQStlRCPa0Nd6CwkNpbQlCEhKUjQSkaAHwFftApQKUClApQK1L6i+kdleGekdNxxma07Y4t0tsIW1YiaW1IieI4dcxKUvnrSkIU2kbK9AHQdXI/Tnn2qxWT1rEI1vuF8xj8fITEvRfdh+NCmSow0uKEL2mH7R37JcHsrAJNldLvSMn531Aj41MxpmDCkfjBmLc27kXnXXYQiF0uM+CgISoS06IWrug7ABBoIBefTZfatC75GsERUWE3cpa7dGvTbj6kRrZPlBiWjwCqK9yhAFAKuPMHkrRSc496YEu1ryBu74taYD9kcaalw05GFymSX4ba3ltFhJTHCJoWl3uFeGQQkKCqCPOemTc7xkQdskK2CyC7QYC3ZdxSuO5GXdbxBW+04lsaU4La0tPJRQPEAHkVLt70b+vbfpB4hcbwLWxaJEGaIbsWPNVLQCWWnQfFLTaVHToB4ck7HZagd0FtUoFKBSgUoFKBSgUoFKBSgUoFKBSgVrpcfREjXKNepT95ek3yVkj16grfdcVEgsu3Vma4hDO+IcUhhKCv4k60CQQry8egllc7Er3ZmuojC1zpS5KJEi3krLvgyUNylnkR6wFPtqLgHPbKSHBpITLYvonZTbs1yLJYmX25MmdfYt8jxnYDikKcakreIeX4gcUOLhQAVKA0OPhoAbSEaHoHXNeGOQXMrhLvpdgblqhrWy+wxCMdxhbalH2VOFL48yFtNnzSFCzuoPo43nMsm6Z3RnMXGhibDLMgSWVOLfW26w4ZDaufJt1YZU2o8tlLpBKk8kOBW9l9BzI7S4/wCH1EMYO2FVqQ4xFWHGVGN4HhpUHEqMcqAeLfIe2pWtKIWMrO9Cs3rIrDOnKxpm3sNsNT7Xbrc8yz4LdxXLMdnk6shpSXShSdhJKQQlKTxAfh9DW9peccOUWyUhExqQ5ClW91Ue9lK5SvGuSQ8C88BJRpSde1GbPlpKMTafQayC2QUR/wCXTDqBisbHyhUV1Oi1AZiKbCkOJV6upTJe4ElIWtRKD3JDYnol0+m9LOmNmxe4XJm7SoHjbkx4yY7YSt5biUJQnSQEJWEDiEj2dhKBpInNApQKUClApQKr/rD0vd6qQsYitXV+zptN7Yurj8Vam31JbbdQUtrHzVHxB3+o/Ggo7OfQddu1wUMWyhOP2hHqyGIT7Tkgjw47jZfUsrCi+la/ES4CFBRUQpKiFDrZV6FmTZFcc3knMretGRS0yiy7APAlMx99suI2UOFKHktgupdGmhoAFAaDkvHoY5ReIv4tk5rAmWzmt5bioTzUh11xmA27sh0gJJhKIA7jxfPts2H0x9HCV026u5LlLGQJcstxjOxoltaj+G5HbV4PhtlQOlIYDJS2CDxS4Qnh7fMKzu3oT5POwu12WPl9nhyoLodVOj215Lsx9DLSETXlLec1KUW1KW42EKJc3yPEheQvXoe5Pd7Xera5ldrdhPMqYhIcguElBurs/wDLBS1NrP5UI0pCkHh3QoHQDE3L0G8luq5pnZrbLr49kZtSvWLWUpfLLcAN+KnkoqQHIK1AKUriXyUhJ5c5M16I97TebdNOUQk+BcnZyViI4Xrek3qVcimGrxAGy83ITGd5AgobHmPZoMp089Fm4dO+lOU4jbb/ABIj97xeBZxKYjL4InMxFsPzCjkNqdKkKJ3v2BsnQqKY76EtztL8SQ9k0QzLfKhu22TGjFtdvabvLs51tgDSWwph5UcBCUp0T7ISSmgkXTj0UbvhOD9QrCcmhQ5eTWSJaGbraIbrLqHWGXm1THgp0lbrpdDiyFJJUVd9nkeJn0TbuL1YJ6LzY7S3BejOiLabe+2i1eFPVLX+L+T6vCL6FeA6SOKkpGkAbRQQd/0DL+5brrFRltqbel21NvM1qE6h6UtNtvMP1qSS4rm+td1bdWoaBLSu3cGuPrJ6JmUojrt2IW9m9W2fMnGJA4tNw7GmR6iRISFyGyhxKoz/ALTaHSRIc9nuQsN1aUClArzx/CHfnqsv6vM/xMmg3Y6H/mV6f/q9b/4ZuptQK4Vwo7inlKYaUp5IQ6SgbcSNjSviO57H40H56hGA0I7WgsOa4D54Ggr9OgO9fjtuivpcDkZlwOKC1hbYPNQ1on4kaHf6hQBbYiUhIisgApIAbHYp+af2aGvhX45bIbqXkrisLDygp0KbB8RQ1oq7dyNDz+FB2ddqUClApQfikhaSlQCgRogjzrqxrRAhNpRHhRmEJX4iUtNJSAvWuQ0PPXbfwoP1u0wWUhLcOOgDjoJaSNcfm+73e74VztMNsI4NIS2nZPFA0Nk7J/aST+2g+6UClApQKUClArrKtcJcz1sxGDLI4+OWk89a1rlretdqD8ctUJ5tCFw460oRwSlTSSEpA1oduw+qvtuDGZWlbcdpC08tKSgAjet/v0N/oFB8uWyG8lQXFYWFlSlBTYOypPFRPb3jsfqr7XCjueLyYaV4qA25yQDzSN6SfiO57H4mg4xaoQCgIkf2iCfySe5CioHy9yiVfpJPvrmjxWYjfhsNNso2VcG0hI2TsnQ+JoOSlApQKUClApQKUClApQKUClApQKUClApQKUClApQKUClApQKUClApQK6tzuLNphrkvIfcbRoFMaO4+vuddkNpUo/sHagwv8v7X/gt7+wZ3+5p/L+1/wCC3v7Bnf7mgfy/tf8Agt7+wZ3+5p/L+1/4Le/sGd/uaB/L+1/4Le/sGd/uafy/tf8Agt7+wZ3+5oH8v7X/AILe/sGd/uafy/tf+C3v7Bnf7mgfy/tf+C3v7Bnf7mn8v7X/AILe/sGd/uaB/L+1/wCC3v7Bnf7mn8v7X/gt7+wZ3+5oH8v7X/gt7+wZ3+5p/L+1/wCC3v7Bnf7mgfy/tf8Agt7+wZ3+5rkj5xbpUhplEa8BbighJcsk1CQSdd1KaASPrJAHvoJBXnj+EOG+tVl/V5n+Jk0G7HQ/8yvT/wDV63/wzdTagUoFKBSgUoFKBSgUoFKBSgUoFKBSgUoFKBSgUoFKBSgUoFKBSgUoFKBSgUoFKBSgUoFKBSgUoFKBSgUoFKBSgUoFKBSgUoFQDr5LNu6OZbOadmNTYlvedheozXojq5QQQwgONKSr2nChOt6O+4NBTmeQrnhV2ukC3vX7I7RhmIqu1zErM7lCdkyXivSy8la1qIRb3CGyoJT45IG1k1jMAyV24ZLNh5Nl7NyRY2bdanHJGbzLVMcUxAZelSBCY2l1SnHXASpYJLWt6GyEbb6r5phWMRod8u9zdujNll9RUPSJCgFRnoDiBb1uE6UGrhITxSewQGBrQqU3e+jGtxsS6kyHrWLE1bblkN2vcm4W43Vb7PgfzlanBGWppqYla0EcC+wSnl4YIS7E8wmw+jWU3JhE5UqXdW7TBZk39+6xC++qPFQuHOPhvrjl10HltKkrDnEpAFSLo71NyTObzDTcEWs2mfY03yMiGy6l6Kw9IWmEHHFurDpdZQpRISjSmz84H2Qg1s6lwlYk5mUvIL1dM5iiddJWN2u4r8KC0wHVepSYo220hKUpaLq2/ELhCgdnt9Xf0mLrYWbk3NvWJiFHYmzY+WJYd/FktMaPHW5EZR6wS69zkaCkOEENO6RyQUgPh/0gc9iY45dJkGxLLjN0Yatsdh9LpfhRkNOL8Xxvmm4nwAgIB4uJPPkk8sXaetmQYy/fLlOucO62SwXMMvKIlKVDgOXhFrSCA+vxlhEKW/4jgUragRoKUCEswfrzmmS9TmMYuOOQrcpE1TUuH4rPrLEdUb1hLqh6yXElvxGWVfkVIWoKUlaeSUjF9Ss3ky+rTsOJfJE+OxOgWxNost8egXi2vFxKnH0wOBbnMKS4ha1OHiltC+IOlbDqXT0lbvKxa33+Miz3+JJjpuxtVnfkMyoAahyZaoz74c8NagtmK2s+yE+KrxGuJG+U+kZmsuz3SRHj2KO3brddLwu5ORxKZdjQ2Y54IbjzHElZefUjkHiCG1eyFApoO/dvSSv0C13K5tsWF12FKdtsnHg4oTIDqZrUMSXn1OJZS0VqW4EOeEVN8SHPna+GPSNyh+TicMw7O1JukoIV3bdckMLmBhlxtluWpSUKSl0lxpUkJ4clBKDyoNkK88fwhp/8tVl7/wDo+z/EyaDdjof+ZXp/+r1v/hm6m1ApQKUClApQKUClApQKUClApQKUClApQKUClApQKUClApQKUClApQKUClApQKUClApQKUClApQKUClApQKUClApQKUClApQKKSFDRAI+BoPwoSd9gdjR2POvgxmirl4aOXx4jdB9KaQsaKUq7a0R7qBpAb4BKQjy467fuoP0oBSBoaHkNeVAgDyAHbXYUAJAJIA2fM686jOVdNrBmsqK9d4z8kMNqZ9XRMeaYebKkqLbrSFhDqNpHsrChrY8iQQkxbSR80H9Ir88JA37Ke/n286D94je9DetbpwTy5aHLWt676oAQka0Bsb91fnhp48eKda1rXuoP0oSQRoHfn286FCSUnQ2nsDryoP2vPH8Id+eqy/q8z/ABMmg19xLrN1AhYpZY0bO8lYjswmW22WrvIShCQ2kBKQF6AAGtVlvlw6jfSBlP21J+/QPlw6jfSBlP21J+/T5cOo30gZT9tSfv0D5cOo30gZT9tSfv0+XDqN9IGU/bUn79A+XDqN9IGU/bUn79Plw6jfSBlP21J+/QPlw6jfSBlP21J+/T5cOo30gZT9tSfv0D5cOo30gZT9tSfv0+XDqN9IGU/bUn79A+XDqN9IGU/bUn79Plw6jfSBlP21J+/QPlw6jfSBlP21J+/T5cOo30gZT9tSfv0D5cOo30gZT9tSfv0+XDqN9IGU/bUn79A+XDqN9IGU/bUn79Plw6jfSBlP21J+/QPlw6jfSBlP21J+/Q9cOo2vzgZT9syfv0D5cOo30gZT9tSfv0+XDqN9IGU/bUn79A+XDqN9IGU/bUn79Plw6jfSBlP21J+/QPlw6jfSBlP21J+/T5cOo30gZT9tSfv0D5cOo30gZT9tSfv0+XDqN9IGU/bUn79A+XDqN9IGU/bUn79Plw6jfSBlP21J+/QPlw6jfSBlP21J+/T5cOo30gZT9tSfv0D5cOo30gZT9tSfv0+XDqN9IGU/bUn79A+XDqN9IGU/bUn79Plw6jfSBlP21J+/QPlw6jfSBlP21J+/T5cOo30gZT9tSfv0A9cOo2/zgZT9syfv0PXDqNof+UDKftmT9+gfLh1G+kDKftqT9+ny4dRvpAyn7ak/foHy4dRvpAyn7ak/fp8uHUb6QMp+2pP36B8uHUb6QMp+2pP36fLh1G+kDKftqT9+gfLh1G+kDKftqT9+ny4dRvpAyn7ak/foHy4dRvpAyn7ak/fp8uHUb6QMp+2pP36B8uHUYj84GU/bMn79Plw6jfSBlP21J+/QPlw6jfSBlP21J+/T5cOo30gZT9tSfv0D5cOo30gZT9tSfv0+XDqN9IGU/bUn79A+XDqN9IGU/bUn79Plw6jfSBlP21J+/QPlw6jfSBlP21J+/T5cOo30gZT9tSfv0D5cOo30gZT9tSfv0+XDqN9IGU/bUn79AHXDqMf/AOIGU/bMn79Plw6jfSBlP21J+/QPlw6jfSBlP21J+/Q9cOo2/wA4GU/bMn79A+XDqN9IGU/bUn79Plw6jfSBlP21J+/QPlw6jfSBlP21J+/T5cOo30gZT9tSfv0D5cOo30gZT9tSfv0HXDqMSP8AygZT9syfv0D5cOo30gZT9tSfv0+XDqN9IGU/bUn79A+XDqN9IGU/bUn79Plw6jfSBlP21J+/QPlw6jfSBlP21J+/T5cOo30gZT9tSfv0D5cOo30gZT9tSfv0+XDqN9IGU/bUn79A+XDqMR+cDKftmT9+ny4dRvpAyn7ak/foHy4dRvpAyn7ak/fp8uHUb6QMp+2pP36B8uHUYj84GU/bMn79Plw6jfSBlP21J+/QPlw6jfSBlP21J+/T5cOo30gZT9tSfv0D5cOo30gZT9tSfv0+XDqMR+cDKftmT9+gfLh1G+kDKftqT9+ny4dRvpAyn7ak/foHy4dRvpAyn7ak/fp8uHUYj84GU/bMn79A+XDqN9IGU/bUn79Plw6jfSBlP21J+/QPlw6jfSBlP21J+/T5cOoxH5wMp+2ZP36B8uHUb6QMp+2pP36pLrb1AyjIcriSLpkt3uUhEJDaXZk911aUhxw8QVKJ1sk6+s0H/9k=)
Figure 2. Hedonic brain circuitry. The schematic figure shows the brain regions for causing and coding fundamental pleasure in rodents and humans. (a) Facial ‘liking’ and ‘disliking’ expressions elicited by sweet and bitter taste are similar in rodents and human infants. (b, d) Pleasure causation has been identified in rodents as arising from interlinked subcortical hedonic hotspots, such as in nucleus accumbens and ventral pallidum, where neural activation may increase ‘liking’ expressions to sweetness. Similar pleasure coding and incentive salience networks have also been identified in humans. (c) The so-called ‘pleasure’ electrodes in rodents and humans are unlikely to have elicited true pleasure but perhaps only incentive salience or ‘wanting’. (d) The cortical localization of pleasure coding might reach an apex in various regions of the orbitofrontal cortex, which differentiate subjective pleasantness from valence processing for aspects of the same stimulus, such as a pleasant food.
- Analogous circuits mediating facial expressions of “liking” and “disliking”
Reward
- A reward reinforces (makes more prevalent/probable) some behavior
- Milner and Olds (Milner, 1989) discovered ‘rewarding’ power of electrical self-stimulation
- (Heath, 1963) studied effects in human patients.
Electrical self-stimulation
“Reward” circuitry in the brain
![[[@nestler2006mesolimbic]](http://dx.doi.org/10.1016/j.biopsych.2005.09.018)](data:image/jpeg;base64,/9j/4AAQSkZJRgABAQEB9AH0AAD//gAuSGFuZG1hZGUgU29mdHdhcmUsIEluYy4gSW1hZ2UgQWxjaGVteSB2MS4xMwr/2wCEAAMCAgMCAgMDAwMEAwMEBQgFBQQEBQoHBwYIDAoMDAsKCwsNDhIQDQ4RDgsLEBYQERMUFRUVDA8XGBYUGBIUFRQBAwQEBQQFCQUFCRQNCw0UFBQUFBQUFBQUFBQUFBQUFBQUFBQUFBQUFBQUFBQUFBQUFBQUFBQUFBQUFBQUFBQUFP/AABEIAOYCBQMBIQACEQEDEQH/xACyAAEAAgIDAQEAAAAAAAAAAAAABgcFCAEDBAIJAQEAAgMBAQAAAAAAAAAAAAAAAQQCAwUGBxAAAQMEAQMDAwIEBAMEBwkAAQIDBAAFBhEHEiExCBNBFCJRMmEVI0JxFjOBkSRSoRc0sfAJGENictHhJSZGY2WCksHxEQABAgQEBAUEAgICAgEFAQABABECAyExBEFR8AUSYXGBkaGxwRMi0eEy8QYUI1IVQjNDYnKCstL/2gAMAwEAAhEDEQA/AP1VpREpREpREpREpREpREpREpREpREpREpRFh5uZY/bpLkaXfbbGkNnS2XpbaFpP7gnYryyOR8SisqdeyiytNIG1LcuDKUp/uSqp5TopIo6gE71c8SsyHI1uzCPlMtHb6fFo714WpXYdKfpUOAq7/pB3vtrddafUkp5n3WOKeTHmjsoUbAlpSx8HocdSpO/woA/kA1JhIuoIXYPURKUvpTxJySo/G7QwkHuB5Mga8/P4NdC+ds0uSlJsnBeZyelAPVdJdtt4BPx98kk/voH+1DCbU35+ihwu1vkPmd1bKjw5aG2FJSVhzNEe6hRHcdIiFJ18nr/ALbrzHmnky3MlV14FyBXSdFVmvtsmA/uAp5pZ/8A4isc2/HyRl+EXdM5zzOWFJsXB2ZTXEthW7pKttvbCj4TtUlR/GyEnX+lff8Aj/mp1n3WuILA39qT7MrNul3Z8jSIS07H/wAWt/t3qWqpXS9y/wApWv3lXDgq6SGmySHLLkVvldSfz0uLaVv9gDXoT6gbsW9q4Z5ICh/SIcA7+3fb/jP9O+u9AHZQjvqFnxkqXJ4f5JZaQNqWm2xHiB+QlqUtR/sAT+1e3FvU9xrlVyRa/wDEQsF7Ukq/g+TRXrRMBB0QGpSG1KI+QnegQfBFYksH3n+FPdWpSpRKURKURKURKURKURK1Z5G9d1pxDLp9ls2NO39qC8uM5McnCMhx5OwpCB7a9gEfqOt9+3zWqZMEsOvT8B4HHx2fFKhjEEMIck17Ucd+wPZXfxFy1Z+ZcTRe7QHGOlZakRH9e6w4ADo6+CCCD8/7gR7JOZb3I5CumG4HiDeW3Oyx2X7vLnXQW6HDW8CppgOe06px1SU9ZSlGkpUnatnVZwRQzACDdcXG4SZgcRMw03+UBI6d+xFV13Pn/wDgPCN7zu8YrdLRcrS47Bcx2X9rr85LwYbZZdAKXUOuqQlDqAQQretgpGFu3qf9vg/Hc8s+Npudxu93i2JVikXERzEmOyvpltuO+2rRbc2D9ncDwN1k4s+9+GipEEM6kPHPMl3yPPb1hWW4o3imQ2y2sXb/AIS6JuMR2O4tbe/dDbZQsKbO0qQPtII7eMdwD6lbdzvJyVpq0uWMW1wSICpD/X/EbYtbiGJyfsT0JcLLn2/doAbV30JAJy3vYNFClFp5943v0S9ybdm9kmsWRhyVPWzMQr2GG99bx0e7Y6VDrG07BAO68LnqY4qbtkW4qz2yCFKkuxGXzKHStxopDh/+BPWglz9AC0kq0oEwGNiPMKSGuu2Hzri8ibepLt6ssfGLdZIV8/jC7holiQ5IQFrbKAEN/wDD/YrrJWSodKekFUlwfkPG+SrW9cMZvEa7xY76oz5ZJC2HUgEocQQFIVopOlAHSgfBBqWe29sjKRUqESlESlESlESlESlESlESlESlESlESlEXw+83GZW664hpptJUtxZ6UpAGySfgaqp776sOJrBMeiKzKLdJTBKXGrGw9cylY7Fs/TIcAXsfoP3ftUURY+T6p7O/HWuyYJyRkLg2ltuPhs+KHFAeOuU20lPfttRFeNSOeuTlEE2XhuyLUodI6LzeynQA7g/Stb2o7HveB42azIbe9+IWN1kjwTmak6PP3IG+/cQ7EN9z/wDp37/9K4d9NDOQKT/jLkXO81YSpSvpJd2RbWFbGtKbt7UYKA8je/8Aao5ooh9x8gAs6LNWv0ycR2eC1Ej8aYqtpvelSbQw+4SfJUtxKlKJ/JJNexv088VtLQtHGeHIWj9Kk2CKCn+38vt5P+9Ycop0WLBTe22uHZ4bcS3xGIMVsdKGIzSW0IAGtBKQAK9NSzBgpulKlEpREpREpRErC5XhWPZ3bTb8ksdtv8HufprnERIbB1rYSsEA/vUgmEuEVVteli1Ym04eOsuynjpwpCW4tvuBmwE61ofSyg6gDXbSOjQ8EV5pV/5x4rSHrtaLRy5YG1KL0nHmzbLy03rfV9K4tTT5GtdKHEqOxpJI7zQsN79+l0O977KyuNOUsc5dxz+M43NXIYbeXGkxpDK2JMN9B0tl9lYC23EnylQB8HwQaltYUIonZKVKJSiJSiJSiJX5dcjenbNcIy+5WkY/dbnAL7piTYkZyQ3IZKlFCtpSodXSfuSe41sjWjVHFSjNAAr0Z33bxX03/B+J4fAz50qfGIecAgkgD7XpXu/gtxvRtxRe+M+PZr2QR1QJ93kJfTAWCFsNJTpPWNnpUdqPT/SNA9wdeLKsNy/AuQOSJdrw6553iGftsPyEY9dY9vultlojJirCVvvMDoUhttaVoc6kKCuw7E2YISJYh3tivH8dxUvHcTnz5H8TFTsKZas6gGL+nnkvIE2Ow5E7ebHicjLJOVOLdvzd1n2lmOwym2wi7I90OLMgKkKAS42noH3lSq8XJ/pmzqFacwxS02p/kTELpktmy5CbhMhMPvvokA3KO4nTKP5iWW1jQCP5ih56t7ySCWPbfs7U0crhFg7Z72LLqTwDn1qVnMvjXjtjiSFe8eRYTZ2bxHeVKkvSW0qnlCFqab+njKkEaV1qKgAD5qVZt6Tb9jdxsIwK9XG4wpWNTcDuX8UdjJNvtTkZz6Rxr222ioMP9B19yylatd9moYPy5eLN7imlslLkm+9+tVD3PTnn+R8fPWaRiNwgXKz4RcrBDcumQxX2HZD8ZDKWYbbIHSyotglT5QRpP2kjqEs5JnJ4Y5itd5RHx+euTx8bO9Z7zdGrcISGnUqL6XHdNrYP2odQglY9tohKh4ka0BLeFtTr65tQgGZst6eG3Vb8Z+nDNsn4kstzh29PS7hmGPwIlwkJYEyRb5smY9EdBClNpWh5GiodlKG/0kDb7i85Tcrtk96yTE7XiDdwfYEOIypt25OobaCFOTXmlKbWd6CEpJ6UJAJJOhi1Ax9vCw0YXval8RCANd3ubqwaUUpSiJSiJSiJSiJSiJSiJSiJSiJVecl8/YPxPIZgXu8B+/SBuLj1qZXNukrt/wCzitBThHj7ukJ7jZG6kAxFhdQSyiMfKecuRVfU2HG8f4xsxT/KVmaF3O5PggfeYsV9ttnX/Kp5Sj8hNeiTxrzReOoy+aINn6lD7bBiDCOhP4SZLz/f9yD/AGoQwLmv9dfCmruhXyj0r2PIpKZPIuTZLyo8ggojZJLQ3AQQrf8A3KK2zHX4T/mNrP2jWqtuxY9asWtjVtstsh2i3s9m4kCOhhpH9kJAA/2qHUrIUoiUoiUoiUoiUoiUoiUoiUoiUoiUoioHm7j26cdXqZzJxww5/iOG0FZDjrCf5GRwkfrBQCNSm0bU26O56ehXUCOm58Qy2057itpyOxTW7hZ7rFblxJLR2lxtaQpJ/wBj3Hwe1Tl+yf6Hj2AZDUrL0qESlESlESlESq9yrnzB8OvblpuF4KpzOg+3EjOyPYJ+FltJAV/7v6hsdu43hFEIA8S0zZsEkPGVMcfyG25XZ410tExqfAkJ6m3mj2P7EeQR4IOiD5qK5/zhhfGd1i2q+XR/+LyWFSW7ZbLdJuMv2UkJLqmYzbi0t7IT1qSEk9gd1IPNCIoajotsJEYBhqu+xczYXlETFpNpv8e4MZO+9FtS2ELUH3mmnHXWz9v8tSEMu9SXOkgoKT93apXdLnEslsl3G4SWoUCGyuRIkvrCG2W0JKlLUo9gkAEknxqpcM6llxarpEvlrh3GA+iVBmMokR32ztLja0hSVD9iCD/rXqqUulRvNJOKxv4KMobtzhk3BuJbRcGEudUtYPQlvYOlkJV3HwDR2R2UkpRFjJ+Qxbde7VanUSlSrkHfYU1FccaT7aQpXuOJSUt9j26yOo9hs1k6IlKIlKIlKIlKIuE7KRvQVruB8GuaFClKIlKIsDmueY7xxYH71lF6hWK1Ma65U54No2SAAN+SSQABskkaqomuXeRuYHEo4vxVvH8dX/8AjDNozrSHUfcOqLABQ854BCnC2k7HY+KEIV7HvTdcswSFcgco5dkwWEh62WqULLb1AEEp9uKEukHQ31OqP4I3U7474XwXiZh1vEMVtdhW9/nSIkZKX3z+XHNda/HyTWRIdxvfm1yVLtZTSlYqEpREpREpREpREpREpREpREpREpREpREpRFwpIWkpIBBGiD81QnCENPFHMmecVMkpx9bDOWY8yo7+nYkOONy46dnfQ2+gLSPgSNfFGJtsZ59BrZL0V+UoiUoiUoiUoiV+bd/ttzteSXK3XsuO3eNKWmW3363HVKKlqGt/rB6wr56v/hIoYwtAIvzo+XbycZrhcUBIgOVfhbWekO3XGNg90mSlLMGbNDsXqBCVkISlxad9+6gQT8lB7nzUOn5nD4Z555mdyS8QMSumWRIEnGMmyCMpVvcQzE9n6ZSkqQFFp/rWW+tKlB4aNbsPDF9KEmpGvo+fSlu1R0cIP+CARU3+FAsOv95za9YbcF220+07nV/Nuu2NWeRbIt7ScckBUxDTzritrfLqOsKIV7QIPmoxPzz/ALV+H+OMUjXu6X27M8XX1q/w4776nk3BMCN7TUrWlF0uNPaSognRBBSvvbJzeur+t3Hp0Y1Ntnq+9/Luywl/zOFc8AttusV8uENUDjuEuzPS8gnpS/NWhwOfw6PG04++l1soUXXVBBCQEBCVbyGYs3/MuOOXs2evV/k5FYsdxa6WN+PcHmkRZblvZXIfabQoJKl7IV2IIUoa7neJBhqa+Af8XFHyZ9FkT4fG96nOciTpXEmV59i8IX0YZM/ww+vquslqNDfkOSBKefkaccQw57TZdDeiok9wV7qK2t8SOtWQy3XsQsPJdkkw5Md2e3b4sJ+EoOrYLzinAx7iQAonp2T0hCVbrKIMCIcnzrdvDJrauKKLtv3eyy/F0rO5vKVpcvOSCJyM5lkhF8gxo1ycnfRB2QC28lTqYzcAMe2ppwIPTtsp2tR6vRxxiLNr4N4RyHNRkT2NZA445nT0ubMfWtxth9MBMhsLUUNJUeg9CRspZ6t6FIjykDelql2yqK0rcQ9vnf76LHY1cMsuUCarjuXkM2azeM6YsS7gt1TyXE2xoQUue4d9ldPR7nfet9ya2J9NTuAIyRDfHzOUvIdsTS7zImrf+hRKCwAJAfIUZ6tOdXQNhKfv1tG8AAAzv1b2DZ50DZBSQ++vf0Wx9KlQlKIlKIlKIlKIlKIoVyhzDi3EFqjy8iuBRJluexAtcRBfnXB7RIajsJ2txZ0ewHb5IFV6q+84crBBsdntvEFgdH/fsiCbleiNH7kxG1ew0fH+Y6sjXdHfQkB9/Dj9eQMXWZwz0w4jjmQs5NfXblyBmDSQEX7LJP1jrB13+nb0Go4J2dNIT51vQGrerEAAU3v9WUlskpUolKIlKIlKIlKIlKIlKIsLeM3xzH0vG6ZBa7aGUlThmTW2vbA8lXUoaA/eoDd/Vhw1ZF9Enk7GFOaBCI1ybkKVvWtBsqJJ32A7moJYgZmgRY0+rjA58gsWCNlWWrSVBS7Fi1wkNJ0dbDvshCgSCAUqPcUjerjAo9xYhZKm+4CuSB9O/mFnftkd8nQ6UvOJ9vez4Kh+Rsd6yY6b257BMnVxQrhFuTIdiSWZTRAIWy4FpII2DsH5FeioRKURKURKURCdDsN/tVEc/tqw3lTh7kJkBLca8qxe5K6ynqi3IJab2PkJkojH9tk/2EPvddFBIzV70opSlESlESlESsBkthxe5vRncgt9olvA+2wu5sNLVvzpBWP3PYfmpEPOeUB1hHDDFC0bN1WdbbS02lCEpQhICUpSNAAfAFdEmRDWr6d91gqUUj2nFDZJPbsfyR2rFgzLMdF6a4ShKAekAbOzoa2fzUpdCkKI2ASO438GuQkJSANADsAPiiGq633WWkD3loQlZDY9wgBRJ0B3+Se2q7KdFJBZKiPIvGFo5NiW9u4v3GBLtz5kwrjaZjkSVGcKChRQ4g70UqIIOwfx2FSCxdQvRguGY9xrZ28dsLYjN9bsxTT0hT0h5bjhU684taitalLV3UonufNSasXdLpSpRKURKURKURKURQTkznLBeIG2BlORRoM2SemLbGUrkz5R8aZjNBTrncgfak+RvzUCTyNzByn9uEYPHwCyOp+3Ic+2ZRSf62bawvr3o70+41ojuk+Klt6W+C4HqEyUn4v9Plh46vEjJZsydmOeTEdMvK7+4Hpakne2mQAERmRs6aaSlIB79R71aNCXpv0UMlKhSlKIlKIlKIlKIlKIlKIla85o9d+febrrx7bciu2PYHicJpWSv2N4RpNxuD/Q4zB98ArbbSx/Mc9spUfeQnqHep5XRnopTY/TBwjiy2bdD43w4S1NlaRLtjEiU6EnutS3QpxZ3ralEnxs+Km0FGF4fGeMIWGyR2Xgy6WAzHSh34QrWgFfse9A5Lw+nlkjPRZy2XiBe45ft06NPZBKS5GdS4kEeRtJIpdbRBvtveg3KFHuEJ5JQ7GltJdbcHyFJUCCP2NQRkjKpJvo44XmP++zx/bLS/oD3bIp22r2DsHcdbZ6gR2V5H5rE3Tg7KuKJSsg4hyK4y1oQBLwvLLvJn2+4pST/kvvrcdhvaUdKSS2SE9aD3NKB2DeHxav7vVC5DPv3U84h5ktvLkC4obttxx7IbO6mNd8fvDHtSre8obSFa2laVAbS4gqSoeD5qwKkhiyXslKhEpREA0kDeyPk1RXreaU16Y8xurPtCTYRFvrKnUFQSuHJakAgDvv+URv96b26CmavRC0uISpCgpKhsKB2CPzXNCGoiUoiUoiUoiV+TfI+XXTPM5u93yH3F3NUtaVRpKifpkpc6RHQlWygNklHR233UNkqB9d/jsqCKbMmRCsIp4/1lXqzrh8VmRQwQww5/G81uV6E8uvGQ8fXi2XKY5Oi2iUhmGt4EqbQpJJb2RvQI2ATsBWiBoAY+zcRYNyt6rucmswxS0ZG5FjWH6V24Q0uuxwqI51Fpw/c2eyTtOjsAg7Hbi8SlQysZNlwgMDlYU0O30XRwkRjkwRG7KK8Bcq55kaMd42sF8jwYYk5G5Bym8tKuj8q1wJzceMlvbiAtW3ikrWT9rIOtqBqRYFzlyTzHktjsEG9Y9isk2q8uT5TMFUhD0qDcBDDkdDjnV7CyeohX3BIP3bII5n3E/cwOnk+fowOVKkXAG3u9/yoqr1b55mmGM3PH5UGNdoeLqu0222uxOzgmUHXkBch911pmPGWI5Uge4XFJUo+Ep6uL36mOUrxjnIuW2i62W027DsbseRN2pduL31i5cJEh+O64VdSWwCrpKdLBKST0jS4IYc0X90O/lQK7p519f7yGd8pZnY8jnYfksqzZiIt3w+5RZc20NpVGTPuKmXEBodSQppTXU273Unt1FX6q9GI8+cki14HnF2vUK5WfIcznYi9jTNuQ2GmmnpqESEPJJWp8GGNj9BCtBCSOqjVYm3Yi4Gjh8ntmykQvUex30/KxHGnqe5UyC14jklzhuqtGXwpUnpkQ4UaFB/4R6RHMB36gvSSj2whxDjZUoErAQEkGRcecjcrZ2eJrRIzyDbZed4w5mTlzasrBMZCGoZFvYQVFK9qlFalq+7oQoJ0dKENS9f16V71DPdRcab3qoe16l8ttuMrzmUxY7vkkbC7q4mc3bw2yHEX9MNt1S0kuCMEBLqmwvw2T3PetoOMnMisuU3rGskzy35rIYhRbgyEwERJ0dLq3kkupaPtlpRa/lkAK+1wKKtA1mXzL+nv4M/Xo0Ehg29/tWVSsVKUoiUoiVwTogAE7/6URRXkLlbEOKLULhl2QwbDFV/lmW5pbp2B0toG1LV3/SkEnv27Gqnl8mckc4lqFxnZZWC449ovZvlMEof6QRv6OAshS1dj9z4Sjv+lY3UgPQGvh7buMqqL2U94p4ExPiNDkq3R37tkkof8dk15dMq5zVfJW8ruE9+yEdKAOwSKsahLqTVKVCJSiJSiJSiJSiJSiJSiJSiJVBeml1dlz3mzGrz1JyxOWu3p5a1ApkwJTSBBdbHwhLTIZI76Wwrv3oz2G97uirP1EO5lH9cHGDuCRrNNyEYxPDTF+ecai9HWvrJU2CvevGgRvW9ea1vzxm8zsF5kGR2+3ysgXyrajPt8FazCcdU2pCmkKcSSUK3ruP6u4I8mgP23OY7NdwzN0NKClVB3vfXNX1w9YL36e+aLrfrnxu5isbLLcIluwHA0fxJIMctl2Y85/LZaCfcQkDY2X1aSNKUu9p3qpsGNvxlZdi2X4NaX1hoXvILUG4LSyNpS4624v29+NrATvyRUFoQN+23e91LVbf6Vxwpsa5w2ZUOQ1KivJC2n2FhaFpPgpUOxH7iu6siCCxS680e1w4kyXLYiMMy5ZSZD7bSUreKU9KStQG1aSABvwO1emsWRKVKJSiJVfeobHV5dwJyPZWw4p6fjtwjthk6WVqjrCdH871RgaFQV7uFMhOX8NYFfVLW4q6WCBNK3P1KLkdtez+/3Vi+a+W3uK7XZGbVY3MoyjILgLZaLO3ITHDz3trcUpbigehtCG1KUrRPgAEmkJMban39fnxUtooKnDvUfOiSbm9yFhcKfJZWlOON2B5yHGUUkApmB5LxIJB2UEbHjXavrH/Tzn9kxu0mNznl8LI0wWm7iqSIt1hPSQhHuKbRJYK0oKwsjSkq0rWxrVSGat/JvUj9eajw36b9Pfx5zZeceyuLx5y2zFs2YSFFNovUVJRbMgQCQCyok+1I0PujqPV8p6h4vChrltgflSlKhEqkuQ/SHgXIuRyL3IFxtMyWorli2PpQiSo76lKStKgCrf3FOt62e/er2Exk3BTPqSjVaJ0iXPh5ZgdWbguBWPjbHGLHj8JMKA0VL6eoqU4tX6lrUe6lE+Sf/CofmHpm41z3LZuTXzG/rL3NQ03JkonSWQ8ltPSgLQ24lKgB27jwTvyaqzYzOjMcYBJL2W6ECACGGwWYvvCODZFZLJaZOORY8GxhSLYi3FcJcJKkdC0MrYUhSEKT2UkEJUNbB1UFyP0o4pkXIOK3JdvhR8TsFhlWePYoqHI5bU6804FtuNqSUp6W1pUn+oOEHYJBwpV1JqGUqu/py40vsiM7Mw22rDEBu1iO0hTTDkRvftsOMoIQ42je0pWlQT8ar1RuBsBiY/erI1jUZNqvVujWq4RitwpkxY7PssNqJVvSW/t35I8k1iQCXUMF77zxFh2Q3V25XGwRZc5wwip9zq2TDdL0U+e3tuEqH763vQrth8WYlAtlst8fH4TcK2XNy8w2Pb2liatbi1PpB8LKnnTv4Ku2u2szESXdTve6LFWXgLjrHb3Ju9tw20RLjIS8hTzccfYHjt4Np8N9f9XQB1aG96FRjlzgReX2PD7XjbeMx7TjiDHYsOQ2VM2AWQ0G2+gpKHmlNpT0jocCVJUQoKGtYkkCjP1RZXi/084vx3gFtxuVDiZC9Htsm1yZ8uE2j6mPIfU++z7aR0pZU4s6aGwAAO+t1K8K4zxXjluYnGbBBspmFCpK4rISt7oBCAtXlQSCQkE6GzrW6hoR/ENvfjVHdSalSiUoiUoi8V5vVvx22v3C6To1tgR0FbsmW6lpttIGySpRAA0KohzmzLuc5bts4agoh48Flp/ka8xyYSQNgqgR1aMxQKSOraWu++o9gZbXfXf5aCVLuN/TjimA3hWRTDKzHOHB/PyjIXBJmb8kNbHRHRs9kNJSO/ffmrWqN/PypcGy4UtKNbUBs6Gz5Nc0ZEpREpRFWHJHqOwfjS6oscm4PXzLHR/IxiwMKn3NzsTv2G9qSnt+pWh+9YaJzDydcmBIj8F3dlhZJaTPyC3MvFP9JUgOK6T+U7Oh+T2qbim/37Z1oovZenOufbjx1l4tt04yzC42NUFEz+P2CELiw0rv1tuIbPWkpI12Cidg66dkZjAPUXxtyhL+hx3MLbLugB67W857ExsgbIUw50rBA/b434qGJYb6ef6UsrHpREpREpREpREqpeaeFJma3S15phl2Ri/JtiaU1b7s4kqjy45V1LgzED/MjrPfwVNqPWjuCCvR0Xj4oy3FuY8mF2veMx7Dy3iKFQLjbZgC5ds9wHu05oe4w6klSHB2IV8HYEwncJYHc03QS8TtckXS5NXeaHo4V9RMb17b6t+Vp0NGpLt336HMZ1Cl1DfU3yrdeObBaYNjKo9zvDrqPrQ2FmO02kFZSCCOslSQAQfKvkbGu2D865Lab/HN6usq/WSQv2p0C5LL7bjSigLIC9+AdjXbRPbSjrlzp8cqdDy28ffXTrnkvP4vFRy54EJpDlrr6eSsrhbJMa4l53m8f4zcoknj7MY6rrjbMF9Lke2zmU9MyGgJ2EIWkJeQnY7h3Q8VtFKQ4uOsNKKXNbSR+R8f6+KtzoI4pMUEssWIDNSlGy09yS7rvS44I2iBcLATJs+BoPOqHgDZACj2AG+nWzrXnyv+1dca9OmQkvSXfaRtSuhIUSB+RraQe47j4I7ECvjJ43xTB44SMXMP2xDmhActegDkuHI6Lt/RlRwPABu2az8e4xpadtvIV2PYnR7ef/P716a+x4XFScbKE6RE8J3vzsuPFCYSxQgnWiP9aAaSBvZHyatrBkqOcj5NaML4/wAjv1/e9iyW63vyZrgJBDKWyV6133oHxSmdlKh/pXslwxv00cWWq6sqjXCHjNvZejuDSmVCOgdB/dI0D/ao2ws5z6zZQJW9AwHFGwlJUPbanXJ5RJ156xHiAb+Eu+fv7iHvXr8oe6velEUezzj3HOUMbfsGVWeLe7Q8QpUaUnfStP6VoUNKQsfCkkKHwRVQN8b8rcKDrwHJFcjYwyARieZSiJzafBTGuWidAa6UPpUO36x5oahFNuJuese5Xel2pLcrHcxtoAueK3pv2J8Q/wDMEns60fh1G0EEdxvVWVS6JX52ct8s5BmmZ3Z+Rc5KIbEt2NHgsyFNtxmg4UghKVD7iE/cojf42BqrEiGCInmy3Z9lnIDr33+IYSViMTMmTIeblAZwDU51cZNXVbNeknkK85ril0g3mU5PdtbjQZlPK63FNuJUelS9/f0qSobPfRH7Gq15H5xm2n1DcjYxe+a3eMrRaoVscs0Rm1w5RkLdZWt7QdYcWshQT9oIP3AD9tc6HljIem+n4+D5vjeGgwuPnSZVIQfJwC3r+1msQ9XN6tmDYVAyfGzP5DuNnXeLhCcmxrQhmL9Qtph1X1C0hLjyEdaWh3GlbKdDefZ9YCMmW+7hWC3bKbdFxyLlEmV9S1FKIj3vAISlRPW+DHcHt7APSfu7DeoknZuztm3qaaVXFIIUMv3qivV/b5Inw/rIGGW+JjE6z3a1qY+pCJz7R+5KwrfupcIUCPtS2sbClBVZLkn1NZHeRjj+J4/crfiknkK34ynKUPsLTLCLgmPLSpggqQwsofaS73JKR2T1JNNej7/PoTR5DGm97Oay+Oet3HshvdubFraasl2uTtqts1F3jPS3HkqWlC3YSSXWW3C2QlR3+pOwOpO+/E/Vnds3s2Gv2vjiULnmTa5VigyruwgPxGmUrkSHVpSr2UoUpCAkgqUVpOk99SQQWO8v7zFNVAqHCxrnq9tjb1uyK8Wu+WCJDteVOTbMp5lYDtrlxo60rASSp0rUUt6WlI61b6tgpuLj3O8pyW6yoGT4FLxFaIrUtiT9c3MjuhZIU0VoCeh5BH3I0Ro7CjSpq1O+/wA5kBwoJy3vrbRT2lFKUoiUoijnIPIuN8V4tJyLK7sxZrPHKUKkPBSipaiEoQhCQVLWokAISConwDVSsZPzNzT1u41bYvEOJuJAaueTQzLvkgEb60QgsNxx8D3VrV8lA8EQWpv1/PVSFkrV6SsKfnMXPNnrrypeWSFty81lfWtMq3vbUXSY7XfX6W99h3J71c8eO1EjtMMNoZZaSENtNpCUoSBoAAdgAO2qZMN76qF+Ttr4ZyHlC9czvYLx1Kn523yZc2LfyDHvKYTVlQmalxbSm+sKWQkk/ak9IeSQdoCam+T8SK5A9YHNkWXw1b+ZpkH+BB6TNv4tKoaja46S4NnqcC1IV2+OgDf3HWRhhLO3ancm/wAPrRQWNt0bTdA5Vj2zi/EOb/U/yhiHJ1vLePYVZbbExjHpFwW3HiwzHHuym9FOylSB95J0F6Pitl/TOvGlcLY+1iGZXLPcdjh1iJe7s4XJDiUOKSUFRQgkIIKBsdgkDwBWA5BSjn1/LWDNR3D1Ukk737DyZWjSpRRDlnk60cPYHcsovPuvMRQltiHGT1SJshaghmOyn+pxxakoSPyfxuqqY475g5pZbk59lP8A2Y2F5KVjFcIkFU/RT3RJuSkjvvsUsNp/ZZ81MLX071/Hd39wIVq8ccRYdxHbHYWI4/CsyH1dcl9lvb8pf/O86drdV+6yTUvqHcAb9ENUqG8jcN4Py5bzDzDFrXkDYT0odlxwX2e4O23RpbZ7DuhQP706IqtXifLXAy1v4lcpHLuFN7WrHMgmBN8ip2naYs1X2yAB1abf6VeB7p3Vs8X8l2Tl7CYGT2Bx4wpXWhceU37ciK8hRQ6w8jZ6HELBSpPwR2JGiY8d57fNQNFK6VKlKURKURKURVfzHxHguYKayfI5knGLvbY5js5Ra7mu2TI7JV1e376FDqR1d+hfUnZOh3NazTeasxwm+rhYhyjNyzHEKI97MLSx7w7aCWpDfQtYH6iVtE67j81XnzRKhd6l2rSg6+Ay0dzWjisTDIhYfyNt6Z9VhI+E8i8u3KVehb7hk0d9hAYnzm/aIH2lam1KcS2UdhpKBs6J0lRAFvcP+lWWuc1eM2bSw0251Jsh6XC723t1aVEa2ojpG96PcAkGgMMZ0XPGKak1OlMgAwZhpZweXKwcc6YJsYLUd8/DQ+FK3LmSc8+nvHkcZ5NeMUjQscyO3QnZ8SS86tEMOMj3AHU9QCUfZoqT0lIOwRqonxjnXJGL8a4xmrCJWY4Vd4DFwciXDrE+3NrR1LCiodRCRodf3JUE76WwrYumSZQEckZ1FWqelHu1O9KLoRyPof8AJJ6uNe2nhqtg8D5Bx7lfHhcrNJTKYI6Hozug6woj9LiATo68HuCO4JB3Wft9tTby6EKKgsg9wN9vyfntof2SKpx4GTPxcrHCkUIItcENXOn5C6UudzyvtqDXsvqVbY8oLJbSlxQ/zEjR7dx3HfzXjC5VqUPdcMqOonvr709+wA8ntvfc/sAOw5WMwkXDZv8Av4GFh/8AUgFARnE3/YXpU+YNqCL6o+nGex3ksjGkty2UuNrSsED9J3rtuu2vSyZsE+XDNllxEHHiqxhMJaK6KBKexG/jdUB6lnhyJkeCcOMEyDkc5F0v7TZICLNEWlx4L0eyXnQ0xo+Q4vX6TW8ByANtUqFfj7zcZlbrriGmm0lS3FnpSkAbJJ+BqqJ9Lso5rcuUeRkul+35Tkq0Wl9KSG37fDYbisuo2kbStTbpB77BHc0CdFfNKhEpRFAOVeD8W5gZgu3mO/DvdsUXLXkNqd+muVtWfKmHwNp2OxB2kg9warhvk/kH0/BuHyhbl5jhTKVBvkHH46lvxW0+DcoSQVJ0nup9nqQdbKEbqQDEwF97/KZK8cYyqzZtYol6x+6w71aJaOtibBeS804PyFJJH7ftVOck+kyz5zkr15t95kWJ2U770mOGA80pzttSB1J6Se5IPUNnYAO95wTIpbmA3HXzoQu1wnikfCp/1YBzAhiHv412+qsjjDi2zcT4+q12hCll1wvSJbwHuvuHypRA8edAdhv+5Pmxni5nGuV83zdM9T72TxrdGXDLISI4iIdSCF7JV1e7vWhrp+fiIouYkhc7F4mbjZ8WImn7oi5/A8KdlD+XvTXF5Lz2HmMG5wrVfG7abTINysjF0YdYDhcbUlt3XtutqW50rBP6yClQqt4Xpnyu3coZDZ7Bk9zxrEXcJtVhVdnIrMpy49DswP8AnQbeSlxJCwkBPuq0k7HTi5Nt0/rx8xWAau999VLLv6QLfKavdrteSP2nFbpbbLANnENDpZVbHW1x3EOlQI2lvpUCO5PVvYrsl+k5TsuJAi51c4OFwcqby6HjjMRnpalCWJbjSnv1KYU8XVhsj7VObJUEgUv/AC3sIu7EfSynEpMKFCzC5x8Rts2RNgWNqMy06lTilkNPSk/zHo6S4opbOj+kLUrWqjOacG3rjTC+H7bhLN+utwwxt+3JyCyqh/XMMuMgKC4kkpZfZcUhHUPcSpBS2odQCqczAMPCn9e9XPRDXvsLx8WekU37j+1J5DduKJj0fKI8u2SJLbshbF4ntydvut/YJCAy2SWh0dZOthIJurjri674jeZF3v8And7zO4KiIgMpnBuPGZYSQer2WkhKnlEbU6rufACR2qDTfYatYf0jurDpREpREpRFRHJrJ5A9TvGOIOJbVa8aiSs0mNOqP855P/CQwlI8lDjrjm/goT+avepIZSS6UqFCjGD8aY3xsb9/hy2ptv8AHbo/ergEuuLD8x4guu6Uo9O9D7U6A14rGScewnia55hyA6yzaJ17Edy8XBbyz9QWW/aZAQVdIIT20kDZPfZqDFyh9PgfjVRFEADFFkqGzS8en/1JZPbk5thshc1BEdi8T0CN9uzptbrD3X0bWRpwdIK1eO5raPH8dtmJ2SHZ7NAj2y1w2w1HiRWwhtpI+AB/v+5O61yp8M+Hmh6eNKHyoD4ZKvInwYiHmgy1CyFcKUEpJJAAGyT8VtZ1ZZ1rhgrJ9T3LTPIkrTvGmISHY+JMdR9u6z0koeuhH6VNo7tMn8+6rt9pOyFQKh/hvPr/AEhCUqUSlEStc8ybV6aOYl51HBRxrmstqNkzKNBu03NWm2bl5AS27pDTx/5uhZ8qolclsZSiJSiJSiL4febjMrddcQ002kqW4s9KUgDZJPwNVSeX+paK7dHMf4+tjma387SHIp3FaPjZUO6gO52NJ7H7xqtUyZDLYG5sN5anTrRVZ876ULCsRsFWHJtgulpMS68k3deUZNMcP8HxGA6sMIWQUJUvpI6UbKQooG1FXT1HyLF4h9OEW1lm/wCZNNXO9LQktWxSE/RwdHqHS0B0dQPfsNA+BslRrwynmP4nNzpkwh9Xs4cUJeH5533lzDU6PkAC9KOfB3V81GuSMRlZ5g92sMHILji02Y0Es3i0udEmKsKCkqSfxtIBHbaSRsb3VwhwuyqfvHpayLL4MSwZbzBkOWYQp9p652C4QIiFXFKNK9hchlCHPZUoDqQeraQE7HcnYNplDDSGmkJbbQkJShA0EgDQAHwKkgb3vNFQ/J3G1z44vb3IfHyER3mkqcutmbSQ1KQNbWEpB8DZKQPP3D7gQq1ePM8t3JGJw73bXAW3U9LrR/Uy6AOpB/tvz4III7Gq8EPJHFABS49jkAK5ePalKH0pkUvI1Hz61UloQTrRH+tWFcPRYmXAXBX78QlDQB9xlI/UNjx57AAjQ157EVkYslExhLjZ7HY0dbBHYg6+a83gAMBipmAtAXjg0Z/uA0Ymg0r0FiMc8AmZ2/C7CQCB379vFUTwgpnMuc+aMy6WpH0lwi4pDloX19LUWOl51Ce50Pekq3ryU/kdvSs4dt+X4WghYzkKTcfUryFd+MLS+5A48x55pvM7oytTbtxeUgOC0skeEKbUhT6wQQlaUD9ZrYK222JZ7dFgQIzUODFaQwxGYQENtNpASlCUjsEgAAAeAKiue9/rJOy9FKIlKIlKItZuacCienN5XMOAMO2OHBmol5lYbdsQrpblK6ZEkxx9okNJUXQ4kAkJWD1bq+Hr8i5NsvwZCHYTobW0+2raXEqSFBQKT4IUD+NA70DuvL/5FjJmCwnNILEkB9KRF6OWo2QD0sytYeATI/uXts09UkFtxz3FBIUCVJJ12Hx8+Cf71rLmXPeT8X+qPKFXSYZXFECNZot1bWlIFmcme8lqd1b7Ne42EObI0HEq1pOza4Li48ZgYJs0ua11Z62/B7FYz4OSYYRvz34KAXHlvJri9gse55ZmLEGfl+ZQZK8UYL811iLJcRGQlCWnOpDaUjv06ABJPYmrKc59s3BuDRrjJuGWXidf74i1xG+SHzaEsuBlTrjhdcZHtx0IQolSUL+7SQCTXfIIaGKmnl5kVue5rRVzTf8Afp5su20etI5eix2vFMXg5HldxvkrH1RIt+R9Ch9iEZhdRLDSkuNKaSoghIUCCOndZJfqsvTibfeI2AB/DJ2Wt4e1dv4wn3xJ+uMJ19THtaDAdS4lKvc6lFI2lIVsQCDXfX9a+Tzm2/R/7WK4y9RN/vlutGN4li0rLMjdReLrL/xBfQyGIrF2kxEJ+oDB61rW2Uto6AlKUaKgEhRxXNOf8iZ23k7GFQMltF3teIRGnbFBcQJka4XKUnSyUL6S7GYiuHfXofUb+aFsjttK+5KG+99G/Kr7kLkq8ccYnKtsC4ckwcutWaY4Lnar9eE3CQ/AkyHdIiuNKKVIeQl1sp2PuQAdaBqwr5yReOX/AFM4dh/sZ3x3Y5+MXhchiS4ba9IebeiFt5r21rBKB1jqOiA6R80I5WbfbI+regM43vpuvxxnCTgt05myrJ8+zu62bja8P+1GnX111pyG1a4slYdbOg6duOkb7HY/vXJ9eTcLGLtcpmPWmXKaxuZkUOJZL6ZhbDDaHDFlksI9l0ocSR0+4CUuJ7FI6pihI/dP66nKmRcY5P8Aj9erfixXeaOQnMlXjMXAbM9kkG1m+XGH/iFwobirdcbjNNrEXa5DnsubSUhCSgfcQsEVxjfqzduUhORM4/NVeMisuLpt1nkXtX8PTLuUqSy2jXtn2ukoKnHUoKlJSB07QNzyk0AfzHxV2alehas13vet22P49u+WXS3TmsxsMKyXSHJ9hDlsmmTEmtltCw80VJQtI2tSClaQepskbBBqVVBDFgipDNy9YvV5xdPbKzHveOXyzPANgpC21w5LW1a2CQh7tseKu+oIzO8t7AO6UoiVUPqjxa6ZTxgUWqO9NdhTGpbkWOCXHEJ2NpHffSVBWtHsnt3ArVNh5oIgNMqHw66KtiYTFJiEN23kfZaRBn3WHkpAW4gkK0gqKdjp8eQN6Gv7a/B/RzAbxbr7htolWu5M3aIIyGxKZVsLUlISrYPcK2DsHuD2NczAiERxEn7i2TPr4vcM4PRlxuFFjGB03u2ZWfqs/U5kk3D/AE58m3q2BRuEHHJ70cpIBS4GF9Ku/wCD3/0rrkE0huvQkgCqkXFGLQ8H4uw/Hbett2DabPEgsONEFK0NsoQFAjzsJ3v53UqrI3UkNRKVCJSiJUG52xH/AB7wrnWOhoPuXKyTIzTZOtuKZUEd/j7unvRhFQ78qovjgTMWuQuEcDyNp1bv8SskOQ4XTtYcLKfcSr/3gvqB/cGp5R+apRKURKqrkD1D49iUpVptAXlORq2hFuthDnQv8LUN6Pn7RtXbwNitccwSwHzWidOhkw8xvkMyVDmuKc+5qcbl8i3dVisJX7iMctp0VJ7aSs/22D19R7nQQathm24twzhs+YxGYtNqhMqfkOjXW50g/qUTtaiToAnuVaHmtcsEPMidz6DRhT3PVaJUoy3nTf5ew6KsuDMXl8hZHN5VydhJmTFFuzxVK6kxGUlSNjtrx2BHklau/UDV91lKhaF2DnSvrtrLZhoWliI3iqfH8Bh4JStytJSiJVCN4jduIua0T8ZiKnYnkjg/ikNtxI+iWN/zAknegSpY18FafhGudjcRJwssTp0YhYi5bMAgVHytMcmObFCZYcg+liroVeFvb+miuOhKihSlgp6Tr8aJ8kf6d/xvlVwmpB/4Hv27dZPnY+B+3+mxXn//ADeLnDnwuEiigyJo/YG/y1CxcdH6EIH3Rh19m7e2pCXozzXX2SSARvt5148/9P7bxwnx4clMplw+w6B7jZ32J/q7A9tfO9aT5AqhjOMS4jLmzYIpc2UXaIUILCIAkVPLF0rqwWyCSWIhLg6KEepXkW68f8Yr/wANdByy+zo1hsnWQdS5Kw2lzXfqDYKnCPwg1ib09ZvRz6bVN2mO7eJNqjhiGysFUi83R9ekdWjtTj769nX/ADHXYCvfAQxfdDXfjft50XPI13veak3p+4uf4j4stVluMlNxySQV3G+3FI/75cn1e5Jd/sVqKU/hKUj4qx6zIAoFN0pUIlKIlKIsRls6xQMauLmTSoESwqYW3NcujqG43tKSQsOKWQnpKSQd9tVr96OL9FyLAr1idtuC7xasNuS7PZ79pbjNwtXR1Q1Ie7BxTbavaVonRa3/AFA1x+K8Ph4nhvoRXcEUsdezOLGq3Spn0oubw3v4WxkC3fR7UpwuOEEdgQkbO+w2f/IqGnhqyy8yzu+XL/7Vj5hbIdqnWuS2Cx7DCX09P5PWJC9jx2H963cPwUPD8PBh4S7Z2cnpl2sFjHHzxGLVVjafRhbcUxnC7Vi+dZJj0nFZdylQ7owmM9JX9af5iV+60tKtJ7bKST3JO/EguvprkZHZozN85GyW93q23Fu6Wi+S2IIkW11KFIKUJRHS2tC0LIUlaTv8jtrpgAMRvqKUPi2gDBa7WWQsPpytdpv+MXybf7zerxZLxNvYkzXGwmRIkRXIpSW0ICG20NunpQ2EgFKSdne6MlcBcgXnkqDZo9pumP4bb88byxRReIzlmU2iWZalstBIlB11Y7sq202t5Sgo9KNQwZq+e3yp0uApclW3A9J9px5i1uY5luR43doTdxjLulvdZ91+PMluS3GVJW2pICHXSW1ABSdeTs79tq9LeOWrAr/ird2vSWbnMjy2rk1LUmdDMdpluOEPHZPQGAfu31Fa9ghRqaPan4bZy6CrwKBdFx9KWP39L8u+3+937IZV5tV5k3uYphLzpt7ynIrAQ20lpDSet0aQgHbilb33qdXviu137lfF8+fkyk3XHrfOt0aOhQ9laJRZK1KBG9j2BrRHnvvQqLW/fizD0QgG+97zXTZuGsetKuQ0OIeuETOpq5t2iSlAtnriMxFto6QCEKbZG9knald/xCXfShZblh9zxm9Zfll/tMmySMeisT5zZEGE8lKVpQEtgOOdKEAOO9agE9iNkmftyDb35nUvADLG8/ce5bds3h3rFLFcJ5esb1pflWe/t255W3OtLUlLySlbHdRC2tOoJXo6XsfGBejzHLLx1Y7PepUxy9RbDZbW7Lt74bTGetzi32H4+07SpLzqyCrYIABT3VuGDE5nx8ncegbLVZOburW4940gcdIu7jFxud6ud3lJlz7nd3w7IkOJbQ0jfSlKEpShtKQlKUjt+STUvooVdc2cPs8t2CF9LcnsfyuyyBcLDf4v+bAlpHYkf1tLH2ONnstClDt2I8PBvML/ACHEuOPZNBbx/kjHChi/WQOdSUqUPslMKIBcjugFSF67fck90mh31P5b0CK06URKUReV21Qn0SEOQ47iZCVIeStpJDqVDSgrt3B+QfNUFkGC5F6fr5JyfBWnbvikhXuXPHlkrWyN/qa1s9IHYHuUfIUjfRqmgkCIVI2b76qniJZYTJf8ofUZjx/auPAeQLNyVjzF3ssgusLADjLmg6wsgHoWASAdH4JBHcEgg17cxxiJm2I3vHZ4Jg3eC/AkAAE+262ptXn9lGshEI4XG95hWYYhHCIoTRVf6S8ql3fiCDjF72jLsHWMWvjKldR+pjIQlLoPlSHWi06lR8hwb7g1c9Z9lndKURKURK6J86PbIMiZLeRHix21OvPOnpS2hI2pRPwAASaIqV9FjUhPpwxl1bTjMCU/OmWtLw06be9MediqWND7iyts/O9g771eNOiUySoDyPzdivGJ+nuU0yrsQnotcPS5B6t6JGwEA6PdRG/jZ0Kwjjhgh5ot9uq0zpsMmAxxmirY2rk/ntaVXNSuP8MdIP0bSj9ZIb7EA60ruN91dAHyhYG1Wtx5xJjHGFvRHsluQh/pCXJzyUqkO6GtqUAND/3UgJGzoDdapcJiImRHsNHA9fGxY2VeVJjii+tOvkNB+ddtMQkDetbNUH6h5UnO8yxLi6A642LksXG5KaOlIjoUejZI0BtDqgf+dpA+RvKdWWRrTzpqPRbMQ30zCM2FtTuuqvaFCYt0NiJGaSzGYbS000gaCEpAAA/YAAV3VuVlkr5ccQ0kqWtKQBslR1ofmsI44YIeaIsBqsgHoF5HbvHQelJU4rwAkfPf/wCR/wBq+PqJ8gp9thDCfkunZHnxr9gP9/nWj5WPjE7GRmRwuXzEXiicQj0r+jR6K0JMMAeafDNcC1uvN/8AEyXHFEfoQrpAOteR/cnevn9hWLzPBoeXYddrEpS46Z0dTSXkKIU2vX2qH7BWjrwfkd63YXgkAmDE46MzZupoIf8A8R8lyellqnTTHCZcAaE73VQL00ZlLu+JS8Yu49u+4u+q3PJIICm0kpQRvuQnpUjfz0A/1VcW9nWvHzXopLmXC+m2VGRHzy4T4eVECQlIA0AOwA+K80m3MSe5bAc3vrT9qvI+R/aq2NwUjiEoycRC41zB1BuD1VqGIyy8OSoTkaNJmepLguFcFuPW2E9eXmka+361ELpZWo9W9+0uToHfbv8AkDtutt/7XvVibTdjrH+M7bAvkSCCdTLnNVJQ1IcGtFLCIq+gDv1uFW+wFV+FTYpmDliO8Lg94Ty1euQ79lnODRnr4Xqr/pXWWpKURKURKURVF6luEHObsNtbFvXbkZDYLmzebUi9Ml+3vvtbBZkt6PU0tKlJJAKk7ChsjRw1r52v/H+ZWDE+VMSt2JM36QYNjv1iuKplrekdPUiI4VtNLYdUAenaSlZGgoE9NSIX/jfe28dVBa6valQpSlESlESlESlESlESlESlEStRebM1w/mPKrcOMEZFfuWrU6/bod5xUqt6LehLg99Mua8wtlUcLT3bUl0KPZKdq3UgPk+79tflCWC+Wee/UDwlZ0Pcq8YsZhAShJN4wtZcUyAoBZfQAfuCT1dQQ2gkHukdhaPDfrD4t5weiwrFkKIV7kdQbs92T9NKWpJIUlAJKXCNbPQpXYio339Gfp7UUGl97/rQXXSilKURUTn/AA/dsHv72c8Zaj3PRM+wpbHsTEb6j0JCk997JRv7iftKTvqnPE3L1r5WtCnGEG3XiOAJlreP8xk/kbAKkb7b0PwQD2qtCPpR8gsbdDR/O/Q9wqEv/gm/SJoajvmPlvZVxyG0vhj1IYlnEMBGPZ84ziWQMJOgJ46lW2Vr5J26wo/hbX4rYSrJL5uV0Du6UooSlESqP9WN3kXHCbTxxanVJvnIdwRYEJaUpLjcFQK57wKe6QiMl37vgqT+akUtvyqjOrntdsi2W2RLfBYRGhRGUR2GEDSW20JCUpH7AACsDnXJOPcc2/6q+XBEcqBLcZH3vO+f0oHcjse/gfJrXFFDAHK1zJkMuExxlVE9lPJvOB9rGI4wbFXRr+KzOoSn09vub1o/kjo14/zBvVT3jbgXGONimVHZXc7yVda7nOPU4VkHZSn9KSdnZA6jvuTWiCD6sQnHuB89+1Opo1OVDFPi+rMtkNOpzf4Vj0q0ughOh2G/2qgeAWnc35P5Dz98lTTss2mERsJLTfSNhJJ1tCGFf3Wr81qibmhB3TfV2VWb90yXC2b+hV6PXSIyQFPpJ/CNqI8+db1+k/6jVecXZUk6iR1PDuPcJATvQOu39xXn8XxuVBH/AK+DH1ZukNQKUJNmPetahdKGS9YywXhut6RaYglXe5w7NFIJK5DyGh20dbVob0CD3+f9sRAzfB58tmM1l9jnTnFBKWUXNlxbihrwnrJ323oeCfmqkPCp2PiEzi8b2aWD9oPe8VaC1BV8pM6GANJHid/hTVppLKOlAAGz2A1X1XrJcqCTAJcsNCKADJViSblKVsRUHlaBxZ6lLJkKVJas+Xs/w+aFb0JI6EIV8AbIjj86909++r8rTLHKYoXer+ddfx2zNWTQxwaH3qijpPYbI+BTXffzW5WbqscoveN33mGx4q64teRWmOnIklCNiOkrVGQSo9gVh15HT5IJPxUd4WeGT8884ZQyFCI1MtuLtnrCkrXBYcddUNdweucpJG/6B2B3XIwMP05uIk6Rcw6CIOaa8z93ei3R1EMXRvLY91eFK661JSiJSiJSiJWuHOtrXgvP2E8tXPFZGW4rbbRJsklVuhmbMsr7r7Trc5DASVrT/LLai396QveiPEEkW3vT+0dlYWJ+prirNnlRrXn1jM9B6V2+ZLTFlIV+Cy70rB7j+n5qy23EutpWhSVoUAUqSdgj8ijh23ohC+qVKJSiJSiJSiJSiJSiJSiJVFei5LVr4IiYwCr6vFLtdLBJS4kJWFsTXglStAAqW2W1k67lZ+d1N9726K9aqzNPTdheWXRy9Rbc3YMlcuNvuarzbm0peLsOQHkEggp2odba1a6lIcIJ7DUZM6K06URKURKp/lXg1V6uictwuScfzOMou+4wstNzexBS5rt1EH9WtK0ArY0U65kAjhbPI0cdnBVfESvqyyIbio7hU1zpzaxknCdyYyKI5Y81xG82m7vW9bRKnfpbhGccW0N6J6Oo9IJ7dwSk7rcRS0pKQVAFR0kE+Trfb/QGplmKOEcwY5jqL5k/DVc1UyZomwc2dj0XNKzW9KURK1nxLKLRmXNmact5BcGImH4h7uIY3IkEhDryVJNxkNjv1KLyAwkpBJDKwPNYkgCtr+X7bX5GMUUMI5ojQLM3HmLMuWH3LdxhZ1xbeCUu5DckdCQnWtthXZJ6gQey1Af0D4kmDenazWKb/GMjlPZdkTmlOS7htbaVDf6UqJJ1vW1E9gPFahCZkXNFYWGvU/Ayua0XOghOKiE2L+AsNev4urbpW9dNfDz7bCQXHEoHgFR1s+K8S73GKgloqfUSEgIT5JHb/T9x+/4OuLjuLYbAfbGeaPKEVJ3TwrZbpcqKZZR3PcsfxjDrzeZcJRgRIinVoQf5jg6SOlO9EKKtDuBrYPfuBieGMZtjHGtnVbre5ZrfLb+pRB99Tq9K7JU4tRJUpSAne/G9fA1xDheIcYJGMP0ZZtDCfuIpcg0fMM7i+QGZJlTQIIXiAubDbUUazbn222TLZOH4Li0/kvNoo6pUC0uNpZtvV9wMqU8pLbOzohAUV9uyewrC3PE+X82iPXHkDkW18T400UqVbcOKTICCdEP3GSnST3A202nz53XqMLgsPgpf0sPCIYRpcmlTn0L1PQLXFGY6xFZ+y+knh8xIsiVisTMHC0Ci45LIcvDjqT9wV1yFLB3vfbQ71kbt6UOG73DdjSeMsYS04tLijGtrcdYUlXUCFNhKh332BAOyD2Jq/wAxFH79e/8A20q9KWWJD0WBZ9HWFtT2kqveZP480pxxvGnsmmKgNuKBHWn7/dGgSAn3Onv4rG8T3WdwRylI4jyO4yJ2PXlT90wi8XB5Tji0bCpNsccUSVONKX1tknam168oNYgWA7b/AAGFShZbCoIUnsCO5GlefNFIC2+kkntrYOj/ANKmxUMqh9U+NuXriaXcI3UmdY32rkw81sLZCT0urBAJ+1ta1eOxSD5AqxMGyFvLcNst4b6tTIjbpC/KVFI6gf3B2P8ASq4aGaQMwPTr4hVoaT4g9wPc76+C5zbL7fgOG3vJrq4WrbaIb02QrXcNtoKlf9BVKWm/c+8vwI17szOM8UWCQymTCYvTC7xcpKFISUF5Da2m2AdklIUtQ7A6O9biHo+/PeXSyK2UYt/HvNmLc6w88vUDEL7Eu0FixXt7HVSGHGmm3StmZ7Lyj/llakqSlatpO9DWqsnhR7CuMOPMnQjM7fchbLxcZ2R3WS4hgxpbshbrgfBO0dIUlAKvKUJI7arlQ8sHEJubwQmzWMQej65ktXVbzWAAa/jfVTrEeV8K5AkLj4zltkv8htPWti3XBp9xKf8AmKUqJ18b15qO2j1NcSX66t2yByTi8m4rX7SYqbqyHCvZHT0lW+rY1rzvtXWi+2hK0qQ5dyth+BSXY+RZHb7M81FTMWiY8GyllTyWUr7/AAXVoR/dQqTTJjFviPSpT7caMwhTrrzywhDaEjalKUewAAJJPiocIzqG4Jzdx/yhcJUHEsysuRTYqet6Pb5iHXEJ3rq6QdlOxrY7VN6CoQhkpUolKIorm2BYTk9ukvZdj9hukJttSnnrxDZdQ2gA7UpTgISACe/xs1SHpJvRZyjO8Xw4rvXDFqkhWP3xZ02xIXpUiBHJ7vMNqJKXB2BUpGyEisnMV90Lb7odVszSsUSlESlESlESlESlESlESteYtxd9PvqEvDN2WhGB8n3BqVAuKz0ogXwR22VxXD4AkNsNqbPytDiT3Kdmfe99HBLYalESlESlESlEVB+snAbbduF8rytFnFwvtgtb8xtDZCDJZQnqdac+FJ6ArW/B3rspQVr5kvOmUZo7jsyRcwt2xkPxJMVJQJLqV7EhaSAk9SEjsUAaKh0gLUBzcSDLJigz9SDfLUP91aBlwsfEcOeaD/2v3DVsRluq3X40zmPyNhduvbCQ048gJkMA79l4DS0fuAfB+QQfmpRV+CLnhEWoXagiEcIiGaVAeYOX7XxLYPqJHTMu0gKTBtwX0qeUPKlH+lsdupR8bA7kgGJkXJCSsZsyGVAY4slrBj/qHzRtN+iMuSLlecjkx48Z4dSxb3XCWQpllKSANqa11aSno6llX3GrT4l9H9pxK12tvKZP8cXb2faj25tRMRreitWylKllStqJOgoqJUFE7qnAP9mFoy48L6aU6NkuTh+fGQgzP4g+Zv5WWwsSGxAjNx4rLceO2OlDTSAhKB+AB2ArrkXKNGB63R21sJBJAPz2/wBf9qnF43D4CX9SfEwyFyewFT+wu9BBFMLQrz/xJ58pEeKvR/qdBA1vt/vr/TtsV8oi3CQU+7IS0lJH2o7lWh5JGtd99vHg9u4rzYxPEuLxAYeH6Mr/ALEfcR0GXQ5g5MHsmGXKvUrsbssdKgpfU8rWtuHf7b2e/wD1+T+a9rbKGgQhCUb76SNV2cBwjDYD74Q8w3iNSb55XPncrTMmxTKGyqX1TzlxeFLwhtQCpT0RlO07OvqEKV2Pn7UnsdfJJry81ZRceG/TW6LCnryJMKDj9nSkBvU2StqHHV8BIS46lWu36f8ASuoIYopkQh0HnXZqwpXTnQ1nRRHQe537ZqX8M8UWjhfjqz4vaI7aDFYQZksDbs6UUj3ZDqj3Wtatkk/kDsAANav/AEgWaMXa8cdcWrgXW82+5zf47f4NghuS5yrfGUkJShps9Wluq/UdAFrz20d9CaDwHsPj0qrZLqp8M9SGdYt6UsJxq2XiTj2c2DNrfh88zbakykQH0OmOpyO+k9P2BI0dKPsHRG+/puXNXNGH2jPbs5ydKu0fj/OI+PJjS7TDbN1YceShSpLiUADtoANhBBKiSe2se5NPHOrvavUjLN1BoTv878VOrvzhy3A5emcJtXtx3KpmcR3Id9MOMHGcbcZMpxSUdHQotpacR1KT3JI3vpB2D9VuFf4x4PyR6CzKVlNhjOXzHZFuR1zGLnGQpyOpkaJJUodBSAepK1J191YTYxKgMcwgDqw/pzRv6WQDlt78/hTvHMoVMxm0Sbk2pi5vw2nJEYIILbxSOtGj4IUenR8EgVnY8lEyM262SkODtvWwfx/cf/1XIw3GcFjcTHhsPE5hq+V2oc9vdbY5EcEAiIuvNf7MxkdhuNpkjcadGdiujW/sWgpV/wBDVS+kq+uXTiZNvkOl2ZZpz8F4KI2k7DmuwHYe50+P6SPiurE4mQtZj8bzvS5XPiDT4T0I9ljfWu708Jojy1ut45Mvtqh35xkbUm3OS20Pf6HaUn9lH5q+kIS2hKEAJSkaCQNACtz5b/O6G4Fq6+JSA5GdSQCCggg/2rVf1h8RYq9wTyDyAzbhEyxrHZUR+bCUpn61pQ6S3IQkhLvT20VjaSkaIA1XFmEDiUIz+nF//UPWl9K5rcA8vxHsVVDUjAZ/LPpfTxZ/h6Rl7K1Lv5xoI2iN9M0mUZfsdursr/MOwrXkbqhbLjU8enO1P3+341buNbrlD1uuWYxrQZGQWnUpR9wulaAlo6CAUpV0jtolQ33YgXIioOlL9nr1vWjrSObI735q7fVE3HyfHOfczSXHoVol4xiltmuLSWiyzIjS3vbI/pK5KFE+CUjW9aq3PWTypjHKfpj5AtOA5XbMtuERmNKuMDHbmy/IRBRLa99R6FHpQEg9RPbp2D2JNREXLxM7ZmuZAqf1e4DCLU+N+XVrlh68AzL08X3mzjuJgFmhXTLUWiS5CuONI6GLbH9oBaZgQpCeopWQA4lSkqV4SVje1NQa23v3dS4NkpUIlKIsTluLW7OMWu+PXdgSbXdYjsKUzvXU04goUAfg6J7/ABVCcHclr4abx7hzkmOuw3qKRacevrrSUW/ImUEoj+06n7UyS0EBTKulRUCQCCKAVoKnz/r2ui2RpREpREpREpREpREpREpRErCZrhNj5Fxe4Y5kltZu9lnt+3IiPg9KxsEEEEFKgQCFAgggEEEbqCHoip7izML5xPnw4mz67P3duUlb+IZRcFfzbpHTorhvr0AqWyD58uI0rWwqr8qX5u+/iv4Q1SlESlESlEXRPgx7pBkQ5bKJMWQ2pl5l1O0uIUNKSR8ggkEfvWlHCXpwi8gYFfozWRPMX3G8lu2POSHP+JbfRGlKS0p0bC/cLRRshXyN9+9aZkqGaGi8897cOqmIwsGJhaK4sdN76WVwdhWdcMZ2qz3O1Lm47eCoOTbepLrDTyEgodV+laNjaTtI3tPc9NbKVhIlmVByHLeg9lGFgjly/pzLj2Xku7sxi1THLcw1KuCGVqjsPOe2hxwJPSlStHpBOgTo6rWiX6a8xz+6v3zN8ht0SXJ/zUMIW/0IHV0tpHUgIAClaAUoAnZ6j3rXiuUSjFHHywipLswuc+g/dlhisNMxRhlwloc9X7aeKy9g9OdjxTJrbeIl4usowOt1bDykNJckEAod6kp2hKPv+0nZJ7kgFKrwt8yXOSppt1tBRva1glzud+CBrsR8f38V80kcaxWM4n9Dh7CGIEDmL2qYqA1YMHenW/akcPlYOQ18zv8AS9S7SqQhQkSVu7I+3wnXyNeD5I/tqvU3BjtEFDKAQSoHXgnyf+p/3P5r2uF4JJlxCfiiZs3/ALHLsMhvVYRziaQ0Gi7ykHXgkeN/FK9GqzJSilUZ6vnCOOLSwEqUJF5ZbKQnYUPaeUQR/wDt7dvOvnVc+tFpcb005TOYQlTlodt12AKw2NRJ8eQe/gaDRNaYW54m6fOp/A91UgrPjrkPlXcw8iXGadR9zbiQtOx8EbFYhrCbAxlz+Ut2eGnJH4ohOXUMj6hTAPUGuvz07768bre7WVpgoNytiPG+KWa95pf8Os1wmGRFmPPO29tx6VLaIbiqUSNqWgqASo/pBPxutbY3Ndsmzbs3eOOMSlWK8XBFxuENqEr3ZD4Ow84slSXXARvqKR4H5FU8RifoxAXN7t0fV8qAmwpRc3FYs4aMQwhybu9unl2V68bcGB3mO5cx3zKV5bcbjB+jsba4DUZFtt6ll1Df2frcAcUkuHRIUQfPayM8vWPYbY3r5kE9Fqt7bjbS31oKwpbq0NIT0gEkqUpKQAO/VVfinDpXFJBkRvDoRQj8jIgu40o3XlTTKPPCvqTYZLfUEgPBSSCoED5Ue418kj8jufyScjEssd+K2t1paHVAlQUo73vf/iB/bQ/ffz3hf+MRSsZMlY6HmgZ4Yg4BemRp2JNGcmqvzcVzQgyyx0XaqwQyeyCkDwlOhrskDXz/AEj/AM6qP2m6YxOvNzslsuJbuFtWn6thrqb9pSvAJI6STv4O/Feim/45wyR94iil9om933XVVv8AamEiE1PZ195HjuPZTZJlovC0XO0ykht6DKHvtOp0nQUlQIV36TXbLuluxuGwl2+sW+MgiOj6rSUhWwEJ76/KQB8/b+e/HEMiE8mA4jG9gC8fxoDr5h1vJLPMl/G/JetVxlMtEJktTAhPdXSUnuDokgADwDv/AKdxrBzmmJ9ofts2NHm291GnIcplKmnUnwFoUCCDofaTrsB2B1Xn+I8Y4lhZsqMzBFEAQCITUODYgAOzGnR9d8uVKihIIp131WXxHC8Zxpkycfx202NUhAStdthNMFxIJIBKEjY2SQD+ar7l3LLJwtbbS1dMNtrnF9zlGNe5bEdHtW511X2PvxwjpUypegtz+kkEgjuPsOEmjESJc2EfyAPmAe65McPKTDorBhYBiSMdftMXHrObFNc+qdgohtGM8skH3CjXSonpSd6+B+K7bDgOMYs+49ZcctFoecbDK3IEFphS2xrSCUJBKew7eO1Wnc8291PmsT7L02TE7HjSn1WezW+1KfUVOmFFQyXFeSVdIGz/AHrK1JJN0SlQiUoiVH894/x3lHFJ2NZVaI18sc1IS/DlJJSrR2FAggpUCAQpJBBAIINEVN+mjJMjazjkHAl36RnOF4e8xDt+VXLX1n1KklTsBxxP2yVMJ6Ap7STtYSrqUFEbCUrY38fmvmiUoiUoiUoiUoiUoiUoiUoijee8cY3yfZBasmtTN0hocDzQWVIcYdAIDja0kKbWATpSSCPzVKpvmdelhAYyBVx5J4oZ37d9abL16sTf9KJKB3lMpGv5yfvSNlQIG6GozpbxbfxVHa6vfE8usmeY9BvuO3WHe7NNbDsedBeS606k/IUDr+4+D2NZehDIlKIlRHljk21cOcf3XL723JetdtShTyYiUqc+9aW06ClJH6lpHn5pRFGmPUhh0688X2+E7KmjkViU/ZZLTX8sBhlLrgd2QUKAVrp1sKBB1o1WPB/IuPYV6meWOLF3ZU28Xm+O5HDQwyBHZ64kdT0dS97LwKSsjWun52FASKhxu23GfR1HdbMPXKJHlsxHZTDcp4EtsLcAWvXnpTvZrTLnf1n5NYeQbrj+HNQ4sK0vqiOyZTHvOPuo0HCE7ASkEkDW963sbAqrOmGGB4DVey/xbgsrjeMilTyeSGEktR7AB2LXfw0Vu+ln1DSubLXcYN5jss321hCnHYwIbfaUSAvp/pVtJBHjxr5AtvMsnx7ErYmdkc+NAhhRSlclWupWj2SPJOh4A3WmOXLxeGMGJAIN7gU8fnouVxvBjg3EJuHhipCaE3Yhw/gVqxnPqUfY5TjXTFSqXijNmkQnLdNbLSHJy32lIkjp+/SUIcR0kp2Vg67bHdxR6lZltyK4TMykSX4j0ZLUeFboaQhpwEE6TsH9O/JJ0P3G+BKwHD5OIlzcPKAbMPnQUfN6M5JegNF4qPjEcyYIQ/JnSp7aeLZ5GlluerfHCgqYx7IZA+CiM2RruCTpZ1opP+gOt6rrT6oJkpQVB4zymWwT0h1Mckb0f+UKHwfn/wADr0QxBj/hLiPg3lr76gLcca7CGAnw+d+a7Tz3mzoHtcQXxKSSAXlrSoEa32DJ7b3okjY1++upXMnKj/UmPxLJC9HpLsoAA67b2Bvuf+nxvYGbNJaGXrc+WTV7070WP+ziIv4ST4lt7svS5m/NstpaY/H1sjFaCUOPz2/sPcDY9zv43/qN68V8S7xz5LS59PY8YhbUn2wtZUUj56v5xB867a8VmDiCKiEeJ8y1PJ1PPjYqiADx3+vRRjMsD5jzy2xmMqdw1uBEfTMS64t1pLLiUq6VEgE6PV0qGyDv8A9WWyfizlzkXFrrYb/lmOSLJdYrsSTEbhh1uQy4ClSVEtAgFJ/pII7aOxs4CXiC5JA8H+R1yKgS8Y784B3uyrqPD5GxDkay8W3vkSfjsaTBbGO3cse/HuZaRpyP7ukqQ+gdJCHCorSOoEnYq0k8E8gyz1TeYLr7iger6dhTaQrx2CXU6Hjt8d/yNZfSmH+Uwv2b5sc6A+LlScLiIv5Tj4Db9vGrMsJmvpuuysXusu6cl3a5Iix3JRanpWqOooT17Wn3FdvtB2BsaPneq1ytvHeV3TE3L/Hx6Y9Z0MqeclhKVI6UdQUdb61aI+7pH9O/jYo4jDxmL/jPMerdBWnhQcxDhwAuVi8FEIoQCYqH0+K0HkDlvVxDfrNeeMLBIs8oO26NDRHK3CkKQppISsOa7BW0knx534NUxn+ZWr1F8z8f4TiMxOQY5i95/wAQ5Pc4TanYbD0VJMOKXx/LU4p9QWUJJIDPcDddWEAgCAltRWnU6Gz08F6WXEIpcMUNab36ZLZOTOYi7C17UBvoSCpXgnwP7GvIm4S5aSY0ZKU/C3Vee+iO3g+f/O685jeKxiacJgIPqTc9Ie5Zn6PR66K7BKDc0wsFyu2ypP8AnzVBPb7WQUfBB7g+Dv8A0/PgirMTyK4zPUHmWJy3w9ZYUFmTEZCEoKCWmAolQSCrZcV8kDQHwNVYOCGZEJvEphmxRUItCOwpm2nmVpnYmKAwQygwfpod/gK4ERGWwnTaNp7g9I3v81icvwmw59jMvHshtMW72SWj23oMpsKbUB47fBGgQRojXbVemk4XD4eHkkyxCOgAWPMTcqqcQ41yziG7SrXYMtl5VhAhrVExu9pD06K6AOlpicpQJaIChp7rKSoDqA7iLx/UrZ7EEN8g4tl/Hb6PtfVcrC9LhpVtY2mTGQ40ewP3bA18AV4XifC5n+R4mKdIjaGX9jmrm8RpWhLPQXc2a/Lmf60AhIqa72/urVwPnjjTOo4ZxvO8euzrR6Fsxp7XuBXbe2+rqHdQ7a7E681I84u+MW/DrlIyqbbo+NrYWiW7cXkIjqbKSFJUpRCdEb+a95h5cMmTBLgNIQAPJhbsqJPMTEfVVP6E3pz/AKRuM1XJ2Q9KFtUjrlElZQl5wN+Se3QEgd9a1rtV81ZIILG6wAYJSoUpSiJSiJVS+oXlS4YNYrfjmJBErkjK3TbsfiqSFpZXoe7NdB3plhB9xRIIJCU/1UZ6b0RnoFKOIeMLZw7x5aMVtRW63Db2/LfV1PS5Cj1PPuK8qWtZUok/n4AAqZUYCgRKURKURKURKURKURKURKURK4UkLSUkAgjRB+aIqKyvgG74VfJ2YcLXOPi18lOfUXLGJgJsd7V/UVtjf0ryh/7dkAkgdYUCaz/F/qEtGd3dzF77Ak4NyFFQDJxe9KSl5Y7j3YzgPRJZOiQtsnQI6gk9qU3qXtT3N6DIIa73u6telESqK9cGP3LKfSxntrs1um3a6SIzIjwrewt551QkNkhKEAlWgCda+N0LtT9+CXWtMjhnMuD/AFY4XJiWG6ZBxbZnrvklrXbbc8+LZ9VCV9VB2lJDY92O0WkHp37gA6lFQqBQuBeerJjNm5wcxu3ScmZyVzL37JGiyjkKzJdQh2MWijpS30AEtABSUg997AmIAliH/vXMm9w2rlxFLCm/bpR+1FNMowx+Zzlml4yrhjNM+yi+Xq23LEb1ED8FqDAAbUGVyt7iKa396CAepPfp0FVNOe/Rrll35AuuQ4i1GusC9PmS/CVJDT8Z9ei4epZSFoKgSlQIUnq10nW6rYmWZkIIDkW63GdtdKle1/xTjEng2NMeI/hHCxOYNCDTSooOtV1WPE819F1lgZe9Etd+Td1/RXWK2pYEYfaphKHd/b1KLoJKVAH2wDrZq+8R5s4v9QEJm13FiH9coBabNkLDfX1Ea20SShZ/dBJAI2BXUlcMIwEM2WHAcRCpY3p0boGFWZ281x3jEvi/FZswj7Ym5XzAAApqWy7XVactcJ41aueONLLGZdtGO5YxdIL4judRTcGmm5MdSS4FaBaZlJKBpJ89lAGrc4f4CZ4hv1ynR725c48qOmOhh6MlC2wCDsrCtK8eOkea5X+vL5/qQ3f4/fhagcHz8OBlQTRMl0bff9q2aVZXRSlESlEVceopKTwnlhVopTFCjvXw4k/Pb4/+o81nOJpAlcW4g4CFE2eHspGhv2Eb1/r2rUA8wnoPnx8vHJVR/wDOR/8Ab8r55Q4ux/mDE3ceyKO67ELzcph+M6WZESQ2rqbfZcT9zbiSNhQ/ceCRVcPcXcy4YVLxHliPk0VK9oteeWlDpCd76fq4vtOfttSF/n477WB9N7I+VaUK5g5Z5WtWDzcfy3ji32oXlRtab/Y7+ibEX1oWpaUtONNvAqbbWACjQKhs/nMYinn5vELbjlowrDsNgwIrUT6/JLsubIeIT0rcEWKgtjqIKtF/+vX5rTC5mVyA2P2wGXMqoaKeToB6k/gKurz6fLph97x2yZxNuWY4ZfpxNyhYyH7ZDMlYIDbrDbylKZTsq0V9JSlW0gIAO1dhttpx+1QrLj0GDjtkZHsxosJtEZsD8NoSANeP0/Pn8V53HT50HLw7BhpseYf7YLmKvUsBmcyarfhIBL5vqWhNOr6e3gLrPMWiM0CVoD7h8rdHUT43/buAf7969tdfA4CTw+V9OVc1JNydSVvmTDMLlcIBCEg+QPG9/wDWqHxRC5Hq8zNxoj249pbbdISeyi3EITvXyNHz/T+51bmt9vcKlOFZYGvwVfNY+4zlpWmLF6VyV7HnXQNeT50e4qhxPExYbDEy6xxfbCNYjbMdz0BKvSoOeLoKrtt9uRBQTvreXsrX+SSSdfjuTXrrfgsLDg8PDJBdrnU5ndhRYxxc8RJUKzDhHjzkFxTmTYLjl/fUnp9+42ph50DRHZaklQ7E+D81DrR6O+HbHd4twi4REUYrnvMQpUh+RCad2SHERXHFMpWnZ6VBAKdnWq6HNV81hTMOrkbbS02lCEpQhICUpSNAAfAFfVYANQIlKlEpREpRErX/ANOUNrk7Lsx5onBUpd3mO2bGluKCkxrNGX7e2xsge++h51RHke38Duvve89W97+VsBSiJSiJSiJSiJSiJSiJSiJSiJSiJUN5R4hxbmKxtW3Jrd9T9M570Kaw4piXBe+HY7yCFtLGh3SR4G9iiKo4/ImY+mSSxbuUJr+X8duLDUTkFLQ+ptxJAS3dG0ADpO9CShISNaWASCdh4M6NdIMeZCkNS4chtLzMhhYW26hQBStKh2KSCCCOxBqHy3vblF30qUSvl5BdaWhLimlKSQFoA2k68jYI3/cGhRUvwtyVf4+Z3/i3kGambmdnBnQLv7CGEXy2LV9j6UIASHGyfbdSkAAhJ1pYrIc78u3Lh2XitxahN3GxzJD0WfGSNSCelK0KZJUB1BKHftPY/kea2Qwc8QhGe/X+2XU4bg4cdioMNFE3MCx6sWfxAdSKe1jfPvGM6IzIRNs12jqZ91A+5pegQrR0QpCtEA68D4Pf8vckx6ZiuRXKx3OM2xcIU0xnm+3SHE9CeoaHdB1vt5CkkjR7e0/x2aBFMw8Xeri16XGT0fKi8hxqRMw8wCOkUJI7HT3VqYPleU3DFsIyS5ZWmdZsVz2xsRbRMlIXKbVJUYTi0rIKwz7cwgJ2QQhfSEhJ3+k1ed4rLglYuOGWGH6r2DuwV/CRxTMPBFEX35bqlK5KuJSiJSiKP8h2H/FGB5FaAlS1zbe+whKPPUpshOv9ddj2/NQf0u5EMk4Vsa1Ohx+KXIzg7/ZpZUhP+ja2/wD6jvWkj/lB6H48uutNFVipPh6g+4/atilblaVCcuOHOuesAw5HW9Dt27zPaSk9I0dtqUfx/KKNf/nf7X0tSUIKlKCUpGyT2AH5rTCzxRHbD8vTJlWk/dHGRq3oFCs5vK42LXW6KgXK4xIrSwmDau8h0909Q0d9ux7bIA3o9xWi1r5CvkCfjU/+ISrr/h9wPW5iS/1gjqClJ2Tv79FBI+FaB7JTXkcFFFNBxx/lNL1ALQA/bCxtZyzAk3JYKlxaZFKjglwuwvoTQ7/Fv0TtF3i360Q7lBcEiJLZS+ytP9SVAEf9KiGbc8ccccPux8mzmwWWW1+uJLuDSX09tge3vr2QOw13+K9oaLqg81qqrsq9ZuGyocBWC3qJkckXBtFwZLLiHI0UDa1htYQo9Q7IV+kknROiK9nphmRMtv8An2XuOxxdblcfaERLoLseOgbQFJHgaUlO/B9vz8CrFMebDBZq1etGplnn+H5pmiPEiT/1q/hb1/KuGZen0T3UMke2lJSlC076lA91DWyR4H42r5PYe6ytMmMHUuF59YAcWsgq3+Dr99//AOaA8JwviZ4txWKCbEOWWYjAKVqz0vSoz71XopsoSpQIzusjSvoqoJSiJSiJSiJSiJSiJVGeh8A+k/jXWtfw066d6/zV+N96IrzpREpREpREpREpREpREpREpREpREpRF0zYUe4xHosphqVGeQW3GHkBaFpI0UqSexBHwa16f4yzL04T5Fy4piHJ8BeUp6Zx468EPQSepS3LW64rpT1E7MdZCN/oKd6qX35OoLkMCrP4q5rxTmO2Ov2CeU3CIfbn2acgsT7e78tPsK0pCh+41+CaneyT48H5/tUIS/8AGqEEnXgdu481zRFU/qB4ln8gWe13/FJDds5GxV83DH56+yVr1p2I7+WX0Atr/HUFeRVN8v8AN+OcvcVXKwXKLKw7kWyli6P4pkbPsS0BvXvOMdymQ2G1O6W2oggEnQ3WcLcwJ6dfRj09mXW4TM+lxCREP+0Pqa+m8lUfC/I+S4BlMc2Bt64iY82yuzjakyzoaSnQOlhPUkLPfsP6SK2e5Z9P2B5NkiuSc0lfwe1wreHbtFWsNNL9sE+466CFABJKSBrq0nv279UYqZgZv1pRYkH9EeId3L2c1K9J/m2AwxxEEQvEKjtQHxDjwyWtOMekO9cwPK5LxRx/DLKmUqVidkvbr31a2gn7ZzrnctKcO1NoUlfQhYUfuUdWEeVPUXw+2prI8bOTW6MjqVNXG+oSEJGiS9H7ga0drSVdzvx36MqdheJjlxf2zS/3Wc2rbJr6XFl8vmQTsIeaTWHTe7eMlxr1+41MJav2OXK2upSD1wHESkb+dhXtqT89tHwfwas+w+qri3IGkqTlca3qO/suaFxjseRtYCSQe2gfg1UxXBsTIBjhHND0/HbRbZOPkzftJY6FWJZ8tsWQte5ar1brm3rfXDlNujWt72kn471lq4cUMUNIgy6IIiDgpSsVKwma5pZOO8XuGQ5DcGbXZ4DZdfkvHQSPwB5JJ7ADuSa0l459SzuGqzZzFbHLRabzcVTbWnIEfT+wFq2VqbQpX29KggAlKtNJJHkVVnxCXyxkWfVrflvAG1FzMZOEjkmUcP5N37P+WWwK+Grfzt9Ll17zfMZNlubLMmNjlvuq7ZDifykpUj/hgh5R6gSepw6V+wquPUN6eMP46wRq7WaVlbU1VwYZHvZfd3gQrewEmVvf272Dv53W2P7ZcR5XLG9Tbo34zvVW5kwiSZkByce6pzj3Ib/i+SIlY7klyhXqQW4UWRLYVe0uNqIBjuB94OlrqCSgoc6kgdwdGt0Ma/xcq2pZzhdleyBpzoU9j6XUx3EhIKVdLn3JV94JR1HeuxI3Xz//ACmZDN4YQ94xlSxOZsRdxWjUqcuCzfrcxY5k+JyPT0CntkEgRNP66f6ATtQHyD+e/wA/7itQea7bjnKPI0yy8UY7MyTLIjobv821LaZtEM+CmQ6tQT9RrZCW9qOvvHgj1HDYJs7hsmGcfu5QdMqGhyF7OaZspx0iXieaDy310/tZvjTibK88gz8Vy3MclxLHccKWv8P2YNwPqUL6l9TktPU4pvQICW1I18qOwRevH/BnHPHVtbYxbErNCb6usykR0uvur3+tbyupa1b+VKJrsyeYSwSP0emnme5WuQD9KGGMVz8N9tLrVnmy5TeWOdbhAskQ3ByH022G3GQFKcDRBecV50A6VJKj2AQk+NisxxdgV/4r9SVpslynR0lzGHLq21HYU4Hll0x3GA6VDoU11MKSSFBXWsdtgjg4yKKVLnYsAkwiJg4qQGAf0YkEWBah52Ekf7OLM2L+IOn7t2d+i2rgY4XAoyE+0g72hOgVf0kkd9AgD53o67arPMR24zYQ0kJH/UnWtk+Se3k1zf8AHOB/+MlfWn1nRX6ClBpvN16bET/qlobLspXtFTSlESlESlESlESlEStdvShk1vwKyS+Fb5cokTLsMmOQI8V95KHrnBWS9GltJJ2sKbXpRSOy0LGh2olFsM082+Fe24hzoUUK6TvpUPIP7/tX3S6JSiJSiJSiJSiJSiJSiJSiJSiJSiJSiKsOWvT9j3Kkhm8tuycYziC2pNty2yqDU+IddgT4db/LTgKSPgHuIVbufrzw7MYsXOUNu2IKy1D5At7J/gs1JVpP1B8wnT22le2/kL7gUrkzbqhOe9FsBGlMzYzb8d5uQw4kKQ60oKSsHwQR2IrsoQhDUSopyRxXinLlgNny2xw71DB62vqWgXI7nYhxpflCwQCFJ0ewp2UwkwkEZLXG7+irJYN1ETGM1tbWOKlqnIkXi0KdvEJYA/kNyWXWvcaURvbgKmyNp6j3EaYwyz41nfv8/wBjvmWylvdUC5XWe7PtLeiFBKIv2taGz2KVK7fpGzVgNMBrV8ybNal+rnqTkvXSJkXHpRw0+P8A54XMBP8A7PWKHprDTUd9zbFkVryi3tzbRcI1yiLAKXYzoWO/514P7GsjWggihXk44IpcRgjDEUIORWDyHBccy5JF7sFsu+xrc6G28QP2KgSKq2++jbiy8oHsWN+zujp05bpjqddJ2PsUVI0Ds/p+avyMficPSXGW0NR5FU5uGlTh98Lqv77/AOj7xyQ42uy5VcoC0JASZ8ZqV061rXT7Z/pA7k/P53WEX6M+R8UdaVifJew2oKCXHpMBJIGhtDanASPz58+N9u5DxuXNDYmSD1DegP5HgudFw8wF5MwjvXe+i+3eN/U7j7YMHKU3FIOg0m5tvK6e3lUhod/37/pI+dnE53zD6juHMYm3/JoMFVqhJSXpEhmKvZJ6UpSGXElSlLKQEpBJ2keSayJ4PiSTWBsrE9rjSlK5VKNjpTANF47+CrI454ayzmeRac353TGkPx2yu0YNHbKINv6vEiUgqV7sojtokpa8DatkQzmz0yzMKVLv2JtP3OxaU69b0kqkQxsfoP6nGwNj5Ukf8w2a8Ri5EE0faA47DPVoiwsGPW6343DGdKcfyh3S29HVj+jbJv4rgNyta5S33LfNKm0qeDgS04kKBQNbSkq6+x3334r3esReuIkJ6gkG5xtlXYf1EDe/kgA+fPjvsYE82GJi0N/1l2FtFgIjHgHFTyn0f8ZqOelbhpEVhGd3mOFypKT/AAlp0Eltk7H1B3/UsHSVf8nf+s1luWvUbjfD3IduseRW26SHbjFfuLK4Lba0dDLqW+k+4tHfagr8Df5OzxMVwiLi0zC4PIxGIvZgzi9HfLOwyV3BRDA4UxtVq6uc65qLZ1628FuuFZJb47F6iTJFukssuLaaAQtTa0pJKXSR30fnXz4rA8CerPi/AOHMKx+FZ7nAMS1RWn24cJlKFSPaR7q9BwFSlKJUSR1K3vXevow4FjIouUQudvYfJ8M6/wD5DDgE83p76e3WhVeeqH1II5bVarbj0mbbcdbQTJjzQGlPPlevvSlRCkhIOvKdk7761gfShl95x3mKw2u2OPM227umLLgBSuhxJbK+sAjp62z1K6gAr7CDvZr1EPDvo8MilRgc/LEbihYkEP2HhlULjHFGZi4YwftoPA07VcvrTut3Dj1g9OvGWR3uzWSZeHLbDfuUsNKQ5cJ/tpK1jrUUhSiEnSdgb7ADdUhnfMFuvOa8DctxX50yzXlK7ZYMYtzSUXCVMmo6HlSVqcCPYaShIKBvTgSrfjXzaGEQjlhpvPPx86Ar08EAlw8sFAs1bPWpdbtnrmFscJ5v/iViO1MkQHFw0rYjuKCQ8rb2ujau/wA9j2qZ5V6rcVxH1B2bieVFlqudxDDarmlbYixn3kuKYYXtQV1rDY0ACT1prIiIFi3m+9OposiNd730uwA77n89gKJPUnsoK7kbH96lACuaVClKURKURKURKURKifIXE2Hcr25MHLsbt9+ZR/lqlsguMne9tuDS0H90kGj5KQWsvBxPwjh/CFvuEHDra7bIs95MiS05LekBx0JCS4fcWo9agB1K8q0N+Bqd1DABgGUJSpRKURKURKURKURKURKURKURKURKURK89xtsS8QJEGdFZmwpCC09Gkthxt1BGilSTsEEeQaIQ6ot70z3DjeQ5P4Uyt7BQVKccxSehU6wPne+lDCj1RNn5jqSBv8AQfB4e9RmYcfwyvkziO+2qNHOpN9xR5u9W1tA0C8QkokpR3JI9kkAfOjqBCwYWG+57DwCMTZXHh+aWHkHHol9xq7w75aJSepmZBdDjav22PB/IPcfNZqpKi6V5LraIN9tz8C4xGJ8J9PS7GkthxtY3vRSdg9wDRZwxGAiKEsQqcvPpVsSZ/8AEMUvl3w64aUC5BkKWkgnYHchegf6QsDudg14lo5049bUltVq5ChIIDY6Ux5JGvGiUBP9ypZ8dz33ZEYmDlmb8yB6jUkm/r4eJ4TikIlcVhaMBhMhFf8A9hmKZA9AFzH9VLGPPpi5ziN7xaSshIcU2HmPHnrIQe+h2CT+od++qmNt9R3G11cS21lkNlwgaTLS4xvf4K0gH/T+/isYpRvDbe9RmAxbTiP8bxcI+pg/+aXlFDfy11Z/gTC1Znj9+UpNtvlunqTvaY0pCyPjfY+P3rMkjYGxv8VqigMNCF5ibKmSTyzYTCeob3StfhZJPPXqEuT13UlODcYXFmNDtRWT9ffFRWpP1ToHboYalMhtJB/mKUvYIAGBAI3vbZuta2BpUotdM/5CwP0/cjXKXarbKlZLc4qTNtkN9DUVIK+pLiurslatqOk/kkgFfUZDjGZYf6pceTapsaZDdt8lqa/a3HR/NCCdfeNhxo70rwe43rY3nBI5YOaFm35aZZNku9M/xydL4WMcf4mvLV2JvpXQZF9VdKEJbQlCAEpSNBIGgBVQcu8MLz/lXjXKf4bab1brGqdDuluu7aVpcjSW0adbSpKklbbjLZAOthau/bRiGOKCLnhNR5+B12aLgcoNDZSWZwLxtOhvxl4DjSW32iyotWlhCwkgjsoIBB7nRHiqMhcP2fhz0tQ4WQY3g688x+1iP9Uu3R5H1YZXpKh1oSpa3GUJJBH6lEfcBs2hiZ8ZDxxHo5c50emWeeuVjB4WXOny5RgoYg7DJw9P0tcskyi25JazAdxTF7e8lYInWa0pgPpV4AV7J0sduyFpPfXbYOpx6bmkccXJWZSbZa4trjDolZVkM0x2IzJ0Fho7CVOFGiNIJJJTtO1V2o8fOlYSPDGP7YutW0NaimRLWuS/0Hiv+IcIwIi4hFHFA1hQuTYCj1fTq9HWxWPcu5Rzvk1qGDWBdv4yafDtxyy+slv+Lsp3/JgR1ALW2sgAvuBKenfSFbqkvTp6YMww/wBQMWDkEN7/ALNuO37tIxR2QhJQ8uY4gtlOlbJQgu7JT2UBrXavNmEgb/I7tVu9vm/8hpve3Ct+w8f5FD9dGW5q9Z3xjUnCo9ujXIdPQ5ITIStTQ+7fVob7j481rnfPSjzByvi2f8g3GVGsmT3u9pyKJjMy3pVcELhJcbgNokpe6GlBpZbA6VDwSoFX24MAK1vT11FDnn2Sh3/e6dt9MAu91yDBrBcb5bHbLe5UFl2dbntdUZ8oHuI7EjsrY/tqs/WRUJSoUpSiJSiJSiJSiJSiJSiJSiJSiJSiJSiJSiJSiJSiJSiJSiJSiJSiJSiKmMq9L9jlX+VkeE3u78Y5NJWlyRKxl4NxJix/VJhqBYeJHYqKQo9tqrHIv3PXGxU1dccsnLdrbSrouNikptFyIBJBcjPEsqJTofy3BtQ7AA6CgFPLdfjoHdAN737jIWn1Z4Mm6M2bMFXDjLIHVBLduzKP9EHTof5UjamHfI/Q4a9Fy9XvDFtuJt//AGjWOfPCikxbU+ZzoI1v7GAs/I76+R+RUEgFv78tOtlAL2WNa9a/DhDapOVu2thYGpVztUyKwnYSQFOuNJQN9Wu5+DVoYjyNinIEcv4zktoyFoDZXbJrcgDsD/Qo/kVLguxtuxqjsWKz0iM1LYUy82h5pY0ptxIUlQ/cGotdOIsIvSSJuI2WQT/UqA2FA9/BA2P1H/epBaytSMVPwp5pEZhPQkeyh939KfGt4lJkKsTkd1J2CxLdTrsR2BUdefjWvjWzvGL9I2IodU5Fu2QQlEH7WpiOkEjW9Fs1tE2IZ78d1XpJX+T8QgAhmGGMCn3Qj3DFedPpemxGEtW/k/LoKE66Q3MUnp18/apPcjQ/H7bO6rXkPhbkHiG5x75iWR5PkFtusxTuQKtKC7cELSwlCJJZLo+qPSyhsjYWOlGgodhImkuOUV7/AP8AoBulKWqXO4/5DImf/LgpZ8GPhc9uq447yq/59cnrbivqDgTLwnSV2K9QDFuTJ6SrSoz2nR5I2BrtvZ0OmzRhnPUcOFGf2SRoqKEuQUDfnQOmf7VMUcuM2Yex97dX+Z/3+BTS8eDMPaI+bOtbebuLeQLNk0m8ZVHauL91ClPXG1oKo6lpbQ2AR0DpPSjQBCd787JAieAZhHwi9S5kXOYuI3gtLZRHjwnJ86WDoqSmK0lbmklCPvUkb6tDZ3VszYYpbCuWfp3o7PoeWx9zP4twscJEYrLIEIgJAiLUY1dwPCxzVr28+pLPrmRZk32DbSoBu5ZCWrK10kElXsj3JH/LrqaB760NVbVn4E5EkwveyXl26NPaUVs2xx5LbY0PDnuIJ1onfQPNU+aD/wBYB0qT856ZMBZ14OLjHDpZ/wCHAw+JJy609PJVhfJ3DsC5qteQeoS43yY0roct8C4Ga6lRSANobDyk/af7d0+PnrhM8CSZe7fi3Ieby0NlRciY5dHd7IAKlBpCRs+CdD58AafXJrABTT850pUmlAkX+T4qEcmDlwSuohHm5p6eZUPxDh66IRKau3D2eZUFNLQjolxLM10dG+oKMlLpcICQOyNEAa7bqZ4qxwZx5cIUvN8Ezy2XKE2lESTyJbp94YghPhMd7b7Cdb1tB33Gz3rCZGYgHt30tRzRmuAXAcUDc7jPGJnFp/1D9sAsNNXLXOeWVQrrZ9ZvBLjTft8r4oEq0lKf4k2O+gQNb7HRHb/5VNuPeZsD5abfXhmYWXJ/p/8ANTbJrbym+yTtSUnYGlJ7ka7itIi5rb2xXnydSplSskSlESlESlESlESlESlESlESlESlESlESlESlESlESlESlESlESlESlESlESlESlEXhvdgtmS29cC726JdYS+6o01hLzaj+6VAg1zabHbrBFEa1wItujDwzEZS0gdteEgDxRF7FJC0lJAII0Qfmqqzb0s8V584qRcMLtsS6dJDd2tLf0E1pXfSkvs9KwQSSNk96gh1LkKKJsHNHCIcXZboOacUb7ptd7cREyCOnY30SxpmToA9nEtqP/ADqPmwOKOdMT5jZls2aW7Dv1u0m6Y3dWjFudtWf6X46vuSN7AWNoVo9KjUkvff7v4Zuopbe9sFYNKIlKIodyDw5hHKzLLeW4vbL6phQUy/KjgvMkHYKHRpaO/wDykVA1+jbixCHBBtd6s61KCkrteTXON7ahvRSESAkeSfHnv5AqCHBGu6700COq65v4r4+4msthmX+bnWWWe63yDZf4JKzGcqN7rylIS8tJd2oJ0SUlWjo9iSdzeyZ5xpwVyxK4wt2NWTA7ZExxu/PXpCo8GKGzILCUOE9JJ6v61E7J153UkiEkmhq/jXXze7ZlGYcui8PJb+XjJZD7/qNxjj23Jhm4xrVHs0MuJhhWjIdclPrLid9I9xKUI760fnG2DiTB+Wpd2TmnMMrmFdiWW7hZ/wCKx4tuglI8PxYnR30Dv3lKB0e3mjhiHfyH7zGbZEF6Gex3v48bg4nyXjS72d23cZ3TFplsgEIci4vIjrajk711JZOk71sb8+andCearu6O6Uoi+S0g6+xPbuO3g73/AOPeq55N9PWCctSWZ97s/s32MD9LfrW8uHcIx+Ch9ohXb8HY/INIvuDGqkkmirq9X3kL0we3dciv0vk7ixLnTOnvwUi92Jrp7SHVNaRKYTr7yG0OJ31nqAVWwVou8HILVDudslsT7dMZRIjSoywtt5tQCkrSodikgggj81IBIffT0WLMvXSoUpXHcJH9R/btRQVzSilKURKURKURKURKURKURKURKURKURKURKURKURKURKURKURKURKURcBIBOt9/ya5pdDVKURKURKgfI3BWCcszIM3KMdYn3OACmJcmXHIsyOCQSESGVIcSNgHQVrYBoahksoSr08ZbjqwcM5tzK0MA6EG/JjXxhKfwkvt+8P9XT/AG8a5RgPPdgK3rfy1jWTHo7RMjxEspKh+HYslspB/dCtHv47ULk313/bnunRdyse9Qd6T/Pzbj/GQSftgY3LuCkAAgDrcltA72CfsGtAD539o4o5b6AF86ySvR2U4rAA337gaOvI/Pj+9QHF6+nyd6KA663OEeR7m6hNz56yVEUBQUizWa2RHF7Gu61MOa1+wHfvXI9KOOXVp1rLMrznOmHd9ca+ZLIRHVvp8sRiy2R9ngp19x7d6yfe6qVW3qL9IWLnjuzWjjbjq2QVSsltS7u3ZmW4rjsFtxXuFahoqCQrfYg/IO6wR9HuO4dzzk8aDYHrHxLesAk2y43ORc1vJblOSBv+ZIdWtJCAFDwkEb1s7oHfLNrePbNr3oKuTMKUG8v03xrtjeM3zJ/SNzJyHlj8afMgWa24LaZTJKmzEgyWg8pKu/UlxzoPUPJQfFTi6el7PuY4ExWMYXH4tgscetWHQktJZvksutu9SfaOy2tCCPcd0vunqA+4CQIooXem8hXM5NalSpNCxvvyVyenziTLJHOlozuZx21xHaLNi/8AAJVtakx1rvEnqSespYPT7aAAQpXckD/Tb6sSx/jv38cntSqjoUoBrfff7H4ooIQnQ7Df7Uoi4WhLiChQCkqBBSobBFa1yoi/R7l8WTBITwff5nsy4RBDeJTnVbQ+0e/TDeWopUjQS0tSCCEKUApc7/VjlYVZ1LLZRC0uISpCgpKhsKB2CPzXNCGoi46gVdOx1Ab18iuaMhCUoiUoiUoiUoiUoiUoiUoiUoiUoiUoiUoiUoiUoiUoiUoiUoiUoiUoiUoiUoiUoiUoiUoiUoiUoiV4b9YbblNmm2i8QI10tc1pTEmFMaS6y82oaUlaFAggj4NEXhbwTG2sTTi6LDbU42loMi0CIj6UIB30+1rp1vv489/NZllhuMy2yyhDTTaQhDaE9KUgDQAA8DXxTqnRfdKIigSnsRv43XAHWgbBSex0fI/2ooIdc1wnsnWyojtsjzRS65J7gfP9q8N+sUDKLJPtF1iNT7bOYXGkxX0BSHW1JKVJIPYgg0UXWuuK5xM9Jsl3DOQHLhK46Q9/93M3cbW8zCjKJ6IVwcG/aLQAQh5WkrQEgkKHfY+23CJdoLEuBJYmQXkBbMiM4FtuJPgpUOxH7ilLDLY9PHXVQ1bL0gHXcgmhBJHca/FFNWXB3sa0fzv4rmilKURKURKURKURKURKURKURKURKURKURKURKURKURKURKURKURKURKURKURKURKURKURKURKURKURKURKURcEgEDv37eK5ojJRSQpOjog9tGigh6LgpCiNgEjuN/BoUJKwrQKgCAojuAdb/wDAf7UdSarrlxGJ8V2NJZbkR3UFDjLyApC0kaIIPYgj4NUlM9NDuHyZU7iPL5vGz8hZecsyWEzrI64SST9G4R7OyTv2FN/nRpdn3vZCLqa5o5E43cQxyZxvJmW/qCVZPgXuXSICe5LkPX1bQHfulDiR+e4BleB+o7i7kyW3Bx7N7PNubpBFpeeEecg9WtKjOBLqT1D+pPn/AEoTy39jtxnp2QPdq737srJSoOhC0LCkEbBT8/g7/FfdSdFFMkpUKUpREpREpREpREpREpREpREpREpREpREpREpREpREpREpREpREpREpREpREpREpREpREpRFwtaUDuf7D5Pb4Fc0ZGSlESlESuN6RtWhod+/YUuoJZEnqSDogkeD5Fc0KkpSiJSiJUczHjbEuRIn02UYxZ8iYAICLpBakBO9dx1pOj2HcfgUQFrKtP/VJxOzOJcw2/Zhx4sHft45f3hH1vehGkF5hI38JbFE8Wcz2MpFo5uZu7aCAlvK8TjSFFOu/UuIuMSfwdD43vvuBDDlTeQdkZHbt6hMWcS5JsWB5/GLZ6mrRJlWOQlY/5Q+ZCFA/G1p/c/NelHP9/t7bTV74Zz6DcXN9LEBiHcGlEa/9szIKE+f6+jxUBwHO7fv01pGW9+bL6lct8mS5ntWjg67/AE+1alXrILbESQOw+1p19ez+Cmh5e5ItEhSr5wndl29KCsycdvcG4LTpOyCy4tlZO+wCArf+28mIob737OpFV2H1EuaTrizkg9R1r+Bo7ePP87+/+x+Nb6JHqU+kYW89xVyYhlsdS1N497qkj9kNuqWr+yUk/tuoqLj2+ChYKSce89YVyfepNlstylNX2Mx9S7aLvbJVtmBrq6S4GZLba1ICiAVJBAJAJBIqwqkhCGSlESlESlESlESlESlESlESlESlESlESlESlESlESlESlESlESlESlESlESlESlESuDvY1o/nfxRFzSiJXCUhKQlIAAGgB8URc1wQSodxrXjXfdERKdFXfezvXft2okqKfuAB2ewO+2+3/SpNVC4V1FQA6enXcnyD2+P964DewkqUSpJ322BvWvH4/Y7pQIzr7pUKUpREpREpREpRF5XbVCfuLFwchx3J7Da2mpSmkl1tCiCtKVa2ASlOwOx6RvxXqojJSiJSiJSiJSiJSiJSiJSiL/2Q==)
- Lateral Hypothalamus (Hyp)
- Medial forebrain bundle (MFB)
- Ventral tegmental area (VTA) in midbrain
- Nucleus accumbens (nAcc)
- Dorsal Raphe Nucleus/Locus Coeruleus (DR/LC)
- Amygdala (Amy)
- Hippocampus (HP)
- Prefrontal cortex (PFC)
![[[@kohls2012social]](http://dx.doi.org/10.1186/1866-1955-4-10)](data:image/jpeg;base64,/9j/4AAQSkZJRgABAQEASABIAAD/2wBDAAMCAgMCAgMDAwMEAwMEBQgFBQQEBQoHBwYIDAoMDAsKCwsNDhIQDQ4RDgsLEBYQERMUFRUVDA8XGBYUGBIUFRT/2wBDAQMEBAUEBQkFBQkUDQsNFBQUFBQUFBQUFBQUFBQUFBQUFBQUFBQUFBQUFBQUFBQUFBQUFBQUFBQUFBQUFBQUFBT/wAARCADuAWYDAREAAhEBAxEB/8QAHgAAAgMAAwEBAQAAAAAAAAAAAAcFBggDBAkCAQr/xABMEAABAwMDAgMFBAgEBAMFCQABAgMEAAURBgchEjEIE0EUIlFh8HGBkaEJFSMyscHR4RYzQlJicoLxF0NTJCUmdKI1NmNzkpWjssP/xAAdAQEAAQUBAQEAAAAAAAAAAAAAAQMEBQYHCAIJ/8QAQhEAAgECBAMECAMGAwgDAAAAAAECAxEEBSExQVHwBhJhcRMigZGhscHRMkLhBxQVI1JiM3KiFhckc4KSwvFDY7L/2gAMAwEAAhEDEQA/APVOgCgCgCgCgCgCgCgCgCgCgCgCgCgCgCgCgCgCgCgCgCgCgCgCgCgCgCgEH4srbdrBD0Puhp2y3DUN60JeRKdtdqZL0qZbpLao0tltscqVhbbmB/6OfSgM7WTZ697fbj7NXOVYLjrHV6RFkXe3TbPNEeBKnTXZU+4RLg2Qyy42p0peaez5iGm0eoyB6C0AUAUAUAUAUAUAUAUAUAUAUAUAUAUAUAUAUAUAUAUAUAUAUAUAUAUAUAUAUAUBF6n1PadF2CdfL9cY1os8FsvSp0twNtMoHdSlHgCgM/a4/SAbW6VurdutLtx1q67G85MvTTLcqKheVANrc8wdKuASMHhQPrQCRifpAN2b7BdbRttZLU+62pCJH6zdcLKyCAoJLQCsHBweD2pxsCBsO6vicukVClbkNF3AB/8AcUEc/YGaA4NY618T7lvSpO5jjRjuh9Jj2uGwVFOcJUUs+8k55SeDxQDs8PPj30zfLA7ZN2b3a9G60tIbZkPzn0MR7nwUh5nOPeJQVLQBhHUkAkUBG6v/AEiKrtZor22O312vs1T+H/8AEQEBgM9J99Cm1OKUSrpwCkcEnPGKA+9C/pA5saBcHNydurlZ5KHB7KnTakzkLb6feLhcU2Uq6uAACCPWgGBofx9bP6uauS7le3NDGEpKSjVvlQi/kEkt/tFdQGME8YJHxoDQ0aS1NjNSGHEusOoDjbiDkKSRkEfIigOWgCgCgKXo/X7uqNea908uEiO3pmZFiofDhJfD0NqQSRj3cF3pxk9s0BdKAKAKAKAKAKAKAKAKAKAKAKAKAKAKAKAKAKAKAKAKAKAKAKAKAKAKA/DnKcYxnnPwoD9oD5ccQ0nqWtKE9sqOBQGJvEF+kptWhJ9ysm3lkOqbtb5aGXblM4tjiACHkoWhfmFxCh08pxkK5xjIGQNKXPX3iu1azcdc3+ZdkxA4yy0elpDTSl9fl9DYSFgH1WCcDvQGvNHeH+16ctCFiGhLYSOSO/xoCi7p3O2aPayAlJ7YA/GgGRsNPjaitiXekFBweodsZNAfe+Wt7XoyxSFuFCClPA9c0BjvaLbVvdzXEnUM9jqj9WUBfY/D+NAbk0ht3EjwUMsspQlCfsoCI1hoqM4ShLYPT2Hc+lAJbcfbC3rtMjzGEqPSSQfs5/nQGaNObo7t7RXq42bbvWVwsUGfJD77SW2pKVLACAQXkrKfdAGEkDj76AeVk3V8RlxhIW9utdUOEDKRBg/H/wDIoBiJ8Zu+Wh9Mwobtj03qmVFYS2u5Ti80/KUB/mLDZCAT6hIA+AoB8+G3xu6a3zujWl7lb5Ol9atw23XYcsJDEx0IKpHsqgtRUhspJyvpOFJ4POALptIoK3t3vV3CrtayP/2mN9cUA4aAKAKAKAKAKAKAKAKAKAKAKAKAKAKA45HmmO75BSHuk9BX+71Y4z8s0B5zadtdzVYNIP3e0bjL1uhi8f8Aistxq4gSYZiyApps8MuOLeMf2X2bJSACOkBVAclk0xu9ZdmNXagurmpmd+dIzLTc4Ml1p6XFnQVQ0R2oyUIAQ7htyR56Ugnz0FZyOmgGVqvw0p0funtpbdOQrreZcHSF2W5eJs2WtqVdoyYnsL8k+Z5fWXC6rpVwrqXwQOAFrYbTuArTtid2ug66g7lI0VeBriRf0zkIfu6oYEcdUo+U5I9s61NKZykIJyenAoC3RrbCSvatW3No13ESiXOVqI3iPdUOGUbBNA832nhTnndIUpHulwo5J6aA5/C5p/cHS24+zdhv8fUciyQ9GTLmi43APqQkzGoLioklauzzEhEpKUr94NrQB2NAbkoAoAoAoAoAoCk7qbw6Y2e0vPvWoLg20IzYUiE2rqkSFqOG20NjklSvdBxgdyQASAMCbg7ka98VpYh6ljsWPSDcpb8ayxElJdT1hTJlKKlBbiAkfulKclRx2wB+ai2Gtlo0yvpjoCkt98DIoCq+GCbE09qWXBUEpWHMAcZ7Djn7KDzN5ziiZowqaSEjo5I+HFAefnibU/InBtKyEhZ4HPw+vx+FAaI8L0FcLRKFlPIRnJ+dAZh8ZOqZlx1MzbEuEJWvGB9fZUID48NWmG7NpKEhLYC1genft8KkGprHEDUDI54z/wB/r+dAVy6w/PlOHsBxyPy+VAJXdxZRHebR3UDj8O1AJLR+3/tN0cmOtBSlLKs447mgHtpXS7L6m2/KBAAwMdx+FASOrNCs+QoJQM45AHegM5blbOw7uy+hbCeQQD04+vX8aApm0e+e7+y24Op5ljvovsdoRoblqvaS5FXiGyhlRDfQrqQ22hCT1YwOc0Bs/wAO/wCkFi67vtn0juJZhpjU01C0Jucbi2yZHmfs2WwVLWgls5y4QnKFcjIFAbCiTGJ8duRGfbksODKHWlhSVD5EcGgOagCgCgCgCgCgCgCgOKNJZmMpejuofZV+642oKSfsIoDloAoAoAoCtbjbh2PajRdy1VqSS5EstuShT7rLC31jrWltIShAKlEqWkYAPegKSnxTbc/4fn3Ny7yoz0G5NWZ60SLXKbuiZzqepmOISmw+pxxOVJAQepIKgcAkAdCd4wdsoFngXA3G6SDLmTLf7DGskx2bHkRGw7KbfjJaLrSm2yFq60j3SD2INAWCw+I3bvVGtIWlbVqNmdd5tjb1HGS004WnYKwFJWlzp6SopUlfRnq6SFYxQFcT4zNo1bRp3LGqD/hFVx/VIlexP+b7V/6fk9HmZ6ffz046fe7c0BLveJ/beLqe3WB+/KZudx1DJ0vGbciPBK7iwlBcZ6unAH7VsBRPSSsAEmgOvD8U+gbpfLRZ7W9erzcbm2+6wxbLHMklDTMxUJ113obIbQl9tSeteBxnOOaAboOaA/aAKAKAKAT/AIjPEXadh9PNpDQu+rLgOi12VpXvuqzjzHCAehtPvHqOASnpByRQGKNP7ZXzdDU8nWus5Bu2pJp6lyXEnpaRlSkstAklDSSpXSgHAB+JJIGg9LaLhQraG/LSl1sY+FRuCra5u4jtPwnv8sjpHwNSDPumdISLbrf22KVeWtfJT9/19tBsbm0u8V6BdS6OQg9x24oDEniIiFF16u46+3PxoDSHhkZErRqGcYPl5+zmgM8+JLaCbP3FjTUtLW0HAchJOO/P9qWBpDZ3Tgj2iK2U46UjOR/D86AdTbIaj+X3V25H19faKEkbNtwU2pQByeePmPr86ECT3A0s/cZhHlkgq9M/HuKA69k0KuFFC/J6TjjCfvp4Auui9NKQ+VqSQB6kemaAmLvaw+6UqGOM9JFAKXcWxiH1+70g+vr86AzZp61Jf17rlso5E2GO2cZhNn1oDt6425i3G1uIejpWCnsUZzx/H1oDq7Obj7ubHEw9HX9UmytRvZItmvKVyYEZPX19TLSVoCFZzzz+8fjQGudgPHexqu/wNHbkW1jS1/XFHTevN6IE19KVKcABH/s46UkjzFnPYEnGQNawZ0a6QmJkOQ1LiPoDrMhhYW24gjIUlQ4II5BFAc9AFAFAFAFAJrxN3fWNpsekv8LLvsa3P39lq/zdMQETblHg+U6ctNKQvILwYSpQQohKlHGMkAYttmgd7EbW7bt2XQV89t2ysib60zLcNvcdujlyW840GOkiSr2RhbZbBwDM4OSKAaLumNS3adu5LGk9V/8AjNd5N0kaL1Q/DkJixIDsE+wtJk5DMboSVNqaXgh49RSThVAPbwq2nSlp0m83pvROotGSjGiC6J1FAkxXJMkNqCiS8f2zgPV1uoyFFSfeVxgB4UAUAtPEftdcN59mNQ6PtM6LbrjcPZ1MSZqFKZSpqS09hYTzg+Xjj40Ao7v4WdcX/Vbm5MrUthZ3Kbv0G7xYjMN42hDEWJIipirJX5qitEp5Re7pUU4RgYIHUHgmm6r1bbdQa3v0a4uzb/dL7qGHZ1yoDbgk29uE1GjrbcS50IQyjrKz+0yrIAOKA+rV4Mr7YNZ23Vtr1ixAmwNUKuTFiZhtptjNqMUQExEYQHgpMJDSR7/lhachA70BRkfo6763pFyxp1hAMVWj27YmGphz2dN9DKYirl3zgxEhvp75JVQF+meC6XfNXXCbdL7DNqm37UV3UiOysSGhcGYyI6m1E4DrLkYOdXbITigK9o7wX600tq/QGpJ1y0rqa5WOHLjzva0S4yfPfvDtwVJYDau4DpT0LynPyoDZlAFAFAFALrf3d+PsjtldNTKZjz57QS3Atj0oRzNfUQA0hRBJVjqVgAnCSccGgMF6H0xftwtUv6w1jMXd9QT1AvPrBCG0js20kkhtsEqIQOMknuSaA1/o3SsVu0DoT74TgYB+B+vuoCo3pT1ouqgMpRnBH5UAv90raqbCL6E4VgkHH20Avdn7uh/UBhy8KIVgds/Xeg2NgyWmrdohxaccoJ+vzpoDz7391Q2q9obKwf2hPfHcj+9AaV8Ll7abtUVHUAlSRjn69KAeWqtDQtRJS8tsKURnPfmgOSw6cRZWQlAACTwc9vrn8KAlnD2AH3Y59ePz/PigPtvCgerH3/dQEVcLXFfWSUAnOcmgPlFtjuBKA2Egn4c0BJRLe1Ea9xIGfgPo5oCGkJSJRyMA898A0BBax0Um9wlqbT73pjvQGZtH7dPjczcpBaUPIuMBKuOMqtzKv50BO6x0yuO2EFsg49RnHagIK1abajMLcWgD7RQC01noGNrS6ezuMIdZHdJQCMff9tAWbRerdzdhy2rSGoJEu1oTHaVZbqtcmOGGlE+UwFkiPlJKcoHAxwekUBvvY3fGx756STdLYfZLlH6W7naHVgvwXscpUOCUkhXSvACwkkeoADHoAoAoDpXq8wNOWeddbpMZt9tgsLkypchYQ2y0hJUtalHgAAEkn4UAstlrpqfcW53LcO7uzbRpu6spj6c008jyy3CSoqE2Sk8h9/IUEHHlthCT75XgBtUAUAUAUAUAUAmd8dydTWXcHbXb/Ssq3Wa56yfnqVe7nFMtERqGwl5SW2QtvrdWVADKsBKVnBwKAzhe/Fzu/OftFk07DgXLU0dGo4qm7Pp56ezepltmsxmSkB8eyMPB09bilqDageSMCgLvb/EfuLq7c3/w4huWXT14uOrbta27tIhKlNwokG2wpK2kt+YgPPKckqSFFSQEJJ6SRQFH1P41txrTo6+SEosDlzsun9SKdlsw1mLNm267RYTchpJcyGloeWoo6j7wPOBQF41jvdvNpW4620ZbWrZrG9aaetE6TfLLZiqU1bZiZJdxbzIAcktKjpIQl0dTboUE5TggaV2t1jF3B2503qSFdI16jXS3sykXCIwuO1I6kg9aWlkqbBOfcUSU9iTigLTQBQBQEfqDUFs0pZJ15vM+Pa7TBZVIlTZTgbaYbSMqWtR4AA5JNAeVyd2NR+KrcuRqm/lLNsguKYtVqjuFceK0P9QPSnrUs5V1kdWF9IOAKAfFkmtWSOhlICTwB/CgHNtdqNMpwNdWEnFATWvdIGWn2lkc9+O+fTmgFVfoBegPtOoJIHb4/X8qAQ1ntTtu16lxtJAUvk/Hv/eiBsOa09cNv1Iweryzx/09/wCH4UQ8WeaviO0vcEXZ2ShKj0OE9OeO9AMLw+bttWy2RmHXehxHBB4zzQG4NuNwmdSQ0oU4FKKRjnvQF4ecS3zQHReWfNwn49vnz/U0B8S5qIyACoAcgH5dqEEMbkJD+QeQcgH1pYEvDQCU9Pbjv6jtQk5ZrxaaJOe3qfl/f6zQFVmTlKldQ+OBx3oCx2eb57ICgDngjPy+hQFK25hx3t1N5epCDi62nBI9P1RHqQT+ptuot4QpaEjOMg9/v/KoApdX6Bl25txtls455xz/AFoCsad25fEkrWyeT3I+HYUBN6l0kzDgKU6nsnurmgMxX/cfUexO51u1bo1RMtlYbmQSrDM+P15Uy4MEDIBAXgqR1Ep7mgNfbL/pENF6zgxo2vmRt5e/IdfkvTnUi1o6XMIQiSspKlqQQrpKB2UPTkDWLLzchpDrSwttYCkqScggjIP4UB90AgrsT4mtfv2Vv9ptRpScBc3Dy3qG6srBEQf740ZaQXfRx1Ib5DbgID9oAoAoAoAoAoAoCsa+2y0rulambbqywwr9DYdD7KJjQUWXACAtCu6FYJGUkHBI9aA+LDtVo/S0mzP2bTVstbtngu223mHGS0IsZxaVuNIAAASpSEqPxIz3oCL1DsJt3quDPiXbR9qnMT7mq8yA4x7y5pbS2qQFDlLhQhKSoEEgY7UB+TNgduJ9nYtT2irKbYxbHbK1DTESlpEJxxDjjASMAIUttCiPUpB70B0T4aNrf8KHTY0LZhZjM/WCo/s/70npKPNUvPUpXQSjJOek9PbigGFabTCsNriW22w2Lfb4jSWI8SM2G2mW0jCUISMBKQAAAO1AdugCgCgPMj9KL4gpeq7o7s1ZrZIaatEmPcLpclLKPMcLIU202EKIW2UPHr6wPeSAAe9AL3wfvRjZkxXceak89Q57d8fcaA1XL0kmclDjRAHcjufyoBg7caYeivtnOekg5+8VAG5dW0ogAOJ6vd54/GpAtZ9piXB9SBgFXGM96Apdx23j2aT7epGOn3gaagumjNVwriwuApQOB0gZ7/RoBL787SJmLfebZ6m159Pr40BjzVG30rTEkyoYKOhXX0jtQD+8PG4ym/JTIcKVowk9XH16UA/r/vCzALQC+D8zQFq03rZi9W0P9Q4BP2cU0QKPrfc9u3yVJDgASruT8OKeAI3SW5DdxfCevIz6U2J3Gzb7yhcdLgUOnGc+v8aEELfdYIaJAV27DP8AOgK23qxiW7gqwO3fPegL9pq4IdSk9Y+Pf4fRoCubdSAjdPeLnJN2tOfn/wC6I+aAaUKUejvnscA+v8vWgIaXrfRjrxjTNQ2lDw/0Llo7/DOcZ7+tY+WY4OMnGVWN/NGEnnmVU5+jnioJ/wCZExBtNquEYPwlx5LJx+0juJWnn5jir2E4VFeDTXgZalWp149+jJSXNNNfAT+9VtfixlBlPH/CK+yqZPuGj3bxd1PvNdfvcAj6+NAcl72oZudvUhyOlTZHIIwMYoC5eGjxWPeHi9RtA7gTnHdCuKKLbd3sKVa1kg+W6o4JZJ8w9R6lhS0j93sBrfd7WF21nqBjarQ1wXBvtwjJlX6+xiCbDa1kp8xKuwlPYUhkemFu9m8EBnaQ0jaNBaXtenbBBattmtjCY0WKyPdbQkYHJ5JPck5JJJJJJNATFAFAFAFAFAFAFAFAFAFAQG4F2lWHQmo7nBWlubCtsmSwtaeoJcQ0pSSR6jIHFALDwgeJi0+KfZi1asiFqPeWkiLercg8xJiUjrGO/QrIWg+qVY7g4Ad1AFAFAKfxLb4f+A+3Dl6iwE3W8y30wbdDU4kJL6welawVAqbSQCsJPVjtjvQGCdL7Be2WWZc7gyhybOddlPHpIBccWVqxnPGVH6FAKm1GRtBuN5PSpEF5zHPHPI5/CiBsPTesWpVpaeaeCk9IPfPzoC+aF3PajTUsvLTgnByfv/nQDxdmNX60BbJClFOe9AJzVUuRYpKnOekK9D9fA03B+ydVN6o026ylX7RAOMHmgEJYNXytN60LLqlIHmYAP2mgNQJeja00wk5Stwo7DHfFAIDcLaxb4fKWO2eMUBnGdbp2g7uXmkqS11cpAxxQHHqHceZc2mlNqVkKGc57/X8KAbe3e55jWUBb+B04JKj8KAqGrNQSdTXlTTDp6RknpP2f3oCRsDNw05JaUesoPYkn0+v40BoDS+qi/bAgqJVjtnOKAhdRKkylLLfV8fxoCjIuMqDNHmZGT9lANTROtUtoR1ue7x7313oD7231Sy7uZuwSsHzbpayOfhamBwaCxMb56kkwduPLhSFNCbJRGdU0eklspUVD7D0gH4jPxxWr9oq06OCtB27zs/KzdvbY5128xdXC5R3aTt35KL8rNte21n4GZzg+gA+A7CuV7dddM82bddcvid2y3u4aclolWqa/bZCSCHIrhbPfPOOCPkc9qrUq1TDy71KTi/B262LrDYqvgp+kw1RwkuKbXy8hq2Tf566Rm7drKMLhGOE/rCMgJfSMd1I7L+0YP21uGB7SVYWhil3lzW/tXE6rlH7QK9Fqnmke/H+qOkl5rZ/B+YwbNtjY9TQE3WyzGLhDWf8AMZOeg47KHdKh8DzW/YfEUsVBVKMrrw+v2Z2vB47DZhRVfCTU4+HyfFPwepF6t0ixZoDiEtjqxxxVwX2xkze7TbHlNNJhi5Xm5PGJbLdnpMh3BJJP+ltCcrWrslI+JAK9wXzwf7ujwoTVaO1eW5Gl7zKDzmoUMqL0aUUJbCXAMkse4lCEgEoKv9ucAelAOaA/aAKAKAKAKAKAKAKAKAqW4e7OjNprWbjrHVFq01DwSly5y0M9ePRKScqPySCaAxjvt+lk2Oi6X1BYtPuXzV8uZDkQ237fA8mOFLbUjJW+UKwCocpSfvoDze8DXivneFLeSLd3VvPaQunRDv8ACaGepjPuvJT6uNE9Q+IKk8dVAf0HWO9wNS2aDdrXLan22cyiTGlMLC23mlgKStJHcEEEfbQHeoDjkSGojDj77iGWWklbjjiglKEgZJJPAAHrQHnHqC6S/Efv/dtUuqkK0/a312+zxHpIfYaQ2vy3H2SngJfLSVnGeAnk0BqC2aSjp06GkIT1hPIAH18PxoDLG+uzIvvmuto6HUZUlSftzQeIlbLr667eH9WXPrS2g9IJ+H30BfdPblCZJbebXknnhVAas2a3Q89LbDyyUKAHJ9e2aAYW4+nU32zqeYHWSnPHNAZPlamnaNvqmFBXlFXag8z41tHi3Vhq7RVJQ/8AvKGec/OgJPa3fM2l1MV14npOFJJyQKAf1s1fa9VMoylJUruOPvoCk7lbHtahYU9DaHWoHICeTQeZnm+bHy7UHUqhrA5PAJ+v70sNypI0FdY6VMsB1BHA4IoC3bcbYXZU/rdYcWSf9QOO/pmlgaNjbPi4w2vMa6VAcqIx9dqgE/aNpfYAlOSB25qQTY27b8vnpPyA9aApOr9pVPpWplrBx6DP3UAq7hZLlp9akltYx6c80Y3KRoHU8iDr7cBSlLHVPgZ+0QGhQeY5rzdEav0XLt5IVJCQ6z1HnzEnI/HkffWJzXCPGYOdOK9bdea++xrHaTLHm2V1cNBXmvWj5x1t7dV7RLHueP7VxnrrrmeT/Prr7n56/wBKdddcyOuuuYfXHpTrrrmF111xJzR+trzoS6ifZZq4ruMONn3mnh/tcR2UPzHoRV5hcZWwVT0lGVnx5PzXH5mVyzNMXlFZV8JPuviuD/zLjx8d7NDuu29OnNU6LuV6ubn6nftcZUm4RQFOq6E499kDKnATgBIHVlQHqCep5Zm1LMY2/DNbr6rmumekOz3abDZ7T7n4K0d4/WPOPxWz5ugbebbXGbMl601XDVC1DcmfJjW1wg/qaDkLTGz285Rwt5Q7rwkHpbFZ3c3Mqe8Wk4b1ukNkDI5STwQR2IPp6fhQD48DPiPc3L0rJ0hqzUDM7XFldLSEPJWmRLhdIDTylqJDrh6HCsp5GASE5GQNVUAUBDayl3mDpG9SdOxWJt/ZhPOW+NKJDLsgIJbQsjnpKgAcc80BijUHjR1Tu5t3HTo5x3S8rUVyiWu1XKzW1253FhbNvTMurnsoQvrQy8tqLkJOCpZJ4BAExojxLT95tV6WTeNxTtVaP8HQrz5TYiMqn3b2p2POZcXKQoFEdxjy1Mo6VZdBJ/doCR283hkattd71Jq/f1nbrUH6+uVrd0nJ/ViGbW0xKcbZa8p9sulwtoQ4XFK97zeBjFAbHoCva93A05tdpWdqXVl5iWGxwk9b82Y50oT8APVSieAkAkngAmgPKHxSfphdRankTLDs1EOm7OOps6jntJXOfHI6mmzlLKT6FXUrsfcPFAed2q9Y33Xd7kXjUd5n366PnLky4yVvuq+1SiT91AQ9AA7jnHz+FAeu/wChq371TqrTmpNsbpClXCwadZTOtt1JyiGHFkGGon0UepaB6AODtjAHpjQGefHPuPI0HsRcolsuEOLeb663bGo8lKVrfjuLCJPQgnJIaUr3hnpyCfSgEF4fpVp05p+HHR0MltsICTgYx2pxsDQFq1Q0XUhKwttR4Ax933UBGbh2xpxsvIQlWR8Pr6xQGUN4tu49/gSFtthLqAVAgcnGfwoBU7V2oszHYUgkONL6Oe/B9aA0bph8WV1pbZKT3z+H1inmDVe3d7RfrGELII6enB5/GgFnvJt3CX5klCAHCCcH480Ble7S7nbrk5ESkqYJwBn0yaA7Fq2znXGSJUVtQWs5IGcc/WP+9AaX2n2yuVvQ27LKkJ/4sg/hQDxiRPJZCCcp7c+tAcE60W+WgpfYbXnuSAPrvQEGrRNhS4V+yN9Wc4Ax9fX2UB2WoVqtycMsNo74wn6+s1AOwbyhKcIScdu2KkAm9dfATg+nHf8AOgOaPdySMj178fL+oqAdpExl8DrSMcd+1SCMvGj7de2ilbSCTwOKAzRE2fVI3I3YTDTn2a520D/qtjC/5mj1BEXK03TTqVIUlY6fh8qbDyKZKf8APkuKWClxRKiCO59T+RzXM+0GWuhV/eqX4ZPXwb+j+fmefO3HZ54Ou8ywy/l1H639sn9Javzunur8daf11182cq666+odv6U666+o666+bDt9fXwp11182OuuvmzmhTZFtlsyory48llQW262cKQoeoNfdOpKnNVIOzW3XXzK1GtUw9SNajJxlF3TW6fh182PvTW6cbVmnnGn20Rr7HR+1Qk4Q+B/5iPw5T6enFdWyfN45hD0dTSoviua+q4eR6U7LdqIZ3S9BWtGvFa8pL+pfVcN1oxPa+mSLk8tpJOCT6+lbHob8J+ftzetO3uJqrTkyRaNRW9fnRZ0dSkLQeMpJBBKFD3VJ7KSSDxQHpp4c92G949pbJfVqWq6ttiFdQqMWAmc2lIfCU8+51klJBIIIoC86n1Na9GaduV9vU5m22i3MLlSpchXS2y2kZUpR+AA+34UAsNmrTftcahl7p6pRNtarjGMTTum31qR+rLapQX5j7ece1PlKHF5yW0htsYKVlQDSh6dtVuWyuJbIcVbHmeUpmOhBb8wgudOBx1EAqx3IGaA68nRmn5iY6X7FbX0xpCpbAchtqDTyj1KcTke6skklQ5JoDlVpezKuz11Nogm5vIDTs0xkec4gYwlS8dRAwOCfSgIzczcjT+0OhLzrDVM9NtsVpjmRJfUMnA4CUj/AFLUSEpSOSSB60B4A+MPxj6t8Wmu3Jtxeetuk4Lyv1Np9C8NxUcgLcx++8od1ntkhOBQGfKAKA/eg+owAcEn0oDvWGyTtSXqBabZFdnXKe+iNFisDqcedWrpShI9SpRAH20B/RJ4MvDRA8LWyFq0qhLb1/kYnXucgD9vMWkdQB9UIACE/JOcZUaAetAYH3cuL/iH31uaQXHdNaWddtcBh2OG1B9KuiWvPdSVLbSBk9kjgZoCla40NP0spK7epbZSAcJ4+uaAse1urrhNWhiWD5qT6dz8aA0s5al3bTiCpIUrp4A+vsoBS6h0WZTTzfR3BHb0oDMGrdLStD6tE1oHyHFe9z8/70A4NE25WoWmiOCoZ+dPIGlttbYLLES2pzKiOxNN9gd3XliXeIxS2SfTOfnTYC4s+ybFwkh2YgDsc4oBmWLQ1qsDIS0ynKcYJHagJsuhtISkBKRwAkfw+vSgOnIuSm/9XPwHx+sUBHvziUn3sDtn6+6gKLrPd3Tmi21uXS6ssYz7q18/GmwsJS+ePXbG1vqaVeEuEHHGcfClwRLX6RXa0Lwm5A4+AJ9P4dqAnbV4/trZ7mBdAkHj3uB99SBj6Y8U23mpelMe+xypfGFEjP5fOoA07TqO1X1pLsKcy+kjIUlXGPrFLAmQVtpBQfT5fP8AofzoCg7aPJG6O8vmjPVdrTyRx/8AZEf6+jQFxvuiLfqCKvqbAWofDH5U2BnjcjaJ60uOPsNFaQerKfT4c18VIQqwdOorp7ooV6FLE0pUa0e9CSs09mhUyIrkdZStJyD3xx3xXJc2ymeXzco603s+Xg/Hk+PndHmXtN2ZrZFVc6d5UJbS5f2y8ddHs+Guhw/ZWA666/XSH18f163O3b8aPrrr7n18f163B6UfXXvHXz/Xrfmhy3YMlp9hZbcQrKVD7OR9hHBHzqrSq1KFRVaTtJbPrhz93nc4bE1sHWjiKEu7OLun4/bdNcVp5tbSekkbgRG7jGSkqSeiQ2Dktr79vge4rsWW46GYYdVV+JaNcn1serMgzmnnmCjiYaSWklyl9nunyJzU+hItktCvNACwMYNZU2OwrPDZuW9tn4noOnl6hZtWldSIdakQpikpbfmjiMG+o+64tSynCMFfug5wMFoDUCAfE1uAVq/bbSaUn+4M5b1Hd2F9/wDjiRXE/MOPp9UsjqAflAFAFAFAePf6Y7xIydSbh23aC0ylIs9gQ3PuyEK4fmuJ6m0KHqG2lBX2un/aKA828ZzgHFAW/bPaTWG8WpGrDorTk/Ul1XgliCyVBtP+5xXAQnt7yiBQHoZsh+hZvd0ajz91NXt2JCgFKs+nkpfkD5LfWOhJ/wCVKxz3oDVj36OfYLafbfUcyBodm8XSNapTrdwvr65joWllZSoJUfLBBAOUoFAZW/RA+E79c3Z7ezUkPMKCpcPTjLyAQ4/jpelAHuEAltJ594rPdINAetVALTxGbhxNtNoNQXSS3IfdfZ/V8VqKMrU+/wDsm/UYSFK6ic8BJxk4FAY28NM6No+1QbfNOHUISlZWSVKOOSSTkkkknPf7aA0dfNI27WURLsbpWsjkd8/GgILT2z7dtuaXC304OeAPr4UA3I7TVviJjkJAxjBoCIutmZktqW0AVKBzg0Blzfiwn2eQUtZWkFQwPlQC0273QTZUqjvK8laCQUkEdvnQDHh76yPbEhiTkA9ge3yoB+bf7mM6ljoZfUnzcev5UAxGkpUnqTgD5UB+lpPSVqNARct/ozgDg9jx8KAVu5e8Vg23gOSrtOaYKRnpzyaAwXvp+kmffTJt2j2SO6PaFE8c9+4qNwYf1tuZqTcCe7KvVzdlqWeroUrISc+lSCtIZed4Slah2yASPy+ygOy1aJ8ghKYj6yeRlCsfwoC4aY2dvupFpKWvJyf/ADO59PjQDGt/hZ1Qy0H4ssoc7goOPvpoC66We3k2ikIdg3CQ+y2oe6Sf60BqHZz9IJPivM27WcF1CuEl7CuPn3PqBQGl9h9w7Hr3X+60+BLQ63NuVqcbSVYJAtTCT+YNAPNn9ilPTkp9DQBLtrF4jqaeSPv+vupYCO3K2U61rehoKuo4ATwckHgY/CqdSnCrBwqK6ZQr0KWJpSo14qUZKzT2aM3PJeN0kONLH6sZWphp3IPnrScLUDj90EdIPYkK+VcwzHKXQpznhYOcb73vZLkt2r3Tly0txfLe2fYKj2XytRw9J1a9SSqSbbvh6Xdfdg0vxSq3785NXhCMI7ykzlx0n4Y9a1Xc4Bv15/frQO2Kl9de0b9ef3Aff9f9qPXrzG/Xn9+tBj7Ea2c0lrZqM4vEC6ARHkk8Bf8A5S/lhXH2KNbHkOMeGxih+Wej8+Hx09pv3YvNZZfmsKUn/Lq+q/P8r9j08myc3j1PKVIeaT1ZB4AH3f2rrJ6aMObsWK8at1SYVifejXWClM2RcI5KVW8clrCh2dURlIByEgq+GVwes3hU1jada7C6Sk2a3rtUWFEFrVDW0215Tkb9i50oQSAkqQSn5KBwM4oBt0AUAUAUB/MjvnuE9uzvHrPWDxUVXq7SZqErUD0NqWShPHHuo6U/dQGi/Ax+jz1B4p5repL68/pvbiM6ULuCUYkXFST7zcUK4wOynSCEngBRBAA9rdpNmNF7F6SY03oewRLDa2gCpLCcuPqAx1uuH3nFn/cok/dQF2oCN1JY2tTaeulnfccZYuEV2I441jrSlxBQSMg8gHIoDp6D0RZ9tdF2TSun4iYVls8RuFEYHPS2hIAyfVRxkk8kkk8mgJ6gMfby3lrejeEW5lxiRp/SRWwh1rqJVOWCmQlWfdPlhCEjA7qXyewAWus9KpgXJHsZDak47H04x2oBg7WakuUGQ0w8pSknA94Y+vSgNBpkhUNLpThRGSKAgLteEx2lOurASgZA7DFLDcoLm+Vqh3D2RT6Cc46VK7ntQHNfY1o13CUtpaPNUkkDg5/hQGftabIM+1PLDPlkn99IH8R9lAIO8W97bfUf7V9S21HCUk9vrFB5Di2113IYucRbZUlJUAByOPrNAbi03cxNsbEhR5U3k/gD/P8AOgOy9LCkn0GOefr50BQ9ytTq03pWdcGSStpsqGPln+lAeIHiD3t1BunrKf7dMc9kbd6UM9Rxj496b6g+NstlZGsA24thxbav+H+tAai2+8FVjuCUP3WOG2u5SoDkevH4UA99MeF3bSwtpCNOxpDgHK1toOfj/pqdmC8MbO6EbZDZ09GZHbKW08f/AE1AIq6eH2zJSZNjShtaRnysD+1AVxFtctb3s0lpSFg47Yz+NQwd1VtZfR7zaVp/l8qncWtoU3V20Vm1K0rzIqG3scLSkA5/D86jZASNha1XstrbU8mwyXixDlxkuJSTyDGbUOx54Vip2Btvw6+My2a5DFq1A4Idx4SVOkgKP30BqSbeIttsM28FRciRIzktwsgKPQhBWrGO5wD/AAzXxOShFyeyLrC4eeLxFPDU/wAU5KKvzk7L4tFI1VqNG4bcDTGm54cReoSJ027RVZ9jti+ykH/1XuW2x3GHF/6ObOpP09qVJ/iV2+UX9Xw95tWAwH8I9LmWZ0/8GThCEvz1lwa4wp/inwfqx/MLjd/aZmFFaFqjNsRI7aWm2WU4ShCRhIHyAxzV/wB1QioJbGo1q9XE1p4itLvTm3Jt7tt3b9rM93VEmwvht1IcRkAg8EelatmGQUMW3UovuT+D80vmvczmOedicHmblXwjVKq9Xp6rfils/GPtTZ2m21uxw+ltflK/19PA+0+nrXPcXl2KwP8AjQsue69/3OHZnkWY5O/+LpNL+pax/wC5eWzs/A+R7wB4wfUfXzrHdde4wG23XViT0vbZV41JaoUNBclvyWkNhIJ56gScfADJPyBq5w0JVK8IQ3bXz/S5kMvoVMRjKNGiryco29+/std+1jw8RumnrNMgwbJEau2sNQyVxrLbXFYS4tI6nZDxBymOyj33F/NKR7y013M9kt3bZVYHh6j7a6NVCLy7jOdUuTOuT6Ql2dJUMuPL+GfRP+lISkcChB++CO+xtC71ax0cqG8V6ijouTMlKkhtsxh0LSpJOcqDySMA/uqz6UBuegCgCgCgP57vAt4UH/FZve1aJgdZ0faUpnX2U0cHyc+4wk+i3VDpB9EhauSmgP6AdPaetuk7FAs1mgsWy1QGUR4sOMgIbZbSMJSkDsABQEhQBQBQBQFU3V1rE2727v2oZsgxWYcY9LqUKX0uKIQ1wkE8rUgcDjOe1AZZ2x0/K0xoJiTcn1yrs+gPzZT56nXn1J6nXFq9VKXkk+pOaArKJ/681AppSiSDj7PuzQDm0ho5trokqTwgeoHyoD711uZC04nyluJSO3T1CgFjrfclNw00+808EpKSRg+mKkGS327pq27Oy4UlfUhZ/dV8z8/r86hLUDF01r/UWiHo7dxLhYJA6znHw54oB9t64Yu9iS66QoqQMKJ+/wDpQGUd2H4WqNcRISHEK6TnANANPQ2iAtyOttOOlQ5x2+uaA13pg+yafZaz09CBn8MVA8gXcAUKSpZynjB71IKpqyGjUtmm29fvJeQUnAyBS423POu6+EXRcXcuQu6XPBLhX7ID/Lq+fwoDR+mNEWLTsZpm3wW2WGxhI6Bk/bQFh9s/bobRgJ+A+Haj11YLRb+hmN1EYVjk+vb6/KgPyRNQolOR+Pp9lAc9qlLjyAUqIT8vr7aAkL1pOFqZkOhCUP47pGPT6/GoBVZeh5kJCvcKgn4D519N8WCvyoCmz0rBSfn9fPNALS1xIr2rNxUy/LS37bBKvMwMJ/V7We9fSjKTtFXGwld4NADTspN609ObYkoJdCGncE9j6GrmGDxNRtQpt23snofPeXFjn8PHi8uWuNt9SaFmBUrUcq3PQYCCoBTjjg8tPKsf7wcnjAqwx9CvRpypTg1JrZ767acNNdeGuzTNl7M1MLDOcJWxc1GlCalJvZKPrP5W01votTUnhk8OOrtrLamRddaqajyCl52xW5pt5g+6APMecSScDH+XgDHBrF4LB1cPrOe/BW+bX2Oh9ue2mU9o6ndwmCu43Sqycoy3vpGLS8fWu+aQ7tQWhE6E60tOSRxkdjWYOMmLd7rA9Z72QEHy85Se3qeKgERo7VarEzlYAbwAUkAg/HIPFQ1dWexDSaaZOr1Xp+7uKeftMdw9z5CehSvjynA/HjtWtZnhcrw9J18RSXhbRt+yxz7tBlvZzA4d4zHYeN9ko3i5PklFpeb4LVkDK1nJ045Ju1pS9aDHbW6lFpCi+QAT0pOepSj2xkAn071zmOIbrp4e1K7srN6ecnd+fyOCU8bN4tSwNsP3mkrSaUU2l603d2/qe3gaj8NFmueoNJq3T1k97frfUUb2ZWQOm2QWnFBuG16ckea6oY63FZ/dQgDruArxxGHhUUu94tWvbS9vG1/aeoslxccfgKVeM3O6a7zVu93W4uVuCk1dLlyPrcu+tFDyHFADBGMfb/er8zZlSPdJlp8SG2cq1TX4Dz2oYkN9UdfSXI7jqUuNq+KVDAI9RigPTegCgCgCgPJj9DFvnY7JfdX7W3BqNDu95dF1tsxWA5LLaOl2OT6lKR5iE/DzaA9ZgQRkcigP2gCgCgCgER4w75a2dtrdp6c64mXf7vFjQ20IKkuLacElaVEfujy2Vnng4A9aAq12bTG0uEDuEFPf4Af1oBIaUkeRrF0LVx15we/f/t3oB/ytXMwLOEIcAV0enoPr+FAZi3xbuV5gvSI7ikqTlQAJoBAafv2rNUIdsyErDYJQVfAfh8qAdG0e090sT6fPbU55igSSCeM0A2dxtrVzdNEoZy505BCcEHHegEOm6ahiSk2NUZzp6unzfTHxoDh1htsrTzrV9ClKkp95XVkZoC57ebmMR2Ee+FYxxx9fH8T94D00fu9FlPojOrCQvj4DOP71OrBZdQXr9WkSkkqjqGT0nt9c1AIRW49kgMLfdkpBT/p/lQGcJrcLVG40u/NjqQBhJJ49KPwBbxLUhvoz8hU2BIWNouSUqV2J/H0/nUAujjYEVPSPQcUBFezq8xRJwCc5+/vQkk4SFKOAMY549OP7/wAKEF2skZQQM9sd6As0SC1JHS4jqzwQRn6+ym2xAst5rMnSWmrlfkNBSYrXmFJ7E5AA/EirvCYd4mvCiuLIk+6rnmLrndWT+v7lIaUfaJbiXH3eokrUlAQn8EpSOPhXq3IexmDo0VOUFd2v11rfTXXCVcRK9ijS9wbhKCgt1WDwc/hW80sgwtLVLUtXVkyKtmpZWjtUWy/wVrYdZeT19KsfA/y/OuFftGySnhqcMZSSTi7eae1lsrO74bt21Mng6jk3Fnu34bNwUbgbU226vup9xgF1xagEpAzlRPoMDv2rgfiZaMXJqMVdv3jIfDM6CzKjOokRnkBxp1s9SVpIyFA9iCPWoumrxJnTnSm6dRWktGno01unya+Gwit7NKR7o35zieQMkn64r68D4Mta3YbjJMKN++4roSAOSTxiqVWpCjCVSbskm35It8RiKeFozr1XaMU23yS1OG1wRbYKGAorIGVqJzlX9K43mWPnmFd1ZbLZcl93u/dsjynn+dVc8xssRPSK0iuUb/N7vxdtkdvPSCcZxzgVi+uuvqa31119T0D2006NNbS2C1ODokMwUKcT6pWr31D09VGu1ZbReHwdKk90l73qz1xkGEeByrD4eS1UVfzer+LM67u265LnvJSlQBJ5xxWS9psBk7dg6g0C/E1haVNi7WF5Nzie0o8xvzWveSVp4yMjnkUB6zaZnPXTTlqmyCkvyIjTzhSnpHUpAJwPTkmgJOgCgCgP5d7fdr5ttrZq4WyZIs2oLLO8xmSwotux321cEfAgjt68igPdbwMePHTviq0xHtF0fj2fcqExmdaSehMwJ/ekRsn3kHuUd0HIORhRA1iCCMjkUB+0AUAUBnDxpaeuNxtOgLxFZDlvsuoBInOqcSnyUOR3WEKAPKsuOoGBk+8T2FAQN6mhdhAOACjI/CgMq6v1dJtes0twEqccUrsPtFAMzTkq8XtDRkJUlB55xj/vQFs1FpRyVaC2EklQwSBwPlQHR2r2abtrq5L7ITk5UpQ+ymnADLk6is+n5SIiEoK84zj6+f40BbYlwhXaAPNCQhY7H4cf3oCgaxtVgschMtyMkKJyFY7fn86AqeodNRNd2paYgAGMdIH8+aAQWoNjLrYn3HYa1Nq79PoMfXrQFPOodQaRuKUzGV9CTwtI/p60BobbzdyFqizCDNdBPSAOrv2oBR+IiDIsthmyre8R0gqHSfkfw/tQH3s5Nau+lIjjbiVuLB6sHnOSP5/l86AZCmPLUlBHKj3oCzWeMWkJUR6fD5fX41ALO+EIjdiTjGKkHDBgqlO9h8vh3oC027TqgUlKSoAfgKAtLMNEVoIOCqgOONe2IL3Q4QO/vYPHx7UBU919S6O1/tje7SzrGxx5EhkhClz2v30nIQQVepTj7cfOquBzChhcRCu5qyfNGwS7N51L1Fgqt/8Aly+x477j2VdrvkgKHudXuqSeoY+R9ftFe6ezmYUswwMK1J3T4878fJ8DQMdha2DxE6GIi4zi7NPRp8n4+BT+2Tj8OwrbCxO85EM62qaA94n3QO5PwH4Y++uV9v6XpMnrq9vV4eDT9ztZ+BeYV/zEb50lt9uI7tNpyYmVJe26Q+z+sLewtZRGbLvvLfaSUqdbxnKsnA74714UxuHxV05y70Pdp4rl1oet+w+c9lvRTo4XC+hxvdl3XJ95ydnZQqSv3Zck0tbfiPQqTH3NiRkpQrRUyOlICENomRQEjsBguADGMVmksQopLu29qOLzl2endz9PF+dOfzUOIpd17/rKLAWmdp2yrSOyoN5WTj4gOMJxS+KX5E78m/sUP3bIZr1cXVj/AJqKf/5qv3293HNa313KauW+x7O4kqSGusLAPxyBzWk9osdNxjg7We8v/FfX3Hnj9pmbUKVf+D4Gt6SGkpSs434qPde1ndvno9j77VonXXX1OD9ddfUs+22nhqXWVuiuN+ZGbcD74xkdCOcH7SEp+/7KzGUYP98xkKbWi1fkvu7I2rsxlf8AFs0pUJK8I+tLyjw9rsvb5m+bCp2XFCnARkZwTj7q7Iertym69TZGUrMwJC/XjvQGJPFXOs7WjL55Sk49le5/6FUB6J6H/wDuXYOcn9Xx8n4/sk0BN0AUAUB/Pl+kb2hf2h8Wuto/keTbL3I/X0BeD0rbkZWsD/ldDqf+mgM5WK/XLS94h3a0TpNsukNwPRpkR1TTrKx2UlSSCCPiKA9OfC1+mJftkaHp7ey3uzkICWkastbQLuMcGRHGOr5rbwf+AnmgPS/bPeTQ+8lkRdtE6ptmpYJGVKgPha2/k43+8g/JQBoC5UAUAqfFDpm56q2Sv8azpQu4RlR56EOOBAUlh9Dqxk9vcQqgM723UDWqtLsPxnOpp1kOJV24IBH34oBYI01FtOqTJkALUpZOSPnn7/SgH/o1+1vsoy2kevA/t8qAYbdmhyGkFCQRigK3rSd+poSmYqcZHoKDzM+XRM2Xew6sKyVev29qA7Oq9bXXTrEdDRV04GcDn0+fzoCx2fUDGt7EG5yulfT+8e/r3oCtp1IdCy1JDuWc4BBzx9v3fxoDtT95IMtHvltfyB/lQFTu18sOo21odS37wwD8fnmgFnftGLt1xEyzS/JPVkJBwO+aArW4t+vrunHoMxpTqCgoCh/3oDPG2++l32s1MYziluRQ6Mtknsfv+2gN46V16xuRDt1yiNhDaxkpTyAefwqEBpod9mjtNpwVHgc/zoC2Wi1ofjoU+cZ/L8KkE7FjQ4SfdQVK9aAl7fK8wduhP5UBXr5qhtMxbcdxLim3C0tKTkpWMZTx689vnXypKX4XcqTpzppOatdXV+K5+RQrtq5283Z2DYIbt8urXDrcdYSywfTznj7qO/7oyr5VbSxHr+jpLvS+C83w+fgbJh8jmqCxmZVP3ei9U5K85/8ALp6OS/ubjD+4ol48P51XudYLpfnIrzUguv3hMJjymf2XQW2091KK1KIK1HJAJwMYrGVMslUrxnJ3T1dttLfP5a7nV8o/aFTyjIMXgsGpJ0+7Gj35d6T73e70npaKja6hFWi2ld3Ev449mGWb0nU1nYBizT0yG2gAlp7HAAHZKgMj0BBHqK9O/s7z5YdPL6stFt5fo/DZvkebcbCVWTqybcnq292+b8zEztsfafKFIUFdifUH+Vek41oSjdMwzTRoHwwbB3LdLUiI6GAhllPnuLcTlKVchsHjuVEfcFHHFcM/aRnUFl08JSl609Fryabv4aWfmZjLY04V4VK8bwTXeSdm1xSfB22Zv/wq+JNS0I0M5t3dpU+LlqQ5YkoeaQAopKnUOFPlgHgkqIryJhcznWfo5023xt+u3vZ6K7S/s3w+TUf4hhcwhGk9YqreMnxSi4p9521/Cvqa/nvpMZI6OgBIwj/b24x8vlWc2XI4U73etzNO/Dr0qM423npSCVH4D+n9a+Z1FSi5ydkrv3FGrVhQpyq1XaMU2/JaszekBCEpHYD54zXDsTiJYmrKtLeTv9vseO8fjamY4qpi6u825e/ZezRB+XzNW3XXXEsOuuuK8DR2ztns+3bWnTqBwRrzqiaIkZtaeUK6FLQ2fhwOf+JaR3NdSybCxy6jF1lapUfu5Lw8fF24Hq79n/ZDF0MrxGNjTvOMfST5xhdKK89e815/0moH5ca1QeVBA6eSTjFbVsbV4Gdt4Y6tQFx2K+PU9OePWgMiXXTZlb27cWu8oZuNmn6khQ5UKWhK2pDS3QlTakngpI4IOc5pxuD1UZZbjsoaaQlpptIShCBhKQOAAB2FAfdAFAFAZC/SQ+EBzxPbTN3LT0ZDmvdMhyRbUcAzWSMuxSfirpCkZ7KTjICiaA8GpsJ+3y3o0llyNIZUW3GXklK0KBwUqB5BBBBHpigOHNASentT3jSN1audjus6z3Jo5RMgSVsOp+xaCCPxoDUG3n6UfxC7fstsuauY1VGR+6zqGEiQr73U9LhP2qPegHzYf03mtIzWL1tnYri7x78C4PxU/goOfxoDUHhB/SNw/GFuHN0I7t+vTy02h+e5KN0EhCkpWhsthHlJPIc759O3NALGHMOz24+pNCyI0qBBtc9xq2JlheVws/sFJUokrT0+71/6ik/PAEhqWSxdZLS2XAQSMEHjt3/L8hQDL0lGFutoeVxjnk0BeLBq9EhYQlR4OO44+3+9AS+pGY8uOHXCMHuD6UAvblHs7J8wKQFJ57igFnuBqyyhlSXHEcDuVCg8hGXHeyNZFusxlKOM4CR35oBc6k3tul/cXHYjPKUfXChn64oCoKuOr1eY6ht4I74JPH59qA6LevNWQXFKLDigk84Sft/h/CgJ6zb/AEqK4hueh1og89SSMfWaAY7e5tq1HbOlxxtZKTnqUPzz99AZK3ygRWL8JURacE8hJFAaO8EG5js9H6hdQXlo5ScHI4/t+dBeyNruRZEd0KdUQrvyRQEPrjUeqSwzC0xNbZdSkOywGMvJZKwMocJwlR5wMdR6ScjHNjiPTStGi/PTh5/obr2ep5RTjUxOcwco6qn61oufdbXeil3nFO12pJJuKad9Gdb9c2e9TFR7fNiyXAkrDTL6XFBAIGT0k/FIyfiPjV2qkJv1Xc1ivgMZhIqWIoygnp60XG75K6Rz621s5ozSsqdFt791n9PlxYUdtSlOuH93PSOEgjJJ9M+uKpV6rpU3KKu+CWt318DIZHllPN8fTwtatGlTbvKcmkox4vXeXCK4u3C9s47XjUE1+ZbdaR75b4U2S7N8xuMppuQ84QVpedHvoScDAHSD2J7A4LCRrTfo6ykk3wW7e93vb76nc+2VTJKEKeY9n6tCpVpwjCzmpOEYL1XTg/VlKz1b7zVk4xvdjls9/j6Sjey29LUKG2nCGGAEIT9gHGfn3NbOoxpruQVkjzriMRXxlWVfEzc5y3cm235t6lUm6/XJmrIdUnPw9e/OKhSad0UD4kmTqyI7EXFE6M8ClbTg6kLB9CMY+P3/AGVd0MTOjJVIOzWz2+SPhq6sQNs8FWn7xLM24IdtMckHmYhCUA8YHWDgfb/Stz/24zPD01FzSXNpX9/Pj1rZ1KdKku/UaivFpfFmjds7Ztjs3a2rZCvVmhoSsLWlElLilLOB1KIJJPbk9gOMAcc/x/aGli6zq4vEKUn4rT2LZeHmYqee5Ph/VnioL/qX0uRsOPtjp+DLl6X1jbbTqxV2lXiPd1IWEl19ZKmHgAOqOUhKCnPoFjCgDWtfvuXL1qdZKV20/Pg9NuBu3+9fKsZUp0cwxEZ4ZU6dJwu1pBWU4tr1aid5KW28ZXjdDK0VvXprX9mBXc7fa7u0ssTLY7ObKmXQBnoUSPMQoEKQscFJGQCCBlaGYYetCM3USv4rW29ua8fqWeZTwGC9FVo4qFShWTlSqJpKcU7Pd6Si9Jx3jLwabrG+lnbh7cXu6J6VqKW20uJOR77iUnBHyJqzzqo6eXVZRe6t72l8jSO1td0cixNSD3SWnKTSdvY2ZI5J47/X9q5B1119Tyx118erja2T2klapktXybGItbSuqOlxJAkLHZWPVAP4kfKtxyHKXiJrFVl6i28Xz8Uvi/bfqvYzsy8dUWY4yP8AKj+FP88lx8YrXzenM5N8tgtdbha/01cFXqFpyOq4fq6wx+tTrzSktOyDJcKeEqUpkcDqIHRn93B2vGYatXqxlJ91Xsvde/tsfoP2M7S5JkOVYnDxoyrS7nfrPRJpyjD0cU9Woqb1dru/NWue5u7+oLTpyFHvbTEe+ttBuZ7G71srcHdxs4Huq/eAIyMkfOs5S7/cXpN/n4+04ZmMcHHF1P3CTlRbvHvK0knwkua2um07X4iBTvotTziZDhQMk5Uo8fb+dVDHbF38KltVu5v2dQttw5lj0owtUjzFhSky3hiP0oIOekIcV1ZBSQnHegN7UAUAUAUAUBg7x5/o1Lf4hJErXO35j2XcEo6pkRzDcW8EDgrI/wAt/wBOvsrsrH7wA8addbf6j201NM09qqzTLBfIiiH4U1otrT3wQPVJxkKGUkcg0BXqAKAKA3l+hn48Vt09P/hiX39f28egNefpDNCm27i6T1ZFluB+7wnLUpgpSlCFR+p1CgRySoPLB9AGxjuaAyVB3dlWO4tMTlKwhXTlR59BQGp9CboQtSWNtKHkkqTjGfX8aAten50e2OqedeAR3HUcD8zQETuHv3brfGWwmSgqAOAFj7KAz5ed273qSWWrc055SuOs5xj8OKAq9+0zd5imX7vPWhpSgFJ6jxk/bQFotW2dnZYQ8I4eOASpQzn8aArd1sUW33tryoaEIBGcIA+Py+sUBdJluYbsqnUspBKTwE/Z+FAVXS9gYnuv+cwkpUTjqSPnQEZqjaa03dxaRES2ojgpTz+VAIPdHafUGhGXJ1pdcXER7xQkEkfnzQCKul/m3bCZLnVgH400GpuL9Grt6Ljcbje3kdXTwjIBHYf1oDam5U236ViuXC6zUQooPSkrOVLV/tQO6j9lV6FCpiJ+joxuypCnKrLuQVxK6d3Yt941TDs0SPMht3OYEuT3FIcf61+6npTyEgYSAecAdu5rM1sgrUaEqnpFdXb0+r5eXtNhl3YwVTEQUu5FKMbtRVt27es76ydpR9Ztu60L3bL9pDaDV1wtEO1zZE+6uGU5MaUytlAS3koUrr6khOFqIUAolZPYpxptOVCjW9FSi25btarT28PqbNjqGbZ/k8M0x+JgqeGj3Yxl3oyak33e7FRs+9+FNO1o22ixV6s8Yn+HNxER59vfTYlDrS+UKCVoKsBWcYwcHmr+FWE7qDTs7Pwf3XI5o1zRf9LTtaeKGfIe2nags2uEjplXO7Orah+bhP7AKQhZLnSoKIxgA8nkZq95viQVDedOtPDvf9OWncB+zuO39t9yK5ZpTjqB5RQFhfmNt4P7RJGPn98eZJYNKW6NfMTn5KBDSOtSiRjFU5TjCLnJ2S1ZTq1IUISqVHaMU23yS3ZNS9eSYyDGsw/V8YDpDgSPOVx3z/p+HqfsrnGP7R160nDC+rHnxf2+ZwHOu3mLxM3Syz+XD+reT9/4fDd+KK5NnSbm4XJkh2Ws/wCp5ZWfzrUp1qlaXeqScn4tv59fTmNfEV8XLv4ibm3xbb+fX04c47ceoA4qnddddfK2Wm3XXXgdRB79vWov1119J666/Tj8hsveb5aS70hHWRz0g5x+NLtq19N+utfcXbxeIlhlg/SS9EpOajd91TaUXJLbvNJJu2qSvsjuNXKWxDdhty30Q3seZHS6oNrwQRlOcHBAPbuBVRVJqLgm7Phd2665FONerGDpRm1GW6u7Pjtty6sXHZm26UvGuIcPVinhEfUG2EpUEsrdJwlDx7hJOACPUgHg1lcqhhKuJjDF3s9uV+T8PL5GydmaOWYjMIUszv3XpH+ly4KXg9Nt3vobri22NBaQ0002y22AhLaEgBIA4AHYYHGPlXYklFKKVkj1XCMYRUYqyWiS4LkiibhXBj/H23EYrQEInzZSiD6NwXU/hl1P5VbVf8WknzfyNryv1MrzKp/ZTj/3VYv/AMRWbv6Ab1PJckNnrCuaur3NWMwblbdRbPaJa3AGlJQT1D0wPzoDZ/gg28mbd+HTTMe4ojfrC4B65rdYT7y2pDq3mQ4ogErS24lJ7gYwDjFAPqgCgCgCgCgCgFzvT4edvfEJYBade6Yh31lAIYkrSUSYxPq08nC0fYDg+oNAece9/wChXuUZ6TP2p1izNYJK0WfUqfLdTznpTIbSUq+HvIT8zQGMNxfAzvrte47+uts745HbJHtdrY9vZIzwetjrx9+KAStys06zyPInw5EF7n3JTKmlfgoUBuX9DRhPitunvJH/AMLyyef/AMePQHrjv9tmndXbC62ZAdNwbCZsHylJSVSGT5jaCVAgJWpIQr5KPwBoDyj3H23W/IXKVCdgSmXFMTojqcORn0+642rBIylQUOCRx3NAVy26kY0KyOmcUu4wEdXr86At1r1Jq7WDXSw4tlhX7qzxj8vrFAcadCSbTeGZd5kOS2VEZySU/XagG7bNMQbhDZct3Qkp7BI+/wBKA6estOF23thxCgUcdiPSgJDSLKZEEske+B6igK/frAFSyUoz73c0BMQYHtVhcaKffSkenyH96A6mlrD0uLPT6nICftoCQmWxlp4rV375/HtQFS1ZEi3C3SGHEIUhSSnBGaA87dzLE3YdYTYrP+UV9SUj0GP7mlwb+8DOpIe2G0E+9XMENISFBtP77qyEhLac+pP4YJ9KucPh6mKqKlT3ZVp05VZqEeJBa+1/ddxdRPXa7PAuqyhmOg/s4yPRCAftGT3JOTXTMHg6WCp+jp78Xxb62RstKlCjHuRJ7aezRY0p3Ut3KmrZACvJHX5fmvEHPIIPuZzkHvj4Vgs8xUFReFvrLfy/X5cC1xGKlh2vRv1t9k7expplh03uTB3T3Hl6fhQmzY7JDU4pLeEtgqcCUo6QOerpJI9enJzXE+1eaTyfL/SYb1ZyfdjotL3bv7rrxtpa5kchwdTO8bKOLnKUNJSu3eTWkbv2teEbpWGupXm8YCgRj5Y+H4V5zliMRUmpzqNyTvdtt301vz0WvguR3KFClRj3IRUVySHL4b9wnNP3OJpFxtCLTJKkQ2220oEdzlWBgD3Ve9x6HGO5rqvZDtBipYlYDFzc1Jeq3q01du7erTXnaySVjnXajIsOsNLHYaKjKNu8ls07LbmvDfW4lv0qu3c/Usrb+/8AszibVbmZsRU8EdDch5bBbbIznKktuYOMe7yRxXa1zOQmYNod0PO083ElF1p9spGHm1JDgGCOknhXoatcVQ/esPOje3eTVzG5lg/4hgq2Evbvxavy5fEcESaxcGQ9HcDrZ7H+R+fNcXxOGq4So6VaNmvj5c0eSsfl+Ky3EPDYqHdkvc/FPiuKfv10OY8896teuuv1x3XXX6n19tOuuv1dddfqU666/V111+pTrrr9XXXX69S53WLZ2GnpboZbdfajIVgnLji+hA4+KiB99VKdKVVtQ4JvhstX56cOncUaFSv3lTX4YuT1W0VdtX3stbK70994210TM1lqOGGwpmAw8h2TM7BtKVZISfVRxgAff2rJ5ZgKuOrxUPwpq78vr0zY+zuR4rOcXD0Uf5cWnKXBK+yfFu2i9vM1vqrcRuGpRS4AclWM12U9WiK1duQi9bjW17zji2WuS4FBWMLfcbQP/pbX+NWyj38VG6/Cm/f/AOn0jaab9B2frSb1rVoRXlThKT+M4beHMi5m7K4iV+Y91J+eKur3ZqxSrDYrz4ldfs6ds8ZL9kivsu36a9lLDEYrypklPJcdShxKQPtJAwagHohabVEsdrh26AwmNBhsojsMo7NtoSEpSPkAAKA7dAFAFAFAFAFAFAFAFAdK42S3XgAT4EWcAMASWUuYH3g0BHWbQWmdOXFyfadO2m1z3EFtcqHBaZdUknJSVJSCRkA4oCeoDO3id8L6d0YU3UOl1IhaxSyAptz/ACbilIAS25n91QSClCgUgFWVZHYDyste1l1u+triq5trYXbZTkaTGKgry3W1dK0ZSSDhSVDIJ7cZoLGo9GGyOQG4qCGnEDBA7g/X8KAsN50SzcrarycPNkZ+dStAUa2Rp+j5fThS4v44+VQBmwFRdWW7ylAIfIwM/H4fwoCHh2B+z3IteSQknA+HpQbaIkp2jpEt5LjbRAPJJoDv2XQjzaVB1ASFAd/woDuxNCphFShg9WSDg/X/AHNRdArmotKupcUr07A5+vjUgV2tG2rPapTrix7qTyO/b+xoDLekvD/dd5twHp62VN21K/eKvUccUsDTO5+38jRFk05ZrZDfcgBtxZWy2pQL2QnBwO4TjGfifnW3ZC6UPSVJNJ6LV8OvcZbAdxd5t6/Q7e3nh9u+oXW5l/bes1oGD5SziTIHwSn/AEA/7lc/AVkMdnFOinCg7y58F9/lz5FevjIwVqerO14k9tJ8jQMxuxOiExFjLQ0yzwEgJOMcfEfzrRak5VZuctWzCOTk7vcc8PaLbXSu0OkdxNFG22+ReIEC0XFu3OIDEuWhKytRSkZMhLinUryeyTkDpJrmvbjAzxeXqrTSvSfefB92zTs342duNuLsjfOx+Op4XGSo1ZWVRJLl3k9Pm+mVLWGpWtJaekXR1PnBooCEf+oVLAwPnjn7q4rluEWNrqi7pNO7XCydm/C9l7bXO+5dgZ5hiY4aLte93ySQwtl7ZM1nrCxSLH564fmJluXCKWgYzWCQ5hwFKve6U9OCffPHBxtnZPLq082vVTXoU2+V9kr+N7re6XtWg9rsR/DsFUwtXSpL1bcfF+SXHbbma411tzpjdCxos+rLFCv9sS8iQIk9oOIDic9KsfEZP4mvQUdlc4Gyraw8O+jtb7SW/bu5RXTZLZHbYtzzZSJEEttlttxpRSQlaUkpBxjBIwa+iDAG7Gx+vvDW/bVXJlOqbLNV5KLpZmlAIcwtQacaJKknpRnrx084yDxVvXw9LEw9HWipLxLLF4LC4+n6HF01OPJrby4r2Ffg7l2YqS3cHTBdz7wcISf7en4ffWtVuzWEqO9OTj8V8Tn2L/Z9lVd3oSlT8mpL3S1+JZ49607KQFIvrAJxkKTx8+yqxU+ys7+pWTXiv1Naqfs2rf8AxYpPzg18mTUO0QpyQW71Cwceijx931zVL/ZbELerH3Mtv93GNvriIW8pddeZMw9FW53Hm35oA+jTWf4mriHZV/nre5fqX1L9m0n/AI+K90Pu+veQO7WmNNo201GmHIcm3diGZsJTyhjz2FB5HujA5LWOc96zuFyHCYSXf1k9d3zVnptxZuOW9isqy6oqvrVJaq8npZqzVlZaptFoX4jbHGtMN63PMx4kllt5llkAYC0hQSEj/mxxWwQjGnFQgrJcEbxTpU6MI06UVGK2S0SIG37gau3RvL1q0tpi9XuYhoPKSiMqOhKOoJ6ut7oRjJHY59e2a+iqfu2vh63c3WuU67NR4WmbTIbHlXO5PpfQ6W1qbU0hDKyo4UFHqPuqB4JBq0oJTc6nN2Xkv1v70bZni/c6OEyu/rU4d+ata1So+81sruMFTi97NNDm0d4AkzI6XNwNZzblITI6/Y7ClMaK42FJISvzErWc4UFYUODxg81dmp6mn9D7f6c20sQs2lrNEsVqDq3/AGSE30I8xZypWPiT3NAWCgCgCgCgCgCgCgCgCgCgCgCgCgPygPL7TFnac1PrRa2+r2jUd1Vx/wDOvf0pcEbqHQL8C6e229ZadB6lJHY96JAt+l9YzIWGpTCkkDCj3B+3mm48C7mFbdWsBTQS1J79JPB+v6VAOjA0TcYk5HlNFGCPeT2PP1+dTsBpx7ey3DaVIaBdCQCSPr1oDruzGmE9ISn7AOR+dAR8icFnpBwPQfX20BxhwrRyfszTyBXtSsOTYTqWseYkcfnQCQe2uvuub4WbgCzbEK97/jH4n50BYNU61a8P8qXbv1bEVbHtNSbhaFpCg7IubDgQqKo5wQsPMlIAB4V3pfgDoX3dzW23mp78xqKbp8W2x6Yb1DNTDgPtOOLUtbPsyVl8pB8xGAspOerGB3pxBa9kd2Zm6GlJbtzdtar5bJzsC4GzPebEKwEqQtpXUolKkLTzk8pV8MUB2N9dRQ9Nba3mZJUlCExnCSfh0HIzQDD8N23rlsuuz2jnLgi6RdPWqbuBNW210IRJuS3G4TWCTkoQ9O971LYOBXy1dBOwxt9fCda903HZMdSbTFiRX324NtR0LmzShXllxRylKEnHupTlXUckYFa1UyLDemliKUFGTWtla719m7u7K747I632X7d4jIkoTXflKUU5S1UIXV+6t3J827Ky01ZZfDHt5b9Dbe257T93nSdPXeK1cG7bOKHfY33EguBtwAKCc5BSrOCDgjnOUwFGNKknCV4vW3J8f/Rhe2mbV80zKpHG0oqtSk4OUbrvRi33bq7V7bNW03Ww46yZz4KAKAhbtonTt/CxdLDbLkF/viXDbd6vt6knNAUtXhf2eW4XDtbo/rJyVfqONnP/AOj50B15XhU2glKCjtzp5kjt7NCSz/8A0xTbYC+X4ANujIdcavGrora3FOJYYvJDbeST0pHRwkZwB8ABQcdTuWDwH7X2W8C4ThetSoS2pHsV+uJkxjnHvFGByMcH0yaA6HhN2S29t2hoSHdFaeXqzRt5nWVV1Nsa9pQ5FkuIZdDhT1BRZ8lYVnPvZoBr7q6gmSvZNFaffLepL6hSTIb5NuhggPy1fDpB6EfFxSfQKxZ15ydqUN5fBcX9vE2zI8JSh3s1xsb0KLWj/PPeFNc7tXlygnxavdLDY4WmbJAtNuYEeBBYRGYaT/pQhISkfgKuoxUIqMdka7isVVxteeJru85ttvxbuzv19FqFAFAFAFAFAFAFAFAFAFAFAFAFAFAFAebWqrDK263c11pmLJ/WLke5LuQdUkIUpExSpKRgKP7pdKAfXoB47UBc7TDGpIyfakJjPDsTjFBuc7e38rrISEOoPqFDP8aBEjbdt5rMlDif2IBz8MYPbvQDRgxwxDaadAUoDHX9fdQHRusZwhXl8A4xkU3BRLlDnrcIbUekdzzxQHSjRZyHMLAUPl6/XzqNATcWE8tGVJx6kVIOpMY6Hh8TwfmeB+FAcSkuRsltORQEBqbRtr1u7anb/aWbkq0zE3CCqQD+wfT+64MEcj4HI45BwMARWotvbFqm5PTbtamJ8t9lhhx10rPmNsv+c0hQBwUpdPXgjn1yOKA79n0pbLBdLndosJmJMuSW/bHmgU+d5SSEFQHu5AURkDPxzQHztnt8vxG7vtKMmCvR+j5bMi6MSW0yBNeICm4xbPHSpPUpSjnHSkYPVwBo/wAN0Zi+ztwddR2W2od9vi7dawynpbTbbcPY2AhPolTjclwY4w7QDroBYaIkDbnW07Q8v9lbLk89c9Ouk4QpKyXJMT5LbWpTiR6tr4/y1VY0n6Go6L2eq+q64G6ZlD+L4GGbUtZ01GFZcVZd2FTykkot8JrX8SGhV8aWFAFAFAFAFAFAFAZxtWqrjoTfjdLSenbWL1dNQKgaggRkOBMWK65H9mkuSnBy0kGMysjBWvzR0g54tqtXu+rDWT4X28XxS+etr2dtmy/J/S0/37Hy9Fhl+a2s2t404/nlzf4Y/mktE3Bt/oEaPanTp803nUt0Wl25XVbYQXVJGENoTz0NIBIQjJxkkkqUomaNH0d23eT3fXDki3zXNP4g4UaMPR0Kd1CF72vu2/zTlvKXHZJJJK3VcGBCgCgCgCgCgCgCgCgCgCgCgCgCgCgCgPwqAUEkgE9hnvQGWvGfoN+Am0bjWtqQ85b8wbs0y2jyxEWer2lw9/2ZQE+vurOe1AZ4iyr3cJSFNOlLSuc5+NNQMO0XmRppoOy5hWvH7uf74pYHMjc2fdZqWo4PT1ckDj1pvoCyz9xBakMtO4W7gdX24FAScXXENaEJfwhSxx1HANNwSrMWNcEFTbgJ+JIoCAvVtl21Sn2UFxIySO5oBd6l3+tWjwpqc0W1g4V9YoDu6b3BteurembbX0rzyQCMj6/l91ATcCSHXQggAj48UBISGlKRhKOnGBkDnn6x93egIeSG4SS7JcSlA5JUcZ+/8KAqlpF6351c7o7RayzGZ6Rdr509TVuaUfkCC6UhZQgkZ6TkgUBpnVlotPhc8OeoBpCEW5sSGtENZHW9NuT6vLYU4o8rUt91Hc8A4GAAKAYm12ho+2e3GmdKRSFs2a3MQvMAx5ikICVLPzUoFR+ZNAWigITWGj7briyLttyQ4EdaXmJDC/LfivJ5Q80scocSeQofYcgkGnUpxqR7sv8A14oyWX5hXyyuq9B67NPWMoveMlxi+K+tmUqPrfUO2qfZNcxXbraWuG9WWuOVoKAO8uOgFTKgBytALZ7+52Fsqs6OlZXX9S+q4ee3kbJLLMFnP83KJKFR70Zuzv8A/XN6TXKLamtvW3d/sWorXqe3Nz7PcYt0hOfuSIbyXUH/AKkkirqE4VFeDujVMThMRgqjo4mm4SXCSafuZI19loFAFAFAVfV25mm9ELaYulyQm4PcMW2MlT8x8+gbYQCtX2gY+JFUKlanS0k9eXH3GawGTY7Mk54en6i3k7RgvOUrRXvKytOudy8pKXdvNOL7q6kOXiSn4DGW4oIPf31/8hqj/Orf2R/1fZfFmZTynJtdMVWXmqMX8JVLcvVj/mRUL1o6z7Qb5bUT7Hbm7fbrw3c9MzXeVLedcaTNYW6skqWvqhvDqUSSXDzVxTpQoxUYKxruOzDFZlW9Pip96VrckktkkrKMVwSSSH4DkZqqY4/aAKAKAKAKAKAKAKAKAKAKAKAKAKAKAKA/CkKIJ7igI3UunLbq/T9ysl4htz7VcY7kSVFdz0utLSUqSSORkH05oDAe4mknNgtw0aTfmrft0xlc2zyXzlamArpU2s9ISVoUSMDPulBPJNAVmbMm325dDJV5I7qB7ck0BedOxGrJBU85jzRnnPr9f3pcWK97Y5fL6lJWS2FcDP19GgJG7TFLvkWMlfSE47UBY06nkWqalLayABnHxoLF2s2u2ZqQ3KCert8qARPim2uj6tsjs+2ICZCQSQnsR3/r+FNAZu2R11M2zuLsSaVpbKveSo8Dk/H76jcDw09vLcty9x7BorR/srd7vLq2mpE0q9nZKWluFThQlRAw2R27kVLBoOP4dt+EM9Lt50M6r0Ul6UP/APA0B9aZ8F+tdU3Z53cvWUVu2NuMrZgaWUoe0pCiXW3lutgpSQEgeXg+8o5GBQGn9vtttM7VaeTY9KWhmzWtLq3vIaUpWVrUVKUVKJUSST3Py7UAvt4h/jPdzazRCPfjtzndV3NKDyGIASI4UPgZb8dQ+Pkn4GgHPQBQBQH5QFJvWzOkrzcF3Fu2mzXVWSq5WR9cCQon1Utkp6/+vNWs8NSm+9az5rR/A2XDdosyw9NUJVPSU/6aiVSPsU07eyx0Bt1q61AJs+5FwUyOQ1fLfHnY/wCtIaWfvUa+VRqR/DUftV/sXP8AF8tr3eKy+N+dOc4fBucfgj9MfdePwmdo+4JA4UuJLiqP24ccAqUsSt3F+/8AUhT7Oz3hWj/1U5f+MT9ETdaV7q7lpC3JPdbUGVJUn7AXUA0tiXxivY39UPSdnaeqp1p+coR+UJHydrb5ekhOpNf3uc1z1RbOlu1sqB9CWwXf/wCWo9BOS/mVG/LT5a/En+N4TD64HAwi+c71X/q9T/QWTSW3mm9CodFis0W3uPf50hCep97nOXHVZWs/NRNVqdGnS1hGz+PvMPj82x2ZtfvdVyS2X5V5RVor2JFiqsYkTnivZVC2hd1Oy2pyVpG5QNSthP8AsiyULf8AxY84ffQDgadQ82lxtQWhQCkqScgg9jQH3QBQBQBQBQBQBQBQBQBQBQBQBQBQBQBQBQBQFL3b2k0/vRo97T2oWXCyVedGlRyEyIb4SUpeaUQQlYClYyCOSCCDQGC9SaJ1nsbNet+rbaVW9MgR4moo6AYk3KOpJA6iptWAoFKwOU8EgjIHDNvi120oS51uEYH19d6A7ukmvYh5ruS4vnnv60B11yFPapBKjkfdj64oCe1ClTT6HOn075xR3669gOuzeUjgqKVDng/1oD4uuqOuA4y4rqSQc559KC3MxvvxFk26a/cIyShCjjKfgT/3oD1x8N3hP0L4b4k+TplE6ddLoAX7rd3UPSvKISQwFpQkBsKT1BOO6jzQDtoAoD8JwM0Amdpca03r3R1ooh2NAfj6Pty8dkREl2UR9smQtB/+XHwoBz0AUAUAUAUAUAUAUAUAUAUBFas05F1hpa8WGcOqFdIb0F8Yzlt1CkK/JRoCq7Aqvg2V0WxqaFIt+oIlrZhzmZYw55zKfKUs/wDMUdYPqFA0Bf6AKAKAKAKAKAKAKAKAKAKAKAKAKAKAKAKAKAKAjNSaatWsLHMs18t0a62qY2WpEOW2HG3UnnBB+YB+0CgMi7v+Cy92idMve2Etl+F0NqRpOYej9p14X5EhSwltARhQQsE5Sr3veAACUv41Ft5d0wNXaduVilhsOEqZU8wU5KQQ831Nn90/6sjgkcigOpar7Hm3NMhp5DqSR7yFAgjigL+opu8NPPUr1xUWBX51qdbJ4xjj7KlggJVudeWc5UM55+/ioulxAut7LHHa0TPW8MlKD39Dg/0qQewEDmDGz38tP8BQHYoAoDNEvxioi7r660x+qIarTZrddF2mf7aC7cJ9tYZdmx1NgEthPngJVznynf8AbQC+t/j+mK2y0hcmNEw2tW3C7pZvtmbmENW+ItyNmWFBPUvzBcIZQCB1F1XJ6TlYDe2j8QWpNx91rzoWXpNFqnaU9rb1NLU64WWnC/i3CMVJHmCRHy+ScdAwO5oB9UAUAUAUAUAUAUAUAUAUAgd+dxb7A3i260BA1Ynb+06ghXOfK1AGY7jzq4yWeiKyZCVNJUQ6pxRKVEpawMckAJHavxEbmbnbgbdWx69XiZb5Fnny5juk7bb/AC7kI15VDbnO+0ZLcd5lIUfIJOVZRxigHD4SdyL/ALtabt2stQ7gRptwvjMp13RDMeK0i0KRI6PKRgCQVNABDhdKsqUDhHAoDRdAFAFAFAFAFAFAFAFAFAFAFAFAFAFAFAFAFAFAFAcUmMzNjOx5DSH47qC2406kKQtJGCkg8EEcYNAZx3T8EOk9UOv3XRbqdC35flACI0VW9SEcFJipUlIKhwVJweAeaAzjOVdNstVPaW1XGFsu7YK2VE/sZjWR+1ZUcBY95PUBnpJ6TyOQJX9ZMyEkqwc8dvl8qAjLhPjRm1q91OB3JxUoFC0NpqL4mt7rbt8x5M+zRU/rDUDKZpYcEBJShQQpPvdXW42MAg46uagHqy0jy2kI/wBqQKA+6AKAS6PB/tW1DgNtaabYnRJUuYbuy4UXCS5KQ+iR58kftHUrTJdBStRH7v8AtGAPxvwf7XtCaUWN5Lsy22i0vviY75qo9tcbciJ6s5yFMtdR7qDaQc4oBiWDb2y6a1fqnU0Bh1u76lXGcuTi3lKS4WGQy10pJwnCBg4xnuaAstAFAFAFAFAFAFAFAFAFAQ+qNHWDW9uTb9R2O23+AlYdEW6RG5LQWOyulYIyMnmgOaLpu0QZzEyNa4UeZHiiCzIajoS43HBBDKVAZDeQD0DjIHHFAde2aJ07Zb7PvdusFrgXq4cTLjGhttyJPOf2jiUhS+QDyT2oCaoAoAoAoAoAoAoAoAoAoAoAoAoAoAoAoAoAoAoAoAoAoBcb47YaI3I0wx/jazLusW3yG3mFxnlsSGllQQOlxtSFAe8Mp6sHAyDgUB57+KnaLVfg028t+qZOu06rgTrym1tRBbCw60FtuuoKlqeX1dKWeknAJJzQFq8NPhyvHiq2r0vr64awFm09cZLyZNpjQ1iU40y+tpaQ+HglCldBwQg4z2NAbr2l2W0psppaDY9NW/y2oja2hNlEOzHUrcLhDjxHUv3lep9B8KAvVAFAFAFAFAFAFAFAFAFAFAFAFAFAFAFAFAFAFAFAFAFAFAFAFAFAf//Z)
What does DA signal?
- Hedonia and anhedonia
- Incentive salience
- Reward prediction error (RPE)
![[[@Hu2016-yw]](https://doi.org/10.1146/annurev-neuro-070815-014106)](data:image/jpeg; charset=UTF-8;base64,/9j/4AAQSkZJRgABAQECWAJYAAD//gAuSGFuZG1hZGUgU29mdHdhcmUsIEluYy4gSW1hZ2UgQWxjaGVteSB2MS4xNAr/2wCEAAMCAgICAgMCAgIDAwMDBAcFBAQEBAkGBwUHCgkLCwoJCgoLDRAOCwwPDAoKDhMODxEREhISCw4UFhQSFRASEhIBAwMDBAQECAUFCBIMCgwSEhISEhISEhISEhISEhISEhISEhISEhISEhISEhISEhISEhISEhISEhISEhISEhISEv/CABEIB5MOugMBIQACEQEDEQH/xAGiAAABBQEBAQEBAQAAAAAAAAAAAQIDBAUGBwgJCgsBAAMBAQEBAQEBAQEAAAAAAAABAgMEBQYHCAkKCxAAAgEDAwIEAwUFBAQAAAF9AQIDAAQRBRIhMUEGE1FhByJxFDKBkaEII0KxwRVS0fAkM2JyggkKFhcYGRolJicoKSo0NTY3ODk6Q0RFRkdISUpTVFVWV1hZWmNkZWZnaGlqc3R1dnd4eXqDhIWGh4iJipKTlJWWl5iZmqKjpKWmp6ipqrKztLW2t7i5usLDxMXGx8jJytLT1NXW19jZ2uHi4+Tl5ufo6erx8vP09fb3+Pn6EQACAQIEBAMEBwUEBAABAncAAQIDEQQFITEGEkFRB2FxEyIygQgUQpGhscEJIzNS8BVictEKFiQ04SXxFxgZGiYnKCkqNTY3ODk6Q0RFRkdISUpTVFVWV1hZWmNkZWZnaGlqc3R1dnd4eXqCg4SFhoeIiYqSk5SVlpeYmZqio6Slpqeoqaqys7S1tre4ubrCw8TFxsfIycrS09TV1tfY2dri4+Tl5ufo6ery8/T19vf4+fr/2gAMAwEAAhEDEQAAAP1VAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA4cPgv14Ps4AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAD8+Q5n9LAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPBA+FuuD9NgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA+fw/nV+0w/dcAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAD+acPLf6qgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA8R/nbDH+kA/oaAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAD5/D+dX7TD91wAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAP5pw8t/qqAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAD8q/ypD9fv0sAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADxH86Q8U3Q+7/uMAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPn8P51fsMPpn5eD6D/AFRDbAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADhPzZD502g/XL2AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAOaD8tfnoPpv8+A81/qqAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAzw/Nn5LDsf0uD6LAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAD5fD8YvEg+rv0mD7WAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAD5E/OAPdvi4PCv33D6zAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAD5/D+dVwfRPEh5V9/B+1gAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAcUH86vmgfS2gH7j96AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAH4IB8ie+hm+KAf1VAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfiWHwt+jIfJnlIf0i+kAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHx8H4vebh7n+mwfoCAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB8mB8jfLgfMn6gh+tIAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfP4fzq/RYf0Oc0H8z+eH9SFgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA/L4PyW/SkP2BAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA8n/mtD3v+icKX8ygcr/VUAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHmX80QfVH6WB86flsH7Gfo8AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfj9+awfup9rAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB+TH5eB9N9aHxv+lIfsCAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHz+H86v2mH7rgfzleFB/UP0YAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB+LAfn5+5gfbwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHyv/P6H3t+2gH804eW/wBVQAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfLP8/YaGwAB+pn6bAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAH4/fmsH7qfawAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfzIcsH9SHwCH4t/pSH7AgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB8/h/Or7AH9EvmwfzoegB/S6AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAH5hh+SP2WH7j6APlAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPOv5mg9L/AKKg47+cgKX9VQAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAedfzOB9I/0BBq1wsAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB+P35rB+6n2sAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAH4K/H4e5+YhzX6Uh+wIAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfP4fzq9qHNVAi/aQP0GAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADkg/ni8fDQlD+h/3YAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPxND4HtA3tQ4L+qoAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAD8gw/NHqA9L8uD+mvdAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPx+/NYP3U+1gAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA5T87AzPtsPkn3QPrgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPOg+GPq4Pmnx8Psb6uAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADnPgwPFuxD9DvQAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAqB8A+Lh9l5QeHfpCFsAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAD5E+VgZ7MH3ZqgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHyv89h9n+ugAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB8KB4nth9k+5h+fnlYdN9yB3X5zAAfXnuAAAAfJIcf9zAfnYHuHzsAAfoOHgnzmH2/6aAB8ufKgHsYfaewB8TB5Z+i4aoAAAAAAAAAAAAAAAAAAAAAAc+H55/bgeoeAh81eih5CAfQgfTv5vB6T9xgADA+RPnENb7uD0jzwPhb6BD6rA/Onz4Op+4w9LA+JvLA/RfVAAAAAAAAAAAAAAAAAAAAAAADmg/Pj9Bw6UD8+fpYPcPmkPNPoEPxd/eIOgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAD8zuZDo/qwPojjw/PPED0n7uDlPh4PsH2sPgz6DD3ADgvEQPQA9rAPmkPNPuEOa/PgPs/4wAAP0ZDTAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPjcPnO2Hu32QFv8AP4PKehD7T9jD85vUA+yfmkOH+zgAOU+Hw8z9ZD7+A+TPBg+8e7Dyr4TDM9jD7mthhfngHrf2iAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfzpB6t3AfB39LYfzufTIHyoH9F/8AMgH2d2AfoZ9QAAAB+ewfjx/T6Hmv82Qfv58Ah8vAfUP67h+GHgwfpj+pgAH4/fJIfffwYDv6HQsfzFAftkH2WAAAAAAAAAAAAAAAAAAAAAADA/ml/REP1M/EsMT6pD5J+MQ+7vssPQ/wwDP/AKeA1QD8OA+XPtkMT9Pg9u/NEPyw9bD+iID+X37eDkfnQP6Xa4fzFAftl9lgAAAAAAAAAAAAAAAAAAAAAAB5SH81f9KgerAfzT/quH6A/lQHyp+wQfzq/wBP4dWAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAH8qv6NhrfnAH7CfSgfzq/rCHw37SH6M/gAH1N/QIH86X6WB+gIH5s/lmHov2iH6uAH5UB8qf0BB5T/ADVh/QJ+fIfJ3QB9I/siG2AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfhQHH/YIfnh7cH7z/AM04fTNgPjb+mUP5mM8P6SvzwD5//dcDyQP57fYw+ia4fsSB/Oh5iH6ufpCHxT+IAfrD+bIff36uB8Tfi2Fv+m0JQAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAD+dIP0B+uQ/nK/pbD+fT9Vw+g/50w/o/wD5wA/Z/wBgD6A6sAAAOMD+Y397w8M/OYP6QgPxTA/azjw/mP8A1HD45/ofAA/H7ykP3X4QP5lf3fDe/EoPrmUP2fAPwYDqPk4Pu/8AYEPgn8qA3v3HD4W80D9nP5pw/cL8+Qx/PA/aL8VQ8c+xg/ZH8SAX5TD7G/acAAAAPx++cw/oF/mPD9jfusPn/wDnVD+n/qw/ICkHwt+1gfZIHzP/AD2h/SP6WGZphsfgAH1X+VIf0v8ApYfy+/qoHGfFYf0a/K4fiV9chL+z4AH8+XqIfJP6LB+rn50h+YfYB+6X5/B5p+zgfzT/ALhB+aeiHOfrSH4t+Xh9vfr8H4N9QHyd93h+wIAAAHlIfzV/0qB6sB/NP92B9g/n8HlP7BB/Or9Zh4T+s4foMB+afyeHinVB++Xn4fibwofq59Fh+C/9GYflaB9LfAIeT/u+H5h/GgesftYHyp8PB5ZbD97/AEAAAAAAAAAAAAAAAAAAAAAAAA/lV/f8Pqv8oQ+Fv3HD+dX+n8Py2+fw/Un8QgvfsSH5R/pYH6AgfkH5eH6LfWISgH5UB8U/sEHn/wCIAf0qerB+FHqwfsCAflcHz54CHpf73h4p+LAcv+u4ei/h6H9LH4xh6X6WHxn4UH9Bn5HB8k+1h+2XwwHyf4oHVfvkHVgAAAAAAAAAAAAAAAAAB+FAerfsCHzv/O6H9HX4Lh+q/sYfgB/TgH81Xtwe2+oB8/8A7rgfjMHh39A4c0wOj80D+bj9Mw+cv30D4p/EsP38/GgPvH9NA/Fbug/Nz99A+ngPmn8fg1fKg/cf3UPxG+cQ+2f2JD+c39dw5n80w/cL8Ew9j+4w5z8yQt/r8HqP4hB6H4OH7afVwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB/OkHi9IP0m/XwP5p9YOR+tg/Y/wDmAD2XYD9ovosAAAAPwF+gQ8J+kA/V8D8UwP2s+BA/Kv8AXcPwt/piD0gD8fvKQ/dcD+XL9mg+dvFA+pvzTD+miUD+dIPo76kD8Uv6eA/md/WYPj/00N7ykP3X/lVD9/8A80w80/ZIPzeoh+z/APNUH7H/AJxh333cH4V/02h3oAAAfLn8/ofvF+Fof069AHz/APzqh/T/ANWH81v6Mh8k+ph+zgHwJ+Nof1G/ksH5kfo+H6o/zGh+6X48B+nf6OB/L76WHI/WYftf+QweKfUwfmn/AE0BKB/NP9vh6x+RYf0xfzIB+ynwGH0tSDyn91w/lV/f8PyoYH7BflMHpH6yB/M/+6ofkb9HB9SfikH9SGwAAAHlIfzV/wBKgerAfzT8+HQcqH0B+wQfzq/0SB8uflWH9PoH5UfA4fu5/PaH7OfCgavswfnH++ofzq/0/h+PwH1X+NAft7kh+Pf9KofjA8PpD8vg/fr8Cg/VX9EwAAAAAAAAAAAAAAAAAAAAAAD+VX9/w+q/zzD8lv31D+dXoAx/6HA5r8KA/Wz84w3f0UD9AQPz++Zw+IPv8PuP5pD1752D8vvQAq+bh/Sp6sH4UerB+wIB+PwfKP7aB/O7+74fl968HVfFgfs/+AAf1VfhQHq3qoflN+24eeflyH9IX4Wh6t3QfA/7uB/Pb+zgfdIAAAAAAAAAAAAAAAAAAfhQHq37AhxX8xQf0GfiKHC5gfqp+qIfy+/sOH4xffIcP+64H4Bh7h+xgfzReZB/RV8sh+fH6ZB+LX9PAfKP4Zh9MeDh+2n2MH8yX6vh8B/UIfriB8qfgAH9CX5Kh7d7WH55/qOH4l/0Xh/Ph+1gcp+VAf0BfzVh+sn2WH87H9B4fP35Vh+9H86ofvZ+cIbv7fgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAH86QfoDEH5nf0vh/NV+g4foN6WHKfzAB/RV9AAAAAAAfm3+fIeM/0MB7wB+KYH7WfhqHzp6mHzj+ugfo4B+P3lIfuv8AGIfhL/SQH4K6odh8tB/Qf9NgfzpB+gP1YH86v9IQfze/oGHTe/B4H4kH7s/y0B+//wCaYerfsCH4Me1h+vn81ofqJ8Fh9V/oCH81f9KgerAAABX/AJlQ9m70P3PA+f8A+dUP6f8AzcP5yPqgOH4wP6aJQ8B/nTD92ftEPwo9WD6O/GgPojycPaP3yD+X39rA9b/nQD+j/wDCANXsA+Wv6Dw+mwP5p/1XD1b8AA/pC/m9D9FtMPpb5rDxL92Q/lo/f8Pyo+qw/Vf+e0PsH9TA/mM/ZUPy1/QEPqz+dUP6f+rAAADykP5q/wClQPVgP5p/1XD9AfyoD5U/YIP51f6Zw+afxLD+o0D8qPlQP6Av5pw/Vf8AOwND6DCX7+D+dP8ApqD8gNMPqv8AKgP6WPzuD8vv6Ug/IPxcPrj8/g/ot/nSD9Af0sAAAAAAAAAAAAAAAAAAAAAAAP5Vf3/D6j/BUNb9XA/nV/pvD8GPr4Prn8KA/py/nCDiv2BD9AQAD8q/jkP2E+aQ9e+dg+VP6Ag8p/mrD+lT1YPwo9WD9gQD8fg8p/dcP5ff2sD8lvTQ9x3Q+vP5/Q/qV/CoPafVQ/P7+i0PzD+FA/on/FgM32IPlT+gIP5p/wBVw/QEAAAAAAAAAAAAAAAAAAPwoD1b9gQ/Nv8AJYP6aP5zA/VfyQPiH+joP5ff3XD4j+BA+q/3XA/IAPiz+icD+YAP6KvyLDyn0gPmf9qQ2/xTD+oL8gw+ZP15D+f36eDzxof0jgfKn4AB/VV+KYHoYfDP2cB+kwfztfsqHLfl2H9AX81Yf0qUA/m6/paD5+/EAP6EP51Q/p//ACpDyn91wAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAD+dIP0B+3A/ml/osD8Jf1XD9AQOU/mAD0LQD9LP0sAAAADzf8AmdD3P+jcAD8UwP2Z/mID9r/tsPxL4QP3vA/H74ID2HwYP1i+vg/nF/pPD1v+coPsb9bwP50g/QH6sD+dX+n8Pxb8JD2P9EQq/hqHs3nwfur+aYerfsCHxd+JYfQPmIf0P/hgH1X+gIfzV/0qB6sAAAB+MYfnj+5IfcIHz/8Azqh/T/8AnIHwn/RkHkX82Qf0JfTAH5LB+Y3tYeXfoYHmmsH7Y/EIfiB/TqH80vtAcl3Afsj/ADph/Sf62H85X2MH63gfzT/quHq34AB/VP8AgWHn/qYfppygfhr7MHn37qh+VH1WH6r/AASH46+4hyn9DQfz5foCH1Z/OqH9P/VgAAB5SH81fpAWv13D8iP1XD9AfyoD5U/YIP57/Xg83/QsP1vA/Kj5UD+gL+acP1XeH5AfUIe2/quH80+sDPrEPqv8qA/pY4IP53/WQ8D/AG/DxT8/g/ot/nSD9Af0sAAAAAAAAAAAAAAAAAAAAAAAP5VffQ5LdD94OcD+dX+n8PzZ+Jg/X/8ACgP6gvhwPw4/akP0BA/nzQPBP2aD7uAPyoD5U/oCDyn+asP6VPVg/Cj1YP2BAPx+Dyn91w/l9/awPL/ywD6b+iQ/SH+a0OtpB9w+qh+f39FoeU/zyh7t88h+8HyQHyp/QEH80/6rh+gIAAAAAAAAAAAAAAAAAAfhQHzp3YeSfsuH3X/NOH6r/Qwfzof0rh/Oh+64e7fzXh9XfuuBwgfgh5OHpfhgf0D/AIBB++H1gH4QbYfd34ah9g/KgfrT5eHzT/QOHzv/ADuh/R57cHyp+AAf1VfimB+mwfz8fRwW/wB5A/CP5jDucYP6Av5qw/pU9TD8FvNA5b9Bw+2f51Q/p/8AypDyn91wAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADxcOw7gPCvSA889ADsAK/g4AHoHoAAAAAHhQdL6gAB5eB3viQe26YcJw4e5geaePhq+7B0HKB5f72B5YFf1sDxcOw6APF/eAl8CDM97DY8BDscQPSOHDQ9IA83Dyf6LDV8fDoOwDw/3ANAAAADig809wDQAx/Fw9480A9TA8ED0jswAPP8AxkOu9wDxzsA7vMDxL2UPD+fDvfaw5fy8PewPLK4etgeH+gBoePh9ARB8+Qh73ph4D2IYnpAef9AHoAHlnm4fRegHi/YB0Hi4e8WAAAAzw8PAPUA8/wDQA7Dz8Of9IDyrog4r6AAA8/58PYPDw9A7APMvKQ9m70OC8xD1DKDoPPw9wAxPncPY/QA4fjw9o8XDsO4AAAAAAAAAAAAAAAAAAAAAAAPmnKD0P2UH44eL+8BheTh675OHvDA8C9QDsAPOvHA9t7sAA8/Dn/YAz/Dw9w0A8n0A9IAPNwz/AFgPB/UA6jycPN/aw7XzcOE9DCvoBx/tAHOB8+e4h2vn4c/7AHh/oAdgAAAAAAAAAAAAAAAAAAHj4eadAHu2qB4eHoHYB4D6yHlnrAdB5ID/AFgABnhQcj7CHVeNB7rbDzzlw9D8EB/sod35IGh6WDPAg9V6sOf8fD6A8vA9QDlPBw9D9gDH+eQ9x88D2Dw8PcNAIvnYOj9rDH8XD3jz8M/1gAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA8D4/p/Sez38gNfPAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPDuz877jvj0fhwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAACqr+ZPqXg+xD0PiwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADK+efO+4+mz0fhwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA+dc7w/136bPc/ImYoboAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB8heb956t7P0eIHV88AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB+WvgftP2D9E+l8Efz59vyf6I/fYAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfmh9FfJ/0p9Sn1n81gAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAGJHT+MP7c/Mf0GH1X85gAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHEfkz8n/Sn7On1n81/nd8rh+3IAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHxHifJf0p97n1v81/hH4MH7T/AFcAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB+aHhfsH0V9S9vyYeh8WAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABmTtNdJCswAAAAAAAAAgVcR32Xon55+X+hfTvt/f8b45/Nx1/Ofa37qAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB8E+4fN/vH0MfSfg4AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABkzv+V/6zfP/ALUH0X4cAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAcl+bvzf7x+p59J+D/hh8ch/URdAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADwOh436n9F4ns/ln8vH63B+V334H63bYAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfKvn/bem+u7+MHT4QAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAeX/ABB83+9emaOXofaR9T/OoAAAAAAAAAYkdPxH97+V+ih7H5iAfAn5Vhw39PYXQAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAD+ZXjA/oK+kwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAD5UD8Y/6Rw8v/ABbDyn9pw+owAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPLfEfF/V/sD86U/0O+Vunwsr6ujr+dvqHbzPk76xnX5v9Xno+YdrzP0D69+d/Z/KvIfszk+m+Pdrg+y4r737/ijy30Pi+w6LXg/Gju75PgT9sQ+pgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA8z6Pk+l6k6/mgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAACBV4x7dy/Qh1/OAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAFHybk+l9n8n6/mv5x/sEPqP8ASwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPEaPk/pXvn5d+t+a+KfteAwPzk/JgP0R/V8NUAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA+aOH7D0X1LbyQ6PFAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADy34V+Z/ftH6vno4HoOjxdHzZVw/3VrwfmX+hXJ9J8iff3d8j538Scv0H3L8FFfoX8d9Hg/XGX3/H/ACn978f0p+WuHt/YP0T6XwX85v649vyfxP2QfrOAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAH5ofRXyf8ASn1KfWfzWAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAYkdP4w/tz8x/QYfVfzmAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAcR+TPyf9Kfs78HfWfzX+eP2mHzJ+44AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfEeJ8l/Sn3v8Ag79b/Nf2f+iYAB5OH4m8OH7afRwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAH5oeF+wfRX1L2/Jh6HxYAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAeW+I+L+r/TXxer0fUOb3un7D1Pz3xryzyP0z6b+Zt/H7DXjru+fTr9H/H1836J/nd0eL+hWF635r8Wfe/l/oJ+WuHt/YPsnpfBfzBf079vyfyj+WIf0TgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB+aH0V8n/Sn1KfWfzWAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAYkdP4w/tz8x/QYfVfzmAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAcR+TPyf9Kfs7+N31n819V9rh+FX9MoAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfEeJ8l/Sn3p/MR9b/Nf9AnuwAABF+bAflH+kgfqjpgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB+aHhfsH0V9S9vyYeh8WAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfNIeregAAAAAAAAAAHGB+cPyMHmnowfV36PB6GAAAAAAAAAAAAAAAAAAAAAAAAB5l4x4/wCpfWn57qvsr4sw9buPpjXzt/8AOXH1v08/NfXzvu7xvfxvIOd8H9m+zfk/0/gP0C8O9X83+Ze2+d/dM77m9f8AMD8teb6D7B430vgvyD/o37fk+a/mHD+mLuwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAD80Por5P+lPqU+s/msAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAxI6fxh/bn5j+gw+q/nMAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA4j8mfk/6U/Z3+dj6z+a/1e+vA/l6/pCD1AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPiPE+S/pT6P/AJ4vrf5r/qAlAAAADxT8Uwx/2vD6AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAD80PC/YPor6l7fkw9D4sAAAAAAAAAAAAAAAAAAAD85vg4P6CvLw/nN/oyD1AAAAAAAAAAAAAAAAAA+Pg676TAAAAA/mnD9V/0BAAAAAAAAAA+VvwzD337S/l36L0zzz8s9L42+YP68+X/AG8+yfueMAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA/Mnwf2L61+KvS+G85/Zzs+WP5+A/Q775AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPzk+gPlf6L+mz6r+dAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADGjo/Hv9nfm/wB7D6f+fAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADjPyv+V/ov9Y/5ffqv50/pv7MP5+P0cD7rAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAD5ozPA/ZvI/hb3/xn97wAAAACuH5k/lgH6c/qSFsAAAAAAAAAAAAAAAAAAAAAAAAAAAAA+L/L/QvX/cenwT4M6/m/hP65D7A9mAAAAAAAAAAAAAAAAAA/NP8AP4P6LfKQ/mr/AKVA8E9WD5h+rg9r5wPhnrg+2PhwPaOxD5O+ng8eeHnX3cGr4SHyp9XB7nnh8U8SH0z+EofUP62B0fw2Ho32WDPicMz8kA/Vz9AQAAAAAAAAPGQ/nf8A1M/FPW9C89/Bva+1/lj4fs5/U+84vyG/oH/tz5H6INEAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfgaH3j+gIH5ch4d+24AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAH5+B+Uv7lB9JgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfJn45h/R+B+SeIH7DgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfjH6WH6ogAAAAAB4SH4pgftZ7sAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfPn8/Afqt8sh8n9wH199fB9Y9KAAAAAAAAAAAAAAAfmn+fwf0W+Uh/NX/SoH4UYod15IH9Mv8+Ye/fMwfoTUD82foAPTvq4Pw7+1g+afsEP0m/AoP0X/M4P6RfxgDzn67D7B/DAPcf2GD8SPrwPjb9TA4f8ww+4/wA3g/af9AQAAAAAAAAPwzDpv1D/AIP+z/ng5f8AuT4797fwm/nj3cv+hfJ/DHz7+g8P7+H6v5oAAAAAY4bAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAGXO038wrj+iL2ZwfPIfgn/TaEoAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABmWY6eE/nEvm+yfmcP6LOjAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAGhjbcdIXzAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADMmOn8zPJL5v2nA+LvyuD+iIAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPOq/D9d+B/wCwXd8j9dgAAAAAABU/LMPzJ/VEP00sAAAAAAAAAAAAAAAAAAAAAAAAAAAB5Dze91fXacP89v2Xr5/6mAUvmEPkH5BDwn6WD7B+wA+gZQAAAAAAAAAAAA/OH85g/o38fD+bX+mIPwa+/A+q/wCdsP6Pf5wg914cPqH9xw/m683D+nf5ED8S/wCooPzp/MgP0J/L4PWPDw/bf8Ow/oU+lgP50g/QH6/D+cL30PN/tQPKfpAP1n/mlD9Wf0BAAAAAAAADK/l6D9+e+/zG/Qv57uJ/0o+A/ff8I/5593K/oYyc3880H9M/cf0/88AAAAAH4/B+wIAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAH5a+B+0+jfkX6XwX9PB2/JjA/mQ/eoPogAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA/ND6K+T/AKU/M3uPrP5r/Zz8Iw6j9twAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADEjp/GH9ufmP6DD6r+cwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADiPyZ+T/AKU8y+3/AKz+a/0LA4r+ZIP6hd0AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA+I/Nvkv6U/IT+n363+a+jAAAAAAAAA+f8A8TQ2/wBrw9oAAAAAAAAAAAAAAAAAAAAAAAAAAD80PC/YPorxDt+T/NH+i/0Pi9MAAAOE+QQ+QfkQD67D68+wA7sAAAAAAAAAA+b/AOeoP2s+Zw+Of6VA/nV/QEPqz+dUP6bf5ug/UT6ID1byQPxS2w/Sf1APwz/eEPzxyg++/wAiQ/ZQD6+/mnD6++7g+rP58g9b/VEP52P13D2b2UPy5+dg/UT8Qw/Zn9AQAAAAAAAA8p/nK5a/ooP8vv0Twjiv1Lzfqz5/+R6sP6c6JPwH/YvL/cv6Q/r35cAAAA5oOO/K4P1Rth6WAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAH5a+B+0+U8H6XwX7uHb8mAfid7KH6mAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAH5oeO/J/0p8V/0dfWfzX1fFB/OZ+uQfewAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABiR0/jD+3PzH9Bh9V/OYAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABxH5L/J/0p8Pf0JfWfzX7cAfzcB+zn1WAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfEfxB8l/Snhf8ARf8AW/zWAAAAAAAAAGYH5Xfm2H6ufpOEoAAAAAAAAAAAAAAAAAAAAAAAAB+aHhfsHp/4v9vyf7n/AFj6HxYAAAAARB8/fH4fIPzMHu318H199NhoAAAAAAAAfn2Hwz2IfqT7gH5A/WAe7fkAH7dfOwfmdmB+onw8H0z0YfC32WH4sfcgYn63h6r+XwfJnah+2/gQflrlB+1/zIH5sfrUHI/m8EX6uB6h+PoaPWh9mfWAAAAAAAAAcF/M7m/6MPj3/Pb7joPq7ioPjuv51+OP6G8L9Uf54/Tj+hL23+svmQAAADw8Pm74OD7x6APuEAAAAAAAAAAAD4pD4pAAAAAAAAAAAAAAAAAK3gftPzz97+l8F+oh2/JgHwT+TwfVYAAAAAAAAB+hgfUAAAAAH5R+XgAAAAAAAAAAAAAAAAAV/Avk/wClP1j+zvrP5rAPkn8Mw+1gAAAAAAAAD6gD9DAAAAAPyj8vAAAAAAAAAAAAAAAAASOmn9kfMf0H94n1X85gB+UYeXgAAAAAAAAH6GB9QAAAAAfl94oAAAAAAAAAAAAAAAAAA3F+T/pT5X/pt+s/mt4B+LAeI+0gAAAAAAAAAfrv2oAAAAHFB+RAAAAAAAAAAAAAAAAAABD4x8l/Sn0V+x/1v81gAAAAAAAAAAfN4fib9Yh+yIAAAAAAflGHl4AAAAAAAAAAAAAAAABX8L9g8l9U7fk/26PQ+LAAAAAAAAOaD5O+QQ+QeHD9Lv1cAAAAAAAAAAAAAAAAAAAD4pD8U/6ggAAAAAAAAAAAAAAAAAAAAAAAAP5sA/T538LfY/kH9gf0/wDO/T3zp+L+t8Ufqh+hcPrP41+dp/TlY/oHxAAAAAA/H4P2BAAAAAAAAAAAAAPmkPmkAAAAAAAAAAAAAAAAA8W8D9p9j/S30vgpTt+TAOc+Fw54AAAAAAAAA+vA9wAAAAA+DOGAAAAAAAAAAAAAAAAAA8f+r/k/6U+3T6z+awA+Tw8DAAAAAAAAAPcQ+uwAAAAPgzhgAAAAAAAAAAAAAAAAK8dPz18d/Mf0H/Qr7R9V/OYB+d354h95gAAAAAAAAH14HuAAAAAHw95oAAAAAAAAAAAAAAAAABneGfJ/0p+xHUfWfzWADA/NYAAAAAAAAAA/QfpQAAAAOaD8+AAAAAAAAAAAAAAAAAADC7X5v94/Q+X6T8HAAAAAAAAAAAAyuJD0sAAAAAAPgwOGAAAAAAAAAAAAAAAAAOY8j9O+kvqTq+b3Tt+VAAAAAAAAAAA8cDmvogAAAAAAAAAAAAAAAAAAAPPA+evsgAAAAAAAAAAAAAAAAAAAAAAAAD84Q/M79yf5J+m5n8/vnOjxr1z9n8n7+77+Rfp/xI+4/wDQn4f9Tj6PAAAAAAPm8PpAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADLna5YcA4AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAMyzHTaL5gAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABoY34Vx0/dn6B3zAH4p+zB+owAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAzJjp2S+YAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAACIPww8W+f3/WX3X/PX7gPnt/H/AMmv6b+e6796v6w+a0AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAD8qw83/ZwAD+bj9nA+qwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA5aLm9zrjp8MAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADh8fS2d++cL5QAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPFAPawAAAAAAAAAAAAAAAAAAAAACv+WwfmR3fibek+f8AxnV5n+kP6b5/6uabAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAD8yfB/YvrX6B9L4YOz5YAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA/OT6A+V/ov6bPqv50AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAMaOj8e/uL5v97/ACA/o6+n/nw4cP5lf6hw2wAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADjPyv+V/ov9gz6r+dAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA+I8T5L+lPvc+t/msAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAD80PC/YPor6l7fkw9D4sAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAD8UwP2sAAAAAAAAAAAAAAAAAAAAAAADH+ZA859BD6Y6AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA/LXwP2n7B+ifS+CDt+TAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPzQ+ivk/6U+pT6z+awAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADEjp/GH9lPmP6D/mE/p1+q/nPq/i4Pyu/oiAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAOI/Jn5P+lP2dPrP5rAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAD4jxPkv6U+9z63+awAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPzQ8L9g+ivqXt+TD0PiwAAAAAAAAAAAAAAAAAAAPn/HDxn7+D55+ZA+vgPn/7jD4Z+tg8Xrh0Hy8Hd/bweT+Xhx314HiPz4HyD1AftYAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAflr4H7T9g/RPpfBB2/JgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAH5ofRXyf9KfUp9Z/NYAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABiR0/jD+3PzH9B/zm/rf9V/Of2H+SYYn7DgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHEfkz8n/Sn7On1n81gAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB8R4nyX9Kfe59b/NYAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAH5oeF+wfRX1L2/Jh6HxYAAAAAAAAAAAAAAAAAAAH5UfmiHtf7CB+HH6CB+b37tB/P7/SkH80/7ZB+ff24Grx4fmz+lYdn+PYerfrIH4Yfo2HwN9mB+1gAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB+WvgftP2D9E+l8EHb8mAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfmh9FfJ/0p9Sn1n81gAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAGJHT+MP7c/Mf0H+NfcfVfzn+rn8/Afo591gAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHEfkz8n/Sn7On1n81gAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB8R4nyX9Kfe59b/ADWAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB+aHhfsH0V9S9vyYeh8WAAAAAAAAAAAAAAAAAAAB+VHx4H9DX5Ih+eHqAeD/wBAofh794h8jfRIfDf9HAfmx8SBg/YIfVf5Jh/TV+WwfHP9CQfig8P2sAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPLef2uw6LXgC+QAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA8z6Pk+l6k6/mgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAACBV4x7dy/Q/AnwH1/OfvV/LuH9IvqAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABR8m5PpfZzr+aAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA/Kj5UD+gL86w/Mz9hAPrsPxr+Kw/Yr8dg9I/oSD+Wr9vw/OP1sPqv8qA/pY+EQ/Gj9rw/Jb6TD9rAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA8f/nSD99fwyD+mUAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAACk2Om+XzAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAZk7WbTgHAAAAAAAAAAAAAAAAAAAAHwp4SH6w1A/LH5eD079rA+V/zVD9v/yJD6I/QEPyx+Yw9p9VD3z4TD9nK4fk94YH0R1wfpMAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfnn4v6r9O+39/wAaHX84AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfBPuHzf7x9DH0n4OAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAZM7/lf+s3z/AO1H8x/0X4d9iYgfuOAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHJfm783+8fqefSfg4AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfEeJ8l/Sn3ufW/zWAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB+aHhfsH0V9S9vyYeh8WAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB+WvgftP2D9E+l8EHb8mAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfmh9FfJ/wBKfUp9Z/NYAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABiR0/jD+3PzH9Bn4UfVfzn8c/riH6WAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABxH5M/J/0p+zp9Z/NYAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfEeJ8l/Sn3ufW/zWAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB+aHhfsH0V9S9vyYeh8WAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB4Hx/T+k9nv5Aa+eAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAeHdn533HfHo/DgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAFVX8yfUvB9ifmH6Hxf5I/0Lh9JgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABlfPPnfcfTZ6Pw4AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAYkePpb5t5oAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHM59mjq1iFYgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB82B+LH9FoXQAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAy8Zg2YTjpsAAAAAAAAAAOQ6/gyDv1AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAD5J+G8z1Tz7xuf6iX9S9sAAAAAAAAAAPmf6Y+P8APD7D0AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA+Z/j/AD/pj5sp/Rnl3oa4/oXl4cb676e/l3cZzjWta82o/Ief7jwn2HoYHoXm5eat8Xm4/wCpfV3D7T0fkr6q+B8y3hffen4Z9IAAAAAAAAAfA/j/AKd9I+z93yQdXzwAAAAAAAAAAAAAAAAAAAAAAAB8lfCeb9VW/t/RDRgAAAAAAAAHw57P89+3++H0P4gAAAAAAAAAAAAAAAAAAAAAAABw/l/wPmfRB996YAAAAAAAAAZc7fmT+pXhfsYe/wDiwAAAAAAAAAAAAAAAAAAAAAAAHgfcfA+Z6Iffen51zge0AAAAAAAAAct+eXz37f8ApsfQ/iAAAAAAAAAAAAAAAAAAAAAAAAAB4x5V7f0P14eJ88AAAAAAAAAAHy1lfPft/wBdH0P4gAAAAAAAAAAAAAAAAAAAAAAAAAHlPR1eEfZvX2h5nkAAAAAAAAAB8OeT+kez++dXzwdnzAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfIHwfl9jp+Rz+meRex0enePcsd52XrdHkXSeByYvvP0PXheU4Tt5Hm49zS9noj9P9Lb5Y+qvF5g+19L4N+4/yzxb/AMXfqftfhN/TeHdgAAAAAAAflr4H7T9g/RPpfBB2/JgAAAAAAAAAAAAAAAAAAAAAAAHwb+Y+P9x3/wBJ9YNaAAAAAAAAMIPyo4L5P+lP1h9Q+s/msAAAAAAAAAAAAAAAAAAAAAAADzTxX8s8X61P1P2gAAAAAAAAAxI6fxh/bn5j+gw+q/nMAAAAAAAAAAAAAAAAAAAAAAAD5Z9L/LPF9bP1P2vyD+Pg/o9AAAAAAAAA4j8mfk/6U/Z0+s/msAAAAAAAAAAAAAAAAAAAAAAAAA+dvDfpPrPvs+b+TAAAAAAAAABgfir9G/Jf0p+kB9b/ADWAAAAAAAAAAAAAAAAAAAAAAAAAHhvV2/L/AOiXf6QeN4AAAAAAAAAAfmh4X7B9FfUvb8mHofFgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB8z/AB/n/THzGj13heaPS7P0HVb8J8Tm7flvPyPVO7Xy/v8AOeTocWfrnnn0HX03d+rv8wfT/gcofZeh5J6t4HLL+SHv9X5ifuuH2mAAAAAAAB4Hx/T+k9nv5Aa+eAAAAAAAAAAAAAAAAAAAAAAAAeSeFzerS+30BTAAAAAAAA+Lvx2D6nzfO+4679l/R+HAAAAAAAAAAAAAAAAAAAAAAAAw+X8Dl9EPf6gAAAAAAAAAqq/mT6l4PsQ9D4sAAAAAAAAAAAAAAAAAAAAAAADgdzwOXoj3+r+ej5sD+n3rQAAAAAAAAyvnnzvuPps9H4cAAAAAAAAAAAAAAAAAAAAAAAAA4zlO30PXji88AAAAAAAAAPjYPyJq+d9x+83sfo/DgAAAAAAAAAAAAAAAAAAAAAAAABymmvB+z67hzcYAAAAAAAAAeHcn0nZ99r5wbeYAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHz18rw/QvhOtev8AhXDlpdvzx6H8s739MeVehro9iHgm78V52/U9no9Qs/Zeh889F+ceRhfSf0/aH13eAH8/AeT/AHgH63gAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHl/48B8zfr+H3j4+H86X9JQehgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAGUH8u/fB+zn2GAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB+BAfYfnQeKfvIAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB80fR3xfnx/wAu/wBp6H6ufFgf0GAAAAAAAAfGPl/oPvHq3Z8yHR4gAAAAAAAAAAAAAAAAAAAAAAAB80fHcH0dZ+u7gtgAAAAFT81A/Kv75D9ZurD5C9W8P9d/Kru/c/Iv2BAAAAAAAAAAAAAAAAAAAAAAADkPPfi/P9xPtPQAAAAAAAAADOnX8/f0T8f9SD2vygAAAAAAAAAAAAAAAAAAAAAAADxfr/i/P7n5f+09D8Tfr4Oz/WEAAAAAAAAOd+H/AA/139Aj3PyIAAAAAAAAAAAAAAAAAAAAAAAAA8p849b3Pp08nwwAAAAAAAADwH+fUP3Zy/D/AF38Uv2q9z8i+xgAAAAAAAAAAAAAAAAAAAAAAAADzjbo8b+q+rrDz/MAAAAAAAAAD5C837z1b2fo8QOr54AAAAAAAAAAAAAAAAAAAAAAAAAPKQ5X2APnT6gC2AAAAAAAAAAAAAeEhrethzPVgAAAAAAAAZQfNP0sGqAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB8G/cf5Z4vz3+GH6n7X9Cv824f1C6AAAAAAAAflr4H7T9g/RPpfBB2/JgAAAAAAAAAAAAAAAAAAAAAAAHwb+Y+P9x3/wBJ9YNaAAAAPmQPxosB+y/0mAB+aH0V8n/Sna/zv/WfzX/Sb6GAAAAAAAAAAAAAAAAAAAAAAAeaeK/lni/Wp+p+0AAAAAAAAAGJHT+MP7c/Mf0GH1X85gAAAAAAAAAAAAAAAAAAAAAAAfLPpf5Z4vrf5h/qfteBfYYfnz+/gAAAAAAAAcR+TPyf9Kfs6fWfzWAAAAAAAAAAAAAAAAAAAAAAAAAfO3hv0n1n32fN/JgAAAAAAAAH4ndqH1BifJf0pm/mD9b/ADX/AEb6AAAAAAAAAAAAAAAAAAAAAAAAAHhvV2/L/wCiXf6QeN4AAAAAAAAAAfmh4X7B9FfUvb8mHofFgAAAAAAAAAAAAAAAAAAAfnN+SwHUB/S0AAH5UB8qfsEH86v9P4dWAAAAAAAAAAAAAfhQHq30MH5t/wBD4ABUD4E+ww9DAAAA8pD+av8ApUD1YAAAGeJBwnZh7hYAAAAAAAAAAAAAAAAAAAAA5/lz849A+c5M3ufb6eX1wTzPzse16n1ugO3T4N+4/wAs8X84vmT9T9r9x/5oA/cz6TAAAAAAAD8tfA/afsH6J9L4IO35MAAAAAAAAAAAAAAAAAAAAAAAA+DfzHx/uO/+k+sGtAAAceH5J/Cwfqz+kIWwAD80Por5P+lPqX8PfrP5r9T/AFvAAAAAAAAAAAAAAAAAAAAAAAPNPFfyzxfrU/U/aAAAAAAAAADEjp/GH9ufmP6DD6r+cwAAAAAAAAAAAAAAAAAAAAAAAPln0v8ALPF9b/Cj9T9r67+vw/nP/qFCwAAAAAAAHEfkz8n/AEp+zp9Z/Nf5rfJgfuy8AAAAAAAAAAAAAAAAAAAAAAA+dvDfpPrPvs+b+TAAAAAAAAPOg/my/o9D54xPkv6U+8v55/rf5r+4f08AAAAAAAAAAAAAAAAAAAAAAAAA8N6u35f/AES7/SDxvAAAAAAAAAAD80PC/YPor6l7fkw9D4sAAAAAAAAAAAAAAAAAAAD80/zuD955gPl4PvH4+D0H4oD5U/YIP51f6RQ+Y/tkPPPnwPu4D4e9pD5b+qQ9z+Hg9h4kPqv4hDH+7A1flkPF/hkPc/0LD58++Q4r4pDY7gPwQ/VkP1B86D4z+jg+k+fD4Wth+Kf9KgerAAB+eIfldznHXofB/Pb3v1B+t5v0nlAAAAAAAAAAAAAAAAAAAM/51+J836Fn+r7vnX6Y+a4/FO9Dx76CR88/RYWD7H0Pg37j/LPF/Ef6b/U/a/Sz8Ow+k/0xAAAAAAAD8tfA/afsH6J9L4IO35MAAAAAAAAAAAAAAAAAAAAAAAA+DfzHx/uO/wDpPrBrQAMD4H/IgPqb9gQ9LAAAD80Por5P+lPqXwT6z+a/58P6VQ7UAAAAAAAAAAAAAAAAAAAAAA808V/LPF+tT9T9oAAAAAAAAAMSOn8Yf25+Y/oMPqv5zAAAAAAAAAAAAAAAAAAAAAAAA+WfS/yzxfW/5j/1P2v3w+gA/mP/AHqD6GAAAAAAADiPyZ+T/pT9nYvrP5r/AJquHD9x/ssAAAAAAAAAAAAAAAAAAAAAAA+dvDfpPrPvs+b+TAAAAAAAAPybDxz9xw+I8T5L+lPvf5p+t/mv8EP6RQ9AAAAAAAAAAAAAAAAAAAAAAAAA8N6u35f/AES7/SDxvAAAAAAAAAAD80PC/YPor6l7fkw9D4sAAAAAAAAAAAAAAAAAAAD80/yuD7R9QD6G/FMP6gv5/Q+qwPlT9gg/nV/ptD+cT9dw+aeED90gP5p+jD175MD+k/8AniCL9HA8x80Cv6CH3j+DQfo5+eAfoP6qH5/f0Ch/OR9Jh9TdGH4sfeIfrN/PKH6P/mqH79flGHn/AKQHxP8A0qB6sAAfjd8Y+Tp+qX1L/n39vKz4/r+a/wArP60+Z+i/29/pb5+UAAAAAAAAAAAAAAAAAA4flfjvP9iPsfQ+Zvpn4/z/ABTvQ+bvVPkeA9e+49K6fTdvzv8AQHyHB/NL+6X1/f8ASH5rB8w/uOAAAAAAAHyL5v3ftvpvV84HR44AAAAAAAAAAAAAAAAAAAAAAAB87/J8P0BY+p7QtgHiP43B5T+xgfZIAAAAHyr6b436n66ez+WfhR7WH62AAAAAAAAAAAAAAAAAAAAAAHK8H8hwezH1/eAAAAAAAAAFCdfhf738n9JD2PzEAAAAAAAAAAAAAAAAAAAAAAADyLqvkODkv50vr+/+oWwH4W/Wgfo8AAAAAAAGB8beN+p/dHxz7P5Z+Of6Uh8M/wBB4AAAAAAAAAAAAAAAAAAAAAAB5l5/6ns/Rp5fjABEHnNQAAAAM/8AnrD95/pMPA6Hjfqf0Wez+Wfig8P2sAAAAAAAAAAAAAAAAAAAAAAAA8/238l+mujqDh84AAAAAAAAAPlXz/tvTfXd/GDp8IAAAAAAAAAAAAAAAAAAAD80/wAxg/TnuQ3fxTD+oL+f0PqsD5U/YIP51f6fw/LHykPDP1hD7hA/mn/U8P0E/lyD9x/xyD9P/wBGQ/mQ6IK9gPtbw8P3n/CUPWfVQ/P79ag/DD+o0JeUD+YD+ioPEfxYD2jx8P1s/H4P3n9jD+a3+lQPVgD4MD8iv3S/lT6T80fHf0Ph/bP8kfw32OP/AGF9TP8AD79If6r+b/Sw9vEAAAAAMcNV4AAAAAAAAAEHy/8AEeb6t0ffr471PzPF5x9AdN+SdbCueofX+gHs9B5eH83X9RAXfm8Pwn/poAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAMf8ALYPzT/SQP1O2wAAAAAAAA+fP5/Q/pa6sAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA+BPgMP33A/Mb58D9vAAAAAAAAAD8KA+kf0uD+Z/8AdIPqYAAAAAAAAAAAAAAAAAAAAAAAAAA+X/xoDzZoAAAAH3sH7GAAAB55/N0H75fSwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfmn+fwf0WgfKn4BB+0/5LB+iYHyp+wQfzq/0/hwX87QXf6bQ0wP5p/q8Ps78MA/pE/A0P1X/QEP56vdg+8emD5u/KgP2f8Ax2D7Z9VD8/v3FD+b39hA+i/oAP5cP1hD6p/BcP2l2w+pf53Q+nvYQ/Jv+lQPVgD+cUP0S+xf87Puv5w6n+h/w37k/ht+A+2f0AcF+Wfkv6+X9MEv714oAAAAB+cPiIeWfteHYAAAAAAAAAEMrm+i4c3c9RuydNy81hN/X1Yb34N4L8txfnX/AED/AFPaVA/l4/o/D1MAAAAAAPlrz/tPYPRenwQ38kAAAAAAAAAAAAAAAAAAAAAAAA8G+Y4/cp/pOs+Prf44+kB+y/uAAAAAAAB80ei+T+lepHrfmp+DYfRv6rgAAAAAAAAAAAAAAAAAAAAAc1xPy3F62fU9oAAAAAAAAAUp0+MPtzzP0IPV/OQAAAAAAAAAAAAAAAAAAAAAAAPLOm+W4vyY7j6nt/VwD5k/EUP6VQAAAAAAAxPkzyf0r6J/mv8AW/Nf6VfQA/LP5hD96gAAAAAAAAAAAAAAAAAAAAADzvhvS9b3083yeM/JEPhv9Xw/R6wAAAAAAAeI0fJ/SvfD1vzU/MEPiD+hgJQAAAAAAAAAAAAAAAAAAAAAA4bXby/6J36Q4+AAAAAAAAAAPmjh+w9F9S28kOjxQAAAAAAAAAAAAAAAAAAAPjH5PD9fgK/48B5/7yHtYHhP6Eh+QH7dBu/zeh9jfr4AB/NP7sGf92B+g34xh92fWAeOflSHCffYfdP5HB5d7sHqfdh8n/r8Hwj8Ah6b+z4fln8qB+7nwYHw9uh+y/lgflT9MB8+fsoHdgcV/MlD/os0/wDLz9E/FzyH/QL4j95fxY/n/wBvk/3j6FZ/nf7I/oh9l/rD5oAAAAqB+Df73h8PcEH6PAAAAAAAAAAAAAAAAAAHwb8uflni9T+tn6n7QB/Px+jIfdwAAAAAAflr4H7T9g/RPpfBB2/JgAAAAAAAAAAAAAAAAAAAAAAAHwb+Y+P9x8T+k+t+PvydrX66/oADwAAAAAAA/ND6K+T/AKU+pT6z+az5pD8E/wClcOrAAAAAAAAAAAAAAAAAAAAA808V/LPF+tT9T9oAAAAAAAAAMSOn8Yf25+Y/oMPqv5zAAAAAAAAAAAAAAAAAAAAAAAA+WfS/yzxfwo/W/wDU/a+wwMr+XoP6V/QwAAAAAADiPyZ+T/pT0/5++s/mv9zwOU/miD+gX6DAAAAAAAAAAAAAAAAAAAAAA+dvDfpPrPvj8/8A5v5P8ivrEP2C9FAAAAAAAAD4jxPkv6U+9z63+azPD+cX9Rw/QMAAAAAAAAAAAAAAAAAAAAAAPDert+X/ANEu/wBIPG8AAAAAAAAAAPzQ8L9g+ivqXt+TD0PiwAAAAAAAAAAAAAAAAAAAAAAAAAAAAD8Xvi4P6Ne9AA/mn/VcP0BAAAAAAAAAAAAAAAAAAAAAAADyn+czkr+iY/y//RCKlKS4paR+Av8ARvhfuf8AR39efLgAAAHmgfk3+14fP/wMH68gAAAAAAAAAAAeKB4oAAAABh/lt+WeL+tX2j+p+0Afkt4oH38AAAAAAHyr4H7T7J6N6XwQdvyYAAAAAAAAAAAB9DB6gAAAAB8o8uAAAABh/mPj+b/lp+k+t9w/rjrXZgAAAAAAAAfmh9FfJ/0p9Sn1n81gH4J+th9dgAAAAAAAAHqAfQwAAAAHyjy4AAAAV6X5Z4v1t2v6n7QAYgfIgAAAAAAUY6fir7j+Y/oMPqv5zAAAAAAAAAAAANsPrsAAAAD5f4kAAAAMxv5Z4v4lf06/qftdWAfz1h9Z/RYAAAAAABh/I/yf9KfJ/wCz/wBZ/Nf12AfkcHhP6IgAAAAAAAAAfXe2AAAABiB8iAAAAAcNwf0n1nyH5v8AN/J/sd9ggAAAAAAAAAfEeJ8l/Sn3ufW/zWAfHP4qh+y7wAAAAAAAAPXg94AAAAAAD5RDlwAAAOQ6u3z3707/AEvUzxvAAAAAAAAAAD80PC/YPor6l7fkw9D4s8kDwQAAAAAAAAAA9Q+hgAAAAAAAAAAAAAAAA+Hvo4PUwAD5E9rD1AAAAAAAAAAAAAAAAAAAAAAAAMT+XgP6Ctz/ADK/QfBfpP1Mxny3T80ex/dcXRfztftnk/0sei/1J84AAAAc0H4q/ueHy58qB+pwAAAAAAAAAAAH5/B+fwAAAAH6IfGP5Z4v7F9r+p+0AfNn5jBxIAAAAAAdv4H7Tz+R6XwQdvyYAAAAAAAAAAAB+oIfYIAAAAH4vePAAAAAfoh+Y+P8Pfpn+k+t9QGtAAAAAAAAAH5ofRXyf9KfUp9Z/NYB4T+eIeAgAAAAAAAAH2CH6ggAAAAfi948AAAAHqvpn5Z4vGfsp+p+0AH5lh+dnooAAAAABLHT6J5t8x/QYfVfzmAAAAAAAAAAAAegB+4QAAAAH4/fPwAAAAfXflX5Z4ur+w/6n7QAfBwfncAAAAAAAFnvPk/6UuftZ9Z/NcoBwgfhwAAAAAAAAAAfuF6AAAAAB5+H4egAAAAdx330n1nof6Y/N/J7AAAAAAAAAAB8R4nyX9Kfe59b/NYAfkgHzYAAAAAAAAAfcIfpCAAAAAAH4vB48AAAB1/V2+g/APf6X2v+n3jeB72AAAAAAAAAfmh4X7B9FfUvb8mHofF1P50w1egAAAAAAAAAA+wf1BAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPwVD2b6e/ir638atf+svmfSfMvF2p/rv+Zeh5F8dfVc39D5+weWAAAAAfhR+tIfmp+iIfRAAAAAAAAAAAAAAAAAAB8s/Tnw3m2j7n0gAAAAAAAAA+HPJ/SPoX17t+VDp8EAAAAAAAAAAAAAAAAAAAAAAAA+WfifO+nLX2foBbAAAAAAAAAPi/wBf8D9m9xPf/GQAAAAAAAAAAAAAAAAAAAAAAAOL+bPhvN/C7+oj7n0tAA/m9D9kvrAAAAAAAzJ2/OP9L/E/Wg938fAAAAAAAAAAAAAAAAAAAAAAAA8N7T4bze+PufSAAAAAAAAAADmfgrwP2b9Gz3/xkAAAAAAAAAAAAAAAAAAAAAAAAA8h8w9j3vq08fwQAAAAAAAAAA+aMzwP2b6qPf8AxkAAAAAAAAAAAAAAAAAAAAAAAAAPMd+nxH3Hq7PzR/O7zvK+o/1BD6mAAAAAAAA+L/L/AEL1/wBx6fBDr+b/ADT/AD8D+iewAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB4oB7WAAAAAAAAAAAAAAAAAAAAAfPgfz5/sd+Be19JeA/y39HyXafX8vvngfxvV+Lf7uf6OfB/Vx0IAAAAA4T8+Q+kfrAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAP5ov3HD6WA4z+ZIP6gOgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADkg/OD81g9h/UEPs6UAAAAAAAADgg/m6/eIPqAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA/FMD9rAAAAAAAAAAAAAAAAAAAAAAD4p/EyT7I+1/wCP/qe94v8ALvR+OPh3+xflf2E/Qb9O88AAAAAAAAAAAAAAAAAAAAAAAAAAADzT0fxeeQ9roAAAAAAAAADxjl+h7zq9vLDTiAAAAAAAAAAAAAAAAAAAAAAAAPNPG5/R5PX3CmAAAAAAAAAeQ9Xw/Xdmd3yIAAAAAAAAAAAAAAAAAAAAAAAGRz3i8/46/SPtdH6YgfGn5Rh/ROAAAAAAFdX8/fRPH9SHb8mAAAAAAAAAAAAAAAAAAAAAAABxex4vPuHtdAAAAAAAAAAAZ3h/D9d9And8iAAAAAAAAAAAAAAAAAAAAAAAAAcpznX3enHJwgAAAAAAAAAB51X4frvTTu+RAAAAAAAAAAAAAAAAAAAAAAAAADm70431bXYOflMcPz5/MgNj9Nw+9tAAAAADyHm97q+z04T8TdfPP2yAAAAAAAAAAAAAAAAAAAPn/HDxn7+D55+ZA+vgPn/7jD4Z+tg8Xrh0Hy8Hd/bweT+Xhx314HiPz4HyD1AftYAAAAAAAAAAAAAAAAAAAAAAHkgfmT8ghxnVh9bfpsHtYAAAAAAAAAAAAAAAAAAAAAAAAAAAAHwb9x/lni3z9T9oAAAAAAAAAPy18D9p+wfon0vgg7fkwAAAAAAAAAAAAAAAAAAAAAAAD4N/MfH+47/6T6wa0AAAAAAAAAfmh9FfJ/0p9Sn1n81gAAAAAAAAAAAAAAAAAAAAAAAeaeK/lni+HfLP6n7X7mAfkxy4fseAAAAAAGJHT+MP7c/Mf0GH1X85gAAAAAAAAAAAAAAAAAAAAAAAfLPpf5Z4vrZ+p+0AAAAAAAAAAHEfkz8n/Sn7On1n81gAAAAAAAAAAAAAAAAAAAAAAAAHzt4b9J9Z99nzfyYAAAAAAAAAAfEeJ8l/Sn3ufW/zWAAAAAAAAAAAAAAAAAAAAAAAAAHhvV2/L/6Jd/pB43gABUD4c/MAOK/S4P0U6AAAAPzQ8L9g+ivqXt+T+ZPwV9D4v+kX0AAAAAAAAAAAAAAAAAAAA/Kj80Q9r/YQPw4/QQPze/doP5/f6Ug/mn/bIPz7+3A1ePD82f0rDs/x7D1b9ZA/DD9Gw+BvswP2sAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA+DfuP8s8W+fqftAAAAAAAAAB+WvgftP2D9E+l8EHb8mAAAAAAAAAAAAAAAAAAAAAAAAfBv5j4/3Hf/AEn1g1oAAAAAAAAA/ND6K+T/AKU+pT6z+awAAAAAAAAAAAAAAAAAAAAAAAPNPFfyzxeq/CL9T9r+mgD+f39EQ+8QAAAAAAxI6fxh/bn5j+gw+q/nMAAAAAAAAAAAAAAAAAAAAAAAD5Z9L/LPF9bP1P2gAAAAAAAAAA4j8mfk/wClP2dPrP5rAAAAAAAAAAAAAAAAAAAAAAAAAPnbw36T6z77Pm/kwAAAAAAAAAA+I8T5L+lPvc+t/msAAAAAAAAAAAAAAAAAAAAAAAAAPDert+X/ANEu/wBIPG8AAABgfIP5gh8/fo+H6U9wAAfmh4X7B9FfTvb8n/O798+h8X+mIAAAAAAAAAAAAAAAAAAAflR8eB/Q1+SIfnh6gHg/9Aofh794h8jfRIfDf9HAfmx8SBg/YIfVf5Jh/TV+WwfHP9CQfig8P2sAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA+DfuP8s8W+fqftAAAAAAAAAB+WvgftP2D9E+l8EHb8mAAAAAAAAAAAAAAAAAAAAAAAAfBv5j4/3Hf/SfWDWgAAAAAAAAD80Por5P+lPqU+s/msAAAAAAAAAAAAAAAAAAAAAAADzTxX8s8X6q/l4/U/a/o99KD+Xj+j0PVQAAAAAAxI6fxh/bn5j+gw+q/nMAAAAAAAAAAAAAAAAAAAAAAAD5Z9L/ACzxfWz9T9oAAAAAAAAAAOI/Jn5P+lP2dPrP5rAAAAAAAAAAAAAAAAAAAAAAAAAPnbw36T6z77Pm/kwAAAAAAAAAA+I8T5L+lPvc+t/msAAAAAAAAAAAAAAAAAAAAAAAAAPDert+X/0S7/SDxvAAAAAAPmz8wA+Qf0BD9NPWwPzQ8L9g+ivBu35P85v6LPQ+LtgAAAAAAAAAAAAAAAAAAB+VHyoH9AX51h+Zn7CAfXYfjX8Vh+xX47B6R/QkH8tX7fh+cfrYfVf5UB/Sx8Ih+NH7Xh+S30mH7WAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfHf1x+eeVcP0P1QAAAAAAAAA/PPxf1X6d9v7/AI0Ov5wAAAAAAAAAAAAAAAAAAAAAAAA+O/gPL+uLn3fphowAAAAAAAAD4J9w+b/ePoY+k/BwAAAAAAAAAAAAAAAAAAAAAAAOC8p/PPK+k/5/f0P1f0R9mD8IP6agAAAAAAAyZ3/K/wDWb5/9qD6L8OAAAAAAAAAAAAAAAAAAAAAAAA+ee9/PPK9JP0P1QAAAAAAAAAA5L83fm/3j9Tz6T8HAAAAAAAAAAAAAAAAAAAAAAAAAPEPIvd+k+zDwvmwAAAAAAAAAA+TMn5v94+yT6T8HAAAAAAAAAAAAAAAAAAAAAAAAADyLo6vn37f7O8PL8YAAAAAAPF/zGD4O+0w/T/wnyP07uvw76vm/3S+ru35UAAAAAAAAAAAAAAAAAAAD4U8JD9Yagflj8vB6d+1gfK/5qh+3/wCRIfRH6Ah+WPzGHtPqoe+fCYfs5XD8nvDA+iOuD9JgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA/JgOa9mD5Z/cwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADzr80A/Ov60Cpuh+4QAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAcl1Xn5PPQ1AAAAAAAAADgcfT6TavkC+cAAAAAAAAAAAAAAAAAAAAAAAA5Lgy6p/dqDAAAAAAAAADh9nm9zfOnwwAAAAAAAAAAAAAAAAAAAAAAAKOX5+XRfGnoa/lH7KH0j+mIAAAAAAARJ+ZepYeyHR4oAAAAAAAAAAAAAAAAAAAAAAAGBe8/LRPQ1AAAAAAAAAACr55ze56adPhgAAAAAAAAAAAAAAAAAAAAAAAAGLlbdHXmPOAAAAAAAAAAHLRc3udcdPhgAAAAAAAAAAAAAAAAAAAAAAAABlVWD2daBniAAAAAAAAAcp+Y2PpeMfsHfP6QXygAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB8G/cf5Z4t8/U/aAAAAAAAAAD8tfA/afsH6J9L4IO35MAAAAAAAAAAAAAAAAAAAAAAAA+DfzHx/uO/+k+sGtAAAAAAAAAH5ofRXyf9KfUp9Z/NYAAAAAAAAAAAAAAAAAAAAAAAHmniv5Z4v1rxn6n7X8yXZh+6v0wAAAAAAAGJHT+MP7c/Mf0GH1X85gAAAAAAAAAAAAAAAAAAAAAAAfLPpf5Z4vrZ+p+0AAAAAAAAAAHEfkz8n/Sn7On1n81gAAAAAAAAAAAAAAAAAAAAAAAAHzt4b9J9Z99nzfyYAAAAAAAAAAfEeJ8l/Sn3ufW/zWAAAAAAAAAAAAAAAAAAAAAAAAAHhvV2/L/6Jd/pB43gAAAAAAAAAB+aHhfsH0V9S9vyYeh8WAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHw79qfmPj3j9O9gAAAAAAAAAPzJ8H9i+tfoH0vhg7PlgAAAAAAAAAAAAAAAAAAAAAAAD4d/NvI+1L36J6wasAAAAAAAAA/OT6A+V/ov6bPqv50AAAAAAAAAAAAAAAAAAAAAAAA858b/MfH+qj9O9j+b3ykP6iNAAAAAAAAMaOj8e/2d+b/ew+n/nwAAAAAAAAAAAAAAAAAAAAAAAD5k9G/MfH9VP072AAAAAAAAAAA4z8r/lf6L/YM+q/nQAAAAAAAAAAAAAAAAAAAAAAAAA+f/FPofqfus+e+WAAAAAAAAAAD4xxflf6L+6D6r+dAAAAAAAAAAAAAAAAAAAAAAAAAA8U6uz5p/QHt9EPI8IAAAAAAAAAPzk8T9a+gPpvs+WDv+OAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPxY8XD+g8AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADzTND10AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAOS6rz8nnoagAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAByXBl1T+7UGAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAUcvz8uiPQ1AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADFytujrzHnAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAD4N+4/yzxb/AI/+p+1+X37OBKAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB8G/mPj/cd/9J9YNaAAAAAAAAAKnFB6AAAAAAAAAAAAAAAAAAAAAAAAAB5p4r+WeL9an6n7QAAAAAAAAAAAAB8Q+aB+lIAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAchpr15nkAAAAAAAAAAAAAAAAAAAAAAHzt4b9J9Z99nzfyYAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfBv3H+WeLf8Azu/U/a/Gb+jQPdgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPg38x8f7jv/pPrBrQAAAAAAAB83h+NHhIfu59ggAAAAAAAAAAAAAAAAAAAAAAAB5p4r+WeL9an6n7QAAAAAAAAAAAAB+Efggf0rgAAAAAAAAAAAAAAAAAAAAAAAcjpp8C+n/W/X632H4Pz4eZ5gAAAAAAAAAfAXsez9+nj+MAAAAAAAAAAAAAAAB4/6p19dw5OQAAAD528N+k+s++z5v5MAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA/FYPz+/ZcP0TAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADlA/Jn4GD9UXh8Pf0LgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB/MVxQf0y+igAAAAAAAAAAAAAAAAAAAAAABxmuvzP9l93f8An/8Af+23znzm2v014h5/n/QvzpK+ivGIz7f5x7e7Z+psOfxXzvr7Ppf5gw593xv0/U+/T5j5f40Pp/qPQfZ/J8j5k6D0fS+nfEfI8fkcb1vX+s+e8LwfnXrPZ9r6CPC8Gj82ej6K/TaXiHn/AG9uv9CcnJ+bfs30P0f0x538/wDOrq6a+2/PnLyfQS8XEAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB/N0HqfowftYAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfB35Bh9J/sUHqGOH81v7Uh9ggAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB4yH87vtwfpv9xgAAAAAAAAAAAAAAAAAAAAAABxmuvwF1/wBj9n7nY8Twsv0DOPBsP1PV+3Pzd5OP7g+Yg+6eZ8TxPjn6k9r2vkH7Au/EPZsMN/47u/v08HwD5G9L0vVfmr0PR+4PDfN8w8O9b1/r3w3g4PTPm/r6vrf5fwx+ou88ryfjH3L1/Y8+10vm371qvj76yiPk37LmPGfKOjp+yvz8xx/Qf4QmPvc8bxgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAD4N+4/wAs8Xmf5kv1P2v3a/IAP6RwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPg38x8f7jv/pPrBrQAAAAB45+NYeOfsOH2mAAfmn8Uh/QEAAAAAAAAAAAAAAAAAAAAAAAHmniv5Z4v1qfqftAAAAAAAAAAAAH5/B8He9hF+vgAAAAAAAAAAAAAAAAAAAAAAAcZrr8z/V/o+l8X9h6fp3uv4+PsOJ5ubzD2nr6/FrHT0+pee8vLU9iS+Z/v3LL4L+6GT/A4/v08bxfF/Lvc9365+AIz+//ADvzfN8I89+j+l+/fAPB8A+Y/c9z6cPP8/1/oPK8n8//AH76L6N/pHl+X8c/fu+/yd7KHzP9ls8ZA9/+ScMK/o3Z2+8Hg+B87eG/SfWffZ838mAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHwb9x/lni/IH5V/qftf0D/y8B/TV3AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfBv5j4/3Hf/AEn1g1oAAADK/L4PzI/RkP1X3QAADC/mkD9zfqsAAAAAAAAAAAAAAAAAAAAAAA808V/LPF+tT9T9oAAAAAAAAAAAA/GYPQPfg/MP+hcAAAAAAAAAAAAAAAAAAAAAAAOM11+Z/svu7/z9+zOvr+M/TOzt9t7X575z8y/1H9f2Pz2+s6rjvIG+e+nm/mz794+Px35s6enr+X6en7uyfl/lvy/+ivsvtO0o+V5HLcv2932N8Z8HB9++Aed5xzfoehxm319f10vzHy/i/wA8fQ/Rdp9McfH8S/ft38neyh47idnZ6UcnH7/geT5P5l/qb73vzHz3zvzt4b9J9Z99nzfyYAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABygfF9QPtHtQAAAAPOg+Bv0+AAAAAAAAA/GMPuz6wAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA+DfuP8s8X8pef/U/a/Yn+eAP1O+0QAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA+DfzHx/uO/wDpPrBrQAAHySH43dqH7Ne8AAAAAfl8Hx9/QEAAAAAAAAAAAAAAAAAAAAAAAeaeK/lni/Wp+p+0AAAAAAAAAAAAfzkB+vPvwfzRf1EBqgAAAAAAAAAAAAAAAAAAAAAAAAADKNVokyAABRbvCQAVG5pUgAAAKdlt4kAAFOSqsEyV247gAkAGe3oCR4p1dfnX1hvsHned87eG/SfWffZ838mAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB51/PKHubA+ef6Gg9bAAAA8fD8y/2UD8JfukPvH8Sw+nv0WAAAAA/mnD9V/wBAQAAAAAAAAAAAAAAAAAAAAPJA887sPYHgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfBv3H+WeL+A/wCjP6n7X3d+PAbf62AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB8G/mPj/cd/9J9YNaADgg/ID4+D9b/0LB4AAAABz4fzT/vAH0wAAAAAAAAAAAAAAAAAAAAAAeaeK/lni/Wp+p+0AAAAAAAAAAAByn8xAf0+9KH81v7aB9PAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAACfJ/o+l9XP87zQD528N+k+s++z5v5MAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAD8k/kEP6JwP52PqsPaflwPEffgl+cQ/ZfMD8o/0XD8VfQA/QH8xg7D9tw5f8oA679sgZ+KAZni4fsD+gIAAAAAAAAAAAAAAAAAAAfAwfld51zV6Bwfia9j+on0mH6JvAAAAAAOfD4eAPcQ+kAAAAAAAAAAAAAAAAAADB0uWP5ef6P+q/U/hQPzj/oHAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADB5Y3HdNgwiD86PybD7R/XcO4AAAAAAAD8sfmEP3sAAAAAAAAAAAAAAAAAAAAAAK1HljXOqwAAAAAAAAAAAPkH8iA/o1A/Eb30P08AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAM6jprvmeQAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAH4W9AH7ZAfibhB7x8Dh+yn4IB+0PxCHXfcYfin/S6H84X6WB+gP8+4fcP6Xh/NJ+t4fF3qwYnFB+xP86ofsJ+gIAAAAAAAAAAAAAAAAAAB+PHw35mn6lfW/wDn/wDbTxfE9fy7+WP9d/MfUn7Y/wBHeC8AAAAAqB5oAdMHYAAAAAAAAAAAAAAAAAAB83h+E/8ATQB5eH83X9RAXQAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAD58/GUOd/ZQPqsAAAAAAAADlP5qA/ez6WAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAD8o+CD9mQPzj+RA/dcAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA5/8AK4PjPEAAAAAAAD63D9VPTQAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAD8mPiQP6NQP5x/uwHfKgfuv/ADAB/RV+fweU/oCH4p/1BB/NP+qgfoJ/OkH3/wDdYfzMfcYbf0WHxj7sH68/zKh+tP6AgAAAAAAAAAAAAAAAAAHwoH4+/uV/LX0f56+MfY8n7R/lH+Jevxv6z+zl+IP6cf1f81+jx7OQAAAAAAAAAAAAAAAAAAAAAAAAAAfFnjn5v5Hzd+4/6R64B/M7+6QfSAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfFn535X2Nd/QPU5z8odH+e36gh+nOgAAAAAAAAAAflQHz1+7gAAAAAAAAAAAAAAAAAAAAAHnvkX5v5H0+fpHrgAAAAAAAAAAAfgX98h99gfPn4JB/TgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAeD+N+/9P9uHgfMHxMH48e3B+qm8AAAAAAAQ/ncHwZ+pIfpjoAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAcJ/PmGk0If6CA/N35UD91/wCYAP6Kvz+Dyn9AQ/FP+oIPwD8hD92vytD5s/cUPyt4wPbvvkPOfxuD6O8BD9hP0BAAAAAAAAAAAAAAAAAAP5zQ+/8A7Q/zv+5/nAr/AOhvw/7k/ht+Ae2f0AcF+P8A5T+3j/S+/wDdvGAAAAAAAAAAAAAAAAAAAAAAAAAAD4N+HvyzxfqL9Mf1P2gD8OPpsP0sAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAD4N/MfH+4/i39J9b8fveNa/Y/1sAAAAAAAAAAAOSD+aL+gcPoMAAAAAAAAAAAAAAAAAAAADzTxX8s8X61P1P2gAAAAAAAAAAK4fy+/0SB7MBX/AJeg/ot9gAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA+dvDfpPrPvvyz5v5P8cPncP2B+6QAAAAAAAAAPnT8bQ5r9iQ+uQAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADKD5hth9N3Q8p5UPdfl8Povz8Mn0EPFPqAOM+ZA+pgPln6bDqPl8Mz6TDq/nEL1gO49AAAAAAAAAAAAAAAAAAA4z+Y6D+izV/y9/RPxT8c/wBCPiP3x/Ez+ffb5T97Oibf873Sv6JvY/6u+aAAAAAAAAAAAAAAAAAAAAPPw8/AAAAA8N/Ij8s8X91vpP8AU/aAPzT+RA/WcAAAAAAAND3AAAAPHw58AAAAAAAAA9QDsAAAAAPF8cAAAADw38x8f4O+ev0n1v2F+49aAAAAAAAAAAAAAPyT8HD9ZAAAAAAAAADsA9QAAAAA8XxwAAAA5zjfyzxfrDpf1P2gAyg8UAAAAAAAA8kD8aP6cgAD8CA/QH72AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA+dvn/6T6zivyz+b+T+/v1kDqAAAAAAAAAAAiD87vyYD65/XwPSAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADyn+czkr+iY/y//RIoNVcggiuMPwH/AKL8L9y/pD+vflwAAAAAAAAAAAAAAAAAAAD80w/NMAAAAD9Zvnz8s8X9aLv6n7QByX4YB5GAAAAAAAGL+w4fdYAB+eP5VhaAAAAAAAAAP13D7hAAAAA/Bjw4AAAAD9ZvzHx/Hv0u/SfW6U1oAAAAAAAAAAAAA4r8Sg8RAAAAAAAAAPuEP13AAAAA/Bjw4AAAAPcPpL8s8X4Q/pF/U/aAD8Gw+cdoAAAAAAAAP16+3gADwwP5yAAAAAAAAAAD+n/qwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPKQ/mrAAAAA919x+k+t8x/Yf5v5L6LAAAAAAAAAAAAAOKD8lfgwMcAAAAAAAAA/SkP2BAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADn/5fEf0KdZ/mF+h+dfCf675nlfqn615f3L6Z/KP0uV/O9/SHg/0o+kf1L84AAAAAAAAAAAAAAAAAAAAAAAAAAHmno/i88h7XQAAAAAAAAAAH5h/P4ft4AAfgx9ph+igAAAAAAAAAAAAAAAAAAAAB5p43P6PJ6+4UwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAMjg/F5/5sP6Z/a6O7AZ/MKH9BvuAAAAAAAAAAAAMDlwAAAAAAAAADqHgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAARBzIAAAAGLl9fd6ycnCAAAAAAAAAAAAAABiBngAAAAAAAAGmGwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB+CAe6++/xd9Z+N/1r/Rng/QXg35J6nx5+wP0nPwPxZ7OX9ER+x+UAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB8yfiKH9KoBnh/L/8A0Zh66AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfzxfqUH2mB4j/AD2h/T3KAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB+bIH6TAAAAAAAAAAAAAAAAAAAAAHziH4Afrr+D+z3fvX8nfSh870eH8r+l+f8Ajl+6n97/ABf1sdMgAAAAAAAAAAAAAAAAAAAAAAAAAB8lfVXwPmWz770wAAAAAAAAAAyv5dw/pa9DA+Vw/FX+lIAAAAAAAAAAAAAAAAAAAAAA+SvhPN+qrf2/ohowAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAOH8v+B8z4D1fvvT/W8D89vh4P3qAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADxjyr2/ofrw8T54AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAD8UwP2sAAAAAAAAAAAAAAAAAAAAAAPhH8YYf279q/xt9V6X53+Wel8dfA/wDZnyf6+fop+p+cAAAAAAAAAAAAAAAAAAAAAAAAAAAB8G/cf5Z4t8/U/aAAAAAAAAAAA/nrD9Nvt4D8oQ4H9nAAAAAAAAAAAAAAAAAAAAAAD4N/MfH+47/6T6wa0AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB5p4r+WeLxf5vfqftf0FAfi36sH6nAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB87eG/SfWffZ838mAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB+dPzyHzj/QuH40/LYfd31eH5V/v+H8/H7DB8Q9cGT+fwd3+04eM/DgeTfteH5TfOgM+mQ/awAAAAAAAAAAAAAAAAAAAAAAPFw/MD5HDzTvQ+sf0+D34AAAAAAAAAAAAAAAAAAAAAAAAAAAAD4N+4/wAs8W+fqftAAAAAAAAAAAfkMB+vIH89Afpp9xgAAAAAAAAAAAAAAAAAAAAAHwb+Y+P9x3/0n1g1oAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADzTxX8s8X2j+bf8AU/a/qF0A/nA/YwPrMAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPnbw36T6z77Pm/kwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPyo/NwP2M5QPzj/VwPw2/oMD+fD93Q/DL9VA+A/16D0X5cD8wftMPov8AGMP2Xxw/LX+hUPxI9eD9rAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPg37j/ACzxb5+p+0AAAAAAAAAAB8PB+Zn9Cgcp/MUH9NvZgAAAAAAAAAAAAAAAAAAAAAHwb+Y+P9x3/wBJ9YNaAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA808V/LPF+tf5oP1P2v3M9jD+Y3+oIOgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAD528N+k+s++z5v5MAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAD8qPlQP6AvyRD4V+rAP1GD8WOjD0PyoPnn+mcP5uvuEPAvUA+q/yoD+lj8uw+Hv6JQ/EDTD9rAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPFvY/m+SY+k6wAAAAAAAAAAPPA/mo/qFD47/LMP6JQAAAAAAAAAAAAAAAAAAAAAA8W+d5PY5voOoKYAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHPcj83yen/h39J1/SftYfk3/AEVAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABwfG9/p+3HB5gAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAH5UfKgf0BfO4fgF9ghb/bkPy7/J4P6Pf57w+rv29D+Y/6pD5p+vg+q/yoD+ljwwP54PpAPD/sYP2sAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAD+ar9ug+C+mD9aQAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPzWD5h9gDlf2RAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPKeVD6AA86+dg9D+iw4zwwPqb55Ds/Uw8s8UD1ukHYeUh9LAeT+Th67jh7WAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB87/AEB8hwWD6/vAAAAAAAAAAAA/EP6AD89P2UD6zAAAAAAAAAAAAAAAAAAAAAAD53+T4foCx9T2hbAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA5Xg/kOD2b5v+v7/wAJ/bA/Qv7xAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADzLz/wBT2fo08vxgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA+DfuP8s8W+fqftAAAAAAAAAAAAfnD+fYeGf1ABqgAAAAAAAAAAAAAAAAAAAAAB8G/mPj/cd/8ASfWDWgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPNPFfyzxfrWp+p+1/LxRD+kX1sAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPnbw36T6z77Pm/kwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA+d/oD5DgsH1/eAAAAAAAAAAAB88/wA7YfW376AAAAAAAAAAAAAAAAAAAAAAAB87/J8P0BY+p7QtgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAcrwfyHB7MfX9/wDPx44H9NQAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHmXn/AKns/Rp5fjAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAFf+XoP1U/TkAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAD8xvMQ/YcAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAwgsaoAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfJf0PzHtPpfmeuHD6AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB8tel/X/EetRfIfb1NAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAKFx8PfeH1PxxF8n9pU0AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAwfjz6/4j7hPkPt6gFsAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADweh9d8R9DnyP24AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAUPh7674j7wPkft6nige4ygAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABnXmaIBGgAAAAAAAAAAH5f8A3X519d/Qvzv1AeP7oAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB+bX0L+p/j31B8n/ln7D+BX9QQdGAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAGLrj+N37V/pP5SfIn5j+ufgx/UKG6AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHFflJ+p/j37IH5Z+w/ld8Uh/ROAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB8U4v6n+PfeB+WfsIAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAYv43fqf49+1Z+WfsP56fjAH1B+0gezAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfnt9h8N2XNfS/Kfd938s/YQmwAAPir6f5H7VPmPrj569jwj6FAPH90AAAAAAAA4rq47HWzR8l+z4XtPpfmeuHD6AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB8tel/X/Eetfln8h9v+UP76B9bAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABQuPh77w+p+OPyp+T+0/Kv9/w+qwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAwfjz6/4j7hPkPt/wT+SQ/qF3QAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA8HofXfEfQ58j9uAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAFD4e+u+I+8D5H7f8WPRgr/AJth+qP6YhbAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAD89Psfhv0L+UvO9T0vM35vdfCvN9X3XwlnnPrHvfNeyeFeB9L7X8pd3n9vF28Hn6ex4X1LR+F/RPNPM/rPi/oL575uz6P7n5j63595X634r6d8x+c+qxOA+j+V+rfgDLbS+1GujPjPvAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA/BgPl79Nw/VEAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA/BAPmb9Qg/UYAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAiD+YJgfur9YAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB/N7+yQfWHzoH4u2w/ar3gAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA/N77T4H3zw/3fnv0B+CPC+h/Qv89I1/Qv8uOvi/R74Cc/pN+enH3/AEFi7YfR/h3ie/5b9C+14HzR790c3g30pjv8U/oRcfCn0P2+fkekeF9H89p9H8t6VQ8f3HZO2HYe9fN/VS/Obn1LuuH0Q5O0AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA+f+6+k+U9D/AJgvm/q/0J8KD97AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAqVHyf9d/QfNRfy+fO/Ufof4oH7yAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABkfM/wBJ8p9X+E/N/V/z9/aIewfqSAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHkVP6T5T2k+b+rAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAACp8n/SfKfXfGfN/V/zJf1AB0AFL8vg/MD9Ng/UTTAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA/O/wCx+F+4Ou+X+vd+eP1HyH6HfnkH6G/noP8AQv8APQP0L/PQP0B+SOPtwPW/c+fPoX5b6/8APT6g+s+M4r6G+d+o+KfvDq4/z09B+p+PTq/M9bvPDebr+ocT576bxTb+n+S7X2v5P7M/L/6f5H67+hfnfqA8f3QAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAD82voX9T/AB7sP57/AMs/Yf6EP57Q/p9lAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAMXXH8bv2r/AEn8p8E/AL8x/XP6Av5+w/p3eAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAcV+Un6n+Pfsh+d35Z+w/F32oHxN+84AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfFOL+p/j33gfln7CAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAGL+N36n+PftX8afln7D+Uf8AROAAAeE/i8HP/tSH0cAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB+en2Pw36Fnx33J+bv2vwPrvzh9b8V+qn56fHfdfoX+egfoX+egfV/mcvyb3z2PDPoX5D7f89PqD6z4zivob536j4p+8Orj/PT7K9DzPjP2j0/J9a+dPF9/6hxPC+h8U9G9/wCb+ffuHHfW/L/p5Prv6F+d+oDx/dAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPza+hf1P8e8U+I/yz9h/eT+YgP6B/cwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADF1x/G79q/wBJ/Kfzs+NvzH9c/df+YIP6F/awAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADivyk/U/x79kPxT/LP2H2b7WD+e/8Ap9B4AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfFOL+p/j33gfln7CAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAGL+N36n+PftX+TH5Z+w8v8AseAAAFcPzW/KYP0W/VkNsAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA80Dq+gAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADPvPQI0KtTja2/Pdz8dtDPDQzwu8rtj1UWG5YTz7VxHYmqV9mfoCpc/0c3V1OfqtNlw22uX6XbF/5f/ZfB/Xf0L879QHj+6AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfm19C/qf49+fHrf5Z+w/qZ+DYfaf6IgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAGLrj+N37V/pP5T+J3tv5j+ufqJ+CYfc36DAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHFflJ+p/j37Ifzcfln7D+zn1MH8wn9CQe3AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB8U4v6n+PfeB+WfsIAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAYv43fqf49+1f8/v5Z+w/oj94gAAAAePh+MHmgftJ9TAAAAAAAAAAAAB8GfFwfuJ50H4L/vQHooAAAAAAAAAAAAAAAAB4IF324AAAAD+acP1X/QEAAAAAAAAAAYHyv8fB5/3AfWf14EoAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB8H/WfF/Rnsni+8Hm+sAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfEfsn335t+Gv7JfA/pP1h+XIeM/tYAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAZemf5p/qB9t+ffza/tJ8L+i/U35XB5j+z4AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABy/wCff335t97/AMyXwP6T/UPth+Cv2+H6FgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB8v5f335t9aHwP6SAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAGX+af335t+mv8ALx8D+k/0e+qgAAAAAM/OwPyQ+9g/WrpQAAAAAAAAAAD80/z+D+i3ykP5q/6VA4r0gPn/AOiw7iuHyp1ofQ3y+HqfQB4D7MHCaAcD9Vg/hA+b/pAO7A+b+JD278Cg+wf1XDqPlEPQ/eAPnkM38Pw/Vf8AQEAAAAAAAAADzf8AEMPFvrz8m9P03z38w9D5G7r+kvA/cj131swAAAAA5QPzTAPpYPsEAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADjA/mS/qADoPlEPxo/pFAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADhA/ma/qFDb+Rw/Hr+j0AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPi78rg/oiA/LPyQP2nAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPnT8Ig/poAAAAAAADy/wDGsPBv2fD6+AAAAAAAAAAPzT/P4P6LfKQ/mr/pUD8KOdCWUP6RfwrCv4SH6yeEh8bfSIc590h+KHvIeF/ouH6Lfz4B9d/HIf0i/kWHyJ9Nh91/iWHqH7RB+SfbB8zfs4HFfjkH1Z8Zh+0/6AgAAAAAAAAcoH88H0b+d936GZ/8U/W+veeeDvifFX695fxn/Q7/AG18j6KAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB5f3ed1/Q8/UGO4AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB5t8r+r4/5k/wBE/lewYQfy9f02h2oAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABC58a8X9Lyvy1/od8z1znA/mA/p1DrQAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAACl5R6vj/nvieV7H7DgfJ/44B/R+AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHFQer4/eHlewAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAEPjXq+P8AInyz5XsfuYAAAAAAAAB8CfkAH2d+vgdgAAAAAAAAB+bf54h/Rl5IH82H9Lofg/8AeIfV388Yf0lfzah9geSB73+24fzgefh/TL8rB+Iv9RYfm9+cAfoZ+ZAfTPyCH7b/AIcB/QF9RgfzpB+gP2SH83H2cHhP1mHmXvAfrp/M+H6v/oCAAAAAAAAB+QAeA/sv/FP1v4G+df1z8x/QD+Fn86+7zX9Cdn5Y7H7L5P7bn3/EAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB+X/3X519d/Qvzv1AeP7oAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB+bXw7+p/j3vH7H/ln7CAfzu/qiH2iAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAYuuP43d5+k/lOV+wn5j+uAH86X61B9hgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHFflJ+p/j3hH6Ofln7D91gc//AC+h/Tf2YAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB8U4v6n+PfeB+WfsIAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAYv43fqf49kfSP5Z+w/piAAAAAAAAAefh+Ovy4H7JfbYAAAAAAAAfJ/4Kh++/ysH5rf02h/O1+gIfVn86of0xfzZB+q/wBEB3eeH4JXg/XzYD8OP6Mg/NHxYPvz8pg/ZRgfUf8ANaH6M/oCHrf88ofSf6WB/OB+wgezelh+Wvn4frf/AD7h+x/6AgAAAAAAABX/AJhQ/a/3D/NT7/8AnOyf9JPgf3Z/Cr+fvcj/AKC+Kq34H0f08dB/SfgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfl/wDdfnX139C/O/UB4/ugAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAH5tfmx+p/j36t/eP5Z+wgH47dAH60gAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAGLrj+N3j36T+U/pH90/mP64AfjX2YfrCAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAcV+S36n+PfBf9IX5Z+w+oAH87Afq99kgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHxTi/qf4994H5Z+wgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABi/jd+p/j3yb+4/5Z+w/SwAAAAAAAAAAfEIfjh9Rh+xvegAAAAAAB+UwfDPYB+s31SH4d/aIfRf4dB/QV8ch+XeaH6+fBYfRfUh8IfoSH4p/SYYn7Lh7t+OwfJfch/QT8iB+T2SH9AHxWH5g/tIHkn5oBF+zQei/iWGh1ofoD9ogAAAAAAAB5l/Nxg/6MT/Lv9F+IfLP6V+f/S74L/MfR5r9HOmT+fn9C4f3m+g/65+YAAAAAAAAAAAAAAAAAAAAPzpD86AAAAAAAAAAAAAAAAAD2H7r86f+znzv1HWnj+6AeefkGHiIAAAAAAAAB+rgfaIAAAAH4d+LAAAAAAAAAAAAAAAAAAes8P8Aqf49+3nqH5Z+wgHxj+TAWAAAAAAAAAD7RD9XAAAAAPw78WAAAAAAAAAAAAAAAAAs64+8/JH6T+U/0NekfmP64AfFn44B2oAAAAAAAAB+rgfaIAAAAH4vfOYAAAAAAAAAAAAAAAAAGj7D+p/j3z7/AEe/ln7CAH5ch+cHcAAAAAAAAAAfvR6KAAAAB50H4LgAAAAAAAAAAAAAAAAAHoNj9T/HvNj8s/YQAAAAAAAAAAAD2kP3EAAAAAAPyB+UAAAAAAAAAAAAAAAAAALPvP6n+PfGv9Mv5Z+w6AAAAAAAAAAAAcZ+QwcZ+74AAAAAAAAAAAAAAAAAAAfFIfin/UEAAAAAAAAAAAAAAAAAAAAAAAAHmn82mD/owf8A5efopz/RPmPo/wBDh058r0n8+v8ASngfvb9Af1v8wAAAAAAAAAAAAAAAAAAAAeUh5UAAAAAAAAAAAAAAAAAfM/2fwfxv9X+D9H+rh5XsgH5DeeB+iIAAAAAAAAB7WHoAAAAAHzpz4AAAAAAAAAAAAAAAAAHzp9FfpH5V9WH5v+qgB4+HmgAAAAAAAAB6AHtYAAAAHzpz4AAAAAAAAAAAAAAAAFLTL5B/Rb9A/NPYD87/AFAAPnQOfAAAAAAAAAPaw9AAAAAA+f8AlAAAAAAAAAAAAAAAAAAMX5b/AEj8q/RP1b83/VQAA+VAAAAAAAAAAD6W1QAAAAMoPmkAAAAAAAAAAAAAAAAAA8apfpH5V7kfm/6qAAAAAAAAAAAAdAH0WAAAAAAeE8UAAAAAAAAAAAAAAAAABS+Qf0j8q/Qv2v8AN/1UAAAAAAAAAAAAAytUAAAAAAAAAAAAAAAAAAAPIg+Q/wBGQAAAAAAAAAAAAAAAAAAAAAAAAi/mKD9oPcP82Pvvzh/PP+zflOs479B4f0D/AEs/ib67zr8FP2Lyv6f9v+kPBAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA+BPgMP33AAP50P1kD7LAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA/IgPnD7Z/mP6H85P2C5a9j8m/nP3fyA+7v6u+b+b+2/SfP8A2nPu+MAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADl9+bzb+ebPX+oKxGhwgfzO/0+h0AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABzWj1ceocvYAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAMFxva9XIHL2AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAEHKdXH2Ry9gAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAYjOrj3jl7AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAZxvVx9qcvYAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB5oHV9AAAAAAAAAAAAAAAAAAAAAAAAHOB/PH7h8B2/p9138AfbBxXy35r/ALX5Hyt/RL/cHyHcAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfl/91+dfXf4JfO/Ufvl9AeP7p8GB+dH9BQAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB+bX0L+p/j31Afln7CAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAYuuP43ftX+k/lIfmP64AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAcV+Un6n+Pfsgfln7CAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHxTi/qf4994H5Z+wgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABi/jd+p/j37Vn5Z+wgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfiAH0N+nIAB/NOH6r/oCAAAAAAAAAAAAHKfNgfXwAAAAAAAAABwX4oB84fW/5f6PecZ+cd/yX7F/Qvh/t16n6WYAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB+X/wB1+dfXf5W/O/UfXf6OeP7p+Kweq/qoAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfm19C/qf499QH5Z+wgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAGLrj+N37V/pP5SH5j+uAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHFflJ+p/j37IH5Z+wgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB8U4v6n+PfeB+WfsIAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAYv43fqf49+1Z+WfsIAAAAAAAAAAAAAAAAAAAAB5TVDh9APojykOV+gA+X/AGsPPurDxL2gPUPiwPUPoAPmLHD6o/m3D9QvVQ+kewD5U9jDxH6pD5e9eD1AD5fA+oPNA5/nAsfQwfk9+eYfvb9AAAAAAAAAAAAHzT8ghyXSh9afUoPAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPy/+6/Ovrv5I+d+o8E/bfx/dP5oA/cT6bAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPza+hf1P8e+oD8s/YQAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADF1x/G79q/0n8pD8x/XAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADivyk/U/wAe/ZA/LP2EAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPinF/U/wAe+8D8s/YQAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAxfxu/U/wAe/as/LP2EAAAAAAAAAAAAAAAAAAAAA/Kj83A+gPlwP208RD5U/oCD+X39rA/P75xDsPNw/pg/BoPqveD8/vpEP2G/nKBoHoH9DQfyq/RoeP8AbB0vkIf0x/k6Hz5ygfdOwH5XevB87f0Gh+SHzuH6Y/rCAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB+X/AN1+dfXeJ879R+Jv9KXj+74v/PEH9PVsAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA/Nr6F/U/x76gPyz9hAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAMXXH8bv2r/SfykPzH9cAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAOK/KT9T/AB79kD8s/YQAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA+KcX9T/AB77wPyz9hAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADF/G79T/AB79qz8s/YQAAAAAAAAAAAAAAAAAAAAD8qPkEP6FfxYDC9tD5U/oCD+X39rA/P70YP1Z/mND+iz8ew+q+xD8oP0RD9X/AObwP1M9TD8Dv6qg/lV/fQPizykP2q/mKD+iz+e0PozmQt/eYfnd/R6H84v6Ih1H5UB/SwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAZd53KU1/Lv/S6q+G/kkP3aAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADNsa42jLYAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAaLH2tcgy2AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAGZOuOyZbAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABSbrjfMtgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAbj647RlsAAAAAAAAAAAAAAAAAAAAAflR8Dh/QJ+IYfQ3oofnN+0ofgl+44fn96sH6rfzAB/RV+PofVf6KB5P+BoftV+RQfqv6sH4Af1VB/Kr+/4fFPlIftZ/MAH9FX4RB9v/AGcG180h+f39FofzpfoCHbfjwH9DHsoAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB/PR+ngfAH1sH6SAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAH5UfnyHZ92H7WVw/C3qQ6D9Jg+L/RQ/Rb+fUP3F/MYPovhA/P73MP2v/EgP0W9GD8Xf6LQ/nS/aIPlHzoP1b/n1D9xfNg/Jbkg/UTQD4u/cQPw7+0Q+s/wlD2j9sgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPy/8Auvzr67+hfnfqPyDZ4/u/nz/Q0HtYAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfm19C/qf499QH5Z+wgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAGLrj+N37V/pP5SH5j+uAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHFflJ+p/j37IH5Z+wgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB8U4v6n+PfeB+WfsIAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAYv43fqf49+1Z+WfsIAAAAAAAAAAAAAAAAAAAAB+VHyoH9AQAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB+X/wB1+dfXf0L879R8OfjB4/u9H/S+AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAH5tfQv6n+PfUB+WfsIAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABi64/jd+1f6T+Uh+Y/rgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABxX5Sfqf49+yB+WfsIAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfFOL+p/j33gfln7CAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAGL+N36n+PftWfln7CAAAAAAAAAAAAAAAAAAAAAfNPmgfcIAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABwfXxdHs4dHnX8zWev6A/tSAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHEbPf5u8cHpAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAETXmnqHb54cPogAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAFXz7v8AN9LOD0gAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA5eLv83rTg9IAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAIvNO/zfUDg9IAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAD8aw+0/sEAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAOXDycAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA94CwAAAAAAAAAAAAAAAAAAeH54AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB6wHUAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAeeB82AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAH2SF0AAAAAAAAAAAAAAAAAA+RMQAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA+kw9DAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAD83g/SED5vA+kAAAAAAA/PkPS/rsDw8Py464P101QAAAAAAD8Y/13DpQAAAAMf4GD9EQAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPiYPlcD7A+wQAAAAAAD8Y/wBdw6UAAOaD8iP2cAAAADzT80w/XcAAAAAAPhkPqP0MAAAAAAAAAAAPgQPs7rQAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAD+f0P6AgPz+A/QEDM0wKgVdUADE/O4PVvtYD8cA/R/3AAAAAKjwsAVIg/Dj9rA6sApA62GPsBF83B9LAY+gFgDM0wAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA/H4Pt73gNbVCKuF3MDTAMwNMD8KP2sDqwAA5QPxT/cwItAADM0w8p/KkP2csBYAx9ALAAAeCB630AGIG2AY4bAAAAY+wGZeCX55D3HVCLJDbAzNMAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA/n1D9igPi71APpv8zA6v2UPtn8uw88+8Q+mfxND3j5XD9OftYPN/wKD6W4cPrPND1uoHH+gB848EH6TUg/LX2sPhn+ioOrA/nK+zg+Yvp4PPPcA/Tn+f0P36/G4Ol9bD6z/MwOr9lD9CQAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA/H4Pt73gN78WAsfo4HwN3oeafsuH5ZtDxH9nA/Ej9Xw/K39yg/Mv2YPs7lA/ng+4w8E/TQPYfytDvu4D7T/AAmD6z+bw/eT58D5G8ED9qvwwD9//BA+Bq4foD+aYdn5SH1N+kgfjB1oedfSAfpYB+KfoAeQ9iHYfOIfvV+MAfqt4eHwZ6gH6Sfm6G94iH7T6AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAH89YfdAHzf9dh8z/TAdX+cIfvp8gh8r8CH0H3AfdP5gh7R9rAfimH6rfP4eU/pIH4cdGH7U/gQH6SceHztwQfsf3ofgr+34dWB/PV/QqH5k+ph9wfgAH9AX8/ofp58/h+owH4zfTAdX+cIfvoAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAH4/Bk+gB92fkmH6+eLh8XfXwfAP6WB+Xv6KB8s/TIfBf7WB+Lf7NB+LX7nhb5QPww/foOK/F0PaPUQ9I/NMP2s/JMP3C/MEPdfqwPh74eD9EfnwPq742D78/O4P0B/NMP0+9pD8Av1kD48/WEPhbzwP0sA/FP8AUQOz/FAP3k/G4P0e/MYP1W/FEP38sB4j+TwffPx4H3D9cAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfz+h/QEB+fwHyT+iIdNoB88+ch+gP5HB7x68H2t+YIe0fawH4ph+q3z+HlP6Qh+JFIP3K/AgP18A7D8cA/bTqw/BX9vw6sD+f3+gIPzT9WD7W/n9D+gL+f0P0n8mD9NAPxI/REOm0A9wAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAD8fg/QH6AA/CgP2s+SQ8M+1ALAfl3+ngHqAfix+1gfLny4Dv1vA5QPxD/ecOU/FYPSPsQPSLAdF+VIftZ+aYerfDIfp588h2H0CH5l8eH7c/j8H6A/mmH6rerB/P7+qgfKP6uB8LeeB+lgH4p/qsHV/imH7r/j8H6A/mmH6rfiSH9BLw+dPzxD9GQPWOoAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA/n9D+gID8/gPa/zDD6h9YB/wALB635iH61fimH1n8uB+h32sB+KYfqt8/h5Tz4fQfBBzNsPDPWw+tuED87vfg+If6BA6sD+f3+gIPzT9WD7W/n9D+gL+f0P3q/DUPpT0MPdfzDD6h9YD7ZAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAD8fg/QH6AA/CgP2svB+IX0yFj9OQ/Fv0gL36Vh+Jf7WB0f82Qfvf7MBygfgB9qB4X+j4dn+TofVPoYfTH5Uh+1n5ph6t8aB6X4CH3t9vB+Dv2yH6E/j8H6A/mmH6rerB/P7++4fh966HjX1QH6WAfin+qwdX+KYfuv+PwfoD+aYfqt8kh8o+6B98/kgHtbA/Um2AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABz4dABngaGYHC+ihY4cOroht54cp1wF0DEDWrhDnh0bA5/ow5fHD0ADmgLuqDwOf6AMyYLvPh0HPh0AHm/Zhq5gcL6KFgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAMcNCwBhBtPBnm4dhtgcEGl1YYW0Hj/5Zh+14AwMrMC30AFIPPPRQmyQ28wJrQef9wEWgGPoBYxw0MwNaUOf6AKlIPzh9wD7GAxNYHYobuOGhmBrShzmOHdgeebwdKAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABXD8Vf16DuAAAAAAAAAAAAAAAAAAAAAPym5QPQP1BB4AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAcoAAAAAAAAAAdBbAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPx4+sQAAD88/0MAAAAA4QPZvqsDPD5U85D076rDj/lQMT6QD24AAAADl/lQOR92D6L8qD5elD6m9LAAACkHwNwgfcPsAAHxNzQfbfRgAfN/UB7QAAc/8mB6r78AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB+f36AgAHzeH4Rf00BKAAAB5SHlX1WAB55/MwH9SWwB+cwfFP73gAAAAAHx4H4+/0igAfjN1QfreAAAB+VAdNw4fnX/RQHqv5UB9LfSwfit+54S+Ah+I36wh9rAHg4fl7+wQdwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfn9+gIAB+RwfmN+94fXYAAAHlIeVfVYAHKfzAB/VUAfn8HxT+64AAAAAB8qB+Pv8ARaAB+P3VB+q4AAAHgIe/AfiH9kh9rfzeh/S48PxI+9A+wwPxT+1g+1gOE/n7D+grtQAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAD8/v0BAAP57Q+Z/v4P2sAAAA8pDyr6rAA5T+YAP6qgD8/g+Kf3XD4M/OgP6CvkQPyI/oyAAPlQPx9/otAA/H7qg/VcAAAAAA/E79Tg86/IUP6IgPyelD9XAPxT+1g+1gPyQ8kD6Uoh+lO2AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB+f36AgAfN/lIfh31Af05aAAAAeUh5V9VgAcp/MAH6/AHzeGJ+64eU/hQH9FvKB/Ol/SwAAfKgfiL+nIAHxP9Wh+q4AAAAB5+H5zfqyHxT+aYf0BAflRyofsCB+Kf2sH2sB/Ol+hIfpD+MIY/7cgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfn9+gIAH5/foCH84XiQfuz9ogAAB5SHlX1WAByn8wAf1VAH5/B8U/uuHlP4UB/Rbygfzpf0sAAHyoH4+/wBFoAH4/dUH6rgAAAAV/wA2w/R+6HzP+PQf0OgfkB3wfqCB+Kf2sH2sB/Nl+5QfQfzuH4G/09AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfn9+gIAH5/foCH5U/lWH3H+44AAAeUh5V9VgAcp/MAH9VQB+fwfFP7rh5Z+EYf0X8kH86n9LAAB8qB+Pv9FoAH4/dUH6rgAAAVw/P77uDb4wOg/nAD+kqwH4QfquH0QB+Kf2sH2sB+Hv2+H3H4yH4kf0OgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfn9+gIHzeHQe4B4f/ADihsf07htgAB5SHlX1WAB4Z/OQH9L/pYH5Zh8bf0EAfzVB+m1sPyL/oFD6QAPgwPyV/pCDbYH4cdWH7OfJgfmH++QAGf+KAcZthU+nA/Ub4BDH9+D4O/YkCIPwo+0Q/QsDxn8fg/Zz81g+xPrAPwz+8Q+2wAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA/P79AQPzuD9EQD+cIPEv25D7xAADykPKvqsDPDw8A94Dj/NwOgD2DyoPJPqAPFPewAOX8nA0A9w8vDjwO46gPGfoYADH8XAAPWA6gD5/xA+mwDzcOHA9g6ADig+ZvoYPSAPBPUA6sAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPz+/QED8Yw+m/Pw8/85DP9ADtvQA+zPVQPKQ8q+qwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAD8/v0BD5vA+kPyoD8rP1TD9VwOU/FMPj79lw/QHykPKvqsAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA/P79AQ/OP9HAPyoD8rP1TD9VwDig/mM0A/pz44PKvqsAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA/P778D86P0cA/KgPys/VMP1XAAP5p/LQ/oE9QDyr6rAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPzj1g8p/TEOg/LMPys/VMP1XAAP5l/PQ/oq6oPKvqsAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA/K77RAA9g/MgPys/VMP1XAPl8P59ftgP3S8pDyr6rAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPz+/QEAA/KgPys/T4P1B4oPmn4pD7L+/Al8pDyr6rAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPz+/QEAA/KgPys+6A+7KgfL/jwfo59jAeUh5V9VgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABx/YAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfn9+gIAB+VAflZ+qYfquAfzxB86fvkH1t5SHlX1WAAAHwIHtH0mAAAAAAAAAAAAAAAAB8nh+Vv1oHW+Lh+mnqoAAAAAAAAAABynwCHyJ+2IdmB5v+YAfXn2GFgAAAAAAAAAAAAAAAAAAAP5rf3XD3sAAAK/5HB8s/QAfspbDh/zjDs+UD9SXgAHyyH5DZIfp199gfIIeGcYH139Uh5l+JQAHQfvEAAAAHyN+ZwcZ+yQfRcQfkX8Vh7d+ygelgGeH4K/uqHSgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfn9+gIAB+VAflZ+qYfquAfkwH5efbwfuZ5SHlX1WAAAH87oe/ftIAAAAAAAAAAAAAAAAAB/L7/RYHq35ph+dP9IQAAAAAAAAAAAAfzy7wfvfdA/KYP1ZAAAAAAAAAAAAAAAAAAAAAzNMAAAAPmf2MOo/AgP1J+uw/DP9UQ97/IYPbv0LAAr/AJch+mPgwfgb/RQHR/hkH9BWeH85v9B4fKnqAdxXD8xP2ZAAACkHyX9gh8M/n8H7z/GgdH9Kh+QfmQfu4AfmsH5Xf0nh1YAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAH5/foCAAflQH5WfqmH6rgH5xh+OPtof0e+Uh5V9VgAAHyv8/B+T39ModKBzXy2H0N8Uh9k+oB5Z5uG383B97caHzz9PA75iD6N9DAAAAD+X3+iwPVviwPxK/qIAYHyf5CH2TdD5k6APq7jA+b/AKzDYAAAD8m/mkPa/wBjwPyoD9VwOf8AjYKX3MHjnjgfZPm4fP312FX5bD07hw+s/IQ+UvTQ+vgPOvj4Ot+oQ7jz8PkHsA+y2h8g/TwdKAAAAAfkQH6hdqH8sv8ATOHZ/HIfmT+/gfNPTB67EGwB+Df6vB4v8HB+74H4cfZgfe0oHyYHKfbwAAAAAHzz8vh+k3KB1YHjn4th/QOB8s5gfjB/RaHVgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfn9+gIAB+VAflZ+qYfquAfHIfg3dD+p3ykPKvqsAAA/JD9ag/BL7WD9HAPzx/LYP1e4cPjb92g/IbykP0O+Lw9j/U4PwT/UEPqv8PQ+7PtYAAAAP5ff28DC/MAP0p++QPzBD7rwg/Af+kgPyL7MP1hzA/O79JAAAAA/Kj9Kg/nn/S8P0W/KgP1Xzw/J79YQ/Mvw8P2x/mYD+j/qw/m9/ewPkr8qw/SPhA+6fhEP13/JAD9dA/Ej9rw+BvoEOq/OQP1s/C0Psv6UD8Bf6HA9WAAAA5IPz+8WD9j7AfzJft6H1h8ch+Uf9EQflz6KH6AgEQfhV+6ofIf40h/SlKH4cfUwfpCAfkMH6s9AAAAAB8rh+YH6+B7WAAfE3GB+hnKB+fn6XB/NP/RaHVgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfn9+gIAB+VAflZ+qYfquAeaB/NAB/SqgeVfVYAB5OH59/oMHxf+fAf0WgeU/hQH9FvJB/Op/SwH5/fP4fsD8TB8WftYH4UfpYH1X+KYfa32sAAAAH8vv7Lh+Wv2+H6rAV/wCd0P0pA+X/ANPgm/AUP6TPiQPrHtQAAAA/Kj9Vw8X/AJ9w/cX5TD9V/jwPgP6+DlPIg/Yn+acP6LerD+dL91wPweD+jcD8K/0bD6548P5j/wCn0PwI+zA/Ru2H5d/WYfXPngW+4D+dL91w9WAAAAzA+fPxYD9D/wBLg+Dvy7D9FPlQN39rwAAA+Bg979+Cv+HAWPsYPyL/AHHD6uAi/HsP2J48OZAADsQ6XzoPzQ+HA/pUeBnh+Vf6wg/8wQ/SvTD+af8AotDqwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPz+/QEDjw8v/OgPzs/RMP0X9AD0AD+bLyIP3k9wDyr6rAAPyuD3h4H40B+7v0gHlP4UB/Rbygfzpf0sB+f3z+H7A/FIfFP7WB+FH6WB9V/imH2t9rAAAAB/L7/RYGL/AD1B+9X02HOfzoB/SkAZgaf4QB+gPAh+jgAAAAH5UfquB8mB+JH12H7X/AwfPn6+AZgaf804f0W9WH86X7rgfhQH9FoH87v6rh9lgfy+/wBIoWvyxD4Y/doPzs+qw+9gOa6UP50v3XD1YAAAAA+Hg+P/ANoQM8D+eIP2a+kwAAD53wQ+pgADC8WD8Tv6MglA+XPLw+8/Bw8hAAD3kPXQGfzRh/R/0AM/PEPvbbDn/wCfIADw/wBoD+hLowAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA/P79AQOPDy8AAPQA9AA/M78hg/X77WDyr6rAA5/wDMkP1WAPyQDz/9rw8p/CgP6LeUD+dL+lgPz++fw/YH4pD4p/awPw9++Q+wfwYD9Eft4Pl/6bCUAA/l9/osD1b8/A/K3+igOt/nAD7x/TsPkn6MDvfkQPxo/bwPpYD5v97DVAAPyo/VcAD86fxrD+qrxEP56P3aD6W+IQ+8/wCb0P366UP5zf33Cx+FAf0WgfkhhB+xnPh+Av8AQ0Hxj9nB8GeCB7H+aofvllB88/bwfzpfuuHq3zOH0XdAAAD8+Q637ZAA/MZgfp4B4/th6L86B5v9UBleOB9RgYn4Zh+unuYAH5LfqCHUAAAAAAGV+JYfuSwPzm+yQ6jxQPW+7AA/mn/otDqwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPxu+kAAAAAAACL8zgl+7A+pvqADPD8s5g/TboAz/AMww+Kf1nDn/AM2A/a/x8Pyb/aQPhzzQP1x/PkPnb9jw+afyTD7h4EPRv0hD+dr+gIPSwD5MD8iP0vD7j2A/HPmg/T7MD8XuBD9Z/v4Bgfiz+1IAH89H7Dh9IAHg4fld+qge6gH5AB+v4H50/lsHa/t0HuH5nB8M/oMHxj9vB1H5iB+y/voYn41h7zCH257AH4MfYwcJ+gIep/jcHxH9dh+0vAh+J36WB9sfzih+5ntwAH5V/NgfX3XB99gfP/z8HUfbIAH5B+oB9ufiWHLAH7Ch9IfMIfP/ANwB6sAAfkT+uwAAAAHk4fhV9jh2v3cHof5yB+doB6WH7xAAH4MfuIHRgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB/9oACAEBAAE/AKqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq+N/wAU9P8Agh8JvE/xY1XSrjU7TwxYtdy2lu4SSYAgYUtwDz3r/h958Kv+iH+K/wDwY2/+Ffstf8FMfAv7U3xYt/hN4f8Ahlruh3c9jPdi7vLyKSMCIAkYXnJzVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVfE7/gsL8Nfhj8SfFnw1v/g74lvbnwnrl7pM1zDfwKk7207ws6g8gMUJAPrX/D7z4Vf9EP8AFf8A4Mbf/Cqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr9pf9tn4CfsrW4tPiJ4jluvEU9v59p4d0uLz7yZCSFcjISJCVI3SMoODjOMV4t/4Lha1JeiPwJ+z/AGUFpGzAy6vrrSyTDjadkcSiMj5sjc+cjkY5+AX/AAWL1X4m/E7wh8LvF/wKtLWXxd4gtdKj1PTddYLa/aZEijYwSQkvh3yT5i/KeBkc1VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX7fn/ACZp8Wf+xfk/9DSq/wCCQ3/J5Wm/9i/qP/oC1VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX7WX/J1Hxl/7KBrv/pfNVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV+2T+0TZfsvfs/+Ivifm3l1raLDw9az8rc6jMCIgRkblQK8zLkEpE4BBxXjXxr4r+I/ivVPHPjnXrvWte1m4a4vr66fc8zn9AAAAFACqoAAAAFQwy3EqW9vE8ssjBURFJZmPAAA6k+lfsUeAfEui/tg/B8+MfBep2NvJ4mhaE6nprxI0sal1K+YoBZWVWGOQQD2qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/b8/wCTNPiz/wBi/J/6GlV/wSG/5PK03/sX9R/9AWqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/ay/5Oo+Mv/ZQNd/9L5qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv+C4XjWVYfhV8Oba5YRu2o6reRFTgsBDFAwPqA1wCPcetV/wRr/Z88GWfw01L9ovV9KtL3xVqOqXGm6TcuC7adaRoiybARhJJHaQMwydgUAgM6mqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv2pv2ufhb+yN4V03xJ8SLXXb6XWppINMsdJsDM9w6BS+6RisUYAdTh3DMNxVX2tj4i/8ABbX4rahezRfCn4Q+F9FsdzLHLrs8+oTsvIV8RNCqN0O07wOmW61rn/BV39t7V5YpNP8AidpmirGpDR2PhyxcSH1PnxSHPbggV4N/4K+/th+G3thr+p+FPFkcO4TDU9EWFpgWzkm1MIUgHAIGOBkMc5/ZR/4KqfCP4+6va+BfiLpSfDzxZdtHDZLcXvn2OpTMQuyKYqpjdmPEcg7gB2NVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVft+f8mafFn/sX5P8A0NKr/glN4g0Lwn+1cnibxNq9ppek6X4W1S4vb27lEUVvEkYLO7HgAAda/aG/4LRR6dq914d/Zn8D2Op2luxQeJPEaSrHccsCYbRSjhcbWV5HBOSGiGOb3/gq7+29davFqUHxO0yzt42Qtp8PhyxMEgU8hi8TSYbocOD6Eda+A3/BaXxPb6lZ6J+0V4B0+806SQJJrnh5WgmgU4G+S2dmWTHJOxk4xhSRz4K8a+E/iP4T0vxz4G1601rQdZt1uLG+tX3JMh/UEEEFSAysCCAQRVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVXxk+OPwr+AHhB/HHxa8YWmg6V5nlQtKGeS5l2lhHFGgLyOQpOFBwAScAE18Yv8AgtprA1KSw+Afwk09bKGQhdT8VyyStcLxyLa3dPL6HkytnI4GCDrv/BWD9tzV/I/s/wCJGk6J5O7f9h8O2b+dnGN3nxyYxg424+8c54x4Y/4K/wD7Y2gCAarqPg/xL5UZR/7U0IR+af77fZnhwR/s4HtX7JXxm8VftC/ALwz8YPF/hO08OXviH7S8djbTSSKIY7iSKOT94qkbxHuA+YFSrA/NgVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVfEf4k+BPhF4M1L4g/EnxNaaD4f0mPfd3tzuIXJwFVVBZ3YkBUUFmJAAJOK+On/BajxTe3txpH7O/wAO7HTrBW2prHiRTPcSjnLJbxsEjOcY3NJxnIBPEX/BVb9uKPVzqT/Fmxlt/OaQae/hvTxAFJOI8iESbR0Hz7uBknk1+zl/wWevrvW7Tw1+054P0y106dki/wCEl8PwyqbUBQC9xakyGQE5ZmiKlRwsTdK/ah1HT9Y/aY+LeraTf297Y3vjrWp7a5t5RJHPG99MyujKSGVlIII4IINVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVeIfEGheE9Cv/E3ibV7TS9J0u3e4vb27lEUVvEgyzux4AAHWv2iP+Czvhnw5qVx4b/Zv8EReJZLaRkfX9f3w2chHGYbeNllkQ8He7RHgjaQQ1eI/+Csf7bOuSxyaZ8Q9H8PqjOTHp3h60cOGxgH7QkpwuDjBB5OSeMfD3/gsj+1H4YurdPHGk+EfGdiqotwJ7E2Ny+1SCySQMI0ZiQWzEw4woXNfsnftzfBv9rbTXtfCtzLoXiyzjMl74a1GRftCIu3dNCy8TRZYDcMMD95VyM1VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX/BQv8Aag8f/sl/BbRfiN8ONI8P6lqWo+KINKli1u3mmhWGS1upSyiKWNg+6BACWIwW46EftT/td/En9rzX9D8R/EnRPDOm3Ph+zktLZNDtp4UdHfeS4lmlJOfQge1V+zl/wUm+Of7MHwzg+FXgDwr4E1DSbe8mu1m1ixu5Zy8pBYFormNcccfL+Jr/AIJvftq/FT9sT/hYn/CzNA8KaZ/wiP8AZf2H+wrS4g8z7T9r8zzPOnlzj7OmMbcZbOcjFVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX/BUn4Nal8Yf2TdV/wCEd0i61PXvCeqWmradZ2cHmzXB3+RKigHJ/dXEj7QCWMagAnGPg/8A8El/2rvifZQax4k03R/h/p8rcL4huGF2yDjctvErspzn5ZTGeM9CCYv+CG2tNo5mm/aTsV1TyWItl8Js0Hm4OF837UG2k4y3l5H909K/ar/Yz+L/AOyNr9nYfECCx1HRtXaQaVrumOz210UwSjBgGilClSUYY67WcAmq/wCCZ37RmqftDfs1WL+K7t7nxR4Lujoup3EhG67REV7ecgd2iZUYnlnidu4qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/b8/5M0+LP/Yvyf8AoaVUN3d20U8FtdSxR3UYinRHKiVAyuFYD7w3IjYPGVB6gV8Fv+CfP7Vnx40CHxb4O+G7WWg3UIltNS1q5Swju1ONpiVz5kisDkOqlCAfmzgH44/sq/H39nBrV/jD8Ob3Q7S+maG0v1liurWdwM7VmhZ0DFckKxDYB44OK/4JKftVal8M/i3D+z94mv5ZfCnj64K6crvldP1TYdjKME4mCrEQP4vLPADGqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr40fFzwh8Cfhh4h+K3ji78rSvD9m87RqwEl1JjEcEe4gGSRyqKCQMsMkDJH7RP7QnxC/aZ+Juo/Ev4hak8k1wxj0+wRyYNMtQxKW8K9lUHk4yzEs2SxNV8Lf+CcX7YXxa0e18RaH8JbjSdIvFLQXevXcWnFwACCIZGExVgQVfy9rDkHFeNf8AgmJ+2r4Khlu3+EDa5aQqCZtF1O2u2YkdFhEgmJGOyY96+AXw9b4T/A/wF8NZokjufDfh2ysboKoAa4jhUTPgMwy0m9jhiMngnrVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX/AAUm/a01f9oz426j4T0DW3b4feCbp7LR7WCQ+TeXCZWa9YZw7M25UboIwuAC7lq+Bn7CH7T/AO0Roq+Kfh78OJY/D8mfJ1jVrhLG3uMZH7kyENMMgqWjVlBBBINfHf8AYw/aP/Zvsl1n4qfDm4s9EeYQx6xZ3Ed5aFj90M8TN5RPQCQIT2FVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV/wVN/bN174tfE3VPgD4I1p4PAPg+6+z6itvJxrGoxN+8eQjrFE/wAiJyN6NIScps8P+Htc8W65YeGPDGkXeqatqlwltZWVpEZZbiVzhURRySSQMV4I/wCCJPxQ1jwsNT8d/GbQfDmuS2olj0q00p9RSOUhj5Us/mxhSPkBZFkGS2NwUFv2rP2Ofiz+yJ4ksdJ+IK6fqGla15p0jWtNkLwXYjK71KsA8cih0JVhj5vlZgCa+H/j3xX8LvG2i/EPwNrEuma94fvEu7G6jP3HU9GHRkYZVkOVZWKkEEiv2dPjTo/7Q3wU8J/F/RrdLVfEFiHurNZRJ9kukJSeHcOoWRXAJAJGCQM4qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv+CoPwE+LP7RXwB0DwT8G/Cf/AAkGtWPjC21Ce1+3W9pst0s7uNn3zyIpw8sYwDn5s4wCR8cP2b/jR+zfqumaJ8aPBn/CPXusW73FlF/aNrd+bGrbWbNvJIF54wxBqvhD+w9+1H8evBkXxB+E/wAL/wC3dAmuJLdLv+2rC1zJGcOuyedH4JHO3Ff8EpP2W/jt+zV/wtH/AIXZ4F/4Rz/hI/7H/sz/AImdpeef5H23zf8Aj3lk27fOj+9jO7jODiqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqvj1/wU4/ZY+Bd7c+H08UXHjfxBbK4fT/DCpdJDIOAktyWWJTuyGCs7pg5XOAfFv/BcHxVPdyp4E+AWlWlsu9YpNW1uS5eTk7HKRxxhOMZQM3PAbvX7UX/BQX4yftbeBdN8AfEjwp4H0+x0rVk1OC40WwuYbjzVili2FpbiUbCsxJAUElVOeMGv+CHfiKWHxJ8WfCJid47yx0y+WQykCJoXuEICY6sJlycjHlgc54qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv2/P+TNPiz/ANi/J/6GlV/wT5+C2gfHj9qzwf4O8W21vdaDZNLqmpWk67lu47ZC6wlcEMryeWrKcAoX74Bhhht4Ut7eJI44lCoiLtCgDAAA6DHavif8NfB3xi+H+ufDLx9pSajoPiC1NveQNwQMhldD/C6OqurdVZVI5FfE3wLqXwv+I3ij4baxJ5l74W1i60yeTZsEjQStGXAycBtu4cngjk9a8J+JtV8FeKtG8ZaDN5WpaDqEF/ZybmXZNDIsiHKkMMMo5BB9COtadqFlq2n2uq6bcpcWl5Ck1vKnSSNgGVh7EEGqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/4Lb/FPUtP8N/Dz4NabeSxWur3FzrGqRr8olEASO3Un+IbpJ2K9MqhwSARX/BHj9lrw/491rXf2jPHGm2mpWPhe8/srQLO4jSVPt3lrJNO6MDgxxyw7D/elY8FAaqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv2sPiJe/Cf9mr4k/EDS7lrfUdK8O3X2CdJNjQ3UieVC4ORysjoeOTjA5qv2FvgRo/7Rn7TXhP4c+JncaCGl1DVlQgNLb28Zk8oZ7SOEjOOQrsRyK07TtP0jT7XSdJsbeysbKFILa1t4hFHBGgCqiIuAqhQAABgAAV4s8K+HvHXhjVfBnizS4dS0XW7OWzv7OXIWeGRSrqSCCMgnkEEdQQea+L/giH4Z/Fnxt8N7e7e6j8KeIr/SUnf70otrh4gxwByQmeg69Kqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv2nPiLe/CT9nn4ifEbSyw1HQvDt3NYMq7tl0YysLEbl+USMhbBzgHHOBU00txK888ryyyMWd3YlmY8kknqTX/BOj41fAf8AZ6+Nd98Vfjc+ob9P0eW30A2lgbkw3MzBZJeCCpEPmR+hEre1f8Pef2NP+g54q/8ABC//AMVX7fX7ev7KX7TH7NusfDvwdqHiC58TwahZ3+irdaOYY1mjmCyEu2dv+jyXAyMEkgZwSDX/AARM8bXGq/Bbx74BuL9Jl8PeIoryCFmYvBHdwBccnAQvbOQAB8xkJzmqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr/AILe/wDJU/hp/wBi/df+lAqv+CQ3/Jmmmf8AYwaj/wChrVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVf8FX/23vEkfii8/Zc+FHiCXT7HTo0/4S6/tHKS3MzqrrZpIp4jVWUyAYJY7CQFdWr4Bf8ABOT9qP8AaI0S08WeGvCdj4e8N36lrTWPEl0bOG4XaGDxxqrzOjArtkWMoc8NwcftQf8ABM7x/wDst/CGX4teJviV4f1mGC8t7SSysLWZW3ykjId8AgEdwM+gqv8AgiH/AMlT+Jf/AGL9r/6UGqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr9vz/kzT4s/9i/J/6GlV/wAEhv8Ak8rTf+xf1H/0Baqv279QstT/AGxPi3c2Fyk8aeJriFmXoJIsRyL9VdGU+4qvhvp97pPw78LaVqNu9vd2Wi2cM8LjDRyJCisp9wQRVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVf8Frvtf8Aw074S3+b9m/4QO28vOdm/wC3Xu/HbONmf+A57VX/AAR91Cyvf2O7e2tblJZbDxNqENyq9YpD5cgU++yRG+jCqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr/gqHDLP+wp8To4InkYLpbFVUkgLqtoWPHYAEn0Aqv+CL2oWVn+1dr1tdXKRy33gW9htlbrLILyykKj32Ru30U1VftZf8nUfGX/ALKBrv8A6XzVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVf8ABT6zu779hb4nwWVrLcSLHpkhSJCxCJqdo7tgdlVWYnoACTwKrQ/DHiXxRLLb+GvD2p6tLAoaVLG0edkB4BIQHA+tf8Kl+Kn/AETPxX/4Jbj/AOIr/hUvxU/6Jn4r/wDBLcf/ABFf8Kl+Kn/RM/Ff/gluP/iK/wCCKXgDxj4R0r4u6x4o8OahpMOp3GjwWq31q8DyNCt40hCuAcATx8+ufSqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr/gt7/yVP4af9i/df8ApQKr/gkN/wAmaaZ/2MGo/wDoa1VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVXx/wDEGoeLPjt8RfE2qlPteqeKtSuJVTO1We5kO1dxJCjOACTgACvCX9g/8JVov/CU/wDIF/tCD+0fv/8AHv5i+b9z5vubvu8+nNadDp9vp9rb6TFbx2McKLbJbqFjWMABQgXgLtxgDjGK/wCCyvxU8IaH+z7pvwmn1KKXxN4m1i3u7exSQGSK1g3l53XqEL7UU/xEtj7rYr/giH/yVP4l/wDYv2v/AKUGqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr9vz/AJM0+LP/AGL8n/oaVX7Ivx7l/Zp/aB8K/Fl7R7zT9Pme31W1ViDNZzoY5cY6sobzFHQtGoPGa+HfxK8A/FzwnaeOPhr4r0/xDod7kQ3llLvXcPvIw6o47owDDuBX7VX7WHwy/ZU+Hl/4n8WazY3HiKS1c6D4cW5UXWpz4ITCDLLCGHzy42qM9WKq3iLxBrPi3X9T8VeI9Rl1DVtZvJr2/u5jl7ieVy8kjY7szEn3NfswfCyf42ftC/D/AOGEVp9ph1vXIFvo+OLOM+bdNg9dsEcrY74xVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV/wAFtPhPd6j4Q8AfGrT0lePRbyfRdRVULBUnUSwyEj7gDQyqSRgmVBkHANf8E2f219D/AGVPHGq+GPiOtx/wgnjBoftt5BG0r6VcR7glx5agtIhVirqvzYCsAxXa3hLxj4S8faBa+KfBHiXTNe0e9Xdb32nXSXEMg74dCRkdx1HetQ1HT9JspdS1W/t7O0t13Sz3EojjjHqzMQAPrVpd2moWkN9YXUVzbXMayQzQuHSRGGVZWHBBBBBHBzVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV+0Z8N5/jB8BvH3wysl3XviLw/d2tiC4QfaTGTBkkgY80JnJAxnkVNDLbyvbzxPHLExV0dSrKw4IIPQ+1fs2fHPX/2b/jV4Y+L2gRvcnRbrF9ZCTYL6zkBSeAnkAtGzbSQQrhWwSor4TfGf4YfHTwnbeNPhX4y0/X9NuI1dvs8n722LZwk0R+eJ8qw2uAeDX7Uv7Unw3/ZW+G95408aanDLqksbR6Lokbg3OpXJB2qqZyIwRl5DwoHdiqnxFrupeKdf1PxNrM/nahq95NeXUmPvyyuXc/izGqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr9oz4bz/GD4DePvhlZLuvfEXh+7tbEFwg+0mMmDJJAx5oTOSBjPIq8s7uwu5rC/tZba5tpGimhmQo8TqcMrKeQQQQQehr/AIJ//tQ2H7Kvx9tfFvih7tvCWu2b6Xry26mRoonZWS4EYPzGORFJwC2xpAoJbB8K+K/DHjrw9Y+LPBniHT9c0XUo/Ms7+wuFnhnXJBKupIOCCD6EEHkGqv8AUdP0q1a91S/t7O3RlVpriURoGZgqgsSBksQAO5IHeqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv8Agt7/AMlT+Gn/AGL91/6UCq/4JDf8maaZ/wBjBqP/AKGtVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV/wVA/Zx1/4K/tI6545jsHPhX4k31xq+m3gbcPtTkPeQtkkqwlkLgHAKyLjoQK+Hf7cv7WXwo8If8IH4F+N2t2Whpbi2gtJ4oLz7LEFKhIHnjd4AAcARlcYGMYGNW1H4kfF7X9X8Va5f674s1eG1kv8AVNQupZLuWKCPG6WV2J2ou5VySACyqOSBVf8ABEP/AJKn8S/+xftf/Sg1VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVft+f8mafFn/sX5P8A0NKq2+C/xCvvg1d/Hux0Nrjwbp+vDQ7y9iyxtroxJKu9ccIQ6Lv6BmVTguu7Q/E/iXwxLLP4a8RanpMsyhZXsbt7dnA5AJQjI9jWoajqGr3suparf3F7d3DbpZ7iVpJJG9WZiST9ahhluJUt7eJ5ZZGCoiKSzMeAAB1J9K/4JW/sUal8FvC0nx5+KGlfZvGPiuzEelaZdW22fRrEsSWfPKyzAIxXAKIFBOWdVqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqviz8LfB3xs+HGvfCvx9YNd6F4itTb3SRttdCGDJIhwcOjqjqSCAyg4PSv2n/ANln4m/sqfEK58E+O7B7iwkYvpGuQQsLXU4Mna6Hna4Aw8RO5T6qVZq0PxP4l8MSyz+GvEWp6TLMoWV7G7e3ZwOQCUIyPY1rnjLxf4ohit/EvivWNWjgYtEl9fSThGIwSodjg4r9hbxXb+NP2PfhHrFs0JS38L2unHyiSN1mPsrde+6A5981VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV/wAFTv2MdZ+EvxI1L4/+BtJluPA3jC8Nxqf2eDC6NqEhG9XwSRHM5Lq+AoZynB2bq0fXNa8PXo1Lw/rF7pl2qlRPZ3DQyAHqNykHB9Ku7vVde1Jrq+urvUdQvJBukldppZnPAyTksTwK+NPwU8XfAfxDpPhDx3D9m12/0O11W6sWjZHsftAZkgcNg+YqBd4wNrll527jVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV/wVK/YM1zQPEmt/tRfCbSftnh/VJGu/FmmW0ZMun3Dcy3wGTuikYlpMcoxLHKsSleA/ix8UfhZdSXvw0+I/iXwrPNjzm0fVZrPzgCDhxGwDjKrw2QcCtV/bq/bC1mwl027/aL8axxTY3Na6ibWQYII2yRbXXkDOCMjIPBIrXPib8RvFOuWnibxT498Qa5q1jcfaLa91TUpbuWKXKneGlZjnKISe+0eleAvFtl4/wDAvhzx3pqotp4j0m11K3CSeYBHPEsi4bA3DDDnAz6Cqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr/gt7/wAlT+Gn/Yv3X/pQKr/gkN/yZppn/Ywaj/6GtVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV8TvhV8OvjP4QuvAfxR8Iaf4j0O85e0vIydj7SokjcEPFIAzASIVZcnBFfFD/giP4V1HULvUvg98aL3RreRt0Gla7pgvAmSMqLmN0YKBuxmNj90FurV4F/4IfWiXdtdfE34+yzWyyf6RY6Fogid0yPuXE0jBSRnrCwBx1rwD+xb8BPhX8HPFXwZ+Hvhb+y7Lxno9zpetaszedqF4k8LRM0k7cnAdiqDCKSSqjcc/wDDkL4Vf9Fw8V/+C63/AMa/ZD/4J3eD/wBkL4iap8QvDXxL13X5dV0WTS5bK9to4owGmhlEnyHlh5OBns7VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVftDfBu0/aD+DfiP4O3/ibUNAtvEkcMU2oWABmjSOeOVlAJwQ4jKMDwVc1/wAOQvhV/wBFw8V/+C63/wAa/Zf/AGPPAv7NPwe174K/2m/jXQvEWrT399FrdhE0cwmt4IXheLBR0KwDIYHO45r40/8ABGL4OeNNUm1r4O+PdT8ANMxd9MuLQ6pZg4Pyx7pEljBO3ku4AzgdMaR/wQ98cz3oj1/9oDQbO02nMtnoUtzIG7DY0sYx77uPQ1+zX/wTY/Zy/Zu1eDxhZ6be+MPFlsyva6v4gKSixcEENbQqojjbKghyGkU52uASKqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr4gfDnwJ8VvC154J+I/hPTPEWh3ykTWV/biVM4IDrnlHGSVdSGU8gg818U/+CK/wT8Ualcal8KviT4g8DrcSbhYXVqur21uP7se545cY/vyuc9+1Tf8EQficsrrb/HTwu8YYhGfS51YjsSAxwcdsn614S/4IdafHLa3Hjv9oe4nj2/6TZ6T4dETBvRJ5J24x3MX4V8CPgl4Q/Z2+F2kfCPwJd6rcaJovmG2bU7gTzZkdpJCWVVHzSO7YCgAscADgVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVatpGla9ptzo2uaZaajYXkZiuLS7gWaKZD1V0YEMD6EYr4z/APBGj4HeO9Xn134VeONY+Hs13M0ktibNdTsY8kHEMTPHJGPvcGVgMgAADB0T/gh340uLpk8R/tCaJY2wjJSSy8Py3Tl8jAKPNEAMZ53HkAY5yP2aP+Ccv7O/7NGpW/ivStLu/FXiy3wYda1zZK1o4z81vEqhIjg43YLDHDDJz+13/wAE6fBX7XnxJ034k+I/iNregXOm6HFpKW1laRSo6RzzyhyX5yTORjp8or/hyF8Kv+i4eK//AAXW/wDjWk2U+naVZafdahLfT2tvHFJdTAB52VQC7Y4yxGT7mqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr47f8ABLr9lb43X914htvD+oeB9fu5JJpr7w3OsMdxKwODLbSK8WNx3Hy1jZjnLc14n/4Ida1FK0ngv9oeyuYnmfbDqnh1oGij/hBkjnfe2MAnYgPXA6Von/BDfX7i0Z/Ef7SGn2NyJCEjsvCz3SFMDBLPcxEHORjaRwDnnA+Hn/BGL9mzw3Pb33j3xb4w8YywyBpLVriPT7SZQFyrJEpmGSG5WYcMB1G4+FPC2g+B/DGk+DPC1h9i0bQ7OKy0+1815BbwRKFjjDOSxCqABknAAqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/bD/YF8H/tieJfD3iTxL4/1jw9L4esZLSKKxtY5RKHfeWJfoe1f8OQvhV/0XDxX/wCC63/xr9lr9nTRP2WfhNb/AAm8P+I73XLS3vp7sXd5CschaUgkYXjAxVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVXx6/wCCs3wr+BvxW1/4Tz/C7xXrN74auDa3t0s9vBG0w5PljcxZMFfmbackjaMZP/D87/q1z/y9/wD7hrwv/wAFw/BV5dbPGv7Pmt6VbeZGBJpevxag+wk+YdkkMAyBjA3YYkglcZP7OX7aH7P/AO1LDLb/AAw8Wsmt2sKzXOgarELW/iQgZYR5KyqpIDNEzqpIyRuXNfGb/gsJ8MPhX8QfFvw4074QeKNa1HwnqV5pcs017BaRT3dtK8T7SPMYRF04cjdtOSgPFf8AD87/AKtc/wDL3/8AuGvCH/Bb/wCGt9Iq+PvgT4l0aPzGDNo+rQamQgX5SBKtvkluCMgAc5J+WvgZ+0n8FP2kNAfX/hB47sdaFsqm9sTmG8sS3QTQPh0BIYBsbGKnazAZrxPrkPhjw1q3iSeBp49JsZrt4kOGcRoXIB7EhcV/w+8+FX/RD/Ff/gxt/wDCv+H3nwq/6If4r/8ABjb/AOFf8PvPhV/0Q/xX/wCDG3/wr9kT9qfw/wDtefDXUviT4b8K6hoFtpuuS6S9tfTJK7vHBBKXBTjBE6jHqpqqqqr9rv8A4KD+HP2QPHmleCvFfwu1vXI9Z0sX9pqFjexxxsPMeN4yrjIdSgJxkYdeckgfs9/8FXPhd8fvi/4e+EFt8N/EHh+68RySw2t9d3cMsSSrE8iqwXBG/ZsBGfmZexJFV4y/4LP/AAh8K+L9c8L2Pwp8R6xb6PqVxZw6jb30KR3iRSMgmRWG4K4UMAeQCM18DP269B+OXwG+JH7QOm/DXWNJ0L4dWt3LJBc3kby38ltaG6ljQqMKQhjGSesnbHP/AA+8+FX/AEQ/xX/4Mbf/AAr/AIfefCr/AKIf4r/8GNv/AIV/w+8+FX/RD/Ff/gxt/wDCv+H3nwq/6If4r/8ABjb/AOFfDHxxafE74a+E/iTYWMtlbeLNDstWhtpmDPAlzAkqoxHBIDgEjjIr9sb9vPwN+x1qvhjRPEXg7VfEl74lt7i4EVjMsP2aOJkVWYuMNvLOAAcjyznGRn/h958Kv+iH+K//AAY2/wDhX7N3x58NftMfB3RPjD4VsLjTrTV2njewupUea0lhmeJkfYSATsDjoSrqcDNVVfth/t8eBf2PNa8PeHfEvgbXfEN94htZLqI2MsUUcUaNsOWc5LE9tuMd+1ah/wAFy9PjvZY9K/ZmuLm0VsQy3HjAQyOPVkWzcKfYMfrWn/8ABcvT5b2GPVf2Zri3tC2JZbfxgJpEHqqNZoGPsWH1r4a/8FiP2VvGd3b6b4ys/Ffga4lyHudRsFurRDk7QJLdnk5AXJMSgFuuAWrwl4x8JePtAtfFPgjxLpmvaPerut77TrpLiGQd8OhIyO46jvVV+0z/AMFRvht+zV8YdX+Dep/DnXfEGo6JDbPd3Nncxwxo80KTKmHGSRHIhyMj5sdQQP8Ah958Kv8Aoh/iv/wY2/8AhXhPxNpXjXwro3jLQZvN03XtPgv7OTcrb4Zo1kQ5UlTlWHIJHoT1qqqqqqqqqqqqqqqqqqqqqqqq+IXimfwN4D8ReNbXRZdXk0DS7i/GnwyCOS6EMbSGNCQQHYKQueMkZIHI/wCH3nwq/wCiH+K//Bjb/wCFfs1/H7wx+038INH+L3hPT7vTrXU5J4ZbC7ZWmtZYZWjZWKkg52hwR/C65wcgV+2D+2D4L/Y88H6F4n8U6De67ceINSaztNOsplilKJGzyTbmG3ap8tSOuZVx0OPBv/BZLwD488X6F4H8PfAvxRJqniHUrfTrFH1KABp55FjjBwCcbmHQGv2pf+CmPgX9ln4sXHwm1/4Za7rl3b2MF2buzvIo4yJQSBhucjFf8PvPhV/0Q/xX/wCDG3/wr/h958Kv+iH+K/8AwY2/+Ff8PvPhV/0Q/wAV/wDgxt/8K/Y9/b68H/tieJfEHhrw14A1jw9L4esY7uWW+uo5RKHfYAoToe/NavqlhoelXmt6pceTZafbyXFzLsLbI0UszYUEnABOACa/4fefCr/oh/iv/wAGNv8A4V+zt/wVX+GH7Qnxh8P/AAetPhxrvh678RNNHa399fQNCsscLyhGwQcv5ZRcZJZlGOeKqqmmht4XnnlSOOJSzu7bQoAySSegxXxs/wCCpX7KPwdvb3QrHxLe+OdatFYG28NRLcQCXnCtcsyxYyOShcr6EjFar/wXK0mG/li0P9mm7u7JceVNd+Llt5G4GcxraSBecj7xyADxnA8J/wDBcHwBezhPHPwC8QaRD5gBfSdbh1FtmOTtkjtxnPbOPcV8Afj/APDv9pb4eRfEz4Yz6m+kvdSWjrqFg9rLHNGFLLhvlcAMvzRsy5JG7crAV+0D8aNA/Z4+DviT4x+JrG4vdP8ADkMTvbW5xJO8syQxopwcFnlQZPAzk4GTX/D7z4Vf9EP8V/8Agxt/8K/Y+/4KCfD/APbB8Va94N8PeDdV8OaloenpfrHf3UMv2mEyCNyoU7vlZowTgj5xkjjNV8Qf+Cz/AMK/CPiTXvDWifBnxXqk2j3E1rFJdX1vaLPNGSpB2+aUQupG75jtwduflH/D87/q1z/y9/8A7hrwn/wW9+GF60I8dfAzxRpClmEx0nVINRKAA7SokFvuJOAQcYBzk9D8Df2jvgz+0f4bPib4ReNrTWYocC7tGVobqzfA+WWBwHXk4DYKtg7WYc1VftM/8FRvht+zV8YdX+Dep/DnXfEGo6JDbPd3Nncxwxo80KTKmHGSRHIhyMj5sdQQP+H3nwq/6If4r/8ABjb/AOFeE/E2leNfCujeMtBm83Tde0+C/s5NytvhmjWRDlSVOVYcgkehPWqqqqqqqqqqqqqqqqqqqqqqqqvid44tPhj8NfFnxJv7GW9tvCeh3urTW0LBXnS2geVkUngFghAJ4ya/4fefCr/oh/iv/wAGNv8A4V8MfHFp8Tvhr4T+JNhYy2Vt4s0Oy1aG2mYM8CXMCSqjEcEgOASOMiqr4nf8Fhfhr8MfiT4s+Gt/8HfEt7c+E9cvdJmuYb+BUne2neFnUHkBihIB9a/ZE/an8P8A7Xnw11L4k+G/CuoaBbabrkukvbX0ySu7xwQSlwU4wROox6qar9sv9t7wp+xp/wAIf/wk3gfVfEX/AAl/27yPsNzHF5H2XyN27f13faFxj+6a/Zw/4Km/D/8AaQ+NHh34L6H8K/EGj3viH7V5V7d3sMkcXkWstwcqvJyISv1Ir9r3x74s+F37M/xD+IPgTVv7M1/QtHe40+78iObyZAygNskVkbgnhlIr/h6P+3b/ANF1/wDLZ0n/AORa+HuqX+u+APDOt6pcefe6ho9pcXMuwLvkeFWZsKABkknAAFVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVeIP+C3/wANbaK7PhX4E+JdRkSQi0XUNWgshKm7GXKLN5Z284AcZ4zj5q/4fnf9Wuf+Xv8A/cNeFf8AguB8OrxkHjj4DeI9JUzEOdK1iHUSI8D5gJEgy2cjbwMAHdzgfAX9vr9l/wDaKvbbQfBPxBTTvEV0q+XoOvRfYLt2bpHHuJjmfrlYXcjBPTmq+L/xQ8P/AAV+GfiH4q+KbTULrSfDVmbu7h0+NHndAQMIrsik8jqwHvWt/wDBcPwVBaI/hv8AZ81u+uTIA8d9r8VogTByQyQyknO0Y2gck54wf+H53/Vrn/l7/wD3DXw9/wCC03wA8QXVvZfEP4feLfCTTKm+6g8rUraFtpL7ypSXaCAFKxMWzkquK8BfEDwT8UvCen+Ofh54o0/xBoOpx77W+sZhIjeqnurqchkYBlYEMAQRX7R/xv0n9m/4L+IvjRreh3esWXh37N5tlayLHJL591FbjazcDBmDHPYGv+H3nwq/6If4r/8ABjb/AOFfsn/tU+Bv2uPhvP4/8G2F3pcun6hJYajpd7IrTWsigMhJX7yOjKQ2MZ3L1U1Xxv8A+CtPw6+CHxY8T/CbVfhJ4j1O78MXzWkt3b30KRzMADuUNyBz3r9kT9qfw/8AtefDXUviT4b8K6hoFtpuuS6S9tfTJK7vHBBKXBTjBE6jHqpqq+NHx2+FH7PPhKDxx8YvFqeHtEub5LCG5aznuS9w6O6oEgR3JKxuc4wApya/4ej/ALCX/Rdf/LZ1b/5Fr/h6P+wl/wBF1/8ALZ1b/wCRa+Cn7Zn7NX7RXiq68E/Bv4k/8JBrVjp76hPa/wBj31pst0kjjZ988KKcPLGMA5+bOMAkVVftl/tveFP2NP8AhD/+Em8D6r4i/wCEv+3eR9huY4vI+y+Ru3b+u77QuMf3TX7OH/BU34f/ALSHxo8O/BfQ/hX4g0e98Q/avKvbu9hkji8i1luDlV5ORCV+pFV8TvHFp8Mfhr4s+JN/Yy3tt4T0O91aa2hYK86W0Dysik8AsEIBPGTX/D7z4Vf9EP8AFf8A4Mbf/CvEP7ReieHv2XYv2oZvDl7NpcvhW08QjSllUTiO4ijkEW77u4CQAnpxX/D7z4Vf9EP8V/8Agxt/8K/4fefCr/oh/iv/AMGNv/hX/D7z4Vf9EP8AFf8A4Mbf/Cv+H3nwq/6If4r/APBjb/4V4Y1yHxP4a0nxJbwNDHq1jDdpE5yyCRA4BI7gNiqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv2qv+Cl/w6/ZY+KLfCjWvhx4j17UobGG7nuLWeGGFRKCVVdxLMcDnIGPfrU3/Bc2FZXW3/Zgd4wxCM/jXaxHYkCyODjtk/WtH/4LkaJPeiPX/wBmy9s7Tacy2fitbmQN2GxrWMY993Hoa+FH/BWv9kn4kXtvpHiHVdd8B30yoobxDZKLZpTjKrPA0iqoJPzyiMYXJxkCtK1bSte0221nQtTtNRsLyMSW13aTLNFMh6MjqSGB9QcVXxg/4LB/Cv4WfEbxV8NrX4R+K9XvPCeqXelXFxJdW9tHNc20rxSbMFz5ZdDhjyQclQeK/wCH53/Vrn/l7/8A3DXhP/guD4AvZwnjn4BeINIh8wAvpOtw6i2zHJ2yR24zntnHuK/Z4/a2+BX7UWlTXnwn8Ypc39kpa90W9j+zX9ooIG9oSfmT5lHmIWTLbd27IFeJ/wDgtB8MPDHiXVvDU/wW8UTy6TfTWjypqEADtE5QsAR0O3Nfsrf8FNPhX+1H8UV+E1j4N1jwtq91YzXOnPqFzFKl68QDPCuzkP5e+QcY2xvyCADX7Zf7b3hT9jT/AIQ//hJvA+q+Iv8AhL/t3kfYbmOLyPsvkbt2/ru+0LjH901+y1/wUx8C/tTfFi3+E3h/4Za7od3PYz3Yu7y8ikjAiAJGF5yc1VV8Sfir8N/g94bl8XfE/wAa6T4b0mLI+0X9wI/MYDOyNfvSP6IgLHsK+I3/AAWj/Z+8OzyWfw68CeK/GDpHIVuJvL0y3dwPkCl98mCc5JjG0YIDZIH/AA/O/wCrXP8Ay9//ALhr4Yf8Fk/2fPGur2eg+MPA3jPwtdX00cMU0VsupwBmIGCIT5xO44ASJifTJAqq/a+/4KHeAP2QfHWj/D/xD4H1jxFqWraSNTP2GdIhBC0skSZLjBLNFJ0PG3nGRn/h958Kv+iH+K//AAY2/wDhXwM+Lvh/49/CTwx8XvC9vLbad4ls/PS3mkR3gdXaOSNmQkErIjqfccgHIFftM/tD+Ff2XfhRe/Fjxho2q6rZW1xHax2mmrGZJJpciPJkZQqZHLckDkK3Std/4Li+FLfyP+EY/Z21W/3bvP8At/iSOz2dNu3ZBLuz82c7cYHXPEP/AAXNhaVFn/ZgdIywDsnjXcyjuQDZDJx2yPqK+FX/AAWJ/Zl8dazDoPjfSfEvgOS4kKx3+oQJdWS/dCh5IWMiEsSMmPYoGWcAnGlatpWvabbazoWp2mo2F5GJLa7tJlmimQ9GR1JDA+oOKrx74w0r4eeBfEfj/XC39neGdJutTvNikt5NvE0r4ABJO1D0Br/h958Kv+iH+K//AAY2/wDhX7Lv/BTv4a/tPfFuz+EOmfD/AFvw7qOo2dxPZ3F9ewSJM8Kb2jCghifLWRuM/cPGMkVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV+37/wAnlfFn/sYJP/QEr9gT9gPwp+2H4U8WeJvE3xD1Xw//AMI/qEFnBBY2ccnmb4y7MzOf90AADGDyc8ftW/8ABI/xH8Ffhzq3xU+FXxCfxbpnh61e81fTNQsltbmC1jXdLPHIrlJQqhmZCEIVTgueK8C+OPFPw08YaR498E6zcaXrmh3S3NldQOUZHXscHlWGVZTwysQcgkV8E/iNb/F/4P8Agv4oW6RJ/wAJRodpfyxRZ2wyyRK0kYySfkcsvU/d6nrX7WX/ACdR8Zf+yga7/wCl81fsHf8ABOjwf+158JdZ+JfiX4laxoEum+IpdIitLGyjkDCO3t5jIWc9T9oxjAxtzk54/bE/4JX+MP2b/AV78WvAfjpPGfhfSmB1W3nsPst7p8TOqLIArOs6Bm+dhsKAg7SoZl+Afxv8Z/s7fFTRPit4Gu3W90qYfaLQyskWoWxI823l29UcD3wQrDlQR4413TfFPwM1/wATaNP51hq/hO5vLST+/FLaM6H8VYGq+EP/AASk/aI+Nfwz8O/FXwr4y+HNrpPiWzF3aQ6hqN7HOiEkYdUtHUHjsxHvX/DlT9qj/ofvhT/4Nb//AOQq/wCCen7L3j/9kv4Laz8OPiNq/h/UtS1HxRPqsUuiXE00KwyWtrEFYyxRsH3QOSApGCvPUCqqqr/gs18Hk8XfALQvi9YWqNfeAdWEV1ISQRY3hWJunBInW1xnoGbBGSD4C8Yap8PPHXhz4gaGU/tLwzq1rqdnvUEedbyrKmQQQRuQdQa8MeI9H8Y+GtJ8XeHrxLrS9bsYb6xnQgiWCVA8bgjIIKsDx61+1z8Vj8Ef2afiJ8TIZmiu9K0WSOwkUuCl5ORb2xyhDAedNHkgggc5HUV8Cf2adX8Mf8E7v+FA6atvY+J/F3gXUY7l79yqQ6jqdtKSJmjViFjaZYyVVjtj43Hr/wAOVP2qP+h++FP/AINb/wD+Qq/ah/Ze8f8A7Jfj+w+HHxG1fw/qWpajo8eqxS6JcTTQrDJNNEFYyxRsH3QOSApGCvPUD9l/9l7x/wDta+P7/wCHHw41fw/p2pabo8mqyy63cTQwmGOaGIqpiilbfunQgFQMBuegP/DlT9qj/ofvhT/4Nb//AOQq+CHgnVfhp8FvAHw4124tJ9S8K+F9N0q8ltHZoZJra1jidoyyqxQshIJVTjGQOlf8FNfitL8Vv2xPGZSZHsPB7J4dsQpJKC1z5wJz1+0vcHjGAQOoJPjj4Y6/4D8NeBfE+rxSrbePNDk1ayD2zx+WiXtza7MsMMSLdZcjjZPGe+T/AMESfi5JcaZ8QPgVf3e4WkkXiDS4iy5CviC6wCd2AVtegKgsc7S3zVVf8Fvf+Sp/DT/sX7r/ANKBX7F37OWk/tT/AB0sfhPrfie70Kym0+6u5bq1t1lk/dJkKoYgDJIyTngEY5yNe/4IfeCptNkTwx8fdbttQHMb3+iRTxH2KpIhGeOc8ehr9qn9ij40/sk6pb/8J7p9vqXhzUZjFp3iPSw8lnPJgsInLKDFNsUt5bDkK5QuFYj9jr9r/wAe/sl/Ee01zSb+7vfCGoXCL4j0Ddvju4MgNJGhYKtwqjKPkcjaxKlgfD2v6L4s8P6Z4p8N6jDqGk6zZxXlhdwnKXEEqB45F9mVlI+tajqFlpOn3Oq6lcpb2llC808r8CONAWZj7AAmvjF8RtS+L/xV8W/FDVl2XPijWLm/MXaFZJCyRjk8IpVRyeFHJr4u/DHX/g18Q9W+G3iiKWPVNH8nz1ltngYebCkq5RwGHyyL169ehr/gk98W5Pib+yPpOgajd+dqXgHUJ9Dl3su9oFCzW7bQchBFMsQJAyYW64JNVVVVVVVVVVVVVVVVVVVVVVVNDDcQvbzxJLHIpV0dchgRggg9QR2r9p/4SP8AAn9oLx58KBFsttB1iVbBSzMfscmJbYktzkwSRE5zyTyetf8ABEX4pxyaT8RvgjeXkKvBcQeIdOt/lDuHUW90/qQvl2Y7gF+2ea/4LJfFY+Mf2ldO+GVpMxtPh9oscc0bFwFvLwLcSEAnbgwm05UZyCCTgBf+CRnweb4jftTw+N721d9L+Hemy6kz5Gw3coMFujDrn55ZBjHMHJ7H9uz/AIJs/HP9qD4/XfxV8AeK/AlhpM+l2tosOsX13FOHiUhiVitpFwc8fN+Ar4v/APBKT9oj4K/DPxD8VfFPjL4c3Wk+GrM3d3Dp+o3rzugIGEV7RFJ5HVgPeq8Pf8Ebv2nfE3h/TPEdh46+F6W2q2cV3Ck2p3wdUkQOoYCzIBwRnBIz3r/gnT+wf8Xv2Q/Gni/xH8SfEfg/UrbX9LhtLZNDvLmZ0dJd5LiWCIAY9Cee1f8ABT34rS/Cr9jvxeLGZI7/AMYNH4dti5OCLnd54wCMk20dwB74JyAQfhZ8Mdf+LfiS78MeG4pXubPQ9U1Z/LtnmJSysprkptQEgyGFYlPTfKg74r4e+NtZ+Gvjzw78Q/Dkvl6p4a1S31G0J6eZDIrqD1BBK4IIIIJBBFeE/E2leNfCujeMtBm83Tde0+C/s5NytvhmjWRDlSVOVYcgkehPWqqv+CiX/BQHxJ8ffF2rfCf4V+IpbP4WaZJ9md7Zih8RSIVLTSnqYA6/u06EAO3LBU+Cv7P3xg/aI8TN4T+D/ge98QXsCh7qSMrFb2aEEhpp5CscQO1sBmBYghQTxWhf8ES/jhd6bHceIvi34J069flra2iubpUHbLlEyfUAY9Ca8cf8EYP2ldBt2u/BfjDwT4pRIwTbC6msrhnychVkj8vGAvJkXknjjJ/Yt+Dl18B/2YvAPw51XS0sNZttNF1rEOFLpe3DGaZXZchmRn8vOSMRgA4Aqv8Agtj8VpdI+HPgT4M2EyBvEepTatqAUncsNqoSJTz913nZunWAcjv4Z+GOv+Kvh540+JGnRStpngb7B/aLLbO6r9rmMUW5wNqfMpxuxnoK/wCCfXxbk+DX7W/w/wBelu/J03WdQXQ9T3MqoYLzEIZ2YgKiStFKTkY8rvyDVfFr/kqnjP8A7GC//wDSh6/Yr/4Jm+Bf2pvgbafFjX/ibruh3dxqV1aG0s7OKSMCJgA2W5yc1+2t/wAEy/Ff7LHg8/FTwl4z/wCEw8HQ3CQ6i01j9lutLMjBImcBmWSNnKpvG0hnUbcHNfs0fHDxJ+zt8a/DHxS8OalLbLp14kepxKCy3di7BbiF1/iDJkj0YIwwyqRWo6hZaTp9zqupXKW9pZQvNPK/AjjQFmY+wAJr4xfEbUvi/wDFXxb8UNWXZc+KNYub8xdoVkkLJGOTwilVHJ4Ucmvi78Mdf+DXxD1b4beKIpY9U0fyfPWW2eBh5sKSrlHAYfLIvXr16Gv+CT3xbk+Jv7I+k6BqN352peAdQn0OXey72gULNbttByEEUyxAkDJhbrgk1VVVVVVVVVVVVVVVVVVVVVVVftZf8mr/ABl/7J/rv/pBNVfsnf8AJq/wa/7J/oX/AKQQ1VftZf8AJ1Hxl/7KBrv/AKXzV/wRU/5NX8U/9lAvP/SCwqv+C5//ADRL/uYP/cdX/BLj/k+34Y/9xb/003lft+f8mafFn/sX5P8A0NKr4T/8kr8G/wDYv2P/AKTpVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVfsif8EsPAv7R/7Pfhf40a/8V9d0m78RNeZsbPT4mjgEF3NbgbmOWJ8ndnjG7GOMn4v/APBFDxxoun3GrfBP4r2XiWWNSyaPrVl9gmbAX5UnV2jdmO4jcsYHALHk14g8PeJ/AviS78OeJdI1DQ9d0e4MdzaXUTQT2sq9ipwVI4P5Gv8AglP+2zr/AMZNKuv2f/ixrb6h4p8OWIudD1O4OZdRsUKo8crk/vJotyYONzpknJRmP7fn/JmnxZ/7F+T/ANDSq8G/8EXPh14n8C6F4in+OHiOG+1bSbe6cJpUJjSWSJWOFLZ2gt03Zx371+2L+xv49/Y68bWHh/xNqtprmh69HLLoet20fki8WIoJVeEsxikTzIyVyy4dcMeQP+CSXx+8RfDv9oyx+EFxqz/8Ir8QlmhmtJnYxwX0cLSQzIucK7+WIScfMHXP3QR/wVH/AOTEvib/ANwn/wBO1nVf8E5v2oU/Zm+P9lN4k1lrTwR4vVdN8RB3PlQjJ8i6ZR3idjlsZEckuOtV+37/AMnlfFn/ALGCT/0BK/4Iqf8AJq/in/soF5/6QWFVX/BbvxOLT4QfDjwX9riX+1fEk+oeQV+d/s1sY9wPov2vB93X0r4VfAv4vfHG71Cx+EvgDVfFE+kxpJepYxhvIVyQhYkgDJVsfQ+lf8MBftlf9G8+Kv8Avyn/AMVX/BKT9mH4/wDwV/aI8Q+Kvir8K9a8NaTdeC7qyhu76NVR52vLJ1jGCeSsbn/gJqqr/guf/wA0S/7mD/3HV/wS4/5Pt+GP/cW/9NN5VftZf8mr/GX/ALJ/rv8A6QTVWl/C/X/jX/wTZ8JfCrwreafa6t4l+FWiWlpNqEjxwI5src5dkV2A47KfpX/DlT9qj/ofvhT/AODW/wD/AJCr9o79nnxp+zB8TJ/hV4+1TRNQ1aCzhu2m0eaWWApKCVAaWONs8c/Lj3NfssfsifEn9rzX9c8OfDXW/DOm3Ph+zju7l9cuZ4UdHfYAhhhlJOfUAe9f8OVP2qP+h++FP/g1v/8A5CrwRol14a8FeH/Dl9JFJc6VpdtaTPCSUZ44lRipIBIypxkD6Cqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv+CvP/J5Wpf9i/p3/oDV+wN+x34f/bE8aeKPDniPxrqHh628O6XHdo1lapK8zvKEAJc4AAB7HJI5GOfFn/BD7w/JppbwL8fdQh1BMkJq2iJLFL6DdHIpT64b6d6/aM/Zg+L/AOy14wj8H/FbQUgF4rvpmq2bNLY6miYDtBKVXJXcu5GCuu5SygMpP/BO/wDbe8Sfs1/EfTPAvjDxBLN8LvEd4sGpW1y5dNIkkOFvIckeWFYgygcMm47Sypiv2sv+TqPjL/2UDXf/AEvmr9g7/gnR4P8A2vPhLrPxL8S/ErWNAl03xFLpEVpY2UcgYR29vMZCznqftGMYGNucnPH7YX/BKvxR+zv4C1D4tfDzx3/wl/hrSNjapaXdmtreWMRwpmBDlZkDkZwFZQw4YBmHwo+KfjT4K/ELQ/id8P8AVXsdb0G6WeBtzBJgD88UoUgtE65R1yNysR3rwR4r03x74K8P+OdGbNh4i0u21G0Oc/up4lkTnv8AKwr4tf8AJVPGf/YwX/8A6UPXgnxl4k+Hfi7R/HXg/VJdO1rQryO7sbmJiDHIhBHTqDjBHQgkHg1+zt8b/DP7Rfwb8NfFvwtKgh1m1H2u1BO6yu0+WeBgecpIGAJ+8u1hkMCf+C5//NEv+5g/9x1f8Ehv+TytN/7F/Uf/AEBaqv2pf2ivCf7Lfwb1f4q+KF+0zQ/6Lo+nBtrajfurGGAH+EHYzM3O1EdgGIAPxv8Ajv8AE/8AaI8c3PxA+KniWXVNRl3JbxD5ILGEsWEEEeSEjUscDknqSSST+zz/AME4P2nP2jNHtfFmh+HLHwx4XvVD2us+I52tY7pMKd0MSq80ilWBV9gjbnD8HH/DkL4p/wDRcfCn/gtuP8a+Bv8AwSk+P/w1/aS+HOs/Emw8L654N03Wo77Ur7SdQM8Mf2dGnjSSKZI5GV5IkjOEIy/PBJqq/bx+K0vxk/ay+IvipZkksbLVn0nTthOz7NZ/6OjLknhzGZPrIeB0HxH+GOv/AAy/4Rf+34pU/wCEr8N2mv2XmWzw5trndsI3D5h8h+YcHtX/AARW+LcniH4S+Mvg1qN3vn8IapHqGnq7KD9lvA25EGdxCTQuzEjg3CjPIAr/AIK8/wDJmmpf9jBp3/obV4N0OHxR4v0Lw1PcPDHq2pW9o8qDLIJJFQsAe4DZrUP+CH/w6lspY9K+PXiO3uyuIZbjRoZo0b1ZFdCw9gw+tftKfs6+O/2XfipffCvx8Lea5ihS6sL62z5OoWjlgk0e7BAyjoQejIwycZP/AARU+OHiS81Txj+zxq2pS3WkWennX9GhkBb7GRNHFcqjdkdpoW2dAwdgMuxNf8FcvitL8Pf2Tbrwrp8yLfePtWt9J6nelsubiZl56EQpGevE3TuPhz8Mdf8Aib/wlH9gRSv/AMIn4bvNfvfLtnl221ts8wnaDsHzj5jwO9fAn4oX/wAFPjJ4N+K2nNKX8M6xBdzRxAFpoAwE0Qzx88RdOo+91HUWl3aahaQ31hdRXNtcxrJDNC4dJEYZVlYcEEEEEcHNVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVft+/8AJ5XxZ/7GCT/0BK/4Ih/8kq+JX/YwWv8A6Tmv2xvj78OfgN8C/FuqeM9fsY9S1DRbq20fSDdKlzqVxJGY0SNOWK7nXe4UhFyx6Yq0s7vULuCwsLWW5ubmRYoYYULvI7HCqqjkkkgADqTX7Mnw01T4Ofs+fD74Y67Ij6p4f0G2t7/Y4dVudm6VVYfeVXZlB7gA96/ay/5Oo+Mv/ZQNd/8AS+av+CKn/Jq/in/soF5/6QWFft5+O/C3w/8A2Q/ijfeKrq3jTV/Dt3o9hFKw3T3l3E0MKovJZgz78AZCxs3AUkV4S8M6r4K/Yt0XwbrsPlaloPwvgsLyPay7JodMWNxhgGGGUjBAPqBVfshf8FCv2Pvhd+zP8PPh947+L/8AZmv6Fo6W9/Z/2BqU3kyBmJXfHbsjcEcqSK/4ej/sJf8ARdf/AC2dW/8AkWvhb8UvAfxq8B6X8TPhlr39s+G9Z877De/ZZrfzfKleGT93MqOuJI3XlRnGRkEGqqqqvjP8ObL4vfCTxh8ML8II/E+i3Vgru20RSSRlUkzhsFX2sDtOCo4PStR0+90nULnStSt3t7uymeGeFxho5EJVlPuCCK/4Jb/FN/ih+xz4Vt7u8ludQ8F3E/h67eTdwICrwKCeoW1mt14yBtxxjA/4LX/FaHSPhf4I+DVlM4u/EerSateBDwttaoUVW56PJOGHHWA8jv8AsY/Bz/hfH7TfgH4dXNn9p0yfVEu9WRofMQ2VsDNOr9gHSMx5PG6RRg5ANV/wWr/5Oo8Lf9k/s/8A0vv6/wCCKn/J1Hin/sn95/6X2FV8YviNpvwg+FPi34oasvmW3hfR7m/MXeZo4yyRjkcuwVRyOWHI61NLq/ibWnmcXGo6pqt0WIRC8txPI3ZRyWZj0A5Jr/gqF+zlbeAv2QvghqNlp1uL74aQ2vhzUJoIlUvHLaLvkcqAGHn22c/3p2P8TGv+CevxYb4Pftc+ANcn1FLTTNZvv7E1MyMFjaC7HlKXY/dVZTFITkY8vk4zVVX/AAW9/wCSp/DT/sX7r/0oFf8ABIb/AJPK03/sX9R/9AWq+Ofwj8N/Hb4S+J/hT4ptIZrPxBp8kEckigm1nxmGdODh45Ajg4PK9CMitR0+90nULnStSt3t7uymeGeFxho5EJVlPuCCK/4JPfEB/HP7Gnh/TZ7nz7jwfql7osrEMCAri4jU5AB2xXMajGRgDnOcf8FNfitD8Kv2O/GhSZkv/F6p4dsQpwHa6z5wJz0+zJcHjOSAOhJH7L/w1n+MH7RHw7+HMNj9ri1jxBareRGESj7JG4kuWKEEMFgSRiCMYBzxmv8AgtB8MIfC/wC0F4b+JljZJDB430Hy7mRUwZryzYRuxPciCS0XnkBR2xj/AIIv/FhvDHx48SfCW+1FIrHxvov2i1hdhmW+syXUKDznyJLpjjqEGQcZFVX7bv7SH7RHhP8Aax+J3hvwr8fPiLo2k6frjxWlhp/im9toLdAi/KkaSBVHPQACoP8Agqj8XPCX7L/g/wCE3gjXNYu/iHA18fEPjPXp/wC0J/Le6meFITMXLuI5EUvJkIqKqg5ymsfH/wCO3iDVzr+t/Gjxxe6kWDC6m8QXLSKQcjad/wAoHYDAHbFf8E8/+CkvxPj+JmhfBH4+eKpfE2g+JbhLHTdc1ObdeaddMAsKyTHmaORwqEyZYM4bfgEGv+ChX/BR+f8AZq1Y/Bz4QWOn6l49lsxNqN/eYlg0NJVzEPKB+e4KkSBXwiq0bESB9o1v47ftXfG/WZtQv/iZ8RvFF5BmQw2V7cvHahtoJSGEhIQ21c7VUEj1r4Q/txftU/AfWreXw78VtdvrOxmHm6D4huZb+ycBmLxmKVsxBizbjEY3yc7gQCP2Sv2nPC37WPwgsviZ4esm029imNlrOlySB2sbxFUugYfeQhlZGIBKsMgEED/gsL8ZPi98MfiT8PbD4a/Fbxh4Ttr3Q7mS5h0PXbmwSdxOAGdYXUMQOMmv+CWvx6+OfxC/ax0/w34++NPjvxLpL6HfyNYax4ju7yAuqLtYxyyMuR2OOKr/AIax/ao/6OY+Kv8A4Wd//wDHa/4Jw+LPFXjr9i/4d+KfG3ibVfEGtXv9p/adR1W9ku7mfZqd0ib5ZCWbCKqjJ4CgdBX/AAVK+PXxz+Hv7WOoeG/AHxp8d+GtJTQ7CRbDR/Ed3ZQK7I25hHFIqgnucc18Lv8AgpJ8UPhD+x9qWjx/EHU/FnxX8R+Mr6Ox1LxDqT6pNoenJZ2WJts5fcGkaURI37vcJWIbaVb9jX42/GP4i/tq/DO88dfFTxXrsmo+IFFyt9q80qSKVc7NhbaEHZAAo6AAV+0r+0f8Pv2W/hhefEzx/NLLGkgt9O022K/aNRuWztijDEDoCzMeFVWPJwD8cv8Agpj+1d8Zdaup9K+IN74B0NmItNJ8L3DWbQpuBG+5XE0j4VQzblU/NtRAxUn4yftJ+G7qDxkfit8S9LudY82SDVTrt9A95kgyMs28GTJYbiCeSM9a/ZR/4K4fFPwV4hsPCn7SOof8Jd4TurgJLrf2ZV1HS0IADYiAFxGp+ZgymTBYhmwqHSdV03XtKstc0a+ivLDUbeO4tLmFtyTROoZHU9wVIIPvX/BR79oX4/eBv2z/AIieFfBPxy+IHh/RbH+zPsum6X4mvLS3g36Zau+yKOQKu52ZjgDJYk8k14L/AOCpvxZ+HP7MNr8N9A13W9a+J1xrF2914r8SXR1M2dkwQxeT55fzJNxdcSAogT7rbxt8TftCfHjxnqsmt+KfjP411O9kyPNuNduG2KWLbEG/CICzEIoCjPAFfsLf8FMPiz4O+JGh/Db47+NLrxZ4M8QXiWbaprNyZbzSZZCqJMbhstJEDgOrk4DFgRghqqqqq/4LYfCr+yPiV4G+MljbYh8RaXLpV86twJ7V98bMMcFo5yoOTkQ9Fxlv+Ca3xTf4VftjeA55byWGw8U3DeHr1I9374Xg2QKQO32kWzc8DbnjGRqOoWWk6fc6rqVylvaWULzTyvwI40BZmPsACa+NvxGn+L/xg8Z/FC4WVP8AhKNcu7+KKXG6GKSVmjjOCfuIVXqfu9T1r/gj/wDBz/hX/wCzJN8RNQs/K1T4i6o92GeHy3+xW5aG3U55I3i4kU8ArMCBg5Nft+f8mafFn/sX5P8A0NKr4T/8kr8G/wDYv2P/AKTpVf8ABbH4rQ6x8RfAnwZsJnK+HNNm1bUAp+UzXTBIlPOdyRwM3TGJxye3/BFj4VR+JPid8QfiVrGkRXelaR4fXRkF1bLJFJLeSBnC7gQSIrdlYD+GbB4fn45fDuX4SfGXxt8MpUcL4Z168sYS+cvDHKwifnnDR7GGezCv+CUXxYb4l/si6NoV/qKXGp+BL6fRJlLASCBSJbclR/CIpVjU458o8kg1Vf8ABRv4rap8If2QfHOtaBNcQaprMMei2k8JAMH2pxHK+7OVIhMuGXkNtxj7whhluJUt7eJ5ZZGCoiKSzMeAAB1J9K/ZN/Z28O/sxfBHQPhno9tbtqUcIuNcv0hVHv75xmSRyM5A4Rck4RFGeK1zxl4P8MSxW/iTxXo+kyzKWiS+vo4GcDgkB2GRX/C2PhX/ANFM8Kf+Dm3/APi60Pxl4P8AE8slv4a8V6Pq0sChpUsb6OdkB4BIRjgfWq/4Ke/FaH4q/tieLxYzNJYeDlj8O2xc8hrXd54wCcAXMlwB7YJwSQP+Cff7M7fED/gnp8YoNQ0VJL34otdxaOZLYHzjYxYs5ASMsEvRLgdAUOCCTUM0tvKk9vK8csTBkdGKsrDkEEdDX7NXxTj+NvwB8BfFI3kNzc+INDt5b94tu0Xir5d0oC8DbOkq44xtxgdKr4tf8lU8Z/8AYwX/AP6UPX/BM39q39nP4Q/stWHgz4l/F3QfD2txa1fTPZXkjLII3cFWwFPBAr/gpN/wUR+DnxW+Eeo/s/8AwR1OXxL/AG3eQf2zrP2V4bWGG3mWYRwmQK0jmWKP5wuzaDhm3cfsX/s0eKf2n/jjofhHS9LaTw9pt1FeeJtQkjJgtLJG3MrH+/KFMaL1LHPCq7LX/BTX4rQ/Cr9jvxoUmZL/AMXqnh2xCnAdrrPnAnPT7MlweM5IA6Ekfsv/AA1n+MH7RHw7+HMNj9ri1jxBareRGESj7JG4kuWKEEMFgSRiCMYBzxmv+C0Hwwh8L/tBeG/iZY2SQweN9B8u5kVMGa8s2EbsT3IgktF55AUdsY/4Iv8AxYbwx8ePEnwlvtRSKx8b6L9otYXYZlvrMl1Cg858iS6Y46hBkHGRVV+05+03+0noH7SfxY0HQf2hfiXp2m6b441m2s7K08WX0MNtDHezKkcaLKFRFUBQoAAAAFeKv+Cq3xes/wBnzwL8Kfhhreq2niu00tovFXi7V5Be3s83mOAsDy7+se1mlbLgsAu0pua6+PXxzvtZXxHefGnx3cassiSLfy+I7tpw6Y2MJDJuyu1cHPGBjpX/AATc/wCCjXxF8V/EXS/2fvj54h/t6LXd0Oga/ePi6iugGZbed8fvhJ91Gb5g21csGG2v+Cgv/BTPVvgX4ovfgh8CINPuPFdrb41jXrlVuI9JldQUjhjyVedQdzeYCikqCrncFv8A4w/tU/GPUrjVp/iJ8S/FVza48w21/d3C2wfOAEjO2IHacAAA4PvXwP8A2/v2pPgVrNneaP8AE/VfEWj2/wAsug+I7qTULSWP+4odi8ODggxMhyOcgsD+zZ+0B4Q/ac+EOjfFrwdDLaQ6hvhvNPncPLYXUZxLC5Xg4OGVsDcjI2F3YH/BYX4yfF74Y/En4e2Hw1+K3jDwnbXuh3MlzDoeu3Ngk7icAM6wuoYgcZNf8Etfj18c/iF+1jp/hvx98afHfiXSX0O/kaw1jxHd3kBdUXaxjlkZcjsccVX/AA1j+1R/0cx8Vf8Aws7/AP8Ajtf8E4fFnirx1+xf8O/FPjbxNqviDWr3+0/tOo6reyXdzPs1O6RN8shLNhFVRk8BQOgr/go9+0L8fvA37Z/xE8K+Cfjl8QPD+i2P9mfZdN0vxNeWlvBv0y1d9kUcgVdzszHAGSxJ5JrSP+ClPxS+Gf7H/h/wP4c+IGoeIPij4g1TUJNQ8Q6xqDaldaNZLIoiUeeXzJJ8wXfkIikhcurD/gnP8Xvit8QP28vh9N45+JfijX21FtUa7GpavPcCYrpV1t3K7EELtXAxgbRjGBX7WH7Vnw+/ZI+G6+O/GkUuo31/cfZdH0W1lVJ9QmxlsE52RovLyEELlRyzqrfGH/gpB+1x8XtXmvR8VdT8HaezfuNM8KTvpscAB4Hmo3nOfUs5z6AcVa/Gn9pXwHqFt4jtvir8SNCu9Th86C8OtXlu15ExD7gxceajEKT1U8da/Y+/4K4eOtD8Q6d4D/ak1CHXfDl3IsKeKUthHeabkBVM6xALPEMDcwUSjczFpOFqGaG4hSe3lSWORQyOjZDAjIII6giv23f2kP2iPCf7WPxO8N+Ffj58RdG0nT9ceK0sNP8AFN7bQW6BF+VI0kCqOegAFQf8FUfi54S/Zf8AB/wm8Ea5rF38Q4Gvj4h8Z69P/aE/lvdTPCkJmLl3EciKXkyEVFVQc5TWPj/8dvEGrnX9b+NHji91IsGF1N4guWkUg5G07/lA7AYA7Yr/AIJ5/wDBSX4nx/EzQvgj8fPFUvibQfEtwljpuuanNuvNOumAWFZJjzNHI4VCZMsGcNvwCDVVVVX7WX/Jq/xl/wCyf67/AOkE1V+yd/yav8Gv+yf6F/6QQ1VftZf8nUfGX/soGu/+l81f8Ev/ANsz9mr9nX4A694J+MfxJ/4R/Wr3xhc6hBa/2PfXe+3ezs41ffbwuoy8MgwSD8ucYIJ/4ej/ALCX/Rdf/LZ1b/5Fr/gq3+1H8CP2lv8AhV3/AApPx3/wkf8Awjn9sf2n/wASy7s/s/n/AGLyv+PiKPdu8mT7ucbecZGf+CXH/J9vwx/7i3/ppvK/b8/5M0+LP/Yvyf8AoaVXxr/4KUftEfESx03wd8PPF2ofD/wnoVnDZWUWh3D2l9dpEiKJbi4Rt4dimdkZVAG2nfgs3hn9oT48eDdVj1vwr8Z/GumXseB5tvrtwu9QwbY434dCVUlGBU4GQa/4Jw/8FFL/AOP95D8DPjTND/wn0Vu8mkatFGI01uKJN0iSIOFuFRWfKgK6qxwpX56r/gsr8Wfip8Lv+FQf8Ky+Jnivwj/af9ufbf7C1m4sPtXl/YPL8zyXXft3vjOcb2x1Nfsqf8FIfiZ8Dm8d6x8TfGPjX4jahqGhrF4astd1+4vbWC+Ey/NIJZCUTYzMSnzNs25G7cvxV/bF/aa+M2szax43+M3iVo5JA8enaffvY2VuV3BdlvCVjBCsV3kFyD8zE5Nfs+ft/ftJfs/a/Y3Vj4/1XxR4dhkxdeHtdvpLq3liLhpFiLljA55IdOhYkhgSD8KviT4b+MXw38OfE/wjcebpPiTT47y3yctHuHzRtjo6MGRh2ZSK/a3/AGofCH7JvwluviJ4it/7S1G4k+yaJpCTCN7+6IJAyeVjUDc7gHavQElQfjL+3n+1f+0L4klS5+I2t6NY6jIsFp4b8Lzy2VuAThYtsTeZOWY/8tGcknAwAqjTvjZ+0/8ACDXI7i1+J3xG8L6m8YkEVxqd3AZo8kAtHIcSJkN94EZB9K/4Jy/8FEtW/aRv3+DXxkjtI/HlnZtc6fqtrEsEWswxhfMDxjhLgcuRGAjLuIVNmG/bd8Q6/wCE/wBk74neI/Cuu6ho2rafobyWl/p9y9vPbuHX5kkQhlPPUEV/w1j+1R/0cx8Vf/Czv/8A47Xwyu7rUPhr4Tvr66lubm50Oykmmmcu8jtAhZmY8kkkkk9Sa/4K7fG/40/DT9pPw3oPw4+L/jXwrps/ge1uZbLRPEF1YwyTG9vVMjJE6qXKoiliM4VR2Ff8Eivjf8afiX+0n4k0H4jfF/xr4q02DwPdXMVlrfiC6voY5lvbJRIqSuyhwruoYDIDMO5r/grt8Q/H/wANP2bPDeu/Djxz4g8K6lP44tbaW80TU5rGaSE2N6xjZ4mVihZEJUnGVU9hX7Gn7cvxK+HXxV1Lxv8AGv41+OPFPh/R/DOoTw6LrHim6uI9QvAiiCFEld13s5ADbSVBLYwDXxv/AG2f2kvjz4kuda8U/FDW9PsGuGls9E0i9ks7KyXnaqxxkbyqsV8x9zkE5bmv2adcht/2UvhV4k8Sawkccfw90e7v7++uNoUDT4nkllkc8DGWZmPqSa/ay/4K5/EfxnrV34T/AGZ7p/Cnhi3ZojrstsrahqBViPMjDgi3iZdpA2+Z3LLnaNV+NP7SvxAS71/Vvir8SNfj0td1zdS61eXEdkrkkZO8rEpIOBwODjpXwT/4KU/tZ/BvWYLq6+JeoeOdH8zN3pPiu4fUBOvI+W4cmeIjcSNr7chdysBtr9mL9pXwD+1T8LbT4l+BWltmEn2XVdMuDmbTbxVVnhY4AcYZWVxwysDhTuVf+CwvxO+JPwx+Gvw+v/hr8Q/EvhO5vdcuYrmbQ9WnsHnQQAhXaFlLAHnB4zX7JX/BRH4mfBHxv4j8X/GDx98QPiNZTeF7m00jRtU8R3F3B/aLTQPDIwmdlQBY5FLgFgrtgHOD8Yf22v2n/jfq82peL/i7rtpaSNmPSNGvJNPsYQDlQIYmAYjsz7n/ANo18Dv25v2l/gLr9rqfhv4naxrGlxTB7nQtcvJL2yuV53LsdiYyc8tGVbIGScV8A/jN4b/aD+EPhn4u+FV8mz8QWYlktTKJGs51JWaBmAGSkisucDOAcAGqqqqqqqqqqqqqqqqqqqqqqqqv+CXH/JiXwx/7i3/p2vKr/gtf8E9HsZvBH7QGjaYlveajNJoOuTxoFFwyxmW0ZsdXCJcKWOSVWMdEFfsa/EB/hf8AtUfC7xmLnyIbfxJbW13Lhjstrlvs85woJP7qaTgAk9O9ft+f8mafFn/sX5P/AENKr4T/APJK/Bv/AGL9j/6TpX/BZf4+/Dnxtc+Cvg54L1+x1vVPD11dX+sz2V0s0dizKsUcDFcjzTiRmXIKALkHfx/wS3+FOv8AxK/a+8K6zp0LLpfghZdZ1W5xgRoiGOJAehZ5pIxt67Q7D7pr/gqP/wAmJfE3/uE/+nazr9m+00K//aI+Ftj4njtJNGuPGmkR6il2QIWt2vIhKJCeAmwtnPGM1+0/8ENX/Z1+Ovi34TapE5h0i+ZtNnIOLqxk+e3lB5yTGyhgCdrhlySpr/glp+05P8evgDH4M8U6p9p8XfDrytNunkI8y6sipFpOf7x2o0TMckmLcxJfJ/b9/wCTyviz/wBjBJ/6Alf8EVP+TV/FP/ZQLz/0gsKqv+C3/icXfxJ+GHgsXcTf2Vod7qHkBfnT7TOke4n0b7Jge6t61/wTj/bV+C37Hmk+O/8AhY2geNdT1PxdcWPlf2JaWs0McNss2N3mzxMHLXD5xuBAXpzn4Yf8FSP2OfibdQ6bL8QLrwfe3GwRw+KLI2aZYkENOpeBMcZLSAfMME4OLS7tNQtIb6wuorm2uY1khmhcOkiMMqysOCCCCCODmqqv+C5//NEv+5g/9x1fsFfFLwH8Ff2svAvxM+Jmvf2N4b0b+0Ptt79lmuPK83T7mGP93Cru2ZJEXhTjOTgAkf8AD0f9hL/ouv8A5bOrf/ItftDf8FHv2L/HXwB+Jngnwr8Zvt2teIPB+rafptr/AMI7qcXn3M1nLHEm97YKu52UZYgDOSQOar9k7/k1f4Nf9k/0L/0ghqv+CvP/ACeVqX/Yv6d/6A1f8EQ/+Sp/Ev8A7F+1/wDSg1VV/wAFhfjJ8Xvhj8Sfh7YfDX4reMPCdte6HcyXMOh67c2CTuJwAzrC6hiBxk1+zJ/wUt+J/wADfA/xLHjfxT4t+InizXobCPwn/wAJFrU9/a6bLH9oE8r+bIWVcSREqmC5jUEgfMvxJ/az/aS+LWsy6343+NPiu6eSQulrbalJaWsB6fu7eIrGnHGQoJ7k1+zT/wAFFP2iP2ffEmnnUvGeq+NfCCSBb3w/rd89wDCSN32eVyzwOAPlwdmSSVOTXg7xboHj7wlo3jfwrfpe6Pr1jDe2NwvSSGVA6nHY4IyOx4r9tL9r3wr+x/8ADKHxTqFgms+JNbme18P6N54i+0yqoMksh+8IYwV3FQTl41437h8VP21/2tP2hvEUsOqfE3xHDFqMx+z+HfDU01laKCwZY1ghbdKFKKVMpkcYzuJJJ0D9oH9qP4L+IfO0r4q/EDw3qa7ZJLS71C4VZQAwQy28xKSABmxvUgZyK/4J1/t+z/tVWF58OPiVZ2ll8RNAsxctcWqCODWLVSqGZUydkqsy+Yg+U7gy7QSif8FHvFnirwN+xf8AETxT4J8Tar4f1qx/sz7LqOl3slpcwb9TtUfZLGQy7kZlOCMhiDwTX/DWP7VH/RzHxV/8LO//APjtV/wVK+PXxz+Hv7WOoeG/AHxp8d+GtJTQ7CRbDR/Ed3ZQK7I25hHFIqgnucc1/wAEevjJ8Xvid8SfiDYfEn4reMPFltZaHbSW0Oua7c36QOZyCyLK7BSRxkV/wWF+J3xJ+GPw1+H1/wDDX4h+JfCdze65cxXM2h6tPYPOggBCu0LKWAPODxmv2L/29viL8KPFXj7x38Zvi34w8a2em+B7j+xNB17xPc3Md9qbXtksKxrK7gOEaViyqWWMSkcZr4ufth/tJfGzxJP4j8a/FvxAoeQtb6bpt7JZWVmuThYoImCjAO3ecuwA3Mx5rwl4r0Xwp8D9C8aeMdbhsNM0vwva3mo393JhYo0tlZ5HY89AT6mv2pP+Ct/xi+IuuXGgfs+3cvgPwjbyYivvJR9Uv9pzud23LAhwuEj+brudgxUX3xk/aT8URSeMtS+K3xL1ePSvLik1WfXb64Fp82UUzFzs+ZuBkctxya+A/wDwU6/ao+DGq2413xvd/EPQFyLjSvE9w1zI+WyWS7bM6OOQMsyAHlDgY+AHx48A/tJfDDS/ip8OryWTTr/Mc9tcJsmsblMeZBKvQOpI5BIIIYEggn/gsr8Wfip8Lv8AhUH/AArL4meK/CP9p/259t/sLWbiw+1eX9g8vzPJdd+3e+M5xvbHU1+yp/wUh+JnwObx3rHxN8Y+NfiNqGoaGsXhqy13X7i9tYL4TL80glkJRNjMxKfM2zbkbty/FX9sX9pr4zazNrHjf4zeJWjkkDx6dp9+9jZW5XcF2W8JWMEKxXeQXIPzMTk1+z5+39+0l+z9r9jdWPj/AFXxR4dhkxdeHtdvpLq3liLhpFiLljA55IdOhYkhgSD8KviT4b+MXw38OfE/wjcebpPiTT47y3yctHuHzRtjo6MGRh2ZSKqqqqqqqqqqqqqqqqqqqv8Agrz/AMnlal/2L+nf+gNX/BEP/kqfxL/7F+1/9KDVft9/BPR/jl+yx440S60xLnV9B02bWtDlCAyw3dqhkCxk5wZEV4j6iQ9Dgiv2IviA/wAUP2S/hb4vmuftFw/h+GyuZSGBkmtC1rKx3AHJeBie2c4yMGv2sv8Ak6j4y/8AZQNd/wDS+av+CKn/ACav4p/7KBef+kFhX/BQv4+/Dn4O/sz+PfD3iXX7E+IvGXh280jRtEW6UXd013G1uZlj5byog7OzkBfk253MoPh7w/rPi3X9M8K+G9Ol1DVtZvIrKwtIRl7ieVwkca+7Myge5r4Q+CJfhn8JfBPw3nu0upfCnh2w0l50ziVra3SIsOBwdmeg618Wv+SqeM/+xgv/AP0oev2kfgNDoH7P3wM/aL8KW9vJofjDw6ulazNbfMsWrWryqC7A4DSRRlNo5DWku7k1/wAEhv2m5/hx8W5vgD4l1TZ4Z8fSGTTUkIC22rqgCkHt50aCM+rLEPWv+C5//NEv+5g/9x1f8Ehv+TytN/7F/Uf/AEBaqv8AgtH8VtU8QfHDwz8IoZriPSPCWii9kiYhUlvLpjufAPzBYo4lBbkFpAAASW/4Jxfs7aP+0d+0vpWh+KrZLnw34XtW1zVrWWESR3kcMkapbuDxteSRNwOcoHGOpEMMNvClvbxJHHEoVERdoUAYAAHQY7VN8U/hhbyvbz/EfwvHLGxV0fWIFZWHBBG/g+1f8LY+Ff8A0Uzwp/4Obf8A+LrT9R0/VrKHUtKv7e9tLhd0M9vKJI5B6qykgj6V+1J8Vofgh+zx4/8Aig0zRz6Jos32FlOD9slxDbDOeAZ5IgT1APAPSvDHhzV/GPiXSfCPh60a61TW76GxsbdFJaWeVwkagDJyWYDj1r/gsV8CtK8F+APg1408O2EMdt4bs/8AhEJ5kgWPdDHCslmmFGFCiK7IUcDccY5z/wAEtfiw3wu/bB8L2F1qKW2l+NoZtAvfMYAO0wD2wGf4jcxQKMYPzkDqQa/4K8/8maal/wBjBp3/AKG1fDfULLSfiJ4W1XUrlLe0stas5p5X4EcaTIzMfYAE1rf/AAUS/Yu0LSrjVrj4+6Dcpbrkw2Uc9zM5JAASONCzHJHbA6kgAkft3/tO6f8AtY/Hu5+ImgaVcafoOmabDpGjR3ShZ3tonkk8yUKSAzSTSnAJwpUZJGa/4I3fs0eKfBmjeIP2jPGOltpy+KrFNN8OwzxlZprLesstzg9IpHSIJ3by2bG0oWr/AILNfFaHxd+0PoXwusZmeDwBoo+0qx4S8vdszgDPTyFtDk4OSeMAE/8ABGn4Jaf4y0T4w+M/FejJc6JrOmp4UVpIAwuI5laS9i3EHjYbXK9DvXI4FeOPCepeAvGmv+BdZGNQ8Oapc6bdDbtxLBK0b8dvmQ1/wTb+LDfF39j7wNf3uopdap4ahfQNQwwLRtaHZCGx/EbY27HPJ35Oc5NV/wAFHvFnirwN+xf8RPFPgnxNqvh/WrH+zPsuo6XeyWlzBv1O1R9ksZDLuRmU4IyGIPBNfAD9t742eBPjR4O8XfFD45/FDxD4T0zVI5NY0yXxPd3S3FvyHBhkl2SYBztbg47da/aT/wCCj37R/wAffEN8mj+N9V8DeEGkkWy0HQb1rU+SwK4uZ49slwWU/MrHy852otfDr9qn9ov4U65Hr/gn4y+K7SdZI3lgm1OW4t7nYcqJYZCySDkjDA8MfU1+wj+1nb/tc/BseKdTtLTT/Fmg3AsPEFlbOdnm7QyTxqeVjlGSAc7WV1yduT428ZeG/h34Q1jx14v1SLT9F0Kzku725lYARxouTj1JxgDqSQBya/ab/wCCqPx/+M2r3GlfC3XL34beEYJpBax6TOYdRukzhHnuVO5G287Iiqgscl8Kwm+If7SfhuOz8bT+OfiXpaXMiyWmrvqd9AJXdSytHMWGSy5YEHkZNfsN/wDBUz4j6H400f4V/tIa+/ibw1rN0tpa+IboKL3S5ZGVUM0nAmg3E7i+ZFDFtxVQlV8Tv2pv2nbD4leLLCw/aP8AihbW1trl7FDDD4wvkSJFncKqqJcAAAAAcCv+CWvjfxp8Qv2TdP8AEfj7xhrfiXVn1y/ia/1jUJbycorqFUySszYHYZwK/wCCu3xD8f8Aw0/Zs8N678OPHPiDwrqU/ji1tpbzRNTmsZpITY3rGNniZWKFkQlScZVT2FfsyftN/tJ6/wDtJ/CbQdd/aF+JepabqPjjRra8srvxZfTQ3MMl7CrxyI0pV0ZSVKkEEEg1+05q2q6B+zZ8WNd0HU7vTtS03wPrNzZ3lpM0M1tNHYzMkkbqQyOrAEMCCCARXw8/ax/aT/4T/wAM/wBu/tMfEv8As3+2LT7Z9r8Z33k+T5y7/M3S7dm3Oc8Yzmv20v8AgpH8X/jT471rwt8KPG2p+E/h1YzS2dnFpF01vNq0allNxPMm2QrKORECECkAhjlj/wAEb/EPiDxN+zD4mv8AxHruoarcx+PLuJJr66ed1QWNiQoZySBkk46ZJ9a/b1/4KgN8EfEepfBX4DW1jqHjHTWWPV9cukE9rpjlSWhiQHEk65TcWyiHKlWYME8Q/tOftYfGHVP7N1T4z/EPXrm9bcumWWqXCxuyAtlLaAhMgAnhOAPavh1+2Z+1d8HdQiHhT43+LYUsWZBpuqXrX9qmSd6/ZrnfGpJZs4UHJzkHkfsGf8FAtB/a5sLrwf4r0u08PfEbSLc3NzYWpc2uoW4Kq09uXLMu1mUNEzMRlSGYFtv7bviHX/Cf7J3xO8R+Fdd1DRtW0/Q3ktL/AE+5e3nt3Dr8ySIQynnqCK+E37Z/7QPhb4qeDPE3jb9oD4oav4d0jxBY3mr6fN4svZ0u7SK4R5omjaUq4eNWUqwIOcHg1+1D/wAFM/j/APHPxNqFn4A8Xax4A8EJMy6fpuk3RtbuWIDAa5uIiHZmGSY1byxnbhsbj4J/aX/aD+HWsx6/4M+NHjDTrxOuNXlljlHPEkTsySDnOHUjPPWv+Ce37Zcv7XPwyv28WWtjY+OfCc0dvrENqSsd3G6kxXSIfuB9rqy5IDISMBlUVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV+37/yeV8Wf+xgk/8AQErQ/GXi/wAMQy2/hrxXrGkxzMGlSxvpLcOwGASEYZOPWtV1bVddv5dV1vVLvUL2fHm3N3M0sj4AUZZiScAAc9gK/wCCQHwo/Zl8WeJ7nxxr2vy6x8WPDuZ7Hw9qVokNvp8e7C3lt87G6cDb85CGFm+5/q5Wr9rL/k6j4y/9lA13/wBL5q+Cn7Zn7Sn7OvhW68E/Bv4k/wDCP6Le6g+oT2v9j2N3vuXjjjZ988LsMpDGMAgfLnGSSfi38efjH8d9XXW/i58RdY8TXMTM0KXc22C3LHJ8qBAscQOeiKo6elf8E9/+CePxC+LXjvQfi78WPC9xoXw70S6ivYYdTtykuvyIQ6RRxNhvIJ2lpWG1h8q7iWKfFn/klfjL/sX77/0neq8IfsY/tS+P/DOneMvBnwS8R6tomrQiayvbeJDHPGTjcuW6cGv+GAv2yv8Ao3nxV/35T/4qv+Ce3w/8Z/C79j7wB4E+IPh660PX9M/tL7Xp90oEkPmaldSJkAnqjo30YVVVVVX/AAVB+DzfCX9rzxTd2lq0el+OFTxFZMxB3NcFhc8jv9pSc4PIDLnOQT/wRK+J0OmePviD8IL69RRrum2+rWEUj7cy2zmOUIO7Mk6EjriInoDX/BVL4qN8TP2wfEemW9y8mn+CbWDQrUFgQGjBknwABgieaVTkknYOcYVf+CJXweSe98ffHrULVGFusfh3SpCSSGbbcXXHQHb9kAPJwzDgH5qr/gtX/wAnUeFv+yf2f/pff1/wRU/5Oo8U/wDZP7z/ANL7Cq/4LJ/FuPwb+zdpnwstZcXvxD1hFlQqpBs7JknkIJ5BE32Tp2JyR0P7IyfDwftLfDu++K3iOx0PwrpWtR6jqN5fKDBi1BnSOQFWBWR4kiIxz5mOOo/av/am/Y6+N/7OHxB+F9n+0D4RlvtZ0WX+z0eaTDXkJE1sDhCQPOij5AJHXBqGaW3lSe3leOWJgyOjFWVhyCCOhr9nD4qW3xv+A/gX4qQXKTS+INFgmvCrKwS7VdlymVAGVmSRTgDleg6Cq/4Le/8AJU/hp/2L91/6UCv+CQ3/ACeVpv8A2L+o/wDoC1VfEPVbDXfH/ibW9KuPPstQ1i7uLaXYV3xvMzK2GAIyCDggGv8AgjJo97pn7JmrXt0qCPVvG17c221skxi1tITn0O+F+PTB71/wWw+LcerfEDwN8E9PlzH4e0+XWNRwikGa5by4kz94FI4XbAwCJl6kfL/wS58W/Bb4bftCXnxO+M/j3SfDVvoGhzro7X/Hm3c5WIlTtbGITMDjB+cc43A/8FQvjv8Asv8A7QP7OUNt8PfjR4Z1jxT4X1y3v7Kyt5XM1zE4aCaNBtx0lWQ5I4h69j+z98Tp/gv8bvA3xThklEfhrXLa6ukiIDS2wcC4jBION8JkTOCRu45q0u7TULSG+sLqK5trmNZIZoXDpIjDKsrDgggggjg5qq/b9/5PK+LP/YwSf+gJX/BJX9kv4a/HDW/FPxW+K2iWPiLTvCU0Fnpmi3kfm28tzIrO808ZG2RVUKFRtykuxYfKuf8Ago9+xf8AAY/sx+JviH8O/hd4U8H+JPBkcd/b3Oi6ZHpy3EIlVZopUgUK+Y2YqWUkMqjKgtVpd3dhdwX9hdS21zbSLJDNC5R4nU5VlYcgggEEdDR+IVpp/wAIP+Fr67HDbW1t4b/tq8TzQiRItt5zje3AAAPzHgdTXjfxhrnxC8Y65478TXX2jVvEGoT397Jk4Msrl2xkkgZOACTgYFfsw/8ABQL9hD9nP4J+F/hloVz4ggvLDT4jrF1D4ecPfXzLuuJnbcS2ZGfaCTtXao+VQK/4KQ/HH9m79oj4geG/iR8CY9QXVprOe28Stdac1oJijIbaTBJDOQ0yscA4VMk8Y/4It/EfUNC+P3if4Zy3bjS/FPh17pYMMQby1lQo3XCjypbgE4yTtGRX/Bb3/kqfw0/7F+6/9KBX/BIb/k8rTf8AsX9R/wDQFqq/4Jcf8mJfDH/uLf8Ap2vK/wCCvP8AyeVqX/Yv6d/6A1fsMfsmyftd/GT/AIQrUtWu9J8M6NZm/wBdvrRVMywhlVYotwKiSRmwGIYKAzbWxtPw1/4J3/shfCXxRoXjnwV8K5bbxH4dkSay1ObXr+VxMq48xo2n8ok5JI2bcngCv+CyPxZ1Lxb+0hYfCuK+/wCJT4D0eHNssmQLy6UTSSMMDBMJtlA5wFzn5iB/wSN/Z28NfGX44a14+8baNb6tovw6sYZ4rO5RJIpNQuGdbcyRsDvVUiuHHHDpGc8YPj7wD4Q+J/hDU/AvjrQrTV9F1e3eC5trmIOMMpG5cg7XGcqw5U4IIIr45fDS6+Dfxi8Z/Cy6keU+GNaubGKV3VmmhRyIpCV4y8exsYGN2CAcgf8ABH/4o3fjr9lZ/B+qX0s954E1yfT4RKxcraSKs8OG2jgNJMgXLFRGOilQP+Co/wDyfb8Tf+4T/wCmmzr/AIJSfsrfD/8AaI+Jvijxd8UNOt9Z0DwDa2pXRZwfLvLu6aURNKOjxItvKShxuZo85UMp/bq/Yk/Z71z9mXxx4k8E/CTwf4R8ReDtHudbsNQ0TSYtNYi2TzpY5BAiiUPFG6hXBAZgRt61XwA8T6h43+A3w28Z6s7Nfa/4R0q/uWeQyMZZrSKRyWbljljyeT3qqqqr/gpV8Hm+Mf7IXjG0srVp9U8JqviLTlUj71qGM3B6k2z3AAHJJGM9Dp2oXuk6ha6rpty9vd2UyTW8qcNHIhDKw9wQDX7Yv7SWnx/8E8NS+Lnh29RJviN4dtLLThBKBsfUEVZkBIbDRxNPkfeBjIypG4eDfCmr+PPF+heB/D0SSap4h1K306xRyQGnnkWOMHAJxuYdAa+HvgnRfhr4D8O/Dzw5F5emeGtLt9OtFI58uGNUUnJJJIXJJJJJJJNV+35/yZp8Wf8AsX5P/Q0qvhP/AMkr8G/9i/Y/+k6VX7Wnxbj+Ov7SHxA+KVtL5llrGsOunOVVS1nAqwWxIXjPkRRZ6855PU/8ExPj3+yv8AP2aY7Dx18ZvDmj+KvEutXWo6nZXbFZ7YKRbwxsQmdvlwCUAk485jxkgf8ABT/xR8H/AIi/tHJ8UPg34+0fxPY+JNFtv7Tewdi0F5ADDh8qBgwpBg5zw2QMDP8AwRe+LUfhf45+JvhHf3vl23jfRxcWcbSKBJeWZZwqqRnJgkuGOCOI+QcArVf8FpbO7uf2U/Ds1tayyx2vjyzkndELCJDZXyBmI+6NzouTxlgOpFfDzVbDQvH/AIZ1vVbjyLLT9YtLi5l2FvLjSZWZsKCTgAnABNV/wWh8GeL/ABR8TvhzP4a8Kaxq0cGg3KyvY2Ek4RjOMAlAcHHY1rGh614evTpuv6Pe6ZdqoY295btDIAeh2sAcGv8AgiH/AMlT+Jf/AGL9r/6UGvjF8RtN+EHwp8W/FDVl8y28L6Pc35i7zNHGWSMcjl2CqORyw5HWtX1W/wBd1W91vVbjz73ULiS4uZdgXfI7FmbCgAZJJwABX7MH7Vf7GPwV/Z6+H/wx/wCGgPCkVxoehwLepvZP9LkHm3JwqDrNJIeeeeSTkn9qq38CxftGfEK4+GXiXT9e8L6jrk9/pd7YFjCYrkify1yAf3ZlMR46oeT1P/BFj4sx+Ifg34t+Dt/fb73wjrAv7KN5FBFndryqLgEhZoZWZsnBnUccZr4tf8lU8Z/9jBf/APpQ9fCH9h79qP49eDIviD8J/hf/AG7oE1xJbpd/21YWuZIzh12Tzo/BI524r4xfs4/HL9n+9hsvjB8NNY8Ni5Ypb3M8ay2s7DOVjuIi0TsAMlVYkDBxgiv2C/8AgpZqfwIfQfgx8UPD+iSfDyS4SBdV0+wjsrnSS7KDcTCFQLpB1kLL5xGW3uRsaGaG4hSe3lSWORQyOjZDAjIII6giv+C2Hxbj1b4geBvgnp8uY/D2ny6xqOEUgzXLeXEmfvApHC7YGARMvUj5f+CXPi34LfDb9oS8+J3xn8e6T4at9A0OddHa/wCPNu5ysRKna2MQmYHGD845xuB/4KhfHf8AZf8A2gf2cobb4e/GjwzrHinwvrlvf2VlbyuZrmJw0E0aDbjpKshyRxD17H9n74nT/Bf43eBvinDJKI/DWuW11dJEQGltg4FxGCQcb4TImcEjdxzVpd2moWkN9YXUVzbXMayQzQuHSRGGVZWHBBBBBHBzVV+1l/ydR8Zf+yga7/6XzV/wSQ/ZA+FXxa8PeJ/jn8V/DemeKodM1b+xdI0fUYBPbQypDHNNPLEwKSkrPEqBshcSEgkqV/4KlfsdfBDQ/wBnC9+Mnw1+G/hnwdrXg28tPtB0TT0sI7y1nnW3aNooVCO4kmicOw3AIw3YOD4N8T6h4J8X6F4z0l2W+0DUre/tmSQxsJYZFkUhl5U5UcjkV8aPiFD8JvhD40+J0ot3PhbQbzUYorh9qTSwws8cecjl3CoBkElgBya1fVdS17VbzXNZvpbzUNRuJLi6uZm3PNK7Fndj3JYkk+pr4K/8FGv2BPgR8MtC+F3ge78RWun6LapG8kXhx42u59o824k+YkvIwLEknrjOAK/4KF/Fr9nv46/Gm2+KPwDhvYf7W01V8QrcWDWomvUdgJwpJyzRlFYjAJjBOWZif+CI3xI1C38dfET4QTXjvZX+kw67bwMCRFLBKsErqc4UstxCG4JPlryNpz/wW9/5Kn8NP+xfuv8A0oFf8Ehv+TytN/7F/Uf/AEBaqv8Aglx/yYl8Mf8AuLf+na8r/gqP/wAn2/E3/uE/+mmzr/gn1+xtbftf/E3VLHxRql7pvg3wnax3OszWO0TzSSsywW6MwIQvslYuVbCxMMAsCPhR+wN+yf8ABHxtpHxI+GvwtfTPE+iLKLPUX1y/uGQyQvDITHJO0ZLRyOOUwN2RggY/4KzfFnUviH+1rq/g77b5ujeALODTLGNJMp5skST3D4wMP5knlnr/AKleew/4I5fs7eGviJ4/8T/GjxnotvqVt4KWC10aC5RJYjezBy8pjIJ3RRou0nABlyMlcr8W/hN4G+N/w+1j4afETRotR0bWLd4nDRqZLdypCzwswISVCdyvj5SAa8feDtU+HnjrxH8P9c2f2l4Z1a60y82EEedbytE+CCQRuQ9Ca/4JX/FG6+Jv7HPhm11K+lvL/wAF3lz4fnllYk7ISslug+UABLae3jAG4YQc5yB+37/yeV8Wf+xgk/8AQEr/AIJK/sl/DX44a34p+K3xW0Sx8Rad4Smgs9M0W8j823luZFZ3mnjI2yKqhQqNuUl2LD5Vz/wUe/Yv+Ax/Zj8TfEP4d/C7wp4P8SeDI47+3udF0yPTluIRKqzRSpAoV8xsxUspIZVGVBarS7u7C7gv7C6ltrm2kWSGaFyjxOpyrKw5BBAII6GvBuuS+J/B+heJJ4Ehl1bTbe7eJCSEaSNXIBPYbsVVVVV+1l/yav8AGX/sn+u/+kE1V+yd/wAmr/Br/sn+hf8ApBDVV+1l/wAnUfGX/soGu/8ApfNXgr4IfGn4l6VLrvw4+EHjXxVpsFw1tLeaJ4fur6GOZVVjGzxIyhwroSpOQGU9xX/DJv7VH/Rs/wAVf/CMv/8A41Xjr4TfFT4XfYf+FmfDPxX4R/tPzPsP9u6LcWH2ry9vmeX5yLv270zjON656iv+CXH/ACfb8Mf+4t/6abyv2/P+TNPiz/2L8n/oaVX7En7NXw3/AGe/gh4XXwx4f08+JNb0e3utd10W4+0300qLIymQ/MIlJASPIUAZxuZif+Cvn7LXgS5+FLftI+EfC9jpfiXRdShj8QXVlbCL+0ba4fy1knC4DSrM8QEhG4hypJAXHwd+IF18J/ix4O+Jlk0vmeF9ctNRZIicypFKrvHgMuQ6hlK7gCGIJANVX/Bc/wD5ol/3MH/uOr9kn4K2X7RP7Rngf4PapqL2Wn67fO1/LGcP9lt4ZLidEODtdo4XRWIIDMCQQDWofsN/sh6j4Ol8EP8As8eB4LGW1+zC6t9HijvkXsy3gHnh/wDb37vUnmvip4Il+GfxP8X/AA3nu0upfCmvX2kvOmcStbTvEWHA4OzPQda/4I4eJ9Q1/wDZDn0m8djD4b8XX+n2gaQsBG0VvcnAP3RvuZOBxkk9Sa/4K4/Fe98f/tZ3/guO8STSvh/ptvptskUu+MzyxrcTvwSA+6VYm6Y8gAjINf8ABNj9oz9lD9mOz8R+NvjD/aTeO9Quha6bNbaO119isBGpYpIG+VpHZgwABxEvJDEV+3P+3P8AsX/tP/s/a14H0uXXbnxZZtHeeGrq40V4ja3Suu4B93CvHvjYHIwwOMqpHwO+JGofCD4xeDPidpt49vL4b1q2u5GUE7oVceahCkEq8ZdGUEZDEZGa/b8/5M0+LP8A2L8n/oaVXwn/AOSV+Df+xfsf/SdK/wCC1f8AydR4W/7J/Z/+l9/X/BFT/k6jxT/2T+8/9L7Cv+C1f/Jq/hb/ALKBZ/8ApBf14T8L6z438VaN4L8N2v2nVtf1CDT7CDO3zZ5pFjjXPbLMo/GvhX/wSO/ZL8IeELPTPiP4a1Dx3r/lq17qd1q13ZIZdo3iGK2ljCR5zgPvYd2Nf8FR/GNn+z/+xTpPwj+HS/2XZa7cWXhe0tkuXZ7fSoIGZ1DPuZxsgihbc2SJTknnPwO+Gl18ZfjF4M+FlpI8Z8T61bWMsqOqtDC7gSyAtxlY97Ywc7cAEnB8BeAfCHww8IaX4F8C6FaaRoukW6QW1tbRBAAqgbmwBuc4yzHliSSSTX/BXP8AZ28NfBv44aL4+8EaNb6VovxEsZp5bO3RI4o9QtmRbgxxoBsVklt3ORy7yHPOB/wRz+KN14P/AGorn4dS30o07x9oc8AtQxCSXlqpuYpCApyVhS7UZKj963JOAf8Agt7/AMkq+Gn/AGMF1/6Tiv2f/hZ/wu741+CvhO2pf2fF4n1iCzuLoDLQwlsysowcuEDbQeC2ASBkjT/2G/2Q9P8AB0PghP2ePA89jFa/ZvtVxo8Ul8692a8I88uf7+/cOxFftGfDC3+C/wAdvHfwssrr7RZ+G9cubWzlLFma3Dkw7iQPn8spu7bs4JHNf8EUfE+oaj8AvGnhS6d5LfRvFXn2rPIzbBPbRbo1B4VQ0RbA7yMe9VVVVVVVVVVVVVVVVVVVVVVVVf8ABLj/AJMS+GP/AHFv/TteVX/BbDVLCH9mzwdoklxi9u/HENxDFsPzRxWN2sjZxgYM0YwTk7uM4OPgVo974h+N3w80DTVRrvUvFWmW1uHbaDJJdRquT2GSOa/b8/5M0+LP/Yvyf+hpVTfEj4iXGnvpM/j7xHLYyQmB7V9VmMbRldpQoWwVK8YxjHFaTFpU+q2UOu3t3Z6a9xGt5cWlqtzNDCWG944meNZHC5IQugYgAsucj9hn4Wfs9fDb4CaNdfs66lFruj+II1ur3xDIB9q1OcZB8/gGMxncghwPL+YEbizN/wAFR/8AkxL4m/8AcJ/9O1nXwm/5Kp4M/wCxgsf/AEoSv+Cxf7Nv/CdfC7TP2hvDWn+ZrXgTFnrPlpl59Llf5WPUnyZnyAOAs8rE4Wv2HPj9/wAM2/tJ+FfiDf3ssGgXEjaZ4gVDgPY3GFdmHUiNxFNgdTCBX/BSbSdP0f8Abb+J9vpdy1xb3N1ZXqyM4bc1xYW07YKgDbulOPbHJ61/wRU/5NX8U/8AZQLz/wBILCqr9uD/AIJz/FD9rr43J8SdP+JPhrQtJ0/Q7bStPtri2mlm2xvLK5faAB+8mfGCeMdMV4n/AOCKHx90/TzdeFPif4H1m5jVma1uftNmz4GQqN5bqWJ4+YqBnlhXxg+DHxM+Avja4+HvxX8J3eg63bxrMsU2GS4hYkLNDIpKSxkqw3KSNyspwysB/wAEgv2s/FNl48i/ZW8Y6tcahoOsWtxceFhKDI2n3UKNNLAp/hheJJnweFdBgAyNX7Zf7b3hT9jT/hD/APhJvA+q+Iv+Ev8At3kfYbmOLyPsvkbt2/ru+0LjH901/wAPvPhV/wBEP8V/+DG3/wAK/Za/4KY+Bf2pvixb/Cbw/wDDLXdDu57Ge7F3eXkUkYEQBIwvOTmv+C5//NEv+5g/9x1eFvCXirx1r1r4V8E+GdV8Qa1e7/sum6XZSXdzPsQu+yKMFmwisxwOApJ4Br/hk39qj/o2f4q/+EZf/wDxqtX/AGY/2k9A0q813Xf2eviXp2m6bbyXN5eXfhK+hhtoY1LPJI7RBURVBJYkAAEmq/ZO/wCTV/g1/wBk/wBC/wDSCGq/4K8/8nlal/2L+nf+gNX/AARD/wCSp/Ev/sX7X/0oNVVf8Fvf+Sp/DT/sX7r/ANKBX7DPwA0P9pX9pLw18NfFVy8WgBZr/VkjdkkuLeBNxhQryC7bEJyCqsxByAD4z/YU/ZN8Y+Br3wOPgN4J0hbmzNvBqWl6JBa3tqwTakq3CKJGdThvmY7iPm3ZNeItEu/DXiDU/Dd/JDJc6VeTWkzwklGeNyjFSQCRlTjIH0Ff8EpvE+oeJP2KfCNrqLvI2h32oafFI8jOzxrcvIgOegUShABwFRRX/BTv4r3vxS/bB8Y2pvEl0zwayaDp8aS7xGIB+/zgkbvtDTZAwRgKRlTX/BN/9qT9kP8AZZ8Da1rHxMbVW+IevagRJd2+itcfZbGNAIoY5NxxuYyu5ULu3IGz5amv+Cg/7Z/7Hn7UvwOm0Twi+tXHjvRryC40G5udGeDaDKqzxtJu4QwtI2DkblXjPI/Y8+I+ofCf9p/4aeM7G7eCOHxFa2t8VDNus7iQQ3C7QRkmKR8DpnHXFf8ABUf/AJMS+Jv/AHCf/TtZ1Vf8Fef+TytS/wCxf07/ANAav+CIf/JU/iX/ANi/a/8ApQa/4Le/8kq+Gn/YwXX/AKTivhd8O9f+LfxF8N/DLwuiNqniXUobG2L8LG0jAF2/2VGWPsDXw/8A+CSv7G/hXwtZ6N4v8Gan411eJQbrWL/Wry0eZyBu2xWsscaJnO1cMwBwXYjNf8FhPiK/ww/Zs8I/BfwnN9gtvFuoC3liWZizafYRoxiGckjzHtSWLZ+XBzuJH7K/wcj/AGgP2hvAvwiuWlWy1/VANQMMixyCzhRprkozcB/Iik2nB5xweh0Dwn4Y8KeG7Twd4Z8P6fpmh2Fv9mttNtbZY4IosY2KgGAOTkY5yc9a/wCCpf7Pnh/4EftJNfeC9Ki0/wAO+ONPGr29rAESK2ufMZLiKONQNiblVwMYHm4BwML/AMEVvijd6J8Z/F/wkur6Uad4n0P+0LaEsWT7ZayKOF2naWhmlJbKg+UoO47cf8Fz/wDmiX/cwf8AuOr9kn4K2X7RP7Rngf4PapqL2Wn67fO1/LGcP9lt4ZLidEODtdo4XRWIIDMCQQDWofsN/sh6j4Ol8EP+zx4HgsZbX7MLq30eKO+RezLeAeeH/wBvfu9Sea+KngiX4Z/E/wAX/Dee7S6l8Ka9faS86ZxK1tO8RYcDg7M9B1r/AII4eJ9Q1/8AZDn0m8djD4b8XX+n2gaQsBG0VvcnAP3RvuZOBxkk9Saqqqqqqqqqqqqqqqqqqqv+CvP/ACeVqX/Yv6d/6A1f8EQ/+Sp/Ev8A7F+1/wDSg1Xxv1Ww0L4LeP8AW9VuPIstP8L6lcXMuwt5caWsjM2FBJwATgAmq/4JoaPe6H+w58LrK/VBLJa31yoVsjy59QuZk/HZIuR2NftZf8nUfGX/ALKBrv8A6XzVonjjxr4atGsPDfjDW9KtnkMjw2WoSwIzkAFiqMAThVGfYelahqOoavey6lqt/cXt3cNulnuJWkkkb1ZmJJP1r/gj58KP2ZdX+3/Eq01+XXvizo0e2TS9UtEg/sWJvlM9ogd/ND7ghnOGX7u2Pf8APXxa/wCSqeM/+xgv/wD0oev2Ufgbov7R/wDwSi8N/CLWGiik1mz1c6ddyLn7Hex6tdvbzcc4WRV3AYLIXXoxrV9K8SeA/FV5omqwXej+IPDuoSW9zFuMc1ndQSFWXKnKujqRkHgiv+Chfxe0T9pn9kv9n345xXjtrcN1qOlatbqVCw3pht/tQZQOCXt0dMYGyUccjH/BIb/k8rTf+xf1H/0Baqv+Cp1nd237dPxGmuLWWKO6j0qSB3QqJUGl2qFlJ+8NyMuRxlSOoNf8ER9VsIfjT8QtDkuMXt34XjuIYth+aOK6RZGzjAwZoxgnJ3cZwcV8Wvhj8Sj8SvGd+Ph54l+zf25fyed/ZM+zZ57ndu24xjnPSq/YD/5M0+Ev/Yvx/wDob1/wWs+LceifCzwZ8F7GX/SvE+qPql7hFO22tV2opzyN8sysCOf3LAkA4P7B178MNE/an8E+Lfi/4s0/w/4b8NXEmqPdXw+RriGNmt1GVbB87y2zjPyHBBwR+3T+0t+yN8ef2WPHXgDw/wDHjwpd679jW/0iFZZC8lzbSLMsafJ9+RUaIZ4zJ1HUaHrWqeG9a0/xFod41pqWlXUd1Z3CAFoZo2Do4ByMhlB544r4XeP9H+K/w28L/EvQWT7B4n0m21CFFlEnlCWNWMZIx8yElWGAQVIIBGK/4K8/8maal/2MGnf+htWk6Vf67qtloelQefe6hcR29tFuC75HYKq5YgDJIGSQK1D/AIJjftzaZZS39z8B7h44F3Mtvr2mzyEf7McdyzMfYAmtKvvG/wAFPiLbahdeH/7J8UeGLwO2n6/oySm3mUdJrW5QjIyCAy5BwwwQCP8Agn5/wUDtf2trXUPAnjrR7HQviFoVqty6Wbt9m1a23bWnhVsmJkJQPGWb76spILLHqOoWWk6fc6rqVylvaWULzTyvwI40BZmPsACa+OXxLuvjL8YvGfxTukeM+J9aub2KJ0VWhhdz5UZC8ZWPYucnO3JJJJP/AATv/aL/AGS/gF+yx4a8JeKPjf4Z0zxJqVxc6prdrMxR4riWTaqthMkrDHCpJJOV4OMAf8FGNY+Ffi39qfxB4/8Ag/410nxLofiuztb+SbTmZktrkR+TNG24D5y0PmnGR++HPYf8ES/izHp/i7x/8EdRvsLrFnBrelxPIqqJYD5VwFGMs7pLAcZ4WAnHUiq/4Kj/APJiXxN/7hP/AKdrOq+A3/BP39mv4RfDLRfCPiP4ReDPGOvw2sZ1jWtb0WDUJLu7K/vWjM6MY4txIRFxhQuctlj/AMFK/wBn3wf+zp+01deG/h/YRaf4d8RaPb63YadFIzrZCR5YZIxu5A823kYLkgK4AwBgf8ESvE+oWnx48eeC4nb7DqnhH7fMokIBltruGOMlehO27k5PIycfeNf8Fr/ive6F8MvAvwd0y8RF8V6lPqWpJHLhzBZqgiR1B+48k+8ZHLW4wflNfsU+LvgF8Pvjxpvjv9ouC4uvDegWst1ZWkWnG9W4vwUEIeMEfKoZ5MnI3RqCOePGP/BUn9g74geFtU8E+Mn8RatoetWr2t/ZXGgSFJonGGBwwIPcEEEEAgggGtXi02HVb2HRrmW40+O4kW0mmTY8kQY7GZexK4JHY1+w98SNQ+LX7Jfwx8b6tePdX8+irZ3dw4Iaaa0ke1kdskkszQEk9ySQADgfFr/kqnjP/sYL/wD9KHr/AIJDf8maaZ/2MGo/+hrX/Bav/k1fwt/2UCz/APSC/r9k7/k6j4Nf9lA0L/0vhr9rL/k1f4y/9k/13/0gmqv2FP8Aglx8IPGHwe0H4v8A7Q2m6h4gv/F1mL7T9ES+ls7a0s5ADA7NAyyPI6YkzvCgSBdpIJN74I+E37DP7NfxE1j4MeE4tA0zRtPv9fWyl1C4uVuNQFqqR5ed5WXeYYEwPlHXHJzq+q6lr2q3muazfS3moajcSXF1czNueaV2LO7HuSxJJ9TX7Af7O3hr4Afs5eE4LbRbeLxP4k02DVNfvSiPPJPOiyeSZFHKRAqiqCV+UkZLEn/gsT+zt4a8SfBeH9obR9Gt7fxL4QvreDVb2JEja70+4dYFEpxukZJmgCcnarycYOV/Z0+KN18Ffjt4E+KVrfS2sfh7XLae8eJipe0LhLmPIVjh4GlQ4UnDnAJr9vz/AJM0+LP/AGL8n/oaVX7Iv7BH7O3w9+BfhceMvhX4R8a+Jtc0231DV9U13SIdRJnljDmOHz0by4kDbAFC7goZhuJr/gqh+zj4F/Z/+POl3Hwz0e00bw94v0cXq6XbbglpcpI0cwjU8JGwEbAA4DM4AUBRX/BG7xPqGjftY3mg27u1p4g8K3kFxEZGCho5IZUk2jgsPLZQT0EjY61VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX7fv/J5XxZ/7GCT/wBASv8AgiH/AMkq+JX/AGMFr/6Tmv8AgsH+y74Jl+Gdv+0l4O8MWmmeItJ1SO38Rz2VsIl1G2uDsWefYvzSpN5SiRiCVlKsWxGB+zb8YdS+Afxz8GfFfTp5Vj0LVInv442wbizY7LmLofvws69DgkHGRUM0NxCk9vKkscihkdGyGBGQQR1BFftZf8nUfGX/ALKBrv8A6XzV/wAEpP2YfgB8a/2dvEPin4q/CvRPEurWvjS6sobu+jZnSBbKydYxgjgNI5+rGvAv7M/7PHwymtrzwF8EvBOi3tp/qb620SAXS/OH/wBeVMhwwUjLcbVxjAxXxZ/5JX4y/wCxfvv/AEneq/YD/wCTNPhL/wBi/H/6G9VVVVVVf8Fqfg83iD4VeD/jXptq73PhLUn03UShAAtLsAo7Z5ISaJFGO854PUfsD/FrTvgr+1h4D8aa7qqWGjPdS2GpzSK7IILiF4ssEBbAdkbp1UZ4zXj7xbe+P/HXiPx3qKut34j1a61KcPJ5jCSeVpGy2BuOWPOBn0FfsNfB5vgd+yv4A8D3lq8GqSaaupaqkhBdbu6JnkRiOMx7xFx2jHJ6mq/4LV/8nUeFv+yf2f8A6X39f8EVP+TqPFP/AGT+8/8AS+wqv+CvnxYbx7+1O3gay1FJ9N+H+kw2AjjYMi3cw8+dsj+LDwxsM8GLGAQa+Bv7K3x8/aUh1mf4KeAH8RR+H2hXUXOo2tmIWmDmMA3Ese8kRvkLnGBnGRn/AIdcft2/9EL/APLm0n/5Krx34G8VfDPxhq/gDxxpD6Xr+g3T2uoWbyJIYZV6jchZWHoVJBBBBIr/AIIsfFmPxD8G/Fvwdv77fe+EdYF/ZRvIoIs7teVRcAkLNDKzNk4M6jjjNV/wW9/5Kn8NP+xfuv8A0oFf8Ehv+TytN/7F/Uf/AEBar/gpT+2N4W+APwj1j4Y6DrCT/EbxppslrY2cGHbT7WXKS3U3PyDZvWPuz4IBVWItLO71C7gsLC1lubm5kWKGGFC7yOxwqqo5JJIAA6k1+yL8FZ/2e/2cfA/wn1AwnVNI0/zNUaLBX7ZO7TTqGH3gskjIG7qq8AcD9sL4sN8bv2mfiH8RYtRS9sL3WprfS5o2BRrG3/cWxXHGDFGjcdSxOSSSfhJ+wx+1R8dvBVv8RPhX8Km1rw9dzSwwXrazYWokeNtrgJPOj4DAjOMZB54qb/gl5+3XBC8z/AlysaliE8R6U7YA7KLokn2AzVf8E4Pi1H8Xv2QPAt/Pe/aNS8NW7aBqIMiu0clphIgxAHLW5t35GfnGS33jVft+/wDJ5XxZ/wCxgk/9ASv+CIf/ACSr4lf9jBa/+k5r9vz/AJM0+LP/AGL8n/oaVXijwxqHjf8AYz1bwXpKs19r/wAMZtPtlSMyMZZtMMa4VeWOWHA5NV8NP+CQfjD4sfD7w98SPCHx/wDB93pHiPT4r21kjsZ22h1BKNzw6NlGU4KspBAIIr/hyF8U/wDouPhT/wAFtx/jX7Ev/BM3xf8Ass/G1Pix4n+J2j65Bb6Tc2kNpYWckbGWXYNzFzjaFD9Oc47Zr/gt9DKvxO+GU5iYRvoN2quVOCwnXIB9QGXI9x61/wAEovE2leHf20/Cttqs3lf23p+oWFtIzKqiZrdpFDFiPveUUAGSWZRjnI8TeJdA8GeHdS8WeKdWt9M0fR7WS6vry4bbHBDGpZ3Y+gANV/wS4/5MS+GP/cW/9O15X/BXn/k8rUv+xf07/wBAav8AgiH/AMlT+Jf/AGL9r/6UGq/4Kz+E9S8O/tr+KNYvh+58UaXpmo2Xy4zElqlqfr+8tZK/4IiePNF0n4h/Ez4cXk8Ueo+JNLsL+yDybTILN5lkRRjDHF2rYyDhCQCAxWv2tvHmi/E/9pr4meOfDc8NxpOp+JLs2FxDJvS5gRzHHMpwOJFRXAxxuxzjNf8ABFXwle6V+z14u8XXTOseveKmitomjwDHb28QMgbPILyOuMDBjPJzx/wVH/5Pt+Jv/cJ/9NNnX/BDH/mtn/cA/wDcjX7WX/Jq/wAZf+yf67/6QTVX7J3/ACav8Gv+yf6F/wCkENVVVVajp9lq2n3OlalbpcWl5C8M8LjiSNgVZT7EEivjt8Lr/wCCnxk8ZfCnUBKX8M6xcWkMkpBaaAMTDKccfPEY36D73QdB8XP2h4vGn7CvwU+CC6n9pv8Awz4g1d9Si3MphSHabPcCuGzHfSqpVsARYOSeP+CRnweb4jftTw+N721d9L+Hemy6kz5Gw3coMFujDrn55ZBjHMHJ7Gq/b8/5M0+LP/Yvyf8AoaVXwn/5JX4N/wCxfsf/AEnSv23viw3wV/ZX+Ifji01FLPUxpL2GlyFhvF3dEQRMgP3mQyeZjB4QkjANQwy3EqW9vE8ssjBURFJZmPAAA6k+lf8ADrj9u3/ohf8A5c2k/wDyVXxi/Yi/ah+AXg4+Pviz8Ln0PQRdR2rXi6vY3YWWTOwFIJnYA7TyRjOBnJFfs/fE6f4L/G7wN8U4ZJRH4a1y2urpIiA0tsHAuIwSDjfCZEzgkbuOatLu01C0hvrC6iuba5jWSGaFw6SIwyrKw4IIIII4Oar9uz4Kah8ff2WvG/gLQbJ7vXo7VdR0aFGIaW6tnEqxqBwWkVXiAbjMgPGARX/BNr9svwh8e/hLofwu8Q67FbfEjwhp6Wd1ZXDCN9St4htjuYOf3n7tVEgHKuCSArKTX/BXn/k8rUv+xf07/wBAav8AgiH/AMlT+Jf/AGL9r/6UGv8AgsZ8WG8E/szWPw60/UUhv/iBrUdvNDuAeSxtsTzFe+BKLVTjtJgnBwfh38PPGXxY8a6T8O/h9ojav4h1uYw2Fks0cRlcKWI3yMqKAqkksQAB1r/h1x+3b/0Qv/y5tJ/+Sq+Of7MHx0/Zrl0aD41+BH8ON4gWdtNJ1C1uxcCHYJcG3lkAK+bHkNg/MK/4JZfFqP4W/tf+G9Pvr37Ppvje3m0C5LSKqmSbD24wQck3EUKDBB+fgnJVq+LX/JVPGf8A2MF//wClD1/wSG/5M00z/sYNR/8AQ1r9vK28E3P7HvxYPj02i2EPhu5e0e5UELfgYstuQcObkwqp9WHI61X7FOpa5q/7JPwkvvESSret4TsUzLGUZ4kiCwuc8ndEqNu/izu71+2F8WG+N37TPxD+IsWope2F7rU1vpc0bAo1jb/uLYrjjBijRuOpYnJJJPwk/YY/ao+O3gq3+Inwr+FTa14eu5pYYL1tZsLUSPG21wEnnR8BgRnGMg88VN/wS8/brgheZ/gS5WNSxCeI9KdsAdlF0ST7AZqv+CcHxaj+L37IHgW/nvftGpeGrdtA1EGRXaOS0wkQYgDlrc278jPzjJb7xqv2sv8Ak6j4y/8AZQNd/wDS+av+CKn/ACav4p/7KBef+kFhX/BUf/kxL4m/9wn/ANO1nVfto+GNQ8Y/snfFjQdKR3u38K3s8MSRtI0rQxmXy1VeSzeXtAHdhVeGf+CM/jXxn4d03xZ4V/aG8Gano+sWsd1Y3lvYXDRzwuoZXU56EEV/w5C+Kf8A0XHwp/4Lbj/Gv2Df+CdPir9kP4qa38SfE3xH0rxB/aXh+TSYLaxtJItnmXEErOxf0+zqAB/eNf8ABb6GVfid8MpzEwjfQbtVcqcFhOuQD6gMuR7j1r/glF4m0rw7+2n4VttVm8r+29P1CwtpGZVUTNbtIoYsR97yigAySzKMc5HibxLoHgzw7qXizxTq1vpmj6PayXV9eXDbY4IY1LO7H0ABqv8Aglx/yYl8Mf8AuLf+na8r/gqP/wAn2/E3/uE/+mmzr/ghj/zWz/uAf+5Gq/4KT+E9S8Jftp/EmDUBlNUvINRtZAuBJFPbxuMf7rFkPuhr/giB480UWnxP+F888MWrGSz1i1jMmXuIAHhmYLjgRsYATnkzrwMZN5d2lhaTX19dRW1tbRtJNNM4RIkUZZmY8AAAkk8Cvjh4xsPiL8afH/xB0pNll4n8UalqlsuScR3F1JKoywU9HHVQfYdK/wCCO/hK98Ofsf8A9sXTOY/FPiq/1K2DR7QI0SG0ODk7hvtH545yMcZP7fv/ACeV8Wf+xgk/9ASv+CIf/JKviV/2MFr/AOk5r9vz/kzT4s/9i/J/6GlV8J/+SV+Df+xfsf8A0nSqqqqv2sv+TV/jL/2T/Xf/AEgmqv2Tv+TV/g1/2T/Qv/SCGqr9rL/k6j4y/wDZQNd/9L5q/wCCKn/Jq/in/soF5/6QWFV/wXP/AOaJf9zB/wC46v8Aglx/yfb8Mf8AuLf+mm8r9vz/AJM0+LP/AGL8n/oaVX7PHi3QPHfwI8AeK/DF+l5p1/4dsjFKvZliVHUjsyurKw7FSO1f8FdPiVoHhD9kPVfA1/Kjap481KzstPh8zDAW9zFdSy4wcqogVD0wZV57H4eeDb/4jeP/AAz8PtKfZe+J9YtNLtmwDtkuJliU4LKOrjqQPcdaqv8Aguf/AM0S/wC5g/8AcdX/AAS4/wCT7fhj/wBxb/003lV+1l/ydR8Zf+yga7/6XzV/wRU/5NX8U/8AZQLz/wBILCv+CknhjUPCv7avxNtb5HK399Bf28hjZFkjntopAVz94KWZCRxuRh2r9j/9g7Vv2w/DWuax4W+LnhzQtS8P3y291pF7bSy3AhZFaO4+TjY7eYg94mz1Ff8ADkL4p/8ARcfCn/gtuP8AGrT/AIIhfEn7VD9u+OPhr7N5i+d5Omz79mfm25OM4zjPGa/b4hln/Y2+LSQRNIw8OysVVSSArKWPHYAEn0Aqv2fPE2leM/gP8O/FOiTeZZan4X06aLLKWTNumUbaSA6nKsMnDAjtX/BZTxLoHiD9rLTbDRNWt72fQfBtnYakkTbja3P2q7m8pvRvLnibHYOK/wCCKn/J1Hin/sn95/6X2Ff8Fq/+TV/C3/ZQLP8A9IL+v2Tv+TqPg1/2UDQv/S+Gq/4LWeE9S1X9n/wd4tsxvttC8UeVdoF5RZ7eQK+ewDRqv1kX3r9knx5ovwx/aa+GfjnxHPDBpOl+JLQ39xNJ5aW0DuI5JmODxGrlyMc7ccZyK/4Ld+PNF1f4h/DP4cWU8Uuo+G9Lv7+9CSbjGLx4FjRhjCnFozYyThwSACpb/gkt4SvfEn7avhrWLVnEfhXSdS1K5Cx7gY2tmtBk5G0b7tOeecDHOR/wW9/5JV8NP+xguv8A0nFfsB/8nlfCb/sYE/8AQHqv2/f+Tyviz/2MEn/oCV/wRD/5JV8Sv+xgtf8A0nNVVVVVVVVVVVVVVVVVVVVVVVVf8EuP+TEvhj/3Fv8A07XlajqOn6Rp9zq2rX9vZWNlC89zc3MoijgjQFnd2YgKoUEkngAZNf8ABSj9rXRf2pfjJZW/gS6ln8E+CreWz0ieSLy/tk0jA3FyqnkI/lxKoODtjUkAsQP+CUXwK1L4sftRab44mts+H/hrH/at/KyZVrlldLSIHs5k3SjPG23ccHFft+f8mafFn/sX5P8A0NKrwN4d8P8Aiz4JeGPDfinQ9P1nSb/w3YxXdhqFqlxBcJ5EfyvG4KsOBwQRX7fv7P2kfs3ftN+I/A3ha1lt/DWoxxatoUMm4+TbTg5iVmA3JHMs0anLHai7mLbq/wCCJ3xnvY9a8cfADU7x5LSe1XxBpMbvgQyIyQ3SqO5cSW7Yzx5THBySP+Co/wDyYl8Tf+4T/wCnazr4Tf8AJVPBn/YwWP8A6UJXiLw/ovizw/qfhXxHp0OoaTrNnLZX9pMMpcQSoUkjb2ZWIP1r9rT4B6l+zT8evE/wquklewtLj7Ro1zIObmwly0D57kL8jH++jjtXxH8f6l8S9X0/xNr0ss+sx6PZaff3Mjbjcm0hW2hfuc/ZobdWJ5Z1dj96v+CKn/Jq/in/ALKBef8ApBYVVVX/AAW88R+A7m2+GnhWK8sZvGNnNeXMsMZDT21i6xqPMxyqvIg2g9fLcjoa/wCCcmj6vrX7avwug0VX8221Ka5mZWKhYIraV5ckdAUVlx3JA71+2X+xB4U/bM/4Q/8A4Sbxxqvh3/hD/t3kfYbaOXz/ALV5G7dv6bfs64x/eNf8OQvhV/0XDxX/AOC63/xr9lr/AIJm+Bf2Wfixb/Fjw/8AE3Xdcu7exntBaXlnFHGRKAC2V5yMV/wXP/5ol/3MH/uOr/glx/yfb8Mf+4t/6abyq/ay/wCTV/jL/wBk/wBd/wDSCaq/ZO/5NX+DX/ZP9C/9IIar/grz/wAnlal/2L+nf+gNX/BEP/kqfxL/AOxftf8A0oNVVf8ABb3/AJKn8NP+xfuv/SgV/wAEhv8Ak8rTf+xf1H/0Bar4tf8AJVPGf/YwX/8A6UPX/BIb/kzTTP8AsYNR/wDQ1r9tzwxqHhH9rv4vaTqass0/i6/v1DxlD5d3KbmM4POPLmTB6EYI4Ir9kr/gnlrf7Xfw+vfG/hD4x+GdKuNL1BrK/wBIurWWS4tTtDRuwX+B1JKsOCVYZyrAf8OQvin/ANFx8Kf+C24/xrwF/wAEXviJ4Y8deHPEurfGrw5PZaTq1rd3MVvp0wleOKVXdULHG4qpAzxnFf8ABUOGWf8AYU+J0cETyMF0tiqqSQF1W0LHjsACT6AVXhPxNpXjXwro3jLQZvN03XtPgv7OTcrb4Zo1kQ5UlTlWHIJHoT1r/gq/4l0DxJ+2Z4jTQNWt7/8AsnTbKwvGgbcsNzHHmSIsOCy7gGAztbKnDKQP+CIf/JU/iX/2L9r/AOlBr/gt7/ySr4af9jBdf+k4r9gP/k8r4Tf9jAn/AKA9V/wXE8J6le+CvhR45iH+gaRqmpabOdv/AC1uooJI+f8Ads5f8iv+CffjzRfhr+2T8L/FPiCeKGx/tSWwklmk8tIjeW0tors2DgBp1JJwOOSoywr/AILQ+PNF8RftF+G/A+lzwz3PhLw2g1FkkyYZ7mRpRCy44IhEMmcnImHTHP8AwRq8JXuuftU6l4ljZ47Tw54VupZXEe4PJLLDEkZORtJDSPnB/wBWR3yP+C5//NEv+5g/9x1f8EuP+T7fhj/3Fv8A003lV+1l/wAnUfGX/soGu/8ApfNX/BFT/k1fxT/2UC8/9ILCqqqqqqqqqqqqqqqqqqqr/grz/wAnlal/2L+nf+gNX/BEP/kqfxL/AOxftf8A0oNV/wAFaP2xvC3h34f6h+y94G1hL3xXrzRDxG0GHTTLIESeS7Z4mlIj+TkiMsWxuTPw+8CeJPih450H4deD7L7XrXiPUIbGxiOQvmSMFBYjO1BnLN0VQSeBXwy8C6b8L/hx4X+G2jyeZZeFtHtdMgkKbDIsESxhyMnBbbuPJ5J5PWv2sv8Ak6j4y/8AZQNd/wDS+av+CKn/ACav4p/7KBef+kFhX/BYr9l3wT4Z8P6B+0V4C8MWmk3lzqg0rxFFp9sIo7gypJJFdOiLtD7o3RpCQWMkQ5NfsTfGe9+A/wC054F8bxXjw6dNqUem6wofar2Nywil3dchAwlA/vRryOor4tf8lU8Z/wDYwX//AKUPX/BLj/kxL4Y/9xb/ANO15X/BYz9mO38H+N9M/aS8I6Z5Wm+LpBY+IliB2x6iifupsdF82JCDjA3Qlj80hJj8f6kfhXcfCy5llmsP+Egi1myQt8ltL9nkhuCB6yr9myTn/ULjHOf+CQ3/ACeVpv8A2L+o/wDoC1Vf8FqfgpqGk/ELwl8fNNsnOla/YjRdSmViRFewF5Id2eFMkLMAF4/0diQDy37Knx6v/wBmn47+F/i3a28t1Z6ZcGLVLSIAvc2Uo2TouSBv2EsuSBvVcnGa+F/xS8A/GfwTpvxD+GviW01vQ9TjDQ3EDco2AWjkXrHIucMjAMp4Ir4s/wDJK/GX/Yv33/pO9V+wH/yZp8Jf+xfj/wDQ3r/gqV8WG+KP7YPiiwtNRS60vwTDDoFlsYEI0I33IOP4hcyTqc8/IAegA+CP7IP7RX7R+iah4i+DHw5bxDp2lXQtbu4OqWdmI5iofYBcSxljtIJ25AyM9a/4dcft2/8ARC//AC5tJ/8AkqvE/hrW/BniXVvB/ibT3sdY0K+msNQtXZWaC4hcxyISpIJVlYZBI44Nf8Ed/i1H44/Zkuvhzd3vmaj8PtYlt1iaRWZbO5JnhbAAIBkNyozn/V8HHyr/AMFef+TNNS/7GDTv/Q2r4Tf8lU8Gf9jBY/8ApQlV/wAFvbbwTF8RvhlPpptB4sn0e+OrIigSmzEsQs2c4yQXF4FyT91unf8A4Jv6lrmmftr/AAuk8PpLJPPqE8E8ccZfdbvazLMSo7LGWbJ4Xbu7V/wUk+LDfCP9j7xxf2Wopa6p4lhTQNPywDSNdnZMFz/ELYXDDHI2ZGMZGh6LqniXWtP8OaHZtd6lql1Ha2duhAaWaRgiIM4GSxA545r/AIdcft2/9EL/APLm0n/5Kr42fsb/ALSX7Onhqz8YfGT4av4f0fUL5bC3uhqtndhrhkeQIRbzSFSUjc5IA+U81+xz8Wo/gh+038PPiLd3v2XTrLWI7fU5TIqKlncAwTsxYEYWOVm5x90YKnDCq/4Kj/8AJiXxN/7hP/p2s6qv+C1f/J1Hhb/sn9n/AOl9/X/BFT/k6jxT/wBk/vP/AEvsK/4LheGNQi8SfCrxmqO9jc2Oo6ezLG2IpY3hkAZugLrKcDqfLb0r9lz9n0/tO/FSL4T23xA0fwnqN5YzT6fNqcbut5NGVJt0C87zGZHGe0TdyK/4chfFP/ouPhT/AMFtx/jX/DkL4p/9Fx8Kf+C24/xr9lH4H3X7N/7P3hL4L32uRaxc+HY7nzr2GIxpK891LcNtU8gAzFRn0r4vwy2/xZ8bW88TRyx+ItQV0dSrKwuHBBHY+1f8EcfE2laz+yZcaDZzf6boHii8hu4mZdw8xIpUcAEnYQ+0EgZZHA6Zr/gtj4l0CD9nzwX4Om1a3XW77xlFf29iW/eyW0NndRyygf3VeeFSfVx74/ZO/wCTqPg1/wBlA0L/ANL4a/ay/wCTV/jL/wBk/wBd/wDSCaq/ZO/5NX+DX/ZP9C/9IIa/bL8J6l43/ZT+KvhvRxuvZ/C95LBHtyZWijMoQD1by9o9yKr9l7x5ovxO/Z0+HHjfQZ4ZLbUfDdmHWKTzBBPHEIp4d2BkxzJJGTgcoa/4K3+PNF8K/sba74Vv54vt/jTVLCwsITJhyYrmO7dwoByAtsQScAF15yQreAvCV74/8deHPAmms63fiPVrXTbcpH5jCSeVY1wuRuOWHGRn1Fft+f8AJmnxZ/7F+T/0NKr4T/8AJK/Bv/Yv2P8A6TpX/Bb3/kqfw0/7F+6/9KBX/BIb/k8rTf8AsX9R/wDQFqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv2/f+Tyviz/2MEn/AKAlf8EQ/wDklXxK/wCxgtf/AEnNftQfC2P41fs9eP8A4YiDzrjW9DnWxTy1f/S4x5tscEHpNHGfXjgg4Ir9g34oN8X/ANkj4beLLm7Se/t9JGl35yN4ns2a2ZnAZsM4iWTnGRIGwoYAftZf8nUfGX/soGu/+l81f8EVP+TV/FP/AGUC8/8ASCwqq+LP/JK/GX/Yv33/AKTvVaB+0h+0R4T0W08N+Ffj58RdG0nT4/KtLDT/ABTe28Fug/hSNJAqjnoABX/DWP7VH/RzHxV/8LO//wDjtf8ABGv4s/FT4o/8Lf8A+FmfEzxX4u/sz+w/sP8Abus3F/8AZfM+3+Z5fnO2zdsTOMZ2rnoKqqqqr9pD4V23xv8AgP46+Fc9uk0viDRZ4bMOisEu1XfbPhiBlZkiYZI5UcjqJoZbeV7eeJ45YmKujqVZWHBBB6H2r9h/4M2/x6/ai8B+ANTsYrvRv7QF/rEM4Jjks7ZTNLG+OcSBBF9ZByOoqq/4LV/8nUeFv+yf2f8A6X39f8EVP+TqPFP/AGT+8/8AS+wrxP4j0fwf4a1bxd4gvEtdL0Oxmvr2dyAIoIULyOSeAAqk8+lfEjxvqnxM+IPiX4ia2zm/8Tatc6jcB3DFXmlZyuQFGBuwMADAGABxX/BIn4Wv4C/ZKtfFl7BsvfHusXOqfPGyOlvGRbQqQQOD5DyKRkFZgc88V/wWS+FB8HftK6d8TLSFhafEHRY5JpGD4a8swtvIAT8uBCLThTnJJIGQW/4JX/F2H4V/teeHtN1C4SLTvHVrL4euGkl2hZZSsluQDwWaeKKMdD+9OPQ1X/Bb3/kqfw0/7F+6/wDSgV8IfjN8SvgL4zi+IPwn8S/2Hr8NvJbpd/Y4LrEcgAddk6OnIA525FeJv+CkX7bni3SpNG1X4/arBBJnc+mafZabMMqV4mtoY5Bwx6MMHB6gEaRofjr4m+JhpugaPrvizxDqLFhb2dvLf3dywHJ2qGdzj61/wT2/4Jjaj8M9fsvjf+0fpVv/AMJJpkyT+HfDomWdNOlXDLdTshKtMrfcQEhCNxJbbs/bZ+LMfwU/ZZ+InjmK++zaiNHlsNKdZFVxeXX7iFkBB3FGkEhXB+WNs4AJFfsl/C1/gv8As1/Dr4a3EHk3mk6HC1/H5bJtu5sz3AwwDD99LJ1APqAeKr9uj4Un4M/tX/EbwZBC6WEmrNqWnEhyptrtRcIqs/LhPNMZbn5o2GSQTX/BEz4uw2Hinx78DdRuEUataxa5pgeTb+9hIiuFUH7zMkkLYHIELHBGSKr9v3/k8r4s/wDYwSf+gJX/AARD/wCSVfEr/sYLX/0nNft+f8mafFn/ALF+T/0NKr4T/wDJK/Bv/Yv2P/pOlf8ABRz9hfxb8CfiLrXxX8BeHbi++GfiG6a78+0hBXRLiVnZ7aRFYssQIykhUIA6pncOfgP+2J+0R+zZC+nfCf4h3FhpE0zTT6RdQR3dnI7BQzeXIDsYhFBZNrYGM4zWq/8ABWz9tPUbCWzs/GXh/TJpMbbq18P27SR4IJ2iUOnIGDlTwTjBwR/wSv8Aj/42+PvwK8Q6j8TPFd1r3inR/FE0Nxc3JO5reSCF4iAAEUbjKoRMAbMkDdk/8FW/2afEnx5+BOn+LPAWjy6r4m+Hl5Lex2MClprmxmQLdJEgBLyAxwSBcjKxOAGYqDp2o6hpGoWuraTf3FlfWUyT21zbymKWCVCGV0ZSCrAgEEcggGr744fta/tPy6T8E774j+M/HDaxdRw2ehG7ZxdyrgqZAMBwu3eWkJC7S5IwTXxa+GXiL4M/EfXvhd4tmsZdY8OXX2a8eykaSEyBQTtZlUkc4zgc1/wS4/5MS+GP/cW/9O15X/BXn/k8rUv+xf07/wBAav8AgiH/AMlT+Jf/AGL9r/6UGq/4KVfsUX/7UfgGx8ZfDqCI/EPwdHJ9jgdgg1azb5ntdx6SBgGjJO0Eupx5m5dF1n4i/BP4hW+s6TPrHhDxl4VviU8yJra6sbhDhkeNwCD1VkcYIJVgQSK+JX/BTL9rv4p+BL74ea948sbLTdVh8jUJtL0yK0uLqEjDxmRRlVf+LZtyMrnaWU/s3/sw/Fb9qLxzb+DvhvoMsltHIh1XV5U22umQFlDSSOSASA2RGpLtg4BwSPg38KfC3wO+Fvhr4TeC4WTSPDVittC8gAed8lpZnxgb5JGeRsADc5wAOK/4Kj/8n2/E3/uE/wDpps6/4IY/81s/7gH/ALka/ay/5NX+Mv8A2T/Xf/SCaq/ZO/5NX+DX/ZP9C/8ASCGqqqqq/wCC0Xwlj8L/ABz8M/FvT7Ly7bxvo5t7yRY1AkvLMqhZmBzkwSW6jcBxHwTgha/4JF/Bm3+HH7Lkfj+8sYo9Z+I2oSX8s2D5n2OEtDbRt2wCs0q4zxcdewqv2/P+TNPiz/2L8n/oaVXwn/5JX4N/7F+x/wDSdK/4LbfFqO20L4ffA2wvf317cTa/qUCyKCscYMFsWXG4hme5xyBmLoxAK/sJfC1/jB+1p8NvCTweZZW+sJql/ujZk+z2YNy6uQDgP5QiycDdIoyM5qv2ufhQfjd+zT8RPhlDC0t3quiySWEahyWvLci4thhMsQZoY8gAkjjB6Gv+Ccvxdh+MX7IXgXUZLhJNS8M2v/CPaigl3sktmBHGWPXc8HkSHPP7zv1NV/wUL/4JkeKW8Ta38eP2cNAbVbHVJhc6v4R0+3LXFtMwPmz2ka8yI7AMYVBZWc7QU+VP+JroGq/8venanptx/tQzW00bfgyOrD2IIrSv26v2wtHsItNs/wBovxrJFDna13qJupDkknMku525PcnAwBwAK+InxK8e/FvxRP41+JPivUPEWuXMaRSXt9LvcoihVUdgAB0AAySepJr/AIIh2d1/wsn4l332aX7N/YdrF52w7N/nk7d3TOOcV/wV/wDizH4+/alXwNp195unfD7R4bBkSRXT7ZN+/nYEDg7XgjZcnDQnociv+CNfwtfxb+0nqvxJuIN1n4C0ORo5PLY7Lu7zBGM42jMP2rqQTjgEbiK/4K7/AApPj79lC48Z2ULvf/D/AFa31IBA7M9tK32eZQq5GB5schY42rExyBkHw9rupeFtf0zxNo0/k6hpF5FeWkmPuSxOHQ/gyivhd4/0f4r/AA28L/EvQWT7B4n0m21CFFlEnlCWNWMZIx8yElWGAQVIIBGK+LX/ACVTxn/2MF//AOlD18Cf+Cgf7Rf7Ofw/h+Gnw01PQYdEgupblEvNLWeTzJCC3zE9OOlfHr9r/wDaG/aV8q1+LHxEur/SraTzLfR7SNLSyjcZwxhiAEjjLAPJuYBiAQDiv2K/2G/iL+1d4zsryfS73Rvh3YzB9W8Qz27LHOqt81tascCWZsFTg4jB3NztV/2vfiBo/wCzh+x/401fw8yaWNK8O/2N4eiilAaCeZBa23l7t24x71fBBysZzwCRX7Jfwtf4L/s1/Dr4a3EHk3mk6HC1/H5bJtu5sz3AwwDD99LJ1APqAeKr9uj4Un4M/tX/ABG8GQQulhJqzalpxIcqba7UXCKrPy4TzTGW5+aNhkkE1/wRM+LsNh4p8e/A3UbhFGrWsWuaYHk2/vYSIrhVB+8zJJC2ByBCxwRkiq/ay/5Oo+Mv/ZQNd/8AS+av+CKn/Jq/in/soF5/6QWFf8FR/wDkxL4m/wDcJ/8ATtZ1U0MNxC9vPEkscilXR1yGBGCCD1BHav29/wBhzxn+zD8QNU8UeHdBu734X6xeNLpOqQxBo9PMjEizmwxKFMhUd8CQAEfMGVfgR+23+0r+zjpsfh74ZfESWLQI5JJBouoW0d5aBn+8UWQFo/m+b92y5bJOctnWP+CtP7aup2RtbLxxoOkylgRc2fh+2aQAdRiZZFwf93PuK/4JkfGvxP8AHL9lix17xv4jute8RaNrl/p2pahduzzTP5guE3EgA4iuYlG35QqqOMED/gq3+zT4k+PPwJ0/xZ4C0eXVfE3w8vJb2OxgUtNc2MyBbpIkAJeQGOCQLkZWJwAzFQdO1HUNI1C11bSb+4sr6ymSe2ubeUxSwSoQyujKQVYEAgjkEA1ffHD9rX9p+XSfgnffEfxn44bWLqOGz0I3bOLuVcFTIBgOF27y0hIXaXJGCa+LXwy8RfBn4j698LvFs1jLrHhy6+zXj2UjSQmQKCdrMqkjnGcDmv8Aglx/yYl8Mf8AuLf+na8r/gqP/wAn2/E3/uE/+mmzr/ghj/zWz/uAf+5Gq/4KgfsO6z+0L4etPjF8KdO+1+OvC1m0F1pka4fWLFSzhY/708bMxVf41ZlGWCA+CfHPxK+BfxCtvFngzV9T8KeLvDl06LJ5XlzW0qkpJFLFICCPvK8bqQeVYHkV8Y/+Cjv7Vnxw8D3vw68WeNbGy0PU18vUIdI09LR7uPvE8gy2w4wVBAYEhsgkV+yj+yL8Uf2sPHdroPhDSLi38N2l1GviDxFJGBb6ZASN5BYgSzbfuQqdzHBO1Qzr8PvAnhv4YeBtB+HXg+y+y6L4c0+GwsYicsI40CgseNznGWbqzEk8mv2/f+Tyviz/ANjBJ/6Alf8ABEP/AJJV8Sv+xgtf/Sc1+35/yZp8Wf8AsX5P/Q0qvhP/AMkr8G/9i/Y/+k6VVVVV+1l/yav8Zf8Asn+u/wDpBNVaT+03+0noGlWWg6D+0L8S9O03TbeO2s7K08WX0MNtDGoVI40WUKiKoChQAAAAK/4ax/ao/wCjmPir/wCFnf8A/wAdr/gkV8Q/H/xL/Zs8Sa78RvHPiDxVqUHji6torzW9Tmvpo4VsbJhGryszBAzuQoOAWY9zX7WX/J1Hxl/7KBrv/pfNX/BFT/k1fxT/ANlAvP8A0gsKr/guf/zRL/uYP/cdX/BLj/k+34Y/9xb/ANNN5X7fn/JmnxZ/7F+T/wBDSvAXgzV/iP468OfDzw89uuqeKNWtdLsTcuUjE9xKsUZdgCQu5xkgHA7GvBf7Q37XX7GWs658LPC/jbW/BF5YXkialoV5aQXUUM527nWK4SSMFlVCJEHzLghipyfiv8ZPij8cvFMnjT4s+ONT8S6u67EmvJBsgTJPlwxKBHCmSTsjVVyScZJr/gk3+xR4ml8W2H7U/wATtCax0axtWfwfaXSAPfTyAp9tKE5ESRl/L3D52dZFICKWqv8Aguf/AM0S/wC5g/8AcdX/AAS4/wCT7fhj/wBxb/003lV+1l/ydR8Zf+yga7/6XzV/wRU/5NX8U/8AZQLz/wBILCv+CpP7EXiP4/6Hpvxk+EmjPqXjXwzata32lQIol1WxDFwUJYZliJchACziQqMsqq3w++JPxP8AgV42/wCEp+HvibVfCniXT/MtpJYP3cijOJIpY2BDDKjKOCMqDjIGLT/grn+2dbWkFtN4n8M3UkUao882gRB5SBguwTaoJ6naAMngAcV/wTu/bo+Pvxq/a507wj8afiVca3Ya/oN/aafpy2kVrbxXEYF0HWO3jVS4jglUPJk7WI3ZIB8ZeFNH8eeD9c8D+IImk0vxDptxp18iEBmgnjaOQDIIyVY9Qa+OfwT8d/s9/E3WfhZ8QtMe21LSpmEM4QiG/tyxEdzCx+9FIBkHqOVYBlZR8Pf2pf2ivhP4ZbwZ8OPjL4o0HQyzslhaXzCKFnJLGMHPlkkknZjJOevNeOvgb8XPD/ws0L9ob4g2ssOl+PtUnTTptRmdr3UW2+a92wYEmNyxw7Nuc5YAqQx/4Iqf8nUeKf8Asn95/wCl9hX/AAWr/wCTV/C3/ZQLP/0gv6/ZO/5Oo+DX/ZQNC/8AS+Gq+O/wa8K/tA/CbxF8I/GIdNP1+18sXEaBpLSZSHimQHjcjqrAd8YPBNftDfs4/FL9mXx7c+A/iboMtswkf+ztTjjY2mqRLj97byEAOMMuV+8hbDBTxXw1/wCCmX7Xfws8CWPw80Hx5Y32m6VD5GnzarpkV1cWsIACRrIwyypj5d+7AwudoVRqF/8AE348/EqW/vH1jxl428WX2W2RtcXN7O3QKijoAAAqgKqqAAFAA/4JwfsY3n7KnwzvNa8eW8Q+InjHy31iJJEmXTIIy3k2qSKSGPzF5GU7SxCgsI1Y/wDBb3/klXw0/wCxguv/AEnFfsB/8nlfCb/sYE/9Aeq/b9/5PK+LP/YwSf8AoCV/wRD/AOSVfEr/ALGC1/8ASc1VVVVVVVVVVVVVVVVVVVVVVVV8Lf29f2svgr4D0v4ZfDL4sf2N4b0bzvsNj/YWn3HlebK80n7yaB3bMkjtyxxnAwAAPi5+1H+0J8eEMHxX+LOu69aFt32B5Rb2m7IOfs8ISLOVBHy8YFfs6fsafH39p3VbWL4d+CruLQZJCtx4l1GJrfTrdVYK+JiMSupIzHFvfvtABI/Zg/Zk+Hn7Knwytvh34Dhe4mkYT6vq06AT6ndbQGlcDO1QBhYwcKvckszft+f8mafFn/sX5P8A0NKr4T/8kr8G/wDYv2P/AKTpX/Bbv4WxtafDj412kGHSSfw9fy+WvIINxbLuAzxtuzgkjngD5s/sCfFBvhJ+1z8N/Ect2kFjqGrJpF+0hAjEF4Ps5ZyWUBUaRJCxPGzODjaf+Co//JiXxN/7hP8A6drOvhN/yVTwZ/2MFj/6UJVf8FXf2YJfjb8Dk+J3hWxSTxT8NVnvGVVO+70xlzcxDA+Zk2JKuegSQAZeq/4Iqf8AJq/in/soF5/6QWFVXx8/a/8AjlZftS/E/wAb/DL4s+K/Dlre+JLmGG3stTlihlgt820Bkgz5bERIvDKdpPHIzV3/AMFNP25L3TW0mb483awvGIy0WiadFLgdxKtuHB4+8Gz71d3XxG+M3jprm5k8QeNfGHiG4GTiW/vr+XbgAAbnchVAAGcAADgV/wAEy/2FdY/Zt0O/+Kvxa01Lb4heIYWtYbATJMNJsNytsJXI86RlDNtZgFCDg7xVVX/Bc/8A5ol/3MH/ALjq/wCCXH/J9vwx/wC4t/6abyq/ay/5NX+Mv/ZP9d/9IJqr9k7/AJNX+DX/AGT/AEL/ANIIar/grz/yeVqX/Yv6d/6A1f8ABEP/AJKn8S/+xftf/Sg1VV/wW9/5Kn8NP+xfuv8A0oFf8Ehv+TytN/7F/Uf/AEBar4tf8lU8Z/8AYwX/AP6UPX/BIb/kzTTP+xg1H/0Na/4Kr/sOeKPitJb/ALRHwe0GXVdf02zFt4j0i0iXzru1iV2S6jG4GSRABGY1DOylNo+Qg/Cv4x/Fn4BeLH8VfCvxlqfhbW0XyJ3gAxIocMY5YnBSRdyAlHUjI5FQ/wDBXb9syOFIn8Q+F5WRQDI+gxhnIHU4IGT7ACv+CYv7aPxx+Ov7R2teD/jh8SLrxBHfeF5pdLia2itoo7iGaEkLFbxpGCYmkJdhn5MA5bB+MXw4034v/Cnxb8LtWfZbeKNHudPMveFpIyqSDg8oxVhweVHBr4pfC/xt8GfHusfDX4h6LLpmu6JcNDcQup2uP4ZI2IG+N1wyuOGUgjrXgv8Aaw/aU+HXg5fAHgb43eLdF0CNZFgsbXUXVbYP94Qt96EZJOEK4YlhgkmviZ8Avil8LvAPgf4ofEXTP7OtfiV9tn0e3uXb7ZJFb+QWuJUI+RJPtKFCTuYAsQFKlv8AgiH/AMlT+Jf/AGL9r/6UGv8Agt7/AMkq+Gn/AGMF1/6Tiv2A/wDk8r4Tf9jAn/oD1X7T/wCz94b/AGnPgtr/AMJPEU/2R9QjE2mX4j3tYXseTDMB3Ab5WUEFkZ1BXdkfGz4GfE39nvx3e/Dz4peGrjStStWYwSlGNvfwgkLPbyEASxNjhh0OVYKwZR4S/wCCo37ZPg7wXa+CtP8AiJY3kdja/ZrXUNR0qG6vIkC7UJkcHzGUYw0gckj5t1aRovxN+O/xFGn6RZ6x4w8ZeKL4u+C09xdzyN80juegycs7EKoySQBmv+Cfn7Isf7JnwbOm6+IZvHHiuSO+8RzRlWWBlUiK0Rl4ZIQz/Nk5eSQg7SoH/Bc//miX/cwf+46v+CXH/J9vwx/7i3/ppvKr9rL/AJOo+Mv/AGUDXf8A0vmr/gip/wAmr+Kf+ygXn/pBYVVVVVVVVVVVVVVVVVVVVf8ABXn/AJPK1L/sX9O/9Aavgh+0f8aP2b9V1PW/gv4z/wCEevdYt1t72X+zrW782NW3KuLiOQLzzkAGvHv/AAUE/bJ+JWiyeHvFPx51v7DNG0UsemW9tpZlRsblZrSKNmB2gYJIwWHRmB8A/DT4l/GTxOnhr4c+Dtb8Va1dSAvDYWzzsu9gPMlYcRpk/NI5CjkkgZNf8E9P+CdVp+zZHF8Wvi3Daah8TbiN47OCGQSwaDC6lGWNhw87oSHkHAVii8Fmev2sv+TqPjL/ANlA13/0vmr/AIIqf8mr+Kf+ygXn/pBYV+3b8LY/jB+yV8SfCUcHmXtvo76pp+2NWf7RZkXKKhIOC/lGMkYO2RhkZzVfstfFBvjR+zr8PPiZPdpc3utaDbtqEiEEG9jXyrnGGbAE8cowTkYwcHIHxa/5Kp4z/wCxgv8A/wBKHr/glx/yYl8Mf+4t/wCna8r41fCfw38dPhR4o+EnixP+Jb4m097V5QgdraThop1B4LxSqki543IM18UPhv4p+EHxC8QfDHxraJb634bvpLO7WMkozKeHQkAlHUq6kgZVgcDNf8Ehv+TytN/7F/Uf/QFqq+N/wZ8E/tAfDDXPhT4+sfP0vWrcosyAebZzDmK4iJ+7JG2GGeDjBBUkH9qj9iP40/spa9cR+KNFuNa8Jll+xeKtPtHNlMGxtWQ8+RLnI8tyCSCVLDmvh58V/id8JNVfW/hf8QfEHhW9l2CeXSdRktftCo25UlCECRM87HBU9xzWr/t0ftea7pV5omq/H/xXPZahbyW9zF9pVd8bqVZcqoIyCRkEGq/Zf8aWXwa/4J5+D/iD4rt3gt/CngiTULi3nbyGcRCR1jG4cM+FVRgklhgHIFa5rWqeJda1DxFrl413qWqXUt1eXDgBpZpGLu5AwMliTxxzX/BL74Wv8MP2OfCD3cHlX3i+SbxBcjy2XIuCBA3zAE5to7c5xjngkYJr/grN8KT8Of2udV8Q2kLjT/Hmm2+swnDsqzYME6bm4Lb4DIVBO0TL0BAH/BH/AOLsPgD9qCTwFqNwkdh8QtJlsULybFW8gzPCTngkqk0YXglpVwc8H/grz/yZpqX/AGMGnf8AobVoesXvh7WtP1/TWQXemXUdzbl13ASRsGXI7jIHFah/wVw/bRvbKW1tvFnhywlkXCXNvoEBkiPqokDLn6qRXizxh4++LfjCbxJ4x1/WPFXiXWJlRrm7me5uJ3OFSNc5OBwqovAGAAAAK/4Jb/sGeJ/hDP8A8ND/ABm0mXS/E1/ZvBoGhXMbR3GmQyDEk9wpI2SuuVERGUUndhm2p/wW0+LMeo+LvAHwR02+yuj2c+t6pEkisplnPlW4YYyrokU5xnlZwcdCf+CZ3wtf4pftjeCI5YPMsfCkkniC8Pls2wWoBhbgED/SGtxliBz1zgGv+CgfwpPxh/ZF+Ifh21heTUdL01tZ08IHLNNZnzyiquS7OiSRhSDzIOhwRX7Fnxdh+OH7L/w+8etcJJfvpKWOpgSb2F5a/uJiw6qXaMyBTztkXkggmv8AgqP/AMmJfE3/ALhP/p2s6qv+C1f/ACdR4W/7J/Z/+l9/X/BFT/k6jxT/ANk/vP8A0vsK/bQ/Zm039qz4Gap8OXuIrPXLWRdQ0C+kj3C3vIwdoPIwkis8bHsH3YJUCvHXgD4n/Ab4gS+FvG+h6r4S8WaDcJKiM/lSwurZjmhlQkMMrlZY2KnAKseteFP+CrH7aPhfR00eb4gaZrgiYmO51bRoJZwuB8pdAu4cE5bLcnnGANO/4Kj/ALWHiH4ieFNR8a/FBLPwvp+vWF3qem6ZpsNpHPBFMrSo7xoZ2R03Bk3kMD93pVf8FSv2afEnwc/aJ1v4mWejyt4L+It42pWd+ilkiv5BuureRsALIZRJKo/iRxgsVfb8L/jX8WfgpqV1q3wn+IWt+F7m/jEd2dOujGtwq52iRPuvt3HG4HGTjGTQ8EftG/tT6H46/aB8U6zqfiHTvAumiXV/EGuXbsoCkbLSA4OXxIXEagKoOWK7l3fsnf8AJ1Hwa/7KBoX/AKXw1+1l/wAmr/GX/sn+u/8ApBNVfsnf8mr/AAa/7J/oX/pBDU0MNxC9vPEkscilXR1yGBGCCD1BHav2/f2EvF37MfjnUfGXhDRbvUfhdrFw01hqEETSLpBdgPslyQP3eGYLG7cOpUZLBgP2ef21/wBob9mDTb/QPhZ4whj0XUpPOl0vUbRLu3SbgebGrcxuQMHaQGGNwJVSvx7/AGkPjB+0x4pg8W/F3xW2q3NjCYLG2iiWC2soycssUSAKpYgbmOWbC5J2jH/BLL9gvxbofiqz/aZ+NPhu40dLCHf4P0m8QCWd5UZGvJozzGqo37tWAZmbzMKFQv8At+f8mafFn/sX5P8A0NKr4T/8kr8G/wDYv2P/AKTpX/Bb3/kqfw0/7F+6/wDSgV/wSG/5PK03/sX9R/8AQFqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqvGH7GX7LXj/AMTaj4y8ZfBLw5q2t6tMZr29uInMk8hABZsN1wBXws+CHwm+CGn32lfCbwJpnhi01KZZruGxQqJpFG0Mck8gcVWr/sL/ALIeu6rea3qvwA8KT3uoXElxcy/ZmXfI7FmbCsAMkk4AAr4ZfCb4cfBnw7J4S+F3hGx8OaPLdPdNZ2YYRmZlVWfBJwSEXp6V4i/Yi/ZO8Wa/qfinxH8CvDOoatrN5LeX93NC5e4nlcvJI3zdWZiT7mvhf8IPhn8FfD8/hX4VeDdP8NaTdXjXk1pZIVR52REaQ5J5KxoP+AiqrUdPstW0+50rUrdLi0vIXhnhccSRsCrKfYgkV/wwH+xp/wBG8+FP+/D/APxVf8MB/saf9G8+FP8Avw//AMVX/DAf7Gn/AEbz4U/78P8A/FV8KPgD8GfgZ/an/Co/h5pPhf8Atvyf7Q+woy/aPJ3+XuyT93zZMf7xqqqqqq1f9hf9kPXdVvNb1X4AeFJ73ULiS4uZfszLvkdizNhWAGSScAAV8L/2YfgB8Fdfn8U/Cr4V6J4a1a6s2spruyjZXeBnR2jOSeC0aH6qKqq+KH7MPwA+Nev2/in4q/CvRPEurWtmtlDd30bM6QK7usYwRwGkc/VjXwv/AGYfgB8Fdfn8U/Cr4V6J4a1a6s2spruyjZXeBnR2jOSeC0aH6qK8TeGtD8ZeHdS8J+JtPS/0jWLWS0vrR2ZVuIJFKvG20glWUlSO4JB4Nf8ADAf7Gn/RvPhT/vw//wAVXhnwzoHgzw7pvhLwrpNvpmj6Pax2tjZ26bY4IY1Cqij0AAqvir8C/hD8cbTT7H4teANJ8UQ6TI8lkl/EW8hnADlSCCMhVz9B6Vof7Dv7JfhrWtP8R+HvgX4c0/VNKuo7uxvLdJElt542DxyIwbKsrKCCOhAqq+LP7NnwL+O11aX3xb+Guk+Jrmwt3t7Wa7Dh4I2OWVWVgRk85HINf8OuP2Ev+iF/+XNq3/yVWhf8E3f2I/Dvn/2f8AtJm+0bd/2/UL2+xtzjb58z7OpztxnjOcDHg74eeAPh1YNpXw+8DeH/AAxZPjdbaNpkNlGcFiMrEqjq7np1Ynuar4lfCv4e/GLw3/wh/wATfC1p4g0X7QlwbG7LeW0iAhWIUjONx4PGcHqBVp+wb+x5Y3UN7afs++FEnt5Fkib7O52spyDgtjqBVV8Tf2XP2e/jN4ij8WfFH4TaD4j1iK1W1S8vIWMghVmZUyCMgF26+tfDz9k39nH4S+KIPGnw1+EmieHtcto5Io72yV0cI67XU/Nggg9CCM4PUA1VeMP2Mv2WvH/ibUfGXjL4JeHNW1vVpjNe3txE5knkIALNhuuAK+FnwQ+E3wQ0++0r4TeBNM8MWmpTLNdw2KFRNIo2hjknkDivGHg/wz4/8M6j4N8ZaNb6tomrQmG9srgExzxkglWA7cCv+GA/2NP+jefCn/fh/wD4qtO0+y0nT7XStNt0t7SyhSG3hQYEcaAKqj2AAFTQw3EL288SSxyKVdHXIYEYIIPUEdq8cfsB/sb/ABEna68R/s/+GYpmkEjvpKy6SWYAjJ+xvFnO4k56nBOSAR4d/wCCcX7EvhaWSbTPgDo87SMjEaje3eoAFc4wtxM4A5OQAA3Gc4FeFfBvg/wJo6eHvA/hTR/D2lxsWSx0qxjtIFbABIjjAUHAA6dhVfEr9gv9kP4ua8/ijxz8D9Hn1SVmaa50+4udMadmxlpBayRiRjgfMwJ688mvhD+zN8AvgLDs+Enwq0Hw9OVKNfRwGa8dSBlWuZS0zLwPlLkZyccmvGH7GX7LXj/xNqPjLxl8EvDmra3q0xmvb24icyTyEAFmw3XAFeAvAHgz4XeErDwJ8PvDtpoegaZ5n2PT7VSI4fMkaR9oJJ5d3Y+7GviP+yj+zp8XvEz+MviX8I9B8Q63JCkL3t5EzSGNBhV4YcAV8LP2cfgb8ENQvtV+E3w00fwxd6lCsN3LYxsrTRqdwU5J4B5qq+K37N/wH+OMMifFb4UeHPEU8kPki+ubIJeRx4PypcptmjHP8LjB5rSv+CZX7DWj38WpWfwGtJJoc7Vu9b1G6jOQQcxy3DI3B7g4OCOQDXhjwl4V8EaLB4b8F+GdJ0DSbbPkWGl2UdpBFnrtjjAUZ9hVePv2Qv2Z/il4svvHfxB+DXh/Xdf1Py/td/dxMZJvLjWNMkMOiIi/QCvhR8Afgz8DP7U/4VH8PNJ8L/235P8AaH2FGX7R5O/y92Sfu+bJj/eNeIvD+i+LPD+p+FfEenQ6hpOs2ctlf2kwylxBKhSSNvZlYg/Wv+GA/wBjT/o3nwp/34f/AOKrw94f0Xwn4f0zwr4b06Gw0nRrOKysLSEYS3giQJHGvsqqoH0qqqqqvin8FfhR8btKs9E+LHgPSfE9lp9wbi1iv4d3kyFSpZSCCMg4Izg8Z6DH/DAf7Gn/AEbz4U/78P8A/FV4e8P6L4T8P6Z4V8N6dDYaTo1nFZWFpCMJbwRIEjjX2VVUD6VVeMPB/hnx/wCGdR8G+MtGt9W0TVoTDe2VwCY54yQSrAduBX/DAf7Gn/RvPhT/AL8P/wDFVp2n2Wk6fa6VptulvaWUKQ28KDAjjQBVUewAAr4m/sufs/fGfxFH4s+Kfws0fxNq8NqtpFd34d2jgVmZY1+bCqGd2wB1YnqTXwy/Zc/Z7+DPiKTxZ8LvhNoPhzWJbV7VryzhYSGFmVmTJJwCUXp6VVXn7Bv7Hl/dTXt1+z74UknuJGkkb7O43MxyTgNgcmvhd8G/hj8FNEuvDfwp8HWPhrS726N3PZ2W4RvOVVDJtJIDFUQEjqFXPQVVV8TP2fvgh8ZvKf4pfCrwz4lng/1Vzf6ejzxjjIWXAdQcDIDYOBnpV5/wTB/YWvrua8m+BEKyTyNI6w+IdUiQEnJ2olyFUc8KoAHQACvBX7CP7H3w/MLeHv2fPCMrwKVjk1W0OqOuX37t120p3AgYbOQOAQOK0/TtP0myh03SrG3s7S3XbDb28QjjjHoqrgAewrxX+xb+yx468Saj4v8AGHwT8P6vrWr3DXF7e3aSPJPI3Vid34Y6AYAwBXwr+Bfwh+B9pqFj8JfAGk+F4dVkSS9SxiK+eyAhCxJJOAzY+p9arxN4Z0Dxn4d1Lwl4p0m31PR9YtZLW+s7hN0c8LqVdGHoQTX/AAwH+xp/0bz4U/78P/8AFV4L8F+Fvh34W0/wV4K0eHStE0qMxWVlCzFIELFtq7iSBknAzgdBgCvFv/BPL9jLxxdS3/iH4E6S9zPcPcTTWl7d2TyyOSWZmgmQtkknByM1/wAOuP2Ev+iF/wDlzat/8lV4N/Yl/ZK8BS29x4b/AGffBgntFjEE99py38sZjwUcPceYwcEA787ieSTUMMNvClvbxJHHEoVERdoUAYAAHQY7V8SvhX8PfjF4b/4Q/wCJvha08QaL9oS4NjdlvLaRAQrEKRnG48HjOD1Aq0/YN/Y8sbqG9tP2ffCiT28iyRN9nc7WU5BwWx1Aqq+Jv7Ln7Pfxm8RR+LPij8JtB8R6xFarapeXkLGQQqzMqZBGQC7dfWvh5+yb+zj8JfFEHjT4a/CTRPD2uW0ckUd7ZK6OEddrqfmwQQehBGcHqAaqvEX7EX7J3izX9T8U+I/gV4Z1DVtZvJby/u5oXL3E8rl5JG+bqzMSfc18L/hB8M/gr4fn8K/Crwbp/hrSbq8a8mtLJCqPOyIjSHJPJWNB/wABFePvAHgz4peEr7wJ8QfDtprugan5f2vT7tSY5vLkWRMgEHh0RvqBX/DAf7Gn/RvPhT/vw/8A8VVXlnaahaTWF9bRXNtcxtFNDMgdJEYYZWU8EEEgg8HNeMf+Cen7F3jq5+161+z54ctpNwbbo7T6QmQoX7lnJEuMAcYxnJ6kmvDH/BOv9inwjKs2k/s/aDOyzLKBqlxc6kNy9BtuZZAV45XG09wa8OeGPDXg/R7fw94R8PaZoml2qhYLHTrRLaCIAAALGgCqMADAHaq+JX7Bf7Ifxc15/FHjn4H6PPqkrM01zp9xc6Y07NjLSC1kjEjHA+ZgT155NfCH9mb4BfAWHZ8JPhVoPh6cqUa+jgM146kDKtcylpmXgfKXIzk45NeMP2Mv2WvH/ibUfGXjL4JeHNW1vVpjNe3txE5knkIALNhuuAK8BeAPBnwu8JWHgT4feHbTQ9A0zzPsen2qkRw+ZI0j7QSTy7ux92NePv2Qv2Z/il4svvHfxB+DXh/Xdf1Py/td/dxMZJvLjWNMkMOiIi/QCvhR8Afgz8DP7U/4VH8PNJ8L/wBt+T/aH2FGX7R5O/y92Sfu+bJj/eNVXxa/Zb/Z4+OrS3HxV+EPhzXr6ZUR9Se18i+KoCFX7VCUmCgE4XfjpxwK0L/gmp+w/wCHNSj1XT/gJp808X3Uv9Vv76I/WKed0bp3U1oHh3w/4T0W08N+FdD0/RtJ0+PyrSw0+1S3gt0H8KRoAqjnoABVeMP2Mv2WvH/ibUfGXjL4JeHNW1vVpjNe3txE5knkIALNhuuAK+FnwQ+E3wQ0++0r4TeBNM8MWmpTLNdw2KFRNIo2hjknkDivGHg/wz4/8M6j4N8ZaNb6tomrQmG9srgExzxkglWA7cCv+GA/2NP+jefCn/fh/wD4qtO0+y0nT7XStNt0t7SyhSG3hQYEcaAKqj2AAFVVVVeIvD+i+LPD+p+FfEenQ6hpOs2ctlf2kwylxBKhSSNvZlYg/Wv+GA/2NP8Ao3nwp/34f/4qv+GA/wBjT/o3nwp/34f/AOKr/hgP9jT/AKN58Kf9+H/+Kr4X/CD4Z/BXw/P4V+FXg3T/AA1pN1eNeTWlkhVHnZERpDknkrGg/wCAivEX7EX7J3izX9T8U+I/gV4Z1DVtZvJby/u5oXL3E8rl5JG+bqzMSfc18L/hB8M/gr4fn8K/Crwbp/hrSbq8a8mtLJCqPOyIjSHJPJWNB/wEVXxX+APwZ+Of9l/8Lc+Hmk+Kf7E87+z/ALejN9m83Z5m3BH3vKjz/uivAX7IX7M/wu8WWHjv4ffBrw/oev6Z5n2O/tYmEkPmRtG+0lj1R3U+zGvGHg/wz4/8M6j4N8ZaNb6tomrQmG9srgExzxkglWA7cCvDP7E/7KHg7xFpvizwz8CfC9jq+j3Ud3YXaWzFreeNgySLuYgMrAMDjggEcivi1+zV8BPjrH/xdj4U+H/EVwIzEt7Pa+Xdom0rtW5j2yqMMSAGABwRyAR4B/YE/Y6+GepJq/hT4C+HzeRSCSKXVGn1UxOMYZPtckoQggEFcYPPWqqviv8AAH4M/HP+y/8Ahbnw80nxT/Ynnf2f9vRm+zebs8zbgj73lR5/3RXgL9kL9mf4XeLLDx38Pvg14f0PX9M8z7Hf2sTCSHzI2jfaSx6o7qfZjVeIv2Iv2TvFmv6n4p8R/ArwzqGrazeS3l/dzQuXuJ5XLySN83VmYk+5r4X/AAg+GfwV8Pz+FfhV4N0/w1pN1eNeTWlkhVHnZERpDknkrGg/4CKr4ofsyfs+fGi6fUfih8HvC+v6jIqo2o3Gnql4VVSqr9oTbLtAY4XdgHBxkAjT/wDgmN+wxpl7Df23wHt3lgbci3GvalPGT/tRyXLKw9iCK+G3wD+CXweQ/wDCrvhT4X8NTMu2S50/TI4riUZY4ebHmOBubAZjgHAwOKr4t/AX4OfHfR10P4ufDvR/E1vErLC93CVntwwwfKmQiSInPVGU9D2rwT/wTx/Yv+H2spr/AId+AeiS3kX3P7WubrVY1POCIruWVARnIO3IOD1Ar4nfAv4Q/GfStO0L4o+ANJ8R6fpEhksba7iOy3YrtJUKRj5QBXwv/Zh+AHwV1+fxT8KvhXonhrVrqzaymu7KNld4GdHaM5J4LRofqor4ofCD4Z/Gvw/b+Ffir4N0/wAS6Ta3i3kNpfIWRJ1R0WQYI5CyOP8AgRrw9+xF+yd4T1/TPFPhv4FeGbDVtGvIrywu4YXD288Th45F+bqrKpHuKqvGHgfwV8QtFbw34+8IaJ4l0l5Fkaw1jT4ryAuv3WMcqsuR2OOK1D/gmN+wxqd7Lf3PwHt0lmbcy2+valbxg/7McdyqqPYACvhV+z/8E/gfbywfCb4YeH/DL3EYinubKzUXE6A5CyTnMkgB5wzEZ5qvip8EPhN8b9PsdK+LHgTTPE9ppkzTWkN8hYQyMNpYYI5I4rwh+xl+y14A8Tad4y8GfBLw5pOt6TMJrK9t4nEkEgGNy5Y84JqvGH7GX7LXj/xNqPjLxl8EvDmra3q0xmvb24icyTyEAFmw3XAFfCz4IfCb4IaffaV8JvAmmeGLTUplmu4bFComkUbQxyTyBxVVVVVVVVVVVVVVVVVVVVVVVVrv/BN39iPxH5H9ofALSYvs+7Z9h1C9sc7sZ3eRMm/oMbs45xjJz/w64/YS/wCiF/8Alzat/wDJVeDv2L/2UfAV7HqXhj4A+DIbuFVWKe501buSPb0ZWm3lW9WGCe5qGGG3hS3t4kjjiUKiIu0KAMAADoMdqrxh4P8ADPj/AMM6j4N8ZaNb6tomrQmG9srgExzxkglWA7cCv+GA/wBjT/o3nwp/34f/AOKrTtPstJ0+10rTbdLe0soUht4UGBHGgCqo9gABXxE+GngL4teFp/BXxJ8Kaf4i0S5kSSSyvot6F0YMrDuCCOoI6kdCah/YI/Y6t5Unt/2fvC8csTBkdIpFZWHIIIbg14+8AeDPil4SvvAnxB8O2mu6Bqfl/a9Pu1Jjm8uRZEyAQeHRG+oFad+wl+yDpOoWuq6b8AfC9vd2UyTW8qQuGjkQhlYfN1BANVNDDcQvbzxJLHIpV0dchgRggg9QR2r/AIYD/Y0/6N58Kf8Afh//AIqvhf8ACD4Z/BXw/P4V+FXg3T/DWk3V415NaWSFUedkRGkOSeSsaD/gIqq+JH7OfwG+ME5vfiZ8IfCniK9O3N9e6XG1zhQAB54AkxgAY3YwB6VN/wAEvP2FJ5Xmf4EoGkYsQniPVUAJ9FF0AB7AYFfDP4EfBj4MxSJ8Kvhd4Z8LyTx+VPcabpscU86biwWSYDzJAGJwGYgcAYAFVVV8V/gD8Gfjn/Zf/C3Ph5pPin+xPO/s/wC3ozfZvN2eZtwR97yo8/7orwF+yF+zP8LvFlh47+H3wa8P6Hr+meZ9jv7WJhJD5kbRvtJY9Ud1PsxqvEXh/RfFnh/U/CviPTodQ0nWbOWyv7SYZS4glQpJG3sysQfrX/DAf7Gn/RvPhT/vw/8A8VXh7w/ovhPw/pnhXw3p0NhpOjWcVlYWkIwlvBEgSONfZVVQPpVfEf8AZR/Z0+L3iZ/GXxL+Eeg+IdbkhSF728iZpDGgwq8MOAK+Fn7OPwN+CGoX2q/Cb4aaP4Yu9ShWG7lsY2Vpo1O4Kck8A81VV8VP2cfgb8b9QsdV+LHw00fxPd6ZC0NpLfRszQxsdxVcEcE818OP2Uf2dPhD4mj8ZfDT4R6D4e1uKF4UvbOJlkEbjDLyx4IAqtR/YS/ZB1bULnVdS+APhe4u7yZ5p5XhfdJIxLMx+bqSSa+HHww+H/wh8Mx+Dfhp4VsfD2iRTPMllZqVjEjnLNgk8nAqvif+yf8As2/GW6uNR+JPwX8L6xqN2rLPqP2IW95ICu35riLbKSB0O7KnkYPNaV/wTK/Ya0e/i1Kz+A1pJNDnat3reo3UZyCDmOW4ZG4PcHBwRyAa+HPwS+D/AMILdrf4XfDLwz4X3x+XLLpmmRQSzLkHEkijfJyB94noPSq+MX7OPwN/aAsobL4wfDTR/EhtVKW9zPG0V1ApzlY7iIrKiknJVWAJwcZAr4f/ALAH7HHwx1ca74T+A2gm+VkeOTVZbjVhEyElWjW7klWNgTncoByAc8DHxX+APwZ+Of8AZf8Awtz4eaT4p/sTzv7P+3ozfZvN2eZtwR97yo8/7or4Wfs4/A34Iahfar8Jvhpo/hi71KFYbuWxjZWmjU7gpyTwDzXxU+CHwm+N+n2OlfFjwJpnie00yZprSG+QsIZGG0sMEckcV4Q/Yy/Za8AeJtO8ZeDPgl4c0nW9JmE1le28TiSCQDG5csecE1VeOfhv8Pfifo48P/EfwPoPijTgxZLXV9Piu40bGNyiRTtbH8QwR61ef8Ewf2Fr67mvJvgRCsk8jSOsPiHVIkBJydqJchVHPCqAB0AAr4Z/BP4RfBmwk034VfDjw/4Xin/17abYJFJPySPMkA3yYJONxOBgDAAFV8V/gD8Gfjn/AGX/AMLc+Hmk+Kf7E87+z/t6M32bzdnmbcEfe8qPP+6K8Bfshfsz/C7xZYeO/h98GvD+h6/pnmfY7+1iYSQ+ZG0b7SWPVHdT7MarxF+xF+yd4s1/U/FPiP4FeGdQ1bWbyW8v7uaFy9xPK5eSRvm6szEn3NfC/wCEHwz+Cvh+fwr8KvBun+GtJurxrya0skKo87IiNIck8lY0H/ARVVVVVVVVVVVVVVVVVVVV8V/2Q/2bfjjrNz4j+Kfwm0nXtWu7dLeW/kkmhn8tPuqJInVlx6gg1/w64/YS/wCiF/8Alzat/wDJVeGf+CeH7FnhIRjSv2ffD8/l3AnX+05bjUsuMcH7TJJlPlHyH5Tzxyc+GfCXhXwVpUeg+DfDOk6DpsWPLstMso7WFMKFGEjAUYVVXp0UDtVV4i/Yi/ZO8Wa/qfinxH8CvDOoatrN5LeX93NC5e4nlcvJI3zdWZiT7mvhf8IPhn8FfD8/hX4VeDdP8NaTdXjXk1pZIVR52REaQ5J5KxoP+Aiqm/YH/Y2nleZ/2ePCIZ2LEJbMgBPooYAD2AwK+Hnw18C/CbwtB4K+HHhq00HQ7WSSSGwtQwjjZ23OVBJxliSfcn1rUf2Ev2QdW1C51XUvgD4XuLu8meaeV4X3SSMSzMfm6kkmvAXgDwZ8LvCVh4E+H3h200PQNM8z7Hp9qpEcPmSNI+0Ek8u7sfdjVfET9kz9m74teKJ/GvxH+D3h/X9duY0jmvruFjJIqKFQEgjOFAH0Ar4cfso/s6fCHxNH4y+Gnwj0Hw9rcULwpe2cTLII3GGXljwQBVVU0MNxC9vPEkscilXR1yGBGCCD1BHavHH7CX7IPxEuLy88T/AHwubnUGD3M+nwvpsjtu3Ft1s0ZDE8swILZOScmrT/AIJg/sLWV3BeQfAiFpLeRZEWbxDqkqEqcjcj3JVhxyrAg9CCK+H/AOzl8A/hU8M/w6+Dfg/QbqDOy9s9HhW5+8W5nKmRsEnGWOBwMAAV408F+FviL4W1DwV400eHVdE1WMRXtlMzBJ0DBtrbSCRlRkZweh4Jr/hgP9jT/o3nwp/34f8A+KrSdJ03QdKstC0ayis7DTreO2tLaFdqQxIoVEUdgFAAHtVfFb9nv4KfHKXTZ/i18N9H8US6Osq2L30RZoBJtLhSCODsXg+n1rwp+xb+yx4G8Sab4v8AB/wT8P6RrWkXC3Fje2qSJJBIvRlIb8COhBIOQa+I/wAMPh/8XvDL+DfiX4VsfEOiSTJM9leKTGZEOVbAI5Fah/wTG/YY1O9lv7n4D26SzNuZbfXtSt4wf9mOO5VVHsABWlf8Eyv2GtHv4tSs/gNaSTQ52rd63qN1Gcgg5jluGRuD3BwcEcgGvh38B/gp8I/n+GPwo8KeGZzGI3udN0mGCeVRnAeULvf7x+8x6mq+In7Jv7OPxb8UT+NPiT8JNE8Ra5cxpFJe3yu7lEUKqj5sAADoABkk9STXwt/Zr+BHwS1W81z4UfC/RPDOoX9v9mubmyiKvJFuDbCSTxuAOPUD0qpoYbiF7eeJJY5FKujrkMCMEEHqCO1f8MB/saf9G8+FP+/D/wDxVfDX4V/D34PeG/8AhD/hl4WtPD+i/aHuBY2pby1kcAMyhidudoyBgZyepNV4+8AeDPil4SvvAnxB8O2mu6Bqfl/a9Pu1Jjm8uRZEyAQeHRG+oFf8MB/saf8ARvPhT/vw/wD8VVfFD9mH4AfGvX7fxT8VfhXoniXVrWzWyhu76NmdIFd3WMYI4DSOfqxr4X/sw/AD4K6/P4p+FXwr0Tw1q11ZtZTXdlGyu8DOjtGck8Fo0P1UVXxD+FHwx+LelJonxP8Ah/4f8VWUO8wRatp0d19nZ12s8RcExvjjchDDsa1D/gmN+wxqd7Lf3PwHt0lmbcy2+valbxg/7McdyqqPYACvAf7H/wCy98Mr+LVvBXwJ8H2OoQbfIvZNNW5nhKgqDHLNvdCQxBKkFu+cVXi3wb4S8faBdeFfG/hrTNe0e9XbcWOo2qXEMg90cEZHY9R2qL/gmf8AsOQ6udbT4C2JuTM02x9Y1BoNxJJHkG4Me3nhdu0cYAxVx8I/hfcfD+6+FK+ANBtvB17C0M+h2dilraNGTkr5UQVQCQDwBXh79iL9k7wnr+meKfDfwK8M2GraNeRXlhdwwuHt54nDxyL83VWVSPcV4i8P6L4s8P6n4V8R6dDqGk6zZy2V/aTDKXEEqFJI29mViD9a/wCGA/2NP+jefCn/AH4f/wCKrw94f0Xwn4f0zwr4b06Gw0nRrOKysLSEYS3giQJHGvsqqoH0qryztNQtJrC+torm2uY2imhmQOkiMMMrKeCCCQQeDmvGP/BOv9inx1qn9sa1+z9oNtPtC7NHuLnSIcAAf6mzlijzwOduep7mvhZ+xb+yx8FtSt9a+HPwT8P2GqWknmWuoXSSahc27/3o5rlpHjPupHHHSq8YeD/DPj/wzqPg3xlo1vq2iatCYb2yuATHPGSCVYDtwK/4YD/Y0/6N58Kf9+H/APiq07T7LSdPtdK023S3tLKFIbeFBgRxoAqqPYAAV8VP2cfgb8b9QsdV+LHw00fxPd6ZC0NpLfRszQxsdxVcEcE818OP2Uf2dPhD4mj8ZfDT4R6D4e1uKF4UvbOJlkEbjDLyx4IAqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqvif8AtnfC74TeOdT+H/iPQfFNzqOleV50tlawPE3mRJKu0vMrfdkAOVHOfrXwk+K3h34z+DovG/hez1G1sZbiSBY7+NElDIcE4R3GPxqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq+Ln7Xvw0+DPjGTwR4o0TxNdX0VvHOZLC2geLa4yMF5kOeOeK+DHxn8LfHTwtdeLfCNhqlpZ2l+9i8eoxRxyGRY43JAR3G3Eq985B49aqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqtW1GDR9LvNWuVdobG3knkWMAsVRSxwCQM4HtX/AA8S+Cf/AELHjf8A8ArX/wCSKqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqvFfiKx8H+FtY8W6nFPLZ6Jp899cRwKGkaOGMuwQEgFiFOASBnuK0n/goD8GtZ1Sy0i18NeM1mvrhII2ksrYKGdgoJxcE4yfSqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv2uv2ifG3wC/4RP/AIQ7S9DvP7d+2/af7Tglk2eT5G3Z5ciYz5rZznoOnOf2TP2mvHvx58Q69pHi/SdAs4dKso54W023mjZmZ9p3eZK4Ix6AVVTTQ28LzzypHHEpZ3dtoUAZJJPQYrwT478F/Erw5beL/h/4p0zxDol2zrBf6dcrPDIyMVcBlJGQykEe1VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVftMfte/Er4M/FGfwR4X0TwzdWMVlBOsl/bTvLucEnlJkGOPSv2U/jP4o+Onw71Dxb4tsNKtLy01qWxjj06KSOMxrDC4JDu53ZlbvjAHHrVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVftp/wDJzHjL/tx/9Ibev2Df+TfLP/sKXf8A6EKqvG3/AAUI/alb4jeNdY8CfHfxLZ6DrmuX1xp9oZ/PitbaSU+VHAs6loUWMIFChSvJwCTn/gkP+0N4t+MPwp8YeDfiL401XxL4j8K6xHcJe6vqM13cvZ3UfyKXlyWCyQzfxHAcAheC1VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVft5/wDJwV3/ANgq0/8AQTX/AATt/wCSJ61/2NNx/wCktrVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVXjn/AJEnxB/2Crn/ANFNVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVXxz/AOSJ/EH/ALFXU/8A0lkrwL/yO/h7/sK23/o1aqv+Cvn7RXxA+Dnh/wCGfhb4W+O9V8NazquqXWqXNxpOoNbzeTaoiJHKEILRO9wx2MCjGHkHbx+w7+278fPFX7X/AMO9M+L/AMZ/EetaBrM0ujzWFzehLaSSaB47YmIFI2fz/J+YgucnGS2DVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVf8ABSr/AJpz/wBxT/20r/gnD/yOvjD/ALBUP/o01Vftv/tk/Ha6/aV+MPg3wd8V/EuleEDqE3h6XRIr1/swjtozaThI2yI/MdZmYpgsXyTwMfs8/tQ/Gb9l/wAUnxN8J/FL2sdwyf2jpF0DNYakqnIWeHIycZAdSsihmCsu41+yJ/wUT+DH7U8Nr4YnnTwf8QGVt/h3ULgEXZUZLWcxAE4287MLINrnaVXeaqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr9vP/k4K7/7BVp/6Ca/4J2/8kT1r/sabj/0ltaqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr/xP4a0q4NnqniLTLOcKCYri7SNgD0OCQcVpuuaJrXmf2NrNjf8Ak48z7NcLLsznGdpOM4P5Gqqqqqqqqqqqqr+9tdNsbjUb2Xy7e1iaWZ9pO1FGWOBkngHgV8OPjt8Kfi7e3mm/DzxX/a1xp8Sy3CfYbiDYhOAcyxqDz2Gaqv2q/gJ8YvGvx88UeJvCvw+1TUtLvPsn2e6hRSkmy0hRsZPZlYfhX7HfgzxT4D+Ctr4e8YaJcaVqKajcyNbXAAYKzDaeCeDVfti/EOH4VfsufE7xu8iLLaeHbm3tS4yBc3A+zwZHcebNHx36VX/BIz4oN4D/AGtrHwnc3aRWHjzSbrS3EhAQTxr9ohbJYYYmBo1+9ky7cZYEVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV+2L8D/iz49+NNz4g8HeBNT1XTm062jW4gQFSyqdw5Pav2IPAPjH4dfCfVdE8b+H7rR76bxFNcRwXKgM0Rt7dQwwTxlGH4VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV4utbi+8J61Y2cTSz3GnXEcUajlmaNgAPqSK/wCGXP2gv+iU63/37X/4qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr4u6VqGu/CfxromkWj3V9qHh2/t7WCMfNLK9u6oo9ySB+NeEP2Zvj1Y+LNEvrz4XazFBb6jbySyNGuEVZFJJ56ACqr/gr78RE8Z/tcz+F7W6d7fwRoNnprRiQmMTyBrqRgM4DbbiNGI/55gH7teDfE+oeCfF+heM9JdlvtA1K3v7ZkkMbCWGRZFIZeVOVHI5FaHq9l4h0TT9f01na01K1jubcuu1jHIoZcjscEcVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX7U/7OGt/tB/8Ix/Y3iWx0n+wPtfmfaYXfzfO8nGNvTHlH8xX7L37L2vfAHXtb1fV/FNhqqarZxwIltA6FCr7snd2qvHHivTfAXgrX/HOstiw8OaXc6jdHO3EUETSPz2+VDWr6rf67qt7req3Hn3uoXElxcy7Au+R2LM2FAAySTgACvjn/wR78OeJPhh4a1D4LX9v4f8f6NoNrb6xZ3UrNZa5cxQKsku7J8iZ3U5ZR5bE5KqSznx/wDDr4hfBzxhceDviF4Z1Pwz4h01ld7W6QxyL3WRGHDKcZDqSD1Br9jX/grV4p8DNa/D39qG8vfEvh5VWO08UIhm1GywAALkdbpOAd/+tBLEmTIC+DfG3hD4ieG7Lxf4F8Safrui6hGJLa9sZxLHICPUdCOhU4IPBANVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVftE/sdeJfjX8SZvHOl+MtM02CWzhgEFxbu7AoCCcrxg5r9mX4J6p8BvAV/wCD9W1y11Wa81eS+Wa3iZFVWhhj24bnOYifxFVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV+0+7x/s/eOmjdlP8AZMgyDjgkAj6EEiv2Xf2XdD+P2h65rOs+K77S/wCyrqO3iitrdX3bkLFiWP0GMevrx40/4J36rpOnf2v8MPiE93qtn+8itL+AW5kZeR5cyH5H4GMrjOMso5r9jf8AaM8fap44l+CPxS1C61K62TjT7q+bddQzw5aSCVj80g2rIQWJZSuOQRtqqqqqqqqqqqvHP/Ik+IP+wVc/+imr/gnD/wAjr4w/7BUP/o01VVVf8FmfiG/hn9mXR/Alne+XceMvEkMc8O5h5tpbI0z9CM4mFqcMCO+AQCP2ZvgDq/7RXjPxB4T0ct5uieEdW1vaqFi8lvbkQJwRgNcSQKTzgN0PSvhl461L4X/Ebwv8SNHj8y98Laxa6nBEX2CRoJVkCE4OA23aeDwTwa0PWtK8S6Jp/iLQ71LvTtUtY7qzuEBAmhkUOjgHBwVIPPrVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVftEfEFviv8d/H/AMRhMkkOv+Ir26tShDBbcysIVBCruAjCLuwCcZPJNa7oOpeG9QGlatD5Nz9ngnMZ6qs0SSoCOx2uuR2ORX/BOf4gt8R/2MvhnqU8yNdaPpraLOikExiyka3iBwqjJhjibpnDDJY5Jqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr/gp58QP+Ff8A7GHjnyGxeeJPs+i22Tx+/mXzc/Mp/wBQk2MZ5xkFc1+xr8P3+KP7VHwu8GC2+0Q3HiS2ubuLLDfbWzfaJxlSCMxQycggjrVfHz9mv4PftL+E28J/FfwpDqCR82eoQ4ivLF/70MwGV91OVb+JTX7YX/BOT4ufsuG68X6SX8YfD5GLf23aw7ZdPUuFRbuLnYfmUeYuUJPVSQtfs3/tYfGb9lrxMuufDLxI66fcTLJqWhXZMljqIAxiSPIw2OA6lXHrjiv2SP8AgoJ8GP2qbK30OG8Twt47SENdeHdQmAMrdCbWU4E698ABwOq45NVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV+1F/yb745/wCwU/8A6Etf8E4f+RI8Yf8AYVh/9FVXw4jsfiJ+3bNrXg0Ry6VHr95ftcwoDG0UavulBXgrI+MN3Mik9TWpajYaPp11q+q3kVrZWMDz3M8zbUijRSzOxPQAAkn2rWf229b8W+Jrjwp+z58J7/xg1uu438yyKrKCVL+Sq7kjyUw7sh5wVBwat/21Pir8PdYtdL+P3wVl0m3uZApvLGKW3wvBLRpKXWbAIyBIO/cYrwh4u8PePPDOn+L/AApqUd9pWpxeZbToCMjJBBB5DBgVKnkEEHpXwx/a48TeJ/jo3wW8ceEdK0WaO6u7I3ENw7E3EIbCjdxhthAPclcZyKrx5+3B/wAIh8cJvhtb+HtMl0DT9WgsL7VpLpg8YyguH2j5R5ZLrgnqnOM4FftM/tX6j8DfFukeDvDnhmx1q9vbL7VcLPM6mMM5SNQFHU7H756cDjPxJ+JsPwj+GNx8QvF2k3N22nwQC7ttOUH99IypgFmACeYwGSSQD0J4Mv7Yv7THiqGPVfhz+zxLJpjk7ZX0q+1IMCFK4ki8tehz05DDp3+Ff7eJ1PxPbeCfjH4NTw5fTXH2d7+3MiRQSnACywSAvGN3BbccZGQACarV/wBqefQf2nJPgdrOkaZZ6HCVEusTTsrx7rEXALD7oG8hSTwBySMGviR/wUR1qHXprP4U+FNKl0qByi3msxyu9zgkb1jR02KRggMSfUDoPGXjHw34A8N3vi3xbqsWn6Zp8e+aaQ/gFUDlmJwAo5JIArVP27/ij4016XS/gd8H/wC0YYNzBbq1n1C5kjBOHaK2ZRFxjI3OAc/Ma0b9svV7q01T4f8Ax3+H1z4N1i/06aO1umgltoWd4iFEkU/zRg5Hz7mHIyB1P/BOH/kdfGH/AGCof/Rpqq8a/tPfAz4d+Jrzwd4x8c/2frGn7PtNt/Zl3Ls3xrIvzRxMpyjqeCevrmvAfxC8H/E/w8nirwNrH9paXJK8Sz/Z5Icuhww2yKrcfSq/4LWfEb+3vjj4L+GVvLvg8JeH3u5PlxsuL2X515UE/ura3bO5h82MAhs/8EQPh5G918T/AIsXdn88UdlolhcbV6MXnuUzjI+7aHAIB7g4Uj9qr4VD4I/tGfEL4YRW3kWei65P/Z0ZbcVspSJrXJwOfIkizgYznHFf8EzvijH8Uv2OfA7y3HmX3hSN/D14N6tsNqQIV4JI/wBGa3OGAPPTGCaqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqvHv7SHwW+GPiB/Cvjnxp/ZuqRxJK1v/Z11NhGHyndHGy846Zr4dfE/wL8WdEm8R/D/AF3+1dOtrprSWb7NLBtmVEcrtlVW+7IhzjHPXg1VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVf3trptjcajey+Xb2sTSzPtJ2ooyxwMk8A8Cv+G0v2Zv+il/+Ua+/wDjNVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVWu61pfhrRNQ8R61dfZ9O0q0lurubYz+XFGhd22qCxwqk4AJPYVY/ti/s4anfW+m2PxH825upVihT+yL1dzsQFGTDgZJHXiq/aw+I0Pwl/Zq+JPj950im0zw7dLZs7bR9rlTybcE7lODNJGMAhjn5cnArwF4SvfH/AI68OeBNNZ1u/EerWum25SPzGEk8qxrhcjccsOMjPqK/4K5/CnTvhv8AtO6fquhWTwab4m8K2EseS7BZLVTZ+WGZmLFYba3JPH3hnJyx/wCCI/xAGpfC74i/C6Zvn0DXLfVYSx5KXkJiZV+Y8K1nkgKADJnJ3YFVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVftK/tK/8ADO//AAjn/FF/8JB/wkH2r/mJfZPI8nyv+mT7t3m+2NvfPH7OH7U//DQet6xo3/CCf2D/AGTapP5n9qfavN3Ntxjykx9cmq/4LffEFIPDXwy+FNvM7Ne313rV3GGICCFFhhJG3DE+fcDhsrtOR8wNf8EZfhzD4n/aU1rx/eQI8Pgvw7K1sxXLJd3TrChBKkAeT9pBwVbkdQWFVU0MNxC9vPEkscilXR1yGBGCCD1BHav2w/8Agkh4O8fJffED9mRLHwp4i2vLP4Yk/dabfvndi3P/AC6OQWAX/Uk7BiIbmPizwf8AEX4L+O5vDni3RtY8JeLPD10r+VMGt7i2lQho5Y3HbgMkiEgjaykgg1+x7/wV41fw4lh8PP2qjcavpcapDa+MLWEyXduAcD7ZEozOoUjMqDzfk5WVnLDwr4r8MeOvD1j4s8GeIdP1zRdSj8yzv7C4WeGdckEq6kg4IIPoQQeQaqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq+PX7Zn/CkPiDN4E/4Vv8A215VpFP9q/tj7NneCcbPJfpjrmv2dvjf/wAL+8E3vjH/AIRj+wvseqyWH2b7b9q3bIopN+7YmM+bjGP4c554qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr9qL/k33xz/2Cn/9CWv2cPFv7SfhzQ9Yg+BvhgapYzXSNfP9iWfZKEwoySMfLXxq+Mf7WD6dF4c+LKax4W0XUHEcxtNKFutyOrASjG8hc5jEgB7+tfsg/Db4R+Ffh3D4r+G+r/2/eaygW/1a4hEU6suCbcxgt5IU4JTJycNuYbTX/BQ74hahofgvw/8AD7S73yh4iuJZ9QWN8O0EGzajDOdjPID05MXXgg/s+fE79mz4Q/CzRfDEHxJ8Pw6lLbpc6vIJGLS3bqDJk7ckKfkHT5VHvXxp+MP7NXxP+GPiDwjqHxJ0C5luLKV7FiC7Q3SoTFIgwPmDY6EZBIzgmv8AgnJ4yvrrTvF3gO8u3kt7J7e+sY2yRHv3pNjnABKxHAHUsa/azsZvg/8AtU6J8UtPRlgv5LPVisWBmSBljmj2qVJDLGC2SN3mtz1x418Y6d4P8B6z47eaKe00rS5r5Csg2zhIyyhTkA7yABzyWHrUXwz1DxL+zp4p+OmpK8+pf8JZCstxImC8JVhK4+UcPPcxAkEjMeMDFfs4+OT8Rfgn4T8ST3a3F59gW2vWyNxnhJicsB0LFN2OOGBHBFaVbf8ADQf7dVzdsv2nR9B1JpnzFuQW9gAiZGT8kkypnPB83pztq9sLHU7V7HUbOC6t5Mb4Z4w6NggjKnIPIB/CvGP7SnwK8B3baf4l+JWlR3Ub7JILXfePE3PDrArlD8pyGxjjPUZ/bE+L3we+M9x4f8QfD9rw61YiW3vpJ7Iw+dAdpjySedrb8f759q/Z91y88SfBHwRq9+265l0W3SRiclyihNx9ztyfc1+1RoeoeKP2v9c8MaTJtvdZvdMsYDu2/PNaW0YBORwd2D7V8Nvhj4N+FHhm28L+DdGtrOGKNRcTpEBLdyAcySt1djz1PAOBgAAf8FDvG+qap4u8M/CjTJGmtra2GoTW8HztLcys8UalRk7lRWwMZxN3yMfB34VeHvg94D07wfoVlBHLFEr6hcRglru5KgSSsx5OSOM9BgAADFftE/DPQfij8KNe0fVrWD7VZ2Ut1p128QZ7WeNd4ZT1AbbtbHVSRX/BOH/kdfGH/YKh/wDRpqq/bT/5OY8Zf9uP/pDb1+wb/wAm+Wf/AGFLv/0IVX7eHxDj+KH7XnxQ8T2979qtYdck021kVlKNFZqtqrIVJBRvJ3Ag/Nu3cEkV/wAEpfhz/wAIB+xp4b1CaPy7vxlqF5rlwmc43uIIjkMw5gtoW4243YI3Ak/8FpfhW3hz44eFfixZ27raeMtFNrcuEGDd2bBSSQc5MMsAwQPuHBPIX/giL8Tnh1/4jfBe5k3LdWcGv2SZb5DE4t7g9Mc+dbdSD8vAbJK1VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVft5/8nBXf/YKtP/QTX/BO3/kietf9jTcf+ktrVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVXjn/kSfEH/AGCrn/0U1VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVfHP/kifxB/7FXU//SWSvAv/ACO/h7/sK23/AKNWq/4LIfEB/C/7K1n4MtbnbP4z8SW1tPFhvntoFe4c5AxxLFb8EgndkZwcf8EwPhzL8Rf2zPBTtA0ll4WW41y8KqSUEEZETfdYAfaJLcEnHB4IYrX/AAW5+Hw1L4XfDr4owr8+ga5caVMFHJS8hEis3ynhWs8AlgAZMYO7I/4I8/EB/Cn7Wg8HyXO2Dxr4fvLJYSGIeaAC6VhgHBCQTDnAwx7kCqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/4KVf805/7in/tpX/BOH/kdfGH/YKh/wDRpqv+CtXj6Pxt+2VrekQSxSw+DdHsdGjkiZWDHYbpxkd1kunQg8gqQelf8EVfh62hfAjxj8RriFEm8VeIltYSFBZ7eziAViQx48y4nXaQCNpPIYYqqr9ob9l34MftQ+Fh4Y+K/hZLuW2V/wCztWtSIb/TWYYLQTYOB0JRg0bFVLK20Y/a4/4J9fGb9la9uNcltH8U+BGmK2viKwgJEK9QLqIZMDdsklCejZ4H7NH7Xvxr/ZU8QnVPhp4i8zSbqQvqPh7UN0thfEgAs0YIKSYVcSoVf5QCSuVP7J37dPwY/ax0eC28P6gmgeNEhL33hW/uA1xFtA3vC+FFxEM/fUAgcsqZxVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV+3n/ycFd/9gq0/wDQTX/BO3/kietf9jTcf+ktrVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVftRf8AJvvjn/sFP/6Etf8ABOH/AJEjxh/2FYf/AEVXxK8CaP8AEzwLrPgfW4I5LfVLR4kdlyYZcfu5V64ZHCsPcV/wT38X6pofxB8S/C3U3aKC8s2u1gkb/V3UDqjqB6lHOTn/AJZDrX/BSLQrhdQ8EeJkZmgkgu7NxswI2UxuvOedwZuMcbD68fDD9ln9mHx58OfDXjCH4erOdW0uCeVhrN7xKUHmKds2AVcMpA4BBr/hiz9mb/omn/lZvv8A49Xw4+BHwp+Ed7eaj8PPCv8AZVxfxLFcP9uuJ96KcgYlkYDn0xX/AAUF8Dx698I7DxnFGTc+FtRXc2OBBcYjf85BB+Xvx8WfjbNq37E3gfRlv2fU/EEg0y93OC7RWLYckk7iWKWxJHUPyecHwR8FLLTv2bbX4MajEsMl7oLwXzLk+XdTqXkcE8nbK5IyP4Rxjiv2Q/ixdfDv4VfFjw9qTvDe+FrWXVrKFyCRNsMTJjacfvUgHOVy5OByT/wTu8CNb+HPEvxO1BN9zq10thayOSX8uMb5W9wzuoyecxHp3/b3+MWteCfCmlfD3wxqEtnd+JxM+oTRZVxaJhTGG7eYzEHBzhCDw3PwC/Yz+FmieC9J8QePtEi8S6/qNtHdS/amY29tvUMIkiB2sACAWcNkgkYBxX7ffgL4feDPCXhSXwh4L8PaFc3WozLK2m6bDavKixjgmNQWAJHXgEiv2Xf+TffAv/YKX/0Jq+MP/KQSw/7Gnw9/6Ls6r9uCC68LftI6P4rvLdpbSWxsruLYCN4hlZXTJGN2UzgZwGUnrVjfWep2NvqWn3Mdxa3cSywSxnKyIwBVgfQgg18StRsdI+HXijU9RuFgtrXRruSWRgTtURNngcn6Dk1/wTh/5HXxh/2Cof8A0aaqv20/+TmPGX/bj/6Q29fsG/8AJvln/wBhS7/9CFfE7xnb/Df4a+LPiJdRebD4W0O91SSPBO5beB5SMDnkJX/E11/Vf+XvUtT1G4/2pprmaRvxZ3Zj7kk18J/A9v8ADL4XeEPhzapEsfhjQ7PTR5TFlYwQrGWBIBOSpOSATnJ5Nf8ABXP4XJ48/ZIvfFltavLf+A9WttTjMYJcwSN9mmGApyoEyyN93Ai3ZwpB/YA+KMfwj/a7+HHiS7uPLsNQ1QaPfFnVEEV6ptw7sxACI8iSE548vPPSqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv28/+Tgrv/sFWn/oJr/gnb/yRPWv+xpuP/SW1qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqrxz/yJPiD/sFXP/opqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq+Of/JE/iD/2Kup/+ksleBf+R38Pf9hW2/8ARq1X/BbD4gR618avA3w2t7nzY/C/h+W9lQBcRTXk2GXON2fLtYTgkjDLjBJz/wAEQPh9HPr/AMT/AIqXNth7OzstFspsqdwld5rhcZyMeRbHkYO7g8HH/BRT4fx/Eb9jT4maatt5txo+l/2zbsCoMRsnW4dhuIH+qjkU98McAnFfs3fEOH4TfH74e/Ea6kRbTQfEVncXhccC281RN9D5ZfB7HBqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv+ClX/NOf+4p/7aV/wTh/5HXxh/2Cof8A0aammht4XnnlSOOJSzu7bQoAySSegxXxo8e3HxU+L3jT4kXNw8zeJdevNQQtuG2OWZmRAGJKqqFVCknaAB2r9hfwA/wz/ZD+FXhSaOWOc+H4tQnimRleKW9ZruRGVuQVe4ZSD0IxVVVVeWdpqFpNYX1tFc21zG0U0MyB0kRhhlZTwQQSCDwc1+2D/wAEjPDfjEXvj79l5bTw/rTYebwpO/l2Nyf4jbuf+Pdz/cP7s9jGK17w/wDEP4O+NpNI17Ttb8I+KtCuMlJA9pc2sinh0YYI6ZDKcEcgkc1+x7/wV4vbFrD4eftVFru3LJDa+MLWH95CuMD7ZEo+ccDMqDd6q3LV4a8TeHfGegWHinwlrljrGj6nCJrO+sp1mhnjPRkdSQRVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX7ef/ACcFd/8AYKtP/QTX/BO3/kietf8AY03H/pLa1VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVXi3/go9+xf4G8Vaz4J8U/Gb7FrXh/UJ9P1G1/4R3U5PIuYJGjlTelsVbDqwypIOMgkc18FPj38Jv2ivCt142+Dfiz/hINFsdQfT57n7BcWmy5SOORk2TxoxwksZyBj5sZyDiqqqqqqqqqqvit8ZPhb8DvC0njT4s+ONM8NaQjbEmvJDvnfBPlwxKDJM+ATsjVmwCcYBr42f8ABbCK2up9L/Z5+FsV3HFJtTV/FbMElAIyVtYHVgD82C0oPKkqDla8Y/8ABTP9tXxldedJ8ZrjR4FYNHa6Pp1taIh2gH5lj8xgcZw7sAScYGAJv2tv2qp5Xmf9pf4phpGLEJ4wvkAJ9FEoAHsBgV4S/wCClf7avhC9N1b/ABtvdUjdlMttq9hbXkbhc4GXjLIPmOdjKTxk8Cvgr/wWw1VLqPTP2g/hbaS2zbV/tXwqzRvHzgl7ad2D5BySsi4I4U5+X4N/tA/Bz9oHQB4i+Efj7TPEECKDcQRSFLm0J7TQPiSM/wC8oB7Ejmqqqqqqqqqqqqqqqqqqqqqqqqqr9pi0ur/4C+NrOxtpbieXS2WOKJC7Mdy8ADk1/wAE89F1jRvBfi2LWNKvLF5NUhKLcwNEWHldQGAyKq88N638E/22jr9loeonQJdc897mGydoY7a9T979xcbI/OcY7eX7Zr44fCPSPjb8O77wNqlx9lld1nsLwR7zaXKZ2uB3BBZGHdXbBBwR4T8bftLfsb3Fz4X1/wAHS6v4WSQyhZY5JLNc8l7e5QYjycEqwPUkqCc1H/wUR8Ra4ktl4X+Bjz3ypvULq73QRQQCSiW6kjnGcjBI+h+BXi7x/wCN/h3aa58TvCr+H/EDXEyT2bWslsNm8mNljkyyjYVGCTypOecD4jeEYvHvgHxD4LlZF/trTZ7VHfpG7oQj8Z+621unav2fPhp4+8YfFjwZ4H8WaLrFvoPhvVZ9RkS6hdIoCoR5EG7jEjwRKQvJ3E+4r9q34c+MPBXxr8YJ4T0PUbjSfG9pHdTNb2rTB1kmSaRSVBwftNuWwecY9cn4GeBG+Gnwi8K+CpU2XGn6epulySBcSEyzAZ5x5jvjpx2HSv23/gR4n+Kvh7RvFXgfTvt+q+HfOSeyjH725gk2H92P4mQqTt6kM2MkAH4cftzfEHwN4csPh14m+GMviDWdMgS1tpWu3tbhkVR5Yki8py7BdvI2lgATkksfj74J/aG+JPhWH40fE7QLyzmkvobHSvDtpayM1pbOkzySvFlmh+aOJct8zFhnGFB/ZfWRPgD4Kgmt54JYNPMUkU8LROjpI6sCrAEcg9sHqMgg18WvDXiK5/b0sdXttA1GWxXxPoDm6S0dogqx2m47wMYGDk54war9q79n6X46+Crc6A8EXibQXebTjL8q3CsPngLfw7sKQx4DKM4BJHgD9qX4zfs2abD8N/ib8OLi/tNNUwafFfM1jNGiEgqkux1ljGRggEYxhiCMXfij4+ftmix8MWHg6fwL8OrmVH1W9eQyG6iG1sLI6IZeRlVRduWG84Ga/wCCe/hvxFoPjXxf/bmgajp2dNiQfa7V4csJTkfMByPSqr9tP/k5jxl/24/+kNvX7Bv/ACb5Z/8AYUu//QhX/BVT4gR+BP2MPFdlHc+TeeLLyz0W0bCnJeUSyrgg9YIJx6jOQQRX7C/gCP4m/tefCrwpNHFLCPEEWoTxTIrJLFZK13IjK2QQyW7KQeucVXxN8C6b8UPhx4o+G2sSeXZeKNHutMnk2bzEs8TRlwMjld24cjkDkda1fStZ8J6/eaJqsEthq2jXklvcxbsPbzxOVZcqeqspGQeor4E/Ei3+MHwY8E/E+3bJ8S6Ha3ky7CmyZox5qYwPuyb144OMjIwaqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv28/+Tgrv/sFWn/oJr/gnb/yRPWv+xpuP/SW1qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqrxz/yJPiD/ALBVz/6Kaqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqvjn/wAkT+IP/Yq6n/6SyV4F/wCR38Pf9hW2/wDRq1X/AAUC+Ib/ABN/bD+J2ure/aLaw1htJtdrMUSOyRbbCAk4BaJ3OOCzswA3V/wSb+HcvgT9jbQ9TuUeOfxnq17rbxvnKhmW3jP0aO1jcezD3rXNHsvEOiahoGoq7WmpWsttOEbaxjkUq2D2OCea8ceE9S8BeNNf8C6yMah4c1S5026G3biWCVo347fMhr9l74hv8WP2dPhv8Qri9+1XmseG7N7+XczbrtYglxyxZjiZJBkknjk5qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr/gpV/zTn/uKf+2lf8E4f+R18Yf9gqH/ANGmv2z/AIgt8Lf2VPih4zhmSK5t/Dtxa2kjkAR3FyBbQtgqwJEkyEKRhiADjOR8LfA178T/AImeE/hxpxcXPifWrTTY2UZKGeZY93sAGySeABk1aWdpYWkNhYW0VtbW0axQwwoESJFGFVVHAAAAAHAqqqqqv2l/2Qfgn+1X4eGl/Erw7s1e1j2ab4hsNsN/Y4JIVZMEPHlmzFIGT5iQA2GH7WX7BHxr/ZPvpdU1qy/4SXwS8m218U6bAwhXL7VW5jyTbSHKcMShLYR3IOP2Vv21PjN+ybr4n8FammqeGruZX1Tw1qLFrW6AyC0Z+9BLhjiROpC71dRtr9lj9tr4J/tY6Kn/AAhesf2X4qt7cS6l4X1GRUvLbHDPH2niBx+8ToGTeI2YLVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV+3n/wAnBXf/AGCrT/0E1/wTt/5InrX/AGNNx/6S2tVVVVVVVVVVVVVVVVVVVVVVf8FofE/iXwx8MPhzP4a8RanpMs2vXKyvY3b27OBACAShGR7Gv2EviR8RNX/a++Fem6r4+8R3tpca8iy29xqs0kcg2PwysxBHsar9qe7u7D9mH4v39hdS21zbeA9bkhmhco8TrYzFWVhyCCAQR0Nf8LZ+Kn/RTfFf/g5uP/i6/ZZu7vUP2YfhBf391Lc3Nz4D0SSaaZy7yO1jCWZmPJJJJJPUmqqqqqqqqqqqqqqqqqqvj9/wVE/Zv+AXjDWfh3qFn4r8R+JtDkaC7tNL05UihnCZCPLO8YxkgFkD4zkA1+xX+2PD+2Roni7xLY+AX8LWPhzUo7K3il1H7XLchlZ/MciNBGdu0bBuwc/Mc1VVVVVftZf8nUfGX/soGu/+l81f8EVP+TV/FP8A2UC8/wDSCwqqqqqqqqqqr9t79vPwP+yHoEWkWVrb+JPiFq0JfTdC8/YltGcgXN0VyViBHCDDSEEAqAzp8YfjZ8Ufj54wm8dfFjxje6/q0i+XG07BYrWMdIoYlwkSZydqAAkljkkk/Aj/AIJs/Hb4vRQ614uiX4f6DKu5LjVrZnu5QehS0BVsf9dGj45Ga8Df8EtP2Y/DESt4qh8ReMJyvz/b9Sa1iz6qtt5bAexdvrVt+wv+yVa6eumxfBDQ2iSMoGlknkkx7yNIXJ9yc+9eNf8Agl7+yz4ltJI/Deka94SufK2xS6dq0twquM4ZluTLuHIBAK5AGCpJNfHj/gmP8bfhhFNrnw4lX4iaJEu50sLYw6hCO+bXc3mDoB5TOx5JRRXw/wDiL8Qvg74wt/GXw78T6l4a8Q6czIl1aOY3XsyOp4ZTjBRgQe4r9g3/AIKE+Gf2rNP/AOEF8bRWPh74mafDveyicrBq8SrlprYMchlwS8JJKj5gWXdsqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv2xfCfinU/2j/F97pvhrVbu3k+xbJoLKSRGxZQA4YAg8gj6iv2HNN1HSvgJZ2eqWFxZzjU7omK4iaNgCwwcEA4r/gt/8QOPhh8KrZv+f3WrwE/7kMGMN/185yv93B+8K/4IrfD99d+Pni/4iT23mWvhXw39mR8sPLubuZQh4OD+6guBg5HOeoBqq/4KZfC1/hb+2N43jig8ux8VyJ4gsz5bLvF0CZm5AB/0hbgZUkcdc5A/4I2fE6Hxb+zNqXw5uL1HvvAmvTRx24fLR2d3+/jYjqA0xux6fIffFVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV+3J4X8S6t8eru70rw7qd5AdLtVEtvZvIuQpyMqCK/wCCf2k6po/wa1i11fTLqxmbxPO6x3MLRMV+zWwyAwBxkEZ9qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq8axvL4N16KJGd30y5CqoySTE2ABX/AAgvjf8A6E7XP/BdL/8AE1VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVfGu3nu/g149tbWF5ppvDGpJHHGpZnY20gAAHJJPGK8E+CPGcXjPQJZfCOtIiapbFmbT5QFAlXJJ29K+IfjGw+HXgDxN8QdVTfZeGNHu9UuVyRmO3haVhlQx6Ieik+x6VeXWpa9qs19dSS3moajcNJI2NzzSu2ScDqSx/M18F/ANt8LPhD4L+G1rAkS+GdBs9PcLt+aSKFVdyVADMzBmZgBuJJ71X/BTX4eR/Dv9s/x7FaWf2ex8RSQa3a/Ko8w3MKtO+FAHNwLj1Jxkkkk1/wAEdfiN/wAJb+ylL4Jnl/0jwN4gurOKPb0t7jF0jZCgcyzXAwSxG3JIBUCqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr9s34C/EH43/8ACH/8IJbWMv8AYv277V9quRDjzfs+zGev+qb6cV+x3+zt8Sfgp4l8Q6p45tdPig1KxjhgNtdiYllfJyAOBiv+C0PxDj8O/s6eG/h7bXvl3ni/xIjyQ7l/e2lpEzycZ3cTSWpyAQO5BIz/AMEqPh8njz9szwvezxPLa+EbG81qdFU4ykfkxEkMCoE08TdwSApGGOKqqqqqtR07T9Y0+50nVrG3vbG9heC5tbmISRzxuCro6NkMrKSCCMEHBr9s3/gkfo+sw6j8Sv2VYE07U9zz3fgyVwttcZGWNlIx/ctnJ8lzsO7CmMKEN3aePPhP42a1uY9b8JeLPDd4MjMlleafcIcggjDxuDggjB6EV+xz/wAFeGnl0/4cftXFA0jeTa+NbWEIATjaL2FAAB1HnRjA+Xcn35K0rVtK17TbbWdC1O01GwvIxJbXdpMs0UyHoyOpIYH1BxVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX7U/7LHxZ+LfxZuPGHg6z0yTTpLCCFWuL0RNuQEN8pHTmv2QvhH4x+DPw01Lwv42htI7661yW8jW2nEq+U0ECA5HfMbcfSqr9pb/AIKseC/2e/j5dfCOw8CP4w0zRLXy9cvrLUVhktb9huEMeQySBFKrICVKuzDOYyp+GX/BWf8Aa+8BXESeI/Emj+ObATF5LfXNNRZNrNllSa38tgeTtLbwvAxtAWvhd/wWw+FOsxxWfxf+FPiDw1c/Khu9GuY9Tt2O0bnZX8qSMFt2FAkIGOTzj4Y/tkfsufGR0t/h78bvDN9dP/q7K6uGsLl/mK/LBciORuVPRehU9GUmqqqqqqqqqqqqqqqqqr/gt7/ySr4af9jBdf8ApOK/YD/5PK+E3/YwJ/6A9V+1l/yav8Zf+yf67/6QTVX7J3/Jq/wa/wCyf6F/6QQ18X/+Crf7O3wV+JniH4VeKvBvxFutW8NXhtLubT9Os3gdwAcoz3aMRyOqj6VpH7bfwBu/2d9E/ab8SeILvwn4Q8QSXEVhFrMKi9mlhnlhaNYYGl3uWgdgELfLgnHONY/4LZ/Am21c2uifCjxxf6crAG7ma2t5GGfmKxeY3GORlgT321+zd+2D8DP2qtNubn4V+JJTqWnxrJfaJqUQt761Q4+Zo9zB0BYKXjZkDcZ5Ga+IHxF8CfCrwteeNviP4t0zw7odipM17f3AiTOCQi55d2wQqKCzHgAk4rxv/wAFpP2dtC1Q6f4L8CeM/FEETMHvmihsYXGFKmIO5kYcsDvRCCvGQcj4N/8ABW/9lr4n6raeHfFMmt/D/UbuRY0m16KP7DvZiF/0mN2CDAUs8qxqu7qQCahmhuIUnt5UljkUMjo2QwIyCCOoIrxt/wAFeP2e/h34u1jwL4w+F3xb07WtCvJLS9tpdIsAY5EODj/TeQcZB6EEEcGv2V/21/g1+19DryfDOLXdOv8Aw40RvNO122iguGikB2TRiKWRXTcrKcNlSBuADKW/aJ/aH+HP7MHw1n+KPxMuL3+zo7qK0trWwiWS5vbiTJWKJXZFLbUdzuZQFRjniof+C0v7LlxKlvb/AA9+LEssjBURNJsCzMeAABe8k+lfG39prwn+z58Frf42/Erwf4rtbCWS2im0iCC2lv7WSf7qSL54iyvRtsrAdia+An/BUD4A/tFfFnQvg34J8I/ECy1rxB9o+yz6rp9pFbp5FvJO+9o7p2GUhYDCnkjOBkj49/8ABUX9mD4Fa/e+Dl1DWPGniCwaWG6tfDsEckNpcJkeVNPI6IDuG1vL8wrzkZGK8Af8Fnv2cfEmrjTPHPg/xb4Qt5GQR6g8KX8CAnDGQRHzFA4PyI5PPTAz4V8V+GPHXh6x8WeDPEOn65oupR+ZZ39hcLPDOuSCVdSQcEEH0IIPINV+0X+2t+z1+y9Gtp8SvGPna7LGZIfD+koLu/dQuQWjDARBsgK0rIrc4Jwcf8PuPgr/AGr5P/CnPGv9m/aNv2j7Ra+d5O77/lb9u/bzs34zxu71+zl+2h+z/wDtSwy2/wAMPFrJrdrCs1zoGqxC1v4kIGWEeSsqqSAzRM6qSMkblzXiDxD4f8J6Ld+I/FOu6fo2k6fGZLu/1C5S3gt0H8TyOQqjkckgV8TP+CwX7KfgfUrzRvC0HivxxcW2VS60mxSGzkcYyPNnkRyM5G5Y2U4yCQQTon/Bbb4F3F0yeI/hF47sbYRkpJZPaXTl8jAKPLEAMZ53HkAY5yPi3+258JPg38BfCX7RfiHRPFt/4W8aXVtb6XFY6dGt2RPBLPFI8U8sYVDHAxB3EnchAIOR+098UPD/AMa/j/44+KvhW01C10nxLqjXdpDqEaRzohVRh1RnUHjsxHvX/BOn9vD4Qfsh+CvF3hv4k+HfGGpXOv6pDd2z6HZ20yIiRbCHMs8RBz6Aj3r9nL9oXwV+0/8ADOD4q+ANM1uw0me8mtFh1iCKKcPEQGJWKSRcHPHzfgK+J/xa+GvwX8LXHjT4peNNM8OaPbKxM95LhpCBnZFGMvK5A4RFZj2BrxZ/wWr/AGf9L1I2fhH4beNddtkyGu5hb2SsexRS7sR7sFPt3r4H/wDBVn9lr4x6zZ+FtX1DVfAWsXnyRDxHHHHaSSddi3KOyLkZwZfLBIwOSoNTTQ28LzzypHHEpZ3dtoUAZJJPQYr4yf8ABW/9lr4Yard+HfC0mt/EHUbSRo2m0KKMWIdWUNi5kdQ4wWKvEsitt6gEGvhL/wAFhP2Z/iDr9t4b8aaV4j8BTXk3lw6hqccc9gudoXzJYmLxksSCWj2KBuZwM4/anu7TUP2nfi/fWF1Fc21z481uSGaFw6SI19MVZWHBBBBBHBzX/BFT/k1fxT/2UC8/9ILCqqqqqqqqqr9rX9pPw3+yt8FtX+J2spDeaiuLXQ9LaQI2oXr8InUHYozI5HIRGxk4B8ZeLvHPxm+IepeMPE13ea/4q8V6iZZmSMySXE8rYWONF6DlUSNRhQFVQAAK/Yg/YH0b4PafY/FD4v6Tbaj4+m2z2VlKBLFoQ6rgcq9x0JfkIQAnQu37U/7dHw0/Zku4vDEmmz+KvF00QmOj2dysK2qHG03EpDeUWByqhWYjkgAqT8CP+Cofwy+K3i6z8F+OvB1z4Eu9TnEFhdyaiL61eRioRJJBHGYizEgEqVHGWGeK/aj/AOCgPw0/Zv1r/hCdP0Wfxl4tjVXutOtbxbaGxU4wJ59rlZCp3CMIxxgttBXP7Ov/AAUt+Gnxu8ZWPw98U+ErvwTreryiHS2lv1vLW5mJwsJl2RskjkgKCmGPy7gSob9s79hfwp+0JpV1438EWttonxDs4JJFmhgRE1sqh2Q3HKgOSFVZicqOG3KF2xS+Nfhb41WaGTVPDXinwzf5VlLW11YXUTfgyMrCv2HP2rtK/a0+DFt4rnNvbeLdFZbLxNYQoUSG5xlZY1JJ8qVRuXk4IZcnaTVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX/BVT4gP47/bP8V2Udz51n4Ss7PRbRsMNoSESyrggYxPPOPQ4yCQa/4IxfD7/hHf2bNd8eXC/wCkeL/EkvlEDg21rGsSZyoOfNNx3YY24wdwqq/4Ld/DCObw/wDDj4z2qbXtLyfQL1sL84lQ3FuOuePJuexHzcleN3/BG/4pv4P/AGm7/wCG9zeSrZeP9DmiigXcVe8tAbiNmxx8sC3YBP8Af684NVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV/wVl/a8T4TfDgfs/8AgbVWj8YeObUtqc0DFW03SixVvmBGHnKtGBz+7EpO0lCdS/af/aF1jwt4V8Gan8YPEtxpHgi8ivdAt3vTmxnhbdDIr/fYxHiPcT5YwE2gYr9hT9rnRP2svhBbateXVvB430BUtPE2nrtQ+dtG25jjBJEMvJBIGGV1/hyaqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr/gqP8RG+H37GXjGG2uUhvPFM1tolsWkClhNIGmUAnLEwRTjA5xk9Aa/Ym+HrfFH9rL4WeD/ACklhfxFBe3MTqCJLe0zdTKQWXgxwOOueeATgGq/4Le/Dn7P4q+Gnxcto932/T7rQ7x842eRIJ4Bgtzu+0XHRRjZyTlQP+CJfxDj0r4seP8A4X3N7sXxFocGpW0bsoV5bSXYQuTneUumbCg5WMkkbBVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV/wAFqviA+u/Hzwh8OoLnzLXwr4bNy6YYeXc3czFxyMH91BbHIyOcdQRX/BED4eOtp8T/AIsXdn8kkllolhcbWHKh57lM4wfvWhwCSO4GVJqqqqqqqv2nv2M/gn+1ZoX2Tx9oX2PXraNl0/xFp6rFeWpxgBmxiWPOD5b5HptPNftWfsLfGv8AZQ1KS68T6b/bnhCSRUtPE+nQsbZy33UlU5MEnGNrcE/dZq/ZU/bo+Nf7KGpR2nhjU/7c8ISSM934X1GVjbOW+88LDJgk4zuXgn7ytX7MX7ZfwT/as0L7V4A137Hr1tGp1Dw7qDLFeWpxklVziWPPHmJkdM7TxVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV+3H+09Zfsq/AfVPG1pcW7eKdUb+zvDNrKm8S3jqT5jL3SJA0jZwpKqucuudE0fxh8XPiDZ6Jpy3Gs+KPF+rLFGXbdJdXdxLjcx9Wdskn1JNa9/wRH+Ht94P0mPw38ZNd0jxXFYxDVZrqziv7C4uhGgkMMY8mSJGkDkbnkIDD05+KP8AwSR/a5+H6T3nhvR9B8d2MbEq2hagFnEeWwWhuBG27CqSqF8bxgthiPG/w3+IPwz1Q6J8RPA+u+Gb8Myi31bT5bR2ICk7RIo3DDIcjjDKehFfC79rT9pP4LxxW3w1+M/iXSbOHb5dg919qtE2qFGLecPEPlVV+70UDoBXwr/4LUfGnw6kNj8Wvhv4c8Y28ahTd6fK+lXbHJ3O5xJExwRhVjjHy8nnI+FX/BW79kr4heTZ+KdW1vwFqEmVMet2JkgLDb92eDzAAcnBcJ91s4+Xd4F+Jvw4+KGmtrHw28e+H/FNlHt8yfSNSiu1iLZwHMbHaeDwcHg+hqqqqqqqqqqqqqqqq/4Le/8AJKvhp/2MF1/6Tiv2A/8Ak8r4Tf8AYwJ/6A9V+1l/yav8Zf8Asn+u/wDpBNVfsnf8mr/Br/sn+hf+kENft+/8nlfFn/sYJP8A0BK+AvwR+O37XHiPSfg/8PZ5b+z8OW8s0balePHp2iQSybpJGwG2b5CMhFZ3I6ELx+0t/wAE6/2g/wBlrwZF8QfG7+Gdb8Pm4S3ub3QL2WcWbucJ5qzRRMAx4DAFckAkEgH9nj4x6z8APjR4T+LWiSy79A1BJLqGPk3Nq3yXEOCQDviZ15IwSDxjNQzQ3EKT28qSxyKGR0bIYEZBBHUEV/wUB/au1f8Aai+OOoz6ffuvgjwtNLp/huzjuC8UkaMQ94cHaXmYbsgcRiNcnbuP7K3/AASMvPjN8KNF+K3xT+JF34Zi8T2aXulaVp1ik0y2z8xTSyM20eZHtdUUEhXXJByo/bQ/ZA8Tfse/EWz8J6nria9omt2rXWj6stuIDOqttkjePc210ymecEOpHUgf8Ecf2ktS8c+BNc/Z78W6tLeaj4MjS+0J7ife501yI3hAJzshkKAdgs6rwFAr/grn+x8PGPhs/tQ+ANM3a14et1i8VQxnBubFBhLkL3eLox7x4PSOv2V/2gNZ/Zk+OHh74taVDLd21hI0Gq2Mb7De2Ug2zRZPGcYZc8B0Q9q/4Kl/tY6L+0R8W9K8HfDnxDDqngTwXZg2t3bPmLUL24RXmmHTIRfLiAYZVklI4ev+CSP7HMPjfxEv7T3xE0dJtD8O3WzwpbzDIudQjb5roqeqwkAIf+evI5jr/grz/wAmaal/2MGnf+htXhzxN4i8H6xB4h8Ka5e6RqlssiwXtlO0M0QdGR9jqQVJRmGRzzXwN/4JGfH74u+BLHx94j8SaD4It9Whjn06w1GOWa7khcbhJIiDEQKlSFLFuTuVMc/tFfs1/FT9l3x3/wAID8U9Kt4rmaHz7C/spTNaahDkr5kLkKSAQQVZVYcZUZGf+CPn7S2t+EPi037N+t37zeGvGi3F1pUThcWepRReYxDHkLLDCylc43rHgAs2f20f2jYf2Wv2f9d+J1vb291rbsmnaDbXD4SW/mDeWWHVlRVeVkGCyxMMrncPhv4E+JH7Unxw0rwTp+pS6t4u8caoxn1DUZy5LENLPczOcsQkaySNjLEIQATgHXv+CIVjB4Jkfw58dLu68WxW+5EudHSOznlAyUGJC6AngMS2OpHYaBr3j34IfEuDW9FvbrQPF3g7VGAeOTa9tcwuUkjYqcEZDIy5IZSQcgmvgN8WtH+O/wAHPCPxc0NEjt/EumpcPCsgcW84yk0OR1McqyIfde1f8FLP2x/En7QHxZ1T4Y+HdVlt/h54K1CS1tbSJyqaleREpJdyf3vm3LGDkKoyMF2r9k79hf4yftdyX2peCjp+j+GdJuBbX2u6ozCFZtoYwxKgLSyBWViBgKHXcRuXOvf8EN9ft9Nkl8MftIafqGoD/VwX/hZ7OJvrIlzKR2/gNftifsZXX7Q/7O3g/wCAnw+8S6f4atvB+qWM9rNe2xdPs1rZT2yRhYgoBxKh4AA2ngcV8b/hXqHwQ+LPif4Tarq1vqd34YvmtJbu3jKRzMADuUNyBz3r9jz9gTxf+2J4a8QeJfDXxA0fw9H4evo7SWK+tZJWlZ03hgU6DHHNfsbfs/6h+yV8Ao/hr4t8W6ZqbadfXd/PqMKGCBI3IYlt5+UKFOSeK/bN/aj8RftXfGbUfG95LcW/hywZ7Tw3pbysUtLQHAfaeBLLgO5HfC8hFx+zx/wRwm+IXwy0jx58Xvife+HtR1+1ivLXR9LsEla0gddyedK7YLspUlFUbOQSxPy/tcfsueK/2SPizL8M/EeqxazaXFml9pGrwweSt9bOWXcY9zGN1dHVkLEjaDkqyk/8EjP2ktS+LnwW1H4TeL9Wlvdf+HMkUdrPcz75bjTZt3kjLHc3lMjx+ip5I7iv+Cxv7T+teCvD+i/s1+Dr64srrxZYnU/EVxEzRs2n+Y0UVurA4KyvHN5g/uxqDkORXwG+BHxC/aQ+JWnfCz4Z2NvPq18rSyTXcpit7OBMb55nAJVFyM4DMSQFBJAPxF/4IpfEfwz8PrvxF4D+MGn+LfE1nbib+wG0Q2K3BC5eOG4M7hnyMLvRA3GSlXlnd2F3NYX9rLbXNtI0U0MyFHidThlZTyCCCCD0Nf8ABFT/AJNX8U/9lAvP/SCwqqqqqqqqqq/4K1fH67+Kv7SE/wAM9Mvd/h34aRmwiRGO2W/kVXupCDjlW2Q/9sSRw1f8Esf2brHxNq+oftB+MNLWe00K5Fn4ciniDI92BuluMMOsQKKjDjeznhoxVfFjx7qPxS+Jfib4h6rdTzz69qc10GnADLGzHy0wCQAqBVABIAUDPFV+z38SLvxP+zL4L+Jfi24nluj4XiudTuHbzJJ3hixLMcAZZzGz4xwWxz1rxf4p1jxz4q1jxl4huPP1PXL6a9u5MnBkkcs2MkkDJwBngYqKWWCVJoJWjkjYMjqcFSOhBHQ18G/FOo+OvhB4G8bawV+3+IPDen6hdbQAPNnto5HwAAANzHsK/wCCqP7NltdaXB+0t4WtYormy8qx8UIoC+fGzLFbXHA5ZSViYkklTEBgIc/8E0f2h5/gJ+01oljqN75fhnx5JFoero8gVI2lcC3nJPA8uUrlj0R5PWqqqqqqqrXPGXg/wxLFb+JPFej6TLMpaJL6+jgZwOCQHYZFVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVe+KvC+m3L2Oo+JdKtbiPG+Ge8jjdcgEZUnIyCD+NR67oc2mS61DrNi+n26O8t2twpijVRliz5wAAMkk8V8TvGc/xI+JPiz4iXUXlT+KNcvdUkjwBsa4neUjA44L1+xr8Po/hf8AsrfC7wYLb7PPb+G7a5u4sqdlzcr9onGVJB/ezScgkHr3qq/b/wDha/xd/ZD+I/hu0g83UNP0s6xYhY2dzLZMLgoiqCS7pG8YGOTJ26j4EfEif4P/ABn8E/E+3fA8Na5a3ky7A++FZB5qYIP3o968cjORg4NWl3aahaQ31hdRXNtcxrJDNC4dJEYZVlYcEEEEEcHNVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVf+J/DWlXBs9U8RaZZzhQTFcXaRsAehwSDitO1XS9YgNzpGp2t9CrlGkt5lkUMACRlSRnBHHuK/aE+OHhP9nP4Q+Ifi74wbzLTRLfNvZq+x765c7YbdDgkF3KgtghV3MflUmvif8SPGnxu+JWufEnxpePqHiHxPfGefywxAY4WOKJSSQiIEjRcnCqo7V8WP+CXXjb4efse6L8Z7SPULr4gafu1HxXoAiLm3sJQu1Y1XJ8y3Ch5PUSSnIEShv2Wv2kPGH7LXxf0r4neFXe4tUYW+taWZNqanYswMsJPZsAMjYO11U4IBU/Dr4g+Efiv4H0X4jeA9XTU9B1+1W5sblUKb0PBBU4KsCCrKQCCCDyKqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqSSOKNpZZFREUlmY4CgdST6V/wnHgn/ocdE/8ABhF/8VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVcTwWsElzczxxQwoXkkkYKqKBkkk8AAd6Txr4NldYovF2iu7nCqt/ESSegA3V/wW/8AHzx6T8MPhZbyy7Li4vdZu03MEzGqQwHHRj+8uOTyvb7xr/gi98PH8RftF+I/iFcWfm2fhDw28ccu1v3V3dyqkfOCvMMd0MEg+gIDYqv+Ct3w8fxv+x5quu2tn59z4L1iz1ZdisXEZc20hAUHIC3JdgcAKhYn5RX/AATz8fP8Of2yvhdqxllEOqawNGmjRmAlF8jWqBgOoEksb4PGUB7VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV8Xfjz8Pfgh/ZP/Cd3V7F/bXnfZPs1qZs+Vs35x0/1i/rXwm/aI+Gvxr1G/0vwNd38s+mwLNOLm0MICsdowSeea/be+IEfxQ/a0+KXi+C5+0QP4gmsraXCgSQ2gW1iYbQOCkCkd8Yzk5Nf8ErPh9H4E/Yw8J3r23k3niy8vNaulypyXlMMTZUnrBBAfUZwQCKqqqqqqqq1bSNK17TbnRtc0y01GwvIzFcWl3As0UyHqrowIYH0IxX7YP/AASC/wCP34gfsnH0kuPBd7cfgxsriQ/Q+VKf75WT7sdWt148+E/jZLm1k1vwl4s8N3hwSJLK80+4Q4IIOHjcHIIOD1Br9jf/AIK56Lrsem/Db9qdotK1L93b2njGBMW1ydu0G9jH+pcsBmZP3ZL5ZYlUsdO1HT9Y0+11bSb+3vbG9hSe2ubeUSRzxuAyujKSGVlIII4IINVVVVVVVVVVVVVVVVVVVVVVVVVVVVfEz9qb4S/CXxS/g7xhe6nHqMcCTMsFk0q7XGR8wPtXwp+Lfg74z+HbjxR4InupbG1vWs5DcwGFhKqI5wD22yLzV5d2lhaTX19dRW1tbRtJNNM4RIkUZZmY8AAAkk8Cv2/P2p7r9qj47X2t6ZJs8H+GfM0zw1CGJ8yBX+e5bP8AHMw39BhBGpyULH/gj3+yV9ktJf2r/HGm4uJ/OsfB8Mq8pHgx3F6Of4syQLkA4EpxhlNVWtaHoniXSrjQ/EWj2Oq6ddrtuLO9t1nhmAIOGRwVYZAOCO1fFX/gmN+xz8VBNc/8Kz/4RDUZsD7b4VuDp+wDdwtvhrYctnPlZ4AzgYr4qf8ABEfxDapLe/BT4y2WoAKTHp3iWza2bIIwPtEO8MSu7rEoyAM4Ylfil/wT/wD2u/hHHLd+JPgpreoWEW4m+0FV1WIIqlmkYW5d4kAU5aRUA/EVpera/wCE9Zi1XRNT1DRtW0+Q+Vc2sz209u4yp2spDKeSD0718Lv+CmP7Y3wteKKP4py+K7GPbmz8UwjUQ+GB5mJE/IBU4lHDHvgj4Y/8FuvD80a2vxn+CuoWjr9698MXqXAf5TnFvcGPb8wUf61uGJ/hw3wu/b//AGQ/i68Vp4b+NeiafqEm0Cw15m0qUuzBRGn2gIkjksAFjZs/gcWl3aahaQ31hdRXNtcxrJDNC4dJEYZVlYcEEEEEcHNVVVVVVVVVVVVV/wAFt9HvZ/gh8P8AX41T7JZeKntpSWwwkmtZGTA9MQSZPbj1r9hXVbDR/wBsL4R3epXHkxSeKLW3VthbMkx8qNcAE8u6jPQZycDJqv2xdUsNH/ZO+MV3qNx5UUngfV7dW2FsyTWkkUa4AJ5d1Gegzk4GTVfs06Pe+Hv2cfhV4f1JUF3pngnR7WcI24CSOyiVsHuMg81+37/yeV8Wf+xgk/8AQEr/AIIgWdonw1+J9+lrCtzNrlnFJMEAd0SByqlupALuQOgLN6mv+Cg1naX37GHxYgvLWG4jXQzIqSoGAdJY3RgD3VlVgexAI5FV4z8Qah4S/Yo1zxVpJQX2i/C24vbYvnHmxaWzrnaQcZUdCD7iq079u39r7SdPtdK034/eKLe0soUht4UmQCONAFVR8vQAAV8V/j98Zfjp/ZX/AAtz4h6r4o/sTzv7P+3urfZvN2eZtwB97yo8/wC6K/4I9XWpW/7YUcNjJKsNz4X1CO8CDIaIGJgG9B5iRn6gVeWdpqFpNYX1tFc21zG0U0MyB0kRhhlZTwQQSCDwc1+194A+Gvww/aO8ceB/hJ4jTWPDWm6ky27oOLSQgGW1DdHELlotw67PXNeCLTwnf+NPD9j481S703wzcapbR6ze2kfmTW1m0qieSNcNudYyzAYOSBweleCNA8J+FfBuh+GvAlpaW3hzTNPht9JhtJPMiW2RAI9jZO4bQPmJJPUk5zX/AAV5/wCTNNS/7GDTv/Q2r4b6fZav8RPC2lalbpcWl5rVnDPC44kjeZFZT7EEiq/4LaeGdKu/gD4F8ZTQ51LS/GAsLeTavyw3NnPJKM43DLWkPAIHHIOAR+x3qt/o/wC1h8HbvTbjyZpPHGkW7NsDZjmu44pFwQeqOwz1GcjBwa/4Lj+INQtvDXwh8KRFPsOo32rXswOd3m26WqR45xjbcyZyCemCOc/Dj4n+P/hD4mj8ZfDTxVe+HtbiheFL2zYLII3GGXkHggCv+G/P2yv+jhvFX/f5P/ia8ReINZ8W6/qfirxHqMuoatrN5Ne393McvcTyuXkkbHdmYk+5r/gmV4l1zQv+Ce666P3z6BJrc2mR3KEx7Y3klC4BBKGUyZwepbkVNNLcSvPPK8ssjFnd2JZmPJJJ6k1/wTh0HQtB/Ys+GMegwxKl7p8t5dOmCZbiS4kaQsR1Ib5eeQFC/wANVX7fv/J5XxZ/7GCT/wBASv8AgiH/AMkq+JX/AGMFr/6Tmv22/EGoeGP2R/i1q2llBc/8IreWys2RsE6eSzAqQQwWQkHsQDz0qv8Ahvz9sr/o4bxV/wB/k/8Aia+Knxu+LPxv1Cx1X4seO9T8T3emQtDaS3zhmhjY7iowBwTzX/BFC61JP2k/GNjFJKNPl8DzSToB8hlS9tBGSfUK8uPq1f8ABVTVb/Uf25PH9neXHmRaZb6Xb2i7APLjOnW8pXIGT88shycn5sdAAP8AghxoOm3Ov/GDxNLDnUNPs9Is4JP7sU73byD8Wt4j/wABqv2rIYbf9qL4xW9vEkccXj7XFREXaFAv5gAAOgx2r/gip/yav4p/7KBef+kFhVVVVVVVVVeLfENp4R8K6z4svl3W2i6fPezLvCZSKNnbk8DhTyeBXifxHq/jHxLq3i7xBeNdaprd9NfX07klpZ5XLyOScnJZiefWvgJ8N7f4Q/Bjwb8N4YFik0PSIYroKqruuWG+4fCkjLStIxwTy3U9ar45/Dq9+Evxg8X/AA6vrBrQ6Hq08NvG2Tm3LboHGSTteFo3BJzhhnmoopZ5UggjaSSRgqIq5LE8AADqa+Anw0k8Gfs6eDfhd4qtmea28Nw2mq27NtKySRZni3KQcBndcg5wM1498Ga18OvGuueA/EVu0OpaBfzWVyjLjLRsV3D1VsAgjIIIIJBzWmaZf61qVpo2k2kl3fX06QW1vEu55ZHYKqqO5JIAHvXwq8JzeAvhd4O8C3MvmTeHNAsdNkfIO5oIEjJ49SlfEbwRpfxK8A+Ivh9rQX7F4h0yexldow/l+YhUOAf4lJDDkEEAgjrWp2GoeG9cutLuJPJvtLu3hdon+5LGxBKsPQrwa+BnjsfFH4L+BPiMbpLiXxJ4dsb+d1ZDiaWBGkU7PlDK5ZWAAwQRgYxVVVVVVVf8Fvf+Sp/DT/sX7r/0oFVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV+2n/ycx4y/wC3H/0ht6T4hv8ADD/gmD8RPE9ve/Zrqa3v9NtZFZg6y3kiWqshUgh187cCD8pXdyARXwS8AP8AFX4x+B/hqkcrp4m8QWWnzGJGYxxSzIsj/LyAqFmJ7BSe1VVTQw3EL288SSxyKVdHXIYEYIIPUEdq+O/w3n+D/wAZ/G3wwnXA8Na5dWcDbw++FZD5T5BP3o9jYPIzg4ORX/BPv4nS/Fn9kH4ceIr29e51HT9NOk3zSOWk82zdoAXJ5LMkcbknk78nkmqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/bz/wCTgrv/ALBVp/6Ca/4J2/8AJE9a/wCxpuP/AElta/4Kd/tgD9on4rf8K88Fap53gHwLcSRWrouF1G/GUmuc/wASDmOM/wB3cw/1lf8ABJv9jxPil44/4aH+IGkpL4V8H3QTRba5hJXUNSXBEozwyQcN3BkKj+FhU0MNxC9vPEkscilXR1yGBGCCD1BHav8Agpd+xo37NXxRbx14JsEj+Hfja6kl06KJAq6Vdkb5bPAwAn3ni4HyZTkxlj/wSy/bQj+Bvjo/BH4h6pFD4E8ZXiva3dzKsaaPqJUKJC5IAilCoj54VlRhtG/dVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVXjn/kSfEH/AGCrn/0U1VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVfHP/kifxB/7FXU//SWSvAv/ACO/h7/sK23/AKNWv+Cr/wAQW8dftmeI9MjmSW18HabZaLbOpBBCx/aJBwoIKzXMqnJY5XrjAH/BFrwBHoP7Pfin4gTRwi58VeJDCjKi7mt7SJVTcw5/1ks+FPA6j7xqq+N3gCP4q/Bzxv8ADVo4nfxN4fvdPhMqKwjllhZY3G7gFXKsD2Kg9q0nVb/QtVstb0q48i90+4juLaXYG8uRGDK2GBBwQDggivAXi2y8f+BfDnjvTVRbTxHpNrqVuEk8wCOeJZFw2BuGGHOBn0FVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX/AAUq/wCac/8AcU/9tK/Y+8dw/C7w98YviTMiSL4W8GTamI3GRI0CvIExuXJYqFA3DJIGRnNQxav4m1pII2uNR1TVboKpdy8txPI2OSTkszHqepNfDfwda/Dz4deFvh/Y7Ps/hnRbPTIdhYjZbwpEMFiWIwg6kn1qqqqqqqqqq/ao/Yl+Cf7WOit/wmej/wBl+Kre3MWm+KNOjVLy2xyqydp4gf8Alm/QM+xo2YtX7VH7F/xk/ZO8SNY+NNL/ALU8OXMhGmeJdPiZrS6XPCvkZhlxjMb987S64Y/sm/t7/Gv9k++i0vRL7/hJfBLybrrwtqU7CFcvuZraTBNtIcvyoKEtl0cgY/Zn/a3+DH7Vvhl9b+GeuumpWSj+09Bv1EV9p5JwC6AkMh4xIhZDnGQwZRVVVVVVVVVVVVVVVVVVVVVVVVVVVft5/wDJwV3/ANgq0/8AQTX/AATt/wCSJ61/2NNx/wCktrX/AAV1/axl+G/w/h/Zy8E6i8PiHxvamXXZ4nINrpJLKYsgghp2Uoc5HlLICPnBH7IH7OGuftR/HPQvhtp8UsejrILzxBfKp22dhGQZTkA4d+I0/wBuRc4GSPDPhnQPBnh3TfCXhXSbfTNH0e1jtbGzt02xwQxqFVFHoABVVVVXxI+BHwY+MFubf4n/AAu8M+JT8u2bUNNjkmTaRjZLjenTHysMjIPBIr4n/wDBGv8AZm8Ww3Fx8Odc8UeBL51YW8cd1/aVnGxHBaKf96wBxwJl4zz3HxT/AOCN37Tfg957n4b6v4Z8f2SybbeKC6Gm3jruxuaO4IhXg5IE7dCOeM/Ej4EfGf4Pzm3+J/wu8S+GgNu2bUNNkjhfcBjZLjY/XHyscHIPIIr4b/Hf4z/B+dZ/hh8UfEvhoDdugsNRkjhfcCDvizsfrn5lODg9QDXws/4LIftN+D3t7X4kaT4Z8f2Sybp5Z7UabeOu7O1ZLcCFeDgEwN0B55z8Lv8Agsp+zX4tjitviToHiXwHeNt8yR4P7TtEyoziSAeacNuH+pGQAepKj4b/AB2+DHxgtxP8MPij4Z8Sn5t0FhqMckybSQd8Wd6dM/MoyMHoQaqqqqqqqqqqr/goR8Cr/wDaC/ZZ8VeE9AsJb3xDo+zWdFt4YBLJNc2+SYkU875ImmjXaQcuOoJVtO1C90nULXVdNuXt7uymSa3lTho5EIZWHuCAa+AP/BXb9njxX4EsD8eNeuPBfi61hWPUAmj3VzaXcoAzLAbdZWRWJJ2vgjBGTwT/AMFFP+Ckngz4/eAP+FH/AAPt9Ql8O6hcRT67q9/bm3N0IZA8UEUZ+YJ5iJIzMFOUUAYyT+y78Ctc/aP+OfhX4U6NbStBqN4smrXKIStnYRkNcTMR0wgIXJG52RcgsKhhht4Ut7eJI44lCoiLtCgDAAA6DHav2/f+Tyviz/2MEn/oCV/wRD/5JV8Sv+xgtf8A0nNft+f8mafFn/sX5P8A0NKrwb4e0/xb8BtD8KasGNjrPhG3srkIBu8uW0VHxuBGdrHqCPUGvit8NvEnwe+JHiP4X+LrfytW8NahLZ3GAQsm0/LIuf4HUq6nurA96/Y8/b+/ZH1z4c6D4G/aG8KeF/Cni7R7VLJ9Um8OJLZ6osSqqTM8cbeTKwyXDAJuUkMAwQar+1t/wTY0ewl1K78b/DSSKHG5bXQftUhyQBtjigZ25IzgHAyTwCa/Zu+InwG+NXhOf4o/Avw7p9rpovJtMN7Ho8djLKY9jONoAcISUOGAJwDjoa/4KQftex/sxfB59D8J6l5fxA8ZxyWuimKRfM0+IDEt4QeRtB2pxy7A8hWr4B/CDxJ+0T8aPDPwq0SSVrzxJqAW6uyC5t4Bl7i4bPXZGrtz1IA6mv2tPgHqX7NPx68T/Cq6SV7C0uPtGjXMg5ubCXLQPnuQvyMf76OO1f8ABIj9rn/hNfCcn7Mnj3WoTrfhi387wq0x2veaev8ArLcE8M8JIKr94xscDbExH/BXn/kzTUv+xg07/wBDatJ1W/0LVbLW9KuPIvdPuI7i2l2BvLkRgythgQcEA4IIr4If8FQf2W/iV4B03WfHXxC0/wAEeJ1t0Gr6PqUcqLBPyGMMu0pLGSpKkMWCldwUnFf8FM/22vCP7U3ibw/4M+FRuJvBnhFppl1K4tjA2p3coVS6I4DpEiLhdwViXfcvC1/wSm+D2r/Ev9rPQvFcVq50T4fwy6rqU+SoEhjeK2jBH8TSuGx3WJ/Sv+CvfwW1T4l/s1W/jvQLZri++HWpf2hcxKoJNhKnl3DAYzlT5LnkAIjkg4GP2b/jQPgB8YND+Jlx4T0zxRYWLPFqOj6hCkkd5bSKVdQWVtjgEMrY4ZRkFdynwT+2p/wTc8baKmrLrHgnQpTxLYa34bW0nhPPB/dFG4GcozDkc54rxF+2n/wTg0K+tNIs9S8H67qN9JHHBbaT4X88MZH2DMxiWFORyGcEAgkYOasND0TStKXQ9L0exs9OVWVbOC3WOEBiSwCKAuCSSRjnJ9a/aN+D2r/AT42+L/hTrFq8R0PUpEs2Ykia0f57eUE9Q0TIfqSDyDX/AAT2/wCCk2hfs3+DG+C/xk0rVb7wpFeSXOjanp6iaXTPNJaWFoiRuiMhaQFfmVpJMhgw261/wVW/Yc0rSrjULD4s3usTwrmOxsvDeoJNOcgYQzQxxg9/mdRx17Voer2XiHRNP1/TWdrTUrWO5ty67WMcihlyOxwRxX/BRzw9qHhv9tX4o2eoBC11qUN7EyA7WjntopUwSBkgPtOOAysMnGa/4Jn/ALcfw1/ZWtvGXg74s2WpppHiGa3vbK/06189oZ41ZHSRNwJVlKFSo4KtnIYFdB8dfDX9uD9m/wAU/wDCA397/wAI94w03UdCFxeWnkywyPE0TOEbdgqXDA4PIBrxl4S1/wAA+LNY8EeKbB7LWNBvprG+t36xzROUce4yDg9xg1+zF/wUJ/Y1+IHgvSdK+O3hfwj4K8bwQ+TfyXHhtDY3siLzPHLHEViDgE7H27WyoLfKTrH7Xf8AwTW0OyN/e+NvhvLGGC7bPw8buTJ/6ZwwM2PfGBX7OvjX4NfFz4dWHxe+DHhux03R9da4hjdNLisp2EE7xMJFTkAtGWAJztZTgZxX/BZ74Pap4d+Nvh/4z2dhcNo3i7SUsrm6EYMcd/bZGwso+UtAYioblvLkwSFIX9hj9rGT9kT4yf8ACaalpN3q3hnWLM6frtjaMomMJZWWWIMQpkRlyFJUMCy7lzuHxG/4K6fsm+HPAlzrvgDxBqfi7xI1rus9DXSbqzInIG1Jppo1jVQT8zIX4U7Q3GfE/iPV/GPiXVvF3iC8a61TW76a+vp3JLSzyuXkck5OSzE8+tf8EVP+TV/FP/ZQLz/0gsKqqqqqqqqr9rL/AJNX+Mv/AGT/AF3/ANIJq+HEUU/xD8LQTRrJHJrVmro65DAzICCO4qq/aR/Y1+EH7TYttS8XwX2leILGBobXWdLZI5inVUlDKwljDcgHBGW2su418B/+CcvwM+CXia18bXNzqni7XtPlWawm1XYsFlKjBkljiRR+8BAwzs2CAQFPNV+0V+xJ8FP2kr0+IvE9lfaN4nEAiXWtJkWOWQKCEEyMGSUDgZID7QFDgAY/Z7/4J+/BH9n/AF638aWz6l4o8TWu421/qxTy7Rjn5oYUUBW24G5ixByQVziqr9qix/s/9pf4qW/2y2ud3jDU5d9vJvUeZcu+0n+8u7aw7MrDtX/BOfVv7b/Yn+FV59jltvL0ua22SjBPk3U0W8ezeXuHswqqqqqq8cfE74a/DG0tr/4k/EPwz4Ttr2QxW02uatBYJO4GSqNKyhiBzgV/w1j+yv8A9HMfCn/ws7D/AOO1/wAFhfid8Nfid8Sfh7ffDX4h+GfFltZaHcxXM2h6tBfpA5nBCu0LMFJHODX/AA1j+yv/ANHMfCn/AMLOw/8AjtaT+03+zZr+q2Wg6D+0L8NNS1LUbiO2s7K08WWU01zNIwVI40WUs7sxChQCSSAKqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv20/+TmPGX/bj/wCkNvX7XXxG/sH9gv4f/DK3l2XHi3xrfXcny5329kg3rypA/e3Nu2dyn5cYILY/4JI/DyPxv+2HpOu3Nn59t4L0e91Zt6qUEhQW0ZIYHJDXIdQMEMgYH5TVVVf8Flfha/hL9pPSviTbQbbPx5ocbSSeWw3XdpiCQZxtOIfsvQ5GeQBgn/giL8U45NJ+I3wRvLyFXguIPEOnW/yh3DqLe6f1IXy7MdwC/bPNVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVft5/8AJwV3/wBgq0/9BNfEP9rcfA79jzWPg/4K1byvG/j/AMQ3SSvDJtk03Sza26Sz5B3K8rAxIcYIExBBQZ/Z4+BPjD9pH4uaD8JfBcLLc6rNm8vTFvj060Tma5k5A2ovQEjcxVAdzqD8K/hj4Q+DPw60D4XeA9P+x6H4cs1trSNgN74yXkcqAGkkcs7tgbndj3qvjP8ACHwZ8ePhnrvwq8e2H2rSNdt/LcqSrwSKQ0cyEEEOjqrDsSMHIJB/aC+BXjb9nD4ra18JvHdti+0uQNbXaIRFf2z8xXER7o69uqsGQ4ZWA/4JZftoP8c/Ah+CPxD1SWbx34Ms1e1u7mVpH1jTwQokLkkmWIlEfPLKyMNx37aqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqvHP/ACJPiD/sFXP/AKKaqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqvjn/AMkT+IP/AGKup/8ApLJXguaG38Y6DPPKkccWp27O7ttCgSKSST0GK+L3juX4pfFbxl8SZkdG8U69e6mI3BBjE8zyBMbmwFDBQNxwABk9a/Ym+HqfC79k34WeD/JeKaPw7Be3MTqQ0dxd5uplILNyJJ3HXHHAAwBVV+2X8P3+F/7VHxR8GG28iG38SXVzaRZY7La5b7RAMsST+6mj5JJPXvX/AATD+IH/AAsD9jDwN5zZvPDf2nRbnB4HkTN5WPmY/wCoeHOcc5wAuKqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv8AgpV/zTn/ALin/tpXxB+IKeBf2Z/ippkczxXXjFNL0W2dSQQWuTcSDhSMNDbSqclRhuucA/sF/DmH4qftffDDwtdQJLaQ60up3SSLujeOyRroo4KsCr+QEIIwd+MjORVVVVVVVVVVXibwz4d8Z6BfeFfFmiWOsaPqcJhvLG9gWaGeM9VZGyCK/bD/AOCQ97YtffEL9lQNd25Z5rnwddTfvIVxk/Y5WPzjg4ikO70ZuFrQvEHxD+DvjaLV9B1HW/CPirQrjAdC9pc2sinlHU4I6YKMMEcEEV+x/wD8Fc/DfjEWXgD9qFrTw/rTZSHxVCnl2Nyf4RcIP+Pdz/fH7s9/LFWl3aahaQ31hdRXNtcxrJDNC4dJEYZVlYcEEEEEcHNVVVVVVVVVVVVVVVVVVVVVVVVVVft5/wDJwV3/ANgq0/8AQTX7L3xh8J/AL9jzx78WvGc+NO8P67dSiBWw93MbW1WG3Tg/PJIUQE8Atk4AJHxa+J3in40fErxF8UvGl69zrHiO+e6nLOWEQPEcSZ5CRxqkajsqKO1f8E4/2UI/2ZPgZbXPibSfI8e+Mdl/r7SqvmWi4/cWeR0EanLDn95JJzjaBVVVVVVV5Z2moWk1hfW0VzbXMbRTQzIHSRGGGVlPBBBIIPBzXxS/YA/ZD+Lry3fiT4KaJp+oSbib7QVbSpS7MWaRhblEkcljlpFbP4DHxP8A+CInh+aNrn4L/GrULR1ztsvE9klwH+VetxbiPb8wY/6puGA/hJb4o/8ABMn9sj4XpPdt8LW8V6fCxUXfhe5XUC5ywG23GLgghd2fKxhlBwSQNV0jX/Cesy6Vremaho2rafIPNtrqB7ae3cYYZVgGU8g8gdjXwu/b/wD2u/hHHFaeG/jXreoWEW0Cx15l1WIIqhRGpuA7xIAoAWNkA/E18K/+C2/iG1SGx+NXwbstQAUB9R8N3jWzZBOT9nm3hiV29JVGQT0YBfhV/wAFOf2OfioIbUfE3/hENRmyfsXiu3On7ANvLXGWthy2MebngnGBmtC8Q+H/ABTpses+GNd0/V7Cb/V3dhcpcRP9HQkHt3qqqqqqqqqv2w/+CT/hH44eJr74m/BTxDY+C/FGps82pabdwE6bfzkZ80eWC9u7Ny5VXVid20NuL+I/+CTn7bOiSxxaZ8PdH8QK7ODJp3iG0QIFxgn7RJEcNk4wCeDnHGfhz/wRv/ag8UapZJ4/1Twv4K0uRUe8mkvP7QuYARyqQw/JI6nggyqvXDnv+yn+yJ8LP2SPBL+GfAkEt/q+o7H1rXr1F+06hIo4Hy8RxKSdkQyFBJJZiztX7V3/AATM/ak+L/7Rfj34l+DNJ8OSaJ4h1Zrmye41hIpDGVUfMpHB4PFf8Ezv2Wvix+yz4F8Z+H/ixZ6ZBd65q0FzaCxvRcgxpFtOSAMHPav2rvhx4m+L37Onj34aeDYreXW/EOkva2SXEoijMhZT8zHoMA81/wAOhv2yv+gJ4U/8Hqf/ABNeAtHvfD3gXw5oGpKgu9M0m1tpwjbgJI4lVsHuMg81+2Z+wD8M/wBry3h8Qz6hL4Y8d6fbrb2mvQRmZZIFLssE8O4K6bpCQww445I+U+OP+CRH7Y/hado/Dmg+GfGUXmBUk0nXIoDtIJ3EXfk4xgAgE8sMZGSPDv8AwSc/bZ1yWSLU/h7o/h9UZAsmo+IbRw4bOSPs8kpwuBnIB5GM84/YA/ZW+IH7IPwj13wR458W6T4gudW1htUjtNHibyrdzBHG4WWUI0hfykHzKqjaP7xr9pT9gH9vX9pf4w658V/FmjeFI3v5PK0+zXXk22NmhIhgXj+FTkn+JmZv4q/4JsfsE+Jv2XZfEfxF+Lq6Y/jTVlGn6fBZTC4SxsRtd28zj55XCggD5ViXk7yF/wCCkH7EmpftW+DNF8Q/De209PiD4auBFBJdz+Sl5YOT5kDN0BVysik9MSAcvXw6/wCCYv7efwp8d6D8R/BNr4Wsdc8O30d5ZTDXlxvQ52uABuRhlWU8MrMp4Jr/AIKp3mt6h+wn9t8TaQml6vNq2lNfWSTLMtvPuO9FdeGUNnDdxgkA8DwFo9l4i8deHPD+pK7Wmp6ta2s4RtrGOSVVbB7HBPNfG7/gjn+0H4Q1y9uPgpfaT498PtIWsYZr2PT9RRCV+SVZikJIDH51kwwQnahISvhV/wAEf/2q/GmswwfEe30T4e6SJCLm6u7+HUZwg28xQ2sjK5O44DyRj5WyR8u79mz9mf4ZfssfDxPh98NbO4ZJpjcajqV6yvdahORjfKygDAUBVVQFUDpksTNDDcQvbzxJLHIpV0dchgRggg9QR2r9pb/gjTaa/rd14q/Zk8VaZoUNy29/DOuPN9ngIXnyLlRI4BIyEdSAW++BgDVf+CVf7cmn38tnZ/CG01OKPG26tfEmnLHJkAnaJZ0fgnByo5BxkYJ8Ef8ABHP9rDxJPAfFV14P8J2zxo8xvNTN1LHkcoEt0dWdeh+cL6Ma0m2v7PSrKz1TUv7QvYLeNLm78kQ/aJAoDSbF4TcQTtHAziv2wf2Jvhl+2B4ZtrXxFO+geKtJUrpPiS1tlllgQnJhlQlfOhJJbYWUq3Ksu5t3xB/4JB/tgeErop4T0rwz44tmkxHJpWsx2zhCWwXS88kKcKuQrOAXABbBI8L/APBJj9tfxBdfZtW8BaJ4ajEkaefqniG1kTDEgvi1eZsLjJ4zyNoY5A+Bng/xL8PPgv4E8AeMb2xvNb8M+HbHTL64spHkhmlt4EiLq0gVmB2ZyygknOB0r/goL/wT6T9rRNN8feAdbsdF8faLam0H24FbXU7UFmWORkUsjqzHa+CMMwI6FdQ/4JXftzWV7La23wbt7+OJsJc2/iTTVjlHqokuFbH1UGv+Cf8A8BvHv7OH7OOnfDX4kpp6a5Hql5dyR2Vz56Ikj/KC2ACcLnjI5HOcgfto/wDBOX4cftYXTeOdI1d/CPxCjhWI6ssLXEGoRouI47iHcOQAFEqYYA8iQKqjxj/wSP8A20fDNz9n0Twn4c8WpuA8/R9fgiTG0HOLwwNjJ29M5B7YJ8Mf8Elf21dflWLVfBGg+G1MyxmTVPEFtIAp6yf6M0x2r3GN3oDX7Cn7NvjT9lb4Fr8LvHPi/TNfv21aa/RtOgZILNJY4g0Cs4VpQJEkbeyqT5mMAAV8Z/gx8PPj/wDDzUvhl8TdDTUtH1FcgjCzWkwB2Twvg+XKmThvcggqzKfjD/wR4/aX8G6vN/wqebR/iFo7N/o7peRabdgE9JI7h1jBHqsjZ9B0r4Hf8Ecfj74w1q1ufjfqOmeAdBjYNdwQXcWo6hKAxBSNYWaFCVX77SHbvU7HIZR+1N/wSd+L2v8Axl1HVf2eNA8Pw+Bm0/T4NPtbrUxBJbmC0igdW3bmcs0RkLnljIc+p/4Jtfs4/Ez9mD4F654A+Ktrp8GrX/iy41OFbK7Fwhge0tIlJYAYO6F+Pp61VVVVVVVVX7QHhjUPG/wG+JHgvSVZr7X/AAjqun2ypGZGMs1pLGuFXljlhwOTWh6vc+H9b0/XrJInuNNuormJZQSpeNgwDAEHGQM4I+taPq2m6/pFjruj3a3VhqNtHc2s6ggSxSKGRhnnBUg8+tftzft3WfwKtZ/hj8Kb62vPiFcRj7TdFVmi0JGGQzqQVecggrGQQoIZxjar/sd/t2+Ef2i7OLwd4xNj4c8f28aqLNpwsOr4X5pLbdg7shiYeWVeQWAYrVV+1j+2b8Pv2Z9AuNOju7bWvHl1BnTdBRyfLJHE1yV/1cQyDtJDP0Xjcy/sp/8ABSTx5afE640P4/6w2seH/Ft+vl3qRBDokz4RdiL/AMu3Chk6r98ZJcPXxf8AFVl47+LXjbxxpq7bTxD4i1DUbdck4jnuHkXqAejDqAfYdK/Yl8Pah4Y/ZH+Emk6oEFz/AMIrZ3DKoI2CdBMqkEAhgsgDDHBBHPWqqqqqr9qf9kT4a/teeH9D8N/EnW/Eum23h+8ku7Z9DuYIXd3TYQ5lhlBGPQDnvX/DlT9lf/ofvir/AODWw/8AkKv+Ci37Inw1/ZD8aeEPDnw11vxLqVtr+lzXdy+uXMEzo6ShAEMMMQAx6g896/4cqfsr/wDQ/fFX/wAGth/8hV8PP+CRP7Nnw08f+GfiNoPjb4lz6l4V1i01Wziu9SsmhkmtpllRZAtorFCyAEBlOM4I61VVVVVVVVVVVVX/AAUh/bV+Kn7Hn/Cu/wDhWWgeFNT/AOEu/tT7b/btpcTeX9m+yeX5fkzxYz9ofOd2cLjHOf8Ah9X+1R/0IXwp/wDBVf8A/wAm1/w+r/ao/wChC+FP/gqv/wD5Nr/h9X+1R/0IXwp/8FV//wDJtf8AD6v9qj/oQvhT/wCCq/8A/k2v+H1f7VH/AEIXwp/8FV//APJtf8Pq/wBqj/oQvhT/AOCq/wD/AJNr/h9X+1R/0IXwp/8ABVf/APybX/D6v9qj/oQvhT/4Kr//AOTa/wCH1f7VH/QhfCn/AMFV/wD/ACbX/D6v9qj/AKEL4U/+Cq//APk2v+H1f7VH/QhfCn/wVX//AMm1/wAPq/2qP+hC+FP/AIKr/wD+Ta/4fV/tUf8AQhfCn/wVX/8A8m1/w+r/AGqP+hC+FP8A4Kr/AP8Ak2v+H1f7VH/QhfCn/wAFV/8A/Jtf8Pq/2qP+hC+FP/gqv/8A5Nr/AIfV/tUf9CF8Kf8AwVX/AP8AJtf8Pq/2qP8AoQvhT/4Kr/8A+Ta/4fV/tUf9CF8Kf/BVf/8AybX/AA+r/ao/6EL4U/8Agqv/AP5Nr/h9X+1R/wBCF8Kf/BVf/wDybX/D6v8Aao/6EL4U/wDgqv8A/wCTa/4fV/tUf9CF8Kf/AAVX/wD8m1/w+r/ao/6EL4U/+Cq//wDk2v8Ah9X+1R/0IXwp/wDBVf8A/wAm1/w+r/ao/wChC+FP/gqv/wD5Nr/h9X+1R/0IXwp/8FV//wDJtTfGvxV+0VIfjH42sNJsta8Qf8fNvpcUkVunkfuE2LI7sMpCpOWPJPQcD9pXxb4n8W+JvDnw7WD7Ra+H7do9KtbaJmkllupAz8ZO52YIoCgcKowTyf8AgiD8Pkg8NfE34rXETs17fWmi2jlSAghRppgDuwxPn2+QVyu0YPzEVVVX/BZH4WR+MP2ZLD4kWtpC174A1yGWWdtoaOzuyLeRVzz80zWhIH9zJHGR+zX+0j4//ZX+I/8Aws/4cWuk3epPp81hLbarHNJbzQyFWIdYpI2bDRowBbGVBxwK/wCH1f7VH/QhfCn/AMFV/wD/ACbX/D6v9qj/AKEL4U/+Cq//APk2v+H1f7VH/QhfCn/wVX//AMm1/wAPq/2qP+hC+FP/AIKr/wD+Ta/4fV/tUf8AQhfCn/wVX/8A8m1/w+r/AGqP+hC+FP8A4Kr/AP8Ak2v+H1f7VH/QhfCn/wAFV/8A/Jtf8Pq/2qP+hC+FP/gqv/8A5Nr/AIfV/tUf9CF8Kf8AwVX/AP8AJtf8Pq/2qP8AoQvhT/4Kr/8A+Ta/4fV/tUf9CF8Kf/BVf/8AybX/AA+r/ao/6EL4U/8Agqv/AP5Nr/h9X+1R/wBCF8Kf/BVf/wDybX/D6v8Aao/6EL4U/wDgqv8A/wCTa/4fV/tUf9CF8Kf/AAVX/wD8m1/wT0/ag8f/ALWvwW1n4jfEbSPD+nalpviifSootEt5oYTDHa2soZhLLK2/dO4JDAYC8dSaqqqqr9p7/gq3+0R8Ffj/AOOPhV4V8G/Dm60nw1qjWlpNqGnXrzugVTl2S7RSeeyge1f8Pq/2qP8AoQvhT/4Kr/8A+Ta/4fV/tUf9CF8Kf/BVf/8AybX/AA+r/ao/6EL4U/8Agqv/AP5Nr/h9X+1R/wBCF8Kf/BVf/wDybX/D6v8Aao/6EL4U/wDgqv8A/wCTa/4fV/tUf9CF8Kf/AAVX/wD8m1/w+r/ao/6EL4U/+Cq//wDk2v8Ah9X+1R/0IXwp/wDBVf8A/wAm1/w+r/ao/wChC+FP/gqv/wD5Nr/h9X+1R/0IXwp/8FV//wDJtf8AD6v9qj/oQvhT/wCCq/8A/k2v+H1f7VH/AEIXwp/8FV//APJtf8Pq/wBqj/oQvhT/AOCq/wD/AJNr/h9X+1R/0IXwp/8ABVf/APybX/D6v9qj/oQvhT/4Kr//AOTa/wCH1f7VH/QhfCn/AMFV/wD/ACbX/D6v9qj/AKEL4U/+Cq//APk2v+H1f7VH/QhfCn/wVX//AMm1/wAPq/2qP+hC+FP/AIKr/wD+Ta/4fV/tUf8AQhfCn/wVX/8A8m1/w+r/AGqP+hC+FP8A4Kr/AP8Ak2v+H1f7VH/QhfCn/wAFV/8A/Jtf8Pq/2qP+hC+FP/gqv/8A5Nr/AIfV/tUf9CF8Kf8AwVX/AP8AJtf8Pq/2qP8AoQvhT/4Kr/8A+Ta/4fV/tUf9CF8Kf/BVf/8AybX/AA+r/ao/6EL4U/8Agqv/AP5Nq4/aF8aftQP/AMLV8faZolhq04+yNDo8EsUASLhSFlkkbJzz82PYV8d/h/428X+Ir7xD4W8Mahq2n+GNAhudYms4TL9hgaaf99Iq5KxjacvjavGSMjP/AATI/ZFh/Z1+DkfjnxfpCR/EHx3ClzfmWPMmnWX3oLQHJ2kgiWQAKS7BWz5Smqqv+CiX7G1j+1L8KJta8K6bEPiP4Tt3n0KZQkbahGPmexkcjJDjcYwSAspBJVWcnwP4x8e/Bfx7pXj7wncXeh+I/DWoFrWaWDBhnj4kjdHGDw210YdGII5r/h9X+1R/0IXwp/8ABVf/APybX/D6v9qj/oQvhT/4Kr//AOTa/wCH1f7VH/QhfCn/AMFV/wD/ACbX/D6v9qj/AKEL4U/+Cq//APk2v+H1f7VH/QhfCn/wVX//AMm1/wAPq/2qP+hC+FP/AIKr/wD+Ta/4fV/tUf8AQhfCn/wVX/8A8m1/w+r/AGqP+hC+FP8A4Kr/AP8Ak2v+H1f7VH/QhfCn/wAFV/8A/Jtf8Pq/2qP+hC+FP/gqv/8A5Nr/AIfV/tUf9CF8Kf8AwVX/AP8AJtf8Pq/2qP8AoQvhT/4Kr/8A+Ta/4fV/tUf9CF8Kf/BVf/8AybX/AA+r/ao/6EL4U/8Agqv/AP5Nr/h9X+1R/wBCF8Kf/BVf/wDybX/BPT/goX8af2tfjTrPw4+I3hnwTp2m6b4Xn1WKXRLK6hmM0d1axBWMtxKuzbO5ICg5C89Qaqqqqr9p7/gq3+0R8Ffj/wCOPhV4V8G/Dm60nw1qjWlpNqGnXrzugVTl2S7RSeeyge1f8Pq/2qP+hC+FP/gqv/8A5Nr/AIfV/tUf9CF8Kf8AwVX/AP8AJtf8Pq/2qP8AoQvhT/4Kr/8A+Ta/4fV/tUf9CF8Kf/BVf/8AybX/AA+r/ao/6EL4U/8Agqv/AP5Nr/h9X+1R/wBCF8Kf/BVf/wDybX/D6v8Aao/6EL4U/wDgqv8A/wCTa/4fV/tUf9CF8Kf/AAVX/wD8m1/w+r/ao/6EL4U/+Cq//wDk2v8Ah9X+1R/0IXwp/wDBVf8A/wAm1/w+r/ao/wChC+FP/gqv/wD5Nr/h9X+1R/0IXwp/8FV//wDJtf8AD6v9qj/oQvhT/wCCq/8A/k2v+H1f7VH/AEIXwp/8FV//APJtf8Pq/wBqj/oQvhT/AOCq/wD/AJNr/h9X+1R/0IXwp/8ABVf/APybX/D6v9qj/oQvhT/4Kr//AOTa/wCH1f7VH/QhfCn/AMFV/wD/ACbX/D6v9qj/AKEL4U/+Cq//APk2v+H1f7VH/QhfCn/wVX//AMm1/wAPq/2qP+hC+FP/AIKr/wD+Ta/4fV/tUf8AQhfCn/wVX/8A8m1/w+r/AGqP+hC+FP8A4Kr/AP8Ak2v+H1f7VH/QhfCn/wAFV/8A/Jtf8Pq/2qP+hC+FP/gqv/8A5Nr/AIfV/tUf9CF8Kf8AwVX/AP8AJtN/wWQ/ad8TL/wjl94G+F6W2qj7JM8Ol3wdUk+RipN4QDgnGQRntVf8E6f28Pi9+15408X+HPiT4d8H6bbeH9Lhu7Z9Ds7mF3d5dhDmWeUEY9AOe9VVVX7T3/BVv9oj4K/H/wAcfCrwr4N+HN1pPhrVGtLSbUNOvXndAqnLsl2ik89lA9q/4fV/tUf9CF8Kf/BVf/8AybX/AA+r/ao/6EL4U/8Agqv/AP5Nr/h9X+1R/wBCF8Kf/BVf/wDybX/D6v8Aao/6EL4U/wDgqv8A/wCTa/4fV/tUf9CF8Kf/AAVX/wD8m1/w+r/ao/6EL4U/+Cq//wDk2v8Ah9X+1R/0IXwp/wDBVf8A/wAm1/w+r/ao/wChC+FP/gqv/wD5Nr/h9X+1R/0IXwp/8FV//wDJtf8AD6v9qj/oQvhT/wCCq/8A/k2v+H1f7VH/AEIXwp/8FV//APJtf8Pq/wBqj/oQvhT/AOCq/wD/AJNr/h9X+1R/0IXwp/8ABVf/APybX/D6v9qj/oQvhT/4Kr//AOTa/wCH1f7VH/QhfCn/AMFV/wD/ACbX/BPT9qDx/wDta/BbWfiN8RtI8P6dqWm+KJ9Kii0S3mhhMMdrayhmEssrb907gkMBgLx1Jqqqqqv2zf8AgqB8fv2df2lPGHwb8E+Efh/faL4f+w/ZZ9V0+8kuX86xgnfe0d0inDysBhRwBnJyT/w+r/ao/wChC+FP/gqv/wD5Nr/h9X+1R/0IXwp/8FV//wDJtf8AD6v9qj/oQvhT/wCCq/8A/k2v+H1f7VH/AEIXwp/8FV//APJtf8Pq/wBqj/oQvhT/AOCq/wD/AJNr/h9X+1R/0IXwp/8ABVf/APybX/D6v9qj/oQvhT/4Kr//AOTa/wCH1f7VH/QhfCn/AMFV/wD/ACbX/D6v9qj/AKEL4U/+Cq//APk2v+H1f7VH/QhfCn/wVX//AMm1/wAPq/2qP+hC+FP/AIKr/wD+Ta/4fV/tUf8AQhfCn/wVX/8A8m1/w+r/AGqP+hC+FP8A4Kr/AP8Ak2v+H1f7VH/QhfCn/wAFV/8A/Jtf8Pq/2qP+hC+FP/gqv/8A5Nr/AIfV/tUf9CF8Kf8AwVX/AP8AJtf8Pq/2qP8AoQvhT/4Kr/8A+Ta/4fV/tUf9CF8Kf/BVf/8AybX/AA+r/ao/6EL4U/8Agqv/AP5Nr/h9X+1R/wBCF8Kf/BVf/wDybX/D6v8Aao/6EL4U/wDgqv8A/wCTa/4fV/tUf9CF8Kf/AAVX/wD8m1/w+r/ao/6EL4U/+Cq//wDk2v8Ah9X+1R/0IXwp/wDBVf8A/wAm1/w+r/ao/wChC+FP/gqv/wD5Nr/h9X+1R/0IXwp/8FV//wDJtf8AD6v9qj/oQvhT/wCCq/8A/k2pP+Cuv7SfxLjb4ca74J+GkGm+KgdKvJbTTb5Jo4bj907RlrtlDhXJBKkA4yD0rxB/yAdS/wCvOX/0A18N/hF4l8afFPwD8OtW0XU9LHjnVtPtbSa5tng8yC6ljUTozIcpskD7wrDHOCKhhht4Ut7eJI44lCoiLtCgDAAA6DHaqqv+CyXwzu9M/ad8N+LdItJbg+OPD8MSwxRl5Jby3laEqqqvzZja2AGWbJPQbRX7Ln/BQD41/skeD9V8D/DnRfCOqaXq+pf2jImu2lzM0U5jSNjH5M8QAZY485B+6K/4fV/tUf8AQhfCn/wVX/8A8m1/w+r/AGqP+hC+FP8A4Kr/AP8Ak2v+H1f7VH/QhfCn/wAFV/8A/Jtf8Pq/2qP+hC+FP/gqv/8A5Nr/AIfV/tUf9CF8Kf8AwVX/AP8AJtf8Pq/2qP8AoQvhT/4Kr/8A+Ta/4fV/tUf9CF8Kf/BVf/8AybX/AA+r/ao/6EL4U/8Agqv/AP5Nr/h9X+1R/wBCF8Kf/BVf/wDybX/D6v8Aao/6EL4U/wDgqv8A/wCTa/4fV/tUf9CF8Kf/AAVX/wD8m1/w+r/ao/6EL4U/+Cq//wDk2v8Ah9X+1R/0IXwp/wDBVf8A/wAm1/w+r/ao/wChC+FP/gqv/wD5Nr/h9X+1R/0IXwp/8FV//wDJtfBDxrqvxL+C3gD4ja7b2kGpeKvC+m6reRWiMkMc1xaxyusYZmYIGcgAsSBjJPWqqqqqr43+NdV+GnwW8f8AxG0G3tJ9S8K+F9S1Wziu0ZoZJra1klRZArKxQsgBAZTjOCOtf8Pq/wBqj/oQvhT/AOCq/wD/AJNr/h9X+1R/0IXwp/8ABVf/APybX/D6v9qj/oQvhT/4Kr//AOTa/wCH1f7VH/QhfCn/AMFV/wD/ACbX/D6v9qj/AKEL4U/+Cq//APk2v+H1f7VH/QhfCn/wVX//AMm1/wAPq/2qP+hC+FP/AIKr/wD+Ta/4fV/tUf8AQhfCn/wVX/8A8m1/w+r/AGqP+hC+FP8A4Kr/AP8Ak2v+H1f7VH/QhfCn/wAFV/8A/Jtf8Pq/2qP+hC+FP/gqv/8A5Nr/AIfV/tUf9CF8Kf8AwVX/AP8AJtf8Pq/2qP8AoQvhT/4Kr/8A+Ta/4fV/tUf9CF8Kf/BVf/8AybX/AA+r/ao/6EL4U/8Agqv/AP5Nr/h9X+1R/wBCF8Kf/BVf/wDybX/D6v8Aao/6EL4U/wDgqv8A/wCTa/4fV/tUf9CF8Kf/AAVX/wD8m1/w+r/ao/6EL4U/+Cq//wDk2v8Ah9X+1R/0IXwp/wDBVf8A/wAm1/w+r/ao/wChC+FP/gqv/wD5Nr/h9X+1R/0IXwp/8FV//wDJtf8AD6v9qj/oQvhT/wCCq/8A/k2v+H1f7VH/AEIXwp/8FV//APJtf8Pq/wBqj/oQvhT/AOCq/wD/AJNr/h9X+1R/0IXwp/8ABVf/APybX/D6v9qj/oQvhT/4Kr//AOTa/wCH1f7VH/QhfCn/AMFV/wD/ACbVz+2r8VP2xNv/AAszQPCmmf8ACI5+w/2FaXEHmfaf9Z5nnTy5x9nTGNuMtnORj9o6z1XVdI0HRtItbu8nu9Qby7S3RpGlcRnGEXJYgFscdz61/wAEW/hle3fx68e+O9UsHiPg3QV014Z4tklvd3c/GQSGVglpcIRtP3jkqcA1VVVVVVVVVVVftafsJfBv9rTSnu9etf8AhHfGcMYWy8UafbK042qQkc6HAuIgSDsJVhjCumTn9pf9kH41/sqeIRpfxK8O+ZpN1Js03xDYbpbC+JBIVZMApJhWzFIFf5SQCuGP/BIb4oftLap8Uz8L/DOt/wBpfDDTbdrvX7bVC0semoVZYvsrZzHI8mP3Y+RgrsRkZFVVVVVVV+09/wAFW/2iPgr8f/HHwq8K+DfhzdaT4a1RrS0m1DTr153QKpy7JdopPPZQPav+H1f7VH/QhfCn/wAFV/8A/Jtf8Pq/2qP+hC+FP/gqv/8A5Nr/AIfV/tUf9CF8Kf8AwVX/AP8AJtf8Pq/2qP8AoQvhT/4Kr/8A+Ta/4fV/tUf9CF8Kf/BVf/8AybX/AA+r/ao/6EL4U/8Agqv/AP5Nr/h9X+1R/wBCF8Kf/BVf/wDybX/D6v8Aao/6EL4U/wDgqv8A/wCTa/4fV/tUf9CF8Kf/AAVX/wD8m1/w+r/ao/6EL4U/+Cq//wDk2v8Ah9X+1R/0IXwp/wDBVf8A/wAm1/w+r/ao/wChC+FP/gqv/wD5Nr/h9X+1R/0IXwp/8FV//wDJtf8AD6v9qj/oQvhT/wCCq/8A/k2v+H1f7VH/AEIXwp/8FV//APJtf8Pq/wBqj/oQvhT/AOCq/wD/AJNr/h9X+1R/0IXwp/8ABVf/APybX/D6v9qj/oQvhT/4Kr//AOTa/wCH1f7VH/QhfCn/AMFV/wD/ACbX/D6v9qj/AKEL4U/+Cq//APk2v+H1f7VH/QhfCn/wVX//AMm1/wAPq/2qP+hC+FP/AIKr/wD+Ta/4fV/tUf8AQhfCn/wVX/8A8m1/w+r/AGqP+hC+FP8A4Kr/AP8Ak2v+H1f7VH/QhfCn/wAFV/8A/Jtf8Pq/2qP+hC+FP/gqv/8A5Nr/AIfV/tUf9CF8Kf8AwVX/AP8AJtXH7QvjT9qB/wDhavj7TNEsNWnH2RodHgligCRcKQsskjZOefmx7Cv2ivGPxX1nwWnwr0S3vZ/AmgzjxDqqWdu7Kl1ODbpLcMMgIq2+EJAAaR8kllx/wSe/ZPT4zfFhvjV4x0x5fCPw8uke1SWI+Vf6rjdEmeMiAFJmHqYQQVcg1VVVVVVVVVeOvhl8OPihpqaP8SPAfh/xTZR7vKg1jTYrtYi2MlBIp2ngcjB4HpXxV/4JI/slfELzrvwrpOt+AtQkwQ+iXxkgLDdndBP5gAORlUKfdXGPm3fFT/git8afDqS33wk+JHhzxjbxqWFpqET6VdscjCIMyRMcE5ZpIx8vTnA+KX7Jf7SfwXjluPiT8GPEuk2cO7zL9LT7VaJtUsc3EBeIfKrH73RSegNeCPiP8Qfhnqg1v4deONd8M34YMbjSdQltHYgMBuMbDcMMwweMMw6E1/wTS/bW/ax+N/x00/4R+OPFWn+KvDFrpd5f6rdahYIl7bQRoqRmOWELuP2h4VJlDZWR+c7NtVVVVVVVVVVVVVVVVVVVX/BXn/kzTUv+xg07/wBDavhN/wAlU8Gf9jBY/wDpQlVVVVVVVVVVVVVVVVVVVVVVVVVX7W/web4DftHePfhfFatBYaXq0kmlqxB/0GYCa25HBIikQHGOQRgEED/gm78bLb4rfs8af4TvLvfr3w+2aTeRsRuNrgmzkAHRfKXyhnkmBz3FftofsOeHv2k7H/hMfCEtnofxBsotiXciFYdUjUYWK428hhwFlAJA+Ugjbt8X+DfHPwo8YT+GPF+jah4d8RaRKjPBLmOWFuGR1ZTyOjK6nB4INfAj/gqb8TvAGmw+G/i5oC+O7CBdsOpC5+zaggz/AMtHIZJwBwMhG7l2r/h6v+zB/Yv9qfYfG/2n/oG/2TF5/wB7b187yunzff6e/FfHH/gq1488W2N14e+C3hVfCNrOrRnV76YXN8VI6xoB5cLckZzIehBU9LKx8cfFHxgtpYwaz4q8Ua7cs21RJeXd7McszE8u7YBYk54BJPBr9i3/AIJ7ab8Gbmx+KXxhS21XxvDl7DTkKy2ujtnh93IlnAGQ33UJO3JAev26fjRafBf9nPxNewXkCa74itn0jR4Xk2u8k42SSKBzmKIvJnpuVQfvV8Gfhnq/xm+K/hL4V6Er/a/E+rQWQkVSfIjdx5kpwCdsce92ODgKTWnafZaTp9rpWm26W9pZQpDbwoMCONAFVR7AACqqqqqqq/4Le/8AJU/hp/2L91/6UCqqqqqqqqqqqqqqv2y/2IPCn7Zn/CH/APCTeONV8O/8If8AbvI+w20cvn/avI3bt/Tb9nXGP7xr/hyF8Kv+i4eK/wDwXW/+Nf8ADkL4Vf8ARcPFf/gut/8AGv8AhyF8Kv8AouHiv/wXW/8AjX/DkL4Vf9Fw8V/+C63/AMa/4chfCr/ouHiv/wAF1v8A41/w5C+FX/RcPFf/AILrf/Gv+HIXwq/6Lh4r/wDBdb/41/w5C+FX/RcPFf8A4Lrf/Gv+HIXwq/6Lh4r/APBdb/41/wAOQvhV/wBFw8V/+C63/wAa/wCHIXwq/wCi4eK//Bdb/wCNf8OQvhV/0XDxX/4Lrf8Axr/hyF8Kv+i4eK//AAXW/wDjX/DkL4Vf9Fw8V/8Agut/8a/4chfCr/ouHiv/AMF1v/jX/DkL4Vf9Fw8V/wDgut/8a/4chfCr/ouHiv8A8F1v/jX/AA5C+FX/AEXDxX/4Lrf/ABr/AIchfCr/AKLh4r/8F1v/AI1/w5C+FX/RcPFf/gut/wDGv+HIXwq/6Lh4r/8ABdb/AONf8OQvhV/0XDxX/wCC63/xr/hyF8Kv+i4eK/8AwXW/+Nf8OQvhV/0XDxX/AOC63/xr/hyF8Kv+i4eK/wDwXW/+Nf8ADkL4Vf8ARcPFf/gut/8AGv8AhyF8Kv8AouHiv/wXW/8AjXxE+B+k/s3+MdQ+C+h65d6xZeHfL8q9u4ljkl8+Jbg5VeBgzFfoBX7Mn7Ang/4sa/4P/ai1Hx/rFhqnhvxHbXcWlQ2sbQStYzpIisx+YBioBx0zUNnaW0k81tbRRSXUgkndECmVwqoGYj7x2oi5PZQOgFVVV8RvAHhn4q+BNd+HHjKze60TxFYyWV7EjlGMbjBKsPusOCD2IBrx9/wRg+AnifxZfa34N8b+IPB2kXPl/ZtEtY/tkdrtjVW2y3DvI25lZzuY4LkDAAFf8OQvhV/0XDxX/wCC63/xr/hyF8Kv+i4eK/8AwXW/+Nf8OQvhV/0XDxX/AOC63/xr/hyF8Kv+i4eK/wDwXW/+Nf8ADkL4Vf8ARcPFf/gut/8AGv8AhyF8Kv8AouHiv/wXW/8AjX/DkL4Vf9Fw8V/+C63/AMa/4chfCr/ouHiv/wAF1v8A41/w5C+FX/RcPFf/AILrf/Gv+HIXwq/6Lh4r/wDBdb/41/w5C+FX/RcPFf8A4Lrf/Gv+HIXwq/6Lh4r/APBdb/41/wAOQvhV/wBFw8V/+C63/wAa/wCHIXwq/wCi4eK//Bdb/wCNf8OQvhV/0XDxX/4Lrf8Axr9kT9ljw/8Ash/DXUvhr4b8Vah4gttS1yXVnub6BInR5IIISgCcYAt1OfVjVVVVVV8b/wDgkt8Ovjf8WPE/xY1X4t+I9Nu/E9813LaW9jC8cLEAYUsckcd6/wCHIXwq/wCi4eK//Bdb/wCNf8OQvhV/0XDxX/4Lrf8Axr/hyF8Kv+i4eK//AAXW/wDjX/DkL4Vf9Fw8V/8Agut/8a/4chfCr/ouHiv/AMF1v/jX/DkL4Vf9Fw8V/wDgut/8a/4chfCr/ouHiv8A8F1v/jX/AA5C+FX/AEXDxX/4Lrf/ABr/AIchfCr/AKLh4r/8F1v/AI1/w5C+FX/RcPFf/gut/wDGv+HIXwq/6Lh4r/8ABdb/AONf8OQvhV/0XDxX/wCC63/xr/hyF8Kv+i4eK/8AwXW/+Nf8OQvhV/0XDxX/AOC63/xr/hyF8Kv+i4eK/wDwXW/+Nf8ADkL4Vf8ARcPFf/gut/8AGv8AhyF8Kv8AouHiv/wXW/8AjX/DkL4Vf9Fw8V/+C63/AMa/4chfCr/ouHiv/wAF1v8A41/w5C+FX/RcPFf/AILrf/Gv+HIXwq/6Lh4r/wDBdb/41/w5C+FX/RcPFf8A4Lrf/Gv+HIXwq/6Lh4r/APBdb/41/wAOQvhV/wBFw8V/+C63/wAa/wCHIXwq/wCi4eK//Bdb/wCNf8OQvhV/0XDxX/4Lrf8Axr/hyF8Kv+i4eK//AAXW/wDjXxO/Z00T9lnxS/wm8P8AiO91y0t4Euxd3kKxyFpRkrheMDFf8E5fhXp9u3in43Lqtwb6/C+HmsSg8tY4NlwJQ3XcTcFSOmFFVVVX7W3/AAT08AftZeItD8Q6p4x1Dwq+i288Xk6XYQstw80nmPK+cfOW6nv1Nf8ADkL4Vf8ARcPFf/gut/8AGv8AhyF8Kv8AouHiv/wXW/8AjX/DkL4Vf9Fw8V/+C63/AMa/4chfCr/ouHiv/wAF1v8A41/w5C+FX/RcPFf/AILrf/Gv+HIXwq/6Lh4r/wDBdb/41/w5C+FX/RcPFf8A4Lrf/Gv+HIXwq/6Lh4r/APBdb/41/wAOQvhV/wBFw8V/+C63/wAa/wCHIXwq/wCi4eK//Bdb/wCNf8OQvhV/0XDxX/4Lrf8Axr/hyF8Kv+i4eK//AAXW/wDjX/DkL4Vf9Fw8V/8Agut/8a/4chfCr/ouHiv/AMF1v/jX/DkL4Vf9Fw8V/wDgut/8a/ZE/wCCdPgr9kP4k6l8SfDfxG1vxBc6locukvbX1pFEiJJPBKXBTnINuox0+Y1VVVVVfG//AIJLfDr43/FjxP8AFjVfi34j0278T3zXctpb2MLxwsQBhSxyRx3r/hyF8Kv+i4eK/wDwXW/+Nf8ADkL4Vf8ARcPFf/gut/8AGv8AhyF8Kv8AouHiv/wXW/8AjX/DkL4Vf9Fw8V/+C63/AMa/4chfCr/ouHiv/wAF1v8A41/w5C+FX/RcPFf/AILrf/Gv+HIXwq/6Lh4r/wDBdb/41/w5C+FX/RcPFf8A4Lrf/Gv+HIXwq/6Lh4r/APBdb/41/wAOQvhV/wBFw8V/+C63/wAa/wCHIXwq/wCi4eK//Bdb/wCNf8OQvhV/0XDxX/4Lrf8Axr/hyF8Kv+i4eK//AAXW/wDjX/DkL4Vf9Fw8V/8Agut/8a/4chfCr/ouHiv/AMF1v/jX/DkL4Vf9Fw8V/wDgut/8a/4chfCr/ouHiv8A8F1v/jX/AA5C+FX/AEXDxX/4Lrf/ABr/AIchfCr/AKLh4r/8F1v/AI1/w5C+FX/RcPFf/gut/wDGv+HIXwq/6Lh4r/8ABdb/AONf8OQvhV/0XDxX/wCC63/xr/hyF8Kv+i4eK/8AwXW/+Nf8OQvhV/0XDxX/AOC63/xr/hyF8Kv+i4eK/wDwXW/+Nf8ADkL4Vf8ARcPFf/gut/8AGtS/4IvfDDwxp114kt/jT4onl0mB7tIn0+AB2jUuASD0O3FV8QtB1z4TfELxt8MYdZu4/wCxdYvNGvhDOUW6FtO8ZDhThxlM4PFfsY/EFvil+yp8LvGc0yS3Nx4dt7W6kUgh7i2BtpmwFUAmSFyVAwpOBnGaqq/4LS/Eefw38DfBfgDT7v7PdeJvEhvGZXAcw2URJCjGR+9ngbcCCCoHIY4/Yr/Ztb9s/wCOl94F8VeMNTsY4NFuNXv9TUie4YRyRRD/AFhO4l5kBzzjntX/AA5C+FX/AEXDxX/4Lrf/ABr/AIchfCr/AKLh4r/8F1v/AI1/w5C+FX/RcPFf/gut/wDGv+HIXwq/6Lh4r/8ABdb/AONf8OQvhV/0XDxX/wCC63/xr/hyF8Kv+i4eK/8AwXW/+Nf8OQvhV/0XDxX/AOC63/xr/hyF8Kv+i4eK/wDwXW/+Nf8ADkL4Vf8ARcPFf/gut/8AGv8AhyF8Kv8AouHiv/wXW/8AjX/DkL4Vf9Fw8V/+C63/AMa/4chfCr/ouHiv/wAF1v8A41/w5C+FX/RcPFf/AILrf/Gv+HIXwq/6Lh4r/wDBdb/41/w5C+FX/RcPFf8A4Lrf/Gv2RP2WPD/7Ifw11L4a+G/FWoeILbUtcl1Z7m+gSJ0eSCCEoAnGALdTn1Y1VVVVVftH/wDBLH4f/tIfGjxF8aNb+KniDR73xD9m82ytbKGSOLyLWK3G1m5ORCGOe5Nf8OQvhV/0XDxX/wCC63/xr/hyF8Kv+i4eK/8AwXW/+Nf8OQvhV/0XDxX/AOC63/xr/hyF8Kv+i4eK/wDwXW/+Nf8ADkL4Vf8ARcPFf/gut/8AGv8AhyF8Kv8AouHiv/wXW/8AjX/DkL4Vf9Fw8V/+C63/AMa/4chfCr/ouHiv/wAF1v8A41/w5C+FX/RcPFf/AILrf/Gv+HIXwq/6Lh4r/wDBdb/41/w5C+FX/RcPFf8A4Lrf/Gv+HIXwq/6Lh4r/APBdb/41/wAOQvhV/wBFw8V/+C63/wAa/wCHIXwq/wCi4eK//Bdb/wCNf8OQvhV/0XDxX/4Lrf8Axr/hyF8Kv+i4eK//AAXW/wDjX/DkL4Vf9Fw8V/8Agut/8a/4chfCr/ouHiv/AMF1v/jX/DkL4Vf9Fw8V/wDgut/8a/4chfCr/ouHiv8A8F1v/jX/AA5C+FX/AEXDxX/4Lrf/ABr/AIchfCr/AKLh4r/8F1v/AI1/w5C+FX/RcPFf/gut/wDGv+HIXwq/6Lh4r/8ABdb/AONf8OQvhV/0XDxX/wCC63/xr/hyF8Kv+i4eK/8AwXW/+Nf8OQvhV/0XDxX/AOC63/xrxL/wR6+Gvwx8Oar8SbD4xeJb258J2U2rQ201hAqTvbIZVRiDkBigBI9a0zQ4fFGpWnhqe4eGPVp0tHlQZZBIwQsAe4DZr4beAdP+HHwz8JfDK2un1G08JaLY6XBcXEYDTC1hSNJGUcBj5atx0PSqqqaGFpVnMSGVFKq5X5lBwSAfQ7VyPYelf8OQvhV/0XDxX/4Lrf8Axr/hyF8Kv+i4eK//AAXW/wDjX/DkL4Vf9Fw8V/8Agut/8a/4chfCr/ouHiv/AMF1v/jX/DkL4Vf9Fw8V/wDgut/8a/4chfCr/ouHiv8A8F1v/jX/AA5C+FX/AEXDxX/4Lrf/ABr/AIchfCr/AKLh4r/8F1v/AI1/w5C+FX/RcPFf/gut/wDGv+HIXwq/6Lh4r/8ABdb/AONf8OQvhV/0XDxX/wCC63/xr/hyF8Kv+i4eK/8AwXW/+Nf8OQvhV/0XDxX/AOC63/xr/hyF8Kv+i4eK/wDwXW/+Nf8ADkL4Vf8ARcPFf/gut/8AGv8AhyF8Kv8AouHiv/wXW/8AjXwx8D2nwx+GvhP4a2F9Le23hPQ7LSYbmZQrzpbQJCrsBwCwQEgcZNVVVVVXxO8D2nxO+Gviz4a399LZW3izQ73SZrmFQzwJcwPCzqDwSA5IB4yK/wCHIXwq/wCi4eK//Bdb/wCNf8OQvhV/0XDxX/4Lrf8Axr/hyF8Kv+i4eK//AAXW/wDjX/DkL4Vf9Fw8V/8Agut/8a/4chfCr/ouHiv/AMF1v/jX/DkL4Vf9Fw8V/wDgut/8a/4chfCr/ouHiv8A8F1v/jX/AA5C+FX/AEXDxX/4Lrf/ABr/AIchfCr/AKLh4r/8F1v/AI1/w5C+FX/RcPFf/gut/wDGv+HIXwq/6Lh4r/8ABdb/AONf8OQvhV/0XDxX/wCC63/xr/hyF8Kv+i4eK/8AwXW/+Nf8OQvhV/0XDxX/AOC63/xr/hyF8Kv+i4eK/wDwXW/+Nf8ADkL4Vf8ARcPFf/gut/8AGv8AhyF8Kv8AouHiv/wXW/8AjX/DkL4Vf9Fw8V/+C63/AMa/4chfCr/ouHiv/wAF1v8A41/w5C+FX/RcPFf/AILrf/Gv+HIXwq/6Lh4r/wDBdb/41/w5C+FX/RcPFf8A4Lrf/Gv+HIXwq/6Lh4r/APBdb/41/wAOQvhV/wBFw8V/+C63/wAa/wCHIXwq/wCi4eK//Bdb/wCNf8OQvhV/0XDxX/4Lrf8Axr/hyF8Kv+i4eK//AAXW/wDjX/DkL4Vf9Fw8V/8Agut/8a8Uf8Ez9X/Z++zf8KdvfEfjz+3t/wDaPnWkUf2LyceVjaed/mydf7lfs1/saeNL74v+HviD8SdL1rwx/wAIHdrqWnI8CFL+UgoY2OSQADnIpbO0S6kvktoluZo0ikmCAO6IWKqW6kAu5A6As3qaqqqqqqqqqqqqrxV4T8MeOvD174S8Z+H9P1zRdSj8u70/ULZZ4Z1yCAyMCDggEehAI5FfAn9nT4Sfs2eG9Q8J/CHwz/ZGnanqD31wsk73EjOwACmVyXZEUBVDE4GeSSSaqqqqqqr43/8ABJb4dfG/4seJ/ixqvxb8R6bd+J75ruW0t7GF44WIAwpY5I471/w5C+FX/RcPFf8A4Lrf/Gv+HIXwq/6Lh4r/APBdb/41/wAOQvhV/wBFw8V/+C63/wAa/wCHIXwq/wCi4eK//Bdb/wCNf8OQvhV/0XDxX/4Lrf8Axr/hyF8Kv+i4eK//AAXW/wDjX/DkL4Vf9Fw8V/8Agut/8a/4chfCr/ouHiv/AMF1v/jX/DkL4Vf9Fw8V/wDgut/8a/4chfCr/ouHiv8A8F1v/jX/AA5C+FX/AEXDxX/4Lrf/ABr/AIchfCr/AKLh4r/8F1v/AI1/w5C+FX/RcPFf/gut/wDGv+HIXwq/6Lh4r/8ABdb/AONf8OQvhV/0XDxX/wCC63/xr/hyF8Kv+i4eK/8AwXW/+Nf8OQvhV/0XDxX/AOC63/xr/hyF8Kv+i4eK/wDwXW/+Nf8ADkL4Vf8ARcPFf/gut/8AGv8AhyF8Kv8AouHiv/wXW/8AjX/DkL4Vf9Fw8V/+C63/AMa/4chfCr/ouHiv/wAF1v8A41/w5C+FX/RcPFf/AILrf/Gv+HIXwq/6Lh4r/wDBdb/41/w5C+FX/RcPFf8A4Lrf/Gv+HIXwq/6Lh4r/APBdb/41/wAOQvhV/wBFw8V/+C63/wAa8Q/sCeP/AILaifA3wv0fxB4x0OJBOmqS28aFpH5dMKccYFfsPfs0658OtR8T/Ezx7p9/pusa1aLoh0e8hTy2tImEwmzkk7mlkQg8YSvBXgPwX8NvD8XhP4f+F9M8PaNBNNNFp+nWywQRvLI0khVFACgszHAAA6AAAAVVVVVVVVVVVVV8Tv2O/wBl/wCMSSH4g/BDwvfXMzBpL21tPsF25BB5uLYxykZHTdjk+pr9mb9hf4K/sn+LPE3i74Yza7PceJbWG1KardJOLKJHZmSFlRTtcmMnfuOYlwRkiqqqqqqqqqqqqqqqqqqqqvjt8Cfh/wDtG/D+b4afEu2vZ9Emuorl0s7kwSeZGSV+YZ456Vof/BJ39kHw9rWn6/pui+KBd6ZdR3NuX1x2AkjYMuRjkZA4qqqqqqqqqqqqqqqqqqqqqqqqqq/4K/8A7K174/8ABVj+0d4K05JdW8GWrW/iGNBh59N3bkmA7mF2cnvskY9Er9lv9oTWv2a/i1p/j7T4Wu9MmX7HrVivJubN2UyBBkDzF2hkJONyjPBNfD74g+D/AIqeD9N8d+A9cg1XRdVi8y3uIj+DIynlHUgqynBUggivjd+zv8Jf2htAj0H4m+F4r5rXcbLUIT5N3ZEjkxSjkA8EocoxCkqcDHxP/wCCR/xA025mvPhF8RNI1yy5ZLPWo2srlBk4QOgeOQ4x8x8sdeBjn/h25+2H/aX2D/hV1t5Pn+X9s/t+w8rbux5mPO37cc42bsfw54r4Z/8ABJP4n6vcx3XxW8e6H4esepttLD310cFgQSwSNM4Ughn4PIBGK+BX7Mvwe/Z10cad8OPDSpfSxeVd6zebZr68XIOJJQBhcgHYgVMgHbmvF3i3w14D8Naj4w8Ya1baTo2kwGe8vLltqRIP1JJIAUZLEgAEkCv2yv2nb/8Aae+Kba7Zpc2nhTRI2tfD1hO2GSMkGSd1BIEsrKCcdFWNcts3H/gjf+y7e2Cap+1D4y0d4hdQvpvhITpgtGSRc3Sg9jtESt3Hm9iDVVVVVVVV+1P+wf8ACD9rzX9D8R/EnxH4w0258P2clpbJod5bQo6O+8lxLBKSc+hA9qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq98KeF9TuXvdR8N6VdXEmN809nG7tgADJIJPAA/CrDTdO0q2FnpdhbWcAJIigiEagnqcKAKqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr/wx4a1W4N3qnh7TLycqAZbi0SRsDoMkE4rTtJ0vR4Da6RptrZQs5do7eFYlLYAJwoAzgAZ9hVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVUkccsbRSoro6lWVhkEHqCPSv+EG8E/wDQn6J/4L4v/ia/4Kr/AA+TwJ+2Z4ovbeJ47XxbY2etQIynGXj8mUgliWBmglbsAWKgYUZ/4Iw/EaHxL+zl4g+Hc86Ne+DfETskatytrdoJIyRuJBMqXX8KrgDGSGNVVf8ABZ34gf8ACR/tJaD4Dt2zb+EPDcfmgnkXNzI0j4wxGPKW36hTnOcjaa/4IgfD94fD/wAT/ipc22UvLyy0Wymyw2mFHmuFxnBz51seRkY4PJqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqri3guoJLW5hjmhlQpJHIoZXUjBBB4II4xSeCvBsTrLF4S0VHQ5VlsIgQR0IO2qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqvLO01C0msL62iuba5jaKaGZA6SIwwysp4IIJBB4Oa/wCCiX7A+tfs2eKbv4nfDrSmuvhbrV1mIQKztoEzni3mJJPlEnEchOOQjYbaX/Zh/a1+JH7L3iCS58NsureHNQlV9V8P3UpSG5IGN8b4Jhl28bwCDgblYKAPgP8Atm/An4/2lrb+HPFcGleIZVUS6Dqsgt7lZDgbY8nbOMnAMZJ6ZAJxVVXxz/aw+Bv7PNs6fEDxhE2sCPfFoWnAXN/LkZH7oECMEdGlKKccNmv2rP2zfiH+1BqaafeR/wBg+DrGfzbDQoJt437QPNnfA81/vY4AQMQByWb9gz9hTxT+1j4wj8Q+IILjS/hnod0o1fUipRr91wxs7Y93YY3OOI1bJyxVW8P+HtC8J6FYeGPDGkWml6TpdultZWVpEIoreJBhURRwAAAMVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV/wW++HMrQ/DL4uWkDGNGu9DvpCpwrELPbAEL1IW6JDN2G0cMa/wCCK3xAfQ/j54v+HU9z5dr4q8N/aUTDHzLmzmUoOBgfup7g5OBxjqQKqq/bL+ID/FH9qj4o+Mzc/aIbjxJdW1pLhhvtrZvs8BwwBGYoY+CAR0r/AIJh/D7/AIV/+xh4GEy4vPEn2nWrnA4/fzN5WPlU/wCoSHOc85wSuKqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq8cfFH4ffDX7F/wnfiyx0X+0fM+yfamI83y9u/GAem9fzFeCvi38NviPdXNj4G8Y6frM9nGJJ47ZiTGpOATkDjNVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV4u+N/wl8B6w3h7xh480zStRSNZGtrhyGCt0PA6GvBnj3wd8RdKl1vwR4htNYsYLg28k9sxKrKFVivPfa6n8RVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVXxr/bM/Zq/Z18VWvgn4x/En/hH9avdPTUILX+x767327ySRq++3hdRl4ZBgkH5c4wQT8FP2zP2av2ivFV14J+DfxJ/wCEg1qx099Qntf7HvrTZbpJHGz754UU4eWMYBz82cYBIqqqqqqqqqqqqqqqqqqqqqqrUdO0/WNPudJ1axt72xvYXgubW5iEkc8bgq6OjZDKykggjBBwa/a3/wCCPX2ue68cfsm3EMRf55/BupXe1c4OfsdzIcDJ2/u5mAGWIkA2oPH/AMM/iH8Ktcfwz8SvBGt+GdUTJFtqli9s0ig43puADoSOGXKnqCRXgj9r39pr4dpFD4W+NXiRIYM+Vb31wNQiQYIwI7gSKBgnjGM89Rmrf/gpl+1xDpJ02TxrpFxcGN1F/JodsJgWzhsKojyuRj5McDIPOfHH7Z37UnxEtjZeJfjX4gFu0flvDprx6Ysi4YFXW1WMOCHIIbOeM5wMeFPBvjb4k+IY/D3grw1rHiTWrtty2un2sl1M+WALFVBOMsMseOeTX7J//BHnXtUvbLxr+1Vdpp2mxskieENPuN1xdDqVubiNsQr0BWIs5BPzxkc+GvDPh3wZoFh4V8JaJY6Po+mQiGzsbKBYYYIx0VEXAAqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/bF8V+KdM/aP8X2Om+JtVtbeL7FshgvJI0XNlAThQcDJJP1NfsOalqOrfASzvNU1C4vJzqd0DLcStI2AwwMkk4qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/bk8UeJdJ+PV3Z6V4i1OzgGl2pEVvdvGoJU5OFIFf8E/tW1TWfg1rFzq+p3V9MniedFkuZmlYKLa2OAWJOMk8e9VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVeNXeLwbr0sTsjpplyVZTggiJsEGv+E68b/wDQ5a5/4MZf/iq/4KqfD6Px3+xh4rvY7bz7zwleWetWi5UbSkohlbJIxiCec+pxgAk1+w78Q4fhZ+1t8LfGNzIiW6a9HY3EjjiKG8VrSRz7Kk7N+FVXxy+IcPwl+DXjb4mSyIG8M6DeX0KuOHmjiYxJzxln2qM8ZIrSdKv9d1Wy0PSoPPvdQuI7e2i3Bd8jsFVcsQBkkDJIFeCPCem+AvBXh/wLowxYeHNLttOtBt24igiWNOO3yoKqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr41zz2vwa8e3NrO8M0PhjUnjkjYqyMLaQggjkEEda8E+NvGc3jPQIpfF+tOj6pbKytfykEGVcgjd0qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr/AIKVf805/wC4p/7aV/wTh/5HXxh/2Cof/Rpqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv28/+Tgrv/sFWn/oJr/gnb/yRPWv+xpuP/SW1qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr/gtX/wAnUeFv+yf2f/pff1/wRU/5Oo8U/wDZP7z/ANL7CqqqqqqqqqqqqqqqqqqqqqqqqvFfg3wf480d/D3jjwpo/iLS5G3PY6rYx3cDMARkxyBlJwSOnc14x/4Ja/sU+L7r7bH8LbjQp2YGRtH1e5gRwFCgeUztGo4B+VVJOSScmpv+CLv7KEuoPeR+LfibFC0xkW0TVrMxouc+WCbQvtA45YtjvnmvCX/BK39inwtem/n+Gd7r0oZWiXV9ZuZY4iM5/do6K4ORkOGHAxjnPgn4e+A/hrosfhz4eeDNE8NaZH9200qwjtY888lUABJJJJPJJJPWqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/bT/5OY8Zf9uP/pDb1+wb/wAm+Wf/AGFLv/0IVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVft5/8nBXf/YKtP8A0E1/wTt/5InrX/Y03H/pLa1VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV45/5EnxB/2Crn/0U1V8TvBdv8SPhr4s+Hd1N5UPijQ73SpHyRsW4geInjngPU0Wr+GdaeB2uNP1TSroqSjlJbeeNscEHKsrL1HQivhP44t/ib8LvCHxFtXiaPxNodnqQ8pSqqZoVkIAJJXBYjBJIxgnIqv+Ct3xDfwR+x5quhWt75Fz401iz0ldjMHMYc3MgBUjAK2xRs5BVypHzA1/wTz8AP8AEb9sr4XaSY5TDpesDWZpERiIhYo10hYj7oMkMaZPGXA71VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVfHP/AJIn8Qf+xV1P/wBJZK8C/wDI7+Hv+wrbf+jVqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr/gpV/zTn/uKf+2lf8E4f+R18Yf9gqH/ANGmqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/bz/wCTgrv/ALBVp/6Ca/4J2/8AJE9a/wCxpuP/AEltaqqqqqqqqqqqqqqqqqqqqqq/au/bV+Ff7Hn/AAi3/CzNA8V6n/wl32z7D/YVpbzeX9m8nzPM86eLGftCYxuzhs44z+zT/wAFAfgj+1LqHiWx8F6X4o0CPwlpo1HUrzxJBa2kEcGSC2+O4kwFAJJbaABnNfEr/gst+zj4R199E8DeFvFHjWCBmWXUreNLK2cjGPK80+Y4+9ksiDgY3A8V+0X+3j+zf+zHOuj+PPF0up+IWyToOgxLeXkYA6yjcqQ9RgSuhbOQCAxFr/wW4+Cz6kkV78HPGsVgZCGniuLWSUJ2IjLqCenG8D3r9nj9rb4FftRaVNefCfxilzf2Slr3Rb2P7Nf2iggb2hJ+ZPmUeYhZMtt3bsgftQftQ+Cf2TvBGn/EH4g+GPFGraPf6kunb9Ct7eZreZo3dPMWWaLCsI3AYZ5ABxkZ8Bf8Fe/2Z/iH468OeANJ8HfEi0vvE2rWumW1xfadYxwQy3EqxK8rLeMVQFwWIUkAE4PSq8cf8FgP2YfAnjTX/A9/4T+IuoXPh3VLnTprvTrCxltrh4JWjaSFzeAtGxUlWIGQQcDpXwg+KXhj43fDPw78VvBgu10bxLZi6tY7tFSaIZKskiozKHVlZSAxGQeTX7Un/BQD4KfskeL9J8D/ABG0Xxbqmqavpv8AaMSaHaW8wigMjxqX86eIgs0b4wD909K/Zh/4KEfB39rTx5ffD74a+DvHdle6bpb6lc3Ws2NrDbpEskceN0VzI28tKuBt5AY54r4/ft2fsz/s26tJ4X+Inj3zfEkMaySaFpNs15dxBlDL5gX5ISysrBZWQsrBgCDmtV/4LcfBWG/li0T4OeNbuyGPKmuri1t5G4Gcxq8gXnI+8cgA8ZwPhF/wV7/Zi+I2s2vh3xdZ+IPAV1dSFEu9XjiksV67d08bkpnGCWQKMjLYyRp2o6frGn2uraTf297Y3sKT21zbyiSOeNwGV0ZSQyspBBHBBBr9qH9qDwB+yX4AsPiN8RtI8Qalpuo6xHpUUWiW8M0yzSQzShmEssahNsDgkMTkrx1I/Zy/4KS/Av8Aag+JkHwq8AeFfHdhq09nNdrNrFjaRQBIgCwLRXMjZOePlx7ivjf/AMFb/wBmz4T+JLnwj4Vstb8f6hYXDQ3lxpAjjsomXIcLPI370hgBlFZCCSH4wfhj44tPid8NfCfxJsLGWytvFmh2WrQ20zBngS5gSVUYjgkBwCRxkV+0X/wUG/Zq/Zm1d/C3jPxLe634lhVWn0Pw9Al3c24YjAlLOkUTYO7Y7q+3B24Zc+Gf+C1/wH1HVY7PxT8LfGuj2UmF+127W955bFgMum9DsALEldzcABTnj/grD8T/AIf/ABi+O/gjx98MvFVj4g0HUfh5ZmC8tWOARf3+UdGAaNxkbkcKynggGv8Agip/ydR4p/7J/ef+l9hVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX7af/JzHjL/ALcf/SG3r9g3/k3yz/7Cl3/6EKqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv28/wDk4K7/AOwVaf8AoJr/AIJ2/wDJE9a/7Gm4/wDSW1qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqrxz/AMiT4g/7BVz/AOimqq/bw+Hkfww/a8+KHhi3s/s1rNrkmpWsaqoRYrxVuVCBQAEXztoAHy7duSQTX/BKH4gp46/Yy8N6Y8zy3Xg7Ur3Rbl3JJJWT7RGOVAAWG5hUYLDCjnOQK/4Le/Eb7R4q+Gnwjtpdv2DT7rXLxNud/nyCCA5K8bfs9x0Y538gYUn/AIIl/DyPVvix4/8Aifc2fmL4d0ODTbaR1Uqkt3LvLLkZ3hLVlypGFkIOd4xVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVfHP/kifxB/7FXU/wD0lkrwL/yO/h7/ALCtt/6NWqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv8AgpV/zTn/ALin/tpX/BOH/kdfGH/YKh/9Gmqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/bz/5OCu/+wVaf+gmv+Cdv/JE9a/7Gm4/9JbWqqqqqqqqqqqqqqqqqqqqqqv+C5//ADRL/uYP/cdXhv4keKfCXhDxX4J0K7S30/xpDbQasQD5ksMEvmrGCDgKXCluOdoHAyD4S/4Jx/treN9AtfEuhfAfU0sbxd0I1HUrLTpiPUw3M0cig9sqM1/wUB/adn/Za/Z+vvFGgGJ/FXiG4/sjQVdwDDNJG7NclepESIzem8xg8NXwK+DPxA/aq+NGn/Drw9qPn654huJbq/1TUZmk8pBmSe5lYks56n1ZiB1Oa+IH/BEmLTfAF5ffDf4zXuseLrK1MsNlqGlR29tqMqocxKwlzAXbG1mLhejdd4+FHxO8afAv4m6H8SPB17cafrfhy+WUJvaLzQrYkglAwSjruR1PUMRX7UWmeH/2tf2B/Euu+Fk8+HX/AAnF4k0YAJJLHNAq3aRA4IWQmNoGxgjc65HNaTqt/oWq2Wt6VceRe6fcR3FtLsDeXIjBlbDAg4IBwQRXxA+NWleG/wBmvWvj/ps0Rs4PB769YYkWRZS1r5sChh8rbmZFB6HcO1Xl3d6hdzX9/dS3NzcyNJNNM5d5HY5ZmY8kkkkk9Sa/4I2/En/hK/2YdQ8A3NxuufA/iCeGGLOdlrcgTofbMzXPHt71/wAFSfHtx46/bR8Z27XDSWfheG00ayD7gUWKBXlXBJAHnyzkbcAgg4yST/wRX+EzeH/hB4v+MGo6eiXPi3VksLCZ1BZrS0U7mU9VVppZFI4yYQSOFNftsf8ABNPxZ+1P+0bYfErwr4y0fwzol7osFtrtzdwPPOLiF2UPFEpAkJhKDDMgHlj5ju+WX/gij8JdG0cX/iP9oXXYVtYVN5ePp1vbQBgBufDudik9AWOMgZPWv2jvgk37PnxZ1X4Zp420fxba2apNZ6xpbgxXUEgyhKhm8t8cMm5tpBwzDBP/AARe+PWt+JvCniv4A+Ir+4u4/Cyx6noTSyKwgtZXKzwgfeCrKUcdRmVvu4AP/Bav/k1fwt/2UCz/APSC/rwD8SPFPwzutZvvCN2lrda5ot1o9xOQd6W1yoWbYQRtZkBXPOAx74I+GP7Bf7XPxj8LW3jX4f8AwV1O80S8VXtbu8vbXThcowyskQupY2kQjkOoKnsa+Onxl8UfsUf8E8vBr31h9i8e23hfRvCtjbsyzLaap9hVZSzxkqfKSC4cEEqzRqMkNmoYvEXjXxKkEbXus69r18FUu7TXF7dTPjkklnd3bqeST714K/4Ij/EXWPBcOreOfjbo/h3xLNCZP7GtdGbUIYWK5SOS586PDZ+VtkbqpBKlxgn4yfB3x78BfiJq3wv+JOkfYNb0iTD7G3xXEbDKTRPxvjdcEHg9iAwIH/BFT/k6jxT/ANk/vP8A0vsKqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/bT/5OY8Zf9uP/AKQ29fsG/wDJvln/ANhS7/8AQhVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV+3n/ycFd/9gq0/9BNf8E7f+SJ61/2NNx/6S2tVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVeOf+RJ8Qf8AYKuf/RTVVf8ABa/4etovxv8ABHxKhiSO38U+HZLF9qgGS4spiXcndnPl3UC8qBhRgnnH/BED4hxrdfE/4T3d788kdlrdhb7lHCl4Ll8ZyfvWg4BA7kZUGv8Agpr8Q4/iJ+2f49ltL37RY+HZINEtfmU+WbaFVnT5SRxcG444IzggHIr/AII6/Dn/AIRL9lKXxtPH/pHjnxBdXcUmf+Xe3xaouAxHEsNwckKTvwQQFJqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqvjn/wAkT+IP/Yq6n/6SyV4F/wCR38Pf9hW2/wDRq1VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV/wUq/5pz/3FP8A20r/AIJw/wDI6+MP+wVD/wCjTVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVft5/8AJwV3/wBgq0/9BNf8E7f+SJ61/wBjTcf+ktrVVVVVVVVVVVVVVVVVVVVVVX/Bc/8A5ol/3MH/ALjq/wCCcfhLQPG/7a3wv0LxLYJeWKX11eiF+hmtbKe5hJHcCWGM474xVf8ABcLxBqE3i/4VeFCUFjaabqN6oXIZ5ZZIUO7nBAWFccZG5uTnj4X/ABf+JnwV1+fxV8KvGeoeGtWurNrKa7smCu8DOjtGcg8Fo0P1UV/w35+2V/0cN4q/7/J/8TWo6he6vqFzqupXL3F3eTPNPK/3pJHJZmPuSSa/4Jl3WpXv7DXwum1WSV51t7+NDKMERJqNysQHsI1QD2xX7S3wsf4J/H3x78LBaS21t4f1y4isEl3bjZs2+1Y7uTugeJsnOd2cnrXxp+P8dz/wR8+Guh22p+ZqPie4t/DM8XnK0sUOnTyM2cfdG20txtPOyZM5BNfCT4Gy+N/2R/jl8W2sGmk8E32gtYFFLsxeaWO5ICncAkUyMdw2bSx6plf+CL3xJ/4Rn48+L/AF5ceVYeKPDZuixOFE9lIGQt2AEU1wcnpx618SfFVz8Svif4p8bqLi4uPFOvXmogPCqyyPcTvJyifKGJf7q8Z4Ffsy/CkfA/8AZ/8AAXwqeFI7rQNFhjv1QJg3jjzLkjZwQZnkOeSc5JYkk/t5/wDBUbx+PHGu/Bj9nHWU0LSdCupbDUfE9qwkudQlTCyLbll/cIjiRPMQsz4DKyrjPgz4T/tP/tbeIrjVvDPh3xn8R9TiZILrV724kuFiZmG1Jbu4bYhy+7DOOCzdASPjX8BPiz+zr4qtfBPxj8J/8I/rV7p6ahBa/bre7327ySRq++3kdRl4ZBgkH5c4wQT/AMEVP+TqPFP/AGT+8/8AS+wr/gtX/wAmr+Fv+ygWf/pBf1+z14X0Xxv8fvhn4L8R2v2nSdf8YaTp9/Bnb5sE13FHIue2VYj8ahhht4Ut7eJI44lCoiLtCgDAAA6DHav+C5Wq38Ok/BvQ4rjFld3GtXE0WwfNJEtkqNnGRgTSDAODu5zgY/4JqaDpviP9uD4W6fqsPmww3l5eIvpLb2FxPEfwkiQ/hVf8FqIYY/2qvDTxRIrS+AbNpCq4Lt9uvhk+pwAPoBX/AARU/wCTqPFP/ZP7z/0vsKqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq8a/sw/Az4ieJrzxj4x8D/wBoaxqGz7Tc/wBpXcW/ZGsa/LHKqjCIo4A6Z65rwH8PfB/wx8PJ4V8DaR/ZulxyvKtv9okmw7HLHdIzNz6Zqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq8e/s3fBb4n+IH8VeOfBn9p6o8SRNP/aN1DlEGFG2ORV4+lfDr4YeBfhNok3hz4f6H/ZWnXF011LD9pln3TMiIW3SszD5Y0GM446cmqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr+ytdTsbjTb2LzLe6iaKZNxXcjDDDI5GQT0r/AIYs/Zm/6Jp/5Wb7/wCPVX/BZn4eP4n/AGZdH8d2dn5lx4M8SQvPNtY+VaXKNC/QHGZjajLEDtnJAP8AwSz8fP4E/bP8H2zyyra+KLe70a58tmGRJCZIwQPvDzoYeDwPvdVFa5q9l4e0TUNf1FnFpplrLczlF3MI41LNgdzgHivGfifUPG/i/XPGerOzX2v6lcX9yzyGRjLNI0jks3LHLHk8nvX7L3w8f4Ufs6fDf4e3Fn9lvNG8N2cd/DtZdt20Qe44YKwzM8hwQDzyM1VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVa7oml+JdE1Dw5rVt9p07VbSW0u4d7J5kUiFHXcpDDKkjIIIzwRVj+x1+zhpl9b6lY/Dny7m0lWWF/7XvTtdSCpwZiDyBweKqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv8Aguf/AM0S/wC5g/8AcdX/AAS4/wCT7fhj/wBxb/003lV/wWd+C2qeNPg54X+MOiWzzN8P76aHUkRRkWd55S+aeMkJLFEOuAJWOO4/ZL+Punfs3/GSw8f6/wCCtP8AFeg3Fu9hrOmXdtHMz2sjIzPD5gKrKjRoyk4zgqSockeFP2yf+CbXi3R01aDxJ4B0wlij2ureHRaTxsADgq8PIwR8ykr1Gcg4m/bD/wCCdk3i/QPAfh658I+INb8RataaZaw6Z4WDxo9zII0ked4liVFZhuw5YA/dNafp2n6TZQ6bpVjb2dpbrtht7eIRxxj0VVwAPYV/wWi+Esfhf45+Gfi3p9l5dt430c295IsagSXlmVQszA5yYJLdRuA4j4JwQuvfFbVNd+CXhD4Ozz3H2Xwrr2ralGAQsbJdxWaopAPzMjwXB3MMgTkA4OB+wz+zXb3P/BOGf4daxZfZ7z4t6XqeoXIuQcRteReTazLuVtv7iK1kVgpwcMATyfhH8QtX+B3xLXxglg6anpVjqlj9lubckpLcWNxalZEJUgAzYbkMADjkYr9gL4SR/Gf9rT4f+F7yLfpunah/bGogsoBhsx52wg8kO6RxkDnEhPGCR4t/t3/hFdZ/4Rb/AJDX9nz/ANm/c/4+PLbyvv8Ay/f2/e49eKr/AIJ7/tS/sueF/wBknwd4V1X4reD/AAfqvh63mi1XTtZ1W3sJ3uDK7vKqO4MofduDLknIGAeB/wAFH/2jvBn7Tf7R0ni34e+bL4f8PaPDodleyIU+3iKaeVp1U8qjPOwXOCVVSQCSB/wRU/5Oo8U/9k/vP/S+wr/gtX/yav4W/wCygWf/AKQX9fsnf8nUfBr/ALKBoX/pfDVf8Fg/g9qnxD/ZqsvHmg2FxeXnw+1YXt1HDGHK2EyGOd8AbsK3kOSOFRXZhgbl+F3xD1/4SfEXw38TPC8iDVPDOpQ31srj5XaNgSjf7LDKn2Y14P8A+Cuf7HuveCYfEXifxHrfhnXPL/0jw9caPcXMyyADcI5oUaF0JJ2szoSBkqhOK/bG/aSvf2q/jrq/xUfTrjTdL8mOw0WwuJvMe1sos7AxHAZmaSRlXgNIwBONx/4Iqf8AJ1Hin/sn95/6X2FVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVftgeAI/ih+y98TvBRjhkmu/Dd1NarKisv2iBPPhzu4H72JPm6r1HIFfDzxjf/AA68f+GfiDpSb73wxrFpqlsuQMyW8yyqMkMOqDqpHself8FA/ihp/g/9iX4heJtM1FHTxJoqaZYPGAftCX7LCdu4EYMMsjeuASMHFfs3fDuH4s/H74e/Di6RHtNe8RWdveK/T7N5qmb6nyw+B3OB3qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq1PXNE0Xy/7Y1mxsfOz5f2m4WLfjGcbiM4yPzFab4g0HWXeLR9csL541y621ykpUepCk4FVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVf8Aifw1pVwbPVPEWmWc4UExXF2kbAHocEg4rTtV0vWIDc6RqdrfQq5RpLeZZFDAAkZUkZwRx7iqqqqqqqqqqqqqqqqqqqqqqv8Agp9+yF8Zf2rf+Faf8KjsdJuP+EW/tb+0Pt9+ttt+0fY/L25B3f6iTPpx61+w9/wTk/aY+Av7Ufgn4sfEHS/D8WgaF9v+1va6ss0g86wuIE2oBk/PKmfQZNVq+k6br2lXmhazZRXlhqNvJbXVtMu5JonUq6MO4Kkgj3r9o3/gjDfXet3fiT9mPxhplrp07PIPDPiGWVTagKSEt7oCQyAnCqsoUqOWlbk1qH/BK79uayvZbW2+DdvfxxNhLm38Saasco9VElwrY+qg18LP+CP/AO1hN4k07WvEHiPwf4LXTriG5S5kuzqE8brl1McMaGNyrKuQ7qvPBbkVX/BRb9lvxN+1T8DrPwz4BgsX8VaDrUN/p32u6FujxlWimQsVIwVcNj5eYxz2a0/4JA/thzXUMN1pnhS3heRVkm/tpX8tSeW2gZOBzjvXhjw5o/g/w1pPhHw9aJa6XodjDY2NuigCKCFAkagDgAKoHHpX7TX/AASr/aL8bfH/AMeeNPhdpfhybwz4h1qbUrE3GqrA6m4PmyKYyPlCyO6gdMAY4r/gmZ+wr8TP2XvFXjXx58YrLSYdW1PT7fTdHGn34udsBkaS534UYy0drtwf4WyOhqv2zv8Agkn4m8XeO9V+Kf7NN9pjLr91Ld6l4X1CYWwt7hyWd7WU/JsdiT5T7BGfusVIVPCv/BJz9tnxFq6aZq/w90fwxbspJ1DVfENpJAhyBgi1kmkzznhCODz0z8Yf+CNnxB0Lwh4MsvgxrNp4o8Rt9sfxXe6jeLYwKxW3+zx20ZBOwEXGWJLHIJxwB/wTg/YX/aG/Zd+PGqePPifouiJo2p+F7nShLZaqk7xyvcW0ykoBkjEDD6sK/wCCk37OPxM/ag+Beh+APhVa6fPq2n+LLfU5lvrsW6CBLS7iYhiDk7pk4+vpXwF/4Ja/tY/D345/Dnx94j0fwymk+GvFml6nftDrSO6wW93HLIVXHzHahwO5qtR07T9Y0+50nVrG3vbG9heC5tbmISRzxuCro6NkMrKSCCMEHBr9qL/gjn4pbxNqHi79mDVtMn0e+maVfC+pzm3lscjJSCdsrIm7OBIUKggFnwWrwH/wSQ/bI8V6zFYeKvCmieC7HzFEt9qet21yAhyWKJaPKzEbQMHaCWXkDcy/tC/8Ei9bs/hP8O/CH7PNxY61r+iXWoTeJ9T1aUWb6nJcJbhJFBZhGkf2cqsQ3YDkli25m/4JtfsJ/H79mD45654++KunaJBpN/4TuNMhay1JbhzO93aSqCoAwNsL8+uPWqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv2q/j18YvBfx88UeGfCvxB1TTdLs/sn2e1hdQke60hdsZHdmY/jX7HfjPxT49+Ctr4g8Ya3carqL6jcxtcXBBYqrAKOOwqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/bF+OHxZ8BfGm58PeDvHmp6VpyadbSLbQOAoZlO48g9a/Yg8e+MfiL8J9V1vxv4hutYvoPEU1vHPckFliFvbsFGAOMux/GqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqvF11cWPhPWr2zmaKe3064kideqMsbEEfQgV/w1H+0F/0VfW/+/i/4VVfH74ep8J/jf49+GsMLx2/hvxFe2VoHUgtbpMwhfBZjho9jDLE4IyT1r9r/wDaBHjr/gnX+zR4PSTNzqnm/bMH5x/Y0ZsB5n7wn94ZRJyDuK7vkxtP/BHn4fv4r/a0Hi+S23QeCvD95erKSwCTTAWqqMHklJ5jg5GFPcA1VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV8XdU1DQ/hP411vSLt7W+0/w7f3FrPHw0UqW7sjD3BANeEP2mPj1f+LNEsbz4o6zLBcajbxyxs64ZWkUEHjoQaqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/4KVf8ANOf+4p/7aV/wTh/5HXxh/wBgqH/0aaqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr9vP8A5OCu/wDsFWn/AKCa/wCCdv8AyRPWv+xpuP8A0ltaqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv20/wDk5jxl/wBuP/pDb1+wb/yb5Z/9hS7/APQhVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV+3n/AMnBXf8A2CrT/wBBNf8ABO3/AJInrX/Y03H/AKS2tVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVeOf8AkSfEH/YKuf8A0U1VVf8ABXb4fDwb+2HqPiGJf3PjbQ7HVVCj5VdENo6j5QAc2ocjLf6zJPzYF74g1nUdI03Qb7UZZtP0jzfsNux+WDzWDSbf95gCa/4Ij/D4ab8LviL8UZl+fXtct9KhDDkJZwmRmX5ejNeYJDEEx4wNuTVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV8c/wDkifxB/wCxV1P/ANJZK8C/8jv4e/7Ctt/6NWqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv+ClX/ADTn/uKf+2lf8E4f+R18Yf8AYKh/9Gmqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/bz/AOTgrv8A7BVp/wCgmv8Agnb/AMkT1r/sabj/ANJbWqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr4n/sY/C74s+OdT+IHiPXvFNvqOq+V50VldQJEvlxJEu0PCzD5YwTknknp0r4SfCnw78GfB0Xgjwve6jdWMVxJOsl/Ijy7nOTyiIMcelVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVXxc/ZC+Gnxn8YyeNvFGueJrW+kt44DHYXMCRBUGAQHhc5555r4MfBjwt8C/C114R8I3+qXdnd373zvqMsckgkaONCAURBtxEvGM5J59KqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqtW06DWNLvNIuXdYb23kgkaMgMFdSpIzkZwfSv+HdnwT/6Gjxv/AOBtr/8AI9VX/Bb34fx3Xgr4afFSC2w+napdaLczAr8wuIhNEpGc8fZpiMDHzNk8jNf8E5/h63w4/Yy+GemTxIt1rGmtrU7qoBkF7I1xEThmGRDJEvXOFGQpyBVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV4r8OWPjHwtrHhHU5Z47PW9Pnsbh4GCyLHLGUYoSCA2GOMgjPY1pP8AwT9+DWj6pZava+JfGbzWNwk8ayXlsVLIwYZAtwcZHqKqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq8TeBfBPjT7N/wmPg7Q9e+x7/s39p6dFdeTuxu2eYp252rnHXaPSvDfw+8BeDZ5rnwf4I0DQ5rhAk0mm6ZDatIoOQGMagkZ7Gqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq1/4YfDTxXqB1fxR8PPDOsXzIEN1f6RBcSlR0G91JwPTNeHfCnhbwfYvpnhHw1pWiWckpme306zjto2kIALlUABYhVGeuAPSqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv+CiXwl1r4zfsj+NvC3hbSb3U9eslt9S0yzsoWmluJYJkdkSNAWdmi81QoBJYjr0rT/2IP2r7q40UX37P/j2ytNZuo4BcP4fuJDahmAMk0aqXiVQckuF4FaHo9l4e0TT9A01XFpplrHbW4dtzCONQq5Pc4A5qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqvvFPhfTG2al4k0u0YMVxPeRxncOo5PX2qDxv4Ku5Vt7Xxhok0rZ2pHqETMcDJwA3oKjkjmjWWGRXR1BVlOQQehB9Kqqqqqqqqqqqq/wCFqeEf+Et/4Qnz7n+0/P8AI2+Qdu/GfvelVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVeDPB2neOviZe6DqtxcQ27S3MjNbkK+VY4ALAgc+1Xv7MfhCS2dNO17WIZyPkeYxyqD7qFUn8xWmaz4w+BfjaPQNUvjLpjMjSxZZopYGY/vEHVWHzdO4IORSsrqHRgwIyCDnNVVVVVVVVVVVX/NyP/cX/APZKqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq+DH/ACWa8/7e/wD0Kq/aHvINf8e6b4f0dPtN9bwLA6oOTLI+Vj+uCp9Pm+tafafYbC2sfM8z7PCse7GN20AZx26V4w+N72uuN4T8BaI2t6qkjROzKxjVxkEADBbBxk5A4PPcaj4/+PfhmN9Z1/wXYvp+0s6xx7xCo6kmORio92zXw7+JOi/EbTpLjT4pLa7tdourSQ5MZI4KsPvKcEZ4PHIFfFH4t+Lvh94vh0uLTtNm0q4ijmjke3kMm3OHGRIFLAgnoMArn1MM0VxCk8Dh45FDIw7gjg18ZPi5qnw9v9O0rQrWxnubiFppxdozBUztTbtZepD5z6CvBHiIeLfCWl+ISEEl5bgzKgwqyj5ZAoyTgMGAyelfFrx5c/D7wwmqadHay309ysMEdwCyngliQpBwAPXqRXw48Sa94q8F23iTXbO2S5uzI8UFrGyfIrFV4djydpOc4wRUvxg+K3ippW8DeAmjswSqzy27ysCpAPz5VM+q4JGfbNN8bPih4OvY4vHfhWNoZeVDQtbsfXa4yp+mD9RXh7X9L8UaNa67o8/m2t0m5CRgqe6sOxByCPWvjR8TNe+HP9j/ANh2lhP/AGh5/m/a43bGzy8bdrL/AHznOe1fED49Xdlqf/CPeAbCO8uVUebcyIZMNtyURFPVeMk9CCMcZr4eeINU1nwJY+IPFJS3u3WZrkunkqipI4BIP3RtUHn614m+P+salrDaB8NNEF827YlxJC0jSkdSkYxgdOW7dQKg+LHxa8JOl74+8FvLpTMBLNHbmJouQM7gSvfgMBk8ZFaNq1jr/wAfLTWdNlMlteakJImIwcFO47HtVVX/AAvP4qf9DV/5I2//AMRWkTy3Wk2VzO+6WW2jd2xjLFQScCq8d+OfCnwz8Hav4+8cauml6DoNq91qF48TyCGJep2IGZj6KoJJIABNWv8AwUW/Ytv30iGw+Omn3M+uXCW9lBDpl88rSOwVVeMQloskgZkCiqqqqqqqqqr9p79p745fDv45eJfB3g7xz/Z+kaf9l+zW39mWsuzfaQyN80kTMcs7Hknr6V+yV8QvF/xP+Dtt4q8cax/aWqSX9xE0/wBnjhyiMAo2xqq8fSqqqqqqqqqqqqqqqqqqqqqqqqqqq/4Xn8VP+hq/8kbf/wCIrSJ5brSbK5nfdLLbRu7YxlioJOBVVVVVVVVVVVV+1v8AtIfGn4Y/GG48K+BvGn9m6XHp9vKtv/Z1rNh2U7jukjZucdM4r9jH4n+Ofiz8LtU8R/EDXf7V1G21+a0im+zRQbYlggcLtiVR96RzkjPPXgVVVVVVVVVVVVVVVVVVVVVVVVVVfGTxJrXhTwTLq+gXv2W7W4jQSeWr4BPIwwI/SvhR8V/H/ibx/pWia3r/ANpsrnzvNi+ywpu2wuw5VARyoPWqqqqqqqqqqqrxXe3Om+F9Y1Gxl8u4tdPnlhfaDtdYyQcHIPIHBr/htL9pj/opf/lGsf8A4zVVVVVVVVVVVVVVVVVVVVVVVVVVXxl+J/jnwp41k0jw/rv2W0W2jcR/Zon+Yg5OWUn9a+B3ivX/ABj4Su9T8R6h9suYtReFH8pI8II4yBhAB1Y/nVV8af2hvg3+zvpWl638ZPG0Xhyy1m8+x2MslpPcebNtLEYhRyoAGSzAKOMkZFfDj9sn9mf4veP4/hf8NPitY+IfEklq90lrZ2Vy0ZiQZY+eYxFkAj5d+faqqqqqqqqqr4ra1qfhr4XeMfEeiXX2fUdK0C+urSbYr+XNHA7o21gQcMoOCCD3FeFP2xf2j9T8U6Npt78R/Nt7rUIIpk/siyXcjSKGGRDkZBPSqqqqqqqqqqqqqqqqqqqqqqqqqqq/ar+IPi/4a/DvT9d8E6x/Z19PrUVtJL9njmzGYZmK4kVh1RTnGePrX7NHx4+K/wAQfijb+HPF/iv7fpz2U8jQ/YbeLLKBg7o41b9aqqqqqqqqqqqq/bf+NPxM+D//AAhf/CufE39k/wBrfb/tn+hQXHm+V9n2f61GxjzH6YznnoMfsU/Hb4rfF3xR4k034h+K/wC1rfT9Piltk+w28GxzJgnMUak8djmqqqqqqqqqqqqqqqqqqqqqqqqqqqr9pfxt4n+H3wun8R+ENU+wail7BGs3kRy4Vidw2urDn6V+z5+0H8X/ABz8X9A8LeKfF/23TL37T58H9n20W/ZbSuvzJGGGGVTwe1VVVVVVVVVVVVftb/tIfGn4Y/GG48K+BvGn9m6XHp9vKtv/AGdazYdlO47pI2bnHTOK/Yx+J/jn4s/C7VPEfxA13+1dRttfmtIpvs0UG2JYIHC7YlUfekc5Izz14FVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVeHP+E1/wCFhX//AAgH/IW8642/6r7m47v9b8teJL39pax0mafWJL6O1VSZHs0ti6KBycwfOoA78Y69q/Z20rwNe3E+piee58S2ylmjulAESHgvEOc8naWJz04Xdz8W/EE3hr4fatqNpLsuWiEEJBwQ0jBSR7gEn8K+B3i74b+CdJu77xBrQt9YvJShH2aaTZCANoBVSOWyTz6elN8c/hQ6lH8UBgRgg2Nwc/8AkOvhxrOlaX8btvhi5dtF1G5mhiWNGQNG6koNrYOFbb15wK/aX8P/AG3wxp/iGIZfTLkxuMf8s5QBnPsyqP8AgRr4Q61/bvw50S6Z90kEH2aTPUGI7Bn6hQfxqPSm+K/xX8WSMoeGysbmG1I+6HVfJiI5HUkvyQM+lfsz6402jar4YnYiSxuBPGrDBCuMMPwZPT+L8v2h7651/wAZaF4J09y0iquEOQpmmcKoPrgKOe24+9aRpdrouk2ej2S4gsoEhjHsqgD+VeJPiT4G8HMbXWdet4p41wLaFTJIOOBtQHb074HSvid8ZvAPjLwrf+G7Wy1WWd9j2tw0CKiyKQQcltwGMqfl6E/Wv2Zb2ebwpqdhI+6O2vt0eSSV3IMj6fLnHqT61+1L/wAyx/29/wDtGvgp4O07w14J0/UY7eM3+rQLcz3G35irjciA9QoUrx65Pev2hfEE2jeATY2su2XVrlbdsHB8vBZ8ex2hT7NX7PvhOy0fwVD4ge1X+0NXLs0rD5liDYVR6Kdu73yM9BiaGG5he3uIklikUq8bruVlIwQQeoI7V4e0O18N/Hi20Oydmt7PVSkO7qEwSAfXAIGe+M1VVWgf8gLTf+vSL/0AVX/BW74hv4I/Y81XQrW98i58aaxZ6SuxmDmMObmQAqRgFbYo2cgq5Uj5gar4EfECP4r/AAV8C/Ehbnz5PEfh+zvZ2IUETPCplUhQACJN6kAAZBxxVVVVVVVVVftp/wDJzHjL/tx/9Ibev2Df+TfLP/sKXf8A6EKqqqqqqqqqqqqqqqqqqqqqqqqqqqtA/wCQFpv/AF6Rf+gCqqqqqqqqqqq8ceO/Bfw08M3njL4geKdM8PaHp6g3F/qNysESZOFGWPLMcBVGSxIABJAr49/HnwD+0f8AEi9+I/w1k1CXQ/KSyhmvbbyGmaEEM6pkkIc8bgreoFaP+378Uv2Rn17wP4M0PSdZtvENhb3lgNTLeXpl4HljlmCJhpfMjWJSpYAGJCP4g37M3xy0X9o/4H+Ffi7o4iik1mzA1G0jbP2O9jOy4h55wsittJwWQo3RhVVVVVVVVVVVVVVVVVVVVVVVVVftC/8AJN5/+vuH+dfAz/kqmh/9vH/pPJVVVVVVVVVVVV45/wCRJ8Qf9gq5/wDRTVVVVVVVVVVVVVVVVVVVVVVVVVVVX7Q3/JR5v+vOH+Rr9mj/AJEO+/7C8n/oqKqr/gtz8Q31D4jfDr4V297+60XR7jVrmFGYAyXUoiQvztJVbVtowCokbkh6/Yh+IEfww/a0+Fvi+a5+zwR+IIbK5lIUiOG7DWsrHIPASdie4GcYODVVVVVVVVVV8c/+SJ/EH/sVdT/9JZK8C/8AI7+Hv+wrbf8Ao1aqqqqqqqqqqqqqqqqqqqqqqqqqqr9uT/kkuk/9jFD/AOk9xX7G3/JbLX/sHXP/AKCKqqqqqqqqqqqppobeF555UjjiUs7u20KAMkknoMV43/at8S/tV/GHxvrkmsXUvgvQ7xIfCOnzxon2S0YurSEKinfN5SSMH3MuQm4hAa/YF+JHhXwl8ZJfBOu3b2+o+NLB4NIJA8uWaDMrRk5yGMYYrxztI4OAaqqqqqqqqqqqqqqqqqqqqqqqqqqr9sn/AJIndf8AYRtv/QjX7KP/ACX7wr/2+f8ApHNVVVVVVVVVVVVX7ef/ACcFd/8AYKtP/QTX/BO3/kietf8AY03H/pLa1VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV8GP+SzXn/b3/wChVWoQ/wDCt/j3EbBvIs5b6NsAceRPgONozwu5gB/sg+lfHTS5dT+GeqfZ1ZntDHcBR3CuN34BSx/CvgN4W8CeLfCt0msaBZXepWN2VkaTJYxsAUY+gyHH/ATX/Cpfht/0J+n/APfB/wAa074beBNJvodS03wxZW91btuilRSCh9RzXjTQY/FHhTVdBZAzXdq6xggHEgGUPPowU9unUV8IfHE3hn4feMrd5Ck2mxi4tVcHiSQeX6cYcR8Z78Y5Nfs4+HDpfgybXJkIm1i4LKT3ijyq/wDj3mfgRWmongD9oqazVfKs9adlTooInAYADPQSgLj24HQV8P7Y+OvjprHipwZLXTJZZI3zwcfuoQf+Ajd/wGvir4pufB/gbUdXsGxeFRDbNj7judu7/gIyRnjIFfA/4Y6F4n02bxr4rj/tSae5dIYZmLKCPvO/99iSeDkd+SePilofh3S/hrrbWmkadaCK12xGO3SPYSwAAwBjJIH41+zB/wAgDWv+vxP/AECv2pf+ZY/7e/8A2jXgD/kQ/Df/AGCLX/0StftM6fNceEdO1CIZS0vtsgx0DocH6ZAH4ivgjq1rq3w10kW7rvsle2mQNko6sev1Uq2P9oVVneW2oftEi6tJBJEdZZVYcg7QVJHtkHmqqv8AhmjwH/0Fte/7/wAX/wAaq0tY7K0gsoWYpBGsalupCjAz+VV/wW++IKXXi/4ZfCm3mdW03TbvWruME4cXEiwwkjbjK/ZrgcMT85yBwTX/AASP+I0vjn9j7TdAu53kufBGtXmjlpGJdoyVuYzyxO0Lc+WvCgCPaB8uTVVVVVVVVXxP/Yx+F3xZ8c6n8QPEeveKbfUdV8rzorK6gSJfLiSJdoeFmHyxgnJPJPTpXwk+FPh34M+DovBHhe91G6sYriSdZL+RHl3OcnlEQY49Kqqqqqqqqqqqqqqqqqqqqqqqqqqr/hmjwH/0Fte/7/xf/GqtLWOytILKFmKQRrGpbqQowM/lVVVVVVVVVVX7XP8AwU7+D37O39o+C/Axh8deP4Yyi2drMPsNhKeP9JnUnJXqYo8scbWMedw+P37TXxm/aZ8TDxL8WfGFxqCwMxsdMhJisdPBABEEAJVCQFBY5dsDcxwK/YP/AGSv2ivjHpFlezaLp/hX4bP5ky67qtvL9ovC6ttNpEGHnKHCkltiFdwD7q/4KifsmzfADxR4Q8ZeHb6/1PwvrenGwe6uwpeC+iZnZGKAAK6PuQYz8jjJ25r/AII6ftI/8IN8UNT/AGePEmobNF8dZu9G8xsLBqkUfzKOw86BMEnktBEoGWqqqqqqqqqqqqqqqqqqqqqqqqqvGXhDTPHGiNoGrT3MVu8iyFrdlV8r05YEY/CvCnwO8JeDtftfEemajq8tzZ7/AC0nljZDuRkOQEB6Me/WqqqqqqqqqqqrVtOg1jS7zSLl3WG9t5IJGjIDBXUqSM5GcH0r/h3Z8E/+ho8b/wDgba//ACPVVVVVVVVVVVVVVVVVVVVVVVVVVXjP4L+FvHOttr2rX+qxXDRLGVt5Y1XC9OGQn9a8D+BtI+H+kS6No1zdzQzXLTs106swYqqkDaqjGEHb1qq/4KOfEb/hZf7ZXxH1CGXdaaHqA0O3Tbjy/sSLBKM7VJzOkzc5xuwCVANQzS28qT28rxyxMGR0YqysOQQR0NfCLx3D8UvhT4N+JMCIi+KdBstTMajAjaeBJGTAZsFSxUjccEYyetVVVVVVVVV4r8OWPjHwtrHhHU5Z47PW9Pnsbh4GCyLHLGUYoSCA2GOMgjPY1pP/AAT9+DWj6pZava+JfGbzWNwk8ayXlsVLIwYZAtwcZHqKqqqqqqqqqqqqqqqqqqqqqqqqqqr4rfCnw78YfDtv4Z8S3uo21rbXq3aPYyIjl1R0AJdGG3Eh7Zzjmvhp+zF4B+FXiiPxb4d1fX7i8jgeER3txC8e1xgnCRKc8etVVVVVVVVVVVX/AAVP/aEh+Cn7M2oeE9J1J7fxR8R2fSNOWJ9rx2o2m9m/3REwi45DXCHsa/ZK8O/FT4h/FC2+E/wk0zSrnVfE3zyy6kknlWsVvHI7SMyMNo2lhz1YoowWGfi78UPHeg/FO50q1A0TU/h74pnOm3EcTxXVrcWtwVjZ9xIEitGpI2jDDp2r9kv9onQ/2ovgdoXxT0s28OoyL9k12yg3BbHUY1UzRAMSdvzK65JOyRMnOaqqqqqqqqqqqqqqqqqqqqqqqqqqr4mfDfQ/ir4Wk8I+Ibu+t7OSZJi9k6JJuQ5HLKwx+FfD79lT4d/DXxfp/jXQtZ8RT32neZ5Ud3cQtGd8bRncFiU/dc4wRziqqqqqqqqqqqqvi5+yF8NPjP4xk8beKNc8TWt9JbxwGOwuYEiCoMAgPC5zzzzXwY+DHhb4F+Frrwj4Rv8AVLuzu797531GWOSQSNHGhAKIg24iXjGck8+lVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVXwl8L+JdN+LV1qWo+HdTtbRvtW24ns3jjOTxhiMc1X7RXgfWdY1XSte0DRr3UHeB7e4W1t2lKbTuQkLk872GcfwitBuJfEHhaym1nT5opbyzVbu2uoSjBiuJFZSBxnPYZH1rWPhn8RfhZ4hfxB8OWuL6yckKsKebIEPPlyx4+ce4B6Z+U4q3+PHxHYpay/DrzbknYVjgnXc2cYC8n8Mmvhf4i+JfiG7v7nxx4e/s2zaJPsi+QYNrgkMNrkvyMHJ44461XxA+HfjOz8c63ZeHtB1OfTtWnDiSC0ZoSrssm0lRhQr8e23066FpFtoGi2OiWmPKsbdIVIXbu2gDOPU4yfc18f/CeuXs+heLfCthd3GpWE3lH7JbtLIoB3xvhQSArBu2MsPx/Z+8HX3hnwrdX+r2E9nfanc5aKeMxusceVXKnkclzz2I+p+IHhNfG3hK/8OCZIZbhQ0ErrkJIrBlPqASMEjnBNeFvEHxT+DslzoU/hS4ubSWQuIpIHZN+Blo5E4II25HI6dDmtY0v4r/FvTLq/wDEGlvpGj2NvLPa6ekZSW5nWNvLAVss3zbRk4GD8vNfs56PrOi6VrNnrWi6hp8jXEboLu1eHeCpHy7gM4xzjpketftI+H9e13/hHf7D0S/1DyPtXm/ZbZ5fLz5WM7QcZwevoa8EW89p4K8P2l1BJDPDpdskkUiFWRhEoIIPIIIwQa8SaBYeKdCvfD+pKTb3sRRiOqHqrD3BAI9wKtNL+LPwQ1W5bSdOfUtLlYF2SBpYJhyFZgvzRtz6jnA+YYo+PfjN8RLZ9F8O+El0hJspPfOjoIxxkB3wBx1ABbB4xWhfDPXvCHxg0q2ttJ1G602zmhLagLR/KYmIF23AEAbywwTxVVVVX/BTX4hx/ET9s/x7LaXv2ix8OyQaJa/Mp8s20KrOnykji4NxxwRnBAORXjf4Jap4L+Bfwz+NN1M8lr8R7rWI7dBjEKWM0UPOM4LO0vJIJ2n5QF3P/wAEQfiClt4v+JvwpuJnZtR0201q0jJOIxbyNDOQNuMt9ptwcsD8gwDyRVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVXx8/aU+D37NPhNvFnxX8Vw2CPxZ6fDiW8vn/uwwg5b3Y4Vf4mFftb/wDBUH4w/tCC+8HfDx7vwB4Em3xPa2l0Rf6jEdyn7ROmNqOp+aFPl5Ks0g5r4N/Aj4s/tA+KR4O+Efgu98Qaiih7gxAJDaRk43zSuQka57sRnoMniv2TP+CUfwo+CwtvF/xpbT/iJ4uEaMtrNbbtL0+TqfLjcZnIPAklUDGCI1PNV+1p8A9N/aW+Anif4VXSRLqF3b/aNGuZB/x7X8WWgfPYFvkY/wBx3HetJ1LxX8KvH9lrFokuleJvB+sRzxLNHh7S8tZgwDKe6yIMj1Ffs/fGTQP2gfg54W+Lnh0okGv2KyT26tuNpcr8k8J90kV1z3AB6GqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqvFvibSvBXhXWfGWuzeVpug6fPf3km5V2QwxtI5yxCjCqeSQPUjrXiLXdS8U6/qfibWZ/O1DV7ya8upMffllcu5/FmNftI/BLVP2dvi/q3wj1mZ57vR7WxkllbGHeezhmfGOMB5GXALAFSNzY3H/glD8QU8dfsZeG9MeZ5brwdqV7oty7kkkrJ9ojHKgALDcwqMFhhRznIFVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV/wU4/aD/4Xt+09rWn6Lqv2vwv4E3aJpOw/I8kZ/0uUckNunDqHHDJHGa/4Iw/s9w6N4K8QftI6/prLf8AiGZ9I0B5U4FlEw+0TIe4edfL7EG2YdGr/gsN+yrL4e8TWn7UHgvSEXSdbaOx8UJbxH9xe4IiumCjAWVQI2Y4G9Uzlpa/4JkftXp+zr8bY/Cfi7VHh8C+O2Sy1AySny7C76W91t5GNx8tzxhJNxJ8sCqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv+Gl/Hf/AECdB/8AAeX/AOO1plzJe6ZaXkoUPPAkjBRgAlQTj25rXNXsvD2iahr+os4tNMtZbmcou5hHGpZsDucA8V4z8T6h438X654z1Z2a+1/Uri/uWeQyMZZpGkclm5Y5Y8nk96/bo/Z7l8J/8Ewvhrpo05LXUvhguj3WqAoQyPcxGC6UB9jANdXSMflLfKMgckf8E0vH0fw+/bP+Htxcywpa67cTaNP5jKuTdQtHEAT0PnGHjqfujk1VVVVVVVVX7Qn7Z3xR+E3xf1/4f+HNB8LXOnaV9m8mW9tZ3mbzLaKVtxSZR96QgYUcY+tfs0fFbxF8Z/hbb+N/FFnp1rfS3s8BjsI3SIKhABAd3OfXmqqqqqqqqqqqqqqqqqqqqqqqqqqq/wCGl/Hf/QJ0H/wHl/8AjtaZcyXumWl5KFDzwJIwUYAJUE49uaqqqqqqqtW1bStB0251nXNTtNOsLOMyXF3dzLDFCg6s7sQFA9ScV+1r/wAFgfD3h37Z4G/ZYtodc1JcxzeLb6Ei0t2GM/ZoHAM5+8N77UBUECVWBHizxh4++LfjCbxJ4x1/WPFXiXWJlRrm7me5uJ3OFSNc5OBwqovAGAAAAK/ZM/4JD+OvH32bxr+0pPdeDtDEiPB4dgwdSvkHJ85uRaoeBg7pD82VjwrH4ZfCr4dfBnwhaeA/hd4Q0/w5odnzHaWcZG99oUySOSXlkIVQZHLM2Bkmq/aY/a9+JXwZ+KM/gjwvonhm6sYrKCdZL+2neXc4JPKTIMcelfsp/GfxR8dPh3qHi3xbYaVaXlprUtjHHp0UkcZjWGFwSHdzuzK3fGAOPX/grt+zvP8ADD4/J8X9Fs9vh34lxmeQxxgLb6lCqrcIcdPMXy5gW5Znlxwhr/gjZ+0j/wAIz431f9mzxLqG3TvFO7UvD/mNxHfxp++iH/XSFA31h9Xqqqqqqqqqqqqqqqqqqqqqqqqr4p+L9S8D+EpNe0mC1muEnjjC3KMy4Y88KQf1r4b/ABx8WeMfGmneG9T0/SIra783e8EMiuNsTuMEuR1UdqqqqqqqqqqqqvEmoz6P4d1TVrVY2msbKaeNZASpZELDIBBxke1f8PEvjZ/0LHgj/wAArr/5Iqqqqqqqqqqqqqqqqqqqqqqqqqqr4qfGfxT4H8WSaDpNhpU1ukEcga5ikZ8sOeVcDH4V8IfHGrfEDw1c6zrNvZwzw3z26rbIyrtCI2SGZjnLnv6VX/BS3x8/w+/Yw+IVxbSyrda7bw6NB5bMuRdSrHKCV6DyTLx0P3TwTX7JPw8f4rftNfDPwJ9j+1W+oeJLR72Hax3WkLia5+6D/wAsY5DkjA6kgZI/4LX/AA9bRfjf4I+JUMSR2/inw7JYvtUAyXFlMS7k7s58u6gXlQMKME84/wCCIHj6OPVfif8ACy4lh33FvZazaJuUPiNnhnOOrD95b8jhT1+8Kqqqqqqqqr4keIr3wf8ADvxR4t0yKCW80TRby+t451LRtJDC7qGAIJUlRkAg47ivDf8AwUB+Mus+ItK0i58NeC1hvr2GCRo7K5DBXcKSM3BGcH0qqqqqqqqqqqqqqqqqqqqqqqqqqqv2j/ir4i+D/gex8S+GbPTrm6udVjtHS+jd0CNFK5ICOpzmNe+ME8enwB/ac8e/FX4hw+EfEOk+H7ezktJpi9lbzJJuQAjlpWGPwqqqqqqqqqqqv25v2gW/Zr/Zt8TeP9OuEj1+7UaXoAZgD9unDBHAPUxoskuO4iPbJr4Z+ANc+KvxD8NfDXwzHv1TxNqlvp9sShZY2lcLvbHIRQSzHsqkngV8bviXrX7Fvw7+F/w1+FGj6HdaZbaXLp4OqW0jMVtI7dVf91JGN7mR2cnOWOfXPwc+Keqfts+HvH3wa+Mnh7Q08P3+ieVKNJglilHmNtDq0skgDIQrqccMoPOMV8d/g14p/Z++LPiL4R+MAj6j4fuvLFxGhWO6hYB4pkB52vGysB2zg8g1/wAEx/2qp/2jPgYvhrxbf+f428AeVYalI7gvfWxB+zXJGOpVTG3Ul4ixI3gCqqqqqqqqqqqqqqqqqqqqqqqqqr4/fEfXPhV8O5vFvh21sbi8ju4YRHexu8e1yQThGU549a+CX7VfxD+JXxP0XwTruj+HYLHUfP8ANktLeZJV8uCSQbS0rDqgByDxn61VVVVVVVVVVVV+0x+178Svgz8UZ/BHhfRPDN1YxWUE6yX9tO8u5wSeUmQY49K/ZT+M/ij46fDvUPFvi2w0q0vLTWpbGOPTopI4zGsMLgkO7ndmVu+MAcetVVVVVVVVVVVVVVVVVVVVVVVVVVVXx0/aj+BH7NX9if8AC7PHf/COf8JH9o/sz/iWXd55/keX5v8Ax7xSbdvnR/exndxnBx8Lf29f2TfjV470v4ZfDL4sf2z4l1nzvsNj/YWoW/m+VC80n7yaBEXEcbtywzjAySBXj/x74S+F3gzVviD471b+zNA0K3Nxf3fkSTeTGCAW2RqztyRwqk1/w9H/AGEv+i6/+Wzq3/yLWk6pYa7pVlrelXHn2WoW8dxbS7Cu+N1DK2CARkEHBANVVVVVVVVVVVVVVfF/9uH9lz4C+M5Ph98WPij/AGHr8NvHcPZ/2Lf3WI5BlG3wQOnIHTOR3r4Iftcfs8/tIarqeifBf4hf8JFe6PbrcXsX9k3lp5UbNtVs3EMYOTxgEmry7tLC0mvr66itra2jaSaaZwiRIoyzMx4AABJJ4FeB/iz8K/id9p/4Vr8TPCniz7Fj7T/Yes29/wCRnpv8l22596qqqqqqqqqvFvinQfA3hXWfG3im/wDsWi+H9Pn1DUbnynk8i3gjaSV9iAs2EVjhQScYAJr/AIej/sJf9F1/8tnVv/kWvCfinQfHXhXRvG3hW/8At2i+INPg1DTbnyni8+3mjWSJ9jgMu5GU4YAjOCAaqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq0D/kBab/16Rf8AoAr/AIKM/EFvhx+xl8S9St5kW61jTV0WBGIBkF7ItvKBlWGRDJK3TOFOCpwR+zR4Aj+Kf7Qfw6+H88cMltrXiSyhu1lRWU24lVpsq2Q37tX+U8Hp3r9pz4dp8Wf2efiJ8OxbNPPrPh27js0SMyN9qWMvAQoBLESpGcDk4wK8G+J9Q8E+L9C8Z6S7LfaBqVvf2zJIY2EsMiyKQy8qcqORyK8Pa7pvinw/pnibRp/OsNXs4ry0k/vxSoHQ/irA1VVVVVVVV+2n/wAnMeMv+3H/ANIbev2Df+TfLP8A7Cl3/wChCqqqqqqqqqqqqqqqqqqqqqqqqqqqrQP+QFpv/XpF/wCgCqqqqqqq/as/4KA/Az9le0uNI1PUv+Eo8b+WTbeGNLmHmKcHBuZsFLZMgA7t0mGBWNhnH7T37a3xz/as1Ly/HviH7F4atrhprDw3pxMVnbn+FnHWeQDgSSEkZbbtDEV+zD+w38eP2qr2G78E6Cml+FlmaO68Taruis4iv3ljwC0z9tsYIDEbigyw/Zb/AGBvgP8AstWUGoaDoieI/GO0faPE+rQh7gHniBOVtl5I+T5iMBmbAxVV+3n/AMnBXf8A2CrT/wBBNf8ABO3/AJInrX/Y03H/AKS2tftnfs8Wn7Tv7PXiX4ZpDCdcWP8AtDw7NIQvk6lCCYvmPCiQF4WbskznrivDPiLxT8MvG+m+KNEmuNJ8Q+GNSjubdnQpJbXMEgIDKeQQy4Kn3Br9n74yaB+0D8HPC3xc8OlEg1+xWSe3VtxtLlfknhPukiuue4APQ1VVVVVVVVVVVVVVVVVVVVVVVftC/wDJN5/+vuH+dfAz/kqmh/8Abx/6TyVVVVVVVVVVVVeOf+RJ8Qf9gq5/9FNVVVVVVVVVVVVVVVVVVVVVVVVVVVftDf8AJR5v+vOH+Rr9mj/kQ77/ALC8n/oqKq/4Le/ECO28FfDT4VQXO59R1S61m5hAX5BbxCGJicZGftMwGDj5WyOBj/gjd8O4fFX7Ud943u0Qx+CvDtxcW5P3hc3BW3XA9PKkuMn6DvX/AAWV8AP4m/Zd03xpbRytL4O8SW80zKjMq29wjwNuxwv714MMeO3VhX/BLv4hv8P/ANs/wVFLe/Z7HxRHdaJefMw8wTQs0KYBAObiO34bI74yARVVVVVVVVXxz/5In8Qf+xV1P/0lkrwL/wAjv4e/7Ctt/wCjVqqqqqqqqqqqqqqqqqqqqqqqqqqqv25P+SS6T/2MUP8A6T3Ffsbf8lstf+wdc/8AoIqqqqqqqqqqqv8AgsF8fv8AhY3x3svg7ol7K2jfDi3KXSBsJLqU4V5Wx/FsjESAnOG8zGMnP/BF74Af29438R/tHa3ZQy2XhmN9G0NnXJW/mRWnlX0KW7hOeouj6V/wUq/5pz/3FP8A20r/AIJw/wDI6+MP+wVD/wCjTX/BXP8AZVh+Jfwtj/aC8HaQ0nijwHCw1dbeLLXmkk5d3wMk25zIDwFjaYnOBj9kX9obWf2Yfjt4f+J2nTynTVkFlr1rGMm706R186PHcjasi/7caV4e1/RfFnh/TPFPhvUYdQ0nWbOK8sLuE5S4glQPHIvsyspH1qqqqqqqqqqqqqqqqqqqqqqqqqq/bJ/5Indf9hG2/wDQjX7KP/JfvCv/AG+f+kc1VVVVVVVVVVVVft5/8nBXf/YKtP8A0E1/wTt/5InrX/Y03H/pLa1VVVVVVVVVVVVVVVVVVVVVV/wWh8T+JfDHww+HM/hrxFqekyza9crK9jdvbs4EAIBKEZHsa/4Wz8VP+im+K/8Awc3H/wAXX/C2fip/0U3xX/4Obj/4uvhZ8U/ifcfE/wAH28/xI8USxSa9Yq6PrE5DAzoCCC/II7VVVVf8Fz/+aJf9zB/7jq/4Jcf8n2/DH/uLf+mm8r9vz/kzT4s/9i/J/wChpVfCf/klfg3/ALF+x/8ASdKqqqqqqqqqqqqqq/4K8/8AJ5Wpf9i/p3/oDV/wRD/5Kn8S/wDsX7X/ANKDX/BZn/hPf+GZdH/4Rvzf+Ec/4SSH/hIvK67NjfZ9/wD0z87bntv8v2r/AIJ5y/E2P9r/AOHCfC6W9W5l1aJdZW3ZgjaTvU3omxwU8oMQG43hMfNtqqqtX1Sw0PSrzW9UuPJstPt5Li5l2FtkaKWZsKCTgAnABNfEP/gtD+zp4djuLb4e+CvGHi+8iz5MksUWm2kvytj947NKvzBRzDwGJ5Iwf2fPirL8cvgv4T+Lc2iJo7eKLH7YLBbgzi2BdgE8zau8gAZbauTzgdKqqqqr9rL/AJNX+Mv/AGT/AF3/ANIJqr9k7/k1f4Nf9k/0L/0ghqqqqqaaG3heeeVI44lLO7ttCgDJJJ6DFeN/23v2R/h488Pib9oHweJ7XeJ4NPvv7RljZG2sjR2wkYODxsxu9q/4ej/sJf8ARdf/AC2dW/8AkWvBX7dv7H3xAMKeHv2hPCMbzqTHHqt0dKdsPsxtuxGdxJGFxkjkAjmtP1HT9WsodS0q/t720uF3Qz28okjkHqrKSCPpVVVVVVVVVVVVVVVVVVVVVV4s8e+FPBUKv4h1aOCR1zHbqC8rjpkIOccHk8e9XH7TvhlZWW18OanJGPutI6Ix45yATj868C/EPQPiFZ3F3oUd5GLVgsqXMOwqTnGCCVPAzwTjIzjNf8Lf8M/8Jt/wgX2HU/7Q+0/Z/M8pPK3YznO/OPwqvHnxD0T4eWdre61a3s6XchjjFqisQQM87mXivDuuWfibQ7PXrCOZLe9i8yNZlAcD3AJHb1rxB4j0Twvpsmq67qEVrbxjq55Y+ijqx9hV5+0v4LhuEis9I1a5i3YklKImBxyoLZPfg7a8G/Erwj47Vk0LUCLqNdz2lwvlyqPXGSGA6EqSBx6iqqq0D/kBab/16Rf+gCv+C3PxAGm/C74dfC6Fvn17XLjVZip5CWcIiVW+bozXmQCpBMecjbg/8Eefh+/iv9rQeL5LbdB4K8P3l6spLAJNMBaqoweSUnmODkYU9wDVftOfDtvhN+0N8RPh4LZIING8RXcdnGkYjAtWkLwEKAAoMTRnA4GcDiv+Cc/xBb4j/sZfDPUp5ka60fTW0WdFIJjFlI1vEDhVGTDHE3TOGGSxyTVVVVVVVV+2n/ycx4y/7cf/AEht6/YN/wCTfLP/ALCl3/6EKqqqqqqqqqqqqqqqqqqqqqqqqqqqtA/5AWm/9ekX/oAqqqqqq+Kfxf8Ahn8EvCc/jf4q+M9P8OaNb8Ge6YlpG/uRRqC8r/7CKze1ftZ/8FcfiD8SRc+Df2cU1DwP4ckjeK41mbauqXYPGY2UkWox0KEyZwQ69K8FeBPiL8ZvGcPhfwN4c1jxX4l1eYv5FrE1xNIzN88sjfwrlstI5CrkliBk1+yR/wAEhPC3hEWPjr9qGW08Sayu2aHwtaSs1jat8rL9okGDcOp4aMfuuCCZVNaVpGlaDptto2haZaadYWcYitrS0gWGKFB0VEUAKB6AYqqqv28/+Tgrv/sFWn/oJr/gnb/yRPWv+xpuP/SW1qv+Ctn7Nv8AwqT49L8WvDun+X4b+JvmXcvlphbfVUx9qU46ebuScEnLNJLgYSv+CMP7RkOkeIPEH7MfiK7RIdcZ9a8OFzgm6jjAuoB3JaGNJVHAUQSnq1VVVVVVVVVVVVVVVVVVVVVVVX7Qv/JN5/8Ar7h/nXwM/wCSqaH/ANvH/pPJVVVVVVVVVVVV45/5EnxB/wBgq5/9FNVVVVVVVVVVVVVVVVVVVVVVVVVVVftDf8lHm/684f5Gv2aP+RDvv+wvJ/6Kiqv+Cu3xAHjP9sPUfDsTZh8E6HY6Uu0/Kzuhu3YfMQTm6CE4X/V4I+XJ/wCCJ/w/fRfgp45+JE9t5cnijxBFZRMS2ZYbOHKsBnGPMupl4AOVOcgDH7Xfw7b4r/sx/EvwHBbJcXWoeHbmSyieMOHuoV86AYIOD5sUeD1BwRyBXw88Y3/w68f+GfiDpSb73wxrFpqlsuQMyW8yyqMkMOqDqpHseladqFlq2n2uq6bcpcWl5Ck1vKnSSNgGVh7EEGqqqqqqqqvjn/yRP4g/9irqf/pLJXgX/kd/D3/YVtv/AEatVX/BWn9r3x38Hte8A/Cz4O+ONT8O+IYWbxBq11YSmNvK+aG2hfB2yI7C4Z4nBX5IiQcivgd/wUn/AGhov2kPBfjr40/FzVdT8Kx3CWGt6aqpb2RtZV8t52toAkZkQsJQ+3dlMD5SVMM0NxCk9vKkscihkdGyGBGQQR1BFVVVVVVVVVVVVVVVVVVVVVVVX7cn/JJdJ/7GKH/0nuK/Y2/5LZa/9g65/wDQRVVVVVVVVVVX7Q/xj0X4AfBbxZ8Wtbli2aBp7yWsMvIubpvkghwCCd8rIvBGASeMZq8uvFfxK8aTX11Jda54m8Vao0kjY3TX15cS5JwOrvI/5tX7LfwR039nf4CeDvhPY28S3Wk6ej6rLGd32i/k+e5k3YywMrOFz0QIvRRX/BSr/mnP/cU/9tK/4Jw/8jr4w/7BUP8A6NNXlnaahaTWF9bRXNtcxtFNDMgdJEYYZWU8EEEgg8HNft8fsv3X7Lfx91Tw3ptpL/wiHiDdqfhm4MZCfZ3Y77fPILwPmMjOSvluQPMAr/gjv+1Vb6x4euv2WPGN/jUdI82/8KSSOT9otmJe4thxwY2JkXkkrI/AEfNVVVVVVVVVVVVVVVVVVVVVVVVV+2T/AMkTuv8AsI23/oRr9lH/AJL94V/7fP8A0jmqqqqqqqqqqqqr9vP/AJOCu/8AsFWn/oJr/gnb/wAkT1r/ALGm4/8ASW1qqqqqqqqqqqqqqqqqqqqqqr/gt7/ySr4af9jBdf8ApOK/Yi8O+H/Fv7WPwx8N+KtD0/WdJv8AXEiu7DULVLiC4TY3yvG4KsOBwQRX/DJv7K//AEbP8Kf/AAjLD/41Vp+yx+zDYXcF/Yfs4/C+2ubaRZIZofB9ijxOpyrKwiyCCAQR0NftHftC+Cv2YPhnP8VfH2ma3qGk295DaNDo8EUs5eUkKQsska445+b8DXh7/gsh+zD4m8QaZ4bsPA3xQS51W8itIXm0uxCK8jhFLEXhIGSM4B+lV8e/+CoHwB/Z1+LOu/Bvxt4R+IF9rXh/7P8AaZ9L0+0kt3863jnTY0l0jHCSqDlRyD1HJ/ZS/bV+Ff7Yn/CUf8Ky0DxXpn/CI/Y/tv8AbtpbweZ9p87y/L8meXOPs75ztxlcZycf8Fz/APmiX/cwf+46v+CXH/J9vwx/7i3/AKabyv2/P+TNPiz/ANi/J/6GlVrHxe0P4C/st6R8WfEuha3q+k+H/DenS3tvo8UUlyI2jiQuqyyRqQu4FvmyACcHFf8AD6v9lf8A6EL4q/8AgqsP/k2vhd8RfDvxe+HPhv4n+EzcDSPE+mw39olyqrLEsig7JArModTlWAYgMpwSOa/al/a0+GP7IvhXR/FfxK0/xBqEWuagbK0tdEtY5piwjZ2dvNkjQIAoB+bdl1wCNxXwx/wWI/Zu8Y+JdJ8I+Hvhv8WLrVNbvobGxt00mwLSzyuEjUAXpOSzAcetVXxD/wCCuv7Nnw08f+Jvhxrvgn4lz6l4V1i70q8ltNNsmhkmtpmidoy12rFCyEglVOMZA6Vqv7bH7P8A4Z+BXhv9oLxr4muPDfh/xdamfR7G/gD6hdEdY0ghaTcw4yQSi7lLMoOa1L/gtt8C4tSWLR/hF47urD5d09y9pBKP72I1lcHHb5xn2r9nL9tD9n/9qWGW3+GHi1k1u1hWa50DVYha38SEDLCPJWVVJAZomdVJGSNy5r4h/wDBXX9mz4aeP/E3w413wT8S59S8K6xd6VeS2mm2TQyTW0zRO0Za7VihZCQSqnGMgdK8Rftx/AHwb8BPDP7QvjHWtQ0XQ/GFuZdF0y5tlfUrxhnMawxu4yMDLbti7l3MNwz+yf8A8FFPh/8Atc/FTWPhl4N+H/iDRf7L0efVY73U5of30MdxDCFMcZba589WxuYDBGT1rVtW0rQdNudZ1zU7TTrCzjMlxd3cywxQoOrO7EBQPUnFfFn/AILAfswfD/V7vQPB1l4j8e3VnMYnu9KgjhsWKnDbJpWDOAc4ZEZGxkMQQT4B/wCCzv7NniTUk07xv4T8YeEI5JAq30lvHfW8anGWk8lvNGPRY34HrxXhXxX4Y8deHrHxZ4M8Q6frmi6lH5lnf2Fws8M65IJV1JBwQQfQgg8g1X7RH/BSH9mX9nPXLjwfrevah4o8TWm5brSfDkKXLWcgJXZPK7pFG+QQU3F1xyoyufB3/Baj9n3WdXj0/wAYfDzxn4dtJWVRfIsN7HFk8tIqurhQP7iufav+CoHjrwd8Tf2nR488AeI7HXdA1jwxp0tlf2cm+OVQrqw9VZWVlZSAysrKwBBA/wCCIf8AyVP4l/8AYv2v/pQa/ay/aw+Ff7KfhXSdV+LHhvxBrem+KLiawS20mzt7ndiPc4lWeWNdhU4xznPIr9mr/goP+xRr/wAR9F+EXwN+A2u+CtS8X3wto3tPC2l6bbNKVzumNtcFiMIBnax4FV8WPjV8KPgZ4bbxZ8WvHmk+GdN+YRPeTfvLhlGSkMS5kmcA52RqzY5xXiz/AILU/s66XcG28I/Dzx3ruyQq080NvZRMuOGQmV3PPZkWvhT/AMFgvgL8SPFOjeCL74dePdI1jXtSjsLEQ2sF9C0kpVYgSkok3NI2wKsbclTnk4/bY/bw+EH7OF3qHwV8ceHfGF9rnibwnLd2lxpVnbS2yJcG4t0DtJPG4IeFicIRgjBJyBX7MX/BVv8AZ2+CvwA8D/CrxT4N+It1q3hrS1tLubT9Os3gdwzHKM92jEcjqo+leHtbtPE3h/TPEdhHKltqtnFdwpMoDqkiB1DAEgHBGcEjPevj1+1B8D/2adFi1f4ueObTS5br/jz02IGe9u+o3RwJlygIILkBAcAsCQDqv/Bbj4Kw38sWifBzxrd2Qx5U11cWtvI3AzmNXkC85H3jkAHjOB+zp/wUG/Zq/aa1ePwt4L8S3ui+JZlZoND8QwJZ3NwFJyIirvFK2Bu2I7PtySuFbFePfiB4J+F3hO/8c/EPxRp/h/QdMj33V9fTCNE9FHdnY4CooLMxAUEkCvG//BaT9nbQtUOn+C/AnjPxRBEzB75oobGFxhSpiDuZGHLA70QgrxkHI17/AIKQfs5ftL/s5fF7wXo2o6n4V8WXHw91sW+keIIkhN6/9nS7ltpUdo5CCcBWKSMASEwDiv2Tv+TV/g1/2T/Qv/SCGqqqr9rz/grV4H+FV7e/D/8AZ8s7Hxp4ntWMd1rU7b9Ks25BWMowa5cEDO0rGMj52IKj4y/tL/HX9oHUPt3xZ+Jesa7GrFobBpvJs4MnP7u3j2xqenO3JCrknArwd8Jfin8RF8zwF8N/E/iKMNtaTS9JmuUQ5x8zIpCjPckAV/wxP+1b/wBEM8Sf9+U/+Krxp8GPi78OY5J/Hvww8U6BbxuyfadR0iaCFirBTtkZQjDLKMqSDuXB5FfCb4//ABo+BWpf2p8JPiTrfhuQyCSSC1n3W8zDHMkD7opPuqPnU8DHSv2Uf+CwPhbxjPYeB/2nNPtPDOrTyCGHxNYRsNOkJACm5jJZoCTwZBmPLZIiUEi0u7TULSG+sLqK5trmNZIZoXDpIjDKsrDgggggjg5qqqqqqqqqqqqqqqqqqqq8Q6zbeHdCv9dulLRWFu8zIDgvtGQo9ycD8a+Gvg+5+MnivUfFXjC4kltIJA08asV812ztiU5yqADtyBgDrkWvhTwvYwLbWXhzTIY16Iloij69K0nQtF0GKWDRNKtLCOaTzJI7aERqWwBnA46AV/zcj/3F/wD2Sq/af/5AGif9fkn/AKBXwl/5Jt4f/wCvMfzNXQn+NfxkfSbydv7F0l5MRgEfuI2AbHQgyNt5PIDD0Aqy0TR9N05dIsNMtYLNVx5CRAIeAOR36Dk18YvD8Hwv8b6P4t8IRR2Yui0yW6riNJYyN4AGMIwdRt9zjg4Gm30Oqada6lbkGK7gSWMhs/KygjkdeDVVWgf8gLTf+vSL/wBAFf8ABYb4gP4r/a0Pg+K53QeCvD9nZNCAwCTTA3TtyOSUnhHGRhV7giv+CI/w+Gm/C74i/FGZfn17XLfSoQw5CWcJkZl+XozXmCQxBMeMDbk1/wAFifh6nhT9rJPGFtC/k+NvDtpeyylSAbiDdasoyxyRHBATgKPnHGck/wDBEf4hx6h8OPiL8K7i9zNousW+rW0LsoJjuojE5TncQrWq7hgBTIvJL1VVVVVVVV+2n/ycx4y/7cf/AEht6/YN/wCTfLP/ALCl3/6EKqqqqqqqqqqqqqqqqqqqqqqqqqqqtA/5AWm/9ekX/oAqqqqqaaG3heeeVI44lLO7ttCgDJJJ6DFftc/8FX/hh8IP7R8CfAw2njvxjHGUbUo5N+k6fKfWRTm5dRg7YzsycFwQyj4xfHH4rfH7xXJ4z+LXjbUPEGonIhWd9sNohx+7ghXCQpwCVRQCeTkkk/sl/wDBLH4w/Hj7H4x+KaXfw98ES4dGu7crqeoJzgwW7j92hwP3kuAQysiyA5HwL/Zy+D37OHhOHwl8JvB1ppcYjC3d+6iS8v3HJknnPzOScnHCrnCqqgAVVVVft5/8nBXf/YKtP/QTX/BO3/kietf9jTcf+ktrVftx/AD/AIaS/Zr8VfD3T7KGfX7aNdT8Ps65KX9vlkVfQyIZYMnoJia8CeNfFPwo8faJ498LXL2Gv+F9SivLRpFI2TRODtdeMqcFWU9QSDwTXwV+K/hv46fCjwv8WvCT/wDEt8Tael0kRcO1tJyssDEcF4pVeNscbkOKqqqqqqqqqqqqqqqqqqqqqqr9oX/km8//AF9w/wA6+Bn/ACVTQ/8At4/9J5KqqqqqqqqqqqvHP/Ik+IP+wVc/+imqqqqqqqqqqqqqqqqqqqqqqqqqqqv2hv8Ako83/XnD/I1+zR/yId9/2F5P/RUVV8fviCnxY+N/j34kwzPJb+JPEV7e2m8klbd5mMK5KqcLHsUZUHAGQK/YF+Hw+Gn7Hnws8OuuJ7rQ01WfcPmD3zNdlW+VTlROEwRxtAy2Mmvj98PU+E/xv8e/DWGF47fw34ivbK0DqQWt0mYQvgsxw0exhlicEZJ61/wT++IcfxN/Y8+GOute/aLmw0ddJutzKXSSyZrbDgE4JWJH55KurEDdVVVVVVVVXxz/AOSJ/EH/ALFXU/8A0lkrwL/yO/h7/sK23/o1aq8u7SwtJr6+uora2to2kmmmcIkSKMszMeAAASSeBX7UPxovf2hfj540+LVy7fZ9a1JxpsbpsMNjEBFbIV6BhCke71bce9eLfBXivwJqFrpXjDQbvSru90+21G3iuU2mW2uYllhlX1DI6n1BypwwIH/BK79oP/hdf7NFn4U1rVftXib4cyJpF6rn941ptJs5Tycgxq0QJwSbdsjuaqqqqqqqqqqqqqqqqqqqqqqr9uT/AJJLpP8A2MUP/pPcV+xt/wAlstf+wdc/+giqqqqqqqqqqv8AgtF+0JLdap4Z/Zq8Pakn2ezUa34iSJzuaZgVtYXx0CoZJSpznzImwNoJ/wCCSf7O9v8AFz9oU/EzxDZ+doPwyjj1CNHjJWbUnJFoCen7sq82Qchoo+CCar/gpV/zTn/uKf8AtpX/AATh/wCR18Yf9gqH/wBGmq/b9/Zbt/2pfgJqOhaRZRN4z8ObtR8MzuSD5y482AkdpowUweA/lsfu18OvH/jD4OfELRfiF4OvG07xD4ZvluLZ5I/uyIcMjqeqsNyMp6gkd6+Afxm8N/tB/CHwz8XfCq+TZ+ILMSyWplEjWc6krNAzADJSRWXOBnAOADVVVVVVVVVVVVVVVVVVVVVVVVVftk/8kTuv+wjbf+hGv2Uf+S/eFf8At8/9I5qqqqqqqqqqqqq/bz/5OCu/+wVaf+gmv+Cdv/JE9a/7Gm4/9JbWqqqqqqqqqqqqqqqqqqqqqqv+C3v/ACSr4af9jBdf+k4r4QfFDX/gr8TPDvxV8K2mn3WreGrwXdpDqEbvA7gEYdUZGI57MPrX/D6v9qj/AKEL4U/+Cq//APk2v2Yf+Crf7RHxr+P/AIH+FXirwb8ObXSfEuqLaXc2n6dexzohVjlGe7dQeB1Uj2r/AIK8/wDJmmpf9jBp3/obV8Jv+SqeDP8AsYLH/wBKEqv+Co//ACfb8Tf+4T/6abOv+CGP/NbP+4B/7ka/4Ln/APNEv+5g/wDcdX/BLj/k+34Y/wDcW/8ATTeV+35/yZp8Wf8AsX5P/Q0qvBvhzR/GPwG0Pwj4gtEutL1vwjb2N7buoIlgltFSRSDkEFWI59a+Nfwu1n4KfFnxZ8KdfGbzwzqk1p5mciaMHMUo9njKOMgHDDIB4H/BGb4qjxd+zlrXwwvLrfeeAdcbyIwuBHZXgM0fOTkmdbzsBjb1Oa/4LWfEb+3vjj4L+GVvLvg8JeH3u5PlxsuL2X515UE/ura3bO5h82MAhs/8EkPgzH8TP2pYPGmqWPn6T8OtPfVGLhShvH/dWykHnILSSqRjDQA57Gq/ay/5Oo+Mv/ZQNd/9L5q/Z3/Z1+Pf7aHiC08CeC757rTPBlikL3+r3biy0S1lmkdYxgMQXkaZljQEsQ5xhWI/aK/4JUfHn4C+Ab/4lafruieNNF0W3e51ZdMSSK5s4V5ebynHzxquWYqcqASRtBI+HXxB8W/CjxxovxG8B6u2m6/oF0tzY3KoG2OOCCpyGVlJVlIwQSDwa+BXxV0344/B7wj8WdJt/s8PibS4rprfdu8iUjEsee+yRXXP+zX7WX/J1Hxl/wCyga7/AOl81adbfGv9qDX/AAn8O/CfhrUPE+o+G9Di0rSNN0y3ZhbWcTkl3JOEBklLPIxVQXHIGAP+CX/7GX7Sn7Ovx+17xt8Y/ht/wj+i3vg+50+C5/tixu99y93ZyKmy3mdhlIZDkgD5cZyRn/gr1+1dq/i74iP+zH4Uv3g8O+FGim154Lg41G+dFkWJwpwUhVl+U/8ALUtkZRcfsNf8E39b/a08O3vxI8U+M38K+DrW6eztXtrVbi61CdF+fYGYLGiFly5DbjlQByw/bt/4J26t+yHo2kePvDfjGXxV4P1W8FhNLcWi29xp9yYyyK4ViJEcRykOAoUqFIyVLf8ABH79pLUvA/xim/Z/8QatK3hzxzG8mlwzT4is9SiRnygJwvnRqyHHLOsIr/gpR+0zqv7Nn7O91N4Pupbfxd4zuDo+j3Ue5TYhkLT3IYYw6RghCDkSSI2CFYV4Z8NeJviD4s03wn4Z0+41fX/EF9Ha2VsrAyXNxK4VRuYgZLEZLEAdSQMmtH/4Ig/EG68HDUNb+PGg2HihoSw0qHRZbi0WTsjXXmq2PVhCcdga+JXw48Z/CLx1rPw2+IOiS6T4h0G48i9tJCDtJUMrKRwyMjK6sOGVlIJBFf8ABEP/AJKn8S/+xftf/Sg1/wAFvf8AklXw0/7GC6/9JxX7Af8AyeV8Jv8AsYE/9Aevij8Q9A+Evw58SfEzxRI40vwzps19cqg+ZxGpIRf9pjhR7kV+0T+0J8Qv2mfibqPxL+IWpPJNcMY9PsEcmDTLUMSlvCvZVB5OMsxLNksTX7MP/BKT4yftAeENL+I3ivxTp/gHwxrduLnTZLmza9vbqFlJjmW3DRhY3+UqWkUlWDAEEZ+Bf/BIW6+Cfx78C/FG7+MWn+LdE8M6gb+6s5tCNhN50cbm3MYMs6ttnELEkqQASCGANf8ABRf9gTxf+0r46Hxu0P4gaPo1l4X8G/ZJrG7tZJJZmt5bq4ZlZeACswUZ7g1XwR/4JK/ET43/AAm8MfFjSvi54c0208T2K3cNpcWMzyQgkjaxU4J47V8Q/iBo/wCzB+zlfeOfFRS+t/Afh2FXjSUQi9njRIoolZs7TLKUQEg4Ljg9K8Y+Lvif+078Z5de1mWXXfGfjrWI4LW3STCtNPII4LaHzGISNSyRopbCqAM8ZrR/+CH+nv4OA1/49XEfit4SSbPRg9jFL2XDOJHUdN3yk9do6V8SvAXjb4A/FnWfAet3EuneJfB2qeWt1ZzFGSSMh4p4nU5XI2SKQQRkdDX7DP7QLftK/s2+GfH+o3CSa/Zq2l6+FYE/boAodyB90yI0cuOMCUdua/4KlftP618a/j/q3ww0y+uIvB3w2vpdMt7TcyLc6hETHdXEiE4LLIJIkP8AcXIx5jV+x1+wv8Tf2xNUv7nw/qlj4c8KaJMkOqa7exNLskYZEUEKkGaXaQxUsigYy4LKG/a6/wCCXHxI/Zk8C/8ACzfDPjSL4geHLHH9syQ6UbC404E4WUxebKHiyQGcMCpIJXaCwr9k7/k1f4Nf9k/0L/0ghqqqv+Ckf/BRe7+J93qv7PvwM1aW28HW0j23iDWoWKPrTqcPbxHqLYEEMf8AlqeP9X/rPgL+zt8TP2jfFyeFfh7pG6KLDX+qXIKWthHkAtI+Dzzwi5ZuwwCR8A/+CdXwE+Dttbap4n0aLx54mWMebe61AslrE+OfJtTlAM4IMm9wRkMM4q2tbaytobKyt4oLe3jEcUMSBFjVRgKqjgAAAADpVSxRTxPBNGskcilXR1yGB4II7ivjp/wT9/Z7+NUE99Z+HIvBniB9zLqmgwJAruTkmaAARyZJyThXP9+v2if2Wfir+zTrw07xxpX2jSbpyun65Zoz2l397C7yPklwpJjb5gORkYJ/4J5f8FDdc/Z/1zS/hH8W9eluvhZdSOkM0kRml0KWQgiRCDu+z7sl4wDjcXUZyGtLu01C0hvrC6iuba5jWSGaFw6SIwyrKw4IIIII4Oaqqqqqqqqqqqqqqqqqqr42wzz/AAu11LYEuI4mODj5VmQt/wCOg1+zJcWz+ENUs0I8+LUi7jHO1o0C/qrVVMyr+0hlmAH9rgDJxyVwKr9p/wD5AGif9fkn/oFfCX/km3h//rzH8zXw68CWnjXxhdeGNa1Oa0kihlYPEu8vIjKCOe2Cxz7e9f8ADMGgf9DRf/8AflK/4Zg0D/oaL/8A78pWh6XHoeiafokMrSpp9rHbq7DBYIoUE/XFVX/DMWu/9DRYf9+HrTrVrLTrWydwzW8KRlgMZKqBn9K/aY8fR/FP9oP4i/ECCWGS21rxJezWjRMrKbcSssOGXIb92qfMOD171/wTs+H8fw5/Y0+Gemm28u41fS/7ZnYlSZTeu1wjHaSP9VJEo74UZAOar/gtt8OJdX+Fnw++KlrA7nw3rVxplyY1JxHeRB1d8KflV7QKCWABlAwS3H/BHn4gP4U/a0Hg+S52weNfD95ZLCQxDzQAXSsMA4ISCYc4GGPcgVVVVVVVVV8d/wBijxR8XfitrnxD03xvpVhb6r9n2W09tIzp5dvHEckccmMn8a/Z3+Euo/BX4aweBtU1e21KeK7mnM8EbIpDkEDB54xVVVVVVVVVVVVVVVVVVVVVVVVVVVf8Mxa7/wBDRYf9+HrTrVrLTrWydwzW8KRlgMZKqBn9Kqqqv2lP2w/gd+yxoD6j8RvE6TazIo+w+HdOZZtQuyehEeR5aYBJkkKpxgEsVU/tZf8ABRf44ftPy3fhu3v38G+A3ZlTw/pc7K13Hzj7ZMMNOSDymFi4U7Nw3H4Afst/G39pvXm0T4TeDLi/gt2xe6tcZgsLHp/rZyNobBBEa7nIyQpAOP2UP+CY/wAEf2dWsvF3iyFPHnjqFUddR1CH/RLCTHzfZrc5XOTxJJucbQV2ZIqqqqqq/aJ/Y68S/Gv4kzeOdL8ZaZpsEtnDAILi3d2BQEE5XjBzX7MvwT1T4DeAr/wfq2uWuqzXmryXyzW8TIqq0MMe3Dc5zET+Iqq/4Krfs2/8KT/aFm8e+H9P8nwv8SPN1K32JhIL8Efa4vbLMso/67ED7tf8EZP2kf7N13XP2YPE2ofuNX8zWPDHmN924RB9qt1zk/NEizKowB5Mx6vVVVVVVVVVVVVVVVVVVVVVVVfEvwdc+O/C0nh+0vYrWR5kkEkilgNp9BXw++BOreDPF+n+JbnXrS5js/M3RRxMrNujZOCf97NVVVVVVVVVVVXiDTX1nQdS0eKVY3vrOWBXYZCl0Kgn25r/AIdw+Nf+ik6J/wCAcv8AjVVVVVVVVVVVVVVVVVVVVVVVVVVXxL+CeqeO/FD6/aa5a2sbQJGI5ImY5UdeK+FXgS7+Hvh240W8v4bt5r1rgPEhUAFEXGD/ALn61+2B4+j+GH7L3xO8aGWFJrPw3dQ2rSsqqLidPIhzu4P72VPl6t0HJFfDzwbf/Ebx/wCGfh9pT7L3xPrFppds2AdslxMsSnBZR1cdSB7jrWnafZaTp9rpWm26W9pZQpDbwoMCONAFVR7AACq/4K9fDx/B37Xd34nhs/LtfG2h2epLIitsaWNTayDOAA/+jozAE/6xWJy2K/4InfEOXW/g147+GVxI8jeFdehvoSwOEhvYiAg9g9pK31c+1VVVVVVVVfEHw3P4z8BeJPB9rdR282uaRd2Mc0ikrG00LRhiByQC2a8P/wDBPPxlo2v6ZrEvxE0WRLG8inZFtJQWCOGIHPfFV/wVS+Okfwf/AGVtX8NaddbNd+I0h0G0VXXcls6lryQqeqeSDCcchrhDX7KPwTuv2hv2gvBnwrjtZZbHU9QWTVmjBHlWMX7y4YsCNv7tWUHI+ZlA5IB/4LHfs4Q698MPDPx28H6JbxTeBFGlasltBtI0yVgIDgcBIZiVCgcC5Y9BX/BOr9oSX9nv9pzw9qGo6klt4Z8VsNE14SuVjWGZh5Ux6hTFMI2LY+55gyAxNVVVVVVVVVVVVVVVVVVVVVVVX7QPwm1H4y+DLPwvpmr22nS2uppeGWeNmUqsUqbQBznMgP4V8Dv2XNf+E/j2HxfqPirT7+GO1lhMMMDqxLgAHJ47VVVVXd3aWNu11fXUNvDHjdJK4RVycDJPA5IFav8AtN/s2aBqt5oOu/tC/DTTtS024ktryyu/FllDNbTRsVeORGlDI6sCpUgEEEGv+Gsf2V/+jmPhT/4Wdh/8dr/hrH9lf/o5j4U/+FnYf/Ha/wCGsf2V/wDo5j4U/wDhZ2H/AMdr/hrH9lf/AKOY+FP/AIWdh/8AHa/4ax/ZX/6OY+FP/hZ2H/x2v+Gsf2V/+jmPhT/4Wdh/8dr/AIax/ZX/AOjmPhT/AOFnYf8Ax2vEX7ZX7Kfhzw/qfiGb9on4c38el2ct09rp3imyubmcRoWKQxJKWkkbGFRRliQBya+M3xS1742/FTxR8V/EypHqPifUpbx4UOVt0Y4jhU4BKxxhEBPJCjPOa/4J8fs+t+zt+zH4a8OarAieIvEC/wBta5hQClxcKpWEkdTFEIozyRuViDgiq/an/Zw1v9oP/hGP7G8S2Ok/2B9r8z7TC7+b53k4xt6Y8o/mK/Ze/Ze174A69rer6v4psNVTVbOOBEtoHQoVfdk7u1VX/BW39lZvhT8WB8evCWnMnhb4h3THUdoGy01ghnkHqBOqtNznLibkDAH/AASP/am/4VZ8VpfgN4u1fyfC/wAQLhf7MWT7tvrJ2JHj089VWI+rLF05qqqqqqqqqqqqqqqqqqqqqqqqqvjl8NL74s+ApfB+nanBYTSXUUwmnQsoCHJGBzXwd/ZN8RfDL4j6R43vvFunXsGm+fugigdWfzIHjGCeOC4P4VVVVVVVVVVVVV+0T+x14l+NfxJm8c6X4y0zTYJbOGAQXFu7sCgIJyvGDmv2ZfgnqnwG8BX/AIP1bXLXVZrzV5L5ZreJkVVaGGPbhuc5iJ/EVVVVVVVVVVVVVVVVVVVVVVV/wW9/5JV8NP8AsYLr/wBJxX7OPwr0/wCN/wAcvBvwm1XVbjTbTxPqS2kt3boHkhBUncobgnjvX/DkL4Vf9Fw8V/8Agut/8a+CP/BJb4dfBD4seGPixpXxb8R6nd+GL4XcNpcWMKRzMARhipyBz2r/AIK8/wDJmmpf9jBp3/obV8Jv+SqeDP8AsYLH/wBKEqv+Co//ACfb8Tf+4T/6abOv+CGP/NbP+4B/7ka/4Ln/APNEv+5g/wDcdX/BLj/k+34Y/wDcW/8ATTeV+35/yZp8Wf8AsX5P/Q0qvhP/AMkr8G/9i/Y/+k6V/wAFp/gWmi+NfCf7Qej2bLb+Iof7G1l1BK/a4VL27njAZ4Q64z0txx1J/wCCRHxWHgH9q+38GXkyJYeP9JuNNJcoqpcxL9ohYsSCCfJkjCjO5plGCcEftqfEb/ha/wC1b8UPG0cvmQT+IJrO0k27fMt7QC1hbG1SMxQIcEZGcEk5J/4JF/Bm3+HH7Lkfj+8sYo9Z+I2oSX8s2D5n2OEtDbRt2wCs0q4zxcdewqv2sv8Ak6j4y/8AZQNd/wDS+av+CKOn2Uf7M/i/VY7dBd3Hjq4hllA+Z447GzZFPsDJIf8AgRq8s7TULSawvraK5trmNopoZkDpIjDDKyngggkEHg5rxxolp4a8a+IPDdhJLJbaVqlzaQvMQXZI5WRSxAAJwozgAZ7Cv+CSeqX+o/sV+HbO8uPMi0zWNSt7VdgHlxm4aUrkDJ+eVzk5PzY6AAftZf8AJ1Hxl/7KBrv/AKXzV/wRN8JaBbfAbxv47hsEGt6l4ubTbi6PLNbW9pbyRIPQB7mYn1yPQVXx/wDEGoeLPjt8RfE2qlPteqeKtSuJVTO1We5kO1dxJCjOACTgACvAX7Xv7THwu8JWHgT4ffGXxBoegaZ5n2PT7WVRHD5kjSPtBUnl3dj7sa+I/wC1d+0X8XvDL+DPiX8Xdd8Q6JJMkz2V5IpjMiHKtgKORX7Hd1qVn+1h8HZtJklSdvHGkRsYhk+U93Gso+hjZwfYmv8AguDql/N4/wDhZoklxmytNHv7iGLYBtklmiWRs4yciGMYJwNvGMnP/BJrQdN1r9tPwzdahD5r6Rpeo3lqD0WX7O0YJHfCysR6HB7VX/BXaGGP9szVniiRWl0HTmkKrgu3lkZPqcAD6AV/wRD/AOSp/Ev/ALF+1/8ASg1/wW9/5JV8NP8AsYLr/wBJxX7Af/J5Xwm/7GBP/QHr/grF4g1DQ/2KfFNnYFFXWtS06yuGOQyx/aUlO0gjBJhVTnIKswxzkeB9M03WvGvh/R9Zk2affapbQXTb9m2J5VVzu/h+UnntVpZ2lhaQ2FhbRW1tbRrFDDCgRIkUYVVUcAAAAAcCq+LP/JK/GX/Yv33/AKTvVfsB/wDJmnwl/wCxfj/9Dev+CyviDUNF/ZIsNNsink694ysbK73Zz5awXNwMYI5328fXIxnjOCPCHi/xN4A8Tad4y8Ga1caTrekzCayvbcgSQSAY3LnPOCa/4b8/bK/6OG8Vf9/k/wDia8feP/GfxS8WX3jv4g+IrrXdf1Py/td/dMDJN5caxpkgDoiIv0Ar/giPdak/wW+IVjLJKbCHxRHJAhHyCVrVBIQfUqkWfotavqt/ruq3ut6rcefe6hcSXFzLsC75HYszYUADJJOAAK/4JS6DpukfsP8AgfULCHZPrd5ql5eN/flW/ngB/wC/cEY/Cv2t4YZ/2VfjJHNEkijwDrbBXXIDLYzEH6ggEehAqv2Tv+TV/g1/2T/Qv/SCGqqv+Csv7VN78GfhHb/BzwXqSQeKPiLDLFeSocyWelD5ZmX+60xPlAkH5fNIwwVh8A/gf4u/aG+JmmfDXwfHsluczX168ZePT7VSBJO4BHA3KAMjczKuQWFfCL4TeCvgl4B0z4eeA9Kis9O0+Mb3C/vLqYgb55W6s7kZJPQYAwoAH7c37dfxM1j4n6x8L/g9411Lwz4a8MXT2V1d6XKbW6v7yJysreeh3rErqVUIVDAFm3AqF/Z9/bg+N/wL8R2Mtx4t1TxR4WWX/TtB1S8adHiJO7yXfc0D5YsCuAW+8GGRXh7XtL8VeH9M8T6HdLc6dq9nFeWcykESQyoHRgRxgqwP41+2X+3f8UfHnxI1vwT8LPGuoeG/BmhXktlby6RctbTam0Z2PM80bbmjZg2xVIUoVLDJOP2Zv27Pi98EvF2nQ+KvFmr+K/BcjiK/0rUrx7hoImKgyW7vlkZMZCA7G+YYBbcPGngvwJ8Z/AN14V8WaXba54c8QWilo3Bw6MAySI3VWHDKwwVIBGK/am/Z2179mn4q33gfUWkutJuN11oeoMCftdozEJuO0DzVxtcAYDDI4IJ/4I/ftXT+MfC11+zF431eW41bwzbteeGZrhgTJpwZRJahjyxidwyKcny2IGFiAFVVVVVVVrnjLwf4Ylit/EnivR9JlmUtEl9fRwM4HBIDsMioZobiFJ7eVJY5FDI6NkMCMggjqCKqqqqqqqqqq1CxtdUsLnTL2PzLe7heGVM43IwIYfkTVvJ4t+APjV/NtjcafcHa3GI7yHPBDfwuP0PqDzbftF/Due3SWdtSt5GHzRPbZKn6qSK+H/xI0j4jRX8+k2lxbrYyKhW4K7yGBIYqpOBwR17Gvjz4a1fwz44t/iBpiv5F1JFIswTiC5iAAB+oRWGep3elab+034ZbTojrGhaml+E/erbIjRFsdVLOCAT2I496+J2t+KvH+nReO9S05tP0SG5Wz02B8neXVmdwcDd/qwCeB0Azg18Jf+SbeH/+vMfzNfFXwj4k8AeN/wDhZvhWBmtZJ/tEjopcQStxIJB2R8nn/aI44zov7SXg28tA2t2N9p90oG5EQTIT/ssMH8wOverb9oLwbqOuafoel2l/M19cxwefKixxoXYAHqSeT6D61VVX7UPxDf4Ufs6fEf4hW179lvNG8N3j2Eu5l23bRFLflSrDMzxjIIPPBzXg3wxqHjfxfoXgvSVZr7X9St9PtlSMyMZZpFjXCryxyw4HJrQ9HsvD2iafoGmq4tNMtY7a3DtuYRxqFXJ7nAHNV/wUU+H8fxG/Y0+JmmrbebcaPpf9s27AqDEbJ1uHYbiB/qo5FPfDHAJxX7OXxAj+FXx8+HnxEnufItdA8SWVzePhT/owmUTj5gQMxFxnqM5GCM1VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVeLfGPhLwDoF14p8b+JdM0HR7Jd1xfajdJbwxj3dyBk9h1Pav2tf+Cwl1eC88D/ALKEHkQHMcvjHUbQiRxwc2lvIBs7jfMpOCcIpw1Xd148+LPjZrm5k1vxb4s8R3gycSXt5qFw5wAANzyOTgADJ6AV+yd/wR51TUpbTxt+1ZdvYWaMHi8H6dcAzTjg/wClXMbfu1IyDHEd5yD5iEYPg3wT4Q+Hfhuy8H+BfDen6FounRiO2sbGARRxgDGcDqTjljkk8kk1VVVVVVVVX7dX7OcP7TX7OviDwTY2iSeJNMX+1PDjkfML2FSRGD2EqF4j2G8HsK+HPjnxF8IviR4e+IWgh4NY8KatBfQxyFkzJDIGMbgYO1sFGXuCQetfCr4k+G/jF8N/DnxP8I3Hm6T4k0+O8t8nLR7h80bY6OjBkYdmUiqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr/gsz8Q38M/sy6P4Es73y7jxl4khjnh3MPNtLZGmfoRnEwtThgR3wCAR/wSz8AP47/bP8H3Dxyva+Fre71m58tGO0RQmOMkj7o86aHk8H7vVhVV/wAFvPh3DdeBfht8WIURZdM1a50S4YfekFxF58QPsv2Wb/vs+1f8Eb/iBH4X/aovPBl1c7YfGfhu6toIsL89zAyXCnOM8RRXHAIB3ZOcDFVVVVVVVVVX/BWT47/8Lb/aeuvBGkap9p0D4a250mBI5N0Zv2Ie9cDGQ4cJAw9bUV/wRW+BUdpovi/9ojWLbM9/J/YWiFkX5Yk2yXMgPJ+Z/JQdMeW/XPHjLwloHj7wlrHgjxTYJe6Pr1jNY31u/SSGVCjj2OCcHscGvjV8J/EnwM+K3ij4SeLE/wCJl4Z1B7V5QhRbiPhop1B5CSxNHIuedrjNf8E3/wBoef8AaH/Zl0a+1298/wATeEJP7D1hnkBknaFFMM5HX95EyZY9XWTHSqqqqqqqqqqqqqqqqqqqqqqqqqqq/aq/4KFfAn9lqO50C/1H/hLPG8cbeX4b0m4QvDJtBUXcvItgcqeQz7SGCMK/aX/bQ+O37VOpBviJ4l+y6DBJvtPDmll4LCBhna5j3EyyAMR5khZhkgYBxVx4X+K3xDu7vxxL4Y8UeIZ9cupru51RdPnuftk7yMZZDIFIZi+7cc9c5r/hUvxU/wCiZ+K//BLcf/EV/wAKl+Kn/RM/Ff8A4Jbj/wCIr/hUvxU/6Jn4r/8ABLcf/EV/wqX4qf8ARM/Ff/gluP8A4iv+FS/FT/omfiv/AMEtx/8AEV/wqX4qf9Ez8V/+CW4/+Ir/AIVL8VP+iZ+K/wDwS3H/AMRX/Cpfip/0TPxX/wCCW4/+Ir9jH/gmDrfj2D4RftA+K9etItAk1C4vPEnhfV9Plguo/sl1OkMaDkSJKYYiwfy8I7Eb+Aaqqqvj/wDBTwt+0P8ACPxF8JfFqolrrdqyW92YRK9hcrzDcICR8yOAcZGRlScMa+IHgTxd8IviBrXw/wDF1lLpviHwxqD2t0gLLtkjbiSNuCUYYdHHDKysOCDX7AX7Udv+1L8BNO1zV72JvGfhzbp3iaBAQfOXPlXAB7TRgPkcB/MUfdqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv+C3v/JKvhp/2MF1/6Tiv2A/+TyvhN/2MCf8AoD1Vf8FPPAGpfED9jDxzHo8cst14f+z6yIo03b4reZWnJ9AsJlfP+xzxk1aXd3YXcF/YXUttc20iyQzQuUeJ1OVZWHIIIBBHQ14X/wCCy37N918PrTWfF+heK7LxWlmpvdGs9PWWNrkKNwhmMgXyy2dpcqQOoz1/aC+NGv8A7RHxk8UfGTxLY29lfeJLpZBaQHKW0McaQwxBsDcVijjUtgbiCcDOK/4IY/8ANbP+4B/7ka/4Ln/80S/7mD/3HV/wS4/5Pt+GP/cW/wDTTeV+35/yZp8Wf+xfk/8AQ0qvhP8A8kr8G/8AYv2P/pOlftjfBBP2hv2cPGnwzt40bVLmxN1o7MCdt9AfNhHHIDMvlk84Dng9K8EeL/Enwv8AHmh+OvDkn2TXfC2qQ31p50ZISeCQOodOMjcuGU9RkHrXwu8Aa/8AGT4oeG/hxosrvqnivVobJJnG8o0rgNK2SMhQWc8jgHmvDHhzR/B/hrSfCPh60S10vQ7GGxsbdFAEUEKBI1AHAAVQOPSqr9rL/k6j4y/9lA13/wBL5q/4Jd/tz+Av2cotf+EHxkv20rwnr98NT0/WI7Z5lsr0xLHKsyxqzlJEihAYAhGQ5GHLJ+0r/wAFSf2efh18N9Vb4P8Ajy08ZeN76zli0WDT4XkgtZ2G1Z53ZQmxCd3ljLPt24AO4Qwy3EqW9vE8ssjBURFJZmPAAA6k+lfsc/CTUvgX+zF8PfhfrUXlappOlmXUIt27yrm4le4mTP8AsyTOvpxxxX7WX/J1Hxl/7KBrv/pfNX/BFT/k1fxT/wBlAvP/AEgsKr/got8FtU+C37WPja2uLZ10vxbfS+IdJm2gLJDdyNI6rgAARzGWPHYIOucn/gnz+3B+z58L/Ao+Df7SXgnR4rHTJpJdG8SjQFvm8uWQvJDcqkbSkq7syuA3ynaQNi7pv2qP+Cb8ELzP46+FhWNSxCaSjtgDsoiJJ9gM1+zn8cf2Sv2hvE3iGH4C6Lo97c+DFtJptSTw2tgCbgS7TB5iJLlTEwYlFAJXBOa/4LR/B3VPF3wg8H/F3RLC4um8D6lPbakIIwRFZ3ip++k4ztSWCJBzgGc8c5HwG+MGv/AH4weFfjB4ZgS4vvDN8J/szvsFzCymOaEtg7RJE8ibsHG7OOK0z/grh+xne+DF8S3/AIq8QWGq+Xufw7JoU73gbIBUSIDbnucmUcDseK/ad+O+r/tK/G/xL8YdWsnsF1mZFstPacyiytYo1jiiBwBnaoLEAAuzNgZr/giH/wAlT+Jf/Yv2v/pQa/4Le/8AJKvhp/2MF1/6Tiv2A/8Ak8r4Tf8AYwJ/6A9ftlfB69+PP7Mvj74ZaRapcatqGm+fpcbHBe7t5FnhUHsWeIJn0Y54JqaGW3le3nieOWJiro6lWVhwQQeh9q/ZW/4K8fCe88DaL4P/AGlLnVdB8SaVZpbXPiJLOS+tdS8tMCaRYQ0qSuFXcAjKXYkFQdq+EP8Ago5+yX8RfiD4Y+GXw78fah4i1rxTeG0thb6Hd28cD7SwMrXEceA2CBtDc9QBzXjjRLrxN4K1/wAOWEkSXOq6Xc2kLzEhFeSJkUsQCQMkZwD9Kmhlt5Xt54njliYq6OpVlYcEEHofav2Lf+CoPwC+Fv7O/hb4W/FqHXdM1zwpC9kJLHTjcQXcAkZopFIbKtsYKwIHzAkcHA/bz+Fs/wC0r+xr4gt/BcEt1qMdnbeItFhMYDzmECUptIJ3vA0qqBg72UZxkH4aeOr/AOGHxA8PfEHTNN0/Ubjw/qEV2tlqFuJre6CMC0UqHqjrlTjBAOQQQCPhR+3b/wAE8PiR4dt9Q1608I+CtYEKNeaTrfh9EMLlRuCTLEY5VDEgEEMQuSq5rxb+2l/wTW8IWQup/EXgfVJZFYxW2k+FzeSOVxkZSAqh+YY3soPODwa+Hy+D7jwfpeueBtEsdL0jXbWHUYIbS2jgDCaNWVmWP5SxXaCQT0HJAFftkfB7VPgZ+0p488B31hcW9mNWmvdIeWMKJ7CdzJA6lQFYBW2EqAA6OuAVIH/BNn/goT4M/Zv0K++DXxoj1CLwpfagb3TNZtIDcf2ZK4USpNEvztE20MDGGZWyNpDZX9vH/gp/8G/GvwW1v4Q/s96zqHiHUPF9ubHUNYNnPYQWVm2POUCZEkkeRd0W3aF2s5JOAGr9k7/k1f4Nf9k/0L/0ghqqr9tr4z3vx4/ac8deN5Lx5tOg1KTTdGUvuVLG2YxRbfQOFMpH96RuT1P/AATJ+B9t8OfgTF8RtU02JPEHjx/tQmYAyR2CnbAmewbDS4B5Ei55GBWu3GrXmt6hd68si6nNdSveiWLynExYl9yYG07s5GBjpgVX7KN3rUf7GHge70ZWudVh8KMbFGG8tKquIlweoyFGPTiqr9myWWf9nT4VzTStJJJ4K0dndzksTZRZJPc1/wAFBvgdF8Zv2edXvdOsGn8Q+DFfV9LMaZkdUX/SIgOpDRBjtHJZEwCeK/Zw+Ll78B/jr4J+LVkz7fDurRy3aJ96W0fMdzGODgvA8qZwcbs1aXdpqFpDfWF1Fc21zGskM0Lh0kRhlWVhwQQQQRwc1VVVVVV/wU7/AOCgPib4M6if2ffgfrKaf4rntVl8Q61FhptKilQNHBCCMJM8bK5k6orKVwzBk+E//BIX4z/GzwxD8Tvjf8af+EW1vxBHHdR2lzp0mtXzRsvytdSPPFsk2hfkBkIBAYqwKgeIv2sf+CV/xttvDV9qlxqHhiaY3MdgZ5DpHiO16O0YP+qmAIBIG+NtuQ6Eb/hB8U/Cfxu+Gfh34q+CLz7Ro3iOzFxAT96JgSskTf7ccivGw/vIaqqqqqqqqqqvbGy1K2ez1CzguoJBh4pow6t9QeDX/Cq/h15/2n/hDtM379+PIG3Oc/d6Y9sYqw03TtKthZ6XYW1nApyIreIRqD9FAHYVeWdnqFrJZX9rDc28q7ZIZow6OPQqeCKi+GPw9guxexeDtLEoYsM24Kgn0U/L9OOO1X+k6Vqtp/Z+qaZaXlsCCILiFZEyOnykEcVaWdnp9rHZWFrDbW8K7Y4YYwiIPQKOAKZVdSjqGBGCCM5q++Gfw/1Gc3V14Q0xpW+8yQCPcc5JIXGTz1PNaR4Q8K6A6y6L4d06zlVdolhtlV8e7YyfxNVVf8NR/wDUjf8AlT/+1V/wV7+KH9gfsh2Pha3Oy4+IGt2VvJD1xbwg3btuKkcSw24xlSd2eQGB/wCCZfw8j+In7Z/gKK7s/tFj4dkn1u5+VT5ZtoWaB8MCOLg2/oRnIIODVVq+lWGu6VeaJqkHn2WoW8lvcxbiu+N1KsuVIIyCRkEGvHHhPUvAXjTX/AusjGoeHNUudNuht24lglaN+O3zIa/ZZ+I0vxb/AGcvhx8Q7udpr3WPDto19I7Fi12iCO4OSzEjzUkxuYtjGec1VVVVVVVfGr9t/wD4U/8AEzWfhz/wrH+1/wCyPI/0z+2/s/m+bBHL9zyGxjzNv3jnGeM4HwG+Lv8Awu/4ew+O/wDhHv7F826lg+yfa/tOPLIGd+xOuemOKqqqqqqqqqqqqqqqqqqqqqqqqqqr/hqP/qRv/Kn/APaqsLr7dYW175ez7RCsmzOdu4A4z+NVX7W//BSr4Lfs1fbvB/h25i8b/EC33xNo9jN+40+Qbh/pc4BVCGUgxLmQdwgIav2hv2rPjb+1Brw1n4r+LXurS3meTT9GtFMFhp+7tFFk8gfLvcs5HVjX7KX/AATv+Of7Uv2TxJb2f/CI+BZpCreJNTtyRMF6m1gyrXHPG4FY8hhvBUiv2av2N/gZ+yvootPhx4a8/W54wt/4h1Iie/u+vG/AESc42RhFOASC2WNVVVVVVV8ev2zP+FIfEGbwJ/wrf+2vKtIp/tX9sfZs7wTjZ5L9Mdc1+zt8b/8Ahf3gm98Y/wDCMf2F9j1WSw+zfbftW7ZFFJv3bExnzcYx/DnPPFVX/BU/9nuX4KftM6h4r0nTUt/C/wAR1fV9OMSFUjuhtF7D/vCVhLgDAW4QdjX/AARk/aR/5Dn7MHibUP8AnprHhjzG+n2q3X/x2YKP+mxqqqqqqqqqqqqqqqqqqqqqqr4h+M/+EC8Nv4h/s37dsmSPyfO8rO44zu2t/KvAvx6/4TXxVY+Gf+EU+x/bPM/f/b/M2bI2f7vljOduOveqqqqqqqqqqqrXdS/sXRNQ1jyfO+wWss/l7tu/YhbGcHGcYziv+HlP/VFv/Lk/+5qqqqqqqqqqqqqqqqqqqqqqqqqqq+Ifxw/4QLxI/h7/AIRj7dshSTzvtvlfeHTbsb+dfDTx7/wsXQZ9b/sn+z/Iu2t/K8/zc4RG3Z2r/fxjHav+C1/xBbW/jf4I+GsMySW/hbw7JeuFIzHcXsxDqRtzny7WBuWIw4wByT/wRA+Hkb3XxP8Aixd2fzxR2WiWFxtXoxee5TOMj7tocAgHuDhSKr/gpL8P3+In7GHxGsrW28280Szi1m3bLfuxaTJLM2ARn9wsw5yBnOMgV+yf8RpfhL+0p8NvH6TtFDpniK1W8ZGKt9llfybgA7lGTDJIMEhTn5sjIqqqqqqqqrx14l/4QvwT4g8Y/Yvtn9g6Vc3/ANm8zy/O8mJpNm7B2524zg4znBrQv+Ci39t63p2jf8Kd8n7fdRW/mf8ACQ7tm9gu7H2YZxnpkV+0V8X7D4B/A/xn8X7+OKX/AIRrS5JrWCYkJcXTER20TFeQHmeJCR03ZqGLxN8QvF6QRtcav4h8S6kFUu4Mt5dzyY5JIG53fqe5r4AfCXTfgV8FvB3wk0rymj8NaXHbzSRDCzXBy88o4H35mkfoD83PNV/wWk/Z8+1WHhf9pPw5pX7yz/4kniSWMdY2O6zlYAdmMsbOeTvhXoBX/BLr9oyX4F/tKad4Y1a7ZfC/xJaHRdRTJ2xXTPiynx6rK5jJJwqXEjHoKqqqqqqqqqqqqqqqqqqqqqqr45fFz/hS/hK08U/8I9/bP2rUUs/I+1/Z9u6OR927Y+f9XjGO/Xivg3+1T/wtzxtF4O/4QP8AsrzLaWb7T/ann42AHG3yl6+uaqviz8Yvhr8DvB1347+KXi2x0LSLRTh53zJOw6RxRjLSueyqCfwya/a1/wCCt3xE+J/2zwV+zxFqHgTwy2Y5NaabZq16vHKMhxaDOR8jM5GDvXJUeAPh18QvjH4wt/B3w88M6n4m8Q6izOlraoZJG7tI7HhVGcs7EAZySK/ZG/4JD+E/BX9nePf2mp7TxPrYj8xfCsHzafZuegnk63LqMZUYjDZH71cMdO07T9I0+10nSbG3srGyhSC2tbeIRRwRoAqoiLgKoUAAAYAAFVVVVVVVX7Sv7Sv/AAzv/wAI5/xRf/CQf8JB9q/5iX2TyPJ8r/pk+7d5vtjb3zx+zh+1P/w0HresaN/wgn9g/wBk2qT+Z/an2rzdzbcY8pMfXJqqr/gsV+ysmsaHZftS+DNOQXmkqlh4sVAS09uWVLa5wOMxsTGx6lXj7Ia/YI/aguv2W/j7pfiPUbuX/hEPEG3TPE1sJCE+zuw2XO3kF4Hw4OMlfMQEeYTVpd2moWkN9YXUVzbXMayQzQuHSRGGVZWHBBBBBHBzVVVVVVVVVVVVVVVVVVVVVVVV8Zfib/wqPwTL4x/sT+1vLuY4fs32nyM7zjO7a3THTFfCj9rn/haHj/SvAv8Awr7+zf7S87/Sv7W87y/LheT7nkrnOzHUYzn2qqqqqqqqqqqqr49ftmf8KQ+IM3gT/hW/9teVaRT/AGr+2Ps2d4Jxs8l+mOua/Z2+N/8Awv7wTe+Mf+EY/sL7Hqslh9m+2/at2yKKTfu2JjPm4xj+HOeeKqqqqqqqqqqqqqqqqqqqqqv+CmX7LXxY/am8C+DPD/wms9MuLvQ9WnubsX16LYCNogowSDk57V+yl/wTM/ak+EP7RfgH4l+MtJ8OR6J4e1Zbm9e31hJZBGFYHaoHJ5HFVWo6dp+safc6Tq1jb3tjewvBc2tzEJI543BV0dGyGVlJBBGCDg1+0z/wR6+KuheKdQ1/9mgWPibwzdzeZaaFeagltfWIYjMQknKxyouThmdX2gAhmG5vhn/wSE/a38Zautr460vQfAOnRsPOu9R1SG9kZcjPlRWjybmwScO0Y4PzDjP7V3/BKLxLe23gDw3+y34W0T+zvD+jywa1qOp3yW95qV20u7zpm25ckZwOFQYVQAAK/wCCYP7IXxl/ZT/4WV/wtyx0m3/4Sj+yf7P+w363O77P9s8zdgDb/r48evPpX/BT79kL4y/tW/8ACtP+FR2Ok3H/AAi39rf2h9vv1ttv2j7H5e3IO7/USZ9OPWv2Hv8AgnJ+0x8Bf2o/BPxY+IOl+H4tA0L7f9re11ZZpB51hcQJtQDJ+eVM+gya/au+HHib4vfs6ePfhp4Nit5db8Q6S9rZJcSiKMyFlPzMegwDzX/Dob9sr/oCeFP/AAep/wDE14C0e98PeBfDmgakqC70zSbW2nCNuAkjiVWwe4yDzVftpx/DdP2qviY3wlv7S88MTa5JNbTWZBgMzqr3IiKkqYxcNMq7fl2gYwMCv+CMn7O8+ueN9c/aT12zxYeG45NJ0FnjB8y8lQfaJFJ6eXC2zI6+eRngiqqv2sv+TqPjL/2UDXf/AEvmr9mD/gnnqv7Wv7Neq/Ez4e+L7TS/GeieLLrTPsOq7hZ31strZyqBIis0Mimabna6tlQdmC1f8Opf24P7Z/sv/hVmn/ZvM2f2l/wkdh5GP7+3zvNx/wBs8+1fsW/8EqPD3wN8SaX8WfjZrtp4o8Y6VIlzpWmWSH+z9LuF5WYs4DTyoQpQ4RUYE4YhWWvj1/wS1/ax+IXxz+Ivj7w3o/hl9J8S+LNU1OwabWkjcwT3cksZZcfKdrrkdjX/AATa/Zx+Jn7MHwL1zwB8VbXT4NWv/FlxqcK2V2LhDA9paRKSwAwd0L8fT1qv2n/2V/hb+1h4EXwZ8RrO4iubBnm0bVrSQpcaZOwALoPuurAAMjgqwA6MFZfib/wRv/ag8J3t0/w91Twv4605Wf7IYLz+zruRRtx5kVxiONmycATOPlOWGRnR/wDglR+3Fqd6LW9+E1jpMZUn7TeeI9PaMEdsQzSNk/7uPcV/wTk/YJ+MX7J3i/XvHfxK8Z+HJY/EOiiyOh6SJLho5PMikSSSd0TDIBMhRAynfnccCvEPh7QvFuhX/hjxNpFpqmk6pbvbXtldxCWK4icYZHU8EEHGK/aT/wCCN3xE0nxDfeIv2aNX0/XtAupJJYfD+p3X2W8seCRDHNIfLnTI2hnaNhuUNuw0lfDb/gkR+114v1mK08caLongTTRIBPeX+qwXrhO5jitXk3HsAzIM9x1r9pj/AIJI+KIvC3w68M/s0xafqbaPZ3a+Jb3VrpbSa9vHaIi4ySRhgGURqMRrEvLFix/4JnfsV/HL9lnxz4z1/wCLFho8FprmkwW1obHUFuWMiSljkADAx3r/AIKZfstfFj9qbwL4M8P/AAms9MuLvQ9WnubsX16LYCNogowSDk57V+yl/wAEzP2pPhD+0X4B+JfjLSfDkeieHtWW5vXt9YSWQRhWB2qByeRxVftq/wDBLnwj+0Zr158UfhTrtj4N8cXil7+C4tidP1ebtJJ5fzQSn+KRVfdjJTcWc+Jv+CT/AO25oOqyadpXw30nxHAmdt/pniKzjhfDEcC5khk5ADcoOGHQ5A/Zn/4Jdftg+E/i54E+JviLTPC/heLwvr1lq1xBqGtiaVkt7pGaIC0EoLOisQNwUg4LKTiq/bX/AOCT/jPx58SNZ+Lf7Ol9pMg8RXD3mp+HL+cWrR3TktI9vJjZsdjuKOV2sWwxBAXSv+CVf7cmo38Vnd/CG00yKTO66u/EmnNHHgEjIind+SAOFPJGcDJHgjRLrw14K8P+HL6SKS50rS7a0meEkozxxKjFSQCRlTjIH0Ffte/8ElfC/wAZPEuo/Er4F+I7Hwh4k1WZ7jUdL1FZX0+9uJHZ5Jg673t2YsSVVGTgYVeSfEP/AASi/be0XUPsWm/DHTNehC5+16f4isUjzkjGLiWJ84AP3cYYc5yB4N/4JA/th+JXtv7f0zwp4Tjl3Gb+09aWZoAGxyLUTAkgZABxyMlTnHwJ+H+s/Cf4MeCfhj4g8QRa3f8AhTQ7XTJL6G38hJRBGEUKvoqqqgnk7cnkmv2zP2Jvh5+2B4RgttUnTQfGOkKRo3iOK2ErwoTloJkyvmwtknbkFGO5SMsG8Zf8El/20vDeryaf4f8AAuj+LLRWYR32ma9aQxsAeCVupInBI5xtOPWv2R/+CQWq+GfF2nfET9prWNJu4tJuBPZ+FdMdp0uJEPyPdT/KNgI3eUgYPldzAbkOo/8ABID9r2DULmDTbDwvd2kczrb3D6wkTSxgkK5TB2kjB25OM4ya+Avg/Wfh78C/hz4A8RpCmreGvCel6ZfrDJvRZ7e0jikCt/ENyHB71VfG/wAWT+Avgt4/8c2jSifw54X1LUYjCQrhoLWSQbSeAcoMZ71o+lXeu6tY6JYBTdahcx28AdtoLuwVcnsMkV4Z8Pab4S8N6T4U0aLy7DRbGGytE2qu2KJAiDCgAfKo4AA9AKr9u79lbxh8Fvin4g8caXol3deBPEV+97aalFDmKzlndna1kIzsKsSELY3Lt5JyB8Cf2f8A4jftCeNLLwj4E0S5khknVb/VWgY2umxHlpJX6DCgkLncxwACTXgfwjpPgDwXoXgbQY9mn+H9OgsLUHr5cUYQE9SSQuSTySSSa/bL/Zd8Yfs//FHW7yHw9OfA+r38txoWpW9vi3jikbeLZipIjePd5eG27gm4DBwP2c/2afiN+0j40tPD/hLR7mPRo51Gra5JCwtdPiGC2XxgylT8sY+ZieyhmGj6RpugaRY6Do9qttYabbR21rApJEUUahUUZycBQBzUsUU8TwTRrJHIpV0dchgeCCO4r4o+FF8B/Ezxd4GRlYeHddvdOBVywPkTvHwSASPl6kA+1fsZ+KLvxl+yh8JtevxKbl/CdjBM805meV4YlhaRmIyS5jLnPQtjJ6mqqqqq1fVLDQ9KvNb1S48my0+3kuLmXYW2RopZmwoJOACcAE1+xxpcv7WX/BQjSvF/jq1S5h1HXr3xTqVtPKZVXyt88MPON6LL5CbcY2DGMDFV/wAFfvhppvjH9km68cyxRLqHgLWLS9gmJw/lXEq2kkQ+U5DNPExGV5iU542n/gid8Q5db+DXjv4ZXEjyN4V16G+hLA4SG9iICD2D2krfVz7VVVVVVVVVVVVVVVVVVVVf8FlviHHq3jv4XfC+1vfMXw74YbUriNGUqkt26oA2DneEtVbDAYWQEE7zj/giH8OftHir4lfFy4j2/YNPtdDs3znf58hnnGA3G37Pb9VOd/BGGBqq/wCCmvw8j+Hf7Z/j2K0s/s9j4ikg1u1+VR5huYVad8KAObgXHqTjJJJJr/gjl8QU8VfsqXHguaZ/tPgrxFdWqRsxYC3nC3KMPlAAMks425JypJxuAqqqqqqqq/bT/wCTmPGX/bj/AOkNvX7Bv/Jvln/2FLv/ANCFVVVVVVVVVVVVVVVVVVVVVVVVVVVWgf8AIC03/r0i/wDQBXxw/aD+EP7OfhM+MPi74ztNFtJNy2ducy3N864ykEK5eQgsuSBtUMCxVcmv2s/+Cqfxe+Of2nwh8JRd/DzwXJG8UwgnB1LUlbg+dMv+qQrgeXFjq253BAX4R/Bf4n/HbxdB4H+FPg7UPEGqy4aRbaL93axkhfNmkPyQxgkAu5AyQM5IB/ZL/wCCS3w1+E7ab46+PF1b+OPFsDLPHpSx/wDEqsHwMKUbm6ZTk7nCpyP3fG4www28KW9vEkccShURF2hQBgAAdBjtVVVVVVVVX7ef/JwV3/2CrT/0E1/wTt/5InrX/Y03H/pLa1VV/wAFBv2bf+Gl/wBm/XPD+jaf9o8WeHM6x4c2JmSS5iU77cY5PnRF4wuQN5jY/cr4VfEnxJ8HviR4c+J/hG48rVvDWoR3lvkkLJtPzRtj+B1LIw7qxHevhV8SfDfxi+G/hz4n+EbjzdJ8SafHeW+Tlo9w+aNsdHRgyMOzKRVVVVVVVVVVVVVVVVVVVVVV+0L/AMk3n/6+4f518DP+SqaH/wBvH/pPJVVVVVVVVVVVV45/5EnxB/2Crn/0U1VVVVVVVVVVVVVVVVVVVVVVVVVVV+0N/wAlHm/684f5Gv2aP+RDvv8AsLyf+ioq/bw+IcfxQ/a8+KHie3vftVrDrkmm2sispRorNVtVZCpIKN5O4EH5t27gkiv+CUPw9TwL+xl4b1J4XiuvGOpXutXKOpBBaT7PGeWIw0NtCwwFGGHGck1Xi3wzpXjXwrrPg3XYfN03XtPnsLyParb4Zo2jcYYFTlWIwQR6g14n8Pah4S8S6t4U1YIL7Rb6ayuQgOPNico+NwBxlT1APqBX7PHxBT4r/Aj4f/Ebznkm1/w7ZXVyXYsy3BhXzlJKruKyB13YAOMjgiqqqqqqqr45/wDJE/iD/wBirqf/AKSyV4F/5Hfw9/2Fbb/0atf8FrfjmkNl4M/Z00a9cSzsfEOuImVHljfDaRkg4YFvtDshHBSJu4r/AIJJ/AtPip+0zH491WzabRvhrajUnOCEN9JuS0ViB1BEsoGRkwdxkGq+K3w28N/GL4b+I/hf4ut/N0nxJp8tncYGWj3D5ZFz0dGCup7MoNfEHwT4k+E/xD17wB4hils9a8LapNZT4BQiSFyodSD0OAysDggggkEGv2Ff2jIf2mv2dfD3jW+u0k8SaYv9l+IkB+YXsKqDIR2EqFJfQbyOxqqqqqqqqqqqqqqqqqqqqqqr9uT/AJJLpP8A2MUP/pPcV+xt/wAlstf+wdc/+gippobeF555UjjiUs7u20KAMkknoMV+1p/wVo+GvwnbUvAvwHtrfxx4tgZoJNUaT/iVWD4OWDrzdMpwNqbU5J8zjafi58Z/if8AHbxdP44+K3jLUPEGqy5WNrmT93axklvKgjHyQxgkkIgAySepJP7Jn/BKv4vfHP7N4v8Aiybv4eeC5I0lgM8AOpakrcjyYW/1SFefMlx1Xajgkr8D/wBnv4Q/s5+Ex4P+EXg200W0k2td3AzLc3zrnDzzNl5CCzYBO1QxChV4qqqqqqqqqv8AgpV/zTn/ALin/tpX/BOH/kdfGH/YKh/9GmqqvEXh/RfFnh/U/CviPTodQ0nWbOWyv7SYZS4glQpJG3sysQfrX7Wv7Out/su/HDXfhZqYuJtPjb7XoV7Pt3X2nSMwhlO3A3fKyNgAb43wMYr/AIJHftUt8VfhbP8AALxfqTS+J/h7aodLaUjN1o4KxxgY5Jt2KRHIACPDgk7sVVVVVVVVVVVVVVVVVVVVVVVX7ZP/ACRO6/7CNt/6Ea/ZR/5L94V/7fP/AEjmqqqqqqqqqqqqr9vP/k4K7/7BVp/6Ca/4J2/8kT1r/sabj/0ltaqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv8AgoP+31+0lcfETx5+zbZvp/g3w9o2qXNhNJpIkF5qlmwIj86ZmO1JIWRisap94glhxXwS+C3j39oD4kaT8MPhzo0t/qmpSDzHC/u7OAEeZcSt0SNAcknqSFGWYA/An4NeFf2fvhN4c+Efg4O+n+H7XyzcSIFkupmJeWZwONzuzMR2zgcAVVV+1l/ydR8Zf+yga7/6XzV/wRU/5NX8U/8AZQLz/wBILCqqqqqqqqqqqqqqqqqqqqqqqqv2sv8Ak1f4y/8AZP8AXf8A0gmr4Z/8lI8Kf9huz/8AR6VVVbWttZwLbWdvFBEmdscaBVXJycAe9VVta21lbQ2VlbxQW9vGI4oYkCLGqjAVVHAAAAAHSqr9qSa2uP2lfio9rZrbIPGWqoY1dmyy3Uis+SScswLEdAWwMACv+Cdlje6f+xT8Kbe/1N7+VtJklWV02FY5LmV0jxk8IjLGD3CA8ZxVVVVVV+05qt/oX7NnxY1vSrjyL3T/AAPrNxbS7A3lyJYzMrYYEHBAOCCK/wCCK8MMv7VXiV5IkdovAN40ZZclG+3WAyPQ4JH0J9ar9vz/AJM0+LP/AGL8n/oaV/wQ/wBQvY/iJ8UdKjuXFpcaLYzSwjo8kc0iox9wJHH/AAI1VVVVVVVVVVVVX7SP7XnwZ/ZT/wCEd/4W5farb/8ACUfav7P+w2DXO77P5XmbsEbf9fHj1yfSv+HvP7Gn/Qc8Vf8Aghf/AOKr/h7z+xp/0HPFX/ghf/4qv+HvP7Gn/Qc8Vf8Aghf/AOKr/h7z+xp/0HPFX/ghf/4qv+HvP7Gn/Qc8Vf8Aghf/AOKr/h7z+xp/0HPFX/ghf/4qv+HvP7Gn/Qc8Vf8Aghf/AOKr/h7z+xp/0HPFX/ghf/4qq/bV+Ldv8a/2kfFfjPTpZW02E2+nWCSqVKRWsCQng8jc6O/1c1/wSS+Hj+CP2PNJ125s/IufGmsXurNvVg5jDi2jJDAYBW2DqBkFXDA/Maqq/wCC33w9aDxJ8Mvitbwoy3tjd6LdyBQGQwus0IJ3ZYHz7jouF2nJ+YCv+CZf7Y3w/wD2U/Evjey+Kl1qcXh7xPY20kL2Vqblku7d2CjaDkBo5pMn/YXPav8Ah7z+xp/0HPFX/ghf/wCKr/h7z+xp/wBBzxV/4IX/APiq/wCHvP7Gn/Qc8Vf+CF//AIqv+HvP7Gn/AEHPFX/ghf8A+Kr/AIe8/saf9BzxV/4IX/8Aiq/4e8/saf8AQc8Vf+CF/wD4qv8Ah7z+xp/0HPFX/ghf/wCKr/h7z+xp/wBBzxV/4IX/APiq/wCHvP7Gn/Qc8Vf+CF//AIqv+HvP7Gn/AEHPFX/ghf8A+Kr/AIe8/saf9BzxV/4IX/8Aiq+Nvxe8GfHr4na18WPh9PdTaBrnkfY3urcwSHyYI4Hyh5HzxP8Ahg96+Af/AAUD/Zz/AGc/h3B8NPiXqmuw63BdTXLpZ6W08flyEFfmB64HSv8Ah7z+xp/0HPFX/ghf/wCKr/h7z+xp/wBBzxV/4IX/APiq/wCHvP7Gn/Qc8Vf+CF//AIqv+HvP7Gn/AEHPFX/ghf8A+Kr/AIe8/saf9BzxV/4IX/8Aiq/4e8/saf8AQc8Vf+CF/wD4qv8Ah7z+xp/0HPFX/ghf/wCKr/h7z+xp/wBBzxV/4IX/APiq/wCHvP7Gn/Qc8Vf+CF//AIqv+HvP7Gn/AEHPFX/ghf8A+Kr/AIe8/saf9BzxV/4IX/8Aiq/4e8/saf8AQc8Vf+CF/wD4qv8Ah7z+xp/0HPFX/ghf/wCKr/h7z+xp/wBBzxV/4IX/APiq/wCHvP7Gn/Qc8Vf+CF//AIqv+HvP7Gn/AEHPFX/ghf8A+Kr/AIe8/saf9BzxV/4IX/8Aiq/4e8/saf8AQc8Vf+CF/wD4qv8Ah7z+xp/0HPFX/ghf/wCKr/h7z+xp/wBBzxV/4IX/APiq/wCHvP7Gn/Qc8Vf+CF//AIqv+HvP7Gn/AEHPFX/ghf8A+Kr9nr9o34Z/tP8Agq98ffCq71C40nT9Uk0yZr60Ns4nSKKVgFJORtmTn1z6VVVVVVfE/wD4KZfstfCHx/rnw08Zav4jj1vw9dG2vUt9HeWMSAAnawPIwRzX/D3n9jT/AKDnir/wQv8A/FV/w95/Y0/6Dnir/wAEL/8AxVf8Pef2NP8AoOeKv/BC/wD8VX/D3n9jT/oOeKv/AAQv/wDFV/w95/Y0/wCg54q/8EL/APxVf8Pef2NP+g54q/8ABC//AMVX/D3n9jT/AKDnir/wQv8A/FV/w95/Y0/6Dnir/wAEL/8AxVV+0P8A8FgvD3g/QB4I/Z48LS6n4jt4BbXOsa5AY7bT5UOxwkAO6Z/lJBJVASpIfla8f/EX4hfGPxhceMfiF4n1PxN4h1FlR7m6cySN2WNFHCqM4CKABngCv2TP+CSXxK+J8tj4z/aFa98C+FXXzF0cJs1e87BWRwRaqepLgvxjYN29fhP8Hfhr8DvB1p4E+FvhKx0LSLRQPLgTMk7DrJLIctK57sxJ/Cqqqqqqqqqr9vP/AJOCu/8AsFWn/oJr/gnb/wAkT1r/ALGm4/8ASW1qqqv+CoH7Nv8AwoP9o++8QaDp/keE/iJ5usaXsTCQXJYfbLcdvllYSBQAFSeNR0Nf8E4f+CiPw6/Z2+GGsfCP43T6qmm2eofbfD11Y2RuSqzZ8+BwCCoDqJFODkyyZIwor/h7z+xp/wBBzxV/4IX/APiq/wCHvP7Gn/Qc8Vf+CF//AIqv+HvP7Gn/AEHPFX/ghf8A+Kr/AIe8/saf9BzxV/4IX/8Aiq/4e8/saf8AQc8Vf+CF/wD4qv8Ah7z+xp/0HPFX/ghf/wCKr/h7z+xp/wBBzxV/4IX/APiq/wCHvP7Gn/Qc8Vf+CF//AIqv+HvP7Gn/AEHPFX/ghf8A+Kr/AIe8/saf9BzxV/4IX/8Aiq/4e8/saf8AQc8Vf+CF/wD4qv8Ah7z+xp/0HPFX/ghf/wCKr/h7z+xp/wBBzxV/4IX/APiq/wCHvP7Gn/Qc8Vf+CF//AIqv+HvP7Gn/AEHPFX/ghf8A+Kr9nr9uv4AftP8AjS98AfCrUtauNW0/S5NTmW+0xrZBAksUTEMScndMnH19Kqqqqqvif/wUy/Za+EPj/XPhp4y1fxHHrfh66Ntepb6O8sYkABO1geRgjmv+HvP7Gn/Qc8Vf+CF//iq/4e8/saf9BzxV/wCCF/8A4qv+HvP7Gn/Qc8Vf+CF//iq/4e8/saf9BzxV/wCCF/8A4qv+HvP7Gn/Qc8Vf+CF//iq/4e8/saf9BzxV/wCCF/8A4qv+HvP7Gn/Qc8Vf+CF//iq1D/goH+zn+0bbH4afDTVNdn1uYi5RLzS2t4/Li5b5iTzg9Ksvi94M+At0nxY+IM91DoGhZ+1va25mkHnDyE2oOT88qZ9Bk18Gfi94M+PXw10f4sfD6e6m0DXPP+xvdW5gkPkzyQPlDyPnif8ADB71VVXjjxfovw98Fa/4+8RySppPhrS7nU79oY97rBbxNLIVUfeO1DgdzX/D3n9jT/oOeKv/AAQv/wDFV/w95/Y0/wCg54q/8EL/APxVf8Pef2NP+g54q/8ABC//AMVX/D3n9jT/AKDnir/wQv8A/FV/w95/Y0/6Dnir/wAEL/8AxVf8Pef2NP8AoOeKv/BC/wD8VX/D3n9jT/oOeKv/AAQv/wDFV/w95/Y0/wCg54q/8EL/APxVf8Pef2NP+g54q/8ABC//AMVX/D3n9jT/AKDnir/wQv8A/FV/w95/Y0/6Dnir/wAEL/8AxVa7/wAFYf2QfEOh6hoGm634oa71K0ltrcPobqDJIhVcnPAyRzVf8Pef2NP+g54q/wDBC/8A8VX/AA95/Y0/6Dnir/wQv/8AFV/w95/Y0/6Dnir/AMEL/wDxVf8AD3n9jT/oOeKv/BC//wAVX/D3n9jT/oOeKv8AwQv/APFV/wAPef2NP+g54q/8EL//ABVf8Pef2NP+g54q/wDBC/8A8VX/AA95/Y0/6Dnir/wQv/8AFV/w95/Y0/6Dnir/AMEL/wDxVf8AD3n9jT/oOeKv/BC//wAVX/D3n9jT/oOeKv8AwQv/APFV/wAPef2NP+g54q/8EL//ABVf8Pef2NP+g54q/wDBC/8A8VX/AA95/Y0/6Dnir/wQv/8AFV/w95/Y0/6Dnir/AMEL/wDxVf8AD3n9jT/oOeKv/BC//wAVX/D3n9jT/oOeKv8AwQv/APFV/wAPef2NP+g54q/8EL//ABVf8Pef2NP+g54q/wDBC/8A8VX/AA95/Y0/6Dnir/wQv/8AFV/w95/Y0/6Dnir/AMEL/wDxVf8AD3n9jT/oOeKv/BC//wAVXgj/AIKlfsm/ELxp4f8AAHhvWfEr6t4l1S20ywWbRXjQzzyrFGGbPyjc65Paqqqqqr4z/wDBRr9mf4C/ErWPhP8AEHVfEEOv6F5H2tLXSXmjHnQRzJtcHn5JUz6HIr/h7z+xp/0HPFX/AIIX/wDiq/4e8/saf9BzxV/4IX/+Kr/h7z+xp/0HPFX/AIIX/wDiq/4e8/saf9BzxV/4IX/+Kr/h7z+xp/0HPFX/AIIX/wDiq/4e8/saf9BzxV/4IX/+Kr/h7z+xp/0HPFX/AIIX/wDiq1D47fD/APaNuP8AhZfw0ub2fRJlFqj3lsbeTzI+G+Uk8cjmpf28Pgl+y1od94G8eXOqnxNeQyapp1vb6e00Mqsnlxh3B+XMkDA+gwaml1fxNrTzOLjUdU1W6LEIheW4nkbso5LMx6Ack18J/A9v8Mvhd4Q+HNqkSx+GNDs9NHlMWVjBCsZYEgE5Kk5IBOcnk1VV/wAFIfh63w6/bM+I9jHCi2ut3ya1bMqhRILuNZpDgMSCJmlU5wSV3YAYV+wL/wAFH/gP8Cf2bdH+EnxbvdbtdU8PaheraCy0w3CSW00xuAxZT18yaUYPPyjtiv8Ah7z+xp/0HPFX/ghf/wCKr/h7z+xp/wBBzxV/4IX/APiq/wCHvP7Gn/Qc8Vf+CF//AIqv+HvP7Gn/AEHPFX/ghf8A+Kr/AIe8/saf9BzxV/4IX/8Aiq/4e8/saf8AQc8Vf+CF/wD4qv8Ah7z+xp/0HPFX/ghf/wCKr/h7z+xp/wBBzxV/4IX/APiq/wCHvP7Gn/Qc8Vf+CF//AIqv+HvP7Gn/AEHPFX/ghf8A+Kr/AIe8/saf9BzxV/4IX/8Aiq8d/wDBUr9k34heB/EPgDw3rPiV9W8S6TdaZYLNorxoZ54mijDNn5RudcntWg6xZeHtc07X9SZxaaZdxXM5RdzCONwzYHc4B4r9qH40Xv7Qvx88afFq5dvs+tak402N02GGxiAitkK9AwhSPd6tuPev+CXnwKf4L/sraJqeq23la748k/t2+DIytHFIoFtGc+kKo54GGkYc4yaqv+Cz/wCzvb6P4l8O/tK+HbPZHr+3R/EQSM4NzEmbWckZ5aFGiOSABBGBkk1/wTd/bQ0X9k/x7r2l/EWbUH8CeKrMG6W0h897S9hyYZlTPRlaSNgvJLRk8JX/AA95/Y0/6Dnir/wQv/8AFV/w95/Y0/6Dnir/AMEL/wDxVf8AD3n9jT/oOeKv/BC//wAVX/D3n9jT/oOeKv8AwQv/APFV/wAPef2NP+g54q/8EL//ABVf8Pef2NP+g54q/wDBC/8A8VX/AA95/Y0/6Dnir/wQv/8AFV/w95/Y0/6Dnir/AMEL/wDxVf8AD3n9jT/oOeKv/BC//wAVX/D3n9jT/oOeKv8AwQv/APFV/wAPef2NP+g54q/8EL//ABVf8Pef2NP+g54q/wDBC/8A8VX/AA95/Y0/6Dnir/wQv/8AFV/w95/Y0/6Dnir/AMEL/wDxVf8AD3n9jT/oOeKv/BC//wAVXgjxfovxC8FeH/H3huSV9J8S6XbanYNNH5bmCeJZYyy/wna65HY1VVVVVeOPF+i/D3wVr/j7xHJKmk+GtLudTv2hj3usFvE0shVR947UOB3Nf8Pef2NP+g54q/8ABC//AMVX/D3n9jT/AKDnir/wQv8A/FV/w95/Y0/6Dnir/wAEL/8AxVf8Pef2NP8AoOeKv/BC/wD8VX/D3n9jT/oOeKv/AAQv/wDFV/w95/Y0/wCg54q/8EL/APxVf8Pef2NP+g54q/8ABC//AMVX/D3n9jT/AKDnir/wQv8A/FV8X/26/gB+0/4ZtvAHwq1LWrjVtPvk1OZb7TGtkECI8TEMScndMnH19K8CftC+Cf2YddPxU8e2eq3mnW9rLax2+m24lllmkX5F5KqgO05ZiAOOpIB/au/4KH/HP9qX7X4buLz/AIRHwLNIGXw1plwSJgvQXU+Fa4552kLHkKdgKg1+zz+yn8bf2oNeOjfCjwm91aW8yRX+s3jGCw0/d3llweQOdiBnI6Ka/ZI/4JqfBb9mr7D4v8RW8Xjf4gW+2RdYv4P3GnyDaf8ARICSqEMoIlbMg7FAStVVVVVVVVVVf8FKv+ac/wDcU/8AbSv+CcP/ACOvjD/sFQ/+jTVVVf8ABTn9lWf9oz4GN4k8JWPn+NvAHm3+mRIgL31sQPtNsDnqVUSL1JeIKAN5I+BPxk8U/s/fFnw58XPBxR9R8P3XmG3kYrHdQsCksL452vGzKT2zkcgVD/wV8/Y2lhSR9X8WxMygmN9CYshx0OGIyOnBI96/4e8/saf9BzxV/wCCF/8A4qv+HvP7Gn/Qc8Vf+CF//iq/4e8/saf9BzxV/wCCF/8A4qv+HvP7Gn/Qc8Vf+CF//iq/4e8/saf9BzxV/wCCF/8A4qv+HvP7Gn/Qc8Vf+CF//iq/4e8/saf9BzxV/wCCF/8A4qv+HvP7Gn/Qc8Vf+CF//iq/4e8/saf9BzxV/wCCF/8A4qv+HvP7Gn/Qc8Vf+CF//iq/4e8/saf9BzxV/wCCF/8A4qv+HvP7Gn/Qc8Vf+CF//iq/4e8/saf9BzxV/wCCF/8A4qv+HvP7Gn/Qc8Vf+CF//iq/Z0/bU+Bv7U2tavoHwm1DWLi70O1S5uxfac1sBG7bRgknJz2qqqqqqqq+J/8AwUy/Za+EPj/XPhp4y1fxHHrfh66Ntepb6O8sYkABO1geRgjmv+HvP7Gn/Qc8Vf8Aghf/AOKr/h7z+xp/0HPFX/ghf/4qv+HvP7Gn/Qc8Vf8Aghf/AOKr/h7z+xp/0HPFX/ghf/4qv+HvP7Gn/Qc8Vf8Aghf/AOKr/h7z+xp/0HPFX/ghf/4qv+HvP7Gn/Qc8Vf8Aghf/AOKr4qf8FA/2c/2jfCUnw0+Gmqa7Prc08dyiXmltbx+XGct8xJ5wRxXw3+L3gz4C+NNO+LHxBnuodA0Lzftb2tuZpB50TwJtQcn55Uz6DJr4F/tL/CD9orw5b+JPht4mSVblpBHY3u23vCqMVaTyC2/ZuVgGxglW9DVVVVVVVVVVVV+3n/ycFd/9gq0/9BNf8E7f+SJ61/2NNx/6S2tVVV+0F+1P8Ff2X7HQ7/4w+J5dLTxFcSw2EcFnJcySeUgaR9kYJ2LujUnHBkT1yP8Ah7z+xp/0HPFX/ghf/wCKr/h7z+xp/wBBzxV/4IX/APiq/wCHvP7Gn/Qc8Vf+CF//AIqv+HvP7Gn/AEHPFX/ghf8A+Kr/AIe8/saf9BzxV/4IX/8Aiq/4e8/saf8AQc8Vf+CF/wD4qv8Ah7z+xp/0HPFX/ghf/wCKr/h7z+xp/wBBzxV/4IX/APiq/wCHvP7Gn/Qc8Vf+CF//AIqv+HvP7Gn/AEHPFX/ghf8A+Kr/AIe8/saf9BzxV/4IX/8Aiq/4e8/saf8AQc8Vf+CF/wD4qv8Ah7z+xp/0HPFX/ghf/wCKr/h7z+xp/wBBzxV/4IX/APiq/wCHvP7Gn/Qc8Vf+CF//AIqv+HvP7Gn/AEHPFX/ghf8A+Kr9nr9uv4AftP8AjS98AfCrUtauNW0/S5NTmW+0xrZBAksUTEMScndMnH19Kqqqqqqqqqqqqqqqqr9pP/gl34N/aY/aGu/jT4h+JeoaHpup2dpFqOl6bpyG4uZYU8outw7FY8xJEozE/Iz2wfgB+zN8HP2ZfCzeFfhN4VSwW4bfe6hcOZ7y+fAG6WU8kcDCLtQHOFGTVVVeOP8Aglr+yb8QvGmv+PvEej+JX1bxLqlzqd+0OtPGhnnlaWQqoHyjc5wO1fs9fs4/DP8AZg8FXvgD4VWuoQaTf6pJqcy312bhzO8UUTEMQMDbCnHrn1qqqqqqqqqqqqqqqqqqqqqqqqrxx4XtPHHgrX/BV+Yfs2v6Xc6fN50AmTZNE0bboyQHGGOVJAPTvWr6Vf6Fqt7oeqweRe6fcSW9zFuDeXIjFWXKkg4IIyCRXwW+IVp8WPhL4R+I1nMsg17SILmYKuNk5UCZMeqyB1OOMrxxXjPxf4e+H/hPV/G/ivUFstH0Ozku72dgTsjRSTgDlmOMBRySQBkkV8DP2/P2e/jcI9NHiL/hEtebj+zNfkS38w/9Mps+W/0yG/2aqpZYoInmmlWOONSzu5wFA6knsK+OP/BQb9nn4MxXWnWXiNfGfiGBmjGl6FIsqpIOoluP9WgB4IBZgcjbkGvgr8V9A+OHwu8PfE/w2vlWuuWgke2Mm9rWZSVlhY4GSjqy5wM4zjBrxX4k0vwZ4W1jxhrcjJp2hafPfXjKASsMMZkcgEgZ2qepFeItcvfE+v6n4k1MqbzVryW7uNuceZI5dsZJOMsepJ96/Zy+H7/Cr4B/Dz4dT23kXWgeG7G2vEyx/wBJEKmc/MSRmUucdBnAwBiqqqqqr4neFx43+GvizwX9liuf7f0O90/yJjtSXzoHj2sewO7B+tf8EhfFR8Jftl2/h67CQzeJ/Duo6V5c0Ll98ey7KjH3GAszktxgEdSKr/gpr4rTwn+xL8R5kltxc6nDaadBHMCRIZ7uFJAuCPmERlYdsrznof8Agh34Slj0/wCLPju4sEMc82mabZ3RzuDItxJOg9sSWxP4VVVVVVVVVVVVVX/BVv8AZb+O37S3/Crv+FJ+Bf8AhI/+Ec/tj+0/+JnaWf2fz/sXlf8AHxLHu3eTJ93ONvOMjP8Aw64/bt/6IX/5c2k//JVf8OuP27f+iF/+XNpP/wAlV/w64/bt/wCiF/8AlzaT/wDJVf8ADrj9u3/ohf8A5c2k/wDyVX/Drj9u3/ohf/lzaT/8lV/w64/bt/6IX/5c2k//ACVX/Drj9u3/AKIX/wCXNpP/AMlV/wAOuP27f+iF/wDlzaT/APJVV4f/AOCTn7VfxM8S+KNV1rTNE8GWs0sl3pFzq2pw3EWoeZKSq/6I8skXykNlk9sZr4J/Di3+EHwf8F/C63eKT/hF9DtNPlliztmljiVZJBkA/O4Zug+90FVVft9fst67+1r8Dofh94Q1DR7DxDpmtW+padc6rJJFACqvHIrPGkjgGOVuApBZVzjqPiv/AMEif2qfBmt2GmfDbR7T4h2U+nrPeaja3tnpsdtcmWVTbqlzcLJJiNInL7QMylRnYSf+HXH7dv8A0Qv/AMubSf8A5Kr/AIdcft2/9EL/APLm0n/5Kr/h1x+3b/0Qv/y5tJ/+Sq/4dcft2/8ARC//AC5tJ/8Akqv+HXH7dv8A0Qv/AMubSf8A5Kr/AIdcft2/9EL/APLm0n/5Kr/h1x+3b/0Qv/y5tJ/+Sq/4dcft2/8ARC//AC5tJ/8Akqv+HXH7dv8A0Qv/AMubSf8A5Kr/AIdcft2/9EL/APLm0n/5Kr/h1x+3b/0Qv/y5tJ/+Sq8OfC3x38FdFt/hl8TNC/sbxLo277bY/aorjyvNYzR/vIXdGzHIjcMcZwcEEDxT+w9+1H8etVb4g/Cf4X/27oE0a26Xf9tWFrmSPh12Tzo/BI524r/h1x+3b/0Qv/y5tJ/+Sq/4dcft2/8ARC//AC5tJ/8Akqv+HXH7dv8A0Qv/AMubSf8A5Kr/AIdcft2/9EL/APLm0n/5Kr/h1x+3b/0Qv/y5tJ/+Sq/4dcft2/8ARC//AC5tJ/8Akqv+HXH7dv8A0Qv/AMubSf8A5Kr/AIdcft2/9EL/APLm0n/5Kr/h1x+3b/0Qv/y5tJ/+Sq/4dcft2/8ARC//AC5tJ/8Akqv+HXH7dv8A0Qv/AMubSf8A5Kr/AIdcft2/9EL/APLm0n/5Kr/h1x+3b/0Qv/y5tJ/+Sq/4dcft2/8ARC//AC5tJ/8Akqv+HXH7dv8A0Qv/AMubSf8A5Kr/AIdcft2/9EL/APLm0n/5Kr/h1x+3b/0Qv/y5tJ/+Sq/4dcft2/8ARC//AC5tJ/8Akqv+HXH7dv8A0Qv/AMubSf8A5Kr/AIdcft2/9EL/APLm0n/5Kr/h1x+3b/0Qv/y5tJ/+Sq/4dcft2/8ARC//AC5tJ/8Akqv+CX/wE+LP7OvwB17wT8Y/Cf8Awj+tXvjC51CC1+3W93vt3s7ONX328jqMvDIMEg/LnGCCaqqqqr9r7/gnr+2B8Uv2mPiH8QfAnwg/tPQNc1h7jT7v+39Ng86MooDbJLhXXkHhlBr/AIdcft2/9EL/APLm0n/5Kr/h1x+3b/0Qv/y5tJ/+Sq/4dcft2/8ARC//AC5tJ/8Akqv+HXH7dv8A0Qv/AMubSf8A5Kr/AIdcft2/9EL/APLm0n/5Kr/h1x+3b/0Qv/y5tJ/+Sq/4dcft2/8ARC//AC5tJ/8Akqv+HXH7dv8A0Qv/AMubSf8A5Kqrj/gll+1n4y+ItrCnhjTNM0/xKLnU5tUvNQja302M3UiLHKYi5aVoxFKFQN8swBIZZAn7Kv8AwT1+BP7LUdrr1hp//CWeN4418zxJq1uheGTaQxtIuRbA5YcFn2naXYVVVVVVVVVVVX7ef/JwV3/2CrT/ANBNf8E7f+SJ61/2NNx/6S2tVVV+2H+yj4b/AGvvhhbfD3Wtc/sG807VIb+w1dLH7TJbFcrIgTemQ8bMpBbAO1iCUAr9rL/gk9408PXfhDSf2TPh1qfiizh02UeINU1HX7SGae68wbCUnliVRtzgRoB6knmv+HXH7dv/AEQv/wAubSf/AJKr/h1x+3b/ANEL/wDLm0n/AOSq/wCHXH7dv/RC/wDy5tJ/+Sq/4dcft2/9EL/8ubSf/kqv+HXH7dv/AEQv/wAubSf/AJKr/h1x+3b/ANEL/wDLm0n/AOSq/wCHXH7dv/RC/wDy5tJ/+Sq/4dcft2/9EL/8ubSf/kqv+HXH7dv/AEQv/wAubSf/AJKr/h1x+3b/ANEL/wDLm0n/AOSq/wCHXH7dv/RC/wDy5tJ/+Sq/4dcft2/9EL/8ubSf/kqv+HXH7dv/AEQv/wAubSf/AJKr/h1x+3b/ANEL/wDLm0n/AOSq/wCHXH7dv/RC/wDy5tJ/+Sq/4Jf/ALGX7Sn7Ovx+17xt8Y/ht/wj+i3vg+50+C5/tixu99y93ZyKmy3mdhlIZDkgD5cZyRmqqqqq/a+/4J6/tgfFL9pj4h/EHwJ8IP7T0DXNYe40+7/t/TYPOjKKA2yS4V15B4ZQa/4dcft2/wDRC/8Ay5tJ/wDkqv8Ah1x+3b/0Qv8A8ubSf/kqv+HXH7dv/RC//Lm0n/5Kr/h1x+3b/wBEL/8ALm0n/wCSq/4dcft2/wDRC/8Ay5tJ/wDkqv8Ah1x+3b/0Qv8A8ubSf/kqv+HXH7dv/RC//Lm0n/5Kr9mn9h79qP4C/FCD4g/Fj4X/ANh6BDZT273f9tWF1iSQAIuyCd35I67cDvX7an/JsvjL/tx/9Lrev+Cd3/BQTxl4N1z4Qfsm3fhjw5F4Nl1K402fU5Fma+lnvbieWDYwkEaDz540IKNlc8gnIqqr/gpf+3t4y+FPjnxR+zH4Y8O+HNU8P+IvAktlrNxdRzLe2l1fRzxkxusmzasDwuFaMklj82MY+CX7Hn7SH7Rvh++8VfBr4aS+INJ028+xXN2dStLNFnCK5jH2iWPcQroTtzjcucZFf8OuP27f+iF/+XNpP/yVX/Drj9u3/ohf/lzaT/8AJVf8OuP27f8Aohf/AJc2k/8AyVX/AA64/bt/6IX/AOXNpP8A8lV/w64/bt/6IX/5c2k//JVf8OuP27f+iF/+XNpP/wAlV/w64/bt/wCiF/8AlzaT/wDJVf8ADrj9u3/ohf8A5c2k/wDyVX/Drj9u3/ohf/lzaT/8lV/w64/bt/6IX/5c2k//ACVVr/wTM/be0O5h1vVPgl5Nlp8i3FzL/wAJJpbbI0O5mwtyScAE4AJqv+HXH7dv/RC//Lm0n/5Kr/h1x+3b/wBEL/8ALm0n/wCSq/4dcft2/wDRC/8Ay5tJ/wDkqv8Ah1x+3b/0Qv8A8ubSf/kqv+HXH7dv/RC//Lm0n/5Kr/h1x+3b/wBEL/8ALm0n/wCSq/4dcft2/wDRC/8Ay5tJ/wDkqv8Ah1x+3b/0Qv8A8ubSf/kqv+HXH7dv/RC//Lm0n/5Kr/h1x+3b/wBEL/8ALm0n/wCSq/4dcft2/wDRC/8Ay5tJ/wDkqv8Ah1x+3b/0Qv8A8ubSf/kqv+HXH7dv/RC//Lm0n/5Kr/h1x+3b/wBEL/8ALm0n/wCSq/4dcft2/wDRC/8Ay5tJ/wDkqv8Ah1x+3b/0Qv8A8ubSf/kqv+HXH7dv/RC//Lm0n/5Kr/h1x+3b/wBEL/8ALm0n/wCSq/4dcft2/wDRC/8Ay5tJ/wDkqv8Ah1x+3b/0Qv8A8ubSf/kqv+HXH7dv/RC//Lm0n/5Kr/h1x+3b/wBEL/8ALm0n/wCSq/Z6/wCCcP7Z/gb4/fDPxt4p+DP2LRfD/jDSdQ1G5/4SHTJPIt4LuKSV9iXJZsIrHCgk4wATVVVVVV+3t+wT+1l8av2svHXxM+GXwo/tnw3rP9n/AGG9/t3T7fzfK0+2hk/dzXCOuJI3XlRnGRkEGv8Ah1x+3b/0Qv8A8ubSf/kqv+HXH7dv/RC//Lm0n/5Kr/h1x+3b/wBEL/8ALm0n/wCSq/4dcft2/wDRC/8Ay5tJ/wDkqv8Ah1x+3b/0Qv8A8ubSf/kqv+HXH7dv/RC//Lm0n/5Kr/h1x+3b/wBEL/8ALm0n/wCSq/Zq+DHxK+Avwut/h98WPDf9h6/Dez3D2f2yC6xHIQUbfA7pyB0zkd6/aq/Yy/aU/aK+Ien+Nvg38Nv+Eg0Wx0WLT57n+2LG02XKTTSMmyeZGOEljOQMfNjOQcfs0f8ABJP472HjTwH8SvirN4f0K10TxJBeax4cu7sT3L21vKsg2SW/mxP5hXaVLqQCec8VVVX7fv8AwTx8b/td/Fjwp4/8F+MfD+hQ6dobaVqram07SbUlklgMMcaFW+aaQNuZMDBG7pX/AA64/bt/6IX/AOXNpP8A8lV/w64/bt/6IX/5c2k//JVf8OuP27f+iF/+XNpP/wAlV/w64/bt/wCiF/8AlzaT/wDJVf8ADrj9u3/ohf8A5c2k/wDyVX/Drj9u3/ohf/lzaT/8lV/w64/bt/6IX/5c2k//ACVX/Drj9u3/AKIX/wCXNpP/AMlV/wAOuP27f+iF/wDlzaT/APJVf8OuP27f+iF/+XNpP/yVX/Drj9u3/ohf/lzaT/8AJVf8OuP27f8Aohf/AJc2k/8AyVWmf8E4f2z/AANqVp428U/Bn7Fovh+dNQ1G5/4SHTJPIt4GEkr7EuSzYRWOFBJxgAmrrSr/AF22l0PSoPPvdQja3totwXfI42quWIAySBkkCvhV/wAEgf2jdV1jwjq3xNsNE0bRp9YVPEWlyaokt3bWSSKXZGgZ438xNwG2QMpPIHWrSztLC0hsLC2itra2jWKGGFAiRIowqqo4AAAAA4FVVfHH4N+EP2gPhXr/AMJPHEcv9la9biNpoMCW2kVg8c0ZYEB0dVYZBHGDkEiv+HXH7dv/AEQv/wAubSf/AJKr/h1x+3b/ANEL/wDLm0n/AOSq/wCHXH7dv/RC/wDy5tJ/+Sq/4dcft2/9EL/8ubSf/kqv+HXH7dv/AEQv/wAubSf/AJKr/h1x+3b/ANEL/wDLm0n/AOSq/wCHXH7dv/RC/wDy5tJ/+Sq/4dcft2/9EL/8ubSf/kqv+HXH7dv/AEQv/wAubSf/AJKr/h1x+3b/ANEL/wDLm0n/AOSq/wCHXH7dv/RC/wDy5tJ/+Sq/4dcft2/9EL/8ubSf/kqv+HXH7dv/AEQv/wAubSf/AJKr/h1x+3b/ANEL/wDLm0n/AOSq/wCHXH7dv/RC/wDy5tJ/+Sq/4dcft2/9EL/8ubSf/kqv2evC2veBvgD8M/BPimw+xa14f8H6Tp+o2vmpJ5FzBZxRypvQlWw6sMqSDjIJHNVVVVVX7Q3hbXvHXwB+JngnwrYfbta8QeD9W0/TbXzUi8+5ms5Y4k3uQq7nZRliAM5JA5r/AIdcft2/9EL/APLm0n/5Kr/h1x+3b/0Qv/y5tJ/+Sq/4dcft2/8ARC//AC5tJ/8Akqv+HXH7dv8A0Qv/AMubSf8A5Kr/AIdcft2/9EL/APLm0n/5Kr/h1x+3b/0Qv/y5tJ/+Sq/4dcft2/8ARC//AC5tJ/8Akqv+HXH7dv8A0Qv/AMubSf8A5Kr4R/sZftKfs6+JLnxt8Y/ht/wj+i3ti+nwXP8AbFjd77l3SRU2W8zsMpDIckAfLjOSM+Mfgl4x/aDsrD4a+CtGl1K5u9TtpryOG7htnjtEkHnyK0zBcqhJA5JPQHpX7Pn/AARjsdK8WXmt/tG+LrfW9Dsb6VNN0XRpJIDqUCuypLdSjDQh1Cv5UTFgGGZAQRXhLwb4S8A6Ba+FfBHhrTNB0eyXbb2OnWqW8MY74RABk9z1Peqqqqqqqqqqq/4KVf8ANOf+4p/7aV/wTh/5HXxh/wBgqH/0aaqqqv29f+CbPxN8T/GLTNS/ZN+BNj/wibaCG1FrXV7O0DalJe3csxK3M6uTskhAwNiqFRcBAq/8OuP27f8Aohf/AJc2k/8AyVX/AA64/bt/6IX/AOXNpP8A8lV/w64/bt/6IX/5c2k//JVf8OuP27f+iF/+XNpP/wAlV/w64/bt/wCiF/8AlzaT/wDJVf8ADrj9u3/ohf8A5c2k/wDyVX/Drj9u3/ohf/lzaT/8lV/w64/bt/6IX/5c2k//ACVX/Drj9u3/AKIX/wCXNpP/AMlV/wAOuP27f+iF/wDlzaT/APJVf8OuP27f+iF/+XNpP/yVX/Drj9u3/ohf/lzaT/8AJVf8OuP27f8Aohf/AJc2k/8AyVX/AA64/bt/6IX/AOXNpP8A8lV/w64/bt/6IX/5c2k//JVf8Esv2R/2hf2b/H/jjW/jR8Pv+EestY0eC3spf7Vs7vzZFmLMuLeaQrxzlgBVVVVVVVV+19/wT1/bA+KX7THxD+IPgT4Qf2noGuaw9xp93/b+mwedGUUBtklwrryDwyg1/wAOuP27f+iF/wDlzaT/APJVf8OuP27f+iF/+XNpP/yVX/Drj9u3/ohf/lzaT/8AJVf8OuP27f8Aohf/AJc2k/8AyVX/AA64/bt/6IX/AOXNpP8A8lV/w64/bt/6IX/5c2k//JVf8OuP27f+iF/+XNpP/wAlV8Lf2Hv2o/gL4sj+IPxY+F/9h6BDBJbvd/21YXWJJBhF2QTu/JHXbgd6+I/wt8d/GrwXqPwy+GWhf2z4l1nyvsNj9qit/N8qVJpP3kzoi4jjduWGcYGSQKvofHXw68Uix1KLXfDHiTw9NtWGdZbK806UHdjadrxMCxOODk571+z3/wAFePj58MpbPRPi9a2/xI8PRKsby3DC21OJRxuW4UFZSAckSoWbAG9eSfgD+3F+zX+0l9m0/wCHvj+G31+eMM3h7WE+xX6HuqoxKTEAZPkvIB3NVVVVVVVVVVft5/8AJwV3/wBgq0/9BNf8E7f+SJ61/wBjTcf+ktrVVWr6rpug6Vea5rN9FZ2GnW8lxdXMzbUhiRSzux7AKCSfav20P2l9S/ap+O2sfERhLBoNrnT/AA5aSDa0FhG7eWzrkgSSFmkcAnDOQCQorwB/wT0/bE+KPgzSPiB4I+Dct/oOuW4uNPupdb0+1M8RJAcRzTo4BxkEqMggjIINf8OuP27f+iF/+XNpP/yVX/Drj9u3/ohf/lzaT/8AJVf8OuP27f8Aohf/AJc2k/8AyVX/AA64/bt/6IX/AOXNpP8A8lV/w64/bt/6IX/5c2k//JVf8OuP27f+iF/+XNpP/wAlV/w64/bt/wCiF/8AlzaT/wDJVf8ADrj9u3/ohf8A5c2k/wDyVX/Drj9u3/ohf/lzaT/8lV/w64/bt/6IX/5c2k//ACVX/Drj9u3/AKIX/wCXNpP/AMlV/wAOuP27f+iF/wDlzaT/APJVf8OuP27f+iF/+XNpP/yVX/Drj9u3/ohf/lzaT/8AJVf8OuP27f8Aohf/AJc2k/8AyVX/AAS//Yy/aU/Z1+P2veNvjH8Nv+Ef0W98H3OnwXP9sWN3vuXu7ORU2W8zsMpDIckAfLjOSM1VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV/wVO+A8vwb/AGptY8R2EbNofxHVtftHMZAS5kci7iLYALCYGTjos6A5OTX/AASn/aItja337N3iS6ijnDzal4ckkkCmUEBri2UdyMNMOpwZSeFFftt/BD4o/tAfBt/AXww8UafpsrXiXF/Y3oZE1SKP5kh81Q2wiQI4BXBZVyygZr4l/Bv4p/B3VjovxN8Cav4fuN7JG93bnyZyOvlTLmOUe6Mw96+HX7SPx4+EsEdn8Pfir4g0mzidXSxW6M1srAjnyJN0fO0A/LyBg5HFf8PF/wBsv/osn/lvaZ/8j18QP2ifjp8VIHs/H/xX8SaxZyffsZb90tmOW5MCYjz8zDO3ODjoAK+F/wADvi38atR/sv4X+AtU1+RWKyTQxiOCEgA4kncrHGcEfeYdR61+wl+zd8S/2avh1qvh34ieLtP1BtZvI7230mxRnj0x9m2T982N7OBECoUKpjOGfdkf8FS/2jLbwx4Hg/Z+8Mah/wATrxN5dzrbROM21grbliODlWlkVTj+4jAjDiv2APgDL+0N+054W8N3unvc+HtCmGs68clV+y27KwjY+kkhjiwMHDkgjGRVVVVVVX7Znw08Y/sO/ttWXxe8JabEdF1LXB4n8Mu6uIJiJVe5s3Oc/K7FGVWz5UsZypfA+DH7ZX7O3xw8CW/jnw58TdB00CFX1HTNX1GG0u9McglkmjdhgDDAOMo20lWIBr/gqN+2Hof7RHibQ/2evgffN4k0PRtSEt3e6aGnTV9SYGOKK22E+ciB2AYAh3f5cgBm/YK/Z1vf2Zf2bdB8B66E/wCEi1OaTV9dVPux3c4UeWPeOKOGMnnLIxHBAFVVVVVVVVVVVVVVVVVVf8M8/Ef/AJ9tP/8AAof4VpVtJaaXZ2kwHmQW6I4BzyFAP8qqqqqqqqqqqqv2kf2SPjD8T/jT4i8c+FbLSpNL1L7L9nae+Ebny7WKNsrjj5kav2Wfhn4p+EvwltvB3jCK2j1GK+uJmW3mEq7XIK/MPpVVVVVVVVVVVVVVVVVVVVVVVVVVVf8ADPPxH/59tP8A/Aof4VpVtJaaXZ2kwHmQW6I4BzyFAP8AKqqqqqqqqqqqr9qf9lj4s/Fv4s3HjDwdZ6ZJp0lhBCrXF6Im3ICG+UjpzX7IXwj8Y/Bn4aal4X8bQ2kd9da5LeRrbTiVfKaCBAcjvmNuPpVVVVVVVVVVVVVVVVVVVVVVVVVV8W/CureMvBsuiaIkTXTXEbgSPsGFPPNfEj9lzxx4o+GvjLw9qFpCy6h4b1KGAW0plk+0NaSiDCKNzfvfL+UckZHevCfibVfBXirRvGWgzeVqWg6hBf2cm5l2TQyLIhypDDDKOQQfQjrXh7XdN8U+H9M8TaNP51hq9nFeWkn9+KVA6H8VYGqqv21PiN/wtf8Aat+KHjaOXzIJ/EE1naSbdvmW9oBawtjapGYoEOCMjOCSck/8Eu/h5H8P/wBjDwVLLZ/Z77xRJda3efKo8wzTMsL8AE5to7f72T2zgACqqqqqqvE9hcar4a1bS7MKZ7yxmhiDHA3MhUZPYZIr/hgz9oL/AKB2h/8AgzX/AAqqqqqqqqqqqqqqqqqqqqqqqqqqr4ufCPxj4y8Yya1okNo1s1vHGDLOEOVHPFfBjwZrfgfwtdaTryQrcS6g86iKTeNpjjUc/VTVVVVVVVVVVVVXxP0DUPFfw08WeF9IWNr7WNCvbO1WRtqmWWB0TJ7DLDmvC/7Dfx60nxLpGqXen6KILO+hmlK6kpIVHBOBj0FVVVVVVVVVVVVVVVVVVVVVVVVVVV+078M/FHxV8A2Hh3wjDbSXlvrEd04nmES+WsMyHBPfLrxX7Ov7OnxK+GnxKg8UeKLXT0sY7OaImG7EjbmAA4Aqqqqqqqqqqqqr9s34C/EH43/8If8A8IJbWMv9i/bvtX2q5EOPN+z7MZ6/6pvpxX7Hf7O3xJ+CniXxDqnjm10+KDUrGOGA212JiWV8nIA4GKqqqqqqqqqqqqqqqqqqqqqqqqqqqv2i/AHiL4l/DSfwv4XigkvnvIZVWeXy12qTnmvgP+zR8Ufh98V9C8X+I7TTk06w+0ec0V4HYb7eSNcKBz8zivjR+zd8D/2hdNTTvi78OdJ194I2jtr2SMxXdqpzkRXCESIMnO0NtJAJBwK/aE/4Iw+NdGlvNf8A2bfGFv4hsFVpE0DXZVtr0HtHFcACGU/9dPJAA5JNeNfAfj74UeKZfC3j3wvrHhjX7Bg7Wl/bPbTJz8si5AypxlXXIPUEjmv2eP8AgqV+0t8EJ7fS/FOs/wDCyPDEe1X07X52NzGgAH7m8AMinAA/eCVAM4UEk1+zx/wUg/Zl/aHnt9CsfE0vhHxNPtVNG8R7LZp5CBkQTBjHLzkBdwc9dgqqqqqqqqqv2p/2WPiz8W/izceMPB1npkmnSWEEKtcXoibcgIb5SOnNfshfCPxj8GfhpqXhfxtDaR311rkt5GttOJV8poIEByO+Y24+lVVf8Fh/2pv+EV8H2P7Mfg3V9mreJ41vfExh+9Bp6tmGAt/CZZE3MAQ2yMA/LLz+xJ+zJqH7VHx40bwLNDcJ4Z09hf8AiW7jRsRWUZBaMMMbXlOIlPUFi2CEIrTtO0/SNPtdJ0mxt7KxsoUgtrW3iEUcEaAKqIi4CqFAAAGAABVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVXxr/AGzP2av2dfFVr4J+MfxJ/wCEf1q909NQgtf7HvrvfbvJJGr77eF1GXhkGCQflzjBBPwU/bM/Zq/aK8VXXgn4N/En/hINasdPfUJ7X+x7602W6SRxs++eFFOHljGAc/NnGASKqqqqqqqqqqqqqqqqqqqqqv24f2W7L9rH4HXvga1kt7bxPpc39oeHLyc7Eju1Vl8uRgCRFIjMrADg7GwSgqWLxr8LfGrQzJqnhrxT4ZvyGVg1tdWF1E34MjKwr9jr9tnwf+0noVt4c8QXNnovxEtIiLzSi2xL4IuWntc/eUgEtHksmD1UBjq+j6Rr+mz6Prul2epWFyu2e1u4FmilGQcMjAgjIB5FeMv+Cfn7JXjWWe7uPhPbaTdz9JtGvJ7ER/PuO2GNxCM5I+5wDgYwMf8ADrz9lL/oD+JP/By/+FeDP2B/2TPBEkN1Y/CDT9UuolAaXWp5tQEhHdopnaLP0QCrKwsdMtIrDTbOC1toF2xQQRiNEHoqjgCv2tP2wPA37MfhWWP7RZ6x43voj/ZWgrNlhnIE9wFO6OEEH0LkFV6My+J/EfjX4u+Pr3xJrc15r3ifxRqG9xFEZJbmeVgEjjjXJ6lUSNRgAKqjAAr/AIJ2fsiQ/ssfBiOfxNYovxA8YLFeeIXL7zaAAmGyUjjESs24jOZHfDMoTFVVVVVVfGb4IfDD9oDwTc+APir4UtNa0ufLQtIm2azlxgTQSj5opBkjcpGQSpyrEHxN/wAEOfCt3qsk3g39orVdL0058u31Pw3HfzL8xxmWO4gU/LtH3ByCeM4H7M3/AATX/Z3/AGbNVh8Xw2F34z8XW0m+11jXlRxYkMCpt4FASNxtU+Yd0gIO1lDFaqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr9qL4eR/Cj9ov4j/D22s/stno3iS8jsIdqrttGlL2/ChVGYXjOAAOeBiv8AgnJ8Rv8AhZf7Gnw31CaXdd6Hp7aHcJtx5f2J2giGdqg5gSFuM43YJJBNVXxo8e23ws+EPjT4kXNwkS+GdBvNQQtt+aSKFmRFDEBmZgqhSRuJA71p1hqniXWrXS7FHu9S1S6SGFXkAaaaRgoBZiBksRyT35NfD3wbYfDrwB4Z+H2lPvsvDGj2ml2zYIzHbwrEpwzMeiDqSfc9aqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq+JXwk+GXxj0B/DHxR8DaP4m05lZVi1C1WRod2MtG/3om4HzIVPA54r9oP8A4It2F19u8Rfs1+PPscn34vDXiHLxnqSsV4uWXsFWRGyT80gHNfFf4H/Fz4G69J4Z+LPw+1jw1fI21PtkH7mfr80My5jmXgjdGzDg88Gv2c/+Chf7Sn7OMsGm6N4vfxR4Xjb5/D/iF3uoFXv5EhPmW55YgIwTccsjV+zn/wAFV/2dfjdLBoHjWd/hp4kmbalrrN0sllMx6CO8CqufaVY+eBmrS7tNQtIb6wuorm2uY1khmhcOkiMMqysOCCCCCODmqqqqqqqqqr47/GTwr+z98JvEXxc8Yl30/wAP2vmC3jYLJdTMQkUKZ43O7KoPbOTwDXxL+IXi740fEjXPiN4un+26/wCKNQe5uBCjbd7nCxxqSSEUbUVcnCqoya/4J6/sqx/stfAm0sNesYo/G/izytR8SSbF8yFyn7qzLAncIFZl4JXe8pHDVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX/Bav/k6jwt/2T+z/APS+/r/gip/ydR4p/wCyf3n/AKX2FVVVVVVVVVVVVVVVVVVVVVVV/wAFA/8Agnto37UWmzfEr4ePFpfxO0uz8uJGEcVvrkafdhnbGRKANscpbAGFb5cMnirwl4++EPja48OeKtJ1Twx4o8P3fzRSBoJ7aaNvldHU+qgrIhIPDKSMGv2fv+Cp3xD8ExWfhr44aO3jPR4VWMatalYtTiQcZcHEdwQAB83lsSSWkY14H/b9/ZQ8d20DwfFaz0S6lZFe01yCSxaFmYgBnceURxklXKqCNxGa/wCGmP2b/wDo4L4bf+FXZf8Ax2vGf7d/7KHgiKU3vxh0rVJomKrBoySagZGHZWhVkx7lgvvXx1/4KweJvEWn3Ph74C+EpPDUc3y/27q7JNeBeP8AVwLuiibgjczS8HgKcERQ+Pvi945WGCPWfFni3xHd4VVD3d3ezt+bMePwA7AV/wAE+P8AgmrafAyfS/jd8bFiv/HxtxJpuihQ0Ph93BBZmBImuNhAyPljJbG87XFVVVVVVVVVVVVVVVVVVVVVVVVVVVVf8NC/Df8A5+7/AP8AAQ1a3EV5aw3cBJjmjDpkYOCMiqqqqqqqqqqqr4hfta/B34Y+L7/wN4qvtUj1TTfL89YbFpEHmRrIuGB5+V1r4afEvwt8W/C0fjDwdNcSadJM8KtcQmJtyHDfKe1VVVVVVVVVVVVVVVVVVVVVVVVVVV/w0L8N/wDn7v8A/wABDVrcRXlrDdwEmOaMOmRg4IyKqqqqqqqqqqqviZ+1N8JfhL4pfwd4wvdTj1GOBJmWCyaVdrjI+YH2r4U/Fvwd8Z/Dtx4o8ET3Utja3rWchuYDCwlVEc4B7bZF5qqqqqqqqqqqqqqqqqqqqqqqqqq8VeKdI8HaQ2ta1JItsjqhMabzlunFeG/jH4J8V61baBo9xdtd3W7yxJblR8qljk/RTX/BYr4c/wDCJftWxeNreP8AceOfD9rdySZ63NuDauuCxPEUNuc4UHfjBIYn/giP8Q49Q+HHxF+Fdxe5m0XWLfVraF2UEx3URicpzuIVrVdwwApkXkl6qv8AgrV4+fwT+xprekwSyxz+MtYsdGjkiZlKjebpxkdmjtHQg8EOQetf8E/fh4/xN/bD+GOhNZ/aLaw1hdWutysUSOyRrnLkA4BaJEweCzqpI3Zqqqqqqqq1K/ttK0661S7ZhBZwPNKVGSFVSTgfQV/w3l+z5/0E9b/8Fjf41VVVVVVVVVVVVVVVVVVVVVVVVVV4q+Lfg3wbqzaJrdxdLdLGrkRwFxhunNeEPGWieONMk1bQJJXt4ZzAxljKHeFVjx6YYVVVVVVVVVVVVV4k17TvCvh3VPFGru62Oj2U15dGNNzCKJC7kDudqnitN/bj+Amraja6VZ6lrJnvJkhiDaawG5iFGTngZIqqqqqqqqqqqqqqqqqqqqqqqqqqqviR8S/C3wq0ODxD4tmuI7O4u1tYzBCZG8xkdhwO2EbmvAH7RXw0+JfiJPC/he71CS+eF5VWe0Ma7VHPOaqqqqqqqqqqqq+Lvx5+HvwQ/sn/AITu6vYv7a877J9mtTNnytm/OOn+sX9a+E37RHw1+Neo3+l+Bru/ln02BZpxc2hhAVjtGCTzzVVVVVVVVVVVVVVVVVVVVVVVVVVVV4/8f+Hfhp4dfxR4olnSxjlSImGIyNubgcCvBP7S/wALviD4nsvCHhy81F9Rv/M8lZrMxqdkbO2Tnj5UNVVV4t8G+EvH2gXXhXxv4a0zXtHvV23FjqNqlxDIPdHBGR2PUdq/aE/4I0/DDxbDea9+z14quPBusOzSJo+qu13pkhxwiOAZ4BnJLEzDoAg618b/ANlb4+fs63r2/wAWPhvqelWfnGKHVY1FxY3B7bLiMlMkEEKSG7EAggfs8/tnftC/sxXUafDPxxK+iCTfN4d1QG706Yk5b9ySDEWOMvC0bnABbHFfs8f8Fhfgr4/t7fRPjtpsvw9107UN9Ekl3ptw5IGQygyQZJ6OpVRyZK0DxD4f8WaLaeI/Cuu6frOk38fmWl/p9ylxBcJ/eSRCVYcdQSKqqqqqviZ+1N8JfhL4pfwd4wvdTj1GOBJmWCyaVdrjI+YH2r4U/Fvwd8Z/Dtx4o8ET3Utja3rWchuYDCwlVEc4B7bZF5qq/wCCtH7V0fxe+KcfwM8F6v8AaPCPgC4JvniZWivdX2lZGBGciFXaEdPnM3UbSf8Agkx+yhH8XvinJ8cvGek/aPCPgC4AsUlVWivdX2q0akHORCrrMenzmHqNwNVVVVVVVVVVVVVVVVVVV+1d+2r8K/2PP+EW/wCFmaB4r1P/AIS77Z9h/sK0t5vL+zeT5nmedPFjP2hMY3Zw2ccZ/Zp/4KA/BH9qXUPEtj4L0vxRoEfhLTRqOpXniSC1tII4MkFt8dxJgKASS20ADOa+JX/BZb9nHwjr76J4G8LeKPGsEDMsupW8aWVs5GMeV5p8xx97JZEHAxuB4r9ov9vH9m/9mOddH8eeLpdT8QtknQdBiW8vIwB1lG5Uh6jAldC2cgEBiLX/AILcfBZ9SSK9+DnjWKwMhDTxXFrJKE7ERl1BPTjeB71+zx+1t8Cv2otKmvPhP4xS5v7JS17ot7H9mv7RQQN7Qk/MnzKPMQsmW27t2QP2oP2ofBP7J3gjT/iD8QfDHijVtHv9SXTt+hW9vM1vM0bunmLLNFhWEbgMM8gA4yM+Av8Agr3+zP8AEPx14c8AaT4O+JFpfeJtWtdMtri+06xjghluJViV5WW8YqgLgsQpIAJwelV44/4LAfsw+BPGmv8Age/8J/EXULnw7qlzp013p1hYy21w8ErRtJC5vAWjYqSrEDIIOB0r4QfFLwx8bvhn4d+K3gwXa6N4lsxdWsd2ipNEMlWSRUZlDqyspAYjIPJr9qT/AIKAfBT9kjxfpPgf4jaL4t1TVNX03+0Yk0O0t5hFAZHjUv508RBZo3xgH7p6V+zD/wAFCPg7+1p48vvh98NfB3juyvdN0t9SubrWbG1ht0iWSOPG6K5kbeWlXA28gMc8V8fv27P2Z/2bdWk8L/ETx75viSGNZJNC0m2a8u4gyhl8wL8kJZWVgsrIWVgwBBzWq/8ABbj4Kw38sWifBzxrd2Qx5U11cWtvI3AzmNXkC85H3jkAHjOB8Iv+Cvf7MXxG1m18O+LrPxB4CurqQol3q8cUlivXbunjclM4wSyBRkZbGSNO1HT9Y0+11bSb+3vbG9hSe2ubeUSRzxuAyujKSGVlIII4IINftQ/tQeAP2S/AFh8RviNpHiDUtN1HWI9Kii0S3hmmWaSGaUMwlljUJtgcEhicleOpH7OX/BSX4F/tQfEyD4VeAPCvjuw1aezmu1m1ixtIoAkQBYFormRsnPHy49xXxv8A+Ct/7Nnwn8SXPhHwrZa34/1CwuGhvLjSBHHZRMuQ4WeRv3pDADKKyEEkPxg/DHxxafE74a+E/iTYWMtlbeLNDstWhtpmDPAlzAkqoxHBIDgEjjIr9ov/AIKDfs1fszau/hbxn4lvdb8SwqrT6H4egS7ubcMRgSlnSKJsHdsd1fbg7cMufDP/AAWv+A+o6rHZ+Kfhb410eykwv2u3a3vPLYsBl03odgBYkrubgAKc8f8ABWH4n/D/AOMXx38EePvhl4qsfEGg6j8PLMwXlqxwCL+/yjowDRuMjcjhWU8EA1/wRU/5Oo8U/wDZP7z/ANL7Cqqqqqqqqqqqqqqqqqqqqqqqr9oH9lb4H/tOaLHpXxa8GxX9zaRsljqts5gvbLPJ8uZecZ52MGQnkqa+Nv8AwRg+L3hue61T4F+M9J8Y6au9odN1Rxp9+BjKoHP7iQ9RuZoh04GTjxr+yF+1F8PDK3iz4BeOLWCBQ0t3Do0t1bIC+wZnhDxgluAN2TlezDP/AAqX4qf9Ez8V/wDgluP/AIivCH7KP7THj3a/hP4C+O76F9225/sKeKDK9R5zqqZ9t2a+DP8AwRm+O/iye1v/AIy+KNE8Cab5g8+yt5Bqd/tAyQBGfIXP3Q3mtjk7TgBv2df2PPgJ+y5ppt/hd4Rxqs8YS817UpPtN/dY/vSEARg90iVEOM7c81VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVoH/IC03/AK9Iv/QBVVVVVVVVVVVV+2n/AMnMeMv+3H/0ht6/YN/5N8s/+wpd/wDoQqqqqqqqqqqqqqqqqqqqqqqqqqqqq0D/AJAWm/8AXpF/6AKqqqqqqqqqqqv28/8Ak4K7/wCwVaf+gmv+Cdv/ACRPWv8Asabj/wBJbWqqqqqqqqqqqqqqqqqqqqqqqqqr9oX/AJJvP/19w/zr4Gf8lU0P/t4/9J5K/wCC2nw8fVvhN4A+J9rZ+Y/h3XJ9NuJEViyRXcW8M2BjYHtVXLEYaQAZ3nH/AAR+8fP4T/a7g8KPLKYfG3h++08RBm2ebCq3auVHGQltKoJ6eYwHWqr/AILf/ECOfX/hh8KrW5w9nZ3utXsOFO4SukNu2cZGPIuR1wd3I4GP+CJ3w7h1v4y+O/iZcIki+FdBhsYVbqk17KSHHfIjtJV9MOfaqqqqqqqrxz/yJPiD/sFXP/opqqqqqqqqqqqqqqqqqqqqqqqqqqqr9ob/AJKPN/15w/yNfs0f8iHff9heT/0VFVVVVVVVVVVVVXxz/wCSJ/EH/sVdT/8ASWSvAv8AyO/h7/sK23/o1aqqqqqqqqqqqqqqqqqqqqqqqqqqr9uT/kkuk/8AYxQ/+k9xX7G3/JbLX/sHXP8A6CKqqqqqqqqqqqq/4KVf805/7in/ALaV/wAE4f8AkdfGH/YKh/8ARpqqqqqqqqqqqqqqqqqqqqqqqqqqqq/bJ/5Indf9hG2/9CNfso/8l+8K/wDb5/6RzVVVVVq2kaVr2m3Oja5plpqNheRmK4tLuBZopkPVXRgQwPoRiv2jP+CR3wC+KsM+ufCKR/hn4jKkrHZxGfTLhgOj2xYGInAG6JlUcko5r9of9hb9o/8AZn8y/wDHvgv+0NAj/wCZg0NmvLEf777VeHnj96iZ7Zr4L/tI/HD9nvUn1H4Q/EbVdASeRZLmyjkEtpdMMYMtu4aNzgY3FdwBIBGTX7Pf/BZ7wVrUNn4f/aR8H3Hh6/ZljfX9Cha5siMcyS2+TNEOOkfnEk8ACvAvxB8DfE/w3beL/h14u0nxHot1xFe6bdLPHuAGUJUna4yAUOGU8EA1VVVV+3n/AMnBXf8A2CrT/wBBNf8ABO3/AJInrX/Y03H/AKS2tV/wUG/ajj/Zd+AOoavod75fjTxRv0zwyqhWMMzL+8uiD2hQlhkMDIYlIwxI+HXgDxh8Y/iFovw88HWbal4h8TXy29skj43SMcs7sc4VQGdmPQAntXwD+DPhv9n34Q+GfhF4VfzrPw/ZiKS6MQja8nYlpp2UE4MkjM2MnGQM4FVVVVVVVVVVVVVVVVVVVX/Bc/8A5ol/3MH/ALjq8N/EjxT4S8IeK/BOhXaW+n+NIbaDViAfMlhgl81YwQcBS4UtxztA4GQfCX/BOP8AbW8b6Ba+JdC+A+ppY3i7oRqOpWWnTEephuZo5FB7ZUZr/goD+07P+y1+z9feKNAMT+KvENx/ZGgq7gGGaSN2a5K9SIkRm9N5jB4avgV8GfiB+1V8aNP+HXh7UfP1zxDcS3V/qmozNJ5SDMk9zKxJZz1PqzEDqc18QP8AgiTFpvgC8vvhv8Zr3WPF1lamWGy1DSo7e21GVUOYlYS5gLtjazFwvRuu8fCj4neNPgX8TdD+JHg69uNP1vw5fLKE3tF5oVsSQSgYJR13I6nqGIr9qLTPD/7Wv7A/iXXfCyefDr/hOLxJowASSWOaBVu0iBwQshMbQNjBG51yOa0nVb/QtVstb0q48i90+4juLaXYG8uRGDK2GBBwQDggiviB8atK8N/s1618f9NmiNnB4PfXrDEiyLKWtfNgUMPlbczIoPQ7h2q8u7vULua/v7qW5ubmRpJppnLvI7HLMzHkkkkknqTX/BG34k/8JX+zDqHgG5uN1z4H8QTwwxZzstbkCdD7Zma549vev+CpPj248dfto+M7drhpLPwvDaaNZB9wKLFAryrgkgDz5ZyNuAQQcZJJ/wCCK/wmbw/8IPF/xg1HT0S58W6slhYTOoLNaWincynqqtNLIpHGTCCRwpr9tj/gmn4s/an/AGjbD4leFfGWj+GdEvdFgttdubuB55xcQuyh4olIEhMJQYZkA8sfMd3yy/8ABFH4S6No4v8AxH+0LrsK2sKm8vH063toAwA3Ph3OxSegLHGQMnrX7R3wSb9nz4s6r8M08baP4ttbNUms9Y0twYrqCQZQlQzeW+OGTc20g4Zhgn/gi98etb8TeFPFfwB8RX9xdx+Flj1PQmlkVhBayuVnhA+8FWUo46jMrfdwAf8AgtX/AMmr+Fv+ygWf/pBf14B+JHin4Z3Ws33hG7S1utc0W60e4nIO9La5ULNsII2syArnnAY98EfDH9gv9rn4x+Frbxr8P/grqd5ol4qva3d5e2unC5RhlZIhdSxtIhHIdQVPY18dPjL4o/Yo/wCCeXg176w+xePbbwvo3hWxt2ZZltNU+wqspZ4yVPlJBcOCCVZo1GSGzUMXiLxr4lSCNr3Wde16+CqXdpri9upnxySSzu7t1PJJ968Ff8ER/iLrHguHVvHPxt0fw74lmhMn9jWujNqEMLFcpHJc+dHhs/K2yN1UglS4wT8ZPg749+AvxE1b4X/EnSPsGt6RJh9jb4riNhlJon43xuuCDwexAYED/gip/wAnUeKf+yf3n/pfYVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVoH/IC03/AK9Iv/QBVVVVVVVVVVVV+2n/AMnMeMv+3H/0ht6/YN/5N8s/+wpd/wDoQqqqqqqqqqqqqqqqqqqqqqqqqqqqq0D/AJAWm/8AXpF/6AKqqqqqqqqqqqv28/8Ak4K7/wCwVaf+gmv+Cdv/ACRPWv8Asabj/wBJbWqqqqqqqqqqqqqqqqqqqqqqqqqr9oX/AJJvP/19w/zr4Gf8lU0P/t4/9J5K/wCChngCP4jfsafFDSBHEZ9L0c6zDI6KTEbF1unKk/dJjhkTI5w5Hev2cviBH8Kvj58PPiJPc+Ra6B4ksrm8fCn/AEYTKJx8wIGYi4z1GcjBGaqv+CnvxA/4WB+2f45EDZtPDf2fRbbJ5HkQr5ufmYf695sYxxjIDZr/AII3/D9/C/7K154zurbbP4z8SXVzBLlvntoFS3UYzjiWK45ABO7BzgYqqqqqqqvHP/Ik+IP+wVc/+imqqqqqqqqqqqqqqqqqqqqqqqqqqqv2hv8Ako83/XnD/I1+zR/yId9/2F5P/RUVVVVVVVVVVVVVfHP/AJIn8Qf+xV1P/wBJZK8C/wDI7+Hv+wrbf+jVqqqqqqqqqqqqqqqqqqqqqqqqqqqv25P+SS6T/wBjFD/6T3Ffsbf8lstf+wdc/wDoIqqqqqqqqqqqqr/gpV/zTn/uKf8AtpX/AATh/wCR18Yf9gqH/wBGmqqqqqqqqqqqqqqqqqqqqqqqqqqqr9sn/kid1/2Ebb/0I1+yj/yX7wr/ANvn/pHNVVVVVVNDDcQvbzxJLHIpV0dchgRggg9QR2r9pL/gl/8As3/H37Vrvh/Sv+Fd+LJ8t/amg2yrbzyH+Kezysb8kklDE7E5Zz0r9oz/AIJrftKfs/Sz6pb+G38ceGEY7NY8PQPOyL2M1uMyRH1OGQf3zXw1+LfxN+DmvJ4n+F3jrWPDWpKylpdPuWjEu3OBIn3ZF5PyuCOTxzX7Pn/BaO/tvsPhz9pPwH9sj+5L4l8PYSQdAGls2wrdyzRuuAPljJ4r4U/G/wCEfxy0CPxL8JviDo/iWxdcuLSf99B04mhbEkLcj5ZFU8jjkVVVX7ef/JwV3/2CrT/0E1/wTt/5InrX/Y03H/pLa1qOo6fpGn3Oratf29lY2ULz3NzcyiKOCNAWd3ZiAqhQSSeABk1+3J+09e/tVfHjVPG1pc3A8LaWv9neGbWVPLMVmhJ8xl7PK5aRs5IDKucIuP8Agj5+yhH4a8LT/tReNdJxq3iCOSz8LRzqpMFiG2y3QU8o8rqUU8Hy1YjKy81VVVVVVVVVVVVVVVVVVVf8Fz/+aJf9zB/7jq/4Jx+EtA8b/trfC/QvEtgl5YpfXV6IX6Ga1sp7mEkdwJYYzjvjFV/wXC8QahN4v+FXhQlBY2mm6jeqFyGeWWSFDu5wQFhXHGRubk54+F/xf+JnwV1+fxV8KvGeoeGtWurNrKa7smCu8DOjtGcg8Fo0P1UV/wAN+ftlf9HDeKv+/wAn/wATWo6he6vqFzqupXL3F3eTPNPK/wB6SRyWZj7kkmv+CZd1qV7+w18LptVkledbe/jQyjBESajcrEB7CNUA9sV+0t8LH+Cfx98e/CwWkttbeH9cuIrBJd242bNvtWO7k7oHibJzndnJ618afj/Hc/8ABHz4a6Hban5mo+J7i38MzxecrSxQ6dPIzZx90bbS3G087JkzkE18JPgbL43/AGR/jl8W2sGmk8E32gtYFFLsxeaWO5ICncAkUyMdw2bSx6plf+CL3xJ/4Rn48+L/AABeXHlWHijw2bosThRPZSBkLdgBFNcHJ6cetfEnxVc/Er4n+KfG6i4uLjxTr15qIDwqssj3E7yconyhiX+6vGeBX7MvwpHwP/Z/8BfCp4UjutA0WGO/VAmDeOPMuSNnBBmeQ55JzkliST+3n/wVG8fjxxrvwY/Zx1lNC0nQrqWw1HxPasJLnUJUwsi25Zf3CI4kTzELM+Aysq4z4M+E/wC0/wDtbeIrjVvDPh3xn8R9TiZILrV724kuFiZmG1Jbu4bYhy+7DOOCzdASPjX8BPiz+zr4qtfBPxj8J/8ACP61e6emoQWv263u99u8kkavvt5HUZeGQYJB+XOMEE/8EVP+TqPFP/ZP7z/0vsK/4LV/8mr+Fv8AsoFn/wCkF/X7PXhfRfG/x++GfgvxHa/adJ1/xhpOn38GdvmwTXcUci57ZViPxqGGG3hS3t4kjjiUKiIu0KAMAADoMdq/4Llarfw6T8G9DiuMWV3ca1cTRbB80kS2So2cZGBNIMA4O7nOBj/gmpoOm+I/24Phbp+qw+bDDeXl4i+ktvYXE8R/CSJD+FV/wWohhj/aq8NPFEitL4Bs2kKrgu326+GT6nAA+gFf8EVP+TqPFP8A2T+8/wDS+wqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqrQP+QFpv/XpF/wCgCqqqqqqqqqqqr9tP/k5jxl/24/8ApDb1+wb/AMm+Wf8A2FLv/wBCFVVVVVVVVVVVVVVVVVVVVVVVVVVVWgf8gLTf+vSL/wBAFVVVVVVVVVVVX7ef/JwV3/2CrT/0E1/wTt/5InrX/Y03H/pLa1VVVVVVVVVVVVVVVVVVVVVVVVVftC/8k3n/AOvuH+dfAz/kqmh/9vH/AKTyVq+lWGu6VeaJqkHn2WoW8lvcxbiu+N1KsuVIIyCRkEGvHHhPUvAXjTX/AALrIxqHhzVLnTbobduJYJWjfjt8yGv2VPiB/wALU/Zs+Gvjx23XGp+G7T7WQePtMcYinx8zHHmo+MknGM85FavqlhoelXmt6pceTZafbyXFzLsLbI0UszYUEnABOACa8ceLNS8e+NNf8c6y2dQ8Rapc6jdHOf3s8rSPz3+ZjX7Knw+/4VZ+zZ8NfAbri40vw3afawBgfaZIxJPj5VOPNd8ZAOMZ5yaqqqqqqqvHP/Ik+IP+wVc/+imqqqqqqqqqqqqqqqqqqqqqqqqqqqv2hv8Ako83/XnD/I1+zR/yId9/2F5P/RUVVVVVVVVVVVVVfHP/AJIn8Qf+xV1P/wBJZK8C/wDI7+Hv+wrbf+jVqqqqqqqqqqqqqqqqqqqqqqqqqqqv25P+SS6T/wBjFD/6T3Ffsbf8lstf+wdc/wDoIqqqqqqqqqqqqr/gpV/zTn/uKf8AtpX/AATh/wCR18Yf9gqH/wBGmqqqqqqqqqqqqqqqqqqqqqqqqqqqr9sn/kid1/2Ebb/0I1+yj/yX7wr/ANvn/pHNVVVVVVVV+0Z/wT6/Zq/aVln1rxP4Sbw/4onYu/iHw8UtLqZupMy7WjnJ4BaRGfAwGWv2iP8Agk/+0Z8IZ7jVvhxZf8LM8OLuZJdIgKX8SgE4ktCSzHjA8oyZ9BkCvDvibxv8MvFMet+Ftc1jwx4h0mZkW4tJ5LS5tpFOGUlSGU5BBU+4Ir9nv/gst8TvCctnoH7Qvha38ZaOirG+saUi2mpxgdXaMkQTnGAFAhPcuelfA39q34AftHWhk+EvxH0/Vb2OMPPpUwa1vYOATuglCuQCcF1DJnoxqq/bz/5OCu/+wVaf+gmv+Cdv/JE9a/7Gm4/9JbWv+Cvf7V7eBPA9v+zZ4I1NF1zxfD53iOWKUFrTTeiwEDlWnbryMRxsCCJQR+xf+zRqX7VPx20b4dq0sGg2uNQ8RXcZ2tBYRuokCNggSSFljQkHDOGIIU1pOk6boOlWWhaNZRWdhp1vHbWltCu1IYkUKiKOwCgAD2qqqqqqqqqqqqqqqqqqqqr/AILn/wDNEv8AuYP/AHHV/wAEuP8Ak+34Y/8AcW/9NN5Vf8FnfgtqnjT4OeF/jDols8zfD++mh1JEUZFneeUvmnjJCSxRDrgCVjjuP2S/j7p37N/xksPH+v8AgrT/ABXoNxbvYazpl3bRzM9rIyMzw+YCqyo0aMpOM4KkqHJHhT9sn/gm14t0dNWg8SeAdMJYo9rq3h0Wk8bAA4KvDyMEfMpK9RnIOJv2w/8AgnZN4v0DwH4eufCPiDW/EWrWmmWsOmeFg8aPcyCNJHneJYlRWYbsOWAP3TWn6dp+k2UOm6VY29naW67Ybe3iEccY9FVcAD2Ff8FovhLH4X+Ofhn4t6fZeXbeN9HNveSLGoEl5ZlULMwOcmCS3UbgOI+CcELr3xW1TXfgl4Q+Ds89x9l8K69q2pRgELGyXcVmqKQD8zI8FwdzDIE5AODgfsM/s129z/wThn+HWsWX2e8+Lel6nqFyLkHEbXkXk2sy7lbb+4itZFYKcHDAE8n4R/ELV/gd8S18YJYOmp6VY6pY/Zbm3JKS3FjcWpWRCVIAM2G5DAA45GK/YC+Ekfxn/a0+H/he8i36bp2of2xqILKAYbMedsIPJDukcZA5xITxgkeLf7d/4RXWf+EW/wCQ1/Z8/wDZv3P+Pjy28r7/AMv39v3uPXiq/wCCe/7Uv7Lnhf8AZJ8HeFdV+K3g/wAH6r4et5otV07WdVt7Cd7gyu7yqjuDKH3bgy5JyBgHgf8ABR/9o7wZ+03+0dJ4t+Hvmy+H/D2jw6HZXsiFPt4imnladVPKozzsFzglVUkAkgf8EVP+TqPFP/ZP7z/0vsK/4LV/8mr+Fv8AsoFn/wCkF/X7J3/J1Hwa/wCygaF/6Xw1X/BYP4Pap8Q/2arLx5oNhcXl58PtWF7dRwxhythMhjnfAG7Ct5DkjhUV2YYG5fhd8Q9f+EnxF8N/EzwvIg1TwzqUN9bK4+V2jYEo3+ywyp9mNeD/APgrn+x7r3gmHxF4n8R634Z1zy/9I8PXGj3FzMsgA3COaFGhdCSdrM6EgZKoTiv2xv2kr39qv466v8VH06403S/JjsNFsLibzHtbKLOwMRwGZmkkZV4DSMATjcf+CKn/ACdR4p/7J/ef+l9hVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV/wqX4kf9Chf/8AfI/xrRoZLfR7G3mQrJHbRq6nqCFAIqqqqqqqqqqqq/ar+Anxi8a/HzxR4m8K/D7VNS0u8+yfZ7qFFKSbLSFGxk9mVh+Ffsd+DPFPgP4K2vh7xholxpWopqNzI1tcABgrMNp4J4NVVVVVVVVVVVVVVVVVVVVVVVVVVV/wqX4kf9Chf/8AfI/xrRoZLfR7G3mQrJHbRq6nqCFAIqqqqqqqqqqqq/bF+B/xZ8e/Gm58QeDvAmp6rpzadbRrcQICpZVO4cntX7EHgHxj8OvhPquieN/D91o99N4imuI4LlQGaI29uoYYJ4yjD8KqqqqqqqqqqqqqqqqqqqqqqqqqvjZoereIvAkum6JYSXl01zEwijHOAeTXwh+HfjbQviJpOq6v4cu7W0g87zJZFAC5hdRnnuSB+NV/wU9+H3/Cv/2z/HPkLi08SfZ9atsjn9/Cvm5+VR/r0mxjPGMktmv+CN/xAfxT+yteeDLq53T+DPEl1bQRYb5LadUuFOcY5lluOASRtycZFf8ABQzx9H8Of2NPihqwli8/VNHOjQxuygym+dbVwoPUiOaR8DnCE9q/Z4+HrfFf47/D/wCHPkpJDr/iKytbkOoYLbmVfOYqWXcBGHO3IJxgckVVVVVVVVV4utbi+8J61Y2cTSz3GnXEcUajlmaNgAPqSK/4Zc/aC/6JTrf/AH7X/wCKqqqqqqqqqqqqqqqqqqqqqqqqqqr42+AfGPiLx1LqWieH7q8tjaxKJY1GMgHI618BfDut+GfB95Ya9pstlcPqTyLHKMEqY4wD9MqR+FVVVVVVVVVVVVfF3StQ134T+NdE0i0e6vtQ8O39vawRj5pZXt3VFHuSQPxrwh+zN8erHxZol9efC7WYoLfUbeSWRo1wirIpJPPQAVVVVVVVVVVVVVVVVVVVVVVVVVVVX7XPgzxR45+G2m6R4R0S41S8h1yKd4YFBZYxBOpbntllH41+y58IviV4N+LFvrfijwdqGnWK2M6GedAFDMBgcHvVVVVVVVVVVVVX7eXwt+IPxL/4Qb/hA/Cl7rX9m/2j9q+zKD5O/wCzbM5I67G/I1+wv8I/iT8OPFfie98c+D7/AEaC806KOB7lQBIwkyQME9qqqqqqqqqqqqqqqqqqqqqqqqqqqqv2pPCniLxn8J7jRPC+kz6lfNfQOsEIyxVScn8K/Zx+DXxR8KfGfw7r/iPwTqNhp1r9q864mQBU3WsqrnB7swH41VVVVVVVVXx5/Y7/AGeP2kYZZvif8O7G41h4fKj12yH2XUIgANv79OXC44WQOoyeOTn9oP8A4I5/GXwJ9u8QfAvXbTx9osPzx6ZOy2mqIvJICnEM20DqrozHhYyeKu7Pxp8NfFbWl9a634V8TaHcDdFKktjeWMy8jIO143HB7EV+zn/wVx+Pnwqlg0P4uonxM8OBsGS8lEGp2ynulwARKBknbKrMeAJEFfs8/t0/s3/tMeXp/gLxp/Z+vv8A8y9rirZ35/3E3MkvHP7p3x3xX7YvwP8Aiz49+NNz4g8HeBNT1XTm062jW4gQFSyqdw5PavgPrz/sifsseNvHfxu0m60KDRdVub9bScqs15m2t0iiiBOC8si+WoJA3HnAya+LXxO8U/Gj4leIvil40vXudY8R3z3U5ZywiB4jiTPISONUjUdlRR2r/gnF+ys37MXwHgbxLpzW/jnxo0eo+IVlA32wCkW9px2iRmJByRJLLzjAFVVVVVVVVVVVVVVVVVVVVf8ABT79kL4y/tW/8K0/4VHY6Tcf8It/a39ofb79bbb9o+x+XtyDu/1EmfTj1r9h7/gnJ+0x8Bf2o/BPxY+IOl+H4tA0L7f9re11ZZpB51hcQJtQDJ+eVM+gyarV9J03XtKvNC1myivLDUbeS2uraZdyTROpV0YdwVJBHvX7Rv8AwRhvrvW7vxJ+zH4w0y106dnkHhnxDLKptQFJCW90BIZAThVWUKVHLStya1D/AIJXftzWV7La23wbt7+OJsJc2/iTTVjlHqokuFbH1UGvhZ/wR/8A2sJvEmna14g8R+D/AAWunXENylzJdnUJ43XLqY4Y0MblWVch3VeeC3Iqv+Ci37Lfib9qn4HWfhnwDBYv4q0HWob/AE77XdC3R4yrRTIWKkYKuGx8vMY57Naf8Egf2w5rqGG60zwpbwvIqyTf20r+WpPLbQMnA5x3rwx4c0fwf4a0nwj4etEtdL0OxhsbG3RQBFBCgSNQBwAFUDj0r9pr/glX+0X42+P/AI88afC7S/Dk3hnxDrU2pWJuNVWB1NwfNkUxkfKFkd1A6YAxxX/BMz9hX4mfsveKvGvjz4xWWkw6tqen2+m6ONPvxc7YDI0lzvwoxlo7Xbg/wtkdDVftnf8ABJPxN4u8d6r8U/2ab7TGXX7qW71LwvqEwthb3Dks72sp+TY7Enyn2CM/dYqQqeFf+CTn7bPiLV00zV/h7o/hi3ZSTqGq+IbSSBDkDBFrJNJnnPCEcHnpn4w/8EbPiDoXhDwZZfBjWbTxR4jb7Y/iu91G8WxgVitv9njtoyCdgIuMsSWOQTjgD/gnB+wv+0N+y78eNU8efE/RdETRtT8L3OlCWy1VJ3jle4tplJQDJGIGH1YV/wAFJv2cfiZ+1B8C9D8AfCq10+fVtP8AFlvqcy312LdBAlpdxMQxByd0ycfX0r4C/wDBLX9rH4e/HP4c+PvEej+GU0nw14s0vU79odaR3WC3u45ZCq4+Y7UOB3NVqOnafrGn3Ok6tY297Y3sLwXNrcxCSOeNwVdHRshlZSQQRgg4NftRf8Ec/FLeJtQ8Xfswatpk+j30zSr4X1Oc28tjkZKQTtlZE3ZwJChUEAs+C1eA/wDgkh+2R4r1mKw8VeFNE8F2PmKJb7U9btrkBDksUS0eVmI2gYO0EsvIG5l/aF/4JF63Z/Cf4d+EP2ebix1rX9EutQm8T6nq0os31OS4S3CSKCzCNI/s5VYhuwHJLFtzN/wTa/YT+P37MHxz1zx98VdO0SDSb/wncaZC1lqS3Dmd7u0lUFQBgbYX59cetVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX/Bb/wCH0cOv/DD4qWttl7yzvdFvZsqNohdJrdcZyc+dcnpgY5PIr/giX4+j0j4x+P8A4azSxRr4l8PwahEXZVLy2U20IueSSl5K2B2QntX/AAW0+Ib6V8JvAHwvtb3y38Ra5PqVxGjMGeK0i2ANg42F7pWwwOWjBBGw1/wR+8AP4s/a7g8VvHKIfBPh++1AShG2ebKq2ioSOASlzKwB6+WxHSqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqvjJ+zx8Fv2gNFfRPi18PdJ19PL8uG6lh8u6thyR5VwmJI8FicKwHJyDk1+0J/wAEX/FWjQ3niL9m3xuniGFWZ08N66UtroLjhYroERStntIsIAHLk142+H3xD+E/iR/D3j7wlrfhfWrOTIg1C0e2kBUgh0LAbh0IdcggggkEGv2RP+Ckn7RXwk1SPwl4m11vH/hGz029uRpuuTs1zCttaSzBILz5pEB8pUAkEqKv3UHWv29/+Cg837Xmj+FfBvg/w/qfhnwxpS/bdUsbuZJGu9RIKr8yfeiiQsEPyljK5ZRtXH/BKP8AZTHxr+MJ+MHiyz3eEfhzcRTwo44vtVyHgjwRgpGB5r85B8oYIc4qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/4Wx8N/8AocNP/wC+z/hUMsVxCk8Dh45FDIw7gjg1VVVVVVVVVVVeKPjz8HfBeuXPhnxV8QdK03VLPZ59rO7B49yB1zgd1ZT+NeEvGXhbx7oy+IPB2t22q6c8jRrcW5JUsv3hz6VVVVVVVVVVVVVVVVVVVVVVVVVVVX/C2Phv/wBDhp//AH2f8KhliuIUngcPHIoZGHcEcGqqqqqqqqqqqrxd8b/hL4D1hvD3jDx5pmlaikayNbXDkMFboeB0NeDPHvg74i6VLrfgjxDaaxYwXBt5J7ZiVWUKrFee+11P4iqqqqqqqqqqqqqqqqqqqqqqqqqrWdc0jw9YnUtav4rS2VgplkOACegrSviF4J13UItK0jxJZ3V3Pu8uGNiWbAJOOPQE1X/BWrwA/jb9jTW9Wgjlln8G6xY6zHHEjMWG82rnA7LHdO5J4AUk9K/4J+/EN/hn+2H8Mdda98i2v9YXSbrczBHjvUa2w4BGQGlR+eAyKxB21/wWK+I3/CW/tWxeCbeX9x4G8P2tnJHt6XNwDdO2SoPMU1uMZYDbnIJYD/giP8PI7D4cfEX4qXFn++1rWLfSbaZ1XIjtYjK4TjcAzXS7ucMY14BSqqqqqqqqru6trG0mvbuZYoLeNpJXboiqMkn2AFf8NRfs+/8ARV9D/wC/jf4VVVVVVVVVVVVVVVVVVVVVVVVVVVrPj3wd4evTputeIbSzulUMYpWIIB6GtD8Q6J4ms3v9B1KG9t45DE0kRyA4AJH5EfnVVVVVVVVVVVVWrapp2h6Vea3q93Ha2On28lxczyHCxRIpZ2PsFBNWn7S/wFv7qGxs/ijoss9xIscUayNlmY4AHHckVVVVVVVVVVVVVVVVVVVVVVVVVVVXivxl4W8DadHq3i3W7bS7OWYQRzTkhWkKlgox3wrH8K8M/Fz4aeM9UXRPC/jLT9SvmRnWCFiWKr1PTtVVVVVVVVVVVVXjj4o/D74a/Yv+E78WWOi/2j5n2T7UxHm+Xt34wD03r+YrwV8W/ht8R7q5sfA3jHT9Zns4xJPHbMSY1JwCcgcZqqqqqqqqqqqqqqqqqqqqqqqqqqqq8TeKvDvg3Sm1vxRq8GnWKuqGeckKGboOPWvD3xl+F3izWLfw/wCHPG2nX+o3W7ybeJyWfapZscdlUn8KqqqqqqqqqqqviT8Kfhv8YvDcvhH4n+C9J8S6TLk/Z7+3EnltjG+NvvRv6MhDDsa/aG/4Jz/AH4XeJ9Rv/hF8bbbw/rB0fUIz4T8TTmRGN1aTRReTcqu6NQZFG2QSE8kyDFfsh/8ABM7xJ+0d4R8f6v4y1+78H3mg3EdhoEyJFd213eBWabzSjHdEq+SA0bc+buBYLg/s7fBDwz+zp8G/DXwj8LRoYdFtR9ruQDuvbt/mnnYnnLyFiAfurtUYCgCqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqtA/5AWm/9ekX/AKAKqqqqqqqqqqqv20/+TmPGX/bj/wCkNvX7Bv8Ayb5Z/wDYUu//AEIVVVVVVVVVVVVVVVVVVVVVVVVVVVVaB/yAtN/69Iv/AEAVVVVVVVVVVVVft5/8nBXf/YKtP/QTX/BO3/kietf9jTcf+ktrVVVVVVVVVVVVVVVVVVVVVVVVVV+0L/yTef8A6+4f518DP+SqaH/28f8ApPJVfGjwDbfFT4Q+NPhtcwJMviXQbzT0DbRtklhZUdSwIVlYqwYg7SAe1adf6p4b1q11Sxd7TUtKukmhZ4wWhmjYEEqwIyGUcEduRX7QHxVn+OPxr8afFma3lt08TaxPdW1vMwZ4LcttgjYjglIlRSRxla/4Jv8Aw+T4dfsZfDixeJ1utbsX1q4d1KmQ3cjTRnBYgAQtCoxgEKGwCxqqqqqqqqrxz/yJPiD/ALBVz/6Kaqqqqqqqqqqqqqqqqqqqqqqqqqqq/aG/5KPN/wBecP8AI1+zR/yId9/2F5P/AEVFVVVVVVVVVVVVXxz/AOSJ/EH/ALFXU/8A0lkrwL/yO/h7/sK23/o1aqqqqqqqqqqqqqqqqqqqqqqqqqqr9uT/AJJLpP8A2MUP/pPcV+xt/wAlstf+wdc/+giqqqqqqqqqqqqv+ClX/NOf+4p/7aV/wTh/5HXxh/2Cof8A0aaqqqqqqqqqqqqqqqqqqqqqqqqqqqv2yf8Akid1/wBhG2/9CNfso/8AJfvCv/b5/wCkc1VVVVVVVVVVVVft5/8AJwV3/wBgq0/9BNf8E7f+SJ61/wBjTcf+ktrVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVf8I9r3/QEv8A/wABn/wrQlZND05HUhltIgQRgg7BVVVVVVVVVVVV+2L4T8U6n+0f4vvdN8Nard28n2LZNBZSSI2LKAHDAEHkEfUV+w5puo6V8BLOz1SwuLOcandExXETRsAWGDggHFVVVVVVVVVVVVVVVVVVVVVVVVVVV/wj2vf9AS//APAZ/wDCtCVk0PTkdSGW0iBBGCDsFVVVVVVVVVVVX7cnhfxLq3x6u7vSvDup3kB0u1US29m8i5CnIyoIr/gn9pOqaP8ABrWLXV9MurGZvE87rHcwtExX7NbDIDAHGQRn2qqqqqqqqqqqqqqqqqqqqqqqqqq+Ptrc3nw7mgs7eWeT7VCQkaFj19BXwT0bV7X4naLPdaVdwxJ5+55IGUD9xIOSRVV+2p8Of+FUftW/FDwTHH5cEHiCa7tI858u3uwLqFc7mJxFOgyTk4yQDkDwn4Z1Xxr4q0bwboMPm6lr2oQWFnHtZt000ixoMKCxyzAYAJ9Aa8PaDpvhbw/pnhjRofJsNIs4rO0j/uRRIEQfgqgVVVVVVVVV41jeXwbr0USM7vplyFVRkkmJsACv+EF8b/8AQna5/wCC6X/4mqqqqqqqqqqqqqqqqqqqqqqqqqqvj7pOq3nxDmms9Mup4/skQDxwMwzg9wK/Zysruy8D3sN7azQOdVkYLLGUJHlRc4Pbg1VVVVVVVVVVVV8a7ee7+DXj21tYXmmm8MakkccalmdjbSAAAckk8YrwT4I8ZxeM9All8I60iJqlsWZtPlAUCVcknb0qqqqqqqqqqqqqqqqqqqqqqqqqqqv21tN1HVPhXpVvplhcXcq+IIWMcETSMF+z3AzgAnGSPzr9kPw9r+m/GW2udR0PULWEafcAyT2rooJUYGSMVVVVVVVVVVVVV/wUX0LW9b/4V9/Y2jX1/wCT/afmfZrZ5dmfsuM7QcZwfyNf8E8/D2v6N4y8Wy6xod/YpJpkIRrm2eIMRKeAWAzVVVVVVVVVVVVVVVVVVVVVVVVVVVV+17YX2pfBm5tdOsp7qY6hbkRwxl2wGOTgc1+y54b8RWHx28MXd9oOo28Ef2vdLNauirm0mAySMDkgfjVVVVVVVVVVVVX7cnhfxLq3x6u7vSvDup3kB0u1US29m8i5CnIyoIr/AIJ/aTqmj/BrWLXV9MurGZvE87rHcwtExX7NbDIDAHGQRn2qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/4LQ/Dx/Dv7Rfhv4hW1n5dn4v8NpHJLtb97d2krJJzjbxDJajAJI7gArn/gmv8O4fiP8AtmfDy0u0RrTQbqTW5t3UNaRtLCQO588Q/QZPaqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr4x/s3fBH9oKfQLn4xeALTxM/hiSaTSxc3E0awGYIJcrG6iQN5ceVcMPlHFfCz9kr9nX4I+N7v4i/Cj4Y2PhrX77TW024uLO6uPLe2aSORk8lpDECXijO4KG+XrgnNVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX/C2Phv8A9Dhp/wD32f8ACoZYriFJ4HDxyKGRh3BHBqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/4Wx8N/+hw0/wD77P8AhUMsVxCk8Dh45FDIw7gjg1VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVrOuaR4esTqWtX8VpbKwUyyHABPQVpXxC8E67qEWlaR4ks7q7n3eXDGxLNgEnHHoCaqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq8V+MvC3gbTo9W8W63baXZyzCCOackK0hUsFGO+FY/hXhn4ufDTxnqi6J4X8ZafqV8yM6wQsSxVep6dqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqrQP+QFpv/XpF/wCgCq/a8+MWpfAD9m3x38WtEWE6poenqunmaLzEW5nmjt4WZe4EkyEjpxzxS/8ABaX9om+XTLO+8E+BNPjF4janeafYXMkrW4kQlYI5bgqr7A6kuWB3DGwjNWl3aahaQ31hdRXNtcxrJDNC4dJEYZVlYcEEEEEcHNVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVWgf8AIC03/r0i/wDQBVVVVVVVVVVVVq2raVoOm3Os65qdpp1hZxmS4u7uZYYoUHVndiAoHqTivhj8fvgz8Z9C1rxN8MfiHpOu6V4cvJbTVLuJ2jjtpIxlmJkC5j2/MJRmNgCVYgE1VVVVVVVVVVVVVVVVVVVVVVVVVftC/wDJN5/+vuH+dfAz/kqmh/8Abx/6TyVVVVVVVVVVVVVVVX/BSD9uH4g/sh3vgDTfhfp3hzUtS8RLfz6hBrMEs0ccEXkrFhYnjbLNJJghx/qyCDnj9if/AIKY/FP9pf8Aads/hh478PeF9F0HWNJvP7MtdNjlWYXUSiYM8kjP5hEUc42gRjnPVcGqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq8IfF34aeP9Tk0bwZ4y0/Vr2GAzyQW7EssYZVLcjpllH4iqqqqqqqqqqqqqqqqqqqqqqqqv25P+SS6T/wBjFD/6T3Ffsbf8lstf+wdc/wDoIqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqtA/wCQFpv/AF6Rf+gCq/4LVfECPQ/gH4Q+HUFz5d14q8SG5dMKfMtrOFi45GR+9ntjkYPGOhIqv2F/Hz/E39kP4VeK5pZZZx4fi0+eWZmZ5ZbJmtJHYtkks9uzEnqTmqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqrQP+QFpv8A16Rf+gCqqqqqqqqqqq/aq/bz+Bn7KdpLpfiPV/7f8ZmMNb+F9MkDXPIUhp35W2QhlIL/ADMDlFfBx+1J+3J8c/2rNSuLXxfrn9keERceZZeF9NYpaQhfuGU/enkHUu/G4kqqAhRaeIdfsNF1Dw3Ya7qFvpOrSQyX9hFdOkF28O7ymljB2uU8x9pYHbvbGMmv+CYH7SP/AAv39m+x8P67qHn+LPh15Wj6pvbMk9uFP2O4PU/PEhjLE5Z4JGPUVVVVVVVVVVVVVVVVVVVVVVVVVX7Qv/JN5/8Ar7h/nXwM/wCSqaH/ANvH/pPJVVVVVVVVVVVVVVVf8FbviHH43/bD1XQrW98+28F6PZ6SuxlKCQobmQAqTkhrko2cEMhUj5Qa/ZU+IH/CrP2kvhr48dsW+l+JLT7Wc4P2aSQRT4+ZRnynfGSBnGeMiqqqqqqqqqqqqqqqqqqqqqqqqq+MF1c2Pwl8bXtjcy29xb+HNQkhmico8brbuQykcgggEEdK+A/wK1b9pS51241T4gXGnvoEdsolubVr55BKZSFGZF2geWx6n71ax+wv8RvCNm+p/DX4pLe6goy0AhfTHfGCoV1kcE5z97aBxz6fsu/tI+O7rx2vwa+K88t1dySTQWt3efJc29zFuLwzE/eztZRn5gwA5yMVVVVVVVVVVVVfsBf8lj1j/sWZ/wD0ptqqqqqqqqqqqqqqqqqqvjx+0n4c+BUtnp2oeHtR1bUtRtnmtY4XSKHCnbh5CSy846I3FaLqP9saNYat5PlfbbWOfy927ZvUNjPGcZxnFVVVVVX7cn/JJdJ/7GKH/wBJ7iv2Nv8Aktlr/wBg65/9BFVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVf8FnviNL4k/aN8P/AA7gnZrPwb4dRnjZjhbu7cySEAMQAYktf4VbIOcgKar/AIIq/EFtd+BHjH4c3EySTeFPES3UIBAaO3vIhtUqFHHmW853EkncRwFGaqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr4nfFT4dfBnwhdeO/ij4v0/w5odnw91eSEb32lhHGgBeWQhWxGgZmwcA1+1n/AMFd/HXj77T4J/ZqguvB2hiR0m8RT4OpXyDgeSvItUPJz80h+XDR4ZT4T8HePvi34wh8N+DdA1jxV4l1iZnW2tIXubidzlnkbGTgcsztwBlmIAJr9lD/AII96BoKWXjf9qS/TWNTVklh8J6fL/okBBzi5nU5nJ+XKJtQYILSBuP+Cv37K+lXPw28PfHn4e+HLSwPgqNNJ1e0sLZYY1053/cOEQAKIpXK4A6T+i1/wT5/aR/4Zp/aP0PX9Z1D7P4T8R40fxHvbCRW8rDZcHsPJlCSFsE7BIo++aqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr45fEOX4t/GXxt8TJZHZfEuvXl9CHBBjhklYxJzzhY9ijPYCpoZbeV7eeJ45YmKujqVZWHBBB6H2r9nL4gP8VfgH8PPiJPc+fda94bsbm8fDD/AEkwqJx8wBOJQ4z0OMjIOaqqqqqqqqqqqqqqqqqqqqqqqq+NX/JG/Hn/AGLOo/8ApNJX/BO3/moH/cM/9uqrxDFbeKf267ceD5UkiTxNZSO0JJBNvHE11yue8U2e3XOBnEjxxxtJI6qqAlmY4AA6k14z/bR8K6d4j/4RT4Z+ENR8e34IUNYSFIpGJwVjIR2fHHIXBJ4Jqy/bb17wzqUOmfGH4Nax4eE5JSZBJG4X5ekMyKWAySSH7gY714V8V+HPHGg2nifwpq0GpaZepuguIsgHHBBBwVYHgqQCDwQK0L9rJr743/8ACl9f8AppMg1OfTzqP9rmZS6BvLIj8lSRIyoBkjG8E9Kr4t/tjaP8LfiXL8PR4Q/tSOz8gX1//aJhFu0gDMBGIn3bUZTweSSOCKR45Y1kidXVhlWU5BB7iv2h/wBpG0+As2iWa+Fv7eudYSZ2iF/9l8hIygDE+W+dxYgdPumr74kWfh74WxfE/wAX6XcWECabDe3dnak3bwGQL8gKgb8FwC2Ao5JIAzWoft2eItYml/4V/wDBy7vbWGTBuLid5WIx0KRJhDnP8bcV8Ov26fDuta1B4b+JPhaXwzcSTGBr5J/MgifdgearBWiA6E/Ng8nAzivF37SVv4S+OmkfBe58KLImqvbRjVG1HZ5bTcKPK8s5+bA+8M57V8U/27dH8M+ILjw98PfDKa4tlKY59RurgxRO65DCJFBLLnHzkjODgEYY2viOxTwlb+LNburfT7U6el3cyyybY4FKBmJY9AM9TXi39vK2k1k6L8LfANxrgEhEdzdSOhnUKSdkKKW98kg4HK88fDb9tXRdb1+18JfFDwnceEdQu5FjhundmtyzHC+YGUNEDkDcdy9yVGcfsBf8lj1j/sWZ/wD0ptqqv+HiX/VH/wDy4f8A7nr/AIeJf9Uf/wDLh/8Auevgb+1y/wAafHaeCk+HR0kG0luGuhq32jYExwV8lepIGc149+IXhD4Z+H5fEvjPWoLCzj4QMcyTvjhI06ux9B2yTgAkap+3drOr3c0Xw3+EV7qFtbt8091MzvtOdu6OJSEJx/fbv6Zr4fftz+ENd1WLw98Q/DNz4VvGlEDXJm863STgHzMqrRDdnqGC45PUhHjljWSJ1dWGVZTkEHuK+L37TEHwl+KOgfDm58GvqUWtQW00l8moCJoFlneI4iMZ3lRHu++uc44618Wv24vCfgvWLjw54F0RfE9zakxzXpuvJtkkBwQhCsZcY5I2j0Jr4d/t8aLq+rRaV8RfCq6LbzybV1GznaaOHOMeZGV3YznLKT1Hy8E1BPBdQR3NtMksUqB45EYMrqRkEEdQR3rx98RPB/wx0B/EnjTWYrC0UlIwRued8EhI1HLMQDx+JwOam/bP8ceJ4Jrr4XfAPXdXsrdv3l64lnVVwchlhjIU8dd56HjvXwp/bI8C/EDXY/CXifR7nwjrc0phhiu5vNhkl3YEXmbVKufR1UZ4znANVrmuaN4Z0i617xBqdvp+n2Ue+e5uHCJGvTkn1JAA6kkAcmtV/bWn1zU7vR/g58JNc8Xtb5AuUWQcYGH8mON32khupU8A9yB4R/bf0ca6nhj4ueBNR8HXYJSW4bfIkTj/AJ6RMiyRjII4D4OM8ZItbq1vrWG9srmK4t7iNZIZYnDpIjDIZSOCCCCCK1v9qLT/AA/8e3+CmqeFVhtYpI1l1ttSCrGHtVn3NEUACgsFJL8AFv8AZr4h/t86FpGqz6V8PPCf9twQNtGpXdyYI5COuyMKWK+hJX6V8IP23vDPjjWrXwv450NfDl7eSCK2vI7gy20kjNhVbIBizkDJ3DPJK1418Rf8If4N17xb9j+1/wBiaZc332fzPL87yYmfZuwduduM4OM9DX7Qnx4/4XzrOk6t/wAIr/Yf9l2rweX9u+0+ZubdnPlpj0xg1ov/AAUB/sfRrDSP+FS+d9htY4PM/t/bv2KFzj7OcZx0r44fFX/hTPgOXxt/YP8AbHlXMUH2X7V9nzvOM79j9MdMV4l/bG8G+Gvhl4d8bzaJLdax4mtnmtdFgus+WEkaNzJMUG1QyMudhJYYAwCR8J/2lPFHxH+GPjHx6vgS1nvfC/zRaba3jIbpQm98OVYhgoYgBTkgDvX7Pf7TemfHi/1fR38NroGoaZDHPFbm/wDtJuYiSruD5aY2NsB6/wCsFV8Wv2sT8OvihH8K/D/w8l8Takwt4yV1MWxNxPgpEq+U+cq0ZySOXxjjJQuY1Miqr4+YKcgHvg8Z/Kqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv2z/iCnxS/ar+KHjOGZ5be48RXFraSOxJe3tiLaFsFVIBjhQhSMqCAc4zXx/8A2eLfwF+wb+z78WYbPyr/AFzUNTbV5UjO2Y3pEtnk/Lgi3tMcqxbJ+faiCv8AgjF8QP8AhHf2kte8B3Df6P4v8Ny+UAeTc2siyJ1YDHlG47Mc7cYG41VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX7Wf/BVz4UfBYXPhD4LLp/xE8XCN1a6hud2l6fJ0HmSIczkHkxxMBjIMimvjV8efit+0N4uk8a/Fnxfd61f/ADLbROdkFlGTnyoIh8saewGTjJJPNfsl/wDBL74zftBtpvjDx/FceAfAc7LKbq9tyt/qEOAwNrAwGFcEYmkwuG3KJANp+Af7Nfwe/Zp8Jr4T+E/hSGwR+bzUJ8S3l8/96aYjLeyjCr/CoqvGXhLQPH3hLWPBHimwS90fXrGaxvrd+kkMqFHHscE4PY4NftBfBrX/ANn74x+KfhH4iDvP4fvmjguGTaLq2YB4JgPR42RsdiSO1f8ABMD9pH/hfv7N9j4f13UPP8WfDrytH1Te2ZJ7cKfsdwep+eJDGWJyzwSMeoqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/bK+IEfwv/AGVvij4z+0/Z57fw3dW1pLhTsublfs8BwwIP72aPggg9O9fDHwXP8SPiT4T+HdpN5U/ijXLLSonyBsa4nSIHnjguOtf8FFfhfp/wl/a/8eeHdE05LPSL+aDU7CNCMbLmBJJMAEkATGZecE7c9CDX/BH34hw+Lv2SIvBzSILnwPr15Y+WBhvJnYXaOfYvcSrz/cPtVVVVVVVVVVVVVVVVVVVVVVVVXxq/5I348/7FnUf/AEmkr9nb4lfGP4d/8JB/wqTwB/wk39ofZv7Q/wCJVc3v2fZ5vl/6hl2bt8n3s528dDXxM/ak/abOnpoXiTQv+EDF6hU3EOkXNlM6NwSrzMzLjB+aPDDnBz0/ZB+EHw+8P+Gl+Jmj+JIfFOtashjkvhGUFj0MkIRvmD5+8zYJGMAA/N+218RdS8FfCuHQtEu5ba88T3ZtZJY32stsq7pQCDn5vkU9trMDjIz+yxc/A74XfDPT7u9+Ivg6HxJrkK3WpyXGr2qTxbwCkBBbcoRduUP8e48ZwPiX4q/Z++J/gvUvB2v/ABP8DzRXkLCCR9ctt1tNtOyVDv4ZSc59Mg5BIP8AwT88a3x1HxL8O7m9L2v2ZdTtIGH+qZWEcrA4/i3xZBP8IIH3q/bQ0W++H/xz8PfFHS0UHUY4LqNs4JurNkB/DYIOa/4SLSf+Ea/4S37R/wAS37D9u83H/LHZv3f9880/hPVPiv4A+J/xz1CKU3dnrNtcRqrFziWR/OQHH3UWWE9gFXPAGD+y/wCNW8d/BDw1qNxcebe2EB0+7yxZg8B2KWJ6s0Yjc9fv9TXxXtx8d/2ybTwXEy3Om6ZcQ6dMY9oxb24Mt0CemQxnXnPOB7Vqml6drWnXOkavZQXlleRNFcW86B0lRhgqwPUEVr3xG+Ffw1gi0jXfFvh/QRboqRWJuI4njXHG2FfmC4I6DHIr9sP4g/A34m2Ok698P/ENvfeJbO7NveeVps8LTWxQ/M0joobYyKAMk4kOOM1+zF4hvPFHwF8HapfsTMlk1qSzZJW3leBSTgdViB9fUnqf2z0vpP2hzHpZl+2NZWQt/KYq/mfw7SOhzjFfCf8AZ9+Hvwx8KWWkDw5pmoar5Cf2jqNxbLK9xNjLkFhlU3Z2qMYGM5OSf2+/Hdzpuj+Hfhlpj+TBf7r29VcKGSMhYUwD93dvJGMZRcdDj9nH4OeH/hT8PdMe305Br2q2cVxqt5In71pHUMYgeoRCdoUYBxkjJNfGL4UeH/i/4KvvDOrWVqb0wSf2Zeyx5eynIBV1b7wXcq7gPvKMV+wF/wAlj1j/ALFmf/0ptqqq/bC/aBvoZz8Evhzc3Emp3hWPV7izJZ/nyBZoACS7ZUtt7ELzuYD9mf4D2vwW8GhtUiil8UauFk1OcEN5Ix8tuhHG1eckfeYk5IC4/aC1DWPjp+0/Z/Ci11WWPSrC9i02AJllhO0NczbM4Lj5x2yI1GRXhDwd4a8B+H7Xwv4T0mDT9Os02xxRryxxyzHqzHHLHknrX7ZXwc8P+Lfh1qPxDs9OSLxH4eiWb7TEm1rm3DASJJj7wVSWBPI24GAxr9h74g3Pi74VTeF9RufNu/ClyLeMlgW+yuN0Wec8ESKOMYQAZwcft9O8Xxm0WSJyrp4agKspwQRc3PIr4L/B3w18G/B1poGkWkL6i8StqeoBcvdz4G45PIQHhV6AY7kk/t3fDHw/L4Os/ibp2n2trq9pfJb3ssabGuoZAQN2B8zKwXBPYtzwBX7IniW78TfATw6187vNpnm2G9znKRORGB7CMov/AAGvjv4isfi/+1Fa/D/xV4oi0fwjoV2LOSW5u0giiVE33D7mbYsjsGjDHnhBjPFaT8U/gPoOm22jaJ8SvAljY2kYjgtrfWrWNI1HZVD4FfttTfCfxHZ6J4/8DeMPDl/4ijvBaXq6VqUM000RjZkkdY2J/dmMKH/2wCThcfAnxldfED4QeFvFmoXX2m8vLAJdTFdpkmiYxSMRgcl0YnAxnpxiq/by8e6nca54f+Emnah9mtJ4Fvr/AHTCOOVnkKRLIc8KnlsxBGPmU9hj4a+JP2evhb4QsPCHhz4o+CVitIgJrj+2rRZLqT+KRyH5YnP0GAOBX7WOrfBH4kfCm/urHx94RvvEWiAT6U1tq9vJOfnUSRKFYswZM/KP4gp7V+w/41vvFXwffRtTvTPP4bv2s4QwOVtiiPGM45wTIoGSQFHbFftI6Dc+K/2udU8K2V4trPrV/plgkzA4jM1rbx5IHJHzcjuK8A/D7wp8M/DVr4V8IaYlpZ2yAMxAMk795JG6sxPf8AAAAP28Phj4f8OanoXj7w/p9rYvrDS22oRQpsEsqgMku0DG4gsGPGcL1yTXwa8S3fjH4UeE/El+7vd3ukwNcO5yXlChXbPuyk/jX/BQj/kcfCf/AGDJv/RgrwX/AMiboP8A2DLf/wBFLX7b/wDyQe7/AOwna/8AoRr9iz4HeHvF+kXXxK8dabFq8VpcGy0ayu18yKLZ88kpQ5DfM+FHQEOcEkEWWiaNpsEltpuk2VrFN/rI4LdY1fjHIA5445rTCf2Y/wBrL7G9yttof2/yXklACjTrnGC2O0YZSSMcxdMcVqN/Z6Vp9zqmoXCw2tnC808rA4REBLMcc8AE1+zDo138av2kNX+KOs2btaaZPNqr9kjnkci3iPU/KNzLz/yx5J71VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVWgf8gLTf8Ar0i/9AFfG7x9H8Kvg543+JTSxK/hjw/e6hCJWVRJLFCzRoN3BLOFUDuWA71pOlX+u6rZaHpUHn3uoXEdvbRbgu+R2CquWIAySBkkCv2//glZN/wT21bwJo+x/wDhXGk6bcaeVi2J5dj5cbnbhyo+z+bgA8HGW25r9jX4gP8AC/8Aao+F3jMXPkQ2/iS2truXDHZbXLfZ5zhQSf3U0nABJ6d6qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqtA/5AWm/wDXpF/6AKqqqqqqqv2iv2tPgd+y7of9p/FPxYkeozQ+bZaDY7Z9Rvl3FcxQlhhchhvcqmQRuzxX7Wv/AAUg+Nf7T32zwrp8v/CF+Apsp/YWnXDNJepx/wAfc/Bl5BOxQiAEAqxG4/AL9mX4zftM+Jv+Ea+E3hC41BYGUX2pzKYrHTwQSDPOQVQkAkLy7YO1Tg1+yX/wS++DH7Piab4w8fxW/j7x7Aqym6vbcNYafMCGBtYGByyEDE0mWyu5RHnaKqq/4LJ/s2/8JN4I0f8AaS8M6fu1Hwtt03xB5acyWMj/ALmU4/55zOV+k/PC1/wTc/aR/wCGdv2j9K/tvUPI8J+NNmj63vbCQ72/cXB7Dy5SMseiPJ61VVVVVVVVVVVVVVVVVVVVVVVV+0L/AMk3n/6+4f518DP+SqaH/wBvH/pPJVVVVVVVVVVVVVVX/BZ34gf8I7+zZoPgO3b/AEjxf4kj80E8G2tY2kfGGBz5pt+oYYznB2mv+CVnw/fx3+2f4TvXtvOs/CVneazdLlhtCQmGJsqRjE88B54OMEEGv+C3vgB7Tx/8NPilFHK6aro91o07hGKRG1mE0YJ6At9rlwOp8tvSv+CI/wAQ3sPiN8RfhXcXv7rWtHt9WtoXZsCS1lMTlOdoLLdLuGMsI15ASqqqqqqqqqqqqqqqqqqqqqqqqvjV/wAkb8ef9izqP/pNJX/BO3/moH/cM/8AbqvG/g3Q/iB4U1Lwh4htUnstSgaJtyBjGxHyyLnoynDA9iBX7FHiPWPBHxi134T6tMqR38c8cluxK7by1Y5KhsH7glBGAThSfu1/wUM0e5n0LwVryA/Z7O7vLaQ7DgPKkTLz0HED8d/wNfCH9lL4E/EL4YeGfGM1pq0tzqWnRtdMmoEL9oUbJsAZAHmK4x26HB4r/hiD4D/9A7Wf/Bi3+FfDD9nP4afCHX5/Evgy11CO9ubNrSQ3F2ZV8tnRzgHvmNea/bf8EJ4m+DT+IoId154XvI7lSqksYZCIpFAHb5kck9BHnPrqHxmkX9hiytku86pcy/8ACMsUfBREJJBAJODaqqnOOX9CM/AL4S6fpn7N9l4J1u12t4p06W41NTHsbN0nGR1DLEY19QU7dB+xp40k+HM3xL8BeJ5XX/hHYJdVeHDKsZtSYro8gYJxCOQD8vTjj9hXwzfeJfFfjD4x68DPcyyNaxTyIW8y4mbzrhgx6MB5fviQ+vP7YHxk1b4U+ALXT/C92bXXPEkzwQXK8NbwooMsiHs/zRqD23kgggV8Av2PfB3iLwfp3xA+K73etX2vwi9is1u3jiSGVdyM7oQ7yMGDk7gASAQcEn9s74Q/DTwB8LtL1nwZ4N0/Sb2bX4beSa3QhmjME7FeSeMop/AV+x7/AMm5eEf+33/0tnr9qv8A5Oq0v/uG/wDoYqv+CgOm3cXxR8P6u8eLa50BYI29XjnlZx+AlT860XVrLxBo1hrumyb7TUbWO5gbIOUkUMpyCR0I6Eirq5t7K1lvLqVY4YI2kkduiqoySfwFfsBf8lj1j/sWZ/8A0ptqqv2nPj5bfBfwj9l0aeCTxVqyFdOhYB/s65w07qT90chcggtjggNX7GnwJuVI+OXjtJZ9Svy8mjrcMXfa4O+6Y7iSz7mA3DOMtzuBFeFnXwP+3dMdfBj+0+J74R4UjP21JRB1x1+0R/nxnjNftD67Y+Hvgh41vL+RVS40a4s4wWxuknQxIB/wJxxX/BPTRbuDw/4z8ROP9Gvby0tY+P44UkZ/0nSv2/f+Sx6P/wBizB/6U3NV+2//AMkHu/8AsJ2v/oRr9iD/AJIPaf8AYTuv/QhWp+BvDup/ti6r4K+IC3H9l6z4luvMWJ/Icm5DyW+CcdXki+oPGciv+GIPgP8A9A7Wf/Bi3+Ff8MQfAf8A6B2s/wDgxb/CvAfgfQfhv4TsPBfhiOZNN07zPIWaQyON8jSNlu/zO1V+27pMWnfHjTtU1eKaTTdR0y1lk8pcEokjpIik4BbC568b19ag/Ym+AF1BHc21pq0sUqB45E1MsrqRkEEDkEd6/wCGIPgP/wBA7Wf/AAYt/hXwt+Dvgr4O6ffaZ4Kgu4odQmWacXFwZSWUYGCenFfFn/k/Cx/7GbQf/RdpVf8ABQj/AJE3wn/2E5v/AEVX7NX/ACQfwV/2DF/9Cav+ChH/ACOPhP8A7Bk3/owV4L/5E3Qf+wZb/wDopa/bf/5IPd/9hO1/9CNfsQf8kHtP+wndf+hCq/b6+HR1Dw/ovxOsIFMulSfYL9hgHyZCTEx9lk3Dr1l6dTXjr49f2v8AsYWGoLff8TzXETQLnEmH3x8TuQAMh4UyccDzwPav2M/h6PBXwbtNXu7fZqHiiX+0JGZcMISAsK9B8uwbx1/1h9aqv25P+SS6T/2MUP8A6T3Ffsbf8lstf+wdc/8AoIqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqtA/5AWm/9ekX/AKAK/wCCtfxETwR+xzrOgw3TxXfjXVrLSITHIVcKJPtMvQ52mO2ZGzwRJg9a/wCCefgB/iN+2V8LtJMcph0vWBrM0iIxEQsUa6QsR90GSGNMnjLgd68ceE9N8e+Ctf8AAusjNh4i0u5066G3P7qeJo347/KxrV9Kv9C1W90PVYPIvdPuJLe5i3BvLkRirLlSQcEEZBIr4G/EOH4t/BrwR8TIpEZvEug2l9MqjAjmkiUypxxlX3Ke2RVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVaB/yAtN/69Iv/AEAVVVVVVV4q8V+GPA3h698WeM/EOn6Joumx+Zd3+oXCwQwLkAFnYgDJIA9SQBya/a4/4LBj/TvAX7Kdp/fhuPGOpW/+8CbKBv8AgJEsw/vDyuj1quq+PPiv4zl1TV77W/FvirXrgb5ZWkvby9lICqB953OAFAHQAAcCv2UP+CPeva81l43/AGpL99H01lSWHwnp8v8Apc4Izi5nU4gA+XKJuc5ILRkc+CPAfgv4aeGbPwZ8P/C+meHtD09SttYafbLBEmTlm2qOWY5LMcliSSSSTVVVV498FaB8SfBGv/D3xTbtPo/iTTbjTr6NG2sYZoyj7W/hbDEhuxwe1fGr4T+JPgZ8VvFHwk8WJ/xMvDOoPavKEKLcR8NFOoPISWJo5FzztcZr/gm5+0j/AMNFfs36V/beofaPFngvZo+t72zJLsX9xcHufMiAyx6uknpVVVVVVVVVVVVVVVVVVVVVVVV+0L/yTef/AK+4f518DP8Akqmh/wDbx/6TyVVVVVVVVVVVVVVV/wAFq/iCmu/Hfwb8ObaZ5IfCnh1rqUFiAlxeSncoUqOfLt4CWBIO4Dgqc/8ABEH4cwtL8Tfi5dQIZUW00OwkC/MoO6e5BJXgHbanCt2O4cKa/wCCvXw8j8Zfsh3fieKz8y68E65Z6ksiKu9YpGNrIM4J2H7QjEAjlFYnC4r/AIJyfEb/AIVp+2V8N9Qmlxaa5qDaHcJtz5n21GgiGQrEYneFuMfdwSFJNVVVVVVVVVVVVVVVVVVVVVVVVfGr/kjfjz/sWdR/9JpK/wCCdv8AzUD/ALhn/t1VfFQf8KR/bHg8Vsxg0+61SDVHdVZSbe4+W5I5GeTOOuD37ivi78NdJ+MXw91DwbeXKRfbEWWyuwnmfZ5l5SQDPI7HB5ViM814O+I/xt/Y91C68H+LvCB1DQJbjfHFM7LC7kH57a4AKgMACVIOMcqrbquv+ChPhZLd2svhvqss4HyJLfRxqT7sFYj8jXwG+J3iH4teDJ/FHiPwlN4fm+3PFbQPFIFlhCqVkVnA3gkspIGMqa8V+HrPxb4Y1bwtqAU22rWU1rLuUnCyIVJwCDxnPBB9xXw88L+JfF3j3QfgHqqmOzg8SzSX1qdylHVVW5J75EVuwHTnOT6JHHHGsUSKqoMKqjAAHYCv2t9F1H4YfG3V9Z0MJDZ+NdJd5FAJB81TFOueoYsnmcHHzjsStfsu+CH8B/BHw5p1zGUvNQhOo3QZNhDz/OoI6gqmxTnuvboP20PhD4i+JngjS9a8Jaa+o6p4anlc2cSs0ssEqr5nlqPvMDHGduMkA454b4Q/ttN4B8FWPgjxp4PudRl0SJbW0ubWcRsYUG1UkRhwyABcg8jGQCMn476v8YvjV4Om+Lfi7w7L4Z8I6NJFDpGmsGP2h5Xw0pLYZuCo8zaqn5QoyXNfscXNvcfs7eF4YZVd7aS8jlUfwMbuVsH/AICyn8a/ar/5Oq0v/uG/+hiq/ae+CMnxq8Bx2ujCBPEOiytcaY8p2iXIxJAWP3Q4CnPTciZIGTXwp/aj8cfACwX4XfEzwPd3VrphZbVZGNtdWqlmJX5gRLHu4U8YGcMwCga98cfil+1JYyfDf4QeBbvRtI1A+RrOtXUxZY4WA3Rs6qFjBXOVyzOpwAOc/sR6X/Yf7QfivRNzn+z9FvLfMi7WOy8t15HY8dKrxd4hg8I+E9a8V3Nu9xFounXF7JChAaRYo2cqCehIXFat8Vm8a/FP/hZHxN0R/EMBnEjaTHem3i8teUgVmWQrED1XGWy3ILE0n/BQ6OONYovg4qqgwqr4gwAB2A+zV8Hv2zf+Fs/EbSPh/wD8K3/sr+1fP/0v+2fP8vy4Hl+55K5z5eOoxnPbFftffs5a746uYfif8PbU3GtWcKxX1hCuJbpFPySxkdZFHBXqVAxyoB8Hft0634W0tfDfxX8B31zrmnhY5LmJxbyS8dZYnUbHxgkjg56L38aeOPjN+2Xqdj4U8GeDX0nwzaziR2kdmgSQAr5txcbAOAWwijPJ4cgY+Evwz0b4SeBNO8FaMxlFsDJc3DgbriduXc49+AOyhRzjNft+/wDJY9H/AOxZg/8ASm5qv23/APkg93/2E7X/ANCNfsQf8kHtP+wndf8AoQr9rj9nPxD45voPil8N4DLrthCqXtlCdktysfKSxNkZkUcberALt5AB8G/t0+J/CtqPDfxY8B3F3qVjGI3uoX+yzswAx5sLrgMRyWBUdPl54n/byvNdvLbSPAHwj1DUL2dgAklwZpOuMJFEhLnn+8OaR1kjWRQcMMgMpU/iDyPoar9qH4ESfGvwdAdDeCLxHojPLYGX5RcKw+eAt/DuwpBPAKjOASR8O/2o/ib8AreL4Z/FbwPd3trpQ8i1E7G2ureNcAKrEFZowMBcY4IwxXaBqH/BQPTJ4FtvDHww1C61CdvLhjuL5VXcQQuAiMzHdtG0YznqK+HXiPVPF/gXRPEmuaNNpOo39mkl3ZTQvE0MvRwFf5guQSM9iOvWviz/AMn4WP8A2M2g/wDou0qv+ChH/Im+E/8AsJzf+iq/Zq/5IP4K/wCwYv8A6E1ft3fDDxB4m0fQ/Hnh3TZr1dDSaDUkhBd0icoUkCjkqrBgxGcbgTwCR8Kv24LO08PeHPBWv+DLu71aCODT/tcN0qxzYxGkjAgkEjG7GeckdcD9t/8A5IPd/wDYTtf/AEI1+xB/yQe0/wCwndf+hCq8f+D9P+IHgrWvBmpD9xq1m8G7ODG5HyOPdWCsPcd68CeDPFPjrxxovwOnlmjt4ddnNzHEMm2JCJcy88ErHbjGe6471ZWVpp1lb6dYW6QW1rEsUMSDCxooAVQOwAAFVX7cn/JJdJ/7GKH/ANJ7iv2Nv+S2Wv8A2Drn/wBBFVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVfEn4q/Df4PeG5fF3xP8a6T4b0mLI+0X9wI/MYDOyNfvSP6IgLHsK8d/8ABZr9mLw7d3Fh4N8M+NfFrwybY7uGyisrWdckFlaaQSjgAgNEOvOK8If8Fpf2bdYkW28XeBfHfh6R5GAmS2t7yBEC5BZllWQEtlcCNgODnBOPhR8avhR8dPDa+LPhJ480nxNpvyiV7Ob95bMwyEmibEkLkDOyRVbHOKqqqqqv2g/jl4Q/Zx+Emu/F3xqJZrHRo1EVpAwE15PI4SKGPPdmYZP8KhmPCmvg5/wWj03xX8TbLw78WPhbY+FvCuq3XkJq1pqj3D6YGbCPOGRRIgyodl2EDLBT92qqqqqqqqqr49f8FSv2sfh78c/iL4A8N6x4ZTSfDXizVNMsFm0VHdYLe7kijDNn5jtRcnua/wCCbX7RvxM/ag+BeuePvird6fPq2n+LLjTIWsrQW6CBLS0lUFQTk7pn5+npVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVaB/yAtN/69Iv/AEAV/wAFvviC0/iT4ZfCm3mQLZWN3rV3GCCzmZ1hhJG3KgeRcDhsNuOR8oNf8ETPh22rfF3x78Ubi2R4PDmgxabA0kYO2e7mD7kJGQwjtXUkdpSDw1V/wUP+Hcvw2/bI+JullH8jWNWOt28jZxIL1RcOR7CSSRPqh7V/wSS+Ib+N/wBjzSdCub3z7nwXrF7pLb2YuIy4uYwSxOQFuQi4wAqBQPlJqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqtA/5AWm/wDXpF/6AKqqqqqv2tf+CkHwT/ZiF54V0+X/AITXx7DlP7C064VY7KTj/j7n5EXBJ2KHfIAKqDuH7R/7Wvxr/ak8QvqvxL8UynSorgzaf4ftGaKwsDgqDHFk7nCkjzH3OcnnBxX7Kf8AwT9+Of7VN1b6vpenf8Iv4J8wC48T6pCfLYAjcLaHIe5fBJGNseVIaRSRn9mP9i74Hfsq6OsfgLw8l74jmh8u+8S6jGst9cZA3qr4/cxEqD5aYBwN24jNVVVVVX/BZv8AZt/tPQdD/ae8M6fm40fZo/ify0627v8A6LcNjj5ZWaFmOSfOhHRK/wCCbn7SP/DO37R+lf23qHkeE/GmzR9b3thId7fuLg9h5cpGWPRHk9aqqqqqqqqqqqqqqqqqqqqqqqv2hf8Akm8//X3D/OvgZ/yVTQ/+3j/0nkqqqqqqqqqqqqqqr9ujx9H8Tf2vPip4rhlilhHiCXT4JYmVklislW0jdSuQQyW6sCOuc1/wSp+HreA/2MvC97cQpFdeLb681qdFUZw8nkxEkMQxMMETdiAQpAKnP7QXw5/4W98DfHnwyjj33HiPw/eWdpzjbcNE3kt95QdsoRsFgDjBOCa8Pa7qXhbX9M8TaNP5OoaReRXlpJj7ksTh0P4Morwn4m0rxr4V0bxloM3m6br2nwX9nJuVt8M0ayIcqSpyrDkEj0J61VVVVVVVVVVVVVVVVVVVVVVVVVVU8EF1BJbXMKSxSoUkjkUMrqRggg9QRxirHw34d0ycXWmaBp1pOAQJILVI2APUZAzVVVVVXGg6Hd3yandaNYzXkZUpcSWytIpX7pDEZGO3pVVVVqOkaTq8aRatplpepGcotxAsgU+oDA4q2tbWyt0tLK3ighiGEiiQIqj0AHAqqqqqtR0TRtY8v+1tJsr7yc+X9pt1k2ZxnG4HGcD8hUEEFtBHbW0KRRRIEjjRQqooGAAB0AHGKqqqq1HRNG1jy/7W0myvvJz5f2m3WTZnGcbgcZwPyFWOnafpluLTTLG3tIQSRFBEI1BPU4HFVVX2nafqlubTUrG3u4SQTFPEJFyOhwcitO0TRtH8z+yNJsrLzceZ9nt1i34zjO0DOMn86qqqqi8PaBb351W30PT470szm5S1RZCzZ3HcBnJycnvk1VV+0h+0Vofwd8P3OkaTfQXPjC9gYWVopDG13DAmlGCABncqt98jHTJH7EPwdvtLsLv4zeKoZTqGuRtFpYuAS4gLZknOTnMjAAE4O0E5Ikqqr9uT/kkuk/8AYxQ/+k9xX7G3/JbLX/sHXP8A6CKqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv2hPjh4T/Zz+EPiH4u+MG8y00S3zb2avse+uXO2G3Q4JBdyoLYIVdzH5VJr9oT9oj4m/tM/EK8+IXxL1p7iaRmWw0+JmFrpkGeIYIyTtUADJ+8x+ZiSSa/Zy/wCCX37Rf7QnhjT/AB7J/ZPgvwxqkfnWV3rhlW4vISuY5obdFJMb5BVnKBl+ZdwK7v2kv+CYn7Qv7Ovha/8AiA8uj+MfC2mKr3l5orSefaR4+aWWB1BCKerIXAXDNtG7b8GPjP8AEL4A/ELTPib8MtcfTtZ01sEMC0N3CSN8EyZG+JwBlfYEEMqsP2Wf2h/Df7UXwW0T4seH4obSe63W2q6ak/mtpt7HjzYGOAehV1yASkiNgbqqqqqr9rL9nuw/ag+BfiD4RXWq/wBl3d/5Vxpt+VLLbXULh42ZQRuQ4KMOcK5I+YAj4Qf8EZ/jZefEWzX4z+IfC+m+DdOvka/fTL2W5uNTgViWS3XYnlhwu3fIVZBIGCOQVqv2vP28f2s5Pjr8Sfh7pPxn1jQfD/hzxVquladaaIsentFbwXLRJmaJVmZtsSnczkglsYBxX/BHHxDr/in9mnxbrPibXdQ1fUJviBeeZd3909xK/wDoFh1dySfzqv27P25v2qdC/aI+IHwr8K/F7U/D3hrQdSksrK20iKK0lSMxgEmdFExb5jzv4OCMHmv+CNHizxV41+H/AMTtd8ZeJtV17UpfEFt5l5qd7JdTNmFmOXkJY5ZmPXqxPc1VVVVVftZf8nUfGX/soGu/+l81f8EVP+TV/FP/AGUC8/8ASCwqqqqqqqqqqqqqqqqqqqqqq8Yftb/sv+A5GtvFXx/8CWlzHIsclqmuQzzxll3KWijZnUFcHcQByOeRmX/gpf8AsOQ6uNDf49WJuTMsIddH1BoNxIAPnC3Me3nlt20dSRg14S/bG/ZV8c3p03w1+0D4Hnu9yqlvPrEVtJKzZwI1lKmQ/Kchc44zjIqGaG4hSe3lSWORQyOjZDAjIII6giqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq0D/AJAWm/8AXpF/6AK/4Ka/EOP4iftn+PZbS9+0WPh2SDRLX5lPlm2hVZ0+UkcXBuOOCM4IByK/4I5fD5PC37Klx4zmif7T408RXV0kjKVBt4AtsigbiCBJFOd2FOWIOdoNV/wW0+HkelfFjwB8T7Wz2L4i0OfTbiRFUK8tpLvDNgZ3lLpVyxOVjAGNhr/giH8Rvs/ir4lfCO4l3fb9Ptdcs0242eRIYJzkLzu+0W/VhjZwDliKqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq0D/kBab/ANekX/oAqqqqr4y/Hb4Tfs/eFf8AhMfi540sdA092KW4mJea6kAyUhiQF5Gx2UHHU4HNftZ/8FXPit8ahc+EPgsuofDzwj5jq11Dc7dV1CPoPMkQ4twRyY4mJycGRhXws+EHxM+N3iu38EfCrwbqHiPWbjkQWqALEv8AfkkYhIk/2nZV96/ZQ/4JGfDj4bJZeMv2iZrHx14lRkmj0WNGOlWTA5CuGwbs8DIdVj5KlHHzGGGG3hS3t4kjjiUKiIu0KAMAADoMdqqqqqqq+IPgTw38T/A2vfDrxfZfa9F8R6fNYXsQOG8uRCpKnna4zlW6qwBHIr41fCfxJ8DPit4o+EnixP8AiZeGdQe1eUIUW4j4aKdQeQksTRyLnna4zX/BPT9oyH9o/wDZq0DWNRu0k8UeF1XRfECZwzTwoojnx6TRGNyQAu8yKPuVVVVVVVVVVVVVVVVVVVVVVVX7Qv8AyTef/r7h/nXwM/5Kpof/AG8f+k8lVVVVVVVVVVVVVXxY8cW/wy+F3i74i3LxLH4Y0O81I+apZWMELSAEAgnJUDAIJzgHJr/ia6/qv/L3qWp6jcf7U01zNI34s7sx9ySa+GPgu3+G/wANfCfw7tJvNh8LaHZaVE+Sdy28CRA888hB1qv2tvh4/wAKv2mviZ4EFn9lt9P8SXb2MO1httJnM1t1A/5YyRnOMHOQSME/8E1/iHL8R/2Mvh5d3cjtd6DayaJNuBwBaSNFCAe48gQ/Q5HaqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqkRZI2jYnDAg7WKn8CORXw9/YV0/TvFk/iL4l+I01yzivJJLbT4t5+0ruJRriQ7SWOQWVRgn+IjIMEEFtBHbW0KRRRIEjjRQqooGAAB0AHGKqq/bk/5JLpP/YxQ/8ApPcV+xt/yWy1/wCwdc/+giqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr/gtz8U9Sm8VfD34I2t5LHYWmny+Ib23H3J5ZZHt7dz7osNyB/wBdmz1FfsF/BDR/2gf2pPBngTxNEkug280mp6tA4BFxb2yGTySD1WR1SNschXYjkVDDDbwpb28SRxxKFREXaFAGAAB0GO1TQw3EL288SSxyKVdHXIYEYIIPUEdq/b0+CGj/ALP37UnjLwJ4ZiSLQZ5k1PSYFAUW1vcoJPJAHRY3Lxrnkqik8mv+CJXxMvbD4l+Pvg/M6Gx1nRY9ag3tgxz2sqwsq88l0ugTweIR0wc/8FePj98ZfgZ/wqf/AIVH8Q9V8L/23/bX9ofYXVftHk/YfL3ZB+75smP941+zJ/wVF+KPwxtPHet/Gjxlrvj/AFGTSYY/C2k30oWA3hmAdpJFXKKIyWPGW27RgnI+JP7f/wC178TtZl1fUPjl4l0KNpC0Vj4bvpNJggHQKogZWYD/AG2Y9ySa/Y4/4KofFnwL470vwl+0T4tuPFfgfU5ktp9TvUDXmjliALgyKN00S9XRtzYyynI2t+1n/wAFaPiz478U33hf9nDWW8IeDLKbZBqyWo/tHVNp5kLSA+RESAVRVV8ffb5jGvg79vH9sLwPrLa5pX7QnjC9lkkDvBrN+dUgb7wIEVz5iKCGPCgdiMFVI/YL/bFsP2v/AIWXOqalYw6b408LSQ2viKxgB8pmkUmK5iznEcuyTCElkaNwcjazf8FMvif4/wDhD+y1f+Mvhp4qvfD2txa1Ywpe2bBZBG7kMvIPBAr4b/t2/tfav8RPC2lal8fvFFxaXmtWcM8LzJiSN5kVlPy9CCRX/BWn43fFj4IfDrwFqvwm8d6n4Yu9S1q4hu5bFwrTRrCGCnIPAPNeIvEGs+Ldf1PxV4j1GXUNW1m8mvb+7mOXuJ5XLySNjuzMSfc18L/2nvj/APBXw/P4V+FXxU1rw1pN1eNezWllIqo87IiNIcg8lY0H/ARXxS/ae+P+jf8ABLb4P/GTS/iprVt4213xpcWWo60kiie5gWXVgI2OMYAt4R0/5ZivGHi/xN4/8Taj4y8Za1catrerTGa9vbggyTyEAFmx3wBXws/aO+OXwQ0++0r4TfEzWPDFpqUyzXcNi6qJpFXaGOQeQOK/4Jm/E/x/8Xv2WrDxl8S/FV74h1uTWr6F728YGQxo4CrwBwBX7e3/AAUX0X9lOeP4a+AtHtPEfxEvrPz3WebFrosbg+XJOF5kkbGRCCh2kOzAFA/jf9u39r/x/qh1XWP2hfGdjJuYrDompPpEKghRjy7Ty1IAUYyCepzlmJ+Df/BSr9rf4RaraS3PxP1Dxpo8Uim60vxPJ9v+0JuJIFw+Z0PzNgq+Pu5DBQB+zN+0V4J/ak+EunfFXwQsttHNI1rqWnTsGl068QKZIHI4bAdGVhjcjo2FJIH7Tf7SHgT9lr4VX/xO8cO1wUYW+l6XDIEm1O7YEpChPQYBZmwdqqxwSAp+Mn/BSr9rf4u6rdy2vxP1DwZo8sjG10vwxJ9g+zpuBA+0R4nc/KuSz4+9gKGIPwl/4KIftcfCXXrbVYvjFrviqxjm33Wl+KLx9UhukO3cheYtJGPlGDG6kHOPvMD8WfHH/Czvip4y+JP2H7F/wlniC/1b7Nu3eR9puHl2Z7434z7V/wAEVP8Ak1fxT/2UC8/9ILCqqqqqqqqqqqqqqqqqqqqv2s/+Csfwt+Dkt94I+B8Vj8QPF8C7XvxIW0ixk7q0iMGuGAxlYiFGcGQMrKPjT+19+0d+0DLMnxO+KusX2nSsSNHtZBZ2CDnA+zxbUYgMVDOGfHVjzXhbwD468dXH2PwR4K17xDPuK+VpWmy3bZCliMRqTnaCfoCaj/ZD/agl0f8AtxPgN40FuFLeW2kyLPwcH9wR5mcjptyRyODmvFfw3+IfgNgnjnwF4i8OszBVGraVNZkkgkD94o5IUkfQ18H/ANqL4/8AwFuIJPhX8Vdd0a0hbP8AZn2kz2L5bcd1tJuiJJzztzyeRk1+yZ/wV28C/Ei6tvA/7R1pp/gfXZdkdtr8Dsul3Tk4Im3km0PIO5i0eNxZo8AGGaG4hSe3lSWORQyOjZDAjIII6giqqqqqqq+IfjXSvhp4A8TfEbXbe7n03wro93qt5FaIrTSQ20LSusYZlUuVQgAsozjJHWv+H1f7K/8A0IXxV/8ABVYf/Jtf8Pq/2V/+hC+Kv/gqsP8A5Nr/AIfV/sr/APQhfFX/AMFVh/8AJtfssft4fCD9rzX9c8OfDXw74w0258P2cd3cvrlnbQo6O+wBDDPKSc+oA96qqqqqqqqqqqqqqqqqqr/hAPAf/Qk6D/4LYv8A4mtT1HTPDujXWrX8i2un6ZavPO6xkiKKNSzEKoJICg8AZ9K8ceLNS8e+NNf8c6y2dQ8Rapc6jdHOf3s8rSPz3+ZjX7LPw5l+En7OXw4+Hd3A0N7o/h20W+jdSpS7dBJcDBVSB5ryYDKGxjPOTVf8Fivhz/wlv7KUXja3j/0jwN4gtbuSTPS3uM2rrgsBzLNbnOGI24wAWI/4Jl/EOP4d/tn+Apbu9+z2PiKSfRLn5lHmG5hZYE+Ygc3At+OScYAJIFVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVf8IB4D/wChJ0H/AMFsX/xNRxxxRrDCioiKFVVGAoHQAdhVVVXl3aWFpNfX11FbW1tG0k00zhEiRRlmZjwAACSTwK/az/4K7+BPAX2nwV+zVBaeMtdMbpN4in3DTrF+g8leDdOOTkbYx8uGkyyj4kfE/wCJXxu8aT+NPiT4q1PxP4h1Bgnn3LbmALErFFGoCxoGY7Y41VRuOAM1+yX/AMEmvij8Wm03xx8dTceBvB8rLKumsANV1CLAIwhBFspzjMo3jB/d4Iavg/8ABP4W/ATwdB4E+E3g+x0DSYm3yLCpaW5lPBlmlbLyuQANzkkABRhQAKqqqqqqqv8AgtF+z3Lc2vhn9pXw9pqEWijRPETxIQ2wsWtZnxxgM0kRY8/NEuSAAP8Agl/+0j/woP8AaPsfD+u6h5HhP4ieVo+qb2wkFyWP2O4Pb5ZWMZYkBUnkY9BVVVVVVVVVVVVVVVVVVVVVVVajpemaxbGy1bTrW9tyQTDcwrIuR0O1gRVj4Q8JaXdR3+meF9ItLmLPlzQWMcbrkYOGABGQSPxqqqqqqqqqqqqqq/4Kr/EFvAf7GXiixt5kiuvFt9Z6LAzEZw8nnSgAqQxMMEq9iASwIKjP7Cnw7b4o/tc/C7wu1sk9vDr0WpXcckYeNoLMG6dXBBBVhDsIPXdjvVV/wWS+HcPhb9qOw8b2iII/Gvh23uJyOGNzblrdsj08qO35+o7V/wAEQ/iBHdeCviV8KprnD6dqlrrNtCQvzC4iMMrA4zx9mhBycfMuByc1VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVrvhvw74os00/xLoOnavaxyCVIL61S4RXAIDBXBAOGIz1wT61o3w98A+HL0an4d8D+H9LvFUqLiy0yGCQKeo3IoOD6VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVf8ABYD7V/w2Nd/aPN8v/hG9P8jfnGzD5257bt3TjOe+a/4ItXdpbftWeIYbi6iikuvAd5FAjuFMri9sXKqD947UZsDspPQGqr/gtLd2lz+1Z4dhtrqKWS18B2cU6I4YxOb2+cKwH3TtdGwezA9CK/4I+6fe3v7YkFza27yx2HhnUJrllHEUZ8uMMfbfIi/VhX/Bc/8A5ol/3MH/ALjq/ZJ+Ctl+0T+0Z4H+D2qai9lp+u3ztfyxnD/ZbeGS4nRDg7XaOF0ViCAzAkEA1+0r+wN+zRrv7OninQfBPwf8KeGdb0PQ7i60TVrGxFvcxXEERePzp1BkmRiu1vMLnDE/eANV/wAE4f2G/gJ/wzv4a+LHxH+Hmg+NPE3jOF72Rtbs0v7ezg8x1hiihlXYDsCszFS29iAxVVx/wVA/Zk8Efs3/AB2sG+GdjDpvhnxjpYv7fSo3dhYTo5jmRN5J8tiFkA3cF3UBVVRX/BGrxbe6J+1TqXhqNXktPEfhW6ilQSbQkkUsMqSEYO4gJImOP9YTnjB/4K8/8maal/2MGnf+htXwm/5Kp4M/7GCx/wDShK8cfDH4a/E60trD4k/D3wz4strKQyW0OuaTBfpA5GCyLKrBSRxkV+05pGlaB+0n8WNB0HTLTTtN03xxrNtZ2VpAsMNtDHezKkcaKAqIqgKFAAAAAr/gkV8EPgt8S/2bPEmu/Eb4QeCfFWpQeOLq2ivNb8P2t9NHCtjZMI1eVGYIGdyFBwCzHuavfgh8FtS8GWHw41D4QeCbrwnpVwbmw0Gfw/avY2kxLkyRW5Ty0cmWU7lUH94/945/bd8O+H/Cf7WPxO8N+FdD0/RtJ0/XHitLDT7VLaC3QIvypGgCqOegAFf8Eevg38IPid8NfiDf/En4VeD/ABZc2WuW0VtNrmhW1+8CGAkqjSoxUE84HGat9H+HXwQ8A6pP4W8I6P4Z8N6Fa3Goz2Gi6fDZQqEQySMsaBEDEKeTjPc96+JnxB8SfFj4geIPiT4uv5bvV/EeoS3t1JI5baXbIRfREXCKowFVVAAAAr9kX9rP/gnF+zR8GfDfg/SvilY23iRtNhfxHqS+E9SNxeXrDfMHlW0y6JIzKgJIVFUZPU/8FJvHP7JvxT+Ivh74jfsxa/Y3l1qlrcR+KLez0O602NZ42Qw3BE0UYd5VkkVig/5YgsctX/BFf4oahovxs8W/CW51F10rxNoJ1CC3YMwN7ayIAVGCEJhmmLHIB8tQckLj/gsp8WL3xZ+0dpnwsttRZtL8CaLF5tqG4W+uh50jn3MBtRz0wf7xr/gmF+yt4U/aX+NOpX/xG05tQ8I+CLFLy8scOI726lcrbwyMuMJhJXI3At5QXBUvj4i/sofs8fE/4fXfw11/4TeGbXSZ7cQ2zaZpcFpNp5Vdscls6IDE6DgY4xlSCpKn4qeCJfhn8T/F/wAN57tLqXwpr19pLzpnErW07xFhwODsz0HWv+CKn/Jq/in/ALKBef8ApBYVVVVVVVVVVVVVVVVVVVVf8FI/+CjOs+NNZ1X9n/4CeIpbDwvYSPba/wCINPusPrLjh7eGSM8Ww5ViD+9OR/q8+Z8FPgL8Tv2g/FP/AAifwz8PtfTQqHvLuVvLtrFDnDzSdFBwcDlmwdoODXwH/wCCafwL+GFpa6p8QrFfH/iIKrStqMf+gwuMEiO26OuRj97vyOy5IqysLHTLSKw02zgtbaBdsUEEYjRB6Ko4Aqrq1tr62lsry3iuLeeNo5YZUDrIrDBVlPBBBwQa+PH/AATp+AHxkim1Pw7pK+AfETL8l9odsqW8jdvNtBiNh1JKeW5J5Y9K/aB/Zq+KP7NviceH/iDpKm0uWb+ztXtNz2l+oClvLcgEMu4BkYAg9iCCf+Cdn/BRPV/gVq+n/Br4yaxcah8ONQmSGxvriUvJ4ckYgBgSebU8bk/g5df4leGaG4hSe3lSWORQyOjZDAjIII6giqqqqqqtX0jStf0q80HXdMtNS03UbeS2vLK7gWaG5hkUq8ciMCroykqVIIIJBr/hk39lf/o2f4U/+EZYf/Gq/wCGTf2V/wDo2f4U/wDhGWH/AMar/hk39lf/AKNn+FP/AIRlh/8AGq8D/Bv4QfDG6ub74a/Crwf4Tub2MRXM2h6FbWDzoDkK7QopYA84PGaqqqqqqqqqqqqqqqqqqqv+CinxAj+HP7GnxM1Jbny7jV9L/sW3QBSZTeutu6jcCP8AVSSMe+FOCDg1+zl8Po/it8fPh58Op7bz7XXvEllbXiZUf6MZlM5+YgHEQc46nGBknFVX7UPw8f4sfs6fEf4e21n9qvNY8N3kdhDtZt12sRe34UMxxMkZwATxwM14N8T6h4J8X6F4z0l2W+0DUre/tmSQxsJYZFkUhl5U5UcjkVoer2XiHRNP1/TWdrTUrWO5ty67WMcihlyOxwRxVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVftPftx/Af9lWyltPG2vvqnikwiS18M6ViW8lDfdaTJCwJ33SEEgHaHOAf2qv28/jn+1ZdS6X4j1f+wPBgkDW/hfTJCtvwVIad+GuXBVSC/wAqkZRUyc/s1/sefHH9qfX0074ceGXh0eJiL7xFqKtDp9mB1BkwfMfkARxhn5yQFDMP2Uf+CdfwM/Zjt7DxFLpcXi/x7DGDL4h1OAMLeTIJNpCcrBjGA4zJjI34Yiqqqqqqqqqr4rfDbw38Yvhv4j+F/i6383SfEmny2dxgZaPcPlkXPR0YK6nsyg18Vvht4k+D3xI8R/C/xdb+Vq3hrUJbO4wCFk2n5ZFz/A6lXU91YHvX/BPn9pH/AIaX/Zv0PxBrOofaPFnhzGj+I97ZkkuYlGy4Pc+dEUkLYA3mRR9w1VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX/Bb/4hxvdfDD4T2l788Ud7rd/b7l6MUgtnwDkfduxyAD2JwwH/AARV+Hya78d/GPxGuInkh8KeHVtYSFICXF5KNrEhhz5dvOu0gg7ieCozVf8ABbD4fvrXwU8DfEi3tvNk8L+IJbKVgWzFDeQ5ZsZxjzLWFeQTllxgE5/4JFfEAeDf2w9O8Oyt+58baHfaUdx+VXRBdo33gAc2pQHDf6zAHzZFVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVePfiB4J+F3hO/8c/EPxRp/h/QdMj33V9fTCNE9FHdnY4CooLMxAUEkCv2o/wDgsZ4o1DVZPDH7K9jFpel28iFvE2rWSy3N0VYFhDbyApHGwG0mQM5BOBGcGvHH7af7Tvi/xpr/AIssPjv8RdBtta1S5vYdJ07xffR22npLKzrBColAWNAwRQAAAor/AIax/ao/6OY+Kv8A4Wd//wDHa/4ax/ao/wCjmPir/wCFnf8A/wAdr/hrH9qj/o5j4q/+Fnf/APx2v+Gsf2qP+jmPir/4Wd//APHa/wCGsf2qP+jmPir/AOFnf/8Ax2v+Gsf2qP8Ao5j4q/8AhZ3/AP8AHa/4ax/ao/6OY+Kv/hZ3/wD8dr/hrH9qj/o5j4q/+Fnf/wDx2v8AhrH9qj/o5j4q/wDhZ3//AMdr/hrH9qj/AKOY+Kv/AIWd/wD/AB2v+Gsf2qP+jmPir/4Wd/8A/Ha/4ax/ao/6OY+Kv/hZ3/8A8dr4N/8ABYv4y+FfG8jfFfQrTxP4IuPLiisYFWO/02ONAgaO4OPtDtjfJ52S7Z2tEDtr4EftI/Bz9pPwsnir4S+MbfVERV+2WEgMN5YOQMpNC3zKQTjcMoxB2sw5qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/wCC3Pwnuotf+HvxztUlktrmzl8OXzbDsheN3ubcZ6ZcS3XHH+q79v2Nfjrb/s4ftG+D/inqbSnRrK4e21hIoy7NZzo0UpCggsUDCQDPLIOvStD1vR/E2iaf4j8Palb6jpeq2sd3Y3lvIHjuIJFDxyIw4KspBBHUEVrmt6P4a0TUPEfiDUrfT9L0q1lu768uZAkdvBGpeSR2PCqqqSSegFfti/HZP2kf2i/GHxUsldNIvLoWujRuCpWxgURQsVPKs6p5jL2aRh0Ff8ESvg9cJcePvj1qFqyRtDH4d0qQlgJAWW4uuPukApaAHk53Djv/AMFz/wDmiX/cwf8AuOr/AIJcf8n2/DH/ALi3/ppvK+LP/JK/GX/Yv33/AKTvVfsB/wDJmnwl/wCxfj/9Dev+C3v/ACVP4af9i/df+lAr/gkN/wAnlab/ANi/qP8A6Atf8Fef+TNNS/7GDTv/AENq+E3/ACVTwZ/2MFj/AOlCVX7WX/J1Hxl/7KBrv/pfNX/BFOaFv2XPFdusqGVPH10zIG+ZQbCxAJHodp/I+lV+37/yeV8Wf+xgk/8AQEr/AIIhTQt8MPiZbrKhlTXrRmQN8ygwMASPQ7T+R9K/aL8Pah4u/Z8+J/hTSQhvtZ8G6vZWwcHHmy2cqLnaCcZYdAT7Gq8Ef8EhP2i/iF4N0Px34Y+I/wAKbjSfEGnw39lIdVvsmKVA65xZkA4IBGTg5Ff8OVP2qP8AofvhT/4Nb/8A+Qq/YN/4Jr/Gn9mL49R/Fb4jeMfBN5ptro91axW+iXN1cTSTS7FG4SwRKqBd5LAschRt5LL/AMFXfD2oaL+29401K8CeTr1jpd7abQc+WtjDbnOQOd9vJ0yMY5zkD/giH470DSfH3xO+HN9dJFq3iTTdPv7BHbb5qWbzrKq+rYu0bHXCscYBIr9ovxHo/jH9oP4n+LvD14l1pet+MtXvrGdCCJYJbyV43BGQQVYHj1r/AIIqf8mr+Kf+ygXn/pBYVVVVVVVVVVVVVVVVVVVX/BVj9rO6+Bvwog+Evgq+8nxd8QreaKW4ilKyadpw+WWVcch5CTGp9BIRyor4E/BbxX8f/iZpPw08Ix7Z75993dsoZLG1UjzbhwSMhAfu5BYkKOWFfA/4H+Af2fvANn8P/h/pvk20P7y7u5QDPfzkANNMw+8xwOOigBQAABX7dn7efxKX4laz8Hvg14lufDWkeGp5bDU9SsX2XV9dL8sqrJjdCsbBkGwhiys27BUL8B/2/fj/APCPxTa3Pifx1rPjbw3PcqdT03XLtr2Vo+AxgmlJkicLkgBthP3lNaHrek+JtE0/xHoF/Fe6ZqtpFd2VzEcrPDIgdHX2ZWBH1r9s79v34p+JfiNrHw++D3i2+8K+FvD11LZNd6ZMYbrUpo2KvKZgA8cYIIVUIyPmYkkBf2a/+CgPxn+FfjfToPiN451fxf4OvbqOPVotYuZL24toScNNBK5LhkDFtmSr7dvGQy/E/wCGHw9+PXw9uvBPjXT4NX0LVollhmhcFomxmO4gkGdrjOVYZBBIOVYg/tM/s+eJf2avinffDzXZ/tto0YutI1JY9i39q5IV9uTtYEMjL2ZTjKlWP/BID9qm98f+Cr79nDxpqSS6t4MtVuPDsjnDz6bu2vCT3MLsgHfZIo6JVVVVVVVVVVVVVVVVVVVVVVVVVVVVX/Cf+O/+h217/wAGUv8A8VX/AAWb+JVzp3wS+F/wwiupDJ4m1CTVbtlc5dLSBUCyfNyrPdhsFSC0Wcgrz/wR5+H7+K/2tB4vktt0Hgrw/eXqyksAk0wFqqjB5JSeY4ORhT3ANVVftMeAI/hb+0H8Rfh/BHCltoviS9htFiRUUW/ms0OFXhf3bJ8o4HTtX/BOz4gR/Eb9jT4Z6kbnzbjR9L/sWdCFBiNk7W6KdoA/1UcTDvhhkk5NVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX/Cf+O/8Aodte/wDBlL/8VWiSSTaLYSyyM7vaxlmY5JJUZJNV8QPiL4E+FXha88bfEfxbpnh3Q7FSZr2/uBEmcEhFzy7tghUUFmPABJxX7W//AAV78UeLxfeBf2Xorvw3o7b4ZvFN3Eq390vzKxtozkW6MOVkP70ZBAiYVo+ifEH4ueMBp2iabrvi/wAUazMWMcMct7d3Uh+8zY3Mx7lj9Sa/ZQ/4I72Vg1l42/asvUvJwqSxeENMuv3UbEcreXCffIyPkhYLlf8AWOpIOgeHfD/hPRbTw34V0PT9G0nT4/KtLDT7VLeC3QfwpGgCqOegAFVVVVVVVVVVV/wWb/Zt/wCQH+094Z0/+5o/ify0/wDAW4bH/AoWY/8ATEV/wSp/aHt/gn+0fD4P1+98jw78SY49JuWaQqkN4GJs5GGcH52eLJ6eeTkAGqqqqqqqqqqqqqqqqqqqqqqvjrqepaR4AmvNJ1G5srgXUSiW3maJwCeRlSDivg14v8War8SdHsNT8U6veW0vn74Z72SRGxBIRlSSDggH6iqqqqqqqqqqqqqr/gqf8Q5fH/7Zniy0EjtaeErW00S03gggRxiWUY7Dz55seowe9f8ABGf4eP4Z/Zl1jx3d2fl3HjLxJM9vNtYebaWyJCnUDOJhdDIJHbIIIFV+378OYfih+x98T9AMCPc6for6vaMVy6y2JFzhPlYhmWFo+ACQ5XK7iR8AviCnwn+N/gL4kzTPHb+G/EVle3W0kFrdJlMy5CscNHvU4UnB4Bqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv2yPEfiLwv8MNM1Dwzr+o6TdSa9FE89jdPbuyGCclSyEErlVOOmQPSv2T/iD498SfF620zxD441/VLM2Fwxt73U5p4ywUYO1mIyKqq/a0/4KjfBv9n9tS8F/Dprfx/48tGaCS2tZ8WGnS4IPnzqCHZGwGhjy2Qys0ZGR8eP2kfjH+0n4pbxV8WvGNxqkiM32OwjHk2dghJISGFflUAHG45dgBuZjzX7Jn/BNj41/tMC28T61DL4F8DTRpNFrWpWbNJfo3INrASpkBHPmEqmDwW6V4I/4Js/sbeCvDNn4ck+EFj4iltVIk1PXJGuLu5YnlpGXaufZFVR2Ar/AIYD/Y0/6N58Kf8Afh//AIqv+GA/2NP+jefCn/fh/wD4qv8AhgP9jT/o3nwp/wB+H/8Aiq/4YD/Y0/6N58Kf9+H/APiq/wCGA/2NP+jefCn/AH4f/wCKr/hgP9jT/o3nwp/34f8A+Kr/AIYD/Y0/6N58Kf8Afh//AIqv+GA/2NP+jefCn/fh/wD4qv8AhgP9jT/o3nwp/wB+H/8Aiq/4YD/Y0/6N58Kf9+H/APiq/wCGA/2NP+jefCn/AH4f/wCKq8/4J8/sYX1pNZTfs9+GVjnjaNzCJonAIwSro4ZTzwykEHkEGv2rP+CQvxB+HwuPGH7N91d+NtAXLTaHcso1S1HJJjwFS5QAAYXbJkgBH5YeE/GHj74S+MIfEng3X9Y8LeJdGmZFubSZ7W4t3GVeNsYODyrI3BGQwIJFfsp/8FiNI1cW/g39qnT4tLu+Fh8WaXbsYJegAubZATGepMkeVOQNiAEnQPEPh/xZotp4j8K67p+s6Tfx+ZaX+n3KXEFwn95JEJVhx1BIqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr44/Bvwh+0B8K9f+EnjiOX+ytetxG00GBLbSKweOaMsCA6OqsMgjjByCRX7UH7KnxS/ZT8ez+EfHulyz6XPIx0bxBBbstpqsQ/iRjkLIARviJ3ISOqlWb4G/tw/tNfs76fFoPw4+JNwuhRMzLo2pQR3tqpZcfIsgLRDIDYjZRkc5yQfjv+23+0r+0fpsnh74m/ESWbQHkjkOi2FtHZ2hZPulljAaT5vm/eM2GwRjC4/Zd/Y/8AjD+1f4rh0jwHoktt4ft7gR6v4luoiLPTl+Ut8xI82XaykQodx3AnauXHwZ+EPgz4EfDPQvhV4BsPsukaFb+WhYkvPIxLSzOSSS7uzMewLYGAAB/wXP8A+aJf9zB/7jq/4Jcf8n2/DH/uLf8AppvK+LP/ACSvxl/2L99/6TvVfsB/8mafCX/sX4//AEN6/wCC3v8AyVP4af8AYv3X/pQK/wCCQ3/J5Wm/9i/qP/oC1/wV5/5M01L/ALGDTv8A0Nq+E3/JVPBn/YwWP/pQlV/wU3+CWufCD9rDxVq9zp+zRPHtw+vaTdxxFY5jMc3KE8jzEnMm4Zzh0YhRIBXwP/ag+O/7N0uqy/Bb4hXHh0a2sa38Qs7e7jn8vOxjHcRyKGG5huABwSM4NfsFfFLx38av2TfAvxM+Jmvf2z4l1n+0Ptt79lit/N8rULmGP93CqIuI40XhRnGTkkmv+CtfwS1z4c/tR6j8R10/Hhz4iW8N9Y3EUREaXMUSQ3MLMesm6MTHtidfQ4+Cf7Rfxp/Zz1e/1z4MeOrjw5d6pCsN6UtYLmOdFJKho50dCQWbB25GTg81/wAE5fjN8Svj1+zPY/EH4seJf7d1+bWL23e7+xwWuY42ARdkCInAPXbn1r/goZ+xZ4k/Zo+KWqeLPDGgyyfDHxJeNcaTe28JEOmSTM7HT3/uFNrbM8NHt5LBwv7OX7en7R37L+lL4Y+H3iexv/DKzPMug63Zi6tUkcgsyFSssYJBJVJFUlmYjcc1qv8AwWe/aw1Gwls7Pwx8NNMlkxturXSLtpI8EE7RLdOnIGDlTwTjBwR/wTL/AGkfHP7S3wP13xH8T9fTVPFOkeJp7W4lWKOEG3aGGSErHEiKi/M6ADJJjLEksa/4KwfseeJvjh4R0n41fDHSX1PxR4LtZLbUdNghDT3+mkmTMWPmd4XLsIh95ZZNvzAK/hDxh4p+H/ibTvGfgnX73Rdc0mYTWV/ZTGKWF8YyCOxBIIPBBIIIJFfED/gpN+2J8V/BM3w11n4hwxWWq2/2O/bStKhtbnUEYBWRnjXK7+jCLYGDFcbSVr4l/DXxh8IvGFx4C8faU2ma9ZWtpcXVlJw9uLm2iuURx1VxHMgZTyrZHav+CKn/ACav4p/7KBef+kFhVVVVVVVVVVVVVVVVVVVft0/GZ/jt+1L478aW9/8AadJtdQbS9GKltgs7X91GyBuQJCrS44+aVjgZwP8AgmD8BrT4efBc/FjV7FR4h8et5kTyRYe306NisKDIyBIwaUkHDK0WeUFVrF7qOpate6jrDs1/dXMkt0XjCEysxLkqAAp3E8YGKr9lrVNdj/Yt8E6npzT3uq2vhA/YlZWmZnjRxCgXksBtRQo7AAVVfs2Syz/s6fCuaaVpJJPBWjs7uclibKLJJ7mv+Ci3wHi+MnwAv/EOmWyt4i8ArLq1g+Pmkt1X/S4c+jRqJAAMl4UHc1+zb8YdS+Afxz8GfFfTp5Vj0LVInv442wbizY7LmLofvws69DgkHGRUM0NxCk9vKkscihkdGyGBGQQR1BFVVVVVVVVVVVVVVVVVVVVVVVVVVVVf8Fa/iCniz9pXS/B9tM/k+CfCWn2UsRJIFxMhumcAqMExzwA4LD5BznIH/BEf4eR2Hw4+IvxUuLP99rWsW+k20zquRHaxGVwnG4Bmul3c4YxrwClVVf8ABYb4fv4U/a0Pi+K22weNfD9netKCxDzQA2rryTghIITgYGGXuSa/4Ij/ABAGpfC74i/C6Zvn0DXLfVYSx5KXkJiZV+Y8K1nkgKADJnJ3YFVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVWgf8gLTf+vSL/0AV+1r/wAFTfg98B/tng74WSWnxC8bxZR1tJw2mae4xkTzof3jjP8Aq4s4KsrtGRg/HX9oz4w/tIeK5vFvxY8ZXeqSGRmtLBGMdnp6ngJbwA7YwBgZ5ZsZZmYlj+yb/wAEz/jb+0dLaeKPFNlceBPAjsrHVNRtytzfJwSLW3bDMCp4lcCP0LkFa/Z6/ZY+Cf7MHhsaB8KfCUVtcyxhL/WbsLNqGoEAczT4BIyu7y1CxqSSqLk1VVVVVVVVVVVV8W/hroHxj+GXib4XeJ4kfTvEumy2UrNHv8lmX5JQMj5kcK45HKjkV488FeKfhR4+1vwF4ot3sNf8L6lLZ3SxsRsmicruRuMqcBlYdQQRwa/YV/aMh/aa/Z18PeNb67STxJpi/wBl+IkB+YXsKqDIR2EqFJfQbyOxqqqqqqqqqqqqqqqqqqqqqq/aF/5JvP8A9fcP86+Bn/JVND/7eP8A0nkqqqqqqqqqqqqqtR1Cy0nT7nVdSuUt7SyheaeV+BHGgLMx9gATXxJ8YXPxE+Inin4gXpf7R4m1q81Obeqht9xM8rZCgKDlz0AHpX7Gvw+j+F/7K3wu8GC2+zz2/hu2ubuLKnZc3K/aJxlSQf3s0nIJB696qtR0+y1bT7nStSt0uLS8heGeFxxJGwKsp9iCRXxD8G3/AMOvH/ib4fao++98Maxd6XctgDMlvM0THCsw6oehI9z1r9j/AMfR/FD9l74YeNBLDJNd+G7WG6aJlZftECeRNjbwP3sT/L1XoeQaqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/bk/5JLpP/AGMUP/pPcV+xt/yWy1/7B1z/AOgiq+P37TPwY/Zn8MjxL8WfGFvp7TKxsdMhIlvtQIIBEEAO5wCVBY4RcjcwyK/a1/4Kc/Gj9oz7Z4R8GyS+AfAcuUbTbG43Xl+nHNzcAA4OCfKj2phireZgNXwU+AHxc/aH8Ur4R+Evgy91u6Ro/tdwibLawRyQJJ5j8sS8NjJydpCgniv2Tf8AglR8I/gbLaeM/izPb/ELxnCyyQpNb40zT3GCpihbmVwQf3knHTCIRk1VVVVVVVVVftSfsJfAn9qqxuL7xTov9h+LzHi38UaVEkd2GVNqCcEYuIxhRtf5towrp1r9qr9gz45/sp3UuqeI9J/t/wAGGQJb+KNMjLW/JUBZ0yWtnJZVAf5WJwjPg4/Zw/a/+Of7LmuR3/w28WSto8kga+8PX5M9heLkFsxE/u3OB+8jKv23YJB/ZQ/4KS/A79pdLLwxq10ngfx3OyQjRNTuV8q9lY4As5+BKT8vyMEkycBWA3GqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqrxR4S8K+N9Fn8N+NPDOk6/pNzjzrDVLKO6glx03RyAqfxFeMf+CWv7FPi+6+2x/C240KdmBkbR9XuYEcBQoHlM7RqOAflVSTkknJrwV/wTE/Yp8FSw3cfwgTW7uFiVm1vU7m7DA9mhMghIGO6Z960XQ9E8N6Vb6H4c0ex0vTrRStvZ2VusEMIzkhEQBVGSTgDvVfHT9lv4EftLf2J/wuzwL/AMJH/wAI59o/sz/iZ3dn9n8/y/N/495Y927yY/vZxt4xk5+Fv7BP7JvwV8d6X8TPhl8KP7G8S6N532G9/t3ULjyvNheGT93NcOjZjkdeVOM5GCARq+lWGu6VeaJqkHn2WoW8lvcxbiu+N1KsuVIIyCRkEGv+HXH7CX/RC/8Ay5tW/wDkqvAHgHwl8LvBmkfD7wJpP9maBoVuLfT7Pz5JvJjBJC75GZ25J5Yk18cP2R/2ef2kNV0zW/jR8Pv+EivdHt3t7KX+1by08qNm3MuLeaMHJ5ywJr4Q/sPfsufAXxnF8QfhP8L/AOw9fht5LdLv+2r+6xHIAHXZPO6cgDnbkV8X/gx8Nfj14Mk+H3xY8N/27oE1xHcPZ/bJ7XMkZyjb4HR+CemcetaT/wAEzP2INC1Wy1vSvgl5F7p9xHcW0v8Awkmqt5ciMGVsNckHBAOCCKr4rfBr4W/HLwtJ4L+LPgjTPEukO29IbyM74HwR5kMqkSQvgkb42VsEjOCam/4JH/sXS6g95H4S8RxQtMZFtE1+cxouc+WCSX2gccsWx3zzXwy+GPgX4N+BdL+Gvw10FNG8N6Ksi2Nis8kwiEkjSv8APKzOxLu7EsxOWr4kfC/4efGDwtP4J+J3g/TPEmiXDB2tL+AOEcAgSIfvRuAzAOpDDJwRmtQ/4JH/ALF17ey3Vt4S8R2McjZS2t9fnMcQ9FMhZsfVifevhJ8Hfhv8CfBVv8O/hT4aTQvD9rNLNFaLczXBEkjbnYyTO7sSfVjgYAwABWraRpWvabc6NrmmWmo2F5GYri0u4FmimQ9VdGBDA+hGK8cf8Ewf2LPHE7Xp+Ev9g3TyBnl0PUri0UgA/KIQ5iUHOflQHgc4yD4d/wCCUf7EuhSvNe/DbUtcYsjRjUdfu8RFc9FhkjDA5GQ4YcDpzn4c/Cj4afCDQm8NfC7wLonhfTXk82W30yySATSYA3yFRmR8ADcxJwAM8VXxh/4J7fsl/G7V5vEniv4WW9jrdw26fUdEuJNPkmOckukREbse7shb3r4KfsHfssfAHVYPEngL4XWkmv2/MWsatPJqFzEwYkPF5pKQuAdu6JUYgYJPOfi5+wx+yt8dvGtx8RPip8Kk1rxDdwxwz3q6zf2hkSNdqApBOiZCgDOM4A54r4KfAT4Tfs6+FbrwT8G/Cf8Awj+i3uoPqE9r9uuLvfcvHHGz755HYZSGMYBA+XOMkk1VVVVVVVVVVVVVVVVVfGrxhc/Dz4N+PPiBYl/tHhnwzqOpw7FUtvt7aSUYDAqTlB1BHrWmabe6zqVpo+mw+dd306QW8e4LvkdgqjJIAySBkkCvCnhrS/BnhbR/B+hxsmnaFp8FhZo5BKwwxrGgOABnao7Cq/bw/Zl8U/BH4wa54qtNInm8E+Kr+W/0vUIo90du8rb5LaTH+rZHZggP3k2kEkMF+DPwV+IHx48b2HgfwBodzeTXE8aXd4IWa302JjgzzuAQkagE88sRtUFiAfAXg7S/h54H8P8AgLRCxsPDumW+n2zOAGdIY1QM2P4iFyT3JNftgfsy+Kf2cvijqdpLpE58IateSzeHtSRMwvAzFlgZhwJYx8rKcE7dwG1hXwA/Z/8AHv7Qnj3TfCPhHRL6Sxkuo11TVUgJg02AnLySOflBCBiqk5Y4ABJrw9oOl+FvD+meGNDtVttO0ezis7OBQAIoYkCIoA4ACqBV1a219bS2V5bxXFvPG0csMqB1kVhgqynggg4INfEzwj/wr/4keK/AXn+d/wAI3rd5pvmbt2/yJ3i3Zwuc7OuB9B0r9jnxbe+Of2VfhT4l1JX+1z+FbGKd3k3tLJDEImkJwOXMZfGON2MnGaqqqqqqqqqqqqqqqqqqqqqviv8AH74M/Az+y/8AhbnxE0nwv/bfnf2f9vdl+0eTs8zbgH7vmx5/3hX/AA35+xp/0cP4U/7/AD//ABNf8N+fsaf9HD+FP+/z/wDxNf8ADfn7Gn/Rw/hT/v8AP/8AE1/w35+xp/0cP4U/7/P/APE1/wAN+fsaf9HD+FP+/wA//wATX/Dfn7Gn/Rw/hT/v8/8A8TX/AA35+xp/0cP4U/7/AD//ABNf8N+fsaf9HD+FP+/z/wDxNV+0R45vfiV8c/HPjW+DBtQ1u4EKOctHBG3lwofdY0Rfwr/gnP8AD1vhx+xl8M9MniRbrWNNbWp3VQDIL2RriInDMMiGSJeucKMhTkCqr/gtz8PhqXwu+HXxRhX59A1y40qYKOSl5CJFZvlPCtZ4BLAAyYwd2R/wS3+P/g/4B/tEXt38RfFdpoPhjxF4fuLK6urwsIkmR45YmJUHB/duozgfvCOpFf8ADfn7Gn/Rw/hT/v8AP/8AE1/w35+xp/0cP4U/7/P/APE1/wAN+fsaf9HD+FP+/wA//wATX/Dfn7Gn/Rw/hT/v8/8A8TX/AA35+xp/0cP4U/7/AD//ABNf8N+fsaf9HD+FP+/z/wDxNf8ADfn7Gn/Rw/hT/v8AP/8AE1/w35+xp/0cP4U/7/P/APE1/wAN+fsaf9HD+FP+/wA//wATX/Dfn7Gn/Rw/hT/v8/8A8TX/AA35+xp/0cP4U/7/AD//ABNf8N+fsaf9HD+FP+/z/wDxNeHv23P2TvFmv6Z4V8N/HbwzqGrazeRWVhaQzOXuJ5XCRxr8vVmZQPc1VVVV4+/a9/Zn+F3iy+8CfEH4y+H9D1/TPL+16fdysJIfMjWRMgKeqOjfRhX/AA35+xp/0cP4U/7/AD//ABNf8N+fsaf9HD+FP+/z/wDxNf8ADfn7Gn/Rw/hT/v8AP/8AE1/w35+xp/0cP4U/7/P/APE1/wAN+fsaf9HD+FP+/wA//wATX/Dfn7Gn/Rw/hT/v8/8A8TX/AA35+xp/0cP4U/7/AD//ABNf8N+fsaf9HD+FP+/z/wDxNf8ADfn7Gn/Rw/hT/v8AP/8AE1/w35+xp/0cP4U/7/P/APE1/wAN+fsaf9HD+FP+/wA//wATX/Dfn7Gn/Rw/hT/v8/8A8TX/AA35+xp/0cP4U/7/AD//ABNf8N+fsaf9HD+FP+/z/wDxNf8ADfn7Gn/Rw/hT/v8AP/8AE18L/i98M/jX4fn8U/Crxnp/iXSbW8aymu7JiyJOqI7RnIHIWRD/AMCFVVVVVXjD9s39lrwB4m1Hwb4y+NvhzSdb0mYw3tlcSuJIJAASrAKecEV/w35+xp/0cP4U/wC/z/8AxNf8N+fsaf8ARw/hT/v8/wD8TX/Dfn7Gn/Rw/hT/AL/P/wDE1/w35+xp/wBHD+FP+/z/APxNf8N+fsaf9HD+FP8Av8//AMTX/Dfn7Gn/AEcP4U/7/P8A/E1/w35+xp/0cP4U/wC/z/8AxNf8N+fsaf8ARw/hT/v8/wD8TVftd/8ABRf44/Gi4vfhd4cv38E+C9Knks3s9LnZLnU1jJjBuZxhipAJ8pNqfN828qrD4H/s9/F79o3xWPB/wi8G3WtXce03dwMRW1ghzh55mwkYIVsAncxUhQzcV+yh/wAErPg58DUsvF3xVW3+IPjaFklRriI/2bp8inIMEBH7xhx88ueVBVUOc1VVVVVVVVVVVVVf8Fm/2bf7N13Q/wBp7wxp/wC41fZo/ify0+7cIn+i3DYyfmiRoWY4A8mEdXr/AIJi/tV6V+zf8arnRPHeuJp3gXxpa+RqdxMx8uyuYgz29wQM8cvGcdpQT90V/wAN+fsaf9HD+FP+/wA//wATX/Dfn7Gn/Rw/hT/v8/8A8TX/AA35+xp/0cP4U/7/AD//ABNf8N+fsaf9HD+FP+/z/wDxNf8ADfn7Gn/Rw/hT/v8AP/8AE1/w35+xp/0cP4U/7/P/APE1/wAN+fsaf9HD+FP+/wA//wATX/Dfn7Gn/Rw/hT/v8/8A8TX/AA35+xp/0cP4U/7/AD//ABNf8N+fsaf9HD+FP+/z/wDxNf8ADfn7Gn/Rw/hT/v8AP/8AE1/w35+xp/0cP4U/7/P/APE1/wAN+fsaf9HD+FP+/wA//wATX/Dfn7Gn/Rw/hT/v8/8A8TX/AA35+xp/0cP4U/7/AD//ABNfC/8Aad+AHxr1+fwr8KviponiXVrWza8mtLKRmdIFdEaQ5A4DSIPqwqqqqqq8Yftm/steAPE2o+DfGXxt8OaTrekzGG9sriVxJBIACVYBTzgiv+G/P2NP+jh/Cn/f5/8A4mv+G/P2NP8Ao4fwp/3+f/4mv+G/P2NP+jh/Cn/f5/8A4mv+G/P2NP8Ao4fwp/3+f/4mv+G/P2NP+jh/Cn/f5/8A4mv+G/P2NP8Ao4fwp/3+f/4mv+G/P2NP+jh/Cn/f5/8A4mvH/wC1b+zn8XvDr+DPhp8XdB8Q63JKkyWVnIzSGNDlmwVHAFeCfH/gz4XeJ7Lx38QfEVpoegaZ5n2u/u2Ijh8yNo0yQCeXdF+rCvBHxK8BfEnwtpPjXwN4r0/WNE12SWLTL2CX5Lt4mkWRY84LFTDLkAf8s2PQE1VVXiLxX4b8J6Vqet+I9btLCy0bT5dRv5ZpAPs9rEpaSZh12KFJJx2r/hvz9jT/AKOH8Kf9/n/+Jr/hvz9jT/o4fwp/3+f/AOJr/hvz9jT/AKOH8Kf9/n/+Jr/hvz9jT/o4fwp/3+f/AOJr/hvz9jT/AKOH8Kf9/n/+Jr/hvz9jT/o4fwp/3+f/AOJr/hvz9jT/AKOH8Kf9/n/+Jr/hvz9jT/o4fwp/3+f/AOJr/hvz9jT/AKOH8Kf9/n/+Jr/hvz9jT/o4fwp/3+f/AOJr/hvz9jT/AKOH8Kf9/n/+Jr/hvz9jT/o4fwp/3+f/AOJr/hvz9jT/AKOH8Kf9/n/+Jr9rT9v39nF/2a/iJYfC/wCM+ia14p1TQ5tO02z0+V/PL3OIDIh+TBjWVpN2fl2Zw2Np+BHw/f4r/GrwL8NltvPj8R+ILOynQlgBC8yiViVIYAR72OCDgHHNVVV/wVM8AP4E/bP8YXCRyra+KLe01m28xGGRJCI5CCfvDzoZuRwPu9VNf8Ea/Hz+Jv2XdS8FXMsrS+DvElxDCrMzKtvcIk67c8L+9efKjjv1Y1VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV+3J/ySXSf+xih/8ASe4r9lHXNE8NfFVvEHiPWLHStL07SLue8vr64WCC3jVAWeSRyFRQOSSQBX7WX/BYTStClu/A/wCyxYW+rXkbNHP4t1GIm2jIyCLWAgeaehEr4TjhHBDDxF4l8ffFzxm+teJNW1jxV4o166VPMmZ7q5upnbCRovJJJIVUUY5AA6Cv2S/+CQvjPx19j8b/ALTN1deE9BbEkPhu0YLqd0Of9exBW2Q/KdvzSEFgREcNXw3+F/w8+EHha38E/DHwfpnhvRLdi62ljAIwzkAGRz96RyFUF2JY4GScVVVVVVVVVVVVXlnaahaTWF9bRXNtcxtFNDMgdJEYYZWU8EEEgg8HNftWf8Ehfh78QhceL/2b7q08E6+2Wm0O5Zjpd0eSTHgM9s5JAwu6PAACJyx+Kfwg+JnwS8Vz+CPir4N1Dw5rNvyYLpAVkX+/FIhKSp/tIzL71+yd/wAFVvi58DYbTwX8WYbj4heDYVEcLzT41PT0AAAimY4lQAH93Jz0xIgGD8EP2ifg3+0X4ZTxT8I/G9jrUIUG5tMmK7smzgrNA2HjOQQCRtbqpYEE1VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV+1l/yav8AGX/sn+u/+kE1fDP/AJKR4U/7Ddn/AOj0qqurW2vraWyvLeK4t542jlhlQOsisMFWU8EEHBBrS9H0jQ7Qafoml2en2ysWEFrAsSAnqdqgDJqr6wsdTtJbDUrOC7tpl2ywTxiRHHoVOQR9a0vR9I0O0Gn6Jpdnp9srFhBawLEgJ6naoAyaqv2qJ7K5/aX+KklhYfY4x4w1NGj80ybpFuXWR8npvcM+Oi7sDgV/wTnstSsf2J/hVBquofbJ20uaVJMY2xPdTPEn/AI2RP8AgNVVVVVVVVVVVVVVVVVVVVVX/BZX4TfFT4o/8Kg/4Vl8M/Ffi7+zP7c+2/2Fotxf/ZfM+weX5nko2zdsfGcZ2tjoa/4ZN/ao/wCjZ/ir/wCEZf8A/wAar/hk39qj/o2f4q/+EZf/APxqv+GTf2qP+jZ/ir/4Rl//APGq/wCGTf2qP+jZ/ir/AOEZf/8Axqv+GTf2qP8Ao2f4q/8AhGX/AP8AGq/4ZN/ao/6Nn+Kv/hGX/wD8ar/hk39qj/o2f4q/+EZf/wDxqv8Ahk39qj/o2f4q/wDhGX//AMaqv2Bv2M4PHvxW+K1p+0x+z1rbeHL2AyaRc6/ot1YJI73EgLW05CNkxtnMbf3W7AjQ9F0rw3omn+HNDs0tNO0q1jtbO3QkiGGNQiIM5OAqgc+lVVfFD4TfDv40+FG8D/FDwvb6/obXUN01lPI6K0sLh4ySjKSARyucMCQQQSD+1n+yJ8ZLL9o3x7ZfCX9mrx23g+31Qx6OdF8JXktoYFRQDE8cZVhwfmBOTkkk5r/hk39qj/o2f4q/+EZf/wDxqv8Ahk39qj/o2f4q/wDhGX//AMar/hk39qj/AKNn+Kv/AIRl/wD/ABqv+GTf2qP+jZ/ir/4Rl/8A/Gq/4ZN/ao/6Nn+Kv/hGX/8A8ar/AIZN/ao/6Nn+Kv8A4Rl//wDGq/4ZN/ao/wCjZ/ir/wCEZf8A/wAar/hk39qj/o2f4q/+EZf/APxqv+GTf2qP+jZ/ir/4Rl//APGq/wCGTf2qP+jZ/ir/AOEZf/8Axqv+GTf2qP8Ao2f4q/8AhGX/AP8AGq/4ZN/ao/6Nn+Kv/hGX/wD8aq88NfGf9njxp4c8T+IPBfiXwN4j0y8h1XRH1zRJLV/Ot5VdJUjuYwJAsirwVZcjBB6V/wAEpv2nfiT+0X4H+IVv8W/GEviPxHoWuW9x9plt0hMdtdQbY41WNFjVA9rMQFGQWbIGRmqqv+CvP7SPjf4O+Gvhz4N+F/jjWPDXiHU9Wm1ae70m7ktpfs9snlrHIyOA8Ujz5MbKysYecbcNqEP7QH7VfjvVvGMfhzxX8R/FUlvbnVLnSdFkvJhHFClvE8q20fHyRIu4jLEZJLEk/wDDJv7VH/Rs/wAVf/CMv/8A41X/AAyb+1R/0bP8Vf8AwjL/AP8AjVf8Mm/tUf8ARs/xV/8ACMv/AP41X/DJv7VH/Rs/xV/8Iy//APjVf8Mm/tUf9Gz/ABV/8Iy//wDjVf8ADJv7VH/Rs/xV/wDCMv8A/wCNV/wyb+1R/wBGz/FX/wAIy/8A/jVf8Mm/tUf9Gz/FX/wjL/8A+NV/wyb+1R/0bP8AFX/wjL//AONV/wAMm/tUf9Gz/FX/AMIy/wD/AI1X/DJv7VH/AEbP8Vf/AAjL/wD+NV/wyb+1R/0bP8Vf/CMv/wD41X/DJv7VH/Rs/wAVf/CMv/8A41X/AAyb+1R/0bP8Vf8AwjL/AP8AjVf8Mm/tUf8ARs/xV/8ACMv/AP41X/BIr4eeP/hp+zZ4k0H4jeBvEHhXUp/HF1cxWWt6ZNYzSQmxslEipKqsULI6hgMZVh2NVVVVVX7bv7N37RHi39rH4neI/CvwE+Ius6Tf648lpf6f4WvbiC4TYo3JIkZVhx1BIr/hk39qj/o2f4q/+EZf/wDxqv8Ahk39qj/o2f4q/wDhGX//AMar/hk39qj/AKNn+Kv/AIRl/wD/ABqv+GTf2qP+jZ/ir/4Rl/8A/Gq/4ZN/ao/6Nn+Kv/hGX/8A8ar/AIZN/ao/6Nn+Kv8A4Rl//wDGq/4ZN/ao/wCjZ/ir/wCEZf8A/wAar/hk39qj/o2f4q/+EZf/APxqq+BX/BKTwp8W7Xwr8W/irf8AiPw9b3c+py+IPC8tq9tc38qapdC3YO+Gghkthbk4Us6/MjLvDj4d/DTwD8JPCdp4G+GvhTT/AA9odjkw2VlFsXcfvOx5Lue7MSx7k1VVVVVVVVVVVVVVXxq+E/hv46fCjxR8JPFif8S3xNp72ryhA7W0nDRTqDwXilVJFzxuQZr9pz/gmt8Y/gPd+F9J8CaD4t+KNzqtjLPql34d8LXM9tYyrJtWIGMSHJXnLbSfQV/wyb+1R/0bP8Vf/CMv/wD41X/DJv7VH/Rs/wAVf/CMv/8A41X/AAyb+1R/0bP8Vf8AwjL/AP8AjVf8Mm/tUf8ARs/xV/8ACMv/AP41X/DJv7VH/Rs/xV/8Iy//APjVf8Mm/tUf9Gz/ABV/8Iy//wDjVf8ADJv7VH/Rs/xV/wDCMv8A/wCNV/wyb+1R/wBGz/FX/wAIy/8A/jVf8Mm/tUf9Gz/FX/wjL/8A+NV/wyb+1R/0bP8AFX/wjL//AONV/wAMm/tUf9Gz/FX/AMIy/wD/AI1X/DJv7VH/AEbP8Vf/AAjL/wD+NV/wyb+1R/0bP8Vf/CMv/wD41X/DJv7VH/Rs/wAVf/CMv/8A41X/AAyb+1R/0bP8Vf8AwjL/AP8AjVf8Eivgh8afhp+0n4k134jfCDxr4V02fwPdW0V5rfh+6sYZJje2TCNXlRVLlUchQc4Vj2NVVVVVX7bv7N37RHi39rH4neI/CvwE+Ius6Tf648lpf6f4WvbiC4TYo3JIkZVhx1BIr/hk39qj/o2f4q/+EZf/APxqv+GTf2qP+jZ/ir/4Rl//APGq/wCGTf2qP+jZ/ir/AOEZf/8Axqv+GTf2qP8Ao2f4q/8AhGX/AP8AGq/4ZN/ao/6Nn+Kv/hGX/wD8ar/hk39qj/o2f4q/+EZf/wDxqv8Ahk39qj/o2f4q/wDhGX//AMar9jr4CfHP4e/Gm18R+Pvgv478NaSmnXMTX+seHLuygV2UBVMksaqCewzzX7YOkarrf7N3jWy0bTLu/uI7e3uXitoGldYYbmKWaQhQSESKN3ZuiqjMcAE1+xx8Rbj4Z/tTfCrxhJfPFb2XiK2s55GLMIbS5cwXGAoJx5c8p2gckn1qqqv+CrvxGbxn+2Z4gsLG+d4fCGk2mhxyIQoA8szTICACQJLqVTuyc7h0wK8IfAT45/ELRV8R+APgv478S6S8jRLf6P4cu7yAuvDKJIo2XI7jPFf8Mm/tUf8ARs/xV/8ACMv/AP41X/DJv7VH/Rs/xV/8Iy//APjVf8Mm/tUf9Gz/ABV/8Iy//wDjVf8ADJv7VH/Rs/xV/wDCMv8A/wCNV/wyb+1R/wBGz/FX/wAIy/8A/jVf8Mm/tUf9Gz/FX/wjL/8A+NV/wyb+1R/0bP8AFX/wjL//AONV/wAMm/tUf9Gz/FX/AMIy/wD/AI1X/DJv7VH/AEbP8Vf/AAjL/wD+NV/wyb+1R/0bP8Vf/CMv/wD41X/DJv7VH/Rs/wAVf/CMv/8A41X/AAyb+1R/0bP8Vf8AwjL/AP8AjVf8Mm/tUf8ARs/xV/8ACMv/AP41X/BPT9kT4b+HvgN8LviR8R/gcmi/FbQptRme61XT5rK/hk+2XkcTTRPtLHyJF2+YpwPLYcqjCqqv2lP2cfgb8UNF1z4lfEH4aaPr/iXQvDN1Dp9/exs7QxxrJMi7c7WCyMzDcD95uxIqqqqqqqqqqqvhP/ySvwb/ANi/Y/8ApOlVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX7WX/Jq/xl/7J/rv/pBNVVVVVV+xx/yU7U/+wDN/6Pgr9pTR73xF8OoPD+mqjXep6zZ2tuHbaDJJJtXJ7DJHNfs//wDBPL9o/wCPnjG98Ox+FJfCWlaHqEljrWta5C0UNpNE+2aKNQMzyrz8qfLkAM6A5r9lz9hL4E/srWNtfeFtF/tzxesZFx4o1WJHuyzJtkEAAxbxnLDanzbWwzv1qqqqqqqqqqqqqqr4ufBf4YfHbwhP4H+K3g7T/EGlS5aNbmL95ayFSvmwSD54ZACQHQg4JGcEiv2sP+CRnxH+GzXnjL9nWa98deGkV5pNFkRTqtkoAJVAuBdjg4CKsnIUI5+Y+JvBXivwYmlt4q0G70o61ZteWMd0mx5YVuJrdmKH5lxNbzJhgDlCcYIJqqqqqqqqqq/4Ih/8lT+Jf/Yv2v8A6UGqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqviH4Tt/HvgDxN4FuhCYfEWj3enSCZSUKzwtGdwHJGG5q8s9R0LVJ7C7jltL/T7hopUJ2vDKjYIyOhDD8xXwn8d2nxQ+GPhb4h2RXy/EOkW96UVdvlu8YLpjLYKtuUjJwR1PWq+MX7Qfwf+AukjVPif43sdKeWMvbWAYy3d12/dQJl2GRgtjaCRuYda+JH/BXnVpJ5bX4QfCezhhVj5V74kuWlaQbhgm3gZQh2huPNblhz8vzf8PUv2ov7W/tHyvBn2ffu+wf2Q/k4/u583zMf8Dz718N/+CvOrRzxWvxf+E9nPCzDzL3w3ctE0Y3HJW3nZg52leDKvKnn5vl+C37Rvwd/aC02a++F/jCDUZrRQ13YSo0F1bAnALxPhtpPG4ZUnvVeK/Eml+DPC2seMNbkZNO0LT5768ZQCVhhjMjkAkDO1T1IrxX4hvfF/ijWPFmpuzXmtX897cF5C5MkshdiWPJOWPJ5Nfs2+Az8MP2ffhz4AktWt7nRPDNjb3cbq6t9pEKmclX+ZSZS52n7ucYGMVVVVVVVVVVVVVVVVVVVVVVVVVVVVf8ADPXw3/59b/8A8CjVrbRWlrDaQA+XBGEQE5OAMD+VVVVVVVVVVVVVf8Fvfh/HdeCvhp8VILbD6dql1otzMCvzC4iE0SkZzx9mmIwMfM2TyM/8Eb/iBH4X/aovPBl1c7YfGfhu6toIsL89zAyXCnOM8RRXHAIB3ZOcDFVVf8FXvic/xE/bD17RoJN1j4H0+10S2ILAMyp58x2kDBE1xImeciNSCRjH/BFf4Zy+HfgX4v8AifeWzQzeMteS2ty2cTWtlGQrj5iMedcXK42g5Q5JBGKqqqqqqqqqqqqqqqqqqqqqv+Gevhv/AM+t/wD+BRq1torS1htIAfLgjCICcnAGB/KqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqvFXhXSPGWkNomtJI9szq5Eb7DlenNap+zh8N9S0TV9Ea2uxHrGl3mnTeZMZB5VzA8EmVPX5JGrV9Kv9C1W90PVYPIvdPuJLe5i3BvLkRirLlSQcEEZBIr4EfECP4r/BXwL8SFufPk8R+H7O9nYhQRM8KmVSFAAIk3qQABkHHFVU00NvC888qRxxKWd3baFAGSST0GK+MfxAuvix8WPGPxMvGl8zxRrl3qKpKTmJJZWdIwCzYCKVULuIUKACQK/Yi+H7/DD9kv4W+D5rb7PcR+H4b25hJYmOa7LXUqncSch52BHQHOMDAqqqqqqqqqqqqqaGG4he3niSWORSro65DAjBBB6gjtX/AAqb4V/9E08Kf+Ca3/8AiK/4VN8K/wDomnhT/wAE1v8A/EV/wqb4V/8ARNPCn/gmt/8A4iv+FTfCv/omnhT/AME1v/8AEV/wqb4V/wDRNPCn/gmt/wD4iv8AhU3wr/6Jp4U/8E1v/wDEV/wqb4V/9E08Kf8Agmt//iK/4VN8K/8AomnhT/wTW/8A8RX/AAqb4V/9E08Kf+Ca3/8AiK/4VN8K/wDomnhT/wAE1v8A/EV/wqb4V/8ARNPCn/gmt/8A4iv+FTfCv/omnhT/AME1v/8AEV/wqb4V/wDRNPCn/gmt/wD4iv8AhU3wr/6Jp4U/8E1v/wDEV/wqb4V/9E08Kf8Agmt//iK/4VN8K/8AomnhT/wTW/8A8RUMMNvClvbxJHHEoVERdoUAYAAHQY7VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVXlnaahaTWF9bRXNtcxtFNDMgdJEYYZWU8EEEgg8HNf8Km+Ff/RNPCn/AIJrf/4iv+FTfCv/AKJp4U/8E1v/APEV/wAKm+Ff/RNPCn/gmt//AIiv+FTfCv8A6Jp4U/8ABNb/APxFf8Km+Ff/AETTwp/4Jrf/AOIr/hU3wr/6Jp4U/wDBNb//ABFf8Km+Ff8A0TTwp/4Jrf8A+Ir/AIVN8K/+iaeFP/BNb/8AxFeLf2fPhF4w02PTLrwVpunJFOJhLpdrHZykhWG0uiglfmzt6ZAPavCX7L3wf8G61Fr2naFLd3EKkRpqEguY1PZgjDG4Y4PUVVVVVVVVVVVVVVVVVWt+B/BXia6S+8R+ENE1W5jjESTX2nxTuqAkhQzqSBkk46ZJ9a/4VN8K/wDomnhT/wAE1v8A/EV/wqb4V/8ARNPCn/gmt/8A4iv+FTfCv/omnhT/AME1v/8AEV/wqb4V/wDRNPCn/gmt/wD4iv8AhU3wr/6Jp4U/8E1v/wDEV/wqb4V/9E08Kf8Agmt//iK/4VN8K/8AomnhT/wTW/8A8RX/AAqb4V/9E08Kf+Ca3/8AiK/4VN8K/wDomnhT/wAE1v8A/EV/wqb4V/8ARNPCn/gmt/8A4iv+FTfCv/omnhT/AME1v/8AEV/wqb4V/wDRNPCn/gmt/wD4iv8AhU3wr/6Jp4U/8E1v/wDEV/wqb4V/9E08Kf8Agmt//iK/4VN8K/8AomnhT/wTW/8A8RWh+DfB/hiWSfw14U0fSZZlCyvY2McDOByASgGR7Gqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/4Kj/AALvfg9+1Xr/AIhtrNk0H4hs2u6fMRkNPIR9sQnAG4T73x2WVM5JzX/BKX9oWC50nUv2cvEuo4ubN5NR8MiQn54my1zbr2G1/wB6B1PmSnotft4/tE/Ez9nT4YWWu/Dfwkt5NrFy9pNrs2JIdGf5ShaHB3tIPMCk4RSnzZyqnWdb8dfFTxc+p6zf6v4n8SaxPjc5e5uLiQnhVUZJ9AqjA6AAV8Fv+CWvxm+INpaa58SdYs/AWm3C7/s08ButQ284BhBVYycD7z5G7lcjFf8ADoj4Of2T5H/C0PGX9p78/adlt5O308ry92ffzPwr4x/8EqvjJ4JtJ9Z+F3iHT/HllCu5rMRfYL7HJO2N2aOQADtIGY9EPStO1Lxn8NfFceo6Xeav4Z8SaLOdkkbSWl1aSAEMOzIcEgg9iQeDX/BP39pH4p/tDfD/AFF/iR4Y+fw68VrF4kjQxR6q23lSnQzLjc7Jhf3ifKuef+CpH7RGm+EPhwnwD0K8Z/EHi5Y7jU/KYj7Jp6SBgGII+aWSMKF5yiSbsBl3fsCfAC7/AGh/2mvCvhuay87QNBuE1nX2ZTs+yW7qxibGD+9fZFwcjzCexqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/4KS/D9/iJ+xh8RrK1tvNvNEs4tZt2y37sWkySzNgEZ/cLMOcgZzjIFfsn/EaX4S/tKfDbx+k7RQ6Z4itVvGRirfZZX8m4AO5RkwySDBIU5+bIyKqq8Ra7pvhbw/qfibWZ/JsNIs5by6k/uRRIXc/gqk14+8Yap8RPHXiP4ga4U/tLxNq11qd5sUAedcStK+AAABuc9AK/ZG+GEvwb/Zm+G/w5u7JrS+0zQYH1C3dCrRXk+Z7lSDzkTSyDnmqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/4KKfD9/hz+2V8TNNW28u31fVP7Zt2BYiUXqLcOw3En/WySqe2VOABiv8Agkf8RpfHP7H2m6BdzvJc+CNavNHLSMS7RkrcxnlidoW58teFAEe0D5cmq/bZ+IKfC79k34p+MBM8U0fh2eytZUJDR3F3i1hcYVuRJOh6Y45IGSPhD4El+KXxW8G/DWB3RvFOvWWmGRSQYxPMkbPkK2AoYknacAZwatLO0sLSGwsLaK2traNYoYYUCJEijCqqjgAAAADgVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV/wUm/bs+P37MHxz0PwB8KtS0SDSb/AMJ2+pzLfaYtw5ne7u4mIYkYG2FOPXPrX/BNr9uz4/ftQfHPXPAHxV1LRJ9J0/wncanCtlpi27idLu0iUlgTkbZn49celVVVVVVVVVVVVVVVVVVVVVVft2/spWX7WPwRu/C+nJbw+MdCZr7wxeTHaqXAA3wOcjCTINhJ4VtjkHZg2tz4z+FPjtLiH7Z4f8V+EtUPyyR7JrG8t5MFWRhwyuhBVhjggjrX7Nv7Tnww/bd+F2pfCr4mWGnweKLrT2tdb0N32Jfxlebm0yd2AQGwDvhcA54Vz+zf+xl8Hv2aFn1HwvZT6x4juWdX13Uwr3CQknbFGAAsS7eGKgFz94kBVWqr9pL9kL4S/tNabG3iuwbTPENooW08QafGq3SIDny3JGJY+uFb7pJKlSTn4xfG/wCCX7BXwh0zwN4dsIP7Th09k8PeHoADLcuAR9ouGGNqNJkvIfmc7toYg48deN/GHxY8b6l438Y6nPq/iDXrkSXExX5pHwFRFUdFChUVQMABQBgCv+Ccn7Icn7LXwea+8W2nl+PfGXlXWuo21jYogbybRWUkHYGZmIOC7sOQoNVVVVVVVVVVVVVVVVVVVVVVVVVVVVf8Lz+Kn/Q1f+SNv/8AEVpE8t1pNlczvullto3dsYyxUEnAqqqqqqqqqqqqrxb4Z0rxr4V1nwbrsPm6br2nz2F5HtVt8M0bRuMMCpyrEYII9Qa8RaDqXhbX9T8MazD5OoaReTWd1H/clico4/BlIr9nj4gp8V/gR8P/AIjec8k2v+HbK6uS7FmW4MK+cpJVdxWQOu7ABxkcEVVf8FNvig3wv/Y28bvaXaQ6h4rWLw/ZhyMSC6bFwuNwJJtVucYycgEjaGI/ZL+Fsfxp/aT+HXw1uIPOs9W1yFr+Py1fdaQ5nuBhgVP7mKTqCPUEcVVVVVVVVVVVVVVVVVVVVVVVX/C8/ip/0NX/AJI2/wD8RWkTy3Wk2VzO+6WW2jd2xjLFQScCqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqvjJ4k1rwp4Jl1fQL37LdrcRoJPLV8AnkYYEfpXwo+K/j/xN4/0rRNb1/7TZXPnebF9lhTdthdhyqAjlQetf8FtvhzDpPxT+H3xUtYEUeI9FuNMuTGuMy2cqurvhR8zJdhQSxJEQGAF5/4Ig/EFLbxf8TfhTcTOzajptprVpGScRi3kaGcgbcZb7Tbg5YH5BgHkiq/4LRfERvDv7PPhn4eWtykc/i/xEJJozIA0lraRl3AXOSBLJbEnoMAH7wr/AIJQ/D1vHX7ZnhvUniSW18Habe61cI6gghY/s8Z5YEFZrmFhgMcqOMZIqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/ar+IPi/4a/DvT9d8E6x/Z19PrUVtJL9njmzGYZmK4kVh1RTnGePrX7NHx4+K/xB+KNv4c8X+K/t+nPZTyND9ht4ssoGDujjVv1qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/4LV/8AJ1Hhb/sn9n/6X39f8EVP+TqPFP8A2T+8/wDS+wqqqqqqqqqqqqqqqqqqqqqqqv8AgoP/AME5dP8A2jYZfiv8HbSx0z4lwKovLd2EEGvxgAASN0SdVACyHhgAj8bWTU9L8e/Cbxo+napZ634R8V+H7lS0ciyWV5YzDDKw+66NgqwIxwQQcEGvgR/wVb8Y+G4odC+PfhxvFFnGu0axpSRwXwx/fiJWKU+4Mfqd1eB/24/2VfH1tHNp3xk0PS5WyGg1yQ6Y8ZABIJn2qeCOVYg8gEkHEXxa+FM9impwfE3wnJZyRCZLhdatzG0ZGQ4YPgrjnPTFeNf22f2V/AtpJc6l8a/DmoukXmJBotz/AGm8h5wo+z7wGJXHzEAZBJAOa+On/BWTUtTtLvQP2fvCM+kmRQsfiHWhG86H+Ix2o3xg8cM7OCDygNX9/wCNvif4ue9v7nWPFPibW5wC7mS8u7yUgBQOrOcAAAdgAOBX/BO3/gmwPg5LZ/G79oDR7efxyjb9E0N2SePRMdJ5CpKvcnquCREMHJc/u6qqqqqqqqqqqqqqqqqqqqqqqqqqqqrQP+QFpv8A16Rf+gCqqqqqqqqqqqqq/wCCkPw9b4dftmfEexjhRbXW75NatmVQokF3Gs0hwGJBEzSqc4JK7sAMK/4JBfERvGf7Itv4XublHufBGvXmmrG0gMggkK3SMRnIXNxIik/88yB92qr/AILefE6V9Q+G3wYs71ljihudf1C3DnDsx+z2rkeoCXYz/tnGOc/8EWvhhH4l+O3iz4o3Sb4fBWhpbwDC/Jc3rsqt1z/qoLgcAj5uSOA1VVVVVVVVVVVVVVVVVVVVVVVoH/IC03/r0i/9AFVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVftC/8k3n/wCvuH+dfAz/AJKpof8A28f+k8lf8Fifh63iz9k1PGFtEnneCfEVpeyylQWFvNutWUZYYBkngJwGPyDjGSP+CZ/xEb4dftmeAZZLlIrPxFNNol0HkCCQXMbLEuScZ88QEDqSAByRVV/wWi+IieIv2hfDPw7tblpIPB/h0STIZCRHdXchdwFzgExR2xJ6nIB4UV/wRC+HaJp/xN+LNzbMWlmtNEspjGQAFVp7hQ2MEnfbEgHIwM/eFVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVftyf8kl0n/sYof/AEnuK/Y2/wCS2Wv/AGDrn/0EVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVf8FePj98ZfgZ/wqf8A4VH8Q9V8L/23/bX9ofYXVftHk/YfL3ZB+75smP8AeNfsi/8ABSL4k+BPD/xQ8d/Hn4i6r42bStHtU8N6HfXSI15fyzlQqHbwAoZ3bBwiMQCcA/Er9vv9rv4oa8+u6j8dPFGhLuYw2PhrUZdItoVOPlCW7KXAwMGQu3Xnk14m8S6B4M8O6l4s8U6tb6Zo+j2sl1fXlw22OCGNSzux9AAa/ac/4K7fGT4havcaF+z/ACN4B8M280iR35jjn1HUY84V3Lqy24IAOyP5gTgyEcVa/tiftYWepJqsP7SnxLadJDIEl8UXcsWT6xNIUI/2SuPav2G/+CrPjLWvGej/AAh/abvrfU4dcultNN8Vpbx28tvcSMqxR3SRhYzESdvmgAqSC+Rudf8Agphrvxe8D/sz3fxN+DPj3VfDGqeE9Utp9QewIzdWcreQyEHPSSWF884CN2JI+Fn/AAUJ/ak034n+D9R8a/HTxFqPh6116xl1azublEjubRZ0M0bsEJCtGGBODgGry7tLC0mvr66itra2jaSaaZwiRIoyzMx4AABJJ4FfEP8A4KIftVa54/8AE2teEfjh4l0vQ7/WLu40ywhlVUtbZ5maKJQVyAqFVAPPFfsC/GXWfjv+yp4K8c+KdWl1HxCkc9hq9xMPnknt5njDsQqgl4xG5IB5fBJIJr/gqZ+2N8avhf8AtC6d8N/g38TtS8N2OkeHYJdTisGCtJeTSSP85OQQIRARwD8zZzxj/gk38Wv2k/j1488a+Lfit8VPEHiLwt4a0uK0gtL5v3T39xIGV1K4DGOKGQFTnHnKTg7c/wDBSb9u/wDai+BPxsufg98N9Q0nwpon9l219ZapFpwuLy+jmXDsWuFaNQsscqDy1yNpJfd8qar+2J+1hrN/LqV3+0p8S45psbltfFF3axjAAG2OKRUXgDOAMnJPJJr4Rf8ABRz9rr4Saza3y/FrVfF2nRSFrnS/FM7alHcqc5UyyEzJ1yCki4IHUZU/sk/tQ+EP2s/hLafETw7b/wBnajbyfZNb0h5hI+n3SgEjI5aNgdyOQNynkAhgP+Crfxf+JnwV/Z28PeKvhV4z1Dw1q1140trKa7smCu8DWV47RnIPBaND9VFfsaf8FCfi7o/xV1LxJ+0H8Z9d1zwhofhnUL2XTrqaP/S7hEHkRRggZldyqKCQMsMkDJHxv/4KO/tV/GXxJc6lY/FDW/BGjC4Z7DSPDF/JYC2jydqvPFtlmO08lzgkZCrwB8CvHdrpf7JXw5+JXxG8UssMHw90rU9a1nU7lpGP+gRSTTyyMSzMTuYsckk9ya/ac/4K0/HH4jeJ73SvgPq0vgLwdBIY7aWOCJ9RvlVsiWWRlbyd2AfLjxgEqzOCa8M/tr/tc+EtVj1nSv2j/iBPcR42pqeuTalCcMG5huWkjPKjqvIyOhIP7U/7Tvin9q/xd4a8e+NtKsbLXNG8Mw6NevZKUiuniuLiXzghJ2FhOMrnAIOMDAr/AIIqf8nUeKf+yf3n/pfYVVVVVVVVVVVVVVVVVVVVVVVVftFfsgfAf9qLR/sPxQ8IIdTiXFpr2nEW2o2nBA2zYO9cE/u5FdM87cgEfGz/AII2fHvwddT3vwW13SfiDphk/cWs0semX6KSMBhM4hbAJywkXO3O0ZCjxj+zD+0Z4AuvsnjD4G+ONNJYKksmhTtDIxUNhJVUo5wRkKxwcg4INV4T/Z7+PHjy7isvBvwZ8a6xJLsINpoNxIiq5AV2cJtRDkfOxCjqTivgn/wR+/aQ+IE9pqPxSutJ+HWiybHlS6mW9v2Rhn5IISUU4wCJJEZcj5SQQP2a/wBiD9n/APZbhS++H/hh73xI0Jin8R6q4uL2QEfMEOAsKnusarkYznrVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVWgf8gLTf+vSL/0AVVVVVVVVVVVVVX/Bbr4dJp/xB+HHxVtbZ8a1pN1pF3IkZ2q1rKsse5sY3Mt1IBnkiI9lr/giN8RE0/4g/Ef4U3N0+Na0m11e0jeQ7Va1laKXaucbmW6jJ4yREOy1Vf8ABRn4oj4r/th/EHVLW++0adoV4uh2ADbljSzQRSBTtXIacTv3HznBYYJ/4JAfDFPBX7KEfjS4gdbzx3rVzf7nY5+zwt9miXb0A3QysDjJEg5I24qqqqqqqqqqqqqqqqqqqqqqqtA/5AWm/wDXpF/6AKqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/aF/5JvP/wBfcP8AOvgZ/wAlU0P/ALeP/SeSv2nPh2nxZ/Z5+Inw7Fs08+s+HbuOzRIzI32pYy8BCgEsRKkZwOTjArwb4n1DwT4v0LxnpLst9oGpW9/bMkhjYSwyLIpDLypyo5HIrw9rum+KfD+meJtGn86w1ezivLST+/FKgdD+KsDVfts/EFvij+1l8U/GAmSWF/EU9jayoQRJb2mLWFxhV4McCHpnnkk5J/4JcfDtvh9+xl4OlubZIbzxTNda3cBYwpYTSFYWJAyxMEUByecYHQCqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv25P+SS6T/wBjFD/6T3Ffsbf8lstf+wdc/wDoIqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/4Ln/APNEv+5g/wDcdXwb+FHin45fFHw18JvBcKPq/iW+W2heQHZAmC0sz4ydkcavI2ATtQ4BPFeEv+COX7KmjaBa2Him98W+IdURf9J1A6iLQSt32xRrhF9ASx9WNf8ABaP4w6p4S+EHg/4Q6JqFxbN441Ke51IwSACWzs1T9zIM52vLPE44wTAeeMH9imf9nLTvjxpviH9qLWrey8GaJay3i211pk1/BqN2pRYreWGKKUsnzNIQyhT5W0n5tp+IH7d//BMz4peAbz4YeOfiNpmp+Gry1Nt9gk8Iapst12FEaH/RP3ToD8jphkIBUggVq9tYWeq3tnpWpf2hZQXEiW135Jh+0RhiFk2Nym4AHaeRnFfA++/4a/8A2B9EtfFep/bbrxz4LuNH1K/lj3ObxFktJbgh1/1gmiaTdtwHGVyMGtR0+90nULnStSt3t7uymeGeFxho5EJVlPuCCK+M37Q5tv8AgmFN8ZF1V/7W8TeALOxWXznE32+9ijtZijMyuzxvJK+8En92XG4DnT/Cmr6p4Y1jxZZRJJY6FNbR3oyd0YnLhHxjG3dHtPOcuvBySP8AgiH8RvtPhT4lfCO4l2/YNQtdcs0253+fGYJzkLxt+z2/VjnfwBhif23viBH8UP2tPil4vguftED+IJrK2lwoEkNoFtYmG0DgpApHfGM5OTX/AASp+EUPww/ZF0HW7iBF1Tx5dTa5dMYsMI3Iit13dWXyYkcDgAytgckn9pY/sd/D7U9K+N/7TGneETq2l2rWmiz61b/bJ3VHEhW2tSG8x1dg25ULJuB3KDXjD/gtL8CvC+7R/hh8IPEuu2djbrHaNcSwaVAxXgIiqJWSMKFwdoPbaAAT+0d8VvB3xt+LGq/ErwT8KrH4eWerqjz6NZXv2mL7QBiSYERxqpc4YqqAZyeSST/wRJ8ZalafGnx98PEeU2GqeF11R183CLLbXUUSnZjklbtvmyMbcYOeP+C1f/Jq/hb/ALKBZ/8ApBf14e8P6z4t1/TPCvhvTpdQ1bWbyKysLSEZe4nlcJHGvuzMoHua+Ff/AARq/Z90PwhZwfFnW/EHibxNJGrX09jffY7WKTaNyQoF3FAcjc5y2AcLnaP+CsPimy+Bf7JHgD9nXwNe3FnY6vNb6ZHE8u+SXStMgQeWzdSfMNmSe+CD96v2dvhDdfHv44eDPhDaXMtsPEuqRwXNxDGXeC2UF7iRVAOSsKSMM/LkckDJHgr9lr9nb4f+C4fh/wCHPg14RGjJCYpYrvSYbp7sMu12neVWaZmUAEuSSMDoAK/4KOfs5eEf2Zv2kLjwj4BR4PDuv6TDrenWTymU2KSyzRNCGIztWSB9uSxCFQWJzX/BFT/k6jxT/wBk/vP/AEvsKqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq0D/kBab/16Rf+gCqqqqqqqqqqqqq/4LBeAI/Fn7Ic/itI4hP4J8QWWoGUou/ypWNoyAnkAvcxMQOpRSelf8E3/iC3w6/bM+HF68yLa63fPotyjkKJBdxtDGMlSQRM0LDGCSoXIDGq+K/j7TfhV8MfFfxK1eWJbXwxo91qDiVtocxRsyoPUswCgDklgByaml1fxNrTzOLjUdU1W6LEIheW4nkbso5LMx6Ack18GPhzZfCL4SeD/hhYBDH4Y0W1sGdW3CWSONVkkzhclnDMTtXJboOlVVVVVVVVVVVVVVVVVVVVVVVWgf8AIC03/r0i/wDQBVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX7Qv/ACTef/r7h/nXwM/5Kpof/bx/6TyVX7Tnw7b4TftDfET4eC2SCDRvEV3HZxpGIwLVpC8BCgAKDE0ZwOBnA4r/AIJz/EFviP8AsZfDPUp5ka60fTW0WdFIJjFlI1vEDhVGTDHE3TOGGSxyT8YviBafCf4TeMfiZeNF5fhfQ7vUVSUjErxRM6RgFlyXYKoXcMlgAQTVpaalr2qwWNpHLeahqNwscSZ3PNK7YAyepLEde5r4e+DbD4deAPDPw+0p99l4Y0e00u2bBGY7eFYlOGZj0QdST7nrVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX7cn/JJdJ/7GKH/0nuK/Y2/5LZa/9g65/wDQRVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV/wXP/AOaJf9zB/wC46v8Aglx/yfb8Mf8AuLf+mm8qv+C4Xh7UIfF/wq8VEI1jd6bqNkpUEsksUkLndxgArMuOcna3Axz+zX+zp4t/ak+JH/CrfA3iXwzo+stp817A2vXU0EVyIiu6KMxRSsZNrF8bQNsbnPGD/wAOVP2qP+h++FP/AINb/wD+Qq/4cqftUf8AQ/fCn/wa3/8A8hV+x58Etf8A2cv2b/Bvwa8U6tp+p6t4fjuzd3On7zAzz3c1xhC4ViF84LkquducDOK/4KVfCx/hV+2N48t4bSWGw8U3C+IbJ5N374Xg3zuCe32kXK8cDbjjGB8U/jpd6z/wTS+DfwlXU3klh8ZavDdESyM7x2WJUic5xtVdUgwhBGI4yMbcV+yp+zvP4z/4Jq/tEeL/ALHuvdauIbiw3Rhm2aKBdF485wW824iyPmPzDvX/AATL+L0Hwc/adTWbyWJbPVPC+r2twJp0gSQRWxvERpHB2bpLONQRzuYDkEg+CPC/iH4xfFHQ/B9rPLc614y1yG0ExQOzTXMwUyEEqOrljkqOuSBzWk6Z4b+HPgqy0awSLTdA8L6XHbwrgKlta28QVRgDgKiDoO1ftHfHrxh+0j8XNf8Aij4uvrhv7Run/s6xeTMenWg+WKCNRwoVFUEgDc25jlmJP7EX/BKjwf8AGj4WaH8avjb4y1iGy8QqbjS9B0Ro4WECylQ9xOyvkSBWPlxhWVSp3hiVX/gpN+zj8M/2YPjnofgD4VWuoQaTf+E7fU5lvrs3Dmd7u7iYhiBgbYU49c+tf8EVP+TqPFP/AGT+8/8AS+wr/gtX/wAmr+Fv+ygWf/pBf1+yd/ydR8Gv+ygaF/6Xw1X/AAXH8Pahc+GvhD4qiCfYdOvtWspiQd3m3CWrx44xjbbSZyQemAecf8E9/HegfDb9sv4X+KfE10ltp/8AaU1g8zttWN7y1mtY2Y/wqHnQkngDJJAyRX/BZXxHo+uftb2GmaZeJNc+H/BtjY6gikEwztPc3AU46HyriFuezCv+CKn/ACdR4p/7J/ef+l9hVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV/wzFoH/Q0X/wD35SrK1WysreyRywt4ljDEYJ2gDP6VVVVVVVVVVVVVX7Q/w+T4r/Ajx/8ADkRPJNr/AIdvbW1CKWZbgwt5LBQy7isgQ7cgHGDwTXhjxBqHhLxLpPirSSgvtFvob22L5x5sTh1ztIOMqOhB9xXhPxNpXjXwro3jLQZvN03XtPgv7OTcrb4Zo1kQ5UlTlWHIJHoT1r/grn8UE8B/skXvhO2u3iv/AB5q1tpcYjJDiCNvtExyGGFIgWNvvZEu3GGJH/BPv4YQ/Fn9r74ceHb2yS50/T9SOrXySJuj8uzRpwHB6qzxxoQeDvweCaqqqqqqqqqqqqqqqqqqqqqqqv8AhmLQP+hov/8AvylWVqtlZW9kjlhbxLGGIwTtAGf0qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqrx14OtfHfh9/D93ey2sbypJ5kagkFfY14N+BOkeDPEln4ltdeu7mWz37YpIlAbcjIckezZqv8AgsT8PU8KftZJ4wtoX8nxt4dtL2WUqQDcQbrVlGWOSI4ICcBR844zkn/giP8AEOPUPhx8RfhXcXuZtF1i31a2hdlBMd1EYnKc7iFa1XcMAKZF5Jev+CsXxAfwN+xp4g023ufIuPGGqWWixMA2SGc3EijAIG6K2kU7sDBPOSK/YC+HEvxQ/bB+GGgCB3t9P1pNXum2kosdiDc4f5WAVmhWPkAEuFyu7Iqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr4yfCbTvjL4XtfC+p6vcadFa36XiywRhmLLHIm3B7YkJ/CvhP+y5oHwn8Xx+L9O8VahfzR28kIhngRVIcYJyOe1VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX/AAXP/wCaJf8Acwf+46v+CXH/ACfb8Mf+4t/6abyq/br/AGXYf2rvgPqHgrTDbweKtImGpeG7qYbVW6RSDC7dQksbMh5wrFHIOwCr/T/iL8E/iE1lf2+seEPGXhW+VtrBra6sbhCGRgeCD91gw4IIIJBBrwr/AMFj/wBrvw7o6aZq9p4B8UXKMSdQ1XRZY52GAMEWs8MeOM8IDyeemNO/4K6/tQeKfiJ4Uk8Ral4X8P8AhmLXrCXWbPRtM8kXNmkymeJpbhp3jV0zuZTkdsDINf8ABbj4Vtd+Hfh58arG3cnT7qfQtQdUBG2VfOt8nOQAY7gdCMuOVOA0moXsunwaVJcu1pbTSTRQnokkiorsPciKMH/dFfspfBy1+Ff7KngH4Ra1pbI0Ph1RrFlcBsi5ug013GwPIHmzyrjjA4wOg+J/g3XPgn8WvGHw5lupY73wzql/o7z7CnnxAyQM4DAHZJESQcDKuCOtf8EhPhM3j39qdfHF7p6T6b8P9JmvjJIoZFu5h5EC4P8AFh5pFOODFnIIFa5o9l4h0TUNA1FXa01K1ltpwjbWMcilWwexwTzXxe+Ffi74J/EjXvhf4406W01bQLx7eTfEyLOgPyTR56xyLtdW6FWBr9nb/gqn8Zf2ffhNp/wkg8F+HPE1joivHpN3fvLFLbxMSwjfyyBIqsxx9044zwMfGz4jfFf44eLbr44fFMXtzN4nupYbW+Nq8Vn+4Cbra2J+XbCssQKAkqHUsSX3H/gip/ydR4p/7J/ef+l9hX/Bav8A5NX8Lf8AZQLP/wBIL+v2Tv8Ak6j4Nf8AZQNC/wDS+Gq/bR/Zxh/al/Z/134YW9zb2utoyahoNzcJlIr+EN5YY9UV1Z4mcZKrKxw2Np8a+CvFfw58V6p4G8c6Dd6Nr2i3DW99Y3SbXhcfoQQQQwJVlIYEgg14P/4Kd/tl+CfBMPgfTPihDeRWkflWmo6npkF5eQIAAF82RT5mMHmUO3PXAAHxA8H/ABSk8PaN8cPiMmoTw/EfUNRex1PUZGafU5LYwm4ny3LIXuFUP0LK4/hNf8EVP+TqPFP/AGT+8/8AS+wqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr9rH4cy/CX9pT4k+AHgaKHTPEV01mrqVb7LK/nW5I2qMmGSM8KFOcrkYNf8E2viA/xE/Yw+HN7c3Pm3miWcui3C4b92LSZ4YVyQM/uFhPGQM4zkGv+C1vxNTXfjF4J+FNnO7Q+FNFlv7lQpUC4vHA2nP3iI7eM5GQPMIzncK/4IifDOWfxL8SPjHdWzJHZ2NvoNjMc4laZ/PuFGG4KiC1Jyp/1gwRhgaqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/4LbfDiXV/hZ8PvipawO58N61caZcmNScR3kQdXfCn5Ve0CglgAZQMEtx/wAEefiA/hT9rQeD5LnbB418P3lksJDEPNABdKwwDghIJhzgYY9yBX/Bb/4gRz6/8MPhVa3OHs7O91q9hwp3CV0ht2zjIx5FyOuDu5HAx/wRP+H8etfGrxz8SJ7bzY/C/h+KyiYlcRTXk2VYDO7Pl2sy8AjDHOCRmqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq+On7LfwI/aW/sT/hdngX/hI/+Ec+0f2Z/wATO7s/s/n+X5v/AB7yx7t3kx/ezjbxjJz8Lf2Cf2Tfgr470v4mfDL4Uf2N4l0bzvsN7/buoXHlebC8Mn7ua4dGzHI68qcZyMEAiq+L37OvwP8Aj3aQ2nxe+GeieJWto2it7m5gKXMCMCGWOeMrKgO4nCsBnDdQCNQ/4JH/ALF17ey3Vt4S8R2McjZS2t9fnMcQ9FMhZsfVifevAf8AwTy/Yz+HF/Fqvh/4EaJdXkW0iXWZp9VG5QQGEd1JIin5iflUc4PVVxXxc+Dvw3+O3gq4+HfxW8NJrvh+6mjmltGuZrc+ZG25GEkLo6kEdmGRkHIJFWn/AATB/YWsruC8g+BELSW8iyIs3iHVJUJU5G5HuSrDjlWBB6EEVXxJ/wCCf37IXxd8b6r8RviB8HotR8Q63Isl/dxazf2gmdUVA3lwTpGCVQZIUbjljkkk/A39mD4Ffs1w6zB8E/AieHF8QNC2pEahdXbXBhDiPJuJZCAvmycLgfMar44/swfAj9pCytbT4xfD2x12WwVls7wSSW11bA9Qk0TK+3POwkqT1Brwr/wSn/Yu8L6umrzfD/UtcMSkR22q6xPLAGyPmKIV3HAIw2V5PGcEfFn9j/8AZu+OGj+GPD3xI+FtjfaX4Lhmg0Gxsru50yDT45REHSOO0kiXaRDEACCF28Yyc/Bv9in9mb9nzxc/jv4P/DiXw9rktnJZSXKa7qFwJIHKs0bRzTujAsiNypwVBGCK+NfwE+E37RXhW18E/GPwn/wkGi2OoJqEFr9uuLTZcpHJGr77eRGOElkGCcfNnGQCPCf/AATh/Yv8DeKtG8beFfgz9i1rw/qEGoabc/8ACQ6nJ5FzBIskT7HuSrYdVOGBBxggiqr42/sqfs/ftFQxj4u/DPTNau4F2w6ihe1vIhjgCeFlkKjrsJK56g14D/4Jd/sYeA9Zi15fhhL4hubeRZII9e1Ga8gjIznMBIilByMrKrjgYA5z8Zv2X/gP+0FomieHPi18PrfWtO8Nsx0q3ivLixFpuVUIT7NJGdu1VG08DaOOK+Cn7GX7NX7Oviq68bfBv4bf8I/rV7p76fPc/wBsX13vt3kjkZNk8zqMvDGcgA/LjOCc1VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX/DTuu/9CvYf9/nrTrpr7TrW9dQrTwpIVHYsoP9aqqqqqqqqqqqqqv+CyHw+j8L/tUWfjO1ttsPjPw3bXM8uV+e5gZ7dxgHPEUVvyQAd2BnBx/wRD+IEd14K+JXwqmucPp2qWus20JC/MLiIwysDjPH2aEHJx8y4HJz+2T8To/jJ+1F8SfiDbyeZa32uS29k+VO+2tlW2gb5QBzFCh7nnkscsf+CXXwwl+Gf7G3hGW9smtr/wAXzXGv3KOhUsJ2CwP7hraK3YH0I7c1VVVVVVVVVVVVVVVVVVVVVVVf8NO67/0K9h/3+etOumvtOtb11CtPCkhUdiyg/wBaqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq+JfjG58C+FpNftLOK6kSZIxHIxUfMcZyK+H3x21bxn4v0/w1c6DaW8d55m6WOViy7Y2fgH/dx+NV/wUU+H8fxG/Y0+JmmrbebcaPpf9s27AqDEbJ1uHYbiB/qo5FPfDHAJxX7OXxAj+FXx8+HnxEnufItdA8SWVzePhT/owmUTj5gQMxFxnqM5GCM1/wAFRPiG/wAQf2z/ABpFFe/aLHwtHa6JZ/Mx8sQwq0yYJIGLiS44XA74yST/AMEb/h+/hf8AZWvPGd1bbZ/GfiS6uYJct89tAqW6jGccSxXHIAJ3YOcDFVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVftA/FnUfg34Ms/E+maTbajLdamlmYp3ZFVWilfcCOc5jA/Gvgd+1Hr/AMWfHsPg/UfC2n2EMlrLMZoZnZgUAIGDxVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVaB/wAgLTf+vSL/ANAFVVVVVVVVVVVVVX/BbH4dy638GvAnxMt0eRvCuvTWMwXOEhvYgS59g9pEv1ce9fsTftG3X7NnjTxx4ittS+zSa14C1axsRISY/wC0FiE1mxG9BkSxBQTk4dlUbnFeDfCmr+PPF+heB/D0SSap4h1K306xRyQGnnkWOMHAJxuYdAa8J+GdK8FeFdG8G6DD5Wm6Dp8FhZx7VXZDDGsaDCgKMKoGAAPQCqqqqqqqqqqqqqqqqqqqqqqqqtA/5AWm/wDXpF/6AKqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/aF/5JvP/wBfcP8AOvgZ/wAlU0P/ALeP/SeSq1fSrDXdKvNE1SDz7LULeS3uYtxXfG6lWXKkEZBIyCDXjjwnqXgLxpr/AIF1kY1Dw5qlzpt0Nu3EsErRvx2+ZDWua3q/ibWtQ8R+INSuNR1TVbqW7vry5kLy3E8jF5JHY8lmYkknqTX7JXw8j+FX7Mvwz8CfY/stxp/hu0e9h2qNt3Mgmufugf8ALaSQ5Iyc5JJyTVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX7cn/JJdJ/7GKH/ANJ7iv2Nv+S2Wv8A2Drn/wBBFVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVf8ADMWu/wDQ0WH/AH4etOtWstOtbJ3DNbwpGWAxkqoGf0qqqqqqqqqqqqqq/wCCgXw8j+Jv7HnxO0JbP7Rc2Gjtq1rtVS6SWTLc5QkHBKxOhA5Kuygjdmq/4JZfDFviT+2R4WupYElsfBlrc6/dqzAECFRFCwz1IuJ7c4GTgH6iqqqqqqqqqqqqqqqqqqqqqqqr/hmLXf8AoaLD/vw9adatZada2TuGa3hSMsBjJVQM/pVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVXxL8HXPjvwtJ4ftL2K1keZJBJIpYDafQV8PvgTq3gzxfp/iW5160uY7PzN0UcTKzbo2Tgn/ezVV/wU1+Hkfw7/bP8exWln9nsfEUkGt2vyqPMNzCrTvhQBzcC49ScZJJJNfs8fD1viv8d/h/8OfJSSHX/EVla3IdQwW3Mq+cxUsu4CMOduQTjA5Iqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/aB+E2o/GXwZZ+F9M1e206W11NLwyzxsylVilTaAOc5kB/Cvgd+y5r/wAJ/HsPi/UfFWn38MdrLCYYYHViXAAOTx2qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqtR0+y1bT7nStSt0uLS8heGeFxxJGwKsp9iCRXxD8G3/wAOvH/ib4fao++98Maxd6XctgDMlvM0THCsw6oehI9z1r/giR8MUsvBHxD+MN1A/m6rqUGiWbsxAEdvH50xA6EM08QyQcGMgEfNmqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr/AIK5fsu/FT4wePvhv42+Efw81jxVey6bd6TqQ02yMn2VYXE8BlfOFVvOuANwABXGSXUV/wAE4f2Lfjh8Nf2v7DxJ8aPhXrfh2z8J6PqF5aXl1biW0mujttVjWeMtEz7biSRRu5EZZcgZqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqy8BeBdMl1WfTfBeg2suvNI2qvBpsUbX7SZMhmIX96Wyd27Oc814b8JeFfBthJpXg/wzpOh2UtxJcyW2m2UdrG80h3SSFYwAXY8lsZJ5NVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVfHP8AbQ8T/CX4p638PtO8F6Xf2+lfZ9lxPcSK7+ZbxynIHHBkI/Cv2e/itqHxo+HEHjfU9Jt9Onlu5oDBA7OoCEAHJ55zVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV+0L+194k+DHxHm8D6X4P03UoIrOGcT3E7oxLgkjA44xX7NPxo1P47eBL7xdq2i2ulzWeryWKw28jOrKsUT7stznMpH4CqvLu0sLSa+vrqK2traNpJppnCJEijLMzHgAAEkngVP4h8P2um2+s3Ou6fFYXfl/Z7uS5RYpvMxs2OTht2RjB5zxVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVWv6i+j6DqOrxRrI9jaSzqjHAYohYA+3Ff8PFfGn/ROdE/8C5aqry7tLC0mvr66itra2jaSaaZwiRIoyzMx4AABJJ4FS+IfD8NpYX02u6eltqskcVhM1ygS7eQZjWJs4csOVC5z2qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqvH/iObwf4E8R+LrW2S4m0PSLq+jhkJCyNDEzhSRyASuK0D/goJ4x1nXtN0iT4e6NGl9dxQM63UpKh3Ckj86qtQ1bStJ+zf2pqdpZ/bLhba2+0TLH50zZ2xpkjc5wcKOTipdW0qHUodGm1O0TULiNpIbRplEsiL95lTOSB3IGBVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV+1F+0XrXwA/wCEZ/sfw5Zat/bv2vzPtMrJ5Xk+TjG3rnzT+Qr9mP8Aac1349a7rWkav4YsNLTSrRJ0e2mdy5Z9uDu7VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVr+ovo+g6jq8UayPY2ks6oxwGKIWAPtxX/DxXxp/0TnRP/AuWqrVdW0rQ7CXVNb1O00+ygx5tzdTLDGmSFGWYgDJIHPcivib+2x+zn8HfjJpXwO+IvjyLR9e1SzFy91Mo+xWG9gsUd1MDiB5AS43gKqLudkDIWtLu01C0hvrC6iuba5jWSGaFw6SIwyrKw4IIIII4Oaqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq17xH4e8LaedW8T69p2kWKsENzf3SW8YY9BucgZPpWheI/D3irT11bwxr2navYsxRbmwukuIyw6gMhIyO4qqqqqqqqqqqqqq/bM/5OU8Yf9uP/AKQwV+wp/wAkAs/+wpd/+hCqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr9uz/kv13/ANgu1/8AQTX/AAT1/wCSLaz/ANjRP/6S2tV/wVE+Icfw/wD2MPGkUV79nvvFElroln8yjzDNMrTJgkE5to7jhcnvjAJGnahe6TqFrqum3L293ZTJNbypw0ciEMrD3BANfDfxha/ET4deFvH9iU+z+JtFs9Th2KwGy4hSUYDAMBhx1APrVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV43/5EvX/APsF3P8A6Kaqqv8Agqp8QI/An7GHiuyjufJvPFl5Z6LaNhTkvKJZVwQesEE49RnIIIrSdVv9C1Wy1vSrjyL3T7iO4tpdgby5EYMrYYEHBAOCCK8BeLbLx/4F8OeO9NVFtPEek2upW4STzAI54lkXDYG4YYc4GfQVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV8b/+SLeP/wDsV9S/9JZK8D/8jr4f/wCwpbf+jVqq/wCC3/xDdbT4YfCa0vfkkkvdbv7fcw5UJBbPgHB+9djkEjsRlgf+CfvxDf4Z/th/DHXWvfItr/WF0m63MwR471GtsOARkBpUfngMisQdtVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX/AAUh/wCad/8AcU/9tK/4J1f8jp4u/wCwXD/6Nqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqrxv/wAiXr//AGC7n/0U1VVf8FpvH0eg/s9+Fvh/DLCLnxV4kEzqzLua3tImZ9q9f9ZLBlhwOh+8Kr9kj/goJ8Zv2Vr230OG8fxT4EaYNdeHb+YkQr0JtZTkwN3wAUJ6rnkfs4fta/BP9qTw9Hqvwz8UwnVYrcTah4fvGWK/sOQpMkWTuQMQPMTchyOcnFVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV+0l4j+JevftR6p8K9A+IWuafYatqGl2NtZjVJ47SJ5oLYBjGrYA3vuOB1yetSfsTfHKCNprH46qbmNS0Ia5u4xvH3csCSozjkA49DVh8cf2jf2X/G1l4U+Ml1eeIdFlJlZLqcXUk8LMQZYLlvnJBGdjnA6ELuBGh6zp/iPRNP8QaTP51jqdrHdW0mMb45FDKce6kVVVVVVfB7Vf2sLn9pa4tvFo8Q/wBkx3szaxbXLSHTYLchwnkbsIBnaYynLbcnI3VVV+3X/wAkAvP+wpa/+hGv2FP+SAWf/YUu/wD0IVVVVVVVVVVVXxb+LnhL4LeF4/FnjH7a1pNdC1hSzhEskkpjdwoBIAyI25JAzjJFfCn4i6d8WfAOmfEDSbC4srTVDP5UFwQZFEczxc7cjkx546Zqq+Of7F/if4tfFPW/iDp3jXS7C31X7Pst57eRnTy7eOI5I45MZP41+z38KNQ+DHw4g8D6nq1vqU8V3NOZ4I2RSHIIGDzxiqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr9oX9kDxJ8aPiPN430vxjpunQS2cMAguLd3YFAQTkcc5r9mn4L6n8CvAl94R1bW7XVJrvV5L5ZreNkVVaKJNuG5zmIn8RVf8Fv8Ax88ek/DD4WW8suy4uL3WbtNzBMxqkMBx0Y/vLjk8r2+8ar/glp4+j8d/sYeD7Z5YXuvC1xd6Nc+WynaYpTJGCB90+TLDweT97owqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq1/TX1nQdR0iKVY3vbSWBXYZCl0Kgn25r/AId0+NP+ijaJ/wCAktVX/Bb/AOIHHww+FVs3/P7rV4Cf9yGDGG/6+c5X+7g/eFV/wTD+IH/CwP2MPA3nNm88N/adFucHgeRM3lY+Zj/qHhznHOcALiqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqvH/hybxj4E8R+EbW5S3m1vSLqxjmkUlY2liZAxA5IBbNaB/wAE+/GOj69puryfELRpEsbuKdkW1lBYI4YgfXFVX/BVr4jf8LA/bK8SafDL5lr4N0+z0O3fbtzsQzyjG1TxNcTLzuztyDtIA0nVb/QtVstb0q48i90+4juLaXYG8uRGDK2GBBwQDggivAXi2y8f+BfDnjvTVRbTxHpNrqVuEk8wCOeJZFw2BuGGHOBn0FVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV+1F+zprXx/wD+EZ/sfxHZaV/YP2vzPtMLP5nneTjG3pjyj+Yr9mP9mPXfgLrutavq/iiw1VNUtEgRLaF0KFX3ZO7tVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVWv6a+s6DqOkRSrG97aSwK7DIUuhUE+3Nf8O6fGn/AEUbRP8AwElqq/4LQ/EN/EX7Rfhv4e2175tn4Q8No8kO5v3V3dys8nGSvMMdqcgA+pIC4/4I1fBXw3418E/FzxN468N2muaHrUlloX2LULQTW04jDTzKyuCjkF7Zh3UgHjINftc/8Ef73R4bzx1+ypNcahbK0k1x4Rv7jdNCmCcWkznMoHQRyneePncnFaXqvjz4UeM4tU0i+1vwn4q0G4OyWJpLK8spQCrDs6HBKkHGQSDwTX7IX/BXzRddjs/AX7Vvk6VqXyxW/jCztsW1yduP9LgjH7lywH7yMeWS/KRKpY6Vq2la9pttrOhanaajYXkYktru0mWaKZD0ZHUkMD6g4qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq+L3/J/lh/2NHh//ANF2lV/wUQg0pvhh4buJlQ6lHrwS3JJ3CFoJTLjtjcsOfoPev2Qnv5P2cvBjaiJfNEFyF81Sp8sXUwjx7bAuPUYNfAv9q3w98dfF134R0jwpqOmTWemvfNNczo6sqyRoVAXnOZQfwqvjr+1b4e+BXi608I6t4U1HVJrvTUvlmt50RVVpJE24bnOYifxFV4C/a68BeMvEfijQdTsLjw/D4UtJ7u6vr2ZTE0cUyxNjbzks6YXkknAyeK8Vf8FEvCenanJaeEPh5f61aRyMourrUBZeYBjDKgjkODz97acYyASQPgh+1J8PPjfL/Y1gZ9G8QojOdKvGBMqryWiccSAA8jhhgnbgZqvC37Vvh7xV8bpvghb+FNRgvoNSvrE3zzoYy1qspZto5w3knH1Farqmm6JptzrGsX8FnY2UTS3FxO4RIkUZLMTwABXi7/gof4L0zUHtPBvgPUddt43K/arq8Firj+8q7JGx/vBT7Cvgd+2D4W+NPiWDwWvhDVtI1ieCSVR5iXFuAi5bMg2sPbKY9xxX7df/ACQC8/7Clr/6Ea/Z0+N/gL4Jfsy2Gq+MNQc3N1ql4LHTrZd9xdsGXO1cgBRkZZiFHAzkgGw/4KNaDLqoh1P4V39vpvmEG4g1VJptmeD5RjRc45xvxnjPevAXj7wp8TPDFr4t8G6rHfafc5XcBteJx96N1PKsMjIPqCMggn4tfEWx+Evw+1X4g6jp09/b6V5O+3hcK7+ZMkQwTxwZAfwpP2x/hhB8LLL4mazb39k2o3M1taaQNktzO8RwxUAgbMFcscAE464zYf8ABRrQZdVEOp/Cu/t9N8wg3EGqpNNszwfKMaLnHON+M8Z71rfx/wDhVoPw2tvitc+J4ptBvQBaGAbpriQ/8skj4PmDB3KcbcHOACavP+CjeiJqQisPhTezWG4AzzausUoXPJ8sRMM47b+fUda+EXxi8F/GrwwPEnhC7cGFvLvLK4AWe0fsHUEjBAyGBIIzzkECviJ+2r8KPBWunwvoEGo+L9SV1Rv7ICNAHJx5YlJ+d+nCBhk4yCCB4f8A2/PCo1NdL+I3w41zws7Hh0k+1CNTjDOrJG4H3vuqx4GAc8aFr2i+KNHtNf8ADuqW2o6dex+Zb3Nu4dJF6cEehBBHUEEHkV8fv2i9E+AH9g/2x4cvdV/t37T5f2aVU8ryfKznd1z5o/I18W/2xPhh8LjbafbpdeINYuLdJ2sbQhPsyOgZPOkPCMQQdoDMByQARnwd/wAFDfBur6tHY+NPAt74es5XVRe2999uWLPVnQRowUcfdDHrx66Xqmm65ptrrGj38F7Y3kSy29xA4dJUYZDKRwQRX7V/7Tmg/F3w2Ph1pvhi/sLnRtf8+S5nmRkkESTREADkZMgP4V8DP20PC/wl+FeifD7UfBeqX9xpX2jfcQ3Eao/mXEkowDzwJAPwr4deNLX4i+B9G8b2VlLaQaxbCeOCVgzRgkjBI47VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVf8FYviBH45/bK1/Tbe58+38H6XZaLEwCgAqhuJFGACcS3Mindk5B5xjGteANc0HwJ4Z+IF9HjTfFVxfQ2DBDhjaGJZPm6ZzKPl6gbSfvCv+CIHxDd7T4n/Ca7vfkikstbsLfc3Vg8Fy+M4H3LQcAE9ycKBVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV/wVX+IKePP2zPFFjbzPLa+EbGz0WB2Y4ykfnSgAqCoE08q45BILA4YY174fa/4c8F+FvHmorD/Zni/wC2f2cVD7j9llEUu7KhfvEY2lvfHSv+CIHxAefw/wDE/wCFVzc4SzvLLWbKHDHcZUeG4bOMDHkWw5OTu4HBxVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVeXdpYWk19fXUVtbW0bSTTTOESJFGWZmPAAAJJPAr4seOJ/ib8UfF/xFuXlaTxNrl5qR81QrKJpmkAIBIXAYDAJAxgHAr4lfD3W/hb4tk8F+I4ni1K3sbG6njdVBjNzaRXIX5WYEATABgcMADxnA/4Jd/EOP4g/sYeCopb37RfeFpLrRLz5lPlmGZmhTAJIxbSW/DYPfGCCaqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/bU+I3/C1/2rfih42jl8yCfxBNZ2km3b5lvaAWsLY2qRmKBDgjIzgknJP/AAS7+Hkfw/8A2MPBUstn9nvvFEl1rd58qjzDNMywvwATm2jt/vZPbOAAK/as/YW+Cf7WGmyXXifTf7D8Xxxqlp4n06FRcoF+6kq8CePttbkD7rLX7VH7Evxr/ZO1lv8AhM9H/tXwrcXBi03xRp0bPaXHdVk728pH/LN+pV9jSKpav2VP26PjX+yhqUdp4Y1P+3PCEkjPd+F9RlY2zlvvPCwyYJOM7l4J+8rV+zH+2j8Dv2q9HV/AXiJLLxHDD5l94a1GRYr63wBvZUz++iBYDzEyBkbtpOKqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr9ovxFa+D/20rvxbfQSzW+iaxo97NFFje6RW9q7BckDJCkDPGauv+ChnwgS3drLwn4wlnA/dpLbW0asfdhOxH5GtZl+M37cPjywls9BfRvC9g5iilIeS009DzI7SEL50xAAwoGcKMKMtXhfw1pHg7w3pnhTQbfydP0m1jtrZDydiKACT3Y4yT3JJ71/wT1/5LTrP/Yrz/8ApVa1X/BQv/ktOi/9ivB/6VXVV8Mvhr/wtz9oW+8D3N/Paabd6leSak0Mmxnt45S7IOoJLKgGQQDg9q0/4VfDPS9Fi8O2PgDw+mnQweStu2nRurIRghtwJbOTktksSSckmvjd4Zs/2Zv2mtN1rwiHg01JbbWrS2TkxQtIyywDkZQmOVQMj5WAz/Ea+EX/ACf5ff8AY0eIP/Rd3X7d/wDbv/Cg5/7I3/Zf7Wtf7U25x9ny23OO3neR14zjviv2E/EnwsPw1g8M6Pc6db+NBLO+rQuAlzdDzHMbpnmRFiCAheFIOQC2TD4W8M22vyeKrbw/p0Wsy27W8moR2yLPJEWDFGcDcy7lBwT1Fft1/wDJALz/ALClr/6Ea/Yf+BXhrxLoTfFfxrZRaw9tcvZ6LZXQEsNsEO55djZG7e7bR0U5bGSCPjD+z/4A+KHgi/0EeGNNstTSB30u+trVIpLa42/L8ygZQkAMp4I9CAR/wTt8X3cPifxT4BeR3tbqwTU4lIysbxSLE5HPBYTJng52DkY5/bM/5Nr8Yf8Abj/6XQV+wf8ABrw14rtdU+KHiuwi1RtLu/7O0q2uVDxQOEDyyFDkE4lQLkYBLHkkFf2ivhZ4d+J/wv1201LTrVtTsLCa40y9dMPbTRqXXDgEhGK7WHcE8ZxX7Gnwz0/4q/EP7N4sl+2+H/CsT6iulT/vIZ7iUpHzG2V2kRqX/veWinI6eK/AvhLxv4Xn8GeJdDtbzSJ4hH9mMYURADCmPH3GX+ErgjtX7Il7qPw2/ag1P4ZNdytDdyX+l3CKAVeS18x1due3kuARn75HQkj9tf4n6t8O/hGNO8PXD2+o+J7r7D9oQ4aGDaWlKnsxACeoDkgggGv2QfA/wy+G/wAN9N8V6jrGg/8ACVeIbcXVxcz3cPm20EgBjgQ5yi7drMOpYnd91Qvxh0D4UfGDwRf+Fdd8QeHJbhoXOnXUl9EHs7jb8kisDleQMgdRkEEHFf8ABPHx9qE0niT4ZXl35lrDEup2ETNzEdwjmxxnaS0JxnAOTjLE1/wUh/5p3/3FP/bSv2VP2f8Awh4f+HmmePfE2lWuueJfFdqmoTXd9Gtx5MUo3oibgdpKMC56licnAAH7ZHwM8F6t8KtS8daFoNhpmt+HVSfzrS2SH7TDuCvHJtAyArFgeuVA6Gv2BfF13r3wfvPDl7I7t4d1R4rdiOBBIokVc5JyHMvYAAr71+37oGg6b8INJvdN0WwtbmXxPCHmgtkjdgba5JBYDJBIB+or9kDwr4Y1L9nTwje6j4c0u6uJPtu+Wezjd2xezgZJBJ4AFWtpa2NvHaWVtFbwRDEcUSBFQegA4FV8Wv20PC/wl+IOq/D7UfBeqX9xpXk77iG4jVH8yFJRgHngSAfhXwY+K2nfGjwPF430vSbjToJbmWAQXDh2BQgE5XjnNVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVXxn/AGvvDXwY8by+B9U8H6lqM8VtHOZ4J0RSHGQMHnjFfAv40aX8dfCN34u0jRbrTIbPUnsWhuZFdmZY43LArxjEoH4VNNDbwvPPKkccSlnd22hQBkkk9Bivi947l+KXxW8ZfEmZHRvFOvXupiNwQYxPM8gTG5sBQwUDccAAZPWv2xP2fP8AhEf+CYHwJ1JdK2ap4Mkt7m9yMm2i1ZJJ7lSWBYH7S9uCAwXOeMBcf8EpfiN/wgH7ZXhvT5pfLtfGWn3mh3D7c43oJ4hjax5mtoV4243ZJ2gg1VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVWs6jHo+kX2ryRmRLG2knZFOCwRSxA/Kv+Hivgr/onOt/+BcVVeXdpYWk19fXUVtbW0bSTTTOESJFGWZmPAAAJJPAr4neM5/iR8SfFnxEuovKn8Ua5e6pJHgDY1xO8pGBxwXr9tv8AZ9Hgr/gmx+z/AKitig1Pwa0H25VCKLcarA1xcjO87iLkQqSpO4ndgDp/wSa+IcPgT9sjQ9LuZEjg8Z6Te6I8jjhWZVuIx9WktY0Huw7Zqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqrxj4jg8H+Edb8XXNs9xDoem3F9JChAaRYY2cqCeASFxWj/8ABQTwbrOr2OkRfD7WY3vrmOBXa6jIUuwUE+3NV+3R4+f4Z/sh/FTxXDLLHOfD8unwSwsyvFLestpG6leQVe4VgR0IzXwX8A3HxU+L3gv4bWsDzN4l16z09wu4bY5ZlV3JUEqqoWZmAO0AntX/AAWc+GyeFv2hfDHjrT9OS20/xV4ZjhLIhUPdWcjRuB8oXCwvajAJI7gArn/giD8Q5ZtE+JvwmuJH8u0urTW7NQCRulVoJyfQ4htvrz6VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVXx+/aL0T4Af2D/bHhy91X+3ftPl/ZpVTyvJ8rOd3XPmj8jXwG/ac0H49avqmkaR4Yv8AS30q2Sd3uZkcOGbbgbaqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq1nUY9H0i+1eSMyJY20k7IpwWCKWIH5V/w8V8Ff9E51v8A8C4qr40ePbb4WfCHxp8SLm4SJfDOg3moIW2/NJFCzIihiAzMwVQpI3Egd606w1TxLrVrpdij3epapdJDCryANNNIwUAsxAyWI5J78mvh74NsPh14A8M/D7Sn32XhjR7TS7ZsEZjt4ViU4ZmPRB1JPuetVWt6HonibR7zw94j0ex1XS9QhaC8sb63WeC4jYYZJI3BVlI4IIINftdf8Ef7KWG/8ffspzPDMqtNP4OvrjcjkAcWc7nKngny5WIJJw6jC1d2njz4T+NmtbmPW/CXizw3eDIzJZXmn3CHIIIw8bg4IIwehFfsf/8ABXz/AI8vh9+1j7x23jSytv8AvkXlvGPqPNiX+4Gj+9JXhrxN4d8Z6BYeKfCWuWOsaPqcIms76ynWaGeM9GR1JBFVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVXxotLW//AG8raxvraK4t7jxJoMc0MqB0kRorQMrKeCCCQQeDVr8GvhBY3Ed3Y/CnwdbzxHMcsWhWyMh9QQmRSRxxRrFEioiLhVUYAA6AD0qv+CfM8EXxr1WOWZEebwzcJErMAXb7RbNgDudqscegJ7VX/BQl43+NWkKjqxTwxAGAOdp+03JwfTgg/jVfsmf8nZ6p/wBxT/0M1X/BQv8A5LTov/Yrwf8ApVdVXwi/5P8AL7/saPEH/ou7rVdK03XNNudH1ixgvbG8iaK4t50DpKjDBVgeCCK+J/8AwT/0e7nl1r4P+JX0idcvHpeolpYgwPAjnGXQDH8Qc5/iFfs6/H/4mfD/AOKcfwF+M95dXyzXo0+Ca9m82exuWP7sCTkyxyFlUZJwGQqQowf26/8AkgF5/wBhS1/9CNfsKf8AJALP/sKXf/oQqv8Agnr/AMlp1n/sV5//AEqta/bM/wCTa/GH/bj/AOl0Ff8ABPX/AJItrP8A2NE//pLa143/AORL1/8A7Bdz/wCimr/gnV/yOni7/sFw/wDo2q+EX/J/l9/2NHiD/wBF3df8FHNKuptI8Ca0m37PaXN9byZznfKsLLjjGMQv39OvOPhR+xR8GPiJ8NfDXjaXxN4vFxq+mxTXKwXdssaT7cSqoMBIUSBgMknA5Nf8O9Pgt/0M/jX/AMDbX/5Hr4N/ssfD34H+J7nxZ4T1jxDd3l3YPZOmo3EMkYjaSNyQEiQ7sxL3xgnj0/4KQ/8ANO/+4p/7aV8EP+SLeAP+xX03/wBJY6/ad/5ID44/7Bbf+hLX/BOr/kSvF3/YUh/9FV/wUL/5Itov/Y0Qf+kt1X7Gf/Jtfg7/ALfv/S6eqr9sz/k5Txh/24/+kMFfsKf8kAs/+wpd/wDoQqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/bs/wCS/Xf/AGC7X/0E1/wT1/5ItrP/AGNE/wD6S2tftvfEB/hh+yX8UvF8Fz9nuI/D81lbS4YmOa7K2sTDaCch51I7A4zgZNfCHwJL8Uvit4N+GsDujeKdestMMikgxieZI2fIVsBQxJO04Azg1+2p8OLf4k/sj/E/wPb2uSPDc13ZW8KEbprMC6gRQrLjMkCKOcDPIIyD8J/HE/wy+KPhD4i2ryrJ4Y1yz1IeUoZmEEyyEAEgHIUjBIBzgnBq0u7TULSG+sLqK5trmNZIZoXDpIjDKsrDgggggjg5qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqvG/8AyJev/wDYLuf/AEU1VX7eHxDf4Yfsh/FDxPb3v2a6m0OTTbWRWYOst4y2qshUgh187cCD8pXdyARXwS8AP8VfjH4H+GqRyunibxBZafMYkZjHFLMiyP8ALyAqFmJ7BSe1ft0eAH+Jv7IfxU8KQxyyzjw/LqEEUKMzyy2TLdxoqrkks9uqgDqTivgv49uPhZ8XvBfxItbh4m8M69Z6g5Xd80cUys6EKQWVkDKygjcGI71DNDcQpPbypLHIoZHRshgRkEEdQRVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV8b/wDki3j/AP7FfUv/AElkrwP/AMjr4f8A+wpbf+jVqv8AgtV8QI9D+AfhD4dQXPl3XirxIbl0wp8y2s4WLjkZH72e2ORg8Y6Eiv8Agkt4Aj8bftlaJq08cUsPg3R77WZI5UVgx2C1QgHusl0jgjkFQe1f8Fp/h3Dr/wAAPCfxGhRDd+EvEX2di3BFteRESY9/Mgt+PTJ7V/wSd+IEfgb9srw/ps9z5Nv4w0u90WViFwSyC4jU5BI3S20ajGDkjnBNVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV/wUh/5p3/3FP/bSv+CdX/I6eLv+wXD/AOjaqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq8b/8iXr/AP2C7n/0U1VX/BWrx8/gn9jTW9Jglljn8ZaxY6NHJEzKVG83TjI7NHaOhB4Icg9a/wCCfvw8f4m/th/DHQms/tFtYawurXW5WKJHZI1zlyAcAtEiYPBZ1Ukbs1VVVftPfsZ/BP8Aas0L7J4+0L7Hr1tGy6f4i09VivLU4wAzYxLHnB8t8j02nmv2sv2CPjX+yffS6prVl/wkvgl5Ntr4p02BhCuX2qtzHkm2kOU4YlCWwjuQcfsxftl/Gv8AZT1z7V4B137ZoNzIp1Dw7qDNJZ3IBySq5zFIRkeYmD0zuHFfsr/t3fA79qvT7ex8Oawmg+MSrGfwtqc6rdfKMs0B4Fwm0E5TkAEsq1VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVXxe/5P8sP+xo8P/wDou0qqr4raF48/ZS/aCfx34dgc6feXs13pdxMu6G7hl5lt5Md13lSDhuFcY+U1N/wUR0u608QaL8K9Rl1mdFSC3e/Vo/ObAxlU3MMngBQW4Hy5yP2gPDfxRt9Y07x/8XUeDXvGKS3Js3h8v7JFGVRExk7flx8h5UbdxJJqv2TP+Ts9U/7in/oZqv8AgoX/AMlp0X/sV4P/AEquqr4Rf8n+X3/Y0eIP/Rd3X7R3x1n+A3hvSdftfDLa299qHkSwNI0KJEEYs3mhWCNu8sAEc5b0pP8Agod8JjYeZJ4P8Wi98skQiG3Me/HA3+aDjpzt49DXwe8N+M/2mP2kx8cr3w6+laBY6pDfTzklo1e2RBBbxvhfNf8AdxbiAMDJIGVU/t1/8kAvP+wpa/8AoRr9hT/kgFn/ANhS7/8AQhVf8E9f+S06z/2K8/8A6VWtftmf8m1+MP8Atx/9LoK/4J6/8kW1n/saJ/8A0lta8b/8iXr/AP2C7n/0U1f8E6v+R08Xf9guH/0bVfCL/k/y+/7GjxB/6Lu6+O/wmsvjT8NtR8FTzRW94xW4026kUkW9ymdrHHOCCyMQCdrtjnFfC747/E39ku+uvhf8SPBlxd6VHM0kVs8nlvAT954JMFZI2ODjpnkEEtm6/wCChnwgS3drLwn4wlnA/dpLbW0asfdhOxH5GvgH8Zo/jn4NufF6eHpdFEOoSWqW0s3mlkVVIkDYXcCWYdAAVYc7cn/gpD/zTv8A7in/ALaV8EP+SLeAP+xX03/0ljr9p3/kgPjj/sFt/wChLX/BOr/kSvF3/YUh/wDRVftm/DzW/iJ8FLm38O2M97f6JfxalHawcvMqK8bhVwdxCSs20YJ28ZPyn9mz9sbwn8J/hxD4A8caDrd0NNnkbT59MihkzHI7SMrh3Qgh2Yg5bIbHG0Z8JeIrXxh4U0XxbYwSw2+t6dBewxS43oksauobBIyAwBxxmq/bM/5OU8Yf9uP/AKQwV+wp/wAkAs/+wpd/+hCqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr9uz/kv13/ANgu1/8AQTX/AAT1/wCSLaz/ANjRP/6S2tf8FpvH0eg/s9+Fvh/DLCLnxV4kEzqzLua3tImZ9q9f9ZLBlhwOh+8K/wCCTvw/j8c/tleH9SntvPt/B+l3usyoSoAKoLeNjkgnEtzGwxk5A4xkiaGG4he3niSWORSro65DAjBBB6gjtXxo8A3Hws+L3jT4bXMDxN4Z16809A275o4pmVHBYAsrIFYMQNwYHvX7DvxDl+Kf7JHwu8Y3MjyXL6DHY3Ejg7pJrNmtJHPuzwM341VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVXjf/AJEvX/8AsF3P/opqqv8AgtX8QW0L4EeDvhzbTIk3irxE11KCQWe3s4juUKVPHmXEBLAgjaByGOP+CSPw8j8b/th6TrtzZ+fbeC9HvdWbeqlBIUFtGSGByQ1yHUDBDIGB+U1eWdpqFpNYX1tFc21zG0U0MyB0kRhhlZTwQQSCDwc18WPA8/wy+KPi/wCHNykqyeGNcvNNPmsGZhBM0YYkAA5Cg5AAOcjg1+xZ8Rv+FsfspfC/xs8vm3E/h+Gzu5Nu3zLm0zaztjaoGZYHOAMDOASME1VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVXxv/AOSLeP8A/sV9S/8ASWSvA/8AyOvh/wD7Clt/6NWq/wCCzvxA/wCEj/aS0HwHbtm38IeG4/NBPIubmRpHxhiMeUtv1CnOc5G01/wRA+H7w+H/AIn/ABUubbKXl5ZaLZTZYbTCjzXC4zg5862PIyMcHk1+298P3+KH7JfxS8HwW32i4fw/Ne20OWBkmtCt1Eo2kHJeBQB0zjORkV8IfHcvwu+K3g34kwI8jeFtestTMagkyCCZJGTAZchgpUjcMg4yM5qGaG4hSe3lSWORQyOjZDAjIII6giqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv+CkP/ADTv/uKf+2lf8E6v+R08Xf8AYLh/9G1VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV43/5EvX/+wXc/+imqq/4Lf/ECOfX/AIYfCq1ucPZ2d7rV7DhTuErpDbtnGRjyLkdcHdyOBj/gid8O4db+Mvjv4mXCJIvhXQYbGFW6pNeykhx3yI7SVfTDn2qqqqrUdO0/WNPudJ1axt72xvYXgubW5iEkc8bgq6OjZDKykggjBBwa/a5/4JCeG/FA1Hx7+y7LFoWszSGaXwndy7bGYn732aQ825J5EbEx84BjUADxF4Z8ffCTxm+i+JNK1jwr4o0G6V/LmR7W5tZkbKSIeCCCAyupxwCD0Nfse/8ABXjV/DiWHw8/aqNxq+lxqkNr4wtYTJd24BwPtkSjM6hSMyoPN+TlZWcsPBvjbwh8RPDdl4v8C+JNP13RdQjElte2M4ljkBHqOhHQqcEHggGqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqtX/AGm/2bNA1W80HXf2hfhpp2pabcSW15ZXfiyyhmtpo2KvHIjShkdWBUqQCCCDXgr4h+APiXpUuu/Djxz4f8VabBcNbS3mianDfQxzKqsY2eJmUOFdCVJyAynuKqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqm8OeHrnUBq1xoWnS3wZXFy9qjShlxtO4jORgYPbAqqrU9K0vWrNtP1jTbW+tX+/BcwrKjfVWBBrQfht8OvCt5/aPhfwD4c0e72kefYaVDbyYxgjcig4wT+daloei615f9saRZX3k58v7TbrLszjONwOM4H5CqtPDnh7T7s39hoWnW102czw2qI5z1+YDPNVqPhzw9rEy3OraFp17MqbFkubVJGCgk4BYE4yTx71UPhzw9b6gdWt9C06O+LM5uUtUWUs2dx3AZycnJ75NTwQXUElrcwpNDKhSSORQyupGCCDwQRxin+EPwmlvxqknwv8JNeiQSC5bRLcybx0bdszkYHNQQQWsEdrawpFDCgSONFCqigYAAHAAHGKv9N07Vbc2mp2FveQEgmK4iEi5HQ4ORVhpunaXbi00uwt7SAEkRQRCNQT1OBgVWneHPD2kTG50nQtOspmQo0lvapExXIJGVAOMgcewq9sbLUrV7LUbOC6t5Mb4Z4w6NggjKnIPIBrTtK0vSIDa6TptrZQsxdo7eFYlLYAJwoAzgAZ9hUkccsbRSoro6kMrDIIPUEeladoGg6O7y6RothZPIuHa2tkiLD0JUDIqofDnh631A6tb6Fp0d8WZzcpaospZs7juAzk5OT3yarWND0TxDYtpmv6PZanaPndb3lus0bZBByrAjoSPoT61YfCH4TaVcC70v4X+ErOcKQJYNEt42APUZVAcUkccUaxRIqIi4VVGAAOgA9K1LQ9F1ry/wC2NIsr7yc+X9pt1l2ZxnG4HGcD8hUEEFrBHa2sKRQwoEjjRQqooGAABwABxirq0tb63e0vbaK4glGHilQOrD0IPBrTtG0jR43i0jSrOySQ5dbaBYgx9SFAzVan8LPhhrWpNrOsfDjwvf6g7b3urnR4JZWbOcl2UknPOc1BBBawR2trCkUMKBI40UKqKBgAAcAAcYqv2zP+TlPGH/bj/wCkMFfsKf8AJALP/sKXf/oQqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/bs/5L9d/wDYLtf/AEE1/wAE9f8Aki2s/wDY0T/+ktrX/BaH4hv4i/aL8N/D22vfNs/CHhtHkh3N+6u7uVnk4yV5hjtTkAH1JAXH/BEDwA8ek/E/4pXEcuy4uLLRrR9jBMxq804z0Y/vLfgcr3+8Kr/grL8O4fAn7ZGuanaokcHjPSbLW4416KzK1tIfq0lrI5z3b0r/AIIrfECPXfgH4v8Ah1Pc+ZdeFfEn2mNMKPKtruFSg4GTmWC4OTk846ACqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq8b/8AIl6//wBgu5/9FNVV/wAFmfiHH4m/aZ0fwJZ3vmW/gzw3DHPDuU+Vd3LtM/QnGYTanDAHvjBBP/BEH4fJB4a+JvxWuInZr2+tNFtHKkBBCjTTAHdhifPt8grldowfmIqv+CrXw5/4QD9srxJqEMfl2vjLT7PXLdM5xvQwSnO5jzNbTNztxuwBtAJ/4IvfEOPxF+zp4j+Htxe+beeEPEjvHDuX91aXcSvHxndzNHdHJAHoSQ2Kqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr43/8AJFvH/wD2K+pf+ksleB/+R18P/wDYUtv/AEatV+2X8QH+KP7VHxR8Zm5+0Q3HiS6trSXDDfbWzfZ4DhgCMxQx8EAjpX/BMP4ff8K//Yw8DCZcXniT7TrVzgcfv5m8rHyqf9QkOc55zglcVeWdpqFpNYX1tFc21zG0U0MyB0kRhhlZTwQQSCDwc18XvAkvwu+K3jL4azO8jeFtevdMEjkkyCCZ4w+dq5DBQQdoyCDgZr9ib4gp8Uf2TfhZ4w855pn8OwWVzK5JaS4tM2sznKryZIHPTHPBIwTVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV/wUh/5p3/ANxT/wBtK/4J1f8AI6eLv+wXD/6Nqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqrxv/AMiXr/8A2C7n/wBFNVV/wU9+IH/CwP2z/HIgbNp4b+z6LbZPI8iFfNz8zD/XvNjGOMZAbNf8Eb/h+/hf9la88Z3Vttn8Z+JLq5gly3z20CpbqMZxxLFccgAndg5wMVVVVVX7Rf7KHwT/AGpPDy6J8VPDHm3ltGUsNasWWC/sMnJ8qUqwxn+B1ZD1K55r9rv/AIJ2fGb9liW58TW8L+Mfh+rLs8RafbkNaBuAt5CCTAc8b8tGdyDcGbYP2b/2sPjN+y14mXXPhl4kddPuJlk1LQrsmSx1EAYxJHkYbHAdSrj1xxX7Jf8AwUK+Cf7UtpZ6Auow+FPHrRgTeHNRuFU3EgBLfZJDgXAwpbaAHABJUAZqqqqqqqqqqqqqv+CtPxu+LHwQ+HXgLVfhN471Pwxd6lrVxDdy2LhWmjWEMFOQeAea/Yy/bN/ak8f/ALUnw28GeMvjb4j1bRNW1pIb2yuJUMc8ZRiVYBenAqv2kfEGs+E/2dvil4q8N6jLYato3gvWL2wu4Th7eeKyleORfdWVSPcV/wAN+ftlf9HDeKv+/wAn/wATX7N/iDWfFv7O3wt8VeI9Rl1DVtZ8F6Re393McvcTy2UTySNjuzMSfc1VVVVVVVVVVVVVVVVVV8Xv26v2VPgbq194b+IHxd0+LXdP3LPpVhbzX9wkgXd5biBGEbngYkKgEjJFfsxfta/Db9rTT/EmtfDHTNdt9O8NXyWUs+rW8cDXLsrMHjRHc7NoUgttbnBUY5qqqqqv2sv+TqPjL/2UDXf/AEvmr/gip/yav4p/7KBef+kFhVVVVVVVVVVVTTQ28LzzypHHEpZ3dtoUAZJJPQYr44/8FMP2T/ghdXmhz+NpfGGvWfyyaZ4YiF5sfONrXBZYFIOdy+YWXByucA+N/wDgt/4qnnni+G/wJ0qzhEbrBPrmrSXLM+Pldo4ljAAPVA5z/eGeP+H1f7VH/QhfCn/wVX//AMm14J/4LfeOba4tY/iN8DNB1GAqFuZdE1SWyZW3DMiJKJgQF3fIWGTj5wK+Cv8AwVP/AGTPi9JHpmr+Krv4fas21Rb+Ko0toJG25YpdIzQhAcjMrRsey1aXdpqFpDfWF1Fc21zGskM0Lh0kRhlWVhwQQQQRwc1VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV+0z+zN8b/iF8b/Eni/wf4J/tDSL/wCy/Zrj+0rWLfstYY2+WSVWGGRhyB0z0r9k/wCH/i74Z/CC28LeN9I/s3VI7+4laDz45sIzDad0bMvP1qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/ax/Zy+M3xM+L1x4o8EeDf7S0x7C3iWf+0LaHLqpDDbJIrcfSv2N/hl43+FPww1Pw74+0T+y9QuNemuooftMU+6JoIEDbomYD5o2GCc8dORX7anxG/4Wv8AtW/FDxtHL5kE/iCaztJNu3zLe0AtYWxtUjMUCHBGRnBJOSf+CXfw8j+H/wCxh4Klls/s994okutbvPlUeYZpmWF+ACc20dv97J7ZwABX/Bb/AOH7z+H/AIYfFS1tsJZ3l7ot7NljuMqJNbrjOBjyLk9MndyeBj/gjF8QP+Ed/aS17wHcN/o/i/w3L5QB5NzayLInVgMeUbjsxztxgbjVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVeKrK51Lwxq+nWUXmXF1YTxQpkLudoyAMngZJHWv+GM/wBpT/om3/lYsf8A49Vfti/EOX4q/tR/E7xvJI7x3XiK5t7UuCG+zWx+zwZHY+VDHx26V/wTL+Hj/Dv9jDwFFd2fkX3iKOfW7n5WHmC5lZoHwwB5thb+oOMgkYNV/wAFv/h472nww+LFpZ/JFJe6Jf3G1urBJ7ZMgYH3Ls4JBPYHDEf8EWfiHLoHx+8V/DiWRxaeLfDv2hQoJBubOUFM+g8ue459cDvVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV8U9F1LxJ8MPF/h3Rbb7TqGqaDfWtpDvVPMlkgdEXcxAGWIGSQBnkivCv7IH7RWm+J9H1G9+Hfl29rfwSyv/a1kdqLIpY4ExJ4B6V8bvH0fwq+Dnjf4lNLEr+GPD97qEIlZVEksULNGg3cEs4VQO5YDvWk6Vf67qtloelQefe6hcR29tFuC75HYKq5YgDJIGSQK8EeE9N8BeCvD/gXRhiw8OaXbadaDbtxFBEsacdvlQVX/BV/4et4F/bM8R6lHEkVr4x02y1q2RVAADR/Z5DwxJLTW0rHIU5bpjBP/BFrx9Hr37Pfin4fzSwtc+FPEhmRVZdy293ErJuUc/6yKfDHg9B901VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV+2z8GfiV8Xv+EM/wCFd+G/7W/sn7f9s/0yCDyvM+z7P9a65z5b9M4xzjIr9i/4GfFP4S+J/Eeo/EHwt/ZVvf2EUVu/223n3uJMkYikYjj1xVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVXiqyudS8Mavp1lF5lxdWE8UKZC7naMgDJ4GSR1r/hjP9pT/AKJt/wCVix/+PVq+qWGh6Vea3qlx5Nlp9vJcXMuwtsjRSzNhQScAE4AJrxx4s1Lx7401/wAc6y2dQ8Rapc6jdHOf3s8rSPz3+ZjX7Knw+/4VZ+zZ8NfAbri40vw3afawBgfaZIxJPj5VOPNd8ZAOMZ5yaqqqqqqmhhuIXt54kljkUq6OuQwIwQQeoI7V+1v/AMEjfAPxE+3eOv2b3tPBviN98s3h6QbNLvG+YkQgDNq5JAAXMQAUBE5avH/w6+IXwc8YXHg74heGdT8M+IdNZXe1ukMci91kRhwynGQ6kg9Qa/Y9/wCCt3jDwC1h8P8A9pp73xX4dDJDB4nT95qVguNubgf8vaDCksf3wG85lJVR8O/iV4B+LnhO08cfDXxXp/iHQ73IhvLKXeu4feRh1Rx3RgGHcCqqqqqqqqqqqr/gt7/ySr4af9jBdf8ApOK/YD/5PK+E3/YwJ/6A9V+1l/yav8Zf+yf67/6QTVX7J3/Jq/wa/wCyf6F/6QQ18QP+ChP7H3wu8Z6t8PvHfxf/ALM1/Qrg29/Z/wBgalN5MgAJXfHbsjcEcqSK8NftB/Bfxb8JbX466T8QtMi8B3izNDreo79PhIileF8i4VGUiSN1AIBOOMgjOsf8FYf2KdK1c6VB4/1jUo1YK17Z6DcmAHOD99VYgeoUg9s18Jfjd8Jvjv4dbxV8I/HemeJtOiZUme1crJbsy7gssTgSRMQc7XVT7cGq1HUdP0jT7nVtWv7eysbKF57m5uZRFHBGgLO7sxAVQoJJPAAya8b/APBUj9i7wTqh0dfidceIJ42ZZn0TS57mGMgKR++KrG4O7gxswBUg4r4N/t9/so/HTVbTw34K+KtpBr17IsVvpOr28unzyyMxVEjMqhJXbbkLGzH5lBAJxVav/wAFMf2I9B1W80LWfjRLZ6hp1xJbXVtN4X1ZXhlRirow+y8EMCCPUV8D/wBpX4H/ALSWn6rqfwV8fW/iSHQ5o4dQVbSe1ltmkBKFo540fawVsNjaSjAHKsB8VPix8O/gl4KvfiH8UvFFvoPh+wZEmu5o3kO52CqqRxqzuxJ+6qk4ycYBI/4ej/sJf9F1/wDLZ1b/AORa8fftB/CT4W/DC0+MnxC8SXeheE7zyPKu7vR7xZQZv9WHthEZ0J9GjBHfFfC39vX9k341eO9L+GXwy+LH9s+JdZ877DY/2FqFv5vlQvNJ+8mgRFxHG7csM4wMkgV8av21v2Yv2fb+TRPiZ8VtPtdaSNm/smxhlv7oEDIV44FfyS3QeaUBz1xzXgD/AIKf/sZfEDVxocfxQbw9cuyLC/iCwksYJCxwf3xBjQDjJkZRzkZwcQzQ3EKT28qSxyKGR0bIYEZBBHUEVXxc+Onwh+A+hR+I/i78QNJ8M2U+/wCz/a5CZbkoAXEMKBpJSARkIrHkeor/AIe2fsV/2r/Z3/CZ+IPI+0eV9u/4R+48nbux5uMeZsx82Nm7H8OeK+Enx0+EPx50KTxH8IfiBpPieyg2faPskhEtsXBKCaFwJIiQDgOqng+hqq+Jn/BRL9jr4ValeaF4g+M+n3+q2WRJZ6LbT6kd4xmPzYEaJXGcFWcYIIOCCK0T/grN+xPq101tfePta0aNYywnvvD10yMQQNoEKSNk5J5AHB5zgHx5+1V8APhp8L9E+NHjH4i29r4L8R3Udrper29lc3kd1K6SOoVYI3cArDKclQPlxnJGf2vvHvhL4pftMfEP4g+BNW/tPQNc1h7jT7vyJIPOjKKA2yRVdeQeGUGv+CWX7XH7PP7N/wAP/HGh/Gj4hf8ACPXusaxBcWUX9k3l35saw7WbNvDIF54wSDXwh+M3w1+PXgyL4g/CfxL/AG7oE1xJbpd/Y57XMkZw67J0R+CRztxWt65onhrR7zxB4j1ix0rS9Ohae8vr64WCC3jUZZ5JHIVFA5JJAFeLP+Cqv7FnhbUjpcHxG1DXXTIkl0nRriWJCO291QN9U3D3r4K/tx/su/tAapbeHPh18VLGTX7pSYtH1GGSwupGAyVjWZVErBcnEZfgE9AcVXxk/b7/AGUfgXqt34b8a/FW0uNesZGin0nSLeW/nikVlV0kMSlInXdkrIyn5WABIxXwl/4KNfsifGXX7bwn4e+KKaXrd7N5VpZa7Zy2H2hvlChJXHlFmZgqpv3sRgKeM/tZf8nUfGX/ALKBrv8A6XzV/wAEVP8Ak1fxT/2UC8/9ILCqqqqqqqqqqvj9+0F8Mv2afh5dfEj4o6w9rYQt5VraW6rJdahORlYLeMsu9yATyQqgFmKqCR+1h/wUP+OP7T17e6HDqtx4Q8CSM6Q+HdMumQXERwMXcq4NwTjO0gRg9FyMn4TfAP4v/HK/aw+F/gPUtbWFwlxdpGI7a3Y44knciNDg52k5IyQDivBH/BIj4l6rbCf4g/Ffw/4ed49yw6bYy6m6khcK+5oFB5cHaWHyjBOcj/h0H8N/+iweJP8AwAg/xrxn/wAEg/G9jbSz/D/4w6NrEqqWjt9V0ySwyd33d6PMCduecAE4GADkfGL9mr42fAa5EfxN8B32m2kjlINRi23FpMc4G2aMlQSBkKxDY6qK/ZX/AG5fjb+ylqlvbeFtZfWvB5maS88LahKTazFhhmjbBaB84bcmASBuDDiv2Zf2n/hl+1X8PI/Hfw7vnjmt2EOraPdMoutLnI+5IoPKsASjj5WAPQhlWqqqqqqviz/wnf8Awqvxl/wqz/kdP+Efv/8AhHP9V/yEfs7/AGb/AF37r/W7P9Z8n97jNf8AG9r/AD/widf8b2v8/wDCJ1/xva/z/wAInX7EP/Dwr/hKvEv/AA2f/wAgX+z4v7F/5A3/AB8eZ8//ACD/AJvuf3+PTmqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq+NHj22+Fnwh8afEi5uEiXwzoN5qCFtvzSRQsyIoYgMzMFUKSNxIHetOsNU8S61a6XYo93qWqXSQwq8gDTTSMFALMQMliOSe/Jr4e+DbD4deAPDPw+0p99l4Y0e00u2bBGY7eFYlOGZj0QdST7nrVf8FQPhzD8Rf2MvGrpAkl74Wa31yyLrkI0EgEzfdYg/Z5JwCMckZIUtX7GHxBT4XftV/C7xnNM8Vvb+Ire1upFYgx29yTbTNgKxIEczkqBlgMDGciqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr48fECP4U/BXx18SGufJk8OeH729t2wpJmSFjEoDAglpNigEEZIzxWh6Pe+Ita0/w/pqo13qd1Ha24dtoMkjBVyewyRzXgjwnpvgLwV4f8C6MMWHhzS7bTrQbduIoIljTjt8qCq/4Kn/DuHx/+xl4suwiNd+Erq01u038ANHJ5Upz2PkTzY9Tgd6/Yh+IEfww/a0+Fvi+a5+zwR+IIbK5lIUiOG7DWsrHIPASdie4GcYODVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVf8ABW74hv4I/Y81XQrW98i58aaxZ6SuxmDmMObmQAqRgFbYo2cgq5Uj5ga/4J5+AH+I37ZXwu0kxymHS9YGszSIjERCxRrpCxH3QZIY0yeMuB3qq/4Lf+AHl0n4YfFK3jl2W9xe6NdvsYpmRUmgGein93ccHlu33TX/AARe+Ib+Hf2i/Efw9uL3y7Pxf4bd44dzfvbu0lV4+M7eIZLo5IJHYgFs1VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV/wUM8fR/Dn9jT4oasJYvP1TRzo0MbsoMpvnW1cKD1IjmkfA5whPav2cvh9H8Vvj58PPh1Pbefa694ksra8TKj/RjMpnPzEA4iDnHU4wMk4qqqqqqqqr4+fs1/B79pfwm3hP4r+FIdQSPmz1CHEV5Yv/ehmAyvupyrfxKa/bD/AOCaPxc/Zqa+8aeDkuPHHw7iZ5DqdrB/pelxAbv9MhXOFUBgZ0zGduW8osqV8A/2lPjD+zT4rXxZ8J/Fctg8nF5p8+ZbO+X+7NCThvZhhl/hYV+yP/wUl+DH7S8Nn4W8Q3Vv4I8fStHCujX9yBDqErAD/RJTgOS3AibEnIADfeqqqqqqqqqqqv8Agt7/AMkq+Gn/AGMF1/6Tiv2A/wDk8r4Tf9jAn/oD1X7WX/Jq/wAZf+yf67/6QTVX7J3/ACav8Gv+yf6F/wCkENft+/8AJ5XxZ/7GCT/0BK8Fv+0X+0jpPg/9mbwFDrHifTvDbXNxo3h+1KxwWzSytJNcSsxVFG6UjzJWCpvwCN53fGr9j/8AaR/Z30e08QfGH4W3uhaXezeRFfJd297AJSCQjSW8kioxCsQGIJwcZxX7IX7Qmtfs0fHjw18RrLUriHRxdJa+ILVHbZd6fIQsyui53lQfMUYOHRT2qv8Agph+2br/AMe/ivqnws8H61e2vw68HXTWS2iSbE1a9iciW6kUfeUONkYYkBU3gKZGA/Zs/wCCWfxx/aH+Htj8UJ/EGieDtC1eMyaSNTjlkubyMMV83ylHyRttJVmOWBDAFWDH9pf9mP4l/sq/EAfD/wCJEFpLJc2/2nT9RsWd7a9hLFd0bMqnIIwykZUkdiCf+CUP7Vev/HX4Vap8NPiHrlxqnizwG0SxXty26W806QYiZ26u8bIyMx5IMZJJJNf8FeP2PGsb3/hqr4eaS7W90yW/jC1t4QFhk4WK9wOz/LHIf72xv4mNfsP/ALTN1+yz8etH8cXl1dnwvf8A+geJLSDLebaP/wAtAg+88TbZF7naVGNxr/gsD+05afEn4jaL8B/BmtRXnhzwbGmoanNbTCSK71KeLMeGUlWEVu4AYH71xKp+7X/BLf8AYyh+PPxCPxh+IujPN4E8F3SNbQTxZh1fUVIZYWB+/FGNruuCGJRDkMwr/grz/wAmaal/2MGnf+htXwz+JfjX4PeNtP8AiL8O9afSfEOlLMLK9SNXaEywvC5AYEE7JGHII5r4U/8ABPr9rv8AaM8PN8VNH8JY0/W5Dcw6n4h1NbeXVDISzTqHJkcMTu8xgA+7KluSPil8JfiP8E/GF14B+KfhK98O69aKHe1ugpDocgSRuhKSISCA6MykgjPBr/gj5+1jrUXiNv2UvGeovdadew3F94Ukmdne2mjXzJ7RckgRGNZJVAA2ssnXeMftN/HvQP2aPgp4i+LuvWj3x0qER2FijbTeXkh2QRFv4VLkFmwSqBiASADqGq/Gj9rj412w1G+1Dxf468Z6gttbK7dGZjhFH3YYIwScDCRopJwATWvf8EX/ANobSvBMmv6f468H6prsFv5z6JA06lyBlo45mTDP2GQoJ7jrXwe+LvxI/Zw+KWn/ABA8Eahd6RruiXBiurWQFBcRhgJrWeMjlG24ZSMggEYZQR8LviHoHxb+HPhv4meF5HbS/Eumw31srjDRiRQSjf7SnKn3Br/gqB+3b4k+IfjbWf2c/hfrcth4L8PXD2evXNrKVfWrxCVliZlPNvG2U2dHdWY7gEx8A/2X/jb+0zrNzo/wg8GS6qmn7ft99LKtva2e7O3zJXIAJ2nCjLHBwDg1r3/BGb9rXSNNkvtP1z4c63On3bKw1i4SV/oZ7aNPzYV+3H+yB49+Lf7Jnw9+AHwVg0+6ufBGqaYEF9dfZkNpaafcWoILljnMkfBJPJ5OM18T/hx4m+EPj/XPhp4yit49b8PXRtr1LeYSxiQAE7WHUYI5r9nT9ir45ftTaJq+v/Caw0e4tNDuktbs32orbMJGXcNoIORjvX/BPz4E+P8A9nP9nOy+GnxLtrGHW4NWvLl0s7kTx+XIwK/MO+B0r/goB+2br37UfxUvdM8Oa1ewfDfw/M1vomnCTbHdspw95Io+80hGVDZ2JgDBLZ+Af/BJf47/ABq+Hmn/ABH1nxPoPgux1qFLjS7PUYpZrqaBgSsrogAjVl2soLFiDkheM/Hz4C/Ev9l/4n3Pw0+I1tDbaxZRx3VreWMjtb3kLZ2XFvIyqzJuVlyVBDIwIBUgf8Ew/wBqbWf2kfgZcaR451eXUvGngO4jstUu5uZLy3kDG1ncjq5VJI2J5YwliSWJr/gq7+1/rPwO8Cab8GfhtrkuneMfG1vJNe3trLsm03TAdhZTjKvM4dFdeVEUhBB2mvhx8NfHXxe8Z6b8Pfht4au9e8Q6tJstLK2ABbAyzMzEKiKASzsQqgEkgDNfEX/gkt+1j8O/h9d+P2h8KeIvsFuLi50bQr+e4v4027n2xvAiSFADlY3Zj/CGq8u7vULua/v7qW5ubmRpJppnLvI7HLMzHkkkkknqTX/BFT/k1fxT/wBlAvP/AEgsKqqqqqqqqqvir8TfCHwZ+HWv/FHx3qH2PQ/Dlm1zdOMb3xgJHGGIDSSOVjRcjc7qM81+1N+0/wDEL9qv4m3XjvxrfPHYW7PDoejxsRBpdqW4RVzy7AKXc8sw7KFVf2Kv+Cd138TLbTPi18cbaez8JXMUV3pGjxybJtVQtuV5j1jt2UAgDDOHBBUYLeNfif8AAP8AZc8G6bZeKdc0PwVoMH7jTtPtrZs4JJPlW0Cs7DJJZlUgEkk85r4Q/tZfs/fHTUTofw4+Itne6uFZhpt1DLZ3LhQSSiTKpkAUEnZuwOuKr4s/tA/Br4GW1vcfFXx/p+gtdrut7d1knuJlDBSyQxK0jKCQCQuB68Gvhb+0d+zx+0tY3fh/wT4v0rX5Hts32h6hatFMYyBvDW86jzUG4KzIHQEgZ5Fftl/8E249Fsr/AOKX7OmnSPaW0bT6l4VXdJIoyS0loeSwAOfJPICnYTkIP2fP2g/iR+zR8SLH4lfDXVfIu4P3d7ZTEm31G3JBeCdARuQ4HPBUgMpBAI/Z++Ongn9pD4UaJ8WPAd1my1SMrcWjuDNp9ynEtvKB0dG9gGUq4yrKTVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVXjP9pn4IfD3xLeeD/F/jf7Bq+n7PtFt/Zt1Ls3orr80cTKcq6ngnr614E+IHhD4meH08U+CNX/tLTHleJZ/s8kOXXhhtkVW4+lVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVXjv9oz4M/DPX38LeN/GX9m6pHEkrQf2dczYRh8p3RxsvOPWv8AgpF+0h4M1v8AYc8RX3w3199RtvFXiC08NG6W1mgMcnF3KmJFU4MMJUnGMPjOen/BP34eP8Tf2w/hjoTWf2i2sNYXVrrcrFEjska5y5AOAWiRMHgs6qSN2aqvHvhKy8f+BfEfgTUmRbTxHpN1ps5ePzAI54mjbK5G4YY8ZGfUVq+lX+hare6HqsHkXun3Elvcxbg3lyIxVlypIOCCMgkV8EvH0fxV+Dngf4lJLE7+JvD9lqEwiZWEcssKNIny8Aq5ZSOxUjtVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV9e2um2VxqN7L5dvaxNLK+0naijLHAyTwD0r/hsz9mv/opP/lHvv/jNf8FcfiNL4G/Y+1LQLSd47nxvrVno4aNirrGC1zIeGBClbby24YESbSPmyP8AgnX8P3+I37ZXwz0023m2+j6p/bM7EsBELJGuEY7SD/rY4lHbLDIIzVV8SfB1r8RPh14p+H97s+z+JtFvNMm3lgNlxC8TZKkMBhz0IPpV5aaloOqzWN1HLZ6hp1w0ciZ2vDKjYIyOhDD8xXwi8dw/FL4U+DfiTAiIvinQbLUzGowI2ngSRkwGbBUsVI3HBGMnrVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVa5rOmeG9E1DxFrV19n0/S7WW6u5tjP5cUalnbaoLHCqTgAk9hVj+1/8As6ale2+nWXxF8y4upVihT+yL1dzsQAMmHAySOtf8FvfiN9o8VfDT4R20u37Bp91rl4m3O/z5BBAcleNv2e46Mc7+QMKT/wAES/h5Hq3xY8f/ABPubPzF8O6HBpttI6qVSW7l3llyM7wlqy5UjCyEHO8Yqv8AgqJ8PI/iD+xh40lis/tF94Wktdbs/lU+WYZlWZ+QSMW0lx93B7ZwSD+xN8QW+F/7WXws8YeckUKeIoLG5lcgCO3u82sznKtwI53PTPHBBwRVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV8SfjN8NfhF/Z3/Cw/Ev8AZP8Aa3m/Y/8AQ55/N8vZv/1SNjHmJ1xnPHQ18Ovjn8K/i1e3enfD7xT/AGrcWEQluE+w3EGxCcA5lRQefTNVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX17a6bZXGo3svl29rE0sr7SdqKMscDJPAPSv+GzP2a/8AopP/AJR77/4zX/BbT4hvpXwm8AfC+1vfLfxFrk+pXEaMwZ4rSLYA2DjYXulbDA5aMEEbDX/BH7wA/iz9ruDxW8coh8E+H77UBKEbZ5sqraKhI4BKXMrAHr5bEdKqqqqqqqqqv2uf+CT/AMMPi/8A2j46+Bn2TwL4xkjLtpscezSdQlHcxqM2zsMDdGNmRkplmY/Fn4O/Er4G+MLvwJ8UvCV7oWr2jECOdMxzqOkkUgysqHsykj8c1+x//wAFX/iJ8IfsXgP4+nUPHXg+PKQ6p5nmatpq9vncgXUYORtkIcBuHKosZ+Fnxf8Ahn8bvCdv43+FXjPT/EejXHAntWIaJv7kkbAPE/8AsOqt7VVVVVVVVVVf8FutPvZfg38O9Vjt3a0t/E00MsoHCSSWzFFPuRG5/wCAmv2DLu0sv2xvhJNeXUMEbeJIIleVwgLuGRFBPdmZVA7kgDk1X7Xd3aWX7KfxjmvLqKCNvAetRK8rhAXeylRFBPdmZVA7kgDk1X7L2n3uk/sz/CPStSt3t7uy8C6LDPC4w0ciWMKsp9wQRX7fv/J5XxZ/7GCT/wBASv8AgiB4f0VPBXxP8VLp0R1aXVLOya7Iy4gWJ3EY/ujc5JxjJ25ztXH/AAUS0TStd/Yu+Kdtq1mlwltpK3UIYkGOaGeOSNwRyCGUfUZByCQa8ReMtS8Efshan8QrR5ZtQ0D4bzapE3m7XeWHTjKDvIOCWUfNg884NV4Y/wCCq37Wfg/w1pPhHw9qPhG10vQ7GGxsbdNAjAighQJGoAOAAqgcelftIftb/GD9quXw/P8AFq50eeXwytyti9hpy2zYn8suHIJ3D90uAemW9TX/AAR21e90z9r/AOxWrII9W8K39tchlyTGHhmGPQ74U59M+teJvDOgeM/DupeEvFOk2+p6PrFrJa31ncJujnhdSrow9CCa/au+C+m/s+fH7xf8J9G8SWmt2GjXn+iXEEm9o4pFEiRS+ksasEcf3lPTOB4I0HTfFXjTw/4X1nxHaeH7DV9UtrO61e7GYdOillVHuJBkfJGrFzyOFPIr4UfCzwX8Ffh5ofwx+H+lJY6JoNqsEC7VDzMB880pUANK7Zd2wNzMT3r/AIK8/wDJmmpf9jBp3/obV4C0ey8ReOvDnh/UldrTU9WtbWcI21jHJKqtg9jgnmrSztLC0hsLC2itra2jWKGGFAiRIowqqo4AAAAA4Ff8FtvAWi3fwg8A/E8wRLq2l+JG0dZBHh5ILm2lmILA8hWtAQCDgu2CuW3fsW+INQ8M/tb/AAf1LTCnnTeMtNsm35x5dzOtvJ0IOfLlbHbOMgjIP/BcLxbe2fgX4VeBI1f7Jq+rajqUpEmAJLSKGNMrjk4vH5yMc8HPHwP+NfjX9nv4iWXxR+Hg0wa/p0M0VrLf2S3KwiVDG7KrdGKMy5HOGI6E1/w95/bK/wCg54U/8EKf/FV4y8Vap478X65431wW41LxDqVxqN4LeERRiaeRpH2IOFXcxwo6DAr/AIJheNL3Sf2ALPWbq3S6j8JTa01tAreWZI0lkuChbBwS8jjdg4BHBxWo6he6vqFzqupXL3F3eTPNPK/3pJHJZmPuSSa/4JoeA9A8C/sZeAW0S1RJ/EUM2salMFw1xczSMNze6xpFGD/diWqr9v3/AJPK+LP/AGMEn/oCV/wRD/5JV8Sv+xgtf/Sc1+2N4tvfA/7KvxW8S6ar/a4PCt9FbukmxopJojEsgODyhkD4xztxkZyKtP8Agrd+2BYWkNhYal4Ptra2jWKGGHw9GiRIowqqoOAAAAAOBX7RX7T/AMU/2pdf0rxN8V7jSZ9Q0azaztpLGwW2/dFy+GwTuwxJHpk+tf8ABE/WL2D9o3xn4fjZPsl54JluZQV+bzIb21VMHsMTyZH09K/4KweJtV179tzxhpWoTb4PDmn6ZYWC7mOyFrKK5IwSQP3tzKcKAOc4yST/AMEP/Buj6j43+Kfjy5hR9S0PTdNsLRmQEpHdyXDykHsf9EjH0Jqv2odPstJ/aY+Lelabbpb2ll461qG3hQYEcaX0yqo9gABX/BFT/k1fxT/2UC8/9ILCqqqqqqqqqr/gsx+0ZL4g8c6N+zT4fu3Fh4XWPVNfVSQJb2aPNvGfaOB9/oTOO61/wT1/ZetPj98TpvE/jHTlufBfg1opr2CVfkv7lsmG3IIwyfIWkH90Kp++KiiigiSCCNY441CoirgKBwAAOgr9qD4uav8AG345eK/HGp6it1bNfy2ukiNgyQ2MTssCKVyCNmGJHDMzN3rw/wCINa8Ka7p/ibw3qc+n6rpVylzZ3UDbXhlQgqyn1BAr4LfEaL4o/Bzwj8TLhVtm1zRYLy6VjhYZSg80Z4+UOHGcDgZ4r47fFrWvjj8V/EXxK1q4nY6teObOGU/8etqpxDCBkgBUCggHBOT1JNeDfGHiP4f+KdL8aeEdVn07WNHuVuLS5hcqyMOxx1UjKsvQqSDwTXgHxbaeP/Anhzx3p8DQW3iPSLXUoInOTGk8SyKpOBkgMB0Ff8FLP2V7T4VeMLf4y+A9IW28LeK7l01G3gXCWGonLkgZ4SYb2AAwrI44BQV/wSM/aRv/AIZ/HQfBLW9Qx4X+Iu5II5GCra6qiZhkBP8Az0VGhKjlmaL+7VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVftmf8AJynjD/tx/wDSGCv2FP8AkgFn/wBhS7/9CFVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX7dn/Jfrv8A7Bdr/wCgmv2tfiBHP8NfBPwqtbnD2euaprV7DhTuEsFnDbtnGRjyLkdcHdyOBj/gid8O4db+Mvjv4mXCJIvhXQYbGFW6pNeykhx3yI7SVfTDn2qqr/goZ4Af4c/tlfFDSRHKIdU1g6zDI6MBKL5FunKk9QJJZEyOMoR2r/gkl8Q38b/seaToVze+fc+C9YvdJbezFxGXFzGCWJyAtyEXGAFQKB8pNVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVeN/8AkS9f/wCwXc/+imqv+C33xBS68X/DL4U28zq2m6bd61dxgnDi4kWGEkbcZX7NcDhifnOQOCf+CJXw5h1f4p/EH4qXUCOPDei2+mWpkXOJbyUuzplT8ypaFSQwIEpGCG4qq/b6+Hw+Gn7YfxS8OxriG61x9VgwPlCXyLdhV+VRhTOUwBgbSMtjJ/4JQ/EFPHX7GXhvTHmeW68Hale6Lcu5JJKyfaIxyoACw3MKjBYYUc5yBVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVXxv8A+SLeP/8AsV9S/wDSWSvA/wDyOvh//sKW3/o1a/4Ka/EOP4iftn+PZbS9+0WPh2SDRLX5lPlm2hVZ0+UkcXBuOOCM4IByK/4I6/Dn/hEv2UpfG08f+keOfEF1dxSZ/wCXe3xaouAxHEsNwckKTvwQQFJqviH4NsPiL4A8TfD7VX2WXifR7vS7lsE4juIWiY4VlPRz0IPuOtXlpqWg6rNY3UctnqGnXDRyJna8MqNgjI6EMPzFfBfx7bfFT4Q+C/iRa3CTL4l0Gz1Byu0bZJYVZ0IUkKysWVlBO0gjtVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV/wUh/5p3/3FP/bSv+CdX/I6eLv+wXD/AOjaqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq8b/8iXr/AP2C7n/0U1V/wWK+I3/CW/tWxeCbeX9x4G8P2tnJHt6XNwDdO2SoPMU1uMZYDbnIJYD/AIIj/DyOw+HHxF+KlxZ/vta1i30m2mdVyI7WIyuE43AM10u7nDGNeAUqqqqqqqqqqr41fAX4UftDeEJPBXxZ8I2mt2PzNbSuNk9lIVx5sEo+aN/cHBxggjiv2w/+CW/xR+ACX3jr4UNe+PPAMCvNMyQhtS0mMH/l4iQASoqkEzRAABXZ0jUAn4K/Hn4rfs9eLo/Gvwm8X3ei3/yrcxId8F7Gpz5U8R+WRPYjIzkEHmv2Rv8AgqL8Ifj+NO8FfEuW08B+PruTyIra4lI0/UJD90QTtwjseBFIQSxCqzkiqqqqqqqqq/4Ke/B29+MX7IniWPSIfO1PwbcReIrWLa5Li3V1mACnr9nlnIyGBIAwMhl8MeI9X8H+JdJ8XeHrxrXVNDvob6xnQkNFPC4eNwRyCGUHj0r4A/8ABRX9mf4zeBLDX/EXxP8AC/gXxAIVGqaLr+rx2JtpwBvEUk5RZkyeGXt1AIIH/BTP/goJ8K/GHwnu/gF8DPFsXiS58SSR/wBuavp7t9mtbWOQP5KSceY8jIoO3K7NwJO7A+CHwo1745fFzwp8JvDMLvfeJdSjtt6j/UQ/emmPX5Y4lkkbg8IeD0rTtPstJ0+10rTbdLe0soUht4UGBHGgCqo9gABX7fv/ACeV8Wf+xgk/9ASv+CIf/JKviV/2MFr/AOk5r9vz/kzT4s/9i/J/6GlVonhKy8f/ALOOn+BNSZFtPEfgmPTZy8fmARz2QjbK5G4YY8ZGfUV4y8Ja/wCAfFmseCPFNg9lrGg301jfW79Y5onKOPcZBwe4wa/Y8v8A/gmX8dPhzoOmeOfhr8PPC/xCs7VLXVNN1if7KL2eNVVp7d5HCyLITuCZLqdwIIXe2q/s4/8ABMzQrCXVdb8L/B7T7KDHm3N3rMMMceSFGWaYAZJA57kV+zd8Kv2PLR5/it+zJ4Q8KDPnaW+taNG7K4DI0saOxwwyqZZcjjGeor9un9qvTf2UPgneeJ7SeGTxfrm+x8MWkibw9zt+aZ1/55xKQxzwTsX+KvAHgvxt8f8A4uaR4L0maXUfE3jXWBG1zOS5aWZy0s8h64UF5HPoGNftBfBrX/2fvjH4p+EfiIO8/h++aOC4ZNourZgHgmA9HjZGx2JI7V/wSp/a4l+OPwtk+DvjbU0l8Z+ALWNYJZZy02paZkJHMQeWaIlYnP8AtREnLmv+CvP/ACZpqX/Ywad/6G1eHtbu/DXiDTPEdhHDJc6VeRXcKTKSjPG4dQwBBIyozgj6ivgh+2D8A/jt4B03xr4d+I3h/T7i4t0e/wBH1DVIYbzTJjkNFNGzBhhlcK+Nrhdykiv+CtH7X/w/+Nmo+Gvg18JfEtv4g0TwxdS6hq2qWU5ktLi8KeXEkLKdkojRpSZBuU+aArDD5/4JkfCu/wDif+2H4KuE077RpnhCSTXdRlaIOsAgQ+Q2Dxk3DQAdxncPu1/wWa+EupeM/gD4d+JukWX2mTwFrDfbdseWhs7tVjeQHPAE0dsCMH72cgKc/s3+N/hb8PvjBofiH40/D638Z+C9zwavpkqlnWN1KiaIbgC8bbXCk4YArlSwZfBPwZ/4JY/EbRU1/wAGaB8JNRs3641BYpIjzxJE8ivGeM4dQcc9DXij4P8A/BKzwbcW9h4h0X4Q299dtGtvYx6kk91MXYqmyCORpGBYEAhSMjFeD/hV8Ovh/wCBV+GXgvwhp+keF1jnjGl20ZWLbMWMoPOTuLtnnvXxh+G2s/B74peKfhfr9vLFe+GtUms280YMiK37uQY7OhVwehDA96/4Jpf8FD/hh8M/hhb/AAA+PGvtoUei3UreHtalhaS3NvKxka3mKAsjLK0hVz8pVwvy7Bu1r9sn9kzQNKuNYvv2k/hvLBarudLLxLa3kxGQPkhhd5HPPRVJ79BWnahZatp9rqum3KXFpeQpNbyp0kjYBlYexBBr/gohpV/o/wC2n8VrPUYPKlk1hLhV3BsxzW8UsbZBPVHU46jODg5Ff8EmP2r/AIL/AAM0zx34B+L3iu38MHWLq21DTdQuw5gnKIySRMVU7GA2MCThtzDgqN3jG7+H/wC11+zf420P4Y+KtM8QaX4r0XUNKs9Rgy8Md2YmVGwduSkhRsccjrWr6TqWg6reaFrNlLZ6hp1xJbXVtMu14ZUYq6MOxDAgj1FfsxS/8Euf2hfBek3eo/DH4beFPGckOzUvDuqXH2WRLhFzIYDJIPOiIBZSpJC/eAKsBrH7O/8AwTH8PWR1LX/Dnwc0y0VgpnvNbghjDHoNzTAZPpX7Nfwv/ZT0nRV+LP7Mng/w5a6f4ihltV1jSoHQXUcM7JIgL87RLEw6YJUHkYNf8FkfhfdeEv2m7D4jR22LDx5ocMgnERUNdWgEEqZyQxWIWpzxgOoxxk/8E3v2tdB/ZV+Ml7L4+urqHwT4vs0s9Xlt4nm+ySxsWguTGmS4TdIpCgsFlYqCflPxG/b/AP2S/h54EufHC/Gvwv4jZLXzrTSNC1SG9vrpyAUiEKMWjYkgHzNgXncRg48feMNU+InjrxH8QNcKf2l4m1a61O82KAPOuJWlfAAAA3OegFf8EVP+TV/FP/ZQLz/0gsKqqqqqqqqrUdQstJ0+51XUrlLe0soXmnlfgRxoCzMfYAE18WPH2pfFX4neK/iVq8sr3XibWLrUHErbjGJZGZUHoFUhQBwAoA4FfsXfCSx+DX7OHhDw7FaLDqOqWaatq7BgzSXdyiu24rwSieXECOMRLyeprxr4Yu/BXjHXvBl+zNdaDqdzYTF49hLwyNG2VycHKnjJx6mq/Zm8E38f7IngjwRr8cljcX/hJIplK4eBbiMkZU9GCyDKnocg1f2N3pl7cabfwNDdWkrQzROOUdThlPuCCKr4CaNf+HPgV8OfD2qw+VfaX4S0u1uY/wC5JHaRow/Agiv2hvhXZ/Gr4LeLfhtcxb5tW06QWTBVJju0+eBhuIHEipnkcZGRnNeHde1zwR4p0vxPotxLY6xoGoRXlpLyjwXEMgdG7EFXUHsQRXw98Y2HxF8AeGfiDpSbLLxPo9pqlsuScR3EKyqMsFPRx1UH2HSqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv2zP+TlPGH/bj/wCkMFfsKf8AJALP/sKXf/oQqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/bs/5L9d/wDYLtf/AEE1+1D/AMj/AKf/ANgeP/0dNX/BG/4fv4X/AGVrzxndW22fxn4kurmCXLfPbQKluoxnHEsVxyACd2DnAxVV/wAFs/h22lfF3wF8Ube2RIPEWgy6bM0cYG6ezmL7nIGSxjukUE9ogBwtf8EQfiC0HiT4m/Cm4mRlvbG01q0jJAZDC7QzkDblgfPtwcthdowPmJqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqrxv/AMiXr/8A2C7n/wBFNVf8FLfH0fxB/bP+INxbSwva6FcQ6NB5bK2DawrHKCV7+cJcjqPunkGv+CO3w9bwp+ya/jC4iTzvG3iK8vYZQoDG3g22qqcMcgSQTnkKfnPGME1Vf8Fr/h62i/G/wR8SoYkjt/FPh2SxfaoBkuLKYl3J3Zz5d1AvKgYUYJ5x/wAEQPH0ceq/E/4WXEsO+4t7LWbRNyh8Rs8M5x1YfvLfkcKev3hVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVfG/wD5It4//wCxX1L/ANJZK0PWLLw9rWn6/qTOLTTLqO5nKLuYRxsGbA7nAPFeOPFmpePfGmv+OdZbOoeItUudRujnP72eVpH57/Mxr9l74eP8KP2dPhv8Pbiz+y3mjeG7OO/h2su27aIPccMFYZmeQ4IB55Gaqq/4KBfDx/hn+2H8TtCWz8i2v9YbVrXarBHjvUW5yhIGQGldCBwGRlBO3Nf8Em/iHL48/Y20PS7mR5J/BmrXuiPI4OWCstxGPosd1Gg9lHvVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV/wUh/5p3/ANxT/wBtK/4J1f8AI6eLv+wXD/6Nqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqrxv/AMiXr/8A2C7n/wBFNVftRfEOP4sftF/Ef4g2179qs9Y8SXj2Eu5W3WiylLflSynEKRjIJHHBxX/BOT4c/wDCtP2NPhvp80eLvXNPbXLh8/6z7a7TxHG5gMQPCvGPu5IBJFVVVVVVVVVVVX7XX/BLf4R/tAS3/jj4Ztb+AvHlyzSzS28ONO1KU4LNcQKPkc4JMsWCSzM6yE5Hxq+AvxW/Z68XSeCviz4Ru9Fv/ma2lcb4L2NTjzYJR8sie4ORnBAPFfse/wDBUj4o/ABLDwJ8Vxe+PPAMCpDCskwbUtJjBx/o8rkCVFUkCGUgAKio8agg/Bv47fCb9oHwqPGPwj8aWPiDT0YJcCIlJrSQjOyaJwHjbHZgM9Rkc1VVVVVVVV+2H/wSH1vVPE198Qv2VBpgtNQZ5rnwddTLai2lxk/Y5WPl7HOcRSFBGejFSFTxH+xH+134Wljg1P8AZx8eztIzqp07RZdQAK4BybcOAORgnAbnGcGvhz/wTY/bI+I+qWVjF8IL3w3aXSo8mo+I5FsIbVGGQ0iNmbI4BRY2cHqo5r9iH9gvwP8Ash6BJq95d2/iT4hatCE1LXPs+xLaM4JtrVTkrECOXOGkIBYKAqJX7Zv7GX7Unj/9qT4keMvBnwS8R6tomra081le28SGOeMoo3LlunBr/gkt8EPix8EPh1480r4seBNT8MXepa1bzWkN8gVpo1hKlhgngHiv2zfB/ibx/wDstfEjwZ4N0a41bW9W0V4bKytwDJPIXU7VB74Br/hgL9sr/o3nxV/35T/4qvhvp97pPw78LaVqNu9vd2Wi2cM8LjDRyJCisp9wQRX7eP8AwTU0f9pnVJPir8LdVsfDvxB8lIryG6QR2WrqgbDSlELpPgovmncCqKpXgMPHH7An7Y3w/naDWP2ffFd+BIEWTQ7UaurZBIYfZDIQMKeSBjIBwSBXh39iP9rvxTLJBpn7OPj2FomRSdR0WXTxlsgYNwEBHByRwvGcZFf8EzvgL8av2avgZr3g741aRY6bLd69JqlhZW12L24hR7eJJFcxF48ZhBVUJOS5PUV+2Z8H/wBuf9qz416n4+uv2efGFtoNnmy8O6cyKRa2aMdrEF8CSQ/O/u2Oiiv+CVn7Dnjr4Q+Idd+OHxu8Iy6Fr0cbaZ4e0y8AM0KMAZ7ogEhdwIiTndjzcgBlJ/4KpfsWeI/jvoeifF/4Q+GbjV/HGhsmnX+n2u0PfaezMVcA43PFIw4/uSOT9wCvgd+zH+3t8Avit4b+LXgz9n/xWNR8P3iymBokCXcJ+Wa3f5j8kkZdCRyN2RggEf8ABVPWJfEX7Cf9u3GkXulTX+raVNLYX0RjntXZiWikH95TlTjg4yCQQa8G6HD4o8X6F4anuHhj1bUre0eVBlkEkioWAPcBs18bv+CaH7WHwd1y9ttM+HOoeO9CikP2LV/DMBvDcx5UAtbJmeN/nXcpQgENtZ1UtXwq/wCCeH7XvxZ1mHS7P4Na34YtmkKT6l4rtZNJgtgNuXZZVErj5h/q43J5wDtbH7Gf7G3gn9j7wDcaDpF//bvibWpFl1zXZLYQtclc+XFGmSUiQE4UsSWZmJ5AXxD4e0LxboV/4Y8TaRaappOqW7217ZXcQliuInGGR1PBBBxiv2lv+CPnxa8Ia3da3+zfInjTw1K26LSr29it9Ssxt+YFpPLimXIOCpV8EDaSNx1X9jr9rDR7+XTbv9mz4lyTQ43Na+F7u6jOQCNskSMjcEZwTg5B5BFeA/8AgnP+2p4zvbSTS/gprGiqypMt3rNxFpogB5ViJXWQMOMqFLqeoFaT/av9lWX9u/ZP7S+zx/bPsm7yfO2jf5e75tm7OM84xmv27f8Agnl4W/axtE8a+DLzT/DPxKsoxGuo3EbC21WJRhYrrYCwK8bZQrMoG0hht2fEH/gn5+2N8N7o22r/AAE8S6pGZNkU/h+3GsJIMsA+LUyMoOwn5wpGVyAWAPhf9h79r3xhdfYtJ/Zy8dwSCSOPOqaRJpiZckA77oRrjjls4XgsQCK+AOleMtB+Bnw+0D4iWD2fijSvDOn2esQPcRzst1FbpHKS8ZKMSyk5Ukc8E9a/4KY/8E+/GH7QGs2fxu+CNlaXfiu1s1s9Z0Z5FgfUoo8+XNHIxCmVRhCrEblC4IKYbUP2P/2rtMvZbC5/Zr+JrywNtZrfwpeTxk/7MkcbKw9wSK/4JofCv4g/B39ljTPB/wATfC114f1r+2L64NjdFfMWN5AFYhScZ2ng84wehFft2f8ABL20+PniG++MHwQ1PT9D8aXvz6rpN6BDY6o4HMqPGhMVw38RbKyEgkodzN4x/YQ/bE8DXP2PWv2dvGdzJuC50ew/tZMlQ337MyrjBHOcZyOoIrwx+w1+2B4vlWDSf2dPHEDNMsIOqaW+mjc3AObnywF9W+6O5Ff8E6Pgr8XvgB+zfb/Dn4y2Gn2Oqwaxc3VlaWl4Lk29rMsb7JXUlPMExnyIyVxtOSSTX7T37NXgH9qn4W3fwz8dCW2YSfatK1O3GZtNvFVlSZRkBxhmVkPDKxGVO1l+MP8AwTf/AGuPhDq81kPhXqfjHT1bEGp+FLd9SjmBOAfKRfOQ+oZBj1I5r4Hf8Ezf2rvjLrVrBqvw+vfAOhswN3q3ii3azaFNxDbLZsTSPhWKrtVT8u50DBj+1N/wTZ+Nvg34y6joXwI+FHiDxL4Kg0/T10/UYFV2ldbSJLhpT8oEjTpK5Cjb84xjoP8AglJ8IPiZ8Ff2dvEPhX4q+DdQ8NatdeNLq9htL5ArvA1lZIsgwTwWjcfVTVVVVVVVVVftQ6he6T+zP8W9V025e3u7LwLrU1vKnDRyJYzMrD3BANeC9KtNd8Y6Dol+rNbahqdvbzBW2ko8iq2D2OCaqv2xf+CdJ+O/jCX4ofCrxHpWg+I79UXVLDUomS0vGXA88SRKzRybRhhsYOQDlTkt+z7/AMEp9fg8R2PiX9oPXNKOlWcvmN4e0uV5nvNpOEmmwojQkAkJuLKcZQ8iKKKCJIII1jjjUKiKuAoHAAA6Cv2t/wDgmrd/Frx1qXxS+DXiLSNI1PWn8/VNH1JGit5rgj55opY1Yq0hwzKykFyzFxuwP2Zv+CW+qeG/F2neOP2gNX0i9ttNcT2/h7TZHlWWZSpQ3EpVQVBBJjXIbC5bG5TVftG6JH4b/aB+JWhW9jJZ29n4t1NLaGQNlYftMnl/e5IKFSCc5BByc5r/AIJ/a2niD9jL4TX6ak18ItBW0815C5U28jwmPJ7J5ewDoAoA4FVVVVVVVVVVVVVVVVVVVVVX/BXj4/fGX4Gf8Kn/AOFR/EPVfC/9t/21/aH2F1X7R5P2Hy92Qfu+bJj/AHjX/Dfn7ZX/AEcN4q/7/J/8TX/Dfn7ZX/Rw3ir/AL/J/wDE1/w35+2V/wBHDeKv+/yf/E1/w35+2V/0cN4q/wC/yf8AxNf8N+ftlf8ARw3ir/v8n/xNf8N+ftlf9HDeKv8Av8n/AMTX/Dfn7ZX/AEcN4q/7/J/8TX/Dfn7ZX/Rw3ir/AL/J/wDE1/w35+2V/wBHDeKv+/yf/E1/w35+2V/0cN4q/wC/yf8AxNf8N+ftlf8ARw3ir/v8n/xNf8N+ftlf9HDeKv8Av8n/AMTX/Dfn7ZX/AEcN4q/7/J/8TX/Dfn7ZX/Rw3ir/AL/J/wDE1/w35+2V/wBHDeKv+/yf/E1/w35+2V/0cN4q/wC/yf8AxNf8N+ftlf8ARw3ir/v8n/xNf8N+ftlf9HDeKv8Av8n/AMTX/Dfn7ZX/AEcN4q/7/J/8TX/Dfn7ZX/Rw3ir/AL/J/wDE1/w35+2V/wBHDeKv+/yf/E1/w35+2V/0cN4q/wC/yf8AxNf8N+ftlf8ARw3ir/v8n/xNf8N+ftlf9HDeKv8Av8n/AMTX/Dfn7ZX/AEcN4q/7/J/8TX/Dfn7ZX/Rw3ir/AL/J/wDE1/w35+2V/wBHDeKv+/yf/E14B8f+M/il4SsPHfxB8RXWu6/qfmfa7+6YGSby5GjTJAHRERfoBXgz9qT4u/C79pn4e+ALT4v3vhz4eP4m0htYs5bqOCyS1kuo/tTSs4wiGPduYkADJyK+Gvxa+Gvxk0S78R/C3xppnibS7G+ewnvNPl8yNLhFR2Td0JCyIcjIww5qqqvjV8f/AIS/s8aDpnif4weLE0DS9X1JdOtrlrWW4BnaN5ACsSswXbE2WxtBIyRmv20f2+PjPD+0t4wT9n/9ojUj4BK2DaR/ZFyjW2DYW5m2EqTnzjLuB5DbgcYr/hvz9sr/AKOG8Vf9/k/+Jr/hvz9sr/o4bxV/3+T/AOJr/hvz9sr/AKOG8Vf9/k/+Jr/hvz9sr/o4bxV/3+T/AOJr/hvz9sr/AKOG8Vf9/k/+Jr/hvz9sr/o4bxV/3+T/AOJr/hvz9sr/AKOG8Vf9/k/+Jr/hvz9sr/o4bxV/3+T/AOJr/hvz9sr/AKOG8Vf9/k/+Jr/hvz9sr/o4bxV/3+T/AOJr/hvz9sr/AKOG8Vf9/k/+Jr/hvz9sr/o4bxV/3+T/AOJr/hvz9sr/AKOG8Vf9/k/+Jr/hvz9sr/o4bxV/3+T/AOJr/hvz9sr/AKOG8Vf9/k/+Jr/glJ8X/iZ8a/2dvEPir4q+M9Q8S6ta+NLqyhu75gzpAtlZOsYwBwGkc/VjVVVVVV+2b+2b+1J4A/ak+JHgzwZ8bfEek6JpOtPDZWVvKgjgjCKdqgqeOTX/AA35+2V/0cN4q/7/ACf/ABNf8N+ftlf9HDeKv+/yf/E1/wAN+ftlf9HDeKv+/wAn/wATX/Dfn7ZX/Rw3ir/v8n/xNf8ADfn7ZX/Rw3ir/v8AJ/8AE1/w35+2V/0cN4q/7/J/8TX/AA35+2V/0cN4q/7/ACf/ABNf8N+ftlf9HDeKv+/yf/E1/wAN+ftlf9HDeKv+/wAn/wATX/Dfn7ZX/Rw3ir/v8n/xNf8ADfn7ZX/Rw3ir/v8AJ/8AE1/w35+2V/0cN4q/7/J/8TX/AA35+2V/0cN4q/7/ACf/ABNf8N+ftlf9HDeKv+/yf/E1/wAN+ftlf9HDeKv+/wAn/wATX/Dfn7ZX/Rw3ir/v8n/xNf8ADfn7ZX/Rw3ir/v8AJ/8AE1/w35+2V/0cN4q/7/J/8TX/AA35+2V/0cN4q/7/ACf/ABNf8N+ftlf9HDeKv+/yf/E1/wAN+ftlf9HDeKv+/wAn/wATX/Dfn7ZX/Rw3ir/v8n/xNf8ADfn7ZX/Rw3ir/v8AJ/8AE1/w35+2V/0cN4q/7/J/8TX/AA35+2V/0cN4q/7/ACf/ABNf8N+ftlf9HDeKv+/yf/E1/wAN+ftlf9HDeKv+/wAn/wATXw4+J/j/AOL3hiPxl8S/FV74h1uSZ4XvbxgZDGhwq8AcAGviN8Avi/8AHj4i3Fr8IvBF14muNB8P29xfwWs0SyRxvcyorKjsrP8AMeQgJAyTgAmv2X/hpP8AB79nf4d/DW+i8q/0Pw/ax6gmQQt2yB7gDCrx5zyYyM4xnJyaqq/4Kofs/eMPj38ANIt/hx4Yvdd8UeHvEUN1bWdmm+SW3kikjmXBYKBlon3EEjy8DG4mvEngL9qb9hDxZofiDU5r34e+J/EOmzG0NlqEE05td6hw/lM6qCwU7Sc8DIBFf8N+ftlf9HDeKv8Av8n/AMTX/Dfn7ZX/AEcN4q/7/J/8TX/Dfn7ZX/Rw3ir/AL/J/wDE1/w35+2V/wBHDeKv+/yf/E1/w35+2V/0cN4q/wC/yf8AxNf8N+ftlf8ARw3ir/v8n/xNf8N+ftlf9HDeKv8Av8n/AMTX/Dfn7ZX/AEcN4q/7/J/8TX/Dfn7ZX/Rw3ir/AL/J/wDE1/w35+2V/wBHDeKv+/yf/E1/w35+2V/0cN4q/wC/yf8AxNf8N+ftlf8ARw3ir/v8n/xNf8N+ftlf9HDeKv8Av8n/AMTX/Dfn7ZX/AEcN4q/7/J/8TX/Dfn7ZX/Rw3ir/AL/J/wDE1/wSk/ae+P8A8a/2iPEPhX4q/FTWvEuk2vgu6vYbS+kVkSdbyyRZBgDkLI4/4Eaqqqqqv2zf2zf2pPAH7UnxI8GeDPjb4j0nRNJ1p4bKyt5UEcEYRTtUFTxya/4b8/bK/wCjhvFX/f5P/ia/4b8/bK/6OG8Vf9/k/wDia/4b8/bK/wCjhvFX/f5P/ia/4b8/bK/6OG8Vf9/k/wDia/4b8/bK/wCjhvFX/f5P/ia/4b8/bK/6OG8Vf9/k/wDia/4b8/bK/wCjhvFX/f5P/ia/4b8/bK/6OG8Vf9/k/wDia/4b8/bK/wCjhvFX/f5P/ia/4b8/bK/6OG8Vf9/k/wDia/4b8/bK/wCjhvFX/f5P/ia/4b8/bK/6OG8Vf9/k/wDia/4b8/bK/wCjhvFX/f5P/ia/4b8/bK/6OG8Vf9/k/wDia/4b8/bK/wCjhvFX/f5P/ia/4b8/bK/6OG8Vf9/k/wDia/4b8/bK/wCjhvFX/f5P/ia/4b8/bK/6OG8Vf9/k/wDia/4b8/bK/wCjhvFX/f5P/ia/4b8/bK/6OG8Vf9/k/wDia/4b8/bK/wCjhvFX/f5P/ia/4b8/bK/6OG8Vf9/k/wDia/4b8/bK/wCjhvFX/f5P/ia/4b8/bK/6OG8Vf9/k/wDia/4b8/bK/wCjhvFX/f5P/ia/4b8/bK/6OG8Vf9/k/wDia079u39r7VtQtdK1L4/eKLi0vJkhnheZMSRsQrKfl6EEiq8B/CL46ftNeI/Eer+BPDGq+N9djkOo63KkyPcO88h3TOHYM5aQsSRnk5OK/Zu+Hcvwm+AHw9+HF0jrd6D4ds7e8D5z9p8pTN9B5hfA7DAqqr/gqp+z544+PfwI0KP4Z+G73XvEnh3xFHcR2NoNzy20kMkcuASBkMYmyegU+taZ4h+On7JvxP1ew0PXdV8C+NNMjfTtTWzuUMiI2x2hcoWRhlY2IyQCo7jj/hvz9sr/AKOG8Vf9/k/+Jr/hvz9sr/o4bxV/3+T/AOJr/hvz9sr/AKOG8Vf9/k/+Jr/hvz9sr/o4bxV/3+T/AOJr/hvz9sr/AKOG8Vf9/k/+Jr/hvz9sr/o4bxV/3+T/AOJr/hvz9sr/AKOG8Vf9/k/+Jr/hvz9sr/o4bxV/3+T/AOJr/hvz9sr/AKOG8Vf9/k/+Jr/hvz9sr/o4bxV/3+T/AOJr/hvz9sr/AKOG8Vf9/k/+Jr/hvz9sr/o4bxV/3+T/AOJr/hvz9sr/AKOG8Vf9/k/+Jr/hvz9sr/o4bxV/3+T/AOJr/hvz9sr/AKOG8Vf9/k/+Jr/glJ8X/iZ8a/2dvEPir4q+M9Q8S6ta+NLqyhu75gzpAtlZOsYwBwGkc/VjVVVVVV/wUK/a9/aY+F37YHj/AMCfD74y+IND0DTP7N+x6fayqI4fM021kfaCpPLu7H3Y1/w35+2V/wBHDeKv+/yf/E1/w35+2V/0cN4q/wC/yf8AxNf8N+ftlf8ARw3ir/v8n/xNf8N+ftlf9HDeKv8Av8n/AMTX/Dfn7ZX/AEcN4q/7/J/8TX/Dfn7ZX/Rw3ir/AL/J/wDE1/w35+2V/wBHDeKv+/yf/E1/w35+2V/0cN4q/wC/yf8AxNf8N+ftlf8ARw3ir/v8n/xNf8N+ftlf9HDeKv8Av8n/AMTX/Dfn7ZX/AEcN4q/7/J/8TX/Dfn7ZX/Rw3ir/AL/J/wDE1/w35+2V/wBHDeKv+/yf/E1/w35+2V/0cN4q/wC/yf8AxNf8N+ftlf8ARw3ir/v8n/xNf8N+ftlf9HDeKv8Av8n/AMTX/Dfn7ZX/AEcN4q/7/J/8TX/Dfn7ZX/Rw3ir/AL/J/wDE1/w35+2V/wBHDeKv+/yf/E1/w35+2V/0cN4q/wC/yf8AxNf8N+ftlf8ARw3ir/v8n/xNf8N+ftlf9HDeKv8Av8n/AMTX/Dfn7ZX/AEcN4q/7/J/8TX/Dfn7ZX/Rw3ir/AL/J/wDE1/w35+2V/wBHDeKv+/yf/E1/w35+2V/0cN4q/wC/yf8AxNf8N+ftlf8ARw3ir/v8n/xNeHv23f2sfFviDTPCviP47eJdQ0nWbyKyv7SaVClxBK4SSNsL0ZWIP1rxF/yL+p/9ec3/AKAa/ZG/Zt8dftHfFjQtL8P+ErrVfDOma5py+KbyNQY7CzllJdnyf4o4psDuVxVVVf8ABUv9i/4zfGz44+EvHvwS+Hl34ifVdD/s/VPs80MKQTW8rFHleV1VN0cwUEkD91Xws/aO+OXwQ0++0r4TfEzWPDFpqUyzXcNi6qJpFXaGOQeQOK/4b8/bK/6OG8Vf9/k/+Jr/AIb8/bK/6OG8Vf8Af5P/AImv+G/P2yv+jhvFX/f5P/ia/wCG/P2yv+jhvFX/AH+T/wCJr/hvz9sr/o4bxV/3+T/4mv8Ahvz9sr/o4bxV/wB/k/8Aia/4b8/bK/6OG8Vf9/k/+Jr/AIb8/bK/6OG8Vf8Af5P/AImv+G/P2yv+jhvFX/f5P/ia/wCG/P2yv+jhvFX/AH+T/wCJr/hvz9sr/o4bxV/3+T/4mv8Ahvz9sr/o4bxV/wB/k/8Aia/4b8/bK/6OG8Vf9/k/+Jr/AIb8/bK/6OG8Vf8Af5P/AImv+G/P2yv+jhvFX/f5P/ia/Zv8Qaz4t/Z2+FvirxHqMuoatrPgvSL2/u5jl7ieWyieSRsd2ZiT7mqqqqqr9pHxBrPhP9nb4peKvDeoy2GraN4L1i9sLuE4e3nispXjkX3VlUj3Ff8ADfn7ZX/Rw3ir/v8AJ/8AE1/w35+2V/0cN4q/7/J/8TX/AA35+2V/0cN4q/7/ACf/ABNf8N+ftlf9HDeKv+/yf/E1/wAN+ftlf9HDeKv+/wAn/wATX/Dfn7ZX/Rw3ir/v8n/xNf8ADfn7ZX/Rw3ir/v8AJ/8AE1/w35+2V/0cN4q/7/J/8TX/AA35+2V/0cN4q/7/ACf/ABNf8N+ftlf9HDeKv+/yf/E1/wAN+ftlf9HDeKv+/wAn/wATX/Dfn7ZX/Rw3ir/v8n/xNf8ADfn7ZX/Rw3ir/v8AJ/8AE1/w35+2V/0cN4q/7/J/8TX/AA35+2V/0cN4q/7/ACf/ABNf8N+ftlf9HDeKv+/yf/E1/wAN+ftlf9HDeKv+/wAn/wATX/Dfn7ZX/Rw3ir/v8n/xNf8ADfn7ZX/Rw3ir/v8AJ/8AE1/w35+2V/0cN4q/7/J/8TX/AA35+2V/0cN4q/7/ACf/ABNf8N+ftlf9HDeKv+/yf/E1/wAN+ftlf9HDeKv+/wAn/wATX/Dfn7ZX/Rw3ir/v8n/xNf8ADfn7ZX/Rw3ir/v8AJ/8AE1/w35+2V/0cN4q/7/J/8TX/AA35+2V/0cN4q/7/ACf/ABNf8N+ftlf9HDeKv+/yf/E18KPj98Zfjp/an/C3PiHqvij+xPJ/s/7e6t9m83f5m3AH3vKjz/uivip8bviz8ENPsdV+E3jvU/DF3qUzQ3cti4Vpo1XcFOQeAea/4b8/bK/6OG8Vf9/k/wDia/4b8/bK/wCjhvFX/f5P/ia/4b8/bK/6OG8Vf9/k/wDia/4b8/bK/wCjhvFX/f5P/ia/4b8/bK/6OG8Vf9/k/wDia/4b8/bK/wCjhvFX/f5P/ia/4b8/bK/6OG8Vf9/k/wDia/4b8/bK/wCjhvFX/f5P/ia/4b8/bK/6OG8Vf9/k/wDia/4b8/bK/wCjhvFX/f5P/ia/4b8/bK/6OG8Vf9/k/wDia/4b8/bK/wCjhvFX/f5P/ia/4b8/bK/6OG8Vf9/k/wDia/4b8/bK/wCjhvFX/f5P/ia/4b8/bK/6OG8Vf9/k/wDia/4b8/bK/wCjhvFX/f5P/ia/4b8/bK/6OG8Vf9/k/wDia/4b8/bK/wCjhvFX/f5P/ia/4b8/bK/6OG8Vf9/k/wDia/4b8/bK/wCjhvFX/f5P/ia/4b8/bK/6OG8Vf9/k/wDia/Yy/bN/ak8f/tSfDbwZ4y+NviPVtE1bWkhvbK4lQxzxlGJVgF6cCqqqqqqq/wCCmf7Vv7Rfwh/akv8AwZ8NPi7rvh7RI9FsZksrORVjEjoSzYKnk4Ff8N+ftlf9HDeKv+/yf/E1/wAN+ftlf9HDeKv+/wAn/wATX/Dfn7ZX/Rw3ir/v8n/xNf8ADfn7ZX/Rw3ir/v8AJ/8AE1/w35+2V/0cN4q/7/J/8TX/AA35+2V/0cN4q/7/ACf/ABNf8N+ftlf9HDeKv+/yf/E1/wAN+ftlf9HDeKv+/wAn/wATX/Dfn7ZX/Rw3ir/v8n/xNf8ADfn7ZX/Rw3ir/v8AJ/8AE1/w35+2V/0cN4q/7/J/8TX/AA35+2V/0cN4q/7/ACf/ABNf8N+ftlf9HDeKv+/yf/E1/wAN+ftlf9HDeKv+/wAn/wATX/Dfn7ZX/Rw3ir/v8n/xNf8ADfn7ZX/Rw3ir/v8AJ/8AE1/w35+2V/0cN4q/7/J/8TX/AA35+2V/0cN4q/7/ACf/ABNf8N+ftlf9HDeKv+/yf/E1/wAN+ftlf9HDeKv+/wAn/wATX/Dfn7ZX/Rw3ir/v8n/xNf8ADfn7ZX/Rw3ir/v8AJ/8AE1/w35+2V/0cN4q/7/J/8TX/AA35+2V/0cN4q/7/ACf/ABNf8N+ftlf9HDeKv+/yf/E1/wAN+ftlf9HDeKv+/wAn/wATX/Dfn7ZX/Rw3ir/v8n/xNad+3b+19q2oWulal8fvFFxaXkyQzwvMmJI2IVlPy9CCRVfB74K/Er49eL18D/C3wzca1qiw/aJ0iHFtb+ZHG00ncIrSpkgE89DXh7QdN8LeH9M8MaND5NhpFnFZ2kf9yKJAiD8FUCqqqqqqqqqqqqviz8Hfhr8cvB134E+KXhKx17SLpSBHOmJIG7SRSDDROOzKQfwzX7YP/BJ/4ifCH7b47+AP9oeOvB8eHm0vy/M1bTV7nYgAuowcfNGA4DcoVRpD/wAEqPhf8TfH37Tlh4i8F+JtY8P6B4PWO/8AEl5ZMyx3UO4+XYyj7rCdlYbW/gSRl5QEVVVVVVVVVVVVVVVVVVVVf8Fef+TNNS/7GDTv/Q2r4Tf8lU8Gf9jBY/8ApQlVVVVVVVVVVVVVVVVVVVVVVVVV+05pV/rv7NnxY0PSoPPvdQ8D6zb20W4LvkexmVVyxAGSQMkgV4Q1e28P+LdE168SR7fTdRt7mVYgC5WORWIUEgZwOMkfWopYp4kmglWSORQyOpyGB6EEdRVV4y+IXgL4d6euqePvGmh+HbR93lzapfx2qyEYyF3sNx5HAyeR61/w2x+yl/0XPw1/39f/AOJrwP8AE/4cfE2xOo/Dzx3oPiOBFBkOmahHcGLIBw6qSUbDDKsARnkCqqv2hdZg8RfHv4ka7azSS29/4t1SaBpchvLa6kKAg9MLgY7dK/YD0gaL+xp8JbIaZ9g8zw/Hc+V5HlbvOd5fMxgZ8zzN+7+Lfu5zmqqqqqqqqqqqqqqqqqqqqqr46fst/Aj9pb+xP+F2eBf+Ej/4Rz7R/Zn/ABM7uz+z+f5fm/8AHvLHu3eTH97ONvGMnP8Aw64/YS/6IX/5c2rf/JVf8OuP2Ev+iF/+XNq3/wAlV/w64/YS/wCiF/8Alzat/wDJVf8ADrj9hL/ohf8A5c2rf/JVf8OuP2Ev+iF/+XNq3/yVX/Drj9hL/ohf/lzat/8AJVf8OuP2Ev8Aohf/AJc2rf8AyVX/AA64/YS/6IX/AOXNq3/yVX/Drj9hL/ohf/lzat/8lV/w64/YS/6IX/5c2rf/ACVX/Drj9hL/AKIX/wCXNq3/AMlV/wAOuP2Ev+iF/wDlzat/8lV/w64/YS/6IX/5c2rf/JVf8OuP2Ev+iF/+XNq3/wAlV/w64/YS/wCiF/8Alzat/wDJVf8ADrj9hL/ohf8A5c2rf/JVf8OuP2Ev+iF/+XNq3/yVX/Drj9hL/ohf/lzat/8AJVf8OuP2Ev8Aohf/AJc2rf8AyVX/AA64/YS/6IX/AOXNq3/yVX/Drj9hL/ohf/lzat/8lV/w64/YS/6IX/5c2rf/ACVX/Drj9hL/AKIX/wCXNq3/AMlV/wAOuP2Ev+iF/wDlzat/8lV/w64/YS/6IX/5c2rf/JVf8OuP2Ev+iF/+XNq3/wAlV/w64/YS/wCiF/8Alzat/wDJVfGH9lrU/hz8RtX8GfBH4N+KI/BWn+T/AGYllY3t/EN8Ecku2Z/MZ/3ryZyxwcjgDA/bf+C3iH4cP4B8ba/4d1HRZvFdheW01rf2zwStPZzjdJsc7gDFc24xtVflyCxLY/4IgfEONbr4n/Ce7vfnkjstbsLfco4UvBcvjOT960HAIHcjKg1VV/wW2+J0Op+Pvh98ILG9RxoWm3GrX8Ub5xLcuI4g47MqQOQODiYHoRX/AASv/ZD+GH7S2v8AjzX/AIzeEZdd8OeG7O1tbO2N9c2aNeTu7Fw8DIzFI4SCu/A85SVOVK/8OuP2Ev8Aohf/AJc2rf8AyVX/AA64/YS/6IX/AOXNq3/yVX/Drj9hL/ohf/lzat/8lV/w64/YS/6IX/5c2rf/ACVX/Drj9hL/AKIX/wCXNq3/AMlV/wAOuP2Ev+iF/wDlzat/8lV/w64/YS/6IX/5c2rf/JVf8OuP2Ev+iF/+XNq3/wAlV/w64/YS/wCiF/8Alzat/wDJVf8ADrj9hL/ohf8A5c2rf/JVf8OuP2Ev+iF/+XNq3/yVX/Drj9hL/ohf/lzat/8AJVf8OuP2Ev8Aohf/AJc2rf8AyVX/AA64/YS/6IX/AOXNq3/yVX/Drj9hL/ohf/lzat/8lV8FPgJ8Jv2dfCt14J+DfhP/AIR/Rb3UH1Ce1+3XF3vuXjjjZ988jsMpDGMAgfLnGSSaqqqqr4gf8E9f2Pvil4z1b4g+O/hB/aev65cG4v7v+39Sg86QgAtsjuFReAOFUCv+HXH7CX/RC/8Ay5tW/wDkqv8Ah1x+wl/0Qv8A8ubVv/kqv+HXH7CX/RC//Lm1b/5Kr/h1x+wl/wBEL/8ALm1b/wCSq/4dcfsJf9EL/wDLm1b/AOSq/wCHXH7CX/RC/wDy5tW/+Sq/4dcfsJf9EL/8ubVv/kqv+HXH7CX/AEQv/wAubVv/AJKr/h1x+wl/0Qv/AMubVv8A5Kr/AIdcfsJf9EL/APLm1b/5Kr/h1x+wl/0Qv/y5tW/+Sq/4dcfsJf8ARC//AC5tW/8Akqv+HXH7CX/RC/8Ay5tW/wDkqv8Ah1x+wl/0Qv8A8ubVv/kqv+HXH7CX/RC//Lm1b/5Kr/h1x+wl/wBEL/8ALm1b/wCSq/4dcfsJf9EL/wDLm1b/AOSq/wCHXH7CX/RC/wDy5tW/+Sq/4dcfsJf9EL/8ubVv/kqv+HXH7CX/AEQv/wAubVv/AJKr/h1x+wl/0Qv/AMubVv8A5Kr/AIdcfsJf9EL/APLm1b/5Kr/h1x+wl/0Qv/y5tW/+Sq/4dcfsJf8ARC//AC5tW/8Akqv+HXH7CX/RC/8Ay5tW/wDkqv8Ah1x+wl/0Qv8A8ubVv/kqv+HXH7CX/RC//Lm1b/5Kr44fsmJ8LvHkvhP4AfBrxLF4SjtYpYlsLW91GPznB8zE0hkYnIGRu49BX7CXwYh8F6Fr/j7xR4G1TRPGV/dnTHm1GK4t5JdPjSOWNRDIQu0SyTHeFyTkFiAAKqqr44fsj/s8/tIarpmt/Gj4ff8ACRXuj2729lL/AGreWnlRs25lxbzRg5POWBNf8OuP2Ev+iF/+XNq3/wAlV/w64/YS/wCiF/8Alzat/wDJVf8ADrj9hL/ohf8A5c2rf/JVf8OuP2Ev+iF/+XNq3/yVX/Drj9hL/ohf/lzat/8AJVf8OuP2Ev8Aohf/AJc2rf8AyVX/AA64/YS/6IX/AOXNq3/yVX/Drj9hL/ohf/lzat/8lV/w64/YS/6IX/5c2rf/ACVX/Drj9hL/AKIX/wCXNq3/AMlV/wAOuP2Ev+iF/wDlzat/8lV/w64/YS/6IX/5c2rf/JVf8OuP2Ev+iF/+XNq3/wAlV/w64/YS/wCiF/8Alzat/wDJVf8ADrj9hL/ohf8A5c2rf/JVfBT9jL9mr9nXxVdeNvg38Nv+Ef1q9099Pnuf7YvrvfbvJHIybJ5nUZeGM5AB+XGcE5qqqqqviB/wT1/Y++KXjPVviD47+EH9p6/rlwbi/u/7f1KDzpCAC2yO4VF4A4VQK/4dcfsJf9EL/wDLm1b/AOSq/wCHXH7CX/RC/wDy5tW/+Sq/4dcfsJf9EL/8ubVv/kqv+HXH7CX/AEQv/wAubVv/AJKr/h1x+wl/0Qv/AMubVv8A5Kr/AIdcfsJf9EL/APLm1b/5Kr/h1x+wl/0Qv/y5tW/+Sq/4dcfsJf8ARC//AC5tW/8Akqv+HXH7CX/RC/8Ay5tW/wDkqv8Ah1x+wl/0Qv8A8ubVv/kqv+HXH7CX/RC//Lm1b/5Kr/h1x+wl/wBEL/8ALm1b/wCSq/4dcfsJf9EL/wDLm1b/AOSq/wCHXH7CX/RC/wDy5tW/+Sq/4dcfsJf9EL/8ubVv/kqv+HXH7CX/AEQv/wAubVv/AJKr/h1x+wl/0Qv/AMubVv8A5Kr/AIdcfsJf9EL/APLm1b/5Kr/h1x+wl/0Qv/y5tW/+Sq/4dcfsJf8ARC//AC5tW/8Akqv+HXH7CX/RC/8Ay5tW/wDkqv8Ah1x+wl/0Qv8A8ubVv/kqv+HXH7CX/RC//Lm1b/5Kr/h1x+wl/wBEL/8ALm1b/wCSq/4dcfsJf9EL/wDLm1b/AOSq/wCHXH7CX/RC/wDy5tW/+Sq1v/gmL+xPpmjX+paD8DHGp2lrJLZFPEWqyMJlUlMKbkhjuA4IOfSv+FIfGn/okHjX/wAJ+6/+Ir4Ifsj/ALPP7N+q6nrfwX+H3/CPXusW6297L/at5d+bGrblXFxNIF55yADVVVV8QP8Agnr+x98UvGerfEHx38IP7T1/XLg3F/d/2/qUHnSEAFtkdwqLwBwqgV/w64/YS/6IX/5c2rf/ACVX/Drj9hL/AKIX/wCXNq3/AMlV/wAOuP2Ev+iF/wDlzat/8lV/w64/YS/6IX/5c2rf/JVf8OuP2Ev+iF/+XNq3/wAlV/w64/YS/wCiF/8Alzat/wDJVf8ADrj9hL/ohf8A5c2rf/JVf8OuP2Ev+iF/+XNq3/yVX/Drj9hL/ohf/lzat/8AJVf8OuP2Ev8Aohf/AJc2rf8AyVX/AA64/YS/6IX/AOXNq3/yVX/Drj9hL/ohf/lzat/8lV/w64/YS/6IX/5c2rf/ACVX/Drj9hL/AKIX/wCXNq3/AMlV/wAOuP2Ev+iF/wDlzat/8lV8FPgJ8Jv2dfCt14J+DfhP/hH9FvdQfUJ7X7dcXe+5eOONn3zyOwykMYwCB8ucZJJqqqqqvil+wT+yb8avHep/Ez4mfCj+2fEus+V9tvf7d1C383yoUhj/AHcNwiLiONF4UZxk5JJr/h1x+wl/0Qv/AMubVv8A5Kr/AIdcfsJf9EL/APLm1b/5Kr/h1x+wl/0Qv/y5tW/+Sq/4dcfsJf8ARC//AC5tW/8Akqv+HXH7CX/RC/8Ay5tW/wDkqv8Ah1x+wl/0Qv8A8ubVv/kqv+HXH7CX/RC//Lm1b/5Kr/h1x+wl/wBEL/8ALm1b/wCSq/4dcfsJf9EL/wDLm1b/AOSq/wCHXH7CX/RC/wDy5tW/+Sq/4dcfsJf9EL/8ubVv/kqv+HXH7CX/AEQv/wAubVv/AJKr/h1x+wl/0Qv/AMubVv8A5Kr/AIdcfsJf9EL/APLm1b/5Kr/h1x+wl/0Qv/y5tW/+Sq/4dcfsJf8ARC//AC5tW/8Akqv+HXH7CX/RC/8Ay5tW/wDkqv8Ah1x+wl/0Qv8A8ubVv/kqv+HXH7CX/RC//Lm1b/5Kr/h1x+wl/wBEL/8ALm1b/wCSq/4dcfsJf9EL/wDLm1b/AOSq/wCHXH7CX/RC/wDy5tW/+Sq/4dcfsJf9EL/8ubVv/kqv+HXH7CX/AEQv/wAubVv/AJKr/h1x+wl/0Qv/AMubVv8A5Kr/AIdcfsJf9EL/APLm1b/5Kr/h1x+wl/0Qv/y5tW/+Sq8a/wDBNP8AY88M+Dde8SeA/gbOvibSdMubvRmg13VLiQXkcTPAVia4YSN5irhCpDHgg5xWi/AL4p6nrNhpuvfB/wAZHTLu6jhvRJol1GphZgHywQFRtJ5yMdcivgX+y38CP2av7b/4Un4F/wCEc/4SP7P/AGn/AMTO7vPP8jzPK/4+JZNu3zpPu4zu5zgYqqqv+HXH7CX/AEQv/wAubVv/AJKr/h1x+wl/0Qv/AMubVv8A5Kr/AIdcfsJf9EL/APLm1b/5Kr/h1x+wl/0Qv/y5tW/+Sq/4dcfsJf8ARC//AC5tW/8Akqv+HXH7CX/RC/8Ay5tW/wDkqv8Ah1x+wl/0Qv8A8ubVv/kqv+HXH7CX/RC//Lm1b/5Kr/h1x+wl/wBEL/8ALm1b/wCSq/4dcfsJf9EL/wDLm1b/AOSq/wCHXH7CX/RC/wDy5tW/+Sq/4dcfsJf9EL/8ubVv/kqv+HXH7CX/AEQv/wAubVv/AJKr/h1x+wl/0Qv/AMubVv8A5Kr/AIdcfsJf9EL/APLm1b/5Kr/h1x+wl/0Qv/y5tW/+Sq8J+FtB8DeFdG8E+FbD7Fovh/T4NP021815PIt4I1jiTe5LNhFUZYknGSSeaqqqqqvFvhbQfHXhXWfBPimw+3aL4g0+fT9RtfNeLz7eaNo5U3oQy7kZhlSCM5BB5r/h1x+wl/0Qv/y5tW/+Sq/4dcfsJf8ARC//AC5tW/8Akqv+HXH7CX/RC/8Ay5tW/wDkqv8Ah1x+wl/0Qv8A8ubVv/kqv+HXH7CX/RC//Lm1b/5Kr/h1x+wl/wBEL/8ALm1b/wCSq/4dcfsJf9EL/wDLm1b/AOSq/wCHXH7CX/RC/wDy5tW/+Sq/4dcfsJf9EL/8ubVv/kqv+HXH7CX/AEQv/wAubVv/AJKr/h1x+wl/0Qv/AMubVv8A5Kr/AIdcfsJf9EL/APLm1b/5Kr/h1x+wl/0Qv/y5tW/+Sq/4dcfsJf8ARC//AC5tW/8Akqv+HXH7CX/RC/8Ay5tW/wDkqv8Ah1x+wl/0Qv8A8ubVv/kqv+HXH7CX/RC//Lm1b/5Kr/h1x+wl/wBEL/8ALm1b/wCSq/4dcfsJf9EL/wDLm1b/AOSq/wCHXH7CX/RC/wDy5tW/+Sq/4dcfsJf9EL/8ubVv/kqv+HXH7CX/AEQv/wAubVv/AJKr/h1x+wl/0Qv/AMubVv8A5Kr/AIdcfsJf9EL/APLm1b/5Kr/h1x+wl/0Qv/y5tW/+Sq/4dcfsJf8ARC//AC5tW/8Akqv+HXH7CX/RC/8Ay5tW/wDkqv8Ah1x+wl/0Qv8A8ubVv/kqv2if2JPh58HP+Ef/AOGY/gtrcP8Aa/2n+2/7NfUNVz5XleRv8x5fL/1k2Mbd3Oc7Rj9nz9jHwl8YNZ1bTf2lfgvr02m6fapLpg1IX+lKJi2G2tG8Rc7exJx1xX/Drj9hL/ohf/lzat/8lV/w64/YS/6IX/5c2rf/ACVX/Drj9hL/AKIX/wCXNq3/AMlV/wAOuP2Ev+iF/wDlzat/8lV/w64/YS/6IX/5c2rf/JVf8OuP2Ev+iF/+XNq3/wAlV/w64/YS/wCiF/8Alzat/wDJVf8ADrj9hL/ohf8A5c2rf/JVf8OuP2Ev+iF/+XNq3/yVX/Drj9hL/ohf/lzat/8AJVf8OuP2Ev8Aohf/AJc2rf8AyVX/AA64/YS/6IX/AOXNq3/yVX/Drj9hL/ohf/lzat/8lV/w64/YS/6IX/5c2rf/ACVX/Drj9hL/AKIX/wCXNq3/AMlV/wAOuP2Ev+iF/wDlzat/8lV/w64/YS/6IX/5c2rf/JVf8OuP2Ev+iF/+XNq3/wAlV/w64/YS/wCiF/8Alzat/wDJVf8ADrj9hL/ohf8A5c2rf/JVf8OuP2Ev+iF/+XNq3/yVXgD/AIJ6/sffC7xnpPxB8CfCD+zNf0K4Fxp93/b+pTeTIAQG2SXDI3BPDKRVVVVVVVfF/wDYe/Zc+PXjOT4g/Fj4X/27r8tvHbvd/wBtX9rmOMYRdkE6JwD1259a/wCHXH7CX/RC/wDy5tW/+Sq/4dcfsJf9EL/8ubVv/kqv+HXH7CX/AEQv/wAubVv/AJKr/h1x+wl/0Qv/AMubVv8A5Kr/AIdcfsJf9EL/APLm1b/5Kr/h1x+wl/0Qv/y5tW/+Sq/4dcfsJf8ARC//AC5tW/8Akqv+HXH7CX/RC/8Ay5tW/wDkqv8Ah1x+wl/0Qv8A8ubVv/kqv+HXH7CX/RC//Lm1b/5Kr/h1x+wl/wBEL/8ALm1b/wCSq/4dcfsJf9EL/wDLm1b/AOSq/wCHXH7CX/RC/wDy5tW/+Sq/4dcfsJf9EL/8ubVv/kqv+HXH7CX/AEQv/wAubVv/AJKr/h1x+wl/0Qv/AMubVv8A5Kr/AIdcfsJf9EL/APLm1b/5Kr/h1x+wl/0Qv/y5tW/+Sq/4dcfsJf8ARC//AC5tW/8Akqv+HXH7CX/RC/8Ay5tW/wDkqv8Ah1x+wl/0Qv8A8ubVv/kqv+HXH7CX/RC//Lm1b/5Kr/h1x+wl/wBEL/8ALm1b/wCSq/4dcfsJf9EL/wDLm1b/AOSq/wCHXH7CX/RC/wDy5tW/+Sq/4dcfsJf9EL/8ubVv/kqv+HXH7CX/AEQv/wAubVv/AJKrW/8AgmL+xPpmjX+paD8DHGp2lrJLZFPEWqyMJlUlMKbkhjuA4IOfSv8AhSHxp/6JB41/8J+6/wDiK+Cn7GX7NX7Oviq68bfBv4bf8I/rV7p76fPc/wBsX13vt3kjkZNk8zqMvDGcgA/LjOCc1VVVVVVVVVVVVVaH4S8K+GJ9TuvDXhnSdJn1q8a91OSxso4GvblgA00xQAySEAAu2ScDmqqqqqqqqqqqqqqqqqqqqv2pf2dNE/am+E1x8Jtf8R3uh2lxfQXZu7OFZJA0RJAw3GDmvDH/AARe+GHhjxLpPiW3+NPiieXSb6G7SJ9PgAdonDgEg9DtxVVVVVVVVVVVVVVVVVVVVVVVVVXlnaahaTWF9bRXNtcxtFNDMgdJEYYZWU8EEEgg8HNfGf4c3vwi+LXjD4YX4cy+GNaurBXdtxljjkKxyZwuQyBWB2rkMOB0r9iL4rwfGD9mjwdrr3sU+p6RaDR9VVSd0dxbAIC/+08XlSnt+9HToL++sdLsbjU9TvYLSztImmuLieQRxwxqMs7MeFUAEkngAV+1l/wU6ura/u/AH7NV1EBau0N54rmiSVZGGQRZo2VK/wDTVwc87VxhzrviHxf8QvEb6v4i1jVPEOu6lKFM91M9zcTuThVBOWJyQAo9gK/4Zw/aG+zfbP8AhQvxF+z+X5nm/wDCLXm3bjO7Pl4xjnNaXrHijwRrov8ARNV1TQNZ0+UqJrWeS1uLeRTyNykMjAj2IIr9lj/gqBf2c1p4H/aTk+1Wh8uG18UW0H7yHAIJu0X74+7+8RQw5JDZyLG+sdUsbfU9MvYLuzu4lmt7iCQSRzRsAVdGGQykEEEcEGv2kfi3Y/A74JeLPiPc3iw3en6e8emKVDGW+kGy3UKeo8xlLcHChiRgGvDHhzV/GPiXSfCPh60a61TW76GxsbdFJaWeVwkagDJyWYDj1rwR4T03wF4K8P8AgXRhiw8OaXbadaDbtxFBEsacdvlQVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX/BZn4eP4n/AGZdH8d2dn5lx4M8SQvPNtY+VaXKNC/QHGZjajLEDtnJAP8AwSz8fP4E/bP8H2zyyra+KLe70a58tmGRJCZIwQPvDzoYeDwPvdVFVVV+3l8UE+L/AO1t8SPFlrdvPYW+rNpdgSTsEFmq2wKAs2FcxNJxjJkLYUsQP+CTPw2/4QL9jzRdbuLfy7zxtql3rMoYfME3C3i59DHbq4HT95nqTVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVf8Fl/2bb/SfF+k/tOeHNP8zTNcji0nxE0aDMN5GpFvM+OSJIl8vceFMKAn51FfsA/tSxfs8fE59B8XXzR+B/F7Rwam7HK2FwuRDdY/ujcUfBHyNuO4xqtf8FLvCfxx8Y/BaC8+Fmp/avCFjuufE+l2KH7TdQjDRzbgT5kCYLNGAMcOdwXKfs0/ss/Eb9p3xWdE8Jwf2fo1lzqmu3MLNbWQxkLxjfK3G2MHJ6nCgsPgH+yh8GP2c9Ljg8C+G1uNYK/6TruoAT3s7YIOHxiJcMRsjCqR1BOSa+PH7L/we/aJ0SbT/HvhiAap5Wy01y0jWK+tD2KyY+ZR/cfcvt3r9qT9kr4h/sv+JRb67F/anhjUZ3TR9egTEdwByI5F/wCWU23koeDhipYAkf8ABLXwn8etF+Ht/rfjjU5Ifh7qsav4b0u/RmuDJkbriEk/urdlBAQghzh12jJk/wCCkf7VFj8ZvG1r8KfA2orc+EvB1y73F1E4aPUdQwUMiMrENHGhZEYYyXlPIKmv+CQH7Nt38Q/jFc/HrXtPl/4R34f5i0+RkIS51SRMBQejeVE5kYdQ0kJ6Gqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq1r4p/DDw3qU2i+IviP4X0vULbb51pe6xBBLHuUMu5GcMMqQRkcgg96/aj0Pw38ef2TviboHhnWtJ1u3vfDt49pcWs8VzAbq3Xz4hvBKgiWJOeqnkcgV8N/GFz8PPiJ4W+IFiX+0eGdas9Th2KpbfbzJKMBgVJyg6gj1rTtQstW0+11XTblLi0vIUmt5U6SRsAysPYgg1VfHf4kW/wf+DHjb4nztz4a0O6vIF2F98yRnykwAfvSbF54GcnAya0nStZ8W6/ZaJpUEuoatrN5Hb20W7L3E8rhVXLHqzMBknqa+Hvg2w+HXgDwz8PtKffZeGNHtNLtmwRmO3hWJThmY9EHUk+561VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVWvfEz4b+FtQOk+J/iD4a0i+VA5tb/VoLeUKeh2OwOD64rw94p8L+L7F9S8J+JNL1q0ilML3GnXkdzGsgAJQshIDYZTjrgj1qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqeeC1gkubmZIoYULySSMFVFAySSeAAO9f8AC7/gt/0V/wAE/wDhQWv/AMXVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVfX1jpljcalqV5BaWlpE0txcTyCOOGNRlnZjwqgAkk8ACoPjR8HbqeO1tfiz4NmmlcJHGmvWzM7E4AAD5JJ4xVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVXiXxv4K8G/Zv+Ev8AGGiaH9s3/Zv7S1CK187bjds8xhuxuXOOm4eteHPH3gTxjNNa+EfG2g65NboHmj07Uorlo1JwCwRiQM8ZNVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVTzwWsElzczJFDCheSSRgqooGSSTwAB3r/hd/wAFv+iv+Cf/AAoLX/4uqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq1f9pv8AZs0DVbzQdd/aF+GmnalptxJbXlld+LLKGa2mjYq8ciNKGR1YFSpAIIINeCviH4A+JelS678OPHPh/wAVabBcNbS3mianDfQxzKqsY2eJmUOFdCVJyAynuKqqqqqqqqqqqqqqqqqqqqqqqq+Ivw98I/FfwPrPw58d6Smp6Dr9q1tfWrMU3ocEEMpBVgQGDDBBAI5Fftcfsr+M/wBkv4qXHgHxHI+o6TdqbjQtaS3aOLUbbPXngSpkLIgJ2kjkhlJ/Ys/4KFXvwZsbP4V/GI3eq+DImCafqaEy3OjJg/u9mCZoc7cKCGjGcbhtQfCu5+Ft94Jsr/4NSeH38LXbyz2zaGIxbF3ctLgR8K28tuXAIbIIBGKqq+J2mfDbVPA+pxfFy00O48KQRifUBrSobVFjYMrvv4GGAI98V+2f/wAFEl8d6XffB74AST2HhuRTb6j4gXMMmoRYwYLdMAxQkcFjhnHy4Vc7/wBnH9nnx7+038UtL+GXgOxlLXMivqeo+Tvi0q0DASXMvIGFB4XILsVUcsK+Cfwe8HfAT4W+H/hN4EtWj0nQLURLJKcy3MpO6WeQ93kcs5xgAtgAKABVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX7Zn/ACcp4w/7cf8A0hgr9hT/AJIBZ/8AYUu//QhXx4+H7/Cn41eOvhs1t5EfhzxBe2VumWIMKTMImBYliGj2MMknBGea/YF+IA+Jn7Hnws8Qu2Z7TQ00qcMfm32LNaFm+ZjlhAHyTzuBwucCq/4LH/E1vCP7L1l8PrSdBc+PNegt542UEtaWv+kuRnkETJajjHDHnsf+CaPwwl+KH7ZHgSJ7Jp7DwtNJr966oWEItV3QufQfaTbrk92HfAqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/bs/5L9d/9gu1/9BNf8E9f+SLaz/2NE/8A6S2tVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVeN/wDkS9f/AOwXc/8Aopqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq+N/8AyRbx/wD9ivqX/pLJXgf/AJHXw/8A9hS2/wDRq1VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV/wUh/5p3/3FP8A20r/AIJ1f8jp4u/7BcP/AKNqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqrxv8A8iXr/wD2C7n/ANFNVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV/wV4+J/wAUNX/ae1L4Za1qep2ng7Q9NsX0fTlndLW8EkIkkujHnaz+a8sW/GQIQOxr/gi58WPip4q0Pxt8M/Eeo3uqeDvC0NpLpEt0xcadLK0ga3jc87GVNwTOEKkgDec1VV+1l/ydR8Zf+yga7/6XzV/wRU/5NX8U/wDZQLz/ANILCqqqqqqqqqqqqq+JXxV+G/wc8N/8Jd8UfGuk+GdINwlsl1qNwIlkmcErGg6u5Cs21QTtVm6KSLT/AIKl/sp+IfiJ4V+Gfw/1bxH4r1LxbrVlpNrc2ejvbW0EtzMkStKboxOFUvk7UY8cDvVVVVVVVVVVVfGz4GfDL9oXwJe/Dz4peGrfVdNuVYwSlFFxYTYIWe3kIJilXPDDqCVYMrMp/az/AOCanxr/AGcbq58ReFrG78eeBhvkXVdNtGaexQHgXUK5KYHWRcoepK/dr4a/GT4p/B3Vl1n4Y+PNX8P3G8PIlrOfJnI6ebC2Y5R7OrD2rwH/AMFavjPocENp4+8C+GvFKRYDXFu0mnXEoydxZl3x55UDbGoGOQc5H/D4v/q3T/y7/wD7jrxv/wAFcfi3q6SweAfht4a8OpJwsl9PLqUsYwMkEeUmc5PKEYOMcZr4qfHn4w/G67hu/il8QdU1/wCzsWgt5nWK3hY5yyQRhY0Yg4JCgkYGcAV+yj/wT5+Of7Ud9Ya3b6NL4X8BSSA3HibU4iiTRhwHFpGcNcPw4BXEYZCGdTwf2d/2bPhV+zD4Eh8DfDDQktw6o2panMA13qkyjHmzydzy2FGFTcQoANVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX7Zn/ACcp4w/7cf8A0hgr9hT/AJIBZ/8AYUu//QhX/BXb4fDwb+2HqPiGJf3PjbQ7HVVCj5VdENo6j5QAc2ocjLf6zJPzYH/BE/4gPrXwU8c/Dae582Twv4givYkIbMUN5DhVBxjHmWszYBJyxzgEZqv+CznxQbxV+0VoPwytLtJLPwLoKtLECC0V7eMJZM4Y4BgS0OCFPU8gqa/4IifC5hD8SPjTeWqbWa38P6dOAC2QPtF0pyuQObM8Ng85HCkVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX7dn/Jfrv/ALBdr/6Ca/4J6/8AJFtZ/wCxon/9JbWqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqvG//Il6/wD9gu5/9FNVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVXxv8A+SLeP/8AsV9S/wDSWSvA/wDyOvh//sKW3/o1aqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/4KQ/8ANO/+4p/7aV/wTq/5HTxd/wBguH/0bVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVXjf/kS9f/7Bdz/6Kaqqqqqqqqqqqqqqqqqqqqqqqr46ftR/Aj9mr+xP+F2eO/8AhHP+Ej+0f2Z/xLLu88/yPL83/j3ik27fOj+9jO7jODj4Iftcfs8/tIarqeifBf4hf8JFe6PbrcXsX9k3lp5UbNtVs3EMYOTxgEmvil/wUd/Y9+Eur3XhzW/i3b6tq9mwWez0G0l1EISSCDNEphDKQQyeZuU8EZr4Mft/fsp/HfV7fwz4L+KFva69dTLDbaVrMD2E9w7EhFiMgCSsxGAqMzdMgZGa+On7UfwI/Zq/sT/hdnjv/hHP+Ej+0f2Z/wASy7vPP8jy/N/494pNu3zo/vYzu4zg4+Fv7ev7Jvxq8d6X8Mvhl8WP7Z8S6z532Gx/sLULfzfKheaT95NAiLiON25YZxgZJAqtX/4KZfsQaFqt7oeq/G7yL3T7iS3uYv8AhG9Vby5EYqy5W2IOCCMgkV8IfjN8Nfj14Mi+IPwn8S/27oE1xJbpd/Y57XMkZw67J0R+CRztxXxf/bh/Zc+AvjOT4ffFj4o/2Hr8NvHcPZ/2Lf3WI5BlG3wQOnIHTOR3r4VftI/BH42+A9T+J/w38f2moeFtFvJLPUNUu7ebToraaOOORw/2pIyAEljO7G35uuQceMf+CqH7F3hDV5NFj+JF7r0sDMks2j6TPPAjA4wJSqrID2ZCyn1r4JftNfAv9oywmvfg98RNP16S0jEl1ZBXgurZSSuZIJQsijcCN2Np7Eggmv2vPip/wTP8QeObn4Z/tYXemXPizwwsKSq2i6qLq1SSNZ44xd2UQJQpMr7BIVBY5Gc1+x98Sv2PvF/hnVvB37IM+mDSPC7QNqdvZaPd2Rja4Mvls73MSNMzeVJ82WICgEgYqaaG3heeeVI44lLO7ttCgDJJJ6DFfEv/AIKVfsb/AAw1J9E1D4sw65qMMmyWDw/Zy6gsf3gSZkHknBXBUOWGRxg5rwl/wVS/Yp8VXpsJ/iZe6DKWVYm1fRbmKOUnOf3iI6oBgZLlR8wxnnGgeIfD/izRbTxH4V13T9Z0m/j8y0v9PuUuILhP7ySISrDjqCRX7WX/ACdR8Zf+yga7/wCl81f8EfvFfhjwN+xt428WeM/EOn6Joum+PLyS7v8AULhYIYF+w6eAWdiAMkgD1JAHJrUv+Cr37E+n64ujR/ETVb2HzFR9RttAujbx5OGY7kDsF6kqhyPu7q+Fnxf+Gfxu8J2/jf4VeM9P8R6NccCe1Yhom/uSRsA8T/7Dqre1V/w9H/YS/wCi6/8Als6t/wDItfEr9o34IfB7wXp3xA+JfxG0zQNE1iFZtOkuRIZr1CqtmG3VTNIQrqSFQldwyBXhT/gqb+xZ4r1waCfibdaM8kgjgutW0i4t7eUk4B8zaRGO+6TYAOpFQ65olxoieJLfWLGXSJLUXaX6XCmBoCu8SiQHaUK/NuzjHOcV48/4Kg/sXeBNUm0Vviq3iC7tZjFN/YOmz3kKkDO5ZwohlXtmN2FfBj9p34B/tDRTt8HfidpPiKe1jMs9mgkt7qKMNsLtbzKkqpuwNxXHI55Gar4l/wDBSr9jf4Yak+iah8WYdc1GGTZLB4fs5dQWP7wJMyDyTgrgqHLDI4wc18NP+ClX7G/xQ1KPRNP+LMOh6jNJshg8QWcunrJ90AiZx5IyWwFLhuDxjmq+OH7R/wAF/wBm/SdM1v40eM/+EestYuHt7KX+zrq782RV3MuLeOQrxzkgCvgp+2Z+zV+0V4quvBPwb+JP/CQa1Y6e+oT2v9j31pst0kjjZ988KKcPLGMA5+bOMAkfGH/goR+yX8EtXm8NeK/inb3+t2zbZ9O0S2k1CSE5wQ7xAxow7ozhh6V8FP2+P2WPj7rMHhbwL8S4YNfuZPLt9J1a2ksZ7hjnAi8wBJSQp+VGZh3AyM1/wVX/AGuP2efjd8FtN+F3ww+IX9teJ9B8cRXGoWP9k3lt5McNreQyN5k0KI2JJEXCsSc5GQCR+z14p0HwN8fvhn428U3/ANi0Xw/4w0nUNRufKeTyLeC7iklfYgLNhFY4UEnGACa+Fv7ev7Jvxq8d6X8Mvhl8WP7Z8S6z532Gx/sLULfzfKheaT95NAiLiON25YZxgZJAqqqqqqqqqqqq+Ov/AATu/ZW+P082reIvAP8Awj+uzyNJLrPhp1sLiRmHzF12tFITgHc8bNkdeTnxr/wQ9haWW4+HP7QDpGWAis9b0LcyjuWnilGTjHAiH1Ff8OQvin/0XHwp/wCC24/xrwh/wQ7umkWfx9+0HEkayMGtdH8Pli6bflImlmG07jyPLYYHXJ+X4K/8Ez/2SvgjqFtrth4GuPFutWbFoNR8VXC37RnOQRAFS3DLxtfyty4BBByTDDDbwpb28SRxxKFREXaFAGAAB0GO1VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVftmf8nKeMP+3H/wBIYK/YU/5IBZ/9hS7/APQhX/Bb34fx3Xgr4afFSC2w+napdaLczAr8wuIhNEpGc8fZpiMDHzNk8jP/AARr8fR+Gv2odS8FXMsIi8YeG7iGFWZVZri3ZJ1255b90k+VHPfopqq/ab+J0vxm/aC+IHxMa9a6g1zXrl7GR3LEWaN5dsufRYEiUdsAYwK/4JvfDCP4W/sc/D+zdMXniSzOv3T4UbzeHzYj8pPSAwLyc/LyFPyiqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/bs/5L9d/9gu1/wDQTX/BPX/ki2s/9jRP/wCktrVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVXjf/AJEvX/8AsF3P/opqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq+N//ACRbx/8A9ivqX/pLJXgf/kdfD/8A2FLb/wBGrVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX/BSH/mnf8A3FP/AG0r/gnV/wAjp4u/7BcP/o2qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqvG/8AyJev/wDYLuf/AEU1VVVVVVVVVVVVVVVVVVVVVVVX/Bc//miX/cwf+46vh78V/iV8OLHxLoHw78SXulp4200aTqqWYxJc25kVvLUj5lLFdp24JV2XoxFaP/wS2/bR1nwcPF8Xwyt7UvCZY9KvNUggvnUdB5TN8rEfwuVbsQDxXiHw9rnhPXL/AMMeJtIu9L1bS7h7a9sruIxS28qHDI6nkEEYxX/BKn9rHWvj38Kr/wCGnj7UXvfFnw/WGNL2Z2eXUNPcERSSMxJeVGVkZuMjyycksT/wXP8A+aJf9zB/7jq/4Jcf8n2/DH/uLf8AppvKr4tf8lU8Z/8AYwX/AP6UPX/BIb/kzTTP+xg1H/0Na/4K8/8AJ5Wpf9i/p3/oDV4H8WfG3xp4Pt/2YPh5c6xqWj+I9e/tEeG9NjLNqF75aIGcLyyqkKtgnYuzefugr8WP2F/2rvgh4Rn8d/Ez4P3umaDasFub631C0vkt8kKGkFtLIY1JIG5gBkgZya+EXxU8XfBP4kaB8T/A2pS2mraBeJPHskZFnQH54ZMdY5FyjL0KsRXgLxhpXxE8C+HPH+hlv7O8TaTa6nZ71IbybiJZUyCAQdrjqBX/AAVH/wCT7fib/wBwn/002df8EMf+a2f9wD/3I1/wWR/aY8U+DrHw/wDs4+DNZuNNXxNpr6n4keAlHuLJpGiht9+PuO8U5cA5IRQflYhv2Sf2SfHf7X3ju/8ABvg3V9M0e30exN5qOo35LLCpO2NVjX5nZm44wAAST0B+IH/BGH9o3w5o/wDaXgXxh4R8YXMauZNPSV7CdyANojMo8tieR87oBxyckj4ceCNK+Gnw+8NfDvRFQWHhnSbbTbcqhUMkMSxhsEscnbk5JOTySea/ay/5Oo+Mv/ZQNd/9L5q+E3gb9oH9oGzt/gL8JdI1jxJYWV9Nrn9jW0iRW0FxJHFA91K7lUU7I4ow0jADJC4Ltu+N37L3x6/Zxl06P4z/AA4vfDqaspNncG4hu4JSM5QTW7yRhxjJQsGxg4wQa/4J+/tCa1+z7+0p4Wvo9SuE8N+J76HSPEForsY5YJn8tJmRc7mhdxIMAtgMo++c1Xw9+Bn7WP7ffiK98W6HYy+If7Bs7bTZ9W1K6S0tLVIYgsVvGThc7RuMcS4BcswXfk/Fn4R/EH4HeOtQ+G/xP8Oy6Nr2m7TLbu6yK6OoZJI3QlXRgeGUkdQcEEDQfj/+0R4u+EOmfsi+F9e1XUvDmo6xvtNGs1eW5u3kKhLQEEkwCQGQQqAu92Y5+Xb8Yf8Agn7+1R8Cvh7/AMLO+IXw9hg0ODH9oSWep292+nbmVU85YnbhmYDcm5QepGRn4UfFPxp8FfiFofxO+H+qvY63oN0s8DbmCTAH54pQpBaJ1yjrkblYjvXgLxhpXxE8C+HPH+hlv7O8TaTa6nZ71IbybiJZUyCAQdrjqBX/AAVp/bN1/XPGtz+y/wDDnWr3T9C0FQPFk0Enl/2ndOoYWxI5MMSEbhnDu5BX92pP7J3/AATq+M/7WHhufx3oupaT4Y8KRXEltBqWqrIWvJkA3CCJBl0UttLkhdwZQSVYD9rT9iz4q/sg6zplr43uNP1fRtc8wabrGmiQwyMmN0bh1BjkAYHbzkZIJwcf8Eef2q9f8SJqX7MnjzXLi/Ok2IvvCclw25obaMhZrTd1KruR0BztXeBhVUD/AILe/wDJKvhp/wBjBdf+k4r4a/Fz4ifB6+1nVPhp4ou9Bv8AXtHm0e8u7Q7ZTaSujSIjdUJMSfMuGGMgg4I0/wD4J2ftpap4Oh8dWXwE1h9LntftUaPeWiXbR9v9EaUXG49k8vceOOa/0uwu/wDlrbXNtJ7o8TqfzBBH4Gv+CdXx+1X9on9l7QfEviW9lvPEXh64k0LWbqVmZ7me3RGWVmb7zvDLCzNzl2bnqB+2H+xX8cvgDLq3xd+Idho8Ph7xF4qmtrF7TUFnlMk/nzx7kAyB5cT5PY4HevA/g/WfiF408P8AgDw2kL6t4l1S20ywWaTy0M88qxRhm/hG51ye1fsPf8E5P2mPgL+1H4J+LHxB0vw/FoGhfb/tb2urLNIPOsLiBNqAZPzypn0GTVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV+2Z/ycp4w/wC3H/0hgr9hT/kgFn/2FLv/ANCFf8FLfAD/ABB/Yw+IVtbRyvdaFbw6zB5aM2BayrJKSF7eSJeeg+8eAa/ZJ+Ib/Cr9pr4Z+O/tv2W30/xJaR3s25httJnENz90j/ljJIME4OcEEZBr9s74ly/CD9lf4mePbS6a2vLPQZbaxnTO6G6uiLaBx8rcrLNGcEY45IGSPhb4Gvfif8TPCfw404uLnxPrVppsbKMlDPMse72ADZJPAAya0nSrDQ9KstE0qDybLT7eO3totxbZGihVXJJJwABkkmqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr9uz/kv13/2C7X/ANBNf8E9f+SLaz/2NE//AKS2tVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVeN/8AkS9f/wCwXc/+imqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr43/8AJFvH/wD2K+pf+ksleB/+R18P/wDYUtv/AEatVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVf8FIf+ad/wDcU/8AbSv+CdX/ACOni7/sFw/+jaqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq8b/wDIl6//ANgu5/8ARTVVVVVVVVVVVVVVVVVVVVVVVVf8Fz/+aJf9zB/7jq/4Js+HNH8VftvfCzTNbtEubeC+u75EdQQJ7WxuLiF+e6ywxsPdRVf8FofAWi+Hv2i/DfjfS4IYLnxZ4bQ6iqRYM09tI0QmZs8kwmGPGBgQjrnj/gjl4g1DSP2uJtJtSht9c8K31tcq+eAjwzKygEDcGiAyQeGYd8j/AILn/wDNEv8AuYP/AHHV/wAEuP8Ak+34Y/8AcW/9NN5VfFr/AJKp4z/7GC//APSh6/4JDf8AJmmmf9jBqP8A6Gtf8Fef+TytS/7F/Tv/AEBq/wCCJvh/Rb/42+O/EN5p0U2o6V4bjjsp3GTbiWdRIVHQFgijdjIGQCAzA/HTRNK8SfBTx9oGt2aXdhf+GdRhuIXJAdGt3BGRgg+hBBBwQQar9giWWf8AY2+ErzStIw8OxKC7EkBWYAfQAAAdgBX/AAVH/wCT7fib/wBwn/002df8EMf+a2f9wD/3I1/wV1/ZG8c/FS20T9oL4a6LLrN54W0ttO17T7YPJcmyWVpYpooxneI2ln3hRuAdW5VWK/Df4ofEL4QeKbfxr8MfGGp+G9bt1KLd2MxjZkJBMbj7siEqpKMCpwMg4r9nv/gs94w0/ULPw/8AtI+D7HV9MdljfX9Ch+z3UIJ5kltyTHKBnpH5ZAHAY9fCnirw9468MaT4z8JarDqWi63ZxXmn3cQIWeGRQyMAQCMgjggEdCAa/ay/5Oo+Mv8A2UDXf/S+av8AgihpOmw/s2eMdcisol1C78cTW09yF+eSKKxtGjQnuFaaUgdi7etf8FV9F0rVf2HPHd9qFmk0+j3Wl3Vi7EgwTHULeEuMd/LmlXns59qhmlt5Unt5XjliYMjoxVlYcggjoaqv+CVmk6bp37DXgC8sLKKCfVLjVbi8dFwZ5RqNxEHb1PlxRrn0QDtX/BbrT7KP4x/DrVY7dBd3HhmaGWUD5njjuWZFPsDJIf8AgRr/AIJH6fZXv7aOhXN1bpLLYaLqU1szDmKQwGMsPfZI6/RjXxu0nTdd+DPjvR9Ysobyyu/DeoRzwSruV1Nu+QRVfsESyz/sbfCV5pWkYeHYlBdiSArMAPoAAAOwArx94tvfH/jrxH471FXW78R6tdalOHk8xhJPK0jZbA3HLHnAz6CvhP8A8FJ/2mvgp8O9E+F3gC98L2eg6BC0VnE+iIzYZ2kdmbI3MzuzE9SSSeTX7Q/7c3x5/ah8H2Hgj4rXug3GnaZqS6jbmy0tbeVZljkj++CTtKytle5CntX/AATr1e90P9tX4U3tgyCWTVpLZt65Hlz200Mn47JGwexwa/4Le/8AJKvhp/2MF1/6Tiv2IPD+i+KP2t/hToviDTor+xl8SW8klvMMpIY8yLuH8Q3IpKng4wQQSKr9uXRNK8P/ALXnxZ0zRrNLW1Hia5mESE4V5SJHIz0Bd2OBwM4AAAFf8ERJZW+EvxItzKxjTxFbsqFjhWNvgkD1IVc/Qelf8Fq/+TV/C3/ZQLP/ANIL+v2Tv+TqPg1/2UDQv/S+Gqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr3wr4Y1K5e91Hw5pd1cSY3yz2cbu2AAMkgk8ACrDTdO0u3Fppdhb2kAJIigiEagnqcDArxb4Z0rxr4V1nwbrsPm6br2nz2F5HtVt8M0bRuMMCpyrEYII9Qa8RaDqXhbX9T8MazD5OoaReTWd1H/clico4/BlIr9n74jf8Le+BvgP4mPLvuPEfh+zu7v5cbbhol85fuqDtlDrkKAcZAwRX/Ba/4nQ6P8JPBHwktL1Fu/EmtSandRI+W+zWkZUBh2VpLhSCepiOOhr/AIJJfC5PiD+1zp/iO8tXksfAmk3OrsxBEfnkLbwqTtI3bpzIqkrnyicnaVNVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV/4Z8N6rcG71Pw/pt5OQAZbi0SRsDoMkE1p2laXpEBtdJ021soWYu0dvCsSlsAE4UAZwAM+wqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqSOOWNopUV0dSGVhkEHqCPSv+EI8F/8AQoaJ/wCC+L/4mqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqeCC6gktbmFJoZUKSRyKGV1IwQQeCCOMUngvwdE6yReE9GR0OVZbCMEEdCDtqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqrUtD0XWvL/tjSLK+8nPl/abdZdmcZxuBxnA/IVp2gaDo7vLpGi2Fk8i4dra2SIsPQlQMiqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqpI45Y2ilRXR1IZWGQQeoI9K/wCEI8F/9Chon/gvi/8Aiaqqqqqqqqqqqqqqqqqqqqqqr/guf/zRL/uYP/cdX7JPxqsv2d/2jPA/xh1TT3vdP0G+db+GMZcWtxDJbzugyNzrHM7KpIBZQCQCa0z9pj9njWfCC+PNO+N3gmTQTb+e142twII12gkOrMGRxkAowDA8EA8V/wAFG/2mdB/ae/aIn17wRdSz+EvDGnx6Ro87b0F4Ed5JbkRt9zfJIyg4BaOOMsAeB/wRP+Fd/qXxO8bfGa707dpui6ONGs7iSIFftVxJHI/lk8h0ihAJHRZ8H71f8FxPCepXvgr4UeOYh/oGkapqWmznb/y1uooJI+f92zl/yK/ZS+Mlt+z9+0R4F+Lt/FcS2Ggal/xMEt1VpTaTRvBcbFYYLeVLJgcZOAGU/MPiL/wU6/ZC8F/D678Z+HvibaeLtQ+zhtO0PTYpkubuVlyiMHjHkD+80gGwAjBbCHxFrd34m8Qan4jv44UudVvJruZIVIRXkcuwUEkgZJxkn61/wSG/5M00z/sYNR/9DWv+CvP/ACeVqX/Yv6d/6A1f8EQ/+Sp/Ev8A7F+1/wDSg18Wf+SV+Mv+xfvv/Sd6r9gP/kzT4S/9i/H/AOhvX/BUf/k+34m/9wn/ANNNnX/BDH/mtn/cA/8AcjXj39oL4M/C3xnoXgH4j/ETR/DWr+I7Wa505dUuBbQypEyKwMz4jViXAVWYFsNjJFfGv9g79kT9ovTZ/FWp+C9J0vUbu3lePxN4amWzZi2SZ38r9zOQed8queMZAzXxD0HQvCvj/wATeF/C/iOLxDo2j6xd2enavCAE1C3imZI7hQCRiRFVxgkfN1Nf8Ed9W8Ral+x/9i1pGFnpXiq/ttILFiDalIZWIySAPPluBwAMg8ZyT+1l/wAnUfGX/soGu/8ApfNX/BFT/k1fxT/2UC8/9ILCv+Co/wDyYl8Tf+4T/wCnazqqr/glx/yYl8Mf+4t/6dryv+C3v/JU/hp/2L91/wClAr/gkN/yeVpv/Yv6j/6AtfFn/klfjL/sX77/ANJ3qv2A/wDkzT4S/wDYvx/+hvX7T3wl1L4HfH3xz8MdQsvs8ekaxP8AYMR7Fls5G8y2kUZOA0LRnGTjJGSQa/YG8R/8E7/iL8ONO+H/AMfvh74G0H4g6QxgbUtclNvBrcZZ2jlWZ3CJKFwjIxGWClc79q3n7M3/AATWsLSa/v8Awd8JLa2to2kmmm1aJEiRRlmZjLgAAEkngV8AvhD+wdqfiybxv+zn4S8CajrPg+48ttW0JjcJaSyxuuElDGNiULfdJxkHjIr/AILe/wDJKvhp/wBjBdf+k4r9gP8A5PK+E3/YwJ/6A9V+37/yeV8Wf+xgk/8AQEr/AIIh/wDJKviV/wBjBa/+k5r/AILP6Vf6j+ydot3ZweZFpnjiyuLptwHlxm0vIg2Ccn55YxgZPzZ6AkfBvxxafDH4veBviTf2Mt7beE/Emn6tNbQsFedLa5jlZFJ4BYIQCeMmvhv+2N+zH8X/ABFpXhD4cfGPRNb1zWo2ksdOiEsc8oWJpW/duispWNGYqwBG05GeKqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv+Cjnw5/4Vp+2V8R9Phjxaa5qA1y3fOfM+2os8pxuYjE7zLzj7uQApAr/gkN8Q4/GX7Idp4YlvfMuvBOuXmmtG7LvWKRhcxtgEnYftDqCQOUZRwua/4K3fFBPiD+1zqHhuyu3ksfAmk22kIoJEfnkG4mYDcRu3TiNmwufKAwdoY/8EUPhhDo/wAJPG/xbu7JFu/EmtJplrK6Zb7NaRhiVPZWkuGBA6mIZ6Cqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/4Ln/80S/7mD/3HV+zR8D7r9pD406F8GNP1yLR7zxBb35tbyaIvGk0FjPcRhwOQjPCqsRkqGJAJGD8RP2Av2v/AIa6vNpWp/AfxRrKJM0cV54dsX1aCdQTiRTbh2VWAyA6qwyAQDxXwF/4Jd/tR/GTV7U+KPCVx8OfDxZTdan4itzFOiE8+XZkiV3A5Ct5a9i65r4CfAvwJ+zj8LdH+FHw8s3j03S1ZpbmYAz31w5zJcTMANzsfwVQqgBVUD9oz4D+E/2lPhDrvwj8XzS21tqsYa2vYRl7K5Q7oplXI3bWAypIDDK5Gc18XP8Agnn+1t8JPEk+hy/BzxB4rsxIRaap4XsJNUguU5w+IVZ4s4+7IqEemCCf2f8A/gmP+1B8ZfEWmr4q8B6n8P8AwtJN/p+reILf7LPDGrYYR2jkTO5AO3Kqh4JYAgn9qj/gnN8Y/DXx28RaD+z18DvFGp+AbKGwj0q9V/tH2lvsMH2iQu7ZLNcecWAAUEkKFUAD/gmb8MPH/wAIf2WrDwZ8S/Ct74e1uLWr6Z7K8ULII3cFWwCeCBX/AAUz/ZR/aL+L/wC1Jf8AjL4afCPXfEOiSaLYwpe2camMyIhDLyw5GRX/AASW/Zx+OXwQ+Inj3Vfix8NNY8MWmp6Lbw2kt9GqiaRZiSowTyBzXxI0+91b4d+KdK023e4u7zRbyG3hQfNJI0Lqqj3JIFf8MBftlf8ARvPir/vyn/xVfsZeD/E3gD9lr4beDPGWjXGk63pOipDe2VwAJIJA7EqwGecEV/wUK/ZB/aY+KX7YHj/x38Pvg34g13QNT/s37Hf2sSmOby9NtY32ksDw6Op9wa/4JEfAD4y/Az/hbH/C3Ph5qvhf+2/7F/s/7eir9o8n7d5m3BP3fNjz/vCv+Con7H/7U3xx+M9v8Tfh74Kh8T+F9M0O206zt9Pv4/tUGJJHkLwSFCSZJTzHv+TaSRghbz9kT9qyyu5rKb9mr4oNJbyNG5h8JXsqEqcEq6RlWHHDKSD1BIr4If8ABKf9qT4n6/Anjnwynw+8PRXSJe32sSobgx9XMFuhLOwXGN2xCTjdw2PhD8KPBnwO+G2g/Cn4fWMtroPh63MNqk0plkcs7SSSOx6u8ju7YAGWOABgD9pH9iL9rHxb+0R8UvFXhv4FeJdQ0nWfGmsXthdwwoUuIJbyV45F+boyspH1r/glJ8IPiZ8Ff2dvEPhX4q+DdQ8NatdeNLq9htL5ArvA1lZIsgwTwWjcfVTX/BQr4f8AjP4pfsfeP/Anw+8PXWu6/qf9m/Y9PtVBkm8vUrWR9oJHREdj7A1/wwF+2V/0bz4q/wC/Kf8AxVV/wwF+2V/0bz4q/wC/Kf8AxVf8E9vh/wCM/hd+x94A8CfEHw9daHr+mf2l9r0+6UCSHzNSupEyAT1R0b6MK/4K1fs4/HL43/ETwFqvwm+GmseJ7TTNFuIbuWxjVhDI0wIU5I5I5r/gmb+yj+0X8If2pLDxl8S/hHrvh7RI9FvoXvbyNVjEjooVeGPJxXxI0+91b4d+KdK023e4u7zRbyG3hQfNJI0Lqqj3JIFf8MBftlf9G8+Kv+/Kf/FV+xl4P8TeAP2Wvht4M8ZaNcaTrek6KkN7ZXAAkgkDsSrAZ5wRX7cv7A3hD9r3SLXxBpepw+HPiBotu0On6oYQYbyMsp8m72qXZFw2xlOUMjHDAlT8Rv8AgnR+2R8Nbi5S/wDgnrGu2sDYju/Dm3VFnXcFDJHCWlAJIOGRWAySAATWifsY/ta+ILtrKw/Zu+IsUiRmQm+8O3FkmAQOHmVFJ5Hyg56nGAcf8Eqv2XP2iP2bk8d3Pxh8M2Og6X4qhsHtLJ9QjuboTQGb5tsLsiKUmO4MdxITgYNf8Fafgh8WPjf8OvAWlfCbwJqfie70zWria7hsUDNDG0IUMckcE8V+xl+xl+1J4A/ak+G3jLxl8EvEek6JpOtJNe3txEgjgjCMCzYY8ciq/bN/Yy/ak8f/ALUnxI8ZeDPgl4j1bRNW1p5rK9t4kMc8ZRRuXLdODX/BJb4IfFj4IfDrx5pXxY8Can4Yu9S1q3mtIb5ArTRrCVLDBPAPFfHz4M+G/wBoP4Q+JvhF4pfybPxBZmKO6EQkaznUhoZ1XIyUkVGxkZwRkAmvi3/wTu/a4+EuvXOky/B7XfFVjHNstdU8L2b6pDdId21wkIaSMfKciRFIOM/eUn/gmv8Asu/tF+Ev2ufBHxD8YfBTxh4d8O6F/aP2++1rSZdPWLzdOuYkwswVnzJIi/KD154BxVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVftmf8AJynjD/tx/wDSGCv2FP8AkgFn/wBhS7/9CFV/wW5+Hj2HxG+HXxUt7P8Ada1o9xpNzMitgSWsolQPxtBZbptvOWEbcAJX/BFr4r2vhnxL8VfAOtXbx6c2gxeIy5RmFulm5inYBQSSVuYsjBJ8sYr4p+Ob34n/ABM8V/EfUA4ufE+tXepSK5yUM8zSbfYAMAAOAABX7F3wuHwc/ZZ+G3gSax+yX0Ohw3eowlcMl5c5uJ1Y7myVklZc5x8owAMAVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVft2f8l+u/wDsF2v/AKCa/wCCev8AyRbWf+xon/8ASW1qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqrxv/wAiXr//AGC7n/0U1VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVfG//ki3j/8A7FfUv/SWSvA//I6+H/8AsKW3/o1aqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/wCCkP8AzTv/ALin/tpX/BOr/kdPF3/YLh/9G1VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV43/wCRL1//ALBdz/6Kaqqqqqqqqqqqqqqqqqqqqqqqr/guf/zRL/uYP/cdX/BLj/k+34Y/9xb/ANNN5VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX7Zn/ACcp4w/7cf8A0hgr9hT/AJIBZ/8AYUu//QhVf8Fg/h3D4u/ZIl8YoiC58D69Z33mHhvJnb7I6D2L3ETH/cHvXw6+I2vfDTUdX1DQZcf254f1LQ7xNqfvLe9tngcZZWxjeG4wflwCM5r9mT4YS/Gb9oL4ffDJLJrqDXNeto76NULEWaN5ly2PRYElY9sDnA5qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv27P+S/Xf/YLtf8A0E1/wT1/5ItrP/Y0T/8ApLa1VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV43/wCRL1//ALBdz/6Kaqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqvjf/wAkW8f/APYr6l/6SyV4H/5HXw//ANhS2/8ARq1VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV/wUh/5p3/ANxT/wBtK/4J1f8AI6eLv+wXD/6Nqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqrxv/AMiXr/8A2C7n/wBFNVVVVVVVVVVVVVVVVVVVVVVVV+2X+xB4U/bM/wCEP/4Sbxxqvh3/AIQ/7d5H2G2jl8/7V5G7dv6bfs64x/eNfs4f8Esfh/8As3/Gjw78aND+KniDWL3w79q8qyu7KGOOXz7WW3OWXkYExb6gVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV4n+BPwg8Z65c+JPFHgDS9S1S82+fdToxeTagRc4PZVUfhXhPwb4X8C6Ovh/whottpenpI0i28AIUM33jz61X7Rvw/f4q/AP4hfDq3tvPute8N31tZJlh/pJhYwH5SCcShDjocYOQSKr/gjH8Lm8VftFa98TLq1SSz8C6CywyEAtFe3jeVHjKnAMCXYJBU9ByCwqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqvFnwT+FHjvWG1/xf4G0zVdReNY2uLhCWKr0HB6CvBvgTwf8PdLl0XwVoFrpFjNObiSC3UhWlKqpbknnaij8BVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV3a299azWV3EssFxG0csbdGVhgg/UGv+GYvgD/0SvRP+/bf41VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVarpen65pd5ourWsd1Y39u9vcwSD5ZY3Uq6n2Kkj8atf2a/gRY3UN7afDHRop7eRZInWNsoynII57ECqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqvG3ww8AfEj7F/wnPhWx1n+zvM+y/aVJ8rft34wR12L+QrwX8Jvhx8Orq5vfA/hCw0ee7jEc72yEGRQcgHJPeqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqru1t761msruJZYLiNo5Y26MrDBB+oNf8MxfAH/AKJXon/ftv8AGqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq8e/wDBHX9ozxR8TPGereEtf8A6Z4cudeu5tFGqapOsslnJMzQllgtnCsEKhh8oDA4BGDX/AATS/ZW8b/st/CbxNpPxO0y0tPFOv+IGmkFtcpcIbSKJEgwyE/xGdsHBG8ZAqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr4j/ABP+H/wh8Mv4y+Jfiqx8PaJFMkL3t4xWMSOcKvAPJxX/AA35+xp/0cP4U/7/AD//ABNf8N+fsaf9HD+FP+/z/wDxNf8ADfn7Gn/Rw/hT/v8AP/8AE1/w35+xp/0cP4U/7/P/APE1/wAN+fsaf9HD+FP+/wA//wATX/Dfn7Gn/Rw/hT/v8/8A8TX/AA35+xp/0cP4U/7/AD//ABNf8N+fsaf9HD+FP+/z/wDxNf8ADfn7Gn/Rw/hT/v8AP/8AE1/w35+xp/0cP4U/7/P/APE1/wAN+fsaf9HD+FP+/wA//wATX/Dfn7Gn/Rw/hT/v8/8A8TX/AA35+xp/0cP4U/7/AD//ABNf8N+fsaf9HD+FP+/z/wDxNf8ADfn7Gn/Rw/hT/v8AP/8AE1/w35+xp/0cP4U/7/P/APE1/wAN+fsaf9HD+FP+/wA//wATX/Dfn7Gn/Rw/hT/v8/8A8TX/AA35+xp/0cP4U/7/AD//ABNf8N+fsaf9HD+FP+/z/wDxNf8ADfn7Gn/Rw/hT/v8AP/8AE1/w35+xp/0cP4U/7/P/APE1/wAN+fsaf9HD+FP+/wA//wATX/Dfn7Gn/Rw/hT/v8/8A8TX/AA35+xp/0cP4U/7/AD//ABNf8N+fsaf9HD+FP+/z/wDxNf8ADfn7Gn/Rw/hT/v8AP/8AE1/w35+xp/0cP4U/7/P/APE1/wAN+fsaf9HD+FP+/wA//wATX/Dfn7Gn/Rw/hT/v8/8A8TX/AA35+xp/0cP4U/7/AD//ABNf8N+fsaf9HD+FP+/z/wDxNf8ADfn7Gn/Rw/hT/v8AP/8AE1/w35+xp/0cP4U/7/P/APE1/wAN+fsaf9HD+FP+/wA//wATX/Dfn7Gn/Rw/hT/v8/8A8TX/AA35+xp/0cP4U/7/AD//ABNf8N+fsaf9HD+FP+/z/wDxNf8ADfn7Gn/Rw/hT/v8AP/8AE1/w35+xp/0cP4U/7/P/APE1/wAN+fsaf9HD+FP+/wA//wATX/Dfn7Gn/Rw/hT/v8/8A8TX/AA35+xp/0cP4U/7/AD//ABNf8N+fsaf9HD+FP+/z/wDxNf8ADfn7Gn/Rw/hT/v8AP/8AE1/w35+xp/0cP4U/7/P/APE1/wAN+fsaf9HD+FP+/wA//wATX/Dfn7Gn/Rw/hT/v8/8A8TX/AA35+xp/0cP4U/7/AD//ABNf8N+fsaf9HD+FP+/z/wDxNf8ADfn7Gn/Rw/hT/v8AP/8AE1/w35+xp/0cP4U/7/P/APE1/wAN+fsaf9HD+FP+/wA//wATX/Dfn7Gn/Rw/hT/v8/8A8TX/AA35+xp/0cP4U/7/AD//ABNf8N+fsaf9HD+FP+/z/wDxNf8ADfn7Gn/Rw/hT/v8AP/8AE1/w35+xp/0cP4U/7/P/APE1/wAN+fsaf9HD+FP+/wA//wATX/Dfn7Gn/Rw/hT/v8/8A8TX/AA35+xp/0cP4U/7/AD//ABNf8N+fsaf9HD+FP+/z/wDxNf8ADfn7Gn/Rw/hT/v8AP/8AE1/w35+xp/0cP4U/7/P/APE1/wAN+fsaf9HD+FP+/wA//wATX/Dfn7Gn/Rw/hT/v8/8A8TX/AA35+xp/0cP4U/7/AD//ABNf8N+fsaf9HD+FP+/z/wDxNf8ADfn7Gn/Rw/hT/v8AP/8AE1/w35+xp/0cP4U/7/P/APE1/wAN+fsaf9HD+FP+/wA//wATX/Dfn7Gn/Rw/hT/v8/8A8TX/AA35+xp/0cP4U/7/AD//ABNf8N+fsaf9HD+FP+/z/wDxNf8ADfn7Gn/Rw/hT/v8AP/8AE1/w35+xp/0cP4U/7/P/APE1/wAN+fsaf9HD+FP+/wA//wATX/Dfn7Gn/Rw/hT/v8/8A8TX/AA35+xp/0cP4U/7/AD//ABNf8N+fsaf9HD+FP+/z/wDxNf8ADfn7Gn/Rw/hT/v8AP/8AE1/w35+xp/0cP4U/7/P/APE1/wAN+fsaf9HD+FP+/wA//wATX/Dfn7Gn/Rw/hT/v8/8A8TX/AA35+xp/0cP4U/7/AD//ABNf8N+fsaf9HD+FP+/z/wDxNf8ADfn7Gn/Rw/hT/v8AP/8AE1/w35+xp/0cP4U/7/P/APE1/wAN+fsaf9HD+FP+/wA//wATX/Dfn7Gn/Rw/hT/v8/8A8TX/AA35+xp/0cP4U/7/AD//ABNf8N+fsaf9HD+FP+/z/wDxNf8ADfn7Gn/Rw/hT/v8AP/8AE1/w35+xp/0cP4U/7/P/APE1/wAN+fsaf9HD+FP+/wA//wATX/Dfn7Gn/Rw/hT/v8/8A8TX/AA35+xp/0cP4U/7/AD//ABNf8N+fsaf9HD+FP+/z/wDxNf8ADfn7Gn/Rw/hT/v8AP/8AE1/w35+xp/0cP4U/7/P/APE1/wAN+fsaf9HD+FP+/wA//wATX/Dfn7Gn/Rw/hT/v8/8A8TX/AA35+xp/0cP4U/7/AD//ABNf8N+fsaf9HD+FP+/z/wDxNf8ADfn7Gn/Rw/hT/v8AP/8AE1/w35+xp/0cP4U/7/P/APE1/wAN+fsaf9HD+FP+/wA//wATX/Dfn7Gn/Rw/hT/v8/8A8TX/AA35+xp/0cP4U/7/AD//ABNf8N+fsaf9HD+FP+/z/wDxNf8ADfn7Gn/Rw/hT/v8AP/8AE1/w35+xp/0cP4U/7/P/APE1/wAN+fsaf9HD+FP+/wA//wATX/Dfn7Gn/Rw/hT/v8/8A8TX/AA35+xp/0cP4U/7/AD//ABNf8N+fsaf9HD+FP+/z/wDxNf8ADfn7Gn/Rw/hT/v8AP/8AE1/w35+xp/0cP4U/7/P/APE1/wAN+fsaf9HD+FP+/wA//wATX/Dfn7Gn/Rw/hT/v8/8A8TX/AA35+xp/0cP4U/7/AD//ABNf8N+fsaf9HD+FP+/z/wDxNf8ADfn7Gn/Rw/hT/v8AP/8AE1/w35+xp/0cP4U/7/P/APE1/wAN+fsaf9HD+FP+/wA//wATX/Dfn7Gn/Rw/hT/v8/8A8TX/AA35+xp/0cP4U/7/AD//ABNf8N+fsaf9HD+FP+/z/wDxNf8ADfn7Gn/Rw/hT/v8AP/8AE1/w35+xp/0cP4U/7/P/APE1/wAN+fsaf9HD+FP+/wA//wATX/Dfn7Gn/Rw/hT/v8/8A8TX/AA35+xp/0cP4U/7/AD//ABNf8N+fsaf9HD+FP+/z/wDxNf8ADfn7Gn/Rw/hT/v8AP/8AE1/w35+xp/0cP4U/7/P/APE1/wAN+fsaf9HD+FP+/wA//wATX/Dfn7Gn/Rw/hT/v8/8A8TX/AA35+xp/0cP4U/7/AD//ABNf8N+fsaf9HD+FP+/z/wDxNf8ADfn7Gn/Rw/hT/v8AP/8AE1/w35+xp/0cP4U/7/P/APE1/wAN+fsaf9HD+FP+/wA//wATX/Dfn7Gn/Rw/hT/v8/8A8TX/AA35+xp/0cP4U/7/AD//ABNf8N+fsaf9HD+FP+/z/wDxNf8ADfn7Gn/Rw/hT/v8AP/8AE1/w35+xp/0cP4U/7/P/APE1/wAN+fsaf9HD+FP+/wA//wATX/Dfn7Gn/Rw/hT/v8/8A8TX/AA35+xp/0cP4U/7/AD//ABNf8N+fsaf9HD+FP+/z/wDxNf8ADfn7Gn/Rw/hT/v8AP/8AE1/w35+xp/0cP4U/7/P/APE1/wAN+fsaf9HD+FP+/wA//wATX/Dfn7Gn/Rw/hT/v8/8A8TX/AA35+xp/0cP4U/7/AD//ABNf8N+fsaf9HD+FP+/z/wDxNf8ADfn7Gn/Rw/hT/v8AP/8AE1/w35+xp/0cP4U/7/P/APE1/wAN+fsaf9HD+FP+/wA//wATX/Dfn7Gn/Rw/hT/v8/8A8TX/AA35+xp/0cP4U/7/AD//ABNf8N+fsaf9HD+FP+/z/wDxNf8ADfn7Gn/Rw/hT/v8AP/8AE1/w35+xp/0cP4U/7/P/APE1/wAN+fsaf9HD+FP+/wA//wATX/Dfn7Gn/Rw/hT/v8/8A8TX/AA35+xp/0cP4U/7/AD//ABNf8N+fsaf9HD+FP+/z/wDxNf8ADfn7Gn/Rw/hT/v8AP/8AE1/w35+xp/0cP4U/7/P/APE1/wAN+fsaf9HD+FP+/wA//wATX/Dfn7Gn/Rw/hT/v8/8A8TX/AA35+xp/0cP4U/7/AD//ABNf8N+fsaf9HD+FP+/z/wDxNf8ADfn7Gn/Rw/hT/v8AP/8AE1/w35+xp/0cP4U/7/P/APE1/wAN+fsaf9HD+FP+/wA//wATX/Dfn7Gn/Rw/hT/v8/8A8TX/AA35+xp/0cP4U/7/AD//ABNf8N+fsaf9HD+FP+/z/wDxNf8ADfn7Gn/Rw/hT/v8AP/8AE1/w35+xp/0cP4U/7/P/APE1/wAN+fsaf9HD+FP+/wA//wATX/Dfn7Gn/Rw/hT/v8/8A8TX/AA35+xp/0cP4U/7/AD//ABNf8N+fsaf9HD+FP+/z/wDxNf8ADfn7Gn/Rw/hT/v8AP/8AE1/w35+xp/0cP4U/7/P/APE1/wAN+fsaf9HD+FP+/wA//wATX/Dfn7Gn/Rw/hT/v8/8A8TX/AA35+xp/0cP4U/7/AD//ABNf8N+fsaf9HD+FP+/z/wDxNf8ADfn7Gn/Rw/hT/v8AP/8AE1/w35+xp/0cP4U/7/P/APE1/wAN+fsaf9HD+FP+/wA//wATX/Dfn7Gn/Rw/hT/v8/8A8TX/AA35+xp/0cP4U/7/AD//ABNf8N+fsaf9HD+FP+/z/wDxNf8ADfn7Gn/Rw/hT/v8AP/8AE1/w35+xp/0cP4U/7/P/APE1/wAN+fsaf9HD+FP+/wA//wATX/Dfn7Gn/Rw/hT/v8/8A8TX/AA35+xp/0cP4U/7/AD//ABNf8N+fsaf9HD+FP+/z/wDxNf8ADfn7Gn/Rw/hT/v8AP/8AE1/w35+xp/0cP4U/7/P/APE1/wAN+fsaf9HD+FP+/wA//wATX/Dfn7Gn/Rw/hT/v8/8A8TX/AA35+xp/0cP4U/7/AD//ABNf8N+fsaf9HD+FP+/z/wDxNf8ADfn7Gn/Rw/hT/v8AP/8AE1p2oWWrafa6rptylxaXkKTW8qdJI2AZWHsQQaqqqqqqqqqqqqqqqqqqrxF+25+yd4T1/U/CviP47eGbDVtGvJbK/tJpnD288TlJI2+XqrKQfcV/w35+xp/0cP4U/wC/z/8AxNf8N+fsaf8ARw/hT/v8/wD8TX/Dfn7Gn/Rw/hT/AL/P/wDE1/w35+xp/wBHD+FP+/z/APxNf8N+fsaf9HD+FP8Av8//AMTX/Dfn7Gn/AEcP4U/7/P8A/E1/w35+xp/0cP4U/wC/z/8AxNf8N+fsaf8ARw/hT/v8/wD8TX/Dfn7Gn/Rw/hT/AL/P/wDE1/w35+xp/wBHD+FP+/z/APxNf8N+fsaf9HD+FP8Av8//AMTX/Dfn7Gn/AEcP4U/7/P8A/E1/w35+xp/0cP4U/wC/z/8AxNf8N+fsaf8ARw/hT/v8/wD8TX/Dfn7Gn/Rw/hT/AL/P/wDE1/w35+xp/wBHD+FP+/z/APxNf8N+fsaf9HD+FP8Av8//AMTX/Dfn7Gn/AEcP4U/7/P8A/E1/w35+xp/0cP4U/wC/z/8AxNf8N+fsaf8ARw/hT/v8/wD8TX/Dfn7Gn/Rw/hT/AL/P/wDE1/w35+xp/wBHD+FP+/z/APxNf8N+fsaf9HD+FP8Av8//AMTX/Dfn7Gn/AEcP4U/7/P8A/E1/w35+xp/0cP4U/wC/z/8AxNf8N+fsaf8ARw/hT/v8/wD8TX/Dfn7Gn/Rw/hT/AL/P/wDE1/w35+xp/wBHD+FP+/z/APxNf8N+fsaf9HD+FP8Av8//AMTX/Dfn7Gn/AEcP4U/7/P8A/E1/w35+xp/0cP4U/wC/z/8AxNf8N+fsaf8ARw/hT/v8/wD8TX/Dfn7Gn/Rw/hT/AL/P/wDE1/w35+xp/wBHD+FP+/z/APxNf8N+fsaf9HD+FP8Av8//AMTX/Dfn7Gn/AEcP4U/7/P8A/E1/w35+xp/0cP4U/wC/z/8AxNf8N+fsaf8ARw/hT/v8/wD8TX/Dfn7Gn/Rw/hT/AL/P/wDE1/w35+xp/wBHD+FP+/z/APxNf8N+fsaf9HD+FP8Av8//AMTX/Dfn7Gn/AEcP4U/7/P8A/E1/w35+xp/0cP4U/wC/z/8AxNf8N+fsaf8ARw/hT/v8/wD8TX/Dfn7Gn/Rw/hT/AL/P/wDE1/w35+xp/wBHD+FP+/z/APxNf8N+fsaf9HD+FP8Av8//AMTX/Dfn7Gn/AEcP4U/7/P8A/E1/w35+xp/0cP4U/wC/z/8AxNf8N+fsaf8ARw/hT/v8/wD8TX/Dfn7Gn/Rw/hT/AL/P/wDE1/w35+xp/wBHD+FP+/z/APxNf8N+fsaf9HD+FP8Av8//AMTX/Dfn7Gn/AEcP4U/7/P8A/E1/w35+xp/0cP4U/wC/z/8AxNf8N+fsaf8ARw/hT/v8/wD8TX/Dfn7Gn/Rw/hT/AL/P/wDE1/w35+xp/wBHD+FP+/z/APxNf8N+fsaf9HD+FP8Av8//AMTX/Dfn7Gn/AEcP4U/7/P8A/E1/w35+xp/0cP4U/wC/z/8AxNf8N+fsaf8ARw/hT/v8/wD8TX/Dfn7Gn/Rw/hT/AL/P/wDE1/w35+xp/wBHD+FP+/z/APxNf8N+fsaf9HD+FP8Av8//AMTX/Dfn7Gn/AEcP4U/7/P8A/E1/w35+xp/0cP4U/wC/z/8AxNf8N+fsaf8ARw/hT/v8/wD8TX/Dfn7Gn/Rw/hT/AL/P/wDE1/w35+xp/wBHD+FP+/z/APxNf8N+fsaf9HD+FP8Av8//AMTX/Dfn7Gn/AEcP4U/7/P8A/E1/w35+xp/0cP4U/wC/z/8AxNf8N+fsaf8ARw/hT/v8/wD8TX/Dfn7Gn/Rw/hT/AL/P/wDE1/w35+xp/wBHD+FP+/z/APxNf8N+fsaf9HD+FP8Av8//AMTX/Dfn7Gn/AEcP4U/7/P8A/E1/w35+xp/0cP4U/wC/z/8AxNf8N+fsaf8ARw/hT/v8/wD8TX/Dfn7Gn/Rw/hT/AL/P/wDE1/w35+xp/wBHD+FP+/z/APxNf8N+fsaf9HD+FP8Av8//AMTX/Dfn7Gn/AEcP4U/7/P8A/E1/w35+xp/0cP4U/wC/z/8AxNf8N+fsaf8ARw/hT/v8/wD8TX/Dfn7Gn/Rw/hT/AL/P/wDE1/w35+xp/wBHD+FP+/z/APxNf8N+fsaf9HD+FP8Av8//AMTX/Dfn7Gn/AEcP4U/7/P8A/E1/w35+xp/0cP4U/wC/z/8AxNf8N+fsaf8ARw/hT/v8/wD8TX/Dfn7Gn/Rw/hT/AL/P/wDE1/w35+xp/wBHD+FP+/z/APxNfDj4n/D/AOL3hmPxl8NPFVj4h0SSZ4UvbNiYzIhwy8gcjiqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq+O3wJ+H/7Rvw/m+GnxLtr2fRJrqK5dLO5MEnmRklfmGeOelf8Ohv2NP8AoB+Kv/B6/wDhX/Dob9jT/oB+Kv8Awev/AIV/w6G/Y0/6Afir/wAHr/4V/wAOhv2NP+gH4q/8Hr/4V/w6G/Y0/wCgH4q/8Hr/AOFf8Ohv2NP+gH4q/wDB6/8AhX/Dob9jT/oB+Kv/AAev/hX/AA6G/Y0/6Afir/wev/hX/Dob9jT/AKAfir/wev8A4V/w6G/Y0/6Afir/AMHr/wCFf8Ohv2NP+gH4q/8AB6/+Ff8ADob9jT/oB+Kv/B6/+Ff8Ohv2NP8AoB+Kv/B6/wDhX/Dob9jT/oB+Kv8Awev/AIV/w6G/Y0/6Afir/wAHr/4V/wAOhv2NP+gH4q/8Hr/4V/w6G/Y0/wCgH4q/8Hr/AOFf8Ohv2NP+gH4q/wDB6/8AhX/Dob9jT/oB+Kv/AAev/hX/AA6G/Y0/6Afir/wev/hX/Dob9jT/AKAfir/wev8A4V/w6G/Y0/6Afir/AMHr/wCFf8Ohv2NP+gH4q/8AB6/+Ff8ADob9jT/oB+Kv/B6/+Ff8Ohv2NP8AoB+Kv/B6/wDhX/Dob9jT/oB+Kv8Awev/AIV/w6G/Y0/6Afir/wAHr/4V/wAOhv2NP+gH4q/8Hr/4V/w6G/Y0/wCgH4q/8Hr/AOFf8Ohv2NP+gH4q/wDB6/8AhX/Dob9jT/oB+Kv/AAev/hX/AA6G/Y0/6Afir/wev/hX/Dob9jT/AKAfir/wev8A4V/w6G/Y0/6Afir/AMHr/wCFf8Ohv2NP+gH4q/8AB6/+Ff8ADob9jT/oB+Kv/B6/+Ff8Ohv2NP8AoB+Kv/B6/wDhX/Dob9jT/oB+Kv8Awev/AIV/w6G/Y0/6Afir/wAHr/4V/wAOhv2NP+gH4q/8Hr/4V/w6G/Y0/wCgH4q/8Hr/AOFf8Ohv2NP+gH4q/wDB6/8AhX/Dob9jT/oB+Kv/AAev/hX/AA6G/Y0/6Afir/wev/hX/Dob9jT/AKAfir/wev8A4V/w6G/Y0/6Afir/AMHr/wCFf8Ohv2NP+gH4q/8AB6/+Ff8ADob9jT/oB+Kv/B6/+Ff8Ohv2NP8AoB+Kv/B6/wDhX/Dob9jT/oB+Kv8Awev/AIV/w6G/Y0/6Afir/wAHr/4V/wAOhv2NP+gH4q/8Hr/4V/w6G/Y0/wCgH4q/8Hr/AOFf8Ohv2NP+gH4q/wDB6/8AhX/Dob9jT/oB+Kv/AAev/hX/AA6G/Y0/6Afir/wev/hX/Dob9jT/AKAfir/wev8A4V/w6G/Y0/6Afir/AMHr/wCFf8Ohv2NP+gH4q/8AB6/+Ff8ADob9jT/oB+Kv/B6/+Ff8Ohv2NP8AoB+Kv/B6/wDhX/Dob9jT/oB+Kv8Awev/AIV/w6G/Y0/6Afir/wAHr/4V/wAOhv2NP+gH4q/8Hr/4V/w6G/Y0/wCgH4q/8Hr/AOFf8Ohv2NP+gH4q/wDB6/8AhX/Dob9jT/oB+Kv/AAev/hX/AA6G/Y0/6Afir/wev/hX/Dob9jT/AKAfir/wev8A4V/w6G/Y0/6Afir/AMHr/wCFf8Ohv2NP+gH4q/8AB6/+Ff8ADob9jT/oB+Kv/B6/+Ff8Ohv2NP8AoB+Kv/B6/wDhX/Dob9jT/oB+Kv8Awev/AIV/w6G/Y0/6Afir/wAHr/4V/wAOhv2NP+gH4q/8Hr/4V/w6G/Y0/wCgH4q/8Hr/AOFf8Ohv2NP+gH4q/wDB6/8AhX/Dob9jT/oB+Kv/AAev/hX/AA6G/Y0/6Afir/wev/hX/Dob9jT/AKAfir/wev8A4V/w6G/Y0/6Afir/AMHr/wCFf8Ohv2NP+gH4q/8AB6/+Ff8ADob9jT/oB+Kv/B6/+Ff8Ohv2NP8AoB+Kv/B6/wDhX/Dob9jT/oB+Kv8Awev/AIV/w6G/Y0/6Afir/wAHr/4V/wAOhv2NP+gH4q/8Hr/4V/w6G/Y0/wCgH4q/8Hr/AOFf8Ohv2NP+gH4q/wDB6/8AhX/Dob9jT/oB+Kv/AAev/hX/AA6G/Y0/6Afir/wev/hX/Dob9jT/AKAfir/wev8A4V/w6G/Y0/6Afir/AMHr/wCFf8Ohv2NP+gH4q/8AB6/+Ff8ADob9jT/oB+Kv/B6/+Ff8Ohv2NP8AoB+Kv/B6/wDhX/Dob9jT/oB+Kv8Awev/AIV/w6G/Y0/6Afir/wAHr/4V/wAOhv2NP+gH4q/8Hr/4V/w6G/Y0/wCgH4q/8Hr/AOFf8Ohv2NP+gH4q/wDB6/8AhX/Dob9jT/oB+Kv/AAev/hX/AA6G/Y0/6Afir/wev/hX/Dob9jT/AKAfir/wev8A4V/w6G/Y0/6Afir/AMHr/wCFf8Ohv2NP+gH4q/8AB6/+Ff8ADob9jT/oB+Kv/B6/+Ff8Ohv2NP8AoB+Kv/B6/wDhX/Dob9jT/oB+Kv8Awev/AIV/w6G/Y0/6Afir/wAHr/4V/wAOhv2NP+gH4q/8Hr/4V/w6G/Y0/wCgH4q/8Hr/AOFf8Ohv2NP+gH4q/wDB6/8AhX/Dob9jT/oB+Kv/AAev/hX/AA6G/Y0/6Afir/wev/hX/Dob9jT/AKAfir/wev8A4V/w6G/Y0/6Afir/AMHr/wCFf8Ohv2NP+gH4q/8AB6/+Ff8ADob9jT/oB+Kv/B6/+Ff8Ohv2NP8AoB+Kv/B6/wDhX/Dob9jT/oB+Kv8Awev/AIV/w6G/Y0/6Afir/wAHr/4V/wAOhv2NP+gH4q/8Hr/4V/w6G/Y0/wCgH4q/8Hr/AOFf8Ohv2NP+gH4q/wDB6/8AhX/Dob9jT/oB+Kv/AAev/hX/AA6G/Y0/6Afir/wev/hX/Dob9jT/AKAfir/wev8A4V/w6G/Y0/6Afir/AMHr/wCFf8Ohv2NP+gH4q/8AB6/+Ff8ADob9jT/oB+Kv/B6/+Ff8Ohv2NP8AoB+Kv/B6/wDhX/Dob9jT/oB+Kv8Awev/AIV/w6G/Y0/6Afir/wAHr/4V/wAOhv2NP+gH4q/8Hr/4V/w6G/Y0/wCgH4q/8Hr/AOFf8Ohv2NP+gH4q/wDB6/8AhX/Dob9jT/oB+Kv/AAev/hX/AA6G/Y0/6Afir/wev/hX/Dob9jT/AKAfir/wev8A4V/w6G/Y0/6Afir/AMHr/wCFf8Ohv2NP+gH4q/8AB6/+Ff8ADob9jT/oB+Kv/B6/+Ff8Ohv2NP8AoB+Kv/B6/wDhX/Dob9jT/oB+Kv8Awev/AIV/w6G/Y0/6Afir/wAHr/4V/wAOhv2NP+gH4q/8Hr/4V/w6G/Y0/wCgH4q/8Hr/AOFf8Ohv2NP+gH4q/wDB6/8AhX/Dob9jT/oB+Kv/AAev/hX/AA6G/Y0/6Afir/wev/hX/Dob9jT/AKAfir/wev8A4V/w6G/Y0/6Afir/AMHr/wCFf8Ohv2NP+gH4q/8AB6/+Ff8ADob9jT/oB+Kv/B6/+Ff8Ohv2NP8AoB+Kv/B6/wDhX/Dob9jT/oB+Kv8Awev/AIV/w6G/Y0/6Afir/wAHr/4V/wAOhv2NP+gH4q/8Hr/4V/w6G/Y0/wCgH4q/8Hr/AOFf8Ohv2NP+gH4q/wDB6/8AhX/Dob9jT/oB+Kv/AAev/hX/AA6G/Y0/6Afir/wev/hX/Dob9jT/AKAfir/wev8A4V/w6G/Y0/6Afir/AMHr/wCFf8Ohv2NP+gH4q/8AB6/+Ff8ADob9jT/oB+Kv/B6/+Ff8Ohv2NP8AoB+Kv/B6/wDhX/Dob9jT/oB+Kv8Awev/AIV/w6G/Y0/6Afir/wAHr/4V/wAOhv2NP+gH4q/8Hr/4V/w6G/Y0/wCgH4q/8Hr/AOFf8Ohv2NP+gH4q/wDB6/8AhX/Dob9jT/oB+Kv/AAev/hX/AA6G/Y0/6Afir/wev/hX/Dob9jT/AKAfir/wev8A4V/w6G/Y0/6Afir/AMHr/wCFf8Ohv2NP+gH4q/8AB6/+Ff8ADob9jT/oB+Kv/B6/+Ff8Ohv2NP8AoB+Kv/B6/wDhX/Dob9jT/oB+Kv8Awev/AIV/w6G/Y0/6Afir/wAHr/4V/wAOhv2NP+gH4q/8Hr/4V/w6G/Y0/wCgH4q/8Hr/AOFf8Ohv2NP+gH4q/wDB6/8AhX/Dob9jT/oB+Kv/AAev/hX/AA6G/Y0/6Afir/wev/hX/Dob9jT/AKAfir/wev8A4V/w6G/Y0/6Afir/AMHr/wCFf8Ohv2NP+gH4q/8AB6/+Ff8ADob9jT/oB+Kv/B6/+Ff8Ohv2NP8AoB+Kv/B6/wDhX/Dob9jT/oB+Kv8Awev/AIV/w6G/Y0/6Afir/wAHr/4V/wAOhv2NP+gH4q/8Hr/4V/w6G/Y0/wCgH4q/8Hr/AOFf8Ohv2NP+gH4q/wDB6/8AhX/Dob9jT/oB+Kv/AAev/hX/AA6G/Y0/6Afir/wev/hX/Dob9jT/AKAfir/wev8A4V/w6G/Y0/6Afir/AMHr/wCFf8Ohv2NP+gH4q/8AB6/+Ff8ADob9jT/oB+Kv/B6/+Ff8Ohv2NP8AoB+Kv/B6/wDhX/Dob9jT/oB+Kv8Awev/AIV/w6G/Y0/6Afir/wAHr/4V/wAOhv2NP+gH4q/8Hr/4V/w6G/Y0/wCgH4q/8Hr/AOFf8Ohv2NP+gH4q/wDB6/8AhX/Dob9jT/oB+Kv/AAev/hX/AA6G/Y0/6Afir/wev/hX/Dob9jT/AKAfir/wev8A4Voej2Xh7RNP0DTVcWmmWsdtbh23MI41Crk9zgDmqqqqqqqqqqqqqqqqqqq8cf8ABLX9k34heNNf8feI9H8Svq3iXVLnU79odaeNDPPK0shVQPlG5zgdq/4dDfsaf9APxV/4PX/wr/h0N+xp/wBAPxV/4PX/AMK/4dDfsaf9APxV/wCD1/8ACv8Ah0N+xp/0A/FX/g9f/Cv+HQ37Gn/QD8Vf+D1/8K/4dDfsaf8AQD8Vf+D1/wDCv+HQ37Gn/QD8Vf8Ag9f/AAr/AIdDfsaf9APxV/4PX/wr/h0N+xp/0A/FX/g9f/Cv+HQ37Gn/AEA/FX/g9f8Awr/h0N+xp/0A/FX/AIPX/wAK/wCHQ37Gn/QD8Vf+D1/8K/4dDfsaf9APxV/4PX/wr/h0N+xp/wBAPxV/4PX/AMK/4dDfsaf9APxV/wCD1/8ACv8Ah0N+xp/0A/FX/g9f/Cv+HQ37Gn/QD8Vf+D1/8K/4dDfsaf8AQD8Vf+D1/wDCv+HQ37Gn/QD8Vf8Ag9f/AAr/AIdDfsaf9APxV/4PX/wr/h0N+xp/0A/FX/g9f/Cv+HQ37Gn/AEA/FX/g9f8Awr/h0N+xp/0A/FX/AIPX/wAK/wCHQ37Gn/QD8Vf+D1/8K/4dDfsaf9APxV/4PX/wr/h0N+xp/wBAPxV/4PX/AMK/4dDfsaf9APxV/wCD1/8ACv8Ah0N+xp/0A/FX/g9f/Cv+HQ37Gn/QD8Vf+D1/8K/4dDfsaf8AQD8Vf+D1/wDCv+HQ37Gn/QD8Vf8Ag9f/AAr/AIdDfsaf9APxV/4PX/wr/h0N+xp/0A/FX/g9f/Cv+HQ37Gn/AEA/FX/g9f8Awr/h0N+xp/0A/FX/AIPX/wAK/wCHQ37Gn/QD8Vf+D1/8K/4dDfsaf9APxV/4PX/wr/h0N+xp/wBAPxV/4PX/AMK/4dDfsaf9APxV/wCD1/8ACv8Ah0N+xp/0A/FX/g9f/Cv+HQ37Gn/QD8Vf+D1/8K/4dDfsaf8AQD8Vf+D1/wDCv+HQ37Gn/QD8Vf8Ag9f/AAr/AIdDfsaf9APxV/4PX/wr/h0N+xp/0A/FX/g9f/Cv+HQ37Gn/AEA/FX/g9f8Awr/h0N+xp/0A/FX/AIPX/wAK/wCHQ37Gn/QD8Vf+D1/8K/4dDfsaf9APxV/4PX/wr/h0N+xp/wBAPxV/4PX/AMK/4dDfsaf9APxV/wCD1/8ACv8Ah0N+xp/0A/FX/g9f/Cv+HQ37Gn/QD8Vf+D1/8K/4dDfsaf8AQD8Vf+D1/wDCv+HQ37Gn/QD8Vf8Ag9f/AAr/AIdDfsaf9APxV/4PX/wr/h0N+xp/0A/FX/g9f/Cv+HQ37Gn/AEA/FX/g9f8Awr/h0N+xp/0A/FX/AIPX/wAK/wCHQ37Gn/QD8Vf+D1/8K/4dDfsaf9APxV/4PX/wr/h0N+xp/wBAPxV/4PX/AMK/4dDfsaf9APxV/wCD1/8ACv8Ah0N+xp/0A/FX/g9f/Cv+HQ37Gn/QD8Vf+D1/8K/4dDfsaf8AQD8Vf+D1/wDCv+HQ37Gn/QD8Vf8Ag9f/AAr/AIdDfsaf9APxV/4PX/wr/h0N+xp/0A/FX/g9f/Cv+HQ37Gn/AEA/FX/g9f8Awr/h0N+xp/0A/FX/AIPX/wAK/wCHQ37Gn/QD8Vf+D1/8K/4dDfsaf9APxV/4PX/wr/h0N+xp/wBAPxV/4PX/AMK/4dDfsaf9APxV/wCD1/8ACv8Ah0N+xp/0A/FX/g9f/Cv+HQ37Gn/QD8Vf+D1/8K/4dDfsaf8AQD8Vf+D1/wDCv+HQ37Gn/QD8Vf8Ag9f/AAr/AIdDfsaf9APxV/4PX/wr/h0N+xp/0A/FX/g9f/Cv+HQ37Gn/AEA/FX/g9f8Awr/h0N+xp/0A/FX/AIPX/wAK/wCHQ37Gn/QD8Vf+D1/8K/4dDfsaf9APxV/4PX/wr/h0N+xp/wBAPxV/4PX/AMK/4dDfsaf9APxV/wCD1/8ACv8Ah0N+xp/0A/FX/g9f/Cv+HQ37Gn/QD8Vf+D1/8K/4dDfsaf8AQD8Vf+D1/wDCv+HQ37Gn/QD8Vf8Ag9f/AAr/AIdDfsaf9APxV/4PX/wr/h0N+xp/0A/FX/g9f/Cv+HQ37Gn/AEA/FX/g9f8Awr4E/An4f/s5/D+H4afDS2vYdEgupblEvLkzyeZIQW+Y9uOlVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVXxc/4Km+P/hv+1Dq3wBsfhX4fvLDTvEkOkpqEt7MsrI7IpcqOMjeeOnFVX7eH7U+v/sh/CDSPiT4c8K6f4gudS8SQaS9tfTvEiJJbXMpcFOcg24GOnzGv2D/2p9f/AGvPhBq/xJ8R+FdP0C503xJPpKW1lO8qOkdtbShyX5yTORjp8oqqqqqqqq/4LC/E74k/DH4a/D6/+GvxD8S+E7m91y5iuZtD1aewedBACFdoWUsAecHjNf8ABLXxv40+IX7Jun+I/H3jDW/EurPrl/E1/rGoS3k5RXUKpklZmwOwzgVVftfftT+E/wBkj4SzfEPXrL+1dUvLgWWh6Os3ltfXLAtgtg7I0VWZnxgYC/edQfBEv/BSb/govdalrugfELUNH8JQXHkTTLqcmi6NFJlWEKxwZe4KbVbJWUpwSwLDdffsYf8ABU39n6EeJfhx8Xr3xEYG86Wy8O+LrmcO2NuXtbxI0mIB6bX9hkV4TtPENh4V0ax8W6pFqeu2+nwR6newxiNLm5WNRLIqgAKGcMwAAAz0HSqqqqqqqr/gqV8evjn8Pf2sdQ8N+APjT478NaSmh2Ei2Gj+I7uygV2RtzCOKRVBPc45r4ZXd1qHw18J319dS3Nzc6HZSTTTOXeR2gQszMeSSSSSepNVVVVVXjLxVo/gTwfrnjjxBK0el+HdNuNRvnQAssEEbSSEAkDO1T1Ir9l7/grJF8e/j9p3wj8TfCyLw3pfiWSSDRb23v2upYpwpaNJxsUEOFK7lHysVyNpLLVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX7fX7fXi/9jzxf4V8NeGvAGj+IY/EOmzXcst9dSRNEySBAAE6gjnmv+H3nxT/6If4U/wDBjcf4V/w+8+Kf/RD/AAp/4Mbj/Cv+Cff7b3ir9sz/AIT3/hJvA+leHf8AhD/7N8j7DcyS+f8AavtO7dv6bfs64x/eNVVVVVVVX/BUr49fHP4e/tY6h4b8AfGnx34a0lNDsJFsNH8R3dlArsjbmEcUiqCe5xzXwyu7rUPhr4Tvr66lubm50Oykmmmcu8jtAhZmY8kkkkk9Saqqqvidd3Vh8NfFl9YXUttc22h3skM0LlHidYHKsrDkEEAgjoa/4Ja/Hr45/EL9rHT/AA34++NPjvxLpL6HfyNYax4ju7yAuqLtYxyyMuR2OOKqqqqr9pzVtV0D9mz4sa7oOp3enalpvgfWbmzvLSZoZraaOxmZJI3UhkdWAIYEEEAiv+CRXxv+NPxL/aT8SaD8Rvi/418VabB4HurmKy1vxBdX0Mcy3tkokVJXZQ4V3UMBkBmHc1VVVVVVVft1f8FJ3/ZP+IGm/Cvwb8PrTxNrkunpqGo3N9fNDBaxyM6xwqiAs0h2bySQFUrgMWOz9n340aB+0R8GvC/xk8M2NxZWPiS1aT7JcHL200cjwzRFsDcFljkUNgBgAcDOKqqqqqqqqqqqq/bR/wCCob/sx/GJ/g74O+Glp4lvdJt7ebWrq91FrdY2mQSLDGqKxz5TxsXY/wAeNpxk/CT4l6B8Zfhl4Z+KfheO4j0zxPpsV9BFcptlh3r80bjpuVsqSCQSuQSCCaqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/ak/5SP+Jv8AsoFp/wCjIaqv+C1f/Jq/hb/soFn/AOkF/X/BFT/k1fxT/wBlAvP/AEgsKqtD8T+GvE8Mk/hrxFpmrRwMFlexu0nCMRwCUJwcdjVVquraVodhLqmt6naafZQY825uplhjTJCjLMQBkkDnuRWheLPCvinz/wDhGfE2lav9l2+f9hvY7jyt2du7YTjO04z1wfSqqtE8b+CvE101h4c8YaJqtzHGZHhstQindUBALFUYkDJAz0yR61/wW9/5JV8NP+xguv8A0nFf8Ehv+TNNM/7GDUf/AENaqv8Agtv4tvb343fD/wACOr/ZNH8KvqURMmQZLu6kjfC44OLKPJyc5HAxz+xB4P0LwR+yN8JNK8P23kw3vhOw1OfIGXuLyFbmZiQBnMkrYzzjAycZqqqqqtV1bStDsJdU1vU7TT7KDHm3N1MsMaZIUZZiAMkgc9yK0/UdP1ayh1LSr+3vbS4XdDPbyiSOQeqspII+lVWq6tpWh2Euqa3qdpp9lBjzbm6mWGNMkKMsxAGSQOe5FaJ4h8P+JrRr7w5run6rbRyGJ5rK5SdFcAEqWQkA4IOPcetf8Fef+TytS/7F/Tv/AEBq+E//ACSvwb/2L9j/AOk6VVVrGuaJ4esjqOv6xY6baKwUz3lwsMYY9BuYgZPpWl6tpWu2EWqaJqdpqFlNnyrm1mWWN8EqdrKSDggg+4NVF4y8Hz62fDMHivR5NYDMp09b6M3AZVLMPLDbshQSRjgDNVeWdpqFpNYX1tFc21zG0U0MyB0kRhhlZTwQQSCDwc18G/8Agnh+y38CfiSvxV8BeDtQXXLWSR9M+3apNcxaZ5iFGEKseflZgGkLsNx5zgiq1zxl4P8ADEsVv4k8V6PpMsylokvr6OBnA4JAdhkVp+o6fq1lDqWlX9ve2lwu6Ge3lEkcg9VZSQR9KqtD8T+GvE8Mk/hrxFpmrRwMFlexu0nCMRwCUJwcdjVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX/AAW9/wCSp/DT/sX7r/0oFfsJ/Df4d6t+yD8K9R1XwF4cvbu40FGlnuNKhkkkbe3LMykk/Wv+FTfCv/omnhT/AME1v/8AEVoXhLwr4X8//hGfDOlaR9p2+f8AYbKO383bnbu2AZxuOM9Mn1qry7tLC0mvr66itra2jaSaaZwiRIoyzMx4AABJJ4FaRrmieIbIajoGsWOp2hYqJ7O4WaMsOo3KSMj0qpPE/hqPW4/DEniLTF1iVd0entdoLhhtLZEedxG0E9OgJqqrWPE/hrw9La2+v+ItM02W+YrapeXaQtOwwCEDEbiNy8D+8PWqr/grz/yeVqX/AGL+nf8AoDV8J/8Aklfg3/sX7H/0nSqqqr4s/wDJK/GX/Yv33/pO9f8ABIb/AJPK03/sX9R/9AWtV1bStDsJdU1vU7TT7KDHm3N1MsMaZIUZZiAMkgc9yK0TxD4f8TWjX3hzXdP1W2jkMTzWVyk6K4AJUshIBwQce49aqq0jxP4a8Qy3VvoHiLTNTlsWC3SWd2kzQMcgBwpO0na3Bx0PpVftZf8AJq/xl/7J/rv/AKQTV/wRU/5Oo8U/9k/vP/S+wrW/EPh/w1aJfeI9d0/SrZ5BEk19cpAjOQSFDOQCcKTj2PpWn6jp+rWUOpaVf297aXC7oZ7eUSRyD1VlJBH0qq1zxl4P8MSxW/iTxXo+kyzKWiS+vo4GcDgkB2GRWn6jp+rWUOpaVf297aXC7oZ7eUSRyD1VlJBH0qqqq/aO/Yj+AH7U+saX4k+Kegag2s6Rbi1g1DTr5raVrcSGQQv1Vk3M5GRkb2wRmvAHgDwf8LfBmkfD3wBocWkeH9Ctxb2FlE7uIkBJ+85LMSSSWYliSSSSSaqtN8b+Cta1JtG0fxhol9qCbt1pbahFLKu372UVieO/HFVVS+MvB8GtjwzN4r0ePWCyqNPe+jFxuZQwHl7t2SpBAx0INVVVVaV4y8H69qEuk6H4r0fUb63VmmtrS+jmljCkKxZFYkAEgHI4JFVrnifw14Yhin8SeItM0mOZisT312kAdgOQC5GTjtVpd2moWkN9YXUVzbXMayQzQuHSRGGVZWHBBBBBHBzVfH/9g79nH9pfxtp3xC+JvhnUH1uxt1tZZ9P1B7UX0CFikcyrwdpdsOu18YBYqqgeFPCvh7wN4Y0nwZ4S0uHTdF0Ozis9Ps4slYIY1CooJJJwAOSST1JJOaq8u7SwtJr6+uora2to2kmmmcIkSKMszMeAAASSeBWieN/BXia6aw8OeMNE1W5jjMjw2WoRTuqAgFiqMSBkgZ6ZI9aqtR8T+GtI1C00nVvEWmWV9fsFtLW5u0iluGJ2gIjEFiSQMAHk4qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv2tdQstJ/wCCh/jDVdSuUt7Sy8dW808r8CONHiZmPsACa/4b8/Y0/wCjh/Cn/f5//ia/4b8/Y0/6OH8Kf9/n/wDia/4Kt/tO/AD41/s7eHvCvwq+KmieJdWtfGlteTWllIzOkC2V4jSHIHAaRB9WFf8ABFT/AJNX8U/9lAvP/SCwr9rL9pnwn+yj8HtQ+JviK3/tC+aRbTRdJWTY+o3jglY92DsRVVndj0VDjLFVPwp+Af7Xn/BT+4l+Knxk+L1xoHw8N1NHZgRl7fejDMVnYI6IFU4QzOdxKDJkZTj4v/8ABHz4hfC7R3+In7NXxe1PWNe0SFp49MlgNhfSMA242txE+N5U4EZC55+c7gtf8Evf24vFvxwTUvgR8bNZe+8a+H7U3OlajcxCObULSMqkkcx43zxll5xudSzNlkdj8Y7Tx1ffCbxjafDDVP7O8XyaHdjQrryw/l3nlN5JwwYff2jODjOcHpWh/wDBJj9qL44tB41/aW+P6aZrd4rNJDePL4hvLYYAVHczJGDxjbHIyqAuCeg/aH/4Ji/HH9kzw3J8cvhZ8Vv+EjtfCv8ApV3faZDLo+p6aoGDcRqsr5RcncyyBlBB2ldxX/gmn+2JrP7U/wALNS0X4gSwv458DyQQajcxpsGo20qt5N0VGAJCY5VdV4BVW+USBR8Wfil4O+Cfw4174p+Pr9rTQvDtqZ7p403O5LBEjRcjLu7IigkAs4GR1rX/AItfth/8FPvi+3w/8KXjadozKZV0G1vpLbSdKtAwXz7thkzMN4BdlZiWIjRQQgH/AARE8aabpEesaF+0np6eJrXbLbQf8I9LBCkysCCLlbguuMZDCLOQOB1r9rH4iftKaF4a0v8AZQ/aYt7i+1f4f6tJe6Xrd3cPNNd2kqFFxMw/0iE4zHIfmX5kbldqf8Ehv+TNNM/7GDUf/Q1qv2jvhXqHxv8Agb4y+E2larb6bd+J9Na0hu7hC8cLFlO5gvJHHav2u/2WNf8A2Q/iTpvw18R+K9P8QXOpaHFqyXNlA8SIkk88IQh+cg25OenzCvhj/wAEeviT8Tvhr4T+JNh8Y/DVlbeLNDstWhtptPnZ4EuYElVGIOCQHAJHGRX7Uv8AwTN8c/ss/Ca4+LGv/E7QdctLe+gtDaWdnLHIWlJAbLHGBiv+CIf/ACSr4lf9jBa/+k5r/grh+xlDrOjy/tVfDTRmOp6cqx+MrS2iz9otgAsd8FHO6PASTAOUKudoicn/AIJb/thN8f8A4Wn4UeOtWafx94DtURpriUNLqumghIrn+8zxkrFITkkmJyxaUgf8FV/2uJfgd8LY/g74I1NIvGfj+1kWeWKcrNpmmZKSTADlWlIaJDx92Ug5QV/wSZ/YyPw98LJ+0t8SdGeLxR4ihKeGrS4idJNO09gQ05U4+ecH5eDiIKQ371gP+CkH7el1+y7odn8NvhdLby/EjxFatcJcyxrLHodru2rcMjArJK7BxGhBUbGZwQFV/hd/wS4+Of7StjF8XP2s/jdren6lrMa3Frp8gOpXwikQMpkkkcJb4G0CFVbaAB8mNtfHr9in9pH/AIJ6H/he/wABPi9qeo+HLK6jN7eWEJtbiyG/EQu7cs8VxDlghYgqS3zIoav2Hv2pLL9rH4HWPji5S3tvE+lzHTvEdlCuxI7tVVvMjUkkRSIysueh3LklCa/bo/bA0j9kH4UJ4gtLW01Pxn4gke28M6XdBvKmkTaZZptpDeVErqSFILM6LldxdfhB+z1+1x/wUx8Taj8RPGvxEuE8OWl00c2t61K72sEuA32extEwuVDglVEaKCctuIDeJf8AgkV8fvgnZP8AEL9nP9od9V8UaZDJKttb2UuhXUwXawjhlSeVXZiv3XKKSFyeeP2mfjf47+P3xKXxl8TtFTTPFmn6bBpGrxiEwGS4ttyPI8JA8lz/ABR9AwbAUEKPhP8A8kr8G/8AYv2P/pOlVX7H37Tur/so+OPFHjvwz4ZTXNf1vwrPomkQysfKhuprm2kWWRR80iqIG+RcFiVGQCSLH/glP+0t+0VaP8UP2l/2hP7K8X6pGJl0+5sG1aSDIx5cjLNFHBgKg2RB1A4424PiCz/aj/4Ji/HhNKsPE1xZq7LdQPayF9M8R2edp3xOCpONyMGXfG3KkfI58YfGr9o//gqP4+Pwt/Z2m1DwB8LNJ0+GXX9QupGiDzvGrslzJFy+JQ0cUKH5ghkbj/V/tE/8Ei/ib8Dvh5qPxP8Ah58T7fxynh2E3uoWaaQ2mXUMEYLSSw/vpRIUUbiMq2AcZIAP/BKL9t/xn8RdXl/Zs+L2u3GtX9vYyXnhrWb2VpbmVIyDLaSucmQhCZEZuQqOpJAQCv8Aglx/yfb8Mf8AuLf+mm8rUdR0/SNPudW1a/t7KxsoXnubm5lEUcEaAs7uzEBVCgkk8ADJr9pL/gof8f8A9qb4o/8ACkP2QzruneH9SmNlp6aTCYtU1tgGLzNJw9tDtBbAKbY1Z5GAyqeD/wDgid8SPE2l/wBufFT4/wCmaJr98xmubay0aTV8O5JbzJ3nh3Pz8xCsMk4Yjk/sqf8ABMj4pfsyftH6N4+X4zW+s+CbKG4e9gsXudMnvpREyQJNAjMjoGkMmDIR8mCCGwf2sv2mfCf7KPwe1D4m+Irf+0L5pFtNF0lZNj6jeOCVj3YOxFVWd2PRUOMsVU/Cn4B/tef8FP7iX4qfGT4vXGgfDw3U0dmBGXt96MMxWdgjogVThDM53EoMmRlOPi//AMEfPiF8LtHf4ifs1fF7U9Y17RIWnj0yWA2F9IwDbja3ET43lTgRkLnn5zuC1/wS9/bi8W/HBNS+BHxs1l77xr4ftTc6VqNzEI5tQtIyqSRzHjfPGWXnG51LM2WR2NVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV/wW9/5Kn8NP+xfuv/SgV+wn8SPh3pP7IPwr03VfH3hyzu7fQUWW3uNVhjkjbe3DKzAg+xr/AIWx8K/+imeFP/Bzb/8AxdaRrmieIbIajoGsWOp2hYqJ7O4WaMsOo3KSMj0r/gpjqFlpP/BQ/SNV1K5S3tLKHQpp5X4EcaOGZj7AAmvHh/az/wCCq/xW8QD4ZN9j+FvhzUGt9N+33b2el2qj/VyS4QvPcOoEhAR2jEgGFUjP7FXwP8Rfshfs1nwN8V/EPh+SfR7y+1O9v9NuJXtYbdvnLF5Y42G1VJPy4GOprx9+0t+1H/wUd+Ol78F/2ZNdvfCXgC0VnZ47o2RezSQI17eTKPNw5dcW6ZABUbWIZ6uf+CHmkjwmI7P9oO7PiZYyxll8PqLN35ITYJt6j7q79zd2287R+zv+0r+0L+wJ+0TZ/s3/ALTXiC7uvA81xFDdLeXBu4dOiuAPKvrOd8MLcNgugO0DzvkEoIFfto/t0fGX4pfHBP2Qv2ONTurS+OoHStR1vTpViub+8AxLDBMcG3ihIbfMpVi0bEMEXMnhn/gine6/4dbWPiv+0He/8JjqEPmT/YtM+1QW05Xo8ssge5AOMn91nBHH3q1Sb9rP/gk/8XNFsJ/FkviD4e6rIWit1d30zV7cODMiwuT9luV3BsqQQWHzOjMD8PvHXhv4n+BtB+Ing+9+16L4j0+G+sZSMN5ciBgGGTtcZ2svVWBB5Ff8Fef+TytS/wCxf07/ANAavH/wa0/9oD9jYfCW+CLLrng20WxmZA3kXiW8clvJz2WVEJ5BIyMjOa/4JSfGC9+C37V0nwx8TXVxY6f48hfRbu1cfJHqULFrZnA53BhNCMZANwc8civ+C1/xgTS/Afgj4GaddJ9p16+k1rU0UHclvbgxwKe215JJD3ObcdO//BKX4Fp8JP2W9O8W6lZtFrnxHmGs3DSAhhaEbbRACB8pjzKDznzzyRgCviz/AMkr8Zf9i/ff+k71+yv+0Bq/7NHxKuviX4Z0NNW1/wDsW60/SLeVS0Qup9qK8gB3Mqjc21eWIVcqGLDSf+CYH7VH7UVunxQ/ap/aDl0PXtT3XEOmXNg2qzWauR8jRLLDDa5AB8qLKqAoIUgqvx5/Y/8A2pP+CeuuW3xY8DeOru58PwXiJb+KfD0kls0LZ+RL23JIQOcrtJlibdsLEvtP/BPP9sdv2tPhVcL4rFvB488JNHba6kKiNLtXB8q7RB90OEYMo4Dq2AFKiv2j/j74O/Zn+Ees/FnxmHng09RFZWMb7ZNQu3yIoEODgsQSWwdqhmwQuK+Hfw6/bH/4Kq6vq/jDxx8UH8K/DO1vngWNVZrGGUFZBb29kjp57IGjJllbPT5ywxXxR/4IweKPCeinxR+z38ZbvUvEenYeDTtTt1sXnPcxXUb4jfHQMuCerrX/AATE/bi+IXj3xbf/ALMP7Qus3t74psYZDoF/qkZS9la2BFxZXJbDvMioZAzguRHNvYkKK/ay/wCTV/jL/wBk/wBd/wDSCav2Nfjx44+AXj/xJqnws8OXGt+OvF3hmTw54ZghtftPl3txeWjiQxZy5WOGTauGBcpuBXdWrf8ABJz9rz42z3PxI+N/xw8Pv4w1PLyQ6hc3F+8QxlYmkVQkYViyiOINGi428cC08T/tV/8ABM745y+En1mXT57aSO8udKW5kn0fxDasSqzBDtDo4R0Em1ZUIYfIykD4XfEPQPi38OfDfxM8LyO2l+JdNhvrZXGGjEiglG/2lOVPuDX/AAUB+HH7VvxQ8FeE/C/7LvjC70SS91h4PEP2TUf7Pf7O0RZJnuBiRYkaNlZIzlzKo2sBxpX/AARD8X6zDNqfjn9peyt9XuZmkn+yeG5NQEjMAWdpZLmJixYtnK88HOSQPGHgj9qn/glR8XNA1/TfFVve6NrLNLC9nPK2ma2keBLb3MDbSHCuvUZXeGR8g7fhB8TPDfx1+Evhn4oaBBnSvFelx3QtpsOYSwxLA/GGKOHjbsSpxkVq1jqH7AH7f0M7WlwmleC/Ey3FvlWZrjRbkEHYdx3MbSZ0zk4cEHkEVaXdpqFpDfWF1Fc21zGskM0Lh0kRhlWVhwQQQQRwc1+3h8a/+FCfss+N/GVld+TrN9ZnSdFIOGF5dfuldeCMxoZJsHg+UR3xX/BFn4FprfjXxX+0HrFmzW/hyH+xtGdgQv2uZQ9w44wWSEouM8C4OR0I/wCC3v8AyVP4af8AYv3X/pQK/YD/AOTNPhL/ANi/H/6G9f8ABav/AJOo8Lf9k/s//S+/rWdV/bK/4Kr+I9Vl+Gupv8PvgxpcwtFhvbqS2gnLKPNWUwqWvZtpJ2f6pAVHyFyz/H7/AIJYftA/s7eB7r4u6D4s0fxVYeG1+2X50fzre9sI0+Y3Kow5SMKGZlfco+bbtVmX/glJ+2347+Lmoan8AfjJ4luNe1jTrH7f4f1e9YyXVxChCzQTyk5lZdyOrNliPMyxwuLy7tLC0mvr66itra2jaSaaZwiRIoyzMx4AABJJ4FftOftt/Hj9s341W3wA/ZbvtY07w1d3z2OmQaXcNa3OuMozJcXEqt8kChHcLlVWMFpOeF8Nf8EQPHl94Y+2+LPj3omk6+Yyy6dZaFJe24baCqtcNNEw+bIJERwBkbulfFrxh+1R+xh4I8Y/softC6xL4s8HeO/Dc0Phm/ju2vEtnRwFa3kkKukeQFkhcfLuRlGPv/8ABEP/AJJV8Sv+xgtf/Sc1Vf8ABav/AJOo8Lf9k/s//S+/o2H7VH/BWnxDqet6P4k/4Vl8D9IvPsdtbSytOJ5VA3looypuZ9ku47ikSghFOdzH9qL/AIJn/GL9kfwzF8ZfAPxEfxbpOiTLJe6hptnJpl9pPIC3ARZZDsDHBdHyuQSNuSvw5/4Kn/HnUv2fF+C2j2Gp+KPjXqOpRaVoOu/Zlnkks5VKh9ijMt2jBUUspD+YHcsyMHT/AII5/Gz4maO/j34u/tG26eO9VX7RdWd1p0uphZHAYpLdmdTuDM4OxGXI+UkGvhJ8b/2hf+CbX7QN58P/ABa93NpWlXixeIfDP2sy2d9bybWFxbZIVZGjKvHKAp5CuMF0rQ9a0rxLomn+ItDvUu9O1S1jurO4QECaGRQ6OAcHBUg8+tf8Fq/+TqPC3/ZP7P8A9L7+v2Tv+TV/g1/2T/Qv/SCGv+Cgn7eum/sn+HoPBvgiK01X4la9bmWzgn+eHSrfO37TOoILEkMI06EqzMcLtb4U/shftlf8FD5Y/i78YPiZe6d4au5v9F1TXxI/mpxuaxsU2RiIbmwV8qMncASd2PEP/BD/AMX6bp/23wN+0fpmoavC26CLUPDkmnRhgCVPnR3EzKdwUcIcAk9QAf2dfh74j+FPwM8EfD3xhr93rWvaLo8MWqXtzfS3jPckbpVWWUl2jV2ZEzgBFUAKAFH7Uf7a3x3/AGo/juv7K37GWrXdhpv2iaxn1TT7kW82ryRgmeX7QcGC2jCPgoQXAJy25UGlf8ESIdT0Ca98c/tFXr+Lb1mmnuLTRfPto5WwW3eZKJJzu3EuTGTkcDBJ0Xx9+1R/wSq+N2keBfH2vXev/DvU5FuHsIpWubLUrEOY3ktBIR9nuUXBKArz5QfchUnw9r+i+LPD+meKfDeow6hpOs2cV5YXcJylxBKgeORfZlZSPrVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV+2XocPij9vrx74anuHhj1bxlFaPKgyyCQxIWAPcBs1/w5C+FX/RcPFf/AILrf/Gv+HIXwq/6Lh4r/wDBdb/41+3j/wAE6fBX7Ifwg0j4k+G/iNrfiC51LxJBpL219aRRIiSW1zKXBTnINuox0+Y1/wAEVP8Ak1fxT/2UC8/9ILCv+C1PxFuNb+O/hL4Z21872Hhjw6LuWEFgEu7qV92QRgnyoYDuGfvEdQRXwo/4KdfsRfCr4Y+FPhrpGseJVtfDGj22noYvD7qHMUaqz9eSzAsSeSWJPJNf8Pef2NP+g54q/wDBC/8A8VXwl+KPg5f+Cmvh34kfCC3uI/DfiX4jJHp8Tp9mZIdSl8iUbMHYoFzJhMfdAXI6ivjZ/wAFSv2Ufg7e3uhWPiW98c61aKwNt4aiW4gEvOFa5ZlixkclC5X0JGK+Kv8AwWc8MeOPCXiTwBY/s3Xd3pPiPR7nTppr/wATrC22eFo3zEls4IAY/wAYz7V/wRR1C9j/AGmPF+lR3Li0uPAtxNLCOjyR31mqMfcCSQf8CNf8FsviFdaL8H/AXw0tZ/LXxTrlxfXACHMkdlEo27s4A33UbYxklVIIwc/8Effhbo/g79lZfiLDYIureP8AVrie4uiwZpLe1le2hj4AwqulwwBycysc4IAr/guB4c0eTwf8LvFxtEGqQalfWInVQGaB443KsepAaMEZ6bmx941/wSG/5M00z/sYNR/9DWqr/gtX/wAnUeFv+yf2f/pff1+yd/yav8Gv+yf6F/6QQ1/wV5/5M01L/sYNO/8AQ2r/AIIh/wDJKviV/wBjBa/+k5rUdO0/WNPudJ1axt72xvYXgubW5iEkc8bgq6OjZDKykggjBBwa/aJ+E/jP/gmf+1zoPxQ+F9rcTeEpbp77w69y7GOWAjbc6dK+SSVRym4/MUdH5bOP2Uvg34u/4KP/ALVviP8AaF+MOnbfBelahHd6nAUZre5dQBbaXGSQSixIpcjOEUbgDMpqGGG3hS3t4kjjiUKiIu0KAMAADoMdq8c/GXwF4o/4KQap8Xfi7qNxc+DNE8dSSMyWz3Bks9PcraoIWAJV/IhBQgAB2yOtf8Pef2NP+g54q/8ABC//AMVXxi/4KffsVfE/4TeMfh1eah4lu4/Eeh3dgsUuhMBvkiZUIJJ2kMVIb+EgEciv+CJHi29s/jd8QPAkav8AZNX8KpqUpEmAJLS6jjTK45OLx+cjHPBzx/wWA+IV14t/a4uPBzT5tfBGh2djHEEKhZJ4/tTtyTklZ4xkYGEUYyCT8Bfhbo/wT+DPg74V6JYJaQeHdJht5UVg2+4I3TyEgAFnmaR2IABLkgAcVX/BWHw5o/h/9tHxNPpFoludY02wvrpUUKrTtAEZwB0LCME+rFieSa+E/wDySvwb/wBi/Y/+k6VVf8E2fDmj+Kv23vhZpmt2iXNvBfXd8iOoIE9rY3FxC/PdZYY2Huoqv+C4OlWE3w/+FmtyQbr201i/t4ZdxG2OWGJpFxnByYYzkjI28Yyc/wDBHnRNK039kJdSsbNIrrVvE19NeSgktK6CONSc9AERRgYHU4ySTNDDcQvbzxJLHIpV0dchgRggg9QR2r9ikDwF/wAFD/BmmeHjshsPFl9pkIl+ciB47i3YE9z5bEZ9ear/AIJcf8n2/DH/ALi3/ppvK/4KwfEm6+Hv7HOu6dp9xLBc+NNUtNCSWIkFUctPKuR2eG2lQ54IcjvX/BEv4R6T/wAI548+Ol7aRTag+oJ4f0+VlUtbpHEk9xtOMjf50GecHZVV/wAFqfiLca38d/CXwztr53sPDHh0XcsILAJd3Ur7sgjBPlQwHcM/eI6givhR/wAFOv2IvhV8MfCnw10jWPEq2vhjR7bT0MXh91DmKNVZ+vJZgWJPJLEnkmv+HvP7Gn/Qc8Vf+CF//iq+EvxR8HL/AMFNfDvxI+EFvcR+G/EvxGSPT4nT7MyQ6lL5Eo2YOxQLmTCY+6AuR1FVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV/wW9/5Kn8NP+xfuv8A0oFfAH/gkP8A8Lz+DPhL4uf8NC/2J/wlGnrd/wBn/wDCJfafs2WI2+Z9sTf067R9K/4cYf8AV0f/AJZH/wB3V+yH+zd/wyn8Gbb4R/8ACZ/8JR9n1C4u/wC0P7O+w7vNYHb5fmyYxjru59BX/BXn/k8rUv8AsX9O/wDQGr9kz4baL8Jf2bfh14H0S3hjW18P2s908YwJ7qaMS3EvPPzSu556Agdq/wCCrvxFuPAH7G3iCwsb5rW68Y6laaHHIhYMVdjNMgIGAGht5VO7AKsw6kA/8E2P2xf2Yf2U/hZ4jsPiTqGsReLfE2tCe4ex0hpwLOKJVgjLgjJDtcNjoPM+tf8AD3n9jT/oOeKv/BC//wAVX/BTf9pr4BftR+JfAfi74OPqU2qaTY3ljrM99p5tmaDfE9qoJyWAZ7o+273rwN8bNQ0L/gm1pnxpup3g1bR/hh5kUru0jS3sFoYYnJVcgyTIh6YXfycAmv8Agm5+0J8Df2cvin4p+KPxtv8AU/t8+kix0hrfT2vH3TSh7iQnPytiJF3dSJGHQmv+HvP7Gn/Qc8Vf+CF//iq/4KK/twfsuftR/Am18H+ALnW7nxTpWuQX2nyXektbiNAjxzDzCTwVflehKqf4RX/BHfxbe+JP2P8A+xrpXEfhXxVf6bbFpNwMbJDdnAwNo33b8c85OecD/grz/wAnlal/2L+nf+gNXwn/AOSV+Df+xfsf/SdK/wCCnXwr1v8AZ4/bDT4r+CpW0uLxdNF4m0i9ttqtbalFIDOQMcOJ1WbOCD5w5JyB8Ffihovxr+EvhP4raA2LPxNpcN2IyMGCQjEsR68pIHQ8kZU4JHNftD6he/t1/wDBR0+A9Gub1dHn16LwzZSx/vnttOs2YXVxGOm35bu4AwAA/Pc1pOk6boOlWWhaNZRWdhp1vHbWltCu1IYkUKiKOwCgAD2qviz/AMkr8Zf9i/ff+k71/wAEuPAegePP2zPBy+IrVLqDw/DdaxBA65VrmCMmBj6FJGSQH+9Gvaq+Nnw0034yfCLxf8LdViieHxNo9xZI0pwIpXQ+VKDtbBSQI4O04Kg4OMV/wSI8cT+Fv2x9L8ORvL5XjLQ9Q02RFUFcxxfawzZIxj7KQCAT82OjEj/gt58Rbi48X/Db4R298ywWOm3OuXdspYCRppPIhduNpKi3uAMHI3tnAIz+zf8A8FI/2L/gd8B/AvwoTV/EUc/h3RYIL42+gv5cl4y77mReejzvK2ep3c1/w95/Y0/6Dnir/wAEL/8AxVeLfjR4B1b/AIKMWHxz+CcN1/wjl/440rU4Ipovsklw8hgN6rDB2eZM1wCcHh885r9rL/k1f4y/9k/13/0gmr/gjf4W0LxH+1rfahrGnw3U/h7wfeahpzSIG8i4+0WsHmDI4PlzyjIwfmqv+C4HhzR5PB/wu8XG0QapBqV9YidVAZoHjjcqx6kBowRnpubH3jX/AASW1e91P9inw1ZXTIY9J1bUra2CrgiM3LTHPqd8z8+mPSvHHjvwX8NPDN54y+IHinTPD2h6eoNxf6jcrBEmThRljyzHAVRksSAASQK+IX/BZn9mrwzqA0/wL4Y8W+MkVmD3sVslhAQCQCnnESEnAODGvDDnOQP22f8AgojZftkfDrRPA8nwRTwnd6JrS6jFqZ8Rfb2MfkyxvCE+zRYDF42Lbj/qgMc5H/BJDUL29/Yu0G2url5Y7DWtShtlbpFGZzIVHtvkdvqxr/gtd8DhLb+Cv2h9G02Vnh3eH9dmTlVQlpbRio+78xuVLng7o1yPlB/4Jq/GBvjH+yF4Ou726afVPCat4d1BmA+9ahRDyOpNs9uSTySTnPU/8FsvjF9u8SeBPgNpl7mLS7eTXtViSbcpml3Q2wZR910RZ2552zg8A5b9iz4FR/s6/s1+DPhxc23layLMX+ulkUOb+4/eTKxXhvLLCEN3WJa/4Le/8lT+Gn/Yv3X/AKUCv2A/+TNPhL/2L8f/AKG9f8Fq/wDk6jwt/wBk/s//AEvv6/Yl8JaB4M/ZI+EemeHbBLWC88I6dqU6r1kubuBLidye5aSVz7DA6CryztNQtJrC+torm2uY2imhmQOkiMMMrKeCCCQQeDmv2KQPAX/BQ/wZpnh47IbDxZfaZCJfnIgeO4t2BPc+WxGfXmv+Ck/xH1D4Zfsa+PtS0a7e31LWYYNHtpFDcC6mSObkEFT5BmwfXHWv+CIvw00251L4k/F69hhkvbGO00XTmzlolk3y3BIK8btluAQ3ZwR0qv8AgsX4c0fVv2SY9bvbRHvtD8TWctlPtAaMyLJE6567WV+R0JVT/CK/4Ih/8kq+JX/YwWv/AKTmqr/gtX/ydR4W/wCyf2f/AKX39fsW+HNH8LfskfB/TNDtEtrebwbpt86KoAM91AtxM/HdpZZGPu1fG/SrDXfgt4/0PVYPPstQ8L6lb3MW4rvje1kVlypBGQSMgg1/wSX0TStX/bR8O3GpWaXEml6TqF1aFycRTCAxh8dCQsjgZzgkHqARX/BbTQdNtPjh4C8RW8Oy91Hwu8Fyw4DrDcuUOPX96wz6BR2r9hrV73XP2P8A4RXt+yGWLwrZ2y7V2jy4E8lPx2Rrk9zzX/Bav/k6jwt/2T+z/wDS+/r9k7/k1f4Nf9k/0L/0ghr4hJL+17/wUVv9C1K8uDp3inx8NKDmQq8WmQTCHK5+6wt4iQv97itO07T9I0+10nSbG3srGyhSC2tbeIRRwRoAqoiLgKoUAAAYAAFV+2x8Rbj4UfsofE/xtY3zWd5b6DLaWdyhYNDcXTLbROpUEhhJMhB6AgE4GTX/AATL/aV/Z7/Zb1rxv4z+MF5qUet6xa22n6S1jpjXLRWwZpLjLDoHdbfj/plz2x/w95/Y0/6Dnir/AMEL/wDxVf8ABS79sn9mn9qn4YeFNO+GE+sXXivw7rxlSW+0s24jsZYHE6hyTy0iWp299ue1f8ErfFt74q/Yp8GW9+rmXQbq/wBNWV5NxljS5d04wNoVJFjA54QHPOBVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVftSf8pH/ABN/2UC0/wDRkNVX/Bav/k1fwt/2UCz/APSC/r/gip/yav4p/wCygXn/AKQWFf8ABZfwbf6H+1JpfiuR/MsvEvhe2eFsAbJIJJYpI8biTgCNtxAH7zAztJr4Z/sf/sN/FL4d+GviP4e+AHg+XTvEul29/bmJZHCCWMMUySDlSSpBAIIIIBBA/wCGA/2NP+jefCn/AH4f/wCKrw9+xF+yd4T1/TPFPhv4FeGbDVtGvIrywu4YXD288Th45F+bqrKpHuK/4K+ftA+J/hJ8ENC+HXgzWJdM1H4jXlxb3lzA7JKdOt0T7RGjAfLvaeBWO4EoXXBDEr/wS3/YZ+DfiT4N2H7RHxV8MaT401HxTJdRaXpurWa3Nnp1vBPJbsWhkBSSV3ic7mBCrs27TuJ+KFp4f8J/BLxpDZW2n6NpOn+G9SkKRIlvBboIJHdsDCqOWYngdSa/4Iqf8nUeKf8Asn95/wCl9hX/AAXA8HXN98Ovhd8QE3/Z9F1q+0yTBXG+7hjlXIzuzixfoMdc4yM/8EivH2m+LP2OtJ8KW0sQvPBOsX+n3MQb58SzNdo5XqARclQehKMOoNV/wXA8R6PH4P8Ahd4RN4japNqV9fC3UgskCRxoXYdQC0gAz12tj7pr/gkN/wAmaaZ/2MGo/wDoa1Vf8Fq/+TqPC3/ZP7P/ANL7+v2Tv+TV/g1/2T/Qv/SCGv8Agrz/AMmaal/2MGnf+htX/BEP/klXxK/7GC1/9JzWr6rpug6Vea5rN9FZ2GnW8lxdXMzbUhiRSzux7AKCSfaviz4y8Y/8FTP209J+H3gm+lsfA2lSS22kzOjlbHTI2ButReNsfvZtqkKQv/LvExG0vX7Jfxo8Yf8ABOv9rLXfhF8XXe18MX18ml+KI9m9YB9611KMDnaEkVzjJaGVvlZwmLS7tNQtIb6wuorm2uY1khmhcOkiMMqysOCCCCCODmvCHgvwF4G/4KRz/Dz45aPp+teF/wDhPNQ0y9h1BvLgnS5aaO1nfaW2DdNBLgnC4wxXBI/4YD/Y0/6N58Kf9+H/APiq/wCGA/2NP+jefCn/AH4f/wCKr4X/ALMPwA+Cuvz+KfhV8K9E8NatdWbWU13ZRsrvAzo7RnJPBaND9VFf8FcPB1z4Z/bR13Wp9+zxboum6nBuKkbEgFmcYJIG6zb72DnPbBPwo8fab8Vfhj4U+JWkSxPa+JtHttQQRNuEZljVmT2KsSpB5BUg8g1X/BWHxHo/iD9tHxNBpF4lwdH02wsbpkIKrOsAdlBHUqJAp9GDA8g18J/+SV+Df+xfsf8A0nSqr/glx/yfb8Mf+4t/6abyq/4Le/8AJKvhp/2MF1/6Tiv+CQ3/ACZppn/Ywaj/AOhrVfsuf8pH/DH/AGUC7/8ARk1V/wAEuP8Ak+34Y/8AcW/9NN5X/BX7wTf+Lf2PbjV7GKWQeEfEljqs6xgcRkS2pJBOcA3anjJGMngEj/giR8TNHuvh18QPg3I6R6pputJr0SswDTwXEMcDlRnJEbW0e7jjzl5OeKr/AILL+Db/AEP9qTS/Fcj+ZZeJfC9s8LYA2SQSSxSR43EnAEbbiAP3mBnaTXwz/Y//AGG/il8O/DXxH8PfADwfLp3iXS7e/tzEsjhBLGGKZJBypJUggEEEEAggf8MB/saf9G8+FP8Avw//AMVXh79iL9k7wnr+meKfDfwK8M2GraNeRXlhdwwuHt54nDxyL83VWVSPcVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVf8ABb3/AJKn8NP+xfuv/SgV+wH/AMmafCX/ALF+P/0N6qv+CvP/ACeVqX/Yv6d/6A1fCf8A5JX4N/7F+x/9J0r/AILDeDb/AMUfsevrdm+IvCPiiw1S6GB80biWzA5YEfPeRngMeOmMkf8ABK34HfstfH74F63b/ET4U+HNf8Y+GNeeK8lu97ztaTRq9vIwGAqllnjA5/1LHPOB/wAMB/saf9G8+FP+/D//ABVf8MB/saf9G8+FP+/D/wDxVftJfB7Rz+xr8QvhF8ONJt9IsrTwjeJpNhbqBGhijaVIV3MAoZk27icLuyc4xX/BJv4e/AD4u/EPxv8ADj40eA9E8Sajc6XbX+hDUi2YxA7rcJGBgEsJ4mIznEWQCAxX/hgP9jT/AKN58Kf9+H/+Kr/hgP8AY0/6N58Kf9+H/wDiq+F/wg+GfwV8Pz+FfhV4N0/w1pN1eNeTWlkhVHnZERpDknkrGg/4CK/4K8/8nlal/wBi/p3/AKA1fCf/AJJX4N/7F+x/9J0r/gq38C0+Ln7Leo+LdNs2l1z4cTHWbdkBLG0A23aEAH5RHiUnjHkDkDIP7Av7XM/w5/YV+NmiX+p+VqPwus3vPDjSMGw2o744EVN4JRb7DPjGPtA5JNf8EU/gg2o+JvGf7QuqRv8AZ9IhHh/SAQCr3EoSa5f1DJGIFHYi4b0FVXxZ/wCSV+Mv+xfvv/Sd6/4JOeINF0L9s/w5DrGoxWjarpd/ZWZlOBLO0O5YwegLBGxnGTgDkgGviv4+034VfDHxX8StXliW18MaPdag4lbaHMUbMqD1LMAoA5JYAcmv+CQ/gG98V/thab4qiguDa+CtFvtQmmTiNWliNoiMcHJIuWIXIJ2E9FIr/gtz4Nv7H40/D34gyPmy1rwvLpcK4HElndPLIc7s8i+j6qBxwTyF/Zv/AGT/ANij4yfATwD8TLL4EeEbttd0G2lu3VXk2XaoEuYy2RuZJ0lRjgcqa/4YD/Y0/wCjefCn/fh//iq079hL9kHSdQtdV034A+F7e7spkmt5UhcNHIhDKw+bqCAa/ay/5NX+Mv8A2T/Xf/SCav8Agip/ydR4p/7J/ef+l9hVf8Fvf+SVfDT/ALGC6/8AScV/wSG/5M00z/sYNR/9DWv+Cofxm8Y/HD9rE/ALStb8rwz4SvLTS9PtGkdLeXUZkTzrmVdud6vMYQcMAkRK/fbd+z1+w7+z3+zx4T0rSNG8A6JrviGyjBu/E2q6ZFNfXMx5Z1dgxhTPSNCAABkscsf+C1t3aJ+zD4SsHuoluZvHltJHCXAd0SxvQ7BepALoCegLLnqK/wCCQ3/Jmmmf9jBqP/oa1+1P8GrT9oD9n3xv8KJrfzbrWNLkbTPnCbL+LEtq249B50ce7kZUsCQCa/4IzfFnUPBvxm8YfALXbZ7ePxPYm8gjnBSW3v7IkPFsOMFopJC2eQYFGOTXwTsLj9vn/gpZefES9jefwtp+rNrc3mRthdNsdkdnEQAuDIUt1YHB+ZycnINf8Fvf+Sp/DT/sX7r/ANKBX7Af/Jmnwl/7F+P/ANDev+C1f/J1Hhb/ALJ/Z/8Apff1+yd/yav8Gv8Asn+hf+kENV+y5/ykf8Mf9lAu/wD0ZNX/AAVB8DXvjn9i7xwNNLG50BrXVxGo4eOCdfNz6ARNI31UV/wRF+JOi28/xJ+Ed7cQxapd/ZNZ09CcNcRoHiuMdvkLQHHU7yegqv8AgsX4j0fSf2SY9DvbxFvtc8TWcVlBkbpDGskjtjrtVU5PQFlH8Qr/AIIh/wDJKviV/wBjBa/+k5qq/wCC1f8AydR4W/7J/Z/+l9/X7J3/ACav8Gv+yf6F/wCkENfFn/klfjL/ALF++/8ASd6/4JDf8nlab/2L+o/+gLVf8Fvf+Sp/DT/sX7r/ANKBX7Af/Jmnwl/7F+P/ANDev+C1f/J1Hhb/ALJ/Z/8Apff1+yd/yav8Gv8Asn+hf+kENNqMf7Lv/BRa41TxVBKLDwj8SJZbhpdu9rCS5YicAHGTbyiQDI5IBxziGaG4hSe3lSWORQyOjZDAjIII6giq/wCCh/g2/wDHf7FnxW0TTX2y22jpqjHAP7uxuIryQcsOqW7Drn0DHAP/AASU+Fv7Ofxul+IPgT4v/DrQfEXiHTltNS0o35ZpWtDvjn2KMYWNzBk5OTOvAxz/AMMB/saf9G8+FP8Avw//AMVX/DAf7Gn/AEbz4U/78P8A/FV8OPhh8P8A4Q+GY/Bvw08K2Ph7RIpnmSys1KxiRzlmwSeTgVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVftSf8pH/E3/ZQLT/0ZDVV/wAFq/8Ak1fwt/2UCz/9IL+v+CKn/Jq/in/soF5/6QWFft4fsg2H7XfwlTQ9NuLSw8Z+HJJLrw5qFyp2K7ACW3kI5WOUIgLYO1kRsHaQfgN+1/8AtQ/8E6dSufg78S/hrd3nh77Q9yvhzXd9pJbu3DyWV0FceW5AJwJIycsoDM7N/wAPv/hr9k3/APCifEv2ny8+X/a0Gzfjpu25xnjO33x2r9h7/goD4v8A2xfin4m8N3XwjTwt4b0jRReWt1DPJfbbgSxo0UtyVjTcwkLIgjBIRzn5ef8Agqb+y14u/aM+Dej698OdM/tPxP4DvJrmHTo42aa9tZ1VZ44QAcybo4XC8ZCMM5wD+xd/wUr179kvwVP8FfiL8Pb3xFoWm31xLYCO7+zXelu7ZltzG6lSnm+Y+DtZXkfJOQB4s/an/aR/4KZXbfAX9n74a3vgnwDdTRReLfEEl0bh4rVpBnzZR5SIpTcfsylnlCsAdu4D/gjHdW+l/tZ+IdP1GT7Nc3Xge+toYZQVZpVvLJ2THYhY3OD/AHTX7RPwM8LftIfB/wAQ/CDxbK9va61Cv2e9jjDyWNzGweGdAcZKuoyuRuUspIDGvCPiX9rH/glr8Y76PUvDCR2uqKkF9bXUck2k6/CgLI8My7cugdirKQ0ZZlZcF0Oq/wDBcHwofDcsmifALVR4ga3HlQ3Wtxm1jmIGcyLHvZFOT91S2APkzkftMfCr9pz4w/CrXP24v2k3fQ7q51LT9N0DQpbRoB/Z8glb91EzFoIUZoygclpC8rsc4Z/+CQ3/ACZppn/Ywaj/AOhrVV/wWr/5Oo8Lf9k/s/8A0vv6/ZO/5NX+DX/ZP9C/9IIa/wCCvP8AyZpqX/Ywad/6G1f8EQ/+SVfEr/sYLX/0nNf8Fdf2x4rW0k/ZS+Heq5uZvLm8ZXULspiTCvFZAjAO4FZJMEjGxT1cD/gmX+yOn7OHwYTxh4rs0Hjvx/DDeaiGQ7tPtMFre0GejAOXkwB877TuEStX/BWj9kb/AIWv8Oo/j94E0WWfxf4JtymqxW4y19pIyzMV7vAxZwRg7GlzuwgH/BIz9sAeMfDY/Ze8fapu1rw9btL4VmkXBubFBl7Yt3eLqo7x5HSOv+Cn/wCwF4k+MN+f2hPglov9o+KobeODxDokCEz6rHGAkdxDz80saBUKdWRV2/Mu1vgL/wAFb/i/8EdHtfhb8f8A4b3Hi1vD6rZC9muG07VbdIxtCXCujCd1UKuWEbnGXZ2JJ1j/AILheBYLIyeH/wBn/Xb273ACK812K1jx3O9YpDn22/iK/Yf/AGjfF/7UvwVk+KHjPwYnhq7/ALauLO3t4oJEhngRI3SWJpGJlU+YVLjA3I4A+XJ/4KNfsXXX7WHw4sNW8Cpbx/ELwgztpCzOsMeoQSsnnW0kh+6cKHRjwGBX5RIzD9l79vT46fsLXEnwR+KPw9vdT8L2F0zvoGqrJYahpDSMWkNuzggIzM0hjZSrMcqybmY+OP8AgsX4y+Itpa+Av2ZfgXqA8aa7Ibazn1GQXzxOwwvkW0S/vZOSQXbapUFlcEgftZ/s3fEr4BXvg/XPi/rr33jL4i6bca5rlvK/my2l29y+9ZJckSuytGzt2kZwCwAY/Cf/AJJX4N/7F+x/9J0qq/4Jcf8AJ9vwx/7i3/ppvKr/AILe/wDJKvhp/wBjBdf+k4r/AIJDf8maaZ/2MGo/+hrVfsuf8pH/AAx/2UC7/wDRk1V/wS4/5Pt+GP8A3Fv/AE03leMvCWgePvCWseCPFNgl7o+vWM1jfW79JIZUKOPY4Jwexwa+M/7MH7Tf/BO/4wr8WfhkdQufD+lXE0ui+K7K2F5HDbSF4xDfKY9iSGIgMGXYS2UOR8vgT/gt7p40CCH4nfAy4bW4lAmuNB1QLbTt3ZYpgWiHQbS8nru7V+zd/wAFT9Y/aS/aV8P/AAl0v4PxeH/C+qx3azX0t7JfXSNHG7QyMUREhR2EcZUh8NIBv5r9vD9kGw/a7+EqaHptxaWHjPw5JJdeHNQuVOxXYAS28hHKxyhEBbB2siNg7SD8Bv2v/wBqH/gnTqVz8HfiX8Nbu88PfaHuV8Oa7vtJLd24eSyugrjy3IBOBJGTllAZnZv+H3/w1+yb/wDhRPiX7T5efL/taDZvx03bc4zxnb747V+w9/wUB8X/ALYvxT8TeG7r4Rp4W8N6RoovLW6hnkvttwJY0aKW5KxpuYSFkQRgkI5z8vNVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV/wW9/5Kn8NP+xfuv8A0oFfsB/8mafCX/sX4/8A0N6qv+CvP/J5Wpf9i/p3/oDV8J/+SV+Df+xfsf8A0nSvHngfwz8S/Bet/D/xlpqahofiGxlsr+2Yld8UilWww5VhnIYYKkAgggGvGfwp/au/4Jc/Gmf4heBkuNQ8MhvIt/EH2BptM1W0dwVtr1VP7lyVAKFlbcpMbEYavDP/AAXB8KS6VH/wmXwC1W31JMLJ/ZmtxzQyYUZceZGjJlt3yfNgY+Y5ONB/4LG618Svin4T+Hvw1/Z9uFtda16ytbm4uL9r+8ktXljFx5NrCiASiPzdpMjLkKzDAK1X7Yn7Evxm/ZJ+My/tEfsxafqbeFre+bVbCbRbUzS+GZQctDLFht1tgkBiChQtHIMff+G3/Bbqe30WKy+LfwS+16pDGA+oaBqYijuG9fs8qkx8dxI2T2Arxj/wW7huLX7B8MP2fribUrhQsM+sazlI5SwwPIhjLSgjIwJEOSOtfDfxLqHjT4deFvGGraTcaXfa7otnf3Nhc25gltZZoUkeJ42JKMpYqVJJBGMnGa/4K8/8nlal/wBi/p3/AKA1fCf/AJJX4N/7F+x/9J0rUdPstW0+50rUrdLi0vIXhnhccSRsCrKfYgkV+0J8L/EX7Pvxx8f/AALtb24kgs9S+ypHGG/020ZkuLQsvJLGNoWxzgkgEjk/sf8AwLH7OX7O3g74WXSRDVrKz+0ay0Tbla/mPmTgNk7grNsDcZVAcDOKqviz/wAkr8Zf9i/ff+k71+yZ+zj4/wDj4PGWtfCDV7iz8ffDu1s9c0CKK4EBuJUn5VHx8kw2q0bZUblwSM7l+Hv/AAWV8V+B9Nn8F/tI/A7UJPGGiyfZbufT3/s+SR0+VxcWky5hlBA3BTtJY4SMAA/tGftt/tBf8FB9QsvgT8KPhdcWGhXd0k66DpZa/vL50ICyXM+1VWFGIfG1EQnc7ttVl/4J7fscx/smfCiQ+J1hl8feLPLuPEEkUiyx2oTd5NrGwAyEVzuIJDSMxBKha/bO/ZY0P9rb4MXfw+ubq307X7GYX3h7VJo2YWl2oIw+3kxSIWRhzjIfazIor4OftF/tXf8ABMvxhefDXx98PbiXw5e3Tzz+H9YVoYLphtV7mwu1DLkhVUuvmRnuu4Araf8ABb/4ayWkD3/wJ8Sw3LRqZo4dWgkRHx8wViqlgDkAlVz1wOlfsZ/8FHfF/wC118c7nwEvwZTw34ZtNBnupbq3upNSkhu1kiEXnT7IkiiZPPABjLM+wAgBs/tZf8mr/GX/ALJ/rv8A6QTV/wAEVP8Ak6jxT/2T+8/9L7Cq/wCC3v8AySr4af8AYwXX/pOK/wCCQ3/Jmmmf9jBqP/oa1/wVY/Za+Ifw/wDjjf8A7S3g7Tbufwt4nkhubi802N9+i38MUaM0pUfuxIyCRZM/fLjghc+Bf+C3EFl4GtrX4ifBS71PxZaW+yW503VUgtb+RUGJCrRloNzZyoEgXqM52j9rHVf2pv2sfhne/tbfFfwzceEPhzoLWtp4U0YK/k3AuZAGnjV2DMCpQtclAsnyInAIX/gj7qFle/sd29ta3KSy2HibUIblV6xSHy5Ap99kiN9GFV/wUG8A+JP2V/2z/EHjD4ftdaNaeMbe41jSbuNj8v26GSG+RWHA/evcfJ/CkiAjaRn/AII2/AtvBHwN1j406xZompfEK+8vT2YAsunWjPGpHGULzmckdGWOJvSq/wCC3v8AyVP4af8AYv3X/pQK/YD/AOTNPhL/ANi/H/6G9f8ABav/AJOo8Lf9k/s//S+/r9k7/k1f4Nf9k/0L/wBIIar9lz/lI/4Y/wCygXf/AKMmrV9J03XtKvNC1myivLDUbeS2uraZdyTROpV0YdwVJBHvX7RP7Iv7Qv7AXxei+NPwRm1W58JWF49zouvWGbmXT4sEtb6ggQADYGViQYpEPJBLIvhz/gt343Tw7b6Vrf7PWmat4kMIiF9Z+IJLaCWcqAG+zeRI2C3OwS85wCOtftT+E/2pvj58KdW/bL/aShuPC+mWl1aab4R8LvavboIZ3IkljhkYvEhCBt7gtMWUghETP/BEP/klXxK/7GC1/wDSc1Vf8Fq/+TqPC3/ZP7P/ANL7+v2Tv+TV/g1/2T/Qv/SCGviz/wAkr8Zf9i/ff+k71/wSG/5PK03/ALF/Uf8A0Bar/gt7/wAlT+Gn/Yv3X/pQK/YD/wCTNPhL/wBi/H/6G9f8Fq/+TqPC3/ZP7P8A9L7+v2Tv+TV/g1/2T/Qv/SCGv+Cmf/BP/wAQ/HyeL45/BawhufGen2Yt9Z0hpRG2rW8YPlvD8uDcKDtIZgHRVA+ZQr/s1/8ABS/44/smWUHwR+MXw+uPEOieG2W0i07UVbS9V0aMY/c5ZDvVQSVjlUNyF3qoUDxZ/wAFwfD6aaU8C/ALUJ9QfID6traRRReh2xxsX+mU+tfs9/EW4+PP7P8A4P8AiF4q0G3trvxRoqNq2nNbMsInwY7hBHIWbyi6vhWLfKRkt1P7Sn7JHx9/YH+NB+O37PUWqzeDbK4lvdL1jT7drr+xYXyHtb1W3/u1VtnmSZSRSMkOSo8B/wDBb+0GixW/xQ+BMratFGokutB1YCC4fncRDMu6ED5cAySZyeRjnxn/AMFtdQ1JF0f4P/s+PLql2qx2k+s6qZh9oYlVQWsCBpRnZjEqkkkYHBrSdQ/tbSrLVPsV1Z/bLeOb7NdxeXNDuUHZIuTtcZwR2INVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV38PPAF/qTaze+BvD9xqDyCRrqXTIXlZx0YuVzngc5qq1vw74f8AE1olh4j0PT9Vto5BIkN9apOiuAQGCuCAcEjPufWtE8O+H/DVo1h4c0PT9KtnkMjw2VqkCM5ABYqgAJwoGfYelV4h8MeGvFun/wBk+KvD2ma1YltxtdQtEuI92CudjgjO1mHToSO9af8Asvfsz6Tew6lpX7O3wxs7u3bdDPb+EbGOSNvVWWIEH6Vp+nafpNlDpulWNvZ2luu2G3t4hHHGPRVXAA9hVeMfgV8EfiJqn9ufED4OeB/E2pbQv2zWPDtrezYAAA3yxs2MADGewrRdD0Tw3pVvofhzR7HS9OtFK29nZW6wQwjOSERAFUZJOAO9ab4H8FaNqTazo/hDRLHUH3brq20+KKU7vvZdVB57881WveHfD/inTZNG8TaHp+r2E3+stL+1S4if6o4IP4ivCfwF+BfgLUhrPgX4L+BPDuoDGLvSfDlpZyjHT5441P61rGh6J4hsjpuv6PY6naFgxt7y3WaMsOh2sCMitI0PRPD1kNN0DR7HTbRWLC3s7dYYwx6naoAyaqtb8D+CvE10l94j8IaJqtzHGIkmvtPindUBJChnUkDJJx0yT61aWdpYWkNhYW0VtbW0axQwwoESJFGFVVHAAAAAHArWND0TxDZHTdf0ex1O0LBjb3lus0ZYdDtYEZFaH4Y8NeGIZLfw14e0zSY5mDSpY2iQB2AwCQgGTj1qb4b/AA7uNQfVp/AXhyW+kmM73L6VCZWlJ3FyxXJYnnPXPNVNDDcQvbzxJLHIpV0dchgRggg9QR2rT/hv8O9JvYdR0rwF4cs7u3bdDPb6VDHJG3qrKoIP0qvGfwr+GHxHRY/iF8OPC/ihVVVVdZ0eC9ACklQPNVuAWYgdiT614e/Z5+APhC6N94T+Bvw/0W5bG6bTvDNpbOdpyPmSMHg8j0NVXjT4Y/DX4kW8dp8RPh74Z8Uwxf6uLWtJgvlTBzwJVYDnmvBnwx+Gvw3t3tPh38PfDPheGXPmRaLpMFirc5OREqg881rng3wf4nlin8SeFNH1aWBSsT31jHOyA8kAuDge1Qww28KW9vEkccShURF2hQBgAAdBjtVVpfw88AaHfxapongbw/p97Bnyrm10yGGRMgqdrKoIyCQfYmq1zwx4a8TwxW/iTw9pmrRwMWiS+tEnCMRgkBwcHFaRoeieHrIaboGj2Om2isWFvZ26wxhj1O1QBk1Vr8PPAFjqSazY+BvD9vqEchkS6i0yFJVc9WDhcg8nnNVpfw88AaHfxapongbw/p97Bnyrm10yGGRMgqdrKoIyCQfYmqrWP2af2cfEV6dS1/8AZ/8Ahvqd2VCme88KWU0hA6Dc0ROB6V4e8JeFfCNobDwn4Z0nRbZsbodOso7ZDtGB8qADgcD0qvEPhjw14t0/+yfFXh7TNasS242uoWiXEe7BXOxwRnazDp0JHetP/Ze/Zn0m9h1LSv2dvhjZ3du26Ge38I2MckbeqssQIP0rT9O0/SbKHTdKsbeztLddsNvbxCOOMeiquAB7CqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqrXPBvg/xPLFP4k8KaPq0sClYnvrGOdkB5IBcHA9q0/TtP0myh03SrG3s7S3XbDb28QjjjHoqrgAewqq1jwF4F8Q3p1LX/Beg6ndlQpnvNNimkIHQbmUnA9Khhht4Ut7eJI44lCoiLtCgDAAA6DHaqmhhuIXt54kljkUq6OuQwIwQQeoI7Vqv7Mf7Nmu38uq63+z18NNQvZ8ebc3fhKylkfACjLNEScAAc9gK8L+B/BXge0+weCvCGiaBbeXHF5Ol6fFaJsjBCLtjUDCgkAdBk4qq8T/s/wDwG8b6gdW8Z/BLwDr98zMxudU8NWl1KWY7mO6SMnJJJPqea8IfCb4V/D0KPAHwz8KeGtm7b/Y+i29lt3fex5SLjPf1qtY8BeBfEN6dS1/wXoOp3ZUKZ7zTYppCB0G5lJwPSoYYbeFLe3iSOOJQqIi7QoAwAAOgx2r9qb9p/wCHn7KnwyuvHfjW+SS/uFeHQ9HjYGfU7oLwirnhFypdzwqnuzKrf8E6/gx4w/a6/au1b9pb4o2jX2jeHdWfWtRupI8Q3erSMZLe3jBP3YziQqMhFjjU4DrmqqaGG4he3niSWORSro65DAjBBB6gjtWh+DfB/hiWSfw14U0fSZZlCyvY2McDOByASgGR7GvG3wm+FfxLEQ+I3wz8KeKvIx5X9t6Lb33l7d2Nvmo2Mb3xjpub1NeDvh54A+HVg2lfD7wN4f8ADFk+N1to2mQ2UZwWIysSqOruenVie5qq1jQ9E8Q2R03X9HsdTtCwY295brNGWHQ7WBGRVp+yx+zDYXcF/Yfs4/C+2ubaRZIZofB9ijxOpyrKwiyCCAQR0NaXpGlaHYRaVommWmn2UGfKtrWBYY0ySTtVQAMkknjqSavLO01C0msL62iuba5jaKaGZA6SIwwysp4IIJBB4Oa0TwP4K8NXTX3hzwhomlXLxmJ5rLT4oHZCQSpZFBIyoOOmQPSq1zwx4a8TwxW/iTw9pmrRwMWiS+tEnCMRgkBwcHFaRoeieHrIaboGj2Om2isWFvZ26wxhj1O1QBk1NDDcQvbzxJLHIpV0dchgRggg9QR2qw/Z0/Z80rX18VaX8Cfh5Z62szTrqUHhezjuRK2dziVYw245OTnJyfWtV0jStdsJdK1vTLTULKbHm211Assb4IIyrAg4IB5HUA1onh3w/wCGrRrDw5oen6VbPIZHhsrVIEZyACxVAAThQM+w9KrXPBvg/wATyxT+JPCmj6tLApWJ76xjnZAeSAXBwPatP07T9JsodN0qxt7O0t12w29vEI44x6Kq4AHsKrXPBvg/xPLFP4k8KaPq0sClYnvrGOdkB5IBcHA9q0/TtP0myh03SrG3s7S3XbDb28QjjjHoqrgAewrW/A/grxNdJfeI/CGiarcxxiJJr7T4p3VASQoZ1JAyScdMk+tWlnaWFpDYWFtFbW1tGsUMMKBEiRRhVVRwAAAABwKq1+HngCx1JNZsfA3h+31COQyJdRaZCkquerBwuQeTzmq0/VtK1b7T/ZWp2l59juHtrn7PMsnkzJjdG+0na4yMqeRkVZ+DfB+navL4g07wpo9rqk7O0t9DYxpPIznLlpANxLHk5PPev+Czv7QPhi38B6D+zloWsRXPiK+1SPVtcghdX+xWkUbeTHKMEq8rypIoBDBYcn5ZF3f8EovhVrPwv/ZB0i8162ltrnxvqk/iKOCVslIJo4ooGAwMB4beOUDLcSA5Gdq1Wt+B/BXia6S+8R+ENE1W5jjESTX2nxTuqAkhQzqSBkk46ZJ9atLO0sLSGwsLaK2traNYoYYUCJEijCqqjgAAAADgVNDDcQvbzxJLHIpV0dchgRggg9QR2rSPAXgXw9ejUtA8F6Dpt2qlRPZ6bFDIAeo3KoOD6VWueDfB/ieWKfxJ4U0fVpYFKxPfWMc7IDyQC4OB7Vp+nafpNlDpulWNvZ2luu2G3t4hHHGPRVXAA9hWt+B/BXia6S+8R+ENE1W5jjESTX2nxTuqAkhQzqSBkk46ZJ9atLO0sLSGwsLaK2traNYoYYUCJEijCqqjgAAAADgVXjH4bfDr4iWv2Hx/4B8OeJrfaF8nWNKhvUwGDAbZVYY3AHHqAa8L/AX4F+B7r7d4K+C/gTQLkyRyGbS/DlpaPvjJKNujjBypJIPYk4qq8Q/s6fs+eLtQ/tbxV8Cfh5rV8V2/adQ8L2dzLjJbG54ycZZjj1JPevCHwx+Gvw9iWDwB8PfDPhqNI2iVNH0mCyCozb2UCJVwC3zEdCeetVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVqNvdXen3NrY37WVzNC6Q3KRq7QOQQHCsCpIODgjBxzXhL/AIJ7/tp/tYfGDUPEH7Res6rollZ3Ette+Jdf3SyzpFK6iOxtjtYxltzJxFEEYsCchW+DHwY+HnwB+HmmfDL4ZaGmnaPpq5JbDTXcxA3zzPgb5XwMt7AABVVRVVVVVVVVVVVVVVVVVVVVVfGn/gm9+2h4c+K/i/4vfs+fHZry68Xatc3901lrE/h/UCZneTY3lkRMqlygIkUcghFBIWX4Mf8ABajxKg0PVPE/i3TbW6ZRJdr42063MIBBzvtp/NAyADs5I4OQTX7Of/BHSSDX4PHf7VXjO312YzfaZfDukzSyJcyN8x+1XbBXY7idyxjk/wDLQg8www28KW9vEkccShURF2hQBgAAdBjtVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX/C2PhX/ANFM8Kf+Dm3/APi6/wCFsfCv/opnhT/wc2//AMXX/C2PhX/0Uzwp/wCDm3/+Lr/hbHwr/wCimeFP/Bzb/wDxdf8AC2PhX/0Uzwp/4Obf/wCLr/hbHwr/AOimeFP/AAc2/wD8XX/C2PhX/wBFM8Kf+Dm3/wDi6/4Wx8K/+imeFP8Awc2//wAXX/C2PhX/ANFM8Kf+Dm3/APi6/wCFsfCv/opnhT/wc2//AMXX/C2PhX/0Uzwp/wCDm3/+Lr/hbHwr/wCimeFP/Bzb/wDxdf8AC2PhX/0Uzwp/4Obf/wCLr/hbHwr/AOimeFP/AAc2/wD8XX/C2PhX/wBFM8Kf+Dm3/wDi6/4Wx8K/+imeFP8Awc2//wAXWl6tpWu2EWqaJqdpqFlNnyrm1mWWN8EqdrKSDggg+4NVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX/AATZ/YT+AH7T/wAC9c8ffFXTtauNW0/xZcaZC1lqTWyCBLS0lUFQDk7pn5+npX/Dob9jT/oB+Kv/AAev/hX/AA6G/Y0/6Afir/wev/hX/Dob9jT/AKAfir/wev8A4V/w6G/Y0/6Afir/AMHr/wCFf8Ohv2NP+gH4q/8AB6/+FfsVfBn9lj9oT9qrxp8JPE9pqreE9S+33XgSU6lJa3TxwTlooGAB3u1oWkbdgj7OfU1/w6G/Y0/6Afir/wAHr/4V/wAOhv2NP+gH4q/8Hr/4V/w6G/Y0/wCgH4q/8Hr/AOFf8Ohv2NP+gH4q/wDB6/8AhX/Dob9jT/oB+Kv/AAev/hX/AA6G/Y0/6Afir/wev/hX/Dob9jT/AKAfir/wev8A4V/w6G/Y0/6Afir/AMHr/wCFfHv/AIJa/sm/D34F/EXx94b0fxKmreGvCeqanYNNrTuiz29pJLGWXHzDci5Hev8Aglx/yYl8Mf8AuLf+na8qq8RatJoHh/U9dg0fUNXk06zluU0/To1e5uzGhYRQq7KpkfG1QzKCSMkDmvif/wAFIPBvwXt4bz4pfs1fHvw5Z3GBFeXvhuyFuzEkBPOF6U38H5N27GDjBFf8Pq/2V/8AoQvir/4KrD/5Nr4af8FEPDPxj0ptd+GP7MPx+8QaWjMg1C18N2QtmZThlWVr0IzA9VBJHcVp122oafa3z2dxaNcQpIbe5ULLCWAOxwCQGGcEAkZHU18VPiZf/DLT7G+0/wCFXjjxy17M0bW/hWzt7iW2AGd8gnniAU9Bgk57V41/4K3/AAN+G+vy+FfiD8E/jd4b1iBQz2Oq+HbO1mCn7rbHvASpxww4PYmof+C0v7LlxKlvb/D34sSyyMFRE0mwLMx4AAF7yT6V8KP2m7z4s6xpGn2/7Nvxo8LWGrrKyax4m0G1sbW3CI7AygXTSqHKbV/dnJdD907qqqqqqviP43u/h94ZfxHZeAvFHjCVJkjGl+HLaGe7YMeXCzSxLtHf5s+xr4kf8FT/AIT/AAf1ePQfil8A/jn4WvplZoItU8OWcAuFBwWiZrzbIoPG5SRnvX/D6v8AZX/6EL4q/wDgqsP/AJNr4Xftlx/F19GuPDX7MPx2tNI1q8ht4tZ1Tw7aWtpEkjKPtDM13vMCh9xdFcYDYyQRVfFz47aj8KNVGnW3wD+KvjmD+z/tkl/4R0m2vYY/mcGHElzHI0oEe7YqNkOmCSSBq3/BZX9mzQdSudG1z4X/ABg07ULOQxXFpd6JZQywuOqujXgKkehFeH/+CxP7O3i3WbTw34V+Evxm1nVr+TyrSw0/QbO4nuH/ALqRpeFmPHQA18JvjPqPxUuru2vPgh8S/AsdtbpMk/i3Tba0S43HGxBFcStvHUhguPrxVVVVWt65onhrR7zxB4j1ix0rS9Ohae8vr64WCC3jUZZ5JHIVFA5JJAFfFr/gsz8AfBupXOkfDHwZ4g8eyW25Re+YumWcrDGNjyK8pH3skxDoMbgcjwz/AMFxfCt3qscPjL9nbVdL0048y40zxJHfzL8wziKSCBT8u4/fHIA4ySP2e/2sfgZ+09pD3/wp8ZQ3V/bxmS80W8Agv7RA23dJCSTsyV+dSyfMBnPFVVfGn/gql8GPAXi2b4bfCHwlrvxc8WxTGFbfQSFs5JACWRLhRI0rDH/LKJ1xn5uMVpv/AAVfj8Ja9ZaP+0v+yx8Q/hLb6gw8i8vI5bgbDkeY0ctvbuVyCDsDng4B6V8PviL4H+K/hGw8d/DnxPY6/oOpKTbX1o+5H2kqwIOCrAggqwBBBBANVVftEf8ABQH9mj9m6e40PxT4z/tzxPb7lfw/oKi7uYnA+7MQRHAc4G2RlbDAhSMmv2SP2j4f2q/g/D8Wrfwe/hqOfUrmzSxe/wDtbYhYAOXEaYLA52449T1qqqvHfxB8DfDDw3ceL/iJ4u0nw5otpxLe6ldLBGGIOEBYjc5wQEGWY8AE1q3/AAVM+D+teJl8F/AT4afEP4u6x5xVl8PaOyxBADlwX/eEcZz5YXbuJYY5+Cvxn/aj+I3iCyh+JH7IKfDnw3OrNNql547t724jHluV22kduHJLqiEO0ZAk3Ddgiqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv+CKn/Jq/in/soF5/6QWFVVV/wUO+MUXwV/ZH8d65b3sUOra/ZnQdJVpmid57wGJ2iZeRJHAZ5l6cw9a/Z/8Aivf/AAM+Nfgr4tae93/xTOsQXN1FauEkuLXdtuYFJ4HmwNLGc9nNWl3aahaQ31hdRXNtcxrJDNC4dJEYZVlYcEEEEEcHNVVVVV+1l/yav8Zf+yf67/6QTV/wS4/5MS+GP/cW/wDTteVVV+0R4T0Tx18BfiH4S8Q2qXFhqXhm/jkVo1cxt5DlJE3AgOjBXVsHayqRyKrwd4T0TwJ4S0bwV4btUttL0KxhsrOJY1QLHEgVeEAUHA5wAM9hVV/wWz8J6Jc/AbwP45ktU/tfTvFy6dDOI13eRcWlxJIpbG7G62jOAQOuQTgj/glP4T0TxT+2j4Um1u1S5XQ7G+1G1ikjV0+0JAyxswYHlDJvUjBV0RgeKqqqqqqr/grD4T0TxB+xR4u1zUrVJLzwzfabe6dKY1LRSvew27YJBIBjncHaQTxzjIP7F/hPRPG/7V3wr8NeIrVLrTrjxNayXFvJGrpOIm83y3VwVZGKBWUjlSRxnNVVf8FrfCeiaV8evBnizT7VIL/XfDJjvzHGqidoJ3CSNgAs+yQIWYn5Y4wMBa/4Ie+E9Ev/ABZ8WPGt1apJqmjWOl2VnK0akxx3L3TS4JG4Em2iHBAIzkHjFVVVVft++OPjR+2R+0Kn7Ff7OtvLf6N4V2y+J5Ip/JtpLvKlnu5c4EFtlAF6mUuAruIwPht/wRJ+GtvosT/GD4u+JdQ1Z4wZY/DaQWcEL9wrTxytIB0yVTPXA6V+0J/wRevtC0S88R/s4ePb3XJbOFpP+Ef8RCMXNztXJWG5iVIy7EEBXRByAXGMnwH48+JHwG+JFn4w8H6lqHhnxZ4ZvGX54zHJDIhKyQzRsOQcMjxuMEZBFfsu/HbTf2lPgX4W+L9haxWc+sW7JqFjHLv+yXcTmOaP1A3oWXdyUZD3qv8Agrf+2hqVreT/ALKPw11SW0CxpL4xv7eXa0qyJuSwBB4QqyvL/eyiZx5it/wTU/ZQ0L9nv4GaX4y1fSYm8e+OrOO/1S8kUNJa20gDwWaH+FFXY7jqZGOSQqBfil8L/BPxn8A6x8NfiHosOqaHrduYbiF1G5D/AAyRsQdkiNhlccqwBFfs1/Hvx3/wTk/aj8Q/B/4g6ncXngmLWm0/xJaRkmILkCLU4YwGYN5RSTaPmeMhD8wXbaXdpqFpDfWF1Fc21zGskM0Lh0kRhlWVhwQQQQRwc1Xxr/Zx+Jfx18T3NtrH7S3i3wf8PmhESeHfB0EenXc5MW2Rp79t7OrMW/deXs28HJOR+2l+zV/wyl8d9S+Flnq91qukPZwahpF7dw+XLNbSgj5wAFJWRJU3JwdmflOVX/gkN/yZppn/AGMGo/8Aoa1VV+15+158PP2Rvh43ibxNImpeItRV49A8PxShZtQmA5Ynny4UyC8hHGQACzKp+Dvwy/aK/wCCr/xc1Hxt8WvHFxpHgfw7Mgu5bSBltbPfkrZafAzFfNKj5pGLlFKtIXLIr/B/4J/C34CeDoPAnwm8H2OgaTE2+RYVLS3Mp4Ms0rZeVyABuckgAKMKABVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX/BFT/k1fxT/ANlAvP8A0gsKqqr/AILZfGKTUPF3gT4DaZeyi20izk17VYkmVopJ5i0NsGUfMskccc7c4yt0D3zVf8EufjlJ8Z/2UNC0vUxEmrfDyQeGrhUZQZILeKM2sgQcqPIeOPJ+88Mh+lVVVVftZf8AJq/xl/7J/rv/AKQTV/wS4/5MS+GP/cW/9O15VVXxZ/5JX4y/7F++/wDSd6qqr/gtX/yav4W/7KBZ/wDpBf1/wSG/5PK03/sX9R/9AWqqqqqqq/4Kj/8AJiXxN/7hP/p2s6/YD/5PK+E3/YwJ/wCgPVVX/Bb3/kqfw0/7F+6/9KBX/BDH/mtn/cA/9yNVVVVV+x/+zjqfwC8J+JtV8dS6PffET4geIrzW/FOpaZI80U0kk8jQxJJJHG7IiOWwyjDyy44OTVf8FifhVovgX9pmw8b6DbQ28fjzQ47y/iRuWvYXaGSQLgBQ0awHgks/mMeTk/8ABGSLV4/2TNWfUmuDbyeNr1tPErkqIPstoDsGeF80S8DHzbj3zXj3xhpXw88C+I/H+uFv7O8M6TdanebFJbybeJpXwACSdqHoDXgwan+0N+0r4ei8aXKXF98RvG1pFqUjs4Ute3iLJyG3Bf3hAwcgYwahhht4Ut7eJI44lCoiLtCgDAAA6DHaq/4LL+BIPDf7Uml+MLKy8uLxd4Xtp7mb5B5t1BJJA3C4biFLYZbOc4BwML/wSr+MMvxV/ZJ0TR9T1BLjVPAd1JoMymQmQQRqr2xIJJCiGRI1PT90QPunFV/wWr/5Oo8Lf9k/s/8A0vv6/wCCQ3/Jmmmf9jBqP/oa1VfHL4w+E/gF8KPEfxa8Zz407w/ZtKIFbD3cx+WG3Tg/PJIUQE8Atk4AJGp638X/ANvD9pyyj1XUkufE/jvVltLNJZG+y6VbFiRGg5KwQR72wMsQrMdzsS3wZ+EPgz4EfDPQvhV4BsPsukaFb+WhYkvPIxLSzOSSS7uzMewLYGAABVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVf8EVP+TV/FP/AGUC8/8ASCwqqqv2tNL8XfG+fxd+3KtxDJ4P8V/ES68NaWsls1tOYLe1jNjLsPDBraIoxB4kgfPLcV/wRo+Lw8G/tE6z8J72fbZfEXRz9nRbfez39iHmiy/8CC3a9z6tsHpVVVVVftZf8mr/ABl/7J/rv/pBNX/BLj/kxL4Y/wDcW/8ATteVVV8Wf+SV+Mv+xfvv/Sd6qqr/AILV/wDJq/hb/soFn/6QX9f8Ehv+TytN/wCxf1H/ANAWq/bK/YX8BeH/AIB+L/iH8GPFHjjwjr/g7SZ9XCp4u1C8gu4bdGkmjkW5ncgmNWwUZcMF4IyD+wl8SPiJq/7X3wr03VfH3iO9tLjXkWW3uNVmkjkGx+GVmII9jVfti/sX+EPiB4D8cfEvwN4o8YeFvHlvp91qkNzZeJbw215PFHvEUlu8pjjRhGUzEE2l9xDY2n9ln4nfErUP2nfhBYX/AMQ/EtzbXPjzRIpoZtWndJEa+hDKylsEEEgg8HNVVV/wVH/5MS+Jv/cJ/wDTtZ1+wH/yeV8Jv+xgT/0B6qq/4Le/8lT+Gn/Yv3X/AKUCv+CGP/NbP+4B/wC5Gqqqqqqq/wCCxXjf/hOP2q9G+HWgpLeT+FPD9rZy20UO6Q3t07T7FC5LZiktcDGck8c5P7IXwQb9nb9nPwV8KryO3Gq6dY+drDwgEPfTsZZ/mH3wruYwx6qi9BgD/gp54sTwn+xL8QmjuUiutXWy022V4y4kMt3CJF44B8kSkE4GQO+Af+CeGgz+I/20/hRp9tBFK8OsPeFZcYC29vLOzDPcLESPcCqr/guVpVhDqvwb1uKDF7d2+tW80u4/NHE1kyLjOBgzSHIGTu5zgY/4IdeJ9t18W/Bc93KfMj0rULWDb8i7TcxzNnsTugHvt9qqv+C1f/J1Hhb/ALJ/Z/8Apff1/wAEhv8AkzTTP+xg1H/0Naqv+C1Pxzvb3xT4S/Z30i9dbDTbUa7rCJwJbiQvHboxznKRrI2MY/fKckjj/gmJ4t+CPwL17xr+0p8cPFtjpUHh+xj0jQLVh9ouru6udzytBboGlLLHEqb8BALhstjJX4lf8Fi9ftLD+3fhV+zD4gl8ODY413xLK9vG6MSuPLhR0XLFMN5xGSRt6GvhT/wW10C/1SPTvjT8HbjSbOabH9p+H777V5CEADfbyhSQDkllcnB4QkYbwV418J/EfwnpfjnwNr1prWg6zbrcWN9avuSZD+oIIIKkBlYEEAgiqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv+CKn/ACav4p/7KBef+kFhVVX/AAUO+MUXwV/ZH8d65b3sUOra/ZnQdJVpmid57wGJ2iZeRJHAZ5l6cw9a1P8AYxk1n/gl/pPwA1TSdQufF2heG28QabavAv2q21xjLem1Vc4yZJ5rTP8AckJ61Xwo+Ieq/CT4neFPifokXnXvhXWLbUooDK0S3HkyK5iZl5COAUb1ViO9eHtf0XxZ4f0zxT4b1GHUNJ1mzivLC7hOUuIJUDxyL7MrKR9aqqqq/ay/5NX+Mv8A2T/Xf/SCav8Aglx/yYl8Mf8AuLf+na8qqr4s/wDJK/GX/Yv33/pO9V/w70/b7/6SEeK//Ch1f/49X/DvT9vv/pIR4r/8KHV//j1f8O9P2+/+khHiv/wodX/+PV+3j+yx+078DvhBpHiz40/tS638StEu/EkFlb6TfarfXKQXLW1y6zhbiRlBCRSJkDP7w84Jr/gkN/yeVpv/AGL+o/8AoC1X7WX/ACav8Zf+yf67/wCkE1fsB/8AJ5Xwm/7GBP8A0B6r4s/8kr8Zf9i/ff8ApO9fsnf8nUfBr/soGhf+l8NVVV/wVH/5MS+Jv/cJ/wDTtZ1+zj4M8X/EP45eDfBPgDxlceFPEOr6ksGn61bzSQyWMpUkSK8RDqcAjKkHmv8Ah3p+33/0kI8V/wDhQ6v/APHq/wCHen7ff/SQjxX/AOFDq/8A8er/AId6ft9/9JCPFf8A4UOr/wDx6v2+/gV8bvgT4v8ACuk/G746an8Tb7VNNmnsbu+v7q6azjWQK0am4diATzgYFf8ABDH/AJrZ/wBwD/3I1VVVVVV8evjV4Q/Z6+FGv/FnxrN/oOiW5aK2WQJJeztxFBHn+N2IHsMk8A1/wTW/Zq8WfHT4r6t+3T8d7V52utWuLzw9FMjxi7v2fL3aowx5EW4pEASA68Y8oZr/AIK8/wDJmmpf9jBp3/obV/wS4/5Pt+GP/cW/9NN5VV/wXP8A+aJf9zB/7jq/4Ih/8lT+Jf8A2L9r/wClBqq/4LV/8nUeFv8Asn9n/wCl9/X/AASG/wCTNNM/7GDUf/Q1qq/b98Xjxx+2V8WdaEm/7N4gk0zPl7P+PJEtMY9vs+M9+vev+CXH7F/wK1b4FeGv2iPHHgux8UeK9durySybVV+0W+nRwXUluojgb93v3QF97KzKSNpXmpoYbiF7eeJJY5FKujrkMCMEEHqCO1f8FXf2M/DfwV8Q6b8cvhXoX9neGPFl5Jb6xp9uuLfT9QI3q0Sgfu45VEh2fdVkIGAyqP8Agk3+1D4i+GfxvsPgVress/gzx9M0UNvO7MljqXlkwyRDnaZSohYAYYtGSfkqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr/gip/yav4p/wCygXn/AKQWFVVfttD/AIaH/bQ+An7Ikf8ApOgaZcN4s8W20n761uIY95SGZU+ZH8m2uIwWIH+np61X7bHwci+BH7UXxA+HthZxW2kx6o19o8UELRxR2V0onhij3dREsghJGRuib0qv+CUXx3tPi3+y5pvgi/1SW58R/DWT+yb5J5Azm0Ys9k4AHEYhBgUHnNq3tVVVVX7WX/Jq/wAZf+yf67/6QTV/wS4/5MS+GP8A3Fv/AE7XlVVfFn/klfjL/sX77/0neqqq/wCC1f8Ayav4W/7KBZ/+kF/X/BIb/k8rTf8AsX9R/wDQFqv2sv8Ak1f4y/8AZP8AXf8A0gmr9gP/AJPK+E3/AGMCf+gPVfFn/klfjL/sX77/ANJ3r9k7/k6j4Nf9lA0L/wBL4aqqr/gqP/yYl8Tf+4T/AOnazr9gP/k8r4Tf9jAn/oD1VV/wW9/5Kn8NP+xfuv8A0oFf8EMf+a2f9wD/ANyNVVVVVWo6jp+kafc6tq1/b2VjZQvPc3NzKIo4I0BZ3dmICqFBJJ4AGTT6bqv/AAVf+NC6nO2q6L+zd8M7wJbo8TQTeKL/AObe6NtBjBTaG+YtHGy4CvMxTQ9E0fw1omn+G/D2m2+n6XpVrHaWNnbxhI7eCNQkcaKOFVVUAAdABVf8FI/Bsvjf9in4m6faWaT3Wm2MGqRMbcytCLW5inldcAlT5McoLDorNnjNf8E5/EP/AAjH7a/wq1LbC3napNZYlfaP9JtZrfIPr+94Hc4Heqr/AILfa21x8Tvhl4bOpI62Og3d2LMSAtCZp1QyFeoD/ZwMng+WcdDX/BDzwldW3hX4r+O5oM22o6hpum28phIw9vHPJKofoeLmElR0wpPUVVf8Fq/+TqPC3/ZP7P8A9L7+v+CQ3/Jmmmf9jBqP/oa1Vfti6Vf6P+1h8YrPUoPJmk8cavcKu4NmOa7kljbIJ6o6nHUZwcHIr/gn9DDB+xl8Jo4IkjU6CrFVXA3NI5Y8dySSfc1X/BQfwDbfEX9jb4o6TNAjy6Xor6xA52homsWW5LKWBwSkTqcYJVmUEbs18FdQvdJ+MngPVdNuXt7uy8TadNbypw0ciXMZVh7ggGqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv+CKn/Jq/in/ALKBef8ApBYVVV/wTwx8ev2if2gP21Ln99Za5rH/AAjPhmcfuGNjCInIlg7P9ni0v5ifveZ6mq/4Lc/CAEfD34+WEH/PXwzqkjT/AO/c2YSP/wADizf7g9Kr/gkH8YpPh7+1F/wr2+vZU0n4jaXLYmIzLHEt7bqZ7eV933jtS4hUDktcjrVVVVX7WX/Jq/xl/wCyf67/AOkE1f8ABLj/AJMS+GP/AHFv/TteVVV8Wf8AklfjL/sX77/0neqqq/4LV/8AJq/hb/soFn/6QX9f8Ehv+TytN/7F/Uf/AEBar9qyGW4/Zc+MVvbxPLLJ4B1xURFJZmNhMAAB1J9K/YHmhg/bI+ErzSpGp8RRKC7YG5lYAfUkgAepFV8Xpobf4TeNZ55Ujji8O37O7ttCgW7kkk9Biv2Tv+TqPg1/2UDQv/S+Gqqq/wCCo/8AyYl8Tf8AuE/+nazr9gP/AJPK+E3/AGMCf+gPVVX/AAW9/wCSp/DT/sX7r/0oFf8ABDH/AJrZ/wBwD/3I1VVVVWo6jp+kafc6tq1/b2VjZQvPc3NzKIo4I0BZ3dmICqFBJJ4AGTXiXxJ47/4Kb+O7r4dfDrUNT8M/s1eGL4R+IvEUSmG48XXEZDfZ7fcOIuhAIwoIkkBYxxjwV4K8J/DnwnpfgbwNoNpo2g6Lbrb2NjaptSFB+pJJJLElmYliSSTVeLfi34K8GeOfB/w11a9uJPEnjia4TSLC2t2lZo4I/MmmkI4jiRdoLMRksAAeceJ/Dmj+MfDWreEfEFol1pet2M1je27qCJYJUKSKQcggqxHPrWu+G/Ff7In7UEWkeIrfzdZ+GHiy2u1Ij2rdrbzJPDMqnoksYjkUH+FxmtO1Cy1bT7XVdNuUuLS8hSa3lTpJGwDKw9iCDVf8FbfHtt41/bI1fSbS4SaPwdotjo29NpXdta5dQVJyVe6ZTnBDKy4+XJ/YF+At7+zt+y/4U8E67Ypa+ItRV9X12MRbGS7ucN5bju8UQhhY88xHHGKqv+C1f/J1Hhb/ALJ/Z/8Apff1/wAEhv8AkzTTP+xg1H/0Naqv+CvfwQv/AIe/tKn4p2llt0P4jWcdwkkaBUS9t40hnjwAMEqsUpJyWaVjnrj/AII//Eyy8Zfslw+BQ9ul94B1q7snhRv3jQXEhu45WGTwzzzIDx/qT6Emv2ztd03w7+yV8YdQ1WfyoZfBeqWaN6y3Fu8EQ/GSVB+Nf8E8fhHf/GL9rrwBpcFpLLp/hzUE8QapIihlhgsmWVd4IPyPMIITx/y1HTqKqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv8Agip/yav4p/7KBef+kFhVV/wUO+MUXwV/ZH8d65b3sUOra/ZnQdJVpmid57wGJ2iZeRJHAZ5l6cw9a/Yn+DknwI/Zc+H3w9v7OW21aLS1vtXinhWOaO9uiZ5opNvUxNIYQTk7Yl9Kr9tj4OSfHf8AZc+IHw9sLOW51aTS2vtHighWSaS9tSJ4Yo933TK0QhJGDtlb1qvA/i/Wfh7408P+PvDckKat4a1S21OwaaPeiz28qyxll/iG5FyO9eCPF+i/ELwV4f8AH3huSV9J8S6XbanYNNH5bmCeJZYyy/wna65HY1VVVftZf8mr/GX/ALJ/rv8A6QTV/wAEuP8AkxL4Y/8AcW/9O15VVX7Y/wAefh58C/gR4x1Hxd4n0y01bUdBvbfRNLmnHn6jdSRNHEkcQ+dl3su5gMIuWJAFV8A/j98N/wBob4faT418BeLNJ1KW6s45NQsba4Bm0+cqPMhliJ3xlWOPmAyNrDIYE1X/AAWe+Pnw+1rwd4V+APhzxFp+reIbLxAdX1mGznWU6X5MEsKRTFThJHNy52H5gI8kKGXd/wAE6PjH4R+B/wC1h4U8WePNUt9M8P3sN1pt7qFxkJaedCwjkYjovmiNWY8KrMxIAzWi65oniXSrfXPDmsWOq6ddqWt7yyuFnhmAOCUdCVYZBHB7Vrmj2XiHRNQ0DUVdrTUrWW2nCNtYxyKVbB7HBPNeO/BnxE/ZV+PNx4f1BJdO8U+AdciurC5kh2iRoZFltrlFyQUcLHIOTwceteCf+C23wrm8IRyfEf4ReK7TxTHb4eHRHt7iynmCnkPLLHJEjEA42yFQcZfGWt9d+O3/AAUT8B+IviL470q7+GnwC0fS7uew8PWd45u/F1xDHuUzz4Qm2SWPJ2KoJ+Qb2UyJ4N8Vav4E8X6F448PSpHqnh3UrfUbF3BIWeCRZIyQCDjco6EV+zh+1d8Qv2ibzTLyH9lrxr4R8L3Nuz3fiHXbuG3hjfYWjFvE4WW6jfAXzUXapPNVVf8ABXD48/Dzw3+zdrfwQHifTLnxj4turFRo8U4kuba2huIrkzyIufLU+SiqXxu3Hbna2P2X/iRpHwg/aI+HfxJ8QNt0nQfEFrNqMmxnMVsXCzSBVBZisbMwUDJIA714P8b+DfiFoUPibwJ4q0nxDpNx/q73TbtLmInAONyEgHBGQeRkZqppobeF555UjjiUs7u20KAMkknoMV/wVs+O3gT4yfH7RtF+Heu2Ouad4M0X7Hc6lZTCWGW7klaSRI3UlXVF8oFgfvFx/DX/AAR0+PPw8+FXxF8ceAvH/ifTNA/4Ti1sDpl3qM4gilubVpwIPMbCKzrcttDEbioUZYgGGaG4hSe3lSWORQyOjZDAjIII6giv+Cnvxr+JHwM/ZeuNf+F11d6fq+t65a6TLq1oSJdLgdJZXmRgCFLGFYdxxjzsghttfsF/tR/Hzwx+1D4E0CHx14l8R6T4w8QW+nazpF/qE13FPHcN5T3BVixDxBzNvGD+7wx2lqqq8Q+INC8J6Ff+JvE2r2ml6Tpdu9xe3t3KIoreJBlndjwAAOtf8FB/+CjWoftGyy/Cj4O3d7pnw0gZTeXDqYLjX5RggyL1SBSBtjPLEB352qngr/gqJ+0/8OfCel+BvAyeCdG0HRbdbexsbXw+ipCg/wCBZJJJJYkszEsSSSa/4e8/tlf9Bzwp/wCCFP8A4qtV/wCCtn7aeo2EtnZ+MvD+mTSY23Vr4ft2kjwQTtEodOQMHKngnGDgj/gkb4d8e/GL4ifED9rH4s+IdQ8Sapb28fhzTdR1KTz3MjBJrgISf3flxi3UBQBidgMDINf8FPf2EtX/AGgNKg+Nvwj05Ljxz4dsTBqOlxofM1uzQlk8vBwZ4svhcZkVtucpGp/4Jpft3fDrU/hXo/7P3xj8W6f4W8WeDo107SpdYvDBHqlqrbYo1kkARJYwUhEJbcwVSoPzBf2nf2/PgJ+zl4Ou7+Lxjo/izxZLC39leHNJ1BLiWaXjaZmj3C3i+YEs+CQG2ByMV+wF+xd8Rf2iPi+n7W/7QmnXEfh7+1jrdrDqMDJJ4jvXbzklVDj/AEVXZX3HKSYCAMpcrVXl3aWFpNfX11FbW1tG0k00zhEiRRlmZjwAACSTwK/4Kz/FbwT8Vv2qkfwLrlpq9r4W8N22i3d3aSiWJrlJ7iaRVccNt+0KhwSAysM5BA/4I7/GH4fXv7PVx8J5/E2n2firRvEFw/8AZlxcrHNcwzhHjliU43gsJEIXJBTnAZc1X7WX7M3hP9q/4Pah8MfEVx9gvlkW70XVlj3vp14gIWTbkbkZWZHU9Vc4IYKw/Z7+JfxP/wCCZX7VF94b+Lnhu7TSbuMWXiOytzvW9syxMN9aMcLJtYFlJxlTLGdjElPDX7TH7PHjDwx/wmXh743eCbnRljaSW6fW4IRbhVDN5quytEVUglXCkA5IFf8ABRT9tQ/tM3tp+yV+zBZ3vi2xvr6KTVL/AEmJ5zrU0WZEtbZFBLwoyiVpMctEpGEQs/8AwT0/Yyj/AGTfhhNc+LI7S4+IPirZNrc0JWRbKNf9XZxyDqEyS5HDOTglVU1X/BTL9pj45ax+1B4r+G6+MfEHhzw34OuIbbTdIsr2W1R8wI/2mQJt8x5PMZlZgdqOApIyW/4JP/Hf4pfG/wCAWsJ8UtU1DXbnwrrh0+y12+kaSa7iaGOTypJGGZJIy3LlixWRAemWqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/wCCKn/Jq/in/soF5/6QWFVX7e8Vp+0p+2B8Cf2KZdXlttEkkm8ReJUjuBsnRY5XWLC/NHOLe1uwrHgfbEbGKqq/bY+DkXwI/ai+IHw9sLOK20mPVGvtHighaOKOyulE8MUe7qIlkEJIyN0TelV/wR++OUXxH/ZxuPhRfCU6t8MLz7OZHZn86yu3lmt2LN3VluItg4VIo/Wqqqr9rL/k1f4y/wDZP9d/9IJq/wCCXH/JiXwx/wC4t/6dryqrxF4f0XxZ4f1Pwr4j06HUNJ1mzlsr+0mGUuIJUKSRt7MrEH61/wAMB/saf9G8+FP+/D//ABVf8MB/saf9G8+FP+/D/wDxVf8ADAf7Gn/RvPhT/vw//wAVWnafZaTp9rpWm26W9pZQpDbwoMCONAFVR7AACvip8EPhN8b9PsdK+LHgTTPE9ppkzTWkN8hYQyMNpYYI5I4r/hgP9jT/AKN58Kf9+H/+Kr/hgP8AY0/6N58Kf9+H/wDiq8Bfshfsz/C7xZYeO/h98GvD+h6/pnmfY7+1iYSQ+ZG0b7SWPVHdT7Mar47fsufAv9pTTYLD4v8AgK01ieyjdLLUEke3u7Td/wA85oyrYz82xiUJAyprwH/wS7/Yw8B6zFry/DCXxDc28iyQR69qM15BGRnOYCRFKDkZWVXHAwBznV9B03WvD954YuodlhfWclnJHD8m2J0KELj7vynA9K+Cn7C/7LnwBu4NY8BfCzT5Nct5PMi1nVi2oXkTgnDRPLkQkA7cxBMgc5OSaqviP8MPh/8AF7wy/g34l+FbHxDokkyTPZXikxmRDlWwCORX/DAf7Gn/AEbz4U/78P8A/FV/wwH+xp/0bz4U/wC/D/8AxVeHv2Iv2TvCev6Z4p8N/ArwzYato15FeWF3DC4e3nicPHIvzdVZVI9xVfFD9mH4AfGvX7fxT8VfhXoniXVrWzWyhu76NmdIFd3WMYI4DSOfqxr/AIYD/Y0/6N58Kf8Afh//AIqv+GA/2NP+jefCn/fh/wD4qvhZ+zj8DfghqF9qvwm+Gmj+GLvUoVhu5bGNlaaNTuCnJPAPNeP/AIf+DPin4P1LwB8QfDtprvh/V41jvbC6UlJQrB1PBBBV1VgwIKsoIIIBr4G/sE/sxfs7+Lz4++HPgWVfEKbxaX+o30t29iroyOsO84TKMylsFipI3YJqqrxh4P8ADPj/AMM6j4N8ZaNb6tomrQmG9srgExzxkglWA7cCv+GA/wBjT/o3nwp/34f/AOKr/hgP9jT/AKN58Kf9+H/+Kr/hgP8AY0/6N58Kf9+H/wDiq/4YD/Y0/wCjefCn/fh//iq+HHww+H/wh8Mx+Dfhp4VsfD2iRTPMllZqVjEjnLNgk8nAqq+Nn7B37LHx/wBVm8SePfhdaR6/ccy6xpM8mn3MrFgS8vlEJM5A27pVdgDgEcY+FX/BN/8AY/8AhHrMPiTRfhVDrOrW0he3uvEF1JqIhPy42wyHycqV3K5QupJIYcYqq+Kn7OPwN+N+oWOq/Fj4aaP4nu9MhaG0lvo2ZoY2O4quCOCea/4YD/Y0/wCjefCn/fh//iq/4YD/AGNP+jefCn/fh/8A4qvAXgDwZ8LvCVh4E+H3h200PQNM8z7Hp9qpEcPmSNI+0Ek8u7sfdjVV8WPgr8KPjp4bbwl8WvAek+JtN+YxJeQ/vLZmGC8MqkSQuQMb42VscZqb/gkf+xdLqD3kfhLxHFC0xkW0TX5zGi5z5YJJfaBxyxbHfPNfB39nH4G/s/2U1l8Hvhpo/hs3KhLi5gjaW6nUYwslxKWldQRkKzEA5OMk1VftBfsPfs4/tM6za+J/id4MlbXrWNYRqum3j2k8sS5xHIVO2QDPBZSwwACBkH4YfCv4e/BjwZY/D74X+FrTQNA0/cYLO3LN8zHLO7uWeRyerOSx4yaqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/4Iqf8mr+Kf8AsoF5/wCkFhVV8I/i5+0n4y/a3+KH7aHwJ/Z/l+KGk61eXmgadPJb/Z0tYI/swt2wDuScWcNurevmvk8kV/w2B/wUe/6MJ/8AJuT/AOKr4r/8FPv2ufgZ/ZX/AAtz9kzSvC/9t+d/Z/2++mX7R5OzzNuM/d82PP8AvCv+H3nxT/6If4U/8GNx/hXi+x+O3/BWvxpP4+8BfDbwzomreANLtdM1Njrbok0E0txLbgLIOqstzkjruXPQV/w6G/bK/wCgJ4U/8Hqf/E1+zv8AAX9vT9gz4qaj4m8MfAH/AITz+3fD5s54LHUTLZruuEdWZkI/ep5DAAjhZjz81eNP+Cvn7Rnw61WbRPHf7MWlaFewXE1u0V/PdQ5kibbIqkjDbTgEqSOR6ivAH/Bav4Ratc6Rp/xD+GXiDQfNtx/aeoWkyXUMMwiJby4xh2QyDaM8gMCehr4N/tSfAX4/QR/8Ku+JOk6tem3hml00y+VdQeajOEaJsHeAj7gudu05qq/ay/5NX+Mv/ZP9d/8ASCav+CXH/JiXwx/7i3/p2vKqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv+CKn/ACav4p/7KBef+kFhVf8ABQ74xRfBX9kfx3rlvexQ6tr9mdB0lWmaJ3nvAYnaJl5EkcBnmXpzD1r/AIJ5fByL4K/sj+A9Dns4odW1+z/t7VmWFonee8AlQSq3IkjgMEDdOYar/guf/wA0S/7mD/3HVX/BDH/mtn/cA/8AcjVV8RvhT8N/i9oL+GPiZ4L0nxHpr4/cX9uJNuHR/lb7y5aNCcEZ2jNf8FCf+Cfw/ZRm0vx38Or3Vda8Da3cTxTtcQbm0WbfmGKSQfeR0YKrHBLRPnllFeFPFfiTwN4k03xh4P1u70jWtIuFuLG9tZCkkEi9GUj8iOhBIOQa/wCCb3/BQwfHqCy+BfxhvcfES0t2/szUiuF16GJCz7sfduERWZuzqpYYIIqv2sv+TV/jL/2T/Xf/AEgmr/glx/yYl8Mf+4t/6dryqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr/gip/wAmr+Kf+ygXn/pBYVX/AAUKkk/aK/a3+A37F1lNLLpMt5/b3ie3jvViSaA7ywyMsk8VnbXrLnr9qTHXNVX/AAXP/wCaJf8Acwf+46q/4IY/81s/7gH/ALkaqq+NXwn8N/HT4UeKPhJ4sT/iW+JtPe1eUIHa2k4aKdQeC8UqpIueNyDNXlnd2F3NYX9rLbXNtI0U0MyFHidThlZTyCCCCD0NeFPFfiTwN4k03xh4P1u70jWtIuFuLG9tZCkkEi9GUj8iOhBIOQa+GPji0+J3w18J/EmwsZbK28WaHZatDbTMGeBLmBJVRiOCQHAJHGRX7WX/ACav8Zf+yf67/wCkE1f8EuP+TEvhj/3Fv/TteVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVf8ABFT/AJNX8U/9lAvP/SCwq8u7SwtJr6+uora2to2kmmmcIkSKMszMeAAASSeBX/BPaOT9or9rf48/toXsUsuky3n9geF55LFYkmgOwK2Dlknis7ayVsdftT564qq/4Ln/APNEv+5g/wDcdVf8EMf+a2f9wD/3I1VVX7WX/J1Hxl/7KBrv/pfNVf8ABMK7u779hb4YTXl1LcSLHqcSvK5YhE1O7RFBPZVVVA6AAAcCv2sv+TV/jL/2T/Xf/SCav+CXH/JiXwx/7i3/AKdryqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr/gjf438FeGv2YfE1h4j8YaJpVy/jy7kSG+1CKB2Q2NiAwV2BIypGemQfSv+Ch/7f/we0T4H+LfhD8KfGun+JvGHiqO60C7SxxPFpsBPlXjSseMtG0kabSfmbcOE5/4JQftW/AD4M/AvxT4A+KvxH0/wvqzeLJNThXUAyJPBNaW0SlGAOSGtn3Dtlf71f8LY+Ff/AEUzwp/4Obf/AOLrS9W0rXbCLVNE1O01Cymz5VzazLLG+CVO1lJBwQQfcGv+C5//ADRL/uYP/cdVf8EMf+a2f9wD/wByNVVV+0jr+i+LP2iPil4p8N6jDqGk6z401i8sLuE5S4glvJXjkX2ZWUj61X7Avw81X4XfsdfC3wlrcu+9OjnUpUMLRND9umkvBEytyHjFwEbP8Smv2sv+TV/jL/2T/Xf/AEgmr/glx/yYl8Mf+4t/6dryqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr/glJ+zD8APjX+zt4h8U/FX4V6J4l1a18aXVlDd30bM6QLZWTrGMEcBpHP1Y1/wAMB/saf9G8+FP+/D//ABVf8MB/saf9G8+FP+/D/wDxVf8ADAf7Gn/RvPhT/vw//wAVXgLwB4M+F3hKw8CfD7w7aaHoGmeZ9j0+1UiOHzJGkfaCSeXd2Puxr/guf/zRL/uYP/cdVf8ABDH/AJrZ/wBwD/3I1VV/wUK/amsP2Y/gJqculav9m8c+LrebT/C0SZEiyfIs10CPu+QkocE8FzGvRjiv+CdH7Cev/H/4jWHxD+JvhKWP4YeH5BNdJfI8P9tTGLfDDEOC8eWjkdvulPlzl+K/ay/5NX+Mv/ZP9d/9IJq/4Jcf8mJfDH/uLf8Ap2vKqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv+CKn/Jq/in/soF5/6QWFVVV/wXP/AOaJf9zB/wC46q/4IneLPCvhf/hc3/CTeJtK0j7T/YPkfb72O383b/aG7bvIzjcucdMj1r/hbHwr/wCimeFP/Bzb/wDxdeNv2v8A9mH4darFonjL44+FNPvZ7cXEcX20S5jLMobMe4dUYYznivi5/wAFXvAdnbX+lfs2/DzxB8TtasLh4Z7mKwmSxt9sqqrlkBZ0kQTFCAPuAng1rP7KX/BRL9tnVT8SviHol0cW6TaYfEF0thClvcs8wS1iOdqDPIIBAKA5wMfs6f8ABIP4M/Djydd+Neof8LB1pd3+hbGg06P/AFq/czukyjxn5iNrpkcGrSztLC0hsLC2itra2jWKGGFAiRIowqqo4AAAAA4FV+1l/wAmr/GX/sn+u/8ApBNX/BLj/kxL4Y/9xb/07XlVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX/AARU/wCTV/FP/ZQLz/0gsKqqr/guf/zRL/uYP/cdVf8ABMH9kL4M/tW/8LK/4W5Y6rcf8Iv/AGT/AGf9hv2ttv2j7Z5m7AO7/UR49OfWv+HQ37Gn/QD8Vf8Ag9f/AArV/wDgj7+yHeaVeWelWvivT72e3kS2u/7YaX7PIVIWTYww204O08HGK+KH7AP7Vv7F39vfFP8AZP8Ainqup6Kunt/aaWP7jURbp5TNuhAKzYcyuNgyqIe7EH4Q/wDBVv8Aat+GlzFF4m8Q2njzTftEk09vrcX76TdFsVFnTDIisFcADru/vV+yV/wUv+D37S2p6Z4B1q2l8H+O72PbHp904a2vZVjjLiCb1Zmk2Rt8xEZ7kCqr9rL/AJNX+Mv/AGT/AF3/ANIJq/4Jcf8AJiXwx/7i3/p2vKqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqrxT8ZPhB4H1y18L+Nfit4P8P6zfbfsunaprttaXE+44XZFI4ZskgDA5NVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVf8EVP+TV/FP/AGUC8/8ASCwqqqv+C5//ADRL/uYP/cdVf8EMf+a2f9wD/wByNVVf8FBP2eLT9mz9prxB4Q0OGGHw5rsa65oEMZA+z2lw7jydo+6I5o5olB5KRoT96rS7u7C7gv7C6ltrm2kWSGaFyjxOpyrKw5BBAII6Gv8Agmt+1Nf/ALTPwEWLxlq/27xz4LuBp+uSvgSXUbZa1uiB03oGQnqzwSNwCKr9rL/k1f4y/wDZP9d/9IJq/wCCXH/JiXwx/wC4t/6dryqqqqv+Cvf7RWpfCr4M6L8KvCGu3eneIfHl4XnnsrnyprewtmRn+ZTvTzJGiUEY3KsoyRkV/wAE8vjjL8fP2VPCXiPVL97vXtBVtD1p3YsxubYKFdmwMs8DQSE+shGTjNVVVVVVVVVVVVVVVVVXxU+Cf7d2n+ELvUvg7+2jd6t4htI3kh0zWvBeiwx3uFyI1mS3xG5IwCwK5IyVGWH/AA9H/bt/6Lr/AOWzpP8A8i18Gbn/AILOfGvw1o/jPS/iroPh7QNetUu9Ov8AXdN0iMT27pvjl8uGzllVWG3G5ATuU42ndXirw5/wWm+HMa69afEzwT8RYbaN5JtO0qy09SwCnqstpauxH3gsbEkgDBztPgX/AILJftA+B/ESeHfjr8KNB1aGxmMOox21vNpOpRNuO7crs8e5cgbPLTIXBYElh+zr+0z8Jf2pPBJ8bfCrWpZo7aQRahpt7GIbzTpSMhJowzAZAJDKzI2DhiVbFVVVVVVVVVVV8UvDvjvxZ4D1Pw/8MviN/wAIJ4luvK+w+IP7Hi1T7FtlRpP9GmISTfGrx8n5d+4cgV+0vp3/AAVl+AugXvjjwl+0RY/ETwvpkLTX0+n+EdLtb20iXlpXtWt23IBgkxvIQNxYKqlq8C/8FX/2yfD/AIw0jWPGPxIt/FGh2t0rahpE+g6fbrdwdHQSQQJIjYJKsGGGAyGGVPwe+Lngf47fDjRfin8OtTa90PXIS8Jlj8uWF1YrJFIn8Lo6spGSMjIJBBNV8VPh58VvGmoWN18PP2gdY+HtvbQslxa2OgadqAunJyHLXcTspA4wpAr9sb9o7/gox+x78R7XwVrv7Stp4g0zVrP7XpGrQeE9Jtzcxg7XEkBgcxOrjGNzAgqQ3JC/s+fDT9vHx98NNI8dfFv9sm70LUNfs4b210nSfBWjym0hkTeomlktvmkKsuVVQEII3NnjTre6tdPtbW9v2vbmGFEmuXjVGncABnKqAqliCcAADPFVVVVVVVVVVVVVVVVVVVVftaaZ8R9K/aS+IsHxYTUP+Ekk8QXUs8l6CGmiaQmF0yAPKMWzy9oChNoUAAAfsIaZ8SNG/ZF+GmmfFlNQTxLBpbiWLUARPFb+fKbSNwQCpW2MC7W5GMHkGqqqqtR1HT9I0+51bVr+3srGyhee5ubmURRwRoCzu7MQFUKCSTwAMmv2g/8AgqN8ePjT4zufhr+xZ4d1i201FkjW/sdFa/1bUgGKGWKLY/kREMm35PNBIJZSdo8c/Fz/AIKG/D64/t34j+P/AI/eFhcTGZW1e+1XT4GYtnCpIVj2542gbe2McV+yh/wVt+KvgnxNZeGP2ktVbxf4RvJkjk1j7Ki3+lKRjzP3Sr9oQEgsGBkxkqxwEOk6rpuvaVZa5o19FeWGo28dxaXMLbkmidQyOp7gqQQfeq+M/wAZ/h58Afh5qXxN+JuuJp2j6auAAA013MQdkEKZG+V8HC+xJIVWYeFf2ov28P8AgoF471PRf2aJ7H4WfD7S5o47/V5RG8luuWdDJOyNI8zhR+7gUAfKHYKxc+LP2Dv2+PBWhHxP8MP2+vGvi7xFZxlk0bVb+8toZyRhlV5rqaNjgtjzEUZAOV6j9lj/AIKqeOtC+IB+Cn7aenxadeR3hsX8SyWYsJdOuVYqY763VQgQtgeYioI8ZZSpLpDNDcQpPbypLHIoZHRshgRkEEdQRVVVftxft3eDP2PvD1vpdtYReIviBrduZdJ0RpjGkMWWUXdywBKxB1YBRhpGVlBUBnT4kf8ABQ39sT4m6hPd6h8b9d0KCVgY7Pw3N/ZMcCgkhVaDbIRyRl3ZiMAkgDHgj9u39r/wBqg1XR/2hfGd9JuBaHXNSfVoWADDHl3XmKAQxzgA9DnKqR+wv/wU+0X9oHWbP4SfGex0/wAN+N7iNI9M1CCXZaa3MODGFb/UTngqmSrkkLtO1Gqv24P+Ch3gf9k2FfBnh2xt/FXxGu4RKmlGbbBpsbAbZLtl+YFgcrEuGYckopVj8OPgh/wUq/ay8P2nxC+LH7Ueq/CDRtSjjuNJ07Q4Gt7poShZHeG0kgKIwk6SytJx8yjamfjd4f8A+CkH7BNqPiToX7Q2sfFXwJBMo1C61hHv/s5dgALiC4eWSGJtiDzIpQAXIypf5v2If28/A/7XugSaReWtv4b+IWkQh9S0Lz96XEYwDc2pOC0RJ5Q5aMkBiwKu1ftL/GTT/wBn/wCBXjL4s3xRpdC012sYWYL594+I7ePns0rIDwSBk4OMV/w9H/bt/wCi6/8Als6T/wDItf8AD0f9u3/ouv8A5bOk/wDyLX/D0f8Abt/6Lr/5bOk//ItfD3/goj/wUG+Jfjzw78PPDnxw8zVPEuqW+nWinwxpOBJNIqKT/owAALZJJAABJIFVVVVV+3b+374Z/ZE0q38L+H9Mt/EXxE1m1aew06ZyLawiztWe62kOVLBtsakF9jfMgwT4w+N3/BUX4t2R8dtP8brfQ3WS5huvDGiX2mWMcK5yQ9pGisigEbmZvu5JJya+H/7f37Znwf1xwnxn8S6q9rcMt5pniuVtUVmBUPE4ud0kfKAEIyMvzYI3Nn9iz9szwZ+2F4Bn1bTrL+xfFmg+XHr+iM5cQM+7ZNC5A3xPsbH8SkFW/hZqvLu0sLSa+vrqK2traNpJppnCJEijLMzHgAAEkngV+11/wV58a6pr9/4G/ZYmt9G0OzmaJvFlxbLPdahjALW8UqlIYidwBdWdhtYeUcitV/bE/aw1m/l1K7/aU+Jcc02Ny2vii7tYxgADbHFIqLwBnAGTknkk18Dv+CrX7Unwq1S1i8beIE+I/h6Nh5+n6zsjuSuCD5d4iGRWORzIJBx93k5/Z8/aD+G/7THw3sfiV8NdV8+1n/d3tlMQtzptyAC8E6Ana4yOejAhlJUg1VV4h8QaF4T0K/8AE3ibV7TS9J0u3e4vb27lEUVvEgyzux4AAHWv2pv+Ch3xy+M3xf1XxJ8M/id4z8D+D7VjaaHpmj63caeXtkY7Z51hdQ00mdxznYCEBIXJ+GV3dah8NfCd9fXUtzc3Oh2Uk00zl3kdoELMzHkkkkknqTVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV/wAEVP8Ak1fxT/2UC8/9ILCqqq/4Ln/80S/7mD/3HVX/AAQx/wCa2f8AcA/9yNVVf8FyPD+i22v/AAf8U2+nQpq2oWesWV1dgfPLBbvaPDGfZGuZyPeRqr/gj98Q/wDhEP2u4PCUsd3NF468P32mokcu2OKaFVvFlkX+LCWsqDuDL6E1X7WX/Jq/xl/7J/rv/pBNX/BLj/kxL4Y/9xb/ANO15VVVVX7c/wAaNS/aU/aB8ZfFXTIpbjwnol5DoGkXKv5kUdunneRhgSB5xiuZwBx8zehJ/wCCNvxzTwR8ctY+C2sXrpp3xCsfM09GyVXUbRXkUDnCB4DOCerNHEvpVVVVVVVVVVVVVVVVVVXxa/5Kp4z/AOxgv/8A0oev2Tv+TV/g1/2T/Qv/AEghqv8Agsj+zZ4e1f4dWH7S3h3SobXX9BvIbDxBNDAR9us5sRwySlQRvilEcas2MrNtLHbGtfsefH/XP2b/AI++F/H+n6nLBpL3kdnr9sJikV1YSsFlEgHB2A+Yuejop7VVVVVVVVVVVVVX/BRHwJ4Z8CftofErwl4C8OW+laTbzWd1FYWURWOFptPt7icqo+6peSR9owqg4ACgAf8ABPz9tfVf2TfiIdJ8R3Et18OPFNxGuvWoVpGsnAKrewqOQ6ggOoB8xABgssZXQ9b0fxNomn+I/D2pW+o6XqtrHd2N5byB47iCRQ8ciMOCrKQQR1BFVX/Bc2GFZfgrOsSCV115WcL8xA/s8gE+g3Hj3PrVVVVVVVVVVVVVVVVVVVVVVqPhjw1rGoWmrat4e0y9vrBg1pc3NokktuwO4FGYEqQQDxjnmqqqqq/bU+HHxZ+PVl4N/Z78DxXuleEfF2pNN458SQTBBaabbbGa1TqfNnLDbldpMeG+Utj4RfBj4Z/AjwZa+AfhV4TtNC0i1ySkWXknckkySysS8jkk/MxOBgDAAA1vQ9E8TaPeeHvEej2Oq6XqELQXljfW6zwXEbDDJJG4KspHBBBBr/go5+zDof7MP7QUuj+Cbd4PCHimxXVdGt2dn+xAsyTW25uWCSIWXOSEkjBJIJP/AASq8S654k/Ys8IJrfzDSLy+sLKUoQZLdLhimSTg7SzICMDCAdQSa/4KvftJal8XP2gr34U6Nq0p8J/DmQ2Ito58xXGpDIuZmUHBdCTAM8r5b9N7Cv2FfhLpvwa/ZT+Hnhmzsvs97qOjwavqu6PY7Xl3Gs0ofk5Kbliz/djXp0Ff8FpfgLZaB4y8K/tEaDYtGvihTo+vukWEN3BGDbSM3d3gV0wcfLarjvX/AASB/ab1L4nfC/U/gR4y1SW71r4fRxPo0sxy0mkthEiz1PkONnOMJLEoGENVVajqFlpOn3Oq6lcpb2llC808r8CONAWZj7AAmv2Yv2RU/bw+IXiP9tT9pq2vW8N+I9WlPhrwyk5jF3bQnyozM6ncII1QRqqlWkeNmJC4Eng7wF4I+Hmjx+H/AAF4R0fw9p0Sqq2umWUdtHhRgZCAZOO55r9qf9gn4GftOeHtTmufDOn+HPG80btY+KNPtRHMs5O4G4VMC4Qnhg+Wwx2sp5rxt4P8XfCD4h6x4J8QpLpniPwnqklrOYZGUxTwvgSRuMEjKhlcdQVYdRX7IXxZuvjl+zP8PPifqVx5+o6to6R6jNgjzby3Zre4fBUY3TQyHGMDPBIwT8evi1o/wI+Dni34ua2iS2/hnTXuEheQILic4SGHJ6GSVo0Huwr9jzw7q/7Wf7c3he9+Kd5ca8+qatNrmvTXIMwlW2iadY3BPETPHFCF6KrAAYGKrxF4f0XxZ4f1Pwr4j06HUNJ1mzlsr+0mGUuIJUKSRt7MrEH614207x7+xN+1brGn+DdZurPXvhz4gk/sq9nj2vcWxGYWkQYBSe2kXevRllYcg18GvijoHxt+FXhb4r+GFePT/E+mxXkcLnLW7sMSQsehaNw6EjglTjiv28vA3jP9rL4i+B/2OfAOpPpum26jxX461domMWnWQZoLRMggSSyN9pKwHBJjjfhVZl+F/wDwTz/Y/wDhVo8GmWHwV0HxHcpD5c+oeKLZNWnuWwoMjCYGNGOwHEaIoy2ANxz/AMMm/sr/APRs/wAKf/CMsP8A41Wo/su/slaTp9zqupfs4fCe3tLKF5p5X8GWAEcaAszH9z0ABNf8E0PA2k/Hn9uPxd8dNP8ACdjo/hbwi15qunaba6bFBbWMt27w2VuI4wI4ykJmZSMndBnk5YVVVVa5f3WlaJqGqWOlXGqXNnayzQ2NuVEt06qSIkLEKGYgKMkDJGSK/ZK/ZCez8Q3v7Vn7Sehxav8AGnxleNqQS7kaePw1C4AhtoUbhZY4wiFjkxhAiEAMXr/gqJ+yL4J+K3wT8R/G7QPD9pZePvBVmdRl1C3hEb6lZQqPPiuCo/ebIVLozZKmMKCFZq/4JM+Jdc0L9tfwtpWkf8eviLS9TsNU+Qn/AEdbV7leQeP31tBycjt1IIr/AIKaeMPHt98N/Cn7Nfwog87xZ8bNYbSYlEmwiyhCvckn+FPniDt0EbPnrX7MH/BOn9nz9nPRLa6vfDFj408ZPCBe69rdqs+1yoDrbQsCkCZ3YIBkIYhnYYA8R+GPDXjHR5/D3i7w9pmuaXdKVnsdStEuYJQRgho3BVhgkYI71/wU0/4J6+FPgnof/C//AIG6a2n+FDdR2+v6CrPImmSSttS5hZiSIXkKoYyfkeRNvyttT/gk78ZNV+G37V2k+CjqMseg/EO3l03ULfexjaeOJ5bWQoAdziRTGDxtE7nIGc1Vft5/F7xn+1F8ZtL/AOCf37Pt67iW6jk8cavblpIbYKQzQy7RxFANjyENzIUi4ZSp+NXgjT/hn8ZPHnw30m7uLqx8KeJtR0m2nucebLHbXMkSs+0AbiqAnAAya+E//JK/Bv8A2L9j/wCk6VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVf8EVP+TV/FP8A2UC8/wDSCwqqqv8Aguf/AM0S/wC5g/8AcdVf8EMf+a2f9wD/ANyNVVf8FtvB+i33wL8A+Pp0lOraN4sbTLVhJhBBd2ksswK9yWsoMHthvWq/Yf8AF+s+B/2vfhDrWgyQrc3Piyy0xzLHvHkXsgtJwB6mGeQA9iQe1V+1l/yav8Zf+yf67/6QTV/wS4/5MS+GP/cW/wDTteVVVVf8FKfj0nwJ/ZY8Rtpt+kPiDxmraBpKiQrIpnRhPKm3kFIRIQ3AVzHzkgH9mn9g2213/gm74l8K6hoyR+O/izYjXbd7lVjeCWAmTSot5zsQgBmOAwF3Mp44rwd4q8U/Cf4g6N4y0QPYeIfCWrQ3lulzCQYbm3lDBZEODjcuGU9RkGvhn4+0L4q/Dvw18SvDMu/TPE2l2+oWwLBmjWWMNsbHAdSSrDsykHpVV4/+Jnw7+FWhP4l+JXjfRPDOmJkC51S9S2WRgM7E3EF3IHCLlj0ANeFf+Co/7P8A8SPjl4Y+CXwu0rxH4hbxHffZF117UWdpGxXKsqSkTMOGB3ImMDGc8fGv4/8Awj/Z48LHxb8WvGljolq6yfZLd33XN86AEpBCPmkblc4GBuBYgHNX/wDwWz+BMGvNZ6f8KPHF1o6zKv25mto5mj43uIfMIyPmwpcZwMlckD9n/wDaT+EP7TnhCTxj8JPEv9oQ2kixX9lcRGC6sJWXISaI8jIzhhlG2ttY7TivjR+1D8Av2e7Jrn4tfE/R9FudqtHpokNxfShvulLWINKVJGN+3aO5Ffsr/t5fDj9rrxz4o8JfDzwl4g0618NWcV0t9qwijN0rts4jjZ9vzZxljkYJwTgftHftzfs7/svSf2X8QfFUt/4h+Unw/oiJdX6Ky7laRC6rECNuPMZc7gRkZI8H/wDBan4BazrkOneL/hx4w8O2E3ynUUMN4sLZHLxowfZjJJUM3AAU5yPBXjXwn8R/Cel+OfA2vWmtaDrNutxY31q+5JkP6ggggqQGVgQQCCK+L/x70H4MS26694D+IeuQTWslzJd+G/Ct1qkFrGn3vOkhUrEcAnDEcc9Km/4K5/sc28r28+seLY5YmKuj6BIrKw4IIJ4PtXwZ+L3gz49fDXR/ix8Pp7qbQNc8/wCxvdW5gkPkzyQPlDyPnif8MHvXxW+KGmfCPw1F4o1bwr4t163kuhbtB4a0KfVbiLKO5leKEFliAjILngFlB+8KvP8Agrb+x/YXc1hf6n4wtrm2kaKaGbw9IjxOpwyspOQQQQQehr4E/Hb4f/tG/D+H4l/DS5vZ9EmupbZHvLYwSeZGQG+Uk8c9aqqqqr4tf8lU8Z/9jBf/APpQ9fsnf8mr/Br/ALJ/oX/pBDVf8FcPiFoHhP8AY71vwdqF5bjU/G+pWNlp1s02JX8i5iupZFQAllVYArHgAyLk5Kq3wz8Baz8U/iH4a+G/h6CWXUfEuqW9hAIot5QyuFL4JAwoJYkkAAEkgAkVU00NvC888qRxxKWd3baFAGSST0GK+K//AAUx/Zt+HmuXHg/whca78TvEkCvusPBtkL5I2VSfnnLLGVBADGMyFd33Tgirf/gs78I9N8Q/2F49+CXxA8O+XgT7lgkmgLAFS0LtGcEEHOc4IIBzXwM/aI+D/wC0h4Wfxb8IPGVvrVrbMsd7blGhubGRhkJNC4DIThsHG1tpKlgM18ZfjD4d+B3hBPGvinRPEup6cbxLWRdB0eXUZYS6sVeRIwSkeVC7zxudF6sK/wCHvP7Gn/Qc8Vf+CF//AIqvBn/BTb9nP4kak+jfDvQviX4p1CKPzJLTRfB1zfSog6sUiDED3xitOvE1LT7XUEt7iBbqFJViuYjFLGGAO10PKsM4IPIPFfFn4u6V8H9KtNY1fwd418QQ3UjoV8MeHbjVntwq7i8qwqTGmP4mwM1a/wDBWL9km/1JNGsbnxrcag8hiS0i8NyvKzjqoQHOeDxivCHiWDxl4Z07xRbaRrGlx6jCJUstYsJLK7gGcbZYZAGjbj7pGaqqqq1zRNH8Tf8ABcDUPDfiDTbfUdL1W1ltL6zuYw8dxBJ4RKSRup4KspIIPUGv+CgX7Eer/snePzrfhuF7r4beJrp/7CuTKZHspMbms5s/NuUZ2Mc70Gclg4H/AAS1/buPw21ey/Zs+LWsIvhPV7ojw7qt3O5/sq6kIAtmzkLBI3IPyhHZieHJWq/4Ln/80S/7mD/3HVVeO/iD4G+GHhu48X/ETxdpPhzRbTiW91K6WCMMQcICxG5zggIMsx4AJrxV/wAFavg5E1xL8K/hR8SPiBp1lM8c+r2Okm2ssKMkqzkyZwQxDxoQGBPpXw0/4LI/sy+MNSj0nxvoXivwS8sm1L27tkvLRV+UDe8LGRSSW/5ZlQFzu5xXh/xBoXi3QrDxN4Y1e01TSdUt0uLK9tJRLFcROMq6MOCCCOa+MH7XngD4Harq1h458B/E0WGiqjXOuWXg67uNMw0SyZW7VfKYKGw3PysrA8qa/wCHvP7Gn/Qc8Vf+CF//AIqtD1ey8Q6Jp+v6aztaalax3NuXXaxjkUMuR2OCOK+N37RnhH4BxQ3fjHwj471CwktzPNqOheGrjUbW0UMFxPNGCkRJIwGIzmv+HvP7Gn/Qc8Vf+CF//iq8EeL9F+IXgrw/4+8NySvpPiXS7bU7Bpo/LcwTxLLGWX+E7XXI7GvH/wATPh38KtCfxL8SvG+ieGdMTIFzql6lssjAZ2JuILuQOEXLHoAa8K/8FR/2f/iR8cvDHwS+F2leI/ELeI777IuuvaiztI2K5VlSUiZhwwO5ExgYznj9qr9tH4N/sj6Laz+P7y71HXtUjZ9M8P6YqvdXCjjzW3MFii3cb2POG2hypA/ZU/4KRfBL9qjxa3w50vS9Y8K+LHheay0/VjGyagiAs4gkRjudEUuyMqnaCV3BWK18d/2sPgF+zdZLN8V/iDY6ffSqGg0m3zc30wPQrAmWC/7bBV961X/gsX4AhsJfEuifs7fEu78MDHlavdRQ28bchDkqZEX95lPvnJA6E4HwW/4KmfsnfGPVIfD9x4j1PwPqk7BIYfFUEdrDM2B924jkeIDJIHmMhOOnIzVfHD9oP4Q/s5+Ez4w+LvjO00W0k3LZ25zLc3zrjKQQrl5CCy5IG1QwLFVyaf8A4LAeCtUjn1XwZ+zZ8UNZ0OPOzUDbxIDtUF9wjMiLtOR988AE4zgeHP8Agr5+xtrmj3upanq/i3w/c2sLyRafqWhM892yjiOM27Sxhmxgb3ReeSvWvgj8U9P+N/wm8MfFjStKuNNtPE9it3DaXDh5IQSRhivBPHaq+M3x8+EP7PvhtfFPxd8c6f4fs5t4tY5mLz3jKBuWGFQXlI3LnaDjcCcA5rWv+Cvvw2ZLnVfAvwD+J3iLw9ZrJ5ustZR28H7stvYENIAgUBssVIycquK+Cn/BV79lj4u6vZeGdav9Y8B6vesI4h4hgRbSSUkAKtzEzKo5+9KIxxjqRmGaG4hSe3lSWORQyOjZDAjIII6giry7tLC0mvr66itra2jaSaaZwiRIoyzMx4AABJJ4FfG//gqh+yj8IGfTNB8T3HxE1hWKm28LhZ7ePjIZrtisJU9P3TSEHqB1r4Y+OLT4nfDXwn8SbCxlsrbxZodlq0NtMwZ4EuYElVGI4JAcAkcZFVVVVVVf8FKfD/iT9q79trwh+zz8INO/tbV/Duhxw6pNGCU095pTLK8zfdWOKFoGJz96TYMsQD8FfhP4b+Bnwo8L/CTwkn/Et8M6elqkpQI1xJy0s7AcB5ZWeRscbnOKrxP4g1Dxd4l1bxVqxQ32s3017clM482Vy743EnGWPUk+pNfBbULLV/g34D1XTblLi0vPDOnTW8qdJI3to2Vh7EEGq/4KzeF7TxB+xP4p1a4MPmeGtU0zUIA8Ac72uktTtJPyHbct8wzxlejEj/gl146u/BH7afgiCO9lgsvEkd5pF8kYJ85Jbd3iQgHp9oit254G3Paq/a5/bz+EX7IM2maD4rsNV1/xPrFv9qtdH0wIrJb7ynnTSOQEQlJFXAYsyEYABYfsqftd/Cz9rzwhfeI/h4NQsb/RJIotY0jUY1WeyeRWKNlCyvG+yTa4POw5CkEDxP4b0bxl4a1bwh4itXudK1yxmsb6BJnhaSCZDHIoeMq6EqxG5SGGcgg814O8IeHPh/4S0bwN4P0xNO0PQLGGx060V2cQwRIERdzksxCqMsxLE8kkkmqr/grVYaVZ/tq+JbjT0QT32k6bNfFZCxMwtljBIJO0+XHEMDHGDjkk/wDBPvwTf/D79jP4V+HdSiljuJNHbUWSUAMovZ5btQQDx8s44PI6EA5Ff8FlfEGoaL+yRYabZFPJ17xlY2V3uzny1gubgYwRzvt4+uRjPGcEf8Ej9QsrL9tHQra6uUjlv9F1KG2VussggMhUe+yN2+imqr/gsZ4XtPD/AO17Hq1sYTJ4l8J2GoThIBGd6yT2o3EH5zttU+Y44wvRQa/4Iy+OrvxH+zFrPg2/vZZn8JeKJ4rSNgdsFrPFHMqqc95muWIGPvZ70tnaJdSXyW0S3M0aRSTBAHdELFVLdSAXcgdAWb1NVX/BUH4wJ8Jf2QvFNpaXSR6p44ZPDtkrAnctwGNzwO32ZJxk8Asuc5AP/BIz4PJ8Ov2V4PG97aomqfEPUpdSZ8neLSImC3Rh0x8ksox2n5PYVVVVVVf8FAPiTovwx/ZB+JmoavcRLJruh3GhWMTnBnnvY2gCqB1IV3f6Rk9BX/BIz9j/AFn4d6LfftI/EnRJbHWfElmLTwzZ3UW2SDT22vJdFScqZiEVMhWCIx5WYVWrfC3wHrnxE0L4saroX2nxV4Zs7iz0m+a5mAtYrgATBYg/lksAAWKlscAgVVftk2Glal+yZ8YbbWFRoE8E6rMgeQoPOjtZHhOQRz5ipgdzgEEHFf8ABMnwTf8Ajb9tP4fraxSm20KS51W9ljA/cxw28mwkE9GmaGPjn95mqr/goH+1zb/stfCVbXw0ftXxB8ZebY+G7RFLGA4Aku2A7R70Cr1aR0GCocr/AME8P2Pm/Zr+HE3jHx9avP8AFHxwq3HiC5uXE0tlGWLLaLJlsnJ3yMD88hGSwjQj9rL/AJOo+Mv/AGUDXf8A0vmr4T/8kr8G/wDYv2P/AKTpVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV/wRU/5NX8U/wDZQLz/ANILCqqq/wCC5/8AzRL/ALmD/wBx1V/wQx/5rZ/3AP8A3I1VV/wVn8H6L4m/Yn8Ua1qiStc+E9U0zU9NKSbQs73SWhLD+IeTdzDHqQe1V4e8Qaz4T1/TPFXhvUZbDVtGvIr2wu4Th7eeJw8ci+6sqke4qv2sv+TV/jL/ANk/13/0gmr/AIJcf8mJfDH/ALi3/p2vKqqqv2pmuP26/wDgo74Z/Z10d2vvBnw7m+yaxJbzsFSNGSXVH3f8s3yqWucf6yNeuRVpZ2lhaQ2FhbRW1tbRrFDDCgRIkUYVVUcAAAAAcCv+ConwKj+C/wC1RrWp6TbeVofj2P8At2xARVWOWRiLmMY9Jld+gwsijnGT/wAEYvjpH4p+EniD4Catdf8AEw8EXhv9KVnUb7C6cs6ovU+XceYzMf8An5Qdqqv20/8AgnP8Kbv4MfEL4y2/irx7rnjrw7otxqkOp+IPEb37PDbBpnhbzAfkESuqqCMYX3z+yv4y1L4fftDeBfGOi+F9Q8SalpmqB7HSLCLzJr64KMsUSjr8zsoOOQMkA9K8Jf8ABN/Wvjn4iPxn/bz8fan4t8WagyyQ+FtHv2t9M0aAsW+xq6/OVAIBELRgNvO+Ut5h/be/4Jd/Anw38DPEvxQ+AOgXXhfXPBunvqdxYPqk91bX1pAGe5ybh3dJBEGdSrYJjC7cvuH/AATB+MOqfCn9rnwppcWoJFo/jlm0HU4JZAqy+cM25GTgOLhYQD1IZlH368f+DoviB4P1LwdN4k8QaAmpRqh1LQNRawvrfDBt0M6coflwSOoJHev+CnP7C/wt/Zw8C+Gvit8MJ9dJ1fXn07WBqupG8aaaWKSaOUMw3bj5M24kkHI6Hr/wTr8T/HWz8T+OPAH7N+hpN458a6TBZ2+t3if6F4etllLTXs7YO0qCoQbXy7D5WOEb4df8EmP2erGKXXvjxqviX4seL9U3S6tqeqavcWsc1wzbmkQQyLKTjgmWWQsSTxkBf+Cm37BvgT9mSx8PfFX4OrdWnhPXNQfTL7S7u7a4Njdsjyw+S7kyNG0ccoIckqYhlm3/AC/8ETPjDql5a+PfgPqmoJLZ2Cxa9pEDyDfDvYQ3QUE5KE/ZjgDCszE8yVX/AAVc8LaF4Z/bP8TyaFp8VmNZ0+x1C7jiQIrXDwhZJAoAALlAzHqzMzHljX/BLj/kxL4Y/wDcW/8ATteVX/BZHwtoXh39rWx1DRtPhtp/EPg+z1DUWjQL59x9ouoPMOByfLgiGTz8tf8ABIb/AJM00z/sYNR/9DWqqqqq+LX/ACVTxn/2MF//AOlD1+zf+25+yd4T/Z2+FvhXxH8dvDNhq2jeC9Isr+0mmcPbzxWUSSRt8vVWUg+4r4t/8Fe/2VvAulXJ+Hd1rHxE1hFxb2tjZS2FsZMsCJJ7hFKqNo+ZI5PvrgH5ivxR+In7TP8AwUO+MJ1Sy8Kar4ivIf3Gl6Ho1u72mjQMeBk/LHu25eWQjcV5IVVVf+CeH/BOO5/Zt1QfGT4w3Nje+PpbV4NO0622zQ6Ikg2yP5v8c7JlCyYVVZ1BffkVX/BQ79pLx38Y/jdo/wCwL8C9eTTxrl9bab4q1FSQJZbjafsxkQkiCONg8wUbmIKHhWVv2e/2a/hP+zN4Hs/Bfwz8OW9u8cKrf6tLChvdTkHLSTygAsSSSF+6o4UAACv2sP2T/hv+1V8N9S8M+JdD0+LxNFZyDw94hMAFzptyATH+8A3mAuR5kXRgScBgrL8CPjV45/Zu+LOj/E7wbNLDqOi3G27sZJHiS9gyBLazAclGAwQejBWHKjHw98baL8SvAfh34heHJvM0zxLpdvqNoT1Ec0auoOcEEBsEEAggggV/wWH/AGZdH+HXjzQfj14I0BLHSfGjPZa4ltEEgi1KJAY5MDhWniDkhQAWt3Y/M5J/4IhePvDdpqXxN+GV0tpBruox2OqWb7QJbuCHzY5VLdxG0sZC/wDTZz61XifxHo/g/wANat4u8QXiWul6HYzX17O5AEUEKF5HJPAAVSefSv8Aglf8I7Lx/wCMviZ+254k8P29peeL/EWoQ+GrXyMLYxSzGa6lizkAFpFgVlOVEUy5Ic1VVVVX/Odr/P8A0KdfFj4WeC/jX8PNb+GPxA0pL/RNdtWgnXapeFiPlmiLAhJUbDo2DtZQa/aw/Ze8cfspfFS98A+KILi60mdnm0DWmg2RaragjDrgkK65CumcqcdmUn/gl7+3mPipotn+zt8YNc3+M9HtyNB1a9ucya5brk+S5b71xEuMHJMiLk/Mjs1f8Fz/APmiX/cwf+46q8Ra/ovhPw/qfinxHqMNhpOjWct5f3cxwlvBEheSRvZVUk/Svg6dc/4KuftV6v4u+KU11F8IPhrtm0/wykhhBSZ2FtFJtJHmSiFmmcNuITYhC7dugeHfD/hPRbTw34V0PT9G0nT4/KtLDT7VLeC3QfwpGgCqOegAFf8ABXj9kr4eaX8P7f8AaT8A+HrHQ9astSis/EENlbiGLUIZywSd1QbfOWXapc43rIdxJVBX/BJf9qXWvhj8Z7X4C+INQebwf8QZjFaxzTNt07UwhMUka8gCbaIWAAyxiYsAhBmhhuIXt54kljkUq6OuQwIwQQeoI7V+0X4c0fwf+0H8T/CPh60S10vQ/GWr2NjbooAighvJUjUAcABVA49K+E//ACSvwb/2L9j/AOk6VX7avhbQvBn7WHxT8OeGtPisdNtvEly9vawoEjgEhEhRFAAVAXIVQMAYA6V+yd/yav8ABr/sn+hf+kENftm/8E5PhPffBfx/8YYvGHjvWPHfhrw/PqcWreI/EUmoNPHaRmV4n3qeDGjhVXaFYrjaMiv2aPiXZfBv47eDvihf6XcanH4bvjdrY25xJdSLG4SJTg4LuVXODjOcHGKb/glp46/aS8Iav8a/2iPipquk/GTxlI2oRWMUYl0/SI2XMNnLG4MvyLhMJIBEMKA+3c37F3/BM39pL4ZftP8Ah/x/8U9J0nRfDfgq8a7S9g1SG6/tRwjrGsCIS6gsVLGVY8LnAJ4r9uv9rKy/ZI+DEvirT7e3v/F2uzGx8N2M/MbT4y88oBBMUSfMQPvMY0yu/cv/AAS//Z0t/wBp7x94v/ap/aEf/hNv7M1AWdnDrKm6W91Ixq8k0wf5WEMTRKkZDKPMBAXy0z9jtPsn2D7LD9m8vyvJ2DZsxjbt6YxxjpX/AAVb/Yb8LfD7So/2lPg34bt9I0p7pLbxXpNjEIre2eQhIbuKNQFjVnxHIAQC8kRC5ZzX/BKn9ufX9M8TaR+yv8VNWe90TUVMHhDUbh8yWFwBlbJmJ5hcBhH1KPtQZVh5dTeNtF/bN/4KmaTpvxUl87wXa+JLjS9M0y4+eE2thHM0ELI3a5mhDSKe87DpitO07T9I0+10nSbG3srGyhSC2tbeIRRwRoAqoiLgKoUAAAYAAFf8FQ/2KPA3xA+FHiD9oHwH4ZtNL8c+E7d9R1WWyiWFdYsk+a4acAfPLGgaQSH5iqFSWGzb+wH/AMmafCX/ALF+P/0N6/af/aA8N/sx/BbX/i14ih+1vp8Yh0zTw+xr+9kyIYQewLfMzAEqiuwB24P7BvwZ1n9vX4y+Kf2tv2onh8U6Tod4tjp2kXEW6ymvAokEHlEkC2t45EPkkEO0yli2JA9pZ2lhaQ2FhbRW1tbRrFDDCgRIkUYVVUcAAAAAcCv+CtPwa0L4VftSHXPCukRWGmeOtHi1eSK3gEUS3nmSRXAUA4JYxpKxwMtMfcn/AII1/Grxd8Qfg34p+GHiia7vbf4e3louk3txIz4tblZdtsCe0TQMQOyyqowFFfH79jX4WftL60mp/FLxD44ubCG1hhi0Kz8Qy2+mxyRtI32gW4+XzmEgVn9I0wAdxb47/DKT4M/Gfxt8KnnluI/C+uXVjbzzbd88CSEQysFJALx7GI7FsHBGK/ZO/wCTV/g1/wBk/wBC/wDSCGqqqqq+JHxc+F/wf0ePXfil4/0HwtYzMyQSapfJbmdgMlYlY7pGA52qCcc4r9pz/gsb4C0bR7jw1+zBp1x4h1i6hkQeI9TtHtrSxJGFeKCVRJM45bEiogwud+WUfsAf8FDPhV8HtV1DQPjP4G+yax4x1A3Gt/EaK4mvLq+mZmKtepIXfYpY8wkKMk+Xksx07UdP1jT7XVtJv7e9sb2FJ7a5t5RJHPG4DK6MpIZWUggjggg1X7QPw0uvg78bvHHwxurP7MPD2uXNtbR7iwNvvJgcEknDRGNhk5wwzzmv+CYvxm034ufsj+E9OW/83WfAsf8Awj+pwMfmiEA/0YgdShtjDhsY3K6jO01X/BWbxD/Yv7E/inTNsJ/t/VNMssu+0jbdJcZUdz/o/T0JPav+CeGlX+s/tp/Ciz02DzpY9Ye4ZdwXEcNvLLI2SR0RGOOpxgZJAqv+Co/7Cvxp+LfxSs/jj8GvDsvimC60uCx1bS4Jo1ubWWJmVJY0Yr5kbK6AhdzKVZj8p+X/AIJW/sa/FD9nWw8VfEb4wad/YuseKre2tbDRfPSSS3t0LO0k+wsFdiyBU3ZUK24ZIC1VXl3aWFpNfX11FbW1tG0k00zhEiRRlmZjwAACSTwK8LeBdQ/4KRf8FAfEniCziuG8GXutfbtUvTE0Bj0S22QQoSo+SaWCKKNe4ZixztY1aWdpYWkNhYW0VtbW0axQwwoESJFGFVVHAAAAAHAr/gqz8NLr4i/sc+IbzTrP7Vd+DdQtdejQMVKpEWimccgHbBPKxByMKcDdiv2Vvi3b/Ar9onwD8Vr6WWPT9C1iM6i0KlnFnKDDclVHLHyZJML3PHerS7tNQtIb6wuorm2uY1khmhcOkiMMqysOCCCCCODmq/4LGeIf7b/a9j0zbCP7A8J2FllHyTuknuMsOx/0jp6YPev+CI+lX8PwV+IWtyQYsrvxRHbwy7h80kVqjSLjORgTRnJGDu4zg4qq/wCCu3i/Wvi5+0n8Nf2ZfB9yk89lDAog81lU6nqU6xxpIoXqsaQMGG7AnYDHIPw98E6L8NfAfh34eeHIvL0zw1pdvp1opHPlwxqik5JJJC5JJJJJJJqqqqqq+MP/AAUx/ZC+EdtNHF8TLfxpqiLmPT/CgGoCTK5H+kKRbgdAf3mQT04OPif/AMFHvE3xo+Ofh34i/FH4UaJ4h8B+Fbjz9L8A3WozC1jlBBFxJKoAnuBjaGlieIKWHlfMSf2Xv2tvhL+1n4Ql8RfDrUJbfUdN2Jq+iXuEutPdlyMgEh4ydwWRflbaRwQVFVVf8FavjVp/wz/ZavPAUF8ia98R7pNOs4doZvssTpLdS4bjaFEcRPJBuFIx94f8Ehf2Wrr4Y/DO9+P/AIy0zyNf+IFukejRyxkSW2kAh1fkAj7Q4WTHIKRQMD8xAr4q/E3wh8Gfh1r/AMUfHeofY9D8OWbXN04xvfGAkcYYgNJI5WNFyNzuozzX7FPws8U/tkfHvVP2/PjppSHRrS+aDwDpMynZE1u+2OZVI2skB3BWB5uBI/BTmv2sv+TqPjL/ANlA13/0vmr4T/8AJK/Bv/Yv2P8A6TpVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV/wRU/5NX8U/9lAvP/SCwqqqv+C5/wDzRL/uYP8A3HVX/BDH/mtn/cA/9yNVVftNeFL/AMdfs5fFDwfpGi/2vqer+D9Ut9PsRGHae6a1kECqDxv83ZtPZsHjFVX7J+rf27+y78ItVbU/7Qmn8D6P9ouTN5rPMLOJZdzZJLhwwbPO4HPOa/ay/wCTV/jL/wBk/wBd/wDSCav+CXH/ACYl8Mf+4t/6dryqqq/ap+N9l+zr8AvGHxYnlt/tmk2JTSoZzxcX0pEdumByw8xlJA/hVicAEj/gk7rHwI+G2g+Mfjl8YfjX8P8ARvG/jC8aztodZ8T2dreQ2aP5kztE8isnnT4JDDkQIRgHJ/4ax/ZX/wCjmPhT/wCFnYf/AB2v+CqXiP8AZz+PX7Ptvrvgb46fDTW/F3gXUBeWNlY+KrGa5urabbHcxRIshZjxFLtHJEHGTgH9hH47/wDDPH7T3g/xvf6p9i0C/uP7J8QO8myP7DckI7yHBOyJxFPgd4RVV+1l/wAmr/GX/sn+u/8ApBNX7Af/ACeV8Jv+xgT/ANAeq+L0MNx8JvGtvPEkscnh2/V0dchgbdwQQeoI7V+yd/ydR8Gv+ygaF/6Xw1X/AAWr/wCTV/C3/ZQLP/0gv6/4Ih/8lT+Jf/Yv2v8A6UGq/wCCvkMMv7G1+8kSO0XiLT2jLLko25xkehwSPoT61/wRU/5Oo8U/9k/vP/S+wqv+CvP/ACeVqX/Yv6d/6A1f8EuP+TEvhj/3Fv8A07XlV/wWr/5Oo8Lf9k/s/wD0vv6/4JDf8maaZ/2MGo/+hrVVVVVfFr/kqnjP/sYL/wD9KHr9nr/gnD+xf46+APwz8beKfgz9u1rxB4P0nUNRuf8AhIdTi8+5ms4pJX2JchV3OzHCgAZwABX7Zf7Knib9kn4v3XgbUGuL7w9qKtd+G9XkQYvbQtjaxACiaM/LIoxztbAV0z/wSK/bJt44o/2T/iLqcUIXzJ/Bl3MSPMZmZ5rFmJxkljJEMDP7xc5Mamq1HULLSdPudV1K5S3tLKF5p5X4EcaAszH2ABNfs33vxy+Mn7Y2m6/8HPFOj+HviZ4p1bVdUstU1eJZra3nkt7mecuGhmBJj80KTG2GZT8p+Yf8Ka/4LK/9HZfCn/wWw/8Ayor/AIU1/wAFlf8Ao7L4U/8Agth/+VFav/wRo/a313Vb3W9V+IvwpnvdQuJLi5l/tK+XfI7FmbC2IAySTgACv2QfhJ43+BH7OPgz4SfEXWNM1TX/AA5Dcw3F1ps0ktu0bXU0kKo0iRsQsLxryo5U9Ryf2s/ghbftFfs9+MvhSY0N/qViZtJkIXMV9CRLAQzfdBdFRjkfI7DOCa/Zv+L2ufs1ftBeFPiatrLDL4b1TytVs5oyrtbPmG6hZSMq/lPIBkEq4Bxla0nVdN17SrLXNGvoryw1G3juLS5hbck0TqGR1PcFSCD71/wVZ+KOpW3wq8Nfs0eBbjzvGfxk1y202C0jfa5tFmTdlgfk8ydreIA8OrSj+E18FfhdovwU+EvhP4U6AM2fhnS4bMSE5M0gGZZT7vIXc8AZY4AHAqqqqr/nO1/n/oU6r9rD9l7wP+1d8Kr3wD4ogt7bVoFebQNaaDfLpV0QMOuCCyNgK6Zwwx3VSPHHgz4ofs0/F+88L6y974d8Y+DdSBhu7V3iZZI2zFcQOQrFGAV0fAypB71+wF+2no/7Wfw3Ww12aGz+Inhi3ii16zLxqb0ABft0KLj927feUKBG7behQt/wXP8A+aJf9zB/7jqr/gq/47l8FfsZeI9PtbpIJ/FepWWjxsWIZg0nnyKvqWjt5AQf4S1f8E6PAP7b3ivwf4x1D9kz4weDPBWnRalBFrEet2ccstzMIyYypayuDtClhjcvJPHOa/4U1/wWV/6Oy+FP/gth/wDlRXxS/ZJ/4Ks/GrwHqfwy+Jn7Svwp1nw3rPlfbbH7OLfzfKlSaP8AeQ6WjriSNG4YZxg5BIr4b/8ABH79qrwJ8RPC3jhPiF8MY28O61Z6iHt9UvjKpgmSTKA2YG75eMkDOKr9rL/k6j4y/wDZQNd/9L5q+E//ACSvwb/2L9j/AOk6VX7fv/J5XxZ/7GCT/wBASv2Tv+TV/g1/2T/Qv/SCGv2sv+TV/jL/ANk/13/0gmr9hbSdN1v9r/4T6fq1lDd2x8SQSGGVdylo8uhI74ZVP4VVf8Fcfive+P8A9rO/8Fx3iSaV8P8ATbfTbZIpd8ZnljW4nfgkB90qxN0x5ABGQa/4JU6PZaZ+w54EvbVXEurXWqXNzubIMg1C4hGPQbIU49cnvVfGL4fWnxY+E3jH4ZXixeX4o0O705XlAxE8sTIkgJVsFGKsG2naVBAJFaHrWqeG9a0/xFod41pqWlXUd1Z3CAFoZo2Do4ByMhlB544r4b+MLX4ifDrwt4/sSn2fxNotnqcOxWA2XEKSjAYBgMOOoB9a/bb+CHjv9l39qLXriKO9srDUdal13wlrMYKrLE8vnL5cn/PWB2EbdDuUNja6k/swf8Fk9POn23hP9qjQ7hbyJQieK9FtQyT4Uc3NquNjHBJeEEEsAIkAJrwP8RfhV8d/Bcms+AfFuj+LPD2owtBO9ncCQbXXDRyr96NipIKOFYZ5Ar4YfDjwz8Ifh/ofw08GxXEeieHrUWtklxKZZBGCT8zHqeTzX/Bbv4lXU/ir4cfB22vdttZ6fPr15bhT+8eWQwQOSRj5RDcAAH+Nsj7tf8EqdHstM/Yc8CXtqriXVrrVLm53NkGQahcQjHoNkKceuT3qvjR+z78Gv2iNAtvDPxk8B2PiSxsZjNaebJJBNbOcbjHNCySIGwoYKwDYGQcCvhD8EfhT8BPC3/CF/CHwVY+G9IaYzyQ27PI80pABeSWRmkkbAAy7E4AHQVX7fv8AyeV8Wf8AsYJP/QEr9k7/AJNX+DX/AGT/AEL/ANIIaqqqqrXPDHhrxPDFb+JPD2matHAxaJL60ScIxGCQHBwcV8b/APgnf+yn8cbC5GofDPT/AAtrM25k1nwxAmnTiQksXdEXy5iSxJMiMT6jgj9rb9k3x7+yL8SB4J8W3UOq6XqEbXGh63BF5ceowAgEmPcxikUkB4yTtJBBZSrH/gi98etX8T+CvFfwB8RXz3I8JNHqehNLIWZLSdis8IHZElCOPe4boABVf8FIv+Cfk/7SNonxf+EltaQ/ETSLMxXlk2Il163jBKJu6C4TlUZsBlIRmAVCvwV+NHx+/YT+L7arbaHqeh6htEWteG9ctZbeLUrcMQFkjbByCGKSrypzglSyt4R/4K/az8UBb+HPhP8Asj+K/EvjC6wq6ZZ6t58MROfmMqQFtgxklkQYBJKgZr/gpL47/aw18eDbP9pLU9E8PJrfn6np/wAPtGkLppEUWI4bm5fLCSeTzZ0++23y3wE37a/4I1fCmXxf+0dq/wAULiFGsfAGiv5bkEst5eBoYwOwBhW7yc54HBycVVVVf8FMv2jvEZ0+x/Y1+BVvca38RviQv2XUrbTnWSW0sJF+aBx/A06E5LYCwh2YhWDV+xf+yd4c/ZH+EcHgy0mt9R8Taoy3XiTV4lZReXPO1UDHIijU7F4GeXIDOwqtR07T9Y0+50nVrG3vbG9heC5tbmISRzxuCro6NkMrKSCCMEHBr9tv/gmt8SPgR4pvfF/wg8N6n4r+HV/M8tulhBJdXWigkHyZ1GWZBnCy85C/PtP3v2Ov+CpHj39m/wAJwfDH4geGZfHfhKxwulN9v+z3mlR/88kdlYSxD+GNtpXOA20BR4e/bt/aw/aatDYfso/spS6ZZ3OIh4w8XXxFjbFhlmVQiLIVHOEeQ8qShBw3xx8Q+KfFPxf8X6v408eP411j+1p7efxAylRqQgbyY5UUgFEMcabVwNq7RgYxX/BMP4Uy/Cr9jvwgb2FI7/xg0niK5Cg4IudvkHJxkm2jtyffjkDJqry7tLC0mvr66itra2jaSaaZwiRIoyzMx4AABJJ4FfsLWt7+1x/wUa8YftH6vBcSaR4emu9Zt/Ok3CMv/oun27EjJKQncMAAG2HQYU144+Mnwg+GN1a2HxJ+K3g/wnc3sZktodc122sHnQHBZFldSwB4yOM1/wANY/sr/wDRzHwp/wDCzsP/AI7X/DWP7K//AEcx8Kf/AAs7D/47X/DWP7K//RzHwp/8LOw/+O1/w1j+yv8A9HMfCn/ws7D/AOO1/wANY/sr/wDRzHwp/wDCzsP/AI7Xgf4nfDX4nWlzf/DX4h+GfFltZSCK5m0PVoL9IHIyFdoWYKSOcGq8Wfs4/ADx1pp0jxb8FvBOp23OxZtDt90ZPUo4UMh91INftyf8EpNK+H3hbWPjH+zVNeyaVpEL3eq+FLyUzvbW6BmkltZmO91RQCYpCzkBiHY4Sv2Xfjprn7OPxz8K/FbRrqVYNOvFi1a2RyFvLCQhbiFgOuUJK5B2uqNglRUM0NxCk9vKkscihkdGyGBGQQR1BFVXiHxBoXhPQr/xN4m1e00vSdLt3uL29u5RFFbxIMs7seAAB1r4f+AfEX/BU39q3UPjl450u9svgb4GmFjpFldI0f8AaMUbFltgVK4eQkzTsCxQMkW4jYwhhht4Ut7eJI44lCoiLtCgDAAA6DHaq+NXiLW/+Cln7VVp+zh8PdUuI/gt8M777T4t1mxuFMeoSoSrNGTgElhJbwkbxzJMAycDwz4Z0DwZ4d03wl4V0m30zR9HtY7Wxs7dNscEMahVRR6AAVX7WX/J1Hxl/wCyga7/AOl81fCf/klfg3/sX7H/ANJ0qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/4Iqf8mr+Kf+ygXn/pBYVVfFD4vfDP4K+H7fxT8VfGen+GtJurxbKG7vmKo87I7rGMA8lY3P8AwE1/w35+xp/0cP4U/wC/z/8AxNf8FePj98Gfjn/wqf8A4VH8RNJ8U/2J/bX9ofYXZvs3m/YfL3ZA+95UmP8AdNV/wSH+P3wZ+Bn/AAtj/hbnxE0nwv8A23/Yv9n/AG92X7R5P27zNuAfu+bHn/eFf8N+fsaf9HD+FP8Av8//AMTXwo+P3wZ+Of8Aan/Co/iJpPin+xPJ/tD7C7N9m83f5e7IH3vKkx/umqr9r/4Yf8Kb/ae+Jfw7h0u002y0/wAQXE2mWdq+6O3sbgi4tEX0xbzRAjscjtVf8EmfF+i+Jv2J/C2i6XJK1z4T1TU9M1EPHtCzvdPdgKf4h5N3Cc+pI7V+1l/yav8AGX/sn+u/+kE1f8EuP+TEvhj/ANxb/wBO15VVVf8ABUzxt4k+P/x++HH7EPw2mhurkXkF7qarlvJv7hWWLzR1UQ2rPMxH8FxntUP/AASD/Y2jhSJ9J8WysigGR9dYM5A6nCgZPsAK/wCHQ37Gn/QD8Vf+D1/8K/4dDfsaf9APxV/4PX/wr9oD4S6l8CvjR4w+Emqea0nhrVJLeCSUYaa3OHglPA+/C0b9APm44r/gnv8AHR/j/wDsreEfEuo3Xna7oMZ0HWmLs7Nc2qqokZj1eSFoJW7bpSO1V+1l/wAmr/GX/sn+u/8ApBNX7Af/ACeV8Jv+xgT/ANAeq+LP/JK/GX/Yv33/AKTvX7J3/J1Hwa/7KBoX/pfDVf8ABav/AJNX8Lf9lAs//SC/r/giH/yVP4l/9i/a/wDpQar/AIK8/wDJmmpf9jBp3/obV/wRU/5Oo8U/9k/vP/S+wqv+CvP/ACeVqX/Yv6d/6A1f8EuP+TEvhj/3Fv8A07XlV/wWr/5Oo8Lf9k/s/wD0vv6/4JDf8maaZ/2MGo/+hrVVVVVfFr/kqnjP/sYL/wD9KHr9k7/k1f4Nf9k/0L/0ghr9sL9l/wAN/tX/AAb1D4e6m8VnrVruu/D2puoH2K9VSE3Hax8p87ZABkqcj5gpHiXw547+C/xFuvD2u2l74f8AFnhLUgGVlMcttcRMGSRCe3CurDggqwyCDX7BH7XmlftYfB62vdVvIY/HvhuOO18T2QCqXkwQl3GqgARzBS2AAEcOnRQzV8SNPvdW+HfinStNt3uLu80W8ht4UHzSSNC6qo9ySBX7JfxPsPgz+0n8OviVq91Lb6bo+uQ/2hNFKYzFay5hnbIBJAikclf4gCvGc1VV4d8TeHfGGkprvhXXLHV9OkmmhS7sp1miaSGVopVDKSCVkjdDjoVIqv8Agqv8AZfg5+05f+MtM094vD3xLWTWbWXJK/bdwF7Hk/xeYyykcgC4XnsP+CS3x9l+Ln7Nq/D7W9Qe5174YzR6Y5cHcdPkDNZEnphVSSEDjCwLn1PwHkn/AGw/+Cj/AIz+P8tt9o8BfBa3fRPDkrOJIp7v95EkiZyrhs3dwCuCm63zzgmqqqqq/wCc7X+f+hTqq/4KBfsR6P8AtY/D8634bhS2+JPhm1f+wrkyCNL2PO5rObPy7XOdjHGxznIUuD8N/iP8VP2YPi/B4s8LTXGgeL/Cd9La3dndwkDcjGOe1uIjjKnDIynBBGQVYAj/AIKaftIeBP2pvhV8A/id4Hdrcu3iC31TS5pA02mXajTS8LkdRghlbA3KynAJKiv+C0Wn3t7+yhoNza27yx2HjqymuWUcRRmzvYwx9t8iL9WFf8ES/ifYaP8AED4g/CLULqUT+JNPtdT01GlPlh7RpEmVUxje6XEZJ44h74GKrxH4m8O+ENJbXfFWuWOk6dHNDC93ezrDEkk0qxRKWYgAtJIiDPUsB3qv2sv+TqPjL/2UDXf/AEvmr4T/APJK/Bv/AGL9j/6TpVft+/8AJ5XxZ/7GCT/0BK/ZO/5NX+DX/ZP9C/8ASCGv2sv+TV/jL/2T/Xf/AEgmr9gP/k8r4Tf9jAn/AKA9VX/BRDSr/R/20/itZ6jB5UsmsJcKu4NmOa3iljbIJ6o6nHUZwcHIr/gkn4rt/Ef7Fnh7R4WhL+FtY1LTpQhJIZrhrr5vQ7bpenbFVXxa/wCSqeM/+xgv/wD0oevgB4Y1DwT8Bvht4L1ZWW+0DwjpWn3KvGY2EsNpFG+VblTlTweR3r4s/B34a/HLwdd+BPil4Ssde0i6UgRzpiSBu0kUgw0TjsykH8M1+0l/wRs8b+GftXiX9mzxJ/wlOnLl/wDhH9WkSC/jHpFN8sU30byz2+Y14M8efGv9lv4nvqPhnUtb8FeL9BuPJvLSaNomyvWG4gcYkQ90cEHIPoa/YU/aysv2uPgxF4qv7e3sfF2hTCx8SWMHEaz4yk8QJJEUqfMAfusJEy2zc3/BbDSr+H9pPwdrckGLK78Dw28Mu4fNJFe3bSLjORgTRnJGDu4zg4/4I3/E+08W/syX/wAOXus6h4D1yaPyDKGKWt2TPE4XAKhpTcjBzkoTnnAqtS+I3g/SviBovwuvNVx4l1/T7vULKySJnJt7ZollkcqCIxmZApfAYhgMkEVX7fv/ACeV8Wf+xgk/9ASv2Tv+TV/g1/2T/Qv/AEghqqqqqqr/AIK/fDTTfGP7JN145liiXUPAWsWl7BMTh/KuJVtJIh8pyGaeJiMrzEpzxtP/AARQ0q/m/aT8Y63FBusrTwPNbzS7gNskt7aMi4zk5EMhyBgbecZGarW/Dvh/xNaJYeI9D0/VbaOQSJDfWqTorgEBgrggHBIz7n1rTdI8P+FtNa00fTNP0iwh3SNFbQJbxJ3ZiFAA6ZJr9uH4+P8AtO/tLeI/HGjPLcaLBIukeHYxuYtZwMVRlXqPNdpJdo6GUjrX/BP/APZpb9mL9nXRvDOuWCQeLtfb+1vEh4Zo7mVQFt8jPEMYSMgErvEjD79VVVX7YP7WOlfs6eGbbwx4UsX8S/FfxgptfB/hm1jM8s1w52JcSxr83kq/bgyMNi4+Zk/Yr/ZM1n4NWmq/GD44alF4n+NnjmQ3Gu6zM3nPYRMFxZRPnbhdo3GMKpIVFykaGqqqv/AXgXVdUbXNU8F6DeakzKzXlxpsUkxZQApLlS2QFAHPGB6V+3h+0Pb/ALNn7OHiPxfaXvleItWjOk+HkWQq/wBsnVgJFIOR5SB5c+qAZGRX7Fv7N2pftRfH3QPh6LWU+HrWQah4luUbZ5GnxMPMAYHIeQlYlIyQ0gbGFYi0s7SwtIbCwtora2to1ihhhQIkSKMKqqOAAAAAOBVV/wAFKvjA3wc/ZC8Y3dldNBqnixV8O6cygfeugwm5PQi2S4II5BAxjqP+COnwmbwT+zNffEXUNPSG/wDiBrUlxDNtAeSxtv3EIbvgSi6YZ7SZAwcmv2i/2K/gb+1NrWka/wDFjT9YuLvQ7V7a0NjqLWwEbtuOQAcnPev+HQ37Gn/QD8Vf+D1/8K/4dDfsaf8AQD8Vf+D1/wDCv+HQ37Gn/QD8Vf8Ag9f/AAr/AIdDfsaf9APxV/4PX/wr/h0N+xp/0A/FX/g9f/Cv2dP2WvhN+yzomr+H/hNZ6nb2muXSXN2L69NyxkRdowSBgY7VVTQw3EL288SSxyKVdHXIYEYIIPUEdq/ac+HNl8JP2hviJ8ONLCDT9C8RXcNgqtu2WpkLQKThfmEbIGwMZBxxg18PdKv9D8AeGdE1SDyb3T9HtLe5i3BtkiQqrLlSQcEEZBIqppobeF555UjjiUs7u20KAMkknoMV8UvF3jb/AIKS/FKT4GfBLXbvTPgH4SvFHjjxZCCsev3CMGFpbMOZUAAxghcnzWBAh3+CvBXhP4c+E9L8DeBtBtNG0HRbdbexsbVNqQoP1JJJJYkszEsSSSar4h2mpX/gDxNY6NHLJqFxo93HaJCcO0rQsECnsdxGPev+GAv2yv8Ao3nxV/35T/4qv+GAv2yv+jefFX/flP8A4qv+GAv2yv8Ao3nxV/35T/4qvEXh/WfCev6n4V8R6dLYato15NZX9pMMPbzxOUkjbHdWUg+4rTv2Ev2vtW0+11XTfgD4ouLS8hSa3lSFMSRsAysPm6EEGvhvp97pPw78LaVqNu9vd2Wi2cM8LjDRyJCisp9wQRVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVf8EVP+TV/FP8A2UC8/wDSCwqq+KHwg+Gfxr8P2/hX4q+DdP8AEuk2t4t5DaXyFkSdUdFkGCOQsjj/AIEa/wCGA/2NP+jefCn/AH4f/wCKr/hgP9jT/o3nwp/34f8A+Kr/AIYD/Y0/6N58Kf8Afh//AIqv+GA/2NP+jefCn/fh/wD4qv8AhgP9jT/o3nwp/wB+H/8Aiq+FHwB+DPwM/tT/AIVH8PNJ8L/235P9ofYUZftHk7/L3ZJ+75smP941Vf8ABYX9lO68W+HLP9qHwZZxG88L2YsvFEKg+ZcWhlUQTqqjkxNI4cnnYynIWKq/4Ij/ABD0q9+FfxC+E4i8vUtH8QR63vaVf38N3bpBhE+9+7ay+Zun75BX7WX/ACav8Zf+yf67/wCkE1f8EuP+TEvhj/3Fv/TteVVV8QfHXhv4YeBte+Ini+9+y6L4c0+a+vZQMsI40LEKMjc5xtVerMQBya/4JV+D9S+Pf7QPxN/bL+IVtDPqcV5LBpuB8kF5d5aZoxjjy7crEvP3ZzkHgiqr/gtf8GE07xN4I+Pel2jiPWIZNC1eRYztWeIGW2JbONzxmcY44gHXnH/BH39oKH4cfHK/+DOv3yRaP8SIUWzeabYkOpW6u0QG44BlRpI8Dl3EK88Cq/ay/wCTV/jL/wBk/wBd/wDSCav2A/8Ak8r4Tf8AYwJ/6A9V8Wf+SV+Mv+xfvv8A0nev2Tv+TqPg1/2UDQv/AEvhqv8AgtX/AMmr+Fv+ygWf/pBf1/wRD/5Kn8S/+xftf/Sg1X/BXn/kzTUv+xg07/0Nq/4Iqf8AJ1Hin/sn95/6X2FV/wAFkPDt3pH7W1rq0z74dd8J2VzCQhAXZJPCyZ6E5i3cdA61/wAEmPGOj+JP2LPDHh/T50a78J6lqNhfRhwWWSS7kulJHUAx3Kfka1HUdP0jT7nVtWv7eysbKF57m5uZRFHBGgLO7sxAVQoJJPAAya/4Kg/HHwV8f/2itP8AFvw7N7c+H9M8Mw6XaapNbNFDqpiu7tpJ7YsAXhEkjxB8YLQvjgAn/gj7qFle/sd29ta3KSy2HibUIblV6xSHy5Ap99kiN9GFV4m/4KU/sdeEviRJ8MtW+Keb22uDa3eoW+nzzWNrOCQY3nVSDggAuoZBuGWGG22l3aahaQ31hdRXNtcxrJDNC4dJEYZVlYcEEEEEcHNVVV8Wv+SqeM/+xgv/AP0oev2Tv+TV/g1/2T/Qv/SCGq/4KnfsTw/GHwXP8fvhvpKf8Jt4VtS2r20EWG1jT41JJwqlpJ4gAVyRmMMvJCAfsvftDeKf2X/jNonxX8MB7mO0Y2+rad5pjXUrCQjzoGI7kKrKSCFkSNiDtxXwo+Kfgv41/DzQ/id8P9VS/wBE121WeBtyl4WI+eKUKSElRso65O1lIqv2zfgXqn7PH7RvjD4f3Wmta6XLfSahoD+UESfTZ3ZoGTACkKMxNtAUPE4AGMV/wT2/4Kb+DtI8HaP8Cv2j9efSptGhFronim6ObeS3TasNtcFVzGyLkCVvlKqNxBGW0jxP4a8Q6OPEGgeItM1PSypYX1ndpNAQBknzFJXAHPWv2uv2/PAnw90Ob4U/AXxHY+NvjB4omj0nRNP0eUXKWNxcERpLJKp8sOCwCxbixcpkBckfAX4Xp8F/gz4O+FpvUvZ/Dukw295doDi6usbp5hnnDytI/PPzc1X/AAVA+AA+OH7L2s6xpFlFL4i+H2/XdPZlw7QRoftcSn/ahDOFwdzxIMcgj9nP9pHx7+zTr3iXXvAeoS29x4k8N3mjyMjf6l5U/c3Cjp5kUqq6t2+YcglT/wAE8/gEP2fP2XvC+h6lb+X4h8SRjXNbDRbHjnuUVkhYEZBiiEUZBz86uRwcCq+MP/BQn9lT4G+Pf+Fa+OfiFKdct5Amox6dp014mmk7cee0akA4JJRNzjByoJAPhTxX4b8c+G9N8X+D9btNX0XV7dbixvbWQPHPGw4ZSPyI6g5BwRVVVf8AOdr/AD/0KdVVf8FQ/wBgsfFTRbv9oj4PaHv8Z6Pbg69pNnbZk1y3XA85Av3riJc5GCZEXA+ZEVqr9tL4LXX7QP7Mvjn4Z6VbJNrF3Yi60gFV3G8t5FmiVSQdpcx+USMHbIwyM5rwN428cfBn4haZ438JXtxonijwvfebbvJDh4JkJVkeNxyCNyMjDkEgjk1+y5/wUX+AP7R2hW1rqfiTT/BPjSOM/bPD+sXiwhioyXtp32pOmAxwMSKFJZAAGPibxZ4V8FaVJrvjLxNpOg6bFnzLzU72O1hTCljl5CFGFVj16KT2Nat+0No37cn7T/gn4H/BC+bUfhr8O9Si8V+M9fMDpDqU1nIrWlrEG274vtGzduGJMFlBWHMlftr+Hbrwv+1z8X9MvH3STeMNQvVOwr8lzM1wnB/2ZVGeh6jg1+zb4x0f4g/s/fDnxhoU6SWmpeGbFwFcP5UiwqskZI/iR1dG91NfEL4i+B/hR4Rv/HfxG8T2OgaDpig3N9ePtRMkKoAGSzMSAFUEkkAAmv2rvHtl8U/2i/HvxE0zSNT0yx8Qas13aW2pQeTcLAyr5bOnO0sm18ZPDDk9a/ZA1Cy1P9lD4OXNhcpPGngXR4WZegkjs4o5F+qujKfcV+1l/wAmr/GX/sn+u/8ApBNX7Af/ACeV8Jv+xgT/ANAeqr/gs1+zhq9p4p0X9prw3pTS6Tf2sWkeI5IVLfZrlCfs88nJIWSMiLdgKDFGD80gz/wRm+P1p4R+JPiL4A+IL3y7XxrGt/ou8gKt/bo3mxjvmWHB5P8Ay7gDlua+IXjbRfhr4D8RfELxFN5emeGtLuNRuz38uGNnYDGSSQuAACSSAAa/Ya+Aur/tT/tP6JYa9YvqehaffDW/Fk88RaOW3STe0UhGBmd8R44OHYj7pqv+Cc/7S1x8V7D4h/CXxr4uvdW8X+CfFWoyW51CZpppdKkuCYmEjEs4jlMkeCTsXygMAqBX/BXn4F+C/F37N1/8an023tvFHgW6tGjvoolWW6tri4itmgkbGXUGVHUE/KVOPvEH/gh7Ya83i/4q6pClwNFXTdOhuGEmI2uTJMYgVz8zBBNzg7Qx5G8Z/wCCyXwIv/HnwW0L4y+HdL+033w+vJE1Tyoxv/s652q0hOdzCOZIflAO1ZZG4AYn9jH9qbXP2SvjNZ/EC2trjUdAvoTY+IdKhdVa7tGIO5N3AljYK6njOCm5VdjXwf8A2gvgz8fNDg134TfETR9fSSHzZLSG4C3dqA20+dbtiWI5wPnUZyCMggn45ftT/Af9nXRLrVvil8RNMsLqBT5WkQTCfULp9oYJHbKS5JDJ8zBUXepZlBzX7A+q+MP2kfiR8Qf24/HejXGkQeI4U8MeCtOc7ktNIt5N82xzzIGuFG5gAvmpNgYwqV+37/yeV8Wf+xgk/wDQEr9k7/k1f4Nf9k/0L/0ghqviJ/wUZ/ZH+F/xFm+F/in4lt/a9jdNaalJZ6fNc2+nTqxR45pEUgMrAhgu7aQQ2CDWh63o/ibRNP8AEfh7UrfUdL1W1ju7G8t5A8dxBIoeORGHBVlIII6giqqqqv8AgsZ8dtC8I/Aa2+BVrfeZ4h8d3kFxPaqAfK0+2lEpkckHG6eKFVA2k7XOcKQf+CVf7MGtfAD4HX3jHxrY3Fj4p+I80F5cWM6tG9nZQq4tYpI2A2y/vZpG74lRSAUNVVf8FU/27NK0bQNV/Zc+EevJc6zqSta+MdQtmJWygP37FHVhmVxlZRghULIfmZgn/BNL/gnPd+EZ9H/aR+POky2+uR7bnwv4dnUo1hkfLd3K9RLg5SI/c+83z4EdV+0J+1l8C/2X7Cxuvi54w+wXeqZNhptrbvc3VyqkBnWNAdqDPLOVXggEtgV8Df2gvhJ+0f4QPjX4ReLYdZsYZBDdxNE8M9nNtB8uWJwGU88HBVsEqWHNV+2N+2N4T/ZY8LWthZ20XiH4i+Jf3Phnw2knzTSM2xZ58HKQB+OxdhtXGGZP2SfhD4G+H+t3/wC0R+0R8Y/CPi344eLVL319LrVtJFoUTLj7Ja4fapC4RnTChR5aYQEv/wALY+Ff/RTPCn/g5t//AIuvE37QfwH8GaVJrfin4zeCdMso8jzbjXbdd7BS2xBvy7kKxCKCxxwDX7P/AO1rL+1P8fvFOm/CKVH+E/gLSUivdUnsjHLrGq3EreX5RZtywJFDIR8gYsfmwCmarxr418J/Dnwnqnjnxzr1po2g6LbtcX19dPtSFB+pJJACgFmYgAEkCv2gfih8Wf8Agpt+1DYeFPhVod3Po9j5lp4ZsJ8xR2FnuBmvrs5ZYi+ELsOyxIAzBd37If7J3gf9kb4ZJ4K8MzPqWs6iyXGv6zKu19QuQuAVXJ8uJASEQdASSSzMxqq/4LE+ONU+I/xl+GH7M3g+F7vUYlF08EcYYzXt9KsFtECMtuCxk4AwROvUgbfhd8P9H+FPw28L/DPQQhsPC+k22nQusQj80RRqhkKjPzOQWbkkliSSTmqqqqqqq8WeKvD3gbwxqvjPxZqsOm6LodnLeX93KCVghjUs7EAEnAB4AJPQAk1+y/8ACzX/ANvP9urXPjDe6Xe2vgmz8TSeI9WmlXb5MCzF7Ow3xgKZXCxocENsSVwSy81NNDbwvPPKkccSlnd22hQBkkk9BivjF8frn9u74kX/AOzh8IviXp/g34M6LIE8c+Npb+K3k1xcnNlY7yN8bYILcqw+Zv3e1Zvhnefs5/B3wPpXw5+HPizwZo+g6ND5Vtaw6xB9Wd2L5d2OWZ2yWJJJya/4Wx8K/wDopnhT/wAHNv8A/F1+0J/wUC/Zs+A3hjUrv/hY+ieKPEsMbpZaDod7HezPcbSUWby2IgTIG5nIIB4BJAPwK1D4iax8HvCOt/FqS0Pi/VdLivdWitbfyI7aWYeZ5CoGb/VK6xk5OShOTmqqv2sv+TqPjL/2UDXf/S+avhP/AMkr8G/9i/Y/+k6VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVfsH/te+IP2Q/hBq/wANfEf7NvxG8QXOpeJJ9WS5srB4kRJLa2hCEOmcg25OenzCv+Hsf/VofxW/8Bv/ALCv+Hsf/VofxW/8Bv8A7Cv+Hsf/AFaH8Vv/AAG/+wr/AIex/wDVofxW/wDAb/7Cv+Hsf/VofxW/8Bv/ALCv+Hsf/VofxW/8Bv8A7Cv+Hsf/AFaH8Vv/AAG/+wr/AIex/wDVofxW/wDAb/7Cv+Hsf/VofxW/8Bv/ALCv+Hsf/VofxW/8Bv8A7Cv+Hsf/AFaH8Vv/AAG/+wq8/wCCrFpqFpNYX37HXxRuba5jaKaGazDpIjDDKymPBBBIIPBzXx68DaD4s8Z3PiT4GfAP4l+FdN1G4lmn0bUtMeaGyyE2pbsibtm7zThvugqB0r9nDxb+0j+zB8UrD4pfD74c+JWuYYzbX9lPotwYtQtHZWkt5MJkBiiEEcqyqe1ePP8AgpZ40+J3wD8X/DXxP+yj47svEfizw3qekvc2FpK1pA9zDLDG4DpvICuhYeoOO1f8EzdI1XQv2IPhtpWt6Zd6fewf2r5ttdwNDJHnVLsjKsARkEHkdCDVV4i1aTQPD+p67Bo+oavJp1nLcpp+nRq9zdmNCwihV2VTI+NqhmUEkZIHNftxeJP28v2sNFtvhz4S/ZU8YeEvA0MiXF5azPFPc6pOuCplZSAkaH7sa5yRvYn5VT9gjVf2x/2QvDmsfDnxX+x54w8Q+F9X1Q6ok2ntFFd2lw0UcUnDttlRkhiwuUKlScndgeAvFlz458JWHim78HeIPCs175m7SNfgihvbbZIyDzEikkQbgocYc/Kyk4JIFeItWk0Dw/qeuwaPqGryadZy3Kafp0avc3ZjQsIoVdlUyPjaoZlBJGSBzX/BQDVf2sv2u/CujfDXwR+x5470Pw7pGsHVHvNUaBrm6lSOWGIeVGxWICOZyfnfJYDgJltJ/YX/AG3NB1Wy1zRvgN4ws9Q064juLS5hjRXhlRgyOp3cEMAQfUV+zf8AtM/tUatFofg39ov9kjxrYapNJBZzeJ9JjtzauzMiG4uYXkQwIAWdzGXxg7UAIUftv/Fb9oT4gfC7xl8DPgh+y/8AEue81qSfSL3xDdxW8Fo9mH2ySW3lzO8yToGUbxH8kmSCflr4U/skftz/AAg+Jfhj4oeGv2d/EsmpeF9Ugv7eKaFfLmMbhjG+HB2OAVbBBwxwRXwW+M3ij4ppLa+K/gL49+Hd9aWqSznX4bY2skhODHBLFKzSEdctGnH5V+1z8bvjvL4I8afCj4Hfsw/EPWdc1K1n0qPxFPBbxafHHLHse4tyszSSMFZtoZEAYAnIG0+E/wBij9t/wV4q0bxloP7P/iqLUtB1CC/s5PLC7JoZFkQ5VwwwyjkEH0I618K/2iPH/jPQLybxz+y78TfCOt6ToM2pXVvLb2k1teTxbf8ARLOXzw0s0m75BIkanBywxk/8FDj+1l+15B4c8F/Dv9lb4i6L4Q0C4e+lOrxwJPfXbJsVmiildEEatKFO9ifNb7vQ/sNfDf8AbS/ZA+Lk/jS8/Zb8Ya54f1rT20/WLCBY0m8verpNES+0yIycBuCruMgkMvgn4g3vjLwTN4xuvht4w8NTw+djQ9btYIr+TyxkbUjmkT5+i5cZ74r/AIKB+JP2qP2qvA2mfCn4W/sl/EXR/DsWoJqGpXmtQW8dzdSRoyxwrHFNIixguWJLEsVTAUA7v2PPgx+21+yh8bdP+Klt+zB4t1iwa1l0/VdPRlga4tZdpbawbG5WSNwGBUlB0OGX4X/EG9+JXh+fXb/4beMPBEkF41sNP8UWsFvcyhURvNVYZpVMZ3lQSwOUbgAAn/gpD+xVq37WPgHRta+Hj2kXjzwfJKbGK5dYo9StZdvm27SEfK4ZEeNmO0Hepx5m5fAOn/tq/sweObhPh/4Y+Jfg3xC0iR3Ntb6NOyXmxj5YeFo2iuEyTtyrqdxxkNz8GP2af25/2ztVsdY/bb8eeK9E+GdpIktz4ZuQukz62UYMsbWVukaxIGRS0kqiQf8ALMZYun/BU74A+N/FH7THgLwT8EPhHrGpafYfD2wsLGy0HR5JILVUvr0BP3a7I1VWUkkgKCCSBzX7Gf8AwTw/bM+GlrfS+IPj5cfCLRtaaKS/0TQJY9Svbho2/ibmCBtvSRGlJHDLitP8FXukfDOH4eWfjTXbu6tdF/s2DxBqdx9o1B2EPlrcyyLs8yboxf5Szc5BOa8TfshftK+FfiRJ8Kb34MeK7nX/ALQYbdLLS5p4btQSBNDKq7HiIVj5mdoCtkja2P2afh94k+FH7P3w++G/i/UftmteHfD9rZ3rhy6xyJGMxK2TuSP/AFanoVQEADgVVfFr9rD9oBvBd/a/BL9jH4sP4ouYXitLnxFaWVtbWTlSFmKxXExlKnB8shAccsKm/YI/bQuJXnn/AGfvFsssjFnd4kLMx5JJL8k1+xx8Z/2nfgz8IdG+Efxp/Y/+JeoR+GLc2ulap4ft7aZ5bcEeXHLFPPGAVBZd6tggINucsfCHiGbxZ4Z07xHc+GtY8Py38IkbS9YiSK7tTnGyVY3dQ3HQMw561X7ff/BNH4g6b8WX8ffs0fD+717w54ukmub3R9NRQdGvMgyKqEgCCQsWQLwhDrhVCA/sreE/+Cl37I+s3U/gD4C+INR0HVJFfU/D+p24e1uGHHmptkDRS7eN6nnC7g4UAfA39oP4mfE7VItB+I37LPj34cXbwtIby+lt7vT0Kj7hmDJJuPQDysHuRX7ZP7GXgH9sLwTa6Rrt7LovibQvNfQtcgTebZnA3xyx5AkiYqhK5DAqCrDkN8Zv2Bf2q/gfqVzbeIfhPqut6bb5K614ct5NSs5E/v7o13xDIxiVIz045GfB37Nn7QPxA1ePQvB/wW8Z6ldyMqkJosyRxZOA0kjKEjXP8TkD3r/gnx/wTZb9njVIvjJ8aZbHUPHohZNL0y2Img0QOCHkMnSScqSuV+VAWAL7gy1U0MNxC9vPEkscilXR1yGBGCCD1BHatA/4J76nF/wUaf4R6h4TuLj4Z2OrNr/2gW7m0fSR+/itHfBGdzJasCQT8xGAVNVVftX/ALKH7Q3gf9obxbpuo/DzxX4j/wCEj8QXd3pGr2Olz3i6ws85dXVkD7pT5i748llZsHOQT/wT2+D3j34G/sqeE/AvxJEtvrnmXV7Jp0j7jpqTzNIsHUgHB3sBgB5HHJBY1VfE/wDaU8eeC73XND8Gfsm/FjxjqWmMY7K5t7ezt9Nv2GOVuDO0ipyfm8gnI+7UXwm/4KIR/tQH9q1/2dtdl8U/282qi1e3AtwpyotsCQN5Qh/dDndtAyScmvht+0n428Z6lo2g+K/2Ufi34RvdRk8u6u7u1sptPsTz8zzi4EhTAHIhzk4x3qqr/gpv/wAE+NQ8Ka7d/tC/Ajwy9x4f1WYN4j0KxjZ5LG8lkx9pgjHJhkZlDIvMbnIGxsR1X7eP/BLn/hdPiHU/jP8AAO6tLDxjqUnnavod7L5VtqcmDumikORFOx25VsRscsSpyW8b/sxftE/DjVDo/jT4J+M9Nn3Msbto80sMxUKW8qZFaOQDcuSjEAnBr4HfsHftP/HnWrWx8PfC7WNF0iVgZ9e1+0k0+yhj3FWdXkAMxBUgpEHYHqAMkfsi/sp+Cf2SPhZF4E8My/2jq9/It1r2tSRBJNQudoHA5KRIMhI8kKCxOWd2av8Agp/+wB46+LXig/tDfBHSP7Z1l7OO38RaFCAtzdeSoSK4gGP3riMBGQndiNNobkD4H+N/2/vgRf3Hgr4J6Z8UNJnnuJPN0GPwzNfIJ1BEhFpPDIqyAKdxCBvkGfujH7NH7Fvx++L/AIu0z46f8FBPFmoeJJNLkN34f8E6hdrNBbTucma4t4/9HiGFQiCMc/KJMBDGf2qf2Zfj/wDG79tf4qWXwu+EniPXFm15mW7S0MFooMY2lrmXbCgO1sbnGcHHSv2T/wBgv9qL4aeD9N8LfEr9rLxB4b8M2chng8KeDJgTA7MZSv2yeM+WDI7mSKONlcsTv71+2r8X/wBonxv8OPGvwP8Agj+yr8Q7yfWludFu/EN/FbxWr2hZo5pLZY5meVZYwQpfy/lkyV42nwb+xd+3N4B8X6F468MfALxRb6x4c1K31HT5ntIpBHcQSLJGxVmIYBkU4Iwe9fAz9or4qfEK70fwz8Uf2VviH4F1i8Vxd6lJHbT6TAyQlyzS+cJVDspRV8psFkBbkkV418FeE/iP4T1TwN450G01rQdZt2t76xuk3JMh/UEEAhgQysAwIIBr9ov/AIJYftA/BXxu/jf9nG1vfGXhuxulvtMezmRNV0tlkDRq0RIMzIduHiBJxkomK+F3/BUX4+eE9FHhb9oP9k7xtqXiPTspNqOmabNYvOewltZIcRvjqVbBPRFr4xw/t5/8FE1Hwy0z4NXXwR+FSXCyalJ4qaW3udQaOTdGZUZFlkA2K6wpFsDgF5T+7Zf2X/2X/hv+yn8N4PAXgK08+7n2y6zrM8YW51W5Ax5j4ztQZISMHCAnqSzNXhf9mr9ujQfiprH7RnwI+GHiW2/s7xZfjT9QieGKSYi4cOBbSuJJoGBKsdhjYblJOCB4R/4KgfHvwlYW+h/H79inx3a66tuJDc6bp91Zi6BJAcW1xDlAdp5EjAkHAGK+Neq/tr/8FMI9K+HfhD4C6h8LvhjbagLy41HxHK9uLwqrCKaRnjR5UAJxFAkgDsCzHarL+zF+zV4B/ZW+Ftp8M/AoluWMn2rVdTuBibUrxlVXmYZIQYVVVBwqqBkncx1HTtP1jT7nSdWsbe9sb2F4Lm1uYhJHPG4Kujo2QyspIIIwQcGv2u/+CSfxH8CavdeMP2aNPuPGHhWdnlfQzMv9o6Zks2yMOR9piACqu0mUkgFWwXMvwK+N0Orjw/N8G/HCaoZhALF/Dt0J/NYgBPL8vduJIAGM8iv2Uf8AglB8Y/ixq9r4j+Oun3vw98HQNHJJaXUW3VNRUkExpCebcbchnlAZSRiN/m2+FPCvh7wN4Y0nwZ4S0uHTdF0Ozis9Ps4slYIY1CooJJJwAOSST1JJOa+NPxm8UfCxIrXwn8BfHvxEvrq1eWA6BDbC1ikBwI55ZZVaMnrlY34/Kvil+yJ+3L8WviR4n+J3iL9njxQmpeKdWudRuIo0DJC00hfy0LuW2ICEUEnCqB2r9iX4s/tBeBPhh4M+B3xs/ZZ+JFpd6K1to9lr9lFBPaLZ7gkct15s6vCsSFQdgkG1MqBwlV8W/wBkH9pPwD8WtT+HurfDTxb4g1OfUpksdTtNLnuY9aUyELcxyKG3CQYc5O5d3zYINfsVfC7xh8GP2Wfh58N/H9w8niDStNd72N33tatPPJOLYnJBMKyrDwSv7v5eMVVV/wAFaf2Zf2nPiZ8SNH+JHgHQNd8b+CrbSYrOPSdHia4m0u6Ej7z9kjy8gkDq3nKrEYKvtVELfBb9rf8A4KD/AACf+ydLsfGHiHTLf92dG8V6Fd38URVuisQs0ePmG1ZFHPI4GPD/AO2x/wAFJfjJbRaT8Kf2NbfRZ75SY9Z1uxu4bRIyrAukly8MRYEAglnHy42NuAr9nz/gmvqt949b4/8A7bHi6L4k+PbmRJ49LaRprK1dNhjMrEKJigXaIQohUZGHGCKr4r/tu/Bb4S6tqXhzUdO8d69rWkyPFc2GieEL6ch1XIVZnjSB93ABWQrk8kDJr4k/Gz/goH+035vgP9nP9nzxB8HNAu7gxT+L/Gco06/WD+8IWXfb5PB8lZ36FWHJr9kb/gmF8LfgDe2nxD+Jl2nj74hRsZVublCbDT5Tuy0ELDMj/N/rZcnIDKsZyTVV/wAFfP2cvjPrnxn0740+G/DGt+J/Ct3odvp7yafayXJ0maKSTMTom4pG5kV1fCqXkZevLf8ABHP9n34ufDSw8cfEzx/oWq+G9I8UW9nbaXpuoQPbyX3lmRzcmJsFUUSBUYr8wkcggD5v2k/HHxW+Hvwh1jxB8Efh3L408ZnZBpmmoflR3ODO4yCyRj5ioI3YAyM5HxN/ZO/4KBfGLxxqvxG+I3wZ8Z6zr+sTGW5uZooxjsqIobCIqgKqKAFAAAAFf8MBftlf9G8+Kv8Avyn/AMVX/DAX7ZX/AEbz4q/78p/8VX/DAX7ZX/RvPir/AL8p/wDFV/wTc/Z21T9nX9mnTtI8W6M2m+LvEl9PqmuW8qDzIHJ8uGEnGcLDHGSvIDvJjIOT4w8W6F4E8M6j4v8AE1xcQ6XpUJmupLezmupFQEDKxQq8jnkcKpPtXiz/AIKIeFN9zo/wX+Avxg+J2urG7W0GneErqztmwyKGllmQSRRkvjf5LYI5AyM+Nf2av28f2+vENteftB3un/CD4ZWt5HLb+FYroXNwVUH96Io8h5cFl3zum0tlY8Eg/s9fsx/B79mDwgPCPwp8N/ZvNw1/qd2wmvdQkCgb5pcDP3c7FCxqSdqrk1VVX7LHgfxh+07/AMFLvEn7QXjTwhqtl4d0S8u9asZNS0+e1WVIgtrpsY3qCZFTyZMcD9w2QB8pqqqqqv2rfA3xK+Jf7O/jnwN8IPED6N4u1XTfL025SbyWcrIjSQCTI2GaNZId5IC+ZkkYzWg+Ef24P2U/FZuvDXhP4oeBtS8ySMvaadcfZr3bgOOFaC5QZQ/xrnafQ18L/wDgpX+3vc2A8P6h+yxd+PNVtozIbqx8NahBO0YOC8scKsnVlGVVAOOMnJ8SfAT/AIKG/t4PYaV+0bfaJ8IPhtHcLLcaFpjbp7xVZSrNCskhd+OPPkVVZdwj6Z+BnwD+Fv7OPgS3+Hnwo8OJpmmxt5tzK7mW4vpyAGnnkPLu2B6KowqhVAUV/wAFH9W/bn+KGtat8Cvgf8IPEC/DoW8KajrOmhWk14yRrI8e8sDHApby2QDLlH3Eq22v+GAv2yv+jefFX/flP/iq/wCGAv2yv+jefFX/AH5T/wCKr/hgL9sr/o3nxV/35T/4qv2bP+CdH7Q3iD47+CdO+Lfwa1rRPBy6olxrV3qEKeT9nhBlaJuW/wBb5YiAxjMgzgciqqv2kf2Iv2sfFv7RHxS8VeG/gV4l1DSdZ8aaxe2F3DChS4glvJXjkX5ujKykfWvhvp97pPw78LaVqNu9vd2Wi2cM8LjDRyJCisp9wQRVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX/9oACAECEQE/AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADG42nhKanNNpu2n9eRRqxrU41I7NXAAAAAAAAAApZlRqYl4dJ8yv6afMAAAAAAAAAJNJXMDmNLGc3s01a2/n832AAAAAAAAAAxmZUcLOMJptvt/w4AAAAAAAAABLMqqzH6tZcvzvtfv+gAAAAAAAAABmmMnhaHtIJN3tqYOtKvh4VJbtAAAAAAAAAAAAAAAAAAAAAAAAAAAHEf+6x/xL8mZd/udP0QAAAAAAAAAGC/5HFT/ALeAAAAAAAAACp8D9Dhr/l78v1AAAAAAAAAA4h/3il/XUAAAAAAAAAAn/wAjz7v/AEkAAAAAAAAAA4h/3Neq/Uyv/c6foAAAAAAAAAAAAAAAAAAAAAAASlGKvJ2IyjJXTuAAA2krlCvTr01Om7ozzD1a+GjGnG7v+jMDCUMNTjJWaSAAAAAAAAAAwmErRzOdVx9131AAAAAAAAACavFpGQ4WtQ9p7SNr2/UAAAAAAAAADO8JWrVqbpxul/mAAAAAAAAAASyybzD61zK3b5WAAAAAAAAAAMzwcsXQ9nF21uYSg6FCNNu7SAAAAAAAAAAAAAAAAAAAAAAAM0wqxWPp0m7Jxf6lLDVMtxkIxleE9PmAABU+B+hkf+4x+f5gAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAV/+RrS/wAL/UzdKU6EFu5L/gmYY94dRhCPNOWyMvzCWIlKlVjyzj0MvxcsTTk5KzTa+4wOO+szqRt8Lt/wfwI5g3Urpr3af3vT/gFPO66qRdanaD2ev57MxuK+rqDtvJL0v1MDmixLqyatGNremu/3Ec8qzleVO1N6X1/PYyP/AHGPz/MxOKo4eCnVdlt1/Qp1IVIKcHdMAAAAAAAAACGOw86zoxl7y6WfT8AAAAAAAAAAbSVzDY2hib+yle2+jX5gAAAAAAAAAYjHYfDyUakrN7aP9AAAAAAAAAACWaWx31Xk+d/K+1v1AAAAAAAAAAMwxv1Sj7Tlvrbe3+Zha/t6MalrXQAAAAAAAAAAAAAAAAAAAAAABms68MdTdBXly/5mXSqV8c5Yp+/FaK1vn/XqPE0YZnOVWSXKkl89WSxNCpmVKVKV7pp/oYCSpVsTHs7/AH3ZkqdOtyv7Ueb8WZRFTjVqP7Un9xnOPws6EqKleX5NPuZxepgaPduP5MzaMaGXunT0WiJ4WCwjoW0tb8DI/wDcY/P8ziP/AHWP+JfkzLv9zp+iAAAAAAAAAAwX/I4qf9vAAAAAAAAABU+B+hw1/wAvfl+oAAAAAAAAAHEP+8Uv66gAAAAAAAAAE/8Akefd/wCkgAAAAAAAAAHEP+5r1X6mV/7nT9AAAAAAAAAAAAAAAAAAAAAAACv/AMjWl/hf6mOXs8woVF1uv6+8jgqFfMa0asb6JrddF2ZRyrB0ZqcIWa83/mZhP2OIr/3ofryldfVa1B9otfcrkZPD5PzLdr83/kYnLqGHy2TUU5WWvW+m3YzH/csP6x/JmfL/AGRPs0VJL2bfSxkf+4x+f5nEf+6x/wAS/JmXf7nT9EAAAAAAAAABgv8AkcVP+3gAAAAAAAAAqfA/Q4a/5e/L9QAAAAAAAAAOIf8AeKX9dQAAAAAAAAACf/I8+7/0kAAAAAAAAAA4h/3Neq/Uyv8A3On6AAAAAAAAAAAAAAAAAAAAAAAFeMnmlJ205X+pnEWlSqJfDJf1+BmkK2HrxxdJXsrNeX9fdoZfmlXGVbeztFLffX1sjN8POpiaPKtHo/S6Zn1OUsOpQV2n+DTRjMJKeX+xjukrfK3+RUxeJxuGdJQsorV97bf8MY+MpYPD2XWP5GOw/wBYw86a3a09ehRxuLr01guS0tm+y9PQyWMo4KKas9fzOI/91j/iX5My7/c6fogAAAAAAAAAMF/yOKn/AG8AAAAAAAAAFT4H6HDX/L35fqAAAAAAAAABxD/vFL+uoAAAAAAAAABP/kefd/6SAAAAAAAAAAcQ/wC5r1X6mV/7nT9AAAAAAAAAAAAAAAAPEPx1q4fEzwHDzi+XR1WubX+4no0v5mmn0VrN5X40cY4LEe1nivaxbu4zjFp+lknH/t1peRwJx3l/FuX/AFjD+7UjZTg3dxb/ADi7e7K2vWzTSAAAAAAAAAAOIYSlhYqKv7y/JmXprCU0+yAAAAAAAAAAwdOazaba094AAAAAAAAAKnwM4cpyj7XmVtv1AAAAAAAAAAz+nOVenyq//DgAAAAAAAAAEsvrvM/rGnL/AMCwAAAAAAAAABm+EqYnD8lPe5gaMqOGhTlukAAAAAAAAAAAAAAAHjfxjLJMi+pYedq2JvHTdQXxvyvdRXq2tUZV4LZrmHC/9twqpTcXONLld5RXXmvo5JXirNNW1V9OCODsXxVm8Mvw8uVWcpSauoxW7tpd3aSV1dtXaV2ngsw8K+MKE51PaUpLVpOKnTbtJNa2cWr2u9VF9SlVp1qcalN3jJJprZp7NeoAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB42Z3LMuLq9NSvCglTXyV5fPmbXyRl2Dhg8HRw0FZQjGK9IpL9DwMyqlhs5zpxWlOapx9FKpf8on0kcNCWCy+v9pSnH5NRb/9JR4L54804Qw6k7yot0n/ANu2cfug4oAAAAAAAAAAlOMVeTsJpq6AAAAAAAAAAVSDlypq4AAAAAAAAAEakJ/C0wAAAAAAAAAJVIR0k0gAAAAAAAAAB5hQWJ+r397/AIFwAAAAAAAAADFYqnhqfPU22KNaFamqkdmAAAAAAAAAAAAAAAGcY147McRi27upOUv/AAKTf6mVY2GNy6hi4u6qQjK/lKKd/wATwJzelis5zmMX/EkqkV5c07v/AMmifSRxkFhsvw1/ecpy+SUV+N/wPo3Y5yoZhhHsnCS+akn/AOkoAAAAAAAAAA4j/wB1j/iX5My7/c6fogAAAAAAAAAMF/yOKn/bwAAAAAAAAAVPgfocNf8AL35fqAAAAAAAAABxD/vFL+uoAAAAAAAAABP/AJHn3f8ApIAAAAAAAAABxD/ua9V+plf+50/QAAAAAAAAAAAAAAAxzawtRr+V/kGA8W89wXDTyKly8tnFT154xe8VrbrZO2i0XRrhTinH8NZnDMME1zK6ae0oveL8nZejSZxlxhj+KczeOxlk0uWMVtGK6K+vVtvq30Vkvo4ytm+NjbemvTSX/B09GAAAAAAAAAAcR/7rH/EvyZl3+50/RAAAAAAAAAAYL/kcVP8At4AAAAAAAAAKnwP0OGv+Xvy/UAAAAAAAAADiH/eKX9dQAAAAAAAAACf/ACPPu/8ASQAAAAAAAAADiH/c16r9TK/9zp+gAAAAAAAAAAAAAABWpxq05U3s01954fcHVOIuI6WX1Ivki3Kp5Ri9VdbNu0U+jdzjvhKtwxndXL53cFrCTXxQez+WsX/eTDgvwhea8HYrMMRG1eqr0b9FHW//AHEa5ddo2a3Po3YeUsbmFZbRjBf+BOT/APbWAAAAAAAAAAcR/wC6x/xL8mZd/udP0QAAAAAAAAAGC/5HFT/t4AAAAAAAAAKnwP0OGv8Al78v1AAAAAAAAAA4h/3il/XUAAAAAAAAAAn/AMjz7v8A0kAAAAAAAAAA4h/3Neq/Uyv/AHOn6AAAAAAAAAAAAAAABWzatwN4i4muk+SNSXNFdadT3reqi4teaRxvwVlnHWUUp06iU7c1KolfSSTs1peMlbTRp2fSz4T+j7jo5gqud1Iexg/hg23O3m0uWL/8Ce1o3ueM/GlHh/I1leDaVavHlSWnJT2bstr/AAx26tfCfR4yuVDh6vjJKzq1LLzjBJX/APAnJfIAAAAAAAAAA4j/AN1j/iX5My7/AHOn6IAAAAAAAAADBf8AI4qf9vAAAAAAAAABU+B+hw1/y9+X6gAAAAAAAAAcQ/7xS/rqAAAAAAAAAATwld5v7Xl93TX/ALdsAAAAAAAAAAZ3QqVsMo01d3X6mX05U8LCMlZpAAAAAAAAAAAAAAAB9IjhqdLG4fOacfcmvZy8pRu4/wDgUbr/ALdPC/xeqcNUll2YRdTDX91r4qbe9k94t6tXTTu1e9jPfHvhjCYRyy+Uq9VrSPLKKT/vOSWi/uqX6rNc0zPiXN5Yiu3Ur1pJJLu3aMYrolokjhHIY5FkeFy2O9OKTts5PWTXrJt/MAAAAAAAAAAlCM1aSuJJKyAAAAAAAAAAVOClzJK4AAAAAAAAAEacIfCkgAAAAAAAAAJU4S+JJgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABxRw7heIcprZbifhmtH1i1rGS807Pz2ejOJ+GMx4dzGeBx0LSWz6SXSUX1T/DZ2aaDwO8N608RDiDMIWhH+FF7yb052v5Uvh7t8yskmwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHi6Cq+x5lzdvxAAAAAAAAAAK1enRjzVJWRTqQqQUoO6YAAAAAAAAAAAAAAAGe8N5TnuH+r5lQjVir2utVfdxatKL800zK/B7g7L8T9Yp4Tmkndc8pSS8lFtp26cyb8xJJWQAAAAAAAAAAcQylHCxcXb3l+TMvbeEpt9kAAAAAAAAABg5zebTi3p7wAAAAAAAAAVPgZw5OU/a8zvt+oAAAAAAAAAGfznGvTUXb/hwAAAAAAAAACf8AyPPu/wDSQAAAAAAAAADiH/c16r9TK/8Ac6foAAAAAAAAAAAAAAAAAAAAAAAAAAAcR/7rH/EvyZl3+50/RAAAAAAAAAAYL/kcVP8At4AAAAAAAAAKnwP0OGv+Xvy/UAAAAAAAAADiH/eKX9dQAAAAAAAAACf/ACPPu/8ASQAAAAAAAAADiH/c16r9TK/9zp+gAAAAAAAAAAAAAAAAAAAAAAAAAABxH/usf8S/JmXf7nT9EAAAAAAAAABgv+RxU/7eAAAAAAAAACp8D9Dhr/l78v1AAAAAAAAAA4h/3il/XUAAAAAAAAAAn/yPPu/9JAAAAAAAAAAOIf8Ac16r9TK/9zp+gAAAAAAAAAAAAAAAAAAAAAAAAAABxH/usf8AEvyZl3+50/RAAAAAAAAAAYL/AJHFT/t4AAAAAAAAAKnwP0OGv+Xvy/UAAAAAAAAADiH/AHil/XUAAAAAAAAAAn/yPPu/9JAAAAAAAAAAOIf9zXqv1Mr/ANzp+gAAAAAAAAAAAAAAAAAAAAAAAAAABicLRxMFCqrrfr+hTpwpwUILRAAAAAAAAAAQwOHhWdaMfefW7/zsAAAAAAAAADSasYbBUMNf2UbX31fT1AAAAAAAAAAxGBw+IkpVI3a21f6MAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAD2kOblur9gAAAAAAAAACUoxV5OwmpK6YAAAAAAAAAAAAAAAAAAAAAAAAAABnlerRw0ZU5Wd/0ZgZynhqcpO7aQAAAAAAAAAGExdaeZzpOV4q+gAAAAAAAAAT0i7GQ4qtX9p7SV7W/UAAAAAAAAADO8XWo1qcacrJ/wCYAAAAAAAAABP/AJHn3f8ApIAAAAAAAAABxD/ua9V+plf+50/QAAAAAAAAAAAAAAAAAAAAAAAAAAA4j/3WP+JfkzLv9zp+iAAAAAAAAAAwX/I4qf8AbwAAAAAAAAAVPgfocNf8vfl+oAAAAAAAAAHEP+8Uv66gAAAAAAAAAE/+R593/pIAAAAAAAAABxD/ALmvVfqZX/udP0AAAAAAAAAAAAAAAAAAAAAAAAAAAMbgqeLpqnNtJO+n9Mo0Y0acacdkrAAAAAAAAAAUsto08S8Qm+Z39NfkAAAAAAAAAEkmrGBy6lg+b2bbvbfy+S7gAAAAAAAAAYzLaOKnGU27rt/wzAAAAAAAAAAHh6Ln7TlXN3tr94AAAAAAAAABUpU6seWcU15q5GEYRUYqyQAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABXxNGhHmqzUV3bS/Mw+Mw2JTdCpGVuzT/IAAAwOY0Mb7T2TfuScXddUAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABxhhYYrEYKhN2UpNO2+ttjM8k/wBXqtLH4So3FNKSe9n6JaPbbR2+WjQAAHCX/MX/ANfZAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAcSf8AIwy//H/8icbSX9lOn1lKKXre/wChnWavLMLDkjz1JNRiu7MozzHvG/UsypqE5K8Wtn1tu1t59LPUyrNK2JxuKw1VJezatZPVO+929duxgs6niM4r4FpcsEmu/S99e77Ec0rTzmWCilyRhzPe97rTe2zXT5lXirOotYt4e1D0e3r+treRm+bSw2Uyx2Hs9ItX2tJpdGuj7mV8QVcfmtWlFWowi7Pu00r326vTtqT4tzKpXniMPR5sPB2eju11d+jtr2XU4Nqxq08TUjs6kmvnYODM6x2YzrLFVOblStolvfskAABnWKq4fDxnSdne34PuYKpKphoTk7tpAAAAAABxrnONy72H1WfLzc19E9uW26fdlJuVOLe9gAAMLjsRPMp0ZS91X0sun4gAAAAABnWdY7D59QwtKpanJwurLrKz1avt5gAATdotoyPG18T7T2sr2tbRLv2AAAAAADLc6x1biGrhJ1L005WVl021tf8AEAAAzjHYjD1qcaUrJ+S7+YAAAAAAGHwdGeAlVcfeV+4AABPHYhZr7BS9ztZfy39QAAAAAAMLCNSvCMtmzNcPSoVlGmrK36sAADOcTVw+G56Ts7r+tTAVZ1cNCc3dtAAAAAAAAAAAAAAAAAAAAcafWPbYP6t/E5ny7b6W30+/QwUcfVzuhDOW+ZXcF7tr/wDbunT1va5nOYYXD5/QlipWhCDa0b95trZJ9EZvnuXYzF4OeFqc041F0ktHZPdIwq9jxNXj/PTUvusv8zJm45vSxP8Az+lWX3Wa/IyZe2zjHV+zjFfJa/kjiLiTLKdCrg23KTTjaK2bXVuy0fa9uxiZufBqb7RX3TSK1OOX8NOMNGqf4yWr+9nD+FhQyqhTS3im/WSu/wAzgujGjRxNKO0ajX3JIPDr+LiPSP6gAAcR/wC6x/xL8mZd/udP0QAAAAAAeI3/ADDf9v8A/tpR/hR9EAABgv8AkcVP+3gAAAAAA4i/5KjC+tP/ANKAAAqfA/Q4a/5e/L9QAAAAAAyf/krK/rMAAA4h/wB4pf11AAAAAAAwv/Isn8wAACf/ACPPu/8ASQAAAAAAMD/vNP1Rnn+8R9P1YAAHEP8Aua9V+plf+50/QAAAAAAAAAAAAAAAAAAADiT/AJGGX/4//kTi9ezhhcSt4VF9z3/JGY4LDV+JacMRBSjOnon3Tb/JFPh3Kqc1ONBJrVeqM+q/U82oYldYVF/4DFyX4jw/1XK8sq/yzi36Tu2ZPXdDKMXjY7ylUmvyX4oyvKcLRyGdeUU5zhKTk9Xqm93tb89St/yRi9F/6cRndNz4eml/In91mZNUjVy6hJfyR/JHB8ozjipR2dSX6BkvDuFymU3QlJ81r3a6eiXcAADG4Kni6apzbSTvp/TKNGNGnGnHZKwAAAAAAZ1w9hc29n7eUlyXtZpb2ve6fYjFRioroAABSy2jTxLxCb5nf01+QAAAAAAY3h3C4vH08bOUlOFrJNW913V9G/XUAACSTVjA5dSwfN7Nt3tv5fJdwAAAAAAw3DuFw+Yyx8JS55X0bVtd+l/xAAAMZltHFTjKbd12/wCGYAAAAAAFPH1IUHRSVnf11+YAAA8sovFfWbvm/Da3b9QAAAAAAKVR05qa3Ri8VPEzU5pXStoAABjMHTxdP2c27b6f0yhQjRpRpx2QAAAAAAAAAAAAAAAAAAAGf4etVx2BlTg2ozu2k3Zaavt8zi3B1MVlNSFKLlJNNJK73XT0ucRYHGYnDUMdhU/bU7O1tdUrq3ddvVeRlHEGb4/F06csPywV+Z2fbu9temr/ABONcDiMThaTw8XKSlbRXdmmnt02TOIcDOeTyo0E3KKjy2WvutbL0MsyyX9gxwc1yuUGnfo5Xe3k2YWWd4jCvKHScYxupSs78q1sntrsrbryuypg8TLhL2Cpy57L3bPm/idt9tfTUhh1VwKo1Fo42fzVmYTEZ9gIyymnSu9VGVnon1T2t1V9n32OEsvq4GliKM4tJTdm1a6SSTXkwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADhTiLFZtOrGvGK5UrWTW9+7fYAADNsbUwlBTppNt219H/AJGEqyrUIVJbtXAAAAAADiziDFZT7H2EYvn5r8yfTl2s13KcnKCb6oAADD5lVqY+WHaXKr976fMAAAAAAM24ixWEzmjgYRi4T5btp396VnbW34AAASdotmUZjVxnP7RJWttfrfzfYAAAAAAMBxFisRnlTASjHkjzWaTvp87fgAAAZpmVXC1YQgk797/5gAAAAAAUcBSqYOVZt3V/TQAAAlmVVZj9WsuX532v3/QAAAAAADDU41a0YPZszLCQw1VQg21a+vzAAAzTGTwtD2kEm721MHWlXw8Kkt2gAAAAAAAAAAAAAAAAAAAAAAADw6/i4j0j+oAAHEf+6x/xL8mZd/udP0QAAAAAAeI3/MN/2/8A+2lH+FH0QAAGC/5HFT/t4AAAAAAOIv8AkqML60//AEoAACp8D9Dhr/l78v1AAAAAADJ/+Ssr+swAADiH/eKX9dQAAAAAAML/AMiyfzAAAJ/8jz7v/SQAAAAAAMD/ALzT9UZ5/vEfT9WAABxD/ua9V+plf+50/QAAAAAAAAAAAAAAAAAAAAADG4yhg6Eq1aVooyni/CZjivq0ISi3e17a211ttp8vPvl2aUcf7T2Sa5JOLvbddrN6FLNKNTHzwST54JNvS1nbbW/XsZ1xPg8rmqdROU3rZdF56/1+Jl+PoY/Dxr0HeL+9eT8/62Dw6/i4j0j+oAAHEf8Ausf8S/JmXf7nT9EAAAAAAHiN/wAw3/b/AP7aUf4UfRAAAYL/AJHFT/t4AAAAAAOIv+SowvrT/wDSgAAKnwP0OGv+Xvy/UAAAAAAMn/5Kyv6zAAAOIf8AeKX9dQAAAAAAML/yLJ/MAAAn/wAjz7v/AEkAAAAAADA/7zT9UZ5/vEfT9WAABxD/ALmvVfqZX/udP0AAAAAAAAAAAAAAAAx+Y4PAUHXxdWNOC3lJqK+9tIyzjzhnM66w+Ex1OVR7R5rNvsk7XfkrsAAAAAOJaX1/MMJlzfutuUvRf8M18yngMJSlGVOnFOO1klY4S/5i/wDr7Iwf/JT4n/BH/wBtOGaMMVUxOPqK8pzaX+FaJfo/Q4epxwmZY3BQ0inGSXbmV38tkHh1/FxHpH9QAAOI/wDdY/4l+TMu/wBzp+iAAAAAADxG/wCYb/t//wBtKP8ACj6IAADBf8jip/28AAAAAAHEX/JUYX1p/wDpQAAFT4H6HDX/AC9+X6gAAAAABk//ACVlf1mAAAcQ/wC8Uv66gAAAAAAYX/kWT+YAABP/AJHn3f8ApIAAAAAAGB/3mn6ozz/eI+n6sAADiH/c16r9TK/9zp+gAAAAAAAAAAAAAAAZ7nWEybLq2PxcrU6abfd9kvNuyS7s4042zPinMJYnFytBfBBP3YLyXd/alu32Vkk2ndHgt4n1sfNZHmtTmqJfu5t6ytvCTe7S1i92rp6pXAAAAAxTtxTQv1ptL1vL9A4St/tf/X2Rg/8Akp8T/gj/AO2mQ8I0cywjrTqtSTaaS7GRcNUspqTnCo5cytqg4Y4bq5ROpKc1Lmtsn0v/AJgAAZpgZYyiqcXazv8Ag/8AMwtF0aEKbd7KwAAAAAAcUcO1c39l7Oajyc2/nb/IhHlgo9gAAKGWTp42WIclZ309QAAAAAAzPhurjM3o46M0lDl0s7+67gAASV4tGVZbPB8/NJO9vwv/AJgAAAAABgeG6uHzmpj3UTUubSzvqAAAZllk8XUhOMkrAAAAAAAUcfGGElQtq7/iAAASyybzD61zK3b5WAAAAAAAw9RUqsZvozMMXHE1VNK1lb8wAAMzwcsXQ9nF21uYSg6FCNNu7SAAAAAAAAAAAAAAAD6RPEzlUw2SUpaL95P1d1BP0XM7eafY4f8ACjJJ8G08vxFCPtqsFKVS15qo1dNS3tC9klZWvde9K/gpwZhs24hrSzCmpwwy1i1dObdkpJ6NK0nZ9Uulzxs4cpcN55hM3yuKpe0u7RVlGpTafMktFe60tum+pw3nNLOsow2YU9qsVK3Z21Xyd18gAAAAOL41sNUw+ZUVd0nZ+j/TdfMy/izA4/E08PRT5pX3VrWV++vyKecwyHNcXSrxbhN8ytbrr1a01t8jhmrXxGdVsRWVnUhzJdldKP4JGHzWPD+Y4jDYmL9nN88WvP8ATp5NGT53h80jOVBO0XbXr5gAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB4k5q8z4rx+IvdKbivSHuK3yVylGEacVT+FJW9Oh4LRprOc/tv7ZfdzVbfqfSPS/svAv+/L/ANJPo+Zq8Tw1VwknrRqO3lGSUl/5NzAAAAAThGcXGSun0ZhMqwOEk5UKSi31S1/4byMVlmDxc4zr0lJrZtf1p5bCoUlU9oormta9tbdr728jGZfhMZFRxFNSttdbGHwtDDU1TowUY9krBw7xL/bEqkfZcnLbre97+S7AAAZjjvqdJVOW93be3R+vYw1b21GNS1rq4AAAAAAcR8R/2P7L91z89+trWt5PuQlzQUu4AAFHNPa4yWG5LWvrft5WAAAAAADMeJfqeaUsB7K/Py63tbmdtrPb1AAAk7RbMszP67ze7y8tut97+S7AAAAAABg+JfrObTy/2VuW/vX7eVuvqAAAZhmn1SpGHJe/nb9GAAAAAABSy/2mFdfm2vpbt8wAACWaWx31Xk+d/K+1v1AAAAAAAoUva1Ywva7sY7CfVaihzX0vtYAADMMb9Uo+05b623t/mYWv7ejGpa10AAAAAAAAAAAAAAAVakacHOWyV/uK9aderKrPeTbfq9Th/jfLqvB1POqtRctOmufXacUk4+rekVu7q26PAfienT4kxWHxM7PFK6v1nFt29WpSt6W3Z9IjiDD4rMMLllGd3RUpTt0lO1k/NJXt2kj6N2JaxWYUO8ab+5yX/twAAAAAAB4dfxcR6R/UAADiP/dY/wCJfkzLv9zp+iAAAAAADxG/5hv+3/8A20o/wo+iAAAwX/I4qf8AbwAAAAAAcRf8lRhfWn/6UAABU+B+hw1/y9+X6gAAAAABk/8AyVlf1mAAAcQ/7xS/rqAAAAAABhf+RZP5gAAE/wDkefd/6SAAAAAABgf95p+qM8/3iPp+rAAA4h/3Neq/Uyv/AHOn6AAAAAAAAAAAAAAAGP8A91q/4X+Qczta+gSk5Ntu7Z9HFP8AtbGu+ns46f8Ab39feAAAAAAAeHX8XEekf1AAA4j/AN1j/iX5My7/AHOn6IAAAAAAPEb/AJhv+3//AG0o/wAKPogAAMF/yOKn/bwAAAAAAcRf8lRhfWn/AOlAAAVPgfocNf8AL35fqAAAAAAGT/8AJWV/WYAABxD/ALxS/rqAAAAAABhf+RZP5gAAE/8Akefd/wCkgAAAAAAYH/eafqjPP94j6fqwAAOIf9zXqv1Mr/3On6AAAAAAAAAAAAAAABi8qrU81nl0I3mqjppealypfecXcH5nwxmDweOj5xkvhmu8X+a3T3Em3ZGdeHub5PkFDOMbHkVWXKoNe8k4tpy7Xs9Hr3tex9G3BvlzHEv/AKdxX/k7f/toAAAAAAB4dfxcR6R/UAADiP8A3WP+JfkzLv8Ac6fogAAAAAA8Rv8AmG/7f/8AbSj/AAo+iAAAwX/I4qf9vAAAAAABxF/yVGF9af8A6UAABU+B+hw1/wAvfl+oAAAAAAZP/wAlZX9ZgAAHEP8AvFL+uoAAAAAAGF/5Fk/mAAAT/wCR593/AKSAAAAAABgf95p+qM8/3iPp+rAAA4h/3Neq/Uyv/c6foAAAAAAAAAAAAAAAHGdLC8L+J0cdi4OVFzjWst9dW135aibt1tbQrYbIuKsri5xhiMPPVdV6p7xktntJO6dncyfwx4SyjErE4PBRVRbOTlOzXVc8pWfmrM8eeMMnjlMsjT58RJxdltTs07yfdq6Ud7O7srX8D8keXcI0qk1aVeUqj9HaMfvjFNeoAAAAAABwhkOMyydV4hL3krWd9rgAAZxg6uKoKFPdO/4MwdKVLDwhLdJIAAAAAAOMMixmaex+rpPl5r3dt+W35FOLjCKfRAAAYbL69PMJV38Lv+IAAAAAAZxkOMxWeUMZTS5IuF9ddJXenoAABNXi0jJcBWwvP7Trb8LgAAAAABl2Q4yhn1XGzS5JOVtdddtAAADNsvr4mrCVPZAAAAAAAUMbRhgpUX8TuAAASy+u8z+sacv/AALAAAAAAAYapGnWhOWyZmeJp4ispQ2tb8WAABm+EqYnD8lPe5gaMqOGhTlukAAAAAAAAAAAAAAAHj3wjPM8nhmmHjephr81t3Te77+60n5JyZwzxrnvDlRyy3EOCbu46OL9Yu6vbS6s7bNGY+NfGWNoOj9ZVNPdwioy+T1a9VZ+Zwnw7iuJc7o4CndupK8pb2jvKT9F33dluzCYWlhcPTw9FWhBKKXZJWS+5AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAE4RnFxkrp6WZ4i+COPwmKnjMhp+0oS1dNfFBvdJfaj2teS2aaXMZd4a8W4+uqNPL6kW+s4OEV6ymkv18jw18N8Lwjg25SVTE1Pjmlol/LG+vKn1dnJ6u2iQAAAAAABlWfYPM3JYdt8tr3Vt/8AhgAAMXi6WFgp1Nr2KVWNWmpx2YAAAAAAZtnmDyvk+sN+9e1lfa1/zRGSlFNdQAAKeYUKld0Iv3lf8AAAAAAAxefYPC42GDqN88rW001dlqAAA2krmDx9HF83snt+v/DAAAAAABQz7B18dLBQb9pG99NNN9QAADFZhQw0oxqPVgAAAAAAQwVWdF1l8K/QAAAeYUFifq9/e/4FwAAAAAAKdOVSahHdmJw1TDzUJ77gAAYrFU8NT56m2xRrQrU1UjswAAAAAAAAAAAAAAAAAAAAAAADw6/i4j0j+oAAHEf+6x/xL8mZd/udP0QAAAAAAeI3/MN/2/8A+2lH+FH0QAAGC/5HFT/t4AAAAAAOIv8AkqML60//AEoAACp8D9Dhr/l78v1AAAAAADJ/+Ssr+swAADiH/eKX9dQAAAAAAML/AMiyfzAAAJ/8jz7v/SQAAAAAAMD/ALzT9UZ5/vEfT9WAABxD/ua9V+plf+50/QAAAAAAAAAAAAAAAAAAAAAAAA8Ov4uI9I/qAABxH/usf8S/JmXf7nT9EAAAAAAHiN/zDf8Ab/8A7aUf4UfRAAAYL/kcVP8At4AAAAAAOIv+SowvrT/9KAAAqfA/Q4a/5e/L9QAAAAAAyf8A5Kyv6zAAAOIf94pf11AAAAAAAwv/ACLJ/MAAAn/yPPu/9JAAAAAAAwP+80/VGef7xH0/VgAAcQ/7mvVfqZX/ALnT9AAAAAAAAAAAAAAAAAAAAAAAADw6/i4j0j+oAAHEf+6x/wAS/JmXf7nT9EAAAAAAHiN/zDf9v/8AtpR/hR9EAABgv+RxU/7eAAAAAADiL/kqML60/wD0oAACp8D9Dhr/AJe/L9QAAAAAAyf/AJKyv6zAAAOIf94pf11AAAAAAAwv/Isn8wAACf8AyPPu/wDSQAAAAAAMD/vNP1Rnn+8R9P1YAAHEP+5r1X6mV/7nT9AAAAAAAAAAAAAAAAAAAAAAAADgfK8ZgqlZ4im43UbX8rgAAZ5h6tfDRjTjd3/RmBhKGGpxkrNJAAAAAABxzlmMxv1f6vTcuXmvbpflt+RSTVOKfZAAAYTCVo5nOq4+676gAAAAABnmV4yvxDh8RTptwi4XfRWld/gAABNXi0jIcLWoe09pG17fqAAAAAAGV5XjKXElXEzptQblZ9NdgAADO8JWrVqbpxul/mAAAAAABh8TRjl8qbl72ugAABPCV3m/teX3dNf+3bAAAAAAAYSUYV4Sk9E0ZvWp1q6lB3Vv1YAAGd0KlbDKNNXd1+pl9OVPCwjJWaQAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAGCzTB41tYeopW3t0AAAr4ilQjzVJWRCcakVKLumAAAAAAGNzPB4Ll+sVFHmva/W2/5oTTSaAAAji6E6jpRleS6AAAAAABXzTB0cRHD1KiU5WsurvovvAAAdkihiqNe/spXsAAAAAAFPNMHVxDw0KidRXuuum4AABWxdCi1GpKzYAAAAAAEcNVlTdRR0XUAAAeLoKr7HmXN2/EAAAAAACMJTkoxV2yrRqUpcs1ZgAAVq9OjHmqSsinUhUgpQd0wAAAAAAAAAAAAAAAAAAAAAAADw6/i4j0j+oAAHEf8Ausf8S/JmXf7nT9EAAAAAAHiN/wAw3/b/AP7aUf4UfRAAAYL/AJHFT/t4AAAAAAOIv+SowvrT/wDSgAAKnwP0OGv+Xvy/UAAAAAAMn/5Kyv6zAAAOIf8AeKX9dQAAAAAAML/yLJ/MAAAn/wAjz7v/AEkAAAAAADA/7zT9UZ5/vEfT9WAABxD/ALmvVfqZX/udP0AAAAAAAAAAAAAAAAAAAAAAAAPD+hVpVK/PFrSO6a7gAAcQwlLCxUVf3l+TMvTWEpp9kAAAAAAHiFQq1fq3s4t259lf+Uor91H0QAAGDpzWbTbWnvAAAAAABxBQqy4mw04xbSdPWzt8QAAFT4GcOU5R9rzK236gAAAAABlFCrHimtNxdrz1s7AAAGf05yr0+VX/AOHAAAAAAAws4LLZq+uoAABOnP8Atrmtpp/6SAAAAAABgmliIN90Z1KMsRHld9P1YAAGfwlLCJRV9V+pliawdNPsAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABgs0weNbWHqKVt7dAAAAAAAAAAMbmeDwXL9YqKPNe1+tt/zQmmk0AAAAAAAAAFfNMHRxEcPUqJTlay6u+i+8AAAAAAAAAAAAAAAAAAAAjhqsqbqKOi6gAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHh1/FxHpH9QAAAAAAAAAPEb/mG/7f/wDbSj/Cj6IAAAAAAAAAOIv+SowvrT/9KAAAAAAAAAAAAAAhRqTjKUVdLfyAAAAAAML/AMiyfzAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPDr+LiPSP6gAAAAAAAAAeI3/ADDf9v8A/tpR/hR9EAAAAAAAAAHEX/JUYX1p/wDpQAAAAAAAAABRSdSK80YrNqVCu6Eoe6ra/K+1jNsLChiLQ+Fq6AAAy/8A3PEei/UAAAAKuElToQqt/Ff8AAwv/Isn8wAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAofxY+qM5/32fy/JGbX5aEXuoop5G5QSlUSm1dL+tf8jD4GVX2kW7Sgm7W7dNzAYJ4uo4J2srmDwrxNdUr2vf8AIwWVrEc0pTtFO39djMcBLCTSvdPZmGyZ1pQ97Rq7028jAZa8TOV3aMep9ReFw+IV7xaVn94f6uf9Pfw/4Ji8vlRxCoQfM35W/wAypkLVN8k7zXQweF+s1PZ3to39xDIJyw/Pze/bb9CnSnUmoRWrK2S8tJunUUpR3X9f0wwOXvEpylLlgt2zH4CWFkrO8Xsyhkkqso+97rint36f8ExuXSw1eNO909mVsn9rh6dHnty31tvf5mGyyWIpxlGW7t6aXv8A8Ar5WoRqyjK/I1pbvbX+uxgcH9aqOHNZJXuAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAUP4sfVFbAYOtinOcrz/luui7bl5YnM0qitZ7eSMRDGSxbrKnLR6aPpsRjGGbSj/Mv0/4Bk37rnm+8Y/e9TBQ9jjK8v5FL+vuKUKtV+zgm/Ix1GrTy6nGqrSTt8tTHYidPA0IQdrr8v8AhytJ0sspwT+Ntv8Ar7jA1ZSy+vTfT9f+GDKsOq2JjfZav5GCrKtjamIe0U2vyX4GBxM44yNRvVvX57mEpKnm04+v46mCxc55gqjfxO3yf+Rg6XLj60kruPM16mWUcVTxcZyg7O97p9e5iIKFacF0bX4lajV+oUqdKLd7t2T+X9eROlVWWNVItOMtLq2j/wCHM0rSWEoU1s4pv7lYxknVy+jOW6bX9fcY3/cMP8zD15UcqlKL1bsvw/Qyluqq1J6uUfxX/DmW/u8NXq+Vvv8A6QAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHh1/FxHpH9QAAAAAAAAAPEb/AJhv+3//AG0o/wAKPogAAAAAAAAA4i/5KjC+tP8A9KAAAAAAAAAAKH8WPqjNZuGYSlHdW/JE1GObwkvtK/4NFfOcXCrKF1o2tjD4ypVx1OrPe6X6GMTw9CSW7qN/JGMSpxxNRfaUbfPcoVng8AqkPim9/JGJrVK2WQlN3fN+jMy/3fD/AOH/ACMeubBYdrs/0/yMvX+x4j0X6hRX1bLpVPtVNF6f1f8AAyn3oV4LdxZhouVaCXdfmUmpZxO3b9EZf/vdP1Rhpyji8Ty72bXyP7dxfdfcVJupNze7dytj62HwlB0rWa/Kxic0xGIpunNqz8jNP4WH/wAK/JFf/kV0v8T/AFMb/uGH+Z/zKv8At79DKanJjIeen3mKp/V8FOH8039y/wCGDC/8iyfzAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPDr+LiPSP6gAAAAAAAAAeI3/MN/wBv/wDtpR/hR9EAAAAAAAAAHEX/ACVGF9af/pQAAAAAAAAABQ/ix9UZz/vs/l+SMdVcPq2IXZfgYjKaeMmq9GdlL+v+HXcrwVDEOMXflZn8488Ix7N/f/wxmFVSy6k+rt+CsYTDxx2AVJStKD/P+vwMxjSjl8adJ3UZWv52d/xZmX+74f8Aw/5GApwxuDeGbtKLuv6+8dGnhsHVoRd5JXfz6f1+pCPPJR7mMyj6woRU7RirJWv890QpxyzHQUpXTWult9PPsQyyjhKrxMp+4tUv6/DuZTVdXMHN7u7Mv/3un6ojiY4fM5yls20/mTyCDm5qpaG/9f5hgYU8bhPq0naUXdf1/XQx+WfVKabndt7GafwsP/hX5Ir/APIrpf4n+pRwyxuXwjGVnBv+vyP+ZV/29+hTm4TjJdHcz+pFypwj2b+8ML/yLJ/MAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA8Ov4uI9I/qAAAAAAAAAB4jf8w3/b//ALaUf4UfRAAAAAAAAABxF/yVGF9af/pQAAAAAAAAAAApySsnoAKTjswE2ndPUeoA5SaSvoAOcnHlvoCbTuiUnJ3buwFKSWjDBYZ4ivGmvn6dTNayq4qTWy0XyDC/8iyfzAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPDr+LiPSP6gAAAAAAAAAeI3/MN/2//wC2lH+FH0QAAAAAAAAAcRf8lRhfWn/6UAAAAAAAAAAAAAAAAAAAYHHvC06iitZWt5bj1DC/8iyfzAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAxeLw+EoyrYiooQjvKTSS9W9EZbxjw/mdZ0MHjadSabXKppt27K92vNXXmAAAAAAABh8DhcM26FKML72SV/uAAAAAAAAAAxOBwuJt7enGdtuZJ272uJJKyAAAAAAAAACpgcLVqqrOlFzWzaTattZ76AAAAAAAAAAAAAAAAAAAACrVFDkUnbtfQAAAAAAAAAAAAAAAAAAAAAAAAAAAAAM5zfCZRgKuOxcuWnTV2/wBF3bdkl1bSOOeO8y4rx7r4mVqUW+SC2iv1k+suvSyslGTi04uzR4L+KFbNGskzSfNWS/dzb1mlvGXeSWqf2knfVXkAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfSI4omnh8joy0f7yf4qC/Ntf4X0OHfCHIq3BtPC4ihF4irBSdS3vxnJcys90o3SstJJap3Z4McFYbOuIKv8AaNPmp4dXcHs5t2SkuqVm2tm0k7q54xcO0eEuIMHm+UQVJT95KKtGM6bWyWiTTXu7PXuzh7OKWc5Vh8wpaRqxUrdm1qvk7p+gAAAAAABwHjsVialdV6sp2UbXbdt+7AAAAAAAAAA49xuKw31f2FSUL817Nq9uXezKLbpxb7IAAAAAAAAAM+x2Kp8R4ajCrJQbhdJtJ3lrdXtr1AAAAAAAAAAAAAAAAAAAAMNRpSy6U3FX11tqAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHiZm0s04sx1e90puC9Ie4reT5b/ADuU4QhCMYbJaeh4K06cc3z/AJd/bJW7JSq20+8+kel/ZeBf9+X/AKSfR+zZ4rhieEk9aFRpf4Ze8v8AyZyAAAAAAAPDr+LiPSP6gAAAAAAAAAeI3/MN/wBv/wDtpR/hR9EAAAAAAAAAHEX/ACVGF9af/pQAAAAAAAAAAAAAAAAAAABhf+RZP5gAAAAAAAAAAAAAAAAAAAAAAAAAAAAFScYQc5PRamIrzr1p1Z/FJtv1buzhzjHAV+D6OdVaiUIU0567SirSj6uStFbu6tujwF4mhHiPF4fEStLFLmXnOLbt81KT+R9IrPaNfG4TLKU03SUpTS6OVlFPs7Ju29pJ9T6N2KaxOYYfo405fc5L8ebt0AAAAAAAPDr+LiPSP6gAAAAAAAAAeI3/ADDf9v8A/tpR/hR9EAAAAAAAAAHEX/JUYX1p/wDpQAAAAAAAAAAAAAAAAAAABhf+RZP5gAAAAAAAAAAAAAAAAAAAAAAAAAAAAGP/AN1q/wCF/kCrVFTdJSfK3e19L97dyE5QkpRdmtvIqVJ1Zuc5Nye7erZ9HGMnm+Naens1p/29v8tfvAAAAAAAPDr+LiPSP6gAAAAAAAAAeI3/ADDf9v8A/tpR/hR9EAAAAAAAAAHEX/JUYX1p/wDpQAAAAAAAAAAAAAAAAAAABhf+RZP5gAAAAAAAAAAAAAAAAAAAAAAAAAAAADSaszMcI8HjKuGlvCUo/wDgLa/Q8NvB/FcT01jsbN0sLfRpe9Pvy30SX8zTV9EnrbO/o8ZLWwrWWV506qWnO1KLf95JJpvunZfyszvJMdk2PqYHHQ5KkHZr8mn1TWqZ9G7BP/hQxT/6dxX/AJM3/wC2gAAAAAAGVZDg8scnh0/ete7vsAAAAAAAAABm2RYPNOT6wm+W9rO29r/kiMVGKS6AAAAAAAAABi8hweKxsMZUT542trpo7rQAAAAAAAAAAAAAAAAAAAAhjasKLop+6wAAAAAAAAAAAAAAAAAAAAAAAAAAAAA4p4MwtfxS/s7FS5KOJmp37qScml2cpqUE+jd/I4145yjgjLacXBOdrU6UbLRK3/bsFtez7JM4Z+kN7fHKjnGHjTpydlOF/d7cyd7ru1a3SLPHjIMsx3DqzdySq0uVRkre/GT+G/Va8y7Wb2bPAzJZZfwjTqzVpV5SqedtIx+9RuvUAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA8cuG69fL6GfYK6rYRpu2/JdO/8A25JJ+Scm9jiribHcR5pUzHGP3paJLaMVtFeS/F3b1bDI8dnnFtDAcJxlenCbkn1UbdfKC5nH/FyrojB4Ojg8NTw1CNoQioxXZRVkvkkAAAAAAAcGZ1jsxnWWKqc3KlbRLe/ZIAAAAAAAAADjXOcbl3sPqs+Xm5r6J7ctt0+7KTcqcW97AAAAAAAAABnWdY7D59QwtKpanJwurLrKz1avt5gAAAAAAAAAAAAAAAAAAAGHwdGeAlVcfeV+4AAAAAAAAAAAAAAAAAAAAAAAAAAAAAVaUKsHTmk4tWaeqae6a7M8RPBPMcuxMsXkdJ1sPLXkWs4eVt5R7NXl0a0u8q4F4kzTEeww2CqN3s24uKX+KUrJfNnhj4a0OEcJKdVqeKqJc0lsl/JG+tr6t2Tk7NrRJAAAAAAAHh1/FxHpH9QAAAAAAAAAPEb/AJhv+3//AG0o/wAKPogAAAAAAAAA4i/5KjC+tP8A9KAAAAAAAAAAAAAAAAAAAAML/wAiyfzAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPDr+LiPSP6gAAAAAAAAAeI3/MN/2/8A+2lH+FH0QAAAAAAAAAcRf8lRhfWn/wClAAAAAAAAAAAAAAAAAAAAGF/5Fk/mAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAeHX8XEekf1AAAAAAAAAA8Rv+Yb/t//ANtKP8KPogAAAAAAAAA4i/5KjC+tP/0oAAAAAAAAAAAAAAAAAAAAwv8AyLJ/MAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAyHhullEpyhNy5rbrtf/MAAAAAAAAADP8Ah2lm/s+ebjyX287f5EIqMVHsAAAAAAAAAGP4bpYzMqeOlNpw5dLae67gAAAAAAAAAAAAAAAAAAAFPHyhhnQS0d/xAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA4Y4kq5vOpGcFHlttfrf/IAAAAAAAAADijiKrlHsvZwUufm36Wt/mQlzQUu4AAAAAAAAAZnxJVwmb0cDGCany663XM7AAAAAAAAAAAAAAAAAAAAFHL41MJKvzaq+noAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAeHX8XEekf1AAAAAAAAAA8Rv+Yb/t/wD9tKP8KPogAAAAAAAAA4i/5KjC+tP/ANKAAAAAAAAAAAAAAAAAAAAML/yLJ/MAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA4Y4bq5ROpKc1Lmtsn0v/mAAAAAAAAABxRw7Vzf2Xs5qPJzb+dv8iEeWCj2AAAAAAAAADM+G6uMzejjozSUOXSzv7ruAAAAAAAAAAAAAAAAAAAAUcfGGElQtq7/iAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABw3k1PNsXKhObilFvTyaX6mZ4SOExlWhF3UW1f0AAAAAAAAAAx3DdLD5NTx6qNyko6W01AAAAAAAAACnFTmo9zijh2llHsvZzcufm36Wt/mAAAAAAAAABw7w3SzTD1as6ji4dl5XAAAAAAAAAAIcN0pZF/aXtHzWeltNJcoAAAAAAAAABwzw3SzeFSU6jjy22Xe/+QAAAAAAAAAAAAAAAAAAAAAAAFOnOrNQgm29ktW/RFSnOnNwmmmt09GvkAAAeH3/Izn/gf/pUTiH/AJGuI/xP8wAAAAAAAAAM5/5JOh6QAAAAAAAAACh/Fj6o8Rv+Yb/t/wD9tAAAAAAAAAA4C/3HEev6AAAAAAAAAAFH/ki36P8A9OMAAAAAAAAAA8Ov4WI9Y/kwAAAAAAAAAAAAAAAAAAAADgqnQnjqkq0FJRg5WaT1Tjqr9Sjjco4ijLCKj7OpZuLst/VfitmipBwk4y3WgBkEMvlhsW8VbmUPdv312878tv8AIOHv+Rrh/wDEvzOIf+RriP8AE/zAADBYSpi8RChT+KTsrmMwzw2IqUJO7g2vudjhvOaeVYuVecXJOLVl6p/oZnio4vGVa8VZSbdvUAAAAAAAAADHcSUsRk1PAKDUoqOvTQAAAAAAAAAKclGal2OKOIqWb+y9nBx5Obfzt/kAAAAAAAAABw7xJSyvD1aU4OTl29LAAAAAAAAAAEOJKUci/s3kfNZ69NZcwAAAAAAAAABwzxJSyiFSM4OXNbbyv/mAAAAAAAAAAAAAAAAAAAAAcGf71iP+vUvzicLOSzihy73/AEd/wMXlMsViMfiISSVKTdu95S2+4ybKp5nilh4SUXZu78jLsorY6jVnSesLadXd2sjMuCsVgsD9Z51JpXlFLZdbO+tuuiMtyqeOp15xkl7OLk/O3Q4f4brZvKXLLlhHd2vr2S0v566FLJ6mV5/h8POSl70WmuzfVdHp5+pW4PxGZ4vE4jnUFzy5bq97P10V9L6+hDL6rx0cHP3ZOSj6Nuxg+BMViFUcqijytpaN81na/kr+voZPwbisfGcqkvZqLcdVdtrfqtPMx2Dq4PETw9X4ou3/AAfRrVGG4GxlXDKpKoo1GrqD3t59n8n52MtyqrjcdHB35ZO616NJt/lYwfA2MxOC9u5qMmrqNt/V30v00fnY4OympiMx9pe3smm09+qt+BxZw/UwVSeMlNNVJuy6q92AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHA1N1cbWgt3TkvvcTK+HlkLnmGOqRfInypX3a80tXslbrcySpKrleZzlu1Fv1bkcDf8jeP+GRwZifq1DG17XcYp/NczX4nCGPxGKxdbDVpuSqRlu769/ubucL/wC64/8A69S/JlOlVqcKS9h0qXnbtb/9l/jsZFVnUzbDc8m7SiteyexnufYmnnrnCTUacrJdNN7rz1uZzRVPiyk19qdN/il+hn2bVo8RKcZaU2kl5ac33638jjTMK0M2jCk+X2aTX+J+836vS5m+Ep4riTDXWlRRk/O1/wBI2MwxWPlnUsZGMvdlpo9k7JejW/e77iw8aPGceXZ3f303f8blPPcTLiJVFN8vPy2vpy35dtttfUhGVDixwg7Jzu/mr/qZ9WqTzHERlJtKcrK/95mR5NUzbESoQmotJvXyaX6mOwksJialCTu4tq/oAAAAAAAAABi+G6uHyyGPdRNStpZ31AAAAAAAAACEeeSj3M/4dq5R7Pnmpc99ulrf5gAAAAAAAAAZLw3VzSjUqwqKPJ3T7XAAAAAAAAAAI8N1ZZR/aXtFy66Wd9JcoAAAAAAAAABkPDdXN4zlCoo8tt0+t/8AIAAAAAAAAAAAAAAAAAAAAA4LbWKrtPX2UvziYnGYnEtOvUlO2123+Zw+m8ozG38sf/bjgZf8K8f8Mjhv/kXZh/g/+SOBv+RvH/DI4X/3XH/9epfkzhziGplVVqS5qct1+q8/Lr9zWMy+hhuIsLVw6/d1XGS7K71t5dfK9jiH/ka4j/E/zM+/5Kqh/ip/mjP/APkeVf8AF/kcbf8AI5qekfyRUko8SYBv/n0vxU0YvjrM6GIqUuSHutraXR2/mMjzStmXElHEVklKzWl7aQl3bKH/ACOI/wDXxf8ApRW/5LBf4l/6SjO/+Rnif8c//SmeH3/Izn/gf/pUTiH/AJGuI/xP8wAAAAAAAAAM5/5JOh6QAAAAAAAAACh/Fj6o8Rv+Yb/t/wD9tAAAAAAAAAA4C/3HEev6AAAAAAAAAAFH/ki36P8A9OMAAAAAAAAAA8Ov4WI9Y/kwAAAAAAAAAAAAAAAAAAAADgz/AHrEf9epfnEOCp0amIrYKrtVi1812+Tb+RkORUclxaVSopVKl1HpaKV2/wAEvI4b/wCRdmH+D/5I4G/5G8f8Mjhf/dcf/wBepfkzLOEK+ZYKGJo1Ertpp30s/K/3O3rqY+pSjn2BwlOV1R5It+a/4FvnocQ/8jXEf4n+Zn3/ACVVD/FT/NGf/wDI8q/4v8jjb/kc1PSP5I4jxbwmaYOuvs04P7m7r5ozXhCWZ4j67gqkeSpq7367tWT33s7WZkGC+p8TQw/Nflclfv7kih/yOI/9fF/6UZhiIYfiv2k3ZKcb+Sslc4uyOrgsVPFOScakm1vfXV3+fn9x4ff8jOf+B/8ApUTiH/ka4j/E/wAwAAAAAAAAAM5/5JOh6QAAAAAAAAACh/Fj6o8Rv+Yb/t//ANtAAAAAAAAAA4C/3HEev6AAAAAAAAAAFH/ki36P/wBOMAAAAAAAAAA8Ov4WI9Y/kwAAAAAAAAAAAAAAAAAAAACFScHeDa9AjKUJKUXZoqYzE1antZ1G5d2239+5GpOKai2k9/MhUnTd4Np+WhGpOCai2r7lDGYnDp+xqON97Nq/3CnJS5k9e5KTk227slWqSmpuTbXW+pKcpS5m7snUnUlzTd35k6k6jTk27dyjj8VRpunSqyjF9E2l9yZCpOEuaLs+5zS5ua+pOcpy5pO78yti8RWjGFSo5KOybbt6X2PD7/kZz/wP/wBKicQ/8jXEf4n+YAAAAAAAAAGc/wDJJ0PSAAAAAAAAAAUP4sfVHiN/zDf9v/8AtoAAAAAAAAAHAX+44j1/QAAAAAAAAAAo/wDJFv0f/pxgAAAAAAAAAHh1/CxHrH8mAAAAAAAAAAAAAAAAAAAAAAAAAAAHB+ZYXL8dOriZ8sXFrZvW8X0T7MznEU8RmFarSd4yk2n5fPUAAAAAAAAADM86wFbh6lhKdS9RKN1Z9N9bW/EAAAAAAAAAKTUakW+jONc5wOY+w+qz5uXmvo1a/LbdLswAAAAAAAAAOEc6wGAwlaniKnK5PTRvp5JgAAAAAAAAAFPOsAuGPqTqfvbPSz/nvva23mAAAAAAAAAAcGZ1gMup1liqnK5NW0b2v2TAAAAAAAAAAAAAAAAOO/F7J+F6rwqi6+IW8ItJR/xS1s2tkk33STRlf0j5Ouo5hgUqb6wlqu7tJWfpePqZJnmX51gYY3AVFOnLZrv1TW6a6p6gAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB4ncZx4XyGeIpv8AfT9ymv7z6vyirv1suouD+JMwyqrn/sJTo3blNtNvX3pWb5mk/ila27vo7ZRk+OzfGQwWBpupVnsl+Orskkt22kurOAM8zPw/4neWZtF06VVpTTaaV/hqJrRpdWnte+qsgAAAAAAAAADL8sxWYVXSw0eaSV7XS00XVrujEYeph6sqNRWlF2a816AAAAAAAAAAVskx1HCRxc6dqbtZ3XXbS9/wAAAAAAAAACMXJpLczLJsbl3L9ahy817ap7Wvs33QAAAAAAAAAGAyTHY+nKphqfMo76pfm0AAAAAAAAAALJMdLBfXVT/dd7rvba99/IAAAAAAAAAAy7JMdmKk8LT5uXfVLf1aAAAAAAAAAAAAAAAA8fc/ljuJVgIy9zDxSt05ppSk/u5V6oyXKKGEyShlzj7kacYNPquWzv663PAbIYYbP81m9XQtTT9Zyvb/AMFr7/M+khltPlwGOive9+DfVrSUfu977zwxz953wrhMTOV6iXJLvzQ92782kpfMAAAAAAAAAA8Pv+RnP/A//SonEP8AyNcR/if5gAAAAAAAAAZz/wAknQ9IAAAAAAAAABQ/ix9UeI3/ADDf9v8A/toAAAAAAAAAHAX+44j1/QAAAAAAAAAAo/8AJFv0f/pxgAAAAAAAAAHh1/CxHrH8mAAAAAAAAAAAAAAABxXmMsxz3GYuTvz1JtenM7L5KyMLXjiKEK0dpJP71c8EcZTrZvnqg7p1VJeacqv/AAPvPpIVYrLsBTvq5zf3RV/zR9HDMHUy3HYJv4Jxkv8At+LT/wDSAAAAAAAAAADw+/5Gc/8AA/8A0qJxD/yNcR/if5gAAAAAAAAAZz/ySdD0gAAAAAAAAAFD+LH1R4jf8w3/AG//AO2gAAAAAAAAAcBf7jiPX9AAAAAAAAAACj/yRb9H/wCnGAAAAAAAAAAeHX8LEesfyYAAAAAAAAAAAAAAAYqpKnQnOO6Tf3IMl8daWA4RjgXTk8ZTj7OD05WrWjNtu94q142d3azs3y+GfHK4Tzp4utFzpTi4zS3tdNSSbSbTXVrRvXU8VOPYcW5rCph4uNCknGCe7bd5Sa1teyVr7Jdbn0cKtszx1O+8Iu3pJq/yv+IAAAAAAAAAB4ff8jOf+B/+lROIf+RriP8AE/zAAAAAAAAAAzn/AJJOh6QAAAAAAAAACh/Fj6o8Rv8AmG/7f/8AbQAAAAAAAAAOAv8AccR6/oAAAAAAAAAAUf8Aki36P/04wAAAAAAAAADw6/hYj1j+TAAAAAAAAAAAAAAADE03VoTpx3aa+9DTTszB4PEYyvHD4am51JaKMU22/JLVmbcEcRZTh/rGNwc4U/5mtF6tXt8w+jhSbzLHVO0Ir75P/IAAAAAAAAAA4HxVDD5jOdaaiuRq7aSvzR7me1IVcyrzg005OzWqevQAAAAAAAAADNsdhanDFGjCrFzShomr6b6XuAAAAAAAAAFFpVIt90ce43C4n6v7CpGdua9mnb4d7MAAAAAAAAADgrHYXD4KvGtVjFt6JtK+nS7AAAAAAAAAAKWOwq4R9h7WPtLP3bq/xt7XvtqAAAAAAAAAAcB47C4alXVerGF3G12lffuwAAAAAAAAAAAAAAAOK8sngeIcZgoxd41ZxS6tcz5fvVrep4V+H2H4XyqOIxEV9aqK85P7K35F2S+13l1aSthM2ybOYVKOGr066StJRlGas9LSSb0fmeLXAC4WzVTwy/2atdw/utW5oP0vePeL6tNn0cculTyvG41rSpOMV/25Fv8A9v8A6sAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAcbZRl+U+J2X5pjY/uK7Tv0VWK5U35J+zk9urezv48eIFWM/8AV7AzsrXqtbu+qp+StrLvdLbmTyDPsdkeY0sfgp8s4P5NdYvumtGjxL4qyTPPDqOPkrus4qmusaifvL/t1KSfdf4kzw14flkfC2EwlRWny80u/NN8zT81dR+QAAAAAAAAABh8LXxMuSjBye9km3bvoVKc6c3Caaa3T0a+QAAAAAAAAAE8DioUVWlSkoPZtO2vnsAAAAAAAAACTbsjE4HFYa3t6coX25k1e29rgAAAAAAAAAUMDisRFyo0pSS3sm7fcAAAAAAAAAALA4p0PbqlLk/ms7dt9t9PUAAAAAAAAAAw+BxWJTdClKdt7Ju33AAAAAAAAAAAAAAAAeJPBseKchqYSFlWj71Nv+ZdH5SV4vtdPWxmUcZHGVVjOb2yk+fmvzc19b31vfe4eD3B+K4izKnUxDbweGlztP4XUfL7qW13yxc/7qSe6AAAAAAAAAADw+/5Gc/8D/8ASonEP/I1xH+J/mAAAAAAAAABnP8AySdD0gAAAAAAAAAFD+LH1R4jf8w3/b//ALaAAAAAAAAABwF/uOI9f0AAAAAAAAAAKP8AyRb9H/6cYAAAAAAAAAB4dfwsR6x/JgAAAAAAAAAAAAAAAHHXhRkvFUvrE70sR/PFLXtzx2lbvdS2XNZWMu+jfgqWIUsZj5VIL7MYKDf/AG85z076X80ZTlGBynBwweBpKnShsl/V231bu29WwAAAAAAAAAA8Pv8AkZz/AMD/APSonEP/ACNcR/if5gAAAAAAAAAZz/ySdD0gAAAAAAAAAFD+LH1R4jf8w3/b/wD7aAAAAAAAAABwF/uOI9f0AAAAAAAAAAKP/JFv0f8A6cYAAAAAAAAAB4dfwsR6x/JgAAAAAAAAAAAAAAAAAAAAAAAAAAB4ff8AIzn/AIH/AOlROIf+RriP8T/MAAAAAAAAADOf+SToekAAAAAAAAAAofxY+qPEb/mG/wC3/wD20AAAAAAAAADgL/ccR6/oAAAAAAAAAAUf+SLfo/8A04wAAAAAAAAADw6/hYj1j+TAAAAAAAAAAAAAAAAAAAAAAAAAAADw+/5Gc/8AA/8A0qJxD/yNcR/if5gAAAAAAAAAZz/ySdD0gAAAAAAAAAFD+LH1R4jf8w3/AG//AO2gAAAAAAAAAcBf7jiPX9AAAAAAAAAACj/yRb9H/wCnGAAAAAAAAAAeHX8LEesfyYAAAAAAAAAAAAAAAAAAAAAAAAAAAU6tSm7wk0/LQlJybbd2AAAAAAAAABKvVlBQcm12u7fcAAAAAAAAABVr1atueTdu7uAAAAAAAAABTr1aaahJr0bQAAAAAAAAAB7eryez5ny9ru33AAAAAAAAAAFOvVpL3JNejaAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPD7/AJGc/wDA/wD0qJxD/wAjXEf4n+YAAAAAAAAAGc/8knQ9IAAAAAAAAABQ/ix9UeI3/MN/2/8A+2gAAAAAAAAAcBf7jiPX9AAAAAAAAAACj/yRb9H/AOnGAAAAAAAAAAeHX8LEesfyYAAAAAAAAAAAAAAAAAAAAAAAAAAAeH3/ACM5/wCB/wDpUTiH/ka4j/E/zAAAAAAAAAAzn/kk6HpAAAAAAAAAAKH8WPqjxG/5hv8At/8A9tAAAAAAAAAA4C/3HEev6AAAAAAAAAAFH/ki36P/ANOMAAAAAAAAAA8Ov4WI9Y/kwAAAAAAAAAAAAAAAAAAAAAAAAAAAweOxODqOph5uMmrXXb+kVq1StUlUqO8nq35gAAAAAAAAAVc0xlXDLDTqNwVrLppsAAAAAAAAACbTTRjczxmN5frFRy5b2v0vv+SAAAAAAAAAAwmaYzCQlChUcU97AAAAAAAAAACzTGLC/VVUfs/5em9/zAAAAAAAAAAMFmmMwSaw9RxvvbyAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAD/9oACAEDEQE/AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADhThPG8S42WDwcoxlGLl7zaVk0uik73kunzM2y2rluOq4Ks0505OLava600uk7fJAAAAAAAAAAZjwDmWByGlnVScHSmotJOXN72104peuoAAAAAAAAAU6bqTjBbt2OL+Bcx4X9j9cnCXtea3I5P4eW97xj/ADK2/UAAAAAAAAADhbgHMuI8LWxGEnCMabs+ZyT2vpaMvxsAAAAAAAAAAUeActnwK8/c5+2s3a8eTSo4bct9l/Nv5aAAAAAAAAAAHh3wvg+I84eCxcpRioOV4tJ3TS6qStr2OK8poZTnWJwNBtwpysm7X+dkl+CAAAAAAAAAAAAAAAAAAAAAAAAAAAPAv/koq3/XmX/pdM47/wCSmx//AF8n+YAAAAAAAAAHFn/JsMH/AIaX5AAAAAAAAABg/wDeafqvzPH7/mXf9xf/AHGAAAAAAAAAB4Jf8iXH+v8A7awAAAAAAAAADC/8mif+GX/p9gAAAAAAAAAHgl/yU0v+vcvzieIv/JU47/G/yQAAAAAAAAAAAAAAAAAAAAAABQw9avLkpQcn2Sbf4FajUozcKkXGS6NWf3MAACEJTkox3ehnGTY7KMXLB42HJUja6unurrWLa28zwhzrL8pzyrXx1VU4OlJJvvzwdvuT+44wxdDF5/jK9CSlCVSTTWzTe4AAAAAAAAAHEnEmU4nw/wALl9KvGVaKp3j1Vlr9wAAAAAAAAAYWUY14Sb0TX5njNxFlWcfUfqFeNTk9pe3S/Jb77P7gAAAAAAAAAPCfiTKcryrGUsbXjTlN6J9fdsAAAAAAAAAAUfEDDQ4KfD3sZc9mua6trUc9t9tAAAAAAAAAADgLimjw3mrxtWm5pwcbJpPVp319DifNoZvm+Ix0IuKqSuk90AAAAAAAAAAAAAAAAAAAAAAAeH3ENTh/gzG5hSpqco1Yqz0WqgunqZjnmC494XxdWrRVPFYVc6tr7urdnvZpNOL0T5X2sAAGD/3mn6r8zxg/5K2v/hh/6QgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAMo/5NxmH/X6H/th4ZuVLB5xiH8EcPJP1afKvV2djgvg2OeutiMVWVHDUVec3bd7JXe/nrbTRto444Iw2TUKOYZbX9thKuilo2pauza0d0n0WqaaVteN+GKOQY2jSoVHOFSnGabVvib009PxOL+EP7BwuArc7k69Pmd1a0rRbivTmRX4KjHB5TKFX97jW1Z6Rj7ySel3rzIx3hFk9TA1YZZj/a4ukm5RvBp26csfeg3sm5PXfy4U4cWeVMXT5mpUqM6kUlfmlGyUfR3OMfDuWRQy6hSk6mIxHMpLS3MuRJR0vvKzbbvvoYjwhy7DUI0sPjVPHQSm6d46pavlh8a8m7pvor6eMH/JW1/8MP8A0hGRcPZlnmJlhsvp881FyavGOiaV7yaW7XmY/A4jA4mphcRHlqQbTV07Nb6ptfcwAAAAAAAAAMXwhnWEyuGaVqNqE7WlzRd+bbRSctfNeoAAAAAAAAAQhKclGO70OIOEs4yH2f8AaNH2ftL8vvRlfltf4ZO1rre24AAAAAAAAAGScIZ1ndCpWwFHnjDST5oq2l/tSTenYAAAAAAAAAApeHntOEP9YvrPRvk5O1Rw+Lm8r/D5eYAAAAAAAAABwTwp/rNmbwPtvZ2i5X5ebZpWtzR79ziLJ/7IzWvgOfn9m7Xta/na7t97AAAAAAAAAAAAAAAAAAAAAAAPDnC5RieD8bSzefJQdVXd7a2jbVedjjqjgco4Rp0eH4p4WvP36ilzNtapPybVnqrW5be8yGQZriuAMHh8toSm61Sc52X8rcY38nZNehRyDNcFwFmOGzGi4ck4ThzebSlbfp+fqcZYeWZ5XkFWO9Sn7K/nFxj+Lb9dbHizUhjsp9rBaYfESpLyXs4u3Xql8zxNqywtXLcDB2dChT2eqk9/R6J3PCzgziLCZvQzSpR5KNnfmaTlGUWvh3tez1S6NXPDH2WD4wzK/wANOnW+6NSH6I8NqtXOONY43Fy5pLnqNvvZpeiV1borJbGG4ixUuJYZtzPndVS+Tlt6W9221tNjxg/5K2v/AIYf+kI8C/8Akoq3/XmX/pdM47/5KbH/APXyf5gAAAAAAAAAcWf8mwwf+Gl+QAAAAAAAAAYP/eafqvzPH7/mXf8AcX/3GAAAAAAAAAB4Jf8AIlx/r/7awAAAAAAAAADC/wDJon/hl/6fYAAAAAAAAAB4Jf8AJTS/69y/OJ4i/wDJU47/ABv8kAAAAAAAAAAAAAAAAAAAAAAAZR/ybjMP+v0P/bDhKTxvBWcYKWqp8lSPk76tfKCv6lbinOMn4FyvEZZV9neVSEvdjLVSlb4ou2ib07maeIvE2Z4SeDxeJ5qc9GuSCvZp7qKe66M4MwazXI8nVr+wxUl8uV1X+KX6mT1HxDlWcUd3KvSqL/uJUcX17f12xOGpZ54oujUV6cJbdLUoXt6OUdvPsZFxxm+dcd0YOtJUOeaVNNqPKoytzJfE9Ltu+u1locD/APJV51/17xH/AKcieDji+I5Um9Z0pxXro/yTMFQqPG06Vve5kred7WPGD/kra/8Ahh/6QjwL/wCSirf9eZf+l0zjv/kpsf8A9fJ/mAAAAAAAAABxZ/ybDB/4aX5AAAAAAAAABg/95p+q/M8fv+Zd/wBxf/cYAAAAAAAAAHgl/wAiXH+v/trAAAAAAAAAAML/AMmif+GX/p9gAAAAAAAAAHgl/wAlNL/r3L84niL/AMlTjv8AG/yQAAAAAAAAAAAAAAAAAAAAAABlVejHw9x9JzXM6sGlfV/Bstzwvr0pSzLA1JKKrUJpXdry2S16+8zw8xGV53k9fhnMp8jlLnpSutJaKyvpdb23kpSWhxv4d5fwzlqm8b7TEOaSjZR92zu+Xmk3ry63stra3PDLOcNg8gzNVZJTpJ1IXtrJ05x0636adzwax2HoZ3Uo4iSjCcL6uy5oSjJa/JnDPEtDC8axzSvK1OdSbk+yqcyu/Jcyb9DA8NcP8J57HMJYlSniJWpRdkoKbvKV7u6UdFLRapa3ucG4ijR4qzl1JpJ066TbVm/aR2ezv0tucIZ2slzvDY+Xwwlr/ha5Zfg2Zpwnw3lOMlxZ9aUqN3UhTVmpVN1aSd2ubXltp1dkzxZr0a/FNepSmpRcYap3XwLqjwL/AOSirf8AXmX/AKXTOO/+Smx//Xyf5gAAAAAAAAAcWf8AJsMH/hpfkAAAAAAAAAGD/wB5p+q/M8fv+Zd/3F/9xgAAAAAAAAAeCX/Ilx/r/wC2sAAAAAAAAAAwv/Jon/hl/wCn2AAAAAAAAAAeCX/JTS/69y/OJ4i/8lTjv8b/ACQAAAAAAAAAAAAAAAGByZSip1/u/wAyplOEnGyjb0Mbgp4WfLLVdGAAAAAAAAAAHgliaGH4grSrTUV7GSu2lrz0+5xvUhU4jx04NNOpKzW24AAAAAAAAAHFONw0/DXCUY1IuaVLS6vou24AAAAAAAAAYRpYin6r8zx2xmGxH9n+xqRlb2t7NO38PsAAAAAAAAAB4M43DUMnx0atSMW3pdpX919wAAAAAAAAAChxtlMPD95G5S9vZq1tNarnv6AAAAAAAAAAHhpxHgcgzp4vGtqHJKOiu7tx6fI4vzPD5nnmJxmHbcJyurqztZdAAAAAAAAAAAAAAAAyjCKtW55LSP59Crm1KGI9jbTa/mYzFRw1Jzlr2Rz08ywrSVn+TGmnZgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAGUUVTwsX1ev9fIqS55uXczmo5UqPmr/gjIZPnnHyRm1FU8VK3XX+vmAAAAAAAAAAUMNWxEuWjBye9km3+BUpzpycJqzW6ejQAAAAAAAAAE8FiYUlVlTkoPq07ffsAAAAAAAAACTbsjEYPE4e3tqbjfa6av6XAAAAAAAAAAoYLE14uVKnKSW9k3b7gAAAAAAAAACHBGbTyL+20o+ws3e+ukuTb1AAAAAAAAAAOHeHMdn+MeEwSTmk5auysrJ/mZtlmIyzG1MHiEueDs7aq/qAAAAAAAAAAAAAAAFKHJTjHsrFWHJUcezsZzScaNLyVvwRkMHzTl6Gfw1hL1QAAAAAAAAAB4F/8AJRVv+vMv/S6Zx3/yU2P/AOvk/wAwAAAAAAAAAOLP+TYYP/DS/IAAAAAAAAAMH/vNP1X5nj9/zLv+4v8A7jAAAAAAAAAA8Ev+RLj/AF/9tYAAAAAAAAABhf8Ak0T/AMMv/T7AAAAAAAAAAPBL/kppf9e5fnE8Rf8Akqcd/jf5IAAAAAAAAAAAAAAAh8SCeWUZ4j2z37dLmJw0MRTcJmFwsMNT5IGfL91D1/QAAAAAAAAAA8C/+Sirf9eZf+l0zjv/AJKbH/8AXyf5gAAAAAAAAAcWf8mwwf8AhpfkAAAAAAAAAGD/AN5p+q/M8fv+Zd/3F/8AcYAAAAAAAAAHgl/yJcf6/wDtrAAAAAAAAAAML/yaJ/4Zf+n2AAAAAAAAAAeCX/JTS/69y/OJ4i/8lTjv8b/JAAAAAAAAAAAAAAACdmmY7FKhh3Nb9PUweJjiaKmt+vqGLzT2eKjCL91b/wBeRn7XJBeoAAAAAAAAAB4F/wDJRVv+vMv/AEumcd/8lNj/APr5P8wAAAAAAAAAOLP+TYYP/DS/IAAAAAAAAAMH/vNP1X5nj9/zLv8AuL/7jAAAAAAAAAA8Ev8AkS4/1/8AbWAAAAAAAAAAYX/k0T/wy/8AT7AAAAAAAAAAPBL/AJKaX/XuX5xPEX/kqcd/jf5IAAAAAAAAAAAAAAAFSjjMBGPW34oweLqYOq01ps0YrO4Onaind9+hlOEdet7SW0fxf9bme1FKvGC6L8wAAAAAAAAADwL/AOSirf8AXmX/AKXTOO/+Smx//Xyf5gAAAAAAAAAcWf8AJsMH/hpfkAAAAAAAAAGD/wB5p+q/M8fv+Zd/3F/9xgAAAAAAAAAeCX/Ilx/r/wC2sAAAAAAAAAAw/EmUx8M3ljrx9vaXudf4zl+WoAAAAAAAAAB4TZvgcrz918bVVOHs5K72u3HT8DjjG4fG8RYvEYealCUm01s1ZAAAAAAAAAAAAAAABkeIThKk91qv6/rczHK1iHz03aX5lHJcROVp6L+uxTp08PS5Y6Jf1cxVb21aVTuAAAAAAAAAAUMTWw8uajNxe102n+BUqTqSc5u7e7e4AAAAAAAAAE8biZ0lRlUk4Lo27aeWwAAAAAAAAAJtPQxGMxOIt7ao5W2u27elwAAAAAAAAAKGNxNCLjSqSinvZtX+4AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAMPXlQqqpHdGHxFOvTU4MM4x8VF0IPXr/l/mAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAR4azaWW/2kqEvYfz9NHy/noAAAAAAAAAAZXk+OzWv9XwVJ1J2vZb2W7/ExuCxGCxE8PiIOM4uzT3TAAAAAAAAAAAAAAAAo16tGXNTlZlTNMVUjyuX3aAAAAAAAAAAAeCWGoYjiCtGtBSXsZOzSevPT7nG9OFPiPHQgkkqkrJaJagAAAAAAAAAcU4LDQ8NcJVjTiptUtbK+3fcAAAAAAAAAMIk8RTXmvzPHbB4bD/2f7GnGN/a3skr/wAPsAAAAAAAAAB4M4LDV8nx0qtOMmnpdJ2919wAAAAAAAAADC/8mif+GX/p9gAAAAAAAAAHgl/yU0v+vcvzieIv/JU47/G/yQAAAAAAAAAAAAAAAAAAAAAAAAAAB4F/8lFW/wCvMv8A0umcd/8AJTY//r5P8wAAAAAAAAAOLP8Ak2GD/wANL8gAAAAAAAAAwf8AvNP1X5nj9/zLv+4v/uMAAAAAAAAADwS/5EuP9f8A21gAAAAAAAAAGF/5NE/8Mv8A0+wAAAAAAAAADwS/5KaX/XuX5xPEX/kqcd/jf5IAAAAAAAAAAAAAAAAAAAAAAAAAAA8C/wDkoq3/AF5l/wCl0zjv/kpsf/18n+YAAAAAAAAAHFn/ACbDB/4aX5AAAAAAAAABg/8AeafqvzPH7/mXf9xf/cYAAAAAAAAAHgl/yJcf6/8AtrAAAAAAAAAAML/yaJ/4Zf8Ap9gAAAAAAAAAHgl/yU0v+vcvzieIv/JU47/G/wAkAAAAAAAAAAAAAAAAAAAAAAAAAAAeBf8AyUVb/rzL/wBLpnHf/JTY/wD6+T/MAAAAAAAAADiz/k2GD/w0vyAAAAAAAAADB/7zT9V+Z4/f8y7/ALi/+4wAAAAAAAAAPBL/AJEuP9f/AG1gAAAAAAAAAGF/5NE/8Mv/AE+wAAAAAAAAADwS/wCSml/17l+cTxF/5KnHf43+SAAAAAAAAAAAAAAAAAAAAAAAAAAAMi4hzLI8TLE5fV5JuLi3aL0bTtaSa3S6XMfjsRjsTUxWIlzVJttuyV293ZWS+SAAAAAAAAAAxfF+dYvK4ZXWrXoQslHlircu2qjzaeb9QAAAAAAAAAhOUJKUd0cQcWZxn/s/7Rre09nfl92Mbc1r/Cle9luAAAAAAAAABknF+dZLQqUMBW5Iz+JcsXfS32otrTsAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB9SxLo+2VOXJ/NZ27b7b6AAAAAAAAAAFDD1q8uSlByfZJt/gVKU6c3Caaa6PR/cAAAAAAAAAAAAAAAAAAAAAAAAAAAeEOS5fm2eVaGOpKpBUpNJ91OCv9zf3nGGEoYTP8Zh6EVGEakkktkk9gAAAAAAAAAOJOGspw/h/hcwpUIqtJU7y6u61+8AAAAAAAAAMLGMq8IvZtfmeM3DuVZR9R+oUI0+f2l7dbclvuu/vAAAAAAAAAA8J+GspzXKsZVxtCNSUHo30924AAAAAAAAABhf8Ak0T/AMMv/T7AAAAAAAAAAPBL/kppf9e5fnE8Rf8Akqcd/jf5IAAAAAAAAAAAAAAAAAAAAAAAAAAA8C/+Sirf9eZf+l0zjv8A5KbH/wDXyf5gAAAAAAAAAcWf8mwwf+Gl+QAAAAAAAAAYP/eafqvzPH7/AJl3/cX/ANxgAAAAAAAAAeCX/Ilx/r/7awAAAAAAAAADC/8AJon/AIZf+n2AAAAAAAAAAeCX/JTS/wCvcvzieIv/ACVOO/xv8kAAAAAAAAAAAAAAAAAAAAAAAAAAAcKcV43hrGyxeDjGUpRcbSTas2n0cXe8V1M2zKrmeOq42skp1JOTSva710u27fNgAAAAAAAAAZjx7mWOyGlklSEFSgopNKXN7u125NevugAAAAAAAABTm6c4zW6d/uOL+Osx4o9j9chCPsua3IpL4uW97yl/KrbdQAAAAAAAAAOFuPcy4dwtbDYSEJRqO75lJva2lpR/FMAAAAAAAAAAWc5ksH9SWIn7H+Tmlyb3+G9t9dt9QAAAAAAAAADBZhjMDV9rhKsqc7WvGTi7drpp2MTia2JqyrV5uc5attttvzb1YAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABKcYq8nYhUhP4WmAAAU6sKl+Xo7AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAZhTVSdOL6srYb6pKNWm9OoAABgf8Al5/iYAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAGM/i0vX/IzJr2FuraMTX9jBWV29EYfE1faeyrRs3sUK0p1Jwl9kp4lzxEqXRHtpPEOmtkrksdiV+85PdMRXcKDqQ8vxKGKlVryivhSHj60pOcI3gjL2pRm13YZfiKlVy53ewAAHhRw9lueZ1Vw2YU+eCpuSV5R1UoK94tPZvyOLMDh8DnmLwuHjy04Tkoq7dknpq2397AAAAAADMMRUpcvI7Xv+gtUgAAOIeEMlwnAmGzSjRtXmqd5c0nfm30cnHXyQAAAAAAYjEVIYmMIvR2/MAADDQjOtCMtm0vxPF7hLJ8h+pf2bR9n7T2nN70pX5eS3xSdrXe1t9QAAAAAApYipLFSpt6K4AAB4XcIZLneWYutj6PPKDtF80lb3b/AGZJPXuAAAAAABhcDh55bOtKPvK+t30+dgAAChwhks/Dl5w6P+02l73NLpVcfh5uX4dNvPcAAAAAADBU4VMRCEldNozrC0cPiIwpKytfr3fcAADwtyDLs7zx4XH0+enySdryWqcbO8Wn1fU40y7DZdn+KwmFjy04Sslduysurbf3sAAAAAAAAAAAAAAAAAAADMefmp8m99CmqssTFYjfptb8DE1acMVFzeiX4mIxNGpUpuDu0/MguXGyXdX/AEMPpXjP+ZyMP72Iqy9EYvGUYxlTvd7aE3fLvu/MklSwdl2/MwtNRoRXkZdFRjOK6NhlW8vkAAB4F/8AJRVv+vMv/S6Zx3/yU2P/AOvk/wAwAAAAAAzb7Hz/AEI/CgAAOLP+TYYP/DS/IAAAAAAMX/vkPl+YAAGD/wB5p+q/M8fv+Zd/3F/9xgAAAAABQ/36XzAAAPBL/kS4/wBf/bWAAAAAABgv+RPP0kAAAYX/AJNE/wDDL/0+wAAAAAAMu/3un6o4j/3qP+FfmwAAPBL/AJKaX/XuX5xPEX/kqcd/jf5IAAAAAAAAAAAAAAAAAAADGfxaXr/kZgrKE+zRWpwnjIqaumhYSgmmooxUvZ14z8n+CucnJQovs1+Jh5OOHqVF1bZQoQjhXJrVpu5L/kXf13MSr4R+hh2pUYNdkYBpqbX8zDD4SFC/K3r3AAA4U4rxvDWNli8HGMpSi42km1ZtPo4u94rqZtmVXM8dVxtZJTqScmle13rpdt2+bAAAAAADEYWFe3M3oJWVgAAMx49zLHZDSySpCCpQUUmlLm93a7cmvX3QAAAAAAqYSFSqqjbuv0AAApzdOcZrdO/3HF/HWY8Uex+uQhH2XNbkUl8XLe95S/lVtuoAAAAAAQwkI1XVTd2AAAcLce5lw7ha2GwkISjUd3zKTe1tLSj+KYAAAAAAFLMqtPDPDpLld+99fmAAAU+Pcyhw5/YChD2Nmr2lz6z59+a2/wDd2+8AAAAAACjVlRqRqR3WpjcbUxdRTmkmlbT+mAABwtxRjOHMc8bhIxlNxcbSTas7Po4u+nczrNq2bY+rjq6SnUd2ldK/ldt/iwAAAAAAAAAAAAAAAAAAAMVCUqtJpXszH03Og1FXZi6VScI1YfEihisRVqJOFl1MypTnTjyK7uYuk3h3GO6tb5FGi/qqpvS6/Mg8TOH1flslu/IlTm8By2d+3XcUFKlyvsQniqS9go37MwFKVKM4tdQAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADBYqddyUktAAAPDfhPBcS5rUweLlKMY03JcrSd1KK6qStaT6dtTiXLaOW5xicFRbcac5RTe9k7a2SV/kgAAAAAAx2KnQ5eVLW+/yFqgAAM84By3A8G0M6pzm6s1BtNx5fe3slFP01AAAAAACvipwxEaaSs7AAAUKaqVYQezaR4n8C5dwv8AVfqU5y9rz353F/DyWtaMf5ne9+gAAAAAAU8VOeJdJpWVwAADw74By3iPL8ViMXOcZU3Zcril8N9bxl+gAAAAAAGHyyjUwEsQ2+ZX9NPkAAAUeActnwK8/c5+2s3a8eTSo4bct9l/Nv5aAAAAAAAYSjGtXhTls3YzbBU8JWVOm201fX1f+QAAHh3wvg+I84eCxcpRioOV4tJ3TS6qStr2OK8poZTnWJwNBtwpysm7X+dkl+CAAAAAAAAAAAAAAAAAAAAAAAAMq3l8gAAPAv8A5KKt/wBeZf8ApdM47/5KbH/9fJ/mAAAAAAGbfY+f6EfhQAAHFn/JsMH/AIaX5AAAAAABi/8AfIfL8wAAMH/vNP1X5nj9/wAy7/uL/wC4wAAAAAAof79L5gAAHgl/yJcf6/8AtrAAAAAAAwX/ACJ5+kgAADC/8mif+GX/AKfYAAAAAAGXf73T9UcR/wC9R/wr82AAB4Jf8lNL/r3L84niL/yVOO/xv8kAAAAAAAAAAAAAAAAAAAAABUqRpxcpPRFDH060+RJopVo1b8vR2FWjKq6a3RicbToOz1ZSqwqwUovQMq3l8gAAPAv/AJKKt/15l/6XTOO/+Smx/wD18n+YAAAAAAZt9j5/oR+FAAAcWf8AJsMH/hpfkAAAAAAGL/3yHy/MAADB/wC80/VfmeP3/Mu/7i/+4wAAAAAAof79L5gAAHgl/wAiXH+v/trAAAAAAAwX/Inn6SAAAML/AMmif+GX/p9gAAAAAAZd/vdP1RxH/vUf8K/NgAAeCX/JTS/69y/OJ4i/8lTjv8b/ACQAAAAAAAAAAAAAAAEKcqj5Yq7KmCxFOPNKDSAAAAADGR9rVp0em7FSppqyWhgf+Xn+JlP/AH2fov0MHFTc6r3b/AwiVOtUprZWf3hlW8vkAAB4F/8AJRVv+vMv/S6Zx3/yU2P/AOvk/wAwAAAAAAzb7Hz/AEI/CgAAOLP+TYYP/DS/IAAAAAAMX/vkPl+YAAGD/wB5p+q/M8fv+Zd/3F/9xgAAAAABQ/36XzAAAPBL/kS4/wBf/bWAAAAAABgv+RPP0kAAAYX/AJNE/wDDL/0+wAAAAAAMu/3un6o4j/3qP+FfmwAAPBL/AJKaX/XuX5xPEX/kqcd/jf5IAAAAAAAAAAAAAAAKNGVaooR3ZhMJTw1Pliter7hm2XRgvbU1p1X6gAAAAE/99j6f5hgf+Xn+JlP/AH2fov0MLgI1qfM5amGwcaDbTvcMHhJUG7u9wAAPD7i2hwzmdTGVqbmpQcLJpO7lF31/wnEOZwzPNcRjYR5VUlKST3V3cAAAAAAMZhHX5bO1riVlYAADOfEDDY/hKhkcaMlOmoLmbVny+W+oAAAAAAVsJKpXjUT2t+AAAFCoqdWM+zT+48R+PMNxV9W9jRcPZc97tO/Ny7W7cv4gAAAAABTwko4h1b73AAAOAvEDDcN4DE4arRlN1HdNNK2luoAAAAAAFDM4U8FLDuOrvr6gAAFHxAw0OCnw97GXPZrmura1HPbfbQAAAAAADC1lRrQqNXs7maY2OMrKcVayt+L/AMwAAOAuKaPDeavG1abmnBxsmk9WnfX0OJ82hm+b4jHQi4qpK6T3QAAAAAAAAAAAAAAAZFh9JVn6L9SvmVZYtzjLRO1ulv8Agmb4qVKglB2cvyMorvEUZUqmtvyZiKLpVZU30AAAAAx6lBwrR+yUsdSqzUI7sWIWFr1IyWj1MFKU8RKcuqv/AJEK6wtacJrR6ow+JhXTcegAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAYCl7PDQj5fnqO7epm9/ZUPT9EZD/En6GeUuXEKS6r8v6QAAAADSasyFClTd4xSJ0adRpyim0csb3tqVKVOorSVyEIwVoqyDCYv6w2uW1gAAOBeEP9aMxngvbey5YOd+Xm2lFWtzR/m3v02M9yv+y8zr4Hn5vZycb2tezte13b0uwAAAAAAxeL9hy6XuLVAAAZt4efUOF6Oe/Web2ig+Tktbm/vczvb/AAq/kAAAAAAFbF+zrRpct72/EAACjT9pUjC+7S+84/4A/wBVPq/+0e19rz/Y5bcvL/ele/N5WsAAAAAAFPF89d0uXYAAA4J8PP8AWbBYjE/WfZ+ydrcnNfS+/NG33MAAAAAACjlftcHLEc9rX0t287/oAAAUvDz2nCH+sX1no3ycnao4fFzeV/h8vMAAAAAADDUfbVo072u7GY4H6nVVPmvdX2t3832AAA4J4U/1mzN4H23s7Rcr8vNs0rW5o9+5xFk/9kZrXwHPz+zdr2tfztd2+9gAAAAAAAAAAAAAACV3YSSSSK+DqLFOilq3p6dzOsO3h4yivh/L+rGRUJRpyqNb7fIz+N4036/oAAAAAAAZVvL5AAAeBf8AyUVb/rzL/wBLpnHf/JTY/wD6+T/MAAAAAAM2+x8/0I/CgAAOLP8Ak2GD/wANL8gAAAAAAxf++Q+X5gAAYP8A3mn6r8zx+/5l3/cX/wBxgAAAAABQ/wB+l8wAADwS/wCRLj/X/wBtYAAAAAAGC/5E8/SQAABhf+TRP/DL/wBPsAAAAAADLv8Ae6fqjiP/AHqP+FfmwAAPBL/kppf9e5fnE8Rf+Spx3+N/kgAAAAAAAAAAAAAACHxILILGffwoeoAAAAAABlW8vkAAB4F/8lFW/wCvMv8A0umcd/8AJTY//r5P8wAAAAAAzb7Hz/Qj8KAAA4s/5Nhg/wDDS/IAAAAAAMX/AL5D5fmAABg/95p+q/M8fv8AmXf9xf8A3GAAAAAAFD/fpfMAAA8Ev+RLj/X/ANtYAAAAAAGD/wCRPP8A7eAAAML/AMmif+GX/p9gAAAAAAZd/vdP1RxH/vUf8K/NgAAeCX/JTS/69y/OJ4i/8lTjv8b/ACQAAAAAAAAAAAAAAARqRdJTe1rmFxVPEU+aD/4AUsdSq1nSg72W5n8v4cfX9AAAAAAADKt5fIAADwL/AOSirf8AXmX/AKXTOO/+Smx//Xyf5gAAAAABm32Pn+hH4UAABxZ/ybDB/wCGl+QAAAAAAYv/AHyHy/MAADB/7zT9V+Z4/f8AMu/7i/8AuMAAAAAAKH+/S+YAAB4Jf8iXH+v/ALawAAAAAAMF/wAiefpIAAAwv/Jon/hl/wCn2AAAAAABl3+90/VHEf8AvUf8K/NgAAeCX/JTS/69y/OJ4i/8lTjv8b/JAAAAAAAAAAAAAAABhHLE5dyRetmv6+QpVsNU0bjJFXMMVVjyynp935GTYWq6qrbRX4mcVVUxTS+zoAAAAAAAYDC1KLlzdQAAPC/ibL+H84qYrHNqEqbirK7u5Qf5JnFGYUMxzrFYug/cqTlJX0dm+wAAAAAAY/DVK3LydL/oJWSAAAz7jbKcbwRh8opSl7aCppprT3d9QAAAAAAxGFqTxMai2VvzAAAw84wrQk9k0/xPFfjPKuI/qf1CTfs+e91b4uS1v/AWAAAAAAFLC1I4l1HtqAAAeGnG2U5DluLw+NlJSqO6sr/ZsAAAAAABhswoU8vlQb953/EAAAocbZTDw/eRuUvb2atbTWq57+gAAAAAAGDqxpYiE5bJozjF0sVXU6eyVvxYAAHhpxHgcgzp4vGtqHJKOiu7tx6fI4vzPD5nnmJxmHbcJyurqztZdAAAAAAAAAAAAAAAAyXFKnVdOT0l+f8Awf8AIxGEo4hWqRv+ZTyjCQlflv6mJrxw9FzfTYlJyk5PdgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAG2xgc3pzioVnaXfoypj8LCN3NP0d/yMwx8sVPTSK2X6gAAAAAABQxNOtfk6AAAcN8MY/iHFywuBSc4xcnd20TS/NozLL62X4yrhK9uem3F21V1owAAAAAAr4mnRtzdTdAAAY7gjNsHk1PN6qj7Gai1rr722gAAAAAAVMTThUVN7sAACnB1JqEd3ocT8GZrw57L6+or2nNy2d/htf8A9KQAAAAAARxNOVR01ugAADh3gjNs/wAPVr4JRcaeju7dL/kAAAAAABTy+vUw7rxtyq/4AAAEOCM2nkX9tpR9hZu99dJcm3qAAAAAABSpSq1FCO70MXg6uFmoVN7XAAA4d4cx2f4x4TBJOaTlq7Kysn+Zm2WYjLMbUweIS54Oztqr+oAAAAAAAAAAAAAAAAAAAAAAABlW8vkAAB4F/wDJRVv+vMv/AEumcd/8lNj/APr5P8wAAAAAAzb7Hz/Qj8KAAA4s/wCTYYP/AA0vyAAAAAADF/75D5fmAABg/wDeafqvzPH7/mXf9xf/AHGAAAAAAFD/AH6XzAAAPBL/AJEuP9f/AG1gAAAAAAYL/kTz9JAAAGF/5NE/8Mv/AE+wAAAAAAMu/wB7p+qOI/8Aeo/4V+bAAA8Ev+Sml/17l+cTxF/5KnHf43+SAAAAAAAAAAAAAAAAAAAAAAAAMq3l8gAAPAv/AJKKt/15l/6XTOO/+Smx/wD18n+YAAAAAAZt9j5/oR+FAAAcWf8AJsMH/hpfkAAAAAAGL/3yHy/MAADB/wC80/VfmeP3/Mu/7i/+4wAAAAAAof79L5gAAHgl/wAiXH+v/trAAAAAAAwX/Inn6SAAAML/AMmif+GX/p9gAAAAAAZd/vdP1RxH/vUf8K/NgAAeCX/JTS/69y/OJ4i/8lTjv8b/ACQAAAAAAAAAAAAAAAAAAAAAAABlW8vkAAB4F/8AJRVv+vMv/S6Zx3/yU2P/AOvk/wAwAAAAAAzb7Hz/AEI/CgAAOLP+TYYP/DS/IAAAAAAMX/vkPl+YAAGD/wB5p+q/M8fv+Zd/3F/9xgAAAAABQ/36XzAAAPBL/kS4/wBf/bWAAAAAABgv+RPP0kAAAYX/AJNE/wDDL/0+wAAAAAAMu/3un6o4j/3qP+FfmwAAPBL/AJKaX/XuX5xPEX/kqcd/jf5IAAAAAAAAAAAAAAAAAAAAAAAAy6jUpuXMrAAAeEOdZflOeVa+OqqnB0pJN9+eDt9yf3HGGLoYvP8AGV6ElKEqkmmtmm9wAAAAAAzKjUqcvIr2v+gtkAABxJxJlOJ8P8Ll9KvGVaKp3j1Vlr9wAAAAAAYmjUlioSS0VvzAAAwsoxrwk3omvzPGbiLKs4+o/UK8anJ7S9ul+S332f3AAAAAABRo1Fi5Ta01AAAPCfiTKcryrGUsbXjTlN6J9fdsAAAAAABhcXQjlk6TkuZ30AAAMPxJlMfDN5Y68fb2l7nX+M5flqAAAAAABgZxhiacpOyTRnmIpV8RGVOV1b9WAAB4TZvgcrz918bVVOHs5K72u3HT8DjjG4fG8RYvEYealCUm01s1ZAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAU61Op8DuAABlOS4/NqzoYGk6k0rtLeyaV/va+8xeErYSvPD148s4tpp7prdAAAAAABUrU6duZ2NwAAMRw1m2HwEcwq0JKjK1pPZ32+8AAAAAAJVqcZKDerAAAjFykox3ZnHDua5RyfX6Dp89+W/W1r29Lr7wAAAAAAVanKfInqAAAZXw1m2a0p1cFQlUjDdrp1/IAAAAAACOErzpOrGN4rqAAAR4azaWW/2kqEvYfz9NHy/noAAAAAABCEpyUYq7ZXw9WjLlqRswAAMryfHZrX+r4Kk6k7Xst7Ld/iY3BYjBYieHxEHGcXZp7pgAAAAAAAAAAAAAAAAAAAAAAAGVby+QAAHgX/yUVb/rzL/0umcd/wDJTY//AK+T/MAAAAAAM2+x8/0I/CgAAOLP+TYYP/DS/IAAAAAAMX/vkPl+YAAGD/3mn6r8zx+/5l3/AHF/9xgAAAAABQ/36XzAAAPBL/kS4/1/9tYAAAAAAGC/5E8/SQAABhf+TRP/AAy/9PsAAAAAADLv97p+qOI/96j/AIV+bAAA8Ev+Sml/17l+cTxF/wCSpx3+N/kgAAAAAAAAAAAAAAAAAAAAAAADK4yTlddgAAPBLE0MPxBWlWmor2MldtLXnp9zjepCpxHjpwaadSVmttwAAAAAAzWLlyWXf9COyAAA4pxuGn4a4SjGpFzSpaXV9F23AAAAAADFRk8ZBpdvzAAAwjSxFP1X5njtjMNiP7P9jUjK3tb2adv4fYAAAAAAKEZfXZO3cAAA8GcbhqGT46NWpGLb0u0r+6+4AAAAAAGDqQWUzi2r+8AAAYbG4ZeE7oe0jz2l7t1f+O3tvtqAAAAAABl7SxdNvujiGcZ4qLi7+6vzYAAHgziKNDiOU6s1Fezlq2kt49zxAqQq8T42cJJpzdmtVsgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAKdanU+B3AAAAAAAAAAqVqdO3M7G4AAAAAAAAASrU4yUG9WAAAAAAAAAAAAAAAAAAAARwledJ1YxvFdQAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADKt5fIAAAAAAAAADNvsfP8AQj8KAAAAAAAAADF/75D5fmAAAAAAAAAAAAABCjUnGUoq6jv5AAAAAAGC/wCRPP0kAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAZVvL5AAAAAAAAAAZt9j5/oR+FAAAAAAAAABi/98h8vzAAAAAAAAAAKKTqRXmjFZvSoV3h50/dVvyvtYzbCwoYi1P4WroAADL/9zxPov1AAAACrhJU6EKrfxX09AAwX/Inn6SAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAACh/Fj6ozr/AH6fy/JGb3UaEXuoop5E5QSlUSm1dR/rX8NDD4CVV1It2lBN2t26bmAwTxdRwTtZXMFhXia8aV7Xv+CMDlSxHNKU7RTt/XYzHL5YSaV7p7Mw2SutKHvaON3pt5GAyx4qcru0Y7s+ovC4bEK94tKz+8P9W/8Ap7+H/BMXl0qOIVCD5m/K3+ZUyBqm+Sd5roYPCfWavs720b+4hw/OWH5+b37Xt+hTpTqTUIrVlbJOWk3TqKUo7r+v6YYDL3iU5ylywW7ZmGXywslZ80XszD5JKrKHve64p3t36f8ABMbl0sNXjTvdPZlfJva4enR57ct9bb3+ZhcsliKcJxlu7bbaXv8A8AxGVqEasoyvyNaW721/rsYHB/WqjhzWSV7gAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAFD+LH1RWy/BVsW5zlef8t10Xbc5pYrNEqitZ7eSMRTxksY6ypy0emj6bfgRjGGbyj/Mv0v8AoZL+655ve8Y/e9TBQ9jjK8v5FL89PwKVOrVapwTfl+pj6FWnltNVVaSdvlqY/Ezp4ChCDtda/L/hyvJ0srpwT+Ntv5f0jA1pSy6vTfT9f+GDKcMq+JjfZav5GCrqvjquIe0U2vyX4GBxU44yNRvVvX57mEpKnm84+v46mBxc55iqjfxO3yf+Rg6XJj60oq7jzNLz6GWUcVTxkZyhKzve6fXuYmChWnBdG1+JWoVv7Po06UW73bsn8tv60J0qqytqpFpxlpdW0f8AwWZrWksJh6S2cU39ysY2Tq5dRnLdNr+vuMb/AMi/D/Mw9eVHKZSi9W7L8P0Mok6qrUXq5R/Ff8OZb+7w1er5W+/+kAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABlW8vkAAAAAAAAABm32Pn+hH4UAAAAAAAAAGL/wB8h8vzAAAAAAAAAAKH8WPqjNpuGYylHdW/JFRRjnEJr7Sv+DRXzrGU6soXWja2MPjalXH06s3rdL9DGJ4ehJLd1G/kjGJU44movtKNvnuYes8Fl6qw+Ob37JGKrVK2VwlN3fN+jMz/AN2w/wDh/wAjHrmwOHa7P9P8jL1/sWJ9F+oUV9Wy2VT7VTRen9X/AAMp96FeC3cGYaLlXhFd1+ZRkpZzO3b9EZd/vdP1RhpyhjMVy72bXyP7dxndfcVajqTc5bt3K2YVsNhKDpWs1+VjE5ricRTdObVn5Ga/wsP/AIF+SK//ACKqX+J/qY3/AJF+H+Z/zKf+3v0MoqezxkPPT7zFU/q+CnD+ab+5f8MGC/5E8/SQAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABlW8vkAAAAAAAAABm32Pn+hH4UAAAAAAAAAGL/3yHy/MAAAAAAAAAAofxY+qM6/36fy/JGPquDw2IXZfgYjKaeNmq9GdlL+v+HXcrwWHxDjF35Xv6HEE4udOMezf3v/AIBmNVSy6l3lb8FYwmHjjsAqKlaUH+f9fgZjGlHL406Tuoytfzs7/izM/wDdsP8A4f8AIwFOGNwbwzlaUXdf1946NPDYOrQjK8krv57IhHnko9zGZP8AWFCKnaMVZK1/nuiFOOWY6ClK6a10tvp59inllDCVXiZT9xapfl/wO5lFV1cwc3u7sy7/AHun6ojiY4fNJyls20/mVOH4Obmqlob/ANP9QwEKeOwn1aTtKLuv6/P5GYZX9Upxk53bexmv8LD/AOBfkiv/AMiql/if6lHCrHZfCMZWcG/6/I/5lP8A29+hTm4TjJdHc4gqRcqcI9m/vDBf8iefpIAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAyreXyAAAAAAAAAAzb7Hz/Qj8KAAAAAAAAADF/wC+Q+X5gAAAAAAAAAACnJKyegApOOzATad09R6gDlJpK+gA5yceW+gJtO6JScndu7AUpJaMMFhnia8aa+fp1M2rKri5NbLRfL/ghgv+RPP0kAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAZVvL5AAAAAAAAAAZt9j5/oR+FAAAAAAAAABi/98h8vzAAAAAAAAAAAAAAAAAAAMDmDwtOoor3pWt5bj1DBf8AInn6SAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAjCU3aKuypha9ON5QaXoAAAAAAABClCHwpIAAAAAAAAACdOE/iSfqAAAAAAAAABKlCUlJpXAAAAAAAAAAAAAAAAAAAAFXqxhyKb5e13b7gAAAAAAAAAAAAAAAAAAAAAAAAAAAAApUpVZqEN2YPBU8NDljv1Y1czbLo017amtOq7efoAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAGR4Ze9Wfov1K+aVlinKMvdTtbpb/gmbYuVKgvZvWXXyMrrvFUJ0qrvb8mV6TpVJU30AAAAAAAMsqTm5c0m9twAAAAAAAAAMzqThycra3/QWyAAAAAAAAADFVJrFwipO2n5gAAAAAAAAAAAAAAAAAAAGEw9GWVTm4py11tr94AAAAAAAAAAAAAAAAAAAAAAAAAAAAAZfSVPCwj5X+/Ud2zN23Soen6IyH+JP0M8pcuIUl1X5af5AAAAAAAGVby+QAAAAAAAAAGbfY+f6EfhQAAAAAAAAAYv/AHyHy/MAAAAAAAAAAAAAAAAAAAAwX/Inn6SAAAAAAAAAAAAAAAAAAAAAAAAAAAAAEruyIxUUkuhiMJNYp0UtW9DOsO3h4Sj9n8v6RkVFxhKo1vovkZ/FONOXr+gAAAAAABlW8vkAAAAAAAAABm32Pn+hH4UAAAAAAAAAGL/3yHy/MAAAAAAAAAAAAAAAAAAAAwX/ACJ5+kgAAAAAAAAAAAAAAAAAAAAAAAAAAAACHxILK9+o0mrCSSsjPreyh6/oAAAAAAAZVvL5AAAAAAAAAAZt9j5/oR+FAAAAAAAAABi/98h8vzAAAAAAAAAAAAAAAAAAAAMF/wAiefpIAAAAAAAAAAAAAAAAAAAAAAAAAAAAAISU4KXczDNI4d8kNZfl6/5FHPKql+8Sa8t/6/q5RqwrQU4O6Zn8/wCHH1/QAAAAAAAoYWnRvydQAAAAAAAAAK+Gp1rc3QtZAAAAAAAAABUwtOdRVHugAAAAAAAAAAAAAAAAAAACnmFenh3Qi1yu/wCIAAAAAAAAAAAAAAAAAAAAAAAAAAAAAYfFyWW88dXFW+7/AIGphMHVxlR66dX/AF1MRkVoXpSu10Zk1apCv7Lo9/K3X9DOKqqYppfZVgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADJ68VN0Z7S/P8A4JhsPChSVOHQK0KOGc8T1a/r79CcnOTk92AAAAAAAZfiKlVy53ewAAAAAAAAAGYYipS5eR2vf9BapAAAAAAAAABiMRUhiYwi9Hb8wAAAAAAAAAAAAAAAAAAADC4HDzy2daUfeV9bvp87AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAm07owOb06kVGs7S79H/kVcZh6cbymjMcfLFSSWkVsv1AAAAAAADKt5fIAAAAAAAAADNvsfP9CPwoAAAAAAAAAMX/vkPl+YAAAAAAAAAAAAAAAAAAABgv8AkTz9JAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAGVby+QAAAAAAAAAGbfY+f6EfhQAAAAAAAAAYv/fIfL8wAAAAAAAAAAAAAAAAAAADBf8iefpIAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAyreXyAAAAAAAAAAzb7Hz/Qj8KAAAAAAAAADF/75D5fmAAAAAAAAAAAAAAAAAAAAYL/kTz9JAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAGGwkaDdne4AAAAAAAAAGKwka9ru1hKysAAAAAAAAAFXCRqVVUvqrfgAAAAAAAAAAAAAAAAAAAAUsznTwjw6irO+vqAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABg8XKu2mrWAAAAAAAAAAxmKdDlsr3uLVAAAAAAAAABWxcqdeNNLe34gAAAAAAAAAAAAAAAAAAAFDLIVcFLEOWqvp6AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAGVby+QAAAAAAAAAGbfY+f6EfhQAAAAAAAAAYv/AHyHy/MAAAAAAAAAAAAAAAAAAAAwX/Inn6SAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAMHhJUG7u9wAAAAAAAAAMZhHX5bO1riVlYAAAAAAAAAK2ElUrxqJ7W/AAAAAAAAAAAAAAAAAAAAAoZnCngpYdx1d9fUAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADivPamTYKOIpwUm5KNn5pv9DKsZLG4GliZKznFO3a6AAAAAAAAAAy/iqtis9qZa6aUYuWt3f3QAAAAAAAAAqy5IOXZHCPE9bO/be0pqPJy7N635v8AIAAAAAAAAADijiqtlGJo0adNSU11b01sAAAAAAAAAAVOKq0eIv7K9muW61u76w5tvwAAAAAAAAAAOLeKq2STpRp01LnT3b6W7eoAAAAAAAAAAAAAAAAAAAAAAAFWrTpQc6kkordvRL1ZSq06sFOnJOL2a1T9GAAAeJn/ACKKf/Xxf+kzOGf+RNhf8EfyAAAAAAAAAAyL/ktMR6zAAAAAAAAADEfwZ+j/ACPC3/mL/wC3P/bwAAAAAAAAAPEf/kY4X0/9uAAAAAAAAAAK/wDyXq9V/wCmkAAAAAAAAAAeKP8AGw3pL80AAAAAAAAAAAAAAAAAAAAAcfVcRDLqUaFRwcqijdNrRxlo7dO6K+AzvheUMa6/tKV0pK729H32TWqfkynONSCnHZq4BxLPM44rArB35XP3rX2038rc17/nYOJv+RNiv8EvyOGf+RNhf8EfyAADH42lgsNPEVfhiru25gsVHF4WniIqynFSS9Vc4ryGpnOCjh6c1FqSld+Sa/UyrBSwWBpYaTu4RSv6IAAAAAAAAADL+Fa2Fz2pmTqJxk5aWd/eAAAAAAAAACrDng490cI8L1sk9t7Sopc/LsmrW5v8wAAAAAAAAAOKOFa2cYmjWp1FFQXVPvcAAAAAAAAAAqcK1pcRf2r7Rct17tnfSHL/AMEAAAAAAAAAA4t4VrZ3OlKnUUeRPdPrbt6AAAAAAAAAAAAAAAAAAAAAHHn+6Yb/AK/Q/KRxgovI8TzdvxurfiYLOY4PDZbhpxblWglfs1GN7/eZ7nNPKcG8TOLkk0rLzM0zqhl9ajTqrSo3r0XKrtsyrj7B4/MPqns3FSdoyb3fS6tpfpqzNc5p5fVw9OUW/ayUF5N2V/xOJuKcPkkIKUeecto3touretvLTUr53SzjhrE4mnFx92SafdLo+q18vQoccYXKsFhML7NzahHms7ct0vLV21totte1TM6Ky+WOh70FBzXmkrmP8RMHhnSUaTlzJSlqly3V7dbu3TRbame8dYPLpU40oe0ckpaOySe2tnq+1jL8dRx2Fp4mi/dkrr/L1T0fmYrxCwNHFOlGnKVNOzmtr+S6r5ryTM1zijgMuljrc8Uk9Oqk0k199zH+IWBwuP8Aqyg5RTtKV9n1sra22eq8rnHWc0sNlfsrc3tk0mtls7/O5wZxLRx9GGBjBp0oRu3s7JRAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADxDqqll9Co9o1Yv7lIzjid8SRhlmXU5Lna5nK2iT7JvRbt36W6nEFKFLN8ppw2i5JeiUEeIf/ACJJf4onHeE+t4jAYe9lObT9Hyp/gcb5dhcHgqGLoQUZUpxtZW07felb/gnGH+95b/1+j+aK1WjT4yj9Ya1pWhfvfp6+8vV23ZxFRp0smxahFK8ZN201a1fzOHuHMJV4djTqQTlVi221rd/DZ9LK1vMyKu6vBlVP7MKq/CT/AFOHMlw8+F3TlFN1Yybfnry/ckrdnqcBZZQqZLOdVc3tW07/AMq91L0VnbtcyTG1MHwpi7PWnKcV5XtqvnK5lmDy2OQwwMpx96Guq+Jq7fqnt2suxLEyr8By5ndxtH7qit+Firw5hI8Luk4JzUOfmsr81ua9999PQqSjieC1UmrtU7K/Szt+hw7h6UMrw04xSbpwu7av3UcQ57TybCxxFSDknJRsvNN/oZdjI43CU8TFWU0nbtcAAAAAAAAADB8VUcVm88tjTalHm1ure6AAAAAAAAAFSShBy7HDfE9HO/a+zpuPJbdrW9+3oAAAAAAAAABn/FVHKK9KjOm5OfZrTWwAAAAAAAAABPiqjHO/7K9m+bTW6trHm238gAAAAAAAAADiPiqjkkqcalNy577NaWt/mAAAAAAAAAAAAAAAAAAAAAcfRUsHhk9V7aH5SMLgMJhU1h6UYX35Ul+RxM0s7yu/80//AG08Q7f2JL/FE4r/AORnln/Xx/8Atp4h/wDIkl/iicYf73lv/X6P5o4p4YpZzRTi+WrD4X+j8vPpv3TwWZYjF8MYujinerSU4O+7stL+fTztfqcM/wDImwv+CP5HDn/JG4j/AA1f/SWcNf8AJPUP8H+ZwD/yIaXrL/0plKDnwrmKX/P1/g6bMF4dZTiMNTre0qe9FPePVX/lOIcnoZVwrXw1BtxTi9bX1nHsl+RiP+RHP/r0/wD0go/8kM/8D/8ASmZB/wAijC/9e4f+ko8TP+RRT/6+L/0mZwz/AMibC/4I/kAAAAAAAAABkX/JaYj1mAAAAAAAAAGI/gz9H+R4W/8AMX/25/7eAAAAAAAAAB4j/wDIxwvp/wC3AAAAAAAAAAFf/kvV6r/00gAAAAAAAAADxR/jYb0l+aAAAAAAAAAAAAAAAAAAAAAOPP8AdMN/1+h+Ug4+p1qeGoY+j8VGafyfV+V0l8ziTiKvn+CbpUnClSs5X1vJvlS+5t+fkcV/8jPLP+vj/wDbTxD/AORJL/FE4w/3vLf+v0fzRm/G+GyvHzwtelJ2SaatrdebWnmr+mhltKrPhzMMbVjyuvzzS8mv87/Kz6nDP/Imwv8Agj+Rw5/yRuI/w1f/AElnDX/JPUP8H+ZwD/yIaXrL/wBKZwtgo43J8dhn9upUXo3GNn8nqZPxvHKcL9QzClL2lL3Va2y2Tu1a211e619eJccsfwlPEqPLzqDt29+JiP8AkRz/AOvT/wDSDLcLPE8GKlBXk4Ssu7TbS+ZwTxDRx+EhhFFqdKKTva2mitrfbe6XzPEz/kUU/wDr4v8A0mZwz/yJsL/gj+QAAAAAAAAAGRf8lpiPWYAAAAAAAAAYj+DP0f5Hhb/zF/8Abn/t4AAAAAAAAAHiP/yMcL6f+3AAAAAAAAAAFf8A5L1eq/8ATSAAAAAAAAAAPFH+NhvSX5oAAAAAAAAAAAAAAAAAAAAAqUqdRJTinbuuoThGcXGSun0ZSwOFpUvY06UVDskkvuWhOlTm05RTa2029CpSp1Y8s4przVydKnNpyinba62fkYnA4XE29vSjO23Mk7elyVOEo8jWnYjCMYqMVZIjQpRg6cYpR7W0+4jThGKjFWXYp0oU48sEkuy0KdKnTTUIpX7K2pXy7B16iqVaMZSWzcU2vm1cnShUhySSa7PYcIuPLbTsQpwhFRgrJdEUcFhqE5TpU4xct2kk3623+Z4mf8iin/18X/pMzhn/AJE2F/wR/IAAAAAAAAADIv8AktMR6zAAAAAAAAADEfwZ+j/I8Lf+Yv8A7c/9vAAAAAAAAAA8R/8AkY4X0/8AbgAAAAAAAAACv/yXq9V/6aQAAAAAAAAAB4o/xsN6S/NAAAAAAAAAAAAAAAAAAAAAAAAAAABxzlOMzPLoUcJDmkpp2ulpyyXVrujI8NVw2WUKNVWlGKTXZpeWgAAAAAAAAAGVZBmNDietjalO1KTnZ3j120Tv+AAAAAAAAAAVouVOUVu0zgLIswyv6x9bp8vNy21Tvbmvs33W4AAAAAAAAAHG2QZjmONw9TC0+aMVrrFW182gAAAAAAAAACrkGYy4uWYKn+5uvevH+RR2vffTYAAAAAAAAAA49yDMc0qUHhKfMoqV9Yq17d2uwAAAAAAAAAAAAAAABg8rq4lc17R7/wCRVyFcvuT18ytRnRm4TVmgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAy/CfWayi9lqx4rDwqKhzWfYq1YUoOc3ZIx1GnjsN7Sk7tbfqgAAAAAAAAAAzPNsHltFVsXPli3a9m9bN9E3smYbE0sTRhWpO8ZK6eq0frqAAAAAAAAABQz/AC6vjZYKnUvVje6tLpvra34gAAAAAAAABKSjFt7IyvPcvzTn+p1ObltfSSte9t0uzAAAAAAAAAAzLP8ALsuqQp4qpyyltpJ36dEwAAAAAAAAACWf5dHH/wBnup++/ltLtfe1tvMAAAAAAAAAAzTP8uytxji6nK5XtpJ7b7JgAAAAAAAAAAAAAAAZLQUMPz9ZfkitVc60p31vczqs5UKa76/h/wAEyGo/fh6MzGgqOJlFbbr5gAAAAAAAAAHiZ/yKKf8A18X/AKTM4Z/5E2F/wR/IAAAAAAAAADIv+S0xHrMAAAAAAAAAMR/Bn6P8jwt/5i/+3P8A28AAAAAAAAADxH/5GOF9P/bgAAAAAAAAACv/AMl6vVf+mkAAAAAAAAAAeKP8bDekvzQAAAAAAAAAAAAAAAGGpqnRhHskSjyya7GcQapUfT9EZCvfm/JGfU0qkJ90193/AA4AAAAAAAAAB4mf8iin/wBfF/6TM4Z/5E2F/wAEfyAAAAAAAAAAyL/ktMR6zAAAAAAAAADEfwZ+j/I8Lf8AmL/7c/8AbwAAAAAAAAAPEf8A5GOF9P8A24AAAAAAAAAAr/8AJer1X/ppAAAAAAAAAAHij/Gw3pL80AAAAAAAAAAAAAAAEUnJIK2TOeK57+49X39PmZhg/rNHlTs1sZbgnhqTUvie5ny/dwfmAAAAAAAAAAeJn/Iop/8AXxf+kzOGf+RNhf8ABH8gAAAAAAAAAMi/5LTEeswAAAAAAAAAxH8Gfo/yPC3/AJi/+3P/AG8AAAAAAAAADxH/AORjhfT/ANuAAAAAAAAAAK//ACXq9V/6aQAAAAAAAAAB4o/xsN6S/NAAAAAAAAAAAAAAABFpNMJyjCPNJ2SKWLoVZcsJpsM/f7uC82AAAAAAAAAAeIeDxGKyunDD03OSmnaKbduWWtl0OHqU6WU4enUi1JQSaas07dUAAAAAAAAABk2XYynxbXrzpSUG52k4tJ321tbUAAAAAAAAAK6bpSS7P8jw3y/F4T6z9YpShfktzRavbmva69AAAAAAAAAAOPsuxmKx+GlQpSmktWot2162QAAAAAAAAABWy7GPjVYhUpezuve5Xy/w0t7W309QAAAAAAAAADxHy7GYurh3h6Up2Ur8sW7bb2TAAAAAAAAAAAAAAAAw1RToQn5IzLHSxNTli/dW3+ZKlVpNOUWu2jRlmN+s0rS+Jb/5mfTTqQh2X5/8MAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAYSrOrl06cH70fy3/zMmwKa9vNen+ZXowrU3Ca0Zl+GrUcdyLpv6f1YzCuq2IlJbbL5AAAAAAAAAAGKxmGwsFOvUUFteTSV+2pSq06sFOnJOL2a1T9GAAAAAAAAABDMcHUrOhCrFzW8VJXVt9L30AAAAAAAAABtJXZhcwwmLv8AV6sZ235ZJ2vtezdr2AAAAAAAAAAxOY4PDSUa9WMG9lKSTfpdgAAAAAAAAADzHBrEfVnVj7T+XmXNtfa99tdttQAAAAAAAAADFZjg8I0sRVjC+3NJK/pdgAAAAAAAAAAAAAAAYDFfVqyl02ZDkcFybdAzXFRoU2o/HJW+X9bAAAAAAAAAAB4mf8iin/18X/pMzhn/AJE2F/wR/IAAAAAAAAADIv8AktMR6zAAAAAAAAADEfwZ+j/I8Lf+Yv8A7c/9vAAAAAAAAAA8R/8AkY4X0/8AbgAAAAAAAAACv/yXq9V/6aQAAAAAAAAAB4o/xsN6S/NAAAAAAAAAAAAAAAAGDzKthlyrWPZ/oVM+m42hCz9b/oirVnVk5Td2wAAAAAAAAAA8TP8AkUU/+vi/9JmcM/8AImwv+CP5AAAAAAAAAAZF/wAlpiPWYAAAAAAAAAYj+DP0f5Hhb/zF/wDbn/t4AAAAAAAAAHiP/wAjHC+n/twAAAAAAAAABX/5L1eq/wDTSAAAAAAAAAAPFH+NhvSX5oAAAAAAAAAAAAAAAAAAAAAAAAAAAPEz/kUU/wDr4v8A0mZwz/yJsL/gj+QAAAAAAAAAGRf8lpiPWYAAAAAAAAAYj+DP0f5Hhb/zF/8Abn/t4AAAAAAAAAHiP/yMcL6f+3AAAAAAAAAAFf8A5L1eq/8ATSAAAAAAAAAAPFH+NhvSX5oAAAAAAAAAAAAAAAAAAAAAAAAAAAPEz/kUU/8Ar4v/AEmZwz/yJsL/AII/kAAAAAAAAABkX/JaYj1mAAAAAAAAAGI/gz9H+R4W/wDMX/25/wC3gAAAAAAAAAeI/wDyMcL6f+3AAAAAAAAAAFf/AJL1eq/9NIAAAAAAAAAA8Uf42G9JfmgAAAAAAAAAAAAAAAAAAAAAAAAAAAq0adWPLUimvNXIwjGKjFWSAAAAAAAAAAjhqMZ88YJS72V/vAAAAAAAAABpNFLD0aV/ZwUb9kl+QAAAAAAAAAFXDUarTnBNrukwAAAAAAAAAB4ai6ntORc3eyv9+4AAAAAAAAABVw1Gtb2kFK3dJgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHiZ/yKKf/AF8X/pMzhn/kTYX/AAR/IAAAAAAAAADIv+S0xHrMAAAAAAAAAMR/Bn6P8jwt/wCYv/tz/wBvAAAAAAAAAA8R/wDkY4X0/wDbgAAAAAAAAACv/wAl6vVf+mkAAAAAAAAAAeKP8bDekvzQAAAAAAAAAAAAAAAAAAAAAAAAAAAeJn/Iop/9fF/6TM4Z/wCRNhf8EfyAAAAAAAAAAyL/AJLTEeswAAAAAAAAAxH8Gfo/yPC3/mL/AO3P/bwAAAAAAAAAPEf/AJGOF9P/AG4AAAAAAAAAAr/8l6vVf+mkAAAAAAAAAAeKP8bDekvzQAAAAAAAAAAAAAAAAAAAAAAAAAAAY7LsLjqapYmCnFO9n32v+LKFCnQpRpUlaMVZLsgAAAAAAAAAKWT4CliniqdJKo73fXXf7wAAAAAAAAAlFSVmYDKMDgOb6rTUOa17dbXt+bAAAAAAAAAAxuT4DHTjPEUlJx2v0AAAAAAAAAAHk+AeL+uOkva/zddrfloAAAAAAAAAAY/J8Bj3F4qkpuO1+lwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA//9k=)
DA and GABA signaling
![[[@Watabe-Uchida2017-gi]](https://doi.org/10.1146/annurev-neuro-072116-031109)](data:image/jpeg; charset=UTF-8;base64,/9j/4AAQSkZJRgABAQECWAJYAAD//gAuSGFuZG1hZGUgU29mdHdhcmUsIEluYy4gSW1hZ2UgQWxjaGVteSB2MS4xNAr/2wCEAAMCAgICAgMCAgIDAwMDBAcFBAQEBAkGBwUHCgkLCwoJCgoLDRAOCwwPDAoKDhMODxEREhISCw4UFhQSFRASEhIBAwMDBAQECAUFCBIMCgwSEhISEhISEhISEhISEhISEhISEhISEhISEhISEhISEhISEhISEhISEhISEhISEhISEv/CABEIDZEMpAMBIQACEQEDEQH/xAGiAAABBQEBAQEBAQAAAAAAAAAAAQIDBAUGBwgJCgsBAAMBAQEBAQEBAQEAAAAAAAABAgMEBQYHCAkKCxAAAgEDAwIEAwUFBAQAAAF9AQIDAAQRBRIhMUEGE1FhByJxFDKBkaEII0KxwRVS0fAkM2JyggkKFhcYGRolJicoKSo0NTY3ODk6Q0RFRkdISUpTVFVWV1hZWmNkZWZnaGlqc3R1dnd4eXqDhIWGh4iJipKTlJWWl5iZmqKjpKWmp6ipqrKztLW2t7i5usLDxMXGx8jJytLT1NXW19jZ2uHi4+Tl5ufo6erx8vP09fb3+Pn6EQACAQIEBAMEBwUEBAABAncAAQIDEQQFITEGEkFRB2FxEyIygQgUQpGhscEJIzNS8BVictEKFiQ04SXxFxgZGiYnKCkqNTY3ODk6Q0RFRkdISUpTVFVWV1hZWmNkZWZnaGlqc3R1dnd4eXqCg4SFhoeIiYqSk5SVlpeYmZqio6Slpqeoqaqys7S1tre4ubrCw8TFxsfIycrS09TV1tfY2dri4+Tl5ufo6ery8/T19vf4+fr/2gAMAwEAAhEDEQAAAP1VAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAydMvk3H/AEn8pu+ref6fvMnwf6OCYAAAAAAAAAAAAAAfLfC/o35Z9vH5z+pgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABHD4Tm/xf6+TufT5+mP1j5fg8D8b+t7voP0/5w8s+d78135l9F1Xc/r3yofTed5fU/Dvs/T7n7T8gHRmAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAcV1cf5E++/vf837/AM6c3V7V+ln53+oSHw/6EAAAAAAAAAAAAAAHzbxX3/5r9jnwH6UAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAVPnD8S+xw+x/HPrWegfo/wA/6Qf0R8D8qUv4V/avR/cf6g/Nz5K+S9Toeg+J9nzP3WH6Xlf1H+a/Kk/8KftXuXo/9lfkgfX+UAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAcV1cf5E/pN+ufif0L87/mX63+bn6Z/pf5P6p8LfJ/aeWeofdfnX3Zxv5H+2fDHsf6X+T/PP0D0c3298/fl36/8AGmd+yfhP0z9i/k/7R8TZn13xHlGn9B819EfJnl+x7L97fHfc/G/B/T/I/df5seV7PtHz59P8j7D98fD/AKFnfB+uPkHtv3X5393T/if78fCf1Xxz/K/ufzxv6D/P/Tdx8r/NfWfJ/QfrP4v994X5F+2fDP6HfUfIec/G/bwfpZ8E+H9D3vz99d8T+pvx/wDAfpPzVr/q3439pe2/jX7v+a/pn6V+UeFdj6nj/obj/jH7x8B8H+2fgX1/9Sfk37QHg/RgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHjvk/wDIf6r9QbX9VfmQelznM+N1/MvrH8qfp3nf1X+pfNL8lY16p6n9f5Xyx69+L/X+r+X/AL58P5Xv/wA4foLvoP8AbvjQ/VvmOP8AJ/wX7fd4z4r2PUfTf3z4fD8S+E9rK9R+S9X03wD9H+f671H3uH5z9I/K/pp+F9PnXtJe345pOZ3PxXsexebfu/xPKVvxD7LD9o9rj7PyT9M+d871f53+++ij+xPyYAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA4rq4/yJ/Sb9c/E/oXA/H/3H8c/vT9p/A/FvON+f9C/zs+d+p9V+r+Lv/L/6y+0+B7H4c5uv9Me5/L/17l/APb+f+PP1y9v5/wCIPE/ovmPt/wDP7g9H2b2Xm6/i/wDWHDo+PvL/AFfG/UH8bvC+j9+9R7OD4w/Uhr508l35/v8A/MbzPX+5/pv4f9BPzM9z57yz7p+s+M+C/pv0vK+mvy68b3f1A+T/ABfewvrbq5PzM/Wjt8/yL89PR8v9kfy++Y+u4r63+s+L9M/OLm6/0j8G+f8Apfmr9fd+f8lOr+3/AD/6n+BfI9v9BuF+Y+u+Vf0s9Lyui7v8x/Ww5O0AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA+coP5A/VvpTy/wDor4Hzjc/n/wC6978e/dfiuG93+C9z5h+mfC7eo+SvovPrN/nv7zT+mv334bT+dPo+Db6P57u8T+s05j9++G4P56/lj9M9awevHivrf3+L5m7D431u1+duzL6p+bfpPP7z237fxfkT2D8P+zu+G6z6B6v+h+D889j+UfT+kfO/03ne35v1vleQe4fn/u+ZP8bs+hPlT7/wvZ+w+38bqj9X+YAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAOK6uP8if0m/XPxP6F8j/Kv2T8sP0l/Vfxv4I+h+zh+9vz7/PP07xP9BfqPkfy//VvbD0/8XfP9P7hTHf4x9N+9/N/GP1O+a+s+RMTs4f0z/Hv5r6r68+kvH938iv1A+z+D+X/L+/zf1B/G7wvo/wBC/avk/s/yK/Sb7/8ANfjHy36X5Ser3ef9l/bv4d/Qh+ZnoeYfpmH5Y9t9x+fesfCrVyH6f5HtPvj4D9K/Mz9aHPkX56ej5f7I/l98x9d6X+iHm+t8K/MH6B+Z/sT4d+Q/t35i/qp+lflP5p/UHpeV9x/jD839V9re3eR7f5rZ/wCt/in319M/hf8ARAeH9CAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfP3Kfxx+tfVXPf1d+YeMZn8q/pv078x/rny2Lp/gP3GD679HweyfJX6V876d6Z9F5/wA59N+I/Ze7/Jv6D4Vyb8d+syPof7/w+6P6m/NOD+ev5Y/TPqnjv2f5Dwf6t/Mvo/lKT8P+yWJn0b4V+1fH957b+lfO/InsH4f9nd8M0n65zP6c/Ofln6C/m39C7/5Z+88TsNX5/u8r+tfW5fEuD+A936m+e/1z5Xhe8/A/uPoGT+1Px8LQAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHFdXH+RP3N+0fgvoXw/wCN71b9YPK9f81eb+0+E/R782Pk/s/QvrbPT8v/ALf+r+N7L87ubq/TD4D4PS9191831vzh/U71vF+RMTs4f0z/AB8+a+q+u/pPx/d/In9QPs/g/l/y/v8AN/UH8bvC+j/Qv2v5P7P8if0m+/8AzX5w4Hq5P0KZ+VfsneX/AB/cPzM+1+AP0zD8se2+4/Pvpb83+Pu/Srvfzz9Otclh0fll9yfrH4x5x8xdvB+yP5ffCfovqH6F+Z6/y5+fP3X51+oPz98j9t8nfsB6Xlfkt9QfQ/M/cf4w/NfVfa30V4n0Nj8+/T8jL/U7HcPgv0gAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA4n50/lz9K9B9L+r8vy/H/OPofob5V+x8j2rrfuPG8azvw37T6g+Sv1D5r0X0X0uf5+6n8d+s9D+fve4vou5+1fH/ADz3X8+/d+7H9hfk/B/PX8sfpn1Tx/7P8h4N9afm/wBD8v8AcfEez63W/pb866P5i+A92b1n5r0fn32X5H1bvhm+f12f1X+ZfJ/oP8h/qvp3zT9d5XtdD6Hg8r+tZfiXB/Ae79Sc9/TX5zzvg382foX070n9MfnYexyAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAcp0c35bUP6C/mXoPavD+i+z+q/GP3jhvz7+9/OPMfXPu/zr9AeC/HP3T8v/Y/27+fuC+mPH9z7r+QPzD9c+OvZP1n8Y85/RL5T7H5pyOnl/QP8uPA+k+nvfPL9j8xP0O+r+M+ePO+zh/Q/8rfH937h9a+a+r/MT74+1+B9D/Pvyfa8k3f0v8n/AEt9C/AP6VPz8+j+WP0DD85eu+v+H+6vh/4r775ji/Y/wv7M+svxX99/OLx39a/FvobxXj7v1N/PX4n770T7h871YfhPTL5u6D9a/FfuP6A/D/6D/Lj6I/R/yz6//KLwfpPrTFqPjva/SPyv7p+hPw3+hg8f3AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA4L5b0vKcX+VP0613X0fn9R477fH9L3P6O/P8AzvyD+bv0H6Q+dvpfOzLH4j9l1XtH9A/C+Ycx8D7n0Wf0/wDm/i/K/wAtfpf0kf1L+acZ4f8Azj+gfR/LfrHy/jP0z8F7nNeJeZ0Y+7+VfT/S/nv9X/mHie//ACz+lzd1+h+Bd8e5tPp6X+lPzzifFf51+/z/AED4/wBb2zzL+hfg/N/pn5P0/I+M+J9n3X5z9ziqd1+de/7q7+wvygKQAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHinp+R+X/6t/pX5T6gfkX7aAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB5t2+f8R/d32fwnbn5/8ApYAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAZR18IAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABqnJ3EeLacdgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABN0yHTIAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHNHp+OAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAdKeZ7BByXGeVqAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAFv0Mw9DMAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADmj0/HAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAOlPM9gg5LjPK1AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAC36GYehmAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABzR6fjgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHSnmewQclxnlagAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABb9DMPQzAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA5o9PxwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADpTzPYIOS4zytQAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAt+hmHoZgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAc0en44AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB0p5nsEHJcZ5WoAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAW/QzD0MwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAOaPT8cAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA6U8z2CDkuM8rUAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAALfoZh6GYAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHNHp+OAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAdKeZ7BByXGeVqAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAFv0Mw9DMAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADmj0/HAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAOlPM9gg5LjPK1AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAC36GYehmAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABzR6fjgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHSnmewQclxnlagAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABb9DMPQzAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA5o9PxwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADpTzPYIOS4zytQAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAt+hmHoZgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAc0en44AAGej84Pmrl7sf6LT/TXc6+AGAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAdKeZ7BByXGeVqAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAFv0Mw9DMAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADmj0/HAAAiD8B/eeL0fP/j6L+/v2H6uINsAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAOlPM9gg5LjPK1AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAC36GYehmAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABzR6fjgAAAcR8YZbeQfm/wAvZ6b/AEcbc4dPKAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAGMjkO3VaHBs2ekEDQAAAAAAAAAAAAAAAAAAcNoTfUlQAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHSnmewQclxnlagAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAByGVovQYM35/3NrRIYAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAW/QzD0MwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAOaPT8cAADxaa/AxPP9T6w+M6nrv6S9+YOnkAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPl7g8Ov0ns6z8E7aNNX6R1wC8wAAAAAAAAAAAAAAAABvwhlv6Rykab/1rpiGmQB8ofV+eoaZAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHSnmewQclxnlagAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADPgnqntN3Re5fCYfQfaZv2s5aAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAC36GYehmAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABzR6fjgAAflVlt+Z/wCzHP1fa/8ANJefTf0ltB08gAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABmI+D/0Dz3DXD88/s3Dq7835QAAAAAAAAAAAAAAAAAA8b8zy6PrA15/zt+4cerwz6kvH409xjT4K+zo16z5zvPuPoao+Qo8t/pTmNcPL+Iy39Z8ya+jvSdeb5HrZdP1Z2G/IDQAAAAAAAAAAAAAAAAB0p5nsEHJcZ5WoAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB4By/ZH1Ocd5v5u9sfo5scdggAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAALfoZh6GYAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHNHp+OAAB8F56fjd6Nw+nueRi6/+lDp5A6eQAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA4RP5m+1I1DTHA+SMt6H3KAa4AAAAAAAAAAAAAAAAAHzXtZdHvRrz/CP03j1fO/23Wf54/ocq/P79ARfnd+hg5fiIn6m63TP8/PcsOve67TD58+xkfBne572LtRzH2peYaYgAAAAAAAAAAAAAAAAHSnmewQclxnlagAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHyN6n1x7MclnmNr4+927o+mTztAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAt+hmHoZgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAc0en44AADQ/Pv575ez6nzbjz39N6zDXIAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAArh+cv6MY9No25gD8+PvnLovGvOAAAAAAAAAAAAAAAAAef8Aylj0/dtPXm/PH7vy6fkD7qc/nD+jyf5+foG1+c36LJv+G3P1j1WmX5+e44dfadNrh8//AGCj8/u0y6fY+225djcrMGAAAAAAAAAAAAAAAAAHSnmewQclxnlagAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHkvzb2x92HFYAfm12R+kpx2AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAFv0Mw9DMAAAAAAAD538GAA7j7JAAAADh/x1D9fPRgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAOaPT8cAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPKvl/Dq2/bWeTY810/wBb6YBpkAAAAAAAAAAAAAAAAAHzPGnmDMer6i9H34/hHdw68z7l0x+IPYpu/wCAh6J75efx/Pl0fSvLa8/Ubznxv6XD42+pJv446DPb6N7/AKOQHIAAAAAAAAAAAAAAAAB0p5nsEHJcZ5WoAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfLW0+U6vbH0t8s40n0Ez3U4bAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAC36GYehmAAAAAAAB8C/DYfGf08H05+uAAAAAeU/wA1Yf0f+1gAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAc0en44AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB0p5nsEHJcZ5WoAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAW/QzD0MwAAAAAAADKD+Wr9/w+ovwJDzL6MD9s/z9D8wfdA/YP8AmrD+hz8iQ/Qn77AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADmj0/HAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAOlPM9gg5LjPK1AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAC36GYehmAAAAAAAAZQfy1fv8Ah9V/JIfOf5Lh/QH+NYfTv6nhB/NWH1b2wfuI8AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAOaPT8cAAAAAAAAAAAAAAAAA8Fy89QAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA9x3byBgAAAAB0p5nsEHJcZ5WoAAAAAAAHO/J+0hogAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAD511X0UZMAAAAAAAAAAAAAAAAAALH2VjQZsAAC36GYehmAAAAAAAAZQfy1fv+Gv+AwfsJ+O4fuh0ofkl5OH79fzeB6l3If0JXQAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA5o9PxwAAAAAAAAAAAAAAAAD8Vft/m7AvIAAAAAAAAAAAAAAAAAPiHPT3u3GwCAAAAAAAAAAAAAAAAAOd82D7DN+YAAAAAAAAAAAAAAAAAAPhP7sz1DTIAAAAAAAAAAAAAAAAAPEJr6D+t9cQuAAAAADpTzPYIOS4zytQAAOCZ3tdHjPs1KwS/AfjXrj61M2AAAAAAAAAAAAAAAAAAH5lfYPqZA0AAAAAAAAAAAAAAAAAHzKz679T8vYJAAAAAAAAAAAAAAAAAA/OfZfowYsAAAAAAAAAAAAAAAAAAPzn/au0HLYAAFv0Mw9DMAAAAAAAAyg/lq/f8ACr+DAfdPwkH7u/EoZ/xsH9BX84If0J/hEH6OfqwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHNHp+OAAAAAAAAAAHF/nXjvnfpiE35utbP6TClNcvyn/VjLYNcQAAAAAAAAAAAAAAAAA/LvLb6y0cdwAAAAAAAAAAAAAAAAAA47xVr9HDq4wAAAAAAAAAAAAAAAAAA/Kf9WMtg1xAAAAAAAAAAAAAAAAAD5di8r63ALgAAAAA6U8z2CDkuM8rUAAPg74f9HP90fxz56+kfmvaf2PODTwHxbon7nOegAAAAAAAAAAAAAAAAAA/HT749nAGgAAAAAAAAAAAAAAAAA+OWfeHtPjbhLAAAAAAAAAAAAAAAAAD8meif1mOegAAAAAAAAAAAAAAAAAA/Jn9ZuiQ56AAAt+hmHoZgAAAAAAAEQfLP0AHV/LIXb4ep+RB5t9OB1nzSH0h5iHEfT4KAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHNHp+OAAAAAAAAAAGJMn88/OuO/I/fLPnDoQ+yDbn/Kf9WMtg1xAAAAAAAAAAAAAAAAAD8u8tvrLRx3AAAAAAAAAAAAAAAAAADjvFWv0cOrjAAAAAAAAAAAAAAAAAAD8p/wBWMtg1xAAAAAAAAAAAAAAAAAD5di8r63ALgAAAAA6U8z2CDkuM8rUAPJ6Xwp5l3x+rX5cYP9ffxkH+wWzy14D4t0T9znPQAAAAAAAAAAAAAAAAAAfjp98ezgDQAAAAAAAAAAAAAAAAAfHLPvD2nxtwlgAAAAAAAAAAAAAAAAB+TPRP6zHPQAAAAAAAAAAAAAAAAAAfkz+s3RIc9AAAW/QzD0MwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAOaPT8cAAAAAAAAAAAD4O7PHo+F/2mDxT4wD9Ojbn/AAb+jOPvCp+4fd+jkCpAAAAAAAAAAAAAAA/LvLb6y0cdwAAAAAAAAAAAAAAAAAA47xVr9HDq4wAAAAAAAAAAAAAAAAAA/Kf9WMtg1xAAAAAAAAAAAAAAAA/Kc5e08zl/dv1v08gXAAAAAB0p5nsEHJcZ5WoBD+J/VH6V/mPov0y+Oof6rfj9L/U/suevAfyr7c/bCw/UXnroTmoAAAAAAAAAAAAAAAD8fvu72cAaAAAAAAAAAAAAAAAAAD5AZ92ezeNuEsAAAAAAAAAAAAAAAAAPyk6J/Vs56AAAAAAAAAAAAAAAA8b/AD964DZfK/8AQdkw47AAAt+hmHoZgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAc0en44AAAAAAAAAAHgM18Qfq7Gn4y/soq+fPnIP0WNuf8p/1Yy2DXEAAAAAAAAAAAAAAAAAOMmvMF5usAOOH2IIAAAAAAAAAAAAAAI/XdMuoN+cAAAAAAAAAAAAAAAAAAMnWQDAAAAAAAAAAAAAAAAAA+XYvK+twC4AAAAAOlPM9gg5LjPK1AKXx1rJ+dfXP7B/lpm/2u/EmT9tV5b8B8W6J+5znoAAAAAAAAAAAAAAAAAAPMfOvUxDaQAAAAAAAAAAAAAAAAAvI+hTxugAAAAAAAAAAAAAAAAAAD5g2X0+YsAAAAAAAAAAAAAAAAAAPyZ/WbokOegAALfoZh6GYAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHNHp+OAAAAAAAAAAHhn4183Z+iHr95cN4tN+X/p016gb835T/AKsZbBriAAAAAAAAAAAAAAAAAHPS/Hzm7ACt+e83+iJUAAAAAAAAAAAAAAAe06ZbBvzgAAAAAAAAAAAAAAAAAB8SfbeewaYgAAAAAAAAAAAAAAAAB8uxeV9bgFwAAAAAdKeZ7BByXGeVqAAB8C7T99fHsv5c+wOifqU478B8W6J+5znoAAAAAAAAAAAAAAAAAAPMfOvUxDaQAAAAAAAAAAAAAAAAAvI+hTxugAAAAAAAAAAAAAAAAAAD4X6Z+6DmoAAAAAAAAAAAAAAAAAAPyZ/WbokOegAALfoZh6GYAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHNHp+OAAAAAAAAAAHK8HNHTD9A8eF1HctA1+U/wCrGWwa4gAAAAAAAAAAAAAAAABz0vx85uwA4P8AGvPb9yptMQAAAAAAAAAAAAAAPadMtg35wAAAAAAAAAAAAAAAAAA+JPtvPYNMQAAAAAAAAAAAAAAAAA+XYvK+twC4AAAAAOlPM9gg5LjPK1AAAAAAAA8B8W6J+5znoAAAAAAAAAAAAAAAAAAPMfOvUxDaQAAAAAAAAAAAAAAAAAvI+hTxugAAAAAAAAAAAAAAAAAAD4X6Z+6DmoAAAAAAAAAAAAAAAAAAPyZ/WbokOegAALfoZh6GYAAAAAAAAAAAAAAAAAAAABWW8bBGwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHNHp+OAAAAAAAAAAAAAAAAAflP8AqxlsGuIAAAAAAAAAAAAAAAAAc9L8fObsAPCPxyy3/Xb6O0yByAAAAAAAAAAAAAHtOmWwb84AAAAAAAAAAAAAAAAAAfEn23nsGmIAAAAAAAAAAAAAAAAAfLsXlfW4BcAAAAAHSnmewQclxnlagAAAAAAAeA+LdE/c5z0AAAAAAAAAAAAAAAAAAHmPnXqYhtIAAAAAAAAAAAAAAAAAXkfQp43QAAAAAAAAAAAAAAAAAAB8L9M/dBzUAAAAAAAAAAAAAAAAAAH5M/rN0SHPQAAFv0Mw9DMAAAAAAAAAAAAAAAAAAAAAb4P7Py/vR431AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHNHp+OAAAAAAAAAAAAAAAAAflP8AqxlsGuIAAAAAAAAAAAAAAAAAc9L8fObsAPk/8q8t/rb9TqgLzAAAAAAAAAAAAAD2nTLYN+cAAAAAAAAAAAAAAAAAAPiT7bz2DTEAAAAAAAAAAAAAAAAAPl2LyvrcAuAAAAADpTzPYIOS4zytQAAAAAAAPAfFuifuc56AAAAAAAAAAAAAAAAAADzHzr1MQ2kAAAAAAAAAAAAAAAAALyPoU8boAAAAAAAAAAAAAAAAAAA+F+mfug5qAAAAAAAAAAAAAAAAAAD8mf1m6JDnoAAC36GYehmAAAAAAAAAAAAAAAAAAAAAUPkL6v8AOPon0PwfsQ4vWAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADmj0/HAAAAAAAAAAAAAAAAAPyn/VjLYNcQAAAAAAAAAAAAAAAAA56X4+c3YAfCvz9nv8APv7pC2zTEAAAAAAAAAAAAAPadMtg35wAAAAAAAAAAAAAAAAAA+Yvp2NAvMAAAAAAAAAAAAAAAAAPl2LyvrcAuAAAAADpTzPYIOS4zytQAAAAAAAPAfFuifuc56AAAAAAAAAAAAAAAAAADzHzr1MQ2kAAAAAAAAAAAAAAAAALyPoU8boAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA/Jn9ZuiQ56AAAt+hmHoZgAAAAAAAAAAAAAAAAAAAAHnXzx9t+T3fqbk9LZPlP0YAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAOaPT8cAAAAAAAAAAAAAAAAA/Kf8AVjLYNcQAAAAAAAAAAAAAAAAA56X4+c3YAfm51Oe3z99bB9immIAAAAAAAAAAAAAe06ZbBvzgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfLsXlfW4BcAAAAAHSnmewQclxnlagAAAAAAAeA+LdE/c5z0AAAAAAAAAAAAAAAAAAHmPnXqYhtIAAAAAAAAAAAAAAAAAXkfQp43QAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB+TP6zdEhz0AABb9DMPQzAAAAAAAAAAAAAAAAAAAAAPmHkf0f8ACjrw+mn/AJx+6AMAAAAAAAAAAAAAAAAAAAAAAAAAAAAA5o9PxwAAAAAAAAAAAAAAAAD8p/1Yy2DXEAAAAAAAAAAAAAAAAAOel+PnN2AH5RfTue0nxuH7CGmIAAAAAAAAAAAAAe06ZbBvzgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfLsXlfW4BcAAAAAHSnmewQclxnlagAAAAAAAeA+LdE/c5z0AAAAAAAAAAAAAAAAAAHmPnXqYhtIAAAAAAAAAAAAAAAAAXkfQp43QAAAAAAAAAAAAAAAAAAB5Da9eIYAAAAAAAAAAAAAAAAAAfkz+s3RIc9AAAW/QzD0MwAAAAAAAAAAAAAAAAAAADy70PF8EP0D8VAPY/afhP2IPK+iAAAAAAAAAAAAAAAAAAAAAAAAAAAAAOaPT8cAAAAAAAAAAAAAAAAA/Kf8AVjLYNcQAAAAAAAAAAAAAAAAA56X4+c3YAfjR+iGe3uP4Vh+1XoGmQCAAAAAAAAAAAAPadMtg35wAAAAAAAAAAAAAAAAAA+b/AKQjQLzAAAAAAAAAAAAAAAAAD5di8r63ALgAAAAA6U8z2CDkuM8rUAAAAAAADwHxbon7nOegAAAAAAAAAAAAAAAAAA8x869TENpAAAAAAAAAAAAAAAAAC8j6FPG6AAAAAAAAAAAAAAAAAAAPhfpn7oOagAAAAAAAAAAAAAAAAAA/Jn9ZuiQ56AAAt+hmHoZgAAAAAAAAAAAAAAAAAAAZHyj9X+cQH0vwQBufRPzP3/Tnyv6KAAAAAAAAAAAAAAAAAAAAAAAAAAAAAc0en44AAAAAAAAAAAAAAAAB+U/6sZbBriAAAAAAAAAAAAAAAAAHPS/Hzm7AD8L/ANhM9vTPy7qfSPvwQVAAAAAAAAAAAAB7TplsG/OAAAAAAAAAAAAAAAAAAHxJ9t57BpiAAAAAAAAAAAAAAAAAHy7F5X1uAXAAAAAB0p5nsEHJcZ5WoAAAAAAAHgPi3RP3Oc9AAAAAAAAAAAAAAAAAAB5j516mIbSAAAAAAAAAAAAAAAAAF5H0KeN0AAAAAAAAAAAAAAAAAAAfC/TP2d45i/Xt+GAAAAAAAAAAAAAAAAAfkz+s3RIc9AAAW/QzD0MwAAAAAAAPyE+OAAAA+tQ/Y0AAAAAA8e8U+7/Hg9X5wA9lofN/e+7Hyf6OAAAAAAAAAAAAAAAAAAAAAAAAAAAAAc0en44AAAAB8IfnVyd36eeH6ZfppJvzAAAAAAAAflP+rGWwa4gAAAAAAAAAAAAAAAABz0vx85uwrB+A/wC9ue2p89Xn+ZX7czal5AAAAAAAAAAAAHtOmWwb84AAAAAAAAAAAAAAAAAAfEn23nsGmIAAAAAAAAAAAAAAAAAfLsXlfW4BcAAAAAHSnmewQclxnlagAAAAAAAeA+LdE/c5z0AAAAAAAAAAAAAAAAAAHmPnXqYhtIAAAAAAAAAAAAAAAAAXkfQp43QAAAAAAAAAAAAAABCHmHjO0+M+MdU+NfO3XHP9pa9C/dnz9PRzh0AAAAAAAAAAAAAAAA/Jn9ZuiQ56AAAt+hmHoZgAAAAAAAeV8mAAAHUB6wAAAAAAfP3mn6F+KB3eMPH9V+D/ACf6T9LHzf3QAAAAAAAAAAAAAAAAAAAAAAAAAAAABzR6fjgAAAHLI/nq+iOH0/sP829M/wB0/ROrhBoAAAAAAPyn/VjLYNcQAAAAAAAAAAAAAAAAA56X4+c3YcGP8W/3gz0I9Mvw6/VfPb3I0xAAAAAAAAAAAAPadMtg35wAAAAAAAAAAAAAAAAAA+JPtvPYNMQAAAAAAAAAAAAAAAAA+XYvK+twC4AAAAAOlPM9gg5LjPK1AD5T0XyH9T9M/YPzzy18Tfa/RP0Qcl+A+LdE/c5z0AAAAAAAAAAAAAAAAAAHmPnXqYhtIAAAAAAAAAAAAAAAAAXkfQp43QAAAAAAAAAAByLPGfGOifGvGeqPGfH+qeg9lk9m9m5b9n8pk/Rm7waflN8Bevj+6/0H5eoQwAAAAAAAAAAAAAA/Jn9ZuiQ56AAAt+hmHoZgAAAAAAAAAAAAAAAAAAAeE+U/f/i4ej4Z3nJ6n0N8/fMfoH0WeD9gAAAAAAAAAAAAAAAAAAAAAAAAAAAAAc0en44AAABzSP5+vqTi9HX+Ch/0f9L2+aDAAAAAAA/Kf9WMtg1xAAAAAAAAAAAAAAAAADnpfj5zdh4Qn+Z/7TzoF5fnlzee36YmmIAAAAAAAAAAAB7TplsG/OAAAAAAAAAAAAAAAAAAHxJ9t57BpiAAAAAAAAAAAAAAAAAHy7F5X1uAXAAAAAB0p5nsEHJcZ5WoAcszp/ww6o/cr8UA/a38Rw/cBeS/AfFuifuc56AAAAAAAAAAAAAAAAAADzHzr1MQ2kAAAAAAAAAAAAAAAAALyPoU8boAAAAAADP8iteNeMdM+M+M9UeM870T6/7NjXs3s3LXs3s3LfYGbADz6l6CSz4G2n8hf2j7o+0Dy9QAAAAAAAAAAAAAA/Jn9ZuiQ56AAAt+hmHoZgAAAAAAAAAAAAAAAAAAAcF84faflIe58iXZ09xi+R/TPUz5/wC0AAAAAAAAAAAAAAAAAAAAAAAAAAAAAOaPT8cAAAAA/PH8qeL0Kf6hp/pad3nAAAAAAAAflP8AqxlsGuIAAAAAAAAAAAAAAAAAc9L8fObsPk9V8hfrbNBefl/4257fupe0xAAAAAAAAAAAAPadMtg35wAAAAAAAAAAAAAAAAAAo3kAwAAAAAAAAAAAAAAAAAPl2LyvrcAuAAAAADpTzPYIOS4zytQA5Vn5Reg+hn92/n3g/wBXfx8T/VHsuevAfCOifuA56NNDgAAAAAAAAAAAAAAAAPMfOvUxDaQAAAAAAAAAAAAAAAAAvI+hTxugAGh554xqvGfGeqPGfGuqfLPQdZ9m9m5r9m9m5a9m9R5qmJYAAAAAAAAB8tfh16WX6d/pfyWGFAAAAAAAAAABDQAGflJ+uHRIc1AAAW/QzD0MwAAAAAAAAAAAAAAAAAAAE+ave+O4k+y/LADp/p/5H9NmPnftwAAAAAAAAAAAAAAAAAAAAAAAAAAAADmj0/HAAAAKgfml+mmWzfzcF+iWvpkDAAAAAAA/Kf8AVjLYNcQAAAAAAAAAAAAAAAAA56X4+c3YfCs35t+mYBUH40Rp93fVNSDkAAAAAAAAAAD2nTLYN+cAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAD5di8r63ALgAAAAA6U8z2CDkuM8rUAAKn4odcfq7+dofrf+Ncv9etrmrwH8uO6PdCkfplz1vnPQAAAAAAAAAAAAAAAHmPnXqYhtIAAAAAAAAAAAAAAAAAUg3PGfN28Y8a6o8Z8a6pm9lD2b2blr2X2flv2Pb56AAAAAAAAAAAwmbvAZep35p5Z47S/CP6r9XL9iLfkbAAAAAAAAAAB5B8G9UBsvkz+gTNhx2AABb9DMPQzAAAAAAAAAAAAAAAAAAAAK1R4l5z91+Ps768/cdD4P9iCNQAAAAAAAAAAAAAAAAAAAAAAAAAAAADmj0/HAAAA4v8lsOj4r+3Ofrz/iNP8ApI7nv8wGgAAAAAA/Kf8AVjLYNcQAAAAAAAAAAAAAAAAA56X4+c3Yfm5N9T99AFQfE8183/rQqC8wAAAAAAAAAAPadMtg35wAAAAAAAAAAAAAAAAAA8z9MmgqQAAAAAAAAAAAAAAAAA+XYvK+twC4AAAAAOlPM9gg5LjPK1AD4v1Wz+cvVH7i/i/k/vP4g0X7MHDp4D4t0T9znPQAAAAAAAAAAAAAAAAAAeY+depiG0gAAAAAAAAAAAAEHnrPL/MGeX+X0eYcmz1n2bGvZfZuWvZvZuW/QDJgAAAAAAAAAAAAAAfC/TP3J8v+Z9v9SHV8UAch+GvbF39xA6g4rAAAAAAAAAAAA/Jn9ZuiQ56AAAt+hmHoZgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAc0en44AAAEX45Y7/EH25zdj/o/XD9Azq4wAAAAAAAPyn/VjLYNcQAAAAAAAAAAAAAAAAA56X4+c3YflFGn079gVIOTmR/ht+4mevVmmIAAAAAAAAAAHtOmWwb84AAAAAAAAAAAAAAAAAAfEn23nsGmIAAAAAAAAAAAAAAAAAfLsXlfW4BcAAAAAHSnmewQclxnlagB5dS8e+jbOl57J/On0totYzfgPi3RP3Oc9AAAAAAAAAAAAAAAAAAB5j516mIbSAAAAAAAAAAABlfkbZ4J0dnqHp8nqHp6PUPVYPo88XoAAAAAAAAAAAAAAAAAAA+F+mfsX4z8b9Y+7z0PykAKAfjX83evj+7nrHlahLAAAAAAAAAAD8mf1m6JDnoAAC36GYehmAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABzR6fjgAAAEYeF89nrWrp/Uc+uIAAAAAAAH5T/qxlsGuIAAAAAAAAAAAAAAAAAc9L8fObsPxojT9EPoepByAfk/8AQOe32+aYgAAAAAAAAAAe06ZbBvzgAAAAAAAAAAAAAAAAAB8SfbeewaYgAAAAAAAAAAAAAAAAB8uxeV9bgFwAAAAAdKeZ7BByXGeVqAAAAAAAB4D4t0T9znPQAAAAAAAAAAAAAAAAAAeY+depiG0gAAAAAAAAAAMZ+UDLP1F6CAEABeR9CnjdAAAAAAAAAAAAAAAAAAAHwv0z9RfCHgfvP6anrfhAAAH5f7z+bX7m9cfTB5uoAAAAAAAAAAfkz+s3RIc9AAAW/QzD0MwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAOaPT8cAAAAgD+eTxzzfWs6o/6VtH0vIAAAAAAAA/Kf9WMtg1xAAAAAAAAAAAAAAAAADnpfj5zdh+F+ev7CemXmDQB8u/AWe37PmmIAAAAAAAAAAHtOmWwb84AAAAAAAAAAAAAAAAAAfEn23nsGmIAAAAAAAAAAAAAAAAAfLsXlfW4BcAAAAAHSnmewQclxnlagAAAAAAAeA+LdE/c5z0AAAAAAAAAAAAAAAAAAHmPnXqYhtIAAAAAAAAAAH55UfMf7Js1iAAAC8j6FPG6AAAAAAAAAAAAAAAAAAAPhfpn3/wDOz5r+mv18Pc/mcAAAD4g/Gr1Mv2L+6eDQMmAAAAAAAAAfkz+s3RIc9AAAW/QzD0MwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAOaPT8cAAAA84T/Ef1Hi9D9Pvxr1y/ebe6OMGAAAAAAB+U/wCrGWwa4gAAAAAAAAAAAAAAAABz0vx85uyt+A+e3726mmIAAU/wtz1/Yf1e8waAAAAAAAAAD2nTLYN+cAAAAAAAAAAAAAAAAAAPiT7bz2DTEAAAAAAAAAAAAAAAAAPl2LyvrcAuAAAAADpTzPYIOS4zytQAAAAAAAPAfFuifuc56AAAAAAAAAAAAAAAAAADzHzr1MQ2kAAAAAAAAAAPkj829D9oO7gBAAAF5H0KeN0AAAAAAAAAAAAAAAAAAAfOuqb+cvzP9Y/sVue//LAVzAAAfOFr8LP0W9PL9SneTsAAAAAAAAAfkz+s3RIc9AAAW/QzD0MwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAOaPT8cAAAAzkfgD+gnJ3/AJa4OPR/SV1Po+QDAAAAAAA/Kf8AVjLYNcQAAAAAAAAAAAAAAAAA56X4+c3Zwf4t57fvAaYgAAfmrN6n6LAFQAAAAAAAAAHtOmWwb84AAAAAAAAAAAAAAAAAAeW+pTQVIAAAAAAAAAAAAAAAAAfLsXlfW4BcAAAAAHSnmewQclxnlagAAAAAAAeA+LdE/c5z0AAAAAAAAAAAAAAAAAAHmPnXqYhtIAAAAAAAAAeIM/IL9e7PcTMAAAALyPoU8boAAAAAAAAAAAAAAAAAAAA+UPgj5z+uv1E9i93+ZA28AAAADy78JO/P379m83pnJYAAAAAAAH5M/rN0SHPQAAFv0Mw9DMAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADmj0/HAAAAMhHyf9bzp8d+nTX0Ma4gAAAAAAAflP+rGWwa4gAAAAAAAAAAAAAAAABz0vx85uzwj8z89v2nNMQAAPFE/ya/cqNJjTIAAAAAAAAAPadMtg35wAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPl2LyvrcAuAAAAADpTzPYIOS4zytQAAAAAAAPAfFuifuc56AAAAAAAAAAAAAAAAAADzHzr1MQ2kAAAAAAAAA4f8X7P0f8ArmQEAAAAF5H0KeN0AAAAAAAAAAAAAAAAAAAAfFnxZ81/Yv379U+3/N4dXxQAAABzbPxB5718f3P7TyNgQAAAAAAH5M/rN0SHPQAAFv0Mw9DMAAAAAAAAAAAAAAAAAAAAAOB7PL744/UAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA5o9PxwAAADFR/O1+y/L3e/nTxdO6kAAAAAAAB+U/6sZbBriAAAAAAAAAAAAAAAAAHPS/Hzm7Pk/5Cz2/W2rpjaAAAD8Tf0lz2+jTTEAAAAAAAAAPadMtg35wAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPl2LyvrcAuAAAAADpTzPYIOS4zytQAAAAAAAPAfFuifuc56AAAAAAAAAAAAAAAAAADzHzr1MQ2kAAAAAAAAymfjd9KWfokZgAAAAAXkfQp43QAAAAAAAAAAAAAJis5zm955znN55zm955zmN5845z4X+zfVPvP2PwL1w1/NwAAAACr+QfTPyB+7m8+0nn6AAAAAAB+TP6zdEhz0AABb9DMPQzAAAAAAAAAAAAAAAAAAAAAIPkn6f8/+kOt8H7EObvAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADmj0/HAAAAM4P5weJ831wT/pw1fT8cAAAAAAAD8p/wBWMtg1xAAAAAAAAAAAAAAAAADnpfj5zdnwr5tnt+k3zZcfTznAAAfBnk2e36mGmIAAAAAAAAAe06ZbBvzgAAAAAAAAAAAAAAAAAB8SfbeewaYgAAAAAAAAAAAAAAAAB8uxeV9bgFwAAAAAdKeZ7BByXGeVqAAAAAAAB4D4t0T9znPQAAAAAAAAAAAAAAAAAAeY+depiG0gAAAAAAAM/KKiD9ZAeSAAAAABeR9CnjdAAAAAAAZ3OWub5zeec5veec5zeeb5zeedfquj6TJ9H0mFdH0eFdJ84D5Wz8d/W3lfTfRflHon2L3/AI/6WedqAAAAAfm7+WnrZft/9XeboGbAAAAAPyZ/WbokOegAALfoZh6GYAAAAAAAHL5TAEAGmPqhAAAAAAAeK+O/e/je79SeP9RbPmvvAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA5o9PxwAAACEPnKXPUA+iZdMgAAAAAAAPyn/VjLYNcQAAAAAAAAAAAAAAAAA56X4+c3Z+bnU57fXvy5S919rcA5AOG/EzPb90trTEAAAAAAAAAPadMtg35wAAAAAAAAAAAAAAAAAA+JPtvPYNMQAAAAAAAAAAAAAAAAA+XYvK+twC4AAAAAOlPM9gg5LjPK1AAAAAAAA8B8W6J+5znoAAAAAAAAAAAAAAAAAAPMfOvUxDaQAAAAAAAPz1+XtD9k9iAEAAAAABeR9C1/G6Od5vaec5veec5zeec5veec5veafQs6Po8K6To8K6PpMK6Po8K6TWxoEAAAfC/TPlf0H8z/T/x5+t/R8x8vfKH1/4F7B9hcl92cGgAAAB8daL8Uf1t9HP9BjytQAAAAD8mf1m6JDnoAAC36GYehmAAAAAAAB8P+FaSDQAe1p/dRnQAAAAAROfj6H9O/n/tfSvnfufTT5L9JAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADmj0/HAAAAAAAAAAAAAAAAAPyn/VjLYNcQAAAAAAAAAAAAAAAAA56X4+c3Z+UX07nt6jyVLjPpkO2KyAD8g40+ufsOpByAAAAAAAAe06ZbBvzgAAAAAAAAAAAAAAAAAB8SfbeewaYgAAAAAAAAAAAAAAAAB8uxeV9bgFwAAAAAdKeZ7BByXGeVqAAAAAAAB4D4t0T9znPQAAAAAAAAAAAAAAAAAAeY+depiG0gAAAAAAfJbPzX/Z+jvSAAKLMPJpZWTRlZNLKy6KWMza6OH0nR4V0fSYV0fR4V0nR4VuvyYAAAAAAAAAAHwv0z4R+gfzn9G/mF+vzjbyva/nf5U+XPa5/efr3i0604dAAAAPBPwm9HP7p/WfmqQ56AAAA/Jn9ZuiQ56AAAt+hmHoZgAAAAAAAAAAAAAAAAAAAETn4+h/Tv5/9b6v5T9H9GPm/uwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA5o9PxwAAABgcDJNEodjPUgAAAAAAAflP+rGWwa4gAAAAAAAAAAAAAAAABz0vx85uz8aP0Qz2xfVLT/AkfWulWYCPkJV8dfsFNBeYAAAAAAAB7TplsG/OAAAAAAAAAAAAAAAAAAHxJ9t57BpiAAAAAAAAAAAAAAAAAHy7F5X1uAXAAAAAB0p5nsEHJcZ5WoAAAAAAAHgPi3RP3Oc9AAAAAAAAAAAAAAAAAAB5j516mIbSAAAAAAHiP5A6H68e6QAgA8v+eN50NajW1ZNbVl63Qw/o3ovF3sEsAAAAAAAAAAAAAAAAAPjTon5I/U757+i/za++U/YD2v51MBr5L+bfbw+jvrbz9ehOOwAAPPWfhV6d6+P7Xbfj7AAAAfkz+s3RIc9AAAW/QzD0MwAAAAAAAAAAAAAAAAAAAADyXxL7n8i6z6Q8f6e2fPfbAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABzR6fjgAAAGWH8xx5nsAHYftz0cv04dfCflzltk/q4Hh/53Teb+tLXVG3P+U/6sZbBriAAAAAAAAAAAAAAAAAHPS/Hzm7Pwv8A2Cz2+e/re1t/PhOH9Og4cGQH4V/tbnt3xpiAAAAAAAAe06ZbBvzgAAAAAAAAAAAAAAAAAB8xfTsaBeYAAAAAAAAAAAAAAAAAfLsXlfW4BcAAAAAHSnmewQclxnlagB8Lar5+5Ptz/ZX8zeS+K7Hpn9Nzz9PAfFuifuc56AAAAAAAAAAAAAAAAAADzHzr1MQ2kAAAAAOHZ+L/AOjln14ZgBW+fdF579L2dsYMAAC8j6FPG6AAAAAAAAAAAAAAAAAAAGJ/jD+yPj/0N+fvtiv6mPa/nUAOUpfIngXt4fT/ANW+brrnJYAAYv4odceYfuta9COHQAAPyZ/WbokOegAALfoZh6GYAAAAAAAAAAAAAAAAAAAAANE4GAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABzR6fjgAAAGSj+f39gObtyvylD6x8Nmv3IOzgOeD8n/2Bx3/AB2/W0fzp5MH6AG3P+Ff1JydwVH2P7TvzBUgAAAAAAAAAAAAAAc9L8frc3Z+A/7j57/OX2PcEDz+UfTp29nKxAPy59Mz2++jTEAAAAAAAA9p0y2DfnAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPzPObrPKVX2f9b78oXAAAAAB0p5nsEHJcZ5WoAAH439M/qL+W9L9k/wAVkfs5d5b8B/PDsj6QAPubF65jQAAAAAAAAAAAAAAAHmPnXqYhtIAAAABlfjdZ9J/omgJDmWfM290T9IaPPQIAAAvI+hTxugAAAAAAAAAAAAAAAAAADnZ6/wAjv2T8r9z+M+ejs+7T2v51AAA4el8feNe5z/Vv1F5W185rACMPya+FPXx/dn3vy9fOPknRBqvh/wDd1AclgAAW/QzD0MwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAOaPT8cAAAA49P+cL9d+PvyvygD92/yi0z/AHsOnjMoPyT/AGBx3/Fz9ox+MfFIfp6bc/5LfqBj0Bpht3mgYAAAAAAAAAAAAAABz0vx/g+bt/Fv9iM9uB+lrzBxz3ygtfp7snmAvnj8zs9v23XTEAAAAAAAA9p0y2DfnAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAOWJZ8zzdr63aC4AAAAAOlPM9gg5LjPK1AADjvyV64/Tr4OR+qX5Bp/qL2/PXgPyh0x9iGVHsEFklgAAAAAAAAAAAAAAAeY+depiG0gAAADGflFBZ+sj4AR5P8AP/RPufsmTUhgAAAF5H0KeN0AAAAAAAAAAAAAAAAAAAfH79T5P/XDyf1/55+UsvX/AEzPa/nUAAAA84tfHPm/t8/1v9K+TvZOegA/PnafyS/azuj6q8z87QKPzR/ZfRBzUAABb9DMPQzAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA5o9PxwAAAAD84/wAv+L0D9UQ+j/Eujl+6TXEyg/JP9gcd/wAff1hH818QH3Ybc/5T/qxlsGuIAAAAAAAAAAAAAAAAAc9L8f8ACOXt/M/9HI26j2O8gcnmCrwn61K0B5x/hxnr+rXuN5g0AAAAAAB7TplsG/OAAAAAAAAAAAAAAAAAAHzR9LxoF5gAAAAAAAAAAAAAAAAB8uxeV9bgFwAAAAAdKeZ7BByXGeVqAHKM/JL9ZuqNX8hkfst+KwftDY5b8B8W6J+5znoAAAAAAAAAAAAAAAAAAPMfOvUxDaQAAAD896Plz9k2bBBT+d9Fxf0zR15iwAAAAAvI+hTxugAAAAAAAAAAAAAAAAAAD8rr9+X9TPJ/SfPvzHw+i/Xk9r+dQAAAAA8n0Xxxyvtc/wBffQ/kbyGFAHyj+IHp5fqX+kXHYYV+TP6zdEhz0AABb9DMPQzAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA5o9PxwAAAACj47NVSa92k0zADOD8y/wBRctvF/wA6Zux+rrnZNsPyn/VjLYNcQAAAAAAAAAAAAAAAAA56X4/8n8vd8hfWEa/RnY6YgIA+d8ydfppayPzwm+e/TAAqAAAAAAAPadMtg35wAAAAAAAAAAAAAAAAAA+JPtvPYNMQAAAAAAAAAAAAAAAAA+XYvK+twC4AAAAAOlPM9gg5LjPK1AD8dumaXYdMfox8S818D65tP6FHBp4D4t0T9znPQAAAAAAAAAAAAAAAAAAeY+depiG0gAAAfJv5qaH7Qd7AcmHzP1fRP0Zd56BAAAAAAXkfQp43QAAAAAAAAAAAAAAAAAAB+Rb+q9L/AEp837OH8a8Ppf2A2vW/BQrnAAAAADxTWfjqp7OH2F7x4+6mNHi9L8Ivr71cv19teRt+TP6zdEhz0AABb9DMPQzAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA5o9PxwAAACiH89/jHnesE1/Thq+n44AAAAAAAH5T/AKsZbBriAAAAAAAAAAAAAAAAAHPS/H/hXk7/ACX2ydftKbTnAACv8nrT1X2Mk8uF+N37qRpeNMgAAAAAAPadMtg35wAAAAAAAAAAAAAAAAAA+JPtvPY/PYX6CyaZAAAAAAAAAAAAAAAAHy7F5X1uAXAAAAAB0p5nsEHJcZ5WoAAAAAAAHgPi3RP3Oc9AAAAAAAAAAAAAAAAAAB5j516mIbSAAB4mz8gP11s92TM8f8H6J989cyakMAAAAAAC8j6FPG6AAAAAAAAAAAAAAAAAAAPxpX2n0f8AoPw+8flJl7X337B3fmIbeCAAAAAACeBbT8fansYfYvs3jdAZPi/ww7Y6H9vkfl7+s2bDnoAAC36GYehmAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABzR6fjgAAAHEJ/wA5n9BHL2dKdPJ3bqkAAAAAAAD8p/1Yy2DXEAAAAAAAAAAAAAAAAAOel+P/AJucnfjfSC0+rC8AAAObH8p/UM6deVl+M/3hnt9UGmIAAAAAAHtOmWwb84AAAAAAAAAAAAAAAAAAfEn23nsfzc5a/bP626ZhrgAAAAAAAAAAAAAAB8uxeV9bgFwAAAAAdKeZ7BByXGeVqAAAAAAAB4D4t0T9znPQAAAAAAAAAAAAAAAAAAeY+depiG0gABxH4vWfoz9foofOVLmfpmjqTFgAAAAAAAF5H0KeN0AAAAAAAAAAAAAAAAAAAfi/n+gfXv3Hzbn51R3+3/U23zodXxYAAAAAAAEXznvPyH2nr4fY3qfi9Bm5v8Y/AfXx92/bbisOWwAALfoZh6GYAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHNHp+OAAAAVw/nm934fS6kvP9dbnXwgAAAAAAAflP+rGWwa4gAAAAAAAAAAAAAAAABz0vx/8AKLj9Dqvrkv3ovAAAADyzxCdfrS9WXxN8457frOaYgAAAAAAe06ZbBvzgAAAAAAAAAAAAAAAAAB8Sfbee2b/Mhjt619zRp+oJ2+eAAAAAAAAAAAAAAB8uxeV9bgFwAAAAAdKeZ7BByXGeVqAAAAAAAB4D4t0T9znPQAAAAAAAAAAAAAAAAAAeY+depiG0gBls/G/6Os/RbjZPmfut5+hrXPQIAAAAAAAALyPoU8boAAAAAAAAAAAAAAAAAAA/O3i/TPUvsCvPPjFaYH3bPUHd+agAAAAAAAAFX5m6J+SvUfWx+xPQ/E3b+Wuq/Kz+pvomQ8/QAAC36GYehmAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABzR6fjgAAAFAPxLl4vRAX7YXu3zwAAAAAAAPyn/AFYy2DXEAAAAAAAAAAAAAAAAAOel+P8A40cfoe3fcLr0grEAAAA+c1dT6WDmvw2i/wBxeq0yAQAAAAAe06ZbBvzgAAAAAAAAAAAAAAAAABBOHkX4K8vZ+/f4Hh+kv6L7YBpkAAAAAAAAAAAAAHy7F5X1uAXAAAAAB0p5nsEHJcZ5WoAAAAAAAHgPi3RP3Oc9AAAAAAAAAAAAAAAAAAB5j516mIbSAN/KSin+sgeN+IbT9CeqYsJYAAAAAAAAAXkfQp43QAAAAAAAAAAAAAAAAAAB+dHn/q/vX07v8yfO7x+VP0z5ftQ9L8fAAAAAAAAAADN+WuqPlf2v08fsDtPE6Px1/YrdBzUAABb9DMPQzAAAAAAAAAAAAAAAAAAAAAAAAAADLwQ7IAAAAAAAAAAAAAAAAAAAAAAAAAAAOaPT8cAAADwH0SL+biNAD7En2wAAAAAAAA/Kf9WMtg1xAAAAAAAAAAAAAAAAADnpfj/4X8Xo/Yv3rRrFZAAAAFUPk316dfXvyeS+g/t5oKgAAAAAPadMtg35wAAAAAAAAAAAAAAAAAAAPlP8s+Xs/fjxvo5fwW/V7l7fu87OAAAAAAAAAAAAAAPl2LyvrcAuAAAAADpTzPYIOS4zytQAAAAAAAPAfFuifuc56AAAAAAAAAAAAAAAAAADzHzr1MQ2kA/Pf5a0P2C8GaxvppnRmLAAAAAAAAAAAvI+hTxugAAAAAAAAAAAAAAAAAAD8r/N/Yvtf3nr+DPPb878yP15839hD1vwkAAAAAAAAAAADF+T+uPmX6D9LL5V/YvzNQ5bAAAt+hmHoZgAAAAAAAAAAAAAAAAAAfDweBfq8Hytmh9dgAHg3dB598fB+oIHBfGAfoYAAAAAAAAAAAAAAAAAAAAAAAABzR6fjgAAAfhd9lcvb+SRz9QB/TBqel5AAAAAAAAH5T/qxlsGuIAAAAAAAAAAAAAAAAAc9L8W/Afi9L9M/s65B5gAAAAHMfKs6/U3g7n4F/Z2WF5gAAAAB7TplsG/OAAAAAAAAAAAAAAAAAAHmfpk18CfKPP1ftSdXH4D+E3J2/sb9ldHKFwAAAAAAAAAAAAfLsXlfW4BcAAAAAHSnmewQclxnlagAAAAAAAeA+LdE/c5z0AAAAAAAAAAAAAAAAAAHmPnXqYhtJ8ms/NT9Z9Dyf0a5+gLGFAgAAAAAAAAAAC8j6FPG6AAAAAAAAAAAAAAAAAAAPxl839t/UP1ft/LSHTxvxr/AGA8f+hto9n+dwAAAAAAAAAAAADmvkfsz/M3+g/VB52oAAFv0Mw9DMAAAAAAAAAAAAAAAAAAD8/g+Lf1JDiMoPqb87g3/wBGA/Cf0sPvf4ODsfuoPS/nQN3z0PAf0rDoPzfDoPSQ+xQAAAAAAAAAAAAAAAAAAA5o9PxwAAADAR8V/cca6BpkAAAAAAAAAflP+rGWwa4gAAAAAAAAAAAAAAAABz0v5j/Eri9L9YfpC4BwAAAAAHlCrxb6oK/Dj9ior1U0xAAAAAD2nTLYN+cAAAAAAAAAAAAAAAAAAPiT7bz2/Lbx7Dp/YbpOrhPmZV+Hn7ac3X9UHXwgAAAAAAAAAAAHy7F5X1uAXAAAAAB0p5nsEHJcZ5WoAAAAAAAHgPi3RP3Oc9AAAAAAAAAAAAAAAAAAB5j516mIbT4n+P2h+g+hrP0V6ZhQSAAAAAAAAAAAAF5H0KeN0AAAAAAAAAAAAAAAAAAAfjN5P7x+svaen+Lhfn/lJ9+eL/RXr57X86gAAAAAAAAAAAAAB+S3609Ehz0AABb9DMPQzAAAAAAAA47FaBoANdHaCoAAAA/P4PT/AM3Q/QmoHxh+tAfPniwM+rAPzYD9h/xUD9dfzpD0Drw9o+QA2vpQPSPycD9wQAAAAAAAAAAAAAAAAAAA5o9PxwAAADNR/P1+m3L3e9nRyexSXAAAAAAAAH5T/qxlsGuIAAAAAAAAAAAAAAAAAc9L+KPym4vT/XL168gcgAAAAAB83snbwD5xh/rd2WuICAAAAPadMtg35wAAAAAAAAAAAAAAAAAA+JPtvPb8Z/PuXt+jfpDSPqTY6fP+Rvxd5e790forp4wqQAAAAAAAAAAPl2LyvrcAuAAAAADpTzPYIOS4zytQAAAAAAAPAfFuifuc56AAAAAAAAAAAAAAAAAADzHzr1MTiN5/F76G2X059MSbxkwAAAAAAAAAAAAAvI+hTxugAAAAAAAAAAAAAAAAAAGfjD4/9B/sho+v+Ag8vzq9x8X+ivqQ9r+dQAAAAAAAAAAAAAA/Jn9ZuiQ56AAAt+hmHoZgAAAAAAAc/nNAAAXw6ITAAAAPz+D2vkA/Pr70D4Q/cUPPPzDD1P6sA+Pg/UH8Pg/Wv86Q9A+lg9W/L4Mn9ag6T8Rg/cIAAAAAAAAAAAAAAAAAAAOaPT8cAAAAzA/mcoeZ7AB/Thq+n44AAAAAAAH5T/qxlsGuIAAAAAAAAAAAAAAAAAc9L/O/4A4fT/YPrNcQEAAAAAAFP5NWnsPyJL+UP1wT9XNMQAAAD2nTLYN+cAAAAAAAAAAAAAAAAAAPiT7bz2/Bj2zl7P1K+TOjl8c+n89/pn4x25PyK/ePHp9vOriAAAAAAAAAAA+XYvK+twC4AAAAAOlPM9gg5LjPK1AAAAAAAA8B8W6J+5znoAAAAAAAAAAAAAAAAAAPMfOvUxzPxx6Z3/0C1XvcuDBAAAAAAAAAAAAAAXkfQp43QAAAAAAAAAAAAAAAAAAHO/kb439E/soez/OwB8Y4fi/0V91HtfzqAAAAAAAAAAAAAAH5M/rN0SHPQAAFv0Mw9DMAAAAAAAAAAAAAAAAAAD8/g9r+lg/EP7mDhvKQ4L9JA8i+RQ/QP5QD9Qfw+D9a/wA6Q9A+lg9W/L4Puv8AOYPZfCw/b4AAAAAAAAAAAAAAAAAAAOaPT8cAAAAjDy2WLAPTn3AAAAAAAAH5T/qxlsGuIAAAAAAAAAAAAAAAAAc9L/LX5N8H9H/Vj0Xo81Dt+WAAAAAAA5Ufyx9JTp8z/nvFfq59B6ZA5AAA9p0y2DfnAAAAAAAAAAAAAAAAAAD4k+289v5yf0z4vY/QM9HwfPvkfHp86+q418H/ADGiv3u9W6uAKQAAAAAAAAAfLsXlfW4BcAAAAAHSnmewQclxnlagBwDPy+/Srqj1b4wxr5l+mOiftE4r8B8W6J+5zno+WtF8UndmAAAAAAAAAAAAAAABzuD0z45xmP0X3l6H4H7ICYAAAAAAAAAAAAAAE6PQji0AAAAAAAAAAAAAAAAAADB+d+b7r9Qj7P8AjwA8G+Sfl/6e+jyPOAAAAAAAAAAAAAAD4L/cv3PxUJ8EAAC36GYehmAAAAAAAAAAAAAAAAAABXB8oQAT84GpfA5IOoiC1RC3XB0gPqhRUPFfisP1XAAAAAAAAAAAAAAAAAAADmj0/HAAAA/GD635uz8sDn6QD+iXS9HygAAAAAAAPyn/AFYy2DXEAAAAAAAAAAAAAAAAAOel/j55/wCB+le2fYPzv7j6BV+3/lEGgAAAAAA818GnW5wir4H/AExk+uTXAAAD2nTLYN+cAAAAAAAAAAAAAAAAAAPLfSpr+Yr9Nvkv3b9Ltn6z8KC48s+RcOrm/B8ej5l/VRx90zdnAAAAAAAAAAHy7F5X1uAXAAAAAB0p5nsEHJcZ5WoAcEz8/PsLpj238aA/aj8Rw/bSTlvwHxbon7nOej5qq/j46ukAAAAAAAAAAAAAAADmuf6ZX2nh+5uHlfooAAAAAAAAAAAAAAABOHfHHoAAAAAAAAAAAAAAAAAAB0GnD9DH1H89gHP/ADt4n6yh5n3oAAAAAAAAAAAAAAfCX7o+9+HhPz4AAFv0Mw9DMAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA4D89g6b9DA1gAAAAAAAAAAAAAAAAAAA5o9PxwAAAPk/wBYz1+TzPQA+/7PRzAAAAAAAAflP+rGWwa4gAAAAAAAAAAAAAAAABz0v8QvdPL+o+p/kn5b+i/sL1D6P8Kx2+j8aAAAAAAAwPO/H1r4X8YZa/eX3vpgpUAB7TplsG/OAAAAAAAAAAAAAAAAAAB5mn/Ot+jXxH9L/T30x735L0p7XwIgeM/I/P2fNnzDzdfe/qxvy/XZ1cQAAAAAAAB8uxeV9bgFwAAAAAdKeZ7BByXGeVqAAH5sfVPVO98Ei/Vb8gE/1I7fnrwH8q+3P2wsP1F566E5qAAAAAAAAAAAAAAAA8x869TENpAAAAAAAAAAAAAAAAAC8j6FPG6AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADxv8/euA2Xyv/Qdkw47AAAt+hmHoZgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAc0en44AAAH5derc/V7/APCbn9XDfnAAAAAAAAAPyn/VjLYNcfJfx85uoMugAAAAAAAAAAAAAACujwD9UY6PpLgY9T5H+ufj/wClPZcT7f8Ak+kXkAAAAAAABy3gM6fm92HP37hpyABtNTFSAAAAAAAAAAAAAAAAAABzyMD9reb6n4I/RDwf1jpOx+x/nNxrgnPxp8d/OnyX9AeH/O+nk+t9h7v5IDQAAAAAAHKJ/qj9b9nnhcAAAAAHSnmewQclxnlagAB+bH1T1T6d+WAfr/8AjEH7GaPLXgPi3RP3Oc9AAAAAAAAAAAAAAAAAAB5j516mIbSAAAAAAAAAAAAAAAAAF5H0KeN0AAAAAAAAAAAAAAAAAAAfD3TP3Cc1AAAAAAAAAAAAAAAAAAB+TP6zdEhz0AABb9DMPQzAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA5o9PxwAAAPx+9l5ez3r8pw/fo6uMAAAAAAAAA/Kf9WMtg1x5n5az0CbAAAAAAAAAAAAAAAyVXnv0JltvDng/kX4z+ovqf3f6H8Tq4vofGRlQAAAAAF5VY+WvB/XPE/Tef3Lh9L+EAB6RUaBriAAAAAAAAAAAAAAAAAAHzT9LRr3H0nvzfnV6R8J/WH0en2P8ALncR9XDxnLcHufnh9RfDf13eh9n824VntflDrd8xNWMxeFta5n2rxJ6xl8QMvoP631xC4AAAAAOlPM9gg5LjPK1AAD82Pqnqn378eUex+c7T+tJwaeA+LdE/c5z0AAAAAAAAAAAAAAAAAAHmPnXqYhtIAAAAAAAAAAAAAAAAAXkfQp43QAAAAAAAAAAAAAAAAAAB8L9M/dBzUAAAAAAAAAAAAAAAAAAH5M/rN0SHPQAAFv0Mw9DMAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADmj0/HAAAA/H75q4vR6bxOL+ni8/2wvdvngAAAAAAAflP+rGWwa4gAAAAAAAAAAAAAAAABz0vx85uwAOI5vd+P/pf47+mvetD7b+TsbOvMHIAATaM3pctzev8Yafw39efVOn+g/xmF8oAe06ZbBvzgAAAAAAAAAAAAAAAAAB8SfbeewaY8L+YvxP9QfqCv1f88Y8eePSfHnmfpSfd2mYe9+TAAAAAAAAAB8uxeV9bgFwAAAAAdKeZ7BByXGeVqAAGbbCdgef+hsBHgPi3RP3Oc9AAAAAAAAAAAAAAAAAAB5j516mIbSAAAAAAAAAAAAAAAAAF5H0KeN0AAAAAAAAAAAAAAAAAAAfC/TP3Qc1AAAAAAAAAAAAAAAAAAB+TP6zdEhz0AABb9DMPQzAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA5o9PxwAAAPhr5j5usJsA/XW518IAAAAAAAH5T/AKsZbBriAAAAAAAAAAAAAAAAAHPS/Hzm7AAA4349+O/pv6M9y9/8c3Iu747Eh0xAV7TjW1np+M/Kfxn9Ve8++fRfibj1fzsAAD2nTLYN+cAAAAAAAAAAAAAAAAAAPiT7bz2DTE+K/E/T+K/QnD1Oe0fd/IuM/ND43+nf1N6X7L+aQ14gAAAAAAAAA+XYvK+twC4AAAAAOlPM9gg5LjPK1AAAAAAAA8B8W6J+5znoAAAAAAAAAAAAAAAAAAPMfOvUxDaQAAAAAAAAAAAAAAAAAvI+hTxugAAAAAAAAAAAAAAAAAAD4X6Z+6DmoAAAAAAAAAAAAAAAAAAPyZ/WbokOegAALfoZh6GYAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHNHp+OAAAAAAAAAAAAAAAAAflP+rGWwa4gAAAAAAAAAAAAAAAABz0vx85uwAADjuf2vj/AOhfjf6f9t1fuf5I1MqNH6IRZrWV84eT+l+QfV3i/qfoZ9n/AC4AAAAe06ZbBvzgAAAAAAAAAAAAAAAAAB8SfbeewaYmVG/5X/oD8R/Unsh91/Kn5y+t/B/1h9fn3n8ngAAAAAAAAAAfLsXlfW4BcAAAAAHSnmewQclxnlagAAAAAAAeA+LdE/c5z0AAAAAAAAAAAAAAAAAAHmPnXqYhtIAAAAAAAAAAAAAAAAAXkfQp43QAAAAAAAAAAAAAAAAAAB8o7z9XGFAAAAAAAAAAAAAAAAAAB+TP6zdEhz0AABb9DMPQzAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA5o9PxwAAAAAAAAAAAAAAAAD8p/1Yy2DXEAAAAAAAAAAAAAAAAAOel+PnN2AAAAcl8d/H/0v9Ae4+7+ST7Xo/Dw5gV8KOv5Pl+B/sj6u2vv/AONg04QAAAD2nTLYN+cAAAAAAAAAAAAAAAAAAPmj6XjQLzAPmv5P+D/rD9PvGfrf55+Df1P8D9evn1387gAAAAAAAAAAfLsXlfW4BcAAAAAHSnmewQclxnlagAAAAAAAeA+LdE/c5z0AAAAAAAAAAAAAAAAAAHmPnXqYhtIAAAAAAAAAAAAAAAAAXkfQp43QAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB+TP6zdEhz0AABb9DMPQzAAAAAAAAAAAAAAAAAAAAAAA4EPHvVg8H68PpIAAAAAAAAAAAAAAAAAAAAAAAAAAAADmj0/HAAAAAAAAAAAAAAAAAPyn/AFYy2DXEAAAAAAAAAAAAAAAAAOel+PnN2AAAAHL4er8c+7fFf1T7wfefyCAeb/Kvxf8AU3sv0T7n5NIex+ZAAAAAB7TplsG/OAAAAAAAAAAAAAAAAAAAAAADPzL+X/dfo75j8n9D+tvo/wCs/nwPS+KAAAAAAAAAAA+XYvK+twC4AAAAAOlPM9gg5LjPK1AAAAAAAA8B8W6J+5znoAAAAAAAAAAAAAAAAAAPMfOvUxDaQAAAAAAAAAAAAAAAAAvI+hTxugAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAD8mf1m6JDnoAAC36GYehmAAAAAAAAAAAAAAAAAAAAAHwoHz38Vh9gXQ+wfkcPXf0OBwAAAAAAAAAAAAAAAAAAAAAAAAAAAc0en44AAAAAAAAAAAAAAAAB+U/wCrGWwa4gAAAAAAAAAAAAAAAABz0vx85uwAAAAA5n46+S/pH27332PzBnz8tfDvqb5/9l9PPtv5TAAAAAAA9p0y2DfnAAAAAAAAAAAAAAAAAAAAAAA8q5PoPzR9H/Mf7n/Tl/6x/nyDQAAAAAAAAAAHy7F5X1uAXAAAAAB0p5nsEHJcZ5WoAAAAAAAHgPi3RP3Oc9AAAAAAAAAAAAAAAAAAB5j516mIbSAAAAAAAAAAAAAAAAAF5H0KeN0AAAAAAAAAAAAAAAAAAAfMGy+nzFgAAAAAAAAAAAAAAAAAB+TP6zdEhz0AABb9DMPQzAAAAAAAAAAAAAAAAAAAAKIfnD8Rh/QPRD5H+wwk5oPzW+0A9+AAAAAAAAAAAAAAAAAAAAAAAAAAADmj0/HAAAAAAAAAAAAAAAAAPyn/AFYy2DXEAAAAAAAAAAAAAAAAAOel+PnN2AAAAABzePp/HXr3w/8AWXE5uXo/WPRfe/x2G3mgAAAAAAe06ZbBvzgAAAAAAAAAAAAAAAAAB80fS8aBeYAAfA/0v8n/AED7CfWfz8AAAAAAAAAAAAfLsXlfW4BcAAAAAHSnmewQclxnlagAAAAAAAeA+LdE/c5z0AAAAAAAAAAAAAAAAAAHmPnXqYhtIAAAAAAAAAAAAAAAAAXkfQp43QAAAAAAAAAAAAAAAAAAB8L9M/dBzUAAAAAAAAAAAAAAAAAAH5M/rN0SHPQAAFv0Mw9DMAAAAAAADnMzm9wFsAGi8elOnwwAAAAD4z/PcP0v93DrgAArh+e/iwfryAAAAAAAAAAAAAAAAAAAAAAAAAAAHNHp+OAAAAAAAAAAAAAAAAAflP8AqxlsGuIAAAAAAAAAAAAAAAAAc9L8fObsAAAAAADn/jj5P+i/TPpT0fipz2/ykAAAAAAAA9p0y2DfnAAAAAAAAAAAAAAAAAAD4k+289g0xAADP0J1CsgAAAAAAAAAAAD5di8r63ALgAAAAA6U8z2CDkuM8rUAAAAAAADwHxbon7nOegAAAAAAAAAAAAAAAAAA8x869TENpAAAAAAAAAAAAAAAAAC8j6FPG6AAAAAAAAAAAAAAAAAAAPhfpn7oOagAAAAAAAAAAAAAAAAAA/Jn9ZuiQ56AAAt+hmHoZgAAAAAAAfn788/Bf2WHH9WAHvfX8x+hp99/GAAADA/Hy2Gh+gIXPXgAAAA+MPkkP2FAAAAAAAAAAAAAAAAAAAAAAAAAAADmj0/HAAAAAAAAAAAAAAAAAPyn/VjLYNcQAAAAAAAAAAAAAAAAA56X4+c3YAAAAAABUna2ViAAAAAAAAAHtOmWwb84AAAAAAAAAAAAAAAAAAfEn23nsGmIAAAAAAAAAAAAAAAAAfLsXlfW4BcAAAAAHSnmewQclxnlagAAAAAAAeA+LdE/c5z0AAAAAAAAAAAAAAAAAAHmPnXqYhtIAAAAAAAAAAAAAAAAAXkfQp43QAAAAAAAAAAAAAAAAAAB8L9M/dBzUAAAAAAAAAAAAAAAAAAH5M/rN0SHPQAAFv0Mw9DMAAAAAAAAAAAAAAAAAIg/KHODw/79DzX9KAsgAAB+SX6ah3AAAAAAAAAAAAAAAAAAAAAAAAAAAAc0en44AAAAAAAAAAAAAAAAB+U/6sZbBriAAAAAAAAAAAAAAAAAHPS/Hzm7AAAAAAAAAAAAAAAAAAAPadMtg35wAAAAAAAAAAAAAAAAAA+JPtvPYNMQAAAAAAAAAAAAAAAAA+XYvK+twC4AAAAAOlPM9gg5LjPK1AAAAAAAA8B8W6J+5znoAAAAAAAAAAAAAAAAAAPMfOvUxDaQAAAAAAAAAAAAAAAAAvI+hTxugAAAAAAAAAAAAAAAAAAD4X6Z+6DmoAAAAAAAAAAAAAAAAAAPyZ/WbokOegAALfoZh6GYAAAAAAAAAAAAAAAAAAAGV8kh8M9wH6884HR+dB6L+a4cB+sgPAAAAAAAAAAAAAAAAAAAAAAAAAAADmj0/HAAAAAAAAAAAAAPOvzbx6Jf1RFpm2H5T/qxlsGuIAAAAAAAAAAAAAAAAAc9L8fObsAAAAAAAAAAAAAAAAAAA9p0y2DfnAAAAAAAAAAAAAAAAAAD4k+289g0xAAAAAAAAAAAAAAAAAD5di8r63ALgAAAAA6U8z2CDkuM8rUAAAA+EtV4H710x93nDp4D4t0T9znPQAAAAAAAAAAAAAAAAAAeY+depiG0gAAAAAAAAAAAAAAAABeR9CnjdAAAAAAAAAAAAAAAAAAAEATgAAAAAAAAAAAAAAAAAAB+TP6zdEhz0AABb9DMPQzAAAAAAAAAAAAAAAAAAAAAOM+EQ8V5APoj5KD3SMP1h1gAAAAAAAAAAAAAAAAAAAAAAAAAAAA5o9PxwAAAAAAAAAAAAD8jf0+x6PnDmw+5zbn/ACG/SjDoDTLtdyoBgAAAAAAAAAAAAAAHKy/J15uwAAAAAAAAAAAAAAAAAAG+x6ZdAb84AAAAAAAAAAAAAAAAAAZukAAAAAAAAAAAAAAAAAB5cRZ84zXUfW9SFwAAAAAdKeZ7BByXGeVqAAABlfjn1R+0P4mB+1k/LfgPi3RP3Oc9AAAAAAAAAAAAAAAAAAB5j516mIbSAAAAAAAAAAAAAAAAAF5H0KeN0AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfkz+s3RIc9AAAW/QzD0MwAAAAAAAAAAAAAAAAAAAAAAGBwvbBOAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAc0en44AAAAAAAAAAAAB+LP7TY9Hi/xUH6em3P8Ah59d8vaFZ/Vvre3OFIAAAAAAAAAAAAAAD8u8tvrLRx3AAAAAAAAAAAAAAAAAADjvFWv0cOrjAAAAAAAAAAAAAAAAAAD8p/1Yy2DXEAAAAAAAAAAAAAAAD89zn6Tx9V9Y/W+3OFwAAAAAdKeZ7BByXGeVqAAAB498F9Ufqr+QKf6j9vz14D+VXbn7aWH6i89dCc1AAAAAAAAAAAAAAAAfkl9tezgDQAAAAAAAAAAAAAAAAAfJrPub1/xtwlgAAAAAAAAAAAAAAAAB+XfRP6iHPQAAAAAAAAAAAAAAAHjf5+9cBsvlf+g7Jhx2AABb9DMPQzAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA5o9PxwAAAAAAAAAAAAD8bv2Bx6PnPxgP0DNuf8p/1Yy2DXEAAAAAAAAAAAAAAAAAPy7y2+stHHcAAAAAAAAAAAAAAAAAAOO8Va/Rw6uMAAAAAAAAAAAAAAAAAAPyn/AFYy2DXEAAAAAAAAAAAAAAAAAPl2LyvrcAuAAAAADpTzPYIOS4zytQAAAOV/KPqj9hvxeH+yd7lrwHxbon7nOegAAAAAAAAAAAAAAAAAA/HT749nAGgAAAAAAAAAAAAAAAAA+OWfeHtPjbhLAAAAAAAAAAAAAAAAAD8meif1mOegAAAAAAAAAAAAAAAAAA/Jn9ZuiQ56AAAt+hmHoZgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAc0en44AAAAAAAAAAAAB8zfDXP1Yn6zueyN+f8AKf8AVjLYNcQAAAAAAAAAAAAAAAAA/LvLb6y0cdwAAAAAAAAAAAAAAAAAA47xVr9HDq4wAAAAAAAAAAAAAAAAAA/Kf9WMtg1xAAAAAAAAAAAAAAAAAD5di8r63ALgAAAAA6U8z2CDkuM8rUAAAA/JHpnombz+p55+ngPi3RP3Oc9AAAAAAAAAAAAAAAAAAB+On3x7OANAAAAAAAAAAAAAAAAAB8cs+8PafG3CWAAAAAAAAAAAAAAAAAH5M9E/rMc9AAAAAAAAAAAAAAAAAAB+TP6zdEhz0AABb9DMPQzAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA5o9PxwAAAAAAAAAAAAAzEXJ2AH4u/dfN1hpkAAAAAAAAAAAAAAAAAHwvnr9AXM9QAAAAAAAAAAAAAAAAAA5zzMPss6OUAAAAAAAAAAAAAAAAAAPgj73z1DTIAAAAAAAAAAAAAAAAAPC5v3T630xC4AAAAAOlPM9gg5LjPK1AAAAAMXaAA8B+UOqPsQyoAAAAAAAAAAAAAAAAAAPyx+0vVxBoAAAAAAAAAAAAAAAAAPlhn2V6z5WwSwAAAAAAAAAAAAAAAAA/NHef0uMKAAAAAAAAAAAAAAAAAAD80f2X0Qc1AAAW/QzD0MwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAOaPT8cAAAAAAAAAAAAAAAAA+c8nLUBgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHv8A0emIMAAAAAOlPM9gg5LjPK1AAAAAAAA5n4o3kNUAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAXfvDCgyYAAFv0Mw9DMAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADmj0/HAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAOlPM9gg5LjPK1AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAC36GYehmAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABzR6fjgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHSnmewQclxnlagAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABb9DMPQzAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA5o9PxwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA87T6raEDAAAAAAAAAAAAAAAAABOaR0wwAAAAAAAAAAAAAAAAAKoeZeny7BSAAAAAAAAAAAAAAAAAAAAAAAAAADpTzPYIOS4zytQAAAAAAAAAAAAAAAAAAAAAAAAAAACMPKvVaUhLAAAAAAAAAAAAAAAAACgFfXAAAAAAAAAAAAAAAAAAAOf5Sl6WSwAAAAAAAAAAAAAAAAAAAAAAAC36GYehmAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABzR6fjgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHwVwfP1eEfsO56E35wAAAAAAAAAAAAAAAAD8yvFeXt/Z86uIAAAAAAAAAAAAAAAAAPz9z1q/NWPR+0J1cQAAAAAAAAAAAAAAAAAAAAAAAAAAdKeZ7BByXGeVqAAAAAAAAAAAAAAAAAAAAAAAAAAAAfj70z7l4TvP7DHn6AAAAAAAAAAAAAAAAAHEfjz2R9gffuNBiwAAAAAAAAAAAAAAACkH5g1/Qz6r9KeagwoAAAAAAAAAAAAAAAAAAAAAAC36GYehmAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABzR6fjgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAH4u/tFj0fEOwH2Mbc4AAAAAAAAAAAAAAAAAfjVjv+yptgAAAAAAAAAAAAAAAAAAB+LOPR+0xtzgAAAAAAAAAAAAAAAAAAAAAAAAAB0p5nsEHJcZ5WoAAAAAAAAAAAAAAAAAAAAAAAAAAAHG/lj1T+wH4xB+w2vy0AAAAAAAAAAAAAAAAAHyvovKvv2gMWAAAAAAAAAAAAAAAAAGaH42/tJ1SHLQAAAAAAAAAAAAAAAAAAAAAAFv0Mw9DMAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADmj0/HAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPxb/aTHo+Q+SD7pNucAAAAAAAAAAAAAAAAAPxqx3/ZU2wAAAAAAAAAAAAAAAAAAD5g+d8Oj9JzfnAAAAAAAAAAAAAAAAAAAAAAAAAADpTzPYIOS4zytQAAAAAAAAAAAAAAAAAAAAAAAAAAAPIPgjqj9Vvx/T/UztOegQAAAAAAAAAAAAAAAAfK+i8q+/aAxYAAAAAAAAAAAAAAAAAflr0T9efR2TCGAAAAAAAAAAAAAAAAAAAAAABb9DMPQzAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA5o9PxwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAD8bv15x6Pzo+uA93NucAAAAAAAAAAAAAAAAAPxqx3/ZU2wAAAAAAAAAAAAAAAAAD5umvj/8AU+NHGuIAAAAAAAAAAAAAAAAAAAAAAAAAAdKeZ7BByXGeVqAAAAAAAAAAAAAAAAAAAAAAAAAAABQ/GTqj9qvxLD9snctgAAAAAAAAAAAAAAAAB8r6Lyr79oDFgAAAAAAAAAAAAAAAAH5xeh9kfbRx2AAAAAAAAAAAAAAAAAAAAAAAAW/QzD0MwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAOaPT8cAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA8i/P3Do9M/RO4DTIAAAAAAAAAAAAAAAAAPz1z1/Qo0yAAAAAAAAAAAAAAAAAD8icd9x2ev6b9D08YMAAAAAAAAAAAAAAAAAAAAAAAAAOlPM9gg5LjPK1AAAAAAAAAAAAAAAAAAAAAAAAAAAAPi7VfNX1n0T9ZnFYAAAAAAAAAAAAAAAAAeI0uH+pqAzYAAAAAAAAAAAAAAAB5sz8pu89HL0P9I+SwwoAAAAAAAAAAAAAAAAAAAAAAC36GYehmAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABzR6fjgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHSnmewQclxnlagAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABb9DMPQzAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA5o9PxwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADpTzPYIOS4zytQAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAt+hmHoZgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAc0en44AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB0p5nsEHJcZ5WoAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAW/QzD0MwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAOaPT8cAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA6U8z2CDkuM8rUAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAALfoZh6GYAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHNHp+OAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAdKeZ7BByXGeVqAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAFv0Mw9DMAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADmj0/HAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAOlPM9gg5LjPK1AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAC36GYehmAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABzR6fjgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHSnmewQclxnlagAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABb9DMPQzAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA5o9PxwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADlfkrPUIs+i6n4Y/TcJDXEAAAAAAAD8+/vbPW8aZHytF9/wC1NBUgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB0p5nsEHJcZ5WoAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB4nS8EOmTpEdp4wn97HNQAAAAAAAHG/JG0/dJjQB+fX3bvOwYUAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAW/QzD0MwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAOaPT8cAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA8n/IXn6vAtHm7PbP1D6OT87v2VuLZtzgAAAAAAfM03+aX7jZahvznzNN/md+5WWob84AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAdKeZ7BByXGeVqAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfDmq/P75H9bH3z6Hzf2l89cl/qYcdgAAAAAAAflF6F35/o2cGgB+UXf9+f6PHBoAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAW/QzD0MwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAOaPT8cAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPxB73j7/2B8A6uL8hP3qx3/C31jLf5l+6g+LPYlX7S/PPZ5/5KUOL0f1y+lO3zz8o4v0D9EWvx0+aOXt/Tn7s6uH+af+kvLfVOnkAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADpTzPYIOS4zytQAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA/ly78/wBu/tLlv8+Pmvrj9IPykkq8V1T9jfCMP9HP0h4b/O/84O2PUf2q567Q47/nX/b/AL89L8W0cH+59Hn35eB+9h5+gAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABb9DMPQzAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA5o9PxwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA/EHvePv/AGB+Xenj/Bz+nDDp/m0/SONfIPmpP9iPwjqP6EfxPqPt3nh/OH771mfie19v+u1P8937eYdX0/1XX5/8637n8/Z60dPGAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHSnmewQclxnlagAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB/Ll35/t39pct/ld8g9+f6+fz5o/fD8Z0fS3tY/zo/oDk/nE/czRfl99eh+pp5ev8wH9MnoZ/nJ+dmq/cH3nzdPM/wAGe2P6QTz9AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAC36GYehmAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABzR6fjgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB+IPe8ff+wPy708f4Of04YdP82n7UqvBvklH7zfy9VP77/gsq+o+/mu3/AFe6OU/FdV9nfbF5/mj+cPL2fdP6y9HL/OZ+8ue3px08YAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAdKeZ7BByXGeVqAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAH8uXfn+3f2ly3+V3yD35/r5/Pmj+p/8BcK+vvfw/ET9zN5/BD703l31hy396nn6fzE/0pehnT/JDJ/CX7odM+r/AIX5v+jU8/QAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAt+hmHoZgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAc0en44AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfiD3vH3/sD8u9PH+Dn9OGHT/Np+1Krwb5JR+838vVT++/531Pgv1Jlv9B/ePTxH5gKp/tJP85PD8On1z9ft+f+b/8ApQi5zp5AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA6U8z2CDkuM8rUAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAP5cu/P8Abv7S5b/K75B78/18/nzR/U/+AuFfX3v4fiJ/S3a/nN9y6p9M/RHiv6AOO/59v2C9DPxjwGT4J/XvZb/wFk/3FODQAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAt+hmHoZgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAc0en44AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAeA6OentvI3n87fWM38te7j53jA+ifkgX0fv1PyhyGW30V67pieNj/ABp/dnHo+T5qj63+QLj5G/YyLDbAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAOlPM9gg5LjPK1AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAD5I0Xvndw/IOK0n2v5rH9e/OMna9IHgX1kzlPkelN9Yh1hjX5wcP35/oJ8yc9enfUGb/ABm+u+yPt44NAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAC36GYehmAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABzR6fjgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfPfu01eKk8bmu265oGgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA6U8z2CDkuM8rUAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAKHg9r6HIYB82fSFp5DAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAALfoZh6GYAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHNHp+OAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAdKeZ7BByXGeVqAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAFv0Mw9DMAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADmj0/HAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAOlPM9gg5LjPK1AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAC36GYehmAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABzR6fjgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHSnmewQclxnlagAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABb9DMPQzAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA5o9PxwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADpTzPYIOS4zytQAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAt+hmHoZgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAc0en44AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB0p5nsEHJcZ5WoAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAW/QzD0MwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAOaPT8cAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA6U8z2CDkuM8rUAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAALfoZh6GYAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHNHp+OAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAdKeZ7AkiEMAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHUgsAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA5o9PxwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADpTzPYAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADmj0/HAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAOlPM9gAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAOaPT8cAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA6M831gAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAG4TAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA5o9PxwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAJprfPO9YAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADmj0/HAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAmmt8871gAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAOaPT8cAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAACaa3zzvWAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA5o9PxwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAJprfPO9YAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADmj0/HAAA8Rmvwi+peL0P2mO/wA0AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAmmt8871gAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAOaPT8cAADwWa/n0+s+H0v3SPQ8sAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAmmt8871gAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAOaPT8cAADwWa/n0+s+H0v2h8E7fP9n9FqAaAAD5r8Sx33/rWp600yAAAAPyjy2/U69riAAAAfkrlv8ArUa4AHgnq0383kafSPk95++FQAABgo8b+glQVIAAAAAAAAAAAAAAAAAAH5m5a/fXcaZg0AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAE01vnnesAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABzR6fjgAAeCzX8+np3n+px+Cnb/AGf3w+2zs88AAPxB+6OXsk/MtP8AfHV6uIGAAAfgLh0/vLq78wAAAB/Phz9X9B50coB+dP1fjv8ALxN/UPyhUfosbYAHL/mDjv8ArN5Lpj8bfpHOgaZAAAAAAAAAAAAAAAAAAAfiDh0frb6jtgDQAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAATTW+ed6wAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHNHp+OAAB4LNfz6bfner+0/nnZwfkB3XJ3/0jS+l5AAAfiD+tvP1eo/i1U/pv81NYn0an2350Ih/RUfsf4Ch+lPxtNfUv3rrh4t+dcar+mgvSP58FX6R/KE19rfYu/N8TfRM18jka/YfzBeX3J+fDPnb9Bs9flH8/wDLb7u/RHfm+Sfu+o8j/OqNOf8A1MH6Kb835+bOHT0vvF5/nEuHR+l/zTvz+4+v1n+b/wCgca7f5uC+8fzNVcv+hk39Bfn5pl01ub+hPzVRv/PWW/7G+o9fCDQAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAATTW+ed6wAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHNHp+OAAB4LNfz6fVvB6f7sHo+V/Ot5H5vrf0f+jej5QNAH4g/obzddT8vFX73/zuuf1M+uNcfwu/bfLaH8Qg/f38E3P339x6Z/ix+jWW3vW30cvz/wDnrh0/s9/N8n+w30hth+GH7NY7/m995teCgfR35yB+nf5NB+22PvzcP+PeHV+3Hn23N+Xn7F5bfg5+0E36Jd6OXQGvwU+yOTu++vxVvL9nON2x/Lr7W5+nyn9Bdcf5/f1bx6PWfz10y/WnM256n4cYdP7+/hG19W/oLUfhv+xGevW/z9J/vj6j08gNAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABNNb553rAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAc0en44AAHgs1/Pp9QcHp/vKz0fK/nA88831/wCl3qPR8kGgD8QfWOPv9a+29+bsf56I0/oX+f8Abn/O/wDZDPU/Fsf6bfjnF/vFsdPJ8TeO8/V9kfnM1e8Xx3/eb+e3XH+hI6OX81fQOfq+aPvMPBQPo785A/Yf8XXOv+rI3/jUn+6XjO3P+Xn6B4dPx3+tWmQa4gH4C/u7z9XNfgcq+sia8z/anTL8XPt2NI/lxr2T32o92/LtrQ+Mefp/o8/Bnfn/AG0drh+J/wC8GW5+IIv1t9R2wBoAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAJprfPO9YAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADmj0/HAAA8Fmv59JfO9b9NPOOjl+Dfp7Hf95z0fKAAD8Qf1t5+r1E6OU/nw5+r+g/zvfm/Fr96MehPwKD9yPw0T/an1Hfm/Mz0PDp+Fv2bBn4tB+8386Af0J7m/L+Pf3Lh1fDv3mHgoH0d+cgfsqdHL8b/IHN1/qb+MLX7seM7c/5efp/j0fkz+6DkN+cA/AX95cOmD8K0/3ud0coB+IPV8fofsF+MGvOv7Rh+Vn1Qq+tP53g/ok/Blr9vdjbn/n6/oPw6Z/wCF+1nqO/ODQAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAATTW+ed6wAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHNHp+OAAB89TX4IfbXF6PyRxcafQX7YdPH6mdPIAAH4n/AKw4dPp5vzH4A4dP7/G/N+Zfz9ydzPqhr9EPw4vLvczLfnf2q1w/NXwjPb0XxhV+7H4Na4ej08t9n9jtub8jfutV4UB9E/nYH6R/mkHI/dUX9ofz+VP1r95ufzF/Zxr8o/CsejvP1C1x9TN+b8FP3Xw6dT877z+SvTOfq+nftzp5Pzx8Mw6P2B/PDbDwz9gpr4u/PDPb2T5qjT+hH8KNub9r+o2w/Of445e30vzOa/Yv1Xs4AaAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAACaa3zzvWAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA5o9PxwAAAACtzKfXjQAAAAAAAAAZ6JLjAAxEbYwxEbYwAbkI2RgAAABlolvgVmU9VAMM5C6AAwAAKdNGwMAAAAMhGlKwAADNRdlYAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAATTW+ed6wAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHNHp+OAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABNNb553rAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAc0en44AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAE01vnnesAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABzR6fjgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAATTW+ed6wAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHNHp+OAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHyn+d3N1/SP0nUfQettzgwAAAAAAAAAAADy75ry1+bMHDp/ZY7OEAAAAAAAAAAAAAACaa3zzvWAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA5o9PxwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA5b42z1+Ovlfj9DI98R9KfSfTyfSfoG/MDQAAAAAUEeBfNeXR82fNnN15H0gn9J/VvTx/RpvzgAAAAAAAAAAAAAATTW+ed6wAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHNHp+OAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB57NfI/15nv8z+ERq75rh/Ova5b+++s64/cfufVwvlqTiU/m75sw6fmv585uv0j6XuPo36Z6eT2huuEqgAAAAAAAAAAAAAAAATTW+ed6wAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHNHp+OAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB8PZbfWXX6ZJ+deW/uffNeh/mlNO+XMejzTkM9em9UDhfNw9U+mLzt+8XOD6q58u+4rz8u4AOP+56nTNecAAAAAAAAAAAAAACaa3zzvWAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA5o9PxwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA8w+X8enT9rHz/lqfX/Wl4hpkAAAAAAAAAAAAAAAAAHzDGnneVj1fS3sO/IFQAeMezzYVAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHzTGnl9jHp+rvlnTKH2aa92N+UAAAAAAAAAAAAAAAAAPgvLf7X3dMT88M9/aPQA7L8y5q58m47+X6eWv053+2Hxrw2PR6F7fcdL+n2/N5N7RcYfKC+dfYct/PvqK8/YDflAAAAAAAAAAAAAAAmmt8871gAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAOaPT8cAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAIQ/PD9Dseic25/wA6P0Px6LBtzgAAAAAAAAAAAAAAAAABwHzDj0fbsO3P+cv6C49Pmfs+mPgHVTfwV9o5b+v/ADFrhsfTbn5iw89vqTjdcOO82z296+fRfUPU64fM+Rj0/U1zblBgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAeZfPGPR9qG3Pj/AAjj0foMbc4AAAAAAAAAAAAAAAAAAHg8X5d9kK/m34Gy25nwrHfyr0+L+lPQ9sPur5g1y/Qk35fzw+6cen4f8Lz1+dPM8N8f6ba9l9o2wz/v7TLTdtzgAAAAAAAAAAAAAAAE01vnnesAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABzR6fjgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABwqfzL9qRqGmPkHylh1/Qv0PpgF5gAAAAAAAAAAAAAAAAHzXtZdHvRrz/CP0xj1fPX23Wf54/oQq/P/AO/g+BvtJPpPlas/QvQWfG/r2W+f2tZ+AfYofIHqs3ubemflX1wSGmQAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfKXpmXR7Ca858qZ6+UfXsaeim/MAAAAAAAAAAAAAAAAAcL8NZbfB3mnH6DfpBz9J7W3P7j1Wke+fGDj6Z45rxr2idO1+ZWui+rHHzl9HtfMXiuevgPzbh0eNeg57eh/qFvzfXB18IAAAAAAAAAAAAAAE01vnnesAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABzR6fjgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABko+Cv0Lz6A15xA/On9Ccei8bc4AAAAAAAAAAAAAAAAB415Zj0fXBtz/nZ9uY9Xyx9vVn+d/6IKvz+/QEX53foiM+CRfbm1ph+ePueHb2nTa8/z/8AYKPz+6TLp3tis/SvXdecKkAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPAebx6PqI25wDL+Acej9EjbnAAAAAAAAAAAAAAAAAPmL4N5+r6Q+k7j2iTXAAAAAAAAAAAAAA5X5xjT5r57Dp/W06uIAAAAAAAAAAAAAACaa3zzvWAAAAAAAAAAAAAAAAAAAAAAD4u+Lgf8ATwff7gAAAAPwrD6o/SwAAAAPgv8AOYP6DQD5FD4C/WwP58v36D8s/uMPsfhA7sAAAAAAAADmj0/HAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAD5U875+z0H0Ss/LrKvy79AqzU1wAAAAAAAAAAAAAAAAAG/B2W/Yc9Gvp30drz/AJu+449XhH6YXl+c31qq8ioJ952d5fNXR57d028+06a8/n/7BR+f31bGvyz6yq9j77fkBoAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAIvg3LfqdaNPbfnuok0Jv67OjkAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAACaa3zzvWAAAAAAAAAAAAAAAAAAAAAAD8fvk8P0l/IUP2X+3gtcmHZ5ocT6OC/z0B9gfo6HW3QTz4Oh6EPzp/L0P6Pc4Os8iD5m+oA/mr/o/D56+jQ+YfymD+klgWrIZd4JwAAAAOaPT8cAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADPRhdSOvz4dU5oAAAAAAAAAAAAAAAAAAADmtNPSGqFVOzoCzq49nAC3qCy4B7dUT3hXuBn6AZ2KPqlaAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAMBirosUmLfGDQAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAATTW+ed6wAAAAAAAAAAAAAAAAAAAAAAfj94oH7o/zih+k3xOGH0Ifqv+G4dR2Qfvx+BIedZgdB/RAH5bfPIeA/tuHl35Eh6V5KH7e8QH5e/wBAQfzV/wBH4fhR+iwfnB5KH2BUDv8A9Sw/m7/pAD2oAAAADmj0/HAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPBIv3suAAAAAAAAAAAAAAAAAAAAAAAACaa3zzvWAAAAAAAAAAAAAAAAAAAAAAD8fvzdDQ+ng/cb8LA6n9sw/Cn2sP2C/mMD9gfzpD6W/WgP5k/2OD2X89A+O/r8Poz8nw/pq/G4PJPuIPy9/oCD+av+j8Pwo/RYOr/ACoD+lj4gD8Zv1eD89P6JQAAAAAOaPT8cAAAAAAAAAAAAAAAAAz4kAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABpPYAAAAAAAAAAAAAAAAAADcxAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfFme/2maYAAAAAAAAAAAAAAAAAABJosAAAAJprfPO9YAAAAAAAAAAAAAAAAAAAAAAPx+8QD95rQH8/ofS362B/P97cH7BfzGB+xX5xB9J/rQH8yn7NB+NH6yB8v6gfS35Ph/TP+Nged/aAfl7/AEBB/NX/AEfh+FH6LB1/5KB/TPTD+aDpw/RP9MgAAAAAOaPT8cAAAAAAAAAAAAAAAAA/Cv6Q4+8KgAAAAAAAAAAAAAAAAAPl2b9ZsRoAgAAAAAAAAAAAAAAAADH4oPo41xAAAAAAAAAAAAAAAAAAD5T+rJsKiCcYCAAAAAAAAAAAAAAA8uT+5frDo5QuAAAAAAAAAAAAAAAAADwn8wefq9wJoAAAAAAAAAAAAAAAAAAPkP3vLcBAAAAAAAAAAAAAAAAAB48n7f2e2ICAAAAAAAAAAAAAAAAAD5Tm/qwqAAAAAAAAAAAAAAAAAAA+WP3ruA35gAAJprfPO9YAAAAAAAAAAAAAAAAAAAAAAPx+8UD97AD+f0Ppb9bA+S/w5Dq/SQ/en8AA5jhA9d/oGD+fTkgb9Qh9M/mCGfzYfu/4uH5e/wBAQfzV/wBH4fhR+iwfWX88AfXf7vB+SX5nh/TB3oAAAAAHNHp+OAAAAAAAAAAGL8IZ6s+/gk+AA677baC8/wAp/wBWMtg1xAAAAAAAAAAAAAAAAAD8u8tvrLRx3AAAAAAAAAAAAAAAAAADjvFWv0cOrjAAAAAAAAAAAAAAAAAAD8p/1Yy2DXHA340C8wAAAAAAAAAAAAAAPl2LyvrcAuAAAAAAAAAAAAAAAAADwXxrLb7fNcQAAAAAAAAAAAAAAAAAA/Ib744+4GgAAAAAAAAAAAAAAAAA+PFX297N1cQVIAAAAAAAAAAAAAAAAAflPlt+rBriAAAAAAAAAAAAAAAAAAH5T/qxlsGuIAAE01vnnesAAAAAAAAAAAAAAAAAAAAAAFNgXwDPCW2BlhyHoYGcFrgw76UKfGh2sITRBh3A3qwVdAM3SChZCzzARcWH4jfQwfsmAAAAAAc0en44AAAAAAAAAAcVgTXz3xOW+H9LC+cPoIPqE35/yn/VjLYNcQAAAAAAAAAAAAAAAAA/LvLb6y0cdwAAAAAAAAAAAAAAAAAA47xVr9HDq4wAAAAAAAAAAAAAAAAAA/Kf9WMtg1x4jt8tw1wAAAAAAAAAAAAAAA+XYvK+twC4AAAAAAAAAAAAAAAAAPBfGstvt81xAAAAAAAAAAAAAAAAAAD8hvvjj7gaAAAAAAAAAAAAAAAAAD48Vfb3s3VxBUgAAAAAAAAAAAAAAAAB+U+W36sGuIAAAAAAAAAAAAAAAAAAflP+rGWwa4gAATTW+ed6wAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHxAHxv+xYbgAAAAABzR6fjgAAAAAAAAAAAfHvNY9HyF+0geG/Hwfpmbc/4N/RnH3hUfcPu/RyhUgAAAAAAAAAAAAAAfl3lt9ZaOO4AAAAAAAAAAAAAAAAAAcd4q1+jh1cYAAAAAAAAAAAAAAAAAAflP8AqxlsGuPnPe8/VZOjlAAAAAAAAAAAAAPynOXsPM1X3b9b9HIFwAAAAAAAAAAAAAAAAAeC/lxz9XtJNH6b6ZdCbYAAAAAAAAAAAAAAAAfkN98cfcDQAAAAAAAAAAAAAAAAAfHir7e9m6uIKkAAAAAAAAAAAAAAAAAPyny2/Vg1xAAAAAAAAAAAAAAAA8e+CMegM9Pl39+qgOjmAAAmmt8871gAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAOaPT8cAAAAAAAAAADzhP8AMH9fctvxn/Z8fgvyqH6Sm3P+U/6sZbBriAAAAAAAAAAAAAAAAAHisVhy8/WAEATgAAAAAAAAAAAAAABS7W8/WTo5gAAAAAAAAAAAAAAAAAA8p9WlhS8067l7d06uIAAAAAAAAAAAAAAD5di8r63ALgAAAAAAAAAAAAAAAAA8F8ay2+3zXEAAAAAAAAAAAAAAAAAAPm3R5usJogCcAAAAAAAAAAAAAAADlw+j9rq4gYAAAAAAAAAAAAAAAAAHwLlt99GuIAAAAAAAAAAAAAAAAAAflP8AqxlsGuIAAE01vnnesAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABzR6fjgAAAAAAAAAB5r+N/P1fpl6fpl8m9hGnz990B7ob835T/qxlsGuIAAAAAAAAAAAAAAAAAc9L8fObsAD5HVfXA5AAAAAAAAAAAAAAA9p0y2DfnAAAAAAAAAAAAAAAAAAD5N+ss9Q0y8w3+P0OwOzzwAAAAAAAAAAAAAAPl2LyvrcAuAAAAAAAAAAAAAAAAADwXxrLb7fNcQAAAAAAAAAAAAAAAAAA864Pn6gizwdPuPQGAIAAAAAAAAAAAAALjXup1cYAAAAAAAAAAAAAAAAAAHxJnt9tmmIAAAAAAAAAAAAAAAAAAflP+rGWwa4gAATTW+ed6wAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHNHp+OAAAAAAAAAAHlng+Wp6YP3z4+qO/+hWgqfyn/AFYy2DXEAAAAAAAAAAAAAAAAAOel+PnN2AHM/kxGv7Gl5AAAAAAAAAAAAAAAe06ZbBvzgAAAAAAAAAAAAAAAAAB8SfbeewmmPlt/g9P0Q7/MAAAAAAAAAAAAAAA+XYvK+twC4AAAAAAAAAAAAAAAAAPBfGstvt81xAAAAAAAAAAAAAAAAAADzrg+fqCLPklV5x9+phUAAAAAAAAAAAAABca91OrjAAAAAAAAAAAAAAAAAAA+JM9vts0xAAAAAAAAAAAAAAAAAAD8p/1Yy2DXEAACaa3zzvWAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA5o9PxwAAAAAAAAAAAAAAAAD8p/wBWMtg1xAAAAAAAAAAAAAAAAADnpfj5zdgB5B+LWW/7e+j6Yg0AAAAAAAAAAAAAHtOmWwb84AAAAAAAAAAAAAAAAAAfEn23nsR3j5g7g9X0yXv8oGAAAAAAAAAAAAAB8uxeV9bgFwAAAAAAAAAAAAAAAAAeC+NZbfb5riAAAAAAAAAAAAAAAAAAHnXB8/UEWfCM38Zft0qUvIAAAAAAAAAAAAALjXup1cYAAAAAAAAAAAAAAAAAAHxJnt9tmmIAAAAAAAAAAAAAAAAAAflP+rGWwa4gAATTW+ed6wAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHNHp+OAAAAAAAAAAAAAAAAAflP+rGWwa4gAAAAAAAAAAAAAAAABz0vx85uwA+ZvyUy3/QD9BLzCoAAAAAAAAAAAAAD2nTLYN+cAAAAAAAAAAAAAAAAAAPiT7bz2K15edXOL0er1+zgCoAAAAAAAAAAAAAD5di8r63ALgAAAAAAAAAAAAAAAAA8F8ay2+3zXEAAAAAAAAAAAAAAAAAAPOuD5+oIs/Nab+JP2QjT3Q1wAAAAAAAAAAAAAC417qdXGAAAAAAAAAAAAAAAAAAB5bNepFSAAAAAAAAAAAAAAAAAAH5T/AKsZbBriAABNNb553rAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAc0en44AAAAAAAAAAAAAAAAB+U/6sZbBriAAAAAAAAAAAAAAAAAHPS/Hzm7AD4t+Est2/t/UuLyAAAAAAAAAAAAAD2nTLYN+cAAAAAAAAAAAAAAAAAAPjn7Gz2KN5cHt8ndJ1fRyhpkAAAAAAAAAAAAAHy7F5X1uAXAAAAAAAAAAAAAAAAAB4L41lt9vmuIAAAAAAAAAAAAAAAAAAedcHz9QRZ+TsaeQ/Rsafpmbc4AAAAAAAAAAAAAXGvdTq4wAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPyn/VjLYNcQAAJprfPO9YAAAAAAAAAAAAAAAAAAAAAIHPN9Xvxhz9wAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAc0en44AAAAAAAAAAAAAAAAB+U/wCrGWwa4gAAAAAAAAAAAAAAAABz0vx85uwA/POLPb5O/UcPezTEAAAAAAAAAAAAAPadMtg35wAAAAAAAAAAAAAAAAAAADNmuM6rDq5zvrzDbnAAAAAAAAAAAAAD5di8r63ALgAAAAAAAAAAAAAAAAA8F8ay2+3zXEAAAAAAAAAAAAAAAAAAPOuD5+oIs/GONPs38650/dS/rzgAAAAAAAAAAAABca91OrjAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA/Kf8AVjLYNcQAAJprfPO9YAAAAAAAAAAAAAAAAAAAAAInPyp9S/SfCWz5r70AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAOaPT8cAAAAAAAAAAAAAAAAA/Kf9WMtg1xAAAAAAAAAAAAAAAAADnpfj5zdgB+XPuGe3H8WH6dGmIAAAAAAAAAAAAAe06ZbBvzgAAAAAAAAAAAAAAAAAAAGRN8r3OW3m/pybjo5QAAAAAAAAAAAAA+XYvK+twC4AAAAAAAAAAAAAAAAAPBfGstvt81xAAAAAAAAAAAAAAAAAADzrg+fqCLPwqz1/Y38sg+6vqu4ByAAAAAAAAAAAAXGvdTq4wAAAAAAAAAAAAAAAAAAPiTPb7bNMQAAAAAAAAAAAAAAAAAA/Kf8AVjLYNcQAAJprfPO9YAAAAAAAAAAAAAAAAAAAAAOL6vO+Z/SPuPyP6EPz39tAAAAABofn952H6OaYfm724foH8dBx/wB6h88+1htgAAAAAAAAAAAAAABzR6fjgAAAAAAAAAAAAAAAAH5T/qxlsGuIAAAAAAAAAAAAAAAAAc9L8fObsAPyA+489vVPxhD91NLTEAAAAAAAAAAAAAPadMtg35wAAAAAAAAAAAAAAAAAA8R9umzERz3eRp5v20aaR08gAAAAAAAAAAAAAfLsXlfW4BcAAAAAAAAAAAAAAAAAHgvjWW32+a4gAAAAAAAAAAAAAAAAAB51wfP1BF0/wJz2/fL45qPmv9akwvMAAAAAAAAAAAAuNe6nVxgAAAAAAAAAAAAAAAAAAfEme322aYgAAAAAAAAAAAAAAAAAB+U/6sZbBriAABNNb553rAAAAAAAAAAAAAAAAAAAAAB82+78hwp9n+VfQHpn59+1hweyAAAAfEgZX08H5n9CH3r8fh7n86B6T9KB8nfowAAAAAAAAAAAAAAAAHNHp+OAAAAAAAAAAAAAAAAAflP+rGWwa4gAAAAAAAAAAAAAAAABz0vx85uwA/Eb9Vs9vY/xwc/bv1lUg5AAAAAAAAAAAAPadMtg35wAAAAAAAAAAAAAAAAAA+JPtvPYwHON3EXxd+NOlOnkAAAAAAAAAAAAAD5di8r63ALgAAAAAAAAAAAAAAAAA8F8ay2+3zXEAAAAAAAAAAAAAAAAAAPOuD5+oIvz38Zc9v3c5rTL8Nf3Dz16w0xAAAAAAAAAAAALjXup1cYAAAAAAAAAAAAAAAAAAHxJnt9tmmIAAAAAAAAAAAAAAAAAAflP+rGWwa4gAATTW+ed6wAAAAAAAHyR5AAAAAB6KH2iAAAAAAeZeAfb/kgez8te+ofmvvd0+U/RwAAAD89g676qD8W9kP28+dQ+S+aDuLodZz4foTZAAAAAAAAAAAAAAAOaPT8cAAAAA8A+CMOn7j5mo+uTXEAAAAAAAD8p/wBWMtg1xAAAAAAAAAAAAAAAAADnpfj5zdgB+Bv7k57dT8YVn8vfrgqC8wAAAAAAAAAAAD2nTLYN+cAAAAAAAAAAAAAAAAAAPiT7bz2ObJodmq57Hz17k6eQAAAAAAAAAAAAAPl2LyvrcAuAAAAAAAAAAAAAAAAADwXxrLb7fNcQAAAAAAAAAAAAAAAAAA864Pn6gi/BPzZz2/aE0x/KH33Pb7hNMQAAAAAAAAAAAC417qdXGAAAAAAAAAAAAAAAAAAB8SZ7fbZpiAAAAAAAAAAAAAAAAAAH5T/qxlsGuIAAE01vnnesAAAAAAAB+c3zYAAAAB7WH6ggAAAACB8k5X6Z+BBph6Bf8T6730+K/VgAAACiH5tVA884gP24+Yw8B+qQ4TzMPIfqEND6lAAAAAAAAAAAAAAAOaPT8cAAAAyEfzoei8Hqfod+aOmX719t18INAAAAAAB+U/6sZbBriAAAAAAAAAAAAAAAAAHPS/Hzm7DmR/hh++mei89pl+GP7fZ7diaYgAAAAAAAAAAAHtOmWwb84AAAAAAAAAAAAAAAAAAfEn23nscuKDr06XAZ6+lr08gAAAAAAAAAAAAAfLsXlfW4BcAAAAAAAAAAAAAAAAAHgvjWW32+a4gAAAAAAAAAAAAAAAAAB51wfP1BF/JPybnt+tBpj8wfn9nt+0BpiAAAAAAAAAAAAXGvdTq4wAAAAAAAAAAAAAAAAAAPmKNPp0vMAAAAAAAAAAAAAAAAAAPyn/VjLYNcQAAJprfPO9YAAAAAAAAAAAAAAAAAAAAytMPkg/TPwIA9u5b5z7r6HPkv0oAAAArB8qfAwfqz8dhs/Nofpt62H56ffQflF7KH1B7SAAAAAAAAAAAAAAAc0en44AAABxCf4TfaXJ39L+YiP6Hu27OAGgAAAAAA/Kf9WMtg1xAAAAAAAAAAAAAAAAADnpfj5zdh5An+UP7ezoF5flP7hnt90GmIAAAAAAAAAAAB7TplsG/OAAAAAAAAAAAAAAAAAAHxJ9t57HJCf1QJ5nnr6Dc35gaAAAAAAAAAAAAPl2LyvrcAuAAAAAAAAAAAAAAAAADwXxrLb7fNcQAAAAAAAAAAAAAAAAAA864Pn6gi/hHz/Pb9LjTGp+Fuev7C+sXmDQAAAAAAAAAAFxr3U6uMAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAD8p/1Yy2DXEAACaa3zzvWAAAAAAAAAAAAAAAAAAAAK1R8fN/Tv5/AX1H5l8f+o+7Hz/2YAAABSD5R+ng3UD5W9HD2MDyX1oPM+AD6LAAAAAAAAAAAAAAADmj0/HAAAAAPxu+C+D0z7XT/AGuPQ8sAAAAAAAD8p/1Yy2DXEAAAAAAAAAAAAAAAAAOel+PnN2HzMq+Fv2CmgvP5m/PHPb9pzTEAAAAAAAAAAAA9p0y2DfnAAAAAAAAAAAAAAAAAADj+wTOOz1t9M0cLNbXQXAXmAAAAAAAAAAAAfLsXlfW4BcAAAAAAAAAAAAAAAAAHgvjWW32+a4gAAAAAAAAAAAAAAAAAB51wfP1BF/mt2me33qaYn5rTel+i4BUAAAAAAAAAAAXGvdTq4wAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPyn/VjLYNcQAAJprfPO9YAAAAAAAAAAAAAAAAAAAAA+dPPv0T8QDs8rtPSfnfufTz5H9LAAAAAAAAAAAAAAAAAAAAAAAAAAAAAOaPT8cAAAAhD5g7fPWjhB9I2NMgAAAAAAAPyn/VjLYNcQAAAAAAAAAAAAAAAAA56X4+c3YfFs14P+pSoLzrfhhnr+vHsF5g0AAAAAAAAAAB7TplsG/OAAAAAAAAAAAAAAAAAAAAcTlvq9DUHMp0uzTDXEAAAAAAAAAAAA+XYvK+twC4AAAAAAAAAAAAAAAAAPBfGstvt81xAAAAAAAAAAAAAAAAAADzrg+fqCL/J36nz2+uzTE8VT/Jn9yo0mNMgAAAAAAAAAALjXup1cYAAAAAAAAAAAAAAAAAAHzRGn0uXmAAAAAAAAAAAAAAAAAAH5T/qxlsGuIAAE01vnnesAAAAAAAAAAAAAAAAAAAAAZ3zr9F8Pxz/rvzT1b3L4b9dU8j6YAAAAAAAAAAAAAAAAAAAAAAAAAAAAA5o9PxwAAAOZ+Jc9fyd/W/m68/8AI8P6LvRezgBoAAAAAAPyn/VjLYNcQAAAAAAAAAAAAAAAAA56X4+c3YfnnNs/Q8AqD83pt36PgFQAAAAAAAAAAB7TplsG/OAAAAAAAAAAAAAAAAAAHOdGmcLltu7t5mY1xfo+W4b8wAAAAAAAAAAAB8uxeV9bgFwAAAAAAAAAAAAAAAAAeC+NZbfb5riAAAAAAAAAAAAAAAAAAHnXB8/UEX+Mf6N57fQJpiAfid+kee30caYgAAAAAAAAAAXGvdTq4wAAAAAAAAAAAAAAAAAAPiTPb7bNMQAAAAAAAAAAAAAAAAAA/Kf9WMtg1xAAAmmt8871gAAAAAAAAAAAAAAAAAAAAAAyLO/HeMOwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAOaPT8cAAADM/nj5uvy30rm63/SGmf7QSd3mgAAAAAAAflP+rGWwa4gAAAAAAAAAAAAAAAABz0vx85uw/LmNPcPtZyFSeOp/kX+50aWDTIAAAAAAAAAAD2nTLYN+cAAAAAAAAAAAAAAAAAAPiT7bz2OAmtLrbyG3HmHpXN1znTyAAAAAAAAAAAAHy7F5X1uAXAAAAAAAAAAAAAAAAAB4L41lt9vmuIAAAAAAAAAAAAAAAAAAedcHz9QRf4Vfsbnt6QaYgHwZ5Jnt+pxpiAAAAAAAAAABca91OrjAAAAAAAAAAAAAAAAAAA+JM9vts0xAAAAAAAAAAAAAAAAAAD8p/1Yy2DXEAACaa3zzvWAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA5o9PxwAAACsHwz9w56QQXntOYAAAAAAAB+U/6sZbBriAAAAAAAAAAAAAAAAAHPS/Hzm7D8gI0+4/pypByAfir+ime30saYgAAAAAAAAAAe06ZbBvzgAAAAAAAAAAAAAAAAAB8Sfbeex55FeI/WNINcOA6Xm69o6eQAAAAAAAAAAAA+XYvK+twC4AAAAAAAAAAAAAAAAAPBfGstvt81xAAAAAAAAAAAAAAAAAADzrg+fqKcX+BP75Z7aBpiAcP+JWe37pbemIAAAAAAAAAAFxr3U6uMAAAAAAAAAAAAAAAAAAD4kz2+2zTEAAAAAAAAAAAAAAAAAAPyn/VjLYNcQAAJprfPO9YAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADmj0/HAAAAAPxP+KvP9QI0/pw1fT8cAAAAAAAD8p/1Yy2DXEAAAAAAAAAAAAAAAAAOel+PnN2H4jRr+q3sdZg0AfCni2e36qGmIAAAAAAAAAAHtOmWwb84AAAAAAAAAAAAAAAAAAfEn23nsebxfxb91quvOjk5NvN19cdPIAAAAAAAAAAAAfLsXlfW4BcAAAAAAAAAAAAAAAAAHgvjWW32+a4gAAAAAAAAAAAAAAAAAB51wfP1Hnuen4y/u5GgaZAB+QkafW32LUg5AAAAAAAAAC417qdXGAAAAAAAAAAAAAAAAAAB8SZ7fbZpiAAAAAAAAAAAAAAAAAAH5T/AKsZbBriAABNNb553rAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAc0en44AAAB5+n+DPoPD6X6N/mZrj+997q4gAAAAAAAPyn/AFYy2DXEAAAAAAAAAAAAAAAAAOel+PnN2H4G56/uT1OmYCAOM/ETPb9z9/TEAAAAAAAAAAD2nTLYN+cAAAAAAAAAAAAAAAAAAPiT7bz2PM4v4T+j43+qTq8/H5bm6/QTp5AAAAAAAAAAAAD5di8r63ALgAAAAAAAAAAAAAAAAA8F8ay2+3zXEAAAAAAAAAAAAAAAAAAPOuD5+o8Ez0/Nn9oZsLzAD5DVfG/7CTQXmAAAAAAAAAFxr3U6uMAAAAAAAAAAAAAAAAAACAJwAAAAAAAAAAAAAAAAAAA/Kf8AVjLYNcQAAJprfPO9YAAAAAAAAAAAAAAAAAAAAAIHPPdRvyBz9oAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAc0en44AAABgI/Bj7C5PQ+VfA89f6Me57/LBoAAAAAAPyn/VjLYNcQAAAAAAAAAAAAAAAAA56X4+c3ZzP4YZ7fvoumIAAH5Fxp9UfZjkKkAAAAAAAAAPadMtg35wAAAAAAAAAAAAAAAAAA8E97izy/Lf4D6nLp/SM7fMh805+v05+/IDAAAAAAAAAAAPl2LyvrcAuAAAAAAAAAAAAAAAAADwXxrLb7fNcQAAAAAAAAAAAAAAAAAA864Pn6j5Jz1+Tf1omgvMAMkPwr/arPb0A0xAAAAAAAAAC417qdXGAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB+U/6sZbBriAABNNb553rAAAAAAAAAAAAAAAAAAAAABE5+V/qT6P4ayfN/dgAAAAAAAAAAAAAAAAAAAAAAAAAAAAABzR6fjgAAADA+bvb4v8w/Xsej9Azp5AAAAAAAAPyn/VjLYNcQAAAAAAAAAAAAAAAAA56X4+c3Z5B+UOe37emmIAAHyUq+Kf2LmgvMAAAAAAAAAPadMtg35wAAAAAAAAAAAAAAAAAAAG+W8/X+fmrl0/oL3vZ5p51jv2GrrgFSAAAAAAAAAAB8uxeV9bgFwAAAAAAAAAAAAAAAAAeC+NZbfb5riAAAAAAAAAAAAAAAAAAHnXB8/UfCOW3n/wClwBUAAB+XfpGe334aYgAAAAAAAABca91OrjAAAAAAAAAAAAAAAAAAA8zmvTCpAAAAAAAAAAAAAAAAAAD8p/1Yy2DXEAACaa3zzvWAAAAAAAAAAAAAAAAAAAAADjerz/mX0z7j8k+gD89/awAAAAAA/N/ng+v/AG8Pzk74PvD5FDkPukPA/YQ2wAAAAAAAAAAAAAAA5o9PxwAAADkU/wCbX+i3l7PSzr4gAAAAAAAAD8p/1Yy2DXEAAAAAAAAAAAAAAAAAOel+PnN2fM3wtnt+wTdMXAABmB+FX7R57eimmIAAAAAAAAAe06ZbBvzgAAAAAAAAAAAAAAAAAAAQ+Uc3X8P+4LXW+t9uQ45Oz1DQaZAAAAAAAAAAAfLsXlfW4BcAAAAAAAAAAAAAAAAAHgvjWW32+a4gAAAAAAAAAAAAAAAAAB51wfP1H5rZbdp95g4qAAA+evzKz2/bldMQAAAAAAAAAuNe6nVxgAAAAAAAAAAAAAAAAAAfEme322aYgAAAAAAAAAAAAAAAAAB+U/6sZbBriAABNNb553rAAAAAAAAAAAAAAAAAAAAAB84e58jwR9p+U/QfpP57+2Bw+wAAAAVQ/Jv9Qw2vy3D7g+Pg+jPmcPRfqAPkX9GwAAAAAAAAAAAAAAAA5o9PxwAAACmH8+vEed6vfDn9/tH0PMAAAAAAAA/Kf9WMtg1xAAAAAAAAAAAAAAAAADnpfj5zdnxb4Pnt+ofgtx9AjgAAPzC7rPb9AzTEAAAAAAAAAPadMtg35wAAAAAAAAAAAAAAAAAA+RvrnPWr49lv8nfZNL4h/TByuDeWD3kWG2AAAAAAAAAAAHy7F5X1uAXAAAAAAAAAAAAAAAAAB4L41lt9vmuIAAAAAAAAAAAAAAAAAAedcHz9R+TuO/1P721V9KpASAR/h1nr+q3ud5g0AAAAAAAAXGvdTq4wAAAAAAAAAAAAAAAAAAPiTPb7bNMQAAAAAAAAAAAAAAAAAA/Kf9WMtg1xAAAmmt8871gAAAAAAAPH+NpAAAAHSh7aSwAAAADzb58+3/Jg9n5S99R/NffbZ8p+jAAAAHnwfkh6CH6lfjQH7efOYfJuAHeyB12KH6AzgAAAAAAAAAAAAAAHNHp+OAAAAVA/NWXm6wD9JrXTyAAAAAAAAflP+rGWwa4gAAAAAAAAAAAAAAAABz0vx85uz882Z7fbfynS+kvTqgCQDwH8vc9v2+dpiAAAAAAAAAHtOmWwb84AAAAAAAAAAAAAAAAAAfEn23ntQ8Uy2+f/AL40y/Mr7sjX0urvyedenYdKnRygAAAAAAAAAAfLsXlfW4BcAAAAAAAAAAAAAAAAAHgvjWW32+a4gAAAAAAAAAAAAAAAAAB51wfP1H4x47/o3fZ4/wDWjN4eYAfnlN83+mIBUAAAAAAAAFxr3U6uMAAAAAAAAAAAAAAAAAAD4kz2+2zTEAAAAAAAAAAAAAAAAAAPyn/VjLYNcQAAJprfPO9YAAAAAAAD5X8ntA0AAB6Aj7AIoAAAAQPkrJ/TPwINMO+0/E+u97Piv1YAAAAAPlIPnn56D9uPmUPnz6dDhvOw8m+nA2PqIAAAAAAAAAAAAAAA5o9PxwAAAAAAAAAAAAAAAAD8p/1Yy2DXEAAAAAAAAAAAAAAAAAOel+PnN2flz7hnt7h5rS5/6vZ0I8gBn4fxr+pPvVZg0AAAAAAAAe06ZbBvzgAAAAAAAAAAAAAAAAAB8Sfbee2X4Xjv5d9s7YfINfPb7GOjj817vm67508gAAAAAAAAAAHy7F5X1uAXAAAAAAAAAAAAAAAAAB4L41lt9vmuIAAAAAAAAAAAAAAAAAAedcHz9R+FWHR+xvkdqvEr+q5qwADy9P8AG391I0vGmQAAAAAAABca91OrjAAAAAAAAAAAAAAAAAAA+JM9vts0xAAAAAAAAAAAAAAAAAAD8p/1Yy2DXEAACaa3zzvWAAAAAAAAAAAAAAAAAAAAMrTD5IP0z8CAPbeW+d+5+iD5H9LAAAAPIQ4v4oD6z8RDuvmEP0m9iD89/vMPyo9yD6L9vAAAAAAAAAAAAAAAOaPT8cAAAAAAAAAAAAAAAAA/Kf8AVjLYNcQAAAAAAAAAAAAAAAAA56X4+c3Z+QH3HntJ6FazvMU/q6esQA/Pib439OQCoAAAAAAAAPadMtg35wAAAAAAAAAAAAAAAAAA+JPtvPbH8Bx3576325/O/gbPf9OZOjj4jU5uvozp5AAAAAAAAAAAPl2LyvrcAuAAAAAAAAAAAAAAAAADwXxrLb7fNcQAAAAAAAAAAAAAAAAAA864Pn6qf4E4dP74fHtz9dfOodN78SDkA/Gj7uz2+qTTEAAAAAAAAuNe6nVxgAAAAAAAAAAAAAAAAAAeWzXqRUgAAAAAAAAAAAAAAAAAB+U/6sZbBriAABNNb553rAAAAAAAAAAAAAAAAAAAAFWo+P2/p38/gL6j8y+P/Ufdj5/7MAAAA44PlX2IPeWh8hemB7mB457GHk3nQfTwAAAAAAAAAAAAAAAc0en44AAABmh/PVJ5/qEoey/rf0cnpR0cx+dOetX9IA86/Nubl/VEWmbYflP+rGWwa4gAAAAAAAAAAAAAAAABz0vx85uz8Rv1Wz28Y+prXRfMRO59BgDg82T/ABi/dWNNE0yAAAAAAAAPadMtg35wAAAAAAAAAAAAAAAAAA+P/sDPbD+d8ei99Lb8p+asa/aHrWmPOZmHR2x08gAAAAAAAAAAHy7F5X1uAXAAAAAAAAAAAAAAAAAB4L41lt9vmuIAAAAAAAAAAAAAAAAAAedcHz9Xnv4y4dP7Z/Mdr7Fy3n8lfQk6eilZAHxP8357frQaYgAAAAAAAFxr3U6uMAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAD8p/1Yy2DXEAACaa3zzvWAAAAAAAAAAAAAAAAAAAAAPnHgf0T8PDs8vuPQvnfuvUj5H9KAAAAAAAAAAAAAAAAAAAAAAAAAAAAAOaPT8cAAAAqh/Ox98cXoZPxmHpPah+zp2+eeZJ/mX+wOW35G/p8P5w5sPuc25/xd+6+brDTL3jv9MgaAAAAAAAAAAAAAAA56X4+c3Z+Bv7WZ7fOv2fcrmuPkn36dPSisgD8cPt/Pb6wNMQAAAAAAAD2nTLYN+cAAAAAAAAAAAAAAAAAAADnvmzk7/QPdOnjPk+a5r7VHn8Llr6UdPIAAAAAAAAAfJBlseFzfun1vpiFwAAAAAAAAAAAAAAAAAeC/EWHT9QFQfUtRpFwAAAAAAAAAAAAAAAB51wfP1eCfmzh0/qNw9r6THlwnzZOv1nt1mAuZ/DbPb9xOr0yAQAAAAAAFxr3U6uMAAAAAAAAAAAAAAAAAACAJwAAAAAAAAAAAAAAAAOL+fM9AVfBH7XKg25wAAJprfPO9YAAAAAAAAAAAAAAAAAAAAAzPnL6P4Xk1+t/NvUvePh/wBcU8f6cAAAAAAAAAAAAAAAAAAAAAAAAAAAAA5o9PxwAAADz9P+eb+gXm7Mr8AEfvB+SdL99Dp4zKD8k/2Bx3/Fn9ph+L/FQfp6bc/4y/fHP1heXtXcaZA0AAAAAAAAAAAAAABz0vx/mebt/DD9pM9vMvp68wccT8zTr9Y71ZgL4u+Yc9v1vNMQAAAAAAAD2nTLYN+cAAAAAAAAAAAAAAAAAAADmvmPk7/bvU+niOJa/O79P8emfzFHotrfmBoAAAAAAAD5dMtjwWa9l+t9MguAAAAAAAAAAAAAAAAADwX4ww6PqcqT6SqNAuAAAAAAAAAAAAAAAAPOuD5+r5J+TcOn7b6217API8OVcF9UjlHB+T8afQP2+0FQAAAAAABca91OrjAAAAAAAAAAAAAAAAAAA+JM9vts0xAAAAAAAAAAAAAAAA5HwuNAmvz9/aRMNsAAAJprfPO9YAAAAAAAAAAAAAAAAAAAAAAMK10+fpnN6AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAc0en44AAAAB+JHxj5/qH3WL7w+TOjm/V06OUyg/JP9gcd/xu/YEfzn4wH6Bm3P8AlP8AqxlsGuIAAAAAAAAAAAAAAAAAc9L8f8g5e38of07jbW9svIHJ4mq85+rBzDjnfwyz2/b/ALDTIBAAAAAAAe06ZbBvzgAAAAAAAAAAAAAAAAAB4j7dN8r8tcvb9O9l18ANfnJ9Z8/Z7RwdZ7+7eYXmAAAAAAAAAHy7F5X1uAXAAAAAAAAAAAAAAAAAB4L41lt9vmuIAAAAAAAAAAAAAAAAAAedcHz9Xwj5/h0+3fSVrrh5DA+W+unX3krI+XVXwF+z80F5gAAAAAAXGvdTq4wAAAAAAAAAAAAAAAAAAPiTPb7bNMQAAAAAAAAAAAAAAAAAA/Kf9WMtg1xAAAmmt8871gAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAOaPT8cAAAAAb4VNHuoOKkAMoPyT/YHHf5m+GpvE/Wdz2Rvz/lP+rGWwa4gAAAAAAAAAAAAAAAABz0vx/wCZuXu+Fvs6Nveu60wAQ0Pl7pZ29+KxPyljT3T7maCoAAAAAAA9p0y2DfnAAAAAAAAAAAAAAAAAAD4k+289uR+T+fq+ztDr88GfL3nvP2fcfLaYV+xTDXEAAAAAAAAAPl2LyvrcAuAAAAAAAAAAAAAAAAADwXxrLb7fNcQAAAAAAAAAAAAAAAAAA864Pn6vzW6LDp7D7SuZR5gGR8lrT6J9CcFMX4W/sPnt6uaYgAAAAAAXGvdTq4wAAAAAAAAAAAAAAAAAAPiTPb7bNMQAAAAAAAAAAAAAAAAAA/Kf9WMtg1xAAAmmt8871gAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAOaPT8cAAAAaH4xfGnB6dUjT+mDU9LyAAAADMRcnYAflP+rGWwa4gAAAAAAAAAAAAAAAABz0vx/4t5O75997nb7Ot6c4AAZfyUtPoP0ZwfMqf56ftLNheQAAAAAAHtOmWwb84AAAAAAAAAAAAAAAAAAfEn23ntxnw5z9f6LHZ5wBy35t4df6fwa8/IeiZbBvzAAAAAAAAAB8uxeV9bgFwAAAAAAAAAAAAAAAAAeC+NZbfb5riAAAAAAAAAAAAAAAAAAHnXB8/V+TvseHV6t9U3mDgAOBH84/WavZHn+aupnt+ixpiAAAAAABca91OrjAAAAAAAAAAAAAAAAAAA+JM9vts0xAAAAAAAAAAAAAAAAAAD8p/wBWMtg1xAAAmmt8871gAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAOaPT8cAAAA89T/nL/AGH4+/pDfm+xJ9sAAAAAAAAPyn/VjLYNcQAAAAAAAAAAAAAAAAA56X4/+efJ38H9JrX61L5wAAOFH81/WSvcrPP8L/17z29eNMQAAAAAAPadMtg35wAAAAAAAAAAAAAAAAAA+JPtvPbiPwU5e39h/pvs88Kg/PLLo+lvc6z8y9My3lOjkAAAAAAAAAD5di8r63ALgAAAAAAAAAAAAAAAAA8F8ay2+3zXEAAAAAAAAAAAAAAAAAAPOuD5+r8Y/qXn6/oD3fTEHIAAeFcTO31PJWPin5NZ7fuVNpiAAAAAABca91OrjAAAAAAAAAAAAAAAAAAA+aI0+ly8wAAAAAAAAAAAAAAAAAA/Kf8AVjLYNcQAAJprfPO9YAAAAAAAAAAAAAAAAAAAAAIHOD0u3KGHYAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHNHp+OAAAARB/Pd99cff1Jrj912N+YAAAAAAAD8p/1Yy2DXEAAAAAAAAAAAAAAAAAOel+P/lzx+hL9fl/QReAAAAHh/Azr9Uy1l+bk2e36OmmIAAAAAAHtOmWwb84AAAAAAAAAAAAAAAAAAfEn23ntwP843J27n7u6Y+/HXxHzZGnkH3lN+e9VnpsHTyAAAAAAAAAB8uxeV9bgFwAAAAAAAAAAAAAAAAAeC+NZbfb5riAAAAAAAAAAAAAAAAAAHnXB8/V+FX3Jz9n1Z6LrzgIAAI/lhX23uoj8TZv9Jfo2oByAAAAAFxr3U6uMAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAD8p/1Yy2DXEAACaa3zzvWAAAAAAAAAAAAAAAAAAAAACNz8sfUf0Xw9g+c+6AAAAAAAAAAAAAAAAAAAAAAAAAAAAAADmj0/HAAAAMwP5wZvO9YBf0cafo+UAAAAAAAB+U/wCrGWwa4gAAAAAAAAAAAAAAAABz0vx/8gOP0PSvuQr1EvEAAABny2r633oXjf5HRp+5tjTEAAAAAAD2nTLYN+cAAAAAAAAAAAAAAAAAAOP7BPwn8D+H0/14/KC8v3p9k6uIwWvzM/TvHqx3ydWdPIAAAAAAAAAHy7F5X1uAXAAAAAAAAAAAAAAAAAB4L41lt9vmuIAAAAAAAAAAAAAAAAAAedeb8/V+BP6O8/b9paevMAgAAMYfyX9Hzp33wYT5N+piYXmAAAAAFxr3U6uMAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAD8p/1Yy2DXEAACaa3zzvWAAAAAAAAAAAAAAAAAAAAADj+nz/mP1H7j8l97Pz79qAAAAAADxHzwPrGiH53ehh9ufKAcl9uB4X6yG2AAAAAAAAAAAAAAAHNHp+OAAABzuwnxgmAd26pAAAAAAAA/Kf9WMtg1xAAAAAAAAAAAAAAAAADnpfj/wCI3H6P1F98s3isgAAADJH8lfRM6ehfioj9F/pSoByAAAAAHtOmWwb84AAAAAAAAAAAAAAAAAAAHyB+U/B6f9C/519fD+a374Y7+mnVx/APvHP2e182l3508gAAAAAAAAAfLsXlfW4BcAAAAAAAAAAAAAAAAAHgvjWW32+a4gAAAAAAAAAAAAAAAAAB514Dzdf4p/pdh1/YBrzAAAAAHnvztOv1pzrz/Ev90412jTEAAAAALjXup1cYAAAAAAAAAAAAAAAAAAHzRGn0uXmAAAAAAAAAAAAAAAAAAH5T/qxlsGuIAAE01vnnesAAAAAAAAAAAAAAAAAAAAAHzp7fyXnx9r+UfRHon55+3BxesAAAAAfn581h+y/5YB9j/H4fUfy4HoP1WHx/+jgAAAAAAAAAAAAAAAAc0en44AAAH4N/ZfJ3fkmc/UAf04avp+OAAAAAAAB+U/6sZbBriAAAAAAAAAAAAAAAAAHPS/H/AMDeL0f0V+3eP64PQ+MAAAAAOC+b51+tPiwnxf8AVRMLzAAAAAD2nTLYN+cAAAAAAAAAAAAAAAAAAOW6lP8APf4/4vR/cg7vN/MH4Y4vQ/ezuezg+e/A8Ov7483S9Od08gAAAAAAAAAfLsXlfW4BcAAAAAAAAAAAAAAAAAHgvjWW32+a4gAAAAAAAAAAAAAAAAAB518kc3X+U36t+X9p7yel8aAAAAAHgyrkfqQf5DfXMX9hmmIAAAAAXGvdTq4wAAAAAAAAAAAAAAAAAAPiTPb7bNMQAAAAAAAAAAAAAAAAAA/Kf9WMtg1xAAAmmt8871gAAAAAAAMSg0AAAAWw3xMAAAAA86+ePtvycPa+T0PqT5n7/YPlf0UAAAAAyvx9D9l/xDD9vPnIPk/GDvgOuyw++ZQAAAAAAAAAAAAAADmj0/HAAAA8135r5L9ejScc+7SaZgAAAAAAAflP+rGWwa4gAAAAAAAAAAAAAAAABz0v55/Cfg9f9Df0E+Y/oGrW+s/nQGgAAADwpVxPuhX4kfuhFbxpiAAAAAHtOmWwb84AAAAAAAAAAAAAAAAAAfEn23nt+V3L83X+vJ2+cfkrlt8vfvHGmn+Ybf6Zcul2ultzhUgAAAAAAAHy7F5X1uAXAAAAAAAAAAAAAAAAAB4L41lt9vmuIAAAAAAAAAAAAAAAAAAedfBvL2fn3+jPx/8AR/2FkfW/zrVKzAAAAIg+Vu9nXzP46l/sEaYgAAAABca91OrjAAAAAAAAAAAAAAAAAAA+JM9vts0xAAAAAAAAAAAAAAAAAAD8p/1Yy2DXEAACaa3zzvWAAAAAAAA4nCqQAAADVDviaAAAAAPkvH/TPwMNOfvNfxPrveT4r9WAAAAAMr8fQ/Zf8Rw/bj5oD52+kA4zgA8r+lw6H6eAAAAAAAAAAAAAAAOaPT8cAAAA5tH84P78cvb7UdfEAAAAAAAAAflP+rGWwa4gAAAAAAAAAAAAAAAABz0v5A/Hvy/pvuD6G+H/AK7+p6H3/wDGGOzTEAAAAjD5Z7Wdvz/9mh/oea4AAAAAB7TplsG/OAAAAAAAAAAAAAAAAAAHxJ9t57fjL23n/QfrIep8yRh+OPkPF6P7rfFPRzeuewRd7ptMQ0yAAAAAAAAPl2LyvrcAuAAAAAAAAAAAAAAAAADwXxrLb7fNcQAAAAAAAAAAAAAAAAAA86/NXl7flD6Y/Pv7J+3vUPs/5bq5HX89XKzAAAAw/k1ae+/jLF/rR9B3kDkAAAAuNe6nVxgAAAAAAAAAAAAAAAAAAfEme322aYgAAAAAAAAAAAAAAAAAB+U/6sZbBriAABNNb553rAAAAAAAAAAAAAAAAAAAAGXph8jn6Z+BAHtfL/O/c/RJ8j+lgAAAAGV+Pofsv+Z4et/Kwfor7SH5+fdQflf78Hu3uoAAAAAAAAAAAAAABzR6fjgAAAFAP54/MPN9baA/pN0/S8kAAAAAAAD8p/1Yy2DXEAAAAAAAAAAAAAAAAAOel/A35o+N9t+q3zr8n/S/2/2/2v8AKN/H04s0vEAAADG+TFp6P+ZUX9E/pTWdgqAAAAD2nTLYN+cAAAAAAAAAAAAAAAAAAPiT7Yz3/CG38v8As/7T9f8ATfkQbedV/FLDoxf1OnT5z+72Y3c1kG2AAAAAAAAB8uxeV9bgFwAAAAAAAAAAAAAAAAAeC+NZbfb5riAAAAAAAAAAAAAAAAAAHnX5Ncnb5X+gXyX9GeW/Z3X4Gpq+9+MUsiohKgAADzofz3rzr+Zv6Ww/rc1wAAAALjXup1cYAAAAAAAAAAAAAAAAAAHxJnt9tmmIAAAAAAAAAAAAAAAAAAflP+rGWwa4gAATTW+ed6wAAAAAAAAAAAAAAAAAAABUrP5Ab+nfgAC+o/Mfj/1H3c+f+zAAAAAIPBw9/iD4v9RD6EA8T9sDxnzIPrMAAAAAAAAAAAAAAAOaPT8cAAAArh8Ry4dAB9sT784AAAAAAAH5T/qxlsGuIAAAAAAAAAAAAAAAAAc9L/M34n8L9F/Zj59w9/I+q1dnb9n8oMQKxeYAAHnw/nX6HnT8teizv9VdfbnAAAAPadMtg35wAAAAAAAAAAAAAAAAAA+JPtTLf+eD2T5L97+1Pu71Plg978mM1H4b9Dx+j7L+jumeV6W4U6OUAAAAAAAA+XYvK+twC4AAAAAAAAAAAAAAAAAPBfGstvt81xAAAAAAAAAAAAAAAAAADzr8YeTu7L9K/J/QviP6I8H9j9Zf9p/LOpp57UcioiKgAAPAeZnbsPydg+4PvW4UqAAALjXup1cYAAAAAAAAAAAAAAAAAAEATgAAAAAAAAAAAAAAAAAAB+U/6sZbBriAABNNb553rAAAAAAAAAAAAAAAAAAAAAHzdwn6L+HB1+Z3nd/OfdeqnyX6UAAAAAAAAAAAAAAAAAAAAAAAAAAAAAc0en44AAAH5o/RWHT+fRlsAfs3e7OAAAAAAAAD8p/1Yy2DXEAAAAAAAAAAAAAAAAAOel/kn5R4n3/6x5nf8t8OfYHyH9K9sfZfzBpbEbUMdwwqAAA8U8WnbtPhiayf1sI7E0yAAA9p0y2DfnAAAAAAAAAAAAAAAAAADwv2iNf5oP07+Z/Y/O/0/wCL7btT7b+YQDC/CLn6ue/WfPf226Lr7fXwA0AAAAAAB8uxeV9bgFwAAAAAAAAAAAAAAAAAeC+NZbfb5riAAAAAAAAAAAAAAAAAAHnX4T8fd9ifbHD9f5b81/Nfuv2/P9n/AC8SONXSz1zsmojKgAh+Z1fIaq0/Or6Xzf6XWNucAAC417qdXGAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB+U/wCrGWwa4gAATTW+ed6wAAAAAAAAAAAAAAAAAAAABk/OH0vwfMH1f5v6b778R+tOPG+qAAAAAAAAAAAAAAAAAAAAAAAAAAAAAOaPT8cAAAD83voTHo+BzHcA/Yi72cAAAAAAAAH5T/qxlsGuIAAAAAAAAAAAAAAAAAc9L/FnuvJ+v/Qw7fnPCfLfiv6p+x1+1/lYeGxfz2x828wcgBlj8l8inX5J87z0/XL1PXmBoAPadMtg35wAAAAAAAAAAAAAAAAAAOHmv5t/6BfH+78iwPn/ANw+9z7r+UgA5JP8e/lfh9L9NvuXbDO6aL6Z/XwAwAAAAAPl2LyvrcAuAAAAAAAAAAAAAAAAADwXxrLb7fNcQAAAAAAAAAAAAAAAAAA86/nu4+/9VPoHm91PjjyP0v1L3bt+UD1PgCUetoZ7ZmVWbCoAOW8fWnjfxZnr6h+q9ZbJeYAXGvdTq4wAAAAAAAAAAAAAAAAAAPLZr1IqQDH8s877Dyryn5X+gMft+X3e77j3Pynuu4938q0js+dAAAAAAAAAPyn/AFYy2DXEAACaa3zzvWAAAAAAAAAAAAAAAAAAAAABBc7b7PN2Dj9MAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAOaPT8cAAAD4O9Ax6PZfl1z93GuIAAAAAAAAB+U/wCrGWwa48j8eY7hnsAAAAAAAAAAAAAABRT/ADY+8eb0vrQ6fMg+IfmP336T9S9n8uDu+TLKe2RtiVrxBo5jD1eKi8f9M81+SFXz/wDot7X5N0hWYAddUWi4AAAAAAAAAAAAAAAAAAA84T/OP+kXL0s78rPnP3b9Nu9+r/n8OjyQCD4sz1/OHy/j9D7m9Ba19mswuAAAAAA8Zl/UH1v08gXAAAAAAAAAAAAAAAAAB4L41lt9vmuJ4rN/PBz9QAAAAAAAAAAAAAAAHPfkbj0fu11L2OKy7vj77j+c/cNM+s/nMAJh693Pag1yAcJh+X+g5vl2fd8J5PZ8j98u6/nwAlDszXEAAAAAAAAAAAAAAAAAADx+b9g6rm+j5Lyn4z+pOJ9G8/631b1b6n8F3+G7PneG4Xwv1bhuP8P9S6Luerwu67j3fyntp/W/PIeB08z5O0PQ+F0NANDQDQ0GfGv7k1mHRzAAATTW+ed6wAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHNHp+OAAAB+P3svL2e9flOH79HVxgAAAAAAAAH5T/qxlsGuPlf5K8/UGWwAAAAAAAAAAAAAAFVPB/RkfpheR5PwfXfO3255H6ZKfTfgoAaSrYpxqkouY+Fvk/wCke9dwfYRMrPzLx76j+dw9D4sAOhanKgAAAAAAAAAAAAAAAAAAPO/RJv239UenjPjHy/0bjP0Bw9MPofx8AAD5Ii/gPH4/S8U0vF/T/Zj2/wAsBoAAAA41P9Ofrfr4QuAAAAAAAAAAAAAAAAADwXxrLb7fNcT4xz3/AD3PP9wAAAAAAAAAAAAAAADNtKv1dOrxwD5V1viv6p+lD7X+VgAALA7pNnmHF9T8hdf8H/YAdHjJ4J6n53xp9J+EAA8N4qAAAAAAAAAAAAAAAAAAAOIVb3hefX+on0J5n6D6h0H0H5BqaXTwStuKGbltm4vP28Zy/lff8xy/k/pnlHHeX+gcr8eez+TfWOv6XxUEDzgr6+fBBpweN/0a9vgB1cIAAE01vnnesAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABzR6fjgAAAfj95zx+h6D8kTX6GmuH6TWunkAAAAAAAA/Kf8AVjLYNcQAAAAAAAAAAAAAAAAA56X4+c3YACB8Z+ufFf1T7ifa/wArAAA/RVN5/Lv+Kfa/iv6q95Pu/wCRAAAAAA9p0y2DfnAAAAAAAAAAAAAAAAAAD4k+289g0xMPPq/Kv9LfjP6Y9DPt/wCXQAAAPHPhD4j+o+7+6u75XrT6n8GAAAAAD5di8r63ALgAAAAAAAAAAAAAAAAA8F8ay2+3zXEAAAAAAAAAAAAAAAAAAPOuD5+oIsAwPiP5D+lftbpfqf5+DbywAAAOA+Rfi/6l+tPRfqf5/Dr+aAAAALjXup1cYAAAAAAAAAAAAAAAAAAHxJnt9t/kDL+RP0c5u7S9wa9YoitVkWNK40tLfmv5mmPhHz3h18R0ee35gfqSl+jZ2+cAAAB+U/6sZbBriAABNNb553rAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAc0en44AAAH5jfFPH6AZ2Afv8AaPoeYAAAAAAAB+U/6sZbBriAAAAAAAAAAAAAAAAAHPS/Hzm7AAA4T5G+N/qD7k0Pr/5nC+cAAMz42+X/AH/1D6M9H4oPX/NQAAAAA9p0y2DfnAAAAAAAAAAAAAAAAAAD4k+289g0xAPjnzv4X+rf0KPuv5SAADH+Kvnv2HwP7e8H9a+il+//AJEAAAAAAA+XYvK+twC4AAAAAAAAAAAAAAAAAPBfGstvt81xAAAAAAAAAAAAAAAAAADzrg+fqCLAD5z8j9K5n6yw9UPe/IAAAOO5/a+OvqL5D+lfVD7f+UAAAAAAuNe6nVxgAAAAAAAAAAAAAAAAAAfEme323V0x/Djr+L0P0l+UNY1frIXp9DXnx8AMDn408z8Wy6fVfoTTHovwih/oD+mOuAa5AAAAflP+rGWwa4gAATTW+ed6wAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHNHp+OAAAAAAAAAAAAAAAAAflP8AqxlsGuIAAAAAAAAAAAAAAAAAc9L8fObsAAA+T/C/Xei+jtfOD2PzMAKc7fHfYfCf179Pr97/AB6AgAAAAA9p0y2DfnAAAAAAAAAAAAAAAAAAD4k+289g0xAMn8p/jv6R/SL0z6f8KDq8M8F8/wCt+GPYvgv62+3+g/Sf4nDTkAAAAAAA+XYvK+twC4AAAAAAAAAAAAAAAAAPBfGstvt81xAAAAAAAAAAAAAAAAAADzrg+fqCLAChPR8N/W3x/wDTPen2X8vAABzPxp8j/Sf0X7N7v5EHo/EgAAAABca91OrjAAAAAAAAAAAAAAAAAAA8zmvTCp5z8EOfq+y/1BvP5/8AlLPflbufRv79Z7/QVHb+868/R8DeX4LfcHH3/qOdvngAAAAH5T/qxlsGuIAAE01vnnesAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABzR6fjgAAAAAAAAAAAAAAAAH5T/qxlsGuIAAAAAAAAAAAAAAAAAc9L8fObsAAA5zH0/if7b+W/oPoD6/+aQCD5F+e/adD6uvlee9+QAAAAAAAe06ZbBvzgAAAAAAAAAAAAAAAAAB8f/YGewaYgB8h+P8Ao3lH6J8n0Jzfvfkvw75P+b/2v91+5/ffyMHf8oAAAAAAAAfLsXlfW4BcAAAAAAAAAAAAAAAAAHgvjWW32+a4gAAAAAAAAAAAAAAAAAB51wfP1BFgAB4j498V/VH2a77X+WAEYmXd8Ze3/E/1b7yfd/yIAAAAAABca91OrjAAAAAAAAAAAAAAAAAAAAADzVP8BP1Z5O77sr9nBX0EAwAAOL/Bfm6/r/8AV3TINcQAAAAD8p/1Yy2DXEAACaa3zzvWAAAAAAAAAAAAAAAAAAAAADlA+SvmgKjQtfYofWoAAAAAAAAAAAAAAAAAAAAAAAAAAAAHNHp+OAAAAAAAAAAAAAAAAAflP+rGWwa4gAAAAAAAAAAAAAAAABz0vx85uwAAAD5txPiv6p+rT7X+VhifyjnfBf2H9dz/AHv8fg8wAAAAAAPadMtg35wAAAAAAAAAAAAAAAAAAAAAAMyN/wAqf0V+K/p/gPibn9n6G+0foPx3VPoPyIAAAAAAAAAPl2LyvrcAuAAAAAAAAAAAAAAAAADwXxrLb7fNcQAAAAAAAAAAAAAAAAAA864Pn6giwAAi+JPmf3n6G9a9f82Dv+PzPjb5f9/9Q+jPR+KD1/zUAAAAAAC417qdXGAAAAAAAAAAAAAAAAAABAE4AB8/fhJyd37e/T/TyBUAAAcqj8Gvp3j9H9cV7fOAAAAAAD8p/wBWMtg1xAAAmmt8871gAAAAAAAAAAAAAAAAAAAAD8o9ANzyYPu36ZCh+OYfYv2yAAAABwQfnn6wH3j4eHwl9qh7r8Ch7n9NB8v/AFAAAAAAAAAAAAAAAAc0en44AAAAAAAAAAAAAAAAB+U/6sZbBriAAAAAAAAAAAAAAAAAHPS/Hzm7AAAAM34c+T/ov697j6P8ST5h837fjvsTl+guH0v4UAAAAAAAAe06ZbBvzgAAAAAAAAAAAAAAAAAAAAAAAfJnx3+c/wBn9V94+h8f64fbfzAAAAAAAAAAAB8uxeV9bgFwAAAAAAAAAAAAAAAAAeC+NZbfb5riAAAAAAAAAAAAAAAAAAHnXB8/UEWAAB5fxfV/NH3B4n6nPT+m/B/jvsPhP69+n1+9/j0BAAAAAAAXGvdTq4wAAAAAAAAAAAAAAAAAAPiTPb7bNMQA+N40/HT98sOn2E6uIAM9HzV+UXN2e9/sLpk42wAAAAAAD8p/1Yy2DXEAACaa3zzvWAAAAAAAAAAAAAAAAAAAA5z8fw7L9Fw9aAAD4+D8+v3FBQAAAPxbD9ffzeD67/MoP2x/EkPtb6HD4N+5g436oAAAAAAAAAAAAAAAOaPT8cAAAAAAAAAAAAAAAAA/Kf8AVjLYNcQAAAAAAAAAAAAAAAAA56X4+c3YAAAAeF+Z995T9k8H1vzr5d4v6p9k6f3P8lBfKAAAAAAAAe06ZbBvzgAAAAAAAAAAAAAAAAAB4X7pNhUAABR+UPF/S/rG57H5wFZgAAAAAAAAAAfLsXlfW4BcAAAAAAAAAAAAAAAAAHgvjWW32+a4gAAAAAAAAAAAAAAAAAB51wfP1BFgAAAfHPp3xX9U+4/Ivqfn+h9XXyvPe/IAAAAAAAAC417qdXGAAAAAAAAAAAAAAAAAAB8SZ7fbZpiAAH5w/nhx+h+/XZ9fB4/8mZbfJfy9y9vqX2pef6dz9vngAAAAAAAflP8AqxlsGuIAAE01vnnesAAAAAAAAUlp+NJ+Zf6BgAAAH3z6/wCY/VR9v/Ix4iHzx8yhf9lD9GwAAAAPzI+3Q9aAAAA/DgP2q+DA5XgA/Xn8iQ9j9lD5S7YNz0gPsEAAAAAAAAAAAAAA5o9PxwAAAAAAAAAAAAAAAAD8p/1Yy2DXEAAAAAAAAAAAAAAAAAOel+PnN2AAAAEKv4h6/wDPP7W4z7M9n8w3D6v+dQAAAAAAAAA9p0y2DfnAAAAAAAAAAAAAAAAAAD4k+289g0xAAAAAAAAAAAAAAAAAD5di8r63ALgAAAAAAAAAAAAAAAAA8F8ay2+3zXEAAAAAAAAAAAAAAAAAAPOuD5+oIsAAADi/jz43+nvQKnF9V9dz/ffx2DzAAAAAAAALjXup1cYAAAAAAAAAAAAAAAAAAHxJnt9tmmIAAH5D47/KuZy9vPfVzX1p9Y9XF6Ga4gAAAAAAAAH5T/qxlsGuIAAE01vnnesAAAAAAAAIGKZeiAAAAaDytmnGfkUHCdoH6Lc+HsfRgAAB8UB8e/ssDwAAAPGA+EfHg++fjUP2H/HgP1X+cg4PJD2r5KD9WgAAAAAAAAAAAAADmj0/HAAAAAAAAAAAAAAAAAPyn/VjLYNcQAAAAAAAAAAAAAAAAA56X4+c3YAAAAAeU/MfxP8AV32Z032H8yht5YAAAAAAAAAe06ZbBvzgAAAAAAAAAAAAAAAAAB8SfbeewaYgAAAAAAAAAAAAAAAAB8uxeV9bgFwAAAAAAAAAAAAAAAAAeC+NZbfb5riAAAAAAAAAAAAAAAAAAHnXB8/UEWAAAAfK/h/rPL/YnN7lw+l/CgAAAAAAAAAuNe6nVxgAAAAAAAAAAAAAAAAAAfEme322aYgAAVg/Pv6Hx3+gJtsAAAAAAAAAAAAPyn/VjLYNcQAAJprfPO9YAAAAAAAAAAAAAAAAAMMOO8oD4f8AMQ/YTWDuAADzn83w+kftUAAAADz4Pkb54D9gPxQD9Cvz4D9raAfAP0WHgnjgfrAAAAAAAAAAAAAAAc0en44AAAAAAAAAAAAAAAAB+U/6sZbBriAAAAAAAAAAAAAAAAAHPS/Hzm7AAAAABOcx9PpDbzAAAAAAAAAAAPadMtg35wAAAAAAAAAAAAAAAAAA+JPtvPYNMQAAAAAAAAAAAAAAAAA+XYvK+twC4AAAAAAAAAAAAAAAAAPBfGstvt81xAAAAAAAAAAAAAAAAAADzrg+fqCLAAAAMTLusadZhfKAAAAAAAAAFxr3U6uMAAAAAAAAAAAAAAAAAAD5ojT6XLzAAAAAAAAAAAAAAAAAAD8p/wBWMtg1xAAAmmt8871gAAAAAAAAAAAAAAAAAAAD4ND4X/QUPO/0GD4hQKPIh+lwAAAAB5EHKfQ4LzQfL/1EHR8QGp0YfNfqIehgAAAAAAAAAAAAAHNHp+OAAAAAAAAAAAAAc7+buW2z+kwpTXL8p/1Yy2DXEAAAAAAAAAAAAAAAAAOel+PnN2AAAAAAAAAAAAAAAAAAAe06ZbBvzgAAAAAAAAAAAAAAAAAB8SfbeewaYgAAAAAAAAAAAAAAAAB8uxeV9bgFwAAAAAAAAAAAAAAAAAeC+NZbfb5riAAAAAAAAAAAAAAAAAAHnXB8/UEWAAAAAAAAAAAAAAAAAFxr3U6uMAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAD8p/1Yy2DXEAACaa3zzvWAAAAAAAAAAAAAAAAAAAAAK3xWFr5sDrvq4M/wClQlAAAAAAAAAAAAAAAAAAAAAAAAAAAAA5o9PxwAAAAAAAAAAAAD8rPvnHo+cOhD7INuf8p/1Yy2DXEAAAAAAAAAAAAAAAAAOel+PnN2AAAAAAAAAAAAAAAAAAAe06ZbBvzgAAAAAAAAAAAAAAAAABy3Upg0AAAAAAAAAAAAAAAAAHy7F5X1uAXAAAAAAAAAAAAAAAAAB4L41lt9vmuIAAAAAAAAAAAAAAAAAAedcHz9QRYAAAAAAAAAAAAAAAAAXGvdTq4wAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPyn/VjLYNcQAAJprfPO9YAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADmj0/HAAAAAAAAAAAAAPxZ/abHo8U+MA/To25/wb+jOPvCp+4fd+jkCpAAAAAAAAAAAAAAA/OjLX6IvY9AAAAAAAAAAAAAAAAAAAct5tS+8zp4wAAAAAAAAAAAAAAAAAA/Nz9I89Q0yAAAAAAAAAAAAAAAA/Kc5e08zl/dv1v08gXAAAAAAAAAAAAAAAAAB4L+XHP1+0k0fpvpj0JtgAAAAAAAAAAAAAAAB+W32RydwCAAAAAAAAAAAAAAAAAD5cVfZ3q3Xwg0AAAAAAAAAAAAAAAAAH5uZa/pGa5AAAAAAAAAAAAAAAAePfBGHQEa/Lv79VmHRzAAATTW+ed6wAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHNHp+OAAAAAAAAAAAAAfjL+ymPR8+fOQfosbc/wCU/wCrGWwa4gAAAAAAAAAAAAAAAAB+XeW31lo47gAAAAAAAAAAAAAAAAABx3irX6OHVxgAAAAAAAAAAAAAAAAAB+U/6sZbBriAAAAAAAAAAAAAAAAAHy7F5X1uAXAAAAAAAAAAAAAAAAAB4L41lt9vmuIAAAAAAAAAAAAAAAAAAfkN98cfcDQAAAAAAAAAAAAAAAAAfHir7e9m6uIKkAAAAAAAAAAAAAAAAAPyny2/Vg1xAAAAAAAAAAAAAAAAAAD8p/1Yy2DXEAACaa3zzvWAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA5o9PxwAAAAAAAAAAAAD4u8W5+ry/9OmvUDfm/Kf8AVjLYNcQAAAAAAAAAAAAAAAAA/LvLb6y0cdwAAAAAAAAAAAAAAAAAA47xVr9HDq4wAAAAAAAAAAAAAAAAAA/Kf9WMtg1xAAAAAAAAAAAAAAAAAD5di8r63ALgAAAAAAAAAAAAAAAAA8F8ay2+3zXEAAAAAAAAAAAAAAAAAAPyG++OPuBoAAAAAAAAAAAAAAAAAPjxV9vezdXEFSAAAAAAAAAAAAAAAAAH5T5bfqwa4gAAAAAAAAAAAAAAAAAB+U/6sZbBriAABNNb553rAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAc0en44AAAAAAAAAAAAAePTXUdy0DX5Hfppj0BpiAAAAAAAAAAAAAAAAAH5s56/UV/HcAAAAAAAAAAAAAAAAAAOT8fZ+gJ08YAAAAAAAAAAAAAAAAAAfmP+nGewaYgAAAAAAAAAAAAAAAAB82Tez9buQuAAAAAAAAAAAAAAAAADwX5yx3+yjTIAAAAAAAAAAAAAAAAAAPyq+4OXtAQAAAAAAAAAAAAAAAAAfJ6r7J9c6uMHIAAAAAAAAAAAAAAAAAfmPlt+nBriAAAAAAAAAAAAAAAAAAH5j/AK6xqGuAAAE01vnnesAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABzR6fjgAAAAAAAAAAAAAAAAHwFgc/UCAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAD7Y9E6OYGgAAAAAAAAAAAAAAAADkvhDLYM9AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAC5+iGuQaZAAATTW+ed6wAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHNHp+OAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABNNb553rAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAc0en44AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAE01vnnesAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABzR6fjgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB4tNd117QNAAAAAAAAAAAAAAAAAEPCp+gDQAAAAAAAAAAAAAAAAAY6PF/a1eqVAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAFUPC/bZrRKkAAAAAAAAAAAAAAAAAOfRD0wAwAAAAAAAAAAAAAAAAA4bhov3B1wAAAAAAAAAAAAAAAAAAAAAAAATTW+ed6wAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHNHp+OAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfnfJz9XzL+vjXVG/MAAAAAAAAAAAAAAAAAfmf4jy9v7PHVxAAAAAAAAAAAAAAAAAB8F56aHxpj0ftedXGAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB+QeO/0/8AJUa/skdPGAAAAAAAAAAAAAAAAAcT+R+HT9Z/ed5BpmAAAAAAAAAAAAAAAAUg/Oefn68X9OLyDXEAAAAAAAAAAAAAAAAAAAAAACaa3zzvWAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA5o9PxwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAD8XP2jx6Pii0H2abc4AAAAAAAAAAAAAAAAAfjVjv+yptgAAAAAAAAAAAAAAAAAAB+LOPR+0xtzgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABzP5UY9H6+/kOL9TOs1xBoAAAAAAAAAAAAAAAA+XYvy77zTDTMAAAAAAAAAAAAAAAAAMZH5F/sbluG2AAAAAAAAAAAAAAAAAAAAAAAE01vnnesAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABzR6fjgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAH4uftHj0fIPLh9zG3OAAAAAAAAAAAAAAAAAH41Y7/ALKm2AAAAAAAAAAAAAAAAAAAfIfluHR+hxvzgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB51+cuPR+r/wCUDn9G/RNMgaAAAAAAAAAAAAAAAAPl2L8u+80w0zAAAAAAAAAAAAAAAAAD8qstvuD3W8wqQAAAAAAAAAAAAAAAAAAAAAAJprfPO9YAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADmj0/HAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPx1/WfHo/OP7UD2s25wAAAAAAAAAAAAAAAAA/GrHf8AZU2wAAAAAAAAAAAAAAAAAD5Ri/nf9NpsNcQAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAh/F7Ho/aX8XQ/Z+XbnAAAAAAAAAAAAAAAAAD5di/LvvNMNMwAAAAAAAAAAAAAAAAD8zvfsOn6zN+YAAAAAAAAAAAAAAAAAAAAAAAAmmt8871gAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAOaPT8cAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA81/PXHo9f+9azDTMAAAAAAAAAAAAAAAAAPg7PT7xNMwAAAAAAAAAAAAAAAAA/KXLZxlv+m/Q9PEDAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA+X4v5I+us9vp035gAAAAAAAAAAAAAAAAA8el8f9IjCpAAAAAAAAAAAAAAAAPOE/wA0tzl7PQP0Y35w0yAAAAAAAAAAAAAAAAAAAAAAAmmt8871gAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAOaPT8cAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAACaa3zzvWAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA5o9PxwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAJprfPO9YAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADmj0/HAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAmmt8871gAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAOaPT8cAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAACaa3zzvWAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA5o9PxwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAJprfPO9YAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADmj0/HAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAmmt8871gAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAOaPT8cAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAACaa3zzvWAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA5o9PxwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAJprfPO9YAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADmj0/HAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPK/yV5+oMtj9EtMvyR/oLavnRygAAAAAAZCPxa/a7HovG/OfA2emh90gGmYAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHxdGnwkcvYejNe9fL1R+z51cYAAAAAAAHzx8WYdP6tG/MAfjr+mWHT6wb8wAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABNNb553rAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAc0en44AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAByPx1lr+dHTcvd94/VHVxfHH6HteZ9dUeS+6h5B14+2KjyylF+wFwfCuenzX+v03znj15+xcg1+Gn9DmPQHRygAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAeGzXz9+O3J3foV9S6ZeseT6ZfXHJXlLgFeo+GC9r03MXhyrvu4aBr8Xvtnm6/sjzjo5eN9wT+APIcOn9Zjp5AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAACaa3zzvWAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA5o9PxwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA/EHvePv/YH5d6eP8HP6cMOn+a/0zDp8u9XDmMQf9Gf56dnn/DFPk7vvP9LevgPx7VfRf2NU/wA8H0byd32V999fB/Nd/RFl0dedPGAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfzF83X+1P2Xth+XfyTzdf6z/hi13vkOPR9C+UOfpT9uunj/CmfDq8J/b+8vow6+L+cX99+bs8H/GiK+hf1j0zyfx/Vf0AHTxgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAE01vnnesAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABzR6fjgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB+IPe8ff+wPy708f4Of04YdP82n7KTfg/ySH7x/zI1H7+/hOq+qcmb8r/oh1wPxGc/d3vtx/Ov91cnd+oHpnd5v87n7hc/X7CdPGAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfzF83X+1P2Xth+XfyTzdf62fgQL+m/8G9M/rb3cPxy/cuNPwO/RHPX5R+sqn9azt87+ZD+lrm7PiL8kc9P0o/Sbo5eA/B7Pb+jI6eMAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAmmt8871gAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAOaPT8cAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPxB73j7/2B+Xenj/Bz+nDDp/m0/alV4N8ko/eb+Xqp/ff8OFX3b9Y3nq/UW2B+OOen079zVHj35IZb7f7gVH82n9C86dodPGAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfzF83X+1P2Xth+XfyTzdf62fgQL+oX8Gdcvrb3kPxW/d3PX8Af2Vmu19J6eT240y/m7/oJ5uzqPmLbm/Gv9Oubt+kfxzvP+gU6eMAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAmmt8871gAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAOaPT8cAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPxd7nk7/wBafmfo4/w3/pNx6P52f2Om/DflQP3I/mfqP3b8ZvP8kLXJ3/oT+o/Z55+fzXiP3nGn4bwcvb+pH2B2ef8Agt/RZGgdHKAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfzZc3X+x/19tz/AJn/ACpz9f6rfhIH9MH4b6Y/VfuQfjj/AEbzX4/fDWPTsfthpl9QHXw/iR9/83X4p+dOeqfuFcfP3gLn9dzp5AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAACaa3zzvWAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA5o9PxwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAGVguAHNo/Dj93cemwb8035656wfoiAaZAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABWB8wAHy18W8/V+u9To5bYH4p/p7z9Xt50coAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABNNb553rAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAc0en44AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAE01vnnesAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABzR6fjgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAATTW+ed6wAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHNHp+OAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABNNb553rAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAc0en44AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAE01vnnesAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABzR6fjgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAATTW+ed6wAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHNHp+OAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABNNb553rAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAc0en44AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAE01vnnesAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABzR6fjgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAATTW+ed6wAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHNHp+OAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABNNb553rAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAc0en44AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAE01vnnesAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABzR6fjgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAATTW+ed6wAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHNHp+OAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABNNb553rAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAc0en44AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAE01vnnesAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABzR6fjgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAATTW+ed6wAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHNHp+OAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABNNb553rAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAc0en44AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAE01vnnesAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABzR6fjgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB5Yn6mNAAAAJ5cn6JjB4v9EpzFSAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAE01vnnesAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABzR6fjgAAAcv8AnRlv812ebq/UX6F7OAKkAPhrPT4Yvc3b6T+uu/KG2AAAAAB476dNahUgAAAAHnWpNdiVIAAAAAfz4c/V/QedHKAAAB+Pl7k7vvv8fan7e/RjXCY0yAAAA4Qmu7KkAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAACaa3zzvWAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA5o9PxwAAMdH8//AIl5/re5dTUfrt7p2+cFSAfDnyNzdn7GaHRxtcw4BPrNMTKzKeQn19Zr8MP2I5+r0qbo5eQup9FntXJApOR+Sv2Hh1fVMW/LakZRtBJxqc/ViP58MOr+g86OUA5qBPrKbX8/X7+c/V84fEYfrle6OXnaSfXjRy6KfYD/AC191w6fs630coAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAE01vnnesAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABzR6fjgAAfBmen42/a/H6H7WO9DywAAAPwD/cLn6u6OjlMFH4t+x8nf4H+mdx7J+HjXvHjmW36F+lb8v4vfUnP1/pb8X9HLyPnGO/3f59rh5/+irX4i/rdGn4y+pY7/o1+f8Ath+ofs2/N+K36h49HAfIovGvuOb+4f58Kn+g86OU+A89PnWlz9f0X0euP5G/a2HT575aj9GNPo5vlx+W3uH6NdHL+RvIYdXS/femP499lz9P6JfYPZwBcAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABNNb553rAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAc0en44AAH5W5bfmZ+uXN1/omd3nAAAAfzm/wBGPP1KdHKfnnnrD+iQcN+JOev7ZfiAP9/PNN+X8df3Vx3/AAt/ZVVZ/EUP1gxry/ML94p0/Eanj0fqF9C9fB+SX2Xj0fUX4rVP6h+zaY/it+oePR7N8/7c/wAwfLfN2fuH/Phef9B50csP8/fP1fr43TL8gf6Ds9f58P6Dw+XfjQP1t/n1F+va6Z/jz+xOPR+eP7M6Yhth+XfvPP1fZZ0coAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAE01vnnesAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABzR6fjgAAfAuen46fT/F6P7oa3oeVHIwAAPwg/Wzm6/czp5D8tctvoD7KqIv598t/3Q/GoP3S5bfm/EL95sej8Lf2VT478fU/v0qew+w9MfzO+Dubt/frqOvz/AMkvsvDq+ovxXqf0/wDadMfxU/UXHo+D+jVe/fnEj90v58Kn+g86OXC/n65uv9Lyp0vuG8v58P6D89vl340D9Yf53w/TkatbAvC/1FqQ1x/Lv3nn6vss6OUAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAACaa3zzvWAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA5o9PxwAAMNH8/8A4t5/rdBtI/aX6m7/ADAuAD5h/Jrl7P1m9Z35+MxE/wAx/wBmI0+GvOpv9H/xqc/uly2/N+IX7zY9H4k/ofNfRv4QtfsH6fthmcMj8uf0Dx6fhz9mLy/K706NPuv8x9Muh+uFX4j/ALk56fjt+yrXzj8Kqv3S/nwqP6Dzo5T8J8On9Ovf9Meb9Dqf58P6D8On5d+NA/W38NXP6P8A0ZpnzNxz+E/7OYdXS+gb8v5zmHT+hm50cgMAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAACaa3zzvWAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA5o9PxwAAAOc+BctvDt/Ho/Qv2Tp4wqQA8YmvhnlObr+gPvjfm+VPjnPb2n7+0yyvjUPunKvL4h+8408I+AI1/S+vrj+feNz9X3F5hrh9H+laZfAH2hnpj/myr+7fXNMvzS6PDp9t+h9efgPgaNfrHjWvvP8APeo/Qg0yONT/ADx4zm7PsH6y35fz3/QhV5d4yH1vyl5fnfw3N2/W/wBe78vhvwbnte/S5zR/M4PtD6X0zCoAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAACaa3zzvWAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA5o9PxwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAJprfPO9YAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADmj0/HAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAmmt8871gAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAOaPT8cAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAACaa3zzvWAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA5o9PxwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAJprfPO9YAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADmj0/HAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAmmt8871gAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAOaPT8cAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAACaa3zzvWAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA5o9PxwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAJprfPO9YAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADmj0/HAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA8JmvbrFT51cmr3nY/byoAD56muP+tUwuAAAAAAAAAAAAAAAAD4Rz0+ovTLgGgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAACPwma96KkAAAAAAAAAAAAAAAAAAAAAAADzP5dy2+7jXEAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAJprfPO9YAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADmj0/HAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA/mL5uv8ApL6no5fyS73m6/o/5TuP1NN+YAPzjz18a/YGaDbAAAAAAAAAAAAAAAAA/AXDp/Vj6i0xC4AAAAAAAA+A/XMej6gNucAAAA8PmvkP9LIsNcgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAyv5j+Xs/qFOrjAAAAAAAAAAAAAAAAAAAAAAAA+Xfyn5ez9+jq4wAAAAAAADlE/y6/XDLYNsAAAAD8vctvuj1q8gpAAAAAAAAAAAAAAAAAAAAABNNb553rAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAc0en44AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAH8xfN1/wBJfU9HL+SXe83X9V/PemX6EfnHU/OX0Lh0e7+I6ZfLPZ5dHuP6PdHIGmX5U9jzdn03xumPwn9D5b/cf526Y/qh+dVT9UVWvm71rPT4G7XLf9TPD+nj8X8Nx6P1N/Li88f5g5+r9f8A6i6+E8Za/NzG5e39kvhfo5fkb37Dp/Sn436OXjvlzDp+r/pbXD8MvUuXt/WPJ6+H87vVcOn9Mfyz1w/Sj5uqY/iPHf54/Tmb+/Ts4AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAyv5j+Xs/qFOrjPk2L+CvQ+br/Tv8+t+f7O5qp8H+/Ufkz+ps3s/It58D5Jj0fpp+YN58D+lc385e21Hd/E4fpb+Y7n7p/OtX5l9vZ6faX5U64+t+kKpPgWL6nwXPX9+jt8+P8789fl/7gx3+hvzNvPE/R1V7b+YV56HlGO/6q/BO2P5nfpFzdX6c/mh08nin6F56Y/mtz9x/leL6v/GPHo+pf111x7035gAAAAAAAAAAAAAAAAAAAAAmmt8871gAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAOaPT8cAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAD+Yvm6+lk5+vnP0+vL1H4g0y+9vzFT/av8Yan7ZGvh39cZr8L/AOgIfrZ1cX8yf6R8PpfYX47aY/uv+M9z+j/4z56f0Lfz9VP3eqOw+09ceU/KCNfqXuBfln+wcafKGEH6Nfg21+2H1Fvy0f5y8ej9Z/ddMvKfyqx3/cj8ebj624BPwX9OQ/Cf+itV+OX1In93fgxUfu7+YNR9GZir5L8jx6P3q+Iunj+Lv2Uz09QOnlAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADK/mP5ez+oU6uPy/+f3l7P3t/P8AvPkuxz29H8nc/LH7hNfiX/QW0fmfeXwj+v2XR8XevtfQ35DI+9fJGelfmhnp/Q//AD3aZ/0c+Lb8vl/4p8vb/S9/Oxef2X97VP4Nfubnr8jfJif79HZwfnl8U8nd+wHT9HJ+Hf6Ec/X6f+LIf0W/zOOf3B+K2u1+yqn8Uv3fjT5G4Fr9CfwYD+hf+f0X0J7JN/ob+BlR+pP2TrhpmmYAAAAAAAAAAAAAAAAAAAAATTW+ed6wAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHNHp+OAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB/MXzdf8ASX1PRy/kl3vN1+o/EAfQvIB+tH496Ze0E341+wNR/Ph+wOev0cdXH/MX/STy9nzT+PSr6IJr9DPh7TL0TKVfO9jPX9l/za35fPcvHp9w9l1x+Rv2sqPwj/RnPT7D/AO4/Vr6i0x8m/n/AMOr+lk6uL8xeB5e39ePzX15/Huxx6dT9RNuf+bT9+stvyn+yme3/ghL+oSb+1Ps7p5P5s/u/k7f15+NOzh+DP2+z0DfnAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADK/mP5ez+oU6uP4//ADN5e398vlDfl/Kv9Y8On88+KjX0T0Go3v0+25z8zxcB+t6r8D7PP19Tvuf1P/FPTLtPY40s8Y1+nH4e1H25+ZeW39JH8+OuP7n4euH40f0N57fLf5UzX79HVx/jF9Ecvb+ip1cX8wX9JPL29p/Mm5/oS/nUm/6hfz335vnL9RI0/Bn+jdP8YPHsd/Qhr9pfzY6OX8wP6QMt+/8A58Kj9lfetucKkAAAAAAAAAAAAAAAAAAAAAJprfPO9YAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADmj0/HAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA/mL5uv+kvqejl/JLvebr9R+IA/Sz8Uk/0x/N1V+ihpl41+wNR/Ph+wOev0cdXH/MX/AEl8vZi/gQz9aL95fV3yFcfjr+6nP1fl3jxp+8n86u3P+n3xDN+5ey1PyT+1NR+WXz7jv9zfkMP9wPqLp4qn86mPR95e6Ndv+Ms3+xH5Sh+lvz/Uan6iXn/Np+/WW35jOV/pv+DVR+o/cXn9A/l+HYfBmHT+4XFdPH+Rf7PY7/QR1cYAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAZX8x/L2fr6VHvf4dxr+xPwzU+ufqPpj/Oz9bc3X9LfkRU/vZ7h1cR+Z8XwX62qvyl89y3+9d3bm+iv5/o09M/SEr8N/2bz0zvzTc/p5+ISr+lb+fG8/3R9G35v5y/1T5e75m8fT/fo7fO+Avzi4+/8AU/1bfm/Jz6Jw6e//AD3D+gD+ZKo/qF/PfbD5y/WGL/nL/bbLfyj41c/pRb2w9N/CnPb7m8Umv2u/A/TL6u/Sao3DXEAAAAAAAAAAAAAAAAAAAAAJprfPO9YAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADmj0/HAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA/OzPX9AdK8vl/bz16Px4PsP51uPJvjjn6vrT3rTLkvqm8/hj6pmu+Ncvzs/QHLXS8GuPi/dw6f0MXo5fgr9EI0+caCf1D873n8jfQGO/Q9rpn5h9dOaP59qp/SIv3b03bA84D4Qdz9X6S/Ne3P8q/R+W/1980bYWfo4X5/fbsXyHwOq+9W3n8MTY9H3J8gaZ/oJ8/Vnzf1YH5z+r57fZRvzAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABW/OzLUM9fsPL2x+JvU8tvuux0cvxf7Vlt33w4H6FmuJ89TWJ9QJ0/gxV5Z9Q56fWPylrj0f0Gz87f0GirH54s37MafavxHef2X1t5+F/GGW/wBSedi+4zbBPi2NPAfrbPX3X4CqKn3uHXfnYL9HPDLjgfrFP4W57Pb9CPiG8vDPes9u3s3n9G/nwL7s8lqfjL9KY06U2wAAAAAAAAAAAAAAAAAAAAACaa3zzvWAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA5o9PxwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAMD8RsOntfn7Dp/e/te3zgaAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAACaa3zzvWAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA5o9PxwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADhE+P9fTuFSAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABNNb553rAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAc0en44AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAE01vnnesAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABzR6fjgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAATTW+ed6wAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHNHp+OAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABNNb553rAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAc0en44AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAE01vnnesAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABzR6fjgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAATTW+ed6wAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHNHp+OAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABNNb553rAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAc0en44AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAeYfL+PTp+1j5/y1Pr/rS8Q0yAAAAAAAAAAAAAAAAAD5hjTzvKx6vpb2HfkCoAPGPZ5sKgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAD5pjTy+xj0/V3yzplD7NNe7G/KAAAAAAAAAAAAAAAAAEIfGFfm7MX7OvPqTbnAMTjZv00qAAAAAAAAAAAAAAAAAAAAAAAAAAAJprfPO9YAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADmj0/HAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAACEPzw/Q7HonNuf86P0Px6LBtzgAAAAAAAAAAAAAAAAABwHzDj0fbsO3P+cv6C49Pmfs+mPgHVTfwV9o5b+v/MWuGx9NufmLDz2+pON1w47zbPb3r59F9Q9Trh8z5GPT9TXNuUGAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB5l88Y9H2obc+P8I49H6DG3OAAAAAAAAAAAAAAAAAAB8uRr1PvRPJ/F0bfV29ePS+MD8+8Sx6vt5duX569hjX0X5hcdJ9M1Pi/ITa9KPzj6zcZPzKPsPodyF5gAAAAAAAAAAAAAATTW+ed6wAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHNHp+OAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHCp/Mv2pGoaY+QfKWHX9C/Q+mAXmAAAAAAAAAAAAAAAAAfNe1l0e9GvP8I/TGPV89fbdZ/nj+hCr8/wD7+D4G+0k+k+Vqz9C9BZ8b+vZb5/a1n4B9ih8geqze5t6Z+VfXBIaZAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB8pemZdHsJrznypnr5R9exp6Kb8wAAAAAAAAAAAAAAAAAfAmW/3Rp6YH54Z9HtHoAem/EgfUPjAfUXwUn+gLteb4F+5MunwpweF/ZQfKvsirC6Nx4h9dh8xfRQeim3OAAAAAAAAAAAAAABNNb553rAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAc0en44AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAZKPgr9C8+gNecQPzp/QnHovG3OAAAAAAAAAAAAAAAAAeNeWY9H1wbc/wCdn25j1fLH29Wf53/ogq/P79ARfnd+iIz4JF9ubWmH54+54dvadNrz/P8A9go/P7pMune2Kz9K9d15wqQAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA8B5vHo+ojbnAMv4Bx6P0SNucAAAAAAAAAAAAAAAAAPjPPb3v1Gs8/4Hz29j7tnp3xMH1R4wL3H5zH9nmvP+d/6IZdHn3zkHGfoCL5L9nDF4sPnf3zPY951w640xAAAAAAAAAAAAAAAmmt8871gAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAOaPT8cAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPlTzvn7PQfRKz8usq/Lv0CrNTXAAAAAAAAAAAAAAAAAAb8HZb9hz0a+nfR2vP+bvuOPV4R+mF5fnN9aqvIqCfedneXzV0ee3dNvPtOmvP5/wDsFH5/fVsa/LPrKr2Pvt+QGgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAi+Dct+p1o09t+e6iTQm/rs6OQAAAAAAAAAAAAAAAAAOeT+D/AGDn7PLvraoj+PA9K5ib+xfg2s/uX4rZ696OLyZs3wP1/U/H/wCgLn5L9nDF4sMrGnTb+qLxmNMgAAAAAAAAAAAAAAJprfPO9YAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADmj0/HAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAz0YXUjr8+HVOaAAAAAAAAAAAAAAAAAAAA5rTT0hqhVTs6As6uPZwAt6gsuAe3VE94V7gZ+gGdij6pWgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADAYq6LFJi3xg0AAAAAAAAAAAAAAAAAADOVT6Ww0YSelELQwB6tlrluiTsc4GvcFn6AU5wawLWAN26ANAAAAAAAAAAAAAAATTW+ed6wAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHNHp+OAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABNNb553rAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAc0en44AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAE01vnnesAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABzR6fjgAAAAAAAAAAAAAAAAH4u+lcncDQAAAAAAAAAAAAAAAAAeMzXcSqgAAAAAAAAAAAAAAAAAAoc8HrJeYAAAAAAAAAAAAAAAAAAeJ+2TQVIAAAAAAAAAAAAAAAAAcYn9+fR/TxhUgAAAAAAAAAAAAAAAAHkH5gc/T6MToAAAAAAAAAAAAAAAAAAHhPpuegMAAAAAAAAAAAAAAAAADgkejdBeYMAAAAAAAAAAAAAAAAADxOa9sKkAAAAAAAAAAAAAAAAAAPHv25vIN+cAACaa3zzvWAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA5o9PxwAAAAAAAAAAAAAAAAD8p/1Yy2DXEAAAAAAAAAAAAAAAAAPy7y2+stHHcAAAAAAAAAAAAAAAAAAOO8Va/Rw6uMAAAAAAAAAAAAAAAAAAPyn/VjLYNcQAAAAAAAAAAAAAAAAA+XYvK+twC4AAAAAAAAAAAAAAAAAPBfGstvt81xAAAAAAAAAAAAAAAAAAD8hvvjj7gaAAAAAAAAAAAAAAAAAD48Vfb3s3VxBUgAAAAAAAAAAAAAAAAB+U+W36sGuIAAAAAAAAAAAAAAAAAAflP8AqxlsGuIAAE01vnnesAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABzR6fjgAAAAAAAAAAfCuemb9+ByH5tTeh+nbVk1x/Kf9WMtg1xAAAAAAAAAAAAAAAAAD8u8tvrLRx3AAAAAAAAAAAAAAAAAADjvFWv0cOrjAAAAAAAAAAAAAAAAAAD8p/wBWMtg1xAAAAAAAAAAAAAAAAAD5di8r63ALgAAAAAAAAAAAAAAAAA8F8ay2+3zXEAAAAAAAAAAAAAAAAAAPyG++OPuBoAAAAAAAAAAAAAAAAAPjxV9vezdXEFSAAAAAAAAAAAAAAAAAH5T5bfqwa4gAAAAAAAAAAAAAAAAAB+U/6sZbBriAABNNb553rAAAAAAAAAAAAAAAAAAAAAABxXlAP9uDbAAAAAPgQPW/qAAAAAPn35oD9GQDzwPBPpYPzm/Q8Pnj3IPRwAAAAAAAAADmj0/HAAAAAAAAAAA8Hmvz2/YHPX8nv0YH84PD7VNuf8df0Q5+oNMfTOuuAaAAAAAAAAAAAAAAA/LvLb6y0cdwAAAAAAAAAAAAAAAAAA47xVr9HDq4wAAAAAAAAAAAAAAAAAA/Kf8AVjLYNcQAAAAAAAAAAAAAAAPBTPQ+fJv0b63vMLgAAAAAAAAAAAAAAAAA8F+Ssej61KzPbqm4VIAAAAAAAAAAAAAAAH5DffHH3A0AAAAAAAAAAAAAAAAAHx4q+3vZuriCpAAAAAAAAAAAAAAAAAD8p8tv1YNcQAAAAAAAAAAAAAAAOc8oiwT/ADo/ZGbDbAAACaa3zzvWAAAAAAAAAAAAAAAAAAAAAAD8fvgAPXfJg/oK9yAAAAA/noD7A/VcAAAAPzp/L0P6WAD40D87v2GD+av+j8Pyc+9Q9M/O0P2fAAAAAAAAA5o9PxwAAAAAAAAAAMoPyT/YHHf8Wf2mH4r8WB+nptz/AImfanL2Bef0X6ptgDQAAAAAAAAAAAAAAHy/F7E/P1AAAAAAAAAAAAAAAAAABk3bj6KOjlAAAAAAAAAAAAAAAAAAD5o+l5sKgAAAAAAAAAAAAAAAD4gMOg8VVfSX1vrgFwAAAAAAAAAAAAAAAAAeC/BGHT9JAj7KvPVNMwAAAAAAAAAAAAAAAPjT0rl7ATAAAAAAAAAAAAAAAAADzAf1H1vVxA0AAAAAAAAAAAAAAAAAHzRF/S5cAAAAAAAAAAAAAAAAcB8v56hF/Dv7ggG/OAABNNb553rAAAAAAAAAAAAAAAAAAAAAAB+P3igfvZ/OUH3iwOu80D9PvyqDxv7bD9BP56A7+wH0V+oofBPw6Ha/rKHxR+VYfcXJB+tfjYfG36Qh/NX/AEfh8A/Vwfmt8rB+n1IPavsoPx7/AFJD2MAAAADmj0/HAAAAAAAAAAAyg/JP9gcd/wAaf2ND548BD9DjbD8p/wBWMtg1xAAAAAAAAAAAAAAAAADnpfj5zdgAAAAAAAAAAAAAAAAAAHtOmWwb84AAAAAAAAAAAAAAAAAAfO30TGgXmAAAAAAAAAAAAAAAAAHy7F5X1uAXAAAAAAAAAAAAAAAAAB4L41lt9vmuIAAAAAAAAAAAAAAAAAAedcHz9QRYAAAAAAAAAAAAAAAAAXGvdTq4wAAAAAAAAAAAAAAAAAAPnaNPokvMAAAAAAAAAAAAAAAAAAPyn/VjLYNcQAAJprfPO9YAAAAAAAAAAAAAAAAAAAAAAPx++Kw+t/gMP6APyXDy/wDVsPlXxMP1K/CAP33/ACTDS/RIPxN/fAOt8YD8qvsoPZPx8D9uPy9D236gD8vf6Ag/mr/o/D8KP0WDzj4AD9y/lgPiH9YQ/FH+moL4AAAAHNHp+OAAAAAAAAAABlB+Sf7A47/Ivy/N8L+qLXoZvzflP+rGWwa4gAAAAAAAAAAAAAAAABz0vx85uwAAAAAAAAAAAAAAAAAAD2nTLYN+cAAAAAAAAAAAAAAAAAAPiT7bz2DTEAAAAAAAAAAAAAAAAAPl2LyvrcAuAAAAAAAAAAAAAAAAADwXxrLb7fNcQAAAAAAAAAAAAAAAAAA864Pn6giwAAAAAAAAAAAAAAAAAuNe6nVxgAAAAAAAAAAAAAAAAAAfEme322aYgAAAAAAAAAAAAAAAAAB+U/6sZbBriAABNNb553rAAAAAAAAAAAAAAAAAAAAAAB+P3yCH6S/WAe1fz+h9J/reH87318H6x/zNB+ovwQH19+rIfzMfq4HwhIHJe7h9Lfl2H9LX5Mh8s/ewfl7/AEBB/NX/AEfh+FH6LB1f5UB/Sx5wH80v3AHWfsqAAAAABzR6fjgAAAAAAAAAANDnOlQcCPd6EQM/Kf8AVjLYNcQAAAAAAAAAAAAAAAAA56X4+c3YAAAAAAAAAAAAAAAAAAB7TplsG/OAAAAAAAAAAAAAAAAAAHxJ9t57BpiAAAAAAAAAAAAAAAAAHy7F5X1uAXAAAAAAAAAAAAAAAAAB4L41lt9vmuIAAAAAAAAAAAAAAAAAAedcHz9QRYAAAAAAAAAAAAAAAAAXGvdTq4wAAAAAAAAAAAAAAAAAAPiTPb7bNMQAAAAAAAAAAAAAAAAAA/Kf9WMtg1xAAAmmt8871gAAAAAAAAAAAAAAAAAAAAAA/H7xQP3sAP5/Q+lv1sD8lPiAP0B/K8P6KvxXDk/sAPzk/owD+dn9Fw+Qfo8Ppb8dA/S74SD9CPQg/L3+gIP5q/6Pw/Cj9Fg9Z/DgP2M/Q4Pwo+Lg/oM+nAAAAAAOaPT8cAAAAAAAAAAAAAAAAA/Kf9WMtg1xAAAAAAAAAAAAAAAAADnpfj5zdgAAAAAAAAAAAAAAAAAAHtOmWwb84AAAAAAAAAAAAAAAAAAfEn23nsGmIAAAAAAAAAAAAAAAAAfLsXlfW4BcAAAAAAAAAAAAAAAAAHgvjWW32+a4gAAAAAAAAAAAAAAAAAB51wfP1BFgAAAAAAAAAAAAAAAABca91OrjAAAAAAAAAAAAAAAAAAA+JM9vts0xAAAAAAAAAAAAAAAAAAD8p/1Yy2DXEAACaa3zzvWAAAAAAAAAAAAAAAAAAAAAAD5X5APtcA+Hg9C+nwrfA4eTfbAfQXwgHZfMYfXf08HyV8wB75ph6N5QHk+wH6GeHh85/cIfnx9+h8Y+9B7x+cAdb9+h+Q3ykH9EqgAAAAAc0en44AAAAAAAAAAAAAAAAB+U/wCrGWwa4gAAAAAAAAAAAAAAAABz0vx85uwAAAAAAAAAAAAAAAAAAD2nTLYN+cAAAAAAAAAAAAAAAAAAPiT7bz2DTEAAAAAAAAAAAAAAAAAPl2LyvrcAuAAAAAAAAAAAAAAAAADwXxrLb7fNcQAAAAAAAAAAAAAAAAAA864Pn6giwAAAAAAAAAAAAAAAAAuNe6nVxgAAAAAAAAAAAAAAAAAAfEme322aYgAAAAAAAAAAAAAAAAAB+U/6sZbBriAABNNb553rAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfnwH5AfvsH0gAAAAAAc0en44AAAAAAAAAAAAAAAAB+U/6sZbBriAAAAAAAAAAAAAAAAAHPS/Hzm7AAAAAAAAAAAAAAAAAAAPadMtg35wAAAAAAAAAAAAAAAAAAytVAMAAAAAAAAAAAAAAAAAD5di8r63ALgAAAAAAAAAAAAAAAAA8F8ay2+3zXEAAAAAAAAAAAAAAAAAAPOuD5+oIsAAAAAAAAAAAAAAAAALjXup1cYAAAAAAAAAAAAAAAAAAGUjVGAAAAAAAAAAAAAAAAAAB+U/6sZbBriAABNNb553rAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAcOC9uAAAAAAAc0en44AAAAAAAAAAAAAAAAB+U/wCrGWwa4gAAAAAAAAAAAAAAAABz0vx85uwAAAAAAAAAAAAAAAAAAD2nTLYN+cAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAD5di8r63ALgAAAAAAAAAAAAAAAAA8F8ay2+3zXEAAAAAAAAAAAAAAAAAAPOuD5+oIsAAAAAAAAAAAAAAAAALjXup1cYAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAH5T/qxlsGuIAAE01vnnesAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABzR6fjgAAAAAAAAAAAAAAAAH5T/qxlsGuIAAAAAAAAAAAAAAAAAc9L8fObsAAAAAAAAAAAAAAAAAAA9p0y2DfnAAAAAAAAAAAAAAAAAADge+VA5AAAAAAAAAAAAAAAAAD5di8r63ALgAAAAAAAAAAAAAAAAA8F8ay2+3zXEAAAAAAAAAAAAAAAAAAPOuD5+oIsAAAAAAAAAAAAAAAAALjXup1cYAAAAAAAAAAAAAAAAAAHAqu+HIAAAAAAAAAAAAAAAAAAflP+rGWwa4gAATTW+ed6wAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHNHp+OAAAAAAAAAAAAAAAAAflP8AqxlsGuIAAAAAAAAAAAAAAAAAc9L8fObsAAAAAAAAAAAAAAAAAAA9p0y2DfnAAAAAAAAAAAAAAAAAAD4k+289g0xAAAAAAAAAAAAAAAAAD5di8r63ALgAAAAAAAAAAAAAAAAA8F8ay2+3zXEAAAAAAAAAAAAAAAAAAPOuD5+oIsAAAAAAAAAAAAAAAAALjXup1cYAAAAAAAAAAAAAAAAAAHxJnt9tmmIAAAAAAAAAAAAAAAAAAflP+rGWwa4gAATTW+ed6wAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHNHp+OAAAAAcp8g56/RfT1PblQAAAAAAAB+U/6sZbBriAAAAAAAAAAAAAAAAAHPS/Hzm7AAAAAAAAAAAAAAAAAAAPadMtg35wAAAAAAAAAAAAAAAAAA+JPtvPYNMQAAAAAAAAAAAAAAAAA+XYvK+twC4AAAAAAAAAAAAAAAAAPBfGstvt81xAAAAAAAAAAAAAAAAAADzrg+fqCLAAAAAAAAAAAAAAAAAC417qdXGAAAAAAAAAAAAAAAAAAB8SZ7fbZpiAAAAAAAAAAAAAAAAAAH5T/qxlsGuIAAE01vnnesAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABzR6fjgAAAFYP55OX871f1Z/MLXH+gzpOvgBgAAAAAAflP+rGWwa4gAAAAAAAAAAAAAAAABz0vx85uwAAAAAAAAAAAAAAAAAAD2nTLYN+cAAAAAAAAAAAAAAAAAAPiT7bz2DTEAAAAAAAAAAAAAAAAAPl2LyvrcAuAAAAAAAAAAAAAAAAADwXxrLb7fNcQAAAAAAAAAAAAAAAAAA864Pn6giwAAAAAAAAAAAAAAAAAuNe6nVxgAAAAAAAAAAAAAAAAAAfEme322aYgAAAAAAAAAAAAAAAAAB+U/6sZbBriAABNNb553rAAAAAAAAAAAAAAAAAAAAAAAUAv4oZnWgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAc0en44AAAB54n+JH2Vyd3c/l4H76ejdfCDQAAAAAAflP+rGWwa4gAAAAAAAAAAAAAAAABz0vx85uwAAAAAAAAAAAAAAAAAAD2nTLYN+cAAAAAAAAAAAAAAAAAAPiT7bz2DTEAAAAAAAAAAAAAAAAAPl2LyvrcAuAAAAAAAAAAAAAAAAADwXxrLb7fNcQAAAAAAAAAAAAAAAAAA864Pn6giwAAAAAAAAAAAAAAAAAuNe6nVxgAAAAAAAAAAAAAAAAAAfEme322aYgAAAAAAAAAAAAAAAAAB+U/wCrGWwa4gAATTW+ed6wAAAAAAAAAAAAAAAAAAAABynyEHyRwQdb6IHQSh+obwAAAAPAQ+FNkP1M+PQ+V/owPrH4AD1H7fD4q+1QAAAAAAAAAAAAAADmj0/HAAAAGh+Anmnn+pB3w/6D3eh5QAAAAAAAH5T/AKsZbBriAAAAAAAAAAAAAAAAAHPS/Hzm7AAAAAAAAAAAAAAAAAAAPadMtg35wAAAAAAAAAAAAAAAAAA8f9gmwqAAAAAAAAAAAAAAAAAA+XYvK+twC4AAAAAAAAAAAAAAAAAPBfGstvt81xAAAAAAAAAAAAAAAAAADzrg+fqCLAAAAAAAAAAAAAAAAAC417qdXGAAAAAAAAAAAAAAAAAAB4/N+wFQAAAAAAAAAAAAAAAAAAH5T/qxlsGuIAAE01vnnesAAAAAAAAAAAAAAAAAAAAeZB+Lv6dB6N8rh9V+2BmfAQcj+nwAAAAH4kB+2H58B6j8CB+3H4kB9U/XYfn39VB3XuYAAAAAAAAAAAAAABzR6fjgAAADQ88+SctqP2QHoC64gAAAAAAAflP+rGWwa4gAAAAAAAAAAAAAAAABz0vx85uwAAAAAAAAAAAAAAAAAAD2nTLYN+cAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAD5di8r63ALgAAAAAAAAAAAAAAAAA8F8ay2+3zXEAAAAAAAAAAAAAAAAAAPOuD5+oIsAAAAAAAAAAAAAAAAALjXup1cYAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAH5T/qxlsGuIAAE01vnnesAAAAAAAB8reT/L/0KHP7gAAHoG3l/YJ9V/OQAAHPB8f/AB2H3n76HagAB8gh+fH7YBqgAAAficH6U/EgdLxgfrj+R4fRHFhwvThB2QfoIAAAAAAAAAAAAAAc0en44AAAGZ43NfhD+9/P10PwMR/RR6B1cYNAAAAAAB+U/wCrGWwa4gAAAAAAAAAAAAAAAABz0vx9ebsVoAAAAAAAAAAAAAAAAAB7TplsG/OAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB8uxeV9bgFwAAAAAAAAAAAAAAAAAeC+NZbfb5riAAAAAAAAAAAAAAAAAAHnXB8/UEWAAAAAAAAAAAAAAAAAFxr3U6uMAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAD8p/1Yy2DXEAACaa3zzvWAAAAAAAA8D8+8L9iDH1QAAOv04Po49/8XAD4eD5m8FD9Mc4PtNQAAA5IPy3/AEsD0kAAADlg+PvlMP0Q/O8P2W/GkP135sPk9oex/HYfqsAAAAAAAAAAAAAAc0en44AAAHM/zR8vYw5uw9ac/wBDeh6PkgwAAAAAAPyn/VjLYNcQAAAAAAAAAAAAAAAAA56X4+zk9GOeegNOIAAAAAAAAAAAAAAAA9p0y2DfnAAAAAAAAAAAAAAAAAAD4r+1M9g0xAAAAAAAAAAAAAAAAAD5di8r63ALgAAAAAAAAAAAAAAAAA8F8ay2+3zXEAAAAAAAAAAAAAAAAAAPOuD5+oIsAAAAAAAAAAAAAAAAALjXup1cYAAAAAAAAAAAAAAAAAAHxXnt9qGmIAAAAAAAAAAAAAAAAAAflP8AqxlsGuIAAE01vnnesAAAAAAAAAAAAAAAAAH5wh5NiB9f8WH3N0IAAAfIAfOv6kgAAAAeCh8Jd6H6Vfj8HrXlQfr5ygfHH0sHwfSD9WQAAAAAAAAAAAAADmj0/HAAAAPO0/zO/TjHd/Z7YTjQAAAAAAAH5T/qxlsGuIAAAAAAAAAAAAAAAAAc9L8f57z/AKHgvXuP6cPT+DAAAAAAAAAAAAAAAAPadMtg35wAAAAAAAAAAAAAAAAAA+JPtvPYNMQAAAAAAAAAAAAAAAAA+XYvK+twC4AAAAAAAAAAAAAAAAAPBfGstvt81xAAAAAAAAAAAAAAAAAADzrg+fqCLAAAAAAAAAAAAAAAAAC417qdXGAAAAAAAAAAAAAAAAAAB8SZ7fbZpiAAAAAAAAAAAAAAAAAAH5T/AKsZbBriAABNNb553rAAAAAAAAAAAAAAAAAAAVA/PT5ED7e+qw9FeFK6Hk/5NB+nf0EAAAABzQWtsBoeeeiAuIGlZDlOgC2AAAAAAAAAAAAAAc0en44AAAAB+dnxDyd1avnr++Oj3+YAAAAAAAB+U/6sZbBriAAAAAAAAAAAAAAAAAHPS/H/AD/yvsfn77I8j9JgPo/xEAAAAAAAAAAAAAAAA9p0y2DfnAAAAAAAAAAAAAAAAAAD4k+289g0xAAAAAAAAAAAAAAAAAD5di8r63ALgAAAAAAAAAAAAAAAAA8F8ay2+3zXEAAAAAAAAAAAAAAAAAAPOuD5+oIsAAAAAAAAAAAAAAAAALjXup1cYAAAAAAAAAAAAAAAAAAHxJnt9tmmIAAAAAAAAAAAAAAAAAAflP8AqxlsGuIAAE01vnnesAAAAAAAAAAAAAAAAAAAAeLh8VcCHl/6GB8WagUvsQPscAAAAAAAAAAAAAAAAAAAAAAAAAAAADmj0/HAAAAOTT/nc7Hz/U+zvi/TL+hmft88AAAAAAAD8p/1Yy2DXEAAAAAAAAAAAAAAAAAOel+P+ReN+gfKf3585+2XT7P+XwAAAAAAAAAAAAAAAD2nTLYN+cAAAAAAAAAAAAAAAAAAPiT7bz2DTEAAAAAAAAAAAAAAAAAPl2LyvrcAuAAAAAAAAAAAAAAAAADwXxrLb7fNcQAAAAAAAAAAAAAAAAAA864Pn6giwAAAAAAAAAAAAAAAAAuNe6nVxgAAAAAAAAAAAAAAAAAAfEme322aYgAAAAAAAAAAAAAAAAAB+U/6sZbBriAABNNb553rAAAAAAAAAAAAAAAAAAAAABzATfM4df8ARQAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABzR6fjgAAAHFp/iP71x99n4gT/oG9C7fPBoAAAAAAPyn/VjLYNcQAAAAAAAAAAAAAAAAA56X4/4H4H6j84/UPyf9Ge0n6F/FwAAAAAAAAAAAAAAAB7TplsG/OAAAAAAAAAAAAAAAAAAHy39SRqF5AAAAAAAAAAAAAAAAAB8uxeV9bgFwAAAAAAAAAAAAAAAAAeC+NZbfb5riAAAAAAAAAAAAAAAAAAHnXB8/UEWAAAAAAAAAAAAAAAAAFxr3U6uMAAAAAAAAAAAAAAAAAAD5bjX6kLyAAAAAAAAAAAAAAAAAAD8p/wBWMtg1xAAAmmt8871gAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAOaPT8cAAAAA/PH6xy3/Fn1/m6v2gk7vOAAAAAAAA/Kf8AVjLYNcQAAAAAAAAAAAAAAAAA56X4/wDMvzf7BytH5X+iPuCj+ifxIDQAAAAAAAAAAAAAAHtOmWwb84AAAAAAAAAAAAAAAAAAAB51zez6KdPjAAAAAAAAAAAAAAAAHy7F5X1uAXAAAAAAAAAAAAAAAAAB4L41lt9vmuIAAAAAAAAAAAAAAAAAAedcHz9QRYAAAAAAAAAAAAAAAAAXGvdTq4wAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPyn/VjLYNcQAAJprfPO9YAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADmj0/HAAAAOOT/AAi/ePn6urOnkAAAAAAAAAPyn/VjLYNcQAAAAAAAAAAAAAAAAA56X4/8j/L/ALh6180+N+n/AG9rfX/zMHT4oAAAAAAAAAAAAAAHtOmWwb84AAAAAAAAAAAAAAAAAAAB8D+B+uffB7/5GAAAAAAAAAAAAAAAAfLsXlfW4BcAAAAAAAAAAAAAAAAAHgvjWW32+a4gAAAAAAAAAAAAAAAAAB51wfP1BFgAAAAAAAAAAAAAAAABca91OrjAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA/Kf8AVjLYNcQAAJprfPO9YAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADmj0/HAAAAK4fgD0nB6XsJpn+utzr4QAAAAAAAPyn/VjLYNcQAAAAAAAAAAAAAAAAA56X4/8UfI/0B9N+Puem957PmWHsfmYAAAAAAAAAAAAAAHtOmWwb84AAAAAAAAAAAAAAAAAAfO30TGh+THyv9BfpJ6H9J+JB0+MAAAAAAAAAAAAAAB8uxeV9bgFwAAAAAAAAAAAAAAAAAeC+NZbfb5riAAAAAAAAAAAAAAAAAAHnXB8/UEWAAAAAAAAAAAAAAAAAFxr3U6uMAAAAAAAAAAAAAAAAAAD52jT6JLzAAAAAAAAAAAAAAAAAAD8p/1Yy2DXEAACaa3zzvWAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAKgfIPrQe380HyV9Th2XySHufogeO+xAAcl4UHO/YgHx2H2D8lh9bgAHkfzoH3OB8/dmHpwHNHp+OAAAAUA/EuXi9EBfthe7fPAAAAAAAA/Kf9WMtg1xAAAAAAAAAAAAAAAAADnpfj/wAFfG/0h9r8/wC1+WfLn3p436ZmH1H4EAAAAAAAAAAAAAAB7TplsG/OAAAAAAAAAAAAAAAAAAHxJ9t57Y35GfHf0v8AaX117v5OHsfnQAAAAAAAAAAAAAAHy7F5X1uAXAAAAAAAAAAAAAAAAAB4L41lt9vmuIAAAAAAAAAAAAAAAAAAedcHz9QRYAAAAAAAAAAAAAAAAAXGvdTq4wAAAAAAAAAAAAAAAAAAPiTPb7bNMQAAAAAAAAAAAAAAAAAA/Kf9WMtg1xAAAmmt8871gAAAAAAAAAAAAAAAAAAAAAAABjTxMAAAAAPyID7P+DQ/UD8rA/Rn81A+/r4fJP6Vh8ufcAAH5i/omH4y/uCB+HwfuB8rB9VYgZ+kHjHx8H6Dd0FP8tQ/VcDmj0/HAAAAAAAAAAAAAAAAAPyn/VjLYNcQAAAAAAAAAAAAAAAAA56X4r8BfEf09+gbvrf5w+F/sv47+m98+y/mEAAAAAAAAAAAAAAD2nTLYN+cAAAAAAAAAAAAAAAAAAPiT7bz28p/Mz4f+rfWv0v9b89D6b8OAAAAAAAAAAAAAAA+XYvK+twC4AAAAAAAAAAAAAAAAAPBfGstvt81xAAAAAAAAAAAAAAAAAADzrg+fqCLAAAAAAAAAAAAAAAAAC417qdXGAAAAAAAAAAAAAAAAAAB8SZ7fbZpiAAAAAAAAAAAAAAAAAAH5T/qxlsGuIAAE01vnnesAAAAAAAAAAAAAAAAAAAAAAAAeV+j4nqh53tgAAAAB+G4fuP8Rh5t44H7E/j4Hq/2KHwh3wesesh7OB+TX6yh+Fn16B+fof0Mfh8H6vfloHvvygH6wfkWH1hxIfr1+SwfrSBzR6fjgAAAAAAAAAAAAAAAAH5T/qxlsGuIAAAAAAAAAAAAAAAAAc9L+ePjr4j+oPuw+u/mv5xs/G/099HVvsf5iBoAAAAAAAAAAAAAPadMtg35wAAAAAAAAAAAAAAAAAA+JPtvPb5r+dfjP6c8h/V2+bfPtP5mAAAAAAAAAAAAAAA+XYvK+twC4AAAAAAAAAAAAAAAAAPBfGstvt81xAAAAAAAAAAAAAAAAAADzrg+fqCLAAAAAAAAAAAAAAAAAC417qdXGAAAAAAAAAAAAAAAAAAB8SZ7fbZpiAAAAAAAAAAAAAAAAAAH5T/qxlsGuIAAE01vnnesAAAAAAAAAAAAAAAAAAAAAAAAfL/0Hxfp3qXn+0Hn+0AAAAfHQfKflgfZvz8H68fkMH6RfIgT4genfNofrUB+Sv61B+G/6ugfkOH7x/h8Hv8A9iB79+HYfqJ8dh+on4eB+4f5Kh+tQHNHp+OAAAAUQ/HSTj7yUPZP0V35tc1xAAAAAAD8p/1Yy2DXEAAAAAAAAAAAAAAAAAOel/LPzv8AEf0/9nH1382cf8g/H/01+gmT9d/NIVmAAAAAAAAAAAAAHtOmWwb84AAAAAAAAAAAAAAAAAAfEn23nt8Xu+P/AKV8w+jZ1+kD7b+XwAAAAAAAAAAAAAAPl2LyvrcAuAAAAAAAAAAAAAAAAADwXxrLb7fNcQAAAAAAAAAAAAAAAAAA864Pn6giwAAAAAAAAAAAAAAAAAuNe6nVxgAAAAAAAAAAAAAAAAAAfEme322aYgAAAAAAAAAAAAAAAAAB+U/6sZbBriAABNNb553rAAAAAAAAAAAAAAAAAAAAAAAAZvyT9X+cSfUVR0R8l+lAAAAAHjf5Xh+0/wCJAftX+K4fuAB+cnuAR/LQfqqB+Sn61h+H37ggfh8H7g/h8H3F50H2H+NQfrt8fB+oP4fB+4P5KB+tYHNHp+OAAAAVg/nH964PTyvKQX7Kc/rOd3nAAAAAAB+U/wCrGWwa4gAAAAAAAAAAAAAAAABz0v478y+I/qD6xPrv5rPiL5r91+s+r9f80Q7vlQAAAAAAAAAAAAA9p0y2DfnAAAAAAAAAAAAAAAAAADK1Ufnl738Z/TeV5BOv6DH238vgAAAAAAAAAAAAAAfLsXlfW4BcAAAAAAAAAAAAAAAAAHgvjWW32+a4gAAAAAAAAAAAAAAAAAB51wfP1BFgAAAAAAAAAAAAAAAABca91OrjAAAAAAAAAAAAAAAAAAAykaowAAAAAAAAAAAAAAAAAAPyn/VjLYNcQAAJprfPO9YAAAAAAAAAAAAAAAAAAAAAAADwvyf7/wDGDpOrzvqGf83/AHYFQAAHzAHgH32HR+HB8O/coe3edBR9OD4H9kD6ZA/Pj6mDw37TA+LA+0/iwPtL4bCP40D9ePJQ+lviwPp74GD9LgOaPT8cAAAA4FP+fT+j7n6qP84QfuB+UQ/3zOnjAAAAAAD8p/1Yy2DXEAAAAAAAAAAAAAAAAAOel/DlD4j+n/pI+u/mw+f/ACv0TK+pOb16B735IAAAAAAAAAAAAAHtOmWwb84AAAAAAAAAAAAAAAAAAAH5a/e3xf8ATfR/l4q/Wyx9p/MoOAAAAAAAAAAAAAA+XYvK+twC4AAAAAAAAAAAAAAAAAPBfGstvt81xAAAAAAAAAAAAAAAAAADzrg+fqCLAAAAAAAAAAAAAAAAAC417qdXGAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB+U/6sZbBriAABNNb553rAAAAAAAAAAAAAAAAAAAAAAAcX1ed80t/Rvw0mT9T9z+H/XQ8j6UAAAAAAAAAAAAAreNB7cAAAHKfmWHP/WofZQAB5F6KGwBzR6fjgAAAFcP56+X8/1YPTxfpl8K78/7aHTxgAAAAAB+U/6sZbBriAAAAAAAAAAAAAAAAAHPS/z+634j+n/dD67+bDnMfT+Kv0P+Y/ecg+t/nMAAAAAAAAAAAAAPadMtg35wAAAAAAAAAAAAAAAAAA867+dPyE/Wn43+ldj8uevwPt/2f3PysO35kAAAAAAAAAAAAAPl2LyvrcAuAAAAAAAAAAAAAAAAADwXxrLb7fNcQAAAAAAAAAAAAAAAAAA864Pn6giwAAAAAAAAAAAAAAAAAuNe6nVxgAAAAAAAAAAAAAAAAAAedTXopUgAAAAAAAAAAAAAAAAAB+U/6sZbBriAABNNb553rAAAAAAAAAAAAAAAAAAAAAAAfNXu/H8QfZ/lZoRr1v0B8j+mXzwPswAAAAAAAAAAAAAAAAAAAAAAAAAA5o9PxwAAAADlfiXLU+3A6jL0y1BgAAAAAAflP+rGWwa40+N5fbDH0wAAAAAAAAAAAAAAMRP4O9z+V/dvVj7D+ZwD4w+l/jP6e72L7P8AmIBAAAAGdHTjHJ9GAAAAAHrOmeoa4AAAAAAAAAAAAAAAAAAB8j/RnN63wH+s3l/oJ8X+n8PR+3p1D3Py0AAAAAj5/Ls5k4fqwAkJsF4AB82EbX1v2/LhcAAAAAAAAAAAAAAAAAHgvjWW32+a4mJn1c4cf0gAAAAAAAAAAAAAAAHC8vlp1tfp+chHIAAAAAAAAAAAA8JBOjznP7haz7/Yzo5AAAAAAAAAAAAAAAAAAAPkeNvrjrOnyNI6vCAAAAAAAASFOBs3zmRxfUhHUTuZSsAD8yP1138IOnzgAAJprfPO9YAAAAAAAAAAAAAAAAAAAAAAD5G+l+DzD6v83Onw7fSet+S/SuvPD+tAAAAAAAAAAAAAAAAAAAAAAAAAADmj0/HAAAAAPzt+NeTuqkafutod3mgAAAAAAAflP+rGWwa4+ffnN8X/AEyHzn7SAAAAAAAAAAAAAABGnT+tve/H+zPr/wCaQDw3i/jf6d+vMn7H+ZAcgAAAHx589+08lP8AI/0kDzAAAALrTxoAAAAAAAAAAAAAAAAAAMTbT7v7g+u/mm95l635/wDFP6deN+qzy/VfgMFaar141rV40r189OU+F/lP6K0JPE/XgckSqAncAPM+jwvu/wCt/wBM/h0LgAAAAAAAAAAAAAAAAA8F8ay2+3zXE+Z/M+4+PT87/soAAAAAAAAAAAAAAACtiZdv351f6P8AwrgVOnyAaAAAAAAAAACQero56szM+v4g5v8AO/7fmWd75UgAAAAAAAAAAAAAAAAAAYiej0t8n33q/pP8Mz2NeaxPea1wr140rV40r141rkXY88w9r4Y6X89/tcNeJsM3GLYA+Xv37+9/kAPa/PAAAJprfPO9YAAAAAAAAAAAAAAAAAAAAAAD5V+i+I50+u/Mj1zzfe1O9+Z++3zx/qAAAAPkj5HD6w+vw+VfkYPvjtw+Ffp8Pf8A5WD6pAPlMOw+Rg/TUAAAAAAPIw8Q+ywAOaPT8cAAAA4ZP+bb9BeH0ulNcP1Nt9XGAAAAAAAB+U/6sZbBriAAAAAAAAAAAAAAAAAHPS/Hzm7AAAx/hr5H+lP0Syvqf53DXjAAAA8r+c/if6r+2Z/tf5YB5gAAAHtOmWwb84AAAAAAAAAAAAAAAAAAfEn23ntHjuatTHo/MP75+N/prr4/rf5rnsVNmxeVixednk47/wA3fp74T+tPsA/RP41AAAAAD5di8r63ALgAAAAAAAAAAAAAAAAA8F8ay2+3zXEAAAAAAAAAAAAAAAAAAPOvi/579ZzfrPHt3cb2fyzOLxAAAAAAAAJR6ujnrn5VRifHfzP733H0/wBvy7rnuflHup1cYAAAAAAAAAAAAAAAAAAHxJnt9q/nl8/+xw/eMFeD3fyOvXjRLAWbF52LF52bF5WJrz4D81Piv6f+3voz6T8UD0viwAAAPyn/AFYy2DXEAACaa3zzvWAAAAAAAAAAAAAAAAAAAAAAA8d9P5/xU+9/GzZz6Ov+hPjf1EPF+rAAAA8y9KD8KP2vD8hP2WD8UPoEP0m/LwPp310PYQD8nQ/WL8Pg/cEA+Ww4z7MD5/8AosPDPXQ46yH5rfrOABzR6fjgAAAFMP5/f2A5ezqDfm9ikuAAAAAAAAPyn/VjLYNcQAAAAAAAAAAAAAAAAA56X4+c3YAAB8e/P/s/vXqfb8jUPT+GAAATPs+Evqz4/wDpj0I+0/lsAAAAAPadMtg35wAAAAAAAAAAAAAAAAAA+JPtvPYNMQD4G9F+J/qD60Ptv5fAAAA4r81/jP6W+pfrj3Py4PX/ADoAAAAAPl2LyvrcAuAAAAAAAAAAAAAAAAADwXxrLb7fNcQAAAAAAAAAAAAAAAAAA868t4/V+HPs34/+k+06D67+ac7IqAcgAAAABMPV0M9s7KrOLKXR8d938J/Xn04777+OS4591OrjAAAAAAAAAAAAAAAAAAA+JM9vtvOo/Kn9Evif6g9VPuP5ZAAAAAAA47H0vzQ+u/hP6v8AqU/Qf4+AAAAAPyn/AFYy2DXEAACaa3zzvWAAAAAAAAAAAAAAAAAAAAAAAjc/P3nH6F+KG/vxfR+9+d/uAY9QAAABEH4jfpIHzv8ApEH4hfU4fS3xOHUage5fVQH5Mh+s34fB+4IHwZyge0/H4Wf0uD8hvuYPnD9Iw/Nn9XQmA5o9PxwAAADMD+ZBnmewAf04avp+OAAAAAAAB+U/6sZbBriAAAAAAAAAAAAAAAAAHPS/Hzm7AAAPG/O+18t+0vN+2wz6P8UAAC58yeH+sY/1OMPc/JgAAAAA9p0y2DfnAAAAAAAAAAAAAAAAAADyb1maCpAPnz5Y+J/qD9Kj7b+XwAAOGw9X82/qn4P+sfrQ/Q/45AAAAAAA+XYvK+twC4AAAAAAAAAAAAAAAAAPBfGstvt81xAAAAAAAAAAAAAAAAAADzrg+fq+eeW+J/qf6vl+z/lzoK+euGmmIAAABOPWvZ7ZuVWcZUZXx38v+/d/9M93yantflpca91OrjAAAAAAAAAAAAAAAAAAA8mmvWSp+SPJPhf6r/RI+6/lQAAAAAAOb/NH5D+jPpP6/wDX/Og9j84AAAAAPyn/AFYy2DXEAACaa3zzvWAAAAAAAAAAAAAAAAAAAAAAAAMTXmp9Jebzn7QAAAABPynD7B1g+TP01D8Sv2tD5S8iDA+5g/PD9WgPyVD9avw+D9wQPxh/XsOj/D8P0g+fgqcAGR+wQfk/+qoWwOaPT8cAAAAjDyyaLAPTn3AAAAAAAAH5T/qxlsGuIAAAAAAAAAAAAAAAAAc9L8fObsAAAoR0fCP6F/I/0fTb9j/NAATD5z4n+K/qT7f1vsf5lDTkAAAAAD2nTLYN+cAAAAAAAAAAAAAAAAAAAAAAMn8mfjP6W/WHb+v/AJzDTlAOE/Nf4v8Apv6v+tPd/Jw9f88AAAAAAA+XYvK+twC4AAAAAAAAAAAAAAAAAPBfGstvt81xAAAAAAAAAAAAAAAAAADzrg+fqzPhf4/+kvtTp/p/wV29p5r+fGwrMALA9a5ntm5VZsKgx8+z4+7/AOF/rj6ZX77+PAEXGvdTq4wAAAAAAAAAAAAAAAAAAAADK/Kb47+kP0o9G+o/CQ6fFAAAAMWOn82PZ/z7+wfto/RP41AAAAAAD8p/1Yy2DXEAACaa3zzvWAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPySzQ+qfqsPxp++w+Rv1/DiPlENiUPnP8AVQD8lQ/Wr8HA/Q28GZ4EHrXm4fp9/OsH7x/nOHoH6TB+Sn61gAc0en44AAAH4wfW/N2flgc/SAf0S6Xo+UAAAAAAAB+U/wCrGWwa4gAAAAAAAAAAAAAAAABz0vx85uwAAAD5M9b+N/p72nO+x/mMHLt6Nvgz2z4z+pvZz7b+UwAAAAAAPadMtg35wAAAAAAAAAAAAAAAAAAAAAAPzc+a/bPpj6L9P4cPS+LOE5/W/Nj64+F/q76qP0L+OwAAAAAAAD5di8r63ALgAAAAAAAAAAAAAAAAA8F8ay2+3zXEAAAAAAAAAAAAAAAAAAPOuD5+o+bfE/UM/wCoo6Q9v8p2rue3PwaZFlGtbjbLzLyYOQDH+Oflv6C9E+me/wCPU9r8tAC417qdXGAAAAAAAAAAAAAAAAAAAAAHxx436Vwn6Bc/rB9D+PgAAAZ/5wfL/u/d/eXT4bj3/wAkAAAAAAD8p/1Yy2DXEAACaa3zzvWAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPnzMA9/DF8FD6fuB5r6GE/zSHufRgfn6H09khh2g+jPDgxPowDx0PYuBDa50Piz9GQAOaPT8cAAAD5P9Yz1+TzPQA+/7PRzAAAAAAAAflP8AqxlsGuIAAAAAAAAAAAAAAAAAc9L8fObsAAAA8p8P+N/p37p5z7D+ZQqN/wCfvH/RvFvsnl+ncfRfiQAAAAAAHtOmWwb84AAAAAAAAAAAAAAAAAAfFf2pnsGmIAfKfk/oHlX6A8n0AfQ/j/nv5v8AxX9O/XX1R9B+Qh6vwAAAAAAAAB8uxeV9bgFwAAAAAAAAAAAAAAAAAeC+NZbfb5riAAAAAAAAAAAAAAAAAAHnXB8/UYnP6Pw99v8Ayv8AQ+6fX/zUaarXx50vWU8vMvJo5ADHz7fjn0r4X+t/pNfvv48AQAXGvdTq4wAAAAAAAAAAAAAAAAAAPivPb7UNMTDz6vyn/Tr43+lu6Ptv5fAAArK/zs0fzv8Asv8AQWb9E/jMHIAAAAAAflP+rGWwa4gAATTW+ed6wAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADAxd0AAAAAAAMYNKcADmj0/HAAAAOOT/np/oQ5+vtzp4wAAAAAAAAD8p/1Yy2DXEAAAAAAAAAAAAAAAAAOel+PnN2AAAAVvg75T+ifv1/0X4bDrdHlZvwH8f/AEv9l9b9T/PwdHjgAAAAAAe06ZbBvzgAAAAAAAAAAAAAAAAAB8SfbeewaYgBxOHqfmL+tPy/7xP579N+Gfm39ffDf1Z9UH6F/HQAAAAAAAAAfLsXlfW4BcAAAAAAAAAAAAAAAAAHgvjWW32+a4gAAAAAAAAAAAAAAAAAB51wfP1BF/MFv4z+o/pQ+z/lwAu7Ma5uZUIOQAAx/jn5b+gvTPpH0filPY/MgAALjXup1cYAAAAAAAAAAAAAAAAAAHxJnt9tmmIB8U4Xwv8AVf3mfdfyoAAEf5/fO/seH+i2fbbPpvw8AAAAAAAD8p/1Yy2DXEAACaa3zzvWAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA5o9PxwAAACqH8/8AX4PT9UKj9sL3b54AAAAAAAH5T/qxlsGuIAAAAAAAAAAAAAAAAAc9L8fObsAAAAPlrw/1jvPoHTgq7fq/nfxD1Hxf9U/Rp9v/ACiAAAAAAAAe06ZbBvzgAAAAAAAAAAAAAAAAAB8SfbeewaYgAB+Wn3T8T/UHQfm2L7J+nfovxoPU+FAAAAAAAAAD5di8r63ALgAAAAAAAAAAAAAAAAA8F8ay2+3zXEAAAAAAAAAAAAAAAAAAPOuD5+oIvm/ij4/+mPubV+s/nINOQAAAAAMPLv8Ajv074f8ArX6RX73+OwAAAC417qdXGAAAAAAAAAAAAAAAAAAB8SZ7fbZpiAcz+WHxf9M/qN1/1387ht5w0PhTzf8AOv7M/SPS/Q/45CsQAAAAAAAPyn/VjLYNcQAAJprfPO9YAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADmj0/HAAAAKQfkNLx9wM/XW518IAAAAAAAH5T/qxlsGuIAAAAAAAAAAAAAAAAAc9L8fObsAAAAPN+P6b5w/QDw/1RfC/X/N/mH7d8j9Otn1H8/gAAAAAAAHtOmWwb84AAAAAAAAAAAAAAAAAAfEn23nsGmIAAfFXA/A/1t5X9lR1/Tp+g/wAfAAAAAAAAAAAfLsXlfW4BcAAAAAAAAAAAAAAAAAHgvjWW32+a4gAAAAAAAAAAAAAAAAAB51wfP1BFnyn4X630n0Rt5gev+bgAAAAGH8cfK/0P6j9I+l8Kp7H5kAAAAXGvdTq4wAAAAAAAAAAAAAAAAAAPiTPb7bNMQA+GfC/Vn/cYg938qPizxP0/wz9KvN+13D63+ewAAAAAAAAD8p/1Yy2DXEAACaa3zzvWAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA5o9PxwAAAPlP1fPX5MM9AD7/ALPRzAAAAAAAAflP+rGWwa4gAAAAAAAAAAAAAAAABz0vx85uwAAAAjVfCP3N8d/Tej8G7+R9P+ne9+RB2/LAAAAAAAAHtOmWwb84AAAAAAAAAAAAAAAAAAfLf1JGoXkAAHkH5rfBf1t92fTn1P4GHqfCgAAAAAAAAAAfLsXlfW4BcAAAAAAAAAAAAAAAAAHgvjWW32+a4gAAAAAAAAAAAAAAAAAB51wfP1BFnIc/s/G/3T85+43j6v8AnUAAAAw8u/449Z+H/rL6JX73+PgAAAAAuNe6nVxgAAAAAAAAAAAAAAAAAAfLca/UheQAcfj6X5c/ql8j/Q/SH2n8zfI/zF+e/wBifpd0/wB7/JIa8IAAAAAAAAB+U/6sZbBriAABNNb553rAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAc0en44AAAH4ufW/N2floc/SAf0S6Xo+UAAAAAAAB+U/6sZbBriAAAAAAAAAAAAAAAAAHPS/Hzm7AAAAAD5o0fjv6doYOXofWZ9t/KQAAAAAAAAAe06ZbBvzgAAAAAAAAAAAAAAAAAAAAAAAQ+A+d9n9CHo/GAAAAAAAAAAAAHy7F5X1uAXAAAAAAAAAAAAAAAAAB4L41lt9vmuIAAAAAAAAAAAAAAAAAAedcHz9QRYB8jd98Z/UfvZ9n/AC4AAABh/HHyv9D+s/RXp/Ah6/5sAAAAAFxr3U6uMAAAAAAAAAAAAAAAAAAAAAAAD4K6H4X+q/tT5h+m/D/jv9LfA/Xe1Ps/5nAAAAAAAAAAD8p/1Yy2DXEAACaa3zzvWAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA5o9PxwAAAPm7ps9PR/NKn28qQAAAAAAAAD8p/1Yy2DXEAAAAAAAAAAAAAAAAAOel+PnN2AAAAAHCfInxv8AULftzTh3D67+bAAAAAAAAAA9p0y2DfnAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA+XYvK+twC4AAAAAAAAAAAAAAAAAPBfGstvt81xAAAAAAAAAAAAAAAAAADzrg+fqCLAOE+Sfjf6f+6bH2P8yA8wA5/L0fjn1z4f+svoo+9/jwAAAAAAC417qdXGAAAAAAAAAAAAAAAAAAAAAAABwf5k/E/1B9o/GvH9J+k3ov3P8qB0+MAAAAAAAAAAH5T/AKsZbBriAABNNb553rAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAc0en44AAAH4/ey8vZ71+U4fv0dXGAAAAAAAAAflP8AqxlsGuIAAAAAAAAAAAAAAAAAc9L8fObsAAAAAE+Gvmf3r3P3P0/gg9P4IAAAAAAAAAPadMtg35wAAAAAAAAAAAAAAAAAA+b/AKQjQLzAAAAAAAAAAAAAAAAAD5di8r63ALgAAAAAAAAAAAAAAAAA8F8ay2+3zXEAAAAAAAAAAAAAAAAAAPOuD5+oIsAPjn579p9Z9t7/AJAPS+FAOf8Ajj5P+i/X/oj1fzwPX/NwAAAAAALjXup1cYAAAAAAAAAAAAAAAAAAHzfGn0gXmAAB+fPz37F4Z+knjfpfqp91/KQAAAAAAAAAAAH5T/qxlsGuIAAE01vnnesAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABzR6fjgAAAfj9rcnd69+eaf7FnRy/RMumQAAAAAAAH5T/qxlsGuIAAAAAAAAAAAAAAAAAc9L8fObsAAAAADyfg+v8AVJO75IHIAAAAAAAAAe06ZbBvzgAAAAAAAAAAAAAAAAAB8SfbeewaYgAAAAAAAAAAAAAAAAB8uxeV9bgFwAAAAAAAAAAAAAAAAAeC+NZbfb5riAAAAAAAAAAAAAAAAAAHnXB8/UEWAHm/H9N8wfcXifq8p9P+BHP5ej8cezfEf1d9An3n8gAAAAAAAAFxr3U6uMAAAAAAAAAAAAAAAAAAD4kz2+2zTEAADzfm9rn/AGjPtDt+YAAAAAAAAAAAA/Kf9WMtg1xAAAmmt8871gAAAAAAAAAAAAAAAAAAOXBenDBYHQgAABw/lYfRgEfNB1IAAAAAAAAAAAAAAAAAAAAAAAAHNHp+OAAAB+P3598HphnqAf04avp+OAAAAAAAB+U/6sZbBriAAAAAAAAAAAAAAAAAHPS/Hzm7AAAAAAAAAAAAAAAAAAAPadMtg35wAAAAAAAAAAAAAAAAAA+JPtvPYNMQAAAAAAAAAAAAAAAAA+XYvK+twC4AAAAAAAAAAAAAAAAAPBfGstvt81xAAAAAAAAAAAAAAAAAADzrg+fqCLABA+Lfc/jP6j9gPs/5c5745+T/AKM9m+gfW/OA9b86AAAAAAAALjXup1cYAAAAAAAAAAAAAAAAAAHxJnt9tmmIAAAAAAAAAAAAAAAAAAflP+rGWwa4gAATTW+ed6wAAAAAAAAAAAAAAAAAAH5/B8U/usHx/lB9rAAHxl7UGF8fB+oIHDfKofcYAAAAAAAAAAAAAAAAAAAAAAAAHNHp+OAAABj5yYIAOndSAAAAAAAA/Kf9WMtg1xAAAAAAAAAAAAAAAAADnpfj5zdgAAAAAAAAAAAAAAAAAAHtOmWwb84AAAAAAAAAAAAAAAAAAfEn23nsGmIAAAAAAAAAAAAAAAAAfLsXlfW4BcAAAAAAAAAAAAAAAAAHgvjWW32+a4gAAAAAAAAAAAAAAAAAB51wfP1BFgAB5V87/Gf1F9t879b/ADj8ce1fEf1b9An3v8fAAAAAAAAABca91OrjAAAAAAAAAAAAAAAAAAA+JM9vts0xAAAAAAAAAAAAAAAAAAD8p/1Yy2DXEAACaa3zzvWAAAAAAAAoQCBgAAEqNEGAAB+fweEe+h3WUHovwyFH9Rg/Fz0sPvf8xA9b+iw+sviYOv8AHg8l/SwO0/LwO40A/UMAAAAAAAAAAAAAAAAAAAOaPT8cAAAAAAAAAAAAAAAAA/Kf9WMtg1xAAAAAAAAAAAAAAAAADnpfj5zdgAAAAAAAAAAAAAAAAAAHtOmWwb84AAAAAAAAAAAAAAAAAAfEn23nsGmIAAAAAAAAAAAAAAAAAfLsXlfW4BcAAAAAAAAAAAAAAAAAHgvjWW32+a4gAAAAAAAAAAAAAAAAAB51wfP1BFgAA34i+a/d/afCvJ/RPbffvpvwcPV/PAAAAAAAAAAuNe6nVxgAAAAAAAAAAAAAAAAAAfEme322aYgAAAAAAAAAAAAAAAAAB+U/6sZbBriAABNNb553rAAAAAAAAeec5UgAAAG0HphNAAAfn8HtX5pB9o7ofCn7dB478Bh3P1YB8RB+rv4aB+uv50h6B7wHrn5yBH+lAdt+KwfuEAAAAAAAAAAAAAAAAAAAHNHp+OAAAAAAAAAAAAAAAAAflP8AqxlsGuIAAAAAAAAAAAAAAAAAc9L8fObsAAAAAAAAAAAAAAAAAAA9p0y2DfnAAAAAAAAAAAAAAAAAACjeQDAAAAAAAAAAAAAAAAAA+XYvK+twC4AAAAAAAAAAAAAAAAAPBfGstvt81xAAAAAAAAAAAAAAAAAADzrg+fqCLAAA8d8/7T5X+kPj/wCmfej9A/jIAAAAAAAAAAC417qdXGAAAAAAAAAAAAAAAAAABRReGAAAAAAAAAAAAAAAAAAB+U/6sZbBriAABNNb553rAAAAAAAAAAAAAAAAAAAfn8HtfaB+KX6ch8Hft4Hi3wyHYfUwSfHwfqD+HwfrX+dIegfSwerfl8Fv9GA7H8ag/cIAAAAAAAAAAAAAAAAAAAOaPT8cAAAAAAAAAAAAAAAAA/Kf9WMtg1xAAAAAAAAAAAAAAAAADnpfj5zdgAAAAAAAAAAAAAAAAAAHtOmWwb84AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHy7F5X1uAXAAAAAAAAAAAAAAAAAB4L41lt9vmuIAAAAAAAAAAAAAAAAAAedcHz9QRYAAESvx72bz/sw9H4kAAAAAAAAAAC417qdXGAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB+U/6sZbBriAABNNb553rAAAAAAAAAAAAAAAAAAAfHweo+4B+cHv4aHwkFj9RA8o/O0P0o8AD7x/K4P0R+Lg7D3EPQPg4PuH8wA9V+dA/bUAAAAAAAAAAAAAAAAAAAOaPT8cAAAAAAAAAAAAAAAAA/Kf8AVjLYNcQAAAAAAAAAAAAAAAAA56X4+c3YAAAAAAAAAAAAAAAAAAB7TplsG/OAAAAAAAAAAAAAAAAAAHnXos0FSAAAAAAAAAAAAAAAAAHy7F5X1uAXAAAAAAAAAAAAAAAAAB4L41lt9vmuIAAAAAAAAAAAAAAAAAAedcHz9QRYAAAAAAAAAAAAAAAAAXGvdTq4wAAAAAAAAAAAAAAAAAAPOpr0UqQAAAAAAAAAAAAAAAAAA/Kf9WMtg1xAAAmmt8871gAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADy/wAFDw70QPvcAAAAAAAAAAAAAAAAAAAOaPT8cAAAAAAAAAAAAAAAAA/Kf9WMtg1xAAAAAAAAAAAAAAAAADnpfj5zdgAAAAAAAAAAAAAAAAAAHtOmWwb84AAAAAAAAAAAAAAAAAAfEn23nsGmIAAAAAAAAAAAAAAAAAfLsXlfW4BcAAAAAAAAAAAAAAAAAHgvjWW32+a4gAAAAAAAAAAAAAAAAAB51wfP1BFgAAAAAAAAAAAAAAAABca91OrjAAAAAAAAAAAAAAAAAAA+JM9vts0xAAAAAAAAAAAAAAAAAAD8p/1Yy2DXEAACaa3zzvWAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAKvzwHV+zAAAAAAAAAAAAAAAAAAAABzR6fjgAAAAAAAAAAAAAAAAH5T/qxlsGuIAAAAAAAAAAAAAAAAAc9L8fObsAAAAAAAAAAAAAAAAAAA9p0y2DfnAAAAAAAAAAAAAAAAAAD4k+289g0xAAAAAAAAAAAAAAAAAD5di8r63ALgAAAAAAAAAAAAAAAAA8F8ay2+3zXEAAAAAAAAAAAAAAAAAAPOuD5+oIsAAAAAAAAAAAAAAAAALjXup1cYAAAAAAAAAAAAAAAAAAHxJnt9tmmIAAAAAAAAAAAAAAAAAAflP+rGWwa4gAATTW+ed6wAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHNHp+OAAAAAAAAAAAAAAAAAflP8AqxlsGuIAAAAAAAAAAAAAAAAAc9L8fObsAAAAAAAAAAAAAAAAAAA9p0y2DfnAAAAAAAAAAAAAAAAAAD4k+289g0xAAAAAAAAAAAAAAAAAD5di8r63ALgAAAAAAAAAAAAAAAAA8F8ay2+3zXEAAAAAAAAAAAAAAAAAAPOuD5+oIsAAAAAAAAAAAAAAAAALjXup1cYAAAAAAAAAAAAAAAAAAHxJnt9tmmIAAAAAAAAAAAAAAAAAAflP+rGWwa4gAATTW+ed6wAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHNHp+OAAAAAAAAAAAAAAAAAflP+rGWwa4gAAAAAAAAAAAAAAAABz0vx85uwAAAAAAAAAAAAAAAAAAD2nTLYN+cAAAAAAAAAAAAAAAAAAPiT7bz2DTEAAAAAAAAAAAAAAAAAPl2LyvrcAuAAAAAAAAAAAAAAAAADwXxrLb7fNcQAAAAAAAAAAAAAAAAAA864Pn6giwAAAAAAAAAAAAAAAAAuNe6nVxgAAAAAAAAAAAAAAAAAAfEme322aYgAAAAAAAAAAAAAAAAAB+U/6sZbBriAABNNb553rAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAc0en44AAAAAAAAAAAAAfLcX03vzQVP5T/qxlsGuIAAAAAAAAAAAAAAAAAc9L8fObsAAAAAAAAAAAAAAAAAAA9p0y2DfnAAAAAAAAAAAAAAAAAADyb1maCpAAAAAAAAAAAAAAAAAD5di8r63ALgAAAAAAAAAAAAAAAAA8F8ay2+3zXEAAAAAAAAAAAAAAAAAAPOuD5+oIsAAAAAAAAAAAAAAAAALjXup1cYAAAAAAAAAAAAAAAAAAHk016yVIAAAAAAAAAAAAAAAAAAflP8AqxlsGuIAAE01vnnesAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABzR6fjgAAAAAAAAAAAAHwWzDp8Q+7Q95N+b8G/ozj7wqPtf6B6OUKkAAAAAAAAAAAAAAD4fz09Ut4dIAAAAAAAAAAAAAAAAAAc/ztT9hnTyAAAAAAAAAAAAAAAAAAB8O/cUaBeYAAAAAAAAAAAAAAAH5TnL2Hmar7t+t+jkC4AAAAAAAAAAAAAAAAAPBfy45+r2kmj9N9MuhNsAAAAAAAAAAAAAAAAPzw+juTtAAAAAAAAAAAAAAAAAAA8DT+uvQ+viBoAAAAAAAAAAAAAAAAAPh3PT7iNMwAAAAAAAAAAAAAAAPHvgjHoDPT5d/fqoDo5gAAJprfPO9YAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADmj0/HAAAAAAAAAAAAAPxd/aDHo8F+Ug/Sk25/yn/VjLYNcQAAAAAAAAAAAAAAAAA/LvLb6y0cdwAAAAAAAAAAAAAAAAAA47xVr9HDq4wAAAAAAAAAAAAAAAAAA/Kf8AVjLYNcQAAAAAAAAAAAAAAAAA+XYvK+twC4AAAAAAAAAAAAAAAAAPBfGstvt81xAAAAAAAAAAAAAAAAAAD8hvvjj7gaAAAAAAAAAAAAAAAAAD48Vfb3s3VxBUgAAAAAAAAAAAAAAAAB+U+W36sGuIAAAAAAAAAAAAAAAAAAflP+rGWwa4gAATTW+ed6wAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHNHp+OAAAAAAAAAAAAAfi7+0WPR4b8gh+mBtz/AJT/AKsZbBriAAAAAAAAAAAAAAAAAH5d5bfWWjjuAAAAAAAAAAAAAAAAAAHHeKtfo4dXGAAAAAAAAAAAAAAAAAAH5T/qxlsGuIAAAAAAAAAAAAAAAAAfLsXlfW4BcAAAAAAAAAAAAAAAAAHgvjWW32+a4gAAAAAAAAAAAAAAAAAB+Q33xx9wNAAAAAAAAAAAAAAAAAB8eKvt72bq4gqQAAAAAAAAAAAAAAAAA/KfLb9WDXEAAAAAAAAAAAAAAAAAAPyn/VjLYNcQAAJprfPO9YAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADmj0/HAAAAAAAAAAAAAPzD+v8ej5h+gA+mzbn/Kf9WMtg1xAAAAAAAAAAAAAAAAAD8u8tvrLRx3AAAAAAAAAAAAAAAAAADjvFWv0cOrjAAAAAAAAAAAAAAAAAAD8p/1Yy2DXEAAAAAAAAAAAAAAAAAPl2LyvrcAuAAAAAAAAAAAAAAAAADwXxrLb7fNcQAAAAAAAAAAAAAAAAAA/Ib744+4GgAAAAAAAAAAAAAAAAA+PFX297N1cQVIAAAAAAAAAAAAAAAAAflPlt+rBriAAAAAAAAAAAAAAAAAAH5T/qxlsGuIAAE01vnnesAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABzR6fjgAAAAAAAAAAAAGZ+d+W3on2/Uhef5UJy9gAAAAAAAAAAAAAAAAAAGAnoqMAAAAAAAAAAAAAAAAAAI6obY5AAAAAAAAAAAAAAAAAADnuhTBoAAAAAAAAAAAAAAAAAM8f6Ae0dHIFSAAAAAAAAAAAAAAAAAedfmvj0apnoAAAAAAAAAAAAAAAAAAHN66oAAAAAAAAAAAAAAAAAADNRq2qQCAAAAAAAAAAAAAAAAADnk+hGgAAAAAAAAAAAAAAAAAAyP1v0xDbEAACaa3zzvWAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA5o9PxwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAJprfPO9YAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADmj0/HAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAmmt8871gAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAOaPT8cAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAACaa3zzvWAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA5o9PxwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADyz8+Mejt/0jqA0yAAAAAAAAAAAAAAAAAPi3z7Dp/RM35gAAAAAAAAAAAAAAAAA+Ds9PDfoTLf7fOnkAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA+Rov5d+oMt/rs6OUAAAAAAAAAAAAAAAAAyfzky2+kfqC4CoAAAAAAAAAAAAAAAAKQfmVx/L2/oD7j0cgVIAAAAAAAAAAAAAAAAAAAAAAE01vnnesAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABzR6fjgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAH44frnj0fnl9SB9Cm3OAAAAAAAAAAAAAAAAAeN/mdh0/sqb8wAAAAAAAAAAAAAAAAAYaNz8R8ej9uDfnAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAACh+OePR+zf4rB+0sm3OAAAAAAAAAAAAAAAAAc/8AHOetL7zEGmYAAAAAAAAAAAAAAAABhon+QP7N5bBtgAAAAAAAAAAAAAAAAAAAAAABNNb553rAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAc0en44AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB+LP7TY9HyJxwfdhtzgAAAAAAAAAAAAAAAAB+NWO/7Km2AAAAAAAAAAAAAAAAAAAYX5E49H7Jm3OAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHkfwhj0fqX+Rwv077bXEGgAAAAAAAAAAAAAAAD5di/LvvNMNMwAAAAAAAAAAAAAAAAA+c5r5n/SSbDTIAAAAAAAAAAAAAAAAAAAAAACaa3zzvWAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA5o9PxwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAD8YP2fx6Ph7pA+vTbnAAAAAAAAAAAAAAAAAD8asd/2VNsAAAAAAAAAAAAAAAAAAzkfkj+mOPR6cb84AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAcb+YmPR+tv45i/XrS1wBgAAAAAAAAAAAAAAAB8uxfl33mmGmYAAAAAAAAAAAAAAAAB5h+ZeHT+vFvfmAAAAAAAAAAAAAAAAAAAAAAAAJprfPO9YAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADmj0/HAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPz75bn6vDP2Gqdo25wAAAAAAAAAAAAAAAAA/GrHf9lTbAAAAAAAAAAAAAAAAAAPxBw6PTPVo0/QHb6eMGAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB+P2O/vvjkafrOdPIAAAAAAAAAAAAAAAAAHy7F+XfeaYaZgAAAAAAAAAAAAAAAHCp/hD918nf6N9z9HGGmYAAAAAAAAAAAAAAAAAAAAAAE01vnnesAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABzR6fjgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB5qn2uqIGAAAAAAAAAAAAAAAAABzKOmGAAAAAAAAAAAAAAAAABwycYn281SAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABAHmPqSbxoAAAAAAAAAAAAAAAAAM9EGuAMAAAAAAAAAAAAAAAAKKOQFVvsmgaAAAAAAAAAAAAAAAAAAAAAAAmmt8871gAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAOaPT8cAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAACaa3zzvWAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA5o9PxwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAJprfPO9YAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADmj0/HAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAmmt8871gAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAOaPT8cAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAACaa3zzvWAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA5o9PxwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAJprfPO9YAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADmj0/HAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAmmt8871gAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAOaPT8cAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAwd5AMAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAIM5GwMAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAmmt8871gAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAOaPT8cAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA8U/DPl7Aw6T9Z9sPxg/pgqb508YAAAAAAHGp/g/8A0CYdN46OU/OjPXmf09TDXEAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAD89c9fzSOL0D1ip+s/lDTL92zr4QAAAAAAAPjz405uv9jjp5AD8Nf1A5uv6KOnkAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAJprfPO9YAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADmj0/HAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAMjxya/JXsuTv/Tjterh+K/0SVfIXqyfzx9itfKPqSf0ZW1x+PqGPR9l3tec/O+NfMv1dT8L+dk/pjstM/58/wCj7DoU6eQAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADhk/PvwB5e39WvrfXDI8ma+pPDHM3HK/pj5BJ+sPR9M+R+Po09S+nXIXH4cfpBzdf1L8k7c/C/ZCr5D+Wctv2MOnkAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAJprfPO9YAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADmj0/HAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAD8Qe94+/wDYH5d6eP8ABz+nDDp/mz9jx6eGxEfQPyw1/SB+VnTx8TmY9Pq36/b8p+NgfVX021/On+lnN2fS31x18H8339AePT3x08YAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB/MXzdf7U/Ze2H5d/JPN1/rX+DDX1L8t5bev2g0v3l6OX+ej7Oy3+J/wBbqj7IOvh/m2/oW5uz48/LqL+5/wBC9sOY/GCNP6Dzp4wAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAACaa3zzvWAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA5o9PxwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA/EHvePv/YH5d6eP8HP6cMOn+bT9pJvwT5LD94/5gqj9+PwZVez0cts/wDoq6OU/EZz92/TFx/Pb6Hx9/65fSPd5v8AO1+4uHV7AdPGAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfzF83X+1P2Xth+XfyTzdf62fgQL+n/8ABjXL6493D8X/ANvM9fwh+msd/HftS4/XQ7vN/mQ/pZ5uz5g/EOL+wP171xwvwgm/6Mjp4wAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAACaa3zzvWAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA5o9PxwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA/EHvePv/YH5d6eP8HP6cMOn+bT9qVXg3ySj95v5eqn99/xbH9nfcV53PdtcD8gs9ffPuOp8y/LPPXH/AHPc/wA039Gk6dKdPGAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfzF83X+1P2Xth+XfyTzdf62fgQL+oX8Gdcvrb3kPxW/f6NP54v2mm/SvSunj9FLz/nM/ejm7H8ptzfhv8ApFy9v0f+S2mf75nTxgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAE01vnnesAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABzR6fjgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB+WPW83X+kPgu/N+Sf7rZb/iP+n815J87B+tv4M1P698teX5Wcfyd/6GfoX18J8TNfIv6VZ6/jPPz9X6l+kdfD+MX9AmegdHMAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfhRh0fp/wDTGmXwZ87Y9H6I/kIH7zfkleX0T62z8wP3Imvz0+AMt5f2BqPoQ6uP8b/rPm68782Jv0v9hLz+KOOT/U46eQAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAmmt8871gAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAOaPT8cAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAACkj8fP2Cy6Lxtznwxnp0n2G0FwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAeJ/FuPR+nxtzgH5M/pHj0ehm3OAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAATTW+ed6wAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHNHp+OAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABNNb553rAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAc0en44AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAE01vnnesAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABzR6fjgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAATTW+ed6wAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHNHp+OAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABNNb553rAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAc0en44AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAE01vnnesAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABzR6fjgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAATTW+ed6wAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHNHp+OAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABNNb553rAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAc0en44AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAE01vnnesAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAg0AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAIUGAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAeTdi1sebADAAAAPWciXoNCjrhhboAAAAHnzPQRAAAEQZeyGHtgoHPgdABj7AHPgdAAAZ4Qa4UKobIAAAAAAAc3OG6AUQnnBlULoEcgQsB04AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB+cX2zc95+fQ88qQAAAP0d8tzqXMZxn0AHhf0KiGwn4R7uxK6LPy/S+mpJbogSYDFDxT6Fa+aPo4LNZPzga9IsJ+W+pNQeagvpoRWEzx5rz76iZ4NnM+iiGAVbQeEe7tAnVtB5NpUvRyXh+SNcD9VM+a9Jr1ClL8V+tGeU8yHnX2CASwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAD8/Pv8ApY3zMHjBcgAAAfo7QyvwOe1T7ZHQYiPPvVWfOf0CDOBD0zwoPSvPWfQ/jCKf0EGxiy/FPoVr5o99D547Vmz5aFX6HDwf1wPNMRr1qun577eHV+FI5P6VZ4BAz6SIZ85NbN9ni/uzPV8KH5jytrvNJHo5LAPnD06lynsiPnL6VD5o+lwOBHh+tCBMAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA+VPcKXnPUB8dlyAAAB+jplfzZpWn4gfS/gclHqmcV9Lo8Iqsvciz6h8GkxLFFnskc76Cjyb6LD5l9DZZ9OR5eBZzWecTNfSnmss45mf3Aes/OyPSPKWel+bB9Kkup8iUvU7Ych9LgcLL8/4G16/pI9HJZwDOM9Ba4X2JHzr9Jh80fS4HzKz6YdLAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADy/wA+pecfZwfn1RuQAAAP0dMr+c+xpdB4eH1T8lh1/Rhx/qYeG9w1Nww/ROAD3rxZHa+uInE/lnubXL++o+bPRmaT5LOazzj1MPPM9r0nyFPqOyD1v5sR9J/GDPs35qR2vqYX/mBnf9kjyft2en+Don82o9h0kcX72ji/lej2D16T562KXqfcTXzR9Li4Xk2ezeSI7voEwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAACDjmZfpgvP+CZ86ZVyAAH6OmVscAwJKoPkArA9QkArWgfAFe+AAmaGmBHTC8AIDZQo2wK4SvB8QSgEQeP8AsrQieZpgUgvAZ10FUILIMhAnBtcLgEUoRvBa4TqAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABjeJtd9oB89/QDOS+SqQNAH6OmVgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHhHyzpP0N6PL+Mve6XtHw4gKQB+jplYAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB8YeY6z9De3w/IfYQ8Y+bqQNAH6OmVnzw17hrpgAAAAAAAeG06Xe+gpggAADjA5D2Jrx72EATAAAAAAAAIg8l9faBPP8AKWvZvFgi9uA8yHxHr7XM8KFb3hHjG0z2MlgHlvLUvZ9SX55fa7QTAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA/OmDWPob2SK6f5vD5/S5AAD9HTK+R8IpexayORxWe6+PI5/2gOO9KH5r6UIE/m36JpfOPtQeY+hg7eTwO3QeDtcf9Ts+dfZg8q9ORg+lB5raDmvTQ8g6pndeNI9YwQ8/9WZF1ssA+eMq59Xy049YL3ap4vizX0O2X82/SLT/AJaZ9SkszA8F9+pfP3vQfNHpbPYiGAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB+cRrH0N2sV5f9Fh8NZ1yAAH6OmV/LnodKpmB6J5EHvXgIeu4iOE+lA+a/pQCmn8lTaT9SfNEv0aUOZGe86EM+dGuwrspcEz1LORy/0cHzvSZ7v4Uj6T+ag76YO68XD6F8dR2HZJnkjW15KzqfREV+IZ1vRo+ePXGekePI6juU/GeDtfRnHSHQB88ek0cFph6XziPZyWAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB+cRrH0N2sV8ifWlK78fgg0Afo7xOV+e+g0vKImfSfzbJ3+mHL98HiXsDPO/YkbPn48zlGvTfDw+gegl/LPbWvXL8OX50a+hvjBn0j4oH0Xop+Gdm1R4wPpP5lR9LfMzPp35eD0PTD0v5wR9Aonf8zawPIaXs11Otvh1GrLZ8y0vp75lD6cZLf80Ne+Vxmoiz4+1fy2c9ms+niGAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB8dmk+udLD+frtL6r8Fl8CWgD7E8vivTnI8wVr07zEdztg8/wDSheD7LOz4JHtfEJ9DzrOg8oEz3APHPYA807UNvzYfpPkYvVPIAqezh4PvM9F5FHp3mAcp64HB0Gew+Eo65zN2sjuhM8va9Q8QCb2APJ4WYvr6HZI+25BHmHQ0vXSWAeOaFL1Ml4eO12gmAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAf/aAAgBAQABPwCqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqrxb4itfB/hTWfFt9BLNb6Jp097NFFje6RRs7BckDJCkDPGa1H/AIKNaHExGkfCq+uRng3OrLAcfRY3qb/go5qrSE2/wmtEj/hV9ZZiPxEI/lWm/wDBRyFmSPV/hK6r/HLba1uPXsjQjt/tf/W8BftsfBDxtcR6ff6neeGbuQhVXV4gkTN7SozKB7uVq3uLe7gjubWeOaGVQ0ckbBldT0II4INVVVVVVVVVVVVVVVVV+13+1B4n+EWq2PgPwBHbQ6td2Yu7rULiIS/Z42ZlRY0PyliUJJYEAY4ycj9mj9sT4heJfiFpvgL4nXlrq1trkpgtr9bVLeaCcglARGFRlYjbjbkEg54INVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVcXFvawPcXU8cMUYy8kjBVUepJ6VrXxq8H6Y7QWLXGpSLxmBNqZ/3mx+gNTftAylsW/hZAo/v3hJP5JxVt+0AhIW88LkD+9Fd5/Qr/WvCHxA0HxqZYdLW5ingQNJFPFggZxnIJHX3z7VVeNfivZeD9ZGjHSpLyQQq7sswTaTnAxg9gD+Nf8NAWf8A0LE3/gUP/ia8E+MLXxrpDapbWxt2jmMTws+4qQAQc4HBBFVWqfHewsrqa0tvDk8zQyMhMlwI8kHB6Bqf9oG5LEx+FogvYNeEn/0AVD+0DIGxceFlI9UvMEfmnNeHvjD4R12VLS4ll02duFF0AEJ9nBwPxxXBHBqq1b456fpuqXenRaDLcLazNEJRcBQ+04yBtPHFf8NAWf8A0LE3/gUP/ia0bVINb0mz1a24ju4VkVc525HI+oPH4VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV8b/+SLeP/wDsV9S/9JZKr4Lfsg+K/jR4NTxtp3izSdNs5LiSBIp45Hk3IQCSAMAc+prxR+wB8YNHtJLvw9q+ga95Y4t4p2t5n47CRQnty47fh4g8O674U1e40DxJpN1puo2j7Zra5iKOh+h7HqD0I5HFfszftOa78HdctvD3iG+nvfBl5KEuLeRi5sCx5mh6kAZyyDhhnjdg1b3EF3bxXVrMksMyB45EbKupGQQR1BFVVVVVVVVVVVVVVVVftWfssal8a76x8ZeDdTtLXX7K1FpNb3hKRXUQZmUhlB2uC7DkYIIGRjn9nH9i3xR4I8c2Hj/4lX+mj+x3Mtjp1pIZmebBCvI2AFC8MAMknGcYwaqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq1bVLHRNNuNV1GYR29shZ27/QDuScAD1NeNvHur+M71jPK0FgjfuLRW+VR2Lf3m9/yxWmaRqetXS2Wk2M13ORnZEhYgep9B7nirb4KeObiIPLBZW5x9yW4BI/75BFXvwY8dWsZeKztrraM7YbgZ/JsV8FfDN9oWmajdarYzWt1cXAjEcyFW2IMggHsSx+uKpiqrkkAAck9q8WawfEHiTUdX3ZW4nYx5/uDhf/AB0CrzTrqxitJblNovIPOiz3Tcy5/NTXwI1oWut3uhyOAt7CJIwf76dh9VJP/Aaqta/5DF9/18yf+hGvAXw/n8dm+EOpx2f2ER53RF92/d7jH3f1rVfgRr9patPpmq2t9Igz5JQxM3suSRn6kU6PG5jkUqykhlYYII68V8EfFF3q+kXOiX8zSvppTyHY5PltkBf+AlfyIHaq8V6wugeG9R1csA1tbsY893PCj/vogV80jd2Zj9STWpafc6VqFxpl4oWe1laOQD1U4OPavgdrIv8AwtLpMkgMmmzkKvpG/wAw/wDHt9VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVfG//AJIt4/8A+xX1L/0lkqv2FP8AkgFn/wBhS7/9CFV+2l8HtK8efC++8bWlki+IPCkBuY50QBpbVeZY3PUqF3OPQqem41X7FXjafxl8CNNtryYy3Ph25l0x3Y8lUCvH+AjkRf8AgNfEH4heE/hf4XufF3jLU1s7C2IUYG55pD92ONerMcHj0BJwASPiR+3p8UPEd1NbfD+2tPC2m5IikMS3N0w9WZwUXPoq5GfvHrTftK/Hp7kXR+K2v7wMAC5wv/fGNv6V8N/28fip4bvYbfx8lr4p0wtiVjCltdIvqjoApx6Mpz/eHWvh98QvCvxR8LWvi/wdqIu7C5ypDDa8Mg+9HIv8LDIyPcEZBBP7UHjLxd8PvgprnjDwRqg0/VNPkttlwYY5dqvcRxt8sispyHxyO9W/7Z37RyTxPcfETzYlcF4/7Isl3DPIz5PGRXx5/bq1S21e58LfBVrRbe1Yxy67NEJjKw6mBGyu0f3mDbuwA5Mn7Svx6lu/trfFbX/MBztW52p/3wBt/SvgJ+3PrD6va+FvjVNbTWlywSLXY4hC0DHp56KApT/aUDb1ORkhHSRFeNwysMqQcgj1FftYftD+LvgTaaLb+FvDdjdS64s+2/vWZ0gaPblRGuMsQ4IJbHB4PNa7+1p+0Hr8rvP8R7y0RsgRWMMdsEHoCihvxJJ96079pj4+aXOLm1+Kuuu6kEC5nFwvH+zIGH6V+zx+27eeJtdsvA/xditIri+dYbPWbdPKRpTwqzJ0XceA64AOMgDLCv2tf2hvjV8Lvi5J4Z8F+MP7M0ptNguIoP7PtZuW3Bm3SRs3JU8E9q+DP7bfiTSNA8Xar8YPEDeIby3itv7CsI7aC1eaUmQSLmKNQF+4WZs7QOAScHxp+198ePGN9JcReM5tDtS2YrPSVFusY9N/Lt+LGvht+2d8ZfBGqQv4g12TxVpO4Cez1EgyFe5SYDerfXcPavjX+3B4a8MeHtNh+Fi2+s61rFmlyZLgHytOR1yBIoPzS/7GQFxknoDqn7Tvx91e9N9c/FPWopCc7LWUW8Y+iRhVx+FfAL9tzxdp/iGz8NfGDUk1XRr2RYV1RoljnsmJwGcqAHj/AL2RuA5ycbSCGAKkEEcV+2R8f/i58Kfidpnh3wD4u/svT7jQYrqSH7Bbz7pWnnQtuljZh8saDGccdOTXwJ/al+PHjP4v+FfC3iXx39s0zUr9YrqD+y7SPzE2njckQYdB0Irxrd6jp/g3Xr/RrgQ39tplzJaysoYJKsTFGwQQcMAcEEV/w2Z+0p/0Un/yj2P/AMZrxP8At16hofwz8Naf4dFrrPjW80yN9W1C4RfItZeQf3aBQZDjdtGFXI68gan+078fdVvTf3HxT1qOQtkJbSi3jHP9yMBce2K+B37cvjLSdctNC+L14msaLcyLG2peSsdxZZ4DtsAEiDvkbscgnG0xSxXESTwSpJHIoZHQ5DAjggjqMV4n8TaD4N0C98T+JtThsNM06Ey3FxKThFHoBySTgBQCSSAASa+K/wC3n8QPEGoT2Hwthi8OaShKx3U0KT3cw/vHcGSMEdgCR/e9P+GkPjx9p+1f8LX8Rb89Pth29Mfd+7+lfBf9u7xbpeq22ifGFotX0mZgjarDbiO5tc8bmVAFkQdwFDdTljwbG9s9TsbfUtOuo7m1uollgmjbcsiMAVZSOoIIOfeqr9p/9pH45fDf41694T8JeNxYaPbx2r2lsNNtZfLD28bPlpImY5cueSevpxXwW/bZ8RaV4f8AF+r/ABi8Qt4guraO1Gh2MdtBbSSyMZRIo8uNfl/1ZZmztC8AlsHxx+2L8dfGd9LLaeLG8PWTN+6s9JQQiMdsycyMfXLY9h0rwR+2D8dvBt/DNc+MJdfslbMtnqyiYSDPI8zHmKeuMNj2PSvg58WfD3xo8DWnjPQAYS7GG8s3YM9pOoG6MkdeCGB4yrA4GSBX7RH7alh8PdTufBHwytbTV9ctWMd7f3GWtrNx1jVQR5jjvyFU8fMcgaz+1F8f9duTc3fxR1iFichLNltVHsFiCivA37Znx08HXsUl/wCJh4jsVIElpqsYk3DpxKAJAcd8kZ5INfBn4yeFfjb4Qj8UeG2aGWJhFf2ErZks5sZKk/xKRyrDgj0IIH7UvjnxV8OPgxq3izwXqv8AZ+q2txbJFceRHNtDzKrfLIrKcgkciv8Ahsz9pT/opP8A5R7H/wCM18GvEGr+LPhR4S8S6/efatS1PSLe4up/LWPzJGQFm2qAoyewAFftkfEzxv8ACn4YaZ4i8A65/ZeoXGvRWsk32aKfdE0E7FdsqsB80aHOM8deTX/DZn7Sn/RSf/KPY/8AxmvC17c6l4Y0fUb2XzLi6sIJZX2hdztGpY4HAySelfG39qT9oDwV8WvFfhXRPHwtdP03U5YrSAaXaSeXFnKLuaIscKR1JPvXwv8A2w/EOi/A3XfHHxL1WPxF4gj1s2WkWvlRWzS5hRsMIlUCNTuYtjPOM8jHif8Aav8Aj54o1N9Rk+IV9piF90drpmLaKIdlAXlh/vFj6k0zKqlmYAKMknjFfG/9vC10HUbnwx8HrGz1Sa3Yxzazd5eDcOCIUBG//fJ28cBgQa1j9qX9oDXJnmuvihqsJY/ds9lqq/QRKv8AnrXhX9sX4/8Ahe6SV/Gn9swAjda6pbJMr4/2gBIPwYV+z3+1D4U+OcLaPNbDRvFFtFvm055NyTKOskLfxAcZU/MPcc1XjT9rf9o7QvGGu6Jb/EURxafqVxbxoNJsmCqkjKACYcngdTXhj9ubVPD3wYjvvEtzF4m8e3OoXEMMUkccEcMKhCssyxKuF+cgKBltp+YYrxH+1X8ffEt693P8R9RsVLEpBpxFrHGPQBACR/vEn3r4XftufFnwdqlvF401E+KtELBZ4blEW4Re7RygAlh6PkHpxnI8MeJdG8ZeHdO8U+Hr1brTtUt1ntpV4yrDuOxHQg8ggg9K+J/xL8MfCTwdd+NPFc7raWxCRwxAGW4lb7scYJALHB74ABJ4Br4g/tvfGnxdfSjw3qUHhXTdx8q2sYlkl29t8zgkn3UIParX9pD48Wcyzw/FfxEzL0Et4ZV/FWyD+VfsY/HD4r/F+61+x8dX1lqNhotvCVvBaLDMZZGYKpKYQrtRyflznHNVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVXx61+TzLDwzBKQmz7TOAepyVQH8mOPcVY2VxqV7b6daJunuZVijU8ZZjgfzrwn4U0zwjpMem6fGpfAM8xX5pX7k+3oOwqqqviRrP8AYfgvUrpH2yyxeRFjruf5cj6Ak/hSI0jrHGpZmOFUDkmvjB4XXS/DPh24hQZ0+IWcpA6/KCD+at+deFdYbw/4j07WAxC206mTHdDw4/FSRSsrqGRgQRkEd6rWv+Qxff8AXzJ/6Ea/Z9/5j3/bt/7VrWNc0jQLN73V7+K2iQZ+duW9lHUn2Fa3fpqus3+pxRmNbu6kmVD/AAhmJx+tfAfQrm2sr/X7iMrHdlYrfIxuC53MPbJA+oNV8eNZ+zaHZaJE+HvZjI4H9xB0P1Zh/wB814F0s6z4v0mw2bla5V5BjqifM36Ka+NOkf2d4zku0TEeoQLMMDjcPlb/ANBB/GvgtrQ0zxgthI+ItShaLB6bx8yn9CP+BVVV8U9c1nw74W/tPQ7wW1wtyis5jV/lOcjDAjritN+LfjltRtVvdfDW/np5ym2hUFNw3chOOM8140+NWoS3Uun+ESkFvGSpu3QM8hHdQeAPqM/So/iL44im89PE96Wz0Z9y/wDfJ4/Svhr8VZfEd0mgeIVjS+ZT5E6DaJiOqkdmx6cH2714u8YaT4N0z7fqTl5HJWC3Q/PK3t6Adz0H1IB1v4w+M9Wmb7Hepp0Gflit0GQO2WOST9MD2q2+I3ji1kEsXia9Yg9JX8wfk2RXw8+Li+ILqPQ/ESRQXsnywTxjakzf3SP4WP5H24zXib4o+O9P8Sarp9nrvlwW19NFEn2WI7VVyAMlcngDrXwh8aeJvFOpahb67qf2pIIFaNfJRNpLYJ+VRVXXxZ+IMdzLGniDCq7AD7JDwAf9yvg/4r1/xVbanJr2ofamtnjER8pE2ghs/dA9B1r4veJdf8L6TY3mg34tmluDHIfKR8jaSOGB9DXgz4n+M9S8U6bYarrqyWk022VWt4kBGD3Cgj868Z/GXWr+9ls/C9x9isYmKrMqgyTY/iyfug9gOfU9h8GNZ1bW9AvZ9X1Ge8kju9qPM5YgbFOM+leNvH2keCbVTdA3F7MuYLVGwzDpuJ/hX3/LPNax8YfG2qSH7Nfx6fFniO2jA/Njk/qKg+I/jm3k82PxNeFs5xIwcfkwIrwj8cJzPHZeLoYzGxwLyFNpX3ZR1HuMY9DUUsU8STwSLJHIoZHU5DA9CD6V8YfFviXwrLpR0HUhbJdLN5g8lH3FdmPvKcfe7V4O+LfiJvEEC+KddD6aI5TMDbxp0jYrjaoOSQAB3JrxN8Z/E2q3Dx6HINMtAfkCqGkYerMc4+gxj1NWXxK8c2Ewmi8R3UuOqzkSqfwbNfDn4hweNrSS3uYkt9StVBljU/K69N698Z4I7ZHrVeO/jNNY3s2j+FEiZoGKS3kg3DcOoQdDj1P5d6uPiJ44uZPMk8T3wY9o5Ng/JcCvD/xQ8fR39tZQ6sL0zyrGsV1GHDEkAcjDd/WscYzVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVfG//AJIt4/8A+xX1L/0lkqv2FP8AkgFn/wBhS7/9CFV8QorefwD4lguwpgk0e7WUMcDaYWByewxVf8E7DOfAXitW3eSNXj2ZHG7yRux742/pX7b/AMUL7xn8Wrjwbb3bf2P4TAt4olPyvcsoaaQj1BIj9tnHU5+Anwmn+NHxL07wWLiS2sirXOo3EYG6K3TG4rn+Ikqg4OC4JGAak/ZQ/Z+fw+fDq/DiwWIptFyrP9pBxjcJs789+uM9q+N/wuuvg78SdV8DTXD3NvbMstlcuoBmgcZRiB3HKn3U9q/Ya+Jt94S+LEXgie5c6T4rjaFoi3yx3KKWjkA9SFZPfcM9K/bM/wCTa/GH/bj/AOl0FV+y1+yFofxM8MRfEb4j3N5/Zd3I66bp1tJ5RuFRirSSP94LuDAKuDxnOMA/Hz9iHwZpngnUPFnwpF9Z6ho9u1xLp01wZ47qJBl9pb5lcKCRyQcYxzmq/Ys+IF346+CNlaanctNe+G7l9Md3OWaNFVoifYI6p/wCvjN8FPC3xw0TTNB8U3N3b2+m6gt2HtCFkcBGVo9xBwG3DJxn5R9af4Rfso/CawSDX/DfgmwXZw/iB4p5JPcG4LEk47fhX7TEf7Imv+B729+HOq+HNP8AFlkEkso9GtmhjuQGAeNljUR8rkg8HIHOCQQSCCpwR0r4HeKrnxv8IPCPie+mMt3eaVD9pkJyXmQbJG/FlY/jX7ff/Jc4P+wDbf8AoyWvhn4A1X4o+O9H8B6LLHFdatOU82QErCiqXkcgdQqKxx3xjvWtf8E//hbL4Rl0/QNY1qHX44D5GoXFwrJJMAcb4wuAhPZcEDHJ5zeWlxYXc1jdxGKe2kaOWNuqspwR+Yr9l39n6L48+Kr6LWL+ez0DQ4kkv5LfAlkeQkRxoSCFzsclsHAXGOQa/aO/Yx8FeDPh1f8Ajr4bT6lDcaIgmvLO5m89Z4MgOynAKsoO7rghTxnmq/Zt8SXHi34FeDNau5zNOdNW3kkJyWaBmhJJ9cx8+9f8FC/+S06L/wBivB/6VXVfsw/8l+8D/wDYUX/0Fq8b/wDIl6//ANgu5/8ARTVX7LH7M8Hx0vNQ1vxLqNzY+G9IdYn+y4Et1ORny1YghQq4LHBPzKB1JH7Rv7F/g3wZ8Pb/AMd/DW61GGbRI/OvLG6m85ZoQQGZTgFWUHcckggHoetfsjeKLnxX+z94Wub2cy3NhFJYOzHJxDIyRj/v2Er/AIKA/FG71DxPpnwm065K2Olwre6iqtjzLiQHy1YeiR/MP+uvsK/ZZ+C9j8a/iUuka6ZRoWk2xvNRETFGmUMFSIMOV3MeSOdqtgg4I1P9m74FapoMnh6T4W+HYIHi8sT21ikNwnowmUCTPuW575r4o+CJfhv8Q9f8DS3DTjR714YpWXBkj6xsR2JQqT9a/YJ8f3Xij4VXvhHULgyz+Fb0RwljkrbTAtGPwdZQPQADtVV+25/ycTr3/XrZ/wDpOlfDfwFq/wAT/HGj+BNCZEu9Wn8sSOMrEgBZ5CB1CorNjvim/wCCf/wePh06amseIP7VMWBqRnQ4kx18rbt25/h6443d68a+E9S8C+LtY8G6uVN3o15JbSso+VyjYDL7MMEexFf8E8PFFza+PPEvg5pz9m1HSlvFQnjzYZFTgepWZs+oX2FftTfE+5+FPwc1XWtLuPJ1bUWXTtOkBwY5ZQcuPdY1kYe6itO0+91vVLXSrCMz3l/cJDCmeXkdgqj8SRXw1/ZX+DvgLwxa6RqHgnRfEOoiMG91DVbFLpppcfMVEgIjXPAVccYzk5J/bX+BPhj4W65o/irwRYLYaVr/AJsc9jHkx29xHtOUz91XVvu9AVOMAgD9ijx/deDfjdp+itcMun+KI2sLmPPBkwWhbHqHAX6O1ftuf8m7a9/19Wf/AKUJVfs8/wDJC/Af/YBtf/RYr/goX/yRbRf+xog/9Jbqq8Ef8iXoH/YLtv8A0UtftPf8l+8b/wDYUb/0Fa8H+FPEnj/xFpvgrwvaSXuoahOVtrfdhQxGWY54UBVyx9F9q0n/AIJyzy6Wj678VEg1F0y0drpPmxRNjkBmlUuPfC1+3H8UL7wH8KovDei3bQah4tne0aRWwy2qLmfb9dyIfaQ0qs7BEBZicAAck18Fv2PPhj4Q8HWMnjzwtaa/4ku4Fkv3vh5scDsMmKNM7cLnG7GSRnIGAP2yv2a/DfwytrH4h/D6yaz0e9uPst9YBiyW0pBKOhJJCttYEdAQMfewPCfijWfBXiXTfFnh67a31HSrhZ4HBI5U8g46qRlSO4JHevBviWz8Z+EdF8XWClbfWbCG7jQnJQSIG2n3GcH3FfFP/kp3i/8A7D19/wCj3r9nb4H3nx28df8ACPG+ew0qwg+06ldooZ0jyAEQHjexOBngAMecYPxE/YH+Hb+DryT4eXusW2v2du0lqLq4EyXbquRG42jBbGAy4wTnBHFV/wAE+vE9zqvws1jwzczmQaHqxNuCf9XFMgbaB2G9ZD9WNf8ABRnVtROpeCtBDutgILq52gkLJLuRckdyqjj03n1r4dp4Pk8daEnxAklTw4b6Mak0W7Ihz833fmx67ecZxzitL+BX7MvjXw/HceH/AAJ4T1PTJkCpdaeFJIA/56xndn15z618KPgv4I+DFlquneCLe6ig1a8+0yLcTeaY8IFVFYjOwYJG4k5Y8+lVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV8YZml+IGoRt0hSFF+nlKf6mvhTBHcfEDSI5BkB5GA91icj9QKr4yeK/EPhq50uPQ9Te1WeOUyBUU7iCuOoPqa/4Wl4+/6GSb/v0n/xNeB/iF4y1XxbpenX+uyzW884WSMxoNwwfQZqvj5rIJ0zw9FJ03XMq/8Ajqf+z18NdH/trxrplsy7o4ZfPkyMjEfzc+xIA/GviHo/9ueDdTskXdKsJliHfcnzAD64x+NV8NNZ/tzwXpty77pYI/Ilz13J8uT9QAfxqta/5DF9/wBfMn/oRqOaaHPkzOmeu1iM1I7ytvkdnb1Y5NeHLrw/Z6lHP4j0y4vrVTzFDN5f4kY+Ye2V+teHNX0TW9IgvPD8sTWYXYiIuzysfwlf4SPT+lV8YNZGr+NbmGOTdFp6LbJj1HLfjuJH4V8BtH+0a3f6065WzgESEj+Nz1H0Ckf8Cr476P8Aa/D1nrMa5ewn2OcdEkGCf++lX8602+m0vULXUrY/vbWZZU+qkEfyqyu4NQsoL62bdFcRLJGfVWAI/Q1VfGv/AJEWX/r5i/nVfD/4P6drGiw654jmnP2td0FvE2zCdmY4zk9QB2x68fFD4d23gyW2vtKnlksbtim2U5aJwM4z3BGcfQ1Y3k+nXtvf2r7ZraVZIyOMMpyP5VBMlzBHcRnKyIGX6EZr4h+JZfFHim7vBLutoHMNqueBGpxkfU5P418H/h/peuW03iPXbVbmJJTFbQOPkJAG5iO/XAB46+1eNfhr4a1bQbo6bo9pZXsETPBJbQiLLAZ2kLgEHGOemaR3idZInKuhyrA4II7ivCGsnxD4Z03WHIMlxAPMwMfOPlb/AMeBrxn/AMjfrn/YSuP/AEY1fAL/AJDGq/8AXsn/AKFVXv8Ax+T/APXVv51+z/8A8eetf9dYf5NXx7/5FvT/APr+/wDZGpWZTuRiCOhBxXg34Pap4n0+PVr+/XT7WYZhBi8x5F9cZGAexP5V4M8I2/w+0e9gfVDdQtIbhnaLZsAQA9zn7ua8Sa7d+JdautZvHJe4clVJ/wBWv8Kj2AwK+E3gCy8V3Fxquso0lhZsEWIMV82QjJBI5wBg++R714k+E/hTWNLkt9N0yHT7xE/cTQrtww6Bh0IPfPPvU8MtvM9vMhWSJirqexHBFfAzxHNqGkXXh+6kLtpzBoCxyfLbPy/QEf8Aj1ftB/8AMA/7ef8A2lSo0jLGilmY4AA5JrQPgboMemRt4gnuZr2RAZBFIESMkdBxzj1PX0rx94QbwXrzaWk7T28sYlt5GGCVJIwccZBBH5HjNfDPUpNM8caTIj4E8wgcZxuD/Lj8yD9QK8XXVxZeFtXvLQkTQ2MzIynBUhDz+HWq+FOj/DvVtAhgubWxutXLN9ojugC+cnG1T/Dtx0/Gl+GPg6DV7XWrLTPstxaSiRVichGYdMqcjjrxiqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr43/8AJFvH/wD2K+pf+kslV+wp/wAkAs/+wpd/+hCq/aq8fWPgD4H+JLia4CXms2r6ZYx5wzyTqUJX/dQu/wDwGq/YX8KT+HPgTBqV1Fsk8QalPfJuXDeWAsS/gfKLD/ez3r4vyXE3xZ8ay3gYTt4ivzIGXaQ32h88due1f8E7I7VviB4qlcL9pXRkWMk87DMu7A9MhP0qv+Chsdqvxa0CSMKLh/DqeZg8lRcTbSR/31z/AIV8BvtP/C7fAP2TO/8A4SSwzj+756bv/Hc1+2Z/ybX4w/7cf/S6Cq/ZhAHwA8DgAf8AILX/ANCanRJEMbqGVhgqwyCK1FLZNQuUsyDAsziLByNuTjB78Yr/AIJzi5/4RjxoWP8Ao/2+18vp9/y33e/TZ/nNftf/ALVWu+ENZm+Ffwz1AWd/BGp1bVIjmSFmGRDEeittILOORuAGCDWlaP4v+IniIWGkWWp+INbv3LFUDTzSnuzHk49WPA7mta/ZD+OHhrwlqPjLxD4fsdPstLtXubiOTUInlWNBkkKhYE47Zqv2S/8Ak3bwX/16zf8ApRJX7ff/ACXOD/sA23/oyWv2JP8Ak4nQf+vW8/8ASd6r4pAD4m+LlUAAa9e4H/bd6/4Jyf8AIE8c/wDX1Zf+gy18cVV/gr4/DAH/AIpfUjgjuLaSq/Yz/wCTa/B3/b9/6XT1/wAFC/8AktOi/wDYrwf+lV1X7MP/ACX7wP8A9hRf/QWrxv8A8iXr/wD2C7n/ANFNVf8ABPX/AJItrP8A2NE//pLa18cAG+C3j8EA/wDFL6l1/wCvaSq/YU/5IBZ/9hS7/wDQhX7VlxdXX7QvjWS7DCRb5EUMD9xYY1Tr22hfb04q2vLyzYtZ3c0DMMExuVJ/Kv7c1r/oMXv/AIEN/jU001xIZp5nlkb7zuxJP4mv+CcSXB1Dx66E+QIdPDjPG4mfbx9A3+TVV+25/wAnE69/162f/pOlfsSf8nE6D/163n/pO9V+1oAP2ifGgAH/AB9Rf+k8dfsC/wDJc5/+wDc/+jIq/wCCjFxdL4U8GWiA/Z5NQuXkODgOsahfboz/AOc0jtGyvG5VlOVIOCCO4r+3Na/6DF7/AOBDf41c6hf3qql3fTzhTlRLIWwfbJr4DpcP8bfAK2pO8eJLAnBx8onTd/47mv23P+Tdte/6+rP/ANKEqv2ef+SF+A/+wDa/+ixX/BQv/ki2i/8AY0Qf+kt1VeCP+RL0D/sF23/opa/ae/5L943/AOwo3/oK1/wT7tLa5+NupTTwq72vhu4khYjlHM9uhI99rsPxNV/wUYkuG8U+DIWDeQun3LJleNxkTdz9Av6etfDmO1m+IXheG+Cm3fWbRZgxwNhmTdn2xmq/bUjtZP2cPFDXAUyRy2RgycEP9riBx77S361X7Kv2n/hnrwT9qzv+wNjP93zX29P9nFfFP/kp3i//ALD19/6Pev8AgnJ/yGvHP/XrZf8AoUtVrYA1m/VQABdSYH/AjX/BOT/kCeOf+vqy/wDQZa/aR+A9l8d/BUelQXUNlrulO02lXcoJRWYAPG+MkI+FyQCQVU4OMHx98MfHfwx1Z9G8b+G7zTJgxEckiZinHrHIPlcfQ/XFeF/GPivwTqS6v4Q8RahpF4v/AC1s52iLAdmwcMPY5Ffsn/tUXXxclbwF47WFPE9tCZba6iQIl/GuN2VHCyDqQOCMkAYNVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVXxu017Txmb3Z8l9bI4bHUqNhH5KPzFeFNYXQPEenaw+SltOrSADnYeGx/wEmopYp4kmgkV45FDIynIYHoR7V4s8BaH4zktpdXe5DWqssfkyBfvYznIPoK+Kfg3SPBuoWNrpD3DJcQs7+c4Y5BxxgCvhx/yPOjf9fI/kar4j6z/bvjPUrtJN0UUpgi542p8uR7Egn8a03VtT0a4N1pN/NaTMhQvE5UlTgkZHbgUfHnjNhtPijUjx0Nw1V8A9Z2zal4ekfh1FzEM9xhX/mn5VWtf8hi+/6+ZP8A0I18BbKzvP7d+12kM+z7Pt8yMNjPmZxn6CtV8FeFNagaC/0KzbIwHSII6/RlwRXjPw6PC3iS90RJTLHAwMTt1KMAwz74OD7ivgtrlxp3i6PShIfs+pxsjoTwHVSytj14I/4FWq6hDpOmXWqXBHl2kLysM4yFGcfpV3czXt1NeXD75Z5GkkY92JyT+ZrTPEevaLE8GkaxdWiSNudYZSoJ6ZOKvvF/ijU7R7HUNfvri3lADxSTFlbByMj6gVXwb1n+1fBcFq75l06RoGyedv3lP0w2P+A1VfGv/kRZf+vmL+dV4N/5E/Qv+wbb/wDota+PIB8H2f8A2Ek/9FyVVvJJF4XilgBMiaeCgHqI+OlUlzcRLsiuJFHorEUby7IwbuY/8DNV8HxIPh9pxc8F5tmT281v65rxn/yN+uf9hK4/9GNXwC/5DGq/9eyf+hVV7/x+T/8AXVv51+z/AP8AHnrX/XWH+TV8e/8AkW9P/wCv7/2RqrTYo4NOtYIUCpHAioo7AKABXjhpF8G62Ys7v7PmHA7bDn9M1XwRWIeCAY8bmu5C+PXj+gFV44WJfGOtrDjaL+bgeu85/XNfAXzP+Eqvdv3P7PbP18xMf1r9oP8A5gH/AG8/+0q0b/kMWP8A18x/+hCq+PwH9saVj/n2f/0KvBv/ACN+hf8AYSt//Ri1NDFcQvbzIHjkUq6sOCCMEflXjn4V614bupbzSraS90tmJR41LPCPRx14/vdPXHSgSpBU4I6Edq8F/FnXfD1zFbavdS6jpxOHSVt0kY9UY88eh4+nWra5gvLaK7tZVkhnjDxuOjKRkEfhVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVfG//ki3j/8A7FfUv/SWSq/ZI+Pnwg+HnwctvDfjPxtbaZqSahcSNbyQSsQrMNpyqEc/WvGH7cfwJ8OWkjaHq194kvFU7ILGzkjUt23SShQB6kbseh6V8b/jt4v+OviGPVvEJjtLCyDLp2mQMTHaq2MnJ+85wMsQM4GAAAK+Cfwf8QfGrxxaeFtHhlSyR1k1O+C/LZwZ+Ziem44IVe59gSNF0fTfD2j2Og6PapbWOnW6W9tCo4jjRQqgfQAV+2X8OrzwN8bNV1RbYrpvihjqNnKBwztjz1z/AHhJuJHo6nvX7N/xci+C/wAUrHxVfRySaVcxNZamkYy3kOVJZR3KsqNjvtI71J8efgvHoP8AwkrfFDw39g8rzAwv0MmPQRA793+zt3Z4xX7QnxUX4x/FPVfGNrE8Wn4W206OQYZbeMYUkdixLOR23Y7V+w78OLzxh8YYPFkkB/szwnE1zNIy/K07qyRJn1yWf/tnX7Zn/JtfjD/tx/8AS6Cq/ZH/AGqPA+ieB7P4ZfEfWU0e50hmTT764B8meFmLBGYZ2MpYjLYXaBzmv2gv2wPh1oHgnUdC+HXiS317xDqdu9vDJZEtFZh1IMzSY2kgE4UEnOM4FV+xV8P7vwN8EbK81OAxXniW5fUyjDDLE6qsQ+hRA/8AwOvjW2oP8YfG7aru+1f8JDfCQN2/fvjHtjGPbFfsDeN/AfhLxb4lsPFeqWWm6jq1rbpptzdyCJCqM5liDsQAWLREA9dnHSv2sfj58PdB+FuueDdM8RWGra74gtGs4rSynWbyUfh5JCpIQBc4B5JI4xkiv2S/+TdvBf8A16zf+lElft9/8lzg/wCwDbf+jJa/Yk/5OJ0H/r1vP/Sd6r4p/wDJTvF//Yevv/R71/wTk/5Anjn/AK+rL/0GWvjf/wAkW8f/APYr6l/6SyVX7Gf/ACbX4O/7fv8A0unr/goX/wAlp0X/ALFeD/0quq/Zh/5L94H/AOwov/oLV43/AORL1/8A7Bdz/wCimqv+Cev/ACRbWf8AsaJ//SW1r43/APJFvH//AGK+pf8ApLJVfsKf8kAs/wDsKXf/AKEK/by+H914c+LieNIrbGn+KrRHEijgXEKrHIvsdojb33H3r9mv42j4GfEH+376zlvNH1G2NpqMER+cIWDLIgJwWUr0PUFhxnI0D9o74E+JLBL+y+Kvhy3R1B8vUL5LKRfYpMVbP4Vrn7SnwF8PQNPffFbw/MqDJWxuhesfosO8mvD+uaZ4o0HTvEmi3Hn2Gq2kd1ayFSpaORQykg8g4I4PSqr9tz/k4nXv+vWz/wDSdK/Yk/5OJ0H/AK9bz/0neq/a1/5OJ8af9fUP/pPFX7Av/Jc5/wDsA3P/AKMir9t34f3Xjb4KT6pplv5t54Xul1HCj5jAFZJgPYKwc+0dabqF3pGo2mrafMYrqynSaCQdUdGDKR9CBXwq/bF+EXj7RbY+JPEVn4W1wIq3dnqMnkxB+haOZvkKE9MkMO47m7+PPwRs4DPN8XfB7KvURa1BK3/fKMT+lfD34u/Dz4qvqa+AfEKasujvGl26QSRqrOG243qNwO1uRxx1r9rTw/P4k/Z68YWdrHvmtraO8XAyQIJkkc4/3Eeq/ZX/AGsfhz4e+Gtj4B+JOtHRr3Qg0VrcyQSSR3MBYsvKBtrLkrggDAUgkkgfth/tK+Gvi/FpvgjwGZLnQ9Muvtk9/LE0X2mcIyKI0YBgqq75LAElugAy1eCP+RL0D/sF23/opa/ae/5L943/AOwo3/oK1/wT1/5LTrP/AGK8/wD6VWtV+318OrzxP8N9M8b6ZbGafwrdP9pCjkWswUO3vtdI/oCx7GopZIJUngkZJI2DIynBUjoRXwX/AGn/AIb/ABK8HWV9rXizSdF1+CFU1Kxv7tLdhKBhnj3kBkY8grnG4A4NftuftC+FPGel2fwv8Bazb6rbw3YutUvrZ98JZAQkSOOHGW3ErkZCjPUDw54e1XxZr+neGdCtWuNQ1O5S3t4wDy7EAZ9B3J7DJrwT4YtfBXg7Q/CFm4eHRdPhtFfbt3+WgUsR6kjJ9zXxT/5Kd4v/AOw9ff8Ao96/4Jyf8hrxz/162X/oUtVrn/Ia1D/r6l/9CNf8E5P+QJ45/wCvqy/9Blr4o/G7wB8HLjRIfHmoT2ceuySxwTRwGVY/LClmcLltvzqMgHkirX4j/Bzx7pDxQ+M/Cms2E64lt5ryGQEejxucj6MK/a50H4P6D8SIYPhFc2BhmtN+pW+nTCW2gn3HAQqSFJXGUBwvHAziv2S4tRl/aI8GDTCwlW6lZyO0Qgk8z8Nm4fjVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV8V/BsvivQFm0+LfqGnkyQqBzIp+8g9zgEe4A70ylTtIwRwQR0rwF8Wb3wtbppOqwPfaen+r2tiSEei54I9jj69qt/jF4AniDy6vLbsR9yS1kJH/fII/Wvi54s0TxXqllNody88dtCyO7RsgJLZ4Bwf0r4cf8AI86N/wBfI/ka8W6wPD/hnUdXDgPb27GMn++eE/8AHiKJLHJOT718MPhz4c1TwnDquvaSl1Pdyu8bO7KVQHaBgEd1J/Gv+FW+AP8AoXIf+/kn/wAVXxW8L2fhfxP5GmW4hsrqBZYUBJCn7rAE89Rn8a+H+s/2D4w0y/ZsRmYRS5OBsf5ST9M5/Cq1r/kMX3/XzJ/6Ea+COvaJof8AbX9sarbWfneR5fnSBN2PMzjPpkfnWqfFDwPplu0za7DcsB8sVt+8Zj6DHA/EivFWvz+Kdfu9cnjEZuHGyPOdigAKPyA/Gvgvok+peMI9SCHyNMjZ3bHG5lKqPryT/wABr42ayNO8If2dG4EupTLHjvsX5mP5hR+NAFiFUZPYVo3wo8HQ6RZRaloUc14sCefI0jgtJgbjwfXNf8Kt8Af9C5D/AN/JP/iq8SaS2ha9f6O2f9EnZFJ7rn5T+Iwa+BWsiz8RXOiyN8moQbkGf448kf8Ajpb8qqvjX/yIsv8A18xfzqvBv/In6F/2Dbf/ANFrXx5/5E+z/wCwkn/ouSq0hVfRbFGAINrGCCOvyivFOhzeHNfvdGmUj7PKRGT/ABIeVP4qRXwp+JGn+GbeTQddZ0tJJTJDcBS3lEgAhgOcHAPHQ59eI/G3g2aMSJ4q0kAjgNeIp/InNN8Q/Bf22306HxBbz3F1KsUSQ5kBZiAOVBA5Pc1XxHsH0/xxrEDrjzLlphx1D/P/AOzV8N/F0Pg7xEL68RmtLiIwz7RkqCQQwHfBA/AmvEPxj8J2GmSS6Lff2hesuIYlidQCehYsBgD06/zp2aRi7NlmOST61+z/AP8AHnrX/XWH+TV8e/8AkW9P/wCv7/2Rqqy/484P+uS/yq+s4dQsriwuBmK5iaJx7MMH9DWr6Xd6LqdzpN8m2e1kKOMdcdx7EYI+tfB3x1p/h6S40HWrhYLW6kEkMznCpJgAhj2BAHPQY968R/ETwvoGmSXi6vaXc+w+RbwTCRpG7fdzge5q6uJby5lu533SzOzucdWJyTXwJ8Py2elXniG4Tb9uYRwAjHyJnLfQscf8Br9oP/mAf9vP/tKtG/5DFj/18x/+hCq+P3/IY0r/AK9n/wDQq8G/8jfoX/YSt/8A0YtXVxFZ2st3O2I4I2dz6ADJ/QVovxI8Ga7GrW+uQQSEcw3TCFwfTng/gTXxTs/AV34eu7+ebT11NUzbSQOolkfsCF5YeuenXiq+HC3C+BdGW5J3/ZgRn+6Sdv8A47iqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq+LWl6hrnwq8Z6LpNq91fX/h6/t7aBB80sr27qqj3JIH41ffA/wCM2nBjefCfxcioMs40WdlH/AgpFf8ACrPid/0TjxR/4J5//ia0b9m/48a9KsNj8KPEUbFtoN7ZmzGfrNtAHv0r4d/8E/PHOrXEV38SvEFloVkDl7WyYXNy47jd/q0yP4sv/u18Ofhj4K+FHh2Pwz4I0eOytQd0rk75biTGC8jnlm/QdAAOKr4w/B7wn8avCUnhbxRE0bI3mWV7Co820lxjcpPUHoyngj0OCPiR+x/8avh/dzGz8NTeJdNVj5V7pEZmLL6tCMyKcdeCPQmk+HfxAlufsUXgXxA9xkjyl0uYvkdRjbmvhh+xf8Y/H15DLr2jv4T0kkGW51RNs2O4SD75b/e2D37V8Lvhd4T+EPhK38IeELRkt42Mk08pDS3Mp+9JIwxknAHoAABgCv2zP+Ta/GH/AG4/+l0FVqH7LfxNuPAXh/4j+DNHn8R6XrVgk8kFkhe5tZOQymIfM6kjIKg9cEDgm1+GHxJvrz+zrL4feJJ7rODCmlTFwfcbciv2f/2H/EF/qtn4r+Mtmlhpluwlj0QuHmuiOQJtpIROmVzuPIO2kjjijWKJFREXCqowAB0AHpX7W37JmveNNdm+J/wwsku9QukH9q6UGCPMyjAmiyQCxUAMnBJGRkk1d/DH4k2N7/Zt58PvEkN1nAgk0qZXP0G3PavhF+xV8TvHlx9v8Z6dc+FNHSNiPtcey6nbb8qrEeU5xlnAx2B6V/wzx8c/+iTeKP8AwWyf4V+y3pGvaB8B/C2h+J9HutL1OxS4jntLmFonj/0mUqSG55Qqfxr9tT4TfEzxt8YYdY8I+Bda1exXRYIjcWdm8qBw8hK5A6gEce9fsjfB74p+EPjno2u+KPh/rul6fDbXSyXN3ZPHGhaBwoJIxySBVfEb4C/GnUviF4o1DT/hd4kuLa61m7lglj092WRGmYqwOOQQQa/YR8AeNvAekeMIfGfhXU9Fe8ubRrdb22aIyhVl3Fc9cZH518XtOvtY+E3jXSNKtJbq9vfDt/BbW8Slnlke3dVVQOpJIAHqa/4Z4+Of/RJvFH/gtk/wr9lPw7rvhT4BeFvD/iXSbrTNStftnnWt1EY5I913My5U8jKsp+hFftw/Cv4keOfixpOreDvA+s6zZw+HYYJJ7O0aVFkFxcMVJA64ZTj3Ffs9fBL4veHvjV4Q1vXfhv4gsbCz1FXnubiwdEiXaeWJHArxfbT3nhPWrS1haWabTrhI40GSzGNgAB6k1/wzx8c/+iTeKP8AwWyf4V+w/wCDPFfgf4TarpHjDw9f6Ney+Ipp47e9gMTtGbe3UOAexKsM+oNfF7Tr7WPhN410jSrSW6vb3w7fwW1vEpZ5ZHt3VVUDqSSAB6mv+GePjn/0SbxR/wCC2T/Cv2OfCviTwb8FLXRPFeh3mk36ajcu1tdwmNwrMMHB5wa+KXww8K/F7wfc+DPFts720zCSGaIhZbWVc7ZIyQcMMkdMEEg8E18TP2MfjL4EvZm0LRX8V6UCfJu9MTdKR23QZLhvZdw96ufh949s5mtrvwPr8Mqfejk0yZWH1BXNaB8BfjR4nlWLRvhf4jkDdJJtPeCP/v5IFX9a/Z/8NeK/B3wd8M+FfG1rHbaxpds0E0STLKEUSN5Y3KSCfL2A4OM1VftdfB74p+L/AI56zrvhf4f67qunzW1qsdza2TyRsVgQMAQMcEEV+yN8Hvin4Q+Oeja74o+H+u6Xp8NtdLJc3dk8caFoHCgkjHJIFV+0z8Fvi14n+OfizXfD3w51/UdPu7mJoLm3sXeOQCCMEqQMHkEfhX7Fnwm+Jngr4wzax4u8C61pFidFuIhcXlm8SFy8RC5I6kA8e1SRxyxtFKiujqQysMgg9QR6V8eP2FNXXUrrxP8ABZYbm0uHaSTQppFieAkknyHYhSnorEEdi3bW/hF8U/Dk5ttb+HPiO0ZW2hpNMl2sf9lgu1uh6E9K034WfE3WpBDpHw78S3jE4xBpMz4PvheK/Yi+EHxY+GWr+IdQ8beGX0jStYsoljWedDK80bnb+7UkqNryctj9au7S2v7WaxvIEnt7iNo5YpFyrqwwVI7gg4xXx2/Y7+IPw/1271LwHoV54j8MTyM9qbKNp7i0U8+XLGPmOORvUEEAE4JxWnfC74lavejTtM+H3iO5uS23yo9LmJB9/l4HqT0rwh+xH4tsvht4m8XeONPZvEA0mY6JoVsfOkE23IaTacF8AhUGcFsnkYr/AIZ4+Of/AESbxR/4LZP8K8DR3kXgnw9FqFo9rdJpVss8Egw0TiJdykeoOR+FftC/BL4veIvjV4v1vQvhv4gv7C71Fnt7mCwd0kXaOVIHI4r9h/4V/EjwP8WNV1bxh4H1nRrOXw7NBHPe2jRI0huLdgoJHUhWOPY1V7Z2mo2c+n39tFc21zE0U0MqB0kRhhlZTwQQSCD618cP2E/Eulajca98G0Gq6XKxc6RNMEuLXuRGzECRPTJDDgfN1rVfhb8S9DuTZ6v8PvEdpMCRtm0uZc49Dt5HI5HrXg79nP41+OrqK30T4daxFFIR/pV/bm0hUeu+XaCMdlyfbmv2b/2T9D+CrDxT4hvIdZ8WyxFBPGp8iyVh8ywg8lj0LkA4OAACc18RvgL8adS+IXijUNP+F3iS4trrWbuWCWPT3ZZEaZirA45BBBr9hH4b+PfAereMJvGfhDVdFS8trRbdr21aISlWl3Bc9cZH51Wsfs+/HCbVr2aH4U+J3R7mRlYadIQQWOCOK/YR8AeNvAekeMIfGfhXU9Fe8ubRrdb22aIyhVl3Fc9cZH51+238GvjB8SfE+ka94P8ADZ1jRNK00xCK1nUzpM0jGRjGcEggRgbd33ecVf8Aw0+I2lzfZ9S8AeI7WUEjZPpUyHjrwVrwx8A/jP4wuIrfQvhpr7iUgLPcWTW0IzjrLIFQcEHrX7Ln7LkXwSin8UeKLu3v/FV/D5JMGWhsoSQSiEgFmJA3NgdMDjJaqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqrxv8ACTSPFMz6np0w0/UH5dgmY5T6so6H3H4g1qvwp8c6U5U6M12g6SWrCQH6D735ipPCvieNtknhzVFYdms5Af5Va+AvGl6QsHhjURnoZIDGPzbFeA/hL4q0zX7HXNWFrax2ku8xGXe7cdBtyP1r4y22u6noVro+iaXdXfnz75zDGW2qg4Bx6kg/8BoeAfGrEKPC+o5PAzbsK0bTYtH0iy0mEgraQJECB12gAn8ar4z+Fr7xBo9jd6TZS3V3ZzlfLiQsxRxycD0Kr+Zr/hAvGn/Qr6l/4DtXhq5vrzw/p8+p20sF4YFE8cqlWDgYYkH1Iz+Na54L8XLql5KPDOpujTyMrJau4I3HnIBr/hGfEn/Qv6l/4CP/AIVbeCfGF2QIPDGpnI4ZrV1H5kAVoXwS8VajIraw0OmQZ+bcwkkI9lU4/MivDXhnSfCmmJpWkQlUB3O7HLyN3Zj618Y9J8UeIfEFvb6ZoV9c2ljb4WSOFmUuxyxB+gUfhXhD4eeJZ/E+mLqmgXsFolwrzPLCVUKvzEE++MfjVV8YfBOtah4lj1fRNJuLtbq3AmMMZba68c46ZXb+Rrw74Y8b6Frthq6eF9TP2SdXYC3blc/MPxGRXBHFV8WtM1DV/B0llpdlNdTm4jIjiQs2AeTgV/wgXjT/AKFfUv8AwHavCtvPaeF9HtbmJo5odPgSSNxgqwjUEEeuRXxm0jVNa8L2trpNhPdzJfo7RwoWIURyAnA7ZI/Ov+EC8af9CvqX/gO1aVG8Wl2cUqFXS3RWUjBBCjIrx/8ADvT/ABvbJMsotdRt1Kwz7cgjrtcdx1+mfqDrHw38aaLIyT6FcTop4ltkMyn3+XkfiBS6HrTsETR71mPAAt2z/KtC+H/jq6vILmy8PXUTRSK6vcr5IBBBB+bH6UOQMjnHSvin8OJvFqR6vooQalbpsaNjtE6DkDPQMD0z68ngVd+GPEdjKYLvQb+JwcANbtz9Djn8K8MfCrxV4huU+0WEunWmfnnuYyhx/soeSf0968ZfDfW7HxHdWnh7w/fXGnxrGIZEhLBv3a5Oe53Zz718EtE1vRIdXh1jSrmz8xoTEZoym/h84z6cfnXxo0bVdb0Gxt9I0+e8kju9zJChYgbGGeO3Sv8AhAvGn/Qr6l/4DtVorJawowwyxqCD2OKr4hfDSy8Zxi+tJVtdUiXaspHyygdFfHP0Pb3rV/h/4x0WRkvNAunVf+WsEZlQ++Vzj8cVbeGvEV7IIrTQtQlfOMJbOcfXjivB/wAE9UvZo7zxWfsdqDn7MjgyyexI4Ufr9OtW1tb2dvFaWkKRQwoEjjUYCgDAAFfG/QNa1z+xf7H0u5vPI+0eZ5MZbZny8Zx0zg/lWleBvGMWqWcsvhnUVRLhCzG3YAAMMmq+NXhzXdb1TTZdI0i6vEit2V2hiLBTu6HFeFfBPi608UaPdXPhzUI4Yb+B5HeBgFUSKSSfTArxnYanqnhbUtN0YRm7uoDGgd9oIPDDP+7mtR8EeLtLcpe+HL9dvVkhMi/99LkfrUHh3xBdP5dtoWoSt/dS1dj+grwb8Gda1K6iu/E0JsbFDloWb97L7YH3R6k8+3eooooYkghRUSNQqKowFA6AVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX7Zn/JtfjD/ALcf/S6Cq/Zi/wCSA+B/+wWv/oTVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV+0b4E1/4mfBnxD4I8LRwSapqX2byFnl8tD5dzFI2W7fKjV/wwn8fv+gdon/gyX/Cvgn4T1jwL8KPDPhDX0iTUdKsVhuFifeoYEng9xzVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX9sS/88U/Ov7Yl/54p+df2xL/AM8U/Ov7Yl/54p+df2xL/wA8U/Ov7Yl/54p+df2xL/zxT86/tiX/AJ4p+df2xL/zxT86/tiX/nin51/bEv8AzxT86/tiX/nin51/bEv/ADxT86/tiX/nin51/bEv/PFPzr+2Jf8Anin51/bEv/PFPzr+2Jf+eKfnX9sS/wDPFPzr+2Jf+eKfnX9sS/8APFPzr+2Jf+eKfnX9sS/88U/Ov7Yl/wCeKfnX9sS/88U/Ov7Yl/54p+df2xL/AM8U/Ov7Yl/54p+df2xL/wA8U/Ov7Yl/54p+df2xL/zxT86/tiX/AJ4p+df2xL/zxT86/tiX/nin51/bEv8AzxT86/tiX/nin51/bEv/ADxT86/tiX/nin51/bEv/PFPzr+2Jf8Anin51/bEv/PFPzr+2Jf+eKfnX9sS/wDPFPzr+2Jf+eKfnX9sS/8APFPzr+2Jf+eKfnX9sS/88U/Ov7Yl/wCeKfnX9sS/88U/Ov7Yl/54p+df2xL/AM8U/Ov7Yl/54p+df2xL/wA8U/Ov7Yl/54p+df2xL/zxT86/tiX/AJ4p+df2xL/zxT86/tiX/nin51/bEv8AzxT86/tiX/nin51/bEv/ADxT86/tiX/nin51/bEv/PFPzr+2Jf8Anin51/bEv/PFPzr+2Jf+eKfnX9sS/wDPFPzr+2Jf+eKfnX9sS/8APFPzr+2Jf+eKfnX9sS/88U/Ov7Yl/wCeKfnX9sS/88U/Ov7Yl/54p+df2xL/AM8U/Ov7Yl/54p+df2xL/wA8U/Ov7Yl/54p+df2xL/zxT86/tiX/AJ4p+df2xL/zxT86/tiX/nin51/bEv8AzxT86/tiX/nin51/bEv/ADxT86/tiX/nin51/bEv/PFPzr+2Jf8Anin51/bEv/PFPzr+2Jf+eKfnX9sS/wDPFPzr+2Jf+eKfnX9sS/8APFPzr+2Jf+eKfnX9sS/88U/Ov7Yl/wCeKfnX9sS/88U/Ov7Yl/54p+df2xL/AM8U/Ov7Yl/54p+df2xL/wA8U/Ov7Yl/54p+df2xL/zxT86/tiX/AJ4p+df2xL/zxT86/tiX/nin51/bEv8AzxT86/tiX/nin51/bEv/ADxT86/tiX/nin51/bEv/PFPzr+2Jf8Anin51/bEv/PFPzr+2Jf+eKfnX9sS/wDPFPzr+2Jf+eKfnX9sS/8APFPzr+2Jf+eKfnX9sS/88U/Ov7Yl/wCeKfnX9sS/88U/Ov7Yl/54p+df2xL/AM8U/Ov7Yl/54p+df2xL/wA8U/Ov7Yl/54p+df2xL/zxT86/tiX/AJ4p+df2xL/zxT86/tiX/nin51/bEv8AzxT86/tiX/nin51/bEv/ADxT86/tiX/nin51/bEv/PFPzr+2Jf8Anin51/bEv/PFPzr+2Jf+eKfnX9sS/wDPFPzr+2Jf+eKfnX9sS/8APFPzr+2Jf+eKfnX9sS/88U/Ov7Yl/wCeKfnX9sS/88U/Ov7Yl/54p+df2xL/AM8U/Ov7Yl/54p+df2xL/wA8U/Ov7Yl/54p+df2xL/zxT86/tiX/AJ4p+df2xL/zxT86/tiX/nin51/bEv8AzxT86/tiX/nin51/bEv/ADxT86/tiX/nin51/bEv/PFPzr+2Jf8Anin51/bEv/PFPzr+2Jf+eKfnX9sS/wDPFPzr+2Jf+eKfnX9sS/8APFPzr+2Jf+eKfnVVUkpRtoAPFeef7orzz/dFeef7orzz/dFeef7orzz/AHRXnn+6K88/3RXnn+6K88/3RXnn+6K88/3RXnn+6K88/wB0V55/uivPP90V55/uivPP90V55/uivPP90V55/uivPP8AdFeef7orzz/dFeef7orzz/dFeef7orzz/dFeef7orzz/AHRXnn+6K88/3RXnn+6K88/3RXnn+6K88/3RXnn+6K88/wB0V55/uivPP90V55/uivPP90V55/uivPP90V55/uivPP8AdFeef7orzz/dFeef7orzz/dFeef7orzz/dFeef7orzz/AHRXnn+6K88/3RXnn+6K88/3RXnn+6K88/3RXnn+6K88/wB0V55/uivPP90V55/uivPP90V55/uivPP90V55/uivPP8AdFeef7orzz/dFeef7orzz/dFeef7orzz/dFeef7orzz/AHRXnn+6K88/3RXnn+6K88/3RXnn+6K88/3RXnn+6K88/wB0V55/uivPP90V55/uivPP90V55/uivPP90V55/uivPP8AdFeef7orzz/dFeef7orzz/dFeef7orzz/dFeef7orzz/AHRXnn+6K88/3RXnn+6K88/3RXnn+6K88/3RXnn+6K88/wB0V55/uivPP90V55/uivPP90V55/uivPP90V55/uivPP8AdFeef7orzz/dFeef7orzz/dFeef7orzz/dFeef7orzz/AHRXnn+6K88/3RXnn+6K88/3RXnn+6K88/3RXnn+6K88/wB0V55/uivPP90V55/uivPP90V55/uivPP90V55/uivPP8AdFeef7orzz/dFeef7orzz/dFeef7orzz/dFeef7oqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqf7/4VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVT/f/Cqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqf7/4VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVT/AH/wqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqn+/wDhVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVP9/wDCqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqf7/wCFVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVU/3/wAKqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqp/v/hVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVP9/8KqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqvEOu6Z4W0DUvE2tTvDp+kWct5dyJC0rJFEhdyEQFmIVT8qgk9ACa+L3/BZnw9p9zPpnwP+Fs+rhCVTVfENwbeNsd1tosuyn1aRDgcrzx4k/wCCrv7YmuyO+l+KPDvh4Nnamm6FC4TOOn2jzTxjuT15zWm/8FQf22LG6W4uvi1a6ii9YLnw7p6oeQeTHAjdsde59iPgh/wWR1kanBpH7Qfw/sZLCVgh1jw2rxyQdt0lvI7CQdyUdSAOFY8V4K8beE/iP4V03xt4G1+01rQ9WhE1ne2r7klXofcEEEFSAVIIIBBFVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVT/f/AAqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqura3vbaWzu4UmgnjaOWORcq6kYII7gjjFT/sM/H/AMT/ABp8Z/C/4ZfDjVNVs/DGu3Nh/ak4FvaLEjny2aeUqmWj2PtBLEMOOa8Of8EaPjtf2QuPE/xL8FaROwBW3t/tF2V9mPloAfpuHvXx/wD+CX3x++B3hG+8fWN/onjLQtLhaa/bSmkS5tIlGXlaGRRlF5JKMxABJAAJFf8ABI/9ofVPB/xZuPgFrWpO/h7xnFJcabDI/wAttqMSbiV9BLEjKfVkj/Gqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqf7/4VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV8U/jT8J/gfoQ8RfFXx3pPhqxkLeT9rl/eXDDlhFEoMkrDOSEVjzXxB/wCCyXwS0K4ltPh38OfE/itoshbi7lj0yCU9ipPmSY/3o1PtWvf8Fn/F2qRTWdj8ANBjs7hZI5YbzW5bjfGwxtJWJO2QeOc9BVfsua3ceHP2k/hZrNs7BrfxhpYcKcFo2uo1dfxRmH41VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV4j8ZeEfB9ut14r8UaVo8TAlGvrxIN+P7u4jJ9hUX7RvwKnn+zJ8U/D4cEjL3QReP9o4H61oniHw/4mshqPhvXdP1W0JwJ7K5SdM+m5CRRIAySOB3rWvj18GPD921hqvxN8Px3CEh4kvVlZCOoYJnafY14Y+I/wAP/GzGLwj400XV5VGWitL1JJFHqUB3D8RVVVVVVVVVVVVVVVVVVVVVa/8AHH4PeGLprHW/iT4ft7lG2vCL1ZHQ+jKhJXp3rwr8Uvhx44k+z+EfHGi6rPjJt7e8RpQPUx53Ae+Kqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqn+/wDhVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVXij4wfCrwXO9p4p+Ifh/TbmM4e2mv4xMv1jBLfpWl/tEfAvWZRBY/Fjwz5jNtVZ9QSAk+g3kZqCeC6gjubWeOaKVQ0ciMGVgehBHUVquraVodhLqmtanaafZwDMtxdTLFGg9SzEAfjT/ALSnwDS7Nkfiz4cMgOMi8BTp/fHy/rWg+JPDviqxGp+GNf07V7MnAuLG6SdM+m5CRmqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr9rn9pDR/2W/gvqXxHvLaO91WWQWOh2DtgXV7IGKBsc7FVWdunyoQDkivih8VfiB8Z/GN748+JXie71vWb1jumnf5YkySI4kHyxxjJwigAelfDX4U/Eb4x+JofB3ww8Hal4i1eYbvs9nFkRrnG+RzhY0yQC7lVGRzXh3/AIJDftY63arcaldeBdAkZMmDUdYkd1PHBNvDKueexxx1rxBot14c17UvDt9JFJc6Xdy2szRElGeNyjFSQDjKnGQPpXwB/wCS7/Df/sbtL/8ASuKqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr9qj9qO+8AXh+Gnw1lSTxLKi/bb1VEn2EOPljRecykEHkYUEcEn5fAH7E3jL4gAeMvjb4x1Czu9QAke0VvtF6wPQyyuSEbH8OGIzzggipv2B/gtJa+RFq/iqOULgTfbYScjuR5WPrwPwr4Bfs6aT8BrjxBNp/iCbVzrTQiN5rcRPBHGG+UkMQxJckkBeg4r9rj4teM/GfxKh/Z+8C3UsFt50FrdJDIUN9dTBSsbsP+Wah1BHTOSc4GPC37AXw3tNIhXxh4j1vUdUZAZ3s5UghRschFKMxGc8k88cCvB37FHhDwF8TND8daD4q1K4stInaZtPv40d2k2ERsJU24CsVOCpzjrVVVVVVVVVVVVVVVVVVVVI8cUbSSOqogJZmOAAOpNfFn42fEb9pL4gn4RfByeaHQJJWiVoXMRvlX788zjlYBgkL3GMgsQo8K/wDBP/4eWemx/wDCZeKda1LUCv7w2TJbQqe4UFWY/Unn0FfFX9hWbw/psvif4OeItSubuwUzDTLwgzuV5/cyxhfmGOFIyf72ev7I37TWseK9Qj+E/wASLx7jVlRv7L1Gc4kuNgy0Mvq4UEhjyQCDzgmqqq+Gf7R/xJ8eftMS/DnUZ9OtdAs7vUYPs1ragNKIFkCF3cs2coCdpUZ7Y4qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqn+/+FVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVcTwWsElzczxxQwoXkkkYKqKBkkk8AAd6+LX7RXxS/aH8en4S/ANr220d3aITWr+TNfqvDzSS8GKDHRcjIPzZJCjwR/wTq8Lw2cVx8R/HGpXt6wDSW+kqkESN3XfIrs498J9K1z/gnj8Ir20ZdB8T+JtNutpCSSyxXEefUpsUnt0YV8MfBGl/Bb4V6X4QbUlntfD1nI9zeGLyxIdzSyybcnALMxxk1YN8Q/25vjNPY3uty6V4c09XuEgJLR6dahgqhUzh533AFj1OT91QoX/gnt8EvsItm1vxYbjZg3AvIQd3rt8rGPb/APXX7PP7M2lfs/6j4jvbLxFLrP8AbXkpbvNbiJ7eJNxKHBIYksDuwPujgd6qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr/gtT4j1CTxD8L/CKu62MNlqF8VB+WSV3iQEj1UIcf759ar/gj7oXhKz/AGadV1/SI7d9b1HxLcRavMBmRfKjj8mJj12hH3gdMyMe5rxZ4p8P+B/DOqeMPFWqQadpGjWkl1e3UzbViiRcsT+A6dScAda8Ya1F4l8W634jggaGPVdRuLpInOSgkkZwCR3AOK+AP/Jd/hv/ANjdpf8A6VxVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVXifXLbwx4b1bxLdjdBpNjNdyjOMrEhc8/Ra/Y98MS/Fb456p8Q/FpF7JpCvqUhkXIe8lfCEg9hl2HoUX0qqr9pfSPEXwc/aWh+JkFgZrS8voNW0+RwfLmaPZ5sRbsdynI6hXU9xXwu+OPw4+L2nRXPhPXoftzJum0u5YR3UBxyDHn5gP7y5X3qqqqqqqqqqqqqqqqqqqqr9rfxlc+DPgVr89hOYrvVdmnQuDggSnEn/kISD6kV+wD4Fs7DwXrXxCngU32q3hsoJCMlIIgpOPTc7HPr5a+lVX7V+hyfCP8AaLg8YeFwLZ78W+twBBhUuBIQ/wD308RY/wC+a8e+KLuz+EHiLxr4duTb3UHhu61CxmKK/luts0kbbWBBwQDggj1FfsZ/Gj4l/FjU/FNt4/8AE39qx6bBbNar9jgg8su0gY/ukXOdo656VXjP9oH4u6T+1db/AA00/wAXeV4bfxJp1m1l9gtmzDKYRIvmGMvzvbndkZ4Ir9ob/hfn9i6T/wAKC/5CH2p/t/8Ax6f6rb8v/Hzx97+7zXw+/wCF1/8AC6Lj/hAf+Sgfarzzv+PX/W/P9o/1v7n+/wBOPTtX7P8A/wANZf8ACZXn/C9/+QB/Zcn2b/kH/wDH15sW3/j2+f7nm9fl/HFfHr9oHwx8C9Ehmv4DqWtagG+wabHJtLgdZHbnZGDgZwSTwAcEjRvHP7bPx0iOueDY/wCw9DmY+TJDFDaQkZ/geXMsg4xlSRnPTpXw4t/21/D3xB0HSPHN5FqXhu6uQt9cuttOkcSrlsumJFYhcAtwWI618QPHvhr4Z+FL3xj4rvfIsbJeijLzOeFjRf4mY8AfiSACRbfHP9qf9obV7u2+DmlJoWi27lHlhSMCIHoJLiUffwQcRgH2714h8U/ts/Am1XxP4q1AazoqOPtDTCG9hUk4AkKgSxg8DIKjJAzk18Bvjx4c+Ofhp9R0+H7Bq9gVTUtNZ9xhJztdWwNyNg4OMggg9Mn9sL4j+OPhb8ONL8R+AtfOl3s+tx2kr/ZYp98bQTORiVWA5jXkDNQftMfGX4l6D4f8C/BLSn1bxWNKil8QazLbRKsExX5gobEKc9WYYydoXIrxR8Sv2yfgNqNprPjzUrieyu5MKLxYbu0mI5MZaPmM8HgFCQDjgV4i/bEvfEnhbw3pfwZ8Mtq3jnxLCxksChmGllSVfcBjccqxUnACgO2BwfG2t/t0fDe0PjvxTq9+tjEwNw0P2S5hhz/z0ijBCr23Yxnvkiv2Z/j2nxy8J3MuqWsFp4g0Z0j1CGEkJIGB2SoCSQG2sCMnBU9iK8Z+L9B8A+F9Q8X+JrwW+naZCZJmAyzdgqjuzEhQO5Iqy+Mf7Uf7R2q3o+DdnB4Y8O2khjNy+wY44DzOpJfBB2xAbcjPYnxF4y/bJ/Z4EPiPxrqVt4l0AyKs7ybLmFSTgKzqqyxk5wD93JHXpXwp+J3h/wCL3gmy8aeHS8cU5MdxbSEF7WZcb42x3GQQe4IPGaqqqqqqqqqqqqqqqqqqqqqp/v8A4VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX7b3ju78FfAm+stOuDFdeJbuPSw6nDLEwZ5cexSNkP8Av1/wT/8AhzYaF8MLn4iz26NqXia6kjimK8pawsUCA9syLITjrhf7tVXifRY/EnhvVfDskvlpqljNas+PuiRChP61+z18SJ/2WvjNq2ifEjSrm3tZUOnaqscRaS2ZWDJMo4Lp346q+4Z4B8K+MvCnjnSY9b8H+IbDV7GQcTWkwkCn0YDlT6g4Iqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr/grh8BdY+JPwd0X4q+GLGS7vvh5PO9/DEmWOnzhPNkwOT5bxRsfRWc9qr4KftGfGb9nfV7nWPhF44utDe/VVvLfy0nt7oLnb5kMishIycNjcMnBGTXxq/bA/aK/aD05NE+KPxJu9Q0iN1ddLtoIrO2LL0Z44VUSEHkF9xB6YrUdM1HSLx9O1bT7myuowpeC5haJ1DKGUlWAIypBHqCDXwKuYLP43fDy7uZAkUHirTHkYj7qi6jJP5Cqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq+O0FxcfBXxzFak+Z/wj96cA4JAhYsPyB4r/AIJ2XVul548sWYefLFp8iDvtU3Ab9XX86qq8beBfCfxF0Cbwz4y0WDUtPmO7y5AQY2AIDowwUYZPIIPJ9a+If7BOsaXI+t/CHxa08kDeZDYai3lTKRyPLnXCls9NwXHdq+Bf7UfxA8A+NIfhX8bpLue1+0rZtc6hkXWnSk4UyOeZI8kZLZIBDAkDBqqqqqqqqqqqqqqqqqqqv2+baef4K6fLDnZb+Ird5ev3fJnX/wBCZa/Ylure5+AGlwwsC9rfXcco9GMpb/0Flqq/4KEXkEnj/wAL6erfvoNGeVx6K8zBf1Rq8VWU2nfsu6xp1wMS2vgGeJxjGGWwIP6iv+Cd/wDyGfG//XrZ/wDoUtV8RP8Ak+i0/wCxw0j/ANCt6r9nz/k8/UP+wnrP8pqrxDAfj9+2TJ4d1mQy6d/bcliItxUfZLMOXVTngOsTnju5NW1tbWdtFZ2cEcMEEYjiijXaqKowAAOgAAGKr/goT4tu5Nd8LeBIp2W2gtH1KaIHh3dzGjEf7IjkA/3zXwW8C6d8Ovhf4e8L2FtHG8NlHJdsoGZbh1DSuSOpLEgegAHQCtT0zT9a0260jVbSK6s72FobiCVcrIjDDKR6EEiv2epLr4T/ALWkvge3un+xPqF9o82Wz5sa7zETjvujjP4npX/BQH/kjWjf9jPB/wCk1zX7EvhnStG+A+ma3ZwKLzX7q5uLyQr8zGOZ4UGeuAsYIHYs3qa/aY0aw1z4EeMra/hWRbfTHuoiVyUkiw6kehyuPoT61/wTy0HTZZvGXiWW3R763FrawSMvMcb+Yzgf7xVM/wC6K1jSrDXtJvNE1SBZ7PULd7e4iYcPG6lWH4gmv2Eri50r43azoiy74pdFuI5B0BaOaLDY9eo/4Ea/4KE+Lbq30nwp4HtpisN7NPfXSg43eWFSIH1GXkPPcD0r4VftcfAb4bfDvQfBdtYeIVfTbJEuWisI8STkZlfPmDO5yx59a8f/ALZPwI8beCNd8JXGn+IpF1bT5rdRJp8eFdkIRv8AWcYbBB7EA1/wTx1y5F54y8NPIzQNFa3caHojgujn6kFP++RVVVVVVVVVVVVVVVVVVVVVVP8Af/Cqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/4KNQXDfDzwpcoT5Eesuj88bmhYrx9Fav2O7u3vP2b/BrW7A+VFcxuO6stzKDn+f0IPeqqvjL+zr8NfjhbI/irTZLfVYI/LttVsmEdxGvUKSQQ65J+VgcZOME5rx9+yb8bfgDJN4/+Ffiy51S0sQXln0xntryGIckvECRIgwMhS3QkqBmv2SP2prz4xifwN45WBPFFhb+dDdRKI01CJSAxKjhZFyCQOCCSAMGqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqSOOaNoZUV0dSrKwyCD1BHpX7Q3/AASL+HPxB1u88W/BTxWPA15eO0suj3FobjTy55PlbWD24JOdo3qOiqowBbf8EZv2iHv/ACrv4kfDmOy3Y86O5vXkx6+WbZRnHbd+Nfs4/wDBKf4OfB7WLPxh8SdZl+Iev2MgltYbi0Ftp9vIDkN5G5jKwPeRivfYDjH7S3/BK+y/aE+NXiL4wWXxs/4RkeIjA8mmf8Iz9sEUkcEcTMJPtMed3l7yNvBY8mtD/wCCMFzoOtafrtl+02PtGnXUdzFnwX/HGwYf8vvqBVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVahYWuqWFzpl7GJLe7heGZDxuRgQw/Imvh3rup/snftD3WmeKYp/7Njd7G+ZVz51nIQ0c6Dv0R/XAZeDmtM1PTdb0+31bR7+C9srqMSQXEEgdJFPQqw4Iqqqv28f7HuvjTYW2jokmojRoEvlhGWMpkk2BgOd+wp+BWtChvbfQ9Ot9ScNeRWkS3DesgQBj+eaqqqqqqqqqqqqqqqqqqr48/D2X4o/CfxB4Ps1U31xbiWx3HH7+Jg6DPbcV2k9gxr9iD4vWXgrxBqXwn8WTiwh1m5Eti1xmMR3oAR4mz0ZwqgZx8yAdWFVq+raXoOl3Wta1fwWVjZRNLcXE77UjQDJJNStd/tY/tQLPY2kraG9zGXDqR5OmwYDM390vzx2eUCvixp8+rfCzxlpVou6a88P30MYx1Zrd1H6kV/wT31mytfGninQZpUW5v9NhmhDNgsInIYAdz+9B+gNV4n13TPEv7a1lq+j3SXFo/jbToo5UOVcxzQxsQe43IcHuOar9n9kj/bSv0kcKzarrKqGOCTiY4HvgH8qq4v0+Bv7ZdxrGvAw2Nv4hnmeQqSEtbwPh/cLHPk+6nvUUsU8STwSpJHIoZHU5DA9CCOoxVf8ABQnwvdw+K/C/jRYWa2utOfT2kAOEeKRpAD2GRM2PXafSvg541074hfDLw74p064ST7RYxpcKpGYp0ULKhA6EMD+GD3rUtRsNI0+51XU7uK2s7KF5rieVtqxRqMsxPYAAmv2fI5/i1+1tN44tbWQWMeoX2sS7lwYo23iIHHfdJGPzr/goD/yRrRv+xng/9Jrmv2O/+TcPCH/b7/6Wz1+0D/yRDxz/ANgK6/8ARZr/AIJ3/wDIG8b/APX1Z/8AoMtV+xR/ycRq3/YLvf8A0dHX/BQ3w1dOng/xfDCzW0f2ixuHxwjna8Y/ECX/AL5r4L+C/gb8RPhd4c8U2nww8F3Es9hHHdk6HbMyXKKFlVsoTneD15IIPev+FKfBr/okngv/AMEFt/8AEV4d8C+CPCEk0vhLwdoeiSXKhZm07TorYygdAxRRkDJ6+tVVVVVVVVVVVVVVVVVVVVVU/wB/8Kqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr9qT4Y3nxY+DOs+HtItzPq1kVv8AToh1kmizlB7shdR7sK/YP+OWl+Gprz4NeLr5bNb67Nxo81w2xROQFkgJP3S21So7tuHVgDVVTMqKWZgAoySTjFfs729tfftorP4HCf2NDrOrS27RL+7Wy2ThOnAUqyKPdh061VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVftVftx/CX9kDUPDum/Evw94t1OXxNDcS2h0O0t5gghKBt/nTxYJ8xcYz36V/w+r/ZX/wChC+Kv/gqsP/k2v+H1f7K//QhfFX/wVWH/AMm1/wAPq/2V/wDoQvir/wCCqw/+Ta+Cf/BU39nz49/FLQPhF4P8H/EOz1jxHM8VrPqWn2cdujJE8h3tHdOwG1COFPOPrVVVVVfHXxlq/wAOfgh8QviF4eW3bVPC/hXU9UsVuULxme3tZJY96ggldyDIBGR3Ff8AD5L9rn/oHfD/AP8ABNN/8fr9nLx/rvxW+AvgD4k+J0tV1fxL4ftL+9W1jMcQlkjDNsUkkLk8Ak1VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVfHX9nrwh8c9KjXUXOm65ZIVstVhjDOg67JF48yPPOMgg8gjJzp/wc/bE+AtzNF8Ob2e/0wuW26bcR3EDk8ZNtNyGIxkhP+BcVF8Tv289UjFjaeCry2lIAE76FHEc9M5k+T9P0r9m7RP2gdNTXtQ+Omom4fUWgexhlu0ke3Khw4CRfu0Ugpwp6r0r4jfD/wDbB8HfErxD45+HGsTajp2sXz3CW9jdpIiR9I0e2n4LBFVcqG6dal+Iv7emuQto1r4Pv7KWRdhuV0WOBh2JDyfIOvUYx1GK+BH7IfiWz8Xw/E/436gl7qkM4uoNPNx9pdrgHIkuJeQxUjIVSwJwScDaaqqqqqqqqqqqqqqqqqqqv2if2PrH4l6jP43+H93baT4im+e7tpgVt75/7+VBMch7nBDHk4JLHTNe/bw+Gtuvh5tB1XWIIRthea0j1LA9pUJYj2Zjj2rUvhj+2L+0LPDYfEKd9E0MOGMd40drAmDkHyIvnkYdi44/vCvgl8C/CPwP8PNpehBrzUbsK2oanMgElyw6AAfcQZOF5xnkk5NEBhtIB46Gvid+y38W/hb8Qm8ffAq3ubqxS4e4shYOoudPLZzEY2/1iYYqMBsrww9bVP24/jDZL4T1mN/DGkXS+Vf309rHZO0Z4bIH7w5GeEABzgkA1efsqeL/AAH8dfBl74F8L3+p+GdFvtLuLzUnmiBkeOVWnk2lwR0J2gcdBmq+Ov7PHxe8I/F65+L/AMHbGe/S7vjqEZstr3FnctkyAxN99WYseAQQxBHr+zp4v/aP8Ua7qP8AwuXwy+m6TFZZtHksVtWafeBjGdx+Xd2xx71+0h+zTpXxxsYNW029i0vxRp0Rjt7uRCY7iPJIilxyACSQwyVyeDmvDMP7bnwNtV8M6T4bu9Z0m3O22gMKajCg7eWyHzEXj7pIA9Bmvh1rf7aHi7x/od/4y0KLRvDFrdK2o25jhtVkjIIPBLSsQDkDpkDNfFD4a+G/i14NvPBnieFjb3GHhmjA8y2mX7sqE9GGSPcEg8E1pXwp/av/AGadZvD8NIR4i0W6fc4tY1niuMdC9uxEiPjGSn03EV4j0n9tH9oS1Twv4h8Pjw9okjj7SksQsIXweDIGLSuARnaARkA44Br4DfAfw58DPDT6dp8/2/V78q2pai0ewzFc7UVcnai5OBnJJJPXj9sn4e+MfiV8MNM0LwPocuq38GvRXMkMciIViWCdS2XIHV1H41+zN4T8Q+B/gf4a8LeKtMfT9VsvtXn2zsrFN91K68qSOVZT1718ZdD1XxL8KPFnh/Q7NrvUNQ0i4gtoFYKZJGQgKCSAMn1r9iv4W+Pvhjpfiu38d+HJtJk1Ce1a2WSWN/MCLIGI2McY3Dr61X7K/wAEvil4B+NWo+J/F3hC407TJrC6jjuJJomDM8qFRhWJ5APaviJ4A8PfE/whqHgvxNAZLK/TAdMB4XBysiE5wykA/mDkEirb4L/tWfs56zdzfCi7fXNGuH3P9jCSJOB0MltISyvgYymcdN1J8WP269dBsNP+GKWMz8LM2j+Tt/4FO+z86+CFn8U7HwBbW/xiuYrjxH9omeR0lRz5bNuUMUAQEZIwuQAF57Cqqqqqqqqqqqqqqqqqqqqqn+/+FVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX7Q37Euj/ErVLnxp8OL+10LXrol7uznQi1vJD1fKgmJz3IBDHkgEknSR+378IYU0Sz07WNX0+3+WGNo4dVTaMAbXG6RVwMBcgD0FN4w/4KGeNEFhp/h280dZPlaQadbWZx3O6flfwweOK8N+FPirL+z7/wAIX4l1y1HjmfRbizbUJbl51WVw6xyPJjcWClcnB+YH7w66bpn7fPwXT+yNLg1HXtMtxthVTFqsZUdNm7MqDsF+XjtWtv8At6/GiyPhe/0O/wBG0y5Bjuh5EWmI6ng73YiQrg4KrkEdjX7Mf7Mem/AbTbjVdVvYdT8VanEI7q6iUiK3iyD5UWcEgkAliASQOBiqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr/gpx+xt8a/2qdd8Aah8JLHSLiLw7aX0V6b7UFtirSvCU2gg54jbPpxV9/wSR/bE06xuNQudF8KiK1iaWQjXEJ2qMnAx6Cq+DP8AwTk/aY+PXw00f4sfD7S/D82ga55/2N7vVlhkPkzyQPlCMj54n/DB71+xx/wTY/ae+Cf7S/gX4o+OdK8OxaFoF3NLePbaus0gVraWMbUAyfmdaqqqqr9rL/k1f4y/9k/13/0gmqv2KP8Ak0X4Qf8AYn6f/wCiVqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqf7/4VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV4t/5FTWf+wfP/AOi2qv8Aglx/yYl8Mf8AuLf+na8rVG1FdMu20dIHvxA5tVnz5Zl2nYGxzt3YzjtXxW/4KNftr/EXX7rSbj4lX/hQpcNCNI8M24sDBICVKCRczscjGGkPStY+NX7X3hqaHX/EHxb+MOlTXLEQ3t5r2pQNIdvO12cEnb6HpX7L3/BV345/DDxJYaL8bdcn8feDJ5lju5LxFOo2KE/NLFMADLgHJSXdnGAyda0XWdJ8SaNYeIdB1CG+03VLWO6srqFtyTwyKGR1PdWUgg+hr/gol/wUk1v4AeJZfgh8Do7J/GUFukus6zdwidNJ8xQ8cUUTfK8xQq5LgoquowxJ2a1+1H+1b8QtVe6v/jp8RtRumzJ5FprdzGiepSGFgiDp91QOlfsQ/tU/tfeIf2jfh/8ADGy+OHinV9O13W4YNSs9cujqimzQ77gL9o3shEKSEFSpGBzX7WX/ACav8Zf+yf67/wCkE1V+zb8QPCnwp/YM+HfxG8b6mlhofh/wHZXV5O3UKsC4VR/E7HCqvVmYAcmvih/wUn/at8cfEDXfFPhb4ra34V0a/vHfTtFsZEEdlbjiOPO35mCgbm/iYk8ZxX/BNzw9+1v8RdMh+O/7Q/xi8WXHhu7iP/COeHrqUR/bwRj7XMNoPlYP7tc/OfmPyhd9VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVT/f/AAqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqrxb/AMiprP8A2D5//RbVX/BLj/kxL4Y/9xb/ANO15VfEX4//ALBf7E3iLW5ZYvD9h421i8nvtTsvD+nC91WaadzJJ50g/wBSCW3COSRFAPyqBXxN/wCCx3wC8a6Bqfgy5/Z28Q+J9A1WJre6tdau7a3WaFgQdyL5wDdMYPB5ByBU5gaeQ2qusO8+WsjBmC54yQACcd8Cv+CY3ii+8VfsS/DqbUZGkn02O808O3eOC8mSID2EYRf+A1+3Ho3iTQ/2vfi5aeKkdbubxXe3UG8dbWaQy2xHA48h4v8A69f8E0P27/gj+zV4T1b4X/Fjw/daSdY1c3yeKLK0+0gqY0QRXKr+92psJUoHH7xvlXknwHP+zp8a9R0z44fDpPBninVNO8xLTxFp8UUl3amSMpJG0gHmISjEFHwcHpX7WX/Jq/xl/wCyf67/AOkE1V+1B+1ldeOPgH8Hv2avBmqEeHfCXhTTZfEDRMQL3UvIUiInukKkcf8APRmzyikf8Eyf2I9H/aS8W3PxR+I72lx4G8HXqRSaT5qtJql5tDrFKgO5IACrMWGH+4MjeVgggtYI7W1hjihhQJHGihVRQMAADgADjFVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV4p/bs/ZJ8F+I9R8JeI/jbo9tqukXL217AlvcTCGVDh0LxxspKkEEAnBBB5Br/h4l+xf/wBF30v/AMALz/4zX/DxL9i//ou+l/8AgBef/Ga/4eJfsX/9F30v/wAALz/4zX/DxL9i/wD6Lvpf/gBef/Ga/wCHiX7F/wD0XfS//AC8/wDjNf8ADxL9i/8A6Lvpf/gBef8Axmv+HiX7F/8A0XfS/wDwAvP/AIzX/DxL9i//AKLvpf8A4AXn/wAZr/h4l+xf/wBF30v/AMALz/4zX/DxL9i//ou+l/8AgBef/Ga/4eJfsX/9F30v/wAALz/4zX/DxL9i/wD6Lvpf/gBef/Ga/wCHiX7F/wD0XfS//AC8/wDjNf8ADxL9i/8A6Lvpf/gBef8Axmv+HiX7F/8A0XfS/wDwAvP/AIzX/DxL9i//AKLvpf8A4AXn/wAZr/h4l+xf/wBF30v/AMALz/4zX/DxL9i//ou+l/8AgBef/Ga/4eJfsX/9F30v/wAALz/4zX/DxL9i/wD6Lvpf/gBef/Ga/wCHiX7F/wD0XfS//AC8/wDjNf8ADxL9i/8A6Lvpf/gBef8Axmv+HiX7F/8A0XfS/wDwAvP/AIzX/DxL9i//AKLvpf8A4AXn/wAZr/h4l+xf/wBF30v/AMALz/4zX/DxL9i//ou+l/8AgBef/Ga/4eJfsX/9F30v/wAALz/4zX/DxL9i/wD6Lvpf/gBef/Ga/wCHiX7F/wD0XfS//AC8/wDjNf8ADxL9i/8A6Lvpf/gBef8Axmv+HiX7F/8A0XfS/wDwAvP/AIzX/DxL9i//AKLvpf8A4AXn/wAZr/h4l+xf/wBF30v/AMALz/4zX/DxL9i//ou+l/8AgBef/Ga/4eJfsX/9F30v/wAALz/4zX/DxL9i/wD6Lvpf/gBef/Ga/wCHiX7F/wD0XfS//AC8/wDjNf8ADxL9i/8A6Lvpf/gBef8Axmv+HiX7F/8A0XfS/wDwAvP/AIzX/DxL9i//AKLvpf8A4AXn/wAZr/h4l+xf/wBF30v/AMALz/4zX/DxL9i//ou+l/8AgBef/Ga/4eJfsX/9F30v/wAALz/4zX/DxL9i/wD6Lvpf/gBef/Ga/wCHiX7F/wD0XfS//AC8/wDjNf8ADxL9i/8A6Lvpf/gBef8Axmv+HiX7F/8A0XfS/wDwAvP/AIzX/DxL9i//AKLvpf8A4AXn/wAZr/h4l+xf/wBF30v/AMALz/4zX/DxL9i//ou+l/8AgBef/Ga/4eJfsX/9F30v/wAALz/4zX/DxL9i/wD6Lvpf/gBef/Ga/wCHiX7F/wD0XfS//AC8/wDjNf8ADxL9i/8A6Lvpf/gBef8Axmv+HiX7F/8A0XfS/wDwAvP/AIzX/DxL9i//AKLvpf8A4AXn/wAZr/h4l+xf/wBF30v/AMALz/4zX/DxL9i//ou+l/8AgBef/Ga/4eJfsX/9F30v/wAALz/4zX/DxL9i/wD6Lvpf/gBef/Ga/wCHiX7F/wD0XfS//AC8/wDjNf8ADxL9i/8A6Lvpf/gBef8Axmv+HiX7F/8A0XfS/wDwAvP/AIzX/DxL9i//AKLvpf8A4AXn/wAZr/h4l+xf/wBF30v/AMALz/4zX/DxL9i//ou+l/8AgBef/Ga/4eJfsX/9F30v/wAALz/4zX/DxL9i/wD6Lvpf/gBef/Ga/wCHiX7F/wD0XfS//AC8/wDjNf8ADxL9i/8A6Lvpf/gBef8Axmv+HiX7F/8A0XfS/wDwAvP/AIzX/DxL9i//AKLvpf8A4AXn/wAZr/h4l+xf/wBF30v/AMALz/4zX/DxL9i//ou+l/8AgBef/Ga/4eJfsX/9F30v/wAALz/4zX/DxL9i/wD6Lvpf/gBef/Ga/wCHiX7F/wD0XfS//AC8/wDjNf8ADxL9i/8A6Lvpf/gBef8Axmv+HiX7F/8A0XfS/wDwAvP/AIzX/DxL9i//AKLvpf8A4AXn/wAZr/h4l+xf/wBF30v/AMALz/4zX/DxL9i//ou+l/8AgBef/Ga/4eJfsX/9F30v/wAALz/4zX/DxL9i/wD6Lvpf/gBef/Ga/wCHiX7F/wD0XfS//AC8/wDjNf8ADxL9i/8A6Lvpf/gBef8Axmv+HiX7F/8A0XfS/wDwAvP/AIzX/DxL9i//AKLvpf8A4AXn/wAZr/h4l+xf/wBF30v/AMALz/4zX/DxL9i//ou+l/8AgBef/Ga/4eJfsX/9F30v/wAALz/4zX/DxL9i/wD6Lvpf/gBef/Ga/wCHiX7F/wD0XfS//AC8/wDjNf8ADxL9i/8A6Lvpf/gBef8Axmv+HiX7F/8A0XfS/wDwAvP/AIzX/DxL9i//AKLvpf8A4AXn/wAZr/h4l+xf/wBF30v/AMALz/4zX/DxL9i//ou+l/8AgBef/Ga/4eJfsX/9F30v/wAALz/4zX/DxL9i/wD6Lvpf/gBef/Ga/wCHiX7F/wD0XfS//AC8/wDjNf8ADxL9i/8A6Lvpf/gBef8Axmv+HiX7F/8A0XfS/wDwAvP/AIzX/DxL9i//AKLvpf8A4AXn/wAZr/h4l+xf/wBF30v/AMALz/4zX/DxL9i//ou+l/8AgBef/Ga/4eJfsX/9F30v/wAALz/4zX/DxL9i/wD6Lvpf/gBef/Ga/wCHiX7F/wD0XfS//AC8/wDjNf8ADxL9i/8A6Lvpf/gBef8Axmv+HiX7F/8A0XfS/wDwAvP/AIzX/DxL9i//AKLvpf8A4AXn/wAZr/h4l+xf/wBF30v/AMALz/4zX/DxL9i//ou+l/8AgBef/Ga/4eJfsX/9F30v/wAALz/4zX/DxL9i/wD6Lvpf/gBef/Ga/wCHiX7F/wD0XfS//AC8/wDjNf8ADxL9i/8A6Lvpf/gBef8Axmv+HiX7F/8A0XfS/wDwAvP/AIzX/DxL9i//AKLvpf8A4AXn/wAZr/h4l+xf/wBF30v/AMALz/4zX/DxL9i//ou+l/8AgBef/Ga/4eJfsX/9F30v/wAALz/4zX/DxL9i/wD6Lvpf/gBef/Ga/wCHiX7F/wD0XfS//AC8/wDjNeCPHHhH4leFNO8ceA/EFprehatEZLO+tW3JKAxVsdwQyspBAIIIIBBqqqqqqqqqqn+/+FVVVVVVVVVXxE8f+F/hX4G1v4i+NL5rTRPD9m93ezJE0jKij+FVBLEnAA9SK/4e5/sb/wDQc8Vf+CJ/8a/4e5/sb/8AQc8Vf+CJ/wDGv+Huf7G//Qc8Vf8Agif/ABr/AIe5/sb/APQc8Vf+CJ/8a/4e5/sb/wDQc8Vf+CJ/8a/4e5/sb/8AQc8Vf+CJ/wDGv+Huf7G//Qc8Vf8Agif/ABr/AIe5/sb/APQc8Vf+CJ/8a/4e5/sb/wDQc8Vf+CJ/8a/4e5/sb/8AQc8Vf+CJ/wDGv+Huf7G//Qc8Vf8Agif/ABr/AIe5/sb/APQc8Vf+CJ/8a/4e5/sb/wDQc8Vf+CJ/8a/4e5/sb/8AQc8Vf+CJ/wDGv+Huf7G//Qc8Vf8Agif/ABr/AIe5/sb/APQc8Vf+CJ/8a/4e5/sb/wDQc8Vf+CJ/8a/4e5/sb/8AQc8Vf+CJ/wDGv+Huf7G//Qc8Vf8Agif/ABr/AIe5/sb/APQc8Vf+CJ/8a/4e5/sb/wDQc8Vf+CJ/8a/4e5/sb/8AQc8Vf+CJ/wDGv+Huf7G//Qc8Vf8Agif/ABr/AIe5/sb/APQc8Vf+CJ/8a/4e5/sb/wDQc8Vf+CJ/8a/4e5/sb/8AQc8Vf+CJ/wDGv+Huf7G//Qc8Vf8Agif/ABr/AIe5/sb/APQc8Vf+CJ/8a/4e5/sb/wDQc8Vf+CJ/8a/4e5/sb/8AQc8Vf+CJ/wDGv+Huf7G//Qc8Vf8Agif/ABr/AIe5/sb/APQc8Vf+CJ/8a/4e5/sb/wDQc8Vf+CJ/8a/4e5/sb/8AQc8Vf+CJ/wDGv+Huf7G//Qc8Vf8Agif/ABr/AIe5/sb/APQc8Vf+CJ/8a/4e5/sb/wDQc8Vf+CJ/8a/4e5/sb/8AQc8Vf+CJ/wDGv+Huf7G//Qc8Vf8Agif/ABr/AIe5/sb/APQc8Vf+CJ/8a/4e5/sb/wDQc8Vf+CJ/8a/4e5/sb/8AQc8Vf+CJ/wDGv+Huf7G//Qc8Vf8Agif/ABr/AIe5/sb/APQc8Vf+CJ/8a/4e5/sb/wDQc8Vf+CJ/8a/4e5/sb/8AQc8Vf+CJ/wDGv+Huf7G//Qc8Vf8Agif/ABr/AIe5/sb/APQc8Vf+CJ/8a/4e5/sb/wDQc8Vf+CJ/8a/4e5/sb/8AQc8Vf+CJ/wDGv+Huf7G//Qc8Vf8Agif/ABr/AIe5/sb/APQc8Vf+CJ/8a/4e5/sb/wDQc8Vf+CJ/8a/4e5/sb/8AQc8Vf+CJ/wDGv+Huf7G//Qc8Vf8Agif/ABr/AIe5/sb/APQc8Vf+CJ/8a/4e5/sb/wDQc8Vf+CJ/8a/4e5/sb/8AQc8Vf+CJ/wDGv+Huf7G//Qc8Vf8Agif/ABr/AIe5/sb/APQc8Vf+CJ/8a/4e5/sb/wDQc8Vf+CJ/8a/4e5/sb/8AQc8Vf+CJ/wDGv+Huf7G//Qc8Vf8Agif/ABr/AIe5/sb/APQc8Vf+CJ/8a/4e5/sb/wDQc8Vf+CJ/8a/4e5/sb/8AQc8Vf+CJ/wDGv+Huf7G//Qc8Vf8Agif/ABr/AIe5/sb/APQc8Vf+CJ/8a/4e5/sb/wDQc8Vf+CJ/8a/4e5/sb/8AQc8Vf+CJ/wDGv+Huf7G//Qc8Vf8Agif/ABr/AIe5/sb/APQc8Vf+CJ/8a/4e5/sb/wDQc8Vf+CJ/8a/4e5/sb/8AQc8Vf+CJ/wDGv+Huf7G//Qc8Vf8Agif/ABr/AIe5/sb/APQc8Vf+CJ/8a/4e5/sb/wDQc8Vf+CJ/8a/4e5/sb/8AQc8Vf+CJ/wDGv+Huf7G//Qc8Vf8Agif/ABr/AIe5/sb/APQc8Vf+CJ/8a/4e5/sb/wDQc8Vf+CJ/8a/4e5/sb/8AQc8Vf+CJ/wDGv+Huf7G//Qc8Vf8Agif/ABr/AIe5/sb/APQc8Vf+CJ/8a/4e5/sb/wDQc8Vf+CJ/8a/4e5/sb/8AQc8Vf+CJ/wDGv+Huf7G//Qc8Vf8Agif/ABr/AIe5/sb/APQc8Vf+CJ/8a/4e5/sb/wDQc8Vf+CJ/8a/4e5/sb/8AQc8Vf+CJ/wDGv+Huf7G//Qc8Vf8Agif/ABr/AIe5/sb/APQc8Vf+CJ/8a/4e5/sb/wDQc8Vf+CJ/8a/4e5/sb/8AQc8Vf+CJ/wDGv2Z/+CkHwn8A+MPE2p/FLxz4xv8ATr+ELp8ckM12EbzCeFZjt+XFf8Pc/wBjf/oOeKv/AARP/jX/AA9z/Y3/AOg54q/8ET/41/w9z/Y3/wCg54q/8ET/AONf8Pc/2N/+g54q/wDBE/8AjX/D3P8AY3/6Dnir/wAET/41/wAPc/2N/wDoOeKv/BE/+Nf8Pc/2N/8AoOeKv/BE/wDjX/D3P9jf/oOeKv8AwRP/AI1/w9z/AGN/+g54q/8ABE/+Nf8AD3P9jf8A6Dnir/wRP/jX/D3P9jf/AKDnir/wRP8A41/w9z/Y3/6Dnir/AMET/wCNf8Pc/wBjf/oOeKv/AARP/jX/AA9z/Y3/AOg54q/8ET/41/w9z/Y3/wCg54q/8ET/AONf8Pc/2N/+g54q/wDBE/8AjX/D3P8AY3/6Dnir/wAET/41/wAPc/2N/wDoOeKv/BE/+Nf8Pc/2N/8AoOeKv/BE/wDjX/D3P9jf/oOeKv8AwRP/AI1/w9z/AGN/+g54q/8ABE/+Nf8AD3P9jf8A6Dnir/wRP/jX/D3P9jf/AKDnir/wRP8A41/w9z/Y3/6Dnir/AMET/wCNf8Pc/wBjf/oOeKv/AARP/jX/AA9z/Y3/AOg54q/8ET/41/w9z/Y3/wCg54q/8ET/AONf8Pc/2N/+g54q/wDBE/8AjX/D3P8AY3/6Dnir/wAET/41/wAPc/2N/wDoOeKv/BE/+Nad/wAFZ/2NtR1C209vFHiG0FzKsZnudEkWOLcQNzkEkKOpODgVVVVVVVVVVVVVVVVV4t/5FTWf+wfP/wCi2qv+CXH/ACYl8Mf+4t/6dryv+ChP7QWs/s3/ALMmu+MfCl2LXxJrFzFo2i3JwTb3E4YtKAerJDHM68H5lXIxmvDGgeJfil4/0nwtp9w1/wCIfF2sQ2cEt3MWae7uZgitI5ySWdwSxyeSa+Gn/BGH9nzQdEt/+FneL/FXinWmjH2lrS4SwtFbHIjjVGkxnu0hzgcDkV470rT9C8b+IdD0kk2WnardW9tufcfLSVlXnvwBzX/BJ/8A5Mj8H/8AYQ1P/wBLJa/bG/YK+Fn7XtjBq2pXcvhrxtp0Hk2PiG0gEpaMEkQ3ERIE0YJJHzKyknDAFlP7R3/BOP8AaS/Zv0u98Wazodl4m8J2ILT63oUpmS3TPDTRMFljHq20oP73Qn9nn9oH4gfs1/E3S/iT8P8AVZYZbWVV1CwMhEGp224eZbzL0KsM4PVThlwQDX7R2vaf4r/Y1+KHijSJfMsdY+GOr3lq/wDejl0yV1PH+ywqtd/Zk+L/AIb+Ami/tH6x4akt/B2u6k1layuCJcbcxzsmOIZCHVXPUp6Mpb9hb9rHVf2TvjPaeIbqeebwbrxSy8T2KAtut93y3CKOssJJYdyC6cb8jRdZ0nxJo1j4g0DUbe/03U7aO5s7q3cPHPE6hkdWHVSpBB96qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr9jn9m/wCGX7Tn7UfxW8KfFS11G4sdLF/fW4s7w27ib7eqZLDORtduK/4dN/sf/wDQF8Uf+Dx/8K/4dN/sf/8AQF8Uf+Dx/wDCv+HTf7H/AP0BfFH/AIPH/wAK/wCHTf7H/wD0BfFH/g8f/Cv+HTf7H/8A0BfFH/g8f/Cv+HTf7H//AEBfFH/g8f8Awr/h03+x/wD9AXxR/wCDx/8ACv8Ah03+x/8A9AXxR/4PH/wr/h03+x//ANAXxR/4PH/wr/h03+x//wBAXxR/4PH/AMK/4dN/sf8A/QF8Uf8Ag8f/AAr/AIdN/sf/APQF8Uf+Dx/8K/4dN/sf/wDQF8Uf+Dx/8K/4dN/sf/8AQF8Uf+Dx/wDCv+HTf7H/AP0BfFH/AIPH/wAK/wCHTf7H/wD0BfFH/g8f/Cv+HTf7H/8A0BfFH/g8f/Cv+HTf7H//AEBfFH/g8f8Awr/h03+x/wD9AXxR/wCDx/8ACv8Ah03+x/8A9AXxR/4PH/wr/h03+x//ANAXxR/4PH/wr/h03+x//wBAXxR/4PH/AMK/4dN/sf8A/QF8Uf8Ag8f/AAr/AIdN/sf/APQF8Uf+Dx/8K/4dN/sf/wDQF8Uf+Dx/8K/4dN/sf/8AQF8Uf+Dx/wDCv+HTf7H/AP0BfFH/AIPH/wAK/wCHTf7H/wD0BfFH/g8f/Cv+HTf7H/8A0BfFH/g8f/Cv+HTf7H//AEBfFH/g8f8Awr/gpb+yb8H/ANlz/hXH/CqLLVLf/hJv7V+3/br5rnd9n+yeXtyBt/1z59ePStI/4J1fs1Xuk2V5PpWvmSe2jd8as45KgntX/DuL9mX/AKBXiD/wbP8A4V/w7i/Zl/6BXiD/AMGz/wCFf8O4v2Zf+gV4g/8ABs/+Ff8ADuL9mX/oFeIP/Bs/+Ff8O4v2Zf8AoFeIP/Bs/wDhX/DuL9mX/oFeIP8AwbP/AIV/w7i/Zl/6BXiD/wAGz/4V/wAO4v2Zf+gV4g/8Gz/4V/w7i/Zl/wCgV4g/8Gz/AOFf8O4v2Zf+gV4g/wDBs/8AhX/DuL9mX/oFeIP/AAbP/hX/AA7i/Zl/6BXiD/wbP/hX/DuL9mX/AKBXiD/wbP8A4V/w7i/Zl/6BXiD/AMGz/wCFf8O4v2Zf+gV4g/8ABs/+Ff8ADuL9mX/oFeIP/Bs/+Ff8O4v2Zf8AoFeIP/Bs/wDhX/DuL9mX/oFeIP8AwbP/AIV/w7i/Zl/6BXiD/wAGz/4V/wAO4v2Zf+gV4g/8Gz/4V/w7i/Zl/wCgV4g/8Gz/AOFf8O4v2Zf+gV4g/wDBs/8AhX/DuL9mX/oFeIP/AAbP/hX/AA7i/Zl/6BXiD/wbP/hX/DuL9mX/AKBXiD/wbP8A4V/w7i/Zl/6BXiD/AMGz/wCFf8O4v2Zf+gV4g/8ABs/+Ff8ADuL9mX/oFeIP/Bs/+Ff8O4v2Zf8AoFeIP/Bs/wDhXxF/4J+fs6eGvh74n8R6Xpmure6Vo13d2xfVGZRJHCzqSMcjKjiv+Cb37G3wT/ad8EeMNd+Klhq9xd6LqsNtaGy1BrcBGi3HIAOTmv8Ah03+x/8A9AXxR/4PH/wr/h03+x//ANAXxR/4PH/wr/h03+x//wBAXxR/4PH/AMK/4dN/sf8A/QF8Uf8Ag8f/AAr/AIdN/sf/APQF8Uf+Dx/8K/4dN/sf/wDQF8Uf+Dx/8K/4dN/sf/8AQF8Uf+Dx/wDCv+HTf7H/AP0BfFH/AIPH/wAK/wCHTf7H/wD0BfFH/g8f/Cv+HTf7H/8A0BfFH/g8f/Cv+HTf7H//AEBfFH/g8f8Awr/h03+x/wD9AXxR/wCDx/8ACv8Ah03+x/8A9AXxR/4PH/wr/h03+x//ANAXxR/4PH/wr/h03+x//wBAXxR/4PH/AMK/4dN/sf8A/QF8Uf8Ag8f/AAr/AIdN/sf/APQF8Uf+Dx/8K/4dN/sf/wDQF8Uf+Dx/8K/4dN/sf/8AQF8Uf+Dx/wDCv+HTf7H/AP0BfFH/AIPH/wAK/wCHTf7H/wD0BfFH/g8f/Cv+HTf7H/8A0BfFH/g8f/Cv+HTf7H//AEBfFH/g8f8Awr/h03+x/wD9AXxR/wCDx/8ACv8Ah03+x/8A9AXxR/4PH/wr/h03+x//ANAXxR/4PH/wr/h03+x//wBAXxR/4PH/AMK/4dN/sf8A/QF8Uf8Ag8f/AAr/AIdN/sf/APQF8Uf+Dx/8K/4dN/sf/wDQF8Uf+Dx/8K+NX7LHwl8Dft+eDf2d9As9TXwfrd1pUV3FLes85W4bEmJMZHt6V/w6b/Y//wCgL4o/8Hj/AOFf8Om/2P8A/oC+KP8AweP/AIV/w6b/AGP/APoC+KP/AAeP/hX/AA6b/Y//AOgL4o/8Hj/4V/w6b/Y//wCgL4o/8Hj/AOFf8Om/2P8A/oC+KP8AweP/AIV/w6b/AGP/APoC+KP/AAeP/hX/AA6b/Y//AOgL4o/8Hj/4V/w6b/Y//wCgL4o/8Hj/AOFf8Om/2P8A/oC+KP8AweP/AIV/w6b/AGP/APoC+KP/AAeP/hX/AA6b/Y//AOgL4o/8Hj/4V/w6b/Y//wCgL4o/8Hj/AOFf8Om/2P8A/oC+KP8AweP/AIV/w6b/AGP/APoC+KP/AAeP/hX/AA6b/Y//AOgL4o/8Hj/4V/w6b/Y//wCgL4o/8Hj/AOFf8Om/2P8A/oC+KP8AweP/AIV/w6b/AGP/APoC+KP/AAeP/hX/AA6b/Y//AOgL4o/8Hj/4V/w6b/Y//wCgL4o/8Hj/AOFf8Om/2P8A/oC+KP8AweP/AIV/w6b/AGP/APoC+KP/AAeP/hX/AA6b/Y//AOgL4o/8Hj/4V/w6b/Y//wCgL4o/8Hj/AOFf8Om/2P8A/oC+KP8AweP/AIV/w6b/AGP/APoC+KP/AAeP/hX/AA6b/Y//AOgL4o/8Hj/4V/w6b/Y//wCgL4o/8Hj/AOFf8Om/2P8A/oC+KP8AweP/AIV+2r/wT0/Zv+Bn7MvjL4p+AdL16LXtE+w/ZHutVaaMebfW8L5Qjn5JW+hwa/4JZ/8AJl/hH/r/ANS/9LJaqqqqqqqqqp/v/hVVVVVeG/j/APAfxn4hg8I+D/jb4C13XbousGl6b4ktLq6lKKzuFijkLttVGY4HAUk8A1Wo6hp+k2M+p6rf29nZ2sbST3FxKI44kAyWZmICgDqTxUn7bf7I0esHQn/aK8DC6DlC39rRmEEf9Ns+XjjruxWnalp2sWEGqaRqFtfWV1GJILm2lEscqHoyspIIPqKqv2+P+TOPix/2L8n/AKGlf8Eyv2RP2cfjl+zhceNPir8LrHxBrUfiW7tFu5rq4jYQpFAVTEcijALt2zzX/DuL9ib/AKIJpX/gwvP/AI9X/DuL9ib/AKIJpX/gwvP/AI9X/DuL9ib/AKIJpX/gwvP/AI9X/DuL9ib/AKIJpX/gwvP/AI9X/DuL9ib/AKIJpX/gwvP/AI9X/DuL9ib/AKIJpX/gwvP/AI9X/DuL9ib/AKIJpX/gwvP/AI9X/DuL9ib/AKIJpX/gwvP/AI9X/DuL9ib/AKIJpX/gwvP/AI9X/DuL9ib/AKIJpX/gwvP/AI9X/DuL9ib/AKIJpX/gwvP/AI9X/DuL9ib/AKIJpX/gwvP/AI9X/DuL9ib/AKIJpX/gwvP/AI9X/DuL9ib/AKIJpX/gwvP/AI9X/DuL9ib/AKIJpX/gwvP/AI9X/DuL9ib/AKIJpX/gwvP/AI9X/DuL9ib/AKIJpX/gwvP/AI9X/DuL9ib/AKIJpX/gwvP/AI9X/DuL9ib/AKIJpX/gwvP/AI9X/DuL9ib/AKIJpX/gwvP/AI9X/DuL9ib/AKIJpX/gwvP/AI9X/DuL9ib/AKIJpX/gwvP/AI9X/DuL9ib/AKIJpX/gwvP/AI9X/DuL9ib/AKIJpX/gwvP/AI9X/DuL9ib/AKIJpX/gwvP/AI9X/DuL9ib/AKIJpX/gwvP/AI9X/DuL9ib/AKIJpX/gwvP/AI9X/DuL9ib/AKIJpX/gwvP/AI9X/DuL9ib/AKIJpX/gwvP/AI9X/DuL9ib/AKIJpX/gwvP/AI9Wt/An4S2v/BTfUfgbb+DLZPA0F5NHHo4ml2Kq6QZwN+/f/rQG+919uK/4Yg/ZW/6I/p3/AIGXX/x2v+GIP2Vv+iP6d/4GXX/x2v8AhiD9lb/oj+nf+Bl1/wDHa/4Yg/ZW/wCiP6d/4GXX/wAdr/hiD9lb/oj+nf8AgZdf/Ha/4Yg/ZW/6I/p3/gZdf/Ha/wCGIP2Vv+iP6d/4GXX/AMdr/hiD9lb/AKI/p3/gZdf/AB2v+GIP2Vv+iP6d/wCBl1/8dr/hiD9lb/oj+nf+Bl1/8dr/AIYg/ZW/6I/p3/gZdf8Ax2v+GIP2Vv8Aoj+nf+Bl1/8AHa/4Yg/ZW/6I/p3/AIGXX/x2v+GIP2Vv+iP6d/4GXX/x2v8AhiD9lb/oj+nf+Bl1/wDHa/4Yg/ZW/wCiP6d/4GXX/wAdr/hiD9lb/oj+nf8AgZdf/Ha/4Yg/ZW/6I/p3/gZdf/Ha/wCGIP2Vv+iP6d/4GXX/AMdr/hiD9lb/AKI/p3/gZdf/AB2v+GIP2Vv+iP6d/wCBl1/8dr/hiD9lb/oj+nf+Bl1/8dr/AIYg/ZW/6I/p3/gZdf8Ax2v+GIP2Vv8Aoj+nf+Bl1/8AHa/4Yg/ZW/6I/p3/AIGXX/x2v+GIP2Vv+iP6d/4GXX/x2v8AhiD9lb/oj+nf+Bl1/wDHa/4Yg/ZW/wCiP6d/4GXX/wAdr/hiD9lb/oj+nf8AgZdf/Ha/4Yg/ZW/6I/p3/gZdf/Ha/wCCgP7OfwV+EPwa0bxL8OPAdromp3PieC0luIriaQtC1tcuUw7sMFo0PTPFfstfsHfskePf2cPht408XfBfTdR1rWvDVnd393Je3StPM8QLOQsoAyT2AFf8O4v2Jv8Aogmlf+DC8/8Aj1f8O4v2Jv8Aogmlf+DC8/8Aj1f8O4v2Jv8Aogmlf+DC8/8Aj1f8O4v2Jv8Aogmlf+DC8/8Aj1f8O4v2Jv8Aogmlf+DC8/8Aj1f8O4v2Jv8Aogmlf+DC8/8Aj1f8O4v2Jv8Aogmlf+DC8/8Aj1f8O4v2Jv8Aogmlf+DC8/8Aj1f8O4v2Jv8Aogmlf+DC8/8Aj1f8O4v2Jv8Aogmlf+DC8/8Aj1f8O4v2Jv8Aogmlf+DC8/8Aj1f8O4v2Jv8Aogmlf+DC8/8Aj1f8O4v2Jv8Aogmlf+DC8/8Aj1f8O4v2Jv8Aogmlf+DC8/8Aj1f8O4v2Jv8Aogmlf+DC8/8Aj1f8O4v2Jv8Aogmlf+DC8/8Aj1f8O4v2Jv8Aogmlf+DC8/8Aj1f8O4v2Jv8Aogmlf+DC8/8Aj1f8O4v2Jv8Aogmlf+DC8/8Aj1f8O4v2Jv8Aogmlf+DC8/8Aj1f8O4v2Jv8Aogmlf+DC8/8Aj1f8O4v2Jv8Aogmlf+DC8/8Aj1f8O4v2Jv8Aogmlf+DC8/8Aj1f8O4v2Jv8Aogmlf+DC8/8Aj1f8O4v2Jv8Aogmlf+DC8/8Aj1f8O4v2Jv8Aogmlf+DC8/8Aj1f8O4v2Jv8Aogmlf+DC8/8Aj1f8O4v2Jv8Aogmlf+DC8/8Aj1f8O4v2Jv8Aogmlf+DC8/8Aj1f8O4v2Jv8Aogmlf+DC8/8Aj1f8Ey/2efgz8cfjD8TfDfxV8C2viDTdDs1fT7eaeaMQMblkyDG6k/KAOSa/4dxfsTf9EE0r/wAGF5/8er/h3F+xN/0QTSv/AAYXn/x6v+HcX7E3/RBNK/8ABhef/Hq/4dxfsTf9EE0r/wAGF5/8er/h3F+xN/0QTSv/AAYXn/x6v+HcX7E3/RBNK/8ABhef/Hq/4dxfsTf9EE0r/wAGF5/8er/h3F+xN/0QTSv/AAYXn/x6v+HcX7E3/RBNK/8ABhef/Hq/4dxfsTf9EE0r/wAGF5/8er/h3F+xN/0QTSv/AAYXn/x6v+HcX7E3/RBNK/8ABhef/Hq/4dxfsTf9EE0r/wAGF5/8er/h3F+xN/0QTSv/AAYXn/x6v+HcX7E3/RBNK/8ABhef/Hq/4dxfsTf9EE0r/wAGF5/8er/h3F+xN/0QTSv/AAYXn/x6v+HcX7E3/RBNK/8ABhef/Hq/4dxfsTf9EE0r/wAGF5/8er/h3F+xN/0QTSv/AAYXn/x6v+HcX7E3/RBNK/8ABhef/Hq/4dxfsTf9EE0r/wAGF5/8er/h3F+xN/0QTSv/AAYXn/x6v+HcX7E3/RBNK/8ABhef/Hq/4dxfsTf9EE0r/wAGF5/8er/h3F+xN/0QTSv/AAYXn/x6v+HcX7E3/RBNK/8ABhef/Hq/4dxfsTf9EE0r/wAGF5/8er/h3F+xN/0QTSv/AAYXn/x6v+HcX7E3/RBNK/8ABhef/Hq/4Ku/s8/Bn9n/AMQfDmz+D3gW18Nw61Zag98kE80vntG8AQnzHbGA7dMdaqqqqqqqqqqqqqqqq8W/8iprP/YPn/8ARbVX/BLj/kxL4Y/9xb/07Xlf8FP/AIK+Ifjb+yfq9l4T0+W/1fwlqMOv21nCpaS4WFJI5lRRyzCGeVgvJJXA5Irw34h1jwj4i0rxZ4cvns9W0W9hvbC5QAtDPE4eNwDkZDKDz6V46/4LG/tGeMPAE/g7RPCvhfw5rF/a/Z7nXdPWZp13LhngR3KxOexO/bnjBwRrejaz4e1a70PxDpt3p+p2UpiurS7iaKWGQdVdW5Vh3B5r/gk5LDL+xL4TSKVHaLUtTWRVbJRvtchwfQ4IOPQj1r4i/wDBVn9oX4b/ALR/xFi8H6loniLwTB4hubXTdJ1iz3xxQwN5IeGSJkkXeIt+CzLlyduSa+Nv/BX34m/F34T6/wDC/TvhR4f8P/8ACT6ZNpuo6gL+W6b7PMhjlESFVCMyMy5YtjPrzXwY+D3jf48/EjRvhh8PtJlvtV1edULKhKWsOR5k8p/hjRTkk/QZJAP7Qfhqx8GfsVfErwdpjs1noXwt1Wwt2f7xjh0uWNSffCiq/Z/+GPhT4z/8E/8AwB8L/G1l9p0bxF4CsrW4UAb4yYVKSoTnDowV1OOGUGvjd8IPFfwF+KfiL4T+NLcpqXh+8aHzQhVLqI8xTp/sSIVcegbB5Br/AIJF/tk7Sn7KfxH1gncXm8GXdw/TgtLYlifYvGP99c/cWqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqvjF8XPBvwL+HWr/ABQ8fXF3FomjIhnNpbNPIzO4RFVR3Z2VQWIUEjJA5rWP+Cpf7THxV1S6tf2Y/wBmqS/sLdihkl0y81u59QxW12JGcYO078ep614c/wCCsv7QPw28UweHf2kfgTaQRBv9Iit7G50fUIkJ5cR3DMrkdlwgOMFh1r4Z/ErwX8YPA2k/Eb4fa1FqmhazD5ttOgwQQSGR1PKurAqynkEEV8QviB4P+FfgzVfiB491yDSdC0WAzXl3MThRkAKAOWZmKqqjJZmAAJNat/wU8/aa+Mvim/0f9k39nw6lplh8zy3OmXGqXfl/wvKIHWOANg/Kd/oGNfB7/gqv410H4hRfDL9rz4XxeEp2nWCfU7Wznsn09mxte5tZizbOQS6kYHIU1bXNte20V5Z3EU8E8YkilicMsikZDKRwQQQQRVV/wS4/5PH+M3/Xhf8A/pyjqqqqqqqqqqqqqqqqqqqqr/gtr/zRj/uPf+4+vD3/ACL+mf8AXnF/6AKqqqqqqqqqqqqqqqqqqqvjL/ySDxz/ANi1qH/pNJX/AARb/wCSX/Eb/sPW3/og1VVVVVVVVVVVVVVVVVVVV+01/wApbvht/wBf+gf+h1VVVVVVVVVVVVVVVVVVVV/wU1/5Mf8AiT/3Cv8A062lf8Es/wDky/wj/wBf+pf+lktVVVVVVVVVU/3/AMKqqqq/4Kd/tb6j8NfC1t+zj8Jbqa5+Inj+MW9wLEF57CymPlhUA5E85JjQD5gu5uCUJ/YO+H+p/Cr/AIKVeD/hvrdxHNqfhy71eyvmjHyLcJpN2JVU5O5Vfcob+IAHAzgV+1t8ZPij+3p+1XbfsmfB/WmtfBumarJYqFcrBdTQbjdX1wV+/HGEk8tehCgr80lTf8Eaf2a38Df2FB4q8Yx+JBb4GvG5jYedj7xttmzy8/wZDY435+av2KfjL8S/2Jv2r7z9kb4saq0vhbVNZGlvE8pMFneS4NrewbvuRzB49w4ysoZuUxVV+3x/yZx8WP8AsX5P/Q0r/gjp/wAmkXf/AGN99/6Jt6qqqqqqqqqqqqqqqqqqqq8Rf8pjtW/7CFx/6Ymqqqqqqqqqqqqqqqqqqqqr/gqT/wAm/eH/APscbb/0jvK/Yr/5NI+EP/Yn6f8A+iVqqqqqqqqqqqqqqqqqqqqr/gjr/wAl8+MX/YPT/wBK2qqqqqqqqqqqqqqqqqqqqr/gt/8A8jT8Jf8AsH6p/wCjLaqqqqqqqqqqqqqqqqry0t7+0msbqPzIbiNo5EyRuVhgjI5HBr/h1x+wl/0Qv/y5tW/+Sq+Fvwt8B/BXwHpfwy+GWhf2N4b0bzvsNj9qmuPK82V5pP3kzu7ZkkduWOM4GAABXxi/4Jpfsk/GfX7jxVq3gW68P6xeuZLu68O3hshcOTks0WGi3E5JYICSSSSa+Cv/AATn/ZQ+Bet2/inw38P31nXbNg9rqPiC5N89uwxho4yBErgjIcJuB6EV8Qf+Ce37H3xT8Z6t8QfHXwfXUtf1y4Nxf3a67qNv50mAC3lxXCouQB91R696+D/wW+GfwD8Gp8P/AITeGzoegR3Mlylmb2e62yyY3sHnd35wOM49uTXxR/4J0fsffFnUp9c134Q2el6pcuXlvNCuJdNLseWZo4mETMTyWKEk855OdP8A+CPv7HVlfG7ubPxlfxFsi1uNc2xjnOMxor47fe/XmvhB+z/8GPgHpEmifCD4d6T4agnAE8lvGzz3GOnmzyFpZMdt7HGT614t8LaD468K6z4J8U2H27RfEGnz6fqNr5rxefbzRtHKm9CGXcjMMqQRnIIPNf8ADrj9hL/ohf8A5c2rf/JVeCfBfhn4deENH8B+DdN/s/Q9As47PTrTznl8mGNQqLvkZnbAAGWJPqa+Nf7Gv7NX7RPiS08X/GH4Yw69rNlZizhvF1K8s38kMzBG+zyxhwGdiCwJGTitI/4Jn/sTeH9Vstd0P4NT2Oo6dcJcWl1b+KtXSSCVGDI6sLrIYEAg+ooKFUKCeBgZOaqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqvGvgvwx8RfCeq+BvGekRapomt2r219aSkgSxsORlSCp6EEEEEAggivDHhXw14K0Kz8MeENBsNG0iwjEdrY2NusEUKjsqqAB/Wv+Cinwo8KfEz9lXxrqGt6bbvqnhLT5NX0m+KDzbWSHDuFbGdsiBkZeh3A9QCP+CLPivWL/AMB/EzwVcyStpujapYXtoGztWS5jmSUKfpbRkj3z3r/gsj8Y9Tu/Eng39n3RriUWkNr/AG3qcURJ+0TSO8VshA7oElbHfzVPYV+y38CdA/Z1+CXhv4caPp8EN9DaRz61cIgDXmoOgM8rHqfm+Vck7UVV6AV/wVr+COjeN/2ff+FvWunxjxB4CuoSblU+eWxnlWKSI+oEkkcgznbtfGNzV/wSs+Lt98Tf2XbXw7rV61xqPgPUZNHDyNl2tdqy25PsqyGIe0I+tVX/AAS4/wCTx/jN/wBeF/8A+nKOqqqqqqqqqqqqqqqqqqqqv+C2v/NGP+49/wC4+vD3/Iv6Z/15xf8AoAqqqqqqqqqqqqqqqqqqqq+Mv/JIPHP/AGLWof8ApNJX/BFv/kl/xG/7D1t/6INVVVVVVVVVVVVVVVVVVVVftNf8pbvht/1/6B/6HVVVVVVVVVVVVVVVVVVVVX/BTX/kx/4k/wDcK/8ATraV/wAEs/8Aky/wj/1/6l/6WS1VVVVVVVVVT/f/AAqqqq/af/aH8J/sw/B/WPij4nKXE1uvkaTp3mbH1G9cHyoVPYcFmbB2orHBxg/8E4P2dfGHxy+Jup/t4ftAbr+91G/ln8ORTx4Fxc52NdheixRAeVCvYrkYEaE/BH/lM/qP/Y3+Jf8A033tfHDxfP8AD74LePvHlpIUuPDnhjUdQhKkA+ZDbSSLjPfKjHvX/BFDwdb6t8WPiN8QrqEzXGh6HbWMUr/Nsa7mZ2PP8RFoRnrgsO5qv+CyPhw+Fv2mvCfjvRi1pc6x4ZgkaaMgN9ptriVRICDkEIYR/wABHvjwVr6eLPBmg+KUKFdZ0y2vBsOVxLEr8EdR81V+3x/yZx8WP+xfk/8AQ0r/AII6f8mkXf8A2N99/wCibeqqqqqqqqqqqqqqqqqqqqvEX/KY7Vv+whcf+mJqqqqqqqqqqqqqqqqqqqqq/wCCpP8Ayb94f/7HG2/9I7yv2K/+TSPhD/2J+n/+iVqqqqqqqqqqqqqqqqqqqqr/AII6/wDJfPjF/wBg9P8A0raqqqqqqqqqqqqqqqqqqqqv+C3/APyNPwl/7B+qf+jLaqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv8AgrD+0do/w/8Agy/wM0e/jk8UePAn2qFGBaz01JAzyN6GR0EagjkeYeNtf8ExfgFqXwT/AGc4NZ8TWT2viDx5cjV7mCVNr29sUC20TDqDsBkIPIMxB5FftsSJ4m/4Kdw+H9SQvaw674asSpbOY5IbR2AB4HMzcfj3qv21bSC9/ZL+LENwu5V8K3kgAOPmRCyn81Ff8ETtRnl0v4vaQx/c21xo06DJ+9It4rcdOkS1VfBLwB+0r8Q/2h/iTpn7MXi240DXra5vptQnh1Y6eZLX7ZjbvH3hvKHb7Z7V/wAMy/8ABW7/AKLdqn/hbvX/AAzL/wAFbv8Aot2qf+Fu9f8ADMv/AAVu/wCi3ap/4W71+xr4K+P3gL4RTaH+0h4mn13xY2sTzJdTamb5haskYjXzD2DK/wAvbPvVVVVVVVVVVVVVVVVVV/wW1/5ox/3Hv/cfXh7/AJF/TP8Arzi/9AFVVVVVVVVVVVVVVVVVVVXxl/5JB45/7FrUP/SaSv8Agi3/AMkv+I3/AGHrb/0Qaqqqqqqqqqqqqqqqqqqqq/aa/wCUt3w2/wCv/QP/AEOqqqqqqqqqqqqqqqqqqv8AhmX/AIK3f9Fu1T/wt3r/AIZl/wCCt3/RbtU/8Ld6/wCGZf8Agrd/0W7VP/C3ev2kfgX/AMFEvBvwW8ReJPjr8Ur/AFjwPZ/Zf7UspfFLXiy7rqJIsxH72Jmib2xntX/BLP8A5Mv8I/8AX/qX/pZLVVVVVVVVVVP9/wDCqqqvby006zn1C/uore2tYmlmmlYKsaKMszE8AAAkmvjH8ffAP7d/7XemWfxP+JNr4L+CXhB5BZveStC11bIy+Y6DBInuSFAOB5cYHBKHdon7bv7EPhrRbDw74f8Ajn4N0/TNLto7WytLd2SO3hjUKiIoXAUKAAPQV8JvjL8LtD/4KpX3xn1bxtp1t4Ik8Ta9cprcjkQGKayu0ifOM4ZpEA46sK+N3xS+Gv7Qf7H3xo1D4P8AjXT/ABPaWXhPVYZprDdIFmSzaXy8YB3FduB/tCv+CHV1bpdfGaxaT9/LHociJg8qpvwxz04Lr+f1qv8AgtpdwSfF74dWCuTND4bnkdcdFe5IU5+qN+VfBbS5tE+DfgPRbnd51h4Z063k3IUO5LaNTkHkcjp2qv2+P+TOPix/2L8n/oaV+yN8QP8Agoj4Y+FMum/st+F73UPBp1aeSSaHRbW6UXZSMSDfKC33RHx0596/4XF/wWf/AOhC1X/wmNP/APia/wCFxf8ABZ//AKELVf8AwmNP/wDia/4XF/wWf/6ELVf/AAmNP/8Aia+HNz4nvfh74XvPG0LReIp9GtJNXjaMRlLswoZgVXhcSFhgcCqqqqqqqqqqqqqqqqqrxJpWpp/wWC1XUX026Fq1/cFZzCwQ/wDEjYfexjrx9aqqqqqqqqqqqqqqqqqqqq/4Ke2N7qHwC0CCws5rmQeMLYlIYy5x9kuxnA+o/Ov2MoJrb9k74SW9zC8UsfhGwV45FKspEK5BB5FVVVVVVVVVVVVVVVVVVVVf8EgtH1bTvjx8X5dQ0u7tUksECNNAyBv9LboSOaqqqqqqqqqqqqqqqqqv2uNc+Ovh34Gazqv7N+nTX3jyK4tRYQRWkdyzRmZBNhJMqcRluvSv+Fxf8Fn/APoQtV/8JjT/AP4mv+Fxf8Fn/wDoQtV/8JjT/wD4mv8AhcX/AAWf/wChC1X/AMJjT/8A4mv20vF37Yfi3U/Csn7XOhXWm3ltb3Q0NZ9Mgs98ZaPzsCEDdgiPr0z71VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVftf/ALXfgb9lLwBNq2o3FrqPi7UYmXQNB8395cydBLIBykCnlm4zjaDuNfsUfspeN/2sfiVN+2D+08z6lpFzfm606wvISBrMyEBG2dFs4iAqp919m3G0HdX/AAUptbn4Xft6aP8AEqa3Ihu7fRtbidVB3/ZnETDp94fZehzwV7EVa3VtfWsN7ZzpNBcRrJFIhyHVhkEexBFf8FBvFdt4P/Y6+Jd9POEe+0xdOhXIy7XMyQ4APX5XYnvgE8YzX/BGDwndWHwm+IHjWWIrDrOvwWURIxu+zQbiR3Izc4z0yD6Gqr/glx/yeP8AGb/rwv8A/wBOUdVVVVVVVVVVVVVVVVVVVVfFXwz4b8Q+FZ5Nf8P6bqb2u3yGvLRJjFukTdt3A7c4GcdcClRUUIigBRgADGKqqr4VfErSPit4VfxNpAVFg1K8sJogSTHJbzvHzkA/Mqq/0cVVVVVVVVVVVVVVVVVPBDcwvbXESSxSoUeN13KykYIIPUEdq+HPh3w/4e8LWcegaFp+mrPGGlWztUhEjDjLBQMnHc1VVVVVVVVVVVVVVVVVVVVdeEvCt9q8XiC98M6TcapCVMV7LZRvOhX7pEhG4Y7c8VVVVVVVVVVVVVVVVVVVVV/wU1/5Mf8AiT/3Cv8A062lf8Es/wDky/wj/wBf+pf+lktVVVVVVVVVU/3/AMKqqrXNHsvEOi3+gairNaalay204RtrFJFKtg9jgnmv+HRn7G//AEA/FX/g9f8Awr/h0Z+xv/0A/FX/AIPX/wAK+Gf7NHwu8Vf8FHrz9mXVrTUW8EQeIdasUhS8Kz+Va2lzLEPNxnO6FMnHPPrXwW/ZP+DvwD+HviL4X+ANN1AaD4pklk1KG+vDcNIZIRC4DEcAooGK/YI8Vzfsc/t0a78GPihdLYW+rvP4ZuLmY+XEtyJVktJ+f4JdgVT0xcBjgAkMVVSzMAAOSeMV+0jqUH7dv/BRvTPA3gWVdV8P213aaDFeQNuRrC1Zpb2dWHGwM90VIOGAUg/NSIkaLHGqqqjCqBgAD0FV+3x/yZx8WP8AsX5P/Q0r/gjp/wAmkXf/AGN99/6Jt6qqqqqqqqqqqqqqqqqqqq+MX/Hzpf8A1zl/mtVVVVVVVVVVVVVVVVVVVVaF/wAhzTv+vuL/ANCFVVVVVVVVVVVVVVVVVVVVV+yf+0B8RfjB438WaD4zurCW10eAPai3tREwPmleSDzwKqqqqqqqqqqqqqqqqqqqq/4Lf/8AI0/CX/sH6p/6Mtqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr9sL9qHRf2UPhLJ48vNJk1bV9QufsGiWPIjlumRmBlYfdjVUZjjk4AGM5Hw9+NHwv8AHvx4u/jX+2hN4w8ZBphcLpekWUEsd5ID8kUvmzxCO3QAARICGGASBkNb/wDBYz9lm0t4rS0+H3xNhhhQJFFHpNgqooGAABeYAAAGK/Zo/b8+Df7Vfju++Hnw88N+MtP1HTtIk1SWXWbO2hhMKSxRFVMVxI2/dOhwVAwDz0B/4Kc/sqa3+0F8K7Dxr8P9Ka/8Y+BWllisoVzLqFlIB50KAcvIpRHRepw6qCzgH9jP/gp98OvBnwr0z4V/tEy6rpup+FLdbKx1eGze6S7toxtjSVUBdJUUBM7SGCgkg5z+2T+1dr37efjDwz+zx+zf4X1i/wBEW/E482LypdUugpVZGXP7q3iRnbc5H3izBdor9mj4IaZ+zt8EvDHwn06aO5m0m13ahdIDi6vJCXnkGedpdmCg8hQo7VVf8EuP+Tx/jN/14X//AKco6qqqqqqqqqqqqqqqqqqqq8ff8ilff9s//Ri1VVWpX0Gl6ddandMBDZwPNISQMKqknk8DgV/wTH+Lcp8U+LfhZrN4M60P7ZsFY4HnqQtwoHqyGNsekTVVVVVVVVVVVVVVVVVXhD/kWdO/64CqqqqqqqqqqqqqqqqqqqqvjF/yfR4R/wCvrS//AEKqqqqqqqqqqqqqqqqqqqqv+Cmv/Jj/AMSf+4V/6dbSv+CWf/Jl/hH/AK/9S/8ASyWqqqqqqqqqqf7/AOFVVVVV8Ov2BfjF4Q/4KCXf7V2peJPBsvhKfXtX1BbKC8uWvxHd2txFGChtxHuDTIWHmYABwTwDX7cn/BPPwr+1iYfG/hbV7bwv8QrGAQC+mhLW2pwr9yO5C/MCv8MqgkAlSrDbtk/Yq/4Kn6xoH/Cp9W+PVuPCbQi0keXxbO0LW+MbCRGZ2j2/LsIwRwRiv2KP2DfA37Iek3WsS6mviXx1q8IivtaaDykghyD9nt0JJVMgFmJ3OQCcABRVft8f8mcfFj/sX5P/AENK/wCCOn/JpF3/ANjfff8Aom3qqqqqqqqqqqqqqqqqqqqr4xf8fOl/9c5f5rVVVVVVVVVVVVVVVVVVVVWhf8hzTv8Ar7i/9CFVVVVVVVVVVVVVVVVVVVVV/wAE/wD/AJKh8QP+vVf/AEe1VVVVVVVVVVVVVVVVVVVVf8Fv/wDkafhL/wBg/VP/AEZbVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVXxB+Fnw4+LGmW2i/EvwRo3iaxs5/tFvbapaLcJFLtK71DA4O1iM+hNf8ADGn7KH/RvHgL/wAEkP8AhX/DGn7KH/RvHgL/AMEkP+FeAPgB8EfhVrM3iH4a/Cvwz4a1O4tWtZbvTNOjt5HhZlZoyygEqWRDj1UelV8Rv2Qv2ZvizrMniLx78GPDuo6rO++e+jga1muG/vSPCUaQ89WJPT0FfDH4G/B/4MWcll8LPhzoXhoTqFnlsbRVmnAOQJJTl3A7bmOKqq/4Jcf8nj/Gb/rwv/8A05R1VVVVVVVVVVVVVVVVVVVV4+/5FK+/7Z/+jFqqqvj3rB8P/A/4gayjlZLTwzqDxEZHz/Z329OnzY5r4J/Ea6+EfxX8L/ES1Z9ujagklyidZLdvknQe7RM6/jVpd2uoWkF/Y3Ec9vcxrJDLG25ZEYZVgR1BBBqqqqqqqqqqqqqqqqrwh/yLOnf9cBVVVVVVVVVVVVVVVVVVVVXxi/5Po8I/9fWl/wDoVVVVVVVVVVVVVVVVVVVVVf8ABTX/AJMf+JP/AHCv/TraV/wSz/5Mv8I/9f8AqX/pZLVVVVVVVVVVP9/8Kqqqqqqqqqv2+P8Akzj4sf8AYvyf+hpX/BHT/k0i7/7G++/9E29VVVVVVVVVVVVVVVVVVVVfGL/j50v/AK5y/wA1qqqqqqqqqqqqqqqqqqqqrQv+Q5p3/X3F/wChCqqqqqqqqqqqqqqqqqqqqq/4J/8A/JUPiB/16r/6Paqqqqqqqqqqqqqqqqqqqqv+C3//ACNPwl/7B+qf+jLaqqqqqqqqqqqqqqqqqqqqqqqqqqqqq1O7bT9Oub9ITN9miaQxjgsFGSB74HHvVhf2eqWMGo2E6zW9xGHide4P+elVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX/AAS4/wCTx/jN/wBeF/8A+nKOqqqqqqqqqqqqqqqqqqqqvH3/ACKV9/2z/wDRi1VVX7cesLov7LHju4DgPcW1vaoDjLebcxIcZ/2WY/hVfsEfFA/En9nnSLG8uTJqfhKQ6Rc7mySkYBgb6eUyL9UNVVVVVVVVVVVVVVVVeEP+RZ07/rgKqqqqqqqqqqqqqqqqqqqq+MX/ACfR4R/6+tL/APQqqqqqqqqqqqqqqqqqqqqq/wCCmv8AyY/8Sf8AuFf+nW0r/gln/wAmX+Ef+v8A1L/0slqqqqqqqqqqn+/+FVVVVVVVVVX7fH/JnHxY/wCxfk/9DSv+COn/ACaRd/8AY333/om3qqqqqqqqqqqqqqqqqqqqr4xf8fOl/wDXOX+a1VVVVVVVVVVVVVVVVVVVVoX/ACHNO/6+4v8A0IVVVVVVVVVVVVVVVVVVVVVX/BP/AP5Kh8QP+vVf/R7VVVVVVVVVVVVVVVVVVVVV/wAFv/8AkafhL/2D9U/9GW1VVVVVVVVVVVVVVVVVVVVVVVVVVVVVLGk0bROMq6lSPY18GPH40a9Xwrq0+LG7f/Rnc8Qynt7K36H6mqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr/glx/yeP8Zv+vC//wDTlHVVVVVVVVVVVVVVVVVVVVXj7/kUr7/tn/6MWqqq/wCClusnTP2cY7BZCP7W8Q2lsQCeQqSy8+2Yh19qSGWVHeOJnWJdzkKSEGQMn0GSB9SPWv8Agmx8UR4P+NV14BvrrZYeNbIxRqxwBeQBpIiT7oZlx3LLVVVVVVVVVVVVVVVVXhD/AJFnTv8ArgKqqqqqqqqqqqqqqqqqqqq+MX/J9HhH/r60v/0Kqqqqqqqqqqqqqqqqqqqqv+Cmv/Jj/wASf+4V/wCnW0r/AIJZ/wDJl/hH/r/1L/0slqqqqqqqqqqn+/8AhVVVVVVVVVV+3x/yZx8WP+xfk/8AQ0r/AII6f8mkXf8A2N99/wCibeqqqqqqqqqqqqqqqqqqqqvjF/x86X/1zl/mtVVVVVVVVVVVVVVVVVVVVaF/yHNO/wCvuL/0IVVVVVVVVVVVVVVVVVVVVVX/AAT/AP8AkqHxA/69V/8AR7VVVVVVVVVVVVVVVVVVVVV/wW//AORp+Ev/AGD9U/8ARltVVVVVVVVVVVVVVVVVVVVVVVVVVVVVa/frpehahqTNj7LaySA5xyqk/wBK6cg8ivhF4/HinS/7G1OfOqWCDLMeZ4xwH+o4B/A96qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr/glx/wAnj/Gb/rwv/wD05R1VVVVVVVVVVVVVVVVVVVV4+/5FK+/7Z/8Aoxaqqr/gqprAg8GeAfD28f6bqd3dbfXyYkXPr/y3/X8v2I/g/bfGR/ip4auY0Ju/Br2dq8g+WK6e4ilgc8HpJbqexwD+Hh7W9d+H3jLTvENgHtNY8O6lHcRLIuDHPDIGww/3lwR9a8E+LNL8e+DtE8a6K4ax1ywhvIOclVkQNtPuM4PuDVVVVVVVVVVVVVVVV4Q/5FnTv+uAqqqqqqqqqqqqqqqqqqqqr4j/AAO+I3iP9qfw98UNJ0iCXw9p89i89w13GrKIj8+EJ3HH05qqqqqqqqqqqqqqqqqqqqr/AIKa/wDJj/xJ/wC4V/6dbSv+CWf/ACZf4R/6/wDUv/SyWqqqqqqqqqqf7/4VVVVVVVVVVft8f8mcfFj/ALF+T/0NK/4I6f8AJpF3/wBjfff+ibeqqqqqqqqqqqqqqqqqqqqvjF/x86X/ANc5f5rVVVVVVVVVVVVVVVVVVVVWhf8AIc07/r7i/wDQhVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV/wW//AORp+Ev/AGD9U/8ARltVVVVVVVVVVVVVVVVVVVVVVVVVVVVVfHHX00vwh/ZUb4n1WURgdwikMx/9BH/AqrRdYvtA1S21fTJvLuLZ9yHsfUH2IyCPevCXiew8X6HBrNgcbxtliJyYpB95T/nkEHvVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX/BLj/k8f4zf9eF/wD+nKOqqqqqqqqqqqqqqqqqqqqvH3/IpX3/AGz/APRi1VVX/BVHWfP8e+BfD2/P2LR7i62+nnTBc9f+mHp279v+CVGjCPS/iL4gdAfPn0+1jY9tizsw6558xPyHvX7e3wu/4Vt+0NrF7Z24j0zxag1e12rgB5CROPr5yu2OwdfrX/BND4o/8JV8IdR+G99cbr3wbe5t1Y8m0uCzpjPJxIJh6AFBVVVVVVVVVVVVVVVV4Q/5FnTv+uAqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv+Cmv/Jj/wASf+4V/wCnW0r/AIJZ/wDJl/hH/r/1L/0slqqqqqqqqqqn+/8AhVVVVVVVVVV+3x/yZx8WP+xfk/8AQ0r/AII6f8mkXf8A2N99/wCibeqqqqqqqqqqqqqqqqqqqqvjF/x86X/1zl/mtVVVVVVVVVVVVVVVVVVVVaF/yHNO/wCvuL/0IVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVf8Fv/APkafhL/ANg/VP8A0ZbVVVVVVVVVVVVVVVVVVVVVVVVVVVVUSAMkjgd6+KHi0eLvFM1xbSbrKzHkWuDwyg8v/wACOT9MVVfDXxxN4K1xXmdm067IS7jHYdnA9V/UEioJobmCO4t5VkilQMjqchgRkEe2Kqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv+CXH/ACeP8Zv+vC//APTlHVVVVVVVVVVVVVVVVVVVVXj7/kUr7/tn/wCjFqqqv+Ck+sf2p+0k1jvz/ZGgWdrjPTJkm9P+m3v/AEH/AATE0Y2PwD1fVZEIfU/E07ISCMxpBAg/8eD9K/4KU/C4eL/gvafECwtt9/4LvRJIwHP2ScrHKPU4cQt7BW9zX7C/xRHww/aH0IXl15Wl+JwdHvdxwo84jyWOeBiZYsnspb1qqqqqqqqqqqqqqqq8If8AIs6d/wBcBVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVf8FNf+TH/iT/3Cv/TraV/wSz/5Mv8ACP8A1/6l/wClktVVVVVVVVVU/wB/8Kqqqqqqqqqv2+P+TOPix/2L8n/oaV/wR0/5NIu/+xvvv/RNvVVVVVVVVVVVVVVVVVVVVXxi/wCPnS/+ucv81qqqqqqqqqqqqqqqqqqqqrQv+Q5p3/X3F/6EKqqqqqqqqqqqqqqqqqqqqr4L/tKeFPjf4h1rw54f0LVrCfQ4w8z3gjCuN5T5drE9R3qqqqqqqqqqqqqqqqqqqqr/AILf/wDI0/CX/sH6p/6Mtqqqqqqqqqqqqqqqqqqqqqqqqqqqqq+NHj5NI09vCmlXH+n3if6SynmGI9vZmH6fUVVVXwT8f+WyeDNXn+ViTYSOeh7x/j1H4juKqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv8Aglx/yeP8Zv8Arwv/AP05R1VVVVVVVVVVVVVVVVVVVV4+/wCRSvv+2f8A6MWqqq/bZ1ka7+1J4+u1fcIb2G1A9PJt4oiOp7of/rV+wLo66T+yv4QkKBZNQkvbmQDHObqVVPH+wq14t8M6X408Lav4Q1qLzLDWrGayuVHXy5UKNj0ODwexrxd4b1r4deNtW8Kaizwap4d1KW2kdAUIkicjevfBKhgfQg18BPiVD8Xvg94V+IKSI0+qaen2wKMBLqPMc649BIj49sHvVVVVVVVVVVVVVVVeEP8AkWdO/wCuAqqqqqqqqqqqqqqqqqqqqr4gftAeO/DH7TGgfCPTYtKOh6lNZJM0tuzTATHD4YMAPbiqqqqqqqqqqqqqqqqqqqqv+Cmv/Jj/AMSf+4V/6dbSv+CWf/Jl/hH/AK/9S/8ASyWqqqqqqqqqqf7/AOFVVVVVVVVVX7fH/JnHxY/7F+T/ANDSv+COn/JpF3/2N99/6Jt6qqqqqqqqqqqqqqqqqqqq+MX/AB86X/1zl/mtVVVVVVVVVVVVVVVVVVVVaF/yHNO/6+4v/QhVVVVVVVVVVVVVVVVVVVVVf8E//wDkqHxA/wCvVf8A0e1VVVVVVVVVVVVVVVVVVVVf8Fv/APkafhL/ANg/VP8A0ZbVVVVVVVVVVVVVVVVVVVVVVVVVVVVeLfEMHhbw7e65MFY28f7tD/G54VfxJH4Zq/vrvVL2fUL6dpri4cvI7dSx61VVXiDQ5/DzaTf2zyLFqFjDdQSZwVfaN+CO4cEj2Ir4c+K/+Ew8LW2ozMPtcJ8m6A/56KBz+IIP447VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX/BLj/k8f4zf9eF//wCnKOqqqqqqqqqqqqqqqqqqqqvH3/IpX3/bP/0YtVVV8ctY/wCEh+NPj3Ww+5b3xLqEsZOeENw+0c4PC4FfszaOdC/Z6+HOnMpVx4aspXUggq0kSyMDnuC5FV/wUu+Fv/CKfF7T/iPYWwSx8ZWX79lXAF5bhUfOOm6Mwn3O4+tf8EuPih51j4q+Dl/cZa3Yazpqk5O1tsVwPYAiAgersaqqqqqqqqqqqqqqrwh/yLOnf9cBVVVVVVVVVVVVVVVVVVVVXxi/5Po8I/8AX1pf/oVVVVVVVVVVVVVVVVVVVVVf8FNf+TH/AIk/9wr/ANOtpX/BLP8A5Mv8I/8AX/qX/pZLVVVVVVVVVVP9/wDCqqqqqqqqqr9vj/kzj4sf9i/J/wChpX/BHT/k0i7/AOxvvv8A0Tb1VVVVVVVVVVVVVVVVVVVV8Yv+PnS/+ucv81qqqqqqqqqqqqqqqqqqqqrQv+Q5p3/X3F/6EKqqqqqqqqqqqqqqqqqqqqr/AIJ//wDJUPiB/wBeq/8Ao9q8SeN/Bfg6ET+L/F+i6HEQCH1LUIrUYPAOXYdwfyrxZ+3t+xx4MDHV/wBoXwncbOv9k3D6p+X2VZM9O1eBPHvg34n+E9O8c/D/AMR2WuaFqsIltL20fcjjuCOqsDwyMAykEEAgiqqqqqqqqqqqqqqqqqqv+C3/APyNPwl/7B+qf+jLaqqqqqqqqqqqqqqqr/gqn+0b8efhV+05a+GPhr8XvFXhrSW8LWdw1lpmpyW8RlaWcM+1TjcQqgn2Ff8ADa/7XP8A0cf8QP8AweTf/FV/w2v+1z/0cf8AED/weTf/ABVf8Nr/ALXP/Rx/xA/8Hk3/AMVX/Da/7XP/AEcf8QP/AAeTf/FV/wANr/tc/wDRx/xA/wDB5N/8VX/Da/7XP/Rx/wAQP/B5N/8AFV/wTB/aU+P/AMUP2q9P8KfEX4x+LPEejvol9K1jqWqSTxF0RdrFWJGR2qqqqqqqv2hNXZLbStBjfiR3uJR/ujav82/IVVVVeLPD51L4K6JqeN9xpUEcqt1PlvhWH6qf+A1+z/q7wa7qGiM/7u7thKoJ/jQ44+oY/lVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVftt/8FLtM+AWvXfwo+EGlWHiHxpaLt1K9vSzWelORkR7VIM0wBBIDBUJAJY7lHiz9v/8AbB8YXsl5ffHPXbAO2Vg0ny7COMDoAIVU4+pJPcmv+CWXxV+I3xf/AGedc8Q/E3xjqXiTU7LxfcWMF3fy+ZIkCWdmypu6kBpHOTzljzXxD/4K/wDifwr8TbrwPF8DrHS7TQddbT9ZlvtVe6n2QzmOcxoiRhHwrAAlgDjOelQTQ3MEdxbyrJFKgZHU5DAjII9sVVVVVVVVVX/BLj/k8f4zf9eF/wD+nKOqqqqqqqqqqqqqqqqqqqqvH3/IpX3/AGz/APRi1VVqV9BpenXWp3TAQ2cDzSEkDCqpJ5PA4FXdzPqF5NeTndNcytI+B1Zjk/qa8LaOnh7wxpGgRqqrplhBbKqgAARxqvGOO1V+3X8LR8Tv2eNdks7XzdU8LEaxZYX5sQg+cvrgwtIcDqyr6V+zZ8Tz8H/jb4U8cy3Bjsba9EGont9llHlykjvhWLD3UUrK6h0YMCMgg5zVVVVVVVVVVVVVVXhD/kWdO/64CqqqqqqqqqqqqqqqqqqqqvjF/wAn0eEf+vrS/wD0Kqqqqqqqqqqqqqqqqqqqqv8Agpr/AMmP/En/ALhX/p1tK/4JZ/8AJl/hH/r/ANS/9LJaqqqqqqqqqp/v/hVVVVVVVVVV+3x/yZx8WP8AsX5P/Q0r/gjp/wAmkXf/AGN99/6Jt6qqqqqqqqqqqqqqqqqqqq+MX/Hzpf8A1zl/mtVVVVVVVVVVVVVVVVVVVVaF/wAhzTv+vuL/ANCFVVVVVVVVVVVVVVVVVeXlnp9s95f3cNtBEPnlmkCKvbkngc4rxZ+1b+zN4HLR+KPj34Es5k+9bjXYJZhnjPlIzPjg84rxX/wVV/Ys8NBksfiJqfiGVBzHpWh3J5xnAaZI0Pbo3f648V/8Fs/hFZeYPA/wY8XauVHyHVL2304Mcd/LM+BnjvxzjtXiz/gtf8ab/evgn4Q+DNHVgQp1Ke51Bl9wUaEZ+ox7V4s/4Kmftp+KA8Vt8TbLQYXJzFpOi2sePo8iO4/Bqb4t/FLzLuWL4jeJIDff8fKwanLCs3zbvmVGAIzzjFW1pq/iDUPIs7a81K+uGzsiRppZGPfAySavP2evj1p/he88baj8FfHFn4f06LzbrVLnw/cxW8Uf98yMgXb0yc4Gea/ZG/bP+J37JPiz7Z4dnbV/CmoShtY8N3MxWG54x5kZwfKmAxhwOcAMGAAHwF/aB+GP7SXgG2+IPwv1wXlm5Ed3aTAJc6fPgEwzxgnY4z2JUjlSwINVVVVVVVVVVVVVVVVVX/Bb/wD5Gn4S/wDYP1T/ANGW1VVVVVVVVVVVVVVVXxM/ZZ/Z6+MniNfF3xQ+E+h+I9ZS2S1W8vY2ZxEhYqnDDgFmP41/wwH+xp/0bz4U/wC/D/8AxVf8MB/saf8ARvPhT/vw/wD8VX/DAf7Gn/RvPhT/AL8P/wDFV/wwH+xp/wBG8+FP+/D/APxVf8MB/saf9G8+FP8Avw//AMVX/DAf7Gn/AEbz4U/78P8A/FV8OP2Uf2dPhD4mj8ZfDT4R6D4e1uKF4UvbOJlkEbjDLyx4IAqqqqqqqvj9Kz+MrWIggR6cmAe+XfJH6D8KqqqCCW6njtreMySyuERAOWYnAAq70GNPA0nhsgMI9LNsMeoj2g/mK+Dcrx/ETS0U8SLMrfTynP8AMCqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqvip4vPw9+F/jDx8qox8NaBfamFcEg/Z4Hl5A5x8navg58L/Gn7T/xv0r4f6dqqtrvi/UJprzUrwFhGMPNcXD4+8Qqu2ONxwMjNft+/sU/Df9kfwj8P5/BviHX9Z1PxDcXkWpXOpSRBGMSQlfKjRBsGXbgsx6c1/wAEbP8Ak2DxN/2Pt3/6QWFftw2lnZ/tc/FaGxVRG3iS4kYKc/O+Gc/99M1fs56rPr37PXwv1y6/1+oeDNIuJMnPzPZxMefqaqqqqqqqqr/glx/yeP8AGb/rwv8A/wBOUdVVVVVVVVVVVVVVVVVVVVePv+RSvv8Atn/6MWqqvj3rB8P/AAP+IGso5WS08M6g8RGR8/2d9vTp82Oa+F2jf8JH8TfCPh4Jv/tPXbK1288+ZOi44IPeqq4t4LqCS1uYklilQpJG65DKRggj0Ir48/DWb4Q/F/xT8PHRxBpV+4s2fOXtnw8Le5MbJn3zX7F/xSHxX/Z68NandXXnapokf9kaiWbc3m24CqzHqS0Ricn1c1VVVVVVVVVVVVVV4Q/5FnTv+uAqqqqqqqqqqqqqqqqqqqqr4xf8n0eEf+vrS/8A0Kqqqqqqqqqqqqqqqqqqqqv+Cmv/ACY/8Sf+4V/6dbSv+CWf/Jl/hH/r/wBS/wDSyWqqqqqqqqqqf7/4VVVVftj/APBQX4f/ALIepWXhC+8Ha14m8V6lYC+trKE/ZLVYSzorPcsG6sjDCI5GDnHGdS/4K8ftMwRHxFF+zVpFp4ewJBNcwXzARk9fP+VOmOduPav2PP8AgpH8Mv2p9Zj+H2raFN4N8cvC0kGnT3IuLfUAgJf7PNhSXABYxsoIHIL7WIr9rf8AbZ+FP7Iuh2zeKFm1zxRqkbPpnh2xlVZpVGR5srnIhh3DbvIYk52q21sT/wDBXH9ptLEeOx+y7YR+CWfK3skN8YyhOAPtmBFnOBnZj2r9kT9tr4W/te6HdnwzDPoXinSIxJqfh69lV5YoyQBNE4wJockKWAUqxAZV3KWqv2+P+TOPix/2L8n/AKGlf8EdP+TSLv8A7G++/wDRNvVVVVVVVVVVVVVVVVVVVVXxi/4+dL/65y/zWqqqqqqqqqqqqqqqqqqqqtC/5Dmnf9fcX/oQqqqqqqqqqqqqqvFfxf8AhL4DMi+N/ij4S8PmL766prVvale3IkcV4r/4KOfsWeEC0d98ddKvpVHEelWdzf7j7NDGydu7D9RXiv8A4LM/sy6QGh8MeEfHmvzAfK4soLWE8f3nm3g5x/BXiz/gt5r0weHwN+z/AGFoQDsn1bXnuM8cExxxR4we285HpXiz/gr7+194hDLot14O8MA52nTNF81l9P8Aj5eYE/h36dq8Wft0/tf+NA661+0L4wiEhO5dMvf7NBz1GLYRjHt0rxD4u8WeLrn7Z4r8UatrVxnPm6jeyXL59cuSa8KfA/4z+PCg8E/CTxlrwfGG03Qrm4XB7lkQgDg8njg14T/4Jsftp+MBHJa/BO902BwCZdWv7ay2A+qSSCT8AuR3rwn/AMEYP2kNXEc3ivxz4E0CJh80aXNxeTJx3VYlT24f1/Hwr/wRE8KQKj+N/j7q18Sfmi0rQ47Tb04DySy5787R16cc+Ff+CRP7HPh7Z/a+keLPE+08/wBq660e7kdfsqw+mOMda/YK/Z7+Bup/Ejxjaa18J/C+sRaXbK1kmq6bHfeQwnIDL5wfDYA+br71ofhrw54YtfsXhrw/puk2+APJsbRIE4zj5UAHc/nVzbW95byWl3BHPBMhSWKRAyupGCCDwQRwQa/b0/4Jcy6d/aXxl/Zh0RpbXL3Gr+D7dCXi5y0tio6r1Jg6rj5MjCD4FfHz4o/s1+P7fx78Mtcewv4T5V5ZzAtb38QPzQ3EeRvQ49ipwVKkAj9kX9tH4X/tb+FTdeHp00fxbp8QbV/DdzOGnt+gMsR482EkgbwOCQGCkjNVVVVVVVVVVVVVVVVf8Fv/APkafhL/ANg/VP8A0ZbVVVVVVVVVVVVVVVVVVVVVVVVVVVVftCWTx6/peokHbPZmIH3RyT/6MFVVV8FotHm8cQRanAJJfKdrMk8LKvOcd/lDY9CK1aeO10q8uZWwkNvI7E9gFJNfA6xa78dxXAXIs7WWQkjpkbP/AGeqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqviZ4RX4gfDfxX4DZ0UeJNDvNNLP0HnwPHk+3z1+z18WNb/ZV/aI0Tx/qehSzXPhTUJ7TV9LYhZGjZXguIx2DqGbbnjcozxmv+Cmv7T/wV/aU8CfDC/wDhN4t/tKawur99QsJ7aS3uLLfHBtEiOAOqsMqWUkHBNfsFftn/AAb/AGWf2T/Flp4x1Z73xbP4su7zSvDltG5mvVazs0RjIFKRRl0cF2PRWwGIAPiPXPFPxb+Imo+Ib2J9Q8ReMNYkuHihUkzXVzKTsQe7vgD6Cvhp4SXwD8OPCngRXRh4c0Sz00FBhT5ECR5Ht8lVVVVVVVVV/wAEuP8Ak8f4zf8AXhf/APpyjqqqqqqqqqqqqqqqqqqqqrx9/wAilff9s/8A0YtVVftx6wui/sseO7gOA9xbW9qgOMt5tzEhxn/ZZj+FfshaMNe/aY+HdiU3eVrKXWMf88FabPQ/88//ANXWqqv+Co/wu8i/8LfGPT7cbLlG0fUmVcfOu6SBj6kr5wJ9EUemP+CYvxROg/EfW/hVqF1i18T2f2qxRm6XduCWCjsWiLk/9clqqqqqqqqqqqqqqrwh/wAizp3/AFwFVVVVVVVVVVVVVVVVVVVVfGL/AJPo8I/9fWl/+hVVVVVVVVVVVVVVVVVVVVV/wU1/5Mf+JP8A3Cv/AE62lf8ABLP/AJMv8I/9f+pf+lktVVVVVVVVVU/3/wAKqqqtf+Ffw48VeMNH8feJfBWkar4h8PwSQ6Xf3lqs0lmsjKzeXuyFbKLhsbhyAQGbM0MNxC9vPEkkcilXR1yGBGCCO4I7V8efBGkfCb/gp7ZeG/gdbxWfleM9EuLGxshtjtbm4NvJJAqpgCPfIw2DAVW29BWo39lpOn3OqajcpBaWcLzTyv0jRASzH2ABNfs0eHpP+ChX7fOr+P8A4mW0l94asmn1u60+4clFs4XWKzsvTaC8O5eNypJ3JNPpemyaadFk0+2bT2gNubRoVMRi27fL2Yxt28bcYxxXxl0tP+CfP/BRSy8Q+CPM07wqb231WC0jyF/sm7JS6tgOcopFwidceWh6rSOkiLJG6srDKsDkEeoqv2+P+TOPix/2L8n/AKGlf8EdP+TSLv8A7G++/wDRNvVVVVVVVVVVVVVVVVVVVVXxi/4+dL/65y/zWqqqqqqqqqqqqqqqqqqqqtC/5Dmnf9fcX/oQqqqqqqqqrW/EXh7w1Zm/8R69p2lWqjme+ukgQYx/E5A7j868V/ts/sk+CzImuftCeCmeL78dhqa37qfQrb7zn2xmvFf/AAVz/Y38Pbv7I1zxV4n29P7K0J4930+1ND+teK/+C3XhC3Vk8D/APWL9jwsmq65HZ7eOpWOOXPOONw+orxZ/wWf/AGjtWDw+E/AngTQYmB2ySW1xeTLxjhmlVOuTynp758W/8FKv20/GBeO4+NV3pkDZ2w6Tp9rZbM+jxxiTt3Y4rxb8cvjT49Mn/CbfF3xlryyklk1HXLi4TnsFdyAPYDArw/4T8U+Lbr7D4V8NarrNxkDydPspLh8ngfKgJ5NeEv2GP2vfGwjbQ/2e/GEayAFH1Oy/s1SD0O65MYx79Oh714T/AOCQn7YHiIRHWrLwh4XDgFxqetCVo+MkEWqTAntwcZ745rwp/wAEQ/EU4jl8c/tAadZkffg0rQXuc+wkkljx9dh+leFf+CMn7M+kKkvifxj481+YH5k+2W9rCRx/CkJcd/4+9eFf+CcH7FnhArJY/AzTL+YdZNVvbq/3c55SaRkHQdFH6mvCnwb+EPgMofA/wr8IeHmjIKHS9Dt7QqQc5BjQc55+vNVVVXw3+A/w0+E2r6lrngfR57O81ZNl08l3JMHG7dwHJA5Paqqv28P+CaGgfHJdQ+LHwPtbTRPiCQ019poCxWuvNnLMTwIrg5J3/dc8NgkuNG1r4n/AX4jx6npNzrHg7xn4XvCCGjaC4tJkPzI6MOR2ZGBVgSCCDiv2Gf8Agor4O/aYsrX4ffEOWy8O/EyCID7Pu8u21rAOZLbJOHwMtCTnuu4A7aqqqqqqqqqqqqqqqv8Agt//AMjT8Jf+wfqn/oy2qqqqqqqqqqqqqqqqqqqqqqqqqqqq+NHhx9d8HyXdtHvuNLf7QoA5KYw4/L5v+A1VVWi6pPomr2er2x/eWcyyKM4zg8j6EcfjXxe8eWNr4Tj0zTLlXudcgVlAPKW7DJY+m4fKPqfSvgL4cex0S68RXEeJNRcJDkf8s0zkj6sT/wB8iqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/b8/4Jvah8YNdvPjX8Bbe1XxXeDfrehSyrCmqOFAE0LNhUmIADKxCv97Ibdv8AG/w1+IXw01N9G+IPgnW/Dt7GxUw6nYyW5J/2dwAYe4yD1rwv4N8XeOdTTRPBXhbV9f1GQgJaaZZSXUpJ4GEjBP6V/wAE+/8AgnB4l8A+KdN+Ov7QGnRWWqaWRP4f8OF1ke3m523NyRlQyjBSMEkNhm2ldtVVVVVVVVVX/BLj/k8f4zf9eF//AOnKOqqqqqqqqqqqqqqqqqqqqvH3/IpX3/bP/wBGLVVX/BS3WTpn7OMdgshH9reIbS2IBPIVJZefbMQ6+1f8E69GGqftQaNe7c/2Rpl9dZx0zCYc9f8Apt7/ANRVV+0x8Lx8Yvgf4q8DwQCS/nsjcaaMci7hPmRAHnG5lCHHZzXw38b6n8M/iB4f8eaUCbrQdQiuhHnb5gRhvjPoGXcp9mNaHrOm+JNE0/xFo1ytxYapaRXVrKvSSKRAyMPqpBqqqqqqqqqqqqqq8If8izp3/XAVVVVVVVVVVVVVVVVVVVVVPoWiXN8mqXGj2Mt7GQUuXt1aRSOmGIyMVVVVVVVVVVVVVVVVVVVVV/wU1/5Mf+JP/cK/9OtpX/BLP/ky/wAI/wDX/qX/AKWS1VVVVVVVVVT/AH/wqqqq+LHizVfAXws8ZeOtC0j+1tS8OeH77UbOwwx+1zQW7yJFhct8zIF4554r4r/8FZv2tk8PGz034P6X8PpbsNGuqXem3M0qnHWIT4j3D/aVx7V/wS9+DvwG8X+OLn48eIvjfZ/EP4uKJLt9JnSWKXS5ZciW5YXAWS6kw2PNUbEL92KkftbaxPoH7Lnxa1W1LCeHwXqqxMpwUdrWRVYfQsD+Ff8ABD3SrWbV/jDrTr/pFpbaNbxnHRJWvGb36wpVf8FstPto/jH8PNURT59x4Zmhc/7MdyxX9ZGr4L6pPrvwd8Ca3dE+dqHhnTriTc247nto2PPfk9ar9vj/AJM4+LH/AGL8n/oaV/wSA13Q9O/ZOurfUNZsbWX/AIS6+YRzXCo2PJt+cE5xxX/CWeFf+hm0r/wNj/xr/hLPCv8A0M2lf+Bsf+Nf8JZ4V/6GbSv/AANj/wAajkjmjWWGRXR1DIynIYHoQfSqqqqqqqqqqqqqqqqqr4xf8fOl/wDXOX+a1VVVVVVVVVVVVVVVVVVVVoX/ACHNO/6+4v8A0IVVVVSyRwxtLLIqIilmZjgKB1JPYV4t/aO/Z98BiRfGPxu8DaRLFnMF1r9ukxwcECPfvJB6gA14s/4Kf/sVeFd8SfFp9auEH+p0nSLqfPOOJDGsfb+9/MV4r/4LVfAfT90fg34W+N9adf4r421gjH2KyStjp1UfSvFn/Bbb4mXgceBfgd4Y0rn5Dq2pz6hjjuIxB3x/nmvFv/BWD9s/xN5i6Z410Pw1HJkFNJ0KA4BGMBrgSsPXIOQe9eLf2vv2pPHAkTxJ+0B46nilz5lvBrU1rC2cZBihZUI46Y4/Gru81nxFqJuL67vNTv7lsF5ZGmllYn1OSSTXhP8AZi/aM8deU3hL4F+OtSimxsuItAuBDycDMpQIBwep7H0rwn/wS1/bU8UiOS4+F9poNvLjbNq2tWseAe5jR3kGMdCuea8J/wDBFH4134ifxr8XPBmjK4BddPguNQdOOmGWEE9BwcdefXwn/wAES/hNZBD44+NXizWCD8w0uwt9OB4/6aefjn/PevCv/BKf9i3w1te/+H+q+IpU+6+q65c9fUrC0an6EEc14V/ZP/Zk8Essnhj4B+BLSZPu3DaFBLMOc8SOrP1A79h6VZWNlptrHY6dZwWtvCMRwwxiNEHso4FVVVVVVVVVVV+2l+wZ8PP2stCk1qzW28PfESyg26dryRfLcBQdsF0FGXi7Bh86dRkZRvib8Lvih+z18RZvBvjzR77w54l0WZJoZEkK5wcxzwSocMuRlXU8EdiCB+wX/wAFQ7LxcumfBr9pbWIrPXfkt9J8WXDhIb/jCx3ZOBHLwAJfuvn5trcvVVVVVVVVVVVVV3eWdhAbm+u4beFSAZJpAijPTk8V/wAJZ4V/6GbSv/A2P/Gv+Es8K/8AQzaV/wCBsf8AjX/CWeFf+hm0r/wNj/xr/gtjqul6p4o+FDaZqVrdiPT9UDmCZZNuZLfGcE46VVVVVVVVVVVVVVVVVVVVVVVVVVVVOiSIY3UMrDBUjIIr4n+AJ/Burtc2kTNpV45Nu+M+WepjP07eo+hqqqvA3gzUvHWtR2iGRbSHb9quDyI0HRQfUgYA/oKsrO10+zhsLKFYoLeMRxIo4VQMAVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVq96dM0q81JYxIbS3klCE43bVJxn8K+Hn/BYTxJ458f+GfBUvwJ020TxBrFpp7XC6+7mETTLGXC+SM43ZxkZxVxb293C1tdQRzROMPHIoZWHuD1q1s7OxgW2sbWG3iXO2OKMIoz1wBxXjn/AILGy+CPGviDwXcfs3rdS6Bqlzp7zr4y2CVoZWjLAfYjgHbnGT1rwh4ksvGfhLRPF+nIyWuuadb30Cv1VJo1dQffDCqqqqqqqqq/4Jcf8nj/ABm/68L/AP8ATlHVVVVVVVVVVVVVVVVVVVVXj7/kUr7/ALZ/+jFqqr/gqprAg8GeAfD28f6bqd3dbfXyYkXPr/y3/X8v+CW2jG4+LnizXymRY+Hfs4JHQzXEbenXEJ7+vXtVVX7Z3wt/4VR+0J4l0q1tfJ0zWZP7W00BcL5U5JZVHosolQeyiv8AgnR8UR47+A0fhG9ufM1LwTdGyYMcsbZ8yQMfQAF4wPSKqqqqqqqqqqqqqrwh/wAizp3/AFwFVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV/wU1/5Mf+JP/cK/9OtpX/BLP/ky/wAI/wDX/qX/AKWS1VVVVVVVVVT/AH/wqqqqrV9H0nX9MuNF13S7TUtPu4zHcWl3As0UynqrowIYexFft+/Cy0/Yj/at8MfEj9n64bw3HqluutafaW5IjsLpJWSaJFz/AKhxtzGflxI6Y24FfEqN/jj+yZ4mOmWP77x18PrmSztyu4rJdWDNGAO5DOuPcV/wRF8U2lp47+Kfgh5kFzq2kafqEcZHzFLWWWNyPYG8TP1FV/wWZ13/AISH9pDwX4I0oG5utN8Lx7oo1y3nXFzLhPclUjOP9oe9eCPD48J+C9A8Kgx/8SbS7ayHlrtX91EqfKB0Hy9Kr9vj/kzj4sf9i/J/6Glfsc/8E1P+GtfhHL8U/wDhdX/CK+VrE+nfYP8AhG/t2fKSNt/mfaY+vmY27eMdea/4ccf9XQ/+WR/93V/w44/6uh/8sj/7ur/hxx/1dD/5ZH/3dXw+8Kf8IH4B8NeBvt/27/hHdHtNO+1eV5Xn+RCse/Zltu7bnbk4zjJ61VVVVVVVVVVVVVVVVVfGL/j50v8A65y/zWqqqqqqqqqqqqqqqqqqqqpfG/grwfq1hceLfGGiaJElxGzPqOoRWyqoYZJLsOPevFn/AAUC/Y08GF11b9oHw3clCQRpJl1PJBxwbVJAefwxz05rxX/wWL/ZQ0ItFoOm+OPEj/wPZ6VHBGee5nlRhxz909R748V/8FvB80Pgf9n3/duNV8Qfzijh+n8deK/+Cxn7Vuuh4vD+l+BvDcZ4je00uWeVfqZ5XUn/AIAPpXiz/goJ+2X4zEq6r8f/ABHarKCCuk+Vpm0Yx8ptkjI475z3znmvFHxG+IXjiRpfGvjzxD4gdmLM2qapNdknjkmRjzwPyFeFvhd8TPHbIngj4deJ/ELSHCDStInuy3bjy1bPNeE/+CeH7Z3jLym034C65ZpIRl9Wlg07YM4JK3EiNxjpjPtyK8Jf8Ebf2pdcMcviTX/A3huHjzEuNRluZgD1wsMTIce7ivCX/BEOyTy5vHf7QM0vTzLbSdAEeOOcSyTN3/6Z9vfjwr/wR5/ZK0FVbXLnxt4lk/iF/q6QpnHYW8cZA78kn3rwr+wN+xv4N2/2R+z34VuNvT+1YX1T8/tTSZ6d68NeBfBHguH7P4O8HaHoUW3bs0zTorUY64xGo44HFVVVVVVVVVVVVeH/AB54G8W3VxY+FPGmhazc2i5uIdP1GK4eEZxllRiVGeOe9V8DvFp8a+AV1mSYyTLq2owyZfcRsvJQuT/ubD+NVVftNfssfCv9qrwO3hL4haYI761V20jW7aNRd6ZKR95GP3kJA3Rn5WwOhCsP2nf2Vfil+yp44bwn4/07z9PumdtH1y2jb7LqcS4+ZCfuuARvjPzKSOoKsf2Df+CnGrfCcaf8If2g9RutV8FqFh0zXWDTXWjKAAscg5MtuMDGAXTtuXCro+saR4h0q013QNUtNS06/hWa0u7SZZYp42GVdHUkMpGCCOKqqqqqqqqqqqv2rv2f/wDhp/4Jav8ABz/hLv8AhGv7VuLab+0f7P8AtnleTMsuPK8yPOdmPvDGc89K/wCHHH/V0P8A5ZH/AN3V/wAOOP8Aq6H/AMsj/wC7q/4ccf8AV0P/AJZH/wB3V+3B+xT/AMMZ6r4S0z/hZn/CYf8ACU291Nv/ALF/s/7N5LRjGPPl37vM9sY754qqqqqqqqqqqqqqqqqqqqqqqqqqqq1LTNP1ixl03U7WO5tpl2vG4yD/AIH36ivF3wK1Wzlku/Ccwvbc5ItpWCyp7Anhh+R+tX/h3X9MkMWo6Le27A4xLbsufocc1aaHrWoSCGx0i8uHPAWK3Zj+grwr8DvEOrSJceIm/su06lOGmYew6L+PI9K0PQdJ8N6dHpejWiQQR9h1Y92Y9ST61VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV8afEvi3wX8IvGXjHwHaWF34g0HQ7vUNPtr+F5YZ5IYmkEbKjIx3BSoww5I5rUP+Cw/7TWpWFzp0/gf4YLHdQvE5TTL4EBgQcf6Z1wa8IeJr/wAFeLdE8ZaVFby3ug6jb39tHcKWjeSGRZFDhSCVJUZAIOO461/w+T/ae/6EX4Xf+Cu//wDkyte/4LA/tVaxpU+nWGi/D7RZ5kKpfWGk3DTQnHVRNcSJn/eQ1eXl7ql9Pf39zNdXd3K0s00rF3lkY5ZmJ5JJJJPcmvgX4b1Twb8EPh74P1xXGpaF4V0yxvBIMMJobWON8jJ53Kaqqqqqqqqr/glx/wAnj/Gb/rwv/wD05R1VVVVVVVVVVVVVVVVVVVV4+/5FK+/7Z/8Aoxaqq/4Ko6z5/j3wL4e35+xaPcXW3086YLnr/wBMPTt37f8ABKjRhHpfxF8QOgPnz6faxse2xZ2Ydc8+Yn5D3qqqv+CnnwuGu/DnQ/itp9tuuvDF39kvnUY/0W4ICsx9FmCKB/02Nf8ABPP4oj4f/H608OX115em+NLc6bIGOFFwPnt2+pcGMf8AXWqqqqqqqqqqqqqrwh/yLOnf9cBVVVVVVVVVVVVVVVVVVVVXiL9oDwJ4Y+KGn/CPUotUOuak8CQtFbq0IMpwuW3Aj34qqqqqqqqqqqqqqqqqqqqr/gpr/wAmP/En/uFf+nW0r/gln/yZf4R/6/8AUv8A0slqqqqqqqqqqn+/+FVVVXj7/gqX8C/hX8efFXwW8faDr1va+G7mO1Gv2CLdQyTeWrSq8QIdAjsUyu/JU8CtX/4Kp/sRabpcl/ZfFS/1WdEytjaeHL9ZnOOgMsKR57cuB7968cax8Tv+CqP7Wtmvg7wxd6V4dsoYbJGk+ddF0tJGZ7i4cfL5rlnYKOrFUBbG6tF0fT/D2jWGgaTAIbLTLWO2togc7I41CqPwAAr4veFfG/8AwTQ/bdtfi74b8PzXXgHWr64uNOSM7Iruwn/4+bAsOFkiLfIDn7kLkHJFX/8AwVQ/YytfA7eLrX4iXt3e/ZvMj0BNIuFvmlx/qTlPKVs8FjJs9GIxn9kz4d+Pv2+f20r/APah8faE1t4Q0PWY9SuicmHzbdV+xafGxGJCgSEycYKKS2DIuar9vj/kzj4sf9i/J/6Glf8ABHT/AJNIu/8Asb77/wBE29VVVVVVVVVVVVVVVVVVVVfGL/j50v8A65y/zWqqqqqqqqqqqqqqqq9vrLTrdrvULyC2gQfNLNIEVfqTwK8RftJ/ADwoWTXPjH4RhkTO6GLVYp5Fx6pGWYflXiL/AIKJfsuaGHWx8WaprjocFNO0iYZOcHBmEan6g4rxD/wVW8A224eE/hPr+pY+7/aN/DZZ+uwTY4xXiH/gqb8XL0Mnhj4feFNLB6Ndme7dR7EPGM9Oo/8AreI/2+/2pvEQeNPiLHpULjmLTtMt4scdnKM4/wC+q8R/HH4zeLw6eJfiv4s1GOQENDPrE5jweoCbtoH4VYabq+vXwtNLsLzUbyUkiKCJppHJPXCgk8n9a8J/sa/tV+N/Lbw9+z944eOQ/JNd6RJZxNzjiScIpGe+a8Jf8Elv2y/EpjOq+FvDvhhXI+bV9difaD3ItvOPvjGfavCn/BEb4gXYRvHPx28PaWeNyaTpE1/9QDI8HsM498dq8Kf8EWf2ftNCSeMPiT451yVAMpavbWMTHGDlfLkbGeRhx+NeFf8AgmR+xV4UVWX4PJq869ZtV1W7uc8Y5QyCP3+73+mPCv7PfwG8DFH8G/BfwPo0iDiWy8P20Mh6dXVNx6DqewoKqqFUAADAAGMVVVVVVVVVVVVVVVVV/wAE/wD/AJKh8QP+vVf/AEe1XM8drbS3UpOyFC7YGTgDJ/lX/BP/AMSyav8ADnxJo9zJumstca46YCrPGvA9t0bn8aqqr4sfCX4f/G/wNqHw6+Jnh231nRNRX54pRh4XAO2WJxzHIuThlwRz2JB/bV/YI+IH7J2tya7p32rxH8Or2bbYa6sXz2pY/LBdheEk6APgJJ1GDlF/Yk/4KBePP2VdUh8KeITd+JPhtdTZutIMm6bTyx+aazLEBT1JiJCOc/dY76+GfxP8B/GPwXp/xA+G3iS01zQtTTdBc27fdYfeR1PKOp4ZGAIPUVVVVVVVVVVVVVVf8Fv/APkafhL/ANg/VP8A0ZbVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVdW1ve20tndRLLDPG0ckbDIZSMEH2IrxP/wAEeP2htOm1bUtK8cfDk6VatNLbLNqN6JjApJXcBaEb9oGRnGe/evCHhm/8a+LdE8G6VNbxXuvajb2FtJcMVjSSaRY1LlQSFBYZIBOOxr/hzX+09/0PXwu/8Gd//wDIdQ/8Eav2mGlRbjx98MUiJ+dk1G+ZgO5ANoM/TIr9mP8A4JOeEvhR4v034h/GDxlD4w1PSJkuLHSbOzMNlFOhyjyM5LTgEAhdqLkchhxVVVVVVVVVV/wS4/5PH+M3/Xhf/wDpyjqqqqqqqqqqqqqqqqqqqqrx9/yKV9/2z/8ARi1VV/wUn1j+1P2kmsd+f7I0CztcZ6ZMk3p/029/6D/gmJoxsfgHq+qyIQ+p+Jp2QkEZjSCBB/48H6VVVXxM8D6d8TPh94h8AaoQtvr2nTWhcjPlMykJIPdW2sPcCpovEHgDxe8LM9hrnhvUipKn5re5gk7e6un6V8KvHun/ABS+G/hv4g6aUEWu6dFctGjZEMhGJI8+qOGQ+6mqqqqqqqqqqqqq8If8izp3/XAVVVVVVVVVVVVVVVVVVVVV8Yv+T6PCP/X1pf8A6FVVVVVVVVVVVVVVVVVVVVX/AAU1/wCTH/iT/wBwr/062lf8Es/+TL/CP/X/AKl/6WS1VVVVVVVVVT/f/Cqqqr4t/sv/ALP3x3Yz/Ff4UaDr955Yj+3vCYLwIBwouYikoUdhuwKsf+CWn7ENlfi+b4SXNyFbclvP4gv2jU5yOBMCQOmGJBHXNeAfhp8PfhXoK+GPht4L0bw1patvNrplmlurvjBdtoBdiAMs2Se5qvHvw78C/FPwzc+DfiL4T0zxFot2QZbLULdZU3Do4zyrDPDDBHUEVbf8EvP2IrXWBq6fB+SQK+9bSXXb94Af90zcj2JI9scV4U8I+FvAvh+z8KeC/DunaHo9gmy1sNPtlghiHU7UUADJySe5JJ61Vft8f8mcfFj/ALF+T/0NK/4I6f8AJpF3/wBjfff+ibeqqqqqqqqqqqqqqqqqqqqvjF/x86X/ANc5f5rVVVVVVVVVVVVVVXirxT4f8E+HNQ8W+KtUh07SdKt2nu7mUnbGi9eBySeAAMkkgAEmvj5+2p8XPit431DUPCHjTxB4U8MDMGn6Zp1/JakwjOHmMTDc7ZJYZIHCjIGTq2ua1r9x9s1zWL3UZ/8Anrd3DTN+bEmvDvwb+Lfi7b/wi/ww8VaqrdHtNHnkQY6ksFwB06mvD37B/wC1N4jCSJ8MZNPhY4Muo6hb2+3nHKGTf7/d/pXh3/glv8Z78JJ4k8beEtJRuqQvPdSL65Hlov5NXh3/AIJTeELcKfFvxe1i/PG5dO0yO0x6gF2l9ucfh6eHv+CcX7MGibP7R0LXdfKDk6jq8ibjjqfs/lfX0/DivD37L37O3hcKdH+DHhMOg+WS601Lp14xw0oZu3r6+prwjpWmaRqmm2mk6da2UAu4sRW8KxqPmHZQBVVVVVVVVVVVVVVVVVVVVVf8E/8A/kqHxA/69V/9HtXxKvxpXw58VaoWA+x6JeTZJxjbC5/pX/BO/XhbeN/FfhkuB/aGlRXQU9zBLt/lOaqqqvEHh7QvFeiXvhvxNo9nqulajC0N3ZXkKyxTxt1VkbII+tft5f8ABM3XPgo2o/Fr4E2N1rHgBS01/pQZprrQl5JIzlpbccfPkug+9kAvX7LH7W/xS/ZO8aDxF4IvPt2jXrqNZ8P3UrC21GMfTPlygZ2ygErnkMpKn9nD9pn4W/tReBIvGvw31hWlhCpqmk3DBbvTJiPuSoD0ODtcZVgDg8ECqqqqqqqqqqqqv+C3/wDyNPwl/wCwfqn/AKMtqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqeCG5gktriNZIpUKOjDIYEYII9MVr37P/7FvwhtoPH3ib4YfDHwlBpVxHNBqt9Z21msEyHchV2x8+5QQBzkcVqX/BST9ibS7p7K5+OVq8iEgm20XUbhOuOHjt2U9Oxr/h5r+w//ANFv/wDLb1X/AORqvv8Agp9+xLawebb/ABfnvHzjyoPDmohvr88Cj9asruHULK3v7YkxXMSyRlhg7WAI4+hqqqqqqqqqv+CXH/J4/wAZv+vC/wD/AE5R1VVVVVVVVVVVVVVVVVVVV4+/5FK+/wC2f/oxaqq/bZ1ka7+1J4+u1fcIb2G1A9PJt4oiOp7of/rV+wLo66T+yv4QkKBZNQkvbmQDHObqVVPH+wq1VVVf8FFvhcfAnx5k8WWNt5em+NbUXyEDCi5TEc6j3JCSH3lr/gmD8UDrPgTxB8Jr+5LXHh66F9YKzf8ALtOcOqjsFlUsfeaqqqqqqqqqqqqq8If8izp3/XAVVVVVVVVVVVVVVVVVVVVV8Yv+T6PCP/X1pf8A6FVVVVVVVVVVVVVVVVVVVVX/AAU1/wCTH/iT/wBwr/062lf8Es/+TL/CP/X/AKl/6WS1VVVVVVVVVT/f/Cqqqqqqqqqr9vj/AJM4+LH/AGL8n/oaV/wR0/5NIu/+xvvv/RNvVVVVVVVVVVVVVVVVVVVVXxi/4+dL/wCucv8ANaqqqqqqqqqqqqqp5obaF7i4mSKKJC0kjttVFAySSegA71+3H+1pL8bvEjfD/wADagy+BtEnOJY2I/ta4XgzN/0zXoi/Vj1AX9h/9kub44+JB488b2LL4F0OcBo3BX+1rhefJX/pmvBdvcKOSSvh34e+APCAVfCXgfw/omzO0adpkNtj1xsUegqqqqrQv+Q5p3/X3F/6EKqqqqqqqqqqqqqqqqqqqqr/AIJ//wDJUPiB/wBeq/8Ao9q/aMvzpvwJ8c3AYjfolxDwcf6xfL/9mr9jHXv7D/aD8PxPJsi1SK5s5Dux96FmUe+XRBiqqqqmVWUqwBBGCCK/bz/4JcQasuo/GT9mHRFhvgGn1bwdaxAJPgZaWyUfdfjJgAw2fkwcI3wi+MXxQ/Z2+INt44+HWuXWia5pshjuIJFPlzqD88FxEeHQ4wVPIPIIYAj9jX9uf4bfta+HVsYJItB8eadAG1Xw9NJywAG6e2Y/62En/gScBhgqzVVVVVVVVVVVVf8ABb//AJGn4S/9g/VP/RltVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX99Z6ZY3GpahcpBa2kTSzyyHCxooyzE+gAJr9q39pbxj+098VtU8aa5qN0mhw3EkXh/SWkPlafaA4QBenmMAGdurMewCgVPoms21oL640m8itiARM9uypg9DuIxzxXgb4eeO/idro8MfDrwdrHiXVTEZTZ6XZvcyLGCAXYIDtQFlBY4ALDnmvDtvNa+HtMtbiMpLDZRI6N1VggBH5iqqqqqqqqq/wCCXH/J4/xm/wCvC/8A/TlHVVVVVVVVVVVVVVVVVVVVXj7/AJFK+/7Z/wDoxaqq+OWsf8JD8afHuth9y3viXUJYyc8Ibh9o5weFwK/Zm0c6F+z18OdOZSrjw1ZSupBBVpIlkYHPcFyKqqqv+Ch/wtHj/wCANz4lsbXzNT8FXA1GIqvzG3PyXC/7oUrIf+uIr9j74oj4S/tA+F9eurrydM1Gf+y9TJOF8i4wm5j/AHUfy5D/ALlVVVVVVVVVVVVV4Q/5FnTv+uAqqqqqqqqqqqqqqqqqqqqr4xf8n0eEf+vrS/8A0Kqqqqqqqqqqqqqqqqqqqqv+Cmv/ACY/8Sf+4V/6dbSv+CWf/Jl/hH/r/wBS/wDSyWqqqqqqqqqqf7/4VVVVVVVVVVft8f8AJnHxY/7F+T/0NK/4I6f8mkXf/Y333/om3qqqqqqqqqqqqqqqqqqqqr4xf8fOl/8AXOX+a1VVVVVVVVVVVVVf8FA/2ujO1/8AAD4Z6qQiMYvE2o28n3iOtmjDsCMSEf7nTeD+y7+zf4h/aO8fxaHb+dZ+HdNZJdc1NV/1ERPEaE8GV8EKOccsQQpFeE/Cnh7wN4a07wh4U0yHT9J0q3WC0tohgRoP1JJySTySSTkmqqqqq0L/AJDmnf8AX3F/6EKqqqqqqqqqqqqqqqqqqqqr/gn/AP8AJUPiB/16r/6Pav2x777D+zn4r2n57j7LCvGfvXUWf/HQ1fCnXj4X+J3hPxD5hVdP1m1lkI7oJV3j8VyPxqqqqqq/bs/4JweFv2jLe9+JnwshtNA+JUcbSTKQI7bXSBwk3ZJuAFl75w+RhkuLf4nfAX4ktBKNZ8G+NfCl6RlWa3ubOZeMgjsQeoyrK3dTz+wj/wAFJvDf7Qcdj8Lvi/cWWg/EcKI7W4BEVrrpGADH2jnPeLo3VOuxaqqqqqqqqqqr/gt//wAjT8Jf+wfqn/oy2qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv2k5L6H9nT4pzaWCbyPwVrDW4AyfMFlLtx+OK/4J+/DXwt8V/2s/A3hXxpp8GoaPHJc309lOm+O5a3t5JY0dTwy70QspyGUEEYJr/grj4aTw/+1LZT21oILTUvCVjLboiBY1CSTQ7VA4AHlDjtn0Ir9kXxd4Y+MP7J3w8vRHZanZyeGbbStTtJY1lj8+3iFvPE6NkYLRt8rDlWB5BBPx4nt/2X/wBtHxTd/A3Uf7Oi8G+JPP0oQylktztV5LY4OTGpd4WUnlQVbPNeAfFtl4/8CeHPHenR7LTxHpFrqUCFt21J4lkUZHXhhVVVVVVVVVf8EuP+Tx/jN/14X/8A6co6qqqqqqqqqqqqqqqqqqqq8ff8ilff9s//AEYtVWpX0Gl6ddandMBDZwPNISQMKqknk8DgVd3M+oXk15Od01zK0j4HVmOT+prwto6eHvDGkaBGqqumWEFsqqAABHGq8Y47VVVVavpVhr2k3uh6rbrcWWoW0lvcwsOJI3Uqyn2IJFfFTwHf/C/4jeI/h9qRZptC1GW2WRhjzYwf3cn0dCrD2YV+y58Uf+Fw/Arwr4xubkTal9kFpqZJy32qD93Ix9C+0SY9HFVVVVVVVVVVVVeEP+RZ07/rgKqqqqqqqqqqqqqqqqqqqq+MX/J9HhH/AK+tL/8AQqqqqqqqqqqqqqqqqqqqqq/4Ka/8mP8AxJ/7hX/p1tK/4JZ/8mX+Ef8Ar/1L/wBLJaqqqqqqqqqp/v8A4VVVVVVVVVVft8f8mcfFj/sX5P8A0NK/4I6f8mkXf/Y333/om3qqqqqqqqqqqqqqqqqqqqr4xf8AHzpf/XOX+a1VVVVVVVVVVVVX7dH7W0fwa0B/hp4B1FT431m3PnTx8/2TbMMeYT2lcZ2DsMscfLu+Evwq8Y/HDx/YeA/CFs1xf6g5ee4kBKWsII8yeVuyrkc9SSAMlgD8Ffg54R+BXw/0/wAAeD7ceTbDfd3boBLfXBA3zSEdzgAD+FQqjgCqqqqqrQv+Q5p3/X3F/wChCqqqqqqqqqqqqqqqqqqqqq/Zn/Zr8V/BHxh4m8ReINe0m/h1uERwx2Zk3IRIX+bcoHQ9q/bzvxZ/AkWxYD7drdtCATjOFkk/9p1XgTXh4q8D+HvEwcP/AGtpVtdEj1kiVj/Oqqqqqr9sP9iH4Z/tbeGS+pRRaH430+AppHiOCAGRADkQzqMGaHOflJyhYlSMsG+NXwQ+J37Onj+48A/EzQ5dK1a0Pm208bFobuLcQs8EnG9CVODwQQQQGBA/YM/4Kji4Om/Bv9p/XFWTK2+keMbl8bySAsV8x4B5wJ+BgDf3cxyRzRrLDIro6hkZTkMD0IPpVVVVVVVVVVX/AAW//wCRp+Ev/YP1T/0ZbVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVXiTQrLxR4d1Tw1qIza6tZTWk4Az8kiFG/RjX7Oup+Kv2cv2w/Dct7oN/eap4N8TS6dq1lY2z3E3lZe1u9iICWIjeQgDrx65r/gr38BNU8c/Dbw/wDG/wAMadLdXXglpbfWI4oyX+wTFSJSOu2KRefQTMx4UkeBPjR8XvhfY32mfDf4n+KPDFpqRzdwaTqs1qkzYxuKowG7HAbqB3rQ9E8R+O/FFj4f0S0utW13Xb1Le2hXMktzcSuABk8lmZup9c18K/Bp+HXwv8H/AA9M4nPhjQLHS/NHR/s8CRbh9dmaqqqqqqqqr/glx/yeP8Zv+vC//wDTlHVVVVVVVVVVVVVVVVVVVVXj7/kUr7/tn/6MWqr496wfD/wP+IGso5WS08M6g8RGR8/2d9vTp82Oa+F2jf8ACR/E3wj4eCb/AO09dsrXbzz5k6Ljgg96qqqqr/gp98Lf7G8eeH/izp9sBbeIrQ2N+yr0uoB8jMfVoiFHtCa/4Jd/FEWeu+J/g9qFxiPUYhq2mqxwPNjxHOo92Qxtj0iaqqqqqqqqqqqqvCH/ACLOnf8AXAVVVVVVVVVVVVVVVVVVVVV4l/Z38E+Kvivpvxh1DVNbj1rS5IHhghniW3YwnK7lMZbnvhh+FVVVVVVVVVVVVVVVVVVVVf8ABTX/AJMf+JP/AHCv/TraV/wSz/5Mv8I/9f8AqX/pZLVVVVVVVVVVP9/8Kqqqqqqqqqv2+P8Akzj4sf8AYvyf+hpX/BHT/k0i7/7G++/9E29VVVVVVVVVVVVVVVVVVVVfGL/j50v/AK5y/wA1qqqqqqqqqqqqr9q39pXQ/wBnHwA+pI1veeKdVV4tD02RvvyAczSAc+UmQT03EhQRnIZvG/xd8eZP23xD4p8TX/A+/LdTyH8gPyCgdgK/ZR/Zm0P9nHwGtjIIL3xXqyJJreooMhnA4hjJ58pMkDoWOWOMgCqqqqqrQv8AkOad/wBfcX/oQqqqqqqqqqqqqqqqqqqqqqq/4KHX5j+HPhjSwx/0jW2mxn/nnC69P+2n61X7Iuvf8JB+z34UleTdLYxS2cg3Z2+VM6qP++Ahx71VVVVVV+0P+zh8Lv2nfAU3gP4m6P50a7n0/UbfCXemzEY82ByDg8DKkFWAAYGv2r/2Pvij+yX4x/sXxfanUvD19Iw0bxFbQlba+Uc7TyfKmAHzRMcjGQWXDH9g/wD4KV+IfgTLp/wp+Nd7e678PCVgsr05mutAHQbf4pbcZGY+WQD5M42N4Z8TeHvGnh7T/FfhPWrTVtH1W3W4sr20lEkU8bDIZWHUfyqqqqqqqqqq/wCC3/8AyNPwl/7B+qf+jLaqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq8YeKtF8C+Etb8b+JLlrfSPD2nXGo38yRtIY4II2kkYKuSxCqTgDJrT/+ClX7B+k317qek+KJLO81KTzL24t/CtzHJcv/AHpGWIFz7nNadfaX4p0C11KCNbnTtXs0lRJo+JIZUBAZT2KtyD618Tv+CTf7Lvj/AFubXvD58R+CZbiQvLZ6JdRm0yeu2KaNzHz0VGVRnAXGMfs7fsJ/s9/sz6j/AMJF4I0G91TxGEZE1zXJ1ubmFWGGWIKqRxZBIJRAxBIJI4qqqqqqqqqq/wCCXH/J4/xm/wCvC/8A/TlHVVVVVVVVVVVVVVVVVVVVXj7/AJFK+/7Z/wDoxaqv249YXRf2WPHdwHAe4tre1QHGW825iQ4z/ssx/Cv2QtGGvftMfDuxKbvK1lLrGP8AngrTZ6H/AJ5//q61VVVVX7YvwuHxa/Z98T6HbW3nanpcP9qaYAMt59uC2F93j8yP/gdfBL4jXPwk+LHhf4iWzPt0bUEkuVT70lu3yToPdomdfxq0u7XULSC/sbiOe3uY1khljbcsiMMqwI6ggg1VVVVVVVVVVVeEP+RZ07/rgKqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr/AIKa/wDJj/xJ/wC4V/6dbSv+CWf/ACZf4R/6/wDUv/SyWqqqqqqqqqqf7/4VVVVVVVVVVft8f8mcfFj/ALF+T/0NK/4I6f8AJpF3/wBjfff+ibeqqqqqqqqqqqqqqqqqqqqvjF/x86X/ANc5f5rVVVVVVVVVVVV8Zvi/4R+B3gDUPH/jG5221oNltbIwEt5OQdkMYPVmwfYAFjwDXxf+LHjH45/EC/8AHfi65M17fvstrWPJjtIQf3cES9lUH6kkk5JJP7Cv7JCfB3QI/ib4/wBOH/CbazB/o9vKuTpFsw+5jtM4++eqjCcfNuqqqqqqrQv+Q5p3/X3F/wChCqqqqqqqqqqqqqqqqqqqqqr/AIKNX3/Ih6Wh/wCf+ZwR/wBcAvP/AH1+lV/wT3143vwx1/w88hZ9M1nzVB/hSaJMD/vqNz+NVVVVVVV8Rfhx4H+LXg7UfAPxF8N2muaDqsRjubO5U4PoysMMjg8q6kMpwQQa/bh/4J6+N/2WtRn8aeEftXiT4aXM2IdR2bp9LLH5YrsKMAZOFlA2scA7WIU/sY/t2/EX9krXl0tvO8Q+AL+fdqXh+SbBiJ4M9qx4jl6ZH3ZAMNg7XX4QfGP4dfHfwLY/ET4YeJLfWNHvRgshxJbSgAtDMnWORcjKn1BGQQTVVVVVVVVX/Bb/AP5Gn4S/9g/VP/RltVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV4Y8cwjxrrHgjUJgDHdu1g7HqD8zR/gSSPbI7Cqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq8a+FNL8eeDde8D62GOneIdMudOuwn3vKniaN8Z77WNfH74AfET9nH4hX3w/8AiFo80Dwyv/Z+oCJhb6nAD8s8DHhlIIyM5Una2CCK/Y1/bK+DPxc+DPhfTdS8eaJoni7RNKgsdW0jUbxLWUSwoEMsQcgSRuFDgpnbu2nBBFf8LU+GH/RSPC//AIOIP/i6/wCFqfDD/opHhf8A8HEH/wAXSOkiK8bhlYZUg5BHqKqqqqqqqqr/AIJcf8nj/Gb/AK8L/wD9OUdVVVVVVVVVVVVVVVVVVVVePv8AkUr7/tn/AOjFqq/4KW6ydM/ZxjsFkI/tbxDaWxAJ5CpLLz7ZiHX2r/gnXow1T9qDRr3bn+yNMvrrOOmYTDnr/wBNvf8AqK07U9P1a3a5066SeNJXiZkPR0Yqw/Mf171VVVV+1F8Lj8H/AI6eKvBlvbmLTluzd6YNuF+yzDzIwvqFDGMn1Q1+wT8UT8Sf2edHsby683U/CTnR7kE87IwDAfp5TIue5Q1VVVVVVVVVVVeEP+RZ07/rgKqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr/AIKa/wDJj/xJ/wC4V/6dbSv+CWf/ACZf4R/6/wDUv/SyWqqqqqqqqqqf7/4VVVVVVVVVVft8f8mcfFj/ALF+T/0NK/4I6f8AJpF3/wBjfff+ibeqqqqqqqqqqqqqqqqqqqqvjF/x86X/ANc5f5rVVVVVVVVVVVeKfFHh/wAFeHNR8WeKdUh07SdKt2nu7mY4WNFHPTkk9ABkkkAAkgV+1P8AtJa/+0f4+bV5TLZ+G9KLw6HprN/qYyRmVwOPNfALHsAqjIXJ/wCCfn7Iv26Wx+P3xK0sG2iIl8M6dcRn9446Xjqf4R/yzB6n5+MKTVVVVVVVaF/yHNO/6+4v/QhVVVVVVVVVVVVVVVOyIhd2CqoySTgAVf8AjjwTpWRqnjHRLPb1+0ahFHjjPdh2rUPj58FtNyLj4neH32jn7PeLP2zx5e6tQ/a3+AdgSieNXu3Ucrb6dcN2zwSgH61qH7cPwctARaWPiS+I+6YbKNQeP9uRTjt0q/8A29/CseRpfw+1W4x0+0XkcPb2D96v/wBvrXJc/wBl/Dawt/T7RqTzY49kTv8A571f/tz/ABbuSVstF8MWi/wlbWZ2HHq0uOvtWoftgfHq9yIPFlrZA9rfTYD29XRjVz+0F8a7uPypfibroGT/AKu5MZ/NcGvHPirxN4qvrefxP4j1PV5oYyI5L+8kuGQE8gFySBwKMZMIlB/i29Pav2f/AI4+Kvgprl9deH4LS7tNTSL7dZ3KnEwjLbSGHKsPMfB5HPINfCv9pz4Z/FExadFqH9i604A/s7UHCGRvSJ/uyfThv9mqqqqqqq1XStM1zTLvRda0+2v7C+geC6tbmISRzxuCrI6nIZSCQQeCDX7en/BMLU/hydS+MX7Oel3Go+FAHuNV8NxKZZ9IA5aSDq0sA5JX70YGfmXJT9mv9qD4pfss+Oo/GXw51UNbzlU1XR7klrTU4gfuSKOjDJ2uuGUk4OCwP7L37WPws/au8Er4n8B6h9m1WzVV1jQbpx9q02Q9mHG+M/wyr8rdOGDKKqqqqqqqv+C3/wDyNPwl/wCwfqn/AKMtqqqqqqqqqqqqqqqqqqqqqqqqqqqqqrUL620yxuNRvJAkFtE0kjHsqjJ/lWoaxeX2uXGvrK0VzNdNcKyNgoxbcMH2r4ceOIPG2hrNKyLqNqAl3EOOezgejY/A5HaqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqvEk0tt4d1S4t5WjlispmR1OCrBCQRXjT47fG34kac+jeP8A4v8AjPxHp0kolax1TXbi5t946MIncoCO2BxVVXhX/kV9H/68IP8A0WtVVVVVVVVV/wAEuP8Ak8f4zf8AXhf/APpyjqqqqqqqqqqqqqqqqqqqqrx9/wAilff9s/8A0YtVX/BVTWBB4M8A+Ht4/wBN1O7utvr5MSLn1/5b/r+X/BLbRjcfFzxZr5TIsfDv2cEjoZriNvTriE9/Xr21rUY9H0e91aXG2ytpJiD32qT/AEr4U/EifwfrzRapcO+l6lJ/pWefKc9JQP5+o9cCo5I5Y1likV0dQVZTkEHoQfSqqqr/AIKi/C37Zonhf4w6dbAyafI2k6k6rz5T5kgYnsquJV+sq1/wTX+KA8IfGm78AX1yEsfGtiY41JwPtcAaSI57ZQzL7ll+lVVVVVVVVVVVXhD/AJFnTv8ArgKqqqqqqqqqqqqqqqqqqqq+MX/J9HhH/r60v/0Kqqqqqqqqqqqqqqqqqqqqv+Cmv/Jj/wASf+4V/wCnW0r/AIJZ/wDJl/hH/r/1L/0slqqqqqqqqqqn+/8AhVVVVVVVVVV+3x/yZx8WP+xfk/8AQ0r/AII6f8mkXf8A2N99/wCibeqqqqqqqqqqqqqqqqqqqqvjF/x86X/1zl/mtVVVVVVVVVVTSwwQvPPKkccalnd22hQBySewxX7cn7Wsvxr8Rv8ADzwJqDL4G0Sc5ljcgavcLx5x/wCmSnIQd/vnqoX9iH9kub45+JR458bWLr4F0O4AkRsr/alwuD5C9/LHBdvQhRySVhghtoY7a2iSKKJAkcaLtVFAwAAOgA7VVVVVVVVaF/yHNO/6+4v/AEIVVVVVVVVVqHiTw7pORquv6dZ7Qc/aLpI8Y6/eI9q1H41/CDS8refE3wyGUHKR6nFIwx6qrE/hWoftUfATTQRL8QYJmGcLb2dxLkj0KoR+uK1D9tb4JWYb7NNrl9tBx9n0/bnHp5jL1rUP28/AUYb+y/BWv3GAdv2h4oc+mdrPj/PWtQ/b8uWDLpXwwiTAO17jVi+fQlViH5ZrUP27viZPldM8LeGrYEEAyxzSsOOoIkUZ/CtQ/bL+Od7n7NrOl2GQcfZ9OjOOO3mbq1D9pn47amGFz8R9QTcDn7PFFB2xx5aLitQ+LXxS1TI1D4keJp1bqj6tNt6YOF3YH5Ve6nqWpNv1HUbm6bOczzM5z68k1Y6JrOqYGmaTe3eenkW7SdPoDVj8HfizqQzZfDPxPIuD8x0mZV49yuPwqy/Ze+PWoD/R/h1drwf9fcwQ9P8AfcVZfsXfG+7H7+x0ez4P+v1BT/6AGqx/YO+Isg/4mXjHw3B1x5Bmm/mi1Y/sBD72pfFH1wkGj/ruMv6Y/GrL9g34dR/8hLxj4jn6/wCoMMX80arL9i74H2n+vsNYvev+v1Bh/wCgBa/Yw8D+E/iD8QfGOl+NNDt9XtdLtgbSK4BxGfOK5wCMnAxzX7Z2g+GfDPxmOheFNDstKs7XSbcNBaxLGpdi7FiB3IZRk88CtG8HQ3/7E2veJTbxmfT/ABpHciX+IR+VFBj/AL6mPHTpXwt8IXvj7x3png3Tb21tbvVTJHBLcsVj3iNmVSQCRuKheh5Irx38NPG/w11P+yvGfh+4sJCT5UrDdFMPVJFyrfQHI7gV8Kv2rviT8OBb6ZqN1/wkeiRYX7HfSHzI19I5uWX6NuUdgK+F37Qnw1+LEaW2iauLLViPm0u+Iinz32c4kH+6SfUCqqqqqqq/bz/4Jf2HxAOo/GL9m7SYLHxO26fVfDEW2KDVGJy0tvnCxTHJJThH7bWzv+HvxF+J/wCz/wDEODxb4I1jUvC/inQ52ikUoY3Uq37yCeJhhlJXDRuMHHI4r9if9vvwB+1fo8XhrVvs3h34j2VvuvdFaTEd4FHzTWjNy6YGWjPzJznco3mqqqqqqq/4Lf8A/I0/CX/sH6p/6MtqqqqqqqqqqqqqqqqvEnxQ+HXg/UBpHirxto+lXpiEgt7u7WN9hyA2Cehwfyr/AIX18Ff+ipeGv/Bgn+Nf8L6+Cv8A0VLw1/4ME/xr/hfXwV/6Kl4a/wDBgn+Nf8L6+Cv/AEVLw1/4ME/xr/hfXwV/6Kl4a/8ABgn+Nf8AC+vgr/0VLw1/4ME/xrw98VPht4u1JdG8MeONG1S+ZGdbe1u1kcqvU4BzgVVVVVVVVV8b/HglY+CtLl+VCGv3B6nqsY+nBPvgdjVeDPEeqeFdbj1nTUaVYF/0mIdJIsgMD+Y57HBrSNVstc0y21bTpfMt7qMPGSMHHoR2I6EetVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV3awX1rNZXSb4Z42jkXJGVYYIyORwatf8AgnJ+xXaQLbQ/AuwZUzgy6pfSNyc8s05J/E1/w7t/Yv8A+iE6X/4H3n/x6v8Ah3b+xf8A9EJ0v/wPvP8A49X/AA7t/Yv/AOiE6X/4H3n/AMeq2toLO2is7aMJFBGI40H8KgYA/IVVVVVVVVVX/BLj/k8f4zf9eF//AOnKOqqqqqqqqqqqqqqqqqqqqvH3/IpX3/bP/wBGLVV/wVR1nz/HvgXw9vz9i0e4utvp50wXPX/ph6du/b/glRowj0v4i+IHQHz59PtY2PbYs7MOuefMT8h718btVGl/Dy+jV9sl88dsn4nLf+Oq1V8CviMJo08D61cfvEH/ABLpXPVe8RPt1X2yOwFVVVXxv+G9r8XPhN4o+HlwiGTV9PdLVnxiO5X54XPsJFQ/QV4d1vXPh74z03xFYK1rq/hzUo7iNJFIMc0Mgbaw/wB5cEfWvBfivS/HfhDRfGmiSB7HXLCG8t/myVWRAwBx3GcEdiCKqqqqqqqqqqrwh/yLOnf9cBVVVVVVVVVVVVVVVVVVVVXxi/5Po8I/9fWl/wDoVVVVVVVVVVVVVVVVVVVVVf8ABTX/AJMf+JP/AHCv/TraV/wSz/5Mv8I/9f8AqX/pZLVVVVVVVVVVP9/8Kqqqqqqqqqv2+P8Akzj4sf8AYvyf+hpX/BHT/k0i7/7G++/9E29VVVVVVVVVVVVVVVVVVVVfGL/j50v/AK5y/wA1qqqqqqqqqqr/AIKCftdeYb74AfDPVMBSYvE+owP19bNGH/kQj/c/viv2X/2cPEX7R3xAi0K1E9n4e04rNrepqnFvFnhEJ4Mr4IUc45YghTXhHwl4e8CeGdN8H+FNMh0/SdJt1gtbaJcBFHf3JOSSeSSSeSaqqqqqqqq0L/kOad/19xf+hCqvdS03TY/M1HULa1TH3p5VjH6mtQ+LHwu0oEaj8R/DNuykgo+qwhsjg/LuzWoftL/AnTdwuPiRpz7c5+zxyz9PTy1bNah+2X8C7PcLbWtTvsZx9n06Rc49PM29a1D9u74Yw7l03wt4luSuQDLHDEp9MYkY4P0rUP2+7RcppPwxlk67XuNWCY9PlWI/zrUP28/Hcuf7K8E6Db9dv2iSWbHp91krUP21/jXegi2k0GwznH2ewJx/38Zulah+1T8e9RysnxAmhU5wtvZW8WM+hWMH8zWofGz4v6oCLv4neJiGzlI9TliU565CsBj2rUPEviPVgRqniDUrzdnP2i7eTOevUnrVtZ3d7J5Nlayzvj7sUZY/kKsPhh8StVAOmfD3xLdAjgw6TM464zkL0zVh+zd8ctSANv8ADbVUyOPtASDvj+NhVh+xz8eL0D7R4csbH/r41KE4/wC/bNVh+wp8VLgBr/xF4YtV9FnmkYfgIwP1qw/YE1OQg6p8TbWHjkQaU0vf1Mi9qsf2CfBsYA1Px5rNxxz5FvHD/Pf2qx/Yi+C9oALifxFe4H/Le+Qf+gRr9Ksf2TvgFYAFfAazvjG6e/uXz+HmY/SrD4FfBrTv+Pb4YeG2OODPp0c3/oYPpVh4Q8J6X/yC/C+kWnH/ACwsY4/5Aegqqqqqq/4J/wD/ACVD4gf9eq/+j2r9se++3/tF+K9pylv9lhXjH3bWLP8A48Wr4ReEm1r9g7W7BYmZ9QsdTu4l4+d4ZHKgf8CgA5r4U68fC/xO8J+IfMKrp+s2sshHdBKu8fiuR+NazoejeItOl0jXtLtdRspxiS3uoRIjfUHIr4q/sQaXfCXVvhPqQsJ+WOlX0heI+0cnLL9G3D3ArxZ4K8X/AA91k6R4r0S80m+iO5BKuA+P4o3HysPdSRXwr/bC+IXgcwaX4tY+KNIQBcXEm26iH+zLzu+j5J6ZFfDT42fDr4r2qv4V12P7aFzJpt0RFcx+vyZ+YD+8pI96qqqqqq/bi/4J4eCv2orC48b+ChZeG/iZbxYjv2Qpb6qFHyxXQUE7ugEwBZQACGUADxJ4Z+JvwF+I8mieILHVfCXjLwveJIoDmGe1mXDRyRup5H3WV1JBBBBINfsGf8FNNF+Ma6b8Ivj1f2ej+OSEt9N1hiIrbXGAwFf+GG4OBxwsjH5dpISqqqqqqq/4Lf8A/I0/CX/sH6p/6Mtqqqqqqqqqqqqqqqqv2yPh54/8UfFyHUvDPgbxBq9oNHgjNxY6ZNcRhw8mV3IpGQCOPcV/wpr4v/8ARKfGP/giuf8A4iv+FNfF/wD6JT4x/wDBFc//ABFf8Ka+L/8A0Snxj/4Irn/4iv8AhTXxf/6JT4x/8EVz/wDEV/wpr4v/APRKfGP/AIIrn/4iv+FNfF//AKJT4x/8EVz/APEV+yH8OfiD4a+MttqniPwJ4h0qzXT7hDcX2lzQRhiowNzqBk+lVVVVVVVV3cLaWs10wyIY2cj1wM/0q9u59QvJ766k3zXEjSSMe7Mck/mar4R6RHrfix9OnBNvLYTrMAP4WXb/ADYH64r4GXN1FpGreH7piW0u+KqD/DuyCB7blJ/Gqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv+CXH/ACeP8Zv+vC//APTlHVVVVVVVVVVVVVVVVVVVVXj7/kUr7/tn/wCjFqq/4KT6x/an7STWO/P9kaBZ2uM9MmSb0/6be/8AQf8ABMTRjY/APV9VkQh9T8TTshIIzGkECD/x4P0r9pLVfk0XQ0fqZLiRfyVT/wCh18Jfh9Z+OodejviUEFqkdvLj/VTM2VbHfAQgj0btnNanpureFtbl0+8V7W/0+YcqcFWHKsp9OhB+hr4WfECHx1oQFy6LqtkAl3GON3pIB6HH4HI9Kqqqv29Phcfht+0PrV5Z23l6Z4sQaxaEDgPKSJxn185ZGx2DrX/BNH4pDxX8INQ+HF/db77wbefuFZsk2lwWdMZ67ZBMPYFB6VVVVVVVVVVVXhD/AJFnTv8ArgKqqqqqqqqqqqqqqqqqqqq+MX/J9HhH/r60v/0Kqqqqqqqqqqqqqqqqqqqqv+Cmv/Jj/wASf+4V/wCnW0r/AIJZ/wDJl/hH/r/1L/0slqqqqqqqqqqn+/8AhVVVVVVVVVV+3x/yZx8WP+xfk/8AQ0r/AII6f8mkXf8A2N99/wCibeqqqqqqqqqqqqqqqqqqqqvjF/x86X/1zl/mtVVVVVVVVVX7c37W8fwX8Pv8N/AOpKfHGs2+ZJ42ydIt2GPMJHSVhnYOoHzn+Hd8J/hZ4w+N3j6w8B+ELY3Go6i5aaeTJjtogR5k8rDOEXPJ6kkAZJAPwT+DXhD4E/D+w8A+ELceXbrvvLxkCy31wQN80nXk44GcKAFHAqqqq1LXNE0YKdX1mxsdwyv2m4WLI6cbiKufil8PLQHzfF2ntj/nnJ5n/oOaufjp8NIARHrktwR2jtJf/ZlFXX7RngaHK29hq9wR0IhRR+r5/Srr9pjTEB+xeE7mX0826WP+St2q6/aX1hwRZeF7OL0824aTH5Be1XP7RXjycEQ2ukW/pst3JH/fTmrr44fEy5BC+IEhB7RWkQ/UqTU3xQ+IkziX/hM9VjdTlWhuDEVIOcgpjFah468basWOq+MtcvS2d32jUJZM85OdzHvX765m/jlllb3ZmY/qSTVh4D8cattGl+DNcvN2Nv2fTpZM56Y2qetaf8APjVqYX7N8MtfTcQB9otDb9eP+Wm3Faf8AsjfHu/Cs/gyK0RiMNcajbj8cByR+Vaf+w78YbsqbvUfDVkv8QlvJGYDPOAkZBOPf8asP2B/E8oX+1PiHpdv6+RZSTY57ZZO1WH7AugRhRqnxIv7jHXyNOSHP0y744qw/YZ+EVsFa81jxNeMOoe6iRT9AsQI/OrD9kH4CWQXzvCVxesvRrjUrj19EdR7dKsP2efgjpoUW3wz0R9vT7RB5/fP/AC0LVYfDv4f6Tt/srwN4es9v3fs+mQx45zxtUd+ahght4hDbxJGi/dRF2gfgKqqqqqqqqqqqq/4J/wD/ACVD4gf9eq/+j2r9oy/GpfHXxzcBg2zW7iHg5/1beX/7LX7MWiW8f7OHhHSLmENDd6bI0iFMblmkkc8e4f8AHOa1rS59E1i+0W6z52n3MlvJlcHcjFTx25FeB9cXxP4K8P8AiRX3f2rpdtdZ9fMiVv61Xifwn4a8aaTJofirRLTVLGUfNDcR7gD/AHlPVW9CMEetfFX9h6RfN1b4SanvHLHSdQlwfpFKePwfH+9WsaH4p8C659g1nT9Q0XVbNwyrIrQyIQeHU+nHDDjuDXwr/bS8Z+FzFpXxDt28S6aoCi6UhLyIeufuy/RsE/3q+H3xW8BfFDTxfeDvEFvduq7prRjsuIP9+M8j0z0PYmqqqqqr9rT9jj4X/tbeEBpfiq3GmeJdOhcaL4itogZ7NjkhHHHmwljkxkjuVKk5r9oD9nb4pfszePJvAvxM0N7SYFn0/UIQWtdRiBGJYJMDcORkcMpOGAPFfsGf8FRbjw4NN+Df7TetyXOlKFg0jxdcNukswAAsV4erx8YE3LLn58r8y2t1a31rDe2VzFcW88YkhlicOkiMMhlI4IIIIIqqqqqq/wCC3/8AyNPwl/7B+qf+jLaqqqqqqqqqqqqqqqqqqqqqqqqqqqqru3W7tZrRmwJo2QnHTIxV9Zz6feT6fdJsmtpWjkU9mU4P6iq+AOoaVa3WsQ3SRR3K26zCdzjESn5x7AEqf/1V8GrYz2mueI1Vli1bUneEMOSiknP5sR+FVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVU0sUETzzypHHGpZ3c7QoA5JPYYqD9ob4AXcKXFr8c/h9NE4yjx+JrNlYeoIk5r/AIX98CP+i2eAv/CktP8A45X/AAv74Ef9Fs8Bf+FJaf8AxyrX45/BK+nW1svjH4HuJnztji8Q2rs2Bk4Akz0BqzvbLUrWO9068hureUZjmhkDo46ZBHBqqqqqqqqqv+CXH/J4/wAZv+vC/wD/AE5R1VVVVVVVVVVVVVVVVVVVV4+/5FK+/wC2f/oxaqv22dZGu/tSePrtX3CG9htQPTybeKIjqe6H/wCtX7AujrpP7K/hCQoFk1CS9uZAMc5upVU8f7CrXxx1Uap8QryJX3JYRR2yn6Dc3/jztX7P+lfYfAzX7r82o3byKSOqrhB+qt+dfGb4cjxZpP8AbmkwZ1bT4zhVHNxEOSn+8OSPxHcY8J+KNS8Ha7b63pr/ADxHEsROBKh+8h9j+hwe1eHtf03xRo1trelTb4LlMgHqh7qR2IPFVVV/wUq+Fo8X/Bmz+INjbb7/AMF3u+RlXJNpOVjkH4OIW9gG9TX7CnxRPwy/aH0KK7uvK0vxQDo96GPygykeS3oCJljGT0Vm9aqqqqqqqqqqrwh/yLOnf9cBVVVVVVVVVVVVVVVVVVVVXxi/5Po8I/8AX1pf/oVVVVVVVVVVVVVVVVVVVVVf8FNf+TH/AIk/9wr/ANOtpX/BLP8A5Mv8I/8AX/qX/pZLVVVVVVVVVVP9/wDCqqqqqqqqqr9vj/kzj4sf9i/J/wChpX/BHT/k0i7/AOxvvv8A0Tb1VVVVVVVVVVVVVVVVVVVV8Yv+PnS/+ucv81qqqqqqqqqv2q/2ldB/Zx8AvqQaC98U6qrRaHprt99x1mkA5ESZBP8AeOFBGciWXxx8XvHpkka98Q+KfE9+AMndLczyNgAdgOg7KoHYCv2Tv2ZdE/Zx8BizmEN54s1hEk1vUE5BYD5YIz1EaZPuxJY9gKqqr4k/GvS/Cnm6P4fMWoasMq5zmK3P+0R95v8AZHTue1alqeteKdVa91C4uNQv7psAkFmY9lUDoPQDgVa/Dzx3e4+z+ENXwehe0dAfxYAVa/Bf4l3eCnhl4x6y3ESY/Atn9Ktv2e/iDcAeaNNts4/1tyTj/vkGrX9mrxG4H23xFpsXTPlI8n16hatf2ZYAAb3xg79MrFZBcevJc/yq1/Zu8HxgG71nV5iOyPGgP/jpP61a/AT4b24Al067ucf89btxn/vnFeHfg98M4tXsIR4Qs5Ua5jVlmLygjcMg7ic1YfCf4XaVtOnfDnwzbsuMOmkwhuOh3bcmrPTdO05PL0+wtrZcY2wxBBj6Cqqqqqqqqqqqqqqqqqqqr9h/wF458JfEbxxfeKvBmu6NbXdsBbzahp0tskx84nCM6gNxzxXxKvzqvxF8VaoWLG81u8myTkndM56/jXwVsBpnwd8D2IADReHbHfg5G4wIWx+JNftL6CPDfx68baaqBRJqj3QUdhcATf8AtSv2SNeHiD9nzwlM0gaWygls5BnO3ypXRR/3wEP41VV4z8A+D/iHpLaL4x0C11O2IOzzVw8R9UcYZD7qRXxU/Yh1zTGm1b4V6j/alqAW/sy9dUuE9kk4Vx7HafdjTL4o8Da+MjUtC1rTpMjIe2ngb9GU18K/23PEGjiLSPilp7azajCjUrRVjuUH+2nCyfUbT67jXgr4g+DPiJpg1bwb4htNTgAHmLG+JISezocMh9mAqqqqqvjn8Bfhl+0X4Cu/h58UNATULCcFra4T5LiwmxhZoJOqOPxBGVYMpIP7YX7FHxL/AGSfFRj1aJ9a8GajMy6N4igiIjl6kRTD/llMFGdp4bBKk4O39hb/AIKN+LP2brqy+HHxMlvPEPw0kfZGgbfc6GWPLwZ+/F1LQ8dSykHIfwX428JfEfwtp3jXwL4hstb0PVYfNs760k3xyrnBwexBBBU4IIIIBBFVVVVV/wAFv/8AkafhL/2D9U/9GW1VVVVVVVVVVVVVVVVVVVVVVVVVVVVV8WPhXdazcv4m8MwCS6Yf6VajgyYH31/2sYyO/Xr1ubS5spmtry2lgmQ4aOVCjKfcHmvA/wAOvEviq7UxRT2OnONs926lQyd1Ufxk+nT1rStLstF0220nTohHb2sYSNfYdz6k9SfU1VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVeKVLeGNXVRkmwnAAH/AEzNVVV8MvjL8VPgzrUev/C7x7rHh27Rw7izuWWKfHaWI5SVeB8rqR7V+wn+2HZftZfDq6m1q0ttN8a+GWjh1uzgbEcwcHZdRKeQjlXBU52spGSCpNVXxU/bZ/a+/aU8W6/8KP2UvhpqvhWw0Kae31fViUa8iERKuZbhsQ2n3W+VS0hI+VyeK/4JF/Ez4keOPjb4ysfGvxC8S+ILaHwsZYodU1ae7RJPtUI3hZGIDYJGevJqv2vv2ntA/ZU+Edz48vrWLUdavZfseg6W7lRd3TAnLkciNFBdjx0CggsK+FX7PP7Zv/BQXT7j4wfEb4233hzwxdzuumC5abypypwTa2cTJGsSkFTISCWX+IhiPEerftjf8Ew/iNoR1rxtN4x8B6rKfKt5LuWbT9QjTHmQ7ZMtaXAUqQV9jmRQy18LfiT4X+MPw70D4m+DLs3Gj+IbJbm2LDDJnh43A6OjhkYdmUiqr/glx/yeP8Zv+vC//wDTlHVVVVVVVVVVVVVVVVVVVVXj7/kUr7/tn/6MWqr45ax/wkPxp8e62H3Le+JdQljJzwhuH2jnB4XAr9mrSl8O/s7fDuym/d7PDNnPKGyNhkhWRs56YLGtd1J9Z1q/1aQkteXMk3PbcxP9a8FaUNE8I6RpRTa9vZxiQYx85GW/8eJqvjj8OP7Iu28YaLb4srp/9MjQcQyn+Ieisfyb6ivhD8Q28GayNP1Gc/2RfsBNnpA/QSD27H2+gpWSRA6MGVhkEHIIqqrxf4X0vxt4U1jwdrcfmWGt2M1ncqBzskQqxGehAOQexxXizw5rXw78a6r4V1Jmg1Xw7qMtrI8eVxLC5Xcp64yuQfTBr4D/ABKg+L/wg8LfEKORGn1WwQ3ip0S5T5J1wOgEivj2xVVVVVVVVVVXhD/kWdO/64CqqqqqqqqqqqqqqqqqqqqviP8AA74jeI/2p/D3xQ0nSIJfD2nz2Lz3DXcasoiPz4QnccfTmqqqqqqqqqqqqqqqqqqqqv8Agpr/AMmP/En/ALhX/p1tK/4JZ/8AJl/hH/r/ANS/9LJaqqqqqqqqqp/v/hVVVV+3J4r/AG89f+LOjfAv9l7RZNM0LWNIF7ceI7ACORTvZJUmupPktgmFIEf7xtwIY52jUP8AglH+2JrNp/wkms/tL6Xe+JwvmKlxq2oy/vOuPtLLuBz32da+A/7Zf7Sv7F3x2HwO/ap1fWNZ8Npcx2uoR6vdm8m02OQjZeW1wxLSQhSG2ZKlcgBWHCOkiLJG6srDKsDkEeor9vb9uv4qX3xWj/ZK/ZSubuPxE90lhq2raaM3cl5Jx9jtmx+62ZG+UEMGyAU2MWuv+CU37W7eHz46H7SUU/j3y/tBsm1C8BMuMlBfFt2/tkoFyfvAfNX/AAT3/bq+KJ+KU37J/wC1Hf3lx4gS5mstI1TUzi8hvYch7K5f/loW2tskYltw2kvvXbVft8f8mcfFj/sX5P8A0NK/4I6f8mkXf/Y333/om3qqqqqqqqqqqqqqqqqqqqr4xf8AHzpf/XOX+a1VVVVVVVV8Zvi/4R+B3gDUPH/jG5221oNltbIwEt5OQdkMYPVmwfYAFjwDXxf+LHjH45/EC/8AHfi65M17fvstrWPJjtIQf3cES9lUH6kkk5JJP7Cn7I6fCDQYvij8QNNH/Ca6xb/6NbTIM6RbsPu47TOPvnqoOzj5s1VVqOo2Gk2Uuo6ldxW1tAu6SWVtqqK+JXxzvtdEui+EJJbPTzlZLo5WWce39xf1PtyK8K+Dtf8AGupDTtCszKwIMszcRwg93bt346nsDXw++FegeA4FuI0F7qrLiS8kXkeoQfwj9T3PYVVVVVoX/Ic07/r7i/8AQhVVVVVVVVVVVVVVVVVVVVVXE8VtbyXMzYjiQuxxnAAyauriS7uZbqXG+Z2dsDAyTk4rw9YHSvD+maWQQbOzihIbqNqAdvpX7fGgnTPjXb6siHZrGiwSlvV0Z4yPwVE/Ov8Agnxr3274Xa74eeTc+l60ZFBP3Y5YkwMf70bn8aqqqvH/AMLfAnxP04ad4z8PwXvlqRDcDKTQf7kg+YfToe4NfFb9ivxf4Z8/V/hxdN4i01AW+xyAJeRj0AGFl/DB7BTWl6v4q8B68L3Sb3UdD1eyYozIWgljPdWHBx6qeD3FfCv9uC5gWDR/izphuFyF/tewjAce8kIwD7lMf7prwv4v8L+NtLTWfCeu2eqWb/8ALS3kDbT6MOqn2IBqqqqrxp4K8JfEXwvqPgrxz4fstb0PVoTDeWN5EJI5V69OxBAIYYKkAgggGv26v+Ccniz9m66vPiP8M47zxD8NJH3yOU33Ohljwk+PvxdAs2B1CsAcM/7H/wC2v8TP2SPFO/RpX1rwbqMytrHhy4lIjl6Aywtz5UwAxuAwwwGBwMfAz48/DL9ovwFafEL4XeIE1CwnAS5t3Gy4sJsfNDPH1Rx+IIwyllIJqqqq/wCC3/8AyNPwl/7B+qf+jLaqqqqqqqqqqqqqqqqqqqqqqqqqqqqqp4opCrPGrFDlSy5wfaqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq8W+JdM8F+FdZ8Y607Lp+hafPf3bIMsIoY2kcgeu1TXiPxt8Rv24f2mNGTxdr3kX/jPXLfTLFGO+DSLaWYKkcacZSNWJPQuQSfmYmvBP/BOf9kLwb4btNBm+Elhr9xBGBPqWryyT3Fy+OXY7gq5/uoqqPSviv/wTT/ZV+IXhG90bw38PLTwfrRhf+z9W0qSRGt5cfKXjLFJUzjKsM4zgqTmv2VPEvgL4afH238B/HH4deGfFXhnWNSGiawup2aTvp0hkMQuYJSNybHOWwcMmeNwUr/wUV/4J6/Db4Y/De7+O/wADdNl0S20aeJdd0PznmgMMrrGs8G8s6MsjpuTJXaxYbdhDf8EqvHF14T/a/wBC0OO4KWvi3TL7TblS2FO2FrlMj1326gf73vVVWr6No+h+EddttF0qzsIp4Ly5ljtYFiWSaUM8kjBQMu7sWZjySSTkmv8AgjH/AMl38bf9iif/AErgqv8AgrV4w1f4g/tQeFvhBptxut9A0q3gggZ+Be3sm529sxi2H/Ac968DeD9G+H3gvQvAnh2AQ6Z4e06CwtEAxiOJAi59yFyT3JzX/BTD4f2Pjz9j7xjPPAj3nhhrfWLF2XPlPFKqyEemYZJl/wCBV/wRw8dXmu/AfxT4EvLgyjwt4g8y1BJ/dw3UQbaPbzI5W+rmqr9nn9q7Tv2Rv2j/AIm+NdR8FXHiVNYmvtPW3hvhamM/bRJvLFHyP3eMY71/w+s8L/8ARvuqf+FFH/8AGK/4fWeF/wDo33VP/Cij/wDjFf8AD6zwv/0b7qn/AIUUf/xiv2Tf2k7H9qv4Wy/E7T/CU/h2KLVZtP8Asc14LliY0jbfuCrwfMxjHaqqqqqqqqqqqqqqqqqrx9/yKV9/2z/9GLValfQaXp11qd0wENnA80hJAwqqSeTwOBV3cz6heTXk53TXMrSPgdWY5P6mvESr4E+Dr6dFtQ6bo0VjEFwNp2LCuMemR+VeD9JGu+KtJ0kruS5u41kGP4NwLf8AjoNVV9Y2mpWc2n30CTW9xGY5Y3GQykYIr4jeBrvwLr72DBpLKfL2cxH309D/ALQ6H8D3r4F/Ef7XDH4I1q4/fQr/AMS+Vz99R1jPuB09uOwzVVX/AAUt+Fp8KfGGw+I1ha7LHxlZDz2VeBeW4VH9hmMwn3O8+tf8EufiiJ9P8U/By/ucvbONY01GOfkbbFcKPQBhCcersfWqqqqqqqqqqvCH/Is6d/1wFVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV/wAPrPC//Rvuqf8AhRR//GK/4fWeF/8Ao33VP/Cij/8AjFf8PrPC/wD0b7qn/hRR/wDxiv2pf+CoOg/tF/AjxP8ABuy+D1/oc3iD7Jtv5daSdYfIu4ZzlBEpORCV6jG7Pav+CWf/ACZf4R/6/wDUv/SyWqqqqqqqqqqf7/4VVVVVX/BattBPx58EJaLF/a48J5vSoG7yvtUvk7vxEuM1YeIdS+Ff7NVt4r8VKJNQ8IeBlvdSEvy7pbWxDy7umPmRs9K/4JB+EJPiX+074x+MfiyQ6jqOgaVNdC4mOXN/fSlWmJ7kxi5B/wCulV/wVS0Of4NftraF8WPB+y0v9W03T9eSRBt231tM0W7j2t4WJ7kn61oWr2viDQ9O16yJ+z6laRXMOf7kiBh+hFV+3x/yZx8WP+xfk/8AQ0r9gj/gop8Ev2WPgbP8MviD4X8b6hqkuvXOoLNo1jayw+XJHEqjMtxG27MbZG3HI59P+H0n7LP/AEIfxU/8FVh/8m1/w+k/ZZ/6EP4qf+Cqw/8Ak2v+H0n7LP8A0IfxU/8ABVYf/JteDvE+n+OPCGh+NNJhuIrLX9Nt9Qto7hQsiRzRrIocKSAwDDIBIz3PWqqqqqqqqqqqqqqqqqr4xf8AHzpf/XOX+a1VVVVVVVeKfFHh/wAFeHNR8WeKdUh07SdKt2nu7mY4WNFHPTkk9ABkkkAAkgV+1P8AtJa/+0f4+bV5TLZ+G9KLw6HprN/qYyRmVwOPNfALHsAqjIXJ/wCCfv7Ig1CSx+PvxM0oG1jIk8M6dcRn94w6Xjqf4R/yzB6n5+MKTVVXjb4geHvAlh9p1a433LqfItIyDJKfp2X1Y8fU8V45+IniHx7eebqc/lWkbEwWcRIjj9z/AHmx/EfwwOK+HHwa1fxmY9U1Uyafo5ORIVxJcD/pmD2/2jx6A81oXh/R/DOmx6TodjHa20XRVHLHuzHqSfU1VVVVVaF/yHNO/wCvuL/0IVVVVVVVVVVVVVVVVVVVVVXxH1AaT8PPFGqbwv2PRbybJbbjZC5znt0614Z08at4k0nStob7ZfQw7Su7O5wvTv16VX/BRXQN9l4K8URx/wCqlurOZtvXcI3QZ/4BJx71/wAE7te+z+M/Fvhgv/x/6ZDdhT3MEmz/ANr1VVVVXxJ+C/w7+K9r5Xi7QkkukUrDf258q5h+jjqP9lgy+1fFT9jn4geCVn1Xwex8UaTHltkEe27iX3iGd/1TJPXaBXhjxf4v+H2s/wBqeFtbvtH1CE7JDExXODyroeGGf4WBHtXwq/bf0+7Fvo/xX037JLwv9r2SFoz7yRDLL7lM/wC6BWh6/ofifTItZ8O6vaalYzD93cWswkQ+oyOhHcdR3qqqqurW1vrWWyvbeK4t542jlhlQOkiMMMrKeCCDgg1+3n/wS4uPDw1D4w/sw6HNdaWA0+reEbdd8lpgEtLZr1ePjJh5ZSfkyp2p+z/+0P8AFH9mbx9B47+GettaTqRHf6fNlrbUYQeYZ48jcOuDwyk5Ug81+yZ+2N8L/wBrbwh/afhS5GmeJdOhU614duZQZ7NjgF0PHmwljgSADsGCk4qv2gvjt4Q/Zt+FupfFvxzp2sX2kaXNBFNBpMMctwzTSrGu1ZJI1wGYE5YcevSv+H0n7LP/AEIfxU/8FVh/8m1/w+k/ZZ/6EP4qf+Cqw/8Ak2v+H0n7LP8A0IfxU/8ABVYf/Jtf8FF/2xfhj+17rXgfUfhrofifTY/DNrexXa65awQs7TNEV2eTNLkARtnOOo61VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVXxj8H3XxD+EXjjwBYuFufEvhvUNMhYsFw89s8SnJ4HLjk14f13xP8N/Gen+I9Hmn0vxB4a1KO4t2dNr21zBICNynuGXBB9wa+En/AAV7+AniXQLVfi3pOteD9eSPF39ms2vrN2A+9EyEyAH+6yfLnG5utfHn/grx8I9F8KXunfASw1XxF4muoWSzv76yNrZWTEcSssmJJCpOQm0AkcsB18J6Jr3j/wAd6N4c0uWe41rxFq0FrbuCWke4nlCq3XJYswPXrX/BRHxHYeGv2NviTcXzjN9YQ2MCFsF5JriOMY+gYtj0U1/wTX0e51j9tP4dLbg7bOS+uZmxnaiWM5/U7V/4EKqq8Vf8ivrH/XhP/wCizX/BGP8A5Lv42/7FE/8ApXBVfthpcr/wVOgN6T5LeJ/C5g3nI8vyLIHHoNwf9ar9s50T9k/4sl3Cj/hEr8Ak45MRA/Wv+CJsN2tj8Ybh932Z5dFWPLcbwL0vgduGTnvx6VVf8E0fDvh/xJ+1/wDGO08RaFp2qQxWV+8cV7apOqN/aKDcAwIBwSM+9f8ACq/hh/0Tjwv/AOCeD/4iv+FV/DD/AKJx4X/8E8H/AMRX/Cq/hh/0Tjwv/wCCeD/4itH0LQ/D1p/Z/h/RrHTLYuXMFnbrCm44y21QBk4HPtVVVVVVVVVVVVVVVVVV4+/5FK+/7Z/+jFqvj3rB8P8AwP8AiBrKOVktPDOoPERkfP8AZ329OnzY5r4XaN/wkfxN8I+Hgm/+09dsrXbzz5k6Ljgg96/aK1YWvhWx0hHw99d7iPVI1yf/AB5kr4A6T/aHjsX7LlNNtZJQT/ebCAfkzH8Kqq8c+DrDxxoE2j3mElHz202MmGQDg/TsR6Gr6y1bwvrUllcrJaahp83UHBR1OQwP5EH6Gvhj4+t/HegrNKyJqdoAl5EOOezgf3Wx+ByO2TVV+3d8Lh8TP2d9cns7bzdT8KkaxZ4HO2IHzx9DC0hx3KrX7NHxPPwf+N/hTxvNcGKwgvRb6kc8fZJv3cpI77VYuAe6ilZXUOjBgRkEHOaqqqqqqqqqvCH/ACLOnf8AXAVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX/Cq/hh/wBE48L/APgng/8AiK/4VX8MP+iceF//AATwf/EV/wAKr+GH/ROPC/8A4J4P/iK/4KReAPAeifsW/EXU9F8E6DYXkP8AZnlXFrpsUUiZ1O1BwyqCMgkfQmv+CWf/ACZf4R/6/wDUv/SyWqqqqqqqqqqf7/4VVVVV8Xviz4H+B3w81j4m/EPVlsNG0aAvIeskznhIo1/ikdsKq+p5wMkfsx/D7xv/AMFFf2ytU/aJ+I2lND4J8P6lFeXUL/PCBDj7HpqE43fKqtIQMEByQDIM/tjJeP8Asn/GBbAt5o8E6qW2nB2C2fzPw2bs+1f8EOdm/wCNOdu7Gg4z1x/xMM/0qv8Agtq6H4q/DaMOpZfD1ySueQDccHH4H8q+BcF3b/BH4e2+obvtUXhXTEn3NuO8WsYbJ5zznmq/b4/5M4+LH/Yvyf8AoaV/wSx/Z0+A/wAVP2ZbnxN8SPhH4V8S6snii8t1vdT0yOeURLFAVTcwJ2gsxx7mv+GK/wBkj/o3LwB/4I4f8K/4Yr/ZI/6Ny8Af+COH/Cv+GK/2SP8Ao3LwB/4I4f8ACtL0vTdD0uz0TR7KGzsNPt0t7W2hQIkMSKFRFA6KFAAHYCqqqqqqqqqqqqqqqqqr4xf8fOl/9c5f5rVVVVVVVTSwwQvPPKkccalnd22hQBySewxX7cn7Wsvxr8Rv8PPAmoMvgbRJzmWNyBq9wvHnH/pkpyEHf756qF/Yh/ZLm+OfiUeOfG1i6+BdDuAJEbK/2pcLg+QvfyxwXb0IUcklYYIbaGO2tokiiiQJHGi7VRQMAADoAO1VVfEn446d4e83RfCrxX2pDKyT53RW5/8AZ2Hp0HfuKnn1vxTq/mzPc6lqN7JgcF3kY9AAP5DpXw2+BFrpvla142ijubofNHYZ3Rxe79mPt0+vYKqqFUAADAAGMVVVVVVVoX/Ic07/AK+4v/QhVVVVVVVVVVVVVVVVVVVVVN+1Rf8Axi+Gvxc0O58JQ6Oui+HrhVmivDMZfNJhwQVXHDe9fBKwGp/GTwNYsAVk8Q2O8E4yonQt+gNV+3XoI1f4C3GohATomqWt1nuAxMH/ALWFfsX68NE/aD0CB5AsWqwXNm5Jx1iZ1H4uiD8aqqqqqq+J/wAAPhp8WEe48QaMLbVCuE1OyxFOMdNxwQ49mBx2xXxV/ZN+JPw6EupaTB/wk2jpk/abGE+dEvrJDyR9VLAdyK8EfEbxv8NtS/tPwb4hutNlJHmxo26ObHZ4zlW/EcdsV8K/22fDWvGDSPibYLod6QFGoW4Z7WQ+rLy0Wf8AgQ7kitN1PTdZsYdT0jULa9tLhd0VxbyiRHHqrKSCKqqqv28/+CZei/GNdR+LvwEsbPR/HJDz6jowAittcYDJZP4Ybg4PPCyMfm2kl68N+Jvib8BfiPHrfh++1Xwl4y8L3jxklDDPazLlZI5EYcjqrIwIIJBBBr9h3/gof4J/ag0+18DeN5LLw38TIIvnsN+y31cKPmltSx+9jJaEksoBILKCR438B+C/iV4dn8I+P/C+m+INFumR5rDULdZoZGRgykq3BwwBHuK/4Yr/AGSP+jcvAH/gjh/wr/hiv9kj/o3LwB/4I4f8K/4Yr/ZI/wCjcvAH/gjh/wAK/wCCvPwb+FPwg8R/DO2+Fvw90HwtFqdjqL3iaVZJbidkeAIXCgZwGbH1NVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX7eX/BNHU/ih4j1D40fs+W9mviDUWabW/DssqwJfzHrPbu2ESVuroxVWOWyGJDeOPhN8UPhpezaf8AEH4e+IfDs8Bw41LTZYF64BDMAGBOMEEg5GDzXgf4TfFD4mXsOn/D34e+IfEU85wg03TZZ1HOCSyghQCDkkgDByRX/BP7/gnJqnwa121+Nfx1gtT4stkJ0XQ4pFnTS2ZSGmmdcq82CQoUlUyWyWI2fti/stT/ALWfw9sPAI+JF54Ug0++N9iGxW6jupQhWPzVLIxVdzkAMOWyc4GP2Gf+CeXiX9ln4s+IPiL428WaHr4fSvsGiNYLKrr5kgaaSRHUBG2xooCs2Q75I4qqrxV/yK+sf9eE/wD6LNf8EY/+S7+Nv+xRP/pXBVf8FcPB2reAf2lPCPxi0qDZFrulQvDMyZX7bYy4YHHXEbWx/E18PfG+h/EzwJ4f+IPhucS6Z4i06G+tiGyQsiBtp/2hnaR2IIr/AIKg/Emw8A/si+JdIe5RNS8Y3FvpFjGSCW3SrLMcdcCGKQZ7Fl9a/wCCQPw6uvC37N+qeN7+1aKXxlr8stszLjzLW3RYUPuPNFxVV/wS4/5PH+M3/Xhf/wDpyjqqqqqqqqqqqqqqqqqqqqrx9/yKV9/2z/8ARi1X7cesLov7LHju4DgPcW1vaoDjLebcxIcZ/wBlmP4V+yFow179pj4d2JTd5WspdYx/zwVps9D/AM8//wBXWv2iNVF34ttNKR8rYWYLD0dySf8Ax0JX7OGk+Vo+ra0y83NwkCEjsi5OP++x+VVVV8aPhx/wlGm/8JDo9vnVbFPnRRzcRDqMd2HUevI9MeD/ABVqPgzXbfW9NbJj+WWInAmjP3kP1x+BAPatB1zTvEmkW2taVN5ltdJuUnqp7qR2IOQR6iqq4t4LqCS1uYklilQpJG65DKRggj0Ir47fDWb4Q/F7xT8O5FbydI1B1s2c5L2z4eBj7mJ0J981+xj8Uv8AhbH7PXhrVbu6E2qaNGdJ1Elst5tuAqsx9WiMTn3c1VVVVVVVVVeEP+RZ07/rgKqqqqqqqqqqqqqqqqqqqq+Ifx4+IHhr9qDw/wDCnS7mxGg6jPYpOj2oaQiU4fD5yKqqqqqqqqqqqqqqqqqqqq/4Ka/8mP8AxJ/7hX/p1tK/4JZ/8mX+Ef8Ar/1L/wBLJaqqqqqqqqqp/v8A4VVVVfFL4peBfgx4F1T4jfEbX4NJ0PSIt888hyzt/DHGo5eRjgKo5JNeINT+On/BXD46xaZ4Zsrrwv8ACbwnOuWuHJisImODNKAdst7IuQqLwo4yFDu3wi+EXgL4GfD/AEr4Z/DfRU03RdJj2xqTuknkPLzSv1eRzyWP0GAAB4+8KWnj3wJ4j8DXzhbbxFpN1p0zFc4SeJo2OPoxr/gkf4yl+E/7U/i74KeMlGnah4h06ewEEvDDUrGUsYT6fu/tX4oB3qv+CkmqS/tF/t7aH8HPB8v2qbTItO8MK0I3gXU0zSytkZ+59oCtnAUxNnGCa0+wtNK0+10uwhEVtZwpDDGOiIoCqB9AAKr9vj/kzj4sf9i/J/6Glf8ABHT/AJNIu/8Asb77/wBE29VVVVVVVVVVVVVVVVVVVVfGL/j50v8A65y/zWqqqqqqq/4KCftdcXvwA+GmqesXifUIT+dmjf8Aowj2TP3xX7L/AOzh4i/aO+IEWhWons/D2nFZtb1NU4t4s8IhPBlfBCjnHLEEKa8I+EvD3gTwzpvg/wAKaZDp+k6TbrBa20S4CKO/uSckk8kkk8k1Varq2maHYS6nq99Fa2sIy8kjYA/xPsOTXxJ+N2peJjLo3hlpbDSzlXlztluR3z/dU+g5Pc84rwd4H8QeONQFjolrlEI865kBEcI9WPr6Acn868BfDLw94CtQbOP7VqDria9lX529Qo/gX2H4k1VVVVVVVaF/yHNO/wCvuL/0IVVVVVVVVVVVVVVVVVVVVVXgW+Fj4D+NhDDfcWlnCgIzndqMef8Ax3dX7KVgdS/aF8F2ygkpeSTYH/TOCR//AGWq/aD0I+JPgj420pV3OdGnmjUcZeJfNUfiUFfCXXv+EX+KPhLxAZNqWGtWskpzj92JV3j8VyPxqqqqqqqq+Kv7NHw0+KYn1C507+yNblBI1OwUIzt6yJ92TtknDejCvir+zH8S/hd5t/JYf23oyZP9o6fGziNR3lT70f1OV/2q+Hnxc+IHwuvRdeDvEU9tEzbpbOQ+Zbzeu6M8ZxxuGGHYivhX+2d4H8XGLSvHsKeGNTYBRcM5e0lb/f6xf8D4H96ra5tr23ju7O4ingmUNHLG4ZXU9CCOCKqqr9tj9gXwD+1fo0viPSTbeHfiPZQbbHWlixHeBQNsN2FGXTAwr8snbcAUPxC+HPxP/Z/+Ic3hLxvo+peF/FOhzrLEwcxupVvknglU4ZSVysiHBxweK/YM/wCCoFh8QDpvwd/aQ1aCx8Tttg0rxNLtig1RiQFiuMYWKY5AD8I/fa2N9VX/AAW//wCRp+Ev/YP1T/0ZbVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVa3ruh+GtMn1rxFrNjpWn2y7p7u9uFgiiHqzuQAPqa1L9tz9kjSrprO6/aD8GPIucm21EXCcEjh49ynp6+h6EV/wAN3fsf/wDRf/C3/f5//ia/4bu/Y/8A+i/+Fv8Av8//AMTVvPDd28V1bSCSKZA8bjoykZBH4VVVXiSCW58O6pbW8bSSy2UyIijJZihAAFf8EpPgN8aPhR8ZPF2s/Ev4X+JPDNjd+GDb29xqmnvbpLL9phbYpYDJ2qTj0Bqv2s/2Z/DX7VPwjvPh5rNyNP1O3lF5omqBNxsrtVIUkfxRsGZHXuGyPmVSPhh43/4KJ/sH2t38K1+Cl94v8MR3LtYq2k3Op2cLMSS1tcWpGxX5Yxvzkk7VYtlfgB+2r/wUQ+KWkeLP2gdAvfAPgrTAVRbmxewS0gYgutpazEyvLJtGZXyuApLEKqHwb4Q8PeAPCej+B/CWnJYaNoVlFZ2NshJ8uKNQqgk8k4HJPJOSck1Vf8EuP+Tx/jN/14X/AP6co6qqqqqqqqqqqqqqqqqqqq8ff8ilff8AbP8A9GLVf8FLdZOmfs4x2CyEf2t4htLYgE8hUll59sxDr7V/wTr0Yap+1Bo17tz/AGRpl9dZx0zCYc9f+m3v/UfEXVRrfjjWtQV9ytdtGjeqp8in8lFfCPSv7J+HmjxMuHuITcMfXzCWB/75K1VVVfHD4c/2HfN4s0a3xp94/wDpMaDiCU9/ZW/Q8dxXwe+Ix8Hat/ZWqTn+yL9wJCx4t5Ogk+nZvbB7UCrKGVgQRwRVV/wVG+Fxg1Lwt8YtPtvkukbR9SdR/wAtF3SQE+5Xzhn/AGFFf8ExPiidC+I2ufCm/uSLbxPZ/a7FGbgXVuCWCj1aIuSf+mS1VVVVVVVVVeEP+RZ07/rgKqqqqqqqqqqqqqqqqqqqq+MX/J9HhH/r60v/ANCqqqqqqqqqqqqqqqqqqqqr/gpr/wAmP/En/uFf+nW0r/gln/yZf4R/6/8AUv8A0slqqqqqqqqqqn+/+FVVVX/BRbwb+2z+0D8ctX8P6f8ACPxpqfw/8K3Rh8Ow6ZpEzWsw2DdckgESSMSw3dFXgY5z8J/iJ/wU8+B/gaw+HXwx/ZqvdG0PTwSkMfgeRnlkb70sjtlpHbAyzEngDoAB/wANP/8ABX3/AKIbqv8A4Qz1+zR4j+Kfi74GeE/Efxs0aTSvG97bStq9nLZG0aFxNIqgxH7mYwhx7571+33+wF8SNY+Jcf7Uv7LEc3/CVpcx3mq6TZzCC5F3FtKXtoSQC52gvHkEsNw3FmFS/ty/8FPdY8Pn4c2f7NOo23ieSH7K+tR+Cb9LlGxgzbGPkI/Q7ivljOdoGMf8E8/+Cf3i/wCE/iub9ov9oh/O8fXfnNpmmy3IuZLFpwRNc3EoJD3DhnXAJCh2JJZsJVft8f8AJnHxY/7F+T/0NK/4I6f8mkXf/Y333/om3qqqqqqqqqqqqqqqqqqqqr4xf8fOl/8AXOX+a1VVVVVV+3L+1tF8FvDsnw58B6ijeONatzvmjfJ0i3YY8046SsM7B1H3zxtDfCv4X+Mvjb4+sPAvhC2N1qepSFpZ5SdlvGD+8nlbkhFByT1JIAySAfgn8GvCHwJ+H9h4B8IW48u3XfeXjIFlvrggb5pOvJxwM4UAKOBVV47+JHh/wFZ77+bz76RcwWUbfO/uf7q+5/DNeNfHviDx1f8A2vWLnEMZPkWsZIjhHsO59Sefw4r4b/BPVfFflav4g8zT9JPzKuMS3A/2Qfur/tH8Aeo0fRdK0DT4tK0axitbWEYSONcfUnuSe5PJqqqqqqqqrQv+Q5p3/X3F/wChCqqqqqqqqqqqqqqqqqqqqqsdQFn4Y+IFtvA+3ahaQ43YziaWTp3/ANX0/HtX7EGni9/aD0m42g/YLG7mztzjMRjz7f6zr7471V9Z2+o2Vxp90u6G5iaKRfVWGD+hrV9NuNF1e80i5yJ7C5kgk4wQyMVP6ivBGujxT4L0DxKHDf2tpdtd5xj/AFkSt/Wqqqqqqqqvip+yf8NPiMs+o6Xajw3rcmWF3YxDypG9ZIchW75K7STySa+KP7PfxK+FEsk+t6Ob3SlPy6pYgywY7b+Mxn2YAZ6E9a+Gfxz+I/wpuEHhnXXk08NmTTLvMts/r8ucoT6oVPvXwr/a9+HXjwRaZ4mkXwvrDYUJdy5tpT/sTYAX6OF6gAmlZJEDowZWGQQcgiqqv2of2TvhV+1d4KPhjx7p/wBm1SzVm0fXbVALrTZD/dP8cZ/ijb5T14YKw/aU/Ze+Kn7LXjiTwd8RdK3W05ZtK1m2UtaalEP4o2PRhxujbDKeowQT+wV/wU71L4bnTfg5+0VqtzqXhMbbfSvEkrNLcaQOixz9WlgHADfejAxhlwE0vVNM1zTLTWtF1G2v7C+gSe1uraUSRzxuAVdGUkMpBBBHBBqv+C3/APyNPwl/7B+qf+jLaqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq1zWdN8OaLf+IdYult7DS7WW6upm6RxRqWdj7BQTX7U37U3xC/ak+IV54n8T6ncwaBbzuNC0JZT5GnQZIX5QcNMVxvk6seBhQqiqrwr/wAivo//AF4Qf+i1qqqqqqqqqv8Aglx/yeP8Zv8Arwv/AP05R1VVVVVVVVVVVVVVVVVVVV4+/wCRSvv+2f8A6MWq/wCCqmsCDwZ4B8Pbx/pup3d1t9fJiRc+v/Lf9fy/4JrwjTfH3j3xsygjRfCciKSOjSTRuO3U+SR+PftZWs+p38FlES011Msak8ksxAH6mrS1hsrSCyt12xW8axoPQKMD9BVVVVqOn2Wq2M+m6hbpPbXMZSWNhwwPWviH4HvPAmvvpspaS0ly9nOR/rE9D/tDoR9D0Ir4GfEYahbJ4L1m4zdW6f6DI5/1kY/5Z/VR09vpVV+018Ll+MXwP8VeCIbYTahLZNcaYAPmF3D+8iAPbcy7CfRz618NvG2pfDP4g+H/AB5pe77VoOoxXQjBx5iqw3xn2Zdyn2JrQ9Z03xJomn+ItGuVuLDVLSK6tZV6SRSIGRh9VINVVVVVVVVV4Q/5FnTv+uAqqqqqqqqqqqqqqqqqqqqr4xf8n0eEf+vrS/8A0KqqH/gomnwy/be+IXwg+KmqRj4d3GrQ2enag4/5AVzHbQxvuIz+4eRXL5+4zFuAWFW9xb3dvFdWk8c0MyB4pI2DK6kZDAjggjBzVVVVVVVVVVVVVVVVVV/wU1/5Mf8AiT/3Cv8A062lf8Es/wDky/wj/wBf+pf+lktVVVVVVVVVU/3/AMKqqqqqqqqqv2+P+TOPix/2L8n/AKGlf8EdP+TSLv8A7G++/wDRNvVVVVVVVVVVVVVVVVVVVVXxi/4+dL/65y/zWqqqqqr9qn9pPQf2cfAD6qxhvPE2qh4dD01m/wBZIBzK46iJMgn1JVeN2RcXHjn4wePWnma98Q+KvE9/6bpbmeQ4AA6AdABwqgDoBX7Jn7Mmi/s5eA1trhYbzxbrCLJrWoKMgMB8sER7Rpk+7MSx7BaOAMk9K+JPx2stIEui+DJIru+GVkvCN0UJ/wBns7e/3R79KVda8UauFUXOpajeye8kkjH/AD+Ar4bfAuw0MRaz4vjivNQGGjtPvRQH3/vt+g9+DWABgdqqqqqqqqqrQv8AkOad/wBfcX/oQqqqqqqqqqqqqqqqqqqqqqr6+aO41rTAW2z6kJSMcZQyj/2oa/4J92AuPjDq98wBFp4dl284IZp4AP0DfpVV+0poI8N/HjxtpgQIH1WS6VQMYE4EwwPTEgr9kjXh4g/Z88JTNIGlsoJbOQZzt8qV0Uf98BD+NVVVVVVVVVJGkqNHIiurAhlYZBHcEV8Vf2O/h743Eup+EFTwtqzZYi3i3Wsp/wBqLI2fVMAdcGviX8EviL8KLpo/FWhSfYt22LUrYGW2l9MOB8pP91gp9q+Fv7RXxL+FTxWml6sdR0dODpd+xkiA/wCmZzuj/wCAkDPUGvhX+1R8NPiUsGnXl6vh7W5CFFjfyAJI3pHLwregB2sfSqqvi/8ABz4dfHfwLffDv4n+G7fWNHvRkK64ktpQCFmhfrHIuThh6kHIJB/bP/YR+Iv7JWvNqiGbxD4Av5ymma/HDgxE8iC6UcRy9gfuyAZXB3Iv7EH/AAUJ8cfst6jb+C/FhuvEnw0uZsz6aW3T6WW+9LaEnAGTloj8rckbWJY/Dr4keBvi34P0/wAe/DnxLZ67oWpxh7e7tmJHurKcMjjoyMAynggGv+C3/wDyNPwl/wCwfqn/AKMtqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr45eGdU8a/BP4g+DdD3f2lr3hXU7Cz2jnzprWSNMf8AAmFeB/DcHivx14f8IahfDT4tY1a2sZ7mQAfZlllWNnOem0MTz6V4B/ZA/Zm+G/h628O+Hvgn4RmjgjVHu9S0mG9urggfekmlVnYk5OM4BJwAOK/4UD8CP+iJ+Av/AAm7T/43X/CgfgR/0RPwF/4Tdp/8bqOOOKNYYUVERQqqowFA6ADsKqqqqqqqqr/glx/yeP8AGb/rwv8A/wBOUdVVVVVVVVVVVVVVVVVVVVePv+RSvv8Atn/6MWq/4Ko6z5/j3wL4e35+xaPcXW3086YLnr/0w9O3ft+w4n9k/Cv4n6zgB9XvtL02Ju4CCeWQD6gqDxXwf0kav8Q9JjZcpau1y/tsBKn/AL621VVVVXj7wVY+OvD8uk3BWO4T57ScjmKQDj/gJ6EenuBVzbat4Y1preYSWeo6dN2OGjdTwQfyIPevhp48tvHegrcuyJqNqAl5COMN2cD+636HI7VVftm/C0fCj9oTxNpNrb+VpmsS/wBracAuAIrglmVR2CyCRAPRRX/BOr4o/wDCefAaPwnfXIk1LwTdGxYE5Y2z5eBj6AAvGPaKqqqqqqqqq8If8izp3/XAVVVVVVVVVVVVVVVVVVVVV8Yv+T6PCP8A19aX/wChVVftEeIv+Ev+PvxI8TrJvTVPFmp3ERz0RrmQoB7BcD8K/wCCcX/BQL/hBZdP+AHxz8Q48NPtt/Det3bcaW3RbaZyeLc8BGP+rPBITGyqqqqqqqqqqqqqqqqq/wCCmv8AyY/8Sf8AuFf+nW0r/gln/wAmX+Ef+v8A1L/0slqqqqqqqqqqn+/+FVVVVVVVVVX7fH/JnHxY/wCxfk/9DSv+COn/ACaRd/8AY333/om3qqqqqqqqqqqqqqqqqqqqr4xf8fOl/wDXOX+a1VVVVXxj+LvhD4IeAdQ8f+Mropa2a7ILeMjzbycg7IYx3ZiPoACTgAmvjD8W/GHxz8f3/jzxfceZeXrbLa2iz5dpCCdkMY/urn6kkk5JJr9hP9kZPhFocPxU+IOmD/hNNXt/9EtZo/m0e3cfdwfuzOp+buoOzj581rWt6T4d06XVdavo7W1hHzO56n0A6kn0HNfEj40at4v83SNEMmn6QcqwziW4H+2R0X/ZH4k14J+H/iDx5ffZtItwlvGcT3coIji/Hu3+yOfoOa8DfDrw94CsvJ0yHzryRcT3kq/vJPYf3V/2R7Zyeaqqqqqqqqqq0L/kOad/19xf+hCqqqqqqqqqqqqqqqqqqqqq1If8TvUjn/l6k/8AQjX/AATpsDLr/jbUwDi3s7SHPYb3kP1/5Z1Vft66CNL+N0Wqony6zo0E7N6ujPER/wB8xp+Yr/gnxr3274Xa74eeTc+l60ZFBP3Y5YkwMf70bn8aqqqqqqqqqqvLO01C1lsb+1hubedCksM0YdHU9QVPBHsa+Kv7FvgzxT5uq/Du5Tw3qJyxtWUvaSn6fei/4DkD+7XxA+FPj34Y332Hxj4euLNWYrDdAb4Jv9yQfKeOcdR3Ar4V/tR/Ev4ZGLT5L7+39FQBfsGoSFjGo7RScsnpjlR/dr4WftIfDT4qrBY6fqg0zWpAAdLvmCSM3pG33ZP+AnPqBVV4l8M+HvGWgX/hXxXo1nq2j6pA0F5Y3kIlinjYcqyngiv28P8Agmn4h+BMmofFb4KWl5rnw83NNeWHM13oI6nd3ltx2k5ZB9/ON5/ZQ/bB+KP7JfjEaz4QuzqXh6+kX+2fDtzMVtr5RxuHXypgB8sijIxghlyp/wCCnH7Rvwv/AGnNK+D/AI++GWsGaJbDU47/AE+4AS602YvbExTICcHrhgSrDkE1VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVWoXsWm2FzqM6s0drC8rhBkkKCTj3wK/ap+I3wX+K/wAV774lfBXwt4g8MR65K11qdhqPkhFu2ILyw+Ux2B2JYqc4bJBw21fhx/wVu/aT8DeFLPwvrmk+FvF72EaxRanqsE63ciAYAlaORVkIAHzFQx6sWJzX/D5z47/9Et8Bf983f/x6v+Hznx3/AOiW+Av++bv/AOPVo17JqWj2OoyoqvdW0crKvQFlBIHtzVVVVVVVVVf8EuP+Tx/jN/14X/8A6co6qqqqqqqqqqqqqqqqqqqq8ff8ilff9s//AEYtV/wUn1j+1P2kmsd+f7I0CztcZ6ZMk3p/029/6D9lVDZfAtbcgh9Q8R3d02QRujWG3iT8mWb8/av2b9J8zU9Y1tl/1ECW6EjqXO5sfTYv51VVVVVfGz4cjxHpx8TaPb51Oxj/AHqIvNxEPbuy9R6jI9K8F+LtQ8Fa9BrVgSyr8k8JOBNGeqn8sg9iBWiazp3iHSrbWdKnEttdJuRu49QR2IOQR6iq/wCCnnwt/t34daF8VtPtwbnwxdm0vmVefstwQFZj6LKEUD/psa/4J5fFEeAPj9a+G7242ab40tjpsgY4UXA+eBvqWBjH/XU1VVVVVVVVXhD/AJFnTv8ArgKqqqqqqqqqqqqqqqqqqqq+MX/J9HhH/r60v/0Kq8SazB4c8O6p4huQDDpdlNdSA8fLGhY/oK/0zVb/APinu7yb8ZHY/wAyTX7T37MPxC/ZZ+IUvgvxpb/abC53S6NrMMZWDU4AfvL12uuQHjJypI6qVZv+Cbf/AAUCMx0v9nT45687zO0dr4U126fO7OFSync85zgRueudhP3Kqqqqqqqqqqqqqqqqr/gpr/yY/wDEn/uFf+nW0r/gln/yZf4R/wCv/Uv/AEslqqqqqqqqqqn+/wDhVVVVVVVVVV+3x/yZx8WP+xfk/wDQ0r/gjp/yaRd/9jfff+ibeqqqqqqqqqqqqqqqqqqqqvjF/wAfOl/9c5f5rVVVVXijxPoHgvw7qHivxTqsGnaTpVu093dTNhY0Uc+5J4AA5JIABJr9qr9pPXv2jvHz6qxms/DOlF4dD01m/wBXGTzM46ea+AT6AKvO3J/4J+/sijU5bL4+fEvSwbSJhJ4Z06dP9c46Xjqf4R/yzB6n5+AFLV4/+J/h/wABW5iuZBd6k65iso2+b2Zz/Cvv1PYHmvF/jXX/ABtqJv8AW7suFJ8mBOI4R6Kv9TyfWvht8DtR8RGLWPFSS2OmnDRwEbZbgf8Asin16nt61pml6dothDpmk2cVrawLtjijXAH/ANf36mqqqqqqqqqqq0L/AJDmnf8AX3F/6EKqqqqqqqqqqqqqqqqqqqqr9ubwp4M8GfEHQtI8H+GtM0cT6Sbq6WzhWLzWeZ1DMB1PyHn61/wTq08R+GvGmqBR/pF9aw524z5aOevf/WdO2feqr/gotoO628FeJ4k+493ZzNj1EboP/HZK/wCCd2vfZ/Gfi3wwX/4/9Mhuwp7mCTZ/7Xqqqqqqqqqqqq1XSNK13T5tJ1rTra/srhds1vcxCSOQehU5Br4qfsRaBrLTat8LtSXR7ogsdNu2Z7dz/sPy0f0O4f7orxn8P/Gvw51T+y/GHh+80u4BPlNIvyS47xyDKuPdSa+Ff7XHxH+H4i0zX5j4o0dMAQ3sp8+Jf9ibk49mDDjAxXwx+PHw2+LEKReGtcWLUtm6TTLzEVwmBzhc4cD1QkDviqlijmjaGVFdHUqyMMhgeoI9K/b0/wCCW63A1H4x/sv6GFly9xq/g62TAfJy0tio6HnJg4GM7OyGaGW3le3niaOWNiro6lWVhwQR2PtVutwtvEt3JG84QCVo0KqWxyVBJIGexJ+tVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVa/aTX+hajY2yhpri0ljjUnGWZCBz9TVp/wAEof2xLmBZZvDPhy1ck5im12IsP++Nw/Wv+HTX7X//AEBfC3/g8T/Cv+HTX7X/AP0BfC3/AIPE/wAK/wCHTX7X/wD0BfC3/g8T/CtCs5tP0PTrC4AEttaRROAcjcqAHH4iqqqqqqqqq/4Jcf8AJ4/xm/68L/8A9OUdVVVVVVVVVVVVVVVVVVVVePv+RSvv+2f/AKMWq/bZ1ka7+1J4+u1fcIb2G1A9PJt4oiOp7of/AK1fs6WzW3wd0AMAGl+0OcD1nkx+mK+Amk/2f4BjvGXD6jcyTcjnAOwf+gE/jVVVVVVXxs+HH/CPX58T6Nb4029f99Go4t5T/JW7eh49K+DfxFPhLVRouq3GNIv35ZjxbyngP7A8A/ge1cEZBzXxM8Dab8Tfh74h8AartFvrunS2pcrnynZfkkA9Vbaw91FSJ4g8AeL2jcPYa54b1IggH5re5gk/mrp+lfCzx5YfFD4ceHPiDpu0Q67p0VyY1OfKkK/vI+/KuGU+61VVVVVVVV4Q/wCRZ07/AK4CqqqqqqqqqqqqqqqqqqqqnsLGWdbqSzgeZcbZGjBYY6YPWq/a718+GP2XPitrCSbJE8I6jDE2M7ZJYGiQ9D/E49vXiv2efDo8W/Hz4beGGj3pqnizTLeQYzhGuow5PsFyT7Cv2g/2ffh5+0p8Orz4dfELT98MmZbC+hAFxp1wAQs0THoRnBB4YEg5Br9oL9n/AOIn7NXxGu/h58QLDy54v3thfwg+RqFuSQs8LdwcYIPKkEHBFf8ABN/9v4fEGDTf2fPjXrcj+Kol8nw9rd1Jn+1UUcW8zH/luAMKx/1g4J3431VVVVVVVVVVVVVVVf8ABTX/AJMf+JP/AHCv/TraV/wSz/5Mv8I/9f8AqX/pZLVVVVVVVVVVP9/8Kqqqqqqqqqv2+P8Akzj4sf8AYvyf+hpX/BHT/k0i7/7G++/9E29VVVVVVVVVVVVVVVVVVVVfGL/j50v/AK5y/wA1qqqqlkjhjaWWRURFLMzHAUDqSewr9uX9raX40+IZPhz4D1F18D6LcHfNG+Bq9wpx5px1iU52DofvnnaF/Yi/ZMn+OnidfG/jSxkTwLodwPNVht/tW4XB+zqf7g4MjDsQo5JKxQ2tlapBBHFBb28YVERQiRoowAAOAAB+FfEn48QWPm6J4HmjnuBlZdQwGSP2jHRj/tHj0z1FpZ634q1cW9rFc6jqN45J5Lu7HqST+pPA718N/ghpnhkw6z4mEV/qi4aOLGYrY+395vc8DsOM1VVVVVVVVVVVVoX/ACHNO/6+4v8A0IVVVVVVVVVVVVVVVVVVVVVX7et+bz46JbFifsOh20IB7ZaR+P8Avuv+CflgLf4N6tesBvu/EM2CD/CsEAHH13VVft26CNX+A82pBATomq211uxyAxaE/hmYfpX7F+vDRP2g9AgeQLFqsFzZuScdYmdR+Log/Gqqqqqqqqqqqqq8Q+GvD/izS5NF8S6NZ6nZSj5oLqESLn1Geh9CORXxW/Yft5/P1j4S6mIXwW/se+clT7RTHkewfPJ+8K8ReFvF3gDWhp3iPSL/AEbUrdt0YlQxtkHh0YcEZ6MpI96+Ff7ZXjvwasGk+N4j4n0tCF82WTbeRL7SHiT6Pyf7wr4dfGH4e/FSz+0eD9finuETdNYzDyriH/ejPOO24ZX3qv8Ags38N/Ang/4m+B/FvhbwvY6Xq3i20v5tbubZPLN9LE8ASR1Hy78O2WABbPJNVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVeKPFHhzwV4fvvFfi7W7PSNH0yIy3l9eSiKKBBwWdjwByOTXw4+Mvwo+MMF9c/Cz4h6D4qi0t0S8fSr1LgW7OCUD7ScEhWx9DVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVftCftw/s+/s0a3beFviH4ivbjXriJZjpek2n2maCJvuvJyqoD1ALbiOQMc18MPjZ8NfjL8O4/ij8OPEkWsaCySeZIiMkkEkYzJFJGwDJIowdpHIKkZDAn/h7J+x//ANBvxR/4I3/xr/h7J+x//wBBvxR/4I3/AMa/4eyfsf8A/Qb8Uf8Agjf/ABr/AIeyfsf/APQb8Uf+CN/8asruHULK3v7YkxXMSyRlhg7WAI4+hqqqqqqqqqv+CXH/ACeP8Zv+vC//APTlHVVVVVVVVVVVVVVVVVVVVXj7/kUr7/tn/wCjFqvjlrH/AAkPxp8e62H3Le+JdQljJzwhuH2jnB4XAr4UWJ074Z+F7UjDDSoHYHsXQMf1avCmlDQ/DOl6Rt2taWkcbjH8YUbj+JzVVVVVVWp6bY6xp8+l6jAs9tcxlJI2HUH/AD17V4+8FX3gbX5dKuC0lu+XtJyP9bH2/EdCPX2Ir4HfEf8Ata0TwbrVxm9tU/0KRzzNGB9z/eUfmv0Oa/4KKfC5vAnx6m8VWdts03xrai+jYD5RcphJ1HvkJIf+utf8EwfiiNa8Ba/8JtQud1z4cuhfWCsettOfnVR6LKCx95hVVVVVVVVXhD/kWdO/64Cqqqqqqqqqqqqqqqqqqqqqr/gp74gOhfsXeN4I3Cy6tPp9lGT73kLtxg9Ujcduuc5Ff8E6/Dn/AAk/7Zvw0smi3x2l7cXzkjIX7PazSqTwf4kUfUjp1qv2o/2YfAP7U3w3uPBfi23S21S2R5dD1lI902mXJHDDoWjYhQ8ecMB2YKy/F/4RfET9nn4lX3w+8d2Mmma7o8qyQzwOdk8ecx3EEgwSjYyG4IIIIDAgf8E7P294Pjjplp8F/izqax/EHTLbFjqE8gA16FBycn/l5VeWX+MAuOjAVVVVVVVVVVVVVVVX/BTX/kx/4k/9wr/062lf8Es/+TL/AAj/ANf+pf8ApZLVVVVVVVVVVP8Af/Cqqqqqqqqqr9vj/kzj4sf9i/J/6Glf8EdP+TSLv/sb77/0Tb1VVVVVVVVVVVVVVVVVVVV8Yv8Aj50v/rnL/Naqqqv+Cgn7XXF78APhpqnrF4n1CE/nZo3/AKMI9kz98V+zD+zl4i/aO+IEXh+y86z0DTts2t6mEyLaHPCKTwZXwQo9i2CFNafZ+A/gz4GsNC06G20XQNFtxBaW6DsOcAdXdjkk8kkkk8k18R/jFrPjVpNM07fp+j5x5Ib55/eQjt/sjj1z1rwL8N/EPj27CadD5FlG2J72Vfkj9QP7zew/EjrXgvwF4e8C2AtNHtg07r+/upADLKfc9h/sjgfrVVVVVVVVVVVVVaF/yHNO/wCvuL/0IVVVVVVVVVVVVVVVVVVVVVX7UPjXQviD8b/EPiTwxfi90txbw28/kvFu8uCNH+VwG4kDjkDp6c1+xHYGz/Z60W4II+3Xl5MM98TNHx/37qq/aE0EeJfgh420kJvc6NPPGvq8K+ao/wC+kFfCXXv+EX+KPhLxAZNqWGtWskpzj92JV3j8VyPxqqqqqqqqqqqqqqrxb4K8J+O9LbRvF+gWeq2hztS4jyYye6MPmQ+6kH3r4qfsP6hZrPq/wo1Q30Yy39k3zhZR7Ry8K30fbwPvE1faf4q8Ca/9mvrbUtC1mwfcoYPbzQt2ZTwR7Ede1fCr9tfxT4eFvo/xKsjr1guF+3w4S7jHqRwsv47T3LGv+CwfxH8F/EzU/hNrPgvXYdQhjsNTWZACkkDF7fCuhwyng9Rg4OCaqqqqqqqqqqqqqqqqqqqqqqqqqqr/AIK7fEPx/wDDT9mzw3rvw48c+IPCupT+OLW2lvNE1OaxmkhNjesY2eJlYoWRCVJxlVPYV/wS0/bi8aX/AI+uf2ffj3441rW5/ExF34Z1XX7+W6uFuTGCbUyzEtsljAeME43AgZMoqv8Agpp4y8X+Af2PvFXibwL4r1jw5rFvfacsOoaTfSWdxGGu41YLJGysAVJBAPIOK/4Ja+N/GnxC/ZN0/wAR+PvGGt+JdWfXL+Jr/WNQlvJyiuoVTJKzNgdhnAqqqv2uv2vfBH7H/g7SfFPjDw3rWuSa7ePaWFrpojXMiJvJkd2GxccZAY5PTvXwE+KyfHP4OeFPi3HoR0ZfFFgLtbBrn7QbcFiNvmbV3dOu0da/b8/5M0+LP/Yvyf8AoaV/wQ6/5FT4t/8AYQ0r/wBF3NVXx7uPiVa/BXxtcfByATeNo9DuToSYUsbnyzs2BvlLjqobgttB4Jr/AIJZa9+2brOr+OU/aK/4TmbwwsMT2E3jCKdLhb4yHekBnAcx7N25R8iEIABk1VVVVVVVVVVVVVVVVVVVVVVVVVVVVVV/wUy+E3xC8G/tTeLPGXiDSL6bQ/Fs0V3pOq+SzQzJ5McZhD9A8ZTYUJzgKcYYZ/4JZfCn4heAf2cfHviLxlpV9pdn4snM2j2N5E0UjRx27K1wEPIWQsFBI+YRA9CpP/CK+J/+hb1T/wAA5P8ACv8AhFfE/wD0Leqf+Acn+Ff8Ir4n/wChb1T/AMA5P8K/4RXxP/0Leqf+Acn+FeF1K+GNIVhgiwgBBGMfu1qqqqqqqqqv+CXH/J4/xm/68L//ANOUdVVVVVVVVVVVVVVVVVVVVePv+RSvv+2f/oxa1K+g0vTrrU7pgIbOB5pCSBhVUk8ngcCry5mv7ya8nbdNcSNI5A6sxyf1NfC7w4tzrfhnw0IwY4ngidQP4IwN3/jqmqqqqqqqr4g+CLLx14fk0ybbHdRZeznI/wBVJ/8AEnoR+PUCpYtW8Ma0Yn8yz1HTp+oOGjdT1H5fQ18OPHVp480BL5Skd9b4jvIAfuPj7wH91uo/Edq/4KIfC0ePvgFceJ7G28zU/BVyNQjKrljbNhLhfYBSsh/641+x38UT8Jv2gvDGuXNz5Om6nN/ZWpEnA8i4IXc3ssgjk/4BVVVVVVVVXhD/AJFnTv8ArgKqqqqqqqqqqqqqqqqqqqq8RftAeBPDHxQ0/wCEepRaodc1J4EhaK3VoQZThctuBHvxVf8ABZLxF9g/Z28LeG432yat4uikYf3o4backf8AfTxn8K/4JC+GxrX7Vlzq7xZXQPCt7dK5XhXeSGAAHBwSsz9xwDz1Bqv2wv2R/Bn7WHw7k0PUEttN8WaYjP4f1xoiWtZOCY5MctC+MMvOOGAyoz448E/ET4DfEq88JeJra88PeK/C96rB4ZijwyKQ0c0UinkEbXV1PQgiv2AP269M/aV8PJ8PfH9zbWHxJ0W2HmDcETW4VHNxEvaQAZkjHTO5flJCVVVVVVVVVVVVVVV/wU1/5Mf+JP8A3Cv/AE62lf8ABLP/AJMv8I/9f+pf+lktVVVVVVVVVU/3/wAKqqqqqqqqqv2+P+TOPix/2L8n/oaV/wAEdP8Ak0i7/wCxvvv/AETb1VVVVVVVVVVVVVVVVVVVV8Yv+PnS/wDrnL/Naqqr9uX9raL4LeHZPhz4D1FG8ca1bnfNG+TpFuwx5px0lYZ2DqPvnjaG+Ffwv8ZfG3x9YeBfCFsbrU9SkLSzyk7LeMH95PK3JCKDknqSQBkkA+FNP+Gf7Jnw6s/hv4QiW/1KJPMuTgLLd3DAbp5yM7c4GF5IUKBwM14p8W694y1I6lrl60z5IjiXhIh/dRew/U9ya+G3wKvNZEWteMo5bSxOGis/uyzD1bui+3U+3BqxsLLTLOLT9OtYre3gXbHFGu1VHsBVVVVVVVVVVVVVVoX/ACHNO/6+4v8A0IVVVVVVVVVVVVVVVVVVVVVWsADV74D/AJ+ZP/QjX7K2njTf2fPBVsFC77Fpsbdv+sleTOP+Bde/Wqq+s7fUbK40+6XdDcxNFIvqrDB/Q1q+m3Gi6veaRc5E9hcyQScYIZGKn9RXgjXR4p8F6B4lDhv7W0u2u84x/rIlb+tVVVVVVVVVVVVVVV44+G/gj4kad/ZnjPw7aalGoIjkddssOe6SDDL+B+tfFX9ibxNoIl1b4Y37a9ZDLGwuCsd1GP8AZbhZP/HT6A1+2tpmo6P4g8O6Xq1hcWV5bRXKzW9xEY5I2zHwynBB+tVVVVVVVVVVVVVVVVVVVVVVVVVVV/wWr/5NX8Lf9lAs/wD0gv6+Pn7Pt/ZfskfAn9sT4aNcQXen6Pbad4iubQlJLeeCdktLwMOQysghLdtsOO5r9hf9qfTv2rfgfYeKbqaCPxbom2w8TWcZA2XIXidV6iOZRvXsDvXJ2E1/wVg/5Mj8Yf8AYQ0z/wBLIq/4JDf8maaZ/wBjBqP/AKGtfFn4t/D34H+Br/4i/E7xJb6LoenAB5pcs0rn7sUaDLSSNg4VQScE9ASPFP8AwWY1zxDrk+lfAT9m7U9etbYlvtOpXTvNImcAm3t0by/qZG644xz8Iv8Agsr4N1TxRD4P/aA+FOoeA5TL5M+qWlw93FbP3M9u0ayxqO+3zCOOOprStV0zXdMtNa0XUba/0+/gSe1uraUSRTxOAyujKSGUgggjgg1+3r+3t/w1vpOjeBf+FU/8Ip/wiOsXM32r+3ft32nK+XjZ9ni2dM9W9PevgD/wV4/4UZ8GfCXwj/4Z7/tv/hFtPWz/ALQ/4S37N9owxO7y/sb7OvTcfrX7XXiv/hPf+Ce3jPxz9g+w/wDCReBoNR+y+b5vkeekUmzfhd23djOBnGcDpX/BL/8AaP8AhX+zD8Gfi94++KesyW9vJqmlw2Njax+bdahN5VyfLhjyMnAyWYqq8ZYZFa1/wW08X22rxX+nfs3W0Xh2Vj5IvdckWe4Ud1lEOxT0OArY6ZPWv2UP2xPhV+134Uuta8CPdadrGkFF1fQr/aLizLZ2uCpKyRMVba49MEKeK+LPjr/hV3wr8ZfEz+y/7T/4RHw/f6v9h8/yPtX2a3eby/M2ts3bNu7a2M5welfsOft7f8Noar4u0v8A4VT/AMIf/wAIrb2s2/8At3+0PtPnNIMY+zxbMeX75z2xz+3B+2ZN+xn4b8L+JF+Ga+MYvEd9PaNGdb/s82zRorgj9xLvBBb+7jA654+Mf/BU34S/CX4deCtZj8MXPiHxz4z8NWGt/wDCLafqCldLF3bpMkdzdbPlPz4AEZdgAxRQy58Nf8Fs9dsfE62XxO/Z8S00lpB5n9m6o4u7aM87hHMgWU4xwTGD1yK+FHxW8CfG7wDpPxL+G+uR6poWsRF4JgpRkYHa8boeUdWBUqehHpg1ruuaL4Y0W98ReItVtdN0vTLd7i8vLqURRW8SDLO7HhVABJJr4rf8FmdBh8VP4S/Z4+D9940xMYodS1Cd7cXZU9YbVEaVlIBILFG9Ur4Jf8FcfD+v+NbD4fftGfCu8+Gd3qLrHBq0lw7WqM3CmdJUR4UJ48zLqM5baoLD9nv/AIKDyfHz9p3xN+zlB8IE0ZPDTalv1s+IvtPnLaTiEEQfZkxvLKceYdvqaqqqqqqqqqqqqqqqqqqqqqqqqqvH/jLS/hx4D8SfELXILqfTvC+kXWqXkVqitM8NvC0rrGGZVLlUIALAZxkjrX/D5P8AZg/6EX4o/wDgrsf/AJMrw9rVr4l0DTPEVlFNFb6pZxXUMcwAdEkQOAwBIBwRnBIz3NVVVVVVVVVVV/wS4/5PH+M3/Xhf/wDpyjqqqqqqqqqqqqqqqqqqqqrx9/yKV9/2z/8ARi18e9YPh/4H/EDWUcrJaeGdQeIjI+f7O+3p0+bHNeEbH+1PFmi6YBn7XqMEOPXdIo/rX7Pmki98ayai65XTrN2U+jvhB/46Xqqqqqqqqr43fDj+3rBvFejW+dRso/8ASY0HM8Q747so/McdgK8EeML/AMEa/DrFkS8f3LmDOBNGeo+vcHsQKP8AYHj3wrLC6x32ka3ZPDNGw4kikUq6MPoSCPrXxR8CX/wx+IniL4f6mS0+hahLah2GPNRW+ST6Mm1v+BV+y98UR8YfgX4V8Zz3Il1FrMWmp/NlhdQ/u5CfTcVDgejiqqqqqqqrwh/yLOnf9cBVVVVVVVVVVVVVVVVVVVVXxi/5Po8I/wDX1pf/AKFVf8FrfEW66+FHhGKT/Vx6pfTpnruNukZ/8dl/Ov8Aglz8T9L+EPj3xh4p1nRJr60u9PtrCSWBgJIFaRnJVTw3Ma5G5e1eCfiJ4M+IumjU/CGvW9+gH7yIHbLCfR4z8y/iMHtkVVftufsX+GP2r/A/naetnpXj/RYmOi6xImBIvJNrORyYmJ4OCY2O4AgsrXdp8RfgZ8SntbhdR8LeM/CGpdj5c1ncRNkMCOCOhBGVZSCMg8/sL/tteH/2q/B39j+IJLPS/iLokQ/tXTEbat5GMD7Xbqf4CThl5KNweGUmqqqqqqqqqqqqqr/gpr/yY/8AEn/uFf8Ap1tK/wCCWf8AyZf4R/6/9S/9LJaqqqqqqqqqp/v/AIVVVVVVVVVVft8f8mcfFj/sX5P/AENK/wCCOn/JpF3/ANjfff8Aom3qqqqqqqqqqqqqqqqqqqqr4xf8fOl/9c5f5rVVX7VP7Seg/s4+AH1VjDeeJtVDw6HprN/rJAOZXHURJkE+pKrxuyL288a/FrxzJeXc13r3ibxJe5ZmOZLiZz+AUDpjhVUdgOPgp4e0T9m7wJJ4b8Gvb33i/WEVtf8AEIXcFYDi3ts/8s0yRvP3mJbHK7dL0nW/FerLY6bbz39/dMWPO5iSeWZj0HPJNfDf4LaT4Q8rV9bMWoawOVOMxW5/2Aerf7R/ADvVVVVVVVVVVVVVVWhf8hzTv+vuL/0IVVVVVVVVVVVVVVVVVVVVVWtADWL4A9LmT/0I18FbAaZ8HfA9iAA0Xh2x34ORuMCFsfiTVVX7Smgjw38ePG2mBAgfVZLpVAxgTgTDA9MSCv2SdcGv/s9+EZ2l3SWlvLaOP7vlTOij/vgKfxqqqqqqqqqqqqqqqqr/AILewQJ4v+FFwkMYlk03U1dwoDMBJb4BPUgZP5mqqqqqqqqqqqqqqqrxb4x1PQdUFjZwWroYQ+ZUYnJJ9GHpX/Cy9e/59LD/AL9v/wDFV/wsvXv+fSw/79v/APFV/wALL17/AJ9LD/v2/wD8VX/Cy9e/59LD/v2//wAVX/Cy9e/59LD/AL9v/wDFV/wsvXv+fSw/79v/APFV4V8aaprurLYXcFokZjZsxowOR9WNVVVVVVf8Fq/+TV/C3/ZQLP8A9IL+v2FvBXhr4kf8E8vAvgLxjpkeoaJr+hX1nfW0g+/HJd3CnB6qwzkMOVIBGCBXgDxF44/4Jb/tsXnhrxPLd3fhG4kFvqBVONV0aZiYbtF6GWMjdgdHjljzgsa/4Kmappuu/sH+JNa0a+hvLDULnSLi1uYXDJNE93CyOpHVSpBB9DX/AASG/wCTNNM/7GDUf/Q1r/gpD498X/tM/ts6J+zL4b1Ix6VoWo2WhafFuJiN/d+WZ7l19VMqxn0WEkY3HPwT+CPw6/Z++H+m/Dn4baDb2Gn2EKrNOIlE99KBhp53ABkkY8knp0GAAB/wUs/Za8JfHT4AeIvHVto9tD448CabNqmm6lHEBLNbwKZJ7VyBl0aMOVU/dkCkYBYN/wAEXPjfrPij4feLfgbr15JcR+Dpob/RWkYsY7a5ZxLCPRVlQOB6zt7V/wAFvf8AklXw0/7GC6/9JxX7Af8AyZp8Jf8AsX4//Q3r9vz/AJM0+LP/AGL8n/oaV/wSL/Zl8C/Gfx74m+JnxH0u31nT/AP2VdM0q6TzIJL248w+dIh4YRrDwpBBZwf4BXjj4f8Agv4leEb/AMB+OvDVhrWg6lAYLixuoQyFSMAr3Rh1VlwykAgggGv2Kk1L9m3/AIKXr8K7PUJXsE17VvC12WIJu7cCUQM2B3kit5Og6YOOcftZf8mr/GX/ALJ/rv8A6QTV/wAEOv8Akavi3/2D9K/9GXNf8Fvf+SVfDT/sYLr/ANJxX/BJv9mbwR4W+B2k/tCa3pVtqnjXxgZja39ynmvptlDI0EcMJOdhbyizMuCQyqeFxX/BQL4BeDPjh+zR41uNY0S1k8Q+FNFutX0LUhEouLea2jaUxLIeQkoQoyk7fmDYyqkf8EQfH2pSR/E/4XXNw8lhCbLWLKIn5YZW3wzn6sFt/wDv371/wWo+OGr6H4X8HfALQ76W3g8RmTVtcCPt86CFwtvEwHVDKJHIPG6FDz2/4J7/ALLXg79nn4D+HNYTRLdvG3i3TINQ13U5I8zgzIJEtlYjKRxqyrtHBYMx5NftRfs4+B/2nfhLrHw+8V6TbSX7W0smh6k0Y83Tb0KfKlRuoG7AZcgMuQetf8EbyW/a6vGZiSfB9/kn/rtbVVVVVVVVVVVVVVVVVVVVVVVVVVeJfD2leLfDmq+FNct/P03WbKayvIs48yGVCjr+KsRX7UP7I/xT/Zf8ZX2k+JtEvL3wy1ww0jxHDbk2t7CT8m5hkRy4wGjY5BzjcuGP7Hv/AAUU+A3ir4Q+HPCvxT8d6f4P8X+HdNhsL1NVYwQXYhQIs8Ux+Q71VSykhgxYAEYJ/wCGy/2UP+jh/AX/AIO4f/iq/wCGy/2UP+jh/AX/AIO4f/iqgmhuYI7i3lWSKVAyOpyGBGQR7Yqqqqqqqqqv+CXH/J4/xm/68L//ANOUdVVVVVVVVVVVVVVVVVVVVePv+RSvv+2f/oxa/bj1hdF/ZY8d3AcB7i2t7VAcZbzbmJDjP+yzH8K+CdgNR+K/hi3xnZfLNj/rmC//ALLX7OWk/Z/D2p6yy4a8uliXI6rGuc/TLkfhVVVVVVVVVfGr4cjwzqZ8RaPb40u/c+Yiji3lPUY7K3UehyOOK+DHxG/4RbUx4f1e4xpV8/yux4t5TwG9lPAPpwfWv+Ckv7P3iK48Sw/Hfwpokl3pclglvr7243NbyR5WOd167DHsQsOF8sZxuFfs2ftQ+Of2c/EYn0qV9T8NXsoOp6JNKRHL0Bkj/wCecoAwGAwcAMCAMfCj4teBvjV4PtfGvgLV1vLKb5JomG2a0lwC0UqfwuMj2IwQSCCaqqqqqq8If8izp3/XAVVVVVVVVVVVVVVVVVVVVV8Yv+T6PCP/AF9aX/6FVf8ABYrxEuqftMaDoMUgMei+EbdXXP3ZZbi4c9/7nl/5xX7IGmOPCGsX8cZZ7vUxCoC5LbI1Ix68yVZ3viXwVrguLO41DRdXsHwSC0E0LdwRwR7g/jXww/bPv7QQ6R8UtPN5HkL/AGrZoFkX3kiGA3uVwf8AZJrwt4w8L+NtLTWfCeuWup2jHBkgfJQ/3WXqjezAGq/b1/Yb0b9qLwv/AMJb4OitdN+JGh27CyuWUImqRDn7LO3r/wA83P3SSD8rEjw34i+IvwK+JVtr2izX/hnxh4TvzhZYjHLazoSrxyRsOn3lZGGCCQRg1+xj+2H4R/aw8AJeK9rpnjfSIlXX9DRz+7OcC4hB5aF+PUoTtYnhmqqqqqqqqqqqqqv+Cmv/ACY/8Sf+4V/6dbSv+CWf/Jl/hH/r/wBS/wDSyWqqqqqqqqqqf7/4VVVVVVVVVVft8f8AJnHxY/7F+T/0NK/4I6f8mkXf/Y333/om3qqqqqqqqqqqqqqqqqqqqr4xf8fOl/8AXOX+a1VfGP4u+EPgh4B1Dx/4yuilrZrsgt4yPNvJyDshjHdmI+gAJOACa+MXxc8X/HLx9f8Aj7xhciS7vG2W9tHny7SEE7IYx2VQfqSSTkkmv2efgz/whNgnjDxHbY169i/cxOvNlEw5Hs7A/N6A7f72fAPww8QePbhZLaM2umo2Jb2RTt46hB/E3t0HcivCPgnw/wCCdOFhodoFZgPOnfmSYjux/oOBVVVVVVVVVVVVVVVVoX/Ic07/AK+4v/QhVVVVVVVVVVVVVVVVVVVVVxPFbW8lzM2I4kLscZwAMmrq4ku7mW6lxvmdnbAwMk5OK8N6edK8O6VpZUqbOyhhwRgjagXp+FVVft66CNL+N0Wqony6zo0E7N6ujPER/wB8xp+Yr/gn1r3274V634fkk3SaXrTOoLfdjliQgY7Dckh/Gqqqqqqqqqqqqqqqqv8Agt//AMjT8Jf+wfqn/oy2qqqqqqqqqqqqqqqq13wZYa/ei+ubqeNxGE2pjGBn1HvX/CstI/6CF3/47/hX/CstI/6CF3/47/hX/CstI/6CF3/47/hX/CstI/6CF3/47/hX/CstI/6CF3/47/hX/CstI/6CF3/47/hWieCtP0K+F/bXdxI4QrtfGOfoKqqqqqq/4LV/8mr+Fv8AsoFn/wCkF/X/AATe/wCTI/hX/wBg+5/9LJ6/4KX/ALJC/tJfBeTxT4S0zzvHvgWKW70oRJ+8v7bAM9pwCWJVd8Y/vqFGA7GvAP7R7/EX/gm58WP2bfFtwbjX/Acem6j4daViXk03+1bUSoCTyYC7D/rnKoAwhr/gkN/yZppn/Ywaj/6GteK7yP4U/wDBXd9T8XzGC2HxIhneaY7FjhvdjRuSeiKk6HPTA7VX7RXirSfA/wABPiJ4r1u4SKz03wxfyOXcLvbyHCICf4mYqoHcsB3r/giH4bv7n4nfErxciv8AYtP0G1sZDjgyTzl1/HFu/wCdf8Fvf+SVfDT/ALGC6/8AScV+wH/yZp8Jf+xfj/8AQ3r9vz/kzT4s/wDYvyf+hpX/AAQ6/wCRU+Lf/YQ0r/0Xc1Xhr/lMHN/2VC9/9Dlr9rL/AJNX+Mv/AGT/AF3/ANIJq/4Idf8AI1fFv/sH6V/6Mua/4Le/8kq+Gn/YwXX/AKTiv+Cb3/Jkfwr/AOwfc/8ApZPX7Qv/ACQH4mf9ifq3/pHLX/BEP/kqfxL/AOxftf8A0oNf8FtNB1G3+P8A4G8Tyh/sF/4OFnASDt82C8neTB9dtxFn8K+EXinSvHHwp8HeMNDnSWx1nQrO7gZW3fK8KtjI7jOCOxBrU9RsdH0261fU7pLazsYHnuJpDhYo0UszH2ABNf8ABG3/AJO5u/8AsT77/wBHW1VVVVVVVVVVVVVVVVVVVVVVVVVXih2j8Nas8blWWxmKkHBBCHkVqPiXxHrAkXVvEGpXombdILm6eTeeuTuJyc1VV4V/5FfR/wDrwg/9FrVVVVVVVVVf8EuP+Tx/jN/14X//AKco6qqqqqqqqqqqqqqqqqqqq8ff8ilff9s//Ri1/wAFLdZOmfs4x2CyEf2t4htLYgE8hUll59sxDr7V+zFYi7+LdhcYz9itLiX6ZQp/7PXwr0kaP8P9FtSm15bcTvkYOZCX5/BgPwqqqqqqqqqtW0qw1zTbjSNTgWa2uoykiHuPUehBwQexArx34MvvA+vzaRdZkhPz2s5GBLGeh+o6Edj7Yr4K/ECDxJpZ8E+IXSa6ghKwecAwuYMYKEHqQOMd1+hr9sH9gmXQ/t3xR+BWlPNp3zTan4cgXc9r3aS1UcmPuYuq/wAOV+Vfgr8b/HnwG8YReLvA2pbC21L6xmy0F9EDkxyL+eGGGUnII5r9n39o3wD+0T4VGteFrn7JqtoijVNGnkBnsnPfj78ZOdsg4PcKwKiqqqqqrwh/yLOnf9cBVVVVVVVVVVVVVVVVVVVVXxi/5Po8I/8AX1pf/oVV/wAFL/ES+I/20PH5ilDwaZ9isYuc48uzh3jqR/rDJ6e4zmv+Cd/gweID4G094d8V7rEt9MuOGSF2LA+xEOPxr4g/CXwF8T7MW/i3Q45541Kw3kR8ueH/AHXHOP8AZOR7V8UP2RPG/hHzdT8Fu3iXTFJPlRx7buIe8f8Ay09Mpyf7orw/4n8VeBNZ/tHw7q17pGo27FXMbFDkHlXU8MMjlWBHqK+GP7aFncCLSvinpwtpOFGqWMRKH3ki5I+qZ/3RWi67oviTTotX8P6ta6jZTD5J7aUSKfUZHf1HUV/wUL/YNs/2hNFuPiv8MbGK3+I+kWv722jUIuvQoOImPGJ1AwjnqMI3G0p8MviX8Rv2fviVZeN/Bd9c6J4k0C5aOSGeIrnB2y288RwSpwVZDg/QgEfsn/tVeBP2q/h1B4o8OTxWWv2EaR6/obyZl0+4I5I7tCxBKSdwCDhlZRVVVVVVVVVVVVX/AAU1/wCTH/iT/wBwr/062lf8Es/+TL/CP/X/AKl/6WS1VVVVVVVVVT/f/Cqqqqqqqqqr9vj/AJM4+LH/AGL8n/oaV/wR0/5NIu/+xvvv/RNvVVVVVVVVVVVVVVVVVVVVXxi/4+dL/wCucv8ANarxN4l0Hwb4f1DxT4n1SDTtK0u3ae7uZmwsaKOT7nsAOSSAMk1+1Z+0rrv7R3j59SDT2fhfSmaLQ9NdvuJ3mkA4818An+6MKCcZP7NnwUa+ltviH4ost0SsG0m0dcmRu0xHoP4R3PPYZ+G/wGuL8Q6143ieC34aPT+Vkk/66Hqo/wBnr64q1tLWxto7Oyt44IIVCxxRqFVAOgAHQVVVVVVVVVVVVVVVVVoX/Ic07/r7i/8AQhVVVVVVVVVVVVVVVVVVVVXxH1AaT8PPFGqbwv2PRbybJbbjZC5znt0614b08at4i0rSyob7Zeww4Iznc4Xp+NVVV/wUW0HdbeCvE8Sfce7s5mx6iN0H/jslf8E7deMHi/xd4XLn/TtNhvAv/XGQoT/5HFVVVVVVVVVVVVVVVVf8Fv8A/kafhL/2D9U/9GW1VVVVVVVVVVVVVVVVVVVVVVVVVVV/wWr/AOTV/C3/AGUCz/8ASC/r/gm9/wAmR/Cv/sH3P/pZPVf8FSv2Vbr4A/FiT4v/AA+s3s/BXxFeUXEVsu2Oxv2G6eAgcCOUZlQdM+YAAEFf8Ehv+TNNM/7GDUf/AENa/wCCoX7CHif45fZfjx8GNKN/4w0eyFtq+jwgLLqltGSY5Yem6eMFht6um0L8yKrfAj/gr9rnww8Kw/Db9pv4Z+I9V8ReHV+yNqlqyRXk2zgLdQTbNsqgAF92W6lQck/tEftffHn/AIKPX9n8A/2d/hRrNl4YkuUmvrZJBLLesrZje8mAEVvAhwdpbbuAJYkKB+xV+ytpX7JPwYtfAS3sGpeIdRnN/wCINRiQqk90yhdkefm8qNVVFzjPzNhS5A/4Kz/A3xV8Yf2brXW/BGj3Oq6r4H1hdSms7aMySyWbRPHOUQcsVLRucfwo1fsk/wDBV2w/Z/8AgjpHwd8e/C/U/EE3hnzYdN1Cy1BIjJA0juscqOvylC20MCcqBwCuW/bY1u38TfsJfEXxHaxPHDqvhFbuNH+8qyeW4B98Gv8Agh1/yKnxb/7CGlf+i7mq8Nf8pg5v+yoXv/octftZf8mr/GX/ALJ/rv8A6QTV/wAEOv8Akavi3/2D9K/9GXNf8Fvf+SVfDT/sYLr/ANJxX/BN7/kyP4V/9g+5/wDSyev2hf8AkgPxM/7E/Vv/AEjlr/giH/yVP4l/9i/a/wDpQa/b0/ZItv2uPg5/wj+kTWtn4y8OzNe+HLy44TzCoEtvI3VY5VCgkdGSNuQpB+An7bX7SX/BPi3PwK+N/wAHtQ1LQLGeRrDT9TlexuLPcxL/AGa42vHNCWJbADAliVcA1qv7UP7Un/BSGzPwV+Bfwjufh34B1rbB4q8WXdy91stCR5iLLsjQbgGUwpueQcblUvj/AII0wTTftbahJFGzrD4MvnkIH3F8+1XJ9ssB+IqqqqqqqqqqqqqqqqqqqqqqqqqqnghuYJLa4jWSKVCjowyGBGCCPTFWv7J37LlpAttD+zl8M2VM4MvhSzkbk55ZoyT+Jr/hlf8AZg/6Nw+F3/hH2P8A8ar/AIZX/Zg/6Nw+F3/hH2P/AMar/hlf9mD/AKNw+F3/AIR9j/8AGqihigiSCCNI441CoiLtCgDAAA6DFVVVVVVVVV/wS4/5PH+M3/Xhf/8Apyjqqqqqqqqqqqqqqqqqqqqrx9/yKV9/2z/9GLX/AAVU1gQeDPAPh7eP9N1O7utvr5MSLn1/5b/r+X7K8UsPjPUtXgID2lhsUldwBaRTyCCDwh4PXnrXwZ+K2jfEfTYtEZYrDxBZQASWY+VLhVGN8P4AZQ8j3HNOjIxR1IYdQRjFVVVVVVVVV8RvA1p478Pvp7bY72DL2cxH3H9D/stjB/A9qB1fwxrWQZbLUdOn+jRup/z7GvBPxh8LeJ7S3t9Q1CHT9UKASwTHYjP3KMeCD2Gc81+2B+wjY+PRe/E/4L2ENn4lbdNqOjJhItTPUyRdo5j3HCueeGyW8HeMvHnwd8bQ+IvDGoXugeIdHmaN1ZCjowOHiljYcg4wyMMeor9ln9rzwh+0TpCaRfeRovjazhzeaSZPluABzNbk8sncryydDkYY1VVVVXhD/kWdO/64CqqqqqqqqqqqqqqqqqqqqvjF/wAn0eEf+vrS/wD0Kq/ao8RHxZ+0r8UteE3mx3Pi/UhA+c5iS5dI+5/gVe+PTiv+CXfg5W8O2XiZ4h5WmaFHFHkjiW4YSEjn+6D/AN91VfEr4GfDz4pQtJr2kLb6ljCalZgRTjjjccEOPZgfbFfFD9lj4heAPO1LR4D4j0dCT59nGfOiX/ppFyR9V3DuSK8GfEDxl8PNS/tLwhr1zp0xI82NDmOXHZ0OVYdeo47Yr4Yftk+HNc8rSviRZLol4QFF/AGe2kPqy8tH/wCPDuSK/wCChn7CWm/G3Rrn9oj4D2kd54tS3E2p6fp5WRNegUY86LacG4VR0GfMAwPmA3fA744fEX9nH4j2XxD+HuotZ6lZExXdpMCYb2AkeZbzpxuQ7R6FSAwIYAj9m79o74e/tO/Dm18feBL3ZKm2LVdLmYefptzjJjkA6g9VccMOeCCBVVVVVVVVVVVV/wAFNf8Akx/4k/8AcK/9OtpX/BLP/ky/wj/1/wCpf+lktVVVVVVVVVU/3/wqqqq+N/x2+GH7O3gh/iB8V/EP9k6T54toCtu80lzcMrMsMaICS7KjkZwMKSSACR8bP+Cxfxg1zUmn+A3gWx8NeF7a4MSahrVqby5u2wSoYAiKElefLG88Z34yK/ZV+IviX4u/s7eAviV4xmt5Na8QaStzevbwiJDIWYHao6DAHFV+3Z/wUP0b9lq4i+G/w/0i08S/Ea9gEr29wzG20mNx+7ecJhpHfgrEpU7TuLAFQ918W/8Ags/H4fPxXbwtfJ4d8v7V/ZqaBpTSCLGcfZdpvcY5xjd61+wb/wAFDtL/AGqJrj4dfEDSrDw78RLGBp44LVmW21WFfvvCHJZJE6tEWY7fmBIDBar9vj/kzj4sf9i/J/6Glf8ABHT/AJNIu/8Asb77/wBE29VVft7ftf8AiT9j/wAIeFvEnhvwfpniGXxBqU1pLFfTvEIgke8MCnUnpzX/AA+2+KX/AERHwr/4Mbj/AAr/AIfbfFL/AKIj4V/8GNx/hX/D7b4pf9ER8K/+DG4/wr/h9t8Uv+iI+Ff/AAY3H+Ff8Ptvil/0RHwr/wCDG4/wr/h9t8Uv+iI+Ff8AwY3H+Ff8Ptvil/0RHwr/AODG4/wr/h9t8Uv+iI+Ff/Bjcf4V/wAPtvil/wBER8K/+DG4/wAK/wCH23xS/wCiI+Ff/Bjcf4V/w+2+KX/REfCv/gxuP8K/4fbfFL/oiPhX/wAGNx/hX/D7b4pf9ER8K/8AgxuP8K/4fbfFL/oiPhX/AMGNx/hX/D7b4pf9ER8K/wDgxuP8K/4fbfFL/oiPhX/wY3H+Ff8AD7b4pf8AREfCv/gxuP8ACv8Ah9t8Uv8AoiPhX/wY3H+Ff8Ptvil/0RHwr/4Mbj/Cv+H23xS/6Ij4V/8ABjcf4V/w+2+KX/REfCv/AIMbj/Cv+H23xS/6Ij4V/wDBjcf4V/w+2+KX/REfCv8A4Mbj/Cv+H23xS/6Ij4V/8GNx/hX/AA+2+KX/AERHwr/4Mbj/AAr/AIfbfFL/AKIj4V/8GNx/hXiv/gsH8SPFcltJcfB3w1AbZWChL+c5zj1HtX/D1b4gf9Ep8Pf+B03+FftHfto/EX9ovQ7HwrqOm2fh/RLWXzriysJXYXko+40jNyVXnC9MnJyQuPgj8Kz421dNc1y1Y6DYSAyIfl+1uOfLB/u9Nx9OOpyPh58UPDXgMpdt8P11G/i4hmfUBGkCjoI08ohTjvkn0xX/AA1z/wBU+/8AKt/9pr/hrn/qn3/lW/8AtNf8Nc/9U+/8q3/2mv8Ahrn/AKp9/wCVb/7TX/DXP/VPv/Kt/wDaa/4a5/6p9/5Vv/tNf8Nc/wDVPv8Ayrf/AGmv+Guf+qff+Vb/AO01/wANc/8AVPv/ACrf/aa/4a5/6p9/5Vv/ALTX/DXP/VPv/Kt/9pr/AIa5/wCqff8AlW/+01/w1z/1T7/yrf8A2mv+Guf+qff+Vb/7TX/DXP8A1T7/AMq3/wBpr/hrn/qn3/lW/wDtNf8ADXP/AFT7/wAq3/2mv+Guf+qff+Vb/wC01/w1z/1T7/yrf/aa/wCGuf8Aqn3/AJVv/tNf8Nc/9U+/8q3/ANpr/hrn/qn3/lW/+01/w1z/ANU+/wDKt/8Aaa/4a5/6p9/5Vv8A7TX/AA1z/wBU+/8AKt/9pr/hrn/qn3/lW/8AtNWH7YX2K+t73/hXe/7PKsm3+18Z2kHGfJ46V/w8c/6o3/5cX/3NX/Dxz/qjf/lxf/c1f8PHP+qN/wDlxf8A3NX/AA8c/wCqN/8Alxf/AHNX/Dxz/qjf/lxf/c1f8PHP+qN/+XF/9zV/w8c/6o3/AOXF/wDc1f8ADxz/AKo3/wCXF/8Ac1f8PHP+qN/+XF/9zV/w8c/6o3/5cX/3NX/Dxz/qjf8A5cX/ANzV/wAPHP8Aqjf/AJcX/wBzV/w8c/6o3/5cX/3NX/Dxz/qjf/lxf/c1f8PHP+qN/wDlxf8A3NX/AA8c/wCqN/8Alxf/AHNX/Dxz/qjf/lxf/c1f8PHP+qN/+XF/9zV/w8c/6o3/AOXF/wDc1f8ADxz/AKo3/wCXF/8Ac1f8PHP+qN/+XF/9zV/w8c/6o3/5cX/3NX/Dxz/qjf8A5cX/ANzV/wAPHP8Aqjf/AJcX/wBzV/w8c/6o3/5cX/3NX/Dxz/qjf/lxf/c1f8PHP+qN/wDlxf8A3NX/AA8c/wCqN/8Alxf/AHNX/Dxz/qjf/lxf/c1f8PHP+qN/+XF/9zV4+/b2k8beCNe8HRfC3+z21vTp7I3X9ueb5QlQoW2fZxu4J4yPqK+EcYk+Kng4NH5iprtm7pu27lWZCRntwDX/AAsr/qC/+TP/ANjX/Cyv+oL/AOTP/wBjX/Cyv+oL/wCTP/2Nf8LK/wCoL/5M/wD2Nfto6wnjD4Ms39lCFtI1S3uhJ5u4gHdER90cHzR+lfAz4wz/AAQ8fx+NYdDGrqLSW2ltDdfZ/MVwP49rYwyqfunOO3Wv+HkX/VGf/Lj/APuav+HkX/VGf/Lj/wDuav8Ah5F/1Rn/AMuP/wC5q/4eRf8AVGf/AC4//uav+HkX/VGf/Lj/APuav+HkX/VGf/Lj/wDuav8Ah5F/1Rn/AMuP/wC5q/4eRf8AVGf/AC4//uav+HkX/VGf/Lj/APuav+HkX/VGf/Lj/wDuav8Ah5F/1Rn/AMuP/wC5q/4eRf8AVGf/AC4//uav+HkX/VGf/Lj/APuav+HkX/VGf/Lj/wDuav8Ah5F/1Rn/AMuP/wC5q/4eRf8AVGf/AC4//uav+HkX/VGf/Lj/APuav+HkX/VGf/Lj/wDuav8Ah5F/1Rn/AMuP/wC5q/4eRf8AVGf/AC4//uav+HkX/VGf/Lj/APuav+HkX/VGf/Lj/wDuav8Ah5F/1Rn/AMuP/wC5q/4eRf8AVGf/AC4//uav+CmH7RH/AA0FrngG8/4Q/wDsH+xbS+j2/wBofavN8x4TnPlptxs985qqqqqqqqqqqqqqqqqqqqqqqqqqqr7TtP1OEW+pWNvdxq25UniEgDYxnBzzgn86trW1soEtbO3ighjGEjiQIq/QDgVV7p9hqUIttRsre6iDbhHPEHXI74Oeas7Gx06AW2n2cFtECSI4Ywi5PU4FV4n+G3w68byJL4z8A+HNfePGxtU0qG6K46YMinHU/nWh+H9A8MadHpHhrRNP0mxiP7u1sbZIIk+iIAB09Kqrv4RfCfUNbbxLffDDwlc6uzF21CbRLd7gsepMhTdnk8571Pa2t1btaXNvFLA67WikQMpHoQeMVY6Vpelq66ZptraCQjeIIVj3Y6ZwOaoaDoa3n9oro1iLvfv88Wy+Zu9d2M596mhhuIXt54kljkUq6OuQwIwQQeoI7VY6PpGll20zS7O0MgAcwQLHux0zgc1faXpmqIkepada3aocos8KyBT6gEHFW1ra2UCWtnbxQQxjCRxIEVfoBwKlijmjaGWNXR1KsjDIYHqCPSrHRdH0t2k0zSrO0ZxhmggWMsPfArxn4z8K/Dzwxf8AjPxvrtro2h6XGJLy+um2xwKWCgsewywH41o+ueDfiBocWqeH9Y0XxJo90A0VxZ3EV5byjqCGUsrV+2L+1d8Pf2XvhLreoXviKwHi+4sJIPDuhxTqbme5dSsb+WDuWJCQzOcABcA7ioP/AARO+DWsW0/jn496pYNBYXVsmgaRM8eDc/vFmuipP8KmO3XI4Lbh1Q1VVVVVVVVVVVVVVVVVVVVVVVVX/BRb9rn453f7RniX4aeFvHuv+E/Dfg+eO0trLSb6Sya5k8tHeeZo2DSEsx2gnaFC4GSSf+CbX7R/xK+N/wAAPGmnfEbWbzVtY8EuYbXWp2PnXEMsDMgkkzl5UZGy55IZM5OSf+GqP2nv+jkPij/4WF//APHa/wCGqP2nv+jkPij/AOFhf/8Ax2v+GqP2nv8Ao5D4o/8AhYX/AP8AHa/4ao/ae/6OQ+KP/hYX/wD8drw1LLceHNKnnlaSSSyhZ3ckliUGST3NVVVVVVVVV/wS4/5PH+M3/Xhf/wDpyjqqqqqqqqqqqqqqqqqqqqrx9/yKV9/2z/8ARi1/wVR1nz/HvgXw9vz9i0e4utvp50wXPX/ph6du/b9h74fal491PxHYaVJEk7RIUeT7o8tXbB9NxZBntnNTQ+IfBfiDy5Bc6Xq+lzAgg7Xiccggj8wRwQe4NfBb4zaT8WNOXRNeMVr4mtY/nVflF0o6yRj1/vL26jjpeadPaHcRvTswH8/Sqqqqqqqqq+KPwhj8bzprOj3MNpqaJtk81TsuAPu7iOQR0zg8Y9K8S+CPFHhKXy9d0iaBCcLMBvjb6OMj8OteF/iX4w8IlItL1V5LVP8Al1uP3kePQA8r/wABIr9or4YfDb9oyxk8TWtlD4Q+IUMXy3yputNU2jAjuNo3K3QLJglRgMSAMSxeL/hl4vCia80PxDodyGSWCbbJBKp4ZHQ4I7hlJBByCQa/ZD/bh0j4wxWfw8+JtxbaX42VRHbXOBHBrGOm3skx7p0Y8r12rVVVVXhD/kWdO/64CqqqqqqqqqqqqqqqqqqqqvEv7O/gnxV8V9N+MOoaprcetaXJA8MEM8S27GE5XcpjLc98MPwrUL620ywudSvJNkFpC8srf3VUEk/kK1jU7nW9WvdZvW3XF/cyXEpyTl3Ys3XnqTX7MPxIufg14m0mCW8aDSri3hsr4bsKgUBUkPX7p69flLVoXiK21aNYpGVLjH3c8P7r/hVVXxQ/Zy+HXxOE19PYDSdakGRqVkoVmb1kT7sn1Pzf7Qr4n/s4/EX4ZGa+msP7X0aME/2jYoWVF9ZE+9H7k5X/AGjXw/8Aix48+GN59p8Ja7LBCzZms5f3lvN/vRnjP+0MN7iv2z/hp4O+K7XXxu+HfhttB8YSsZfE2i2432upE/eu7fukueXjIwwO4Evu3/A/9oD4rfs6+LP+Ew+FPieTTLqVFjvLd0Etvexg52TRHhh1weGXJ2kE5r9mv/gq98I/if8AZPDPxqtovh/4jkCoL1pGk0u6k6EiQ5a3yecS5UD/AJaE1Z3lnqNpDfafdw3NtcRiSGaFw6SKRkMrDggjuKqqqqqqqqqqr/gpr/yY/wDEn/uFf+nW0r/gln/yZf4R/wCv/Uv/AEslqqqqqqqqqqn+/wDhVVVV8afgd8N/2gvB8XgP4p6K+qaJFqEF8bZJ3hLSQkkAshDBSCynBBwx5HWv+Cw3gvwj8PvgX8KvCXgbw1pug6NY67dLb2On2ywRR/6OMkKoAyepJ5J5JJr9gf8A5M4+E/8A2L8f/ob14k17T/C3h3VPE+rS+XY6PZTXly+cbY4kLuf++VNf8E7vDFx+1h+3Hr/xs+JlumoHRhc+JZoZv3kf2ySVUtowD/DFvLIOg8hBjHFV+2/oz/sff8FCNO+Kvga1NjaX1xZ+Kre3tlCqd8jR3kQ7YkeKfK8cTY4GKtbmC9tYby1kEkM8YeNwMblIyD+VV+3x/wAmcfFj/sX5P/Q0r/gjp/yaRd/9jfff+ibeqqv23/2O5f2xfCnhrw1F48TwwfD2oS3ZmbT/ALV52+PZtxvTGOuea/4cgXf/AEcRD/4TZ/8Aj9f8OQLv/o4iH/wmz/8AH6/4cgXf/RxEP/hNn/4/X/DkC7/6OIh/8Js//H6/4cgXf/RxEP8A4TZ/+P1/w5Au/wDo4iH/AMJs/wDx+v8AhyBd/wDRxEP/AITZ/wDj9f8ADkC7/wCjiIf/AAmz/wDH6/4cgXf/AEcRD/4TZ/8Aj9f8OQLv/o4iH/wmz/8AH6/4cgXf/RxEP/hNn/4/X/DkC7/6OIh/8Js//H6/4cgXf/RxEP8A4TZ/+P1/w5Au/wDo4iH/AMJs/wDx+v8AhyBd/wDRxEP/AITZ/wDj9f8ADkC7/wCjiIf/AAmz/wDH6/4cgXf/AEcRD/4TZ/8Aj9f8OQLv/o4iH/wmz/8AH6/4cgXf/RxEP/hNn/4/X/DkC7/6OIh/8Js//H6/4cgXf/RxEP8A4TZ/+P1/w5Au/wDo4iH/AMJs/wDx+v8AhyBd/wDRxEP/AITZ/wDj9f8ADkC7/wCjiIf/AAmz/wDH6/4cgXf/AEcRD/4TZ/8Aj9f8OQLv/o4iH/wmz/8AH68Xf8EdJvCcltHJ8eEuftKsQV8Pldu3H/Tc+tf8Ooj/ANFqH/gj/wDt1R/8Eo1DqZPjRuQEblGiYJHfB804rR/2MhoOl22j6R4utLe0tIwkUa2DcAdz8/JPUk8knNf8Mk6h/wBDxbf+ADf/ABdf8Mk6h/0PFt/4AN/8XX/DJOof9Dxbf+ADf/F1/wAMk6h/0PFt/wCADf8Axdf8Mk6h/wBDxbf+ADf/ABdf8Mk6h/0PFt/4AN/8XX/DJOof9Dxbf+ADf/F1/wAMk6h/0PFt/wCADf8Axdf8Mk6h/wBDxbf+ADf/ABdf8Mk6h/0PFt/4AN/8XX/DJOof9Dxbf+ADf/F1/wAMk6h/0PFt/wCADf8Axdf8Mk6h/wBDxbf+ADf/ABdf8Mk6h/0PFt/4AN/8XX/DJOof9Dxbf+ADf/F1/wAMk6h/0PFt/wCADf8Axdf8Mk6h/wBDxbf+ADf/ABdf8Mk6h/0PFt/4AN/8XX/DJOof9Dxbf+ADf/F1/wAMk6h/0PFt/wCADf8Axdf8Mk6h/wBDxbf+ADf/ABdf8Mk6h/0PFt/4AN/8XX/DJOof9Dxbf+ADf/F1/wAMk6h/0PFt/wCADf8Axdf8Mk6h/wBDxbf+ADf/ABdf8Mk6h/0PFt/4AN/8XX/DJOof9Dxbf+ADf/F1Y/sgajfXtvZL46tlM8qxhjp7HGSBn7/vX/DufWv+iq2X/gob/wCO1/w7n1r/AKKrZf8Agob/AOO1/wAO59a/6KrZf+Chv/jtf8O59a/6KrZf+Chv/jtf8O59a/6KrZf+Chv/AI7X/DufWv8Aoqtl/wCChv8A47X/AA7n1r/oqtl/4KG/+O1/w7n1r/oqtl/4KG/+O1/w7n1r/oqtl/4KG/8Ajtf8O59a/wCiq2X/AIKG/wDjtf8ADufWv+iq2X/gob/47X/DufWv+iq2X/gob/47X/DufWv+iq2X/gob/wCO1/w7n1r/AKKrZf8Agob/AOO1/wAO59a/6KrZf+Chv/jtf8O59a/6KrZf+Chv/jtf8O59a/6KrZf+Chv/AI7X/DufWv8Aoqtl/wCChv8A47X/AA7n1r/oqtl/4KG/+O1/w7n1r/oqtl/4KG/+O1/w7n1r/oqtl/4KG/8Ajtf8O59a/wCiq2X/AIKG/wDjtf8ADufWv+iq2X/gob/47X/DufWv+iq2X/gob/47X/DufWv+iq2X/gob/wCO1/w7n1r/AKKrZf8Agob/AOO1/wAO59a/6KrZf+Chv/jtf8O59a/6KrZf+Chv/jtf8O59a/6KrZf+Chv/AI7X/DufWv8Aoqtl/wCChv8A47X/AA7n1r/oqtl/4KG/+O14C/YN1Pwb4x0nxRcfEi0vE024EpgXS2QuQDjkyHHOO1f8K2n/AOgvH/35P+Nf8K2n/wCgvH/35P8AjX/Ctp/+gvH/AN+T/jX/AAraf/oLx/8Afk/418QPgVJ458G6r4TbX4rc6jBsWVrYsI2BDK2NwzggcU//AATk1pjkfFax/wDBQ3/x2v8Ah3FrX/RV7H/wUN/8dr/h3FrX/RV7H/wUN/8AHa/4dxa1/wBFXsf/AAUN/wDHa/4dxa1/0Vex/wDBQ3/x2v8Ah3FrX/RV7H/wUN/8dr/h3FrX/RV7H/wUN/8AHa/4dxa1/wBFXsf/AAUN/wDHa/4dxa1/0Vex/wDBQ3/x2v8Ah3FrX/RV7H/wUN/8dr/h3FrX/RV7H/wUN/8AHa/4dxa1/wBFXsf/AAUN/wDHa/4dxa1/0Vex/wDBQ3/x2v8Ah3FrX/RV7H/wUN/8dr/h3FrX/RV7H/wUN/8AHa/4dxa1/wBFXsf/AAUN/wDHa/4dxa1/0Vex/wDBQ3/x2v8Ah3FrX/RV7H/wUN/8dr/h3FrX/RV7H/wUN/8AHa/4dxa1/wBFXsf/AAUN/wDHa/4dxa1/0Vex/wDBQ3/x2v8Ah3FrX/RV7H/wUN/8dr/h3FrX/RV7H/wUN/8AHa/4dxa1/wBFXsf/AAUN/wDHa/4dxa1/0Vex/wDBQ3/x2v8AgpR+zzefADW/AdneeKYdaOs2l9IrR2hg8ry2hGDlmznf7dKqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr47/BLwZ+0T8MNW+Evj+XU49F1gxGZ9Ouvs8ytHIsiMpIZTh1U4ZWXIGQcVrX/BE+706+kn+HH7TN/p1rMcGC90ImQAE4zJFOofr/AHB3PfFfDH/gix8NNH1qPXPjL8XNb8aqsglk0+xsv7MimPdZZTJLK6k55Qxt7ivCfhLwz4E8N6d4P8G6FZaNoukwCCysLOERRQIOyqPfJJ6kkk5JJqqqqqqqqqqqqqqqqqqqqqqqqqv2n/8AgnN8HP2nPGq/EfUNb1nwx4jlhSG/udNEbx3yoAqNIjqfnCgKGBHAAIOBXwI/Zp+G37O3wtl+FXw+hvBZ3hlkv7+7dXubyeRAjSuwAXIUKAAAAFHHUn/hzH8CP+ipePf++rT/AOM1/wAOY/gR/wBFS8e/99Wn/wAZr/hzH8CP+ipePf8Avq0/+M1/w5j+BH/RUvHv/fVp/wDGa06yj03TrXToXZ0tYUiVm6kKoAJ/Kqqqqqqqqq/4Jcf8nj/Gb/rwv/8A05R1VVVVVVVVVVVVVVVVVVVV4+/5FK+/7Z/+jFr/AIKT6x/an7STWO/P9kaBZ2uM9MmSb0/6be/9B/wS48NeX4P8U+K5YsNJfC2hcjqpVCw/NF/Ovi38JNN+I+m/aLby7XW7VCLa5IwHH/POTHVT2PVSc+oNzba/4O15oJhc6ZqumzcFWKPE46EEfgQRwQc18CPjvZfEeyTw54jlit/ElvHyMBVvlA5dB2bH3l/EcZC3ujRyZktCEb+4eh+npUsUkLmOVCrDsRVVVVVVVVU0MNxE0E8SSRupVkdchh3BHpXir4GeDtfD3Glxto923RrdcxE+8Z4/75K14p+DvjXwwHnFh/aVon/LezBfA9Sn3h+WPevit8IPD/xP0wx3aC01e2Qi0vlT5k/2HH8SZ7dR1GOc+LPCPiT4fa++ja7avaXlu2+KVCdsgB4kjbjI469QeuCCK/Y+/b2j1L7D8LvjvrCpd/LDpniS4cKs3ZYro9A3QCU8H+PByzAhgCpBBHBFVVVXhD/kWdO/64Cqqqqqqqqqqqqqqqqqqqqq/aa8Qnwn+zn8TvEUblZbHwjqbwkf89fs0gT/AMeK14XsTqniXSNMVdxu76GED13OB/Wvir8MNf8AhL4qPhbxAu53toriCYIVWWN16jP91gyH3U1+y/8AE9vF3hMeGNSuydY8PKqoxbDS2/RG+q/cP0U9TWheKFuNtnqThZOiSngN9fQ1VUQCMEA8dDXxP/ZZ+HfxAMupaTAPDusOCftFlEBFI3rJFwD9V2k9ya+JfwK+InwtkabXtINxpu7CalZ5lgPpuOMofZgM9s18RP2evA3jrzb6zgGi6q4J+02kYCSN6yR8BvcjBPrXxA+Dfjj4dO82r6b9o08NhL+1y8R9N3dD7MB7Zr9nb9tD48/sz3UUHgTxU174fDlpvDuqlrixkyfmKpkNCx67oypJxnI4r9mr/gpX8Bvj0tn4e8S36+AvF822Madq04FtdSHjFvc8K2TgBX2OScAN1qqqqqqqqqqr/gpr/wAmP/En/uFf+nW0r/gln/yZf4R/6/8AUv8A0slqqqqqqqqqqn+/+FVVVVf8Ftv+SWfDb/sYLr/0nFfsD/8AJnHwn/7F+P8A9Dev2w9QuNM/ZR+L13akeZ/whWqRgnPAe2dCRg9QGJHviv8Agh1aW73XxmvWjzPFHocaNk8Kxvywx05KL+X1qv8AgtpbwL8WvhxdLGBNJ4cuEZ+5VbgkD8CzfnXwLuri/wDgj8Pb27x59x4V0ySTC4+ZrWMnjtyar9vj/kzj4sf9i/J/6GlfsjfGn9vr4f8Awpl0H9mj4aXuveEW1aeZ7qHwy1+oumSMSL5g9FWP5e2fev8Ahp//AIK+/wDRDdV/8IZ6/wCGn/8Agr7/ANEN1X/whnr/AIaf/wCCvv8A0Q3Vf/CGevhzf+JNV+HvhfVPGVo1rr95o1pNqsDQ+UYrpoUMylP4cOWG3t0qqqqqqqqqqqqqqqqqq+MX/Hzpf/XOX+a1VVVVVVVVVVVVVVVVVVVVoX/Ic07/AK+4v/QhVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVftceLfjX4H+Bms+I/2e/D82teN7e4tVsrKLTzes6NMiykRDriMsc9sZr/AIaf/wCCvv8A0Q3Vf/CGev8Ahp//AIK+/wDRDdV/8IZ6/wCGn/8Agr7/ANEN1X/whnr9tL4l/tZ/EjU/Cs/7VXgq68O3dhb3S6Kk+iHTvORmj80gH7+CsfPbPvVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX/AAVc+PHxo+FHxk8I6L8NPij4k8M2N34YFxPbaZqD26Sy/aZl3sFIydqqM+gFf8Enfiz8TPiz8O/Heo/Ezx5rXie6sNat4rWbVLx7hoUMJJVSxOATziv21fiz4w+Bn7MvjL4p+AZ7WLXtE+w/ZHuoBNGPNvreF8oevySt9Dg1+yH/AMFFv2lfjN+0f4J+GXjbVfD8uia7dTRXiW2krFIVW3lkG1gePmRaqqqqqqqqqqr/AIJcf8nj/Gb/AK8L/wD9OUdVVftYePPFPww/Zy8feP8AwTqS2Gu6HpD3FjctAkwikDKM7HBVuCeCCK/4egftsf8ARW7X/wAJ3Tv/AIxX/D0D9tj/AKK3a/8AhO6d/wDGK/4egftsf9Fbtf8AwndO/wDjFf8AD0D9tj/ordr/AOE7p3/xiv8Ah6B+2x/0Vu1/8J3Tv/jFf8PQP22P+it2v/hO6d/8Yr/h6B+2x/0Vu1/8J3Tv/jFf8PQP22P+it2v/hO6d/8AGK/4egftsf8ARW7X/wAJ3Tv/AIxX/D0D9tj/AKK3a/8AhO6d/wDGK/4egftsf9Fbtf8AwndO/wDjFf8AD0D9tj/ordr/AOE7p3/xiv8Ah6B+2x/0Vu1/8J3Tv/jFf8PQP22P+it2v/hO6d/8Yr/h6B+2x/0Vu1/8J3Tv/jFf8PQP22P+it2v/hO6d/8AGK/4egftsf8ARW7X/wAJ3Tv/AIxX/D0D9tj/AKK3a/8AhO6d/wDGK/4egftsf9Fbtf8AwndO/wDjFf8AD0D9tj/ordr/AOE7p3/xiv8Ah6B+2x/0Vu1/8J3Tv/jFf8PQP22P+it2v/hO6d/8Yr/h6B+2x/0Vu1/8J3Tv/jFf8PQP22P+it2v/hO6d/8AGK/4egftsf8ARW7X/wAJ3Tv/AIxX/D0D9tj/AKK3a/8AhO6d/wDGK1D/AIKYftm6pZvYX3xWtZYJMb0/4R6wXOCCORDnqBXxE+Ifi34reL77x3451NdQ1rURGLidYUhDeXGsa4RAFGFRRwB69Sa/4J8eGk0D9mDw/elAJtbu7u9k9f8AXNEv/jsSn8ar4q/CbSPiRphkRY7XWrdP9FvNvXH8D45KH8weR3Bv7DxB4K8QNaXSXGm6rpswKlWKvG45VlYfgQR1618DP2gbL4gQxeGfFMsVp4iiTCP91L4AcsvZX7lfxHGQLi1guk2TID6HuPpV7pU9sC8f7yMdwOR9RVVVVVVVVVV4p+Gng7xcHk1TSkS6b/l6t/3cufUkcN/wIGvi3+xbcfELSm0mzWLVYdxNtOxEFxaMeNysflI9RnnH3a+NXwY8TfA/xpd+D/EF3Y6gsD4h1DT5fNgm45Ab+F16Mh5Ug9RgnwX+2l+0n4B8NWXhLw78RpP7N05PLtUvLG3uniQdEEkqM20DgAnAGAMAAV/w8C/as/6KLa/+CSz/APjVf8PAv2rP+ii2v/gks/8A41X/AA8C/as/6KLa/wDgks//AI1X/DwL9qz/AKKLa/8Agks//jVf8PAv2rP+ii2v/gks/wD41Vn/AMFNP20LC1jsrT4r2qQwrtRf+Ee084H1MGa/4egftsf9Fbtf/Cd07/4xX/D0D9tj/ordr/4Tunf/ABiv+HoH7bH/AEVu1/8ACd07/wCMV/w9A/bY/wCit2v/AITunf8Axiv+HoH7bH/RW7X/AMJ3Tv8A4xX/AA9A/bY/6K3a/wDhO6d/8Yr/AIegftsf9Fbtf/Cd07/4xX/D0D9tj/ordr/4Tunf/GK/4egftsf9Fbtf/Cd07/4xX/D0D9tj/ordr/4Tunf/ABiv+HoH7bH/AEVu1/8ACd07/wCMV/w9A/bY/wCit2v/AITunf8Axiv+HoH7bH/RW7X/AMJ3Tv8A4xX/AA9A/bY/6K3a/wDhO6d/8Yr/AIegftsf9Fbtf/Cd07/4xX/D0D9tj/ordr/4Tunf/GK/4egftsf9Fbtf/Cd07/4xX/D0D9tj/ordr/4Tunf/ABiv+HoH7bH/AEVu1/8ACd07/wCMV/w9A/bY/wCit2v/AITunf8Axiv+HoH7bH/RW7X/AMJ3Tv8A4xX/AA9A/bY/6K3a/wDhO6d/8Yr/AIegftsf9Fbtf/Cd07/4xX/D0D9tj/ordr/4Tunf/GK/4egftsf9Fbtf/Cd07/4xX/D0D9tj/ordr/4Tunf/ABiv+HoH7bH/AEVu1/8ACd07/wCMV/w9A/bY/wCit2v/AITunf8Axiv+HoH7bH/RW7X/AMJ3Tv8A4xX/AA9A/bY/6K3a/wDhO6d/8Yr/AIegftsf9Fbtf/Cd07/4xXxB/wCCgX7V/wAUvBerfD7xt8S4L7Q9bt/s99bJollCZY8gkb44Qy8gdCK/Yr8ED4iftV/DLwq8Qlik12O6nQj70VsrXEg/74hav2svg8fij8On1PSLXzNf8Nh7mzCj5p4sfvYfckKGA67lA/iNfDrxtffD3xhp3imxLMLWTFxCDjzoTw6fiOnocHtVncWup6VY65plyl1p2pQLPaXCfdkRhkfQ84IPIPFaD4ne022eoMXh6K/Up/iKjkjljWSJw6sMqQcg1VEgDJI4HetV8VadEWtIrZb1GUrICcIQeCOhz/Kv2ide+BPhbX49Cs/hQW1SWEy3Uml6ubIW277gCeU8eSMnGwYGOua1PXdMundLDSZorZwQYrm5WclT2JCKD37V8Qv2fvCficyaj4UhTQdQbkxR820h906p/wAB4/2TXi7wF4p8D3X2bxBpbwoxIiuE+aKX/dYcfgcH1FfC/wDbr/ar+D3hqLwf4H+Ll7Fo9vgW1rf2lvfi3UfwxtcRuyJ/sqQo9K/4egftsf8ARW7X/wAJ3Tv/AIxX/D0D9tj/AKK3a/8AhO6d/wDGK/4egftsf9Fbtf8AwndO/wDjFf8AD0D9tj/ordr/AOE7p3/xiv8Ah6B+2x/0Vu1/8J3Tv/jFf8PQP22P+it2v/hO6d/8Yr/h6B+2x/0Vu1/8J3Tv/jFf8PQP22P+it2v/hO6d/8AGK/4egftsf8ARW7X/wAJ3Tv/AIxX/D0D9tj/AKK3a/8AhO6d/wDGK/4egftsf9Fbtf8AwndO/wDjFf8AD0D9tj/ordr/AOE7p3/xiv8Ah6B+2x/0Vu1/8J3Tv/jFf8PQP22P+it2v/hO6d/8Yr4pft4ftRfGjwHqfw0+IvxEg1Pw9rHlfbLRdFs7cyeVKkyfPHErjDxoeCM4weCa/wCCWf8AyZf4R/6/9S/9LJaqqqqqqqqqp/v/AIVVVVV/wW2/5JZ8Nv8AsYLr/wBJxX7A/wDyZx8J/wDsX4//AEN6+MnhCf4hfCDxz4BtMef4l8N6hpkWTj5p7aSIc9uXFf8ABFTxnBovxd+Inw4vJDBca7odvexxyfLuezmZGXB/iAumOPRW9Kr/AIK8a5L8Q/2tvC3wx8Nt9rvtK0GzsDAGPF7dzu6pjsTG9uf+BCvDmiweG/Dul+HbaUyRaVZQ2sbsMFljQIDjnstV+3x/yZx8WP8AsX5P/Q0r/gjp/wAmkXf/AGN99/6Jt6qqqqqqqqqqqqqqqqqqqq+MX/Hzpf8A1zl/mtVVVVVVVVVVVVVVVVVVVVaF/wAhzTv+vuL/ANCFVVVVVVVVVVVVVVVVVVVVV+wPf3158TvH0d3ezzKlqu1ZJCwX9+emaqqqqqqqqqqqqqqqqqqqq/4Lf/8AI0/CX/sH6p/6Mtqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/4LOf8l38Ef8AYoj/ANK56/4It/8AJL/iN/2Hrb/0Qa/4Ka/8mP8AxJ/7hX/p1tK/4J2/8nofDD/r/uf/AEjnqqqqqqqqqqqv+CXH/J4/xm/68L//ANOUdVVfEv4eeG/iz4D1r4ceL4p5NG1+1NteJBKYnKEgnaw6HgV/w6b/AGP/APoC+KP/AAeP/hX/AA6b/Y//AOgL4o/8Hj/4V/w6b/Y//wCgL4o/8Hj/AOFf8Om/2P8A/oC+KP8AweP/AIV/w6b/AGP/APoC+KP/AAeP/hX/AA6b/Y//AOgL4o/8Hj/4V/w6b/Y//wCgL4o/8Hj/AOFf8Om/2P8A/oC+KP8AweP/AIV/w6b/AGP/APoC+KP/AAeP/hX/AA6b/Y//AOgL4o/8Hj/4V/w6b/Y//wCgL4o/8Hj/AOFf8Om/2P8A/oC+KP8AweP/AIV/w6b/AGP/APoC+KP/AAeP/hX/AA6b/Y//AOgL4o/8Hj/4V/w6b/Y//wCgL4o/8Hj/AOFf8Om/2P8A/oC+KP8AweP/AIV/w6b/AGP/APoC+KP/AAeP/hX/AA6b/Y//AOgL4o/8Hj/4V/w6b/Y//wCgL4o/8Hj/AOFf8Om/2P8A/oC+KP8AweP/AIV/w6b/AGP/APoC+KP/AAeP/hX/AA6b/Y//AOgL4o/8Hj/4V/w6b/Y//wCgL4o/8Hj/AOFf8Om/2P8A/oC+KP8AweP/AIV/w6b/AGP/APoC+KP/AAeP/hX/AA6b/Y//AOgL4o/8Hj/4V4r/AOCVn7JOkaBdajZaN4mE0OzaW1pyOXUHjHoa/wCHcX7Mv/QK8Qf+DZ/8K8CeCtB+HPg/SvA3hiKWPS9GtxBapNIZHCDnlj1PNVXxV+FOk/ErSxzHa6xbKfsl4V/8cfHJQ/mDyO4Os6Nr3gvXpNN1KGax1KwkBUqxUqRyrow7dCCK+Bfx/TxXbxeGfFFxGmswqBG7EKLxR3X0f1Xv1HcC3uYLpN8Mgb1HQir3SIbjMkBEcnpjg1PbzWz+XMhU9vf6VVVVVVVSqzkKgJJ6ACrfRrqYAyARL/tdfyq8XQvD2nzatrF5DBa2yb5Z7lwqIB39K+Mv7TGo+JxP4Z8ASS6fpByk16MpNdDuF7xof++iOuMkVpXwju/jN5nhJNFW9tJMfaJZQVjth2cv/Cw5xjk8471pX/BNr9na10+GDVYtfvbtV/ezLqLRBm74UDgfn9a/4dxfsy/9ArxB/wCDZ/8ACv8Ah3F+zL/0CvEH/g2f/Cv+HcX7Mv8A0CvEH/g2f/Cv+HcX7Mv/AECvEH/g2f8Awr/h3F+zL/0CvEH/AINn/wAK8P8A/BKj9kbUtEs7650bxOZZogzY1twM/TFf8Om/2P8A/oC+KP8AweP/AIV/w6b/AGP/APoC+KP/AAeP/hX/AA6b/Y//AOgL4o/8Hj/4V/w6b/Y//wCgL4o/8Hj/AOFf8Om/2P8A/oC+KP8AweP/AIV/w6b/AGP/APoC+KP/AAeP/hX/AA6b/Y//AOgL4o/8Hj/4V/w6b/Y//wCgL4o/8Hj/AOFf8Om/2P8A/oC+KP8AweP/AIV/w6b/AGP/APoC+KP/AAeP/hX/AA6b/Y//AOgL4o/8Hj/4V/w6b/Y//wCgL4o/8Hj/AOFf8Om/2P8A/oC+KP8AweP/AIV/w6b/AGP/APoC+KP/AAeP/hX/AA6b/Y//AOgL4o/8Hj/4V/w6b/Y//wCgL4o/8Hj/AOFf8Om/2P8A/oC+KP8AweP/AIV/w6b/AGP/APoC+KP/AAeP/hX/AA6b/Y//AOgL4o/8Hj/4V/w6b/Y//wCgL4o/8Hj/AOFf8Om/2P8A/oC+KP8AweP/AIV/w6b/AGP/APoC+KP/AAeP/hX/AA6b/Y//AOgL4o/8Hj/4V/w6b/Y//wCgL4o/8Hj/AOFf8Om/2P8A/oC+KP8AweP/AIV/w6b/AGP/APoC+KP/AAeP/hX/AA6b/Y//AOgL4o/8Hj/4V/w6b/Y//wCgL4o/8Hj/AOFf8Om/2P8A/oC+KP8AweP/AIV/w6b/AGP/APoC+KP/AAeP/hXxD/YT+AHhr9qDw/8ACnS9O1saBqM9ik6PqTNIRKcPh8ZFf8Om/wBj/wD6Avij/wAHj/4V8Gf+Cfn7OfwF+IWn/E74faVrkWuaZHNHbvd6m06KJY2jf5SOu12H41X7XPwd/wCFafEFte0e12aB4lZ7i3Cr8sE/WWL2GSGUejYH3TX7E3xXt7yK5+CviaYSRvvudFMh6HlpYR6d5B/wP2rW/D9xpTmWPMtsT8r45X2NaLr9xpLiNiZbcn5oyenuPSrS8tr6Bbi1lDo3cdvYjtWo6pZaZD5t1KASPlQcs30Fav4hvNUJiU+Tb9o1PX6nv/KvF+vWngzwfqnjTVflsdMhLkk482Q8JGvqzMQo9M5OBzWr6prfjfxNcapdiS81TWLrOyNSS8jnCoo9OQoHpgVpP/BPzWLqygm1n4mW1jcPCrSwQ6SZ/LkIG5N3mrkA5Ge/oK/4d6f9Ve/8t/8A+6Kvf+Cc9nqVrJY6h8VIbm3lXbJFL4cDqw9wbipv+COnwfuJGmk+IGqxu5yUgstiA/7IMrED2ya/4c3fBz/oouuf+Ao/+OVH/wAEb/gwHUy/EPXmQH5lW2Ckj2O84/Kv+HOfwD/6HfxR+aV/w5z+Af8A0O/ij80q3/4I7fs7ru+1+L/F8vTb5c0aY9c5Vs9vSv8Ahzz+zX/0NHjX/wADIv8A43X/AA55/Zr/AOho8a/+BkX/AMbpP+CPv7KwRRJrfj9nA+YrqsABPfA+znFf8Ofv2U/+g18Qf/Bvb/8AyPUX/BIH9lGORXfVPH0oHVH1eEA/lAD+tf8ADoz9kn08a/8Ag5X/AONV/wAOjP2SfTxr/wCDlf8A41Vv/wAElP2RIVKyWHi64JPBk1sjH02oK/4dN/sf/wDQF8Uf+Dx/8K/bV/4J6fs3/Az9mXxl8U/AOl69Fr2ifYfsj3WqtNGPNvreF8oRz8krfQ4Nf8Es/wDky/wj/wBf+pf+lktVVVVVVVVVU/3/AMKqqqq/4Lbf8ks+G3/YwXX/AKTiv2B/+TOPhP8A9i/H/wChvVftm/Aj4rfsT/tOwftg/BPTGuPC17qz6k7RwM8Wm3M2Rc2tyF+7BNvkCsNoAkKAqyqTf/8ABbD4Sf8ACDtdaZ8IPFx8Ym2+Wwnlt/7PSfHQ3Ik8xkB7+SCR2FfsAfs1/FX9or9oGb9tj4+6fNFp0eotqmmC7tzF/at//wAsZIUPIt4PlKN03Rxqu4K2Kr9vj/kzj4sf9i/J/wChpX/BHT/k0i7/AOxvvv8A0Tb1VVVVVVVVVVVVVVVVVVVV8Yv+PnS/+ucv81qqqqqqqqqqqqqqqqqqqqrQv+Q5p3/X3F/6EKqqqqqqqqqqqqqqqqqqqqr/AIJ//wDJUPiB/wBeq/8Ao9qqqqqqqqqqqqqqqqqqqqq/4Lf/API0/CX/ALB+qf8Aoy2qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr/gs5/wAl38Ef9iiP/Suev2fP2yPjZ+zFo2raD8K7/SILXWrpLm6F7p63BMirtGCSMDHavjD/AMFC/wBo/wCOnw41f4WePdU0CXQtb8n7WlrpSwyHypo5kw4OR88SZ9sjvXwp+J3ir4M/EHRvib4Imto9c0KV5LN7mESxhmjaM5U9flc1/wAPZP2v/wDoNeFv/BGn/wAVX/D2T9r/AP6DXhb/AMEaf/FVoV3NqGh6df3BBlubSKRyBgbmQE4H1NVVVVVVVVV/wS4/5PH+M3/Xhf8A/pyjqqqqqqqqqqqqqqqqqqqqrx9/yKV9/wBs/wD0YtVVVXxR+F2kfEnSPJl2W2qWyn7HebeVP9xvVD6duo751vRNc8H65LpWqwS2WoWUgIwcEEcq6sOo6EEV8D/jgPFKQ+GvEd2sOuxLiCcnaL1QP0fHUd+o7gWesxy4jusI3Zux/wAKlhhuY9kqK6kcZ/pV7o8sOZLYmRP7uOR/jRHaqqqqoLO5uTiGIkepGB+dW+hKMNcy5/2U/wAahtre3XbBEq8dQOfzr4g/FHwh8NtLN/4h1EGZgfs9nCQ007Dsq9h6k4A9eRXxS+MXir4p3+/U5haaZC5Nrp0LHy4/RmP8b4/iPvgDOK+FnwP1vx88Wrar5mnaFnPnlcSXAHURA9u248DtnBFeHfDWh+E9Ki0bw/p8VpaxdFQcse7MerMfU1VVVV4Q/wCRZ07/AK4CqqqqqqqqqqqqqqqqqqqqvjF/yfR4R/6+tL/9Cqqr40/DDT/i58PdS8IXQjS6dfO0+dx/qLlAdjfQ5Kn/AGWNW8/iLwB4sSeIy6dregX2QCMNDPE/II9iMEdDXwn+Imk/Fv4faZ4wsViBvIvLvLYHP2edeJIyPQHkZ6qVPete8LvahrvTVLw9Wj6lPp6irDU73TJDJZzbdwwykZB/CpJLq/uN8jPPNIcDuSfYVpHhEfLcar+EIP8AM/0r9tn4tx+IPEsHwq8P3C/2Z4ebff8AlHCy3ZHCccERqcezMwP3a/Yk+EP/AAk/iub4na1a7tN8PP5dgHXiW8Izu/7ZqQf950PY1VVVVVVVVVVVV/wU1/5Mf+JP/cK/9OtpX/BLP/ky/wAI/wDX/qX/AKWS1VVVVVVVVVT/AH/wqqqqrXPDHhvxPDFb+JPD2matHAxaJL60ScIxGCQHBwcVYadp+k2UOm6VY29naW67Ybe3iEaRj0VVwAPYVU8EF1BJa3MKTQyoUkjkUMrqRggg8EEcYq2/Zz/Z7stYHiGy+BHw8g1UPvF9F4Xs0nDf3vMEe7PA5zQVVUKoAAGAAMYqq/b4/wCTOPix/wBi/J/6Glf8EdP+TSLv/sb77/0Tb1VVVVVVVVVVVVVVVVVVVV8Yv+PnS/8ArnL/ADWqqqqqqqqqqqqqqqqqqqqtC/5Dmnf9fcX/AKEKqqqqqqqqqqqqqqqqqqqqr/gn/wD8lQ+IH/Xqv/o9qqqqqqqqqqqqqqqqqqqqq/4Lf/8AI0/CX/sH6p/6Mtqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/b1/YK+I/7WXxH8PeM/BnjHw3pFro+if2fLFqhnDu/nSSbl8tGG3DgcnOQa/4cx/Hf/oqXgL/AL6u/wD4zX/DmP47/wDRUvAX/fV3/wDGa/4cx/Hf/oqXgL/vq7/+M1/w5j+O/wD0VLwF/wB9Xf8A8Zr/AIcx/Hf/AKKl4C/76u//AIzWjWUmm6PY6dK6u9rbRxMy9CVUAke3FVVVVVVVVV/wS4/5PH+M3/Xhf/8Apyjqqqqqqqqqqqqqqqqqqqqrx9/yKV9/2z/9GLVVVVXxM+GejfEnRjZ3gWDULcE2d4Fy0Teh9UPcfj1rxD4e17wTr0mkatBJZ39m4ZGRiM4PyujDqDjII/Qivgr8cE8VCLwp4tuI49XAC21yflW7x2PYSfo3162eoz2hCg70/uN/T0q1vbe8XMT/ADAcqeCKvdMguwXX5JP7wHX61dWc9o22ZOOzDoaqlVnIVFLE9ABVvo11MAZMRL/tdfyq30mzgwSnmMO7/wCFSSRQpukdUUepxVxrcCfLboZD6ngV8Xf2hbHwes2iaNLHqGtAFTEp/dWx/wCmhHJP+xnPqRWsazr/AIx1ptQ1W6uNR1G7cKvBZmJPCqo6DnAUCvhT+zrHbGHxB8QYEkl4aHSzyqehlPc/7HT1z0CRxxxrFEiqqDCqowAB2AqqqqqvCH/Is6d/1wFVVVVVVVVVVVVVVVVVVVVfGL/k+jwj/wBfWl/+hVVVX7cfwd+w38Hxf0G1/cXpW21lUXhJQMRTfRgNhPTIXu1fsdfGL/hX/jweD9Zu9mh+J5Eiy5+W3uukb+wbOw/VSeFqtX8Jw3swuLF1gdj+8Uj5T6ke9aXoljpSYgTfKRhpWHJ+noK+O/xStfhD8ONR8Ub4zqMg+zaXC4z5ly4O3I7hQC5HopHetD0bxB8QfFtro1h5l9rGuXm0NISTJI7ZZ2PpyWJ9Mmvhz4F0n4a+CtJ8FaKN0GmQBGlK4M0h5eQj1ZiTjtnHaqqqqqqqqqqqqv8Agpr/AMmP/En/ALhX/p1tK/4JZ/8AJl/hH/r/ANS/9LJaqqqqqqqqqp/v/hVVVVVVVVVV+3x/yZx8WP8AsX5P/Q0r/gjp/wAmkXf/AGN99/6Jt6qqqqqqqqqqqqqqqqqqqq+MX/Hzpf8A1zl/mtVVVVVVVVVVVVVVVVVVVVaF/wAhzTv+vuL/ANCFVVVVVVVVVVVVVVVVVVVVV/wT/wD+SofED/r1X/0e1VVVVVVVVVVVVVVVVVVVVf8ABb//AJGn4S/9g/VP/RltVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV/wS4/5PH+M3/Xhf/8Apyjqqqqqqqqqqqqqqqqqqqqrx9/yKV9/2z/9GLVVVVVfEn4Z6H8SdIFnfj7PfQAm0vEXLRH0Pqp7j+RrxR4X13wPrsui6zA1vd2zbo3Una65+V0buDjr/Igivgr8cYfEUcHhPxhdrHqy4S1unOBd+ise0n/oX16qzIwdGIYdCDjFWetA4iux7BwP5imWG4iwQsiMPqDV3ojbt9mwIJ+4x6fjVvoSABrmXcf7q8frUNtBbrthiVfoOauNRtLb5XlDMP4V5NXGtzyDbboIx6nk1fX0VvBLfajeJHFCpeSWZ9qoo6kk8AV8V/2iLjUfO8PeAJ3t7blJtSHyvL6iIdVX/a6ntjqfC3hHxD441dNJ0Cye5uHOZHPCRDuzt2H+Rk18MPg14f8Ah3At7Jt1DWnXEl46cR5HKxD+Eds9T7DgVVVVVVeEP+RZ07/rgKqqqqqqqqqqqqqqqqqqqq+MX/J9HhH/AK+tL/8AQqqqrxV4Z0jxn4c1Hwrr1sJ7DVLdoJ0PXDDqD2YHBB7EA18SfAer/DLxvqngvWATPp02I5QMCaM8xyL7MpB9jkdRX7LnxgX4s/DiBdTuvM8QaCFtNSDH5peP3c3/AANQc/7St2xVV+1t8Xj8TPiPLo+lXYk0HwyXtbTaflmmyPOl98soUHptQEfeNfsM/CEQW118YNbtf3k4e00YOvRBxNMPqf3YPs/rVVVVVVVVVVVVVf8ABTX/AJMf+JP/AHCv/TraV/wSz/5Mv8I/9f8AqX/pZLVVVVVVVVVVP9/8Kqqqqqqqqqv2+P8Akzj4sf8AYvyf+hpX/BHT/k0i7/7G++/9E29VVVVVVVVVVVVVVVVVVVVfGL/j50v/AK5y/wA1qqqqqqqqqqqqqqqqqqqqrQv+Q5p3/X3F/wChCqqqqqqqqqqqqqqqqqqqqq/Y/wDgh8UPhZ488Ya1488L/wBmWeq24S0l+2wTeYfNLdInYjg55Aqqqqqqqqqqqqqqqqqqqqr/AILf/wDI0/CX/sH6p/6Mtqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv+CXH/J4/xm/68L//ANOUdVVVVVVVVVVVVVVVVVVVVePv+RSvv+2f/oxaqqqqqviP8ONE+I+iHTtRUQ3cIJs7xVy8D/1U8ZXv7EAjxV4T17wRrUmi65aPb3EJzG4ztlXPDo3dTjr26HBBFfBD43rrqweDvGF2BqQASzvJDj7T6I5/v+h/i+vWrW9ntGzE+V7oehq31S0nj3PII2Ucqx/lVxrcCfLboZD6ngVcahd3I2vLhf7q8Cq8X+NPD3gbSW1fxDfrDHyIohzJMw/hRe5/QdyBXxN+MHiH4jXBtnY2OkRtmGxjfhsdGkP8TfoO3qfhh8GvEHxFnS9kDWGio2JLx05kx1WIfxHtnoPrwfCng/w/4K0lNG8PWK28K8ux5eVu7O3Un+XbAqqqqqqqvCH/ACLOnf8AXAVVVVVVVVVVVVVVVVVVVVV8Q/gN8QPEv7UHh/4raXbWLaDp09i87vchZAIjl8J1NVVVX7afweHjLwYvxF0W13av4Zib7UFHM9lnLfjGSXHsX68V8Avivc/B/wCI1h4jZ3bS7g/ZtUhXJ327EZYAdWQgMPUjHc1aXdrqFpBfWNxHPb3EayQyxtuWRGGVYEdQQQc1+1f8Xf8AhV3w2lstKuvL17xEHtLHa2GiTH72Yf7qkKD2Z1PY18Kvh5qXxT8eaT4K0zcn22XNzMBn7PAvMkh+ig4z1JA71oOh6X4a0Wx8PaLapb2OnW6QW8Sj7qKMD8eOT3PNVVVVVVVVVVVVV/wU1/5Mf+JP/cK/9OtpX/BLP/ky/wAI/wDX/qX/AKWS1VVVVVVVVVT/AH/wqqqqqqqqqq/b4/5M4+LH/Yvyf+hpX/BHT/k0i7/7G++/9E29VVVVVVVVVVVVVVVVVVVVfGL/AI+dL/65y/zWqqqqqqqqqqqqqqqqqqqqtC/5Dmnf9fcX/oQqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/4Lf/8AI0/CX/sH6p/6Mtqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr4r/Hv4M/Ayxg1D4tfEnQ/DKXQJtor25/fTgdTHCuZHA7lVIGR61pv/AAUy/Yb1a7Fla/HqzSTIGbnRdRt05OPvyW6r39ffpVn+0r+zzqPhW78b2Hxw8DXGhWCBrq+j163aODPQP8+VYngKeSeAM14j/wCCr37Fegai+nW3j/VtZ8tirT6dodw0WR6NIqbh7rkH1r4Wf8FEv2Qfi/rNp4a8NfFu2sdZvpBHb2Os2c2ntI5xtVZJVETMSdoUOSTwByM1VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX/AAS4/wCTx/jN/wBeF/8A+nKOqqqqqqqqqqqqqqqqqqqqvH3/ACKV9/2z/wDRi1VVVVVV8Qfh9ofxE0NtJ1VPLmjy1pdKuXt39R6g8ZXv7EAjxf4Q13wJrsuia3AYp4juilTOyVOzoe4OPqDwcEV8EfjkdVaDwd41vf8ATSQllfStjz/SNz/e9D/F0PPWqqvij8atB+HsL6dZmPUdcK/JaK3yw56NKR0Hfb1PHQHNeJfE/iHxtrDarrt7LeXcp2xqBwgzwiKOg56Dv7nNfCn9nWW68nxB8QYGjh4aHTCcM3oZfQf7PX1x0MFvb2sEdrawxwwwoEjjjUKqKBgAAcAAdqqqqqqqqvCH/Is6d/1wFVVVVVVVVVVVVVVVVVVVVVVVVSxRTxPBNGskcilXR1yGB4II7iv2kPhDL8IfiPdabZwOND1PN1pMhHHlk/NFn1Rvl9cbSfvV+xZ8arfVvCd18N/FGppHdeHIGuLGaeQKGsh95ST/AM8j69FYdlNfH74qT/F74kah4kSRxplv/oulxNxst0JwcdixJc+hbHYV+xd8If8AhC/BD+PtZtQur+J0UwB1w0FmOUHtvPzn1AT0qqqqqqqqqqqqqq/4Ka/8mP8AxJ/7hX/p1tK/4JZ/8mX+Ef8Ar/1L/wBLJaqqqqqqqqqp/v8A4VVVVVVVVVVft8f8mcfFj/sX5P8A0NK/4I6f8mkXf/Y333/om3qqqqqqqqqqqqqqqqqqqqr4xf8AHzpf/XOX+a1VVVVVVVVVVVVVVVVVVVVoX/Ic07/r7i/9CFVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX/Bb/AP5Gn4S/9g/VP/RltVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV+0h/wVr+CHwW8Q33gnwBoF78RNe0yZ4LyS1ultLCCVThk+0FXMjA8HYhXj72ah/4K6ftXa+n9p+E/2Z9GuNNk/wBS6WOoXY6f89EZVP4AV8TPihr/AMSvj9d/Ff8Aan8HeJr+HWLsyXOj2l02kyQ24/1cFu88Uu2KNSoC7csOrBmL18GfAH/BIz9oWG18L2z+Lfhv4mnARLfW9deBpn6fu53MluxJwAp2MSeF7Cb/AIIueKb345ajo+nePhp/wmgSGay1m6MdzqV0GUF4BEm1Q6sGHmMFXBVgrHKjwb/wSz/Yq8JaRFp178LrjxJdKoEuoaxq9080pHcrE6Rr/wABRa/4Ka/sN/sz/AP4NWfxU+FWk3fhTWJdcgsE00ahNdW98JEkdwFmZ2RlEZYFWC4BG3kEfsV/8FOvgXqHwz8EfCj4z+NdU0XxnptjHp1zrGtW22yu3RtkTG5V32kx7N0kwQbgxJ5BMckc0aSwyK6OoZGU5DA9CD6VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV/wS4/5PH+M3/Xhf/8Apyjqqqqqqqqqqqqqqqqqqqqrx9/yKV9/2z/9GLVVVVVVVePvh/oHxD0Y6VrMW2SPLWt0g/eW7kdR6jpleh/Ijxt4J1zwFrkmia3BtdfmgmUHZOnZ1Pp7dQeDXwT+OqXi23g3xteEXORHZahI3Evokh/vdg3fvzyalkihiaaaRUSNSzMxwFA6knsK+K/7RSjzvD3w8uMnlJtUx+YhH/s5/Dsa0TQfEHjPWl03SLSfUL+6Ysxzk8nl3Y9BzyTXws+B2ieAo4tW1bytS13GfOK5jtj6Rg9/9o8+mORVVVVVVVVVeEP+RZ07/rgKqqqqqqqqqqqqqqqqqqqqqqqqr9pD4Qw/F/4cXWmWcCHXNMzdaTIRz5gHzRZ9JF+X0ztJ+7UU+oaXcSrBNPaT7JIJQpMbbWBR0PfBBKkHsSDX7Onwml+L3xKsdFuYWOj2GLvVXxgeSpH7vPq7YX1AJP8ADUUUUESQQRrHHGoVEVcBQOAAB0FVVVVVVVVVVVVVX/BTX/kx/wCJP/cK/wDTraV/wSz/AOTL/CP/AF/6l/6WS1VVVVVVVVVT/f8Awqqqqqqqqqq/b4/5M4+LH/Yvyf8AoaV/wR0/5NIu/wDsb77/ANE29VVVVVVVVVVVVVVVVVVVVfGL/j50v/rnL/Naqqqqqqqqqqqqqqqqqqqq0L/kOad/19xf+hCqqqqqqqqqqqqqqqqqqqqq/ZP/AGgPiL8YPG/izQfGd1YS2ujwB7UW9qImB80ryQeeBVVVVVVVVVVVVVVVVVVVVX/Bb/8A5Gn4S/8AYP1T/wBGW1VVVVVVVVVVVVVVVVVVVVVVVVVVVVa7rmi+GNFvfEXiLVbXTdL0y3e4vLy6lEUVvEgyzux4VQASSa+On/BZf4TWUWr+EfhD8Pdf8Uxz281qutz340iMFkKiaBTHLIwBII3rGcjoOtf8E+/jt8LPgl8YEHxN+Ctv45/4SKS3stMvIrRbu+0q4Mm1Wt4ZDsfeXAOMSDA2t1VgqqoVQAAMAAYxWu+H9A8UaXNonibRNP1fTrhds1nf2yXEUg9GRwQR9RXxN/4JVfso+P8AxhpPjHRfDVz4Qez1CK41HTNHcJY6nErhnhaBsiIMBtzEUABPyk8i3trezt4rS0gjhggQJFFGgVUUDAVQOAAABiq8cfDL4bfE6zttO+JPw+8NeLLWylMttBrmkwX6QuRgsiyqwViOMjnFf8FXv2bv2WfhJ8GNK8ZeCfAejeEfGuoa5HbadDo6/ZkvIQjNPut1+TYoCHeFBVmQZ+bB/YHu/El/+xx8KLnxZNcy37aAgV7g5c24kcW/Pp5Aix7Yqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr/AIJcf8nj/Gb/AK8L/wD9OUdVVVVVVVVVVVVVVVVVVVVePv8AkUr7/tn/AOjFqqqqqqqq8eeA9D+IOhvo2sxYZctbXKr89u/95fb1HQj8CPHHgXXvAGsto+uW+M5a3nTmOdM/eU/zB5HevhH+0Fbabph8P+P7uUraQk2l9tLs6qP9W4HJbsG79D6n4pfGzXfiDI+mWBk07Q1b5bZW+efHQykdfXaOB7kZr4bfCbxH8R7wNaRm00uJsT38qfKPVUH8bew4HcjivBXgPw54B0pdL0CzCFgDPcOMyzsO7N/IdB2FVVVVVVVVVXhD/kWdO/64CqqqqqqqqqqqqqqqqqqqqviH8ePiB4a/ag8P/CnS7mxGg6jPYpOj2oaQiU4fD5yKqqqqr9tL4Of8Id4yT4h6FabdI8TSn7SqLxBe4y34SAFx/tB+nFfsv/CL/hU/w1t49SthHr2t7bvU8j5oyR+7hP8AuKcH/aZqqqqqqqqqqqqqqq/4Ka/8mP8AxJ/7hX/p1tK/4JZ/8mX+Ef8Ar/1L/wBLJaqqqqqqqqqp/v8A4VVVVVVVVVVft8f8mcfFj/sX5P8A0NK/4I6f8mkXf/Y333/om3qqqqqqqqqqqqqqqqqqqqr4xf8AHzpf/XOX+a1VVVVVVVVVVVVVVVVVVVVoX/Ic07/r7i/9CFVVVVVVVVVVVVVVVVVVVVV/wT//AOSofED/AK9V/wDR7VVVVVVVVVVVVVVVVVVVVV/wW/8A+Rp+Ev8A2D9U/wDRltVVVVVVVVVVVVVVVV4g+JXw58J340rxV4/8N6NemMSC21DVYbaQocgNtdgcHB5xjg1/wvD4K/8ARYPBP/hQWv8A8XX/AAvD4K/9Fg8E/wDhQWv/AMXX/C8Pgr/0WDwT/wCFBa//ABdf8Lw+Cv8A0WDwT/4UFr/8XX/C8Pgr/wBFg8E/+FBa/wDxdf8AC8Pgr/0WDwT/AOFBa/8AxdaD8Tfht4q1BdI8L/ELw1rF8yF1tbDVoLiUqOpCIxOB3OKqqqqqqr9tT/gpN4H/AGUfECfDjQPC7eMfG5t0nu7P7Z9mt9NRwCnnOFZjIykMI1H3SCSoZc/ta/8ABTDx5+058Hv+FOal8OYvB0jatFc6rPaam8i3sUQYrbtC0alAJNjnLNkxrwMc/sM/sq/BP4UfAPwP4i0fwZo2p+I/EmgWmp6lr13aJPcTyXMKysqSMCUiUOFVFwCACcsSTqvwg+FWuazpPiPVvhx4autW0K6W60y/k0uLz7OZfuvHJt3KR7H09KqqqtRj1CXT7mLSbq3tr5oXFtNcwGeOKUg7WeNXQuoOCVDqSONy5yPjJ/wSi8Z/H7xVeePvir+1zqut+IrlCkUjeFFjtrVMkpFDALoiOJc/dB5OSSSST+x58QPjZ+xN+2TB+xl8UfEcuseE9cu1srSLzHlghknTfaXVqG5jVztR0Hygs2csmaqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/wCCXH/J4/xm/wCvC/8A/TlHVVVVVVVVVVVVVVVVVVVVXj7/AJFK+/7Z/wDoxaqqqqqqqq8beCdD8e6HLoetwblPzQzKPngfs6n19uhHBrx34D1z4f64+i6xDuVstbXCKdlwn95ff1HUH8DXwp/Z5vNcEOv+O4ZbTTzhobDlJZx6v3Rfb7x9uCbGwstMs4dP061itra3QJFDEgVUA7ADpVVVVVVVVVVXhD/kWdO/64CqqqqqqqqqqqqqqqqqqqqvjF/yfR4R/wCvrS//AEKqqqqq1vQNF8SWaafrumwX1vHcRXCxTLuAkicOjfgyg+/Q5BIqqqqqqqqqqqqqqqv+Cmv/ACY/8Sf+4V/6dbSv+CWf/Jl/hH/r/wBS/wDSyWqqqqqqqqqqf7/4VVVVVVVVVVft8f8AJnHxY/7F+T/0NK/4I6f8mkXf/Y333/om3qqqqqqqqqqqqqqqqqqqqr4xf8fOl/8AXOX+a1VVVVVVVVVVVVVVVVVVVVoX/Ic07/r7i/8AQhVVVVVVVVVVVVVVVVVVVVVf8E//APkqHxA/69V/9HtVVVVVVVVVVVVVVVVVVVVX/Bb/AP5Gn4S/9g/VP/RltVVVVVVVVVVVVVVVV+3N4I8aeIfjVDf+H/CGtanbDQ7dDNZ6fLMgYPLkblUjPI496/4VX8T/APonHij/AME8/wD8RX/Cq/if/wBE48Uf+Cef/wCIr/hVfxP/AOiceKP/AATz/wDxFf8ACq/if/0TjxR/4J5//iK/4VX8T/8AonHij/wTz/8AxFf8Kr+J/wD0TjxR/wCCef8A+Ir9ifwL420D46Wuo674O1zTrVdMulM93p0sKAlRgbmUDJqqqqqp2KQSOsscZVCQ8gyq8dSMjj8R9ak8f/8ABU79qn4g+NPDvw58dWOn2/gnUpLG/i8M6vaabawSK7qmx95uJEk8pijszKQDyBmrH9jf/gq38YAfEXjb45XXhm8jgRY7PVPG1xA+3HAEdgskaMRjO7aSevOa/Ym/Y++Onhf9uuXxD+1R4T1XVb3RtDutYsvEGpTtqNtqV4jwQRyC6bcJJVWYuFch1KK2AVFftKfsifBb9qbw1LpHxE8ORR6vHEV0/wAQWUax31i3bbJj50z1jfKn0BwR8FtA/wCGbfgV4I+GvxM8aafe3WivFoFhfRI6/b91w0VlGkZy3mGHygyDcFKuclF3VVVVVXxW/wCCrf7Nvwn1rW/COq6H47u/Eug3Ulpc6WujC3ZZkOOWmkUBT1DDOV5AORn9lXRfip+3t+3Un7WniPww+h+DfCl9FdBlYmKJrePbaWUUhA86TcFkkYAADeSF3opqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/4Jcf8AJ4/xm/68L/8A9OUdVVVVVVVVVVVVVVVVVVVVePv+RSvv+2f/AKMWqqqqqqqqqv8ASNK1R7d9S062umtJfNtzNEHMTgY3Lnofeqqqqqqqqqqqrwh/yLOnf9cBVVVVVVVVVVVVVVVVVVVVXxi/5Po8I/8AX1pf/oVVVVVVVVVVVVVVVVVVVVVf8FNf+TH/AIk/9wr/ANOtpX/BLP8A5Mv8I/8AX/qX/pZLVVVVVVVVVVP9/wDCqqqqqqqqqr9vj/kzj4sf9i/J/wChpX/BHT/k0i7/AOxvvv8A0Tb1VVVVVVVVVVVVVVVVVVVV8Yv+PnS/+ucv81qqqqqqqqqqqqqqqqqqqqrQv+Q5p3/X3F/6EKqqqqqqqqqqqqqqqqqqqqr/AIJ//wDJUPiB/wBeq/8Ao9qqqqqqqqqqqqqqqqqqqqq/4Lf/API0/CX/ALB+qf8Aoy2qqqqqqqqqqqqqqqqqqqqqqqqqqvLS11C0msL63juLe4jaOaKRdyyIwwykHqCCQRXjv/gk1+0b4E8e61dfsr/Gqz0bwn4hZonhuNbvNMuYbdmz5E5t0YTxLnGc5OOUz11H9gT/AIKR/BPW18a/Bn44P4m1e+sYINQay8RSW0r+TGEjhdbwrHNGigJGWPAx8qCv2sNH/wCCj/gTwnpfiH9qHxf4xj8Oa1dvata23iKJ7YSAbgk0dm5hVmXdsLA/dbHTFL/wV9/ZCsvAsGsWn/CUnVI7Vdnh2LSCJo3C4EfmkiHAPG4P05x2r9hjxz8TP26f2ota/ab+KMa2/hb4Z2z2nhPQ4iTbWF3dhhlc8SSrAH8yQjcWkiI2hVUahqem6TDHPqmoW9pHLPFbxvPIEDyyOEjQE9WZ2VQOpJAHWqqqqv8Agtj8Pfh/pXiTwD8TNLlsrXxhrUdxZ6nbIVEl5bQhDDO6dTsJaPeRyCq5wgFfAySC4+CvgK8t9HsNKF34bsLhrKwtVtoIHkt0dljiQBUXcx+UAYqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr/glx/yeP8Zv+vC//wDTlHVVVVVVVVVVVVVVVVVVVVXj7/kUr7/tn/6MWqqqqqqqqqqqqqqqqqqqqq8If8izp3/XAVVVVVVVVVVVVVVVVVVVVV8Yv+T6PCP/AF9aX/6FVVVVVVVVVVVVVVVVVVVVX/BTX/kx/wCJP/cK/wDTraV/wSz/AOTL/CP/AF/6l/6WS1VVVVVVVVVT/f8Awqqqqqqqqqq/b4/5M4+LH/Yvyf8AoaV/wR0/5NIu/wDsb77/ANE29VVVVVVVVVVVVVVVVVVVVfGL/j50v/rnL/Naqqqqqqqqqqqqqqqqqqqq0L/kOad/19xf+hCqqqqqqqqqqqqqqqqqqqqq/wCCf/8AyVD4gf8AXqv/AKPaqqqqqqqqqqqqqqqqqqqqv+C3/wDyNPwl/wCwfqn/AKMtqqqqqqqqqqqqqqqqqqqqqqqqqqqqvFHhTwx430C88K+MfD+na5o+oR+XdWGoWy3EMy5zhkYEHkA9OCAa+K//AAS5/ZQ1nwF4tHw2+D1npXjO80i7Gh3Z1q/8mC9aJhCxiacxBd+3gpgdccV+xF+3XY/sK6B4w+B3xi+E3iF7n/hIJL1/sgSK7tbnyo4ZIZopSvygQIQQerNwQQa8Dftq+Kf22v29/hDoU0Mnhj4f6Hq8t5puhy3QfzrqC1nlS4nYLh5SUVUXBCZwpyzMa+HvxE8FfFfwnZ+Ofh74htdb0O+eVLe8tmyjtHI0bj1yHRh+FV8Pfj/8Kvid4v8AFHw98MeJ4f8AhKvBuoz2Wr6Lc4huoTFIU81UJ/eQtgMHXIwyhtrZUV/wWL03x54F8P8Agr42fD/4o+KPDhmvjoGpafput3FrHcFo5J4ZhHG4UMoimVmxkho/Sv2SP+CXbfGOx8JftEftC/FCfxFpfiC0t9Wi0WEyyz3iOodEurmU7gOgZUBJBIDr1qGGG3hS3t4kjjiUKiIu0KAMAADoMdqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq+Pvxz8Efs5/DDVPij48uJBZWOIra1hwZr65fPlwRA9WbBPoqhmPCk14X+O//BSv9trUdT8RfA6WPwf4RsLgxIbNoLS2ikABEX2mVTLPIFKltvyjIJVNwB8P/tuftm/sd/E/T/AX7X+kXHiHw/eEF5ZoYTcfZ87TPaXMOEn29SkhJP3SYycjwz4l0Lxn4c0zxZ4X1OHUdI1i0jurG6hbKTRSKGVh9QRVV/wS4/5PH+M3/Xhf/wDpyjqqqqqqqqqqqqqqqqqqqqrx9/yKV9/2z/8ARi1VVVVVVVVVVVVVVVVVVVVXhD/kWdO/64CqqqqqqqqqqqqqqqqqqqqvjF/yfR4R/wCvrS//AEKqqqqqqqqqqqqqqqqqqqqv+Cmv/Jj/AMSf+4V/6dbSv+CWf/Jl/hH/AK/9S/8ASyWqqqqqqqqqqf7/AOFVVVVVVft7/wDBRS4/Z91yP4K/BTTbbW/iLcon2yeaIzxaV5oHlIsa8y3DBlYKflUFSQ27aLrwz/wWgXw+fiofFWrmfy/tR0SKfTzchCM4+xBNhOOfKwX7bd3FfsAf8FEr39ojWZvgz8aLGy0n4hWsTyWNzbxmCLVljH72MxE/u7hACxUcMoYgLtINV+3x/wAmcfFj/sX5P/Q0r/gjp/yaRd/9jfff+ibeqqqqqqqqqqqqqqqqqqqqvjF/x86X/wBc5f5rVVVVVVVVVVVVVVVVVVVVWhf8hzTv+vuL/wBCFVVVVVVVVVVVVVVVVVVVVVb6fYWjtJaWUELOPmaOIKT9cVVVVVVVVVVVVVVVVVVVVV/wW/8A+Rp+Ev8A2D9U/wDRltVVVVVVVVVVVVVVVVVVVVVVVVVVVVVeNPgp8GviTfR6n8RPhJ4L8U3kKbI7jWtAtr6RF9A0qMQPav26v+CePi/UPFeg/H39jjSLDQfEfhuKFZtA0cRabhrc5huLMLtjWRQArR/LuCrjJyG139pj/grT408KXXwdu/gl4ngvNStmsbrWYfAtzbXbxyAozefgW8eQGHmKqgZJBBAI+G3xG/bl/wCCYBbRPGHwwe88Dau6Xk9ler9osVndFDGK8gLLDNwqMpJUlc7Tw1Wf/Bb/AOG0mk+dqHwI8Sw6ntGLaHV4JYc85/elFbH3ednc+nPgr4P/ALSf/BQL48+OP2gfhDoUHgWV7o6jDqUl7cWttBOAkaW8F3HHue42AsWUKM5J2bgKtdG/4Lc/DtP7D0zUdc1S3QBY7mW50XVi4XjPmXG+Qdf4sE960j/gn1+3h+1b4w0vXP2wfiHPo+h2DbjFe6nDeXCRsQWFtbWxMETMAAWYqRxkNt214S8K6F4G8K6P4K8MWK2ekaDYQ2NhbqSRFBEgRFyeThVAyetVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV/wWE+I+s+KvjL4M+B+kPJJaaLpq3rW6Njzr67kKKCOhKxxptJ6ea/qa+DHww0P4MfCrwv8LvD0ESWnh3TYrYvGm3z5QMyzH/akkLuT6sa/4Kn/AAq074g/spav4o+xJJq/gW7g1OymCfOsbSLDcJu6hTHJvI6ExKewr/gkP8Sr3xf+zZf+CNSuWll8E65LbWu45K2s6iZB+EjT/QYHQVVf8E6/iJ8Pvhv+1x8X9V+Injvw94Wsrm1v4YLnWtThsY5ZP7RQ7FaVlBbAJwOcAmv+GqP2YP8Ao5D4Xf8AhYWP/wAdr/hqj9mD/o5D4Xf+FhY//Ha/4ao/Zg/6OQ+F3/hYWP8A8drwh468EfEPSTr3gDxlofibTFmaE3uj6hFewiRQCyb4mZdwDKSM5GR61VVVVVVVVVVVVVVVVVfErU9N0/wndrf6jbWxl2eWJpVTdiRM4yecZH50CGAKkEEcEVVVVVVVVVVVVVVVVVVVVSOkaNJI6qqAlmY4AA6k14E1HT9R8L2Laff29yI4gHMMofafQ4PFVVVVVVVVVVVVVVVVVVVVXFx4cTU44bqfTRqBK+WkjIJc9sA/N9Kqqqqqqqqqqqqqqqqqq/4ao/Zg/wCjkPhd/wCFhY//AB2v+GqP2YP+jkPhd/4WFj/8dr/hqj9mD/o5D4Xf+FhY/wDx2v8Agof8fvgR42/Y7+IHhjwZ8bPAWv6zef2b9m07TPElpdXE2zUrV22RRyFmwisxwOApPQGv+CWf/Jl/hH/r/wBS/wDSyWqqqqqqqqqqf7/4VVVVVVeLPEVj4P8ACus+LdSBNpomnz31wAcHy4o2dsfgpr/gmP4Vu/2jf20fEfxz+IcaahcaDFc+IHMo3r/aVzMFhwD0CB5nT+6Ykx0GK/4KD6NN+y5+37pnxd8D2ws/7Sax8VwxQAIjXAlZLlemP3jwOzg5z5xz1qwvrXU7C21KylElvdwrLC4H3kYAg/iCKr9vj/kzj4sf9i/J/wChpX/BHT/k0i7/AOxvvv8A0Tb1VVVVVVVVVVVVVVVVVVVV8Yv+PnS/+ucv81qqqqqqqqqqqqqqqqqqqqrQv+Q5p3/X3F/6EKqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv+C3/APyNPwl/7B+qf+jLaqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqmhhuIXt54kljkUq6OuQwIwQQeoI7VdfAD4D3t4dRvPgn4CnuySxnl8N2jSZPU7jHnsKsNPsNLsodO0uyt7S1t0CQwQRCNI1HQKowAPYVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX7ZBfUv+CpMVhfwh7dfEvhiFFZcB42gsifryzCq/bQhin/ZO+LKTRpIo8J3zAMuQCsRIP1BAI9CK/wCCJs121j8Ybdy32ZJdFaPK4G8i9D4PfhU47cetVX7L/wCyv4O/ay/aW+KHgzxn4i1nSLXR3vr+KXTDGHd/twj2t5isNuHJ4GcgV/w5j+BH/RUvHv8A31af/Ga/4cx/Aj/oqXj3/vq0/wDjNf8ADmP4Ef8ARUvHv/fVp/8AGa/Zl/Zx8Lfst/DiT4aeENd1XVrGXU5r8z6kYzKHkVFK/u1UbQIx2zyaqqqqqqqqqqqqqqqqqv8Agtr/AM0Y/wC49/7j68Pf8i/pn/XnF/6AKqqqqqqqqqqqqqqqqqqqvjL/AMkg8c/9i1qH/pNJX/BFv/kl/wARv+w9bf8Aog1VVVVVVVVVVVVVVVVVVVV+01/ylu+G3/X/AKB/6HVVVVVVVVVVVVVVVVVVX/DmP4Ef9FS8e/8AfVp/8Zr/AIcx/Aj/AKKl49/76tP/AIzX/DmP4Ef9FS8e/wDfVp/8Zr9rj/gmV8Jf2ev2evFfxg8MePvFupanoH2PyLW/Nv5MnnXkEDbtkatwsrEYI5Ar/gln/wAmX+Ef+v8A1L/0slqqqqqqqqqqn+/+FVVVVVX7Y1zc2n7J/wAYJbQZc+CdVjI25+VrZ1f/AMdJ+lf8EOreB7j4z3TRgzRpoSI56hWN+WH4lV/Kq/4Lawwr8V/hvOsSCV/DtyrOF+YgXHAJ9BuPHufWvgXNd3PwR+HtxqBb7VL4V0xp9ybTvNrGWyO3OeKr9vj/AJM4+LH/AGL8n/oaV+yN/wAE2tb/AGsvhTL8UdO+LNj4bii1afTvsc2jvcsTEkbF94lXg+ZjGO1f8OQPFP8A0cRpX/hNyf8Ax+v+HIHin/o4jSv/AAm5P/j9f8OQPFP/AEcRpX/hNyf/AB+vhz4Vk8C/D3wv4IlvVvJPD2jWmnNcLHsExghSMuFycA7c4ycZqqqqqqqqqqqqqqqqqq8UePvHVz/wVq1PwZceNdel8Pw31wsekvqUrWqD+xWfAhLbAN4DdOvPWqqqqqqqqqqqqqqqqqqqqv8AgpP4p8T+EPgZoOqeEvEmqaLet4uto2udOvJLaQp9kuzt3IQcZVTjpkA9q/Y/1TU9c/ZZ+FWsa1qV1qF/eeFLGW4urqZpZZnMSks7sSWJPUk5qqqqqqqqqqqqqqqqqqqqr/gk18RfiF40+OHxZ0/xj488Q67a2VirW0Gp6pNdJAftTDKLIxC8ccdqqqqqqqqqqqqqqqqqqv2ufgDeftOfAzWfg9YeJodAm1W4tZVv5rU3Cx+TMkhGwMpOdmOvGa/4cgeKf+jiNK/8JuT/AOP1/wAOQPFP/RxGlf8AhNyf/H6/4cgeKf8Ao4jSv/Cbk/8Aj9ftpfsW6n+xvqfhXTNS+IFr4oPii3upUeDTmtPI8lowQQXfdnzPbGKqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv+Cp2jan8Mf2yfDnxWs4GZNT03T9Tt5AWUNcWcpjZM54IWKE8YxvHetA1zS/FGg6b4l0S6S507VrOK7s5kORJFIgdGBHYqwP41/wUi8cWfgf9jrx401wiXWvRQaRZxsxUyvPModRjuIVmbHopr/gjb4HudG+Bfi3x1dQtH/wkviIQW5bP7yG1hUBh2x5ksq/VT7VVf8ABLj/AJPH+M3/AF4X/wD6co6qqqqqqqqqqqqqqqqqqqq/4La/80Y/7j3/ALj68Pf8i/pn/XnF/wCgCqqqqqqqqqqqqqqqqqqqr4y/8kg8c/8AYtah/wCk0lf8EW/+SX/Eb/sPW3/og1VVVVVVVVVVVVVVVVVVVV+01/ylu+G3/X/oH/odVVVVVVVVVVVVVVVVVVVVf8FNf+TH/iT/ANwr/wBOtpX/AASz/wCTL/CP/X/qX/pZLVVVVVVVVVVP9/8ACqqqqqr4s+Dm+Inwr8ZfD9GUN4m8P3+mAsSADcW7xckEED56/wCCMHjVPDHxv8ffC7VnNpdeINEjnjhmJQtcWUxBj2k8uEuJWxjICN05qv8AgrJrM/xV/bK8M/Cfwsy3V/pekWGkCFWLEX13O8iqQOmUmt+AM8/QDQdHtfD2had4fsc/Z9MtIrWHccnZGgVf0Aqv2+P+TOPix/2L8n/oaV/wR0/5NIu/+xvvv/RNvVVVVVVVVVVVVVVVVVVVVXiL/lMdq3/YQuP/AExNVVVVVVVVVVVVVVVVVVVVX/BUn/k37w//ANjjbf8ApHeV+xX/AMmkfCH/ALE/T/8A0StVVVVVVVVVVVVVVVVVVVVf8Edf+S+fGL/sHp/6VtVVVVVVVVVVVVVVVVVVVVX/AAW//wCRp+Ev/YP1T/0ZbVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVft7/soP8AtVfCFNN8Otbw+M/C8z3ugSTMEWYsoEtszH7qyhU56B40JIGa/Zz/AOCh/wARv2QvDh+AHx/+FGtagPDRaLTRJKbS9so9x/cusikSRA52MCMLwNy7dvxN+Iv7Qv8AwVQ+JeieC/h74En8P+BdDuCwaVmltbF2GHuru4ChWk2ZCRqMgFgoJZmPwe+Fvhr4KfDHw58LPCMZGmeHbFbeORlAed/vSTOBxvkcs7Y4yxqq/wCCXH/J4/xm/wCvC/8A/TlHVVVVVVVVVVVVVVVVVVVVX/BbX/mjH/ce/wDcfXh7/kX9M/684v8A0AVVVVVVVVVVVVVVVVVVVVfGX/kkHjn/ALFrUP8A0mkr/gi3/wAkv+I3/Yetv/RBqqqqqqqqqqqqqqqqqqqqr9pr/lLd8Nv+v/QP/Q6qqqqqqqqqqqqqqqqqqqq/4Ka/8mP/ABJ/7hX/AKdbSv8Agln/AMmX+Ef+v/Uv/SyWqqqqqqqqqqf7/wCFVVVVVVftyfsvfF/9mX9oRP2zf2ctNubnSpNTbVr5LK3aVtIvGz9o86NeTazhpNzDgeY6NtBQtef8FtYJ/BXk6T8B518ZSW/lp5urB7FZyMb8BBIy558vgn7u/wDir/gnt+yF8WPiL8Z5v2zP2ldOvLe4a8l1DSLTU7cxXGoXsmcXbREDy4Yw2YxgZYIVAVQWqv2+P+TOPix/2L8n/oaV/wAEdP8Ak0i7/wCxvvv/AETb1VVVVVVVVVVVVVVVVVVVV4i/5THat/2ELj/0xNVVVVVVVVVVVVVVVVVVVVX/AAVJ/wCTfvD/AP2ONt/6R3lfsV/8mkfCH/sT9P8A/RK1VVVVVVVVVVVVVVVVVVVV/wAEdf8Akvnxi/7B6f8ApW1VVVVVVVVVVVVVVVVVVVVf8Fv/APkafhL/ANg/VP8A0ZbVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVrXhnw34ljjh8ReH9N1VITmNb20ScIfUBwcVY2FjplpFYabZwWltCu2KCCMRog9Ao4Aqqr9iv8AaH+FX7OH7UvxX8U/FnXLjTNO1NL6ytpIbKW5LTfb1fBWNSQNqMcniv8Ah6Z+xf8A9FF1T/wn7z/43X/D0z9i/wD6KLqn/hP3n/xuv+Hpn7F//RRdU/8ACfvP/jdf8PTP2L/+ii6p/wCE/ef/ABuv+Hpn7F//AEUXVP8Awn7z/wCN1/w9M/Yv/wCii6p/4T95/wDG6/4emfsX/wDRRdU/8J+8/wDjdf8AD0z9i/8A6KLqn/hP3n/xuv8Ah6Z+xf8A9FF1T/wn7z/43X/D0z9i/wD6KLqn/hP3n/xuv+Hpn7F//RRdU/8ACfvP/jdf8PTP2L/+ii6p/wCE/ef/ABuv+Hpn7F//AEUXVP8Awn7z/wCN1/w9M/Yv/wCii6p/4T95/wDG6/4emfsX/wDRRdU/8J+8/wDjdf8AD0z9i/8A6KLqn/hP3n/xuv8Ah6Z+xf8A9FF1T/wn7z/43X/D0z9i/wD6KLqn/hP3n/xuv+Hpn7F//RRdU/8ACfvP/jdf8PTP2L/+ii6p/wCE/ef/ABuv+Hpn7F//AEUXVP8Awn7z/wCN1/w9M/Yv/wCii6p/4T95/wDG6/4emfsX/wDRRdU/8J+8/wDjdf8AD0z9i/8A6KLqn/hP3n/xuv8Ah6Z+xf8A9FF1T/wn7z/43X/D0z9i/wD6KLqn/hP3n/xuv+Hpn7F//RRdU/8ACfvP/jdf8PTP2L/+ii6p/wCE/ef/ABuv+Hpn7F//AEUXVP8Awn7z/wCN1/w9M/Yv/wCii6p/4T95/wDG6/4KdftTfBf9pj/hWv8AwqDxHdar/wAI3/a39oefp81r5fn/AGPy8eYo3Z8mTp0xz1FaP/wUC/Zds9IsbSfxrfiWC2jRwNGuThgoB/g9q/4eFfssf9DvqH/gluv/AIiv+HhX7LH/AEO+of8Agluv/iK/4eFfssf9DvqH/gluv/iK/wCHhX7LH/Q76h/4Jbr/AOIr/h4V+yx/0O+of+CW6/8AiK/4eFfssf8AQ76h/wCCW6/+Ir/h4V+yx/0O+of+CW6/+Ir/AIeFfssf9DvqH/gluv8A4iv+HhX7LH/Q76h/4Jbr/wCIr/h4V+yx/wBDvqH/AIJbr/4iv+HhX7LH/Q76h/4Jbr/4iv8Ah4V+yx/0O+of+CW6/wDiK/4eFfssf9DvqH/gluv/AIiv+HhX7LH/AEO+of8Agluv/iK/4eFfssf9DvqH/gluv/iK/wCHhX7LH/Q76h/4Jbr/AOIr/h4V+yx/0O+of+CW6/8AiK/4eFfssf8AQ76h/wCCW6/+Ir/h4V+yx/0O+of+CW6/+Ir/AIeFfssf9DvqH/gluv8A4iv+HhX7LH/Q76h/4Jbr/wCIr/h4V+yx/wBDvqH/AIJbr/4iv+HhX7LH/Q76h/4Jbr/4iv8Ah4V+yx/0O+of+CW6/wDiK/4eFfssf9DvqH/gluv/AIiv+HhX7LH/AEO+of8Agluv/iK/4eFfssf9DvqH/gluv/iK/wCHhX7LH/Q76h/4Jbr/AOIr/h4V+yx/0O+of+CW6/8AiK+JP7eP7NHiP4deKfD2k+Mr6W+1PRby1to20i5UNLJC6qCSmBkkcniv+CaP7XfwK/Zt8C+MtD+Lfie70y81jVoLizSDTZ7kPGsW0kmNSBz2Nf8AD0z9i/8A6KLqn/hP3n/xuv8Ah6Z+xf8A9FF1T/wn7z/43X/D0z9i/wD6KLqn/hP3n/xuv+Hpn7F//RRdU/8ACfvP/jdf8PTP2L/+ii6p/wCE/ef/ABuv+Hpn7F//AEUXVP8Awn7z/wCN1/w9M/Yv/wCii6p/4T95/wDG6/4emfsX/wDRRdU/8J+8/wDjdf8AD0z9i/8A6KLqn/hP3n/xuv8Ah6Z+xf8A9FF1T/wn7z/43X/D0z9i/wD6KLqn/hP3n/xuv+Hpn7F//RRdU/8ACfvP/jdf8PTP2L/+ii6p/wCE/ef/ABuv+Hpn7F//AEUXVP8Awn7z/wCN1/w9M/Yv/wCii6p/4T95/wDG6/4emfsX/wDRRdU/8J+8/wDjdf8AD0z9i/8A6KLqn/hP3n/xuv8Ah6Z+xf8A9FF1T/wn7z/43X/D0z9i/wD6KLqn/hP3n/xuv+Hpn7F//RRdU/8ACfvP/jdf8PTP2L/+ii6p/wCE/ef/ABuv+Hpn7F//AEUXVP8Awn7z/wCN1/w9M/Yv/wCii6p/4T95/wDG6/4emfsX/wDRRdU/8J+8/wDjdf8AD0z9i/8A6KLqn/hP3n/xuv8Ah6Z+xf8A9FF1T/wn7z/43X/D0z9i/wD6KLqn/hP3n/xuv+Hpn7F//RRdU/8ACfvP/jdf8PTP2L/+ii6p/wCE/ef/ABuv+Hpn7F//AEUXVP8Awn7z/wCN18b/ANpz4O+Nv+Cg/gr4/eHfENzP4L0a70mS8vXsJo3RbdsykRMoc4HoOe1f8PTP2L/+ii6p/wCE/ef/ABuv+Hpn7F//AEUXVP8Awn7z/wCN1/w9M/Yv/wCii6p/4T95/wDG6/4emfsX/wDRRdU/8J+8/wDjdf8AD0z9i/8A6KLqn/hP3n/xuv8Ah6Z+xf8A9FF1T/wn7z/43X/D0z9i/wD6KLqn/hP3n/xuv+Hpn7F//RRdU/8ACfvP/jdf8PTP2L/+ii6p/wCE/ef/ABuv+Hpn7F//AEUXVP8Awn7z/wCN1/w9M/Yv/wCii6p/4T95/wDG6/4emfsX/wDRRdU/8J+8/wDjdf8AD0z9i/8A6KLqn/hP3n/xuv8Ah6Z+xf8A9FF1T/wn7z/43X/D0z9i/wD6KLqn/hP3n/xuv+Hpn7F//RRdU/8ACfvP/jdf8PTP2L/+ii6p/wCE/ef/ABuv+Hpn7F//AEUXVP8Awn7z/wCN1/w9M/Yv/wCii6p/4T95/wDG6/4emfsX/wDRRdU/8J+8/wDjdf8AD0z9i/8A6KLqn/hP3n/xuv8Ah6Z+xf8A9FF1T/wn7z/43X/D0z9i/wD6KLqn/hP3n/xuv+Hpn7F//RRdU/8ACfvP/jdf8PTP2L/+ii6p/wCE/ef/ABuv+Hpn7F//AEUXVP8Awn7z/wCN1/w9M/Yv/wCii6p/4T95/wDG6/4emfsX/wDRRdU/8J+8/wDjdf8AD0z9i/8A6KLqn/hP3n/xuv8Ah6Z+xf8A9FF1T/wn7z/43X7b/wC3z+zH8bP2XvGvwx+HnjS/v/EGs/YPsdvLo9zAr+Vf28z5d0CjCRueTzjHev8Agln/AMmX+Ef+v/Uv/SyWqqqqqqqqqqf7/wCFVVVVVVVB4I8F22q/27beENFi1MtuN4mnxLNu9d4Xdn8aqq/b4/5M4+LH/Yvyf+hpX/BLH9ov4D/Cv9mW58MfEj4u+FfDWrN4ovLhbLU9SjglMTRQBX2sc7SVYZ9jX/Dan7JH/RxvgD/weQ//ABVf8Nqfskf9HG+AP/B5D/8AFV/w2p+yR/0cb4A/8HkP/wAVX/Dan7JH/RxvgD/weQ//ABVf8Nqfskf9HG+AP/B5D/8AFV/w2p+yR/0cb4A/8HkP/wAVX/Dan7JH/RxvgD/weQ//ABVf8Nqfskf9HG+AP/B5D/8AFV/w2p+yR/0cb4A/8HkP/wAVX/Dan7JH/RxvgD/weQ//ABVf8Nqfskf9HG+AP/B5D/8AFV/w2p+yR/0cb4A/8HkP/wAVX/Dan7JH/RxvgD/weQ//ABVf8Nqfskf9HG+AP/B5D/8AFV/w2p+yR/0cb4A/8HkP/wAVX/Dan7JH/RxvgD/weQ//ABVf8Nqfskf9HG+AP/B5D/8AFV/w2p+yR/0cb4A/8HkP/wAVX/Dan7JH/RxvgD/weQ//ABVf8Nqfskf9HG+AP/B5D/8AFV/w2p+yR/0cb4A/8HkP/wAVX/Dan7JH/RxvgD/weQ//ABVf8Nqfskf9HG+AP/B5D/8AFV/w2p+yR/0cb4A/8HkP/wAVX/Dan7JH/RxvgD/weQ//ABVf8Nqfskf9HG+AP/B5D/8AFV/w2p+yR/0cb4A/8HkP/wAVX/Dan7JH/RxvgD/weQ//ABVf8Nqfskf9HG+AP/B5D/8AFV/w2p+yR/0cb4A/8HkP/wAVWu/Ff4Z3H/BVDU/ipB470V/B8t7O6a4LxTaMp0ZogRJnbgyfJ9eK/wCGpf2cf+i3eDf/AAbRf41/w1L+zj/0W7wb/wCDaL/Gv+Gpf2cf+i3eDf8AwbRf41/w1L+zj/0W7wb/AODaL/Gv+Gpf2cf+i3eDf/BtF/jX/DUv7OP/AEW7wb/4Nov8a/4al/Zx/wCi3eDf/BtF/jX/AA1L+zj/ANFu8G/+DaL/ABr/AIal/Zx/6Ld4N/8ABtF/jX/DUv7OP/RbvBv/AINov8a/4al/Zx/6Ld4N/wDBtF/jX/DUv7OP/RbvBv8A4Nov8a/4al/Zx/6Ld4N/8G0X+Nf8NS/s4/8ARbvBv/g2i/xr/hqX9nH/AKLd4N/8G0X+Nf8ADUv7OP8A0W7wb/4Nov8AGv8AhqX9nH/ot3g3/wAG0X+Nf8NS/s4/9Fu8G/8Ag2i/xr/hqX9nH/ot3g3/AMG0X+Nf8NS/s4/9Fu8G/wDg2i/xr/hqX9nH/ot3g3/wbRf41/w1L+zj/wBFu8G/+DaL/Gv+Gpf2cf8Aot3g3/wbRf41/wANS/s4/wDRbvBv/g2i/wAa/wCGpf2cf+i3eDf/AAbRf41/w1L+zj/0W7wb/wCDaL/Gv+Gpf2cf+i3eDf8AwbRf41/w1L+zj/0W7wb/AODaL/Gv+Gpf2cf+i3eDf/BtF/jX/DUv7OP/AEW7wb/4Nov8a/4KLfGf4TfEb4JaJofgP4i6Br+oQeKre4ktrC+SaRIha3SlyqnIUM6jPqw9a/ZN/ay/Zl8Jfsy/DDwx4m+O/grTNW0vwvZW97ZXWsRRy28qxKGR1JyCCMEGv+G1P2SP+jjfAH/g8h/+Kr/htT9kj/o43wB/4PIf/iq/4bU/ZI/6ON8Af+DyH/4qv+G1P2SP+jjfAH/g8h/+Kr/htT9kj/o43wB/4PIf/iq/4bU/ZI/6ON8Af+DyH/4qv+G1P2SP+jjfAH/g8h/+Kr/htT9kj/o43wB/4PIf/iq/4bU/ZI/6ON8Af+DyH/4qv+G1P2SP+jjfAH/g8h/+Kr/htT9kj/o43wB/4PIf/iq/4bU/ZI/6ON8Af+DyH/4qv+G1P2SP+jjfAH/g8h/+Kr/htT9kj/o43wB/4PIf/iq/4bU/ZI/6ON8Af+DyH/4qv+G1P2SP+jjfAH/g8h/+Kr/htT9kj/o43wB/4PIf/iq/4bU/ZI/6ON8Af+DyH/4qv+G1P2SP+jjfAH/g8h/+Kr/htT9kj/o43wB/4PIf/iq/4bU/ZI/6ON8Af+DyH/4qv+G1P2SP+jjfAH/g8h/+Kr/htT9kj/o43wB/4PIf/iq/4bU/ZI/6ON8Af+DyH/4qv+G1P2SP+jjfAH/g8h/+Kr/htT9kj/o43wB/4PIf/iq/4bU/ZI/6ON8Af+DyH/4qv+G1P2SP+jjfAH/g8h/+Kr/htT9kj/o43wB/4PIf/iq/4bU/ZI/6ON8Af+DyH/4qv+CWfxj+FPwr+NHxT1r4kfELQfDdhqtkqWNzqd6kCXDfambCFjz8pB+lf8Nqfskf9HG+AP8AweQ//FV/w2p+yR/0cb4A/wDB5D/8VX/Dan7JH/RxvgD/AMHkP/xVf8Nqfskf9HG+AP8AweQ//FV/w2p+yR/0cb4A/wDB5D/8VX/Dan7JH/RxvgD/AMHkP/xVf8Nqfskf9HG+AP8AweQ//FV/w2p+yR/0cb4A/wDB5D/8VX/Dan7JH/RxvgD/AMHkP/xVf8Nqfskf9HG+AP8AweQ//FV/w2p+yR/0cb4A/wDB5D/8VX/Dan7JH/RxvgD/AMHkP/xVf8Nqfskf9HG+AP8AweQ//FV/w2p+yR/0cb4A/wDB5D/8VX/Dan7JH/RxvgD/AMHkP/xVf8Nqfskf9HG+AP8AweQ//FV/w2p+yR/0cb4A/wDB5D/8VX/Dan7JH/RxvgD/AMHkP/xVf8Nqfskf9HG+AP8AweQ//FV/w2p+yR/0cb4A/wDB5D/8VX/Dan7JH/RxvgD/AMHkP/xVf8Nqfskf9HG+AP8AweQ//FV/w2p+yR/0cb4A/wDB5D/8VX/Dan7JH/RxvgD/AMHkP/xVf8Nqfskf9HG+AP8AweQ//FV/w2p+yR/0cb4A/wDB5D/8VX/Dan7JH/RxvgD/AMHkP/xVf8Nqfskf9HG+AP8AweQ//FV/w2p+yR/0cb4A/wDB5D/8VX/Dan7JH/RxvgD/AMHkP/xVf8FefjH8Kfi/4j+Gdz8LfiFoPiqLTLHUUvH0q9S4FuzvAUD7ScZCtj6Gqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqrxb/wAE8/2OfHHibVPGHiT4MQT6trN091ezQ6zqFsskznLuI4p1RckknaoGST3r/h2V+w//ANER/wDLk1X/AOSa/wCHZX7D/wD0RH/y5NV/+Sa/4dlfsP8A/REf/Lk1X/5Jr/h2V+w//wBER/8ALk1X/wCSa/4dlfsP/wDREf8Ay5NV/wDkmv8Ah2V+w/8A9ER/8uTVf/kmv+HZX7D/AP0RH/y5NV/+Sa/4dlfsP/8AREf/AC5NV/8Akmv+HZX7D/8A0RH/AMuTVf8A5Jr/AIdlfsP/APREf/Lk1X/5Jr/h2V+w/wD9ER/8uTVf/kmv+HZX7D//AERH/wAuTVf/AJJr/h2V+w//ANER/wDLk1X/AOSa/wCHZX7D/wD0RH/y5NV/+Sa/4dlfsP8A/REf/Lk1X/5Jr/h2V+w//wBER/8ALk1X/wCSa/4dlfsP/wDREf8Ay5NV/wDkmv8Ah2V+w/8A9ER/8uTVf/kmv+HZX7D/AP0RH/y5NV/+Sa/4dlfsP/8AREf/AC5NV/8Akmv+HZX7D/8A0RH/AMuTVf8A5Jr/AIdlfsP/APREf/Lk1X/5Jr/h2V+w/wD9ER/8uTVf/kmv+HZX7D//AERH/wAuTVf/AJJr/h2V+w//ANER/wDLk1X/AOSa/wCHZX7D/wD0RH/y5NV/+Sa/4dlfsP8A/REf/Lk1X/5Jr/h2V+w//wBER/8ALk1X/wCSa/4dlfsP/wDREf8Ay5NV/wDkmv8Ah2V+w/8A9ER/8uTVf/kmv+HZX7D/AP0RH/y5NV/+Sa/4dlfsP/8AREf/AC5NV/8Akmv+HZX7D/8A0RH/AMuTVf8A5Jr/AIdlfsP/APREf/Lk1X/5Jr/h2V+w/wD9ER/8uTVf/kmv+HZX7D//AERH/wAuTVf/AJJr/h2V+w//ANER/wDLk1X/AOSa/wCHZX7D/wD0RH/y5NV/+Sa/4dlfsP8A/REf/Lk1X/5Jr/h2V+w//wBER/8ALk1X/wCSa/4dlfsP/wDREf8Ay5NV/wDkmv8Ah2V+w/8A9ER/8uTVf/kmv+HZX7D/AP0RH/y5NV/+Sa/4dlfsP/8AREf/AC5NV/8Akmv+HZX7D/8A0RH/AMuTVf8A5Jr/AIdlfsP/APREf/Lk1X/5Jr/h2V+w/wD9ER/8uTVf/kmv+HZX7D//AERH/wAuTVf/AJJr/h2V+w//ANER/wDLk1X/AOSa/wCHZX7D/wD0RH/y5NV/+Sa/4dlfsP8A/REf/Lk1X/5Jr/h2V+w//wBER/8ALk1X/wCSa/4dlfsP/wDREf8Ay5NV/wDkmv8Ah2V+w/8A9ER/8uTVf/kmv+HZX7D/AP0RH/y5NV/+Sa/4dlfsP/8AREf/AC5NV/8Akmv+HZX7D/8A0RH/AMuTVf8A5Jr/AIdlfsP/APREf/Lk1X/5Jr/h2V+w/wD9ER/8uTVf/kmv+HZX7D//AERH/wAuTVf/AJJr/h2V+w//ANER/wDLk1X/AOSa/wCHZX7D/wD0RH/y5NV/+Sa/4dlfsP8A/REf/Lk1X/5Jr/h2V+w//wBER/8ALk1X/wCSa/4dlfsP/wDREf8Ay5NV/wDkmv8Ah2V+w/8A9ER/8uTVf/kmv+HZX7D/AP0RH/y5NV/+Sa/4dlfsP/8AREf/AC5NV/8Akmv+HZX7D/8A0RH/AMuTVf8A5Jr/AIdlfsP/APREf/Lk1X/5Jr/h2V+w/wD9ER/8uTVf/kmv+HZX7D//AERH/wAuTVf/AJJr/h2V+w//ANER/wDLk1X/AOSa/wCHZX7D/wD0RH/y5NV/+Sa/4dlfsP8A/REf/Lk1X/5Jr/h2V+w//wBER/8ALk1X/wCSa/4dlfsP/wDREf8Ay5NV/wDkmv8Ah2V+w/8A9ER/8uTVf/kmv+HZX7D/AP0RH/y5NV/+Sa/4dlfsP/8AREf/AC5NV/8Akmv+HZX7D/8A0RH/AMuTVf8A5Jr/AIdlfsP/APREf/Lk1X/5Jr/h2V+w/wD9ER/8uTVf/kmv+HZX7D//AERH/wAuTVf/AJJr/h2V+w//ANER/wDLk1X/AOSa/wCHZX7D/wD0RH/y5NV/+Sa/4dlfsP8A/REf/Lk1X/5Jr/h2V+w//wBER/8ALk1X/wCSa/4dlfsP/wDREf8Ay5NV/wDkmv8Ah2V+w/8A9ER/8uTVf/kmv+HZX7D/AP0RH/y5NV/+Sa/4dlfsP/8AREf/AC5NV/8Akmv+HZX7D/8A0RH/AMuTVf8A5Jr/AIdlfsP/APREf/Lk1X/5Jr/h2V+w/wD9ER/8uTVf/kmv+HZX7D//AERH/wAuTVf/AJJr/h2V+w//ANER/wDLk1X/AOSa/wCHZX7D/wD0RH/y5NV/+Sa/4dlfsP8A/REf/Lk1X/5Jr/h2V+w//wBER/8ALk1X/wCSa/4dlfsP/wDREf8Ay5NV/wDkmv8Ah2V+w/8A9ER/8uTVf/kmv+HZX7D/AP0RH/y5NV/+Sa/4dlfsP/8AREf/AC5NV/8Akmv+HZX7D/8A0RH/AMuTVf8A5Jr/AIdlfsP/APREf/Lk1X/5Jr/h2V+w/wD9ER/8uTVf/kmv+HZX7D//AERH/wAuTVf/AJJr/h2V+w//ANER/wDLk1X/AOSa/wCHZX7D/wD0RH/y5NV/+Sa/4dlfsP8A/REf/Lk1X/5Jr/h2V+w//wBER/8ALk1X/wCSa/4dlfsP/wDREf8Ay5NV/wDkmv8Ah2V+w/8A9ER/8uTVf/kmv+HZX7D/AP0RH/y5NV/+Sa/4dlfsP/8AREf/AC5NV/8Akmv+HZX7D/8A0RH/AMuTVf8A5Jr/AIdlfsP/APREf/Lk1X/5Jr/h2V+w/wD9ER/8uTVf/kmv+HZX7D//AERH/wAuTVf/AJJr/h2V+w//ANER/wDLk1X/AOSa/wCHZX7D/wD0RH/y5NV/+Sa/4dlfsP8A/REf/Lk1X/5Jr/h2V+w//wBER/8ALk1X/wCSa/4dlfsP/wDREf8Ay5NV/wDkmvh78PPBfwp8G6Z8P/h5oFvovh/SI2js7KAsyxhmLMSzEszMzMxZiSSSSSTVVVVVVVVVVP8Af/Cqqqqqqqqqr4mfDvwz8W/AGvfDXxjBNLoviKye0vFglMUgRu6sOjAgEHkZHIPSv+HLX7LP/Q+/FT/waWH/AMhV/wAOWv2Wf+h9+Kn/AINLD/5Cr/hy1+yz/wBD78VP/BpYf/IVf8OWv2Wf+h9+Kn/g0sP/AJCr/hy1+yz/AND78VP/AAaWH/yFX/Dlr9ln/offip/4NLD/AOQq/wCHLX7LP/Q+/FT/AMGlh/8AIVf8OWv2Wf8Aoffip/4NLD/5Cr/hy1+yz/0PvxU/8Glh/wDIVf8ADlr9ln/offip/wCDSw/+Qq/4ctfss/8AQ+/FT/waWH/yFX/Dlr9ln/offip/4NLD/wCQq/4ctfss/wDQ+/FT/wAGlh/8hV/w5a/ZZ/6H34qf+DSw/wDkKv8Ahy1+yz/0PvxU/wDBpYf/ACFX/Dlr9ln/AKH34qf+DSw/+Qq/4ctfss/9D78VP/BpYf8AyFX/AA5a/ZZ/6H34qf8Ag0sP/kKv+HLX7LP/AEPvxU/8Glh/8hV/w5a/ZZ/6H34qf+DSw/8AkKv+HLX7LP8A0PvxU/8ABpYf/IVf8OWv2Wf+h9+Kn/g0sP8A5Cr/AIctfss/9D78VP8AwaWH/wAhV/w5a/ZZ/wCh9+Kn/g0sP/kKv+HLX7LP/Q+/FT/waWH/AMhV/wAOWv2Wf+h9+Kn/AINLD/5Cr/hy1+yz/wBD78VP/BpYf/IVf8OWv2Wf+h9+Kn/g0sP/AJCr/hy1+yz/AND78VP/AAaWH/yFX/Dlr9ln/offip/4NLD/AOQq/wCHLX7LP/Q+/FT/AMGlh/8AIVf8OWv2Wf8Aoffip/4NLD/5Cr/hy1+yz/0PvxU/8Glh/wDIVf8ADlr9ln/offip/wCDSw/+Qq/4ctfss/8AQ+/FT/waWH/yFX/Dlr9ln/offip/4NLD/wCQq/4ctfss/wDQ+/FT/wAGlh/8hV/w5a/ZZ/6H34qf+DSw/wDkKv8Ahy1+yz/0PvxU/wDBpYf/ACFX/Dlr9ln/AKH34qf+DSw/+Qq/4ctfss/9D78VP/BpYf8AyFX/AA5a/ZZ/6H34qf8Ag0sP/kKv+HLX7LP/AEPvxU/8Glh/8hV/w5a/ZZ/6H34qf+DSw/8AkKv+HLX7LP8A0PvxU/8ABpYf/IVf8OWv2Wf+h9+Kn/g0sP8A5Cr/AIctfss/9D78VP8AwaWH/wAhV/w5a/ZZ/wCh9+Kn/g0sP/kKv+HLX7LP/Q+/FT/waWH/AMhV/wAOWv2Wf+h9+Kn/AINLD/5Cr/hy1+yz/wBD78VP/BpYf/IVf8OWv2Wf+h9+Kn/g0sP/AJCr/hy1+yz/AND78VP/AAaWH/yFX/Dlr9ln/offip/4NLD/AOQq/wCHLX7LP/Q+/FT/AMGlh/8AIVf8OWv2Wf8Aoffip/4NLD/5Cr/hy1+yz/0PvxU/8Glh/wDIVf8ADlr9ln/offip/wCDSw/+Qq/4ctfss/8AQ+/FT/waWH/yFX/Dlr9ln/offip/4NLD/wCQq/4ctfss/wDQ+/FT/wAGlh/8hV/w5a/ZZ/6H34qf+DSw/wDkKv8Ahy1+yz/0PvxU/wDBpYf/ACFX/Dlr9ln/AKH34qf+DSw/+Qq/4ctfss/9D78VP/BpYf8AyFX/AA5a/ZZ/6H34qf8Ag0sP/kKv+HLX7LP/AEPvxU/8Glh/8hV/w5a/ZZ/6H34qf+DSw/8AkKv+HLX7LP8A0PvxU/8ABpYf/IVf8OWv2Wf+h9+Kn/g0sP8A5Cr/AIctfss/9D78VP8AwaWH/wAhV/w5a/ZZ/wCh9+Kn/g0sP/kKv+HLX7LP/Q+/FT/waWH/AMhV/wAOWv2Wf+h9+Kn/AINLD/5Cr/hy1+yz/wBD78VP/BpYf/IVf8OWv2Wf+h9+Kn/g0sP/AJCr/hy1+yz/AND78VP/AAaWH/yFX/Dlr9ln/offip/4NLD/AOQq/wCHLX7LP/Q+/FT/AMGlh/8AIVf8OWv2Wf8Aoffip/4NLD/5Cr/hy1+yz/0PvxU/8Glh/wDIVf8ADlr9ln/offip/wCDSw/+Qq/4ctfss/8AQ+/FT/waWH/yFX/Dlr9ln/offip/4NLD/wCQq/4ctfss/wDQ+/FT/wAGlh/8hV/w5a/ZZ/6H34qf+DSw/wDkKv8Ahy1+yz/0PvxU/wDBpYf/ACFX/Dlr9ln/AKH34qf+DSw/+Qq/4ctfss/9D78VP/BpYf8AyFX/AA5a/ZZ/6H34qf8Ag0sP/kKv+HLX7LP/AEPvxU/8Glh/8hV/w5a/ZZ/6H34qf+DSw/8AkKv+HLX7LP8A0PvxU/8ABpYf/IVf8OWv2Wf+h9+Kn/g0sP8A5Cr/AIctfss/9D78VP8AwaWH/wAhV/w5a/ZZ/wCh9+Kn/g0sP/kKv+HLX7LP/Q+/FT/waWH/AMhV/wAOWv2Wf+h9+Kn/AINLD/5Cr/hy1+yz/wBD78VP/BpYf/IVf8OWv2Wf+h9+Kn/g0sP/AJCr/hy1+yz/AND78VP/AAaWH/yFX/Dlr9ln/offip/4NLD/AOQq/wCHLX7LP/Q+/FT/AMGlh/8AIVf8OWv2Wf8Aoffip/4NLD/5Cr/hy1+yz/0PvxU/8Glh/wDIVf8ADlr9ln/offip/wCDSw/+Qq/4ctfss/8AQ+/FT/waWH/yFX/Dlr9ln/offip/4NLD/wCQq/4ctfss/wDQ+/FT/wAGlh/8hV/w5a/ZZ/6H34qf+DSw/wDkKv8Ahy1+yz/0PvxU/wDBpYf/ACFX/Dlr9ln/AKH34qf+DSw/+Qq/4ctfss/9D78VP/BpYf8AyFX/AA5a/ZZ/6H34qf8Ag0sP/kKv+HLX7LP/AEPvxU/8Glh/8hV/w5a/ZZ/6H34qf+DSw/8AkKv+HLX7LP8A0PvxU/8ABpYf/IVf8OWv2Wf+h9+Kn/g0sP8A5Cr/AIctfss/9D78VP8AwaWH/wAhV/w5a/ZZ/wCh9+Kn/g0sP/kKv+HLX7LP/Q+/FT/waWH/AMhV/wAOWv2Wf+h9+Kn/AINLD/5Cr/hy1+yz/wBD78VP/BpYf/IVf8OWv2Wf+h9+Kn/g0sP/AJCr/hy1+yz/AND78VP/AAaWH/yFWjf8Eav2U9J1az1O58TfEjVYrWZZHsb3VLTybgKc7JPLtUfaeh2spx3FVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVU/wB/8Kqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqp/v8A4VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV41/aJ+Anw51NtD8dfGXwboepJ9+yvdagjnT/ejLblH1Arwn418G+PtJXXvA3i3RvEWmudq3mlX0d3ET3G+NiM+2aqqqqqqqqqqqqqqqqqqqmKqpZmAAHJPGK0H4m/DXxXqsmg+FviH4a1jU4UZ5LKw1aC4mRVIDEojFgASATjgkVVVVVVVVVVVVVVVVVVVVanqmmaLYTarrGo2tjZWyb5rm6mWKOJfVmYgAe5oftYfsvtqY0df2hvh0bonaFHiW127vTfv257YznPHWrDULDVrKHUtLvre8tLhA8M9vIJI5FPRlZSQR7iqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqp/v/hVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVzcW9pbyXd3PHDBAheWWRgqooGSxJ4AAGSTUn7Wv7LkWsf2BJ+0R8Olvg5jMZ8SWoAcdVLb9oORjGc5468VbXVrfW0V3ZXMVxBMgeKWJwyup6EEcEH1qqqqqqqqqqqqqqqqqqqq17xDoHhbS5db8T67p+kadb4828v7lLeGPJwNzuQoySBya8MeMfCHjjT5NV8F+KtH1+yimMElzpd9HdRpIAGKFo2IDAMpx1wwPeqqqqqqqqqqqqqqqqqqqrxj8Q/h/8O7FNT+IHjrw/wCGbOQ7UuNZ1OGyjY+gaVlBrwj+01+zr4+1JNF8GfHLwNq+oynbFZW2vW7TyH/Zj37m/AGqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv8AgqH+2L4h+CHh3Tvgz8MNWfTvFniq0a51DUoW2zadp5LIPKI5SWVlcBxyixsRgsrL+z7/AMEjfB/iD4fWfiz9ovxP4nXxXrkIupNN0u5jhGnCT5gkryRyNJNggseFDErhsbj8VfAPxY/4JY/tB6D4s+HXiu71nwlrytJbi5/dpqcEbKJ7K6RTtLoHQiQAffV1CkFR8PfHOg/E7wJ4f+Ifhecy6V4j06G+tC3DBJUDBWHZhnaR2II7VVVVVVVVVVVVVVVVVVVXxe8Cf8FB/wBsn4seLfhfLeJ4C+Fmha3daeLtFksrO/gjkKrIQCZrwugVtufKDddhr/gkxY/2Z+2Fq2m+b5v2TwzqMO/bt3bbiAZxzjOOlVVVVVVVVVVVVVVVVVVVX7SH7Cv7Qv7U37RWsXHjL4xyaZ8JrNYJNGhb980RaMeZDFaoVTcHDZmkOSGX7+CBrn/BGL4IT6A8Hhr4q+OLPWvL/d3V99lubbf6mFIo2x7eZX7K/wAWPi1+wp+1cf2dPiJqbSeGdQ1uHTNXsTKz20bT7RBqFvuxsBEkbscDdGSGG5V21VVVVVVVVVVVVVVVVVVVVVVVVVVVVVT/AH/wqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr9sj47fFb9tv8Aadg/Y8+COrNbeFbLVX06URytHFqFxDuNzdXDLy0EISTanIOzcAzMoE3/AARZ+A7+Bv7Kg+JXjJPFYt+NYYwNbefjqbXYD5ef4PNDf7dfsM/Hf4nfsfftP3X7HXxk1OSbw5f6x/ZUcMsxePTr6TH2ae3J+7DPujyvAxKr8EMDVVVVVVVVVVVVVVVVVVXxw8TeNfBXwe8ZeMfhzo9tq3iXQ9Fub3TbG5ieRLmWKMvs2RkMxIUgKpBJwO9ftH/A39t34ofBPxJ+1Z+1T4qm0rTvD4tX0rwveExyYuLqG3BjtI8R2oAmyS/71tmGHO6v+CLf/JrPij/sf7z/ANILCqqqqqqqqqqqqqqqqqqq1y01G/0W/sdG1U6Zf3FrLHa3ohWU2srKQkoRsq21iG2kYOMHivh//wAEnfin8Y9VuPiN+2N8adWbWr+QmSy02dby6K5OC1zLujjHpGiMACOVxtH7RH/BG7w94b8A6p4q/Z+8d+I9S1jSLV7n+xNeEM7agqLlo4pYY49khAO0FSGOASudw/4JHftb+MPiPHq/7O/xI1ubV7vw/po1Hw9f3UheY2iyLHLbu55fYZIymckKXH3VUCqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq+JCH47/8ABV9NC1ZGubG38b2lg1uw3L9n09UEiY5+VhBIT/vk8VX/AAV28IWuv/srReI2izc+GPEdpcxyAchJQ8DL9CZUP1UV/wAEofFdz4l/ZA0rTbm480+Gtbv9OjJbJVS63AU8np9o4HGBjtiqqqqqqqqqqqqqqqqqqqr/AIJW/wDJ63iL/sAap/6UwVVVVVVVVVVVVVVVVVVVVVX7Z0cHxp/4KUW/gvwSfPuf7X0fRZZ7c/8ALdBH5zZA48rcysT08o9hVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVU/3/wqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq+NPi+f4ffBzx349tWIn8N+GtR1KLHXfBbSSLj3yor/gip4Nh1r4v/EP4i3kRnn0LQoLOOWTLbHvJixbJ/iItWGeuC3qar/gsZ4cbwh+1D4V8f6G72l3rHhu3naePhhdW1xIokB65CeSOnG0cnt4O19PFfhDQ/FEYQLrGm294oQ8ASxq/HJ4+b1P1qqqqqqqqqqqqqqqqqqqv+CoX/JivxM/7hX/AKdbOv8Agi3/AMms+KP+x/vP/SCwqqqqqqqqqqqqqqqqqqqqrxN4i0fwf4b1XxZ4hvUtNL0Wymvb2dzgRQxIXdj9FUmv+CP/AIZ1HxR+1b4h8dWVm1rpeieHruScIfkR7maNYoTgdwJGHA/1R+lVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVXwutjoP/AAVzvItQcIZfiRrjqSpH+v8AtTRj8fMUenPpzVf8FU7qC3/Yz8TQyybXudT02OIYJ3MLpGx7fKrHn0r/AII8abNZfsr6xcy526h41vJ48qR8otbSP8eYzVVVVVVVVVVVVVVVVVVVV/wSt/5PW8Rf9gDVP/SmCqqqqqqqqqqqqqqqqqqqqv26P2yvD/7LHw+lstGvLW8+IevQMmh6aSHNsDlTeTL2jQg4B++w2jgOV/4JcfsneILa+n/a3+LcU8msa4kreG47wlpnWfPn38hPO6UMyoTyVZ2OQ6mqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqp/v/hVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV+2Dptxq37KfxesrUZlPgrVJFXBJbZbO5AA7kLge5Ff8ABDq5gW5+M9m0gE0sehOiY6qpvwx/Asv51X/BbS6t3+Lvw5sUkzPF4bnkdMHhWuCFOenJRvyr4J6bNo3wZ8BaPcn99Y+GNNgkypX5kto1PB5HI6VVVVVVVVVVVVVVVVVVVX/BUL/kxX4mf9wr/wBOtnX/AARb/wCTWfFH/Y/3n/pBYVVVVVVVVVVVVVVVVVVVVV/wU0/bGuviDqafsa/ACeTWr/Vb2O08SXOmnzGuJ/MATTYiPvHeFMpHGQEz/rFr9hP9lO1/ZQ+Ctv4Z1MwXHi/XpFv/ABJdRHcvn7cJAjd44l+UHuxdhgNgVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX/BQPw/rH7Nv7eGgfHzTNPc6brN5YeIbQRjaks9q0aXUGT/ABN5au3tcD148H+LvDvj7wrpPjXwlqcWoaPrdnHd2VzGciSN1BB9jzgg8ggg8iv+Cynxm0iLwr4R+AumX8cuqXV//beqRRtlreGON4oFf08xpJGA6/ugeARn9h34TX3wW/Zb8CeCdas2tdYaxa/1OKRcPHcXMjTFHHZkV1jP+5VVVVVVVVVVVVVVVVVVVV/wSt/5PW8Rf9gDVP8A0pgqqqqqqqqqqqqqqqqqqqq/bO/bR8F/sm+D1ULb61451eJjouhGTAx0+0XGOUhUgjsXI2rjDMn7If7HPj/9qnx8P2u/2s7ia907U7lbzTdHvIMHWAo/du6HiOzUBQkeP3gA6J9+OOOKNYYUVERQqqowFA6ADsKqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqn+/+FVVVVVVVVVVVVVVVVVVVVVVVVVVVVVXiHQtN8UaBqXhnWIfOsNWs5bO6j/vxSoUcfirEV/wTx8Vz/smftw698Evibcrp41s3HhmaaX93GL1JVe1k5/hlKFEPQ/aEOcc1X7VWpRftwf8FGdL+HPgaY6potrdWnhyO7t2yptbdmlvp1IONiF7ohgfmVAR1FRxRwxrDCioiKFRFGAoHQAdhVVVVVVVVVVVVVVVVVVVf8FQv+TFfiZ/3Cv/AE62df8ABFv/AJNZ8Uf9j/ef+kFhVVVVVVVVVVVVVVVVVVVV+3r/AMFC9Z0vXZ/2Y/2WZZtW8a6jL/Z2qazpqmaSzmc7PslnszuuSTtZxnyycL8+TH/wT8/4J96d+zTp6fE/4nLa6r8S9UtgFQKJI9AjcfNFE/O+ZgcPKOMZRfl3M9VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX7Tv7NHgL9qb4azeAPGge0uYHNxpGrwRhp9NuQMB1BxvQjh4yQGHcMFZfDn7H/wDwU5/Z4hufBPwP+JVndeGpZneL7BrEKwpu6sIbtQYnOct5YPIPJ4J/Zo/4JleKrT4mR/HP9rbxjbeLvEEdyt7DpSXMl4JrlcFJbueQDzNhAxEoKnC5YqChqqqqqqqqqqqqqqqqqqqq/Yh/YR+Nv7PP7RurfFXx7d+GZNFvtLvbWJdPv5Jpt800bplWjUAYQ559Kqqqqqqqqqqqqqqqqqqqvi7/AMEy/wBs34qfFLxF8R9Z8W+EtRvNW1OW4hubvWphKkW8+SoAhwgRAqhV4UAAcCo/2Jv+CpMUaww/tOakiIoVVX4jaoAoHQAbeBX/AAxV/wAFTf8Ao6DVP/Dj6p/8TXw80vX9D+H/AIZ0TxXftfa5p+j2lvqV007TGe5SFVlcyN8z7nDHceTnJ61VVVVVVVVVVVVVVVVVVVVVVVVVVVVVT/f/AAqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr9u3/gnXon7U9zF8R/h/q9l4Z+ItpCsMs90jC11aJRhFnKAskiAALKqsdvyspAUpN+zP/wAFeda8Pn4T6n8VGTwzJD9kku5vE8OHgxgq0yKbpkKkgg5JGQQRxX7Dv7AfhL9kewufE2tapB4l+IGqweRc6pHCUhsoCQTBbK3zYJALSNgtgcKAQaqqqqqqqqqqqqqqqqqqr9tT4OeLv2gP2ZfGXwi8CS6fHrmvfYfsrX8zRQjyb63nfcyqxHyRNjg84+tf8E6/2aviJ+yx8Eta+HvxMn0eXVNQ8VXGqQnS7lp4/Je1tYhlmRSG3QPxjpjnniqqqqqqqqqqqqqqqqqqv2kPC3xN8cfA7xf4R+Devx6L4x1axFvpl/JcG3EBaRRIfMVWZCYvMAZRkEggg4I8K/8ABKb9ubwLrcXibwR488MeHtYhVlj1DSvE91aXCBhhgJI4QwBHB55r/hiL/gq3/wBHTar/AOHK1X/4mv2e/wBkv/go14I+NXhDxZ8Vv2h9R1zwjpmorLq2nSePdRvFuYQpBUwyDY/JHDcVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVT/f8Awqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqn+/8AhVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVP9/8ACqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqf7/4VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVT/f/Cqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqf7/4VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVT/AH/wqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq+KXxH8PfCH4e678S/FaXb6R4etDc3a2kQklKAgHapIBPI7iv+Hvn7KH/AED/AB7/AOCeH/4/X/D3z9lD/oH+Pf8AwTw//H6/4e+fsof9A/x7/wCCeH/4/X/D3z9lD/oH+Pf/AATw/wDx+vGX7Rvw+8E/s7r+0tqwv/8AhF5tCtNXtoPKAuZkuljNvEEzgSM0sa9cAkknAJrwR/wWettS8d29j48+DkWkeE7u5WNryy1Vri5sYyceY6mNRKB1IXYcZxk8G3uILu3iurWZJYZkDxuhyGUjIIPoRVVVVVVVVVV47/4K8+BfD/xK1TwB4G+EOp+Mbez1I2FjqVrrCRLqMgYJmJPKfKs+QpydwweM4GiXWpX+i2F7rOlrpuoXFtHJdWSziYW0pUFo/MAAfaSRuwM4ziqqv2w/24rj9kLX9B0/Wfg9e+I9J8RWsklpqlvq626+dGwEkLIYmwyho2znkP7Gv2Uv2o/Bv7WHw4k8deGLCTSb2wvHtNU0ie4WWWzkHKHcANyOhDBsAZ3Dqpqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqp/v/hVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX7TX7XPwp/ZOsNA1D4o2+uyxeJJZ4rMaVZpOQ0IQvvDOmBiRcde9f8Pi/wBkj/oH+P8A/wAE0P8A8fr/AIfF/skf9A/x/wD+CaH/AOP1/wAPi/2SP+gf4/8A/BND/wDH6+G3/BU79mX4qeP/AA/8N/DFj41XVvEuoRWFk11pUUcQlkYKu9hMSFyeTg1+2r+2j4V/Y58IaTqN/wCHZvEfiLxJLNHpGlR3H2dGEQXzZZZdrbEXzIxgKSxYAYG5l/Y2/wCCo+mftHfEq1+EPj/wDB4W1zVkkOj3lletPb3UiIXaF1ZQ0bbFYqcsGIx8pIzVVVVVVVVVfGP4r+E/gd8MvEHxU8bXXlaT4es2nlVSA875CxwpngvI5VFHqwr9mn/gp637Tvxd0n4TeEv2f9Ss5L1JJ73UJdfSSOwtoxl5nUQAkZKqBkZZ1GRnNVVV8Xv+CtFp8Efiv4g+E/jj9njV4r7w7qBtriaHX4yJYuCk8amEZV42V1BIyGGSK8HeLvDvj/wnpHjbwlqcWoaNrllHeWNzGeJIpFDKfY4PIPIOQeRVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX7VngDxP8U/2dfHvw88F2Ud3rmu6S9vYwSTLEryFlOC7EKvAPJNf8Osv20P+id6X/4UFn/8cr4y/Bf4g/APxtL8PfibpUGn65DbR3DwQ3UdwojkGVO+MleQOma+Bf7PHxU/aQ8Q3/hX4TaHb6nqOmWX2y5jnvYrYLFvVMhpGAPzOowOea/4dZftof8ARO9L/wDCgs//AI5Xif8AZk1L4qfsN6B+zX4pvU0TXrbwdo9i84ImS0v7OGA4JU4dPMhKEjOVJI7V4H/4JE/tJar47g0Xx3c+HdE8NQ3C/bNZttRFyZYQfmNvEBvLkdBIEHr6VpOl2WiaVZaLpsfl2mn28dvboSTtRFCqM9+AKqqqqqqqqr/gol8brj4HfsveI9R0e9a213xOy6FpUiNho3nVvNcHqCsCTEMOjbenWv8Aglr8G7f4p/tQ6f4h1azE+leArNtZkDqCrXIYJbKfcSP5o/64/nVVX/BRT4OW/wAY/wBlXxZDDZ+dq/hOE69pjKm51e2UmVQByd0BmXHclfQV/wAEvfjdc/Cf9pzSvCt5eMmhfENBo95EWwv2k5NpJju3m/ux6Cdqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqf7/4VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV/wVB/Zc+Mv7Tmg/D6w+D3h611WbQLu/kvln1CG18tZUhCEGRl3ZKN06Yr/AIdSftsf9E40r/wobP8A+OVe2k9heT2FyoWa2kaORQc4ZTg8/UV8F/2EP2lf2gfA0XxF+F/g6w1HQ5rmW2SebV7e3YyRkBxskcNwSOcV+zP/AME2v2tvht+0H8PPH/i7wHptrovh/wARWl7fzJrlrK0cMcgZmCLIS3A6AZr/AIKN/sUeJv2tfC3hrV/h5q9haeK/B7XIgtdQdo4b+Cfyy6bwDsdTEpUkbTuYEjgj9hD/AIJj/F34YfGjR/jL8cpNL0i38LO8+naRaXq3U91clCiNI0eUSNdxbhixZQCAM1VVVVVVVVVf8FpPjlcTar4Q/Z30a+dbe3h/t7W0RuJJGLRWsbY5+VVmcqeP3kZ7Cv8AgjJ8F7fw98KPE3xx1KyX+0fFuoHTtOlZQStjbffKnqA87OGHfyFqqqv+C1PwYtrTUfBHx90qz2NfB9B1eRUwGdA01qxxxuK/aFJPOEQdBx/wRn+Odz4o+GniX4Ea3etLceDbhdQ0cSNk/Yrlm8yNR/djnBb63A9Kqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv+Csv/J3+o/8AYA0//wBAav8AgjH/AMl38bf9iif/AErgqv28viJ40+E/7KHjn4gfDzXpdG8QaV/Z/wBjvoo0dovM1C2ifCuGU5SR15Hev+HiX7aH/Rd9U/8AACz/APjNeHrma80DTLu5k8yaezieRiMbmKAk/nXjL9vz9s3QfF+uaFH8dtRC6dqVxbKEsLTAEcjLxmHOOO9fHL/gqx8VdZ8GeF/BXwb1P+yL2Lw9Yf8ACSeJJbdHurnUGt4zcpArLtjRZC6l9uWIJXaoBbS/2yf2rtH1hddtP2iPH0lyr7xHda5NcwZzn/USs0WOOm3HbpXgL/gr7b2n7PmoX/j7w5DqHxX0yZLOytbeMw2mqK6krdyY/wBWE2kSRr1Yps2hz5fxA/bs/az+I2szavqPxx8T6OskhaOz8P38mlQQjnCKsBUkAHGWLE8Eknmv2Uv+Cm3xk+GHjHTtE+NXizUPGvgi8lWG9k1FvPvtPU8efHMfnk28FkctkA42k5q1urW/tYb2yuI57e4jWSGWNtyujDIYEdQQQc1+2T+2d4J/ZK8JQyXVsmueMtYjY6NoSzbNyg4M8zAEpCp46ZdhtXozL8Q/+Cgf7X3xU1eSQfFnWNCiuHK2+m+GSdPSIHOFRov3rderOx6c8CvAv7d37Xfwz1hLq1+NfiXVDbvtmsfEdy2pxvg8owuNzLzkEqVYdiK/Yp/bP8M/tb+DrppbCLRPGmgqg1nSEkLRlWyFuICeTExBBB5RuCSCrNVf8FpfHU1x4v8Ahv8ADOGcLFYabdavPEG5dp5BDGzD2FvKAf8Aaav+CLvg+G0+F/xD8f8AlDzdV16DTA5IJC2sAlwO45uxn149K1fVdM0HSb3XNZvobPT9Ot5Li7uZm2pDEilndj2UKCSfQV+0x/wVZ+MXjzxDf6F8BtRPgvwlDI0Vverbo+oXyA/6xncEQhsAhUAYdCx6VH+1z+1PFff2gn7RnxHMuc7W8S3TR/8Afsvs/Sv+CWXx+/aB+POj+Np/i54w/t/RvDrWdtptxcWcUdwZpBI0imSNV3hVVM7wT8457VqFhZ6rYXOl6jbpPa3kLwzxOOJEYFWU+xBIrXLPVPhX8TtQsNPuWj1Lwhr0sUEpIJWa1nIVsjg/MgPFeFfEFp4t8L6P4q08YttZsIL2Abt2EljV1578MKqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqn+/+FVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVXiz/AJGnWf8AsIT/APoxq/4JGf8AJm+mf9h/Uf8A0Nar/grJ+0d8bP2ff+FV/wDCnPH114a/t7+2P7R8i3hl+0eT9i8rPmI2NvmydMfe57V+xB+2/wDtU/FL9qn4f+AfHvxh1DV9A1e8njvbKWztUWZVtZnAJSIMPmVTwR0r9u3x146+Gf7J/j7x38NfET6H4j0eCzls79I0dogb2BZAFdWU7o2deR/F2618D/8Agpb+0jpXxb8K6j8X/jRqupeC7fUUbXLX+z7bMttzvAEcQYnHIAIyQBkV+0L/AMFP/wBpT4w6/ex+B/F198PPCu9lstO0Sbybry+gaW6XEhcjrsKKOw4yfhV/wUG/a1+FPiG21mD4ya/4otY5Q1zpnie+k1SC5QdUJmYyID6xsp96/aP/AOCwhufAGgab+zjpYsfE2t6YtxrOoalCJhocp4a3hRhtlkBUnzGUptKnaSx2Xn7Zv7Wd9q/9tzftH/ENbjfv2Q+ILiKHOc/6lGEeOem3Ff8ABN7/AIKKeLPjL4nh+Anx2vob7xJcW7yaBryxLC1/5Slnt51XCmQIrMrgDcEYN82C1ft1f8FQo/gv4hvfg/8AAK307V/FensYtY1u6Hm22lyg4MEcYwJZhzuJOxD8pDNuCa/+2d+2L43urjVLv9oH4ghogZJv7J1SXT4ox6lLXy0VfwAr4E/8FSP2nvhPrlsfGniqX4h+HDIPtena2wafb3MV0B5it6bt68fd718HPi74J+O3w30X4pfD7UGutG1uDfGJFCywOpKyRSKCdrowKkZIyMgkEE1X7d/jyb4j/te/FLX5JxLHa6/LpduVbK+VZAWqle2CId3HUsT3zX7Fvg2HwF+yf8KfDkMKxMPC9peTICDtmuU+0S9OD+8mbn+dftO/tF+Df2XPhJqPxS8YI92YnW10zTYnCSajeOCY4VJ4UYVmZsHaiMcEgA/F7/gpX+1v8VtWuLi0+Jd34N0tmb7Ppnho/YhCpJwDOv75zjAyXxkZAXNeHv21v2t/DF2t5pn7Rvj6aRW3BdR1uW/TPulwXUj2IxX7G/jb4l/Er9mjwJ8QPi5fW934n8QWDXlxNBarbq8Tyv5DbEwoLQ+WxwAMtwBX/BTvwVb+M/2LvHTtCHutANpqtoxIHltFcRiQ8/8ATF5h+P4V/wAEs/HUvgn9s3wjZmYx2vii0vNIujuxkPC0sY98zQw8f/qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/4Ky/8nf6j/2ANP8A/QGr/gjH/wAl38bf9iif/SuCq/4Ka/8AJj/xJ/7hX/p1tKrwr/yK+j/9eEH/AKLWvir/AMlQ8Yf9h6+/9HvX/BMb9iv4O+O/hG/xv+LHhKx8V32s309tpdlqCebbWlvC3lsxizteRpA/LA7Qq7cEkn/gqh+y98O/gR4x8JeNPhboMOh6T4xiuo7vTLbIgt7m3MR3xqfuB1mHyj5QYyQBmv2V/g5Z/tAftBeC/hHqN9NaWOu3rm9lhIEgt4YXnmCE8BjHE6gkHBIOD0Pxb/4J3fsy+LPhHq/g7wb8J9F0DW4dOkGjarZRstzDdKh8ppJMlplLABg5bIJ6HBFfsN+Jrvxd+yN8LNYvnZ5k8PxWZZurC2ZrdSeTnIiHPev2v/i9qfxw/aL8beOb28aa0/tOWy0pC2VhsoGMcKqOgyq7zjqzse5Nf8E6/i/+xP8As9fCK31/xz8RtEsfiVr8sz6pLcadcSz2UIkZYrdJFiIVSiq7BTyz8k7Vx/wUw+LP7H3x68BaX4y+FfjzR9U+ImkajHCz21jcQy3lgysHR2eNQ2xvLZdx+UbwPvGv+Cd/xGvvhv8Atd+Abi1nK23iG8Oh3sW/aJo7oeWgPribyXA7lBVV/wAFdNTkv/2tBaOXxp3haxt03PkYLzScDsMyHj1ye9f8ElLOC2/ZEtpogd114jv5JMn+IeWnH4ItftyWHiLU/wBkb4p2fhVJHvz4fldljOGa3Rla4A9f3Ky8d+neq/Z2/bt/Y4+K3gzSfh3ctongO4W2jtv+EY1y1jt7IEAJshlI8h1zgKGKuQR8vWvAHwy+HHwxstQs/hr4O0fw7Zaxem/uoNLtlgilnZEQuEX5VyqIMKAOM4ySTX7YFnDZftVfFyC3BCt4y1OQ5Ofme4d2/VjX7JWpSat+y58JL2UMH/4QvSo2LPuLFLWNCxPvtz+NVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVT/AH/wqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq8Wf8jTrP/YQn/wDRjV/wSM/5M30z/sP6j/6GtV/wXH/5op/3H/8A3HV/wTi/5PY+Fv8A2ELn/wBI56/4KO/8mTfFL/sH23/pZBVfBj/gm7+zD4A+Fem+EPGnwr0XxXr01kv9tavqUZlmmuWX94YmzmFASQoj2kAAklssf2s/g7YfAH9ovxz8JNImml07QtQBsGnbdILaaJJ4VZv4mEcqAnjJGcCv+CVX7Jnw5/aF8Y+K/HHxW0pNZ0TwWlrFa6RKSIbu5uPNIeUA5ZI1izs6MXGchSD/AMFTP2Lvgx4C+DFr8avhJ4H03wpqGialBaapb6ZEYbe6tpiUVjEPlV1kKYYAZDsDnC4/Zy8T3ngv9oD4b+KrB2EumeKtOlIX+NBcIHTqOGUsp5HBr9rL4s3XwN/Zx8ffFHTZFTUNG0lhp7sAQl3MywQMQeoEsqHHfGKt92t61F/a2r+U1/dD7TfXRaTYXb55XIyzYyWPUnmvhB+13/wTn+Bnw/0z4b/Dv4u6Bp2l6dAsblNKuxJdyBQGmmYQfvJHxlmP6AAD/goJffs6+IPj3J4y/Zp13Tb/AEDxDpsd1qcOn2slvFbaj5kiyhUdF2h1WNztGNzse9f8ES/iTfywfEf4Q3c5ezgNrrdjGX/1btmG4IHuFt+nTbz1qq+JOpya58RfFOtSl99/rV5cN5j7my8zscnueeteBrODT/BHh7T7YERW2lW0UYY5O1YlA/QV/wAFvNP8STeCvhVqdokh0G21PUor4g/KLp44Db5Hrsjucfj61+yr8VvBPwR+PXhX4m/ELwavijQdGnka5sPLSRxviZFmRJPkZ42YOoYgZUcqcEeBvjf+xn+2loy+HbDUPCHjCaa3O/QNdsEW9hXHzbYJ13fL/fjyAeQ3Q1oGg6P4W0HTfDHh6wisdL0ezis7G1iBCQQRIEjRc9lVQB9K/a7s4L79lT4wwTglV8CazKMHHzJZyuv6qK/Y61KTSf2r/g/dRKxZ/G2lQEK+04luo4zz6Yc8d+neqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr/grL/wAnf6j/ANgDT/8A0Bq/4Ix/8l38bf8AYon/ANK4Kr/gpr/yY/8AEn/uFf8Ap1tKrwr/AMivo/8A14Qf+i1r4q/8lQ8Yf9h6+/8AR71/wSz/AOTL/CP/AF/6l/6WS1/wWt/5Ff4Uf9f+qf8Aou3r/gmV/wAnwfDX/uK/+mq7qq/4J2/8mX/DD/rwuf8A0snrxJo994f8Q6poGph/tmm3s1tcCQYbzI3KtkeuQa+BX/BNb42/tDfDDSfiv4D8cfDyLSdXMypBqGoXkdxA8UrRukipauqnKZGGOQVPev8AhzX+09/0PXwu/wDBnf8A/wAh18Df+CT37QPw4+M/gX4heJ/Gfw9n0rwv4isdUuorDUbxp3S3nSXage0VSTsAwWA9x1qq/wCCv2jNpv7Vllf+UVXVfCVlOGySGKzXER+n+rHH0Pev+CROrQ6l+yY9lFjdpXiq+tpME/eMcEvP4SipI45o2hlRXR1KsrDIIPUEelftO/8ABIm28Ra1feM/2bNf07Rzdu0svhjVNyW8bk5b7NMoYop7RsuB2cLgD4u/A74r/AfxEvhb4s+Cb/w/fSqzW5nCvFcqMAtFKhZJAMjJVjjIBxX/AASX/aj8YRePx+zT4s1mfUtB1SxmuPDy3EhdtPuIU8x4YyeRE0SyNt6KyAgDc1V+1bq8Wu/tOfFfVICDFL4y1QREAjci3Uiq2DyMhQce9fsw6K3h79m34V6LJD5U1p4M0pJkyTiX7JGX68/eLcfyqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqf7/4VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVeLP8AkadZ/wCwhP8A+jGr/gkZ/wAmb6Z/2H9R/wDQ1qv+C4//ADRT/uP/APuOr/gnF/yex8Lf+whc/wDpHPX/AAUd/wCTJvil/wBg+2/9LIKqv+CoX/J9PxM/7hX/AKabOv8AgiD/AMir8Wv+whpf/ou5r/gq3/yZN4v/AOwhpn/pZFXwn/5Kn4N/7GCx/wDShK/4KhaNfaz+xH8QBp4dnsn0+6kjQZLxpfQb/wAFUlz/ALtWFql9fW1lJeQWizyrG1xcEiOEMQC7lQTtGcnAJwOhpP8Agi7+1FIiyR+P/hSysMqw1W/II9R/oVf8OWv2pf8AoffhX/4NL/8A+Qq/4J4/sC/Fj9kr4h+KPGnxJ8SeEtSj1jRV0+0j0O8uJmVvOSRi4mt48DCLggnqeO9VXxc0VvDnxW8aeHniMTaX4hv7Uxkk7DHcOmMnnjFfDPV4fEHw28J69b7fK1LQ7O5TBJGJIEYYz7Gvi18JfAfxw8A6p8NfiRoaapoeqxgSxklHidTlJY3HKOpAIYfjkEg/Hv8A4I9fG3wRc3msfBHWbLx7oiFnhsZpFstSjTrtKuRFKQOMq6lj0QZxWp6Z4q8A+J5tL1ay1Tw94h0S62ywTRva3VlOh7g4ZHBA9CK/4Jm/tOeJf2kfgPPH4+vjfeK/Bl8NNvr18b76FkDwTvj+MjejH+Ix7jyxr9s3WItD/ZM+L97KVxJ4M1K1GQTzPbvCOnvIPb14r9iXRG8Qftc/CKwSDzTF4ssbrbkjAt5BMW49BHn0454zVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV/wAFZf8Ak7/Uf+wBp/8A6A1f8EY/+S7+Nv8AsUT/AOlcFV/wU1/5Mf8AiT/3Cv8A062lV4V/5FfR/wDrwg/9FrXxV/5Kh4w/7D19/wCj3r/gln/yZf4R/wCv/Uv/AEslr/gtb/yK/wAKP+v/AFT/ANF29f8ABMr/AJPg+Gv/AHFf/TVd1Vf8E7f+TL/hh/14XP8A6WT1/wAFP/2R/EPwx+Keq/HXwno8tz4K8ZXRub6WCPI0zUZOZVkx0SV8yK543Oy8EDd+zb+2L8bP2Wby6X4caxaXOj6jIJb3Q9UhM9pNIBjzAAyvG+MAsjLkBd2Qoxcf8Fofi89kY7X4PeD47vaAJZLq5dM9zsDA468buPU1/wAE6/23vid+1J4+8deGvim2ixTWWm21/pFppVmbeOCNZGjn5ZndsmSD7zHGPeqr/gtL8PJntPhv8WLW2zFFJdaJezbejMBPbrn6JcnHtX/BFnx3DJo/xK+GM86rLBc2er2sZPLh1aGYgei+XAP+Biv2xPixffBL9mjx78RdHv8A7Hq1lpht9LnADNFd3DrBC6qcglHlD4II+XkYzXw2/wCCyPxn8O2EVh8TPhz4e8YtCoX7Za3DaVcS+pfaskef92NR7V+2d+234i/bCvfDsd54HsvC2keGRcNaWkV4byWSSbyw7PKUTIAiUBQg6knPGP8Agkj8C/E3ir47r8b59Pmg8OeCbS5jhvHUhbi9uIWgESf3tsUsrMedvyA/eBrxh4m07wV4S1vxlq77bDQdOuL+6bOMRQxtI5/75U14N8Pa58afi5o3hiJmk1bxr4git2dBnEtzOAz85wAXLEnoASasbG00yxt9NsIFhtrSJYYYl6IigBQPYAAVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVP9/wDCqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqrxZ/yNOs/9hCf/wBGNX/BIz/kzfTP+w/qP/oa1X/Bcf8A5op/3H//AHHV/wAE4v8Ak9j4W/8AYQuf/SOev+Cjv/Jk3xS/7B9t/wClkFVX/BUL/k+n4mf9wr/002df8EQf+RV+LX/YQ0v/ANF3Nf8ABVv/AJMm8X/9hDTP/SyKvhP/AMlT8G/9jBY/+lCV498FaD8SPBGvfD/xRbefpPiLTZ9PvYxwTFKhRip7MAcg9iAe1ftI/s6fED9mP4m6h8OvHenyBY3d9K1IRFYdUtd2EniPI5GNy5yjZU8iv2ff+Co/7RfwI8LWHgO6j0bxr4f0uIQ2MOtJILm1iH3Y0njYEoBwA4fAwAQAAPEX/Bar48X1mYPDHwu8D6VOykGe5NzdlfdVEiDP1yPav2FPjt4g/aM/Zq8OfEjxjdwT+I5Li8tNWaCFYk82K4cJtVeADCYjj1Pfqar/AIKP/Dmb4b/tjfEG1+zeVaeILxdctHC4Eq3aCSRh/wBtzMp91Nf8E/PHsPxE/Y7+GOqpOrzaZo66ROoOTG1kxtwG9ykSN9GB71/wU9/bB+JP7MbfDfS/hHrltZa1q13d32oRz2qXCS2sKJGsUisM7XeZjlSDmLqK8Of8FvPGtrpnk+Lv2f8ARNT1HZgT6br0tjFu9fKeKZse2/8AGvj78Zdf/aF+L/iT4xeJ7CzsdR8RTxu9raAiOBIokhjQZ5OI40BY8k5Pev8Agkl8BvE/wk+Aep+NPGOnTafqHxBv47y1tJlKuljFGVgdlP3S5eVgMfdKHvx/wVk8fReDf2Odc0RZwlz4x1Wx0qDB+bAl+0yY9ilsynt83uK/4JEfDibxj+1lb+L5LbfZ+B9Fu79pGHyiaZfs0a/UiaRh/wBcyeoFVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVXxl/Ya/Zw+PnjaX4hfE3whfajrk1tHbvPDq1xbqY4xhRsjcL0PXFfAv8AY5+Af7OHiG/8U/CbwteaZqOp2X2K5kn1Oe5DRb1fAWRiB8yKcjmq+K/wr8F/GzwBqnwx+IenS3/h/WfJ+2W8Vw8DP5UyTJh0IYYeNDwecY71/wAOsv2L/wDoneqf+FBef/HKs7SCxs4LG2XbFbxLHGpOcKowOfoK1j/gmP8Asea9q17repfD/U5LzULmS4uHGvXahpHYsxAEmByTxXwi+EPgT4F+BLL4b/DfTJrDQdPklkggluXnZWkcu53uSxyzHvxXx5/Zj+Dv7Stno9j8XfD9zqkWgyTSWKwX81r5bShQ5JjYbshF69MV8KP2Bf2Y/gp4/wBK+Jvw88GX9j4g0bzvsdxLrFzOqebC8L5R3KnKSOORxnPaq/4dZfsX/wDRO9U/8KC8/wDjlfDL4beEfhB4E0n4b+A7CSy0HRInjsoJZ3mZFZ2c5dyWPzO3U961HTtO1iwuNK1axt72yu4miuLa5iEkcqMMMrK2QykcEEYr4if8EuP2Q/H9/Nqln4S1XwlcXBLSf8I7qJgj3EHlYZVkjTHHyoqrwOOudL/4I8/srWFwJrvxD8RdSQEfubrVrZV4/wCuVsjc/Wvgl+yf8AP2d55tQ+E3w9tdJ1K6tzb3GpSzy3V1LEWVinmSsxVSyKSq4UlQccCqr9rv4HJ+0R+z74r+Gdukf9q3FsLrRpHOAl7Cd8Qz2DEGMnssjGv2E/jKf2c/2pvD+s+JpG07Sr+WTQfEAnBjNvDMwUs+fuiOZInbI6RnvXj34e+CPij4YuvBnxD8L6d4g0W8x51lfQiRCw+6w7qwPIYYIPQivFX/AAST/ZF8RTyTaVZ+L/DSyHKxaVrW9U5HT7Skx7Y5J6n2x4L/AOCTv7IvhO8hvdU0vxR4qMLBli1rV8IxByNy2yQ7h0GDwQOQec+GfC/hvwXoNn4X8I6FYaNpGnRCK1sbG3WGKFR2VFAA9fcnNf8ABVj43W3w0/Zun+H9hfKmu/ES4FhDGp+dbOMrJdSf7pHlxH/rt+I/4JF/AKfxv8YL/wCOGs2LHRfAsLQ6fI6/LLqUybQBng+XCzse4aSI1VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVU/wB/8Kqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr3/AIJX/sW6heT31z8OtUaa4kaSQjxBeDLMck48z1NfBf4JfDv9n7wNF8OvhfpM2naHDcy3KQTXUlwwkkILnfIS3JA4zVftAfso/BL9p/8AsH/hcXhy61X/AIRr7T/Z3kajNa+V5/lebnymXdnyY+vTHHU18L/+CeH7K3wc8e6R8S/APgjULLX9DkeSynl1q6mVGaNkJKO5U/K7Dkd6+KPwz8H/ABj8Bav8NPHthJe6BrkSx3tvHO8DOqurjDoQy/Ming1/w6k/Ym/6Jxqv/hQ3n/xyq+Lf/BPv9lz44fELVfij8RfBWoX/AIi1ryftlxFrNzAr+VCkKYRHCjEcSDgc4z3r4A/sufBn9mSz1mw+D3h660qHX5IZL5Z9QmuvMaIMEIMjNtwHbp1zXxi+DvgL48+Ar34afEvS5tQ0DUJIpJ7eK5kt2ZonDoQ8ZDDDKD1rRv8Agl1+xpoOr2Ou6Z8PdUjvNOuY7m3dtfvGCyRsGU4MmDyBwar4lfCj4bfGLw3J4R+KHgrSvEulOdwt7+3D+U2Mb42+9G+ONyEMPWvE3/BHj9kXXrx7nSp/HXhyNmyLfTNajdFHPANzDM2PqxPA5614Y/4I/wD7IGgTxy6rF418SKhy0Wqa2I1f2P2aOE/kRXwx+FHw5+DHhWLwT8LfCFh4d0WKQyi0tEIDyEANI7MSzuQqgsxJOBzxVV/wWS/Z9uPFnw90D9oPw/ZmS88Gk6drQRcs1hM48qQ+0czEY/6eCTwtf8EXfjta203i79nTWr1Ukun/ALe0JXbG9gqx3UYz32rA4UdhKexr43fsvfAb9oy3t4vi/wDDqw1y4s4jFa3weS2urdSc7VniZX27iTsJK5JyOTWq/wDBGv8AZO1G5E9nr/xG0xASfJtdXtmU+3722dv1718I/wDgm1+yR8HtWg8Q6X8PJPEGrWrh7e78RXTX3ksOhWI4hyDghihII4Iqv+Cx3xytfG/xm0L4MaHfLNY+AbJpdR2Hj+0LkKzIccHZCsP0aRx2Nf8ABJ39n2f4Rfs8t8QtfsGt9f8AiTNHflZFw0enxgraqR23BpJfpKvTFVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV48/YD/AGRfiZ4w1Xx74z+D8F7rmt3BuL+5h1i/tRNKQAzmOGdEDHGSQoySSckknRNG0/w7oth4f0pJlstNto7a2Wa4ed1jRQqhpJCzucAZZiSepJNVVfGX9kL9nn9oLxFa+Kvi/wCApvEGp2NoLS2kbW762SKIMzbVjhnRBlmJJ25PGScDHwt+E/w8+Cng21+H/wAL/DMGhaDZySSRWscskpLu252aSRmd2JPVmJwAOgAFVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVT/AH/wqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqrxD4f0XxZoOo+F/EemwahpWrWslre2k67knhkUq6MPQqSK+H3/BP79kf4U+M9J+IXw++FU2jeIdEm86xvYvEepuYmKlT8r3JVlKsylWBUgkEEE1VVWvf8E5v2NPFPii/8Z+I/g82pa1ql9JfXt1deI9UkNxO7l3d1NztOWJJBGOcYxUEEFrBHa2sKRQwoEjjRQqooGAABwABxiqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqp/v/hVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVP8Af/Cqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqf7/AOFVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVU/3/AMKqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqp/v/AIVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVT/f/AAqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqKK3JAJ9xXlp/dH5V5af3R+VeWn90flXlp/dH5V5af3R+VeWn90flXlp/dH5V5af3R+VeWn90flXlp/dH5V5af3R+VeWn90flXlp/dH5V5af3R+VeWn90flXlp/dH5V5af3R+VeWn90flXlp/dH5V5af3R+VeWn90flXlp/dH5V5af3R+VeWn90flXlp/dH5V5af3R+VeWn90flXlp/dH5V5af3R+VeWn90flXlp/dH5V5af3R+VeWn90flXlp/dH5V5af3R+VeWn90flXlp/dH5V5af3R+VeWn90flXlp/dH5V5af3R+VeWn90flXlp/dH5V5af3R+VeWn90flXlp/dH5V5af3R+VeWn90flXlp/dH5V5af3R+VeWn90flXlp/dH5V5af3R+VeWn90flXlp/dH5V5af3R+VeWn90flXlp/dH5V5af3R+VeWn90flXlp/dH5V5af3R+VeWn90flXlp/dH5V5af3R+VeWn90flXlp/dH5V5af3R+VeWn90flXlp/dH5V5af3R+VeWn90flXlp/dH5V5af3R+VeWn90flXlp/dH5V5af3R+VeWn90flXlp/dH5V5af3R+VeWn90flXlp/dH5V5af3R+VeWn90flXlp/dH5V5af3R+VeWn90flXlp/dH5V5af3R+VeWn90flXlp/dH5V5af3R+VeWn90flXlp/dH5V5af3R+VeWn90flXlp/dH5V5af3R+VeWn90flXlp/dH5V5af3R+VeWn90flXlp/dH5V5af3R+VeWn90flXlp/dH5V5af3R+VeWn90flXlp/dH5V5af3R+VeWn90flXlp/dH5V5af3R+VeWn90flXlp/dH5V5af3R+VeWn90flXlp/dH5V5af3R+VeWn90flXlp/dH5V5af3R+VeWn90flXlp/dH5V5af3R+VeWn90flXlp/dH5V5af3R+VeWn90flXlp/dH5V5af3R+VeWn90flXlp/dH5V5af3R+VeWn90flXlp/dH5V5af3R+VeWn90flXlp/dH5V5af3R+VeWn90flXlp/dH5V5af3R+VeWn90flXlp/dH5V5af3R+VeWn90flXlp/dH5V5af3R+VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVebF/wA9U/OvNi/56p+debF/z1T8682L/nqn515sX/PVPzrzYv8Anqn515sX/PVPzrzYv+eqfnXmxf8APVPzrzYv+eqfnXmxf89U/OvNi/56p+debF/z1T8682L/AJ6p+debF/z1T8682L/nqn515sX/AD1T8682L/nqn515sX/PVPzrzYv+eqfnXmxf89U/OvNi/wCeqfnXmxf89U/OvNi/56p+debF/wA9U/OvNi/56p+debF/z1T8682L/nqn515sX/PVPzrzYv8Anqn515sX/PVPzrzYv+eqfnXmxf8APVPzrzYv+eqfnXmxf89U/OvNi/56p+debF/z1T8682L/AJ6p+debF/z1T8682L/nqn515sX/AD1T8682L/nqn515sX/PVPzrzYv+eqfnXmxf89U/OvNi/wCeqfnXmxf89U/OvNi/56p+debF/wA9U/OvNi/56p+debF/z1T8682L/nqn515sX/PVPzrzYv8Anqn515sX/PVPzrzYv+eqfnXmxf8APVPzrzYv+eqfnXmxf89U/OvNi/56p+debF/z1T8682L/AJ6p+debF/z1T8682L/nqn515sX/AD1T8682L/nqn515sX/PVPzrzYv+eqfnXmxf89U/OvNi/wCeqfnXmxf89U/OvNi/56p+debF/wA9U/OvNi/56p+debF/z1T8682L/nqn515sX/PVPzrzYv8Anqn515sX/PVPzrzYv+eqfnXmxf8APVPzrzYv+eqfnXmxf89U/OvNi/56p+debF/z1T8682L/AJ6p+debF/z1T8682L/nqn515sX/AD1T8682L/nqn515sX/PVPzrzYv+eqfnXmxf89U/OvNi/wCeqfnXmxf89U/OvNi/56p+debF/wA9U/OvNi/56p+debF/z1T8682L/nqn515sX/PVPzrzYv8Anqn515sX/PVPzrzYv+eqfnXmxf8APVPzrzYv+eqfnXmxf89U/OvNi/56p+debF/z1T8682L/AJ6p+debF/z1T8682L/nqn515sX/AD1T8682L/nqn515sX/PVPzrzYv+eqfnXmxf89U/OvNi/wCeqfnXmxf89U/OvNi/56p+debF/wA9U/OvNi/56p+debF/z1T8682L/nqn515sX/PVPzrzYv8Anqn515sX/PVPzrzYv+eqfnXmxf8APVPzrzYv+eqfnXmxf89U/OvNi/56p+debF/z1T8682L/AJ6p+debF/z1T8682L/nqn515sX/AD1T8682L/nqn515sX/PVPzrzYv+eqfnXmxf89U/OvNi/wCeqfnXmxf89U/OvNi/56p+debF/wA9U/OvNi/56p+debF/z1T8682L/nqn515sX/PVPzrzYv8Anqn515sX/PVPzrzYv+eqfnXmxf8APVPzoSRscCRSewzVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVZ/8fUX++KqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqrP/j6i/3xVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVZ/8fUX++KqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqrP8A4+ov98VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVftsaxq2gfso/EzWdB1W707ULTQ3e3u7SdoZYW3r8yupBU+4Nf8L++O//AEWzx7/4Ul3/APHK/wCCaXxb+K3iz9rzwtofin4neLNZ06ax1EyWeoa1cXETlbSQqSjuVOCARkcGqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqrP/j6i/wB8VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVft3/8AJn/xU/7AEn/oaVX/AASz/wCT0PCP/XhqX/pHLVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVn/x9Rf74qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/bv/5M/wDip/2AJP8A0NKr/gln/wAnoeEf+vDUv/SOWviT8YfhZ8HdLTWPih4/0TwzazEiA6hdrE85HURpndIR6KCauv8AgqB+xVbX5sR8V7qZFcK1xF4fvzGPU/6ncQPYH2zXwq/aI+B/xwiZ/hV8TtC8QzRpvktILjZcxr/eaBwsqr7lQKqqqqr9oL9v74Bfs2eMpPh/49PiS712G1juXttL00SgJICV+d3Rcnb0z3Hvi8/4LM/s9JLt0/4a/ESaLb96a3s4mz9BcNx+NeFf+Cvn7K2vXEdrrum+N/De7G+4vtKjmhXPXm3lkcgY/ud6+GvxY+G3xj8Op4r+GHjXS/EelsQrTWU4YwtjOyRDh4nxztcA+1VVVVVVfte/8FFv2lfgz+0f42+GXgjVfD8eiaFdQxWaXOkrLIFa3ikOWJ5+ZzWhXc2oaHp1/cEGW5tIpHIGBuZATgfU1VVVVVf8PIf2m/8AhqT/AIVN/avh7+wP+E//ALE2f2QvmfZft/kY35zu2d/Xmqqq/aH/AG2vgH+zHrVh4Y+JGuahLrd/ALhNO0uyNxLFAWKiWTJVVUlWABbccEgEV8OPiL4O+LfgjSPiJ4A1mPVNB1uDzbO5RSu4AlWVlYAqysrKykAggg9K/wCCjf7SPxO/Zm+FvhnxZ8LbvToNQ1TXxZXDXtmLhTF9nlfgEjB3IvNf8E5P2kfid+018LfEviz4pXenT6hpWvmyt2srMW6iL7PE/IBOTudua+IvxE8G/CfwVqvxC+IGuRaRoGiw+beXcis2wFgqgKoLMzMyqFAJJIAHNfs8/tufAT9pzXtR8K/DfWdSj1nTYDcGx1Sz+zSTwBgrSx4LBlBZcjIYbgSMVVVVVXjvxz4T+Gfg/VfHvjnWodK0LRLc3F9eShmESDA4VQWYkkAKASSQACTX7P8A+3N+z9+0r4qvvBPw61zUotbs4nnjtNUsjbNdwoQGkiOSGAyDtJDY524BIqqqqqqqqqqqqqqqqqqqqqv29f29f2g/2ev2g7z4cfDjUtDh0aHSrS5RLvTFnffIpLfMT046V8DPFuseP/gl8PfHfiGSJ9V8R+FdM1K+aKPYhnntY5JCq/wjcxwO1VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVWf/AB9Rf74qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/bv/wCTP/ip/wBgCT/0NKr9m/456l+zl8UIPitomkw6lqmnadeQWMFwSIhPPA0SO+OSql9xUYLYxkZyPH/xC8a/FPxXfeN/iD4lvdd1vUXLT3d3JuY+iqOiIOgRQFUcAAVWh67rfhnV7TxB4b1i90vVLCUS2t7ZTtBNA46MjqQyn3Br/gnT+3LcftHaJN8MfifdwL8QtAthKl0AIxrVqMAyhRgCZCQHUcEEMBjcFqqqq/4Ky/8AJ3+o/wDYA0//ANAav2cf2Af2RfEnwF+HXjDxD8HrbUtZ8ReE9L1HUbq51S9Yy3E1rHJIwUTBUBZzwoA6cV8V/wDglB+y5440i5TwDpep+AtZMbfZ7qxvpbuDzOxkgnd8r/so0f1rw54j+Mv/AAT7/aaubCS4aLVfDN8kOrWUM7C11myYBgD/AHkkiYMhIyhIOAy4HhTxLpHjXwto3jHw/cefpmu6fBf2UpGN8M0ayI3GeqsDVVVVVV/wUS/5PQ+J/wD1/wBt/wCkcFeFf+RX0f8A68IP/Ra1VVVVVf8AN93/AHVv/wBy1VVV+3v/AME6/in+0F8Yk+Lnwn1zRHOpWNva6lYatdPA0MkQ2LJGwRgUKBcrwQQSN27A/ZD+AVz+zR8BtA+FOpa3Fq2p2bzXOoXUG4QmeaQuyxBgDsUEKCQC2CxA3YH/AAWc/wCSEeCP+xuH/pJPX/BGP/khHjb/ALG4/wDpJBX7XnwCuf2l/gNr3wp0zW4tK1O7eG50+6n3eSs8MgdVlCgnYwBUkAlchsHGD+wR/wAE6/in+z98YW+LvxY1zREOm2Nxa6bp+lXTztNJKNhkkYoqhAm4heSSwJ27cGqqvin4k1Dwb8MPF/i/SFha/wBD0G+vrUTKWQywwO6bgCCVyoyMjiv2HP2y/wBo79oH9r/wxoHxK+I1xdaHPaahI2kWlvFaWuUtJGXKRqC+GAI3liCAc8Cq/aq+B8v7RvwG8UfCK01lNKvNYiiksruQExpPDMk0YkAydjNGFbAJAYkAkCv2Ff8Agm98WPgP8cbT4wfFbXtBji8P29zHptlpN3JcPczTRPCXkJRQsYjkcgcsW25AxVVVVVVVVVVVVVVVVVVVVVV/wVl/5O/1H/sAaf8A+gNX7K//ACbB8H/+xC0T/wBIIaqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqs/+PqL/fFVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX7d//Jn/AMVP+wBJ/wChpVeA/BHiL4leNNE8AeEbE3es6/fRWVlDnAMkjAAsf4VGclugAJPSvgj/AMEvf2aPhr4Xtbbx74Wj8e+JJIR9u1HU5JBDvx8whgVgqJnoW3P6tziv2tP+CWHwt8S+C9T8X/s6aFJ4a8XadC9xHo0Vw8lnqoUEtEqOWMUpAAQqQmeCo3b1dHjcxyKVZSQVYYIIr4DfFTU/gl8Y/CPxT0mV1k8PanFPOidZrcnbPF9HiaRP+BVbXNve20V3azLLDPGHjdTkMpGQR7EVVVVf8FZf+Tv9R/7AGn/+gNX7K/8AybB8H/8AsQtE/wDSCGq/4LAvo7/tV6cumGI3KeDrIagEPIn+0XJG738ow/hiv2JrfULb9kn4Txalu84+F7R13dfLZd0f4bClfBv/AIKT+PviX+1RY/s86v8AC/RNNtLjXL7TZb2O7lM0f2dZjnaeMkxAEdsmv2qf+Cs9p8P/ABZf+AP2evD+j+JJ9MdobvxDqTvJZmYcMsEcbKZQp48wuFJHAZcMf2CP2hfHf7TPwKk+InxEttIh1aDXbmwA0y3eGNo40iZSVZ3+b94RkEDAHHUn9or9qD4Sfsv+FYvEnxN1qRZ73cum6TZIJby/dQNwjQkAKMjLsVUZAJyQD42/4LIfGbXtUey+FHwm8NaVbSMVhGpmfUrlgOhHltEoJAyRtbHqcZrwb/wWP+OOg6slp8UfhX4V1e0jIE0eni40y6XJzks7ypwCMDYM4685r9m79qz4RftS+Gpdb+G+rypf2IX+0tFvlEV5Yk9C6AkMhwcOpKnpkEECv+CiX/J6HxP/AOv+2/8ASOCv2kf+CjXwy/Zl0vTfAehaW3jLxzFptubjTobjybfTt0SlftE2G+cghhGoJx94pkE3v/BYv9p6fUvtVl4T+HdtaqfltW026kBGT95vtIYnGBkbRx0Ffssf8FVfA3xj8R2Pw9+L3h2DwV4g1KRYLDUILky6fdzMQFjJYboGYnC7iynuwJANftU/tS+Df2T/AADa+N/F+haxrB1O8+xWFpp8YxJPsZ8SSMQsa7UY55JwcKcHHiP/AILB/tFeJr2SP4b/AAo8J6Zaqd3lzQXOpzoM8ZdXjXHb/V/lXgz/AILG/HXQdZS2+J3ww8KaxYxvieKxS4027X6M7ypxxwUGfXnI+Anx++HH7SPw/tviJ8NdUeezdzDd2lwoS4sJwAWhmQE7WAIIwSCCCCQc1X/N93/dW/8A3LV+1V/wVO8AfBTxDf8Aw8+FWgReN/E2myNBf3UlwYbCxmUkNHuUFp3UjDBdqg8b8ggRf8Fiv2n11MXk3hT4dSWpIzZjTLpVA4zhvtJYH3JIyemOK/ZE/wCCkXw4/aW1iD4f+J9HPgzxvOp+yWclz59rqRUEkQS4Uh8Any3AOPulsHFft0ft8+L/ANkr4h6B4L8O+ANH1+HWNF/tB5726kiaNvOkj2gJxjCA/jX7Mvxc1H48/Arwl8W9W0i20u78RW0sstnbSM8cRSeSPCluTwgPPrX/AAWc/wCSEeCP+xuH/pJPX/BGP/khHjb/ALG4/wDpJBX7Q3xPv/gv8E/F/wAU9L0u31K78Nac11FaXDlEmIYDDFeQOe1fsOf8FB/GP7WXxZ1b4c+Ifh7o2g2+neHZtVS5srqWR2eO4t4ghDcYInJz6qKr9qH/AIKyeFfhf4m1DwB8EfC1r4v1XS5nt7zWL6dksIpkO1kjRMPcAEEFgyLx8pYHNWv/AAV5/at065j1bWvAXgWbTLhvlibSbuBCDziOT7QecdCd30NfsV/taT/tdeAtW8Wz/Du58Ly6HfLYzt9rFzbXMxQO3lPtVgVUoWVl48xcM3OP20f+CkPjb4ffEH4mfs4WXw20O701LOTShqUt3Ks2y5s13PtHy5HnHA6cCv2bfjtq37Nvxb0v4t6HoNprN5pcFxElndytHG4miaMkleeA2a+HH/BX34j+OPiH4X8F3Xwf8NW0Ov61aafJPHfzlolmmSMsARgkBs4NfHP4hXnwm+DnjL4madp0N/deGNFudQhtZ3KpM0SFgrEcgHHav2Kf+CinjT9qf4xTfDLX/hzomh20Wi3GoC6s7uWRy0bxKFw3GD5h/Kq/aI/aj+D/AOzD4aTX/ibr7JdXSt/Z2kWaiW9v2A58uPIwvYu5VASAWyQD8Qf+CznxU1C8mi+F/wAKfDGjWOSscmtTTX85XH3v3bQqp6HGGA6ZPWvDn/BY/wDaN0+6U+JPBHgLWLTdl0itLm1lxxwriZlHfqh61+z18WLz46fBrwv8Wb7wm/hqTxJatcLpr3f2kxIJGVW8wKu4MFDj5Rwwzzmqr4i/8FJ/H3gb9rGb9nRPhfokthH4otNHXUJbuVZjFO8QEu0cZ2yZA6dK/bL/AOCm/h/9n/xNc/DD4UaFp/i3xdYZTVLm7mYWWmS/88WEZDTSj+JVZQnQsW3Kv/BOj9rX4oftWaF44vvibZaBBN4cvLOOzOlWkkGVmSUsHDyPnBjGMY6nOeMfH79on4W/s1eCm8bfE/XGtoZWaKwsbdPNutQlAz5cMeRk4xliQq5G5hkV4s/4K9fHrxnrkum/BD4N6Ja2oyY47uC41W8ZAfvERNGq5HUbWxnr3rwP/wAFhfjN4X19NI+N/wAI9EvbaJ9tyumxz6bewg9SUmeRWIB4XCZx1HWvgx8afh58fvANj8R/hnrY1DSrzKOjrsmtJlA3wzJ/BIuRkcgggglSCf26v24fiR+yH4x8NaVovw50TXtE8S6bJNBeXlzLFItxFJtmjwvBAV4Wz/tkdsn9lD49R/tLfAzQPiw+m2+m32oPcQX9hBIZFtZ4ZmQqCeSCoRx7OKr9pT/gq14j+Dnxv8V/C3wV8ONB17TfDN2tmb+6vZY3knWNfPUqowNkpdP+AZ714F8YaR8QvBOgePNAk36d4i0y31C0JOT5c0auoOO4DAH3qSSOKNpZZFREUlmY4CgdST2FfDT/AIKm/Ej4v/HzRvhB4I+EmgTWGv8AiH7DaX0l5MZVsvMO64ZRxlYVaQgcfKa+MfxW8NfA/wCGeu/FTxdbajcaT4fgWa5i0+386dwzqihVyB95lySQAMkkAE146/4LJfFPXNTk0/4O/B7QrCByywNrEk2o3DgA/OEhaJVbHOPnAx1NaX/wV8/ah8O6nGnjLwD4KvbdtrPbSaddWcxTkfI/nEDPqUYcfWv2S/2z/hn+1roFzL4bgm0PxPpUYfVPD93MJJYUJwJYnAAmiyQu/CkHAZV3Lmvjj/wUn8ffCT9qTUP2frX4X6Jd2Vnq9jZJf3F3KkzR3McL79o46TceoxX7aH/BSrw1+znr0vwz+GuiWXi3xrbL/wATFrmdlstKYjISTZ80suMZRWXaCMtn5a/4JyfthfFX9rFfiGPibYeHbc+Fm002J0m0kgyLn7VvD75HyB9nTb0Iy2SeMftAftFfC79mnwQ3jf4m6y0EUrGKwsLdRJdahMBny4Y8jPGMsSFXIyRkV47/AOCyXxp1rV2tfhZ8L/C2jWMkmyBdTE+o3T8gLyjxICf7uxuuAeMn4X/8FjPHula9b6T8ffhRpklgzIs93oCS2lzbqQMyGCd3WU98Boxzx6V/wUl8f+EPin+0bF4/8B63Bq2ha14Z06ezuosgMuxgQQeVZSCrKQCrAggEV+yv/wAmwfB//sQtE/8ASCGqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqrP/j6i/3xVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV+3f/yZ/wDFT/sASf8AoaVX/BLuytLz9tLwW9zAkptrTUpYtwzscWUwDD3wTVV+1/4W0/wZ+1F8UPDukxJFZweJrySCJBhYklcyBFHYKH2ge1V+zfqU+tfs7fC3WLpiZ77wXpE8hZix3PZRMeTyeT1NVVVX/BWX/k7/AFH/ALAGn/8AoDV+zP8Atf8A7L+k/s8fDPw9rHx08I6dqei+ENKsL+0vtQW3kguIbSJJEKvgnDKRkZHHBNfGT/gp/wDst/DLRrl/Cnis+PddVD9m03RY38pn6DzLllEapkclS7AchTxXgnwX8Yf+ChX7Tt5qN0jm88Q363Wu6lFCxttFsVwoxnOFSNBHGjHLEKCeS1eHtB0rwtoGmeF9CtFtdN0ezis7OBekUMSBEUewVQK+J/jDxF4D/aW+IPinwnqcun6ra+KdbjguomKvF5s08TMpHIba7YPY4Nfsb/8ABNr4R/Dz4f6R4t+Nngmw8WeONXtUuLuz1iEXFppgf5hAsDDYzqNodnDHcCFwOthoPwz+CHgrVrvw34Y0Pwj4b0uGfU76DSdPjs4Iwke6WXy4wq52RjJxztq7v/iT/wAFB/2t7a1uL+SG78X6k0NokrF49G0yIM+1V6YihVmIGN75P3nJr4Ifs9/Cb9nnwpbeFPhh4TtNPWOJUur9o1a8v2GMyTzY3OSRnHCjooUAAfHn9nL4T/tG+ELrwp8SfDFrdSSQslnqkcSre6e/O14ZcErgnO3lW6MCCRWha18SP2B/2r7iJLppNT8E6qbTUI4iUj1awfaxUqSfkmhZHUHJUlG+8oI0TWdN8R6Lp/iHRrpbiw1S1jurWZekkUihkYexUg/jX/BRL/k9D4n/APX/AG3/AKRwV/wT/wD2GPB1r4J039ob47aJB4t8beM4xqtnDrEQuYtPhly8cpR8h55FZZC7glNygBWDE+JfA/gzxnobeGfF3hPR9a0l4jEbG/so54dmMYCMCAMAfpX/AAUe/Y80f9mbx/pvij4e200fgbxgZTaWzsZP7NukwZLfcckoVZWTcScBxzsyf+Cbnx41P47fs0aZL4lv5bzxD4QuW0TUbiZ90lyI0VoZWPUkxOiljyzI5PWvif8ACT4dfGfw9B4U+J3ha11/Sba+ivo7S5LBPPizsY7SMjDMCDwQSCCCRXhzwt4Y8H6XFofhHw5peiadAMRWenWaW0KfREAUflX7eP7OvgT43fALxdquqaFZjxR4Y0a51LRtVWEC4ikt4zL5W/GTHIEKFTx8wPUA1/wSG+Jup+Fv2j734cC6Y6X420aYPb7sL9qtQZo5MeojE6/8D9qr403upab+0X481HRbme31C18a6jLaTW7FZI5VvZCjIRyGDAEY5ziv2L/2Bvhv8BPBWl+JPHvhfTvEPxIv4Uub++1C3S4GlyMAfItg2VTZ90yj5mO45CkKPiH8Kvhv8WdDm8N/EnwTo/iLT54yhivrVZGQeqP96Nh1DKQQeQRX7aH7N+pfsf8Ax5TSPCuqX66JeKmr+F9S8wrPCoc/IZFx+9ikTG4YONjcFsD9kn4zv+0B+zz4N+J94y/2nf2Rg1VQAMXkDGKZsD7oZ0Lgdlda/wCCzn/Jd/BH/Yoj/wBK56/4J2/8mX/DD/rwuf8A0snr/gs5/wAkI8Ef9jcP/SSev+CMf/JCPG3/AGNx/wDSSCv27/8Akz/4qf8AYAk/9DSv+CNn/Jz3ib/sQrv/ANLrCv2s/E2veDf2Zvib4l8LtImqWPhi9a2liba8BMZUyqexQEuP92v+CVngf4XeOf2mmt/iRZWGoXGmaHNe6DY3wDRzXqSRDcEbiR0jaRwpBxgtjKZF/pmm6rYS6Vqmn215ZzpsltriJZI5F9GUggj2IrwT8PfA3w10qfQvh/4U0zw9p1zdyXklnp1usEXnSY3uEXAXOBwABxX7RHgXwRe/Bz4ka5eeDtDn1L/hFdTk+2S6dE029bSTa28ruyMDBzxgV/wTH0fSNd/bD8Kabrel2moWkljqJe3u4FljYi0kIJVgQcEA1b/DP4b2k8V1afD7w1DPC4eKSPSYFZGByGBC5BBAOa/bL/5NQ+LX/Yo6h/6Jav8AgkH/AMnX3f8A2KN9/wCjrevHvjPRPhz4I17x94knMWl+HdNnv7th18uJC7BR3YhcAdyQO9aTZ/Ff/gob+1UkF9qHlan4ouneWZgZYNE06IFsKOPkij+VRxvcjJ3OTXwK/ZP+Bf7POg2eleAPAunf2jAi+frd5bpPf3UgHLtMwyuTk7U2queAK8W+BPBHj7TX0fxx4P0XxDYyLta21SwjuoyP91wRXh7w/ovhTQNN8L+HNOhsNK0i0itLG0hGEghjQIiKPQKoH4VVft467qPhf9u/x54m0aYRX+ka3Y3drIRnZLFbW7ocezKK/YK/YC8D+LvBNl+0V+0Zo58Xa/4yZ9S0/TtTYyW6QSnctxOn/LaWXJk+fKhXU43cjwZ8MPhj8MIr0fDv4f8AhnwnFfbGvF0bSobBJtgbaziJVDEBm5PTJr4meJ/Fv/BRH9tm08L6Nq0sei6lqjaboeeU0/SYNzyTheBvaNJJiDyWYLnAXHwg+DPw4+BXgqy8BfDPw3baVptpGokdUBmu3A5lnkxmSQ85Y/QYAAH7U/7MPgH9p34aan4X8Q6NZr4gitJDoOtGICfT7kDKYfGfKLYDp0Kk9Dgj/glf8Zdc+F37TEfwp1G4kj0bx2kthd2sh+WG+hR3gkx2bKvF7+bz0GP+CtHwsPjr9mE+NLK3D33gPVYb4sPvfZZT5Eygem54nPtGa/4IvfEg3fhX4hfCO6nGdOvrfWbJCckrMnkzY9ADDB+L18UvHumfC34beJ/iPrBH2Tw1pNxfyKTjzPKjLBB7sQFA7kitC+F/ib4tfCv4uftB6jcXFzceEtQ0+5vpjz9plv7mRZWPrhirHHTIzwa/4JRfFb/hYH7Llt4QvbppdR8BalNpriQ5Y27nzoG/3QJHjA7CL6V+3H8Tj8JP2VviF4qtrow39xpTabYMpwwnuyIFZfdRIX/4BX/BHP4TjxH8ZPE3xbv7Pfa+DtKFpZSMMbby7JXcvqRDHMp9PNHqK8Y+EfD3j/wnrHgjxZpyX+ja7Yy2V/bMxXzYZFKuAykFTgnDAgg4IIIzXw/+FPw0+FGkJoXw18C6J4bskQKY9OskhMmAOXYDdI3AyzEk9zXxd+Dnw8+Ofgm/8B/Ejw5a6pp17C6RvJEpmtHIwJYXIJjkU4IYenORkV+y94m1v9nD9tfw3p8d+T/Zni5vDeqlTtWe3knNrMSO4Gd4B/iRT2zVf8FINSu9F/b08e6xp8my6sLjR54G/uummWbKfzAr9gv9hLwl8UfDK/tPftK6c/ivWfGV1NqGnaZfuTAUeRibq4QY815G3MqsSgVgSCSNvgj4UfC74ZNfP8Nvhx4Y8KHUxGL0aJpEFiLny93l+YIlXdt8x8Z6bj61+1F8VPGX7aH7V8ul+Gbhr60u9YXw/wCELPcRFHb+b5aSYGceYx8125xu9FAH7M/7J/wp/Zh8HWeieD9Bs7jX2gUar4hmtx9rv5cDed5yyR5HyxA7V9yST8YvgZ8Lvj14TuvB3xO8JWWrWs8TJDcNEoubNiOJIJcbo3B5yOD0IIJB/aI+DWq/s/fGfxT8I9WuftTaDebba6K7ftNtIiyQS4HALROhIGcNkdq/ZX/5Ng+D/wD2IWif+kENVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVWf8Ax9Rf74qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/bv/AOTP/ip/2AJP/Q0qv+CZ/iLSfDn7Z3gOXV7pLeLUBe2MUjnA86W0lWJfqz7UHuwqppYoInnnlSOONSzu52hQBySewxX7R3j+z+Knx68f/ELTZTJY654hu7iycjBa38wiE/8AfsJUcck0iwwozu5CqqjJJPQAV8LfC8ngj4Y+EPBUqIkmgaDZaeyoAFBhgSMgAcY+Wqqqr/grL/yd/qP/AGANP/8AQGr4a/8ABIOx+KHwv8FfEix+P8+kf8JV4csNVlspfC4uvIe4t0lZFcXUeVBfAyueK8Cf8EaPg1ot5Fd/EH4oeJvE6REE21nbRabFLgDIfmV9p5+6ynpz6/C74Q/DT4KeGI/B3wt8G6d4e0pG3vFaxndM+AN8sjEvK+ABuck4AGeKrU9OtNX/AG3brSb+IS2178VXgmjYZDI+qFWH4gmq/b61e70P9jr4p3tk5WSTRfszEHHyTzRwuPxWRhX/AARu0Wxvv2jPE+sXMYefTfCE32bI+40l1bqzD32gr9GNVX/BYLR9P0z9qvTb2zQLNq3g2yuboj+KQXF1CCf+AQxj8K/Yl1K81X9kn4T3V82ZE8MWsC9fuRL5adf9hFr/AIKJf8nofE//AK/7b/0jgrwfBDbeEdDtreNY4otNt0RFGAoEagAD0xVf8FdNJsdQ/ZMF9cqpm0zxPYzWxIGQzLLGf/HZG/Kv+CJ19dS6Z8X9Mdv9Gt59GmjHPDut4H9ukaV+1d+1v8Ov2TvBcOv+LFk1TW9ULpouhW0gWa9dQNzFjkRxLldzkHGQAGJAq2/a5/4KO/th6xeaf8CdJvNJ0mKTy5I/DdlHbwWxxlVlv7g5R9vOBImeSF6AeJP2Nv8AgpjqXhnVPEnjj42akLWzs557qzvvHt3M3lBC0ihE3pggEbQQO3Sv+CZX/J8Hw1/7iv8A6aruqnghuf26JLa4jWSKX4slHRhkMDquCCPTFVX/AAWp0mxl8FfC/XGVftltql/bRtgZMckUTN79Yl/Ov+CO99dXn7LGs29w2Y7HxteQwA54Q2tm5/8AHnav+Czn/Jd/BH/Yoj/0rnr/AIJ2/wDJl/ww/wCvC5/9LJ6/4LOf8kI8Ef8AY3D/ANJJ6/4Ix/8AJCPG3/Y3H/0kgr9u/wD5M/8Aip/2AJP/AENK/wCCNn/Jz3ib/sQrv/0usK1zRdK8S6Lf+HNdsYr3TdUtZbW8tpRlZoZFKOjexUkH61+0z/wTp+O/7O3i+fx58E7XWvE3hS0uftWmajozOdT0nByokSPEm5O00YIwNx2E4HwL/wCCs/x1+HEtvoHxh0u28faTbkRSTzAWepQgcf61RskIHUSJuYjlxya+CHxz+G/7Q3gK0+Inwx1v7dp07GOaGVQlxZTDG6GaPJ2OMg4yQQQQSpBPx+/5IR8SP+xR1X/0kkr/AIJZ/wDJ6HhH/rw1L/0jlqv2y/8Ak1D4tf8AYo6h/wCiWr/gkH/ydfd/9ijff+jrev8Agptrd5on7Fvjz7C5SS/ewtGcHBVHvYd4/FQV+jV/wRa0LTrn4k/EjxJLEhvtP0O0tYHI5WOaZmkx9TBH+VVVVX/BRL/k9D4n/wDX/bf+kcFeA9OtNI8DeHdJ0+IRW1lpNrBBGBgIiRKqgfQAV+0TrN14d/Z++JviCwYrc6X4N1a6hIOCHjs5WXB7cgV+zXrf7QPh34lprP7M9lqdz4zt7CbaNO0mLUZUt22rI3lSxyLjlQW25GevNf8AC6v+Cx//AEBvHv8A4QFh/wDIlf8AC6v+Cx//AEBvHv8A4QFh/wDIlfs8/syftXWX7UHw88e+Kvgv4ysI4/GthqGrajdaM8MUcf2tHuJGO0Ko27z0AGeMV8SfBGmfEv4e+Jfh5rKg2XiTSbnTpiRnas0bJuHoRuyD1BANf8E4PFWqfBj9trS/BniDdZPrBvvDWqRM2NkwyUQjuftEEa/jX/BXj4qjwb+znYfDizn23vj7VkikUNg/ZLUrNKR/20+zLj0Y1+w1+zNa6j/wT51bwbrdqkN78XbK/vZGmTmETR+TaP8AQLFFMvu+a/4JKfEW++HP7S2u/B/XjLaDxfp0ts1rIduzULItIoYeoj+1L9SK/wCCz/xONn4U8A/B2yuiG1O9n1q/jU4ISFfJgz6qzSzHHTMYPYY/4JkfCdfhh+yZ4bv7q1EWp+NZZNeuyV5KTYW356kG3jhbHQF29cn41fGfwD8Afh5qPxL+I+q/Y9K08BESNd011M33IYUyN8jYOBwAAWJCqSPFP/BRn9sr9pbxjN4K/Zh8GzaHbsWMNvpGnLqF/wCTnG+eeRWjiHT5lVApONx4NQ/si/8ABVjx7EbvxR8Ytc0gyfObW/8AH8yqre0dqzoDyelWnh/xL4S/aVh8K+M9Q+3+ING8crZardee0/n3cV9smfzG+Z9zqx3Nyc5PWq/4Ka/8nwfEn/uFf+mq0r4M6daaR8HvAuk6fEIray8NadBBGBgIiW0aqB9ABXxj1a60D4ReONdsWxc6b4b1C5hIOMPHbOy8/UCv+CY2j2WsftpeBDepvWxi1C5jQ9DItlNtJ+hIb6gVVf8ABWRVH7YGpFQMnQdPzx1+Q1+yv/ybB8H/APsQtE/9IIaqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqs/wDj6i/3xVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV+3f8A8mf/ABU/7AEn/oaVWnajf6RqFrq2k309nfWUyT21zBIY5IZEIZXRhyrAgEEcgivgx/wWTfTPDVvo/wAdfhte6tqtpEFOsaDNEhvCOAZLd9qo2OSVfBPRVr9q7/gqj4t+NXhC/wDhr8JvC0/g3QdWiaDUtQuboS313Aww0KhAFhRgSGwWZhxlQSDX/BO79nm9+Pf7ReiT32ntJ4X8GTR6xrUrJmNvKbMEB7EyShQV7osh7VVVVV/wVl/5O/1H/sAaf/6A1fsr/wDJsHwf/wCxC0T/ANIIaqq/5vu/7q3/AO5aq/aI+HU/xc+BXjv4bWSq17r+g3VtZBm2j7T5ZMJJ9PMCZ9q/4J5fGXTvgB+1To1/4xuBpmk67DN4f1aa5Hl/ZBMylGfP3VWeKHcTgKu4npQKsoZWBBHBFSSRxRtLLIqIikszHAUDqSewr9tv4qr+05+1nreoeAYn1Sya6t9A8PLCdxvRGRGrJ6iWZnZenDrnnNfCHwKnww+FPg74cLKkp8MaFZ6c8iDiR4YVRnH+8yk/jX/BRL/k9D4n/wDX/bf+kcFfsy/Frw/8bvgV4O+IXh68hlF5pcMV9DG3NreRoEnhYdQVcNjPVSrDhgar/gsp8aNHXw54S+AWl3qTanLfDXNVjRgTbxIjxQK3oXMkjY9IwejCv+CQPwwv/CP7PWseP9Ttnhl8b60ZLQMpG+0tl8pH/GU3H4AHvX/BUDxjrPir9sfxfpupTSG18NW9lpunQuT+6h+zRzNgejSzSt9GFfAbwH4U+Gnwa8HeC/BVtbx6Tp+j24ieBQBcM0YZ5iRwzSMS5buWJr9oPxZofgb4GePfFXiK8S2sLDw9eGR2ONzNEyIg9WZ2VQO5YDvX/BMr/k+D4a/9xX/01XdV8RfE0ngn9q7xP4zhtxPJoHxDu79ImOBIYdQaQL+O3FeCfGfhr4i+ENH8deD9Ui1DRdds47uyuYzw8bjIyP4WHQqeVIIOCDVf8Fg/jRo/jX4s+G/hHoN6lyngS0ml1N42BVby68smI47pHFGT6GQjqDX/AATd+GF/8L/2R/CFrq9s9vqPiMza3cROpBUXDZh4PTMCwkjjBJr/AILOf8l38Ef9iiP/AErnr/gnb/yZf8MP+vC5/wDSyev+Czn/ACQjwR/2Nw/9JJ6/4Ix/8kI8bf8AY3H/ANJIK/bv/wCTP/ip/wBgCT/0NK/4I2f8nPeJv+xCu/8A0usKqv8Ago3+y/8ACj4i/Avxh8WJvD1hpnjTwppr6hb61bxCKW5WLloJtuPNVlBClslWIIIGQf8Agjb4v16w+PfirwPbSzPo+seF3vLqBSdqT29xCsUpHQYWeRP+2gr48QTXXwO+IltbxtJLL4U1RERRksxtZAAB9a/4JdXdta/tpeC47iZUa4tNSiiDfxP9imOPyU1X7Zf/ACah8Wv+xR1D/wBEtX/BIP8A5Ovu/wDsUb7/ANHW9ftnfDK/+MH7LnxF8BaRbvPqN1pBurKGMEvNPayJcxxqBzl2hVQP9qv+CZXx80D4G/tFxWvjC/Sy0HxrYnSLi6lbbHazmRHgkc54XepQk8KJcnABNVVVX/BRL/k9D4n/APX/AG3/AKRwV4V/5FfR/wDrwg/9FrXjrwvbeOPBHiHwVeSbINf0q50+VsZ2rNE0ZOPo1fsWfET/AIZe/bD0ZviI39k2trfXXh7xAZm2C037oSznsscyxsxP8KGo5I5o1lhkV0dQVZTkEHoQfSqqv+Cgnha//Z6/bpl+Ifh+HyV1K8sfF2mkcfvxIDKcjubmCVvXDD61/wAFAvE0/wC1N+2f4J+EHgi9M9mllpemWcgGUWW/2XDzfQRTw7vTyjnpXh3QNK8KeHtL8LaFbC303R7KKzs4R0jhiQIij6KoFftfaRcfsn/8FD7f4l6PCYdOu9ZsvFluiKf3kcsv+lp9GlS5XA/hcdK/bYmu/wBp/wD4KGQ/DTQLxrm0+16X4asriA7/AC4SFknkHbCST3DemFzWkaTp2g6RY6FpFslvY6dbR21rCg4jijUKij2CgCv+C0njHWZvGvw6+HwmkTSbXSrjUzGCQss8kvlZPYlViOPTzG/vV/wS78B+FPCn7IfhfxDoVtbtqXiua6vdXvEUb5pUuZYUQkZOI0jVQucA7jgFjU0sUETzzypHHGpZ3c7QoA5JPYYrxP4m0jxt+2Jq3jLQLjz9M174lzX9lLjG+GbUjIjfirA1X/BTX/k+D4k/9wr/ANNVpXwr/wCSYeD/APsA2P8A6ISvEWiWniXw9qfhy+/49tVsprSbjPySIUbj6E18APF95+yn+1z4e1rxjE9s3grxJNpuuKFJMUR8y1umA77UeRgO+B9asb6y1Sxt9T027hurS7iWa3nhcOksbAFWVhwVIIII6g1X/BQL4taH8Zv2qvGPiXwvdRXWjae8WlWVzEwZbhbZAjyKw4ZWkEhVhwV2nvX7K/8AybB8H/8AsQtE/wDSCGqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqrP/j6i/wB8VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVf8FATqB/Y5+J0elxySXEumRRKkce9mVrmFWAGD1Umv+EV8T/8AQt6p/wCAcn+Ffsm/sK237Tn7LHjS+libQfG2l+JmXw/f3duY0k2WsTPbzcbjE5kXkDKMAwyNyt8Vvgn8VPgh4hl8MfFLwPqegXiOyxvcwnybjH8UMoykq/7SEiq/Z2/YW/aA/aM1K0l0Lwlc6D4ZlYGfxFrEDQWyR9zEDhpz2AjBGcbmUc1+zj+zp8P/ANmP4b2vw88BWzyZbz9S1KcDz9RuSAGlkx0GAAqDhQAOTkmqqqr/AIKtaFreoftc6hcWGjX1zEdBsAJIbd3XIRs8gV+y5DLB+zL8IoJ42jkj8CaKro6lSpFjCCCOxqqr/hHfEH/Dc/2r+wtQ8n/hbG/zPsr7dv8Aaud2cYxjnNVX/BQP/gm94g8Y+JdS+Of7PWki+1DVJGuPEHhuMqjzTHlrm2zgMznJePOSxLLksVHwl/4KO/tV/s0afD8L/Hfh+LX7XSEEUFj4rtJ7fULKMZCxiTKvtHQCRXIACggAAfFj9vX9rT9r+xm+EngTwmdP07Vl8q60rwnYz3F1exnrHLLlm8s9woQEZDZGa/4J+f8ABOXU/hFrVn8bvjxaW48VWw3aHoKSLMumErgzzsuVabBIVVJVM7iS2AlftQ+D7j4gf8FGNb0vUvDOo3Ojat4202wuwsMgWWE/Z4ZMOo4BAbkHjNePPh3+13/wTQ+IOr+Ivgtd6hrPwy1SZpo5XtGvbJo/4UvYhjyZkGF81Sm8DhuWRfFP/BYP9pnXNDk0vQ/Dngfw9eTRlW1G0sZppYjj70azSugPX7yvX7On7Df7QX7XfxBHxI+Lq65pPhnULoXWseIdbV0u9TGeUtkf5nLAbQ+BGgHBJUIfC3hjQfBXhrS/B/hbTYtP0jRbOOzsbWIHbDDGoVFGeTgAcnk9TX/BUr9i3xx4s8Wn9o74UeHbrXTc2cUHibTbKEy3KNCoSO6SNQWdPKVEcDJURq2NpYr8Bv8AgqT8cvgd4DsPhrqvhvRPF2n6JELfTptSeWG6tol4WFnU4dFAwoK7gMDJAAGmp+2R/wAFQvEOkWHjLTz4J+EFldJc3M1paSW1pIBjJiMhZrufaSF5MaE5IXPP7Afh/X/AX7c3gOLV9DvkSx1LUbCWWS2eNQz2dzADnBA+Zhx+Ge9V8OPg9afHP9uW/wDBfjLw/q8Ph/xV4p1sTyJE8bQqwuZEdXxjKsEYZ4JHOQcVceIP23/+CY/iC/8AC1gn9t+AJLlpLKa9sZLzR7kFuHVlZWtZT/HGHUk8kONrH4gf8FZ/2pPH+kN4Z8Kad4a8JTXo8r7Xo1lLLeEtxtjaWR1UnIAKpuB5BBr9jT/gnH8TPjF4ys/in+0NomoaN4PS5+2zWerb0v8AX5Cd2GRvnSJicvI+GYHCg7i6xQxQRJBBGkccahURF2hQBgAAdBiv+Cx+javqXxz8FS6dpV5dInhMKzQwM4B+1z8EgGv+Ce1rc2f7Gvwytru3kgmjsLgNHKhVl/0ufqDyK/4LH6dqGpfAvwVDp1jcXTp4sBZYYmcgfZJ+SBniv+COGnahpvwL8aw6jY3Fq7eLCVWeJoyR9kg5APav257ee7/ZF+KVtawSTSvoLhI41LMx3r0A5Nf8EeNE1nTf2mvEs+o6TeWsbeBLtQ89uyAn7dY8ZI64Br9sHXfip4e/Zy8Z3vwV0PWNU8Yy2a2+nppCF7mDzJFSSaML8xZIy7DYC24AjuR4N/4Kz/tQ/DiBvCvxO8HaF4kvrECN5dTspdOvVYf89RGQpP8A2zU+pNfHD9vD9pn9svST8G/C3giKz0vU2jNzo3hqynu7q/2sCFkf5mKBwp2qq8gZJFf8E2/2Ltf/AGa/DOq+PvibbwweN/FcMcH2FHEh0uzU7vKZ1JUyO+1mAJA2IAc7qv7G01OxuNNv4FmtruJoZom6OjDDA+xBIr42fBT4zfsQ/HKK/totR08aNqn2vwt4ljgzBeIrbo2DYKF9oxJEenIIKkE/s5f8FSv2hfix8bPAfwu8U+FvA66Z4g1eCxvbjTtNuUuij/KWBa5ZF55J2YABwBX7YsE1z+yr8V7e2heWWTwnfqiIpZmPktwAOtf8Ej9D1rTv2qbq41DSL22i/wCETvVDzW7IM+db8ZIqv28P+CaHi5fFOrfGT9nPQv7X03VJXutU8L2if6RaTNy8lqmf3sbMSfKX5lJwoZThfhP/AMFEP2tv2b7WP4faxNBrdlpQWCPSfF+nymeyReNiuGjmXAGArlguAAABitb/AOCtH7Wvj1V8OfD/AMIeFdJ1G5G2KTSdInvrvd/sJLJIh7cGNq/Ze8QfEPxV+z74F1v4saVqmneL5tKVNWg1S2NvcmaNmj8ySMgFWkCCTBA+/wBBVV/wUK8Pa/eftk/Ey5s9D1CeJ7+32SR2rsrf6HB0IGK8LqV8MaQrDBFhACCMY/drVf8ABQr/AIJ16t8XtZuvjh8CbKGTxXcKDrmhF1hXUtqgCeBjhRNgAMrEB8ZBDZD/AAt/bs/a/wD2Q4IvhZ4s0VrzT9MQRW2ieMtNnjnsYxwFhfMcgTAAAYugA+UCtU/4LN/Gu7txB4f+Efgy1unOFe5kurkZPTCK6H9a/wCCenx2/ar+N3iHxprPx+8NanbeHriztZfD91Jon9nWkTq8gljhJUNLvDqxYl8eXjI4FV/wWK+EF34q+Gfg34q6Jp0t1e+GtTk0+7SCMu5trpdysQOyyQqP+2xr/gk78H9f8Z/tB6t8XvGmn3zxeB9HWO0mvo3DG7nTyIsFhyEt45h7fJ7VX/BYj4N3Pir4aeEPizoWly3WoeGtSfTrwW8Jdza3K7lZsc7UliUD3nNf8EnvhHr/AIy/aL1v4weM7C/ceD9LZobi9jcM99d5iQ5briFbj6ZWq/4KX/si+If2jvh9pXjP4cWQvPGXgrzjHYAhX1KzkCmSJCesisisgJGcuBksK/Zw/bi+PP7GcWqfDOTw1b6hpSXTSTeH/ENvNbzadcEfP5ZBV4i3BZGBGRkAEknVf2mv23/+ChcEnwj+Fnge08M+FdTxDrN9p0UyQCLjetzevkLGc5McYDuAVw4JB+KnwR1f9m39pp/AsVvqWt2nhDVdOma/NmyLcgxwTsw2ghVJc464GM5NV/wUs8P69ffts/Ee6stEv7iF/wCy9skVs7qcaXaA4IGOoIr4XI0fwy8IxupVl0GyBVhggiBKr/go3/wT68Q/FfWrj49fA3TI7zxG8CjX9BQhJNR2KFW4gzwZQgAZON4UFcvkP8FP27f2pf2S7M/DF4IbvS9LkKr4e8WadMH0/J+ZIyGjliGc/ISVByQvJzqP7Xv7eX7cof4WfCzwtBpOmX7CDUZfDNlNAixt94XV7K7+UmDyFZNw+XDZwf20P2Q5/wBmWf4d+ENIt7jW9RvvDr3OualaxO8c999ocOEGMrGqGNVyASBuIBJA/ZSlM37MHwkVoZYnh8E6RC8cqbWV47SJGBHblTVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVn/wAfUX++KqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqtR0zTdYs5NO1bT7a9tZhiSC5iWRHHurAg1pfwf8AhJod+NU0T4W+EdPvQdwubXQ7eGQHOc7lQHrzVVVVVVVVVVVVrHh7QPEUC23iDQ9P1OJPux3lqkyjp0DAjsPyFaXo2j6Haix0TSrPT7cdIbWBYUH/AAFQBVVVQ+CPBdtqf9tW3hDRItQDbhdpp8Sy5HfeF3Z4Heqqr3wR4L1PURq+o+ENEur8EEXU+nxPLkdDvKk/rVVVSRxzRtDKiujqVZWGQQeoI9K0zwZ4O0W9bUtG8KaNYXj53XFtYRxSHPXLKoPOT+dVVVVVVa14W8MeJFRfEPhzS9UEf3Be2aT7evTcDjqfzNaRoGheH7b7HoGi2Gmwf88rS2SFe56KAOpP51Vahp2natZyafqljb3lrMu2SC4iEiOPQqcgj61o3hPwt4cLt4d8NaVpZkGHNlZRwbunXYBnoPyFVVVrPhvw74jiWDxBoOnanGn3UvLVJgv0DA4rSPD2geH4fs+gaHp+mxYx5dpapCvUnooHck/jVVVVVaro2j67amx1vSrPULc9YbqBZUP/AAFgRWkeDfCHh+XztB8K6Ppsmc77SxjhOcYzlQOxIqqqqqqrWvCPhPxLJHL4i8MaTqrwrtja9so5yg9AXBwKtrW2s7eO1s7eKCGJdsccaBVUegA4AqqqqrWPDXhzxEqJr+gabqYiOUF5aJNtPqNwOKtrW2s4EtrO3igijGEjjQKq/QDgVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVWf/H1F/viqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqz/4+ov98VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVWf/H1F/viqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqz/AOPqL/fFVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV/wUY/aW8f8A7M3wV0/XfhnFBFrviHWF06LUri3WZLBfKeRnCMCrSHywFDArjccHFeB/+CsX7XPhWVB4h1rw54whHBTVtHjhbHs1r5PPuc++a8A/8FpPCF0I4Pih8FtX04jAe50HUY7wN6kRTCLb9N7fWvAH/BRz9j34gmKC3+LdroN3JgG28QW0un7M+srjyfykOPyrw54s8K+MtPGq+EPE2k65ZHGLnTb2O5jOenzRkjpVVVVVVVVVVVVVVXj/APai/Z1+Fwlj8dfGjwnplxBnzLP+0kmuRjr+4jLSf+O18QP+CvP7MXhgyW/gvTvFXjKdciOW1sBZW7Y9WuCsgB/65mviB/wWZ+LurGS3+Gvws8MeHYmyFm1O4l1KZR2K7fJQH6qw7c9a/Zy/4KW/tUa38fPCOh+O/FVt4m0HxLrVrpl1pI0i2g8tZ5VjDwtDGrh135ALEHGD2Iqqqqqqqqqqqqqqqqqz/wCPqL/fFVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV8Svhf4A+MXhC78B/EvwvZ6/od6VaW0uQwwynKujqQ6OOzKQRk8814+/wCCPX7OfiIyXHgbxR4s8JTtnZELhL+3T/gEo8w/9/K/aC/4JleLvgdaHUbf41eC9YicE21ndJNY390OmUgVZVxngkuFHrWsfC/x/oeTf+Fb4ovV4E89QPUlM4H1rR9d8R+E9TXUvD+s6lo2owH5Z7O4e3ljPsykMK8Af8FDf2v/AIdiKGw+MmpazaxgA2+vxx6lvA7GSYGUfg4rwB/wWh+IFgsVv8T/AIOaFrIHyvc6Lfy2DAf3vLkEwY+25QT6dK8Af8FaP2UPFxjg8T3niXwZOwwx1XSzPFu9A9sZTj3ZV98V4B/aB+B3xSES/D34teFddml4W2tNUiafPoYSRIPxUVVVVVVVVVrevaF4b099W8Ra1YaXZRcPc3tykEa/VnIArx9/wUJ/ZA+Hgki1D40aVq9zHwLfQkk1IufQPCrRj8XArx9/wWg+HNgZIPhj8HvEGtMMhZ9avotPTP8AeCR+cWHsSp+leP8A/grX+1Z4uEtv4Xn8MeDLdgVQ6ZpgnmA93uWkGe2VVfoDzXxA/aG+OnxUEsfxC+LnirXYJQQ1pdapKbfnriEERgfRRXgn4TfFH4lzCD4efDrxL4lbdtJ0rSproKf9oopC/jivAP8AwSx/a+8beTNqnhHSPCNrLgibXdVjUhfUxQebIp9mUH+dfD//AIIs6XH5dx8VPjbdT5x5ln4f0xYdvrieYtn/AL9D19q+CH7AP7MnwD1+y8YeEPBt1qHiPTgfsurazfPdSwsRguqfLEr4P3ggI7EZNVVVVVVVVVVVVVVVVVZ/8fUX++Kqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqvjt8WLL4M/Dq+8Xyxxz3xIt9NtpDgTXLg7Qe+0AM7dOFOOSK/Z9+A2sftH65f/Fj4uavf3elNdFDukKy6jKByob+CJBhflx/dXbtOLP4B/BKwsV0+D4UeFWiVdoabS4ppMYx/rHBcn3zmv2qv2H/AAFe+Gr3x38PfC9qZdPRpr/R5YvPjkhAyzw7slGXGdgOCPu4IAb9nj9jf9lX9ofS9T8H+JfDmqeHfFemJ9oh1DRNUeJru3JwSYpfMiDRsVB2oMhl6nJPxA/4Is3qmW5+FXxuglB/1Vl4g0wx7f8AeuIWbP4RCvH/APwTJ/bC8B+ZND8ObfxPaRdbnQNQiuc/SJikx/COvFvw+8eeAbv+z/HXgnXvDt1kqIdV02W0ckegkUHtXgH9qj9o74YeUngf41+LdPghGI7R9Se4tl/7YSlo+392tM/4KxftTx+Gb3w14juPD2sPeQGMaqtkbK+gJ/ijeBljB/7Z/Qiv2UP+Cnvw/wDh/BrGn/GrTPGNxdazdxzLqUMq6iI9qbT5hkdZMf7oY+1eAf27f2SPiQI00D45eHrSeTAFvrMjaW4b+6PtIQMe3ykj0zVhqFhqtnFqOl31veWs67op4JBIkg9VZSQR9KlubaF44priJHlOI1dwC59h36iqrx58bPg98L42b4ifFHwt4ddRkQ6jqsMErcZwsbNvY47AE14+/wCCrf7I3g0SRaFruv8AjG4TICaLpTIm73e5MS4913fjXj7/AILSeKrkyW/wu+CulWAAxHda9qMl2W9zFCI9v03mvH//AAUf/bC+IHmQS/FmfQLR+ltoFrFYbPpKq+d+chrxH4s8W+NdR/tTxd4m1fXr9uPtGpXsl1Kc/wC05Jr4efshftNfFWKG58DfBPxPeWtx/qby4tPsVvJ2ys05SMj33V4B/wCCO37QviDyrjx34w8JeE7dh88Syyahcp/wCNVjP/f2vhx/wR6/Z802NLrxt8RvE3jGeI4dLN4tPtm9QUUSSDt0lFeAP2LP2Vfhm0U3hT4G+GBcwYMd1qNqdRmQ/wB5ZLkyMp9wRVvb29pBHa2kEcMMShY441CqgHQADgCrm7tLRA93dRQqTgGRwoJ/GlZXUMjAgjII71VVVVVVVVVVVVVVVVVWf/H1F/viqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr/goV4knm8TeE/B6SkQ2thLfOgP3mlfy1JHsIWx/vGvhF4XtvBfwv8AC/hi2iCfYdLhEgAxmVlDSN+Lsx/GqZVdSjAEEYII618L4P8AhVX7Zcfh3Tz5NnH4gutOSIE4aCbesSnHoGjP1Ar9v67urT4a+HXtLqWFjrgBMblSR5EnpX7IM81z+zr4SmuJnlkb7bl3YsT/AKbOOpr4h6fYan4E8QWWpWUF3bvplxuiniEitiNsZByK/ZW/ZW+Avxy8Z6/ofxE+H9rdQQaQZ4Ws5pLJ4pfOQbwYWXJwT97I56V+0P8A8ElfBXg/QPEXxB+HXxN1LT9K0u1+0RaXqNqt4+RgMolUpxknGQSB3NeDP+CfHx/+J2hanr/wyt9D8QRaXMIZLX+0FtJ3YruG0TbY8Y9XFfEX9mP9oP4TRT3XxC+DvinSLO1BM18+nvLaoB1JuI90WP8AgVeE/iD498A3JvPAvjfXvDs7HLS6VqUtozcY5MbAnirP4/8AxShuGutS8RtrDucu2pJ5zMfUvw5/Fq8Df8FB/wBoX4ZWd5ZeBNS02wW9gWI+fE90sBBzuijkcxqe2SjccV8QP2vf2nfigZE8Z/HDxVdQTZ8y1tb02Nu2fWG32R49tteHvC/i3x1rA0fwn4d1fxBqs+XFrp1pJdzycjLbEBY8kc46mtT/AGM/2lvD2gWvinxb8K9U8PaTdzCGO51XbblXIJAaMnzEyAfvKOlfs4f8EqV+MvhG08c+J/jbaWNpJK8U+n6TpbTzRSIfmRpZGQKdpVh8jcMK+Lf7G3wH/Z1+MWg6D/Z2s+LNJtbezvr+LV7wFroec3mpiERqFZU2gds8k9a+HP7Ed58VLBPiFJY+Gfhtp2pp5ml6ZpWgruW3P3GKq0fDDBDMzOwwT2rUrT4s/sY/EyxRNZa70y5ImVIZGFrqkAIDqyHhXA49VJBBIIJl0b4tfto6jqOsWfiibwh8NrW4a30+B42drsr1dolZRI3qWbapO1ckMa+KXwU+J/7KGp6b418LeNZZ7KebyY9Ss0a2ZJcFvLmiLMCrAHAJYHBBA7/BP4kRfFn4ZaL42EaRXN1EY72FOkdxGSkgA7AkbgPRhX7V/wC0Rqnwut7LwJ4CIfxZrUe8SrH5rWcJO1Sqc5kdgQoIOME45FeHf2HvEvjawTxN8YfiTqQ169UPJboPtLwZ52vK7Hc3PIUYB6E1r+n/ABb/AGJfGOlXeneJpPEHhDUnbFu5aOGcLjzI3jJYRSgHKupOevI3KPDPiHS/F3h3TfE+izebY6rax3NuxGDsdQRkdiM4I7Gqqqqqqqqqqqqqqqqqs/8Aj6i/3xVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVfHP4++Efgdokd1qwN/rF4p+waXDIFeXH8bHnZGDxuwcngA84sPHH7Y/7RryX/AILkudB0FmZY5bFxp8CdjtnJ82QjvtJwc8Cj+zD+2BZodRtfjKrXONxSLxPfCQk9RkoBn15xX7NGlftAaRFr+n/HDUrm4jtpIU0sTyxTs+QxkcSplmH3AAx4OelftR/tQ3XwouYvAngSCG48UXMSyTzzR+YlkjfdAT+KRuoB4AwSDkCtP+Bn7Z3xGs4/Eeu/Eu40Z7kCSO01DXLi3ZQwznybdGWPtlSFI6EDFfCbwL+194G+J2h6V4t8W3mqeFZ5WN/dSX41CLy0Qts/e/vYyxAUEAcn61VVVVVVVVVVVVVVVVVVVV+0T+14PAWsTfDv4YWcOqeJIn8m6u5F82K0kPHlog/1kvOCOinAIY5Asvgx+2t8TLddc8Q/ES60MzgOlrf6zNaH2PkWysqHBPBCkdwK8QRftj/s3oNf1DxPe63okLDzpmu21O0UZxh1lHmRKePmATrwcmv2ev2i/D/xz0eSB4I9M8S6fGGvtO37gy9POiJ5KEkAjqpIBzkMaqqrwx+1B4P8ZfF7/hUPh/Q9Ua6hluobi+uAkcSvArFtigszAlCMnbVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV+0F+2Dpvw31KbwT8PrK31vxHEdlzPKS1vZv02YU5kkHdQQFPBJOVFj4F/bh+MES6vqnizUfDtpdDdGl1qB0xdp6ZhgG8DH95c/Wrj9nz9tHwlD9t0H4pzanInzfZrPxLcHcR/szqiE8DrXwjXx0PhvoR+Jly03iZ7ctfs0aRsrFiVUhPlyEKg46kE18Tv2ofif8AFHx63wv/AGd45Y4TK0CX1uime8K/ekV2+WGEYyG4OOSRnaD+zN+2HKn9sP8AGPF6y7jGfFF6JgcYxuCbc44+9j3r9mbTP2hNJfxBpnxv1K5uLezMKaZ9peOdpSdxdxMuSy4CDDEnJPTvVVVVVVVVVVVVVVVVVVVf8FBtLmi+JfhzWmB8q60L7OpI43RTyMf0lWvAusW3iHwR4f12zcNDqGl286Eejxqf61VeGnj8eftw/a9MYSQjxTPOkiHho7be24HngiLPvntmv+Cg3/JM/Dn/AGHR/wCiJK/Y7/5Nw8If9vv/AKWz18W/HPgnwX4I1qbxj4w0TQo5dNuFjbUtQitQ5MbAAb2GSTxX7NP7aHwJ+Afi3Xdf8X63qGoJcaV9nhg0iyNw8knmo2AWKp0U9WFfH/8A4K6aJ4/8Ga14C+HnwcvY7bV7cwnUta1NY3jXIJ/0eJWBPH/PXj0NeHf23/2kPBOjahoHgDxxH4Zs9TmEtyNPsYjIzKMDEkiu68f3SK8a/FX4nfEm4+1fEL4ieJPEsmcg6tqk10F/3Q7EKPYYAr4b/Ab4z/GCZ4fhl8MPEfiJYSomnsrB2hhLZ2h5SAiZwcbmGcH0rwB/wSQ/ap8WNFN4rXwx4Nt2wZBqOpfaZgP9lLYSKT7F1+teLv8Agi98VbCwSfwP8Y/C+tXQjzLBqVjPpwDY5VHQzbvQEhffFat+zb8atM8VXfgu08BahrOrWlxPAbfR4/tzyNDu8zYkeXYAIx+70Ga1HS/E/gzWBa6rp2qaFqtoQ4juIZLWeI9jhgGX61Z/tU/tFWvhyfwhP8YfEmpaLchQ9hqt2dQiAVgy7BPv2YIByuDX7NP/AAUp+In7PsGo6XqHgTR/FOmalNHNJF9qksZUdQQSrgOo3AgH5P4RXxi/bp+GXx4+I+n+LtQ8Nax4bt1sYLW6hnK3KrsdixDx4YjD/wB0Hivh1+1n+zB8SIILbwJ8a/CM7lQkFjPfLY3GAMALBPskwAOy8V/wUKitpvCHg69VUd11CdY5Ac/K0ak49jtX8q/Zht4Lb4BeCoreNUU6dvIA/iaRmY/iST+NftmWdvc/s6+Jppowz2ktnJCSPusbqJCR/wABdh+NfsBSyS/BjVEd8iLxJOqDHQfZ7c/zJ/Oviv4z1j/hrfVPE1l4fk8QXmia0qWem+Wz+abZQqAKgJwCm/gepr/hrv4/f9G73v8A4BXn/wARXxu+LHxn+N/hK38J6v8AAvVdPS2vku47iDT7p3DKjrjBToQ5r9ky017T/gJ4b03xFpt1Y3dq11GIbqFopAn2iQrlWwRwcDjoBVVVVVVVVVVVVVVVVVWf/H1F/viqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqry7trC0mvryZYoLaNpJZG6IqjJJ+gFfD/AEq+/a1/aQutY8UNMdGiL3lxAWP7qyjYLFbqe2SyA4xnLt1qysbLTbODT9OtYba1toxHDBCgRI1AwFVRwABxiqr4xXyeDf2yZfEXjSF3sLXXrG9kZk3brVViKsB/EFVcY9VIqxv7HVLKDUdMvILu1uYxJDPDIHSRSMhlYcEEdxVVVVVVVVVVVVVVVVVVVVXx4+IEnww+E/iHxhaOovba28qxyM/v5GEcZx32lgxHopr9hL4XW3iHV9X+MHiOI3kunXBtdOaf5/8ASWUNLMSeSwVlAP8AtseoBqryztNQtJtPv7aK5trmNopoZUDJIjDDKwPBBBIIPrXjHT7j9lX9puG90J5U0m2uY7u3jJJMlhNkSRH1wPMQE90Dda8XeJoPCvgvWvGQt/tkOj6XcX/lI+3zliiaTaG5xkLjOO9fs/8A7SFj8erzWrOz8Jz6OdGihdmkvBN5nmFhgYVcY2frVeIP2stO0H42xfBd/BNxNNLq9ppw1AXwVQZzGA+zZnA8zpnnHWvj98d7T4EaPpWr3fhuXWBqly8Cxx3QhMZVd2clWzXw5+M1r4E+N1z8XJdAlvIp7q9nFitwEYfaN+Bv2kfLv9Ocdq+A/wC1Vp/xz8X3fhK08F3GkPaaa96Z5L4TBgskabdoRcf60HOe3vXi7xj4X8B6FP4l8X61baZp1sPnmnbGT2VQMlmODhQCT2FeJv8AgoH4fhvnsfA/w71DV13FY57u7Ftvx3WNVckH3IPt2r4YfttWvjzxnpXgXVvhlqGm3+qXCW8b296LgIx6sysiFVHJPUgDvVzcW9nby3d3cRwwQIXllkYKqKBksSeAABkk14+/bx8C6DqMmj+A/DV54pmjcxi5M32WB2zgbDtZ3Gf9lQex5zWg/wDBQKwTUEs/HPwxvtNhYjM1neec6qe/luiZ+u78K8I+L/Dfjzw/a+J/CWrQ6jpt4pMU0WRyOCpB5VgeCCARXx6+NEHwM8KWPiq68OS6xFeagtkYo7kQlGaN3ByVbPEZHSvGP7YPw/8ABvgjw/4kurGe81nxFp8d7BottOrPAjDrLJjCDORnBJIOFwDjw9/wUM0641RYfFPw2ms9PdsGey1ETyRDPdGRA3H+0OnTmvHn7Qvwx8AeBtO8e3+tfbrPWot+kwWa7pr7gE7FONoXI3FsBTweSAY/+CiCHUwJfhWRp27BK6vmYD1x5WCfbP496+GfxN8J/FrwrB4t8H3jy2sjGOWKZdktvKAN0ci5OGAIPBIIIIJBqWWKCJ555UjjjUs7ucBQOpJ7DFePf25PB2jazJ4d+HPha98Y3aMU8+KbyYHYf88yFdpAD3CgHsSOa8O/t6adDrEek/Ev4a6l4eRyM3EMxmaMHozROiNt9wSfQeuk6tpev6Xa61ot/Be2N7EJbe4hfckiEZBBFVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV+0d8SpvhV8I9Z8S6fKE1OZRZ6cT/DPLwGHui7n99mK/Ya+D1jrj33xm8VW4vZ4btrfSVuBvxKADLcHPVssFU9iHPXBFV4xtdRvvCGuWWjsRfz6bcR2pHUStGwTH/AiK/YV8T+HfDXxX1HSPEE0Vnfaxp5tbCWfCfvRIrGHJ6MwHA7lMdSBVVVVVVVVVVVVVVVVVVVVVftg/CG9+KPwzGoaBaNc634Zka6tYUGWniYATRqO5IVWA6koB3r9jX9ozRLHRYPhB471OKwltHb+xry5cJG6MdxgdiflYMTtzwQdvBABBDAFSDxxX7Uv7Z3wn+B/ha90iy8f6LceMLyJ4rWzt7tJ5LMngzSqhJXb2UjLNjjAJHwH/bj+CHwL8Q6r401PQ/EfinWWtTbaZDZwRwwrvOZJHklYMpO1VGEbhnz2z+0z/wAFN/GPx+0i08OaL8MtJ8NafYXv2mKSe+kvp2IRkGSFjUcMTjae3NXH7Xv7S8nha18E2Hxl8RaTodkJBBZaRc/2eFDuzuC0AR3BZ2OGY9cdKurvWfEWovdXt1eanf3By8krtNLIfcnJNfCH9lP49fHTWX0P4eeAbi5ngh8+d72eKySKLcF3kzMpIyw4UE89Kuf+CR/xh8M+BNZ8cfED4h+GdPGj2Ml2bDS45b6VwgyyszCJV4BOQW/Gv2Jv2Df2cPihFrsvxMj1jX9R0eWFksm1FrSF4XDfMRDtkPzKQSH4yPUUP2Pv2e9B8Ea14X8AfCHwlod5qOmzWsGoppiSXMTvGVVvPcNKcHB+92r9kb4n2vwW+Jms+CfiBJ/ZFprBW0upLk7Vs7yFmCeYeir80ilugJUk4BIjkjmjWWGRXR1DIynIYHoQfShJG0hhWRTIqhmTPIBzg49Dg/ka+K0Go/s9/tY/8JxJZyPp8+q/2vCUGPPt5yftCLngMN0qY7cHoaa0+HHxh8J2l9eaVofizQNQj82Bb2zjuoXB65SQEAjGCCMggg4Ir9tz9mD9jXwt4W2aN8MNM0bxvqrr9iXRbmW0FvEGy8rW6N5QUgFRlMknjgNXw2/4JN33xZ+HEfjvR/ihL4dnvZ3/ALPstS0v7Qs8C8CRpEdCmWDY+RsgA/xV8b/2I/jP8DfGQ8Fap/Yuv3UkCTwyaRdsVdHZlXiZY2DZU5AB+prx58EvjD8L3ZPiJ8L/ABR4dVTgTajpU0ETc4ysjLtYZ4yCRWm+PPG2kWcWnaX4u1i3s4H3x2qXsghVsYyI87entXwg/wCCn37T3wj0Sw8Lpd+G/Euj6bGIre21bSwrRxj+ESW7RsTz1bcfrXj3/grJo/xd+Eus/Dzxj8H7vRtR1IW4W+03VFuoSY7iOViY3RGQEIRjc3J61+wf+2P+zXoPw91Hwj4r+KmmeHNXuddkuo7fWFe0QxNBAgbznHlD5kcY35GMnAIJ+LHiXSPh5+0/Y/GHwnrGna/4f1G7h1S3utLvEuY50KhLhFdGK7vv8Z6Op714Y8UeH/GehWniTwvqtvqOnXsYeGeF9wPqD6MOhU8g5BwakeOKNpJHVUQEszHAAHUmqqqqqqqqqqqqqqqqqqz/AOPqL/fFVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVfHW6nsvgv45uLY4kHh+8VSMgrmFgSMdCAc1/wTstrZ7zx5dsqm4ii0+OMnqFY3Bb9UX9Kqq/aA/Zy8M/HTTYbiS5GleIbCMpZ6ksW/cnJ8qVeCyZJI5ypJI6kFdE/av/AGVmkm01bufw9A5eQ24+36ew6lmT70IPdiIyfWvgB+154d+LV9B4R8UWEWheJZVxAqSE216wGSIyeUbj7jZz2Yniqqqqqqqqqqqqqqqqqqqr9vm4ng+CunxQk7LjxFbpL1+6IZ2/9CVa/Ylt7eH4AaVJABvnvrt5cd2EpXn/AICq1Vf8FCYLZfH3he5QL9ofR3RznnaszFfwyzfrXiV7iX9ljVZLsHz2+H8xlDLtO77Ac5HbntX/AATv/wCQz43/AOvWz/8AQpar4if8n0Wn/Y4aR/6Fb1c2dpeKqXdrDOFOVEkYbB9s18ALW1m/bJ1C1lt4nhGp6wBGyArwJscdKt9O0+0cyWljbwuRgtHEFOPTj6Cv2ivEWv8Ax8/aMtPhJo98Y9O0zUBpdqpzsSXj7TOyjqVIYf7sY6ZNfDj4U+BvhVokOi+DtCt7UpGFnuzGDPcsOrSSdWJPOOgzgADirnSNJvL611O70y0nvLEsbW4kgVpICwKsUYjK5BIOMZBNft8fEy/0Tw9o/wANNIumhOuB7rUijYZoEICR/wC6z7if+uYHQmv2V/gjoHw1+HuleILrS4ZPE+t2q3V3eSxgyQLIoKwoeqqFIyB1bJPGAPil8LPCfxb8KXfhjxPp8Uhkjb7Ld+WDLZy4+WSNuoIPUdCMg8Gv2N/F2u/Dn426j8INYlIt9UluLSeAt8sN7bBjvH1EbofXK+gr/goD/wAka0b/ALGeD/0mua/Yk+D+gL4Ih+LWv2UWpaxqsskWnSXKiT7HbwsYvkBztYsjjPZQoGBnP7Xnwu8OeM/hLrXiabTYF1zw7bG7tL1UAkCIQZI2bqylN3B6HBr9iX4a6T8Rtev/ABH40j/tWx8HQxQ6XYXR8yGOSZ5HLbDkYUhmxjG6TPUV8UPhj4a+KXgu+8Ja5ptrIZbd1sp3iG6zm24SSM4ypBx06jIOQSK/YC8R6hpvxG8ReCZ3Zba+0s3LRMSds8EqqMDoPllfJ/2RX7dfxJvvCfw70/wXpF00Fz4rmkS5dDg/ZYgvmLkdNzOgPqu4dzX7MHhD4Z/C34b6XetrWhL4j1q0judTuZLyPzVLgMIck5VUBAK9NwJ71+0Fo/wz+J/ww1vS77XdBm1GzspbnS5/tcRkgnRdyhTnIDFQrDuD9K/4J+eNr7UfDfiPwFe3Bkh0eaK7sVZslEm3iRR6KGQN9XPrVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVf8ABQi6nT4f+GLFW/cy6w8jjn7yQsF9ujt/nNfsj21tbfs8eEFtlUCSK4dyO7G5lyT/AC/DFVVftBfsYQ+NtWu/G/wvu7XTtWumMt3pk42QXEh5Mkbj/VuepBG0k5yvOdM+Of7T/wCznd2+h/EXSLzUNMRvLjh1lDIrgdRDdqTuIAwPmcAdq+C/xx8G/G/QX1Tw5I9tfWe1b/TbgjzbZj0PHDIcHDDrjnBBAqqqqqqqqqqqqqqqqqqr4z/Gr4cfADwLcfEP4oa+ul6RBKsEZEZkluZ3yViijXl3IVjgdArMcBSR4+/4LQfDiwEkHwy+D3iDWXHypPrV7Fp6Z/vBY/OJHsSp9cV8Y/2wPHnxZ8V3nimw8MeGfB73hJlh0a1c+axJJkfzncbznkqqA9cZyTrXxb+KfiPSzoWufEbxJe6WRj+z5dTlNuB6CLdsA/CvBPwm+KPxLmEHw8+HXiXxK27aTpWlTXQU/wC0UUhfxxXgH/glj+19428mbVPCOkeEbWXBE2u6rGpC+pig82RT7MoP86+H/wDwRZ0uPy7j4qfG26nzjzLPw/piw7fXE8xbP/foevtWs/8ABInQNF1JpfAWuaZq1nuzH/wkcsySxr6N5SNG56c7Fz6CvgP+wV4P+GOr23iTxbcabqlzZOHtdOsLIQ2sbjkO/QyYPIBCjI5zX7PP7M2u/BTxzrPim/8AEmnaha6pZPbpBbwOjRkyo4PPGMKR+Valp1jq+nXWk6lbJcWl7A8FxC44kjdSrKfYgkVrn7CfxD8P+IpdT+EnxIs7S2YsIXu7iezuYUP8BeFW38YBPy59K+CHgXxV8OvAFt4Z8Z+J/wC3tUjuJZZLwySSZDtkLuk+Y49/Wvjl+yj4H+M123iGC7k0DxEUCvfW8IkS4A4Hmx5G4gcBgQcYByAANP8A2Qf2lPDif2R4V+N0FhpOCPLt9YvrYY5/5ZIpX9e5r9nj4A+JPg5fa3rXiXx+/iC91+OJbiPynwrRlirGR2LOcOw5A618Wfg94L+M3h0aB4us33QMXs72AhZ7RyMEoxB4PGVIIOBxkAhf2Mvjv4LuJ7f4W/GqOysJnycX93prN/vLCHBPA5zXgH9hm3XXl8UfGPxnJ4nud4kezhL7JnHeWZzvce2F+vare2t7O3itLSCOGCBAkUUaBVRQMBVA4AAAGK+PP7LGv/F/4mWfjvTfFen6fBa2kEBt54HZiY3ZiQRxzuqSOOaNoZUV0dSrKwyCD1BHpX7TX7C/wx+PFnp76F4P8IaBqkFw73l/FpSwTXKMuArSQhWbB55Nap/wRv8Agvqnhy1jsfH/AIl0TxAkO24ngMd1ZvJ/eEUiiTHt5lfED/gjd8dNCMk/w8+IPhTxVAmdsd0sum3D+mEIkjH4yCviB+xN+1Z8MzI/ij4G+Jnt4c77rTLYalCoH8Re2MgVfdsfnQfVNEvWRXurC8gbDAFopEPoehBrwT+0l8c/h3I0nhD4na9YbxiTyrx1Mn1YEMfzqH9tn4leI3t7T4oarrWvWQdRKRq8zsFBzlUlZ1J69wM1+z1/wUo/Z4+OPiDRvhzDc+IfD/ibUttvaRa5aIqXs+PupLE7rubHAfZknA5IBqqqqqqqqqqqqqqqqqz/AOPqL/fFVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVeLNBh8VeFdZ8LzuFj1fT57N2I6CWNkJ/Jq/Y78ZH4WfGy/8AA3iw/Yf7ZD6ZMJTgQ3kT/IGPuQ6fVxVVVMqupRgCCMEEda/a/wDBekfCn402OreBETSjf2kWqRxWw2C1uVlcFowOFBMasAOAc4wK0S9l1PRdP1KeLypbq1jleMjGxmUEj8M1VVVVVVVVVVVVVVVVVVX7XHg668Z/ArXoLCEy3WkmPUYkVckiI5kx/wBszIfwr9gH4g2N34Y1r4Z3dyq31hdG/tI2bBkgkCq4Uf7LqCf+ugqq/aN1eT47ftMW3g/wxcC5gt5YNEtZYvmXIcmaTjsrvJk/3Uz0r4uWMdv8FPGmmWEGEi8LX8MESjoBauFAH5Cv+CeE0S6/40tjIPMeztGVe5CvICf/AB4fnVfEF45P257Vo3VgPGOkjIOeQ1uCPzGKr9nz/k8/UP8AsJ6z/Kaq+HFwnhP9t9zrsmw/8JTqUG+Q43POJ44jz/eaRD+NVX/BQWwu4/id4d1N932a40EQx56b455S/wCkifpXgrVbLXfB2hazp0ivbX2mwTQkHPytGpH86r4cvH4x/bcfUtDk8y1bxLf3SzJyGijErbs+jAf+PCv+CgP/ACRrRv8AsZ4P/Sa5r9jv/k3Dwh/2+/8ApbPX7QP/ACRDxz/2Arr/ANFmv+Cd/wDyBvG//X1Z/wDoMtV+xR/ycRq3/YLvf/R0df8ABQ+yul13wXqDMxtpLS7iQdg6vGW/Ehl/L2rwf+wz8LfFnhLRfE9v428RlNW0+G6GxoMDzEDED5D0ziv+HfPwz/6HTxN+cH/xuvgl+zb4T+Bmpanqnh7W9V1CbVIEhkF6Y8IqsTxsUcknv6VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX7bXgy68V/BG41GwhMk/hy9j1BlUcmIBo5PwCybj7Ia/YO+I1hrXw8ufhvc3KrqXh24eaCEnBe1lbduUd8SM4PpuX1qqqvE3hjQPGOh3XhzxPpVvqOnXsZSa3nTII9R3BHUMMEHkEGv2blvPh3+1qfBekXkk9ouoalpFwQf8AXwxCUqxx6NCj/hVVVVVVVVVVVVVVVVVVVft//ss+Jf2qvg/Y+G/BOsWtn4h8P6oNQsob2Vo7e7HlvG8TFQdrYYFWIIBGDgMWHgf/AII5/tB65Ksnjjxr4P8ADFrkgiKWW/nHuEVVTH/bTPtXgD/gjh8BtCWOf4g+PfFniu5Q/NHb+Vpts/1RRJJ+UorwB+xZ+yr8M2im8KfA3wwLmDBjutRtTqMyH+8slyZGU+4Iq3t7e0gjtbSCOGGJQsccahVQDoABwBVVVVVVVVVVVVVVXjb4VfDH4lW/2X4hfDzw34lj24UatpcN0V91LqSp9xgivH3/AAS2/ZA8bmSbTvBuq+E7mUczaFqkiAH1Ec3mxj6BQK8f/wDBFm/TzLn4WfG+3l/552XiDSzHj6zws2f+/Qr9nz/glL8efC3xw8L+KPiTrHhqx8O+GNWt9SmuNOvnuJLswSLIkUSFFK7mUAs23aMkAkAGqqqqqqqqqqqqqqqqrP8A4+ov98VVVVVVVVVVVVVVVVVVVVVVVVV+3x+374h/Y78R+GPC/h34b6d4im8RafLeNc3uoPCsOyQJtCIhLZ5Odw7ccVf/APBa/wDaUkKf2Z8NvhnAADvE9nfTZ9MYulx+taV/wWw/aPhuA2t/DP4bXcGRmO1tb23b3+ZrmQdPav2fP+CwfwX+J2r2nhb4ueGbn4c6jduI4dQkvBeaazdB5ku1Hhyf7yFB/E4HNRyRzRpLDIro6hkZTkMD0IPpVVVVVV+39+0X+0H4M/bA+JPhjwf8d/iHoej2F9bra6fpvii8tbeAG0hYhI45AqgsScADkk96/wCCNfxZ+KnxR/4W/wD8LM+Jnivxd/Zn9h/Yf7d1m4v/ALL5n2/zPL8522btiZxjO1c9BVVVVVf8FfviR8RPhn8EfBmrfDfx94j8KX114qEE9zomqzWEssf2WZtjNEyllyoODxkA1+zJ+03+0nr/AO0n8JtB139oX4l6lpuo+ONGtryyu/Fl9NDcwyXsKvHIjSlXRlJUqQQQSDVVX/BSr9qjxx+y38F9M1H4aLbw+JfFOqHT7bULiETLYRrGzySqjAq0nCqoYFfmJIOMV+x3/wAFJf2lLT49eFPCnxP8d3PjLwz4u1m30y9tb+3jMlq1xKsazwuihlKMwJTlSuRtzgj9pzVtV0D9mz4sa7oOp3enalpvgfWbmzvLSZoZraaOxmZJI3UhkdWAIYEEEAiv+Gsf2qP+jmPir/4Wd/8A/Ha/ZD1zWvE37Lvws8QeI9YvdV1TUPCtjPeX19cNPPcStEpZ5JHJZmJ5JJJNfts/8FHP2k4v2gvGHgD4YeN5/BnhzwVrVxpNvBYQR+bdS20hikmlkdSzbnRiFGFC7RgnLH/gmZ+1j45/aj+EutR/E1obrxP4Ov4rW41KGBYRfwyoWjkdFAUSApIrbQAQFOASarXfjz8E/DPiPTvBuufFjwna+INWvobGx0htWha7nuJXEcaLAGL5LsozjAJGSKqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr9qn9lPUPHeoS/Ev4ZwRnXSoOo6buEf2zaMCWM8AS4ABBIDAAg7vveBf2zvid8MMeDvi54QudXkshs8y6LWd9GBx8+5SJOnUgE8ksauP8AgoR8P1tt1r4E8QSXGOI5JYUXP+8GJ/Sv2dP2g7348P4hmufCR0S30t4fsmJWmEyuH3Zk2qCylVyABgMOteMv2zPGnwy+KfiDwz4r+HpuNCgvnj07eHs7gwr8vmKWBWRWKlhwPvdfTUP+ChPghLB30rwBrk97t+SK4niij3Y7upY4z/s14H8A/Er9rn4rx/Ebx1pr2fhmJ4/Pm8to4TBGeLa33ZLE85OcDczHkgFVVVCIAABgADGKqqqqqqqqqqqqqqqqqqqSOOWNopEV0YEMrDIIPUEV8Zv2d/iF8DPG4+KnwUjvZdIgmNxEtkhkm0wnO6N0GS8OMjOCNpIbpk+Ff+Cg+k/2ckXjrwFeJqEYxJJpcytHIfUJIQU+m5vrXxD/AGyvHXxThfwJ8F/BupWM+oqY2uYyZ71kPBEaRjEfHVssRngrjNfspfswXPwtJ8fePI4m8UXUJjtrRWDjTo2HzZYZBlYcEjgDIBO41qFha6pYXOmXsYkt7uF4ZkPG5GBDD8ia0658e/sb/Gqaa60qS7tQJIAJMxxarZMQQyOAQGGEPGdrDBHUVqv7dx8R2ceg/C74a6td+JtQHlWqXJWRY5COCqR5aXBzx8vqfSovBfizwH+0z4JsvH2ox3WvX+v6XqOoSBt2yWa5R2DN0LA5yRxnpxzVeOLjxD+zT+1PeeNJtFkns5NTub60RmKJeWlzu3KrkHlRIVzzhlGa+Cn7TfhP45a9feH/AA/oOq6fNp9n9qdrzZhl3quBtY92HXFftgfs5eJdZ17/AIXD8NrGe7vNif2pZ2gPnh4xhLiIDljtVQQvI2ggHJx8OP29xpelxaN8WfCt/dX9oPLe/wBMCB5ivGXhcqFfjkhsE/wivCn7bXhTx38QdA8EeGfBurJDrF2LeS8vnRGhyDtIjjL5ydoJLDAJPNftOfA8/G3wIllpTRR+INGka40t5TtVyQBJCT2DgLz2ZVzxmvgv+014o/Z6ik+Fnxc8JatLY6c5+zIECXVkCSSoVyFkjJyVO4Y5wSCMfFP9uB/GWiSeD/g34X1qDUtWQ2/2y6jXz4w3BEMUTPlyOA2cjqBnBH7IH7OWqfDC1uPHvji1WDxBqkHk21kwBaxgJBbcegkcheB90DGckgf8FAf+SNaN/wBjPB/6TXNfsd/8m4eEP+33/wBLZ6/aB/5Ih45/7AV1/wCizX/BO/8A5A3jf/r6s/8A0GWq/Yo/5OI1b/sF3v8A6Ojr9o34Mx/Gv4dzaDaPFDrVhJ9q0qaThRKAQUY9QrqSD6HaecYr4S/tJ+Ov2b0f4WfFPwXfXFjYSN9nidvKubQMxJCFvlljJyV5A5OGIwBe/wDBQP4Tpbs2neEvFk84HypNBbxKfqwmYj8q+CXxRPxi8A2/jZtCk0hp7maL7M8hkwEb5WDlV3AqV5AxnI7VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV1a219ay2V5bxz29xG0csUihldWGCpB4IIOCK+Kf7OHxR+A3jT/hZHwTbULvSoJGlt2s1MtxYA/ejkjwTJFgkZwQV4YDqfCH/BQV7e1Sy+IXgB5LuIbZLnS5wocjg/uZPunr/H+ArWf+ChfhOK3J8P8Aw61a6mxwLy7jt1B+qhye1eHfibr/AIn+Av8Aws+18N3C63Lo9xcQ6ZFA7M1wgdVVUI3FWZQR6qQea8Jft/X2nR/2X8Tfh7Ib23+SWfTpfKYsODuhk+6fX5+vYV43/b+ivtLfTvhp4IvI9TuV8uO61J1IhY8ArFGTvb0yQM9QelfsdfADxTo2uz/GX4i2dxa39xHIum2t2pE5aX/WXEgPKkgsoB5O5iQOCaqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqz/4+ov98VVVVVVVVVVVVVVVVVVVVVVVVV/wW9/5Kn8NP+xfuv8A0oFfsOfArwb+0h+0f4e+E/j671W30XUba8mnfTJkhmJht3kUBnRwASoB4zjOCOteOf8Agiz+zzqmg3EPgDx3410HWljb7LcX1xBe2+/t5kQiRiOP4XXqevFfE74d+JPhJ8QvEPwz8XwRx6x4a1CWyuxExZGZGxvQkAlGGGUkDIYHAr/gkH8c9b+KP7Peo/D3xNqEl7f/AA4v47K0lkOWGnzIWt0JPJ2Mk6D0RUHarrxH4esp2tbzXtOgmjOHjlukRl+oJyKs76x1GAXOn3kF1ESQJIZA65HXkVp/xp+DereL0+HulfFrwZe+KpGdU0O3162kvmZEZ3At1cyEqqMxG3gKSeAarxB4l8N+E9ObV/FPiDTdGsYzhrrULtLeJfq7kAfnWmftLfs463fjStG/aA+G9/ek4FtbeK7KWTOQPurKT1IHTqaR0kRZI3VlYZVgcgj1FV/wUi/5Pc+Kf/YQtv8A0igr/ghj/wA1s/7gH/uRrxP8efgb4J1M6J4z+M/gXQdRD7DZ6p4itbWYN0xskkDZ9sVo2uaJ4j02HWPD2sWOqWE4zDdWdws8Ug9VdCQfwNU7pGhkkdVVQSzMcAAdTWuftE/s++GJza+JPjt8PdJmB2mO+8T2cDA+mHkBrwl8RPh94/he48CeOvD3iSKJQzvpOpw3iqD0JMbNgGq/4Laf8kB8C/8AY4D/ANI56/ZO/wCTqPg1/wBlA0L/ANL4a8WeOfBPgOwGqeOfGOh+HbJshbnVtQis4zjr80jKOOK8GfFz4UfEh3j+HfxP8J+KWiBMi6LrVvfFAOpIidsVX7T/AMAfhR+0l8MpPh18WrlrKyN0tzp+oQ3KQT2V0qsFkjZwVJ2s6lSCCGPsR+zR/wAEyP2ePgV8TtN+JWo/FSTxzrOkXQl0O0uDBbwW0+R5chjRmaWVTypyFDYO3IBH7WX/ACav8Zf+yf67/wCkE1V+xR/yaL8IP+xP0/8A9ErX7UH/AATP/Z8+PnxN1D4l2fxSl8D6/qs2/WreAwXEFzKAA0gidlaKRsfMdxBPO3JYn9lr9nr4Sfs0/DYfD74TXZ1CF7k3Op6nPcJPcXtyygb5GQBVAVVVUAAAHclmP/BZHxeLT9mHQbPw54p8m5m8c20F3FZXu13i+w325HCHJXcFyDxkCv2U5obf9qL4OzzypHHF4+0Nnd22hQL+EkknoMVbeI/D17OlrZ69p08znCRxXSMzfQA5NP4q8LxuY5PEmlqykhla8jBBHXjNWOq6Xqiu2mala3YjIDmCZZNuemcE46UxVVLMwAA5J4xX/CW+FP8AoZ9J/wDA2P8Axqyv7DUoftGnX1vdRBiu+GQOuR1GQTzVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVrHh7QPEMAtdf0PT9ThHSO8tkmUfgwIqz+EXwn0+YXGn/DDwlbSjo8OiW6MMc9QmetQwQW0KW9tCkUcYwiIoUKPQAdK1TRtI1y2NlrWlWeoW56xXUCyr+TAirT4S/CrT7kXth8MvCdtcA5EsOi26PnOeoTPUClRERY41CqowqgYAHtVVVVVVVVVVVVVVVVVVVVVaz8NPhx4iuTe+IPh/4b1O4JyZbzSYZnJ+rKTWi+GfDfhqA23hzw/pulREcx2VokCnHsgFVWs6DoXiOzOneINFsNUtWOTBeWyTIT/usCK0DwX4O8KBl8LeE9G0cOMMLCwjt8/XYoq68P6De3i6jeaJYT3aEFZ5bZGkUjphiMjHaq1vw74f8TWX9m+I9D0/VbQnP2e+tUnTPrtcEV4f8GeD/CauvhXwpo2jCQYcafYR224dedijPNVrvw/8B+KJvtHiXwToGry/89L7TYrhuOnLqTWi+FvDHhtWTw74c0vSw4wwsrNIMj32AVWu+FvDHiiBbbxL4c0vV4lB2x31mk6jPXAcGtA8EeC/CjM3hbwhomjs4wxsNPityfrsUVWoaXpmrwLbarp1rexK29Y7iFZFDAEZAYHnBIz71Z2Nlp1sllp1pBbW8edkUMYRVycnAHA5JP41c21veQPa3cEc0MqlXjkQMrA9QQeCK07RtH0dXTSNKs7ISkFxbwLFuI6Z2gZqrLw74f0y4N5puh6fa3DAgywWqRsQeoyBntVa14c8PeJbYWXiLQtO1W3GcRXtqk6jPXhwRVh8J/hZpM4utK+GnhWzmXpJb6NbxsPxVAaVVVQiAAAYAAxiqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqtc8AeBPE8vneJPBWg6tIer32mxTn83U1pHw6+Hvh6VZ9A8CeHtNlQ5V7PS4YWU+oKqMVWu+CvBvig58TeEtG1c4x/p9hHccf8AA1NaH8P/AAF4YmFx4a8EaBpMo6PY6ZFA35ooNVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVWf/AB9Rf74qqqqqqqqqqqqqqqqqqqqqqqqr/gt7/wAlT+Gn/Yv3X/pQK/4JP/8AJ7ng/wD7B+p/+kUteMvG3g/4d+Hbvxb468TaboOjWCF7i9v7hYY0AGcZY8k44Uck8AGv2rvizpXx0/aK8efFXQoHi0zXtVZrAOpVmt40WKJ2U8hmSNWI7FiK/wCCIngfUdL+G3xL+IdzBLHa+IdYstPtmdcCT7HFK7svqM3YGemVI6g1/wAFPrS3s/26fifDbR7EaTTJCMk/M+l2jMefVmJ/GtN/a88aeDf2RLH9mrwBrt9pTanr1/e+ILy3kaOR7ORIVitI3HKo7CVpAPvDap4Zwf8Aglx/yfb8Mf8AuLf+mm8r9v39tXT/ANkX4eW0Hh+C21Hx/wCJ1kTQ7OcborZFwJLqYAglFyAq/wAbHHQMR42+IXxi/aI8dRaj4y8Ra/418Tatc+VaQOXuJC8jDEVvCvCAnAEcagdABXxG/Y9/ab+EvhEeO/iH8GPEOj6CAplvpIlkS3DEBTMI2YwglgP3gXkgdeK/ZJ/br+MP7K/iWxis9bvvEHgYyhdR8L3lyWhMZPzPb7s+RKOoK4BIAYMOnw88e+F/in4G0P4ieC9RF9oniGyju7KbGCUcZww/hZTlWXqGBB6V/wAFIv8Ak9z4p/8AYQtv/SKCvhN8dfi/8NfCHi34XfCXVb7TpfiRPYQ38umK4vp1t/PCW8LJ8yiRrkhgvzNtC5wzA+PvgL8c/hnpkPiH4lfCPxl4bsLsjZe6votxbRs7dFLuoAc/3SQ3tX7N37Unxa/Zd8aW3ir4c+IJ1sWnRtU0SaUmz1OMEbkkTkBiowJAN69j1B+EXxP8MfGr4ZeG/ip4OnMmk+JbBLqBXILwk8PE+ON8bhkYDgMpr/grF4N/aI8W/tH6D4G8DXXjXxR4e8U6DFdaf4a0szz28U8LtFOfs8eV4HlO0jDjzeSAK0//AIJb/twajpR1Rfg5Hb/Jujt7jXrCOZxjP3TN8p7Ybaa1TTPit8AfiLLpuoRa/wCB/Gnhy4GQkj2d1aPgFWV1IOCpBDKcMpBBIOa/4Jqftha1+1N8LNR0f4gXMU3jrwVLDDqVykaxjULeUN5NxsUABz5bq4UYyobjeAP+C2n/ACQHwL/2OA/9I56+HfjXUvhr8QPDHxF0WC3n1DwrrNpqtpFcAmN5beZZUVwCDtLIAcEcVr3gz9rn9qK51H416n4E+Inj5bqR2l1mDR7m6gGCcxw7EKBE5HlxjagwMAVpWreIfB+uwatompahous6XPuhubaV7e4tZVOMqykMjA5HYiv+CZH7a2s/tM+C9S+HvxN1CO48f+D4kle82qjatYsdonKrgeZG21HIAB3xt1Zsf8FtP+SA+Bf+xwH/AKRz1+yd/wAnUfBr/soGhf8ApfDX7WX/ACav8Zf+yf67/wCkE1V+xR/yaL8IP+xP0/8A9ErX7a//ACdz8X/+xw1D/wBHNX/BEP8A5JV8Sv8AsYLX/wBJzX7UPw4+Ifhj4xfEHxN4k8B+ItJ0e/8AGWpraahfaXNBb3Be5mdAkjqFYsilhgnIBI4Fadp2oaxqFrpOk2Nxe317MkFta28RklnlchVREXJZmJAAAySQK/4J8/Bf4xeGf2x/hnrviT4TeMtK020v7hri8vtBuYIYQbSYAs7oFUEkDk9SK/4Ki/Ao/Bn9qjW9X0yy8nQvHyf27YlV+VZpGIukz0yJg74HRZUr/gjZ8V18G/tHat8M7278uz8f6KyQxlsB7y03TRZ+kJuh9TX7ZPxJ/wCFSfst/Evx1FceTdWmgT21lJnBS6ucW8BH0llQ/hQUsQqgkk4AAr9jP4NL8Bf2Z/Afw5nt/K1O30xbvVgww3224JmnU+u13MYz/Ci1VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX7PH7R2t/GrxV4i8Pap4bsdNj0WEPHJBMzmQ+YV5B6cCqqqqqqqqqqqqqqqqqqqqqqqqqqrP8A4+ov98VVVVVVVVVVVVVVVVVVVVVVVVV/wW9/5Kn8NP8AsX7r/wBKBWmarqmi3sepaNqV1YXkJPl3FtM0UicYOGUgjir/AFrxn45vre21PV9a8QXmClvHcXEt3J6kICSfU4Ffs3/8Ezf2kfjvrFldeI/Cl98P/CTMDdaxr1oYJinGfItXKyyMR0JCp/t9AfhH8KvBvwR+HGhfCzwBp/2TRPD9qILdWOXlYktJLIRjdI7szscDLMeAOK/4Kj/8n2/E3/uE/wDpps6/4Ja/su+Bf2jvjJrOp/E3TU1Xw34J0+O6fS5GKpe3UrlYVkwQWjUJIxXoxVQcqSDoPww+GnhWa1ufC/w88M6PNYqVtZLDSILdoAVKkIUUFQVJHGOCR3r/AIKi+Pr7x3+2d40tp7hpLPwvHa6PYoSf3aRQq8gGfWaSY/j361/wRV+FuieKPi943+KOq2MN1ceCtKtrfTjKoP2ee9aUGVfRhHbypn0kb1rX9B0bxToWoeGfEOnQahperWslpe2k67knhkUq6MO4Kkg/Wviz4MHw4+KnjL4eLKZR4X8QX+lh26t9nuHizx67K/4Iw+Or/wAQ/s1694Lv5nlXwn4mlSyyeI7e4iSXYP8Atr57f8Dr/gpF/wAnufFP/sIW3/pFBX/BEz4aeFvEfxE+IvxM1exjudX8IWNha6U0iBhbm8Nx5si56PttgoPXa7jua8Z+DfDPxC8Kar4I8ZaPb6romtWr2t7Z3CblljYYI9iOCCOQQCMEA18VPBT/AA2+J/i/4dSXBnfwrr19pTStjLm3neItxxzszX/BGXxZe67+yzq/hy9lkdfDfi26gtA33Uglhgm2jn/npJMT/vD3r9rD9vD4KfslpDpPimW617xfd25mtPD+mbTMIzwJJpG+WCMkcE5Y4JVWAOPGn/Bav9oLVrtx4F+HHgjw9ZFiUW8W41CdR2Bk8yJD/wB+x+FftAftCfEP9pjxxD8Q/idJpcmswadHYCSwsltleKN3ZSyjOW/eEbj2CjtX/BFDUruL9pTxjo8cpFrc+Bp55EyeXjvrNUOOnAlf8/rX/BbT/kgPgX/scB/6Rz18DfBumfEb41/D74e608i6f4n8U6bpd20ZwwiuLqOJyp7Ha5rRtG0nw9pFnoOg6db2GnadAlvaWtvGI44IkAVUVRwAAAAK/wCCzfwZ8OeBfjJ4T+KXh3T4LF/iBYXQ1KKFAqzXlo0QecgfxOlxEGPcpnqSa/4JZeLL3wv+2x4HtbeWRbfX4L/TbxE/jja0lkUHnoJYomP+7X/BbT/kgPgX/scB/wCkc9fsnf8AJ1Hwa/7KBoX/AKXw1+1l/wAmr/GX/sn+u/8ApBNVfsUf8mi/CD/sT9P/APRK1+2v/wAnc/F//scNQ/8ARzV/wRD/AOSVfEr/ALGC1/8ASc1/wWr/AOTV/C3/AGUCz/8ASC/r9k7/AJOo+DX/AGUDQv8A0vhqv+CtvwKHxU/Znk8f6VZebrfw2ujqSFEyzWMmEul9gAI5SewgPrXwV+I978H/AIu+Dvihp5cyeGNatr5406yxpIDJH9HTcp9mr/gsr8WrG2/Z18E+CdC1KOaPx7rK36yRtlbiytog+R7GSe2YH2r9gL4Mj45/tW+BvC15bGbSdKvP7Z1YFNy/ZrTEmxh/dkkEUR/661VVVVVVVVVVVVVVVVVVVVVVVVc+IdAs5mtrvXNPgmj4aOW5RWX6gnIr/hKvC/8A0Mml/wDgZH/jX/CVeF/+hk0v/wADI/8AGv8AhKvC/wD0Mml/+Bkf+Nf8JV4X/wChk0v/AMDI/wDGv+Eq8L/9DJpf/gZH/jX/AAlXhf8A6GTS/wDwMj/xr/hKvC//AEMml/8AgZH/AI1/wlXhf/oZNL/8DI/8a/4Srwv/ANDJpf8A4GR/41/wlXhf/oZNL/8AAyP/ABr/AISrwv8A9DJpf/gZH/jX/CVeF/8AoZNL/wDAyP8Axr/hKvC//QyaX/4GR/41/wAJV4X/AOhk0v8A8DI/8a/4Srwv/wBDJpf/AIGR/wCNf8JV4X/6GTS//AyP/Gv+Eq8L/wDQyaX/AOBkf+Nf8JV4X/6GTS//AAMj/wAa/wCEq8L/APQyaX/4GR/41/wlXhf/AKGTS/8AwMj/AMa/4Srwv/0Mml/+Bkf+Nf8ACVeF/wDoZNL/APAyP/Gv+Eq8L/8AQyaX/wCBkf8AjX/CVeF/+hk0v/wMj/xr/hKvC/8A0Mml/wDgZH/jX/CVeF/+hk0v/wADI/8AGv8AhKvC/wD0Mml/+Bkf+Nf8JV4X/wChk0v/AMDI/wDGv+Eq8L/9DJpf/gZH/jX/AAlXhf8A6GTS/wDwMj/xr/hKvC//AEMml/8AgZH/AI1/wlXhf/oZNL/8DI/8a/4Srwv/ANDJpf8A4GR/41/wlXhf/oZNL/8AAyP/ABr/AISrwv8A9DJpf/gZH/jX/CVeF/8AoZNL/wDAyP8Axr/hKvC//QyaX/4GR/41/wAJV4X/AOhk0v8A8DI/8a/4Srwv/wBDJpf/AIGR/wCNf8JV4X/6GTS//AyP/Gv+Eq8L/wDQyaX/AOBkf+Nf8JV4X/6GTS//AAMj/wAa/wCEq8L/APQyaX/4GR/41/wlXhf/AKGTS/8AwMj/AMa/4Srwv/0Mml/+Bkf+Nf8ACVeF/wDoZNL/APAyP/Gv+Eq8L/8AQyaX/wCBkf8AjX/CVeF/+hk0v/wMj/xr/hKvC/8A0Mml/wDgZH/jX/CVeF/+hk0v/wADI/8AGv8AhKvC/wD0Mml/+Bkf+Nf8JV4X/wChk0v/AMDI/wDGv+Eq8L/9DJpf/gZH/jX/AAlXhf8A6GTS/wDwMj/xr/hKvC//AEMml/8AgZH/AI1/wlXhf/oZNL/8DI/8a/4Srwv/ANDJpf8A4GR/41/wlXhf/oZNL/8AAyP/ABr/AISrwv8A9DJpf/gZH/jX/CVeF/8AoZNL/wDAyP8Axr/hKvC//QyaX/4GR/41/wAJV4X/AOhk0v8A8DI/8a/4Srwv/wBDJpf/AIGR/wCNf8JV4X/6GTS//AyP/Gv+Eq8L/wDQyaX/AOBkf+Nf8JV4X/6GTS//AAMj/wAa/wCEq8L/APQyaX/4GR/41/wlXhf/AKGTS/8AwMj/AMa/4Srwv/0Mml/+Bkf+Nf8ACVeF/wDoZNL/APAyP/Gv+Eq8L/8AQyaX/wCBkf8AjX/CVeF/+hk0v/wMj/xr/hKvC/8A0Mml/wDgZH/jX/CVeF/+hk0v/wADI/8AGv8AhKvC/wD0Mml/+Bkf+Nf8JV4X/wChk0v/AMDI/wDGv+Eq8L/9DJpf/gZH/jX/AAlXhf8A6GTS/wDwMj/xr/hKvC//AEMml/8AgZH/AI1/wlXhf/oZNL/8DI/8a/4Srwv/ANDJpf8A4GR/41/wlXhf/oZNL/8AAyP/ABr/AISrwv8A9DJpf/gZH/jX/CVeF/8AoZNL/wDAyP8Axr/hKvC//QyaX/4GR/41/wAJV4X/AOhk0v8A8DI/8a/4Srwv/wBDJpf/AIGR/wCNf8JV4X/6GTS//AyP/Gv+Eq8L/wDQyaX/AOBkf+Nf8JV4X/6GTS//AAMj/wAa/wCEq8L/APQyaX/4GR/41/wlXhf/AKGTS/8AwMj/AMa/4Srwv/0Mml/+Bkf+Nf8ACVeF/wDoZNL/APAyP/Gv+Eq8L/8AQyaX/wCBkf8AjX/CVeF/+hk0v/wMj/xr/hKvC/8A0Mml/wDgZH/jX/CVeF/+hk0v/wADI/8AGv8AhKvC/wD0Mml/+Bkf+Nf8JV4X/wChk0v/AMDI/wDGv+Eq8L/9DJpf/gZH/jX/AAlXhf8A6GTS/wDwMj/xr/hKvC//AEMml/8AgZH/AI1/wlXhf/oZNL/8DI/8a/4Srwv/ANDJpf8A4GR/41/wlXhf/oZNL/8AAyP/ABr/AISrwv8A9DJpf/gZH/jX/CVeF/8AoZNL/wDAyP8Axr/hKvC//QyaX/4GR/41/wAJV4X/AOhk0v8A8DI/8a/4Srwv/wBDJpf/AIGR/wCNf8JV4X/6GTS//AyP/Gv+Eq8L/wDQyaX/AOBkf+Nf8JV4X/6GTS//AAMj/wAa/wCEq8L/APQyaX/4GR/41/wlXhf/AKGTS/8AwMj/AMa/4Srwv/0Mml/+Bkf+Nf8ACVeF/wDoZNL/APAyP/Gv+Eq8L/8AQyaX/wCBkf8AjX/CVeF/+hk0v/wMj/xr/hKvC/8A0Mml/wDgZH/jX/CVeF/+hk0v/wADI/8AGv8AhKvC/wD0Mml/+Bkf+Nf8JV4X/wChk0v/AMDI/wDGv+Eq8L/9DJpf/gZH/jUUsVxEk8EqSRyKGR0OQwI4II6jFVVVVVVVVVVVVVVVVVVVTukaF3cKqjJLHAAHev8AhKvC/wD0Mml/+Bkf+Nf8JV4X/wChk0v/AMDI/wDGv+Eq8L/9DJpf/gZH/jX/AAlXhf8A6GTS/wDwMj/xr/hKvC//AEMml/8AgZH/AI1/wlXhf/oZNL/8DI/8a/4Srwv/ANDJpf8A4GR/41/wlXhf/oZNL/8AAyP/ABr/AISrwv8A9DJpf/gZH/jX/CVeF/8AoZNL/wDAyP8Axr/hKvC//QyaX/4GR/41/wAJV4X/AOhk0v8A8DI/8a/4Srwv/wBDJpf/AIGR/wCNf8JV4X/6GTS//AyP/Gv+Eq8L/wDQyaX/AOBkf+Nf8JV4X/6GTS//AAMj/wAa/wCEq8L/APQyaX/4GR/41/wlXhf/AKGTS/8AwMj/AMa/4Srwv/0Mml/+Bkf+Nf8ACVeF/wDoZNL/APAyP/Gv+Eq8L/8AQyaX/wCBkf8AjX/CVeF/+hk0v/wMj/xr/hKvC/8A0Mml/wDgZH/jX/CVeF/+hk0v/wADI/8AGv8AhKvC/wD0Mml/+Bkf+Nf8JV4X/wChk0v/AMDI/wDGv+Eq8L/9DJpf/gZH/jX/AAlXhf8A6GTS/wDwMj/xr/hKvC//AEMml/8AgZH/AI1/wlXhf/oZNL/8DI/8a/4Srwv/ANDJpf8A4GR/41/wlXhf/oZNL/8AAyP/ABr/AISrwv8A9DJpf/gZH/jX/CVeF/8AoZNL/wDAyP8Axr/hKvC//QyaX/4GR/41/wAJV4X/AOhk0v8A8DI/8a/4Srwv/wBDJpf/AIGR/wCNf8JV4X/6GTS//AyP/Gv+Eq8L/wDQyaX/AOBkf+Nf8JV4X/6GTS//AAMj/wAa/wCEq8L/APQyaX/4GR/41/wlXhf/AKGTS/8AwMj/AMa/4Srwv/0Mml/+Bkf+Nf8ACVeF/wDoZNL/APAyP/Gv+Eq8L/8AQyaX/wCBkf8AjX/CVeF/+hk0v/wMj/xr/hKvC/8A0Mml/wDgZH/jX/CVeF/+hk0v/wADI/8AGv8AhKvC/wD0Mml/+Bkf+Nf8JV4X/wChk0v/AMDI/wDGv+Eq8L/9DJpf/gZH/jX/AAlXhf8A6GTS/wDwMj/xr/hKvC//AEMml/8AgZH/AI1/wlXhf/oZNL/8DI/8a/4Srwv/ANDJpf8A4GR/41/wlXhf/oZNL/8AAyP/ABr/AISrwv8A9DJpf/gZH/jX/CVeF/8AoZNL/wDAyP8Axr/hKvC//QyaX/4GR/41/wAJV4X/AOhk0v8A8DI/8a/4Srwv/wBDJpf/AIGR/wCNf8JV4X/6GTS//AyP/Gv+Eq8L/wDQyaX/AOBkf+Nf8JV4X/6GTS//AAMj/wAa/wCEq8L/APQyaX/4GR/41/wlXhf/AKGTS/8AwMj/AMa/4Srwv/0Mml/+Bkf+Nf8ACVeF/wDoZNL/APAyP/Gv+Eq8L/8AQyaX/wCBkf8AjX/CVeF/+hk0v/wMj/xr/hKvC/8A0Mml/wDgZH/jX/CVeF/+hk0v/wADI/8AGv8AhKvC/wD0Mml/+Bkf+Nf8JV4X/wChk0v/AMDI/wDGv+Eq8L/9DJpf/gZH/jX/AAlXhf8A6GTS/wDwMj/xr/hKvC//AEMml/8AgZH/AI1/wlXhf/oZNL/8DI/8a/4Srwv/ANDJpf8A4GR/41/wlXhf/oZNL/8AAyP/ABr/AISrwv8A9DJpf/gZH/jX/CVeF/8AoZNL/wDAyP8Axr/hKvC//QyaX/4GR/41/wAJV4X/AOhk0v8A8DI/8a/4Srwv/wBDJpf/AIGR/wCNf8JV4X/6GTS//AyP/Gv+Eq8L/wDQyaX/AOBkf+Nf8JV4X/6GTS//AAMj/wAa/wCEq8L/APQyaX/4GR/41/wlXhf/AKGTS/8AwMj/AMa/4Srwv/0Mml/+Bkf+Nf8ACVeF/wDoZNL/APAyP/Gv+Eq8L/8AQyaX/wCBkf8AjX/CVeF/+hk0v/wMj/xr9hbXNEtfiT46e61ixhVrVdpkuFUH9+emTX/CVeF/+hk0v/wMj/xr/hKvC/8A0Mml/wDgZH/jX/CVeF/+hk0v/wADI/8AGv8AhKvC/wD0Mml/+Bkf+Nf8JV4X/wChk0v/AMDI/wDGv+Eq8L/9DJpf/gZH/jX/AAlXhf8A6GTS/wDwMj/xr/hKvC//AEMml/8AgZH/AI1/wlXhf/oZNL/8DI/8a/4Srwv/ANDJpf8A4GR/41/wlXhf/oZNL/8AAyP/ABr/AISrwv8A9DJpf/gZH/jX/CVeF/8AoZNL/wDAyP8Axr/hKvC//QyaX/4GR/41/wAJV4X/AOhk0v8A8DI/8a/4Srwv/wBDJpf/AIGR/wCNf8JV4X/6GTS//AyP/Gv+Eq8L/wDQyaX/AOBkf+Nf8JV4X/6GTS//AAMj/wAa/wCEq8L/APQyaX/4GR/41/wlXhf/AKGTS/8AwMj/AMa/4Srwv/0Mml/+Bkf+Nf8ACVeF/wDoZNL/APAyP/Gv+Eq8L/8AQyaX/wCBkf8AjX/CVeF/+hk0v/wMj/xr/hKvC/8A0Mml/wDgZH/jX/CVeF/+hk0v/wADI/8AGv8AhKvC/wD0Mml/+Bkf+Nf8JV4X/wChk0v/AMDI/wDGv+Eq8L/9DJpf/gZH/jUPiPw7cypb22v6dLLIdqIl0jMx9AAeaqqqqqrP/j6i/wB8VVVVVVVVVVVVVVVVVVVVVVVVV/wW9/5Kn8NP+xfuv/SgV/wSntre8/bV8JW11BHNFJp2ph45EDKw+xy9QetWOlaXpauumaba2gkI3iCFY92OmcDmqr/gqP8A8n2/E3/uE/8Apps6/wCCHX/I1fFv/sH6V/6Muar/AIKfeBb/AMD/ALaPjqS5gZLXxGLXV7F2H+tjlgRXYcngTRzL/wAB7dK/4IsfFXQ/Cvxf8a/C3V76K1n8b6XbT6cZWC+fcWTSnyV9WMdxK4HpG3tXiPxDofhLQNR8UeJdUt9O0nSbWS6vbu4bakEUalndj6AAmvit4zPxI+KPjH4iNEYj4o1++1QxsACn2id5cYHpvr/gjR8P7/wx+zJq/jTUbZ4v+Ew8STTWZYEeZbW8aQhh2/1qzjgfw9fT/gpF/wAnufFP/sIW3/pFBX/BFD4n+EvDHxF+IXwx1vUobTV/Gdnp9xpCyuEFy1mbnzYlz1fbchwvXbG57V448beFvhx4R1bxz401m30vRNEtXuby6ncKqIoycerHoFHJJAGSa+J/jSb4k/Evxb8RLmExTeKddvdVkjOPka4neUjjjgvjiv8AgkR4Pn8A/sd3fjTV4HRPE+v32rQkL8xtoY47cYGf79tKR0zu9MV8UviL4i+LvxF8RfEzxZdvcat4k1GW8uC77tm9vljX0RF2oo4AVQAABX/BNP8AYd/Z11/9nzw58bviB4L0nxx4i8UNcybdXiF3aWEcVxJCIUgbMbN+63MzqWDEgYA5/wCCvuifDTwr+0H4a8KfDnw94f0X+z/CsTalZaPYxWqRzPcTMnmLEoG8x7DzztKnoRX/AARU/wCTqPFP/ZP7z/0vsK/4Laf8kB8C/wDY4D/0jnr4G+MtM+HPxr+H3xC1pZH0/wAMeKdN1S7WMZYxW91HK+0eu1DWg69ovinRLDxJ4c1W11LS9Tt0uLO8tpBJFPE4BV0YcEEEHNf8FlvjT4c+IHxn8LfC/wAN6nb36/D7T7n+0JYHDrFe3bRmSHcOrJHbw5HYsRwQRX/BJzwHe+Mf2zfDetQwGS08H6bf6rdkg4AMDW0fPr5txGf+An3r/gtp/wAkB8C/9jgP/SOev2Tv+TqPg1/2UDQv/S+Gv2sv+TV/jL/2T/Xf/SCaq/Yo/wCTRfhB/wBifp//AKJWv21/+Tufi/8A9jhqH/o5q/4Ih/8AJKviV/2MFr/6Tmv+C08E037KnhuSKNnWHx9ZvIQPuL9hvlyfbLAfiK/Zv17SvCv7RHwt8T65dpbabo/jTSLy8nc4EUMV5E7sSewVSaBVlDKwII4IrXNF0vxLomoeHdbs47zTtUtZbW8t5BlZoZFKOhHoVJB+tftAfCTVPgR8aPF/wl1be0nhzVJIIJXGDPbn54Jcdt8TRvj/AGq/aC/aCv8A43eEvhD4eupJj/wrzwXHok4l5LXCTyrvB7g26Wg+qmv+CKnwXOj+A/GXx31S0K3HiG8XR9Kd1wfstvh5nU/3XlZVPvb1VVVVVVVVVVVVVVVVVVVVVVVV8Fv2Xr79tP8AaZ+KXhvUviTJ4cutPu7/AFSW/l0v+0GuG+2+WVK+dFt/1mc5PTGK/wCHJX/VzX/ll/8A3dX/AA5K/wCrmv8Ayy//ALur/hyV/wBXNf8All//AHdX/Dkr/q5r/wAsv/7ur/hyV/1c1/5Zf/3dX/Dkr/q5r/yy/wD7ur/hyV/1c1/5Zf8A93V/w5K/6ua/8sv/AO7q/wCHJX/VzX/ll/8A3dX/AA5K/wCrmv8Ayy//ALur/hyV/wBXNf8All//AHdX/Dkr/q5r/wAsv/7ur/hyV/1c1/5Zf/3dX/Dkr/q5r/yy/wD7ur/hyV/1c1/5Zf8A93V/w5K/6ua/8sv/AO7q/wCHJX/VzX/ll/8A3dX/AA5K/wCrmv8Ayy//ALur/hyV/wBXNf8All//AHdX/Dkr/q5r/wAsv/7ur/hyV/1c1/5Zf/3dX/Dkr/q5r/yy/wD7ur/hyV/1c1/5Zf8A93V/w5K/6ua/8sv/AO7q/wCHJX/VzX/ll/8A3dX/AA5K/wCrmv8Ayy//ALur/hyV/wBXNf8All//AHdX/Dkr/q5r/wAsv/7ur/hyV/1c1/5Zf/3dX/Dkr/q5r/yy/wD7ur9tv9iT/hjj/hDP+Lm/8Jf/AMJd9v8A+YL/AGf9l+zfZ/8ApvLv3faP9nG3vnjTv+CVn9oafa33/C+Nn2iFJNn/AAi2du5QcZ+189a/4dQ/9V8/8tX/AO66/wCHUP8A1Xz/AMtX/wC66/4dQ/8AVfP/AC1f/uuv+HUP/VfP/LV/+66/4dQ/9V8/8tX/AO66/wCHUP8A1Xz/AMtX/wC66/4dQ/8AVfP/AC1f/uuv+HUP/VfP/LV/+66/4dQ/9V8/8tX/AO66/wCHUP8A1Xz/AMtX/wC66/4dQ/8AVfP/AC1f/uuv+HUP/VfP/LV/+66/4dQ/9V8/8tX/AO66/wCHUP8A1Xz/AMtX/wC66/4dQ/8AVfP/AC1f/uuv+HUP/VfP/LV/+66/4dQ/9V8/8tX/AO66/wCHUP8A1Xz/AMtX/wC66/4dQ/8AVfP/AC1f/uuv+HUP/VfP/LV/+66/4dQ/9V8/8tX/AO66/wCHUP8A1Xz/AMtX/wC66/4dQ/8AVfP/AC1f/uuv+HUP/VfP/LV/+66/4dQ/9V8/8tX/AO66/wCHUP8A1Xz/AMtX/wC66/4dQ/8AVfP/AC1f/uuv+HUP/VfP/LV/+66/4dQ/9V8/8tX/AO668Z/8Ewf+ER8H654r/wCF4fa/7F024vfs/wDwjPl+b5UbPt3fajtztxnBxnoa/Yt/YJ/4bA8L+I/En/C1/wDhEv8AhH7+K08j+wft/n749+7d9oi246YwfrX/AA5K/wCrmv8Ayy//ALur/hyV/wBXNf8All//AHdX/Dkr/q5r/wAsv/7ur/hyV/1c1/5Zf/3dX/Dkr/q5r/yy/wD7ur/hyV/1c1/5Zf8A93V/w5K/6ua/8sv/AO7q/wCHJX/VzX/ll/8A3dX/AA5K/wCrmv8Ayy//ALur/hyV/wBXNf8All//AHdX/Dkr/q5r/wAsv/7ur/hyV/1c1/5Zf/3dX/Dkr/q5r/yy/wD7ur/hyV/1c1/5Zf8A93V/w5K/6ua/8sv/AO7q/wCHJX/VzX/ll/8A3dX/AA5K/wCrmv8Ayy//ALur/hyV/wBXNf8All//AHdX/Dkr/q5r/wAsv/7ur/hyV/1c1/5Zf/3dX/Dkr/q5r/yy/wD7ur/hyV/1c1/5Zf8A93V/w5K/6ua/8sv/AO7q/wCHJX/VzX/ll/8A3dX/AA5K/wCrmv8Ayy//ALur/hyV/wBXNf8All//AHdX/Dkr/q5r/wAsv/7ur/hyV/1c1/5Zf/3dX/Dkr/q5r/yy/wD7ur/hyV/1c1/5Zf8A93V8Tf2Mv+Fc/tb+Gv2WP+Fk/wBo/wDCQ3Gnw/27/Y3k+R9qOM/Z/Obdt9PMGfav+HJX/VzX/ll//d1f8OSv+rmv/LL/APu6v+HJX/VzX/ll/wD3dUf/AARTSW5mtR+0zhoQpP8AxRfUH/t99q/4clf9XNf+WX/93V/w5K/6ua/8sv8A+7q/4clf9XNf+WX/APd1f8OSv+rmv/LL/wDu6v8AhyV/1c1/5Zf/AN3V/wAOSv8Aq5r/AMsv/wC7q/4clf8AVzX/AJZf/wB3V/w5K/6ua/8ALL/+7q/4clf9XNf+WX/93V/w5K/6ua/8sv8A+7q/4clf9XNf+WX/APd1f8OSv+rmv/LL/wDu6v8AhyV/1c1/5Zf/AN3V/wAOSv8Aq5r/AMsv/wC7q/4clf8AVzX/AJZf/wB3V/w5K/6ua/8ALL/+7q/4clf9XNf+WX/93V/w5K/6ua/8sv8A+7q/4clf9XNf+WX/APd1f8OSv+rmv/LL/wDu6v8AhyV/1c1/5Zf/AN3V/wAOSv8Aq5r/AMsv/wC7q/4clf8AVzX/AJZf/wB3V/w5K/6ua/8ALL/+7q/4clf9XNf+WX/93V/w5K/6ua/8sv8A+7q/ab/4Jb/8M5fA/wAS/Gb/AIXp/wAJD/wj32X/AIlv/CMfZPP8+6ht/wDW/apNu3zt33TnbjjOR/wS9vry+/Yu8Fi8upJ/s91qMMXmMWKIt5LhR7DPA7VVVVVVVVVVVVVVVVVVVVft0XE9r+yH8VJbaeSJ/wDhHpV3IxU4ZlDDI7EEg+oNfsg/8E4P+GrvhPL8T/8Ahcv/AAi/lavPp/2H/hHft2fLSNt/mfaY+vmYxt4x15r/AIclf9XNf+WX/wDd1f8ADkr/AKua/wDLL/8Au6v+HJX/AFc1/wCWX/8Ad1f8OSv+rmv/ACy//u6v+HJX/VzX/ll//d1f8OSv+rmv/LL/APu6v+HJX/VzX/ll/wD3dX/Dkr/q5r/yy/8A7ur/AIclf9XNf+WX/wDd1f8ADkr/AKua/wDLL/8Au6v+HJX/AFc1/wCWX/8Ad1f8OSv+rmv/ACy//u6v+HJX/VzX/ll//d1f8OSv+rmv/LL/APu6v+HJX/VzX/ll/wD3dX/Dkr/q5r/yy/8A7ur/AIclf9XNf+WX/wDd1f8ADkr/AKua/wDLL/8Au6v+HJX/AFc1/wCWX/8Ad1f8OSv+rmv/ACy//u6v+HJX/VzX/ll//d1f8OSv+rmv/LL/APu6v+HJX/VzX/ll/wD3dX/Dkr/q5r/yy/8A7ur/AIclf9XNf+WX/wDd1f8ADkr/AKua/wDLL/8Au6v+HJX/AFc1/wCWX/8Ad1f8OSv+rmv/ACy//u6v+HJX/VzX/ll//d1f8OSv+rmv/LL/APu6r79kj7H+2Ddfsn/8LB3/AGa4kh/t/wDsnG7ZYm6z9n87jONmPM9/av8Ah1D/ANV8/wDLV/8Auuv+HUP/AFXz/wAtX/7rr/h1D/1Xz/y1f/uuv+HUP/VfP/LV/wDuuv8Ah1D/ANV8/wDLV/8Auuv+HUP/AFXz/wAtX/7rr/h1D/1Xz/y1f/uuv+HUP/VfP/LV/wDuuv8Ah1D/ANV8/wDLV/8Auuv+HUP/AFXz/wAtX/7rr/h1D/1Xz/y1f/uuv+HUP/VfP/LV/wDuuv8Ah1D/ANV8/wDLV/8Auuv+HUP/AFXz/wAtX/7rr/h1D/1Xz/y1f/uuv+HUP/VfP/LV/wDuuv8Ah1D/ANV8/wDLV/8Auuv+HUP/AFXz/wAtX/7rr/h1D/1Xz/y1f/uuv+HUP/VfP/LV/wDuuv8Ah1D/ANV8/wDLV/8Auuv+HUP/AFXz/wAtX/7rr/h1D/1Xz/y1f/uuv+HUP/VfP/LV/wDuuv8Ah1D/ANV8/wDLV/8Auuv+HUP/AFXz/wAtX/7rr/h1D/1Xz/y1f/uuv+HUP/VfP/LV/wDuuv8Ah1D/ANV8/wDLV/8Auuv+HUP/AFXz/wAtX/7rr9qL9iT/AIZr+H+n+Ov+Fm/8JH9u1iPTfsv9i/Y9m+GaTfv8+TOPJxtwPvZzxg/Br/gkd/wt34T+Evif/wANBf2V/wAJRpFvqH2H/hE/P+zeagbZ5n2td+M4ztGfQV/w5K/6ua/8sv8A+7q/4clf9XNf+WX/APd1f8OSv+rmv/LL/wDu6v8AhyV/1c1/5Zf/AN3V/wAOSv8Aq5r/AMsv/wC7q/4clf8AVzX/AJZf/wB3V/w5K/6ua/8ALL/+7q/4clf9XNf+WX/93V/w5K/6ua/8sv8A+7q/4clf9XNf+WX/APd1f8OSv+rmv/LL/wDu6v8AhyV/1c1/5Zf/AN3V/wAOSv8Aq5r/AMsv/wC7q/4clf8AVzX/AJZf/wB3V/w5K/6ua/8ALL/+7q/4clf9XNf+WX/93V/w5K/6ua/8sv8A+7q/4clf9XNf+WX/APd1f8OSv+rmv/LL/wDu6v8AhyV/1c1/5Zf/AN3V/wAOSv8Aq5r/AMsv/wC7q/4clf8AVzX/AJZf/wB3V/w5K/6ua/8ALL/+7q/4clf9XNf+WX/93V/w5K/6ua/8sv8A+7q/4clf9XNf+WX/APd1f8OSv+rmv/LL/wDu6v8AhyV/1c1/5Zf/AN3V/wAOSv8Aq5r/AMsv/wC7q/4clf8AVzX/AJZf/wB3V+yF+xl/w1b478X+Cf8AhZP/AAi//CKW4l+0/wBjfbftOZTHjZ50ezpnqfT3r/hyV/1c1/5Zf/3dX/Dkr/q5r/yy/wD7ur/hyV/1c1/5Zf8A93V/w5K/6ua/8sv/AO7q/wCHJX/VzX/ll/8A3dX/AA5K/wCrmv8Ayy//ALur/hyV/wBXNf8All//AHdX/Dkr/q5r/wAsv/7ur/hyV/1c1/5Zf/3dX/Dkr/q5r/yy/wD7ur/hyV/1c1/5Zf8A93V/w5K/6ua/8sv/AO7q/wCHJX/VzX/ll/8A3dX/AA5K/wCrmv8Ayy//ALur/hyV/wBXNf8All//AHdX/Dkr/q5r/wAsv/7ur/hyV/1c1/5Zf/3dX/Dkr/q5r/yy/wD7ur/hyV/1c1/5Zf8A93V/w5K/6ua/8sv/AO7q/wCHJX/VzX/ll/8A3dX/AA5K/wCrmv8Ayy//ALur/hyV/wBXNf8All//AHdX/Dkr/q5r/wAsv/7ur/hyV/1c1/5Zf/3dX/Dkr/q5r/yy/wD7ur/hyV/1c1/5Zf8A93V/w5K/6ua/8sv/AO7q/wCHJX/VzX/ll/8A3dX/AA5K/wCrmv8Ayy//ALur9sv9j26/Yr1/walh8UZPE1zr0dxdQ3MWknTXs3t3i2lcTykklwQcrjb3zxVVVVVWf/H1F/viqqqqqqqqqqqqqqqqqqqqqqqqv+C3v/JU/hp/2L91/wClAr/gk/8A8nueD/8AsH6n/wCkUtVVf8FR/wDk+34m/wDcJ/8ATTZ1/wAEOv8Akavi3/2D9K/9GXNV/wAFBf2JLX9rjwHaal4VntdP+IPhdJDpFxP8sd7C3L2krD7oJAZGOdrZ6B2NeMvAXxY+AvjZNJ8ZeHtf8GeJ9JnEtu0qvbTRuhBWWGVT8wBwVkRiOhBr4j/tdftLfFzwqngj4i/GbxFrWhIRvsZZxGk+Onm7ApmwcH94W5wetfsif8E+/jH+0/r+n6pf6JqHhf4eiQPfeIry3MXnxDqloj8zO2CAwBRTyxyAreBfBPhn4beDdG8A+DdMTT9E0Cyjs7G2TnZGi4GT1LHqWPJJJPJr/go1cwXf7bHxVltpA6rqkMZIGMMlpCrD8CpH4VL+y58WrP8AZ+8N/tReFrC61Lw5e3l1Fdy6fGxn0eS3mKLLJtyRG23Ik42spBxlS3jH4xfF34i6fb6T8QPir4v8T2VoQ1vbazrlzexwkDAKpK7BcAkcCv2Qv2Jfin+1j4utItL0y70fwRBN/wATfxNPARDEin5o4CeJpj0CrkKSC2BXgrwX4b+Hvg3RvAPhLTY7LRNBsIrGxthzshjQKoJPLHA5J5JJJ5Nftqfsk+Nv2Wfixq2l3WiXj+CtSvJJvDWsiItBPbsxZYWccCaMfKynBONwG1ga+E/7V37RnwQ0C48JfCj4sa1oOkXkxlexh2SxiRhgsiyK3lseMlME4Gegr4w/Bv47+FvDPh/43fGnTtWif4j3t41rcazLI1/dNCImaeZZPmCyed8hY5YIxwF2lv8Agi9fW9p+1dr1vMTvvfAt7DFgfxC8sn5/4CjV/wAFtrmBPgV4Bs2kAml8Ws6Jjqq2koY/gXX86/Z1/Z48Y/tM+MNW8A+Aprca5YaDdarawTkKt00JQeTuJ+Vn34BPG7GcDLCP4h/tB/BldR+GNt4++IHglLeZ01DQIdWu9ORZD94SQK6jJ9xzXgnwJ43+KPii18I+AvDOp+Itd1B/3NnZQNNK5zyxx0UZyWOAOpIFf8E+P2MIv2SfhpdTeKJba88e+LPKm1uaE70s0QHy7SNv4ghZizDhmbuFU1/wW0/5ID4F/wCxwH/pHPX7J3/J1Hwa/wCygaF/6Xw1+1l/yav8Zf8Asn+u/wDpBNVfsUf8mi/CD/sT9P8A/RK1+2v/AMnc/F//ALHDUP8A0c1f8EQ/+SVfEr/sYLX/ANJzX7cPwAv/ANpX9m3xR8NdBEP9vgR3+iGVtqm7gbcqEngeYm+LJ4HmZPSvFHhbxJ4K1+98K+L9Bv8ARtY02UxXdjfW7QzQOOoZGAIr/gmP8SfiLq37ZXws8Hap4/8AEd5oMCamkWlXGqzSWsappV2VCwligAIGABxgVX/Ba34KWum+IfBfx/0qGOM6xG2havjA3zRAy28mOpJj85SewiQV4e0DV/Fmv6Z4W8PWMl7qusXkVnY20Y+aeeVwkaL7szAfjXwF+E+lfAv4NeD/AISaP5bQ+GdLitpZYwQJ7j7082P+mkrSP9Wqqqqqqqqqqqqqqqqqq8ZeNPCXw98N3vi/xx4isNE0bTo99ze3swijjHbk9STgADkkgAEmviV/wWS+DXh3UZdP+Gfw41/xlHCSBe3VyulQSn1j3JJJt6feRD7V8NP+Cyfwh8SarbaZ8Tfhnrng2G4cIb+1vV1WCDJ+9IAkUm3HXYjn2PWtE1vR/Euj2XiDw9qlrqWmajAlxaXdrKJIp4nGVdGHBUgggitU1PTdE0261nWdQt7GwsYXnurq5lEccEaDLO7HAVQASSeABXxW/wCCxvwj8Ka5Po/wu+HGreOLe2co2oz340q3mx/FEGikkZfdkQ+2Oa/Z3/4Kp/Bb41eJ7HwP4y8PX3gDXNUlWGxa7u1u7KaVuFj+0BUKMx4G9ApOBuyQKqq/4Jcf8nj/ABm/68L/AP8ATlHVVVVVVVVVVVVVVVVVVVVX/BbX/mjH/ce/9x9eHv8AkX9M/wCvOL/0AVVVVVVVVVVVVVVVVVVVVfGX/kkHjn/sWtQ/9JpK/wCCLf8AyS/4jf8AYetv/RBqqqqqqqqqqqqqqqqqqqqr9pr/AJS3fDb/AK/9A/8AQ6qqgugni+5tieJYVAHuFB/lmqqqqqqqqqqqqqqqqqr/AIKa/wDJj/xJ/wC4V/6dbSv+CWf/ACZf4R/6/wDUv/SyWqqqqqqqqqqqqqqqqqqqr9u//kz/AOKn/YAk/wDQ0r/gkH/yahd/9jdff+ibeqqqqqqqqqqqqqqqqqqqqtb/AOUweqf9f9x/6Y2qqqqqqqqqqqqqqqqqqqqr/gqJ/wAkA0D/ALHG2/8ASO8r9jT/AJNQ+Ev/AGKNh/6JWqqqqqqqqqqqqqqqqqqqqv8AgkL/AMl2+L3/AF4J/wClbVVVVVVVVVVVVVVVVVVVVV/wWt/5Gf4Uf9eGqf8Aoy3qqqqqqs/+PqL/AHxVVVVVVVVVVVVVVVVVVVVVVVVV9ouj6o6SalpVndsgwjTwLIVHtkHFWnh7QLCdbqw0TT7aZQQskNsiMM9cEDNVVXfh3w/fztdX2hafcTvjdJLao7NgYGSRnoAKsdH0jSy7aZpdnaGQAOYIFj3Y6ZwOaqvEXhXwv4v086T4s8N6XrdkTk22o2cdzGT67XBH6Von7Of7PXhm+Gp+G/gP8O9KvFO4XFj4Xs4JAeedyRg9z+dVVx4a8OXkz3N1oGmzTSHLySWiMzH1JIyatbGxsrYWdlZwQQDOIoowijPXgcVdfs6fs+Xusf8ACRXnwJ+Hk+q7i326XwvZtPuOcnzDHuzye/c1a2trZW0VnZW8UEEKBIookCKijgAAcAY7VWsaNo/iHTZtH17SrPUrC5XbNa3kCzRSj0ZGBBHsRXh39n74C+D9VXXfCPwR8A6JqaPvW807w1aW0yt6h0jDA8nnPer7S9M1REj1LTrW7VDlFnhWQKfUAg4qz0DQtOmFzp+i2FtMAQJIbZEbB68gZq+0vTNURI9S061u1Q5RZ4VkCn1AIOKstB0PTZvtGnaNY2su0rvhtlRsdxkDOOK8afCf4WfEny/+FifDTwr4p8ldsf8AbWjW99sGc4Hmo2BnmvCXgDwJ4AsjpvgPwVoPhu0YANb6RpsVnGcDA+WJVHAqr7TNN1SNYdSsLa7RG3Kk8SyAH1AIPNQ+GPDVvKk9v4e0yOWJgyOloisrDkEEDg1NDDcQvbzxJLHIpV0dchgRggg9QR2r/hEvCn/Qs6T/AOAUf+FQQQW0KW1tDHFFGu1I0UKqgdgB0FT+GfDd1M9xc+H9NllkYs7yWiMzHuSSMk1Y6Xpmlo0emada2iucssEKxhj6kAVXjT4W/DH4kxxxfET4c+F/FKQrtjXWtHgvggznAEqtgZ5+teEfhH8KPh/MLnwF8MfCfhuYJsEmkaLb2bbf7uYkU45PHvX7Rn7V3wV/ZY0Sy1b4ueILu0m1ZJjpWn2djJcXGoNDs8xY8DYCPMTJkdFG4c81+21+2t4w/bC8a2t1PZPofg7QS66Jonm7ypbG+4nYcPMwAH91FGF6szf8Eg/2V7nxx8RJf2kfF2lt/wAI/wCDnaDQRMny3mpMuGkUHqsKNnP/AD0dCDlDiqqqqqqqqqqqqqqqqqr4zfCbwr8dPhjr/wAKfGaTf2Vr9t5MskG0SwOrB45YywIDo6qwyCMivhP+y98A/gnokGi/D34YaHZtEgWS/ntFuLy4OMFpJ5AXbPJxkKMnAA4r/go7+yd8LPGvwD8U/FLQfB+laP4y8I2p1KPUbG1S3e7hQgzxT7QPMHl72UtlgyjBwWB/4I1/EvWfEnwm8afDLU72S4tvBup29xp6yEnyIbxZSY19F8yCR8esjetf8FiPj9qWg6F4b/Z48O6g9ufEER1bxAI22mS1SQpbRH1VpUkcj1hTsTX7Dn7KfhD9nD4P6HNJoFq3jnXLGK71/U5YQ1wssihjbK/VY4wQm0YDFSxGWr/grP8AsueD3+HMf7Rvgzw9Z6ZrWjX0Vv4ha0gES3ttOwjSaQLwZEmaNd3UiU5J2qK/4JyfHLU/jp+zFot/4j1BrzxB4WuZNE1K4kfdJOYVVoZGzySYZIgWOdzKxz1xVf8ABLj/AJPH+M3/AF4X/wD6co6qqqqqqqqqqqqqqqqqqqq/4La/80Y/7j3/ALj68Pf8i/pn/XnF/wCgCqqqqqqqqqqqqqqqqqqqr4y/8kg8c/8AYtah/wCk0lf8EW/+SX/Eb/sPW3/og1VVVVVVVVVVVVVVVVVVVV+01/ylu+G3/X/oH/odVVX90Lfxabktwkygn2wAf0qqqqqqqqqqqqqqqqqq/wCCmv8AyY/8Sf8AuFf+nW0r/gln/wAmX+Ef+v8A1L/0slqqqqqqqqqqqqqqqqqqqq/bv/5M/wDip/2AJP8A0NK/4JB/8moXf/Y3X3/om3qqqqqqqqqqqqqqqqqqqqrW/wDlMHqn/X/cf+mNqqqqqqqqqqqqqqqqqqqqq/4Kif8AJANA/wCxxtv/AEjvK/Y0/wCTUPhL/wBijYf+iVqqqqqqqqqqqqqqqqqqqqr/AIJC/wDJdvi9/wBeCf8ApW1VVVVVVVVVVVVVVVVVVVVf8Frf+Rn+FH/Xhqn/AKMt6qqqqqrP/j6i/wB8VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX/BWb4DeLPjT8AtC1D4e+EtT8ReJfC/iKKSKy02ze6nktZ0aKYKiAtgP5DE9AqEnjkfsyf8EhvjD8QNWs9f/aCz4D8LIyvJp8c0cuqXq8HaqruS3B5BaQlx/wA8znI8D+CPCnw28I6T4D8D6Jb6RoWiWy21jZQAhYo19zkkk5JYkliSSSSTVVVVVVVVVVVVVVVVVVVV/wAFQvjjo3ws/Zo1fwSl4h8RfEEf2XYWwb5hb7la5mI/uCP93/vSr1Ga/wCCSPwZ1P4efs/ah8Q9dtGt7z4hagt1axuCCbGBSkDkHpudp2HqjIe9ft5vH48/4KS23g3UnZ7WDUfD2klZBkLHMlvIwAB6f6Qx7ck/Wq/bg0u21j9kb4r2lyAUj8NXFwMrn5oQJV/8eQV/wRR1iafw78WfD7M3lWV7pV0oIGA0yXKnHfpAv6e9VXwS+K37Q/wj/aH+JOtfs4fD648Xa5eXN9b31rDoVxqhhtvtm7eY4CGUb1QbjxzjvX/Dan/BU3/o2DVP/Db6p/8AFV/w2p/wVN/6Ng1T/wANvqn/AMVX/Dan/BU3/o2DVP8Aw2+qf/FV+xr8Sfjn8VfhFN4n/aE8ET+FfFCaxPbpYTaNPpjG2VIyj+VOS2CzON3Q49jVVVVVVVVVVVVVVVVVV/wW1/5ox/3Hv/cfXh7/AJF/TP8Arzi/9AFVVVVVVVVVVVVVVVVVVVXxl/5JB45/7FrUP/SaSv8Agi3/AMkv+I3/AGHrb/0Qaqqqqqqqqqqqqqqqqqqqq/aa/wCUt3w2/wCv/QP/AEOqqtcbfq92SRxKw/KtNuPten29xnJeME/XHP61VVVVVVVVVVVVVVX/AA2p/wAFTf8Ao2DVP/Db6p/8VX/Dan/BU3/o2DVP/Db6p/8AFV/w2p/wVN/6Ng1T/wANvqn/AMVX7SP7T/7fXxF+C3iLwb8a/gRf+HfBmofZf7T1OXwTf2CwbLqKSLM8pKJulSNeeu7A5Ir/AIJZ/wDJl/hH/r/1L/0slqqqqqqqqqqqqqqqqqqqq/bv/wCTP/ip/wBgCT/0NK/ZN/aL/bb+FXwtl8Mfs9/Ba98VeF21Wa4e+h8IXupgXLJGHTzYCFGFVDt6jPvX/Dan/BU3/o2DVP8Aw2+qf/FV/wANqf8ABU3/AKNg1T/w2+qf/FV/w2p/wVN/6Ng1T/w2+qf/ABVfDzVNf1z4f+Gda8V2DWOuaho9pcalatA0JguXhVpUMbfMm1yw2nkYwaqqqqqqqqqqqqqqqqqtb/5TB6p/1/3H/pjaqqqqqqqqqqqqqqqqqqqqv+Con/JANA/7HG2/9I7yv2NP+TUPhL/2KNh/6JWqqqqqqqqqqqqqqqqqqqqv+CQv/Jdvi9/14J/6VtVVVVVVVVVVVVVVVVVV+1j48+L3w1+CGseLvgX4Sm8S+MLW4tUs9Oi0ubUGkR5kWUiGEh2whY5HTGTwK/4bU/4Km/8ARsGqf+G31T/4qv8AhtT/AIKm/wDRsGqf+G31T/4qv+G1P+Cpv/RsGqf+G31T/wCKr9sn4x/tOfGDUvC1z+0p8NLnwhc6ZBcppCT+HbrSvtCO0ZlIE5JfBVOV6Z561VVVVVWf/H1F/viqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr4+/HjwD+zl8NtR+JPj/AFAR21quyzs42An1C4IOyCFSeWbH0UAscAE18EfhF8Uv+Cmn7QF58dvjNDLY/DjSLoQvBFIyRtFGd0em2p4JGGBlkGD8zHIZ1xp+n2Gk2FtpWl2cNpZ2cKw29vBGESGNAFVFUcKoAAAHAAr/AIKDRR/Dj/gorp/jzUYhFa3M+g60XfIDxweVEzfT/RWHGPu/WgVZQysCCOCK/bx1638N/sffFTUbl0VJdBezBbpuuJEgUfXdKAPev+CK/hue18A/E3xe0GIdS1ixsUl/vNbwyOy/gLpT/wACqq/4Jcf8nj/Gb/rwv/8A05R1VVVVVVVVVVVVVVVVVVVV+05+zn8Gvj14dsL74r+Df7dn8M+Z/Zb/ANoXNr9n+0PCJeIJEDbhFH97ONvGMnNvbw2lvFaW6bYoUCRrknCgYA556Cqqqsr6y1K3F3p93FcwlmUSROGUlWKsAR6MCPqKqqqqqqqqqqqqqqqq1rR9O8Q6PfaBq9v9osdStpLa6h3snmRSKVdcqQRlSRkEEetfs5/AP4S/AbwdLY/Cjwn/AGHDrjpc36fbri586VV2hszSOV44wuBVVVVVVVVVVVVVVVVVVVVXif8AZa+BHjL4t6b8dfEngX7Z450eS3kstV/tO7j8poDmI+SkoiO33Q575qqrU2D6ldsB1nc4/wCBGvCFwJtJ8gnmCQqB7Hn+pqqqqqqqqqqqqqqqqqv+Cmv/ACY/8Sf+4V/6dbSv+CWf/Jl/hH/r/wBS/wDSyWqqqqqqqqqqqqqqqqqqqr9u/wD5M/8Aip/2AJP/AENK/wCCQf8Ayahd/wDY3X3/AKJt6qqqqqqqqqqqqqqqqqqqq+I/7NnwU0z4zW/x6sfBfl+O7xpZJtV/tG6bcxgFuT5JlMQ/dErwnv15qqqr6+stNtmvNQu4raBCqtJK4VVLEKMk9MkgfjVVVVVVVVVVVVVVVVVePvg58N/jjY6Z4O+KPhz+29Ih1OK6S2+2T22JQGjDboXRvuyOMZxznqBjwZ4O8OfD7wnpHgbwfp32DRNCs47PT7Xznl8mGNdqrvkZmbAAGWJPqaqqqqqqqqqqqqqqqqqqqq/4J2eA/CfhL4rfES+8PaV9lnu7RVmbz5JNw+0MejMQOfSqqqqqqqqqqqqqqqqqqqqv+C1v/Iz/AAo/68NU/wDRlvVVVVVVZ/8AH1F/viqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/aC+P3w//AGbPhve/Ej4hXzJbwnybGyhwZ9QuSCUgiB6sdpJJ4UAscAUfixon7av7QX/Cb/tZ/GC18EeB9MbdBpcQnl8q33cWlqsaNh2wPMncAnrz8qjwb+29+wJ8PPDGn+C/A/xY8P6LomlReVZ2Nppl4kcS5ycDyeSSSSTkkkkkkk18J/2q/wBn346eI7jwj8J/iVZeIdXtLF76a1htZ4ikCuiM+ZI1GA0sYxnPzfWv+Cun7OWsePvAmi/HXwhpUt7f+Co5LXW4oULOdOc7lmx1KwybicDhZmY8ISP2Jv8AgoF8HfHvwf0Xwx8XPiPonhTxp4aso7K+bXr6Oyi1BI1CpcRzSFUZnUDcmQwbdxtKk/8ABS79sXwp8bbDR/2bPgJqR8WJd6nFPqt9pameO7mU7be0tyufOJdg5K5GVjCkncB+xZ8BJv2cf2d/DXw81SNF12VW1DXNrBgL2fDOgIJB8tQkWRwfLz3qq/4Jcf8AJ4/xm/68L/8A9OUdVVVVVVVVVVVVVVVVVVVVePv+RSvv+2f/AKMWqqqr/gnt8aT8RfCvi/whqF2ZL3Rddnv7UOxybS8leXjPJxL5pP8Avr071VVVVVVVVVVVVVVVV4Q/5FnTv+uAqqqqqqqqqqqqqqqqqqqqr4ofFD4g6P8Ate+GfAul+K7230C7uNPWewRgI5A5+fIxnmqqrpg9zK4OcyMefrXgq42Xc9qW/wBZGGA9wf8A69VVVVVVVVVVVVVVVVV/wU1/5Mf+JP8A3Cv/AE62lf8ABLP/AJMv8I/9f+pf+lktVVVVVVVVVVVVVVVVVVVX7d//ACZ/8VP+wBJ/6Glf8Eg/+TULv/sbr7/0Tb1VVVVVVVVVVVVVVVVVVVV8Vv8AX6b/ALkn81qqqv25Nfm8Ofsu+NL20uXhupUtIIHjYqwZ7uFSQR0IXce3SvgH8Sofi/8AB3wr8QUdTPqlgv2xV4CXUZMc4A7DzEfHtiqqqqqqqqqqqqqqqqtF/wCQxYf9fMf/AKEKqqqqqqqqqqqqqqqqqqqqr9g//kpfjz/r1X/0eaqqqqqqqqqqqqqqqqqqqq/4LW/8jP8ACj/rw1T/ANGW9VVVVVVn/wAfUX++Kqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr9oH9m74Y/tNeGdO8JfFK01CfT9Kvvttutldm3YS7GTJIByNrtxX/Dpv9j/AP6Avij/AMHj/wCFf8Om/wBj/wD6Avij/wAHj/4V8Af2IPgP+zT4xvPHfwu07WYNVvtMk02Zr3UWuEMDyRSMApHB3Qpz9fWpoYriJ4J40kjkUq6OuQwIwQR3GK+JX/BKr9k74ia5Pr9lpniPwdLcyGSa38OX8cMDMc52xTRSrGOfuoFUYGAK+AP7CP7OP7OOqR+JfBHhS41HxFChWLW9buPtdzCCCD5YAWOIkEgsiKxBIJwcVVV/wS4/5PH+M3/Xhf8A/pyjqqqqqqqqqqqqqqqqqqqqrx9/yKV9/wBs/wD0YtVVV8TtYXw98NfFniBnCjTNDvbkscceXA7Z547V+xF8UT8Lf2h/DtxdXXlaZ4hY6Pf5OBtnIEbH0CzCIk9gD061VVVVVVVVVVVVVVVVXhD/AJFnTv8ArgKqqqqqqqqqqqqqqqqqqqq+MX/J9HhH/r60v/0KqokAZJ6DvTEsSxPJPNaBcC11e1kJ4Z9h/wCBcf1qqqqqqqqqqqqqqqqqv+Cmv/Jj/wASf+4V/wCnW0r/AIJZ/wDJl/hH/r/1L/0slqqqqqqqqqqqqqqqqqqqq/bv/wCTP/ip/wBgCT/0NK/4JB/8moXf/Y3X3/om3qqqqqqqqqqqqqqqqqqqqr4rf6/Tf9yT+a1VVX/BTPWV079nez05XG/VfElrBtyMlVimkJx6Axr+Y9a/4Jc/FES2fin4OahdfNAy6xpiMf4TtiuAPYEQkD/aY+uaqqqqqqqqqqqqqqqtF/5DFh/18x/+hCqqqqqqqqqqqqqqqqqqqqq/YP8A+Sl+PP8Ar1X/ANHmqqqqqqqqqqqqqqqqqqqqv+C1v/Iz/Cj/AK8NU/8ARlvVVVVVVZ/8fUX++Kqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv8Aglx/yeP8Zv8Arwv/AP05R1VVVVVVVVVVVVVVVVVVVV4+/wCRSvv+2f8A6MWqqq/a61g6F+zR8RL1ZCpk0SW1yCR/ryIccevmYpHeN1kicq6HKspwQR3Ffs8fE2P4w/Bfwr4+aZHvL6xWPUApHy3URMc3HbLoxA9GHrVVVVVVVVVVVVVVVVXhD/kWdO/64CqqqqqqqqqqqqqqqqqqqqvjF/yfR4R/6+tL/wDQqqrlttvKw5whOPwpF3sqA9TimDwSlc4eNvyIq2mW5torhOkqBh+Iqqqqqqqqqqqqqqqqr/gpr/yY/wDEn/uFf+nW0r/gln/yZf4R/wCv/Uv/AEslqqqqqqqqqqqqqqqqqqqq/bv/AOTP/ip/2AJP/Q0r/gkH/wAmoXf/AGN19/6Jt6qqqqqqqqqqqqqqqqqqqq+K3+v03/ck/mtVVV/wVV1gweFfh94fDnF7qF7c7ecHyY4lz6f8t/1r9m34nt8H/jb4U8cyT+XZW16sGo88fZZf3cxI74RiwHqo6daVldQ6MGBGQQc5qqqqqqqqqqqqqqqq0X/kMWH/AF8x/wDoQqqqqqqqqqqqqqqqqqqqqqv2D/8Akpfjz/r1X/0eaqqqqqqqqqqqqqqqqqqqq/4LW/8AIz/Cj/rw1T/0Zb1VVVVVWf8Ax9Rf74qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/4Jcf8nj/Gb/rwv/8A05R1VVVVVVVVVVVVVVVVVVVV4+/5FK+/7Z/+jFqqqv8AgolrA0z9l7W7HeB/a2o2Nrg98TrNj/yD7dKr/glz8UhnxT8G9Quuw1jTEZv92K4UZ/7YsAP9s+tVVVVVVVVVVVVVVVVeEP8AkWdO/wCuAqqqqqqqqqqqqqqqqqqqqr4xf8n0eEf+vrS//QqqtTYJpt0392Bz/wCOmrJd95AgxzKo5+teIrb7NrFygHDtvH/Auf55rwtcfaNGiUnJhJQ/hyP0Iqqqqqqqqqqqqqqqqr/gpr/yY/8AEn/uFf8Ap1tK/wCCWf8AyZf4R/6/9S/9LJaqqqqqqqqqqqqqqqqqqqv27/8Akz/4qf8AYAk/9DSv+CQf/JqF3/2N19/6Jt6qqqqqqqqqqqqqqqqqqqq+K3+v03/ck/mtVVV/wVP1hZ/iT4K8PhwTZaHLcleMjzpyufX/AJYfpVfsWfFH/ha37PPhrUbq483U9Dj/ALI1Ak5Pm24ARiepLRGJyT3Y1VVVVVVVVVVVVVVVaL/yGLD/AK+Y/wD0IVVVVVVVVVVVVVVVVVVVVVXwl/Z38E/BrXNW1/wvqmt3VxrMYSdL+eJ0UBy3yhI0I5Pcmqqqqqqqqqqqqqqqqqqqqv8Agtb/AMjP8KP+vDVP/RlvVVVVVVZ/8fUX++Kqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv+CXH/ACeP8Zv+vC//APTlHVVVVVVVVVVVVVVVVVVVVXj7/kUr7/tn/wCjFqqqv+CpOsm3+D/hXQFfBvvEYnIHBIit5Rjr0zKO3UDpXwd+El38V9O8fNYK7XfhbwrNrFqicmSSKeDcmOpzCZsf7W2v2ePibJ8H/jP4V8fGdktLC+WPUAM/NaSfu5uB1IjZiB/eAPao5I5o0lhkV0dQyMpyGB6EH0qqqqqqqqqqqqqqqq8If8izp3/XAVVVVVVVVVVVVVVVVVVVVV8WfCnii9/bS8K69ZeG9Un0yG500yXsVnI0KBW+YlwNox3yeKqtbbbpF2QcfuWH5jFaUgfU7NMZzOmR/wACFeNrYLc290o++hQ/gf8A69eCbjm5tCewdR+h/pVVVVVVVVVVVVVVVVX/AAU1/wCTH/iT/wBwr/062lf8Es/+TL/CP/X/AKl/6WS1VVVVVVVVVVVVVVVVVVVft3/8mf8AxU/7AEn/AKGlf8Eg/wDk1C7/AOxuvv8A0Tb1VVVVVVVVVVVVVVVVVVVV8Vv9fpv+5J/Naqqr/gpDrB1T9pe5sTIW/sjRLO1AJPy5DTYH/f7P41q3wkvLH4A+HvjRArvBqXiO90u6JHEYWKJoMc9ytzk8dAK/4JjfFEaB8Sta+FeoXW218U2f2myRj/y924JIUerRGQn/AK5LVVVVVVVVVVVVVVVVov8AyGLD/r5j/wDQhVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV/wWt/5Gf4Uf8AXhqn/oy3qqqqqqs/+PqL/fFVVVVVVVVVVVVVVVVVVVVVVVV9BJdWVxbQymOSWJlR1OCpIwCD2xXw58bQeNtCW5dkXULYBLuIcYbHDAf3W/xHaqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/wCCXH/J4/xm/wCvC/8A/TlHVVVVVVVVVVVVVVVVVVVVXj7/AJFK+/7Z/wDoxaqqr/gqxrO/UPhz4eR8eTDqF1Iueu8wKp6dtj9+9f8ABKzQkutS+JGtXEIeOO1sLQB1yriRp2cY6HiNcj3HrX7Q3wyk+D/xm8VfD8Rstrp98z2BbndayASQnPc+W6g/7QPpX7EXxRHxS/Z48O3F1debqfh1Do9/k5O6AARsfUtCYiT6k9etVVVVVVVVVVVVVVVXhD/kWdO/64CqqqqqqqqqqqqqqqqqqqqqqvETbdEuiCPuAc+5FaEofWLQZ/5ag/lXi+28/SDKBkwOG/A8H+deGLj7PrMIJ4lBQ/iOP1Aqqqqqqqqqqqqqqqqr/gpr/wAmP/En/uFf+nW0r/gln/yZf4R/6/8AUv8A0slqqqqqqqqqqqqqqqqqqqq/bv8A+TP/AIqf9gCT/wBDSv8AgkH/AMmoXf8A2N19/wCibeqqqqqqqqqqqqqqqqqqqqvit/r9N/3JP5rVVVftoawNd/ah+IN6HDeVqKWuf+uEEcOOP+ueK+EXwXHxH/4J0W3gVbbfqeq2N7qNkAvP2pLuWWDHGRuCIp9mPUV8OvGuqfDPx/oHjrSgwvNA1GK6EZO3zAjDfGfZl3KfYmtC1rTfEuh6d4j0a5FxYaraRXVpKBgSRSIHRvxUg1VVVVVVVVVVVVVVWi/8hiw/6+Y//QhVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV/wWt/5Gf4Uf9eGqf+jLeqqqqqqz/wCPqL/fFVVVVVVVVVVVVVVVVVVVVVVVV5cx2dpNeSEbII2dsnHAGT/KvBviu+8Ha7Bq9mSyD5LiHOBLGeq/XjIPYgVpOq2Ouabb6rps4lt7mMPG39COxHQjsaqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/wCCXH/J4/xm/wCvC/8A/TlHVVVVVVVVVVVVVVVVVVVVXj7/AJFK+/7Z/wDoxaqqr/gp7rIvvjxoukRvlNM8Mwhh6SSTzse/93y/T+Vf8EtNGWD4TeLtf2ANe+IhbbsDJENvG31x++P6+9f8FR/hbh/Cvxj0+3+8G0bUmUdxult2OP8AtspJ9EFf8Eyvij/wjnxT1b4YX9yFtPFtl51orH/l7twWAA/2ojLn/cX8Kqqqqqqqqqqqqqqrwh/yLOnf9cBVVVVVVVVVVVVVVVVVVVVVVXipgNDuBjqVH/jw/wAK8NLv1y1APdj+SmtQthd2M9tjJkjKj644qCVreeOdPvROGH1BpHWRFdDkMMg1VVVVVVVVVVVVVVVf8FNf+TH/AIk/9wr/ANOtpX/BLP8A5Mv8I/8AX/qX/pZLVVVVVVVVVVVVVVVVVVVV+3f/AMmf/FT/ALAEn/oaV/wSD/5NQu/+xuvv/RNvVVVVVVVVVVVVVVVVVVVVXxW/1+m/7kn81qqqvjPrJ8RfGDxxr2/cNQ8R386n2e4cgDk8YIHWv2b9GGgfs/8Aw60soFdPDNi8i+jvCruOCf4mPNfto/C3/hVH7QviXS7W2EOma1J/a+mgLtXypySyqOwWUSoB6KK/4Jz/ABQ/4Tr4Dr4QvbjfqPgm7NkwJ+Y20mZIGPsMyRj2iFVVVVVVVVVVVVVVVov/ACGLD/r5j/8AQhVVVVVVVVVVVVVVVVVVVVVfsH/8lL8ef9eq/wDo81VVVVVVVVVVVVVVVVVVVV/wWt/5Gf4Uf9eGqf8Aoy3qqqqqqs/+PqL/AHxVVVVVVVVVVVVVVVVVVVVVVVV8XteTQ/BF5GsmJ9QH2WIeu77/AOGwN+JFV8HvH48OakNA1WfGm3z/ACO54glPAPsp4B9OD61VVVVVVVSyRwxtLLIqIilmZjgKB1JPYV8e/wDgr18MfAviiTwH8DPA938TdVilMD38V39nsTLnG2Aqjvcc5GVVVPBVmFeGf+CzXiXw94jttK+O37N97oljckM8+n3ckdxDH0Lrb3CKJec/8tE+tfC/4peA/jP4H0z4i/DbxFb61oOqx7re5iyCpHDI6n5kdTwyMAQeteMPGHhf4f8AhfU/GnjTXbTR9D0e3a4vr66fZHDGvUk9yeAAMkkgAEkCvHX/AAWVvNW8UT+H/wBnn4Aah4otLYki91GeQS3CDgsttAjMi57s5OCMqp4r9mv/AIK5fDz4p+MLP4cfGfwW/wAO9avbgWttqBvfPsGnzgJKXVHtyW+Ubgyg/eZar9pP/gojpPwo+J0fwF+C3ww1X4r/ABILBLrTdNlZIbJyobYzIkjSSBSCyqoVB95gQQPh7/wU31jQfijp3wg/a6+BGp/CTU9X2fYtVmuHe1JdtqNIjopSIsCvnK8ihvvBQGYV+2j+1hbfsqfD7TNU0fQY/EvjPxPqUen+HtBLsGvH3L5jkICxVQVGAMl5EHfjwTc+LL3who1548sLCx8RXFlFJqdpYuzwW07KC8aM3LBSSu44zjOBnFVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVf8EuP+Tx/jN/14X/AP6co6qqqqqqqqqqqqqqqqqqqq8ff8ilff8AbP8A9GLVVVft+6x/a/7U/i2JX3R6dFZWsZOe1rEzDH+87D9e9f8ABOrRv7M/Zg0i92Y/tfU766yc84lMOev/AExx26V+0X8MU+MPwW8VeAo4VkvL2xaTT9w6XUX7yHntl1VSfRjXgPxfq3w38d6H420pSt/4f1GK6SNsruMbglG74YAqR6E14c1/TPFnh3S/FGiT+dp+r2cV3aSf3opEDoePZhVVVVVVVVVVVVVVVeEP+RZ07/rgKqqqqqqqqqqqqqqqqqqqq8Y/tJ/8Il8dNI+Cv/CF/a/7VltY/wC0v7S8vyvOOM+V5Rzj/eGfaqrxewXRiufvSKP8/lXhRN2tQkDO1WP04x/Wq1i2+y6pcwAYCyEqPY8j9CK8PXAutGtnJyUTYf8AgPH8gKqqqqqqqqqqqqqqqv8Agpr/AMmP/En/ALhX/p1tK/4JZ/8AJl/hH/r/ANS/9LJaqqqqqqqqqqqqqqqqqqqv27/+TP8A4qf9gCT/ANDSv+CQf/JqF3/2N19/6Jt6qqqqqqqqqqqqqqqqqqqq+K3+v03/AHJP5rVVWs6lDo2kX2rzgGOxtpJ3BOOEUsfXsPSpHudQu2dg01xcyEkKuWd2PYDuSelaBpSaHoOm6JGQV0+zit1x0wiBf6V/wU6+Fo1/4b6J8VNPtd134Xu/st66rybS4IALHuFlCADt5rV/wT1+KI+H/wC0BZeHr668vTfGdudMlDHCi4zvt2+pcGMf9dTVVVVVVVVVVVVVVVaL/wAhiw/6+Y//AEIVVVVVVVVVVVVVVVVVVVVVX7B//JS/Hn/Xqv8A6PNVVVVVVVVVVVVVVVVVVVVf8Frf+Rn+FH/Xhqn/AKMt6qqqqqrP/j6i/wB8VVVVVVVVVVVVVVVVVVVVVVVVfF7xinirxGbWylD2GmbooSOjv/G/0JAA9gD3qq+Dfj8a/p48NarcZ1GyT9y7nmeIfzZeh9Rg+tVVVVVVV/wVq+OOs/CX9mdfCnhi/ks9V+IOo/2VJNGSrx2Ko0lztI6FgI4j/syt0r/gk5+zR4O+HvwD0n443+j29z408crNOt9NGGksbFZGjihiP8IcJ5jEYLbwDkIK/aS/Z98EftLfCjWfhp4y021kkureRtKv5It0mmXm0+VcRsOQVbGQDhlypyCRX/BHH4s+IPBXxp8X/s4+IriWOy1m0mvLe0kbIt9StGCShR0BeLfuPfyEr/gtB8dtXu9f8Jfs0+HLuX7KLdda1qGHJNzM7tHawkDk7Qkj7ecmSM9VFfsifs0+E/2XvgzongbR9KtU16a1in8R6iqgy318VBkJcclFYlUXoFA7kk/8Fj/2bfDOpfDew/aR8OaLb2niDRL+Kx16eCIJ9ttJjsjklwPmeOXy0DHnbKQSQqgf8E/f2ir74n/sWWXjfxPdS6jrPgKC70vVZXctJcGziEkbMTyWa3eHLHJLZJPJr/gjRoC+NNf+MHx98Vsmo+KNS1CC1+2y5aRDMZbi6bJ/56OYiec/J+f/AAWY8CaNr37MukeOJrWL+1fC3iSBba5K/OILhHSWIH0ZlhYj1jFfslePLjxb+yd8NPHXinUVEzeE7VtQvLlwu4wxBJJnYnAz5ZYnpyTx0r9muzvf26/2xNf/AGufEdtM/wANfhhcHSfAFpPGVSe5XkXG0/xKG885GQ8sIz+6xVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVf8EuP+Tx/jN/14X/AP6co6qqqqqqqqqqqqqqqqqqqq8ff8ilff8AbP8A9GLVVVftO6yuvftD/EbUUcOo8SXkCMCCGWKUxggjqCEFfsf6MNC/Zl+HdiE2+Zo63WMY/wBe7TZ6Dr5n/wCvrVftt/C4fCz9ofxFZ2lt5Wma+w1jTwBgbJyTIAOwWYSqB6AV/wAE4PiiPG3wOfwTfXXmaj4JuzbBWOW+yS5khJ74B81B7Riqqqqqqqqqqqqqqq8If8izp3/XAVVVVVVVVVVVVVVVVVVVVV8Yv+T6PCP/AF9aX/6FVV40bGlRqCPmnHH/AAE//WrweobV8/3YWP8AKq8Z2wj1GO4UcTR8/UcfyxXgq4D2txaE8xuGA+ox/Sqqqqqqqqqqqqqqqr/gpr/yY/8AEn/uFf8Ap1tK/wCCWf8AyZf4R/6/9S/9LJaqqqqqqqqqqqqqqqqqqqv27/8Akz/4qf8AYAk/9DSv+CQf/JqF3/2N19/6Jt6qqqqqqqqqqqqqqqqqqqq+K3+v03/ck/mtVVftDayPD/wG+IWrh9rweGb8RH0kaB1TuP4itfCXR18RfFXwX4fZQw1PxDYWpU458y4Re/Heq+JPgfTfiX4A8QeAdWIFtruny2rOVz5TMpCyAeqttYe4FXEPiDwB4vkt3Mmn654b1IqSpw9vcwSY491dP0r4UePrD4p/Dbw38QtOKCLXdOiuHjQ8RSkYkj+qOGU+61VVVVVVVVVVVVVVov8AyGLD/r5j/wDQhVVVVVVVVVVVVVVVVVVVVVfsH/8AJS/Hn/Xqv/o81VVVVVVVVVVVVVVVVVVVV/wWt/5Gf4Uf9eGqf+jLeqqqqqqz/wCPqL/fFVVVVVVVVVX7Un/BR74WfspfEuL4X+MvA3irV7+XS4dQFxpgtzEEkZ1C/vJFO4GM54xyK/4fafAH/olHxA/75tP/AI9X/D7T4A/9Eo+IH/fNp/8AHq/4fafAH/olHxA/75tP/j1f8PtPgD/0Sj4gf982n/x6v+H2nwB/6JR8QP8Avm0/+PV/w+0+AP8A0Sj4gf8AfNp/8er/AIfafAH/AKJR8QP++bT/AOPV/wAPtPgD/wBEo+IH/fNp/wDHq/Z6/wCCo/wg/aN+MHh/4M+Fvh94w0zVPEP2nyLrUBbCGPyLaW4bdslZuVhYDA6kdqqqqqqqr4y+Pn0CwHhvSZsX99GTNIp5hiPHHozcj2GT3FVVaVdanpNxHr2mSNFJYzJtlX+FiCQCO4IVuOmMg9a8FeK7Txl4fg1i2ASQ/JcRA58qQdR9OhHsRVVVVVVX/BZ74Yaz4s+AXhr4iaPZyXMfgnW2/tAICfJtrpBGZT7CVIF/7aDtmv8AglV+0N4L+JH7N2gfCk61ZweMPAsclldabJKqSz23mO8M8adWTYyoxGcMhzjcM/Fj4vfDr4H+C77x98TvFNnoukWMbMXnkAknYDIihT70kjdAigk1/wAE9PEcnj3/AIKU6R440q2eytNb1PxBqUls8gJjhntLt1jJHDEM6flmv2qZV8df8FbbDQtRZzZt448L6WqMAdkW2yVwPYsztz/eqv8Agodptrqv7FnxWtbtSUTRlnUAD78U8Uidc/xIv/1utf8ABFdovEfwh+LvgfUUZrCTU7YyDcDkXFtJG4AI67Yh1znjjjn/AIJkfELTP2Vvjh8U/wBlT426pa+HNTu7+I6bc6jKttBNc25kRlDOdv7+KSKSM5wwXAJLKK/4K5/G7w74x8HeEf2YPhpqdv4l8Y+JfEMFzc2GlzC4eJFDRwwsEPEks0qbVPOIySBlSf2rdb8VfDj4C/B7/gnJ8Jp47j4i+OtGsdI1ZrdwBbWKIEuZGxyqTSLLk8jyo5s9Qa8eftM+BP2DND8F/safs+/Di9+I/wARLaxijTSrNigWaUGRppyiszTSktL5SgYRgSyrtzB/wUp+OPwV8b6L4c/bR/ZpfwNo3iCTFtrWlyvIkCggMxTMizBNyl1Rw6g52kkA6Zqena3plprOj30F7YX8CT2tzA4eOaJ1DI6sOCpUggjqDVVVVVVVVVVVVVVVVVVVVVV+1F+2r8G/2VLGG28ZXlzq3iW+h82x8P6aFa4kTOBJIWIWKPII3NycHarYIHiz/gs/8Yru9kbwL8I/Bul2m790mrTXN/IF92jeAZ/Dj3r9gr9pnxl+1b8G9U+IXjnQ9G0vUdN8RTaUItJSVIXSO2tpd+JHdgS07DG7GAPcnxH/AMFRf2UNA8X2/gy21vxFqt1JqKWFxc22jvDBZuZBG7Sm4MTBUOSxVWOAcA1VVVVVVVVVf8EuP+Tx/jN/14X/AP6co6qqqqqqqqqqqqqqqqqqqq8ff8ilff8AbP8A9GLVVVeLdYPiLxVrPiAuWOp6hPcljnJ8yRmzzz3r4V6MPDvwv8H+Hgu3+y9AsbXbjGPLgRfU+nqar/gpt8Lj4j+F2j/E+wty134SvDDdsq/8ulwVXJ/3ZViA9N7V/wAE/Piifh5+0HpuiXlz5emeMoTpU4Y8Cdjut2x/eMiiMf8AXU1VVVVVVVVVVVVVVXhD/kWdO/64CqqqqqqqqqqqqqqqqqqqqvjF/wAn0eEf+vrS/wD0Kqrxs3+hW6DvKT+leC1zqUzZ6QEY/wCBD/Cq8Z23m6fFcKMmGTB+hH+IFeEbgQ6sIS3E0ZXHuOf6VVVVVVVVVVVVVVVVf8FNf+TH/iT/ANwr/wBOtpX/AASz/wCTL/CP/X/qX/pZLVVVVVVVVVVVVVVVVVVVV+3f/wAmf/FT/sASf+hpX/BIP/k1C7/7G6+/9E29VVVVVVVVVVVVVVVVVVVVfFb/AF+m/wC5J/Naqq/bq1j+xf2V/HEqPiS6itbVBzz5l1ErD/vgsfw96/Y30c67+078PbJVLGLVPtOAD/ywiebPH/XOqr/gov8AC0+Bfjw/i2xthHpvja0F6hVcKLqPEc6j3JEchPrLX/BMD4o/2x4I8Q/CS/uM3Hh+5F/p6seTbznEiqPRZV3H3mqqqqqqqqqqqqqqrRf+QxYf9fMf/oQqqqqqqqqqqqqqqqqqqqqqv2D/APkpfjz/AK9V/wDR5qqqqqqqqqqqqqqqqqqqqr/gtb/yM/wo/wCvDVP/AEZb1VVVVVWf/H1F/viqqqqqqqqqr9vz/gnN8bv2qvjpB8Tvh54p8D6fpcWg22nmHWb66hn82OSVmIWK3kXbiRcHdnrwK/4cqftUf9D98Kf/AAa3/wD8hV/w5U/ao/6H74U/+DW//wDkKv8Ahyp+1R/0P3wp/wDBrf8A/wAhV/w5U/ao/wCh++FP/g1v/wD5Cr/hyp+1R/0P3wp/8Gt//wDIVf8ADlT9qj/ofvhT/wCDW/8A/kKv+HKn7VH/AEP3wp/8Gt//APIVf8OVP2qP+h++FP8A4Nb/AP8AkKv2M/8Agl98fv2df2lPB3xj8beLvh/faL4f+3fabfS7+8kuX86xngTYslqinDyqTlhwD1PBqqqqqqdlRS7MAFGST2rxVrcviTxDf61KxP2mZjGG/hQcKPwUAVVV8LtCXxPpvifQyAWmskeHI6SqxKH254+hNfAfXJLDxPcaBK5Eeowkqh7Sx5P/AKDvz9B6VVVVVVVruhaL4n0W98OeItLtdT0vUrd7e8s7qISRTxOMMjqeCpBIINfFP/gi9oVx4nk8UfAP4yXvhKMzGW30zUrR7n7Ix7RXMbrIFHIAZWbGMsetRf8ABMz4cfBDw/q37QP7XXxm1X4j6f4M05759I2PbwTsg+SF5ZJHklDuVVUHl5ZlByCVP/BHP4e6x45+PXj39oDVLFYrHS7CW1jaOLy4jfXsokZYwOMJFG4KjoJU6ZGf24Vj+Ef/AAVK034haki2+nHX/DfiBGdML5UItlkbryN9tKTyOcj3qv8AgpV4mtvC/wCxP8S7ieRRJqFpbWECEZLvPdRIQOR0Us3sFPXpX/BE3wtcaf8AAzx14wmhCJrPihbWJimC629shJz3XdOw9iGr9oX9i/8AZ6/aekg1D4o+C/M1m1i8qDWdPna0vEjHRC68SKMnAkVgMnGMmvBH7JH7H/7C/hzXPjna+GpPtHhnT5bqbXtauTe3NuiqQVgBwiSPnYNihmL7ckHFf8E8Ph94o+Mfjrxl+318XbAprnjq5ltPCdnLlhp2modhaPI6EIsKtwSsbtz5lf8ABMdYfjJ+2j8c/jx4kU3eqWrzmxNx8zW32y6kA2dduyGDyhzwrFRwa/4Km+DdJ8W/sVeNr2/gje68NzWOp6fIwz5Uq3UcTEe5imlX/gVf8EyPFeo+Lf2Jvh1carM8txpsV5pwdznMUF3LHEB7LEI1/wCA1VVVVVVVVVVVVVVVVVVVVXjDxLYeC/CWt+MdUDGy0LTri/uAg5McMbSNj3wprXdV+J/7UPxrudSFrc6/4z8c6qfJtojks7nCRrnhI0QBRnCoickAZr9qD9ij4h/sn+GvCWtfELxHoV9d+KpbiNbPS2kkFqYljYhpHVQx/eAcDGQeSOa/4I2f8mweJv8Asfbv/wBILCv2ztBs/Df7V3xW0qwVVh/4Si8nVVXaFMzmUqB2AMhH4dulfBHxHP4y+C/gDxdcymSbXPC+m30jsMFmmtY3J/Nqqqqqqqqqr/glx/yeP8Zv+vC//wDTlHVVVVVVVVVVVVVVVVVVVVXj7/kUr7/tn/6MWqqvidrC+Hvhr4s8QM4UaZod7cljjjy4HbPPHavDOkN4g8SaToEYJbUr6G2UDOcyOF7A+voaVFRQiKAFGAAMYqviF4L0z4i+Bte8CawB9k13T5rORiu4x71IDgf3lOGHoQK1PT9f8AeL7rS7ovY634c1J4pCp+aC4gkwSPo6/pXwg+INl8V/hf4Z+IdiEVdc06OeWNekU2NsqD/dkV1/CqqqqqqqqqqqqqqvCH/Is6d/1wFVVVVVVVVVVVVVVVVVVVVfGL/k+jwj/wBfWl/+hVVeOGAjs09S5/LH+NeCVzc3TjHEaj9f/rVWtWwu9KuYAMkxkqPccj+VafcfZL6C5Bx5cgJ+mef0rgjg1VVVVVVVVVVVVVVX/BTX/kx/4k/9wr/062lf8Es/+TL/AAj/ANf+pf8ApZLVVVVVVVVVVVVVVVVVVVV+3f8A8mf/ABU/7AEn/oaV/wAEg/8Ak1C7/wCxuvv/AETb1VVVVVVVVVVVVVVVVVVVV8Vv9fpv+5J/Naqq/wCCmesrp37O9npyuN+q+JLWDbkZKrFNITj0BjX8x61/wTl0ZdU/ab029KBjpGk3t0CQPlzH5OR/3+x+NVX/AAUM+Fp+IHwBu/EVja+bqfgu4GpRlVyxt8bLhfoEIkP/AFyr9kD4oj4SftAeF/EF1c+VpmoT/wBmamScL5FxhNzeyP5cn/bOqqqqqqqqqqqqqqtF/wCQxYf9fMf/AKEKqqqqqqqqqqqqqqqqqqqqr9lL4HfEb4V+NvFeteNNIgtLTVYAlq8d3HKWPmluQpJHB71VVVVVVVVVVVVVVVVVVVV/wWt/5Gf4Uf8AXhqn/oy3qqqqqqs/+PqL/fFVVVVVVVVVVVVVVVVVVVVVVeLZmtvCus3CH5otPnZecciNjVVVfs96XLFYatrEikJcSRwxkjGdgJY/+PD8qsLX+xvjgLaBGCjVX2qo6LJk/lhvyqqqqqqrVNSsdG0y71jU7gQWdjA89xKwJEcaKWZsDJ4AJ4rxf/wVL/Yo8KaY97afFO48Q3KrlLHSNGunlk+jSpHGD/vOtfGP4+/tIf8ABUzx9Z/B74LeCLrRfA2nzrPLavMTEhGQt1qNwBtUAZKRAHByB5jYNfs0fs9eD/2YfhFpHwp8Hk3C2YM2o6g8YSTUbt8ebO4BOM4CquTtVVXJxmv+CwH7LutfErwRo/x+8D6RJfar4ItpLXXIIELSPphJkEoA5IgcyMwHRZWY8Ia/Yz/4Kd/BLxN8IdF8L/Hrx5B4W8a+HLOOzu7jUkk8nVUjUKlykqhhvZQu9W2nfuKgqeP+Chn7X1r+2V4n8K/s0fs0WmoeJtLXVVmkube3dDq9/tZI0iRgG8mNXkJdgASd3Cxhj+yl8C7T9nD4BeEvhJDLHPe6VaGTVLiPpPeysZJ2B6lQ7FVzztVfSq/bb8Ra3+1n+0T4Q/YH+HeozR6LZzx618RdQtuRbQJh0hJz1VGVsHIMk0HQqa8N+HdE8I+HtM8KeG9OhsNK0e0itLG1iXCwwxqFRAPQKAK/Z28UaR+wZ/wUL+JPwz+LN1H4f8I+O3l/snVLg+VapHJOZ7GV34UR7HkhZvupJkEgKxH/AAVT/ae+HMv7PsvwN8AeMdM8SeKvH17aRfYtGvFu5ILSOZJi7eUTjzHjjjVTy4dsAgGv2M/hDqXwJ/Zh+H/wx1uEQ6tpumGfUos5MV1cSvcTRk9yjysmeny8cVVVVVVVVVVVVVVVVVVVVVXxx8L3vjf4KfEDwXpsbSXev+FtS0+3RBy0k1rJGoHB5yw7V+w98WfCvwP/AGovBXxB8ckQ6LaT3FreXLRljZrcW8kHnYHICGQFsAnbuwCa/wCCyWu6J4m8BfB/XfDmsWWqabeXepyW15ZzrNFMhjtsMrqSCPcGv+CWPxI8EfCb9jHxv47+IXiK00bRdM8b3kk9xcPjOLCxwqL1d2PCooJYkAA5r40/Ea4+L3xb8X/E64ga3PibWbm+jgc5MEbuTHGT32ptX8K+CHhe78EfBX4f+C9QieO60Dwtpunzo/3leG1jjYHgc5U9hVVVVVVVVVf8EuP+Tx/jN/14X/8A6co6qqqqqqqqqqqqqqqqqqqq8ff8ilff9s//AEYtVVftdawdC/Zo+Il6shUyaJLa5BI/15EOOPXzMV+zTow1/wDaD+HOmlA6nxLZSup7pHMsjDr/AHUNVVf8FHvhaPBHxyXxpYWvl6d42sxdEquFF3FiOcD6jynPqZDX/BL34ojU/CfiT4Q6hdbrjRrganpyMeTby4WVVHosgVvrNVVVVVVVVVVVVVVXhD/kWdO/64CqqqqqqqqqqqqqqqqqqqqvjF/yfR4R/wCvrS//AEKqrxw2ZbRM9Fc4/L/CvA6A/bHx/cAP/fX/ANaqIBGDz7Gr63+yXk9r/wA8pGUZ9Aa0i4+16Xaz5yTGAT7jg/qKqqqqqqqqqqqqqqr/AIKa/wDJj/xJ/wC4V/6dbSv+CWf/ACZf4R/6/wDUv/SyWqqqqqqqqqqqqqqqqqqqr9u//kz/AOKn/YAk/wDQ0r/gkH/yahd/9jdff+ibeqqqqqqqqqqqqqqqqqqqqvit/r9N/wByT+a1VV/wVV1gweFfh94fDnF7qF7c7ecHyY4lz6f8t/1r/glno3n/ABS8Y+IAmfsXh9LXdzx506Nj/wAgfpVVq+lafrulXmiarbJc2WoW8lvcwuOJI3Uqyn2IJH418VfAN/8AC74keI/h7qRdptC1GW2WR1wZYwcxyY9HQqw9mFfstfFEfGD4E+FfGFxc+dqQtBZ6mWOW+1Qfu5GPu+0SfRxVVVVVVVVVVVVVVov/ACGLD/r5j/8AQhVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV/wAFrf8AkZ/hR/14ap/6Mt6qqqqqrP8A4+ov98VVVVVVVVVVVVVVVVVVVVVVVqlmNR0y70/dj7TA8WfTcpH9amikgleCZCkkbFWUjBBHBFVVfDG+sNQ8DaVLp8EUKxw+XJHGMBZFOGOPc/N/wKtDQeIvjtcXluQ8NrdSyMw9I02A/wDfQX86qqqqqq1zR7LxDomoaBqKu1pqVrLbThG2sY5FKtg9jgnmvDH/AASp/Yo8OXYvLj4bahrbocomqa3dOinj+CN0VunRgRz9MeC/Afgn4caDD4X+H/hLSPDuk2/+rstMs0togcAFiqAAscDLHk9zVMqspVgCCMEEV8Rv+CY/7G/xJ12fxHefDKTQ766kMlwdCv5bKKRj1PkqTGnXPyKtfAz9kP8AZ4/ZweS7+E3w3sdN1OaMxzarcSPd3jqeqiaUsyKcDKptU4GRxVV8Gf2YPhT8CvE/i7xt4Ms9SuPEXjm6+063qmqXhup7hy7OcMQNql3ZiB1OM9Bivjj+zb8FP2j9EttD+MXgSz16OxZmsrku8FzaE43eXNEyuoOBlc7WwMg4FfBT/gn9+yt8BPEMHjDwR8OVufEFo2611PV7uS+ltm7NErnZGw7Oqhveqqqqqqqqqqqqqqqqqqqqqqv+CiX7BXiv4a+MtZ+Nvwo0GfVfA+t3El5qNpZQF5NCnb5pCyLz9nZizK4GEyVbACsxnmaBbYzOYlYusZY7QxABIHqQAM+wppZWiWAysY0YsqEnAJABIHqQo/Ielf8ABOz9iPxX8Z/iDovxc8daDNY/Dvw7dJeRvdxFP7anjbMcUQI+eIOoMj/dIBQZJO2qqqqqqqqq/wCCXH/J4/xm/wCvC/8A/TlHVVVVVVVVVVVVVVVVVVVVXj7/AJFK+/7Z/wDoxaqq/wCCiWsDTP2Xtbsd4H9rajY2uD3xOs2P/IPt0r9gfRjrH7VHg9yhaPT0vLqTAPG21lCn2+dk61VV/wAFBfhaPiH+z5qGuWVv5mp+DJhqkBVckwAbbhfYCMmQ/wDXIV+yX8UT8Ivj54W8T3F15Om3VyNP1MscL9mnwjM3sjFJPrGKqqqqqqqqqqqqqq8If8izp3/XAVVVVVVVVVVVVVVVVVVVVVq3wi+HOueM7X4har4XguPENk0bW980sgZDH9whQ23j3FVXjZs31umRxDn9T/hXglR9mumB5LqMfQVVeLrbydXaQDiZA349D/KvBtwJdNe3J5hkOB7Hn+eaqqqqqqqqqqqqqqr/AIKa/wDJj/xJ/wC4V/6dbSv+CWf/ACZf4R/6/wDUv/SyWqqqqqqqqqqqqqqqqqqqr9u//kz/AOKn/YAk/wDQ0r/gkH/yahd/9jdff+ibeqqqqqqqqqqqqqqqqqqqqvit/r9N/wByT+a1VV/wVP1hZ/iT4K8PhwTZaHLcleMjzpyufX/lh+lf8EqdGEWg/EPxCUybq7sLVWI6eUkzEDj/AKarnnsKqq/4Kf8AwvGj+O/D3xY0+2At/ENqbG/dVx/pMA+RmPq0TBR7Q1/wS7+KIste8T/B7ULrEepQjVdNRjgedHhJ1HqzIY2x6RMaqqqqqqqqqqqqqtF/5DFh/wBfMf8A6EKqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv+C1v/Iz/Cj/AK8NU/8ARlvVVVVVVZ/8fUX++Kqqqqqqqqqqqqqqqqqqqqqqr4zeEZdA8Syavbxf6DqrGRWA4SX+NT9T834+1VVeD/iTfeEPDWq6LZxs0924a0lzxAxGHb64C49xzXwO8JS6VpE3iW/jK3OpgCEMORCO/wDwI8/QA96qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqvLqCxs5765fbFbxNJIQM4VRknH0FeHP8Agpb+yF4s8Q6X4V0Px9qU2o6xew2VnG2hXaB5pXCICxjwAWYDJ4FeP/2KP2U/idezan4v+B/hya8uCWmubGN9PllY9WZ7Zoyx9yc14M/YO/ZB8A3sWo+HvgR4fe4hO6N9TM2p7GByCBdPIAQeh6jitU/4KMfsV+F9Tu/DOpfGAWV3pE72dxajw1qf7iSJijJ8ttj5SpHHHFaff2eq2FtqmnXCT2t5Cs0EqdJEYAqw9iCDVVVVVVVVVf8ABLj/AJPH+M3/AF4X/wD6co6qqqqqqqqqqqqqqqqqqqq8ff8AIpX3/bP/ANGLVVX/AAVJ1k2/wf8ACugK+DfeIxOQOCRFbyjHXpmUduoHSv8AgmLo63/x+1XVJEBXTPDNw6E44d54EH/jpfn/ABqqrUdPstX0+50rUrZLi0vIXhuIXGVkjdSrKfYgkV8Xvh7e/Cn4neJfh3flmfQ9QkgikcYMsOd0UmO2+Nkb/gVfsm/FH/hbvwD8K+KLm483Ure2/s/UiTlvtMHyMzdeXULJ/wBtBVVVVVVVVVVVVVV4Q/5FnTv+uAqqqqqqqqqqqqqqqqqqqqqqrxmwbVUAH3YFH6mvBS/8S2Zs9ZyPyUVVeNrYNb212o5Rih/EZH8j+deDLjy9QltieJY8j6j/AOsTVVVVVVVVVVVVVVVf8FNf+TH/AIk/9wr/ANOtpX/BLP8A5Mv8I/8AX/qX/pZLVVVVVVVVVVVVVVVVVVVV+3f/AMmf/FT/ALAEn/oaV/wSD/5NQu/+xuvv/RNvVVVVVVVVVVVVVVVVVVVVXxW/1+m/7kn81qqr/gpDrB1T9pe5sTIW/sjRLO1AJPy5DTYH/f7P41/wTI0Yaf8As+6jqbr8+qeJbmRWxj5EhgjA6/3lf061VV+2H8Lv+Fs/s++KNCtoPN1LTIP7U00AZPn24LbV93j8yMf79fBP4jXXwj+K/hf4iWrPt0bUEkuUTrJbt8k6D3aJnX8atLu11C0gv7G4jnt7mNZIZY23LIjDKsCOoIINVVVVVVVVVVVVVov/ACGLD/r5j/8AQhVVVVVVVVVVVVVVVVVVVVVfstfHj4gfFrxp4p0Pxhc2MlrpEAe2FvaiIg+aV5IPPAqqqqqqqqqqqqqqqqqqqqr/AILW/wDIz/Cj/rw1T/0Zb1VVVVVWf/H1F/viqqqqqqqqqqqqqqqqqqqqqqq8QeH9M8T6VNo+rQ+ZBMOo4ZGHRlPYivGnwv8AEXhGaSYW732nAkpdwpnA/wBtRyp/T3qooZbiRYIImkkc4VEUkk+wFfD34L3t3PFq/i+3NvaoQyWTcPKR/fH8K+3U+1KiIixxqFVRhVAwAPaqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr4m+PdN+Fnw98RfEjWdM1LUNP8M6dNqF5b6dGjztDEpaQorsikhQW5YcA1rf8AwWH/AGZdS0a/06DwP8TlkurWSJC+mWIALKQM4vOnNfDDxNYeCviV4S8ZarFPLZaDrtlf3MduoaR44Z0kYIGIBYhTgEgZxyOtf8Pk/wBmD/oRfij/AOCux/8Akyte/wCCzP7P8GlTy+GPhp8Qb7UlQ+RBfwWdpC7Y43SJcSsozjohrxb4kv8Axn4q1nxfqqxLe65qE99crEu1BJNIzttHOBljgV+zkt+v7PPwuXVYvLvh4M0gXMeMbZPscW8ce+aqqqqqqqqr/glx/wAnj/Gb/rwv/wD05R1VVVVVVVVVVVVVVVVVVVV4+/5FK+/7Z/8Aoxaqq/4Ksazv1D4c+HkfHkw6hdSLnrvMCqenbY/fvX/BKjRzJq3xF19lIEFtp9qjEHB3tOzY7ceWv5iqqq/4Kg/C06X4u8OfF3T7XFvrdudN1B1XgXEI3RMx9WjLAe0Nf8EvfiiNN8VeJPhDqFziHWYBqenKx4FxEAsqgerxlW+kNVVVVVVVVVVVVVXhD/kWdO/64CqqqqqqqqqqqqqqqqqqqqtQ+IngTSvEkHg7UvF2lW2uXRRYdPluVWaQv90BM5Oe1VXi5g2suoP3Y1H6V4PTbpBOPvTMf0A/pVV4jtvtOjXCgZZF3j2xyf0zWi3AtdVtZicASAE+gPB/Q1VVVVVVVVVVVVVVX/BTX/kx/wCJP/cK/wDTraV/wSz/AOTL/CP/AF/6l/6WS1VVVVVVVVVVVVVVVVVVVft3/wDJn/xU/wCwBJ/6Glf8Eg/+TULv/sbr7/0Tb1VVVVVVVVVVVVVVVVVVVV8Vv9fpv+5J/Naqq/bQ1ga7+1D8Qb0OG8rUUtc/9cII4ccf9c8V+wbox0b9lfwWHTEt6Lu6fjrvupdp6D+AJ/8Aqqqqv2o/hcfg/wDHTxV4Nt7bydO+1m70sAYX7LN+8jVfUIGMefVDX7BHxQPxJ/Z50ixvLkyan4SkOkXO5skpGAYG+nlMi/VDVVVVVVVVVVVVVaL/AMhiw/6+Y/8A0IVVVVVVVVVVVVVVVVVVVVVX7B//ACUvx5/16r/6PNVVVVVVVVVVVVVVVVVVVVf8Frf+Rn+FH/Xhqn/oy3qqqqqqs/8Aj6i/3xVVVVVVVVVVVVVVVVVVVVVVVVV74P8ACeoyGa98N6ZNITy7Wqbj9TjNadoOh6QP+JTo9lZnGCYLdUJ+pA5qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqvE/h/T/FnhvVfCurRiSx1ixmsrlCM7o5UKOP++WNa5+yb+0xoeoahZS/AD4jTxafNLG13D4UvWhdUJBkVxFgoQMhs4xzWn6ff6tf22laVZT3t7eTLDbW1vEZJJpGIVURVyWYkgAAZJIr/hlf9p7/AKNw+KP/AIR9/wD/ABqof2Uf2obiVII/2cficGc4BfwleoAfdjEAB7mv2Yf+CXPxt+IXjDTNZ+NnhqXwZ4LtZ0mvYbyVRe36KcmGOJSWi3YwXk27QcgMeKgggtoI7a2hSKKJAkcaKFVFAwAAOgA4xVVVVVVVVVf8EuP+Tx/jN/14X/8A6co6qqqqqqqqqqqqqqqqqqqq8ff8ilff9s//AEYtVVf8FPdZF98eNF0iN8ppnhmEMPSSSedj3/u+X6fyr/glpoywfCbxdr+wBr3xELbdgZIht42+uP3x/X3qqqv2t/haPi78AvFPhm2tvO1O0tjqOlhVy32m3y6qo9XUPH/20r4PfEK9+FHxQ8M/EOxZt2iahHNKinBkhPyzJ/wKNnX/AIFWn39nqthbapp1wk9reQrNBKnSRGAKsPYgg1VVVVVVVVVVVVXhD/kWdO/64CqqqqqqqqqqqqqqqqqqqqvjF/yfR4R/6+tL/wDQqqvFLbtcuACDtCgf98ivCHiDTmvT4T3Fb6OyW+CnGHiaV48jvwUGf94e9VUsaSxtE4yrqQR7Gpont5nhY4eNip+oNWVwLuyguQf9ZGGPsSOaqqqqqqqqqqqqqq/4Ka/8mP8AxJ/7hX/p1tK/4JZ/8mX+Ef8Ar/1L/wBLJaqqqqqqqqqqqqqqqqqqqv27/wDkz/4qf9gCT/0NK/4JB/8AJqF3/wBjdff+ibeqqqqqqqqqqqqqqqqqqqqvit/r9N/3JP5rVVXxn1k+IvjB4417fuGoeI7+dT7PcOQByeMEDrX7N+jDQP2f/h1pZQK6eGbF5F9HeFXccE/xMeaqqr/gqL8LReaH4Y+MOnWuZdPlOk6k6rz5T5kgY+yuJR9ZRX/BNj4ojwf8arrwDfXWyw8a2RijVjgC8gDSREn3QzLjuWWqqqqqqqqqqqqq0X/kMWH/AF8x/wDoQqqqqqqqqqqqqqqqqqqqqqv2D/8Akpfjz/r1X/0eaqqqqqqqqqqqqqqqqqqqq/4LW/8AIz/Cj/rw1T/0Zb1VVVVVWf8Ax9Rf74qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqtRso9S06506V2VLqF4mZeoDKQSPzrwZ/wAEifgn4J8YaF4z0/4l+N57rQNTt7+CKY2uyR4ZFkVWxEDtJUA4IOKvNb0XTpfs+oaxZW0u3ISa4VGx64J6V/wlXhf/AKGTS/8AwMj/AMavvHXgjTYPtOo+MtDtYQceZPqEUa5+pbHao5I5o1lhkV0dQVZTkEHoQfSqqqqqqqqq/wCCXH/J4/xm/wCvC/8A/TlHVVVVVVVVVVVVVVVVVVVVXj7/AJFK+/7Z/wDoxaqq/b91j+1/2p/FsSvuj06KytYyc9rWJmGP952H696/4J1aN/Zn7MGkXuzH9r6nfXWTnnEphz1/6Y47dKqqqv2r/hcPhF8evFXhW1txDps10b/TFVcKLaf51VR6IS0f/AK/4J/fFBfiJ+z1pmjXdyJNT8GynSZ1J+byVAa3bH93ymVB7xNVVVVVVVVVVVVVeEP+RZ07/rgKqqqqqqqqqqqqqqqqqqqq+MX/ACfR4R/6+tL/APQqqvELbtauj/t4/ICtZ+IB8I/tReF5JLnFpJpcOlXecABJ5ZGGfYO8bHPpVVXia2+z6zOAOJMOPxHP65rwlcCfR1jJyYHZP6/1qqqqqqqqqqqqqqr/AIKa/wDJj/xJ/wC4V/6dbSv+CWf/ACZf4R/6/wDUv/SyWqqqqqqqqqqqqqqqqqqqr9u//kz/AOKn/YAk/wDQ0r/gkH/yahd/9jdff+ibeqqqqqqqqqqqqqqqqqqqqvit/r9N/wByT+a1VazqUOjaRfavOAY7G2kncE44RSx9ew9Kke51C7Z2DTXFzISQq5Z3Y9gO5J6VoGlJoeg6bokZBXT7OK3XHTCIF/pVVVfG74cW3xc+E3ij4dXATfrOnvHbO/SO4XDwufZZVQn6V4e1vXfh94y07xDYB7TWPDupR3ESyLgxzwyBsMP95cEfWvBPizS/Hvg7RPGuiuGsdcsIbyDnJVZEDbT7jOD7g1VVVVVVVVVVVVov/IYsP+vmP/0IVVVVVVVVVVVVVVVVVVVVVX7B/wDyUvx5/wBeq/8Ao81VVVVVVVVVVVVVVVVVVVV/wWt/5Gf4Uf8AXhqn/oy3qqqqqqs/+PqL/fFVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX7bv8AwUK+Jfxc8d6z4G+E/jK+8PfD3Srl7W3fS52t5tY2Ha08sqkP5bEHbGCF2lSwLdJpZbiV555WkkkYs7uSSxPUk9zVV4V/5FfR/wDrwg/9FrVVVVVVVVVf8EuP+Tx/jN/14X//AKco6qqqqqqqqqqqqqqqqqqqq8ff8ilff9s//Ri1VV+07rK69+0P8RtRRw6jxJeQIwIIZYpTGCCOoIQV+x/ow0L9mX4d2ITb5mjrdYxj/Xu02eg6+Z/+vrVVVV/wVE+F41Hwv4Z+L2n24M+kTtpeosByYJcvCx9lkDj6zCv+Cb3xS/4Qr43y+B766EeneNrM24DNhRdwhpISfqvmoB3MgqqqqqqqqqqqqqvCH/Is6d/1wFVVVVVVVVVVVVVVVVVVVVfGL/k+jwj/ANfWl/8AoVVWssH1a8I7TOOfY1+0HfNd/FzWnR+LdbeJNpHy7YI88j/aya+EnjJPH/w40HxSZhJPdWirdYPSdPkk47fOpP0Iqq8b2wD212o6qUJ+nI/ma8E3G2e4tC33kDqD7cH+Yqqqqqqqqqqqqqqr/gpr/wAmP/En/uFf+nW0r/gln/yZf4R/6/8AUv8A0slqqqqqqqqqqqqqqqqqqqq/bv8A+TP/AIqf9gCT/wBDSv8AgkH/AMmoXf8A2N19/wCibeqqqqqqqqqqqqqqqqqqqqvit/r9N/3JP5rVV+0NrI8P/Ab4hauH2vB4ZvxEfSRoHVO4/iK18JdHXxF8VfBfh9lDDU/ENhalTjnzLhF78d6qqqq/b2+F3/Ctv2htYvbO3EemeLUGr2u1cAPISJx9fOV2x2Dr9a/4JofFH/hKvhDqPw3vrjde+Db3NurHk2lwWdMZ5OJBMPQAoKqqqqqqqqqqqq0X/kMWH/XzH/6EKqqqqqqqqqqqqqqqqqqqqr9g/wD5KX48/wCvVf8A0eaqqqqqqqqqqqqqqqqqqqq/4LW/8jP8KP8Arw1T/wBGW9VVVVVVn/x9Rf74qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv2hdbu/DPwC+JfiSwdlutJ8H6tdwFW2kPHZyupBwccqOxr9kX4JWX7RH7QvhH4VavdzW2lalPLNqUsLBZBbQRPNIqE5wziPYDg4Lg4wDX/BRj4J+A/gJ+0Mngj4a+HU0bQJfD1nd21stxLMSzGRHdnlZmJLRnuR/Kvgn+xd+x98Yv2cfA3iK7+D+jPP4i8LWMt1qFpLLDcLctAnmsJEYYdZQ+e2Qcgjir15P2Gf2xNV0iKysPFemeENY+zz2+pWUU/wBv0+TZIAQ6kJN5TIQ642uOOMg6FrGmeIdD07X9FuEn0/UrSK5tJU6SRSIGRh7FSDVVVVVVVVVf8EuP+Tx/jN/14X//AKco6qqqqqqqqqqqqqqqqqqqq8ff8ilff9s//Ri1VV4t1g+IvFWs+IC5Y6nqE9yWOcnzJGbPPPevhXow8O/C/wAH+Hgu3+y9AsbXbjGPLgRfU+nqaqqqvjL8PLP4sfCzxP8ADu8Cf8TvTpIYHfpFOPmhkP8AuyKjfhWkalr3w+8Y2erWoksta8OakkyK+Q0NxBIDg49GXBrwF4w0z4heCNC8caO3+h67p8N5EpOSgkQMVPupJB9waqqqqqqqqqqqq8If8izp3/XAVVVVVVVVVVVVVVVVVVVVV8Yv+T6PCP8A19aX/wChVVai2+/uXznMznP4mvilejUPiN4kuFbcBqU0anOchGKgjHbC1+xD40Etlrvw+uZstA41C0Unna2ElA9gRGf+BGqrxXbfaNGkYDJhYOP5H9DXhy4+zazbEnh22EfXgfriqqqqqqqqqqqqqqv+Cmv/ACY/8Sf+4V/6dbSv+CWf/Jl/hH/r/wBS/wDSyWqqqqqqqqqqqqqqqqqqqr9u/wD5M/8Aip/2AJP/AENK/wCCQf8Ayahd/wDY3X3/AKJt6qqqqqqqqqqqqqqqqqqqq+K3+v03/ck/mtVX7dWsf2L+yv44lR8SXUVraoOefMuolYf98Fj+HvX7G+jnXf2nfh7ZKpYxap9pwAf+WETzZ4/651VVVV/wUp+Fw8X/AAXtPiBYW2+/8F3okkYDn7JOVjlHqcOIW9gre5r9hf4oj4YftD6ELy68rS/E4Oj3u44UecR5LHPAxMsWT2Ut61VVVVVVVVVVVVov/IYsP+vmP/0IVVVVVVVVVVVVVVVVVVVVVUFhY2rM9rZwQsw+YxxhSfriqqqqqqqqqqqqqqqqqqqqv+C1v/Iz/Cj/AK8NU/8ARlvVVVVVVZ/8fUX++Kqqqqqqqqqqqqqqqqqqqqqqqr+GW5sbi3t5milkiZUkU4KMRgEfQ18PfGtt420JLzKJfW+Eu4R/C/8AeA/ut1H4jtVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVXxB8Mjxr4C8SeDS6r/b2kXdhuboPOhaPJ9vmr9iDx1F8Fv2wPAWs+KCbGCDV5dJ1FZjsEH2mOS1JkJ+6EeRWJ7bDX/BZz4R38z+Bfjlp1m0ttBFJoOqSqn+p+ZprbJHO0lrkZPAO0dWr9n3/AIKFftCfs3+BH+G/gyXw9quhK8j2MGt2Uk509pCWfyWjkjIBYltrbl3EnHJz448aeKvid4z1bxz4w1OXVNe167a5vLhlAMkjHoFHAA4AUDAAAAAFfAnw3qvg34H/AA88Ia6jLqeh+FNMsbxXGCJobWONwffcpqqqqqqqqqv+CXH/ACeP8Zv+vC//APTlHVVVVVVVVVVVVVVVVVVVVXj7/kUr7/tn/wCjFqq+J2sL4e+GvizxAzhRpmh3tyWOOPLgds88dq8M6Q3iDxJpOgRgltSvobZQM5zI4XsD6+hpUVFCIoAUYAAxiqqqqv2/vhaPhz+0Jqmq2Nr5emeMIhq1uVXCiVyVuB9fNVnPtIK/4Jl/FEeJfhTqvwxv7rdeeEb3zbRGOD9kuCzAD12yiXPpvUelVVVVVVVVVVVVeEP+RZ07/rgKqqqqqqqqqqqqqqqqqqqq8b/s4634s/aC0X4y2/iSxgs9LltHeyeFjI/knJww45qquG3zyPkcuTx9a8VStceKNYnYANJfzsQOxMhr4FeND4C+Kmga5JMEtHuBa3hY4Hky/IxPsuQ31UVVXcC3VrLbEj97Gy8+4pGeCVXXh42yM9iKhlWeFJkPyuoYfiKqqqqqqqqqqqqqv+Cmv/Jj/wASf+4V/wCnW0r/AIJZ/wDJl/hH/r/1L/0slqqqqqqqqqqqqqqqqqqqq/bv/wCTP/ip/wBgCT/0NK/4JB/8moXf/Y3X3/om3qqqqqqqqqqqqqqqqqqqqr4rf6/Tf9yT+a1Vf8FM9ZXTv2d7PTlcb9V8SWsG3IyVWKaQnHoDGv5j1r/gnLoy6p+03pt6UDHSNJvboEgfLmPycj/v9j8aqqqq8W+GdL8aeFtX8Ia1F5lhrVjNZXKjr5cqFGx6HB4PY14u8N618OvG2reFNRZ4NU8O6lLbSOgKESRORvXvglQwPoQa+AnxKh+L3we8K/EFJEafVNPT7YFGAl1HmOdcegkR8e2D3qqqqqqqqqqqq0X/AJDFh/18x/8AoQqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/wCC1v8AyM/wo/68NU/9GW9VVVVVVn/x9Rf74qqqqqqqqqqqqqqqqqqqqqqqqu7iOztJruUgJBGzsTxwBk/yrwV4tvfBmuw6ta5eI/Jcw5wJYz1H17g+taZqVlrOn2+qadOs1tcxh43HcH+R7EdjVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVNLFBE888qRxxqWd3O0KAOST2GKT9jf9iuTx/rfxM1fwT4a1rXNf1CW/ujqmofabcTSMWcrbs/lYLEtgqeTXjzwD4K+LHgfUvAXjbRrXWfDuuW3lXNs/wB104KsrDlWUhWVlIKkAgggGvid/wAEYvHUWtzTfBr4raDd6RLITDb+Jlmtp7dOymSCORZT/tbI/pX7Kv8AwSk8OfCLxfp3xI+M3iy08W6xo863Gm6VYwMljBMvKSyM+HmKtgqu1FBAJ3dBVVVVVVVVVf8ABLj/AJPH+M3/AF4X/wD6co6qqqqqqqqqqqqqqqqqqqq8ff8AIpX3/bP/ANGLVV+11rB0L9mj4iXqyFTJoktrkEj/AF5EOOPXzMV+zTow1/8AaD+HOmlA6nxLZSup7pHMsjDr/dQ1VVVVX/BSP4Xf8Jp8EIfHNjb79Q8E3gnYhcsbSYrHMB34byXPYBGr9iD4o/8ACrv2h/D091ciLTPEROj3+44G2cr5bH0xMsRJ9M+pqqqqqqqqqqqqrwh/yLOnf9cBVVVVVVVVVVVVVVVVVVVVVU7BULYztGcVRRtU1Py7cYa7nxGG7Fm4zj61498KXPgfxnrPhK6LFtLvHiRmGC6Zyj/8CUqfxr4GeNB4++Fmga9LP5t2tsLa8JOW86L5GJ92wG/4EKqtetvs2r3UQHBcsPoef614auPtOjW5Jy0YKH8Dx+mKqqqqqqqqqqqqqv8Agpr/AMmP/En/ALhX/p1tK/4JZ/8AJl/hH/r/ANS/9LJaqqqqqqqqqqqqqqqqqqqv27/+TP8A4qf9gCT/ANDSv+CQf/JqF3/2N19/6Jt6qqqqqqqqqqqqqqqqqqqq+K3+v03/AHJP5rVV/wAFVdYMHhX4feHw5xe6he3O3nB8mOJc+n/Lf9a/4JZ6N5/xS8Y+IAmfsXh9LXdzx506Nj/yB+lVVVVV/wAFLvhb/wAIp8XtP+I9hbBLHxlZfv2VcAXluFR846bozCfc7j61/wAEuPih51j4q+Dl/cZa3Yazpqk5O1tsVwPYAiAgersaqqqqqqqqqqqtF/5DFh/18x/+hCqqqqqqqqqqqqqqqqqqqqq+Fn7QHgT4wa1qmg+E4tUS50eMPcG8t1jUjdt+Uhjnke1VVVVVVVVVVVVVVVVVVVVf8Frf+Rn+FH/Xhqn/AKMt6qqqqqrP/j6i/wB8VVVVVVVVVVVVVVVVVVVVVVVVfFvXk0LwRfBZMT34+yxD13/e/wDHA36VXwb8f/2DqA8N6rPjT71/3LseIJTx17K3Q+hwfWqqqqqqqqviN/wWZ8I+AfiD4n8CRfArU9TXw5rN3pq3qa+kQuhBM0fmhDASobZu2knGcZr/AIfi+FP+jdtV/wDCkj/+MV+xn+1xpv7YngTW/HGm+B7nwwmi6v8A2c1tPfrdGU+Ukm8META/eYxjt1rxN4m8PeDPD2oeK/Fes2mlaPpNu9xe3t3KI4oIlGWZmPQYr4j/APBaTwpa+JJtC+CPwT1XxhaQuQupahfmx88Dq0dukUj7T2LlTjGVGeP2Zv8AgrT8JPjb4rsPh98RfClz8PNf1OZbexnmvxd2FxMcBUM2yNomZuAGXbnA35IzX7TH/BRLwb8DviFB8FPhz8PNb+KnxIlZVl0LRnKLbMyhljd1jkZpSpDeWkbYH3ivAr4X/wDBT6ym+J9h8H/2nPgfr/wa13VCi2V1qkzvbOznEZkEkUTRIxBUSAOmRyyjJFftj/tW6T+yf8ObHxJH4fPiXxNr+ox6foGgRzNG99KSDIcqrMFVT2U5ZkX+LNeCb7xRqvg/RtT8baLa6Pr93ZRS6jp1tcGeOzmZQWiEhA37Sdu7ABIJHFVVVVVVVVVVVVVVVVVVVVVXxf8ABtx8RvhL42+HtncJBceJ/Dt/pcUr8CNri3eIMevALg14i8Pa34S17UPC/iXTJ9O1bSbmS1vbO4TbJBNGxV0YdiCCK/Zs8eeHfiX8BPAfjDwvfR3Nld6DaxttbcYZo41jlibHRkkRlI9Vqqqqqqqqqqqv+CXH/J4/xm/68L//ANOUdVVVVVVVVVVVVVVVVVVVVePv+RSvv+2f/oxaqv8AgolrA0z9l7W7HeB/a2o2Nrg98TrNj/yD7dK/YH0Y6x+1R4PcoWj09Ly6kwDxttZQp9vnZOtUjpIu6N1YAkZBzyDgj8xiqqqrxP4d0vxd4b1TwrrUAm0/WLKWzuoz/FHIhRh+RNeOPCesfDjxxrPgzVHaPUfD+oy2skifLlo3IDrzwDgMD6EV+z18TY/jD8GPCnj8yq11qFgqX4XjbdRkxzDHYeYjEA9iD3qqqqqqqqqqqq8If8izp3/XAVVVVVVVVVVVVVVVVVVVVVVXjBLSZ8/djY/pWqTi2027uTIU8qB33DquFJzXw2sBqnxE8LabjIudZtIj16NMoPT2r9tjwUNN8XaV44tYiItZtjb3JA486HGCT6lCoH/XM1+xB40EV3r3w+upjidV1CzUnjcuElH1IMR/4CaqvGttsvYLkDiWMqfqp/wIrwTcZjubQnoQ6j68H+Qqqqqqqqqqqqqqq/4Ka/8AJj/xJ/7hX/p1tK/4JZ/8mX+Ef+v/AFL/ANLJaqqqqqqqqqqqqqqqqqqqv27/APkz/wCKn/YAk/8AQ0r/AIJB/wDJqF3/ANjdff8Aom3qqqqqqqqqqqqqqqqqqqqr4rf6/Tf9yT+a1Vf8FT9YWf4k+CvD4cE2Why3JXjI86crn1/5YfpX/BKnRhFoPxD8QlMm6u7C1ViOnlJMxA4/6arnnsKd0jQvI6qo6ljgCqqqqv26/haPid+zxrslna+bqnhYjWLLC/NiEHzl9cGFpDgdWVfSv2bPiefg/wDG3wp45luDHY216INRPb7LKPLlJHfCsWHuopWV1DowYEZBBzmqqqqqqqqqqq0X/kMWH/XzH/6EKqqqqqqqqqqqqqqqqqqqqr9g/wD5KX48/wCvVf8A0eaqqqqqqqqqqqqqqqqqqqq/4LW/8jP8KP8Arw1T/wBGW9VVVVVVn/x9Rf74qqqqqqqqqqqqqqqqqqqqqqqq+MPjCPxP4k+w2M++x0sGKMg8PJ/Gw9uAB9M96qvg94/HiPTR4f1WfOpWKfu2Y8zxDgH3YdD68H1qqqqqqqtV1K00bS7zWL+UR21jbvPM5IG1EUsx546A9a/4I5+B9F+Jnjr4xfEnxp4d03VWVbKKNb61S4US3MtxLKVDqcEeUmTx94de3/CpvhX/ANE08Kf+Ca3/APiK0Tw34d8M272nhvQNN0qCV98kVjaJArNjG4hAATgAZ9q/4LQfHzVjqHhX9mfw3ezLb3Fuuta9FCT/AKSzOUtITjqAY5JCp6kxHqBX7G/7LnhD9lz4O6N4X07RrUeKb2zin8S6oEDTXd4ygum/r5SMSiL0AGcbmYn/AILFfsw+E38B2P7S3hHQrXT9c0/UY7LxG9tCsYv4J8rHPLtHzSpKETceSsuCTtUD9gj9o69+LH7GWn/ELxXdyajrXgm3utM1mZmJe4ezjDozNyS7W7QFmOSWYnvX/BHHQD8QPF3xg/aK8ZNHqXirUdQitlvZV3PG1w0lxdMCckb28nv0UjvX/BZr4e6Jr/7N2i/EGSzgGseFfEUMUF0V+f7Nco6SxA+hdYWx/wBM/wA/2TvHt345/ZY+GnjzxLf5urnwraPqF3cuF3yRRBJZXYnAyUZiT654r4A29x+3t+2hrX7UGu2zTfCz4RXH9l+B7aZf3d3er8wuNp/iGRcHgEF7ZedhqqqqqqqqqqqqqqqqqqqqqqrWb2TTdHvtRhRXe1tpJVVuhKqSAfbiv2kf2q9V/ac1M+IPGHwi+H2j69lV/tvRbW8gvZI1wAkpa5aObgBdzxlgAApUcV8M/jr8Y/g01wPhb8Stf8NR3h3XEFjeMkUzYwGaPlGbHAYjI9a/4bv/AGv/APov/ij/AL/J/wDE1/w3f+1//wBF/wDFH/f5P/ia8OzzXfh7TLq4kMks1lE7u3VmKAk/nVVVVVVVVVf8EuP+Tx/jN/14X/8A6co6qqqqqqqqqqqqqqqqqqqq8ff8ilff9s//AEYtVX/BUnWTb/B/wroCvg33iMTkDgkRW8ox16ZlHbqB0r/gmLo63/x+1XVJEBXTPDNw6E44d54EH/jpfn/Gr+7i0+xuL+c/u7aJpH+ijJ/QV8G/ihLY6xJ4e8QXWbTU52kglc8QzuckeysT+B+pNVVVVf8ABTP4XDwz8V9K+Jen222z8X2Xl3TAcfbLcKjE+m6Iw49SrH1x/wAEufiiCnin4N6jdcgrrGmIx/3YrhRn/tiwA/2z61VVVVVVVVVVVeEP+RZ07/rgKqqqqqqqqqqqqqqqqqqqq+KfxE8d6V+2J4Y8Hab4u1S20O6uNOWbT4rllhkDn5wUBwc96rVW2aXdsCARA+M+u014wnNt4R1u5DKDFptw4LdBiNjzX7PNh/aPxq8JW4XdsvvOxz/yzRnzx/u1+0t4L/4Tb4Q6zbwQCS80pRqFr8uSGiyXA9zGXH1Ir4ReM28AfEjQfFJl2QWt2qXXvA/yScd/kYke4FAqyhlYEEcEVXjG283S1nUcwSAn6Hj+eK8KXHkaxGhPEysh/mP1Aqqqqqqqqqqqqqq/4Ka/8mP/ABJ/7hX/AKdbSv8Agln/AMmX+Ef+v/Uv/SyWqqqqqqqqqqqqqqqqqqqr9u//AJM/+Kn/AGAJP/Q0r/gkH/yahd/9jdff+ibeqqqqqqqqqqqqqqqqqqqqvit/r9N/3JP5rVV/wUh1g6p+0vc2JkLf2RolnagEn5chpsD/AL/Z/Gv+CZGjDT/2fdR1N1+fVPEtzIrYx8iQwRgdf7yv6da+OOrHTPh7dwo+2S/mjt0Pfk7j/wCOoR+NfBv4jDxbpX9iatcZ1ewQZZjzcRDgP/vDgH8D3OKqqq4t4LqCS1uYklilQpJG65DKRggj0Ir48/DWb4Q/F/xT8PHRxBpV+4s2fOXtnw8Le5MbJn3zX7F3xRPxX/Z68Nand3XnapokZ0jUSWy3m24AVmPUs0Ricn1Y1VVVVVVVVVVWi/8AIYsP+vmP/wBCFVVVVVVVVVVVVVVVVVVVVV+wf/yUvx5/16r/AOjzVVVVVVVVVVVVVVVVVVVVX/Ba3/kZ/hR/14ap/wCjLeqqqqqqz/4+ov8AfFVVVVVVVVVXxW/af8AfB/xOnhLxNpHiC6vHtEuQ9jbwvHtYsAMvKpz8p7enNf8ADefwf/6Fzxj/AOAdt/8AH6/4bz+D/wD0LnjH/wAA7b/4/X/Defwf/wChc8Y/+Adt/wDH6/4bz+D/AP0LnjH/AMA7b/4/X/Defwf/AOhc8Y/+Adt/8fr/AIbz+D//AELnjH/wDtv/AI/X/Defwf8A+hc8Y/8AgHbf/H6/4bz+D/8A0LnjH/wDtv8A4/Xw5/a2+G/xP8Z6d4G0DRfEkF/qXm+TJeW0CRL5cTSNuKzMR8qEDAPOPrVVVVVVVfGPx83hzTh4f0qbGo38Z3up5giPBI9GPIHpyfSqqtIvtT0a7i13THaOSzlXbIOgYg4B9iFbj0zXg7xRZ+MNAt9ZtAFZhsnizkxSD7y/yI9iKqqqqqq/aFg8TXfwG+Itj4K0u61HxDeeFtRt9KtbUZklupLd0iC8j+Nl7ivgP8NP+Cpv7L/hTVNO+EHwfvdKstZuEvL0yWlheTysECqNjuzDAz8oUHJOfbwV/wAFaf2jfhD4xi8GftYfBlWjSQfavL02XSNTgjJwZBFKfLlAwcDEYb+/3r4b/EbwZ8XPA2j/ABG+H2uQ6toGu2wns7qLI3DOGVlPKurBlZTgqykEAg1+1hInj/8A4K02Ph3UXdrJ/GvhjSlRxnZEVslcAA9CzyN/wKq/4KIaXa6v+xX8VrS7UMkejpcKCoPzxXEUqHn/AGkX6V/wRbaLxL8HPi94E1BWNjLqduZBkci5tpI3xx12xD9K/wCCV/jux/Zz+M3xT/ZS+L2oW+g69cajEdN+2uII7i5tzJHJGjNjJljeKSP+8qkjORn/AILC/F/RtT8BeFf2aPB90mseM/E3iC3uZ9MsnEs0USBkijZFyQ8s0qBFPJ8tuOlftT+JPFfwi/Zu+EH/AAT4+Fkkdx8TfH+i2Oi6glswH2W0CKl1I23O1ZpRKu7keWs5JyAa8YftI/C3/gnj4O8F/skfCDwLqPxI+IcNlEF0bTW8tpZ5iWae4dUdvNlcs4iVS2wr91ShOlf8FPfiF8MvHWkeEf2wf2Z9X+Gmn683+ia1FLJJHEuQGZo2T94q7lLmNyygj5DkCrG+stTsbfUtNu4rq0u4llgnhcOkqMAVZSOCCCCCPWqqqqqqqqqqqqqqqqqqqqq1Wx/tTS7zTfN8v7XbvDv2527lIzjv16Va/wDBFDQUgVb39oi/lmGdzxeGEjU88YU3LEcY7mv+HKXhf/o4LVP/AAnI/wD4/X/DlLwv/wBHBap/4Tkf/wAfr/hyl4X/AOjgtU/8JyP/AOP1pdiumaZaaasvmC0gSIORjdtUDOPwqqqqqqqqqv8Aglx/yeP8Zv8Arwv/AP05R1VVVVVVVVVVVVVVVVVVVV4+/wCRSvv+2f8A6MWqr/gqxrO/UPhz4eR8eTDqF1Iueu8wKp6dtj9+9f8ABKjRzJq3xF19lIEFtp9qjEHB3tOzY7ceWv5ivjDq39kfDzVXRsSXSC2Qeu8gN/47uqvgv8Rj4p0z/hHtXuN2q2CfI7nm4iHAb3YcA+vB9aqqqv24vhd/wtH9nfxBDaW5l1Pw2BrNiFXLFoAfNUAckmFpQB3O2v2dfic/wf8AjR4V8etOY7Oyvlj1DGebSX93NwOuEZmA9QKR0lRZInV1YZVlOQR6iqqqqqqqqqqq8If8izp3/XAVVVVVVVVVVVVVVVVVVVVV8Yv+T6PCP/X1pf8A6FVa623R7sj/AJ5EV8TZvI+HPiZ9u7Ok3K4zj70bD+tfsj2AvPjdpdxtz9htLqYcHjMTR/8As/epI45Y2ilRXR1IZWGQQeoI9K+KXg6TwB8Qdd8JOpEdhdsLcn+KFvmiP4oy/jX7OvjT/hOfhHod/NcebeWEf2C7JOT5kWFBJ9WTYx/3qrVLb7Xp1xbAZLxnaPft+uKtJza3UNyvWJw2PoaBVlDKQQRxVVVVVVVVVVVVVf8ABTX/AJMf+JP/AHCv/TraV/wSz/5Mv8I/9f8AqX/pZLVVVVVVVVVVVVVVVVVVVV+3f/yZ/wDFT/sASf8AoaV/wSD/AOTULv8A7G6+/wDRNvVVVVVVVVVVVVVVVVVVVVXxW/1+m/7kn81qq/bQ1ga7+1D8Qb0OG8rUUtc/9cII4ccf9c8V+wbox0b9lfwWHTEt6Lu6fjrvupdp6D+AJ/8Aqr9pLVsvo2hRv0ElzIv1wqH9HrRNZ1Dw9qttrOlTmK6tX3I3Y+oI7gjgj0NeC/F2neNtAg1qwIVj8k8JOTDIOqn+YPcEVVVVf8FR/hd5F/4W+Men242XKNo+pMq4+dd0kDH1JXzgT6Io9Mf8ExfiidB+I+t/CrULrFr4ns/tVijN0u7cEsFHYtEXJ/65LVVVVVVVVVVVaL/yGLD/AK+Y/wD0IVVVVVVVVVVVVVVVVVVVVVX7B/8AyUvx5/16r/6PNVVVVVVVVVVVVVVVVVVVVf8ABa3/AJGf4Uf9eGqf+jLeqqqqqqz/AOPqL/fFVVVVVVVVVX7S37NPxF+LfxFj8VeFZ9HSyTTYrYi7uWjferOTwEPGGHev+GGvjT/z9eGv/A6T/wCN1/ww18af+frw1/4HSf8Axuv+GGvjT/z9eGv/AAOk/wDjdf8ADDXxp/5+vDX/AIHSf/G6/wCGGvjT/wA/Xhr/AMDpP/jdf8MNfGn/AJ+vDX/gdJ/8br/hhr40/wDP14a/8DpP/jdf8MNfGn/n68Nf+B0n/wAbr4C/sp/E/wCGvxY0Lxr4iuNDbT9O+0ecLa7d5Pnt5I1wCgB+Zx36VVVVVVUzKqlmYAAZJPGK8Wa7L4m8RX+tSuSLiYmMH+FBwo/BQKqq+GWgr4n0vxPohQNJLZJJBkciVGJQj054+hNfAbXZLLxJceH5XPlahCWRT2lTnp/u7s/QVVVVVVVVf8FLvgt4R+K/7KXjDXdX062/tzwRYSaxo+omP97bmLDSxhhzskjDKV6Z2t1UEf8ABEPxprWp/D74neAbuaWTTNA1WwvrIOSRG93HMsqr6D/RUOB3YnvX7a6R/Cn/AIKoad481KJbfTv+Ej8Na8jSDCmKIWokfOeR5lvLnp3Haq/4KTeI7bwz+xP8Tbmd0D3tlb2MKsM72nuoY8AZHIVmPtjPPSv+CJfhiex+CPj3xdLBsTVvE6WkTlcFxb2yMSPUZuCPrn8P2kP2Gf2ef2pbuLWviP4au7TxBBEIU13RrgWt2Yx0RyVZJAO3mIxXoCATXw3/AGJf2Qf2INJ1b45Pp2o39z4VsJbx9e8RXK3c9lGiEsYI0SOMSMPlBVN5J2gjcQf+CfXgbxN8d/ib4y/b/wDizp8keo+K7iXT/BVlMSV0/TkPls8eeMbVEIYAZ2zMf9Zmv+CaiRfG79t343/H/wATqLzUtPec6cZyWa1+13Lomz02W8BhHPCtiv8Agql4H0jxh+xd4x1O+toXvfC9xZanp0rrzDILmOJyD1GYppV/EV/wTN8X6j4y/Yn+HF3q0zy3OmQXWmb2/wCeVtdSxQgeywrGv/Aaqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv8Aglx/yeP8Zv8Arwv/AP05R1VVVVVVVVVVVVVVVVVVVV4+/wCRSvv+2f8A6MWqr/gp7rIvvjxoukRvlNM8Mwhh6SSTzse/93y/T+Vf8EtNGWD4TeLtf2ANe+IhbbsDJENvG31x++P6+9ftI6sI9M0fQ0bmed7hwOwRdo/Pefyr4f8Aw/ufHkesR2k3lzWFoHgzwrSlvlVj2BVXH1we1WF9q3hfWo721aS0v9PmPDDBRlOCpH5givA3jGw8caBDq9mQko+S5gzkwyAcj6dwfQ1VVU0MVxE8E8aSRyKVdHXIYEYII7jFftCfDKX4P/GXxV8PzEy22n3zNYFh961kAkhOe58tlB9wR2r9iP4of8LT/Z38OXV1ceZqXh5Do1/k5O+AKI2J7loTExPqTVVVVVVVVVVVeEP+RZ07/rgKqqqqqqqqqqqqqqqqqqqq+MX/ACfR4R/6+tL/APQqrxKwXQ7on+6o/NhXxnmEHwu8ROWIzabOP9pgv9a/YksPP+JmragyZW10R1BweGeaLHP0Vqr9t7wUttq+h+PrWHC3sTWN2wGB5ifNGT7lS4+iCv2JPGgsfEes+A7qX93qcAu7UE8CWLhwPdkYH/tnVVqdt9k1C4tgMBJCFHt2/TFaFcfatItZSckR7T9V4/pVVVVVVVVVVVVVf8FNf+TH/iT/ANwr/wBOtpX/AASz/wCTL/CP/X/qX/pZLVVVVVVVVVVVVVVVVVVVV+3f/wAmf/FT/sASf+hpX/BIP/k1C7/7G6+/9E29VVVVVVVVVVVVVVVVVVVVfFb/AF+m/wC5J/NaqvjPrJ8RfGDxxr2/cNQ8R386n2e4cgDk8YIHWv2b9GGgfs//AA60soFdPDNi8i+jvCruOCf4mPNfG3Vf7U+Id/Gr7o7FEtkP+6uW/wDHman+G1/L8Obbx5ZF5cyyfaIcfdiDbQ474BVs+xB6A18NfHlz4E19LpmZ9PucJeQjuvZgP7y9fzHerS6tr+1ivbOdJoJ0DxyIchlIyCKqqr9pj4Xj4xfA/wAVeB4IBJfz2RuNNGORdwnzIgDzjcyhDjs5r4b+N9T+GfxA8P8AjzSgTdaDqEV0I87fMCMN8Z9Ay7lPsxrQ9Z03xJomn+ItGuVuLDVLSK6tZV6SRSIGRh9VINVVVVVVVVVVaL/yGLD/AK+Y/wD0IVVVVVVVVVVVVVVVVVVVVVX7B/8AyUvx5/16r/6PNVVVVVVVVVVVVVVVVVVVVf8ABa3/AJGf4Uf9eGqf+jLeqqqqqqz/AOPqL/fFVVVVVVVVVVVVVVVVVVVVVVeLJ2tfC2s3MZ+aHT53XBxyI2NVVV+z3pcsdlq+sOvyTyRwRkjqUBLf+hLWm2v9i/G5bWBSqrqrhFUdFfPH0w1VVVVVVftMftPfDj9lPwXpnjr4lJqM1hqmrxaZFDp0SSz73R3MgRmXKKsZLc5GRjJIB0L/AIKNfsV+IdLj1S2+PWjWquuWgv7e4tZkPcFJIwcjpkZB7E1/wUF/4KReCfjF4Duf2ev2b3v9di8STRwavrQspIlnjEgItrWNwJGZ3VQzFQCp2ru3Ej/gmn+zDrP7NXwAVfGlk9p4v8Z3Q1TVbWQYeyTYFgt2/wBpUyzA8h5GXtX/AAWL/Zk1rx54Q0T9ojwZpUl7e+DLV7HxBFCm6QaaWMkc+BzshkaXdjOFmLHCoxr9jP8A4KVfBLx78IdF0P41/EXTPCXjnw/Zx2epNrM/kRal5ahVuo5W+VmdQC6ZBDlsDbgn/gpF+2DpP7V2teGP2Zf2bftfiuwGrJPdXVlA+NVv8GOGCAEBmjTe5LEbWYqRwgY/sj/AmH9m79nzwj8J3kjm1LT7Uz6vPHyJb6ZjJPg91VmKKe6otV+3X4p139qL48eDv+Cf/wANNSkjs5Z4tY+IV9bsCLS0j2yJE3uqFZdp4LyWw9a8KeFtA8D+GNJ8G+FdNh0/R9Es4rOxtYlwsMMahVUfgBz361+y34m0b9h//goV8TvhF8Vb6Hw/4c8aySrpGpXTeVb7XnM9i7ucKqNFJJGW6LJ8pIwcf8FYP2ivAuk/s5XXwY8N+J7DVvFXj67tIY7HT7lZ5YbSOZJ3lYITtVmjSNQeX8w4BCsR+xR8I9U+Bv7LXw9+G+vW32fV7HTWudRhIw0VzcyvcSRt/tI0pQ/7npVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV/wAEuP8Ak8f4zf8AXhf/APpyjqqqqqqqqqqqqqqqqqqqqrx9/wAilff9s/8A0YtVX7fusf2v+1P4tiV90enRWVrGTntaxMwx/vOw/XvX/BOrRv7M/Zg0i92Y/tfU766yc84lMOev/THHbpXx71Uah49ks0fK6daxw4H94jef/QwPwr9njSfsnhG71R0w9/eEKcdUQAD/AMeL18dPhx9rhfxtolv++hX/AImESD76jpKB6gcH257HPw58c3XgTX0vgWksZ8R3kIP3k/vAf3h1H4jvVle2mpWcN/YXCT29wgeKRDkMpHBFVVV/wVH+FoWXwr8Y9OtfvhtH1N1XuN0luxx7ecpJ9EHpX/BMr4ojw58UtW+GF/c7bTxbZ+daKx4+124ZsD03RGXPrsWqqqqqqqqqqq8If8izp3/XAVVVVVVVVVVVVVVVVVVVVV8Yv+T6PCP/AF9aX/6FVeK226JMoP3mUfqP8K+P85g+EmulZArOIEGe+Z48j8s1+wtYB73xhqbKf3UVpCpIPO4yk4P/AABfzFV+0B4LHjv4Ta9pEMHm3ltB9sswBlvNi+YBfdlDJ/wKvhx4um8B+OtE8XQkkabdq8qjq0R+WRR9ULD8ahmhuYY7i3kWSOVAyOpyGBGQR7YqvGNsItVEwHE0YP4jj+QFeC7jzLGa1JyYpMgexH/1jVVVVVVVVVVVVVf8FNf+TH/iT/3Cv/TraV/wSz/5Mv8ACP8A1/6l/wClktVVVVVVVVVVVVVVVVVVVX7d/wDyZ/8AFT/sASf+hpX/AASD/wCTULv/ALG6+/8ARNvVVVVVVVVVVVVVVVVVVVVXxW/1+m/7kn81qtZ1KHRtIvtXnAMdjbSTuCccIpY+vYelSPc6hds7Bpri5kJIVcs7sewHck9K0TT4PDnhyw0ouqw6XZRw7uwWNAufpgVrOoyavq97qsud95cSTEHsWYn+teE9Fj0fwlpmhzRKfIskjmRhkFivz5HoSTXxa+HkngnWvtNjGx0m+YtbNjPlN3jP07eo+hr4G/Ef+zrhPBetXH+i3D/6DI5/1Uh/g+jHp7/Xiqqq/bO+Fv8Awqj9oTxLpVra+Tpmsyf2tpoC4XypySyqPRZRKg9lFf8ABOj4ojx38Bo/CN7c+ZqXgm6NkwY5Y2z5kgY+gALxgekVVVVVVVVVVVaL/wAhiw/6+Y//AEIVVVVVVVVVVVVVVVVVVVVVXwl/Z38E/BrXNW1/wvqmt3VxrMYSdL+eJ0UBy3yhI0I5Pcmqqqqqqqqqqqqqqqqqqqqv+C1v/Iz/AAo/68NU/wDRlvVVVVVVZ/8AH1F/viqqqqqqqqqqqqqqqqqqqqqqtVsv7R0u80/cB9pt3iye25SP61LG8MjQyKVdGKsp7Edaqq+GN1p154F0mTTLeOBFh2SRoOkinDn15YE8+taMg8Q/Hee6tzvitrqV2YDgCNNgP0LBR+NVVVVVVfFH4PfC/wCNnhs+Evit4H0rxNpe/fHDfQ7mhfGN8TjDxNgkbkIOD1rXP+CQH7HGrXrXVhp3jDRomJItbHXC8a+wMySP+bGvgd+w5+zH+zzqSa/8OfhpajXYwfL1jU5XvrqLPBMbSkiI4JBMYUkHBzmqlijmjaGWNXR1KsjDIYHqCPSviN/wSp/Y6+Ieuz+IIvB2q+Fri6kMk8Ph7UTbQMx67YXV0jH+zGFUY6da+AX7FX7OX7NV0dX+GHgKOPXHjMb61qM7Xd5tPDKrucRAjgiMKD3zVV8E/wBlb4Y/Anxj4z+Ifhm41vVvE/jy6+0azq2tXSXE7kuzsqFEQIhdyxUDHC9lAFfH79ln4H/tN6RaaX8XvBkWpzacW+wahBK1vd2mfvBJUIO045RsqTg4yAR8EP8AgnD+yr8BvEtr4z8NeDLvWtesHEllf6/dm8a0cHIeOPCxq4OCH2blIBBB5qqqqqqqqqqqqqqqqqqqqqq8SyyweHNVnglaOSOymZHQlSpCHBBHQ1B+2J+1ZbQpBH+0T8QGVBgGTX53b8WZiT+Jr/hsv9q//o4fx7/4O5v/AIqv+Gy/2r/+jh/Hv/g7m/8Aiqtf21P2s7OdbiH9oXxwzpnAl1aSVeRjlWJB/EV8Jf8Agq3+1F4C1C3TxxqmmeP9HVgJrbU7SO2uAnfy54FUhveRZB7dMfs/fH/4e/tKfDmz+I/w7v3e2kbyL2znG2fT7kKC8EqjowDAgjIYEEEg1VV8fv8AgqnNJr1x8MP2TvAGoeKvEYkkt21S+0+VkV0yG+z2i4lkIwTuk2BSvKsK/wCCZn7XH7Qn7Q/xa8U+Gfi/8QRr2mad4dN5a240mztfLm+0RJu3QRIx+V2GCSOar4+/HPwR+zn8MNU+KPjy4kFlY4itrWHBmvrl8+XBED1ZsE+iqGY8KTXhf47/APBSv9trUdT8RfA6WPwf4RsLgxIbNoLS2ikABEX2mVTLPIFKltvyjIJVNwB8P/tuftm/sd/E/T/AX7X+kXHiHw/eEF5ZoYTcfZ87TPaXMOEn29SkhJP3SYycjwz4l0Lxn4c0zxZ4X1OHUdI1i0jurG6hbKTRSKGVh9QRVV/wS4/5PH+M3/Xhf/8Apyjqqqqqqqqqqqqqqqqqqqqrx9/yKV9/2z/9GLVV+07rK69+0P8AEbUUcOo8SXkCMCCGWKUxggjqCEFfsf6MNC/Zl+HdiE2+Zo63WMY/17tNnoOvmf8A6+teK9V/t3xNqmrh9y3V3I6H/ZLHaPywK+HWkjRfA2i6eV2utosjjHR3+dv1Y06JIhjkUMrAgqwyCK+Lvw6fwXrH27Toj/ZF8xMJA/1L9TGf5j2+hr4HfEb+yLxPB2s3H+hXb/6HI5/1Mp/h/wB1j+TfUmqqq/aN+GMfxh+CvirwGsIkvLyxaXT8jkXUREkOD2y6Kp9mNeAvGGr/AA28d6H420kMl/4f1GK6SNiV3GNwWjbvhgCpHoSK8O69pfivw9pnifRLgT6fq9nFd2kv9+KRA6H8VIqqqqqqqqqqrwh/yLOnf9cBVVVVVVVVVVVVVVVVVVVVXxW8DeNdS/bL8LeKNO8H63daNb3GnGbUYdPle3jCN8xaULtGO+TxVeMG26PtBHzSqOfx/wAK/aQlMfwsvEAB826gU57fOD/Sv2HbDy/BXiLU9v8Ax8aokOcdfLiU9f8Atp+tVXxp8FnwB8Tte8NxxhLaO6M1mAOPIk+dAPoGCn3U1+y/40HjL4QaUk9x5l7omdPuMnnEePLP/fspz6g1XjW2D2UFyBzFIVP0I/8ArCvB1x5WqNbluJoyAPcc/wAs1VVVVVVVVVVVVX/BTX/kx/4k/wDcK/8ATraV/wAEs/8Aky/wj/1/6l/6WS1VVVVVVVVVVVVVVVVVVVft3/8AJn/xU/7AEn/oaV/wSD/5NQu/+xuvv/RNvVVVVVVVVVVVVVVVVVVVVXxW/wBfpv8AuSfzWq/aG1keH/gN8QtXD7Xg8M34iPpI0DqncfxFa+Eujr4i+Kvgvw+yhhqfiGwtSpxz5lwi9+O9fE/Vf7G8A61eK+Ha2MKeuZCE4/76z+FeCtJ/tzxdpGlFdyz3cYkGP4Act/46DVeJvDmm+K9EudD1SPdDcLwwHzRsOjL6EH/Doa8T+HNS8Ia7caJqK7ZrdspIvAkX+F19iPy6dq+D3xFXxjpH9k6nOP7X09AJCTzPGOBIPfoG98HvVVVf8FPPhcNd+HOh/FbT7bddeGLv7JfOox/otwQFZj6LMEUD/psa/wCCefxRHw/+P1p4cvrry9N8aW502QMcKLgfPbt9S4MY/wCutVVVVVVVVVVaL/yGLD/r5j/9CFVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX/Ba3/kZ/hR/14ap/6Mt6qqqqqrP/AI+ov98VVVVVVVVVVVVVVVVVVVVVVVXxj8JS+H/FEupwRf6FqrGaNgOFkP31/P5vofaqqvBnxLuvB/hnVdGtome4uXD2b/wwsRh2P4BSB69a+BnhSbTtLn8U36MLjUhtgDjkRA5Lf8CPP0UHvVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVWqNp66Zdtq0kSWSwObl5W2qse07yx7DGcmvisPh38VfjYvhn9mf4Xf8I/oeo6hHpfh/Tlv7m6n1GR5BHHLI1zIxR5GYYQbVUEA5ILHwT/wRc0eTw3aTfET40X0Wuyxhrq30jTkNvAxHKK8jbpMf3sLn0FfFf/gjTLpPhG91j4Q/Fa71jW7KF5Y9K1bT0iF6VGfLSVG+RzjA3KQSRkqOa/ZT+F3wd+L3xWt/hN8Y/EfiPwxda84tNF1Cw8ry4745Cw3EciFsO2FUqRhsKRhty/tmf8E9vG37Kem2/jjSvESeLfBV1ci2e/W1NvPYSsPkWeMMw2tggSKcEjBCllDf8Eh/irqPhT9ou7+GEl639l+OdJmAti3ym7tVM0cgHqIluB77vaqqtF+B/wAJvhPo3jHVvh94E0vR9R8QpfXep30UO64uZJi8jhpWy+zexIjztXgACv8AgjH/AMl38bf9iif/AErgqv8AgsJ8R9Z8VfGXwZ8D9IeSS00XTVvWt0bHnX13IUUEdCVjjTaT081/U18GPhhofwY+FXhf4XeHoIktPDumxWxeNNvnygZlmP8AtSSF3J9WNf8ABU/4Vad8Qf2UtX8UfYkk1fwLdwanZTBPnWNpFhuE3dQpjk3kdCYlPYV/wSH+JV74v/Zsv/BGpXLSy+CdcltrXcclbWdRMg/CRp/oMDoKqv2K/wBof4Vfs4ftS/FfxT8WdcuNM07U0vrK2khspbktN9vV8FY1JA2oxyeK/wCHpn7F/wD0UXVP/CfvP/jdf8PTP2L/APoouqf+E/ef/G6/4emfsX/9FF1T/wAJ+8/+N18FPjl8N/2hfBr+PPhbrE+paNHeyWTTTWklswmRVZhtkAOMOvOMc1VVVVVVVVVVVVVVVVVePv8AkUr7/tn/AOjFqq8W6wfEXirWfEBcsdT1Ce5LHOT5kjNnnnvWix/8ID+z9o9iv7t9G8K2tpGOm11t0jXuf4sd/wAa8PaW2t69p2jqM/bLqOI44wGYAn8qVVVQiAAKMAAYxVeItA03xRo1zomqRb4LlMEjqh7MvoQea8WeGNS8H67caHqS/vITmOQDAlQ/ddfY/ocjtXwb+Iw8W6V/YmrXGdXsEGWY83EQ4D/7w4B/A9ziqqv22vhd/wAKs/aH8R2drbiLTPED/wBsaeAuBsnJMigdgswlUD0A6V/wTh+KA8bfAxvBV9ciTUfBN4bUKTljay5khY/Q+ag9BGKqqqqqqqqqq8If8izp3/XAVVVVVVVVVVVVVVVVVVVVVVeNWH9mwpjrOD+Smv2opVj+HNshBJl1WJR7Hy5D/Sv2N7AWfwbW4C4+3arcTfdxnASP8f8AV1VftweCwRoHxBto+fm067IH1kiP/o0E/Sv2K/Gi6R451HwXczYh1618yBSf+W8OWwPrGZCf90VWvWwutHuogMkR7h9V5/pWl3AtNRtrgtgJINx9uh/Sqqqqqqqqqqq/4emfsX/9FF1T/wAJ+8/+N1/w9M/Yv/6KLqn/AIT95/8AG6/4emfsX/8ARRdU/wDCfvP/AI3X7b/7fP7Mfxs/Ze8a/DH4eeNL+/8AEGs/YPsdvLo9zAr+Vf28z5d0CjCRueTzjHev+CWf/Jl/hH/r/wBS/wDSyWqqqqqqqqqqqqqqqqqqqr9u/wD5M/8Aip/2AJP/AENK/wCCd37bX7O37PfwBuPAXxS8W3um6zJ4hurxYYdKuLkeU8cKqd0aEdUbjOeK/wCHpn7F/wD0UXVP/CfvP/jdf8PTP2L/APoouqf+E/ef/G6/4emfsX/9FF1T/wAJ+8/+N14c1/S/Fnh3S/FOhztNp2sWUN5ZyMhQvFKgdCVPIyrA4PIqqqqqqqqqqqqqqqqqq+K3+v03/ck/mtV+3VrH9i/sr+OJUfEl1Fa2qDnnzLqJWH/fBY/h71+xvo5139p34e2SqWMWqfacAH/lhE82eP8ArnX7Rmqi28M6do6Ph727MhHqsa8/q6/lX7P2lfbvHLagy5XTrSRwcdGbCD9Gb8qqvix8PIvHGiedZRqNWsVLWz9PMHeMn0Pb0P1NaPq+q+Fdbh1Swdre9sZT8rAjkcMrD0PIIrwd4r07xnoFvrmnsB5g2zRE5MMg+8p/p6gg96qq+JngfTviZ8PvEPgDVCFt9e06a0LkZ8pmUhJB7q21h7gVNF4g8AeL3hZnsNc8N6kVJU/Nb3MEnb3V0/SvhV490/4pfDfw38QdNKCLXdOiuWjRsiGQjEkefVHDIfdTVVVVVVVVVVov/IYsP+vmP/0IVVVVVVVVVVVVVVVVVVVVVUF9Y3bNHa3kEzIPmWOQMR9cVVVVVVVVVVVVVVVVVVfF74u+BPgX4EvfiR8SNUmsNB0+SKOeeK2edlaRwiYRAWOWYDpxX/D0z9i//oouqf8AhP3n/wAbr/h6Z+xf/wBFF1T/AMJ+8/8Ajdf8PTP2L/8Aoouqf+E/ef8Axuv+Cmn7Tnwd/aV1zwDffCLxDc6pFoNpfR3xnsJrXy2leEpgSKN2QjdOmKqqqqqrP/j6i/3xVVVVVVVVVVVVVVVVVVVVVVVeJPDmmeKtIm0bVYt8MoyrDho2HRlPYj/EdCa8Z/DXxF4Omd57druwB+S8hQlcf7Q/gP149CapEaRljjUszHCqBkk+1fD34Nahqc8Wr+LLd7SyQhktXGJJsf3h/Cv15Pt1pI4441iiRVVBhVUYAA7AVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV+0Lp2q6x8AviXpOhKzale+D9WgswoOTM9nKqYxk/eI7V8IPiDP8J/ip4S+JltYpfP4X1m21D7K7bROIpFYpntkAjPbOa+En7UHwH+N2gWuveAPiVoty08e6TT7m7S3vbY45WWByHUjnnBU4yCRzXx5/a6+Bv7PnhS917xZ440q71OGFmstCsbxJb2+kxlUWNSSoJIy7AKoOSag8Q+IdW8fx+K9MULrt5rAvbcQ/Li5abeu3nj5yMc/jX/BQdNJk/Y0+KC6ysbW40uIoJASPOFzCYTx380Jj3xX/AATiiupv21fhklnu8wXV4x2tg7BYXBf8NobI9Kqq8Vf8ivrH/XhP/wCizX/BGP8A5Lv42/7FE/8ApXBVftkF9S/4KkxWF/CHt18S+GIUVlwHjaCyJ+vLMKr9tCGKf9k74spNGkijwnfMAy5AKxEg/UEAj0Ir/gibNdtY/GG3ct9mSXRWjyuBvIvQ+D34VOO3HrVV+w98AfhJ+0N+1T8WfDHxg8Jf2/pumx393awfb7i08ub7eqbt0EiMfldhgkjnpX/Dsr9h/wD6Ij/5cmq//JNf8Oyv2H/+iI/+XJqv/wAk1/w7K/Yf/wCiI/8Alyar/wDJNfCD4KfDL4CeE38DfCbwz/YWiSXb3bWv2ye6zM6qrNvnd25CKMZxx061VVVVVVVVVVVVVVVVVePv+RSvv+2f/oxar4nawvh74a+LPEDOFGmaHe3JY448uB2zzx2rwzpDeIPEmk6BGCW1K+htlAznMjhewPr6Gv2gdUXT/A8OlQkKb+6RNgGPkQbj+oSvgXpI1P4g2s7LuTT4JLhhjvjYP1cH8Kqq+Kfw+h8daEfsqIuq2QLWkh43esZPocfgcH1rTNR1bwtrcWoWbva3+nzHhlwVYcMrD06gj614L8Xaf410CDWrBgrEbJ4c5MMgHKn+YPcEVVV/wU4+Fw8RfC/R/ihYWwa78J3vkXjgc/ZLghcn12yiIDPTzGr/AIJ9/FH/AIV5+0Fp2hXtyI9M8ZQnSpgxwBOTut29yZFEY/66mqqqqqqqqqqvCH/Is6d/1wFVVVVVVVVVVVVVVVVVVVVVXjdgLW1TPWQnH0H/ANev2rJwvg7SLXzOZNT37cddsbjP/j361+zDYDT/AIGeGIyuGmjnmYlcZ3zyMP0IFVXxn8GDx98Mdf8ADccIkuZbRpbQY58+P548emWUL9Ca8G+JbzwX4t0nxTZAmbSryOfZ03hT8yH2ZcqfrWn31pqlhbanYTLLbXcKzQyDoyMAVI+oIplV1KMMgjBBq6gNtcy2zHmJyvPscVplwLvTre4zkvGM/XHP61VVVVVVVVVV/wAOyv2H/wDoiP8A5cmq/wDyTX/Dsr9h/wD6Ij/5cmq//JNf8Oyv2H/+iI/+XJqv/wAk1+3R+wv+yv8ABv8AZX8b/Ej4b/Cz+yPEekfYPsV7/beoXHlebf28T/JLO6HMcjrypxnIwQCP+CWf/Jl/hH/r/wBS/wDSyWqqqqqqqqqqqqqqqqqqqr9u/wD5M/8Aip/2AJP/AENK/wCCcX7Gn7Nnx7/Z7uPHHxY+G/8Abutx+IrqzW6/ti+tcQpHCVXZBMi8F25xnnrX/Dsr9h//AKIj/wCXJqv/AMk1/wAOyv2H/wDoiP8A5cmq/wDyTX/Dsr9h/wD6Ij/5cmq//JNaBoWk+FtB03wxoNr9m03SLOKzsoN7P5UMSBEXcxLHCqBkkk9yaqqqqqqqqqqqqqqqqqvit/r9N/3JP5rVf8FM9ZXTv2d7PTlcb9V8SWsG3IyVWKaQnHoDGv5j1r/gnLoy6p+03pt6UDHSNJvboEgfLmPycj/v9j8a/aG1X7Z4xt9MR8pp9moK+juSx/8AHdlfs46UIdC1XWWX5rq5WFSR2jXPH4v+lVVfHb4c4L+ONFt+P+YjEi/gJQP0b8D6mvhj4+uPAmurLMzPpl2Ql5EOcDs4H95c/iMjvkW1zb3ttFd2kySwzIHjkQ5DKRkEH0Iqqr/got8Lj4E+PMniyxtvL03xrai+QgYUXKYjnUe5ISQ+8tf8EwfigdZ8CeIPhNf3Ja48PXQvrBWb/l2nOHVR2CyqWPvNVVVVVVVVVVov/IYsP+vmP/0IVVVVVVVVVVVVVVVVVVVVVX7B/wDyUvx5/wBeq/8Ao81VVVVVVVVVVVVVVVVVfFT4UeAPjZ4Lu/h58TdA/tnw/fSRyXFn9qmtt7RuHQ74XRxhlB4YZxzX/Dsr9h//AKIj/wCXJqv/AMk1/wAOyv2H/wDoiP8A5cmq/wDyTX/Dsr9h/wD6Ij/5cmq//JNf8FQv2bvgt+zrr3w+svg34L/4R+HXLO/kv1/tG6u/OaN4QhzPI5XAdvu4znmqqqqqqz/4+ov98VVVVVVVVVVVVVVVVVVVVVVVVEBhtIB46Gr3wJ4N1GQzXfhnTnkY5ZhAFJPuRjNaZ4Z8O6KwfSdEsbVwMb4oFVv++sZqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr9vv9g/xZ8B/GGq/Ev4daDcal8NdWne5DWkJc6CzHLQzAfdhBPySdMYViGALVX/AATd/Yk8W/E74haF8c/H+hz6b4F8NXUd9pwuoija1dRndF5akfNArhWZz8rY2DOWK/8ABRj4WfHP41/AmP4b/BLw5a6s19qUdxrMcl/HbSPBB86RxiQqrFpdjcsMeWBzu4/4Jjfsk/F7wD+0bqvjr4s/DvWvDMHhTRpY7JtQtGiW5urk+WPKf7siiITbipIBZfWqqvFX/Ir6x/14T/8Aos1/wRj/AOS7+Nv+xRP/AKVwVX/BU7RtT+GP7ZPhz4rWcDMmp6bp+p28gLKGuLOUxsmc8ELFCeMY3jvWga5pfijQdN8S6JdJc6dq1nFd2cyHIkikQOjAjsVYH8a/4KReOLPwP+x148aa4RLrXooNIs42YqZXnmUOox3EKzNj0U1/wRt8D3OjfAvxb46uoWj/AOEl8RCC3LZ/eQ2sKgMO2PMllX6qfaqr/glx/wAnj/Gb/rwv/wD05R1VVVVVVVVVVVVVVVVVVVV4+/5FK+/7Z/8Aoxar9rrWDoX7NHxEvVkKmTRJbXIJH+vIhxx6+Ziv2adGGv8A7Qfw500oHU+JbKV1PdI5lkYdf7qGv2jtVFx4g0zRkfIs7VpWA7NI2Mfkg/Ov2btJ22+s646/feO2jOOmAWb/ANCSqqq+Ovw481H8caLBl0H/ABMYkHUDgSge3Ab2wexNfDXx5c+BNeS6Znk0+5wl5CO69mA/vL1H4jvVpd2uoWkN9ZTpNBPGHikQ5DKRkEfhVV8RPBemfEbwJr3gTWFBtNd0+a0diMmMupCuPdThh7gVqNj4g+H/AIvudNuDJYa54c1JonK8NBcwSYJGe4df0r4RfEGx+K3wx8NfEPT9gTXNOjnkjU5EU2Nssf8AwCRXX/gNVVVVVVVVVXhD/kWdO/64CqqqqqqqqqqqqqqqqqqqqvGP7Sf/AAiXx00j4K/8IX9r/tWW1j/tL+0vL8rzjjPleUc4/wB4Z9qrxw3FmgI/jJH/AHzX7WMxXSfDtuCMPcTtg9flVB/7NXwcsP7M+E/g+zKhWGi2rsu3GGaNWI/Mmqqv2gvBR8C/FjXdKihCWl1Mb2zAGB5UvzYHsrbl/wCA1+yb40/4Sv4SWmmXE4e88PStYyAn5vLHzRHHYbGCD/cNV4qthBrMjAYEyhxx7YP6g14PuBNpRgJ5gkIA9jz/AFNVVVVVVVVVVVVf8FNf+TH/AIk/9wr/ANOtpX/BLP8A5Mv8I/8AX/qX/pZLVVVVVVVVVVVVVVVVVVVV+3f/AMmf/FT/ALAEn/oaV/wSD/5NQu/+xuvv/RNvVVVVVVVVVVVVVVVVVVVVXxW/1+m/7kn81qv+CqusGDwr8PvD4c4vdQvbnbzg+THEufT/AJb/AK1/wSz0bz/il4x8QBM/YvD6Wu7njzp0bH/kD9K+IGq/23411nUg+5ZLt1jPqiHav/jqivhPpI0f4faNblcPNB9ofPUmQlx+hA/CqqpoYriF4J41kjkUq6OMhgRggj0xXxW+HsvgfW/Ms42bSb5i1q/Xyz3jJ9R29R9DXwL+IwtJk8Ea1cfuZm/4l8jn7jnrH9CeR78dxVVX/BQ/4Wjx/wDAG58S2Nr5mp+CrgajEVX5jbn5Lhf90KVkP/XEV+x98UR8Jf2gfC+vXV15OmajP/ZepknC+RcYTcx/uo/lyH/cqqqqqqqqqq0X/kMWH/XzH/6EKqqqqqqqqqqqqqqqqqqqqr9g/wD5KX48/wCvVf8A0eaqqqqqqqqqqqqqqqqqqqq/4LW/8jP8KP8Arw1T/wBGW9VVVVVVn/x9Rf74qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqdEkQxuoZWGCrDIIrxx+wv+yR8RL2bUfE3wK8O/aZzull00S6YztnJY/ZXjySep6nnPU14H/YX/ZI+Hd7DqPhj4FeHTc253RS6kJdSaNs5DD7U8mCD0PUcYxikRI0WONQqqMKoGAAPSqqqrxV/wAivrH/AF4T/wDos1/wRj/5Lv42/wCxRP8A6VwVX7e/7KD/ALVXwhTTfDrW8PjPwvM97oEkzBFmLKBLbMx+6soVOegeNCSBmv2c/wDgof8AEb9kLw4fgB8f/hRrWoDw0Wi00SSm0vbKPcf3LrIpEkQOdjAjC8Dcu3b8TfiL+0L/AMFUPiXongv4e+BJ/D/gXQ7gsGlZpbWxdhh7q7uAoVpNmQkajIBYKCWZj8Hvhb4a+Cnwx8OfCzwjGRpnh2xW3jkZQHnf70kzgcb5HLO2OMsaqv8Aglx/yeP8Zv8Arwv/AP05R1VVVVVVVVVVVVVVVVVVVV4+/wCRSvv+2f8A6MWq/wCCiWsDTP2Xtbsd4H9rajY2uD3xOs2P/IPt0r9gfRjrH7VHg9yhaPT0vLqTAPG21lCn2+dk618VNV/tn4gazcq+5IrgwJ6YjATj2ypP418FtJ/sr4eacWXbJel7lxj+8cKf++QtVVVJHHLG0UqK6OpVlYZBB6gj0r4t/Dx/BOtfabCNjpN+xa3OM+S3eMn27eo+hr4G/EYabcJ4L1m4xa3D/wCgyOf9VIeqH2Y9Pf61VV/wUc+Fw8D/AB0HjGwtvL07xtZi7BAwouosRzqPqPKkJ9ZTX/BL74ojVPCHiP4RahdZuNEuBqWnIx5NvMdsqqPRZApPvNVVVVVVVVVV4Q/5FnTv+uAqqqqqqqqqqqqqqqqqqqqr4xf8n0eEf+vrS/8A0Kq8btm4tU9EY/qP8K/a0m3XHhi32/cS6bOeuTEP/Zf1rw9YDStA0zSwu37HZxQ7cYxtQDp26VVV+274KF1ouiePrWImWxlNjdED/lm+WjJ9AGDD6yCv2NvGg8P/ABKm8L3MxW28SWpjQE4HnxZdCf8AgPmD6sKrxvbZS1u1HQlGP6j+RrwVcBLye1J/1kYYfVT/APXqqqqqqqqqqqqr/gpr/wAmP/En/uFf+nW0r/gln/yZf4R/6/8AUv8A0slqqqqqqqqqqqqqqqqqqqq/bv8A+TP/AIqf9gCT/wBDSv8AgkH/AMmoXf8A2N19/wCibeqqqqqqqqqqqqqqqqqqqqvit/r9N/3JP5rVf8FT9YWf4k+CvD4cE2Why3JXjI86crn1/wCWH6V/wTPiXw98P/it44YDcptYo2I/ihinfA47mVe/p0rTrKbVdStdOgOZrydIkJ5+ZmAH6mra3itLaK0gXbHCgRF9ABgfoKqqrxV4Z03xfodxoWppmOZco4HzROPuuvuP15HQ14i0DVPCWuXGjaihjubV/ldcgOP4XU+hHP8A9evhD8RU8aaP/Z2pTD+2NPQCbJwZ06CQe/ZvfnuKqtX0qw17Sb3Q9Vt1uLLULaS3uYWHEkbqVZT7EEivip4Dv/hf8RvEfw+1Is02hajLbLIwx5sYP7uT6OhVh7MK/Zc+KP8AwuH4FeFfGNzcibUvsgtNTJOW+1Qfu5GPoX2iTHo4qqqqqqqqqtF/5DFh/wBfMf8A6EKqqqqqqqqqqqqqqqqqqqqr9g//AJKX48/69V/9Hmqqqqqqqqqqqqqqqqqqqqv+C1v/ACM/wo/68NU/9GW9VVVVVVn/AMfUX++KqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqSSOKNpZZFREUlmY4CgdST2Fftg/8ABTP4q/EnxhqfhH4FeK73wj4HsJXt4L3Tn8m91XaSpnMw+eJG6oiFTggsSThdS8f+O9aumvtY8ba9fXLZ3TXOpTSueSTlmYnqSfqTX/CVeJ/+hk1T/wADJP8AGv8AhKvE/wD0Mmqf+Bkn+NeFyW8MaQzNkmwgyT/1zWqqqqqrWvDPhvxLHHD4i8P6bqqQnMa3tok4Q+oDg4qxsLHTLSKw02zgtLaFdsUEEYjRB6BRwBVVX/BLj/k8f4zf9eF//wCnKOqqqqqqqqqqqqqqqqqqqqvH3/IpX3/bP/0YtV/wVJ1k2/wf8K6Ar4N94jE5A4JEVvKMdemZR26gdK/4JtWsMPxq8Q+J7lFMOgeEbu5yccMZYR/6Dv5/xom51K9zzJcXUv4s7H/E1pOnx6VpVnpcONlnbpCuB2VQP6VVVVeJvDmm+K9EudD1SPdDcLwwHzRsOjL6EH/Doa8UeG9T8Ia7caJqSlZrdspIowJF/hdfY/p06ivg78RR4w0j+yNUnB1fT0AcseZ4+gf69AffB71Vf8FB/hd/wsL9n2/12yg36l4MnGqQkDkwAbLhfp5beYf+uQr9kn4pf8Ki+PvhbxLc3Qh027uf7O1Ms21fs0+EZmP91GKSf9sxVVVVVVVVVV4Q/wCRZ07/AK4CqqqqqqqqqqqqqqqqqqqqvjF/yfR4R/6+tL/9CqvGrA6jAg6iDP6n/Cvi58WPC3xF/aL/AOFMWhki1fwvd21rKkpBS7WdYZC8YP8AdMhRgf7oOSDxVVXxR8HxePvh9rvhN0VpL6zcW5b+GZfmiP4OqmtA1jUfCXiTT9ds1aO90i8SdEcFSHjYHaR+GCPrWh6xZeIdEsNe06Tfa6jax3EDeqOoYfoRXie2+06NPgZaLDj8Dz+ma0G4Frq9rITwX2H/AIFx/Wqqqqqqqqqqqqv+Cmv/ACY/8Sf+4V/6dbSv+CWf/Jl/hH/r/wBS/wDSyWqqqqqqqqqqqqqqqqqqqr9u/wD5M/8Aip/2AJP/AENK/wCCQf8Ayahd/wDY3X3/AKJt6qqqqqqqqqqqqqqqqqqqq+K3+v03/ck/mtV/wUh1g6p+0vc2JkLf2RolnagEn5chpsD/AL/Z/Gv2P4x4e/Y91i5JxN4l8WyIhxg+WkUIx16Zifnj730r4NaT/a3xD0sMm6O0LXD8dNinaf8AvrbVVVVXxb+HkfjbRPtdhEP7XsELW5AwZV6mM/XqPQ+mTWg65qnhPW4NX052iurSTlGBGezIw9CMgivCXijTvGOhW+t6a/yyjEkZOWicfeU/T9Rg96qv+Cn3wt/sbx54f+LOn2wFt4itDY37KvS6gHyMx9WiIUe0Jr/gl38URZ674n+D2oXGI9RiGraarHA82PEc6j3ZDG2PSJqqqqqqqqqrRf8AkMWH/XzH/wChCqqqqqqqqqqqqqqqqqqqqq/YP/5KX48/69V/9Hmqqqqqqqqqqqqqqqqqqqqv+C1v/Iz/AAo/68NU/wDRlvVVVVVVZ/8AH1F/viqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/aIXVm/Z/8AiaugBzqZ8HasLIIPm877HLsx77sV4Z8Paj4u8SaT4U0ZEe/1m+hsrVZG2qZZXCICew3MOa8A/wDBID9nLQ/D9tb+Pdc8UeJtaMa/a7qK8Wyg8zHPlRIpZV/3mY+/av8Ah03+x/8A9AXxR/4PH/wr/h03+x//ANAXxR/4PH/wqys4dPsrewtgRFbRLFGGOTtUAD9BVVVVVVVVVf8ABLj/AJPH+M3/AF4X/wD6co6qqqqqqqqqqqqqqqqqqqq8ff8AIpX3/bP/ANGLVf8ABVjWd+ofDnw8j48mHULqRc9d5gVT07bH796/YQQ6d4R+K+v8q8tvpmmwsc4bzZJnkH/fEX6ivhZpQ1j4gaLaldyx3AnbI4xGC/PtlQPxqqqqqviv8PovHGhmWzjUatYqWtX6eYO8ZPoe3ofqa0fV9V8K63Dqlg7W97Yyn5WBHI4ZWHoeQRXg7xXp3jTQbfW9OYDeNs0ROTDIPvKf6HuCD3qtS06x1jTrrSNStkuLS9geC4hfpJG6lWU+xBIr4ufD6++FXxM8S/DzUAxk0PUJII3cYMsWcxSf8DjKN/wKv2UPigfi98BfCviq5uTNqUNqLDU2ZssbmD927N7uAsn/AAMVVVVVVVVVXhD/AJFnTv8ArgKqqqqqqqqqqqqqqqqqqqq+MX/J9HhH/r60v/0Kq8Ytu1cLnO2FR9OTX7QXxB1qy/ay8c+P/C+pyWepaT4wun0+7iPKNbzlI3HYjEY4PBBwetfsRfto+Gf2sPBJt9QFrpPj7RIlGs6Qj4Ey8D7VbgnJiYkAjkxsdpJBVmqqr9pnwUfBXxe1iKGIJaau39o2uBgYlJ3gDtiQSDHoBX7HvjM+JPhcfD1zPvuvDl00ABOT5D/PGT7ZLqPZBU8KXEEkD/dkQqfoRimV4JSp4eNsfQirWdbq2iuFxiVAw/EVVVVVVVVVVVVf8FNf+TH/AIk/9wr/ANOtpX/BLP8A5Mv8I/8AX/qX/pZLVVVVVVVVVVVVVVVVVVVV+3f/AMmf/FT/ALAEn/oaV/wSD/5NQu/+xuvv/RNvVVVVVVVVVVVVVVVVVVVVXxW/1+m/7kn81qv20NYGu/tQ/EG9DhvK1FLXP/XCCOHHH/XPFfB6I6T+zJ8MtCPyy3EN/qU6/wB7zbyVYz0H8Cf5GK/Zu0oPfazrbp/qoo7eNiP7xLN/6Cv51VVVVXx1+HHku/jfRLf5HP8AxMYkH3WPSUD0PRvfB7k18LfH83gbXVNw7Npd4Ql3GOdvpIB6j9RkelQTw3UEdzbSpLFKgeN1OQykZBB9CKr9sX4XD4tfs++J9DtrbztT0uH+1NMAGW8+3BbC+7x+ZH/wOvgl8Rrn4SfFjwv8RLZn26NqCSXKp96S3b5J0Hu0TOv41aXdrqFpBf2NxHPb3MayQyxtuWRGGVYEdQQQaqqqqqqqq0X/AJDFh/18x/8AoQqqqqqqqqqqqqqqqqqqqqqv2WvgN8QPhL408U654wtrGO11aAJbG3uRKSRKW5A6cGqqqqqqqqqqqqqqqqqqqqv+C1v/ACM/wo/68NU/9GW9VVVVVVn/AMfUX++Kqqqqqqqqqqqqqqqqqqqqqqqr6KaeyngtpmileJljdeCjEcEfjXgHxlaeNtBj1CMol3DhLuEH/Vv6j/ZPUfl2NVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVXUlvFbSy3bIsCRsZTJ90KBzn2xX7S/hr4WfCz42yeIv2cPijpXiHw5LfDUtIOntJ5ujyBw6wMXUBgjY2MC2VA3YYGvhv/AMFmPhbJ4Us0+Lfw18WW3iOKMJdN4fit7m1nYDBkXzpomTPXYQ2M43HrX/D5P9mD/oRfij/4K7H/AOTK/wCHyf7MH/Qi/FH/AMFdj/8AJlafexalYW2owKyx3UKyoHGCAwBGcd8Gqqqqqqqqq/4Jcf8AJ4/xm/68L/8A9OUdVVVVVVVVVVVVVVVVVVVVePv+RSvv+2f/AKMWq/4Ke6yL748aLpEb5TTPDMIYekkk87Hv/d8v0/lX7I0Ytvg7qjoBu1DxK/mcDOILaLZ74/0iT26+9fs5aSLjxDqWsMuVs7QRKSOjSN1/JD+dVVVVVXx1+HGQ/jfRLf8A7CMSD8pQP0b8D6mvhj49n8C68s0ru+m3ZCXkQ547OB6rn8Rkd6trm3vbaK7tJklhmQPHIhyGUjIIPoRVf8FQfhaNK8ZeHPi5p1riDXbY6dqLquALiEZiZv8AaeIlfpD27/8ABLz4ojTvFHiX4QahdYh1iAappyMcATxYSZR6s0ZRvpCaqqqqqqqqrwh/yLOnf9cBVVVVVVVVVVVVVVVVVVVVXxi/5Po8I/8AX1pf/oVV4ynih1W4mnkVI4YgXZjgKAuSTXiXWJPEXiPVdflBD6nezXLA9QZHLH19a8DeN/iB8DviLYeMPCt5eaB4o8NXhKrLGY3ikU7XiljbGVI3I6MOQSCK/Y9/a48G/tYfDxNb09rfTfFmlosev6GJctayHgSx55aF8ZVucHKk5Uk1Vftr+Chqfg/SvHFrFmbRbk29wQP+WM2ACT7Oqgf9dDX7IvjMeGPitFo1zNttfEVu1owJ4Eo+eM/XIKj/AH6rxFbC21i5QDh23j8Rk/rmvCtwLjRolJy0LFD/ADH6EVVVVVVVVVVVVf8ABTX/AJMf+JP/AHCv/TraV/wSz/5Mv8I/9f8AqX/pZLVVVVVVVVVVVVVVVVVVVV+3f/yZ/wDFT/sASf8AoaV/wSD/AOTULv8A7G6+/wDRNvVVVVVVVVVVVVVVVVVVVVXxW/1+m/7kn81qvjPrJ8RfGDxxr2/cNQ8R386n2e4cgDk8YIHWvCcf2bwF4O0wcCx8NadEV/uObdHkHGf+WjvXwI0kad8P4Lplw+o3Ek5yOcA7B+GEz+NVVVVVPBDdQSW1xEksUqFHRxkMpGCCPQivil8P5vA2ulbZGbS7wl7SQ87fWMn1H6jB9a+BfxH8l4/A+tXHyOf+JdK5+6x6xE+h/h98juKqv2ovhcfg/wDHTxV4Mt7cxact2bvTBtwv2WYeZGF9QoYxk+qGv2CfiifiT+zzo9jeXXm6n4Sc6PcgnnZGAYD9PKZFz3KGqqqqqqqqtF/5DFh/18x/+hCqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr/gtb/yM/wAKP+vDVP8A0Zb1VVVVVWf/AB9Rf74qqqqqqqqqqqqqqqqqqqqqqqquriO0tpbqU4SGNnY+wGT/ACrwN4wvPBeuxanAWe3f5LqEH/WR55/EdQfX2JrT9Qs9VsYNS0+dZre4jDxuvcH/AD07VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVeKEaTw1q0calmaxmCqBkk7D0q1+Bnxsv4FubH4O+N7iJshZIvD106nHXBCYr/hQPx3/wCiJ+Pf/Cbu/wD43X/Cgfjv/wBET8e/+E3d/wDxuv8AhQPx3/6In49/8Ju7/wDjdeGY5IvDekwyoyOljCrKwwVIQZBHaqqqqqqqqq/4Jcf8nj/Gb/rwv/8A05R1VVVVVVVVVVVVVVVVVVVV4+/5FK+/7Z/+jFqv2/dY/tf9qfxbEr7o9OisrWMnPa1iZhj/AHnYfr3r9mS2e3+EOnSN925uriVRz/z0KE/+OfpX7Pmkiy8FS6m6/PqN2zKcdUQBR/48HqqqqqqmhiuIXgnjWSORSro4yGBGCCPTFfFb4ey+B9b8yzjZtJvmLWr9fLPeMn1Hb1H0NfAz4j/Ypk8E61cfuJm/4l8rn/VuesZ9mPT347jFftc/C4fFz4A+KfDdtb+bqVlbHUdMAXLfaLfLhV93UPH/AMD7V8HPiHefCf4peGPiJZFi2iahHNMinBlgPyzRj03Rs6/8CqwvrPVLG31LT7lLi1u4VlglQ5WRGAKsPYgg1VVVVVVVV4Q/5FnTv+uAqqqqqqqqqqqqqqqqqqqqrVvhF8Odc8Z2vxC1XwvBceIbJo2t75pZAyGP7hChtvHuKr9pXXx4Z+GfxG8SCXy20zw3fToc4O5LRiuORySAAMjkivhx4eHi74h+FvChj8wazrVnZbP73mzImP8Ax6v+Cif7BEHxz0u6+M3wm05IviBplsTfafDEANehQdOOftKqMKf4wAh6KR8Ifi38Qv2eviVYfEHwJfPpuu6NK0csFxGdkyZxLbzxnBKNjBXgggEEMAR+y3+094C/am+G9t408J3EdrqlsqRa5ory7ptMuCOVPQtG2CUkxhgD0YMoqvHfha18b+DdZ8J3YXZqdnJCrMM7HI+R/qrBW+oq0udU8L69DdxBrbUdIvFdQw5imifIz9GWvCfiG08W+GNK8T2JHkapZx3CLnO3eoJU+4JIPuK8bWwW4trpR99Ch/A5/rXgm4+a5tCeoDqP0P8ASqqqqqqqqqqqr/gpr/yY/wDEn/uFf+nW0r/gln/yZf4R/wCv/Uv/AEslqqqqqqqqqqqqqqqqqqqq/bv/AOTP/ip/2AJP/Q0r/gkH/wAmoXf/AGN19/6Jt6qqqqqqqqqqqqqqqqqqqq+K3+v03/ck/mtazqUOjaRfavOAY7G2kncE44RSx9ew9KlkuL65eaQtLPPIWYgcsxPPA9Sasrdre0t7Qnc0Uapx3IAFeGdKGh+HdM0cAA2dpHE3uwUBj+JzVVVVVV4t8L6d4x0K40PUl+WUZjkAy0Tj7rj6fqMjvWv6HqnhTW59I1BTFdWcnDKSM91dT6EYINfCT4hx+NtFFpfSqNXsEC3Cngyr0EgHv0PofQEVX/BUX4W/bNE8L/GHTrYGTT5G0nUnVefKfMkDE9lVxKv1lWv+Ca/xQHhD403fgC+uQlj41sTHGpOB9rgDSRHPbKGZfcsv0qqqqqqqqrRf+QxYf9fMf/oQqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/4LW/8jP8ACj/rw1T/ANGW9VVVVVVn/wAfUX++Kqqqqqqqqqqqqqqqqqqqqqqqvixrq6F4Hv2WTE16v2WIepfhv/HQxqvgx4/Gi3y+FtWnxY3j/wCjOx4hlPb2Vv0P1Jqqqqqqqqr9on9sj4A/swWBb4meM4zrLx77bQNNAudQnGMg+UCBGp7PKUU84Nfshf8ABQjwX+2B441rwP4Z+HutaBNoulHUHnv7qKRZEEqR7QE5BzID+FVruuaL4Y0W98ReItVtdN0vTLd7i8vLqURRW8SDLO7HhVABJJr4p/8ABav4S+GNdm0n4V/CvWfG9nbuUOpXeojSIZsfxRKYpZCv++qH2r9mb/gqx8EPj74osPAHinQb/wCH/iXVJVhsI766S6srqVsBYluQqFXJyAHjUE4AO4har9pv/god8Mv2e/Gtv8JfD3hTW/iL8Q52QN4f0NcG3LgFEkk2sfMYEERojtjltuVz8Iv+CoPg/wAT/Euy+EPx5+D/AIn+DHibU2RLOPXmZoC7nEayNJFDJFvPCs0ezPVhVfte/tVeHf2S/hrb+NNQ0B/EmtatqEdhougQ3f2eS/lY5fD7JCqomSSEbkovVga8E6r4j17wfo2teL/DUfh3Wr+yinvtIS9+1ixlZQWhMuxN5XOCQoGQcZHJqqqqqqqqqqqqqqqqqqqqrxj8Rfh78O7WC9+IHjzw74Zt7ltkEus6pDZJK3HCmVlBPI4HrVpqmmahpsWsWGo2tzYTxCaK6hmV4njIyHVwcFcc5ziv+F/fAj/otngL/wAKS0/+OV/wv74Ef9Fs8Bf+FJaf/HK/4X98CP8AotngL/wpLT/45X/C/vgR/wBFs8Bf+FJaf/HKjkjmjWWGRXR1BVlOQQehB9Kqqqqqqqqr/glx/wAnj/Gb/rwv/wD05R1VVVVVVVVVVVVVVVVVVVV4+/5FK+/7Z/8Aoxar9p3WV179of4jaijh1HiS8gRgQQyxSmMEEdQQgr4I2A074T+GLcDG6xE2P+ujF/8A2avAeknQ/BmjaWybXis0Mi46Ow3N/wCPE1VVVVVV4p8Nab4u0O40PU0zFOvyOB80Tj7rr7g/4dDXiPw/qfhLXLjRtRUx3Fq/yuvAcfwup9CMH/69fCH4ip400f8As7Uph/bGnoBNk4M6dBIPfs3vz3FV+1b8Lf8AhUPx58VeE7a2EWmy3RvtLCrhfs0/zoq+yEtH9YzX7AHxRHxG/Z70zSby436n4PlOk3AY8mJAGgbHp5bKn1jaqqqqqqqqvCH/ACLOnf8AXAVVVVVVVVVVVVVVVVVVVVVVf8FC/EB8P/sy/E6+SUh7i1islAOC3n3EULAcj+Fzn2B4PSv2H/Dy+J/2ufhRprKD5PiS3vcHj/j2zcev/TL/APXVf8FIP2AR8RLbUv2gvgro5Piq3j83X9DtYedWRfvXEKj/AJeFHLL/AMtAMj5xh/2fvj/8Qv2aviPZ/ET4fXwjuIR5N/YT58jULckF4JVHY4BBHKsARgiv2e/2g/h5+0r8OrP4h/D7UQ8bgR6hp8rD7RptxgFoZVHQjs3Rhgjg1VftWeCz4S+LuoXsEW2019BfwkDje3Eo+vmKzfRhX7GHjT+2/h7eeELmfdceHro+UpPIgmyy/XDiT6ZFeLrYT6QZQMmBw39P614YuBb6zACeJcofxHH64qqqqqqqqqqqq/4Ka/8AJj/xJ/7hX/p1tK/4JZ/8mX+Ef+v/AFL/ANLJaqqqqqqqqqqqqqqqqqqqv27/APkz/wCKn/YAk/8AQ0r/AIJB/wDJqF3/ANjdff8Aom3qqqqqqqqqqqqqqqqqqqqr4rf6/Tf9yT+a1+0NrI8P/Ab4hauH2vB4ZvxEfSRoHVO4/iK14LsRqfjHQtNIB+16nbw4PfdIo/rXgDSf7b8a6LppXcsl2jSLjqiHc3/jqmqqqqqqq+L/AMOk8aaP/aOmwj+2NPQmHAwZ06mM+/ce/wBTXh3X9U8Ja5b6zprmO5tX+ZGBAcdGRh6EcH/GvCviXTfF2h22uaY/7udcPGT80Tj7yH3B/Pg9DXxv+G9r8XPhN4o+HlwiGTV9PdLVnxiO5X54XPsJFQ/QV4c1zW/h/wCMtN8R6eGttW8O6lHcxLIMFJoZAwDD/eXBH1rwX4r0vx34Q0Xxpokgex1ywhvLf5slVkQMAcdxnBHYgiqqqqqqqrRf+QxYf9fMf/oQqqqqqqqqqqqqqqqqqqqqqv2Wvjx8QPi1408U6H4wubGS10iAPbC3tREQfNK8kHngVVVVVVVVVVVVVVVVVVVVV/wWt/5Gf4Uf9eGqf+jLeqqqqqqz/wCPqL/fFVVVVVVVVVVVVVVVVVVVVVVVXxl8Xp4j8R/2ZZTB7HSt0alTw8p++304Cj6E96rpyDyK+EPj8eJ9L/sTVJwdUsEADMeZ4xwG+o4B/A9zVVVVVVVV4S/4JO/CHw14h8Z/FH4va/feO7y5u9QvdL0yV3itbeMu7wmdt3mXEoXYTlgmSwKuMMf+CJf/ACX7x1/2J5/9K4Kr/gs9+0Hq1q3hj9mbwzfyRw6hbjWfEKQk5uF8wpaQHHUbo5JCvciI9hX7Ff7Jngv9l34R6LpcGg2beNdRso5/EervErTy3LqGeFX6iKMnYqjAO3cRlia/4LAfsqeDdL8Haf8AtK+AfDtnpGqWupJZeJVsoBCl3HNnyrlwuB5iyBULdWEq5Pyiv2Ev2kLr4wfsdaZ8SvF169/rXhC2utP16Zn3PPLZoHEjHqXeBoXYkcs5Pev+CPnh7/hZnj34v/tL+NympeKr2/jtobuVQzQvcmWe6Zc5Klv3KjGMKGHQ4H/BZz4a6Fr37PGh/Ew2UK634W8QRW8d1tw7WlyjrJFnqR5iwsPTafU1+yn4+vfiF+zD8NfH3iK933t/4XtJNQuZm2+ZNHEEllY9tzIzE+9fBiOX9v79trVP2hdUja5+EnwVuP7P8HQyR/ur/UAQwnwevzYuCeCB9lBHBqqqqqqqqqqqqqqqqqqqqqq/4KW6v431P9sLxraeMp7sw6cbeHRoZS3lxWPkI0flA8BWLMxx/Gz981/wSX1jxvqH7NPxG0zW5bqXw3pt/ImiNMWZY3a2LXMcZPAQExNtHAaRj1Y1VVXhX/kV9H/68IP/AEWtVVVVVVVVV/wS4/5PH+M3/Xhf/wDpyjqqqqqqqqqqqqqqqqqqqqrx9/yKV9/2z/8ARi1Xi3WD4i8Vaz4gLljqeoT3JY5yfMkZs8896/Z+8EX/AIq07RPC9iYZH0u0tBNAz7XNuAqyOoPB28ZGc/MMA84K7flIxjjGMYqqqqqqqvi38O4/G+i/arCJRq9ghNu3TzV6mMn37Z6H0BNaHrWqeFNbg1bT3aG7s5OUcEZ7MjD0IyCK8JeKNO8Y6Fb65prjbKuJIicmKQfeQ/T9Rg96/wCConwuGo+GvDPxf0+2zPpMx0rUXUcmCTLwsfZZA4+swr/gm98UR4K+N8vge/uvL07xtZm3VWbCi7hzJCT9V85AO5kWqqqqqqqqvCH/ACLOnf8AXAVVVVVVVVVVVVVVVVVVVVVqfxQ+H2j+K7bwLqfiuxt9fu2jWCwdiJJC/wBzAx3qv+CruvnS/wBnmfTlk51vxXbWpAPVVWab16ZhX8cV/wAEp/Dv9t/tj6BqW3P9gaRqN71xjdAbf1H/AD8e/wBO4qv+Ck3/AAT/ADcjVP2jfgdokj3LM1z4q0G1izuGCXvYFHOeMyIAc5LjGGz+y9+074//AGWviNb+M/CF01zplyyRa3o0khEOpW4PKkfwyKCSkmMqT3UsrfBj4y+Afj58PdN+JXw41cXulagpVkcbZrSZQN8EyZOyRcjI6EEEEqwJr9s7wWNc+Hdp4ut483Hh26/eED/lhMQjfk4iPsM1+yt40Xwh8XtOtrmfZaa8jafLk8b3wYvx8xUX/gRq/txd2U9tjJkjZRn1xxUErW88cyfeicMPqDSOssayIcqy5Bqqqqqqqqqqqv8Agpr/AMmP/En/ALhX/p1tK/4JZ/8AJl/hH/r/ANS/9LJaqqqqqqqqqqqqqqqqqqqv27/+TP8A4qf9gCT/ANDSv+CQf/JqF3/2N19/6Jt6qqqqqqqqqqqqqqqqqqqq+K3+v03/AHJP5rX7dWsf2L+yv44lR8SXUVraoOefMuolYf8AfBY/h718G7Vrv4n+HY487o7vzRjt5al//Za/Z98DX93d2vj9Gt7nTvssqK0T5eC4yFZHU8g7SxzyCCDnnFVVVVVVVXx1+HP2SZ/G2i2/7iZv+JhGg+454EgHoeh98HucfCn4hS+B9bEV5IzaTfMFuk6+We0gHqO/qPoKiliuIUnglWSORQyOpyGBHBB9MV+3p8Lj8Nv2h9avLO28vTPFiDWLQgcB5SROM+vnLI2Owda/4Jo/FIeK/hBqHw4v7rffeDbz9wrNkm0uCzpjPXbIJh7AoPSqqqqqqqrRf+QxYf8AXzH/AOhCqqqqqqqqqqqqqqqqqqqqq/YP/wCSl+PP+vVf/R5qqqqqqqqqqqqqqqqqqqqr/gtb/wAjP8KP+vDVP/RlvVVVVVVZ/wDH1F/viqqqqqqqqqrWfF+k6HdiyvVnMhQONiAjB/H2r/hY/h7+5d/9+h/jX/Cx/D39y7/79D/Gv+Fj+Hv7l3/36H+Nf8LH8Pf3Lv8A79D/ABr/AIWP4e/uXf8A36H+Nf8ACx/D39y7/wC/Q/xr/hY/h7+5d/8Afof41/wsfw9/cu/+/Q/xrS/GujavfRadaLcCWXO3fGAOASe/oKqqqqqqr4v+P28L6YNE0uXGp38Z+YHmCI8FvqeQPxPaqqtF1LU9Cvode0t2jks5VxIBxkg/KfYgMMdxmvCXiay8XaFb61ZfL5g2yxk5MUg+8p/z0IPeqqqqqqq8W/8AIqaz/wBg+f8A9FtX/BEv/kv3jr/sTz/6VwVX7W7x/ET/AIKyWPhjUXdrB/GXhjSFSVc7ImWzWQYB6F5JDwR97sar/gonpVtrP7FXxWs7oApHpEdwMrn5obmKVf8Ax5BX/BF5o/FHwW+L3w/vy5spdTgLjAxi6tXifHfOIR+nvX/BJnxpbfA/4t/FT9lT4nXUejeJpdRjbToLkiIXNzbGSKeNC2NzMrROgGdyqzDjr/wWO+K2kN8MPC37PHh64/tLxh4q16C7bS7T97MttGHVAUXLBpJpIwgx82x8ZxX7T3ivxd8Dv2WPhF+wh8Mh5/xU+IeiWOh3ENtIAbW3KKl5ISB8qyymSMMR9zzmzlM14j/aF+DP/BNn4deC/wBl7wL4U1T4hfEAWcbjQ9IO2W7uZiS808gV2RpZN2yNUdwuwbQu0nw5/wAFTdT8JeO9L8E/tW/s1eKPhHDrbD7Hq13JLLEqkgb3jkhiby1JG50LkZ5UYq1urW+tYb2yuYri3uI1khlicOkiMMhlI4IIIIIqqqqqqqqqqqqqqqqqqqqr4sfs3/Av453Fne/Ff4Z6P4hu7BPLt7qeNo50TOdnmRlXKZydpOMk8cmrDw78NvhJ8PJ9I03SdG8L+DdBsJpbiGKNba1tbdVZ5ZHxgAbQzMx5PJJ71/wjf/BJH/n5+DP/AINU/wDjlWP7D37GmqWNvqWn/AjwncWt3EssEqROVkRgCrA7uhBBr/hhD9j/AP6IB4W/78v/APFV/wAMIfsf/wDRAPC3/fl//iqt7eG1t4rW2jCRQoEjQdFUDAA/AVVVVVVVVVX/AAS4/wCTx/jN/wBeF/8A+nKOqqqqqqqqqqqqqqqqqqqqvH3/ACKV9/2z/wDRi18TtYXw98NfFniBnCjTNDvbkscceXA7Z547VptmdQ1G0sF5NzOkQH+8wH9a8F+KtR8DeJ9O8T6Q2J9PlDBM4EidGQ+zKSPxrR7/AETxroNj4j0qUPbahAssMg+8AR0YeoOQR2IIq7064tDll3J2cD/OKqqqqqqr46/DgxSP440WD5HP/EwiRfut0EoHoeje+D3Jr4XfECbwLrgNw7Ppd4Ql3GOdvpIB6j9RkelfF7wFpvxj+EniPwI00Tw+INMdLWbOVWXAeCT3CyKjfhWjapr3w/8AGFlrNosllrXhzUkmjWRSrQ3EEgOGHqGXBFeA/F+mfEHwToXjfRnBs9d0+G8hGclRIgbafcEkH3Bqqqqqqqq8If8AIs6d/wBcBVVVVVVVVVVVVVVVVVVVVXxi/wCT6PCP/X1pf/oVV/wWA1/ytA8AeGVf/j+1XULxkB6eSkaAn/v+f1r/AIIweHjdfGfx74rC8ad4XSyJ9PtFzG/r/wBO3p/9eqr/AIKP/wDBP0+C5tQ/aB+Bugu3h+ZnuPEuiWyZ/sxictdQKOfIJJLoP9WfmHyEiP8AY/8A2t/Gf7KHxDj1vTpLjUfCmpuqeINDEmFuoxwJI88LMmcq3GeVJwxr4Z/ErwX8YPA2k/Eb4fa1FqmhazD5ttOgwQQSGR1PKurAqynkEEV4p8PWXizw1qfhnUVBt9TtJLeQ4+6GUjI9xwR7iry11Twtr81nKWttS0i7ZGI6xyxvjj6MteAvFVv448F6L4ttgoXU7NJXRTkRyYw6f8BYMv4VrNt9l1S5gA4EhKjHY8j9DXh64+1aNbOTkouw/wDAeP5AVVVVVVVVVVVX/BTX/kx/4k/9wr/062lf8Es/+TL/AAj/ANf+pf8ApZLVVVVVVVVVVVVVVVVVVVV+3f8A8mf/ABU/7AEn/oaV/wAEg/8Ak1C7/wCxuvv/AETb1VVVVVVVVVVVVVVVVVVVV8Vv9fpv+5J/Na/4KZ6yunfs72enK436r4ktYNuRkqsU0hOPQGNfzHrX7O1mLr4mW02AfslpNKMjpldn/s9fswfEkeEPGX/CL6nc7dL8QMsYLHiK5HEbewb7h9ypPSrvS7W6ywGx/wC8o/mKutNubUFmTcg/iX/PFVVVVVVXNtb3ttLZ3UKSwzIUkjcZDKRggj0Ir4n+ALjwJrphhVn0y7JezlPOB3Qn+8ufxGD34+BXxGxs8D61c/8AYOkc/nET+q/iPQV/wUq+Fo8X/Bmz+INjbb7/AMF3u+RlXJNpOVjkH4OIW9gG9TX7CnxRPwy/aH0KK7uvK0vxQDo96GPygykeS3oCJljGT0Vm9aqqqqqqqtF/5DFh/wBfMf8A6EKqqqqqqqqqqqqqqqqqqqqr9g//AJKX48/69V/9Hmqqqqqqqqqqqqqqqqqqqqv+C1v/ACM/wo/68NU/9GW9VVVVVVn/AMfUX++KqqqqqqqqqvGPhPV9b1ZbywSIxiFU+d9pyCf8a/4V54k/55W//f2v+FeeJP8Anlb/APf2v+FeeJP+eVv/AN/a/wCFeeJP+eVv/wB/a/4V54k/55W//f2v+FeeJP8Anlb/APf2v+FeeJP+eVv/AN/a/wCFeeJP+eVv/wB/a8L+DNb0nXbXULtIRFFv3bZMnlCBx9SKqqqqqqYqqlmYAAck9q8Ya9L4n8SX+syOSs8p8oH+GMcKPyA/GqqvhroK+J9J8T6KEDSvZJLBxyJUYlcfXJH0NfATXpLPxBc+HpZD5N/CXjUnpInoPdd2foKqqqqqqrxb/wAiprP/AGD5/wD0W1f8ES/+S/eOv+xPP/pXBVftnRxfC7/gqvpvjbUYlg07/hJ/DOuK0hIUxRi1ErZJ6eZBL9Onaq/4KReILbw3+xP8T7qd0Bu7CCyiVv4mnuoYsD3wxP4Zr/giT4Znsvgr8QPF0lvtj1XxNHZxyHgv9ntlY/gDcdfXPpx+0z+wL+z7+1PqaeKPG2m6no3iiKJYv7d0K4W3uJUUYRZQ6PHIAAACV3AcBgK+GP7B37KX7HKan8fPEF1rHiHUvDFpLqEmu+J7pLg2CxoWaSGJERPMwPlLBnBwFIJr9gXwb4i/aL+MHjL/AIKA/FPTmik1u4l0zwJYT5YafYx5jaRO3CjydwxlvtDY+ev+CcUcXx6/bp+Nf7QvitRfahpTznS/PG42n2md4otmeR5dtAYR6K2K/wCCq/w/0bxp+xn4r1q9tIH1Hwjc2epadO6/NExuY4ZQDjI3RSyDHQnbnpkf8E1PGeo+OP2KvhvfatPJNd6bbXOll37x21zLFCBz0EKRL9Qaqqqqqqqqqqqqqqqqqqqqr4leDLf4j/DnxV8Pbu5NvB4n0S80uWYDJjW4heIsB7B818Svhx4w+EnjjV/h3490eXTdb0S5aC5gcHBx910P8UbDDKw4ZSCOtfshfGvwV8cPgL4R1/wprFpNeWGk21nq9gkwMthdRRhJI5EzuUFkJUkDcpVhwaqqqqqqqqqqr/glx/yeP8Zv+vC//wDTlHVVVVVVVVVVVVVVVVVVVVXj7/kUr7/tn/6MWv2utYOhfs0fES9WQqZNEltcgkf68iHHHr5mK+G1mL/4geHbYjIOpQswPcK4Y/oKOn3o08ap9mf7IZjCJsfL5gAO364INfsp/FE6dPN8OdXmLQTlrjTiTyj4zJGPYgbgPUN61FNDcx74nV1Iq80aKXL2xEb/AN09D/hU8Etu/lzIVb3qqqqqqnghuoJLa4iSWKVCjo4yGUjBBHoRXxT+H8vgbXSLVWbS70l7SQ87fWMn1GfxGD64+BXxGEDx+B9auPkdv+JdK5+6x6xH69V98juK/b9+Fv8Awrn9oTVNVsrYR6Z4wjGrWxVcASuStwufXzVZz7SLX7A/7XfhfwtoVj8B/iRdrpsSXMh0LVZpMQDzXLG3lJ+587MVc/Kd2DtwM1VVVVVVXhD/AJFnTv8ArgKqqqqqqqqqqqqqqqqqqqq+MX/J9HhH/r60v/0Kq/4K26+Lz4u+DPDavkWHh57vb6Ge4df/AG3Ff8EbvGOm+GvE/jTw1qEaRv4uNrHa3DH/AJbWqyuI/Qblnc/VQO9VVTwQXUEltcwpLFKhSSORQyupGCCD1BHGK/4KKfsBzfBjUbv40/BvRHfwBePv1TToAWOhTs2MqvUWzEjB52MdpwCtfsSftn+J/wBlHxwIb97zVfAOsyga1o8bgmNuALqAHgSqAMjIEijaSMKy+DfGPhj4heFNK8b+DNZt9V0TWrVLmxvIDlZY2HBweQR0KkAqQQQCCK/a98Fnwz8VZNct4dtn4jt1uUIHAmXCSj65Csf+ulfsT+MzqXhDVvA91PmXRrkXFsrHnyZc7gB6B1Yn/roK8Z23l6jFcAcTR4P1H/1sV4KuN1tcWhPKOHAPuMH+VVVVVVVVVVVV/wAFNf8Akx/4k/8AcK/9OtpX/BLP/ky/wj/1/wCpf+lktVVVVVVVVVVVVVVVVVVVX7d//Jn/AMVP+wBJ/wChpX/BIP8A5NQu/wDsbr7/ANE29VVVVVVVVVVVVVVVVVVVVfFb/X6b/uSfzWv+CqusGDwr8PvD4c4vdQvbnbzg+THEufT/AJb/AK1+y7Z+Z4o1jUAP9Tp4iz/vyKf/AGSpoLuxlQTRyQS7ElTcCp2sAysPYggg+9fBH4rnx34MtrjU5BLqNhi3viPvFwOJCO+4YOfXcO1Q3EFym6GRXHt2/CrvSLa4+aMeU/qo4P4VdWFzaH94mV7MORVVVVVV4y8J6d400CfQ9QGN43QygZMMgHyuP5EdwSO9azpGq+Fdbm0u/Vre9sZeGUkcjlWU+h4INeFtb0b43fDrV/BHifa097p8tjqUYABkikQp5qj1wfwb8K8WeHNa+HfjXVfCupM0Gq+HdRltZHjyuJYXK7lPXGVyD6YNfsq/ta+Ev2htBh0e8mj0zxtp9uDf6bIwH2kKAGuIOgZCTyvVCcHjDGqqqqqqtF/5DFh/18x/+hCqqqqqqqqqqqqqqqqqqqqq/YP/AOSl+PP+vVf/AEeaqqqqqqqqqqqqqqqqqqqq/wCC1v8AyM/wo/68NU/9GW9VVVVVVn/x9Rf74qqqqqqqqqqqqqqqqqqqqqqrxXcNaeF9Yuoyd0NhO644ORGT/Sqqq/Z60yRLTWNXdTsleOCM47qCW/8AQlrSrYaH8bVtYAVVdVkVAo6K+cD6YbFVVVVVVV4t/wCRU1n/ALB8/wD6Lav+CJf/ACX7x1/2J5/9K4Kr/gsl+zVrPjDw3oP7RvhDSZb248K2zab4iSFCzpYFzJDPgfwRSPKGIzgTAnCqSP2M/wDgov8ABL4p/CHRdP8Ai58UNA8I+O9Cs47TWE8QajHYx3zxqFF1FLKVR/MA3MgO5WLDG3azf8FN/wBsLw7+0heeHP2Y/wBni6l8W2/9rx3F/e6YhlTUb3Bjt7W3x/rVBkZiwypYpgnaTX7HXwGH7N37O3hH4WXWxtXtLY3WsyIwYPfTsZJgCOCqFhGpHVY1Peq/b48YeIP2kPjL4N/4J+fC7U2j/te4i1Tx9fQfN9hso8SJE/YEKBMVOMsbcZ+YivBng/w78PvCOj+BvCWnR2GjaDZRWdjbRjAjijUKo9zgZJ6kkk9a/ZB8RaR+xl/wUH+KfwS+J19BoOj+MJpYtGv7x/KhkzOZ7Alz8oEkMrrnOBJhM5r/AIK2fHTwp4R/ZpvvhFZ61a3Hinx9dWsEFhBMHnitI5knknKg5CN5Sxgn7xk4zgkfsO/CbVPgl+yn8PPh7r9p9m1e201rvUYWTa8U91K9w8b8feTzfLPpsx2qqqqqqqqqqqqqqqqqqqqqtfuprHQtRvrZws1vaSyRkjOGVCQcfUV8af2rPjf+0LBFbfFvxFpOti1P+iynw5p8FxbLnOyO4jgWZUJ6rvwe+a0jXdb8PXX27QNZvtNudpXzrO4eF8HqNykHHtX/AAtT4of9FI8Uf+Dif/4uv+FqfFD/AKKR4o/8HE//AMXXhh2k8NaS7uWZrGEkscknYOaqqqqqqqqr/glx/wAnj/Gb/rwv/wD05R1VVVVVVVVVVVVVVVVVVVV4+/5FK+/7Z/8Aoxa/4KJawNM/Ze1ux3gf2tqNja4PfE6zY/8AIPt0r4DWZvPilo5IytuJpW46YiYD9SK+HPw0tfE3wEOiXaLHcavLLewSMP8AVTA7Y2+hCDP+yx9aik1bwrrqyJvtNS0q6yARzHKjd/xFfD/xpB4x8M2HibTZBG1xHiaJTnypRw6H6HpnqCD3qz1mKbEdziN/73Y/4VLDDcx7JUV1I4z/AEq80WSPL2pMi/3T1H+NEEHaRgjsaqqqqrxb4X07xjoVxoepL8sozHIBlonH3XH0/UZHetf0PVPCetz6RqCtFdWknDKSM91dT6EYINat4J+H37Wvw4fwV8RbUDXdKQm21GEBbi2Y4AniJ7HADoflOBkD5SPj1+z54+/Z78WN4c8YWYms7gs2m6tAh+z30YxypP3XGRuQ8qT3BDH9kD9u6+8BGw+GPxm1Ca98NDbDp2sPl5tMHAVJe7wjseWQccrgLYX9jqtjb6lpd7BeWl1EstvcQSCSOVGGVZWGQykEEEcHNVVVVVVeEP8AkWdO/wCuAqqqqqqqqqqqqqqqqqqqqr4xf8n0eEf+vrS//QqmbZC7g42qTn04r/gpZrg1b9q7XNPD5/sXS7C0we26BZ/Qf89/f69h+yxPqHhnwbbeIdKuXtb5NXe7tp0OGR02BWH0KV8J/iDYfFL4f6P40sSitewAXMKn/Uzr8sieuAwOM9QQe9VVappem63pt1o2s2FvfWF9C8F1a3MQkjmjcbWR1OQykEgg8EGv+Cgn7CGo/s5a/P8AEv4cWMtz8M9WuQFQMZH0Od+kEhPJiZs+XIfZGO7az/sGftyax+y74o/4RPxjNd6l8N9bnBvbVCXfS5jx9qgXvx99B94AEfMoB/am0XRviT8FbL4g+Fr+21S30wx39neWkgljuLSYBWZGHBUgxvkdkr9mrxovgn4vaNcXE3l2eqMdPuecDbLgIT7CQRk+wNeM7bzdOiuFGTDJg+wI/wAQK8I3Hk6usRPE0bL+I5/pVVVVVVVVVVVf8FNf+TH/AIk/9wr/ANOtpX/BLP8A5Mv8I/8AX/qX/pZLVVVVVVVVVVVVVVVVVVVV+3f/AMmf/FT/ALAEn/oaV/wSD/5NQu/+xuvv/RNvVVVVVVVVVVVVVVVVVVVVXxW/1+m/7kn81r/gqfrCz/EnwV4fDgmy0OW5K8ZHnTlc+v8Ayw/Sv2TtJlu4tbkt498t5c21tEMdWG7gfi4/Sv2gPhfCvhHTfEWiwZl8PWsdpcKi/ftlAVWx/sn9GPpXwY8fHwF4ygnuptumahi3vgTwqk/LJ/wE8/TcO9RSvGRLDIQccFTVrrZACXa5/wBtR/MVHLDcR7onV1I7VdaNbzgtAfKf0A4/KrmyuLU4lj47MOQaqqqqr4xfDpfGGj/2tpcIOr6ehMYA5njHJj+vce+R3rwx4j1PwhrtvrWmuVmt2w8bcCRf4kYeh/Tg9RX7R/7KHgr9qPwyvxM+HTW2keNTD/rWASK/KgAw3IAyJFxhZBzjAO5dpW9sfHfwi8cG1vIdR8NeKfDt2GAJMU1tKuCrAjtjBBGQQQRkGv2RP24NF+MUVn8PfiVcW+l+NkQR29wcRwaxgAZTskx7x9D1Xuq1VVVVVaL/AMhiw/6+Y/8A0IVVVVVVVVVVVVVVVVVVVVVX7B//ACUvx5/16r/6PNVVVVVVVVVVVVVVVVVVVVf8Frf+Rn+FH/Xhqn/oy3qqqqqqs/8Aj6i/3xVVVVVVVVVVVVVVVVVVVVVVWr2X9paVeacCB9qt5Iuf9pSP61JG8TtFIpV0JDKRyCOtVVfDCbTLjwLpL6VAkMYh2yIv/PUHDk9+WBP4itJUa/8AHma4tyGitruRmZegEaFc/iwA/GqqqqqqqvrODUbG40+5BMVzE0UgBwdrDBx+Br9nf9h/4Dfsu+J9S8X/AAo07WbfUdVsPsNy19qLXKmLzFfAUgYO5F5qrq1tb61lsr23iuLe4jaOWGVA6SIwwysp4IIJBBr4jf8ABI39kLx/rs+u6bYeKfBrXMhkktPDmpRx2+49dsc8UwjGf4U2qOgAHFfs8fsIfs3fsy6iPEPw98ITXniNY2jXXdZuPtd3GrDDCM4VIsgkExopIJBJBxVV8D/2TPhz8CfHvjb4paNrniXxH4t8f3Jm1bVvEFzDNMoLl2ji8mKJY4yxBK4P3EHAUAV+0Z+yL8DP2ptOs7X4r+F3nv8ATVK2Gr2Extry2UnJRZBkMhOTscMoJJAB5r4Hf8Exf2VvgT4otPGml6JrXirW9OlEthdeJrxLkWkgOVkSKKOOMuDghmVipAIwRmqqqqqqqqqqqqqqqqqqqqqrULKLUrC506dmWO6heJyhwQGBBx74NWv/AAR9/ZUt4Fhl1v4hXLrnMsur24Zue+23A9uBX/DoP9lD/oI+Pf8AwcQ//GK/4dB/sof9BHx7/wCDiH/4xX/DoP8AZQ/6CPj3/wAHEP8A8Yqxs4dOsbfT7csYrWJYk3HJwoAGfwFVVVVVVVVV/wAEuP8Ak8f4zf8AXhf/APpyjqqqqqqqqqqqqqqqqqqqqrx9/wAilff9s/8A0Ytf8FSdZNv8H/CugK+DfeIxOQOCRFbyjHXpmUduoHSv2ZLBrv4gXMypuMGmvsAGTuZ0UY/AmvDWkpoPh3TNEQDFhZxQcdyqgE/pX7TXgD7DqEPj3TYMQXpEN+FH3ZQPkf8A4EBg+6ju1fs5eP8A/hHPEx8K6hPjT9bYLGWPEdwOFP8AwIfKffb6VVpqNxaEKrbk/uN/T0q01C3vBiNsPjlG61d6fb3gy67X7OBzV3p1xaHLLuTs4H+cVVVVVXxf+Ha+NNG+36bCP7XsEJhwMGZOpj+vce/1NeHdf1TwlrlvrOmuY7m1f5kYEBx0ZGHoRwf8a1vR/hf+0X4Am8NeJrC11TT76INNaO4E9nIOjqR8yOp6MOvuCQf2n/2SvGn7OmtG8XzdZ8H3kpWw1hIsbD2inA4STnAP3XxkY5Vf2Tf2z/EvwGvYPCPi17nWfAlxL81tu3TaYWPMkGf4epMXAJ5GCTnwn4t8NeO/Dtj4s8IazbarpGpRCW1uoGyrqfY8qQeCpAIIIIBBFVVVVVXhD/kWdO/64CqqqqqqqqqqqqqqqqqqqqvjF/yfR4R/6+tL/wDQqv22WNwwxlYmIz9K/bD1weIf2oPiZfh93leILi0zjH/HuRBjoP8Annj/AB618FrEWHwv0GHbgvA0pyMZ3yM38iK/Yh+LB8NeL5/hnq1yF07xEfMsizYEV4q9B6eYgx/vIg71VVXiTw3oHjDQL/wt4o0i11TSNUt3t7yyuohJHPGwwysp6giv27v2Ite/ZZ8XHxH4bjm1D4c69dONKvDln0+Q5b7JOTk7gM7HP31Un7wYD9g/9ttfg003wM+L9zLffC7xQXtmmkcs2hPNlXlQc/uG3EyIBkHLrzuV9Usn0XV7mxhvY5zZzssVzbyBklCn5ZI2BIKsMMrA4IIINeBfFCfEn4UaX4mVlaa/sA04XtOnEgH/AG0RqsLj7LewXIP+rkBP0zz+lcEcGqqqqqqqqqqv+Cmv/Jj/AMSf+4V/6dbSv+CWf/Jl/hH/AK/9S/8ASyWqqqqqqqqqqqqqqqqqqqr9u/8A5M/+Kn/YAk/9DSv+CQf/ACahd/8AY3X3/om3qqqqqqqqqqqqqqqqqqqqr4rf6/Tf9yT+a1/wUh1g6p+0vc2JkLf2RolnagEn5chpsD/v9n8a/YB8MjVb7ShJHuV9bku2JH8MMasM+25MfjV1a299ay2V3Ck0E8bRyxuMh1IwQR6EHFfEvwTceAfF95oD7mt8+bZyN/y0hYnafqMFT7g1+zz4+HirwmPD1/OG1HQ1WP5jzJB0RvfGNp+g9aqKaWBw8MhVh6Va62rYS7XB/vqP5ileK4j3IyuhH1FXeiwy5e3by2/u44/+tVxaT2zbZoyPQ9jVVVVXxz+HH9nXD+M9Ft/9FuH/ANOiQf6qQ/8ALQD0Y9ff68fCb4hyeCNa+zX0rHSL5gtyvXym6CQD26H1H0FftE/sz/Dn9qDwql2l1a2viG1iI0rX7UCTb38uQKcSRE9jypJKkZIPxM+F/j34KeM5/CPjbSptM1SzYSQTIT5c6Z+WaGQY3KccEcggg4IIH7H37eseqfYfhb8ddWWO9+WDS/Elw+Fn6BYrk9n9JTw38WD8zAhgCpBBHBFVVVVVov8AyGLD/r5j/wDQhVVVVVVVVVVVVVVVVVVVVVQWFjasz2tnBCzD5jHGFJ+uKqqqqqqqqqqqqqqqqqqqq/4LW/8AIz/Cj/rw1T/0Zb1VVVVVWf8Ax9Rf74qqqqqqqqqqqqqqqqqqqqqqqvjB4Tk8O+Kpr6CLFlqjGaIgcK5++v5nP0YVVV4J+Js/g/wtqujwxtJczOHsWIysTMMOx+gCkDua+BfhWey0+58WagrefqI2Qb+pjByW/wCBMB/3znvVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV+29/wUz+I/wAI/jBqfwe+CulaLCvhopHqmq6lbm5ee4ZFdo4k3BVRQwUkgksDjAGT+xb+2FeftSfBXXvFesaNZaX4u8JM0GqW9ruNvKTEXinjViWVW2sChYkFDzgiv+Hvn7V//QP8Bf8Agnm/+P1/w98/av8A+gf4C/8ABPN/8fr/AIe+ftX/APQP8Bf+Ceb/AOP1/wAPfP2r/wDoH+Av/BPN/wDH60S8l1HRdP1C42+bc2scrhRgbmUE49uaqqqqqqqqr/glx/yeP8Zv+vC//wDTlHVVVVVVVVVVVVVVVVVVVVXj7/kUr7/tn/6MWv8AgqxrO/UPhz4eR8eTDqF1Iueu8wKp6dtj9+9fsB+Gv+Ei+KDI8e5EuLXcCOGVDJK4/KP9arxJ4f0/xToV74f1SPfbXsJjbjlT2Ye4OCPcV4i0LU/CPiG80LUAYrzT5ym5cjOOVde+CMMD6EV8IvHcfj/wbbajNIDqNr/o98vfzFH3voww3pkkdqoEggqcEdKs9ZkjxHdDev8AeHUf41FNDcx74nV1Iq80aKXL2xEb/wB09D/hU8Etu/lzIVb3qqqqr4u/B/UNU1JvE3hCyWaS4BN5aqwUl/76g8HPcevPOTTJq2g3+1hd6fe256ENFJGf0Iq2+Ll7qmiXPhP4h6JY+K9DvovKura8jG6RD2JwQ3Y8jOQOR1r9o79nXSfAdxN4z+F15c33hKZt0lldj/StJJx8rnpJGSfldckdGxwzfs1ftR+OP2cvEXmabI2p+Gb6UHU9FlkISToDLEf+WcoAxu6HADA4GPhX8WPA3xn8H2vjXwDrCXtjONssbYWa0lwC0UyZOxxkcdDkEEggmqqqqvCH/Is6d/1wFVVVVVVVVVVVVVVVVVVVVeNP2br3xZ8e9G+M8fiyC2h0qW1c6e1mWaTyTkgPuAGfpxWsMF0m8OM/uHH6V491z/hKPHPiLxKXDnVtWurvcD18yVnz/wCPV+xT8B9P+J+gahpWsxBbTSfC8NrDKV5hvZUAikx32+XISPp61qNh4g8BeK5rC58yw1rQL4qSpw0M8T8Mp9ioIPfg18HviPYfFf4eaT4xs2RZ7iIR30K/8sLlQBIn0zyPVSp71VVXjjwP4T+JPhPU/A3jnQrXWND1iAwXlncrlZFPcY5VgQCGGCpAIIIBr9tL9jfxX+yb46W3Ek+reCtakdtC1lo+Tjk28+BhZlHpgOPmAHzKvwQ/aBfwPAnhbxg1xdaKD/o06DfJZ56jHePvgcjsDnFfsCfFbQvGnhfWfC2ka5bajbWsi3tm0T7sI+FlUjquGCHBAOZDWq6bcabdPFLCyoWPlsRww7YP0rSvFF7YBYbgm4gHGGPzL9D/AENWGp2WpxeZaTBiB8yHhl+oqqqqqqqqqq/4Ka/8mP8AxJ/7hX/p1tK/4JZ/8mX+Ef8Ar/1L/wBLJaqqqqqqqqqqqqqqqqqqqv27/wDkz/4qf9gCT/0NK/4JB/8AJqF3/wBjdff+ibeqqqqqqqqqqqqqqqqqqqqvit/r9N/3JP5rX7aGsDXf2ofiDehw3lailrn/AK4QRw44/wCueK/4JweGdnhEa9JHzBZuoJ/vTTuQR/wCP9fpVftAeAP+Ew8IHVtPg36nogaaIKOZIv8AlonvwAw91x3r4d+MrrwH4tsfENuWaKNtl1Epx5sLffX+RHuBVleWupWUGoWMyzW9zEskMi9GVhkEfUEVVQ3E1u2+GQqfbvVrrUb4S6XYf7w6f/Wr91PH/DIjD6g1d6JHJ89q2w/3T0/+tU9tNbtsmjKntkdfpVVVXdpbX9rLZXkKTQToUkjcZDKRgg147+D3iTw1fXNzpGnTX+k7i0MkI3vGvXDqOeOmcY4zx0rRfEWveGrr7Voeq3FlLn5hG2A2P7ynhvoRXxF1TwR8efCB8F/Gbw0sxjBNhrWnKEurCUjHmIDwQeNy52sAMrwCPi58JNc+EniNtIvruHUtOnJbTtUt1Kx3cfqVPzRuOjI3IPQlSGP7H/7dl/8ADw2Pwy+Md/NfeFxth0/V3zJNpY4ASTu8AHT+JB0yuAun6hYatYW+qaXewXdndxLLb3EEgdJUYZVlYcEEEEEVVVVVov8AyGLD/r5j/wDQhVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV/wWt/5Gf4Uf8AXhqn/oy3qqqqqqs/+PqL/fFVVVVVVVVVVVVVVVVVVVVVVV4o8M6Z4t0eXRtUjyj8xyAfNE46Mvv/AEJHevGPw98Q+Dbhxe2zT2Wf3d5EhKMO2f7p9j+GetUAWIVRknoK+Hnwe1HWp4tV8T20lnpyEMsEgKyXHtjqqn16nt1yIoooIkggjVI41CoijAUDgADsKqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv21/+CYvjr40/GDUPi98GfEfh+3PiIRvq2mavNJb+VcKioZYnSNwyuFDMrYIbcQSGwv7Gn7Hkn7LXwY1zwbqGu2WreK/FTvNqt5bqy26N5RjihjLAM0aZY7ioJLtwBgV/wAOa/2nv+h6+F3/AIM7/wD+Q6/4c1/tPf8AQ9fC7/wZ3/8A8h1/w5r/AGnv+h6+F3/gzv8A/wCQ6/4c1/tPf9D18Lv/AAZ3/wD8h1otlLpujWGnTsjSWtrHE5QkglVAOPbiqqqqqqqqq/4Jcf8AJ4/xm/68L/8A9OUdVVVVVVVVVVVVVVVVVVVVePv+RSvv+2f/AKMWv+Cnusi++PGi6RG+U0zwzCGHpJJPOx7/AN3y/T+Vf8EvPDIubnxL4oePK2jeSrY6OVTH/jrSVVftM/D/APtDTYfHmmQZuLECG+Cjloiflf8A4CTg+zei18EfH3/CC+Mofts+zS9Txb3mTgJk/JIf90nk+hagQRkHPpVVDPLbuHhkKn271Z6zFNiO5xG/97sf8KlhhuY9kqK6kcZ/pV5oskeXtSZF/unqP8aIIO0jBHY1VVVa94Y8P+J7b7JrulW94gGFMifMn+6w5X8DXir9nVG33Xg/Vdp6i0vOR/wFx/Ij8a8SeENf8NyNZeI9Fmt1kygMibo5PUBhlW49DXxu/Zzk0zz/ABb8PbNpLTl7rTI1LND6tEOpXqSvUduOB8Evjn49+Ani+LxX4H1Lar7Uv9PmJNvfRA5KSL+eGGGXPB5OfgB+0T4B/aH8Jrr3hS6FtqVsqjU9HnkBnsZCO/TehP3ZAMH2IKiqqqrwh/yLOnf9cBVVVVVVVVVVVVVVVVVVVVVfHLxAPCfwY8c+KfN2f2R4dvrvdnG3y4HbP6Vp1m2oaha2Eed1zMkS49WIH9a/Ys8EL4T+CtpqktuI7nxFcyXhBXBWIfu4l+m1N4/66V+3X8K/7P1iw+LGk22INS22eqbV+7Oq/upD/vIpUnoPLXu1fsSfFT/hFPHU3w+1S626b4nx9m3HiO8UfLj03rlfchKqqqvip8LfBPxo8Bat8N/iDo8eo6NrEBjlQgb4m/hljYg7JEOGVuxAr9qD/gl58X/gjaah4z+HV0PHfg+zVppWhj8vULKIDJaWHpIoHVoyT1JRRXwy+K/xI+CniyLxj8M/Fl/4e1q2BQy25GHXPKSRsCsi5AyrgjjpX7OX/BXDwb4sitfB/wC0x4fi0G+kKxL4g02JpLKU9N00PLwn1Zd65JOEAqys/D/i3RLXxX8P9fsNb0i/j8y1uLO5WeKZfWORSQw/Gla6sZ8o0kM0Z91IrSvGCttg1VcHoJlH8x/hUUsU8aywyK6MPlZTkGqqqqqqqqv+Cmv/ACY/8Sf+4V/6dbSv+CWf/Jl/hH/r/wBS/wDSyWqqqqqqqqqqqqqqqqqqqr9u/wD5M/8Aip/2AJP/AENK/wCCQf8Ayahd/wDY3X3/AKJt6qqqqqqqqqqqqqqqqqqqq+K3+v03/ck/mtfGfWT4i+MHjjXt+4ah4jv51Ps9w5AHJ4wQOtfsPeGv7B/Z/wBCu5I9smpxJLnH8IjVcfg4eqIBGDz7Gvjf8P8A/hBPGMpsoNulapuns8DATn54x/uk8D+6Vr9mbx+L/TZvAWpT5nsQZrAseWiJ+ZP+Ak5Hs3otVVVb3U9q26CQj1HY/hVrrUMoCXI8tvUdP/rU6RTx7WCupHfkVd6GrAvaPg/3G/oamglgfy5oyrehFVVV4p+Gng7xcHk1TSkS6b/l6t/3cufUkcN/wIGvFX7P3iLTA1z4auo9VgHIibEUyj8Ttb8wfavGvgix13T7nwp4z0NzHIPnguIzG8bdmXPKsOxFfF34La58ML43Me++0Kd8W16F5Q9klxwre/Ruo7gfsm/tmeJvgHfQ+E/FDXGs+BLiX95aBt02nMx+aWAnt3MRIBPI2kknwh4w8MePvDlj4t8Ha1barpOoxCS3ubd9ysO4I6qwPBU4IIIIBFVVVWi/8hiw/wCvmP8A9CFVVVVVVVVVVVVVVVVVVVVV8Jf2iPBPxm1zVvD/AIX0vW7W40aMPO1/BEiMN5X5Skjk8juBVVVa/wCL/CfhWHz/ABP4n0rSUxkG9vEgz9NxGa8Tftg/Ajw4GSHxRPrM6ZzDplm8n5O21D+DV4l/4KA2a7ovB3w7mk/uz6neBMfWOMH/ANDrxL+2h8c9f3JY6xp2hxNxs06xXOP96Xew+oIrSP2sfj7o7AR+P5bpM8peWkE2fxZNw69iK0f9vH4sWQWPV9B8N6ig6t5EsMh/FXK/+O960f8A4KCaXJtTxB8NLqHH3pLPUllz7hWRcfTdWj/txfBLUgq3517SiR8xurAOAf8AtkznH4Vo/wC0h8C9dCmx+J2ix7un2yVrT/0cFx1rSfEvhzX0D6F4g03UVxkG0ukmH/jpPpVVVVVVVVVVVf8ABa3/AJGf4Uf9eGqf+jLeqqqqqqz/AOPqL/fFVVVVVVVVVVVVVVVVVVVVVVVTokiGORVZWGCrDINXvw58DahIZbnwxY7mOSY4/Kyf+A4rS/B/hbRZBNpWgWNvKOkiwguPox5qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv+CgX7eHxe/ZR+JXh3wb8O/Dvg/UbLV9CF/PJrVnczSLJ58keFMU8YC4QcEE5zzX/BPn9rD4i/tZeDfFniH4i6N4b0650HU4bW2TRbaaFGR4txLiWWQk59CB7V+0j8a7X9nX4LeIvjJe+H5dch8PfZd1hFciBpvPuorcYcqwGDKG6HOMd81+z/8A8FT9A+Pfxh8NfCKz+DWoaNN4jnkiS+l1tJlh2QvJkoIVJzsx1HWqqqqqqqqqqq/4Jcf8nj/Gb/rwv/8A05R1VV8WfiRonwf+HGv/ABM8RxXMumeHbM3VyltGHkKggfKpIBPI7iv+HwP7OH/QE8Yf+CyP/wCPV/w+B/Zw/wCgJ4w/8Fkf/wAer/h8D+zh/wBATxh/4LI//j1f8Pgf2cP+gJ4w/wDBZH/8er/h8D+zh/0BPGH/AILI/wD49X/D4H9nD/oCeMP/AAWR/wDx6v8Ah8D+zh/0BPGH/gsj/wDj1f8AD4H9nD/oCeMP/BZH/wDHq/4fA/s4f9ATxh/4LI//AI9X/D4H9nD/AKAnjD/wWR//AB6v+HwP7OH/AEBPGH/gsj/+PV/w+B/Zw/6AnjD/AMFkf/x6v+HwP7OH/QE8Yf8Agsj/APj1f8Pgf2cP+gJ4w/8ABZH/APHq/wCHwP7OH/QE8Yf+CyP/AOPV/wAPgf2cP+gJ4w/8Fkf/AMer/h8D+zh/0BPGH/gsj/8Aj1f8Pgf2cP8AoCeMP/BZH/8AHq/4fA/s4f8AQE8Yf+CyP/49X/D4H9nD/oCeMP8AwWR//Hq/4fA/s4f9ATxh/wCCyP8A+PV/w+B/Zw/6AnjD/wAFkf8A8er/AIfA/s4f9ATxh/4LI/8A49X/AA+B/Zw/6AnjD/wWR/8Ax6v+HwP7OH/QE8Yf+CyP/wCPV/w+B/Zw/wCgJ4w/8Fkf/wAerW/+Ctn7NmuaXNpcuk+Mo1m25ZdMjJGGB/57e1ftafFzw78b/jjrPj3wgt8uiz29rBZC+iEcu2OBA+5QSB+8345PGD3wP+CavhkaT+z9J4hePbJrurzupx96OI+WD/30HFVV7ZWuo2c+n30CzW9zE0UsbDh1YYIP1Br4i+DLrwH4tvfD0+5oo232srDHmwt91v5g+4Nfs/8Aj/8A4TDwguk38+/U9ECwybjzJF/yzf34BU+65PWqqqtNRuLQhVbcn9xv6elWmoW94MRth8co3WrvT7e8GXXa/ZwOau9OuLQ5ZdydnA/ziqqq8c/EXwx8PLOC78Q3Mga5crDBCm+R8dSBkcDjJPqPWv8Ahpz4b/8APHWf/AVf/i6uv2kvhbfW72t7YapcQyDDxy2SOrD3BfBrxT4g+Bush7jQl13SLkg4VLVXhJ91L5H4EAelfG74D6J4gjuvF3gGWOLVclp9NWIot3zgunQI+OSOjex6+BfHvjb4TeLrbxZ4M1i60bWtOfbuXK7gCN0ciHhkOACjDBr4e/8ABTb4OXXhDT3+JmjeJLLxIse2+j0yxjmtncfxxlpQwDddpHHTJ6n/AIeW/swf88fHH/gpi/8Aj1f8PLf2YP8Anj44/wDBTF/8er/h5b+zB/zx8cf+CmL/AOPV/wAPLf2YP+ePjj/wUxf/AB6v+Hlv7MH/ADx8cf8Agpi/+PVpX/BXH9m3StOt9Oj0jxjItum0M2mRgn/yNX/D4H9nD/oCeMP/AAWR/wDx6v8Ah8D+zh/0BPGH/gsj/wDj1f8AD4H9nD/oCeMP/BZH/wDHq/4fA/s4f9ATxh/4LI//AI9X/D4H9nD/AKAnjD/wWR//AB6v+HwP7OH/AEBPGH/gsj/+PV/w+B/Zw/6AnjD/AMFkf/x6v+HwP7OH/QE8Yf8Agsj/APj1f8Pgf2cP+gJ4w/8ABZH/APHq/wCHwP7OH/QE8Yf+CyP/AOPV/wAPgf2cP+gJ4w/8Fkf/AMer/h8D+zh/0BPGH/gsj/8Aj1f8Pgf2cP8AoCeMP/BZH/8AHq/4fA/s4f8AQE8Yf+CyP/49X/D4H9nD/oCeMP8AwWR//Hq/4fA/s4f9ATxh/wCCyP8A+PV/w+B/Zw/6AnjD/wAFkf8A8er/AIfA/s4f9ATxh/4LI/8A49X/AA+B/Zw/6AnjD/wWR/8Ax6v+HwP7OH/QE8Yf+CyP/wCPV/w+B/Zw/wCgJ4w/8Fkf/wAer/h8D+zh/wBATxh/4LI//j1f8Pgf2cP+gJ4w/wDBZH/8er/h8D+zh/0BPGH/AILI/wD49X/D4H9nD/oCeMP/AAWR/wDx6v8Ah8D+zh/0BPGH/gsj/wDj1f8AD4H9nD/oCeMP/BZH/wDHq/4fA/s4f9ATxh/4LI//AI9X/D4H9nD/AKAnjD/wWR//AB6v+HwP7OH/AEBPGH/gsj/+PV/w+B/Zw/6AnjD/AMFkf/x6v2jf+CovwU+KPwK8bfDnwhpfieHV/EmkS2Fu9zYpHGPMwrbmEpIGwt2NfBXwve+Nfi94M8J6bEJLrVNctYIUP8TGVcA+1aBotj4b0LTvDumJstNLtIrWBT2SNAq/oBXj7wbpfxC8G6t4M1hQbbVbZoi23JifqkgHqrBWHuBWt6Rr3gLxZd6NemSy1fQr0xl42KtHLG3DoevUAg/Q18EPiVbfFn4a6T4uR0+2tH5Goxrx5VygAkGOwPDgf3XWqqqq/sbTU7G402/gWe2uomimibo6MMMD7EEivjx/wS6+FHxSjuNS8JavPoesMpMcsyhxnsC6gEr0++HOOhFfHv8AYC/aO+AgutV1TwbP4g8O24ZzrOixm5jjjH8UqLl4h7sMD1r4R/H34v8AwK1U6t8LPHuqaG0jZntoZybe46f6yE5R+g5IyOxFeDP+Cut/eeHltPil4EtrvV4sKLyyj2bxjk8MAOf4duB6ntpX/BVH9nm4sIpNY0TxfaXmMSRQ2MUqA/7LeaCR9QK07/grB+zppb7rS38ahSfmjbSoirfh51f8Pgf2cP8AoCeMP/BZH/8AHq/4fA/s4f8AQE8Yf+CyP/49X/D4H9nD/oCeMP8AwWR//Hq/4fA/s4f9ATxh/wCCyP8A+PV/w+B/Zw/6AnjD/wAFkf8A8er/AIfA/s4f9ATxh/4LI/8A49X/AA+B/Zw/6AnjD/wWR/8Ax6v+HwP7OH/QE8Yf+CyP/wCPV/w+B/Zw/wCgJ4w/8Fkf/wAer/h8D+zh/wBATxh/4LI//j1f8Pgf2cP+gJ4w/wDBZH/8er/h8D+zh/0BPGH/AILI/wD49X7Yf/BSH4KfH39nLxd8JfCOl+JIdW137F9ne8sUjiHk3sE7bmEjEfLE2OOuK/4JZ/8AJl/hH/r/ANS/9LJaqqqqqqqqqqqqqqqqqqqv27/+TP8A4qf9gCT/ANDSv+CQf/JqF3/2N19/6Jt6qq/ac/ao8AfsraDoviDx9Z6rcW+uXclrbiwt1lYOibjuBdcDFf8AD4H9nD/oCeMP/BZH/wDHq/4fA/s4f9ATxh/4LI//AI9X/D4H9nD/AKAnjD/wWR//AB6v+HwP7OH/AEBPGH/gsj/+PV/w+B/Zw/6AnjD/AMFkf/x6v+HwP7OH/QE8Yf8Agsj/APj1f8Pgf2cP+gJ4w/8ABZH/APHq/wCHwP7OH/QE8Yf+CyP/AOPV/wAPgf2cP+gJ4w/8Fkf/AMer/h8D+zh/0BPGH/gsj/8Aj1f8Pgf2cP8AoCeMP/BZH/8AHq/4fA/s4f8AQE8Yf+CyP/49X/D4H9nD/oCeMP8AwWR//Hq/4fA/s4f9ATxh/wCCyP8A+PV/w+B/Zw/6AnjD/wAFkf8A8er/AIfA/s4f9ATxh/4LI/8A49X/AA+B/Zw/6AnjD/wWR/8Ax6v+HwP7OH/QE8Yf+CyP/wCPV/w+B/Zw/wCgJ4w/8Fkf/wAer/h8D+zh/wBATxh/4LI//j1f8Pgf2cP+gJ4w/wDBZH/8er/h8D+zh/0BPGH/AILI/wD49X/D4H9nD/oCeMP/AAWR/wDx6v8Ah8D+zh/0BPGH/gsj/wDj1f8AD4H9nD/oCeMP/BZH/wDHq/4fA/s4f9ATxh/4LI//AI9Xiz/gqt+zZ4kSKX+y/GSS20b+Wg02IByccE+acdBzippZ725eaVjLNO5ZjjlmJ54Hqa+FvhxfCPw38M+GwoVrDS4I3A/v7AW/8eJqq+LngOPx/wCDbnTYY1Oo2oM9gx4/eKPu59GGV/EHtXh3XdT8I+IbPXNPJivNPnDBGyM44ZGHXBGVI9Ca8N6/p/inQrLxBpcm+2vYRIgPVT3U+4OQfcVVVVW15cWpzDJx3U8g/hVrrFvPhJv3T+/Q/jUkUNxHskRXU9Min0K3ZiUmdR2HXFf2DD/z8P8AlX9gw/8APw/5V/YMP/Pw/wCVeMfij8MPAusNoHiLxNJHfogeSGG2eUxgjIDFQQCRg4POCD3Ff8NBfBX/AKGm9/8ABdL/APE1rfxg/Z48SWv2PXNUlvI8fKJdMlyvurbcqfcEV4g0X9l/X4bmxl8a6v8A2fdoUltJtLeZCD1HK5x9cn3r9pP9n7wt8OtTOv8Awj8TXXiPwtMpaVJ7N4p9Nb+6+R88Z7OOR0YcBm/Zy/ae8ffs7eIFm0S7e+8OXkytqeizMTFMOAXj/wCecu0YDDrgBsgDCf8ABS/9mJ41Z7bxwjFclTpUJ2n0/wBdX/Dy39mD/nj44/8ABTF/8er/AIeW/swf88fHH/gpi/8Aj1f8PLf2YP8Anj44/wDBTF/8er/h5b+zB/zx8cf+CmL/AOPVaf8ABTX9mK0u4LtLfxuzQSK4U6TFyQc/89vav+HwP7OH/QE8Yf8Agsj/APj1f8Pgf2cP+gJ4w/8ABZH/APHq/wCHwP7OH/QE8Yf+CyP/AOPV/wAPgf2cP+gJ4w/8Fkf/AMer/h8D+zh/0BPGH/gsj/8Aj1f8Pgf2cP8AoCeMP/BZH/8AHq/4fA/s4f8AQE8Yf+CyP/49X/D4H9nD/oCeMP8AwWR//Hq/4fA/s4f9ATxh/wCCyP8A+PV/w+B/Zw/6AnjD/wAFkf8A8er/AIfA/s4f9ATxh/4LI/8A49X/AA+B/Zw/6AnjD/wWR/8Ax6v+HwP7OH/QE8Yf+CyP/wCPV/w+B/Zw/wCgJ4w/8Fkf/wAer/h8D+zh/wBATxh/4LI//j1f8Pgf2cP+gJ4w/wDBZH/8er/h8D+zh/0BPGH/AILI/wD49X/D4H9nD/oCeMP/AAWR/wDx6v8Ah8D+zh/0BPGH/gsj/wDj1f8AD4H9nD/oCeMP/BZH/wDHq/4fA/s4f9ATxh/4LI//AI9X/D4H9nD/AKAnjD/wWR//AB6v+HwP7OH/AEBPGH/gsj/+PV/w+B/Zw/6AnjD/AMFkf/x6v+HwP7OH/QE8Yf8Agsj/APj1f8Pgf2cP+gJ4w/8ABZH/APHq/wCHwP7OH/QE8Yf+CyP/AOPV/wAPgf2cP+gJ4w/8Fkf/AMer/h8D+zh/0BPGH/gsj/8Aj1f8Pgf2cP8AoCeMP/BZH/8AHq/4fA/s4f8AQE8Yf+CyP/49XwW/bz/ZV+CviHWvEOl3Pj3UpNajCSR3GjwoI8OWyCs3viv+HwP7OH/QE8Yf+CyP/wCPVoX/AAUy+FPjLQ31TwR4T1nU5omKSQ3UkdpsbsGwZCARyDgj9a8S/t4fFDU90XhvQND0WM/ddke6lX/gTEL/AOOV4l/aA+NHi0Oms/EjWTG/3orWb7IjexWEKCPYirSy1fXr77PY2l3qN5Kc7Io2mkf3wMk14a/Zi+OnikI9l8Pb+0ibrLqJWzCj12ylWP4A14a/YE8bXpSTxZ420fTEIBKWcL3bj1B3eWAfoTXhv9hb4RaTsk1/UNc1yUfeSS4FvEfosYDD/vutX/Y1+AWpqRbeGLzTWI+/aalNn8pGcfp2rWv2AfAtxuPh7x1rliSOBdwxXQB/4CI+K1j/AIJ/+NYCw8P+PtEvQD8v2y3ltc/Xb5mOK1n9jP496UCbXw3Y6oqjlrPUYv0EhQn8q1n4H/GHQcnU/hn4jRF+9JHp8kyD6sgIH51c2l9plz5F5bT2lxGc7JUMbqR3weR0rR/ir8TfD4VdE+IXiKzRRxHFqcqp+K7sH8q0f9rf4+6PtUeOTexr/wAs7yyglz/wLYG/XvWj/t7/ABNtdqa14V8Oagi9TEktu5+p3sP/AB2tH/4KB6DMFXxB8N7+2wMM1nqCT59wGVMfTP41p/7b/wADr2Pfcza9YNj7lxp+4/8AkNmH60/7avwLA/d6vqTem7T5B/Q0/wC2v8FxxHqF43+9aSj/ANkNfF//AIKSfBz4X+EbrWrQTavrDRsNN0pYpImu5e2XZcIgyCzc4HQEkAxf8Fo/ioIUWf4MeFHlCgOyX1woLY5IHOB7ZP1qD/gtV40WFVuvgLokkoHzNHrsqKfoDESPzq1/4LYaqkIW9/ZytJZcn5ovFbRjH0Nq386tP+C11pJGTf8A7PEsLg/KsPiYSAj6m2XH5VZ/8FqPCkm/+0PgXqsOMbPJ1iOXPrnKLjt61af8FofhrI5F/wDB7xLCgX5TDdQSEn6FlwPxq1/4LN/BZ5Qt78MfGsUePvRR2sjZ+hnX+dQf8Fkf2e5plik8D+PYFbrJJp9qVX67bon8hX/BQ79rT4eftW6z4I1D4f2WrW8fhy1vIroahbLCSZWiK7drtn/VnPTtVVVVVVZ/8fUX++Kqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/4LOf8AJd/BH/Yoj/0rnr/gi3/yS/4jf9h62/8ARBr/AIKa/wDJj/xJ/wC4V/6dbSv+Cdv/ACeh8MP+v+5/9I56qqqqqqqqqqr/AIJcf8nj/Gb/AK8L/wD9OUdVVftSfDzxJ8Wf2e/HXw48HxW8ms6/pTW1klxKIkLllPzMegwDX/Dpr9r/AP6Avhb/AMHif4V/w6a/a/8A+gL4W/8AB4n+Ff8ADpr9r/8A6Avhb/weJ/hX/Dpr9r//AKAvhb/weJ/hX/Dpr9r/AP6Avhb/AMHif4V/w6a/a/8A+gL4W/8AB4n+Ff8ADpr9r/8A6Avhb/weJ/hX/Dpr9r//AKAvhb/weJ/hX/Dpr9r/AP6Avhb/AMHif4V/w6a/a/8A+gL4W/8AB4n+Ff8ADpr9r/8A6Avhb/weJ/hX/Dpr9r//AKAvhb/weJ/hX/Dpr9r/AP6Avhb/AMHif4V/w6a/a/8A+gL4W/8AB4n+Ff8ADpr9r/8A6Avhb/weJ/hX/Dpr9r//AKAvhb/weJ/hX/Dpr9r/AP6Avhb/AMHif4V/w6a/a/8A+gL4W/8AB4n+Ff8ADpr9r/8A6Avhb/weJ/hX/Dpr9r//AKAvhb/weJ/hX/Dpr9r/AP6Avhb/AMHif4V/w6a/a/8A+gL4W/8AB4n+Ff8ADpr9r/8A6Avhb/weJ/hX/Dpr9r//AKAvhb/weJ/hX/Dpr9r/AP6Avhb/AMHif4V/w6a/a/8A+gL4W/8AB4n+Fap/wSs/a20ixk1G80fwwIYcbiutITyQBxj1Ir/h3D+0z/0CvD3/AINl/wAK/Zu+HOp/CX4H+E/h9rcUEepaVaOLxYZA6iWSV5HwwAyMueaqq/aG8ADxX4T/ALe0+HdqWhq0gCjmSD+NfcjAYfQ+tfDTxrceAfF9nr0ZZrfPlXka/wDLSFiNw+owGHuBVrdW19aw3tnMk0FxGskUinIdWGQR7EEVVVQJBBU4I6VZ6zJHiO6G9f7w6j/GopobmPfE6upFT6NazPvQmP1C9P8A61LodmBy0jfU0uk2C9IMn3YmviR458LfC/w3Lr2rwxSSnKWdoMCS5k7Kueg7luw/AHxl4v1zx94iuPEOtzebc3LYjiQHZCn8MaL2Uen4nJJNWf7MHxZvLKC8bTbC3E8YcRz3io6ZGcMOx9u1P+zB8UU4aPR/p/aC0/7NHxNXjytLJ9rwH+lf8M2fE7/n107/AMCx/hXxX/YF8e+O4n1jR7TSbLXEX/WG7AS5xjAfA4OBgN+eR0/4dw/tM/8AQK8Pf+DZf8K/4dw/tM/9Arw9/wCDZf8ACv8Ah3D+0z/0CvD3/g2X/Cv+HcP7TP8A0CvD3/g2X/Cv+HcP7TP/AECvD3/g2X/Cv+HcP7TP/QK8Pf8Ag2X/AAqw/wCCU/7XGpWcV9a6N4YMUy7k3a2gOPpiv+HTX7X/AP0BfC3/AIPE/wAK/wCHTX7X/wD0BfC3/g8T/Cv+HTX7X/8A0BfC3/g8T/Cv+HTX7X//AEBfC3/g8T/Cv+HTX7X/AP0BfC3/AIPE/wAK/wCHTX7X/wD0BfC3/g8T/Cv+HTX7X/8A0BfC3/g8T/Cv+HTX7X//AEBfC3/g8T/Cv+HTX7X/AP0BfC3/AIPE/wAK/wCHTX7X/wD0BfC3/g8T/Cv+HTX7X/8A0BfC3/g8T/Cv+HTX7X//AEBfC3/g8T/Cv+HTX7X/AP0BfC3/AIPE/wAK/wCHTX7X/wD0BfC3/g8T/Cv+HTX7X/8A0BfC3/g8T/Cv+HTX7X//AEBfC3/g8T/Cv+HTX7X/AP0BfC3/AIPE/wAK/wCHTX7X/wD0BfC3/g8T/Cv+HTX7X/8A0BfC3/g8T/Cv+HTX7X//AEBfC3/g8T/Cv+HTX7X/AP0BfC3/AIPE/wAK/wCHTX7X/wD0BfC3/g8T/Cv+HTX7X/8A0BfC3/g8T/Cv+HTX7X//AEBfC3/g8T/Cv+HTX7X/AP0BfC3/AIPE/wAK/wCHTX7X/wD0BfC3/g8T/Cv+HTX7X/8A0BfC3/g8T/Cv+HTX7X//AEBfC3/g8T/Cv+HTX7X/AP0BfC3/AIPE/wAK/wCHTX7X/wD0BfC3/g8T/Ctf/YS+P/hv4k2Hwo1PTtEXX9ReFIETUlaMmXhMvjiv+HTX7X//AEBfC3/g8T/Cv2Mv+Ccfx9+D/wC0h4R+JnxQ0nQF0Dw+1zNILbU1ncytbSpCQoAPEjo2f9mqr9uv4Trb3Nj8XNHtvluStlq4UdHA/cyn6gFCfZB3r9i74sHwX4/PgbVLkLpXipljjLtxDdqD5ZHpvzsPqSnpVVVVVV+0p/wTX+Anx8N34g0CxXwH4vuMudU0m3HkXD+s9rlUfPUshRyTksa1b/gkh+1nY6lc2enQ+EtRtYpCsN3Hq/lrMvZgrqGXI7EcfrWt/wDBLb9qfw3pdxrWvw+DdPsLVN01xc+II40Qe5I/DHc1P+zh8SYpniSPTJQjFRIl38r47jIBwfcA14Z/ZK+NvjTWIPD/AIV8OQ6pqFwf3cFvOGOO7HjCqO7HAHcil/4JN/tgMoJ0TwuCR0OuJkfpX/Dpr9r/AP6Avhb/AMHif4V/w6a/a/8A+gL4W/8AB4n+Ff8ADpr9r/8A6Avhb/weJ/hX/Dpr9r//AKAvhb/weJ/hX/Dpr9r/AP6Avhb/AMHif4V/w6a/a/8A+gL4W/8AB4n+Ff8ADpr9r/8A6Avhb/weJ/hX/Dpr9r//AKAvhb/weJ/hX/Dpr9r/AP6Avhb/AMHif4V/w6a/a/8A+gL4W/8AB4n+FfGH/gnn+0f8DPhxq/xT8e6ZoEWhaJ5P2t7XVVmkHmzRwphAMn55Uz6DJr/gln/yZf4R/wCv/Uv/AEslqqqqqqqqqqqqqqqqqqqq/bv/AOTP/ip/2AJP/Q0r/gkH/wAmoXf/AGN19/6Jt6qq/wCCmP7N/wAXP2kPAvgzQ/hF4dg1a80fVp7i7SW+htdkbRBQQZWUHnsK/wCHW/7af/RMtP8A/Chsf/jtf8Ot/wBtP/omWn/+FDY//Ha/4db/ALaf/RMtP/8AChsf/jtf8Ot/20/+iZaf/wCFDY//AB2v+HW/7af/AETLT/8AwobH/wCO1/w63/bT/wCiZaf/AOFDY/8Ax2v+HW/7af8A0TLT/wDwobH/AOO1/wAOt/20/wDomWn/APhQ2P8A8dr/AIdb/tp/9Ey0/wD8KGx/+O1/w63/AG0/+iZaf/4UNj/8dr/h1v8Atp/9Ey0//wAKGx/+O1/w63/bT/6Jlp//AIUNj/8AHa/4db/tp/8ARMtP/wDChsf/AI7X/Drf9tP/AKJlp/8A4UNj/wDHa/4db/tp/wDRMtP/APChsf8A47X/AA63/bT/AOiZaf8A+FDY/wDx2v8Ah1v+2n/0TLT/APwobH/47X/Drf8AbT/6Jlp//hQ2P/x2v+HW/wC2n/0TLT//AAobH/47X/Drf9tP/omWn/8AhQ2P/wAdr/h1v+2n/wBEy0//AMKGx/8Ajtf8Ot/20/8AomWn/wDhQ2P/AMdr/h1v+2n/ANEy0/8A8KGx/wDjtf8ADrf9tP8A6Jlp/wD4UNj/APHa/wCHW/7af/RMtP8A/Chsf/jtf8Ot/wBtP/omWn/+FDY//Ha1b/gmx+15obRJqnw90+EyglB/btm2cdfuyH1FeDf+CfP7RsPi/Q5vEfg3T4tJj1K3a/f+1rZ9sAkUyHaHJPy54A5qqqv2jvAH/CO+JV8V6fBiw1tiZcDiO56sP+BD5vru9K/Zm8fmx1GXwFqU+IL0mawLH7soHzp9GUZHup7tVVVVVW1/c2hAikyv9xuRX9vPjH2Zf++q/t5/+fZf++q/t5/+fZf++q+NPx6i8AaW2kaPHFJ4gvYj5A3BhaqePNYev91T1I54HNlY+IPGmvi1tI7jU9W1KYsdzbnlc8szMT9SSTgdTX/CgPi5/wBCl/5P2/8A8cr/AIUB8XP+hS/8n7f/AOOV/wAKA+Ln/Qpf+T9v/wDHKk/Z9+LMqNHJ4PV1YEMrX1sQR3BHmV8R/wDgnr8Y7vUf7U8AeEbeKOdiZrGXUrZFiPrGd/3f9k9O3HA/4d6/tT/9CRp//g6tf/i6/wCHev7U/wD0JGn/APg6tf8A4uv+Hev7U/8A0JGn/wDg6tf/AIuv+Hev7U//AEJGn/8Ag6tf/i6/4d6/tT/9CRp//g6tf/i6h/4J3/tV3M0dvD4G09pJWCov9tWoyScD/lpX/Drf9tP/AKJlp/8A4UNj/wDHa/4db/tp/wDRMtP/APChsf8A47X/AA63/bT/AOiZaf8A+FDY/wDx2v8Ah1v+2n/0TLT/APwobH/47X/Drf8AbT/6Jlp//hQ2P/x2v+HW/wC2n/0TLT//AAobH/47X/Drf9tP/omWn/8AhQ2P/wAdr/h1v+2n/wBEy0//AMKGx/8Ajtf8Ot/20/8AomWn/wDhQ2P/AMdr/h1v+2n/ANEy0/8A8KGx/wDjtf8ADrf9tP8A6Jlp/wD4UNj/APHa/wCHW/7af/RMtP8A/Chsf/jtf8Ot/wBtP/omWn/+FDY//Ha/4db/ALaf/RMtP/8AChsf/jtf8Ot/20/+iZaf/wCFDY//AB2v+HW/7af/AETLT/8AwobH/wCO1/w63/bT/wCiZaf/AOFDY/8Ax2v+HW/7af8A0TLT/wDwobH/AOO1/wAOt/20/wDomWn/APhQ2P8A8dr/AIdb/tp/9Ey0/wD8KGx/+O1/w63/AG0/+iZaf/4UNj/8dr/h1v8Atp/9Ey0//wAKGx/+O1/w63/bT/6Jlp//AIUNj/8AHa/4db/tp/8ARMtP/wDChsf/AI7X/Drf9tP/AKJlp/8A4UNj/wDHa/4db/tp/wDRMtP/APChsf8A47X/AA63/bT/AOiZaf8A+FDY/wDx2v8Ah1v+2n/0TLT/APwobH/47X/Drf8AbT/6Jlp//hQ2P/x2v+HW/wC2n/0TLT//AAobH/47X/Drf9tP/omWn/8AhQ2P/wAdrwH+w/8AtEfE7U7/AEbwP4X0/UrzS03XUf8AasEPljdtzmRlB5GOCa8Y/wDBOH9r/wADeFtU8Ya58Lo207R7V7m7+yavaXMqxIMuyxpIWbABJCgnAPFeAPHWq/D/AF+LWNOYyQthLq2LYWePPIPofQ9j7ZB+C/wx+A3xS8OaZ4ptPjF/aMWoRK4s41jsZYn/AIomWQsdynKnHpwcEGtB/Z7+EGhKkkHhC3vZAP8AWX0jXG7/AICxK/kK0K9Hhq2FlollZWlsP+WENska/koFW/jZuFu7EH1aNsfof8atvFOjXAAa4aFj2kQj9RkVDc21yu62uI5RjqjA/wAqp3SNd0jqoHdjirjxBo1tw9/GxHZPn/lmrjxpYoMW1rLKR3YhB/WrjxlqUvEEUMI7cbj+v+FarcS65CbfVyl3EesUqBl/75xitS+D/wAL9Zyt54B0VmYYLQ2awsfxQA1qX7J3wr1TJs/D+qWBbJzaXUhx9BJvH/661H9hzT7kkaJ4j1u1/u/a7JZsfl5ftWo/sK/E6ME6NrWk3YGcC5325P6MM/jXj74LeNPhvP8AZPEUmjNODhobXVYZZU+sYbeB/wABr+yr/wD54f8Ajw/xr4i+MtN+G2hnVtbwZpcraWqyDfcOOwHYDjLdB9SAfFPifW/HWvy6zqshmurlgkUUYJEa9FRF9PbuTnqTXgf/AIJb/tI+JvDFjr+teFbnSZr+ITR2clxaLJFGwBXzFedWR8dUIBXocHID/wDBJj9oYsTHbELngF7Ikf8Ak3Un/BJr9pQORDYwsvYtPaKfy+0mpf8Agk7+1EGxBpFk646ve2qn8vPNTf8ABKD9rAECDQNLcY536pbLj8pTU3/BKP8Aa+AHkeF9Dc99+tW64/JjU3/BKj9smNQyeDNClOcbU163BHvywqT/AIJY/tnom5Ph7pMhH8K+ILQH9ZBXx2/Zl+Mf7Nl3o9j8XvDdvpE2uxyyWKxX8N15ixFQ5JiZtuC69cZzVVVVVVZ/8fUX++Kqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/4LOf8AJd/BH/Yoj/0rnr9iD9vHSP2Q/CniXw3qPw2u/Eza/qEV2ssGpraiEJHs2kGN85654r9qX/gqDoP7RfwI8T/Buy+D1/oc3iD7Jtv5daSdYfIu4ZzlBEpORCV6jG7Pav2bvi9bfAT43eFfi7d6HJrMXhy4llaxjuBA02+GSPAcqwXG/PQ9K/4fWeF/+jfdU/8ACij/APjFf8PrPC//AEb7qn/hRR//ABitLvV1PTLTUlj8sXUCShCc7dyg4z+NVVVVVVVVV/wS4/5PH+M3/Xhf/wDpyjqqqqqqqqqqqqqqqqqqqqrx9/yKV9/2z/8ARi1VVVMoYbSAQRggivjR4BPgLxlPb2kO3TNQzcWJA4VSfmj/AOAnj6bT3r9mjx//AGro8vgbUZ83WmKZLMseXgJ5X6qx/Jh6VVVVVDPLbuHhkKn270uvSBAGt1LY5O7H6U+u3JGEijH1BNeM/iPZ+CdCm17W7xYoY/ljREBeZz0RAepOD9ACTwCa+IXxB8QfEnX313X7jJUbLa3U/Jbx54Vf6nqT+Q+A3wX/ALMW38c+LLT/AExl36fZypzB6SsD/H6DtnPXGDz3qqqqqqqvCH/Is6d/1wFVVVVVVVVVVVVVVVVVVVVfGL/k+jwj/wBfWl/+hVVVXjfwjpXj7wjqvg7Wk3Wmq2rQuQMmMnlXH+0rBWHuBXinw5rvw98X3/hrVN1tqmiXZjZ4yRhlOVdD1wRhgfQg18B/ifbfFv4aaX4o81DqMafZtTjXjy7lAN/HYMCHA9HFVVVVVXxp/ac8BfCCKbS1nXW/EarhNMtpB+6OODM/IjHTjljkcYOa+KXxn8e/F/VDf+LdXY20bk2unQEpbW3+6meT/tNlj64r4Kfsm+Ofin9n1zXFk8PeG3AZbqeLM1yv/TGM4OD/AH2wvORu6V8OvhZ4H+FWjDRvBmixWqsB59y43z3LD+KSTq3fjgDPAFVVVVVVVX/BTX/kx/4k/wDcK/8ATraV/wAEs/8Aky/wj/1/6l/6WS1VVVVVVVVVVVVVVVVVVVft3/8AJn/xU/7AEn/oaV/wSD/5NQu/+xuvv/RNvVVVVVVVVVVVVVVVVVVVVXxW/wBfpv8AuSfzWqqqrx34RsvHPha+8N3m1TcR5hlIz5Uo5Rx9D19QSO9TRat4W11on32mpaVdY46xyxt1H0Ir4d+MrXx54SsfENuUWWRdl1Ep/wBVMv31+nQj2Iqqqqqqr4tfFXTvhxo5SForjWrpD9ktWOcdvMfHRR+p4HciWTXPGGvGRzPqOq6nP6bnlkY9B/h0A9AK+EHwlsfhzpX2q9WK4127T/SbgDIiX/nmh9B3Pc+wGKqqqqqq0X/kMWH/AF8x/wDoQqqqqqqqqqqqqqqqqqqqqqv2D/8Akpfjz/r1X/0ear/gpb+wn/wrPU7z9oL4Q6MR4R1Kfdr2l20ZI0i4c/69FA+W3duo6I7AD5WAX4D/ABXPw28VQ22tXk6+GtRlVdRSKLzWg7eeiZGWXuMjcOOuCLzQPij8OtGsPFvh3X7i68L6nElxp+t6JetNYXUbjKNleBnPRwD7Vov7SnxK0zal/PYaqg4P2m2CNj2Me38yDXw6+O/gTxy0Om6lqMfh7VpOBDfviCRvRJunPowXngZqfw9q8C7jZtIpGQ0R35/KissD4IaN19QQRSa1qyR+Umoz7fdySPx61LNPcNummeRvV2JNW+kandY8ixmYHoSuB+Z4q38HapKAZnhhHcFtx/T/ABq38E2igG5vJZD6IoX/ABq38O6NbgFLFHI7yZb9DxUcUUShIo1RR0CjFeI/FfhnwhYnUfFGv2Ol24Bw91Ose4jsoPLH2GTXjj9tLwRo/mWngjSLrXrgZC3E2baAH1GQXb6bV+teOP2kfi1478y3uvEj6XZSZH2TSwbdMehYEuw9ixFeHPCPirxnffYPDGg3+q3JPzC3hZ9ue7N0Ue5IFfFj4c6b+zv8OpPiT8dvE1to6SZj0zQbFluL/U7jGRCnOxB3aTLBByQSQD4+8c638RvEs/iDWJCWkOy2twcrbxZO2NfpnrjkknvX/BOX/gn1D4HtdO+P3xz0AnxPJtn8PaFeRcaWmAVuZkP/AC8HqqH/AFQwT85Hl1VVVVX/AAWt/wCRn+FH/Xhqn/oy3qqqqqqs/wDj6i/3xVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX/BU/9nH45fGf4w+E9d+Fvw01jxJp9j4aFtcXFlGrLHL9pmbYckc7WU/jX/DCH7X/AP0QDxR/35T/AOKr/hhD9r//AKIB4o/78p/8VX/DCH7X/wD0QDxR/wB+U/8Aiq/4YQ/a/wD+iAeKP+/Kf/FV/wAMIftf/wDRAPFH/flP/iq8O281r4e0y1uIyksNlEjo3VWCAEfmKqqqqqqqqr/glx/yeP8AGb/rwv8A/wBOUdVVVVVVVVVVVVVVVVVVVVePv+RSvv8Atn/6MWqqqqvjL4CXx94MuLS1iDanY5uLE45Lgcp/wIcY9dp7V4X8Q6j4O8R2XiDT8rc2E24ox27x0ZD7EEg/WtA1vT/Eui2WvaXL5lrewiSMnqM9QfcHII9RVVVVVeI/EWkeFNGude1u6EFraplierHsqjuxPAFfEr4jav8AEfXW1G8ZobKElbKz3ZWFPX3Y45P4dAK+A3wWN01v468XWhEKkSabaSD/AFh6iVx/d6FR369MZqqqqqqqq8If8izp3/XAVVVVVVVVVVVVVVVVVVVVV8Yv+T6PCP8A19aX/wChVVVVft2/CvI0/wCLek2392x1bav/AH5lP6oT/uCv2NPip/wg3xJXwnqd1s0nxXttjuPEd0M+S3/AiTH7l1J6VVVVV4r8XeGvA+iz+IfFmtW2mafbj55p3xk9lUdWY9lAJPYV8bf20fEPisXHhz4XCfQ9JbKPqLHbd3A77Mf6lT7fN7rkivBngTxn8TdfGh+EdHudVv5TvlK/djBPLyOeFGe7Ec+5r4J/sdeEPAH2fxB47+z+I9eUBkiePNpaN/sof9Yw/vMMeigjNAAAKoHA4FVVVVVVVVf8FNf+TH/iT/3Cv/TraV/wSz/5Mv8ACP8A1/6l/wClktVVVVVVVVVVVVVVVVVVVX7d/wDyZ/8AFT/sASf+hpX/AASD/wCTULv/ALG6+/8ARNvVVVVVVVVVVVVVVVVVVVVXxW/1+m/7kn81qqqqr9pzwB5NxB8QNNg+SbbBqAUdHAxHIfqBtP0X1r9nnx//AMIt4rHh6/nxp2tssfzHiKfoje2c7T9R6VVVVVVfE34k6T8N9CN/dYnv7gFbK0B5lcDqfRRxk/h1NaxrGu+NNek1LUZpr/Ur+UABVJLE8KiqO3QACvgr8HrfwHYrrutxLLr13H8wIBFmp6op/vf3m/AcZJqqqqqqqtF/5DFh/wBfMf8A6EKqqqqqqqqqqqqqqqqqqqqr9g//AJKX48/69V/9Hmq1TS9N1vTbrRtZsLe+sL6F4Lq1uYhJHNG42sjqchlIJBB4INft+/sT6j+y/wCNP+Ep8HWtxdfDfxDO39nTtlzpk5yTZysSScAEo7feXIOWVif+CdP7cSfBPWE+Cfxb1ETfDjXpyLa5ujvXRLiTqcEH/R5Dw69FY7+Mvu+JX7JPgHxrC+r+CJY/DuoyjeggXfaTZ55jH3M8cpgDrtNfET4O/ED4X3GzxVobpas+2K/g/e28p7YcdCf7rAN7V8M/2gfiN8MHitdM1U6hpKcHTL5jJEB/sHO6P/gJxnqDXw3/AGmfhl8SxDpeqOmh6tIMCz1FlMcjekcp+VvTB2sewNXGg+H4gbm4tYolXqWcoo+vOKn8X/DjQQRdeKvDlht6mW/hiI7c5YfSr/48fBvTsi4+JGhvtznyLkT9PTZn/wCvV9+1X8C7MFU8YyXTrnKwafcH8iUA/Wr79tD4Q2pK29n4ivMHAMNnGo6f7ci/Sr39uTwonGneBdWn/wCu1zHF9em6vG/7YvxL8SK9r4ZhtPDVq4IzAPPnx3BkcYH1VVPvV7f+IPFmq/aL+8v9Y1K5baGld55ZD6DOSfpXgf8AZQ+LHjAR3Oo6bF4esn5MupErIR7RDLZ9m2/WvBH7H3wx8NCO58Rm68S3iYJ+0nyYAR6RIeR7MzD2r42/Gf4V/spfCq68b+J4bfTtLs/3On6XYQrHJe3BB2QQoMDcdpyTwoBJOBX7SP7SHxE/ae+Ilx488eXuyKPdFpWlQuTb6Zb5yI4wepOAWc8seTgAAf8ABOD/AIJ8nQl0/wDaD+PHh0f2mQlx4X0G8j5tB1W7uIz/AMtOhjQ/c+8Ru27Kqqqqqv8Agtb/AMjP8KP+vDVP/RlvVVVVVVZ/8fUX++Kqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv+CXH/ACeP8Zv+vC//APTlHVVVVVVVVVVVVVVVVVVVVXj7/kUr7/tn/wCjFqqqqqv2ifh+PC/ikeI9Og26drbM5CjiK4HLr7BvvD3LelfsyePxbXU/gDUrj93cEz6eWPRwPnjH1A3Aeob1qqqqrV9X03QdNuNX1e8jtbS1QvLLIcBR/UnoAOSTivix8UtS+JOsbhut9Js2Is7XP4eY/qxH5DgdyfgX8GD4llh8Y+KrUHSYmzaWsi/8fbD+Ij/nmD2/iPtnKqqqEQAADAAGMVVVVVVVVV4Q/wCRZ07/AK4CqqqqqqqqqqqqqqqqqqqqvjF/yfR4R/6+tL/9Cqqqq8WeGdK8Z+GtT8Ka3D5tjqtq9vMo6gMMbh6MDgg9iAa8aeFNY+HnjHU/Ceqlo7/RrtojImV3YOUkXuAy7WHsRX7PHxQj+LPwv0zX7idX1W0H2PVF7i4QDLY/21Kv/wACx2qqqvjb+1r4J+GH2jQPDjReIfEkYKmCKT/R7Vv+msg6kHqi89iVr4hfE7xt8VNbbW/GWtS3suSIIF+WG3U/wxxjhR09z1JJ5r4J/sceLfHot/EHj1p/DuguA6QlMXd0vbap/wBWp/vMM9MKQc14K8B+Efh3okfh/wAG6Hb6bZxgbhGuXlbH3nY/M7e5Jqqqqqqqqqq/4Ka/8mP/ABJ/7hX/AKdbSv8Agln/AMmX+Ef+v/Uv/SyWqqqqqqqqqqqqqqqqqqqr9u//AJM/+Kn/AGAJP/Q0r/gkH/yahd/9jdff+ibeqqqqqqqqqqqqqqqqqqqqvit/r9N/3JP5rVVVVWu6Lp/iPRrzQtUi8y1vYWikXvgjqPQjgg9iBXizw1qHg3xHe+HdRBE9lKVVwMCRequPYjB/Gvgz4+Xx94NgubqYPqdhi3vgTyzAfLJ/wIc/XcO1VVVVeP8Ax5o3w90CTWdVkDyHK2tqrYe4k/uj0HcnsPwB8WeKta8ca9PruszmW4uGwka52xL/AAog7Af/AFzySa+BvwYXwpBH4s8U2oOtSrm2gfn7GhHUj/noQefQcdc1VVVVVVVVov8AyGLD/r5j/wDQhVVVVVVVVVVVVVVVVVVVVVfCz9n7wJ8H9a1TXvCc2qPc6xGEuBeXCyKAG3fKAoxyaqviJ8PfCHxW8E6v8PPHejQ6poWt25gu7aUdRkFWU9VdWCsrDlWUEciv2vv2U/F37KPxMl8Lap51/wCHNSLz+HtZMeFvIAeUcgACZMqHUeoYcMK/4Jjft1iL+zP2ZvjDrQ2Erb+ENXuZANvZbCRienQRE+vl/wBwC7s7TULWSyvrWG5t5lKyQzRh0cHqCp4I9jXxO/Y88HeJll1PwDOvh7UmyfszAvaSH/d5aP8A4DkD+7Xj74VeO/hnd/ZfF2gTW0Ttthu0HmQTH/ZkHGcc7TgjuBU089wVaed5Sq7VLsTgDsM9qsfDviDVAv8AZmhahd7sbfItXkznpjAPWrL4P/FbUQGtPhx4kZSOGbTJUU/QsoFWX7NHxx1ADyPAFynGf39zBD7fxuKs/wBj340XXE9hpNp1/wBdfqf/AEANVl+xB8Q5Mf2h4r8OwDJz5LTSkDHui968I/sQ+HbKcXPjXxZdaoqkEW1lD9mQ+u5yWYj6bfrXhH4deBvAdv8AZ/CHhew03K4aSKPMrj/akbLt+JNV8cfjZ4D/AGfPhxqfxM+IWpfZ9PsF2w28ZBnvZz9yCFSRudiOnQAFiQqkj9p79p74hftTfEKXxp40ufs1hbbotG0WGQtBpkBP3V6bnbALyEZYgdFCqv8AwTg/4J8SmXTP2h/jvoCeWFW48L+H72I5J4KXs6HoMcxow5yHOMLmqqqqqqv+C1v/ACM/wo/68NU/9GW9VVVVVVn/AMfUX++Kqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv+CXH/J4/xm/68L//ANOUdVVVVVVVVVVVVVVVVVVVVePv+RSvv+2f/oxaqqqqr4geDrPx34UvfDtyVV5U3W0pH+qmX7jfTPB9ifWkbV/CuuhlMlnqelXXfhopY2/oRXgPxdZ+OfCtj4jsyqm4jxPED/qpRw6/genqCD3qqqr28tNOs5r++uEgt7eMySyOcBFAySfwr4x/Fy7+IepHTtMkkh0G0f8AcREbTOw/5aOP5DsPcmvgl8HJvG94niLxBC8ehWr/ACqRg3jg8qP9gY+Y/gO5EUMNvClvbxJHHGoVERdoUAYAAHQY7VVVVVVVVVXhD/kWdO/64Cqqqqqqqqqqqqqqqqqqqqvit4G8a6l+2X4W8Uad4P1u60a3uNOM2ow6fK9vGEb5i0oXaMd8niqqqqv26vhOtzYWXxc0e2/e2u2y1cKOsZP7qU/RjsJ77kHav2QPiwfh58S4vD+pXITRvFJS0n3NgRT5Pkye3zMUPs+T0qq8d/ELwf8ADXQ38QeMtbgsLVQRGrHMk7D+GNBy7ew6dTgc18bf2wPGHxEFx4f8FCbw74efKMyvi7ul/wBtwfkUj+BfcFmBxXw4+FXjj4r6yNG8F6M90yY8+5k+SC2B7ySdB7DknHANfBT9lHwN8K1g1vWUj8QeJEAYXc8f7q2b/pjGehH99st6bc4qqqqqqqqqqqv+Cmv/ACY/8Sf+4V/6dbSv+CWf/Jl/hH/r/wBS/wDSyWqqqqqqqqqqqqqqqqqqqr9u/wD5M/8Aip/2AJP/AENK/wCCQf8Ayahd/wDY3X3/AKJt6qqqqqqqqqqqqqqqqqqqq+K3+v03/ck/mtVVVVV+0t4AGr6JF4302DN3pa+XdhRy8BPDf8BY/kxPavg149bwF4ygurmYrpl9i3vlJ4Ck/K+PVTz9Nw70rJIgdGDKwyCDkEVVVXjHxfongfQpte1242Qx/LGi8vM5HCIO5OD9OSeAa8d+Oda+IOvSa5rEgXjZb26H5II88KPX3Pc/lXwH+C401Lfxx4ttP9Ldd9hZyp/qB2lcH+P0HbOeuMVVVVVVVVVov/IYsP8Ar5j/APQhVVVVVVVVVVVVVVVVVVVVVVVV+0J8A/Av7SPwy1L4aeO7QeTcr5ljfIgM2nXSg+XPEfUE4I6MpZTwTXxz+Cfj39nP4nal8N/HNo1vqOmyCS0u4siO9gJPl3ELd1YD6qQVOGUgf8E4P254/jjoNv8ABf4p6uv/AAsDRbbFheTv82u2yDlie86KBvHV1G/nD4q+sLDVLV7HUrKC7tpRiSGeMSIw91OQasvD+g6YQdN0SwtSuMeRbJHjHToKqqqqr4v/ABd8C/Az4f6p8SfiJrCWGk6XETjIMtzKQdkMS5G+RyMBfqSQASP2q/2q/iB+1Z8QH8U+KZmsdFsS8eh6HFKWh0+En8N8rYBeQjJIAGFCqP8AgnJ/wT2n8QXGk/tC/HXRAujx7bnw34evIcm+OMpd3CN0iHDIh++cMfkwHqqqqqqqv+C1v/Iz/Cj/AK8NU/8ARlvVVVVVVZ/8fUX++KqqqqqqqqqqqqqqqqqqqqqqqvFHxZ+FfgedrTxp8TPCmgTopZotU1m3tGAAySRI4OMc1+2Z/wAFNvhv8DPCOn23wK8Q+EfiN4s1wyCF7HVo76x0xE25kuDbuSzEsAsYZScMSQAA2mfFb/gtX480+Lxf4Z0XV7HS9RQTWsH/AAjuj24CEZBVLqPztpBGC2c+pq+/4KTf8FAf2d9StLL9on4O2U1rdMwjbWdBm0t7nb94QzxERMRkZwj4yPWtJ+O3/BWX9rxj4l+EOiXvhnwtcMWtX0y0ttLtCoOMpdXZ8yb0Ox2Gc8DkDWdJ/wCC0nwjspPGVxrWvazaWSNJcxQXWm6uQijnNv8AO7AjP3FJGM5HBr/gnp+2vcfte+BNXtfFulWum+NvCDQJqq2YK295FKH8q4jUklCTG6smSAQCCA4Aqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv+CXH/J4/xm/68L//ANOUdVVVVVVVVVVVVVVVVVVVVePv+RSvv+2f/oxaqqqqqv2mvAH2O+h8fabD+5vCIL8KPuyAfI//AAIDB9wPWv2dPH//AAjXij/hF9Qn26frbBU3HAjuOiH/AIF936lfSqqppYbeF555UjjjUs7u20KAOST2GK+N3xjl8b3b+HPD8zR6FayHc4JBvXB+8f8AYGPlH4nsB8HPhHd/ETUxqGoo8Og2cmLiUHaZmAz5aH8snsD6kVY2NnptlBp2n20cFtbRiOKKNcKigYAAqqqqqqqqqq8If8izp3/XAVVVVVVVVVVVVVVVVVVVVVVVVVXibw7pfi7w9qPhjWoBNY6pbPbzp32sMZHoR1B7EA18QPBWr/DfxrqvgzWOLvSrgoJFBUSpwUkX2ZSrD61+zj8V4fiv8L7DV7y7V9Y0xRZ6sGOD5qAYkPs64bPTJYdq+Nv7ZXhXwR9o8O/DvyPEOuICj3QbdZ2rf7wP71hxwvy+rZBFeLfGXjD4k+IG1rxTq93q+p3LBE3c4yeEjReFGTwqgDJ96+Cf7FOueI/s3iP4rtNpGmMA8elRnbdTj/pof+WSnjjl+o+U814a8L+HfB2jQeH/AAto9tpun2wxHb26bQPUnuzHuxyT3NVVVVVVVVVVVX/BTX/kx/4k/wDcK/8ATraV/wAEs/8Aky/wj/1/6l/6WS1VVVVVVVVVVVVVVVVVVVft3/8AJn/xU/7AEn/oaV/wSD/5NQu/+xuvv/RNvVVVVVVVVVVVVVVVVVVVVXxW/wBfpv8AuSfzWqqqqqubaC8tpbS6iWWGZCkkbDIZSMEH2Ir4m+CJ/APi+80JgxtSfNspGH34W+7z3IwVPupr9nfx/wD8JT4U/wCEd1C43ajoaiMFjzJB/A34fdP0XPWqqvEfiLSPCmjXOva3dCC1tUyxPVj2VR3YngCviV8RtX+I+utqN4zQ2UJK2VnuysKevuxxyfw6AV8B/gsblrfxz4usyIlIk02zkXG89RK49OhUd+vTGaqqqqqqqqrRf+QxYf8AXzH/AOhCqqqqqqqqqqqqqqqqqqqqqgvrG7Zo7W8gmZB8yxyBiPriqqq/bL/ZJ8LftYfDV9DuBb2Hi3R1ebw7q7rjyJSOYpCASYZMAMByCFYAlcHVtK+IvwL+JUumahHqHhfxl4Q1EHKsY5rS4jIZXVh1H3WVhkMCCMg1+wv+2Vof7VngD7LrEttY/EHw/Cq65pyYQTrwBdwLknymJAYfwOcHgoWqqqqqvif8TvA/wc8D6n8Q/iHrsGlaJpMW+aaQ5Z2/hjjUcu7HAVRySa/a5/a38d/tYePW1zWpJNO8M6Y7poOhJJmOzjP8b44eZgBuf8BhQBX/AATr/wCCes/j650r4+/HHRmj8LwOlz4f0K5jwdWYHK3E6n/l3yAVQ/63qf3f+sCqqhVAAAwABjFVVVVVVVX/AAWt/wCRn+FH/Xhqn/oy3qqqqqqs/wDj6i/3xVVVVVVVVVVVVVVVVVVVVVVVf8FD/wBpr47+Ov2nLT9jb4NeNP8AhDtOeSysrq8TUP7P/tC7ukST95dAhkgVZFQxr95g4IclVHgn/giJZy20d78Uvj5dTX8oDXFvomkgKrkgtieZyX78mNeoOO1a/wD8EQPhlcwuvhb46+J9PlI+RtQ0qC8CnHOQjRZ7dxWp/sZf8FF/2LXk8Ufs9/Ea88VaFaEySWWgXDyFlB6y6ZOCkhPpGJWGT0r4A6Zq37YH7O/gzxf+2B8GvDz6vBqbajpunz20gjcRho4rqS3kPyM6tITExZCCr4GQqwww28KW9vEkccShURF2hQBgAAdBjtXiYeIT4b1UeERYnXfsM39li/dkt/tWw+V5rKrMI9+3cQrEDOAelad/wTb/AOChf7N6Xnjv4OfEPR31JYd1zbeEfEVzFPdonzbGjmhhScdSEYtk9ATiv+CZ/wC2p4t/al8JeIPCPxSFtJ4z8G+Q8l/DEsP9p20pcCRolAVZEZNrbQFO9CAOaqqqqqvjX8cvhj+z14GuPiD8VfEsOk6XC3lwrjfNdzEErDDGOXcgHgdACSQASNc/4LJ+OvE2qXa/BP8AZhvdV0mzfBub67lnmZTnaXjt4ysJIHTe/wBa/Zg/4Ky/Cr42+KrT4d/E7wtL8O/EWoTC3sJ5r4XNhdTHgRtKVRoXY8KrKVJ435IBr9qb9r74SfsleFYNc+IN7PearqW5dJ0Kw2tdXrKOWAJASMHAaRuBkAZJANx/wWb+K8iN4qsP2XIP+ETWUr9ok1O4YYBxg3IhEYbOB93rxX7JH7bvwl/a90e7HhIXOieJ9JjV9S8Pag6maJDwJYmXiaLPylgAVONyruXd4m8S6B4M8O6l4s8U6tb6Zo+j2sl1fXlw22OCGNSzux9AAaP/AAU3+MXxU1bVrj9lf9kHxJ478K6FOUuNZnklU3AXkqkccbBHIIKpvd8EEoOg/Y8/bs+H37W0ep+H4dCu/CHjnQkL6j4cvpvNYRhgplhk2r5iqxCsCqsrEZGCCaH7XHiTxj+2Yf2XfhL4R0zWNI8NWX2nxn4iuJ5Maa45aGNUG1n+aKPk8O7Aj921VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX/BLj/k8f4zf9eF/wD+nKOqqqqqqqqqqqqqqqqqqqqvH3/IpX3/AGz/APRi1VVVVVeI9B0/xRoV74f1OPfbX0JjfjlfRh7ggEe4FeJdA1Lwh4ivNB1DKXWnzFdy8bscq6+xGCPqK+EHjxPH/g22v55FOo2f+j3y9/MA4fHowwfrkdqpiAMkgADkntXx1+NH/CSSS+DvCl2DpMZxd3Kf8vTA/dU/88wQOf4j7dfhT8LdS+JOs+WS9vpNowN5dbf/ABxOxcj8hyewOkaRpug6bb6Po9nHa2lqmyKJBgKP6k9STyScmqqqqqqqqqqvCH/Is6d/1wFVVVVVVVVVVVVVVVVVVVVVVVVVX7dXwqOpaNYfFfSLXNxpgFnqmxeTAx/dSH/dclSf+mi9lrTPFPiPRdK1HRNH1u8s7HV1Rb+CCUotyEztD46gbjx05r4VfBDx/wDGHURa+FdKK2UbbbnU7kFLaD1BbB3NyPlUE85wBzXwZ/Zq8AfB6GLULe3Gr+IdmJNVuoxuQnqIU5EQ9xljnliOKqqqqqqqqqqqqr/gpr/yY/8AEn/uFf8Ap1tK/wCCWf8AyZf4R/6/9S/9LJaqqqqqqqqqqqqqqqqqqqv27/8Akz/4qf8AYAk/9DSv+CQf/JqF3/2N19/6Jt6qqqqqqqqqqqqqqqqqqqq+K3+v03/ck/mtVVVVVXx/8ADxh4PfVLCANqeihposDmSLH7xPyG4e64718PvGN14E8WWPiK23PHC+y5iH/LWFuHX645HuAasb601Oxt9RsJ0mtrqJZIZFOQ6sMgj8DVavq+m6Dptxq+r3kdraWqF5ZZDgKP6k9AByScV8V/ijqPxJ1ncN1vpNoxFna5/8iP6sR+Q4Hcn4F/Bg+JZYfGPiq1B0mJs2lrIv/H2w/iI/55g9v4j7ZyqqqhEAAAwABjFVVVVVVVVVWi/8hiw/6+Y//QhVVVVVVVVVVVVVVVVVVVVVfsH/APJS/Hn/AF6r/wCjzVVVV/wUJ/Ygs/2lPCTeP/AFhDD8SfD9sRb4AQaxbrk/ZXOQA4yTG57/ACnAbK/DT4kfEL4A/Euw8ceD7u50XxJ4dumV4p4ypyCVlgmjOCVYbkZDg/QgEfsu/tLeCP2pfhhaePfCkiWt/Dtg1rSHl3S6bdYyUPQsh5KPgBh6EMoqqqtd8Q6B4X0yXWfE2uafpGnwD97d39ylvEn1dyAOh6mviX/wUs/ZC+Gwlt1+I7eK72HP+i+GrU3u7H92YlYDn/rpX7Y/7YnjX9rHxyby6efS/BukysNB0LfxEOnnTYJDzsOp5Cg7V4yW/wCCen/BPSf4uT2Hxt+NukSQ+CIXEuj6POhVtcYHiSQdRbAjp/y0/wBz70cccUawwoqIihVVRgKB0AHYVVVVVVVVV/wWt/5Gf4Uf9eGqf+jLeqqqqqqz/wCPqL/fFVVVVVVVVVVVVVVVVVVVVVWpfEfwBpUWqPeeMtGD6LazXV/Cl7G8tvFEheRmjBLAKqknjtV34y/a/wD+CqnxU1/Qvhx4hl8I/DXRXG+zkvpLaxs4HLCL7R5QLXNxIEY7SCAQ2Ni81q//AAQ78aQ6L5+gftB6Jeattb/Rbzw/LbQbuw85ZpGwe58vj0NfsqfCzxr8GPgH4S+HvxF8YXviXxJp1oTqN7c3j3QSR2LeTG7/ADGKJSsa+y5wM4FVVV+29+3n4f8A2ffD+p+BPhrY3Xij4k3dtJDDFYxGW30R2XCzXLgEb1yGWEfMcDdsBBP/AATC/ah+F37MnxM8SaH8YdM1LT7jxw1nZxa7IAY9M2NISJ0IDqjtIhMgJxtUlQMsEdJEWSN1ZWGVYHII9RVVVVVft1+JfEH7W3/BQXQ/2cLTWJ7fw7omr2nh61EfSB5djX11tPBdcsvusCepr4a/DPwN8IPBWmfD34deHrXRtD0mER29tAmMkAZd26vI2Ms7ZLHkmv8Agsn+zZ4Ws/DOiftLeE9Ht7DWhqiaZ4ia2j2fbUkRjBcPjjejR+WW+8wlUE4QY/YB+Nt98cv2T/B3jTxNqJuNa0yCTS9YuZX5ea1YoJXY9WeIRSMT3c18NrB/+Ck//BRG81bxnJcXfgu1lnvntN7J5eiWbBLa3GOVEjvCHIIOZ5GGGNQeHPD1toCeFLbQdOi0WO1+yJpqWqLbLAF2+UIgNoTbxtxjHGK/aG0Fv+CeX/BQfTPGHw5jl0/wxLPba3aWELFVOm3DtHd2Yxxs3JcIowdq+WeSM1/wWC+I13oH7JWn6NoF8/keNvEVpaXEkTYWW0WKW4IyOoLxQ8dxmv2Lvh9o3wz/AGVvhh4a0a0hh83w1Z3940Q/1t1cxLPO5PU5kkbBPYAcAYHxWtIfgp/wWR8Gax4TRbOPxxJZSX8EC4SRr2KS0mLLwPmdPNPX5zu61+2n+0lYfsufAXWviAkkUniG7H9n+G7R13effyqdhK90jAaVgeoTbnLCv+CeX7N2o/Af4MHxL46Sab4i/EWca14mubr5p0eTLRwO3UlA7M2f+WkkntVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV/wAEuP8Ak8f4zf8AXhf/APpyjqqqqqqqqqqqqqqqqqqqqrx9/wAilff9s/8A0YtVVVVVVftMfD86lpkPjvTIM3GnqIr4KOXhJ+V/+Ak4Ps3oK+Cfj7/hBPGUL3k+3S9Sxb3oJ4QE/LIf90n8i1AgjIOfSvjz8aTcNceBfCN4RGpMeo3kbff7GJCO3Zj36dM5+G3w41f4j66mnWStDZQkNe3ZXKwp7erHHA/HoCa8N+HNI8J6LbaBodsIbS1XCjqWPdmPck8k1VVVVVVVVVVXhD/kWdO/64CqqqqqqqqqqqqqqqqqqqqvGn7SF74T+PejfBePwnBcw6rLaodQa8KtH5xwSE2kHH15qqqqqtd0XTfEmi33h/WLZbix1G3e3uIm/iR1KsPbg18NP2FLSz1651L4m6st7ptrdOtlp9o5U3UasQrzOMFQygHYvPPLDBFaTo+laBptvo+h6dbWFjaoEgt7eIRpGo7BRwKqqqqqqqqqqqqqv+Cmv/Jj/wASf+4V/wCnW0r/AIJZ/wDJl/hH/r/1L/0slqqqqqqqqqqqqqqqqqqqq/bv/wCTP/ip/wBgCT/0NK/4JB/8moXf/Y3X3/om3qqqqqqqqqqqqqqqqqqqqr4rf6/Tf9yT+a1VVVVVV8cfh/8A8IL4xkexg2aXqu6e0wMBDn54x/ukj8GWv2ZfH327T5/AWpT5msgZrAsfvRE/On/AScj2Y+lXt5aadZzX99cJBb28Zklkc4CKBkk/hXxj+Ll38Q9SOnaZJJDoNo/7iIjaZ2H/AC0cfyHYe5NfBL4OTeN7xPEXiCF49CtX+VSMG8cHlR/sDHzH8B3Iihht4Ut7eJI441CoiLtCgDAAA6DHaqqqqqqqqqqtF/5DFh/18x/+hCqqqqqqqqqqqqqqqqqqqqq/YP8A+Sl+PP8Ar1X/ANHmqqqqv+CmX7CP/Ca2WoftG/B3RgfEFlEZfE2kW0XOoxKObqJVHMyjJcfxqMj5lIf9mP8AaQ8b/sv/ABPsviD4QmNxatiDWNKkciLUrUn5o2x0YfeR/wCFgOoLKfhB8W/BHxy+HmkfEz4e6qL3SNXhDKGwJbaQffhlUE7ZEPysOncEggmvib+13+zT8IDLB49+MvhyzvIciSwtrn7bdKR2MEAeRemOVAr4m/8ABZb4U6L5ln8KPhnr3iadQVW61SdNNgz2ZQokkcexCH6da+Jv/BVH9rL4geZa6D4h0jwTZOSBFoVgPNK9gZpzI4PuhT8OleLPHfj74kaqupeN/GGveJtRc7Um1O/lvJck9FLsT+Ar4ZfsRftT/FtYbjwl8GddisZvuX+qxDTYCv8AeV7gpvX/AHN3pX7PH/BH7UtH8VaZ4p/aI8W6LqOmWTiZ/DujGWQXbDlUmndUwmfvKinI4DDOasrGy02yt9N020htbS1iWKC3gjEaRIoAVVUcKoAAAHAAqqqqqqqqqv8Agtb/AMjP8KP+vDVP/RlvVVVVVVZ/8fUX++Kqqqqqqqqqq1wkaJqBU4ItZcH/AICa/tzWv+gxe/8AgQ3+Nf25rX/QYvf/AAIb/Gv7c1r/AKDF7/4EN/jX9ua1/wBBi9/8CG/xr+3Na/6DF7/4EN/jX9ua1/0GL3/wIb/Gv7c1r/oMXv8A4EN/jX9ua1/0GL3/AMCG/wAa/tzWv+gxe/8AgQ3+Nf8ABPS7u7z4ZeI3u7qWdl10gGVyxA8iP1qqr48ftn/s4fs33Y0j4o/EW2tdaaNXXRrKF7y82sMqWjjB8sEDIMhUHsa1r/gsx+yrY6K9/o2g+PNTvhJsjsP7MhgJGPvl2m2he3GWz2xzX7X3/BVqT45/Ci1+G3wL0XxV4O1DWLopr11O8Ykktgo2wW8kTlvncnedqnaoUZDsBef8EUviTH8M7HX9E+L2lt46Nos13oVzYNDapIVy0KXSux3A/LuMYVj/AHRzX/BIPSPFXwsHxe+AnxH8L3Ph7xVoOq2Woy2t3HtklimjeIspHDxgwKQykqfNBBOaqqqqqv8Agttp3wxT4f8AgHU5odPT4hSa08ds6KBcS6UIJDNvxyyLN9n27uhZ9vVq/ZJl1yf9lz4TTeJGnbUX8G6WZjOMSH/Rk2lvfbtznn15zVVVVVftTLqn7Jv/AAU1/wCFra1pc0mjy+JbXxPbsq5N3ZzEfafLz/EG+0Rj0ZQemK8EeOfCHxJ8L6f408CeIrHW9F1OFZbW8s5hIjggHBx91hnBU4KnggEV/wAFe/2h/hlpfwOufgBa65bal4z1+/tZpLC1mV202CCVZTJPjOwsVVVQ4Y7i3RTX/BOvVtT8K/8ABMz41eJhO+7Tp/El7ZFWBKCLSIDxngfOjHHvnvX/AAQ/0m2n+InxR1xh/pFnotjbR8fwyzSM36wpVf8ABcLTbWLx98K9XRf9IudH1CCQ4H3I5omX36yv3/LnP7Z3w48Q/HT/AIJh/DjxdomnzXeq+GdA0DxLNBEvmSPAdPCXGDgZCpcGVsY4i/Cv2Cf2o/hf8T/2YvB1ve+NtF07XvBmiQaVrtjfX8cM0BtY1iFwyswPlyIiuH+7liM5UgeA9Rg/bM/4KuH4oeBR/aXgP4ZRRk6qqlopFtoXSJlJ4/eXbuyY6opYDg1pd7p/7e37beofETWbuB/gV+zsH+yTTygWuo6gp3NMSTtMZeLzC2SvlW8W4DzTV/8A8FIPjf8AGXxZreifsUfs0XHjzRPDz7LnX9VkeKKY842xhowm4AlVaQuw52Lgiv2Sv+Cg2l/Hvx7qPwO+KngG5+HHxP0tpVGkXUrNHemIEyom9VaOVFBYxsDlQWDEAgVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV/wS4/5PH+M3/Xhf8A/pyjqqqqqqqqqqqqqqqqqqqqrx9/yKV9/wBs/wD0YtVVVVVVXlna6haTWF7Ak1vcRtHLG4yHVhgg+xBr4j+C7nwF4tvfD8wZoEbzLSVh/rYW+6fr2PuDVz8fNfb4bWngyz8yLUUQ29xqG7DGAYChe+4j5S3oM9TkeBPA2tfEDXotD0ZAON9xO4+SCPux/kB3P514O8HaJ4G0KDQdDtwkcYzJIR88z45dj3J/TgDgVVVVVVVVVVVVeEP+RZ07/rgKqqqqqqqqqqqqqqqqqqqq+MX/ACfR4R/6+tL/APQqqqqqqqqqqqqqqqqqqqqq/wCCmv8AyY/8Sf8AuFf+nW0r/gln/wAmX+Ef+v8A1L/0slqqqqqqqqqqqqqqqqqqqq/bv/5M/wDip/2AJP8A0NK/4JB/8moXf/Y3X3/om3qqqqqqqqqqqqqqqqqqqqr4rf6/Tf8Ack/mtVVVVVVfFrwJH8QPBtzpUUam/t/39i54xKo+7n0YZX8Qe1aBrWp+EfENprViWhvdOn3BXBByOGRh6EZUj3NfGH41XPxBWLRtFjmstGjCvLG5w9xJgH5sfwqeg9eT2A+Dnwju/iJqY1DUUeHQbOTFxKDtMzAZ8tD+WT2B9SKsbGz02yg07T7aOC2toxHFFGuFRQMAAVVVVVVVVVVVWi/8hiw/6+Y//QhVVVVVVVVVVVVVVVVVVVVVfsH/APJS/Hn/AF6r/wCjzVVVVVf8FL/2Ez8OtRvf2hfhBouPCuozmTxFpVtHxpM7t/x8RqOkDseVAAjYjHysAn7Pn7XHxt/Zji1u1+FXiG3t7TXo1+02l9bC5hSVfuzxo3CyAZXPQgjcDtXb8S/2pf2ifjB50fxE+MPiXVbafPmWS3ZtrVs9f9Hh2Rf+O18Pvgx8WvizdLZ/DX4b+IvEjF9jSafp0ksUZ/25ANiD3YgV8M/+CRn7TXjEQ3Xjm78OeBrRiN8d5d/bboA91jt9yHjs0in9cfDP/gj3+z34XENz8R/FHiTxvdJjzIRINMtJPX93ETKP+/1fDf8AZ6+BvwhjjHw1+FPhrQZo12i7ttPQ3LD/AGp2Bkb8WNVVVVVVVVVVV/wWt/5Gf4Uf9eGqf+jLeqqqqqqz/wCPqL/fFVVVVVVVVVVMqupRgGBGCCOtf8IP4K/6E/RP/BfF/wDE1/wg/gr/AKE/RP8AwXxf/E1/wg/gr/oT9E/8F8X/AMTX/CD+Cv8AoT9E/wDBfF/8TX/CD+Cv+hP0T/wXxf8AxNf8IP4K/wChP0T/AMF8X/xNf8IP4K/6E/RP/BfF/wDE1/wg/gr/AKE/RP8AwXxf/E1/wg/gr/oT9E/8F8X/AMTWnaRpOjxNBpGmWllG7bmS2gWIMcYyQoHOAKqq/wCCoGseGfDH7ZPgbxN8afgUmveBYNKjhkuLa/mt5NctQxZ03RlAs0EjSYXd8yOu7AYFdP8A2tP+CbXh7xgniHRP2F7m7ja9ihkXU7qO4iSyWM7pUtJHkhM2/A8s8MMsZAeK+M/x4/4JL/ES0s/Eum/DDxNoPiHTriO6tz4U8Ow6bJI6sH2TR71t5FJADE/NydrDOa/ZQ/aQs/2qvhY/xW0zwhc+HbB9WuLK0trm8W4klSIJ+9baoCEszDZ82NucnPH7W37Vvwt/ZW0e317VNLtte+Imr2zWnhvQrZR9tvgzj5WcAtFb+YFJJ4YrhQzDA+HUHjODwLoa/ES+t7rxPJZpLq7W0QjhS5cbpI4lH/LNGYouSSVUEkkkmqqqr/gppoX7Rnh/4YP8avgB8W/E+gQ+GYMeI9G066MazWm7/j6jwNyvGT8+CAU542Hd+wD+xjZftl6jcftGfHz4o33iyy0nWTZ3mi3VxNcXl/PGiSKLq4kbcISsi/KuSwyNyYNQww28KW9vEkccShURF2hQBgAAdBjtVVVVVftP/snfCb9rHwbF4X+I9hPDe6ezSaVrNiVS7092GDsYghkbA3RsCrYB4IVha/8ABGf4o+G9Vms/BP7VZ03RLpj5zxaXcW8xGDgNFHcbXOMDJYfQdK/bn/Zt+CH7FXwf0X4YeHNVuvFXxN8d3ou9V1zUFVXg02AklIoQSIVkn8sgkszeS4LEDA/Za/Z/1fwp/wAE54vhJqenumueL/CGqTXdtJH8wl1GKVkjZcjDLHLEhUkcqenb/giR4mtbD4wfEXwXO6pc6r4dgvIlYYJ+zXGxgOev+kg4x2zxg5r/AILceJLe++MPw58GwuHuNL8OT3kiBeQLm4KLznr/AKMeMfzr4Q+FJPBnwg8E+B7yBFfQvDdhp0sXl7QDDbJGV284Hy4xXxA/4JWfsd+P/E83ik+DNV8PTXMxluLTQtRa2tZWPXERDLGD/dj2AdgK/a+1LwH+xp8AtP8A2Z/2VvBlvpPjf4u3f9k6VZ6eWku3STbFNcySMTI8hDrCjM2QZMqcRkD9pf4Z237Fn/BLzVfhT4UuB/auqC1sta1G2yv2u6u5kN45PBKNGjwgHny9gPQ1/wAEzPAukeCP2MPADabbRJc+IIZ9Wv5lGDPNNM+Gb3WJYk+iD61/wUYt4vhd/wAFBfgZ8XPDa/ZtR1FtNa+8r5TcNb33lsW9d8DrEefuqBVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVf8EuP+Tx/jN/14X//AKco6qqqqqqqqqqqqqqqqqqqq8ff8ilff9s//Ri1VVVVVVX7QngAeLPCR1zT4N2p6IrSqFHMkP8Ay0X8MBh9CO9eEvCes+Ndct9A0O3Mk8x+ZyPlhTu7Hso/+sMkgV4A8BaN8PdAj0XSl8yQ/Nc3LKA9xJ6n0A6Adh+JNVVVVVVVVVVVXhD/AJFnTv8ArgKqqqqqqqqqqqqqqqqqqqq+MX/J9HhH/r60v/0Kqqqqqqqqqqqqqqqqqqqqv+Cmv/Jj/wASf+4V/wCnW0r/AIJZ/wDJl/hH/r/1L/0slqqqqqqqqqqqqqqqqqqqq/bv/wCTP/ip/wBgCT/0NK/4JB/8moXf/Y3X3/om3qqqqqqqqqqqqqqqqqqqqr4rf6/Tf9yT+a1VVVVVVX7R/gAeHvEi+LNOg22GtMfOCjiO5xlv++gN313V8KfhbqXxJ1nyyXt9JtGBvLrb/wCOJ2LkfkOT2B0jSNN0HTbfR9Hs47W0tU2RRIMBR/UnqSeSTk1VVVVVVVVVVVaL/wAhiw/6+Y//AEIVVVVVVVVVVVVVVVVVVVVVX7B//JS/Hn/Xqv8A6PNVVVVVqemadrWm3Wj6vY297Y30DwXVrcRiSOeJ1KujqchlZSQQeCDV1/wRs+EmofETV9fn+Juv2fhK7ujLYaBYWkaTWiEAtGbqQuGUNuC/ushduWY5NfDL9gL9kv4VeVPovwg0vVr6LB+26/u1OQsOjBZi0aH3RF9atbS1sbaOzsbaK3ghXbHFEgRUA7ADgCqqqqqqqqqqqqq/4LW/8jP8KP8Arw1T/wBGW9VVVVVVn/x9Rf74qqqqqqqqqqqqqqqqqqqq8Y+BfBPxE0R/Dfj7whoviTSpGDNY6tYR3cJYdG2SAjI7HGRUn7MP7OMnhObwKPgT4DTQbhleWwi8P20cbyKCFkO1AfMAJw/3hk8814i/4JnfsZa1oGq6RpnwcsdGvNQs5oINRtbu5aWzd0IWaNXkZNyEhhkEZHTFaX+xh/wVB/ZcvtU8J/s4eMpNY8MXk7Okula1Z28MmcDzWtr51EMpUKGKbugG9gAa/ZK8BeMvGH/BSPRPDX7TniG81DxZ4Z1S5ur7+1b/AO1SXF9ZxNJBEkhLBlWRUdQvylUwvBFftE/tC/Dv9mX4Z3/xL+Il+Vt4P3VjYwkG41G5I+SCFT1Y4ySeFALHAFfA7xprnxK+Dfgr4i+JLC3sdR8VaHa6rNaW+dluLmNZVjGeTtV1XJ5JHaqqq/aC+M1p+z78K9W+LOpeENa8R6doZR7610hY2nihZgrTYdlBRSQW54GT0BI/aH/4LB+HfiX8L/EXwx+E/wAH9XS+8W6ZPpUl7rc8RFvHcRtFIUgi3+a+1iFBYDJBIYDaf+CU37Pvjf4Gfs+X2o/EPS7nSdY8a6t/aKaZcoUltLZY1ji8xTyrth32nkKyZwcgVVVVVXxi+P8A8HvgBpmn6x8YPG9r4bs9VnaCzlngmlEsiruZQI0YjA55xXxv/wCCw37O3grR7m2+DdtqfxB11kK2zm0l0+xjfH3pHmVZWAOPlSP5um5etfslfso/GX9tr42J+1h+1Tb3J8JtcLeW1veQmIayyEGG3ghP3LJR1bo4G0bizuoVVUKoAAGAAMYr48eFfG3/AATd/bqtPjZ4Y0Ge48Da7qU99p6x5WG5tLjP2yw3dFePe2wHOAIXOecD/gpt+xc3gceNj8XYgfs3m/2ObGb+0A+P9T5O37+flznZnndt+avgd4W8a/8ABSb9uq6+NviPw9cWngPQtRgvb5ZPmitbS2x9ksN2MNJIUXeBjhpXGOBVeINe0bwroOo+J/EWpQ2GlaRaS3d9dzHCQQxqWd2PoFBJ+lfsW6BrP7Xn7Sfi39vT4gadNH4f0qaTRPhzp90nEEKAq0wGTyqOwJGVMs82CDGMft7fBzV/jp+yl468DeGrE3eux2seo6XAq7nlntZVm8tBz87ojxqPVxzX/BLv9q74Yal+zrpXwd8beNtK0DxZ4Da4tpLXV75LVrm0MryRyx+YQGCB/LZRyvlgkAMK+KGuab+3T/wU38BaH8L7xdc8FfC9LabU9Vtm8y1kS1uDc3EiuOCjyNFbKw4ZsEEqQ1VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV/wS4/5PH+M3/Xhf8A/pyjqqqqqqqqqqqqqqqqqqqqrx9/yKV9/wBs/wD0YtVVVVVVUyhhtIBBGCCK8H/D/wAMeBVvR4esvKfUJ2lmdjluSSqD0Rc4A/meaqqqqqqqqqqqqrwh/wAizp3/AFwFVVVVVVVVVVVVVVVVVVVVfGL/AJPo8I/9fWl/+hVVVVVVVVVVVVVVVVVVVVV/wU1/5Mf+JP8A3Cv/AE62lf8ABLP/AJMv8I/9f+pf+lktVVVVVVVVVVVVVVVVVVVX7d//ACZ/8VP+wBJ/6Glf8Eg/+TULv/sbr7/0Tb1VVVVVVVVVVVVVVVVVVVV8Vv8AX6b/ALkn81qqqqqqqvGXhPTPG/hy78N6ruWK5UbZFHzRODlWHuCPxGR3rw34c0jwnottoGh2whtLVcKOpY92Y9yTyTVVVVVVVVVVVVWi/wDIYsP+vmP/ANCFVVVVVVVVVVVVVVVVVVVVV+y18BviB8JfGninXPGFtYx2urQBLY29yJSSJS3IHTg1VVVVVVVVVVVVVVVVVVVV/wAFrf8AkZ/hR/14ap/6Mt6qqqqqrP8A4+ov98VVVVVVVVVVVVVVVVVVVVVVV+2v/wAEyx+0N8QG+Nfwf8cWvhLxtOsX2+G8R1tbySJQsc4liBkhlCqikhWDbQcAglv2sf2A/wBq74Q/DOX4z/GP4kaf48t9LvIrWf7Hqt9qUtlBLkCZ3uYk2x+YI0IGeZF98fskf8FCv2YPEXwL8I6L4u+JWj+C/EPhvRbXTtR0zWZvsw3wRLF5kLsNkiPs3AKSwBwQCK0b/grH8O5v2v8AV/CGpa/Cnwam0+HTtN1w27BYtQjLM923y7xDIXMPIIAiifCgua8OeJ/DfjHRrfxD4S8QabrWl3a7re90+6S4hlHqroSp/A1/wUn/AGrfid+yd42+FHiz4Z+JLGaPUE1KLXfDN8Vkhv4FNsYpCg+eMgmVRKpBzgcjcp+GP/BYz9l/xZpsH/CxLHxJ4G1PaPtEc1k2oWwbjPlywAuwHq0Sn2r9or/gqz+ybL8L/FHhXwFfar491LXdIubCG2j0meztSZojGfOe5WNggDHIVGJxjHOR/wAEY/ghe6R8PPFfxh8XeE7QJr+oW8Xhu7vLGMzrHAsgnlhcrvVHeRVyDgmE+mTVVVVVV+0X+y18Jv2ptE0jw/8AFiz1O4tNDunubQWN6bZhIy7TkgHIx2r4af8ABPv9kD4T38Gr+GPgrpN1qVu26O71iWXU3VuzKtw7orDjBVRjr15oKqqFUAADAAGMVXxE+GngH4t+FbnwT8SvCem+ItEuyGksr6EOoYfddT1Rxk4ZSGGeCKj/AOCTX7FCayNU/wCEC1prcSbhpzeILryMf3c7/Mx/wPPvXgD4c+BPhV4XtfBfw48J6Z4d0SzyYrKwtxEm49XbHLOcDLNlieSTVfGb4Q+Ffjt8PNS+F/je41SPQtXMf22PTrs20kyo4cIXAJ2llXIHXGDwSD8Ofh74T+E/gXRPhx4F0tdP0Lw/Zpa2VuDuIRerMx5Z2JLMx5ZmJPJqvjB/wTt/ZN+N3iq48b+LfhubPXb2Uy313o97LY/bHPJaSNDsLE5JcKGYkksa+C/7P3wd/Z68OyeGPg/4FsPD1pcMHupI90s90wzhpZpC0kmMnAZiFycAA1VVVVVVVVVVVVVVVVVVVVVVVVVVVVXxC+IHg/4V+DNV+IHj3XINJ0LRYDNeXcxOFGQAoA5ZmYqqqMlmYAAk1q3/AAU8/aa+Mvim/wBH/ZN/Z8OpaZYfM8tzplxql35f8LyiB1jgDYPynf6BjXwe/wCCq/jXQfiFF8Mv2vPhfF4SnadYJ9TtbOeyfT2bG17m1mLNs5BLqRgchTVtc217bRXlncRTwTxiSKWJwyyKRkMpHBBBBBFVX/BLj/k8f4zf9eF//wCnKOqqqqqqqqqqqqqqqqqqqqvH3/IpX3/bP/0YtVVVVVVVVVVVVVVVVVVVVV4Q/wCRZ07/AK4CqqqqqqqqqqqqqqqqqqqqvjF/yfR4R/6+tL/9Cqqqqqqqqqqqqqqqqqqqqr/gpr/yY/8AEn/uFf8Ap1tK/wCCWf8AyZf4R/6/9S/9LJaqqqqqqqqqqqqqqqqqqqv27/8Akz/4qf8AYAk/9DSv+CQf/JqF3/2N19/6Jt6qqqqqqqqqqqqqqqqqqqq+K3+v03/ck/mtVVVVVVVVVVVVVVVVVVVVaL/yGLD/AK+Y/wD0IVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVf8Frf+Rn+FH/Xhqn/AKMt6qqqqqrP/j6i/wB8VVVVVVVVVVVVVVVVVVVVVVVWqaXpmt6bdaNrOnWt/YX0LwXVpdQrLFPGwwyOjAhlIJBBGCDXj3/gkD+yJ4116XXNLXxl4QSdiz2Gg6pELYMepVbiGZlH+yrBRngAYrUv+CSH7IV38O38EafoviCy1MSGWLxKNTMl+H24AYEeSycDKeWB1wQTmtR/4I1fGrwhqc8vwg/aVsIbWc4L3EF1pkxTrtbyGkDY454z1wOlfBr/AII53MHjiz8b/tJ/FyPxdHZzrNJpFhFLIt+VOVWa5nIby+BuQJkgkbl6nx3/AME8/wBjT4i3cuoa98B9DtruYsWl0eWfS/mOctstpI0Jyc8qRnrmvBf/AATZ/Yr8C38Oq6d8ENP1G7hfcrazfXOoxnByMwzSNER9U+tWtra2NrDZWVvFb29vGscMMSBEjRRgKqjgAAAADpVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV/wWR+Mep3fiTwb+z7o1xKLSG1/tvU4oiT9omkd4rZCB3QJK2O/mqewr9lv4E6B+zr8EvDfw40fT4Ib6G0jn1q4RAGvNQdAZ5WPU/N8q5J2oqr0Ar/grX8EdG8b/s+/8LetdPjHiDwFdQk3Kp88tjPKsUkR9QJJI5BnO3a+Mbmr/glZ8Xb74m/su2vh3Wr1rjUfAeoyaOHkbLta7VltyfZVkMQ9oR9aqv8Aglx/yeP8Zv8Arwv/AP05R1VVVVVVVVVVVVVVVVVVVV4+/wCRSvv+2f8A6MWqqqqqqqqqqqqqqqqqqqqq8If8izp3/XAVVVVVVVVVVVVVVVVVVVVVqfwu+H2s+K7bxzqfhSxudfs2jaC/dT5kZj+4Qc9qqqqqqqqqqqqqqqqqqqqq/wCCmv8AyY/8Sf8AuFf+nW0r/gln/wAmX+Ef+v8A1L/0slqqqqqqqqqqqqqqqqqqqq/bv/5M/wDip/2AJP8A0NK/4JB/8moXf/Y3X3/om3qqqqqqqqqqqqqqqqqqqqr4rf6/Tf8Ack/mtVVVVVVVVVVVVVVVVVVVVaL/AMhiw/6+Y/8A0IVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVf8Frf+Rn+FH/AF4ap/6Mt6qqqqqrP/j6i/3xVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV+2xInib/AIKdw+H9SQvaw674asSpbOY5IbR2AB4HMzcfj3qv21bSC9/ZL+LENwu5V8K3kgAOPmRCyn81Ff8ABE7UZ5dL+L2kMf3NtcaNOgyfvSLeK3HTpEtVXwS/Z6+KP7R37Q/xJ8LfCrx/b+E9Q0y5vry4uZ7qe3EsX2zZsBhBY/M6nB44r/h1v+2P/wBHNaX/AODrUv8A4iv+HW/7Y/8A0c1pf/g61L/4iv8Ah1v+2P8A9HNaX/4OtS/+Ir9jX4HfET9n34RTeA/id41g8U6xJrE94t9DczTgROkaqm6YBuCjHGMc1VVVVVVVVVVVVVVVVVf8Fevi38T/AIXwfCu1+HXj3W/DkWsf2z/aCabePALryjYmPeFPzbS74z03H1rQ5ZLjRNPnmkLySWsTOzHJJKDJNVVVVVVVVVVVVVVVVVVVXxVv73Svhf4w1TTLuS2vLPQL6a3niYq8UiwOVZSOhBAINf8ABJX4o/Eb4ofDbxzc/ETxvrHiOXTNZt4bN9Su2nMEZhJKpuJwCecVVVVVVVVVVVVVVVVVVVVV+0F8bPi74e/4Ka+AfhlofxI8QWPhO/vdFS60WC+dLWYStiQNGDghu/rVVVVVVVVVVVVVVVVVVX/Drf8AbH/6Oa0v/wAHWpf/ABFf8Ot/2x/+jmtL/wDB1qX/AMRX/Drf9sf/AKOa0v8A8HWpf/EV+0j+wT+0r8Ffgt4i+JnxA+Olh4h0DRvsv2vTYtTvpmn826ihTCyqEO15Ebk/w5HOK/4JZ/8AJl/hH/r/ANS/9LJaqqqqqqqqqqqqqqqqqqqv27/+TP8A4qf9gCT/ANDSv2Tf2KP2gv2g/hbL47+GHxlsvC+jx6rNZtYzajeQMZUSNmfbCpXBDqM9eK/4db/tj/8ARzWl/wDg61L/AOIr/h1v+2P/ANHNaX/4OtS/+Ir/AIdb/tj/APRzWl/+DrUv/iK+Hmgan4U+H/hnwtrV+t9qGj6PaWd3cqzMJ5YoVR3Bb5iGZScnnnmqqqqqqqqqqqqqqqqqrxJ8Yfine/8ABUi++F938Qddm8I295OkWiPeubVFGjmXAjzgDzAH+vNVVVVVVVVVVVVVVVVVVVVf8FDfiD44+GvwX0PxB8P/ABZqfh/Un8VW8DXen3LQSGM2t0xXcpzjciHHqor9lTxBrfiv9mv4Z+JfEurXOp6rqfhiyuLy8upDJLPK0SlndjySTySaqqqqqqqqqqqqqqqqqqqq/wCCYHxs+LvxR+MnxQ0b4i/EjxB4jsdLslayt9SvnnS3b7Sy5QMTj5QB9Kqqqqqqqqqqqqqqqqqv2sfhF44+OfwQ1j4cfDrxZD4c12/uLWSDUZZ5YVjWKZHcbogXGVUjgd+a/wCHW/7Y/wD0c1pf/g61L/4iv+HW/wC2P/0c1pf/AIOtS/8AiK/4db/tj/8ARzWl/wDg61L/AOIr9sn9mX4u/s2al4Wsfix8SbXxdNr0FzJYvBeXNx9nWJow4PnqCMl16enNVVVVVVn/AMfUX++Kqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv+ClNrc/C79vTR/iVNbkQ3dvo2txOqg7/ALM4iYdPvD7L0OeCvYirW6tr61hvbOdJoLiNZIpEOQ6sMgj2IIr/AIKDeK7bwf8AsdfEu+nnCPfaYunQrkZdrmZIcAHr8rsT3wCeMZr/AIIweE7qw+E3xA8ayxFYdZ1+CyiJGN32aDcSO5GbnGemQfQ1Vf8ABLj/AJPH+M3/AF4X/wD6co6qqqqqqqqqqqqqqqqqqqq/4La/80Y/7j3/ALj68Pf8i/pn/XnF/wCgCqqqqqqqqqqqqqqqqqqqr4y/8kg8c/8AYtah/wCk0lf8EW/+SX/Eb/sPW3/og1VVVVVVVVVVVVVVVVVVVV+01/ylu+G3/X/oH/odVVVVVVVVVVVVVVVVVVVVf8FNf+TH/iT/ANwr/wBOtpX/AASz/wCTL/CP/X/qX/pZLVVVVVVVVVVVVVVVVVVVV+3f/wAmf/FT/sASf+hpX/BIP/k1C7/7G6+/9E29VVVVVVVVVVVVVVVVVVVVa3/ymD1T/r/uP/TG1VVVVVVVVVVVVVVVVVVVVf8ABUT/AJIBoH/Y423/AKR3lfsaf8mofCX/ALFGw/8ARK1VVVVVVVVVVVVVVVVVVVV/wSF/5Lt8Xv8ArwT/ANK2qqqqqqqqqqqqqqqqqqqqr/gtb/yM/wAKP+vDVP8A0Zb1VVVVVWf/AB9Rf74qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/4Kc/sqa3+0F8K7Dxr8P9Ka/8Y+BWllisoVzLqFlIB50KAcvIpRHRepw6qCzgH9jP/gp98OvBnwr0z4V/tEy6rpup+FLdbKx1eGze6S7toxtjSVUBdJUUBM7SGCgkg5z+2T+1dr37efjDwz+zx+zf4X1i/wBEW/E482LypdUugpVZGXP7q3iRnbc5H3izBdor9mj4IaZ+zt8EvDHwn06aO5m0m13ahdIDi6vJCXnkGedpdmCg8hQo7VVf8EuP+Tx/jN/14X//AKco6qqqqqqqqqqqqqqqqqqqq/4La/8ANGP+49/7j68Pf8i/pn/XnF/6AKqqqqqqqqqqqqqqqqqqqvjL/wAkg8c/9i1qH/pNJX/BFv8A5Jf8Rv8AsPW3/og1VVVVVVVVVVVVVVVVVVVV+01/ylu+G3/X/oH/AKHVVVVVVVVVVVVVVVVVVVVX/BTX/kx/4k/9wr/062lf8Es/+TL/AAj/ANf+pf8ApZLVVVVVVVVVVVVVVVVVVVV+3f8A8mf/ABU/7AEn/oaV/wAEg/8Ak1C7/wCxuvv/AETb1VVVVVVVVVVVVVVVVVVVVrf/ACmD1T/r/uP/AExtVVVVVVVVVVVVVVVVVVVVX/BUT/kgGgf9jjbf+kd5X7Gn/JqHwl/7FGw/9ErVVVVVVVVVVVVVVVVVVVVX/BIX/ku3xe/68E/9K2qqqqqqqqqqqqqqqqqqqqr/AILW/wDIz/Cj/rw1T/0Zb1VVVVVWf/H1F/viqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqviN+yF+zN8WdZk8RePfgx4d1HVZ33z30cDWs1w396R4SjSHnqxJ6egr4Y/A34P8AwYs5LL4WfDnQvDQnULPLY2irNOAcgSSnLuB23McVVV/wTe8Z+DvBP7Xfxh1Dxn4s0bQbWe0v4op9Uv47VJH/ALRQ7VaRgCcAnA5wDX/C/vgR/wBFs8Bf+FJaf/HK/wCF/fAj/otngL/wpLT/AOOV/wAL++BH/RbPAX/hSWn/AMcr/hf3wI/6LZ4C/wDCktP/AI5X/C/vgR/0WzwF/wCFJaf/AByv+F/fAj/otngL/wAKS0/+OV/wv74Ef9Fs8Bf+FJaf/HK/4X98CP8AotngL/wpLT/45X/C/vgR/wBFs8Bf+FJaf/HK/wCF/fAj/otngL/wpLT/AOOV/wAL++BH/RbPAX/hSWn/AMcr/hf3wI/6LZ4C/wDCktP/AI5X/C/vgR/0WzwF/wCFJaf/AByv+F/fAj/otngL/wAKS0/+OV/wv74Ef9Fs8Bf+FJaf/HK/4X98CP8AotngL/wpLT/45X/C/vgR/wBFs8Bf+FJaf/HK/wCF/fAj/otngL/wpLT/AOOV/wAL++BH/RbPAX/hSWn/AMcr/hf3wI/6LZ4C/wDCktP/AI5X/C/vgR/0WzwF/wCFJaf/AByv+F/fAj/otngL/wAKS0/+OV/wv74Ef9Fs8Bf+FJaf/HK/4X98CP8AotngL/wpLT/45X/C/vgR/wBFs8Bf+FJaf/HK/wCF/fAj/otngL/wpLT/AOOV/wAL++BH/RbPAX/hSWn/AMcr/hf3wI/6LZ4C/wDCktP/AI5X/C/vgR/0WzwF/wCFJaf/AByv+F/fAj/otngL/wAKS0/+OV/wWM8f+A/HX/Cov+EJ8baB4h+w/wBt/af7L1KK78jf9g2b/LY7d21sZ67T6GtB+MPwjj0LTo5Pip4QV0tIgytrluCCEGQRvr/hcnwg/wCireDv/B7bf/F1/wALk+EH/RVvB3/g9tv/AIuv+FyfCD/oq3g7/wAHtt/8XX/C5PhB/wBFW8Hf+D22/wDi6/4XJ8IP+ireDv8Awe23/wAXX/C5PhB/0Vbwd/4Pbb/4uv8Ahcnwg/6Kt4O/8Htt/wDF1/wuT4Qf9FW8Hf8Ag9tv/i6/4XJ8IP8Aoq3g7/we23/xdf8AC5PhB/0Vbwd/4Pbb/wCLr/hcnwg/6Kt4O/8AB7bf/F1/wuT4Qf8ARVvB3/g9tv8A4uv+FyfCD/oq3g7/AMHtt/8AF1/wuT4Qf9FW8Hf+D22/+Lr/AIXJ8IP+ireDv/B7bf8Axdf8Lk+EH/RVvB3/AIPbb/4uv+FyfCD/AKKt4O/8Htt/8XX/AAuT4Qf9FW8Hf+D22/8Ai6/4XJ8IP+ireDv/AAe23/xdf8Lk+EH/AEVbwd/4Pbb/AOLr/hcnwg/6Kt4O/wDB7bf/ABdf8Lk+EH/RVvB3/g9tv/i6/wCFyfCD/oq3g7/we23/AMXX/C5PhB/0Vbwd/wCD22/+Lr/hcnwg/wCireDv/B7bf/F1/wALk+EH/RVvB3/g9tv/AIuv+FyfCD/oq3g7/wAHtt/8XX/C5PhB/wBFW8Hf+D22/wDi6/4XJ8IP+ireDv8Awe23/wAXXxb+LfwpvfhT40srL4neE7i4n8PX8cUUWtW7PI5t3AVQHySSQABX/BIL4j/DzwR8NviBaeNPHvh3QJ7nXLd4ItU1SG0aVRAQWUSMCRnjIr/hf3wI/wCi2eAv/CktP/jlf8L++BH/AEWzwF/4Ulp/8cr/AIX98CP+i2eAv/CktP8A45X/AAv74Ef9Fs8Bf+FJaf8Axyv+F/fAj/otngL/AMKS0/8Ajlf8L++BH/RbPAX/AIUlp/8AHK/4X98CP+i2eAv/AApLT/45X/C/vgR/0WzwF/4Ulp/8cr/hf3wI/wCi2eAv/CktP/jlf8L++BH/AEWzwF/4Ulp/8cr/AIX98CP+i2eAv/CktP8A45X/AAv74Ef9Fs8Bf+FJaf8Axyv+F/fAj/otngL/AMKS0/8Ajlf8L++BH/RbPAX/AIUlp/8AHK/4X98CP+i2eAv/AApLT/45X/C/vgR/0WzwF/4Ulp/8cr/hf3wI/wCi2eAv/CktP/jlf8L++BH/AEWzwF/4Ulp/8cr/AIX98CP+i2eAv/CktP8A45X/AAv74Ef9Fs8Bf+FJaf8Axyv+F/fAj/otngL/AMKS0/8Ajlf8L++BH/RbPAX/AIUlp/8AHK/4X98CP+i2eAv/AApLT/45X/C/vgR/0WzwF/4Ulp/8cr/hf3wI/wCi2eAv/CktP/jlf8L++BH/AEWzwF/4Ulp/8cr/AIX98CP+i2eAv/CktP8A45X/AAv74Ef9Fs8Bf+FJaf8Axyv+F/fAj/otngL/AMKS0/8Ajlf8L++BH/RbPAX/AIUlp/8AHK/aM8d+B9V/4Kl/D3xhpnjPQrzQbW+0Np9Ug1GKS1iCN85aUMUUL3yeO9f8L++BH/RbPAX/AIUlp/8AHK/4X98CP+i2eAv/AApLT/45X/C/vgR/0WzwF/4Ulp/8cr/hf3wI/wCi2eAv/CktP/jlf8L++BH/AEWzwF/4Ulp/8cr/AIX98CP+i2eAv/CktP8A45X/AAv74Ef9Fs8Bf+FJaf8Axyv+F/fAj/otngL/AMKS0/8Ajlf8L++BH/RbPAX/AIUlp/8AHK/4X98CP+i2eAv/AApLT/45X/C/vgR/0WzwF/4Ulp/8cr/hf3wI/wCi2eAv/CktP/jlf8L++BH/AEWzwF/4Ulp/8cr/AIX98CP+i2eAv/CktP8A45X/AAv74Ef9Fs8Bf+FJaf8Axyv+F/fAj/otngL/AMKS0/8Ajlf8L++BH/RbPAX/AIUlp/8AHK/4X98CP+i2eAv/AApLT/45X/C/vgR/0WzwF/4Ulp/8cr/hf3wI/wCi2eAv/CktP/jlf8L++BH/AEWzwF/4Ulp/8cr/AIX98CP+i2eAv/CktP8A45X/AAv74Ef9Fs8Bf+FJaf8Axyv+F/fAj/otngL/AMKS0/8Ajlf8L++BH/RbPAX/AIUlp/8AHK/4X98CP+i2eAv/AApLT/45X/C/vgR/0WzwF/4Ulp/8cr/hf3wI/wCi2eAv/CktP/jlf8L++BH/AEWzwF/4Ulp/8cr/AIX98CP+i2eAv/CktP8A45X/AAUY+L3wl8U/safELQfDHxR8I6vqdz/ZnkWVjrdvcTS7dTtWbaiOWOFVicDgAntX/BLP/ky/wj/1/wCpf+lktVVVVVVVVVVVVVVVVVVVX7d//Jn/AMVP+wBJ/wChpX/BKf4p/DDwb+zBdaR4v+I/hfQ78+KbyUWupaxBaylDFAA2x3B2kg84xwa/4X98CP8AotngL/wpLT/45X/C/vgR/wBFs8Bf+FJaf/HK/wCF/fAj/otngL/wpLT/AOOV/wAL++BH/RbPAX/hSWn/AMcr/hf3wI/6LZ4C/wDCktP/AI5X/C/vgR/0WzwF/wCFJaf/AByv+F/fAj/otngL/wAKS0/+OV/wv74Ef9Fs8Bf+FJaf/HK/4X98CP8AotngL/wpLT/45X/C/vgR/wBFs8Bf+FJaf/HK/wCF/fAj/otngL/wpLT/AOOV/wAL++BH/RbPAX/hSWn/AMcr/hf3wI/6LZ4C/wDCktP/AI5X/C/vgR/0WzwF/wCFJaf/AByv+F/fAj/otngL/wAKS0/+OV/wv74Ef9Fs8Bf+FJaf/HK/4X98CP8AotngL/wpLT/45X/C/vgR/wBFs8Bf+FJaf/HK/wCF/fAj/otngL/wpLT/AOOV/wAL++BH/RbPAX/hSWn/AMcr/hf3wI/6LZ4C/wDCktP/AI5X/C/vgR/0WzwF/wCFJaf/AByv+F/fAj/otngL/wAKS0/+OV/wv74Ef9Fs8Bf+FJaf/HK/4X98CP8AotngL/wpLT/45X/C/vgR/wBFs8Bf+FJaf/HK/wCF/fAj/otngL/wpLT/AOOV/wAL++BH/RbPAX/hSWn/AMcr/hf3wI/6LZ4C/wDCktP/AI5X/C/vgR/0WzwF/wCFJaf/ABytY8a+DZP+Cr+peMo/Fuit4fa+nK6qL+I2pB0VkBE27Zjf8vXrx1r/AIXJ8IP+ireDv/B7bf8Axdf8Lk+EH/RVvB3/AIPbb/4uv+FyfCD/AKKt4O/8Htt/8XX/AAuT4Qf9FW8Hf+D22/8Ai6/4XJ8IP+ireDv/AAe23/xdf8Lk+EH/AEVbwd/4Pbb/AOLr/hcnwg/6Kt4O/wDB7bf/ABdf8Lk+EH/RVvB3/g9tv/i6/wCFyfCD/oq3g7/we23/AMXX/C5PhB/0Vbwd/wCD22/+Lr/hcnwg/wCireDv/B7bf/F1/wALk+EH/RVvB3/g9tv/AIuv+FyfCD/oq3g7/wAHtt/8XX/C5PhB/wBFW8Hf+D22/wDi6/4XJ8IP+ireDv8Awe23/wAXX/C5PhB/0Vbwd/4Pbb/4uv8Ahcnwg/6Kt4O/8Htt/wDF1/wuT4Qf9FW8Hf8Ag9tv/i6/4XJ8IP8Aoq3g7/we23/xdf8AC5PhB/0Vbwd/4Pbb/wCLr/hcnwg/6Kt4O/8AB7bf/F1/wuT4Qf8ARVvB3/g9tv8A4uv+FyfCD/oq3g7/AMHtt/8AF1/wuT4Qf9FW8Hf+D22/+Lr/AIXJ8IP+ireDv/B7bf8Axdf8Lk+EH/RVvB3/AIPbb/4uv+FyfCD/AKKt4O/8Htt/8XX/AAuT4Qf9FW8Hf+D22/8Ai6/4XJ8IP+ireDv/AAe23/xdf8Lk+EH/AEVbwd/4Pbb/AOLr/gpL4/8AAfiv4F6Hp3hfxtoOsXcfiy3leCw1KK4dUFpdgsVRiQuWUZ6ZI9a/ZH+NXwa0T9mD4W6RrPxb8GWF/Z+FbGK4tbrXraKWF1iUFXRnBUg9QRmv+F/fAj/otngL/wAKS0/+OV/wv74Ef9Fs8Bf+FJaf/HK/4X98CP8AotngL/wpLT/45X/C/vgR/wBFs8Bf+FJaf/HK/wCF/fAj/otngL/wpLT/AOOV/wAL++BH/RbPAX/hSWn/AMcr/hf3wI/6LZ4C/wDCktP/AI5X/C/vgR/0WzwF/wCFJaf/AByv+F/fAj/otngL/wAKS0/+OV/wv74Ef9Fs8Bf+FJaf/HK/4X98CP8AotngL/wpLT/45X/C/vgR/wBFs8Bf+FJaf/HK/wCF/fAj/otngL/wpLT/AOOV/wAL++BH/RbPAX/hSWn/AMcr/hf3wI/6LZ4C/wDCktP/AI5X/C/vgR/0WzwF/wCFJaf/AByv+F/fAj/otngL/wAKS0/+OV/wv74Ef9Fs8Bf+FJaf/HK/4X98CP8AotngL/wpLT/45X/C/vgR/wBFs8Bf+FJaf/HK/wCF/fAj/otngL/wpLT/AOOV/wAL++BH/RbPAX/hSWn/AMcr/hf3wI/6LZ4C/wDCktP/AI5X/C/vgR/0WzwF/wCFJaf/AByv+F/fAj/otngL/wAKS0/+OV/wv74Ef9Fs8Bf+FJaf/HK/4X98CP8AotngL/wpLT/45X/C/vgR/wBFs8Bf+FJaf/HK/wCF/fAj/otngL/wpLT/AOOV/wAL++BH/RbPAX/hSWn/AMcr/glR478D+DfjX8VdQ8X+M9C0O1vLFVtp9S1GK1SY/amOEZ2AbjnjtX/C/vgR/wBFs8Bf+FJaf/HK/wCF/fAj/otngL/wpLT/AOOV/wAL++BH/RbPAX/hSWn/AMcr/hf3wI/6LZ4C/wDCktP/AI5X/C/vgR/0WzwF/wCFJaf/AByv+F/fAj/otngL/wAKS0/+OV/wv74Ef9Fs8Bf+FJaf/HK/4X98CP8AotngL/wpLT/45X/C/vgR/wBFs8Bf+FJaf/HK/wCF/fAj/otngL/wpLT/AOOV/wAL++BH/RbPAX/hSWn/AMcr/hf3wI/6LZ4C/wDCktP/AI5X/C/vgR/0WzwF/wCFJaf/AByv+F/fAj/otngL/wAKS0/+OV/wv74Ef9Fs8Bf+FJaf/HK/4X98CP8AotngL/wpLT/45X/C/vgR/wBFs8Bf+FJaf/HK/wCF/fAj/otngL/wpLT/AOOV/wAL++BH/RbPAX/hSWn/AMcr/hf3wI/6LZ4C/wDCktP/AI5X/C/vgR/0WzwF/wCFJaf/AByv+F/fAj/otngL/wAKS0/+OV/wv74Ef9Fs8Bf+FJaf/HK/4X98CP8AotngL/wpLT/45X/C/vgR/wBFs8Bf+FJaf/HK/wCF/fAj/otngL/wpLT/AOOV/wAL++BH/RbPAX/hSWn/AMcr/hf3wI/6LZ4C/wDCktP/AI5X/C/vgR/0WzwF/wCFJaf/AByv+F/fAj/otngL/wAKS0/+OV/wWF8d+B/HPiT4YS+CvGeheIEs7HUluG0vUYrsQlnt9oYxsducHGeuDVVVVVVZ/wDH1F/viqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqrx7/AMEf/hJ4z8a654usfil4n0qPWr+a8NktrBMtu0rl2VWIB2gsQM8gYyT1r/hy38L/APotXij/AMF8H+Nf8OW/hf8A9Fq8Uf8Agvg/xr/hy38L/wDotXij/wAF8H+Nf8OW/hf/ANFq8Uf+C+D/ABr/AIct/C//AKLV4o/8F8H+Nf8ADlv4X/8ARavFH/gvg/xr/hy38L/+i1eKP/BfB/jX/Dlv4X/9Fq8Uf+C+D/Gv+HLfwv8A+i1eKP8AwXwf41/w5b+F/wD0WrxR/wCC+D/Gv+HLfwv/AOi1eKP/AAXwf41/w5b+F/8A0WrxR/4L4P8AGv8Ahy38L/8AotXij/wXwf41/wAOW/hf/wBFq8Uf+C+D/Gv+HLfwv/6LV4o/8F8H+Nf8OW/hf/0WrxR/4L4P8a/4ct/C/wD6LV4o/wDBfB/jX/Dlv4X/APRavFH/AIL4P8a/4ct/C/8A6LV4o/8ABfB/jX/Dlv4X/wDRavFH/gvg/wAa/wCHLfwv/wCi1eKP/BfB/jX/AA5b+F//AEWrxR/4L4P8a/4ct/C//otXij/wXwf41/w5b+F//RavFH/gvg/xr/hy38L/APotXij/AMF8H+Nf8OW/hf8A9Fq8Uf8Agvg/xr/hy38L/wDotXij/wAF8H+Nf8OW/hf/ANFq8Uf+C+D/ABr/AIct/C//AKLV4o/8F8H+Nf8ADlv4X/8ARavFH/gvg/xr/hy38L/+i1eKP/BfB/jX/Dlv4X/9Fq8Uf+C+D/Gv+HLfwv8A+i1eKP8AwXwf41/w5b+F/wD0WrxR/wCC+D/Gv+HLfwv/AOi1eKP/AAXwf41/w5b+F/8A0WrxR/4L4P8AGv8Ahy38L/8AotXij/wXwf41/wAOW/hf/wBFq8Uf+C+D/Gv+HLfwv/6LV4o/8F8H+Nf8OW/hf/0WrxR/4L4P8a/4ct/C/wD6LV4o/wDBfB/jX/Dlv4X/APRavFH/AIL4P8a/4ct/C/8A6LV4o/8ABfB/jX/Dlv4X/wDRavFH/gvg/wAa/wCHLfwv/wCi1eKP/BfB/jX/AA5b+F//AEWrxR/4L4P8a/4ct/C//otXij/wXwf41/w5b+F//RavFH/gvg/xr/hy38L/APotXij/AMF8H+Nf8OW/hf8A9Fq8Uf8Agvg/xr/hy38L/wDotXij/wAF8H+Nf8OW/hf/ANFq8Uf+C+D/ABr/AIct/C//AKLV4o/8F8H+Nf8ADlv4X/8ARavFH/gvg/xr/hy38L/+i1eKP/BfB/jX/Dlv4X/9Fq8Uf+C+D/Gv+HLfwv8A+i1eKP8AwXwf41/w5b+F/wD0WrxR/wCC+D/Gv+HLfwv/AOi1eKP/AAXwf41/w5b+F/8A0WrxR/4L4P8AGv8Ahy38L/8AotXij/wXwf41/wAOW/hf/wBFq8Uf+C+D/Gv+HLfwv/6LV4o/8F8H+Nf8OW/hf/0WrxR/4L4P8a/4ct/C/wD6LV4o/wDBfB/jX/Dlv4X/APRavFH/AIL4P8a/4ct/C/8A6LV4o/8ABfB/jX/Dlv4X/wDRavFH/gvg/wAa/wCHLfwv/wCi1eKP/BfB/jX/AA5b+F//AEWrxR/4L4P8a/4ct/C//otXij/wXwf41/w5b+F//RavFH/gvg/xr/hy38L/APotXij/AMF8H+Nf8OW/hf8A9Fq8Uf8Agvg/xr/hy38L/wDotXij/wAF8H+Nf8OW/hf/ANFq8Uf+C+D/ABr/AIct/C//AKLV4o/8F8H+Nf8ADlv4X/8ARavFH/gvg/xr/hy38L/+i1eKP/BfB/jX/Dlv4X/9Fq8Uf+C+D/Gv+HLfwv8A+i1eKP8AwXwf41/w5b+F/wD0WrxR/wCC+D/Gv+HLfwv/AOi1eKP/AAXwf41/w5b+F/8A0WrxR/4L4P8AGv8Ahy38L/8AotXij/wXwf41/wAOW/hf/wBFq8Uf+C+D/Gv+HLfwv/6LV4o/8F8H+Nf8OW/hf/0WrxR/4L4P8a/4ct/C/wD6LV4o/wDBfB/jX/Dlv4X/APRavFH/AIL4P8a/4ct/C/8A6LV4o/8ABfB/jX/Dlv4X/wDRavFH/gvg/wAa/wCHLfwv/wCi1eKP/BfB/jX/AA5b+F//AEWrxR/4L4P8a/4ct/C//otXij/wXwf41/w5b+F//RavFH/gvg/xr/hy38L/APotXij/AMF8H+Nf8OW/hf8A9Fq8Uf8Agvg/xr/hy38L/wDotXij/wAF8H+Nf8OW/hf/ANFq8Uf+C+D/ABr/AIct/C//AKLV4o/8F8H+Nf8ADlv4X/8ARavFH/gvg/xr/hy38L/+i1eKP/BfB/jX/Dlv4X/9Fq8Uf+C+D/Gv+HLfwv8A+i1eKP8AwXwf41/w5b+F/wD0WrxR/wCC+D/Gv+HLfwv/AOi1eKP/AAXwf41/w5b+F/8A0WrxR/4L4P8AGv8Ahy38L/8AotXij/wXwf41/wAOW/hf/wBFq8Uf+C+D/Gv+HLfwv/6LV4o/8F8H+Nf8OW/hf/0WrxR/4L4P8a/4ct/C/wD6LV4o/wDBfB/jX/Dlv4X/APRavFH/AIL4P8a/4ct/C/8A6LV4o/8ABfB/jX/Dlv4X/wDRavFH/gvg/wAa/wCHLfwv/wCi1eKP/BfB/jX/AA5b+F//AEWrxR/4L4P8a/4ct/C//otXij/wXwf41/w5b+F//RavFH/gvg/xr/hy38L/APotXij/AMF8H+Nf8OW/hf8A9Fq8Uf8Agvg/xr/hy38L/wDotXij/wAF8H+Nf8OW/hf/ANFq8Uf+C+D/ABr/AIct/C//AKLV4o/8F8H+NfAD4JeGP2dvhRovwk8IXt7e6fo4lb7VespmnkllaR3baAB8zkAAcAAc9TVVVVVVVVVVVVVVVVVVVfFj4a6D8Yvht4i+F/ia4u4NM8SWD2dxLaOEmjDdGQsCNwIBGQRxyDX/AA5b+F//AEWrxR/4L4P8a/4ct/C//otXij/wXwf41/w5b+F//RavFH/gvg/xr/hy38L/APotXij/AMF8H+Nf8OW/hf8A9Fq8Uf8Agvg/xr/hy38L/wDotXij/wAF8H+Nf8OW/hf/ANFq8Uf+C+D/ABr/AIct/C//AKLV4o/8F8H+Nf8ADlv4X/8ARavFH/gvg/xr/hy38L/+i1eKP/BfB/jX/Dlv4X/9Fq8Uf+C+D/Gv+HLfwv8A+i1eKP8AwXwf41/w5b+F/wD0WrxR/wCC+D/Gv+HLfwv/AOi1eKP/AAXwf41/w5b+F/8A0WrxR/4L4P8AGv8Ahy38L/8AotXij/wXwf41/wAOW/hf/wBFq8Uf+C+D/Gv+HLfwv/6LV4o/8F8H+Nf8OW/hf/0WrxR/4L4P8a/4ct/C/wD6LV4o/wDBfB/jX/Dlv4X/APRavFH/AIL4P8a/4ct/C/8A6LV4o/8ABfB/jX/Dlv4X/wDRavFH/gvg/wAa/wCHLfwv/wCi1eKP/BfB/jX/AA5b+F//AEWrxR/4L4P8a/4ct/C//otXij/wXwf41/w5b+F//RavFH/gvg/xr/hy38L/APotXij/AMF8H+Nf8OW/hf8A9Fq8Uf8Agvg/xr/hy38L/wDotXij/wAF8H+Nf8OW/hf/ANFq8Uf+C+D/ABr/AIct/C//AKLV4o/8F8H+Nf8ADlv4X/8ARavFH/gvg/xr/hy38L/+i1eKP/BfB/jX/Dlv4X/9Fq8Uf+C+D/Gv+HLfwv8A+i1eKP8AwXwf41/w5b+F/wD0WrxR/wCC+D/Gv+HLfwv/AOi1eKP/AAXwf41/w5b+F/8A0WrxR/4L4P8AGv8Ahy38L/8AotXij/wXwf41/wAOW/hf/wBFq8Uf+C+D/Gv+HLfwv/6LV4o/8F8H+Nf8OW/hf/0WrxR/4L4P8a/4ct/C/wD6LV4o/wDBfB/jX/Dlv4X/APRavFH/AIL4P8a/4ct/C/8A6LV4o/8ABfB/jX/Dlv4X/wDRavFH/gvg/wAa/wCHLfwv/wCi1eKP/BfB/jX/AA5b+F//AEWrxR/4L4P8a/4ct/C//otXij/wXwf41/w5b+F//RavFH/gvg/xr/hy38L/APotXij/AMF8H+Nf8OW/hf8A9Fq8Uf8Agvg/xr/hy38L/wDotXij/wAF8H+Nf8OW/hf/ANFq8Uf+C+D/ABr/AIct/C//AKLV4o/8F8H+Nf8ADlv4X/8ARavFH/gvg/xr/hy38L/+i1eKP/BfB/jX/Dlv4X/9Fq8Uf+C+D/Gv+HLfwv8A+i1eKP8AwXwf41/w5b+F/wD0WrxR/wCC+D/Gv+HLfwv/AOi1eKP/AAXwf41/w5b+F/8A0WrxR/4L4P8AGv8Ahy38L/8AotXij/wXwf41/wAOW/hf/wBFq8Uf+C+D/Gv+HLfwv/6LV4o/8F8H+Nf8OW/hf/0WrxR/4L4P8a/4ct/C/wD6LV4o/wDBfB/jX/Dlv4X/APRavFH/AIL4P8a/4ct/C/8A6LV4o/8ABfB/jX/Dlv4X/wDRavFH/gvg/wAa/wCHLfwv/wCi1eKP/BfB/jX/AA5b+F//AEWrxR/4L4P8a/4ct/C//otXij/wXwf41/w5b+F//RavFH/gvg/xr/hy38L/APotXij/AMF8H+Nf8OW/hf8A9Fq8Uf8Agvg/xr/hy38L/wDotXij/wAF8H+Nf8OW/hf/ANFq8Uf+C+D/ABr/AIct/C//AKLV4o/8F8H+Nf8ADlv4X/8ARavFH/gvg/xr/hy38L/+i1eKP/BfB/jX/Dlv4X/9Fq8Uf+C+D/Gv+HLfwv8A+i1eKP8AwXwf41/w5b+F/wD0WrxR/wCC+D/Gv+HLfwv/AOi1eKP/AAXwf41/w5b+F/8A0WrxR/4L4P8AGv8Ahy38L/8AotXij/wXwf41/wAOW/hf/wBFq8Uf+C+D/Gv+HLfwv/6LV4o/8F8H+Nf8OW/hf/0WrxR/4L4P8a/4ct/C/wD6LV4o/wDBfB/jX/Dlv4X/APRavFH/AIL4P8a/4ct/C/8A6LV4o/8ABfB/jX/Dlv4X/wDRavFH/gvg/wAa/wCHLfwv/wCi1eKP/BfB/jX/AA5b+F//AEWrxR/4L4P8a/4ct/C//otXij/wXwf41/w5b+F//RavFH/gvg/xr/hy38L/APotXij/AMF8H+Nf8OW/hf8A9Fq8Uf8Agvg/xr/hy38L/wDotXij/wAF8H+Nf8OW/hf/ANFq8Uf+C+D/ABr/AIct/C//AKLV4o/8F8H+Nf8ADlv4X/8ARavFH/gvg/xr/hy38L/+i1eKP/BfB/jX/Dlv4X/9Fq8Uf+C+D/Gv+HLfwv8A+i1eKP8AwXwf41/w5b+F/wD0WrxR/wCC+D/Gv+HLfwv/AOi1eKP/AAXwf41/w5b+F/8A0WrxR/4L4P8AGv8Ahy38L/8AotXij/wXwf41/wAOW/hf/wBFq8Uf+C+D/Gv+HLfwv/6LV4o/8F8H+Nf8OW/hf/0WrxR/4L4P8a/4ct/C/wD6LV4o/wDBfB/jX/Dlv4X/APRavFH/AIL4P8a/4ct/C/8A6LV4o/8ABfB/jX/Dlv4X/wDRavFH/gvg/wAa/wCHLfwv/wCi1eKP/BfB/jX/AA5b+F//AEWrxR/4L4P8a/4ct/C//otXij/wXwf41/w5b+F//RavFH/gvg/xr/hy38L/APotXij/AMF8H+Nf8OW/hf8A9Fq8Uf8Agvg/xrR/+CMnwestVs7vVvix4q1CzgmV57RbWCEzoDkpvAJUHpkDNVVVVVVn/wAfUX++KqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqrP/j6i/wB8VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVWf/H1F/viqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqvHf7Z37LPw11mXw74w+N3hu11O2cxz2tvK948DjqsggV9jDHKtgivhx8XPhh8YNJfW/hh480XxNZwkLM+n3aytAx6CRAd0ZOOjAGqqqqqqqqqqqqqqqqqqqq8vLPT7Sa+v7uG2treMyTTTOESNQMlmY8AAdSa8E/tA/A/4leIJPCnw9+LPhbxHrEMTzPZaZqcdzII0IDPhCflBZRnpyKqqqqqqqqqqqqqqqqqqqvFfjHwj4E0aXxF428UaToGlwf6y91O8S1hT2LuQM+1D9vj9jttTGkD4++HPPJwHPnCL/v7s8vHvurw14q8MeNNGg8ReDvEema5pV0MwX2nXaXMMn+66EqfzqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqtU1TTNE0641fWdRtbCxs4mluLq6mWKKFFGSzuxAVQOpJxV1+3r+x5aaudDl+P3hprgPs3xNLJDn/rsqGPHHXdivD/iLw/4s0e18Q+FtcsNY0u9TfbXthcpPDMvqjoSrDjsaqqqqqqqqqqqqqqqqqqqvG3xA8C/DXRG8R/EHxho/hzS1cJ9r1S9S2jLnooZyMsfQc14A+Jnw9+K2jTeIfhr4z0jxNplvctay3emXSzxpMqqzRllJAYK6HHow9aqqqqqqqqqqqqqqqqqqqviP8cvg58II42+J3xM8O+G3mGYoL+/SOaUeqRZ3sPcAgV4T/be/ZL8b6imkeH/AI8+FzdyHbHHeztY727BTcKgJPYA5NI6SIrxuGVhlSDkEeoqqqqqqqqqqqqqqqqqqqqqqqqqqs/+PqL/AHxVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV/wVX/AGtvEXw30zTv2e/hnq81jrfiazN1rt7aviaCyYskduhHKtKVcsRhgigdJK/Z2/4JL/CGy+HGn6l+0Bb6rrPi/VLdZrqzttQe1g0ssMiFfLwXkUHDMxK7hhRgZP7RXwc8d/8ABM34/eF/if8ABbxPqFx4b1jzGsWvGyZAjL9o0+72ALIjKyEHAJzkANHur4WfETQ/i58N/DfxM8Nsf7O8SabDewozAtDvUFo2I/iRtyH3U1VVVVVVVVVVVVVVVVVVXxU/Zq/bo/bH+MPijwx8TfGn/CH/AAn0TXbm300+T5UF3bRykRSxWiMHuGKbW8yZtoJO1uMV/wAEm7FNN/bF1jTY5C62vhrUYlZhgkLcQDP6VVVVVVVVVVVVVVVVVVVVfHj/AIJ0+OP2l/2lNZ8c/En4zajD8OkigOj6fE/nXUGY1E0EKsPKt0DqW37XLbhlSctWuf8ABIP9lu/0B9N0XU/Gel6iI8Raj/aSTsH7F42j2sPUAL7Eda/Z98afFD/gn/8AtmN8EvEetNdeHdQ1q20zW7dNwt7uC42i3vo1P3XVZEfjnG9CTVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVfthfF34j/tuftU2f7Jfwo1jyPCumau2nqqyEQXV1AGN1ezbfvxxBJAg5GIyy8virf/AIJGfssR+CV8O3MnimbXfI2t4gGpFJvNxy4hwYgueilSccbifmr9mv4ifEL/AIJ+ftg3v7O3j3W3uvBmsatDY3ysStuROF+y6lECSIztePzME/LuU5aNStVVVVVVVVVVVVVVVVVVfGvUviBovwi8Yaz8KbSK68X6fo1zcaPbyWxuPPuEQssaxggu7Y2qO7EZBHFftB/slftS6v8ABHxX+1h+1Z8Q5jrekR2n2DQJ3W4mC3F3BCQwjIhtUAmLCOME5GCFOa/4I2f8mweJv+x9u/8A0gsKqqqqqqqqqqqqqqqqqqrW7C51XRr/AEyy1SfTbi7tZIYb23CmS2dlIEiBgRuUkMMgjIGRXwq/4JHR+Ibu58bftVfFTWfEHiHUJ2lntdJuy28/3p7qZWeViMZChcY+8wr9oL/gkR8MW8B6nrvwC1fX7HxNpts89tpeoXIu7fUdi58kHaHSRsYVslckAgA7h/wSI/aY8WeJLrWP2cfGOqzajaaRph1Lw5NOxZ7WJJESa23HkpmVGQH7uHHTAFVVVVVVVVVVVVVVVVVVVVVVVVVZ/wDH1F/viqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqrxcG+N3/AAVnXTtUb7VbWvj2C1aEkFfJ01VDJjOApFs2R6s3cmq/4K4eFbXXv2Tm12SENP4b8RWV1FJ0KiTfAw+h84ceoB7V/wAEmvFE/iH9kOx0ye4Mo8OeIL/T4wX3FFJS42+3/HwTj3qqqqqqqqqqqqqqqqqqqq/4JW/8nreIv+wBqn/pTBVVVVVVVVVVVVVVVVVVVVVftbJB+0P/AMFLIPA3gsfbAutaXoc00HOGgCfanPtF+9DenlGqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqvid4nfwR8NfFnjSJlD6Bod7qClsYBhgeQZzxj5e/Ff8EbvC6678dfG/jzUA1zPo/h4QpLKdxEt1OpLknksVhcZ/2jVf8ABZrwxBpnxm8B+NLWPyp9X8PSWsjocFmtpywbjnIFwBn0A9K+EviWXxr8KfBfjGaYSya74esL93Dbtxmt0kJz3zu61VVVVVVVVVVVVVVVVVVX/BTX/kx/4k/9wr/062lf8EbP+TYPE3/Y+3f/AKQWFVVVVVVVVVVVVVVVVVVVVeM/FmieAvCOs+NvEl2ttpWg2E17eSscbYokLtj3wOB3OK/4JF+ENU8W/tNeJfiSLNotO0HQ7gzOg+RLi6lVY4vxRZz/ANs6qqqqqqqqqqqqqqqqqqqqqqqqqrP/AI+ov98VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVfDa2Hhr/grndxajIU874j6y6koR/x8/aWjGPfzVGffNV/wVVvLe1/Y18SQTPh7vVNOiiGOrC5Rz/46jV/wR70qbT/ANlXVLuQtt1Pxne3EeUx8otrWLg9xmI8/h2qqqqqqqqqqqqqqqqqqqq/4JW/8nreIv8AsAap/wClMFVVVVVVVVVVVVVVVVVVVVft+/tt6N+zP4Km8G+DNSt7r4l69bFbC3Uh/wCyoWBBu5RyARz5aH7zc4Kqc/8ABL39j3WPA9nJ+0x8V7KdfFHiO3YaDa3gJmtbWXmS6kyc+bMDxnkIST/rCBVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV+0dpc+ufs8/FDRLXPnX/gzV7ePC7juezlUYHfk9K/4Iq3lunjT4o6e0n7+bS9PlRcdVSWYMfzdfzqv+C1V5byeNfhdpyPmeDStQldcdFeWEKfzjav2c9Km0P8AZ6+F+iXBYy6f4M0i3fcm07ks4lOR25HSqqqqqqqqqqqqqqqqqqq/4Ka/8mP/ABJ/7hX/AKdbSv8AgjZ/ybB4m/7H27/9ILCqqqqqqqqqqqqqqqqqqqqv+ChP7WmrftA+LLP9j39m8y6/DfahHb6xdae28ardq+VtYm6GGNlDvJnaWXqEQlv2N/2ZNI/ZY+DVj4HjlhvPEF+/23xBfxg4uLtlA2pnny41ARemcFsAuaqqqqqqqqqqqqqqqqqqqqqqqqqrP/j6i/3xVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV/wAFHfCmv/s8ftreG/2idBsSbPW7mx1u0YfKjXti0STwk+4jiduxEx6818O/H/hX4qeB9F+IXgrVIr/RtdtEubWZGzww5Vh/C6nKsp5VlIPIr/gsZ8cdGvrHwp+zv4f1GO71OG+Gta3DA2422I2jtomwfvOJZXKHkARt/EDX7Gnwivvgd+zP4F+Hms23kavbWButTiI+aO6uHaeRG90Mnl/8Aqqqqqqqqqqqqqqqqqqqq/4JW/8AJ63iL/sAap/6UwVVVVVVVVVVVVVVVVVVVVftx/tz+GP2VvDn/COeHDZ6z8R9Wg3afpjtujsYzwLm5AIIXrtTILkdgCR+xT+w/wCMPi54ui/a4/a1nu9U1DVboajpWjalGC962Mx3NyhGFiA2+XAABhVyAgCtVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV3aW19azWN5Ck0FxG0csbjIdWGCCPQg4r9lvXh+wz+35q/w6+IN3/Z+iXU9x4eury5bZGLaZ0ls7ps8BWKW5LHG1ZGJIwa3ps37xtxnOeMetftV64v7cn7fej/Dj4c3g1LRbOW38PW17btujaCF3lvbpSMgopechx95Y1IzkVbW1vZ20VnaxLHDBGI441GAqgYAHtgVVVVVVVVVVVVVVVVVVVX/BTX/kx/4k/wDcK/8ATraV/wAEbP8Ak2DxN/2Pt3/6QWFVVVVVVVVVVVVVVVVVVVX7bX7ePivx74rb9lb9kZ7jVtW1WY6dqet6Wd8s8rcNbWbA4AADB5+gGdpAUuf2F/2FfDv7LGgHxV4nkt9Z+I2sWypfXqoGi02M8tbWxIzjON8nBcqOABg1VVVVVVVVVVVVVVVVVVVVVVVVVWf/AB9Rf74qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/aC+AHw+/aU+HF58N/iHZO9tKwmsr2DAuNPuVBCzRMQcMASCDwykg8GrD/gnH+3V8Gri80L4C/tH2dr4dvZS5Ftrl7pLOcAb5IER0VyMDKux469K/ZY/4Jh2vwz8eQfGP9oHxrB478X2twLu1tYvMltYbvO77TJLNiS4kDcruVQCNxDHG2qqqqqqqqqqqqqqqqqqqr9jb/gn78Sv2bfj/AKp8XPFXjTwzqmnX+m3lqltp5n85WmmjdSd8argBCDz3FVVVVVVVVVVVVVVVVVVVXxI/4JN/tF/EL4jeIfiBf/GTwjc3etapNeLPdz3jzKGclAW8o4KrtUAHACgDgCv+HW/7Y/8A0c1pf/g61L/4iv8Ah1v+2P8A9HNaX/4OtS/+Ir4eaBqfhT4f+GfC2tX632oaPo9pZ3dyrMwnlihVHcFvmIZlJyeeeaqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/bN/YV8D/ta6da6zHqY8NeN9Jh8my1pLfzVnhyT5FwgILoCSVYHKFiRkEqYv8Agn3/AMFFv7EX4Xt+0lYJ4NEYtvIXxZqJtxbY/wBWIfJztxx5fC9unNfsb/sK+Af2S9PudYTUj4l8bapD5N7rctv5SwxZBMFvHklEyAWJJZyATgAKKqqqqqqqqqqqqqqqqqqr9rj4Ma9+0N+z14r+D/hjVbDTdT1/7H5F1f7xDH5N5BO27YGblYmAwOpFfsH/ALMni39lH4Qav8O/GWv6Rq97qPiSfVI59MMhjWN7a2iCnzFU7swMemMEc9aqqqqqqqqqqqqqqqqqqr9ov4f+Mvir8EvFvw6+H/iaPw/r2v2QtbbUZJZIlhBkXzAWjBYBow6cD+KtI/4I/wD7SPh+7/tDQPjV4P0262FPPs7q9hfaeo3LEDg4HFf8Ot/2x/8Ao5rS/wDwdal/8RXwA/4J6/tQ/C34zeEviD4x+Puna1ouhagtxe6fHqt/I1wgUgqFkQKeo4PFVVVVVVVVVVVVVVVVVVVVVVVVVVn/AMfUX++KqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqrP/AI+ov98VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVWf/AB9Rf74qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqs/+PqL/AHxVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVZ/8fUX++KqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqrP/j6i/3xVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVZ/8fUX++KqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqrP8A4+ov98VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVWf8Ax9Rf74qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/ak+IfiT4Tfs9+OviP4Plt49Z0DSmubJriISoHDKPmU9Rgniv+Hsn7X/8A0GvC3/gjT/4qv+Hsn7X/AP0GvC3/AII0/wDiq/4eyftf/wDQa8Lf+CNP/iq/4eyftf8A/Qa8Lf8AgjT/AOKr40ftGfETwP8A8E99P/aE0ZoD4y1Xwjol0bpbYGK2ur9bdZJxGcqApmYqpBXdsBBGRXgj9tL9p7wP47t/H9t8aPFeq3aXKy3NnqurTXVpeKDzHJC7FdhGVGACoPylcAjQdUOuaFp2tNZzWhv7SK4NvMMPDvQNsYeozg+4qqqqqqqqqvF/irQvAvhXV/Gnie/Sz0jQrGW9vZ26RxRIWY47nAOB3PFaZ+2V+2H+0r+0TbeFvhp8UNb8Nw+M9dEGmaVZMnlaZak4BPy/MI4VLu38RVmxzitE019G0Ww0iTUbvUHsraOFru7ffNcFVAMkjd2bGSfU1VV/wVH8SftEfCHR/Dfxh+DfxS1/QtBeT+ytcsLOUeXFM254JwCCRuAdGPAysfdjX/BMP9sDxF8e/C2tfDL4pa+2qeNPDJ+2W99OQJdQsJGwSwAGWikIUnjKyR9wSaqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr/AIKT/tT/ABa/Zf0PwHf/AApvdMgl8QXV9Fem+shcArEkJTaCRjl2r/h7J+1//wBBrwt/4I0/+Kr/AIeyftf/APQa8Lf+CNP/AIqv+Hsn7X//AEGvC3/gjT/4qv2dP+Clv7UPxM+PHgH4feKNW8OyaR4h1+0sr1YdHWNzFJIFbawPBwetf8FYf2kPiv8ABfw14K8EfC/xDfeGz4uN9LqGrWLGK48u38kLDDKPmjyZtzMhDDCAEBmB/wCCc37Wvx0tv2jPDPww8R+OfEHi3w34ulltLmx1W9lvWtXETus8LSEtHtZMsAdpUsSMgEVVVVVVVVVX7dH7Sa/syfAbU/FWkzxf8JTrL/2b4eifB23LqS0xXnKxIGfkYLBFP3q/4JvfE/8Aar/aO+Nk1542+NPii/8ABfg21F5qsDzKEup5Mrb27ELnDFXcjusLD+Kqqq/bg+Nn7X/7Mn7RWr+FdH+OfidfDeoldW8OiSRJFFpKx/cncpz5ciyR4bJKopP3q/Zc+PGlftJfBHw78U9PSGC7vYjBqtnG2RaXsfyzR+oXOGXPOx0PeqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqrP/j6i/3xVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV8WvhnoPxl+G/iD4XeJ7u/tdK8R2ZtbqawkRJ0QkHKM6uoPA6qR7V/wAOa/2YP+h6+KP/AIM7H/5Dr9t/4AeDf2afjvd/C7wJqes3+lQaZa3SzavNHLOXlUlgWjjjXHHHy/ia/wCCf37Lfw+/au+JXiLwb8RNY8Q6dZaRoRv4JNFuIYZGk8+OPDGWKQFcOeAAc45r/hzX+zB/0PXxR/8ABnY//Idax4H+DvgL9n+3+F/xJ1PTV+HWh+H7TQru58S30VvE9rGkdvEZ5j5aK7ERjcNnzkbcHFeB/wBlb/gln4H8dweOYvj/AOEta+yXC3FrpGr/ABA02exhdTlfkUq8iggfLI7g9wailiuIknglSSORQyOhyGBHBBHUYqb9qP8AZlgleCf9ov4YRyRsVdH8XWKlSOCCPN4Na78QvAXhfwonjrxH410LTPDkkKSx6td6hFFayI4yjLKzbWDAgrgndkYzWl/t4fsf6xrC6DZ/H3wyt0z7A1zJJbQ5zj/XSKseOeu7HerW6tb+1hvrG5iuLe4jWSGaJw6SIwyrKw4IIIII4NeP/wBsb9l/4X6vL4f8a/Gzw3ZanbP5dxZwztdywP3WRIA5Q8dGwRx6ivhl8avhN8Z9Ol1T4WfELRPEsFvjz1sboPJBnp5kZw8ee24Cqr4jftR/s7fCW+k0r4hfGPwxpGowf62wa+EtzH/vQx7pF6HqvNeAP2vf2ZPijqcOieB/jX4Yv9RuDtgspbr7LNM2cbY45gjOfZQT3qqr/grv8WrjwR+zxpvw50y7MN54/wBVEM+1sFrK22yygY55kNup7FWYHrX/AARv+F0PiH4w+LfirfWwki8H6RHaWbMv3bm8ZhvU+oihlU47S89RVeOPiL4A+Gekf298Q/GuieG9PyQtzql9HbI7f3VLkbm6cDJ5qP8A4KAfsbzX39nJ8e9AEoJG5op1j/7+GPZ+teCviL4A+JWl/wBt/DzxtoXiWwGA1xpOoR3SIT2Yxsdp4PBweD6V+1B8L4fjL+z748+HL2yT3Gq6LObFWTdi7jHm27Y68SpGeK/Yx+Lk3wT/AGl/AnjZrxoNPbUksNV+YhWs7n9zKWHcKHEgB/ijU9qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv2of2Qfhp+1rYeHtO+I+ueJtOj8NTTy2h0S5ghLmYIG3+bDLkARrjGOp61/w5r/Zg/6Hr4o/+DOx/wDkOtbsotN1m/06BmaO1uZIkLnJIViBnHfAr9iH/gnT8Ev2l/gPafFDx34o8cWOqz6ndWrQ6Te2sUASJgFIWS3kbPPPzY+lfDP/AIJVfs8/Cr4heHfiT4d8Y/ES41PwzqMN/aQ3uoWbwvJEwZQ6paqxXI5AYH3FftZeCf2W/iR4P0/wZ+034s8NaBb3M0k+i3Wp69DpVzHKgUSPbPKw3YDoGXDL8y7h92v2T/gZ+wJ8EfHMV38GvjH4Y8ZeONQjkt7Ca88X2Go3qqVJkS3ig2AEqpyyoW27hnBIPifxV4X8E6Hc+J/GfiTS9B0ey2fadQ1S8jtbeDe4Rd8khCrl2VRk8lgOprQf2iP2fvFWsWnh3wv8dfh7rGq30nl2tjYeJ7O4nnf+6kaSFmPHQCvif8efgz8FreOf4p/EvQPDjTIXht7y7UXEyjq0cK5kcdsqpr4bftafs2/F7V08P/Dz4x+HdU1WU7YbBpzbXExxnEccwRpOBn5Qa1XVdL0PTrjV9a1K10+ws4zJcXV1MsUUKDqzOxAUD1JxV3+3r+x5Zat/Ys3x+8NNcb9m+JpZYc5x/rkQx4467sV4c8TeHPGOiWniXwlr2n6zpN9Hvtb6wuVnhmXplXQkEZGODVXNzbWdvJd3lxFBBCheSWVwioo6kk8Ae9eJf25/2RfCd62n6v8AH3ws86MVYWM73wUjqC0CuB09a+G/xv8Ag/8AGGGWX4YfErw94la3GZodPvkklhHTLx53qD2JABqqr/grp8Wrjxp+0Za/DO1ui2meANLjiaINlftl0qzSv6f6s26+xQ1/wSp+F0PgH9lLTfE89sE1HxzqNxqk7MuHEKsYIFz/AHdkRkH/AF1PrVfEr9oz4E/B6cWfxL+LHhvQLwpvFlc3ym5K/wB4QrmTHvtxXh/9uz9kHxPdLZaZ8fvC0cjNtBv5nsVz/vTqi/rWnalpus2EGqaRqFtfWdygeC4tpVljlU9GVlJBHuK/4LH/AAvh1/4MeF/iraWyG98J6ybO4kCYP2W6XByR1AliiAB6eYcdTn/gjT8XJtM8c+Mvgjf3jfZNbsV1nTo3Y7UuYCI5go/vPHIhPtAPSqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqrP/j6i/3xVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVf8ABWX/AJO/1H/sAaf/AOgNX/BGP/ku/jb/ALFE/wDpXBVf8FNf+TH/AIk/9wr/ANOtpVeFf+RX0f8A68IP/Ra18SrW3sfiN4qsrVNkNvrd5HGmSdqrM4Ayeegq71T9pX9sa+0DwnoOg6/4ui8DeH7PTbDTdOheSCwgggSLzX/hWSUx7mdzlmO0HCqo8ceAvGnw08SXPg/4geF9S8P61Z486yv7doZFBGVYA9VI5DDII5BNeC/2zfjz4E+A+q/s+eGPFlzbaLqd0rwXaSN9qsYCGE1tBJnMccjFCQuMYfH+savF37M37QngXwevxB8Z/B7xVpGgMqu9/eac6LEH+6ZAfmjBJAy4HJA6mvg/8XPG/wADfiFpPxK+H2rPY6tpUobGT5dzESN8Mq/xRuOCPxGCAR8OvGul/ErwB4b+IWiDFh4l0m21G3Utkok0SuFPuN2D05B6V/wU4/bq8SeCNXuP2cfg5rUumaktuj+JtatJSk8HmKGW0hccoxQqzuOcMqgj5q/Zy/Y4+OX7Vs+oXvw706xi0ywm8u91rWLpoLZJmG7ZuVXd3wQSFVsAjOMjP7SP7I/xl/ZY1SxsvibpVlLY6puFhq+lztPaXDL95AzKrK4BB2uqkjkZAOP+CV37ZXirxvqMv7N/xS1yfVru2smufDGpXcpknaOLmW0dySX2p86E8hUdc4CAVX/BZjxTLqHx48G+D0l3QaN4VF1tyflluLmUMMf7sERz7j0r/gjf4bi079nTxN4lZALjWPFsse4d4obaAL/4+8v6V8ZfiXpnwb+FPir4pavAbi28M6XNefZw20zuq/u4gexdyq57bq+MHxj+IXx28cX/AMQPiT4iuNU1O9c+WrufKtI85WGFM4jjXso9yckkmx/4JS/tfX/hiDxCnh7w7DcXEKyrpM+sLHdpkZ2sCvlq3Ygvwetf8Eq/2ZviH8DbD4j+IPiv4NvPD+u6hf22mW0F4q5MMCNI8kbKSrxu06DepKkxEA5Bqvjl4bi8G/Gv4geEIECxaJ4o1KyjUcgLFcyIMe2FFfB/xTJ46+EngjxtNJvk8QeHNP1B2JPzGa3SQnkA/wAXcCqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqvFX/ACM+sf8AX/P/AOjDX/BJv/kz/Tf+w9qH/oa1X/BbX/mjH/ce/wDcfX/BO3/k9D4Yf9f9z/6Rz1/wUbtYLz9iv4mw3Cb1Wzs5AMkfMl9bsp49GUGvhj8Qdd+E/wAQNB+JHhjyjqvh28S7sxMCUMi9AwBBK+ozyMjvUHwY/ap/aLj1X40W/wAPfGnjVdRmkmu9bFm832p1yG8s/wDLQLt2hYwQuAoAwBUUt5pt4k8Es1rd2soZHQmOSKRTwQRyrAj6giviL+0l+09+2Ta+Dvg7K2q+IJdLslt49K0mF3k1e4jBJu7hVz5kgRRkn5V2sw27mJ+JvwX+K3wZv7XTPin4A1nwzcX0ZktRf2xRZ1GAxR+VbGRkA5GRnGRX/BMj9pHxH8I/j5ovwyu9Wkfwf8QLxdPubGV8pDfSDbbTxj+FzJsiOMBlfnJVcXl5aafZz399cxW9tbRNLNNKwVY0UZZmJ4AABJNftwftxeMv2m/GN94c8Oard6X8NdMuGj03TIpGj/tEK3F1cj+NmwGVDxGMADduZvhF/wAEx/2nvjF4EtfiFpln4c0DTtRgFxp0Ou38kE95EwyrokcUm1WB48wpkc9CCfEPh/4ufsz/ABWm0fUjqng/xt4WuQVltpzHLE2Mq8ciHDI6kEEEqyt3Br9hT9p+T9qX4I2/ifXFgh8V6Fcf2dr8UKhFkmVQyToo+6siEHHQMHA4UVVftPeKZfG37RnxM8TyS+Yt74q1AwkknEKzukQ59EVR+Hav2d/DcXg/4BfDjwvEgX+zPCumwPz1cW0e9vxbJ/Gv+ClP7T3iH9nL4LWdh4DvWs/Fvja6ksbC9X71jbxoGuJ0/wCmgDRovoZN2cqM/Df4c/Eb9oL4mWXgjwfbT674q8RTyyb7q5wZGCtJLLLK56BVZixOT7kgHxr/AMEsf2v/AAfpb6raeEtG8SpFGXkh0XVUkmUDriOURs59k3E9ga/Yy+F118HP2YPh54E1PT5LLU4NIW71G3mjKSw3VyzXEsbg8hkaUoQem3HQV+3x4bi8V/sefFLTJUDC30U3y54wbWVLgH/yFX/BPnxTJ4R/bG+GV+km1bzU30+RecMLmCSAA4B7yKfqB061VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVZ/8fUX++Kqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr/grL/wAnf6j/ANgDT/8A0Bq/4Ix/8l38bf8AYon/ANK4Kr/gpr/yY/8AEn/uFf8Ap1tKrwr/AMivo/8A14Qf+i1r4q/8lQ8Yf9h6+/8AR71/wSd8LaHov7I2ma9p1hHHf+IdYvrjUJ9o3zNHMYUBPXaqRrgdASx/iNf8FrPDelr/AMKq8XR26LqMv9pWM0oXmSJfIdFJ64VmkIH/AE0av+Cd3hnRPF37Zvwz0fxBYx3dol5dXgikUEebbWU9xCxB/uyxI34V4v8AC+k+NvCes+Dddto7jTtcsJ7G7ikTcrxSoUYEd+GNV/wT3u7i+/Y0+F011L5jrpcsQJGMKlzMijj0VQPwr4peNL34kfErxT4/1C4ea48RaxdX7M7EkCWVmAGegAIAHGAAOK/Z2/4KY+LP2cPhJo/wl8MfCLw5fWulNNJJez3kyS3UksrSM7heM/MF+igdq/am/wCCiPin9qn4ZR/DXxP8LdA0mKDU4dQt722u5ZJIZY1dflDccrI6n2NfsY+Ibvwx+1h8JdSsZHSSbxZY2TFDg7LmUW7j6FJWB9iaqv8AgrXLcSftd3STsxSLw5YLCGHAXDk49txb8c1/wSb/AOTP9N/7D2of+hrX7TXwqu/jf8A/HHws024SG/17SXjsXkban2lCJIQ57KZEQMewJNeJfDXiDwdr9/4V8VaPd6Vq+lztBeWV3EY5YJFOCrKen+TXwE/4K6fFvwDZ2Hhr4xeGrXx1pVqiw/2jFN9k1JUGBudsGOYhR3VGY9XySa+An7RHwr/aU8Gf8Jr8Ldea7ghcRXtlcx+VdWEpGdk0eTtJHRgSrYO1jg1X7Zf/ACdf8Wv+xu1D/wBHNX7HrXL/ALKnwjN2ZC48G6aF8wHOwW6BPw27ce2Kqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq8Vf8jPrH/X/P8A+jDX/BJv/kz/AE3/ALD2of8Aoa1X/BbX/mjH/ce/9x9f8E7f+T0Phh/1/wBz/wCkc9f8FEv+TL/if/14W3/pZBVeE/C2h+CPC+k+DvDNhHZaTollFZ2VvGoAjijUKo49gK/4KJeG9L8K/tn/ABO0rR7dIbeW+tr1kVdo825s4LiU4HrJK5/Gv+CKvhnRZR8UvGEtjHJq0B06xguGUFoYH895FU9QHZIy3r5a+lf8FZfC+k63+yNqGt3ttG154e1uxubKUplkaSTyHAPUArKcjocD0BHwfu7iw+Lfgi+s5fLntvEenyROBnay3EZBwfcCv+Ch/jS98Cfsc/EbU9MuHiu7+yh0yNkYqdt1cRwSjI6fupJPr071p1xb2eoWt3dWSXkMEyPJbSMVWZQQShI5AIGMj1qL/gs/8SoIkgg+CPhSOONQqIl/OoUAYAAA4GK/av8A2mtT/au+IGn/ABE1vwXpfh2/stJTTpEsJnkFwqSSOrsW/iHmlfoB6Cv+CLfiG7t/iX8RvCayP9lv9Ctr1kz8u+CfYDj1xcNVV41luZ/GWvTXjM1xJqdy0pcYYuZGzn3zXhX/AJFfR/8Arwg/9FrX/BW74C+K/in8I/DvxG8H6fPqVx8PLm7kvrOBC7/YrlYvNmVRkt5bQRE+ilm6Ka+GfxL8bfB7xvpfxF+HeuSaTr2jyF7W5RQ4+ZSrKysCrKykqVIwQTXwI/4LHadfXFpoP7Q/gRNOD4R/EHh/dJGp6bpLViXA7ko7H0SvDniPQPGGgaf4o8Laxa6ppGqW63FleWsgkjnjYZVlYdQRX7VH/JsHxf8A+xC1v/0gmr9k5rlP2o/hCbQyb/8AhOdHB2A52G8iD9O23OfbNVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVWf/H1F/viqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/4Ky/8AJ3+o/wDYA0//ANAav+CMf/Jd/G3/AGKJ/wDSuCq/4Ka/8mP/ABJ/7hX/AKdbSq8K/wDIr6P/ANeEH/ota+Kv/JUPGH/Yevv/AEe9f8Es/wDky/wj/wBf+pf+lktf8Frf+RX+FH/X/qn/AKLt6/4Jlf8AJ8Hw1/7iv/pqu6qv+Cdv/Jl/ww/68Ln/ANLJ6+NHgG++Fvxb8YfDvULZoZfD2tXVoqsu3dGkhEbgf3WTaw9Qwr9gH9nj9h79pT4aW+i+N9Dmf4naQ8q6rZnXbm2e8iLs0VxDGrgMojKq20fKykkAMuf+HWX7F/8A0TvVP/CgvP8A45XgL/gnn+yD4B8d6Z4t8HeEbmLxF4WvIL63Da/czG2lVt0TPG0hGCVJAYYODVV/wWR8Nz6f+0T4X8TBG+zax4SiiDHvLDcz7wPbbJF+df8ABHXxHHqn7NGveH3n3T6L4uuAIy+dsUtvbupx2Bfzfbgn1rVdV0vQtLvNb1vUbaw07Trd7i7u7mURRW8SKWeR3YgKqqCSTwACa+NH7K/7Of7U+mW+s+OfClhqtzLbqLHxDpVx5N0I/wCHbPGcSpycK+9BkkDvX7eP7E9j+yHrXh668O+OJNe0HxWbkWcN7Esd5atB5e4SFcLIpEq4dQvOQVHBP/BKPxtrvhv9rrRvC+mzzf2f4t0y+tNRhUkoVit3uI3YdMh4QAeo3kfxGq+P/iKLxh8dviL4qgmEsWr+K9TuonBJBSS6kZcZ7bSMe2K+CvhmfwV8GvAXg26jaObQfDGnWEiN1VobaOMg/itVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVeKv+Rn1j/r/AJ//AEYa/wCCTf8AyZ/pv/Ye1D/0Nar/AILa/wDNGP8AuPf+4+v+Cdv/ACeh8MP+v+5/9I56/wCCiX/Jl/xP/wCvC2/9LIKqv+Cmv/J8HxJ/7hX/AKarSv8Agin/AMiv8Vv+v/S//RdxX/BU3/ky/wAW/wDX/pv/AKWRV8K/+SoeD/8AsPWP/o9K/b88A33xI/ZC+JHh/SrZp72201NSgRF3Mfsk0dw4UdyUidQOp3VYTWtvfW9xe2Yu7eKVWltzIY/NQEFk3DlcjIyORmvhB+wr/wAE7fjr4HsPH/w28OXmqabexKXRfEd2JrSQj5opk83McinIKn6jIIJb/gln+xcqlm+HmpgAck+ILwY/8iV+z7+yf+z5+z9f6h4p+CmhyWsuuWwtri5Oqy3qSxo+cKXdgMMOcelVX7Rfhufwf8fviP4YuEZW03xVqUSbuSyC5fY30K7T+NfAjxHH4w+CHw+8VRz+b/a3hfTrpmL7jue2jZsnrkEkHPOc5rWPE3hvw9Lp9vr/AIg07TZdVufstgl5dJCbqfaW8uMMRvfarHaMnAJ7V8dv+CaP7M3xsuLzXbLQJ/BPiK7LO+oeHyIopZDzuktmBiOSSSUCMxPLV+0H8HL/APZ/+Mnib4P6lr1prU/hy4jj+32qFEnSSJJUO0k7G2SKGXJ2sGGTjNf8EavG2u638GfGXgjUZ5ptP8Na5HJpxkJKxLcxlpIk9AHjL49ZSe9ft3eIovC37IHxU1OWYRibQJLEMSRk3TrbgceplA/Gv2AfDM/iz9sT4X6dBGz/AGXVzfuRxtW2hknJJ9P3WPfIHeqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqrP8A4+ov98VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX/BU7wL438QftZ6hqOg+Dtc1K1OhWCie006WZCwRsjcqkZHpX/BH3wZ4v8N/HDxndeIvCmsaXDL4UKRyXthJArN9qgOAXABOATiq/4KRaTqmufsW/EXS9F026v7yb+zPKtrWFpZHxqdqThVBJwASfYE1/wqr4of8ARN/FH/gnn/8AiK8MI0fhrSY3Uqy2MIKsMEEIK+J/ww+Jdx8S/Fs8Hw88TSRya7esjppE5DAzvggheQRX/BMfR9W0L9jzwppmt6Xd6feR32ol7e7gaGRQbuQglWAIyMGv+CyXhfxL4m8NfC6Pw34d1PVWt77UjKtlZvOYwUt8FtgOM4PX0r/gm98P/HmiftpfDrVNa8E69YWcH9p+ZcXWmzRRpnTLpRlmUAZJA57kVX/Cqvih/wBE38Uf+Cef/wCIr/gn9pmo6P8AsefDXTNX0+5sryCxuBLb3MLRSRk3cxGVYAjgg8+tf8FCP+Cfl3+0Rcj4u/CIWkHj22t1h1CwncQx6zDGMIQ54WdQAgLEKyhQSu0GvG3wz+KXwe1wWHjvwZ4g8K6nay/umvbSS2O4cho3wAw4yGUkHqDR+NHxu1OBdGPxZ8cXcMoEa2h166kVsYwoTeQegwMV/wAEg/B/xj8I+O/HV/4s8AeJtL8MeJNHhlGqalYywRXN5DN8iqZADIxSec7hkDByQSM1X/BYn4UzeKfgl4b+Kun2xkuPBOrNDdsq/ctLwKhYn2mjt1A/6aGv+CM3xMh0j4leOPhPe3AVfEelQ6lZB2wPOtXKuqj+80dxuPtD7V+3Xp3xF1v9lbx34c+FfhfUNf1/WbOOzW0sE3y/Z3lQXBVM5f8Ac+Yu1cklhwea0bx5+0B8Bb6TSND8W+PPANzuPm2UV3daaWIxnfFlQe3BHpWoat8cv2ivFEB1G+8Z/EXXwohgV2uNTnjVjwiD5iqk54GB1r/gmv8AsI+K/gZf3Hxs+MVjFY+KL+xNrpGj7xI+nQyEGSWUqSBK4AUKCdqlg3LEL+0N8S7b4OfA7xv8TLi4ET6Dos81qT/FcldkC/8AApmjX8a/ZP8AhXcfGv8AaM8B/D8WzXFre6xFPqQIyBZwnzrgk9sxxuATxlgO9VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV4o+FvxNk8S6s8fw58TsrX0xUjR5yCN55Hy1/wSx0TWdA/ZL07Ttd0m9067XXL9jBd27QyAF1wdrAHBqv+CzXhTxR4n/4U/8A8I14b1TVvs39ued9hs5J/K3fYNu7YDjO04z1wfSv+Cf3w98faP8Ath/DXUtX8D+ILK0gvrgy3Fzpk0UcYNpMASzKAOSBz61/wUB0zUdZ/Y8+JWmaRp9zfXk9jbiK3toWlkci7hJ2qoJPAJ4r/hVXxQ/6Jv4o/wDBPP8A/EVX/BSL4f8AjzXP20viLqmi+Cdev7Ob+zPKuLXTZpY3xplqpwyqQcEEH3Br/gjd4X8S+GfDXxRj8R+HdT0pp77TTCt9ZvAZAEuMld4GcZHT1r/gpxo+ra9+x54q0zQ9Lu9RvJL7TilvawNNIwF3GSQqgk4AzXww+GHxLg+JfhKef4eeJo449dsmd30idQoE6ZJO3gYp0SRDG6hlYYKsMgiv20v+CYnj7wZ4o1P4h/s7+HJvEXhC/ke5k0GyXdeaSzElo4ohzNCM/Jsy6j5Sp272tr7x/wDDHXZFsr3xB4T1qAbJBFJNYXCc9Djaw5HQ1L4z+N3xRdfD0vizxx4ueQbVsWv7q/LZ7CPc2enpX/BN/RPiL4W/ZQ8OeEfiZ4S1fw9qWjXl5FbWuq27QTtbSTGZHKN8yjMzKAwBG3GMYqq/4Kv/AApm8BftRXPjO2timm+PdOh1CJwuFFxEognQe/7uOQ/9dq/4JdfEuH4hfskeH9IkuA9/4MvLjRroFsttVvNhOOoXypo1HbKH04/4LA+D/jH45s/AVn4K+Hmv614X0RLy71K90yza6SO4kMaIJFjyyBURjuIC/vcZzwNF/aS/aS8D2T+HtD+N3xA0i2jXZ9ij166jWHj+FC/yH6AHpXw6+Bvx8/aM8Ss3gjwV4j8U3+ozl7rVZkdoQ7H5pJ7qT5F9SWbJ9zX7Gf7MVj+yp8GrXwHJfQajr1/cNf67fQqQkt0yquyPOD5aKqquQM4LYBYgf8Fhvibb+GvgFoXwzt7kC/8AGetrJJF/etLQeY5/7+vbfr6V/wAEbfhVPrfxW8W/F+8tSbLwzpS6daSMuAbq6YFip7lYomB9POX1qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqz/wCPqL/fFVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVTQxXETwTxpJHIpV0dchgRggjuMVZaPpGmsz6bpdnas4wxggWMke+BVVXxH8A+Hfip4C1/4c+LLX7RpHiKwlsrpAPmVXXG9T2dThlPZlB7V4R+En7S/wCyJ+0zY6/pnwp8Wa1deBNcwbnS9GuJrfU7Ugq/lyIpGyaB2HXK78EAgitE1e08QaLYa7YJOlvqNtHcRJcQtFIquoYB0bDIwBwVIBByDV1aWt9A1re20VxC+N0cqB1ODkZB46gVa2lrZQLa2VtFBCmdscSBFXJycAcDkmq/4KzeK/iZ4k8KeH/gP8NPAHijXINRmGq69d6XpFxcxLHGSILcvGpXJfdIy9R5cZ/ir/gk/wDsp6/8NdG1v46/Efw7eaTr2uodM0axv7doZ7azVwZpXjYBlMkiKACAQsRPSQVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVe6dp2pRrDqNjb3SIcqs8QcA+oBzVrZ2djAttY2sNvEudscUYRRnrgDiqqv+CiP7NFx+0d8BblfDOni58Y+D5G1LQ1UfPcALie2U/9NEAIHd44xkDJr/glvqvxj+CXxpu/A/jP4W+NNO8K+PIFglurvQrqKGzvYtzQSMzJhVYNJGTxy6EnC1V7pWl6i0b6hptrdNF/qzNCrlPpkcdBVV/wUHb44/tHftI6jP4Z+EHjm78N+GVGiaHJH4fujHcqjnzJ1OzBEkrNhu6LH6V+xv8As+Qfs0fAPQPh1OsTa3KGv9emjIIkvpQC4BHBVFVIgR1EYPc1VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVZ/8fUX++KqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqrP8A4+ov98VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVWf8Ax9Rf74qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqs/8Aj6i/3xVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVZ/8AH1F/viqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqz/4+ov8AfFVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVn/x9Rf74qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqs/+PqL/fFVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVn/x9Rf74qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqs/wDj6i/3xVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVZ/wDH1F/viqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqz/wCPqL/fFVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVn/wAfUX++KqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqrP/j6i/wB8VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVWf/H1F/viqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqz/4+ov98VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVWf/H1F/viqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqtG/al/Zw8R+IbTwjoPxu8G3+tX1yLW2sLfVonmlmJ2iNVByWJ4xVVVVVUxVVLMwAA5J4xWs/tTfs0+H9VOh618fvh/aXyv5ckEniK2DRNnGH+f5P+BY9a0bW9F8SaXb634d1iy1TTrpN9vd2Vws8Uq+quhIYe4NfED4nfDv4U6IfEfxJ8baN4b07JVJ9SvEgEjAZ2oGOXbH8Kgn2rTf+Cif7F2raj/ZVp8d9LSYMF3XOn3ltFk/9NZIVTHvuwKtbq1v7WG+sbmK4t7iNZIZonDpIjDKsrDgggggjg1VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVWf/AB9Rf74qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr4j/E3wD8IvCd343+JHimx0HRbIfvLm6fG5j0RFGWdz2RQWPYV8Zv8AgsvFb3c+l/AP4YxXUUbEJq/iaRgsmOCVtYWDAHqC0oPqorXf+Cpn7aGrz+dp/wARtL0Vc5EVj4fs3Ue37+OQ4/GvDn/BVX9snQ50l1PxrofiFVPMWpaBborex+zrEfyIr9h39ovxp+1D8FX+JfjjwtpeiXiavNYRLpzyGK5SKOMmULISU+d3XG5vudaqqqq/a1/4KhfD/wCB2qX3w++E+m23jbxjYu0V5PJKV07TpR1R3U7pnB4KIQAcguGBUJ+03/wUY/ag1CceAdf8eXlusu1ovBunvZQWhI4VprdVKj3kkJ96HwB/4Km2KjWIU+LscijeJIvGDmYE99q3O/PPPGeua/Za+Nn/AAUJ0D9o3wL8GfiRq3i6Ow17Ugt7aeMdJaQyWcQMtw8c8qCQsIkkwyuVyRnNVVVVVVVXxj/a8/Zz+Al7Lo/xN+KOmafq8Mau2lW6yXd2oYZTdDCrMm4YILhRgg5wc14W8Rab4w8MaR4t0ZpDYa3YQX1qZU2v5Usaum4djtYZHaqqqqqqq+Ln7Q/wT+A9tBcfFr4j6T4da5iaW3tp3aS4nReCyQRhpHAPGVU88V8I/iv4N+OHw90v4n/D+7uLnQNYa4WzmuLdoHkEM8kDNsbkAvE2M4OMcDpVVVVVVVV+y9/yet8Pf+x9tv8A0pqqqqqq/wCCoX7X/izxn8S9S/Z18Aa5c2XhXw232XWls5Sp1e9IBkjkK8mKIkR+X0Lhyc4Xb8OP+CNnxG8UeCbTxD47+LWneE9avbdZk0aPRXvTBuXIjmlMsexxkBgquAc8nFfAX4b6L+xd+yvFoHirU7W5j8Gaff6rrmoWiELcsGknkdQ2CSE2oAeflUUbn42f8FF/2nIdPuNTzqOuTym3SZ2NpoOnJliFUdERMD1kcjJLPmm/4ItfDM6LFbp8bPE41cAeZdHTYDA3riHO4fjIatbW3sbWGytIhHBbxrHEg6KqjAA/ACqqqqqr4r/HT4QfA3TLbVfiz8QdJ8NQ3ok+yJdyky3Xl7fM8qJQXk270ztU43LnqK+DPxq+H/x+8GD4gfDPUri/0NryW1iuZ7V7cyPEQGIRwGC5PGQD7VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVn/wAfUX++KqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqvGHizw/wCA/CureNPFepRWGj6JZyXd7cyHAjijUsx9zgcAck4A5NftaftVeN/2q/iRceKNduJrPw9YSSReH9E3/u7C3J4LAcNM4Cl37nAGFVQK+FH7D/7Unxq0iDxF4E+EuoyaPcpvg1HUJorCGZezRmd0MinplAw/WvEv/BNX9s/wzZNqEvwek1GGNSXGm6raXUgx2Eay72P+6pr9iD4bX3wn/ZU+HXg/V9Mn0/UxpX22+tbiJopoZ7qRrhkkRgGV180KVIBUrg9Kqqqv+CoX7WmqfAz4e2Xwq+H+rNZ+MPG0MjTXcL7ZdO04Ha8iEcq8rZjVuwWQjDBTX/BOT9g/Tf2gbiT4wfFq1kfwJpN0YLLTAWjOs3Kj5tzAgiBCQDtOWb5cgKwOheH9C8LaRa+H/DOjWOk6ZYxiO2srK3WCGFB0VEQAKPYCqZEcqWUEocqSOhxjI/An86qta/aC+AnhvUDpPiL43+ANLvlbYbW98S2kEobjjY0gOeRxjvWgeJvDfizT11bwr4h03WbFjtW50+7S4jJ9AyEiqmligieeeVI441LO7naFAHJJ7DFWWs6PqUjQ6dq1ndOi5ZYZ1cgepAJ4rxP4w8I+CtP/ALW8ZeKtH0Gxzt+06nfR2sefTdIwFeD/AIw/CP4h3LWXgD4p+EPE1wmd0Wj65b3rrgZORE7HpzVXuo6dpsay6jf29qjHarTyiME+gJNf8FQ7yzv/ANsnxZc2N3DcRNYaaFkicOpxZxZwRxXwF8TeG4vgX8OYZfEOmo6eE9LVla7QFSLSPIIzxVreWd/AtzY3cNxE2QskTh1OOuCOKrxf8YPhJ8PrgWfj74peEfDU5AxFrGt29kxyMjiV1PIrwh8Rfh78Qrd7rwD488O+JYYgN8mkapDeKmemTEzAVVz4h0Czma2u9c0+CaPho5blFZfqCcioZ4LmFLi2mSWKRco6MGDD1BHUVV7rWjabKINR1eytZGXcEnnVCR0zgnpwa/tXS/sP9p/2la/Y/wDn485fL64+9nHXj61/wWY1LTdT+Jnw6fTdQtrtU0G5DGCVXCnzx1wTiv8Agmpr+g2X7E3w4tb3W7CCZP7U3Ry3KIy51S7IyCcjgirLUtN1NGfTdQtrtUOGMEquFPvgnFXmt6Lp0v2fUNYsraXbkJNcKjY9cE9KhnguYUuLaZJYpFyjowYMPUEdRVXmt6Lp0v2fUNYsraXbkJNcKjY9cE9KhnguYUuLaZJYpFyjowYMPUEdRVeK/jN8HvAlwbPxx8WPB3h64BwYtW122tGB+kjg9q8IfFn4VfEKUweAfib4U8SyIpZk0fWbe8IA4JxE7ccVVfsvf8nrfD3/ALH22/8ASmqqq8YfEv4cfD2NJvH3xA8N+GkkXKNrGqw2QYZxkGVlyM8V4P8Ai18KfiHK1v4A+J3hPxNLGpZk0fWre9ZQOpIidsCqvdZ0fTZFh1HVrO1dlyqzzrGSPUAnpXwjurX4g/tveGNW8Tyo0OtfEqG9vjM4Kvuv/NcMeBg8gnjrX/CVeF/+hk0v/wADI/8AGv8Agop410uy/Yw+JbaNrthNdT2tnbCOK6VmZJr63ifAByfkdq/4I2roNn8YfHeuard2ttcW/hqO3t5J5VT5ZLlGcDPvEn5VZ63ouoy/Z9P1iyupQudkNwrtgd8A9Krxf8Rfh78PreO68e+PPDvhqGUHy5NY1SGzVsdcGVlBrwh8YPhJ8Qrg2fgH4peEfEs6g5i0jW7e8YYGTxE7HpVVXi34pfDHwAdvjv4j+F/Dh2g41fWILPg9D+8deK8L/Gv4M+OLpLDwV8XPBfiC5kbakOl6/bXTsfQLG5JPSri4t7WFri6njhijGXkkYKqj1JPSv+C1Gq6Xqn/CnP7N1K1u/K/t3f5Eyybc/wBn4zgnGcH8q/4JTa7oen/si6dbX+s2NtKNdvz5c1wqNjevOCc1Y6rpeqb/AOzdStbvysb/ACJlk256ZwTjOD+VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVWf8Ax9Rf74qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/wCCwPxRu/CP7P8Aovw5025aKbxzrO262nG+0tQJHX8ZWtj9AfWq/wCCXP7Lvh347/FPVPHnxA0mLUvC/gOOGRbGdcxXt9KWMKuOjxosbuy9CfLBBUkFESNFjjUKqjCqBgAD0qqqqqv+ChPxCuviN+158Qryaffb6FqH9iWkYOREloBE4H1lWVj7sa+Bnw00/wCDvwd8HfDHTbdIk8O6RBbS7QB5k4UGaQ44y8pdyR1LE1VV488b+Gfhr4M1nx94y1NLDRdBs5Lu9uH52ogzgDqzE4CqOWYgDk18Z/2qf2lv29viivwr+FcGr2Xh3U52i0vwtp0/k+bCOs17KCA/A3NvPlR446Fm8Mf8EVvGd7oiXPjH48aPpOrNHlrPT9BkvolbHTznmhPpzs9fx+MP7PH7Uv8AwTz8WWPjfQPF9zbaddzeVZeJfD87rBMwyRBcxMMBiAT5cgZGGcFtrY/YL/bNtP2r/Al1Y+JYrTT/AB74ZVF1e1g+WO7ibhLqJTyFJBVlydrY6B1r9ouzh1D9nv4n2FwCYrnwZq8UgBwdrWcoOPwNfseftPzfsqeIfHHjWx0yPUNV1TwnJp+j28ykxfbWurdkeUgg+WkazMQMbiqrkZyPDH/BPj9sX9rhv+Fy/Grx1baLca1GJrY+IpJZbxom5XbbRrtt4sHhCUIH8AFftI/sP/Hv9kM2PjbVLq21HQxcqtt4i0GeRfsk/VBICFeFyR8rcrnADZOK/wCCZH7ZHiH9oDwxqfwr+J2pG/8AGXhK1S4t9Rk/1mp2G4IXkP8AFLG5RWbjcJEJy24n9vX9lfxh+1l8OPD3gzwZ4i0bSLrR9b+3yy6oZAjp5Eke1fLVjnLg8jGAa/aH+BfiH9nD4qal8JvFOr6dqeo6ZDBLJc6fv8phNEsigb1VuAwByOteDP8AgkT8bPG/g7QvGen/ABL8EQWuvaZb38EUxut8aTRrIqtiIjIDAHBxmv2OvgX4h/Zw+AeifCbxTq+nanqOmXN3LJc6fv8AKYTTvIoG9VbgMAcjrX/BQz9v/wAa6f41u/2b/wBnbVbqyvrOcWeu61p2Wu5Lpjt+x2pUbkZSQrOvzl/lXbtO74Uf8EfPiJ450GHxb8afiqvhjVdTj899Mg086hcxM/P7+VpUUSeqjfyfvcGv2jv2APjj+xpbRfGn4bfEKfW9F0eZWk1nSUk06/0oscK7ort+7JKrvVzyfmVQa/4J1ftmXf7Tnga98JePp4v+E/8ACUaG8lRQg1S1Y7UuQoACuG+SQAYBKMMb9q/8FT/2QP8AhZXg9v2hfh/pRk8U+FrXbrltBHltR09efNwOskAyfUx7hn5FFf8ABJr9rfI/4Zb8fap/z0uPCFzM31eayz/31In/AG0XP3Fr4t/FHwn8Ffhxr3xP8b3v2fSNAtGnlx9+Z8hY4kHd3cqij1YdBzXgbwp8Uv8AgpB+1hd3+s3ElqmrT/bNXuozvi0TS4yFWOLPHyqVjQfxO25urNX7Qf7MsHxC/ZN1X9mb4Uf2X4cgktLC10v7VvEFvHbXcEx3FAzEssTc4JLNk9Sa/ar/AGSvGn7JXiDQvDvjTxJomsT69ZSXUD6WZSsao+whvMRTnPpXwC/4JlfFn9ob4SaD8YPDHj7wjpum6/8AaPItb83Amj8m4kgbdsjZeWiYjB6EV+wN+yX40/ZK8H+KvDnjTxJomsT69qUV1A+lmUrGqR7CG8xFOc+lf8FfP+Tr7T/sUbL/ANHXFfsaf8mofCX/ALFGw/8ARK1X/BXz/k6+0/7FGy/9HXFfsaf8mofCX/sUbD/0Stf8FAfhp+0f8Uvhp4f8N/s4+JNTsb641oQa1aWd+liJ7N4m/eSTkhgkbIAUVvmEhyrEKK8K/wDBFnxxqWnC68efHrR9J1F/meDTdDl1JAT1zJJNASf+A1+1B/wTy+MP7JGjQ/Ffw340j8TeH9NuY/N1fTYZLC70yQsBHI8Ydtiliqh0kOGIBxkE/wDBMn9sLxH+0J4Q1X4bfE3Uxf8AjHwfFHLDqDnEup2LHaHk/vSRvhWfjcJIyctuJr9l7/k9b4e/9j7bf+lNVVf8FHf29tX+Bcv/AApL4OX0UPjW8tlm1bVsBzo8LjKJGpBHnupDZP3EKkAlwV+CH/BLr4z/ALQujxfFv44/Eu68MSeIR9qjjv7WTUdUulYZWWbzJE8vcCCAzM2OoXNftC/8Es/iv8BPDdx8VPhL8QT4utfDyG8uooLR9P1GzRPmM0QV3EgQAsSrKwA4VsGv+CZn7cniH4yNN8CfjDrD6h4r0+1Nzomr3D5l1O3T/WRSn+KZBhg/JddxblCW/b1/YK+I/wC1l8R/D3jPwZ4x8N6Ra6Pon9nyxaoZw7v50km5fLRhtw4HJzkGvh98MdW+I3xX0T4R6XqFnbalrmsppUNzcbvJjkeTYGbaC23PoM1/w5j+O/8A0VLwF/31d/8Axmvj9/wTK+LP7PXwk174weJvH3hHUtN0D7P59rYm486TzriKBdu+NV4aVSckcA1+yp+yV40/a18Qa74d8F+JNE0efQLKO6nfVDKFkV32AL5aMc59cV+w7/wTw+J37Lfxom+Jfi/xt4X1axl0S408W+mmcyh5HiYN88ajaBGe+eRX/BRn9vDUf2e7eH4RfCW6hHjzVbUT3mosqyDRbdvuFUIKmZ8EgNkKo3EHctfAv/gmd8bv2m9Jj+Mnxu+Jl34cTxEBdQPqVvJqWp30bcrLIHkTy1YYKlmJI52gEE/H3/glD8Uvgz4auviT8IPiD/wmMegxG8uLNLJrDUIFjG5pINruJSoBbAKNxhQx6/8ABMr9ujxL8UL/AP4Z9+MuuS6p4ghtmm8O6zdSbpr6ONcyW0zHl5FUF1c5LKH3HKgtX7cf/BSzxp4r8Ual8Iv2cPEE2j+HbKZrS88Q6e5F3qso+VhbyDmKINkBk+Z8ZDBTg/Cf/glH+0h8WtPj8ZfEjxDp3goan++8rVzJeajJu53yRLwpOejyBweqivGn/BGj4zaNpz33gP4peGPEVzCm4Wl1by6c8hH8KNmRc9Mbio9SK139nnxJrf7Fa/s322uRJ4guPCUGmTX+pTvKn2vahld2G5ipcPjGcAgDgV+1l+xj47/ZD/4RX/hNvFOgaz/wln2z7L/ZZmPlfZvI37/MRevnrjGehziv2bv+CcfxS/aa+GMHxS8JeOfCul6fPeT2q2+om4EoaIgE/JGwwc8c1/wT8/Yx8d/sh/8ACe/8Jt4p0DWf+Es/s37L/ZZmPlfZvtO/f5iL189cYz0OcVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVZ/8fUX++Kqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv+C2BuzrXwjV9/2UWur+VkfLv3Wm/HvjZn8Kr/gi/d6fJ8HvH9hEY/t0PiWKSYADd5b2yCPPfGUkx+PvVVVVVV8dJJNK/ao+IUus3OZLP4g6m11Ly2St/Jvb1PQn1oFWUMrAgjgiqqv+CyfxMv8Aw78IPB3wusLh4l8Y6tNdXuwkebBZLGfLb/ZMs8L4PeIelf8ABHT4O6Xovwr8R/Gy+sYm1jxJqTabZTsuWisrcKWCn+HfMzbh38lPSq+N3wq0P43fCfxP8LvEEEL2+v6dLBFJKufs8+MwzD0KSBHH+7X/AAT68c6t8J/2yfBNtNI1sNX1CTw9qVuXxvFxmJUbHXE4ibHqgr4/f8kI+JH/AGKOq/8ApJJX/BPb4eaF8Tf2uvAWgeJbaO5060uJ9Rlt5FysrW0DzRqR3HmIhIPBAI71X7S3grS/iJ+z78RPB2rwxyQX/hy82eYuRHMkTSRSdDykiIw9Cor/AIJa67eaR+2l4O0+1kYR61Y6nZ3ADYDItlLOARjkb4EOOOQD2xVf8FTf+T0PFv8A14ab/wCkcVfAH/khHw3/AOxR0v8A9JIq+NPjtvhf8IPGvxGiRXm8NaBe6hDGykh5YoWdFIHYsFH41+wZ4w+EegftOWvxS/aG8Yw6fZaJbXOqW93fxS3H2rU2ZVjLBFZi48ySUMRgNGDnOK/4eJfsX/8ARd9L/wDAC8/+M140/bo/Yc8deD9c8E618ctIl0/XtOnsLpH027YGOWMo3Hk88Ma/4JoeNbvwZ+2P4IjhuGjtfEC3WlXiDP7xJYHZFOO3nRwtzx8vbqHRJEMbqGVhgqwyCK/4KA/svax+yb8arH4p/C/z9M8J6/qH2/Qri0yh0bUI2EjW4Yfdww8yL/ZyvPlkn9rH9tTxv+2do/w2+FnhnQbyCVIoDqum2y5/tTXJD5Q8pRkmMZzGp5zMwOdoNfsTfsr6R+yv8H7Xw7PFBP4u1oJd+JL5Pm8yfB2wo3/POIEqOxJduN5FV/wWl/5Kf8OP+wDdf+jxX/BMr/kx/wCGv/cV/wDTrd1X/BXz/k6+0/7FGy/9HXFfsaf8mofCX/sUbD/0StV/wV8/5OvtP+xRsv8A0dcV+xp/yah8Jf8AsUbD/wBErVfFL9vH9lH4Q3k+k+KPi7pl3qlvlXsNHSTUZVcdUYwhkRuOjsuK/aA/4Kj/ALLnxK+EPjf4X6b4f8eXk/ibQL7T7W5bSbcQwzyRMkUjbrlW2q5R+FJ46Z4r/glDrN1pf7YejWNsxEer6JqNrOAcZQRecM+vzQpVfsvf8nrfD3/sfbb/ANKaqru6gsrWa9upNkNvG0kjYJ2qoyTgc9BXwy8f+Bvit+2VYfFT9oPXIdP8M6r4ln1nWJrtHmjRE3zQ25VAzFCyRQ7QCApHYGv+HiX7F/8A0XfS/wDwAvP/AIzT/wDBQ/8AYskQxv8AHXSWVhgq2n3hBH/fmvhV4w8P/DT9tzQ/FHw31RZfDWnfEMxabdQxtGJ9LkvDFlVxuUPbORgjIDYI7VX7L3/J63w9/wCx9tv/AEpqv+Cmv/Jj/wASf+4V/wCnW0r/AIIt/wDJT/iN/wBgG2/9Hmq0v4g+B/jZ+2gnxN+N+uxWnhDW/Fj6jqk11HJIi2MbForcqoL7DHHHCBjIBFD/AIKIfsXKoVfjtpQAGABp94Mf+Qa/4eJfsX/9F30v/wAALz/4zWneNvC3w7/bTh8ffCzVo5vC+mfEL7ZpM8ETRJJp7XmRGFwGCGFihGASCeBnFf8ABQT4oah8Jv2TPHWvaLcPDqep2yaRZyIcGNrpxFIwP8JWJpWBHIYDp1H/AASO+B2ifEj42a18S/ElhFe2fw7s4JbKGVAyi/uGcQyEHg7EhmYejbG6gVVV/wAFtf8AmjH/AHHv/cfX/BJv/kz/AE3/ALD2of8Aoa1VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVn/x9Rf74qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/4LD/DG78VfAXw/wDEjT7YyyeCNa23ZC/6q1u1WNmz/wBdktl/4FVf8E/P2r7L9ln4uz3Piwzt4M8WQR2Wt+ShdrZkYmG6CjlvLLOCo52SOQCwArwt4r8MeONAs/FPg7xBYa1pGoR+Za31jcLNFKvqGUkexHUHg1UM8FzGJreZJUOQGRgwODg8j3GKqqqv+CmHwvvfhr+1v4rvHt2XT/GCx65YSEcOJl2zfiJ0m464we9fsV/GzTPjz+zh4P8AF1vfpPqthYR6Zrke7LxX1uipIWHbeAso/wBmRfeqqv8Agtdo12Lv4TeIFQtamPVbV2A+44Nqyg/UFsf7pr/gk74h0/Wf2QNM0uzlVp9B1zULS6UNkq7SCcZHb5Jkqrq5t7O2lvLudIYII2eWSRtqoqjJJPYACv2aYpfiN+274B1HSYXUX3xBt9WVNu4rFHd/aWz06Ihye3XFfH7/AJIR8SP+xR1X/wBJJK/4JZ/8noeEf+vDUv8A0jlqvip/yTDxf/2Ab7/0Q9f8Eyv+T4Phr/3Ff/TVd1X/AAVN/wCT0PFv/Xhpv/pHFXwB/wCSEfDf/sUdL/8ASSKv22dKutZ/ZK+K9lZJulTwxdTkAE/LEvmN0/2UNfsD/s4/DH9qP4u6r8N/iZrniHS4oNAl1CwfRriGF5ZY5oUZGMsUgI2SM2AoPyk54wf+HNf7MH/Q9fFH/wAGdj/8h1/w5r/Zg/6Hr4o/+DOx/wDkOvhB/wAEu/2fPgt8StA+KXhnxV4+vtV8OXJuLS31LULR4GfYyjesdsjEDdkYYcgduKr/AIK5ftQaHBoEX7LnhuOyv9TvJIL/AMRXDoJDpyIRJBCh/glfCuxGCIyByJTjwNqHxQ/Y0+N/gj4h+I/BxtdTs7e11q1sr+NSLywuojnaedjNG7pn70bg5GVIr4Y/Ejwl8X/AOifEnwNqS32i69arcW0o4ZezI4/hdGDIy9mUjtVf8Fpf+Sn/AA4/7AN1/wCjxX/BMr/kx/4a/wDcV/8ATrd1X/BX5GX9q6yLKQG8IWRUkYyPOuBkfiDX7F08Nz+yb8JpLeQOq+FLJCR2ZYwrD8CCPwqv+Cvbo/7WFsquCU8JWIYA9D5twefwI/Ov2NP+TUPhL/2KNh/6JWv+Cqn7YvjC18ZXP7M3w412fStNsLSNvFF1aSlJbyWZA62u8crEsbIWAPzlyp4Ug/sqf8Em/AuoeCtH8e/tFX+p6hqer2sd0nhyxuDawWkcihlSeRf3jybSMhGQKSR82M18Vf2Ff2SfBXwU8cal4c+CGiQXml+GtQubS5llnuJYpY7Vyjh5ZGbIKg8nrz3Nf8Es/wDk9Dwj/wBeGpf+kctV+y9/yet8Pf8Asfbb/wBKaqvHul3Ot+BvEWi2S7ri/wBJureEYJy7xMo6c9SK/ZC+EXgb46/tC+F/hJ8RtU1fTdI1/wC1RNcaZLFFOk0dvJJGoaVHUBmjC/dJO4DjOR/w5r/Zg/6Hr4o/+DOx/wDkOv8AhzX+zB/0PXxR/wDBnY//ACHXhn/gkR+zR4W8SaT4ns/GPxJubjSL6G8ihudSsjFI0ThwrhbRWKkqAcEHB4I61X7L3/J63w9/7H22/wDSmq/4Ka/8mP8AxJ/7hX/p1tK/4It/8lP+I3/YBtv/AEea1myk1LR77ToXVXuraSJWboCykAn25r9lz4U+EPi7+0T4S+EfxMvtX0rSNcvJ7O7k06SOG6imEEpiVTKjqCZljUgqeCQOcV/w5r/Zg/6Hr4o/+DOx/wDkOv8AhzX+zB/0PXxR/wDBnY//ACHWk/8ABHz9mPSNVstWi8Z/Eud7K4jnWKbU7Io5RgwVgLQHacYOCDjuK/4K2aPd6n+yJc3tshaPSfEdhczkLnah8yEE+nzTIPxr/gin4isVPxW8JSOq3jjTL2FS3Mka/aEcgeilo+f9sdO9VX/BbX/mjH/ce/8AcfX/AASb/wCTP9N/7D2of+hrVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVWf/AB9Rf74qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq8ceC/DnxG8H6x4E8Xael9o2u2Ulpe27cb43XBwezDqCOQQCOlftW/ssePP2V/iLceFfEdtNd6FeSO+g62sZEWoQDB69FlUModOx5GVKk14O+JPxF+HU8l18PvH/AIj8MTTf6yTRtVmsmfjHJiZSeCR+NeIPj/8AHjxbYtpnir42+PdZtGUq1vqHiS7uIyDjIKvIRg4H5Cv+CZviz/hK/wBjTwMksga40VrzTZsAjHl3MhjH/fp4/wDPFVVVX7e37IkP7Vfwtii8Oi2t/HPhhnuNBuJjsWcMB5tq7dlkCqQTwrqpJALV+zn+0j8YP2HfivqVs+jXKwif7J4n8K6jugE5jJHoTHMmTtcA9TkMpIPwl/4KN/snfFbT4JJfiVZ+D9TZR52neJ2FgYjjn98x8lhnIyHzxyBmrj9ov9ny1tGvrn47fD2O3VdxlfxPZhQOxz5n0rQv23v2ZPFvxO0L4Q+DPibaeIPEPiCWWK2GnQyS28ZSJ5PnnwI/mEZUBSxJI4AOa/b8/Zyvv2k/2fNR8OeG7ZJvFOgXC6toaHgzzRqyvAD/ANNI3dRnjfsJ6ZH7AP7Xj/skfEnVfCXxJtb+Hwb4hmWDWYTAxm0q7iJVZ/KPzcZZJEHzYwcEoFPhf4yfCXxtoS+JfCfxL8M6rpbR7zc22qRMsYxn5/myhA6hsEYOcV/wUN/4KD+ANN+H2r/BH4HeKrPxFr3iO3ez1bV9MuBLbabauMSIkq5WSV1JT5CQgLEkNgV/wSN/Zg1m68U3H7TXi7Snt9L0+3lsvDAniKm5nkGya5TP8CR74gw4LSOM/Ia+P3/JCPiR/wBijqv/AKSSV/wSz/5PQ8I/9eGpf+kctV8VP+SYeL/+wDff+iHr/gmV/wAnwfDX/uK/+mq7qv8AgrX4WvdC/a1udbnhkFv4j8P2N3BIeVby1aBgDjqDDyOeoPcV+x1+1p8D/Gf7OvgeHV/id4X0PXNB0W10vVdO1TVYLSaKe3iWIvskYEo+zerDIw2M5BAjn8J/EXwnIbHUNO17w9rtnJCZrS4WeC7gcFHCuhKspBIyDWu6N8Uf+Cd37WcN5DaSTT+Gb9p9NlnBSHW9Lk3L94DGJImZGxnZID3SvgZ+2n+zx8fNAtNR8M/ELStM1eWNftOg6vdpaXttIRygRyPNAP8AHHuXkcg8VrnxN+G3hm0N/wCJPiF4a0q2VSTNfatBAgAxk7nYDHI/MV8Mfjf8J/jQdYPwr8cad4lj0C5S2v5rFmeOORl3KFcgK4IH3kJXqM5Bx+2F+0zoP7LXwc1DxvdNDc6/fBrPw7pznP2q8ZeGYdfLjHzueOAFByy1+wL+zrrv7Xf7QOofFf4qGfVvDeg3/wDamvXN2u4arfuxeO3OeCGbLuMEbF2nG9TX/BRf9k8ftI/CFte8J6aknjzwYkl1pWxcPfQYzNaZ7lgAyA9HUDgOxr/gl9+1y/wc8f8A/Ci/H2o+V4R8YXgFjNcPtXS9SbCqTn7sc2FRs8BgjcDeTX/BarwteprPwu8axwyPaS2uoafLIPuxyK0MiA8cFg8mOedh4GOf+CXv7UPwitP2cbD4TeMvHug+G9c8H3t2qQatqEVn9qt553uFljMjAOA8zoQORtGeCM+GPGPhHxvp8mq+C/FWj6/ZQzGCS50y+juo0lABKFo2IDAMpI6gMPWv+Cz3wr1L+2vAfxrs7N5bFrOTQb+ZV4gdHae3B/3hJcY/3Pev+CZv7cHwu8P/AAptPgJ8YPGVh4Z1Dw/cS/2Jf6pMILW6tZXMnltM3yo6O7jDkAqUAzggfFr9t39mP4P+G7jXdZ+Lfh/WLpIma20rQ7+LULu6YA7VVImbYGIxvcqnqeK/aU+JPj/47fES8+PnjXQn0y18ZzSDR485jW2ttsIjjPBcIAFL4AZw/fIH7Gn/ACah8Jf+xRsP/RK1/wAFH/CWt+E/2xfHx1iKXy9ang1KwmfpNbywoAV9lZXj+sZr4D/tL/Cf49fDrSfG/hbxdpEc1zaob/TJLxEuNOuNo8yGSMncu1sgHGGGGBIINf8ABQL9uH4d+EPhhrnwW+FniWz8UeOvGNpJpTx6ROtymmQzDy5TI8ZI85kZkSMHeGYMcADd/wAEs/8Ak9Dwj/14al/6Ry1X7L3/ACet8Pf+x9tv/Smqqv2yvg545/Y6/arbx34QimstJvtZ/wCEh8JaikWYkbzRK1ue2YpCUKE8xlCeHr9mr/goF8Bvj/4Zsjqni7SfB/jARAX+haveLb/vehNvJJhZkJyQFO8D7yitR+IHgLSLM6hq3jfQLK1Ucz3OpRRIOCfvMwHQE/QV4B/aC+CvxV8Van4L+GvxJ0XxNqujWy3N7Hpc/wBojijZtoIlXMbfNwQrEjjOMjNfsvf8nrfD3/sfbb/0pqv+Cmv/ACY/8Sf+4V/6dbSv+CLf/JT/AIjf9gG2/wDR5qv28vgh40/ZY/ajm+JvhKKey0XXtZ/4SHw1qkMeEtrsSiaSDPRXilyQv9wofUD9l/8A4KF/A74+eGLC28TeK9J8G+N1iVL/AEbVLtbZJZRwXtpZCFlVuoQHevIIwAxvvH/gPTbP+0NR8baDa2oGfPn1KKNMYzncWx0Ga8EftDfBH4m+NL74ffDv4maH4l1zTbI3t1BpVx9pSKEOsZbzUzGSGZQVDEjIyBmvjj8LNK+N3wi8V/CnWJBFB4k02S2SYru8ibhoZcd9kio+P9mvhZ4/+Kn7CH7SrX2paM8Gs+Grl9O17SJWKJqFm5UyRhsfddQkkb4IyI2wRwfg7+2z+zT8bdFttR8N/FHRtMv5YwZtH1u7jsbyBu6mORgHwSBujLLz1rxx+1B+zr8ONOfUvGXxq8IWKIu4Qpqkc87j/YhiLSP/AMBU14e17SvFWgaZ4n0K6Fzp2r2cV5ZzBSPMhlQOjYPIyrA8+tf8FqfC17eeCfhf40ihka10nVNQsJXH3Ve5ihdAeOpFo+OR0PXt/wAEof2mfhV4f+Deq/B7x5430Xw1rGj6vNe2X9q30dpHeW0yoT5byEBnV1fcvXaynkZx4W8ceCfHVtcXngjxjoniGCzl8m4l0rUIrtYZNobYxjYhW2spwecEHvVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVn/wAfUX++Kqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq+IHw68C/FXwtdeCviL4W0/wAQaJej97Z3sW9dw6Op6o4zw6kMDyCK+LH/AARp+H2t3k2p/Bz4n6l4ZDksumavaf2hCp7KkqskiL/veYfer3/gjR+0ik5TTviH8NZ4QOHnvb2Jv++RasP1rw1/wRc+Kd1NEvjH4z+FdNiIXzG0yxuL5h/ewJPJz7ZIz7V+yr+zR4f/AGU/hlJ8NfDvinVdfiudSk1Ge51BUTEzxxoyxog+SPESnaSxyWOecCqqqr9oL9kP4E/tMWar8S/CYOrQxeXba5pzi2voF7ASYIdRkkLIrqCc4rxt/wAEV9RWeSf4cfHO2lhYnyrXW9IaNkGOAZonYNzgE+WvrjtVh/wRf+Mss+zVPi74Lt4cfft4LmZs5H8JRB0z39PXI/Z7/wCCTfg34PeO/D/xK8T/ABe1zXtZ8OX0V/ZQ6dZJp0AmjYMok3NK8iZHIBTcODwSDX7UX/BOr4JftL6nP4xEl14P8ZTj99rGmRK6XhAABuIGIWQgD7ylHPGWIAFal/wRc+LEWoiLR/jL4SurDdgz3NlcQS7c9fLUOM47b+vfvXwR/wCCPvwr8Ganba/8Y/Gt345mt2DrpVta/YLIsO0h3tJKvHTKA9CCOK0zS9N0TTbXRtGsLexsLGFILW1t4hHHBGgCqiKuAqgAAAcACvH/AIW/4TrwH4k8E/b/ALF/wkGkXWn/AGnyvN8jzoWj37MjdjdnGRnGMjrX7LP/AATF/wCGZ/jRpHxe/wCF3/8ACSf2Vb3MP9nf8Iz9j8zzoWjz5n2qTGN2cbTnGOOtV4q0P/hJ/C+seG/tX2b+1bCe087Zv8rzIym7bkZxuzjIz6iv2Zf+CW//AAzl8cPDXxm/4Xp/wkP/AAj32r/iW/8ACMfZPP8APtZrf/W/apNu3zt33TnbjjORX7WX7IXw7/a18IWuieKrmfSNb0hnfR9ctYhJLaF8b0ZCQJIm2qSmQcqCGHeb/gi98YRqLRQfGDwa9gGws721yspGOpjCkA57b/xr9nX4S3XwK+CnhT4SXviT+3pvDdm0Dah9nMAm3SO4AQsxUKHCjnoo6dB8fP2cPhJ+0r4UXwn8VPDv21LYu1hf27+Td6fIwwXhkGcZwMqwZG2jcpwK8d/8EWteS+lm+GPxssJ7NjmK317TXikjGejSwlg/HOQi88Y71oX/AARa+JtzMq+JvjZ4YsIt+Gaw02e7YLxyA5iyevGR2554/ZA/Yv8ACH7IOl67B4d8Z634h1DxKLf+0ZrxY4YMweZsMUKglP8AWtnc7Z46V+1j/wAE7vGH7WHxL/4TrxF+0YNI02ytxa6PokfhQzx6fF1b5/ti73dssz7QT8oxhVA/Z4+BPhL9nD4TaL8KvCBM8OnIZLy+ePZJf3b4Ms7jJwWIwFydqhVyQoNV8fv+CSvhT4v/ABY1v4meDfiy3gmDX5vtV1pS+HvtyJdNzLJG/wBoi2q7fPtwcMWwcEAfCXwn4u8CfDrQ/B3jnx3/AMJnrGj2otpdcawNm94qcI0kZkk+fYFDNuO4gscEmvj98BvAH7SHw2vvhn8Q7OR7K4YTWt1AQs9hcqCEniYggMNzDBBBDMCCCa8Qf8EXfifDrEsfhX4yeFrvS9/7qXULOe3nC57ogkXIH+1z7V+xR+zJq37J/wAIbv4b6140h8S3N9rk2qGeC0aCO3EkMMfkqGZiwBhLbvlyXPHGT8RPh34M+LHgzU/h/wCP9Ct9X0LV4fKurWYHnuGUjlXUgFWBBUgEEYr4j/8ABF7Wv7YluPhH8YrBtMlcmO08Q2bpLAueFMsIYScY52J9K+Dn/BGvQNI1i11j44fEz+3rWBlZ9G0S2a2jmI6q9w53lM8EKiEj+IZ4/ay/4J2+G/2k4vA9h4V8b2fw803wNps2n2tla+Hhdo8LFCirieLYE2HjnO8n6/A34Zy/Br4ReFPhXN4hbXW8MabHYjUGtvs5uAmcHy977BjAxuPTrX7Vn7Hfwv8A2s/DlrYeLjPpOv6UGGla9ZIpntg3WN1PEsROCUOCDyrKSSbv/gi78Wk1TybH4yeEZdO3YFxLZ3Ec2318oBlz7b/xr9lf/gmt8If2c9Wt/G3iDUJPHXjC3H+jXt9aLDbWLH+KC3y2H7b3ZiOq7ea/Zw/4JcH9nX45+HvjNYfHX+2U0CS5K6W/hjyDMk1vLAVMv2psECXOdhyV6DtXwu/4JOf8K2+Nfh74w/8AC/f7R/sHXo9W/s7/AIRTyfO2S7/L837W23PTdtP0qqr4sfCD4c/HDwbc+Afif4Yttb0a5YP5UpKPDIM7ZInUho3GSAykHBI6Eg/En/gi7BLfS3fwh+MxgtG3GOw8RWG94z2BuISNw7f6oY960r/gi78XJptuufGPwhaRZGGtLS4uGxzn5WEft379q/Y//wCCfHhD9kvxNe+OrT4ja34m8QX+mvp0zPbR2dp5LvHIcQgu27dEuCZCAM8d6r4Xf8EnP+FbfGvw98Yf+F+/2j/YOvR6t/Z3/CKeT52yXf5fm/a2256btp+lV+038Ef+Gjvgf4l+DP8Awk//AAj3/CQ/Zf8AiZfYvtfkeRdQ3H+q3pu3eTt+8Mbs84wf2Lf2Cf8Ahj/xP4j8Sf8AC1/+Et/4SCwitPI/sH7B5GyTfu3faJd2emMD61XxP+Fnw/8AjN4NvPAPxL8M2uu6HfYMltPlSjj7skbqQ0bjJwykEZPPJr4mf8EXtPuL+W9+D/xjks7VyTHp/iGx85oz6faISuRn1iyB3NaX/wAEXvi9NPt1r4w+D7WHI+e1tbm4bHf5WVB+v5V+yH/wTr8I/sp+MJPiInxK1rxL4ilsJLFsWsdlaeTIVZgYgZHY5jQgmTAx06Yr9pX9jj4K/tS6bEnj/R5rPXLOMpY69pjLDeQL12EkFZI8/wADg4ydu0nNeKv+CLPj63vH/wCEH+Nvh+/tS/yf2rpk1o6rzwfLMoJHAzxnrgdK8G/8EWPEct3FJ8QfjlpttbLgyw6NpLzu/qqySugX/eKN9K+GHgDTPhV8O/Dnw10bUb+/sPDOmw6fa3F+6vM8UShU3lVUZCgDgAYAr4yfCDwP8d/h1q3wx+IWmm70jVowC0bbJbaVTlJom52ujAEHkHkEFSQfFv8AwRd+IkOsTDwJ8Y/Dl5pRYmE6vZzW06L2DCMSKxHTIIz1wM4H7DX7JOt/sjeBtf8AC+ueO7bxHNr2pJe7bWzeCK2KxhCBuYliwAycL0A5qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqz/4+ov98VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVWf/H1F/viqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqz/AOPqL/fFVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVn/AMfUX++KqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqrP/AI+ov98VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVWf/AB9Rf74qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqs/+PqL/AHxVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVZ/8fUX++Kqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqrj9uf9kW1nktbj49+F0lhco6NM+VYHBH3fUVp9/Z6rYW2qadcJPa3kKzQSp0kRgCrD2IINV8T/wBob4J/Be/stL+KfxI0jw1d6jC01pFeuymZFO0sMA8A8V8L/jh8JfjXb6hdfCnx5pniaLSXjS9eycsIGcMUDZA6hW/Kvif8UfAPwa8GX3xA+JXiS30TQtPAE1zMGYlmOFREUFncnoqgk+nBr4Cftlfs/ftJ6peeH/hf4wkn1mwiM8mm31o9rO0IIBkQMMOoJAO0kjIyBkZqqqv2xf2u9N/ZD8M+HvEeo+BrnxMuv30losMF+tqYSib9xJR856Y4r9j79v7R/wBrnxxrPgrTvhleeGn0bSTqDXE+qLdCQeakewKI1x9/Oc9qqqqqqqqqqqqqqqqqqqr9oH/gqfoHwE+MPiX4RXfwa1DWZvDk8cTX0etpAs2+FJMhDCxXG/HU9K/Zu+Ndr+0X8FvDnxksvD8uhw+IPtW2wluRO0PkXUtucuFUHJhLdBjdjtmqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq4nhtbeS6uJAkUKF3duiqBkn8hX/Dd37H/AP0X/wALf9/n/wDiaqqqqqqqqqqqqqqqqqqqqqqqqqv2kfjXa/s6/BbxF8ZL3w/LrkPh77LusIrkQNN591FbjDlWAwZQ3Q5xjvmv2f8A/gqfoHx7+MPhr4RWfwa1DRpvEc8kSX0utpMsOyF5MlBCpOdmOo61VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVWf/H1F/viqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqrxV/wAjPrH/AF/z/wDow18K/wDkmHg//sA2P/ohKr/gtL/yU/4cf9gG6/8AR4r/AIIp/wDIr/Fb/r/0v/0XcV/wUN/Zu8cftN/Aq38K/Dm6gOvaFrMeqW9hcTiGO/CxSxtFuPyq+JcqzYGQQSN2R/wTt/YN+P8A8LvjxY/GL4s6D/wiem+HbW6jt7OS9imm1GWaFodpWF2CxqJGcliMsqYB5Iqqqv8AgtJ/yS/4cf8AYeuf/RAr/gjH/wAl38bf9iif/SuCqqqqqqqqqqqqqqqqqqq/4KJf8nofE/8A6/7b/wBI4K/4Jlf8mP8Aw1/7iv8A6dbuqqqqqqqqqqv28/8Agob8Sf2Y/iZF8Kfh74K8O3dxPo8N+dU1ZpZtvmvIu1YY2TkeXnJYjnpxz/wTx+OfxH/aH+A978Qvihq9vf6v/wAJJdWkbQWkdukUCRQlUVUAyAXY5bLc8npVVVVVVX7Yv7QmqfsxfBK9+KejeG7XXLuC+t7SO0up2ijzKSN5Kgk4x04z6iv2Jv2+/j5+01+05ZeCvGk2gab4bk0m8uG0zStO2KzxqChMkjPJxnswB9O1VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV4q/5FfWP+vCf/0Waqqqqqqqqqqqqqqqqqqqqqqqqqq/4Ka/8mP/ABJ/7hX/AKdbSv8Agnb/AMnofDD/AK/7n/0jnqqqqqqqqqqr4teNpfhp8KvGfxHg09L+Twp4fv8AVUtHkMYuDbW7yiMsAdobZjODjPQ14C/4KlftFfGD49/DrwRaWHhjwv4f17xfpdheW9jZGeea2mu4o5EaWZmA3IzDcioRnIweaqqqqqqr9rf/AIKifGf4W/Fvxb8IPhz4P8MWCeHL5rP+1b5JbuaXCg71TciIeejBxX7KHjzxR8UP2c/AXxA8a6it9rmu6StzfXCwJCJJC7DIRAFUYAGABVVVVVVVVVVVVVVVVVVVVVVVVVn/AMfUX++KqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqvFX/Iz6x/1/z/8Aow18K/8AkmHg/wD7ANj/AOiEqv8AgtL/AMlP+HH/AGAbr/0eK/4Ip/8AIr/Fb/r/ANL/APRdxX7cX7R3in9lv4LwfEvwhoelatfy63b6ebfUhIYgkiSsW/dspyDGO+OTX7Ff/BRv4pftNfHG1+FvizwP4V0vT59Nubo3GnC4EoaJQQBvkYYOeeKYqqlmYAAck8Yr9pz/AIK46H4I1678F/s7eHtN8U3Fi7RXHiHU3c2JkBwRBHGVaZev7wuqkj5Q6kMdD/4K8ftVadqyX2rWngzVrPd+8spdKeJSp7K8cgYHHQkn3Br4if8ABXWyX4QeGfG/wr8KaR/wlV3qMlnr/h7W5JJTYhYgyyxPEyeZGxJAYgHggqCK/YD/AGtvH37W3hvxd4g8b+GNB0ZdAvre1tBpfmjzS6Oz7xI7dMJjGOpr/gtJ/wAkv+HH/Yeuf/RAr/gmx8c/h9+zt4z+IvxK+I+ptb6fa+E/Kt7eEBp764a6hKQQoSNzttJ6gABmJABI+In/AAWH+P2uazI/w58J+FvDGjq/7iK6ge/uWH/TSQsqc8cKgx6mv2Tf+CsCfEXxfp3w4/aC0DSNBu9WmW3sPEGmM8Vr5zHCJPFIzGMMSB5gbaCRlVXLCqrwH/wVp+OPib4qeHPh5q/w18DQRar4gtdLupbcXRaNZLhYnZMy4yASRkelftff8FWbv4eeM9S+GP7PWjaRqtzo0z22peIdSV5oROpKvHbxKV37CMeaxKk5wpADH9g344+Ov2h/2e7H4jfEWWwk1qXU7u1d7O28hGSNgFO3J5wa/a+/4KL/AA2/Zi1GXwJoelnxh46SMNNp0NwIbfTtygp9olwSGIIYRqCSOpQFSdW/4K9/tV32pC80/T/A2nWyuStpFpMsileMBmeYsenUEck9sAfszf8ABXTSvGniKx8FftDeGNN8NS6hIsMHiLS5HWyjkPCi4ikZmiUnjzA7AEjIVQWHx28Z+Lvh/wDBfxf8Qfh9pdhq2teHtIl1G1tLwO0VwsI8yRcIQxJjV9oB5bHXpXwc/wCCunxR8b/Ffwf4L8ZfD7wbp+i6/rVrp97d2v2lZLdJpFj8wFpWHylgTkHgGq/bE/4KceOf2f8A47av8JPh54N8L6zaaFbWwvLrUTOz/aZIxKygxyKu1UeMYwSG3AnsP2O/2hJf2nfgRo/xR1CxsrDV5bm5s9UsrMsYreeKQgBdxLYaIxPgnjfX7aX7Rt7+y58DLz4l6NpthqOsyaja2GmWt+X8mWWRizhthVjiGOZhgjkDtmv2LP8AgoT8cf2p/jda/DjVPh54R0/RLbTri/1a8sVufNgijUKm3fKVyZpIV5B4Jr9tv9uq2/ZCk0TQ7b4bX3iPWPEVpLcWc8t0LWyjCMFYFwGd3BKkoFAwy/Nzx4h/4K+ftUatcF9H03wPokIb5I7fSpZWxzwzSytk9OQB06CvhR/wWS+KGm6vbWvxl+H2ga3o7MqzXWiLJZ3cQ7uFd3jkI67cR5/vCvht8R/B3xd8DaR8RfAOsR6noet24mtZ1GD1wyMp5V1YFWU8ggg9K/by/ab8bfso/CrRPiF4K8PaJrEuo+II9Lnh1TzdqK9vPKGURspzmAjk96/Zs/4Kct46+G3xR+J/x30TRNA03wEumLYw6KkrTajPd/agIFWR23OTbrjoACzMQoJHh7/grB8fviT8cPCXh7Q/Dvhbw74Y1nxBZ2L2H2d7ucwTTojb52YZfaxwVRB04NfEL4g+DvhV4M1Tx/4+1230jQtGgM13dTHhR0CqByzsxCqoBLMQACTXxh/4LIfEPUNZubD4HeANG0nR43ZIb/Xke6up17P5aOqRf7p8ztz2r4Y/8FjPjVousW8XxX8E+G/EujFgLh9NiexvFGeWVt7RtgfwlBnGNw61+2N8RfCvxd/aS8Z/EnwTevdaJ4gktbm0eSPY4BtIQysvZlYMrDnBU8nrX/BMr/kx/wCGv/cV/wDTrd1VftK/tX/CT9lnwzFrXxE1SWbUb9X/ALL0WyAe7vmXqVUkBEBIy7EAZxycA+P/APgsd8edbvZk+HngTwl4Z08k+ULtJdQuVHvIWSM/9+68K/8ABX79p/Rr5JfEejeC/EFnu/ewy6dJbPt4+48UgCnjqVbqeOmNAvNS1HQdN1DWNOFhqFzZxS3VoHLi3lZAXjDEAnaxIzgZx0FftP8A/BV74f8Awn1y98C/Bnw/D451ywkeG71Ka5MWnW0qnBVCoLXBBBB2lF9HbkC7/wCCuf7WtzqH2yH/AIQq1h3Z+yRaM5jxnpl5WfHb72f518AP+CxFjrWr23h39onwVZ6LDcsEGv6CJGggPrLbuXcL6sjMR/d71pmp6brem2us6NqFvfWF9Ck9rdW8okjmjcAq6MuQykEEEcEGv2/f23PiR+yP4h8H6Z4M8JeG9YtPEtlcyyPqnnF45IXQEL5bqNuJF655rwX/AMFV/Dtt+zbL8XPifoGnyeMbjX7rStK8M6I7xfafJhgkMjvKXMcY89dz/N1AVSQceIv+Cvn7UmqasbzQtN8F6LZK+Y7OPTJJ8rngO8khJOOCV2/QV+xn/wAFPtL+Oviqx+FXxj0HTvDXirU2EWlahYO4stRmPSHY5ZoZG/hyzK54ypKhv2uPjPr37PX7PXiv4weGNLsNS1PQPsfkWt+H8mTzryCBt2xlbhZWIwRyBX7Tf7R3in9qT4jxfEvxfoelaTfxaZFp4t9NEgi2Rs7BvnZjkmQ98cCv2Zf+Ch3xO/Zb+HEnw08IeCfC+rWMupzX5n1ITmUPIqKV/dyKNoEY7Z5NfsWftA+Jf2mvgbafFLxZoumaXqE+pXVq1vpwcRBYmABG9mOTnnmv2tv24vhb+ydYQadrMM3iHxfqEPnWPh+zmEb+XkgSzyEEQxkqQDhmYg4UgMR4n/4LA/tPavqDTeHdD8E6FZq4MUCafLcPtGeHeSUhs55KqvQYxzn4D/8ABYzVbjXbTQ/2h/A2mxabcyLG2ueHkkRrTJxvlt3ZzIozklGBABwjHim1h/Evg4698PNX0m9k1LTjcaLeysZrOZnj3QyMYyC0RJUnaQSOhrxp/wAFaf2mvh34u1XwN4y+DHgjTtZ0O7e1vrWRbsNG6nBwfOwQRyGGQQQRkGvgX8Y/C3x++FXh/wCKvhCYfY9atg0tuz7ns7hfllgf/aRwV9xgjgg1+3l+3/L+ypq+g+BfAOiaP4g8V6hEbzUYdQdzFY2vKx5EbKfMkYEgZ4VCSPmU1+wn+13+0N+1rrmrat4m+HvhPQ/A2hRmK41GzjuPNubxgCkEW+Ur8qne5wcDaP4wR+25+3p8R/jJb+Mv2etd8HeG7LRtI8TzRRXtoJxcOLS4dELbpCvIHOF+mK/Zu/aB8S/sy/E6D4peEtF0zVdQgsprVbfUQ/lFZQAT8jKcjHHNfsO/8FDvid+1J8aJvhp4v8E+F9JsYtEuL8T6aJxLvjeJQvzyMNpEh7Z4FVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVeKv+RX1j/rwn/8ARZqqqv2uP+CiHwr/AGX72TwXYWEnjHxyIw8mkWtwIYbEMAVNzNhtjEHcI1VmIwTsDKx8R/8ABXr9qfV71ptE0/wVodsCdkEGlyTHH+00sjZPuAv0r4Q/8FlfHVlqsFj8cfhzo+p6U7bZL/w8HtbmEE/eMUrukuPQGP6+vwz+Jvgb4w+CtN+IPw68QW+saHqke6C4iyCpHDI6nlHU8MrAEHrX7UH/AAUy+Pf7PPx28V/CRfht4LurTRblTYXNwl1vuLaWNZYnYrKFLbHAbAA3BhxjFfsmfHOf9o/4CeGvizf2NnY6lqYniv7O0LGO3nhmeMhdxJAIVWAJPDiv24P2nbz9lL4NR+PdD0zTtT1zUdWg07TrO/L+W7MrySMwQhsLHG3OQAxXPXB/Yg/4KPeMf2mPjJJ8KviF4S8N6ILnSJ7rTJtMMwea4iKFoiJHYYMXmvxjHl+9V4y/4LF/FrT/ABprmk+D/hx4LvtGttUuINLnnF00s9usrLEzbZQCzIFJwAMngV4MuvEl/wCD9DvvGVla2ev3Gm28mqW1qD5UF00amVE3EnarlgMknAqv+CgP7YXxB/ZGsfA+oeB/C/h/WYvE0t9FeDVfOzE0IgMezy3XgiSTOc9Fx3r4S/8ABTu0vv2bfFHx1+M/h7SrLUNM186Pomi6K0iNqk5t0lVcyM+wDcxaToqjoWKq37P/APwU3+P/AMbv2nPBvgTUNK8LaR4W8RaqLWWwtbJ3kSIqxH753JL8D5gAD/dHSvHfjvwj8MvCOpeOvHevW2j6FpEPm3l5cEhY1yABgZLMSQoUAkkgAEmvjH/wWV8XXGrz6f8AAj4c6VZ6VExWPUfEgee4nAP3hDE6LFn0LSfh0HhH/gsH+0to2oxyeK/DvgzxFYb8zQfYpLSUr6JIkhCn3ZG69DX7LH7W3w0/aw8Hza94MeXTtY0wqmr6FeSKbiyZvusCOJImwdsgAzgghSCB+2/+398bP2UvjLH4B0TwB4T1PQ9R0mDUNNvL5LgyurFo5FYpIq5WSN+AAdpXPXJ/YV/an1b9rL4T6l408S6Ppek63pGtSafc2enl/L2CKOSOQB2ZhuDsOvVDX7UPxqT9nn4EeK/i0lta3V3otsgsLW5JCT3MsixRKQpDFdzgkAg7QTkYyP2Xv+CqXxA+Mnx28KfC34g+CPCekaR4luXs/tlgbgSxztGxgA3yMCHlCJjH8dV+0h/wVa+JHwn+OfjL4Z+BPAnhHVNH8M6ibCK6v/tBmeWNVWcNskVeJhIowBwBnnJr4B/Ebxv8QvgD4Y+KnxB8N29lruuaQdUk0zSY3I8p9zwLGsjEl2h8s4LfeYjivif/AMFm/Hd1fT2nwf8AhNo2l2akpHdeIppLudx/e8qFo1jb23SDjv0qx/4K6ftaWl+bydfBN7ESSLSfRnEY5zgFJVfGOPvdPfmv2N/+Ckvg39pTXoPhr430CPwj43njZrJI7gy2eplQWZYmYBo5AoJ8ts5AJDE8VXxa/wCCtPxx+HHxU8Z/Dy3+Gvga4i8L+IL/AEuOWQXRaRbe4eIMcSgZITJwBX7a/wDwU3g+BPiy7+E3wY0TTPEHinTcpq+paiXa006Xj9yqIVMsoByx3BUOAdx3Kv8AwTm/aT+Jn7Tfwq8R+KvijPpk2o6Tr5sYHsbT7ODF5EUnzKCQTlzzxxiv2xP+Cgnw6/ZXmHg/T9N/4S3x3LEsh0iG58mKxjYZV7mQBipIIZYwCxGCdoKsb/8A4K9/tWXWqi+tNP8AAtnbKTixTSZXjI7ZZpi/5MOfyr4u/wDBSTSv2lv2R/iF8KPiD4Vt/Dnja5g0+WxlsJGey1Ly9StXkVFcl4XCKzbWZwQjHcDhT/wTt/5PQ+GH/X/c/wDpHPVVcXFvaW8l1dTxwwwoXkkkYKqKBksSeAABnJr9or/grz4L8FatdeFfgF4Vt/GN1asUk13UJmisN4OD5UaYedf9rdGPTcDmrz/grh+1vc3v2qCXwZaxbs/ZYdFJjAz0y8jPjt97Nf8ABPT9tb4p/tZ3finR/H/g3QLJfCtpbSPqmk+bEsskzOERonZ8ErFI24OB8vC88fH/APaK+Fv7NPgk+N/ifrTW8MrmKwsLZBJdahKBny4Y8jJxjLEhVyMkZFfEj/gsj8adav5Yfhd8P/DHhrTckRPqIk1C6I6Alg0ca567djYOBk9/B3/BYP8AaU0W+jbxb4c8G+JLHdmWI2clnMR6JJG5VfqUav2Vf2zvhR+1ho07eEpZtI8S6bEJNS8P3zgzwKTjzI2HE0WcDeuCCRuClgD4kvr3TPDuqalpsUMl3aWU0tukxIRpFQlQ2OdpIGcdq+C//BW34rfEH4ueDPAfizwJ4I0vSPEWuWmn3l7CLlXgSaVYywLSlRjcOWGB1PFftHf8Ff7rSPEd54V/Zv8AC2lajZWUjRP4i1tJJEuiDgtbwIyEJxw7sd2fuDv8P/8AgsH+0PoWtQy/EDw14W8UaQXH2iCG2awuAvfy5VZlB/3kb8K+Bnxv8BftDfDfTfib8O9QafTr3Mc0Ey7ZrK4UDzIJV52uuR0yCCGBKsCf+ChX7enxH+HHjv4hfsxaV4O8N3Wg6r4d/s+XULgT/akS+ssSMu2QJlfNO3K44Gc18O/Gd/8ADf4geGPiJpVrb3V74W1m01S2guM+XLJbzJKqvtIO0lADgg4PWvgP/wAFW/jJ8WPjR4J+Gms/DrwZaWPibW7bT7ie1W582JJXClk3SkZAPGQRWsavpXh/SbzXdc1C3sdO063e4u7q4kCRwRIpZ3ZjwFCgkk9MV8fv+CxK6XrV34d/Z28EWGp2tq7RjX9f8zy7gjjdDbRsjBe4Z3BORlB30X/gr7+1Pp+p/a9V0vwPqto0mXtJNLliAXuqMkoI+rbu3Wv2Pv26/hx+1nZXGjW9i3hnxrpsPm3ehXNwJfOj4Bmt5MDzEBIDAqGUkZGCGP7d37TXx6/ZX0vRvHXgDwL4Y8Q+D70/ZdRnv47gzWF2SSm4xyKPLkXhTjhlIJ+Za/Yq/wCCmOo/tDfFNvhV8VfDegeHb7VLYtoNzpzSqlxOmWeB/MdvmZMshBAyhXksteM/F/h34f8AhLV/G/izUo7HR9Cspby9uHPEccalmwO5OMADkkgDk0v/AAWF+O3iTxYNC8C/BrwpdjU9Q+z6PZSLdS3UoeTbDG2yUBpCCoO0AE9AK/a5/at+M37KvwF8DeO9U8K+Er/xjruoR2etWWJzZ2sjQSSssREm5tpQJuLENgnAyAPjD8TdW+M/xO8RfFLXbCzsdQ8SXhuri3tA3lRsQBhdxJxx3Jr4Pf8ABU/4w/Bn4Y+HfhboPw98G32n+G7IWtvcXa3PmyKCTltsoGeewFfAf4gan8WPgv4J+Jes2VraX/ibRLa/uLe13eVE8qBiqbiTtBPGSTVVVVVVVVVVVVVVVVVVVVVVVVZ/8fUX++KqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqvFX/ACM+sf8AX/P/AOjDVv4o+OEdvFHaeIvHKwqgESx3d0FCgcYAOMYxX/CVfHf/AKGXx7/4GXf+NeKNS8aanPBJ401DWrudEIgbVJZZGVc87fMJIGfSv+CKf/Ir/Fb/AK/9L/8ARdxX/BXz/k1C0/7G6y/9E3Ff8Em/+Tv9O/7AGof+gLX/AAVN+NuqfCX9muTw74bv5LTWPHt8NJE0TlHitAhe5ZSP7yqsR9pj6V+wx+zRbftR/HWz8Ga7LPD4Z0i0fU9dkgbY726MqrErdmkkdFyOQu5h92rX9lD9maz8ML4Og+AvgU6WsPk+XJocEkjAd2lZTIz8A7yxbPOc81+35+zXon7MXx6m8K+EJJT4Z13T49V0iKZy72sbu8bwFyctseNsE87WXJJyT/wRb/5Jf8Rv+w9bf+iDX/BaT/kl/wAOP+w9c/8AogV+wn+zLp/7U3xyh8GeIrye28N6NYvqmstbtsllhR0RYUbnaXeRAT2UMRzivEf/AAT2/ZB8Q+DpPBsXwY0jSkMDRwajp5eK9gYjiQTkl3YHn94XB7gjiviz8PdR+E3xN8U/DPVphPdeGNWuNPacJtE4ikKrIBk4DqAwHowr9kvx7f8AxP8A2afhv421a6NzqF/4ft1vJ2bJlniHlSuT6s8bE+5NVWs6jf6R44vtW0q8mtL2y1WSe2uIXKPDIkpZXUjkEEAg9iK/Yr/4Jg+AviB8MdJ+L37QU+q383ii3W80zRLW5a1SK1fmOWaRfnd5Fw4ClQFYZ3E4XxnZ/Dz9gv8AZS8X6r8MdIez0zw5bTXWnWl1dPcBr64ZY4gzSEsVMzx5GemcV+yn8K4/2pP2mNK0D4j+IJpLDUrmfVvEt/cXO2W4jTMkuZCc75ZCqFs5HmE9RXh/w/8ABbwp4VXwP4b03wfp+gLCITpsCwLA6AYwydH9y2STyetf8FLPgb8P/gx8ebe8+FsdhbeHfF2mDUFsbKRDFZXQkZJo41X7kZxG4HQGRgAAoA/4Jr/FC6+NH7IujWXidzfXPhe4n8N3jzDPnxRIjRA+uLeaFDnrtJ718a/h/f8AwT+Nni34epLLHL4V1yeC0mPDNEkmYJf+BR7G/Gvh18SNI8afB3w58Wrm8ig0/VvD1vq9xKeFhV4FlkyB02/MCO2DXiZ/E/7R3xO+JPxJtYpGnkTUvE93GyljDbLKGK8ZwEV1UdgFAr/gi98Twl18QfgxeXP+sjg17T4s9CpEFyfx3Wv5Gv8AgtL8Rd118OPhJa3H+rjudbvYs9dxEFu2P+A3I/Gv+CNfwoGjfDXxj8Y7+3AuPEmpJplgzLyLe2XdIyn+68su0+8FfFv9nr4PfHe68P3PxZ8F23iIeGJpptOhuZHWNHlCh9yqwDghF+VsrkA44rxH47/Yy+BtrL4V17XPhT4SWJfKl0dFs4ZAuMYa3QbsYJHK96/4KJ3X7KGv+PtB8Zfsv61o0zarDcx+I7HSLOS2toZ42j8qZUZFUGUPIDs4Ji3HliT/AMEXPH2pah4R+I/wyvblns9GvbLU7BGJOw3CyxzAdgM28Rx6sx9a/wCCyn/JsHhj/sfbT/0gvq+Afwt8bfHz4jaN8DPB+pPbL4mv45royFjBCLeOUm5kQfeMcTz7eh+cqCN9fDX/AIJd/stfDqTQtXbSte1rxDoVzDeRard6rJGzXETK6v5URWMKHUEKQeOCW61/wWM+Nmp6r8QPDfwE0u+dNJ0OxXVtTiRyBNeTFliDjv5cS7h/13PpX/BMT9irwV8dE1f4y/F3SRq3hvRL4WGl6RIzLFe3aorySTAYLRorxgLnDMzBuFIb4ifsWfsv/ErwlP4R1T4L+FdLjeAxW19oulQ6fdWZx8rRSxKCMHB2nKnHzKRkV8a/hfqnwV+LPir4VaxcC4ufDWpy2guAmz7RGDmOULzgPGUbHbdX/BMr/kx/4a/9xX/063dVrms6b4c0W/8AEOsXS29hpdrLdXUzdI4o1LOx9goJr4+/GbxX+0T8Xdd+JviSWaS51i6K2NpncLO2BxBboB2VcDjqxLHlia/ZO/4JofBn4Y+CNK1z4z+DNP8AGXjm9hSe+i1VBc2enuwybeOA/u325AZ3DZYEjA4rXf2Nf2VPEMcCXX7P/ge0a1mSaKXTNHi06RXVgynfbhCRkDgkg9CCCRX/AAU8+O2q/Bf9m250vwxqElnr3jm8Gj288LFZILcoz3EinsdiiPI5BlBHTI/Yq/ZzT9p/49aT8PNSmng0C0gk1LXZoG2yLZxFQVQ84Z5Hijz23lucYrR/2bP2fdB8LL4K0v4K+C49FEIie0k0WCUSrjGZGdS0jHAyzEsTySTX/BSD9lPQv2aPi3p9/wCArNrXwZ40tpbnTbVnL/YriJlFxApJJKDfE65OcSbf4cn/AII8fHPVvFXgXxN8DfEOoPcnwi0d9ohkfLJaTMwliH+ykoDD/ruR0AFf8Frf+Rn+FH/Xhqn/AKMt6/4Jz/sleHv2o/ifq1z4+86Twd4MtoZ9StYJTE97NMzCCAspDKhEUrMykHCYBG7cPih/wTw/ZY+IPgO98JaR8LNF8Lagbcrp2saRb+RcWkwXCOxB/fDONyvncCehwwvrXXvAfi2eyaaSx1rw7qLRmSJiGguYJMZU9irrwfatIsfB/wC0b8CPD0vj7w9Za1oXjbQdO1K70+dD5UvmJFcpkZBwHCsPoK/4Ka/C34e/CH9o628J/DPwnY+HtIfw1aXLWdmpVDK0s4Z8EnkhVH4V/wAEyv2YvgD8X/2cLjxX8S/hZoniHV08S3dst3eRsXESxQFU4I4BZvzqz0D4X/s0/CfW7rwl4Zs/D/hbwzY3erXFnZDaoEcZklYZJ5IStKfxX+2B+07Znxbrnkan8QvEK/bbyR9y2NuzfNs3H7kMKkKp4AjUcV8L/AHwI+DfhC18D/Dqx8M6Vplvb+S4SWIyXQxhmmkJ3Ss3csTmv+CsHwG+F/gLxP4W+K3wstdI0+LxS1xa61Yaa0awrcxBGjnWNOFMis4fAAzGp6sxP/BHr4tan4x+CXiT4XatcyXDeAtTiayZz/q7S8EjrGPpLFcH6OB0Ff8ABVb9kMePPCrftH+ANK3+IvDduE8R20EeWvrBRxPgdXhHU94s5P7sA/8ABPP9tOz/AGXdf8Q+HPH1xdz+CNcspbsQQq0jW+oQxkxmNR084KImPTPlEkKhNaPpHxW/bi/aWkiib7T4m8c6m89zOwLQ6dbjqx9IYYlCgdcKqjJIz8GfhF4P+BXw00P4XeBrPydL0S3EYkcDzLmU8yTyEdXdiWPbnAwAAP8Agpz+zj8DfA37POufE3wj8NNH0vxTe+IbZ7jVII2E0jTSs0pJJx8xJJ4r/gm78NPAfxZ/absfB/xH8L2Wv6NJo17M1ldqShkRQVbAI5FfDj9mL4A/CHxC3iv4Z/CzRPD2rvbPbNd2cbK5iYqWTknglV/KqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqvFX/ACK+sf8AXhP/AOizVVX7TfxdX4EfATxr8Vo1RrrQ9MY2CSLlWu5WWK3DDuvmyR59s14F8KeMfj/8YNI8JxalLfeI/G+trHNfXbmRmlnkzLPIepxlnY9cA18Iv2RP2fPgx4PtfCXhr4Z6DetFCEu9T1PTorq8v2x8zyyupJycnYMIucAAV/wVO/Y08DfDTSNN+Pnwj8N2uhWF1fCx8Q6XZRiO3SSQEw3EUY+WMEqyMq4XLRkAEsT/AMEjPjnq3g744XPwSvtQdtA8dWssttbO/wAsOo28ZkWRc9N8MciNjG4iPP3QK/4LOfCwWHjLwL8ZLGD93q9jLo1+yrgCWBvNhJ92SWUfSIV/wRi+IC6n8MfHnwxnuAZdB1mHUoEbr5d1FsYD2DW2T6F/ev8Agsv8TxrPxQ8F/CSyud0PhrSZNSvFU8faLp9qq3uscCsPQTe5x8LNT1v9k79q/wAKap4kdrafwlrdo2pnaUP2SdF87jqM28z/AJ1+1V8Rl+FP7N/xD8fw3IiuNO0C4WykDY23My+TAf8Av7JHX7CnwoHxj/ao8B+GLm3EunWGoDVtRDLlfItB5xVv9l2RI/8AtoKqv+C1v/Ir/Cj/AK/9U/8ARdvX7K3wF8YftUfE7SfgvpOuzafo1uZ9U1G4ZWkj0+ACNJpljzgyPthjHTJKZIAzXwn/AOCcf7MHwc8SaB418L6Brdx4j8OTLPa6le6vK7tKBjc8a7YzkE8BAOa/4LC/HbU9a+I2ifs/6RfSR6P4cs01LVYkcgT3swPlBx3EcO1l9529Aa/4Jgfs2/CHwv8ACLT/AI5ePLbQdQ8YeKHlksBqTRudKtEkaNBGjkhXk2Fy/Xa6Dj5s/tf/AAS+DX7Q/wAIPEmmaufDreJrTTZp9C1dZoVubW6jQtGokzkxsVCOhOCpPQgEfsFfFrU/g/8AtUeBdVtLmRLHXtRi0PVIlPEtvdusXzDuEkMcn1jH0r/gsl8LBrvwl8I/Fuyg3XPhXVXsLsqv/LtdqMMx9FlhjUe8p9a/4Iw/EBdO+I/j/wCGFxcALrekQanbo3/PS1lMbhfcrcg49E9q/wCCzfxPGm+AfAvwgsrnE2ualNq16inBENunlxBvZnmYgesPbAzd6P4t+CfinwV4uVjBqMlnp/ibSJChXCl/Mhb1+9H1HpUPxR8NXHwgT41Rzf8AEgfw3/b4ctgi1+z+fkn/AHK+HnhfXv2ivj7ovhq5mZtT8e+JV+2zqCSn2ibfPL34VS7n2Fadp1jpGnWuk6ZbJb2llCkFvCgwscaKFVR7AACtH/Zi/ZK+Cy6p41n+HHgzS2urqW6vNY15Y5vLeRyzYluSREuTgKpUYA4r46/GL/gm78RvDGo+CviR8Qvh5fpcxNBHeabCLuezcg7ZYJrdHKMp5DA4PQ5DYPh7xFqngPxjp3irwrqe3UPD+pR3en3aAr+8hkDRuBnIGVBxXh7WIPEWgaZ4gtk2xanZxXMak5wsiBgOg7H0FftUf8nPfF//ALH3W/8A0umr9hr/AIJ46J+0R4Vf9oP9ofWNYvrLxFeTy6fp0Ny0UuoESMJbq5mOXIeQPgKQTjcWwQK0L4a/B39hv4I+OvEPw30Gex0bTLO51y6tbi+luDPPFBgKGkLEbvLRcDjJ+tT3Pjf46fFRZLy7k1bxZ451xIxJKxzPd3MoVR3wNzgADoMAdK+A/wCwX+zp8FPBljolz8N/D/irXvs6jU9a1vTo72W5mI+coJQwijzwEQDgDO45Y/8ABTv9iD4Z+B/h4f2gfg94XtPDR0y8ig1/S9Pi8q0khmcRxzxxj5YmWVo0KoApDg4BU7v+Cdv/ACeh8MP+v+5/9I56qv8Agr1+0Hq3gf4f6F8C/C2otbXXjZZbnWnibDiwiIVYvULLITnHUQsp4Yiv2BP2Pov2sPiPf/8ACUXVzZ+CvCkUU2sSWzBJbmSQt5NsjEHbu2OWbHCqQMFlNaD+xx+yl4c0mLRtO/Z38ATQQjCvf6DBfTH6zTq8jfixr4X/AAK+EfwVk1qT4VeBtP8ADS+IZo5tQisgyxyvGpVCEJKoAGPyqAOScZJNft5/HbVfjz+0l4p1R9Qkl0Lw7eS6PoVvuPlx28DlGkUesrq0hPX5lB4UAf8ABOn9hz4ZeGPg/oPxj+J3gzTPEXi7xdapqFouqWyXMOmWcnzQeVG4KiR49sjORuG8KMAHd+2D+wz8Kfjr8MdZm8JeA9G0Xx9p1m8+iajptmlq9xMi5W3m2ACRHxsy2ShYEdCD8E/i14m+BPxU8O/FDwrcSR32g3qySwhtouYc4mgf/ZdCyH0znqAav9Y0/wARfD248QaTOJrLU9Ga6tpB/HHJDuVvxBBqv2Zv+Cb/AMBPhn8NtKX4n/DzSvGHjO/tI5dYudXi+0xW8zAM0EMbfIqofl3Y3NgknBCj/gqD+yD4B+AGs+G/iR8J9J/sjw/4plmtb3S0dmhs7tFDq0W4kqsilzszhTGcYBAH/BF7x9f2/j34g/C2S6ZrK/0iHWYoWJxHJBMsLso6DcLiMH12L6V+2P8As4/A3xV8Jfit8WvEPw00e/8AGFl4L1O5t9YljYzxy29jIYXBzjKbFxx2Ffs76Do/ir9oH4Y+F/EOnxX+lav4x0mzvrSYZSeCW8iSRG9mViD9a8M/sbfsu+DfEOneK/C3wU8O6dq+k3KXNldwxOHglQ5V1+bqCK/4LE/HbU/DfhDwx8BPD99JbnxRv1LWzG5UvaRMFhiOOqPKHY+8C+pr/glN+zb8K/EGjar8ffipbaNqN3a6kdP8PafqjI0cBjRGkuvLc4ZizhEJHymNz1II+MXwy+BHx08DXngL4gQ+HbyzuLVobW4EkIn09tpCSW79Y3Q8jHHYggkH4feMfEP7M/7QWneKNNvPM1DwJ4heK4Nuw23UcUjRTxg5wVkj8xOvR+vevG/gvwh8XPAOqeCfFlhHqfh7xJYmG4iYY3xuAVZT1VgdrK3VWAI5FftA/Bfx5+yP8dbvwfPf3dvd6Ldpf+HtahBia5tw+63uYyOjArhgPuujDJxmv2yP+ChE/wC0R8AvAvw08PtLY6hqFut347SONo42uYW2xwJ2aNmU3BAyBmEZyrCv+CUP7H6wxRftR/EPS/3kivF4PtJ0+6vKSXpB7n5kj9t7c5Rh8Tvg98MfjPpFroPxS8Gad4k0+xuftNvb3qFljl2ld4wRztZh+NftheEvDfgT9pv4ieD/AAfo9vpWjaVrLw2VlACEgQIp2qD25Nfsffse/sx+O/2Y/h14v8X/AAY8ParrOq6Mk15eXETGSaQswLNhuuAK8M+GdB8G+HtO8J+FtLg07SNJtktrKzhBCQRIMKij0AGKqqqqqqqqqqqqqqqqqqqqqqqrP/j6i/3xVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV4q/wCRn1j/AK/5/wD0Ya+Ff/JMPB//AGAbH/0QlV/wWl/5Kf8ADj/sA3X/AKPFf8EU/wDkV/it/wBf+l/+i7iv+Cvn/JqFp/2N1l/6JuK/4JN/8nf6d/2ANQ/9AWv+C2FtfvpHwhu4i32KK51iObB48xlszHn3wkn61/wRZ1LSoviL8StHmeP+0rnRLOe3UgbjDHM6ykd8bpYc/Ue1V/wWblib4+eCoFlQyJ4PVmQHkA3dwASPQ7T+R9K/4It/8kv+I3/Yetv/AEQa/wCC0n/JL/hx/wBh65/9ECv+CMf/ACXfxt/2KJ/9K4Kr9u//AJO/+Kn/AGHn/wDQEr/gnb/yZf8ADD/rwuf/AEsnqq8Vf8jPrH/X/P8A+jDXwmhhg+FfgyCCJI44/D9iqIi7QoFugAAHQY7V/wAFWYr6T9jfX2tCfKj1fTmucHH7vz1Az/wMpXwM+BXjz9onxyPh18N4tPk1k2ct2sd7dC3Rkj27sMc5PzA49AfSv+HTX7X/AP0BfC3/AIPE/wAK/wCHTX7X/wD0BfC3/g8T/Cv+Ccv7NnxJ/Zl+E/iPwr8UIdPh1TVfETXkEVlci4UQ/Z4UDFh3LK4x2wD3r/gsF8LB4U+P+i/EyztQlp450YCeQKBvvLTbFJ0/6Ytbda8FftENoX/BITVokvmbV7a6uvBcDFsEm5l8wrntts7hsf7g+tf8EkfgXYePPDfxi8WeIrYf2drmjnwjFJt3bo7hC92v4L9mOO+e2K/Yg8T6r8Bf23PCekeIG+ySjXJ/DOrwk4AeYtbbSTjAWcxtzj7nNf8ABSfxzN8Qv2x/GcNtK1xb+HzbaLZooJK+TEvmKB/13ebj3r9mj4WR/BT4B+Bfhj9nSK50TR4Vv1TobyQeZcsPYzPIfxr/AIKS/t2+PtS+Imufs+/CbxHdaF4e8PSGz1y+sZTFcaldAYmi8xSGSFCfLKjG5lfJK4Ffssf8ExPiX+0T4QsPiZ4q8X2vgrwxquZNPaSza7vL2MEgyLFuQIjEfKzNkj5gpUgt+3d+w/4Y/ZC0XwZqGgeO9U8RTeJZ7qGcXlokKxeSsRBUKSeTIeCeMCv+CKf/ACM/xW/68NL/APRlxX/BZT/k2Dwx/wBj7af+kF9X/BIaGKX9rGd5I0ZovCd80ZZclD5sAyPQ4JH0Jqv+CoMV8n7bHjtrsnypLfTGtcn/AJZ/2fbg4/4GHr/glLd6Zc/sc6FDYSRNPa6xqMd6E6rKZywDe/lvGfoRVf8ABQ+SOX9s74ntFIrgajApKnPItIQR+BBFf8Eyv+TH/hr/ANxX/wBOt3VftapqL/sufFpNJJ+0HwXquAAMlPssm/Ge+zd7+nNfB6+0vTPi54H1LW5I00608R6fLdtJ90RLcRlyfbaDVV/wWxhv20n4QzxbvsSXGsLN6eYVszH+iyfrX/BGC501PjL49spXj+3y+GI5IFI+YxLcxiQj23PFn6iq/wCC1V3pqeCPhdYSlPt82q6hJACOfKSKESY9tzxfpX/BGezv5P2h/GGoRF/sUPguWKYAnb5j3lqY89s4STGeeuO9f8Frf+Rn+FH/AF4ap/6Mt6/4Ilf81m/7gP8A7kKr4/f8l3+JH/Y3ar/6VyV+yv8A8mwfB/8A7ELRP/SCGv8Agr5/ydfaf9ijZf8Ao64r/gkH/wAmoXf/AGN19/6Jt6/bdivpv2SPiumnE+aPDN0zYOP3YGZP/HA1fC/4b+JPi/4+0X4a+DxaNrWvTmCyW6nEMbOFLYLnpkKQPfA71/w6a/a//wCgL4W/8Hif4V/w6a/a/wD+gL4W/wDB4n+Ff8E1/wBj/wCMP7MGq+PtS+K1tpVuNft7CKwSxvRc7/KacyFiMbcb0x65PpXjjxX4X8C+DtZ8YeNdQt7PQdHsZbnUJpxlFhVSWGP4iRwF6sSAASa+IWr+GPEHjzxDrvgrw+2haBqGp3E+maW0m82Vu8hMcWe+1SB+Ff8ABHr4pfDfwt8S/Evw48QaZbWvirxbbRnRdXkPzTJCGaSyGfulhiQYxu8sgkkIKr/grJ/yZ/qP/Ye0/wD9Dav+CTf/ACd/p3/YA1D/ANAWqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq8Vf8ivrH/XhP/6LNVVf8FWIb+X9jfxA9nu8qLVtOa6x/wA8/tCgZ9t5j/Sv+Cb1zp1p+2v8MpdUeNYWub6NDIOPNfT7lYvx8woB74qv+Col3p1t+xT44gvSnnXVzpkVnuHPmi/gc49/LST8M1/wT8s7+/8A2yfhdBphfzl1WSVthIPlpbyvJ07bFbPbGc8V/wAFJPhYPil+yP4vW2tRNqHhNU1+z+UEp9myZyP+3dp+lf8ABIzx03hj9qhvCctwRB4w8PXdmkWeGmh23Kt9QkMw+jGvEkj/ALYf/BQ6W1iH2zTfFHjZLVc850q1YIWx0/49Lctjpnv3r/gr98NW8LftIab8Qbe322njfQopJJduN91a/uJB74hFt+f5/tdftCSeN/8Agml8HIU1HzNS8YXFtZank/NMNNR452I9TcRwP+PuK/4Iw/Cjy7Px78br+0XMrxaBpkpXkBcT3WPYk2vTupqq/wCC1v8AyK/wo/6/9U/9F29f8EXIYW+KnxEnMSGVPD9uquV+ZQbjkA+h2jI9h6VX/BSOK+i/bX+JY1AkyNcWLISc/uzp9ts/8c2ivhf/AME4/wBpb4w+ANF+JXgnTvDlxomvQGazkm1dI3KhipDLjghlII9q/wCHTX7X/wD0BfC3/g8T/Cvh/wD8Erv2rtD8e+Gtb1fTPDMNjp+r2txdSJrCOyRJMrOQuPmIUHjvX7TvwtX40/s/eO/holsJ7rWNGmFghAP+lxjzbc8/9No46/4J7+OX+H37YXw4vpJ2jg1XUm0e4Q8eZ9rjaBFP0leNvqor/gon4t1T47/tu6j4I8Nn7U+kz2XhXSoy2A0+4b146f6TPKvfgD6D/grf8DLDwH4c+Dvirw7bf8S7Q9G/4RGSTZt2x2yK9ovHqv2k47Y7840D9oSRv+CQWsaYdR/4nGn358GqzH5mSWdJtuPT7HKyD/c9jX/BHz4Uf8JX8eNc+Kd9aLJZ+BtIMdtIy/cvbvMaEf8AbFLkH03D1r9qH486V+zZ8FPEHxW1G0W9uLCNYdNsWbaLu8lO2JCey5O5iOQitjnAqfXP2i/27vjXYaDf63d+JfEmtTuLO2mmMVlpkIBZyifdghRQScDJx/Ex58Hf8EV7AadHL8QPjrOb50/eW+j6OBFE3HAklclx152L1HHHPjDRIvDXi3W/DkE7TR6VqNxaJI4wXEcjIGIHchc18K/+SYeD/wDsA2P/AKISv2qP+Tnvi/8A9j7rf/pdNX7GEMUP7J3wmSGNEU+E7FiFXAyYgSfqSST7mv8AgoBFfTfsb/FJNOJ80aQrNg4/dieIyf8Ajgav2JrvTLL9rX4UT6tJEkB8TWqKZOnms22L8fMKY98VX/BTiSNP2IPiOjyKrOdLVATgsf7UtDgevAJ/Cv8Agnb/AMnofDD/AK/7n/0jnqq/4LFJqI/ai0Rrwk258F2n2TgY2farvd07793XnkdsV/wRevtLk+EfxB0yGSM6jB4jhluFH3hE9uoiJ9t0cuPoaqtXhv7fVb231Xd9tiuJFuQ3XzAxDZ/HNfBS5029+DPgK80d42sJ/DGnSWpjHymI20ZQj22kVXjq703UPG/iG/0cobC51W6ktSgwPKaVimB6bSK+DVnf6d+yh4F0/VS5vrX4eafFc+YSW8xdPjD5zznIPXmqr/gs5/yQjwR/2Nw/9JJ6/wCCNn/Jz3ib/sQrv/0usK/ao/5Ng+L/AP2IWt/+kE1fsr/8nPfB/wD7H3RP/S6Gq/4LERXyftTaO90SYn8F2ZtsnOEF1dggenz7+Pf3r4GfsIfH79orwKPiH8M7HQbjSPtktmTdaosEiyx7SwKkEjhlP0Nf8Omv2v8A/oC+Fv8AweJ/hX/Dpr9r/wD6Avhb/wAHif4V4M0afw74O0Lw9dSK82maZb2sjL0LRxqpI9siv+Cy3xC+GF1aeDfhkmnQ33j2xmfUGvY3AbTbF1KmF8dTM4Vgp6CLd/EM6Xc2llqdpeX2npf20E6STWkjsizorAshZeQGAIyORnivgN8SfAPxc+EPhfx18M0hg8PX2nxpa2UahPsHljY1syrwpiKlMDj5eMgg1X7d/wDyd/8AFT/sPP8A+gJX7CP/ACZ/8Kv+wAn/AKG1VVVVVVVVVVVVVVVVVVVVVVVVWf8Ax9Rf74qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq8Vf8jPrH/X/P/wCjDXwr/wCSYeD/APsA2P8A6ISq/wCC0v8AyU/4cf8AYBuv/R4r/gin/wAiv8Vv+v8A0v8A9F3Ff8FfP+TULT/sbrL/ANE3Ff8ABJv/AJO/07/sAah/6Atft+/s6ah+0l+z1qXhrwzbJN4o0C5XVtDjJ2meaNWV4QfWSJ3UA8b9mcYyPgl8Y/iB+zJ8XLD4heFomtdZ0OaS3vtOvY2RZ4z8s1tOnBAOMEcFWUEYKjEf/BZ74KN4ZF1N8JvGy+IfKz9gWS1a08z+79p8wPt/2vJz7V+0j8XfiX+0J4/l+NvxD0x7KHxAGg0eJEZbeK2tzt8qEty6oWO5u7s54OQP+CLf/JL/AIjf9h62/wDRBr/gtJ/yS/4cf9h65/8ARAr/AIIx/wDJd/G3/Yon/wBK4Kr9u/8A5O/+Kn/Yef8A9ASv+Cdv/Jl/ww/68Ln/ANLJ6qvFX/Iz6x/1/wA//ow18K/+SYeD/wDsA2P/AKISv2hPhLZ/HX4KeMPhNeSxxHxFprw200i5WG5UiSCQjuFlSNvwrwZ4q+KX7JPx3tteh05tJ8YeCNReK5sL2M7X4KSROARujkjYjIPKsGU9DXw2/wCCsf7Kfi3QoLvx1rGr+BtVEY+02V7pc97GJOMiOW2STevXBZUPHIFeMP8Agq1+x34bsXudC8Wa74rnCZS20vQ7iFi3pm6WFR9c9PXpXhTxHp3jHwto3i7SH3WOt6fBfWrE5zFNGroeP9lhX/BWL4Xf8J3+y1N4vs4N9/4E1WDUQVXLG3kPkTKPb97HIfaKh8R9bHwob4Qqf+JO3iEa4w39bgW5gHGP7hPOe9fsA/Cc/CD9lHwNod3arDqesWh1rUflwxluz5ihh/eWEwxn/cr/AIKR+A734L/tm6t4p0BfsaeIvsvibTZFH3J2OJWz6/aYZXx/tD1r9k3w1eftSft2aT4g1q0Ah1HxLdeK9WjALqiRytdFD/stKY4/+B1X7XnhHXfBH7T/AMUNC8RRzC7bxRfXiSSg7pobiZp4Zeeu+ORG/Gv2Qv8AgoP+zNZ/s8eDvCXj/wAd2vhHXvBuh22l3lleW0xEwto1iWaJkRhJvVAxUfMGJGMYJ/4KLftieHf2qPHOh6T8Pbe6Xwf4PjnWzurqMxSahPMU8ybyzyiARIqBvm+8SBu2j/gin/yM/wAVv+vDS/8A0ZcV/wAFlP8Ak2Dwx/2Ptp/6QX1f8Eg/+Tr7v/sUb7/0db1X/BYj9n/WJNW8P/tGeHtNafTxZro/iBokJ+zurs1tM+P4WDtGWOACkY/iFfsEftzP+ydq2qeGPGGkXuseBfEMyz3MNkV+0afcqu3z4lYgSBlCq6Fl4VSDlSrfEj/grr+zf4f8KTXvw5tde8VeIJIT9ksZLBrKGOTHHnSSYwo77A5PTjqPiNrnjPxj4svviF47in/tTxhPLqzzywmJbnzZXzJGOnl71dRjgbcDpX/BMr/kx/4a/wDcV/8ATrd1Wr6VYa7pV5omq263FlqFvJb3MLdJI3Uqyn2IJFftK/ALxX+zb8W9a+Gfia2mMNtM0uk3zphdQsmY+VMp6ElRhgPuuGXtX7Jv/BWLw3oHgvTfh9+0jZaq15pEC21r4lsIftP2mFAAv2mPIcSAAAyLu39SAck+Jv8AgrP+yRotmJdE1TxN4guGICw2mjvCFJOMs0xTCjqcbjjoCeK/4KA/s73v7Sv7O19pHhOEXfiXw9OusaFHGQTduiMrwKf+mkbsF7FwmeOa/Zy+OPif9mH41aP8TNL0+SeXSZZLbU9LlYwm6t3GyaBsglW7gkHa6KSDjFaT/wAFOv2NNT8LL4kufifPp0/k75NJutHuvtcb4/1eEjZGbkDKuV/2uDj9tn9qi8/aw+LreLbOwn03wzo1ubHQbGcgyLDuLNNKASBJIxyQMhQEXLbdx/4Jbfszax8EPg7e+PvG2mSWPif4gPFcfZJ02yWdhGD5COp5V3LvIR2DICAykV/wWt/5Gf4Uf9eGqf8Aoy3r/giV/wA1m/7gP/uQqvj9/wAl3+JH/Y3ar/6VyV+yv/ybB8H/APsQtE/9IIa/4LJ+Eb/TP2gfCnjJoJPsGu+FUt4pW+609tcS+Yq/RJoD/wAD96/4J1/t7fDT9m/wDrvwt+Ldrq0dhPqjanpmoWFsLjDSRokkLoCCB+6VlYZzuYHGBnwB8Qfht+1p8E7zXPDkd9ceEvFtrfaVKt7b+TJLEd9vN8uTgH5sZ7HkV8R/AXxL/ZL+PEugXrzad4j8G6rHeaVfiPatwsb77e5jzwyOFBxz3U8ggfBP/grd+z/4s8N2cXxnfUPA/iKKELfNHp817ZTSAcvCYQ8iqSM7XT5c43NjdXiL/gqb+xfolibvTPiHqviCUKSLXTvD94khx2BuI4k5/wB764r4MfFbw98cvhf4e+K3hWC5g0vxFbGaCG52+bFtdkZH2kjcGRgcEjI61/wVY/a7Xx14mf8AZs8A6nu0Hw3db/ElxDJ8t7fp0t8jqkJ+8D1l7fuwT/wTM/Yo8PeL/hP4o+K/xb0Jbm18eabcaLolvKnzRWTZSe6TI+V2cbUbqBGSOHFfGH4Y+Pf2WfjlqPgy8vbix1zwpqSXOlapBmMyorCS2uoiOmQFb/ZbIPINfsZ/tMaV+1J8FtN8al7eHxJp+LLxHYxHHkXajl1XqI5BiReuNxXJKGv+CnXhG/8AFn7G/jF9Mgkmm0Oez1Ixx9THHcIJWPssbu56cL+B/ZA+Pdt+zX8fPDvxT1PTZ9Q0m1E1rqltb4817aaMozJnALKSrgEjds25Gcj4A/txfAr9pfxrc+Afhdda5PqllpMupz/bdONvGkMcsMRG4scsWnTAAxgNyOM1VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVeKv8AkV9Y/wCvCf8A9Fmq/wCHznwI/wCiW+Pf++bT/wCPV/w+c+BH/RLfHv8A3zaf/Hq0LxZ4G/4KFfspeJIdJ0zUtE0nxXDdaZENSRDLbXEJBjmIjZgQsojfGedvatU0r4j/ALOvxd+w6laz6D4y8DavHKoYcxXELh0dT0dDhWBHDKQeQa+EP/BUv9l7xz4Ns9T+IPixvA/iJYgNQ0u8sridFkA+ZoZYo2V0J6Zw2Oqiv+Ci37c+h/tNXel/Dj4XLdjwNoF0byS9uYjC+qXe0oriM/MkaIzhdwDEuxIGBX/BIf8AZm1g65f/ALTXizTJLbT4LaTTvDAmTabmR/luLlQedqqDEG6MXkH8Farpdhrel3mi6pbJcWd/bvb3ELjIkjdSrKfYgkU9z4q/ZP8A2jNUi01mbWvAOtX1jDIx2GTaJIRIDjoyNuHHQiv+COHwnPiD4ueKvi9f2qva+EtKWxsndel3dE5ZT6rDFKp9ph61/wAFdfhavjL9mu0+IFpbB73wHrEU7yYyVtLkiCUAe8jWzH2Q1rfxH1vXvhr4W+GV827T/CWoajd2LZ5X7YLfemPQNblv+Bmv2KfhSPg1+y/4A8GTw+Xfvpa6hqIZcMLm6JnkVvdDII/ogqq/4LW/8iv8KP8Ar/1T/wBF29f8EW/+Sn/Eb/sA23/o81X/AAWA/Z01mDxPpP7SXhvTJLjSry0j0vxC0Mefss8ZIgnfHRXQiLceAY0GcuK/YK/4KLWn7OXh3/hUXxY0m/1LwWlw8+mXunqJLjS3kYtIhjZgHiZyX4IZWZzht2Av/BTv9iNrUTn4ySq+zd5B8OanvBx93It9ue3XHvXwB/bq+CX7S3xK1T4afDBdcluNK0ptRN7fWi20NxGskcbCJSxkJBlUncq/jVftYeD7z9nH9s3xTHpFsI49J8Sx69pCgbU8qV1u4lX2Qt5efVDX/BOnwbqPx/8A23G+J3iG2WeLRJ73xTqJk+YG6kciHn+8J51kH/XI1/wUR+Fq/FX9kjxxY29sJtQ8OW667YnGSj2h3ykDuTB56gerVbfEfW7f4Uaj8Iw27SL/AMQWmtgE48ueG3uIDgf7SzjP/XMV/wAEsvhSPhz+ylpXiC7h2aj46vptYm3LhlhJEUC/QxxCQf8AXU1/wVq8I674o/ZLfUNFjmki8NeJLPUtQSMEk2+yaAkgdVD3EbH0257Zr9gr4/eEf2cP2iNN8d+PIZv7AvLC402+uoYjK9ksu0iYIvLAMihgMnazEAkAH41/8FPf2Zvh94IutT+H3jOHxv4luLVjpemWEEoQyEYVp5GVVjQEgsud+Oi1qeo3msajdatqM5mu72d555G6vI7FmP4kmvhX/wAkw8H/APYBsf8A0QlftUf8nPfF/wD7H3W//S6av2NP+TUPhL/2KNh/6JWviD4L0n4j+BPEPw/1xc6f4j0u40+5O3JVJo2QsB6jdke4FfELwJ47+AfxS1HwZ4jhm0rxJ4U1EbJUBX542DRTxE9UYBXVu4INfA//AIK6/BXXPBtjbfHG21fw34ptYFjvZ7Sxa7tLx1GDLH5fzoWIJKMuFJwGYc1+3Z+3i37WWnW/wh+C3hjWI/CmnvJqmo3FzABc6h9mieQt5aFvLgiRZJCScnaGIXZz/wAE7f8Ak9D4Yf8AX/c/+kc9VX/BWn9m3XPif8ONH+M3gzTpL3VPAaTJqttCm6SXTpMM0oA5Pkuu4gfwSSMfu1+yZ+1R4y/ZP+JP/CZ+HbVNU0nUYlttb0eWXy0voAcja2DskQ5Kvg4yQQQxB8P/APBWL9kLV9Hi1DVdd8R6JdsmZLC70WWSRGxyN0O9D7Hd+Vfs1fth/Cv9qzUfFdl8MrbWYovCRtfOl1O3WA3In83a0aBmO0eSQS2DkjgcE/8ABR79mzWPgV8fdX8TWWnyHwh47u5tU0q6VP3cc8jb7i2J7MjsSB/cZMdDj/gn/wD8FHvh38Pfhvp/wS+P2q3OkRaADFomvfZ5LmJrcsSsEyxqzoUJ2qwBXbgHbsy37Y3/AAU9+EUHwv1fwJ+z54jl8S+JvEVm9m2qRWstvb6XFIpWSQNKql5dpwoUEAncW+Xa37Hv7N2v/tOfGnR/BdpYz/8ACPWU0d34kvlUhLWyVssu7tJJgxoOu5s4wrEeIreC08IanaWsSRQw6bMkcaDARRGQAB2AAqq/4LOf8kI8Ef8AY3D/ANJJ6/4I2f8AJz3ib/sQrv8A9LrCvjZ4Vu/HfwZ8e+CNPRnuvEPhjUtOgVCAS89tJGoBPfLCvAnivUvhx4/8O+ObC3Dah4W1i11GGGX5cy28yyKrenzIAeK+Ef8AwUu/Zy+Mfinwx4D8Pp4mtvEniaZLeKxudNAWCZlJKvKH2kDB5Gc8cV/wV0/Z01n4gfD/AET44+EdMkvL7wQktvrUMMe6RtPkIYTYHJWGQMSB0WVmOApNfsH/ALdF7+ybquoeGPFOj3OteA9fuFuLy3tCPtNjchQhnhDEK+5FVWQkZ2oQw24bTv8Agp/+xNfWUV1c/Fy50+WRctbXPh3UWkjPoxjgdc/RiK+F/wDwUT/Z4+M/xh0X4M/Dm513UdQ1uO4aDUZ7D7LagxQvKU/eMJCxWNsDZj3r9pb4/eFf2afhFq/xQ8TlZ3tlEGl2G/a+oXjg+VCvscFmPO1FdsHGK8E+Gfib+2f+0db6Xd373viXxzqrT6jfupKWsX3pZSP4Y4olIVfRVUdhX/BS/wDYj0Dwr8JfC/xT+EGgiC28AaZDo+tWsSDdLYLxFdNj7zo7EO3JIkDHAQ1/wTF/azb4H/FAfCjxlqgj8EeOblEDzOQmm6iQFim9AsmFifp/yzYkBDmv+Cj/AIRv/CP7ZHxAS7gkWHV57fUrSR+k0c1vGSy+wkEifVD161+xZ/wUu+C/ws+AegfCv4vQ65Y6t4WWS1t7qysftMV3bmRnjPykFGUNsIIwdobPzEDwL4v0n4h+CPD3j/QFnXTPEulW2p2S3CBJBDcRLKm9QSA21xkZOD3qqqqqqqqqqqqqqqqqqqqqqqqs/wDj6i/3xVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVqf/BGj4Y6nqN1qL/GfxOjXU7ylRp8GFLMTgc+9eGtEi8NeHNK8OQTtNHpVlDaJI4wXEaBAxA7kLVftcfsJeEv2ufEegeIvEfjzV9Ak0CyktIorK2jlEod95Yl+h7V+yJ+x14Z/ZD03xNpvhvxlqfiBfE09tLK19bpEYTCsgAXZ1z5h6+lftT/ALN2h/tT/DKL4Za/4lvtDtotUh1AXVnCsjlo0kULhuMHzD+Vfsx/8E3vBH7MXxUt/inoPxJ1zWruCxntBaXdpFGhEoALZXnIxVfH79gn9m/9ozUpPEXjDwtc6R4hmP77WtBnW0uZ+MZkBVo5D0+Z0LcAZxxXgn/gkL+y94Y1ePVfEWq+MfFccT5Ww1C/jgt2HbeII0kP4OAfSv8AgrDrngyH44+GPhV4CtrCz0vwB4WhsnsLGJY4rGWSR5PJVVHGIjAcf7XrnP8AwRz8KXGkfs5eIfE91Ds/t/xVN9nJX78MMEKBs+nmGUY7bT68ftcfsj+HP2ufDmgeHPEfi/UtAj0C9kuopLKBJWlZ02EEP0GK/ZL/AGBfB/7JXjTWPGnhzx/rGvz6xpf9nvBe2scSxr5qSbgU5zlAMe9V8af+CVPw++NPxU8S/FTVPit4h0678SXrXUtpBZQukJIAwpPJHHevgP8ACPTfgP8ACTw58JNI1i51S08OwSRRXdzGqSSh5XkJIXgcuRx6VVan/wAEaPhjqeo3Wov8Z/E6NdTvKVGnwYUsxOBz714a0SLw14c0rw5BO00elWUNokjjBcRoEDEDuQtV+0l+xZ8C/wBqGFbzx7oM1j4hgi8q28QaU6wXiIOiOSGWVB2V1bGTtK5JrWv+CKN19tZvDv7QsRtGJKJe+Gj5iDsCyXGG+uF+leDf+CK/hC1uIpviB8dNX1OHIMlto+jR2Le4Eskk357B9K+H/gnRvhr4G0H4feHZLp9L8OafDYWRupfMkEMSBEDNgZIUAdO1fEDwXpHxH8C+Ifh/ry7tO8R6XcafdYGSsc0bISPcBsg9iBWn/wDBGH4TWt/bXN58XvE93BDMry27WMKCZQQSm4HIyOMjpmoIILaCO2toUiiiQJHGihVRQMAADoAOMV+15+xD4I/a8ufDV/4i8U6j4evfDUdxElxY26SNcRylDsff2UoSP99vWv2SP2CvAX7JPiXXvFugeLtU8Ranrdilist9bxxfZoQ+91XZ13ssROf+eY9ar9qn9iD4Q/tXwW2o+Kje6J4n0+HybPXdO2+aIgSRFKjArLGCSQOGBPDAEg6X/wAEUIE1XdrX7RLy6ajjCWvhcRzSr3G5rlljPTnD/QV4o/4Jh/s+6n8E7f4M+FH1Hw/s1iDVbvX9qXV/eyxRSxKJXYAbNsz4RQqg8hckmv2SP2E/Cv7I3iTXvEPhvx9q2vnX7GO1mhvrWOIR7H3BlKH3IwR3r9rH9mHQf2svhzp3w58Q+KL/AEG207W4tVS5soUkdnjhmiCENxgicnPqor9ln/gnX4K/ZY+JkvxM8P8AxG1rXLqXS5tPNreWkUaBZGjYtlecjyxx71Wt6Jo/iXR73w94g0y11LTNRge3u7O6iEkU8TjDI6ngqQSCDXxZ/wCCN3ww8T6xcax8JfiXqfg2Gdi/9l3tj/acEZ/uxP5kciL0++ZD156AfCP/AII5/CjwnrVvrXxX+Iup+N47ZlkXTLaxGmW0jDHyykSSSOvX7rRk8dsg/tK/8E2Pht+0b4607xnL411TwpHpeiW+kW2maXYwm3ihgZymwHG0AOF2jgbRX7OPwP079nL4Q6P8INH8RXut2WiyXDwXV5GqORNO8xXavAAaRqqvjh+z38Jf2i/Cw8J/FbwrBqkEJZrO6QmK6sXIwXhlHzITgZHKtgbgcV4x/wCCK2h3F88/w/8Ajxe2NoW+S11fQ1unUe80csYPb/lmK8I/8EVfD8F1HN49+PWoX1sD+8ttI0NLV8Z7TSyyjkf9M+PetB0eDw9oWnaBa3E88OmWkVtHLOwaR1jQKGYgAFiBkkADPYV+0p/wTv8AgH+0jqc/i29tLzwp4snGZtY0UIn2tuzXELArKf8AaG1zxliBipf+CJ+om/2Q/tG232IniR/CreYBj+59qwfT7w9fav2ef+CZH7PvwK1i18Xast7468SWTCS2utZRBbWsg6PFbL8u4dQZC5UgEYIzVftefsP+FP2vdR8M6j4j8c6roDeGYLmKFbK2jlEwmaMktv6Y8sdPWv2QP2LfC/7H/wDwlv8AwjXjXVPEH/CW/YvO+3W8cXkfZvP27dnXd9oOc/3RVeOP+CQXw28ceNNf8aXXxg8S20+v6pc6hJBHYQFYmmlaQqCTkgFsc18NvBVr8Nvhz4V+HNjfS3lt4V0Sz0qG5mUK8yW8KRK7AcAkICQOMmv2if2cPhn+074EPgT4k2E5jgm+0aff2cgjubCfaV8yNiCOQSCrAqeMjIBF9/wRQvG1Nhpn7REK6cz/ACmfwwWmRfQgXIViB3yufQV+zD8BrX9mn4M6N8IbPxNLr66VLcStfyWotjK00rSHEYZtoBbAG49OtfH/APZf+DX7TGgR6J8U/C4uri0VhYarav5F7YluvlSgHg8EowZCQCVJArxP/wAEU7WS+eXwZ8f5YbNmOy31Tw+JZIx7yxzKGP8AwBa8L/8ABFTSYrhJfGvx+u7qAH57fS/D6wMR7SyTOB/3xx718O/gBo/wc+Bg+Bvwp8V61pFrb21zFYatdsl3dWjzuzvKvyqhYO7MBtChjnBHFJ/wRk+F01+t5qfxp8XXYebzLgNZwB5snLfOckE8888nNaBoOj+FtC07wx4e0+Kx0vSbSO0srWIYSCGNQiIvsFAH4V+1v+w58OP2uJtB1bxFrV94f1vQUeCPUrCFHee2Y7vJkDdVV8sp/hLP/eNfss/sAaZ+yj49uPGfgz4xeINRtdRtDa6lpN3ZRLBeJ1jY7TkOjcq3UAsOjGtT03T9Z0260fVrKC8sb6B4Lm2nQOk0bqVdGU8FSpIIPUGvip/wRo8Ea9r8+r/CT4rXfhfT7iQsNJ1LTTqCQZ7RzCVH2joA4c+rcc/sW/8ABPFP2SPH+pfEe7+Kx8UX2p6HJpRtI9G+xxwq80MpfcZpC5zAo6L941VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVqdimp6bdaa8hRbqB4iwHKhlIyPzr/hy38L/wDotXij/wAF8H+Nf8OW/hf/ANFq8Uf+C+D/ABr/AIct/C//AKLV4o/8F8H+Nfsx/s96N+zH8K7f4V6D4hvdatbe9nuhd3cSxyEykEjC8YGK/aO/Y++CP7UOnRR/EbQJYNYtE2Weu6Y6wXsC/wB3eVZXTk/I6sBkkYPNa7/wRRkbUGfwz+0Kq2LMdkd94b3SxL2BdLgBz152r9K+Df8AwSG+BngLVrbXPiX4p1X4hXFqwZLKW2Gn2LsOhkiR3dwDg7TJtOMEEHFWGn2Gk2FvpelWVvZ2dpEsVvbW8QjjhRRhVRVwFUAAADgVX7Rf/BMX4bftD/FrV/i3ffEDWvD97rUcAurSys4njaSKJYvMy3OWVFz789zX7K37MPhL9lH4c3Pw+8K6vd6v9u1KTULvULyJUlmkZEQAheAqrGoA9ST3NfFH4faN8V/hz4l+GviAsun+JdLnsJpFUFohIhUSKDxuUkMPdRXh3/gjb8JNF8QaXrGofFXxHqlrYXkU89jNYwKl0iOGaJiOQGAKk+9VVftefsdeGf2vdN8M6b4j8ZanoC+GZ7mWFrK3SUzGZYwQ2/pjyx09a/ZH/YS8JfsjeI9f8ReHPHmr6/Jr9lHaSRXttHEIgj7wwKdSenNVrOjaR4i0m70HX9LtNS02/haC7s7uFZYp42GGR0YEMpBwQRg18Xv+CPfwW8Z6nPrHws8b6v4Ce4fc1i9qNTs4/URqzpIoPoZGA7DHFWf/AARQ1t7hVv8A9oixig/iaHww8jfgDcqP1r9lf/gnZ8Mf2W/F6/ETSPGviTX/ABJ9jktDLcmOC2Ebgb8QopbkgH5nbGBVftY/8E+fAH7V/jrTPH+t+M9V8O6jp+lrp8n2C1jkFxGsjuhbd3BkYZ9Megr9kT9i3wR+yHbeJB4a8Sahr974me38+7voEjeKOEPtjUL0BMjE+vHoKv7Gz1SxuNM1C3S4tbuJoZ4ZBlZEYYZSPQgkVN/wRd+Fbyu8Xxm8URoWJVPsEB2jsM55rwx4c0nwf4Z0jwjoNv5Gm6JYw2NlF/zzhhQIi/gqgVq+k6Zr2lXmh61YQX2n6hA9vdWs8YeOaJ1KujKeCpBII96+LH/BGv4eeI9budY+EnxRv/CVtcSFxpOoaf8A2jDDk/djlEkbqgHQP5h9W718Ff8AgkJ8GvAGuQeIPil4xv8A4hyWrh4dPaxGnWTMDn97GJJHlA443hTzlSDivir/AMElPhd8T/iP4i+IS/EvXdEPiHUJLxtPs7CEw27OcsqZ/hzk47ZrwP4Yj8E+CfD/AIMhvXvE0DSrbT1uJF2tMIYljDkZOCduce9fEr/gkf8ADn4lfEXxV8Rb74u+I7O58U63earNbRWMLJC9xM8rIpJyQC5AJ9K+E3w9svhN8MvC/wAMtO1Ga/tvDGlwafDdToFeZYkChmA4BOOgqv2mv2OPgz+1VpkCeP8ATbmy13T4jHp+vaa6xXduuc+WxIKyR552ODjLbSpYmn/4InodWzH+0aRpn3gD4VzN977n/H1t+7/H6/w18KP+Ce3wL+D3wy8X+A/DK6jc6v430C70XU/E18UlvVguIWjdYQFCRKN24KByVXcWwMfBH/glh4D+B/xW8N/FfRPi14jvr3w3d/aIrWayhRJsoyFWKnIBDEHFVTKrKVYAgjBBFfHj/glX+z78XdWuvE/g66vvh7rF4xeZdKhSWxkc5Jc2zY2HPaN0X25zS/8ABE/WTeeW/wC0XZC13ECUeFmL47HZ9qxn23fjX7IP7C3g39kS81nWtE8d654i1XXrWO2vGuoooLbYjblKRKCwbJPJkIweneviL8NvAnxb8JXngb4jeGLLXtEv1xNaXceQGHR0YYZHGeHUhgehFfET/gjB4D1XUJr34XfGPVtAtnJZbDV9MXUQmf4VlR4mCjtuVj0yT1rwN/wRZ8K2d/HdfEf446lqtopy9no+jpZMcH/ntJJLwRj+AY9e9fCL4LfDH4EeEYfBPwr8J2miaYh3yiMF5bmTGDJLIxLSOfVicDAGAABqdimp6bdaa8hRbqB4iwHKhlIyPzr/AIct/C//AKLV4o/8F8H+NV+1p+yp4e/a18FaP4L8ReK9Q0GHR9U/tCOeygSRnbynj2kPwBhyfwr9k/8A4J8eDf2TviLqPxF8O/ELWdeudR0SXSmtr21iiREeaGUuCnOQYAMdPmNV+0n/AMEpvhj8afF2o/EH4f8AjG58Ca1q873OoWwsRe2VxMxJeRY96NEzMSWwxXJyFGTX7On/AAShl+Cnxb8NfFjWvjcmryeGr37VDp1roBgE5CkDdK07bRycgIeg5qSOOaNoZUV0dSrKwyCD1BHpXx0/4JOfAL4pardeI/h/ql98OdTuyzSQafbLc6fvJyWFsxUp/uxuijsopf8AgihrxuQj/tEWAt9+DIPDDl9vrt+04zjtn8a/Zz/4JbfDD4C+O9E+J178R/EniPxFoE3nWZSKKxtQ5BUlohvcjaSMeZjk5z2/ax/Yds/2t/EOk6n4s+LmvaNpWhW7R2GkWNnE0McjnMkzFjlnbCjPQBQB3J/ZJ/YS+HH7JOqa74i0DX9R8R63rcCWovtQhSNrW3U7mjjCdnYIWJ6+WmMYOdY0jS/EGkXug63YQ32nalbSW13azpuSeKRSrowPVWUkEehrUv8AgjD8I7nULq4074u+KbO1lmdoLY2cMnkoSSqbjy2BgZPXGa+GnhLVfAfgLRPBmteLbzxPdaNaLanVr2IJPdInCNJgkFwoUFurEZPJNftXfsW/Cr9rTS7JvFst5o/iPSImj03XLAKZYo2OTFIjcSx552nBUk7WXc2YP+CJ+pG+KXX7RlqLME4ePwqxkYf7pusDP1OPevhn4Jt/hr8N/Cnw5tL+S+h8K6HZ6VFdSIEadbeBIg5UEgFggJHbNVVVVVVVVVVVVVVVVVVVVVVVVn/x9Rf74qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqrx/4kufBvgXxF4ustKudUuNF0q5vYbG2haWW6eKJnWNEQFmZioUADPNfDb/gpl+138LA+l3fi+z8U20LsBZ+J7E3DQtnkeYjRzYHTazkDGABXjT/gr7+074l0aXSfD2i+DPC800ZU6hY2Ms1xGSMZTzpXjHXujcgc18FfgD8bv2vPiRJb+G7LUNVuNQvTLrniTUN729oznc8txMc5c5JC5LMegr4M/Cnw38D/AIW+G/hT4T3tp3h2yW3SWQAPcSElpZmA4DPIzuQOAWOOKqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqs/+PqL/AHxVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVeN/gR8E/iVcG9+IHwl8IeIbs/8vWo6NBPMPpIylh0HetI/Yr/ZM0O4+02P7Pfgh5Mgj7XpSXQBGeiy7h39PT0FaTo+kaDp0OkaFpdnp1jbLthtbSBYYox6KigAD6Cqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqs/8Aj6i/3xVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVZ/8AH1F/viqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqz/4+ov8AfFVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVn/x9Rf74qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqs/+PqL/fFVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVn/x9Rf74qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqs/wDj6i/3xVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVfHP4++Efgdokd1qwN/rF4p+waXDIFeXH8bHnZGDxuwcngA84sPHH7Y/wC0a8l/4LkudB0FmZY5bFxp8CdjtnJ82QjvtJwc8Cj+zD+2BZodRtfjKrXONxSLxPfCQk9RkoBn15xX7NGlftAaRFr+n/HDUrm4jtpIU0sTyxTs+QxkcSplmH3AAx4OelftR/tQ3XwouYvAngSCG48UXMSyTzzR+YlkjfdAT+KRuoB4AwSDkCtP+Bn7Z3xGs4/Eeu/Eu40Z7kCSO01DXLi3ZQwznybdGWPtlSFI6EDFfCbwL+194G+J2h6V4t8W3mqeFZ5WN/dSX41CLy0Qts/e/vYyxAUEAcn61VVVVVVVVVVVVVVVVVVVV+0T+14PAWsTfDv4YWcOqeJIn8m6u5F82K0kPHlog/1kvOCOinAIY5Asvgx+2t8TLddc8Q/ES60MzgOlrf6zNaH2PkWysqHBPBCkdwK8QRftj/s3oNf1DxPe63okLDzpmu21O0UZxh1lHmRKePmATrwcmv2ev2i/D/xz0eSB4I9M8S6fGGvtO37gy9POiJ5KEkAjqpIBzkMaqqrwx+1B4P8AGXxe/wCFQ+H9D1RrqGW6huL64CRxK8CsW2KCzMCUIydtVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX7QX7YOm/DfUpvBPw+srfW/EcR2XM8pLW9m/TZhTmSQd1BAU8Ek5UWPgX9uH4wRLq+qeLNR8O2l0N0aXWoHTF2npmGAbwMf3lz9auP2fP20fCUP23QfinNqcifN9ms/EtwdxH+zOqITwOtfCNfHQ+G+hH4mXLTeJnty1+zRpGysWJVSE+XIQqDjqQTXxO/ah+J/xR8et8L/ANneOWOEytAl9bopnvCv3pFdvlhhGMhuDjkkZ2g/szfthyp/bD/GPF6y7jGfFF6JgcYxuCbc44+9j3r9mbTP2hNJfxBpnxv1K5uLezMKaZ9peOdpSdxdxMuSy4CDDEnJPTvVVVVVVVVVVVVVVVVVVVXl3aafaTX9/dRW9tbRtJNNK4RI0UZZmJ4AABJJ6V8Qf2t/iV8TvFf/AAr/APZy0m4VJGZI75bdZLq6A+86h/lhj/2mGcYJKdAP2av2xdZiGral8XvIuyu7yJ/E14ZEPXaPLRkHJ6BsUPjP+1D+zVr9tpXxRjudd0qViEXUpROtyoxuMN2MtuxjhicZ5Xmvht8SPC/xW8J2vi/wneGW1n+WWJxiS2lAG6OQdmGR7EEEEgg1VVXjTxbpPgTwrqfjDXBObHSbczziBN7lR2UEjJ5HUivgj8dNE+OljrGqaBot9p9rpV0kCm8Zd8u5d2SqkhfTGTVVVVVVVVVVVVVVVVVVVVVVVVVVVVVWf/H1F/viqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqry7trC0mvryZYoLaNpJZG6IqjJJ+gFfD/Sr79rX9pC61jxQ0x0aIveXEBY/urKNgsVup7ZLIDjGcu3WrKxstNs4NP061htrW2jEcMEKBEjUDAVVHAAHGKqvjFfJ4N/bJl8ReNIXewtdesb2RmTdutVWIqwH8QVVxj1UirG/sdUsoNR0y8gu7W5jEkM8MgdJFIyGVhwQR3FVVVVVVVVVVVVVVVVVVVVfHj4gSfDD4T+IfGFo6i9trbyrHIz+/kYRxnHfaWDEeimv2EvhdbeIdX1f4weI4jeS6dcG105p/n/0llDSzEnksFZQD/tseoBqryztNQtJtPv7aK5trmNopoZUDJIjDDKwPBBBIIPrXjHT7j9lX9puG90J5U0m2uY7u3jJJMlhNkSRH1wPMQE90Dda8XeJoPCvgvWvGQt/tkOj6XcX/AJSPt85Yomk2hucZC4zjvX7P/wC0hY/Hq81qzs/Cc+jnRooXZpLwTeZ5hYYGFXGNn61XiD9rLTtB+NsXwXfwTcTTS6vaacNQF8FUGcxgPs2ZwPM6Z5x1r4/fHe0+BGj6Vq934bl1gapcvAscd0ITGVXdnJVs18OfjNa+BPjdc/FyXQJbyKe6vZxYrcBGH2jfgb9pHy7/AE5x2r4D/tVaf8c/F934StPBdxpD2mmvemeS+EwYLJGm3aEXH+tBznt714u8Y+F/AehT+JfF+tW2madbD55p2xk9lUDJZjg4UAk9hXib/goH4fhvnsfA/wAO9Q1ddxWOe7uxbb8d1jVXJB9yD7dq+GH7bVr488Z6V4F1b4Zahpt/qlwlvG9vei4CMerMrIhVRyT1IA71c3FvZ28t3d3EcMECF5ZZGCqigZLEngAAZJNePv28fAug6jJo/gPw1eeKZo3MYuTN9lgds4Gw7Wdxn/ZUHsec1oP/AAUCsE1BLPxz8Mb7TYWIzNZ3nnOqnv5bomfru/CvCPi/w3488P2vifwlq0Oo6beKTFNFkcjgqQeVYHgggEV8evjRB8DPClj4quvDkusRXmoLZGKO5EJRmjdwclWzxGR0rxj+2D8P/Bvgjw/4kurGe81nxFp8d7BottOrPAjDrLJjCDORnBJIOFwDjw9/wUM0641RYfFPw2ms9PdsGey1ETyRDPdGRA3H+0OnTmvHn7Qvwx8AeBtO8e3+tfbrPWot+kwWa7pr7gE7FONoXI3FsBTweSAY/wDgogh1MCX4VkaduwSur5mA9ceVgn2z+Pevhn8TfCfxa8KweLfB948trIxjlimXZLbygDdHIuThgCDwSCCCCQalligieeeVI441LO7nAUDqSewxXj39uTwdo2syeHfhz4WvfGN2jFPPim8mB2H/ADzIV2kAPcKAexI5rw7+3pp0OsR6T8S/hrqXh5HIzcQzGZowejNE6I233BJ9B66Tq2l6/pdrrWi38F7Y3sQlt7iF9ySIRkEEVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX7R3xKm+FXwj1nxLp8oTU5lFnpxP8M8vAYe6Luf32Yr9hr4PWOuPffGbxVbi9nhu2t9JW4G/EoAMtwc9WywVT2Ic9cEVXjG11G+8Ia5ZaOxF/PptxHakdRK0bBMf8CIr9hXxP4d8NfFfUdI8QTRWd9rGnm1sJZ8J+9EisYcnozAcDuUx1IFVVVVVVVVVVVVVVVVVVVVV+3l8R7vw34D0zwDpdwYpvE8ztdshwfs0O0lPbc7J9QrDvX7Hnwn0/4f/Cux8SXFqv8AbniqFby5mZfmSBuYYweoXYQxH95z6DFfFH4c6D8VvBOo+C9fhRoruMm3mKZa1nAOyVPQqT+IJB4Jr9i7xfrHw/8AjTqHwr1hzHFrBmtZoCciK9ttxBHpwsin1JX0FfHv43WnwK8N6f4iu/D0usLf332URR3IhKHYzbslWz93GPevhH8RIPix8PNJ8f22lPpseqedttXlEpj8uZ4vvADOfLz071rOoro+j32rNEZRY20k5jBwW2KWxntnFfAT9qKw+O3iPUPDtp4On0dtPsftRlkvRMHG9U24CLj72c+1ftaftI2OiJ4q+Bz+E55p7uwjjGoi8CqvmIr58vbnjOOtfs4/tM2HwI0TWNIu/CM+sNql0k6vHeCHy9qbcYKtmvgd8Xrb42+Cn8Y2uhyaSiX0lr9nknExyiqd24KOu/pjtXxZ+Ovw6+DNkkvi/Vi19Ohe2021USXMw9QuQFXII3MVGQRnPFXX/BQae4u2GgfCCe5tITl2l1UhyvqQsLBOh7tX7P8A+0lpvx6l1OytPCN/o9zpMMck5edZ4fnJAUOAp3HaeCo4Brx1498J/Dfw7P4o8Y6xFp9hAdoZslpHPREUcsxweB6E9ATWtf8ABQG2mvntvBHwtvdRgQkia8vfKdl9fLjR8dv4jXw3/bp8A+K9Ug0PxroVz4UuZ22LcyXAuLYN0w77VaPJ7lcDuR1pWSRFeNwysMqQcgj1FeP/ANpWy8A/GPRPhFd+EZbh9als0TURfKixi4l8vcU2k/KcnqM47V8WP24fBXgXWrnw34P0N/FF3ZuY57lboQWyOOqq4VjIQeDgAehNfCL9t7wl8QPEFp4V8WeHZPDd7fyiK1uBdC4t5JCcKrNtUoSSAOCM9SK+N37WfgT4Oai/hqCyn8Q+IIlBmsreURR22RkCWUg7WIIO0KxxycZGfBf/AAUD0DUtWisfHHgebSLOVgv26zvPtIiz3aMop2+pUk+xqyvbPUrK31HT7mO4tbqJZYJo23LIjAFWUjqCCDmvGnjbwv8AD3w9c+KfF+rw6fp1qPmkkySzHoiKOWY44UDNal+3jcalezQfD34P6prFrC3NxPcFXI944432546tXwh/bN8DfEnW7fwp4h0i48LazdP5dus84lt5ZOgQSYUq5PADKATxnJANVVVVVVVVVVVVVVVVVZ/8fUX++Kqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq+Ot1PZfBfxzcWxxIPD94qkZBXMLAkY6EA5r/gnZbWz3njy7ZVNxFFp8cZPUKxuC36ov6VVV+0B+zl4Z+Ommw3ElyNK8Q2EZSz1JYt+5OT5Uq8FkySRzlSSR1IK6J+1f+ys0k2mrdz+HoHLyG3H2/T2HUsyfehB7sRGT618AP2vPDvxavoPCPiiwi0LxLKuIFSQm2vWAyRGTyjcfcbOezE8VVVVVVVVVVVVVVVVVVVVft83E8HwV0+KEnZceIrdJev3RDO3/oSrX7Etvbw/ADSpIAN899dvLjuwlK8/8BVaqv8AgoTBbL4+8L3KBftD6O6Oc87VmYr+GWb9a8SvcS/ssarJdg+e3w/mMoZdp3fYDnI7c9q/4J3/APIZ8b/9etn/AOhS1XxE/wCT6LT/ALHDSP8A0K3q5s7S8VUu7WGcKcqJIw2D7Zr4AWtrN+2TqFrLbxPCNT1gCNkBXgTY46Vb6dp9o5ktLG3hcjBaOIKcenH0FftFeItf+Pn7Rlp8JNHvjHp2magNLtVOdiS8faZ2UdSpDD/djHTJr4cfCnwN8KtEh0XwdoVvalIws92YwZ7lh1aSTqxJ5x0GcAAcVc6RpN5fWup3emWk95Ylja3EkCtJAWBVijEZXIJBxjIJr9vj4mX+ieHtH+GmkXTQnXA91qRRsM0CEBI/91n3E/8AXMDoTX7K/wAEdA+Gvw90rxBdaXDJ4n1u1W6u7yWMGSBZFBWFD1VQpGQOrZJ4wB8UvhZ4T+LfhS78MeJ9PikMkbfZbvywZbOXHyyRt1BB6joRkHg1+xv4u134c/G3UfhBrEpFvqktxaTwFvlhvbYMd4+ojdD65X0Ff8FAf+SNaN/2M8H/AKTXNfsSfB/QF8EQ/FrX7KLUtY1WWSLTpLlRJ9jt4WMXyA52sWRxnsoUDAzn9rz4XeHPGfwl1rxNNpsC654dtjd2l6qASBEIMkbN1ZSm7g9Dg1+xL8NdJ+I2vX/iPxpH/atj4Ohih0uwuj5kMckzyOW2HIwpDNjGN0meor4ofDHw18UvBd94S1zTbWQy27rZTvEN1nNtwkkZxlSDjp1GQcgkV+wF4j1DTfiN4i8Ezuy219pZuWiYk7Z4JVUYHQfLK+T/ALIr9uv4k33hP4d6f4L0i6aC58VzSJcuhwfssQXzFyOm5nQH1XcO5r9mDwh8M/hb8N9LvW1rQl8R61aR3Op3Ml5H5qlwGEOScqqAgFem4E96/aC0f4Z/E/4Ya3pd9rugzajZ2Utzpc/2uIyQTou5QpzkBioVh3B+lf8ABPzxtfaj4b8R+Ar24MkOjzRXdirNkok28SKPRQyBvq59aqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr/goRdTp8P/AAxYq37mXWHkcc/eSFgvt0dv85r9ke2trb9njwgtsqgSRXDuR3Y3MuSf5fhiqqv2gv2MIfG2rXfjf4X3drp2rXTGW70ycbILiQ8mSNx/q3PUgjaSc5XnOmfHP9p/9nO7t9D+IukXmoaYjeXHDrKGRXA6iG7UncQBgfM4A7V8F/jj4N+N+gvqnhyR7a+s9q3+m3BHm2zHoeOGQ4OGHXHOCCBVVVVVVVVVVVVVVVVVVVf8FA7id/ixoNoxPkxeHY3Tr95ricN+irXhi3t7Tw1pNpaACCGxhSIDoFCADH4AVVSRraftzhNLUfN4xBYIN332zL/6E+fTmv8AgoN/yTPw5/2HR/6Ikr9jv/k3Dwh/2+/+ls9eNf8AkTde/wCwZc/+imr/AIJ9f8lM8Rf9gI/+j46/aZsLF/gb41u2s4GnGmNiUxjcMMvfrX/BP2wsLzwb4ra7soJ2XU4QpkjDEfuu2a8R6xpPgfwpq3iWe3SKy0eymvJo4lC7ljQsQAO5C4/KvgF4Eu/2ovjNrPjT4kTveafYkXeowqxUSs5IhtwRysYCnoc7Y8d8jR9E0bw9p0WkaBpVpp1lAMRW1rAsUaD2VQAKtdI0mxu7q/sdMtLe6vipup4oFR5yudpdgMtjJxnpk+tfH7Wtb+P37TFr8LtPvnTTdN1EaTbIpysTKf8ASpiO7Aq/4RqK8DeAvCnw48PW3hnwhpEFjZ26BSUQB5mAwXkYcu57k1+2d8E9B8UeAL74k6RpkNv4g0BRNcTRRhTd2+QHEmPvFR8wY8gKR3r9hn4j3/jD4aXnhLVrhp7nwncJDDIzZb7NKGMSnv8AKUkUf7IUdq/blF0fj3Yixd1uf7GtPJZG2sH82XaQexzjmvg58H/C3we8IWWgaLp9ub8QqdQ1Dyx5t1MR87Fuu3Odq9AMe5P7eHww8PeF9W0Hx/4b0+DT5dYeWC/jt0EavKgVklCjoxBYEjrtB6kk/sh/DHRtN+H1n8UNZgXU/Ffip5b251K6HmzIrOwCq5yRuA3MepZjnOBj9ub4X6Bqnw3f4lWmmW8Gs6JcwrcXUcYV57eRxHtcj72HdCCenzY61+xJ4ovPEnwKtbS9mMraDqM+nxsxJOwBJVH4CYKPQACv2n/EVz8Xv2i9G+Dx1kWWhaRcw200jSBESSQK88xJ43Kh2jPAKn+8a8Jz/CrwNoFr4Z8Kax4e0/TrNAscMV7EMnHLMd2WY45Y5JPU1+3T4a8B6jo2k/EjwxqOkvrkd8LS+NlcRl7iJkYo7hTklDGFB64fBOAMfs9+Nbz4hfBnwt4o1Kfzb2a0MN1ITkvLC7RMx92Kbv8AgVVVVVVVVVVVVVVVVVVZ/wDH1F/viqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqvFmgw+KvCus+F53Cx6vp89m7EdBLGyE/k1fsd+Mj8LPjZf+BvFh+w/2yH0yYSnAhvIn+QMfch0+riqqqZVdSjAEEYII61+1/wCC9I+FPxpsdW8CImlG/tItUjithsFrcrK4LRgcKCY1YAcA5xgVol7Lqei6fqU8XlS3VrHK8ZGNjMoJH4Zqqqqqqqqqqqqqqqqqqqv2uPB114z+BWvQWEJlutJMeoxIq5JERzJj/tmZD+FfsA/EGxu/DGtfDO7uVW+sLo39pGzYMkEgVXCj/ZdQT/10FVX7RuryfHb9pi28H+GLgXMFvLBolrLF8y5DkzScdld5Mn+6melfFyxjt/gp400ywgwkXha/hgiUdALVwoA/IV/wTwmiXX/GlsZB5j2doyr3IV5AT/48PzqviC8cn7c9q0bqwHjHSRkHPIa3BH5jFV+z5/yefqH/AGE9Z/lNVfDi4Twn+2+512TYf+Ep1KDfIcbnnE8cR5/vNIh/Gqr/AIKC2F3H8TvDupvu+zXGgiGPPTfHPKX/AEkT9K8FarZa74O0LWdOkV7a+02CaEg5+Vo1I/nVfDl4/GP7bj6locnmWreJb+6WZOQ0UYlbdn0YD/x4V/wUB/5I1o3/AGM8H/pNc1+x3/ybh4Q/7ff/AEtnr9oH/kiHjn/sBXX/AKLNf8E7/wDkDeN/+vqz/wDQZar9ij/k4jVv+wXe/wDo6Ov+Ch9ldLrvgvUGZjbSWl3Eg7B1eMt+JDL+XtXg/wDYZ+Fvizwlovie38beIymrafDdDY0GB5iBiB8h6ZxX/Dvn4Z/9Dp4m/OD/AON18Ev2bfCfwM1LU9U8Pa3quoTapAkMgvTHhFVieNijkk9/Sqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/ba8GXXiv4I3Go2EJkn8OXseoMqjkxANHJ+AWTcfZDX7B3xGsNa+Hlz8N7m5VdS8O3DzQQk4L2srbtyjviRnB9Ny+tVVV4m8MaB4x0O68OeJ9Kt9R069jKTW86ZBHqO4I6hhgg8gg1+zct58O/2tT4L0i8kntF1DUtIuCD/r4YhKVY49GhR/wqqqqqqqqqqqqqqqqqqqq/4KE+DrqSPwt4+toS0EQl066cL9wn95F+f738h61+zX8QbH4jfBzw9qUNysl7p1qmn6gm7LJPCoUlh23KFf6OKrxBruleF9DvvEWuXiW1hptu89xK5wFRRk/U9gO5wK/ZY07Ufix+03d/ES+t/wBzZzXesXWQSqSSlljQHsQ0mQPSM+lf8FBYnb4X+HpQhKLryqWxwCYJcD9D+VfsbzRS/s5+FY45AzQteo4/un7ZMcfkQfxrxy8cXgnxBJI6oiaVclmY4AAibJJr/gn1/wAlM8Rf9gI/+j46/aX/AOSC+Nf+wW3/AKEtf8E9P+RM8Wf9hOH/ANFV+0JY3Wo/A/xva2W4y/2LcOAvUqi7mH4qpFf8E8NWsVPjXQWlVbyQWdzGhPLxr5qsR9Cyf99Cqr4bzL4U/bgZdck2sfFGpQB3P33nE6RH/gTSJj6iq/aC1Wy0b4IeOLu/lVI5NCurZSxxmSaMxIPxd1Ff8E79MugPHGsNuW2b7FAnHDuPOZvyBX/vqv21/wDk4jSf+wXZf+jpKr/goX/yJnhL/sJzf+iq/Zo/5IL4J/7Ba/8AoTV+2L/ybh4u/wC3L/0tgr/gn/8A8ka1j/sZ5/8A0mtq8eeCdO8Vftf6t4I8Xalc2VprPiBo2uIiPMQTLuhC7sjndGOexr/h3z8M/wDodPE35wf/ABuv+HfPwz/6HTxN+cH/AMbr4V/DbSPhL4IsfAuh3t1d2tk8rrNdFTI5kkZznaAON2Onaqqqqqqqqqqqqqqqqqs/+PqL/fFVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV+1T+ynqHjvUJfiX8M4IzrpUHUdN3CP7ZtGBLGeAJcAAgkBgAQd33vAv7Z3xO+GGPB3xc8IXOryWQ2eZdFrO+jA4+fcpEnTqQCeSWNXH/AAUI+H622618CeIJLjHEcksKLn/eDE/pX7On7Qd78eH8QzXPhI6Jb6W8P2TErTCZXD7sybVBZSq5AAwGHWvGX7ZnjT4ZfFPxB4Z8V/D03GhQXzx6dvD2dwYV+XzFLArIrFSw4H3uvpqH/BQnwQlg76V4A1ye92/JFcTxRR7sd3UscZ/2a8D+AfiV+1z8V4/iN46017PwzE8fnzeW0cJgjPFtb7sliecnOBuZjyQCqqqhEAAAwABjFVVVVVVVVVVVVVVVVVVVJHHLG0UiK6MCGVhkEHqCK+M37O/xC+BnjcfFT4KR3sukQTG4iWyQyTaYTndG6DJeHGRnBG0kN0yfCv8AwUH0n+zki8deArxNQjGJJNLmVo5D6hJCCn03N9a+If7ZXjr4pwv4E+C/g3UrGfUVMbXMZM96yHgiNIxiPjq2WIzwVxmv2Uv2YLn4Wk+PvHkcTeKLqEx21orBxp0bD5ssMgysOCRwBkAnca1CwtdUsLnTL2MSW93C8MyHjcjAhh+RNadc+Pf2N/jVNNdaVJd2oEkAEmY4tVsmIIZHAIDDCHjO1hgjqK1X9u4+I7OPQfhd8NdWu/E2oDyrVLkrIschHBVI8tLg54+X1PpUXgvxZ4D/AGmfBNl4+1GO616/1/S9R1CQNu2SzXKOwZuhYHOSOM9OOarxxceIf2af2p7zxpNosk9nJqdzfWiMxRLy0ud25Vcg8qJCuecMozXwU/ab8J/HLXr7w/4f0HVdPm0+z+1O15swy71XA2se7Driv2wP2cvEus69/wALh+G1jPd3mxP7Us7QHzw8YwlxEByx2qoIXkbQQDk4+HH7e40vS4tG+LPhW/ur+0Hlvf6YEDzFeMvC5UK/HJDYJ/hFeFP22vCnjv4g6B4I8M+DdWSHWLsW8l5fOiNDkHaRHGXzk7QSWGASea/ac+B5+NvgRLLSmij8QaNI1xpbynarkgCSEnsHAXnsyrnjNfBf9prxR+z1FJ8LPi54S1aWx05z9mQIEurIEklQrkLJGTkqdwxzgkEY+Kf7cD+MtEk8H/BvwvrUGpasht/tl1GvnxhuCIYomfLkcBs5HUDOCP2QP2ctU+GFrcePfHFqsHiDVIPJtrJgC1jASC249BI5C8D7oGM5JA/4KA/8ka0b/sZ4P/Sa5r9jv/k3Dwh/2+/+ls9ftA/8kQ8c/wDYCuv/AEWa/wCCd/8AyBvG/wD19Wf/AKDLVfsUf8nEat/2C73/ANHR1+0b8GY/jX8O5tBtHih1qwk+1aVNJwolAIKMeoV1JB9DtPOMV8Jf2k/HX7N6P8LPin4LvrixsJG+zxO3lXNoGYkhC3yyxk5K8gcnDEYAvf8AgoH8J0t2bTvCXiyecD5Umgt4lP1YTMR+VfBL4on4xeAbfxs2hSaQ09zNF9meQyYCN8rByq7gVK8gYzkdqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq6tba+tZbK8t457e4jaOWKRQyurDBUg8EEHBFfFP9nD4o/Abxp/wsj4JtqF3pUEjS27WamW4sAfvRyR4JkiwSM4IK8MB1PhD/goK9vapZfELwA8l3ENslzpc4UORwf3Mn3T1/j/AAFaz/wUL8JxW5Ph/wCHWrXU2OBeXcduoP1UOT2rw78Tdf8AE/wF/wCFn2vhu4XW5dHuLiHTIoHZmuEDqqqhG4qzKCPVSDzXhL9v6+06P+y/ib8PZDe2/wAks+nS+UxYcHdDJ90+vz9ewrxv+39FfaW+nfDTwReR6ncr5cd1qTqRCx4BWKMne3pkgZ6g9K/Y6+AHinRtdn+MvxFs7i1v7iORdNtbtSJy0v8ArLiQHlSQWUA8ncxIHBNVVVVVVVVVVVVVVVVVVV4/8DaD8SfCGpeC/EkBksdSi2MV4eJgcrIp7MrAEduOcjIoaZ8cP2LvG1xqdlZNqXh66bY85jdrK/jydu/af3Uo7ZOQc43KTmz/AOChPgZ7FX1DwBrkV5t5ihnikj3Y7OSpxnvtrxz8VfjV+15qEfgnwF4SnsfDyTAzQQuWQkHh7qcgKAOoTA57MQK+AnwT0f4H+C10G1mS81W9YTapfBMedKBgKvcIoyFB9SeCxr9pH4X3Xxb+E+p+GNLVW1W3dLzTlY4DTR5+XJ4G5S65PALA18AP2ldY/Z6GpeAfGvhe+u9NF20jW2fJubCfADja/UHaMqSuDznk14i+O/j39qhm+E/wd8KXmjaVqACa3q94wYxW5+8rbflRSMgjcWf7oxk5/YFgNn8V/FFlI6l4dFdDjvtuIwSPavit4Tn8dfDXxN4Qs9v2rVNMmhttzbR5xUmPJ7DeFzX7Onx9l/Zx1XxD4a8ZeF9Qntr2VPPt0xHcWlxFuUjY+Acg4IJBG0V8KfiVoXxq8DL4s0zS7i3sbqWa2e2vApYhTtO4AkYIPT3rx/8ADP4n/sl/Ez/hYXgC3mufDyyMbW7EbSxCFzzbXQByOwySN2AwIYfLof8AwUG+H82mq/iXwR4gtdQCfNHYmG4iLd8M7oQP+AnrXwG/aR074763r+n6d4YudKt9Ihhkt5J5hI04YuH3ADCEYTAyc5PPFftgfs7+JtV19fjL8NbOe5vkVDqlpaA+erxgBLiIDliAqggcjapAPJHw/wD2+tGs9Ch074p+FtYfWbRfLlutLjiZbhhxuaN3Ty29QMjOcYBwPir8ZfiF+1rq9p8Ofhh4RvYNFimWWSFmBeVuiy3Dj5Y0XsuSM85JwB8DfhLp/wAGPh7ZeD7adLm8LNcahdKu0T3D43Ed9oAVRnsozzmv21/+TiNJ/wCwXZf+jpKr/goX/wAiZ4S/7Cc3/oqv2aP+SC+Cf+wWv/oTV+2L/wAm4eLv+3L/ANLYK/4J/wD/ACRrWP8AsZ5//Sa2r9sP9nzxH4o1K2+Lnw3sprjWLKNU1G1tc+dII+Y54wOWdRhSBzgLgcGvh9+3zbafpcWkfFfwjqUmpWa+VLe6WqEzMvGXhkZNjcc4bGc4C9K1z/goH4IEQi8IeAfEGoXcmFjjvnithuJwP9W0pPbjAz0461aXMd7aQ3cKuEnjV1DqVYAjIyD0PtVVVVVVVVVVVVVVVVVWf/H1F/viqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqrWPD2geIYBa6/oen6nCOkd5bJMo/BgRVn8IvhPp8wuNP+GHhK2lHR4dEt0YY56hM9ahggtoUt7aFIo4xhERQoUegA6VqmjaRrlsbLWtKs9Qtz1iuoFlX8mBFWnwl+FWn3IvbD4ZeE7a4ByJYdFt0fOc9QmeoFKiIixxqFVRhVAwAPaqqqqqqqqqqqqqqqqqqqqq1n4afDjxFcm98QfD/wAN6ncE5Mt5pMMzk/VlJrRfDPhvw1Abbw54f03SoiOY7K0SBTj2QCqrWdB0LxHZnTvEGi2GqWrHJgvLZJkJ/wB1gRWgeC/B3hQMvhbwno2jhxhhYWEdvn67FFXXh/Qb28XUbzRLCe7QgrPLbI0ikdMMRkY7VWt+HfD/AImsv7N8R6Hp+q2hOfs99apOmfXa4Irw/wCDPB/hNXXwr4U0bRhIMONPsI7bcOvOxRnmq134f+A/FE32jxL4J0DV5f8AnpfabFcNx05dSa0Xwt4Y8NqyeHfDml6WHGGFlZpBke+wCq13wt4Y8UQLbeJfDml6vEoO2O+s0nUZ64Dg1oHgjwX4UZm8LeENE0dnGGNhp8VuT9diiq1DS9M1eBbbVdOtb2JW3rHcQrIoYAjIDA84JGferOxstOtkstOtILa3jzsihjCKuTk4A4HJJ/Grm2t7yB7W7gjmhlUq8ciBlYHqCDwRWnaNo+jq6aRpVnZCUguLeBYtxHTO0DNVZeHfD+mXBvNN0PT7W4YEGWC1SNiD1GQM9qrWvDnh7xLbCy8RaFp2q24ziK9tUnUZ68OCKsPhP8LNJnF1pXw08K2cy9JLfRreNh+KoDSqqqEQAADAAGMVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVrngDwJ4nl87xJ4K0HVpD1e+02Kc/m6mtI+HXw98PSrPoHgTw9psqHKvZ6XDCyn1BVRiq13wV4N8UHPibwlo2rnGP8AT7CO44/4GprQ/h/4C8MTC48NeCNA0mUdHsdMigb80UGqqqqqqqqqqqqqqqqqqqqqliiniaGaNJEcEMjrkEehFTfCL4T3F19un+GHhKS5J3GZ9Ety+fXdszVlYWGmWqWWm2UFrbxjCQwRiNF+ijgVVeIPAfgbxZKk/inwZoWsyRjCPqGnRXBX6F1OK0zSdK0W0Ww0bTbSwtkOVgtoViQfRVAFWPh/QdLuGu9M0SwtJ3Uq0sFskbMCQSCQM4yAfwqtf+H/AID8WTrdeKfBOgazOgwsmoabFcMo9AXUkVpulaXotmmnaNptrY2sedkFtCsSLnrhVAAplV1KMAQRggjrVx8Kfhdd3n9oXXw28KzXX/PeTRoGf/vopnufzqw07T9LtVstLsbe0t0+7DBEI0X6KMAVWt/Dj4eeJrk3viPwH4d1W4PWa+0uGdz+LqTWlaNo+hWi2Gh6VZ6fbKcrBawLCgPsqgCqvvDvh/U7hbzUtD0+7uFACyz2qSMAOgBIJqtR0fSdYjSLVtLtL1IzlFuYFkCn1AYHFW1ra2VulpZW8UEMQwkUSBFUegA4FXtjZalbPZahaQXVvJjfFNGHVsHIyDweQDWn6XpmkwG20rTrWziZi7R28KxqWwBnCgc4AGfYVWu/D/wF4pm+0+JvBGgavMOkl/pkVw3Ax1dSelaH4E8D+F383wz4N0LSXx96x06K3P5oo9T+dVVVVVVVVVVVVVVVVVVn/wAfUX++KqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqrP/j6i/wB8VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVWf/H1F/viqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqrw78QP29P2oPj38QfCnwh+NWvWs+j315cmxGutp9tbWyXPlIkar8oA3IMYyepJOSf+GZf+Ct3/AEW7VP8Awt3r/hmX/grd/wBFu1T/AMLd6/4Zl/4K3f8ARbtU/wDC3ev+GZf+Ct3/AEW7VP8Awt3r/hmX/grd/wBFu1T/AMLd6/4Zl/4K3f8ARbtU/wDC3ev+GZf+Ct3/AEW7VP8Awt3r/hmX/grd/wBFu1T/AMLd6/4Zl/4K3f8ARbtU/wDC3ev+GZf+Ct3/AEW7VP8Awt3r/hmX/grd/wBFu1T/AMLd6/4Zl/4K3f8ARbtU/wDC3ev+GZf+Ct3/AEW7VP8Awt3r/hmX/grd/wBFu1T/AMLd6/4Zl/4K3f8ARbtU/wDC3ev+GZf+Ct3/AEW7VP8Awt3r/hmX/grd/wBFu1T/AMLd6/4Zl/4K3f8ARbtU/wDC3ev+GZf+Ct3/AEW7VP8Awt3r/hmX/grd/wBFu1T/AMLd6/4Zl/4K3f8ARbtU/wDC3ev+GZf+Ct3/AEW7VP8Awt3r/hmX/grd/wBFu1T/AMLd6/4Zl/4K3f8ARbtU/wDC3ev+GZf+Ct3/AEW7VP8Awt3r/hmX/grd/wBFu1T/AMLd6/4Zl/4K3f8ARbtU/wDC3ev+GZf+Ct3/AEW7VP8Awt3r/hmX/grd/wBFu1T/AMLd6/4Zl/4K3f8ARbtU/wDC3ev2idZ/4KHfsuf8I/8A8LX+PfiiD/hJvtP2D7D4pkud32fyvM3YI2/66PHrz6Va/DX/AIKh3trDeQfHTXTHPGrpnxjIDgjI/Sv+FWf8FSP+i565/wCFi9f8Ks/4Kkf9Fz1z/wALF6/4VZ/wVI/6Lnrn/hYvX/CrP+CpH/Rc9c/8LF6/4VZ/wVI/6Lnrn/hYvX/CrP8AgqR/0XPXP/Cxev8AhVn/AAVI/wCi565/4WL1/wAKs/4Kkf8ARc9c/wDCxev+FWf8FSP+i565/wCFi9f8Ks/4Kkf9Fz1z/wALF6/4VZ/wVI/6Lnrn/hYvX/CrP+CpH/Rc9c/8LF6/4VZ/wVI/6Lnrn/hYvX/CrP8AgqR/0XPXP/Cxev8AhVn/AAVI/wCi565/4WL1/wAKs/4Kkf8ARc9c/wDCxev+FWf8FSP+i565/wCFi9f8Ks/4Kkf9Fz1z/wALF6/4VZ/wVI/6Lnrn/hYvX/CrP+CpH/Rc9c/8LF6/4VZ/wVI/6Lnrn/hYvX/CrP8AgqR/0XPXP/Cxev8AhVn/AAVI/wCi565/4WL1/wAKs/4Kkf8ARc9c/wDCxev+FWf8FSP+i565/wCFi9f8Ks/4Kkf9Fz1z/wALF6/4VZ/wVI/6Lnrn/hYvX/CrP+CpH/Rc9c/8LF6/4VZ/wVI/6Lnrn/hYvXiDwP8A8FOfDWg6l4j1T466+tnpVnLd3BTxfIzCONC7YHc4U8V+z43/AAUa/ad0bVtd+Ffx58TT2ui3SW10b3xXJbkSMu4YBJyMV/wzL/wVu/6Ldqn/AIW71/wzL/wVu/6Ldqn/AIW71/wzL/wVu/6Ldqn/AIW71/wzL/wVu/6Ldqn/AIW71/wzL/wVu/6Ldqn/AIW71/wzL/wVu/6Ldqn/AIW71/wzL/wVu/6Ldqn/AIW71/wzL/wVu/6Ldqn/AIW71/wzL/wVu/6Ldqn/AIW71/wzL/wVu/6Ldqn/AIW71/wzL/wVu/6Ldqn/AIW71/wzL/wVu/6Ldqn/AIW71/wzL/wVu/6Ldqn/AIW71/wzL/wVu/6Ldqn/AIW71/wzL/wVu/6Ldqn/AIW71/wzL/wVu/6Ldqn/AIW71/wzL/wVu/6Ldqn/AIW71/wzL/wVu/6Ldqn/AIW71/wzL/wVu/6Ldqn/AIW71/wzL/wVu/6Ldqn/AIW71/wzL/wVu/6Ldqn/AIW71/wzL/wVu/6Ldqn/AIW71/wzL/wVu/6Ldqn/AIW71/wzL/wVu/6Ldqn/AIW71/wzL/wVu/6Ldqn/AIW71/wzL/wVu/6Ldqn/AIW71/wzL/wVu/6Ldqn/AIW71/wzL/wVu/6Ldqn/AIW71/wzL/wVu/6Ldqn/AIW71/wzL/wVu/6Ldqn/AIW714w8Sf8ABQjwN8ctI/Z31/49+Kk8Ya3LaxWkUXiiR4C1wcR5kzge/pX/AAzL/wAFbv8Aot2qf+Fu9f8ADMv/AAVu/wCi3ap/4W71/wAMy/8ABW7/AKLdqn/hbvX/AAzL/wAFbv8Aot2qf+Fu9f8ADMv/AAVu/wCi3ap/4W71/wAMy/8ABW7/AKLdqn/hbvX/AAzL/wAFbv8Aot2qf+Fu9f8ADMv/AAVu/wCi3ap/4W71/wAMy/8ABW7/AKLdqn/hbvX/AAzL/wAFbv8Aot2qf+Fu9f8ADMv/AAVu/wCi3ap/4W71/wAMy/8ABW7/AKLdqn/hbvX/AAzL/wAFbv8Aot2qf+Fu9f8ADMv/AAVu/wCi3ap/4W71/wAMy/8ABW7/AKLdqn/hbvX/AAzL/wAFbv8Aot2qf+Fu9f8ADMv/AAVu/wCi3ap/4W71/wAMy/8ABW7/AKLdqn/hbvX/AAzL/wAFbv8Aot2qf+Fu9f8ADMv/AAVu/wCi3ap/4W71/wAMy/8ABW7/AKLdqn/hbvX/AAzL/wAFbv8Aot2qf+Fu9f8ADMv/AAVu/wCi3ap/4W71/wAMy/8ABW7/AKLdqn/hbvX/AAzL/wAFbv8Aot2qf+Fu9f8ADMv/AAVu/wCi3ap/4W71/wAMy/8ABW7/AKLdqn/hbvX/AAzL/wAFbv8Aot2qf+Fu9f8ADMv/AAVu/wCi3ap/4W71/wAMy/8ABW7/AKLdqn/hbvXxh+H3/BT74GfDjV/in49+OmvRaFonk/a3tfGEk0g82aOFMIOT88qZ9Bk1/wAE+PiX41+LX7KXhHxh8Qtdm1nW2lvLaa/nA82dYrmRELkfeYIFUt1OATkkk1VVVVVVVVVVVVVVVVVVX7XXjTxL8O/2aPiL408G6o+na1pWiSyWV2ihmgckLvUEEZAY4OODg18CNA/4KWftHeCH+IPwz+O/iKfSI76SyZrzxbJbv5saozfKSeMOvNf8My/8Fbv+i3ap/wCFu9f8My/8Fbv+i3ap/wCFu9f8My/8Fbv+i3ap/wCFu9f8My/8Fbv+i3ap/wCFu9f8My/8Fbv+i3ap/wCFu9f8My/8Fbv+i3ap/wCFu9f8My/8Fbv+i3ap/wCFu9f8My/8Fbv+i3ap/wCFu9f8My/8Fbv+i3ap/wCFu9f8My/8Fbv+i3ap/wCFu9f8My/8Fbv+i3ap/wCFu9f8My/8Fbv+i3ap/wCFu9f8My/8Fbv+i3ap/wCFu9f8My/8Fbv+i3ap/wCFu9f8My/8Fbv+i3ap/wCFu9f8My/8Fbv+i3ap/wCFu9f8My/8Fbv+i3ap/wCFu9f8My/8Fbv+i3ap/wCFu9f8My/8Fbv+i3ap/wCFu9f8My/8Fbv+i3ap/wCFu9f8My/8Fbv+i3ap/wCFu9f8My/8Fbv+i3ap/wCFu9f8My/8Fbv+i3ap/wCFu9f8My/8Fbv+i3ap/wCFu9f8My/8Fbv+i3ap/wCFu9f8My/8Fbv+i3ap/wCFu9f8My/8Fbv+i3ap/wCFu9f8My/8Fbv+i3ap/wCFu9f8My/8Fbv+i3ap/wCFu9f8My/8Fbv+i3ap/wCFu9XPjb9v+1+PM37N8vx98Wf8JnbyvE8Q8TymDctsbg/vM4/1Y9OvFf8ACrP+CpH/AEXPXP8AwsXr/hVn/BUj/oueuf8AhYvX/CrP+CpH/Rc9c/8ACxev+FWf8FSP+i565/4WL1/wqz/gqR/0XPXP/Cxev+FWf8FSP+i565/4WL1/wqz/AIKkf9Fz1z/wsXr/AIVZ/wAFSP8Aoueuf+Fi9f8ACrP+CpH/AEXPXP8AwsXr/hVn/BUj/oueuf8AhYvX/CrP+CpH/Rc9c/8ACxev+FWf8FSP+i565/4WL1/wqz/gqR/0XPXP/Cxev+FWf8FSP+i565/4WL1/wqz/AIKkf9Fz1z/wsXr/AIVZ/wAFSP8Aoueuf+Fi9f8ACrP+CpH/AEXPXP8AwsXr/hVn/BUj/oueuf8AhYvX/CrP+CpH/Rc9c/8ACxev+FWf8FSP+i565/4WL1/wqz/gqR/0XPXP/Cxev+FWf8FSP+i565/4WL1/wqz/AIKkf9Fz1z/wsXr/AIVZ/wAFSP8Aoueuf+Fi9f8ACrP+CpH/AEXPXP8AwsXr/hVn/BUj/oueuf8AhYvX/CrP+CpH/Rc9c/8ACxev+FWf8FSP+i565/4WL1/wqz/gqR/0XPXP/Cxev+FWf8FSP+i565/4WL18ZtZ/4KHfAfwva+L/AIgfHvxRFp15fpYxNa+KZJm85o5HAKg8DbE/P09a8AfB3/gqh8TPBGh/EHwt8ctak0jxDYRXtk0/jJ43MUihlLL2OD0r/hmX/grd/wBFu1T/AMLd6/4Zl/4K3f8ARbtU/wDC3ev+GZf+Ct3/AEW7VP8Awt3r/hmX/grd/wBFu1T/AMLd6/4Zl/4K3f8ARbtU/wDC3ev+GZf+Ct3/AEW7VP8Awt3r/hmX/grd/wBFu1T/AMLd6/4Zl/4K3f8ARbtU/wDC3ev+GZf+Ct3/AEW7VP8Awt3r/hmX/grd/wBFu1T/AMLd6/4Zl/4K3f8ARbtU/wDC3ev+GZf+Ct3/AEW7VP8Awt3r/hmX/grd/wBFu1T/AMLd6/4Zl/4K3f8ARbtU/wDC3ev+GZf+Ct3/AEW7VP8Awt3r/hmX/grd/wBFu1T/AMLd6/4Zl/4K3f8ARbtU/wDC3ev+GZf+Ct3/AEW7VP8Awt3r/hmX/grd/wBFu1T/AMLd6/4Zl/4K3f8ARbtU/wDC3ev+GZf+Ct3/AEW7VP8Awt3r/hmX/grd/wBFu1T/AMLd6/4Zl/4K3f8ARbtU/wDC3ev+GZf+Ct3/AEW7VP8Awt3r/hmX/grd/wBFu1T/AMLd6/4Zl/4K3f8ARbtU/wDC3ev+GZf+Ct3/AEW7VP8Awt3r/hmX/grd/wBFu1T/AMLd6/4Zl/4K3f8ARbtU/wDC3ev+GZf+Ct3/AEW7VP8Awt3r4E+JP+ChH7R3iXXfCfwz+PfiqfUPDkQkvlvPFEluqqXKfKSTn5hX/DMv/BW7/ot2qf8AhbvX/DMv/BW7/ot2qf8AhbvX/DMv/BW7/ot2qf8AhbvX/DMv/BW7/ot2qf8AhbvX/DMv/BW7/ot2qf8AhbvX/DMv/BW7/ot2qf8AhbvX/DMv/BW7/ot2qf8AhbvX/DMv/BW7/ot2qf8AhbvX/DMv/BW7/ot2qf8AhbvX/DMv/BW7/ot2qf8AhbvX/DMv/BW7/ot2qf8AhbvX/DMv/BW7/ot2qf8AhbvX/DMv/BW7/ot2qf8AhbvX/DMv/BW7/ot2qf8AhbvX/DMv/BW7/ot2qf8AhbvX/DMv/BW7/ot2qf8AhbvX/DMv/BW7/ot2qf8AhbvX/DMv/BW7/ot2qf8AhbvX/DMv/BW7/ot2qf8AhbvX/DMv/BW7/ot2qf8AhbvX/DMv/BW7/ot2qf8AhbvX/DMv/BW7/ot2qf8AhbvX/DMv/BW7/ot2qf8AhbvX/DMv/BW7/ot2qf8AhbvX/DMv/BW7/ot2qf8AhbvX/DMv/BW7/ot2qf8AhbvX/DMv/BW7/ot2qf8AhbvX/DMv/BW7/ot2qf8AhbvX/DMv/BW7/ot2qf8AhbvX/DMv/BW7/ot2qf8AhbvXxx8a/wDBQ39kfxB4Zn+J/wAc9fNzqpkubCFfEDahDKIGTcJUb5SpLqNrAggmqqqqqqz/AOPqL/fFVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX/BLj/k8f4zf9eF//AOnKOqqqqqqqqqqqqqqqqqqqqv8Agtr/AM0Y/wC49/7j68Pf8i/pn/XnF/6AKqqqqqqqqqqqqqqqqqqqvjL/AMkg8c/9i1qH/pNJX/BFv/kl/wARv+w9bf8Aog1VVVVVVVVVVVVVVVVVVVV+01/ylu+G3/X/AKB/6HVVVVVVVVVVVVVVVVVVVVX/AAU1/wCTH/iT/wBwr/062lf8Es/+TL/CP/X/AKl/6WS1VVVVVVVVVVVVVVVVVVVft3/8mf8AxU/7AEn/AKGlf8Eg/wDk1C7/AOxuvv8A0Tb1VVVVVVVVVVVVVVVVVVVVrf8AymD1T/r/ALj/ANMbVVVVVVVVVVVVVVVVVVVVV/wVE/5IBoH/AGONt/6R3lfsaf8AJqHwl/7FGw/9ErVVVVVVVVVVVVVVVVVVVVX/AASF/wCS7fF7/rwT/wBK2qqqqqqqqqqqqqqqqqqqqr/gtb/yM/wo/wCvDVP/AEZb1VVVVVWf/H1F/viqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr9o//gqt8KPhncz+EPgpp4+InihXaAzxs0enW0ucAeYBuuDn+GL5T/fBr/gnl+3F8Z/2pfif4k8IfEq08NQ2GlaCb63/ALLsZIH83z4kwS0j5Xa54x1xzVfFj4qeCfgp4A1b4lfELVhYaJo8W+Zwu55GJCpHGv8AE7sQoHqeSBkiP9vf9u39pvxDqUf7KXwkhsNB0x8GRLFLyRAegnubgiAOwGQiqDjPLYzXw4/4KY/Hv4N/Eq3+GX7anw7+xW8rotxqUWmtZ3lmjHAuDGuY7iHIOTGAcbipcgIdN1LT9Z0201jSL2C8sb6BJ7W4gcOk0bqGV1YcFSpBBHUGqr/glx/yeP8AGb/rwv8A/wBOUdVVVVVVVVVVVVVVVVVVVVf8Ftf+aMf9x7/3H14e/wCRf0z/AK84v/QBVVVVVVVVVVVVVVVVVVVV8Zf+SQeOf+xa1D/0mkr/AIIt/wDJL/iN/wBh62/9EGqqqqqqqqqqqqqqqqqqqqv2mv8AlLd8Nv8Ar/0D/wBDqqqqqqqqqqqqqqqqqqqqr/gpr/yY/wDEn/uFf+nW0r/gln/yZf4R/wCv/Uv/AEslqqqqqqqqqqqqqqqqqqqq/bv/AOTP/ip/2AJP/Q0r/gkH/wAmoXf/AGN19/6Jt6qqqqqqqqqqqqqqqqqqqq1v/lMHqn/X/cf+mNqqqqqqqqqqqqqqqqqqqqq/4Kif8kA0D/scbb/0jvK/Y0/5NQ+Ev/Yo2H/olaqqqqqqqqqqqqqqqqqqqq/4JC/8l2+L3/Xgn/pW1VVVVVVVVVVVVVVVVVVVVf8ABa3/AJGf4Uf9eGqf+jLeqqqqqqz/AOPqL/fFVVVVVVVVVVVVVVVVVVVVVVVVfET42fB74Rm2T4ofFHwt4VlvUL2sOr6rDayXCggM0aOwZwCQCQCBnmr/AP4KIfsV6cUW4/aC8PuXBx5ENxPjHrsjOPxrSv8AgoX+xbrNwLWz/aD8OxvkAG7Se1Xn/aljVf14rwj458FfEHSV17wH4w0XxHprHaLzSb+O7izjON8bEZwRxmqqqqqqv2iP+CsXh79n34z+J/g7efBTUdam8NXEcL38WuJAs++FJMhDCxXHmY6npX7E37cek/tof8Jn/Zfw6u/Cv/CHfYN/n6kt39p+1faMYxGm3b9nPrnd2xzVVVVX7Zv7XGm/seeBNE8cal4HufFCa1q/9nLbQX62piPlPJvLFHyP3eMYHXrXws/4LG+Gvih8T/B/w0t/gPqeny+LdesdIS7fxAkgt2uZ0hEhUQjcF35xkZx1FVVfHb48/DT9nD4fXPxJ+KetPYaVDKsEMcMRlnvJ2BKQwoPvOQrHkgAKSSACR+zx/wAFPv2df2ifiBb/AAy0uz8SeF9b1FymlLrtvCkV+wGRGjxSuFlIBwrYB4AZiQK+KnjmH4YfDDxf8S59OfUI/CWg32rvaJL5bXAtoHmMYYg7SwTGcHGc4Nf8PxfCn/Ru2q/+FJH/APGK+DXxHg+MPwo8JfFK20p9Mi8VaRb6illJMJWtxKgbYXAG4jOM4FftGf8ABTf9nb9nHx3P8NNWt/EXijxBp7KNTh0K2ieOwYgNseSWRAZNpB2puxnDFSCK+AX7QPwz/aW+Htv8SfhZq8t3pskzW9zb3MflXFjcKAWhmQE7XAZTwSCGBBIINVVVVVVVVVVVVVVVVVVVVVVV4P8A2R/gJ8Fv+Ev8ceCvA1v/AMJJrI1C7m1a9/0ieHzi7mKEkYhjXdtAQAlQNxY81/wRj/5Lv42/7FE/+lcFV/wWP+LOp6z4+8GfATSJ5Hs9NshrF7BHk+ddzs8UKkd2SNHI/wCvg9e37OvwZ0H4A/Brwx8L9Ds4Im0qxj/tCaNADd3rKDPMx7lnzjJOFCqOFFf8FYPg3pXj79mm4+I0Vgja78P7yK6guFUeY1pNIsM8Wf7vzxyH/rl7nP8AwSb+Kd94/wD2YR4T1a6ea78C6tLpsLOcn7K6rNDz6KZJEA7KijtVV+wL8Yfhj8Gf2sPi5rvxS8aad4b0+9gv7a3uL1yqyS/2gjbBgHnarH8K/wCG7v2P/wDov/hb/v8AP/8AE1/w3d+x/wD9F/8AC3/f5/8A4mv+G7v2P/8Aov8A4W/7/P8A/E18M/iz8N/jLoM/if4XeMLDxHpVrdtZzXVkxZEnVEdkOQOQsiH/AIEKqqqqqqqqqqqqqqqqqv8Agtr/AM0Y/wC49/7j68Pf8i/pn/XnF/6AKqqqqqqqqqqqqqqqqqqqvjL/AMkg8c/9i1qH/pNJX/BFv/kl/wARv+w9bf8Aog1VVVVVVVVVVVVVVVVVVVV+01/ylu+G3/X/AKB/6HVVVVVVVVVVVVVVVVVVX/Dd37H/AP0X/wALf9/n/wDia/4bu/Y//wCi/wDhb/v8/wD8TX/Dd37H/wD0X/wt/wB/n/8Aia/b8/az/Zv+Jv7JPjvwP4B+MGga3r2pf2d9ksLWVjJN5eo20j4BUdERmPsDX/BLP/ky/wAI/wDX/qX/AKWS1VVVVVVVVVVVVVVVVVVVft3/APJn/wAVP+wBJ/6Glf8ABMr9pz4A/CD9nC48J/Ev4qaJ4e1d/Et3crZ3kjK5iaKAK+ADwSrD8K/4bu/Y/wD+i/8Ahb/v8/8A8TX/AA3d+x//ANF/8Lf9/n/+Jr/hu79j/wD6L/4W/wC/z/8AxNaPq2m+INIsdd0a8ju9P1G2jubS4jOVlikUMjj2KkH8aqqqqqqqqqqqqqqqqqtb/wCUweqf9f8Acf8Apjaqqqqqqqqqqqqqqqqqqqqv+Con/JANA/7HG2/9I7yv2NP+TUPhL/2KNh/6JWqqqqqqqqqqqqqqqqqqqqv+CQv/ACXb4vf9eCf+lbVVVVVVVVVVVVVVVVVVeP8A4h+CfhZ4YuPGfxC8SWehaJaOiT3t2xEcbOwVAcA9WIH41/w3d+x//wBF/wDC3/f5/wD4mv8Ahu79j/8A6L/4W/7/AD//ABNf8N3fsf8A/Rf/AAt/3+f/AOJr/gq78cPhL8a/EHw4uvhT480zxNFpNlqCXr2TlhAzvAUDZA6hW/Kqqqqqqz/4+ov98VVVVVVVVVVVVVVVVVVVVVVVVV/wW9/5Kn8NP+xfuv8A0oFfCv4T/ED42+NrH4c/C/w6+ueItRWRrayW4ig3iNC7nfKyooCqTyw6V45/4J5/tmfDrQbjxL4n+BOrf2faxtJO+nXtpqTxov3mMdrLI4UDknGAAT0Br4V/F/4k/BLxba+OPhb4vv8AQNXtmH722kwkyjqksZysqHurgiv2Nv2l9M/at+Bul/EyG1istYhlbT9esYidltfRqpfZkk7GV0kXJJAcAkkGqqqqqr/gpF/ye58U/wDsIW3/AKRQV/wQx/5rZ/3AP/cjVVVVVf8ABbT/AJID4F/7HAf+kc9fsnf8nUfBr/soGhf+l8NVVf8ABUX9mr4j/tG/A/SIvhZZtqmt+E9XN+dIWRUa9haJkfy9xAMi5UhSRkFwMkgH9jH9g79pbxH+0H4O8QeKPhn4m8EaD4S1611TUNU1qwk08gWsqShIBIA0juVCqygqM7icCv2sv+TV/jL/ANk/13/0gmqv2KP+TRfhB/2J+n/+iVr9ub9hf9pPSv2jfGvjLwl8MvEvjXw94y1251awv9EsJL8x/aZDK0MqRBniMbOUBYBSqgg+n/BLH9mL4lfs5/CHxBffFS0k0rWPGWoxXUeivIHaxgijKo0gGQsrlmJXJwqpnDZVf27f2qNZ/ZC+DmnfEjQPCNl4ivdU8QQ6RHb3ly0McPmW9xN5p2glsfZwNuVzuzkY5+GX/BUP9q74z/tD/DLwde654f8ADnh/XvGmkWOoafo2koBcW0t5EkkZknMsgDKxBKsp57VVVVVVVVVVVVVVVVVVVVVVeKv+RX1j/rwn/wDRZr/gjH/yXfxt/wBiif8A0rgqv20JRr3/AAVCi0bUU8y2h8Q+GbQJuPMbwWbsPbmVulV+2nbQXf7JnxZiuIw6r4UvXAPGGWMsp/AgGv8AgibezyWHxg01iPJgm0WVBjnc4vQ36RrVV+yL+zJ8OP2pP2nvip4Q+Jc+sxWOkm+vrc6ZdLA/m/blj+YsjZXa54x1xX/DoP8AZQ/6CPj3/wAHEP8A8Yr/AIdB/sof9BHx7/4OIf8A4xX/AA6D/ZQ/6CPj3/wcQ/8Axiv2dP2cPh7+y/4IvfAHw1n1eXTL/VZNTlOp3Kzyec8UUTYZUUBdsKcY65554qqqqqqqqqqqqqqqqqv24v2QfDf7Uui+GtT1/wAYaloj+DPtfkJZ26SC4+1Nbq27d02+QMY/vGrC0SwsbexRy628KxhiMEhQBn9KqqqqqqqqqqqqqqqqqqqvFnh+HxZ4V1nwrcXDwRazp89k8qDLRrLGyFgD3AbNfsW/sreH/wBlvwNqumaB4r1DXF8TXUd7K15AkZhZE2bV29RjnmqqqqqqqqqqqqqqqqqqqqviJ+w/4U+In7UHh/8AagvfHOq2mqeH57GWLS4raNoZTanKgsfmG7vVVVVVVVVVVVVVVVVVVX/DoP8AZQ/6CPj3/wAHEP8A8Yr/AIdB/sof9BHx7/4OIf8A4xX/AA6D/ZQ/6CPj3/wcQ/8Axiv2y/8AgnF+z38BP2bPGHxY8DXvi2TW9C+xfZVv9SjlhPnX0ED7lWJSfklbHI5xX/BLP/ky/wAI/wDX/qX/AKWS1VVVVVVVVVVVVVVVVVVVft3/APJn/wAVP+wBJ/6GlfsDfsC/Az9pf4GT/EX4i3fiiPVYteubFV02/jhi8qOOJl+Vo2OcyNzn0r/h0H+yh/0EfHv/AIOIf/jFf8Og/wBlD/oI+Pf/AAcQ/wDxiv8Ah0H+yh/0EfHv/g4h/wDjFeFPDeneDfC2jeENIaZrHQ9PgsbUzNucxQxrGm4gDJ2qMnA5qqqqqqqqqqqqqqqqqq8efsb+GdF/atH7U0XjPU5dV1CWaZtKa3QQITZ/ZcB/vfdbd9aqqqqqqqqqqqqqqqqqqqq+P/7PGi/tNeFdJ+Hmu+Ir3Rbe31qG9FxaRLI5ZY5YtuG4xiYn8BXwm+Htl8Jvhl4X+GWnajNf23hjS4NPhup0CvMsSBQzAcAnHQVVVVVVVVVVVVVVVVVVVVV+y1+w/wCFP2WfGninxp4f8c6rrk/imARTQXltHGsIEpkypXk8nHNVVVVVVVVVVVVVVVVVXxz+Cng39oX4b3/wt8eS6jHo2pSwyzNp86wzBopFkXDMrADcozx0r/h0H+yh/wBBHx7/AODiH/4xX/DoP9lD/oI+Pf8AwcQ//GK/4dB/sof9BHx7/wCDiH/4xX/BRr9kz4W/sq614G0/4Y3GuSxeIrW9lvP7Uu0nIaJoQuzaiYGJGz17VVVVVVWf/H1F/viqqqqqqqqqqqqqqqqqqqqqqqqv+C3v/JU/hp/2L91/6UCv+CT/APye54P/AOwfqf8A6RS1X7dmi+FfD/7X3xV0nwZHbx6XF4gkYRQY2RzuqvcIoHChZmlG0cDGO1f8EOLjV38N/F60mDf2XHfaS9sSePPZLoS4Gf7qw9vT8P2wv+CoP7RP7O37R/jL4OeFvCHw6vNJ8PS2v2SfUdPvJZ3Sa0hnG9kukUn97jhR+PWvDv8AwVck0f8AZIt/jR8SvDGi3XxA1jXr3SdF0HR/MtrefyEiczyeZJI6xIJkDEElmKqAMll/Yt/4KL/tM/tC/teeDvh1441vQ7fwtr7agbnS9P0mOJVEVhczRhZG3SjDxIeXOcY7mviT8SfBHwh8E6p8RPiL4httG0DRofMurucnAyQFVVHLOzEKqgEsSAASa+Nv/Bafx9qOq3OmfAD4faTo+kxuyRan4hVrq7nXs4hR1jhP+yxl+orTP+Ct/wC2jYX4vLrxb4c1GIHP2S60CBYzyDjMYR+gx97ofXmv2R/+Cs3gn41eItP+G/xp8PWfgnxLqUghsNStrgtpt7Mfuxnf81uzE4UMzqTxuBIBr/gpF/ye58U/+whbf+kUFf8ABPn9sXwR+x54C+MPiHX9Mn1rX9efRYdB0aGTyvtbxi+MjvIQfLjQSJuOGOXUAHOR4q/4K/ftga7qzXuhah4V8OWYkJjsrPRlmXbngM85dicdSCueeB0H7Kv/AAWHfxL4jsfA37Teg6VpSahMkFt4o0lWhghdiAPtULM21STzKjAL3QDLBHSRFkjdWVhlWByCPUV+3P8A8FL9f/ZZ+Iknwi8H/CSLUtaWwhvV1fWbspaPHKDjy4YvnkAZSpYumGRhgjDVrn/BXb9szVpzLp/iDwtoq7siKx0GN1Ht+/Mhx+NfC3/gs5+0D4d1a3j+KnhTwz4v0csouDawNp94o6Eo6kx574MfJ4ytfAv44/D79or4bab8Ufhpqb3Wk6gWjeKdNk9nOmPMgmTJ2yLkZGSCCGBKsCf+C2n/ACQHwL/2OA/9I56/Zz8R6L4P/aE+F/i7xJfx2Wk6H4z0i+v7qQHbBBDeRPI5xzhVUn8K+O//AAWb+KOsa/eaX+z94V0jw/oEMjJb6nrFubu+uVHAk8vcIoQeuwiQ9Pm6ivhh/wAFj/2l/C+tQP8AErS/DfjbRy4+0w/Yxp9zs7+VLD8in/ejb8Otfs+/tAfDr9pf4bWXxN+GuoyTWM7tBdWtwoS4sLhQN8MygnawBB4JBDKwJBBr/goj+1b8RP2R/hh4b8a/DfRvDmpX2sa9/Z88et2000ax+RJJlRFLEQ2YwMkkYJ4718EP+Cuv7SfxL+NPgD4ca74J+GkGm+KvFGm6VeS2mm3yTRw3F1HE7RlrtlDhXJBKkA4yD0r9rL/k1f4y/wDZP9d/9IJqr9ij/k0X4Qf9ifp//ola/aH/AOCtH7Rvwk+Onjv4Y+G/Bfw3udL8L69dafZzX2m3rzvHFIVUyMl2qlsDkhQPYV/wTp/a7+JP7Xngrxd4j+JOieGdNufD+qQ2lsmh208KOjxbyXEs0pJz6ED2r9vT9uP4s/H3UPEHwE8YeHvCNn4f8HeNrmWwudNtLiO7ka1NzbR+Y0k7oQUlYttRcsARgcH4eeNdV+Gnj/wz8RtBt7SfUvCusWmq2cV2jNDJNbTLKiyBWVihZACAynGcEda/ZK/4Km/tB/Hv9onwX8IvGHg/4eWej+I7qaK6n03T7yO4QJbySDY0l06g7kA5U8Z+tftN/wDBUD9rX9nf46+L/hDd+BfhjNBoV+w0+5n0m/D3NnIBJbyHF4AWMTJuwMBtw7V/wTr/AOCg3j39rXxr4s8A/FPQ/Cul6npemRajpI0O2ngE0SyeXcCQTTSkkGSDbtxgFs54x8U/HNl8Mfhn4r+I+o7Db+F9Fu9SkVzgOIIWk2/U7QABySQK/wCH1f7VH/QhfCn/AMFV/wD/ACbX7IvxL+J/xl/Z+8K/FT4uaToml654nhe9is9ItpYIY7RnIt2xLLIxLxhZM7sYccDFVVVVVVVVVVVVVVVVVXir/kV9Y/68J/8A0Wa/4Ix/8l38bf8AYon/ANK4Kr/gpzY3vwu/bj0L4niCRor2y0rWYWTHzPayeUyjPG4fZ1OD/eHrWnahY6vp1rqumXSXNpewpNbzIcrJG6hlYexBB/Gv+CiPjG08F/sdfEa6nuAk2qWMel2ybgGle5mSMqM9cIzsR/dU1/wRl8IXWm/Brxx41nRlj13xDHaQBhgMttACWHtuuGH1U1Vf8EuP+Tx/jN/14X//AKco6qqqqqqqqqqqqqqqqqqqq8ff8ilff9s//Ri1VVVVVVVVVVVVVVVVVVVVXhD/AJFnTv8ArgKqqqqqqqqqqqqqqqqqqqq8e/tEeNfC37SOhfB3TtL0STRtTms0mnmglNwomOG2sJAox2yp/Gqqqqqqqqqqqqqqqqqqqqv+Cmv/ACY/8Sf+4V/6dbSv+CWf/Jl/hH/r/wBS/wDSyWqqqqqqqqqqqqqqqqqqqr9u/wD5M/8Aip/2AJP/AENK/wCCQf8Ayahd/wDY3X3/AKJt6qqqqqqqqqqqqqqqqqqqq+K3+v03/ck/mtVVVVVVVVVVVVVVVVVVVVaL/wAhiw/6+Y//AEIVVVVVVVVVVVVVVVVVVVVVX7NP7RHjX4zeL/Evh/xRpeiWtvo0Ie3awglR2PmFfmLyODwOwFVVVVVVVVVVVVVVVVVVVVf8Frf+Rn+FH/Xhqn/oy3qqqqqqs/8Aj6i/3xVVVVVVVVVVVVVVVVVVVVVVVVX/AAW9/wCSp/DT/sX7r/0oFfsu/H/UP2YvjNo/xi0zw3b69PpMNxF9gnuGgWQTQtEfnAYggNnoelfE7/gtD8c/FWgT6N8Ovh94d8F3NzEY21Jp31G4hzn5oQ6pGrdOXRx149La18SeN/EiWtnb6jruva3eYSOJHuLm9uJG7AZZ3Zj7kk1/wT4/Zivv2Xf2fLHw14mijXxb4huTq2vKhDfZ5nVVS3DDr5caKDjI3lyCQQa/4Kj/APJ9vxN/7hP/AKabOv2cf2f/AIm/tV/ETTvhH4DuUH2eGW7nuL6V/sul2wKiWZgASMsUXCjLMyj3H7Ln/BKnS/2a/jP4V+NI+Nd14gvvDi3W+w/sFbWOcz2k1ucP57lQvnbuhztxxnI/4LGfH3WPF/xts/gLp2oMnh/wNaw3F5bI/wAs+o3EYk3tjrsgeNVB+6Xk/vV/wT4/ZR079rD43Hw/4smuYvCHhqy/tHW/s7FHuV3hI7ZXH3DIxJJ67EfGDgj4yf8ABLv9ljx58Nb/AMMeAPh3ZeDfEcNo/wDY2sWU8xaGcL8nnBnbzoyQA27LYJIIbmtX0q/0LVbzRNVtmt77T7iS3uYXGGjkRirKfcEEV/wTS+PGrfHv9lnRdR8Tag974g8KXcmg6lcysS9wYVR4ZGJ5LGGWIFj95lY9c1/wUi/5Pc+Kf/YQtv8A0igr/gnz+x1a/tefFLUbDxRqV3YeDvCdrFdazJZkLNO0jFYbdGIIQvskJbBwsbAckEfG7/gkR+zb4n+H99a/BjRL7wf4utbUtp102rXF3BdTKOEnSd3wr9C0e3aSDggFTqOn3uk6hc6VqdrJbXllM8NxBIu1opEJVlI7EEEGv+CXnxd1T4u/sh+HW127a61LwfdzeHp53OWdLdUeDPuIJoVz32571+0T+xP8Af2ivH+h/FX4zw3s8fhfS5LSS1S++x21zAJDIpnkXEgVC0hG11++cmvEnjb/AII+fC61k8M6rpvwbutgMTmx8ODXJQQMEefBDMwPHXfwe+a/a5H7N0/xiudT/ZXvbqTwTqNlHcG1ntp4RYXZZxLDGJxvKYVHHUDzCoOFwP8AgiN8QdQtviL8RPhS927WOo6JFrUUDElY5beZIXZR0BZblAe52L1xx/wW0/5ID4F/7HAf+kc9eFfDWr+NPE+j+DvD1t9p1XXb+CwsYc48yeaRY41z2yzAfjXw5/4JEfso+HfAlvofxA0TVfFviR4B9s1ttWubTbKR83kRROqKgPADq5wBkmv25/2T7j9kb4yt4MsNRudT8M6zaf2hoN7cKBKYSxVopCAAZI2XBIABBRsDdgf8EdPi7qng/wDaUufhS92x0jx/pUy/ZieBeWkbTxyD3ES3Cn13D0Ff8FtP+SA+Bf8AscB/6Rz1+yd/ydR8Gv8AsoGhf+l8NftZf8mr/GX/ALJ/rv8A6QTVX7FH/Jovwg/7E/T/AP0Stftr/wDJ3Pxf/wCxw1D/ANHNX/BEP/klXxK/7GC1/wDSc1/wVQ/Y2+CfwR+GUXxs8CWGrxeJ/FfjsRajJc6g00TLcwXlxLtjIwv7yNcegyK+Avg/RfiF8c/hz4A8RpK+k+JfFml6ZfrDJ5bmCe7jikCsPuna5wexr4S/8E2P2YPgn8RdF+KPgXSfEMWu6BK8tm9zq7zRhmjaM7kI5+V2r/gtX8CjPY+D/wBorRbIbrZjoOuMic7G3S2shx2DechJ/vRj0r9gr4sH4NftZfD3xTcXYg0691NdJ1It937PdjyCzf7KM6Sf9sx16V/wVm+JP/CBfsea1olvceXeeNtUtNGhKn5gm43EuB6GO3ZCen7z1Ir4LfDPU/jL8WvCPws0gSfaPE+r29kZEGTDG7jzZfoke9z7Ka0DQ9L8MaFpvhrQ7RLXTtJtIrSzgQYEUMSBEUewVQKqqqqqqqqqqqqqqqqqvFX/ACK+sf8AXhP/AOizX/BGP/ku/jb/ALFE/wDpXBVf8FHv2TtU/aW+E1prHgazSfxv4KkludNtz8pv7dwPPtgf752I6Z43Jt43kj9kb/gp3o3wW+Hlt8Ff2i/CviX7V4QX7Dp1/Y2qvMsMZ2rbXMMjoyPEBsBGcgAEAqWb9pj9o/4j/wDBSP4h+HPgb8APBGqw+GbK6+0pFeAK802Chu7soWSCGJGYAZP325ZmVR+z/wDBrQv2f/g94Z+E3h+QTxaFZhbi62bDd3LkvPMRzjfIzEAk7QQM4FVX/BLj/k8f4zf9eF//AOnKOqqqqqqqqqqqqqqqqqqqqvH3/IpX3/bP/wBGLVVVVVVVVVVVVVVVVVVVVVeEP+RZ07/rgKqqqqqqqqqqqqqqqqqqqq+MX/J9HhH/AK+tL/8AQqqqqqqqqqqqqqqqqqqqqq/4Ka/8mP8AxJ/7hX/p1tK/4JZ/8mX+Ef8Ar/1L/wBLJaqqqqqqqqqqqqqqqqqqqv27/wDkz/4qf9gCT/0NK/4JB/8AJqF3/wBjdff+ibeqqqqqqqqqqqqqqqqqqqqvit/r9N/3JP5rVVVVVVVVVVVVVVVVVVVVWi/8hiw/6+Y//QhVVVVVVVVVVVVVVVVVVVVVfsH/APJS/Hn/AF6r/wCjzVVVVVVVVVVVVVVVVVVVVX/Ba3/kZ/hR/wBeGqf+jLeqqqqqqz/4+ov98VVVVVVVVVVVVVVVVVVVVVVVVV/wW9/5Kn8NP+xfuv8A0oFfsbfALQ/2mvj5ovwf8Q69faPZ6raXkrXlkivJG0MDyLgNwQSoB9jWif8ABEv4E21yJPEPxa8dahCDnyrRbW1JGDwWaOTvjt0z65HwH/Y3/Z0/ZuYX3wt+HVna6wY2R9avXa7vmVhhgJpCTGpAAKx7VOORVf8ABUf/AJPt+Jv/AHCf/TTZ1/wQ7hhbxf8AFmcxIZU03TFVyvzKDJcZAPodq5HsPSq/4KJWuoWn7anxWh1Pf5zauki7ySfLe2iaP8PLKY9sV/wQ2vdMj1n4xabK0X9oT2ujSwKQN5iRrwSY743SRZ7cjPaq/aWvtM1T9o34q6norRtp93421ma0MYAUxNeylNoHAG0iv+CJFjfx/BP4g6lIH+xT+KY4Yc5x5iWsZkx2ziSPPfpntX/BSL/k9z4p/wDYQtv/AEigr/ghlJGJPjXCZF8xl0FlQnkgf2hkgegyPzFV+0lqWl6z+0T8U9X0NkOnX3jXWJ7MooC+S97KyYA4A2kdK/4IpabeW/7N3i/U5lKwXfjWZIQQQW2WdruYdsZYDI7qR2r9vf8AbZ8cftL/ABL1jw1oniC5s/hrod7JbaRpdtMUivxG2Ptc4B/eM5G5QeEUgAZ3M37J3/BNv4y/tU+F0+IdprOk+E/B8s0kNrqWoI80t60bFXMMKY3KrBlLMyjcCBnBx+2z+yGf2OPHug+Bm+IQ8XHW9GGom5Gk/YPJ/fSR+Xt86Xd/q87sjr04r/gip/ydR4p/7J/ef+l9hX/BbT/kgPgX/scB/wCkc9fspSRw/tSfByWWRURPH+hszMcBQL+HJJ7Cq/4LkalpcuvfB/R4WT+0raz1ie4AUbvJke0WLJ643RTY/H1Nf8Ey9NvNT/bg+GMdmpzb3F9PK2DhUSwuC2SM4zjaM8ZYDvX/AAW0/wCSA+Bf+xwH/pHPX7J3/J1Hwa/7KBoX/pfDX7WX/Jq/xl/7J/rv/pBNVfsUf8mi/CD/ALE/T/8A0Stftr/8nc/F/wD7HDUP/RzV/wAEQ/8AklXxK/7GC1/9JzX/AAWr/wCTV/C3/ZQLP/0gv6/ZO/5Oo+DX/ZQNC/8AS+Gq/aL+D+m/Hz4IeMfhJqSxj/hINMkitJZBkQXS4e3l/wCATJG30FappmqeHtYu9G1S2lstR0y5eC4hf5XgmjYqyn0IYEfUV/wUi/aTPxz+G/7PNpFfK81x4OOu6uiNkG9mYWzg+6SWlwB3w5z1Ff8ABGD4LnxR8Y/Enxs1K0Y2fgnTvsenyMuAb27DKxU9ysCyggdPOX1qqqqqqqqqqqqqqqqqqqSOOaNoZUV0dSrKwyCD1BHpXhj4afDjwTdy6h4M+H/hvQbqeLypZ9M0mG1eRMg7WaNQSuQDg8ZAqq8dfAL4H/E6+Gq/EL4R+EfEN+oAF5qGjwzT4AwB5hXdt9s46eleC/h34A+G+mHRfh74J0Lw1YsQXttJ06K0RyO7CNRuPJ5PPJ9aqq/4Jcf8nj/Gb/rwv/8A05R1VVVVVVVVVVVVVVVVVVVV4+/5FK+/7Z/+jFqqqqqqqqqqqqqqqqqqqqqvCH/Is6d/1wFVVVVVVVVVVVVVVVVVVVVfGL/k+jwj/wBfWl/+hVVVVVVVVVVVVVVVVVVVVV/wU1/5Mf8AiT/3Cv8A062lf8Es/wDky/wj/wBf+pf+lktVVVVVVVVVVVVVVVVVVVX7d/8AyZ/8VP8AsASf+hpX/BIP/k1C7/7G6+/9E29VVVVVVVVVVVVVVVVVVVVfFb/X6b/uSfzWqqqqqqqqqqqqqqqqqqqqtF/5DFh/18x/+hCqqqqqqqqqqqqqqqqqqqqq/YP/AOSl+PP+vVf/AEeaqqqqqqqqqqqqqqqqqqqq/wCC1v8AyM/wo/68NU/9GW9VVVVVVn/x9Rf74qqqqqqqqqqqqqqqqqqqqqqqqr/gt7/yVP4af9i/df8ApQK/4JP/APJ7ng//ALB+p/8ApFLVVX/BUf8A5Pt+Jv8A3Cf/AE02df8ABDr/AJGr4t/9g/Sv/RlzVf8ABYz9lzXY/E9l+1H4R0qS60q7tYtO8UeShZrWaP5YLlwOiOm2It0UxoP4xX7KP7S/iz9lH4vWXxQ8MWceowGB7LVtLlk8tb+zcqXj34JRgyI6tg4ZBkEZB+NP/BaPwjq3w2vdI+B/w98T6f4v1K2eBNQ1oW8cGmMy486MRySGZ16qGCDOCcgbT4c8OeI/G/iOw8L+F9JvNY1vWLlYLOztozLLcSucBVA5JJ/xr9jL9n0fszfs8eGPhfdtFJrUaPe65NEcrJfTHdIAR1CDbED3WMHvX/BSL/k9z4p/9hC2/wDSKCv2X/2lvib+yL8Qrb4k+DbFLm01S1a3vtNvldbbVbYPggMOQyuh2yLnawIOQWU/Gn/gtB4y8a+Ar7wr8KfhSvg3V9Utmgl1q51o3slmrqQxt0WKPEgz8sjE4PO3PTQdB1rxTrdh4b8OaXc6lqup3CW1nZ20RklnlchVRFHJJJAxX7KH7P5+AH7M3hn4N3c6JqsOnyPq9xAQf9NuCzzFWH3gjPsVu6xrXjTwjrvgDxfrXgbxPZPaavoF/NY3sDjBSWJyjD6ZHB7jBr9i7/gqj8NvgH8BNI+D3xT8B+KL278MmePTL3Qo7eVLmGSV5QJRLLGY2VpCuV3AgA9c5/bF/ak179rf4wzfEjU9IXR9Os7NNP0bTFl8w21qjMw3twGkZpHZiAOoHRQa/wCCKn/J1Hin/sn95/6X2Ff8FtP+SA+Bf+xwH/pHPWljV4bn+1NFF2s+mbbr7RbBt1ttZdsm5fuYYr83GCR618Ov+C2firQ/A9tpHxG+C9v4l8S2cCxnVbPWvsMd6QMb5IjC+xzjLbTtJJwqjiv2i/2g/Hv7TvxRv/in8Q5rdb24iS2tLO1DLBYWyZ2QxBiTtBZmJJyWdj3r/gjP+zZrdnfa7+014p0uS2srizbSPDQmjwbjc6m5uEz/AAjy1iVhwS0o/hr/AILaf8kB8C/9jgP/AEjnr9k7/k6j4Nf9lA0L/wBL4a/ay/5NX+Mv/ZP9d/8ASCaq/Yo/5NF+EH/Yn6f/AOiVr9tf/k7n4v8A/Y4ah/6Oav8AgiH/AMkq+JX/AGMFr/6Tmv8Agrn4D1Hxr+x5f6npto9w3hDXrLWJVQEssQElu7AA9FFzk9cAE9sjwx4i1bwf4l0nxboNz5Gp6JfQ31lLjPlzQuHRsezKDX7Jf/BVXWv2kfjT4L+B+rfBWx0W819LoXusW+vNLGGgsprgmO3aEFQzQhcGVsBupxVf8FZPgX/wqf8AaeuvG2l2nl6J8Sbc6tCVXCreKQl2g9y+yU/9d6nu7u5it4Li6lljtYzFAjuWESFmcqoP3RudmwOMsT1Jr/gnL8D2+Bf7KPhPSdRszBrnidDr+rqylWEtyqmNGB5BSBYEI/vK3TNVVVVVVVVVVVVVVVVVVVVVVVVf8EuP+Tx/jN/14X//AKco6qqqqqqqqqqqqqqqqqqqq8ff8ilff9s//Ri1VVVVVVVVVVVVVVVVVVVVXhD/AJFnTv8ArgKqqqqqqqqqqqqqqqqqqqq+MX/J9HhH/r60v/0Kqqqqqqqqqqqqqqqqqqqqv+Cmv/Jj/wASf+4V/wCnW0r/AIJZ/wDJl/hH/r/1L/0slqqqqqqqqqqqqqqqqqqqq/bv/wCTP/ip/wBgCT/0NK/4JB/8moXf/Y3X3/om3qqqqqqqqqqqqqqqqqqqqr4rf6/Tf9yT+a1VVVVVVVVVVVVVVVVVVVVov/IYsP8Ar5j/APQhVVVVVVVVVVVVVVVVVVVVVfsH/wDJS/Hn/Xqv/o81VVVVVVVVVVVVVVVVVVVV/wAFrf8AkZ/hR/14ap/6Mt6qqqqqrP8A4+ov98VVVVVVVVVVVVVVVVVVVVVVVVV+2H+wL4P/AGxPEvh7xJ4l8f6x4el8PWMlpFFY2scolDvvLEv0Pav2Zf8Agl/4C/Zk+MGlfGDQfijr+tXmlQXMKWd3ZRRxuJoWjJJXngNn8Kqq/aP/AOCWPw//AGkPjR4i+NGt/FTxBo974h+zebZWtlDJHF5FrFbjazcnIhDHPcmv2Of2EvCX7HGpeKNS8M+PNX8RN4ogtoZkvraOIQiFpCCuzrnzDnPoKrUtM03WdOudI1ewtr6xvYWhubW5iWWKaNhhkdGBDKQSCCMEGvjR/wAEbvgL4/1W41z4X+LdX+HdxdMWeyjtl1GxQnr5cTukiDrx5u0dgAMVon/BDif7cG8SftHRmzRuY7HwwRJIOf4nucIenOGr9mn9hn9n79lgf2l4A8OT6h4kkjMcviLWZFuL0qeqxkKqQqckERqpYYDFsVXx2/4JO+APjx8XPE3xd1z4v+JLC+8S3YuJbWCxhdIcIqKqk8kBUA5pP+CZnwG1H9nTR/2fPFVzqOqf8I/d3d3pniWKOO21C2luJC74IBVlPyqUYEEIvQqCLr/ghvK2sYs/2kkXSixOZfCpadBzhcC6Cseg3cdScdq/ZW/4J8fAr9lW7XxPoNveeJfGJjaP+39XCmS3Vhhlt4lAWEEZBPzOQSN+Diq/a3/4JwfBv9qrWG8cNqN54O8atEIptY0+BZo70KMIbmAlRIygABlZGwACSAoHhv8A4IdQx6skni/9ohp9MRxvh03w55U0q9wHknZYz052v9K+J/8AwSh/Z58b/Dvwh8OvCF7qfg6DwncXc5v7aNLm61OS4WISPcO+CzfuI8YwFHCgDiv2T/8Agm74N/ZN+KMvxQ8M/E7XNcuJ9Km02S0vbOKNDHI8bk5Q5yGiXrkfzH7YP7ImgftheEtB8IeI/G2qeHrXQtRa+VrG3SUzOYzGAwfpgM3T1r9nD/glx8KP2e/HN/4wm8Y6h4yt9T0S60i50rWNOh8iSG42iQnGeqqVx6Ma+K//AARP+H+v65cat8IPi5qHhSzncuNJ1TTf7RjiyfuxzCSNwo7Bw59W718Gf+CMPwf8G6za678XfiDqfj37MwkGl29kNMtJGH8MuJJJJFB9Gjz34yK0rStM0PTLTRdF0+2sNPsIEgtbW2iEUUESAKqIq4CqAAABwAK/a/8A2SvDn7YHgjRvBHiTxdqPh+DRdV/tGOexgSVpG8p49pD8AYkJ/Cvhj/wR6+Gvwx+JPhP4k2Hxi8S3tz4T1yy1aG2msIFSd7adJVRiDkBigBI9a+J3ge0+J3w18WfDW/vpbK28WaHe6TNcwqGeBLmB4WdQeCQHJAPGRX/DkL4Vf9Fw8V/+C63/AMa+EPw5sfhD8LvCvwu03Up9QtfC2lQadDdToEeZYkChmA4BOOgr4vf8EiPhx8X/AIo+KfijqXxf8SafdeKdVn1Ga1gsIWSFpXLFVJOSBnvX7Hv7Hnhr9jzw14g8NeGvGOp+IYvEN9Hdyy31ukTRMibAAE6gjnmtY0jS/EGk3mg63p9vf6dqNu9vd2txGHjnidSroyngqQSCD618W/8Agil8OPEet3OsfB74raj4QtZ5C66RqWnf2lDDk/djl82ORUA6B/Mb1bvX7Jn/AASpt/2avjP4f+NeqfGyTxHfeH0ufI0230AWcZae2lt23SGeQsAsxIwq8gVX/BY39o/4YeP7zw18DfBk9rrWteE9RlvdX1S3kV47F2jMf2NWGdzn70g6KUQcncF/YP8A2dLj9pX9ozw54SvdONx4Z0eVdU8RsyEx/Y4WBMTH/ps+2L1w5P8ACaCqqhVAAAwABjFVVVVVVVVVVVVVVVVVVVVVVVVf8EuP+Tx/jN/14X//AKco6qqqqqqqqqqqqqqqqqqqq8ff8ilff9s//Ri1VVVVVVVVVVVVVVVVVVVVXhD/AJFnTv8ArgKqqqqqqqqqqqqqqqqqqqq+MX/J9HhH/r60v/0Kqqqqqqqqqqqqqqqqqqqqv+Cmv/Jj/wASf+4V/wCnW0r/AIJZ/wDJl/hH/r/1L/0slqqqqqqqqqqqqqqqqqqqq/bv/wCTP/ip/wBgCT/0NK/4JB/8moXf/Y3X3/om3qqqqqqqqqqqqqqqqqqqqr4rf6/Tf9yT+a1VVVVVVVVVVVVVVVVVVVVov/IYsP8Ar5j/APQhVVVVVVVVVVVVVVVVVVVVVfsH/wDJS/Hn/Xqv/o81VVVVVVVVVVVVVVVVVVVV/wAFrf8AkZ/hR/14ap/6Mt6qqqqqrP8A4+ov98VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX/BY3xD8WvB3wf8IeIfh94813QdDu9Wn0nX7bTLpoBeCaLzIfMZMNtAgmUjO0+Zg54r4cfDLx78X/ABfY+A/hr4Wv/EGu6g2IbS0j3EDjLufuoi5BZ2IVRySBX7DX7IOifsi/CgaFPLb6h4x18pc+JNSiGVeVRhIIiQD5MYLAZGWZnbjcFFVVVVVVVVVVVVVVVVVVVVVVVVf8EuP+Tx/jN/14X/8A6co6qqqqqqqqqqqqqqqqqqqq8ff8ilff9s//AEYtVVVVVVVVVVVVVVVVVVVVV4Q/5FnTv+uAqqqqqqqqqqqqqqqqqqqqq68KeFr3VotevPDelT6nCVMV7LZxtMhX7pDkbhjtzxVVVVVVVVVVVVVVVVVVVVX/AAU1/wCTH/iT/wBwr/062lf8Es/+TL/CP/X/AKl/6WS1VVVVVVVVVVVVVVVVVVVft3/8mf8AxU/7AEn/AKGlf8Eg/wDk1C7/AOxuvv8A0Tb1VVVVVVVVVVVVVVVVVVVV8Vv9fpv+5J/Naqqqqqqqqqqqqqqqqqqqq0X/AJDFh/18x/8AoQqqqqqqqqqqqqqqqqqqqqqtI8KeFvD88t1oPhvStNmuBiaS0s44WkGc/MVAJ59aqqqqqqqqqqqqqqqqqqqq/wCC1v8AyM/wo/68NU/9GW9VVVVVVn/x9Rf74qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqvjJ8Fvhx8fvBEvw6+KmgnV9BnuYbiS2W5kt2MkTblIeNlde4OCOCR3r4WfBL4SfBDRm0D4TfD7RfDFnLjzhY2wWS4KjAMspy8pA7uxNVVVVVVVVVVVVVVVVVVVVVVVVVX/AAS4/wCTx/jN/wBeF/8A+nKOqqqqqqqqqqqqqqqqqqqqvH3/ACKV9/2z/wDRi1VVVVVVVVVVVVVVVVVVVVXhD/kWdO/64Cqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/4Ka/8mP/ABJ/7hX/AKdbSv8Agln/AMmX+Ef+v/Uv/SyWqqqqqqqqqqqqqqqqqqqr9u//AJM/+Kn/AGAJP/Q0r/gkH/yahd/9jdff+ibeqqqqqqqqqqqqqqqqqqqqvit/r9N/3JP5rVVVVVVVVVVVVVVVVVVVVWi/8hiw/wCvmP8A9CFVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX/Ba3/kZ/hR/14ap/wCjLeqqqqqqz/4+ov8AfFVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX/BLj/k8f4zf9eF/wD+nKOqqqqqqqqqqqqqqqqqqqqvH3/IpX3/AGz/APRi1VVVVVVVVVVVVVVVVVVVVXhD/kWdO/64Cqqqqqqqqqqqqqqqqqqqqtc+OHw48O/EGy+F2q6xPH4h1B4kt7cWkjKxl4T5wNoz7niqqqqqqqqqqqqqqqqqqqqv+Cmv/Jj/AMSf+4V/6dbSv+CWf/Jl/hH/AK/9S/8ASyWqqqqqqqqqqqqqqqqqqqr9u/8A5M/+Kn/YAk/9DSv+CQf/ACahd/8AY3X3/om3qqqqqqqqqqqqqqqqqqqqr4rf6/Tf9yT+a1VVVVVVVVVVVVVVVVVVVVov/IYsP+vmP/0IVVVVVVVVVVVVVVVVVVVVVXw8+OHw4+Kmq6jovgvWJ7u70pN90klpJCEG7bwWAB5Haqqqqqqqqqqqqqqqqqqqqv8Agtb/AMjP8KP+vDVP/RlvVVVVVVZ/8fUX++Kqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv+CXH/ACeP8Zv+vC//APTlHVVVVVVVVVVVVVVVVVVVVXj7/kUr7/tn/wCjFqqqqqqqqqqqqqqqqqqqqqvCH/Is6d/1wFVVVVVVVVVVVVVVVVVVVVfGL/k+jwj/ANfWl/8AoVVVVVVVVVVVVVVVVVVVVVf8FNf+TH/iT/3Cv/TraV/wSz/5Mv8ACP8A1/6l/wClktVVVVVVVVVVVVVVVVVVVX7d/wDyZ/8AFT/sASf+hpX/AASD/wCTULv/ALG6+/8ARNvVVVVVVVVVVVVVVVVVVVVXxW/1+m/7kn81qqqqqqqqqqqqqqqqqqqqrRf+QxYf9fMf/oQqqqqqqqqqqqqqqqqqqqqqv2D/APkpfjz/AK9V/wDR5qqqqqqqqqqqqqqqqqqqqr/gtb/yM/wo/wCvDVP/AEZb1VVVVVWf/H1F/viqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr4k/FT4c/B7w1L4v8Aib4x0zw7pMR2/aL2baZHxnZGgy0j4BO1AWODxXiz/gsF+y/od7JY+HtC8ceJFjbC3drp0NvBIPVfOlST8DGK/Zj/AGmPBH7Vnw+u/iJ4D0bXNMstP1V9LuINXiijk89IYZWK+VI4KYnUAkgkg/KOM698b/gz4X1m08OeIviv4R07V765jtrbT7jWYEuJZXYKqrEW3EliB0qqqqqqqqqq/wCCXH/J4/xm/wCvC/8A/TlHVVVVVVVVVVVVVVVVVVVVXj7/AJFK+/7Z/wDoxaqqqqqqqqqqqqqqqqqqqqrwh/yLOnf9cBVVVVVVVVVVVVVVVVVVVVXxi/5Po8I/9fWl/wDoVVVVVVVVVVVVVVVVVVVVVf8ABTX/AJMf+JP/AHCv/TraV/wSz/5Mv8I/9f8AqX/pZLVVVVVVVVVVVVVVVVVVVV+3f/yZ/wDFT/sASf8AoaV/wSD/AOTULv8A7G6+/wDRNvVVVVVVVVVVVVVVVVVVVVXxW/1+m/7kn81qqqqqqqqqqqqqqqqqqqqrRf8AkMWH/XzH/wChCqqqqqqqqqqqqqqqqqqqqq/YP/5KX48/69V/9Hmqqqqqqqqqqqqqqqqqqqqv+C1v/Iz/AAo/68NU/wDRlvVVVVVVZ/8AH1F/viqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq1PUbHR9NutX1O5S3s7GB57iZ+kcaKWZj7AAmv2pf2jPGf7Ufxc1Dxrrd1cnTUne38PaUCSlhabsIiqP42ADO3VmPoFA+IXwI+MPwm0LRfEfxL+Hur+GrHxCZF01tSh8iSYoFL/umO9cB1+8o61/wRs/5Ng8Tf8AY+3f/pBYV+0/4XX4aftNfEbw7ortax6T4qvHsDEdphjMxkiAIPBVWUZ9u3Svht4o/wCE5+HPhXxrkf8AFQaJZ6hxjH76FJOMcfxduKqqqqqqqqr/AIJcf8nj/Gb/AK8L/wD9OUdVVVVVVVVVVVVVVVVVVVVePv8AkUr7/tn/AOjFqqqqqqqqqqqqqqqqqqqqqvCH/Is6d/1wFVVVVVVVVVVVVVVVVVVVVfGL/k+jwj/19aX/AOhVVVVVVVVVVVVVVVVVVVVV/wAFNf8Akx/4k/8AcK/9OtpX/BLP/ky/wj/1/wCpf+lktVVVVVVVVVVVVVVVVVVVX7d//Jn/AMVP+wBJ/wChpX/BIP8A5NQu/wDsbr7/ANE29VVVVVVVVVVVVVVVVVVVVfFb/X6b/uSfzWqqqqqqqqqqqqqqqqqqqqtF/wCQxYf9fMf/AKEKqqqqqqqqqqqqqqqqqqqqr9g//kpfjz/r1X/0eaqqqqqqqqqqqqqqqqqqqq/4LW/8jP8ACj/rw1T/ANGW9VVVVVVn/wAfUX++Kqqqqqqqqqqqqqqqqqqqqqqqqq88QaBYTm1v9c0+2mUDMc1yiMM9OCc1Xifxt4L8FWyXnjLxfougwScJLqd/Faq3IHBkYA8kfmK0D4vfCXxXerpnhb4o+EdYvGIC29hrdvcSEnphUcnmqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr9oqz1DUf2ffidp+kNi/ufBurxWpGeJWs5QnTJ6kdq/4J4TeCof2xfhw3jo2/2I3sy2huQPLF8beQWuc9/OMYT/bKV/wWt/5Ff4Uf9f8Aqn/ou3r/AII53Fvafss+K7q6njhhh8dXjySSMFVFFhYksSeAABnJr9p34gaX8VP2hPiF8QNCcPpms6/cy2MgBHmW4cpE+OxZFViPU1+zfpdzon7O3wt0W9XFxYeC9It5RgjDpZRK3XnqDVVVVVVVVVf8EuP+Tx/jN/14X/8A6co6qqqqqqqqqqqqqqqqqqqq8ff8ilff9s//AEYtVVVVVVVVVVVVVVVVVVVVV4Q/5FnTv+uAqqqqqqqqqqqqqqqqqqqqr4xf8n0eEf8Ar60v/wBCqqqqqqqqqqqqqqqqqqqqr/gpr/yY/wDEn/uFf+nW0r/gln/yZf4R/wCv/Uv/AEslqqqqqqqqqqqqqqqqqqqq/bv/AOTP/ip/2AJP/Q0r/gkH/wAmoXf/AGN19/6Jt6qqqqqqqqqqqqqqqqqqqq+K3+v03/ck/mtVVVVVVVVVVVVVVVVVVVVaL/yGLD/r5j/9CFVVVVVVVVVVVVVVVVVVVVV+wf8A8lL8ef8AXqv/AKPNVVVVVVVVVVVVVVVVVVVVf8Frf+Rn+FH/AF4ap/6Mt6qqqqqrP/j6i/3xVVVVVVVVVVVVVVVVVVVVVVVfEn4q/Df4PeG5PF3xQ8a6T4a0mNtgudQuBH5j4zsjX70j4BO1QTweK8W/8FjP2RfD1zJa6Hb+OfFAUHZcaboyQxMe3/HzLE4B4/g79K/bL/4Kta18a/B9h4F/Z/i8T+BbG5Mh1+9nkSG8uV4EcMMkMjFIyNxfBDN8q527g3wE/Ye+BP7RVrax2P7dnhq28Z6gN8uhzeG5hM0zdVR7qeBp23E5KIc9a+Kf7Fn7bf7Fk2gat8FPit4i8Q2muaoNPtoPCEt5FMJyrOgntBlDGwR+SWUFfmxkZ+H3/BIP4z/FmU+P/wBqP423Gn65qQ8ye0iLaxfZPaa5kkCKw54TzBz1rxr/AMEQtJTRZ7j4cfHe+/teGMtBBrGkIYZ3HRTJE4aMf7W18elf8ErP23oH8O638Gf2gvizpNomim3bwrf+ItWSGSWNy6yWgmlYbwhWMopJIDlR8oUCGaG4hSe3lSWORQyOjZDAjIII6giqqqqqr9sD9sb4efsgeB7fXvE1vJrHiDWGePRNBt5RHJdsgG53Y58uFcqGfBOWAAJPGgftE/8ABXr9pTTD8Q/gj4Kt9B8L3DsbL7Lpum28U0eSAY31NmeYDGC6fKSD06D4N/8ABUT42/Cb4pL8Hf25PBC6d++SK41dNPNld6dv+7LLCn7uaA8HdEFIBLDfwtW9xb3lvFd2lxHPBMgeKWNgyupGQykcEEYOa/bw/wCChXh39kuC38EeEdKtPEnxF1K38+OyuJGFtpkDZCzXG3DMWIO2JSpIyxZRt3H4u/8ABaXU/Dw+LFj4UvofDrxfaU02LQNK8wxYyMWjg3pGMHGN2K/YK/4KN2n7TmqS/Cn4qaRYeHPiJbRNJai13JbauiDMnlo5LRzIAS0ZY5ALDADBfjB8VPCfwR+GfiD4qeN7podH8O2bXE4QAvK2QqRID1d3ZUUerCvh98Z/+CoP7Yml3nxZ+Bdx4J+GngZbqRdIttTgjlbUliYho1klgmaQ5+VpNsKFlIUghgP2Jv25vHnxS+JOu/sy/tJ+Frbw78U/Dxm8uS2i8qK/EP8ArY2TLBZVX5wyEo6ZYbcDdXw+/aG+Mf7SP7buveEPhF4xbSvgp8LIhb+IZ4dOtpv7cvwzL5SzSxs6K0gZcxsuY4GYEF1IqqqqqqqqqqqqqqqqqqqqqSOOaNoZUV0dSrKwyCD1BHpX7a/7Knif9lv4t31pBp1x/wAIZrN1JceGdTUEoYid3kM3aWLO0g4JADDhq+Inx++Mfxb8N6D4S+JfxA1LxJp3hlpG0tdQ2SywGRUV8zFfNfIjQYdmxjjGTnRfjh8WfDfwv1L4MeH/ABzqGneDdZvZLzUdLtdsS3crpHG/mOo8xlKQxgoW2fL05Of2Nv2Z/EP7Tvxl0nwtb6dOfDGm3Ed14lvwhEdtaK2THuGMSS7SiAc5JbGEbEcccUawwoqIihVVRgKB0AHYVVVVVVVVVX/BLj/k8f4zf9eF/wD+nKOqqqqqqqqqqqqqqqqqqqqvH3/IpX3/AGz/APRi1VVVVVVVVVVVVVVVVVVVVXhD/kWdO/64CqqqqqqqqqqqqqqqqqqqqvFf7OOieK/jLpfxluPEl9Be6XJbOlkkKmNzCcjLHnmqqqqqqqqqqqqqqqqqqqqv+Cmv/Jj/AMSf+4V/6dbSv+CWf/Jl/hH/AK/9S/8ASyWqqqqqqqqqqqqqqqqqqqr9u/8A5M/+Kn/YAk/9DSv+CQf/ACahd/8AY3X3/om3qqqqqqqqqqqqqqqqqqqqr4rf6/Tf9yT+a1VVVVVVVVVVVVVVVVVVVVov/IYsP+vmP/0IVVVVVVVVVVVVVVVVVVVVVXwW/Zx0T4K+Ida8Q6X4kvtSk1qMJJHcQqgjw5bI2/XFVVVVVVVVVVVVVVVVVVVVf8Frf+Rn+FH/AF4ap/6Mt6qqqqqrP/j6i/3xVVVVVVVVVVVVVVVVVVVVVVXxm/aW+BP7PlpDc/GD4maT4ekuEL29pIzz3Uy5xuS3iDSsueNwXA7mv2x/2ivhd+1H+2Bp3iHX/GfiGf4Paf8AYrGCWwtDHc2tpsVrp4oZQAJGmMnzEElQnDbQtfAD9jf/AIJ8694D03xf8J/ht4U8b6NeJiPVtRd9UaZwRuDrOSI3BADJsQryCoyRXiD9iH9kLxLC8Go/s5eA4VcYJ0/Ro7BhxjgwBCOvavjl/wAEZfg54ptLjUvgP4q1LwTqoDNFp2oyvqGnv6JuY+fH7uXk/wB2v2Ovg/8AET4GfAPQfAHxV8dXninxLbtJLdTz3j3SWgY/JbQu/wAxijQKBnuWxhcAV4n0GDxT4a1bwxcX15ZRavYzWclzZyCOeBZUKF42IIVwGyCQQCBwa8ff8EXf2dbzw9eSeB/iH4z8ParFAzQXOo3NveWiMBndLH5SMV9SJFwOea/4Is/F3xvrul+PPg1rmqXGp6B4Zjtb3RmkcutiZXlWWJCeQjlVdV4AIc4+Y1VVVVVftITD9qj/AIKm2nw08UXjy+HbLxPZeHFtyzLssrYhrmNT1Bd/tBB7GQHkCrDT7HSrC20vTLOG1s7OFYbe3hjCJDGoCqiqOAoAAAHAAr/gs38ING8SfAnQ/jHb2KLrng/V47SW5VcF7G5yrIxHXEwhK54G58feNf8ABMv4uy+NP2KPD2q+KNQy3gc3ekXVzIcbYLX54s+yW7xL/wAAr9hvRZP2z/8AgoLq/wAXviNaLfWunyXXiiezuPnQFJEisoPdYjJDtB4IgAORmq/4KLeGLj9lb9u/RvjL8O7cWA1g2fim3SEbIxeJMyXKY7iRot7g8H7QR0OB/wAFivG0l7+yX4MbQpZBpvirxPZzSOeC8AtJ5UVh7tsb6pX7K3h7TfC37M/wr0LSYEit7fwfpjEKu3fI9tG8jkerOzMfdjX7RcA8Jf8ABYv4X6x4di2Xmt/2S98Yxy3mrLayE4/6YIAc9hzxX/BRP9pDU/gX8GY/CHgGWWX4jfEqc6J4at7Ynz4y+1ZrhAOdyiRUQjkSSxnkA1+xl+zdpn7LnwH0P4dJDA+uzr9u8Q3cYB+0X8ijzMMOqoAsa/7KA9Saqqqqqqqqqqqqqqqqqqqqp3SNC7uFVRkljgADvXjO9+AXxZ0GbwH421rwV4l0zU2WNtOur+3mEjnhSg3ZD5PylcMCeCDXj/8A4I5/AHxDezX3gPxt4q8JiUkrZu0eoW8PsgkCyY/3pGPvXgz/AII0fBTSb2K68b/E7xX4hiiOTbWsMOnxyc9GOJG244+VlPuK+Gfg34J/BTwvD4H+Gdl4d8O6VbMSbe3uEDyP0LyuzF5H4wWcluAM8UCrKGVgQRwRVVVVVVVVVf8ABLj/AJPH+M3/AF4X/wD6co6qqqqqqqqqqqqqqqqqqqq8ff8AIpX3/bP/ANGLVVVVVVVVVVVVVVVVVVVVVeEP+RZ07/rgKqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr/gpr/yY/8AEn/uFf8Ap1tK/wCCWf8AyZf4R/6/9S/9LJaqqqqqqqqqqqqqqqqqqqv27/8Akz/4qf8AYAk/9DSv+CQf/JqF3/2N19/6Jt6qqqqqqqqqqqqqqqqqqqq+K3+v03/ck/mtVVVVVVVVVVVVVVVVVVVVaL/yGLD/AK+Y/wD0IVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVf8Frf+Rn+FH/Xhqn/AKMt6qqqqqrP/j6i/wB8VVVVVVVVVVftL/ta+NPgl8RY/Bnh/wAN6Jf2z6bFdGW8Eu/c7OCPlcDHyDtX/DxP4of9CR4W/Kf/AOOV/wAPE/ih/wBCR4W/Kf8A+OV/w8T+KH/QkeFvyn/+OV/w8T+KH/QkeFvyn/8Ajlf8PE/ih/0JHhb8p/8A45X/AA8T+KH/AEJHhb8p/wD45X/DxP4of9CR4W/Kf/45X/DxP4of9CR4W/Kf/wCOV8BP2zvHnxZ+LOhfD7WvC2gWdnqv2jzJrUTeYnl28koxucjkxgHI6E1VVVVXj/4g+CvhZ4S1Dx38QvEtloWg6XGHur67fakYJAAAGSzEkAKASSQACTV//wAFf/2RVGq22l6h4nkuLO1mexmuNGeO3vZkQmOMFS0ib2AXcyADOTgV+xP+yxc/8FDPiB43/aB/aP8AF+rX+nWmpJFNBaTeVJf3TLv8oOc+TBFH5ahEwcMoBULzq3/BLP8AYg1TRV0i3+EM+nSRoyx3tpr18J0J6tl5mVz6b1YDsAK+DPwj8H/Aj4YeH/hN4Dgmj0bw9a+TC9wVaWdixaSWQqFBkd2Z2IAGWOABgCqqq/bX0L9t74t6NqPwp/Z48L6LoHhbUIHttW1y/wBZhS81KJ1w8UKDd5ERBKlj+8b0QZ3fDjxH+1T/AMEnfiLAnxA8B2E/hPxu8J1KKIxXEepRwFh+4uk+aOaMSvhHOPnyUIIavCfijRPG/hXR/Gfhu8W70nXrCC/sJwMebBMgdG/FWBqqqqqv+CgPgjxv+yj+3Rp/7SGgaU82ja3rFr4h0ufaVikuotn2q0dwDhmZGY8fcnHUg18L/wBv79kr4oeErbxRD8bPC3hyZ4Va60vxFqkWm3VrJjLRlJmXzCDxuj3KexNf8FM/27/hr8YPhle/Ab4GTnxZaNcQXviTxBBC/wBls4Ypk8uOJiBvLTGIGT7g4UFi/wAv7AcF94X/AOCW3xu8RQt+9uI/E9/bblIAMekRRjockboTzx+ma/4IeWlu/jT4r3rR5ni0vTY0bJ4VpZywx05KL+X1qv8AguJbwL4y+E90sYE0mmamjP3KrLAQPwLN+dfHD4DeJf2n/wDgmV8O9A8NWou/Fel+EPD+vaZaAbftNzFp6LJEvozRTTKo6bioOAcj9i//AIKT/Arwd8BdE+Fv7QHiO68G+LPh7YjSJILnS7mUXcFsPLi2CJHKyKipG0b4bchIGDx+zVHqn7X37dHiX9uXXtLl0D4WeAY5G0271NBDG4htmhiBY5XKLvuZCGIjbaM4Ir4D+INM/aU/aC+IX/BRP4uO9l8LfhPbT2XgmG6iwAkCMzXAQ9XVGaTHJ864UKcxAV4C+NH/AAUQ/bzu9d8bfs++KfDfwj+HWmXjWmnvf26Sy3sigHaZTBM7uAy7ygjjG4KNxBr9mD9tP48eF/2ipv2Pv2z7KwbxZcvs0XxFZQRwLdyFS6K4jCxvHKg/duiIQ3yMuSdlVVVVVVVVVVVVVVVVVVV4l8S+H/B2g3vifxVrNppWk6bEZru9u5RHFAg6szHgD3NeJP2xf2Vbnw7qltb/ALQfgSSWWymREXWoSWYoQABmvgrqenaJ8ZPAWs6vew2djYeJtOuLq4mfYkMSXMbO7E9FCgkn0Ff8Nl/sof8ARw/gL/wdw/8AxVa9+3L+yN4e0qfV7z4/eErmK3QsYrC9F5M+B0WKLc7HjsK+IfiKx8YeP/E3i3TLFrKz1vWLu9t7ZgAYI5ZmdUOOOAwHHHFfs+azdeI/gH8NPEF80jXOp+D9KupjIcsXktImbJ7nJOaqqqqqqqqr/glx/wAnj/Gb/rwv/wD05R1VVVVVVVVVVVVVVVVVVVV4+/5FK+/7Z/8AoxaqNBJIsefvECpomhlaFuqEiqqqqqqqqqqqqqqqqqqq8If8izp3/XAVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX/BTX/kx/wCJP/cK/wDTraV/wSz/AOTL/CP/AF/6l/6WS1VVVVVVVVVVVVVVVVVVVft3/wDJn/xU/wCwBJ/6Glf8Eg/+TULv/sbr7/0Tb1VVVVVVVVVVVVVVVVVVVV8Vv9fpv+5J/Naqqqqqqqqqqqqqqqqqqqq0X/kMWH/XzH/6EKqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv+C1v/Iz/AAo/68NU/wDRlvVVVVVVZ/8AH1F/viqqqqqqqqqr43/sjeG/jh4zTxnqvi/UtMnSyjtBBbwI64RmIbLc5O/9K/4dzeCf+ij63/4CRV/w7m8E/wDRR9b/APASKv8Ah3N4J/6KPrf/AICRV/w7m8E/9FH1v/wEir/h3N4J/wCij63/AOAkVf8ADubwT/0UfW//AAEir/h3N4J/6KPrf/gJFX/DubwT/wBFH1v/AMBIq+Ev7Fnhf4SfEHSfiDpvjXVL+40rztltPbxqj+ZC8RyRzwJCfwqqqq/bk/4KI+Kv2WPH2n/Cnwb8GJdd1vVbCK8tNS1G5ItZ1dmTbFFFl5GDoVILIQR0IIJX/gpR/wAFE/HE8ngTwl+zxp9v4pRo5ZBpvhDUZZ4YWGVJhllkChhzvYYx09a/a0+Nf7b/AMXLHwr8Kf2mPCOo+HohqYl06O48Py6at/cOoiVmIGyXbl8bBwZH9gL7/glR+yDqHwysvAL+DLy21WztVj/4Si0vZI7+SfaA0zZLRtuOTsZCgzgAV+wB+zB8U/2QPGXxK+F3imSLW/B2tNaan4c8QWo2pK6b45o5oySYpirQHbypCMVZsHarpIu6N1YAkZBzyDg/qMVVVVV4v+L3wl+H10bDx78UfCPhu5EInMOr63b2biMkgPtldTtJUjPTINf8FVf2ufhJ8fPDvhT4CfBPVV8Z6ha+Ik1C7v8ATImliEohkgitoGA/fs5uCTsyMqoBJJA/Zs8C638Mv2ffhz8PvEqxpq/h/wAM2Nnfohysc6QqJFB7gNkZ74zVVVVVfEr4XfD34xeEbrwJ8TvCen+ItCvCDJZ3kZIVhnDowIaNxk4dCGGTgitQ/wCCN37Il7rX9q2194/sbbcT/ZtvrMRgx6ZkgaXH/bTPHWv+Ck9p8DPgJ4T8Ofscfs5+EbOyvrvUItY8UvbM1zeTuqMlpBNKxZ5GYyvJ5ZOF/dlQA9fs/fs53Pw3/Yr0r9nzUNkOq3vha7ttTJbIjvL5JHmG4dQjzMgPogr/AIIyeKz4S/aK8bfDPWg1pd634fYpDIcE3NnOuY8f3gkkx+iGq/4LOeKR4s/aL8EfDXRGa8vNF8PLvgjOStzd3DbY8f3ikcJ+jrXw+8MDwT4B8NeDBKJf7A0e008OP4vJhWPP47a8Z/s7fAL4ja2viXx78FvBHiHVgQWvtS0G3uJpMDADu6EuAOzZFf8ABQ74gavqcHgv9gf4A2tvZeJviZJHFqEFhEIotL0dWO4FUA2I+x2YL0ihlBGHGf8AgoB4F0j9nj/gm3N8IPh5E8Gj2Mml6XLIBh5l+0rLLK5H8UsqbmxgEyEdDiv+Cc2i6Zof7Ffwtt9LjQJc6ZLdSsuMtLLcSvISR1O5iPYADtX/AAU/hOhftv8A7P3i7QUP9uE6eFCDLMYNT3Q8L8xO6Rx156Cqqqqqqqqqqqqqqqqqqqr4neC7T4kfDbxV8Pb0gQeJdFu9NdiPuiaFo93sRuznsRV9ZXWnXk+nX0DQ3NrK0U0TjBR1OGU+4IIqqr9m/wDZj+Jv7TPjuw8K+CdCvBpZuEGra41ufsumw5G93c4UvtyVjzuYjA7kaBoem+GNB03w1o0Hk6fpNnFaWkWc7IokCIufZVAqqqqqqqqqv+CXH/J4/wAZv+vC/wD/AE5R1VVVVVVVVVVVVVVVVVVVV4+/5FK+/wC2f/oxaqe7XT7eXUGxi0iaY5/2FLf0rSNct/FvhvR/F1ooWPVbNJWQfwPj5l/A5H4VVVVVVVVVVVVVVVVVVVXhD/kWdO/64Cqqqqqqqqqqqqqqqqqqqqvi/fXsf7cPhK1ivZ1ha50zMayEKctzx0qqqqqqqqqqqqqqqqqqqqr/AIKa/wDJj/xJ/wC4V/6dbSv+CWf/ACZf4R/6/wDUv/SyWqqqqqqqqqqqqqqqqqqqr9u//kz/AOKn/YAk/wDQ0r/gkH/yahd/9jdff+ibeqqqqqqqqqqqqqqqqqqqqvit/r9N/wByT+a1VVVVVVVVVVVVVVVVVVVVov8AyGLD/r5j/wDQhVVVVVVVVVVVVVVVVVVVVVfsLX17d/Enx1HdXs8ypartWSQsB+/PTNVVVVVVVVVVVVVVVVVVVVf8Frf+Rn+FH/Xhqn/oy3qqqqqqs/8Aj6i/3xVVVVVVVVVVVVVVVVVVVV/wUE+C37Y2k/Hzwx+09+znfa54mtdBt1SHRrP/AEiTSpQAsoS1J/ewzqqlggJ3bsgcNWofH3/gsbNon/CwJfhjrtrpdjqUd01mnhOJJtnlFTAbUj7S8BBLMdpKtg71wBXj7/gp1+1v4r8I3ui+Iv2YPCJgtFE19NqPhe/uYbVo8OJfLlk2xshAcMxO3APvX/BPb9sjT/jJ8GL7V/jd8a/Dk3jmHWLl7yzvZ7XTntbXCCLZEAgMWATvAb5iQWzX7WP/AAUSXV/E+lfs0fsc65Z+IfHXi2/h0yTxNZuJ7XS2lkCbYGwVlk5yZRlI15yWzs+HngrTvhz4F0LwLpdxPc2+iWMdsLm4YtLcuq/PNIxyWkkbc7McksxJOTVVVVX/AAUx/Zg8N/Hn9n3W/GsNpFB4x+HmnXGq6XfBQGlt4lMlxbOf4kZEYqOzhSOCwP8AwRo+EXwW8T+D9e+LGp+C4r34heGdda0t9Su3aVbW3eBGR4Yz8iSEmVS+CwA4IDEVVVVVVfta/tfeCP2QPDmg+JPG/hnXNag8QXslpBHpQiLRsibyW8x1GMelfEr/AIK//Fj4qMfAH7LXwVvbLWtTUx299cA6pfcjkwWkSbQ44IZjIPVe9fsH/wDBOXxpofjyH9pr9q1ze+LXuG1DS9EvZftU8N2zBhe3cm4gzA5ZY/m2sQzEOu1a/bx/Z4+Kn7Jn7ScP7anwLsHm0G61b+1bt4bdpF0m+fIuEuFHP2e43P8APkDMrp8vyFh/wW3+EJ8EC7b4PeLx4w+zc6eJrc6f52On2nf5mzPfyc+1fsMfs7fFr9rb9pOT9tT4+aZJBoFvqY1Wy+0W7RJqt4gAtkt0bn7NbhUIckg+UiZY7ytfFb4meFfg18OPEPxR8a3ht9G8N2L3Vyy43PjhI0BIBd3Koo7swHev+Ccvwz8UfErxF4w/bx+MNkD4p+JdzJF4dgk+YadpasEzHnkBhGkSEgHy4QckSmv2sfgmf2iP2efGvwjt5Yor/WbAPpssvCpeQus0GT/CpkjVWI/hZuvSv2Gv27fBX7L3gC7/AGXv2sINa8Fa14EvbiOxnuNNmuQ0UkjSmB1hV3DB3co+CjIy4YADPgAa3/wUZ/4KAaP8edH8OanafCP4Vm3Wyvr6Dy1untXaaGMDlTJJcyeYy9VhUBsHaDVVVVVVVVVVVVVVVVVVVV4g+APwZ1my1I/8Kg8DPf30Uv8ApM3h+2LGVwfnZvLJzuOSevevh5/wSF/aE8NeP/DPiLxJ4r+GF/pGlaxaXV/arfXkpngjmV5ECPaBW3KpG1iAc4JAr/hQPwI/6In4C/8ACbtP/jdQ/Af4HW0yXFt8GfAsUsZyjp4dtVZSO4IjyKsbCx0y0isNNs4LS2hXbFBBGI0QegUcAVVVVVVVVVVf8EuP+Tx/jN/14X//AKco6qqqqqqqqqqqqqqqqqqqq8ff8ilff9s//Ri1XxDvRp3w/wDEt4DgppFyqn0Zo2RT+bCv2U/E41nwRqvgueXNxo1x9ptlP/PKTOQPo4Yn/fFVVVVVVVVVVVVVVVVVVV4Q/wCRZ07/AK4CqqqqqqqqqqqqqqqqqqqqvjF/yfR4R/6+tL/9Cqqqqqqqqqqqqqqqqqqqqr/gpr/yY/8AEn/uFf8Ap1tK/wCCWf8AyZf4R/6/9S/9LJaqqqqqqqqqqqqqqqqqqqv27/8Akz/4qf8AYAk/9DSv+CQf/JqF3/2N19/6Jt6qqqqqqqqqqqqqqqqqqqq+K3+v03/ck/mtVVVVVVVVVVVVVVVVVVVVaL/yGLD/AK+Y/wD0IVVVVVVVVVVVVVVVVVVVVVX7B/8AyUvx5/16r/6PNVVVVVVVVVVVVVVVVVVVVf8ABa3/AJGf4Uf9eGqf+jLeqqqqqqz/AOPqL/fFVVVVVVVVVVVVVVVVVVVVVq+k6Zr2lXmha1YQX2n6jbyW13azoHjnidSroyngqykgg9Qa+I3/AARX+CviXxDc6v8AD34meIfCFjcyF/7LltE1GK3z/DEzMjhR2Ds596+Dmi/D39gv/gouND+M97djw74UluYrHWbizZ9q3Fqfs120cYYlSsmDsB2sxP8ACcftYf8ABVP4L/Dz4e3dh+z74xsPGXjrUU8qxeC3ke000HrcSu6hXIGdsalstjcAvX9mxNb/AOGfPhvceJtSuNQ1q88L6fd6ndXDbpJrqaBJZnY8ZJkdjQngadrVZozNGiu8YYblViQpI6gEqwB74PpVSa1o0Wrw+HpdWs01S4t3uIbJplE0kSEK8ipncVUsoJAwCy56iq/an1j4veGfgR4r8XfA2/sYPFvh6zbUbeG8sxcpdwwjfNCFP8ZjDlcdXCjoSa0f46f8FIf+Cguiav8ADXwTfR3fh1hHba+umQWulwLHLuws87ESbGVXzGrHeARtbpX7EP7K9r+yR8FIPAFzqkOqeIdTvH1HXb2AMInuXVUCRBufLRERQSAWIZsLu2iqqqqq8cfDH4a/E60trD4k/D3wz4strKQyW0OuaTBfpA5GCyLKrBSRxkV4Y8D+CvBFqbHwX4Q0TQLY9YdL0+K1Q/8AAY1AqqlijmjaGWNXR1KsjDIYHqCPSk/Zw/Z4i1keIovgL8Ok1YP5gv18LWYnDj+LzPL3Z4HOaREjRY41VVUYVQMAAegqvGHgfwV8QtFbw34+8IaJ4l0l5Fkaw1jT4ryAuv3WMcqsuR2OOK0zS9M0TTLTRdF061sNPsIEt7S0tYViit4kUKkaIoAVVUABQAAAAKrxn8JPhT8R57e5+IXwy8J+KJrQYt5NZ0W3vWiwc/KZUYrz6VpOj6RoGm2+jaDpdnp1haJst7S0gWGKJfRUUAKPYCqqqqqqqqqqqqqqqqqqqqq/bT/4KiXHwk8Y6n8IvgRpGm6nrmjym31fXNRBlt7WcffhhiUje6HhmY7VYFdrYyNS/wCCkP7aep3TXUnxvu4N2dsdtpNjEiDPQBYfwycn1Jr/AIeJftof9F31T/wAs/8A4zV9/wAFBP2ydQg+zz/HrXFXOcwW9tC3/fSRg/rXhyaW58O6XcXErSSy2ULO7HJZigJJqqqqqqqqqv8Aglx/yeP8Zv8Arwv/AP05R1VVVVVVVVVVVVVVVVVVVV4+/wCRSvv+2f8A6MWq+PV6LL4S6/hgHnWCFM+pnjz/AOOhq/Z68X/8If8AFLSpppglpqZNhc5OBtkxtJ9MSBD9Aav7cW108QHGcr9DVVVVVVVVVVVVVVVVVVXhD/kWdO/64CqqqqqqqqqqqqqqqqqqqqvjF/yfR4R/6+tL/wDQqqqqqqqqqqqqqqqqqqqqq/4Ka/8AJj/xJ/7hX/p1tK/4JZ/8mX+Ef+v/AFL/ANLJaqqqqqqqqqqqqqqqqqqqv27/APkz/wCKn/YAk/8AQ0r/AIJB/wDJqF3/ANjdff8Aom3qqqqqqqqqqqqqqqqqqqqr4rf6/Tf9yT+a1VVVVVVVVVVVVVVVVVVVVov/ACGLD/r5j/8AQhVVVVVVVVVVVVVVVVVVVVVfsH/8lL8ef9eq/wDo81VVVVVVVVVVVVVVVVVVVV/wWt/5Gf4Uf9eGqf8Aoy3qqqqqqs/+PqL/AHxVVVVVVVVVVVVVVVVVVVVVVX7Q37H/AMBP2obe1b4r+DvtOp2EZis9XsZ2tbyBMk7BIv30ySQjhlBJIAJzX7Tn/BIb4faF8GNT1j9m228R6l420y4S7S01PUFnN9bKrCW3iUKiiTlXXOSTHtH3q/Z+/wCCt+ufBP4e6d8Hvjb8HtT1PVfBtsmmQXltdC0uDHCPLSK4glTKyIqhS2cnHKg5J0j/AIKT/GTS/wBqPUv2r5/Bd1J4O1uFNAm0Brh/sy2cQ3xwJc7Nv2hGZ5t23rLJ8oV+PAX/AAVX/Yy8aaUl5q3xAv8AwlebAZLDW9JnEiHvh4Fkjb8Gz7dcf8FJv2v/AIa/Gzx18KZ/2W/G+tah4s8H3V6Y9Y0m2uLNlkuTbiOKFmVJJH3RH7q7fmwCdxA0v/gpP+3n8AbdfD3x/wDgYdW+xrtN9rOiXOl3EoXqxmjAhcYB+ZY+euTXjT/gp/8AtZftH+Hr/wCHHwO+BCWEmvQSWU9xpNrc6vdoki7XETBVSM4LZYodoORtI3V/wTL/AGSPFX7Lnwl1e/8AiNDDbeL/ABvdQ3N9Yxsr/YLeFWEELuuQ0gMkrNgkDeAOhJqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr4t+LZfAHwp8Z+O7dlEvhvw9f6khZcgNBbvIMjByPk9D9K+Efwy8X/tA/FvQvhr4cuFl13xVfshubpiQnDSTTyHkkKiyO2MkhT3r9rj9nM/stfFs/Cs+J5PEGzSra9+3vY/ZBIZQ24Km9/lDKRncehrwX/wSH8GfET4P+G/Hvh341ara6p4k8P2ep26XOmRy2yvPAkoU7WVtnzYz1A5weleB/h/4B+F37SJ+Ff7VWiX/APY2mao2l6y+m6h5H2R94C3IfaTJCBhiBtJRtwOQAbG3trSxt7SyI+zwxKkWG3fKAAOe/AFVVVVVVVVV/wAEuP8Ak8f4zf8AXhf/APpyjqqqqqqqqqqqqqqqqqqqqrx9/wAilff9s/8A0YtV+0/eG3+G0Fsjc3WrQoR6qI5WP6haR3jdZInKuhyrKcEEdxXhLxInjnwDoXjBGUy3dqoucdpV+WQfQOrVVVVVVVVVVVVVVVVVVV4Q/wCRZ07/AK4CqqqqqqqqqqqqqqqqqqqqvjF/yfR4R/6+tL/9Cqqqqqqqqqqqqqqqqqqqqr/gpr/yY/8AEn/uFf8Ap1tK/wCCWf8AyZf4R/6/9S/9LJaqqqqqqqqqqqqqqqqqqqv27/8Akz/4qf8AYAk/9DSv+CQf/JqF3/2N19/6Jt6qqqqqqqqqqqqqqqqqqqq+K3+v03/ck/mtVVVVVVVVVVVVVVVVVVVVaL/yGLD/AK+Y/wD0IVVVVVVVVVVVVVVVVVVVVVX7B/8AyUvx5/16r/6PNVVVVVVVVVVVVVVVVVVVVf8ABa3/AJGf4Uf9eGqf+jLeqqqqqqz/AOPqL/fFVVVVVVVVVVVVVVVVVVVVVVVXif4W/DHxveJqPjP4c+F9fu4k2JPqmjwXUir6BpFJA9q1P4d+ANa8Jv4C1bwRoN54akj8t9Hn06J7Qr6eSV2Yzz0614t/4JWfsV+KtRfU4fhtfaHJId0kek6xcQxMfaNmZUHsgUcdOtfBT9gD9lb4Ca/B4t8EfDdLnxBaPvtdU1e6kvpbZuzRBzsjYdnVQ3vVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVfGvw1N4z+DXjzwfbQtLNrnhjUbCONc5dpraSMAY55LYr/gmz4o0/wr+2b8PrjVJkig1GS708O5xiWe0lSID3aQxr/wACr/gs58P7vTvit4G+J0Vu32PW9Ck0uRwOBPazNJz6EpcqBnqEOOhr9kX/AIKmeEvgz8FtN+FXxc8G+I9WuvDMLW+j3+jiGQT2+SY4pllkTZszsDLuyoXjIOfj/wDFq7+O/wAZPFfxbvNKj01/Ed8Z47NH3+RGqrHGhbjcwRFycDJycDOK/Z3n1W6/Z++GNzrpkOpy+DdJe88w5bzjZxF8++7Oaqqqqqqqqr/glx/yeP8AGb/rwv8A/wBOUdVVVVVVVVVVVVVVVVVVVVePv+RSvv8Atn/6MWq/axvQmmeGdOVs+bPcysPTasYH/obVtJGQOBxnFfsh+KVv9H1zwBdS5eBhe2oJ6K2FfHsGCH6uaZSpKsOQcEVVVVVVVVVVVVVVVVVVeEP+RZ07/rgKqqqqqqqqqqqqqqqqqqqq+JXwW+JfiD9q7w58S9I8NfaPDdhPYNcXv2yBNgiPzny2cOcey89s1VVVVVVVVVVVVVVVVVVVV/wU1/5Mf+JP/cK/9OtpX/BLP/ky/wAI/wDX/qX/AKWS1VVVVVVVVVVVVVVVVVVVft3/APJn/wAVP+wBJ/6Glf8ABIP/AJNQu/8Asbr7/wBE29VVVVVVVVVVVVVVVVVVVVfFb/X6b/uSfzWqqqqqqqqqqqqqqqqqqqqtF/5DFh/18x/+hCqqqqqqqqqqqqqqqqqqqqq/ZL+C3xL+GPjjxbrHjjw1/ZtpqcAS0k+2QTeYfNLdI3YjjnkCqqqqqqqqqqqqqqqqqqqqv+C1v/Iz/Cj/AK8NU/8ARlvVVVVVVZ/8fUX++KqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqrD/AIJGaEnxl1T4kn4yaloelLr76poen6FYJHc2A83zYgLiQsqmNsAfu2+6DX7UP7OXhb9qH4RX/wAMfEt01lceYt1pWppGHewvEBCS7eNykMyMvGVdsEHBHxO/YF/ax+F+tzaRd/BzXvEcCSFYNQ8NWcmqQXCjo6+SpdAfSRUPtX7KX/BMP4x/EbxjpviD43+F7nwd4JsLhJby21D93e6kqkEwRwg741bhWd9uATt3EcQQQW0EdtbQpFFEgSONFCqigYAAHQAcYqqqqqqqqqv+CXH/ACeP8Zv+vC//APTlHVVVVVVVVVVVVVVVVVVVVXj7/kUr7/tn/wCjFqv2rb3zPFWiaaGyINNM2PQvKy/yjFeFPBf9v/B/xVrcMIa5029gmjIHOyJG8wfTZKW/4CK+Cni4eCfiXomryziO1kn+zXZJwPKk+Uk+ykhv+A1rFv5N2XUfLIMj696qqqqqqqqqqqqqqqqqvCH/ACLOnf8AXAVVVVVVVVVVVVVVVVVVVVVVUfiUkPx7PwrlmXyrjwyl/DntOs7hlHuY8N9Fqqqqqqqqqqqqqqqqqqr/AIKa/wDJj/xJ/wC4V/6dbSv+CWf/ACZf4R/6/wDUv/SyWqqqqqqqqqqqqqqqqqqqr9u//kz/AOKn/YAk/wDQ0r/gkH/yahd/9jdff+ibeqqqqqqqqqqqqqqqqqqqqvit/r9N/wByT+a1VVVVVVVVVVVVVVVVVVVVov8AyGLD/r5j/wDQhVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV/wWt/5Gf4Uf8AXhqn/oy3qqqqqqs/+PqL/fFVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV8ZE8WyfCDxzH4Anmh8UN4b1AaJJAcSJe/ZpPIZevzCTYRx1pv27f2wVJVvj94pBHBBlQY/8dr4Ua/H4s+Fvg/xPHqY1EatoNldfa927zzJAjF8++c/jVVVVVVVVVVVf8EuP+Tx/jN/14X/AP6co6qqqqqqqqqqqqqqqqqqqq8ff8ilff8AbP8A9GLVftJ3v2r4pXVruz9hs7eLHplBJ/7Ur9nDSYH+Fc0N1EHh1K9nMit0ZSqxkfkmK8W6BP4W8Tan4duAd1hcvGpYY3KD8rfiuD+NfDTxOfHvws0XxBJIJLyKDyro5582P5HJ+uA30Iqqqqqqqqqqqqqqqqqq8If8izp3/XAVVVVVVVVVVVVVVVVVVVVVVV8UviWfDP7Zx8XC4CWmiaha2kzZ+7B5KRz9fZ5f881VVVVVVVVVVVVVVVVVV/wU1/5Mf+JP/cK/9OtpX/BLP/ky/wAI/wDX/qX/AKWS1VVVVVVVVVVVVVVVVVVVft3/APJn/wAVP+wBJ/6Glf8ABIP/AJNQu/8Asbr7/wBE29VVVVVVVVVVVVVVVVVVVVfFb/X6b/uSfzWqqqqqqqqqqqqqqqqqqqqtF/5DFh/18x/+hCqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr/gtb/wAjP8KP+vDVP/RlvVVVVVVZ/wDH1F/viqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq1C9i02wudRnVmjtYXlcIMkhQSce+BX7WPxA/Zb+LXjDUfiH8DvDHjXwpquq3TT3+majaW32K4kY5eZPLmZoGJJYoA6knjZX7Of8AwUY+P/7OPhOHwBoo0TxL4atN32Kx1u3kdrLcSxEUkbowUsSdrbgMnAGa/wCHznx3/wCiW+Av++bv/wCPV/w+c+O//RLfAX/fN3/8erRr2TUtHsdRlRVe6to5WVegLKCQPbmqqqqqqqqq/wCCXH/J4/xm/wCvC/8A/TlHVVVVVVVVVVVVVVVVVVVVXj7/AJFK+/7Z/wDoxar4zXov/ij4kmDA+XemH8YwI/8A2WvgbZCx+FWgRbQDJFJKTjrvldh+hFftR+FxY+I9P8V28f7vU4DDOQP+WseMEn3Qgf8AADX7HnixFuNb8B3UnyzoL61UnjIwko+pBjP/AAE1cQmCZ4W6ocVVVVVVVVVVVVVVVVVXhD/kWdO/64CqqqqqqqqqqqqqqqqqqqqvHv7RHjXwt+0joXwd07S9Ek0bU5rNJp5oJTcKJjhtrCQKMdsqfxqq+Jes/wDCR/ETxPrwfcL/AFi6mTjGFaVio/LAr9n/AMZDx58HfC+vyS77kWS210SckzQ/u3J+pTd/wIVVVVVVVVVVVVVVVVVVf8FNf+TH/iT/ANwr/wBOtpX/AASz/wCTL/CP/X/qX/pZLVVVVVVVVVVVVVVVVVVVV+3f/wAmf/FT/sASf+hpX/BIP/k1C7/7G6+/9E29VVVVVVVVVVVVVVVVVVVVfFb/AF+m/wC5J/Naqqqqqqqqqqqqqqqqqqqq0X/kMWH/AF8x/wDoQqqqqqqqqqqqqqqqqqqqqqv2af2iPGvxm8X+JfD/AIo0vRLW30aEPbtYQSo7HzCvzF5HB4HYCqqqqqqqqqqqqqqqqqqqqv8Agtb/AMjP8KP+vDVP/RlvVVVVVVZ/8fUX++Kqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq1bVtK0HTLrWtc1O007T7GJpbm7u5lhigjUZZ3diAqgckkgCvG//AAVl/Y28G6tLo9l4r13xO0DlJLjRNJaSEMOu15WjDj/aTcp7EivgH+2d+zn+0rO+l/C34gQT61Chd9Gv4ms7zaBksscgHmKB1aMsF4yRmq+InxK8A/CTwpdeN/iV4t03w7odnxLeX0wRdxzhFHV3ODhFBY9gauv+Cvv7G8GsHTIdU8X3NsH2jUYtCYQEf3sM4lx/wDPPSvhR8Y/hh8cvCkfjb4UeM9P8R6Q7+W01q5DQyAAmOWNgHifBB2uAcEHGCDTFVUszAADknjFfEn/gqT+x98NvE03hObxtqPiK7tJ/IuptB083VvAwJDfviVSQDHJjL9eM84+CX7QPwh/aL8LP4u+EHjK212yt5BFdRhGhntJCMhZYnAdCQDgkYbBIJFV4l/ao+GXh39oHQ/2ZYrfW9X8b63ai6aHTLRZodPhIZt9zIXXywEQuRgnaVOPmUGqr4tfFDwl8Fvhvr/xR8c332XRfDtm1zcMMb5DwEiQHq7uVRR3ZgK/YT/aP+NX7VPg3Wfip8QPBHh3wz4Te9a08Ox2SztcXpRj5sjPI5UxqcRgqvzOsn3dmD8c/+Ckfgf4F/tCj9nrVvhtrmqaiZ7CEajb3cSRZuljZTtb5vl8wZ+lVX7SX/BSPwP8As2/GtPgprfw21zWb54LWUXtrdxRxgT9BtbnjvU00NvC888qRxxKWd3baFAGSST0GK/Z8/wCClfwM/aN+NE3wW8JaP4hsL2dbhtGv9QijWHUxApeTaquXQlEd1DDlVOdp+Wqqq/a2+IXin4Ufs2/ED4jeCL6Oz13QNIe5sZ5IFmWOQMoyUcFW4J4Ir/glz+1L8Z/2ntA+IV/8YvEVrqs3h+8sIrAwafDa+WsqTF8iJV3ZKL16Yqq/bX+M3xo/Z9+D8vxa+EPhPw/4kh0OcNr1nqcU7PFathRPF5TrwjEbwQflbdkBDn9m749eE/2lvg9oPxa8JERR6lEY76xaQM+n3icTQOfVW5BIG5GRsAMKqqqqqqrX7Sa/0LUbG2UNNcWkscak4yzIQOfqatf+CUH7YdxbrNL4b8N2ztnMUuuxFl577dw9+D3r/h01+1//ANAXwt/4PE/wr/h01+1//wBAXwt/4PE/wr/h01+1/wD9AXwt/wCDxP8ACtCs5tP0PTrC4AEttaRROAcjcqAHH4iqqqqqqqqq/wCCXH/J4/xm/wCvC/8A/TlHVVVVVVVVVVVVVVVVVVVVXj7/AJFK+/7Z/wDoxaALEKOp6V4tvhqnirWdSD7vteoTyg+u6Rjn9a8B2I03wP4fsMYMGmW6tn+95a5/XNfHHwv/AMJT8ONThhiD3Onr9st+MnMfLY9yhcfUivhd4tbwN4+0XxKZSkNtcqtzjnML/LJx3+ViR7gVrcKlo7uMhlkGMjofQ/lVVVVVVVVVVVVVVVVVeEP+RZ07/rgKqqqqqqqqqqqqqqqqqqqq+MX/ACfR4R/6+tL/APQqrxnrP/COeDtd8Q7sf2XplxdZ9PLjZv6VX7AfjLz9I8S+ALifLWsyajaoTztcCOXHsCkfHq59aqqqqqqqqqqqqqqqqqv+Cmv/ACY/8Sf+4V/6dbSv+CWf/Jl/hH/r/wBS/wDSyWqqqqqqqqqqqqqqqqqqqr9u/wD5M/8Aip/2AJP/AENK/wCCQf8Ayahd/wDY3X3/AKJt6qqqqqqqqqqqqqqqqqqqq+K3+v03/ck/mtVVVVVVVVVVVVVVVVVVVVaL/wAhiw/6+Y//AEIVVVVVVVVVVVVVVVVVVVVVX7B//JS/Hn/Xqv8A6PNVVVVVVVVVVVVVVVVVVVVf8Frf+Rn+FH/Xhqn/AKMt6qqqqqrP/j6i/wB8VVVVVVVVVVVVVVVVVVVVVVVVVVVDPDcIXgkDhXZCR2ZSQR+YIqqqqqqqq/4KhfHDx38cP2idB/Yt+Gl+6afbX9jaXsEchRNQ1W6KGNZWHWKJJY+OgcuTkquPg1/wTZ/ZT+Fngiy8Oa58MdG8aaz5K/2lrWu2v2mS6mx8zIjErCmeAiAYGMljlj/wUc/ZB079kHxT4U/aK/ZzuL7wzpF3qoga3trhidG1JVMkLwOxLeXIscp2kkKUIztcKv7KXxuh/aL/AGfvBvxbKRRXur2OzUoYxhYryFjFOAOoUyIzKD/Cy9etfHXxV4z/AOCkn7ddp8D/AA34hntPAehalPY2DxAvDb2ltn7XqBXOGkl2MEJxwYUOOSbb/gnN+xpb+Bl8Bn4I6PNALfym1KVpDqDNjmQ3QbzA5PPBCjoFA4rwPqnin/gmV+3xN4Ck125uPAuqXttb37TthL7SLk5huHUcGW3Lt8wA+aOQDCuc/wDBVH4y6z8IP2TtTt/DWotZ6p411KHQUnjfbJHDKkkk5XByMxQtGT283sSDX7Af7Lfwx+GX7MPhC8v/AARo+oeIPGuiwaprl7fWCTTT/akEqwMXBIjjjZE2fdypOMsSfC2h2n7Gf/BWSy8BeAV/s3wV8RVii/sqNmWJIbyJtiKM4xHeR5T0T5RjJr47/GPwv8APhL4k+Lfi6TNj4fs2lSANte7nJCwwJ/tSSMiA9BuyeATX/BNn4O+Kb/TvE37ZHxij87x/8YLhrm2MiYNnphYNGqAklVk2oVXtFHCB3qqr/grv8RPEPjnxn8Lv2PfBE5N94pvoL++hVsCaWaY2tlGxHbf57EH/AKZnsK+GPw98O/Cb4eeHfhp4Ttlh0nw3p0NjbKFwWCKAXb1ZjlmPdmJ71/wUH/5SYJ/2EPDv/oq3qq/4Km/8n6Qf9g/R/wCdOiSIY5FVlYEMrDII78V8A/2O/wBjT4QfGK7+JXwgu7O68UyLOLKyGvpdppiSArKLeIHco2krlyxCkgEZNV4t8b+CvAOmf21468YaJ4c08Ej7Xq2oRWcWQMn55GUdOeteC/iz8K/iUZV+HXxM8KeKjAMyjRdZt77yx0yfKdsfjT+NfBqa0PDT+LdFXV94Qaeb+IXG4jIXy927OMHGK/b8/wCTNPiz/wBi/J/6Glf8EOv+RU+Lf/YQ0r/0Xc1qfx7+BWi6/wD8InrPxq8B2Gth9n9mXXiO0iud3p5TSBs+2KR0kRZI3VlYZVgcgj1Fa5omk+JdE1Dw5r1hFfaZqtrLaXtrMuUnhkUo6MO4ZSQfrX/BOTXtX/Zo/bO+J/7GOv38j6XqF1dSaOspwXuLUGSKRV7GayJdv+uSdcZFVVVVVVVVVVVVVVVVVVV/wS4/5PH+M3/Xhf8A/pyjqqqqqqqqqqqqqqqqqqqqrx9/yKV9/wBs/wD0Ytane/2bpl3qIbH2WB5cnttUn+lW8El1cRW0Qy8rhFB9ScCoIY7eCO3iHyRIFUH0AwKdEkQxyKGVgQVYZBFfEDw03hDxlq3h7YVjtblvI3d4m+ZD/wB8kV8EPE//AAnXwi0ueWTfeafGbKfud8IAUn3KbCfdqqqqqqqqqqqqqqqqqrwh/wAizp3/AFwFVVVVVVVVVVVVVVVVVVVVfGL/AJPo8I/9fWl/+hVX7UusjQ/gJ4uuQ2HuLVLVRjr50qRn/wAdYn8KihlnYxwRPIwVmKqpJAUEseOwAJPoBX7KvjP/AIQv43+H5pZtlrq7tptxk4yJsBM+3miM/hVVVVVVVVVVVVVVVVVV/wAFNf8Akx/4k/8AcK/9OtpX/BLP/ky/wj/1/wCpf+lktVVVVVVVVVVVVVVVVVVVX7d//Jn/AMVP+wBJ/wChpX/BIP8A5NQu/wDsbr7/ANE29VVVVVVVVVVVVVVVVVVVVfFb/X6b/uSfzWqqqqqqqqqqqqqqqqqqqqtF/wCQxYf9fMf/AKEKqqqqqqqqqqqqqqqqqqqqr9g//kpfjz/r1X/0eaqqqqqqqqqqqqqqqqqqqq/4LW/8jP8ACj/rw1T/ANGW9VVVVVVn/wAfUX++Kqqqqqqqqqqqqqqqqqqqqqqqqqql8bx+D/itqejX8wXStTaFyScC3mMajf8AQ4+b8D254IyDmqqqqqqq+JWrQ/BP/grTP4o+IM5h0638f299NdTnYkNpdBGSUk8bI45lJPpGeh4COkiLJG6srDKsDkEeor/gr5rvhPT/ANkG90TW9RtY9W1XW7IaLbO/7yaWOQNKyKOSFi35J4G4Dqy5/wCCZfjbVfCf/BOr4peLGmeE+EtR166sZSWGxYdMt59wPPSQv0HXtnNf8ER/C9rffGH4h+MZY0abSPDkFnEW6r9puAzED/t2Az7+9V/wW88O2tr8V/ht4rjjjFxqXh65s5GA+Yrb3Adc+3+ktj8a/wCCj+m6t8Uv+Cc/wp+KNosty1gNC1e/cA4EN1YNGXIPOfNnhHP945r9jTx7onxJ/ZY+F/iTQ7yGdU8M2VjdLGf9TdW0SwTxkdRiSNhz2we9fEaWD45/8Fk/CmleGCLy28AC1XULiEZSJrKGS5k3EH+GaRYT0+f5fr+0ddXP7d37ZOg/sl+HrqWT4afCqf8Atbx7dQSYS4u0+U24I/iXcIB0ZWlnOD5dfF7/AIKdfsj/AAD1x/h5FqOr+JrzRQLWe18J2EU8FkUG3yvMkkiiJUAAqjNtI2nBBA/Z0/a7+Bf7U2n3lx8J/FMk9/pihr/SL+A215bKTgOYzkMhOBvQsoJAJBOKqo1b4uf8Fryt+hlsPCl2THGwP7r7FpfyEcf8/OG5/vdema/4KV63b+Gf+ChupeJLuKSWDSX0O7ljjxudY7eByFzxkhe9f8PtPgD/ANEo+IH/AHzaf/Hq/wCH2nwB/wCiUfED/vm0/wDj1ftaftEeF/2o/wBqKw+K/hDRNU0nT50060W21IRiUNEwDE+WzDBzxzVf8Es/+T9J/wDsH6x/Ov2tf2idH/Zc+BuufFbULaO9voNtpo1hI20Xl9LkRIeQdowztg52RtjnFfs3fsofGf8A4KU+K9X+PPx9+JWq2fheO8a3S7RA811IMFreyjb93BDGCAW2lQSAFY7yvxJ/4JCW3w40kfEP9kz4u+M7Dx74d/0vTYNXu4CbqROfLjngjh8lyOAWDKT8rbQxYfsn/EvxN8a/+ClXgb4jePbCGHxBqupn+0olh8sfaYNMeJn2EfIxaLftx8pOB0r9vz/kzT4s/wDYvyf+hpX7COh/tF/GCLxP+zh8CvEb+EdJ8Tz29/4w8Txlg9lYwq8axKVIbMjSn5FYM+0AlUEhrx//AMESfC8Hgm5m+Gfxh1q48V21uzwQ6vaQi0vJAOI/kw0IY8biXxnkGv8Agk5+058QfCXxfm/ZJ+I+oX0+l38d1Ho1pfOzSaPf2qs8tumT8kbJHLlOgdFwBubNftlq3wm/4Kw/Cbx/YBof+Ejk0G4vJY85dXuXsJhzxnyIgMehHIzVVVVVVVVVVVVVVVVVVVV/wS4/5PH+M3/Xhf8A/pyjqqqqqqqqqqqqqqqqqqqqrx9/yKV9/wBs/wD0YtfE+9/s/wCHXiS4DYP9lzxg+hdCo/8AQq8A2Q1Lxx4fsSARNqdurZ/u+Yuf0zVV+1R4X8q90nxhAnyzobO4OMfMuWQ/UguP+Aiv2P8Axctj4k1TwXczYj1SAXFspP8Ay1j+8B7lDn/gFalbi3u3QD5W+Zfoaqqqqqqqqqqqqqqqq8If8izp3/XAVVVVVVVVVVVVVVVVVVVVV8Yv+T6PCP8A19aX/wChVX7eGsix+Eum6PG/7zUtaj3AjrHHHIx/8e2V+yP4HsvH3xNv9J1GMNap4evhIxXOwSoLcke+JzV9aap4W1+4sJma21LSLxo3KHBimifBwfZlrwF4pt/HHgnQ/F1sV26tYRXDKv8AA7KN6/8AAWyPwqqqqqqqqqqqqqqqqqv+Cmv/ACY/8Sf+4V/6dbSv+CWf/Jl/hH/r/wBS/wDSyWqqqqqqqqqqqqqqqqqqqr9u/wD5M/8Aip/2AJP/AENK/wCCQf8Ayahd/wDY3X3/AKJt6qqqqqqqqqqqqqqqqqqqq+K3+v03/ck/mtVVVVVVVVVVVVVVVVVVVVaL/wAhiw/6+Y//AEIVVVVVVVVVVVVVVVVVVVVVX7B//JS/Hn/Xqv8A6PNVVVVVVVVVVVVVVVVVVVVf8Frf+Rn+FH/Xhqn/AKMt6qqqqqrP/j6i/wB8VVVVVVVVVVVVVVVVVVVVVVVVVVV8VL9NS8f6xNG2VilEI9jGoQ/qpr4MePxrNkvhXVZ831nH/ozseZoh/D/vL+o+hqqqqqqqv2+v+Cf2k/tb2Fn4y8HarZ6D8RNGtvs1vdXat9m1G3BLLBOVBZCrMSsgBI3MCrAgr4B8E/8ABZ74L6db/DDwdbT6ho9onk6fNeX+kahFbxKMARzXDl0QAALG+AAAAozg/ttfAL4vfDz4WWfxe/a/+Lj+Lfin4t1FNO0DRYLrfDpVqg825m+UKmRiOPy4lEamfdliQV/Y8+Eeo6R/wSy1/wAP3NrLFf8Aj3wt4gvxF0YrdQSxQMM8fNCkTDt8wr/gh/qlrD8Qfilojt/pN1o1hcRjP8EU0itx16zJVf8ABcDVLab4h/C3REI8+00W+uHGf4ZZo1Xj6wtXwo+G2g6p+y14H+E3jbRI9R0p/Aum6VqNheKGEiLZRxsrYxzx1GCCMjGKtv8AgmN8bfhRquq2H7MH7YviPwV4S1uZnm0qeKUvb5wCweKRVeQAACQLG2ABnjNeJfh38L/+CU/7Ovir4g+HNcufFfxZ8agaZYazqUYE15eSZYeXEC2yGMhp3UsxdlVWckpgeB/EP7BP/BNzxp42u7maD4s/EKOGXWL+Vj9pgu72QRrGXPzb4IZZWzziYyMODX/BKj4DeEvAH7MGifEmbRLOXxV4+8+9vr+SENKLbzXjhgDnny9iByBgFpGznANftQ+HdK/ZC/4KWfC34l/DGyi0LR/G81pLq9naDy4WM1y1tfKI14CtEySYxjzCWAyBVV+zSh0//gsV8R7a9KxS3F/r5iViMvuHmLj6p830qv8AgorY2ep/8FI5dN1C2juLW6u9AinhkXcsiNDbhlI7ggkEV/wxR+yL/wBG4fD/AP8ABFD/APE1/wAMUfsi/wDRuHw//wDBFD/8TX/BRL4d+Bfhf+2ra+FPh14S0vw5o8dppUqWOm2ywRK7H5mCrxk45NV/wSz/AOT9J/8AsH6x/Ov+C42v39t4W+EfhaJn+xahqGq3kwDYXzII7ZEyO5xcyYPbn1r9gfw/pfhv9jj4TWGkQqkVx4dhvJMADdNcFppScdy8jVXwKhht/wDgr9fW9vEkccXxO8SqiIu0KA16AAB0GO1ft+f8mafFn/sX5P8A0NK/4IcwQDw38XblYY/Oa+0lGk2jcVCXRAz1wCTx7mq8Foumf8Fgp004fZwfihqORHx/rHm3/nubP1NV/wAFTkOo/tyfA/R7MrJeSWGmBYsgH59VlVOTwMlSPwqqqqqqqtcvJdO0W/1C32mW1tZZUDDI3KpIz7cVB/wWP/ahhhSKTwb8Mp2UYMkml3oZvc7bsD8gK/4fJ/tPf9CL8Lv/AAV3/wD8mV/w+T/ae/6EX4Xf+Cu//wDkyrX/AILK/tKpOrXvw/8AhnLCM7kisL6NjxxhjdsBzjsa+Ev/AAWZ8L6pqFvpfxp+FdxocEjBX1bQ7s3ccee7W7qrhR1JV3Povr4S8W+GfHnhvT/F/g3XLPWNG1WBZ7O9tZA8cqHuD2PYg4IIIIBBqqqqqqqqr/glx/yeP8Zv+vC//wDTlHVVVVVVVVVVVVVVVVVVVVXj7/kUr7/tn/6MWv2gr0Wnwo1iNWw1y0ESn/tqhP6KRXwLsjffFXQY8ZEckkpPpsidh+oA/Gqr4t+F/wDhL/h/q2kxRB7lIfPthjnzI/mAHuQCv/Aq8DeJp/BvjDSPFFuWzp10kjqvBePOHX8ULD8a1MwX9hb6naSrLE6BkdeQyMMgj26fnVVVVVVVVVVVVVVVVXhD/kWdO/64CqqqqqqqqqqqqqqqqqqqqvjF/wAn0eEf+vrS/wD0Kq/4KCa0JNU8G+HUf/UW91dSL672jRT+Hlv+df8ABPvRPM1Dxl4idP8AUwWtpExA53mR3HrxsT8xX7Yng0eE/jdqd5BBstfEEMeoRYHG5gVl59TIjMf94etfsMeMv7d+FV14Vnl3T+G79kRc5xBNmRD/AN9+aPwqqqqqqqqqqqqqqqqqv+Cmv/Jj/wASf+4V/wCnW0r/AIJZ/wDJl/hH/r/1L/0slqqqqqqqqqqqqqqqqqqqq/bv/wCTP/ip/wBgCT/0NK/4JB/8moXf/Y3X3/om3qqqqqqqqqqqqqqqqqqqqr4rf6/Tf9yT+a1VVVVVVVVVVVVVVVVVVVVov/IYsP8Ar5j/APQhVVVVVVVVVVVVVVVVVVVVVfsH/wDJS/Hn/Xqv/o81VVVVVVVVVVVVVVVVVVVV/wAFrf8AkZ/hR/14ap/6Mt6qqqqqrP8A4+ov98VVVVVVVVVVVVVVVVVVVVVVVVVVeJNbt/DmhXut3JG20hLBT/E3RV/EkD8auJ5bqeS5ncvLK5d2Pdick1YX93pd7BqNhO0NxbuHjdeqkV4F8YWnjXQYtSh2pcx/JdQg/wCrk749j1H/ANY1VVVVVVXxv/a9/Z4/Z80e51H4i/EvSY72BCYtFsLhbrULhscKluhLDJGNz7UBIywHNaXYfFv/AIK0/tVw67qml3OhfDfw2RHMUyYtK05XDGFZMYe7nzyevO7GyPA0rRdJ0TRrPw7pNhDbabp9qlrbWqLhIoUUIqAf3QoAx6V8O9Sm/wCCbX/BRG+0rxhFPa+DbmeexkufLY+bol44e3uFx94RskJcLn5oZFGSKHjPwg3hQeOl8U6SfDZtPtY1cXkf2TyMZ83zs7NmOd2cV8S9Sk/4KTf8FEbDSPBMc934NtJYLCO7aIgR6LZuXuLhuAVWR3mKBsHM0anBOKREjRY41VVUYVQMAAegpiqqWZgAByTxivACt/wUF/blu/ipcg3fwa+A832Xw8rLmDVdT3ZEwBOGDOglJH8EVuGA3nP/AAU98Ban4+/Yw8cRaNbyT3egm21cRRrktFBMpmJ9lhMj/wDAK/4JgfEfRPiB+xx4KsNPvIX1Dwos+kanbowLQSRyu0eR1G6F4m/E+lftzPD8ev8AgpR8GPgr4UZL6bw01h/bHlZY22bg3dyDg/wWkaSdR97HFVXxZK/AD/gsp4X8Z36/ZtL8aXdlJEzHYhW/tG053Y9MC4DsSeBjJ6VX/BQf/lJgn/YQ8O/+ireqr/gqb/yfpB/2D9H/AJ1X/BLP/k/Sf/sH6x/Ov+CtXwF1v4v/ALOEPjHwrZPeat8Ob59Tlt403PJYNHsudo9UAjlP+zE3U4r/AIJYftsfDe5+EWm/s9fE7xfp3h/xJ4Ykkh0WXU7lYItTtJHLoiSNhfNjZynlk5KhCu75tvx5/au+CP7PPgm98W+NfHOkvcxW7vp+j216kl3qMoB2xxRqS2CwALkbVzliBX7EfifWvG//AAUd8J+M/ElsbfV9f8U6pqF/CylTHPNBdSSLg8jDMRg1+35/yZp8Wf8AsX5P/Q0r/gh1/wAip8W/+whpX/ou5qvDX/KYOb/sqF7/AOhy1XxnX/hf3/BYzwh4U00i7tPA17pqSvnKBbCI6hMp9hKzxkHq2R35qqqqqqrU7FNT026015Ci3UDxFgOVDKRkfnXx48CfDj4e/E3UPAnwp8f33jex0uU2s2pyaatqk1yrEOsAWRzIgPyh+NxBxlcMfBP/AASr/a28aeG7TxHNonh7w6L2MSR2OtakYblUIyC8caPsJH8LEMOhAPFfFb/gmd+1T8JvCV5411Dw7o+v6bpsLz339h3/ANoltolGWkMbqjMoGSdgYgAkgAE1+zx+zv4h/aW8W3HgPwZ418I6P4gS2aeysNeu57d9SCgs625jhkVnVVyVYqcZIyFYr8fP2X/jT+zVq9tpnxW8JtYwX5b7BqNtKtxaXe0DcEkXowyMowVh1xgg1/wR4+O+rab45179nvV755dI1izk1bR45HyLe7i2iZEHpJEdx7Aw5x8xNVVVVVVVVf8ABLj/AJPH+M3/AF4X/wD6co6qqqqqqqqqqqqqqqqqqqq8ff8AIpX3/bP/ANGLX7Ul6IPANjZKwDXOqJkZ6qschP67a/ZlshdfElpyoP2PTppASOhJRP8A2eqqvit4XHhDx9q+jxRbLbzjNajGB5T/ADKB7DO3/gNfs4eKl8Y/CeysrmXdc6OW0+XJ52oAYz9NhUfVTTo0bmNh8ykgiqqqqqqqqqqqqqqqrwh/yLOnf9cBVVVVVVVVVVVVVVVVVVVVV14U8LXurRa9eeG9Kn1OEqYr2WzjaZCv3SHI3DHbniq/bd1oar8cp7AOD/Y+l21rj+6WDTf+1hX7COiiw+Eeoau6ASanrUpVvWOOONR/48Hr9vbwZ/aXgrQvHFtFmXRr1rW4IXnyZhwSfQPGo+slfsReMh4d+L58O3E2228S2MluFJwPOj/eIT+CyKPd6qqqqqqqqqqqqqqqqr/gpr/yY/8AEn/uFf8Ap1tK/wCCWf8AyZf4R/6/9S/9LJaqqqqqqqqqqqqqqqqqqqv27/8Akz/4qf8AYAk/9DSv+CQf/JqF3/2N19/6Jt6qqqqqqqqqqqqqqqqqqqq+K3+v03/ck/mtVVVVVVVVVVVVVVVVVVVVaL/yGLD/AK+Y/wD0IVVVVVVVVVVVVVVVVVVVVVWkeFPC3h+eW60Hw3pWmzXAxNJaWccLSDOfmKgE8+tVVVVVVVVVVVVVVVVVVVVf8Frf+Rn+FH/Xhqn/AKMt6qqqqqrP/j6i/wB8VVVVVVVVVVVVVVVVVVVVVVVVVVfHLxquoXieEdOm3Q2bb7tlPDS9l/4COvufaqrwH4yu/BWux6jFuktZPkuoQf8AWJ6j/aHUfl3NWN9aanZQ6hYTrNb3CB43XowNVVVVVX7dv7HGpftkeEvC/hnTPHlr4Xbw7qMt200+ntdCYPHs2gB0xjrnmvhp/wAEUvg74fv4L74o/FPxD4wSE7msrGzTSYJf9lyHlk24/uupz3HSvh98OPAnwo8KWfgf4b+FdO8PaFYLiCysYRGgJ6ux6u5xlnYlmPJJNV+1N+x/8I/2tvC9tonxCtLmz1XS9x0rXNPKpdWRbG5QWBDxtgbkYY7jacMI/wDgiRrIvBpkv7T7nw8Jt5hXw0wkI6n5PtWwN23c+uO1fsvfshfCD9kzwvPofw40+4udT1LadV1zUGWS7vSvRSygBIwSdsagAZycsSxr9oT4f+OPit8IPEXw4+H3jeDwlqniG2Nm+sSWrTtb278TBFVlIdk3IGyNu4kcgV+zN8AfDP7M3wZ0D4SeGpFuf7OjMuo35i2Nf3knM07DnGTwoJO1FRcnbmryztNQtJrC+torm2uY2imhmQOkiMMMrKeCCCQQeDmvEv8AwSx1Hwr471Txp+yp+0r4p+ENvrjEXulWccssaoSTsjkjniby1JJVHDkE8MMV+yf+wj8Ov2XNT1Pxw/iPVfG/xB1xHTUfE2rjEhV2DSCKPcxTewDMzO7sR97HFVX/AAV++AWq+M/hboXx/wDBlvP/AG98Nrg/bntgRJ/Z8jKfNGOcwyqj5H3VeRicLX7JXx6039pP4BeFfilazxHUbq1FtrUCYH2fUIgFnTA6At86j+46nvXx8/4Jq6H8dv2jh+0Pd/Fu+0e5W40+b+yo9HWZP9EWNQPMMqn5vL/u8Z71VftOf8E1dD/aW+OSfG2++Ld9oM8dvaQjTotHW4X/AEfod5lU/N9OKr9mP/gmrof7NXxyf422Pxbvtene3u4f7Ol0dbdR9o6neJWPH05plVlKsAQRggivjp/wSM/Zz+LGvXXirwXqOrfDzUr2RpLi30uNJ7F3bJLLbvgxnP8ADG6p6KOtfA3/AIJF/s7fCbxDaeL/ABvrGrfEK/09xJBa6pFHb2CuDkO0C5MhBH3Xdk9VNfsbW1l8Vf8Agq1q/jfw2PtmhxeKPE2vJNF937NJ9qWGTI7F54fbkDvXx3+FVv8AHH4QeKvhJda1JpEXiewa0e+jgEzQAkHcEJG7p0yK/Yr/AGKtL/Y00vxXpmmfEG68UjxTcWszvPpy2nkeSsgAAEj7s+Z7YxVad/wTV0PTv2tn/atX4t30l23iabXP7FOjqEDSFj5Xm+bnA3fe29ulfGr4reG/gd8KfE/xX8VyquneG9PkuTGW2m4l6RQqf70khSMdsuM8V/wSW+EHiTxbr/j39tL4kQs+reNb65ttIkkTHm+ZP5t7cKD/AAmULEpHTy5R0xVVVVVVVftAavq/h/4DfEjXvD0kiappvhHVbmxaIkOs8dpK0ZXHOdwGMV+z94s8NeA/jn8P/GvjGMvoeheJLG9v8R+YVhinRmYL/FtA3Y74xWk6tpWvaZa61oepWuoafexLNbXVrKJYpo2GVZGUkMpHQivGPi7wx4C8Lap4y8Z6xa6XomkWz3F9d3LbUijUc59SeAFHJJAAJIFaB8QJ/B3xesvid4Ihk059K8QLqemwIcGJVn8xIzg8jbhSOhGR0Nf8FGfCOieLv2OviCusW8byaNaxalYysoLQXEMyEMpPQspeMn+7Iw71/wAE4b64sP21PhnLaybTJc3kLg9Cr2NwrD8j+eKqqqqqqqq/4Jcf8nj/ABm/68L/AP8ATlHVVVVVVVVVVVVVVVVVVVVXj7/kUr7/ALZ/+jFr9rG9Ih8N6arcM1zK4+gjC/zav2UbLzNY8Qajj/UWsMWf99mP/tOqqv2qfC4ktdJ8YwR/NExsrggfwnLRn6Ahx/wIV+yR4uGj+O7rwtcSYh162/dg/wDPaIFl/NDJ+OK1q38q6EqjiUZ/Edf6VVVVVVVVVVVVVVVVeEP+RZ07/rgKqqqqqqqqqqqqqqqqqqqqqv2gtbHiH41+M9SEm9Rq01ujeqwnyhj2xGK/Ze0UaF8BfCFqUAeeza6Y9z50jyA/98uK+MHg0fED4YeJPCKxCSa/sJPsykf8t0+eL/x9Vrwj4ivPB3ivSPFNjn7RpF9Fcoucbijhip9jjB+taffWuq6fbanYyiW3u4UmhcfxIwBU/iCKqqqqqqqqqqqqqqqq/wCCmv8AyY/8Sf8AuFf+nW0r/gln/wAmX+Ef+v8A1L/0slqqqqqqqqqqqqqqqqqqqq/bv/5M/wDip/2AJP8A0NK/4JB/8moXf/Y3X3/om3qqqqqqqqqqqqqqqqqqqqr4rf6/Tf8Ack/mtVVVVVVVVVVVVVVVVVVVVaL/AMhiw/6+Y/8A0IVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVf8Frf+Rn+FH/AF4ap/6Mt6qqqqqrP/j6i/3xVVVVVVVVVVVVVVVVVVVVVVVVVfFPxufBugAWTr/aV8THbZ52YHzPj2yPxI96kkkmkaWWRndySzMckk9STVVY2c2o3tvp9tt825lWKPccDcxAGT2GTXwV8bTaZqP/AAherSFYLhz9l38GKXunsG5/H61VVVVVVVVVVVVVVVappmna3pl3o2r2MF7YX8D291bTxh45onUq6Mp4KlSQQeoNfs4/si/DD9lefxCnwp1TxMmneJZUmudK1DUFubaGRCdrxLsDKwVtpJY7lC7slQRVVVV8V7rx1ZfDHxXdfDDTF1Hxemj3X9hWzSRxiS9MbCHLSEIAH2k7iBgVp/8AwUo/bm+CluPDvx+/Zhm1W7tB5Q1CbTLrSnuGGPmZ0R4ZPrGqqcj8fiD+3J+3b+11oF38Lfgh+z5qfhyx1yE21/eaZaXFxP5T/K6m8kCRW6NypYgH0cc5/wCCdf7Cs37JvhnU/Fvj25s774g+KIUhuvsp3xaXaqQwtkf+NmcBpGHykogGQu5qqq/aS/Zk8CftT+FNP8EfEbXfEtpothefbDaaPepbLcyhdqGXdG5YKC2BwMsSckDHg/wh4a8AeFdJ8E+DtIg0zRNDtI7Sws4RhYYkGFHPJOByTkkkkkkk1VVVVVVWoafZatYXOl6lbR3NpeQvDPDIuVkjYFWUjuCCQfrX7Wv7Mfi79lz4rX/g7WbO4l0C8lkm8O6syfJf2mePmHHmoCFdeoPP3WUn4dftE/HX4R2f9mfDX4teJ/D9gWZvsNpqLi33Hq3kklNx9dua+JXx7+NPxiSGH4n/ABQ8R+JLe3bfDa31+7wRNjG5Ys7FbHcDNfsK/s1eIv2jPjpodvHpcreE/Dd7BqHiO+ZD5SQxtvEG7+/MV2ADkAs2MKa/4KreL9f0X9lu98H+HPD2r6jL4s1CCC8uLOyklisrWB1nkeWRVKxgtHGgDEbgzY4U1/wSW+H974q/autvF6W7mz8FaNeXks2DtWSeM2qIT6ss8hAPZG9Kqqqqqqqq/wCCXH/J4/xm/wCvC/8A/TlHVVVVVVVVVVVVVVVVVVVVXj7/AJFK+/7Z/wDoxa/anvRL410uwVgRb6YHPsXkf+ij86/ZTsvL8O65qOP9fepFn/cTP/tSqqviJ4ZXxh4K1bw+qBpri3Jt89pV+ZPp8wH4Zrw5rd74W8Rafr9kStzpl0kyKeMlWBKn2OMH6013Z+IdAtdZ02QS291AlzA/95GUEH8jVVVVVVVVVVVVVVVV4Q/5FnTv+uAqqqqqqqqqqqqqqqqqqqqrX/jx8PvDfxIsPhTqdzerr+ovCkCJaloyZThMv2q8uoLGznvrlwsVvE0kjHjCqMk/kK1S/n1jVLvVLglp724eZ++WdiT+prwfow8OeEdE8PKuBpem29qB6eXGq9vpVftC+DB4E+MfifQYo9ls14bq1GMARTASKB7Lv2/8Br9kbxkPGHwO0WKWXfdaEz6ZP7CLBj/8hNGPwNVVVVVVVVVVVVVVVVf8FNf+TH/iT/3Cv/TraV/wSz/5Mv8ACP8A1/6l/wClktVVVVVVVVVVVVVVVVVVVX7d/wDyZ/8AFT/sASf+hpX/AASD/wCTULv/ALG6+/8ARNvVVVVVVVVVVVVVVVVVVVVXxW/1+m/7kn81qqqqqqqqqqqqqqqqqqqqrRf+QxYf9fMf/oQqqqqqqqqqqqqqqqqqqqqqvhp8ePh98WtX1PQ/B9zeyXWkpvuRcWpiAG7bwT15FVVVVVVVVVVVVVVVVVVVVf8ABa3/AJGf4Uf9eGqf+jLeqqqqqqz/AOPqL/fFVVVVVVVVVVVVVVVVVVVVVVVVV8YtbfWPHF5CHzDp4FtEM9NvLf8AjxP5CqqvDpkHiDTDCMuLyLaPfeMV8XdGfwz47fULA+Ut5tvIWUY2SZ+bHvuUt+IrQdTTW9EsNXQAC8tklIH8JZQSPwPFVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVfEn4WfDv4w+F5/BnxN8I6d4h0ec7jbXkedjDgPG4w0bgE4dCGGTzXjj/gjZ8DdavZrzwL8RfFfhpJTlbW4SLUIYueiZCPtx/edjnnPavA//BGz4G6Lew3njr4i+K/EyRHJtbdItPhl56PgO+3H911Oec9q+HHww+H3wh8LW3gr4aeE9P8AD2i2vKWtnHt3MQAXdjlpHOBl3JY4GTVaZ4W8MaJqF9q2i+HNLsL7U9v225tbNIpbrbnb5jKAXxubG7ONx9TVVVVVVVVX/BLj/k8f4zf9eF//AOnKOqqtX1fS9A0y41jWtQgsrG0TfPcTuESNfUk9BX/C+/gp/wBFT8Nf+DGP/Gv+F9/BT/oqfhr/AMGMf+Nf8L7+Cn/RU/DX/gxj/wAa/wCF9/BT/oqfhr/wYx/41/wvv4Kf9FT8Nf8Agxj/AMa/4X38FP8Aoqfhr/wYx/41/wAL7+Cn/RU/DX/gxj/xr/hffwU/6Kn4a/8ABjH/AI1/wvv4Kf8ARU/DX/gxj/xr/hffwU/6Kn4a/wDBjH/jX/C+/gp/0VPw1/4MY/8AGv8AhffwU/6Kn4a/8GMf+Nf8L7+Cn/RU/DX/AIMY/wDGv+F9/BT/AKKn4a/8GMf+Nf8AC+/gp/0VPw1/4MY/8a/4X38FP+ip+Gv/AAYx/wCNf8L7+Cn/AEVPw1/4MY/8a/4X38FP+ip+Gv8AwYx/41/wvv4Kf9FT8Nf+DGP/ABr/AIX38FP+ip+Gv/BjH/jX/C+/gp/0VPw1/wCDGP8Axr/hffwU/wCip+Gv/BjH/jX/AAvv4Kf9FT8Nf+DGP/Gv+F9/BT/oqfhr/wAGMf8AjX/C+/gp/wBFT8Nf+DGP/Gv+F9/BT/oqfhr/AMGMf+NeNfjj8Hb3wzeWtn8TPDs0z7NqJfoScOpOBn0FfHfxDp3iX4jXl7pN/DeWkVvDFFNC+5GwgJwc9ixH1r9mmyFr8MknA/4/L+aU/htT/wBkqqqvjb4X/wCEV+I2p20UWy2vm+2W/GBtk5bHsHDj8K/ZY8W/8JH8NF0S5lD3OgTtbEE8mFvmjJ9uWUf7lXcBt7h4T/CePp2qqqqqqqta8Q6F4ctkvNe1e00+GR9iSXEoQM2CcDPfANf8LU+G/wD0PGjf+Ba/41/wtT4b/wDQ8aN/4Fr/AI1/wtT4b/8AQ8aN/wCBa/41/wALU+G//Q8aN/4Fr/jX/C1Phv8A9Dxo3/gWv+Nf8LU+G/8A0PGjf+Ba/wCNf8LU+G//AEPGjf8AgWv+Nf8AC1Phv/0PGjf+Ba/41/wtT4b/APQ8aN/4Fr/jX/C1Phv/ANDxo3/gWv8AjX/C1Phv/wBDxo3/AIFr/jXhf46fBq18PWNvcfE7w5HLHCAyPqCAg/TNf8L7+Cn/AEVPw1/4MY/8a/4X38FP+ip+Gv8AwYx/41/wvv4Kf9FT8Nf+DGP/ABr/AIX38FP+ip+Gv/BjH/jX/C+/gp/0VPw1/wCDGP8Axr/hffwU/wCip+Gv/BjH/jX/AAvv4Kf9FT8Nf+DGP/Gv+F9/BT/oqfhr/wAGMf8AjX/C+/gp/wBFT8Nf+DGP/Gv+F9/BT/oqfhr/AMGMf+Nf8L7+Cn/RU/DX/gxj/wAa/wCF9/BT/oqfhr/wYx/41/wvv4Kf9FT8Nf8Agxj/AMa/4X38FP8Aoqfhr/wYx/41/wAL7+Cn/RU/DX/gxj/xr/hffwU/6Kn4a/8ABjH/AI1/wvv4Kf8ARU/DX/gxj/xr/hffwU/6Kn4a/wDBjH/jX/C+/gp/0VPw1/4MY/8AGv8AhffwU/6Kn4a/8GMf+Nf8L7+Cn/RU/DX/AIMY/wDGv+F9/BT/AKKn4a/8GMf+Nf8AC+/gp/0VPw1/4MY/8a/4X38FP+ip+Gv/AAYx/wCNf8L7+Cn/AEVPw1/4MY/8a/4X38FP+ip+Gv8AwYx/41/wvv4Kf9FT8Nf+DGP/ABr/AIX38FP+ip+Gv/BjH/jX/C+/gp/0VPw1/wCDGP8Axr/hffwU/wCip+Gv/BjH/jXxT8b+D9V/bE8MeMdN8S6dc6Ha3GnGbUIp1aGMIfnJccDHevi18e/hW3ww8VQ6B8RNBvdSn0i5htILe9R5HkeMou0A5JywP4V8MdFHiP4j+FtBZdy3+sWsLjOPlaVQx/LNVX7fvgwxaj4Z+IFtCNtxE+m3TgYwykyRZ9yGl/75H4fsDeMhZeKfEHgO5mAj1O0S9tlY4/eRHa4HuVcH6R1VVVVVVVSXFvFxLPGmP7zAV4o+I3gTwVZx33irxZpumwSyeXG004+ZsZwAMk8Cv+GkfgV/0U7Rv+/jf4V/w0j8Cv8Aop2jf9/G/wAK/wCGkfgV/wBFO0b/AL+N/hX/AA0j8Cv+inaN/wB/G/wr/hpH4Ff9FO0b/v43+FW/7RPwOunMcXxQ0EEDP7y58sfm2B+Ff8L7+Cn/AEVPw1/4MY/8a/4X38FP+ip+Gv8AwYx/41/wvv4Kf9FT8Nf+DGP/ABr/AIX38FP+ip+Gv/BjH/jX/C+/gp/0VPw1/wCDGP8Axr/gox8Xfhf4o/Y0+IWheHPH2h6lqNz/AGZ5NrbXqSSSbdTtWbCg5OFUn6A1/wAEs/8Aky/wj/1/6l/6WS1VVVVVVVVVVVVVVVVVVVft3/8AJn/xU/7AEn/oaV/wSD/5NQu/+xuvv/RNvVVXijxt4Q8E28F14u8S6fo8Ny5SF7ydYhIwGSAT1OK/4X38FP8Aoqfhr/wYx/41/wAL7+Cn/RU/DX/gxj/xr/hffwU/6Kn4a/8ABjH/AI1/wvv4Kf8ARU/DX/gxj/xr/hffwU/6Kn4a/wDBjH/jX/C+/gp/0VPw1/4MY/8AGv8AhffwU/6Kn4a/8GMf+Nf8L7+Cn/RU/DX/AIMY/wDGv+F9/BT/AKKn4a/8GMf+Nf8AC+/gp/0VPw1/4MY/8a/4X38FP+ip+Gv/AAYx/wCNf8L7+Cn/AEVPw1/4MY/8a/4X38FP+ip+Gv8AwYx/41/wvv4Kf9FT8Nf+DGP/ABr/AIX38FP+ip+Gv/BjH/jX/C+/gp/0VPw1/wCDGP8Axr/hffwU/wCip+Gv/BjH/jX/AAvv4Kf9FT8Nf+DGP/Gv+F9/BT/oqfhr/wAGMf8AjX/C+/gp/wBFT8Nf+DGP/Gv+F9/BT/oqfhr/AMGMf+Nf8L7+Cn/RU/DX/gxj/wAa/wCF9/BT/oqfhr/wYx/41/wvv4Kf9FT8Nf8Agxj/AMa/4X38FP8Aoqfhr/wYx/41/wAL7+Cn/RU/DX/gxj/xr4k/Gb4TalNYHT/iLoFwI0cMY75G25K46H2rSfHvgnXtQh0nR/FelXd5cHbFBHdKWc4zgDPPANLo183BVF+rf4VdWc9o4SVRz0YdDVVVVVVVVVVVVVVUcEs52wxs5HYDOK+wXv8Az7Sf9819gvf+faT/AL5rWr2z8O6e+qa9dxWFpGQGnuHEagk4Aye5Nf8AC1Phv/0PGjf+Ba/41/wtT4b/APQ8aN/4Fr/jWk/Ff4Zw6pZSy+OtFVEuELMbtQAAwyetf8L7+Cn/AEVPw1/4MY/8a/4X38FP+ip+Gv8AwYx/41/wvv4Kf9FT8Nf+DGP/ABr/AIX38FP+ip+Gv/BjH/jX/C+/gp/0VPw1/wCDGP8Axr/hffwU/wCip+Gv/BjH/jX/AAvv4Kf9FT8Nf+DGP/Gv+F9/BT/oqfhr/wAGMf8AjX/C+/gp/wBFT8Nf+DGP/Gv+F9/BT/oqfhr/AMGMf+Nf8L7+Cn/RU/DX/gxj/wAa/wCF9/BT/oqfhr/wYx/41/wvv4Kf9FT8Nf8Agxj/AMa/4X38FP8Aoqfhr/wYx/41/wAL7+Cn/RU/DX/gxj/xr/hffwU/6Kn4a/8ABjH/AI1/wvv4Kf8ARU/DX/gxj/xr/hffwU/6Kn4a/wDBjH/jX/C+/gp/0VPw1/4MY/8AGv8AhffwU/6Kn4a/8GMf+Nf8L7+Cn/RU/DX/AIMY/wDGv+F9/BT/AKKn4a/8GMf+Nf8AC+/gp/0VPw1/4MY/8a/4X38FP+ip+Gv/AAYx/wCNf8L7+Cn/AEVPw1/4MY/8a/4X38FP+ip+Gv8AwYx/41/wvv4Kf9FT8Nf+DGP/ABr/AIX38FP+ip+Gv/BjH/jX/C+/gp/0VPw1/wCDGP8Axr/hffwU/wCip+Gv/BjH/jX/AAvv4Kf9FT8Nf+DGP/Gv2NPG/g/wZ4/8Zah4s8S6dpNte24W3lvJ1iWU+cThSevHNf8AC+/gp/0VPw1/4MY/8a8P+JPD/izTE1nwzrVlqljISqXFpMsqEg4IyD1B7VVVVVVVVVVEgDJPT1qS9soh+9u4Ux/ekAp9c0dDg6jAfo+f5V/b2jf9BGH868S/GP4XeDruKw8T+ONM0+5mj8xIppCGKZxuwB0yCPwNf8NI/Ar/AKKdo3/fxv8ACv8AhpH4Ff8ARTtG/wC/jf4V/wANI/Ar/op2jf8Afxv8K/4aR+BX/RTtG/7+N/hVv+0L8ELpS0XxR8PjacHzLsR/kGxX/C+/gp/0VPw1/wCDGP8Axr/hffwU/wCip+Gv/BjH/jX/AAvv4Kf9FT8Nf+DGP/Gv+F9/BT/oqfhr/wAGMf8AjX/BYTx14N8ceJPhjL4P8T6brKWljqSztZ3CyiIs9vgHHTOD+VVVVVVVn/x9Rf74qqqqqqqqqqqqqqqqqqqqqqqqqvE8jTeJdWlcHc99MSG65LnrVVXw10eXWvG+k20aFlguFuJTjgLGdxz7HAH41+0NZI+maPqIA3xTyRZ9mUH/ANk/WvhJK8/w70d5DyEkUZ9FlcD9AKqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/a+/4KqW3wr8W6j8MPgLoOl+INW0mR7fUtc1Jne0t51JV4oYkKmVlPBcsFDAgBxzWpf8FQf22L+6a4tfi1a6ehziC28O6eyLyTwZIHbvjr2Hua/4ea/twf8ARb//AC29K/8Akav+Hmv7cH/Rb/8Ay29K/wDkatAup77QdNvbqTfNPZxSSNgDLMgJOBwOTVVVVVVVVVf8EuP+Tx/jN/14X/8A6co6qq+P+l6lrXwZ8W6Vo+n3N9e3OnMkNtbQtLJI24cKqgkn2Ar/AIUx8Yf+iUeMf/BFc/8AxFf8KY+MP/RKPGP/AIIrn/4iv+FMfGH/AKJR4x/8EVz/APEV/wAKY+MP/RKPGP8A4Irn/wCIr/hTHxh/6JR4x/8ABFc//EV/wpj4w/8ARKPGP/giuf8A4iv+FMfGH/olHjH/AMEVz/8AEV/wpj4w/wDRKPGP/giuf/iK/wCFMfGH/olHjH/wRXP/AMRX/CmPjD/0Sjxj/wCCK5/+Ir/hTHxh/wCiUeMf/BFc/wDxFf8ACmPjD/0Sjxj/AOCK5/8AiK/4Ux8Yf+iUeMf/AARXP/xFf8KY+MP/AESjxj/4Irn/AOIr/hTHxh/6JR4x/wDBFc//ABFf8KY+MP8A0Sjxj/4Irn/4iv8AhTHxh/6JR4x/8EVz/wDEV/wpj4w/9Eo8Y/8Agiuf/iK/4Ux8Yf8AolHjH/wRXP8A8RX/AApj4w/9Eo8Y/wDgiuf/AIiv+FMfGH/olHjH/wAEVz/8RX/CmPjD/wBEo8Y/+CK5/wDiK/4Ux8Yf+iUeMf8AwRXP/wARX/CmPjD/ANEo8Y/+CK5/+Ir/AIUx8Yf+iUeMf/BFc/8AxFf8KY+MP/RKPGP/AIIrn/4irj4Q/Fm0ha4uvhf4thiT7zyaJcKozxySnriv+FdfEH/oRPEP/grm/wDia+D2j3ehfDTQtLvrWW2uI4XaSGZCjozyM5BU8g/N0qqqv2pPC4vfD+neLLePMmnTGCcgf8s5OhPsGAH/AAOv2V/Fw8O/EpNFuJdtrr8BtiCcASr80Z+vDKP9+tdt8GO5Udflb+lVVVVVVKpchVGSa+OXxEXx54teDTbjfo+lFobQqflmbPzy/wDAiAB/sgdCTVj4I8Z6naR3+m+Edau7aYZjmg0+WRHHTIYKQelf8K6+IP8A0IniH/wVzf8AxNf8K6+IP/QieIf/AAVzf/E1/wAK6+IP/QieIf8AwVzf/E1/wrr4g/8AQieIf/BXN/8AE1/wrr4g/wDQieIf/BXN/wDE1/wrr4g/9CJ4h/8ABXN/8TX/AArr4g/9CJ4h/wDBXN/8TX/CuviD/wBCJ4h/8Fc3/wATX/CuviD/ANCJ4h/8Fc3/AMTX/CuviD/0IniH/wAFc3/xNQ/B74t3MSz2/wALfF0sbjKumh3DAj2ISv8AhTHxh/6JR4x/8EVz/wDEV/wpj4w/9Eo8Y/8Agiuf/iK/4Ux8Yf8AolHjH/wRXP8A8RX/AApj4w/9Eo8Y/wDgiuf/AIiv+FMfGH/olHjH/wAEVz/8RX/CmPjD/wBEo8Y/+CK5/wDiK/4Ux8Yf+iUeMf8AwRXP/wARX/CmPjD/ANEo8Y/+CK5/+Ir/AIUx8Yf+iUeMf/BFc/8AxFf8KY+MP/RKPGP/AIIrn/4iv+FMfGH/AKJR4x/8EVz/APEV/wAKY+MP/RKPGP8A4Irn/wCIr/hTHxh/6JR4x/8ABFc//EV/wpj4w/8ARKPGP/giuf8A4iv+FMfGH/olHjH/AMEVz/8AEV/wpj4w/wDRKPGP/giuf/iK/wCFMfGH/olHjH/wRXP/AMRX/CmPjD/0Sjxj/wCCK5/+Ir/hTHxh/wCiUeMf/BFc/wDxFf8ACmPjD/0Sjxj/AOCK5/8AiK/4Ux8Yf+iUeMf/AARXP/xFf8KY+MP/AESjxj/4Irn/AOIr/hTHxh/6JR4x/wDBFc//ABFf8KY+MP8A0Sjxj/4Irn/4iv8AhTHxh/6JR4x/8EVz/wDEV/wpj4w/9Eo8Y/8Agiuf/iK/4Ux8Yf8AolHjH/wRXP8A8RX/AApj4w/9Eo8Y/wDgiuf/AIiv+FMfGH/olHjH/wAEVz/8RX/CmPjD/wBEo8Y/+CK5/wDiKvfCPivTtdh8Lah4Y1a21q4KiHTprKRLiQt90LERuOe2BzX/AApj4w/9Eo8Y/wDgiuf/AIivgR8LfHml/FrQr7xJ4J1/SrWyaWZ5r3TZrdVIibZ8zqBncVq08S6vaKEFz5yjoso3fr1pPGt8P9ZaQN/u5X+pr9plU8f/AAZ1zTTpuLnTkXULZxJna0Jy3GO8ZkH418E/Gg+HvxV8NeK5HK29pfKl1g/8sJAY5PrhHY49QKTxdozj5pJU/wB6P/DNJ4j0SThdQT/gSsv8xTa5o4GTqUH4PmpPE+hoMC93H0WNj/SpPGOkoMIlw/phAP5mpPG1sB+6sJG9Nzhf8ak8bXBGIrCNfTc5P+FSeMdWfhEt09MIf6mpPE+tycC92+yxqP6VJrWrSjDalcfg5X+VSXNzKMS3Ej/7zk18QPiF4b+G+gya74huwvBFtbIR5ty4/hQflk9B1NfEb4i+IfiZ4hk13XZ8KuVtbVCfLto/7qj19T1J/AC3+BXxlu7eK6tvhh4keKZA8bDTpPmUjIPT0r/hQfxr/wCiW+Jf/BdJ/hX/AAoP41/9Et8S/wDguk/wr/hQfxr/AOiW+Jf/AAXSf4V/woP41/8ARLfEv/guk/wq4+BfxntkDyfCvxUwJwBHpM0h/JVJr/hTHxh/6JR4x/8ABFc//EV/wpj4w/8ARKPGP/giuf8A4iv+FMfGH/olHjH/AMEVz/8AEV/wpj4w/wDRKPGP/giuf/iK/wCFMfGH/olHjH/wRXP/AMRX7SPwy+JHh74LeItY1/4e+JdNsLf7L5t1eaTPDFHuuolG52UAZYgDJ6kDvX/BLP8A5Mv8I/8AX/qX/pZLVVVVVVVVVVVVVVVVVVVV+3f/AMmf/FT/ALAEn/oaV/wSD/5NQu/+xuvv/RNvVVX7cPhLxV4t8KeGbbwp4Z1XWZbfUJWlj0+ykuGjUx4BYICQM9zX/CmPjD/0Sjxj/wCCK5/+Ir/hTHxh/wCiUeMf/BFc/wDxFf8ACmPjD/0Sjxj/AOCK5/8AiK/4Ux8Yf+iUeMf/AARXP/xFf8KY+MP/AESjxj/4Irn/AOIr/hTHxh/6JR4x/wDBFc//ABFf8KY+MP8A0Sjxj/4Irn/4iv8AhTHxh/6JR4x/8EVz/wDEV/wpj4w/9Eo8Y/8Agiuf/iK/4Ux8Yf8AolHjH/wRXP8A8RX/AApj4w/9Eo8Y/wDgiuf/AIiv+FMfGH/olHjH/wAEVz/8RX/CmPjD/wBEo8Y/+CK5/wDiK/4Ux8Yf+iUeMf8AwRXP/wARX/CmPjD/ANEo8Y/+CK5/+Ir/AIUx8Yf+iUeMf/BFc/8AxFf8KY+MP/RKPGP/AIIrn/4iv+FMfGH/AKJR4x/8EVz/APEV/wAKY+MP/RKPGP8A4Irn/wCIr/hTHxh/6JR4x/8ABFc//EV/wpj4w/8ARKPGP/giuf8A4iv+FMfGH/olHjH/AMEVz/8AEV/wpj4w/wDRKPGP/giuf/iK/wCFMfGH/olHjH/wRXP/AMRX/CmPjD/0Sjxj/wCCK5/+Ir/hTHxh/wCiUeMf/BFc/wDxFXfwq+KFgVW++G/im3Lg7RNo86Zx1xlKt59W8NazFcw+fY6lplwHUOhSSGVGyMg8ggjoa+H3xE0jx14R03xHBMqS3MQFzEAf3Mw4dfoD09iD3qWGG5iMcgDqw/yRV/p0lm25TuiJ4bHT61VVVVVVVVVVVVBbT3LbIIyx7nsKtdDRcPdPuP8AcXgfnSpDBHtRVRVHpgCvtln/AM/cP/fYrxJ4w8N+EtGuNd17V7e2tLZcsxcFmPZVHVmPYCvi/wDF7W/itrfnTF7XSLRj9hsd3CDpvfHVyO/boO5Np4F8bahaxXth4O1u5t5l3RTQ6dK6OPUELgiv+FdfEH/oRPEP/grm/wDiaT4bfESV1ii8A+I3djhVXSpiST2A21/wpj4w/wDRKPGP/giuf/iK/wCFMfGH/olHjH/wRXP/AMRX/CmPjD/0Sjxj/wCCK5/+Ir/hTHxh/wCiUeMf/BFc/wDxFf8ACmPjD/0Sjxj/AOCK5/8AiK/4Ux8Yf+iUeMf/AARXP/xFf8KY+MP/AESjxj/4Irn/AOIr/hTHxh/6JR4x/wDBFc//ABFf8KY+MP8A0Sjxj/4Irn/4iv8AhTHxh/6JR4x/8EVz/wDEV/wpj4w/9Eo8Y/8Agiuf/iK/4Ux8Yf8AolHjH/wRXP8A8RX/AApj4w/9Eo8Y/wDgiuf/AIiv+FMfGH/olHjH/wAEVz/8RX/CmPjD/wBEo8Y/+CK5/wDiK/4Ux8Yf+iUeMf8AwRXP/wARX/CmPjD/ANEo8Y/+CK5/+Ir/AIUx8Yf+iUeMf/BFc/8AxFf8KY+MP/RKPGP/AIIrn/4iv+FMfGH/AKJR4x/8EVz/APEV/wAKY+MP/RKPGP8A4Irn/wCIr/hTHxh/6JR4x/8ABFc//EV/wpj4w/8ARKPGP/giuf8A4iv+FMfGH/olHjH/AMEVz/8AEV/wpj4w/wDRKPGP/giuf/iK/wCFMfGH/olHjH/wRXP/AMRX/CmPjD/0Sjxj/wCCK5/+Ir/hTHxh/wCiUeMf/BFc/wDxFf8ACmPjD/0Sjxj/AOCK5/8AiK/4Ux8Yf+iUeMf/AARXP/xFf8KY+MP/AESjxj/4Irn/AOIrQfCPivxXcz2Phfwxq2sXFoMzw2FlJcPEM4yyoCV545rVPhb8TdE0+bVda+HPiewsrZd01zdaPPFHGM4yzMoAGSOtfA7406x8I/EAYXE8miXrgXttG+Cp6ean+0B2/iHB7EaH8Q5tW0q21XSr+11GzuoxJBOFyHU9Dxj9eR3pPG10BiSxib6MR/jSeN4TxJpzD/dkz/QUnjPTG4eC4U/7oP8AWk8V6I/DXDp/vRn+maTxBosnC6jF/wACyv8AOn17RkGW1GE/Q5/lUnirRE4W6Z/92Nv6ipPGemKMRwXDn/dA/rUnjeMDEWnMfTdJj+lSeNr1hiKzgX/eJP8AhUni/WHGFaFP92P/ABzUniTW5ODfsP8AdRR/IVJqupy8PqNwfbzSKklll5klZ/8AeOaSCaXiKJ3/AN1SaTSNUkGU064I9fKIr40fE+x+D2grc6jbGTVr5WGnWTfKZCOrt6IuRk9+g9RqOoeJvH/iZru7e61bWdVnCqqIXeV24VEUfgAoHHAFf8KD+Nf/AES3xL/4LpP8K/4UH8a/+iW+Jf8AwXSf4V/woP41/wDRLfEv/guk/wAKb4DfGlTg/CzxP+GmyH+lT/BH4yQMEf4U+LSSMjZo07/qqmv+FMfGH/olHjH/AMEVz/8AEV/wpj4w/wDRKPGP/giuf/iK/wCFMfGH/olHjH/wRXP/AMRX/CmPjD/0Sjxj/wCCK5/+Ir9sjwd4t8I6n4Xi8V+FtX0V7mC5MC6jYyWxlCtHkqHAzjIzj1FVVVVVVn/x9Rf74qqqqqqqqqqqqqqqqqqqqqqqqqviHpr6V421m0dcZu3lQf7L/OP0YVVV+z7qNlHqep6VJDGLmeFZYpSvzbVOGXPp8ynHsa/aFvoV0rSdM3AyyXDy49FVcfzb9K8A6bJpPgvR7CZdsiWis6nqrN8xH4E4qqqqqq/a0/4KR/Bj9ljW5/AT6bqPjDxxbxJJNo1i4gitA6h0+0XDAhCysCFRZGwQSACCb7/gsR+0fDEniZP2a9GtvDT8rNOl6Qy5OMXHyp267ex/D9jv/gpH8Lf2rNY/4QPUNFl8F+N/KMlvpd1eLcQ6gqjL/Z5tqlnUAsY2UNtyRuCsRX7Y3/BQj4V/skNF4Yl0+Xxb44uofOi0KzuVhW1jP3XuZSG8oMOVUKzHrgKQ1Sf8Fd/2mfD8Np4w8YfstWVv4PvGUwXLQ31oJkbpsupA0bEjOCE59K/ZY/az+Fv7WvgeTxX8P7ie01DTnWLWNEvSoutOkYHbuAJDxtglJF4bBBwysq/GP4veBfgR8OdY+KPxF1Q2WiaNEGkMa75Z3YhUiiTI3SOxCgZA5ySACRYf8FFf2wviFod38VvhB+xZPqXw3sWlY3lzcyyXF1EmQXiKBd2CrbvLjlCkEZyCa/ZA/bH+Hf7YHgq513wxay6N4g0Zkj1rQbmYSSWZfOx0cAeZE21gHwpypBA71+z9+1r42/aR/aL8f+FPAPhvQz8IvAT/AGR/Ej+a1zqV5jaFhIYR7CyyNnB+RUPBkGKqv+Ci/wC1dJ+y78DZG8MXix+N/GDyafoHI3WoCgz3eO/lKygf9NJI8gjNfsT+Afix4J+A+i3vxy8eeI/E3jbxHGupaiNZv5Lg6aJFBjtUDfcKJjf3MjPyQFxrX7e/7Qtj/wAFCo/2b7fU9D/4QxvG9to5iOlqZ/szsgYeZnO7DHnFVVVVVVVX7VPw8+I3xK+Cmu6N8IPHet+FPGtnH9t0S80u/e1M08YJFvIVIzHICyc8KxVv4a/4Jn/taax+0T8Lb3wN8SL2WT4h+AHS11KS5G2a/tjlYrhwefMBVo5OM7lVicyYFVVVVVVV8dfEmqeDfgh8QvF+hs41LQvCup31mUOGE0NrI6Y687lFW9veaneRWtrFLc3V1KEjjQF3ldjgADqSSfzNeAf+CNfxf8QeH7bVfHfxO8P+Fr+4jVzpsNnJfvBkfdkcMihx0IUsP9o1/wAOUvE//RwWl/8AhOSf/H6/4cpeJ/8Ao4LS/wDwnJP/AI/Wk2J0zSrLTWlEhtLeOIuBjdtUDOPwqqqqqqqqqv8Aglx/yeP8Zv8Arwv/AP05R1VVVVVVVVVVVVVVVVVVVV4+/wCRSvv+2f8A6MWqqqqvGHh6DxX4X1Pw7PtxfWzxozDIR8ZRvwYKfwqzutR8Oa1Be25a3v8AS7pXTPBjljbI/IitH1Wz8YeFrHXLBgYdStEniH93coOD7g8GiMcHqKqqqqrGeBX7RnxC/wCEK8LDwnplwF1nX4iJSp+a2tTwx9i/Kj23dOK+Gfga6+IPiy10KLclsv728mUf6qFev4nhR7ke9WVlaabZQafYQJDb20axwxqMBFUYAH4Cqqqqqqq8If8AIs6d/wBcBVVVVVVVVVVVVVVVVVVVVXxi/wCT6PCP/X1pf/oVVPb29zH5VxCkq/3XXNXPhDSJ2Lx+bD7I3H65qTwRbEYiv5F9NyA/4VdeAmuIZLd72KWOVCjq8ZAYEYIPJrx/4UufA3jbW/CF3zJpN9Lbhj/GqsdrD2ZcH8a+C91qnj74W+HvE9ui3DzWginbzFDebETG5IJyMshP0IPepNA1iMfNp8p/3Ru/lS6TqjNtXTrnI7GFhik8Oa1KPlsH/wCBEL/M0nhHWXHzJEn+9J/hmo/BN83Et5Av+6Cf8Kj8DrwZdSJ9lix/Wo/BWmr/AKy5uG9gQP6UnhPREADQO+P70h/pik8P6NGMLp8R/wB4Fv518VfiT4H+DXhWXxL4ghtw7ZSysokVZbuXH3EHYdCW6KPwB+J/xO8UfFnxRP4n8T3QLHKWtrGSIrSLPEcY7D1PUnk1+yx+yv8AZvsfxN+Jum/vvll0nSZ0/wBX3WaZT/F0KoenU84Aqqqqqqqv+Cmv/Jj/AMSf+4V/6dbSv+CWf/Jl/hH/AK/9S/8ASyWqqqqqqqqqqqqqqqqqqqr9u/8A5M/+Kn/YAk/9DSv+CQf/ACahd/8AY3X3/om3qqqqqqqqqqqqqqqqqqqqr4rf6/Tf9yT+a1+054DNlqVv480+L9zfEQXoUfdlA+R/+BKMfVR61+zb48/sHxI/hG/uAtlrJ/cbjgJcgYGP98Db7kLWn6m9qRFIS0Xp3X6UrQ3MWVKujD65rUdKa3zNbgtH3Xuv/wBaqqqqqqqqqqoLS4ujtgjLY6noB+NWuiRIA1y/mN/dHAoLFBHhQqIo+gFXWtQR/JbL5jep4Aq5vLi7OZpCR2UcAfhXibxLo3hHRp9d128WC1txyerO3ZVHdj2H9K+JnxL1n4k6yby8YwWFuSLOzDZWJfU+rHufw6V8Efgi/iZ4fFvi22K6Qh3Wtq4wbsj+Jh/zz/8AQvp1SOOONYokVVQYVVGAAOwFVov/ACGLD/r5j/8AQhVVVVVVVVVVVVVVVVVVVVVfsH/8lL8ef9eq/wDo81eWdpqNpNYX9tFc21xG0c0MqB0kRhgqwPBBHBBr9p/9nS6+Emst4m8M28k3hLUpj5R+8bCViT5LHqV/use3B5GW/Zh+LsnhPxXY+BvEV07eHtauVhRy/wDx4zucK4zwELEBhxjO7sQZPA8R/wBVqLD2aLP9ak8E3YH7q+hb/eUj/GpPB2rrwrW7/wC65/qKk8L62n/Llu/3ZFP9ak0TV4x82nXB/wB2Mn+VR6Pqsv3NOuPYmMj+dR+GNbkAxZED1Z1H9aTwdq7j5mt0/wB5z/QGk8EXR4kvol/3UJ/wqPwRAP8AW6g7f7sYX+pqPwZpS43y3Dn0LAfyFR+FtDTraFyP70jf40miaRGMLp0Bx/eTd/OktLSMYitol/3UAqvjP8YvDnwY8JSeINYIub2bMenaesgWS6lx+iDgs2OB6kgHxr4z8U/E7xZc+JfEV1JfanqEu1I0BIjGfkijXnCjOAB+pJJ/Zc/Zng+GtjF438b2Mcviu6TMELYYaZGw+6O3mkZ3N2B2j+Imqqqqqr/gtb/yM/wo/wCvDVP/AEZb1VVVVVWf/H1F/viqqqqqqqqqqqqqqqqqqqqqqqqq+PPhGWeO38X2UW7yVEF4AOi5+R/zJU/Vaqq8H6+3hfxLYa4AzJbS/vVXq0ZGGA98E/jitN/tD4xfERL27tymnW2GeM8iKBTkIT6sf5k9BWABgdqqqqqqtb/Zq+B/if4vR/HPxL8PNL1fxjBp8VjBeX0XnpCkbMyusTZQS/PjzMbgFUAgDmeCC6gktbmGOaGVCkkcihldSMEEHggjjFf8FC/h7pP7JP7a2neJ/g9bRaHHc21j4r02ztF8uKxuPtEqOiKOFQyWzNtHygPtAAAFeOviLo3gL4Xa78VtXjkGmaDoc+rzROQjtHFCZdnf5iBtA55IHNf8E4PhZL+2H+1d4w+Pnxqgj8QQeHJBq91BcgvFPqVxI32ZGU5BijWKVlTOB5Ua4K5FeIfD2heLNDvvDPibSLTVNK1OBre8sruISRTxsMMrKeCCK+Esl7+wh/wU1f4cafqFxF4Xu9fTRZo5XJE2magEa1Mh/iMXmwOW7tE3TJFf8FrvFWrXVl8IvhNYXTxWmt6je310rMRG0sYhhgJA67RPN24yMdTXg7wnongPwlovgnw1Zpa6VoNhDY2UCjASKJAij8lFfsz20Pwg/wCCvPxG+HHhcG20bxCupLJaRHEaCSGO/UBegCuCq+gJA4Jr/gpB+0Nrfwu+Flj8Hvhi8tz8S/i1cf2Noltat+/ggkKpNMuOVY71iQ5B3Sbgf3Zr9k79nrRP2Yvgb4f+FmmCGW/t4vtOtXsa4+238gBmk9doICLnoiKKqq+L0h/a1/4K1+G/hlev9q8KfDaaOKWEjMbLZxG7uQy/7dwRAT3AX0qvE3/KYOH/ALKhZf8AocVVVfG3/gpP+yf8DNYu/DOseNrrxJrtgzJdab4btftrwOvBRpSyQhwQQU8zcpHzAV4A/wCCu37H/jfVI9K1XUfFfg4yyBI7jxBpKiEk9CXtpJtgzxlsAdSQOa0nVtK17TLXWtC1O01HT72JZbW7tJlminjYZV0dSQykcgg4NfET9vX9nz4VfGhPgL451LXNO8TyXdpbKzaYzW2bkIYpPOBxsxIMt/DhgeQar9o39sz4Gfsr6hoelfFjWNRhvPEEM01pBYWTXLCOMqpZwCNoJbA9dreleEfFOieOfCmjeNfDV4LrSdf0+C/sJwMeZBNGrxtjtlWBxXx8/aA+G37NfgMfEX4pajdWmkNfRWUZtbYzySTSBiqqg6/Kjk+ymvgf8bPBP7Qvw8s/ih8OzqL6DfzzQ20t9am3eUxOUchSSdu9WXPqprxE/wDwyJ/wV7sb3T3+x+HPitPE1xCDhXj1RjE5Y9Ao1CMyegC49aqqqqqqqtZ0jTvEGj32g6vbLc2OpW0ltdQt0kikUq6n2Kkj8a+OPwP+IH7Ff7QNlDrGlzXdlo2sRan4b1OVCINVt4ZlkjO4DAcYVZEHKt6gqT8N/wDgoJ+yh8RPCln4il+Luh+G7qaMfadL1y6FncWsmPmQh8BwP7yEqfXtX/DZf7KH/Rw/gL/wdw//ABVf8Nl/sof9HD+Av/B3D/8AFVBNDcwR3FvKskUqBkdTkMCMgj2xVVVVVVVVVf8ABLj/AJPH+M3/AF4X/wD6co6qqqqqqqqqqqqqqqqqqqq8ff8AIpX3/bP/ANGLVVVVVftAeFx4c+I15PBFsttXUXkWBxubiT8d4Y/8CFfskeLv7Y8CXXhW4lBn0G5PlAnnyZSWH5OJPwxWqW4gvGCj5X+Zf61VVVVWqarpXhHw9feMNfk8uy06IyY7ueige5YgD3NeMvFerePfFN94l1Ri9zfzZWNSWEa9EjX2AwB/9evgp8O18A+Eo/tsIGr6mFmvSRynHyx/8BBOf9ot2xVVVVVVVV4Q/wCRZ07/AK4CqqqqqqqqqqqqqqqqqqqqvjF/yfR4R/6+tL/9Cqqqq/bq8GjRPilY+LIIdsPiTTwXfH3p4MRt/wCQzD+dfsCeMhdeH/EfgK4mG+wuUv7ZSeSkg2SY9gyIfq9VVVVVXxY+LHhX4P8AhWXxL4luNztlLKyjYCW8lxwijsOmW6KPwB+J/wAT/FXxc8VTeKPFF1vkb5LW1jJ8q1izxHGOw9T1J5Nfsv8A7KMWmR2fxI+KOm7r/KzaZpE68W/dZZlPV+hCH7vU88LVVVVVVVV/wU1/5Mf+JP8A3Cv/AE62lf8ABLP/AJMv8I/9f+pf+lktVVVVVVVVVVVVVVVVVVVX7d//ACZ/8VP+wBJ/6Glf8Eg/+TULv/sbr7/0Tb1VVVVVVVVVVVVVVVVVVVV8Vv8AX6b/ALkn81rxV4csfFvh2/8ADuoqDDewlN2MlG/hce4IB/CtX0vVPCmvXGlXZaC/0y4KFkJG11PDKfTgEH6Gvhh41h8feDrLXdyC7C+VeRrxsmX73HYHhgPRhVlfS2b5Q7kP3kNW9xDdRCSFsjuD1H1rUdJ3ZntF5/iQf0/wojHB6iqqqqqqqq3s7i6OIYyR/ePAFW2iQRgNcN5jeg4AomKGPqqIo+gFXWtxJ8lqvmN/ePAq4up7pszSlvQdAPwqvE3iXRvCOjT67rt4sFrbjk9WduyqO7HsP6V8TPiXrPxJ1k3l4xgsLckWdmGysS+p9WPc/h0r4K/AyXxE1t4t8YW5TSgd9rZuMNdejMO0ft/F9OqRxxxrFEiqqDCqowAB2Aqq0X/kMWH/AF8x/wDoQqqqqqqqqqqqqqqqqqqqqqv2D/8Akpfjz/r1X/0earxDoGkeKtDvfDmu2Ud3YahA0M8Mi5DKR+hHUHsQD2r4jeCdS+HHjjWPBepBvO0q6aNJCMebH1jkHsyFW/Gv2avif/wtT4U6Zqt7dCXV9NH2HUwT8xljAw5/30KtnpksO1VVVVVVVVV8VPij4Z+EfhK58VeJLgfIClpaK2JLubHyxoP5noBkmvib8SvE3xZ8W3PizxNcbppjst7dCfLtYh92OMHoB+ZJJPJNfso/syL4SgtfiX8QdPRtbnRZNLsJk5sFIyJHB/5akHgH7n+8flqqqqqqr/gtb/yM/wAKP+vDVP8A0Zb1VVVVVWf/AB9Rf74qqqqqqqqqqqqqqqqqqqqqqqqqrm2t7y3ktLqFJoZkKSRuuQwPBBFfED4Qar4fnk1Hw9BLfaYxLbEG6S39iOrL/tD8fUkY4PUVXhTwR4g8Y3Qg0mzPkhsS3Mg2xxfU9z7DmvB3g/S/BekLpmnDfI3zXE7DDTP6n0HoO35mqqqqqqqurq1sbWW9vbmK3t7eNpJZZXCJGijLMxPAAAJJPSvHE0v/AAUh/wCCjFvYeFIZLrwXZXMNoboISqaLZNumnJ/hEztJszjmeNTzX/BSrWZ9C/Yf+KV7bFt8llZ2p2nB2z39vC34bZDX/BEbSrWH4MfEPW0X/SbrxPFbyHH8EVsjL79Znqv+Ctky+Fv22rDxJpysLlPD+l3zENtJkjllVcEdOIl5r/gs18IfEfin4W+C/jJ4XsZ528AahcR6k1vHl4La6EW2diOdqSwIM9jLnpk18JP23P2e/iR8FrD4t6n8U/DGhiLTkm1ywvdSjin024C/vYmiJ3k7gwTCnzBgrnIr9hK7Pxg/a9+NX7dviSNtI8AaLDqBt9RvsqsQbbtBJzzFZREuB93eg6EV+zn4h0v4z/Fn4kf8FOPj0z6X4C8FRTad4FtryPP2e3iyplVMnMgD7QBkNPcyhcFBjR/29f24/j1DqPjr9lb9krS9R8AadcvEl5r1wWubtU+95YFxArPjqkQm2k4yxHP7Ff7dvh39q3+1/BfiHwxJ4N+I3hpWbUtBmkZhLGjCN5Yiyqw2yEK8bDchZRls5qq/4JeMvj39uf44/FK5kWeSa31OVHPHzXeppJuAIyOIyO2AxGOeK8Tf8pg4f+yoWX/ocVVX/BWb9qbxF8EPhVpHwv8Ah/qs2neJPiCZ1uL+3fbLZ6dEFEuxgco8rSKgYdFEmMHBH/BPD/gnN8JrL4S6D8Zvjp4KsfFfibxVbJf2Gm6rB51pp1lIA0IMDZSWR0xIxcEKHCgAqzN+0r/wTf8A2dfjR8PtR03wL8N/DfgTxdBbs2j6poWnR2EYnUEqk8UIVJI2OFYlSwByDkc/8Ejv2h/GXgr4s6p+yL4/u7s2N6Lt9Gsrl9zaZqNtue5gT+6jokzEA4DxZAy7E/8ABbH4Vyab4t+Hvxy0uBo/7QtZdEv54wV2SwsZrckj+Jlkn564iHpx8AviPD8X/gl4G+JsUyyP4j0G1u7jaPuTtGPOT6rIHX8K/bTtdf8A2w/2x/i3YeGLl5dM+DXg6/eNYxuymmJuuFA/vNdzSLxyQAOtf8EmPi0PiP8Asl6b4YvLtZNS8A6jPpEqlsuYCfOgYjsuyUxj/rifSv8AgtZ8R7vWfFHw0+Amhs086Ry6zdWsY3NJLM/2e1AA/i+S447+YK+Anwtsfgn8F/BnwpsBGR4a0eC1meMYE1wFzPLj/blLv9Wr/gtVaS+GPiX8F/iPp4Vb77LfRxsT/FaT28qZ4zjNwe/5d7K7g1Czgv7Vy0NzEskZIxlWGQcduDVVVVVVVeKHaPw1qzxuVZbGYqQcEEIeRV3eXd/Mbm+u5riZhgySuXY46cnmqqvCv/Ir6P8A9eEH/otaqqqqqqqqr/glx/yeP8Zv+vC//wDTlHVVVVVVVVVVVVVVVVVVVVXj7/kUr7/tn/6MWqqqqq/ac8LjVfBlv4jhTM+i3HzkD/llJhW/8eCfrX7NXi4eFfilYW88u211tDYSgnjc5BjP13qo/wCBGtbtxJbCZR80R5+h/wAiqqqqtNsDeSbnH7pDz7+1ftT/ABRXxDrifD/RLgHTtFkzeMh4luQMbfogyP8AeLegr9nT4cDxJrp8X6rb7tO0iQeQrLxNcdR+CcN9SvvVVVVVVVVXhD/kWdO/64CqqqqqqqqqqqqqqqqqqqqvFH7OnhbxX8XtM+Md5rmqRanpclu8VrEY/JYwnK5ypbnvzVVVV+234M/4ST4OnX7eMNc+Gr2O5yBz5T/u5APbLIx/3K/ZN8Zf8Ib8cNCMs2y11otpk/OM+bjyx/39WOqqqqq+Lfxf8JfBzw02v+JbkvNLuSxsIiPNu5APuqOyjjLHgZHcgH4n/E/xV8XPFU3ijxRdb5G+S1tYyfKtYs8RxjsPU9SeTX7LH7LA0QWfxM+JmnA6jgS6VpU6f8evdZpVP/LTuqn7vU/NgLVVVVVVVVX/AAU1/wCTH/iT/wBwr/062lf8Es/+TL/CP/X/AKl/6WS1VVVVVVVVVVVVVVVVVVVft3/8mf8AxU/7AEn/AKGlf8Eg/wDk1C7/AOxuvv8A0Tb1VVVVVVVVVVVVVVVVVVVV8Vv9fpv+5J/Nar9qDwMoNp4+0+AAki2v9o6n/lm5/wDQSf8AdFfs4+NR4d8Ynw7dy4s9dCxrk8JOufLP45K+5K+lVbXMtrIJIW57g9DVnfQ3seUOHA+ZD1FajpaXIM0GFl7js1OjRsY3Uhl4II6VVVVVVW9jc3RHlR8f3jwKttFt4gGnPmt6HgflTtFBHlmVFUd+AKutbjXKWqbz/ebgflU9zPctumlLHsD0H4VVeJvEujeEdGn13XbxYLW3HJ6s7dlUd2PYf0r4mfEvWfiTrJvLxjBYW5Is7MNlYl9T6se5/DpXwQ+B7a60HjDxhaEaaCHs7ORcG5PZ3H/PP0H8X06qqqoRAAAMAAYxVVWi/wDIYsP+vmP/ANCFVVVVVVVVVVVVVVVVVVVVV8Hf2dPC3wX17WPEGg65ql9NrUYSZLwx7UAct8u1Qep71Vft3fC832laZ8VtKtQZdPxZaoVXkxMf3Uh/3XJUnr+8XsK/Y6+KC+AfijHoGpXRj0nxSq2coZsKlwD+4c/iSn/bTPaqqqqqqqqviD4/8MfDPwvdeLPFd+tvaWwwiDmSeQj5Y41/iY46duScAEj4wfF3xL8ZfFkniTX3EMEQMdhYxtmO0iznaPVj1ZjyT6AAD9kz9mMWy2PxX+IdgDMwEujaZPH/AKv+7cSA9+6Dtw3XGKqqqqqqr/gtb/yM/wAKP+vDVP8A0Zb1VVVVVWf/AB9Rf74qqqqqqqqqqqqqqqqqqqqqqqqqqrVvBHhLXJDNqnh+ymlb70nl7XP1ZcE1a/DPwFZyiWHwxZsw6CUGQfkxIqKGG3iWC3iSONBhURdoUewFVVVVVVUxVVLMwAA5J4xX/BQP9tfxN8cvFy/sZfsrPJrf9tXQ07WtS0yTLanMThrKFxhRAMHzZc7WAYZEasX/AGF/2M/D37Inw1+xXb2up+OtfVJfEOqxLlcjJW2hJAIhjz1PLtliBlVX9v7wnceNP2Nfivotpb+dJFoTX4TbuOLSVLkkD1AhJ/Cv+CIPiq0ufh78TvA4mQXWn6zZ6j5ZGGKTwtHkeoBtufTIz1FV/wAFFYx8Xf8AgpDpfw30sJdTJLoOgFPL3LvmKSFSOjAfahn8Qelajp2n6vYXGlatY297ZXkTQ3FtcRCSOaNhhkdWyGUgkEEYINax/wAEsf2KNZ8SnxG/wuurVXlMkunWes3UNo5JyRsV8ov+yjKB2Ar9t+9t9RuvAH/BNT9mLR7Dw/N4wmjm1+PTYRHDpWlqxkw4Xn5tjzSc7isQB3edz/wVZ0zSPgT+xv8ADf4B/DyCSw8PHWYbUoDzNBawO+JCMbmeZ0lY45dc1+zR4M0n4e/s9fDjwdosCRW2neGbFTtH+slaFXlkPu8jO592NeMoI/hb/wAFptBvfDiiCLxY0DXtvENisbrT2hlyO+XXzT6sc1Vf8EkAnhz9q/41eCLxj9vhsLjhhtOLfUBG/wAvUfNKv0qvE3/KYOH/ALKhZf8AocVVX/BbGPVB+0Z4LllDf2c3gmJbckDHnC9uvNx3ztMP6V8N59Lu/h14WutDaNtOm0WzezMf3TCYUKbfbbiq+FCpqv8AwWBuG8IsjRn4la1JuiXKsifaWuCMZ4KrKc9O+QOa/wCClHwn/wCFt/sfeNrO1tvO1HwvEniCwwm4q1plpSB6m3M6/wDAu/Sv+Cb37SWn+Ef+Cf8A431XWblGm+Dz6jJFDIc+ZFIhubdev8c7yxgeoFf8EdfhPc6t4F+JPx18b27X1z491E6Yktzgm5hTc90/HUSSzbT7wnp3/wCCbF5e/s3/ALcnxS/ZW1m4dbXVftNvZeZ964msJHlt5B6B7SSd/wDvmvCFl/w1x/wV41TWp1e98PeAtXkueBuSODSQsMJHbY92sbc9fMPrVf8ABcK+TUNf+DHhu0jMl7DBrEvlqcswmezRAB15MTfWtJsP7L0my0vzvN+x28cO/bt3bVAzjnGcdKqqqqqqqeCG5gktriNZIpUKOjDIYEYII9MVa/sm/st2cC28P7OXw0ZVzgy+FbOVv++mjJ/Wv+GV/wBmD/o3D4Xf+EfY/wDxqv8Ahlf9mD/o3D4Xf+EfY/8Axqv+GV/2YP8Ao3D4Xf8AhH2P/wAaqKGKCJIII0jjjUKiIu0KAMAADoMVVVVVVVVVX/BLj/k8f4zf9eF//wCnKOqqqqqqqqqqqqqqqqqqqqvH3/IpX3/bP/0YtVVVVVr+j23iDRL/AEO8GYb+2eB+OgZSMj3Gc1dW+o+HdZltZGaC+0y5KEqcGOWNsZH0IrwV4jtfHHgvSvEkJUrqdmryKP4JMYdfwYMPwqWNopGib7yEg1VVVpayXkwij6fxH0FfHD4kW/ws8DSPp8qLq+oBrfTkPJDY+aUj0UHP1KjvXh3QdV8Z+IrTQ9P3TXuoTYLuScZ5Z2PXAGWJ9q8LeGtN8I+H7Lw7pMe23s4goYjBdv4nPuTkn61VVVVVVVV4Q/5FnTv+uAqqqqqqqqqqqqqqqqqqqqqqqqrxb4ds/F3hbVvC1+AbfVrGa1kJH3RIhXI9xnI9CKnh1Twtr0lvIWttS0i8KtjrFNE+D+IZa8E+Jbfxn4O0TxZa7RHq9hDchVP3C6AlfwJI/Cqqqr4wfGDwt8GvCz+IPEEwmupQyafp8bgS3coHQeijI3N0A9SQD8R/iP4p+Kvim48VeKr0zXMx2wwpkR20YPyxxr2UZ+pJJOSSa/ZY/ZYGiCz+JnxM04HUcCXStKnT/j17rNKp/wCWndVP3ep+bAWqqqqqqqqq/wCCmv8AyY/8Sf8AuFf+nW0r/gln/wAmX+Ef+v8A1L/0slqqqqqqqqqqqqqqqqqqqq/bv/5M/wDip/2AJP8A0NK/4JB/8moXf/Y3X3/om3qqqqqqqqqqqqqqqqqqqqr4rf6/Tf8Ack/mtV4m8P2PirQL7w9qS5t76Fo2OMlD2Ye4IBHuBWqadqfhfXLjTLrdb3+m3BRipwVdDwyn8AQfoa+G/i+Hxz4N07xArqbiSPy7tVGNky8OMdgTyPYiqjlkhcSROVZehFWGpR3aiOQhZQOnr9KvtPivUzwsgHytj+dTwS28hilXDD9aqqqrawubsjy48L/ebgVbaNbQ4aU+a49RwPwqSSKCPdI6oo9eKutcUApaJk/3mH8hU081w2+aUufftVVXirxVongzRZtd167ENtDwABl5GPREHdj6ficAE18TPiXrPxJ1k3l4xgsLckWdmGysS+p9WPc/h0r4IfA9tdaDxh4wtCNNBD2dnIuDcns7j/nn6D+L6dVVVUIgAAGAAMYqqqtF/wCQxYf9fMf/AKEKqqqqqqqqqqqqqqqqqqqqqqrxV4b0zxh4b1PwrrMXmWWq2sltMB1CuuMj0I6g9iAa8aeFNY+HvjHVPCWrZjvtHumiLqCu7HKSL3AZSrD2Ir4AfE6P4tfDDS/E0sqnUoV+yamo423MYG447BgVcDsHAqqqqqqqvGHi/wAPeA/Dt54q8U6jHZadYpukkbkk9lUdWYnAAHJJr46fG7xD8a/FDajeu9to1m7LpenZ4gjJ+82ODIwA3H8BwBX7J37Mf9uvZ/FL4h6ew06JxLpGmzJj7URyJ5Af+WefuqfvdT8uN1VVVVVVVX/Ba3/kZ/hR/wBeGqf+jLeqqqqqqz/4+ov98VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX/BTX/godqep6pr/AOzF8FNQubCx0+4m07xdrSZilupUYpNZQ9CsSsGWR+rkFR8mTJ+w5+0Z+wF+yR4UGqarrmva78RtVhA1XW18PSFLZT/y7Wu4hliHdsBpDywACqv/AA95/Y0/6Dnir/wQv/8AFV4P8UeEPjV8MtM8WaTC994Z8Y6Ss8UV3CUM1rPH910PTKtgj3rwxf8Ajf8A4JS/toXsXiXRtR1PwHrCS26SxDnVdIeQNHNEThTcQsqbkOPmDrkK4evH3/BVn9kDwv4EufE/hTx3P4r1k25ax0K1065t5ppSPlSR5YgkS5xuYk4GSA3AP/BNb4H+P/2j/wBpTWP20fitp7jSbHUrq/s53jKR6hq0pYAQqesUAZjkHCssSjOGxXxs+LnhX4EfCvxH8WPGU4XTfD1k05iDhXuZfuxQIT/HJIVQe7DPGa/4Jt/CPxRr8fin9tf4xwibx38Xbh5tOEin/QdK3AoI9xJVJNiBRziKGHBwxr/gq58Edd+MP7Ls2q+FNNlvtX8C6pHrIt4Yy8stqI3iuFQDk7VkEp9oj9K/Ys/bl+A3jT9nDwvF43+Kvhvw14k8IaNBp2t2et6lFZylreMRidBIw81JFVWymcM204PFfszNcftlf8FM/FH7T/h+0uT4A8CKy2F3NGyJOy2ptLZQDhg0mZbnaR8oXDYyAarwaW/Zi/4LF6tpF8wtdG+JV7cJHKflEq6oguIwo44+2qsWPVTjPGa+NPjvw/8AC/8A4Klax8RPFcs8ej+HPiDb3180ERkdYozGzFVHU4HSv+HvP7Gn/Qc8Vf8Aghf/AOKrQ/8AgrD+yD4h1rT/AA/put+KGu9Suo7W3D6G6gySMFXJzwMkc1/wVK/ZK179ov4Tab41+HeltqHjPwE800NjEuZdRsZQvnwoP4pFKJIi9TtdVBZwD+wF/wAFMvAfw8+Hun/Af9pS+u9Bfwsgs9G102cs8Zt1JC21wkas8bx8IrBSu0ANtK5b9p//AIKzfA3wd4A1DTf2ffEjeMPGmoW7RWFxFYyw2mmMwx58jTIvmFc7lRQwJADFRX/BJH9kbxhoutXn7VPxS0u5tJdRs5LfwxBegiecTH9/euG+YBlBRC3Lh3bGCrHUdPsdX0+50rU7WO5s7yF4biCRcrJG4KspHcEEg/WvGeo+Mf2cbv45fsmWkd1NH4i16006cquWuIrC7kkt3AHJ81XjYYHIbkc4r9mX4SQfAr4A+BfhTHEiXGg6PEl8UAAe8f8AeXLDHZpnkP41/wAFLLXV/wBmb9tT4Y/tbeGtPM0WpxxPdxgbfPuLLEU0ZcdPMtZIkGf7rdQMD/gi98Lbm18A+O/jxrsLyah4s1VdOs7mY5d4YAXmcH0eaXB94O3ev2rF/wCGm/8Agqx8P/hDpmbnTfBj6da6iEG5NkDNqF4c9AfLbyiegZAME9aqqqqqq/4KLftc/HO7/aM8S/DTwt491/wn4b8Hzx2ltZaTfSWTXMnlo7zzNGwaQlmO0E7QoXAyST/wTa/aP+JXxv8AgB40074jazeatrHglzDa61Ox864hlgZkEkmcvKjI2XPJDJnJyT/w1R+09/0ch8Uf/Cwv/wD47X/DVH7T3/RyHxR/8LC//wDjtf8ADVH7T3/RyHxR/wDCwv8A/wCO1/w1R+09/wBHIfFH/wALC/8A/jteGpZbjw5pU88rSSSWULO7kksSgySe5qqqqqqqqqv+CXH/ACeP8Zv+vC//APTlHVVVVVVVVVVVVVVVVVVVVXj7/kUr7/tn/wCjFqqqqqq/aR8L/wBh+Pjq8EW221uETAgYHmr8sg+vCsf9+v2P/Fxv/DeqeDLmXMmlzi4tlPXypPvAewcZ/wCB1rdvsuFnUcSDn6j/ACKqqiieaRYolyzHAFKLLRNOmu7u4jiht42luJnOAqqMsxPYAA18YfiPc/E7xpda5udbCHMGnwtxshU8EjszfeP1x0Ar9nL4c/8ACP6GfGWqwY1DVowLZWHMNv1B+r8H6BfU1VVVVVVVVXhD/kWdO/64Cqqqqqqqqqqqqqqqqqqqqqqqqq/a88Gf8Ih8b9Xmhi2WuvImpQ4XGTJkS/iZVkP4iv2HfGQ8QfCSbwxPNuufDV88SqTkiCXMiH/voyj/AIDVVVfGD4weFvg14WfxB4gmE11KGTT9PjcCW7lA6D0UZG5ugHqSAfiN8RfFfxY8Vz+KPFF2bi6nOyCCMHy7ePPyxRr2UZ+pJJOSSa/Zd/ZWh8MxWvxF+JmmJJrLgSabpc6ZFiOokkU/8te4U/c6/e+7VVVVVVVVVV/wU1/5Mf8AiT/3Cv8A062lf8Es/wDky/wj/wBf+pf+lktVVVVVVVVVVVVVVVVVVVX7d/8AyZ/8VP8AsASf+hpX/BIP/k1C7/7G6+/9E29VVVVVVVVVVVVVVVVVVVVfFb/X6b/uSfzWqr9qDwT9l1Gz8dWUf7u8xbXmB0kUfI34qCP+AD1r9mfxsdI8STeD7yXFrrA3wbjgJOo/9mUEfVVqqBKkMrYI6Edq07VVlAguWAforHvV1aQ3kXlyjkfdYdRV3ZzWcmyQZB+6w6Gqq2065usFE2r/AHm4FW2kWsGGcea47sOB+FSzQwJulkVAOmautcyClon/AAJh/IVNNLO2+WQufc1VVXivxXongzRJ9e167ENvCMKo5eV+yIO7H0/E4AJr4j/EbWviPrR1HUXMNpCStnZq2VgX+rHjLd/YAAfBD4HtrrQeMPGFoRpoIezs5Fwbk9ncf88/QfxfTqqqqhEAAAwABjFVVVWi/wDIYsP+vmP/ANCFVVVVVVVVVVVVVVVVVVVVVVVX7d/wuRTpnxa0q35YrYaqFXqcEwyn8AUJP+wK/Yr+KLeDviOfBOo3G3S/FQESBmwI7tc+UR/vZKY7lk9Kqqqqqq8Qa/o/hbRLzxFr9/HZ6dp8JluJ5OiKPpyT2AHJJAHWv2hvj9rHxr8RGO2eez8M6e5/s6wZsFj0M0gHBcjPHIUHA6kn9lf9mR/H9zD8QPH2nuvhq3bdZWkg2/2k4PUjr5QI5/vHjoDSoiIscahVUYVQMAD2qqqqqqqqr/gtb/yM/wAKP+vDVP8A0Zb1VVVVVWf/AB9Rf74qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqvE/wCwD+x94z8S6t4w8TfBHTL/AFjXL6a+1C6e+u1ae4mcySSELKACzMxwABzxiv8Ah27+xH/0QLSf/Bhe/wDx6v8Ah27+xH/0QLSf/Bhe/wDx6vCHhHw54C8LaV4K8IaXHp2i6JaJaWFojMywQoMKgLEsQAAOSTXxR+D3wv8AjZ4bPhL4reB9K8TaXv3xw30O5oXxjfE4w8TYJG5CDg9a0P8A4Je/sRaDrK61B8G/tbxuXitr7Wr24gQ56GN5iHAHGH3D1yea0fRtH8O6Va6F4f0qz0zTbGJYbWzs4FghgjUYCIigKqgdAAAKr46/s9fDD9pDw1Y+D/ixpt/qWjWF6L2Ozt9RmtEeZVKqz+Uyl8BmwDwNxNWGn2OlWFtpemWcNrZ2cKw29vDGESGNQFVFUcBQAAAOABVeN/8AgnF+xl8QfFEvi/XvgtZxahcymW6XTdQurCG4YnJLQwSogJJJJUKSTyTXgD4c+BPhV4XtfBfw48J6Z4d0SzyYrKwtxEm49XbHLOcDLNlieSTVV/wVo/Zz1zxp4D0T9pL4dQyr4p+GLb71rdT5z6eHEglXAJJt5f3nYBXlY/dr9kn9oTRf2nPgV4d+J+nTQjUpYRaa5aRn/jz1CNQJkI7Akh1/2JEPevHH7Cf7JnxI8Wan458bfBnTdV13WZzPfXkl7dI00hABYhJQo4A6AV/w7d/Yj/6IFpP/AIML3/49Wmf8E7/2MdG1K01jS/gVpcF5YzpPbyi/vCY5EYMrAGbHBAPNV8Yf2Jf2XPjvqkuv/Ej4Q6VeazMD5up2Ukthcyt/eke3dDK3vJu7elfDb/gnn+x58K9Ui13w18FtMutSgcPFc6xcTakY2HIKpcO6KQehCgj1qq8Vfsgfs1+N/iaPjJ4q+E+l6j4xF3b3Z1SSadWaa3CCJ2RXCMVEaDlSDt5zVfGD4DfCL4/aNY+H/i/4Is/Elhpt0bq0iuJZI/JlKlSwaNlblSQRnHTjgV8Ovhx4I+Evg3Tvh98OfD1vofh7ShILOwgLMsW+RpHOWJYku7MSSeTX7Q3xt8Mfs7/B7xJ8WvFMqNBotoTa2pfa17dN8sMC98u5UZwdo3MeFJr/AIJRfAnxLqL+LP2zfirE83ib4iXNwNJeePaxgkmMlzdBSOPNlAVcY+SNsZVxVVVVVVVftP8A/BOb4OftOeNV+I+oa3rPhjxHLCkN/c6aI3jvlQBUaRHU/OFAUMCOAAQcCvgR+zT8Nv2dvhbL8Kvh9DeCzvDLJf3926vc3k8iBGldgAuQoUAAAAKOOpP/AA5j+BH/AEVLx7/31af/ABmv+HMfwI/6Kl49/wC+rT/4zX/DmP4Ef9FS8e/99Wn/AMZr/hzH8CP+ipePf++rT/4zWnWUem6da6dC7OlrCkSs3UhVABP5VVVVVVVVVX/BLj/k8f4zf9eF/wD+nKOqqqqqqqqqqqqqqqqqqqqvH3/IpX3/AGz/APRi1VVVVVftH+Fxrvw/fVYI91zokwnXA5MbfLIPpyrH/cr4A+Lv+EO+KWj3csm21v3+w3OTj5JSACfYOEY/StUt/tFm4UZZPmX8P/rVVAEkADJ7CtM08Wsfmygeaw7/AMI9K/av+KRsbNPhnotzie6VZdUdTykfVIvq3DH2A7Ma+DHw8b4geLI47uInStOxNfN2Yfwx/wDAiPyDUiJGixxqFVRhVAwAB6VVVVVVVVVXhD/kWdO/64Cqqqqqqqqqqqqqqqqqqqqvi/fXsf7cPhK1ivZ1ha50zMayEKctzx0qqqqr9vjwYL/wl4f8dW0IMulXb2dywHPlTDKk+wePA95K/Yd8ZHQPi1N4YnmC23iWxeJVJwDPFmRD/wB8iUfVqqq+M/xo8LfBbwyda1txdX9wCunabHIFkunHXnnagyNzkYGR1JAPxG+Iviv4seK5/FHii7NxdTnZBBGD5dvHn5Yo17KM/UkknJJNfstfssp4VS0+JHxI08NrbASabpsy5FiO0kg/56+i/wAHX733aqqqqqqqqqq/4Ka/8mP/ABJ/7hX/AKdbSv8Agln/AMmX+Ef+v/Uv/SyWqqqqqqqqqqqqqqqqqqqr9u//AJM/+Kn/AGAJP/Q0r/gkH/yahd/9jdff+ibeqqqqqqqqqqqqqqqqqqqqvit/r9N/3JP5rVV408MWnjLwvqPhu7CgXkJWN2H+rkHKN+DAGnTVPDOtlG32mo6XdY9Giljb+hFeCPFVp418Laf4ks9o+1xAyxg/6qQcOv4MD9Rg96qq0/VjGBBdNleiue319qlhhuYjHIA6sP8AJFSaHP52yJ1MfZm7fhVrpNrb4Zh5rjuw6fQVPcQW6b5pFUds/wBBV1rbMNlomP8AaYc/gKllkmbfLIXb1Jqqqq8V+K9E8GaJPr2vXYht4RhVHLyv2RB3Y+n4nABNfEf4ja18R9aOo6i5htISVs7NWysC/wBWPGW7+wAA+CfwKGrpB4w8a2hFnw9lYuMef6PIP7nov8XU8dVVVUIgAAGAAMYqqqqrRf8AkMWH/XzH/wChCqqqqqqqqqqqqqqqqqqqqq/YWvr27+JPjqO6vZ5lS1XaskhYD9+emaqq8b+EdK8feEdV8Ha0m601W1aFyBkxk8q4/wBpWCsPcCtd0bXPAniq80S/32mq6JeNGzISCkkbcMp9MgEH0wa+CnxGt/ir8NdG8XpJGbuWHydQjTjy7lOJBjsCfmA/usKqqqqrUdQsNJsLjVNUvIbW0tImlnnmcIkaKMlmJ4AAFftK/tF3/wAY9Y/sTQZJ7Twnp0h+zwsSrXjjjzpB/wCgqeg56k4/Zg/ZtuvitqUXi7xZayQ+EbKUjaSUbUZF6xoeoQH7zf8AARzkraWdpYWkFhYW0VvbW0axQwxIESNFGFVQOAAAAB2qqqqqqqqqv+C1v/Iz/Cj/AK8NU/8ARlvVVVVVVZ/8fUX++KqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqeCC6gktbmGOaGVCkkcihldSMEEHggjjFeFfAfgbwJBPa+B/BmheHobpg88eladFaLKwGAWEagMQOMmqqqqqqqqvFPgvwd45sItL8a+E9G8QWcMwmittUsI7uNJACocLIpAbDMM9cE+tWNjY6ZY2+maZZwWlpaRLDb28EYjjhjUAKiqOFUAAADgAVVVVVVVVfGTRPEfib4QeOfDfg67mtde1Xw3qFppc8EpikiupLaRImVwQVYOVIIIwecirr46/tAWVzLZ3nxk+IMFxA7RyxS+IbxHjdTgqwMmQQRgg9K+CHifTvGnwb8D+KtK1L7fban4fspluC+5nJhXduPJ3BshgeQQQeaqqqqqqqqqqr/AIJcf8nj/Gb/AK8L/wD9OUdVVVVVVVVVVVVVVVVVVVVePv8AkUr7/tn/AOjFqqqqqq1GwtdV0+50y9j3293C8Mq+qsMEfka1/R7vw3r19ol02LjTrl4WYcZKtgMPY4BH1r4XeLE8c/D/AETxIZQ81zaqtz7TJ8knHb5lJ+hFXtv9muXiA4Byv0qtI0/gXcy/9c1I/WviX49034ceEbzxLflXkiXZawE4M0x+6v0zyfQAmr+91rxj4hlvbp5L7VdWusnA+aSVzwAPqQAO3FfDDwJbfD7wla6IgR7xx5t7KP45iBnHsOFHsM9SaqqqqqqqqqvCH/Is6d/1wFVVVVVVVVVVVVVVVVVVVVfGL/k+jwj/ANfWl/8AoVVVVVfGrwb/AMJ/8KvE3hWOLzJ7uwdrVcZzPH+8i/8AH0UfjXgjxNceC/GGi+LLTJk0i+iuQo/jCMCV/EAj8asry21Gyg1CymWW3uYllidejKwBBH1BFV8aPjR4X+C3hdtZ1mQXOoXAZdO05HxJdSD/ANBQZG5u3uSAfHnjvxX8U/Fdx4n8T3j3moXjBIokB2Qpn5Yol5wozwOpJJOSST+y1+yynhVLT4kfEjTw2tsBJpumzLkWI7SSD/nr6L/B1+992qqqqqqqqqqq/wCCmv8AyY/8Sf8AuFf+nW0r/gln/wAmX+Ef+v8A1L/0slqqqqqqqqqqqqqqqqqqqq/bv/5M/wDip/2AJP8A0NK/4JB/8moXf/Y3X3/om3qqqqqqqqqqqqqqqqqqqqr4rf6/Tf8Ack/mtVVftN+CRpPiG28Y2URFvq48u5wOFnQDB/4Eo/NWNfsweNjZ6pd+Br2fEN8DcWQY9JVHzqPqoz/wA+tVVVp+pvakRSEtF6d1+lSXtpHEJmnXaw+XHU/hV1rbvlLVdo/vMOfyqSR5W3yOWY9yaqqqq8V+K9E8GaJPr2vXYht4RhVHLyv2RB3Y+n4nABNfEf4ja18R9aOo6i5htISVs7NWysC/1Y8Zbv7AAD4H/A7+0vs/jPxnZ/6IMPY2Mq/670kcf3PRf4up46gADA7dBVVVVVaL/wAhiw/6+Y//AEIVVVVVVVVVVVVVVVVVVVVVX7B//JS/Hn/Xqv8A6PNVVV+3d8L2stX034raVajyNQAstUKr0mUfupD/ALyArnt5a+tfsP8AxQXwx44ufh5qlztsPEyhrTc3yx3aA4HtvTK+5VBVVVVVzcW9pbyXd3PHDDCheSSRgqooGSxJ4AAGSTX7Uf7Sc/xR1CTwV4PuXi8KWM3zyqxU6lIp4dh/zzBGVU9fvHnAX9m79njUvjNro1XV0ktvCmmzAXs4yrXLDnyIz6kY3N/CD6kVpOk6ZoWmWujaNYw2djZRCK3t4U2pGgGAABVVVVVVVVVV/wAFrf8AkZ/hR/14ap/6Mt6qqqqqrP8A4+ov98VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVWqXq6Zpl3qTR+YLSB5SgON21ScZ/Cv2qfj78Gv2hvFF7488J/AW88C+J9Qn8y+vbfxKtxbX7E/NLLa/ZVxK3dkkUEklg5JNfAv9tn9oz9nXRm8MfDnxzjQS7Omk6lbJd28LsSWaION0WSSSEIBJJIJOa/4eyftf/wDQa8Lf+CNP/iq/4eyftf8A/Qa8Lf8AgjT/AOKrQrubUND06/uCDLc2kUjkDA3MgJwPqaqqqqqqqqr/AIJcf8nj/Gb/AK8L/wD9OUdVVVVVVVVVVVVVVVVVVVVePv8AkUr7/tn/AOjFqqqqqqr9p3wuNL8Y23iS3ixDrFviQgf8to8KfplNn5Gv2OvFxktda8DXMwzEwvrRSecHCSgewIjP/AjWu2+VjuVHT5W/pWmaebqTzZR+6Q/99H0qeaK2hMshAVRwP6Cv2hfifJ4/8Wtp1jchtJ0dmigCn5ZZOjv7jjA9hkfer9mn4cC5nb4g6tBmOEtFpqOv3n6PL+H3R77vQVVVVVVVVVVV4Q/5FnTv+uAqqqqqqqqqqqqqqqqqqqqr4xf8n0eEf+vrS/8A0Kqqqqq+Pvgw+Avi/wCJ/DscIjtlvmuLRQMAQzfvEA9gHC/VTX7JvjIeMvgfoRlmEl1ogbTJ+fu+Tjyx/wB+jHXxo+NHhf4LeF21nWZBc6hcBl07TkfEl1IP/QUGRubt7kgHx3478WfFTxZP4m8TXb3uoXjhIoo1O2Jc/LFEnOFGeB1JJJySSf2XP2XIvA8Vt8Q/iHYrJ4ikUPYWEi5GnA9HYd5v/QP97pVVVVVVVVVVVV/wU1/5Mf8AiT/3Cv8A062lf8Es/wDky/wj/wBf+pf+lktVVVVVVVVVVVVVVVVVVVX7d/8AyZ/8VP8AsASf+hpX/BIP/k1C7/7G6+/9E29VVVVVVVVVVVVVVVVVVVVfFb/X6b/uSfzWqqviJ4Rh8ceD9R8OuE86aLdbOw+5MvKH25GD7E1YXmp+F9dgvoA1vf6XdBgrDBSRG5BH1GCK8L+IbLxZ4d0/xFp5/c30AkCk5KHoyn3DAg/SqqqqqqqqvGPjDQ/A+hza7rtz5cUfEca8vM/ZEHcn9OpwBXxE+I2u/EfWP7R1VxFbQ5W0s4ydkCn+bHjLd/YYA+B/wO/tL7P4z8Z2f+iDD2NjKv8ArvSRx/c9F/i6njqAAMDt0FVVVVVWi/8AIYsP+vmP/wBCFVVVVVVVVVVVVVVVVVVVVV+wf/yUvx5/16r/AOjzVVVfEnwPp/xJ8C6z4J1IhYtUtjGkhXPlSj5o3x/suFP4VdW2veBfFcltLvsdZ0C/IJHWGeF+o+jLmvhP8QLH4o/D7R/GtkFRr6AC5hU/6mdflkT1wGBx6jB71VVTsiIXdwqqMkk4AHrX7Vv7TTeNri5+G3gG/U+HoH2399E3/IQdT9xT/wA8gR1/jIz93Gf2f/gLrXxt8RmIvJZeHtPZTqV+E5/65R54MhH4KOT2B8O+HdE8J6JZ+HPDmnQ2OnWEQjt7eJcBAP1JJySTySSTyaqqqqqqqqqq/wCC1v8AyM/wo/68NU/9GW9VVVVVVn/x9Rf74qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqtbspdS0a/wBOgZFkurWSJC5IALKQM47c1a/8EZf2gXgVr34mfD2KbnckU15Io54wxgUnjHYV/wAOY/jv/wBFS8Bf99Xf/wAZr/hzH8d/+ipeAv8Avq7/APjNf8OY/jv/ANFS8Bf99Xf/AMZrRrKTTdHsdOldXe1to4mZehKqASPbiqqqqqqqqq/4Jcf8nj/Gb/rwv/8A05R1VVVVVVVVVVVVVVVVVVVV4+/5FK+/7Z/+jFqqqqqqr4/+Fx4k+HF7NFHuudIYXkWBzhciT8NhY/UCvg54uHgj4kaJrssuy2FwILrJwPKk+Rif93O76qKuIEuYWgc8MPypUit4Qi4VEHftX7SHxUPhrw6dC0q52alqyNHFtOGhh6PJ6gn7o/E9q8AeDL7x74ps/DtluVJG33MwGfJiH3m/LgepIHetL0ux0XTbbSNMt1htbOJYoY1/hUDA/wD11VVVVVVVVVV4Q/5FnTv+uAqqqqqqqqqqqqqqqqqqqqr4xf8AJ9HhH/r60v8A9Cqqqqqv2/PBq22t+GvHttFxe28mn3TAYAaM74yfch3H0QV+zd+0NY/BPQfF1nqdpNfNfRRT6XarwrXK5VgzfwqVZST6R4HJFeOfHXi34peKZvEnii+kvtRu2EccaKdsa5+WKJB0UZ4A6kknJJJ/Zc/Zci8DxW3xD+IdisniKRQ9hYSLkacD0dh3m/8AQP8Ae6VVVVVVVVVVVVV/wU1/5Mf+JP8A3Cv/AE62lf8ABLP/AJMv8I/9f+pf+lktVVVVVVVVVVVVVVVVVVVX7d//ACZ/8VP+wBJ/6Glf8Eg/+TULv/sbr7/0Tb1VVVVVVVVVVVVVVVVVVVV8Vv8AX6b/ALkn81qqqv2kfBI8P+Ll8SWMO2z1wF3wOFuF+/8A99DDe5LelfsveNtr3vgO+uOGzdWAY9/+WiD8MMB7MaqqqqqqqvGPjDQ/A+hza7rtz5cUfEca8vM/ZEHcn9OpwBXxB+IOt/ETW21XVZPLhjytpaK2Ut09B6k8Zbv7AAD4H/A7+0vs/jPxnZ/6IMPY2Mq/670kcf3PRf4up46gADA7dBVVVVVVWi/8hiw/6+Y//QhVVVVVVVVVVVVVVVVVVVVVfsH/APJS/Hn/AF6r/wCjzVVVV+3P8Lk0LxTY/E7Srfbaa8Ps9+FXhLpF+Vj2G+MfnGx71+wv8UW0fxPe/C3U7j/Q9bButPDNgJcovzqP9+Nc/WMetVVV+1j+06NTN58LPhzqYNlhodZ1GE/689GgjYfwdQzD73QcZ3fBH4K+IfjV4rTRtND2umWpD6lqJjyttH6D1dsEKv1J4BNeC/Bnh74f+GbHwn4XsVtdPsI9qLj5nP8AE7n+JmPJPcmqqqqqqqqqqq/4LW/8jP8ACj/rw1T/ANGW9VVVVVVn/wAfUX++Kqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/af8A+CivwZ/Zj8Xr8PNS0rWPE/iaOJJb2z0zy1SxVwGUSyOww7KQwQA8EElcjPwN/aR+Gv7Q3wtf4rfDu6u5LG1Msd9Y3UYjubKeNA7QyKCV3bSpBDFSGHPXH/D5z4Ef9Et8e/8AfNp/8er/AIfOfAj/AKJb49/75tP/AI9X/D5z4Ef9Et8e/wDfNp/8er/h858CP+iW+Pf++bT/AOPVp17HqWnWuowoypdQpKqt1AZQQD+dVVVVVVVVV/wS4/5PH+M3/Xhf/wDpyjqqqqqqqqqqqqqqqqqqqqrx9/yKV9/2z/8ARi1VVVVVVPBDcwSW08ayRyoUdGGQwIwQfwrxl4dm8J+KdT8OT5JsblkQn+JOqN+KlT+NfBTxf/wm3wz0XV5ZvMu4oPs12SckyxfKSfdgA3/Aq8Y+KNP0PSrvUb+5EVjYxGSeT1x2Hqc4AHckV448Xah458TXniPUCQ074hizkQxj7qD6D8zk96+BHw5Hgjwsuo6jBt1fV1WScMOYY/4I/bg5PucdhVVVVVVVVVVVeEP+RZ07/rgKqqqqqqqqqqqqqqqqqqqq+JXwW+JfiD9q7w58S9I8NfaPDdhPYNcXv2yBNgiPzny2cOcey89s1VVVVX7V3gz/AITP4H69FDFvutHVdTt+OhhyZD/36Mo/Gre2uLy4itLSCSaeZwkUUaFmdicAADkknAxX7MH7LMfgNbfx/wDESyjl8RkbrGxfDrpw/vtjIMp/Jfr0qqqqqqqqqqqqqv8Agpr/AMmP/En/ALhX/p1tK/4JZ/8AJl/hH/r/ANS/9LJaqqqqqqqqqqqqqqqqqqqv27/+TP8A4qf9gCT/ANDSv+CQf/JqF3/2N19/6Jt6qqqqqqqqqqqqqqqqqqqq+K3+v03/AHJP5rVVVfFXwWvjvwTf6JEim8RfPsiR0mTkD23DK59Grw7rl/4U8QWOu2I23OnTiQI3GcH5lPsRkH61omr2XiDR7LW9Ok3219AssZPUBhnB9x0I9aqqqqqq8Y+MND8D6HNruu3PlxR8Rxry8z9kQdyf06nAFfEH4g638RNbbVdVk8uGPK2lorZS3T0HqTxlu/sAAPgf8DRcC38Z+NbLMfD2FhKv3/SSQHt3Cnr1PHUAAYHboKqqqqqqrRf+QxYf9fMf/oQqqqqqqqqqqqqqqqqqqqqqv2S/gt8S/hj448W6x448Nf2baanAEtJPtkE3mHzS3SN2I455Aqqqq+Lfw+sfij8PdZ8F3gQPe25NrKw/1NwvzRv9AwGfUEjvVnd694G8UxXkHmWGs6DfBgGGGgnifkEezLgivh142034jeCNH8aaUy+TqlqsjIDnypBxJGfdXDL+FVX7WP7Tp0pbv4WfDnU2W/y0WsalC2Ps46NBGw/j7Mw+70HOdvwk+E3ib4w+LIPDPh6ApCpV769ZMx2cOeXb1PXavVj6DJHw5+HXhn4W+FLTwj4VtTHa243SSOcyXEpxukc92OPoBgDAAFVVVVVVVVVVVX/Ba3/kZ/hR/wBeGqf+jLeqqqqqqz/4+ov98VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV/wAFFv2TPjjaftJeJfiL4Y8A+IvFfhzxhNHeWl9pWny3pgk8tEeCURhjGVZTtyMMpXBJDAf8E1P2dPiX8FvgB421L4i6HfaRqvjWQy2eiXMZWeCGKBlRnj+8kkjO3yEbgFTOCcD/AIZX/ae/6Nw+KP8A4R9//wDGq/4ZX/ae/wCjcPij/wCEff8A/wAar/hlf9p7/o3D4o/+Eff/APxqv+GV/wBp7/o3D4o/+Eff/wDxqvDUMsHhzSoJ42jkjsoVdHUqVIQAgjsaqqqqqqqqr/glx/yeP8Zv+vC//wDTlHVVVVVVVVVVVVVVVVVVVVXj7/kUr7/tn/6MWqqqqqqq/al8Li013TfFlvFiPUITb3DAf8tI+VJ9ypx/wCv2UfGktjPrXgh5cJeILy2BOMOuFkx7lSn4Ia/aV+I39oX6+ANIuc29kwk1BlP35eqx+4Ucn/aI7rX7Pvw5/wCEu8SjxBqdvu0rRnDEMOJp+qJ7gcMfwB61VVVVVVVVVVVeEP8AkWdO/wCuAqqqqqqqqqqqqqqqqqqqqqqqqqqvLS2v7SaxvIllguI2jljboysMEH6g1+z7+yppHwr1S58W+J5INU1xLiVNNwN0dnCGIVxnrKy4JP8ACDgdyaqqqqqqqqqqqqqv+Cmv/Jj/AMSf+4V/6dbSv+CWf/Jl/hH/AK/9S/8ASyWqqqqqqqqqqqqqqqqqqqr9u/8A5M/+Kn/YAk/9DSv+CQf/ACahd/8AY3X3/om3qqqqqqqqqqqqqqqqqqqqr4rf6/Tf9yT+a1VVVftCeCR4V8bvqlnBssNcBuI8DhZc/vV/Mhv+B1+y/wCNhc6feeBL6fMtoTc2QY9YyfnUfRiG/wCBH0qqqqqq8Y+MND8D6HNruu3PlxR8Rxry8z9kQdyf06nAFfEH4g638RNbbVdVk8uGPK2lorZS3T0HqTxlu/sAAPgd8DvP+z+NPGln+74ksLCVfvekkg9O4U9ep461VVVVVVVaL/yGLD/r5j/9CFVVVVVVVVVVVVVVVVVVVVVVVVVX7b/wvPhjx3b/ABC0y1C6d4lXbclFwI7tBhs+m9AG9yHNfsJfFBbLU9S+FGqXOI7/ADe6WGbpKq/vYx9UAYDp8jetV+1b+0yvgi2ufht4Bvj/AMJFPHtv76Jv+Qch/gU/89SO/wDADnqRj4dfDrxV8V/Flv4W8L2pnu7gl555CfLt48jdLI3ZRke5JAGSQK+E/wAKPDHwf8Jw+GPDcG5ziS9vJB+8u5sYLt6DsF6AfiTVVVVVVVVVVVV/wWt/5Gf4Uf8AXhqn/oy3qqqqqqs/+PqL/fFVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVftN/t4fCD9lHxbpXg34ieHfGGo3ur6d9vgk0Wztpo1j8x48MZZ4yGyh4AIxjmv2Yv2sPhz+1loOteIfhzo3iPTrbQbtLW5TWraGF2d03AoIpZARj1I57V8RPiJ4L+E/g3UPiB8Q9ei0bw/pXl/bL6WN3WLzJFiTKoGY5eRF4HevAf7a37LnxP8AFuneA/Afxd0/Vte1Z2jsrKOzuUaZlQuQC8YUfKrHkjpVVVVVVVVVVVf8EuP+Tx/jN/14X/8A6co6qqqqqqqqqqqqqqqqqqqq8ff8ilff9s//AEYtVVVVVVV8a/C48VfDnVLWNN1xZJ9stxjndGCSB7lN4/GvC/iTU/COuW3iHR5FW7tN3llhkfMpU5HcYJrStM1fxdr8GmWYe61DU7jALtks7HLMx/Mk/U14L8J6f4J8NWXhzTQCltH+8k24Msh+85+p/IYHaqqqqqqqqqqqrwh/yLOnf9cBVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVf8ABTX/AJMf+JP/AHCv/TraV/wSz/5Mv8I/9f8AqX/pZLVVVVVVVVVVVVVVVVVVVV+3f/yZ/wDFT/sASf8AoaV/wSD/AOTULv8A7G6+/wDRNvVVVVVVVVVVVVVVVVVVVVXxW/1+m/7kn81qqqq+NHgkeOPAt5aW8W6/sQbqzwMkuoOVH+8uRj1I9K8G+Jrvwb4n0/xJZZL2UwZkzjzEPDr+Kkj8a0+/tNVsLbU7CdZra6iWWGRejKwyD+Rqqqqrxt420PwFocut63cYVflhhU/PO/ZVHr79AOTXjzx7rvxB1p9X1mchFJFtaqx8u3T0UevTJ6k/gB8Dvgd5/wBn8aeNLP8Ad8SWFhKv3vSSQencKevU8daqqqqqqqq0X/kMWH/XzH/6EKqqqqqqqqqqqqqqqqqqqqqqqqqvjf8ADaD4r/DXV/CBCi8kj87T5GOPLuU5jOewJypPoxrw9rmt+AvFtjr1gGttU0O9WVUkUjbJG3KMPTgqR6Eivjb+1/omk+AdNj+G92k/iDxJp6zBwwb+yUdeS/8A02ByAvQEbjxgN4R8JeKfid4utvDmgQSahq+qTMxeVyeTy8kjnOAOSWP6k18Evgv4c+C3hSPRtLSO51O5VX1PUSmHuZB2HcIuSFXt16kk1VVVVVVVVVVVV/wWt/5Gf4Uf9eGqf+jLeqqqqqqz/wCPqL/fFVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVf8FnP+S7+CP8AsUR/6Vz1/wAEW/8Akl/xG/7D1t/6INf8FNf+TH/iT/3Cv/TraV/wTt/5PQ+GH/X/AHP/AKRz1VVVVVVVVVVX/BLj/k8f4zf9eF//AOnKOqqqqqqqqqqqqqqqqqqqqvH3/IpX3/bP/wBGLVVVVVVVMoYbSAQRggiviP4YPg/xtq2gKm2GC4LW/H/LJvmT/wAdIH1Br9mz4cDTNNbx5q1vi7v0KWCuvMcPd/qxHH+yP9qqqqqqqqqqqqqvCH/Is6d/1wFVVVVVVVVVVVVVVVVVVVVfED9oDx34Y/aY0D4R6bFpR0PUprJJmlt2aYCY4fDBgB7cVVVVVVVVVVVVVVVVVVVVV/wU1/5Mf+JP/cK/9OtpX/BLP/ky/wAI/wDX/qX/AKWS1VVVVVVVVVVVVVVVVVVVft3/APJn/wAVP+wBJ/6Glf8ABIP/AJNQu/8Asbr7/wBE29VVVVVVVVVVVVVVVVVVVVfFb/X6b/uSfzWqqqqvjr4JPg3x1cvawbNP1XN1a4GApJ+dB9Gzx2BWv2ZPGo1Xw/ceDLyYm50kmW23HloGPI/4Cx/Jh6VVVVeNvG2h+AtDl1vW7jCr8sMKn5537Ko9ffoBya8eeO9c+IOuPrOszYVcrbWyn5LdP7q+/qepP4AfBL4FOzweMfHFjhVw9jp8q/e7iSQenop+p9DVVVVVVVVVaL/yGLD/AK+Y/wD0IVVVVVVVVVVVVVVVVVVVVVX7MP7QHjv4weMfE+g+LItKS20eAPbm0t2jbPmFfmJY54HtVVVVVftpfC5fBXxJXxdplts0zxWrTttXCx3S484f8Cyr+5ZvSvDfhvWvF+u2Xhrw5p8l7qOoSiKCGMZJJ7n0AGSSeAASeBXwB+A+h/BPwyIB5N74hvkB1LUFT7x6+VHnkRr+G48nsBVVVVVVVVVVVVVf8Frf+Rn+FH/Xhqn/AKMt6qqqqqrP/j6i/wB8VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV/wWc/5Lv4I/7FEf8ApXPX/BLf9pr4FfAfwB430j4t/EOz8O3mqaxBPZxT208pljWHaWHlowHPHOK/by/bN/Zj+LP7KHjn4f8Aw8+LNhrPiDVP7P8AsdjFZ3KNL5eoW0r4LxqowkbtyR0r9inx54R+GP7UXgHx3471qLSdB0m8ne9vZY3dYVa2lQEhAWOWZRwD1r/h4l+xf/0XfS//AAAvP/jNf8PEv2L/APou+l/+AF5/8Zq2uYLy2iu7aQSRTxh42AxuUjIP5VVVVVVVVVX/AAS4/wCTx/jN/wBeF/8A+nKOqqqqqqqqqqqqqqqqqqqqvH3/ACKV9/2z/wDRi1VVVVVVVfFD4Pjx7468P6uFC2So0eqsDgmNCGQD3bcy57DB7VDBDbQR21tEkcUSBI0RcBVAwAAOgxVVVVVVVVVVVVXhD/kWdO/64CqqqqqqqqqqqqqqqqqqqqvjF/yfR4R/6+tL/wDQqqqqqqqqqqqqqqqqqqqqq/4Ka/8AJj/xJ/7hX/p1tK/4JZ/8mX+Ef+v/AFL/ANLJaqqqqqqqqqqqqqqqqqqqv27/APkz/wCKn/YAk/8AQ0r/AIJB/wDJqF3/ANjdff8Aom3qqqqqqqqqqqqqqqqqqqqr4rf6/Tf9yT+a1VVVV8efBC+MPAs9zaw79Q0bddW2BksoH7xB9VGcdyq18PfFs/gfxfp3iGFm8uCXbcoP44W4cY+nI9wDVtcQXltFd2syywzIHjdTkMpGQR7EVVV428baH4C0OXW9buMKvywwqfnnfsqj19+gHJrx5471z4g64+s6zNhVyttbKfkt0/ur7+p6k/gB8Dvgd5P2fxp40s/3nElhYSr9z0kkHr3Cnp1PPSqqqqqqqqqrRf8AkMWH/XzH/wChCqqqqqqqqqqqqqqqqqqqqq/YP/5KX48/69V/9Hmqqqqq/aB+GMfxZ+F+q+G4IFfU4F+16WxwCLmMHaATwN4LISegfPav2cP2edM+DGhHUdUEN54q1KIfbLkKCtsvXyYj125xuP8AEQOwAFVVVVVVVVVVVVVX/Ba3/kZ/hR/14ap/6Mt6qqqqqrP/AI+ov98VVVVVVVVVVVVVVVVVVVVVXxD+KPw5+EmiQeI/id420fwvpdzdLaQ3mqXa28bzsrOIwzEAsVjc49FPpXw++J3w6+LGiSeJPhl430XxRpcNw1tJd6VeJcRxzKFYxsVJwwVlODzhge9V448eeCvhp4cn8XfEDxTpvh7RbV0Sa/1G4WCGNnYKoLNwCWIA9zXgT4h+Bfih4fTxX8OvFul+I9HkleJL7TblZ4i6cMoZSRkdxVVVVXxv+Ken/BD4TeJ/ixqulXGp2nhixa7ltLdwkkwBAwpbgHnvX7HP7aHhf9sjTPFGpeGfBeqeHl8Lz20MyX1xHKZjMshBXZ0x5Z6+tVVzcW9nby3d3cRwwQIXllkYKqKBksSeAABkk14D+Kfwy+KdreXvw0+IPh3xVBp0/kXcmj6nFeLBJzhXMbHaTgkZ6jkZFVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVf8FnP+S7+CP8AsUR/6Vz1VVVeFf8AkV9H/wCvCD/0WtVVVVVVVVV/wS4/5PH+M3/Xhf8A/pyjqqqqqqqqqqqqqqqqqqqqrx9/yKV9/wBs/wD0YtVVVVVVVVVVVVVVVVVVVVV4Q/5FnTv+uAqqqqqqqqqqqqqqqqqqqqr4xf8AJ9HhH/r60v8A9Cqqqqqqqqqqqqqqqqqqqqr/AIKa/wDJj/xJ/wC4V/6dbSv+CWf/ACZf4R/6/wDUv/SyWqqqqqqqqqqqqqqqqqqqr9u//kz/AOKn/YAk/wDQ0r/gkH/yahd/9jdff+ibeqqqqqqqqqqqqqqqqqqqqvit/r9N/wByT+a1VVVUQCMHn2NfGLwSfA3jm90+CLbY3Z+02WBgCNyflH+6wK/QA96/Zt8bf8JB4RfwzeS5vNCIRMnl4GzsP/ASCvsNvrVV438caB4B0V9Y1y5A4Igt1P7y4f8AuqPyyeg71488d658QdcfWdZmwq5W2tlPyW6f3V9/U9SfwA+B3wO8n7P408aWf7ziSwsJV+56SSD17hT06nnpVVVVVVVVVVWi/wDIYsP+vmP/ANCFVVVVVVVVVVVVVVVVVVVVV+wf/wAlL8ef9eq/+jzVVVVVVVVVVVVVVVVVVVVX/Ba3/kZ/hR/14ap/6Mt6qqqqqrP/AI+ov98VVVVVVVVVVVVVVVVVVVVVX/Bav/k1fwt/2UCz/wDSC/r9gT9ofxB+xt8bNH0H4i3Rt/h98TtOs57uR2bybdZhm3vlyB/q33xSEcYEn3ti0jpIiyRurKwyrA5BHqK/4Kwf8mR+MP8AsIaZ/wClkVf8Ehv+TNNM/wCxg1H/ANDWqqq/4Kb/ALU3xY/Zd+G/hTVPhJd6XZ6h4i1Wa1uLu8shcvCiRbgY1Y7AcnncrdK/Y98c+KviZ+zH8O/HvjjV31TXtb0dbi/vHiSMzSF2BbagVR0HAAFft+f8mafFn/sX5P8A0NK/4Idf8ip8W/8AsIaV/wCi7mqr45fDWT4yfB3xj8K4dcl0eTxTo1xp6X0a7vIMiFQSuRuXsy5GVJGRmv8Agnd+wN8Qf2Q/EPi3xZ8Q/HOiapda9ZxWNtY6I80kCxpJvMsjSxxkvwAAFwAW5OeKqqqqqqqqqqqqqqqqqqqqqqqqqqqqqta8G+EPEk8d14i8K6Pqk8SbI5L2xjnZFznALAkDJJx71/wqv4Yf9E48L/8Agng/+Ir/AIVX8MP+iceF/wDwTwf/ABFf8Kr+GH/ROPC//gng/wDiK/4VX8MP+iceF/8AwTwf/EV/wqv4Yf8AROPC/wD4J4P/AIikRI0WONQqqMKoGAAPSqqqqqqqqq/4Jcf8nj/Gb/rwv/8A05R1VVVVVVVVVVVVVVVVVVVV4+/5FK+/7Z/+jFqqqqqqqqqqqqqqqqqqqqqvCH/Is6d/1wFVVVVVVVVVVVVVVVVVVVVfGL/k+jwj/wBfWl/+hVVVVVVVVVVVVVVVVVVVVV/wU1/5Mf8AiT/3Cv8A062lf8Es/wDky/wj/wBf+pf+lktVVVVVVVVVVVVVVVVVVVX7d/8AyZ/8VP8AsASf+hpX/BIP/k1C7/7G6+/9E29VVVVVVVVVVVVVVVVVVVVfFb/X6b/uSfzWqqqqq/aI8EjxP4JbWrODff6ETOu0ctCceYPwADf8BPrXws8Zv4F8a2GtPIy2jN5N6o7wtgMcd8cNj1UUjxyxrJE6urDKspyCD3FePPHeh/D7Q31nWZtzHK21sp+e4f8Aur7ep6AfgK8beNtc8e65Lret3G5j8sMKn5IE7Ko9PfqTya+B3wO8n7P408aWf7ziSwsJV+56SSD17hT06nnpVVVVVVVVVVVaL/yGLD/r5j/9CFVVVVVVVVVVVVVVVVVVVVV+wf8A8lL8ef8AXqv/AKPNVVVVVVVVVVVVVVVVVVVVf8Frf+Rn+FH/AF4ap/6Mt6qqqqqrP/j6i/3xVVVVVVVVVVfa7o+mzC2vtQihk27grnBx6/pX/CWeG/8AoMW/5mv+Es8N/wDQYt/zNf8ACWeG/wDoMW/5mv8AhLPDf/QYt/zNf8JZ4b/6DFv+Zr/hLPDf/QYt/wAzX/CWeG/+gxb/AJmv+Es8N/8AQYt/zNWniHRL+4S0s9ShlmfO1FPJwMn9BVVVVVf8Fq/+TV/C3/ZQLP8A9IL+vil+zO3x4/4JkfCb4zeEtH3eLfhxoM5lSIbnvNLjuJROvQZMezzlHYeaBktX/BKH9ro/GL4aH4F+ONV83xh4EtVFhLM436jpYIVD6s8JKxt/smI5JLGv+CsH/JkfjD/sIaZ/6WRV/wAEhv8AkzTTP+xg1H/0Na/bX/bT8GfsfeB4L26s01zxjriuuhaGJtgfbgNPMwyUhUkdOXPyrj5mXwXc/wDBWn9srSV+JPhrx4fA/he9Jk0sx3S6JbzJk/6lY1a4kT0eQsD2Y4rXv2lP+CkH7A/ivSIv2g5v+E48I38pjia/ljuoL0L95Yr5F82KXaCQsufXYwzXwJ+N3gT9on4Y6R8Vfh3fPNpeqIVeGZQs1nOhxJBKoJ2up47gghgSrAn9vXw7+3To2k6NcftW679u8Nzaxcjw/H9qspdkm3J4twGH7vH3v518AfBn/BWXUPgz4SvPgl4s+zeBZdPU6FD9v0tNlvuOBtlXeOc/e5r9rqHxTb/8E9vGdv45m83xJF4GgXWH3K268CRCc5T5TmTd93j04r9gr9qbxl8BvBnjL4efBvwDL4y+KPxD1axh8P6eYmeGFIop/NnkCkFtu9cLlRjczMFU5+KPg/8A4LH+D9Iuvi94h8d67NHp8TXV7Y6Hq9pJ9kjUZYmzhAjcKM5CK/TPbNf8E4P+Cg+tftM3N78I/i8lmnjrTLM3llqNrCIY9Xt1IEm6MfKkyFlJCYVlJIVdhz+0X4k1vwZ+z58T/GHhjUHsdY0Lwbq99p90iqzQXENnLJHIAwIJVlU4II45r/gk7+018df2gvEPxHs/jF8Q7vxLDotlp8lgk9tBF5DSPOHI8pFzkIvXPSv+Crvx6+MXwB8B+Ate+D3j298NXep6vc216beCGUToIlZciVGwQQcYx945zxi1/be/a6/aa8OeEvg5+xxos9/4i0vwppr+OfGl5bwR7dQe2T7QEaYCGEeYJRuKl3YN5agJub4o+Mv+Cqn7Gl3afET4j+P9a1HRLm6WNrm4votY055Dk+TIhyYdwyBgJn+FsgY/Yu/as0T9rn4PQ+PLfT49L1/Tbg2Ov6ZGxZLa5Chg0ZPJidWDKT0+ZSSVJP7Qnx68Cfs1/CzVfit8QLmQWNhtitrSDBmv7l8+XBED1ZiCc9FVWY8KTXhr49f8FLP2/PEep3vwO1NvBHhDT7gxl9OuF0+0syfmWN7sgzzy7du4JkDIOxA2Cfip/wAFI/2DrrTvGH7REy/E/wCF73McGrTw3aX0loHIG5ZyqTRv0CmUGJjhchmBH7Bf7YHxp+P/AO2l4q8Ka58U9R1nwAlpq1/o2ny2cEK+Styi22cRiQbY5BwTnOM96qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/4Jcf8nj/ABm/68L/AP8ATlHVVVVVVVVVVVVVVVVVVVVXj7/kUr7/ALZ/+jFqqqqqqqqqqqqqqqqqqqqqvCH/ACLOnf8AXAVVVVVVVVVVVVVVVVVVVVV8Yv8Ak+jwj/19aX/6FVVVVVVVVVVVVVVVVVVVVX/BTX/kx/4k/wDcK/8ATraV/wAEs/8Aky/wj/1/6l/6WS1VVVVVVVVVVVVVVVVVVVft3/8AJn/xU/7AEn/oaV/wSD/5NQu/+xuvv/RNvVVVVVVVVVVVVVVVVVVVVXxW/wBfpv8AuSfzWqqqqqkjjljaKRFdWBDKwyCD1BFfFDwY/gTxpf6Cqn7Lu86zY/xQtyv1xypPqpr4a/HDRNH+Fhk8SXfmahoZFrDbK37y6XH7rA9MAqT22Z7gHxt411zx9rkmt63PudvlggXOyBOyKPT36k8mvgn8CfsZtvGPja1BnwJLLT5F/wBV6SSD+93C9up54FVVVVVVVVVVVWi/8hiw/wCvmP8A9CFVVVVVVVVVVVVVVVVVVVVV+wf/AMlL8ef9eq/+jzVVVVVVVVVVVVVVVVVVVVX/AAWt/wCRn+FH/Xhqn/oy3qqqqqqs/wDj6i/3xVVVVVVVVVV480bVb/W1nstOnnjECruSMkZya/4RnxD/ANAa7/79Gv8AhGfEP/QGu/8Av0a/4RnxD/0Brv8A79Gv+EZ8Q/8AQGu/+/Rr/hGfEP8A0Brv/v0a/wCEZ8Q/9Aa7/wC/Rr/hGfEP/QGu/wDv0a/4RnxD/wBAa7/79GvB2haxZ+I7O5utMuIok37neMgDKMP54qqqqqv+C1f/ACav4W/7KBZ/+kF/X/BOBEk/Yg+Fsciqytp10GVhkEfbJ88V+1z8J/GP/BO/9rvRfjN8IITa+GtVvZNU8PqAfJj5xd6bJg52bXK47xyrg7lJH7fPxU8MfHf/AIJpaj8WPA8zT6Rrr6Vchc7mtm+3RJJFJjgPHIDG3bcK/wCCQ3/Jmmmf9jBqP/oa1+02kv7Uv/BUZPhprt5LJokXiex8NrEJCDBZW+37UqHsWb7S/wDvPVhp9jpVhbaXplnDa2dnCsNvbwxhEhjUBVRVHAUAAADgAV+2V8KNE+M37MvxB8GaxZxTypolxfaa7gZgvbeNpYHU/wAPzoFJHVWYdCa/4Ih/EHUk134lfCme7Z7CW0tdatYGJxFKjmGZl7AurwA9z5a+hr/gt7/ySr4af9jBdf8ApOK/YD/5M0+Ev/Yvx/8Aob1+35/yZp8Wf+xfk/8AQ0r/AIIgeDPD1zd/FL4gXNhHLrViun6daXDqC0EEvnSSqp6je0UWf9wVXwQ061+FX/BWlPDvhOJLbT7L4g6tp9tbxrsWK2mFxH5YA4wqSYH+6OlftZf8mr/GX/sn+u/+kE1f8EOv+Rq+Lf8A2D9K/wDRlzX/AAW9/wCSVfDT/sYLr/0nFf8ABLfwT4f8I/sXeCdR0ayjivPEz3epanOFw1xObmSJSf8AdiiiQey+5r9rPwrpHjX9mL4qeHdbto57abwlqMqiRNwjmigeWGQD1SSNHHuor/gh9rN7F43+Knh1JP8AQ7rStPunQ/8APSKWVFI/CZs/hX/Bbr4h6hc+P/h38J4rorY6fo82tzQjgSSzzNAjN67Vt5APTe3rX7Mnwu0X4M/ALwL8OtEtIoE0zRbc3TRgfvrqRA88pI6l5Wds+9eNPCOheP8AwhrXgbxPZJd6Tr9hNY3sDjIeKVCjD2ODwexwa/4I2/8AJ3N3/wBifff+jraqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv8Aglx/yeP8Zv8Arwv/AP05R1VVVVVVVVVVVVVVVVVVVV4+/wCRSvv+2f8A6MWqqqqqqqqqqqqqqqqqqqqq8If8izp3/XAVVVVVVVVVVVVVVVVVVVVVPoWiXN8mqXGj2Mt7GQUuXt1aRSOmGIyMVVVVVVVVVVVVVVVVVVVVV/wU1/5Mf+JP/cK/9OtpX/BLP/ky/wAI/wDX/qX/AKWS1VVVVVVVVVVVVVVVVVVVft3/APJn/wAVP+wBJ/6Glf8ABIP/AJNQu/8Asbr7/wBE29VVVVVVVVVVVVVVVVVVVVfFb/X6b/uSfzWqqqqqr9pTwSde8KR+KLKDdeaISZcDlrdvvfXacN7DdQGeByewr4H/AAOGnC38Z+M7PN3w9jYyr/qPSRx/e9F/h6nnpVVVVVVVVVVVVWi/8hiw/wCvmP8A9CFVVVVVVVVVVVVVVVVVVVVVYaFomlSyT6Xo9jZySjEjwW6xl/qVAzVVVVVVVVVVVVVVVVVVVVX/AAWt/wCRn+FH/Xhqn/oy3qqqqqqs/wDj6i/3xVVVVVVVVVVVVVVVVVVVVVf8Fq/+TV/C3/ZQLP8A9IL+v+Cb3/Jkfwr/AOwfc/8ApZPX7WP7Onh79qL4Ja58LtYEUF/Kn2nRL91ybC/jB8qT12nJRwOqOwGDgjwn8XPFHwu/Z/8Ajx+xZ8Tkn0yW9kgutOt7jJNnqllqFs9xAOwWWKBiD03RDGd9f8Ehv+TNNM/7GDUf/Q1r9qF7v9lf/gp//wALN1iylTR28T2XiVHCFjcWU5U3JTOckN9pT2ZeO1aRq+l+INJstd0PULe/07UbdLi0urdw8c8TqGR1YcFSCCD71+218ZNC+B37MnjrxZq19FDeXukz6Xo8LEbri+uY2jiVVP3tpJkYf3I2Pav+CInw01Jbj4kfGC7s3jsHittDsJyvE0mTNcAH/ZAtv++/av8Agt4jn4UfDWQKSq+IbkFscAm34/kfyr/gn5cW91+xj8J5LaZJVXQghZTkBlldWH1DAg+4r9vz/kzT4s/9i/J/6Glf8EOv+RU+Lf8A2ENK/wDRdzVeGv8AlMHN/wBlQvf/AEOWv2sv+TV/jL/2T/Xf/SCav+CHX/I1fFv/ALB+lf8Aoy5r/gt7/wAkq+Gn/YwXX/pOK/4Jvf8AJkfwr/7B9z/6WT1+0L/yQH4mf9ifq3/pHLX/AARD/wCSp/Ev/sX7X/0oNf8ABbn4calb+Pvh58XYrR30+/0eXQ551BKxSwTPPGrdgWW4lI9fLb0r9k34x6D8eP2e/BXxB0S/iuJZtLhttTiQjNtfQoqXETL/AAkOCRwMqysOGFfFn4l+GPg58NvEXxO8YX8VrpXh2wkupmkbb5jAYSNfVncqigclmA71/wAEbf8Ak7m7/wCxPvv/AEdbVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV/wS4/5PH+M3/Xhf8A/pyjqqqqqqqqqqqqqqqqqqqqrx9/yKV9/wBs/wD0YtVVVVVVVVVVVVVVVVVVVVV4Q/5FnTv+uAqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv+Cmv/Jj/AMSf+4V/6dbSv+CWf/Jl/hH/AK/9S/8ASyWqqqqqqqqqqqqqqqqqqqr9u/8A5M/+Kn/YAk/9DSv+CQf/ACahd/8AY3X3/om3qqqqqqqqqqqqqqqqqqqqr4rf6/Tf9yT+a1VVVVVXdrb31rNZXcSywTxtHLGw4ZWGCD7EGvhv+z7p/hbxFda/r8kd99luW/sqEjIRAfllf1fGMDoDz1xiqqqqqqqqqqqqq0X/AJDFh/18x/8AoQqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/wCC1v8AyM/wo/68NU/9GW9VVVVVVn/x9Rf74qqqqqqqqqqqqqqqqqqqqqv+CoPwE+LP7RXwB0DwT8G/Cf8AwkGtWPjC21Ce1+3W9pst0s7uNn3zyIpw8sYwDn5s4wCR+xN8N/Gnwi/Za8AfDj4h6N/ZPiLRLOeO+svtEU/ks1zK6jfEzI2VdT8rHrVf8FNP+CfXjf46eKtJ+MvwA8LQ6p4qulFl4i00XkFobpETEN0HnkRNyqoiYZyR5eB8rV/wTl+DHxK+Av7M9j8Pvix4b/sPX4dYvbh7P7ZBdYjkYFG3wO6cgdM5Hev21P2K/BP7YPgm3sb28XQ/GGhq7aHriw7/AC92C0EyjBeFiAeuUPzL/EreBvh//wAFa/2PdPk+HXw60AeKPClvKwsUtza6rapkli0KuVnhUnJ2sqrkk7cnNP8AsS/t/wD7bXjbS/En7WnihPCfh2wLeTDcSW7SW8TEbltrK2JVHbABeYq2FBJbCg/CP4S+Bfgb8PNH+GHw50gafoeiw7IUZt8krk7nlkb+KR2JZj6ngAYA/br/AGXZf2svgRdeANGv7Wx8R6XfR6poVxdZEX2mNXQxyFckI8cjrnBwSrYO3FfCLwb/AMFdv2cdHf4P/DLwXq0OjRXMj2sMsOm31rbu5LO0M8pZUViSxUsF3EnALNn9pjwJ8R/ir+yJ4u8A6ToqX/jfXvDKQfYUuIoVkvCEMih3YRqNwbksB71/wSr/AGZfjf8As2+H/iJY/GrwR/wjs+u3lhJp6f2la3fnLEk4c5t5ZAuC6/exnPHeq0P9ij9puz/4KRyfHy5+GezwG3jy61Qat/bNif8ARXZysnkifzeQw+XZu56V+0N4W17x18AfiZ4J8K2H27WvEHg/VtP02181IvPuZrOWOJN7kKu52UZYgDOSQOa/4JV/so/H79m3xB8RL741eAf+Edh12zsI9Pb+1LO785onnLjFvLIVwHX72M5471/wVO/Zw+NH7SHw/wDA+h/BfwZ/wkV7o+sT3F7F/aNraeVG0IVWzcSRg5PGASa/Ym+G/jT4RfsteAPhx8Q9G/snxFolnPHfWX2iKfyWa5ldRviZkbKup+Vj1r4yaBq3iv4QeOfC2gWn2rVNY8N6hZ2UHmKnmzS20iIu5iFXLMBkkAZ5IFf8Esv2R/2hf2b/AB/441v40fD7/hHrLWNHgt7KX+1bO782RZizLi3mkK8c5YAV8d/gf4D/AGivhjq3wq+Ilg8+l6moaOaEhZrOdeY54WIO11P4EEqQVYg2P7BH/BRX9k3xZfX37Kvj6LW9MvHB3WGo21oLgDhTc2d8fILgE4OZMDODzivCH7E37Yf7RniTSvEv/BQH4wh/B+gzLdt4Vs7qBRdsnJEq2ipbxLgYaQb32lgCmd1f8EYdKuL39qvW9QjBEOneCrx5GK8Za5tUVc9ick/8BNVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVf8EuP+Tx/jN/14X/AP6co6qqqqqqqqqqqqqqqqqqqq8ff8ilff8AbP8A9GLVVVVVVVVVVVVVVVVVVVVVeEP+RZ07/rgKqqqqqqqqqqqqqqqqqqqq1/48fD7w38SLD4U6nc3q6/qLwpAiWpaMmU4TL9qqqqqqqqqqqqqqqqqqqqq/4Ka/8mP/ABJ/7hX/AKdbSv8Agln/AMmX+Ef+v/Uv/SyWqqqqqqqqqqqqqqqqqqqr9u//AJM/+Kn/AGAJP/Q0r/gkH/yahd/9jdff+ibeqqqqqqqqqqqqqqqqqqqqvit/r9N/3JP5rVVVVVVVVVVVVVVVVVVVVWi/8hiw/wCvmP8A9CFVVVVVVVVVVVVVVVVVVVVV8NPjx8Pvi1q+p6H4Pub2S60lN9yLi1MQA3beCevIqqqqqqqqqqqqqqqqqqqqr/gtb/yM/wAKP+vDVP8A0Zb1VVVVVWf/AB9Rf74qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv2lfjxoX7NPwZ134w+INMk1ODRjAkdhFMInupZZkiVFYggH59xODwprwd/wVp/Yt8S6Wl9r3jfW/CdyVBey1TQLqZ1PcBrVJkOPXdX7Y3/AAVh+GGtfDbW/hl+zXPq2t634jtXsX1+Syks4LKCUFZGhWULK8xUlVyihSwbJIwf+CTn7J/in4GfDnW/ij8SdFn0rxN468lLTTrqMpPY6fFuK+YpwUeV3LFDyFSPOCWAqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/4Jcf8nj/Gb/rwv/8A05R1VVVVVVVVVVVVVVVVVVVV4+/5FK+/7Z/+jFqqqqqqqqqqqqqqqqqqqqqvCH/Is6d/1wFVVVVVVVVVVVVVVVVVVVVfGL/k+jwj/wBfWl/+hVVVVVVVVVVVVVVVVVVVVV/wU1/5Mf8AiT/3Cv8A062lf8Es/wDky/wj/wBf+pf+lktVVVVVVVVVVVVVVVVVVVX7d/8AyZ/8VP8AsASf+hpX/BIP/k1C7/7G6+/9E29VVVVVVVVVVVVVVVVVVVVfFb/X6b/uSfzWqqqqqqqqqqqqqqqqqqqqtF/5DFh/18x/+hCqqqqqqqqqqqqqqqqqqqqq/YP/AOSl+PP+vVf/AEeaqqqqqqqqqqqqqqqqqqqq/wCC1v8AyM/wo/68NU/9GW9VVVVVVn/x9Rf74qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqtU0rS9c0640fWtNtdQsbuMx3FrdQrLFKh6qyMCGB9CK8Uf8ABOX9ifxfem/1X4AaLBKzEldLu7vTY8n0jtpY1A56Y/lXwt/Y2/Ze+C2pQ618N/gt4e03U7Zg1vqE8b3tzAw6GOa4aR0PupBNVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV/wS4/5PH+M3/Xhf8A/pyjqqqqqqqqqqqqqqqqqqqqrx9/yKV9/wBs/wD0YtVVVVVVVVVVVVVVVVVVVVV4Q/5FnTv+uAqqqqqqqqqqqqqqqqqqqqr4xf8AJ9HhH/r60v8A9Cqqqqqqqqqqqqqqqqqqqqr/AIKa/wDJj/xJ/wC4V/6dbSv+CWf/ACZf4R/6/wDUv/SyWqqqqqqqqqqqqqqqqqqqr9u//kz/AOKn/YAk/wDQ0r/gkH/yahd/9jdff+ibeqqqqqqqqqqqqqqqqqqqqvit/r9N/wByT+a1VVVVVVVVVVVVVVVVVVVVov8AyGLD/r5j/wDQhVVVVVVVVVVVVVVVVVVVVVfsH/8AJS/Hn/Xqv/o81VVVVVVVVVVVVVVVVVVVV/wWt/5Gf4Uf9eGqf+jLeqqqqqqz/wCPqL/fFVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX/BLj/k8f4zf9eF//wCnKOqqqqqqqqqqqqqqqqqqqqvH3/IpX3/bP/0YtVVVVVVVVVVVVVVVVVVVVV4Q/wCRZ07/AK4CqqqqqqqqqqqqqqqqqqqqvjF/yfR4R/6+tL/9Cqqqqqqqqqqqqqqqqqqqqr/gpr/yY/8AEn/uFf8Ap1tK/wCCWf8AyZf4R/6/9S/9LJaqqqqqqqqqqqqqqqqqqqv27/8Akz/4qf8AYAk/9DSv+CQf/JqF3/2N19/6Jt6qqqqqqqqqqqqqqqqqqqq+K3+v03/ck/mtVVVVVVVVVVVVVVVVVVVVaL/yGLD/AK+Y/wD0IVVVVVVVVVVVVVVVVVVVVVX7B/8AyUvx5/16r/6PNVVVVVVVVVVVVVVVVVVVVf8ABa3/AJGf4Uf9eGqf+jLeqqqqqqz/AOPqL/fFVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX/BLj/k8f4zf9eF//AOnKOqqqqqqqqqqqqqqqqqqqqvH3/IpX3/bP/wBGLVVVVVVVVVVVVVVVVVVVVVeEP+RZ07/rgKqqqqqqqqqqqqqqqqqqqq+MX/J9HhH/AK+tL/8AQqqqqqqqqqqqqqqqqqqqqq/4Ka/8mP8AxJ/7hX/p1tK/4JZ/8mX+Ef8Ar/1L/wBLJaqqqqqqqqqqqqqqqqqqqv27/wDkz/4qf9gCT/0NK/4JB/8AJqF3/wBjdff+ibeqqqqqqqqqqqqqqqqqqqqvit/r9N/3JP5rVVVVVVVVVVVVVVVVVVVVWi/8hiw/6+Y//QhVVVVVVVVVVVVVVVVVVVVVfsH/APJS/Hn/AF6r/wCjzVVVVVVVVVVVVVVVVVVVVX/Ba3/kZ/hR/wBeGqf+jLeqqqqqqz/4+ov98VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV8Vv+Ck/wCyd8J9cn8NXnje78R6laOY7mLw9Zm8SFh1BmJWIkcghXJBBBwa+BP7c37N37RGqx+GvAnjZrbxBKpMWjavbmzuZgBk+XnKSkAElUZiACSMDNVVf8EuP+Tx/jN/14X/AP6co6qqqqqqqqqqqqqqqqqqqq8ff8ilff8AbP8A9GLVVVVVVVVVVVVVVVVVVVVVeEP+RZ07/rgKqqqqqqqqqqqqqqqqqqqq8Ufs6eFvFfxe0z4x3muapFqelyW7xWsRj8ljCcrnKlue/NVVVVVVVVVVVVVVVVVVVVf8FNf+TH/iT/3Cv/TraV/wSz/5Mv8ACP8A1/6l/wClktVVVVVVVVVVVVVVVVVVVX7d/wDyZ/8AFT/sASf+hpX/AASD/wCTULv/ALG6+/8ARNvVVVVVVVVVVVVVVVVVVVVXxW/1+m/7kn81qqqqqqqqqqqqqqqqqqqqrRf+QxYf9fMf/oQqqqqqqqqqqqqqqqqqqqqqvg7+zp4W+C+vax4g0HXNUvptajCTJeGPagDlvl2qD1Peqqqqqqqqqqqqqqqqqqqqv+C1v/Iz/Cj/AK8NU/8ARlvVVVVVVZ/8fUX++Kqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv8Agq7+03rXwm+G+l/B/wADatJY6947SVtQuoHKS22mx4VlUjBUzOdmR/Ckg4yDX7EH/BOr4QeHvhHovjz42eBrHxV4u8UWUd81rqsJkg0uCVQ0cKwn5TJtKl2YEhiVGACW/wCCkn7FvhL4Dabo37RXwAsJ/DNva6pFDqljYzuq2M5O63u7c5zFiRQpCnAZoyoHzV+xJ8fLn9o/9nbw74/1iSNtfti+m65sXaDeQYDPgcDzEaOXA4HmYHSqr4JX37V9h+0P8SZP2RkuH8SNc3w1IQ21lMfsf2zuLsFAPM8vlfm/DNf8JH/wWw/599U/8FXh7/43X/CR/wDBbD/n31T/AMFXh7/43X/CR/8ABbD/AJ99U/8ABV4e/wDjdfsU3v7UV/8ACrVJv2tUnXxeviCZbIT29nCfsH2e38vi0ATHmmfk/N68Yqqqqqqqqqqqqqqqqqq/4KaftV/E/wDZq0/wDpvw6g0OWLxd/aX9of2laPMR9mNoY9m1128zPnOc8dK0m5kvtKsrybb5k9vHI4UYGSoJx+dVVVVVVVVVVVVVVVVVVVXxC1y98L+APEviXTBEbzSNHu7u3Eq7lMkULOu4AjIyoyOK/wCCbf7THxG/aV+HvijUPiJDo8cvhzUobOzGnWrQgxtEWO4M7ZOe/FVVVVVVVVVVVVVVVVVVVVfGX9sr4u+Av28PB/7N2h23h9vCeu3WlxXTz2TvdAXLYk2yCQAe3ynHvVVVVVVVVVVVVVVVVVVX/CR/8FsP+ffVP/BV4e/+N1/wkf8AwWw/599U/wDBV4e/+N1/wkf/AAWw/wCffVP/AAVeHv8A43X7SOtf8FRLz4LeIrb9ouG/X4eN9l/tgy6fo8S8XURhy1sglH78Q/dP14zX/BLP/ky/wj/1/wCpf+lktVVVVVVVVVVVVVVVVVVVX7d//Jn/AMVP+wBJ/wChpX7Jur/8FG7H4WywfsqRXreC/wC1ZjIYbHSph9s2R+Z810pk+75fH3fTvX/CR/8ABbD/AJ99U/8ABV4e/wDjdf8ACR/8FsP+ffVP/BV4e/8Ajdf8JH/wWw/599U/8FXh7/43Xw8fxTL8P/DMnjgMPEjaPaHWAyop+2GFfOyI/kH7zdwvy+nFVVVVVVVVVVVVVVVVVXi/9rX4p6x/wUAn/Zpu7fQh4RsLmeGJ0tHF1tGmm55k34z5ij+Hpx71VVVVVVVVVVVVVVVVVVVV+2Z8efHH7Onw00bx58P4tLk1KfxHBZsuo27TR+WYJ5DhVZTndCnOema/Z68d638UfgZ4E+IviVLVdV8R6Da314trGUiEskYZtikkhcngEmqqqqqqqqqqqqqqqqqqqqv2B/2yvi7+0v8AE7x94Q+Itt4fjsfDNqstkdNsnhcsZzH85aRsjaB2HNVVVVVVVVVVVVVVVVVX7WN18ebP4Iaxcfs0rM3j5bi1FgIobaVvL85POwtyDH/q93UZ9Oa/4SP/AILYf8++qf8Agq8Pf/G6/wCEj/4LYf8APvqn/gq8Pf8Axuv+Ej/4LYf8++qf+Crw9/8AG6/bJ1D9srUNS8LH9sCO5W8SC5Ghefa2EP7vdH52PsagHny/v8+neqqqqqqz/wCPqL/fFVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX/BRSRvib/wUJsfh9dzeZDbHQ9DVSxGxJ9kxHTjm7Y8Z61HHHFGsMKKiIoVVUYCgdAB2Fft5eH7XxN+x/wDFPT7qNHWDQnvVDjo9s6TqR75iFf8ABFbxBJc+Cfij4UMuY9O1TT71UwflNxFMhPpz9mH5fSqr/glx/wAnj/Gb/rwv/wD05R1VVVVVVVVVVVVVVVVVVVV/wW1/5ox/3Hv/AHH14e/5F/TP+vOL/wBAFVVVVVVVVVVVVVVVVVVVXxl/5JB45/7FrUP/AEmkr/gi3/yS/wCI3/Yetv8A0Qaqqqqqqqqqqqqqqqqqqqq/aa/5S3fDb/r/ANA/9Dqqqqqqqqqqqqqqqqqqqqr/AIKa/wDJj/xJ/wC4V/6dbSv+CWf/ACZf4R/6/wDUv/SyWqqqqqqqqqqqqqqqqqqqr9u//kz/AOKn/YAk/wDQ0r/gkH/yahd/9jdff+ibeqqqqqqqqqqqqqqqqqqqqtb/AOUweqf9f9x/6Y2qqqqqqqqqqqqqqqqqqqqr/gqJ/wAkA0D/ALHG2/8ASO8r9jT/AJNQ+Ev/AGKNh/6JWqqqqqqqqqqqqqqqqqqqqv8AgkL/AMl2+L3/AF4J/wClbVVVVVVVVVVVVVVVVVVVVV/wWt/5Gf4Uf9eGqf8Aoy3qqqqqqs/+PqL/AHxVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV+3LBH4P8A+CmNp4o1Hctrc6v4c1JmYgDy4o7aNiPQfuG6981X7cGqW2kfsjfFe7uSAj+Gri3GTj5pQIl/8ecV/wAEU9Dlt/C/xW8SGOQR31/plorH7paCO4cge4FwufqKqv8Aglx/yeP8Zv8Arwv/AP05R1VVVVVVVVVVVVVVVVVVVV/wW1/5ox/3Hv8A3H14e/5F/TP+vOL/ANAFVVVVVVVVVVVVVVVVVVVXxl/5JB45/wCxa1D/ANJpK/4It/8AJL/iN/2Hrb/0Qaqqqqqqqqqqqqqqqqqqqq/aa/5S3fDb/r/0D/0Oqqqqqqqqqqqqqqqqqqqqv+Cmv/Jj/wASf+4V/wCnW0r/AIJZ/wDJl/hH/r/1L/0slqqqqqqqqqqqqqqqqqqqq/bv/wCTP/ip/wBgCT/0NK/4JB/8moXf/Y3X3/om3qqqqqqqqqqqqqqqqqqqqrW/+Uweqf8AX/cf+mNqqqqqqqqqqqqqqqqqqqqq/wCCon/JANA/7HG2/wDSO8r9jT/k1D4S/wDYo2H/AKJWqqqqqqqqqqqqqqqqqqqqv+CQv/Jdvi9/14J/6VtVVVVVVVVVVVVVVVVVVVVX/Ba3/kZ/hR/14ap/6Mt6qqqqqrP/AI+ov98VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVf8FkvgbqV9B4T/aE0Wxea306H+w9cZEz5CGRpLWRsdFLvMhY8BmjH8Qr9jD9pjwz+0t8GNE1yDW7eXxZpdnFa+JNPMgE8F0ihWlKZz5cpG9W5HzFc5VgP+Ct/7THhbT/hxH+zd4Y1qC91/XLyG51+GCQOLG0hcSRxy4+7I8qxsF6hYySAGXP/AATt+Bup/An9mHQdI8R2TWmveJZ31zU7eRCr27zqgjjYHkMsMcQZT91tw7VVf8EuP+Tx/jN/14X/AP6co6qqqqqqqqqqqqqqqqqqqq/4La/80Y/7j3/uPrw9/wAi/pn/AF5xf+gCqqqqqqqqqqqqqqqqqqqr4y/8kg8c/wDYtah/6TSV/wAEW/8Akl/xG/7D1t/6INVVVVVVVVVVVVVVVVVVVVftNf8AKW74bf8AX/oH/odVVVVVVVVVVVVVVVVVVVVf8FNf+TH/AIk/9wr/ANOtpX/BLP8A5Mv8I/8AX/qX/pZLVVVVVVVVVVVVVVVVVVVV+3f/AMmf/FT/ALAEn/oaV/wSD/5NQu/+xuvv/RNvVVVVVVVVVVVVVVVVVVVVWt/8pg9U/wCv+4/9MbVVVVVVVVVVVVVVVVVVVVV/wVE/5IBoH/Y423/pHeV+xp/yah8Jf+xRsP8A0StVVVVVVVVVVVVVVVVVVVVf8Ehf+S7fF7/rwT/0raqqqqqqqqqqqqqqqqqqqqv+C1v/ACM/wo/68NU/9GW9VVVVVVn/AMfUX++KqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqvEvhrQPGXh/UPCvinSLXVNI1W3e3vbK5jDxzxsMMrD0x+VePv+CN3h6bxFNrXwc+NupeF7OZ2KafqGnm7a3Vskqk6SxsVHAAZScdWJ6/s4f8ABK74S/BjxRZePviB4nuviF4g06UT2cdzZLa2MEwORIYd0jSupxgu+3PO3OCKqvE3/BLX9q3Sfif4r8XfB/41+GNEsNb1G5mt7hdb1DTrt7eSUyBJhBbsMgkZAcglQeOg/wCHdP8AwUV/6Om0v/wt9a/+R6/4d0/8FFf+jptL/wDC31r/AOR6/wCHdP8AwUV/6Om0v/wt9a/+R6/4d0/8FFf+jptL/wDC31r/AOR6/wCHdP8AwUV/6Om0v/wt9a/+R6/4d0/8FFf+jptL/wDC31r/AOR6/wCHdP8AwUV/6Om0v/wt9a/+R6/4d0/8FFf+jptL/wDC31r/AOR6/wCHdP8AwUV/6Om0v/wt9a/+R6/4d0/8FFf+jptL/wDC31r/AOR6/wCHdP8AwUV/6Om0v/wt9a/+R6/4d0/8FFf+jptL/wDC31r/AOR6/wCHdP8AwUV/6Om0v/wt9a/+R6/4d0/8FFf+jptL/wDC31r/AOR6/wCHdP8AwUV/6Om0v/wt9a/+R6/4d0/8FFf+jptL/wDC31r/AOR6/wCHdP8AwUV/6Om0v/wt9a/+R6/4d0/8FFf+jptL/wDC31r/AOR6/wCHdP8AwUV/6Om0v/wt9a/+R6/4d0/8FFf+jptL/wDC31r/AOR6/wCHdP8AwUV/6Om0v/wt9a/+R6/4d0/8FFf+jptL/wDC31r/AOR6/wCHdP8AwUV/6Om0v/wt9a/+R6/4d0/8FFf+jptL/wDC31r/AOR6/wCHdP8AwUV/6Om0v/wt9a/+R6/4d0/8FFf+jptL/wDC31r/AOR6/wCHdP8AwUV/6Om0v/wt9a/+R6/4d0/8FFf+jptL/wDC31r/AOR6/wCHdP8AwUV/6Om0v/wt9a/+R6/4d0/8FFf+jptL/wDC31r/AOR68U/8Eqv22vHX2X/hNvjh4M8Q/Yd/2X+1fEuqXfkb9u/Z5lqdu7aucddoz0pP+CZv7fcaLFF+0d4dVUGFVfF+rgADsB9mr/h2h+39/wBHI+H/APwsNX/+Rq/4doft/f8ARyPh/wD8LDV//kav+HaH7f3/AEcj4f8A/Cw1f/5Gr/h2h+39/wBHI+H/APwsNX/+Rq/4doft/f8ARyPh/wD8LDV//kav+HaH7f3/AEcj4f8A/Cw1f/5Gr/h2h+39/wBHI+H/APwsNX/+Rq/4doft/f8ARyPh/wD8LDV//kav+HaH7f3/AEcj4f8A/Cw1f/5Gr/h2h+39/wBHI+H/APwsNX/+Rq/4doft/f8ARyPh/wD8LDV//kav+HaH7f3/AEcj4f8A/Cw1f/5Gr/h2h+39/wBHI+H/APwsNX/+Rq/4doft/f8ARyPh/wD8LDV//kav+HaH7f3/AEcj4f8A/Cw1f/5Gr/h2h+39/wBHI+H/APwsNX/+Rq/4doft/f8ARyPh/wD8LDV//kav+HaH7f3/AEcj4f8A/Cw1f/5Gr/h2h+39/wBHI+H/APwsNX/+Rq/4doft/f8ARyPh/wD8LDV//kav+HaH7f3/AEcj4f8A/Cw1f/5Gr/h2h+39/wBHI+H/APwsNX/+Rq/4doft/f8ARyPh/wD8LDV//kav+HaH7f3/AEcj4f8A/Cw1f/5Gr/h2h+39/wBHI+H/APwsNX/+Rq/4doft/f8ARyPh/wD8LDV//kav+HaH7f3/AEcj4f8A/Cw1f/5Gr/h2h+39/wBHI+H/APwsNX/+Rq/4doft/f8ARyPh/wD8LDV//karr/gmP+3rfW0tleftEeGri3njaOWGXxdq7rIrDBVlNtggg4INeGP+CWX7cPgi3mtPBfx38IaBDcuHni0vxRqtosrAYDMI7UAnHGTX/Dun/gor/wBHTaX/AOFvrX/yPX/Dun/gor/0dNpf/hb61/8AI9f8O6f+Civ/AEdNpf8A4W+tf/I9f8O6f+Civ/R02l/+FvrX/wAj1/w7p/4KK/8AR02l/wDhb61/8j1/w7p/4KK/9HTaX/4W+tf/ACPX/Dun/gor/wBHTaX/AOFvrX/yPX/Dun/gor/0dNpf/hb61/8AI9f8O6f+Civ/AEdNpf8A4W+tf/I9f8O6f+Civ/R02l/+FvrX/wAj1/w7p/4KK/8AR02l/wDhb61/8j1/w7p/4KK/9HTaX/4W+tf/ACPX/Dun/gor/wBHTaX/AOFvrX/yPX/Dun/gor/0dNpf/hb61/8AI9f8O6f+Civ/AEdNpf8A4W+tf/I9f8O6f+Civ/R02l/+FvrX/wAj1/w7p/4KK/8AR02l/wDhb61/8j1/w7p/4KK/9HTaX/4W+tf/ACPX/Dun/gor/wBHTaX/AOFvrX/yPX/Dun/gor/0dNpf/hb61/8AI9f8O6f+Civ/AEdNpf8A4W+tf/I9f8O6f+Civ/R02l/+FvrX/wAj1/w7p/4KK/8AR02l/wDhb61/8j1/w7p/4KK/9HTaX/4W+tf/ACPX/Dun/gor/wBHTaX/AOFvrX/yPX/Dun/gor/0dNpf/hb61/8AI9f8O6f+Civ/AEdNpf8A4W+tf/I9f8O6f+Civ/R02l/+FvrX/wAj1/w7p/4KK/8AR02l/wDhb61/8j1/w7p/4KK/9HTaX/4W+tf/ACPV/wD8Epv21tV8RQeMNT+Nvgq8161KGDVJ/EmqSXURT7hWU2pddvbB47V/w7p/4KK/9HTaX/4W+tf/ACPX/Dun/gor/wBHTaX/AOFvrX/yPX/Dun/gor/0dNpf/hb61/8AI9f8O6f+Civ/AEdNpf8A4W+tf/I9f8O6f+Civ/R02l/+FvrX/wAj1/w7p/4KK/8AR02l/wDhb61/8j1/w7p/4KK/9HTaX/4W+tf/ACPX/Dun/gor/wBHTaX/AOFvrX/yPX/Dun/gor/0dNpf/hb61/8AI9f8O6f+Civ/AEdNpf8A4W+tf/I9f8O6f+Civ/R02l/+FvrX/wAj1/w7p/4KK/8AR02l/wDhb61/8j1/w7p/4KK/9HTaX/4W+tf/ACPX/Dun/gor/wBHTaX/AOFvrX/yPX/Dun/gor/0dNpf/hb61/8AI9f8O6f+Civ/AEdNpf8A4W+tf/I9f8O6f+Civ/R02l/+FvrX/wAj1/w7p/4KK/8AR02l/wDhb61/8j1/w7p/4KK/9HTaX/4W+tf/ACPX/Dun/gor/wBHTaX/AOFvrX/yPX/Dun/gor/0dNpf/hb61/8AI9f8O6f+Civ/AEdNpf8A4W+tf/I9f8O6f+Civ/R02l/+FvrX/wAj1/w7p/4KK/8AR02l/wDhb61/8j1/w7p/4KK/9HTaX/4W+tf/ACPX/Dun/gor/wBHTaX/AOFvrX/yPX/Dun/gor/0dNpf/hb61/8AI9f8O6f+Civ/AEdNpf8A4W+tf/I9f8O6f+Civ/R02l/+FvrX/wAj1/w7p/4KK/8AR02l/wDhb61/8j1rv/BMr9vjxTpM+g+Jv2i/Der6bc7fPsr/AMXavcQy7WDLuR7YqcMqkZHBAPav2P8A4Fax+zf+z94b+E3iHWbPVNW003E17cWQbyPNmneUrGWAYqocLuIBOCcDOBVVVVVVVVVVVVVVVVVVVftEfDDUPjR8EPGfws0jVLfT77xJpUlrbXNyjNHHIcFd4XnaSoBIBIBzg4wfDf8AwS+/bt8G6cdI8IftA+FtDsTIZTa6b4r1a1iLkAFtkdqBkgDnGeBX/Dun/gor/wBHTaX/AOFvrX/yPX/Dun/gor/0dNpf/hb61/8AI9f8O6f+Civ/AEdNpf8A4W+tf/I9f8O6f+Civ/R02l/+FvrX/wAj1/w7p/4KK/8AR02l/wDhb61/8j1/w7p/4KK/9HTaX/4W+tf/ACPX/Dun/gor/wBHTaX/AOFvrX/yPX/Dun/gor/0dNpf/hb61/8AI9f8O6f+Civ/AEdNpf8A4W+tf/I9f8O6f+Civ/R02l/+FvrX/wAj1/w7p/4KK/8AR02l/wDhb61/8j1/w7p/4KK/9HTaX/4W+tf/ACPX/Dun/gor/wBHTaX/AOFvrX/yPX/Dun/gor/0dNpf/hb61/8AI9f8O6f+Civ/AEdNpf8A4W+tf/I9f8O6f+Civ/R02l/+FvrX/wAj1/w7p/4KK/8AR02l/wDhb61/8j1/w7p/4KK/9HTaX/4W+tf/ACPX/Dun/gor/wBHTaX/AOFvrX/yPX/Dun/gor/0dNpf/hb61/8AI9f8O6f+Civ/AEdNpf8A4W+tf/I9f8O6f+Civ/R02l/+FvrX/wAj1/w7p/4KK/8AR02l/wDhb61/8j1/w7p/4KK/9HTaX/4W+tf/ACPX/Dun/gor/wBHTaX/AOFvrX/yPX/Dun/gor/0dNpf/hb61/8AI9f8O6f+Civ/AEdNpf8A4W+tf/I9f8O6f+Civ/R02l/+FvrX/wAj1/w7p/4KK/8AR02l/wDhb61/8j1/w7p/4KK/9HTaX/4W+tf/ACPT/wDBKD9tCTxI3jJ/jP4HbxAxJbVT4j1M3RJTYSZvsm/7ny9enHSv+HaH7f3/AEcj4f8A/Cw1f/5Gr/h2h+39/wBHI+H/APwsNX/+Rq/4doft/f8ARyPh/wD8LDV//kav+HaH7f3/AEcj4f8A/Cw1f/5Gr/h2h+39/wBHI+H/APwsNX/+Rq/4doft/f8ARyPh/wD8LDV//kav+HaH7f3/AEcj4f8A/Cw1f/5Gr/h2h+39/wBHI+H/APwsNX/+Rq/4doft/f8ARyPh/wD8LDV//kav+HaH7f3/AEcj4f8A/Cw1f/5Gr/h2h+39/wBHI+H/APwsNX/+Rq/4doft/f8ARyPh/wD8LDV//kav+HaH7f3/AEcj4f8A/Cw1f/5Gr/h2h+39/wBHI+H/APwsNX/+Rq/4doft/f8ARyPh/wD8LDV//kav+HaH7f3/AEcj4f8A/Cw1f/5Gr/h2h+39/wBHI+H/APwsNX/+Rq/4doft/f8ARyPh/wD8LDV//kav+HaH7f3/AEcj4f8A/Cw1f/5Gr/h2h+39/wBHI+H/APwsNX/+Rq/4doft/f8ARyPh/wD8LDV//kav+HaH7f3/AEcj4f8A/Cw1f/5Gr/h2h+39/wBHI+H/APwsNX/+Rq/4doft/f8ARyPh/wD8LDV//kav+HaH7f3/AEcj4f8A/Cw1f/5Gr/h2h+39/wBHI+H/APwsNX/+Rq/4doft/f8ARyPh/wD8LDV//kav+HaH7f3/AEcj4f8A/Cw1f/5Gr/h2h+39/wBHI+H/APwsNX/+Rq/4doft/f8ARyPh/wD8LDV//katf/4JVftteK7NNO8UfHDwZrFrHKJUgv8AxLqlwiuAQGCvakBsMwz1wT61pn/BNX/goHomnW2kaL+0roNhY2cQit7W18ZaxFFCijAVEW2AVQOgAxX/AA7p/wCCiv8A0dNpf/hb61/8j1/w7p/4KK/9HTaX/wCFvrX/AMj1/wAO6f8Agor/ANHTaX/4W+tf/I9f8O6f+Civ/R02l/8Ahb61/wDI9f8ADun/AIKK/wDR02l/+FvrX/yPX/Dun/gor/0dNpf/AIW+tf8AyPX/AA7p/wCCiv8A0dNpf/hb61/8j1/w7p/4KK/9HTaX/wCFvrX/AMj1/wAO6f8Agor/ANHTaX/4W+tf/I9f8O6f+Civ/R02l/8Ahb61/wDI9f8ADun/AIKK/wDR02l/+FvrX/yPX/Dun/gor/0dNpf/AIW+tf8AyPX/AA7p/wCCiv8A0dNpf/hb61/8j1/w7p/4KK/9HTaX/wCFvrX/AMj1/wAO6f8Agor/ANHTaX/4W+tf/I9f8O6f+Civ/R02l/8Ahb61/wDI9f8ADun/AIKK/wDR02l/+FvrX/yPX/Dun/gor/0dNpf/AIW+tf8AyPX/AA7p/wCCiv8A0dNpf/hb61/8j1/w7p/4KK/9HTaX/wCFvrX/AMj1/wAO6f8Agor/ANHTaX/4W+tf/I9f8O6f+Civ/R02l/8Ahb61/wDI9f8ADun/AIKK/wDR02l/+FvrX/yPX/Dun/gor/0dNpf/AIW+tf8AyPX/AA7p/wCCiv8A0dNpf/hb61/8j1/w7p/4KK/9HTaX/wCFvrX/AMj1/wAO6f8Agor/ANHTaX/4W+tf/I9f8O6f+Civ/R02l/8Ahb61/wDI9f8ADun/AIKK/wDR02l/+FvrX/yPX/Dun/gor/0dNpf/AIW+tf8AyPXhv/glN+2t4Nu7m/8ACHxt8FaHdXi7bmfTfEmqWrzDOcO0dqCwzzz3r/h3T/wUV/6Om0v/AMLfWv8A5Hr/AId0/wDBRX/o6bS//C31r/5Hr/h3T/wUV/6Om0v/AMLfWv8A5Hr/AId0/wDBRX/o6bS//C31r/5Hr/h3T/wUV/6Om0v/AMLfWv8A5Hr/AId0/wDBRX/o6bS//C31r/5Hr/h3T/wUV/6Om0v/AMLfWv8A5Hr/AId0/wDBRX/o6bS//C31r/5Hr/h3T/wUV/6Om0v/AMLfWv8A5Hr/AId0/wDBRX/o6bS//C31r/5Hr/h3T/wUV/6Om0v/AMLfWv8A5Hr/AId0/wDBRX/o6bS//C31r/5Hr/h3T/wUV/6Om0v/AMLfWv8A5Hr/AId0/wDBRX/o6bS//C31r/5Hr/h3T/wUV/6Om0v/AMLfWv8A5Hr/AId0/wDBRX/o6bS//C31r/5Hr/h3T/wUV/6Om0v/AMLfWv8A5Hr/AId0/wDBRX/o6bS//C31r/5Hr/h3T/wUV/6Om0v/AMLfWv8A5Hr/AId0/wDBRX/o6bS//C31r/5Hr/h3T/wUV/6Om0v/AMLfWv8A5Hr/AId0/wDBRX/o6bS//C31r/5Hr/h3T/wUV/6Om0v/AMLfWv8A5Hr/AId0/wDBRX/o6bS//C31r/5Hr/h3T/wUV/6Om0v/AMLfWv8A5Hr/AId0/wDBRX/o6bS//C31r/5Hr/h3T/wUV/6Om0v/AMLfWv8A5Hr/AId0/wDBRX/o6bS//C31r/5Hr/h3T/wUV/6Om0v/AMLfWv8A5Hr/AId0/wDBRX/o6bS//C31r/5HrVf+CUf7Ynj3VtMb4n/HXwprFnayhGnu9e1LUJraFmHmGFJrcAtgZ27lBIGSOtVVVVVVn/x9Rf74qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqs/+PqL/fFVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVn/x9Rf74qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqs/wDj6i/3xVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV+0d+0T4A/Zk+G118RfHlxJIN/2fTdOgI8/UboqSsSZ4HCksx4VQTycA6P8Atl/8FKP2jze+Lf2fvhXbad4YtpmSJrPS4p0OOqG4uztmkHG7ywuOPlGa/Zz/AOCnXji0+J0XwT/bA8HweGdTluRaLrAtHsWs7hsbFvIHJCo2QPNXaFypK7SXWqqqqqqqqqqqqqqqqqqq/aZ/4Kf/AAc+Cl5feC/h3at4/wDGVpM9tNb2shisrKZTtKyT4PmMG/giDcggspr9g79vn45ftL/Hq7+HPxF03wvaaVFol1fKmm6fLDKksckSqu55X+UCRhgjPTmqqqqqqqqqqqqqqqqqqqr9q7/gpB4l+FvxQvvgF8DPhFfeJvG9i6wzTahazPH5rosirBaxYluAUYHduQdwGHNa3+07/wAFZ/DVo/jXW/hXqEGjwr5ktsfByvHGg6l1UGZFHUkkY7kV+xH/AMFItI/aR11Phf8AE3RbDw144mjL6c9k7/Y9V2qWdEVyWikCgtsLMGAODkYqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv22v2/vDH7K4h8E+F9It/E/xBv4BMlhLIy2+nRNnZLcFfmYsR8sSkEjJLKNu65/aT/4K0x6Afiq/wvli8NeX9qNovhiIhYcZyYcm6CAc7j2GScc1+w/+374d/asS58F+KdItfDfj/ToDcNZQSs1tqMAwGlty2WUqSN0TFiAQwZhu2VVVVVVVVVVVVVVVVVVV4r8V+G/A3hzUPF3i/WrXSdG0qAz3l7dPsjhQdST+Qx1JIA61+0L/AMFf7HT7i50D9mjwhFrItuZvEWuQSLb7dwBMVspVypJADyMmCcbDkV/wT2/aO+IX7UPwX1nx/wDEmHSItT0/xRPpkK6ZbNBH5KWtrKCVZ2O7dM/OemOKqqqqqqqqqqqqqqqqqqrW9VtdB0a/1y9jnkt9OtZLiVLeFpZGRFLMERcszYBwoGScAV4j/wCCmv7Svxu8SXXhz9kH4C3NzY2zgG9udNl1K62/wvIsZENsDgjDF/Zq139uX/gpJ8AJrfWvjZ8OIpNImkEe7WfDX2e3cn+BZ7YoqvjOASfocV+yb+1f4E/ay8AP4p8MQtpms6W6w63ok0u+SwlYHaQ2BvicKxV8DOGBAKkCqqqqqqqqqqqqqqqqqqqqqqqqqs/+PqL/AHxVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV/wUP8T63+0f8Atx+Hv2fNIv2TT9Eu7DQLTaNyJdXjRPcT4z1HmRo3QYg/E+CfBfhr4deEdI8C+DtLh07RdDtEtbK2iXAREGOfVjySx5Ykk5JNf8FlPg3pV34N8I/HbTbBE1TT9Q/sTU5o1Aae3lR5IS57iN43Uf8AXb6Y/YH+Kd98X/2UfAniTWLp7jVLC0bSr6WQ5aSS1cxK7HqWaNY2JPUsaqqqqqqqqqqqqqqqqqqrwL+yP8BPh98Stf8AjBo3ge3uvGHiLVJ9Sm1S/wD9Ie1lmcu4t1I2wjczHKjd8xyxFf8ABK3/AJPW8Rf9gDVP/SmCqqqqqqqqqqqqqqqqqqqqz8EeDtP8V6h46svDOmQ+I9Vgjt73VUtlFzPFGMIjSY3FR6ZxVftk+HrD4I/8FEYdW+G1otpI2s6RrcFnagRiO6kaN5FUAcB5FZiMf8tDxjiqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqtf1rT/Dehaj4i1aXy7LSrSW6uXAztjjQu5/JTX7B/hib9rz9uDXfjH8TLSPUINKNx4juLadfMiNwZFjtIcH+GPcGUHjFuoqv2wdHH7HH/AAUC074leBrU2Gn3N1aeJ7e2txtXy5XeO8hUDACyNHcDaMALJjpireeG7t4rq2kEkUyB43HRlIyCPwqqqqqqqqqqqqqqqqqqr4g+B9C+JngXxB8PPEyStpXiTTZ9PvPJYLIscqFGZCQQHAOQSDggHFftn/s/fCD9n7/gn98SfDnwo8FWOjROuk/abkKZLm7YapafNNM2Xf1wTtGeABxX/BGz/k2DxN/2Pt3/AOkFhVVVVVVVVVVVVVVVVVVVV4O8EeDvh7oUXhjwL4Z03QdKhZnSz0+2WCMMxyzYUDLE9SeT3rxz4K8N/EfwdrHgPxfpsV/o2u2clpeW8ig7kcYyMg4YHDK3VWAI5Ff8EktU1Xw9+1nrHhWxna5sL/w7ewXZQ4Q+TNEySkc9xtHp5h55qqqqqqqqqqqqqqqqqqqqqqqqqqs/+PqL/fFVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVXwylHiL/grjePqSb/K+JOtKo3Hj7ObkRn8PKXjpx6VX/BVK2gn/Yy8Tyyxhnt9S014if4W+1IufyZh+Nf8Edb2e7/Za1y3lIK2fje8iiwP4TaWb8/i7VVVVVVVVVVVVVVVVVVVVf8ABK3/AJPW8Rf9gDVP/SmCqqqqqqqqqqqqqqqqqqqqvHPjbwv8N/CGreO/Gmrw6Zomh2r3N7dSniNFHYdWYnACjliQBkkV+zN4c8R/t4ft56l8eNd0qaDwvoGqxa1dq/KwRwYXT7QnoWPkx7h0IjkPeqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv2udRn0r9lr4tXlqcSjwZqkanJBXfbOhII6EBsj3r/gibZ28l58Yb9kzPDFokSNnorm+LD8Si/lVf8ABaO2gX4sfDy7WMCaTw7PG7eqrcEqPwLN+dfBG9n1P4LeANRuSDNdeF9NlkwMDc1rGTj8TVVVVVVVVVVVVVVVVVVVf8FNf+TH/iT/ANwr/wBOtpX/AARs/wCTYPE3/Y+3f/pBYVVVVVVVVVVVVVVVVVVVVV+3X+0zpP7NPwN1TU7fUEXxd4jhl0/w3ahvnM7Lh7jHUJCrbyem7YvG8V/wR+/Z41Hw14c179onxJYvby+JYf7L0BZFwXs0kDTzY/uvLGir0/1LHowNVVVVVVVVVVVVVVVVVVVVVVVVVVn/AMfUX++KqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqvHrj4F/8ABWVNZ1h/s9ldeN7a8adiAog1FF3yZ/ur9ofP+6e9V/wV58Y2mgfstW3hhrgC68T+JLWCOEMAWjhV5nfHdQ0cYPu61/wSu8IXXhb9jzQL67RkfxJql9qaIwwQhl8lT9CsAYezVVVVVVVVVVVVVVVVVVVVf8Erf+T1vEX/AGANU/8ASmCqqqqqqqqqqqqqqqqqqqrxD4h0Hwlod94m8T6xZ6VpOmQNPeXt3MIooI1GSzMeAK+P/wAa/ip/wUo+Nll8APgJDcwfDrTbsSNdSwvHHKE4bULsjlYlyfLjOCcjjewC/s7/ALP/AIG/Zq+GOnfDTwNAXjg/e39/KgE2o3TAB55MdCcABeiqFUdM1VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV8ZvCVx4/wDg/wCOfAlou648ReG9Q06EZx889s8a/qwr/gjD4wt9K+KXxF+Ht1KYrjW9Ftr6KN/l3G0mZGHPO4C7zj0DHtxX/BXDW5PHX7U/hX4ceHz9qvdL0C1tDArAkXd1cSMqYHQlGgPP94fU+GtFi8OeHNK8OwymSPS7KG1R2HLCNAgP44qqqqqqqqqqqqqqqqqqqv8Agpr/AMmP/En/ALhX/p1tK/4I2f8AJsHib/sfbv8A9ILCqqqqqqqqqqqqqqqqqqqr9oX9on4a/s0+ALjx38RNWWP5WTTtNiYG51OcDiKFD16jLH5VBySK+BfwY+LX/BTD443Px5+OstxY/D7SZliWKFWjinjRyV06zPZBk+ZIDuBY873yNJ0nTNB0qz0PRbCCy0/T4Et7W1gjCRwxIoVUVRwFAAAFVVVVVVVVVVVVVVVVVVVVVVVVVVn/AMfUX++Kqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv+Cp37IPiL4t6Lpvx2+GGkS6j4j8L2bWusafbRlpr2xDF0kiUcs8TM+VAyyvx9wA/s/8A/BXnwFpfw40/w/8AH7w54mfxTo9utvJqOk28VxHqQQYErq8iGOUgDcOVJywIztHxB8U/F3/gqt+0PpGgeBvDl3ofgXw4PLjluF3x6TbSMDNdXDD5TPIEULEp52KoPDufBXg/QPh94P0XwL4Wsxa6RoFhDY2UOclYokCLk9zgZJ7kk96qqqqqqqqqqqqqqqqqqqv+CVv/ACet4i/7AGqf+lMFVVVVVVVVVVVVVVVVVVVX/BQL9trxR+0H43v/AIW6DJd6J8PPDeovAbQH97qc8TlWuJwDgqCD5cecDhj8xG39m/8A4KKfs1/sw/Du38BeAvgT4oeV9suq6rNe24uNTuAMGSQjoByFQcKOBkkk/wDD6T4X/wDRFfFH/gwg/wAK8EeJ7fxx4K8P+NLS1ktodf0u21CKCRgWiWaJZApI4JAYA1VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV+1n8Hfib+wv+1JbftVfCXSTceENR1Z7+NljZoLSafIubG4x9yOXfJsPA2uAvzR5q+/4LJ/AlfBLanp3w98YyeKTbfJpMyQLbrPjo1yJCfLz/EI9xGPkHb9hP4BfE39qL9oq4/bN+NunOmi22ptqdi08JRdTv14gWBW5+z2+EIbJGYkQFsPtqqqqqqqqqqqqqqqqqqqv+Cmv/Jj/wASf+4V/wCnW0r/AII2f8mweJv+x9u//SCwqqqqqqqqqqqqqqqqqqqvjj8WdH+Bfwm8S/FnXtPub+z8OWfnta2xAkndmVEQE8KC7qC3OAScHGK8f/tFyftD/HSD4oftJHV9U0CGX5dC0WYQiC1U5W0hZziJD/FJy7cn7x3Dw5/wWB+CPg/QNP8AC3hb9n7XtL0jS7dbeysrW8t444I1GFVVA4GK+C//AAVV+H3xp+Knhr4V6X8KfEOnXfiS9FpFd3F7C6QkgncwXkjjtVVVVVVVVVVVVVVVVVVVVVVVVVVZ/wDH1F/viqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqvFv7NX7PXjzV38QeMfgl4I1fVJWLS3t1ocDzSk93fbufp/ET39TXhrwn4W8F6RF4f8ABvhrStC0yD/VWWmWUdrCnAHyxxgKOAO3aqqqqqqqqqqqqqqqqqqqqvD/AMMPhp4T1R9c8K/Dzwzo2pSIyPeafpEFtMysQWBdFDEEgEjPOBVVVVVVVVVVVVVVVVVVVVcfAj4H3k8l1dfBrwNNPM5eSSTw7aszsTksSY8kk8kmv+FA/Aj/AKIn4C/8Ju0/+N1/woH4Ef8ARE/AX/hN2n/xurOzs9Ps4NP0+1htrW2iWKCCGMIkSKMKqqOAoAAAHAFVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX9hY6pZTadqVnBd2lzGY5oJ4xIkikYKspyCCOxq2/ZS/Zks9XGu2v7P3w9jvlfesi+HLYbW/vKuzaDxnIGc81FDFBEkEEaRxxqFREXaFAGAAB0GKqqqqqqqqqqqqqqqqqqqtd8P6D4p0mfQfE2iWGr6bc7fPsr+2S4hl2sGXcjgqcMqkZHBAPavDPg/wl4KsH0rwb4X0jQbKWYzSW2mWMdrG8hAUuVjABYhVGcZwo9KqqqqqqqqqqqqqqqqqqrWtD0TxJpc+h+ItHstU066ULPZ3tus8UoBBAZGBVhkA8jqBX/CgfgR/0RPwF/wCE3af/ABuv+FA/Aj/oifgL/wAJu0/+N1pPwY+D2galb6zoPwo8G6bqFo++3u7TQraGWFv7yuqAqfcGqqqqqqqqqqqqqqqqqqqqqqqqqqz/AOPqL/fFVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVn/AMfUX++KqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqrP/AI+ov98VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVWf/AB9Rf74qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqs/+PqL/AHxVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVZ/8fUX++KqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqrP/j6i/3xVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVpvj7wJrWuXHhjR/Gug32sWeRcadbalFLcQ46741YsuPcVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVXt7ZabaS3+o3kNrbW6F5Zp5BGkajqzMeAPc14a8Y+EfGlm+oeDvFWj67axtsefTb6O6RW9C0bEA+1VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVZ/8fUX++Kqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv20df17wr+yv8AErxD4X1u/wBI1Ww0R5LW+sLl7ee3cOvzJIhDKfcEV/w1R+09/wBHIfFH/wALC/8A/jtf8NUftPf9HIfFH/wsL/8A+O1/w1R+09/0ch8Uf/Cwv/8A47X/AA1R+09/0ch8Uf8AwsL/AP8AjtfHLxf8T0/4Jb6d4w8Lazq1z4lvPh/oE2pams7y3jQyxWv2yYyZLFjG0pdychS7E5Ga8Gat4s0Hxbo+r+A7y/tfEdrexPpctgW89bjcPL8sLyWLYAHfOO9aDLqs+hadNrsCQ6k9pE15EhyqTFBvAPoGyKqqqqqqqqr4xfE7Qfgv8LvE3xS8Stmw8N6fJdPGG2mdxxHEp7NJIUQe7Cvhb4Z8a/ttftX6fY+KtQkudS8ba015rd2oOLe0Qb5tn91UhTZGucDCLxWiaJpPhvRbDw7oNhDY6bpdtHa2drCu1IIo1CoijsAoA/Cqqv8AgsB+z7b+I/h/o/7Q2g2I/tPwtImna0yJzLYzPiJ2P/TOZgo9pznoK/4JAftIXV7Fq/7M/inUTILSJ9U8MmVuVTd/pNsvtlhKo95T0xVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV/wAFd/if8Svhp4c+Gc/w4+InibwrJf3uordvomrT2JuAqQbQ5iZdwG44z0yfWv8Ahqj9p7/o5D4o/wDhYX//AMdr/hqj9p7/AKOQ+KP/AIWF/wD/AB2v+GqP2nv+jkPij/4WF/8A/Ha/ZN/aO/aG8SftM/DDQPEPx5+ImqaZf+KLKG7sr3xReTw3EbSqGR42kKspHBBBBr/gtFrXjuz8KfDjRdOnvIvCGoXd82piEsIpruMQm3WXHHCmZkU9SHP8PH/BMnW/iDpn7XfhLT/A8t41nqSXEWv28THypLEQuzNKOmEcIyk/x7QPvYNVVVVVVVVV/wAFOP2kLr4F/AY+FvC+om28VfEB5NOs5I2xJbWiqPtUy9wdrpGD1BlDDla/4JH/ALPtt8Qfi5qXxp8RWCT6T4AjRNOWVCRJqUwOxx2PlRhm56NJEw5FVVV/wUw+Ai/Aj9pGfxN4Xt2stB8dK2s6cYQY1trvf/pMSEYwVkKyADAVZkA6V+wf+0VJ+0n+z3o/ijWbtZfE+iOdK1/oC9zEqlZseksbI5OANxcD7tVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVWf8Ax9Rf74qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq8W+EvDfjzw3qHg/wAX6Pb6ro2qwmG8srgEpMhIJVgO3Ar/AIYQ/Y//AOiAeFv+/L//ABVf8FIvhp4D+E37Td94P+HHhey0DRo9GspksrRSqCR1JZsEnk4Ff8EsPg98MfjP8YfFmg/FLwZp3iTT7Hw0bm3t71Cyxy/aYV3jBHO1mH41/wAMIfsf/wDRAPC3/fl//iq+I/jb4Zfs1fBa48S+I7FtP8EeEbG0sRaWdsZ/It2eO1hjWPuo3xrj0rwt+2P/AMEuPBHi0+O/CPwxg0nXt5eO9tfBwR4GPVoh0iPugU8n1NWV3DqFlb39sSYrmJZIywwdrAEcfQ1ff8FWf2RdPvJ7C41rxOJbaVonA0RyNynBxz6ivil+2R+z98HPAegePvHPjMW8HinTYdR0bToITNfXsEqB1ZYV5VcMMsxVQeN2eK0v/gsd+zZe6wtjqHgvx/p9i77VvpLK2kCDPDOiTlgMZJ27j7GvCPxf+GPjv4eRfFfwv430m88IyWzXDas1wIoYEQZfzS+DEVwdyvtKkYIFfED/AIK5fsveD9Zm0Xw5Y+LPGH2eQo19pljHFatjOdjTyI7c9wm09QSMZ/Zy/bz/AGff2mtUHhjwdrN/o/iYxl49E1yBbeedVGWMLKzpLgAkqrbgASVABNVqWpabo2n3GraxqFtY2NnE0txc3MqxRwooyzuzEBVAGSTwK+JX/BVz9lHwDqU+kaLqHiHxpcQEo0ug2Cm3DjPHmzvGGXj7yBwcjGa8Af8ABW/9ljxjqcOk+IovFfg5pTtF3q2npLbAk4AL28kjjtksgUZ5OMmtF1vRfEukWmv+HdXs9U0y/iE1peWc6zQzxkcMjqSGU+oNVX/BZT4oT6D8J/B3wnsLrY/ivVZb29VTy1vaKu1GH90yzI31i9jX/BF/4cW994s+IXxXvLZWfSrG20ixdlzhp2aWYj0IEMQz1w5Hc1Xx7/bU/Z4/ZwuDpHxD8aiXXhGHGh6XCbu82kZBZVwsWRgjzGTI6ZqP/gsv+zq195Uvw6+IqWmTiYWtmX9iU+04/wDHq/Z6/a1+CX7T9vfH4VeILye+0mJJNQ0++sZLea1VyQpbIKMCVI+RmxjmvjP8PrP4r/CXxh8Nr2JHTxHot1ZJuONkjxkRuD2KvtYHsQK/Z6+JN78Dvj14L+IxLQHw7rcTXyNkH7OT5dynsTE8i/jQKsoZWBBHBFVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV8Ufgf8JfjXb6fa/FXwJpniaLSXkeyS+QsIGcKHK4I6hV/Kv8AhhD9j/8A6IB4W/78v/8AFV4jt4bXxBqdrbRhIobyZI0HRVDkAD8BX/BN79lv9nv4s/sx2Pi/4j/CjQ9f1mTWb2F7y7jYuURgFXhhwBXhn9jb9l3wb4h07xX4W+Cnh3TtX0m5S5sruGJw8EqHKuvzdQRX7Wv7QP7OfwZ0HSPC37RujnV9I8ZC5FvYyaONRhl+zeUXLoeFIM0ZU9c8jGK/Ze/al/YJPjzT/hb+zt4MPh/XfFUrRI8Ph37MbgojSbZJySxUBGIBJAPQDNfGH4s+D/gZ8OdX+Kfj2e6i0LRPJ+1vawGaQebNHCmEHJ+eVPoMmvhv/wAFJP2Yfit470T4c+EtY199Z1+6W1sludJaKMyN0BYnjpXx8/4KLfs1/ADWrvwlq2u6h4m8R2LGO60vw/brcNbSD+CWV3SJWB4ZQxZcHK54r4W/8FX/ANl/4i6/b+G9cHiLwVPdyiOG71y2iFoWPADSxSP5fPVnVVGQS2M4+LPxt+FnwO8IN44+J/jKx0XSTxA7sZJLpiMhIY0y8rEc4UHA5OACau/+CyP7N0OrfY7XwN8QrixV9rXgsrVcjP3lQ3GSOh5wfYV8Dv2gvhP+0Z4TPi/4UeKItUtoGEd5bOhiubGQjISaJvmQnBweVbBKkgZqvib8Wvhr8GfDMni/4oeM9N8O6VG2xZryTDSv/cjjXLyPjnagJwCcYBrxL/wWI/Zn0m9az0Hwr4711EYj7VFYwQROPVfMmD/mi18D/wDgot+zB8d9at/C+i+KL7w3rt4+y003xJbLaPct2VJEd4Sx4ATzNxJAANVVf8FUPihP8QP2sNX8Ow3Xmab4HsbfSbZVPy+YUE07Y/veZKUJ/wCmQ9K/4JofDm3+Hn7IPg+b7Osd74qafW7xguPMMz7Yj/34jgH4VJJHFG0ssioiKSzMcBQOpJ7Cvi1/wVC/ZS+FmqXGg2niDVvGmoWrNHMvhq0SeGNwSMefI8cbDI6xs4rw/wD8Fi/2adTultdc8IePtHVmwLh7G3njUerbJy/5Ka+GPxP8DfGXwRp3xF+HGuDV/D+qh/st2IJIdxRyjgpIqsCGVlII6iv+CvPw4h8Vfsz2njyG2DXngjXIJmmxyltcnyJF+hla3P8AwEV/wR4+KM/hr476/wDC64nxY+NNGM0SE/8AL3aEumB/1xe4z9F9Kqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqs/wDj6i/3xVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVf8FZf+Tv8AUf8AsAaf/wCgNX/BGP8A5Lv42/7FE/8ApXBVf8FNf+TH/iT/ANwr/wBOtpVeFf8AkV9H/wCvCD/0WtfFJEj+Jvi6ONQqrr16FUDAAE79K+BH7K/7RX7a93cat4avLJ9P8N2drpTa1r928NtbpBAscFrH5aO7FYkQYVCFBBYjcCf2hv2bvif+zF42XwN8TrC0We4g+0WN9YzGa1vos4LxOVVuGBBVlVhxkYIJ8H+LPixq/hyT4C+CdX1i60nxfrFtK3h6zYlb+9X5IvlHJJLLlejFIyQTGpX4mf8ABMH9p/4W/DK++KGsQeGNRtdKszd6jpumajJNeWcKrukdlaJY2CKCW8t24BIyBXh/X9a8Ka7p/ibw3qc+n6rpVzHc2V3A214JUYMrqfUEA18D/iIvxb+Dvgv4mBI45PEmh2t7PHH92KZ4wZUHsr7l/Cv+Cpn7W+v+PfiRqP7PHg3WJLfwh4UmWLWVgkwNT1BcM6uRyY4WwgQ8eYrMc4Tb+xZ/wTr8R/tT+Hbj4i+JfFreFfCENy1taSxWguLnUJU/1nlqWVURScbznLAgDgkftt/sC6/+yVa6X4v0nxWfE/g/WLo2a3ctr9nnsrnazrFKoZlYMiMVcYyVYFRwW/4JKftKa74Z+Jx/Z18Q6s8/hvxTDNPo0U8hIsb+NTIyx5OFWVFkyvd1QjlmzVf8Fjtfn1D9pHw54f3N9n0nwhAyqf8AnpLc3BYj2KrGPwr/AII9+H00v9l7VdZaLEus+LrqUOR1jjgt41HXoGR/Tqa/aZ+KV18FPgF45+KGnIj32g6RJJYh13KLl8Rwlhg5USOhI7gHp1rXdc1jxPrN74i8Q6pc6jqmpXD3F5d3Mhkknlc5Z2Y8kkknNaV/wRamv/CVpfXH7Qlumt3Fskrpb+H/AD7QOyglUk88M65OBJgZHO3tX/BPr9jvxR+yXonjm28b6lpGo6t4h1SAW91pru6SWUEZ8sneqsrF5psrjsOTwar9pLw+nhT9ob4m+G4otkWneL9UhhXGP3YupNh6nGV2n8a+AGvz+K/gP8N/FF1Izz6v4S0u7lZjkl5LSN2yfXJNVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVeKv+Rn1j/r/n/9GGv+CTf/ACZ/pv8A2HtQ/wDQ1qv+C2v/ADRj/uPf+4+v+Cdv/J6Hww/6/wC5/wDSOev+CiiI/wCxd8TldQwFjbEAjPIvICD+deHPEeueENdsfE3hnU5tO1XTZhNZ3cBxJBIOjqezDqD2PNfCv/gmT+1B8ZPh5bfFHTY/Dek2mrQfa9Ptdb1GWK7v42G5ZFVInVQ/UGRkzkHoc14n8M694M8Ran4S8UaXNp2saPdSWl9aTDDwTRsVdTjjgg8jg9RxXwz8CftGftr+KvD3wx0TVbrXz4R0cWtnNqlyUtNGsUbjewB2jJVRgM7YUYIUY/aa/Yo+Nf7KVvpep/EWLRtQ0jWJTBb6rot089uJwpbyX8yON1cqCwyuCAcE7Tj/AIJ+/GPWPg7+1J4LmtNQeLSvFOoRaHq9uWxHNDcuI0Zh/sStHID22nsSD4r8T6L4K8L6v4x8R3q2mlaHYzXt7O3SOGJC7t+Cqa/ac/aQ8c/tP/E6+8eeLbyaOxjd4tE0rzMxaZa7vljUDgsQAXfGWbnoAB8Hf+COeqeKfh/ZeI/iv8T7nw1r2qWyzR6RZaYs/wBg3KCqzOzjc4z8yKAARjcetftJfs/eL/2Y/ivqHwu8W3UF3LbRpdWGoW6lEvbVyfLmVTypyrKV52srDJABP/BMP9pPXPj18ELrw3421VtQ8UeArmOyuLuaQvNd2kik28shPJf5JYy3fygSSSTVV+0Dr8/ir47/ABF8SXLMZNS8ValPhuqhrmQhe/AGBj2r4L+H08J/BzwJ4Wjj8saP4a0+zCYxjyraNPU/3fWv+Cu/xu8R/Dn4NeHvhn4Y1Cawl+Id3cx6hPCxVnsbZYzLCCOgd54Q3IyoZTkMa/Zu+CF7+0b8ZfD/AMINP8SWegya20xa/uYzKIUiieV9sYILuVQgLkZJGSBkjxr/AMEWNdttLe4+Hnx0stR1BIzttNY0VrSORv8ArrHJIVB/3D9a/Zv+Fr/BT4EeBvhdcCH7XoGjwxXxhOUN2w33BU91MryEHuDX7bvh9PE37JHxX014vMEPhm6vQMdDbDzweo6GLP4d+lfsJa/P4a/a/wDhVqNtIyvNr8dmSpwdtyjQMPoVlIqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqz/4+ov98VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX/AAVl/wCTv9R/7AGn/wDoDV/wRj/5Lv42/wCxRP8A6VwVX/BTX/kx/wCJP/cK/wDTraVXhX/kV9H/AOvCD/0WtfFX/kqHjD/sPX3/AKPev+CVtpbW/wCxn4YlghRHudT1KSZgMF2F065PvtVR9AK/4LXWNo2i/CbUjAv2pLrVYVk6HYVtSR7jKg+3Pqa/4Jp2lteftufDSG6hSVFk1KQKwyAyaZdMp+oZQR7itQsLTVLC50y+iEttdwvDNG3RkYEMPxBNV/wTwd5P2MPhgXcsRp9wASc8C7nA/QV4s8QXfi7xTrPirUHZrrWdQnvZyxyS8sjOxJ78sa+G/wC3b+1X8IvBOmfDr4dfFNdI8PaMjrZWS6Dp83lB5Gkf55IGdsu7HLE9a+Lf7aX7THx28IN4D+KvxKGuaE9zHcm1Oi2Nt+9jztYPDAjjGT3xzzX7IF5f2P7VPwjm0wMZm8Z6ZEwXOfLe5RJOh6bGb29ciqr/AIK12s9v+13dSyptW58OWEkRyDuUB1z7fMrDn0r/AIJNOj/sgaequCU1/UAwB6Hep5/Aj86+M3wx0j40fCrxT8K9cm8i08S6ZLaGcR7zbyEZjlC9yjhHA9VFfG/4C/E/9nnxpc+B/id4bn0+6jZvst0qlra/iBwJYJcYdDx7qeGCkED4F/t5ftLfACG00jwt46fV/D1mAseha4hvLZEB+5GSRJEvXiN1HPSv2Nv21/BH7XPh68itdO/4R/xhokatqmiST+aPLPAngfA3xFuDwChIB4Ks1ftkSRy/tW/FpopFcDxfqKkqc8idgR+BBFfsfWk1n+yp8I4bhQHbwbpkgAOfle3RlP5MKqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq8Vf8AIz6x/wBf8/8A6MNf8Em/+TP9N/7D2of+hrVf8Ftf+aMf9x7/ANx9f8E7f+T0Phh/1/3P/pHPX/BRL/ky/wCJ/wD14W3/AKWQVVraW1jaw2NnCkMFvGscUaDARVGAAPQAAV/wUpsbTT/22viXb2UCxRtLp8pVe7yabau7fizMfxr/AIIpWtsNB+LF6IU89rvSozJj5toS5IH0yTX/AAVWsLS8/Y18R3FxEHkstV06aAn+BzcqhI/4C7D8a+FDtH8UvBrxuVZdfsSpBwQROnIr/gpf4gu/D37F3j97B2WXUBZWRcHG1JbyESfUFNy/8CrTr+60nULXVLCRUubOZJoWaNXCuhBUlWBBGQOCCD3r/h5r+3B/0W//AMtvSv8A5Gr4z/Hv4s/tC+ILLxV8X/FY1/VdOshZW1x/Z9taFIA7OE2wRoD8zsckE89a/wCCL15qCfF34g6fErfYpvDcMsx5wJUuFEffHR5Ooz1x3zVeNbWex8Y69ZXSbJrfU7mORMg7WWRgRkcdRXhN0k8K6M8bhlbT4CpByCPLXkV/wUc/ZU179pv4RWFz4DgiuPGPgy5lu9MtZGCfbYZFUT26sSArtsjZcnBMYBxnIurTxZ4A8Tta3lvqvhzxDolyNySLJaXVlOhyOOHjcHB7EV8CP+Cq/wC0N8Mbi00v4kXEHxF8Px4SRdQIhv0T1S5UZdv+uquT6jrXwc+L/gf47/DrSfid8PNTN3pGrRkqsi7JbaVTiSGVcna6MCCOQeCCVIJ/askji/Zf+L7SSKoPgPWlBY45NjMAPxJA/Gv2TbSa9/aj+EMNuoZ18caPIQTj5UvImY/kpqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqz/4+ov8AfFVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV/wVl/5O/1H/sAaf8A+gNX/BGP/ku/jb/sUT/6VwVX/BTX/kx/4k/9wr/062lV4V/5FfR/+vCD/wBFrXxV/wCSoeMP+w9ff+j3r/gln/yZf4R/6/8AUv8A0slr/gtb/wAiv8KP+v8A1T/0Xb1/wTK/5Pg+Gv8A3Ff/AE1XdVX/AATt/wCTL/hh/wBeFz/6WT1+1P8ACjU/gr+0D44+H2o2ssUVnq802nvID++s5WMlvID3zGy564YMOoNf8E+P2kf2R4fh5Z/B79oPwb4H0rX9ImkGn+INb0S3ki1GGSRnCyztGfLdCxXMhClQuGzxVxf/ALAVrZnULi9/Z/jtQobzmfSAmD0OenORj1zXwb1f9jT4o+I9UPwQ0T4davq3g+S3lurrSNAhQ2byF/KeOcRAMcxPzGxxgZxkZqv+CzfhGXT/AI2eB/G6w7YNb8MtZBgMB5bW4dmP123UY+mK/wCCNXiiPUPgD4u8JNIGn0bxW1wBkZWK4toQowOfvwynJ65x2rx3400D4ceCtd8f+KbloNI8O6fNf3siJuYRRIXbav8AExAwB3JA71onxr/ZK/ac8Lf2TD418B+MtNvFDSaPqzQmVT2L2twBIh54JQc9DX/BSv4Ffs2/Bfxf4bf4D6vaQX2spcvrPh+01H7ZHYBPL8qQZZni3lpBsYkHZ8oUA5/4JX3Ouwftm+FodIDm1udN1KPVNoOBbi0kZc47ecsHXviq+LHiP/hYHxd8ZeLrFXnPiTxJfX0IUb2fz7h3GMdSd46Cvhp4VXwL8OPCnghQoHh7RLPTgFAAHkQJHxjjHy9uKqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq8Vf8AIz6x/wBf8/8A6MNf8Em/+TP9N/7D2of+hrVf8Ftf+aMf9x7/ANx9f8E7f+T0Phh/1/3P/pHPX/BRL/ky/wCJ/wD14W3/AKWQVVf8FNf+T4PiT/3Cv/TVaV/wRT/5Ff4rf9f+l/8Aou4r/gqb/wAmX+Lf+v8A03/0sir4V/8AJUPB/wD2HrH/ANHpX7YHws1D40/sz/EH4caPA8+pajpJmsIE4aa5t5EuIYx/vSQovp81Wlxc6TqMN2sKC4sp1cRzxBgHRs4ZG4IyOQR7GvgB+0T/AME+/jb4VsLzVfCXwn8G+JmhUahomuaTZWhhmHDCKWVFSZCeVKndjG5VOQPEPij/AIJ5+FrM3uuX/wABYECkhEj0yWRwOu2NAXb8Aa+C0PwK1/wrafEn4F+HvDNvo2vxMIdR0jRksDdJHIyEECNHwHRhhh1Gaqv2q/CMvgX9pX4neF5IfKS18U37264x+4kmaSI8esbofxr9mfxRH40/Z3+GnihJA7X/AIU055iCDiUW6LIOMDhww6Dp0FfGf9oX4Tfs+2uhX/xZ8TjRLTxDfmytLhrd5VWQIXLOEBKoMAFsYBZc4GSPHPhj9jz9qPw8bvxZN8O/G1msJVNTg1GB57ZOchLmJxJEOuQGHI5HFftLeC/ht8O/jp4v8F/CHxR/wkPhLS7xU03UPPWfeDEjSIJE+VxHIzx7h97ZnvX/AARduddk+FXxDs7kP/Y0PiC3ezJBx9oa3xOB2+6tvn6iv+ChHiiLwl+xx8TL93w95pkenxqCAWNzPHAQM+0hJ9ga/wCCc3hCTxl+2R8O7YQ74NLup9TnbaCIxb28kiE5B/5aCMZ9WHTrVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVn/wAfUX++Kqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr/goZ+xV+018dP2j73x78LPhn/behS6RZ26Xf8AbNjbZkjUhxsmnR+CRzjFf8Eyv2R/2hf2eviz4p8TfGD4ff2BpupeHTZ2s/8AatndeZN9oifbtgmdh8qMckAcdar9uj4Y+OPjJ+yv43+G/wAN9D/tfxHq/wBg+xWX2mK383yr+3lf55WVFxHG55YZxgZJAr/h2V+3B/0RH/y5NK/+Sa0C1nstB02yuk2TW9nFHIuQdrKgBGRx1FfED/gm7+2lrnjzxJrWl/Bjz7O/1e6uLeT/AISLTF3xvKzK2DcgjIIOCAa/YG+FHj/4Kfsx+Hfh58TdA/sbxBYXd9JcWf2qG42LJcyOh3wu6HKsDw3GecV/wVC/Zu+NP7Reg/D6y+Dfgv8A4SGbQ7u/kv1/tG1tPJWRIQhzPIgbJRvu5xjnFfsL/sL/ALVHwb/ao8EfEj4kfCz+yPDukf2h9tvf7bsLjyvN0+4iT5Ip3c5kkReFOM5OACRX/Dsr9uD/AKIj/wCXJpX/AMk1+xl8O/GPwn/Zj8CfDz4gaP8A2X4g0aznjvrP7RHP5TNcyuBviZkb5WU8Metftk/sR+Bf2ttBtrqa+Hh/xpo8RTTNcjg8zdHyfs86ZG+IsSQc7kJJHBZW+In/AATY/a++H1/NbR/DFvE9nGT5d/4fuku45gATlYyVmHA6NGM9BWl/sPftc6xcC1tP2ffGKPwM3VgbZef9qUqP1r/gmf8Asf8A7RH7OXjTxJ4t+K2maTouj69oy2v9mLqKXN0blJkeOQiHdGECecD8+cuOOuKr/grL8H5viH+zWnjnSrTztS+H2pLfttUljZSjyrgAegJhkJ7LEa/4I+/FSHwj8fdc+GV/cpFbeO9H/wBHB6yXlmWkjX/vy90fqBX7Yvwq+IHxv/Z28VfC34Z6hplnrWuJAgfUJnijaJJkkkjDKrEM4TZyMfMckdR4q/4J9ftieEZ5IL74Ga1fLGeJdKkhv1cZABHkux7jggHrkDBx4L/4J5ftheN7yG1tPgtqmkxSMA9zrUsVgkIzgswkYOQMdFVjjnByK/Ya/YW0T9kzSb3xFr+rW+u+Pdctlgvr23Rlt7ODIY28G7DMpZVLOwBYqvCgc/tjfFaH4L/s0ePvHIuhDex6TJZ6ac4Y3dx+5hIHU7XkDkDshPGM1+wV8ILj40ftTeCdBNr52m6JeLreqllyq21oyyYYf3Xk8qL/ALaCqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq8Qf8ABNP9tm+17Ur21+Cm+Ge7lkjb/hI9LGVZyQcG5yODX/BPT4PfEb4Gfs32XgH4p+Hf7E16HV7y4e0+1w3OI5GBRt8LunIHTOR3qv8AgqR+zJ8cP2jv+FZf8KZ8Ef8ACQ/8I9/a39pf8TK1tPI8/wCx+V/x8Spu3eTJ93ONvOMjP7GX7Bv7V/wn/ac8CfEP4gfCn+y/D+jXc8l9ef25p8/lK1tKgOyKdnb5mUcKetftnfDvxj8Wf2Y/Hfw8+H+j/wBq+INZs4Y7Gz+0Rweay3MTsN8rKi/KjHlh0r/h2V+3B/0RH/y5NK/+Sar9uj9hf9qj4yftUeN/iR8N/hZ/a/h3V/sH2K9/tuwt/N8rT7eJ/klnV1xJG68qM4yMgg1/wS+/Zu+NP7Oug/EGy+Mngv8A4R+bXLuwksF/tG1u/OWNJg5zbyOFwXX72M5471+318KPH/xs/Zj8RfDz4Y6B/bPiC+u7GS3s/tUNtvWO5jdzvmdEGFUnlhnHFeAP+Cbv7aWiePPDetap8GPJs7DV7W4uJP8AhItMbZGkqszYW5JOACcAE1X7Z3/BLi1+LniTUfir8BdT03Q/EWpO0+qaHegx2l/OTl5opFB8mRjnKldjMc5T5ifEv7BX7YHha7ezv/gL4kumVtok02NL9G9w0DOMcV4a/YA/bE8Vzxwaf8CNetd5xv1NobBVA6kmd09Pqe2civ2FPgv8S/gB+zxpXwx+KVzpcmp2F9czW0Wn3DTpbwTP5nlsxVRvEjyk7cryME1Vf8FgPg/N4S+OukfFuxtMad460xY7mVVOPt1oqxtk9BmE2+Omdrehr/gkn8VIfG/7Mh8BXFyjaj4B1Wa0Mf8AF9luGM8Ln2LvOg9oq/4KZfsnfHz9peTwfqXwmg0jVNP8LW10JNJmvxa3Ms87R5dDJiIrtiQfM6kHPXJrVP2G/wBrrSLkWl3+z94vkckgG1svtK/99RFl/WvhP/wS3/ar+IuqwR+KPC1t4E0cuBPqGs3MbOq/xbLeJmkZsdAwRT/eHOPgB8CvBX7OXwv0v4W+BYpWs7DdLcXU+DLe3L48yeTHG5iAMDhVCqOAK/4LL/FaHS/h14M+DNjdD7Zr2pPq18in5lt7dSkYYf3XklJHvB+f/BGX4QXEmp+N/jtqNpi3hgTQNLkZfvuxWa5Iz02hbcZH99h9aqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqrP/j6i/wB8VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVa3o2leI9Gv/D2uWMV7pup2slreW0q5SaGRSrow9CpIP1q7/wCCa37XXwe+OI8X/A7SNP1bTvDOtpfeHdUm1e3geWNHDxiWN3Vs4+RxgBsNjKkVol3qN/othe6xpTaZf3FtHJdWTSrKbaUqC8e9CVbacjIODjNVVf8ABSb4H/tP/tIReGPh58H/AAhbXfhLS2bUdRuJ9Wt7Y3N6QyRpsdw22NC5zjBMv+zX/BOj9jjV/wBmDwNq+v8AxFs7VPHniiYJdRwzLOtjZxE+XCrrkEsxaRipwcxjqlVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX7YP7O1h+078DNZ+HBaCDWYyL7QbuUcW99EDsyeyurPGx5wshPUCv2CP2UP2xv2X/jfDr3ibwHZL4P1+1aw19YdctZDGn3op1QSZZo5AOmTsdwASaqqr9rn9i39tf8Aab/aA134iN4E0230V5VsNDS4161H2XT4iRGWUOSCxZ5WUAkNIwGeK+AHwZ8Pfs//AAh8N/Cfw2RLBolrtuLkpta7uXJeaZv952YgdhgdAKqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqs/8Aj6i/3xVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVZ/8AH1F/viqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqz/4+ov8AfFVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVn/x9Rf74qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqs/+PqL/fFVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVn/x9Rf74qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqs/wDj6i/3xVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVZ/wDH1F/viqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqz/wCPqL/fFVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVsX+6PyrYv90flWxf7o/Kti/wB0flWxf7o/Kti/3R+VbF/uj8q2L/dH5VsX+6PyrYv90flWxf7o/Kti/wB0flWxf7o/Kti/3R+VbF/uj8q2L/dH5VsX+6PyrYv90flWxf7o/Kti/wB0flWxf7o/Kti/3R+VbF/uj8q2L/dH5VsX+6PyrYv90flWxf7o/Kti/wB0flWxf7o/Kti/3R+VbF/uj8q2L/dH5VsX+6PyrYv90flWxf7o/Kti/wB0flWxf7o/Kti/3R+VbF/uj8q2L/dH5VsX+6PyrYv90flWxf7o/Kti/wB0flWxf7o/Kti/3R+VbF/uj8q2L/dH5VsX+6PyrYv90flWxf7o/Kti/wB0flWxf7o/Kti/3R+VbF/uj8q2L/dH5VsX+6PyrYv90flWxf7o/Kti/wB0flWxf7o/Kti/3R+VbF/uj8q2L/dH5VsX+6PyrYv90flWxf7o/Kti/wB0flWxf7o/Kti/3R+VbF/uj8q2L/dH5VsX+6PyrYv90flWxf7o/Kti/wB0flWxf7o/Kti/3R+VbF/uj8q2L/dH5VsX+6PyrYv90flWxf7o/Kti/wB0flWxf7o/Kti/3R+VbF/uj8q2L/dH5VsX+6PyrYv90flWxf7o/Kti/wB0flWxf7o/Kti/3R+VbF/uj8q2L/dH5VsX+6PyrYv90flWxf7o/Kti/wB0flWxf7o/Kti/3R+VbF/uj8q2L/dH5VsX+6PyrYv90flWxf7o/Kti/wB0flWxf7o/Kti/3R+VbF/uj8q2L/dH5VsX+6PyrYv90flWxf7o/Kti/wB0flWxf7o/Kti/3R+VbF/uj8q2L/dH5VsX+6PyrYv90flWxf7o/Kti/wB0flWxf7o/Kti/3R+VbF/uj8q2L/dH5VsX+6PyrYv90flWxf7o/Kti/wB0flWxf7o/Kti/3R+VbF/uj8q2L/dH5VsX+6PyrYv90flWxf7o/Kti/wB0flWxf7o/Kti/3R+VbF/uj8q2L/dH5VsX+6PyrYv90flWxf7o/Kti/wB0flWxf7o/Kti/3R+VbF/uj8q2L/dH5VsX+6PyrYv90flWxf7o/Kti/wB0flWxf7o/Kti/3R+VbF/uj8q2L/dH5VsX+6PyrYv90flWxf7o/Kti/wB0flWxf7o/Kti/3R+VbF/uj8q2L/dH5VsX+6PyrYv90flWxf7o/Kti/wB0flWxf7o/Kti/3R+VbF/uj8q2L/dH5VsX+6PyrYv90flWxf7o/Kti/wB0flWxf7o/Kti/3R+VbF/uj8q2L/dH5VsX+6PyrYv90flWxf7o/Kti/wB0flWxf7o/Kti/3R+VbF/uj8q2L/dH5VsX+6PyrYv90flWxf7o/Kti/wB0flWxf7o/Kti/3R+VbF/uj8q2L/dH5VsX+6PyrYv90flWxf7o/Kti/wB0flWxf7o/Kti/3R+VbF/uj8q2L/dH5VsX+6PyrYv90flWxf7o/Kti/wB0flWxf7o/Kti/3R+VbF/uj8q2L/dH5VsX+6PyrYv90flWxf7o/Kti/wB0flWxf7o/Kti/3R+VbF/uj8q2L/dH5VsX+6PyrYv90flWxf7o/Kti/wB0flWxf7o/Kti/3R+VbF/uj8q2L/dH5VsX+6PyrYv90flWxf7o/Kti/wB0flWxf7o/Kti/3R+VbF/uj8q2L/dH5VsX+6PyrYv90flWxf7o/Kti/wB0flWxf7o/Kti/3R+VbF/uj8q2L/dH5VsX+6PyrYv90flWxf7o/Kti/wB0flWxf7o/Kti/3R+VbF/uj8q2L/dH5VsX+6PyrYv90flWxf7o/Kti/wB0flWxf7o/Kti/3R+VbF/uj8q2L/dH5VsX+6PyrYv90flWxf7o/Kti/wB0flWxf7o/Kti/3R+VbF/uj8q2L/dH5VsX+6PyrYv90flWxf7o/Kti/wB0flWxf7o/Kti/3R+VbF/uj8q2L/dH5VsX+6PyrYv90flWxf7o/Kti/wB0flWxf7o/Kti/3R+VbF/uj8q2L/dH5VsX+6PyrYv90flWxf7o/Kti/wB0flWxf7o/Kti/3R+VbF/uj8q2L/dH5VsX+6PyrYv90flWxf7o/Kti/wB0flWxf7o/Kti/3R+VbF/uj8q2L/dH5VsX+6PyrYv90flWxf7o/Kti/wB0flWxf7o/Kti/3R+VbF/uj8q2L/dH5VsX+6PyrYv90flWxf7o/Kti/wB0flWxf7o/Kti/3R+VbF/uj8q2r2UflVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVf8NO/Cr/AJ/NS/8AAM/414F+Ivhv4jWt1eeGnunhs5FjkaaEx/MRnA9eP5ivEGuWXhrRbzXtRWU2tjEZJvKj3sFHUgew5+lf8NO/Cr/n81L/AMAz/jX/AA078Kv+fzUv/AM/41/w078Kv+fzUv8AwDP+Nf8ADTvwq/5/NS/8Az/jX/DTvwq/5/NS/wDAM/41/wANO/Cr/n81L/wDP+Nf8NO/Cr/n81L/AMAz/jX/AA078Kv+fzUv/AM/41/w078Kv+fzUv8AwDP+NV4j8XeGvCMENz4l1iDT4rhykTzEgMwGSBj2qwvrPVLG31LT7hJ7a6iWWGVDkOjDII9iCK1PU9P0bT59U1S7jtrS2QvLNIcKijua8OeLvDPi+Ca48Na1bahHbsElaFs7CRkA/hVaX468Ia3rM/h3SdftbrUrXf51vGxLJsO1s9uDxVVVVVVrHxm0HRviJb/DefS79764lhiWdAnlgyAEE854zzxVVVVVXd1b2NrNe3cyxQW8bSSu3RFUZJP0Arw54x8L+L455PDOt2+oralRMYSTsJzjP1warxJ448JeEHgi8S69a6e1yGMImJG8LjOMfUfnSOksayRsGVhlSO4NVXjzxnYeAPDNx4n1K1nuYLZ0Vo4MbyWYKMZIHevAnjOw8f8Ahm38T6Zaz28Fw7qsc+N4KMVOcEjtVVH4v8LTa+3hWLX7FtXjXLWYmHmDjOMeuOcdcc1VePPGdh4A8M3HifUrWe5gtnRWjgxvJZgoxkgd68CeM7Dx/wCGbfxPplrPbwXDuqxz43goxU5wSO1VVVr3iDRPDGntquv6lDY2isEMspwAx6CvDni3w14ut5bnw1rVtqEdu4SVoWzsYjIB/Cq1zXdH8N6bJq+u6jDZWcRAeaU4AJIA/UivDnjHwv4vjnl8M63baitsQsxhYnYTnGfrg1VVVVVVVV8QPHOnfDzw4/iTU7S4uYElSIxwAbstwDyQMV4I8W2PjvwvZeKtOtp7e3vvM2Rz43rskZDnBI6qTVVWo67oekFV1bWbGyLfdFzcLFn6biKs76x1GAXOn3kF1CeBJDIHU/iOKqaaG3iaaeVI40GWd22hR7mrDWtG1YsulavZXhj++Le4WTb9dpOKqvtNt532b7RF5v8Azz3jd0z069KqW8s4H8ua7hjbH3XkAP5V/aWm/wDQQtv+/q/40uoWEjBEvrdmY4CiUEk/nVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV8A/DA8MfDDSo5IttxqSm9n4wSZMFf8AyGEH4VqNhbapp9zpl4m+C7heGVfVWBBH5GvEOi3Ph3XdQ0G8H7/T7mSBzjGSrEZHscZ/Gqqqqqqv2ktA/tv4W3tzGm6XSZ47tABzgHY34bXJ/Cv2c9c/tv4VabEzhpdMlktH9trblH/fDpX7UPiD+yfhwNIikxLrF2kJAOD5afOx+mVUfjX7Nj3fhL4h+Jvh/qLbZGhD4YYy8LY4HYlZCfwrxPrMPhzw5qevTFdun2ks+D3KqSB+JAH41+ydoUs6+IPGt4S8txItpHIw5J/1kp/EmP8AKpZYoYzLNIqIo5ZjgD8atNS07UQx0/ULa52fe8mUPj64NV8UPHvi7TPjd4W8H6drk1tpNzc2DTW8Sqvmb7jawZgNxBAAxnFU7pGheR1VVGSWOAKtNT0zUGZLDUbW5ZB8yxTK5X64NV45/wCTqNL/AOv2w/8AQEq7vbKxj829vILdP70sgQfmajkjmjWSGRXRhlWU5B+lVc3VrZwme7uYoI16vK4UD8TVreWd9F59jdw3Eecb4nDjP1FVXjf/AJEvX/8AsF3P/opq/ZD/AOQb4m/672//AKDJVfte/wDIR8M/9cLn/wBCjrTf+Qba/wDXBP8A0EUxVVySAAOSe1W+p6beSNDaajbTSISGSOVWII6ggGv2k/8Akkeqf9d7f/0atfs3FV+EWlkkACa4yT2/etVrqOnX5ZbG/trgx8OIpQ+364PFVY/AXR7L4oSfElNcum33L3S2JjAxM+dxL5yVyxIXA9M4qru+sbFVa9vYLcOcKZZAmT7Zr9pBkk+EOpujBlaa3IIOQR5q1+zb/wAkj0v/AK73H/o1qmmht4mmnlSNFHzM7bQPqatNR0/UFZtPv7e5CHDGGUPg++DVV+1PciD4Yxxb8faNUhjxjOcK7Y9vu1+z9fXPgT4kHwfqEjLD4h0u3nQNx+9MKzJ+QeRfriq/aN1GfxL4g8K/CjTZiJNSu0nutv8ACGbYhPsB5jEH0Br9kWR4j4s06YqrxPanbnnP70N/IVVVUGraVdTm1tdTtJplzmOOZWYY68A5qv2ifHvi7wv4g0DRfD2uTWFrfRl7hYVUM5DgfexuAx2BFVVXOq6VZyrBd6nawSt91JZlVj9ATQKsoKkEEcEV+07/AMkqn/6/YP5mv2d/+SOeH/8At5/9KZaqvG9h4h1TwpqVh4T1P7Bq0sBFrPkDa2RkZwcZGRnqM5HStN/ZU0S7hN34y8Xaxf6nN800ts6oN3fmRXZvqcfQV4v8N+Jf2bvFuna34Y1ua602/Jwkny+aEI3RSqODwww2B1yMEVYapZ6hpFtrcUgS1ubZbhWfjCMobJ9ODVodd/aX8fXsE+qXVj4R0k7hFEcfKSQnB48x8E5IO0A+nPxL+AVn4H0N/HHw31TUrS90YedLG0+W8tfvOjAAggckdCM9O/wW+IMnxF8EwanfMv8AaVm5tr4KMBnUAhwP9pSD6ZyB0r4sJ8XtSvtP8P8Aw3eC0tLyFzeagxCtAQQMbznaCCMbQW4OOlfDjw7deFP2lLLQL7VG1G5tZJ/OunBBldrN2YnJJ6t1J561XxF+AGj/ABF8Sv4lvfEF5aSvCkXlRRKwAUYzk1/wyN4c/wChv1L/AL8JXw5+H9jqPxwi0nwzqE93pXhu9W5lvZFAL+QwPGOMNIAo9RzVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVeDfDsvi3xXpPhuEnN/dJE7Acqmfnb8FBP4Uq29nbBFCRQwJgDoEUD9AAK8EeLLHxx4Zs/EunDEV1vGzPKlXKkH06Z/Gv2pPC39j+PIfEEEW2DXLYMxAwPOjwrf+O+WfqTVVVVVVXiDSIdf0LUdDuMeXqFrJbsSOgdSuf1r9lDUprG+8TeDr07JoWSdYyeVZSY5P18sV8Yw3jf44eD/AAFF+9t7LZNcoO25t8gP/bKNT+NeNmPgn9p7RteJ8u31nyN5/hxIpt2z9CAx/Ov2m/EA0f4ZTadHJiXWLqO3AHXaD5jH6fIB/wACr4I+HP8AhGPhhodlIm2e5g+1zZGDulO8Z9wpVfwrxj8B7bx/4sudc8S+L9WfTW2fZtOhfAgIQBsFtwAJBOAo6nmvij8CbT4caC3jz4f69qdtcaU6NKksw3BSwXcjKAQQSMg9Rn0wfAGu/wDC3fhL/wATaVo59Ss59Pv3iABDlSjOvYEqQ2MYBOK8cfCrRvC/xX0PwBZ6jeTWeqSWiyTS7fMXzZSjYwMcAcZFfDv4EeHPhvr7eIdK1jUbqZrdoDHcFNuGKkn5VBz8orx78Eo/iL4oOq674v1SPSVhRY9NgbASQZDMC2VXIx/DnOea+JH7POleDfDlx4y8B63qlte6OnnskswJKL95kZQCrAc++O1fAzx1eePvANvqOqSeZqFlM1pdSYx5jKAQ+PUqy598/SvjPqOpaT+0F/aOjWy3GoQPaNaxMMh5fLTYMd+ccVbfs22niKA6t8SfFusanrl0u6aSGZVSEn+FQytkDp2HoBWoL4s/Zn8c2cdpqs+oeG9QO/yXGFnjBw6leiyrkHcOuR2JWvG3jrTfCHge68aZW4iSBXtUBx5zvgRj6EkE+gya8D/CnV/jVbf8LB+KHiG/aG8d/sNnbsECoDjIyCETIICgZOMk88/EP4Z6n8Cvs3xA+G2v3y20cyxXlvcMGxk/Lu2gB4yeCCMgkYPPHg/xJa+MPC+meJrRdiahbrIUznY3Rlz7MCPwqvG//Il6/wD9gu5/9FNX7If/ACDfE3/Xe3/9Bkqv2vf+Qj4Z/wCuFz/6FHWo67YeGPCEniDU5NttYWQlkx1OFGAPcnAHuRXh3SfHf7SmrXmra/r02l+GrOby1toM7c9QiL0ZgMEu2eowOwu/2UfAjQKNL13XLS6j5SZpY5OfUgIP0Ir9oyGO3+DuoW8QOyKS2VQTk4Eqgc18JvAviv4p+CrfRL7X5tH8J6ZNInlWw/eX0rMXYnPG1QwHORntnOPid8G9T+DcFp458FeJb1ooJ1jkZsJLAx6NlcBkJGCCO4HIJx8MvFzeOvA2leJpkVLi5iK3CqMASoxR8DsCVJA9CK0D/k7eX/r9u/8A0lkr44fEqX4b+EvtGmlDq2ouYLLcMiPAy0hHfaMcerLnjNeAP2f5fH2mReN/ibr+pzXOqoJoIY5Rv8tuVZ3YN1ByFGMDHPYfGH4a698LPC8tt4f8Q3l94U1SZI7i0uyGa2lDB0YYwOdpGQB6HOQa/Zt/5JHpf/Xe4/8ARrV4q+AVp468WXmveKfF+rzWEjg2mnxOALcbQGAZtwALA8BRwetfFP4JQ/C3Rh8QPh7r+p20mmyp56STDeoZgoZGUD+IqCpyCD7Yr4WeMJPHfgTSvEdwFF1NGY7kKMDzUJViB2BI3Y7A1X7W1zt8G6NZgt+91Pfjt8sTD/2avjVpN74Sj+HvxF0sDz7OytreRwMAyRKrx5/3hvH0WtJ1ay1rR7PXLKUG1vbdJ42PHyMoYZ9ODXwkLfEz44eIPiNLl7LSgy2ZbtuBji4/65q5PoSK/ZgdrXxx4v01nQtsBIHUlJWGR7fN+oqqrWtW8S/tAfEG+8EaFrMumeE9ILC6lhz/AKQAdpLDI3bmB2qeABnGRWufsq+GI9KeXwlreq2ur26braS4mVkdwOA21QVyR94HjOcHpX7P3xB1bxroep+EfFVxJJqujgRmaT/WSRNlfm9WUjBJ65HU5NfGP4VaN8ONf0bSNK1G8uo9SjLSNcbcqd4XjaB2rwR+zv4X8C+KLLxVp2t6pcXFj5myOcx7G3xshzhQejE1Xjjw9qHirwxeaFpWu3GkXNyFCXkBIaMBgW6EEgqCMZHWof2UvARtWW+1vXLi7cEvcCaNfmPcLsP6k/WvhlqniD4TfF+T4Tanqz3ukXb+Xb7idqMyb4nVf4SeFYDjJ74Br9p3/klU/wD1+wfzNfs7/wDJHPD/AP28/wDpTLVVVftT6wmt6x4e8A6Qv2nURIZHiQ5IeTCRp9TycehB714+ifwb8D9R02Gb5tP0NbJXXP8AcWLI796/ZQ0+KDwBqGoADzbrVHUkf3UjTA/Mt+dalZRalp11p0wBjuoXicEZGGUg/wA6/ZDvnW+8SaYWJR4reZRjoVLg/wDoQ/KqsP8Ak75v+u8v/pC1VXxt8eDwF4Eu7y1n2alfZtbEA8q7Dlx/urk/XHrX7N/gYeFvAya3eRFdQ18i4fcOVhGfKX8QS3/A/aqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/ZS8MDUfGF/4nmiJj0i12RMR0llyMj/AIAHH/AhXxw8Sf8ACMfDHWruOTbPdQ/ZIOx3S/KSPcKWP4V+yX4l+0aRrPhKaTLWky3cAJ/hcbXA9gVU/wDA6/aY8Mf298NpdShi33GizrcqR18s/I4+mGDH/dqqqqqqqtLj/wCEG/aouLQDZba48mD0yJ08z/0aMfhXweQeNPjf4z8ev+8gsWaC2c/7TbIyP+2URH/Aq/ay0eUaV4f8WWhKS2F29uzqORvAdD+BjP8A31Xxj1QfE/xf8PPC1mx8nUrSC6lCH7guSu7PuqIT+NRxxwxrDEiqiKFVQMAAdAK1T4t/EL4leL7nwZ8H1tbO1tN3nanOoYlQwUvlgQqknAAUsevHb4g/Cr4kad4I1bXfEfxi1K/S1ty82nosghmGR8pPmAY+qV+yx/yTGT/sKTf+gR18Xv8Ak5Lwh/1303/0pNV4q+L/AI58X+N5/hz8IYLeOW1Z0uNSmUNjYQHYbgVVAeM4Yk4x1GfFvwm+KMfhDWda8R/GfUZvsmnXE89jAknlTqsZLRk+Yo2sBjlMcniv2Sv+RK1j/sKH/wBFJXjtVf8Aan0sMAf9OsDgjuFSq/a4Nt/wiuhqxX7R/aDeWD12+Wd34Z2fpXxka/i/Z58CwzkgN9jEnY8Wr7Afw/UV4N8G/tD3XhLR7nw58RNFttLlsYntIGY5ijKgqp/cHkDg8nnPJrXvhR+0V4n0mfQtd+IOhXdjc7fNhZ2UNtYMORAD1UH8K0H4X+LdL+D8Hw+h8WDS9WimZ1v7CSQiNTMX2qfkbkHB6dT1r/hRnxX/AOi863/3+uP/AI7Vn4Y1rR/hdd+F9W16fW9RGn3cbXsxZnmL7yudxJ4DBeT2r9kK6gNv4mst/wC+D20gU91xIMj8f5iq/a2lS68QeGtLt/nuRbSt5Y64d1C/mUNftGJc2nwcu7WFtyrLbRzEcZUOvP8A30Fr9m9LRfhFpJttvmNLcGfH9/znAz/wEJVftJ/8kj1T/rvb/wDo1a/Zt/5JHpf/AF3uP/RrV+0SAfg54g46fZsf+BMVfsxkn4VW4z0vZ8fmK0D/AJO3l/6/bv8A9JZK/a8a4Or+G42z5Atpyn+8WTd+gWrJbZLOBLLb9nWJRFt6bcDGPbGK/aT2/wDCpNTzjPn2+M/9dVr9m4qvwi0skgATXGSe371quviv8R/it4quvC3whNtp2n2efN1KdQSyZxvJYHaD2UKW7+uPiX8LPiJpPgTVfEHiP4v6lqcdtErTacFkEM2XUYP7zGATnlOw/D9mL/klVv8A9fs/8xVftfXG208LWgJ/eSXTnn+6Ih0/4FXxd8HDxF8J9Q0WKFTc6faLcWyqM4eEZwv1UMv/AAKvCnxMey/Zr1e3a4xf6c7aZBlvmKzcqR9FaTH/AFzr9nnwn/wi/wANLCaaPbdawTezZHOHAEY+mwKcerGvgN/oHx28Y6WVWMBL1QhOSCl0gwPwz+VVWsyTxaRfS2h/fpbSGLBx8wU4/XFfA7QPihrUesN8NvFWnaQYjCLxbk4aTO/YR+7fgYf061/wg37UP/RTtE/76P8A8j18HPg7418B+NNT8U+KdY0u7Go2kkbi0lkZmleVHLEMijHyt+dftTf8jr4V/wCuB/8ARoqq+LvxQs/hf4fjvzbLd6heu0dlbs20MwHzMx67VyM45OQOM5GieGv2gPiPYwa9rXxBHhmzvEElvbWkW2RY2AIO1NpGQcgM5Prirnw1q3hP9oLw5o+teKbvxBcreWjm9ugwcgtwvzO5wPrX7Tv/ACSqf/r9g/ma/Z3/AOSOeH/+3n/0plqqr4ufFnSfhloxZWiutZuVxZ2Rfnv+8cDkIMfieB3I+B3wu1eTU5Pit8QRJLrV+zSWkM64aIMOZWHZiDgL/CPwx8YtOk1T4X+JbSJNzDT3lC/9c8P/AOy1+yhfRT/D6/sAw8211RyV77WjjIP5hh+FapfRaXpl3qczAR2kDzOT0AVST/Kv2Q9PdrrxLqrJ8iR28Kse5Jckfov5iqsP+Tvm/wCu8v8A6QtVV4weT43fHW08J2shl0LQWKTlT8uxCDO2f9pgIwfZTSRxxRrFEioiLhVUYAA6AD0qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv2cvDB8O/DKyupottxrEjXj5HO1sLH+GxVP/AjX7W/iQNLofhCGT7ivezr7nKR/wApPzFfALxL/wAIz8UNJeSTbBqRNjN7iTAT8PMEZ/CtV0211jTLvSL5N9vewPBKp7qylT+hrXdHuvD+tX+hXoxPp9zJBJxjJRiMj2OM1VVVVVV+1FZXmheJPC3xA0v5Li3byvMx914nEkefruf8q/Zk8PDR/hnFqUiYm1i6kuCSOdiny1H0+Qn/AIFXxy0E+IfhZr1rGm6W2t/tUfHIMRDnH1VWH41+zTp934p+IR8Q6gfMj8O6YsMJIzsYr5aD/vgSH61LGJYmiLEB1IypwRn0r4beJZf2f/HesaJ430u6EF1GIjNDHknax2SKCRuQgnoc9O4Ir4k/Fyb4n+FdV8OfDjRb+azhtXuNV1G4QRJDBEN7KvJ5bbjnBPQA5JH7K0iSfDOdEbJj1WZWHodkZ/kRX7SNhq3hv4ieHfiRa2TzWlqsGXx8qzQylwrHtuBGM9cGvh78bPCfxK1V9E0O01KG7htDcyC5iVVVQyqQCGOTlx26V4c1i5+AHxc1U+LNMupbC+WWJJ40y0kTSB0lTJAboARnjJ7jFeNvjMfiToGoeDvhfo2oXct3auL69nj8qK2g2kvzk8lQV5x14ya/ZJkRvB2swhvnXU8kegMS4/ka+K+j634g/aHl07wxNGmrJFFPaFmxiSG2EoHsTswM8ZIzxWmftOeGrS0Nl420PVdK1q1Gy6tkttylx125YEfRsY9T1q+bxP8AtM+OrN7XTLiw8L6YxQyuOIkJBcluhlYADaM4wOwJPxG+H9n448CXPhC38u2ZI1NixB2wyR/c/DHyn2Jr4f8Axg1j4OQf8K++Jvh3UEhsmb7JNEgLopJJABIEiZJIYHjOOeMah+1L8OoIsabaavqFw3EcSWwQFuwJZuPwBqwu11Cwtr5I3RbiFZAjjDLuAOCOx5qq13RvFX7PHxFn8X6Hpb3vhu+ZxtXITy3OfJcgHYykDaT1wOvzCpf2sPAK2Hnw6Nrb3W3iBoo1AbHdt5498Z9q+H/hXxb8YviRF8U/GOnNZ6RZyLJaxSKQJNnMccYPJQH5mbGCcjuceMPDNl4y8Maj4Yv2Kw38Bj3gZMbdVYD2YA/hXgrxl4u/Z11a78LeNPD9xPo91N5iPD03YAMkLH5XDKBlSQRgdDkG+/an+HUNsr6dZaxe3D8LAtuqYPuS38s1+0bLHcfB7UZ4jlZJLZlyMcGVa/Zt/wCSR6X/ANd7j/0a1ftEf8kc8Qf9u3/pTFX7MX/JKrf/AK/Z/wCYrQP+Tt5f+v27/wDSWSvj58Nrr4h+Eo20eISatpMjTW0Z4MqkYeMHsThSPdQO9fDv9o2Dwno0PhD4i6PqSXmlIIEmiiBfaowqyIxUhgMDPOeM9yfij438RfGXwve3nhzQrrT/AAnoI+0z3V0uHu5QdqqAMjjcSQCcdSegP7PVst98FrOyMpTz2uoyy9V3SMMj86+FfjBvgJ4u1vw7470q6iS8EaPJDHuKmMttdQSNyMHJyPavih8V7n4peEtU0L4e6LfPpdrD9o1bUrlBEixx/PsXk8khfQnHTGSP2YJEf4WRIjZMd/OrD0PB/kRVftSbb7xl4R0kBGLI3yseu+VV5Hp8v86ZVZdpAIIwQRV98PtQj+L0/wAJrVpE0281eObyxnHkBWdXHusUjj65qKKKCJIIUVI41CooGAoAwAK+G+NP/ae8R2wG37TNfcN1O5vM4/LP0r9oL4oeJfAI0TTvCU8SXupNK0m6ASsFXYFABzySxHTtWgRarDoenxa5dC41FbZPtcoQKGl2jeQFwAM56VWpaX4q/Zy8f3fibR9IfUPCmokhwgIVIy2RGzYOx1P3SeCPqcQftS/DGW1E8qavFJjmFrUFgfqGx+tfDf4j6b8TNMu9W0vTb2zgtrkwoLpQGkG0HcMEjuRgE4x71+1f4e1RxoXi+wt5JILASQ3DquRCSytGT6AncM9M4Hevh98ffCnxA1Sx8O2mn6jbapcwszpJGvlIyLuYBg2SODjj64qv2q/CGsa1ouk+JNLt5bmHSDMt3GgLGNJNmJMegKYJ9x2BrQP2ofBCeHLRNSsNUGqQwJHJbQQBhJIAB8jbsYJ9cGrq/wDE2pfHvwv4j8X6Y2lzazeWk9paO2WhgMnlxqw6gkoScgcnOBnFftO/8kqn/wCv2D+Zr9nf/kjnh/8A7ef/AEplrwF8c/Gnib4tP4P1LR7ePT5Zp4vJWJhLaiMMQWbPJ+UA5A5PGOlV8W/iGnwz8IPry2f2q5nmFtaRnhfNZWYF++0BGPHJ4HGcjwj8QNAh8Xz+OfiLpt/4i1MyiSBN6LErDozA9ccBVGAMfTH/AA1z4c/6FDUv+/6V4R8RWXj7wjZ+IEsWitdUhfNvMQx27ipBxwc4P51plxrX7M/j+9i1HTrq88K6ucJNGudygkoQTx5iAkFSRkHPoa+JPx2g+IGiv4E+Gek6leXusDyppGg2kRn7yqoJJz0JOABn1yPg78Pj8OfBVvo9yUbULlzcXzLyBKwA2g9woAX0JBPeqsP+Tvm/67y/+kLVXxo8dr4A8CXmpW0wXUbsfZrAA8iVgfnH+6oLfUAd6/Zl8DHw/wCEH8VX0f8Ap2vkOm7qluudn/fRJb3BX0qvj/8AFLxP8NrXSI/DdnBu1B5PMuZ4i6ps24QDOMncevYceo8A+Ib3xZ4M0jxFqVkLW6v7ZZJYlBAB6ZUHkKcZHsRyaqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqrCxtdMsLbTbKMR29pCsUKD+FFAAH5AV4r+DPgDxtrD694i0ye4vHRULrdyIAqjAAAOBVv+zp8KbWeO5ttFuo5YXDxuL6XKsDkEfN6iq8UfAr4c+L9Xude1bS7gX12waaWG5dNxChc7c4HAGePevE37Jnh+4tpJfCPiC8tLoDKRX2JYmPplQGUe/wA30rxP4X1vwfrM+g+ILFra8gPKnkMp6Mp6FT6iqqqqqmt4LhQk8McoByA6g4P40kccaCOJFVVGAqjAH4Uyq6lGAYEYII61Da21vn7PbxR7uuxAufyqtR0nStXiWDVdMtL2NTlUuIVkAPqAwNW1hY2VsLKys4ILcAgRRRhFHrwOKht4LdSlvDHGCckIoXn8KnghuYWguIkljcYZHXcGHuDVhoujaUWbS9JsrMv9429usefrtAq/0zTdVg+zanp9teRZz5dxEsi59cEGrLTtP023+y6dY29rCOkcMQjX8hxUNtbW4It4I489QiBc/lX2W2877R9ni83/AJ6bBu6Y69elahoeiasyPqmj2V40f3DcW6yFfpkHFRQw28SwW8SRxoMKiLtCj2Aqr2wsNRh+z6hZQXUR6xzRB1/I1YaBoWlMH0vRbC0YZwYLZIz7/dAqqplV1KMAwIwQR1qLwt4ZhuPtkHh3S0uP+eq2aBvzxnsKqrm1tryE293bxTxt1SRAwP4GrHw9oGmSebpmh6faPkndBapGcnryBUsMU6eXNGkinqrrkflUUMUKCOGNEUdFVcAfhUsUU0ZiljV1PVWXIP4VFDDAnlwRJGv91VwPyFC0tRN9oW2iEv8Af2Dd+fWqv9B0LVZUn1PRrC8kj+49xbJIV+hIOKFvbiAWywx+UF2iPaNoHpj0qKGGBPLgiSNf7qrgfkK1HR9I1iNYtW0uzvUQ5VbiBZAD7BgagsbK1thZWtpBDAAQIo4wqgd+BxUUEFumy3hSNSclUUKM/hVS2trO6yTW8Ujr91mQEj6VVULW1WXz1t4hKf4wg3fnWveAfEPjb472Wv6vpTxeHPD8EfkTSMuLiRPnAC5z/rH5yMEJ9KqmVXUowDAjBBHWh4U8Li4N0vhvSxOesn2OPcfxxmlVVUIgACjAAGMU6JIhjkVWVgQysMgjvxVloOh6bM1xpujWNrK2dzw2yxsc9ckDNVUHhzw9a3n9oWuhadDdE5M8dqivn13AZqS1tpnWSW3id1+6zICR9DUsMM6eXNEki/3XXI/Ko4ooYxFDGqKvRVXAH4VFpmmwXkuoQafbR3c4xLOkSq8gHZmAyfxqpoILhAk8KSKDkK6gjNf2Zpv/AD4W3/fpf8K/szTf+fC2/wC/S/4VHFHEgjiRUVeiqMAfhVxbW93C1vdQRzRsMMkiBgfqDWn6No+kqU0nSrOyDdRbwLHn/vkCqoWtqJvtAt4vN/v7Bu9OvXpVTW1tcgC4gjl29A6BsfnSoiIERQqqMBQMACqvtO0/VIPsupWNvdwhgwjniEi5HQ4ORmlVVUIgACjAAGMVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVeMvE9p4M8L6j4nvYzJFp8Jfyw20yMSAqg9ssQPxr/hrzTf+hGuf/A9f/iK+E/xktfircalb2+gy6cdOSNiXuBJv3lh2UY+7+tfGXxZrHgnwBfeItBkiW8t5YlQyx71w0iqePoasv2pviXbSh7mLR7tM8pJasvHfBVhivhN8VtN+Kejz3MNobLULFlW7tS+4LuB2sp4ypwfcEH2J/ai8HW2seCF8VwwD7bokq7nA5aB2Csp9cMVPt83qaqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv2sPE32LwvpnhWCXEmp3JmmAP/ACziHAPsWZT/AMBqv2Q/+Qj4m/64W3/oUlftJ/8AJI9U/wCu9v8A+jVqv2R4rlvE2uzID9nSwRZDjjeX+X9A9fHKaCD4TeJHnI2m1CjP95nUL+pFVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX7RfiX/AISL4n39vFKWg0hFsoxnjcvL8f77MPwFV+yH/wAhHxN/1wtv/QpK+JXgn/hYfhG68K/2n9g+0vG3n+R5u3Y4b7u5c5xjrVn+yFp6SA6h47uJkzysOniI4+pdv5V4E+H3hr4daQdI8OW7gSNvnnmbdLOwGAWIA7dAAAOeOTX7UXxIsJrOL4d6PdpNL5om1Jo2yI9v3Yjjvn5iO2F9aqqqqqvi78fPFOheLb3Q/AxtjaaNGEvp5IBIPOJAIz2AJC/UGvB+sSeIfCWi67MVMuoWEM8m0YAdkBYAfUmqqqqqqqqq+J3xX+INl8TIPhx8PorB7iSGPPnxbmMrKXIyW2hdm09PWtT1v9qTwnaPrWoWWlara243zRxRRuVUdSVTaxH0zivhD8XNP+KWlzsbQWWqWG0XVsH3KQejoeu0kHg8g+vBNVVVVV8YPGN74E8Aah4h0toxfRtHHbeYu5dzOoOR3+Xcfwr4A/F7WPiI+qaR4oktzqNoEngMUXl74Tw3Hs23n/bFUPib4p1D4+/8K70ya2/sW2H+kHyAX+WDe3zdvnIWqqqqqqqqqqqupZILWWaCAzyRxsyRA4LkDgA9s9K+B/xM8eeP9V1q38U6NFb2tmAYpI7dovKk3EGI7ic8DPqMc9RVVXiHXtL8L6Ld6/rNyIbOyiLysevsAO5JwAO5Ir4X/tA3PxH8Xp4ZfwxFYxvDJKswui7DaOBjaOv1okAZJ6eteIv2hbi61+Xwt8LvCk3iW8hLB7gbjFkcEqFGSoPG4lR9QQa0D9om9sfEEXhn4o+EZfDtxNgJcfMEXJwCysMhM5+YEj9TQKsoKkEEcEVXxN+NB8Ca3b+FdJ8JX+taxdQCaKOPKoVJIBXaGZjlTkADp1q+/aC+KXhzZqHiv4UPaacxA3PHNBjPbewIB9iK8AeP9B+I2grruhO6hX8ueCUASQSAZKkD2III4I/ED4r/ABV134eahYWWkeDZdaS7haR5Edx5ZBxj5Vav+GmPGn/RJrn/AL+y/wDxurj9qrxHaTx2t18NvJmlx5ccl06s2TgYBjyeeK8WeJLLwh4dvvEuoW9xPb2Ee+SO3QM5GQOASB35yeBk0fj58UdehN/4O+EN3NY4yk0kU04cexRVBPTgZr4dftE6f4r1yPwn4o0R9E1WaTyocuWjeT+4QQCjHoAc5PcEgVVVXxF+PGkeD9XHhTw/pE/iDXyQhtbckLG56KSASzdPlUfUirn4+/EbwrJDd+PvhXNZ6bO4Ami3oUB7ZbKlsfwkrn2rwx4m0Xxholt4g0C7FxaXK5U4wykdVYdmB4Iqvi98X9O+FtjaqtomoanetmK0MuzEY+87HBwM4A45OfQ4+E3xBm+JfhZvEU+lpYMt28HlJKZB8oU5zgf3qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq+vLjUr241C7k8ye6laWVj/ABMxyT+Zqv2Q/wDkI+Jv+uFt/wChSV8VvH8vw28O2viJLFLuNtQignjJIPlsGLFf9rC8Z4rRtY03xBpVrrWkXSXFneRCSGRe4P8AI9iOx4r4++O/i94W16bQn1kWej3ys9lPYw+U0sfdS/LBlzg4I7HuKdmkYu7lmY5JJySfWqqqqq+KnjmH4e+Cr7xBuQ3ePJso2/jmb7vHcDBYj0U1b/D64sP2dNf8VanGZNW1uWC+LyDLiFZl2kk9yGd89wwzX7P+of2l8JNBdny8CSwMPTZKwH/ju2vif4+i+G3hV/Esmm/b2E6Qpb+f5W4sf72GxgAnp2rVP2gLG30vSotG8L3mseItUso7r+ybJjL9nV1DDe4XOcEHAXODzgEE+Gf2kryTxVB4X8e+DpNCe5lWNZGZ0aFmOF8xHAIUn+Lt1xVeHf2kfD2qaZrmta3pR0yz0mWOK32XXnyXzvvwqJtXBwmTyQM8nHNax+0p4w0iSLUbz4UXtno8rYjlvDJEZR/suU25I5xz/WvB/ivSvG/hyz8S6M7G3u0J2OMNGwOGVgO4II/Xoa8U/GP/AIR74maV8N7Tw59ul1EwCS6N55fkGRiD8mxs7VAbqM5xx1qm+MfmfF0fCmz8OecVbEl/9s2hcQ+a37vYen3fvDmqqvhww8XftM+IfEAbfHpv2po3x1C4t1x9VOa1LUbHR9PuNU1K6S3tbSNpJpXOAigZJr9lrTLm+8ZeI/FdrbvDpvkNAikYXfJKrhR2yqp+G4etfEL4m+F/htpy3mu3DPcTg/ZrOEAyzEdSB2UdyePqeKu/2hPiFaWo8QTfB69h0QAMZpWlX5D0bf5eADxzjHvWk/FTS/Efw4v/AIg+HrF7n+zreWSaxlkEbq8a7mQsA2PlwQcHII45xXwn+JcXxS8PXGuJpH9mvbXbW7QG487oisG3bV67sYx2rxh4n0/wZ4a1DxNqRzBYQl9gODI3RUHuzED8a+FPxE1H4maJPr9x4X/si0SbyrZjd+cZyPvkDYuADgZ5ycjjFVX7WuqtF4X0PQYyS99ftNtH8QiTGPzlFeJraT4JfGjQNcU+Xp11Z26z7fu7BGIZhx1I2h/qRSsrqGRgwIyCO9fADPij4ueNPGpbzIx5ixnsBNMSmP8AgMRH0rx38QvDPw60n+1fEV2VMhK29tEN0s7DqFX8sk4AyMnkVcftCfEF7M+IbH4P339hqu/7RKZfmQD7+8JgL74IHrXw2+I+i/EzQTrOkxyW8kMnlXNrKQWhfGeo6qR0Pf2wRTfGPzPi6PhTZ+HPOKtiS/8Atm0LiHzW/d7D0+794c1Xhj4x/wDCUfE3U/h3aeHNkelmcSX/ANs3Z8pghPl7OMsQPvV8W/iwvwtt9LdNB/tWbVJXRIhdeSV2hcn7jZ5YDHvXjX9oeLSfED+E/Bnhx9e1C3fy7iUSERRyA4KrtUlsHgngA+teKfFmheC9Fk13xJfpbW0WBnBLSOeioo5JPp+JwATQ/aF8a+IBJe+BvhLqWoabESPtDh3L468IpAPsC1fC/wCOWifES+fQLzTpdH1yJWP2SV9wk2/e2NgHIxkqQCPfBx4l8R6P4S0S68Qa7diCztEy7YySScBQO5JIAHvSftCeOdfEmoeCvhJqF/pcTEeewkcvjrjYuAfYFscV4E8S3vjDwvZ+INQ0OXSJrneDaSvuZNrFcnIB5xnBAr4kfGfwh8NgLS/le+1R13JYWxBcDsXJ4QH35PYGm/aE+JJT+1E+DOof2Xjd5hSfG318zy9v44rxB42Phv4ey+OtQ0lkkhso7h7FpdrK77QIy2ODuYAnFfDDx3L8R/CqeJ5NF/sxZJ3jjh+0eduC4G7dtXvkYx261VV4+1a/+OPxFtvhl4bnYeH9Hm8zVLqM8OynDtnuB91R3Yk8jBHwwtLew/aV1fT7OJY7e0N3DDGBwiIAqqPYAAV8fvE8/hb4Y6lNZymO51ArZQuDgr5n38e+wP8AQ4r9mrwnaaF8ObfWvIUXutu00shHzbFYrGufTA3fVjX7Tfhe01r4cy655S/bNEmSWOTHzFHYI659PmDf8BFfAzxZ/bfwn07UtVuhu0uOS3uJnbosXQk/9c9uTXgL4yeDPiPqd5o/h+S8S5s0MgW5hCedGCAXTBPGSODg8jivs1v9o+1iCPz9mzzdg3bc5xnrjPOK8eXeg2Xg3WJvE0kS6cbORZhJjDAqQFHqxOAB1ziv2RdN1KKx8RarIrLY3EkEURI4eRA5bH0Dr+ftVTSxQRPPNIqRxqWdmOAoA5JPpivAEMnxn+OF/wCO72NpNG0FgbNXGB8pIgXHY5DSH3HvU8EF1A9tcwxzRSqVeORQysDwQQeoI7UFVVCqAABgADpXxsgs/EPx10XTfCDJLqv+jx3TwDOydZCctjuqBSfQAZ6HFVXijVW0Lwzq2uRqGbTrCe4UEdSkZb+lfsp6Pb6peeIfGmpAXOpCVYo5pBuZS+5pGz6tkc/X1rX9E0/xJot7oOqQrLa30LRSKwzjI4I9wcEHsQDX7KurXlhrfiPwPcy7kiX7QibuEdH8uQge+5M/7orxV4m0rwf4fvPEeszbLWyj3MB1c9FVfUk4A+tRaLrnjvwz4z+NviyMo0tlJDpMJ+6in5GKg/wqpKA9yWPXmv2WP+SYyf8AYUm/9Ajqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr9kP/kI+Jv+uFt/6FJX7U//ACTGL/sKQ/8AoD1+zt8Wh4U1RfBuv3WNI1GT/R5HPFrMf5K3Q+hwfWviL4D0r4i+GLjw9qICSEb7W425NvKB8rD27EdwT9a8Q6BqnhfWrvw/rVsYLyykKSoensQe4IwQe4INVVVVV8XvGOg+Ovi1YeE9b1lLPwxoExS8mO4iSQcy42gnPAjHHByehrx38WvhLrfgLW/DuneLLMyXGmyw2sKwSKN4Q7FHyYAyFHtX7KOoi5+H19p7Pl7PVH2j0Ro0I/XdX7XGrGLQdA0BGBN1dyXDKOv7tAo/9Gn8q+EHw6svh74TtbZrdTq13EkmoTsMsXx/q8/3U+6B7E96/amtYdR8a+FdMsl26hcQGMuq/NhpQI/fht/51471caD4K13WA4V7TTpnjz/fCHaPxbAr9l74c2WryXPjvWbdZ47GfydPikGVEwALSY7kAqAfUnuBj48NZp8JfETXqKyeQgQEZw5kQIR/wIiv2Y7Oa2+FVvNKxK3d7PJECOighOPxRq8MY8X/ALVWp6kCZItIec4PQeTGIBj23EH61XwM/wCKr+OHivxgcyQxC4eFjzt8yUBB/wB8BhVVrWoxaPo99q8xGyxtpJ2z0wilj/KvgxpXxgvF1XxJ8OZ7FDPKIbua62Esw+fADA/3gTj2rxVqXjq/8Z2HhD48a9qdjpc7oSlksSxMpOA/y/KVB6khivPHatD0Dw94H0BdL0azjsdOskZyBk9BlmY8ljxyTzXgfxX4Q8bfFLU/iF8TNatLa3syP7Ls7tiQOT5YC8jCKMn1Zs+tfEL9on4dp4d1DSvD93JrV7e20kEccdu6xqXUjLlwMjnoM56cdv2ffh9qmifDnUbbxJbyWz+IHc/ZpVw0cRj2Asp6E8nB7Yr9ky6ltJ/FXh254lgkhkC+hBdH/ktfHnWdQ+IXjnR/g54Zn3bZlkvmXkLIRn5sdo48sR7+orw9oWneGdDsvD+lReXa2EKxRDucdSfcnJJ7kmqr4z58U/Hjwd4TQeZFbeQ0y56bpS0n/kNFNftLeDv+Ek+Hz6xbQ77zQJPtC4HJhOBKPpjax/3K+G/xGOpfAW+1ie4zf+HdPntpCepaOPMR/FSgz65r9k/SBa+CNS1hlw9/qJRTjqkaLg/99M9ab4s8I/EX4x3vir4g61aw6FpW4abbXLHZKFbEY29weXYeuByK8W/tIfDTSdMuE0a/bWrwxssVvDbsEJI43M4A2+uMn2r9l7wVrXhrw5qet6zbS2p1qSL7PBKu1vLjDYcjqNxc9eyg9CK+BhHiv44eK/F5JeGIXDwsedvmSgJ/44GFXt1BYWc99cNtit4mkkPoqjJ/QV+yraTarr3irxhdKTLLsjDnnLSO0j8/8BX86/aiurvWPHnhjwtphL3aQBoVU4/ezS7VA9D+7X9K8AeBdG+H/h2DRNKhTzdoa7uMfPcy4+Z2PXrnA7DivGrXHxn+PMHgaSd/7E0ORkmRGI4jAMx9mZh5ee3B9as7K0060hsLC2it7e3QJFFGoVUUcAADoK+NNtF4X+PXhrXtFTZc3htZ5kTjzHExQ9P7yqFPrz61+09omsax8OFm0qN5Y9OvkubuNBk+UEdS3HJClgSPTJ7V8B/iX4S8Q+E9J8J2tzHZ6vplmkElnJ8pl2LgyIejbsFiByCTkY5PjvxPD4M8Iar4nlVX+wW5aNGOA8hwqKfqxUfjX7PHgRPFM978WfGKjUb66unFn9oXcAwPzzYPGc/Kv93afbFftTaubD4appqSANqeoRRMvqihpCf++kT86+DukDQ/hf4bsNu1msVnYEYIaXMh/V6qvj58SrjwzpcPgzwzKX8Q66BHGI2w0ETHbu9mY5VfxPGBXwi8D6F8NPC0enm/s5NTu8S6hOsqndJjhQf7q9B+J718Prm2T9pzX53uIxGZr3DlgAefWv2sJTN8P9Jkt5FeA6uuWUggt5UuP/Zq+EgQfDHwwI9uP7Mh6eu0Z/WvjWVHwq8S7iP+PI9fXcK/Zcs0vvhZqVlfxebbXOqTxmNjw0bQxBh9D81fD74J+EPhvq93reiy31xc3MZiQ3UisIYyQSq4UdSBycnA+ufHGj65r/he90jw3rbaRqM/l+TeKSDFtkVm+7zyoK/jXj34MfFu1hXXtb1weNbOwPmSWct3OW2gfNhMg4x/cbNfBH4ieGPG/hv+ztC0iHRptJVUl06MjagPR0PGVJzkkZz1znJr9pbx4fDXg0eGbCfGoa/mIhTyluP9Yf8AgWQvuC3pXwY8C/8ACA+A7LS7mLbqF1/pN9kciVwPl/4CoVfqCe9V47+FXxc8Q6tqd7oPxPltLG6kLW9gbmaJUXH3SUzgfhXwz1q0+CfjIeFfiD4OtrO/vCEXXFkLtsY4ByxI8sngldpGPmBI48T+K/D/AIN0k654k1D7JZK6oZfKeT5m6DCAn9K/4aI+Dn/Q4/8AlPuf/jdf8NEfBz/ocf8Ayn3P/wAbrQvFvgv4oaNqNv4f1Q6hZlGtroiCSIjepBHzqOxNeEdd139mzxnqGieKtKubrRtRICzwrjzQhOyWPJwThiGXIIz1451D9o7SNZQaN8NdB1PW9dul220T2+yOM/3n5zgcE9B6kda/ZXtLm58feIdYuZhK0Vg0cso+6zyTK2fx8smvFuqv8fPiTD4O03URB4S0CTzLy5EgUTsDhmU9CTyideNzdDiviv8A2LYfCDXdJ0qS0igt9O8qCCJxhVXAAA+gr9ly6tYvhq8MtzEkh1SbCM4BPyx9qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/ZD/5CPib/rhbf+hSV+1P/wAkxi/7CkP/AKA9V+zt8WR4t0lfB2vXWdY02L9xI7c3UI757svAPqMHnmvj/wDCVfHWiHxBodsDr2mRnaqrzdRDkx+7Dkr75HfhlKnaRgjggjpVVVV8YfHSfD/wNe6vDKov5x9nsFPXzmBw2P8AZALfhjvXwP8Agt4cvvBcfiLxvocWo32sv9oiFyCTHD/CevVuWz6EVL8E/hXJG0a+CtOUspAYIcj3HNfsl3Ulpf8AirQLg4kTyJAvoVMiv/Na+M4/4Sv4++FfCqhXitvsyTKefvSF34/3AvFV4p/4q79qnStMALx6O0ABHQeVGbg/huJH1r9pXVhpnwpvrZXAbUrmG2U5wfvbzj8IyK+AukDR/hPoMTKBJcxPcuQOvmOzKf8AvkqPwr9qrV/sXw8tdLRvn1LUEVh6oisx/wDHglfCvSP7D+HHhzTSpV006J5FI5DuN7D/AL6Y1+yzNHqXjTxTqt42L+aANtY/Nh5S0n/jwTP1r4vfECx+H/g68vWuFGo3cTw6fCD8zyEY3Af3VzuJ+g6kV+yRpAg8Na5rhTBu71LcE9xEm7j8ZaXxl4Uk8QHwpH4hsG1cA5sxMDIMDJGPXHOOuOar486uNG+FGvSq+JLmFbZBnGfMcKw/75LH8K/Zr0gaZ8KbG4KgNqVzNcsMYP3vLBP4Rj9K/a5sI5PD/h/U9i74bySEN3w6A4+nyCrl7vX/AIPSSWTGS61Hw0TEQclne24x+JFfs3eCvh7410rVoPEujW9/qdlcq6CWRlIhZQBgBhkblbPHcetaL8PvA3huRbnRPCml2cycrMlsvmL9HI3frVaP4ktfhN8ZviBeXSjyltLqSCFjgPI8kckS/Q7sfQ+1fs3+C7w2t98UfEW6XVNfkcwPIPmERbLv7F2/RR614g8T+HfCtol94j1m10+GR9iNPIF3t6AdT+FWl3a39rDe2NzHcW86B4pY2DK6kZBBHUEVXg8DxZ+1Nrmr8smjLOFyeAY0W34/Fif1q7tbe+tZrK7iWWC4jaOWNujKwwQfqDV7dXfwxHxC+Gt1M+28jijty3G/bOhVsdt0Lsfyr4TaDcad8FtJ0y2+W5u9NkmQnj5ptzr/AOhj8q/Zv8HeBfGFxrmk+L9HivNQtfKkt4pndGCfMHwAR0OzOemRWj/DfwD4fmW50fwhpVvOhyswtVZ1PsxyR+BrxLeTWHhvVNQtcmW2sZpY9p53KhIx+Ir9kOK0Gm+JZ1dTctPbq655CBX2n6Elvyr9oLx/Y+EvA95osVwrarrcDW8EKn5ljYYkkI6gbcgH1I9Dj9lzSBYfDM6gyfNqd/LKGPdVxGB9Mo35mkB8X/tXkkB7fSJDx12+TBj/ANG/zqvhTcJ4d/aP8S6drL+XcX73sMDSHG92mWVTz/eRSR65HrVzc29nbyXd3PHDDCheSSRgqooGSST0AFaO7/G39oJfEFnG0mhaA8bpKy4HlwkmP/vuXLAHnaTxwaIBGCM+xr9pDwboHgXVdF8W+EFXSb+7ncvFbHYodNpEqKPukE4OOORxnOfjs2q6r8CjfTRbZ2js57yMD7uSu4fgxH5V+zrqVhf/AAm0e3spEMli00NxGDyj+azc/UMG/Gq/anuJtZ8T+E/BlrIPNlDPtAz800ixoSP+AN+Zq7vdI8NaQbrULyCxsLGIK0szhFjUYA5P4D3NaJr2i+JdPTVdA1O3v7RyVWWF9wyOoPofY1Xjj4AeDfH/AIin8TazqetQ3VwiIyW00aoAqhRgNGx6D1r/AIZN+HP/AEGvEf8A4Ew//Gq8K/Cvw9rvxl1T4d3d5qKadYyXKxyxyIJiIz8uSVK/X5RXxQ+FtppfwKm8JeHmubpNAY3tubhg0jASM8mSoAyFkkxgdh9a/Zt8VWmvfDe10hZ1N7ojtBPHnkIWLRtj0KnH1U1+054ptdG+HcmgCdftmtzJHHHn5vLRg7tj0+VV/wCBV8CvDU3hb4YaRaXkXl3N0rXcykYIMhyufQ7NgPvWh+L/AAt4mmubbw/r9jqEtmcTpbzBynvx29+lJNDK8kUUyO8J2yKrZKEgEAjtwQfxFX17Z6bZzahf3MdvbW8ZeWWRtqooGSSfSv2a1fUvit4i1vSYWi0s205xtwFEkymND6HCk/8AATUjxxRtLK6oiKSzMcAAdST6V4XDfHT463HiS5RpNB0DDwKw+UojEQqR23vlyPZhVSzQwhPOmRN7BE3tjcx6Ae/tVftaappd9qnh7QbJln1O2WZ5kj5ZFk2BFOO5Kk469PWrLw/aah4V03RfFOm2uomK1hWeK7hWZTIqAElWBGc55r/hWvw5/wChA8N/+CqH/wCJr/hWvw5/6EDw3/4Kof8A4mtI8PaB4fikg0DQ9P02OVt0iWdskIcjuQoGTV5ZWeoW7Wt/aQ3MLfejmjDqfqDxXxk+JHhT4ceHr3wt4RhsIdd1GMxeTZRKn2VWGC77Rw2CdoPOSD0r4FfDObwn8PriDW4pINQ8QKXukA2vDGVKonPRgCTyOCxHav8Ahk34c/8AQa8R/wDgTD/8ar4hfs3eBvCfgrV/Eem6rrslzp9sZIknniKEgjqBGDjn1FfAL4N+F/GGhW3jbUr/AFSO+sNU/dRwSxrEfK2MuQUJ6nnkfhVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVf8MxfFT/n003/AMDB/hX/AAzF8VP+fTTf/Awf4V+z78K/Fvw5vNam8Sw2qLfRQrD5MwkyVL5zjp94V8c/BOu+P/BSaF4djhe6W+jmIlk2DaqsDz+Ir/hmL4qf8+mm/wDgYP8ACtG/Z6+M3h/VbXWtIWwt7yzkEkMi3gypH4cg9COhBxWjzancaVaza1Zx2t+0Q+0wxyb1R++09xnp7V8Yv2ddU8QeJG8ReA4rVRflnvbaWQRhJe7r/vckj1ye/H/DMXxU/wCfTTf/AAMH+Ff8MxfFT/n003/wMH+Ff8MxfFT/AJ9NN/8AAwf4V/wzF8VP+fTTf/Awf4VXxN+D1/8AE3xLpd/qPiRING03aDp6wEtJlsyHdu4LABc44AFRQxQRJBBGqRxqFRFGAoAwAB6Yqvh98HLnwJ491rxdHr0Vxa6qsyraC2KGIPKHX5txzgDHQdatfg7cr8YZPipfa9FOpZjFZi3IKfufKX5t2OBz061XhT4O3OgfFDVviPqGvx3p1AzmG2WAoYTI4I+bcchUBXoK+MfwvvvilpmnaXa67Hp0VnO00m+AyeY23auMMMYBb860XTIdF0aw0aAgx2FrHbpgY4RQo47cCvjF8Ir34qzaQI/EKafb6Z5u6MwGQyF9uTnIxgJjv1qKKOGJIYlCpGoVVHYDpXiX9nDW4vFk3in4c+Lxoz3ErSGNi8bQFjlgjpklSf4SBxxzVr+z+8ml6nd+IvFtzrfiXUbJ7WPUbxWkS0Vxhtilic7SRnPfgDJr4XeA/wDhXHhGHwy18l5Ik0ksk6RGMOWPpk9AAPwrS/2e7qy+LD+Pp/EaSWIv3vo4AjecXZi2xj0wCevOQMYGar4ufDy9+JnhqDw7Z61HpypdrPI7wGTeFVgFwCMcsD+ArwloEfhbwvpXhyKUSjTrSOAyBdvmMqgM2O2Tk496+Lvw3f4oeGbfQYtUSwkt71blZmh8wcI6lcZHXf69q8H6JceGvCuleHrq8W6k020S385UKhwg2g4JOOAK8Ufs53KeI5PFXw08VP4eupWLNB8yqjH72x0OVU/3SCOfTim+B/xP8Rotr43+L95LZ4xJbWm8iQehyVHpyVNRoY41jMjOVUDc3U47mviJ+zzH4/8AHR8WP4hS0tpxCLm2FsWdwgCnDbhglQB04q0tLawtIbGyhSG3t41jijQYCKowAB6AACvjb8IL74pxaS+m6zFZT6a0gKTqxR1fbkjHQjYO3Oe2K8FeGYvBvhTTPDEV21yNPgEZmYYLtnJOOcDJOB2GKr4WfCO4+H2t65r+o65Hqd1rLAhlgMfl/MzPnJOckr6dKr4u/AWP4ma9beILHXItMuEt/JuA1sZPN2nKtww5wSPoB6Vp1jDpmnWum2/+qtIEhTjHyqoA/QV44/Z5Op+JH8Y+APEsnh/U5ZDLIihgnmH7zqyncme4wRyenSj8FPi1r0Ys/F3xiuTZniSG18xt49DygP4g1Y6bDZaTb6Q0jXEVvbrAWmwTIqqFy2OMkDnirn9mvxToPiGbVPhx48Ok20zEBWaSOSJCc7NyZ3ge+Og+tXH7O0E/hnVLS68UT6j4j1gRrPrF8hlaNFdWKou7IB2gElice3FeAvCq+CfB+meFhcrcGwiKtKqbA7FixOMnGSxr4e/B258G+O9a8c6hrsV/Pq4mxGluU8syyiRjksc9AO1V8VfgdpXxEvItf07Un0jXYFAFyibllC/d3AEEMOzA5HvgYk+BPxT8RQppPjT4tTXGlqRuiiaSVpAP7wbaCfdt2K8GeB/DngHRk0Tw3Z+TCDukkc7pJn7s7dz+QHYAV42+FHxB1PxXeeLPBfxMuNLe9VA9m4dI12qFHKkgjAzyvUnnmtA/Z8vL3xBB4n+KXi+bxLc2+DHasG8rI5AYsclM87QFGeuckVqWnWWr6fcaVqNuk9rdxNFNE44dWGCPypv2dPGHhXVJr34Y/EWXToJz80M5dCAM4DFMiTGeMqOteAvhh450TxLF4o8Z/Ei61qSGF40s8OYhuGCQWbA/BR9a8R/B258R/FbTfiLda9ELbTWhMdkbYlv3WWHzbsffOelfFfwHN8R/B0/hu11EWU5lSaKR1JQsv8LAc4OfwODzivg/8NpPhh4Wk0S51Jb25ubpriZ0UqisVVdqg84wo5PXNVVeG/g3c6D8VdR+JD6/FNHfPOwtBblSnmH+9u5x9KZVZdpAIIwQRWu/s76hp3iCXxJ8K/GEvh6ackvbHcEXPJCspzsz/CQQPyA8Ofs8z3HiCPxR8TvFc3iW8hKmOBg3l5HIDFjllB/hAA9c5xWp2Meqabd6ZJK8aXcDws6HDKGUgke/NfB34E33wz8SX2v3/iCK8Elu1tBFBGyZUsrFnz3+UfKMjnOa8ZfALWtT8UX3jHwf8Q7/AEm/v5d8qNuXHsJI2BCgDABB471P+z58SvE5S18cfFaa6slYEwq8s4OPRXKqD74OK8D+A/Dvw90VdF8O2pRC2+aaQ7pJ3xjcx/oMAelePfD+q+K/Cl/4c0fVk02bUI/Ke5aIvtjP3wACPvDK/QmvhP8ADS1+GHht9HS7S8u7idpbm5WLZvPRQBk4AUDjPUk96r4nfDq2+Jnh6PQrnWLrT/JuBPHJCoYbwCBuU9R8x4BHOOaX4HfGexjGnaX8ZLgWI+Vd1zcRso9lBOPoDXw9/Z30PwnqyeJvEWqy6/q8b+ZG8qbY45Ou/BJLMD0JPXnGcEVVV45+E/xS8WeJ7+8sPibNpmi3LKYbOO4m+QbVBBRSF6gnr3r4f/s9eDvBN4ms300uuapGd0c90gWONv7yx88+7FsdsVVeOPDb+L/CWqeGYrtbZtQgMQmZNwTkc4yM9K+E3w+m+GnhZvDk+qJfs128/mpEYx8wUYxk/wB39aqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr/2gAIAQIRAT8AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAApU3VqRgt20vvKXh07fvMRr5R/W/6GZcBYvD03Uw81Ut0tZ/LV3/AaadmAAAAYThPC1cgeJd/auLknfa2yttqlr1uwAAAAAAAAAAAAAAAAAAAAAAKk4U4Oc3ZLVt6JJdTO/GfA4as6WAoOql9pvlT9FZtrzdjhPxVwmdYyGCrUHTqS+Gz5otpX7Jrbs13aDO/F7D5bmNbBfVXP2cnG/MldrfTlfUyPNYZrl1HGwVlUSdt7d1fyZxrxpS4Zo0pyouo6jaSTstLbuz79iPjjU5/ewS5f8ev/pH6HC/FeA4hwjr4V2cdJRe8X5+T6Pr6ppGK8a8NRrzpRwjai2r861s7X+HqYTEQxOHp14bTSa9GrgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABgv96p/4l+aM6zenleF9vOLkrpWXn/wxknEeDzVNUrqa3T39V3Rx1lccNjY4imrRqXv/AIlv9+j9bgAAAGF4to0sjeEaftUnFdrPr8l07/gAAAAAAAAAAAAAAAAAAAAAAHjJxJPC4SnldGVpVfen/hWiXpJ3v/htszhzgbOc/g6uEglBac0nZX7Ldv5Ky6nBHAOcZNxNQq4yn7kVNqSd435Wrd09dLpX6bDskZhOpj8XicWtnJyf/b0v82eE2L9vwxShf+HKUfx5vykeOP8Au+C9Z/lEyTwvxmb5HDMsPXXNJStBp/Zk425r9baaW79zwmzCrhOJYUL2jVUoteicl87q3zZmmKWEwNbEfyRlL7k3+gqFSVGVbomk/VptfkzgLF/WuGsFUvtBR/8AAPd/QAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAMF/vVP8AxL80ce/8ipf4l+TOH8ZLCZnRqJ9Un6PR/mcfUFPLIz6xkvxTX+RwlwzTzFvEYn+HF2S25n/kv62Mx4PyzE0HCnTUJdGv1XVfj5mDwKjmtPCYiP21GS+dnsYvgnATxdFUoctNKTlq3e1rLVu19duiM74RwNfBy+q01GpFaW626PvfvvcyLLqeYZhTw1SVk77b6Ju3zsQ4XyiFP2aoJrzvf79zifJFleM5KfwSV4/qvl+TRR4UwWLyWlUo07VZKPvXl1a5na9tr9PQqcH5VLCewjTs7aS+1fvf9NjhvhHD0sP7TG01Kbvo9kvTu9/u8zi/JKeW4uLoq0Jq6XZrdemz+ZgeG8rq5GsTKlepyN3vLdJ62vb8DgzJcBmNKs8TT5nFq2rW9+zRk3B+CoxnPEU1KTbsnso30+durOL8ip5bioyoK1Od7Ls1uvTZo4R4dhmVSVauv3cdLd329F19UZvwhgMThZRw9NQqJaNaa9n6/eiUXFtPdHB+W4TH4+VLEx5oqLaV2tbx7NdGzB8G4GGPq1Zw/dqyjF3a+FXbu23re1/PyONOHMPhKccXhY8qvaSW3k127P5HDuSvNcYqTdoLWT8uy83/AMHoV+Espq0HRVJR7Nbrzv1+ZQyz2WcQwWIV1zqL6XV189UcZZFl+X4anPDU+VuVnq3082zhTh3LMdlqrYilzSu1e8lt6NHCeU4LH42tSxEOZRV1q11t0aMr4SwEcVXq1Kd4qVoxd2kkld6766anDWQvNsU4ydqcdZP8kvNlXhPJ50fZKil5q9/v/wA7mcZXUy3GSw83e2z7p7P/AD8zG8G4Cth6SwsOWUmru8naNrvRtr/gmL4PyurhHRp01GVtJdb93387lHA1quLWFS99y5fnexg+Esqw9BU5UlN9W93/AJfI4lwOHwWZ1KGHVoq2m9rpP9QAAAAAAAAAAAAAAPEfEVMw4sr04a2cYR+SS/8ASmzKMso5ZgKWDor3YRS9e79W9X5sOIsX9UyfFV76xpza9VF2/E4ayz6zw/m1a2sI07f+B8z/AAieCGL5sFi8Nf4ZRl/4Emv/AG08cf8Ad8F6z/KJ4dYzD4PgvD4ivNRhFVG2+iVSZ4bUZY/jCGIgvdjzzfkmml+MkeImL+rcMYyd948v/gTUfyZhcs9pwViMXb4a8PuUWvzmjwdxftuHPZX/AIc5L77S/Vh4hcaZ3k/EEMNha/LScYtrlg9209XFvp3OJ/E3OMZmUpYCu6dGDtFLql1l3vvZ6JaW3b4E4lln+TwxNS3tItxnb+ZW1+aafk7roeJ3H+Kyyssty6fLUtecuqvtFX2bWrfRNWOAvEfMqWZ08JmNZ1KVR8t5auLezvva+jT0W6tbXxT4jzPJMHh6mX1eSUpNPSLurf3kzw+zfGZrkFLFYyfPUbld2S2k0tEktvI4E464gzPiOlg8XiOam+e65YLaMmtVFPdLqcV8Z53l/F8MBRr8tByp3XLDaVubVxvrr106WOFuN8wz3jLk9o1h7T5YLayWjfdvfXbZaHiV4jYrAYp5ZlkuWUfjnu03ryxvotN3vrZWaZwp4oZrgcZCOYVXVoSdpc2so3+0nvp2d1baz1OPM2xeXcO18bganLOPJZ2T0c4rZpp3T7EvFXOFkXsnWviZTkua0U4wUY2skkrttpOztZ+VvDLj7Ma+ZRy3MKrqRqX5XLdSSva+7TSa12drdTxB4tlw9liqUUnWqPljfZd5NdbL8Wr6XMu8QeJ4Y2M44pzba92WsXd7W6fK3kAAAAAAAAAAAAAAAAAAAAAAAAAAAAAGC/3qn/iX5o49/wCRUv8AEvyZh7qtC3dfmcbW/sap6x/NGXuOV5BCpb4Yc3zav+bscF5/i8ZiqlDEz5rrmV+lnql5a7dLHEGFVLijDTX25U38+a36I4mzeeWYB1afxtpK/d9fkk/mcJ53VzTCSdb44uz810f6fIx1WeV55UqUlrCTaXTX/gMyPF8Q4rG08RecqbavfSNr62W2i7HiHSTwtGp1UmvvX/AMmrxw+RUq0towv9yucJ8QV81jVVdJSi01bs76fKxmfFNXDZ3TwUUuS8VLveXX5Jr8TxCpJ4GlPtK33p/5GW/8k0v+vcvyZ4dfwsR6x/JmP4rq0M8hg4pezTUX3u7a/K/5niDSUsupz6qa/FP/ACRlk/7L4ZVaPxcrl85bfmjgbOMViq1WhiKjnpzK7vbWz+Wq02RxLh1QzavBfzX+/X9TgD/kaS/wP84nFfE1fK69KjQSbau79r2S+dmcScmKyKrNbOKkvvTOAsPGll1TEP7T/CK/zbOG+I8bic65as24VL6X0Wjasum1vPqcQYdU+J8LNfacH81K35JHiH/udH/F+hwN/wAiiP8AikcCf8jOv/hf/pSOIuJ6eUShTjDnnLW17WXfZ76nAuFjSyr2nWbb+7T9GZNxTisRnnLOd6U20l0X8tvPa/e54iYWNqFdb6xf5r9TFY1YLK3iGr8sU/nbT8ThLPq2a0ant0uaDW2mjvb8mYTDwjxjKPnJ/Nxb/U43x2ZYOpSnh5uMLbr+bz+WyfmY7GVcbiJV6vxS3+6wAAAAAAAAAAAAAAFa1bj1qXXFW/8AKtjxCzPF5bw9XxWEnyVIuNnppeUU97rZmS+IPEtfMsPSqYtuMpxTVo6pySfTseKGL+r8L4mz1lyxXzkr/hc8NMq+scI5ira1eeK+UNPxbPBXF8mcV6Dek6d/nGS/Rs8cf93wXrP8ojqTcFC7t26Hgtjssj7bCqNsS9bt/FFdI9rX1Wt73vZWXjNi/ZZBCinrOol8km/zSMpyrn8Ma6trJSn/AOAT/wAoHgfi7xxmGb/kkv8AyZP9A8ZP+Sij/wBe4/nIzXw3w2D4Qhmak/bpRnJdLSt7tunKmne+6fdW8DsQ3DG0W9FyNfPmT/JHDmDpcUcazniVzU3Kc2u8V8K9PhT8tDxbyDCZVmVCvg6apxqRekVZKUWtUlorprbqr9TxZxf1zIMuxP8APaX3wT/U8KP+SWoes/8A0pnhh/yV1D/uJ/6RI8Wm1xRUa/lh+R4dcA1Ms9jnWJrWk4t8ltoyWl3fe2rVtNjhjALibiiMcTrGpKU5+msmvntptc8WuFsFlOKoYjBU1CFRNNLZSjbVdrp/hfqY7Hyxvhb7STu1GEf/AACrGK/BI8NuDMNxFia0sW37OmlonZuUr217JJ/gU8FLI+LaeHUr+yrRSfdKStf1W5431JvHYSn0UZP5tq/5I4V4S4YzPI8HXpUk5xUW5LSXPGzkpfO6aeltujAAAAAAAAAAAAAAAAAAAAAAAAAAAAADBf71T/xL80ce/wDIqX+JfkzKMO8Rj6NJdZL89fwOPaqhlSj/ADSS/Bv9DP8ATh6py/yx/Q4Hv/a8bdpfkcUW/t7Aesf/AEtHiF/uFL/F+jPDq98Sunu/+3FTKqeY8V1ac1eEbSfnaMdPm39xjc5wGAqU6FSVpSskkvkttkeIX/Iup/41/wCkyKP/ACSr/wCvT/Jnh3/Hr+i/NnEP/JSS/wAUPyieIP8AyLKf+Nf+kyMt/wCSaX/XuX5M8Ov4WI9Y/kzOf+Shn/jX5o49/wCRUv8AEvyZlUIvLqEd1yR/JEacIv3UkcZ2/tqtb+7/AOko4A/5Gkv8D/OJx/8A8jSP+BfnIxn/ACTH/cJf+ko4LS/sWl6y/wDSmKjTTuoq/ocTW/t7Aeq/9KR4h/7nR/xfocDf8iiP+KRwJ/yM6/8Ahf8A6Ujjpt5u12ijhvTh+ny/yy/NmR3/ALUw1v54/wDpSPEK39n0u/P+jOJf+Seqf4Y/nE8OvixP/bv/ALcZzjngeJ5Yha8rj93Kk/wITw2PwykrTpzXya/r7jiLKlluYToR+HePo/8ALb5AAAAAAAAAAAAAAAcSz/s3jGvWf2K3P/5Mp/qZ3k2EzzLpYOu37OdneLs9GmrOz7GOynD5TxnDBYdvkhVppX1eri/LueNeL5Mow+HT+Od/lGL/AFkjA8SZvgKPsMLiZwhvZSaV3voeGWL+r8UYV9Jc0fvi7fjY8cf93wXrP8onh/kWW5jwfh6eKoRmpc97pX/iTW+6a6O90ZBGWS8Z0qEJfBW9nfunJwf3pnjhi7zweGT2U5P58qX5Mp8SZvTwn1OGJmqVmuXmdrPdW7O+p4N4v2XEEqXSdOS+aal+SYeMn/JRR/69x/ORxT/yRVb/AK9R/JHgs3GWPcd1GP8A7cQnODvF2fkVKtSpbnk36niAn/qfk9/5Y/8AptHhR/yS1D1n/wClM8MP+Suof9xP/SJHi3/yU9X/AAw/IzFcmUVlDpTlb/wFng6oviN3/wCfcrffH9DxuUf7Nwr687/9JMJf/iFle/8AMv8A07A8D/8Ad8b6w/KRxl/yXNX/AK+Q/KJ4tcMV80y6ni8NHmqUb3S3cXa9u7TSdu1zhPifFZBmEMRSk+RtKceko9dO66Po/K5TqRqQU4O6augAAAAAAAAAAAAAAAAAAAAAAAAAAAADBf71T/xL80cYYDE43LlSw8eaXMnbys+5wlwrWwdb63i1aS+Fb2vu3by0seIOOjPEUsLF/Cm36vb8F+JhoLM8gjCL1nTt80rfmjgfJsVQxdSvXg4qK5VdW1bV7eiX4nEOIVXijDQX2HTXzcr/AJNHF+VVcwy7loq84tSS77pr7n+BwZlFbAYOUq8eWc3t1SW1/wAThqvDEZ5jai+Xonb9EZxlOLxPE8Wovlbi720SSV9fVP5+p4hf8i6n/jX/AKTIo/8AJKv/AK9P8meHf8ev6L82cRf8lJP/ABQ/KJ4g/wDIsp/41/6TIy3/AJJpf9e5fkzw6/hYj1j+TM5/5KGf+Nfmjj3/AJFS/wAS/JmAorNOHIUU9ZQ5fnHT80f6uZs6vsvq8r+mn/gW34lSnKnNwlutPuOAP+RpL/A/zicf/wDI0j/gX5yMZ/yTH/cJf+ko4FrRrZTKk/sya+T1/VmO4YzLD4mVKNGUlfRpNpro7rb5mDw9XAZvRp11aUZwuvmn+RxflNbMcAoUFecWml33T/O/yOHMtnl+W06FT4tW/V9PlscCf8jOv/hf/pSOOf8Akby/wo4IrxrZRGH8ja+93/UyThvFUc9UZwahTbd7aNL4bPrfT8TxExC5aFBb6v8AJL9TiX/knqn+GP5xPDr4sT/27/7ccQ4GtjeIatCirydrfKCf6HBdLMsLmLoThKMLPmTTSv0fr+aPEHl/tGnbfkX5sAAAAAAAAAAAAAADxjymWGzyOMS92tFa/wB6Putfdy/ecDeKWXUMuhgs1m4SpqylZyTitr8qbTS02s973MzzPDZnxrHF4aV6c6tOzs1ezitnr0PG/F82OwmGv8MZS/8AAnb/ANtOGPDzh+vkuFq4nDKVSUIybblvJX6PzMdQp5Hxi401aFKsml2jzJpfdoeOP+74L1n+UTgXjDJMq4ToQxWIipwU7xveXxyaXLvqmrdNexw5KpnPGFCtbWdb2jXa0ud/ckzxhxft+I/ZL/l3CMfvvL/25GE8NeGo4enGrhE5JK7vLV21fxdzg5vK+MqFN/ZqOH33h+oeMn/JRR/69x/ORxT/AMkVW/69R/JHgek6+NXlD85HCuaQ4X4pf1nSEZTpy8le1/k0n6bFXjXh6nGL+uQlzNJKMlJ3e2kbtfNK3U8bKLllGHqrpUt98X/kcBeIGXZNw7Ww+Il+9g5OEbP3rrRXWi9697201PCxt8V4ZvtP/wBIkeLf/JT1f8MPyHSjVw/s5bNW+9HBeYx4f4npvFPljGUoT8r3jd+Sdm/JHjFxDhMfiMPhMLUU1TTcnF3V5WsrrS6S19TGYGWD8LOSS1ahL/wKrGS/Bo8D/wDd8b6w/KRxl/yXNX/r5D8omY8R5Zl2LpYXF1VCVRNxb0WltG9k3fS+/qeLFLK4Z3GWAcbyjefLa3Nd66aJtWv8n1OHlNZPhFPf2cL+vKgAAAAAAAAAAAAAAAAAAAAAAAAAAAADCSjHEU5N6Jr8yGcZdNXjXg/+3l/mZtxjl+DptUZqpPolqvm9rempisTVxVaVaq7yk7s4M4jo4WLweJlaLd4t7Lun2XVfMzPiHAYGg6kqik+iTTb+7p57GAxVTFZ1Rr1H70qkW/8AwJGYZpQwLpuu7Rm7X6J2vr5aWOIOJcLgcJL2VRSqSVopO9vN26L8ThfNoZdmMalR+5JWflfr8ml8jNeI8DgsK6qqKUn8KTTv93Tu/wBTjfM8Di8BCNCrGUlNOyd9LS1/IpZtgFw37B1o8/s2rX1vZ6WOBsfhcJWrOvUUU0rXdurM8xVCtn0q1OacOaOq20UbnG+aYLF5fCFCrGTU07J305ZamAzfAQ4fVGVaKnyNWvrez0OBcxwmEp11XqKN3G13buZriKNTO51YSTjzp36W01OM81wOKy1U6FWMpcydk79GcGcRUcGnhMTK0G7p9E+qfk/wfqVs2wFGnzzrRS9V+Hf5GY1KdXGVZ037rlJr0bdjgrGYfC5jKdeaiuVq7dtbo41xmHxWYxnQmpLlSunfW7MVm+Alw97BVo8/s0rX1vZaWOFM9jleKaq/w56Pyts/l19RZrgHT9oq0eXvzL/M4oxmHxWazr4eV07a+aSWn3GVcQ4LGYONaVRRkl7ybSs+vy7GX8V4DEVKznVUYp2jd2urb/N3+Vjg7HYXC5jWnWqKMWnZt2v7yOMMVQxOZupRmpRstVqcJZ9DLMS4Vn+7nv5NbP8AR/8AAK+d5dRoe2lWjy+TTv6JaszzNZ5njZV3otkuyW3+b82Z/m+ArZHOjTrRc7R0T10aOBcwwuElX9vUUb8tru17cxDNMDT4pniZ1FyPaS1V+VL7t0Vs4y+lS9rKtHl9U/utv8jPs0/tLHzxC0WyXktv8/mAAAAAAAAAAAAAABxfwvh+IctlhKj5ZLWMv5Zf5PZr9UjNOB+IMurOnVwspecU5Rfo0n9zs/I4Q8Pc/wARmFDEVKDpU4SjJufuvRp2UfibdtNLd2eJuQZ3mfEM6mGws5wjGMU1FtPS7s/VtGEw8cPh6dGO0Ul9yseJfCWbYriGriMHhpzjNRd4xbV0lFrT0PFHLM0zTKsvqUcPOc0m5JRbcW4x3SV1rco8L53XmoU8HUb/AMEv8tPmeGvh9XyWUsfj0lWatGO/Knu21pzPbTZdddOIOGM5zPjCdd4WfspVYrm5XblTUb37WV79g4k4WzqhxVWxeFws5wVRTTUW09VJ2fr+IeKvDmb5hnsa2Ew06keSKvGLaveWmhxFgsTX4Sq4alByqOklypa3stLdzwgyPMstrYt4yhKmpKFuZNXs5Xtc8UeAsZVxss0y+m5qfxxSu00rcyS1aa3tqnd7PTKeF88xuJjHDYabaa1cWkvVuyX3nG2QSzzJK2Ep/Ho4/wCJapfPVfMyjgnOswzCOD9hKDv7zlFpRXVtv8F16HCPCmZZZxqp/VprDxlVSk07cvLNRd/PS3qeJvDWcY/iGpWwuGnODjHVRbWi1KSapxT7HifwFjPr0s0wFNzhPWairuMurstWnu+zvfRnCHh/mWc4yHtqUqdBP3pNNXXaN929tNFu+z8QcurYnhfEYXB03KVoKMYq7spx2S7JfceEGTZhltDFrGUZU3JwtzJq9lK9rnFXDOc4jjCpiaOFnKm5wfMou1ko3d/keMOQ5tjcXRxWHoudKMLPlV2ndt3S1ta2u3exwnwHmecY6EalGUKKfvSkmlZbpX3b2023ehGMYxUYrRAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAGSf8jPDf44f+lI8Qv8AcKX+L9GAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAZbXhQxtGtP4Yyi36Jps4u4iwOZ4WFPDttqV9Vbo0AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfV49z6vHufV49z6vHufV49z6vHufV49z6vHufV49z6vHufV49z6vHufV49z6vHufV49z6vHufV49z6vHufV49z6vHufV49z6vHufV49z6vHufV49z6vHufV49z6vHufV49z6vHufV49z6vHufV49z6vHufV49z6vHufV49z6vHufV49z6vHufV49z6vHufV49z6vHufV49z6vHufV49z6vHufV49z6vHufV49z6vHufV49z6vHufV49z6vHufV49z6vHufV49z6vHufV49z6vHufV49z6vHufV49z6vHufV49z6vHufV49z6vHufV49z6vHufV49z6vHufV49z6vHufV49z6vHufV49z6vHufV49z6vHufV49wr1HThzI+vS7H16XY+vS7H16XY+vS7H16XY+vS7H16XY+vS7H16XY+vS7H16XY+vS7H16XY+vS7H16XY+vS7H16XY+vS7H16XY+vS7H16XY+vS7H16XY+vS7H16XY+vS7H16XY+vS7H16XY+vS7H16XY+vS7H16XY+vS7H16XY+vS7H16XY+vS7H16XY+vS7H16XY+vS7H16XY+vS7H16XY+vS7H16XY+vS7H16XY+vS7H16XY+vS7H16XY+vS7H16XY+vS7H16XY+vS7H16XY+vS7H16XY+vS7H16XY+vS7H16XY+vS7H16XY+vS7H16XY+vS7H16XY+vS7H16XYw9d1b3WwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAYz+EAAAAAAAAAAAAAAAAAAAAAAAABgftfIAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAMZ/CAAAAAAAAAAAAAAAAAAAAAAAAAwP2vkAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAGM/hAAAAAAAAAAAAAAAAAAAAAAAAAYH7XyAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADGfwgAAAAAAAAAAAAAAAAAAAAAAAAMD9r5AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABjP4QAAAAAAAAAAAAAAAAAAAAAAAAGB+18gAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAxn8IAAAAAAAAAAAAAAAAAAAAAAAADA/a+QAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAYz+EAAAAAAAAAAAAAAAAAAAAAAAABgftfIAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAMZ/CAAAAAAAAAAAAAAAAAAAAAAAAAwP2vkAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAGM/hAAAAAAAAAAAAAAAAAAAAAAAAAYH7XyAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADGfwgAAAAAAAAAAAAAAAAAAAAAAAAMD9r5AAAAAAAAAAAAAAAATrt/CRryT1NGgAAAAAAAAAAAAAAAAAAAAAAAAAMZ/CAAAAAAAAAAAAAAAAAAAAAAAAAwP2vkAAAAAAAAAAAAAAAAqMpE4ODsyi7wAAAAAAAAAAAAAAAAAAAAAAAAAAxn8IAAAAAAAAAAAAAAAAAAAAAAAADA/a+QAAAAAAAAAAAAAAAE6sY6dSpU57GH2YAAAAAAAAADskKUXsxtJaiaa0AAAAAOaLdrgAAAAAAAABjP4QAAAAAAAAAAqNRq6iWbdidKcVdoAAAAAAAAAAAAADA/a+QAAAAAAAAAAAAAAASdo3NZPzJ03FK5h9mAAAAAAAAABJOpK3RCpRTTRy+0k79CEFFWQAAAAFVu1lux0FbTcpy5ogQk5IAAAAAAAAxn8IAAAAAAAAADC04yk5S2RLHS5tFoYdq86r6E8XKcHFrcAAAAAAAAAAAAADA/a+QAAAAAAAAAAAAAAAVvgZQtzmI6Iw+zAAAAAAAAAB7FJe7cINKTiAAAAAPWogho5IptuKbE5NSsU+e/kTlJvliRclH3jmqNcy2HK8LolObV1sSnKWsdiMk43OabXMthNNXAAAAAMZ/CAAAAAAAAAAXu4V+bDCx56c4dSUWnZgAAAAAAAAAAAAAYH7XyAAAAAAAAAAAAAAACqrwaE2ndEpOTuzD7MAAAAAAAAAJbFL4EFSmpbblOTkrPdAAAAAf8vPkC+KRS+BFPeXqUtmU9bsq/AxKysL+G0JL2dvIpJciFpSZ7WFrFL4EAAAAAYz+EAAAAAAAAABP/AHWPr/mFOpKElJFbkr0udboAAAAAAAAAAAAADA/a+QAAAAAAAAAAAAAAAEsPd6MdBctrlOnydQAAAAAAAAAKWicewEPikAAAABPSaYU9U33KXwIpatlLZlLqisrwE01cX8NsXwL0KXwItekxRi0tBJJaAAAAAGM/hAAAAAAAAAAUl7TDyit0BhvgqegAAAAAAAAAAAAAGB+18gAAAAAAAAAAAAAAAAAAAAAAAAAAnB35okPaN67D9onZFOPKtdwAAAAJw5o2LVJLlIxSVkcs07LYpw5bkVKMrdCcZKXNEipSi+Y5aiXKhwtDlQk+SxBNRSZCPu2YvaR0Qr2VwAAAADGfwgAAAAAAAAAKFb2U79BrCN81/wCvQpOhLmjLq9CtUpQp+zp9QAAAAAAAAAAAAAMD9r5AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABjP4QAAAAAAAAAAAAAAAAAAAAAAAAGB+18gAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAxn8IAAAAAAAAAAAAAAAAAAAAAAAADA/a+QAAAAAAAAAAAAAAAAAAA6sO57WHc9rDue1h3Paw7ntYdz2sO57WHc9rDue1h3Paw7ntYdz2sO57WHc9rDue1h3Paw7ntYdz2sO57WHc9rDue1h3Paw7ntYdz2sO57WHc9rDue1h3Paw7ntYdz2sO57WHc9rDue1h3Paw7ntYdz2sO57WHc9rDue1h3Paw7ntYdz2sO57WHc9rDue1h3Paw7ntYdz2sO57WHc9rDue1h3Paw7ntYdz2sO57WHc9rDue1h3Paw7ntYdz2sO57WHc9rDuaNAAGM/hAABToTqK8T6nVPqdU+p1T6nVPqdU+p1T6nVPqdU+p1T6nVPqdU+p1T6nVPqdU+p1T6nVPqdU+p1T6nVPqdU+p1T6nVPqdU+p1T6nVPqdU+p1T6nVPqdU+p1T6nVPqdU+p1T6nVPqdU+p1T6nVPqdU+p1T6nVPqdU+p1T6nVPqdU+p1T6nVIYWShJNavY+p1T6nVPqdU+p1T6nVPqdU+p1T6nVPqdU+p1T6nVPqdU+p1T6nVPqdUq0ZU7cwBgftfIAAAAAAAAAAAAAAAAAAAhSjKN2ewgewgewgewgewgewgewgewgewgewgewgewgewgewgVKUYxbRyROSJyROSJyROSJyROSJyROSJyROSJyROSJyRIwi5JHsIHsIHsIHsIHsIHsIHsIHsIHsIHsIHsIHsIHsIHsIHsIEKUW2ewgewgewgewgewgewgewgewgewgewgewgewgewgewgSowUWyHwIAAxn8IAUW1ojcnTlHdBTnKFBuL6n1mr/MfWav8x9Zq/wAx9Zq/zH1mr/MfWav8x9Zq/wAx9Zq/zH1mr/MfWav8x9Zq/wAx9Zq/zH1mr/MfWav8x9Zq/wAxGTcV6HMzmZzM5mczOZnMzmZzM5mczOZnMzmYm2yeIqqTSZ9Zq/zH1mr/ADH1mr/MfWav8x9Zq/zH1mr/ADH1mr/MfWav8x9Zq/zH1mr/ADH1mr/MfWav8x9Zq/zH1mr/ADFOtUdObb1Vj6zV/mPrNX+Y+s1f5j6zV/mPrNX+Y+s1f5j6zV/mPrNX+Y+s1f5j6zV/mPrNX+Y+s1f5j6zV/mPrNX+Y+s1f5ivJypwb31AMD9r5AAAAAAAAAAAAAAAAABUqOOiWpTqOWj3Kk3GyW5TnzLXcKXwAAAAAVv4bAAAAAIfGgAAAACnvL1AAAAAn8DIfAgADGfwgClFRoPu0zCxXM5PoRquvCUZb7oF/u79QAAAACPwR9AAAAAFuVPjYAAAAFL+FP5AAAAAFX+DD5gGB+18gAAAAAAAAAAAAAAAAAsrjVqqI61G+xtV9UFL4AAAAAK38NgAAAAEPjQAAAABT3l6gAAAAT+BkPgQABjP4QEIuUlFDknKaWyVilpQmzB/xRqzsL/d36gAAAAEfgj6AAAAALcqfGwAAAAKX8KfyAAAAAKv8GHzAMD9r5AAAAAAAAAAAAAAAAAAfFV9Cl8UiX8WIRjUa91nJV7kU+VXAAAAK38NgAAAAEPjQAAAABT3l6gAAAByVe5KFW2rIfAgADGfwgMLF3cl0KNKolO63RGLVCae5hP4qJatmHUHSfPtc5ML3MRGikvZsAAAAI/BH0AAAAAW5U+NgAAAAUv4U/kAAAAFCNBx996nJhe5ilBQgo7agGB+18gAAAAAAAAAAAAAAAAAlJRjcpRtG73ZT+KSHrVQUvgAAAAAlrFgAAAAFPcAAAAAsgAAAAJ/AyHwIAAxn8ICnWnTXulHETmpX6IpSdSM092YRWk5dkC/3d+oAAAABh/4SAAAAAlsAAAAAU6cZU5N7qwAAAABV/gw+YBgftfIAAAAAAAAAAAAAAAAAK0lzJPYjWjJ2RJqFS72ZT96TkFL4AAAAAJfCwHugAAACnuAAAAAQ3YAAAAE/gZD4EAAYz+EAUqigpJ9UUans5qRWr03DlprcF/u79QAAAADD/wAJAAAAAS2AAAAAKP8ABqfIAAAAAq/wYfMAwP2vkAAAAAAAAAAAAAAAAADhF7oUIrZEoqSsxJJWQUvgAAAAAl8LAnugAAACnuAAAAAQ3YAAAAE/gZD4EAAYz+EAAC/3d+oAAAABh/4SAAAAAlsAAAAAUf4NT5AAAAAFX+DD5gGB+18gAAAAAPZv2fP0vYAAAAAAAAAAAAApfAAAAABL4WBU3FqgAAAKe4AAAABDdgAAAAT+BkPgQABjP4QAAL/d36gAAAAGH/hIAAAACWwAAAABR/g1PkAAAAAVf4MPmAYH7XyAAAAAAy/D+3wleK3VmvVXAAAAAAAAAAAAAKXwAAAAAS+FgT3IaoAAACnuAAAAAQ3YAAAAE/gZD4EAAYz+EAAC/wB3fqAAAAAYf+EgAAAAJbAAAAAFH+DU+QAAAABV/gw+YBgftfIAAAAADhyPuVH6fqZjh/YYmUFt09GAAAAAAAAAAAABS+AAAAACXwsBq8iHYAAACnuAAAAARi02AAAABP4GQ+BAAGM/hAAAv93fqAAAAAYf+EgAAAAJbAAAAAAAAAABV/gw+YBgftfIAAAAADIafLhL923+n6HEWHvGNZdNH+gAAAAAAAAAAAAFL4AAAAAJfCwPti0kAAABT3AAAAAAAAAAJ/AyHwIAAxn8IAAF/u79QAAAADD/AMJAAAAAS2AAAAAAAAAACr/Bh8wDA/a+QAAAAAGDo+xw8KfZfj1MZh1XoSp91+PQaadmAAAAAAAAAAAAFL4AAAAAJfCwF8RLSQAAAFPcAAAAAAAAAAn8DIfAgADGfwgAAX+7v1AAAAAMP/CQAAAAEtgAAAACNNyi5LoAAAAAVf4MPmAYH7XyAAAAADK8P7fFQj0Wr+X9WAznD+xxcrbS1/z/ABAAAAAAAAAAAACl8AAAAAEvhYEfiZU6G6AAACnuAAAAARbbYAAAAE/gZD4EAAYz+EAAC/3d+oAAAABh/wCEgAAAAJbAAAAAFH+DU+QAAAABV/gw+YBgftfIAAAAAOHaKUJ1e+n3AZ3FVsLCst07fo/xQAAAAAAAAAAAAUvgAAAAAl8LAhuyauiOsQAAAp7gAAAAEN2AAAABP4GQ+BAAGM/hAAAv93fqAAAAAYf+EgAAAAJbAAAAAFH+DU+RClOfwoaadmAAAAFX+DD5gGB+18gAAAAAyWHLgoed3+IDaq5dXj/LJ/ncAAAAAAAAAKtV3sh1HGCYAAUvgAAAAAl8LAh1GrohsAAAFPcAAAAAhuwAAAAJ/AyHwIAAxn8IAAF/u79QAAAADD/wkAAAABLYAAAAIYepPZEMD/MyFGEFZIcorRsxlGz50AAAAFX+DD5gGB+18gAAAAAyxWwdP0BtJXMunz4TE+jf4MAAAAAAAAAewoXg5FT+FEWwABS+AAAAACXwsHsQ2COjsAAAFPcAAAAAhuwAAAAJ/AyHwIAAxn8IClTdSSiirhFGHNF3sU6bqSUUVsKow5ou9gX+7v1AAAAAMP8AwkAAAABLYAAAUXJ2RDCVJb6EMFBfE7kaUIfCidanDdk8bH7KI15zpzbe1htsozVanZlSm4ScWAAAAVf4MPmAYH7XyAAAAADKWpYKn6fqGYcywtTl3szLXy4LEPyt+DAAAAAAAAAGroUkoOJOScYxRbQAApfAAAAABL4WD2IbBtIAAAKe4AAAABDdgAAAAT+BkPgQABjP4QCbWxhHrJdLGG92M59kYX3nKHdAv93fqAAAAAYf+EgAAAAJbAAEac5/CiGCm/idiGEpR31EoxWmhPE0o9bk8a38KJ1qk92ApyUXFdQoVfZzT6GLpKcOZboAAAAq/wAGHzAMD9r5AAAAABkFRSwvL2b/AMwqQU4OL6jg8PlUlLeb/r8gAAAAAAAAArU2ndbFKDlK4AAUvgAAAAAl8LCWxDYJ7oAAAKe4AAAABYAAAACfwMh8CAAMZ/CASu0idNUqT5NblP8A3eZhP4qJLVkVfDv1OVjTAAAADD/wkAAAABLYAhh6k9kQwP8AMyGGpR2RKUYrV2J4ymttSeMqPbQlOUt3cAADB1eaPI+hiKXs56bAAAFmzlZV/hQ+YBgftfIAAAAAMgxCp13Te0vzQHEGIUqkaMfs7+r/AOB+YAAAAAAAAAU6nOipU5EtDcAApfAAAAABL4WEvhIfCE1oLVIAAAp7gAAAAAAAAAE/gZD4EAAYz+EAYSXv8vRkF+5mjCK9VD1ZQq+zouVr6n17+7+JXxHtUla1gAAAAw/8JAAAAANXRDCVJdLEMDFfE7kaUIfCidWnDdk8bFfCrk8VVl1sNt7gAAKLaukOXvpeT/QITcJKSKsI16V18hoAACjifZx5bXPr3938TFT54Ql6gGB+18gAAAAAjJwkpReqMBmlLEwSbtLqv8jHZlRw0HreXRf10KlSVSbnJ3bAAAAAAAAAKtRwasQqOC0J1HPcWwABS+AAAAACXwsJ/CR+EJK6IbAAAFPcAAAAATTAAAAAn8DIfAgADGfwgKVF1G0ihRlSvOfQwzUnKD6op05UYylL0QL/AHd+oAAAABh/4SAAAAtc5WcqNCeJpR6k8c/sonXqT3YAAABR/g1PkVJWr016/oBg6tnyPqYylyy5l1AAACr/AAYfMAwP2vkAAAAAAAAAAAAAAABVpudrEKbmtCNBp6sAAKXwAAAAAS+FhPYjsg3IdgAACnuAAAAAQ3YAAAAE/gZD4EAAYz+EBGUou8XqVK06itJibTuipWnUVpMF/u79QAAAADD/AMJAAABFXByQ5Mk7oAAAAAo/wanyMRK2KpfMDYhKNelZk4OMnFgAAFX+DD5gGB+18gAAAAAAAAAAAAAAABuED20O4AAUvgAAAAAl8LCewtkAtJAAAFPcAAAAAhuwAAAAJ/AyHwIAAxn8IAAF/u79QAAAADD/AMJAAABsjcCWwAAAABR/g1PkYyVsVT/rqAYer7OfkzGUuaPOugAABV/gw+YBgftfIAAAAAAAAAAAAAAAAk+Z3LNuwAAUvgAAAAAl8LCpsLYCWkkAAAU9wAAAACG7AAAAAn8DIfAgADGfwgAAX+7v1AAAAAMP/CQAAAldkuwBLYAAAAAo/wAGp8jMZWxEfJL8wAwtVThyvdFel7OdgAAKv8GHzAMD9r5AAAAAAAAAAAAAAAAPYSXsm/Mq25YyRugACl8AAAAAEvhYVOgBUWgtUAABT3AAAAAIbsAAAACfwMh8CAAMZ/CAABf7u/UAAAAAw/8ACQAAAtEPUAlsAAAAAQqKMJR7mZ61/kRd4pgFObhJSRXpqtTvHfoAABV/gw+YBgftfIAAAAAAAAAAAAAAADcir05Iu3ZCWgABS+AAAAACXwsJ7oAmvdI6xAAAp7gAAAACilcAAAACfwMh8CAAMZ/CAABf7u/UAAAAAw/8JAAAJXJbABLYAAAAAMx1xD+RQd6MX5IAMHV+w/kYujyS5lswAAq/wYfMAwP2vkAAAAAAAAAAAAAAAAyM4y2FRipXAACl8AAAAAEvhYS+JADV0Q2AAAp7gAAAAAAAAAE/gZD4EAAYz+EAAC/3d+oAAAABh/4SAAAih6sAJbAAAAABmGuIl8vyMG74eIAJtO6Fy16X9bkouLswACr/AAYfMAwP2vkAAAAABhsP7anUtvFX+5/5AAAAAAAAAANXRRtGTTOaPcAAKXwAAAAAS+FhL4kAEdJWAAAp7gAAAAAAAAAE/gZD4EAAYz+EAAC/3d+oAAAABh/4SAAAeiAAlsAAAAouWyuRwtaX2SOXze7RHL4Ld3I4SjHoZnBSxlVLp+iRl1BzwUZx6Xv94AGFq8k7PZmNo/bQABV/gw+YBgftfIAAAAADh2kn7ST8l/mYzDuhXlT7P8On4AAAAAAAAAANt7gAAUvgAAAAAl8LB/GMAekgAAKe4AAAABDdgAAAAT+BkPgQABjP4QAAL/d36gAAAAGH/hIAAIq7JasACWwABGlOWyI4KtLpYjlz+1IjgKS31I4elHaKNEiWIpR3kSx9Jbakswk/hiRxNSdObvtYq3niKrb6M4drReHdN73f5IxeFt78AAMPUVWnyvcq03Tm4gAVf4MPmAYH7XyAACzLMsyzAAMow/scJFPd6/f/AMCxxFQSnGqvR/oAAAAAAAAAB7KHY9lDsAAFL4AAAAAJfCw+2MWwE90AABT3AAAAAIbsAAAACfwMh8CAAMZ/CAABf7u/UAAAAAw/8JAAAtEAAS2LX2I4erLaJHAVXvZEcuj1kRwVFdLkaUI7JEpxjuyWLorqSzCC+FXJY+o9kkSxVaX2hty3YBR/g1PkUVzYmr6S/MymTUZW6GGxKqrle5i8Ly+/DYAKNR05pmJpKpT5o7oACr/Bh8wDA/a+QAAJqxdF0NqwACV2kKKikl0M5kpQqx7KL/FoAAAAAAAAAAAACl8AAAAAEvhYL4h6sQE1oR1SAACnuAAAAAQ3YAAAAE/gZD4EAAYz+EAAC/3d+oAAAABh/wCEgABK7JdgLXOSTPZM9kjkj2FGMVoiVanHeSJY2itnclmC+zEljqr20JV6kt5AAAFH+DU+RgVfETfr+ZlekpxE3F3W5hsQqsbPcxeF5Pejt+QAYOrdcjMVS5J6bMAKv8GHzAMD9r5AAAAAAnZpkWpJNGaKUZYhvZ8qX5/oAAAAAAAAABzR7nNHuAAFL4AAAAAJfCwj8TOowJK6IbAABT3AAAAAIbsAAAACfwMh8CAAMZ/CAABf7u/UAAAAAw/8JAAEVoPUCEbK45RXUdRDqPoSnJrclOUt3cAAAACk0qU1fsZbrVmzA+7ipx9fzCMnFpp6mHrxrRs9zFYX2b5o7fkARk4tNElGvS0GmnZgFX+DD5gGB+18gAAAAAMnxka+HUG/ejp/kzPsZCpNUoO9t/X/AIAAAAAAAAAAAUJu/KFWTSSjuyi24IqXlLkTKTabi+gUvgAAAAAl8LCG7EPYWwEN7AABT3AAAAAIxabAAAAAn8DIfAgADGfwgKCg5+/sc1OvFxSszD01OeuyJyp14PlWqBf7u/UAAAAAw/8ACQAFrktEBCN2VJaWAJbAAAAABlWrm/T9Sj7uOkvUCE3CScXqUa0a8PPqjFYZ0ndbAGEq8suV7MxlKz50AVf4MPmAYH7XyAAAAAA22AAAAAAAAAAZSiptyZUhGMk0hxUKqsBR+BENZyY9Kq9AjWUVax9YXYi+ZJgAAAEvhYQ6iDqBtIAAKe4AAAAAAAAAH1hdiVdNNWIfAgADGfwgDCfxUUtKdRowf8S3dDRRpupRcV3PqNTuirh5U0rsAAAAMP8AwkABFEtWCV3YdoxHqAS2AAAAAJ6RZlS0n8iXu5gv66AEJyhJSjuUqsK8PzRicM6T02AKM1WpWZUg4ScWUsNOpG6Z9Rqd0YmDhThF+YBgftfIAAAAAAAAAAAAAAABq6KVRR0ZVmptJFX+JECj8CKW8vUl/FiFKKcTlj2AAAACXwsexDYWwM3CWjQAAU9wAAAAAAAAAOWPYnFcr0IfAgADGfwgDDrkhKp8in/u8zCfxUS3ZCTjQbT6nt6n8zJVJS3dwAAAAw/8JAAbICEbK5OXMwAlsAAAAASpqVCpK+yMqXuS9TFe7jYP0/MAKdSVOXNEpzhXpmIw7pS8gKFX2c79DF0lOHMt0RqTirJnt6n8zK7cqUG33AMD9r5AAAAAAAAAAAAAAAAFam07rYo023foSp3mpXAo/AiGk5IetVeSCl8AAAAAEvhY9iPwi2B6oQVFoLVAAFPcAAAAAi22wAAAAJ/AyHwIAAxn8IBRcmkjEtRiqS6FHWlNGEXvt9kPUX+7v1AAAAAMP/CQAJXZLsEI3dypLogAJbAAAAAE9MJW9DK1+6fr+iMx0rQl/W4AFKrKnLmiRlTr0/Ir0JUpW6AYSrzR5H0MRS9nOy2Cr/Bh8wDA/a+QAAAAAAAAAAAAAAABKUVuz2sO4BSTULMqKUZc0SnF6ye7Cl8AAAAAEvhZLYj8IBswmrxIaoAAp7gAAAAEN2AAAABP4GQ+BAAGM/hAYVwU7yZKnhpO7l+JSnGnVfbYqeypQag7tgv93fqAAAAAYf8AhIAErIeold2G1GI9QAJbAAAAAFXTB1vQyxfuPmZstIP1/Qi7xTAAo1pUpXR+7xFPyK9CVKVmEJuElJFWEa9K6+Q0Vf4MPmAYH7XyAAAAAAAAAAAAAAAAJS5ncAACl8AAAAAEvhZL4RbIAZ0GrohsAAU9wAAAACG7KkuWwAAAAT+BkPgQABjP4QAAL/d36gAAAAGH/hIASuS2CEbInLmYABLYAAAAAr/7lW9F+Zly/cL5mar93F+ZQd6UX5IAAoVpUpXWw1Tr0/IrUZUpcrDB1bPkfUxlHllzLZlX+DD5gGB+18gAAAAAAAAAAAAAAAGrooRTu2csewAAUvgAAAAAl8LJ/CR2QAII6SAAKe4AAAABDdmI2RSlzR9AAAACfwMh8CAAMZ/CAABf7u/UAAAAAw/8JAEUPVlON3cqS6IAAJbAAAAAGLdsDV+X5mXr/Z4/P8zMleh8zBu+HiAAFCvKlLyJwp16ZVpSpy5WbbEJRr0rMxEXGnCL6XAMD9r5AAAAAAAAAAAAAAAACqcsm4n1iXYAAKXwAAAAAS+Fk9iOyABg9JAAFPcAAAAAhuzEdCD5J2YAAABP4GQ+BAAGM/hAAAv93fqAAAAAYf8AhIAeiEruxJqMR6gABLYAAAAAzCTWGkr72MCrYeP9dTHq+Hl/XUy93w6+YAAGHxDpPyKlOFen+RUpypy5ZGGq+znrszH/AGfn+gBgftfIAAAAAAAAAAAAAAAA9hM9hMAAKXwAAAAAS+Fk9iOwAPYWxPobgAU9wAAAAAr6ySMRDaSKcuaIAAAE/gZD4EAAYz+EAAC/3d+oAAAABh/4SAirslqyEeVEpczAAAlsAAAAAZn/AAPmYNWoQ9DFq9CXoZY70WvMAAAw2IdJ2exWpQrw0+TJwlCXK9ys26UL+YBgftfIAAAAAAAAAAAAAAABTi5cvU9vA9vAAAKXwAAAAAS+FlTYjsAB1JrQjqgAKe4AAAAAVdaiJRUlYpPlnysAAACfwMh8CAAMZ/CAABf7u/UAAAAAw/8ACQC0RCN3cqS6IAAAlsAAAAAZon7Bev6Mwy/cw9EVVenJeTMqfuSQAAAYXEum+V7FehGtDTfoYiDjThF76gGB+18gAAAAAAAAAAAAAAAIRftWWZZgABS+AAAAACXwsqdBbAAMeqIbAAU9wAAAABNMnrVQV42akiEuaKYAAAT+BkPgQABjP4QAAL/d36gAAAAGH/hIErsau7Emox0AAACWwAAAABmj/cJef6MoK1KK8kNXRlekpxAAAAwuJ9m+WW35GYWbjbz/AEAMD9r5AAAAAAAAAAAAAAAAPYhUU9gAAKXwAAAAAS+Fk90LYAB6oRDewAFPcAAAAAhuyTtVuJprQnFSjYoStLlYAAAT+BkPgQABjP4QAAL/AHd+oAAAABh/4SCK0ErIlLmYAAAS2AAAAAM1fuRXmU1aCXkGB93Ezj6/mAAAAVf4UPmAYH7XyAACyLIsiyAAAAAAAAAAAANym1CbTOeHcAAKXwAAAAAS+Fk90dAADZktJG6ACnuAAAAAQ3ZPWoyE+VmjRWjyy5kRkpRTAAAJ/AyHwIAAxn8IAAF/u79QAAAADD/wkbkUVJdEAAABLYAAAAAzb7C9f0ErJIKPu46S9QAAACr/AAYfMAwP2vkAAFy5cuAAAAAAAAAAAAD1AAApfAAAAABL4WS+IAAGSV0QfQAKe4AAAABDdm9RlSnfVEJ8uhUipxsUJa8oAABP4GQ+BAAGM/hAAAv93fqAAAAAYf8AhIihvliAAAAS2AAAAAM01nBAS93MF/XQAAAAq/wYfMAwP2vkAAAAAAAAAAAAAAAAckOxyQ7AABS+AAAAACXwszabhhZyi9V/mjAYuOKw8ai36+oAAIkrPQTugCnuAAAAAQ3ZGpFV4xe7vb5BUp31RTnbRlVcs7oTUldAAAT+BkPgQABjP4QEYuTSRXw8adNdyjS9pNRKtGlKDlT6Av8Ad36hSgpzUT6jHufUY9z6jHufUY9z6jHufUY9z6jHufUY9z6jHufUY9z6jHufUY9z6jHuQpKEVG5jMTHC0HUavbp3JcR8z/hfj/wD/WL/AKd/j/wD/WL/AKd/j/wD/WL/AKd/j/wD/WL/AKd/j/wD/WL/AKd/j/wD/WL/AKd/j/wD/WL/AKd/j/wD/WL/AKd/j/wD/WL/AKd/j/wD/WL/AKd/j/wD/WL/AKd/j/wD/WL/AKd/j/wB8Q3X8P8AH/gH9rf3Px/4B/a39z8f+Af2t/c/H/gH9rf3Px/4B/a39z8f+Af2t/c/H/gH9rf3Px/4B/a39z8f+Af2t/c/H/gH9rf3Px/4B/a39z8f+Af2t/c/H/gH9rf3Px/4B/a39z8f+Af2t/c/H/gFXEfWa0NLdPxP7P8A734f8E/s/wDvfh/wTNYfVcZF76Jn9r/3Px/4B/a/9z8f+Af2v/c/H/gH9r/3Px/4B/a/9z8f+Af2v/c/H/gH9r/3Px/4B/a/9z8f+Af2v/c/H/gH9r/3Px/4B/a/9z8f+Af2v/c/H/gEKvtsLSna17/mAYH7XyAAAAAAAAAAAAAAAAhNuo0c8u5zy7gABS+AAAAACXwszWlKth5U47yaX4oyPFSw+JdCel9PRr+rfcboAAaHqrEXZ2AKe4AAAAAopXM2xPsK9Cd9m/u0T/BkJKUbhUp9UN3jYpVOXR7AAAT+BkPgQABjP4QEZOLTRUbeHi33KGlOcvIwms3HugX+7v1CE3CSkuh9eqfyo+vVP5UfXqn8qPr1T+VH16p/Kj69U/lR9eqfyo+vVP5UfXqn8qPr1T+VH16p/Kj69U/lR9eqfyohVqzinZfiZhgq2MjGLkkkf6vT/wCfi+7/AIJ/q9P/AJ+L7v8Agn+r0/8An4vu/wCCf6vT/wCfi+7/AIJ/q9P/AJ+L7v8Agn+r0/8An4vu/wCCf6vT/wCfi+7/AIJ/q9P/AJ+L7v8Agn+r0/8An4vu/wCCf6vT/wCfi+7/AIJ/q9P/AJ+L7v8Agn+r0/8An4vu/wCCf6vT/wCfi+7/AII+Hp2/ifh/wT+yX/P+B/ZL/n/A/sl/z/gf2S/5/wAD+yX/AD/gf2S/5/wP7Jf8/wCB/ZL/AJ/wP7Jf8/4H9kv+f8D+yX/P+B/ZL/n/AAP7Jf8AP+B/ZL/n/A/sl/z/AIFHLXTqRnzbNPY+vx/lPr8f5TNKH1ypGadrKx/ZEv5/wP7Il/P+B/ZEv5/wP7Il/P8Agf2RL+f8D+yJfz/gf2RL+f8AA/siX8/4H9kS/n/A/siX8/4H9kS/n/A/siX8/wCBCl7LC0oXva/5gGB+18gAAAAAAAAAAAAAAAFTSlzH1ePc+rx7gABS+AAAAACXwsqrmqQXn+SZxDhHSrLEQ2l+a/zMuxixOHjU67P1AABokiLugKe4AAAABLY4hpSnCM1tH9f+GMoxPtcNCTfSz9VoBUp9USiUqtvdYAAT+BkPgQABjP4QBP8A3ePqU/8Ad5mE/ioluzDxhKk1N6XPZYbv+JiIUopcjAAAADD/AMJAAAAAS2AAAAAAAAAAoQoyj771PZYbv+JilFQgo7agGB+18gAAAAAAAAAAAAAAAFJ+1tcnNqaSAAApfAFacotJM9tPue2n3PbT7ntp9z20+57afc9tPue2n3PbT7ntp9z20+57afcdWbW4lzST7GNwscTQlSfXbyfQyXEyw2JdCel9PRr+rEX0AABpWKdSM1eDujmZzMVSS2Z7afc9tPue2n3PbT7ntp9z20+57afc9tPue2n3PbT7ntp9z20+57afc9tPue2n3HVm1a5isL9YwlWHVrT1Wq/E4brc1OdLtr9+n+X3lKfNGwTmooxvEMaNaVOFO9tL3/Sxl+cQxU+SS5ZdPMVWaVrntp9z20+57afc9tPue2n3PbT7jqzatch8CAAMZ/CAJ/7vH1KWtGaMIv3noPUX+7v1AAAAAMP/AAkAAAABLYAAAAApL91P5AAAAAFX+DD5gGB+18gAAAAAAAAAAAAAAAF/GKv8WIAAFL4AnTjPc9hA9hA9hA9hA9hA9hA9hA9hA9hA9hA9hA9hAdCCQopbBxDg3TqrEQ2e/r/wTL8UsVho1Ouz9f8Ag7idwAG0jO8w9hS5Iv3pfgu/+Rw5SreylKT9x7Lz6s5InJEhSjJnsIHsIHsIHsIHsIHsIHsIHsIHsIHsIHsIHsIHsIHsIHsIEaMW2RgoqyMKlgs5dPo218nqv0NacxyVrleraMpvZJv7jLcrnmU5ycrW3dr6v5oXCjTTVbX/AA//AGwsBiUkvbf+Sr/MWBr9av8A5Kv8xYKrfWr+CPqU/wDn4/uX+R9T/vv8P8hYVJfE393+QsNFdWSowUWyHwIAAxn8IAn/ALvH1MLJczi+uh7L2EJNvV6IF/u79QAAAADD/wAJAAAAAS2AAAAAKP8ABqfIAAAAAq/wYfMAwP2vkAAAAAAAAAAAAAAABOTjUbRKbk0z28wAApfAAAAABL4WBjMNHE0JUpdfz6GS4mWFxbo1NE9H69P8hqzE7oAbSMRXhRpyqTeiKVOrmeM167+S/r8SnTjTgoQVkgKe4AAAABDdhxRRdOvTrx66fNbf15EJrEUI1Y9UmXbVjPqvsMA49ZNL9X+Rw7h/ZYFSe8m3+i/IAACfwMh8CAAMZ/CALu1glKUt3cF/u79QAAAADD/wkAAAABLYAAAAAo/wanyAAAAAKv8ABh8wDA/a+QAAAAAAAAAAAAAAAE6LlJu59Xl3Pq8u4AAUvgAAAAAl8LAOI8HyVI4iPXR+vR/12MrxixeGUn8S0fr/AME2ZuDl2Erme4/29X2FP4Y/i/8AgGUZesJQ1+J7/wCXyAKe4AAAABDdhxBh/bYGTW8dfu3/AAucNYj2mD5HvF2+W4qSUrnE1V1cTTw8en5v+vxKNKNKlGnHZJL7gAAJ/AyHwIAAxn8IAAF/u79QAAAADD/wkAAAABLYAAAAAo/wanyAAAAAKv8ABh8wDA/a+QAAAAAAAAAAAAAAAAAABS+AAAAACXwsAxWGhiKMqUtmZRiJYLGOjU0T0fr0f9dxq6E7DdxRM6zD6rQ5YP35beXmcP5f7Sf1ia0W3r3+X5+gAU9wAAAACG7CpCNSDg9nocPTeGx88PLrdfOP/AuGD/23OnU3SbfyWi/QAAAn8DIfAgADGfwgAAX+7v1AAAAAMP8AwkAAAABLYAAAAApziqc03q7AAAAAFX+DD5gGB+18gAAAAAAAAAAAAAAAAAACl8AAAAAEvhYAcR4LlksRFb6P16P9DKcZ9aw0ZN+8tH6/8EkriiVq0KNN1JvRCVXNMb6/gl/X3lKlClTVOCskAFPcAAAAAjFpsDOE8Hmsa62dn+j/AK8zMcSqGDnVT2Wnq9EcK4e1OpWfXRfLf9AAACfwMh8CAAMZ/CAABf7u/UAAAAAw/wDCQAAAAEtgAAAAAAAAAAq/wYfMAwP2vkAAAAAAAAAAAAAAAAAAAUvgAAAAAl8LADE0IV6MqUtmjKq8sDjXSqaJ6P16P+ujAz/MPa1Pq9N6Lfzf/A/MybL/AKrQvJe/Lfy8v66gAU9wAAAAADijD8+GjVW8X+D/AODYzDMPa5VRp31ej/7d0/HRmUYf2GCpw62v9+oAABP4GQ+BAAGM/hAAAv8Ad36gAAAAGH/hIAAAACWwAAAAAAAAAAVf4MPmAYH7XyAAAAAAAAAAAAAAAAAAAKXwAAAAAS+FgAcR4KzWIivJ/o/0+4yjG/WsMm37y0f+fzM4zBYSh7r96Wi/z/rqZDl/tqvt57R283/wPzAAKe4AAAAABjsOsRhp0u6/Hp+JgqDr4iFHu/8Ah/wLKwAABP4GQ+BAAGM/hAAAv93fqAAAAAYf+EgAAAAJbAAAAAFOnGVOTe6sAAAAAVf4MPmAYH7XyAAAAAAAAAAAAAAAAAAAKXwAAAAAS+FgAYihCvSlTls0ZfiJZdjHCrts/wBH/XRk5Vc0xunXbyX9fiUKEKNKNOC0QABT3AAAAAIttsAy/L/Z5vVlbSOq/wC3v+BdAAABP4GQ+BAAGM/hAAAv93fqAAAAAYf+EgAAAAJbAAAAAFH+DU+QAAAABV/gw+YBgftfIAAJYvDxbjKok15o+vYX/n7H71/mfXsL/wA/Y/ev8yOMw0moxqJv1QAAAAAAAAAAAAAAAUvgAAAAAl8LAAOJMFdRxEV5P9H+n3GSZd9Wo88178vwXb/MAAKe4AAAABDdgChFScktWAAAE/gZD4EAAYz+EAAC/wB3fqAAAAAYf+EgAAAAJbAAAAAFH+DU+QAAAABV/gw+YBgftfIAAMxw1aWKqNQbV+zPqtf+R/cz6rX/AJH9zMBhq0cVTbg7XXRgAAAAAAAAAAAAAABS+AAAAACXwsAAlCM1aSugAAKe4AAAABDdgAAAAT+BkPgQABjP4QAAL/d36gAAAAGH/hIAAAACWwAAAABR/g1PkAAAAAVf4MPmAYH7XyAAAAAAAAAAAAAAAAAAAKXwAAAAAS+FgAAAAFPcAAAAAhuwAAAAJ/AyHwIAAxn8IAAF/u79QAAAADD/AMJAAAAAS2AAAAAKP8Gp8gAAAACr/Bh8wDA/a+QAAAAAAAAAAAAAAAAABUlK6jHcpybvGW6Cl8AAAAAEvhYAAAABT3AAAAAIbsAAAACfwMh8CAAMZ/CAChS9pNLoVKdKcG6e6Bf7u/UAAAAAw/8ACQAAAAEtgAAAAAAAAAAq/wAGHzAMD9r5AAAAAAAAAAAAAAAAAAENZyZtV+QUpRUbNnPDuaNAAAAEvhYAAAABT3AAAAAAAAAA54dyc4uL1IfAgADGfwgAoaU5yMJrNx7oYv8Ad36gAAAAGH/hIAAAACWwAAAAAAAAAAVf4MPmAYH7XyAAAAAAAAAAAAAAAAAAKW8vUl/FiEKMZK59Xj3EkkkAAAAVv4bAAAAAIfGgAAAACnvL1AAAAD6vHuSoRUW7kPgQABjP4QAU/wDd5mE/ioluzD1FTpOVup9eX8piMQqqStsAAAAEElCNuwAAAAC1ZUXvMAAAACkk6U/kAAAAFDEqnHltc+vL+UxU+eEJeoBgftfIAAAAAAAAAAAAAAAAAAp/HJD1qr0Cl8AAAAAFb+GwAAAACHxoAAAAAp7y9QAAAAJ/AyHwIAAxn8IAKWtGaMIv3l+yHqL/AHd+oAAAABH4I+gAAAAC3KnxsAAAACl/Cn8gAAAACr/Bh8wDA/a+QAAAAAAAAAAAAAAAAABUbhPmKV5NyYUvgAAAAArfw2AAAAAQ+NAAAAAFPeXqAAAABP4GQ+BAAGM/hABhZJT5Xs9B01Qpyd9XoC/3d+oAAAABH4I+gAAAAC3KnxsAAAACl/Cn8gAAAACr/Bh8wDA/a+QAAAAAAAAAAAAAAAAABa4FOpGMbNntodz20O57aHc9tDue2h3PbQ7ntodz20O57aHc9tDue2h3PbQ7ntodz20O5VqwlBpM54nPE54nPE54nPE54nPE54nPE54nPE54nPE54kZxUkz20O57aHc9tDue2h3PbQ7ntodz20O57aHc9tDue2h3PbQ7ntodz20O57aHc9tDuQqwTep7aHc9tDue2h3PbQ7ntodz20O57aHc9tDue2h3PbQ7ntodz20O57aHc9tDuTqwcWkyHwIAAxn8IABtsIRcqDSV9T2FT+VnsKn8rPYVP5Wewqfys9hU/lZ7Cp/Kz2FT+VnsKn8rPYVP5Wewqfys9hU/lZ7Cp/Kz2FT+VnsKn8rPYVP5WRT5UvIsyzLMsyzLMsyzLMsyzLMsyzEncnRqOTtE9hU/lZ7Cp/Kz2FT+VnsKn8rPYVP5Wewqfys9hU/lZ7Cp/Kz2FT+VnsKn8rPYVP5Wewqfys9hU/lZ7Cp/KynSmqc1bsewqfys9hU/lZ7Cp/Kz2FT+VnsKn8rPYVP5Wewqfys9hU/lZ7Cp/Kz2FT+VnsKn8rPYVP5Wewqfys9hU/lZ7Cp/KyvFqlBPzAMD9r5AAAAAAAAAAAAAAAAAAADowfQ9jT7HsafY9jT7HsafY9jT7HsafY9jT7HsafY9jT7HsafY9jT7HsafY9jT7HsafY9jT7HsafY9jT7HsafY9jT7HsafY9jT7HsafY9jT7HsafY9jT7HsafY9jT7HsafY9jT7HsafY9jT7HsafY9jT7HsafY9jT7HsafY9jT7HsafY9jT7HsafY9jT7HsafY9jT7HsafY9jT7HsafY9jT7HsafY9jT7HsafY9jT7HsafY9jT7HsafY9jT7HsafY9jT7HsafY9jT7HsafY9jT7HsafYskrAAGM/hAABSxEqasj67U7I+u1OyPrtTsj67U7I+u1OyPrtTsj67U7I+u1OyPrtTsj67U7I+u1OyPrtTsj67U7I+u1OyPrtTsj67U7I+u1OyPrtTsj67U7I+u1OyPrtTsj67U7I+u1OyPrtTsj67U7I+u1OyPrtTsj67U7I+u1OyPrtTsj67U7I+u1OyPrtTsj67U7I+u1OyPrtTsj67U7I+u1OyPrtTsj67U7I+u1OyPrtTsj67U7I+u1OyPrtTsj67U7I+u1OyPrtTsj67U7I+u1OyPrtTsj67U7I+u1OyPrtTsj67U7I+u1OyPrtTsj67U7I+u1OyPrtTsj67U7I+u1OyK1aVW1+gBgftfIAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAMZ/CAAAAAAAAAAAAAAAAAAAAAAAAAwP2vkAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAGM/hAAAAAAAAAAAAAAAAAAAAAAAAAYH7XyAAAAAAAAAAAAAAAAAAAAAAAAG0lqaNAAAAANoAAAAAUk9mAAAAAAAGM/hAAAAAAAAShKO6AAAAAtfYaa0YAAAACpzauloAAAAAAGB+18gAAAAAAAAAAAAAAAAAAAAAAACUfaTs9kUvdbj2AAAAAalzpsAAAACpFyVkSh7NpoAAAAAAAMZ/CAAAAAAADCxTk5PZCq+3hKLWu6AAAAAwrhHV73sYj+LIAAAAD64lJJL3TEwUajSAAAAAAMD9r5AAAAAAAAAAAAAAAAAAAAAAAAFLXmfmf8vfkAAAABU+KIAAAABX1ikAAAAAAAGM/hAAAAAAABS0oTZhP4thrUAAAAKX8SPqjEfxZAAAAAWvoYzWqAAAAAAYH7XyAAAAAAAAAAAAAAAAAAAAAAAAKOz9R/wAX5AAAAAVPiiAAAAAS96ol2AAAAAAADGfwgAAAAAAAp/7vMwn8VEtWwAAAApfxI+qMR/FkAAAABhoc1RdkVZ883IAAAAAAwP2vkAAAAAAAAAAAAAAAAAAAAAAAAU9JSRHWo/IAAAACp8UQAAAAJyUY3KUbRu92AAAAAAAGM/hAAAAAAABQ96nOPkYRWk5dkAAAABS/iR9UYj+LIAAAACX7qjbrIAAAAAAMD9r5AAAAAAAAAAAAAAAAAAAAAAAAFSEm+aO5ThyrXcAAAACcW5JroAAAABVg5pJHs6vc6AAAAAAAGM/hAAAAAAABRqezmmVa1Pkcaa3AAAAAg1Gab6FWSnNtAAAABGykr7EsTQlur/IrzpzkuRWAAAAAAMD9r5AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABjP4QAAAAAAAAAAAAAAAAAAAAAAAAGB+18gAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAxn8IAAAAAAAAAAAAAAAAAAAAAAAADA/a+QAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAYz+EAAAAAAAAAAAAAAAAAAAAAAAABgftfIAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAMZ/CAAAAAAAAAAAAAAAAAAAAAAAAAwP2vkAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAGM/hAAAAAAAAAAAAAAAAAAAAAAAAAYH7XyAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADGfwgAAAAAAAAAAAAAAAAAAAAAAAAMD9r5AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABjP4QAAAAAAAAAAAAAAAAAAAAAAAAGB+18gAAAAAAAAAAAAAAAAAAAAAAAAAACdSMXZnt4EasZJ26FOpzoAAITU27BOXLG5FqSTAAAAAAAAAADGfwgAAAAAAAAAAhBzkoo+pVPIeDqJX0I4ZypOdwAAp0OeDlfYCrQcIqV73AAAAAAAAADA/a+QAAAAAAAAAAAAAAAAAAAAAAAAAABWpyk1Y5G5cvUp0pJNMpU3FO4ABN2i2UF7gTV4tFB3hYAAAAAAAAAAMZ/CAAAAAAAAAAChJQqJsp1YVL8r2KmIppON9SGIjGi4dQAAl7mFXmBL38KvIAAAAAAAAAMD9r5AAAAAAAAAAAAAAAAAAAAAAAAAAAAv4xN2i7FGcpXuz20knqQrNbkaslK7ZOSjG57Wd73IyUoplb4GUlaCJ1ZN6MptygmzD9UAAAAAAAAAAGM/hAAAAAAAAAAAYH7XyK/8SXqU6NN0OZrXUw1GE6butRYamoctijhYxj7yuzEU1TnZbGHw0FBOSu2YuhGFpR2DFaUYr+tjCUIyXNIxlOEYppalDXDyXr+QAAAAAAAAAYH7XyAAAAAAAAAAAAAAAAAAAAAAAAAAABfxifwMw/UpRUp6k4L2nKivBRasVn7iHKHs+W5h9Ylf4CP8P5FGUU2mWSWhR+NgAAAAAAAAABjP4QAAAAAAAAAAGB+18iv/ABJepS/3b5MwX8N+phq0pzabJ1pLEKN9DFxUqkF3MXCc0lFFVN4f3twxvwxMOubD2XmTU4+7IwmtKQAAAAAAAAAYH7XyAAAAAAAAAAAAAAAAAAAAAAAAAAABfxifwMw/UofGyf8AFXyMT0K3wRFQk1e5SpuCsyv8BT+BFWk46rYou8CjrJgAAAAAAAAABjP4QAAAAAAAAAAGB+18iv8AxJepS/3b5MwX8N+pgv4j9Cp/vXzRif4sCtXjStcrYuE4OKTDG6xiYauqfuy2MZFSp83YwulKT/rYAAAAAAAAAMD9r5AAAAAAAAAAAAAAAAAAAAAAAAAAAAv4xP4GYfqUPjZP+KvkYnoVI3pIjWcVYpyco3KyvBlN/urkqzlGxRVoGHWjYAAAAAAAAAAYz+EAAAAAAAAAABgftfIr/wASXqUv92+TMF/DfqYL+I/Qqf7180Y12lFk4QrwRiMNGnTunqGJ96hF/wBbFPCxqU072ZirQoqJR0w0n6gAAAAAAAABgftfIAAAAAAAAAAAAAAAAAAAAAAAAAAAPZxvfqNJqxGEY7EacYu6HTi3fqShGW5ZJWHQg2KKSsiSvFooWcWj2ELlR2gyirQAAAAAAAAAADGfwgAAAAAAAAAAKdWdO/KyUnJ3Yq81DkT0KdedNWiynUlB3iOpJz5+pUqyqW5mUsROmrIq151FZgvfwtuxTxNSCsipVlUd5Mqe5hlHv/w4AAAAAAAAAYH7XyAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAUIp3QSipKzEklZAAAAAAAAAABjP4QAAAAAAAAAAAAABGpOKaT3AnUnNJSewAAAAAAAAAYH7XyAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADGfwgAAAAAAAAAAAAAAAAAAAAAAAAMD9r5AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABjP4QAAAAAAAAAAAAAAAAAAAAAAAAGB+18gAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAxn8IAAAAAAAAAAAAAAAAAAAAAAAADA/a+QAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAYz+EAAAAAAAAAAAAAAAAAAAAAAAABgftfIAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAMZ/CAAAAAAAAAAAAAAAAAAAAAAAAAwP2vkAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAGM/hAAAAAAAAAAAAAAAAAAAAAAAAAYH7XyAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAACUYyVmrnsKX8qPYUv5Uewpfyo9hS/lR7Cl/Kj2FL+VHsKX8qPYUv5Uewpfyo9hS/lR7Cl/Kj2FL+VHsKX8qPYUv5Uewpfyo9hS/lR7Cl/Kj2FL+VHsKX8qPYUv5Uewpfyo9hS/lR7Cl/Kj2FL+VHsKX8qPYUv5Uewpfyo9hS/lR7Cl/Kj2FL+VHsKX8qPYUv5Uewpfyo9hS/lR7Cl/Kj2FL+VHsKX8qPYUv5Uewpfyo9hS/lR7Cl/Kj2FL+VHsKX8qPYUv5Uewpfyo9hS/lR7Cl/Kj2FL+VHsKX8qPYUv5Uewpfyo9hS/lR7Cl/Kj2FL+VHsKX8qPYUv5Uewpfyo9hS/lR7Cl/Kj2FL+VHsKX8qPYUv5Uewpfyo9hS/lR7Cl/Kj2FL+VHsKX8qPYUv5Uewpfyo9hS/lR7Cl/Kj2FL+VHsKX8qPYUv5URpxj8KsAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHK+xyvscr7HK+xyvscr7HK+xyvscr7HK+xyvscr7HK+xyvscr7HK+xyvscr7HK+xyvscr7HK+xyvscr7HK+xyvscr7HK+xyvscr7HK+xyvscr7HK+xyvscr7HK+xyvscr7HK+xyvscr7HK+xyvscr7HK+xyvscr7HK+xyvscr7HK+xyvscr7HK+xyvscr7HK+xyvscr7HK+xyvscr7HK+xyvscr7HK+xyvscr7HK+xyvscr7HK+xyvscr7HK+xyvsAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAEvhYAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAS+FgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABL4WAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAEvhYAAAAAAAAAAAAAAAS+FkJSclqAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAEvhYAAAAAAAAAAAAAAAS+FkPiQAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAS+FgAAAAAAAAAAAAAABL4WQ+JEqkY6NiaaugCVaKdiFSM9gArScY3QAVZuNrAKSbsjnftOUckmkAOyRGSkroAAAAAqycY3QtUAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABL4WAAAAAAAAAAAAAAAEvhZF2aY3dlCVpWAIwjKcrlSCg1KJuAYj4EAFfoBGm1NyP8Al8SptzUgKkmpRSJK6aKUHGNmAAAAAV/gFsgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAJfCwAAAAAAAAAAAAAAAl8LLX0HQjy2W4nZpgFP+JIr/AOTioruRqc07LYq1eV2W5Tq8zs9zEfAipWs7IpzU0VKri7Ip1ebR7lfoTrtOyITU43FO83E/5fEp2molSaginVcnZoqVG5+hCo5RbKcueNyrV5dFuUqrm7MHVsn5EZ3hzM9vK+2hOdocyIu6TIVOaTXYqTcWkupVqcui3KdVydmVJ8qR7W9TlRVq8ui3J1OeFnuLZAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAS+FgAAAAAAAAAAAAAABL4WQ+JBLdi2QEJJVJXZVkptRiV9ErEIKMSiuZuTKys1JGI+BFOCUSmuWo4lle5NWqJoxGyIwSjYpLlm4kP4rP+XxP+JEqWdSKZZFVLniNJRdij8BRjzNyZZBGKlUaZWWiiuo4Jx5SHvUmim/3aKOkl5k9akUWV7lZJSizEfCinBKKfUpJSk5GIirJi2QAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAEvhYAAAAAAAAAAAAAAAS+FkPiQS1ZbQD2fPN6kKUYFfoT+FlD4Cv8BW/hoH/GRUnK/LHccZKUeZlfoC/jMh/FZ/y+J/xIlaDkrrdFKpzaPcq/HEl8LKf8J/Mo/AgKXxyK9+ZW3OasUYtRaZB2pMtyuAtar8irNq0Y7k4zVnJmI+BHQofAV/gFsgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAJfCwAAAAAAAAAAAAAAAl8LB17xt1IR5pJAFP8AiSCv0GrqxQd42MQ/dSK6tBITukxa1vQp6zkVfjiYjZGjRDWq2Q/is/5fE/4kQkkqqaKvxxJfCykr02ig04WApfFIraSiwJac0fMrK0U+xS1lJkdarK+yMR8CN0UNmjEP3UhbIAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAACXwsAAAAAAAAAAAAAAAJ/CynDmdhpp2ZQpte8wCmvfkFdbBO9OV1syF6kuZ7Ir6pIc5U/dKMGld7sm3Tm2upaTkpMrK7ih1JwXKUYcsddyK/est+9Jr95Eq1JQlpsU7znzPoV07JroQm5wdyh8BLmpyutilUlN6hSXvSKsOaNhVpRVim24pslG9VFVXgyivcKl4T5kT5pLmZiFeKFsTvTndEuacXJi2QAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAEvhYAAAAAAAAAAAAAAAA0mAABuBZMGkwLIALXAsg3ACyABpMA3AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAACXwsAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAJfCwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAl8LAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAACXwsAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAKtWUZWRGvF76GjAAACdWMT28m1YAAAAl8LAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAACdKMycOV2uRnKOzI109xNNaAAE60VtqTqykRpylsQoRjqwAAACXwsAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAJOyuU4Jq73JU489u5Og1tqJyiyNZ3VyMoyWjBtJak66WxKcpbsjRlIjRjHzAAAAAl8LAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAACr8DErIq7xYv4j9CpFSkkydG0rIacWKbTuhuUnqRpNysxU024oSl8BTTjJxJJzlboindNxfQAAAAl8LAAAAAAAAAAAAAAAAAAAAAAAAAACcm3yxIRmnqyXNOVo9CCklZgAAAAVJcsRxqJc1yMlKNwIyUloAAAAAAAAAAAAAAAAAATlyq5yVHrcnJqKS3IxmnqwAAAAKy9w3RV1shfxH6FWSUkTqtyuizk+7Fh5Nbji0RqNNPcpzi5MX8Rj/iL0FOSbsiDbndoAAAAl8LAAAAAAAAAAAAAAAAAAAAAAAAAACkrpvuFPdoAAAAAlrNIKel12Iy5o3PaKz8inUWxOoo6dSE+ZDrK+w5JRuSrJOxOqouwmmrjrK+2huAAAAAAAAAAAAAAAS1qJdgnpOIAAAABNXi0U5KUUVKkedeRKo27oUXJ2QqKUkmSSjKLQU7XcSVKMp2RKDV/IjNxehGqpTTZTau0AAAAEvhYAAAAAAAAAAAAAAAAAAAAAAAAABLYpfAgqRafNEhJSVwAAAAP+XnyBfFIpfAinvIpbMgk5NlR2gxQXJYX8JoUV7OxTinATtSEo8lim7wQAAAAAAAAAAAAAAAv4jCpDmVinN35ZbgAAAAVKqhp1HJtkacpbH1fTfUjStYqQcrW3HDmjZkack9WTptu6epCHKiMbNslRUncnRlHbUTa2KVRXswAAACXwsAAAAAAAAAAAAAAAAAAAAAAAAAAauij8NgKatKSAAAAAelRBHXmZS+BFPeRS2ZS2ZW+A3QtYSF8C9Cl8CP8Al0z2EWtyMVFWQAAAAAAAAAAAAAAA9KgFRe9FgAAAAVKLlK6I0Yx8wAAAJU4y3Pq7TVmAAAAS+FgAAAAAAAAAAAAAAAAAAAAAAAAABK9OV+jI1VJpI9ta6aKSdm31AAAAAqxbV1uh1rqyWpCPLGxGfInEpK10QlaTiN8kn2ZdVIuwqto26jjy07C+BehS+BEFenYjVUVZid0mAAAAAAAAAAAAAAAVYtq63R7dW21JVOW2gpe0krbIAAAAAAAAAAAAACXwsAAAAAAAAAAAAAAAAAAAAAAAAAALXEkthxTAAAAADlQWVyxZXGk0WRZXuAWHFMAAAAAAAAAAAAAAADlVxpNFkloAAAAAAAAAAAAABL4WAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA5WaQAAAAAAS+FgAAAAAAAAAAAAAAAAAAGhoaGhoaGhoaGhoaGhoaGhoaGhoaGhoaGhoaGhoaGhoaGhoaGhoaGhoaGhoaGhoaGhoaGhoaGhoaGhoaGhoaAAAAABoaGhoaGhoaGhoaGhoaGhoaGhoaGhoaGhoaGhoaGhoaGhoaGhoaGhoaGhoaGhoT+KJoaGhoaGhoaGhoaGhoaGgAS+FgAAAAAAAAAAAAAAAAAAEaPOr3Pq3mfVvM+reZ9W8z6t5n1bzPq3mfVvM+reZ9W8z6t5n1bzPq3mfVvMnQ5Y3uez8z2fmez8z2fmez8z2fmez8z2fmez8z2fmez8z2fmez8z2fmez8xU7ySufVvM+reZ9W8z6t5n1bzPq3mfVvM+reZ9W8z6t5n1bzPq3mfVvM+reZ9W8yNDmvrsfVvM+r36n1bzPq3mfVvM+reZ9W8z6t5n1bzPq3mfVvM+reZ9W8z6t5ksPZN3IfAgAAAAKsOeaVz6t5n1bzPq3mfVvM+reZ9W8z6t5n1bzPq3mfVvM+reZ9W8z6t5n1bzPq3mcmrVz2fmez8z2fmez8z2fmez8z2fmez8z2fmez8z2fmez8z2fmez8xwstxYe63Pq3mfVvM+reZ9W8z6t5n1bzPq3mfVvM+reZ9W8z6t5n1bzPq3mfVvMlRs0r7n1bzPq3mfVvM+reZ9W8z6t5n1bzPq3mfVvM+reZ9W8z6t5n1bzPq3mfVvMpR5ZNAEvhYAAAAAAAAAAAAAAAAAVKigU6imipUUEQmpq6Cl8AAAAAFb+GwAAAACHxoAAAAAp7y9QhqmAAAAT+BkPgQAAAAD/iL0AAAAAOr9QAAAAJbEfhQAAAAE/jiAAAAAR+OXyAJfCwAAAAAAAAAAAAAAAAAcU3caSqq3US5qjfYS5ajXcKXwAAAAAVv4bAAAAAIfGgAAAACnvL1Cnu/UAAAAn8DIfAgAAAAH/EXoAAAAAdX6gAAAAS2I/CgAAAAJ/HEAAAAAj8cvkAS+FgAAAAAAAAAAAAAAAAAPWr6FP4pEv4qCM5pWSPaVe34EW3FXAAAAK38NgAAAAEPjQAAAABT3l6hSfvMAAAD2lXt+BKpUaaaIfAgAAAAKrkprlPaVe34FOU5fEgAAAA6v1AAAAAlsR+FAAAAAT+OIAAAAVJzTtFHtKvb8Ck25SvuAS+FgAAAAAAAAAAAAAAAAA2krlJNpyfUp/FIlrVQUvgAAAAAmk4tAAAAAFNJyAAAAAFFLYKXxgAAAE/gZD4EAAAAA/4i9AAAAACaSk7AAAAAWT0LaAAAAAT+KIAAAABH45fIAl8LAAAAAAAAAAAAAAAAACtJNqNxVIPRMuo1HfqRtKo2gpfAAAAABL4WA90AAAAU9wAAAACDbbCn8aAAAAJ/AyHwIAAAAB/xF6AAAAAFT4gexugAAAFuAAAAAT+KIAAAABH45fIAl8LAAAAAAAAAAAAAAAAACVKMndioxTuicIzVmRioqyCl8AAAAAEvhYEt0AAAAU9wAAAACG7CL95AAAAE/gZD4EAAAAA/wCIvQAAAAAqfEEtiOsQAAAFuAAAAAT+KIAAAABH45fIAl8LAAAAAAAAAAAAAAAAAAAKXwAAAAAS+FgVNzdAAAAU9wAAAACG7A3QAAAE/gZD4EAAAAA/4i9AAAAACp8QS+EhsAAAAtwAAAACfxRAAAAAI/HL5AEvhYAAAAAAAAAAAAAAAAAABS+AAAAACXwsCe5HWIAAAFPcAAAAAhux7FvcIaxQAAAE/gZD4EAAAAA/4i9AAAAACp8QT+EhuAAAAtwAAAABxTafYAAAAAj8cvkAS+FgAAAAAAAAAAAAAAAAAAFL4AAAAAJfCwJK8iGwAAAFPcAAAAAgtWS2Er02UtYIAAACfwMh8CAAAAAf8RegAAAABU+IJ7C0aAAAAW4AAAAAAAAAAR+OXyAJfCwAAAAAItJpmLoOhXlT7P8A4b8AAAAAAAAAAAAAKXwAAAAAS+FgfbI6OwAAAFPcAAAAAJfCyCvTZR+EAAACfwMh8CAAAAAf8RegAAAABU+IKmxLSwAAAC3AAAAAAAAAACPxy+QBL4WAAAAAAldpHEWHtONZddH+gAAAAAAAAAAAAFL4AAAAAJfCwF8Q9JAAAAU9wAAAAAn8LKKvBlDZoAAACfwMh8CAAAAAf8RegAAAABU+IKmxNe6LVIAAAFuAAAAAT+KIAAAABH45fIAl8LAAAAAAwNP2mJpx80Zlh/b4WUFvuvVf1YAAAAAAAAAAAACl8AAAAAEvhYEfiZPdAAAAU9wAAAABSvfyJ/Cyj8JS0m0AAABP4GQ+BAAAAAP+IvQAAAAAqfEFToSV0Q2AAABbgAAAAE/iiAAAAAR+OXyAJfCwAAAAAMgoc+Jc/wCVfi9P8wzPD+wxUorbdfP+rAAAAAAAAAAAABS+AAAAACXwsCO7JrQjqkAAAFPcAAAAAhuyp8DKPwC0qgAAAT+BkPgQAAAAD/iL0AAAAAKnxBPdG5DsAAAC3AAAAAJ/FEAAAAAj8cvkAS+FgAAAAAZHh1Swqk95a/5f15hxDQUoRqrdaP8AP+vUAAAAAAAAAKlbldke0Sim+oAAUvgAAAAAl8LAh1JK6IbAAABT3AAAAAIbsqfAyj8BLSqgAAAJ/AyHwIAAAAB/xF6AAAAAFT4gl8SBaSAAABbgAAAAE/iiAAAAAR+OXyAJfCwAAAAA3KVNU6cYLokgxtqlPE0+1n+C/wAgAAAAAAAABnK5JyKn8OItUAAUvgAAAAAl8LAgEOwAAAU9wAAAACG7KvwMpfAitpJMAAACfwMh8CAAAAAf8RegAAAABU+IJfEglpJAAAAtwAAAACUW5JgAAAAEfjl8gCXwsAAAAAKKvUivNAe0U8Xio/3fyVgAAAAAAAAB6oi17JoqNckULYAApfAAAAABL4WD2IbAtJAAABT3AAAAAIbsq/Ayl8CK60QtUgAAAn8DIfAgAAAAH/EXoAAAAAVPiB/GFRaC1QAAAtwAAAAAAAAAAj8cvkAS+FgAAAABF8sk+xo1oGGjKnVxTlulJfiAAAAAAAAAFWPLIpx5pJAABS+AAAAACXwsJbENglpJAAABT3AAAAAEktit8BT+BFZXiU9YIAAAJ/AyHwIAAAAB/wARegAAAABU+IPthNe6R1iAAALcAAAAAAAAAAI/HL5AEvhYAAAAAGX1lWwsJeX4rQM1pxw9CrJPWo1+H9P7wAAAAAAAAAvCou57lNAABS+AAAAACXwsJbENgqLQWqAAAKe4AAAAAVvgKfwIqq8GUdYAAABP4GQ+BAAAAAP+IvQAAAAAqfEC+IGrohsAAALcAAAAAk2pJAAAAAEfjl8gCXwsAAAAADIsdGnJ0JvR7ev/AARtJGcY5YmtaD92O36sAAAAAAAAAnUjDRlKpGCdyrUU7WFsAAUvgAAAAAl8LCfwkfhCauiOqAAAKe4AAAABdMrfCQ+FEleLKHwtAAABP4GQ+BAAAAAP+IvQAAAAAqfEEfiYEdJWAAAFuAAAAAT+KIAAAABH45fIAl8LAAAAAAJYqvKHI5trtdgAAAAAAAABXi3awot7IjSk3sAAFL4AAAAAJfCwnsR2QNXRDYAAAp7gAAAAEN2V/hIfCgo7tAAABP4GQ+BAAAAAP+IvQAAAAAqfEEN2A9JAAAC3AAAAAJ/FEAAAAAj8cvkAS+FgAAAAAAAAAAAAAAAEKagtC6AACl8AAAAAEvhYT2FsgI6SsAAAU9wAAAACG7K/wopO8bBDSo0AAAE/gZD4EAAAAA/4i9AAAAACp8QQAnugAABbgAAAAE/iiAAAAAR+OXyAJfCwAAAAAAAAAAAAAAACpJykAABS+AAAAACXwsKmwtgHpIAAAp7gAAAAEN2V9kUnq0G1UAAAJ/AyHwIAAAAB/wARegAAAABU+IexDYCa0I6pAAALcAAAAAn8UQAAAACPxy+QBL4WAAAAAAAAAAAAAAAA9jlXs7lSMYpSQAAUvgAAAAAl8LCp0AJ9AAACnuAAAAAQ3ZX2RTfvBU0qIAAAJ/AyHwIAAAAB/wARegAAAABU+IexDYCSuiGwAAC3AAAAAAAAAACPxy+QBL4WAAAAABFpNMxdB0K0qb6MAAAAAAAAAHqiKvSaJTcopdhbAAFL4AAAAAJfCwn0AJrQjqkAABT3AAAAAIxs2V9kR0aCvumbgAAE/gZD4EAAAAA/4i9AAAAACp8RLYhsAQ3sAAAtwAAAAAAAAAAj8cvkAS+FgAAAAAJXaRxFh7TjWXXR/L+vwAAAAAAAAAAUIpOwqFpb6AABS+AAAAACXwsJ7oAkrohsAABT3AAAAACv0LaC1SK691EdYoAAAn8DIfAgAAAAH/EXoAAAAAVPiJfCQ+EA2kAAAtwAAAABtJpAAAAAEfjl8gCXwsAAAAADBU/aYmnHzRmeH9vhZQW+69V/VgAAAAAAAAAaujD6NoAACl8AAAAAEvhYS+JABDewAAFPcAAAAAK/QSvBlPWKKyvApawQAABP4GQ+BAAAAAP+IvQAAAAAqfET+Ej8IBLRoAABbgAAAAE/iiAAAAAR+OXyAJfCwAAAAAMgo8+J5/5V+L0/zDMsN7DEygtt16P+rAAAAAAAAAA5tyuj2k+4AAUvgAAAAAl8LB/EMA2kAABT3AAAAAINtsr7ogrwZR2aKivFlHWIAABP4GQ+BAAAAAP+IvQAAAAAqfET2F8J0AqLQWqAABbgAAAAE/iiAAAAAR+OXyAJfCwAA5WzlZys5WOLAAyPD+zwvO95a/5f15hxDQUoRqrdaP8ANf15gAAAAAAAAB9XXc+rruAAFL4AAAAAJfCw+2M6AT0aAAAp7gAAAAEN2V/iRS+Ep6SaGrqxQ2aAAAJ/AyHwIAAAAB/xF6AAAAAFT4ipsfZEBNXRDVAAAtwAAAACfxRAAAAAI/HL5AEvhYAAcyscyOZHMhyTQAFGmqdOMF0SQY61Sliafaz/AAX+QAAAAAAAAAAAAUvgAAAAAl8LBfEPViAqLQWqAACnuAAAAAQ3ZW+JFL4TaoFLSTQAABP4GQ+BAAAAAP8AiL0AAAAAKnxFTYewA1dENgAAW4AAAABP4ogAAAAEfjl8gCXwsAAAAAKKvUivNAe0U8Xio9OX8lYAAAAAAAAAAAAKXwAAAAAS+FhH4mdR7gTV0Q1QAAU9wAAAACG7K3xIp/AirpJMI6VWAAAT+BkPgQAAAAD/AIi9AAAAACp8RU6DGtBbAR0kAAC3AAAAAHFNp9gAAAACPxy+QBL4WAAAAAEHyyT7GjQYVShVxUpbpS/MAAAAAAAAAPaT7ntJ9ylUctGFSfKtNylJyjdlSUrqMdynJu8ZboKXwAAAAAS+FhHdiHsLYGrogAAFPcAAAAAgtWVvjIfCistER1iiWlVAAAE/gZD4EAAAAA/4i9AAAAACp8RPdDBAPSQAALcAAAAAAAAAAI/HL5AEvhYAAAAAGX1lWwsJeX4rQM1pxw1GrJPWo1+H9MAAAAAAAAAIU1Nu2xKkoySb0Yo8lRIGkyj8CIazkzar8gp1IxjZs9tDuJpq6AAAAJfCwh1ECAjpIAAKe4AAAAAVfjIfCiorxKWsSrpJMAAD20O5OrBxaTIfAgAAAAJyUaib7HtodyM4y2AAAACp8RL4kdQYFToboAAW4AAAAAAAAAEqkYuzZ7aHcptSlJoAl8LAAAAAAyHHRg3Qm9Ht69vmGc41Ymtywfux/PqwAAAAAAAAB7FCS1RXkm0kVf4kQKPwIpby9SX8WIU6cJRu0exp9hJJWQAAABL4WENmLYGA9JAABT3AAAAACr8ZH4UNXVijs0V1ohaoAAPY0+xOlBRdkQ+BAAAAATipVEn2PY0+xGEY7AAAABU+Il8SOoPYWwTWhHVAAC3AAAAAJ/FEAAAAJU4yep7Gn2KcUpSS8gCXwsAAAAAAljcROHI5tr1AAAAAAAAAAqQ5ZWKceaSJwk6ifQCj8CKfxyQ9aq9ApfAAAAABL4WPYjsLYHsIKnQ3AAKe4AAAAApXv5FX42LYIaTaKyvEhrFAAAT+BkPgQAAAAD/iL0AAAAAKnxD+MQHUJK6IbAAC3AAAAAJ/FEAAAAAj8cvkAS+FgAAAAAAAAAAAAAAAEoxkrMjGMVZAFH4EVG4T5ileTcmFL4AAAAAJfCyWxH4ToB1Ca0I6oAAp7gAAAAEN2VPjYD0qFVXgylrBAAAT+BkPgQAAAAD/AIi9AAAAACp8R9sQDN0EN7AAC3AAAAAJ/FEAAAAAj8cvkAS+FgAAAAAAAAAAAAAAAFSo5MuwAtcCl8AAAAAEvhZLYjsAM3RJXRDYAAp7gAAAAEN2TtzgVdGmS1iyh8IAAE/gZD4EAAAAA/4i9AAAAACp8QviYtgGINpgAC3AAAAAJ/FEAAAAAj8cvkAS+FgAAAAAAAAAAAAAAAPYpU1O9z2EAAApfAAAAABL4WT+EjsgBiCGjsAAU9wAAAACG7KvxMjJSimFZe6R1iih1QAAE/gZD4EAAAAA/wCIvQAAAAAqfELdi2AHuE9GmAALcAAAAAlFuSYAAAABH45fIAl8LAAAAAAjZNXMVQdCtKm+jAAAAAAAAAAhUjBtdD6xAAAKXwAAAAAS+Fk9iOyAB7htIAAp7gAAAAEN2VfiZQlZ8oVFeLKTvEp6VGgAAJ/AyHwIAAAAB/xF6AAAAAFT4iO7OgA9hbFRaC1QAC3AAAAAAAAAACPxy+QBL4WAAAAAAldpI4hw/LUjWXXR+q/4H5AAAAAAAAAAckuxyS7AABS+AAAAACXwsnsR2AB7C2J7oAAp7gAAAACSWxPWZVXLO6ItSSaGroo9UbVQAAJ/AyHwIAAAAB/xF6AAAAAFT4iIACJK6IaoABbgAAAAAAAAABH45fIAl8LAAAAAAwVP2mJpx80Zph/b4WUVutV6oAAAAAAAAALrY5o9zmj3AACl8AAAAAEvhZU2I7AAIqLQWqAAp7gAAAAA9anzKsOaNihL7IR0qNFTSogAAJ/AyHwIAAAAB/xF6AAAAAFTdkAAGPVEOwAC3AAAAAJNqSQAAAABH45fIAl8LAAAAAAyGjz4rn/lX56BmOH9hiZQW269GAAAAAAAAAQv7VgAAFL4AAAAAJfCyp0MsxHt8NGXXb7gAGS1iQ2AAp7gAAAAF0xa1EFRck7oTTV0T0qIr7pgAAT+BkPgQAAAAD/iL0AAAAAKu7IYhLGexfWN/wAWAA9hbC0kAAtwAAAACfxRAAAAAI/HL5AEvhYAAWLFixYADIsP7PDc73l+XQOIqCcY1l00f5r+vMAAAAAAAAA5UncUKb2PZQ7AABS+AAAAACXwsrOyucN19J0vn+j/AEN0AD2N1YgABT3AAAAAIbshrUQVoc0ShK65SstmVtYpkdYoAAJ/AyHwIAAAAB/xF6AAAAAFXdmOxHss1py8kvvv/mJ30AA6ktJAALcAAAAAn8UQAAAACPxy+QBL4WAAF0XRdF0NoACjTVOlGC6JIMdarSxNPtZ/gv8AIAAAAAAAAAauihpJoAACl8AAAAAEvhZjHalJ+T/Iyio6OMhf7Wn37fjYTswAOotJAAU9wAAAACG7J140pRb6tL79AJfu6hV1iS1pFPWCAACfwMh8CAAAAAf8RegAAAABW3Znl3jJeVvyMJV9th4VO6/Hr+IncAGT2QndAAtwAAAACfxRAAAAAI/HL5AEvhYAAAAAUlepFeaA51PF4qPTl/JWAAAAAAAAACUnJ3LvuAAFL4AAAAAJfCzMnbDVH/df5GZYd0aOHqx35V961/UpVFWpRqLqkxO2gAMmrq5F3QAU9wAAAACG7M+m44W6et0Yeqq1GNRdUn94V4XjfsU5c1NrsLWm0UdYAABP4GQ+BAAAAAP+IvQAAAAArbsxOG9vLFvquW3yV/0OHK/PhpU39l/g/wDg3JRsJ3AGWurEXZ2ABbgAAAAE/iiAAAAAR+OXyAJfCwAAAAAg+WSfY3QYVSjWxUpbpS/MAAAAAAAAAPq8D2EAAApfAAAAABL4WZs7YSp6Gc4bny9pbxs/u0f4HD9f2mDUesXb9SSIvoAMVnofCwAp7gAAAAEftHET/wBnivP9GcOV/a4FR6xbX6/qG5/DnYp9UUNmgAAn8DIfAgAAAAH/ABF6AAAAAFfcy6ClKu31k19yRk7eFzGVB9br7tUDVhO4B1JrqRd0ALcAAAAAAAAAAI/HL5AEvhYAAAAAGArKthYT8vxWjDNKUcNRrT61Gv6/MAAAAAAAAAjVvNxPbyPbyAACl8AAAAAEvhZmMeak493FffJFSnGpBwezVjh+bo4qdCXX81/TCSsJ3AGiSuiDurAFPcAAAAA5bJmY4dYmvRovZt/gjhiq6depQlv+q0f5gV4Xjcov3kJuEhNNXQABP4GQ+BAAAAAP+IvQAAAAAr7mXR/dSfeUvza/QzqLw2YQrrrZ/dv+FhNSSaBqwncBq50sJ2YAtwAAAAAAAAAAj8cvkAS+FgAAAAAZDjYxboTe+3r2DOsbHEVlCD92P59QAAAAAAAACNK0+Y+ry7n1eXcAAKXwAAAAAS+FmIjzSivNfhr+gY//AGTNo1ejs/v0f6gNWYndADQ9GLVAU9wAAAACXwshDnx1N/yxk/vsip/seeJ9JP8A9K3/ABAaTVicXCRpVj5opz5XZgAE/gZD4EAAAAA/4i9AAAAACvuYKNqEfPX79TiOhz4ZVFvF/g/+DYyev7bBQfVafd/wLBuNWE7gNEloQd1YBbgAAAADkk0gMZmmFwq/eT17LV/16mM4nrVPdoLlXd6v/JfiYfiPG0tJNSXmv1VjD8U4eelWLi/vX+f4GHx+FxH8Kon5dfu3AAAj8cvkAS+FgAAAAAEsfipw5HUdgAAAAAAAAAVRupykqijJRAAApfAFSryOx9YfY+sPsfWH2PrD7H1h9j6w+x9YfY+sPsfWH2PrD7H1h9j6w+w67atYa5pxfZ/ow4moc1KFVdHb7/8AhvxMur+3wlOfW34rRg1dGzN0GIrwoUnUnsj/AFjwfZ/d/wAEfEGDk0tfu/4JGWmhznORq8p9YfY+sPsfWH2PrD7H1h9j6w+x9YfY+sPsfWH2PrD7H1h9j6w+x9YfY+sPsfWH2HXbVrGGhetKXkl+L/4BxVRalTrLzX6r9TC1lXoQqLqkwKlNTQ1KDJTUtSNZxVj6w+x9YfY+sPsfWH2PrD7Eq7atYh8CAAAAAf8AEXoE5csbn1h9j6w+x9YfY+sPsfWH2PrD7H1h9j6w+x9YfY+sPsfWH2PrD7H1h9ipPmdyjHlpxXZIxlBV8POn3T+/ocM1/wCJRfr+j/QBq41Y5jmKudYSlNwlLVeTHn2Bf2vwZhsXSxEeek7o5znPaH1h9j6w+x9YfY+sPsfWH2PrD7H1h9j6w+x9YfY+sPsfWH2PrD7H1h9j6w+x9YfYda7TtsY3M1haLqONzGZ/jMRpF8sey/z3/IwuBxOKlalFvz6feYPhaCtLESv5L/P/AIYxHDODqfw24v71+Ov4mI4YxdPWm1Jfc/x/zK+Fr0HapBr1Rh82xtDSFR27PX8zD8VTWlanfzWn4P8AzFxJgGr3a+RSz3C1qip03dvyZ9YfY+sPsfWH2PrD7FKXNJsAl8LAAAAAAAAAAAAAAAAX8Yq/xYgAAUvgCtTlJqx7CZ7CZ7CZ7CZ7CZ7CZ7CZ7CZ7CZ7CZ7CZ7CY6M7EY2YZnQ9vhJw62/FanDVfmozpPo7/f/wAMA1dF2jmZxFi3Jxw8Xtq/0/r0FkONaTsvvP8AV/G9l95ltDEU6Cp1t1t6HIzkYqUnsewmewmewmewmewmewmewmewmewmewmewmewmewmewmewmKjJ7FKDitdzP6HtsBPvHX7t/wucN1/aYJQ6xbX6/qATgpqzJ0nCLk3oiGdYWc1CF23otD2Ez2Ez2Ez2Ez2Ex0ZJXIfAgAAAAH/ABF6BVTcHZXZy1f5H+H+Zy1f5H+H+Zy1f5H+H+Zy1f5H+H+Zy1f5H+H+Zy1f5H+H+Zy1f5H+H+Zy1f5H+H+Zy1f5H+H+Zy1f5H+H+Zy1f5H+H+Zy1f5H+H+Zy1f5H+H+Y+e9nB/h/mBT/wBjzi3Rv/0r/ggDVzlRmGIWGw0qnXp6mHy/FYqLnTjf5rf5s/sPH/8APv8AFf5mVYLH4Wr71P3Xvqvv3Lz/AJH+H+Zef8j/AA/zL1P5H+H+Zy1f5H+H+Zy1f5H+H+Zy1f5H+H+Zy1f5H+H+Zy1f5H+H+Zy1f5H+H+Zy1f5H+H+Zy1f5H+H+Zy1f5H+H+Zy1f5H+H+Zy1f5H+H+Zy1f5H+H+Zy1f5H+H+Zy1f5H+H+Zy1f5H+H+Y1VTS5H+H+ZiMP7ajKnNWv6ffo2YXIcNSs6nvPz2+7/Mp1FCKiloiM4y2CVWKKk3NWexWyfCVvsWflp/wPwK3C83rRl8n/n/wCeRY+LtyX+a/zMmyyph5SqVlaWy/pE55gptRw91095IVTHPfD/8Ak0f8xSxT3oP74/8AyQvbv/l0/vj/APJGHTV7qzAJfCwAAAAAAAAAAAAAAAJS5arZOopSUux9YXYAAKXwAAAAAS+FgGX/AOyZrKl0d1+q/QAcUytOFKm5y2SuZXRljse6s9lq/wBF/XRAAU9wAAAACG7CrTjUpyg9mmvvOGqjo4qph5b/AKp/8EAOIs2528NSei3fn2+XXzOH8p9jBYiqveey7Lv6v8gAJ/AyHwIAAAAB/wARegAAAABU+IDiSi41KdaPp92qMPVVajGouqTADiHFOtXjh4a2/N/1+JgcKsNh40lut/XqAC3AAAAAJ/FErR5o+hFXQop9SUJR3HKTVrkaUpEaMV5miROpfRCjdX6FKPNK4AR+OXyAJfCwAAAAAAAAAAAAAAAKlOTk7I9lPseyn2AACl8AAAAAEvhYBn8HQxkK8ev5r+kQnGcFKOz1AOJMXywjh4vV6v06fj+RkuD+r4VX+KWr/T8AAKe4AAAABDdgYn/Y87U+jaf/AIFo/wAbgGfZqsLT9lTfvy/Bd/8AL7zIMp+s1Pb1V7i/F/5L+uoABP4GQ+BAAAAAP+IvQAAAAAqfEBnlD2uClbeOv3b/AIXOHq/tMJydYu36gGLxEcPQlVl0RkeHlicW689eXX5v+r/cAAtwAAAACfxRBfu6hUp21RCatZnLCOo5xXUdWPQlNyIUurK8tFFFKHLEAI/HL5AEvhYAAAAAAAAAAAAAAAAAABS+AAAAACXwsA4hoe0wnMt4u/6GR1/a4KK6x0/y/CwE5RhFyk9EYeLzLMeaW279FsvyQABT3AAAAAIbsDiuhpTrLzX6r9TBV/b4aFTul9/UMyx9PBUHUlv0Xd/1uYLC1s0xbcn5yfb+tkUqUKVNQgrJbAAE/gZD4EAAAAA/4i9AAAAACp8QE4RnBxez0Mhm6GMnQl1uvmv6YBxJi7yjh4vbV/p/XoZXg/quFjB7vV+v9aAALcAAAAAcU2mGIjdXKUuaBUp21Ra4oSfQVKTI01EbSVyCdSd2ABH45fIAl8LAAAAAAAAAAAAAAAAAAAKXwAAAAAS+FgGIoqtSlTfVNHDdVwq1KEvX7tGBxDi/ZUFRi9Zfl/wTIMH7HDe0e8vy6f5gAFPcAAAAAgtWBntD22An3Wv3b/hc4Zr+0wfI94v8Hr/mVqsKNN1JuyRi8TXzTFpRW+iXZf1qzL8BTwdBU479X3YAAT+BkPgQAAAAD/iL0AAAAAKnxAGY/wCyZrGqtnZ/owK1WNKnKctkrmV0pY7HurPZav8ARf10QAAtwAAAAAGk1YpPlnysLAGIlpylKHLEACPxy+QBL4WAAAAAAAAAAAAAAAAAAAUvgAAAAAl8LACr/secKXRv/wBK3/EHZIm3meY2Xw3/APJV/n+bFFRSS6AAFPcAAAAAAnCM4OL2ehw5N0MbUw8vP74v/hzPs1+tVPY0n7i/F/5dvvMiypYSl7Sovfl+C7f5gAAT+BkPgQAAAAD/AIi9AAAAACp8QBxLQ5qMKq6O33/8MZZX9vhKc+trfNaBxJi+WEcPF6vV+nT8fyMlwf1fCq/xS1f6fgAALcAAAAAArxs1JEJKUUwBtJEV7Sd2AAR+OXyAJfCwAAAAAAAAAAAAAAAAAACl8AAAAAEvhYAcTUP4dZen6r9TB1vb4eFTuvx6me4v2GFcE/elp8uv+XzOHcHyUXXe8tF6L/N/kAAFPcAAAAAAM+pyw+YSlB25lf71Z/qcO5TzNYqqtF8K/X/IAAAn8DIfAgAAAAH/ABF6AAAAAFT4gDMaHt8JOHW34rU4ar3pzpPo7/f/AMMTlGEXKT0RhoPMsx5pbbv0Wy/JAAAtwAAAACfxRAqR5o2KEvsgFaqmuVFGHLEAAj8cvkAS+FgAAAAAAAAAAAAAAAAAAFL4AAAAAJfCwAzih7bBTS3Wv3f8A4cr8+GdPrF/g/8Ag3MfUlmGYKnB6Xsv1ZTpxpwUI7LQAAKe4AAAABGV7gGZZTHG1qUpPSN7+a7EYxjFRitEAAAT+BkPgQAAAAD/AIi9AAAAACp8QAYH/ZM2dLo219+q/Q4hxfsqCoxesvy/4JkGD9jhvaPeX5dP8wAAW4AAAABP4ogFRck7oTTV0Tqxj6k6kpblOi5avYAAI/HL5AEvhYAAS2Zzy7nPLuc8u5zy7mRyk8ZFN9H+QAAAAAAAAAAAAAAFL4AAAAAJfCwAaUlZlPETy+rWprdpr8d/uOG8HpLES9F+v+X3gAAU9wAAAACG7AAAAAn8DIfAgAAAAH/EXoAAAAAVPiADiGm6OKp149fzX9IlKWaZguz/AAS/r72KKikl0AABbgAAAAE/iiAVo80fQ55cvLchSlIhSjH1AAAj8cvkAS+FgAAewpfyr7j2FL+Vfcewpfyr7j2FL+VfcRpU4u8YpP0AAAAAAAAAAAAAAApfAAAAABL4WABnmDlLHRUFrO337f5GHoRoUo047JAAAU9wAAAACG7AAAAAn8DIfAgAAAAH/EXoAAAAAVPiADiCh7TBuS3i7/ocO4Pkouu95aL0X+b/ACAAAW4AAAABP4ogBGjFO4AAAR+OXyAJfCwAAAAAAAAAAAAAAAAAACl8AAAAAEvhYAE6FOdSNRrWN7fMAAAp7gAAAAEN2AAAABP4GQ+BAAAAAP8AiL0AAAAAKnxABWpRq05U5bNNFOnGnBQjstAAAFuAAAAASi3JMAAAAAj8cvkAS+FgAAAAAAAAAAAAAAAAAFSbjZLcpz5lruFL4AAAAAJfCwAAAACnuAAAAAQ3YAAAAE/gZD4EAAAAA/4i9AAAAACp8QAAAAC3AAAAAAAAAACPxy+QBL4WAAAAAAAAAAAAAAAAAAR1qN9jar6oKXwAAAAAS+FgAAAAFPcAAAAAskAAAABP4GQ+BAAAAAP+IvQAAAAAqfEAAAAAtwAAAAAAAAAAj8cvkAS+FgAAAAAAAAAAAAAAAAAFL4pEv4sQjSlJXTPYT7kVaKQAAABVdoOwAAAABB++kAAAAAQbbYAAAAewn3JUZJXuQ+BAAAAAVYuU0kewn3KdNx3YAAAAN3bAAAAAewtUgAAAAJtqUUAAAAFSnKTumewn3KMXGTTAJfCwAAAAAAAAAAAAAAAAACn8UkPWqgpfAAAAABW/hsAAAAAh8aAAAAAKe8vUAAAACfwMh8CAAAAAf8RegAAAAB1fqAAAABLYj8KAAAAAn8cQAAAACPxy+QBL4WAAAAAAAAAAAAAAAAAASahUu9mU/ek5BS+AAAAACt/DYAAAABD40AAAAAU95eoAAAAE/gZD4EAAAAA/4i9AAAAADq/UAAAACWxH4UAAAABP44gAAAAEfjl8gCXwsAAAAAAAAAAAAAAAAAAlFSVmJJKyCk0o7nNHuc0e5zR7nNHuc0e5zR7nNHuc0e5zR7nNHuc0e5zR7nNHuc0e5WknB2ZdF0XRdF0XRdF0XRdF0XRdF0XRBrnRzR7nNHuc0e5zR7nNHuc0e5zR7nNHuc0e5zR7nNHuc0e5zR7nNHuc0e5CSu9epzR7nNHuc0e5zR7nNHuc0e5zR7nNHuc0e5zR7nNHuc0e5zR7nNHuTlFxepD4EAAAABJpVF6HNHuc0e5zR7nNHuc0e5zR7nNHuc0e5zR7nNHuc0e5zR7nNHuc0e5zR7l1d+pdF0XRdF0XRdF0XRdF0XRdF0SasRlGy1OaPc5o9zmj3OaPc5o9zmj3OaPc5o9zmj3OaPc5o9zmj3OaPc5o9ybTlHU5o9zmj3OaPc5o9zmj3OaPc5o9zmj3OaPc5o9zmj3OaPc5o9zmj3OaPchZzlbyAJfCwAAAAAAAAAAAAAAAAAAB4dN7n1ddz6uu59XXc+rrufV13Pq67n1ddz6uu59XXc+rrufV13Pq67n1ddz6uu59XXc+rrufV13Pq67n1ddz6uu59XXc+rrufV13Pq67n1ddz6uu59XXc+rrufV13Pq67n1ddz6uu59XXc+rrufV13Pq67n1ddz6uu59XXc+rrufV13Pq67n1ddz6uu59XXc+rrufV13Pq67n1ddz6uu59XXc+rrufV13Pq67n1ddz6uu59XXc+rrufV13Pq67n1ddz6uu4kkkgAAAAKlJTe59XXc+rrufV13Pq67n1ddz6uu59XXc+rrufV13Pq67n1ddz6uu59XXc+rrufV13Pq67n1ddz6uu59XXc+rrufV13Pq67n1ddz6uu59XXc+rrufV13Pq67n1ddz6uu59XXc+rrufV13Pq67n1ddz6uu59XXc+rrufV13Pq67n1ddz6uu59XXc+rrufV13Pq67n1ddz6uu59XXc+rrufV13Pq67n1ddz6uu59XXc+rrufV13Pq67n1ddz6uu59XXc+rruU6agAS+FgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABL4WAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAEvhYAAAAAAAAAAAAAAAAAAAAAAABKSitWJprQAAAABtLQAAAAB2sKcZbMAAAAAAAAAAAAAAAFKMtmAAAAA7JGjWgAAAADnFO1wAAAAAAl8LAAAAAAAAAAAAAAAAAAAAAAAAHFVJu+yKXutxAAAAAcZc6bAAAAAqQ51a5KCptNAAAAAAAAAAAAAAAFZ+6kupKCptNAAAAAVoykvIp/AgAAAAPYLl31KTvBAAAAAAEvhYAAAAAAAAAAAAAAAAAAAAAAABS15n5n/L35AAAAAVPiiAAAAAVtYpAAAAAAAAAAAAAAAE9ZxRXXuC1QAAAAT+BlP4EAAAAA9iivcAAAAAAJfCwAAAAAAAAAAAAAAAAAAAAAAACjs/Uf8AF+QAAAAFT4ogAAAAD96ol2AAAAAAAAAAAAAAAJ/xIlb4GLYAAAAJ/Ayn8CAAAAAqu0bLdkY2ikAAAAAAS+FgAAAAAAAAAAAAAAAAAAAAAAAFPSUkR1qNgAAAAVPiiAAAABOSjG5TjaOu7AAAAAAAAAAAAAAAKujiytqku4AAAAE/gZT+BAAAAAR9+d+iAAAAAACXwsAAAAAAAAAAAAAAAAAAAAAAAAqU23eL1KcOVWAAAAAlDmafYAAAACrTc0rM9hPudAAAAAAAAAAAAAAAJxUo2IUpJ3k9gAAAAJK8WiMbRSAAAACSurHsJdynBxVmwAAAAACXwsAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAJfCwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAl8LAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAACXwsAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAJfCwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAl8LAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAACXwsAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAJfCwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAl8LAAAAAAAAAAAAAAAAAAAAAAAAAAAK1SUWkj28yFSUk79ChJyTTAAGUZOV2wq35HYpy5opgAAAAAAAAAAAAAAAAAAAAFRuMW0e3mKtNtInNqrYAAJybqKKApyfO4tgAAAAAAAABL4WAAAAAAAAAAAAAAAAAAAAAAAAAAATpRm9RQTnykacYXKcYJe6zmi9mKUZbMU4t2T1Dnje1wqO0GUV7iJTitGx2lEw7vFoAAAAAAAAAAAAAAAAAAAAAlFSVirTULWIUYtJjjBzvfUcktLjnG9r6kpRjuzclKMd2JprQIa1WNpLUUoy2ZLSsgAAAAAAAAAl8LAAAAAAAAAAAAAAAAAAAAAAAAAAABfxifwMw/UV3dITcWWcWirO0Lo9nLl5ilLmgiv8AAQ0poUJVG2inG0Eihu0AAAAAAAAAAAAAAAAAAAAAGJ2RT+BE/wCN9xX0mmSUt2S5pe8UHeHoNSqydig2pOLCjrJlZuU1FFKm4y1Kv8SIAAAAAAAAAS+FgAAAAAAAAAAAAAAAAAAAAAAAAAAAv4xP4GYfqUVeZNL2piVsVn7kR1Xy8tjD/Cyv8BFXppeRCbgxNSV0UfjYAAAAAAAAAAAAAAAAAAAAAYnZFP4ET/jfcYj4kYhLkRFL2JS+CRCo4bEJc1RMMP1Kyancp1FNeZW+KIAAAAAAAAAS+FgAAAAAAAAAAAAAAAAAAAAAAAAAAAv4xP4GYfqUPjZP+KvkYnoVl7kSFKnKKZGEYqyK69wp/AitTTXMihrAoayYAAAAAAAAAAAAAAAAAAAAAYnZFP4ET/jfcYj4kYj4ER/g/JlFXhJFGMZNpipQTugofE0ThGasykrVLFXWpFAAAAAAAAABL4WAAAAAAAAAAAAAAAAAAAAAAAAAAACT9qT+FmHW5RT52TT9qjEK9iUOamke8tCkmoK5VV4Mp60tC8muUhHkgYde62AAAAAAAAAAAAAAAAAAAAAGIV0in8CJp+1+4xCbkjEK8URX7ow60ZUg4S0KF3K4Q0qtFVyU20UIO/MyWtZAAAAAAAAABL4WAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADV1YoppNMKt+R2KUbQSAAAAAAAAAAAAAAAAAAAAAAAAAnFqomgKcW5tsAAAAAAAAAJfCwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAl8LAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAACXwsAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAJfCwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAl8LAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAACXwsAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAJfCwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAl8LAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAACXwsAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAJfCwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAl8LAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAACXwsAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAJfCwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAl8LAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAACXwsAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAJfCwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAl8LAAAAAAAAAAAAAAAAAAAAAAAAG0t2ADaW7G1a5GcZbMAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAACXwsAAAAAAAAAAAAAAACpVUNOp7afci+aKYFSajG5+9nqik5OOoACkm7IADmTduoAFbeIAOLqzeuxVTtGCIU1DYAOZXt1AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAJfCwAAAAAAAAAAAAAAAZKXM7ipTavYirRSAmueoo9gG0tzcLq9rhT+OQXV7XAI/xWANpITTWhW3iBdXtcKGzJ/wAVBdXtcG0kJprQX8ZgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAEvhYAAAAAAAAAAAAAAAVXaDKceaSQAUtZyYPYUHVbdyneEuRlZ8skyVKVuZivUpkaTk2r7E24U0uo6LUb31H+8pFKScPQpxcot9xUm5OJUjy0kiPwop/xJE6TnLfQgnCfKVt4hUlyxuOg+W99TWpTsQpOa0Y6TUlE1p0z2Dcb31KUuaKGnVlpsiFJxvqeyblykI8sUgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAJfCwAAAAAAAAAAAAAAArfw2UfjAChvIKrtBlJWgirpOLKyvOKJK6KHwFP45FVrmjcdWDW5Q+EleLlFdSMeWKRH+Kyv8BH4UU/4kiUlFakpqU42K28Qr2vFM9rDuULWaKHwsn/ABUV2tEe1h3KbtGVigrQBfxmAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAS+FgAAAAAAAAAAAAAABVV4Mg+WSZo0AU9KkkFb4GU9YIrbxRV+OI9jD/AU/jkVYqU0mfV4dyEFBWRVd53XQVmiP8AFZX+Aj8KKf8AEkcvtKjvsicIxnGxW3iFZJyij6vDuQpqC0KHwsn/ABUV0m4o+rw7jpqMHYo/AgX8ZgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAEvhYAAAAAAAAAAAAAAANXRKPK7HM0rXIaxQFVOM1JCqRa3G1OLSKMk42G1OqrdCr8cR7GH+Ag0qkkyv0kugpRavccly3IQvTfmUZXhbsJpVWVleDsQqR5FdlF3m2UmlOSJyUqisVt4hXXuproRmpK4mmtCh8LJ/xUV17t10IzUlc0ktCjJK8WKSbsmXSramjQAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAEvhYAAAAAAAAAAAAAAAFSkp+p9Xl3IxskgJRUlZjw76Mp01BWJ0OZ3TKdNQRKHNJO+w9UU4citcqUeZ3TOVOPKx4d30Y6V4cqYkkkiNPlk2mVaXO7ojG0bH1fXfQULSbKlHmd0z2KVrPYnDma12BpNWHh9dGQgoxsU4citcdO81K40mrDw+ujIQUY2KlFSd0U6XJ1KtLn1RCPLFIAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAACXwsAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAJfCwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAl8LAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAACXwsAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAJfCwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAl8LAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAACXwsAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAJfCwAAAAAAAAAAAAAAAAAAAAAAAlKMd2HNG9r6jlFO3UCcuWNyEuaNwAAAAqT5FcTukwAAAAAAAAAAAAAAAAAAJSjHdgAAAAABKooyUQAAAAAAAAAAJfCwAAAAAAAAAAAAAAAAAAAAAAAxOyFsL+MSpt1EwKvwMo/AgAAAAxHwIh8CAABTbqcoASdotlKTlG7AAAAAAAAAAAAAAMT0AAAAAACr/FiAAD2KU3NO4AVZuMdCOqQAAAAAEvhYAAAAAAAAAAAAAAAAAAAAAAAYnZC2F/GJVGpqJUnyIjXdnclLmpXFV5YJLcpz54h7d8rfUdS0FJka7b1KlRxaRVnyRujm9zmFVfJzMVeV9SUrRuh1nZW3KtTkVluTqc8LPch8CJNRV2OvK5UmoRI13fUqS5Y3ROu7KxGpzQuKo1LmOdKHMxV3fUqzcUrE5pQ5idRun6lKo01EAAAAAAAAAAAAAAxPQJyUY3FXlfUqT5Ypocvc5inPnjcp1HNsqS5Y3Q6z0SKtTkVluUqvM7MVRufKTqcslEqz5FoVKjjFdynWbdmVZuCVhVLz5UVKzTshz55xYVJqCKdWUpWZVq8ui3KdbmdmVJuLSJV3zabFSdoXRTqOOiKtTkXmU6zk7MdRqfKyrUcbJFeTb5SjUctOwAAAAAEvhYAAAAAAAAAAAAAAAAAAAAAAAYnZHvFH40T/ioqrmqKJXhFRuj/lyUacXG7KGkmgpwU5WZVS5oxK8VyehV1hFlfVJF/wByU4p00mVnDlt1FrRKNNSV2Nc9WxXgkk0Q+BGIeiRGlFRtYq+9UUStBct10G70SlTi4alDW6Iwi6jRW6RRWiuTToW56RzOUVEqRXJ6FCMXG/UAAAAAAAAAAAAAAxPQK+rUSrTjyadD4qPoN/uSi7Rkuxh17pW+BlGmpK7FaVV36FWymmh6VkVtZvyK+rihxi9ys4tpxMRrFFOCjHm6lGKk22Sio1VYJrnq8ooRWyKaU6jbK0VGSaMR0KlKKp+aIJSpalCMWrsaUqvoV7JpoxEdEyP7yon2MQlZMhGKSaAAAAAAl8LAAAAAAAAAAAAAAAAAAAAAAADE7IWwv4xP+Kh/xv67Ff4D/lyUfgRS/iMMPuysm5pHsJlSLVJLsL3pRXkX/d28yq+SFkOlaHM2L+CUPgKf8VmI+BEPgRX+JA/4xW+Bn/Lko/w0UPiZD+KyurySPYTKa5YJMptc6KnwMofAAAAAAAAAAAAAAAYnoFX+Kip8DKSvTZvBLzHo5opK0EVvgZR+BDg5TaQqE7lVWnFjV1NnxTiV221FFSlyJalf4UP4PkYf4WVf4sQj/GYYfdmI2RX2iVfgZS/hmH+FkoOVRpHsJlWypsw7V2jEfCiPwoAAAAACXwsAAAAAAAAAAAAAAAAAAAAAAAMTshbC/jE/4qK14zUkVaqmrI/5clH4EUv4jDD7srqzUke3hYqK8GUNZDXv8vmYhe6ipUi6dkL+CUPgJPkq3K1RTskQ+BGIjdJka8eXXcre7NSK1SMo2RKNqNij/DRQ+JjkoVW2V9UpIVeDRVl7nqThaEWN81O/kUJpLlAAAAAAAAAAAAAAMT0PrEexW1SkipVi4WW5SjaCKavNLsV9JepFWikVvgZR+BEnyVLiqwbSTMQvdTKavTfmUNZehVdqiZXmpNJFf4UJXiilNQbTHLmqJhVvGpcjVjJpIi1Co7lRqpNJGJ6FX4GUVemUZqOjJvkqXPawbSTK75mookuSojEL3UUpqSsAAAAABL4WAAAAAAAAAAAAAAAAAAAAAAAFSmp21D2a5+a5KmnJSuSipKzKkIwhoRjzU0iEFGNiNNRk3cIU1DqSipKzPq6vuWVrFOmoHslz8xKKkrEaCSeoqaUOW5CCirFSmpo9guW1xKySGk1Zn1dX3JwUo2ZGgk7tk480bEI8sbEKai27lSipu9zkTjys+rq+5OkpJK+xKClGxCHLGwqCUrpgAAAAAAAAAAAAAFSmp21Pq67nInHlFh0nqwVJKXMTpKbTCceaNiEVGNipTU0QoKLvcnBSjYjFRjYhSUCpTU0ewjy2uTpqaSuJWVipRUne57FaW6BOCmrMhQUXe5UpKfqU6Sh6lSmp21JR5o2IQUY2HQXNdMqU1NWI4dJ3ueyXPzXKlNTsSipRsynS5OoAAAAAEvhYAAAAAAAAAAAAAAAAAAAAAAAAFabi1YlKVRpCVlYAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAACXwsAAAAAAAAAAAAAAAAAAAAAAAABpNaijFbIAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAl8LAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAACXwsAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAJfCwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAl8LAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAACXwsAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAJfCwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAl8LAAAAAAAAAAAAAAAAAAAAAAAAAACcm3yxIRmnqyXNOVo9CCklZgAAAAVJcsRxqJc1yMlKNwIyUloAAAAAAAAAAAAAAAAAATlyq5yVHrcnJqKS3IxmnqwAAAAB89R6PQpyd+V7gOSukAAAAAAAS+FgAAAAAAAAAAAAAAAAAAAAAAAAABSV033Cnu0AAAAAS1mkFPS67EZc0bntFZ+RTqLYnUUdOpCfMh1lfYcko3JVknYnVUXYTTVx1lfbQ3AAAAAAAAAAAAAAAJa1EuwT0nEAAAAAqO0GQVopE9JxZzXlyjlZpEqiU79jmXLcjV5naxOoo6EJqSJVEkz2qcbkJqaJ1FHTqQkpIAAAAl8LAAAAAAAAAAAAAAAAAAAAAAAAAAJbFL4EFSLT5okJKSuAAAAB/y8+QL4pFL4EU95FLZkEnJsqO0GKC5LC/hNCivZ2KcU4CdqQlHksU3eCAAAAAAAAAAAAAAAF/EYVIcysU5u/LLcAAAACt8AVegv4j9CfxxH/ERUV5KJZEFeTZa1T1IJObYor2jLWqEbOTZG3O7AAAAEvhYAAAAAAAAAAAAAAAAAAAAAAAAAA1dFH4bAU1aUkAAAAA9KiCOvMyl8CKe8ilsylsyt8BuhawkL4F6FL4Ef8umewi1uRioqyAAAAAAAAAAAAAAAHpUAqL3osAAAACqrwZF3iiprKKF/EfoT+OI/wCIiX8RBS2ZL+IiHxSF/EY/4i9CNNSbuRpqLugAAACXwsAAAAAAAAAAAAAAAAAAAAAAAAAAJXpyv0ZGqpNJHtrXTRSTs2+oAAAAFWLaut0OtdWS1IR5Y2Iz5E4lJWuiErScRvkk+zLqpF2FVtG3UceWnYXwL0KXwIgr07EaqirMTukwAAAAAAAAAAAAAACrFtXW6Pbq22pKpy20FL2klbZAAAAANJqxGfJ7rIXlLmZUfLNSLuU0yo+WSZU1ipIjVUmkN8kn2ZD3pORD4pC/iMf8Reg3ySfZkJqewAAABL4WAAAAAAAAAAAAAAAAAAAAAAAAAAFriSWw4pgAAAAByoLK5YsrjSaLIsr3ALDimAAAAAAAAAAAAAAAByq40miyS0AAAAAGk9waTRZDSaCyWw0mgsWRZDSaLJLQAAAAl8LAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAACXwsAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAJfCwAAAAAAAAAAAAAAAAAABKrLVM5Kvc5Kvc5Kvc5Kvc5Kvc5Kvc5Kvc5Kvc5Kvc5Kvc5Kvc5Kvc5Kvc5KvckqsVdsvU7l6ncvU7l6ncvU7l6ncvU7l6ncvU7l6ncvU7l6ncvU7l6ncvU7i9o3a5yVe5yVe5yVe5yVe5yVe5yVe5yVe5yVe5yVe5yVe5yVe5yVe5yVe5yVe5yVe4lVezOSr3OSr3OSr3OSr3OSr3OSr3OSr3OSr3OSr3OSr3OSr3OSr3OSr3OSr3HGqle5HWKAAAAAq87klFnJV7nJV7nJV7nJV7nJV7nJV7nJV7nJV7nJV7nJV7nJV7nJV7nJV7nJV7nJV7l6ncvU7l6ncvU7l6ncvU7l6ncvU7l6ncvU7l6ncvU7l6ncvU7l6ncbqJbnLVa3OSr3OSr3OSr3OSr3OSr3OSr3OSr3OSr3OSr3OSr3OSr3OSr3OSr3OSr3GqqaVzkq9zkq9zkq9zkq9zkq9zkq9zkq9zkq9zkq9zkq9zkq9zkq9zkq9zkq9zkq9ylzXakwCXwsAAAAAAAAAAAAAAAAAAApfAAAAABW/hsAAAAAh8aAAAAAKe8vUAAAACfwMh8CAAAAAf8RegAAAAB1fqAAAABLYj8KAAAAAn8cQAAAACPxy+QBL4WAAAAAAAAAAAAAAAAAFSaiinJyjdlSbTUY7lObkrPdBS+AAAAACt/DYAAAABD40AAAAAU95eoAAAAE/gZD4EAAAAA/4i9AAAAADq/UAAAACWxH4UAAAABP44gAAAAEfjl8gCXwsAAAAAAAAAAAAAAAAAHFNoo/AiOtRs2q+qCnOKjZs9rDuJprQAAAArfw2AAAAAQ+NAAAAAFPeXqAAAAHtYdydSDi1ch8CAAAAAk0qiv2Paw7ilGWzAAAADq/UAAAACWxH4UAAAABP44gAAABKcY6NntYdyDTnJryAJfCwAAAAAAAAAAAAAAAAAKPwIpby9SX8WIQpRlG7PYQEklZAAAAFSPNFoAAAAApxTlcAAAAAjFK4AAAAewgSoQUWyHwIAAAACcFKaT7HsIEKcY7AAAABKNpMAAAAC19BKysAAAABKKbT7AAAABOlGbuz2ECnFRlJIAl8LAAAAAAAAAAAAAAAAAAo/Ain8UkPWqgpfAAAAABL4WAAAAAU9wAAAACMrtgAAAAT+BkPgQAAAAD/AIi9AAAAACp8QAAAAC3AAAAAJSs0gAAAACPxy+QBL4WAAAAAAAAAAAAAAAAABR+BE3yTv0ZT96TkFL4AAAAAJfCwAAAACnuAAAAAQ3YAAAAE/gZD4EAAAAA/4i9AAAAACp8QAAAAC3AAAAAJ/FEAAAAAj8cvkAS+FgAAAAAAAAAAAAAAAAAWSWg0mrMsktApfAAAAABL4WAAAAAU9wAAAACG7AAAAAn8DIfAgAAAAH/EXoAAAAAVPiAAAAAW4AAAABP4ogAAAAEfjl8gCXwsAAAAAAAAAAAAAAAAAAApfAAAAABL4WAAAAAU9wAAAACG7AAAAAn8DIfAgAAAAH/EXoAAAAAVPiAAAAAW4AAAABP4ogAAAAEfjl8gCXwsAAAAAAAAAAAAAAAAAAApfAAAAABL4WAAAAAU9wAAAACG7AAAAAn8DIfAgAAAAH/EXoAAAAAVPiAAAAAW4AAAABP4ogAAAAEfjl8gCXwsAAAAAAAAAAAAAAAAAAApfAAAAABL4WAAAAAU9wAAAACyAAAAAn8DIfAgAAAAH/EXoAAAAAVPiAAAAAW4AAAABZAAAAAEfjl8gCXwsAAAAAAAAAAAAAAAAAAApfAAAAABL4WAAAAAU9wAAAAAAAAACfwMh8CAAAAAf8RegAAAABU+IAAAABbgAAAAAAAAABH45fIAl8LAAAAAAAAAAAAAAAAAAAKXwAAAAAS+FgAAAAFPcAAAAATT2AAAAAn8DIfAgAAAAH/EXoAAAAAVPiAAAAAW4AAAAA2k0gAAAACPxy+QBL4WAAAAAAAAAAAAAAAAAAAUvgAAAAAl8LAAAAAKe4AAAABDdgAAAAT+BkPgQAAAAD/iL0AAAAAKnxAAAAALcAAAAAn8UQAAAACPxy+QBL4WAAAAAAAAAAAAAAAATqRjuKSaTAACl8AAAAAEvhYAAAABT3AAAAAIbsAAAACfwMh8CAAAAAf8RegAAAABU+IAAAABbgAAAAE/iiAAAAAR+OXyAJfCwAAAAAAAAAAAAAAACV5Nsqfw4m6AAKXwAAAAAS+FgAAAAFPcAAAAAhuwAAAAJ/AyHwIAAAAB/wARegAAAABU+IAAAABbgAAAAE/iiAAAAAR+OXyAJfCwAAAAAAAAAAAAAAAHsRt7JlT+HFC2QABS+AAAAACXwsAAAAAp7gAAAAEN2AAAABP4GQ+BAAAAAP8AiL0AAAAAKnxAAAAALcAAAAAn8UQAAAACPxy+QBL4WAAAAAAAAAAAAAAAASTi3EinNpAABS+AAAAACXwsAAAAAp7gAAAACja4AAAAE/gZD4EAAAAA/wCIvQAAAAAqfEAAAAAtwAAAABxTafYAAAAAj8cvkAS+FgAAAAAAAAAAAAAAAE6cZkKUYAABS+AAAAACXwsAAAAAp7gAAAAAAAAAE/gZD4EAAAAA/wCIvQAAAAAqfEAAAAAtwAAAAAAAAAAj8cvkAS+FgABi88rUa8qairJ26/5n+sWI/lX4/wCZ/rFiP5V+P+Z/rFiP5V+P+Zh8+r1a0IOK1aXXq/UAAAAAAAAAAAByit2UJRSd2V5RaSTFsAAUvgAAAAAl8LAAAAAKe4AAAAAAAAABP4GQ+BAAAAAP+IvQAAAAAqfEAAAAAtwAAAAAAAAAAj8cvkAS+FgABiMip1qsqjm1c/1cpfzv7j/Vyl/O/uP9XKX87+4o5BSpVIzU3o0/uAAAAAAAAAAAAxC2CMXJ2QAAUvgAAAAAl8LJu0WyjNVKcZLqgAAAAp7gAAAAEN2AAAABP4GQ+BAAAAAP+IvQAAAAAqfEAAAAAtwAAAACXxRAAAAAI/HL5AEvhYAAAAAAAAAAAAAAADSZTpcq1EktgAApfAAAAABL4WYl2oTfZP8AIyep7TBU32Vvu0AAAACnuAAAAAQ3YAAAAE/gZD4EAAAAA/4i9AAAAACp8QAAAAC3AAAAAJ/FEAAAAAj8cvkAS+FgAAAAAAAAAAAAAAAFStK9ke1n3AACl8AAAAAEvhZmDthKr/uv8jhupzYaUOzAAAACnuAAAAAQ3YAAAAE/gZD4EAAAAA/4i9AAAAACp8QAAAAC3AAAAAJ/FEAAAAAj8cvkAS+FgAAAAAAAAAAAAAAAPY5bx5iVJRs+gAAUvgAAAAAl8LM2dsFU9DhqparOHdX+7/hwAAAAp7gAAAAEN2AAAABP4GQ+BAAAAAP+IvQAAAAAqfEAAAAAtwAAAACfxRAAAAAI/HL5AEvhYAAAAAAAAAAAAAAAD1QlekydRSgkLYAApfAAAAABL4WZ27YCp8vzRklTkxsPO6/AAAAAKe4AAAABFNNgAAAAT+BkPgQAAAAD/iL0AAAAAKnxAAAAALcAAAAAkm5IAAAAAj8cvkAS+FgAAAAAAAAAAAAAAAEaSSa7kaMnKzAACl8AAAAAEvhZxA7YGS80KPsJ4ep3Sf8A5M/0FsAAAAU9wAAAAAxFb2XJ5tL7wAAAAn8DIfAgAAAAH/EXoAAAAAVPiAAAAAW4AAAAAAAAAAR+OXyAJfCwAAAAAAAAAAAAAAAHsUJOTd2AABS+AAAAACXws4jdsGl/eX5MzOjbL8NPyt96uYSp7XDwn3S/IAAAAp7gAAAABxBW9lSpS7TT+64AAAAT+BkPgQAAAAD/AIi9AAAAACp8QAAAAC3AAAAAAAAAACPxy+QBL4WAAAAAAAAAAAAAAAAc6jK8T6xLsAAFL4AAAAAJfCziZ/7PBef6GY0ebKIf3VF/kv1Miqc+Ciu11+IAAABT3AAAAAIyu2cWS/dUo+bMJU9rh6c+6T/AAAAAn8DIfAgAAAAH/EXoAAAAAVPiAAAAAW4AAAABKVmkAAAAAR+OXyAJfCwAAAAAAAAAAAAAAAD2Ez2EwAApfAAAAABL4WcTv3Ka83+hXo8+BlT/ALv6HDVS9KcOzv8Af/wwAAABT3AAAAAIbs4sledKPk/0/wAjIqntMvp+V19zAAAAJ/AyHwIAAAAB/wARegAAAABU+IAAAABbgAAAAE/iiAAAAAR+OXyAJfCwAAAAAAs2rgAAAAAAAAAAABS+AAAAACXws4n1nSj6/oWVrGSfucdUo+v4MAAAAp7gAAAAEN2cUu+LhFfy/qzhapzYSUOz/NIAAAAn8DIfAgAAAAH/ABF6AAAAAFT4gAAAAFuAAAAAT+KIAAAABH45fIAl8LAAAAAADL6Ht3UprfldvVNf8MAAAAAAAAAAAAFL4AAAAAJfCzP/AHsZRj/W4T/cZ15N/mv8zdAAAAU9wAAAACG7Mygq2dQg/L/M4Wk41atJ+X4XX6gAAAE/gZD4EAAAAA/4i9AAAAACp8QAAAAC3AAAAAJ/FEAAAAAj8cvkAS+FgAAAAABw7G+Ik+y/VGaYf2GKlFbPVfMAAAAAAAAA+sSPrEinUU15gAFL4AAAAAJfCzNvezWjH/D/AOlMOIF7LF0qy7fk/wDgkWnHQAAACnuAAAAAQ3Z/E4h9P0iZb+5zupT783+f6AAAAE/gZD4EAAAAA/4i9AAAAACp8QAAAAC3AAAAAJ/FEAAAAAj8cvkAS+FgAAAAABw5StCpPvZfd/w5xDh+alGqumj9H/X4gAAAAAAAACpuTaiOk1JRZTTjUSYABS+AAAAACXwsxvvZ1TXa3+YcTUr0IT7O33r/AIBl1T2mFpy8kAAABT3AAAAALIwHv55Ufbm/yMb+4z2E+7X4rlAAAAJ/AyHwIAAAAB/xF6AAAAAFT4gAAAAFuAAAAAWQAAAABH45fIAl8LAAAAAADKqHscJBdXr9/wDwDE0FXoypvqiUXFuL3QAAAAAAAAPYw9rNGIeqKv8AEiAAUvgAAAAAl8LJ+9nqXb/5EM7pe0wM/LX7mcP1OfBqPZtfr+oAAAFPcAAAAAMh9/Mq0/X8ZI4nTp4mjWXb8nf9RNSSaAAAAn8DIfAgAAAAH/EXoAAAAAVPiAAAAAW4AAAAAAAAAAR+OXyAJfCwAAAAADBUPb4iFPu/w6llYk1FNvoZ3QVPFNraWv8AmAAAAAAAAA7wlZEbzkietVAAFL4AAAAAJfCyh72eSfa/5WDEU/a0ZQ7pr8Dhqp/Eh6MAAACnuAAAAAcys/I4VXNUrT9PxucVU74aE+zt96/4Bl1T2mDpS/ur8gAAAJ/AyHwIAAAAB/xF6AAAAAFT4gAAAAFuAAAAANpNIAAAAAj8cvkAS+FgAAAAAHD9NSxLl2X+QVLcj9DNH7XB0KvW1vy/yAAAAAAAAAqUozIUowAACl8AV8Zh6DUak0n5s/tXA/8AP2P3n9q4H/n7H7z+1cD/AM/Y/ef2rgf+fsfvP7VwP/P2P3n9q4H/AJ+x+8/tXA/8/Y/ef2rgf+fsfvP7VwP/AD9j95/auB/5+x+8/tXA/wDP2P3n9q4H/n7H7yWaYJp/vY/eZXKNXNqk4u6978wMt/cZpUp97r8br8AAKk4U4uUnZI/tLB/8/V95/aWD/wCfq+8/tLB/8/V95/aWD/5+r7z+0sH/AM/V95TzPBJ/xY/ef2rgf+fsfvP7VwP/AD9j95/auB/5+x+8/tXA/wDP2P3n9q4H/n7H7z+1cD/z9j95/auB/wCfsfvP7VwP/P2P3n9q4H/n7H7z+1cD/wA/Y/ef2rgf+fsfvP7VwP8Az9j95/auB/5+x+8/tXA/8/Y/ef2rgf8An7H7yrmmD9lUtVV7O2vkcKR/dVJeaOIKfPl8/Kz/ABOHqnPl8F2uvxv+oAFWtTpQcpyskf2xgP8An6j+2MB/z9Qs4wD09qj+1cD/AM/Y/ef2rgf+fsfvHmWDmuWNVNvzIfAgAAAAH/EXoFWpCnBzm7JH9q4H/n7H7z+1cD/z9j95/auB/wCfsfvP7VwP/P2P3n9q4H/n7H7z+1cD/wA/Y/ef2rgf+fsfvP7VwP8Az9j95/auB/5+x+8/tXA/8/Y/ef2rgf8An7H7z+1cD/z9j95/auB/5+x+8eY4Sc7RqJt+ZdMAAAC6KuMw9KXLUmk/NizLB3/ir7z+1cD/AM/Y/ef2rgf+fsfvP7VwP/P2P3n9q4H/AJ+x+8/tXA/8/Y/ef2rgf+fsfvP7VwP/AD9j95/auB/5+x+8/tXA/wDP2P3n9q4H/n7H7z+1cD/z9j95/auB/wCfsfvP7VwP/P2P3n9q4H/n7H7z+1cD/wA/Y/eSzTBOS/ex+8o43DV5ctOom/JgAA2ke0h3KmaYKnJwlUSaP7YwH/P1CzfAN29qj+1cD/z9j95/auB/5+x+8oVYVW5wd0+oBL4WAAAAAAcOJe0qeiDFzUKE5dk/yK7vlNLyk/1AAAAAAAAAqVne0T20+4AAUvgDiXC162Ig6cHJW6Jvq+x/Z2M/59S/8Bf+R/Z2M/59S/8AAX/kf2djP+fUv/AX/kf2djP+fUv/AAF/5H9nYz/n1L/wF/5H9nYz/n1L/wABf+R/Z2M/59S/8Bf+R/Z2M/59S/8AAX/kf2djP+fUv/AX/kf2djP+fUv/AAF/5H9nYz/n1L/wF/5H9nYz/n1L/wABf+R/Z2M/59S/8Bf+Rw/ha1LESlUg1p1TXVAY39xnMZ97fjoLYAOI8RyYZU1vJ/gv6QsDipJNU5NejPqGL/59S/8AAX/kfUMX/wA+pf8AgL/yPqGL/wCfUv8AwF/5H1DF/wDPqX/gL/yP7Pxb2pS/8Bf+R/Z2M/59S/8AAX/kf2djP+fUv/AX/kf2djP+fUv/AAF/5H9nYz/n1L/wF/5H9nYz/n1L/wABf+R/Z2M/59S/8Bf+R/Z2M/59S/8AAX/kf2djP+fUv/AX/kf2djP+fUv/AAF/5H9nYz/n1L/wF/5H9nYz/n1L/wABf+R/Z2M/59S/8Bf+R/Z2M/59S/8AAX/kf2djP+fUv/AX/kf2djP+fUv/AAF/5H9nYx/8upf+Av8AyOH6VWhh5KcWm3s1bojHS9rhakGt0/yOF66VCdN9Hf71/wAA9pB9R1YIdfsh1pMcpPqcQ4zRYeL83+i/X7j+y8a1/Cl9zP7Kxv8Az6l9zHleNS/hS+5n9nYz/n1L/wABf+R/Z2M/59S/8Bf+Rg8Bi44inKVKSSa6PuQ+BAAAAAP+IvQM5pzqYGpGCu30Xqj+zsZ/z6l/4C/8j+zsZ/z6l/4C/wDI/s7Gf8+pf+Av/I/s7Gf8+pf+Av8AyP7Oxn/PqX/gL/yP7Oxn/PqX/gL/AMj+zsZ/z6l/4C/8j+zsZ/z6l/4C/wDI/s7Gf8+pf+Av/I/s7Gf8+pf+Av8AyP7Oxn/PqX/gL/yP7Oxn/PqX/gL/AMj+zsZ/z6l/4C/8iph69BqU4OPa6aKFRVKUai6q4tUAAAN3Kk404OctEivKtja8pxi230SvZfI/s/F/8+pf+Av/ACP7Oxn/AD6l/wCAv/I/s7Gf8+pf+Av/ACP7Oxn/AD6l/wCAv/I/s7Gf8+pf+Av/ACP7Oxn/AD6l/wCAv/I/s7Gf8+pf+Av/ACP7Oxn/AD6l/wCAv/I/s7Gf8+pf+Av/ACP7Oxn/AD6l/wCAv/I/s7Gf8+pf+Av/ACP7Oxn/AD6l/wCAv/I/s7Gf8+pf+Av/ACP7Oxn/AD6l/wCAv/I/s7Gf8+pf+Av/ACP7Oxn/AD6l/wCAv/I/s7Gf8+pf+Av/ACMPKtgcTCc4uLXRprTqQxHNFNaoVePYVWD6jqQXUdddEOtJ7DqSfU1ZjK31ehKrJbf0iGGxOKcpwg5d7Lqf2Vjf+fUvuY8sxqX8KX3M/s7Gf8+pf+Av/I/s7Gf8+pf+Av8AyMipzp4OMZqzXR6PdgEvhYAAAAABw7JKtOPl+oZvFywU0v61MR7uWUo923+YAAAAAAAAPYp03M+reYAAUvgAAAAAl8LAOJYONWlVW+q+7X9SlNTgpLqAGbN4vMY0Y7Ky/wA/68hRSSSACnuAAAAAQ3ZOEZbksOmtzh+Lhi6tHrb8nb9R05dhUpMVDuxUYmIqUsPRlVlskZVh55jj3VqapO7/AEX9dEABP4GQ+BAAAAAP+IvQAAAAA4hoe1wkmt42f9fIyCt7XBKPWLa/UWjsAAA3cUbHEeM5YLDxer1fp0/ryMiwXsMMpte9LX5dP8/mLcAAAAAn8UTiTBe2w3tYrWH5df8AM4eq/WMJyN6xdvl0/wAh0H0Y6Ul0FTk+gqDfUVCPVipwXQsjijGc044aPTV+vT+vMyrBLCYWNPru/V/1YAI/HL5AEvhYAAAAABk9ZUsZC+z0+/8A4IV6ftaUod00Zzy05U8PF/AvxYAAAAAAAAFJqDcWc8O4AAUvgAAAAAl8LAOI6XNhFLs/6/Qymp7TB035W+7QAq1I06bm9kjIabr4ydeXS7+b/wCBcACnuAAAAAQ3YGG/cZ9KPdv8VcAOKMddxw0X5v8ARfr9xkuAWEwqTXvS1f8Al8vzAAn8DIfAgAAAAH/EXoAAAAAYimqkZQezVvwOHajpYipQl/TRJCd0AAN3FGxVqRpwc5PRGFpSzPHuU9t36dv0LC3AAAAAJ/FEnCM4uL2Zw7J0MdUw78/vi/8AhwADEV4UKUqktkrmTUJY/MHXqapPmfr0X9dgAI/HL5AEvhYAAAAAAm001uZfjI4qipLfqvMxFeFCm6k3ojEVpV6sqkt2AAAAAAAAAAABS+AAAAACXwsAzan7TBVF5X+7X9Dh2pzYZx7MAz/Eezwjit5af5mQYf2eDUnvJ3/RAAU9wAAAABRSuBm37nOaVTvyv8bP8ADE4iGHoyqz2SMpw88wx7rVNUnd/ov66IAAn8DIfAgAAAAH/EXoAAAAAVPiK6+q5ypdG1+Oj/G49TZgAxu4lYOI8ZywWHi9Xq/Tp/XkZHgvq+GUmvelr/l/XmC3AAAAAHFNp9gxn+yZ3Gp0bT+/R/qAAcUY20Y4aL31f6f16GR4L6rhIp/FLV/5fJfiABH45fIAl8LAAAAAACjXqUZc1OVmV8XWr29pK9gAAAAAAAADkh2OSHYAAKXwAAAAAS+FgFSCnBxfU4dk4ValJ7/5afqA9jPakq+LhQj0/N/0ilTjTpxhHZKwAFPcAAAAAA4rg1KlUXmvyKc1UhGS6q4HFGOvKOGi9tX+i/X7jJcD9UwkU17z1f8Al8gACfwMh8CAAAAAf8RegAAAABU+I4mpWdOqvNfqv1MPVVajCouqTJK6Iu6AY3cSsFWpGlTc5PRGEpyzLHuc9t36LZfoAtwAAAAAOK6GlOsvNfqv1MHW9vh4VO6TACpUjTg5yeiVzA05ZnmTqT2vd+i2X5IAAj8cvkAS+FgAAAAAAAAAAAAAAAQm3UaOeXc55dwAApfAAAAABL4WAGGXsM5lDu3+OotUE2ktTKovF5lKs9ld/ov68gACnuAAAAAAcUU+bBxl2f6Myip7TA0n5W+7T9AxWIhh6Mqstkv6+8yfDyx+Odarqlq/Xov66IAAJ/AyHwIAAAAB/wARegAAAABU+Iz2j7TBS7xs/wCvkZBW9pglH+Vtfr+oNWZuh2SG7iVgOI8ZywWHi99X+hkeC+r4ZSa96Wv+X9eYC3AAAAAAz+h7XAT7x1+7/gXOGq/tMFyfytr9f1ADifG+zoKhF6y39F/m/wAjh7BfV8IpyXvT1+XT/P5gAEfjl8gCXwsAAAAAAAAAAAAAAABU0pcx9Xj3Pq8e4AAUvgAAAAAl8LADNv3OaU6nez/G35EdgznEeywk2t3ovn/wLnDmH5MM6j3k/wAF/wAG4ABT3AAAAAIbsDPKftMvqLsr/c7nDVTnwKj/ACtr9f1DifHc0lhYPbV+vRfr9xk+B+qYWMWveer9f+AAAE/gZD4EAAAAA/4i9AAAAACp8RWpqrTlB9U0cN1HCrUovff7tH+YNXQnYeolYCrVjSpucnojBUpZlj3Oe279Oi/QAW4AAAABL4ohVpxqU5QezTX3nDFR08RUoS3t+Kdv1AJNJXbIqWbZnf7N/uiv8/zYkkrIAAj8cvkAS+FgAAAAAAAAAAAAAAAPYoNtu7AAApfAAAAABL4WAHE9PSnUXmjC1PaUoz7pMlscRVnKpCjH1+/RGFoKjQhTXRWAAKe4AAAABDdgYmn7WhOHdNfejhSpeFWHaz/P/IxeJhhqEqstkv6+8ybDTx2Odapqk7v16L+uiAAAn8DIfAgAAAAH/EXoAAAAAVPiCP8AsudW6Sf/AKV/wQJLUSSAOI8ZywWHi99X+hkmC+r4ZNr3pav9EALcAAAAAn8UQKn+yZ6n0k//AEpf5gHEeN9hhfZRfvT0+XX/ACOGsF7LDutJay29F/n/AJAABH45fIAl8LAAAAAAAAAAAAAAAAHO0rxPbzAACl8AAAAAEvhYAcQUufBN/wArT/T9TJKnPg4eV19xLV2MOvrubOfRO/yWi/QAAKe4AAAABDdgGRfuczq0fVfczifG8044aD21fr0X9d0ZRgVhMLGD+J6v1/4GwAAE/gZD4EAAAAA/4i9AAAAACp8QcR03Tr060f6tr+pSmqlOM1s1cACtVhSpyqS2SMDSlmOPc6m279Oi/T0ABbgAAAAE/iiBxVRcalKsvT7tV+ph6qrUY1F1Sf3gYqUs1zNQi/dvZei3f5shCMIqMVotAAAj8cvkAS+FgAAAAAAAAAAAAAAAH1eXc+ry7gABS+AAAAACXwsAMwpe0wlSPkzhupehOHZ3+9f8AzCv7DCzqdbaer0OGsPanOs+ui+W/wDXkAAFPcAAAAAhuwDFV1gc5lVa03++P+ZkmGnjsc69TVLV+vT+vIAAAn8DIfAgAAAAH/EXoAAAAAVPiDiGj7TB838rT/T9TJK3tcFDutPu/wCBYADiTGWisPF76v8AT/P7jJMF9XwybXvS1f6IAFuAAAAAT+KIHEVD2uAk+sWn+n5M4dr+1wEV1i2v1/Jhn2N+rYRqL96Wi/V/ccL4LlpyxEt3ovTr97/IAACPxy+QBL4WAAAAAAAAAAAAAAAAqicuU+sR7H1iPYAAKXwAAAAAS+FgA0mrGRfusXUovz/BnEte0IUV11f5L9TL8P7DCwp9UtfV6sAAKe4AAAABFNNgHFFJrFxkuq/FN/8AAMowP1TCxg/ier9f+BsAAAT+BkPgQAAAAD/iL0AAAAAKnxBiqPtqE6fdNHDNbSpSfk/0f6ABWqwpU5VJbJGAoyzHHupU23f6L+ugAC3AAAAAJJuSAxFFVqMqb6pr7zhaq4zqUXvv92j/AEDNa0swzFUaeyfKv1f9dEUaMKNKNOGyVgAAI/HL5AEvhYAAAAAAAAAAAAAAAEINVGzkl2OSXYAAKXwAAAAAS+FgAJewzu3Rv81/mVv9tzdR3SdvlHf79fvAAAp7gAAAAABisBHEYmlVltC/36W/zAAACfwMh8CAAAAAf8RegAAAABU+IDCf7LnEodG2vv1X6ABxJjLRWHi99X+n+f3GS4L6thVde9LV/ogAFuAAAAAAFP8A2TPWukn/AOlf8EzjG/VMJKafvPRer/y3OF8FdyxMl5L9X+n3gAAEfjl8gCXwsAAAAAAAAAAAAAAAD2icuUcop2uAABS+AAAAACXwsADiDmo4ynWjvZfen/wxw1QcpTrP0/V/oAABT3AAAAAAAAAAJ/AyHwIAAAAB/wARegAAAABU+IDPouhjadePl96f/DEWpJNAV60KNKVSWyRl9GWYY51Km27/AEX9dAABbgAAAAABxPTdLE0q8d/1Tv8AqZ1jHj8VClS1Ssl6v+rGEw0cNQjSj0X/AA7+YAABH45fIAl8LAAAAAAAAAAAAAAAAX8Yq/xYgAAUvgAAAAAl8LAA4mpXo059nb71/wAAyjD+xwcF1ev3/wDAAAAp7gAAAAEW22AAAABP4GQ+BAAAAAP+IvQAAAAAqfEBxHR58Kpr7L/B/wBIyit7XBU32Vvu0A4kxmiw8X5v9F+v3GTYL6thlzL3pav9EAALcAAAAAk2pJAHEtD2mC5+sWn+n6nDGC9pWeIltHRer/yX5gAABH45fIAl8LAAAAAAAAAAAAAAAAulVbZUlF1E0z2sO4AAUvgAAAAAl8LAAzHCfWqHs/Nfnr+FxJJWQAAFPcAAAAAhuwAAAAJ/AyHwIAAAAB/xF6AAAAAFT4gMwo+2wtSHdP71qjhmtelOn2d/v/4YK9aFGlKpLZIy2jLH451amy1f6L+uiAABbgAAAAE/iiAYyh7fDzp900ZfhFhcNCkt1v69QAAAj8cvkAS+FgAAAAAAAAAAAAAAAVfjYAABS+AAAAACXwsAAAAAp7gAAAAEN2AAAABP4GQ+BAAAAAP+IvQAAAAAqfEAZX/s2aTo9HdfqvwQcR43RYeL83+i/X7jJ8F9Wwquvelq/wDL5AAAtwAAAACfxRAAAAAI/HL5AEvhYAAAAAAAAAAAAAAADimcsexyx7AABS+AAAAACXwsAAAAAp7gAAAAEN2AAAABP4GQ+BAAAAAP+IvQAAAAAqfEAZx/s2ZQrLrZ/do/wMRXhRpSqS2SMsoSx+OdSpqlq/0X9dAAAFuAAAAAT+KIAAAABH45fIAl8LAACxYsWLAAAAAAAAAAAAAAAFL4AAAAAJfCwAAAACnuAAAAAQ3YAAAAE/gZD4EAAAAA/wCIvQAAAAAqfEAcS0eahCoujt9//DGZZl7bCUqMX0TfqtP+D9xlGC+q4ZJ/E9X/AJfIAABbgAAAAE/iiAAAAAR+OXyAJfCwAA0saGhoO1gAAAAAAAAAAAAAACl8AAAAAEvhYAAAABT3AAAAALAAAAAT+BkPgQAAAAD/AIi9AAAAACp8QBmtH22DqR8r/dqZJgvrGJTa92Or/QAAAW4AAAAAAAAAAR+OXyAJfCwAAAAAAAAAAAAAAAAAACl8AAAAAEvhYAAAABT3AAAAAAAAAAJ/AyHwIAAAAB/xF6AAAAAFT4gBpNWZluAjhKPJ1bu/69AAABbgAAAAAAAAABH45fIAl8LAAAAAAAAAAAAAAAAAAAKXwAAAAAS+FgAAAAFPcAAAAATTAAAAAn8DIfAgAAAAH/EXoAAAAAVPiAAAAAW4AAAAA2k0gAAAACPxy+QBL4WAAAAAAAAAAAAAAAAAAAUvgAAAAAl8LAAAAAKe4AAAABDdgAAAAT+BkPgQAAAAD/iL0AAAAAKnxAAAAALcAAAAAn8UQAAAACPxy+QBL4WAAAAAAAAAAAAAAAAAAAUvgAAAAAl8LAAAAAKe4AAAABDdgAAAAT+BkPgQAAAAD/iL0AAAAAKnxAAAAALcAAAAAn8UQAAAACPxy+QBL4WAAAAAAAAAAAAAAAAAAAUvgAAAAAl8LAAAAAKe4AAAABDdgAAAAT+BkPgQAAAAD/iL0AAAAAKnxAAAAALcAAAAAn8UQAAAACPxy+QBL4WAAAAAAAAAAAAAAAAAAAUvgAAAAAl8LAAAAAKe4AAAABDdgAAAAT+BkPgQAAAAD/iL0AAAAAKnxAAAAALcAAAAAn8UQAAAACPxy+QBL4WAAAAAAAAAAAAAAAAAATnGK1IyUldBS+AAAAACXwsAAAAAp7gAAAACilcAAAACfwMh8CAAAAAf8RegAAAABU+IAAAABbgAAAADim0+wAAAABH45fIAl8LAAAAAAAAAAAAAAAAAAGlOpr0IJRqOKCPtbe7sfvv6sRvyq+4AAABVdoNoAAAAAg/eSAAAAAIybb8gAAAA/ff1Yl7azuQ+BAAAAAVebnXLufvv6sU+f7QAAAAN3kwAAAAG7IWqAAAAAlJqSXcAAAAKntL+6fvv6sUubmlzbgEvhYAAAAAAAAAAAAAAAAABT1lIl/FQUvgAAAAArfw2AAAAAQ+NAAAAAFPeXqAAAABP4GQ+BAAAAAP+IvQAAAAA6v1AAAAAlsR+FAAAAAT+OIAAAABH45fIAl8LAAAAAAAAAAAAAAAAAAKfxSJfxUFL4AAAAAK38NgAAAAEPjQAAAABT3l6gAAAAT+BkPgQAAAAD/iL0AAAAAOr9QAAAAJbEfhQAAAAE/jiAAAAAR+OXyAJfCwAAAAAAAAAAAAAAAAACOlRruL3qr8gpfAAAAABW/hsAAAAAh8aAAAAAKe8vUAAAACfwMh8CAAAAAf8RegAAAAB1fqAAAABLYj8KAAAAAn8cQAAAACPxy+QBL4WAAAAAAAAAAAAAAAAAAVKamQgoKwexn0Z7Kp3PZVO57Kp3PZVO57Kp3PZVO57Kp3PZVO57Kp3PZVO57Kp3PZVO57Kp3PZVO46NRrc9hPuewn3PYT7nsJ9z2E+57Cfc9hPuewn3PYT7nsJ9z2E+57Cfc9hPuewn3PYT7nsJ9z2VTueyqdz2VTueyqdz2VTueyqdz2VTueyqdz2VTueyqdz2VTueyqdz2VTueyqdz2VTuexqdz2VTueyqdz2VTueyqdz2VTueyqdz2VTueyqdz2VTueyqdz2VTueyqdz2VTueyqdx0aj6kVaKQAAAAFSnKTTiz2VTueyqdz2VTueyqdz2VTueyqdz2VTueyqdz2VTueyqdz2VTueyqdz2VTueyqdz2VTuewn3PYT7nsJ9z2E+57Cfc9hPuewn3PYT7nsJ9z2E+57Cfc9hPuewn3PYT7nsJ9z2E+57Kp3PZVO57Kp3PZVO57Kp3PZVO57Kp3PZVO57Kp3PZVO57Kp3PZVO57Kp3PZVO57Kp3PY1H1PZVO57Kp3PZVO57Kp3PZVO57Kp3PZVO57Kp3PZVO57Kp3PZVO57Kp3PZVO57Kp3PZVO5Tg43uwCXwsAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAJfCwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAl8LAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAACXwsAAAAAAAAAAAAAAAAAAAAAAAAqTlfljuU58y13AAAAAdW81FAAAAAVJ8qI1JKSUgAAAAAAAAAAAAAACclGNyFSXNaXUAAAACpUUF5kHzRTAAAAAdWb1S0ItSSaAAAAAAJfCwAAAAAAAAAAAAAAAAAAAAAAACnrKTFpVaAAAAAnFKUbAAAAAWTK/wAKYAAAAAAAAAAAAAABU1lFFb4U+wAAAAFSKab8in8CAAAAAsrWKHwAAAAAAEvhYAAAAAAAAAAAAAAAAAAAAAAABR2fqP8Air0AAAAAqfFEAAAAAq6yjEAAAAAAAAAAAAAAAl/EiV/gFsAAAAE/gZT+BAAAAATdotlJWggAAAAACXwsAAAAAAAAAAAAAAAAAAAAAAAApfaXmb1fkAAAABU+KIAAAABT96TkAAAAAAAAAAAAAAAT/iRK/wAAAAAAE/gZT+BAAAAAVPekoIAAAAAAJfCwAAAAAAAAAAAAAAAAAAAAAAABy9nN32ZS95uXcAAAACp8UQAAAAK8rRt3FXilZI3QAAAAAAAAAAAAAAFZOya6DkqkkkAAAABP4GU/gQAAAAN2VyFVJttashNTQAAAAABL4WAAAAAAAAAAAAAAAAAAAAAAAANJrUsrAAAAANJgAAAANJnLHsAAAAAAAAAAAAAAACilsAAAABuWQAAAAHLHsJJbAAAAAAEvhYAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAS+FgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABL4WAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAEvhYAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAS+FgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABL4WAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAEvhYAAAAAAAAAAAAAAAAAAAAAAAAAAABowAAAAAAAAAAAAAAAAAAAAAAAAANAAAAAAAAAAAAACXwsAAAAAAAAAAAAAAAAAAAAAAAAAAAryaasznl3KcpNS16GHvZgADdkUFdNhVV4MpO8EAAAAAAAAAAAAAAAAAAAAAVW1B2OeXcjOTa1J39qgAAn71VLsBT92o4gAAAAAAAABL4WAAAAAAAAAAAAAAAAAAAAAAAAAAAShGW6FFe0scsYK6KcoSXuiqxafkQqRnsKtFuwe3hewVXaDKK9xEq0YuwmpR0MPs0AAAAAAAAAAAAAAAAAAAAANJqzK8IxSsQpxcU7DnDnt1JVIxaTHWgnYnVjF2YmmronUjHRkZKSugp61Wyc4xWpCpGeiJaVkAAAAAAAAAEvhYAAAAAAAAAAAAAAAAAAAAAAAAAAAL+MT+BmH6ii5OyGnF26k4OD1KsrQ9T2XucxRd4Ff4CGlNFOnz31IR5YpFH4mgAAAAAAAAAAAAAAAAAAAAAxOyKfwIn/G+4xHxIlTcUmzklKPMUH7pGHtJNlK8ZuIUNZMq+9UsU6XI9yt8cQAAAAAAAAAl8LAAAAAAAAAAAAAAAAAAAAAAAAAAABfxifwMw/UofGyf8X7jE9Ct8ET2kmrXMP8LK/wEFemkJyhIjJSimUfjYAAAAAAAAAAAAAAAAAAAAAYnZFP4ET/AI33GI+JGI+BEf4JS+CRGco7Mptuomww/Urxalco1ObR7lb4ogAAAAAAAABL4WAAAAAAAAAAAAAAAAAAAAAAAAAAAC/jE/gZh+pQ+Nk/4q+RiehVXuRZTcHFXI8rWhXXuFN+4ityyhdFD4Chq2wAAAAAAAAAAAAAAAAAAAAAxOyKfwIn/G+4xHxIxHwIj/B+TKKvGSKLinZi5G7LcKPxNDcX7rKSSqWRV1qRQAAAAAAAAAS+FgAAAAAAAAAAAAAAAAAAAAAAAAAAAqcvaX6E1eLSKMJRvcpU5Rk2yVOTqXWxXhKVrDheHKx0pp2sU4uMbMqK8Gil71OyPZTvawoqEDDr3QAAAAAAAAAAAAAAAAAAAAArQlK1iCaikSpydS/QrU5SasVouUbIjFqnbqUYSincqUXe6KNOSd2EdKrRVpycroo03HVj1regAAAAAAAAAS+FgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAUoON0wqJuLSIR5YpAAAAAAAAAAAAAAAAAAAAAAAAATg3NSQEINSbYAAAAAAAAAS+FgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABL4WAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAEvhYAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAS+FgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABL4WAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAEvhYAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAS+FgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABL4WAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAEvhYAAAAAAAAAAAAAAXZdl2XZdl2XZdl2XZdl2XZdl2XZdl2XZdl2XZdl2XZdl2XZdl2XZdl2XZdl2XZdl2XZdl2XZdl2XZdl2XZdl2XZdl2XZdl2XZdl2XZdl2XZdl2XZdl2XZdl2XZdl2XZdl2XZdl2XZdl2XZdl2XZdl2XZdl2XZdl2XZdl2XZdl2XZdl2XZdl2XZdl2XZdl2XZdl2XZdl2XZdl2XZdl2XZdl2XZdl2XZdl2XZdl2XZdl2XZdl2XZdl2XZdl2XZdl2XZdl2XZdl2XYAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADi7CVzlZys5WcrNwGgBK4AAW0CxuMNwtoAAW0AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAACWxEAFufaHuS3B7IIgBLceyD7IN3FuPcFuS3PsgDSsgPsgnZEtUjZ2LK5I5tCQAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAEhbgAdTexfUeqG9BPoJal76C0Q9VcJbj2Q3bRC2Hqhq7RdN2L2Y9ULceqHsCN0N6Be6PsgdEPcluPoJXY9WAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA9WP4gAHsLZCSY7ND2RojqxbmlhtW0G0yW52HufZPsnUW5ZNjs0R3Fqje4K1i6toPZCVzTZH2QSuPce5LcfQiAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAALWRLcAB/CL4Raqw1ZDV0NXdzqWsLVWGrA1ce4432Oh9ke6Lam90NWRHciLZjSsLsONkWuhblrH2SSSIrQcbFru49WPofZErsYAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAXG7ikAAAAAAAAAMAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAFG6La2JRViOqAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAACOw/iJbEdgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAWw9xyuhLQASuMAAshqwAldjVmNAAANK4WuhK41YSuNKxyprccRxVgOUZbQAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABq7ElcAFsNXZy6CVxqyGhqwS3GrscS2thoW5LoNXdhrQascuthocdBK5a7sOKGg6WPsm8Rqx0CRHcCW5Jaj2AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfxEtGAFnofaFqxbH2SRLcNLiNkdTcjubo6stZEtz7QtXcjuWvEWwtx7iV2b3HsfZG7seq0ErserNGrIs2WaGrj3PsgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAP4iSuiOqAWo3qdSJ9kfwjV2S1Ytz7R0Nx7FhbETozcavI6sWwiO49iO5YTsJ3N0fZLC2YgjuXLi1ZfUvoAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHMi6ucyE7M5kcyE7BcvoX0sN3RzFxHUTG0xu6L6WOgnYTG09jmLidhu6si6sJl0X0sCYnYbuX1G7ovpYHJW0AvpoF9LAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAf//aAAgBAxEBPwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAACvVVKlKo9opv7it4oq9qWG085foo/qZV4j4LE1VTxVN079b3Xz0TX3PzYmmroAAAAx3GeLo8SLCRt7FSUGrb3sm776N6LayAAAAAAAAAAAAAAAAAAAAAAAUXJpJXZg+EK1SClXny+Vrv56r9TNOGKuDoutCalFb6Wf63DB8KVMRh4Vvapcyva3/BMbhXhcROi3dxdjJ8nlmM5RU+VRt0vuPgyNtK2v+H/gmZZXXwFXkqbPZ9H/AF1QU+Dqk4Rl7VK67f8ABKtN06koPdNr7gAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAzD/AHSr/hl+TMhySrnGL+rU5KLs3d+Vu3qcQcLY3JnGVW0oPRSW1+z7P+r7nh5m8sXl8sNUd5UrJf4Xt91mvSwAAABjOCa1fiFY1SXsm1J97q2lvNrfs38wAAAAAAAAAAAAAAAAAAAAAA4Sy+NWrLEzWkdF69/kvzMwzrB4FqNWXvdlqzOc8wmMy2caMvebWj0e9/0ChGNClTpeSX3L/gHFNLkzKT/mSf4W/Q4M/iVvRfqYziSjhcY8NUg7K2t+6T2+ZxRh41culPrFpr77fqYal7WtCn3aX3sc4qah1s391v8AMzyl7PMa0fO/36/qAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAGYf7pV/wy/Jnhx/yOX/gl+aOJsDHG5TXpNXfK2vVar8UeG9dwzaVPpKD+9NP/ADONeLKmVqOFwr/eyV29+VdNO789u2qMs43zbCYhTq1HUh1i+3k+j/DujH5hzZPVxmFl/wAu3KL/AO3brR/kzB8f5jTwNf21TmqtxULqKtfm5nZJXtZb9WulzIONsww+Nj9bqudOTtK/S/VdrdtrHEeZ1ctyyriqUVKUbWvtq0r/ACuVOL87nV9p9YafZWS+7b70cI5+84wPPU/iQdpfo/n+aZX4yx+Dz6rSr1L0YOfu2j0T5Ve197dfUpccZxHGrEyqXjfWH2bdkunrv5nFXG2JrYn2WAqONONtVo2/XstrevkcEZ/VzXBSjXd6lN2b7p7P10a+VzMOKs3pcQvCQrWp+0Ubcsdm1pe1/wATjzP8yyurQWEqcqknfSL2t3T7me8cY/EShTwtRxjFK7WjlKyu79r6WWn6cEcQ1c2wcoV3epTaTfdPZ+ujT/4JxtxRUyqlGhhn+9nrf+Vd7d29F6MyTjbMsLi4yxNVzpt+8nrZd12t22ZGSlFNO6ZxxmmMy3LoVsJPllzpN2T0tLun1SMdx3mE8to0adT967uckkn8TslZJLS17Lt5nAfFGJxtWWCxc+aSV4t76bp9+667nFOfRyfAOqlectIrz7vyS/RdTD8aZ1SxKrOs5d4v4X5W6fKxic29tkdTH4Z2fs3JdbNJ/LR6djgTiHMszxVWni6vMoxutIrW/kkcZcUZtl+aOhhqvLCyduWL39U2ca5zj8uy+hWwtTllJ2eifS/VMzjjXMZYPD0aVW03G85KybbbstNtNXbucWcRLJsGpQSdSWkU/wAW/Jfm0UuNM7p1/be3b8nbl+7p8rGR5vSzXAwxNPS+jXZrdf5eVjAcd5jQxNb63U54xUuWNoq8rpLVJP18jB8cZvSxir1arlG+seluyXTya+dyvmFClgnjG/cUeb5Wvp6mP40zjE4h1Y1nBdIx0SX6/M4UzDE4/KaWIxLvJ31ta9m1f8AAAAAAAAAAAAAAAOH6caGVwk+qbf8AXpYxeInia8qs95O4YCl7XF0od5L8zMcR7PHYWF93L8rfqcZUrVqVTumvuf8AwTgz+JW9F+pn9GdbN6lOmrt8tv8AwFHEM1Qylwb1dl+X6JmQUvaZlSXZ3+5X/Qq4i2cU6V94P8/+AcW0uTH8380U/wA1+gZDk+DxeAdSpC8rtbvt5My7hzCUsOlWgpTa1v08l2sZ3lywOLdOPwvVen/AOHMipYmH1jEK8dku/n+hnnD+GlhpVcPDllFX02aW+hw1l+GxlWpGvHmSStq1+TRn2Eo4XHSpUVaKS017eZneSYDDZfKrSp2kra3fVruzLMoweIyp1pwvO0tbvdXtpe34GZ5NQwWUX5b1NLvrfrbyOHuH6VeksRiFdPZfq/0M04awtei3QjyzW1tn5NfqZJhaWIx8KNaN073Wq2TfTXoLhjCPG8yjamktLvWV33d9FY4jyPDwwzxFCPK47pbNenkZFlSx+J5Z/BHV/ovmYjIctlRcXTSSW60f3/5gAAAAAAAAAAAAAAAAAAAAAAAAAAAABmH+6Vf8MvyZ4cf8jl/4JfmjEpOhNPs/yOAL/wBu07dpf+kszRSzfiWdK/xVOX0SfLf7lc494awWBwdLEYSCjZ8rS63Wjfnpv1vqcNYt1uEMVTb+CNRfLlv+rOE8khm+ZKhUfuJOUrdlZW+ba+Rxpw/RyjGxjQv7Oaur62a0a/J/My+jDOOHqVKs9JwSb66Wv+KOIcDwxhMvqYW0IVFF8ttZ3S0u1eWr7nhhWaxlel0cU/udv1M+w0sVxFWoQ3nUsvVtI404Zw+Tui8O24yTTv3VtfnfYyrg+ji+HqmPk37S0nHtaPS3m019x4Y1WsfWp94X+5pfqZr/AMlY/wDr7H80eKP8bDekvzRl3BlHE8PTx0m/atSlHtaN9LedvxXmeGlVxzSpT6OD+9Nf5szeH9scWPDyfu8yj8o/F+T+88Q8jweEoUcRhqahrytJWT0utuuj13fU4UxLxGS4abevLb/wH3f0PEr/AJE8P8a/KRwbwlh83w9WviG0k+WNu9rt/K60OFVPCcRUab3UnF/c4niRiZV8zpYaP2Y/jJ/5JHFfC2AwmQc1GmlOly+9bWWqTu+t73126HDWJdXhDF02/gVRL0cb/m2eGP8Av1f/AA/qjxD/AOR3L/DE8Rv+RRh/8S/9JZwvwjVzuE6sqnJCOl7Xbdtt1srHiLi5Vs49lfSnFL5vV/mjPeDsHhuH+anC1Wmk3Lq/5r+W9u1jwwxb/wBowzemkl+T/QwmXvMM3WFTtzzav2V9X9xxrw5h8nr0vq7fJNPfXVWv+aMbiZz4FhJPpGPyU0v0seH+XZTjqVaniYKVS6snvy23Xz3a20MvwFHA4aGGo/DHa/rcAAAAAAAAAAAAAACHu5Hp/wA+v/bTIsNSxGPhSqq8XfT5MxmQ5dDD1JRpK6Ta1fb1OG6XtMyp+V3+H+ZxFieTNcP/AHbP/wAm/wAkcYUubCQn2l+af+RwZ/Erei/U5Y3vbU4woYl8lS96a0t2fd97/h+fCNLnxzl/LF/jZf5mKxNuI4eVo/ev+CcZ0taNRea/L/ghwl/yL3/if5Iw3EFStmrw1lyXaXe66/O35HGdNXoz/wAS/IzCrLLcnjGm7SSSXq93+ZwtjquKw84VZczi93q7P/hmcL0vZY7EU/5dPudjij/kZT9F+SOJP+RVP/t380cLq+Wx9X+Zn+eRxPPg6cbq9r36p9jMq39nZY3T3ikl+X/BOF8yrYqlOnWldxtq97P/ACt+JRoKjxLyrZtv74tnEOb1MBTgqVuaV9+y3/MlWWNyuU7fFB/fb/M4NivYVX1uvyMzzTMcNjKsJSdndJPaz2t8uvfcAAAAAAAAAAAAAAAAAAAAAAAAAAAAAMw/3Sr/AIZfkzw4/wCRy/8ABL80Z3iY4bLa9VvaMvvtp+J4cUXPOHL+WDf4pfqcN68TU+b+eX5M8Qknkc7/AM0fzOEL/wCrmY9uWX/pDPDL/kY1v8H/ALcjxSS5cL39/wD9tKecVMs4No1KbtOV4x8rylr8kn87GAyHMcypVMTSjeMbtybtru99W/6Z4Zf8jOr/AIH/AOlRK/8AyWK/6/L/ANKR4of7th/WX5I4Z/5JWH+Gf5yPDP8A5G1T/r2//SoGa/8AJWP/AK+x/NHij/Gw3pL80ZF/yTNP/r2/yZ4cf8jl/wCCX5ozmcv7TxEr2fPP/wBKZOrOatKTZwJf+wKH/b3/AKXI8Sv+RPD/ABr8pHht/wAief8Ajf5RMD/yV3/caX/pTOPm3n1XyUf/AElEq1SSs5N/M4TT/wBW8x9Jf+kM8Mf9+r/4f1R4h/8AI7l/hieI3/Iow/8AiX/pLPDuKWSK3WUv0OK9eJavN/NH8onEKTyjFX/59z/9JZ4ZX/tKr/g/9uicKf8AJT0/8U//AEmR4o/Bhf8At/8A9tMiy6OY8IwwrfxRlbyfM2n96RUp4vLcW4yvCpB+jT8v61RwvnDzXLIYiXx7S9V/mrP5gAAAAAAAAAAAAAAZfH6xlEILrC34WMHi6uDrqrT+JX3+4o4qeKyh1p7uMtvmcH0ubFTn2jb73/wCvl+Erz56lNSfdo4jpe0y2p5Wf3NfocGfxK3ov1M9xuIw+bVJUptWt1/uroY5rGZRKbW8Ob52ucGUvdrVPRfnf9CWX4SVX2rppy3vbW6OLaXPgFL+WS/VBwl/yL3/AIn+SMt/5HEf8T/U4wSaoJ93+hKKkrNXFCMdkZFb+1cXbu//AEpnFH/Iyn6L8kcSf8iqf/bv5o4W/wCRbH1f5mHfNi4X/mX5nFt/7P0/mX6nBt/rFT0/Uq2/1lp+n/trOM/4lH0f6GU/8iWP+GX6nC2ZQw2IlSqO0Z9fNf5/5GaZbSx2HdOS97o+z/y7kouLae6AAAAAAAAAAAAAAAAAAAAAAAAAAAAAMw/3Sr/hl+TOB8ywuAzN1sTPljytX13uu1zjXjHD46h9SwTvB6ylqr22Svra+rfkrHhnl0qeGq4uS+NpL0W/3t2+Ri5vKeJZVJLSFS//AG63f8Ys8Qs9weIwNLDYeopuT5nZ3skna/a7f4HDOFdLhDFVH9uNRr0UeX80zgnOKOW5opV3aE04t9tmn96t8zj3O8PmWOhDDy5oU1a/Rt728tEjizDTw3D2X0n0Wvq43/VmR5zgcLwlJOa50pq19XKTdtN9mvlfseGX/Izq/wCB/wDpUSv/AMliv+vy/wDSkeKH+7Yf1l+SOGP+SVh/hn+cjwz/AORtU/69v/0qBmv/ACVj/wCvsfzR4o/xsN6S/NGRf8kzT/69v8meHH/I5f8Agl+aMxrPKOKZ12tIz5reUtXb5MfFOSqiq31mNu19f/Afi+VinUjUhGcXo1dfM8Sv+RPD/GvykeG3/Inn/jf5RMD/AMld/wBxpf8ApTPEWhKhnMay+1FP5q6/RGX8W5TisLGtKvGDtrFtJp9VZ6v5bmOxNDMskr1MPLmjKE0n8mvzRwTnOHyzMnPEStCUWm+2zT016W+ZxVmtPM80qYil8GiXola/z3PEb/kUYf8AxL/0lnh5/wAiSP8AikeIOGlRzuVT+eMWvkuX9DiDinB1+HXKnUTqVYpct9U38V1ura/h3PDDCvmxGIe2kV+Lf6HCn/JT0/8AFP8A9JkeKPwYX1n/AO2nDOPoYDhijiK7tCN7/ObX6nHtbKsXlUa9OcZVLrlaabt1WmtrfczwzUv7Lq9ud/8ApMf+AAAAAAAAAAAAAAABwnilUwTpX1g/wev53M64arzxDrYVXUtWrpWfXfSxh8NUw+TulUVpKMr/AInBtK1CrU7tL7l/wTMs+x0MZVhTqWim0tF007FGcsblKcndyg0/W1vzODP4lb0X6mdZTjMVmk3Sptp216fCupmCjhMpnC+kYcv4WRwnS5Mv5v5m3+n6FXiHMHUk41Xa7tov8jNksTlE5LrFS+6zDhL/AJF7/wAT/JGW/wDI4j/if6nGf8Oj6v8AQzPDPMssXs92lJf16Ecnx8m17KSt3Vl97ODppYqpHvH8mv8AMzzIq+Lx8KlNe60k32t/wDiZf8JdT5fmjhb/AJFsfV/mczjU5lumZxh3jstkqWraTX5/ijhPAVaFOpVqxacrJJ6PT+vwKVZVuJbro2vui0cZ/wASj6P9DKP+RLH/AAy/Uw+X4jEUpVKUeZR3tvr5HDEsS8G1Wvo7K+9rL8DH2eLq225n+bAAAAAAAAAAAAAAAAAAAAAAAAAAAAAMbCU8NUjFXbi1+BUyPM6b5ZYeaf8Ahl/kZNwLmeOqp14OlT6uSs/lHe/rZGDwlHCYeFCirRirJf1+Jx5wtXxk1jsJHmklaSW7S2a7vo12sZTwxmOY4hUo03FdZNNJff18tzM8HSweQV8PSXuwpSS+UX/TMsyjEZjGqsOuaUFzW6tXSdvPW/8AwThrhTF5hjI+2puNKLvJtNXt0V+r28tzjDJZ5plcqVJXnF80V3a0t80387GT8K5hmGLVF05QivibTVl8+vZfoeH+U5hgsyqTxFGUIuDV2mteaOn5lbJcxfFSxCoS9n7VPms7Wutb9jxDy7F42hQjhqbm03eyvbRHD2Dr0eHIUKkGp8s1ZrXVytoeH+T4/B5nOpiKMoRcGrtNa80dPwMyyPMqnErrxoScPaRd7O1rrW54iZXjcbVw7w1KU7KV7K9tjJ8LWpcPwoTi1NQas972eljgPJswwmaupiKMox5WrtNK90cecLVse443CR5pxVpRW7XRru126q1tihkmZV6ns6dCTl/henr2+ZllKrSwNGnVXvxjFP1SSf4nH2BxOMyuNPDwc5c6dkru1panAOBxODyuVPEQcJc7dmrO1o6mDyPMo8TfWHQlye1bvZ2tzPW/Y4y4dlm+DXsf4sLuPnfdfOyt5olk2Yqr7J0J83blf+RwhgMRhMmp4fFRtJc2nk23r9+xnPC+PwWOlQhSlKLfutJu66bde67mZ8GZjhqVBU6LnJxblZXs77adlb53OOcvxeMyvDwoU3KSabSV2vdZwPg8RhMojSrwcZc0tGrM414cnm2EjOgv3tO9vNPdfqv+CYfh/M69f2EKEubzTVvVvRL1OHsmhlOAhhk7veT7ye/y6LyRw3keZUOIadarQkoKUtWnbVSseIuWYzGwwyw1JztzXsr2vylTKMwq8HQwkKb9ot4vR2U29n12dijkeZVq3sYUJc3blat632+Zw5lCyrLqeGbvJat+b3+7ZeSAAAAAAAAAAAAAAAyrMp4DEKrHVbNd0YbOcBiIKUaqXk3Z/czNc+wNPDzpxnzSaastd1bfY4dx2Dw2AjGpVSk2203r2/JFWbqVJTfVtnDuaYWlgI06tRRab0b87/qcN4jDYbFV4yqJLpd2Ts3sTzLBQjd1Y/ejiLPYYxKhQ+Bat93/AJGBzHB4bKYwVVcyi9L63d3b1uGX5lg55ZClUqpPl5Wm/kHDOYYShgnCrUUXzPRv0MBWpwzWNSUko8zd/vOK8bhsRCkqM1Kzez9DhvO6MaKw1eVmtm9rdjFZlgqNNupUVvW7+5amT45YLGQqv4dn6P8Aq5i84wdDDurzp9knq/6/AzXM8Picn5faJ1Go3V9b3V9Dh3McJQwEYVKiTu9G/Mlq2cN53R9gsNXlZx2b2a7eqM2z3D4Si+SSlN7Ja/N+X5mRYiFPMqdWrKy1u35pnFeLw+InSdGalZPb5GWZjg6eUxpyqJS5XpfXqcJ47C0aU6VSaUm766aWXUzTO8NhKDcZpz6JO+vn5DbbuwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADiD/AJFGK/69z/8ASWeGX/Ixrf4P/bkAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAZthp4nL69Cn8UoSS9WmkcFcL5hlOMqVcSkk42VnfW6f6AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAH1p9j60+x9afY+tPsfWn2PrT7H1p9j60+x9afY+tPsfWn2PrT7H1p9j60+x9afY+tPsfWn2PrT7H1p9j60+x9afY+tPsfWn2PrT7H1p9j60+x9afY+tPsfWn2PrT7H1p9j60+x9afY+tPsfWn2PrT7H1p9j60+x9afY+tPsfWn2PrT7H1p9j60+x9afY+tPsfWn2PrT7H1p9j60+x9afY+tPsfWn2PrT7H1p9j60+x9afY+tPsfWn2PrT7H1p9j60+x9afY+tPsfWn2PrT7H1p9j60+x9afY+tPsfWn2PrT7H1p9j60+x9afY+tPsfWn2PrT7H1p9j60+x9afY+tPsfWn2CKuz2a7ns13PZruezXc9mu57Ndz2a7ns13PZruezXc9mu57Ndz2a7ns13PZruezXc9mu57Ndz2a7ns13PZruezXc9mu57Ndz2a7ns13PZruezXc9mu57Ndz2a7ns13PZruezXc9mu57Ndz2a7ns13PZruezXc9mu57Ndz2a7ns13PZruezXc9mu57Ndz2a7ns13PZruezXc9mu57Ndz2a7ns13PZruezXc9mu57Ndz2a7ns13PZruezXc9mu57Ndz2a7ns13PZruezXc9mu57Ndz2a7ns13JRsAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAENwAAAAAAAAAAAAAAAAAAAAAAAAKnQAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAIbgAAAAAAAAAAAAAAAAAAAAAAAAVOgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAQ3AAAAAAAAAAAAAAAAAAAAAAAAAqdAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAhuAAAAAAAAAAAAAAAAAAAAAAAABU6AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABDcAAAAAAAAAAAAAAAAAAAAAAAACp0AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAACG4AAAAAAAAAAAAAAAAAAAAAAAAFToAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAENwAAAAAAAAAAAAAAAAAAAAAAAAKnQAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAIbgAAAAAAAAAAAAAAAAAAAAAAAAVOgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAQ3AAAAAAAAAAAAAAAAAAAAAAAAAqdAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAhuAAAAAAAAAAAAAAAAAAAAAAAABU6AAAAAAAAAAAAAAABa5DCxS94nhoNe7uNWdgAAAAAAAAAAAAAAAAAAAAAAAAAIbgAAAAAAAAAAAAAAAAAAAAAAAAVOgAAAAAAAAAAAAAAAEsRCKXcp1FNXRiY2mAAAAAAAAAAAAAAAAAAAAAAAAABDcAAAAAAAAAAAAAAAAAAAAAAAACp0AAAAAAAAAAAAAAAAp0JT16FKkqaepi1qgAAAAAAAAAEm9ESpyitURhKWyHFp2YAAAAA6c0r2AAAAAAAAACG4AAAAAAAAABdBdMAAAAAAAAAAAAACp0AAAAAAAAAAAAAAACEeaSRpGPkiFWM20jF7oAAAAAAAAACLVKmn1ZKvKUXFjn7Kmkt2VajqNMAAAAChFNuT2QsS3LVaFeCjOyArQUWkuwAAAAAAABDcAAAAAAAAACT0sjkViWyQoJMAAAAAAAAAAAAACp0AAAAAAAAAAAAAAADDq9RGJ/hmFW7MXugAAAAAAAAASu7GJfv27BVi5U4yQAAAAAtKD82FX3owZWiozaRKFOLjfaxX9nbXexSpwUeeZUUJSSh1OSinyPcVNKrysjSpJ8r3ZClTjpPdk6dp8qOSinydSUeVtAAAAAENwAAAAAAAAAN5BLRpm4AAAAAAAAAAAAAFToAAAAAAAAAAAAAAAFFpTRKKkrMjGMVZGL3QAAAAAAAAAR+JGI/iMKNbk0exXgoyutmAAAAA/wCAvUJfBAxH8Rlf4Y+hifiXoYjRRj5GHV6iJO8myetaLJNut8yu/wB4ySvXQ8PNyuYhfvGAAAAAQ3AAAAAAAAAA+0DSaFeLsAAAAAAAAAAAAABU6AAAAAAAAAAAAAAAAQxS5bSWp9YfPexWq+0toAAAAAAAAABiFdqS6gVf4cAAAAAKa5qUolrlaycY9jEr94zEKyivIxPxL0MQr8r8jDu1REo2lYn/ABorsS/i/Mr/AMRjdq6JTmpNXY23uAAAAAQ3AAAAAAAAAAekkBLdAAAAAAAAAAAAAAVOgAAAAAAAAAAAAAAAAAAAAAAAAAAUqkXHkmVFRUXy7i9hJJsrVFOWmwAAAAFKpySuXoRfMicnKVxzpSipS3RWmppMnKnOmnfVFOcJQ5JlRwjJcnQ5qLfP1FUTq8zG06l/MrNSm2itNOalFj9jU95sny8ztsAAAAAQ3AAAAAAAAAAlG6Pe2GpaWEm3dgAAAAAAAAAAAAAVOgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAQ3AAAAAAAAAAAAAAAAAAAAAAAAAqdAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAhuAAAAAAAAAAAAAAAAAAAAAAAABU6AAAAAAAAAAAAAAAAAAAHsKjWx7Cp2PYVOx7Cp2PYVOx7Cp2PYVOx7Cp2PYVOx7Cp2PYVOx7Cp2PYVOx7Cp2PYVOx7Cp2PYVOx7Cp2PYVOx7Cp2PYVOx7Cp2PYVOx7Cp2PYVOx7Cp2PYVOx7Cp2PYVOx7Cp2PYVOx7Cp2PYVOx7Cp2PYVOx7Cp2PYVOx7Cp2PYVOx7Cp2PYVOx7Cp2PYVOx7Cp2PYVOx7Cp2PYVOx7Cp2PYVOx7Cp2PYVOx7Cp2PYVOx7Cp2PYVOx7Cp2PYVOx7Cp2PYVOx7Cp2PYVOx7Cp2PYVOw1Z2AAIbgAA5JHOjnRzo50c6OdHOjnRzo50c6OdHOjnRzo50c6OdHOjnRzo50c6OdHOjnRzo50c6OdHOjnRzo50c6OdHOjnRzo50c6OdHOjnRzo50OSbRzo50c6OdHOjnRzo50c6OdHOjnRzo50c6E09gCp0AAAAAAAAAAAAAAAAAAAKtaUZWR9ZqH1mofWah9ZqH1mofWah9ZqH1mofWah9ZqH1mofWah9ZqH1moUq85TSY6kj2kj2kj2kj2kj2kj2kj2kj2kj2kj2kj2kj2kj2kj2kidSSg2j6zUPrNQ+s1D6zUPrNQ+s1D6zUPrNQ+s1D6zUPrNQ+s1D6zUPrNQ+s1CpWnFRt1R9ZqH1mofWah9ZqH1mofWah9ZqH1mofWah9ZqH1mofWah9ZqH1moQxE3JIq/GwACG4BoF0waTkcsTlicsTlicsTlicsTlicsTlicsTlicsTlicsR7gAAAACirHLE5YnLE5YnLE5YnLE5YnLE5YnLE5YnLE5Yjiro5YnLE5YnLE5YnLE5YnLE5YnLE5YnLE5YnLE5YiSTYBU6AAAAAAAAAAAAAAAAABSoqSvJ6FaioJOL0KVJSTcnoitSUGrbMMR8YAAAAGH/iIYAAAAFT+EwAAAACt8MfQAAAACn8a9Sr8bAAIbgEtZE9rDXK1YH8QAAAABLcAAAAAWwAAAAEt0AAAAALdgFToAAAAAAAAAAAAAAAAAF3awneg/InpSiu4/eoejDEfGAAAABh/4iGAAAABU/hMAAAAArfDH0AAAAAp/GvUq/GwACG4D2LaIfxInsD+IAAAAAluAAAAALYAAAACW6AAAAAFuwCp0AAAAAAAAAAAAAAAAAAfu0PUrfDD0I/wZBOVJO0lqe0odvwJtOTtsAAAAYf8AiIYAAAAFT+EwAAAACt8MfQAAAAPaUO34EZ0W1ZalX42AAQ3An2JNOw7OSJ/CdCV76F5EbvcAAAAJbgAAAAC2AAAAAe6AAAACV76F5Ebtu4BU6AAAAAAAAAAAAAAAAABCDnJJFeScrLZFbWEGR/gP1DEfGAAAABD4kBbQAAAArfCAAAAAPUAAAACn8a9Sr8bAAIbgOKZKKVhpKxPVWB/EAAAAAT3AAAAAAAAAAG9UAAAAALdgFToAAAAAAAAAAAAAAAAAGHg+RyW5LDzirtkY+0pWW6Kq5Kah1DEfGAAAABH4kBFXiwAAACt8KAAAAAK3wx9AAAAAKfxr1KvxsAAhuANXsSV1YUXfUH8QAAAABPcAAAAAAAAAAlugAAAABbsAqdAAAAAAAAAAAAAAAAAAU5JWTHUm1ZsjJxd0yTcndhiPjAAAAAj8SApq6YAAABW+FAAAAAFb4Y+gAAAAFP416lX42AAQ3AAB/EAAAAAT3AAAAAAAAAAJboAAAAAW7AKnQAAAAAD2sfa+y62v+j+7T7wAAAAAAAAAAAADEfGAAAABH4kBT2JKzAAAArfCgAAAACt8MfQAAAACn8a9Sr8bAAIbgAA/iAAAAAJ7gAAAAAAAAAEt0AAAAALdgFToAAAAABneNWEzHB1G9HzRfo+X8nqAAAAAAAAAAAAAYj4wAAAAI/EgKfwlRagAAAVvhQAAAABW+GPoAAAABT+NepV+NgAENwAAfxAAAAAE9wAAAAAAAAACW6AAAAAFuwCp0AAAAAA46nerQj2Tf32/yMkxv1zAU6retrP1Wj+/f5gAAAAAAAAAAAAYj4wAAAAI/EgI6QKq2AAAArfCgAAAACpJSUUuiAAAAAp/GvUq/GwACG4AAP4gAAAACe4AAAAAAAAAAAAAAALdgFToAAAAABxlWVTMuRfZil+v6nBGNtOphW9/eX5P9PuAAAAAAAAAAAAAxHxgAAAAR+JAP+GS1gAAABW+FAAAAAAAAAAFP416lX42AAQ3AAB/EAAAAAT3AAAAAAAAAAAAAAABbsAqdAAAAAAM0xX1rG1ay2bdvTp+BleMeDxlOuuj19Nn+AmpJNAAAAAAAAAAAABiPjAAAAAj8SAl8CI6wsAAABW+FAAAAAAAAAAFP416lX42AAQ3AAB/EAAAAAT3AAAAAAAAAAGwAAAABbsAqdAAAAAA4gxqwmXVJp6tWXq/8ld/IDhfG/Wsugm/eh7r+W34WAAAAAAAAAAAADEfGAAAABH4kBP4EUuw1Z2AAACt8KAAAAAKkFFRt1AAAAAp/GvUq/GwACG4AAP4gAAAACe4AAAAAAAAABLdAAAAAC3YBU6AAAAABxxim6tLDp7Jyfz0X5P7wOFJvC5hVwr2krr5ar8GAAAAAAAAAAAAGI+MAAAACPxICp8KKfxFRe8AAAFb4UAAAAAVvhj6AAAAAU/jXqVfjYABDcAAH8QAAAABPcAAAAAAAAAAluhtIAAAABbsAqdAAAAAA4rq+0zWou1l+C/UBReHzvCT/mhH8YuP6IAAAAAAAAAKNCPKnLc9ip1JLsAAGI+MAAAACPxICrsiOjRVWqAAACt8KAAAAAK3wx9AAAAAKfxr1KvxsAAhuAAD+IAAAAAnuAAAAAAAAADkkOfYbbLXIPoAAAAC3YBU6AAAAABxA75nW9QUXJpLcz2l7LMsBbul9zj/AJgAAAAAAAACV3YlO01BFP8AjSJbsAAxHxgAAAAR+JAtyr0CprFMAAAK3woAAAAArfDH0AAAAAp/GvUq/GwACG4DdkKV3YbshSu7A/iAAAAAJ7gAAAAAAAA5oc30G2zlbPZjik0ElZ6Cd0AAAALdgFToAAAAAHEkOXNay8/zSYZKqbzGgqiuuZf8D8TP4qea4GK6Nv8AFP8AQAAAAAAAAAWjJQbqRkiELSlJj1YABiPjAAAAAj8SCO6Ku6DemAAAFb4UAAAAAVvhj6AAAAAU/jXqVfjYABDcDcnsiWtkT2uD+IAAAAAnuAAAAAAANpDmhzbNWcjYqaFFIBpMJK6IOzsAAAALdgFToAAAAAHGeHcMwU+kor71p/kFGo6VSM1umn9x7WON4ihyO8acfxa/+2X3AAAAAAAAABQqKUUuqK9RRjbqAAGI+MAAAACPxII/EipuFPVWAAACt8KAAAAAHqAAAABT+NepV+NgAENwBPmeo/iRP4QfxGgAAAAE9wAAAAABySHPsOTZa4oNigiyAAAmupF3QAAAaC3YBU6AAAAABxngXWwca8VrB/g9/wAbAcFYFwoTxMt5uy9Fv97/ACAAAAAAAAACtS9m1qUqXtG9Rq2gABiPjAAAAAj8SCHxIqfEFPcmtWAAAVvhQAAAAAAAAABT+NepV+NgAENwCew/iRPYJK7sez8yMbAAAABPcAAAAAckhz7DbYotipnIgAAAo4XnyytWW8Z018pKp/kgauhe6wAACUbns/MirNoAqdAAAAAAqU4VIOE1dPRozrh7EYCq3CLlTez/AEfZ/mZRkOJx9VLlcYdW1+Xd/wBMoUKdGlGlTVopWQAAAAAAAABQoxmncqUY1LXKdKMNiWrYABiPjAAAAAj8SCn8RU+IIfEiotQAACt8KAAAAAJQcUvMAAAACn8a9Sr8bAAIbgNpEmnoiW1xtSaSB/EAAAAAT3AAAAAFFsVM5UgAAACW6Mnw3tOFszqfyyoP8Zr9QJxurkHdWAAABbsAqdAAAAAAAAAAAAAAAAKFVQvcqVYwtcniU42igAAxHxgAAAAR+JBT3J/EwKq0QAABW+FAAAAAFb4Y+gAAAAFP416lX42AAQ3AaTFFLY3FFLYH8QAAAABPcAAAAAAAAAAlujhXDe14Kzj/ALhv/wABfMANcrN0AAALdgFToAAAAAAAAAAAAAAAAKNSp5n1ep2AADEfGAAAABH4kFPcluwJawAAAK3woAAAAArfDH0AAAAAp/GvUq/GwACG4AAP4gAAAACe4AAAAAAAAABLdHh9hfbcE5su6n96p3QBJXRB9AAABbsAqdAAAAAAAAAAAAAAAACMVGKSG0kMAAxHxgAAAAR+JBSHuBDWFgAACt8KAAAAAK3wx9AAAAAKfxr1KvxsAAhuAAD+IAAAAAnuAAAAAAAAAAS3R4SYb23CeLh/POa/8pwQATVncTugAAFuwCp0AAAAAAAAAAAAAAAAjuhv98l5FFe/KI1Z2AAMR8YAAAAEfiQUuowKXYkrOwAAFb4UAAAAAVvhj6AAAAAU/jXqVfjYABDcAAH8QAAAABPcAAAAAAAAAAau0eDcOXhq/epJ/hFfoY2j7HE1KX8smvudgBpNWIuzsAAAt2AVOgAAAAAAAAAAAAAAABUaVWLLJaj1YABiPjAAAAAj8SCnswCn8RUXvAAAVvhQAAAABKbkkuwAAAAFP416lX42AAQ3AAB/EAAAAAT3AAAAAAAAAADwjhy8LUX3lN/+TNfocV0fY57jafarP/0p2/AAJx6kXdAAAt2AVOgAAAAAAAAAAAAAAAFr6E6c4/ESrzlHlAADEfGAAAABH4kEPhYAtGiqtgAAK3woAAAAAAAAAAp/GvUq/GwACG4AAP4gAAAACe4AAAAAAAAAAeFkOXhLCefP/wCnJHiLR9lxRjY/3r/ek/1ADc+Fm6AAFuwCp0AAAAAAx+MWGrULvSUuX71p+KQAAAAAAAAAC0ZiFzwTicsuwAAYj4wAAAAI/Egj8DACesUwAAK3woAAAAAAAAAAp/GvUq/GwACG4AAP4gAAAACe4AAAAAAAADsOce46iHUfQc5PqeH9VYbhXL0/taf+BSkzxbp+z4rrv+aMH/5Kl+gAE43RB9AABbsAqdAAAAAAOOMRy+wpxeur/JL9TLMYsZg6ddfaWvrs/wAQAAAAAAAAAFFJWQMAAxHxgAAAAR+JAv4ZFXTAFrAAACt8KAAAAAK3wx9AAAAAKfxr1KvxsAAhuAAD+IAAAAAnuAAAAAAANpDnEdVdB1Gxyb6m5ySfQ9mz2S7jhFNGW1fqvC+TW+1Ogv8AwJ/8E8b8O4cQUqi2lSj96lJflYhO+jAAkrMTugAFuwCp0AAA0NDQAA4nxv1rMp2ekfdXy3/G5wTjG6U8NLp7y9Nn+NvvAAAAAAAAAA9vU7nt6ncAAMR8YAAAAEfiQP8AhkNiaswKeqsNAABW+FAAAAAFb4Y+gAAAAFP416lX42AAQ3AAB/EAAAAAT3AAAAAByiuo6kR1ew6kmNt9SzexySZ7J9z2SOSJZAEt0cQ1lhOEcll2nh/whf8AQ8c6Fsbg6veMl9zT/UnC2xCd9GAEldEXZ2AAW7AKnQAAGixYsABJqMW+xOTnJye7OF4OFTDzX2nUXySi1+KYAAAAAAAAAAAAGI+MAAAACPxIJfAhaRJq6uBT3Jr3gAArfCgAAAACt8MfQAAAACn8a9Sr8bAAIbgAA/iAAAAAJ7gAAAXRdFy49TlbPZyZ7LzPZxOVIAAAlujxOk6PCOWwW6dP8KbPG+KrYPAYhbXn/wCTKL/QauicOUhO+jACcepF3QALdgFToAAAAAEoqUWicHCTi90cPyhUp4KMd4+0b/L9UAAAAAAAAAB7OXY5JdgAAxHxgAAAAR+JBP4ES+EjrGw0EPiRVWoAAVvhQAAAABW+GPoAAAABT+NepV+NgAENwAAfxAAAAAE9wAAAZYsWLIskAAAAA1qjxj/d5Jl9Lz/KKX6nib/tPBeXYnzpv/wKm3/kDSaJR5WQnzaADV0fCwAW7AKnQAAAAADijKp4TGSqpe5N3T83uvv/AAODssqUKMsRVVnLRJ9u/wA3+QAAAAAAAAAAYmmnHmW4UYRk25bIxCSnoUuWEOdldRklOPUMR8YAAAAEfiQVPhRPYg9Sa1AqapMAAK3woAAAAAqSUlFLogAAAAKfxr1KvxsAAhuBK9tCziSdkWcWD+IAAAAAnuAAAMSAAAAAABRcmkjxzajQwFNd6n4KH+ZxF/tPhjhqn8ip/g+QBpNWJRcWRkpIAnG6IPSwAt2AVOgAAAAADSaswAAAAAAAAALXK83TSjEozlODV9SMnUou+4GJ/iFXSnBC1oPyYVMPzu9z6o+5OPLJoAAAAj8SCpuip0CXvRAesAAArfCgAAAAAAAAAPqj7kcK1JO5V+NgAENwCfwj3RPYJNJnOhSTAAAACe4AABuAAAAAABgIe0xdKHeSX4o8dZ3rYGHZVH97h/kYX/avCmcOsU//ACWrzfkANJqxKLiyMuZAElyvQTuhySOdEXdsAqdAAAAAAAAAAAAAAAAFo0V6Tmk1uUafs4u5R/gy/roBif4hW+GHoR/gyCu2p6M5pdx6gAAAEfiQtyeskT3CD0sNWdghrGwAAVvhQAAAAAAAAAHNLuU5NzWpV+NgAENwCWrSH8SJ/CdBpORyoskAAAAE9wAAHqWAAAAAAAyKPPm2Fh3qQX/kyPHKd8wwke0H+Mv+AcE/7V4dY+l/Kqq+6Cl+oANJqw04sjJSQEldEHZ2GkzlQkrsAqdAAAAAAAAAAAAAAAAChVUopdSvUUY2W5GqlTcLAYn+IVdacWLSh6sMR8YAAAAEfiRHdEviRPcIuzKi6hS7ElZgAFb4UAAAAAVIKKjbqgAAAAKfxr1KvxsAAhuAR11JbonsD+IAAAAAnuAADEgAAAAAADhSHPxBgI/9Paf/AKWjxvnfO8PDtST++cv8jwh/2jh3MML3b/8AJoW/QACUU1YacWRkpICa6kXdAt2AVOgAAAAAAAAAAAAAAABGEpbI9jU7AGIalPRlJxlDkkytKNlCOyDEfGAAAABH4kR+JD+Ie4HxRCHxFRe8AAVvhQAAAABW+GPoAAAABT+NepV+NgAENwJ3a0LyJK6FeT1B/EAAAAAT3AADcAAAAAAAOB4c/E2BX/TyL+53PGmfNxHBdqUV/wCTTf6ngVW1x1J/9O3/AOlp/oYyj7HEVKX8ra+52AAlFSRrFkZKSBpNWF7rBbsAqdAAAAAAAAAAAAAAAALXIxUYpIGAAYj4wAAAAI/EiHxIe7AIPoSVmLRlVbAAFb4UAAAAAVvhj6FKnz3XkAAAAFP416lX42AAQ3AAB/EAAAAAT3AAHqWAAAAAAADw6hz8VYJf3m/ui2eMM+biea7Qgvwv+p4H1rZviaXenf7pJfqcVUfY57jafarP/wBKYABKKkjWLE1JBON1cg7qwt2AVOgAAAAAAAAAAAAAAAC0aMVNqyTOeXcAAMR8YAAAAEfiRT+Il1AE7MmroJ6wAAK3woAAAAArfDH0MKtWYiHLP1AAAAKfxr1KvxsAAhuAAD+IAAAAAnuAAxIAAAAAAADwthz8XYPy53/5TkeLE+bivErsoL/ySL/U8G63s+JVH+aEl+T/AEPESj7HijGx/vX+9J/qAAEoqSE3Fiaa0BrlZHVsAqdAAAAAAAAAAAAAAAACVJTglLdH1WHcAAMR8YAAAAEfiRT3J7MAI6xsNC1gAAVvhQAAAABW+GPoYRaMqpVKd0AAAAU/jXqVfjYABDcAAH8QAAAABPcADcAAAAAAAA8I6fNxTRfaM3/5K1+p4mz5+K8Y/OK+6EUeF1b2XFeE8+ZffCX62PFqj7PiqvL+ZQf/AJKl+gAAEoqSE3Fiaa0JK6IAFToAAAAAAAAAAAAAAAAfWaZ9ZpjAAMR8YAAAAEfiRT3KmwAQepNalPZoaAArfCgAAAAAw2kGzCz+yVocs7AAAAU/jXqVfjYABDcAAH8QAAAABPcAHqWAAAAAAAAPBuHNxLftTk/yX6niBPn4nxr/AL7X3JI4Grex4kwMv+nkV97t+p410eTiCnP+alH71KS/yAAAJwUkJuLE01oLdgFToAAAAAAAAAAAAAAAA6clFS6H1aofVqgAAYj4wAAAAI/EikVAAJaxKe5Ne8ABW+FAAAAABR0oy+ZGTjJNFeKnT5kAAABT+NepV+NgAENwAAfxAAAAAE9wBiQAAAAAAAAeCcL8QVX2pS/9LgcaT5+Isc/+ns190mjI63sc0w1X+WcH90kzxzo2xuDq94yX3NP9QAAAnDmRGTixWbYBU6AAAAAAAAAAAAAAAAVJJ0UXRdDAAMR8YAAAAEfiRS6lQACDurC0kVVqABW+FAAAAAEoOKXmQ0oMMNO8eVlWHJJoAAAKfxr1KvxsAAhuAAD+IAAAAAnuAbgAAAAAAAAeB9O+bYmfanb75L/I4mn7TO8ZLvVqP/yZkJOElJbo8cIKrg8BiFtef/kyi/0AAAAnC60KewBU6AAAAAAAAAAAAAAAAJXdipRlBK4AAGI+MAAAACPxIp7E9wAIuzJrqVNUmABW+FAAAAAFb4Y+hCPNQsSi07Mpy5ZJmJgpQ5l0AAACn8a9Sr8bAAIbgAA/iAAAAAJ7gPUAAAAAAAAA8DIXxeMl2jFfe3/kZvPnzCvLvOX5sPE7/aeDMuxK703/AOBU2/8AIAAAAW7AKnQAAGy5cuAAAAAAAAAAAAFZe1ppxPZT7AABiPjAAAAAj8SKfwk9wAD4okbSjYatoAFb4UAAAAAVvhj6FJfu0VaSmtNxpp2Zh5KUOVlSHLJoAAAp/GvUq/GwACG4AAP4gAAAACe4PYSAAAAAAAAAPAmH+/y/69r/ANLMRP2lWUu7bDiP/afDLC1P5FT/AAfIAAAALdgFToAAFrlkWRZAAAAAAAAAAAABZAwADEfGAAAABH4kU1eOg9WAAQfQT5ZFSPUAK3woAAAAArfDH0FpSXoUatvdexVpKautynJwmYqF0pIAAAp/GvUq/GwACG4AAP4gAAAACe4bsAAAAAAAAAPBP93l+Pq+cfwjJ/qBhv8AavCmUesU/wDyWrf8gAAABbsAqdAAAAAAAAAAAAAAAAD2k+57SfdgABiPjAAAAAj8SPDrCU8XxHhKNSPNGTd0+q5ZXX3HGXDdXh/OKuClrFO8X3i9n+j80wABOzJq60IPmViUbOwBW+FAAAAAFb4Y+hHA1auBq4iC0p8t/JSdk/vsvmFGtb3WVqKmrrcotTp8r6EouLaAAAp/GvUq/GwACG4D2IyuyTshN3swfxA3ZHOznZzs52c7OdnOznZzs52c7OdnOxu7OFeHK3EGa08BSly8125WvypK7drq/bdatH/EBf8AqYf+Uv8A7of8QF/6mH/lL/7of8QF/wCph/5S/wDuh/xAX/qYf+Uv/uh/xAX/AKmH/lL/AO6H/EBf+ph/5S/+6H/EBf8AqYf+Uv8A7of8QF/6mH/lL/7of8QF/wCph/5S/wDuh/xAX/qYf+Uv/uh/xAX/AKmH/lL/AO6H/EBf+ph/5S/+6H/EBf8AqYf+Uv8A7of8QF/6mH/lL/7of8QI/wCph/5S/wDuh/xAj/qYf+Uv/uh/xAj/AKmH/lL/AO6H/ECP+ph/5S/+6H/ECP8AqYf+Uv8A7of8QI/6mH/lL/7of8QI/wCph/5S/wDuh/xAj/qYf+Uv/uh/xAj/AKmH/lL/AO6H/ECP+ph/5S/+6H/ECP8AqYf+Uv8A7of8QI/6mH/lL/7of8QI/wCph/5S/wDuh/xAj/qYf+Uv/uh/xAj/AKmH/lL/AO6HD3B64VyXGU/b+15lKV+XltaL0tzSue18j2vkeGOCWdcE4jAOXLzSnC9r2uou9rq9r3tdep/xAX/qYf8AlL/7of8AEBf+ph/5S/8Auh/xAX/qYf8AlL/7of8AEBf+ph/5S/8Auh/xAX/qYf8AlL/7of8AEBf+ph/5S/8Auh/xAX/qYf8AlL/7of8AEBf+ph/5S/8Auh/xAX/qYf8AlL/7of8AEBf+ph/5S/8Auh/xAX/qYf8AlL/7of8AEBf+ph/5S/8AuhxRkX9h5xXy72nP7O3vW5b3ipbXdt7bgFToAAAAAAAAAAAAAAABUpxVFPqezh2R7OHZDAAMR8YAAAAEfiR4eY6ll+e0sbVV40o1JNelOR4u8P0s5yOnm+E950lzXX2qctb/AC0kuy5hqwABGSasWs7oqRugCt8KAAAAAJSckl2PDPI1m+U5xhbXcqcEv8Xvyj/5NFFSm4SsFGt9lnLaXMivR51dbgAAU/jXqVfjYABDcBq6F8RLVpE9gfxBBU3JKpe3luezy/vP8D2eX95/gezy/vP8D2eX95/gezy/vP8AA9nl/ef4Hs8v7z/A9nl/ef4Hs8v7z/A9nl/ef4Hs8v7z/A9nl/ef4Hs8v7z/AAGsuTt7/wD5KcEcaZbwxVq1o4eVSc0ldySslq0tHu7X9Ef8R3w3/QC//A1/8if8R3w3/QC//A1/8if8R3w3/QC//A1/8if8R3w3/QC//A1/8if8R3w3/QC//A1/8if8R3w3/QC//A1/8if8R3w3/QC//A1/8if8R3w3/QC//A1/8if8R3w3/QC//A1/8if8R3w3/QC//A1/8if8R3w3/QC//A1/8if8R3w3/QC//A1/8if8R3w3/QC//A1/8if8R3w3/QE//A1/8if8R0w//QE//A1/8if8R0w//QE//A1/8if8R0w//QE//A1/8if8R0w//QE//A1/8if8R0w//QE//A1/8if8R0w//QE//A1/8if8R0w//QE//A1/8if8R0w//QE//A1/8if8R0w//QE//A1/8if8R0w//QE//A1/8if8R0w//QE//A1/8if8R0w//QE//A1/8if8R0w//QE//A1/8if8R0w//QE//A1/8if8R0w//QE//A1/8iZn400MZga2Gjg2nOMo351pdNX+Hpc9l5nsvM4A8RqfC+Bq4WdB1OaXNdStbRLs+x/xHjD/APQC/wDwNf8AyJ/xHjD/APQC/wDwNf8AyJ/xHjD/APQC/wDwNf8AyJ/xHjD/APQC/wDwNf8AyJ/xHjD/APQC/wDwNf8AyJ/xHjD/APQC/wDwNf8AyJ/xHjD/APQC/wDwNf8AyJ/xHjD/APQC/wDwNf8AyJ/xHjD/APQC/wDwNf8AyJ/xHjD/APQC/wDwNf8AyJ/xHjD/APQC/wDwNf8AyJ/xHjD/APQC/wDwNf8AyJxVnkc8zmvmEYciqW0ve1oqO9l27AFToAAAAAAAAAAAAAAABKrJwUT61LsfWpdgAAxHxgAAAAR+JGW1PZUMTJb+zt/4FKMX+DZ4McSQzHKqmT4l3lSWifWnLdefK3Z+TSON+GpZDnNbB29z4oPvF7fd8L80xgAEZX0YicbMCt8KAAAAAIq7R4H4+jQrV8LL4q2q/wC4aV1907/JniHkv9m8Q4vDxVlzOUf8MveSXpe3yAo1r+6xMr0L+9EAAKfxr1KvxsAAhuAfaH8SJ/CdCV76F5EbvcAAAAJ7gAAAAAAAAAAAAAASvfQvIjdt3AKnQAAAAAAAAAAAAAAACUI+wTtqU6UZU3J7oAADEfGGHpQnF3R9Xp9j6vT7H1en2Pq9PsfV6fY+r0+x9Xp9j6vT7H1en2Pq9PsfV6fY+r0+x7Cmuh7TkpTivtK34p/ocLZ/WyLNqOPpfZeq7xekl81t2dn0PFXIKOe5DDNsH70qS5019qnJJv7l7y7K/cnG+oAAQk9jG4GvhanscRBxlZOz0dmk196aPZxPZxJUoSWqPq9PsfV6fY+r0+x9Xp9j6vT7H1en2Pq9PsfV6fY+r0+x9Xp9j6vT7H1en2Pq9PsfV6fY+r0+wqFNPRHCue/2TnmBxDdowm+b/DNKMvwueOuVcmKwuYRXxJwfrHVfem/uK9PllddQp03N6HDHgxPNMroY6vjOT2i5uX2d2k9ve51urP4etvM438LcVw/hli8PU9tSXxO1nF9G1d+69r9HvuOhTbvY+r0+x9Xp9j6vT7H1en2Pq9PsfV6fYVCmndIq/GwACG4B9oe6J7A/iAAAAAJ7gAAAAAAAAAD3QAAAAAt2AVOgAAAAAAAAAAAAAAAEv93RR/gy+f5AAAYj4whVlBWR9ZqH1mofWah9ZqH1mofWah9ZqH1mofWah9ZqH1mofWagsRNtIlJyQeC3E8Mbl88lxDvKmm436wb1X/brf3SS6HHHDcsgzurhEv3b96D7xe33fC/NXJxtqAAo3PCngtZ1mP1vERvQotN32lLdR9FvLysup4447LJZhRoUY3xMV78r7R3jFrvrfyVu+ntJHtJFStKK0PrNQ+s1D6zUPrNQ+s1D6zUPrNQ+s1D6zUPrNQ+s1D6zUPrNQ+s1D6zUKlacVG3VE5ubuzP3/rR4Zwxa1qU4Rn/29T92b+aUvvNKsBQbly9TLcBPFYqjhKXxVJRivWTS/U4246wvBOFwuHhQ9o5JqMebltGCitXaXdJadCr9ICFWm6c8sTi1Zp1dGnumvZFXiHh6c5SWWNJt6e2lp5fAVM9yVv3Mvt61ZP8A9tRPPMrcfdwKT86k3/kf21gv+gOP/gU//kh5vSvphYffU/8AlhUzOM3dUYr05/1myeOco2UEvS/6tkMRNySKvxsAAhuAfaJrQvzNA/iAAAAAJ7gAAAAAAAAAEt0AAAAALdgFToAAAAAAAAAAAAAAABCCnSSZGnGMXFH1amMAAxHxgAAAAR+JAcNZ5WyTNaGPpbweq7xekl81deT1PFPI6PEPDkM0wfvSpLni11g0nJfdaXya6iakiUbMAjC5k+U4nNcdSwWFjec3ZeXdvyS1fkjMMXl3AXC6jTV3BWinvOo+r9XdvslZbIx2Nr43EzxOIlzTm2231b/r5AVvhQAAAABW+GPoHgVmVPGZVjMpraqL5rd4zVpL0uv/ACYzLA1MrzKvgam9OUo/c7X+ZyxTueD+Wf2lxVTqtXhQjKfz+GP4yuvQ8Z83+v8AFE6MXeNGKgvX4pfjKz9AAAKfxr1KvxsAAhuABZA/iAAAAAJ7gAAAAAAAAAEt0AAAAALdgFToAAAAAAAAAAAAAAABDERjFKx9ah2PrUOwAAYj4wAAAAI/EgDwS4mjicHUyXEO7heUL9Yt+8vk3f8A7efRHiBww+Hs8qYeC/dS96H+F9P+3XdfJPqNKSGrBGHclJI8IuDo5Vl7zbGK1WqtL/Zp7/Jy3flbbU8SeMpcR5s3Sf7ineMF37y/7e/BWW9wCt8KAAAAAK3wx9A8IM4/s7iqhGTtGsnTfz1j/wCTKK+Z43ZP9T4k+tRXu14qX/b0fdf4JN+pKs3BRPA7AU8BkmMzetopO1/7tNXb+9v7jM8dUx+NrYup8VSUpP1k2/1AAAp/GvUq/GwACG4AAP4gAAAACe4AAAAAAAAABLdAAAAAC3YBU6AAAAAAAAAAAAAAAAAAAGI+MAAAACPxIA4ezuvkuZ0cfQ+KDvbutmvmro8Sslo8UcM08ywXvTpr2ke7g1eS9ba23vG3UjKxKN0RikSlbY8LuDHxBmntsRG+Ho2cu0n0j895eWnVHjLxmsFhf7Fwkv3lRe+19mH8vrLr/d/xABW+FAAAAAFb4Y+gYTE1MLiKdek7Sg1Jeqd1+J4xYannPCeFzigrqDjL0hUST/8AJuQOJf8AjGPDOng1pUqRjD/t6p71RfdzIAAAp/GvUq/GwACG4AAP4gAAAACe4AAAAAAAAAA1doAAAAAW7AKnQAAAAAAAAAAAAAAAAAAAxHxgAAAAR+JAB4IcUKrh6mS15axvKF+sX8Ufk9beb6I8RuGP7Az2pRpq1Kfvw9H0/wC3Xdeln1Iy6EpX0RleW4nM8ZSweGjzVJuyX6+iWrfRak5ZdwBwtpryL0dSo/8AN+toryMyzHEZji6mLxMuapNtt+b/AEWyXRaABW+FAAAAAFSSkopdEB4cyjxJwDWyqbvKCnT+9c0H8m7L/CcG5LLM+IsLgZx3muZeUfekvuTPH7N+fFYTLYv4U5v1k+WP3JP7wAACn8a9Sr8bAAIbgAA/iAAAAAJ7gAAAAAAAAAAAAAAAt2AVOgAAAAAAAAAAAAAAAAAABiPjAAAAAj8SADI83r5RmNHHUH71Np+q6r0aun5M8Qsnw/FnC0Mwwa5pwj7SHdxa96Prbp/NFIDwc4MWAwf9s4uNqlRe5f7MO/k5f+k27s8T+M3xBmrp0Jf7PSuo/wB59Z/PZeSW12ABW+FAAAAAAHgPnH1fOq2Ak9K0Lr/FDX/0ly+44S4P+o8fZni+X3IpOPa9b3nb0tKJ4i5v/avE+Mrp3ipcq9Ie7p62v8wAACn8a9Sr8bAAIbgAA/iAAAAAJ7gAAAAAAAAAAAAAAAt2AVOgAAAAAAAAAAAAAAAAAABiPjAAAAAj8SAA8EOKXKNTJK8tryp+n2o/+3Jf4jxL4VeQZ5OFNWo1bzh5JvWP/br09LPqeGnBr4izVOtH/Z6VnPz7R/7etr5J9bHjBxlHK8AsowkrVaq9632YbW8nLb/DfugACt8KAAAAAAOFc3eUZ3hcdfSE03/h2l98W0cTZpTyrJ8VmKteEG0+7s+Vf+BNfeNuTu3qAAAU/jXqVfjYABDcAAH8QAAAABPcAAAAAAAAAAb1QAAAAAt2AVOgAAAAAAAAAAAAAAAAAABiPjAAAAAj8SAAybNa+VZhRxuHfv02mvPun5NXT8mcZ5LR434YpYnAa1NJ076b6Si307P+9FGFoZdwDws3N35FdvZzqPovV6LtFa7MzfNcTmuOq43EyvObu/0S8ktEuiQABW+FAAAAAFSCio26oA4w4v8Arnh1l9Hm/eVWoy9KOjv6vkfzAAACn8a9Sr8bAAIbgAA/iAAAAAJ7gAAAAAAAAAEt0AAAAALdgFToAAGH4YzvEUo1aODqyhLVNU5tNd00rM/1Q4h/6AK3/gqf/wAif6ocQ/8AQBW/8FT/APkSvwvnlClKrVwVWMYq7bpzSS7ttWSAAAAAAAAAAAAAAADEfGAAAABH4kAAeB/FFnUyWtLe86f/ALdH/wBuS/xM8VeNP7dzP6rhpXw9FtLtKWzl6dI+Wq+IAAK3woAAAAArfDH0AJV6sqUaLleMW2l2btf77L7gAAAp/GvUq/GwACG4AAP4gAAAACe4AAAAAAAAABLdAAAAAC3YBU6AABwPnuVUeHMFTqYmnGSgk05xTXqm9D/WPJv+gun/AOBx/wAz/WPJv+gun/4HH/M4zz7Kq3D2Np08VTlJ05JJTi23bokwAAAAAAAAAAAAAAAxHxgAAAAR+JAAFDEVcPUVSjJxkr6p2eqs/vWgAAFb4UAAAAAVvhj6AAAAAU/jXqVfjYABDcAAH8QAAAABPcAAAAAAAAAAlugAAAABbsAqdAAAAAAAAAAAAAAAAAAADEfGAAAABH4kAAAAAVvhQAAAABW+GPoAAAABT+NepV+NgAENwAAfxAAAAAE9wAAAAAAAAACW6AAAAAFuwCp0AAAAAAAAAAAAAAAAAAKUIcrnLYrQirSjswxHxgAAAAR+JAAAAAFb4UAAAAAVvhj6AAAAAU/jXqVfjYABDcAJOyE2nqD+IAAAAAnuAAAAAAAAAAAAAAAC3YBU6AAAAAAAAAAAAAAAAAAFTSlFG9D0YV4Sc9Eezn2Y01oAAAAR+JAAAAAFb4QAAAABgAAAB7OfZkKc1NaFX42AAQ3ACWrSJ7A/iAAAAAJ7gAAAAAAAAAAAAAAAt2AVOgAAAAAAAAAAAAAAAAABX+GHoR/gyCpXlCVkj61LsSlzO4AAABh/4iGAAAABU/hMAAAAArfDH0AAAAD61LsRxMpSSsVfjYABDcAH8SJ/CdCSuz2fmRjYAAAAJbgAAAAC2AAAAAe6AAAACUbs9n5kVZtAFToAAAAAAAAAAAAAAAAAAVdacGLSg/UMR8YAAAAGH/iIYAAAAFT+EwAAAACt8MfQAAAACn8a9Sr8bAAIbgBLdE9gfxAAAAAEtwAAAABbAAAAAS3QAAAAAt2AVOgAAAAAAAAAAAAAAAAABTSqU+TqivaMVBdAxHxgAAAAYf8AiIYAAAAFT+EwAAAACt8MfQAAAACn8a9Sr8bAAIbgBJXRfmaB/EAAAAAS3AAAAAFsAAAABLdAAAAAC3YBU6AAAAAAAAAAAAAAAAAAG2wFajOcro+r1Ox9Xqdj6vU7H1ep2Pq9TsfV6nY+r1Ox9Xqdj6vU7H1ep2Pq9TsfV6nY+r1Ox9XqdilRnGabQ6cj2cj2cj2cj2cj2cj2cj2cj2cj2cj2cj2cj2cj2cj2cidOTptI+r1Ox9Xqdj6vU7H1ep2Pq9TsfV6nY+r1Ox9Xqdj6vU7H1ep2Pq9TsfV6nY+r1Ox9Xqdj6vU7FSjOSil0R9Xqdj6vU7H1ep2Pq9TsfV6nY+r1Ox9Xqdj6vU7H1ep2Pq9TsfV6nY+r1Ox9Xqdj6vU7EKE1JOxV+NgAENwAAduY5l3OZdzmXc5l3OZdzmXc5l3OZdzmXc5l3OZdzmXc5l3OZdzmXce4AAAAAmrbnMu5zLucy7nMu5zLucy7nMu5zLucy7nMu5zLucy7nMu5zLuNptHMu5zLucy7nMu5zLucy7nMu5zLucy7nMu5zLucy7nMu5zLucy7is2wCp0AAAAAAAAAAAAAAAAAAAPb1Lbn1ip3PrFTufWKnc+sVO59Yqdz6xU7n1ip3PrFTufWKnc+sVO59Yqdz6xU7n1ip3PrFTufWKnc+sVO59Yqdz6xU7n1ip3PrFTufWKnc+sVO59Yqdz6xU7n1ip3PrFTufWKnc+sVO59Yqdz6xU7n1ip3PrFTufWKnc+sVO59Yqdz6xU7n1ip3PrFTufWKnc+sVO59Yqdz6xU7n1ip3PrFTufWKnc+sVO59Yqdz6xU7n1ip3PrFTufWKnc+sVO59Yqdz6xU7n1ip3PrFTufWKnc+sVO59Yqdz6xU7n1ip3PrFTuNtu4ABDcAAHFM9mj2aPZo9mj2aPZo9mj2aPZo9mj2aPZo9mj2aPZo9mj2aPZo9mj2aPZo9mj2aPZo9mj2aPZo9mj2aPZo9mj2aPZo9mj2aPZo9mj2aPZo9mj2aPZo9mj2aPZo9mj2aPZo9mj2aPZo9mj2aPZo9mj2aPZo9mj2aPZo9mj2aFFIAqdAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAhuAAAAAAAAAAAAAAAAAAAAAAAABU6AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABDcAAAAAAAAAAAAAAAAAAAAAAAACp0AAAAAAAAAAAAAAAAAAAAAAAAFFy0Q01oAAAABZtaAAAAAbkoSjugAAAAAAAhuAAAAAAABdMAAAAAAAAAAugAAAAAAqdAAAAAAAAAAAAAAAAAAAAAAAACMvZUk1uyvaUYzXUAAAAC8PZuMewAAAAFKag7sjU9tFxYAAAAAAAQ3AAAAAAACW1i3K0AAAABO7I7IAAAADk08yLugAAAAACp0AAAAAAAAAAAAAAAAAAAAAAAAK+0V5D1ofMAAAACl8EwAAAADD/E35AAAAAAABDcAAAAAAAHuiewAAAAEtiOyAAAAAIbAAAAAAFToAAAAAAAAAAAAAAAAAAAAAAAAYjdegv4D9QAAAAKXwTAAAAAI+5Sb6sAAAAAAAIbgAAAAAAA/iRP4ToAAAAEtiOyAAAAAk9BKyAAAAAAKnQAAAAAAAAAAAAAAAAAAAAAAAArawiyelGK7gAAAAUvgmAAAABCHPJIryTlZbIAAAAAAAIbgAAAAAABLdE9rAAAAAS2I7IAAAADdgAAAAABU6AAAAAAAAAAAAAAAAAAAAAAAAFKpDl5Z7FaopvTZAAAAAQmoxku4AAAAFCpGDdz2tH+X8B2bAAAAAAAIbgAAAAAABJXVhRd9QAAAAGroSsgAAAAexyyIppagAAAAAFToAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAENwAAAAAAAAAAAAAAAAAAAAAAAAKnQAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAIbgAAAAAAAAAAAAAAAAAAAAAAAAVOgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAQ3AAAAAAAAAAAAAAAAAAAAAAAAAqdAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAhuAAAAAAAAAAAAAAAAAAAAAAAABU6AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABDcAAAAAAAAAAAAAAAAAAAAAAAACp0AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAACG4AAAAAAAAAAAAAAAAAAAAAAAAFToAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAENwAAAAAAAAAAAAAAAAAAAAAAAAKnQAAAAAAAAAAAAAAAAAAAAAAAAAAAhRlNXR9VmSoSjZPqVqPs7agABUpciWoU4c8rXJx5ZNAAAAAAAAAABDcAAAAAAAAAAHojnRzpjlrYAAG7OwCdwAAAAAAAAAqdAAAAAAAAAAAAAAAAAAAAAAAAAAAChVhCLTY5x5eboVa0XZroV6qnawABTjzTSMU7zsFN2mmYqNp3AAAAAAAAAACG4AAAAAAAAAASV0OLQoscbyuAAG8gNpAAAAAAAAABU6AAAAAAAAAAAAAAAAAAAAAAAAAAAA/wCB8iCTkkYmnGKXKh4eDasirQjJK2hKhBxskQg5yUT2NO1rFSHJJow6vURX1qMhQhGNmrsrRUZtIxSuosAAAAAAAAAAIbgAAAAAAAAAAVNkR2Q2+axJtM5nclJvYi7olJt2RF30COrZOVtEQbZL4kAAAAAAAAAFToAAAAAAAAAAAAAAAAAAAAAAAAAAAD/gfIp/GvUxeyK03GnoQm3S5jDzck7lCK9pI5antea2hil7yMN8ZL+N8zEQlJLl6Dbb1MR/DiAAAAAAAAAAENwAAAAAAAAAAKmyI7IfxFTcnFJHKuUhsyDSFbm0CHUl8QrMnugAAAAAAAAAqdAAAAAAAAAAAAAAAAAAAAAAAAAAAAf8D5FP416mL2Rif4aKf8D5MwnUofHMliIJ2sV6iqNWMN8ZW+NlGspqz3MRFKehifgigAAAAAAAAAAhuAAAAAAAAAABU2RHZD+IqblTYXwkdmKNxQaYQ3ZKNyG9ie6AAAAAAAAACp0AAAAAAAAAAAAAAAAAAAAAAAAAAAB/wPkU/jXqYvZGJ/hop/wPkzCdSlJKs13KmHjN3K0FCdkUHaoirG9axHDxjPmTMQ+apZGL6IAAAAAAAAAAIbgAAAAAAAAAAVNkR2Q/iKm5U2F8JDYu4sjK7CPxMcrMjqyWskAAAAAAAAAFToAAAAAAAAAAAAAAAAAAAAAAAAAAADqzceW+gnZ3ROpKe5OrKas2KrJR5U9CFSUPhG23cWJmlYlJyd2QdpJmJTU1JH1mdrFJOVRGJd527AAAAAAAAAABDcAAAAAAAAAABpMtoOKbuOKY0miytYSS2HFMUUg2kOKYkktBayAAAAAAAAACp0AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAlOUkk3sEZOLuhtt3YAAAAAAAAAAQ3AAAAAAAAAAAAAAGkwEkgAAAAAAAAAqdAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAhuAAAAAAAAAAAAAAAAAAAAAAAABU6AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABDcAAAAAAAAAAAAAAAAAAAAAAAACp0AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAACG4AAAAAAAAAAAAAAAAAAAAAAAAFToAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAENwAAAAAAAAAAAAAAAAAAAAAAAAKnQAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAIbgAAAAAAAAAAAAAAAAAAAAAAAAVOgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAQ3AAAAAAAAAAAAAAAAAAAAAAAAAqdAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAALvuXfcu+5d9y77l33LvuXfcu+5d9y77l33LvuXfcu+5d9y77l33LvuXfcu+5d9y77l33LvuXfcu+5d9y77l33LvuXfcu+5d9y77l33LvuXfcu+5d9y77l33LvuXfcu+5d9y77l33LvuXfcu+5d9y77l33LvuXfcu+5d9y77l33LvuXfcu+5d9y77l33LvuXfcu+5d9y77l33LvuXfcbuAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHNHuc0e5zR7nNHuc0e5zR7nNHuc0e5zR7nNHuc0e5zR7nNHuc0e5zR7nNHuc0e5zR7nNHuc0e5zR7nNHuc0e5zR7nNHuc0e5zR7nNHuc0e5zR7nNHuc0e5zR7nNHuc0e5zR7nNHuc0e5zR7nNHuc0e5zR7nNHuc0e5zR7nNHuc0e5zR7nNHuc0e5zR7nNHuc0e5zR7nNHuc0e5zR7nNHuc0e5zR7nNHuc0e5zR7nNHuc0e5zR7nNHuc0e5zR7nNHuc0e5zR7nNHuc0e5zR7nNHucy7gAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABD4kAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAEPiQAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAQ+JAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABD4kAAAAAAAAAAAAAAAENZIqQjyPQAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAIfEgAAAAAAAAAAAAAAAh8SKvwP0AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAACHxIAAAAAAAAAAAAAAAIfEir8D9CNKcldIlFxdmARw8pK5UoygtQAoU4zlZjAKNOM73AdOSipdD2cfZc3UVOTi5LZACV3ZE4ODswAAAACjCM5WZJJNoAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAACHxIAAAAAAAAAAAAAAAIfEiUeaLQkkrIxMFKF+qAJzlGlGzKVR1E4yGrAGF+N+gwDDfaAnVUqaj1P+YchVSpuPUClCLhJvoRdpJlaanK6AAAAAMN8ZL4mAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABD4kAAAAAAAAAAAAAAAEPiQ7JCxEue72JK6sAVv4UTDfGKmpuT7E6KjTu9yjQ51d7Fahyq8djC/G/Qp4ZSjeRVpckrFGgpq7ZWoKCutjDfaIYZON5FWm4SsSpJU1K+5/zDkaSlTcr7FKl7R2K1CMY3TKdJKna+5VoqEkk9yrTUJWRRoc6u9ivRjBXTBYdNpJk6f7zkifVY2tfUhT5p8rJx5ZNFSlyxT7lKmpptvYo0VNXb0K2HUY3iUqam2h0UqXM9yjQU1d7EKPJO62JfEwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAIfEgAAAAAAAAAAAAAAAh8SKvwP0COsUS3YFSDlSjZFGm6cXKRhtWypUlOW+hiHypQRh3zRcGYVe+yrUcp6dCq+eipHM7WKb5qMl2MLuydRyne5XtOnGRU/gRP+Ycp/wAGRS0pSa3Lu1ii37KRFtyRiVepYxEuVKCLu1gqT5KSa3KGrcn0FNqfMVFy1kytH960Yizg/Ip6UZM5naxQd4SiYX436Fao5SaT0K75YKCMLNu8SXxMAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAACHxIAAAAAAAAAAAAAAAIfEiorwYJWSQ9WB7T2dOLsVa0pq3Qw32iGskYn+IYb4yh/FkMX+7so0o8vPPY5oSpy5TDfaB64dFT+BE/5hyn/BkYeai7PqV6Kjqtij/CkQ+JFXWsvkYj+IwK/wAEDDqLhK5yYf8ApmIlFyTiypG9aJfmU0S0oLzKNKMlzS2ISpyuoowvxv0N5GK+NehhvjJfEwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAIfEgAAAAAAAAAAAAAAAh8SBYZKd76FR8sGwCt/CiGG+0RdmmYpe8mYVe82YZ3qMkrNofu0F5lbSnFFFfupGFV2xq2hUXLQiip/Aif8w5T/AIMgg+ag0+hR/hSIfEiu7VUzFRtO4Ff4IFBc0JRAjZ8svIw7vJruivpGK8ielBJGGW5hfjfoPcxSu0zCq8myXxMAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAACHxIAAAAAAAAAAAAAAAIfEirNRjcTTV0Ymon7qAKv8KIYb7QQ5a0Enuipy0ocsd2YbST9BU4VbSMRNNqK6EIxq00n0PdUJQj0Rh3ZSZ7KnUfOYialKy6FS3sYn/MOU/4MijShOGu5V5adPkW7MM1dxfUqU4wnFIxOsyHLWhZ7orUoQirbhX+CBRnySuSw8JO6ZVjFStEjJewZRdqiMS05lJRqU+R9CChB8iML8b9CW5T5atPle6I8lOSgifxMAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAACHxIAAAAAAAAAAAAAAAATaVkwAAB6hdoLtbAXYAbbD1B6gPUAu7WADbYAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAACHxIAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAIfEgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAh8SAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAACHxIAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAKNGE4XZPDSW2o1YAAAIUJzPq0FF9wAAACHxIAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAKdaUNEU588b2J04zWqJ4Vr4WSi07MAC1yGGlLfQhRhHYnVhDdlTESkrLQAAAAh8SAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAACEeaSRWqtS5YuyRGrJ0rrdEMTF6PQajNdyWHXI1EnCUd0Ci5OyIYVv4iFOMdkTrwjpuypXnLTY3AAAACHxIAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAMOr1ESd22UNVJeRL+AvUoyapyaIVk4czE4zWmpKEZKzFGMVoTqqMOZakqvLBSZLk0qle04KaISVKCdtWV0pJTXUAAAAh8SAAAAAAAAAAAAAAAAAAAAAAAAAAClTgo88ypOlKDstSChTgpSV2ys4NpxAAAAApU+eVhSoyfJYqQ5JNATg4NJgAAAAAAAAAAAAAAAAAFKnzyse0oL3bFOEZTb6IqTpSg7LUAAAADD/AMRDVnYobSfkS/gL1KMHKnJIp0VGHK9R8sV2Q8VBOyLp6E6UZRcVoV6cuSKWtiX8CPqL+A/UlShJRu7aFWMY01FO+oAAABD4kAAAAAAAAAAAAAAAAAAAAAAAAAAbmIdmoroFdXjF+QAAAABD3aMmFdc3K+5UhyS5T2OsVfcr0brmvsilRdTXoVaXI0r3FhW476kYNy5SOGbW5Tw7krt2JRadhYVuO+o0AAAAAAAAAAAAAAAQ92i33Ckr05JAAAAAFKXLNMrw5ZspUn7N92RpRUVF6kmoq7JVuam5R6FNupCSb1CtdxjJEarjTUpainFpeZOnGasyVFxpOK1K6bjGQAAAAQ+JAAAAAAAAAAAAAAAAAAAAAAAAAAEfiRiP4jCjOMo8kirT5JWAAAAAf8BeoS/hwMR/EZX2j6GI+JehWfLCMEUVzTSJ1G6nMSS9umupKb9sV5tVNOhOKddDcnU5iurVGAAAAAAAAAAAAAAAS/gL1ClU5JXK1KNueOwAAAAFKg5q/QUUkluTqwhuxYpX1WhOvzJoo1VC6ewqnLPmRUrQlFpLVlKsox5ZK6KtXnemxOalGK7EcQ4xSIV4S8hpNWK9K8Vy9AAAACHxIAAAAAAAAAAAAAAAAAAAAAAAAAAWjMSvfv3ArPmhFgAAAARXNQa7BU0UImI/iMr7R9DE/EvQxG69DDfxBrUl/FgiX8X5lf8AiMf8deg8TJO1ipNzldgAAAAAAAAAAAAAACXNQ9GBRd4SQAAAAFKvGELPcniJy20AAAAhVlDZn1mLi7rUAAAAh8SAAAAAAAAAAAAAAAAAAAAAAAAAACCjWpqL3RPDuMG2z6tzJNMxDV1FdAAAAAKE0nyvZiwyUrt6FWfPO6J0lUanfQxLT5WitDnipp9BRVamknqhxdGabHh1KfNfQ51OsmiX8X5lf+IytLlqpk8Pzy5ovRk48smgAAAAAAAAAAAAAAChNJ8r2Y8K76PQhQU7pPVEoKlTavqwAAAAAAAAAAAAAIfEgAAAAAAAAAAAAAAAAAAAAAAAAAAvbYcnLdilJKyYAAAAAOUmrXDmdrXG7nM7WuJtbMbb3OZ2tcB6jdxSklZMAAAAAAAAAAAAAAADnla1xNrYbb3YAAAAAAAAAAAAAEPiQAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAEYKUW+wAAAAAAQ+JAAAAAAAAAAAAAAAAAAAFmWZZlmWZZlmWZZlmWZZlmWZZlmWZZlmWZZlmWZZlmWZZlmWZZlmWZZlmWZZlmWZZlmWZZlmWZZlmWZZlmWZZlmWZZlmWZZlmWZZlmWZZgAAAAFrlmWZZlmWZZlmWZZlmWZZlmWZZlmWZZlmWZZlmWZZlmWZZlmWZZlmWZZlmWZZlmWZZlmWZZlmWZZlmU1+7kWZZlmWZZlmWZZlmWZZlmWZZlmWAIfEgAAAAAAAAAAAAAAAAAACdfkfLY+t+R9b8j635H1vyPrfkfW/I+t+R9b8j635H1vyPrfkfW/I+t+R9b8iniOeSjY9r5HtfI9r5HtfI9r5HtfI9r5HtfI9r5HtfI9r5HtfI9r5HtfI9r5EqtoOVj635H1vyPrfkfW/I+t+R9b8j635H1vyPrfkfW/I+t+R9b8j635H1vyPrfkTxHKlpufW/IeKt0PrfkfW/I+t+R9b8j635H1vyPrfkfW/I+t+R9b8j635H1vyI4nmaVir8bAAAAAoz5KbfmfW/I+t+R9b8j635H1vyPrfkfW/I+t+R9b8j635H1vyPrfkfW/I+t+R9b8h1NFoe18j2vke18j2vke18j2vke18j2vke18j2vke18j2vke18j2vkKpd2sSxVnax9b8j635H1vyPrfkfW/I+t+R9b8j635H1vyPrfkfW/I+t+R9b8j635EcRzRbtsfW/I+t+R9b8j635H1vyPrfkfW/I+t+R9b8j635H1vyPrfkfW/I+t+R9b8ivLnjF+oBD4kAAAAAAAAAAAAAAAAAFKi6hVoumUqTqMq03B2DEfGAAAABh/wCIhgAAAAVP4TAAAAAK3wx9AqK1vRAAAAFP416lX42AAAAAv4L9QAAAAB/DH0AAAAAj8SJ/EwAAAAKf8OQAAAABU/hw+YBD4kAAAAAAAAAAAAAAAAAHPJR5UXcqDv0Je7RSXUk+ain2DEfGAAAABh/4iGAAAABU/hMAAAAArfDH0CstI+gAAABT+NepV+NgAAAAL+C/UAAAAAfwx9AAAAAI/EifxMAAAACn/DkAAAAAVP4cPmAQ+JAAAAAAAAAAAAAAAAAAbUPVlb4IehH+BIJ06UneT1PY0f5vxJpKTSAAAAMP/EQwAAAAKn8JgAAAAFb4Y+gV17kQAAAPY0f5vxI0qKas9fUq/GwAAAAKMYyptSfU9jR/m/ErQpxS5XcAAAAH8MfQAAAACPxIn8TAAAAAp/w5AAAABSp05R956nsaP834mIUVGKjtqAQ+JAAAAAAAAAAAAAAAAABGLk0kYhq6guhW1hD0I/wGGI+MAAAACDakmgAAAACq2o2AAAAAJSckk+gV1ekgAAACn8a9Sr8bAAAAAX8F+oAAAABBtxVwAAAAL22HqAAAABT/AIcgAAAACp/Dh8wCHxIAAAAAAAAAAAAAAAAAMPBpOa36EqVS12jlc6KtuiouSko9QxHxgAAAAR+JAJXiwAAACt8KAAAAAKsUoxt2Cqr02AAAAU/jXqVfjYAAAAC/gv1AAAAAKfwoI7oas7AAAAPYAAAAAp/w5AAAAAFT+HD5gEPiQAAAAAAAAAAAAAAAAAQrSirIlXnJWZTqSg9Cc3N3YYj4wAAAAI/EgIK8WAAAAVvhQAAAABW+GPoE1eLQAAABT+NepV+NgAAAAL+C/UAAAAAp/Cgj8SKi94AAAB7AAAAAFP8AhyAAAAAKn8OHzAIfEgAAAAAAAAAAAAAAAAAADEfGAAAABH4kBT2GrOwAAAFb4UAAAAAVvhj6C3BqzsAAABT+NepV+NgAAAAL+C/UAAAAAp/Cgh8SKq1QAAAD2AAAAAKf8OQAAAABU/hw+YBD4kAAAAAAAAAAAAAAAAAAAYj4wAAAAI/EgKfwlRe8AAABW+FAAAAAFb4Y+gt0N++kVFabAAAAp/GvUq/GwAAAAF/BfqAAAAAU/hQU/iKq0AAAAewAAAAAptRaXUAAAAAqfw4fMAh8SAAAAAAAAAAAAAAAAAAAMR8YAAAAEfiQENIlVaoAAACt8KAAAAAKrTjG3Yj8SJP96iurTYAAAFP416lX42AAAAAv4L9QAAAACn8KCnuS1iwAAAHsAAAAAAAAAAFT+HD5gEPiQAAAAAE4uUWkZbi1jMJTrr7S/Hr+IAAAAAAAAAAAAGI+MAAAACPxID/l2VNYpgAAAVvhQAAAAAQ+JFV2qxMSveQAAAFP416lX42AAAAAv4L9QAAAACn8KCnuR1ugAAAHsAAAAAAAAAAFT+HD5gEPiQAAAAAE5KMW+xwRjealUwreq1Xo9H+NvvAAAAAAAAAAAAAxHxgAAAAR+JAP4DeAAAAFb4UAAAAAFP40Yh2qIxS2AAAAp/GvUq/GwAAAAF/BfqAAAAAU/hQUupB+8ya1YAAAPYAAAAAp/wAOQAAAABU/hw+YBD4kAAAAABm9ZUcBWn2i/vtZfiZFjfqePp1G9L2fo9Pw3+QAAAAAAAAAAAAGI+MAAAACPxICfwIp6qwAAAFb4UAAAAAThypa7lP40Yn4yvrTTAAAAp/GvUq/GwAAAAF/BfqAAAAAU/hQUupB+8VFqAAAD2AAAAAKf8OQAAAABU/hw+YBD4kAAAAABxnilSwCpJ6zf4LV/jYMhxv1zL6dRvVKz9Vp+O/zAAAAAAAAAAAADEfGAAAABH4kBU+FFPcmrSYAAAVvhQAAAABW+GPoUvjRiPjJa0EAAABT+NepV+NgAAAAL+C/UAAAAAp/Cgp7MKqukAAAD2AAAAAKf8OQAAAABU/hw+YBD4kAAAAABxdjfrGYOmnpBW+e7/y+QcE4twqzw0tpLmXqtH9/6AAAAAAAAABSw/Ors9i3Nxj0AADEfGAAAABH4kBV2RD4kVFqAAAFb4UAAAAAVvhj6FH40Yj4yGtFoAAACn8a9Sr8bAAAAAX8F+oAAAABT+FBD4GEtYAAAA9gAAAACn/DkAAAAAVP4cPmAQ+JAAAAAA7WMTWdavOo95Nv72GVXoVsBXX2nKP/AJM1/wC3AAAAAAAAACV9ByjFqBT/AIsiS1YABiPjAAAAAj8SBFXoFVXSAAACt8KAAAAAK3wx9Cj/ABEV/jZh9YNAAAAU/jXqVfjYAAAAC/gv1AAAAAKfwoI/AENYWAAAB7AAAAAEJJQku4AAAABU/hw+YBD4kAAAAAGKdqE35P8AIB0XTyzL5dVP823+gAAAAAAAAAtGicX7aLKcf3kmS1bAAMR8YAAAAEfiQLdFXdA9YAAABW+FAAAAAFb4Y+hQ/iIr/wARmG3aJK0mgAAAp/GvUq/GwAAAAF/BfqAAAAAU/hQL+GFLqiSs7AAAD2AAAAAAAAAACp/Dh8wCHxIAAAAAKkOeDj3Q007MMwlCth8ujBaSlB28kv0uAAAAAAAAAFKanBMqzUYNgABiPjAAAAAj8SCO6Ku4Q1jYAAAK3woAAAAAcm0r9Ch8aK38RmHfvlZWmwAAAp/GvUq/GwAAAAF/BfqAAAAAU/hQP+GFP4ioveAAAHsAAAAAAAAAAFT+HD5gEPiQAAAAAGeYV4bMKtPzbXo9V+YcOVp47FYeDXu0Iy+9uy/C33MAAAAAAAAAGp0n2LVKr7gABiPjAAAAAj8SCHxIqfEFLsSVmAAAVvhQAAAAAYf4yt8bKOk0Yhe+AAAFP416lX42AAAAAv4L9QAAAACn8KCXwIFo0VVsAAAPYAAAAAhBOEn2AAAAAKn8OHzAIfEgAAAAAOMMnlXgsXSV5RVmvLv8vy9BJt2W5wxlEsBheaovfnq/JdF/n6gAAAAAAAAFOjKauivRlUasUaLhe5LdgAGI+MAAAACPxIIfEVPiCn8RUXvAAAFb4UAAAAAOLRh/jKvxsg7SRiVqgAAAp/GvUq/GwAAAAF/BfqAAAAAU/hQT+BAT1imAAAPYAAAAAp/w5AAAAAFT+HD5gEPiQAAAAABDLsHCr7WNKKl3srgAAAAAAAABhpRV02OUVuydaEY6MAAMR8YAAAAEfiQU/iJ/EC0aKq2AAAK3woAAAAArfDH0MN8ZVXvsMSrpMAAAKfxr1KvxsAAAABfwX6gAAAAFP4UFTZALWAAAA9gAAAACn/DkAAAAAVP4cPmAQ+JAAAAAAAAAAAAAAAAFSo6lizAADEfGAAAABH4kFPcluwJ6xTAAAK3woAAAAArfDH0MN8TMQtUwq60UwAAAp/GvUq/GwAAAAF/BfqAAAAAU/hQVegFPVNDQAAD2AAAAAKf8OQAAAABU/hw+YBD4kAAAAAAAAAAAAAAAAU4KEUgYABiPjAAAAAj8SCnuS3YC1gAAAVvhQAAAABW+GPoYb4mYle6mG9AAAAKfxr1KvxsAAAABfwX6gAAAAFP4ULcq9AKe5Ne8AAA9gAAAACn/AA5AAAAAFT+HD5gEPiQAAAAAAAAAAAAAAAEdWhzftFHyKU5TcotjAAMR8YAAAAEfiQUuowKeqsNAAAVvhQAAAABW+GPoYbdldXpsKWtJoAAAKfxr1KvxsAAAABfwX6gAAAAFP4UR3RV3Ah8SKq1AAAewAAAAAAAAAAVP4cPmAQ+JAAAAAATTlFpPUy7FxxeEp119pfj1/EAAAAAAAAAFoyelaLI04xk2upLVsAAxHxgAAAAR+JBT2YBT3Jr3gAAK3woAAAAAnNSSXYw3UmrxaDDapoaAAAKfxr1KvxsAAAABfwX6gAAAAFP4UR+JFTcAqapMAAB7AAAAAAAAAABU/hw+YBD4kAAAAABKSjFt9DgnG89GeFb1jqvR7/c/zAAAAAAAAAAlObabexLEpxslqAAGI+MAAAACPxIIfCwCPxIqrUAACt8KAAAAADDdS+tiatJowz95oqK02AAAU/jXqVfjYAAAAC/gv1AAAAAKfwoh8SKnxAD1gAAA9gAAAABQck2ugAAAABU/hw+YBD4kAAAAABm1ZUcDWn2i/vtp+JkON+p5hTqN6N2fo9Pw3+QAAAAAAAAAJ2aMVrFMAADEfGAAAABH4kEfgYAVNUmAABW+FAAAAABhtmSdqiRXVpsw/wAZXVpsAAAp/GvUq/GwAAAAF/BfqAAAAAU/hRT+In8QBDWNgAAHsAAAAAU/4cgAAAACp/Dh8wCHxIAAAAADjPFKll6pJ6zf4LV/jYMix31zL6dRvW1n6rT8d/mAAAAAAAAACppR5Xqexp9hgAGI+MAAAACPxIF8AldMAesAAAK3woAAAAAqxSjG3Yw3wsrO1SJiVqmUnaaMSvfAAAKfxr1KvxsAAAABfwX6gAAAAFP4UU9x6zJKzApdiSswAAewAAAABT/hyAAAAAKn8OHzAIfEgAAuXRdF0XQAHF+N+sZh7JPSCt893/l8g4JxjjUnhpbSXMvlo/68gAAAAAAAAA+tvsfW32AADEfGAAAABH4kH/LshsSVmBDWNgAAK3woAAAAArfDH0MN8LMT8ZX96mmRdmjFLYAAAp/GvUq/GwAAAAF/BfqAAAAAU/hRT3FrMmtLgQ+IqL3gAAewAAAABT/hyAAAAAKn8OHzAIfEgAAauWLFiwAMxVZ1686r+02/vYZT+4r4Csvtc0f/ACZr/wBuAAAAAAAAAAAADEfGAAAABH4kD+AWkSa0Ap72JKzAACt8KAAAAAK3wx9DDfAzEfGfFQDEawTAAAKfxr1KvxsAAAABfwX6gAAAAFP4UUupHWTNJKwC0ZVWwAAPYAAAAAp/w5AAAAAFT+HD5gEPiQAAAAAYl8tCbXRP8gPYunluXz6qf5yv+gAAAAAAAAAAAAYj4wAAAAI/Egn8CH8AveiBD4ioveAACt8KAAAAAK3wx9DD/AV/4jKHvQaCetBAAAFP416lX42AAAAAv4L9QAAAACn8KKXUh1IvUmrMCesAAAewAAAAAptRaXUAAAAAqfw4fMAh8SAAAAACrDng490NNOwZg4VcNl8IKycoNLyS/S4AAAAAAAAAeyh2PZU+xiKKirxClT535IrwjGdkUoQ5XOWxWhFWlHZhiPjAAAAAj8SCp8KJ7Ig9bE1qC0ZVWwAAVvhQAAAABVacY27GH+ArazZhnq0VFabRHWgwAACn8a9Sr8bAAAAAX8F+oAAAABT+FFPZkNgmroBawAAB7AAAAAAAAAABU/hw+YBD4kAAAAABnWFeGzCrT8216PVfgw4erzx2Kw1NrSjGT+bdl+FvuYAAAAAAAAAVKrppLdkKznBtLVEpKpRbBNpWRif4hU0pRRvQ9GFajOcro+r1OxKLTswAAACPxIKnRFToEleIE9YAABW+FAAAAABh/gKus2UHaaMQrTKGsGgAAPq9TsQoTUk7FX42AAAABTg50ml3Pq9TsTpSgtQAAAAp/CiHwMXwBDVWGrMKezQ0AAPYAAAAAAAAACFKU1dH1ep2K0XGEU/MAh8SAAAAAA4xyiVaCxdJXcVaXp3+XXy9C19EcL5TLA4VzqK056vyXRfqwAAAAAAAACO6MVBuzRhoNJtlH+FL+ugGJ/iFf4YehH+DIK1WcZWTPrFTuSbk7sAAAAj8SET3RPcIO6sNWYR1gAAFb4UAAAAAFD+GiprJkXaSZilszDdUSVm0AAH1ip3IV6jklcq/GwAAAAIScaTa7n1ip3J1JTWrAAAACn8KI/AP4Ai7MmtQp7k17wAA9gAAAACn/DkAAAAEas4qyZ9YqdytJyhFvzAIfEgAAAAACnlWBp1fawpRUu9kAAAAAAAAABTmpwTK01GDZCcVRavqBif4hV1pwYtKD9QxHxgAAAAR+JC3RL4kT3CLsyaCntYaswACt8KAAAAAJw5Utdyh/DRLWTCt71JMwz98qq02AABT+NepV+NgAAAAL+C/UAAAAAp/ChfwyfwoB+9EI6NFVagAD2AAAAAKf8OQAAAABU/hw+YBD4kAAAAAAAAAAAAAAAARnKOzJTlPdgGJ/iFNKpT5OqK9oxUF0DEfGAAAABH4kR3Q/iJbgP3ohT3Jr3gACt8KAAAAAK3wx9Cj/DQ9WEfeoWKHxoxC98AACn8a9Sr8bAAAAAX8F+oAAAABT+FH2Cp0Ag+hJWYVNUmAAPYAAAAAp/w5AAAAAFT+HD5gEPiQAAAAAAAAAAAAAAABTpRhFLqWQwDbYDEfGAAAABH4kQ+JD3HqBB9CSsyOjRVWoABW+FAAAAAFb4Y+hS/hoDD6xaIaTRiV7yAAAp/GvUq/GwAAAAF/BfqAAAAAU/hR9lFTcBOzKi6hvTAAHsAAAAAU/wCHIAAAAAqfw4fMAh8SAAAAAAAAAAAAAAAAjuivVcEkj6zUAADEfGAAAABH4kQ+IfUATsya0uFTWKYABW+FAAAAAFb4Y+hRXuIqQ5ZNBhn71iqrTZiVdJgAAU/jXqVfjYAAAAC/gv1AAAAAKfwo6InuAL3ohT1jYAAewAAAABCSUJLuAAAAAVP4cPmAQ+JAAAAAATTcWk9TAYuOLwtOvH7Sv8+v3PQAAAAAAAAACpSdSK7n1WfdAABiPjAAAAAj8SKfxE9mAC96Ng3pgAFb4UAAAAAVvhj6FL4EYqF4qSCi7TRiV75W1pJgAAU/jXqVfjYAAAAC/gv1AAAAAKfwoexLcAg9bE1ZlPexJWYAD2AAAAAAAAAACp/Dh8wCHxIAAAAACTUYtvocE43noTw0nrF3Xo9/x/MAAAAAAAAADnj3OeHcYABiPjAAAAAj8SKe5U2ACD1sTVmU9Y2AAK3woAAAAAcm0r9CnpBehQkp0+Vk4uMmhOzTMSrpMetAAACn8a9Sr8bAAAAAX8F+oAAAABT+FDHqALQmroh8SKi1AAewAAAAAAAAAAVP4cPmAQ+JAAAAAAZrWVHA1p9ov77aHD+N+qZhTm3o9H6P/J2fyAAAAAAAAACztc5Jdjkl2AADEfGAAAABH4kU9ypsABNXRT3sSVmABW+FAAAAAAtKfyKM+WaMVDaSCp71FMpa0WgAAKfxr1KvxsAAAABfwX6gAAAAFL4US2AAjqrGzKq2AAewAAAABCCcJPsAAAAAVP4cPmAQ+JAAAAAAcZYpUsv9knrNpfJav9PvDI8b9cwFOq3raz9Vo/v3+YAAAAAAAABUS9ivkAwADEfGAAAABH4kUupx7kv9k55Vw6Vou0l6SSb+53XyAAhqrEdJFRagAVvhQAAAAA4tJXJaUn6BTaqU7MlFp2ZT96i0YbWLQwAAp/GvUq/GwAAAAF/BfqAAAAAUdkTyXn4WWZxWsa0oP0cItfc0/vAAi7MmtSWsAAHsAAAAAU/4cgAAAACp/Dh8wCHxIAANC6Loui6AA4wxvt8f7JPSmrfN6v8ARfIOCcY4zqYaWzXMvlo/0+4AAAAAAAAA5pNKNyU6sd2e2qdwAAxHxgAAAAR+JGEp+0qRh3aX3njtlC58JmEVunTfy96P5yGrABF2ZNWaZVWiAArfCgAAAACt8MfQq6UmFCfLO3cxULNSRhndNGH0m0VFaTQAAU/jXqVfjYAAAAC/gv1AAAAAKOyOEMl/tHw4xtBK8nKcl6wUGvv5bfMnHqgAH70SOsLAAPYAAAAAp/w5AAAAAFT+HD5gEPiQAANFixYsABi6zr4idV/abf3sMovQxGBrL7XNH/yZr/24AAAAAAAAATszErmgmgAAMR8YAAAAEfiRkFL2uZYeHecF98keJuX0804XxSp6ype+vJw+L/yXmGlJDQAP3ojXNAACt8KAAAAAK3wx9DL8rqZj7WnDeMJz/wDAIuT/ACAjarSsUNJ2Iq1ZorK02AAFP416lX42AAAAAv4L9QAAAAChsjwijCHC1CLesnN272lZ/ocUZX/ZedYrBWsoTaX+HeP/AJK0Sj1QAQeliGjaJKzsAD2AAAAAKf8ADkAAAAAVP4cPmAQ+JAAAAABiXy0JtdE/yA9i6eW5fPqp/nK/6AAAAAAAAABGEYqyOWPYYABiPjAAAAAj8SOC6XtM/wADH/p7T/8ASkcC5zDM8xzrLqrulVm0u8ZXg/l7q/8AAjNMDPLsfWwdTenKUfudr/MlG6ACD6EXZ2KkbMAK3woAAAAAq7R9DwdwsMRxFKE1ePs53Xk7Rf5mcZfPLswr4Oe9OUo/+Atr8Qw07St3KkeWqpLqVNKsWYle+AAFP416lX42AAAAAv4L9QAAAADD7IyDO/7Jw3DdNu0ajrKXpOfKr+jlf5HjllP1fO6ONitK0LP/ABQ0f/kriRldE49gCLsyTs0ycbq6AB7AAAAAFP8AhyAAAAAKn8OHzAIfEgAAAAAqw56co90xqzsGYOFTDZfCmtHKDXol/wAEAAAAAAAAAPrMz6zMAAMR8YAAAAEfiR4d01PiXBJ9Jp/dd/oeF+fOhxipyfu4hzi/WT5l/wCTJL5njPlP1PiV14r3a0VL5r3X+Sb9SMuhOPVACdmTXVDtOI0AVvhQAAAABU2h6HgbSvnlefak198of5HjRlP1LimpVivdrRjP5/C/xjd+obGlWndblb7MjFLVAABT+NepV+NgAAAAL+C/UAAAAAw+yOM8RKlhcopQdnChGXo5Tk/0R4mU45/wRh81prWPJP0UlyyXybV/QIyuiUbbAC96JTfQnGzAHsAAAAAAAAAAFT+HD5gEPiQAAAAAGc4V4bH1aXZu3o9V+DDIK8sdisLStpRjJv1bsvwt+IAAAAAAAABOilTUup9VgfVYDAAMR8YAAAAEfiRwTV9jmsa38kK0v/AaU3+hgsXUweJp4mk/ehJSXqndfkeM2Ep5nw9hM3oaqLT/AO3KiTv96j94RldE42AIu6IqzKkeoBW+FAAAAADnzOK7Hh/m08my3NMyh8VOFNL1lOx47YCGMyrBZrR1ina/92aUov0938QMNOz5TERvBkoxqQJRcXZgAFP416lX42AAAAAv4L9QAAAADD7HGlXmxmHpf8+6FCP/AJSjJ/jI8LqkM94MxGU1HrHnh6KavF/KTdvQq0p0pypzVmm015rc2E00SjbYCDS0GuqJLmQA9gAAAAAAAAAAqfw4fMAh8SAAAAAA4yymVSKxlJXcVaXp0fy6/LsHCuUywWFdSorTnr6Lov1YAAAAAAAABOspU1HqfWodmfWodmAAGI+MAAAACPxIySp7GOIqdqU1/wCBLk/9uDhD/jJfDqrgHrOEZQXrH3of+2r5AJqSJRsBsRkmaNWGrOwFb4UAAAAAQ1kijiPYcK42PWrVox+UVUk/xS/Awf8AxkfhXKnvOlBrzvRd0vnGK+8BNp3RCcakSzoyt0ZVpqautxqwAFP416lX42AAAAAv4L9QAAAADD/CcUVPaZtWX8to/wDgEVH9DwPzb6tntTBSelaGn+KGq/8AJeY8Tsp/s3ijFQStGb51/wBv6v8A8muvkGwmmiUbARl0YipGzuA9gAAAABQcotroBw5wHn+ftPB0HyP7cvdh9739IpvyOG/AvK8Jy1c1qOtP+VXjD/5KX3xXdGceC/C+OvKjCVGX9x6f+Ay5l8lYzjwGziheWArwqrs7wl+sfvkjOOEs8yi/17Czgl1teP8A4Erx/EAACp/Dh8wCHxIAAAAABpNWKeTZfTq+1hRipen5LZAAAAAAAAAA6UVS5upCjzQcr7AAAYj4wpUVUV7n1Rdz6ou59UXc+qLufVF3Pqi7n1Rdz6ou59UXc+qLufVF3Pqi7iwqT3KNT2dCtH+aKX/k8X+geBGbezx2Jy+T0nFTXrF2f3qX4HHOU/2VxFi8KlaKk2v8MveX3JpAnY0khqwZLk+KzjH08DhVepN2V9tFdt+SSuz/AIgjxN/NT/8AAn/8iT8F+J4Qcr03ZbKTu/JXitWVKTUrSVmj2SPZInQUla59UXc+qLufVF3Pqi7n1Rdz6ou59UXc+qLufVF3Pqi7n1Rdz6ou59UXc+qLufVF3I4ZJp3MdiLZXToJ7zk38oxS/OR4BZlGpQxuWVNVpNLvdcsvyj95n+WSyzNMRgn/AMu5yj8k2k/mrMClVcGJxqR01I0+VWTKmHU3c+qLufVF3Pqi7n1Rdz6ou5HDJNO5V+NgAAAAL+C/UKceeSR9UXc+qLufVF3Pqi7n1Rdz6ou59UXc+qLufVF3Pqi7n1Rdz6ou59UXcpUuVWWpmdVVsdWqL7UpP72zhrNXlWcYbG30pzTfpe0vvV0ePGVJrCZlBd4N/wDk0f8A24BOwmpI9mj2aMv8LOJcfhKeKoUFyTSavKKdns7N31WovCDixf8ALhf+Bx/zOIOHMxyPELDY+nyyautU01to1ddD2SPZIdJNH1Rdz6ou59UXc+qLufVF3Pqi7n1Rdz6ou59UXc+qLufVF3Pqi7n1Rdz6ou59UXcjh0otX3OEuD58RZlHA06yptpu8k3ot0kt3bW10rJ6nDfhHw5k/LUqU/b1V9qeqT8o/CvndruZ7xVkuQ0ubHV4w00jvJ+kVd/hbzOJPHmvU5qWTUeVfzz1fyitF83L0Mn8cuIsI1HGRhXj5rll98dPvizJ/HLh7F2jjIzoS81zR++Ov3xRlmfZVmsObBYiFRf3ZJteq3XzRnHh5wzm13iMJHmf2orkfreNr/O5nHgDhp3lluLcf7s1df8AgUbNL/t1lbwU4sp1HGNOEkuqmrP77P70jNfCziHK8HPGYynGNOG75o97LS93dux9Uj3Pqke59Uj3Pqi7mIhyxjH1AIfEgAAAAAAAAAAAAAAAJf7uij/Bl8/yAAAxHxhQqxhFpn1mmfWaZ9Zpn1mmfWaZ9Zpn1mmfWaZ9Zpn1mmfWaZ9ZpixFN6E5pxsg4Dzb+y+I8JiW7R5lF+kvdf3J3+R475T7LMcNj4rSpFxfrF3X3qX4ARdhpSRyI8EOG1TpVc4qrWXuQ9F8T+bsvkyr4xcLU5uPtJO2mkXb5D8Z+Ff55/8AgDOPMxyXH5vPGZVJ8tTWScbWl1a8pb+rfke0ie0iSrQitT6zTPrNM+s0z6zTPrNM+s0z6zTPrNM+s0z6zTPrNM+s0z6zTPrNM+s0yVaEUr9SvV52knojwizb+z+K8Om7Rqp03/28rr/yZRPG3KfqfE7xEVpWhGXzXuv/ANJTfqAU6jg7ooSeIqxpUouUpNJJK7beiS82Ynwm4iwuHliK6hCEU225rRJXd/QeIp9z6zTPrNM+s0z6zTFXg2kir8bAAAAAX8F+oYVQdaKlJRXd3svuTf4HscJ/0FQ+6p/8rPY4T/oKh91T/wCVnscJ/wBBUPuqf/Kz2OE/6CofdU/+VnscJ/0FQ+6p/wDKz2OE/wCgqH3VP/lZ7HCf9BUPuqf/ACs9jhP+gqH3VP8A5Wexwn/QVD7qn/ys9jhP+gqH3VP/AJWexwn/AEFQ+6p/8rPY4T/oKh91T/5Wexwn/QVD7qn/AMrPYYeK5o4mDa8qn/yAGNtxL4ZKpvUpwT8+ak7S+bin94AnY5zgzIZ59ndDApe63eT7RWsvvWi82jOuMuHuHqkMLjq6pycbqKjJ+7svhi7LSyvbY/4i1wd/0Gf+U6v/AMgeI/E/B/EmWqNDGJV6bvBuFRXT+KLfJs1r6pdLn1XDf9BMPuqf/IH1XDf9BMPuqf8AyB9Vw3/QTD7qn/yB7HCf9BUPuqf/ACs9jhP+gqH3VP8A5Wexwn/QVD7qn/ys9jhP+gqH3VP/AJWexwn/AEFQ+6p/8rPY4T/oKh91T/5Wexwn/QVD7qn/AMrPY4T/AKCofdU/+VnscJ/0FQ+6p/8AKz2OE/6CofdU/wDlZ7HCf9BUPuqf/Kz2OE/6CofdU/8AlZ7HCf8AQVD7qn/ys9jhP+gqH3VP/lZ7HCf9BUPuqf8AysWHwrTaxUNPKp/8rMpzN5TmVHGYWopum09OZLzi+ZJ6rR6bM4i8Ys8zG9PB2oU/7us//Ant/wBupPzMSp4io6tSTcnu222/VvUnSlHctchh5S30KEfYyU4NqS2ezXoZT4m8TZZZRxLnFdKnvfi/e+5oynx6oaRzLDW86bv/AOSy/wDkmUfFvhCrTU3inFvo4VLr7otfczxV4+wOdUaOByypz0k+aTtKN3slaST01b0tquqMJlHCFXC051s49nUaTlH2FSST6q6te219uxVybhZfw86i/WhWX6Mq5Vkqv7PNab9addflSZVwWAhflx9KXpGt+tJGOUFJKElJd1ez27pP70AQ+JAAAAAAAAAAAAAAAARhz0UiFLlg433Pqj7gABiPjAAAAAj8SAONUuJPDylmK1nBQqP1XuT+68n8gBSsZZgcRmOMpYPDq86klFer/RbvyOP8xocJ8IQy7CO0pr2ce9re/L7r3a+1JMACt8KAAAAAK3wx9AwOLqYPFUsTT+KElJesWmvyPG/CU8xyHBZvR1Se/wDdqRTT++K+8APBvw++q0o55j4fvJL93F/Zi/t+sl8PaOvXTxh8Qf7RrvJsBP8Acwfvtfbkun+GL++WuyTYAU/jXqVfjYAAAAC/gv1AAAAAKfwoDwMzKGIwOMyurqk1JLupLlkvTRfeZ3ls8tzKvgpb05Sj9zsn81qAHgxw/DLsoq5xifddS9m+lOO7+bTb8opnF2f1M9zmvjpbSdop9IrSK+7fzbYAPYAAAAAp/wAORh58s7dyTSaRJ2RGcZLRihFbInWhHqTryltoWcmU6CWstyU0morcrz5YW6sAKn8OHzAIfEgAAAAAAAAAAAAAAAKVSCgk2e2p9z21PuMAAxHxgAAAAR+JAHg7iqea8NYvKKzuouS/7cqJ/rzfeYzC1MLiKmHqK0oNxfqnZ/kAeB3DXtsVVzirHSn7kP8AE17zXpHT/t59jxU4l/tniCpGnK9Kj7kfNr4n85aeaSAArfCgAAAACt8MfQDJP+Mj8LqmG3nThKPzpPmgv/AVFfMA8KPD959jPr2Mj/s1J7P7clry/wCFby+S6u3i5x+slwn9l4GVsRUWrX2IP8pS2XZa6e6AAU/jXqVfjYAAAAC/gv1AAAAAKfwoDwmzb+z+KaCbtGqnTf8A29rH/wAmUTxpyn6pxH9ZivdrRUvmvdf4JP5gHDuS1s5zShgKW9R2v2W8n8kmzxazqjkfDtPKMJ7rqpQSXSnFK/36R802AA9gAAAACn/DkD/e0vMo1ub3XuVqX2o7jnUmrCozfQWGk92QpxgtCrX6RMNC95srz55gBU/hw+YBD4kAAAAAAAAAAAAAAAAAAAYj4wAAAAI/EgDwXzb6nxIsNJ+7Wi4/Ne8vya+Z4tZT/Z/FFaUVaNVKa+ekv/JkwMPh6uJrQo0o805NJLu27JfNmeV6XA3BSoUJfvFHki+9SV3KXy96S9Eh6gAVvhQAAAABW+GPoB4AZom8Zl0nuozS/wDJZfnE4myt5XnOKwVtKc5Jel9PvVg4L4SxXE2aQwdHSC1nL+WPV+r2S6vyTa4mz7LOBeH4Qw8Umly0od33fWy3k+r63aMwx+Jx+KqYrEzcqk222+rf9aLZLQAAp/GvUq/GwAAAAF/BfqAAAAAU/hQGFxNTC16dek7Sg1Jeqd0eMOGp5twzhM3oq6i0/wDt2ol+vKAeBvDahSrZzWWsvchfsvil83aPykjxB4k/t/Pq2Jg704+7D/DHr/287y+YAD2AAAAAFJpNLqGFnZ8pXhyz0KNXm0e47JDqwXUeIgtipWc9OgouTSRVap07IACp/Dh8wCHxIAAAAAAAAAAAAAAAAAAAxHxgAAAAR+JAGTZjPLcxoYyG9OUZfc07fPY8c8uhicBg80papNxb7qS5ov00f3geC3DX1/NpZjVjeFBaec3t/wCAq78nynjJxL/aWd/UqUr08PePrN/F92kfJp9wACt8KAAAAAKrTjG3YDwpzb+zuK8K27RqN03/ANvKy/8AJuU8csp+q8RxxUVpWgn/ANvR91/go/eZdl2JzHF08JhY81Sbsl5/5dW9ktWcO5JlfAnD0p15pcq5qk+spdl1t9mK/Vs4y4rxfEuaTxlfSO0I9Ix6L16t9X5WSAAKfxr1KvxsAAAABfwX6gAAAAFP4UAcEW4j8Pa2XPWcFOmvVe/D7rxXyGmnZhlmXV8xxlLB0FedRqK9X+i3fZHH+YUOE+EIZdhHaU17OPe1vfl917vpKSYAA9gAAAAAItxaaK8VOnzIG7gGGhd8xiJ809OgAFT+HD5gEPiQAAAAAAAAAAAAAAAAAABiPjAAAAAj8SADAf8AGS+GUqW9SnBrz5qTvH5uKX3hGLk1GKu2YWFLgPghylb2qjd+dWey80nZf4Y3KtWdWpKpN3k222+re7AAK3woAAAAAAwuIqYavCvTdpQaa9U7o8Z8NTzbhnB5vQV1Fp/9u1Yr9VFfM8JuAFkeD/tLHRtiKi0T+xB628pPeXZaaa38VuP3xBjfqWEl/s1J6f35bOXoto+V31sgAAp/GvUq/GwAAAAF/BfqAAAAAU/hQB4E5t7LMsTgJPSpFSXrB209VK/yOPsp/sviTF4ZK0eZyXpL3l917fIPA7hr22Kq5xVjpT9yH+Jr3mvSOn/bz7HipxKs6z+cacr0qPuR82vifzlpfqkgAB7AAAAAAGGknHlZUhyyaAEm3ZE2qVOyAAKn8OHzAIfEgAAAAAAAAAAAAAAAAAADEfGAAAABH4kAHgPmqf1vLZPtNL/yWX/tpxPlTyrOcTgraQm0v8N7x++Njwj4a/tbPo4ipG9LD2m/8X2F965vSJ438S/Wswp5TSl7tH3pec5LT/wGP4yYAAVvhQAAAAAAeE+LoZxwhQo14qfspcrT11hJThp5Jxt5o8ZfEH2FOWR4CfvyX72S6J/YXm/tdlp1dgAAKfxr1KvxsAAAABfwX6gAAAAFP4UAcEZt/ZfEOExTdkppP/DL3Zfg2zx4yn2eNwuYRWk4uD9Yu6+9Sf3GHw9XE1oUaUeacmkl3bdkvmzPMRS4G4KVChL94o8kX3qSu5S+XvSXokPUAAewAAAABT/hyApT5JJmKhdKSAtcoUWnzSMRPmlZdAACp/Dh8wCHxIAAAAAAAAAAAAAAAAAAAxHxgAAAAR+JAB4Z5t/ZvFGFm3aM3yP/ALf0X/k1n8jxxyn6tnlLGxWlaGv+KGj/APJXE4RwdDgzgyWNxKtUcfaS7tte5D8o+TbMdjK2NxNTE1nec25N+bd2AAFb4UAAAAAThypeYBwT4g1eGcrx1Ckr1KnL7PtGWqlJ/K1l1aV9LlatUrVJVKknKUm22923u36gAAFP416lX42AAAAAv4L9QAAAACn8KADi3/jJfDmlj1rOEYz/AO3o+5P/ANufyPBbhr6/m0sxqxvCgtPOb2/8BV35PlPGTiX+0s7+pUpXp4e8fWb+L7tI+TT7gAA9gAAAACn/AA5AFJqpS5WSTTsU8PKW+iIUowWhVrxirLVgABU/hw+YBD4kAAFH+JH1R9Tw/wDz7X3I+p4f/n2vuR9Tw/8Az7X3I+p4f/n2vuR4u4ejDhaq4wSfNDov5gAAAAAAAAAAAAAAMR8YAAAAEfiQAU6k6c1ODs07p9mjHZLhOMcvyzG1LcsXGo13Tj70PnKyfkmeOXEt5Ucmoy29+f8A7avzbXnFgAAVvhQAAAABW+GPoAAAABT+NepV+NgAAAAL+C/UAAAAAp/CgA8GMXTzLh/F5RWd1FtW/uVE0196l95So0eAODJXadWKbv8AzVZaL1S0X+GNyrVnVqSqTd5Nttvq3uwAAewAAAABT/hyAMPPlnbuezjzc1tSpWhD1KlaU9OgAAFT+HD5gEPiQAAH9r5j/wBBE/8AwKX+Z/a+Y/8AQRP/AMCl/mf2vmP/AEET/wDApf5n9r5j/wBBE/8AwKX+ZWzDGV4clWrKS7OTa+5sAAAAAAAAAAAAAADEfGAAAABH4kAB4T8S0KfCNWWJlZYVyv8A4X76+9tpLysZ3mtbNsxrY6t8VSTfp2S8krJeSAAArfCgAAAACt8MfQAAAACn8a9Sr8bAAAAAX8F+oAAAABT+FAB4NZt9S4ljQk/drRcfmveX5WXqeN/Ev1rMKeU0pe7R96XnOS0/8Bj+MmAAA9gAAAACn/DkAE8RKSstAAAAqfw4fMAh8SAAAAAAAAAAAAAAAAAAAMR8YAAAAEfiQAGGzfFYbA18FSlanW5eZd+Rtr8WAAAVvhQAAAABW+GPoAAAABT+NepV+NgAAAAL+C/UAAAAAp/CgAyzH1cvxtHF0vipyUl6p3+59THYytjcTUxNZ3nNuTfm3dgAAPYAAAAAhJKEl3AAAAAKn8OHzAIfEgAAAAAAAAAAAAAAAAAClSUk3J6IrUlBq2zDEfGAAAABH4kAAAAAVvhQAAAABW+GPoAAAABT+NepV+NgAAAAL+C/UAAAAAp/CgAAAAHsAAAAAAAAAAFT+HD5gEPiQAAAAAAAAAAAAAAAAABPSlFdx+9Q9GGI+MAAAACPxIAAAAArfCgAAAABybSuAAAABT+NepV+NgAAAAL+C/UAAAAAp/CgAAAAHsAAAAAAAAAAFT+HD5gEPiQAAAAAAAAAAAAAAAAABW+GHoR/gyCdaEXZo+sU+xNqUm0AAAAUEnUSGAAAABUSdNgAAAAFWKUY27AAAAB9Yp9iNeDaSRV+NgAAAAUZqNNt9z6xT7FapGaVkAAAAFkoq3YAAAACKTaJpczAAAAAhFOEgAAAApVYQjZo+sU+xiJKUYteYBD4kAAAAAAAAAAAAAAAAAAVtYQZH+A/UMR8YAAAAGH/AIiGAAAABU/hMAAAAArfDH0AAAAAp/GvUq/GwAAAAF/BfqAAAAAP4Y+gAAAAEfiRP4mAAAABT/hyAAAAAKn8OHzAIfEgAAAAAAAAAAAAAAAAACMfaUrLdFVclNQ6hiPjAAAAAw/8RDAAAAAqfwmAAAAAVvhj6AAAAAU/jXqVfjYAAAAC/gv1AAAAAH8MfQAAAACPxIn8TAAAAAp/w5AAAAAFT+HD5gEPiQAAAAAAAAAAAAAAAAABGTi7pkm5O7CvGTnojkl2OSXY5Jdjkl2OSXY5Jdjkl2OSXY5Jdjkl2OSXY5Jdjkl2OSXYoQkqi0HFnKzlZys5WcrOVnKzlZys5WcrOVnKzlZOLdNqxyS7HJLsckuxyS7HJLsckuxyS7HJLsckuxyS7HJLsckuxyS7HJLsckuxVhJxjZdDkl2OSXY5Jdjkl2OSXY5Jdjkl2OSXY5Jdjkl2OSXY5Jdjkl2OSXYpwlzrQq/GwAAAAIpui7dzkl2OSXY5Jdjkl2OSXY5Jdjkl2OSXY5Jdjkl2OSXY5Jdjkl2OSXY5Jdhp2XocrOVnKzlZys5WcrOVnKzlZys5WcrOVkU7onCTk9Dkl2OSXY5Jdjkl2OSXY5Jdjkl2OSXY5Jdjkl2OSXY5Jdjkl2OSXYpwlyS0OSXY5Jdjkl2OSXY5Jdjkl2OSXY5Jdjkl2OSXY5Jdjkl2OSXY5Jdjkl2KiapwXqAQ+JAAAAAAAAAAAAAAAAAAACxTtsfWn2PrT7H1p9j60+x9afY+tPsfWn2PrT7H1p9j60+x9afY+tPsfWn2PrT7H1p9j60+x9afY+tPsfWn2PrT7H1p9j60+x9afY+tPsfWn2PrT7H1p9j60+x9afY+tPsfWn2PrT7H1p9j60+x9afY+tPsfWn2PrT7H1p9j60+x9afY+tPsfWn2PrT7H1p9j60+x9afY+tPsfWn2PrT7H1p9j60+x9afY+tPsfWn2PrT7H1p9j60+x9afY+tPsfWn2PrT7EnzNsAAAACnWcFZI+tPsfWn2PrT7H1p9j60+x9afY+tPsfWn2PrT7H1p9j60+x9afY+tPsfWn2PrT7H1p9j60+x9afY+tPsfWn2PrT7H1p9j60+x9afY+tPsfWn2PrT7H1p9j60+x9afY+tPsfWn2PrT7H1p9j60+x9afY+tPsfWn2PrT7H1p9j60+x9afY+tPsfWn2PrT7H1p9j60+x9afY+tPsfWn2PrT7H1p9j60+x9afY+tPsfWn2PrT7H1p9j60+x9afY+tPsfWn2KtV1LAEPiQAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAQ+JAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABD4kAAAAAAAAAAAAAAAAAAAAAAAARhKTskNNOzAAAAAUW1dAAAAAJX0RKnKK1QAAAAAAAAAAAAAABuSpyitUAAAAAk27IaadmAAAAAqU2rpAAAAAABD4kAAAAAAAAAAAAAAAAAAAAAAAAKTp0lbdle0oxmAAAABzRdNxj0QAAAAFKag2yFR1YuMgAAAAAAAAAAAAAACglzOT6EZurGUWAAAABQlGPqVv4jAAAAA+svm0WhXilN2AAAAAAIfEgAAAAAAAAAAAAAAAAAAAAAAACvtFeRvQ+YAAAAFL4JgAAAAGH0k35AAAAAAAAAAAAAAAFPSlJmH/iIaswAAAAp/GvUrfxGAAAAAld2MTrMAAAAAAh8SAAAAAAAAAAAAAAAAAAAAAAAAMRvH0F/A+YAAAAFL4JgAAAACXJRb6sAAAAAAAAAAAAAAAhrRkYf+IiWrYAAAAU/jXqVv4jAAAAAoQTld7InLmk2AAAAAAQ+JAAAAAAAAAAAAAAAAAAAAAAAAFXWnFk9KMV3AAAAApfBMAAAACnDnkkV5qUrLZAAAAAAAAAAAAAAAFHWMo+Rh1aTl2QAAAAFP416lb+IwAAAAJfu6fL1YAAAAAAQ+JAAAAAAAAAAAAAAAAAAAAAAAAFKrGMeWSuirU55eQAAAAEJqMZLuAAAABRqRg3c+sU+w9WAAAAAAAAAAAAAABTnySTKlaLjaK3AAAAAi0pJlSSlJtAAAABFpNMeJg+hWnGbVkAAAAAAQ+JAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABD4kAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAEPiQAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAQ+JAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABD4kAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAEPiQAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAQ+JAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABD4kAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAEPiQAAAAAAAAAAAAAAAAAAAAAAAAAABQpRnFtn1amVKMIuNurMTCKs0AAWvoYiMY2SCiouaTK0FGbSAAAAAAAAAAAAAAAAAAAAAKUVKaTPq1Mlh6aTIU4ujfqAAEIRVJyaAqwj7OMkgAAAAAAAAAh8SAAAAAAAAAAAAAAAAAAAAAAAAAAAKdaUFZDm/Zc3UnWlOyKspytzIcJLoShKO6HTko3aD2U7XsFFXqIxLvUI05yV0hXjJGKXvJgAAAAAAAAAAAAAAAAAAAAEJOMk0UKsql7lTESTcRSqezsloKMmrpHs5ON7aEacpbIatoRhKWyJRadmFT3aCQouTsiUJR3Qvew7XYAAAAAAAAAIfEgAAAAAAAAAAAAAAAAAAAAAAAAAAAf8D5FP416mL2Q+Wyb6EoxmkXjNaalGnzVLPoe0jz8nUrwUZuxhleoVVzVWiU4U0kytJSm2jE6xiwAAAAAAAAAAAAAAAAAAAAAwnUrfGyn/A+TMMr02hOL0XQioR91GJjaenUvGjBJmJipQUkGJ0hFGHSjT5ivVjOCsUNaUkAAAAAAAAAEPiQAAAAAAAAAAAAAAAAAAAAAAAAAAAP+B8in8a9TF7IxP8ADRTf7j5MwmzRQXvyPYLn5rmLXvIw3xk3aq35lWmqkSUXF2ZiP4cQAAAAAAAAAAAAAAAAAAAAAwnUrfGyn/A+TML8DMM7zZNv25XV5xKtJVLXZUilRaDFbRMPaVOxWo8j02MP8EgAAAAAAAAAh8SAAAAAAAAAAAAAAAAAAAAAAAAAAAB/wPkU/jXqYvZGJ/hop/wPkzCdSg/fkipWqRk1cnOU3qYb+IVl+8Zh6rvyMxSXOYnSMUAAAAAAAAAAAAAAAAAAAAAGE6lb42U/4HyZhfgZhfjfoT/j/NFd2nExE5xScSVaclZsMTrCLITlB3RXtKlcoaUpMAAAAAAAAAIfEgAAAAAAAAAAAAAAAAAAAAAAAAAAAbXsLX6FP40YpppWMS06aINewt6mFaVyM+Sq2WjJX3MRZ1HYou1RFVJVlfyLR+IqSVSpoYt3aQAAAAAAAAAAAAAAAAAAAAAYVpXKus2U2vYW9TDNKDMM0pv0J29v9ximm1YpVFUj5mJUVCyCr71CLKKjKmluYma5eVEdMOwAAAAAAAAAh8SAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABOzuYlxlZphRSc1cryUpuwAAAAAAAAAAAAAAAAAAAAAAAAEGpUXFsCrJKlGKYAAAAAAAAAQ+JAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABD4kAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAEPiQAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAQ+JAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABD4kAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAEPiQAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAQ+JAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABD4kAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAEPiQAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAQ+JAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABD4kAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAEPiQAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAQ+JAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABD4kAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAEPiQAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAQ+JAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABD4kAAAAAAAAAAAAAAAAAAAAAAAAKLlsgAUXLZHK27dSdOUd0AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABD4kAAAAAAAAAAAAAAABRouer2Pq9O2xOKjJpAUqbnKx+5hpoYhQUlygAOElFNgAOElHm6AAYf4ZAApKjTWmrKLTcpsq1XUsAHJLl5ugAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABD4kAAAAAAAAAAAAAAACV9CMVGKSHWpp2uTd5NgQfs6PN1Y9QUXLZDVixytq9grL91EOWVr2LXCxUX7iJYLCi3ohprRmH+GQHLK17G5iVqkQX7iRY5ZWvYEm9EOLWjJL/Z0AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAQ+JAAAAAAAAAAAAAAABRV6iKsuWDYAV9KcUCV3ZEpxoxUUtSso1Ic6KEeaDRGtBy5ESSp1SdaMYp9ynFVKjfQWIjKXLbQX7utYrU7VLLqVpKMoxXQlWioKRSkpVm0T+JlX+DEp1owha2pVaq0uZdDD/DIKMOeaR9YjzcttB2pVfIqVowaTFWi4OXY0rVV2PrEVLltoV4KM7IXLRgr7lSvGdvIdaKgpFWSnJtAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAQ+JAAAAAAAAAAAAAAABh/4iMR/DYAYnaIUVeoiu71GUNacolB2pyZHSSMV8a9Ct/CiUE3CVtz2FRdDFL30RSmoz7FSXNJsqfwImG+Mn8TKv8GJGEpuyI03ClJMw/wAMgw6dpNbnsKnYxS1Rit0Q/gS/rsYZP3mtz2FTsVo3nC5iXeduwS/3dAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAEPiQAAAAAAAAAAAAAAAUXaoipHmg0NW0AK3vUosMP/ERVXvsw/wyZR/hSFujFfGvQrfwolCTjSk0fWp9kVKjm7soxSp8r6jVnYqfwImG+Mn8TKv8GJzeypK27Kc5SpyuzD/DIMO7QlY+tT7IqVXUtcxW6IfwJf12MO7RlY+tT7IVVzqRuYhfvGEv93QAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABD4kAAAAAAAAAAAAAAACdmRkpRTQ4RbvYqq02BQtOm4MlRnF2sJOnNNmJpvm5l1EvZ0XfdlH+FIW6MV8a9CpFypRsYezvB9SVOSdrCptzUWVaiVVeRiYWnfuOLlQVihpUSZUoydR2W5iElTSRWi5U4tFODjRdzD/DIMM1zOPcnTlGViUXF2Zit0Q/gS/rsYZrmt3J05RlYs4SVzEU3K0ojhJK7RyuVBJDVtAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAIfEgAAAAAAAAAAAAAAAKVZw06DxULaInLmk2BGTi7oWKjbVFWp7R3KeJ5Y2aKtV1GQqcsXG24irU9o72KVdQjZnO1LmR9aVttRVrT52iT5m2VKvPFK2xRr8isyc7z5kPFe7otSVTmgo9iliOSNmPEN303KdTkTVtwTad0LFK2q1Kk+eVyrU9o1oRqWpuFtxNp3QsUrarUqT55XKWI5VZlasqiskUa3IrMqSUpNgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAQ+JAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABD4kAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAEPiQAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAQ+JAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABD4kAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAEPiQAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAQ+JAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABD4kAAAAAAAAAAAAAAAAAAAAAAAEYSlsg5JcvNbQVOTi5LYCEOeSRUhyStcAAAAKVP2jtclGzaAAAAAAAAAAAAAAAAAAAjCUtkAAAAAAEKPNByvsAAAAAAAAAABD4kAAAAAAAAAAAAAAAAAAAAAAAGE6ktyX+7kKsVSceoFD+IjE/xAAAAAwvxv0Knxv1AACVKKpKXUAIR5pJFeChKyAAAAAAAAAAAAAAMJswAAAAAAo/wZfP8gAASu0ivTjCyQAUaanKzJpKTSAAAAAAh8SAAAAAAAAAAAAAAAAAAAAAAADCdSW5L/dyNFSpuVyjS9o/IlhY3ViMVGskiVFTm29itS5JWD6suZK+h7LmqOKJ4WPL7u5SoxnFu+qKNNTlZns/3nKSor2igmPCwtpuQheaixYeN3fZFGjz6vYhRUJ3WxU+N+pGLk0kfVYWsUqXPK3QnhY8vu7lOCnPlZHDRu7k6XLUUejJUk4KJ7K9TkRLCx5dNyjSjO6e5ClzT5SFGKqWT2K9JSTkAAAAAAAAAAAAAAYTZhTpucrIlhY203KVJSk4sVO9TlK1NQlZFakoJWZTgpySYsOveb6FGjz6vYrUFFXiSpRVNSTKdFTg5FGmqjsynRU5NX0RVw8VG8SjSVRtMlRUaXM9ylh1KN5Cp8lOS9fyClSdR+RVoQjC6KNBTV3sVsOoq8SlSU0/IjhY8uu5SpXqcr6Fakpq76FGl7R+RWw6jG8RUYyp8yepRoqabZh4JLmRiKSjquoAAAAAEPiQAAAAAAAAAAAAAAAAAAAAAAAYTqWiYi3snYp/wH8yj7lJyMPUk5WbP+Yj+uxiKsoySTMVZwTCtU5I3W5Rb5ZT6mHlLns+pR0qSiYZWcjl/2gqzaqtooKpzXew1bEGIquOiE+ShdbmGqOV02VPjfqYWOrZOtNyumUfdpORh6jc7N7ijbEf12K9WUZ2T2MVtFonOSopp6mH+1JlCUvaa9S/JXOVRlKZSnL2m+5iZyUrJ6AAAAAAAAAAAAAAGE2YYZJRcilVk6mr3GuWuvMUf9oZXXNKL7mKd5JFD+IjEVXF8qJXjQSXUo3lTcWL3sO0UElTS7mHXKpMUpLRPcw8ZqLUjCr3mVqjlLkWxiZuKUUQm5UXfzCm+SjzDqTas2VW4UkkUJc8HFmF6lOtKVS19GVW41tDEzlFpJivGhpuzDXcXFmFlq4k17Kk0YWTbaKs5SbTYAAAAAEPiQAAAAAAAAAAAAAAAAAAAAAAAYTqS3Jf7uU/4D+Yv93/ruYb+If8AMR/XYxP8QxH8KIYr4UYdpU2z6zTKc06za6kvdhJ+Za9W/kUYqdRtntk6nIiX+8GJ+Mq/wI/IwvxsqfG/Uw3wMF/u5Q/iIf8AvCMR/EZifgiVP4CMM0oM+s0yrJTndFZN02il8aMV8YAAAAAAAAAAAAABhNmFH+C/mU/jRWdqsRq1Ry8hLmjBmId6jKH8RGJ/iCnGFOLY8RTsUNackJ8rghrlpz9TDxSi5spVfaN6GG+Ji/i/MxfxIo/wZfP8gn/u6DF/CjCdTDbyKP8AERX/AIqMXuiE4xpJs+s0yim6isYtOyZhfiZP4mAAAAABD4kAAAAAAAAAAAAAAAAAAAAAAAGE6ktyX+7lP+A/mYe0qbiyjQ5Hds/5iP67GJ/iGI/hRDFfCjDWlFxHhp3sUtKiMTpCwn+65vIwtuZop0ZKo2yX+8GJ+MgvaUbFCk4XvuVPjfqYWSu0Tw0nLTYoWlTcShRlGV2RlzV7mI/iMxPwRORzopIw+jlBjw072RRp3qW7EKilUlES5aqXmYmk2+ZAAAAAAAAAAAAAAGE2Z9Vl3MOrc0GUqEozu9kYiV6mnQqtezbMPrBeRN3k2UP4iMT/ABGRj7Sil1HQmk2zCv3miq7VY+RidIepRXNRaRh6bincw3xMlpNlan7SKcRQ5KTQUbTpcpKhOKbZOPtaStuU4ulTbe5hepR/iIxGlUxFJzSa6FNKpR5R4eaTbMMlGLkyP7yi11ML8bRWpOLv3AAAAAAh8SAAAAAAAAAAAAAAAAAAAAAAAClVdO9kPUdVunyWI1WoctiE3B3RSqSqVFfoTny1WypPnlcnVc4qNtgq1XUSViE3F3Q8Vpoi+tyrVdSx7Z+z5CE3CV0SxLdrIdVufPYqT55XKVV02PEtyvYk+ZtkZOLuj61psQqOEroniXJWSsQnySuVJc8rlSq5pKxSruCta5zvn5kfWtNiFdxbdtyE3GXMVJ88rksRzQ5bAAAAAAAAAAAAAAFKq6d7I+tPse0anzIlim1ZIJVnKCiU6zgmkEJcskypPnlcpVXBlTE80bJFObhK6Jzc5XKtZ1EtClVdNn1mXNexTquDbSG7u5TxDhG1rnt5a36hTqOD0KmI542SKVZw06FWs5q3QpVXTvZEJcskypPnlc+svk5WinUcHoTxN1ZI9s/Z8iRSqunexGbjLmRWre0SVgAAAAAIfEgAAAAAAAAAAAAAAAAAAAAAAAAKFOM4u5GEKUWyTu7gAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAQ+JAAAAAAAAAAAAAAAAAAAAAAAAAJtbMlOUt2AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAQ+JAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABD4kAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAEPiQAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAQ+JAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABD4kAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAEPiQAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAQ+JAAAAAAAAAAAAAAAAAAAAAAAAAABSpwUeeZUnSlB2WpBQpwUpK7ZWcG04gAAAAUqfPKwpUZPksVIckmgJwcGkwAAAAAAAAAAAAAAAAAClT55WPaUF7tinCMpt9EVJ0pQdlqAAAABuWp0klJXZWpxspx2AUG4OXYAAAAAAAh8SAAAAAAAAAAAAAAAAAAAAAAAAAANzEOzUV0Curxi/IAAAAAh7tGTCuublfcqQ5Jcp7HWKvuV6N1zX2RSoupr0KtLkaV7iwrcd9SMG5cpHDNrcp4dyV27EotOwsK3HfUaAAAAAAAAAAAAAAAIe7Rb7hSV6ckgAAAACjG9RIqy5ptlL3qUojp2gpXI0+aLlfYhRvStfc9m3PkRUw/LG9ylQc1e+hVpODsRw7lbUeHfPylWi6bRSouaveyKtNwdgAAACHxIAAAAAAAAAAAAAAAAAAAAAAAAAAj8SMR/EYUZxlHkkVafJKwAAAAD/gL1CX8OBiP4jK+0fQxHxL0Kz5YRgiiuaaROo3U5iSXt011JTftivNqpp0JxTrobk6nMV1aowAAAAAAAAAAAAAACX8BeoUqnJK5WpRtzx2AAAAAw38Qe5h/tehL+AvUp/wpEf4LKPuwlIu2VnywjFF+ah6FWVqUUTm3QQ3zUPRlRNQjFE7ujG/QAAAAh8SAAAAAAAAAAAAAAAAAAAAAAAAAAFozEr379wKz5oRYAAAAEVzUGuwVNFCJiP4jK+0fQxPxL0MRuvQw38Qa1JfxYIl/F+ZX/iMf8deg8TJO1ipNzldgAAAAAAAAAAAAAACXNQ9GBRd4SQAAAAFB2qIqRtJoo6QkyX8BepT/hSI/wAF+pDWjIMRuvQh/BkVvggS/gx9RfwH6lSs4JJLoVKzmrMAAAAh8SAAAAAAAAAAAAAAAAAAAAAAAAAACCjWpqL3RPDuMG2z6tzJNMxDV1FdAAAAAKE0nyvZiwyUrt6FWfPO6J0lUanfQxLT5WitDnipp9BRVamknqhxdGabHh1KfNfQ51OsmiX8X5lf+IytLlqpk8Pzy5ovRk48smgAAAAAAAAAAAAAAChNJ8r2Y8K76PQhQU7pPVEoKlTavqwAAAAE7NNE6Sq2lFlblhBU0UoqpTcOo4qFOUepRjz05RKVoycH1J4dxi3cUfa01Z6orWhBQW5W+CBL+DH1F/AfqKKq00k9UVaTgldgAAAEPiQAAAAAAAAAAAAAAAAAAAAAAAAAAXtsOTluxSklZMAAAAAHKTVrhzO1rjdzmdrXE2tmNt7nM7WuA9Ru4pSSsmAAAAAAAAAAAAAAABzyta4m1sNt7sAAAAAUmtmPUTa2ZdibWzHqOTa1Ym1sx6jdy7tYu7WE2tmNt7sAAAAh8SAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAACHxIAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAIfEgAAAAAAAAAAAAAAAAAACTox0aPaUO34HtKHb8D2lDt+B7Sh2/A9pQ7fge0odvwPaUO34HtKHb8D2lDt+B7Sh2/A9pQ7fge0odvwPaUO34HtKHb8CLoydki1LsWpdi1LsWpdi1LsWpdi1LsWpdi1LsWpdi1LsWpdi1LsWpdi1LsNUlG9j2lDt+B7Sh2/A9pQ7fge0odvwPaUO34HtKHb8D2lDt+B7Sh2/A9pQ7fge0odvwPaUO34HtKHb8D2lDt+B7Sh2/A9pQ7fgSdGKV1ue0odvwPaUO34HtKHb8D2lDt+B7Sh2/A9pQ7fge0odvwPaUO34HtKHb8D2lDt+B7Sh2/A9pQ7fge0odvwPaUO34CnQbSS/AqJKTSAAAAAoqCg3JHtKHb8D2lDt+B7Sh2/A9pQ7fge0odvwPaUO34HtKHb8D2lDt+B7Sh2/A9pQ7fge0odvwPaUO34HtKHb8D2lDt+B7Sh2/A5aVloWpdi1LsWpdi1LsWpdi1LsWpdi1LsWpdi1LsWpdi1LsWpdi1LsKNJ6WHOgna34HtKHb8D2lDt+B7Sh2/A9pQ7fge0odvwPaUO34HtKHb8D2lDt+B7Sh2/A9pQ7fge0odvwPaUO34HtKHb8D2lDt+AnRabS2PaUO34HtKHb8D2lDt+B7Sh2/A9pQ7fge0odvwPaUO34HtKHb8D2lDt+B7Sh2/A9pQ7fge0odvwPaUO34HtKHb8D2lDt+BXUGouKAIfEgAAAAAAAAAAAAAAAAAADEfGAAAABh/wCIhgAAAAVP4TAAAAAK3wx9AAAAAKfxr1KvxsAAAABfwX6gAAAAD+GPoAAAABH4kT+JgAAAAU/4cgAAAACp/Dh8wCHxIAAAAAAAAAAAAAAAAAKVN1HYrQUJWRSpxacpbFamo2cdmGI+MAAAADD/AMRDAAAAAqfwmAAAAAVvhj6AAAAAU/jXqVfjYAAAAC/gv1AAAAAH8MfQAAAACPxIn8TAAAAAp/w5AAAAAFT+HD5gEPiQAAAAAAAAAAAAAAAAARk0rLqYn+IVNKUUb0PRhWpzlK6R7Gp2GmnZgAAAGH/iIYAAAAFT+EwAAAACt8MfQAAAAPY1OxCjNSTsVfjYAAAAEIuVJpdz2NTsSpyjugAAAAfwx9AAAAAI/EifxMAAAACn/DkAAAAEacpK6R7Gp2KsWoRT8wCHxIAAAAAAAAAAAAAAAAADE/xCt8MPQj/BkFWtKErI+szJScndgAAAFKXLNMAAAAAqytCwAAAABOfMkuwAAAAfWZkMRNySKvxsAAAACnNwpNrufWZlSrKaswAAAAjLmigAAAAL21G7u4AAAAEZ2i13AAAACFaUFZH1mZWk5Qi35gEPiQAAAAAAAAAAAAAAAAAGJ/iFbWEGLSg/UMR8YAAAAEfiQAAAABW+FAAAAAFSCio26gAAAAU/jXqVfjYAAAAC/gv1AAAAAKfwoAAAAB7AAAAAEYKUZPsAAAAAVP4cPmAQ+JAAAAAAAAAAAAAAAAAAYn+IQj7SlyrdFZKEFAMR8YAAAAEfiQAAAABW+FAAAAAFb4Y+gAAAAFP416lX42AAAAAv4L9QAAAACn8KAAAAAewAAAABT/hyAAAAAKn8OHzAIfEgAAAAAAAAAAAAAAAAAG29xNrZjbe4Yj4wAAAAI/EgAAAACt8KAAAAAK3wx9AAAAAKfxr1KvxsAAAABfwX6gAAAAFP4UAAAAA9gAAAACn/AA5AAAAAFT+HD5gEPiQAAAAAAAAAAAAAAAAAABiPjAAAAAj8SAAAAAK3woAAAAArfDH0AAAAAp/GvUq/GwAAAAF/BfqAAAAAU/hQAAAAD2AAAAAKf8OQAAAABU/hw+YBD4kAAAAAAAAAAAAAAAAAAAYj4wAAAAI/EgAAAACt8KAAAAAK3wx9AAAAAKfxr1KvxsAAAABfwX6gAAAAFP4UAAAAA9gAAAACn/DkAAAAAVP4cPmAQ+JAAAAAAAAAAAAAAAAAAAGI+MAAAACPxIAAAAArfCgAAAABtsAAAACn8a9Sr8bAAAAAX8F+oAAAABT+FAAAAAPYAAAAAu7AAAAAFT+HD5gEPiQAAAAAAAAAAAAAAAAAABiPjAAAAAj8SAAAAAK3woAAAAAAAAAAp/GvUq/GwAAAAF/BfqAAAAAU/hQAAAAD2AAAAAAAAAACp/Dh8wCHxIAAAAAAAAAAAAAAAAAAAxHxgAAAAR+JAAAAAFb4UAAAAASi0lfqAAAABT+NepV+NgAAAAL+C/UAAAAAp/CgAAAAHsAAAAAKLabXQAAAAAqfw4fMAh8SAAAAAAAAAAAAAAAAAAAMR8YAAAAEfiQAAAABW+FAAAAAFb4Y+gAAAAFP416lX42AAAAAv4L9QAAAACn8KAAAAAewAAAABT/hyAAAAAKn8OHzAIfEgAAAAAAAAAAAAAAACnRlNaDpyUnEAAMR8YAAAAEfiQAAAABW+FAAAAAFb4Y+gAAAAFP416lX42AAAAAv4L9QAAAACn8KAAAAAewAAAABT/hyAAAAAKn8OHzAIfEgAAAAAAAAAAAAAAANxcsUolP+LJDVnYAAxHxgAAAAR+JAAAAAFb4UAAAAAVvhj6AAAAAU/jXqVfjYAAAAC/gv1AAAAAKfwoAAAAB7AAAAAFP+HIAAAAAqfw4fMAh8SAAAAAAAAAAAAAAAAjuid/bxKa/eyZLVsAAxHxgAAAAR+JAAAAAFb4UAAAAAVvhj6AAAAAU/jXqVfjYAAAAC/gv1AAAAAKfwoAAAAB7AAAAAFP8AhyAAAAAKn8OHzAIfEgAAAAAAAAAAAAAAACPLOKkStCLYAAYj4wAAAAI/EgAAAACt8KAAAAAJz5kl2AAAAAp/GvUq/GwAAAAF/BfqAAAAAU/hQAAAAD2AAAAAIztFruAAAAAVP4cPmAQ+JAAAAAAAAAAAAAAAAFOrKnoVK0qisAAGI+MAAAACPxIAAAAArfCgAAAAAAAAACn8a9Sr8bAAAAAX8F+oAAAABT+FAAAAAPYAAAAAAAAAAKn8OHzAIfEgAA4b8IMqzTKMPjaleopVIptLlsr9ro/4gZk3/QTU/wDJf/kT/iBmTf8AQTU/8l/+RP8AiBmTf9BNT/yX/wCRM88GspwGV4nF08RUcqcJySfLZuMW1f3dtAAAAAAAAAAAAIwlLZGJhKTVkYaEo3uiW7AAMR8YAAAAEfiQNAAAABW+FAAAAAAAAAAFP416lX42AAAAAv4L9QAAAACn8KAAAAAewAAAAAAAAAAVP4cPmAQ+JAABknjDjsry6jgYYaMlTioptu7sf8R1zD/oEh/4FI/4jrmH/QJD/wACkf8AEdcw/wCgSH/gUjNfGfHZhgK+DlhYJVISg3d3XMmr/iAAAAAAAAAAABhXo0EpKKux6gAGI+MAAAACPxIoU/aVYw7tL7zMsJLCYyrhpbwlKP3O36AAAABW+FAAAAAFX4Y+gAAAAFP416lX42AAAAAv4L9QAAAACn8KAAAAAewAAAABT/hyAAAAAKn8OHzAIfEgAAAAAAAAAAAAAAAE2tmVq3O1yjk3uwAAxHxgAAAAR+JGR0va5phqfecF98keJuB+qcVYyFtJS5v/AAJKT/FsAAAAK3woAAAAArfDH0AAAAAp/GvUq/GwAAAAF/BfqAAAAAU/hQAAAAD2AAAAAKf8OQAAAABU/hw+YBD4kAAAAAAAAAAAAAAAAUsPHlTluewp9hgAGI+MAAAACPxI4Npe04iwEf8Ap7T/APSkeOeB9lnlHEJaVKa++Laf4OIAAAAVvhQAAAABW+GPoAAAABT+NepV+NgAAAAL+C/UAAAAAp/CgAAAAHsAAAAAU/4cgAAAACp/Dh8wCHxIAAAAAAAAAAAAAAABK7SHJcyiQqud0t0AAGI+MAAAACPxI8O6ftOKcCv76f3Jv9Dx5wPPl2ExVvgnKP8A4Er/APtgAAAAVvhQAAAABW+GPoAAAABT+NepV+NgAAAAL+C/UAAAAAp/CgAAAAHsAAAAAU/4cgAAAACp/Dh8wCHxIAAAAAAAAAAAAAAABbolpWXoQpcs3IluwADEfGAAAABH4keFFL2nF+DXbnf3U5HizgViuFMQ1q4OMl8pJP8ABsAAAAK3woAAAAAqSUlFLogAAAAKfxr1KvxsAAAABfwX6gAAAAFP4UAAAAA9gAAAACEkoSXcAAAAAqfw4fMAh8SAAAAAAAAAAAAAAAAJ1nKSdtiVeCjdMAAMR8YAAAAEfiR4N0ufiuk/5Yzf/ktv1KlT+2MJnOAerjKUEvWlC3/k1/uJLUAAAArfCgAAAAAybLPryxCS1p05T/8AAbP8gAAAAp/GvUq/GwAAAAF/BfqAAAAAU/hQAAAAD2AAAAAAAAAACp/Dh8wCHxIAAAAAAAAAAAAAAABWujEwjFKyAAAxHxgAAAAR+JHgfS5+JZv+WlJ/+TQX6nAeZKXGGd4VveXMv+3G4v8ANHEeB+o5vicKlpCcor0Uml+AAAABW+FAAAAAB4QZZ/aGYY+ja/Nh6kfnJxX+YAAAAU/jXqVfjYAAAAC/gv1AAAAAKfwoAAAAB7AAAAAAAAAABU/hw+YBD4kAAAAAAAAAAAAAAAAOnzQUZH1WHcAAMR8YAAAAEfiR4D0r5ziZ9qdvvlH/ACOCcy9n4k4nXSrOvH8XJfjFI8W8D9V4qxDS0nyyXzik/wAUwAAACt8KAAAAAKkFFRt1Po/UObG46r2jBfe2/wD204jwX1LOMVhv5Kk4/dJoAAAAp/GvUq/GwAAAAF/BfqAAAAAU/hQAAAAD2AAAAAIwUoyfYAAAAAqfw4fMAh8SAAAAAAAAAAAAAAAAPrNM+s0xgAGI+MAAAACPxI8A6V8Tjp9owX3uX+RlOZfVuLaWMb0Va79HPX8Gzx5wPJmGExSXxwcf/AXf/wBvAAAAK3woAAAAArfDH0Po+0bYbH1e8oL7lJ/qeLGC+q8XYxJaScZL/t6Kb/G4AAABT+NepV+NgAAAAL+C/UAAAAAp/CgAAAAHsAAAAAU/4cgAAAACp/Dh8wCHxIAAAAAAG1e3UAAAAAAAAAAAAMR8YAAAAEfiR4CxUMNmFV7Xh+Cm/wBSU5OblfXc8Wksz4SwOZR1d4v5Thd/ikAAAAVvhQAAAABW+GPoeAtJQ4fxFV6Xqv7lCH+Z484L2Wf0a6Wk6a++MpJ/g0AAAAU/jXqVfjYAAAAC/gv1AAAAAKfwoAAAAB7AAAAAFP8AhyAAAAAKn8OHzAIfEgAAAAAAzvGLCKhWb0U0n6NNP/P5AAAAAAAAAAAABiPjAAAAAj8SPBz9zw1mNfs3/wCSwv8AqGH/AOFbwpcd5U4v5eznf/0lfiNagAAAVvhQAAAABW+GPocE4iWXeGWJxUd/3jXrpH9Dx8w8K+By/HQ2vJX8pKMl/wCksAAAAp/GvUq/GwAAAAF/BfqAAAAAU/hQAAAAD2AAAAAKf8OQAAAABU/hw+YBD4kAAAAAAHG80sDTj3l+Sf8AmcP4365l1Obeq0fqv81Z/MAAAAAAAAA+qw7n1WHcrUXD0AAMR8YAAAAEfiR4c/uPDzMqv/X5/dSiv0DwanHMOG8dlkn1a+VSFvziytBwm4yVmgAAAK3woAAAAArfDH0H/sng95yX/pVf/JnG3/Cn4YYPFbumqTfql7N/iwAAACn8a9Sr8bAAAAAX8F+oAAAABT+FAAAAAPYAAAAAp/w5AAAAAFT+HD5gEPiQAAAAAAcc106lGiuib+/RfkzgnG8leeGb0krr1W/3r8gAAAAAAAACVVRinIVWMouS6FW06TaAAMR8YAAAAEfiRwx+48LcXP8AmVX8fdDwIx3s82xOFvpOCl84yS/KTONsD9S4gxlBLRVJW9G7r8GgAAAK3woAAAAAbbOLv9l8K8FT/nVL8VznDX/Cp4U4jD7unGov/AX7RfoAAAAU/jXqVfjYAAAAC/gv1AAAAAKfwoAAAAB7AAAAAF3YAAAAAqfw4fMAh8SAAAAAADiTFrE5lUknovdXy0/O5gMXLCYqnXj9l3+XX70QnGcVKLumAAAAAAAAEdWjFp3TMKrplHWjJeoABiPjAAAAAj8SMPah4RSl3T/Gvb8g8K8d9U4swrb0nzRf/b0Wl+NjxnwP1fiedT/n5CMvuXJ/7aAAABW+FAAAAAB4tf7NwTlmF7On/wCS0mv1PAupHGZLmGXSenMn8qkXH/20qU5U5uEt1p9wAAAFP416lX42AAAAAv4L9QAAAACn8KAAAAAewAAAAAAAAAAVP4cPmAQ+JAAAAAAGbYz6pgqlZbpaer0X4jbbuyEHOSit2cKYt4jLowlvB8v3bfhp8gAAAAAAAAFy1Iolywg7FOyoMAAxHxgAAAAR+JGc/uPCejD+ZQ/GpzBk2N+o5lh8Vf8AhzjL/wABaf6HjzgU5YLFxW6nF/KzX5yAAAArfCgAAAABUpOUYrd7Hj9JUsHl2Hjtef8A5KoL9TwCxnJnGKw1/jp83zjJL8pM40wX1PiLHUOiqTt6OTa/BoAAACn8a9Sr8bAAAAAX8F+oAAAABT+FAAAAAPYAAAAAUG02ugAAAABU/hw+YBD4kAAAAAAca1nDL4wX2pL8E3/kFC/tY27o4eXsMzxmH6Xuvvf6NAAAAAAAAAU60oadCrWlPToAAGI+MMs4azjNKTq4HDTqRTs3GLaT3tp5H+oPFH/QBV/8AZ/qDxR/0AVf/AGf6g8Uf9AFX/wBn+oPFH/QBV/8AZ/qDxR/0AVf/AGf6g8Uf9AFX/wBn+oPFH/QBV/8AZ/qDxR/0AVf/AGf6g8Uf9AFX/wBn+oPFH/QBV/8AZ/qDxR/0AVf/AGf6g8Uf9AFX/wBkeAeKLr/AGCr/wCAM4+oVcD4c4HDVYuMv3Sae6fI20/NNagcff8ACr4fYPHbuKpSb83Hkl/5MxrUAMJg8RjK8aGHg5zlokldt+SP9ROJv+gCp/4Az/UTib/oAqf+AM/1E4m/6AKn/gDP9ROJv+gCp/4Az/UTib/oAqf+AMq8BcTtaYCp/wCAM/1B4o/6AKv/AIAz/UHij/oAq/8AgDP9QeKP+gCr/wCAM/1B4o/6AKv/AIAz/UHij/oAq/8AgDP9QeKP+gCr/wCAM/1B4o/6AKv/AIAz/UHij/oAq/8AgDP9QeKP+gCr/wCAM/1B4o/6AKv/AIAz/UHij/oAq/8AgDP9QeKP+gCr/wCAM/1B4o/6AKv/AIAz/UHij/oAq/8AgDP9QeKP+gCr/wCAMy3gLiN4/De0wNRR5o3bi7Jcyu36I+kDW5sdgaXaM397S/8AbTwgxn1bi7DJvSalF/OLa/FI8ZMF9X4uryS0qRhL/wAlUX+MWABgcvxWPxEMPhabnOWiSWrP+IbcWf8AQDP7l/mf8Q24s/6AZ/cv8yXhxxXGN3gJ/df8mf6g8Uf9AFX/AMAZ/qDxR/0AVf8AwBlTgniPDwdargqkYRV23FpJLVt+SW5V+NgAAAAL+C/UMFgcTjq8cPhoOdSW0Urt9dF6H+oPFH/QBV/8AZ/qDxR/0AVf/AGf6g8Uf9AFX/wBn+oPFH/QBV/8AZ/qDxR/0AVf/AGf6g8Uf9AFX/wBn+oPFH/QBV/8AZ/qDxR/0AVf/AGf6g8Uf9AFX/wBn+oPFH/QBV/8AZ/qDxR/0AVf/AGf6g8Uf9AFX/wBn+oPFH/QBV/8AZPgniOhRdSrgqijFNtuL0S1bfyHFpAAABa5ysy7hjOcyoutg8LOpBO14xbV+34ofAfE1v8AcKn/AIAz/UHij/oAq/8AgDP9QeKP+gCr/wCAM/1B4o/6AKv/AIAz/UHij/oAq/8AgDP9QeKP+gCr/wCAM/1B4o/6AKv/AIAz/UHij/oAq/8AgDP9QeKP+gCr/wCAM/1B4o/6AKv/AIAz/UHij/oAq/8AgDP9QeKP+gCr/wCAM/1B4o/6AKv/AIAz/UHij/oAq/8AgDP9QeKP+gCr/wCAM/1B4o/6AKv/AIAyHAXE6hJfUKn/AIAzM+F86yyiq2Nws6cG7XlFpX7X+QAAJN7Hsp9jBcCcSY3DwxGHwc5QlqmluvvP+IbcWf8AQDP7l/mT8OeK4Ru8BP5K/wCCP9QeKP8AoAq/+AM/1B4o/wCgCr/4AzMsBicDUWHxVNwqR3i1Zq9nqvRpgEPiQAAAAABx0/3NFeb/ACQZdSdXGUoLrJfmYSPLxJXt1gn/AOkoAAAAAAAACnh48qctz6vT7DAAMR8YeCWfZTl+T4injMVTpSdS6U5xi2uWOqTa0P8AXPhv/oYUf/BsP/kj/XPhv/oYUf8AwbD/AOSP9c+G/wDoYUf/AAbD/wCSP9c+G/8AoYUf/BsP/kj/AFz4b/6GFH/wbD/5I/1z4b/6GFH/AMGw/wDkj/XPhv8A6GFH/wAGw/8Akj/XPhv/AKGFH/wbD/5I/wBc+G/+hhR/8Gw/+SP9c+G/+hhR/wDBsP8A5I/1z4b/AOhhR/8ABsP/AJI/1z4b/wChhR/8Gw/+SP8AXPht/wDMwo/+DYf/ACR40cQZXj8loUcHiYVZe0TahOMmkoyV2k27agcM/wDCt4X18Nu6cai+cX7RfmkTWoAeCGTfWs8qY2S92hHT/FPRf+S8xV4ryClUlTqY6lGUW006kE01o01fRrqj/XHhz/oYUf8AwbD/AOSP9ceHP+hhR/8ABsP/AJI/1x4c/wChhR/8Gw/+SP8AXHhz/oYUf/BsP/kh8Y8OL/mYUf8AwbD/AOSP9c+G/wDoYUf/AAbD/wCSP9c+G/8AoYUf/BsP/kj/AFz4b/6GFH/wbD/5I/1z4b/6GFH/AMGw/wDkj/XPhv8A6GFH/wAGw/8Akj/XPhv/AKGFH/wbD/5I/wBc+G/+hhR/8Gw/+SP9c+G/+hhR/wDBsP8A5I/1z4b/AOhhR/8ABsP/AJI/1z4b/wChhR/8Gw/+SP8AXPhv/oYUf/BsP/kj/XPhv/oYUf8AwbD/AOSP9c+G/wDoYUf/AAbD/wCSP9c+G/8AoYUf/BsP/kj/AFz4b/6GFH/wbD/5IfGXDa/5mFH/AMGw/wDkjxmzPA5pnVGWFqxqQjTSvGSkr80m1dNra2hwnUWCz7BYhPSNSDfpzK/4XPHzLpf2phMUvtQcf/AZX/8Abx0ZroLDzYsL3YsPBChFbI8EuF23UzqtHvCn/wC3SX/pKf8AiRLjzhiLcXj6V1/fX+Z/r7wv/wBDCl/4Gv8AMjx3wxJpLMKX/gcV+bP9c+G/+hhR/wDBsP8A5I/1z4b/AOhhR/8ABsP/AJI4p4s4fr5FjaVLHUpSlSqJJVINtuDSSSldtvRIq/GwAAAAF/BfqHhti8PhOKsHWxFRQhFu8pNJL3ZLVvRH+ufDf/Qwo/8Ag2H/AMkf658N/wDQwo/+DYf/ACR/rnw3/wBDCj/4Nh/8kf658N/9DCj/AODYf/JH+ufDf/Qwo/8Ag2H/AMkf658N/wDQwo/+DYf/ACR/rnw3/wBDCj/4Nh/8kf658N/9DCj/AODYf/JH+ufDf/Qwo/8Ag2H/AMkf658N/wDQwo/+DYf/ACR/rnw3/wBDCj/4Nh/8kf658N/9DCj/AODYf/JH+ufDf/Qwo/8Ag2H/AMkYXN8ozaM6OGxFOtpqozjLR6a8rej2M3wEsBj6+DnvTlKP3O34klZgAAWuKKSMFhKuMxNPDUFzTm0ku7eiMqw+V8K5PQwlevCnGKtzSkoqUt5PVrVu7t29D/XHhz/oYUf/AAbD/wCSP9c+G/8AoYUf/BsP/kj/AFz4b/6GFH/wbD/5I/1z4b/6GFH/AMGw/wDkj/XPhv8A6GFH/wAGw/8Akj/XPhv/AKGFH/wbD/5I/wBc+G/+hhR/8Gw/+SP9c+G/+hhR/wDBsP8A5I/1z4b/AOhhR/8ABsP/AJI/1z4b/wChhR/8Gw/+SP8AXPhv/oYUf/BsP/kj/XPhv/oYUf8AwbD/AOSP9c+G/wDoYUf/AAbD/wCSP9c+G/8AoYUf/BsP/kj/AFz4b/6GFH/wbD/5I/1z4b/6GFH/AMGw/wDkj/XLhtrTMKP/AINh/wDJGdYbLOLMkxGEw1aFVSVlKMlJRmtY3cW7Wdrrs/MxGAlRqypTTjKLaa7NaNDwz6MdCa6CozfQWGb3YsNBbipQWyNEjhvJ553m1DL6T1m9X2S1k/kk2YrOcjyGnSwuIrwopRSjGUkvdWml+i2P9feF/wDoYUv/AANf5kOOuGJuyzCl85xX5s/1z4b/AOhhR/8ABsP/AJI/1z4b/wChhR/8Gw/+SPFLGYbGcT4mvhqkZwbjaUWmnaEVo1dPVW9QCHxIAAAAAA44p3wlKfaVvvX/AAA4ZqRhmtFyXVr700vxMGlPP8RJfZjFffZ/oAAAAAAAACV2irVVNI+trsAAGI+MAAAACPxIA8CsTCvgMdgJ7Xi7eUk4v/0lGPw0sNiJ0Z7wbi/VOwAeHNKHDvBFbNKq96SlU17JcsF82tP8RVqTqzlUm7ybbb83uAFb4UAAAAAVvhj6EKsobEMW4tO2qPGdwxnD2AzJbOS+6pDm/wDbT2kGtx16a6jxK6IeIm9jJstxWb5hRwNDWdSSS8u7fkldvyRx/m2H4O4RjgcE7TlH2cO9re9P1trf+aSYAFP416lX42AAAAAv4L9QAAAADwezX6jxNRg37tVSg/nqv/JopfM8ZMr+pcTyrRXu1oxl817r/wDSbv1Jq6AAC1xRSRKV9EeCHC/t8VUzmtH3afuw/wATXvP5Rdv+3n2PF3ij+188eGpSvSoXivOX23965f8At2/UewAAAABT/hyPBPiX+zs7eX1Zfu8QrLymvh+9Xj5trseNGTxyziBYmmvdrx5v+3lpL79JPzbFiYvcVWD6jqwXUeJj0HiZPZDrTfUbb3PArhpUcLWzqutZ3hC/SK+J/Nq3/br7niBxM+Ic+rYuLvTXuw/wR2+93l6sAKn8OHzAIfEgAAAAADifCvEZZUS3j733b/hcMJX9hiKdX+Vp/c7nC3NXhWxk1rUl+C2/Nr5AAAAAAAAAV4upGMonsp9gAAxHxgAAAAR+JAHgfjvY8Q1KDelSm184tNfhc8R8D9T4nxtPvPm/8DSl+oBgMHUxmKp4akrynJRXq3ZfieMGMpZRwxhsnoO3Pyxt/cppfrygAVvhQAAAABW+GPoBnf8AwqeEtGstXTjD/wAkn7N/crgB4E8K2jVzuvHe8Kf/ALfL/wBtT/xI8TuK3xBn1SVOV6NK8IdmlvL/ALeevpbsABT+NepV+NgAAAAL+C/UAAAAAyzGVMFiaOJp/FTkpL1i7/oeNmDp4/JMHmtHVJ2v/dqRTT++K+8g7qxNWYACVxRSRKV9EYHBV8diqeFoK85tRS827HEOOw/AvCEcPhn+8S5Id3NpuU/lrL1suo227vcewAAAABT/AIcjD4iph60K1J2lFpp9mndP7zxnoU814XwWb0lezi/SNSN/zUQADJ8rr5rj6OBoK86klFeV+volq/JHiVmlDhThGGWYN2lOKpR78qXvy9WtG+8rgAVP4cPmAQ+JAAAAAAEoqUXF6pmdZVPL8VKm17r1i+6/zWzMHg6uLrxo0ldv+r+iMHhYYXDwoQ2irf8AB+YAAAAAAAAAMAAxHxgAAAAR+JAHh1jvqfFOBqX3mo/+Bpw/9uPG/A+xz+FdLSpTX3xbT/CwB4O5N9e4jhWkvdopzfr8Mfxd/keMmc/XuJJUIu8aEVH5v3pfnZ+gAFb4UAAAAASm5JLsBwB/wp+G2Nwe7gqsV68qmvxYBkeT4jOMxo4DDK86jsvLu35JXb8keIWbYfhDhKGXYJ2nOPs4d7W9+Xrbr/NJMAAp/GvUq/GwAAAAF/BfqAAAAAU/hRlD/wBYPDCpQes6cJR+dJ80V/4CooWg0pIAEriikiUr6IPBDhf2+KqZzWj7tP3Yf4mvefyi7f8Abz7HizxR/bOeyoUpXpULwXZy+2/vVvSKfUHsAAAAARm1FruHDf8AxkPhfVwm86cZx+cHzwX3cqAAPAjhjnrVc6rR0j7kPV/E/krRXq+x4p8T/wBu8Q1XTd6VL3IdnZ+9L/t6V7PtYACp/Dh8wCHxIAAAAAAMVg6GKp+zrQUl5/1oYPLcJg01QpqN/v8Av3AAAAAAAAAPaT7ntJ92AAGI+MAAAACPxIAwmIlhsRTrQ3g016p3PHHDQxGX4HH09rtX8ppSX/pLGCV2eD+CpZVw5ic3rqyld3/uU0/15vuRmGNq47F1cVVfvVJOT9W7sACt8KAAAAAAPAHExqUMfg57e5K3qpJ/kjHYZ4XFVKEt4ScfudgPArhXkpVM7rx1leFP0XxS+b91eku54ncU/wCsGf1KlOV6NP3IdmlvL/t53fpbsAAU/jXqVfjYAAAAC/gv1AAAAAKfwo8B8xU6WNy6eq92aXr7svyiZ5lzy7M8Rg3/AMu5yj8k2k/miD6E49QEriSSJSuGX4CvmGLp4TDq85tRS83p93fsjiTG0OBuDo4bCv8AeW5IPq5y1lP5ay7Xsuo9QewAAAAAHgBmacsZl0nuozS/8ll+cTiPK3leb4nBdKc5RXono/mrMAMFg62MxNPDUI805tRS7tuyOKcXR4G4JjhMM7VOX2ce7lK7nP8A9Kl62QABU/hw+YBD4kAAAAAAAAAAAAAAABUpxVFPqezh2R7OHZDAAMR8YAAAAEfiQAcQ/wDCt4X4fEbunGm/nF+zb/O5JahQpyqTUYq7eiXmeIdSPDnAtLK6b96ajT+73pv52af+IAArfCgAAAAADwIxvsuIqlBvSpTf3pxf5XPEXBfVOKsfStvNy/8AA/e/9uDIsnxGcZlRwGH+KpJL0XVvySu35I8SM5ocJcKQy3AvlnNezh3UUvfl620vvzSTAACn8a9Sr8bAAAAAX8F+oAAAABT+FHhHmf1LiqjFvSqpQfzV1/5NFHjJln1PiidVLSrGM/nblf4xv8wTUkNW0EriSSJSuB4H8L+1r1M5rR0heEP8TXvP5LT5vseLPFH9s57KhSlelQvBdnL7b+9W9Ip9QHsAAAAAB4SZp9Q4sw137tS8H/28tP8AyZRPG7K1hOJ3XitK0Iy+a91/+kp/MAPA3hj65mdTNqsbwo6R/wAbW/8A27H8ZJ9Dxj4n/tfPnhaUr0sPeK85fbf3pR/7d8wACp/Dh8wCHxIAAAAAAAAAAAAAAACVWTgon1qXY+tS7AABiPjAAAAAj8SADw7/AOFXw+xmB6x9rFfOPMv/ACZlRah4YZN/aXEuGhJXjT/eP/t3Vf8Ak3KvmeOOcfWc6pYGL92jHX/FPV/+SqIABW+FAAAAAFX4Y+gHhbjPqnF2ClfSUnH/AMCi4r8WjxvwXsOKnUt/Epwl914f+2h4F8KKlQqZ5XWsrwp37L4pfNrlXo+54lcU/wCsOfVa1OV6UPch6LeX/bzu/Sy6AABT+NepV+NgAAAAL+C/UAAAAAp/CjLMdPA46jiob05Rkv8At1p/oeOmBhiMBgszpapNxv3U0pR+Xuv7wi7MlG4kkiUrgZdgK+YYulhKCvObUUvN6fd37I4qzChwPwhDC4R2qW5Id3J6yn+cu3M0uo9QHsAAAAAU/wCHIMDi6mDxVLE0/ihJSXrFpr8jx0wdPG5Ngs1o6pO1/wC7UjzJ/wDkq+8Ap051ZqEFeTdklu2yvKl4f8Dcqa9so2XnVn180nr/AIYk5ynJyk7tgAFT+HD5gEPiQAAAAAAAAAAAAAAACtdGKiklZAAAYj4wAAAAI/EgA8BMavaY3BvqoSXyun+cTPcC8FmOIwtv4c5R+5tfoRV2eB+VwoYLF5pV0TfKm+iiuaT9NV9xxBmss1zXEY2X/LyTa8lfRfJWQABW+FAAAAAFb4Y+gGR4z6lmmGxN/wCHOEv/AAGSf6H0gsFbEYHFLqpxfycWv/SmcP5LXzrM6GX0PiqO1+y3b9Ert+h4mZ1h+FeFoZbgnyzqR9nBdVFK0pfdpffmlcAACn8a9Sr8bAAAAAX8F+oAAAABT+FBW/4X/C1S3nSh9zouz++EX94EZaEpXAPA/hf2tepnNaOkLwh/ia95/JafN9jxW4o/trPZU6Ur0qF4R7N/al83p5pJgD2AAAAAKf8ADkBhP+Mh8KpU950oNeadF3S+cEvvAPBfhj+089+u1Venh7S9Zv4V8tZeTS7njfxP9fzeOWUpfu6G/nN7/wDgKsvJuQAAVP4cPmAQ+JAAAAAAAAAAAAAAAAHs04cstT6tTGAAYj4wAAAAI/EgA8Gcd9X4phTv/FhOP3Lm/wDbTxVwP1XinFJLSfLJfOKb/G5FWRnb/wBVvDiOGWlScFD/ALeqXc/uTlb0QAAVvhQAAAABW+GPoAeK/wDwpcD5fmC1d6cn6Tpu/wCNjwL4WVHDVc7rrWd4wv0ivil82rf9uvozxG4ofEOfVcRB3pQ9yH+Fdf8At53l80ugAAFP416lX42AAAAAv4L9QAAAACn8KDwQxlPGZTjcrq6pO9vKceV/+k/iY/BzweLq4afxU5Si/WLaf5AAZZl1fMsbSweHV51Gor5/ot32RxZj8PwTwhHCYN2nb2cO/M/in67y7czS6gA9gAAAACn/AA5AeAmYQrYPG5ZU1V1NLupLll+UfvM4y+WXZhXwct6c5R/8BbX6Ba+iMiw9HgPgh4ivH96o88k+tSVlGPy92L9GzE4iria069WXNOTbbfVt3b+bAACp/Dh8wCHxIAAAAAAAAAAAAAAAA+tQ7H1qHYAAMR8YAAAAEfiQAcGY76lxDgq99FUin6N2f4NnjrgeTNcNif54OPzjK/5SRwRk/wDa3EGFwjV4uScv8Mfef3pW+Z475z7TF4bLYPSCc5estF80k/8AwIAAK3woAAAAArfDH0AMgyqfFPhrQwEZWk7Rv25Ku/8A4AjxQzrD8L8LQyvBe7KovZxXVQStJ+ttL73lcAAAp/GvUq/GwAAAAF/BfqAAAAAU/hQeC+Z/VeJlQb0rQlH5r3l/6S18zxYyz6jxViLK0alpr/t5a/8AkykAB4H8L89SpnVaOkbwp+v2pfJe6vWXY8VuKP7az2VOlK9KheEezf2pfN6eaSYAPYAAAAAp/wAOQHg1mn1LiulTb0rRlB/dzL8YpfM8ZcrWC4rqzirKrGM193K/xi38w8JuGP7b4hpzqRvSoe/LtdfCvnLW3VJnjvxP7fF0smoy92n78/8AE17q+Udf+3l2AAAqfw4fMAh8SAAAAAAAAAAAAAAAAlRkoKR9Vl3Pqsu4AAYj4wAAAAI/EgAjJxkpReqPGGMcx4bwOZwXVP5VIX/NI8Csn58RiszktIpQj6v3pfclH7zjXOf7Xz/FYtO8XJqP+GPux+9JP1YAAVvhQAAAABUkpKKXRAHgRjoz4dr0ZP8Ah1G/ROMX+aZ4i8UviLPauJg70o+7D/Cuv/bzvL5pdAAACn8a9Sr8bAAAAAX8F+oAAAABT+FBw9mX9m5thsZ0pzi36Jq/3q6PHrLU5YPHx6qUH/6VH85ABlmXV8yxtLB4dXnUaivn+i3fZHGWY0OC+EYYLBu02vZw73a96fru7/zNAAPYAAAAAhJKEl3AyfMJZdmNDGQ3pzjL/wABaf4nj5l8K2EwWZU9Vdwb7qSUo/lL7w4By7D8H8HTzHFq05x9pLva3uR9bW0/mk0ZpmNfMsbVxld3nUk5P1bvp5LZeQAAFT+HD5gEPiQAAAAAAAAAAAAAAAFSpF0Ur6ntId0e0h3QwADEfGAAAABH4kABUf8Aa3hSnvKnBfL2c7f+kr8TLv8AjFfDZ1tqk4OXnzVdI/NJxv6AAAVvhQAAAAAAZBxbWyjJMwwFH4sRyJPslzc/3pqPzv0AAACn8a9Sr8bAAAAAX8F+oAAAABT+FAcSf8L/AIZ0sUtZ04wl84Pkm/u5mAHgfwvz1KmdVo6RvCn6/al8l7q9ZdjxS4o/tzPZqlK9KjeEezt8Uvm+vVJAAPYAAAAAAMb/AMZD4VRqLWdKCfnek7N/OKf3nhxwz/b/ABBRw81enD35/wCGLWn/AG87R+dzx34mUadHJKMt/fn6L4F99215RYAABU/hw+YBD4kAAAAAAAAAAAAAAAA6UlDmFTlJXSAAAxHxgAAAAR+JAAeDTpZlwxi8srax5pJ/4akUv0keOubRpYfCZVT0vebXZL3Y/LWX3AAAVvhQAAAAAAAAABT+NepV+NgAAAAL+C/UAAAAAp/CgPB+rDNeF8ZlNV6Jyj6RqR/zUitSnRqSpzVpRbTXmtGBlOWV8zx1LBYdXnUaS+fV+SWr8jjXMsPwbwlHB4N2m17OHe7XvT9bXd/5mu4AA9gAAAAAA8DcXTx2SY3KauqTv/27Ujytf+Sv7zwz4ahwnkWJx2Ye7NuUpO20Kd0vvs5ed0cRZ1WzrNK+PrfFUk3bstkvkrL5AAAFT+HD5gEPiQAAAAAAAAAAAAAAAEv93RR/gy+f5AAAYj4wAAAAI/EgAPAbHcmZYvCX+OCl/wCAO3/t54mZz/anE2JqRd4wfJH0ho/vld/MAACt8KAAAAAKkFFRt1AAAAAp/GvUq/GwAAAAF/BfqAAAAAU/hQHgfmf1fPqmFb0qwdv8UXdf+S8x4lZZ/Z/FOMppaSlzr/t9cz/FtfIDwO4Xu6mdVo7XhT/9ul/7an/iR4o8UrPc9mqUr0aPuQ7P+aXzf3pIAAewAAAABCClCT7AHgjmn1XidYdvStCUfmveX4Ra+Z46cTfVcup5RRl79b3peUIvRf8Ab0v/AElrqAAAFT+HD5gEPiQAAAAAAAAAAAAAAAHK5UEkUqclSaa1PYVOwAAYj4wAAAAI/EgAOCeIv7BzX652hUXzcHyr5zUSUpTk5Sd2wAAK3woAAAAArfDH0AAAAAp/GvUq/GwAAAAF/BfqAAAAAU/hQHBmZ/2bxBg8VeyjNJ/4Ze7L/wAlbPHjLPZ4/C46K+OLg/WLuvvUvwDKcsr5njqWCw6vOo0l8+r8ktX5HHOZ4fg/hKGAwbtOUfZw77e/P1td3/mkgAAewAAAABT/AIcgDhvM3lecYXG30pzi36Jq/wB6ujjHiGef51Xx8vhk7RXaK0ivu1fm2AAAFT+HD5gEPiQAAAAAAAAAAAAAAAFH+GgGAAYj4wAAAAI/EgAAAACt8KAAAAAK3wx9AAAAAKfxr1KvxsAAAABfwX6gAAAAFP4UAeINs+8P8PmS1lBU5v1a5JL73r6B4H8LNupnVePeFP8A9ukv/SU/8SPE7ij+3c9m6cr0qXuQ7O28v+3n17JAAA9gAAAACn/DkAAAAAVP4cPmAQ+JAAAAAAAAAAAAAAAAcztuc0u5zS7gABiPjAAAAAj8SAAAAAK3woAAAAArfDH0AAAAAp/GvUq/GwAAAAF/BfqAAAAAU/hQB4aNZ5wNicrk7yj7SC8uZc0X/wCBN29DJ8pxGaZhSwFBe/OSj6d2/JK7fkjjzNcPwhwnHA4N2nKPs4d9vel621v/ADSTAAAewAAAABT/AIcgAAAACp/Dh8wCHxIAANC6Loui6AAAAAAAAAAAAAAAMR8YAAAAEfiQAAAABW+FAAAAAFb4Y+gAAAAFP416lX42AAAAAv4L9QAAAACn8KAPAjM/ZZpicE3pUgpL1g7flJ/ccEcCLLOIcwzGrGy55Rpf4Ze82vk1FPykeJnFP9v55OVJ3o0vch2aT1l/28/wsAAA9gAAAACn/DkAAAAAVP4cPmAQ+JAAA0WLFiwAAAAAAAAAAAAAABiPjAAAAAj8SAAAAAK3woAAAAAeoAAAAFP416lX42AAAAAv4L9QAAAACn8KAPDzM/7O4nwdZuycuV+k/d/C9/keKvFH9i5FKnSlatWvCPdL7UvktPJtMAAAewAAAAAAAAAAVP4cPmAQ+JAAAAAAAAAAAAAAAAAAAGI+MAAAACPxIAAAAArfCgAAAAAAAAACn8a9Sr8bAAAAAX8F+oAAAABT+FAEJypzU4uzWq9UcecXVOJczWI2hGKjFdtLyfzlf5WXQAAAewAAAAAAAAAAVP4cPmAQ+JAAAAAAAAAAAAAAAAAAAGI+MAAAACPxIAAAAArfCgAAAACUHFK/UAAAACn8a9Sr8bAAAAAX8F+oAAAABT+FAAAAAPYAAAAAUG02ugAAAABU/hw+YBD4kAAAAAAAAAAAAAAAAAAAYj4wAAAAI/EgAAAACt8KAAAAAK3wx9AAAAAKfxr1KvxsAAAABfwX6gAAAAFP4UAAAAA9gAAAACn/AA5AAAAAFT+HD5gEPiQAAAAAAAAAAAAAAAAAABiPjAAAAAj8SAAAAAK3woAAAAArfDH0AAAAAp/GvUq/GwAAAAF/BfqAAAAAU/hQAAAAD2AAAAAKf8OQAAAABU/hw+YBD4kAAAAAAAAAAAAAAAAAAAYj4wAAAAI/EgAAAACt8KAAAAAK3wx9AAAAAKfxr1KvxsAAAABfwX6gAAAAFP4UAAAAA9gAAAACn/DkAAAAAVP4cPmAQ+JAAAAAAAAAAAAAAAAAAAGI+MAAAACPxIAAAAArfCgAAAACt8MfQAAAACn8a9Sr8bAAAAAX8F+oAAAABT+FAAAAAPYAAAAAp/w5AAAAAFT+HD5gEPiQAAAAAAAAAAAAAAAAABCnKbsicHB2YYj4wAAAAI/EgAAAACt8KAAAAAJTckl2AAAAAp/GvUq/GwAAAAF/BfqAAAAAU/hQAAAAD2AAAAAIzai13AAAAAKn8OHzAIfEgAAAAAAAAAAAAAAAAAC7hRVt2VHz0lJ7oJqjf3ty2H/q5Pl5ny7AAAAFGKlNIAAAAAqRTptgAAAAFSCio26gAAABbD/1ciqF1bf5lX42AAAABR5PZvm2uWw/9XKypWXIAAAACilFW7AAAAAJJuxJJNoAAAACEFKEn2AAAACkqXL7+5bD/wBXMRy8seXbUAh8SAAAAAAAAAAAAAAAAAAK3wQXkR1oSDEfGAAAABh/4iGAAAABU/hMAAAAArfDH0AAAAAp/GvUq/GwAAAAF/BfqAAAAAP4Y+gAAAAEfiRP4mAAAABT/hyAAAAAKn8OHzAIfEgAAAAAAAAAAAAAAAAACt8EPQj/AAZBiPjAAAAAw/8AEQwAAAAKn8JgAAAAFb4Y+gAAAAFP416lX42AAAAAv4L9QAAAAB/DH0AAAAAj8SJ/EwAAAAKf8OQAAAABU/hw+YBD4kAAAAAAAAAAAAAAAAAASXNRT7Ely0Uu4Yj4wAAAAMP/ABEMAAAACp/CYAAAABW+GPoAAAABT+NepV+NgAAAAL+C/UAAAAAfwx9AAAAAI/EifxMAAAACn/DkAAAAAVP4cPmAQ+JAAAAAAAAAAAAAAAAAAFKq4FWo6juHt6TWqPb0f5fwPb0f5fwPb0f5fwPb0f5fwPb0f5fwPb0f5fwPb0f5fwPb0f5fwPb0f5fwPb0f5fwPb0f5fwPb0f5fwPb0f5fwPb0f5fwFXpLZH1mn2PrFPsfWKfY+sU+x9Yp9j6xT7H1in2PrFPsfWKfY+sU+x9Yp9j6xT7H1in2PrFPsfWKfYeIptWse3o/y/ge3o/y/ge3o/wAv4Ht6P8v4Ht6P8v4Ht6P8v4Ht6P8AL+B7ej/L+B7ej/L+B7ej/L+B7ej/AC/ge3o/y/ge3o/y/ge3o/y/ge3o/wAv4DxFJ9D29H+X8D29H+X8D29H+X8D29H+X8D29H+X8D29H+X8D29H+X8D29H+X8D29H+X8D29H+X8D29H+X8D29H+X8D29H+X8D29H+X8D29LsTalJsAAAAClVhGLUke3o/y/ge3o/wAv4Ht6P8v4Ht6P8v4Ht6P8v4Ht6P8AL+B7ej/L+B7ej/L+B7ej/L+B7ej/AC/ge3o/y/ge3o/y/ge3o/y/ge3o/wAv4Ht6P8v4H1mn2PrFPsfWKfY+sU+x9Yp9j6xT7H1in2PrFPsfWKfY+sU+x9Yp9j6xT7H1in2PrFPsfWKfY+s0+x7ek+h7ej/L+B7ej/L+B7ej/L+B7ej/AC/ge3o/y/ge3o/y/ge3o/y/ge3o/wAv4Ht6P8v4Ht6P8v4Ht6P8v4Ht6P8AL+B7ej/L+B7ej/L+B7elbY9vR/l/A9vR/l/A9vR/l/A9vR/l/A9vR/l/A9vR/l/A9vR/l/A9vR/l/A9vR/l/A9vR/l/A9vR/l/A9vR/l/A9vR/l/A9vR/l/A9vR/l/ArVIzsorYAh8SAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAACHxIAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAIfEgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAh8SAAAAAAAAAAAAAAAAAAAAAAAAKVOLjzT2K1NQemwAAAACoWpuT3AAAAApU1OVrk6MHBuD2AAAAAAAAAAAAAAAKcOeSRUpQcW4PYAAAACjR535FSKjJpAAAABuKhTXut6k4uMmgAAAAACHxIAAAAAAAAAAAAAAAAAAAAAAAAraQjEl71FPsAAAABCTlCV+wAAAAFzDfE15AAAAAAAAAAAAAAAFHSEpGH1k490AAAABTk7qPmVv4jAAAAAu73MT8YAAAAABD4kAAAAAAAAAAAAAAAAAAAAAAAAYjePoL+A/UAAAACl8EwAAAACirRlIAAAAAAAAAAAAAAAh/BkYf+IiW7AAAACn8a9St/EYAAAAEI80kiu+aowAAAAACHxIAAAAAAAAAAAAAAAAAAAAAAAAr6qL8h6UPmAAAABS+CYAAAAFrla0IKCAAAAAAAAAAAAAAAKetKSMMvfHqAAAAFP416lb+IwAAAAKS5IOb+RuAAAAAAQ+JAAAAAAAAAAAAAAAAAAAAAAAAEYe1pJLdFe0UoLoAAAABS+CYAAAAGGgpSv2JYaUndsas7AAAAAAAAAAAAAAAUGuZxfUVP2UG29QAAAAKfxr1K38RgAAAAldpFSg5JJPRFWk4O1wAAAAACHxIAAAAAAAAAAAAAAAAAAAAAAAAUnHZj1AAAAAu0tAAAAAE2tmc8u4AAAAAAAAAAAAAAASlKW7AAAAAHqAAAABzy7jbe7AAAAAAIfEgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAh8SAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAACHxIAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAIfEgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAh8SAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAACHxIAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAIfEgAAAAAAAAAAAAAAAAAAAAAAAAAAABpoAAAAAAAAAAAAAAAAAAAAAAAAACzsAAAAAAAAAAAAAQ+JAAAAAAAAAAAAAAAAAAAAAAAAAAAGGhGUXdHs4dkVoRTjZdTFpaAACV2kYppWigoO1RGIjaowAAAAAAAAAAAAAAAAAAAACik6iTPZw7IlThyvRFNL2H3gABD3aDfcCt71KMgAAAAAAAAAh8SAAAAAAAAAAAAAAAAAAAAAAAAAAAIzlFWTHJ+xvfWw5Tm0myrCatzMdCaaXcqUZQsSw81G4fVptXGiir1EYh3qMhh5yVxxcJWfQxa1TAAAAAAAAAAAAAAAAAAAAAE2ndGGnKV7sq1JqTVyNOo6d09CNKUo3QsPNxuU6Epq6JRcXZlOjKauidOUHZhV0oxRCnKbsipRlBXZD3qDQAAAAAAAAAQ+JAAAAAAAAAAAAAAAAAAAAAAAAAAAA/4HyKfxr1MXsiUlGN2JxlFMhONRaFCCdT0PbL2nIYmKUzDL94VFzVWvMq1VTS0KslOTaMTrCIAAAAAAAAAAAAAAAAAAAAAYTqVvjZT/gfJmF1gyNSMm0uhzxUlAxUffVic1SgkivadJSQYn4YooWjSuVayqRSsUNackAAAAAAAAAEPiQAAAAAAAAAAAAAAAAAAAAAAAAAAAP+B8in8a9TF7IxP8NFP+B8mYTqUPjmexhzXtqYv4kYb4yo7VG/McY1IlSDhKxiP4cQAAAAAAAAAAAAAAAAAAAAAwnUrfGyn/A+TML8DML8b9Cf8f7ivrUgTpxnuirFKk0gxW0TDSThy9jEUVHVGH+CQAAAAAAAAAQ+JAAAAAAAAAAAAAAAAAAAAAAAAAAAA/4HyKfxr1MXsjE/w0U/4HyZhOpQf7ySKsaim7XJ8ydpbmH/AIiKyvUaKHPGfKzFfGYrSMUAAAAAAAAAAAAAAAAAAAAAGE6lb42U/wCB8mYX4GYX436E/wCP80Yh2nFmIjJpOJJVOXW9gxOsIsSnFKSK+tLXco6UpMAAAAAAAAAIfEgAAAAAAAAAAAAAAAAAAAAAAAAAAAdWDpct9bEGlJMxFSM7crK9WEoJJkKsFR5b66mHqRhe7FPlqcyFWptXuVpRlO6KTtNFa0aqbPbU7XuSl7SoYp3kkAAAAAAAAAAAAAAAAAAAAAGHqRhe7KjUptohVgqXLfXUoVYRjZsoTjGV2yU4urzX0MROM7crKVeLjaT1MRUhKNkwn71BPsUasFBK5iKqkuWIvdw4AAAAAAAAAQ+JAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAV5xmk1uFJxU02VZc82wAAAAAAAAAAAAAAAAAAAAAAAAIVI+zcWBVqRcFGIAAAAAAAAAQ+JAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABD4kAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAEPiQAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAQ+JAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABD4kAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAEPiQAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAQ+JAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABD4kAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAEPiQAAAAAAAAAAAAAAWRZFkWRZFkWRZFkWRZFkWRZFkWRZFkWRZFkWRZFkWRZFkWRZFkWRZFkWRZFkWRZFkWRZFkWRZFkWRZFkWRZFkWRZFkWRZFkWRZFkWRZFkWRZFkWRZFkWRZFkWRZFkWRZFkWRZFkWRZFkWRZFkWRZFkWRZFkWRZFkWRZFkWRZFkWRZFkWRZFkWRZFkWRZFkWRZFkWRZFkWRZFkWRZFkWRZFkWRZFkWRZFkWRZFkWRZFkWRZFkWRZFkWRZFkWRZFkWRZFkWRZFkWQAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAF7jLoui6LoAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABDAB7HQWwtgBgAtgOoD2OgPYWx1AAOoNXEWudBFhAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAIewAHQ6FtAsND2LD1AWwWuPcC2ha6B7ADCwWsdQDoLYGdAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAOhugABjOobnQewyxYWwdDqdToPY6A9hgMswDqBbQ6C2BgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAD2FsAB1HuM3DyOhuM3DYLnU6hfQNx7DGDL3Oo9jc6m4y9zyA6j2AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA3LDQAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADZ0ExgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAzoIYAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAM6FhgD2AADU3AGdAAAAGM3GDZe5cC4AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAdB7AAzoXGbgB0AHsHQDoFxnQAOp1AB7ALYWwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB0FqgA0Og9hnUA6DNzoDOp0Nw6D2GdQewPYYdS2gPYDQ0DodQAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAOghgFkdBnUOh0HsdBgDGB0Ogxj2Oo9gauNB1AYPYLDLaFtQAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAsdCzHqixYeoWALFgGrlixYGrjVyxYsNXLFhq5YAauWLaFgLAWAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAD//2Q==)
Figure 1. Firing patterns of identified dopamine and GABA neurons in VTA. (a) VTA neurons were recorded while mice performed an odor-outcome association task in which different odors predicted different outcomes (see legend on right). Odors were presented for 1 s (gray shading), and outcomes were presented after a 1-s delay. Neuron types were identified based on their optogenetic responses. Dopamine neurons (left) showed phasic excitations to reward-predictive cues and reward. GABA neurons (right) showed sustained activation during the delay. Data from Cohen et al. (2012). (b) Reward expectation modulates dopamine neuron firing. The plot on the left shows when outcome was presented, and the right-hand plot shows when outcome was omitted. Different odors predicted reward with different probabilities. Higher reward probability increased cue responses but suppressed reward responses. Data from Tian & Uchida (2015). Also see Fiorillo et al. (2003) and Matsumoto & Hikosaka (2009a,b). (c) Reward context-dependent modulation of dopamine responses to air puff–predictive cues and air puff. The task conditions during recording differed only in the probability of reward. Dopamine neurons showed both excitation and inhibition in high-reward contexts (left) but only inhibition in low-reward contexts (right). The response in reward trials is not shown. Data from Matsumoto et al. (2016). Abbreviations: CS, conditioned stimulus; VTA, ventral tegmental area.
Expectation modulates DA signaling
![[[@Watabe-Uchida2017-gi]](https://doi.org/10.1146/annurev-neuro-072116-031109)](data:image/jpeg; charset=UTF-8;base64,/9j/4AAQSkZJRgABAQECWAJYAAD//gAuSGFuZG1hZGUgU29mdHdhcmUsIEluYy4gSW1hZ2UgQWxjaGVteSB2MS4xNAr/2wCEAAMCAgICAgMCAgIDAwMDBAcFBAQEBAkGBwUHCgkLCwoJCgoLDRAOCwwPDAoKDhMODxEREhISCw4UFhQSFRASEhIBAwMDBAQECAUFCBIMCgwSEhISEhISEhISEhISEhISEhISEhISEhISEhISEhISEhISEhISEhISEhISEhISEhISEv/CABEIC30KXgMBIQACEQEDEQH/xAGiAAABBQEBAQEBAQAAAAAAAAAAAQIDBAUGBwgJCgsBAAMBAQEBAQEBAQEAAAAAAAABAgMEBQYHCAkKCxAAAgEDAwIEAwUFBAQAAAF9AQIDAAQRBRIhMUEGE1FhByJxFDKBkaEII0KxwRVS0fAkM2JyggkKFhcYGRolJicoKSo0NTY3ODk6Q0RFRkdISUpTVFVWV1hZWmNkZWZnaGlqc3R1dnd4eXqDhIWGh4iJipKTlJWWl5iZmqKjpKWmp6ipqrKztLW2t7i5usLDxMXGx8jJytLT1NXW19jZ2uHi4+Tl5ufo6erx8vP09fb3+Pn6EQACAQIEBAMEBwUEBAABAncAAQIDEQQFITEGEkFRB2FxEyIygQgUQpGhscEJIzNS8BVictEKFiQ04SXxFxgZGiYnKCkqNTY3ODk6Q0RFRkdISUpTVFVWV1hZWmNkZWZnaGlqc3R1dnd4eXqCg4SFhoeIiYqSk5SVlpeYmZqio6Slpqeoqaqys7S1tre4ubrCw8TFxsfIycrS09TV1tfY2dri4+Tl5ufo6ery8/T19vf4+fr/2gAMAwEAAhEDEQAAAP1VAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA+JuCD7z6MAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA/no+bA/pU9WAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAP56PmwP6VPVgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADn/AMuw+a84PqL9Tw2AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA/no+bA/Zj4jCb9PQ+qwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAM/+foPpbyAPjP9LQ/X0AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAP56PmwO398D5N0w/o+9YAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA8sD4e8SD89veg/owAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAD+ej5sD+gn6hD8bfznD9dv00AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAD5n/AAFDqPsAPz/9oD+kIAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAP56PmwP6VPVg/OD8dA/Uf9YwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA/Gr86g/d36dD+YX2gP6QgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA/no+bA/cT7dD8J/jYP25+8QAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA/N38dw9JmDzj1sP6RwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA/ni8TDM1g5v3gP6HtQAAAAAAAAAAAAAAAAAAAAAAAg/OoOb9ED7P60Pmv1MPQQAD5j9kDuADywOQ+ggAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAACt8BB5R90h7oAAH5Oh+sQAAAAAAAAAAAAHxV9qgAAAAAAAAAAAAAAAAAAABH+d4eFfYoefYgfpAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAH59SB7F8Mh6Z+iQdOAAAAAAAAAAAAAAAAAAAAAAAGV+EAftj4qH5WftuHlHrAekAAHyr7WHfgHkgcZ9HAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAH5o9IH2OB6GAAH4Jh+9gAB+IP7fAAAAAAAB+X4fYfvofC33SAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAGV+HofuuB8ZfPQdj9RB+Yv7bh5t+c4dh9jh8n+GB9edgHkv2AH5H7odV+swfj5ph4l+nAfUoAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAH5q8+H6f2w+OM0Ptn8cg/Xv8JQ+kvFg+o/eA/E77kD7x/OAPRvqgPkfzMP2f8ANQ/Kfpw+6PyLD2j9WQ/MD9nw/LHz4OT/AFzD5p+Vgx+vD9cAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADK/D0P3XA+dfgEOx+wQ+Mfs4PmH6HD5A+7A/Lv9Kg+mvj4PFPNg+7fogPx2/QsPyF/dcOQ/KEP2uAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAh/O4Phj9Yw8vyQ/QH8KA/cL8KA/a3vg/AL98g/B/8AewPmn87w/aasHyv+cofo5+cgfsj3IH5Jh92fQAfgn+vAfnV+xIfO/wAEB79kB+jH4GB+9FsAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADK/D0P3XA/Mj0UPGvsEO0/OAOC/bsPyW+7A9S/MkE+pA8U8AD9YvSQ/LT6lD8uf3sDlPxoD9wgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADjvxaD7v54P0I/CMP3H/CgP3jsh+E37hB+FX72B80/Hwfqv8Ai4H2/wDMofTv5nh+9U4H5Jh92fQAfgn+oQfL/wCpQea/lEH1NrB91/hQH7haoAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAGV/PiH6aeBhxv7LB+V/wBgh9LfhQH0x+n4fkn92B5GwPjv7iDxT3oPz4+1Q/Pz91w/CP8AewOU/GgP3CAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA+GK4eB+gh9gfjmH3Z+Yof0W/iMHuXowfN37Uh+EH6DB6H8fB+q/4Rh+hP52B+kPlwfPn1oH0j8RByX6ah+Hn7sB+Ef6Gh8VfokHkmsH3X+FAfuFqgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAR/NAVfYQ9EA8/6QNzzoOm3w876wPMPPw+msgMvuA8s8mD6Z0w8j9cCv5mHqgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHMfO4d/7gB4wGb14ehebhP5GH07YDifFg+g+bD0DlQ8D9sDQ6oPD+CD6UuB8v8A0kHn3rgY3zSHs3pIcfIHWeTh6pIAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAACfGofDXiwM9hD7W+3gnAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAD5B+Bw+y/XQf5SHyB9WB96AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAih5/2IePeFB9wSAoAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAGdGABatgAAAB5f4aH2EAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfhf+j4fh2gfrv9/gAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfz0fNgAH6ffrYAAAAH5/fkmH9NoAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB+LHzgGl+kIeR/q+AAAAB8ah5mBYugAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHOBzP2wHuQAAAN/O0McAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADMDlPrgPrIAAAAAAAAAAAADO+QA/CX+pUOM+Ag5X7tD6C+SA+QPrMOB/JMP6RPzoD7O9yAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAIpQ8+/NsPvPyYOm+pQAAAAPxvD9OgK1MAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA3w1PzgD9VwAAAyvydD9NwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAANEOs/HQP2OAAAAAAAAAAAAAPj4PwW/qqDzT5CD5u+KQ/o8/mQD9b/o8PIvyTD7G44P3QsAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHzF+PAfWv2UH01bAAAAAAPyTD9bAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAOH+Ag/TUAAAMr8vQ/VcAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA/JIP1tAAAAAAAAAAAAAPj4PwW/qqD5R/E4PX/lUP6i/wyD5m/XQND8Xwg/bkPu8AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADCD5M+NQ+Zfdg+8/ssAAAAD8kw/WwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADz/4TD9LAAAAyvy9D9VwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAD8kw/WwAAAAAAAAAAAAA+Pg/Bb+qoPxr8ND7O/I0P6f8A5XD43+dQ/ST8gw/Un8vA/o39AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAb88h5F9ygAAAAfkmH62AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAef/CYfpYAAAGV+XofquAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfkmH62AAAAAAAAAAAAAHhofm1+y4eL/lWHunmAfqT+UoZ/wCkIJ8TB+s35Gh9Lfc4AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAflP9sB+TfyyH9AH0UAAAAB+SYfrYAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABUD5c+fw+bv0QD6vA8/8AhMP0sAAADK/L0P1XAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAxQ+Y/AQ+YfssP0HAPyTD9bAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPwl9lD8+/0XCf9WwAAAAPyTD9bAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAMnwIPmfwIPlPy0PZ/pEPpz6WD3wDz/AOEw/SwAAAMr8vQ/VcAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADJ+dw+ZfAQ+W/Mg9s+lA+pPfA+hwD8kw/WwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAD44/GAP1IyA1f0gAAAAA/JMP1sAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAOG+ZQ+TfmUPEvSg+lfdQ+nvoMNcAAAPP/hMP0sAAADK/L0P1XAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPP8A5kD5V+Ww8Q9fD6U93D6a+jAsAAAAfkmH62AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAM+Oww/r4I+pAAAAA/JMP1sAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADyr5mD5m+RA8x+ig+kvqgPo7vgAAAAAADz/4TD9LAAAAyvy9D9VwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA8i+bA+W/kwPOPpEPpH6nD6P7YAAAAAAAPyTD9bAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAhmcgqAAAAAAAAAAAAAAAAAAAAAAD8uvzmD0f7cA/TsAAAAD8kw/WwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABgePfKwfKPzeHF/QIfVH10H0ZoAAAAAAAAAAef/AAmH6WAAABlfl6H6rgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAeLfKYfJ/zsHK/QQfU31qH0beAAAAAAAAAAPyTD9bAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA8z7z0/A0DzPfAAAAAAAAAAAAAAAAAAAAAAD8I/0xD8k/skM/8AV4AAAAD8kw/WwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAwvkQPl347Dy36sD7E+jA+jpgAAAAAAAAAAAA8/wDhMP0sAAADK/L0P1XAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAMD5CD5a+RQ83+mw+zPpAPoWcAAAAAAAAAAAAD8kw/WwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPFuZ+z/LPo8+M/UwAAAAAAAAAAAAAAAAAAAAAA+BfyFDFaH78fTAAAAAH5Jh+tgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAR/PwfFfyKHjHtwfU/1gH1JogAAAAAAAAAAAAAAHn/wmH6WAAABlfl6H6rgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABB8+B8Y/IAeN+4B9WfVwfUGiAAAAAAAAAAAAAAAH5Jh+tgAAAAAAAAAAAAHJwB2fPhT6wAAAAAAAPjYPr+0AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAANrAWQcAAAHzHx/wCk/g3ufrPxH62Hm+8AAAAAAAAAAAAAAAAAAAAAZnz0Hzx9Lh2vqYAAAAH5Jh+tgAAAAAAAAAAAAAAAAAAAAAAAAAAAABzAfLfxQHxnVD7i+yw+quhAAAAAAAAAAAAAAAAAA8/+Ew/SwAAAMr8vQ/VcAAAAAAAAAAAAAAAAAAAAAAAAAAAAA5QPmD4kD46jD7R+1w+pOiAAAAAAAAAAAAAAAAAAPyTD9bAAAAAAAAAAAAAPhT8+A/e35ZDz/wC6wAAAAAAA/FMP2X1QAAAAAAAAAAAAAAAAAAAAAAAAAAAA889D7vIDh9cAAAAAp841kVWpdkNvUVPEABxXX5vzQfpH4RN9P/Ofc9OfH/qAAAAAAAAAAAAAAAAAAAAAB+P32wHpn4nB9xfpYAAAAB+SYfrYAAAAAAAAAAAAAAAAAAAAAAAAAAAHnnxcHwd82B7/APaIfZnvgAAAAAAAAAAAAAAAAAAAAef/AAmH6WAAABlfl6H6rgAAAAAAAAAAAAAAAAAAAAAAAAAABwnxCHw98wh9DfZAfZX0AAAAAAAAAAAAAAAAAAAAAB+SYfrYAAAAAAAAAAAAB8KfPwene5h5/wCth8J5wfp36kCflYFTpw/TX8lQ1visP6FvzuDhog/WH8cQ7z9VQAAAAAAAAAAAAAAAAAAAAAAAwNuX5Z+m/q/znqj479QAADm+Aa4/nbVbdFubiNzbT29RU8QAQfKn03wOKfW/mhv49X1LY/NP3wJsAAAAAAAAAAAAAAAAAAAD4g/GoMb63D9iPTgAAAAPyTD9bAAAAAAAAAAAAAAAAAAAAAAAAADzAPhf4pDwP69D7N+4w3AAAAAAAAAAAAAAAAAAAAAADz/4TD9LAAAAyvy9D9VwAAAAAAAAAAAAAAAAAAAAAAAADzcPg/4mDwL7CD7E+7g2gAAAAAAAAAAAAAAAAAAAAAA/JMP1sAAAAAAAAAAAAA+FPQA/Pj7DCT4rD9tvOw/M39fw+cflQP05/HQPrrxgP04/DsP1y/MYP2L/ADHD6k/KIP3xAAAAAAAAAAAAAAAAAAAAAAAA+cPd+P4Ltvq/zj6Ud+b/ALsVR/JviOk1N1re3g3d5G5uI3NtPb1FTxAB4/4n97+NB63zZ0WHX6B7V8T+th5P0YAAAAAAAAAAAAAAAAAAfJfxkB5eHyB+hAfcH1QAAAAB+SYfrYAAAAAAAAAAAAAAAAAAAAAAAHKfAwfBXhwfa324H1T14AAAAAAAAAAAAAAAAAAAAAAAAef/AAmH6WAAABlfl6H6rgAAAAAAAAAAAAAAAAAAAAAABzHwcH5++Fh9rfcIfVXWgAAAAAAAAAAAAAAAAAAAAAAAAfkmH62AAAAAAAAAAAAAfCnoAewfh8H6Q/FQftX5cH5+frSHz18aB+qH4xB9v/OYfqV+Eofsb+V4fs7+Vofan5dh+4QAAAAAAAAAAAAAAAAAAAAAAAHyJ9N8Bnezex8v6tofCfsIRsZQef8AC0uTxqWrvC3NxG5tp7eoqeIAPlbnP0z8CDfjPX/M+g4f6l8H7EPnfuAAAAAAAAAAAAAAAAAAA+GPz6AAAP0V+1wAAAAPyTD9bAAAAAAAAAAAAAAAAAAAAADED5K/P8PkH6wD9DfrQN0AAAAAAAAAAAAAAAAAAAAAAAAAA8/+Ew/SwAAAMr8vQ/VcAAAAAAAAAAAAAAAAAAAAAyA+R/gQPjz6kD9FProNsAAAAAAAAAAAAAAAAAAAAAAIYrkOq8X07Z7vlH5Jh+tgAAAAAAAAAAAAHyt1we9/AAerdkH5+Ww/T3sgPzHDlvaA++PyoCnmB+svwSHk3cB+lP5eB+oYAAAAAAAAAAAAAAAAAAAAAAAHifrfN+P+pfWfm3vR+e/toAAABEHNYDWZE1qbYbeoqeIAPkCl+o/z0FZn0l4P2Pkf0t5H0weD9kAAAAAAAAAAAAAAAAAAHO+Pha82AA9t9UAAAAA/JMP1sAAAAAAAAAAAAAAAAAAAA8TD89vgUPevs8PuvuwAAAAAAAAAAAAAAAAAAAAAAAAAAAA8/wDhMP0sAAADK/L0P1XAAAAAAAAAAAAAAAAAAAA8XD8+PgwPafs0Pu3vwAAAAAAAAAAAAAAAAAAAAAIOc8n0OG5n8s++4nm/y/7zd9H+p8D38/pD8UPyTD9bAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAACCdwCsAAAAAAACnAyW4hwAB8sc1+mfgQb8Z9efM/f+EfQ/ne6Hh/WgAAAAAAAAAAAAAAAAAB8Qfl+HufwCAB+v36WAAAAAfkmH62AAAAAAAAAAAAAAAAAAEXx8H5w/LYfcv6JB9IgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHn/wmH6WAAABlfl6H6rgAAAAAAAAAAAAAAAAADPkoPzb+VQ+2P0QD6eAAAAAAAAAAAAAAAAAAAAAxeH+P+j5DivyX9E5K98N9V23Zfpnw3U9l+o/B6R9N4YAfkmH62AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHknh33n44Hr/ADJ6Bxer0/svxv6kHl/RAAAAAAAAAAAAAAAAAAAQhNyweaexhogAAAAH5Jh+tgAAAAAAAAAAAAAAAAHHfnmH52aIfpH95h1IAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAef/CYfpYAAAGV+XofquAAAAAAAAAAAAAAAABy358B+c6B+kP30HXAAAAAAAAAAAAAAAAAAA1GFwPxP1PD8R+M/peN2fkel2fd/rv5z2Wl+n/Ch0YgAAAH5Jh+tgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAz52934/z8+2/Jjt+T0vo+b84/dQVAAAAAAAAAAAAAAAAAAAAfFX4ehmekB/QJ68AAAAB+SYfrYAAAAAAAAAAAAAAAHmf5lB+fX14H6O/VYOAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA8/8AhMP0sAAADK/L0P1XAAAAAAAAAAAAAAADz780A/O76qD9H/rUHgAAAAAAAAAAAAAAAACI5Hj/AIH6/wA+478a/S6XeY6d32/7P+Z9ZJ+g/HBSAAAAAAA/JMP1sAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABA4fm/e+N6HuvO91xxeqAAAAAAAAAAAAAAAAAAAAAfhH70H6J/kQHuX6qAAAAAfkmH62AAAAAAAAAAAAAAHif5iB8Pfeofpv64AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAef8AwmH6WAAABlfl6H6rgAAAAAAAAAAAAAHkP5jh8G/cQfp77SAAAAAAAAAAAAAAAAABj+ffG/S+a8J+DfrWh2PVh6B2H7J+adYv3nyQMAAAAAAAAAD8kw/WwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAD81PBQ+4/zSD6K+pA+r9IAAAPyTD9bAAAAAAAAAAAAAPOvyYD4r/RQP0r9BAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADz/4TD9LAAAAyvy9D9VwAAAAAAAAAAAAOL/KEPg39Bw/TD00AAAAAAAAAAAAAAAAM/g/lve4vzP8V/T8rt/D9X0fu/3H8r6g+++RAAAAAAAAAAAAAD8kw/WwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAD8U/gEAAA/pq9AAAAD8kw/WwAAAAAAAAAAAOe/O4Pyz+9Q/VXtAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAIuX8n0M3b8n0ekPrPnvP/hMP0sAAADK/L0P1XAAAAAAAAAAADE+AA/KP7RD9ZPQQAAAAAAAAAAAAAAA5rz+zznzv8S/UeM7H4n6j0Dvf2r8w7CT9H+KCkAAAAAAAAAAAAAAB+SYfrYAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABWshzPnwezgAAAAH5Jh+tgAAAAAAAAAAnw2H44/Qofrf7+AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAc7wdeZzHx30uLi/F/TcrifBfW5HQeb3X/pP9d/OuqP2b8z8/8AhMP0sAAADK/L0P1XAAAAAAAAAAD45D8Y/Zg/WH6bAAAAAAAAAAAAAAER595p+SfonDZn5H+i+gehfp3wvomn+xfmwdGIAAAAAAAAAAAAAAAAAfkmH62AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfid+iofi54oH7y/YoAAAAH5Jh+tgAAAAAAAAB48H4q8KH7IfWoAAAAAAAAAAAAAAAAAAAAAAAAAABz3O/JfQ43K/D/VY+F8X9Pyk/wAf9HudR9d87t7X2vzHUbH3XynQy/U+AFyAef8AwmH6WAAABlfl6H6rgAAAAAAAAHmAfjJ4mH6+/bIAAAAAAAAAAAAAFfz35r2/O/LvwL9d1/WPY87uey/b/wAslPb8sAAAAAAAAAAAAAAAAAAAA/JMP1sAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA/DnLDwT9QQ8y/WEAAAAD8kw/WwAAAAAAACuH5e/m2H6m/osFsAAAAAAAAAAAAAAAAAAAAAAERyXMfDfV8xzf539ljcn8N9Xc6bfLa6b7n5Xc2vtfl+rv/cfKvOrAAAAAAAPP/hMP0sAAADK/L0P1XAAAAAAAAYH5x/k4H6p/o2GoAAAAAAAAAAABBFec+a/jv6T5p1f5f956B6z+5/lO8foXxwAAAAAAAAAAAAAAAAAAAAAAH5Jh+tgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHgH5Mh+knnodD9xAAAAAfkmH62AAAAAAAB5d+H4Q/t2HrgAAAAAAAAAAAAAAAAAAAAHL8d8N9Xg8b+cfacxm/n32Gl1Xt+X0/U/o/xWx1X3XymofS+IAAAAAAAAAAAB5/8Jh+lgAAAZX5eh+q4AAAAAAAcP8AiYHm/wC3Ie9AAAAAAAAAAAAcb5PoeV+Wfz1+x9H6T7XmeqbP7t+Th184AAAAAAAAAAAAAAAAAAAAAAAB+SYfrYAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHhvKB9NPAQPnmAPo4AAAD8kw/WwAAAAAAPj4PxK/RkP1UsgAAAAAAAAAAAAAAAAAZnH/M+5zHH/AJj91xmb+a/b2eu9Xg6Xsf0v4frNP9F+M0z2fMAAAAAAAAAAAAAAAA8/+Ew/SwAAAMr8vQ/VcAAAAAAPmYPw0+2A/XnTAAAAAAAAAAAMfyn4P6zymj+D/rXqfq37T+YdSfpfxAAAAAAAAAAAAAAAAAAAAAAAAAAAH5Jh+tgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAeZfjWHzb6cEnmAe4/siHtwAAAH5Jh+tgAAAAAB+an5Zh+3X16AAAAAAAAAAAAAAAFTLTkOO/Ovs+N438o/QcvrPK7+l639J+J7jX/T/hNVfd8kGAAAAAAAAAAAAAAAAAAB5/8ACYfpYAAAGV+XofquAAAAAAfBX47B+yf3WAAAAAAAAAAAcP4v+P8A6P5/3n5n9z7n1/8ASv4g49fzgAAAAAAAAAAAAAAAAAAAAAAAAAAAD8kw/WwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADnvIAh9ZDrQAAAAD8kw/WwAAAACp+OofJP7xh7QAAAAAAAAAAAAAYPF1cTw/5T+gcRzn5N+hdN2P0/g9l2P6x+fdRJ938mFIAAAAAAAAAAAAAAAAAAAAAAPP/AITD9LAAAAyvy9D9VwAAAAG/lGH57/uyH0aAAAAAAAAAVMr8s8e/Bf1vO9f4Oz1fa/oL8dDs5wAAAAAAAAAAAAAAAAAAAAAAAAAAAAA/JMP1sAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPyTD9bAAAAMMPw158P3U7IAAAAAAAAAAAOT8vv4nhvyP8AROPwvy77z0HtP0T43te1/XfzmyfQeQAAAAAAAAAAAAAAAAAAAAAAAAAAHn/wmH6WAAABlfl6H6rgAAAVA/FjwIP3j9MAAAAAAAAAxuTo8X8j/m/9r7H2H63530h/7Z+XhSAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA/JMP1sAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPngOV+qwp6QAAAAH5Jh+tgAAAYf4Lh237dhpgAAAAAAAAGZzbec8D+PfpPExfmP3Xbd9+k/EdT2f6t+fuPZ80AAAAAAAAAAAAAAAAAAAAAAAAAAAAA8/+Ew/SwAAAMr8vQ/VcAAAKv4bhxv7vB0wAAAAAAAHO8HX4l5f/Nn7d6h7X+p/A9Ifp/wgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfkmH62AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfnH+S4e2/a4aX6XAAAAAfkmH62AABUD8JNgP3EtAAAAAAAA1PkeD/O/svNuM/Df1XvPR/vvke1679i/NnHr+cAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB5/8Jh+lgAAAZX5eh+q4AAIH4teOB++GyAAAAAAAHOef1+D+efzR+4+u+0/sf5tsH6J8YAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB+SYfrYAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB+EP6Yh+Sv22HMfrGAAAAB+SYfrYAAH41h4d++QaYAAAAAAnG+R6PmXlH8/fsFnvc79D9D/dPyi2fUeEAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHn/wmH6WAAABlfl6H6rgAB+XAfBv9B4dUAAAAAAFTK/CfIv5o/cfZfav1/8AOL5978kAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfkmH62AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfm5+awcXGH7n/WoAAAAH5Jh+tgAB+e35bB/Q/2YAAAAAGH5J+ffYeUH4V+sek+s/s/5j1h+n/CgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHn/AMJh+lgAAAZX5eh+q4AB8gfiUH9DnrAAAAAAAyX4n4v/ADh+1dv9C/ffJbZ+p/AgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB+SYfrYAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABQ8T+C+u9U6f6z54T0OL51vfhf6v6t0/7J+ah6PEAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAnxQHzx9Th9cAAAAAH5Jh+tgB4oH89X7+B9CAAAANT828b/Ef1Hj/TvivqPWu2/ov8XU9nzQAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA8/8AhMP0sAAADK/L0P1XAODD+cz9qg+xQAAAAADgfnz8O/VLn0N2c3Xn7L+aAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfkmH62AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAZflfxH1PW937vkmJ3cvnXsHgeuH1fz55T8z7jPWZYfUeEAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfnF9Wh+cHykH7rezAAAAAfkmH62AR/z5B9qfp6AAAFPHTyXxn+ff2G369vl6td/e/wAjDbMAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADz/wCEw/SwAAAMr8vQ/VcA/D4Ox/ZAAAAAADO+e/zb7bhfevh/qvU3f0B+PgwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAD8kw/WwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA8m+Y93e7vt5Tnuvn8gzfxX9P+gdH9x/K+f8a/PvsPerf6J8YGsAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAH4P/AFiH5R/eQejfqmAAAAB+SYfrYB8EfmEH9FVoAAzefbxTx/8Am39s7f2j7X5jvV/ZfzQAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPP/AITD9LAAAAyvy9D9VwPlv8Nw/pH6UAAAADz/AMP1fm30P+fv1/6Kl/pv8NCpAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPyTD9bAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADyb5j3d7u+3lD2vMPOfA9fM9Z59jyKa4v6R8f0Q/QfjwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAD4D/FUP1F2w0v00AAAAA/JMP1syw/m6/ZsPr4AiivH/Cf50/Z/RvcPvPk+lP1L4IAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADz/wCEw/SwAAAMr8vQ/VdA/np/QMP0UAAAAYn4P5F/NX7f9H+k/uv5OH0PjgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB+SYfrYAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAeTfMe7vd328vz36T8L9ZzvB+N6Xvu5+wfm+JwvzXuebfTnleg4/R/igAAAAAAAAAAAAAAAAAAAAAAAAAAAKHPdvldgcXqgAAAAAAAAAAAAAAAAAAAAAAc2EFELnWgAAAAfkmH62fm8HxJ+/gAHCfN/4X+ra/0H38nYn7L+aAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHn/wAJh+lgAAAZX5eh+q/xoH5Bf0ehbAAACj8xflv3tP6d5OjaP1386AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA/JMP1sAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAK+dvlaz72OmBtc2856XEYXF1atjpwDSAAAAAAAAAAAAAAAAAAAAAAAAAAAAKfyv8AT/n/ANTXvm/uwjUAAAAAAAAAAAAAAAAAAAAA+BPxiDNA/X79LAAAAAPyTD9X/wCa0P25+pgi8O+I+p8b+ivzL7n1A/oT8dAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADz/4TD9LAAAAyvy9D9V/56w/Rb7+AAADn+Hq+Wez/nT9o+jZv6W/DwuQAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPyTD9bAAAAAAAAAAAAAAAAAAAo4axacsOnEAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABAjkBQAAA4r5o+1/KPQvorxvqA8P68AAAAAAAAAAAAAAAAAAAAA/Cb1wPucD1X0EAAAAD8kw+i/y/D+iHG5Oj5esfzR+4fSm9/Sv4iHbygAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB5/wDCYfpYAAAGV+XofaP4Mh/S7fAAAOL+W/wj9Y9p9v8A0b4sPsfmwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAD8kw/WwAAAAAAAAAAAAAA5Px38t+9uxfOe19Bv8A3T8pPPfD9XI9Z5dw+n8IAAADxvB/Gf0z6CP2b8zAAAAAAAAAAAzefa3PrmFyAAAAAAAAAAHN9J0cQc/aAAAAAFYMLPajtM2dRN4gAAPnTz79G/DD3nj9P1M+F/YAAAAAAAAAAAAAAAAAAAAAD80PjQPrkD61+jAAAAAPyTDxX67D7++Tfxb9O9S97+x+bcfcfKgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAef/CYfpYAAAGV+Xoc7IH66gAB5/wCH6vzH9G/i36d6ef0L+OAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB+SYfrYAAAAAAAAAAAAAAfMf0v+cfay+TfQePb6jj6fK6vwP13V+i/oPx/j8nxX0/tHnf6B8hQ9A4OvbPtPmPnJn4D+vfRU373+RhcgAAAAAAAABSx0+SPRf52/ZvX+g/oH8fDu5QAAAAAAAADwXpPqvzn1Y+V/RgAAMEOH5alhR1O3uI3NtPb1FTxAAedd3kfPB+i/hpOq+ouj/NP3sMOwAAAAAAAAAAAAAAAAAAAD8nfjQAD9OvvMAAAAD8hw/Oz+jPDXw71X+ef2T3Q/o38WAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPP/hMP0sAAADK/L0Phr9mA+pAADhvlz8E/W/pz0D9e/OQ93ygAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA/JMP1sAAAAAAAAAAAAAAPmT6b/Ovszl/s/mfNOV/GP036W8q/Tfhn8P8H9Z7/z/AOlfEefZv5f956P336n8CYXdy+QdD+W/e6XpP3/yAfQeOAeXeXfif6gHzftgAAAAAByeD8X9OvoPocXt/d/01+Gh7XmAAAAAAAc7vx/LFv8ASfwj6c6L82/dyth2fOvndrialTvbwbu8jc3Ebm2nt6ip4gAg+UfpvgMg+t/NSzN9T9M/HfqSnz/2gAAAAAAAAAAAAAAAAAAAB80h4X9Vh6+AAAAAfz8Bwf7N/mn3Gz9H9nMH3/yAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB5/8Jh+lgAAAZX5Ah8Bf1ABoAHNed2fJf0f+H/qnpR/QP4+AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB+SYfrYAAAAAAAAAAAAAAfL307+bfbyeU/R+JZ8v/Pvsfp3zn9I+JzPO/zH7r6dzf2X808Hl/D/ANT9v1v278tPEfmPdr3vjfpeU+k/ovGD9B+PAOW5f4n6gPN7QAAAAAA8U4/8L/V+09g/Q/jfULH7r+UBpAAAAAAAHjnqfPeLdz9p+Venei/C/r4cPr8twDXDcZSwtWlu7yNzcRubae3qKniADzn55+4/Iw9v5IuTp6L6X8j+l9gfOfdAAAAAAAAAAAAAAAAAAAAfBH4nB12IH9DnvIAAAAH804fQP6Lfz/8Ar/11P++/kQXIAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB5/8Jh+lgAAAZX87IeifvuABR+Q/x/8AR/ZPZvs/mQ+t+eAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA/JMP1sAAAAAAAAAAAAAAOI8g/JP0Tag8n0PoHxP7/5DJr/nP2va2vr/AJvha3xn03s/k/2Pzftmr+g/HHzT8P8AVfSx+hfHfNn0X+dfZ2D9F+MAAAAAAAAADA4erxT1388+y60/UfgwAAAAAAAAA4Ts8v5u3/0H8V997D88/bg5fQAACHk2uYx2qukLc2x7eoqeIAPFvHPv/wAXD1Pnjp+ft9eqfKfpHqR8/wDaAAAAAAAAAAAAAAAAAAAAfhD+iIfZv5VhH+rAAAAAH83uWn6Z/bn4d+qdmfu35OAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHn/wmH6WAAABlfy1B+qX6sAAfL9z+d/2b6UP6I/GQAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAD8kw/WwAAAAAAAAAAAAAAAI5JYUmPlhSastSkAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHMHoeJ055/tgAAAAAAmcFJGrmim8AA8j8P+8/HA9f5k9r8n6Tfj+e+39GPD+uAAAAAAAAAAAAAAAAAAAA/HzwUPq/4PD9Nf0JAAAAA/Dn532f0e+qPl/eD9K+IAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADz/4TD9LAAAAyv5ag/df7TA8p+Z9zwz6/+H+qtH69+cgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAH5Jh+tgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABgfKn1/wCYtPpPhDr+b0Nn3X5P9Ivng/YgAAAAAAAAAAAAAAAAAAAcl+OoeAfWwfrpcAAAAA/EL5v2/wBcfTKkPovGAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADz/wCEw/SwAAAMr+WoP6OvcAo/Gv4z+mfUHcfpvwwez5gAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB+SYfrYAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHi/q/OeNn334yAe8eqfnv7aHne4AAAAAAAAAAAAAAAAAAABl/nUHj31cH24AAAAAfiH837X7XXve8oNsgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA8/+Ew/SwAAAPPf5lw/Zno6J/TfwH9d+mj98/IvifJoAPqv0OQ+ag8mKD2hH0MI8t+Y2Aze+0EECPhgoAPsnpZD5NDgyg+hke0CPD/AWAzvPrKQ5oPjYoAPueeQ+L2YIw+pEelCPnLx5gM9Z+lZDzwPlQoNYPtgkPhFmt81h+tggAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPOvPPqvzqT0zPbuD5n78AAAAAAAAAAAAAAAAAAAAA/ID4MD6M+VQ/bL7eAAAAA/DPyfR+7D1/OPWw+hRHnPzKwA3PsQCBHxMMAPr3o0HyuHDjD3cPaRHjvgLADuPqhBzgfIQwA+2Z0Hx2zDAPpMPTRHgPjrAD1T6MQcAHy8MNUPssQfD7AA+rw7ER8y+csAPcPcEHkgfJ/PM/SwQAAGV+SYfr5mcnR8c/X34z+mdAfuP5WAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAH5Jh+tgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAH4g/bQfcn5VBa/UsAAAAD8jg/WMAmCwBHVAAdcAAogAFxwFUIwCwEwENcACS0A0KYABeAKYNALISgV4QAJbIEYVQBQugFEAAtg8CrGABPOBCHnHxAH6WAAABlfl6H6r/PXwn1kH0ZFB998iAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAH5Jh+tgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAH5j/AJsB9D/LYe8daH7V9YAAAH5Jh+tgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHn/wmH6WAAABlfl6H6lfFv4t+nfX/Qfrf52HdygAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB+SYfrYAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB+fXzaAAAfrj0wAAAfkmH62AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAef/CYfpYAAAGV+XoffPmv4X+rfVB+6flIAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAH5Jh+tgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHzH62Hy98zB+rHRgAAAAfkmH62AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAef/CYfpYAAAGV+XofV/qP4d+qern7j+VgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfkmH62AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfhB+kgfhP74H3T+lwAAAAH5Jh+tgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHn/AMJh+lgAAAZX5cy/evsn8T/Udg/bvy0AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAD8kw/WwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAD8kPzfD6z+xA4T9UwAAAAPyTD9bAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPP8A4TD9LAAAAyvyJ8j0fsT7H/PfsQ/Wfz0AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAD8kw/WwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAACh8fh9OeZhu+xAAAAAfkmH62AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAef/AAmH6WAAABlfin8h9H+mX0f53YH6B8gAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB+SYfrYAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB8z/AIch6P8ApUGx9qgAAAAfkmH62AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAef/CYfpYAAAGV+I/xX0/64eu9nMH1PggAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfkmH62AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfhf8AVofBf3uHnX6ygAAAAfkmH62AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAef8AwmH6WAAABlfiP8V9P+zPae95Iev5wAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAH5Jh+tgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAH47c2Hy11ofov+hwAAAAH5JB+toAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHn/AMJh+lgAAAYf4pfFfUftftfU/Ph2c4AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAH5Jh+tgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHJ/j6Hzx9Uh+veiAAAAB+JQfWgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHLsD9LAAAAxPwE+e9j73s+75QaQAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB4yH6+AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfLQfKP6Ehz/xcHun2OAAAAB5mGAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAeiB04AAAHn4ZIAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB1odSAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHL/AMywe3cgHl3qQfsZ9igAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB85/kKH9A34Bh+gv3uAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB87fgQHv3hIeo9KH6Q/cIAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAcd+YgAAB9m/T4AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAGPzod0AAAEMwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAGPzod0AAAEMwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHgH4qhkelB+r30wAAAAAAAAAAAAAB+f35Jh/TaAAAH4GfYwfpaAAAAAAJ+AAftv6SAAAfzpfsiH04B+GH6oh78AAAAAAAAAAAAAAAAAAAHgH4qhkelB+r30wAAAAAAAAAAAAAB+f35Jh/TaAAAH4GfYwfpaAAAAAAJ+AAftv6SAAAfzpfsiH04B+GH6oh78AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB8qfgAH9J/5Rh5V/QYHmXzeH01fDwz6nD579TDC4kPV/mALv1qHPeMBsdwGh8rB45+aAf02gHzAHGfWofHnsYXeEA7sPcPn0PL/AK5DZA+bfyVD9/PmQPW7YeHfUwfyq/vSH2CB8BfMwfssAAAAAAAAAAAAAAAAAAAHyp+AAf0n/lGHlX9BgeZfN4fTV8PDPqcPnv1MMLiQ9X+YAu/Woc94wGx3AaHysHjn5oB/TaAfMAcZ9ah8eexhd4QDuw9w+fQ8v+uQ2QPm38lQ/fz5kD1u2Hh31MH8qv70h9ggfAXzMH7LAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfKn4AB+3v5sB9N/oGH4I/bwfCn72h/Oz/TGH85f6lB4zaD6c+fg+RvqUPvD8Dw6D9qw/Gf0MPPMUP6bQPz5/KoPsX9Tw/C/7ACb5KD5f/SkPefzMD2zkA/foD82vKA/Xz+ZIP1s6sPw9/qYD+VX96Q+wQPBPxSD+igAAAAAAAAAAAAAAAAAAAPlT8AA/b382A+m/wBAw/BH7eD4U/e0P52f6Yw/nL/UoPGbQfTnz8HyN9Sh94fgeHQftWH4z+hh55ih/TaB+fP5VB9i/qeH4X/YATfJQfL/AOlIe8/mYHtnIB+/QH5teUB+vn8yQfrZ1Yfh7/UwH8qv70h9ggeCfikH9FAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHyp+AAfe/xEH78fGIfn79Ih8g/ueH5GfdofA/s4eZ/reFb8rA859HD9f/AOesP6VfKQ/AD+o0PhL8nA/ptA+F/wAVA+/v1yD8C/sAP1X/ACRD48/oUD8C+dDv/kkP6j9sPyuhD9V/5kg/Wzqw/D3+pgP5Vf3pD7BA8p/AMP6UgAAAAAAAAAAAAAAAAAAA+VPwAD73+Ig/fj4xD8/fpEPkH9zw/Iz7tD4H9nDzP9bwrflYHnPo4fr/APz1h/Sr5SH4Af1Gh8Jfk4H9NoHwv+Kgff365B+Bf2AH6r/kiHx5/QoH4F86Hf8AySH9R+2H5XQh+q/8yQfrZ1Yfh7/UwH8qv70h9ggeU/gGH9KQAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHyp+AAf1K/iAG99BB+c/7VBX+jA/Kz4TD9NfzPDI/pgD+dH79DlvlEP2B/nrD+n/AMzD+cz96w+OfhcP6bQOF87D8FP1ZD8+PsAPbvxUD9uvfg/HucP1Oxw+oHh+c3hofsX/ADJB+gW+H5W/1HB/LL+twfSPqocT+Ogf0LgAAAAAAAAAAAAAAAAAAB8qfgAH9Sv4gBvfQQfnP+1QV/owPys+Ew/TX8zwyP6YA/nR+/Q5b5RD9gf56w/p/wDMw/nM/esPjn4XD+m0DhfOw/BT9WQ/Pj7AD278VA/br34Px7nD9TscPqB4fnN4aH7F/wAyQfoFvh+Vv9Rwfyy/rcH0j6qHE/joH9C4AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHzp+Lof0OfDwfAf7mh+MHy0Hff0NB86fjUH7s/luHQfr4H5f/nEH0PtB+oX4mB/QV0Yfkl8KB9X+VB+/wH5vfmSHp/7ch+Tf06Hyn4oB9Oh+kn4xh5D9kB+zwHhf4rB/Q7/MkHRfUgfN/wCyofjoAfouFXy8P1wAAAAAAAAAAAAAAAAAAAD50/F0P6HPh4PgP9zQ/GD5aDvv6Gg+dPxqD92fy3DoP18D8v8A84g+h9oP1C/EwP6CujD8kvhQPq/yoP3+A/N78yQ9P/bkPyb+nQ+U/FAPp0P0k/GMPIfsgP2eA8L/ABWD+h3+ZIOi+pA+b/2VD8dAD9Fwq+Xh+uAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAH4wB+qv88oftp9YAAAAB+Nf6fh6qAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAH4wB+qv88oftp9YAAAAB+Nf6fh6qAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfnkH1X7EAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfnkH1X7EAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHAfPIaX0qG8AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAflKH5b1wP6FA+lgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPkX8Dw9m/aANn3kNIAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAMsPhbPD6V94AAAAAAAAAA/Lz9QwAAAA4Thw9zAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPyD/ADRD9nv0MAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA8/D8Zf1gD8rv1HD6EAAAAAAAAA/BP97AAAD8YP2fDx/wAvD6wAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPyM/MgP1q/UAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADz8PyT/AGsD88elDJ+aQx/1XDyX5CDmcYP2X/NYPvv0MPx+/YEPwT/esPyi4cPrbfD8Z/vwPtfwoPtT8kgm78P1P/KIDw4P0y+nAPEA+A/OQ/Wj8wg/XGQPyS/ScPzKoB+h1APiPuA90+XQ5r9jA+dfzcDt5A/Xn84Q8qjD9c5AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA+Yv58g2v0rDf/QcO/AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADz8PwG+vA83/XAPyG/c4PJPyoD7c+aQ/Wz86A3vmEP1B9WD8E/wB7A/BP97A5L5sD8wP3/D8E/wB7A+VPmkM76iD6w/HkPv8A/JoP2xqh+Q37fAAeN/AwdldD0G8HnXzKH6PaYflT+iQfGf7GhF86B+W/6sB+TX7mg78Cw/bf8nw/Uj87A+3vqAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAD8+g/J7zsLn9AIfToAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAefh+Q/7OBueIB+df7BhS/CoP0M86D9G/j8PEvGA/Tr1oPwR/e4PwT/ZIPyX/ScPyi/fgPwT/ewPlT5pDyX9RA9X/NEPbfzUD97MoPw9/dcD84Q53rw4/wDRIPx42Q/Xj8NA+7wO7lDxT9LA/Fr7HD5G/QEPyj/fID8Cw/Vj8zw+ywPrT00AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABObDbtAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB5+H5J/tYBCH4M/pIHyf9BB0H5qB+qn5rB+uHyOHmHp4fFH76h+Cf6nB+ff2IH5mf0Ih+An6uBT+aQ+i/ggPvj8ww/dL8Mg/ezKD8Pf3XA/HUPqb5mDuf1ED8WejD9g/wA0w536vDq/Mg8U/SwPwH/VUPyn/WMPjLdDW+HA/f38KQ/TOiH2LIAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAGeHkntYAGD8kh6t78HxT5GHrvvQetxB8i+qAz24PBPew+V+kDS9yDyXyEPo3mg9Q8QDxX6wDqfBA97iDw73YDCD5J99BvrAeb2A9AA+Z/OA+mtUMT0sPNPn4Pc+sDrPnoOy/H8P3U5QPk/qg+rJQAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPj3hQ+/AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA+MPkcOK/QkPrMAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAMMNwDD3AAAAAAAAAAAAAAAAAAAAA8Y7QOzAAAAAAAAAAAAAPmUPpoAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPOw8j2A9w2AAAAAAAAAAAA5kPmVAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAq4YfY3QgAAAAAAAAAAAAAAAAAAAAAAB+HwfuCB+H37ggAAAAAAAAAAAAAAH57/oQAAAAH5p/SwfSwAAAAAAAAAAAAB+HofuEAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAcwH4ZZ4eo5gePfcQfqKAAAAAAAAAAHwcGP68AAfncH6IgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHy92Ae4gAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAH53foiABZ8SD1D6/AAAAAAAAAAAAAAAAAAAAAAAD8Pg/cED8Pv2SD80MsPfvvQPzg4gPrrxAPsv4vD7P/ADgC5IH6nfCweP8AuAfZ/wCYQUPrQPx2/RcPsL8/Ad+ogfFXggeE/qCH0t5IHwP7gD/FQ9joh9efNQdz1Aea+RB5N+gIeb/NofV3y2GJ8ah/RCAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAYYYXcgcMHSefh6qAAAAgfn5OH378oh8g9QH6hqB8KB7X9AAAH5Jh+tgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB8U+gB9LAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHx8H5R/owH6CgHx8Gt9VgAAAAAAAAAAAAAAB8Fh+TYAAAGmH9IoB+HwfuCB+H37Ah+R37Yh+Fv6kB+ev2mH58fscH43/AEqH6QfhSH71/mGHuf56B+if5tB9v92H2YB+Kf7WB+L32gHzp7eHxj+3YflR9gh9LfP4fnl+twfiF+jYfmp+mwcZ4cEG+H3hRD4u8/D3CoHvnygH6f8A4JB+9oAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfikH6Ffg4H0D/QKAAAAAB81/CQfsDhBu/kmH6D+ygfCge1/QAAB+SYfrYAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfFPoAfSwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB+NH7LgAHx8Gt9VgAAAAAAAAAAAAAABygefgAAAPD2gA/EAP2ylD8S/18D8vv2BD8Pf0QD5a+1w2u9D8UPrIP0R/FIP3C/OUOn+RQ/S2YPkn6OD6iA/FH9rg/Eb9MQsdeH5T/tSH5d/VwfS3z+Hx9+hofjt+noO9vD8e/YQ4WAP0w/KEP1H/ADYD6A9AC/4uH6NfhKH7tAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB+KAfO/3uHxB/Q+AAAAAByn5Uh+wICfikH7K7IHwoHnnrAHoYfSf53B+iIAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB8v9gHuAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAEX5YB4Z6UH1nTA+ew+r/qsAAAAAAAAAAAAAAAAAAAAAAAPl4PgO4H3r7MH54fqKH5CfriH5F9IH0/88h98fnQH6HfhEH23yIfr1+XYZPuYfXv5Ih6T+hAfB3RB9EfBAd3+lYfm5ywc7+igfQPiYfLn6Hh+X/Nh7X98h+P36Qh4j5uH3j+Owe8ZYe49sH0v+OIdv40H7ivAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAArBzdQPi37zD4f+9wAAAAAOU/KkP2BA/Lf6YD6wAPhQKHsgHXh7OAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB5j5OHyJ9HB9p+Wh+Wf6EAfNIfQP1WAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABlfjQH7WAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAGQH4kZAfVv2UHtwAAAAAByn5Uh+wP5eB80/VofZ/roHwoHtf0AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAcF4OHzl83B4XxgVPWg7b9LA+yfNw+B/1AA+Pg1vqsAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAACHw0PeAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAyg+U/jQPEv2TD2YAAAACPnw6TkQzQOs1APhQPa/oAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAQPM/Bg8T8EDzLmA7T3AKH0MH0J2oehWwg/KEOPuB+r3VgfHwa31WAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfMIe5fm6H0B9jBYAAAAAAAAAAD4UD2v6AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAEDifLg8Q+bQ8Q5IPT/eA9x9uD03sgAAAAAAD8k/1sAAPj4Nb6rAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPwdD61+Cwxv6OgAAAAAAAAAAA+FA9r+gAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADO8mD5d8WDxnzEO99RD3H6ED37qAAAAAAAAAAD4+D5J9yD9BQD4+DW+qwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAOPDB9OAAAAAAAAAAAAAA8l7YOkAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA/OsPyH/AHkD8iP6HwAAAAAAAAAAA+FA9r+gAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAwQ8k+dw+YfBQX1QPq32oPa+/B4AAAAAAAAAAAAAAAAB8fBrfVYAAAAAAAAAAAAAAAAAAAAAAAfgKH0d3IfpmAAAAAAAAAfHQfTvUAAAB8qB4p+iwAAAAAAAAAAAAAH5ffYIfQAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAFQPi314Or9UAAAAAAAAAAAD4UD2v6AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADyn5lDxL5MDyf2IPXfoUPpn6AC2AAAAAAAAAAAAAAAAAAAAAfHwa31WAAAAAAAAAAAAAAAAAAAAAAAH4fB+4IHyd2gcj2ofL/AC4c5+poeK/F4egfocHw74KH1Z+UQfS36xB+a+KH6TeaB8H4ge4/osDPzKDQ+qg+L/QA980Q2vEg+wfhsOp+eQ+qPWQ+JPSw0fnMPCf1UD6AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA/IgPkX7wD4f/AKIQAAAAAAAAAAA+FA9r+gAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPOw/HDrw/aQAAAAA4UPCflsPmfwUOY+oA9/8AocPrHtgAAAAAAAAAAAAAAAAAAAAAAAAPj4Nb6rAAAAAAAAAAAAAAADkvL7/PT8x+6AAAka9rP2H83PwFD6O9bD7+/GMOj/YsPwD/AHoD5H87D5I/TAPzu+8g+Q/1zA/JMP0s+Fw6jfDxDwkP23+KQ479FgP55w/oR/D4P0w/O4P0T+Og3+BD7D5kPrz5XD89P1UD84f2CD8Yv25D8hPvsPoAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAD8Mw+hvOg+P8A+ksAAAAAAAAAAAPhQPa/oAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAD4OD5hhD7q+qwAIPngPC/jsPnDhg9f+rg+kvooPUIuLp139nODRz3B13dTbMOnAAAAAAAAAAY+WFIAAAAAAAA+Pg1vqsAAAAAAAAAAAAAAAOJ8f0vMD8h/RgAAJKn6BP3T8pPw+D9wQG/jQHR/rsH4X/ugHzh8tB8n/pyBlB4n+lgH5Jh+ln5tB7334em/kmH7lfIQeTfosB+HofuF+DYfpyB9G/kyHYagecfrWH4tfqsH5X/p2Hx9+hAflR+yAflb9pB9AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfPgfiLwYfrP+iQAAAAAAAAAAAfCge1/QAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAQfkOH6/gfjb98h8ffPAfOEwfS31sH1J7aF0Ail/NXbfiH6n0/e/q35+Ht+WeP/ACvv9L3focZT9Xg889P8H1g+j8UAAAA898L1uph0z4/0/l6A+j8UAAAA888H1vPer/OvtPWj9j/NT4+DW+qwAAAAAAAAAAAAAAAAAAAAAAA/C8PZLYe9dQHiX08H5EfVweVfrYHxn4sHd/piH45dwH2J5YHy/wDpiH5b+pB91fLIfP2qHvf3+B+M4fsx+eYeLd2H6dfmkHpvuIfmd+0Afi17UHhn6dB8ofpGH5ETh5v+oIe7gAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfI4fWWIH5e/rGAAAAAAAAAAAHwoHtf0AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAFP8AG4P2diD+erjg+rvrEPfPcAAAAAM/DX5v+mvz37Gbzj6nwOk1OnHzOL4P63nV+f8AX9Byvsfm/H/oH5r3OKz7n0bgvc8rR9f97yPLuX+P+k7zw7h6ul9j9vy+Z7L3/I8cb+e/Y+0cJ+g/H5fM/B/W+57P7B+bmQz5099/MfuulP1j89Pj4Nb6rAAAAAAAAAAAAAAAAAAAAAAAAAAAAD8Pv3BAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA/IIPGfEQ+gf3QAAAAAAAAAAAD4UD2v6AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAj8qD4j+aQ+UOtD6ivh+jH0IAAAAAAAAHDePfkX6L6vzfpcXZ731Xz/AIh0v539noen/oPx/wAvfRnyP0XjHvvteX8n/Vfj+mzzv6v5/wAW+m/iPqfEvof6DxjxDr5/SoO3m88538y+5+gq/wCwfm/hmp+PfpHoL/0T43gvb4rG8K8b0/XvNk/QO5+y+aPj72fM1vqsAAAAAAAAAAAAAAAAAAAAAAAAAAAAPmX6aAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADzcPkZgfnnlh+nf3SAAAAAAAAAAAHwoHtf0AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAGAHyp8fh8ac4H199Yh9S9AH5rgfrYAAAAAAAAANR8x93+HfqvU736Z8N4t0vwH1+76H+j/FfMX0P8d9L4v796/m/Mn03xdXzh7Zy9HhPueOnlfvXv+QeIa5+lQdvN55yv5d959Jwftn5d8+bX4p+oeon6p8D5d7t5Xf5P0/Pt2Mf3fyfhHvfxX058ffb/AC+t9VgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfOofk6AAdd+3wAAAAAAAAAAAfCge1/QAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHjfxSHyX8rB719hh9jfSIPAAAAAAAAAAAAy+bfw5v4j+o+h9J+l/EeB9Z+afbpv/AEPj8pH4nqdj6Z+mfD/M3t3xX0/lf0H9B4/iLPlvf5L6d9zyfBT5n3e/b9v8t5b7R4fq8V1vp8Pkrvhfq/UeS+0+Y3YdJyPFvzn7PH9b8P1fcPMf2n8x6T0T3PKPj73vJ1vqsAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADED5F6wPJwNj7pAAAAAAAAAAAD4UD2v6AAAAAAAAAAAAAAAAAAAAAAAAAAAAG/OIfGfxMHnH2cH1z9kB1oAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAQRXMc98V9PznH/nv2V2Hi6eB9p5OjU2/wBM+H7fC+w+b6Xm9I+efdfyz77tz9l/ND4+DW+qwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPNg/K/1YPlYDrv2+AAAAAAAAAAAD4UD2v6AAAAAAAAAAAAAAAAAAAAAAAAAAYHzN8bB8MUg+z/tMPqu4AAAAAAAAAAAAAAAAAAAAAAAAAAAABXzrF5j5T6HleZ/N/teS5383+23t32PN6jqv0T43rNr9F+L1T3vJAAA888H1vLe7/M/ufWj9q/MD4+DW+qwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADmA/OPED9CvVAAAAAAAAAAAA+FA9r+gAAAAAAAAAAAAAAAAAAAAAAAA4UPhT83wqfaIfa31MDwAAAAAAAAAAAAAAAAAAAAAAAAAAajA5n5L6LnOa/P/ALDh8X8x+6l6zs5er6T9K+J67qf0v4e0e35YAAAAAAAAAAHx8Gt9VgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAH4rh5f6iHzb/RyAAAAAAAAAAAHwoHtf0AAAAAAAAAAAAAAAAAAAAAABgfAwfAngIfW/6Mh9fPAAAAAAAAAAAAAAAAAAAAAAAAAAOc8/s4jkvyv77j+U/LvvbXUb49H0f6D8f2fQ/pfw+sfReMAAAAAAAAAAAAAAB8fBrfVYAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB+Ogfan0qH4Ef0MgAAAAAAAAAAB8KB7X9AAAAAAAAAAAAAAAAAAAAFL8dQMgPiv6UD9EfsQNkAAAAAAAAAAAAAAAAAAAAAAACOXz3H/F/T8Py/wCTfoXKS/E/Ud51n6X8P2HXfqfwWqfT+EAAAAAAAAAAAAAFXK83V5Oic9DkAD4+DW+qwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAD8bA/OgAA/b/7rAAAAAAAAAAPhQPa/oAAAAAAAAAAAAAAAAAAAhD4H+MQ6nwcP299eAAAAAAAAAAAAAAKmWnHnx/0fdn2vzAAAAAAYPF1cRx/5X99wvMfkv6HqdD6HH6D1365+d9VsfonxgdfOAAAAAAAAACBh8p8l9C7nvn/Y9Yv/AKN8WfJn5v8Abc3Q/Cf1i99I/o3xfo5+/wD5AAHx8Gt9VgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHAAniYd12QdVrAAAAAAAAAAHwoHtf0AAAAAAAAAAAAAAAAABgB+fX5vB6X+/QH5Hh+rumAAAAAAAAAAAAAAeafO+z4fF/PP7J9L9T/S34eHpcQAzjvG9Lz7kPxv8AS+So/nn2Pd9V+g/H991369+c3j6HxwAAAAAAGS3lLK8y+K+n5/n/AMy+5s/TP33yJB+g/H/FfVfyl/QLt76LxfoHQ/of8bMDDV2pF2T0eIAAD4+DW+qwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAD8Mw/Q78Pw9i/ocAAAAAAACEPzO5gP1eAPhQPa/oAAAAAAAAAAAAAAAAMIPy6/NYPun7gD4w/YULn43B+w9oAAAAAAAAAAAAABPnb4H676KZ978l88/RPwf1mRxvpcXC8F+NfpfL7fx30vo3Y/r/wCcdd036f8ACOPT4QAAAGY3D1P2bk4fh6vEM7+fP2LA6zg6/qs/qv8AAOc8N/LfvOg6HDaf037f5YPovGAAAAAAAAAAA+Pg1vqsAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA/GYPjv9PA/P7+iIAAAAAAAPzYD0/6aDvgD4UD2v6AAAAAAAAAAAAAAAMgPzW/MkPpz9gQ9ZA8I/CEOg/UoPusAAAAAAAAAAAAAAZ8//CfWfQZ938n8sc//AC5+84vYcHX2/ffsP5v1939L+HDbMAqYnk+hzfa+X36p9V4Hivj387fs2Nb+D+t9r9u/pT8QOQ9zyuc0/k/oeg2PsPm5z0eMAAAAAAAAAAAAAAAA+Pg1vqsAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABgP58PlX6/D5J+vQkAAAAAAD8GA+3vlwP0i+igPhQPmX0QD2QPt0AAAAAAAAAAGB8IfkqHtn62B9CAAAAAAAAAAAAAAAAAAFTzv5f3uE8w/Cf1f0Wj6vB1PoP3fyfaSfpnwxQy05fJ+L+n9Hm+4+VPkv8y+54zX/Df1Xovb/1n8978/afzHkz5j3dzQ+g8cN8gAAAAAAAAAAAAAAAAAAAPgTzkDxkPvz6rAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA4APxK+sA9Szg8Boh+x4AAAAAAfiQH7bfLAeI/osB8KB6R7wBeDdAAAAAAAAAAPnb8cw5z9eA+wAAAAAAAAAAAAAAAAAI+V8n0fOOW/G/0rzjs/hPq/Quw/Z/zOtjVO96J9d86VfZ8z4uq/x7/SPS7H0/hfRm5/Sn4iZNToTb5BcgAAAHzF+dfZ9PQ+V+g9L679Q+D8l9m4eoPrfnjw3436X3I+y+aOV8zuwvR+LpD6DyAAAAAAAAOeqgfJYdp9VgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAcT8aB5BqB9F/XAWQAAAAAA+PQ+PudD9Zu/A+FA9r+gAAAAAAAAAAADkfydD4T/VYP0ctAAAAAAAAAAAAAAABQw18984/Hv0jzm7+UfoPddB+j/Fe2W/3P8p5f5O/Ev1Klrfl/3fS/VX7t+UKfpnw2RpcfTKdnMAAAAAAAAAHzJ+dfZ/Tbf0T435g+k/zL7ni6/ocfpOX9p8xgZXyX0Xo3j3ThpdtFZ3R+z5vkMv579j69xH3vyNXmPjfpvZp/0z4YLkAAAA+Pg1vqsAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAD4UD2v6AAAAAAAAAAAb+RQeXfN4fvl7MAAAAAAAAAAAAAAAc15p+e/Y+f8AGfh36p2Wh7vk8lH877WV6L1c/wBRH9Xfz9Di+d2b6+hxgwAAAAAAAAAAAAAPmT86+z+mz9F+M8G9I/NPuPJOr8H1uo4/6Hx++8P8/r9l4Xu5faeL+s+e5rmPzf7b2bI+++R4Wp+e/Y9vX+o8Gh677Hmh9V4AAAAB8fBrfVYAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABGHyj6OHonTgAAAAAAAAAAB8KB7X9AAAAAAAAAAQh+T30GH3J5uH5+fqWAAAAAAAAAAAABgcPV535j+JfqPCH5T+gd/9P/v35Dewf0b4zk+l+b9roJfr/nApAAAAAAAAAAAAAAAAAB8yfnX2f02fovxnzJ9CfmH3fhXvvVz+FRfG/TfRPzJ9l8z9NePez5vMenfKfQc1wfjel9OQftn5d85634V+sewUP1j89889v8/sD7H5sAAAA+Pg1vqsAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA/KAPhL9Cw+IP6IAAAAAAAAAAAA+FA9r+gAAAAAAAAA8s/E0PI/2aD7H5APzW/VgAAAAAAAAABvK+V38hh/m/23n/Kfk/6F1/R/TeH13Wfpnw3VXPvvkQ1zAAAAAAAAAAAAAAAAAAAAAD5K/L/u/Uub8f0u6777/wCQ8C+jPK9D5z6/5b3/AF35k+i8X3TjOzn4T1r5z2uatdnNzFP5v2vT/PPo/F9gofYfN+ee3+f2B9j82AAAAfHwa31WAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfhYH1T5qHxP/SYAAAAAAAAAAAHwoHtf0AAAAAAAAHz2H4N/oOH6m/lADebD9W++AAAAAAAAA8V+Q+j8Hq/zD+63vRvp/C9D9K/e/yS2fV/PgAAAAAAAAAAAAAAAAAAAAAAAABSx0i0ZpU3xr2YuCQH1QtZGGuonRi2YM+bm3tV+jCZKUc6YXIAAAB8fBrfVYAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB8yh+GfFB+s/6aAAAAAAAAAAAB8KB7X9AAAAAAAAHwJ+Pgfrx+gwAAAAAAAAHHeT6Hn3H/jf6X6H0v23y3l/lv5J+idB679F43qtr94/JQ1gAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPj4Nb6rAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPGA9O+Fw9z+jgAAAAAAAAAAA+FA9r+gAAAAAAD8+g/Jf95w+hgAAAAAAAAPnnzX+Z/3Hucfoy4zY+R+h9Q9T/dvyfpz9I+KAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPj4Nb6rAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPwhD7p/KoNT+jYAAAAAAAAAAAPhQPa/oAAAAACoH48ekB5B+mYe7AAAAAZXAfIfR+Y9l+a/b+4H7z+S8p5Z+afceOp+B/rnQfSv69+c9YftP5iAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHx8Gt9VgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAH5Yh+Vv7mh+VH9FQAAAAAAAAAAAfCge1/QAAAAAHywHI/aoUfygD9bQAAAPE/j/AKTwnuP5/wD1/wBH9P8A3v8AI4vBPH9Lh+//ADP7nD+nf0n4mx86d/H9HL+gfHgwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA+Pg1vqsAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAQPIN8OD99AAAAAAAAAAAD4UD2v6AAAAADM/BgPrL9SA8V+Kg/T8A57g68Xu+ToMr0eO3a0z57w/81+48w9b/MPu/cNX+lfxD58+g/jvpDwXSPcrH2XzQaQAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfHwa31WAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfl6H5Lgd7/TaAAAAAAAAAAAHwoHtf0AAAeEh+fjg5fywP0G+dw6f9Og0+K8f0fDeI/mv9v8AbPcv3r8kD7L5rG8G/M/uPMvXvy/7z2rX/pH8TDr5/KeX/If0a27DX3E/ZfzQAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPj4Nb6rAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPxDD79+vwAAAAAAAAAAAAPhQPa/oAAA/GQP2Qrh/Pr+xYfSQABx3y1+Ffq/t3rn6D8dcPtPmYvGfjfpfDfVvxP9Q900/6V/EA6cQAqZaRaEsN8gAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA+Pg1vqsAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA/JAPgD0EO1/fgAAAAAAAAAAAPhQPa/oAAA/IEP1+A/IT9ewMLi6d07eYr53YNIOE8X0/nPU/nL9p+iup/pT8RD0+EAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA+Pg1vqsAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA+ew4CMNj6vAAAAAAAAAAAD4UD2v6AAQPx1rB2NUPaP0ER474L/OX7T9d9F+/wD5CHfyEEV4B5b/ADP+4/Qnqn7r+UB9N4YAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfHwa31WAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfLwfhdz4fr7+ioAAAAAAAAAAAfCge1/QAHigfLf6Hhz35PB+vXzV+bfbc19L/P+x1Z+0/mIBxfjf5L+h9hy3l9/tXT/AK7+dB6PEAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHx8Gt9VgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAH4Xh96+sh+JH9HIAAAAAAAAAAAfCge1/QAHFB+Xf68B5j+eAfoj5R+KfqH01a/ZvzINYPLPmvc+afrD8k/Q+5h/ffyLwj334v6cPt/lgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAKYeaerhzHjAe96YAB8fBrfVYAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB+Oocl6SHyF/RWAAAAAAAAAAAHwoHtf0AAB8nfmkHO/u+HQgABFFfOnnv8y/ufuMH6J8Z7Rx/6J8bxXsvj+iH1vzwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAQxhgdQCgAAAAHxUzzr9Ew/OP6+D5W/Q4AQfHwa31WAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAcoH5U8YH6de9AAAAAAAAAAAB8KB7X9AAAH4nh7/+mPNuHTgAUPmH8s++Pp1Gh5l9l81xmr+c/ae0P/ZvzMKQAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAeVfCzPaucD73EfMHNMLgfXAjwvlWeJ/WIeH/ZIfm5+kYAg+Pg1vqsAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA8TD8ksQP0/+tQAAAAAAAAAAA+FA9r+gAAGfzt+b2/qJ9s/lv31o/ZPzQyOXo+Uuy/nH9p+jJv6V/EAuQAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA8r+fmfav5sB+k4gAADMD8y/q1nyh9Ys5n6+R+b36QgCD4+DW+qwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAD8MQ9R9sD8tf6WwAAAAAAAAAAA+FA9r+gAAPOvzA/OvtP0Q909LhD7P5njvnH8V/T5vrj1/NU/U/ggAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPK/AWfaX5sh+kwj5g5phcD64rI4IZ8PffYfCH6CB+fn6NACD4+DW+qwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAD8qw6L3oPyM/ZUPY+zAAAAAAPIQ/OC+H6y2QPhQPa/oAADwPwb8W/TvuHU/YPzcxYvxb3/4/6Tg+d4+j2I/SPigAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAOY4QPYvnVn0UIAAAAAPAvfQ8a8JZ9V9ggAPj4Nb6rAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPxwD42AAP2K+zgAAAAAD8YA/YD4mDr/ALJA+FA9r+gAAON8C/IP0f6vP1/84PA/iPqe59D9nzE8A8T1PoA+5+VAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA+Pg1vqsAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA8YD0/4fD2r6OAAAAAAAD8Qg/b35nD5z/SQD4UDwf00DtA+2LQHkgYvgH4J+t9T716vBxvpfp8Jw/6n8CDOj9NQY4eTjB4e2iDxYIBh6ijfA855UAZ2PfoOZDzUYaIeviIPFgBh6+jRA8rwwBnf9ig4sODGHQI9QAyvJABh7ah4HjtFgB6b0aDz0ORGHXI9CA5zzIAZa9mQAeHjAD1vVQeXhz4w7xHy1wQHhgfb31WAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfhCH3T+VQan9GwAAAAAAAfiWH7YfGgY/3OB8KBl+xgWmfQ1xB4lFfOvuH4J+uXu3/Z/wAy6/T7uTgOA6sQDp/V0GIHjgwkD3oQeEhXGHrYdGI8w5BgB2/o6DlA8sGGoHtQiv4SAMPakagHkGAwA9H7dBwgeeDDow9bEZHjAAw96RIB4jnsAPV+nQeaBxgw7MPSxHMeUMALnuSADwEYAez66DyQOcGHoYV8xGD8hB9DfVYAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB+WIflb+5oflR/RUAAAAAAAHx2Hx/wAkH7IdCB8KB7X9AAAHlnzXt8bT/Kf0Hk/r36fwQ/VPgQAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA+Pg1vqsAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAaHxd7iE/rYAAAAAAAAZOiEoB8KB7X9AAAHzz8J9Z9DH3fyfzz9DfCfWB938mAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHx8Gt9VgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAH5WB+fH6Mh8R/0OgAAAAAAAAAAB8KB7X9AAAZ3zp+Zfc/Tifpnw/zt9FfB/Wh978iAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHx8Gt9VgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAH4Uh9c+aB8N/wBJgAAAAAAAAAAAfCge1/QAEEV85ej/AI9+kcPgfP8Ase7dd+4flgepwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB8fBrfVYAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB8sB+GHJB+rv6jAAAAAAAAAAAB8KB7X9AAcr5ndyfq3mdx8/eZ3fQJ9z8qAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB8fBrfVYAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHl/zUH2X46Hzv+ggAAAAAAAAAAAfCge1/QAEEV85fMn5x9r9rRYa+vH6j8GAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB8sdkHuoAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB+cf6OAAHx8Gt9VgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfzf9QHNeYh9mft+AAAAAAAAAAAHwoHtf0AABn/h18T9R+wnqPpcQfSeIAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB8U+gB9LAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAH5J/rYAAfHwa31WAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABQD+eb+igP5t/wCh0OrAAAAAAAAAAAA+FA9r+gAAIvws+N+m/dCx9d84GkAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfFPoAfSwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB+Sf62AAHx8Gt9VgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAMD+WKwGdoh339LQAAAAAAAAAAAfnmG77aGDhBh/ml8x7v6in0/hAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHzN1we1gAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAH59foKAAfOofUH1WAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB5fCABZ9NAAAAAAAAAAADjw+Z+cDe3QlAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAD6K6EAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA5jxEPo+YAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAMviw9IA8F7IPSAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAoB88evh29APnT6BDZAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAM4PxW+0w+e/wBawAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAQPyRzQ/YH8+wwfnQP1j9KAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPjcKH56h+4H5Dh9i/EYftgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfK4eK/okH4h/t4AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHKflSH7A/iIH7d/JQeXfoMAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAH5BB+if5sB+yP46B+rXYgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfHAcn95h+In7dgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAByn5Uh+wP4iB+3fzCHgP6OAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHhXwIH6U/mkH66fkUH6X+jAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAecB+Y36zh+N37VAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAByn5Uh+wP49B+ofweH1B9JAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHhofnd+vITfiEH7e/iKH7dvAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA+HfAQ/Qr1YAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAMf4+D7Z4wPzW9TD9CQAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA/OgPKHB+g3Fh8Y/aQfSwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfkIHk3oofqL6UAAAAAAAAAAAHzmH4s/wBGQAAB+VfpofoOAAAAAfOAfOX6OAAHh4fnp+vYH5oh9Ye/AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAH804fr18WB0n7FB8gPD60+NA+yPlUPqT5fD3X53DnfsgOK5MOR+pg+MeGD8uP6WAOBD5I+jg8Y7kM3kAPrsPOvm4Pr7swPxQD9Ib4anUB43GH40f0hAfLQfBP7NgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAH804fWvzuH6ueAB4jxIfdf5dh+0v4DB+8X4dB/R/+fYfLvYB9cfkCH6IOD5Z78Pnv+lgD+eMPpT7lD89vaw1PjEPDf3zD8NPvsPiz+h0D+cgP6IPx/D0D6BD8vv1sD8aP6QgOTD8Bv6JQAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAD+acP0R/OwP2s/KQOU6APtDz8POeEDs8EP1w/DwPWOTD9FvgsP31/nyD9JPTw/EH+lgD8zQ/Mb9YA+Xvaw++P56w/bvy4Px493DnP6JgP5tA/pE/G8O++gg/L/9bA/Gj+kIDCD+ef8Ao1AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAP5pw/cL4aDJ9AD47+sQ+3+HD8Hv3qD8HP15DqfytD2/gA/Rb4UD97PycD4p9NDxP+lgD8pA8H6wIvaw+O/NA+pP0CD8QPvoPU/0cA/BMP2P/ADFD5R9LDwv9rQ/DT61D2f76D8o/3SAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPmkPccUPO/oIPmvkw+mNkPmT6oD5q9yDo/lEPRcAPS+ED3uIPlL1sPPPpoD5vDkfqsPJ9sPNM8D6LDmvnkPXPbwPgMOJ8aDc/SgPIPoIPBAOl+PQ9l+4wAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAIg5f4pDufsoAAAOQDrHgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAwPHPZgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAACkHwl+YQdF/QoCgAAAAAAAAAAAAAAAAAAAAAAAAH58hofewAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAGEH55fmaHon6gB9pygAAAAAAAAAAAAAAAAAAAAAAAAAH5Jhq/quAAAAAAAAAAAAAAAAAAAAAAAAAAAFYLCgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHFh+bn5vB9Hfp+H1WAAAAAAAAAAAAAAAAAAAAAAAAAAAB+SYav6rgAAAAAAAAAAAAAAAAAAAAAAAAAAB+eYfnF+nwfcdgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPLA/M38/g+yv0+D34AAAAAAAAAAAAAAAAAAAAAAAAAAAAA/JMNX9VwAAAAAAAAAAAAAAAAAAAAAAAAAAArh8PfmAHL/pwH6A64AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAch8sB6H9JgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHhAfmF8WB99fpqHrAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfkmGr+q4AAAAAAAAAAAAAAAAAAAAAAAAAAAACB8ifmCHhf6UB+jXWAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHzT/PWH2n+64AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHy6H5f/Mofo5+k4dyAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB+SYav6rgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHzl+YAfJH6HB+l3o4AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHzT/PWH2n+64AAAAAAAAAAAAAABzeoGgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAR/GQfmD5aH6X/oiHQgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAH5Jhq/quAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHjX5ih8Lfcofp17SAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAeQh+aXIB7r+TQfaf7rgAAB8kh+bcIfpj24fGP6jAAAAAAAH4/fawfVYAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAFL4ND8w4w/Tz70C+AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAH5Jhq/quAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB51+agfnf9Xh+n/0kAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHBfzjhxX0OHh+QH2n+64AAAfzph/QnshSpB5v7EAwMvXAAz5gtAfj99rB9VgYG+AAAGKG0AMDG3AAAAAAMzTBmOG2AAZGoDwKtcNIAAAAAAAAAAAAAAAAAAAAAAwPzyD8zPSg/UD7RCQAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAD8kw1f1XAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAOW/OYPzU9tD9QPr8FAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPzJ/IsP0h/YkPhT8QA+0/3XAAAD+eoP1D+zwf4GH5ufswH89HtIeR/oCH6IfnyHxJ3geDfuuH5i/awe5fjaHo3n4ft1bA/EQOw8+DF/eMPHfyUD0GMP2T/n4D98tUP5+P3yD8gcgPYvqgPy39HDmv22D80fCAi8YD99PJQ/Jn0cP1z7MAAAAAAAAAAAAAAAAAAAAOI/NsPzh+kw/T/6nAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPyTDV/VcAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAMr4AD8x+gD9P/ALkCyAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB+OH5yB+3H3kHzT/PWH2n+64AAAcCH5bfNwftNRD80/2sD+aH+lcMf+fsP6Fv54w/oinD+eT92w/L77WD5H9kD6U/M0Puf62A/D0P0e+pA/CX9cg/K79Ag9M/IUP2F/EgP6AtUP56P6Ag/H36fD74/nuD94uvD8o/bQ8t3Q/ST8pg9w5gPHf1jC+AAAAAAAAAAAAAAAAAAAHlX5lh8A/Z4fp772AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAH5Jhq/quAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABX+Ig/MDkQ/TX9BQ2AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA/L38lw/RX9lQ/PX8Xw+0/3XAAAAA+aQ/Mz9SQ/NP9rA/nS/otA/nSD9+/w1D9/APwQ/a8Py++1g/PP1IO5A+yPZAPw9D9V/aw/FP9LA/HL7YCyB93fhMH7/AOwH89H9AQfj7+gIfS3854f0YAfAfEhQ9WD7r/NMOr+9w/N34QD9x/VAAAAAAAAAAAAAAAAAAPBfzCD4w++w/Tb1UAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAD8kw1f1XAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAE+SQ/L/wQP0o/R0OtAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPOP5ww5j14OR48PtD92gAABPmQPoH4XD5/wD0DD80/wBrA/nS/otA/nSD+i3+fQP1Z3A/CD+jUPy++1g8c8mD9XuZDrwD8PQ/Vf2sPxT/AEsD4c9LD9FeTDr/AMOQ++/VQ/n9/pjD8ff0BD6W/D4PvT6yD8Kv10D469WD7r/NMOr+yQ7D8ng+jPt0AAAAAAAAAAAAAAAAPln8vw+bP0eD9JO3AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPyTDV/VcAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAD51D8v/kcP0K/S4PSgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADwsPzWrh+k3wIHsv6KgAAFP8tg8a9uD9MeSD5W+8Q/Mb9OQPzGD9OeED88fRQ+Bv3WD5N95D1787Q+Z+2D9bbIHwEH2D3gfB31SHdfmiHh3sYfqV5OH5a+uh3X3wHxb9Ih6pzgfmF58H3f8AWQfJfYB9CfK4bHMB8GeuB+oN0AAAAAAAAAAAAAAjD4v/AC/DzT9MQ/Q/fAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAD8kw1f1XAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPHQ/Mf4RD7h/TsPbAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPmgOr8kD80f31AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAoB8F/mKDv07D7yvAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfkmGr+q4AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAefB+af52B9V/qAH0qAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHziHyP1wfd/UgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABzwfnh+ZweofqAH2bIAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABQD5O+Uw1fpYPrK+AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHMB+c/5qB7P+n4fYTgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADhg/Nn84w+mP1AD6jAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAD8L/ksJvq4P3QAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAMsPgP8xg2v09D7ntgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAeG2g9o8nD8yvgUPtP8ATwPeAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPkf8DQP3zD63AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAACAPiT8wA4v9NQ/QfZAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADykP58f2QD4Y+NA/QD9Mw9TAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAGfzcgf0jA8AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABA+TPzAD5//AElD9H+vAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA4L+fIPH+uD9I/0kDtQAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPzJBf01AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPnn8wA+QP0HD9MfTAPDQ9yAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAOWD+f/5+A6UP2L++gAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAOMA7MAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPH/wAyA+Dvt4P07/KAP0Q+vgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPHw8s6UN/ow6DqwUAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAOA/NUPzphCr+/AfS4AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABSD8i/zWA60P6BvdwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPKg+dOfDneeDpf0dBQAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAOQD8+/lgOH60PpP9BA9OAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA/EcPW/pMO05gPnv5DD9/QAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAGgvCh2/xeHu3rgOAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA8e/FYPvX8jQ63+lcHgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAACB+bGMAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADLwfpfOAAAAAAAedfztBJ+vIflT/AEShvAAAAedBF6UB5oGt2oAHxD8pBvgeH+sBsAAAAAAAAAAAAAAAAAAAAAAAAAfO/wBEAAAAAAAAAAAAAAAAAAAAAAAAAAfO/wBEA1Q4L9Jg+hAADkuKD2IDiM0PSQDzAPyC9gAA8X70OrAAAAAAAAAAAAAAAAAAAAAAAAAAPmv6UAAAAAAAAAAAAAAAAAAAAAAAAAA8O9PDogPI/wBag9eAAPlLzQPt3bD5Z4kPtl4EQfkP+qQAH59fWoeoAAAAAAAAAAAAAAAAAAAAAAAAAflV+qoAAAAAAAAAAAAAAAAAAAAAAAAAH5VfqqD/AM5Q/QfuwAAAAABAX4MDyj9OA/Jr9dwAAAA8s+KA8X+0Qb8wh5t+qIesgfnT9gB6sB+af3YHoAAAAAAAAAAAAAAAAAAAAAAAAAH5J/rYAAAAAAAAAAAAAAAAAAAAAAAAAB+Sf62AAfH2sH1WAUg/MhQn+zA/NzYD7R+qgPFA+af0BAA/P76WD2sAAAAAAAAAAAAAAAAAAAAAAAAAA/JP9bAAAAAAAAAAAAAAAAAAAAAAAAAAPzp+wA9WA+FPaw+gACkHyRrh8T/c4fKProYX6GAAfkn+tgAH50/YAerAAAAAAAAAAAAAAAAAAAAAAAAAfkn+tgAAAAAAAAAAAAAAAAAAAAAAAAcb+eAfM37tAfl8H6LdWAAAAAAfiiGR9lB9h9wAAAAAAAfOPy6Hz9+1AfO/zMH6VAfnT9gB6sB+af3YHoAAAAAAAAAAAAAAAAAAAAAAAAAH5J/rYAAAAAAAAAAAAAAAAAAAAAAAAAB+Sf62AAfH2sH1WAAGSH43ejB+hHZh+SH7KgeKB80/oCAB+f30sHtYAAAAAAAAAAAAAAAAAAAAAAAAAB+Sf62AAAAAAAAAAAAAAAAAAAAAAAAAAfnT9gB6sB8Ke1h9AAABwwfln0QfoX6iH4o/tqAB+Sf62AAfnT9gB6sAAAAAAAAAAAAAAAAAAAAAAAAB+Sf62AAAAAAAAAAAAAAAAAAAAAAAABB+QwS/r2B+Xwfot1YAAAAAAgeDfGofHTA+wv0RDuQAAAKofjR+tYfj1+3YeHfEwfqUB+dPgIdQB85/twHoAAAAAAAAAAAAAAAAAAAAAAAAAH5J/rYAAAAAAAAAAAAAAAAAAAAAAAAAB+SfqAeXgeS/o2H1WAAH5Rh9rfn0H6t9UH4oftqB4oH5QfRIHUBsfSwe1gAAAAAAAAAAAAAAAAAAAAAAAAAH5J/rYAAAAAAAAAAAAAAAAAAAAAAAAAB+dPmIdGB4F+nwfQAABV/G4P1W/NsP0Z9KD8Uv2zAA/no+qwAPsT3gOwAAAAAAAAAAAAAAAAAAAAAAAAA+Mvs0AAAAAAAAAAAAAAAAAAAAAAAAD8kwAD5oD92OrAAAAAAAAA4wPj36MD2kAAAKv5Fh+g/u4fkX+kofLfTh9mAfnT9gB6sBzW6FgAAAAAAAAAAAAAAAAAAAAAAAADkutAAAAAAAAAAAAAAAAAAAAAAAAAA5LrQAPj7WD6rAAPzN9lD7N/PMPVevD5Q/TcDxQPk77xAmC7eC0AAAAAAAAAAAAAAAAAAAAAAAAAAc70QAAAAAAAAAAAAAAAAAAAAAAAAAGLmgoHxH9Dh9AAFEPxj+6Q988KDxX1cPPP0QAA/JP9bAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPDPx8D7M/ScH/l8H6LdWAAAAAAAcZ+T4foN+BgfYX7fAAAAB8yfBodD9Eh9Y/mSHXfpsEgH50/YAerAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfH2sH1WAeaB+WPWh6J+jAfl3GH6e7YHigfNP6AgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHwp7WH0AAcEHxMB2n3EHxFgB98SAAfkn+tgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAH45/sAHxZ0wfXP5fB+i3VgAAAAAAHh/4nh2HsAfGf8ASCG2AAAAAAAAAAfnT9gB6sAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB8fawfVYAAAAAAAHigfNP6AgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHwp7WH0AAAAAAAAAB+Sf62AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAnjgfjz+wAfE/1sH0Z+Xwfot1YAAAAAABF+CoeR/0BB/Pv/SWEwAAAAAAAAAB+dP2AHqwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHx9rB9VgAAAAAAAeKB80/oCAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfCntYfQAAAAAAAAAH5J/rYAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABU8qD45+cw+RaofU3mgfrd9Wgfl8H6LdWAAAAAAAHEB8dfVYaHoIAAAAAAAAAAB+dP2AHqwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHx9rB9VgAAAAAAAeKB80/oCAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfCntYfQAAAAAAAAAH5J/rYAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAQ+HB8/+Ch+cu6H1N9RB9Ze4g/8kw/WwA/L4P0W6sAAAAAAAxv5vQyP2aD8bf6YAtAAAAAAAAAAH50/YAerAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfH2sH1WAAAAAAAB4oHzT+gIAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB8Ke1h9AAAAAAAAAAfkn+tgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHnYfIHyIHivugfRX0MHsnoAAABifi8H6zerAfl8H6LdWAAAAAAAeHfkIH0B9FB+Mn9Lwb4AAAAAAAAAB+dP2AHqwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHx9rB9VgAAAAAAAeKB80/oCAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfCntYfQAAAAAAAAAH5J/rYAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAnigfHXyYHAd2H159bB7sAAAAAAYP5/wDyvv8AuH1dvifl99H436LdWAAAAAAAVP50Q4yMPav6BgAAAAAAAAAAD86fsAPVgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPj7WD6rAAAAAAAA8UD5p/QEAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA+FPaw+gAAAAAAAAAPyT/WwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAMQPlj42D5h9pD66+ww9DAAAAAAAAAPL/AJz2rXo2kH5fe/5H6LdWAAAAAAAVeZD4ceH077mAAAAAAAAAAAfnT9gB6sAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB8fawfVYAAAAAAAHigfNP6AgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHwp7WH0AAAAAAAAAB+Sf62AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAYgfL/AMdh81zB9N/ZwfS90AAAAAAAAAAAy/n/APNft3+/Naf5ffpPxP6LdWAAAAAAAef/AI/h+1vwaH5Jf05AAAAAAAAAAAH50/YAerAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfH2sH1WAAAAAAAB4oHzT+gIAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB8Ke1h9AAAAAAAAAAfkn+tgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA0Pnv4pD5c8tD7T+zg+mtYAAAAAAAAAAAAADwb2n4L61PGA94/L7735L9FurAAAAAAAMf8Hw8Mxg/Vr9KgAAAAAAAAAAD86fsAPVgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPj7WD6rAAAAAAAA8UD5p/QEAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAKAfEfr4fRgHwp7WH0AAAAAAAAAB+Sf62AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABS+PQ+OvjQG/ZYfU32uGyAAAAAAAAAAAAAAAHjW3+ZfcpkB7F+X36b8N+i3VgAAAAAH5VBkMD4U9XD6i/WEAAAAAAAAAAA/On7AD1YAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAD4+1g+qwAAAAAAAPFA+af0BAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAACuH44fpyHrwHwp7WH0AAAAAAAAAB+Sf62AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAFIPkL4eD409PD7d+vg+i1AAAAAAAAAAAAAAAAAATwn4r6i97Rtkv5ffX/Ofot1YAAAAAB+Doc+AAaH75AAAAAAAAAAAH50/YAerAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfH2sH1WAAAAAAAB4oHzT+gIAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAflR+oQaoHwp7WH0AAAAAAAAAB+Sf62AAAAAAAAAAAAAAAAAAAAAAAAAAAABQ+YQ+J/hAPTPukPtP2wAAAAAAAAAAAAAAAAAA5Hyu/iuY/K/v+D5r8o/QfoD2v8ApD8TPy++w+c/RbqwAAAAADw4PRvn8ACX6mAAAAAAAAAAAPzp+wA9WAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA+PtYPqsAAAAAAADxQPmn9AQAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA/Kj9Qg1QPhT2sPoAAAAAAAAAD8k/wBbAAAAAAAAAAAAAAAAAAAAAAAAAAAD5/8Az7D4K9ED7I+9w92AAAAAAAAAAAAAAAAAp8p4PrcNxH5D+jce/wDPvse+6f8ASfiO32/074WfW9zyj8vurD9FurAAAAAAP52Q/QL8eQANL+pgAAAAAAAAAAA/On7AD1YAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAD4+1g+qwAAAAAAAPFA+af0BAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADhw/FH9Aw+4wPhT2sPoAAAAAAAAAD8k/wBbAAAAAAAAAAAAAAAAAAAAAAAAAOBD4N+BQ+f/ANCwufc4fVgAAAAAAAAAAAAAAAV+O8L1uF4n8e/R+H1/z/7Dte5/U/ge76D9b/O1PQ5AAAPy+D9FurAAAAAAM4L2aHmWqHcaoAAAAAAAAAAB+dP2AHqwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHx9rB9VgAAAAAAAeKB80/oCAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAH49B+n/wAYh7P9Wh8Ke1h9AAAAAAAAAAfkn+tgAAAAAAAAAAAAAAAAAAAAAAAny0H5q/IYfY330HyV+voR/ksH64gAAAAAAAAAAAAQ8X4Xq+c8D+H/AKpy/R/H/R912363+ed/qfrP56HTgAAAAAH5fB+i3VgAAAAAAEP4Qh8cqH6e/rYAAAAAAAAAAAfnT9gB6sAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB8fawfVYAAAAAAAHigfNP6AgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAEXAh5D8yB5N87B4z6CH9FHxyG99hh8Ke1h9AAAAAAAAAAfkn+tgAAAAAAAAAAAAAAAAAAAAAHIfnCH56Afo9+hQdgB8zfnOCfpIH0cAAAAAAAAAAN5Xy+/gfOfxb9O5Db+C+t9D9D/ZfzTsrn6h8IGuYAAAAAAAB+Xwfot1YAAAAAAB4f/PqH7188H4M/wBP4a4AAAAAAAAAB+dP2AHqwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHx9rB9VgAAAAAAAeKB80/oCAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAVQ8j8mDzXxEPC/Eg803w9E+qw9U+iA9xyYr8Xfq/4z6b9T3fbfL/CntYfQAAAAAAAAAH5J/rYAAAAAAAAAAAAAAAAAAAAHg35oh8N/Xgfp59HBKAAHn4P70AAAAAAAADO59vL/AD/8V/T+CufmX3Pf+i/rf532m1+sfnwdnMAAAAAAAAAAB+Xwfot1YAAAAAABl/zyB+sGYH5C/teHoXtYAAAAAAAAAH50/YAerAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfH2sH1WAAAAAAAB4oHzT+gIAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABB4kHinjQeQeaB4R62Ho/tIe7e3h6r0AAAANR8+e2/nX2mwfo3xfwp7WH0AAAAAAAAAB+Sf62AAAAAAAAAAAAAAAAAACB8efmYHz7+igfo96OAAAAAAAAAAAABFL4Pz/8r+/4Hmfxv9K7/wBM/Sfie73/ANj/ADUO7lAAAAAAAAAAAAAD8vg/RbqwAAAAAADz/wDmVAAAPv79rAAAAAAAAAAPzp+wA9WAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA+PtYPqsAAAAAAADxQPmn9AQAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA4IPnb57D5h40Ge2B7f66HuHpgdSAAAAAAABTx0+a/YPyL9G9AP2P8ANfhT2sPoAAAAAAAAAD8k/wBbAAAAAAAAAAAAAAAAAGB8W/lmHLfpsH3ztgAAAAAHlfzXucH9IeJ6YfoHyAAAQ8N4Hr+Xeafz7+wavsfu+V0ncfsv5pOe/wCQAAAAAAAAAAAAAAAAfl8H6LdWAAAAAAAR5IS+UB3u0Fi6AAAAAAAAAB+dP2AHqwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHx9rB9VgAAAAAAAeKB80/oCAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHFB4T4oHyz5iGR2wfRvs4fWfogAAAAAAAAAAAAHz58L9X6353ht23bfU+B8Ke1+x5v0AAAAAAAAAB+Sf62AAAAAAAAAAAAAAGSFHoA+HPyxCn+pYfd1gAAAAAAAOO8n0PO/dfB9YPtPmTI5OjzbgPxj9M8+3vzf7b071D93/ACfpD9D+NAAAAAAAAAAAAAAAAAAA/L4P0W6sAAAAAAA81/F0P0e/CMPof+iMAAAAAAAAAAA/On7AD1YAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAD4+1g+qwAAAAAAAPFA+af0BAAAAAAAAAAAAAAAAAAAAAAAAAAAAD5r/NcP1K9yAYHlPz2HzR4sHkXdh9AeuB9f+oBqAAAAAAAAAAAAAAAAfPHwf1v0H5/7nlM9HF8Ke1/QeR9AAAAAAAAAAfkn+tgAAAAAAAAAAAAAHgP5xB1vxcFj9Vg+3JQAAAAAAAA43zD8w+76bN5d/ZfOvs/mfJuE/nj9l9C7/7z5L0ba/ZfzMOznAAAAAAAAAAAAAAAAAAAAPy+D9FurAAAAAAAPIvwVDX+/wAPzl/osDfAAAAAAAAAAPzp+wA9WAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA+PtYPqsAAAAAAADxQPmn9AQAAAAAAAAAAAAAAAAAAAAAAAAAAAA/PD43D6l5YPmT5xC776H037gH0J7WAAAAAAAAAAAAAAAAAAAUvMvkfovDPX/wAh/RvbHf0X+MfCntbX0AAAAAAAAAB+Sf62AAAAAAAAAAAAAAflF+jwd/8Ai4H7OTgAAAAAAAAAeP8AefC/WdJ8h+N6XNdV+O/pPq3qX9B/jt8+y+aAAAAAAAAAAAAAAAAAAAAAAA/L4P0W6sAAAAAAAPyQDyT9og/D396AcAAAAAAAAAAfnT9gB6sAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAUg827MOjA+PtYPqsAAAAAAADxQPmn9AQAAAAAAAAAAAAAAAAAAAAAAAAAE8WD5t+OQ92/UwPx0cH1P8ASoemgAAAAAAAAAAAAAAAANy+ToxuZ+Q+j5PB/OftONyvzz7JOi1zrfY/7B+cTn69+c/CntYfQAAAAAAAAAH5J/rYAAAAAAAAAAAAGP8AkQHwv9oBofaQfYoAAAAAAAAByfl9/i3Dfzt+zdx2f1/znpmv+yfmodXOAAAAAAAAAAAAAAAAAAAAAAAAfl8H6LdWAAAAAAfGQfU/TAAZXxmH3CAAAAAAAAAAfnT9gB6sAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAcoH4d/r8HuIHx9rB9VgAAAAAAAeKB80/oCAAAAAAAAAAAAAAAAAAAAAAAHAB8Z/MYfKvPh9L/VQeC/oqH50ftSDwAAAAAAAAAAAAACPh/C9XC5v89+x5Lmfzb7bHn+d9ra6L6nwet2P0H47utf9I+JvQepw/P3ffkv6H6Wfr35z8Ke1h9AAAAAAAAAAfkn+tgAAAAAAAAAAAz83g/Mr6XD9h+7AAAAAAAAAA5rzH88+y8wyvw39V9X7P9a/O8H2v6z58Pr/AJwAAAAAAAAAAAAAAAAAAAAAAAAAPy+D9FurAAAAAAPzlD8uupDtOaDzj9AQ/W0AAAAAAAAAA/On7AD1YAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAq53+Qv62eb27B63n/H2sH1WAAAAAAAB4oHzT+gIAAAAAAAAAAAAAAAAAAAAHMB8tfKofDNUPoj6yD6a+jQlA+a/wAa/I9H9t/ce/kDoxAAAAAAAAAKXK+B6/I4/wCdfZ8vhfCfWc30Hgev1PQfoPx/Vdb+j/FbVz7D5sNswAAAAAA+FPaw+gAAAAAAAAAPyT/WwAAAAAAAAAA84D8afLQ/Zf6YAAAAAAAAAiiuF8f/ACD9H4L134f6nv8AvP3z8jk4K55717g6w+q8AAAAAAAAAAAAAAAAAAAAAAAAAAPy+D9FurAAAAAAAIvGQ4Tqg9s0AAAAAAAAAAAD86fsAPVgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADwfHT8S/1I8b0/uQ9/yPj7WD6rAAAAAAAA8UD5p/QEAAAAAAAAAAAAAAAAAAA82+Fg+KPnQPof6fD7D+pwkAAADx/wCIfmvb+9fSfpfEAAAAAABMXi6eW5f8++x5Hjfy77zF6PwfW7Db/R/i+g6L7f5bptD7j5UN8gAAAAAAAAAA+FPaw+gAAAAAAAAAPyT/AFsAAAAAAAAAA+QfxLD74/V4NwAAAAAAADI5d/GvIP5x/atr1D0OP2S7/QX48G2YAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfl8H6LdWAAAAAAAAAAAAAAAAAAAAAB+dP2AHqwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABh+cgvzqHL/M4fsj+YIfoB6wHx9rB9VgAAAAAAAeKB80/oCAAAAAAAAAAAAAAMDxfeD03xoPgX42Dx37KD7k+sg6oAAAAAAzPnf8y+6+jb36P8SG2YARy8XlvlPoPPeR/Hv0jl3/ABH1PT9T9v8AL9Z2P6f8J1Uv3nyQXIAAAAAAAAAAAAAAfCntYfQAAAAAAAAAH5J/rYAAAAAAAHigef8Aw+Hw1+1IfY4AAAAAAAHI+X3+Meafzd+2ene1fqXwPUH6n8EAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfl8H6LdWAAAAAAAGKHy77+HYXQAAAAAAAAAAD86fsAPVgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA4nGDz7wMLHjIcXyofNNoOD64Ifew73Aw1/br8wdsvuv2jLT4+1ts/qsAAAAAAADxQPmn9AQAAAAAAAAAAAAADzgP52vUw3Pn8Puj7cD6i1wAAAAAAAgivnL1b8l/Q/KfpP2/LZyv1nz3J8f8AmH3fLcf+Yfd3N+49E7P9h/OOo2f0X4sOvnAAAAAAAAAAAAAAAAAAD4U9rD6AAAAAAAAAA/JP9bAAAAAAADx78qQ2PBA/d/uwAAAAAABPOfn/AGPFOZ/nD9q9g9i/cPyzWPv/AJAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPy+D9FurAAAAAAAM/wDnRDy39uA/Hn+lgJgAAAAAAAAAD86fsAPVgAAAAAAAAAAAAAAAAAAAAAAAAAAqB5p1wdQAAAAAAABAD/nAOU8oD5ush8oXQ801An+sQj9cD0CMPrKYO/7MAA+Cfhfq/wA7PuX0eL786j1/O+Ptb0uL6rAAAAAAAA8UD5p/QEAAAAAAAAAAAADOD5K/M8P3J+cA+Lv1tDoAAAAAAAAADK5d/JPcPl/e+esr8y+58+vfBfXdV2v6H8b0/afqPwemfUeCAAAAAAAAAAAAAAAAAAAAAAHwp7WH0AAAAAAAAAB+Sf62AAAAAAAH5f8A3sHpX5EB+u4AAAAAA3zD5r2/DG/zr+z+1eq/0H+PTn1nzwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAH5fB+i3VgAAAAAAHhv5AB9C/SQfjJ/SkHRgAAAAAAAAAH50/YAerAAAAAAAAAAAAAAAAAAAAAAAAAAB5sH88X60h9+gABCC/Ngct4GGJ89hR+YQ5/wBDD16YPtvKD6b2Q9G9BAAAAAAAAA8+8L1fyV/UX5P6H2Y/Sfifj7WD6rAAAAAAAA8UD5p/QEAAAAAAAKAYfRBZAPFw+R/y7Du++D9nPm0Hfb4AAAAAAAAVfPfmfc87478Q/U8DvfJ9Da6L9A+P7Ts/1T4EPX84AAAAAAAAAAAAAAAAAAAAAAAAA+FPaw+gAAAAAAAAAPyT/WwAAAAAA+ZQ/B37LDqvpYPvEAAADmfO7OUX4n6nw4/G/wBK919L/on8ZU+i8YAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA/L4P0W6sAAAAAAAzP5xQRwdD/RWAAAAAAAAAAAfnT9gB6sAAAAAAAAAAAAAAAAAAAAAAAAAAH42h92fCwfZmQHg/igcJ8uhF6cHovvQb/sYdl9Ch6BoBZAAAAAAAApY6cpJ8r7/O4Hyv0Ccj857VLmPC9X8vP2n/oH8f8AdD2fN+PtYPqsAAAAAAADxQPmn9AQAAAAAAA8RD8av29D0b5xD8y/kMPuX70D6l4kP5wPqAP3TcAAAAABk8F8j9F59xv4n+oc71/gev3/AKH+1/mHVeH+jx++n3vyIAAAAAAAAAAAAAAAAAAAAAAAAAAAHwp7WH0AAAAAAAAAB+Sf62AAAAAAfOf4Ih+0PsIebfXYTAAAAeZfO+z8w7f8ufvPvnqf9P8A4Sp9N4YAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfl8H6LdWAAAAAAAHmYfnw8P0P9BAAAAAAAAAAAPzp+wA9WAAAAAAAAAAAAAAAAAAAAAAADnPAA868iD5ivh+6XwQH5gbofUXrYej/R4bfUA8AAAAAAI5fHS/KfQcXR+N+lyuW+U+gOe+e9nMPA9fone/5Gx1P1Hgp1P0fi6uJ9D4/jHtumfq5938p8fawfVYAAAAAAAHigfNP6AgAAAAAAFf8gQdth4r8zh+jn6Ph2AA0OB0g64AAAE5Lyu/zLjvxH9S4v0j5v2+v7L9g/OOxm/RPjAueM8B/Gv0v6K6f9U+BD0eIAAAAAAAAAAAAAAAAAAAAAAAAAAPhT2sPoAAAAAAAAAD8k/1sAAAADnPKg/Ab9fw+8gAAAADD4+n5+xP5z/aO50Pr/nPbz9p/MQAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPy+D9FurAAAAAAAOa/M8Pu/8Og+vv1tAAAAAAAAAAAPzp+wA9WAAAAAAAAAAAAAAAAAAAA83D5I84D5r8ADzn3EF+pA+gqwfHP0iH6hPAAAAAAM7n1j84+V+gp8b+f/AGFDL+S+h5Sf5P6DWn9rzOg6H67513d/V/Prv/UeDqn0Pj6J38gAAAAAAfH2sH1WAAAAAAAB4oHzT+gIAAAAAAB4X82h+g346h2f6bh0AAAAAAAZPLvxXlP5B+j8TN+Wfe+kdz+ufnfoGh+s/nob5AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHwp7WH0AAAAAAAAAB+Sf62AAABEH4IfJIfrp+mwAAAADUeTfP387fs/W+6fReL3fz79d879BH3XygAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAH5fB+i3VgAAAAAAHhf41h6NXD5j/pRDVAAAAAAAAAAPzp+wA9WAAAAAAAAAAAAAPjYPlD3kPv8Aqh88+GB89+EB84WQ9i+lg+ifbw9z6EAA4fK/xe+//wA/+w+2j9F+MKuV5/CfJ/QpxHxf09HlPh/qq2T837b+39Hi3t79A+P6N31vzveV/s/mtVe/jBgAAAAAAAAAAAHx9rB9VgAAAAAAAeKB80/oCAAAAAAVg/Mr4VD7z86D9egAAAAACv5d8p9B5HyX87fs3Zehfa/L+kdN+3flwejxAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfCntYfQAAAAAAAAAH5J/rYBwYfMv0MHqH5jB7dwQbH3gAAAAHJfOX4z+mM+kkbfz57/k6m34Xq+yH7L+aAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfl8H6LdWAAAAAAAUf50w81/oeD+f/wDpbC2AAAAAAAAAAfnT9gB6sAAAAAAAAAAAABB+PYfsX+WofOfyqHZ+ih9I/Twe2+yhOAAAABl+K/n32GryXx/0kvJ/Oe1Tw/lPoLHZejxWut+z+Z2Oq+8+St7f1HhXD1/OAAAAAAAAAAAAAAAAAAD4+1g+qwAAAAAAAPFA+af0BAAAAAA8b/EILH6lB+cv7dB14AAAFHHX5rofzd+18b2Pher7D6R/Rn4xZPqPBAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAD4U9rD6AAAAAAAAAA/JP9bAOVD8d/16DuPxgD9huTD8//ANRQAABvh/xn03ivu/5J+iexP/oz8WMfm38B+jPg/rLR+j/FgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfl8H6LdWAAAAAAAFQOf1g4b04AAAAAAAAAAA/On7AD1YAAAAAAAAAAOdD5a+SA+V90P2Z/NQPo/6uD14AAAADP5P532eZ5L82+25vP+E+twPSpf0x5r/Q/wCNcL3/AMP9Vtb36B8fdPZ80AAAAAAAAAAAAAAAAAAAAAAAD4+1g+qwAAAAAAAPFA+af0BAAAAD4V+Eg+Xf07D9OeRDxz3UOhAAG8F4Xq+X+Zfz3+xe58t9l836n3/6n8CHueUAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB8Ke1h9AAAAAAAAAAfkn+tgAH5J/rYHkH87ofTO4H7P9aAAZvPt8y0P5o/cPpvoP6Q/FA7+Q53g6+X7zx/SvH0/hAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfl8H6LdWAAAAAAAUPwOD5VA0v6mAAAAAAAAAAAD86fsAPVgAAAAAAAAPCvlEPl741D3P6iD669hD5V+ow9lAAKmR5ffx3I/mv2/G5P5p9vh73leh1Xa/pXxG9r/AHvyXUeUbZcgv5h939Ez/vv5EFyAAAAAAAAAAAAAAAAAAAAAAAAAfH2sH1WAAAAAAAB4oHzT+gIAHgvzGH3nXD8lfMQ9+/WUAAAAIvHPj/pPIT8A/XvT8L7n5X6U539n/MuA9h8P1Q+r+fAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA+FPaw+gAAAAAAAAAPyT/WwD4CD5LQPMfoEPp2sH3WAAcb8wfhn6r6P9CfZ/MzH6B8gAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfl8H6LdWAAAAAAAeI/z2h+3O4Ev1MAAAAAAAAAAAfnT9gB6sAAAAAAHxOHy/81B4J9mh9M/YIewgAABR8/8Alve5LjPyT9E48/PvsO4639G+M6Lp/wBD+M6mf7n5ULkAKOOnj3d/nv2PaH6R8UAAAAAAAAAAAAAAAAAAAAAAAAAAHx9rB9VgAAAAAAAeKB80/oCAB4H+QofuB+a4fbH0MH5DfryAAAUPHvhvqvIN38N/VfoPr/6W/EDz7n18fX8O/VfozU/oX8cDpwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA+FPaw+gAAAAAAAAAPyT/WwD8gA/X/4ED8iP6TA6YAAPJPl/e+dPfPxL9R9qP6V/EAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA/L4P0W6sAAAAAAAyv5zQ+/d4LX6RAAAAAAAAAAAH50/YAerAAAAAHkoflD0ofsn+XgfYvvwAABy/j35j91JwX559lzvYfN+32vSfqnwHXdb+m/DB6nAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB8fawfVYAAAAAAAHigfNP6AgcYH5MfoEH5S+Bh9re7hgfqABjcnRx3Kfmv2/i/oH5Z997L3H9H/ioez5gxPyLb/N/tfRT9J+JAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPhT2sPoAAAAAAAAAD8k/1sA/HIPqf49B37TAAAHh3Pfhn6t7N439B4/0gfqfwIAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfl8H6LdWAAAAAAAcP/PgEoGl/RgAAAAAAAAAAAfnT9gB6sAAAHkvzGHwj8wh+gXJI/Sn83w/YDpGZvm3x/wBJwvnP4Z+qy+q9fP8AQPjv7b+Xdv1fseaHp8IAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfH2sH1WAAAAAAAB4oHzT+gIHjYfIH6Rh+MH6Nh80fn4H9CdsD53+D+t5vD/ABb9O+oOo/pP8RD0uIAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPhT2sPoAAAAAAAAAD8k/1sA5wPwJ+yw5X9eAADxH476Txb3T8l/RPY/nb9O+E+iT735IAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPy+D9FurAAAAAAADED482Q+vLIAAAAAAAAAAB+dP2AHqwGH4AHruOH5hfeIfln+4QdX8jfDfVcL2HTj6NwX5J+h8v1/y3v+od1+6/lHUfO7W5ynxH1P0rJ+5/lIUgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA+PtYPqsAAAAAAADxQPmn9AQPLw/ALUD9Gf0gAAMLi6vnX0j8Y/TPX/nv7n5X6EPu/kwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPhT2sPoAAAAAAAAAD8k/1sAA/FP4/D+if0UADxjxX+df2f6y8n/R/iub9R+d9n0o/ZfzQAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPy+D9FurAAAAAAAIf54w8HhD7i/b4AAAAAAAAAAA/On7AD1YD8XPsQPib44D9ff0CD8eP2HDH+evy/7z6Z8E9bzqfcfL+96Jf/W/zsN8ivndG1ydFw7+QAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAD4+1g+qwAAAAAAAPFA+af0BA+QQ/JnxEP6V+7ARHjHhP83/tnr2X6/nej+f+z5n0afsP5uAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB8Ke1h9AAAAAAAAAAfkn+tgHjIfgl+mofo8AeTfMe74F9Z/nH2vRn7d+WgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAH5fB+i3VgAAAAAAHh/4gB/QzkB/N5/SuGyAAAAAAAAAAfnT9gB6tywfh9+pQfO/oIeG8aH037H+a/b+T9n+Kfp/Z9/+wfnHoB+q/AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB8fawfVYAAAAAAAHigfNP6AgfjWH7J/liH6E99nfg+L+DfrXJfT/p8PUcN+o/B+LfQPwP1vQn6v+fgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfCntYfQAAAAAAAAAH5J/rYAB/MP/Q4HrgHnfy5/PX7H9d9Z+4/lQepwgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAH5fB+i3VgAAAAAAHIfzYh+keEHwP/TSEwAAAAAB5L+aAR/rgG6B+dP2AHq34+h+gvxoHk/7OS/n9fwz9W8j9T+T+h931f6Y/Dg6cAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPj7WD6rAAAAaH5482H6S3wPFA+af0BA/FYP1e/PHyfQ/SXC8/s3O17uX56+hfnPaD7v5MAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPhT2sPoAAAAADz/nw9gAAPyT/WwDgw/mq/qOB+bz7fHfov8ANH7h9In9P/hIAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfl8H6LdWAAAAAAAHwwH5b3A/Wr65AAAAAAAPxu/WYPmngA/QAD86fsAPVvx0D9g/jkLHpHw31XXeoet5/z79BeR6IfdfKAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHx9rB9VgAAAfDgQd8Hgv6TAeKB80/oCB835X5D9J/jX6ZyFXyu/S7r6XxOM97+k8QPtvlwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA+FPaw+gAAAAPFg/NXsw+mfscAD8k/1sA+Vw/HP98vm/a6XwT5L6KDrvl/evex/pvwwfWfPAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAH5fB+i3VgAAAAAAB4aHwdsh+hffgAAAAAAH4h/t4Hifw4H6ngfnT5MHXeDh8vfX/wA17f3bU/B/1no+2+w+c8a9j+0+Y6L0D7n5Uxw81GEweuCDyAGDD0pGwB5/zoAzru0Qc0HBDDSD1ARW8mAGHqyLgHmeSAM7rqEHHBxww30ehgZXmQAw9iQAeTVmAHo+2g4IOaGHyijjgPGfvMPrQAAAD8ew/Uzrg/Fb9swPFA+JvtkDmPN7u49a8n0PnHjfxr9L9nofrv513dr6/wCc6Maby4AB0oSgc9WAA2dEDKDKALgbwFfnQADqgAOZjAA3rgGKGeAaIbIFLCAAl6UBA5YAA6SYDACoAawagGbjgAW98DyUM4D44+sg+gAAAAPyvD7m9XD8av2oAA/I79YwMUPFIfzX7f0f5l/PPsvsbpv3z8i+ePeflvfuO+6+UKgMALATAQQAASWgGhTAALwBSBAC0EgFaIACawBGFUAcFwAogAFwHAVYwALEwEIVwCQLQDaYABeA/L4P0N7QAAAAAAA5D+aYPY8gE/o7BwAAAAAB+Lv7Fh80eSB+hIH5094HfjPMfq38j/Q/F/pPrw43lvvfkcBPT4ew9cA5wPFhhZD6DEHzuEYw9pR0YHkfHsAPQvSkHFh5OMNoPchFLwAAYe9o0QPEsBgB6n3KDzkPORh1gewCMDxIAYfRiADwCkwA9l6dB5QHFDDpg7sR8idcH1wAAAB+RwfpV1YfkD+zQHigflv9LAZ/l9/rPbfzl+08j7z+h/G+HH6/+cm+H1KIp/I7AA+rA2BHzNyjAD3T1BB46Hjow7cPo0RzXzAwAPsQAR8m5rAD6N7dB4KHmow9QD3QRwXzwwA2PqwCFHyCMAPqXfQfNgccMPaA9cEeU+IsAOx+k0HxEHG3g+bP0UD6AAAAA/IsP0t9GD8Rv26AA/nM9zAGfVnun4h+pcx6l4PrYkvfycvyX67+dfYm59B458oM44A+hQ9bEeH+HsAO/wDqFByofJIwA+4BB8aMygD6hDvxHzn5WwA9b+hUHnIfMow3A+xBEHxMwAPr0OjEfK/DsAPd/aUHjoeAjDuA+qBHOfITAD4m9LQ75pD9PfsVgIAAAAAPKPwGD+k+qH8w39NwawAAAAAB85/nWEH6/B1IH50/YAerNT+ffoT4X6vxX05G4fefJgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfH2sH1WAAAB8wh8d9EHvf2KB4oHzT+gIHzb+ffYfQHguWne+n/R+KH0figAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHwp7WH0AAAAB8IBtesh8bfqKAB+Sf62AV878I9/8Ai/p/CvXsdOG8t+S+i+oj9r/LwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA/L4P0W6sAAAAAAAwv5xA9yzQ8D+sg+qf0ZAAAAAAAAAA/On7AD1bH5Oj496X+Wf3v0T3H9/wDyEPsvmgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPj7WD6rAAAAA+fawfRYB4oHzT+gIHzj8B9f9G+DdGPsmt9b86HVgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfCntYfQAAAAFcPzFzw/TbcAA/JP9bAI5fyx7l+OfpXkn0d9D49ev8AafMax6HGAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAH5fB+i3VgAAAAAAHK/jwEoAB9QfoYAAAAAAAAAB+dP2AHq3zv8AB/W+8eZdfN0nc+v5we15gAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHx9rB9VgAAAAAAAeKB80/oCB574PrfMvtv4V+r+3n9J/iQAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfCntYfQAAAAAAAAAH5J/rYA1P5v8m/lf99+yei/pr8MDv5AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPy+D9FurAAAAAAAKfyaH03+XAfUX2QAAAAAAAAAAAfnT9gB6t87/B/W++eZ+hx7neejxB7flgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfH2sH1WAAAAAAAB4oHzT+gIGRy7/Ff2d+KfqG6fuv5QAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHwp7WH0AAAAAAAAAB+Sf62AeXfN+3w/Ifiv6f7v6D+4/lQe75QAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAH5fB+i3VgAAAAAAHhv4/h9MfDgcb/TMFkAAAAAAAAAA/On7AD1bE4+nxDovyD9H9oP2v8ALwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAD4+1g+qwAAAAAAAPFA+af0BA57g6/k/wC0fzH7oP1386AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAD4U9rD6AAAAAAAAAA/JP9bAPn74b6v6B5P6XwuW9V83uD6bwwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPy+D9FurAAAAAAAOV/mtDE/fAPxU/o7DXAAAAAAAAAAPzp+wA9WAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA+PtYPqsAAAAAAADxQPmn9AQAMbZ5OgOvnAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAD4U9rD6AAAAAAAAAA/JP8AWwDxr5L6GjgfnH2vum9+z/mYdvKAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB+Xwfot1YAAAAAAAeEhwH1SHxF90gAAAAAAAAAAH50/YAerAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfH2sH1WAAAAAAAB4oHzT+gIAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB8Ke1h9AAAAAAAAAAfkn+tgCI+I/rP8L/V+qP3X8oAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPy+D9FurAAAAAAAD8fQ/O79wA+Jf2tAAAAAAAAAAAPzp+wA9WA8i9JDUAAAAAAAAAAAAAAAAAAAAAAAAA8N9yAAAAAAAAAAAAAAAAAAAAAAAAAA8N9yAA+PtYPqsAAAAAAADxQPmn9AQAPJ+zDpQAAAAAAAAAAAAAAAAAAAAAAAAADwL30AAAAAAAAAAAAAAAAAAAAAAAAADx/wBDDdA+FPaw+gAAAAAAAAAPyT/WwCvFfCP1d+O/pPpB+y/mgAAAAAAAAAAAAAAAAAAAAAAAHknrYAAAAAAAAAAAAAAAAAAAAAAAAAHknrYH5fB+i3VgAAAAAAHj34Yh759uh+Ov9HodGAAAAAAAAAAfnT9gB6sB+af3YHoAAAAAAAAAAAAAAAAAAAAAAAAAH5J/rYAAAAAAAAAAAAAAAAAAAAAAAAAB+Sf62AAfH2sH1WAAAAAAAB4oHzT+gIAH5/fSwe1gAAAAAAAAAAAAAAAAAAAAAAAAAH5J/rYAAAAAAAAAAAAAAAAAAAAAAAAAB+dP2AHqwHwp7WH0AAAAAAAAAB+Sf62AAfnT9gB6sAAAAAAAAAAAAAAAAAAAAAAAAB+Sf62AAAAAAAAAAAAAAAAAAAAAAAAAAfkn+tgH5fB+i3VgAAAAAAHF/wA8YezdaHyJ/TmEoAAAAAAAAAB+dP2AHqwH5p/dgegAAAAAAAAAAAAAAAAAAAAAAAAAfkn+tgAAAAAAAAAAAAAAAAAAAAAAAAAH5J/rYAB8fawfVYAAAAAAAHhYfPX3yAB+f30sHtYAAAAAAAAAAAAAAAAAAAAAAAAAB+Sf62AAAAAAAAAAAAAAAAAAAAAAAAAAfnT9gB6sB8Ke1h9AAAAAAAAAAfkn+tgAH50/YAerAAAAAAAAAAAAAAAAAAAAAAAAAfkn+tgAAAAAAAAAAAAAAAAAAAAAAAAAH5J/rYB+Xwfot1YAAAAAAAfDoflw8P1f+xwAAAAAAAAAAD85/sMPVQPzT+7A9AAAAAAAAAAAAAAAAAAAAAAAAAA/JP8AWwAAAAAAAAAAAAAAAAAAAAAAAAAD8k/1sAA+PtYPqsAAAAAAAD5eDlPswAD8/vpYPawAAAAAAAAAAAAAAAAAAAAAAAAAD8k/1sAAAAAAAAAAAAAAAAAAAAAAAAAA/On7AD1YD4U9rD6AAAAAAAAAqB+VH6xAAfnT9gB6sAAAAAAAAAAAAAAAAAAAAAAAAB+Sf62AAAAAAAAAAAAAAAAAAAAAAAAAAfkn+tgH5fB+i3VgAAAAAGYH56d6H2t8kh8n/riAAAAAAAAAAAfGXyeGiBw/WBcAAAAAAAAAAAAAAAAAAAAAAAAA879EAAAAAAAAAAAAAAAAAAAAAAAAAA879EAA5v8AR0PcwAAAAAAAPLw/LrpQkA5fYDQAAAAAAAAAAAAAAAAAAAAAAAAAAOG7kAAAAAAAAAAAAAAAAAAAAAAAAADlN4LoHNfqIHpoAAAAAAAGUH4y+hAoHnPYhqgAAAAAAAAAAAAAAAAAAAAAAAAeSetgAAAAAAAAAAAAAAAAAAAAAAAAAeSetgeYh+z24AAAAAAflwH5oWA7TxgPqL9/wAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPwzD9Au1D8S/wCh4OrAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPwzDyvWDwz6RC/8Au6AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB8LB8zgAFz9dQAAAAAAAAAAAAOGD8z9oP1KlAAAAAAAAAAAAAAAAAAAAAAOeD8zPsgPoz5jD4s+gw+7AAAAAAAAAAAAAAAAAAAAAD4jD5q9eD9E/i8PmD63D64AAAAAAAAAAAA44PzJ0Q/U2yAAAAAAAAAAAAAAAAAAAAAAAHzMHxh+tAM/MMOL/TEPQgAAAAAAAAAAAAAAAAAAAACkH5ccYH6Ie0B+XeKH6i9QAAAAAAAAAAAHHB+ZOiH6m2QAAAAAAAAAAAAAAAAAAAAADng/Mz7ID6M+bw+IPdg++QAAAAAAAAAAAAAAAAAAAAD4uD5g9QD9IPjwPlP6qD7FAAAAAAAAAAAAAAAAAAAAAAAA/HoP1I+Qw637MAAAAAAAAAAAAAAAAAAAAAAOQD4f7kPtb8Pw/b78XA/YToQAAAAAAAAAAAAAAAAAAAAMIN38RA/Yz8hQ/an8OQ/cYAAAAAAAAAAAD8ig/Sj5oB/3CAAAAAAAAAAAAAAAAAAAAAAAHi4flz+1wfGXLh9bfluH7DgAAAAAAAAAAAAAAAAAAAAAGEgfjV9Sh6z3gfFn6mAAAAAAAAAAAB+RQfpR80A/wC4QAAAAAAAAAAAAAAAAAAAAADkA+H+5D7W/EEP26/GwP1o68AAAAAAAAAAAAAAAAAAAADCDd/FYP1y/IMP2z/D0P2+kAAAAAAAAAAAAAAAAAAAAAAAD8QQ/b756D5L/TkAAAAAAAAAAAAAAAAAAAAAAA+KQPrb8gg/Z38uQ+zveAAAAAAAAAAAAAAAAAAAAAAKf4nh+o3xaH6kfjIH68dGAAAAAAAAAAAH4gh+33h4fEP6nAAAAAAAAAAAAAAAAAAAAAAAAB+Kf7WB+Wf20HuX4gh+3wAAAAAAAAAAAAAAAAAAAAAAfNYfL/k4fq31gfiV+3QAAAAAAAAAAAfiCH7feHh8Q/qcAAAAAAAAAAAAAAAAAAAAAAAfFIH2T+LYftj+Z4fUX0cAAAAAAAAAAAAAAAAAAAAABX/EsP1Y+Bg/V38eQ/U7uAAAAAAAAAAAAAAAAAAAAAAAA/EwP2w+VQ8k/QgAAAAAAAAAAAAAAAAAAAAAAA+KQPs78WQ/a78kQ/Sf0cAAAAAAAAAAAAAAAAAAAACAPyL/AEFDrvzZD9bvxSD9ppwAAAAAAAAAAA/FIP2k+cA+ev0dAAAAAAAAAAAAAAAAAAAAAAAAD8U/2sD84foYPpD8Wg/a0AAAAAAAAAAAAAAAAAAAAAA8r/LoP2O/KsP0I6oPyk/YgAAAAAAAAAAAPxSD9pPnAPnr9HQAAAAAAAAAAAAAAAAAAAAAAD4pA+1fxKD9tvynD789cAAAAAAAAAAAAAAAAAAAAAjD8kvu4PT/AMsQ/YL8XQ/Y/TAAAAAAAAAAAAAAAAAAAAAAAD4oD508xD9de7AAAAAAAAAAAAAAAAAAAAAAAPikD7W/IwPX/nYP2dAAAAAAAAAAAAAAAAAAAAAPxmDP99D70/JYPo3yQP1vAAAAAAAAAAAA+Og+V+AD9ZfRwAAAAAAAAAAAAAAAAAAAAAAAA/FP9rA85/JQPSPqQPsUAAAAAAAAAAAAAAAAAAAAA4IPwF++Q9L+gw/NLWD73+kAAAAAAAAAAAA+Og+V+AD9ZfRwAAAAAAAAAAAAAAAAAAAAAAD4pA+1vyaD035wD9plAAAAAAAAAAAAAAAAAAAAA/HwON+iQ+6/ypD6F8WD9ewAAAAAAAAAAAAAAAAAAAAAAAAPO+lDfAAAAAAAAAAAAAAAAAAAAAAADLA1GB5R6mE4AAAAAAAAAAAAAAAAAAAABxIVQO0eHl/rAKAAAAAAAAAAAAB59vB0YAAAAAAAAAAAAAAAAAAAAAAAAHL9QBzIZHegAAAAAAAAAAAAAAAAAAAAAFEONAudoHHsDswAAAAAAAAAAAAPPt4OjAAAAAAAAAAAAAAAAAAAAAAADLA1EDyf1ALIAAAAAAAAAAAAAAAAAAAABxYUwOynDyz1cHgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAcGHl8ge1bwAAAAAAAAAAAAAB8MB2f1oAAAUfiIPuScAAAAAAgD5O+uAAADk/Mw94A4nysPooAAAAAAAAAAAAAAAAAAAAAAAAAAAAAaHifMB1vs4KAAAAAAAAAAAAAcuH5zfoeHZAAB4mHL/SgAAAAAAB5JWD2QAAPHw3PRAPnIPWuzAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA5HyUIfYw6sAAAAAAAAAAAAAA+NqofaYAAIHw39lBtgAAAAAB8hB9duAAOfDx/3wJOdDwP6eAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPxTD499XDxP8AoOD3UAAAAAAAAAAAAA/BMPa/2BAAAPOfwZD99+1AAAAAAPgoPmv9iQAAD4U/GgP6ggMv+cYP6LegAAAAAAAAAAAAAAAAAAAAAAAAAAAAAyg/lq+ig86+gw/eIAAAAAAAAAAAAA8pD+av+j8PawAA/NgPJv2AAAAAAAAP58/2aD3IAAP5/Q+lv1sA+Cg+aP2MAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA/Kj80A+hPm0P3e+tgAAAAAAAAAAAAAPxTyQ/cIAAM4P56/20D3EAAAAAA+SvzrD9yQAA+VA/AD+pUNWMP5zP3xD0YAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA/FMMn9wg/mc/VAPs/wDKQOZ/U8PmmMPrz8ug/Zf8ig+5vkYPmn2oP1j+DgwfAg/Y78ng8n8QD64/YED5RD82fXQ/TT8lg/Wn8lgAP00wQ/M3sw/WrqwPwnD9GfVA/L79nw/PO6HQ/jQH9QQB+HofdH20AAAAAAAAAAAAAAAAAAAAAAAAAAAABlB/LV+/4c9+FIf1NfGgfn37gH6k/jqH6jfL4avuofnR+m4flPyofoT9lh+LP0cHU+6h+WnUB+fH9H4e1wB+VnzyH6UcMHnnvIfBwHa/rmH5j/Lofb36AAeeB/Pd/SoH5zXQ+8fxuD9T/wAWg+lv1sA5IP5vf6bwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPyo+Pw/oX/nyD6k/UQPyj8tD9LuODw/9KA/HH9hw/OP3oPJvkEPUP1nD5X8dD57/aEPzC+fA849ZD9wgPnX8vw6X9mA/GD9QQ/OblwPv4PRPyyCf9Yw9NA/Gf6ZD68/JYP13+Qw88+iQ/AD+pUNUD8fvZw/RoAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA/FMPkf1YOc/oED8PvdQ6L44D9FPzoD9CvxzD+gr+fUP6YvEA8n/IMP2L+YQ+GP1ICx+Y4fst+N4fb/wCwIH84gfUn6HBvfzgB/SZ8uB8OfP4f0H/zqh+yHwaHt365gfzeh+7/AJuH4Af1Kh+KWsH2D+NAf1BAH5Nh0P6hgAAAAAAAAAAAAAAAAAAAAAAAAAAAAZQfy1fTgeU/o4H3P/OqH7qfkIH6VfnOH6K/nOG39nh8yftIHyp8gh8G/wBQwfytfVwfq3+LYfTftYfkp/R+Htfy2H4A/u2HvX5lh4p+vgfLn5JB98/Qgfmn+vYfg7/RsHsHyoH5Yfv0H4e6wfsD/MAH9Cn5Jh9LfrYAB/ML/SaHWgAAAAAAAAAAAAAAAAAAAAAAAAAAAAflR+bAfQ/jYf0Dfl2HNfVgflD+3AfiB+2wfg7+7Qfih+8wVfIQ/JP9LAg/KYP019sD8cv2vD8nfdw/cID8BewD9NPoEP5av3/DgfjkPiT+iAPwl/RAPFuUD9yAP5/f1fDuP5wA/pU/NQPFP1BD8AP6lQ1QPzb8gD9gwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAD8Uw82+zA/Jb+lgP5w+yDreoD9qv5rg+x6QbHlgfvj/ADoB9P8AymH6beKB55+7Qfi9yAfuh+B4e3/sCB8BB+R3sofsv/OOH9KnmYfgv++wdB/OSH0ZOH1d+tIH824fvT5mH4Af1Jh+KmsH2D+NAf1BAH5PBt/qMAAAAAAAAAAAAAAAAAAAAAAAAAAAABlB/LV+xYfD/pIfa/4hB9CgfoD54HjPlIebfRAfa2oH5B/X4fn9/UqH8tX7zh9cfywB++HsAfzX/wBH4e14ofip8XB+yHggeKfvYH4q+QB+/f5Bh8JewAfsOH0h8nB+XX77B+HuoH7B/wAwAf0KfkmH0t+tgAH8w/8ASSHXgAAAAAAAAAAAAAAAAAAAAAAAAAAAAflR8PB+qH4hh/RR+LYcT34H72B/NB9nhz2wHy3/AEsB/PB6yHif2sHV/CIf0X/k8Hyf/QiH4n5ofuEB8X/joHUf0Ah/MP8Av+Hon88wftN9eB/Ln7yG17WH7WAfz7/rSHafzgB/Sl+aweKfqCH4Af1Khqgfm940H7DAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAH4phk/twH8wP7Gh8XcQH3L6aH2j/ADsBwv7VB+D36qh9qfziB+uH5dh+qXigef8A7rh+ff46h+kv5yB+gf7Agfm0HOflYH9B/wDPCH9EP4Nh331cH3T+GYfYfvYfU30AB+CofpvYD8AP0UD4r9UD9K/xND7xA/Wn8iw+0vuUAAAAAAAAAAAAAAAAAAAAAAAAAAAADKD+Wr9/w8v/AB0D+iz+csP0n7UPunzgP59/0yDx743D+lr8rw+W/poPzk/qVD+Wr96Q+wf55Qt+oh8J/wBH4e1+MB8i/Eoeq9KHin3GH4vfeoemfRAfjX+oIJ+moHlYfgT/AElB+Hvy+H17+fwftN+focZ9Eh7X+jofzW/06goAAAAAAAAAAAAAAAAAAAAAAAAAAAAH5UfH4f0L/wA4YfdnMB8jfoeHVfe4fhb8hh/QL/PgH3/+sgfzG/qaH59/eQdX8Jh/Rb8eB+EX6YB8CfVIfuEB+eeUH47/ANIAfzffviH495Afaf2aH5I+nh9Z+7B9VAfir9bh9EfzgB97/MoSfsiH4G/ewWP1BD88/Ug/SUAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA+RAq/YofD3bh9Gfn2Hn/1WH1v8mhzP2KH5zfcIel/BAed+qh61zIYf2cDfgMOI9yDV+rgPzyDy77CD234CD7P+MAAPsXVD4Jph91e1AfnYHj/ANkB+FX6nB9O/Kwfa/x0AB95/wA9If0R9YAAAAAAAAAAAAAAAAAAAAAAAAAAAABWD83PuENz4CD7780D4k0Q/Qfug/N37JDkPDw+/eGD85/dA8w/RkPzm+2A9U88D89Po4PHvu0O08uD4C1g/QjwcOQ6UPBQNP8ARkPjb5jDv/0hCUD+df8AcoPybwA+5toOv5APNAPS3B8cftMAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB80+Vh91fIgZX2GHwL5CH0N9vh85+HB9+/nMH1T7gHxV4UHtvahueMB95gfEHjQfRWeH2UB8JeGB9S/WQfm59nh8YAH1AHq35/BzX2aH0eB8WfD4fqR/OAH6/fTIfHn6EB8QgH3CH4O/uqHrAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAGcHxN6wH4J/0/AAAABwHgYfXYAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHgDw/NP0EP1nAAAA+Tg9n9KAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAACP4ND6i/mpD+pnRAAADmg+T/ALVAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAKocV34AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAc2gdKAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAEfBh3coAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHwx9JXPe/GCYVIAAAH3P5tFdJUDj/AF4PF/agBPk+sAAPm5r6REwAAM0PNfXGvF/YgmE/Nxr0gT8W9paPN0z0gAA4BnNeyC84yWeuksAAOT6wAAA8v0GvQBPL8Sa0vbw8r5lnt+JL869ra8j5Zlj31AJgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfnL+gFztfDKfEFyAAAB+jvIZX5/SpZPrwcjdDm/YUfJX0CxvJh7X80h3OgHrXjCGe1guKn4p9CtfNH0cHluKzouCC974jzX0QPFOeZ6REjm/bwteKB599WM+eazPo8hnnjXE+tM+Ufdmer4cPybrqWbpB6OS68ofOvr9LyT2IPLvVA8K+mUFFPwT6JaBMAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAD4m+xKXBcUHzSXIAAAH6OplfzVp2qmiHtGfL8S7hrgvpdHP8AjTNrzhn1Z86ydFksoduFDtkeX/QofNHqrOS9vR5fkhdtM8u22ex+fSafVh4l3DPWvAET9sziMRn0oQ4vkul7J56B9LgRS/NPFLXsWkj0clkYfMXstLmPYkfMf06HzR9LgeBB6/vJgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHgGzS84+gg+DS5AAAA/R0yv5t27Wz5gH1X8ro9kww4j6ZR8q+6M4Lg2fVfzpJtazO815b0D5p+oKXy37EHL+1I8vAs5rPONZntnnEmn5Oz2TDD2L5vR778as+wvBA980Jb/AJcpfQ92X8+/S7S/P4en+HM9J0g67Zlx/OFL3Oyn4f6815n7qj5o+lwp/PLPpLMl3ZgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA5vxmlw32GHx751SyRoAAP0dMr856Fq/wAeq+Gha7IMtgYMrOmy0YGyz1bw9DvawnE/MOOpegegJ+J12vR3okrBzPogeHbbO884Ra7IO28xD07ywPU/Lw4X6FCXzIOX9GZT55HrnIp+Z6VLubKPOvaw4zyVlj2hHB4LPb7Evy/1BrjNAfR+ai6noUwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAoBhdKHyV49pINAH6OmVgAAAAAAAAAAAAAAAAAAAAAAAAAAAAABzvRAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB5r8a3P0F6En8ZfSVL0r4jQFIA/R0ysAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAD5d8E0n6G9Hl/HP1DS6b4xQFIA/R0yswA2pQAAAAAAAAwMhrs3pgAAAFQKmsGLtAAAAAAAAAAAGRrgAEQSwgyyAgZ2kEUYFkKFsJAABtMLwDUB4AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfAvOax9DfSWdS8Cn8q8VrAAAfo6ZXD8b0vp7sJaQBYAr2AAAAA+ZPU6XnP0citZTAAA8ha86+o2fOf0YitZTAACKUK1gFrBZAqWg819LYCPMfG7X1B86IT6BDd8zT5GRrsOJDyr64Dynzpn1oQxgfMnRWum9Wk+Ye8o9iIYAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfnEax9DdxFavDB8ylyAAH6OmV+JatJaIczx7X1n8hp9b6aHmv0oj5s+kwET+UPTLVrARFaZT7NEfqKD54CH29njGyEMIYn0mj5rs0T9AjP5hnqPCo9j8ODf6oKXqyDIT8axLXdbslvgmQeyI8T69nqXOy/Kvd2vK/P2fShLMcPDPoVr5i9LZjXw9iJYAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfnEax9DdrFfL/wBzs+U/KWgaAP0dq5X8pdzcmcP6H+dkes8ujTsh5f8ARbPnr6OQc2n4zjXP0R8tp+h+vI8Pho+lCGfOjXs/zqz1fxBnpvpKPIPcw8gwGfTPzTJ9L/MzPcvBQ9Yrh6j4Uj1bfTPm9rW8spep92nZ45Hs8afLeOUvpD5vR9CXE835tpe8cAg7wOGy2ZnLtdJnj+oyGAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAH5xGsfQ3axXyJ6ba+uvhuXzhUgH6O+EZ16r0SfzU619J/NsnSekM8j9xR8q9hS6nGT978kR0fDh6D4Iz0T2RHyT9CM57sEdL86h9FfHbPqX5zD0j0hHzJs0vVfM0/pT5kR1oze8sD2rx0PU/NA9cpI9EEz5opfQXzqEnvAeZXwy2B6z46H0DxsvhvPrX1PLDAPLPNqX0Pqy+K5Kl7ESwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA4Ea3bifLDXZZqfPjQB30ScoEQEsQLIEUoUrIOgCywHNBYQjuBHIFecFjCSMJIQrXwo2wdGErQo6AVXhPSCZ4CBKARBLXAsBWkCpoAxQcBmaIOAAM/QAARoPAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA//aAAgBAQABPwCqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq+O3/AAVU+DnwC+LPiL4Q+JPh34y1HUvDc8cNxc2ItvJkLxJICu+UNjDgcgcg1/w+0+AP/RKPiB/3zaf/AB6vhv420/4m/Drwt8SNJtLi1sfFei2erW0FzjzYo7mFJVV9pI3BXAOCRnvVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV/wUi/5Pc+Kf8A2ELb/wBIoKr9k7/k1f4Nf9k/0L/0ghqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/4KRf8nufFP/sIW3/pFBVfsnf8mr/Br/sn+hf+kENVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVXxB8caD8MvA2vfEPxQ1yNI8N6fNf3zW1u00iwxIWcqi8nABPoOpwMmvi//AMFtNZlurjT/AIEfCSzgtlJWLU/FE7SyOMnn7NAyhDjB5lb3FeI/+Cp/7bviB3Fv8WbXR4X6wadoNkgGCDw7xO46f3q0j/gpv+2/o90LlPjhPdrkbobzRrCZGA7cw5H1Ug+9fs/f8Fotfj1S10H9pLwPYT6dKyode8OxvFLb+ry2zMwkHOSYyhAHCseK8H+MPC/xA8L6Z408Fa7aaxoesW63FjfWr745o26EHseoIOCCCCAQRVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX/BSL/k9z4p/wDYQtv/AEigqvEP/BRX4c/sn/sufBnwXpGmL4v8fz/DnQZDpMVyIodORtOhKPdSAEqTkERAbmXklAVY+N/+CrP7aXjC9efTPiFpvhW1Zyy2ei6LbhF9AHnWWXA/3+e+a8Af8FYP2y/Bl/FPrfjfSvGFkjfPZa1o8ChhnnElusUgPpliB6dq/Y2/4KC/C79raI+GRaN4V8e2sBln0G6nEi3SKPmltZcDzFHUqQHUZOCo3VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVeItB0vxV4f1Pwvrdstxp2r2ctneQt0khlQo6/irEV4I/4J2/tTfEjxx4g8JeE/h3cR2Ph7V7rTbjXdVb7DZM8ErRMyO/Moyv/ACyV8VoH/BEH4n3Nisnij46eF9PvMDdDYaXPeRg98O7RE/8AfP5V+05/wS2+OH7O3g29+I+m65pPjfwzpaGTUptOieC5soh1leB85jHGWRmKjJICgtVf8EZ/2itU0jx5rH7NfiDU3l0jXrWTU9Ailcn7NeQjM8cY7LJFukI6AwZHLnNVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX/AAUi/wCT3Pin/wBhC2/9IoKr4VfBv4t/tAeK08I/CzwfqfijVxEu9YcBLeJQFVpZXISJAAFBdlHAA7CvGf8AwSz/AGz/AAb4fl8Qn4b2muR26F5rXRtTiurlQB/DCCGkPbbGGPtVzbXFncS2l3BJDPA5SWKRCrIwOCpB5BBGCDXhDxd4k8BeKNK8aeD9Xn0vW9Eu0urG8gbDwyocqR2I45B4IyDkE1+yx8ddP/aS+A/hT4t2kcEF3qtr5ep2sJO21vYiUnjAPIXepK552sp5zmqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr44/tPfAr9nLTY774u/EPT9GmuEL2unrunvLkDvHBGGcrkY3EBQepFfED/gt18PtOuZbX4YfBLXNcjViqXWs6pHpwOP4hHGkxIPGAWU4POOlePf8Agsr8WvG3hrWPCS/BvwXb6dren3FjdLPPc3DeXMjI2CGUfdY9QQf0qv2CdbufD/7ZHwlv7RmDy+I4rQ7Tg7bhWhb8Nsh/+tVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX/AAUi/wCT3Pin/wBhC2/9IoKr/gm/8FPDvwd/ZQ8FXmm2MI1fxvpsHiDVrwKDJcNcoJIVLddscLooXoDuPVmzX/BZb4BaF4H+Jfhn42eFtLhs4vHMU9vrKQJtV7+DaROQP45Y5MEjqYSx5Yk1/wAEQvG11qHw9+Jvw7mlcwaJq9lqUCs3AN3FJG+0df8AlzUnt8w9aqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr9vr9rOL9k34LSa7oohn8Z+JJXsPDlvKAyxy7cyXLqfvJEpU7f4maNTwxI8aeNfFvxG8T6h408c+Ib3W9c1WYzXd9eSmSSVj79gBwFGAoAAAAAr4Lfs+/GH9ofxE/hf4QeBr7xBd26q11LHtit7RTnDTTOQkYODgMQWwcAkVoX/AARZ/aa1G1Fxrfjn4d6U7KCIDe3U8inuG22+0fgxqv2KP+TufhB/2OGn/wDo5aqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/4KRf8nufFP/sIW3/pFBVfsnf8mr/Br/sn+hf+kENV/wAFuLuyT4JfD6wd1+1zeKpJIlI5KJayByD6ZdPzFV/wQ3064fWPjDqo2iCG20eA5zks7XbDHbgIc/UetVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV/wW18Qalc/HvwJ4UlaT7Bp3hD7bAC3yiW4u50kwPXbbRZ/D05r/gklpvhGz/Yw8P3vhxLf+07/AFfUZNfaPBY3a3DIgfvn7MttgHsQe9fF74o+Fvgt8NPEPxP8ZX8Nrpfh+xkuX8yQIZnA+SFM9Xkfair3ZgKr9ij/AJO5+EH/AGOGn/8Ao5aqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/4KRf8nufFP8A7CFt/wCkUFV/wS5/a08F/FX4I+HvgnrevW1n478EWY0+PT55Aj39jEMQSwAn59ke2N1HIKbiAGFalqWm6Pp9xqur6hbWNlZxNLcXNzKsUcKKMszMxAVQBkk8Cv8Agp7+1h4e/aW+MWn6H8PdRN94N8CW81rZXi8JfXUrKbieP1jIjiRT3EZYcMKr/gjt8KbzwP8Asy3vj3VbQw3Xj7W5Lq2LLhmsrdRDESOvMi3DDsVZSOuTVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX/BZT9nbW/G/gPw78fPCemyXk/gpZbLXo4Y9ziwlYMk+BzsikDBsZwJ9xwqsRXwX/aU+On7O95d3nwa+I+o+HP7Qx9rt40juLecgEBngmV42YAkBiuRng18Yv2mPj/8AtFT2kPxb+Jer+JUt5d1rYlUgt0kPG5beFUjD4ONwXPOM1rGj6r4f1a90DXdOudP1LTbmS2vLS5iMctvNGxV43U8qysCCDyCDX7GN1b2f7WvwemuZNiN400yMHBPzPcIqjj1ZgPxqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr/gozdW95+2v8VZbaTeq6pDGTgj5ktYVYc+jKRXw38Aa/8VPHmhfDjwqsDax4ivUsrFZ32I0znChm52gnAz261q2k+IPB/iC60XWbC90fWtHumiuLedGhntZ42wVIOCrKw+oIrxH8Uvib4x09NI8W/EbxPrdjGRstdR1ee5jXGMYR3IHQduwqv2O/2SvG37WfxQtPDGj2tzaeF9PlSXxHrYj/AHdjb55VSeDM4BVE5OcsRtViPC3hjQ/BXhnSfB3hjT47HSNDsorKwtYx8sMMSBEUfRVAqqqqqqqqqqqqqqqqqqqqqqqq1C9h02wudRuAxitYXlcKMnaoJOB64Fa1/wAFtvgXBLt8O/CLx3fR9mvXtLU9B2SWXvkdfQ98BP8AguL4SLqJP2d9XC5+YjxHGSB3wPIrwL/wWa/Zk8RXMdl4y8M+M/CbO2GuZbOO9t4xnqTC5k6YPEZ/x+GXxa+Gfxn8Np4u+FnjfSfE2lM2xp7CcOYnxnZInDRvjB2uAcEHHNV+17+3Z8Lf2Pm0TS/Fmjavr+u6/G81tpumhFMcCMFaWR3ICgnIUAEkqegGa+A3xw8EftGfC3R/iz8Ppbr+ydXV18i8jEc9rLGxSSKRQSAysD0JBGCCQQaqqr9rv9vn4U/sgaronhjxToes+INf1u3N2mn6aI1+z2ocoJZHkYAbmVwqgHJRs7cAn4KfGLwX8fvhfoXxa8AXFxJouvQs8KXMYjmgdHaOSKRQSA6OjKcEg4yCQQTVV+0n+0X4B/Zc+F9z8UviEL2ezS5js7OysY1ee9uZAxSJNxCj5UdiSQAqMeTgH9kX9tD4aftiaDrV/wCCtM1TRtV8OSxJqel6kELxrKG8qVHQkOjbHHYgqcgAqTVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVapqemaJpt1rGs6ja2FhYwvPdXV1MsUUEaDLO7sQFUAEkkgACvjN/wAFlPgT4E1ifQ/hZ4O1f4iSWxKvfrcjTLJ2HBEcjo8jgY+95YU8EFhzWk/8FyLwXW3Xf2b4WtmYfNaeKirxjucNbEMenGVr9k/9rv4d/te+E9T8TeAtG17SpNDnjt9RtdVtlTy5XUsBHIjMkgwp6EMOMqNwzVVVft+/t+/tPfAX9p7xF8M/hn400/T9A0+zsZLe3m0e2uGVpbaN3O90LHLMTyeKqqqqqqqqqqqqqvBv/BVv9nbxv8d7X4K6ZpfiGK21PVBpWm+JJY4xaXNyz7EO0NvSJ3wquR/EpZVGSKqqqqqqqqqqqqqqqqqqqquba3vLeW0u4I54JkKSxSIGV1IwVIPBBHBBr4//APBG74VePtYvPE/wU8ZT+Abm6YyNo89n9s08OeoiwyyQKTzjLgdFVRgDTv8AgiL8YpdQ8rV/jR4NtrHd/rra0uZ5cZ6+WyoM4ycb+vHvX7L3/BMP4Dfs5axZ+NdVnuvHnjCxYSWuparAsVvZyDBEkFsCwVwRkM7OynlStfHX/gkT4S+Nfxe8VfFdfjRqOhN4o1Br2TT4tCSdYZGA34fzl3ZYFug+9+NfDv8A4I16D8OfiB4Y+IWm/tB6nPd+F9ZtNUt4m8OooeS3mWVVJE/AJQDNVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVfHf/AIJHeHPjj8X/ABV8Wrv44ajpE3ie/a7exj0FJlgJUDaHMwLdOuBXwD/4JI+F/gX8YvC3xbi+Nepa3J4YvftUdhJoaW6ztsZQC4mYqAWB6HOMd6/as/4J7/Az9qy5bxPrcN34Z8ZiNYx4g0pVDzqowq3ETfLMAMAH5XAAG/aMVr3/AARD+LVvfPH4X+NvhHULMMdkt/YXFnIR2JRPNAPXjca+E3/BE3wdpWoQan8aPi7e6/BE+5tK0Ox+xJJjs87s7lT3Cqh9G7j4b/DH4f8Awg8I2fgT4ZeE9P8AD2hWIPk2dnHtBY4y7sctI5wMu5LHuTVVVVVVVVVVVVVVVVVVVVVVVVXi3/kVNZ/7B8//AKLav+CXPgLwR8R/2sdN8N/EDwjo/iXSf7FvpjYatZR3cDSIi7WMcgKkjPGRTfs5/s9NAbVvgR8OzCU2GM+F7PbtxjGPLxjHGK+MX/BMr9kb4tabcpZfDi38E6vIreRqXhj/AELyWPIzbj9wy5xkFAcZAK5zXjvwd+0X/wAEwP2hrS50LxEwEyCfTtRgRhY+IbEON8U0ZPUHCvGTuQlWU8o5/Z2+Ofhb9pD4P+H/AIueElaC31iAi5s5H3PZXKHbNAx4ztcEBsDcu1sAGv2yv2Cfh/8Atiz6FrWs+KdR8M+INAie2h1Czt0nWa3ZtxikjYjOG3FSGGC7ZBzx+z18B/Bv7Nnwo0f4SeBpLuew0vzJJLq7YNNdzyMWklfAABJOAAMAADtmqqq/bJ/4J9/Dz9sPWtD8W6v4s1Pwx4h0S1+w/bbS3S4W5tN7SCN42I5VnkKsGGN7ZDcY+BHwV8H/ALPPwp0H4ReBvtL6VoUThZ7pw01xK7tJJK5AA3M7McAADIAwAKqq/ag/Zs8E/tV/Cq4+Fnje7vLGEXcd9YX9mFMtldRhlWRQwIYbZJEKnqrtgg4I/Y3/AGJPAX7HGia5beHPEOoeIta8SSRHUdUvIVhzHFu8uOONSQigu5OSSSeTgACqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/4LS/G7XfDPgfwd8DfD99La2/i2SfUNbMbFTNb27IIYSe6NIzOR6wp71/wTg/YN+DGlfBDw38Zvid4K0jxf4q8YWw1C2GrWyXdvp1q/wDqUjifKFymHZyNwLYGNvPiT9nb4BeMNPOl+Jvgp4H1K2KlVSfQLZig/wBk7MqfcEGvgp8AvhX+zv4e1Lwn8IvDX9iaRquqSanNa/aJJws7xxxttaRmYLtiXCkkDnGBxVVVV/wVg/5Pc8Yf9g/TP/SKKqqq/aO+L3xU17/go8ngTV/iP4juvDek/EnTLex0Z9Tl+x28YuIOFg3bB7nGT3qqqqqqqqr/AILNfFj4n+Ar/wCG/hvwN8QvEPh7TNbsdSbUrbS9RltFvCjwBRJ5ZBcAM3B45r9gmaW4/Y6+FM88ryyyaCjO7sSzMZHJJJ6k1Xgr/gkl8DfBPx5s/jFY+LNcn0jStVXVNN8MPDGsNvOj+ZGpmHzPEjgEJtBwoDMwzmqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqvFv/Iqaz/2D5//AEW1f8Ehv+TytN/7F/Uf/QFqq/4KyfCvTviD+yJrPilrVX1XwHfW+qWUoX5wjSLBOmeu0xylyOhMSntX/BEH4gX8tt8TvhXcyu9nA9nrFmmfljkffDOfqwS3/wC+TX/BYb41/F34O/8ACo/+FV/EjxB4U/tf+2/t/wDZN89t9p8r7D5e/aRu2+Y+M9Nx9a/YM8XeKPH/AOyJ8OfFvjXxBf63rWpWNw13f3s7SzTsLqZQWc8khVUfQCtc/ad/aU/ZU/bdbwR8Tfjb4z1vwl4W8WJHe2mp6pLcR3WkTEFZGQkhm+zSq464cD0qOSOaNJYZFdHUMjKchgehB9K/aV+Llr8B/gN42+LE8iCXw/pMklkr4xJdviO3Q57NM8Y/Gv8AglN49/aW+Pfxp1jxR8Q/jV411vwh4K0zdPZXuqyywXV5c7o4EdScMAizSd8Micc5r/gr/wDHL4xfB/xJ8Mrb4WfEzxF4Vi1Ox1J7xNKv3txcMjwBC4UjJAZsfU1+xV4o8R+Nv2VPhn4r8Xa3eaxrOp6Ikt5fXkpllncuwLOx5J4HWv8Agpd+3/45+DfieD9nr4DXQs/Fc9tFPrOsxxLNNZCYZitoEYECV0KuXIJCum35jldG/wCCen/BS3x7psHjvxF8bDpOsToJVs9b8b6g1/H/ABAExJIiHnp5gwTziv8Agnp4K/bZ8Daj448OftW+J9a1DStMS1j0FdTvo9SNzI5cyyx3eWkZFVY12M3Bf7qkGv8AgpT488afDT9kXxR4u+H/AIp1Lw9rdrfaekF/p9w0E0avdRq4DLyAVJB9jX/BIj4vfFH4v/Djx/qXxR8f654qutP1u3htZtVvHuGhQwElVLE4BPOK+Ofxr8Dfs9fDHWPir8Qb14dL0mMbYYgDNdzNxHBEpI3O54AyAOSSACReftQ/t/f8FBfiBfeFPgLLqvhXQLY7nstAvjYQ6fAxIVru/wDld2IU/KCA2G2x9RWsf8E8/wDgpb8P9Pk8Z+FvjSdW1SJDM1poXje/jvWP3iMzJEjNntvOSO/Gf2M/+CovxD8PePLb4IftdTvNDJef2dF4hvLYW15pd0G2CK9XChoww2mQqHQ5LlhkrX7T/wC1Z8W/gz/wUnuPD158bPFem/DfRfFGgT6ppi3sr2sWnyW9nNdp5C53KUkl+UAk545rV/jD/wAFCv8AgoZrWq3f7OMeofD/AOF9ldPa213DqH9liYA9ZrpT5sshUgvHDlEBAIJOW+KnhH/go5+w7f2njrxH8RvF0GmT3e1NXsvEMmqafNOedk8UhIy2MgTRgN2yQcf8E/8A9sZP2u/hZd3niG2tLHxv4WmjttdtbYFYplcEw3MakkqsgVwVzw8bditVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX/BaD4FeIvF3gbwj8cPDeny3kHg1riy1xIkLNDbTtGYpiAOEWRWVj281T0BNf8E4/wDgo/8ADPw58OdD/Z8+O+sp4cutAU2uh+ILo4s7i3LEpDO//LBkB2h2+QqBllI+bSdY0jxBpltrWg6rZ6lp95GJLa7tJ1mimQ9GR1JDD3Bqv+CqfxK+IHwq/ZitfE/w18Zat4a1dvFNnbte6ZdNbymJopyyblOdpKqSPYV/wSX+KPxF+Lf7OPiLxH8TvG2seKNUtvG11aQ3mqXbXEscC2Vk4jDMSQoaRzj1Y+tV/wAFPv2lPj/8L/2q7/wp8OvjH4s8OaPHoljKljpuqSQRK7I25gqnGTjmvDM8114b0m5uJWkllsYXd2OSzFAST7k1/wAFYP8Ak9zxh/2D9M/9Ioq+Iv7Sf7aH7efjzXfBP7FsOpeGfhtoNybaXxDBcjTnvGHSWS7OHjDD5lhh+cKwLg5+X4nfsmf8FJf2c9KuPiqvj3xJqdvpqtc6hf8Ahvxdc3Mtqi/MzyxsUkdOCWIV1AyWwM1/wTk/4KQ+IvjT4jt/gR8eru1m8UXELHQdfSNYP7SKKWaCdFAUTbQSrKAHCkEbsF/2+vAf7WPj3wf4Vs/2Ttf1PStWtdSmfVnsdbTTWeAx4QFmddw3du1eNfDHxy0/9pCbwl4w1S8m+K6+I7e2ku5NRWWb+0mkQRP9oDEbgxT584HXPFaN+z1/wWIh1ixm1L4i+KWtI7mNp1bx5CwMYYbuPO54zxX7f/ijxZ4K/ZA+IvirwN4o1Hw9rmmW1pLaajp85hnhIvYNwVhyNylkPsxr4Sf8FKfi98Ov2UdW0ufxhf8AjL4seJPGM1roN1q8rX8un2YtbXfJsbO7EjYiQjDPI5wwUqf2Ifg1/wAFBrr9p/wP8bPjza+PH8IEX7ajNr/iFcosthcLErWLzeai+a8WE8oBTg4AXj9vjWf21Ley8GeF/wBj+xMkviKe6g1m7trWI3FntWNom86c+VBGwMoLkAhgoDZYCp/+CcH/AAUa8a2n/CQeMf2oLUau372K1vfGOqTvEx/hLrGyIRgfc3AcVoH7U37bv/BPr4w23w/+Peraz4o0T5J7nSdY1E6gl7ZsSomsrtyzp0bAB2hgQ6ZBA8FeMPD/AMQvB2ieO/Cl6LvR/EGnw39hPjBeGVA6EjscEZB5ByD0r9qf9qD4f/sofDGf4geNWe8u53Nvo2jwOFm1K6xkIpPCIB8zueFXszFVa1+On/BSD9v/AMUahb/CTUdd0jQLWbbLb+Hbz+x7CwVuiTXW5XlbbyVZ2J5IQDij/wAE8f8Agpl4TDeIfDXxakl1OMEqmmePLqG4c9SA7+WvJA6sBnH1r4UeE/2tZv2EtV8PeO/EuszfGvUNJ1JNPkurqGC6sZi7pap9oTALbVR/MZicv97AFftj/D79r7wDqXheH9rTxFqmq3d5BctohvtdTUjHGrR+dtKu2zJMfBxnA9K+CPwP/wCCpPiP4TeGNb+DfjrxHZ+CryxV9Fgt/GcVrHHBk4CxGUFBnPGBX7I/hz4y+E/2efCfh/8AaC1K6v8Ax/afbP7XuLrUFvZH3Xk7Q5mUsHxA0Q68AAdq+DX7TX7Qutf8FFbP4c6t8Z/F134Yf4iX9k2kTapI1sbdJ5gsXlk42AKoA6cCq/bT/wCCofjlvHt38Bf2SYw97Bef2ddeI4LcXlxeXZbYYLGLDLgMdvmEMzN9wKAGbS/2AP8Agpn8VtOTxl40+M8uk6hOnnJZeIvG1810u7kKFgSVI+g+Tcu3IGBggP8AtEf8FBP+CenxDsfDfxi1fVPEmi3WZIrDX79tTstShUgMbW7JaSJlz0BG0speMggH9nv49eBP2lfhZpXxW+H9zIbG/wB0VzaT4E1hcpjzIJQOjKSDnoysrDhga/4K1/Er4j/Cb9nzwx4s+GHj3XPC2py+MoLGafSrxrdpoXs7tyrFeSN0KHHSvBX/AAUL/aU8bfBD4b/s8/AA6t4y+M+sW98fEGv3UP2m5tR9tn8lEMo8vcLYRM88mURSvO7cU8efsY/8FUrbTZPiLqPxH8Sa/qsCvO9lpvjqeS+h/vCNN6oTgcJExJAAAJwK/wCCe/8AwUe+KI+KOkfAD9ozxDca5Ya7cjTtL1nUlxfWF8TtjhnkOGlR3/d5cF1dly23OKqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq8W/8iprP/YPn/wDRbV/wSG/5PK03/sX9R/8AQFqq/wCCh2pWek/sWfFa6vZQkb6MsCkkD55Z4o0HPqzqPxr/AIIi6RNP8YfiLrqk+VZ+GobZht43S3KsOc8cQtxjn8Of+C5//NEv+5g/9x1f8E3v+TI/hX/2D7n/ANLJ6/4LW/BD7Lqvgv8AaF0ey/d3qNoOtSIuAJEDS2rn1LL56k+kaDPTH/BOv4y/8Lt/ZK8Fazd3Hm6t4dgOgaoS2T51oFRGJ7s8BgkOe7n61/wWt+MzaR4G8F/AjS7tRP4gu31nVUVvmFvB8kCsP7ryu7fW3Ff8Ezfgafgl+yj4bbU7Iwa740J1/Uw64ZROo+zoc8jbbrFlT0Znr/guN/yNXwj/AOwfqv8A6Mtq/YD/AOTNPhL/ANi/H/6G9f8ABTHwb45+CX7cZ+NsumSXWma5d6dreh3VwjNBNLaRwo9uzdMq8AynUI6HuK/Z3/4KG/s0/tDWVhZWPjW18LeKrpVWTw9rswtphKRykMrYjnBOcbDuIGSq9Kr/AIKwf8mR+MP+whpn/pZFX/BEP/klXxK/7GC1/wDSc1/wW2+KOpXXjnwB8Fra7dNO0/Sn166hVvllnmlkgiLD+8iQTY9BMfWv2Bfgno3wO/ZZ8D6NZWEcWq6/psOta1N5YWSa6uo1kIc9/LRkiHtGPeq/4LSfBXR/CvxL8H/GrQrBIH8aWs9lrBiQKr3VqI/Llb1d4pAn0gH4/sGfEq++LX7Ivw08YatdG41EaUdPvJHfc7yWkr2xdz/eYQhznn5s96/4KZWB1T/goD8QtMWURm7uNGhDkZ27tLshnH418PPAXhn4XeBdC+HXg3T0stF8PWMdnZwqOQiLjcx7sxyzMeSxJPJr9qHwJo/xL/Z1+I3gvW7aKaC/8N3pj81AwinjiaSGQZB5SREcHHBUV/wRd8QXun/tPeIfD8cjGz1fwdcGWMPgeZFc27I5HcgNIv8AwM1VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVXtlZ6lZz6dqFpDdWt1E0U8E0YdJUYYZWU8FSCQQeCDX7SP8AwR0+Gvj++vPFXwB8Sr4E1S4ZpG0S8iafTJHPaMr+8tgTyQPMUdFRRWreH/24f+CbPi+3vlu9W8O6fdXH7q4tJ/tuias2OVdeYy5VT8rqkoAyMda/YZ/bW8O/theBLu4uNPg0Txr4e8tNc0mJyYyHzsuLfcSxibaRg5KMNpJBVm/4LJf8mi2n/Y4WP/om5r/gip/yav4p/wCygXn/AKQWFV/wV5/5PK1L/sX9O/8AQGrwl/yKmi/9g+D/ANFrX/BWD/k9zxh/2D9M/wDSKKvgL8IfDvwI+EPhb4VeGrOCG30HT44p3iTH2m5KgzTt6tJIWYn39AKZVZSrAEEYIIr9t7wIP2UP24dT1L4bwDS7a01Ky8T6DHGuxbdnKzFUHQIs6SqoHAUAdq0bVLXXdHsdbsW3W+oW0dxCc5yjqGXp7EV8eP8AlKRe/wDZUNN/9KLeq/4KRf8AJkfxT/7B9t/6WQV/wRr+C+g+P/jh4k+KHiGwhvU+HmnQNp8U0e5Yr66Z1jm543JHBPj0ZgwwVBqtS1LTdH0+41XV9QtrGys4mluLm5lWKOFFGWZmYgKoAySeBXxE/wCCon7Gfw9u5dNX4mTeJ7yA4ePw7p8l4n/AZyFhcf7rmv8AgpB+2f8AA79ryx8F/wDCs/Dniux1bwpc3qT3WsWNvAk9vMI8BDHPIxw0QIDKuNx6EkV/wS31y71z9iD4fC9d3k099RtFd2yWRL6fZ9AqlVA9FFf8FS/HniL4z/tpH4U6RKZ7fwslloWk224hGurlY5JXxzhjJMsZPpEvpXwR+D3hD4C/C7w/8KvBFkkGm6HarGZNuHupjzLPIe7u5LE++BgAAVX/AAXG/wCRq+Ef/YP1X/0ZbV+wH/yZp8Jf+xfj/wDQ3qvgR/ylIsf+yoal/wClE9ftw/FHUvg1+yh8SPH2h3b22qW2k/ZLCeNsPDPdSpbJIn+0jTBx6bc1/wAEXfgno3iv4k+L/jVr1hHcv4LtoLLRhLGGEd1ciTzJl9HSKPb9JzVft+fBXR/jh+yv450W8sEn1TQNNm1vRZQgMkV3axtIAhPQyIHiPtIa/wCCJfxKvrH4mePfhDPdE2Gr6Kms28TvhUntpkhfaP7zpcLnHaIelf8ABav/AJNX8Lf9lAs//SC/r/gix8NPDWl/BDxV8V1s0k8Q674gk0x7lly0VpbxROsansDJK7NjGcJn7oqv+CmPha1+Fn7cHijVfCATT31BrLXYRAuzybqSNWkcYA+ZpkeQn1cnNaTff2rpNlqfleV9rt45vL3btm5QcZ4zjNVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVeLf+RU1n/sHz/wDotq/4JDf8nlab/wBi/qP/AKAtVX/BZf8AaP0fT/Bmk/szeHtRWbWNWuotU8QJGci2tY8mCJ/RpJNsmOoESk8OM/8ABH74H6h8Nf2d9Q+JOu2D2uo/Ei/S6t1kTa50+3Vkt2IPOGZ53X1V1I4Oa/4Ln/8ANEv+5g/9x1f8E3v+TI/hX/2D7n/0snr9rz4Kx/tB/s5+N/hdFbpLqV/pzT6QWwNt9ARLb8/whpEVCf7rt64r/gi18X5vD3xO8ZfAjV7h44PEdiNT0+GUY2XlqdsqAddzxPk57W/bv8S7a5/b0/4Kcv4Qt5DeeGLLWhpZZDlE0jTsm4ZWGMCVknZT/enUelQww28KW9vEkccShURF2hQBgAAdBjtX/Bcb/kavhH/2D9V/9GW1fsB/8mafCX/sX4//AEN6+KXwl+HHxs8H3PgP4peEbHxDol0QzW10pBjcdJI3Uh43GSA6EMASM4Jr9p3/AII3DQ9E1Pxr+zV4svdQayiec+FtZAeaVVBLLbXCgbm4wsci895M9f8Agkh+2J4+1fxyP2ZPiJ4gu9b0u70+WfwzcXkplmsZIFLvbB2+ZojEHZQSdhjwuA3H/BWD/kyPxh/2ENM/9LIq/wCCIf8AySr4lf8AYwWv/pOa/wCC0nhu9079prw34kdGNnrHg+BIpCuB5sNzcB0B74V4z/wOv2a/F+mePv2e/hv4v0iZXt9S8L2DkD/lnIIEWRD7q6sp91NV/wAFwPGWkjSPhb8PYrhJNTa5vtUmiB+aGELHFGxHo7GQD/rk1f8ABMfwve+Fv2JPhzBqEbR3Gox3uobG7RzXkzxEexiMbf8AAq/4KN/8pF/Gn/YQ0L/022VV8Wf+SV+Mv+xfvv8A0nev+CNv/J3N3/2J99/6Otqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr4wfDPwh8Yvhn4i+G/jmxtrnR9bsJIZvPUEQNtykyk/daNgHVuCCoOa/wCCQ2uaxpX7ZGm6bpry/ZdZ0DULbUFUEgxLGJlLe3mRR8+pHrX/AAWNt5p/2QoZIoyy2/i2weQj+FfLnXJ/FlH41/wRRvrWT9mXxdpiSg3EHju4lkTH3Uexsgp/Exv+VV/wV0ura4/bO1mGCeOR7bQtOjmVWyY2MW7B9DtZTj0YeteEv+RU0X/sHwf+i1r/AIKwf8nueMP+wfpn/pFFWm6jY6xptrq+l3SXNnewJPbzIcrJG6hlYexBB/Gq/wCCsPiK38c/tral4f8AD6m8utE0nTtIZIRuMlwymbYuOrD7QqkdmBHUGvCGiN4b8JaJ4ceTe2ladb2hbOcmONUz/wCO18eP+UpF7/2VDTf/AEot6r/gpF/yZH8U/wDsH23/AKWQV/wQx/5rZ/3AP/cjVf8ABSz9oD4kftA/tNJ+yh4E1KRPDukaxaaPb6dFKUTVNWlKKXmx94JLIIlU5C7Cw5avgf8A8Es/2V/hX4fsovGHg2L4geI1iX7bqets7QvJwWEdsG8tEz0DBmxwWPNf8Fg/g18Hvhd8NPh5dfDT4V+D/Cl1ea3cxXE2i6Jb2Mk8awAhXaJFLgHnknmv+CT/APyZH4P/AOwhqf8A6WS1+2jK/wAJf+CkHiLxTrMEjW2m+LdM1zhOZIStvcfLg88ZX6g9Ksb2z1Oxt9S065juLW6iWWCaNtyyIwBVlPcEEHNVX/Bcb/kavhH/ANg/Vf8A0ZbV+wH/AMmafCX/ALF+P/0N6r4Ef8pSLH/sqGpf+lE9f8FP/Dd74l/Yj+IcenI0k2nCxviirnMcV5C0h9gse9s/7Nf8EP8Axfpj6L8UvAMkypqEd1YanEh6yxMskTkeyssef+ug/Cv2lvGWk/D79nr4j+MNZuEittN8MX7DeceZK0LJFGP9p5GRB7sK/wCCLPhe91L9pPxR4pWNvsWieEJopJB0E09zAI1P1WOY/wDAa/4LV/8AJq/hb/soFn/6QX9f8Ebf+TRbv/scL7/0TbVX/BXn/k8rUv8AsX9O/wDQGrwl/wAipov/AGD4P/Ra1VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV4t/5FTWf+wfP/AOi2r9gv9oHwP+zN+0LY/FD4hWmr3GjxaZdWjrpduk0qvKoCttd0G0Y55z7Gp/8Agst+yRFC8sej/EOZkUkRpo0AZj6DdcAZ+pFfGz/gtF4m8R2U3hr9nT4YS6NdXg8qDWdcdbm5QtwPKtY8oJM4wWeQE9VNfsmf8E3fjF+0T47T41ftURazpfhu9uhqF1FqsjLqfiF2O7DAnfDExxud8MVOEHO9bDT7HSrC20vTLOG1s7OFYbe3hjCJDGoCqiqOAoAAAHAAr/guf/zRL/uYP/cdX/BN7/kyP4V/9g+5/wDSyeq/bI0PxR+xL+3tqHxI+H0a2i6nLJ4i0N8EIFvEljuEHoBK1woA6KV4AIr/AIIsfA97Pw/4x/aK1u2JuNYm/sPRpJF+byYysl1ICeoeTyVznrC+ar/guN/yNXwj/wCwfqv/AKMtq/YD/wCTNPhL/wBi/H/6G9eLf+CrvwG+HXxz8Y/B3x7oWu2tl4Y1L7DBr+nqt5DPIiIJxJECHjKTeYny+ZkJn5c4HxS/4K5/sn+FfCF5qPw48Ral428QG3b7DpsGj3VpH5xHy+dJcJGFQHG4puOOgNf8EifgV4y8d/tDSftE6np08Ph7wjDeeXfvEUS81C5ieExR9A22OaVmxkL8gI+YEf8ABWD/AJMj8Yf9hDTP/SyKv+CIf/JKviV/2MFr/wCk5r/go3+yPeftU/BmE+D4I38c+DZZb3Q0cqn2tHUCe13NwvmBEKkkDfGgJAJNfsPf8FA/EX7HMl98Cvjj4T1u58J2l/Ji3EPl6h4fuWb96nkyld0ZfLNGSpVizDJJU+Nf+CwX7JXh/wAOyal4Tn8TeKtVMRMGmwaW9p+8xwJJZtqoueCy7yOSFPGfAXw8+PX/AAVG/aZuvHPia3ntNB+0Rx6xqsMRFnodgnKWkBbhpNpO1OSzO0jcFmrw9oGj+FPD+meFvD1jHZaXo1nFZ2NtGMLBBEgSNF9gqgfhX/BRv/lIv40/7CGhf+m2yqviz/ySvxl/2L99/wCk71/wRt/5O5u/+xPvv/R1tVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV+2/8A8FFdb/ZF+N3hXwFpngKx8TaRd6G2oazFLcPbTbpJmSEQzAMqlRC5ZSjZEi8r1rwx/wAFkv2UNZsUl17S/HGgXQUeZDPpcc67u4V4pWyPchfoK/ax/wCCvelePPAmr/DT9nbwnrenvr1tJZ3fiHWNkEsMLgq4toY3ch2UkCRmBXnC5wR/wSQ/Y78W/DOHVP2iPihoM+k6nr1gLDw5p93GUnjs3KyS3LoeU8wrGqAgNtVz0cZ/bL+B91+0T+zZ41+Fuk+WNYvrNbnSC5Cg3lvIs0SEnhQ7J5ZY9BIT2r9h79sLXv2G/iX4h8P+OvCeo3fhzWJVtvEOlKojvbG5ty4WSNXIG9dzqyMV3AjkFRXxK/4LN/ADSPC0svwr8J+KPEviSeH/AES2v7RbK2hkI48597McHHyop3dNwzkftS+C/jdpPjHTfiZ8f0eDxX8UbF/EEtpPEYp7aJpnijSSMj918sQ2x9VTaCAQQPCDpJ4S0SSN1ZW063KsDkEeWvIr/go74VuPiP8A8FGr/wCHukXcaX3iC50HSo32mTyZbi3t0Xcq8n/WKcdSCPWv2bf+CifiT9kWJ/2Wv2wPA+uRz+ByNP0/VbCJZZ4Ldf8AVRyxsyiWIJjy5oycpsG1vvV8aP8Agsz8F9G8J3cHwP8ADeu+IvE9xCy2cuqWYtLK1c8B5cuZJMcHYqgNjBdetf8ABOP9lv4g/tK/HhP2ovi3Dd3PhnRdYfVpNQvgQ2uasH3oqA/ejSQh3b7uVCDOTtr/AIKS+DfFnwP/AG5NU+I9rYyQW2t3ll4i0G7ZWMc0kaxeYN2fvLPG2VByAyHADCvhd/wVo/Zo+I934T8N3Fh4q0nxP4mv7TTm0+SwWWK1uZ5FiyZw4Volds78BtozsB+Wv+CkX/JkfxT/AOwfbf8ApZBX/BDH/mtn/cA/9yNV+3v4H8cfsx/t13Hxft9LZ7PU/ENv4t8O3kqkw3Mscsc0kZbpuSdWBXqFKHowrw7/AMFcf2OtU8FweItd8Ua3outGANP4ffRbmedJMfMiSohhYZyAxdcjBIXoP27v2gviX+2RDD8a9J8Capovwe8H6kND0Wa6ADS3lwjSPJJglTIy24DBCVjAjUsS+W/4JNXEE/7E3hSKKUM0Gp6mkgH8Lfa5GwfwZT+Nf8FXf2K/E/xk03T/AI+fCfRJNU8S+HLE2etaVaxFp7+yVmdJYlHMksZZwVGWZGGPuBT+xX/wVVvfgb4R0/4P/Hfw9qviHw7oyi30rVtPZWvdPhXhbeSOQqJY06KdysijbhgFC61/wWJ/ZD03TXvNOXxtq1wB8lpb6Msbk47tLIqgevP4Gv2Y/j7on7TnwZ0b4w6Do8ukQ6rNdQvp004mktmhneLDsoAyyor4xwHA561/wWy+Fuua74D+H3xZ0jTpbix8L3l5YatJGpbyUuhCYXYD7qB4HXdjG6VATyK/Ya/4Kg/Bv4R/APRfhB8Z7LX7PUfCayQWN9Y2gu4r22aRnQEBgyOm8pgjaQqndkkD4W/Efw58YPh34f8Aid4RNz/Y/iSyS7sxdRhJQjdnUEgMMEEZNfAj/lKRY/8AZUNS/wDSievFXhnRPGvhjV/B3iSyS80nXLGaxvrd+ksEqFJFP1ViK8VeEPjx/wAEuP2obfxTo9nPd6UksselajPGws/EOmuRvgkZeBIAE3J1R1RwCNhPg3/gsD+yVr/hqLVPFVz4l8MasIsz6XNpT3REgHKxyw5V1yMBm2E5BIXkD9tj9vXxb+2xqOmfAb4DeD9di8K3d7GVsjBv1DX7pTmMNFEWCxKfmWMFskB2I2gL/wAE8/2SZ/2Ufgu1h4oSBvG/iyZL7xA0RDi32qRDahhwwiDOSRwXkkwSMGv+C1f/ACav4W/7KBZ/+kF/X/BG3/k0W7/7HC+/9E21V/wV5/5PK1L/ALF/Tv8A0Bq8Jf8AIqaL/wBg+D/0WtVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVXNvBeW8tpcxrJFMhSRG6MpGCPyrVf+Can7D+ssWu/gJp8ZLl8Wuq39rz9Ip149ulWn/BML9haynW4g+BELOoIAm8QanKvP8AsvckH8q+Hf7OvwH+Ekwuvhr8IPCfh67H/L5ZaVElweMYM2DIR143dz6mqr4r/AH4M/HP+y/+FufDzSfFP9ied/Z/29Gb7N5uzzNuCPveVHn/AHRXgnwP4S+G/hXT/BHgXQrXRtC0pGSzsLZSI4FZixCgknlmY/jVfFX9nf4I/HG5069+LPw10bxPcaTHJHZSX0RZoVcgsAQRwSoODXgTwF4N+GPhSx8D/D/w7Z6HoOmBxaWFom2OLe7O2B7szE+5NV8Vf2efgn8cbjTrr4tfDjSPFE2kJIli99GzGBZCpcLgjqVXP0FeEPB/hnwB4Z07wb4N0a30nRNJhENlZW4IjgjBztUHPHJr4rfsa/svfG7UZdZ+JXwX0DU9TuG3T6hbrJY3U7esk1s0cjn/AHmNeGv+CYv7Efhi/XUoPgvFqEyNuRdT1a8uo19vKeUow/3lNaLoeieGtJtdB8OaPY6VpljGIrWysrdYIYEHRURAFUewAFfEL4ceBvix4VufBHxG8M2ev6FdvG89hdqTHIyMGQkAg8MAfwr4WfBD4TfBDT77SvhN4E0zwxaalMs13DYoVE0ijaGOSeQOKr4wfsufs+fHxhP8WvhRoev3ioI1v3ia3vAg6KLmEpLtGT8u7Arw/wD8Evf2IvD2orqUfwdOoSIcxx6hrV7cRKf+uZl2sPZga8NeF/DPgzRLXw14P8Paboek2SbLaw060S2ghX0SNAFUfQVXjX9kn9m34j+NLr4i+OPg/oGs+Jb14nuNSuYmMsjRIkcZJDAfKkaKPZRVajp9lq2n3OlalbpcWl5C8M8LjiSNgVZT7EEivhp+yz+z18G/EbeLvhf8J9D8Oay9s9q15ZRsrmJipZOWPBKqfwqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq+IHwu+G/xX0ceH/iX4F0LxRp6EtHb6rYR3KxMRgsm8Eo3+0uD71rn/BLf9iDW7r7YPg9Jp8jEl1sddvokbp/B5xVcY/hA6nOa+GP7En7KXwd1KHWvh/8EfD9nqVswe3vbwS6hPAwOQ0cly8jI2e6kGqr4zfscfs0ftAaidc+Kvwm0vVdXKbDqcEktldOAMLvlt3RpNoAADlgOmMV8Kf2Ff2T/grrFt4j8AfBnSYNYtGV4NQ1CabUZoZAB+8ja5eTym4zlAuMnGM18U/2bfgT8btUsta+K/ww0XxNf6dbm3tbi+iLPFEWLFAQRxuJOPc1p2n2Wk6fa6VptulvaWUKQ28KDAjjQBVUewAArXP2YfgB4m+JkXxk174V6Je+Noby2vI9aljYzrPbBBBIDnGUEUYHH8Ir4x/s4fAz9oGyhsvjD8NNH8SG1Qpb3MyNDdQKTkrHcRFZUUnkqrAE9a8Kf8EyP2J/COrJrNr8GYdSniYNFHq2p3d7CpBzzDJIY3B4yHVun1zp+nafpFhb6XpNjb2VnaRiK3treIRxxIowFVVwFAHAA4qvi18FPhT8dvDP/CIfFvwNpviXS1fzIortCHgfGN8UqESRNjjcjKcEjODXgr/gmr+xn4B8UWXjHQPhK7appl5Hd2El1rd9OtrLG25GVGmKtghT84b7o968b+B/CXxI8K3/AII8c6Fa6zoWqIqXlhcqTHMqsGAYAg8Mqn8K+FHwB+DPwM/tT/hUfw80nwv/AG35P9ofYUZftHk7/L3ZJ+75smP941XxJ+Ffw4+MPhqXwh8T/BmleJdIlbd9mv7cSCN+zo33o2/2lII9a0f/AIJifsQaJq51m3+CUVzIrh4oLzWb64gjI6jy3mKsD3Dhh9K8XfAv4O+O/Aln8MPFPw30G88JadKklnoq2aw2tsyBghjjj2hMBm4GB8xr4bfCz4e/B7w0PB/wy8K2Xh7RRcPcCytAwjEj43MAScZwKr4tfsTfssfHDUp9c+Ivwa0W81a5JafUrMy6fczOf45JLZo2lb3fd+VaB/wS7/Yh0C7W9X4OHUZUYMg1DW76ZFx6xmbYw9mBrwh4K8H/AA/0G38LeBPC2k+HtHtc+TYaZZpbQoT1IRABknknGSeTWtaJo3iTSLzw/wCIdKs9T0zUIGgu7K8gWaG4iYYZHRgQykHBBGDWpf8ABLn9h7VNYbV5fg00PmMWktbbXb+GBic5IRZhsHPRCoGBx1rwV4L8LfDrwnpfgbwVo8WlaHotutvYWUTMywxjooLEk/UkmtJ/ZJ/Zt0Hx6nxR0f4P6Ba+K0v31BdVjiYTC5clnlzuxuJZj071Xi/wV4P+IOg3Hhbx14W0nxDo91jzrDU7NLmFyOhKOCMg8g4yDyK1f/glv+xBrGpnU2+D0loWbdJBaa7fRROc8/IJsKO2F2ivhF+zb8CfgNA8Xwj+F2h+HJZk8uW7ggMl1Kn917iQtKy8ZwWIzzVfFD4QfDP41+H7fwr8VfBun+JdJtbxbyG0vkLIk6o6LIMEchZHH/AjXw0+FHw5+DfhxvCPwv8ACNh4c0d7l7lrOyUqhlYKGfBJ5IVR+FV8R/2Uf2dPi94mfxl8S/hHoPiHW5IUhe9vImaQxoMKvDDgCre3gtLeK0to1jihQJGg6KoGAB+AqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqndI0MkjqqqCWZjgADqa/aA/4Kpfsz/BW4udB8M6jcfEXxBbko1toMim0icZ4ku2+TqMHyxIQeor4if8FmP2lvEtzLH4A8NeEfBtiSfKxbPqF0v+9JKRG34RL/hq/wDwUe/ba1tma8+P+rxlmBItLGztOQMcCKFcD279TWlf8FGf22NGYPZ/tAa1IQ24C7tLW6GceksTce3Svh//AMFkP2pfDEsMXjXTPCPjK1U/vTc2Bsrhh/svAyop+sZ+lfAb/grj+zh8VZ7bQ/iLFe/DTWp2Cg6pKLjT2Y9hdoBsHXJlSNRx83PFjfWOp2UGo6beQXdpcxrJBPBIJElRhkMrDIIIwQRxVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX/BU+Hxff/slajoPgeLWLnU9a1ywshZaUsjzXqO53Q7I/mkDYGUwQcdK+Bn/AAR7/aC+I9rb618U9Z034b6ZOgdbe5i+3agQemYEZUTI7PIGHdeteAv+COn7KHhiGJ/F9z4t8ZXIwZRe6n9khYgchUtlR1B9C5PvWlf8E6/2KdGt/s1n+z9oMi4Azdz3N03H+1LKx/XnvWpf8E8P2LNWt/st1+z74fROebaW4t25GPvRyK3f1968df8ABH39kPxRFIfC9r4r8HTEfuv7N1hrmNT7rdCViPbcD718bP8AgjR8bvBkNzq3wc8XaT49sogWWxnT+zb/ABjOFVmaJyOmfMUnjC84H/BIPRPiD4R+CvjfwN8RdN13Sr/QfFzwxaVq8csT2StbQuVSOT7iszM3AAJYnnOaqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqKqxBIBIPGR0qviN+0B8FPhBrmj+G/ij8TtA8LX+vRySaemq3Yt0lSMqHYyNhEGXUZcjPOM4OND8QaB4o0yLWvDOuafq2nzj91d2Nyk8Un+66Eg/ga/aT/bK+BH7Ktna/8LR8R3D6tqEZlstD0uAXN7PGDguELKqJkEBpGUEggEkHHw3/AOCwP7KvjrxFD4d8QWvivwWtzLsh1HWrOJrQZ4XzHglkaPJ6krtHUsBzVpd2moWkN9YXUVzbXMayQzQuHSRGGVZWHBBBBBHBzW1c7gBkjBOKqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqrrxH4esp2tbzXtOgmjOHjlukRl+oJyK/wCEt8Kf9DPpP/gbH/jX/CW+FP8AoZ9J/wDA2P8Axr/hLfCn/Qz6T/4Gx/41Y6xpGqF10zVbO7MYG8QTrJtz0zgnHSqqqqq/aG/aN+Gf7MHgqy8ffFW71CDSb/VI9MhaytDcOZ3illUFQRgbYX59ceteFv8Agqr+yP4y8T6P4Q0TWvE76jrl/BY2iyaI6KZZpFjQE54G5hk9qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr/gsB4ou9e/bCutGmkcw+G/DthZQoRgAOrXBI55yZzzx0x2zX7Bui+D/gB+wX4V8XarNFaWLaHceKdcvAoBfzA0zO3TJWFY4x7RqK+N3xb8TfHb4q+Jfiv4uuHfUPEN884iMhdbaLpFAhP8EcYRB7KK8e/A/4v/C7RNF8R/ET4b6/4e0zxDEJNMu9QsnhjuQVDYBI4baQdpw2DnFf8EcvjVrPxD+AmufDHxBfveXHw61GKKxeRstHYXKu0MZ7kLJFOBnou1RgKKqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/wCCkX/J7nxT/wCwhbf+kUFVVf8ABDr/AJGr4t/9g/Sv/RlzVVVVVf8ABav/AJNX8Lf9lAs//SC/r9nr/kv3wz/7HDSf/SuKqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/4LNfCvU/DX7QmifFSO1Y6T4z0SOEzhMAXlqSjoSP+mTQEZ5OW/u1c/trfH66/Zzh/ZdfxJaL4MhUQ7ktAt29sH3i2aYHmLdjjG4gbSxX5a/YG+A0v7Qn7TvhLwteaWbzw/o9wNX1/em6MWluQ2x/9mWTy4sf9NPY1/wWm1nSLT9mXwzod0scmoah4ygktFb7yLFa3PmOPpvRT/10Ff8ABDfT71tQ+MWqBnW0WHRoSCCA8hN4RjsSoBz3G8etVVVVVXxr/wCCq37M/wAEfiHrXwx1TTPGniHV/D87W19Loun2728VwvDxb5p4yWVsqxCkAg8nFf8AD7T4A/8ARKPiB/3zaf8Ax6v+H2nwB/6JR8QP++bT/wCPVY/8FsP2cJJGXUvhn8SYEC/K0FrZSkn3BuVwPfNf8Pq/2V/+hC+Kv/gqsP8A5Nr/AIfV/sr/APQhfFX/AMFVh/8AJtf8Pq/2V/8AoQvir/4KrD/5Nr/h9X+yv/0IXxV/8FVh/wDJtf8AD6v9lf8A6EL4q/8AgqsP/k2v+H1f7K//AEIXxV/8FVh/8m1/w+r/AGV/+hC+Kv8A4KrD/wCTa/4fV/sr/wDQhfFX/wAFVh/8m1/w+r/ZX/6EL4q/+Cqw/wDk2v8Ah9X+yv8A9CF8Vf8AwVWH/wAm1/w+r/ZX/wChC+Kv/gqsP/k2v+H1f7K//QhfFX/wVWH/AMm1/wAPq/2V/wDoQvir/wCCqw/+Ta/4fV/sr/8AQhfFX/wVWH/ybX/D6v8AZX/6EL4q/wDgqsP/AJNr/h9X+yv/ANCF8Vf/AAVWH/ybX/D6v9lf/oQvir/4KrD/AOTa/wCH1f7K/wD0IXxV/wDBVYf/ACbX/D6v9lf/AKEL4q/+Cqw/+Ta/4fV/sr/9CF8Vf/BVYf8AybX/AA+r/ZX/AOhC+Kv/AIKrD/5Nr/h9X+yv/wBCF8Vf/BVYf/Jtf8Pq/wBlf/oQvir/AOCqw/8Ak2v+H1f7K/8A0IXxV/8ABVYf/Jtf8Pq/2V/+hC+Kv/gqsP8A5Nr/AIfV/sr/APQhfFX/AMFVh/8AJtf8Pq/2V/8AoQvir/4KrD/5Nr/h9X+yv/0IXxV/8FVh/wDJtf8AD6v9lf8A6EL4q/8AgqsP/k2v+H1f7K//AEIXxV/8FVh/8m1/w+r/AGV/+hC+Kv8A4KrD/wCTa/4fV/sr/wDQhfFX/wAFVh/8m1/w+r/ZX/6EL4q/+Cqw/wDk2v8Ah9X+yv8A9CF8Vf8AwVWH/wAm1/w+r/ZX/wChC+Kv/gqsP/k2v+H1f7K//QhfFX/wVWH/AMm1/wAPq/2V/wDoQvir/wCCqw/+Ta/4fV/sr/8AQhfFX/wVWH/ybX/D6v8AZX/6EL4q/wDgqsP/AJNr/h9X+yv/ANCF8Vf/AAVWH/ybX/D6v9lf/oQvir/4KrD/AOTa/wCH1f7K/wD0IXxV/wDBVYf/ACbX/D6v9lf/AKEL4q/+Cqw/+Ta/4fV/sr/9CF8Vf/BVYf8AybX/AA+r/ZX/AOhC+Kv/AIKrD/5Nr/h9X+yv/wBCF8Vf/BVYf/Jtf8Pq/wBlf/oQvir/AOCqw/8Ak2v+H1f7K/8A0IXxV/8ABVYf/Jtf8Pq/2V/+hC+Kv/gqsP8A5Nr/AIfV/sr/APQhfFX/AMFVh/8AJtf8Pq/2V/8AoQvir/4KrD/5Nr/h9X+yv/0IXxV/8FVh/wDJtf8AD6v9lf8A6EL4q/8AgqsP/k2v+H1f7K//AEIXxV/8FVh/8m1/w+r/AGV/+hC+Kv8A4KrD/wCTa/4fV/sr/wDQhfFX/wAFVh/8m1/w+r/ZX/6EL4q/+Cqw/wDk2v8Ah9X+yv8A9CF8Vf8AwVWH/wAm1/w+r/ZX/wChC+Kv/gqsP/k2v+H1f7K//QhfFX/wVWH/AMm1/wAPq/2V/wDoQvir/wCCqw/+Ta/4fV/sr/8AQhfFX/wVWH/ybX/D6v8AZX/6EL4q/wDgqsP/AJNr/h9X+yv/ANCF8Vf/AAVWH/ybX/D6v9lf/oQvir/4KrD/AOTa/wCH1f7K/wD0IXxV/wDBVYf/ACbX/D6v9lf/AKEL4q/+Cqw/+Ta/4fV/sr/9CF8Vf/BVYf8AybX/AA+r/ZX/AOhC+Kv/AIKrD/5Nr/h9X+yv/wBCF8Vf/BVYf/Jtf8Pq/wBlf/oQvir/AOCqw/8Ak2v+H1f7K/8A0IXxV/8ABVYf/Jtf8Pq/2V/+hC+Kv/gqsP8A5Nr/AIfV/sr/APQhfFX/AMFVh/8AJtf8Pq/2V/8AoQvir/4KrD/5Nr/h9X+yv/0IXxV/8FVh/wDJta3/AMFufg3Arf8ACOfBnxnfEbtovbq2tc88Z2NLjI69ce9H/guTo320Rr+zXem028ynxYvmA4/ufZcY991fsmftcfDr9r3wNeeLPBFnfaXfaPcrbatpF8VMto7LuRgykh43Aba3BJVgQMVVVVVLJHDG0ssioiKWZmOAoHUk9hXiz/gtX8CdI1680zwv8M/F2vWFtM8cWomSG1W5CkgOiMSwRgMjcFbBGVB4r/h958Kv+iH+K/8AwY2/+Ff8PvPhV/0Q/wAV/wDgxt/8K/4fefCr/oh/iv8A8GNv/hX/AA+8+FX/AEQ/xX/4Mbf/AAr/AIfefCr/AKIf4r/8GNv/AIV/w+8+FX/RD/Ff/gxt/wDCv+H3nwq/6If4r/8ABjb/AOFf8PvPhV/0Q/xX/wCDG3/wr/h958Kv+iH+K/8AwY2/+Ff8PvPhV/0Q/wAV/wDgxt/8K/4fefCr/oh/iv8A8GNv/hX/AA+8+FX/AEQ/xX/4Mbf/AAr/AIfefCr/AKIf4r/8GNv/AIV/w+8+FX/RD/Ff/gxt/wDCv+H3nwq/6If4r/8ABjb/AOFf8PvPhV/0Q/xX/wCDG3/wr/h958Kv+iH+K/8AwY2/+Ff8PvPhV/0Q/wAV/wDgxt/8K/4fefCr/oh/iv8A8GNv/hX/AA+8+FX/AEQ/xX/4Mbf/AAr/AIfefCr/AKIf4r/8GNv/AIV/w+8+FX/RD/Ff/gxt/wDCv+H3nwq/6If4r/8ABjb/AOFf8PvPhV/0Q/xX/wCDG3/wr/h958Kv+iH+K/8AwY2/+Ff8PvPhV/0Q/wAV/wDgxt/8K/4fefCr/oh/iv8A8GNv/hX/AA+8+FX/AEQ/xX/4Mbf/AAr/AIfefCr/AKIf4r/8GNv/AIV/w+8+FX/RD/Ff/gxt/wDCv+H3nwq/6If4r/8ABjb/AOFf8PvPhV/0Q/xX/wCDG3/wr/h958Kv+iH+K/8AwY2/+Ff8PvPhV/0Q/wAV/wDgxt/8K/4fefCr/oh/iv8A8GNv/hX/AA+8+FX/AEQ/xX/4Mbf/AAr/AIfefCr/AKIf4r/8GNv/AIV/w+8+FX/RD/Ff/gxt/wDCv+H3nwq/6If4r/8ABjb/AOFf8PvPhV/0Q/xX/wCDG3/wr/h958Kv+iH+K/8AwY2/+Ff8PvPhV/0Q/wAV/wDgxt/8K/4fefCr/oh/iv8A8GNv/hX/AA+8+FX/AEQ/xX/4Mbf/AAr/AIfefCr/AKIf4r/8GNv/AIV/w+8+FX/RD/Ff/gxt/wDCv+H3nwq/6If4r/8ABjb/AOFf8PvPhV/0Q/xX/wCDG3/wr/h958Kv+iH+K/8AwY2/+Ff8PvPhV/0Q/wAV/wDgxt/8K/4fefCr/oh/iv8A8GNv/hX/AA+8+FX/AEQ/xX/4Mbf/AAr/AIfefCr/AKIf4r/8GNv/AIV/w+8+FX/RD/Ff/gxt/wDCv+H3nwq/6If4r/8ABjb/AOFf8PvPhV/0Q/xX/wCDG3/wr/h958Kv+iH+K/8AwY2/+Ff8PvPhV/0Q/wAV/wDgxt/8K/4fefCr/oh/iv8A8GNv/hX/AA+8+FX/AEQ/xX/4Mbf/AAr/AIfefCr/AKIf4r/8GNv/AIV/w+8+FX/RD/Ff/gxt/wDCv+H3nwq/6If4r/8ABjb/AOFf8PvPhV/0Q/xX/wCDG3/wr/h958Kv+iH+K/8AwY2/+Ff8PvPhV/0Q/wAV/wDgxt/8K/4fefCr/oh/iv8A8GNv/hX/AA+8+FX/AEQ/xX/4Mbf/AAr/AIfefCr/AKIf4r/8GNv/AIV/w+8+FX/RD/Ff/gxt/wDCv+H3nwq/6If4r/8ABjb/AOFf8PvPhV/0Q/xX/wCDG3/wr/h958Kv+iH+K/8AwY2/+Ff8PvPhV/0Q/wAV/wDgxt/8K/4fefCr/oh/iv8A8GNv/hX/AA+8+FX/AEQ/xX/4Mbf/AAr/AIfefCr/AKIf4r/8GNv/AIV/w+8+FX/RD/Ff/gxt/wDCv+H3nwq/6If4r/8ABjb/AOFaz/wXE8IQL/xT37PGr3rbRxeeIo7XnPP3YJOMY5/l1qX/AILjeJmkYw/s56WqbjsVvE0jEDtk/Zxk++BX7En/AAUD8HftjXWr+Fj4PuPCXi3Q7QXsunPfC8iubbeI2likCIfldowysgx5iYLc4qqqqqqqqqqqqqrrw54evZ2urzQdOnmc/PJLaozN9SRk1/wVV8PaBYfsVeLrqw0TT7aZdQ0wLJDbIjDN5FnBAzVf8Il4U/6FnSf/AACj/wAK+MPxV+FX7M3w81H4o+OIP7K0W0eOKZ9N00yySyO22NAsY7scAsQoJ5IzXjf/AILgeHba7ktvhx8A9Q1C2BOy71rW0tHP1hijlH/kStC/4LheMIbtT4m/Z+0a7tSwytjr0lu6jnJBeKQE9DjA6Ed8j9ln9t/4I/tZ2U1v4E1G60vxJYw+be+HtVVY7qNAQDIm0lZY8kDcpyMruCkgVX7Vf/BSj4G/sw6zceBxb3vjPxpbL+/0jS5UjismIBC3Nw2RGxBztVXYcZVQQTN/wXB8fNqHmW/wC8PpY7uIX1uZpcf9dBGBn32V+xb/AMFFvDv7YHiu/wDAEfwt1fwxr2m6W+ozOL1L2z8lJI4z+92xurFpVwuw8Z54r/gtX/yav4W/7KBZ/wDpBf1+z1/yX74Z/wDY4aT/AOlcVfFD4leEfg98Ptd+JnjrUlsdE8P2b3N1KfvNjhUQfxO7FUVe7MB3pv8AguL4TyQv7O+rEZ4J8SRj/wBoV+yb+0X4o/af8BSfEzUPg/d+B9AuJNmkSXuqC5l1ILkPKiCJNsQI2hiTuIbAwMmqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqquLq1tVRrq5ihEkixoZHC7nY4VRnqScADvVfHX4EfDb9o34eXnwz+KOjG+0q5cSwyxN5c9lOoISeB8HZIAzDOCCGZSCrEHXP+CHmuHxCV8NftA2I0N5eHvtBc3MKZ6bUl2SMB3ygPoK/ZR/ZD+GH7I3gufw14FW41DVdUZJNY1u9UfaL51GFGBxHGuW2xjONxJLEkn9vj9g7xh+2NqPh/WdC+L1toCeGLKWGz0a90xpbeSaVwZJjMkm5GYJEuNjcIPU1/wT4/ZI179kb4S6v4Y8aajpOoeJte1qS8vLjTJHkgEKKscCKzojHhWflRgyEdqqqqqqv2XPht8P/ip/wVF+Nvhn4keDtI8TaTHrHim4Wy1S0W4iEq6qAr7WBGQGYZ9zX/DJv7K//Rs/wp/8Iyw/+NV/wyb+yv8A9Gz/AAp/8Iyw/wDjVX37Hv7J+ookdx+zX8M1CHI8jwtaQH8SkYJ/Gv8Ahij9kX/o3D4f/wDgih/+Jr/hij9kX/o3D4f/APgih/8Aia/4Yo/ZF/6Nw+H/AP4Iof8A4mv+GKP2Rf8Ao3D4f/8Agih/+Jr/AIYo/ZF/6Nw+H/8A4Iof/ia/4Yo/ZF/6Nw+H/wD4Iof/AImv+GKP2Rf+jcPh/wD+CKH/AOJr/hij9kX/AKNw+H//AIIof/ia/wCGKP2Rf+jcPh//AOCKH/4mv+GKP2Rf+jcPh/8A+CKH/wCJr/hij9kX/o3D4f8A/gih/wDia/4Yo/ZF/wCjcPh//wCCKH/4mv8Ahij9kX/o3D4f/wDgih/+Jr/hij9kX/o3D4f/APgih/8Aia/4Yo/ZF/6Nw+H/AP4Iof8A4mv+GKP2Rf8Ao3D4f/8Agih/+Jr/AIYo/ZF/6Nw+H/8A4Iof/ia/4Yo/ZF/6Nw+H/wD4Iof/AImv+GKP2Rf+jcPh/wD+CKH/AOJr/hij9kX/AKNw+H//AIIof/ia/wCGKP2Rf+jcPh//AOCKH/4mv+GKP2Rf+jcPh/8A+CKH/wCJr/hij9kX/o3D4f8A/gih/wDia/4Yo/ZF/wCjcPh//wCCKH/4mv8Ahij9kX/o3D4f/wDgih/+Jr/hij9kX/o3D4f/APgih/8Aia/4Yo/ZF/6Nw+H/AP4Iof8A4mv+GKP2Rf8Ao3D4f/8Agih/+Jr/AIYo/ZF/6Nw+H/8A4Iof/ia/4Yo/ZF/6Nw+H/wD4Iof/AImv+GKP2Rf+jcPh/wD+CKH/AOJr/hij9kX/AKNw+H//AIIof/ia/wCGKP2Rf+jcPh//AOCKH/4mv+GKP2Rf+jcPh/8A+CKH/wCJr/hij9kX/o3D4f8A/gih/wDia/4Yo/ZF/wCjcPh//wCCKH/4mv8Ahij9kX/o3D4f/wDgih/+Jr/hij9kX/o3D4f/APgih/8Aia/4Yo/ZF/6Nw+H/AP4Iof8A4mv+GKP2Rf8Ao3D4f/8Agih/+Jr/AIYo/ZF/6Nw+H/8A4Iof/ia/4Yo/ZF/6Nw+H/wD4Iof/AImv+GKP2Rf+jcPh/wD+CKH/AOJr/hij9kX/AKNw+H//AIIof/ia/wCGKP2Rf+jcPh//AOCKH/4mv+GKP2Rf+jcPh/8A+CKH/wCJr/hij9kX/o3D4f8A/gih/wDia/4Yo/ZF/wCjcPh//wCCKH/4mv8Ahij9kX/o3D4f/wDgih/+Jr/hij9kX/o3D4f/APgih/8Aia/4Yo/ZF/6Nw+H/AP4Iof8A4mv+GKP2Rf8Ao3D4f/8Agih/+Jr/AIYo/ZF/6Nw+H/8A4Iof/ia/4Yo/ZF/6Nw+H/wD4Iof/AImv+GKP2Rf+jcPh/wD+CKH/AOJr/hij9kX/AKNw+H//AIIof/ia/wCGKP2Rf+jcPh//AOCKH/4mv+GKP2Rf+jcPh/8A+CKH/wCJr/hij9kX/o3D4f8A/gih/wDia/4Yo/ZF/wCjcPh//wCCKH/4mv8Ahij9kX/o3D4f/wDgih/+Jr/hij9kX/o3D4f/APgih/8Aia/4Yo/ZF/6Nw+H/AP4Iof8A4mv+GKP2Rf8Ao3D4f/8Agih/+Jr/AIYo/ZF/6Nw+H/8A4Iof/ia/4Yo/ZF/6Nw+H/wD4Iof/AImv+GKP2Rf+jcPh/wD+CKH/AOJr/hij9kX/AKNw+H//AIIof/ia/wCGKP2Rf+jcPh//AOCKH/4mv+GKP2Rf+jcPh/8A+CKH/wCJr/hij9kX/o3D4f8A/gih/wDia/4Yo/ZF/wCjcPh//wCCKH/4mv8Ahij9kX/o3D4f/wDgih/+Jr/hij9kX/o3D4f/APgih/8Aia/4Yo/ZF/6Nw+H/AP4Iof8A4mv+GKP2Rf8Ao3D4f/8Agih/+JrRP2Yv2bfDZRtB/Z/+HNjJHt2yweF7NZMr90l/L3Eg9yc14r+HPw6h8Ea3Yp4E8OJafYZ3aEaXCE3CM/MV24zwOfav+CGP/NbP+4B/7kaqqqq8W/8AIqaz/wBg+f8A9FtX/BFz4eeAPHv/AAuL/hOfA3h/xF9h/sL7L/a2mQ3fkb/t+/Z5itt3bFzjGdoz0Ff8M9fAH/ohvw//APCZtP8A43X/AAz18Af+iG/D/wD8Jm0/+N1/wz18Af8Aohvw/wD/AAmbT/43X/DPXwB/6Ib8P/8AwmbT/wCN1/wz18Af+iG/D/8A8Jm0/wDjdf8ADPXwB/6Ib8P/APwmbT/43X/DPXwB/wCiG/D/AP8ACZtP/jdf8M9fAH/ohvw//wDCZtP/AI3X/DPXwB/6Ib8P/wDwmbT/AON1/wAM9fAH/ohvw/8A/CZtP/jdf8M9fAH/AKIb8P8A/wAJm0/+N1/wz18Af+iG/D//AMJm0/8Ajdf8M9fAH/ohvw//APCZtP8A43X/AAz18Af+iG/D/wD8Jm0/+N1/wz18Af8Aohvw/wD/AAmbT/43X/DPXwB/6Ib8P/8AwmbT/wCN1/wz18Af+iG/D/8A8Jm0/wDjdf8ADPXwB/6Ib8P/APwmbT/43X/DPXwB/wCiG/D/AP8ACZtP/jdf8M9fAH/ohvw//wDCZtP/AI3X/DPXwB/6Ib8P/wDwmbT/AON1/wAM9fAH/ohvw/8A/CZtP/jdf8M9fAH/AKIb8P8A/wAJm0/+N1/wz18Af+iG/D//AMJm0/8Ajdf8M9fAH/ohvw//APCZtP8A43X/AAz18Af+iG/D/wD8Jm0/+N1/wz18Af8Aohvw/wD/AAmbT/43X/DPXwB/6Ib8P/8AwmbT/wCN1/wz18Af+iG/D/8A8Jm0/wDjdf8ADPXwB/6Ib8P/APwmbT/43X/DPXwB/wCiG/D/AP8ACZtP/jdf8M9fAH/ohvw//wDCZtP/AI3X/DPXwB/6Ib8P/wDwmbT/AON1/wAM9fAH/ohvw/8A/CZtP/jdf8M9fAH/AKIb8P8A/wAJm0/+N1/wz18Af+iG/D//AMJm0/8Ajdf8M9fAH/ohvw//APCZtP8A43X/AAz18Af+iG/D/wD8Jm0/+N1/wz18Af8Aohvw/wD/AAmbT/43X/DPXwB/6Ib8P/8AwmbT/wCN1/wz18Af+iG/D/8A8Jm0/wDjdf8ADPXwB/6Ib8P/APwmbT/43X/DPXwB/wCiG/D/AP8ACZtP/jdf8M9fAH/ohvw//wDCZtP/AI3X/DPXwB/6Ib8P/wDwmbT/AON1/wAM9fAH/ohvw/8A/CZtP/jdf8M9fAH/AKIb8P8A/wAJm0/+N1/wz18Af+iG/D//AMJm0/8Ajdf8M9fAH/ohvw//APCZtP8A43X/AAz18Af+iG/D/wD8Jm0/+N1/wz18Af8Aohvw/wD/AAmbT/43X/DPXwB/6Ib8P/8AwmbT/wCN1/wz18Af+iG/D/8A8Jm0/wDjdf8ADPXwB/6Ib8P/APwmbT/43X/DPXwB/wCiG/D/AP8ACZtP/jdf8M9fAH/ohvw//wDCZtP/AI3X/DPXwB/6Ib8P/wDwmbT/AON1/wAM9fAH/ohvw/8A/CZtP/jdf8M9fAH/AKIb8P8A/wAJm0/+N1/wz18Af+iG/D//AMJm0/8Ajdf8M9fAH/ohvw//APCZtP8A43X/AAz18Af+iG/D/wD8Jm0/+N1/wz18Af8Aohvw/wD/AAmbT/43X/DPXwB/6Ib8P/8AwmbT/wCN1/wz18Af+iG/D/8A8Jm0/wDjdf8ADPXwB/6Ib8P/APwmbT/43X/DPXwB/wCiG/D/AP8ACZtP/jdf8M9fAH/ohvw//wDCZtP/AI3X/DPXwB/6Ib8P/wDwmbT/AON1/wAM9fAH/ohvw/8A/CZtP/jdf8M9fAH/AKIb8P8A/wAJm0/+N1/wz18Af+iG/D//AMJm0/8Ajdf8M9fAH/ohvw//APCZtP8A43X/AAz18Af+iG/D/wD8Jm0/+N1/wz18Af8Aohvw/wD/AAmbT/43X/DPXwB/6Ib8P/8AwmbT/wCN1/wz18Af+iG/D/8A8Jm0/wDjdf8ADPXwB/6Ib8P/APwmbT/43X/DPXwB/wCiG/D/AP8ACZtP/jdaP8HvhH4dl8/w/wDCzwhpkmc77PQ7eA5xjOVQdiRUMMNvClvbxJHHEoVERdoUAYAAHQY7V+xxpF9rX/BRz482vga1Kwo/iN7eKBxbhLca1AFABIAABUbf8Kqqqqqqqqqqqqqq/wCCsH/JkfjD/sIaZ/6WRVVftJ/Bu1/aB+BfjL4P3FzBbSeItOaK0uJ0LJBcoyyQSMBzhZUjJxzgGvAf/BHr9l7wFoI1L4v+Mtf8T3MKBru5mvl0myTA5KonzqM55aU9ulftzfshfsTfD74Zah46/Z6+Nnhy38T6M0LSeFD4vttQfUIWlWNzDGzmYSLuLnlgVRxgHmvgL8Wdc+Bvxh8J/FTw/ezQT6BqcU0yxEjz7cnbPCR3V4i6EejV8Y/G1z8P/g144+I+jqlzceG/DGoaraqAHWR4LaSVPqCUH1zWr6tqevarea5rV/PfahqNxJc3dzO5eSeV2LO7MeSzMSST1Jr9mj/gmL+xz8YvgF4W8bnxj4i8R6prelwzajqOmaxHGtndsimWBYhGwjMbEoVkDMMc1+x5+wb4P/Y68UeNdd8M+Mr/AMRQ+KIbSCzGo2qRz2EURkZ0aRDtl3s6HIRMbBx3r/gtX/yav4W/7KBZ/wDpBf1+z1/yX74Z/wDY4aT/AOlcVf8ABV/9sRviz4+f9nzwFqZbwj4LvG/teeGT5NT1NflZeOscGWQdi+88hUNf8E6/2DdQ/ac8VJ8RfiHYT23wx0C5An3BkOt3C8m2jYEERjjzJB0+6PmJKadp1hpGn2uk6VZQWdlZQpBbW0EYjjhjQBVRVHCqAAABwAKqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv+CkPw78c/FL9kjxV4Q+HPhq+17XJbuwmhsbJN80ixXUbuUXqxCqTgc8cV4O/ay/bT/Zj1UeHIviD4z0J7QYbQvE9u88cajjaLe7VjGOMfIFPvXwq/4La+LLIQWHxq+D+naqgwJNR8OXbWkgHr9nm3q5xjpIgz6dB8Kf8Agpf+x58WDBaQfFCPwrqMxwLHxRAdOK+mZiTBn2EpNaXq2la7p8Oq6JqdpqFlcLuhubWZZY5B6qykgj6VVVVVVVX7D/8Aylh+OH/YQ8V/+nVaqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqvjrey6Z8EfiFqVuqNLaeFdTlRXBILLayEZx24r/AIIf6fDH8PfilqisfNuNZsYWBHAEcMhH/ow1VVVV4t/5FTWf+wfP/wCi2r/ghj/zWz/uAf8AuRqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr/glai3n7cPxt1W6Jlu/7O1MGViSx36rCXz9Sqn8Kqqqqqqqqqqqqqq/4Kwf8mR+MP8AsIaZ/wClkVVX/BQb9tBf2RvhvYx+GLW1v/HXixpYdFhuPmitEjA826kUfeCl0CpxuZu4VhWveNP2g/2rviFZ6drmv+J/iD4q1e4K2NiXabDEZYQwrhIUABJCKqqAScAE144/4Jm/tSfDT4Ua18XfHukeHdG0zw/ZG7vbKTV1mulQEAgLCHQtyON/41Wj6Xp+ueAbHRdWtUurG/0eO3uYJB8ssbwhXU+xUkfjX7YP/BOL4x/s7+JdS13wV4c1Hxh8O5ZXlsdT0+BriawiJyI7uNQWQqOPNxsYYOVJKL8Pvij8R/hNra+I/hn451vwzqQwGn0y9e3MgHRXCnDryflYEc9K/wCCdP8AwUg134++I4/gd8cTZ/8ACYy27y6LrNtEsC6qI13SQyxr8qzBAzgoArKrDClRv/4LV/8AJq/hb/soFn/6QX9aNrGp+HtYsdf0S9ktNR0y5jubS4jOGhljYMjr7hgD+FXf2tp2kvvNM8uJGaXO59w3Bjnk5BBz3zmv+CUP7U3h/wCLPwZtfgdqUVjp3iz4d2axR21vEsK3+nbsJcKi4G9WYJKe7Mrk5kOKqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq8YeA/BHxD0ltB8e+D9F8R6a2SbTVbCO7iyRjO2QEA4718Vv+CTH7I/xFM174b0LV/Aeoy/N5ug3xMJb3gn8xAv8Asx7Bx9c/Fb/gi38cfDYnvfhN4+8O+NLVCTHaXitpd2w7ABi8RI7kyJ9OcDUPBX7aH7HepPqMmk/Eb4cYc772yeaKynYcEGaImCXoONzdvavhV/wWH/af8EmCz+IFl4d8f2MYAke8tfsN2QOmJYMID6lomJx65Nfss/8ABT34P/tN+ONI+FUfg7xH4X8Y6ysxtbafy7q0k8qF5nC3CENkJE5+aNR0wSTVVVVVV+w//wApYfjh/wBhDxX/AOnVaqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv2hf8AkgPxM/7E/Vv/AEjlr/giH/ySr4lf9jBa/wDpOaqqqq8W/wDIqaz/ANg+f/0W1f8ABDH/AJrZ/wBwD/3I1VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV/wSk/5PS+Nn/YP1D/ANOcdVVVVVVVVVVVVVVf8FYP+TI/GH/YQ0z/ANLIqqv+C2ula7D8ePAet3Cyf2Nd+ETb2hI+X7RFdzNPj32TW+fwr/gm9+0F8OP2bv2jk8Z/FMyQaHqmiXGlNqCW7T/2dJJJDIs5RQXK4hKHaCQJDwRmv+Cgn/BR/wCAvjH4D678Hvgl4kPi7WPF8SWt3eR2c0Ntp9tvVpCWlRS8jBdqqoIG4sSNoVq/4STTfBnwr/4TDWZNlhoXh/7ddNnG2KG38xz/AN8qa+E3/BWn9kv4hafAPGOvan4A1dgBLZ6xZSTQ7+M7LiBXUr/tOIzx0Ff8FTPit+xh8UtL0DU/gZcaHq3xFfVTJq2r6NZNCkliYn3LPJtVZpTIYip+ZlCuCQCAf+Ce+g+IfEH7ZXwsg8Nxymaz1pby5eNciO2iRnnLHsDGGXJ7sB1Ir/gtX/yav4W/7KBZ/wDpBf18G9A0nxX8XvA3hfXbX7TpuseJNPs7yDeyeZDLcxo65UgjKsRkEEZr/grX+x9pd/8ADnSfj78MfD9vZzeBrKHTNZsbOEIp0tcLBIqqP+WBIU/9M3ycCOvgv8XfGPwI+Jug/FTwJemDVtCuhKqMxEdzGeJIZAOsboSrD0PGCAa+A3xq8H/tDfCjQPiz4JnzYa1bhpbZ3DSWVwvEtvJj+NHBHoRhhwwJqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqlijmjaGWNXR1KsjDIYHqCPSviZ+wx+x58clu7nWvhN4dj1B3YS6l4fP9nzrL3Z2tyqu/P/AC0De46V8E/+CWngr9nn9o7wr8c/hz8T9YudM8Pve+boesWcc0siz2c9uNtzGUA2mYHBjOQOoPWqqqqq/Yf/AOUsPxw/7CHiv/06rVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV+0L/yQH4mf9ifq3/pHLX/AARD/wCSVfEr/sYLX/0nNVVVVeLf+RU1n/sHz/8Aotq/4IY/81s/7gH/ALkaqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/4JSf8npfGz/sH6h/6c46qqqqqqqqqqqqqq/bR+AniP9pj9nzXPhD4T1nTdL1LVLq0liudR3iFBDOkjA7FZskKQOOtf8OS/j9/0Vf4f/8AfV5/8Zqv2rf2V/AH7Wvw0bwD40ll0+8s5vtOjazbRh5tOuMYLBSQHRhw8ZIDDByGVWXxT/wRt/ar0jWXsvDeq+C9f08uRDepqT2x254LxyR5U46hS49zXwE/4IyQ6RY6hrXx/wDGmn6pqs2nzw6bo+j+YbW1uJImWOeaZwrSFGYMEVAu5BlnGVr/AIcl/H7/AKKv8P8A/vq8/wDjNfEL4Q6n8Tv2d9S+CWreK30i+1vw2mk3uq2cIm2MYlSVlRtu5WwwIO0lWPTt40/4IuftI6NdynwX438D+I7Fc+U0txPY3D49Y2jZFzx/y0P9a8Kf8EaP2p9avI08S694G8PWm7Eskuoy3MoX1RIoiGPsWXp1FfsdfsI/C79kDTLq/wBGvJ/EnjLVYRDqPiG8gETeUCD5MEQLCGIsAxG5mYgbmIVQv7f37Lfi/wDa4+Dej/DfwV4h0fR77TfE0GrST6oZBG0cdtcxFR5asdxM6npjAPNfDD/gjp8cvA3xL8JeNtQ+J/gW4tfD+u2WoTwwm73yJBOkjKuYQNxCkDJAzWsaRpfiDSL3QdbsYb3TtStpLa7tZk3JPFIpV0Yd1ZSQR6GvGv8AwRP+J8/i7WZ/AHxQ8IweG5L6V9Jh1Jrk3MVsWJjSUrEVLquASDzjPGcV+wT+xh+0f+x94o1ax8QfETwdrvgbxDHvvtNtJboS290i4juIQ8QXcR8jgkbl2nOUAqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv+CoP7Z/xy+Df7Rvh/wR8F/iFf+GoNC8Px3N6kEUckdzc3EjH95HKrJIFjji27gcF39TX7CnxI+L3xg/Zq8NfE34032n3OueIJJ5rZrSxFr/oiyGOIyKpKl28tnyoUbXUYyCT/AMFAP26vHX7QfxK1vwR4K8V3en/DHQ7qSzsLOwuGjTV9h2tdTlSPNDsCUU/KqbeNxYnwD8RvHnws8SW3i/4c+LtU8O6xaMDFd6fctE3B+62OHU91YFSOCCK/YS/agP7VvwGsfG2rxwQeKNIuG0zxDBCoVDcoqsJkXPCSIyPjoGLqMhc1VVVVVfsP/wDKWH44f9hDxX/6dVqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqtV1jSNDs21DW9Vs9PtYwd891OsKLgEnLMQBwCfoDXin/gqH+xR4V1i70OX4tPqc9kxSSXS9JurqB2HURzLHskH+0pKnPWviP/AMFtvhrpjzWvwp+Duv68y5WO71q9j06PP94JGJmZfYlD9K1j/gsl+1lf6wL/AEvTPAmmWaN8thHpMsqMPRneYuT6lSv0Ffsnf8FGvgn+1PqVn4ItbfUPCvju5id10K+UzJceWheQwXCDa4VVJw4jbg4UgZqq/aF/5ID8TP8AsT9W/wDSOWv+CIf/ACSr4lf9jBa/+k5qqqqrxb/yKms/9g+f/wBFtX/BDH/mtn/cA/8AcjVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVa5428F+GFdvEni/RdJEXDm+v4oNny7ud7DHy8/TmviZ/wVH/Y++Gfih/Cc3ji/wDEl1byCO6uPD1j9stoGPX99uVJMd/LL46dQQPiT/wW3+H+nSS2nwl+DOt64RlUvNcv49PQH+8IohMzDPYsh+nSvFv/AAV9/bC8Qao15oOqeFfDNrk+XZ2GipMoHbc1wZGJ46ggZJ4HAH7HP/BVrwX8Xrjw18KPjNo9/ovjvUGg0+31SztzcWer3TYRSUiXdbvIxHy7TGCSdyjAFVX/AASk/wCT0vjZ/wBg/UP/AE5x1VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVf8FdvD9/o/7ZmraldxOsOuaFp13asy4DRrF5BIPf54HFaL/wAFMfht4P8A2AtO+EPhT+1rf4o2nhz/AIR2KBbQrFbfKYjeib7pAiO4D73mYBUL81fBf4X6x8avix4U+FOgsUvPE+qQ2YlCbvIjZv3kpHcJGHc+ymv+Cn37GP7PfwI/Z28LeNfhT4Ph8P6vp2u22kyzrcyO+owyW8zHzQzENLuhVt4AON3bGP8AghxrN59p+L/h5izWvl6RdICeI5M3atgerDb/AN8Cqqqqqr9h/wD5Sw/HD/sIeK//AE6rVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV4m8XeE/BWltrfjLxRpOg6cjbWu9TvY7WIH0LyEDPB7149/4KMfsZ/D62eW/wDjho+sTqSqW2gpJqbSEdg0Ksg+rMB7149/4LdfC3S7qOH4afBPxL4jhBImm1jU4dJx7oI1uCw6fe2n2r4j/wDBZ39onxFq/m/Dbwf4V8H6UgGy3uI21S4Y453zNsUj0CxKR3Jr4j/tYftI/FjVbrVfG/xp8W3YupGc2UOqS21pFnqI7eJljQdBgKOgzmrq7vdSuDcXt1PdTvgF5XLsew5PNfCj9in9qf43afPq3w3+C+t31jb4zd3rw6bDIT2jkunjWU+oQtjIzjIr4b/8EZf2kfFCx3XxC8UeFPBVu334GnbUbpP+AQ4iP/f2vCn/AARQ/Z+sdDNp41+J3jvWNVfGb3TmttPjTB/gieKYjPQ7nbjpjrX7NX7EPwB/ZUkutS+Geg3tzrt7B9nuNc1i5FzePDkExqVVUjUkAkIi7iBnO0Yqv2hf+SA/Ez/sT9W/9I5a/wCCIf8AySr4lf8AYwWv/pOaqqqq8W/8iprP/YPn/wDRbV/wQx/5rZ/3AP8A3I1VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVeKPF3hPwRo8viHxp4o0nQNKgOJb7VL2O0gj4z80khCjgHqe1eN/+ClX7FvgeCVp/jRZazcxqSlrolnPfNKR2V0Tyx0/icD35r4jf8FufAlnC8Hwl+C+uapMSQtx4gvorFE4+95cPnFuccbl+vavij/wWQ/aX8X28Fp8OdI8OeAVSMedcW9sNRuJJO5BuFMar6L5ZI7sa+Kn7Un7Q3xs1F9S+Jnxd8Q6uzoqfZkufstqoXpttoAkKngEkICTyc1aWV5qE62thaTXMzA7Y4Yy7HHXAHNfCb9iP9qX41XUsHgr4Oa8kEERke91a3Om2xxxtWW42K7Z42qSR1IA5r4cf8EYP2iPErxz/ABG8Z+FPBtq2N0cbvqd0nr8ke2M/9/a+H/8AwRg/Zx8P2RHj/wAY+L/Ft+y7S8c0enQL05WNAzg9esjDB6d6+BP7A/7MX7O3iUeNPh94Ekl8Qxqy2+p6rePeS2oYYbygx2xkgkFlUNgkZwSKqq/4JSf8npfGz/sH6h/6c46qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv+Cjv7El/wDtYeB9M8RfD9rWL4geEhILCO4cRR6nbPgvbM54RgwDRsx2glgcByy65+zb+0J4c8QnwprPwQ8cwat5vlpajQLh2lbOB5e1CJAexUkHsa/4JgfsCeKvgvqUvx8+NuijTfFNxaNbeH9GmIaXTYpBiSeUD7kzqSgTOVVn3cthf+CzknxT8VaJ4H8B+EPhx4o1Twzpkk2s6vq9lpc09pHcYMMEbSIpVWVDOx3HpIuO9f8ABE/4c3+g/CLx78Sr22eFPFWt29la7wR5kdlG+XA9PMuZFz6ofSqqqqqr9h//AJSw/HD/ALCHiv8A9Oq1VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVXxA+OvwW+FKOfiR8VvCnhuRBnyNS1aGGZv92ItvY47AE18RP8Agrh+x/4IeW20DXPEPjW5jyNuiaUyR7vTzLkxAj3XcPTNfGb/AILUfErxFaf2X8D/AIbaf4Q+dt+qavcjUrh1wMbIgiRxEHOd3mg8dK+KH/BQz9r74tWNtpmvfGTVNKtbZMNF4fxpXnnu0jW+1n7DaTt44AOSfEvjn4gfEO5tF8YeMfEPia4hPl2g1LUJr103EfKnmMxGTjgda8F/sVftZfEF4R4Z/Z98aNHcf6qe+0xtPhccciW48tMc9c46+hr4ff8ABGP9pPxHJDN498WeD/CFo8eZEFxJqF0jY4Xy41EZ9CfN47Zr4Y/8ESPBGnXj3fxg+Mmq65AFxFZaDYJp+GwOWllMpYdflCr0Bz1FfBr/AIJy/slfBSWa+0n4Z2/ibUZgV+2+KgmpvGvokbr5SH/aVA2DgkitI8H+EvDxDaB4X0jTSpJBs7GOHBIwT8oHUcVVVVV+0L/yQH4mf9ifq3/pHLX/AARD/wCSVfEr/sYLX/0nNVVVVeLf+RU1n/sHz/8Aotq/4IY/81s/7gH/ALkaqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr4iftBfA34Sh1+JPxb8KeHZkGfst9qsSTt/uw7vMb8FNfET/grz+yH4N8yDwvqPiXxvcrlV/sjSWgi3D1e6MXHuqt7Zr4wf8FqfiT4i0250j4L/DLT/CEkg2x6vqd4NSuEH95IfLWNW7YbzB/T4g/8FAf2xvihoR8NeLfjtrJ09my8emW1tpTSezvaRROy/wCyxI9q1TxR4p8QQRWOteI9V1KGOVpIobq8kmVXb7zBWJAY9z1NeCP2Gv2ufiJb2134X+APitre7VWgnv7UadHIrdGD3JjG09d2cY5zivh9/wAEXf2ifEP2a5+IHjjwf4RtpSPOhjkl1G7hHf5EVYmPTgTfiK8Af8ER/hxo2vR3/wASPjbrPifS4m3f2fp2jJpRkx/C8pmmO08Z2hT6EdR8Kv8AgnX+yL8HvEH/AAlPhr4VQahqiMxt5tbupNRW2B7RxykxggcBipYf3utWOnafpduLXTLG3tIQciOCIRrn1wOOwqqqqqv+CUn/ACel8bP+wfqH/pzjqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqmmht4XnnlSOOJSzu7bQoAySSegxXxg/wCCrX7Jfwq1KfQtL17V/HWo2zmOVfDVqs0EbD1uJXjjce8Zcfrj9n7/AIKsfDD9oD4u6D8H9K+F/ijSb/xFPJDZ3dxcQSRKUieQmQKwK/LHjjdyfxrxt8Tfhv8ADW0W/wDiJ8QPDnhi3cEpJrGqQ2avjrtMjDP4V8N/iV8Oviv4cPir4X+KtM8QaL9qlgN5p8m+IzKcyDI6nLZJ75z3qqqqqq/Yf/5Sw/HD/sIeK/8A06rVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVXjn9qv9mv4bSS23jX46eCtNuoOJLNtYiluF+sMbNJ/wCO18Zv+CwX7M/gKw8v4Ww6p8StWLlfJtopdMtY8dS888W7ntsjcHB5HGfiV/wWy+Kmv6R9g+Fvwf0LwjdyKRJe6jqb6u6e8aiKFVP+8HHtXxC/bT/as+KXnR+Mfjx4smt58+baWN6dPt3B7GG2EaEexFeGPBfjj4h6sdL8F+E9d8TanI25rfS7CW9mYk9dsasxJOa+Hn/BMn9sz4iMkifCeTw3aN1ufEV5HY7frESZvyjrwV/wRB8dXLWsvxF+Omg6eu8G6g0XS5bw7e4SSVouTwMlOM5wcYPgP/gkv+x34J1i31m/8Oa/4rktirJb67qhkg3rzuaOFYw4PdX3KehGK8Lfs/fAXwLq8XiDwR8EfAPh/VIRiO90rw1aWk6fSSOMMPzqqqqqqqq/aF/5ID8TP+xP1b/0jlr/AIIh/wDJKviV/wBjBa/+k5qqqqrxb/yKms/9g+f/ANFtX/BDH/mtn/cA/wDcjVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVXj/APa5/Zi+GE0lp43+Ovg6wu4SVls49TS5uYyP70MJeRfxWvix/wAFhv2YfB+jXEnwwi1vx/rGMW0EVlLptsTzzJLcIHVRj+GNicjjqR4u/wCC1/xv1XRrix8IfCzwjoN9KxEd9PNNemFD02oSilx6tlf9mviP+2V+1J8WJZ28a/HPxXcQXBPmWVnfNY2pHp5Fvsjxjjla8J+A/HvxG1M6X4G8Ha94m1B2ybfStPlvJST3Kxqx5r4e/wDBLz9s74gGCWT4YReGLOYcXXiHUIrXZ/vQqXnH/fuvBv8AwQ98VXNvFN8Qvj/pOnzAjzLbRtDkvFI7gSyyxEfUxn6V4J/4JP8A7GfhS10wav4H1PxPfWCL511qmsXAF3IP4niidI8Z/hC7cYBzznQfgf8ABbwtdWt/4a+EXgvS7uyVVtriz0G2hlhCgAbXVAwwAO9VVVVVVVVf8EpP+T0vjZ/2D9Q/9OcdVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVC8t2vG08SDz0iEhQ8ZUkjI9eQfpx6iqqqqqqqqqqqqqqqqqqqqqqqqq/4LG/tReIvDi6P+zL4K1aWxj1iw/tLxPNbyFXnt3dkhtSR0Riju6/xDyx0LBvhd+zJ8efjToOqeKvhn8M9U1jRNFjle91MmO2tYvLTfIommZEZ1XBKKS3I45Gf+Cb3/J7nwr/AOwhc/8ApFPX/Bb/AMHWlx4J+GPxASNVubHVbzTHcDl0miSVQfYG3bHpuPrX/BD/AMZXN54K+KXw+muT5GlapYanbws38VzFLHIVGc/8usWfqPWqqqqqr9h//lLD8cP+wh4r/wDTqtVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVNNDbwvPPKkccSlnd22hQBkkk9BivHH7Y37LXw90DUPEXiD48+C5YNMfy57fTdXhv7nzD0jWCBnkLHHTbwMk4AJHj3/gtP+z3olnPH8Pvh/wCM/E+oIp8kXccOnWrnOBmQvJIOmf8AVHqPfHjn/gsl+1P4jnkTwhpXg3wla7v3Qt9Oa7nUejPO7Ix9xGv0r4rftg/tN/G20bTviV8Z/EOqWDgiSwhlWytZM/34LdY42/FTjJ9a8EfCD4r/ABMkWL4dfDPxT4mLPszpOkT3Sg98tGpAA7k4A714A/4Je/tn+PLq2Sb4Wp4Zsp2Ae917UYLZYR6tErNN+UZrwj/wQ8VNSgm8eftA+bp6gGe20jQfLlf1VZZJWC/7xRvpXw0/4JufscfDG0t47b4P6f4lvIR8974mY6k8x9Wjk/cj6LGo9q0Lw74f8LabHo3hjQ9P0iwh/wBXaWFqlvEn0RAAOg7VVVVVVVVVVV+0L/yQH4mf9ifq3/pHLX/BEP8A5JV8Sv8AsYLX/wBJzVVVVXi3/kVNZ/7B8/8A6Lav+CGP/NbP+4B/7kaqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqvFv7aH7KPgltTh1/4/8AgpLnR1f7Va2+qx3M6sgyyLHEWZn4xsUE54xnivGv/Ba74DaUJ4fAnww8aeIZosiN71rfToZTzyGDyuB05KA8njjn4h/8Fhv2sPFdzKPBY8LeCbTkQrY6YLyZR/tvcmRWb3CKPaviZ+15+018YbGbSviJ8a/E+qadc/6+wS6+y20vs0EISNhx0K18P/gn8YfixIY/hn8L/FPicK215dL0ma4jjP8AtyKpVB7sRXgv/glf+2p4waJ7n4Z2fhy2l24uNa1i3iwD3McbvKMdwUz+teEf+CHlmI7C58eftBzM/kj7fZaRoAAEmeRFcSTHK44y0QJPOB0r4U/8E3/2PvhNaQpZ/Cax8T38WN2oeKManJIR0JjceSp/3I1rRdB0Pw3p0Wj+HdGsdLsYRiK1srZYIk+iIAB+VVVVVVVVVVVV/wAEpP8Ak9L42f8AYP1D/wBOcdVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVfGDWb3wrqHh3xPppHnW0s0UiE4EqMFJRvb5fwPNaBrmn+JdIttZ0yXfDcJkA9UPdT6EHiqqqqqqqqqqqqqqqqqqqqqqqqv+CumnajZftnazc3zMYb/AELTprMHPEQi8s49vMjk/Wv2XtV8Eat/wTp8PN4Ce1Nna/D+e3vUgwPL1BLZxeBwOjGfzGOeTuB75r/gm9/ye58K/wDsIXP/AKRT1/wW1+Knh+bSvh/8FrG9guNYivJtb1CJHy1pGI/Jg3Dt5m+Ygekee4r/AIIcaPdNffF/XypFskOkWykj77k3bHB9gBn/AHhVVVVVV+w//wApYfjh/wBhDxX/AOnVaqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqrxT498DeB9LvNb8Z+MtE0LT9OCG7utSv47aO3352b2dgF3YOM9ccV8Sf+Cuv7IvgS+l0zQdQ8TeN54iVMuhaYBAGGePMuHi3Dj7yBhyMZr4of8FtPiFf3ctt8GvhHoWkWQG1LrxDPJezv/teXC0Sxn2LOO+e1fGz9tL9pb9oCeZfiD8U9V/syaIwto2mSmxsShGCrQRELJnnmTceSM44rQvAfjjxTbfbPDPgzXdXg83yvNsdOluF38fLlFI3YI468j1r4N/8ABKX9qz4uaBY+Kb3TNF8EadfEmOPxNNNb3flg43/Z0id1zjgPsJGD0INfDP8A4IpfBzSLK3n+LHxO8T+I9RABlh0hItOtc91+ZZZGA6ZDIT1wM4Hw7/4Jy/sa/DS4hv8ASfgrpmrX0OCLjXppdTyR0byp2aIHvwg5qzsrPTrSKw0+0htraBAkUMMYRI1HQKo4A9hVVVVVVVVVVVVVftC/8kB+Jn/Yn6t/6Ry1/wAEQ/8AklXxK/7GC1/9JzVVVVXi3/kVNZ/7B8//AKLav+CGP/NbP+4B/wC5Gqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq8QeP/AvhRb8+JvGOi6W2l6dJqV6l3fRxPb2cZAe4dSciNSygvjALAZ5FfFj/AIK6fso/Dy9m0jwrc6/4+vIgQZNEs1jtVccbTNOybh/tRq6+9fFb/gth8StXZrP4M/CrQ/D0GCDea7O+oTt/tKkflJGenDeYOvrx8av22f2nPj9Gln8Q/irqT6ZGTs0vTQun2v8AwKOEL5h5ODJuIzwa8FfCf4p/EpLiT4c/DTxV4qW0kWO4bRdGuL4QuwJVW8pG2khWIB67T6V8Df8Agld+1R8ZtOl1zWNEs/h3p8UojUeLEntLufg5MdssbOAOATJ5YO75S2Dj4T/8EWPg5oNot18YviJr/izUCObfS1XTbRPUHPmSOR/e3J/u14F/4J4fsZ/D2aO60b4EaHfXEZyJdaebVMn12XLug6DooFWGn2GlWUGm6XZW9paW0Yjgt4IhHHEgGAqquAAB2FVVVVVVVVVVVVVX/BKT/k9L42f9g/UP/TnHVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX7Q96jS6LpqkF0WaVh3AO0D+TV8J/Hx8Jav/AGdqMx/sq+YCXJ4gfoJPp2Ptz2oFWUMrAgjgiqqqqqqqqqqqqqqqqqqqqqqqr/gqz+x54i+PHgrS/i/8M9Jk1Lxb4Kt5YbzT4FzLqGnElyIx1eSJ9zKg5YSSAZbaD8Ofjf8AF74PwapYfDf4g6zoFrrMLwalZ2058i6VlKHzIWyjHaSAxGRngivAnjzxd8MfFmn+OvAWuz6Pr2lM7WV9Aql4GZGRiu4EZ2sw6cZrUtT8b/FHxi1/qt9rHinxR4gu1UyzPJd3d9O5CquTlnY/KoHPYCv+CfP7M17+y/8As86d4Y8SwRx+LPEE51bX0Vt3kTyKqpb5BIPlxqinHyl95GQc1VVVVVfsP/8AKWH44f8AYQ8V/wDp1Wqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqri4t7SB7m6uI4Yoxl5JGCqo9ST0r42ft7fst/AW5stO8Y/Eq2v9Qvk8yOy0NP7RkSPkb3MRKoMjADMCewIBI+L3/BbWcXE+n/Aj4QRGFCRFqnim4JL+/2WBht+vnHPoK+Jf/BSH9sf4nrNbX3xhvtBsZQQLTw7CmmhAeoEsYExH1kNaH4h+K3ijS/EHgrQJ9e12HxNc21/rNrBFJeS3ctuZBFLJgMx2md+fVua+D//AATY/a7+MkIv7L4aSeFdNI+W98Vs2mBvTbEymZgRyGEZU+tfC3/giNrktwtz8bPjNY28Cjmy8LWrzM/uJ7hUC9+PKP4V8Jv+CXP7H/wpv01Z/At140vYlxHJ4tuFv4xxgk24RIGJ/wBqM47Yrw/4c8PeEtHtvDvhTQdO0bSrJNltY6fapbQQL1wkaAKo9gKqqqqqqqqqqqqqqqv2hf8AkgPxM/7E/Vv/AEjlr/giH/ySr4lf9jBa/wDpOaqqqq8W/wDIqaz/ANg+f/0W1f8ABDH/AJrZ/wBwD/3I1VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVfX1jptpLf6jeQWttAu6WaeQRpGPUseAPrXxv/b9/ZZ+BOjLqOu/E3TfEl9I5SLR/C91Dqd4xGM7lSQJEOesrIDyASQRXxe/4LbM4ex+A3wfKgr8mpeLJ+QcdPstu5HB7+cfpXxJ/wCCjX7Y/wATvNg1H4z6nolnJkC08OommBAeoEkIEpH+85rwff8AxZ8T+I9TtfBP/CS6/r3iOwms9QisYpr66vreTaJEcKGd1O1M+4FfCP8A4Jq/tf8AxduQsXwvufB9kMh77xcH0xEP/XJlM5+qxEe9fDj/AIIh6g19Dc/F743262aN+9svDenlpJBn+GefAQ49Ymr4Z/8ABL39jj4Z6la60nw7ufFF/Ztvhl8R3z3ke71aAbYX+jRkVoPhzw94V0yPRfC+g6do+nwk+XaWFqlvEmeuEQAD8qqqqqqqqqqqqqqqqq/4JSf8npfGz/sH6h/6c46qqqqqqqqqqqqqviZ8V/hz8G/Di+Lvih4vsPDmjvcpareXrFUMrBiqcA8kKx/Cvhd8ZvhX8bNGutf+FHjrSvE9hY3P2a5msJdwhl2htrA4I+VgenP4Gq8feP8AwZ8LvCV947+IPiK00PQNM8v7Xf3bERw+ZIsaZIBPLui/VhXw0+K/w5+MnhxvF3wv8X2HiPR0uXtWvLJiyCVApZOQOQGU/jVVVVVVVVX7ZP8AwUb0T9kD4n6Z8NNS+FN74nl1LQYtXW7g1dbUIJJ54fL2mJ8kfZyc5/i6cVpV8uqaXZ6msZjF3bpMEJzt3KDjP41VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVSSRRLulkVB6scU+q2CnatwHI7IM/r0pL9JeI4z/wACNIxdcnFVVVVfFjXk8QeN72WF90Nni1iPsmd34bi1V8FfH41G2Xwfq0+bq3T/AEJ2PMkY6p9VHT2+lVVVVVVVVVVVVVVVVVVVVVVVUfibw9N4luPBsWs2ja5aWMV/Np4lHnJbSvJHHKV67GeGVc9Moa+Pf/BOz9lv9obVLjxJ4n8G3Gg+Irty9zrPhycWU87Hq0iFXikYnku0ZY+tRf8ABEf4JLqHmT/GHxu9jniFIbVZf+/mwj1/gr4F/sT/ALL/AOyhFP4v8JeGYU1Syt3e48TeIbsT3FvEF+dhI22OBdudzIqZGckivgn+0l8Lf2h77xZF8KNXk1jTvCF9FY3OqLHtt7qZkLHyCeXRcAb8AMclcrhjVVVVVfsP/wDKWH44f9hDxX/6dVqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqrxv8UPhr8NLQX3xE+IPhzwxAylkk1fVIbQOB/d8xhu6dBXxW/wCCsH7Inw8sbtfDHiq/8d6vApEVlotjIsbv2BuJlSPb6shfA6AniviJ/wAFpP2g/EKzWvw78B+EfCNvICI5p1l1K6i9MOxSI/jEe1fFf4+/Gf45ao2rfFj4la54klLbkhu7ki3hP/TKBMRRD2RQK8C/BL4wfE6+tdN+H3ww8T6/NeOqQmx0uWSPnBBaQLtVcEEsxCgHJIHNfBv/AIIv/GXxPL9t+NPjjR/BNkrcWenqNUu5MHkEqyxRgjo29z6rXw2/4JM/seeAvKuNb8Ma342vIsES6/qj7A3c+VbiJCP9lw34nmvBPw68AfDTSBoPw78FaH4a04Yza6Tp8dqjEd2EYG49eTk8n1qqqqqqqqqqqqqqqqqqr9oX/kgPxM/7E/Vv/SOWv+CIf/JKviV/2MFr/wCk5qqqqrxb/wAiprP/AGD5/wD0W1f8EMf+a2f9wD/3I1VVVVVVVVVVVVVVVVVVVVVVVVVVVVVV45+LHwu+GNt9r+I/xG8M+GIipZTq+qw2hcf7IkYFj7DJNfEn/grH+x54EjuYNC8V6v40v4FYLBoemSBGfHA86fy0IJx8ylgM98Yr4kf8FoP2hvEU9zbfDfwX4S8I2Ls3kSTxSajdxjPGXdliJxj/AJZda+Lf7Qvxt+O9+uofFv4ma34kaNi0Vvcz7LeEnqY7dAsUZ/3VHb0rwl+z98dvHrwr4L+DXjXWlnICS2Wg3EsfOOS4TaByOSQBnrXwV/4IzfGzxc9vqfxm8X6R4G0140kaztB/aN/kjJRlUrFGe27e+D/Ca+G3/BKD9jrwB5U+reEdX8aXkWCJ/EGpu67u/wC6g8qMj2ZW/rXgr4cfD74baYNG+HngfQfDNjgZt9J06K0RsdyI1GTyeTzzVVVVVVVVVVVVVVVVVVVf8EpP+T0vjZ/2D9Q/9OcdVVVVVVVVVVVVVX/BZL/k0W0/7HCx/wDRNzX7FHx48UfsT/Gbwv4g8XvKvw++JulQSals3GN7RpXjS7Ve7280cyngkqJQudwNWl3aahaQ31hdRXNtcxrJDNC4dJEYZVlYcEEEEEcHNf8ABUf/AJMS+Jv/AHCf/TtZ1/wRt/5NFu/+xwvv/RNtVVVVVVVVX/Bav/k6jwt/2T+z/wDS+/rwl/yKmi/9g+D/ANFrVVVVVVVVVVVVVVVVVVVVVVVVVVVVVeB/H0F14o1nwZfXALwX0/8AZ8jH76B2zH/wHnH+z9KqqqqqqqtQ1nR9KXdqeqWtrxkCaZUJ+gJq9+KnhC2JS2u5r1hxiCI4z9WwPyqf4rTznbp2kxx+jTSFv0GP503i7X7/AIkvzGp/hiUJj8Rz+tQSyzPvllZ2PUscmrPp+FWnQVF9yqqqr4p+N08H6A0VpKP7TvgY7ZQeYx/FJ+Hb3I96JLHJOSfWqtLu5sLqG9sp2hngcPHIpwVYdCK+H/jS18baEl6pRL2DCXcIP3X9QP7p6j8R2qqqqqqqqqqqqqqqqqqqqqqqv+CrXxT8e/Bf9t/wJ8Rfhr4judF17S/AVm0FxCchlN/f7o3U5V42HDIwII6iv2JP28/AX7W3h5NEvvs3h/4i6db7tS0IyHbcqoAa5tSeWiJPKElozwcjDt+0b+1N8HP2XPCn/CSfFDxGsNzcI39m6Pa4lvdRYdRFHn7oOMuxCLkZIJAP7XP7f/xk/atv59Gurt/C/gRJM2vhqwuCUlAOVe6k4M79DggIMAhQck/8EQ/+SVfEr/sYLX/0nNVVVVVX7D//AClh+OH/AGEPFf8A6dVqqqqqqqqqqqqqqqqqqqqqqqqqqqqr4w/tCfBb4BaQNY+LnxG0fw6kkZeC2uJt9zcgcHyrdMyyc8fKpx3xXj7/AILVfAbRo7m3+Hnw38YeJbqIMIZb3ydOtpWwcENukkCk46xg4PSvjx/wUo/an+Oc9xZr44l8F6BISE0nwy7Wfyekk4PnSZGAQWCHn5RnFWlj4q8c659nsLPVfEGs3rZ2QxyXdxM3TOBlmPSvh5/wTm/bK+JIim034Jato9rIQGuPEDx6WIwf4jHOyykf7qE18I/+CJehJpdte/HX4t38uoud02neFokjiiH90XE6Mz+58pcdBnrXwW/YV/Za+AtxBqngX4V6fNrVvgpq+rltQulb++jSkiJuOsap1Pqaqqqqqqqqqqqqqqqqqqqqr9oX/kgPxM/7E/Vv/SOWv+CIf/JKviV/2MFr/wCk5qqqqrxb/wAiprP/AGD5/wD0W1f8EMf+a2f9wD/3I1VVVVVVVVVVVVVVVVVVVVVVVVVVVV8Wvjp8IfgVoX/CRfFr4gaR4btGB8lbubM1wR1EUK5klPsik18QP+C1vwR0O6ntPh18LvFPirychLm9uItMhmOOCvErhT6sgPtXx/8A+Cmn7UPx0e40yy8Wf8IL4dlyF0vw27W7uvI/e3OfOfIOCAyof7laXo/inxtrQsNE0vVdf1e7JYQ2sEl1PKe52qCzdq+Hn/BNv9sv4j+XNZ/BnUNBtXxuuPEU0embM+sUpE35IcV8FP8AgihoyWKah+0L8T7ua9ZgRpnhMrHFGB2e4njJfPQhY1xzhj1Hwe/YN/ZT+B88GpeDPhJpdzq1u26PVdZ3ajco4/jRpiwiPvGq9/U1VVVVVVVVVVVVVVVVVVVVVf8ABKT/AJPS+Nn/AGD9Q/8ATnHVVVVVVVVVVVVVV/wWS/5NFtP+xwsf/RNzVz+zNJ+0r/wSm+GfjXwrpCyeNvh7bapc6ekSbpL20j1C5S4txgZJKxiVV5+dNo++TX/BIz9rpvH3g1/2avHmqmTxB4UtzN4cnnky13pwwDACeS0BIwP+ebAAYjJr/gqP/wAmJfE3/uE/+nazr/gjb/yaLd/9jhff+ibav22f2z/CP7H/AIBg1Gezj1rxhru+PQdFMmwSFcb55iOVhTIzjlmIUY+Zl8B6D/wVZ/bd0cfE3Sfi9L4A8MX7F9LddWm0C3nQE/6hLNHndARgPLkN2ZsHHjT4n/8ABS//AIJ661pOr/Fbxo/j7wdeXIh8+/vn1eyu3xkxfaZVW5gkKhiu7bnBOHCkV+zZ+0P4G/ag+FOnfFPwI8sUM7tb39hOQZdOu0AMkDkcEgMrBhwyspwM4FO6RoZJHVVUEszHAAHU1+0F/wAFIPjl8bvi6P2fP2E9PMpluHtY9dgt457rU3TPmSQGXMUFuAGPmsMlRv3Rirz9jv8A4K26ZZN41tf2oLi/1RE84aLD48vzIzY+4IpI1tc8dN233r9jH/gpT8Rm+KMf7N37YFp9l8Qy3/8AZllrktmtnNDfb9i215EgVBubCrIqrg43Ahi6zTQ28LzzypHHEpZ3dtoUAZJJPQYr49f8FEf2if2kPjAfgL+xBb3Nrp73D21vqlhErXurbPvziSUbbW3GMhvlbA3M4DbBa/srf8Fd/hzZHx9oP7SJ8Q6nAom/sGXxhdahI55JQQ3sX2Untjfjpiv21f2hPF37RvxJ0HxD8RfB7eGfGPhfw3H4f8QWPlNEovILy6dmWN8tGCsyZRiSrBhkjFeEv+RU0X/sHwf+i1r9tP8A4KU+ONF+Jb/s3fsjafHqni5L0abfa0loLx1vS237LZwkFXkVvlZ3DKGDKF43VF+x9/wVv8TWK+M9T/aeuNJ1ORPNOiv47voJFbrsMdtGbUHnGA233xXwH/4KNftAfs9fF9vgN+3NaTz2sNwltPq1zbRpeaUWA8uVmiAS5tyCCXGWwdwZsbTDNDcQpPbypLHIoZHRshgRkEEdQRXw7/4KE/E/4QftHfGC4+LHxJ1vxR4Y8Kxa1b6F4eu7pFF1eJfpDaxKduRhSxYgEqiu2DjFfAKH/gqB8ePjP4G+OPju58T6L8OZdftLy70yLVo9ItX08SAuq2IkWSWLZz+8VjICDlzzVVVVVVVVVVVVVVVVVVVVVVVVXjfxXaeDvD8+rTlWmxstoif9ZIRwPp3PsKjv7yK+XU4rl1ull81ZQcMHznd9c18O/G1v420JLpiqX9tiO7iHZuzAf3WxkfiO1VVVVa98R/APhksmueL9KtZFzmFrlWk/74XLfpWsftR/DuzLR6Lb6lqzj7rpD5MZ/F8MP++a1L9pzxPqBKaJoVhYIejTM07j6H5R+hq9+JnjvXiRqHie82t1jgbyVx6YTGfxq1d5G3yOWY9SxyTVl2qz7VadvpVn/SrPp+FWnQVF9yqqqv72102xn1C8lEcFtG0kjHsoGTXi/wAT3ni/XrjWbskBzthiJyIox91R/X3JNVVeCfF154M12HVbYs8J+S5hBx5sZ6j6jqD6/jWnahZ6tYW+pWEwlt7mMPG47g/56VVVVVVVVVVVVVVVVVVVVVVV/wAFDP8Agnte/tY3ll8UPAPitdP8caHpC6dFp9/xZ6hbxySyqgcDdDLumk+c7lPygheWrXvDfxf/AGb/AImx2Ou6drfgfxt4buVngLZgngcH5JY3U4dDg4dSVYdCQa8efEDxt8UfFN942+IXijUPEGu6i+64vb6YyO3oo7KoHARQFUYAAAxX7Kv/AAS1+Nnx7+x+K/iHHP8ADzwXKQ4nv7Y/2hep/wBMLZsFVI6SSbRggqHHFfAD9nD4T/sy+C/+EH+E+gPY2s0glvbq4mM1zfTAY8yVz1bHZQqjsAKqqqqqv2H/APlLD8cP+wh4r/8ATqtVVVVVVVVVVVVVVVVVVVVVVVVVVfGb9pv4Dfs+2Quvi58TdH0KZ03Q2DSGe8mHqltEGlZe27btGRkivix/wWz8EaaZ9P8Agr8JNT1uQArHqPiC6WyiDf3hBFvd19i8Z+lfEv8A4KU/tkfEy+nnl+Lt34Zs5CTHYeGol0+OAeiyLmYj/fkY+9RQ/Eb4weMRFCniPxr4r1mXgATaje3knf8AvSOfzr4af8Egf2sPHWm22r+Jj4W8EQTgMbbWb95LtVI4PlW6SKD0+V3UjPIBGK+AX/BHn4F/DuWDW/jFrd38R9ViIdbRojY6dGfQxKxeXB/vPtOOU5xXhLwL4I8A6aujeBfB+ieHbBAAtrpWnxWkYx0+WNQKqqqqqqqqqqqqqqqqqqqqqqr9oX/kgPxM/wCxP1b/ANI5a/4Ih/8AJKviV/2MFr/6TmqqqqvFv/Iqaz/2D5//AEW1f8EMf+a2f9wD/wByNVVVVVVVVVVVVVVVVVVVVVVVVVVfGL9o34H/AAC03+0vi38StH8P7l3RWksplupx6x28YaVxx1CkDuRXxZ/4LZeCdNaaw+Cnwk1PWpBlU1HxBdLZRA/3hDFvd1+rxn6V8Vv+CkX7YXxYvHluPi3f+FbMnMdh4UJ0tIvpIh85v+BSNUs/j74peKIkmm1/xd4j1JliiVmmv7u5boqr953PoBmvhH/wSI/al+I2mx6z4v8A7B8AWkozHBrM7S3jDsfJhDBO/DsrD0r4G/8ABG34H+BLyLWvjH4q1H4i3cR3JYLCdMsVPBG9EkaSTBHeRVPdTXhPwT4N8A6RF4f8DeFNH8P6bCoWOz0yxjtYlA9EjAH/AOuqqqqqqqqqqqqqqqqqqqqqqqr/AIJSf8npfGz/ALB+of8Apzjqqqqqqqqqqqqqq/4LJf8AJotp/wBjhY/+ibmv+CXH/JiXwx/7i3/p2vK/b5+Bniv9iP8Aae0T9ov4LJ/Zmg65qZ1PSXjjzFp+oqd1xaMo48qQFmCcAo7oBhCa/a9+NPhz9pL/AIJaeLPiv4OQ+TrFppT3Vmrb3sbiPVbT7RC2Bn92ytzgZUBuhr/gjb/yaLd/9jhff+ibav2zXn/aR/4Kc2vwq1e+lOjR6/pHhiJQ5Bgtj5Rudueh8yW4YY7kdetabpun6Pptro+k2cNpZWMCQW1vCgRIY0UKqKo4CgAADsBX7THwv0X4y/APx18OdctIp49V0S4+zGQf6i5RC8Eo9CkqI34elf8ABEfx5qNr8UPiH8LzdO1hqegRawsJJKpLbTpCWUdiVugD0ztXrgYr/goj8QdQ+Gn7G3xK1/R7t7e/u9Pi0u3eMlWH2ueO3kKsPukRySEHjkdc4r/giT8L9Gh8CePvjRNaxy6reaumgW0zAFreGGGOeVV7gO1xFn18pfQ1X/Baf4a6V4S+MPgP4saJCtnfeL9MuYL14TtaSexaHbMSOd/l3ESZ9Il9K/aQ+Outyf8ABLuf4tx3zLrni7wPpUE0gBBM1+IIbnBAGCFlmIPHQY7V/wAESfhfosPgPx98Z5rSKTVrzWF0C2nYZaCGGGKeVV9A7TxE+vlL6VX/AAWP8KaJ4c/a3tNS0izjgn8SeELLUdQKKB5twJ7m33nHfy7eIZ9q+KHjm4+GX7Mfif4h2MgW78OeCbi+tSRnM8VmWiHQ9XCj055r/gi38N9N8Y/GXx78XdeX7dqHhHTbeGzkuDvZLi/eXfPk87/Lt5Uz6St61X/Bbb4X6LN4D8A/GeC0ij1az1htAuJ1GGnhmhlniVvUI0EpHp5retf8E5fHuo/EX9jH4a6zrF09xfWNjNpcruSSVtLiSCLJ7nyo4+f1NfBL4baF8W/+CnB8DeJraG50uf4h61eXVvMm5J1tHurry2Xur+QFIPBDEGqqqqqqqqqqqqqqqqqqqqqqqqq+MniqTxB4rl02J/8ARNIZoI1B6yf8tG/MY+iiq8EXOp+EbODx/aM72aX5sr2EdHjKq359efXbUckc0ayxOGR1BUjuD3qq1SXUYNNuptHs4bu+jhY20E0xhSWQD5VZwrbQTgZwcelfEb46ftU6NPNbTfDoeGrePIFzbae16B7+cS8R/KtY+KHxF8Wsy+I/Gmr3iMTmF7llj9/3a4UflVn/AA1ZdBVn2qz7VZdvpVl2qz7VadvpVn/SrPp+FWnQVF9yqqq+PXiySCC38I2UuDOomuyD/CD8i/iQSfoKqqrRPDo1vwrrmoW6ZutHaGbA6tE28SflhW/A18BfFjrNP4PvJcoymez3HoR99R9R82PZvWqqqqqqqqqqqqqqqqqqqqq/b/8A20PE/wCxxo/gvUvDPgrS/ETeKLm8hmS+uJIhCIViIK7OufMOc+gr/h958U/+iH+FP/Bjcf4V/wAPvPin/wBEP8Kf+DG4/wAK+PX/AAUn0z9pXw1/wjXxa/Ze8GamIVYWWoR6lcQ3lgx/ihmUbl5AJU5VsDcrDivCHjLXPh/4y0vx14LuzY6pod8l5p0ssaT+TIjZQsrLtfGBnK4PpVj/AMFuPi9DY28N/wDBnwjc3UcSrNMl7cRiRwBuYLztBOTjJxnqa/YF/bC8S/tieD/FXiXxJ4O0zw9L4e1KK0iisbh5RKHj3liX6HtVVVVVV+w//wApYfjh/wBhDxX/AOnVaqqqqqqqqqqqqqqqqqqqqqqqq+InxW+Gfwk0Y+IPib480PwxYYbZLqd6kHmkDJWNWO6Rv9lQSfSvj5/wWc+GnhlZ9F/Z78HXPjHUFLKusawr2VgnoyRcTTD2YQ9jk9K+Kv8AwUW/bA+Lbzw6p8YNR0DT5icWHhoDS40U9V8yLEzL2w8jccdzXhnwh8Qfit4mGkeEfDuu+LNevnLGGytpby4lJ6s20FvcseB1Jr4R/wDBHD9o7xvHb6h8S9e8P/D6ylxvhmf+0r1FPfyoWEf4GYH1Ar4Pf8Ekv2VPhvEt14y07VfiHqm0bpdauDDbo3qlvCVGOOkjSdetfD34P/Cr4S2T6f8ADH4c+HPC8MoAmGlabFbNNju7IAzn3Ymqqqqqqqqqqqqqqqqqqqqqqqqq/aF/5ID8TP8AsT9W/wDSOWv+CIf/ACSr4lf9jBa/+k5qqqqrxb/yKms/9g+f/wBFtX/BDH/mtn/cA/8AcjVVVVVVVVVVVVVVVVVVVVVVVVXj/wCKHw4+FWjN4g+JPjnQ/DOnqDifVL5LcOR2QMQXbkfKoJOelfHv/gst8I/CUVxo/wAA/DF5441MZVNU1BHsNPQ/3grATS+m3bGO4Y18WP8Agon+178XpLiLVfi9qOgadOTjTvDR/suJFPVN8WJXXBxh5GyOua8PeF/HnxQ8SDSfCuga74r1+/cv9nsbWW9uZm7sQoZj7k/jXwc/4I7ftG+P7a21X4k61ofw7sJsEwXWb++VTzu8iIiMcfwtKrA9QK+Df/BJT9lf4ZtBqPi+w1T4h6rHyX1uby7VW9Vtotqkf7Mhk6/l4F+EXwq+GEJg+G/w18L+F1ZNj/2RpMNoZBx94xqCx4HJyT3qqqqqqqqqqqqqqqqqqqqqqqqqq/4JSf8AJ6Xxs/7B+of+nOOqqqqqqqqqqqqqr/gsl/yaLaf9jhY/+ibmv+CXH/JiXwx/7i3/AKdryv2jvgT4V/aR+D3iD4S+K0VItUg3WV55YZ7C7TmGdPdW6gY3KWXOGNeFPiH4z/Z58DfHz9jP4qh9Oj1yzbyoZAWW11a0ljlVkzj93cwx7Q2PmxARwTn/AII2/wDJot3/ANjhff8Aom2r9uuy1f8AZo/4KQ2vxjfTJX0y51jSvFNiMcXUcZiFzGD0yZYZlx1AZTxkGvCnirw7458M6X4x8Javb6po2s2qXVjeW77kmicZVgfoenUHg1+2B8bNA+AP7O/jLx7rF9FFd/2bNZaPAzfNdX8yMkEajqfmO5sdERj0Ff8ABEf4XanJ4l+IfxoubRo9Pt7CLQLKdl4mlkkWedVP+wIrcn/rov4V+3/8M9T+LX7IPxH8J6FZvdanFpyajZwxjLu9pNHcMqjuzJE6gdy2Bziv+CKHxp0G30rxt+z/AKpqEFtqc9+uv6RFI4VrwNEkNyqZ6sghgbA5IZjjCk1X/BXz4r2Xxg/aE8IfBHwITrF54LgktJktRvZtTvpIg1suPvMFitwcdGYrwVNftR/AHUU/4Js6n8FNKgF1qfg3wbppTyxzLJpvkSzMoHVnWCXjHJb1r/gid8Y9Cj0Dxx8A9Rv4bfVDqK+INLhc4a7R4kgudvqU8mA4znDkgYVjVf8ABW/4meGfiP8Atbz2Xhi9ju08GaBbaFezRNuQ3Uc080qgjuhuBG3oyMO1fEbwHL8Uv2bfEPw3tgv2jxN4Mn063LNtCyy2hSNs9sOVPPHFf8EfPizp/wAKvj94u+DHjWQaRc+NrWK3t0u/3TDUrKSTbbEH7rss04APVkCjJYCq/wCC13xs0G603wX+z5pV9Fc6na351/V40bJtAInhtlb0Z1mnbHUAKejCv2Dfhdqfwe/ZI+HHgrXLR7bVBprX99DIu145buZ7ko4/vIJVQjtsx2r9j3/lLUv/AGOHiz/0l1Gqqqqqqqqqqqqqqqqqqqqqqqqq8SRSw+ItUhnP71L2ZXz6hzmqt7Swj/Z6lZWQmTMjseP3n2gAfjgAe/414djki8P6ZFMCJEsogwPUEIM1VVWseEvC3iFWTXfDel6iHHP2q0SU/mwNa5+zL8I9ZLSW2iT6VK38dhcMmPojblH4CtW/ZJngJk8NeL45B/DFfW5XH/A0J/8AQa1D4F/EvQyWbQRexr/y0s5Vlz9F4b9KbTdR0yYW+pWFxaSj/lnPE0bfkRmrLt9Ksu1WfarTt9Ks/wClWfT8KtOgqL7lVVV8RdUbWPG2sXZcsq3LRR5/ux/IMfgufxqqqv2f7JLseIY54w8EkMMTqR94NvyPy/nWlifwV8RIImc503UvKdjxuTdtY/ipP51VVVVVVVVVVVVVVVVVVVVX/BUb9lr4z/tPaB8PbD4O+HbXVZvD95fyX6z6hDa+WsqQhCDIy7slG6dMV/w6f/bc/wCib6T/AOFFZ/8Axyv+HT/7bn/RN9J/8KKz/wDjlf8ADp/9tz/om+k/+FFZ/wDxyv8Ah0/+25/0TfSf/Cis/wD45X/Dp/8Abc/6JvpP/hRWf/xyv+CYH7Mvxg/Zk8BeNdB+MGgWul3mtavBc2aQX8V0HjWHaxJjZgOexqqqqqq/Yf8A+UsPxw/7CHiv/wBOq1VVVVVVVVVVVVVVVVVVVVVV4w8ceCvh7osniPx74u0bw5pUR2ve6rfR2kIY9BvkIGTjp1NfG/8A4KvfstfCmGSy8H6zdfEbWgh2W2hDFqrdvMunwgB9YxIR3FfGj/grh+1F8S2n0/wLdaX8OtIkJVY9JiE92U7BrmUEg9PmiWM1qGp+PPij4nSfVNR1/wAXeI9ScIj3E01/d3LckKCSzuevHNfAv/glJ+1F8XGttS8X6Pb/AA40OQgvca+pF2U7lLNf3m4f3ZTFn1r4P/8ABJr9k/4a21vc+LdD1H4g6xGoMlzrdyyW+/uUtoSqbT/dkMmPWvB/gPwR8PdJXQfAXg/RfDmmrgi00qwjtIsgYztjAGcd6qqqqqqqqqqqqqqqqqqqqqqqqqqr9oX/AJID8TP+xP1b/wBI5a/4Ih/8kq+JX/YwWv8A6TmqqqqvFv8AyKms/wDYPn/9FtX/AAQx/wCa2f8AcA/9yNVVVVVVVVVVVVVVVVVVVVVVeK/F/hTwLodx4m8a+JtL0HSLQZnv9Tu0toYvTc7kAZ+vNfGH/grD+yd8NdPdfCPiG++IOr4YR2eiWzpErA4BkuJgqBTg8p5h74wQa+NX/BXX9pz4lG4074ftpfw40iUFVTTEF1elT2a5lHB/2o0jNavrnj74p+KBe67q+v8Ai7xHqThFlup5b+7uGJJCgsWdjknj3r4Hf8Er/wBqn4wG21LxB4bh+HuhykFrvxJuiuCvGdlouZd2D0kEYP8Aer4Kf8Emf2WfhhawXXjbSr34ja2gBe51mRorVWHXZaxMF2n+7KZfrXhHwN4K+H+kroXgPwhovhzTU5Wz0qwjtIgfXZGoH6VVVVVVVVVVVVVVVVVVVVVVVVXd5Z2MJuL67ht4l6vLIEUfia1D4y/DTTpvs83imGVwcE28Ukyj33IpGPoa0nV9N13ToNV0i8S5tbhA0ciZGQfUHkH2OCKqq/4JSf8AJ6Xxs/7B+of+nOOqqqqqqqqqqqqqr/gpD8BviX+0Z+z1b/D/AOFGjW+p63H4jtb1oJ7yO2UQxxzKzbpCBnLrxnPNfsKfCfxv8Df2VPA/wt+I2mw2HiLRP7Q+2W8Vwk6p5uoXEyYdCVOY5UPB4zjtVf8ABS79gPxN+0feaH8VfgnpFlN43tQLDV7Sa5jtVv7QAmOXe5C+ZGfl55ZHAz+7AP8AwTe+A3xL/Zz/AGerj4f/ABX0a303W5PEd1erBDeR3K+S8cKq26MkZyjcZzxX7Xf7Inw+/a8+HyeFfFMr6Xrels8uha7BEHlsJWADAqSPMifCh0JGcAgggEeEP2eP+Cr/AOx+l34N+B1/b+IvCbXDPCljd2N3a7m53pBe7ZYWPVgihS2cluCT+wd+31+2L4x0zxF+1/4+j8OaJYkmO3luLeeWCNiCy21paHyI3YAAu7K3C7g+3FfCH4ReA/gX8PdJ+GPw30ZdO0TSItsaE7pJnPLyyv1eR2yWb1PGAABVftQf8ErfGFv8Rn+OH7G/iS38PauLz7eNBa6NibS5zkvYzjhAW58t9qrkgMFwgnj/AOC3XifTm8BXcN1pySR+TPqiTaLayqnQkXMTbgeR80fz8cHrX7EX/BMWx+BPieD4y/HLXLLxX49hczafa2++W00yZuTOZHAaefnhioVCSRuO1wyqylWAIIwQRX7TX/BKr4peFviRN8ZP2MNcW1drtr230KLURpt5pc7Ek/Y7glU8vJOFZ0KAhcsOmkeBv+C03xVtv+Ff+J/GN74W0SdRBdapeXOlWTxp0J86zU3THGclMk9zyK/aP/4JMfETSvCnw98Ofs/aNB4s1WygvpfF2u32oQ2Ul9dytEYyElk4RQrhVBOByxLMSfDNtdWfhvSbO+gMFzBYwpNEWDbHCAMuRkHBBGRxX7b3/BL5PjV4vuvjV8AtbsvDfjW7k+0alp1yzQ22oXA5+0RyICYJyQCTja7YYlWLM1mP+C3ejacvgKG1u7lVTyItUml0SeUJ03G5diScAnc5L89c4r9lj/glj4jtPiGnxz/bB8UW/ivxD9r+3Joa3L3qzXOcia9nf/WkNz5a5UkDLMMoa/Z0/YY/aQ+Hf/BQRfj74q8IWNt4LHiPX737YmrW8j+TdwXiQt5SuXyWmjyMZGecYNVVVVVVVVVVVVVVVVVVVVVVVVV8YPhpqUupS+K9AtHuYrgZu4Ily6OBguAOSDxnHQ5PToylSVYYI4II6V8NfCvinxLp0Gkamklt4ZhuxdyCRCpuWAGEXPJXjnt171gAYHaqqqqqq5tbW8hMF3bxTxnqkiBgfwNaj8L/AANqRLNoUNvIejWxMWPwX5f0q7+C1ghLaTrM0folxGH/AFGMflU3w38S2BzHFBdKO8MnP5Nihpt/YkLeWU0JA/jjIqz/AKVZ9Pwq06CovuVVVWtjGs34PUXUn/oRqqqvg/4UuPDPhUTXy7bvUnE7oRgouAEU++Mn23Y7V8YLYQfEXURbg7pRE4CjncY16fiM/jVVVVVVVVVVVVVVVVVVVVVfEH4h+CfhT4P1Hx98RPElnoWgaTF5l3e3TEKgzgAAAszEkAKoLMSAAScV8Cf27v2Zv2jPFU3gb4beO5H1+NXeCw1Gyks5LyNBlnh3gB8DJKg7wASVwCa+PP7bn7Ov7NXi+z8C/Fzxbe6XrF9pqajBDBpVxdBrd5JI1bdGjAHdDIMHnjPcV/w9g/Yj/wCikar/AOE7e/8Axuv+HsH7Ef8A0UjVf/Cdvf8A43X/AA9g/Yj/AOikar/4Tt7/APG6+AX7TXwf/ab0fVdd+D/iC51Wz0W5S2vHnsJrUpIy7gAJFUnjuKqqqqqv2H/+UsPxw/7CHiv/ANOq1VVVVVVVVVVVVVVVVVVVVXxJ/bT/AGV/hJqGo6N46+N3hu01PSQReafbzteXMLggFDFAHYPkj5MZHUgDmvib/wAFs/HVzqGrWPwh+EuiWOnljHpt/rtxJcT7enmPDGUUMRyE3EKcZLd/jB8cvi3+0H4rPiz4r+NNR8Rakx2W8crbYbZSf9XBCgCRL7KoyeTkkmvC3/BMX9qPx3ongPWPDPg17ePxdpk13qD62f7PXRHS6liVZ1f94Q8KwyrtQsRIcDCk18FP+CLXwx8Ppbar8d/H+peLL1SrSaVo4NhZA90aU5mlU/3lMJ9q+FvwI+DfwS07+zPhR8NtB8MxsgSSWys1WeYDp5kxzJJ06uxNVVVVVVVVVVVVVVVVVVVVVVVVVVVVV+0L/wAkB+Jn/Yn6t/6Ry1/wRD/5JV8Sv+xgtf8A0nNVVVVeLf8AkVNZ/wCwfP8A+i2r/ghj/wA1s/7gH/uRqqqqqqqqqqqqqqqqqqqqqviv+2X+zD8Edbn8MfEz4xaLpWs20e+fTo1lu7iEbdwDxwI7IxBBCsATkYHNfG7/AILUeNU8X3mm/s+eAPDp8N2+YoNS8S288tzdnHMqxRSxiJc9FbecAE4ztHxw/aP+NH7Ruvx+IPi946vdbkt8i0tDiG1sweoigQBEJGAWxubAyTivhn/wTU/aR+Lfwn0T4heGPCNzY3+q6/JZtYa2Bp4TT/s8UkWoAy4YxM7SJ8oZjhSoxyfgj/wRZ+HmhLbat8fPiFfeJ7xQGk0jQ1NlZq3dGmbM0q+6iE/1+FXwA+CvwP08ab8J/hnoPhtdmx57S0BuJh/00nbMsn/A2NVVVVVVVVVVVVVVVVVVVVVVXl9Y6fD9ov7yC2iBwXmkCL+ZrXPil4B8Pqpv/E1o7OMqlsTOxHrhM4/HFa/+0n4fs5mg8P6JcakAOJpZPs659hgsR9QP61dftMeJXBFl4d0yLrjzWeTHp0K1rfxW+IGuXrXkvie+tAT8sNlM1vGg9AFPP1OT71eX2pavc+ff3tze3Dcb5pGkc/icmtK+Gfj3W4TPp3hW+aMDIaVBCD9C5Gfwr4UfC/4l6L4gh1C8uZtD0+Nw1xGJ1c3IH8GwEjnplunbmqqv+CUn/J6Xxs/7B+of+nOOqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqk0+wlmFxLZW7yjpI0QLD8etVVVVVVVVVRAYbSAeOhqbRtLnOXsYgT3Vdp/SjoFqnMErp7HkVFYSw8Blb9KjUquCOaqqr4kaU+j+ONYtWXCyXLTJx/DJ84x/31j8Kqqrw/qsOu6HYavARtu7dXwP4SRyPwOR+Fa+q+MPjR9kt8PEb+OE4GfliAEh9/uMaqqqqqqqqqqqqqqqqqqqqr/grv8OPH3xB/Zat7rwPY3eow+GPEUGp6xZWsZd2tBDNGZdq5LCN5UYjGApZzgJmv+CfPw58f+P8A9rT4dXHgWyvDH4c1211TV72KM+XaWUL75hI44USIrRAH7xcAZzX/AAWr/wCTqPC3/ZP7P/0vv6qq/wCCIf8AySr4lf8AYwWv/pOaqqqqqv2H/wDlLD8cP+wh4r/9Oq1VVVVVVVVVVVVVVVVVVV1dWtlbSXl7cxQQQoXkllcIqKOpJPAGO5r4/f8ABU39mH4LLdaT4a1yT4i+IoQVSx8PyK1srjPEl4cxgZBB8vzGHGVr44/8FVf2qPi491pvhnxBb/DzQp8qtp4fXbclO2+7bMu73j8sH0rwR8OviN8V9dbQvh54L1/xXqzDzZbfS7CW8lVSwBkcICVXJGWbAGeTXwB/4I5/Gjx2o1f45a7bfD3TGiDQ2UGzUNQkYkEBkRvLiXaTyXLA4BQc4/Z4/wCCfn7NH7Nt5b+IPCXhGTWvE9uo2a9r0ou7mJv70S4EULdfmjRWwcFiKqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr9oX/AJID8TP+xP1b/wBI5a/4Ih/8kq+JX/YwWv8A6TmqqqqvFv8AyKms/wDYPn/9FtX/AAQx/wCa2f8AcA/9yNVVVVVVVVVVVVVVVVVVVNNDbwvPPKkccSlnd22hQBkkk9Bivj3/AMFPP2W/gilxpmmeKT498QwllGm+G3WeNHHGJLnPlKM5BCl2GPu18c/+Cr/7UXxZkuNO8GavbfDjQ5chLbQvmuyvbfduN4Yf3ohF9K8MeEfiN8Y/GJ0bwnoWueL/ABPqsjzvFbRSXdzOxOXlc8seTlnb1yTXwD/4I9fHjx/cW2rfGjUbL4eaGw3Pbb0vdSkGMjbGhMceeM733Ln7hPFfAL/gnL+y7+z7d2mvaN4Ok8TeJLMh4tZ8RSC7licfxRxALFGQeQypvH96qqqqqqqqqqqqqqqqqqqqqrxD4z8LeFrdp9d1u2ttn/LMvukP0RcsfyrWP2h/Atla+Zpa3mpTkkCJYjEB7lm6D6A1qv7TOqXFmY9G8L29nck/6ye5M6qPZQq8/U4+tap8ZviTqqtHJ4llt0bPy2saQkfRlG79au77UtVnV728ubyZjhTLI0jEn0zk1pPhDxRrt0LPSdAvbiXdtIWEgKf9pjwv4kVof7Pvj3UrsRavb2+k24HzSyTJMf8AgKoxz+JFW37MemJj7Z4tuZfXyrRY/wCbNXh74EeANBkM89nNqspXA+3MHVfXCgAfnmtJ8M+HdCB/sXQ7CyJ6tBbqhP1IGTVVVV/wSk/5PS+Nn/YP1D/05x1VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVXx48Ive2Vv4ssYS0lmvk3QUf8syflb8CSD/ALw9Kqqrwp8VLvwt4LvPD8EbPeeafsMp+7Crg7z+BGQPVvavgP4TmLXHjK+Q/OGhtNw5bn53/Tbn/eqqqqqqqqqqqqqqqqqqqqqqtrKzsldbK0hgEshkkEUYTe56scdScDmq8d/FT4ZfC+yXUPiR8Q/Dfhe3kUmN9Y1SG08zHXZ5jAsfYZNWH7c37H+pX39nW/7RngdJQ23dcaosEecgf6x8JjJ656c9BmtA8R+HvFmlQ654W17TtZ024H7m8sLpLiGT/ddCVP4Gqqqqqqv2H/8AlLD8cP8AsIeK/wD06rVVVVVVVVVVVVVVVVVV8T/jF8LPgtoDeJvip480fwzp4B2Pf3IR5iMZWKMZeVuR8qKx9q+PP/BabwzpZuNE/Z0+H8mtXCkqmueIg0Ft06x2yESOOnLtGRj7pr42/tZftCftD3Dn4qfE3VdTsDJvj0mFxa2MRHTFvFtQkDgMwLepOTXwJ/Yx/aO/aMlgm+G3w3v30eVsNruor9j09B3ImfAkx3WMOw9K+An/AARm+FXhSG21n4/+LLvxpqgCs+k6Yz2OnxnupcYmmHH3gYuCcr3r4efCz4b/AAl0JfDXwy8D6L4Z01QN0GmWaQCQgYDOVGXbr8zEk55NVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVftC/8kB+Jn/Yn6t/6Ry1/wAEQ/8AklXxK/7GC1/9JzVVVVXi3/kVNZ/7B8//AKLav+CGP/NbP+4B/wC5Gqqqqqqqqqqqqqqqqqr4i/FL4c/CPw9J4q+JvjbR/DWlRkgXGo3SwiRgM7UBOXb0VQSewr49/wDBaLwLoRudE/Z38DT+JrpMqmt66r2lnn+8luMTSL0++YT7evxv/bA/aL/aIlli+J/xO1O80yRyy6NaN9ksE5yB5EWFcjsz7m9+TXwO/ZC/aH/aKuYh8L/hrqV3prsFfWbtPslhEM4JM8mFbHUqm5vRTX7P/wDwRj+HnhsW+uftEeMJvFt8AGbRNHaSzsUPdXm4mlHuvlde9fDf4RfC/wCD2i/8I98LvAWh+GLEgeZHptmsJmI6NI4G6Rv9pyT71VVVVVVVVVVVVVVVVVVVSSRxRtLLIqIgJZmOAAO5Naj4+8E6XbNdXvirTFROoS5WRj9FUkn8BWs/tI+E7MvHoulX+ouv3WfECN9Cct+a1rP7RnjS+ymkWdhpqfwsEM0g/Fvl/wDHa1j4g+NteBXVPFF/Kh6xrKY0P/AVwP0q1sL7UHKWNnPcuOoijLkflWmfBH4lamkMq+H/ALNFMAQ9zOke0f7SZ3j6bc1afsyai0sf27xXbLHgGTyrZmbPcDJH5/pWmfAT4caeiC40y4v5EHL3Fy4yfXCFR+GK0rwz4d0NVXR9DsbPaMBobdUb8SBk/jVVVVVVVf8ABKT/AJPS+Nn/AGD9Q/8ATnHVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVTwQ3UEltcRLJFKpV0cZDAjBBHpXxI+Ft/4UuZdT0mGS50dyWDAbmt/9lvb0b8+etVXw8+GWp+MrqO8u43ttIjb95MwwZcdVT19Ceg+vFWdna6faQ2NjAkMECBI41GAqgYAFVVVVVVVVVVVVVVVVVVVVVVf8FJ/239R/Za8H6d4I+G89v8A8LD8WQvLbTyoJBpNmp2m5KHhnZgyxggrlHJzt2t4q8W+KPHWvXfinxn4i1HXNYv5C9zfahctPNKx7lmJP4dqr4F/tF/F/wDZx8WQeLfhT4wu9NdZVe7095Gez1BR1juIchZFIyM8MucqynBH7K37Rfhj9qX4M6R8VPDsS2lxMWtdX07fubT72MDzIie4wyup7o6k4JIFVVVVV+w//wApYfjh/wBhDxX/AOnVaqqqqqqqqqqqqqqqq+Nf7SfwQ/Z50c6v8W/iHpeiM0Ze3sGk828uh0/dW6ZkcZ43BdozyQOa/aI/4LM+O/En2jQP2cfCieFbE7kGuazGlzfOOQGjg+aKE9D83m/hXiPxV8SfjH4u/tTxRrmv+MfEmpPsR7maW9uZiSSEQHLY5OEUYHYCvgF/wSR/aO+KwttY+I/2X4aaFKA2dUi8/UHU/wB20VgUPYiV42Hoa+BP/BM/9lX4HLb6g3gz/hNtfhwTqnicLd7W65jt8CFMHodhcf3j1qOKOGNIYY1REUKiKMBQOgA7Cqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr9oX/kgPxM/wCxP1b/ANI5a/4Ih/8AJKviV/2MFr/6TmqqqqvFv/Iqaz/2D5//AEW1f8EMf+a2f9wD/wByNVVVVVVVVVVVVVVVVfGH4/fBv4BaGNf+LnxB0rw7byKTBDcSF7i5x1EUCAySH/dU474r9on/AILO+JNV+1+HP2afB66Jb5KJ4k12NJ7lh/eiteY4+RkGQyZB5RTXjb4gfE341eLf7d8deKNc8XeIb+QRxvdzPcysWPyxxJztGTgIgAHAAr4Af8EoP2lfjClrrXjizg+Gvh+bDGXW4ma/dPVLMEMp9pmi9Rnv8CP+CYP7K/wTNrquoeFX8eeILcK39o+JdtxGjjqY7UAQqM8jcrsuB83eoIILWCO1tYY4oYUCRxooVUUDAAA4AA4xVVVVVVVVVVVVVVVVVVTsiIXdwqqMkk4AHrWr/Fv4daK5iu/FFrJIpwUtg05z9UBA/E1rf7SXhe1hZdC0i+vp8fKZsQx/nkt+GPxrV/2g/iDqDMLCay01D90QW4dgPq+7+QrW/G/i7xJEbfW/EV7dwltxheUiPPrsGB+lWega5qPl/wBn6NfXPmnEfk2zvu+mBzWg/s/+PdYjWe9itNKjPIF1JlyPZVBx9DitH/Zo0OAq+u+Iru8I6pbxLCPpk7iR+VaR8I/h1oxVrbwvazOP47rM5z64ckD8BUFtb2sSwWsEcMa/dSNAoH0Aqqqqqqqqqq/4JSf8npfGz/sH6h/6c46qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqmVXUowBBGCCOta98G/BOuStcR2kunTNklrNgik/wC6QVH4AUf2edMMmR4mudn90265/PP9K0L4LeCtGkS4uLebUpk5Bu2BQH/cAAP0OajjjijWKJFREGFVRgAD0FVVVVVVVVVVVVVVVVVVVVVVV/wVzl1eT9s/Wk1J5DbxaHpy6eHOQIPJydvoPNM34k1/wS6/YO+EfxM+Go/aB+Mvh+HxQ19qE1toekXbFrSGOFtjzSxg/vXaQOoR8qAucEsCL/8AZc/Zp1PSm0S8/Z++HLWLJsEK+GLRAgxgbdsYKkdiMEdq/wCCn/7Fvgj9mTxP4e8c/CiGWy8KeMHnhfSZJWlGm3UQRiI3cljHIrEhWJKlG5wVA/4Ic65q0kPxd8NMZH0yBtJu0yflhmcXSNx6usa/9+6qqqqqv2H/APlLD8cP+wh4r/8ATqtVVVVVVVVVVVVVVV8fv2x/2ef2arOYfEzx9arrCR7otA04i61GbIyuIVPyAjo0hRP9qv2if+Cwnxm+Ia3fh34H6RD8PtElBQag5Fzqkq9CQ5/dwZHZFZlPIkq2tPiR8Y/GRitYPEfjXxXrEuW2ibUb27c4GSfmdz05Oa/Z2/4I3/FDxmbXxB+0J4iTwTpEihzpGnOlzqcgPZm+aGA4wc/vCOhUGvgV+yl8BP2cNOS0+FPw90/T77yvLn1edftF/cjvvuHy+Cedi7UHZRVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV+0L/yQH4mf9ifq3/pHLX/BEP8A5JV8Sv8AsYLX/wBJzVVVVXi3/kVNZ/7B8/8A6Lav+CGP/NbP+4B/7kaqqqqqqqqqqqqqqr47/tYfAL9m6w8/4r/EGx0++eMvb6Rb5ub64442wJlgD0DvtTPVhX7Q/wDwWQ+KnjUXOgfs/wDhyLwNpThk/ta/CXepSqeAVXBigJHUASEHkOKubv4kfGPxkJLmfxH418V6xLhdxm1G9u3POAPmdz14Ga/Z1/4I9/GT4hrZ+IvjlrEXw+0SUhzpsYW51SVOuCgPlwZHdyzKesdfAP8AY7/Z6/Zss0Hwx+H9nFqoXEuuagBdahLkYP75hlAR1WMIp9Kqqqqqqqqqqqqqqqqqq13xj4W8MjGu69Z2bkZEckg3keoQZY/lXiT9ovwlpgEfh20n1mQ9WObeNfxZdxP/AAHHvV/8f/iHerPHDdWVosoIXybfmMH+6SSc+9ahrut6uS2qaxe3hY5PnztJ/M1oPgPxj4nQS6H4evLmInAl2bIz/wADbC/rWl/s8eP75lN8NP09f4vOuN7D6BAwz+NaJ+zd4Ts41bXNUvtRmA+ZUIgj/IZb/wAerTfhP8OdKKm18J2TsvQ3Cmf/ANDJpI44o1iiRURBhVUYAA6ACqqqqqqqqqqqqv8AglJ/yel8bP8AsH6h/wCnOOqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr/gsF8CfBnjjR9G+K2mePPCek+OfDOnmG50bVNatrG41XTC7sjQrM6l3jk83ag5fe4XLKqn/gnZ/wUc8Lfs3eEZ/gx8ZNM1Obwub2S70nVtPjEz6e0mDJFJFkFoiwLhlywZm4IbKv/wAFYf2JFQsvxG1dyASFHh28yfYZjr/god+3BZ/te+LNE0rwVo97pfgrwmJjYi+CrcX1xLtDzyKpIRQqKqLkkAsScttX/gnv+1R+xh+yX8HLjSfFfxQvLjxn4ouxe641toF48VvtUrDbo/lDeEUsSf78j4JUA1/w9g/Yj/6KRqv/AITt7/8AG68NeIdK8X+G9K8V6FO02m61Yw3tnIyFC8MqB0YqeQSrA4PIqqqqr9h//lLD8cP+wh4r/wDTqtVVVVVVVVVVVVVXxt/aC+EP7O/hZ/Fvxa8aWWi2xVvstszeZc3rj+CCFcvI3IzgYXOWKjJr9qH/AIK5/Fz4nveeFfgNBP8AD3w0zFP7S3htWu09fMGVtgeDiPLgj/WYJFeDvAnxQ+N/jL+w/BXhzXfGPiXUpDLKlvE91M5J+aWVznAycs7kAdSa/Z5/4Iv6/qQtfEH7SvjVNJgO1z4d8PyLNcH1Wa5IMaHjBEYkyDw4NfB/4AfBv4BaGfD/AMIvh/pXh23kUCeWCMvcXOOhlncmSQ/7zHHbFVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV+0L/yQH4mf9ifq3/pHLX/BEP8A5JV8Sv8AsYLX/wBJzVVVVXi3/kVNZ/7B8/8A6Lav+CGP/NbP+4B/7kaqqqqqqqqqqqqqviz8Zfhf8DPCk3jX4reM9P8AD2lRZCSXLkvO4GdkUa5eV8fwoCfav2o/+CwPxC8dG88Jfs4abN4M0NwY2168VX1O4Hcxr80dsCMjje/QhkPA0XQPiX8avGx0/QtN8Q+NfFmtTGSRYklvru5c/ekdvmY9cl2OB1JFfs8f8EZPGniJLTxD+0f4uXwxZuNzaBojpcXxH92SchoYj7KJeO4PT4K/s1/A/wDZ50gaV8JPh5peiO8QjuL8R+beXQHP724fMjjPO0naOwA4qqqqqqqqqqqqqqqqqtY8QaFoEIn1vV7SxQj5TPMELfQHk/hWtftBfD/TNyWE15qkg4H2eAqufcvt49wDVx+0nqVzLeLb6LBZxG1cWhLGVxNxtLHgbcZ4x361r3j/AMZeJpWk1fxFeSqTxCkhjjH0RcD9K03SdV1q5FppOnXN7Of+WcERc/kK0b4EfETVpkW50uLToW6y3UyjA/3VJbP4Von7Nfh63gceINbvL2Zj8n2ZRAqD8dxJ/KvDHw08GeErfytL0eKSQtua4uVEshI6fMRx9BgVgAYHaqqqqqqqqqqqqqqq/wCCUn/J6Xxs/wCwfqH/AKc46qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv+C1f/ACdR4W/7J/Z/+l9/VVVV+z1/yQH4Z/8AYn6T/wCkcVVVVVfsP/8AKWH44f8AYQ8V/wDp1WqqqqqqqqqqqqvHfxE8B/DDw/N4p+InjDSPDmkwcPd6ldpboTj7qliNzHsoyT2FftTf8FiPCOg2c/hT9luyXxBqkqMr+JtTtXitLUnjMEDhXmcdcuFQEDiQEgeL/GnxR+Ovjptd8Xa3rnjLxVrMwjjaQvczzMT8sUUag4HOFjQADoABX7LH/BHzxt40W08YftLalceEtHbbJF4dsXVtRuFxkedJ8yW4PHy4d/vAiMgGvhT8GPhb8DfDCeD/AIT+CNM8N6WpDSR2kXzzuBjfLI2XlfHG52Jx3qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq6u7Wyga5vLmKCJB80krhFH1Jq4+LHw4tp/s0ni+xZ84zGxkX/vpQR+ta18cvh1o8QaLVpNRkPSKziLn82wv65rwJ450/4gaTLq+m2N1bRQ3BgIuAoLMFUkjBPGGFVX7Qv/JAfiZ/2J+rf+kctf8ABEP/AJJV8Sv+xgtf/Sc1VVVV4t/5FTWf+wfP/wCi2r/ghj/zWz/uAf8AuRqqqqqqqqqqqqrxj438G/DzQLjxT488VaT4e0e1/wBdfandpbQoT0G9yBk9h1Pav2l/+Cw/ww8Iabc+H/2brA+Mtffci6xf28lvp1qf7yo22Wc9RjCL0O5hwfib8WPit8fvGp8U/EjxVqvinX71xDAJSX2bj8sUESjbGuTwiKBk9Mmv2Vf+CRvxO+KC2XjD4/3V34C8NShZY9KSMHVrtc9GVgVtQR3kDOO8Y618GfgD8IP2ffDY8LfCPwPp+g2rhftM0SF7i7ZRw00zZeQ9fvEgZ4AHFVVVVVVVVVVVVVVVVaz8SPAmgO0WqeKLFJU4aKN/NdT6FUyQa8SftKaZbkweFdEku2H/AC3u28tPwQZJH1K14l+Nvj3xNbCza/i02Ldlhp4aEt7FtxbHtnvzTHVNcvgC11qF5OcDO6WRz+pNaN8EviNrIV/7D+wxt/HeyCLH1Xlv/Ha8P/s0WyFZfFHiF5eOYLFNoz/vtnI/4CK8PfB74f8AhxxNbaGl3OOkt6fOI+gPyj6gVBbW9rGIbWCOJB0VECgfgKqqqqqqqqqqqqqqqqqv+CUn/J6Xxs/7B+of+nOOqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqp7bTDqMdzJBam/8phHIyL5vlgjOD12gsM9vmHrVfEH4m/Dz4UaA3if4leNdG8M6UrbBdaneJArv2RdxyzH+6uT7V8O/2yv2W/ixr8fhbwD8bvDepavM+yCyedraW4b+7EsyoZD7Jmqqqqqqv2H/APlLD8cP+wh4r/8ATqtVVVVVVVVVVU7pGhkkdVVQSzMcAAdTX7Wf/BVD4OfBKx1Dwp8Ib2x8f+Ok3RKLeQvplhJ0LTTqQJSD/wAs4ickEMyGvit8Y/iv+0D4zfxZ8TPFepeJdau5NlukhJSEMeIoIV+WNc4wiADPqea+A3/BLj9qP40SWOqa54aTwB4cuSGfUfEA8ufy+CTHaA+axwRgOI1P94V+zD+xH8C/2VdNjk8D6B/aXiaSLZeeJdTUS3k2fvKh+7Ch6bIwMgDcWIzVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVa58QvBPhu6NjrPiO0t7lfvQ5Lsv1CgkfjV38ZfhraW32k+KIZePlSKJ3c/gBx+OKuv2lPBsbFbTR9XnwDgskaAn/vonH4VfftOdV03wh9Hnvf/AGUJ9O/+NX37SXjCeJ47HSdLti33XKvIy/TLYz9RV18YfiTdymWTxXcoeyxIkYH4KBWteMPFHiJVTW9fvbxFGAkkp2j/AID0/SgCxCqMk9BirL4feONStvtll4U1OWHHD/ZmG76Z6/hWnfCP4i6pOLeHwtdw+r3IEKj8WI/Svhl4RuPBPg+00K9eF7tXkkuGhYlSzMcYJAPC7R07VVftC/8AJAfiZ/2J+rf+kctf8EQ/+SVfEr/sYLX/ANJzVVVVXi3/AJFTWf8AsHz/APotq/4IY/8ANbP+4B/7kaqqqqqqqqqqqv2pP+CnPwE/Z/t9Q8OeEtTh8e+N4A0aaZpk261tZen+kXIyoweqJufIwQucj47/ALRPxd/aU8ZSeMfip4pn1K4LMLKwiJjtNPQ/8s7eHJCLwBnlmxlmY818Cf8AgmT+1T8cI7PV5vCEfgnw/dEMNT8SlrVmj67o7YAzNkYKkoqtkfNjJH7Kn/BP74Gfss2tvq+maYvifxqE/feJNUgVpY2xyLaPlbdev3cuQcF2FVVVVVVVVVVVVVVVI8caGSR1VVHLMcAV4s+OHgfwyXtra7bVrxOPKsyGVT7yfd/LJHpWsftJ+KbtWTRtGsLBT0aQtO6/Q/KP0rWviX488QI8Wp+KL1opBhoon8lGHoVTAI+taL4X8ReJHaPQdFvL7YcO0MRZUPu3QfjXh79njxpqjJJrctrpEBPzB3E0uPZVOPzYVYfs4eCLba19qGq3jAfMplWNT+AXI/OtF8M+HvDkQh0LRbOxGwIzQwhWcDpubqx9ySaqqqqqqqqqqqqqqqqqqqq/4JSf8npfGz/sH6h/6c46qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv8AgsF8TNdvf2sdK8O6XrFzaJ4J8PWy2xt5mjaG4uGaaSRSDlWZGhHHZBX/AAT4XxXo37HXhDxd8UvHmta7qGsWs+s3N9rmoyXTWtqzHykWSUlliWBEbGcAs2OK/bA/aX8T/tS/GjWfHmqajdf2Db3Elv4b02RiEsLENhAE6B3ADuepY4zgKBHJJDIksMjJIjBkZTgqR0IPY1/wS9/aU1/9oX9nttP8capJqHirwLeDTL28mbdLeW5QNbTSHu5XfGSeWMRYkljVVVVVV+w//wApYfjh/wBhDxX/AOnVaqqqqqqqqqq+Pf7WfwH/AGcNHvr34k+PtNh1W1tTPD4ftrhJdSu8j5Fjtwdw3HADNtTuWABI/av/AOCg/wAcP2o7260aXVZfCngZmKweG9MuGVJk7G6kGGuGPcHCDAwgPJ/Z6/Y/+Pf7TupJB8L/AAVPJpKTeVda9f5ttPtTxu3TEfOwyCUjDvgg7a/Y4/4J2fCj9la3g8Uam0XjD4hOn7zXLq2Cx2ORylpEc+WOxkJLtzyqnYKqqqqqqqqqqqqqqqqqqqqqqqqqqOAMk9K1j4heCNBcxap4osIpFODEsvmOv1VckflV58evhnbJug1i4uzj7sNnID/4+FFXX7Sng2NSLPRtYmYZxvSNAfx3k/pWo/tMeIJHP9k+G9PgX+EXEjzH/wAdKVdftD/EK4BER0y3znHlWpOP++mNaj8YPiTqaNHP4ruY1btbqkBH0KAH9au729v5fPvrya5kPV5pC5/M1pOi6tr94un6Lp895csCRHEhY4HUn0FQfCz4iXEwgj8IaiGPQyRbF/76bAqx/Z0+IF3EslxLpVkTjMc1wzMP++FYfrVn+zHqkgH9oeLLWH1ENq0v82Wrb9mbQEK/bPE2oSgdRFEkefpnditP+Bvw0sIBE+hNdv3luLhyx/BSAPwArS/hn4B0acXOn+FbFZVOVeRDKVPqN5OPwqOztInEkVrCjDoyxgEVVVVftC/8kB+Jn/Yn6t/6Ry1/wRD/AOSVfEr/ALGC1/8ASc1VVVV4t/5FTWf+wfP/AOi2r/ghj/zWz/uAf+5GqqqqqqqqqqvjV+0z8C/2erAXfxa+I+laJPJA01tpzSebeXKjPMcCZdgSMbsbc8Eiv2wP+Ck3xf8A2lL2+8K+Er278F/DxmaOPSbSbZc6hH2a8lU5bP8AzyUiMZwd5G4/AX9lz44ftK60dI+Evge71KCGRUvNUm/cWNln/nrO3yg452DLkA4U1+xv/wAExfhn+zfd2vj74g3dr448fQEPbXElvix0pxzm3iblpAf+Wz8jAKqhyTVVVVVVVVVVVVVVf6lp2l25utTv7e0hXrJPKI1H4k1r3x+8A6PIbeymutVkHU2kWEB/3mIz9RmvFX7RfiHU0a18L2MelREYM0hEsp+mRtX8j9RWp61rGtzG41fVbq9kJzunmZ/yyeK8L/Drxh4wkUaLo0pgPW5mHlxKPXcev0GT7Vov7M1msaP4i8SzSOR88VlEEAPoHbOf++RWkfBj4caOFaPw7FdyL1e8czZ+qk7f0q1s7Swt0tLG1ht4UHyRRRhFX6AcCqqqqqqqqqqqqqqqqqqqqqqv+CUn/J6Xxs/7B+of+nOOqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr/gsP4H1Lw5+1kPFk9vJ9h8W+H7S4t5iPlZ4QYJEB9VEaEj0kU96b/gpd4xh/Yzg/ZX0zwTHZ6omlf2HL4kTUDhtN5UosGwFZGiPlE7yMZbGTgfsq/BW4/aF/aB8F/ChYpWstW1FW1R48gxWMQMlw24fdPlIwB/vFR1Ir/gsf4O8A6J+y34NksdB07T73R/FFrp+ii2t1jMFsbW4LwIAPlixGh2jABVPx/4IbPf/wBqfGOKPJsvs+imXpxJuvdnv03/AJfSqqqqqr9h/wD5Sw/HD/sIeK//AE6rVVVVVVVVVajqOn6RYXGq6tf29lZWcTS3FzcSiOOGNRlndmICqACSTwBX7bH/AAVos7OLUPhf+ynqC3F0d0N940KZji7MtirD5z1HnsMDBKBsrIPhj4E8cftCfF7SPClrHrniDWPEmqRf2jdokt5cLHJKomuZWwzYUMWZ2yB1NfA//gjh4B8E/FXVfEvxb8UJ408JWE6t4d0gI0DXIIyWvsYB2HgJGdr43NgZjOkaPpHh/S7XRNB0uz03TrKIRW1naQLDFAg6KiKAFUegGKqqqqqqqqqqqqqqqqqqqqqqqndI0MkjqqqCWZjgADqau/i38OLKd7efxbZl0OD5YaQfgVBBrUf2jPAtpKYbK11S+AP+sjhVFP03MD+la1+0zdsSnh3wzCgHSS9lL5/4AmMf99Grr9oX4iXC7YpdOtz6xWuf/Qiau/jJ8TL6MxTeLJ1BGMxQxxH80UGr3UL/AFKY3Go3091KerzyF2/MkmtH8MeIvEL+XomiXt6e5hgZlX6noPxq2+CXxNumCjwy0Y7tLcxLj8N2atv2cvH0+PNudIt+OfMuHOP++UNaV+zLCEDa54pcv3jtIMAf8CY8/lVt+zf4FiAM+oaxOcchpkUfkEz+tWPwN+GdkQx0Brl16Ge5kb9AwH6Vp/hrw7pMPkaXoVhapj7sVsq5+uBzUVvBACIIY489QqgZ/Kqqqqqqqqv2hf8AkgPxM/7E/Vv/AEjlr/giH/ySr4lf9jBa/wDpOaqqqq8W/wDIqaz/ANg+f/0W1f8ABDH/AJrZ/wBwD/3I1VVVVVVVVU00NvC888qRxxKWd3baFAGSST0GK/bR/wCCsXhTwFb3/wAOP2Zb6z8R+J8NDc+JtolsNOPQ+QDxcyjs3MQOD+85ULH8Tfjl8QCI49f8a+MvEdyWIVZL28vJSMk4GWOACfQAdgK+GP8AwRp+H1l428N+LPHni/UtS8Lx6Jaz6l4blTyp5NT2J5sbzJ/y7lt52rh+Qu7A3Hwx4W8NeCtAsvCvg/QbDRtH02IRWljY26wwwIOyooAFVVVVVVVVVVVVVfX9jplq97qN7BawRjLyzSBFX6k1r/7QXgTSS0OmvdatKvA+zx7Ez7s2PzANeJ/2h/F2rq1voVvBo0J/iQ+bKf8AgTDA/Bc+9X2o6rrV19o1G+ur65c4DzSNI5P45NeHfg74/wDEmx4NEaygb/lvfHyVHvtPzEfRTWg/szsJVl8TeI1MY+9DZRnJ/wCBt0/75rw94D8I+FoFh0XQbWFlHMzJvlb6u2T+HSqqqqqqqqqqqqqqqqqqqqqqqqq/4JSf8npfGz/sH6h/6c46qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv2yv2QfBv7YHw3i8K6zff2P4h0aVrjQdbSAStaSMAHjdcgtFIAu5QQcqjdVwdc/4JL/ALauleITo2neBtD1mz83YNWs/EFqlvtz9/bM8c2O+PLz7V+wF+wPpn7I2j33ivxdqVlrnxD163WC6u7VT5GnW2Qxt4CwDNudVZ3IG7agAAXLf8FS/wBmj9p39pM+EbL4P+GtN1nwz4agnubizXVIre7mvJCFJ2TbEKpGg24fJMjjHAz/AMEo/wBm3x18AvhB4p1T4neGbrQPEnirXAGsLpdskdrapsiZgDxukknI7FSpBINVVVVVX7D/APylh+OH/YQ8V/8Ap1Wqqqqqqqqv2g/2kvhL+zJ4Kfxr8VPEa2iSBlsNOgAkvNSlUf6uCLILHkZYkIuQWZQc1+15+3x8YP2r9Tn0m8u5PDXgSKXNn4aspzskAOVe6cY8+TgHkBFP3VByT+yd+wR8av2rr6HU9GsT4c8EpLtuvE+owN5JwcMttHwbiQYPCkICMM6kjP7N/wCy58I/2WvBa+Efhjoeye4Ctqer3WJLzUpAPvSyADgc4RQEXJwMkk1VVVVVVVVVVVVVVVVVVVVWqfE/wBo121jqHimzSeMkOiEybSOoOwHB9jWqftHeDbO7Ntp+n6hfxo2DOqrGpHqoY5P4gVqn7TZKlNF8KDP8Ml1c/wDsqj/2atW+PHxG1QssGqW+no38FrbqPyZtzD86ufHXjW8bfc+LtYc9gb2QAfQA4FXWq6pfKUvdSurgE5IlmZ+fxNaN4G8X+IrY3mi+Hr27gBx5qR4Un2J6/hWj/AL4iarH5s9nZ6apGV+2T4J/BAxH4gVpP7MupPIG13xNbRIOqWkLSE/i23H1wat/2b/AkRDTX+szkDkNOig/kgP61b/Av4YwEM3h95iMY8y7lPT2DAVpnhXwzosPkaToNharjB8u3UE/U4yfxrAAwO1VVVVVVVVVVVVV+0L/AMkB+Jn/AGJ+rf8ApHLX/BEP/klXxK/7GC1/9JzVVVVXi3/kVNZ/7B8//otq/wCCGP8AzWz/ALgH/uRqqqqqqqqr4x/Gr4Z/APwTdeP/AIp+KbXRdJtvlQyHdLdS4yIoYx80khwcKo6ZJwASP2zv+CjvxP8A2n7q78H+FJLvwh8ONxRNJilAuNSUdHvJF656+Sp2DPO8gNX7Ln7F/wAaf2sNc+z+BNHWw8O2suzUfEeoKyWdr3KqRzLJgj92mTyNxUHdX7Kf7GXwh/ZK8MnT/BVgdT8R3sQXVfEl9Ev2q76EouM+TDkAiNTjgFizDdVVVVVVVVVVVVRwBknpXiX4ueAvC++K71tLq5Trb2Y858+hI+VT7EivEv7SWs3Ya38LaRDYoRgT3J82T6heFH47q13xN4h8T3P2nXtXub6TPyiV8qv+6o4UewArw18HPHvieNLi20j7HbP92e9bygR6heWI9wMVoH7NOnwsk3ibxBJc45aC0j8tfpvbJI/AV4f8G+F/C0SxaDolralRjzFTMjfVzlj+Jqqqqqqqqqqqqqqqqqqqqqqqqqqqv+CUn/J6Xxs/7B+of+nOOqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv2xf2jL39lj4PD4t2fhmDxBHa6xaWl1YSXBgMkMrEMUkAO1xgEEqw7Y5yPgZ/wUa/ZU+OlrbwWfxDtvCeuSgB9G8TuthKG6YSVj5MuT0COW9VGcVDNDcQpPbypLHIoZHRshgRkEEdQRVUxVVLMwAA5J4xXxt/b6/ZY+BFtOnib4oafrGrwhgui+H5F1G7Zx/AwjOyI8f8tWSv2Kf2tpf2wvCni3xxF4OTw5p2i69/Zun2zXJnmkjEEcnmStgKGJf7qjC4xlutVVVVV+w/8A8pYfjh/2EPFf/p1Wqqqqqqqr9tv/AIKKfD39lizuPBfhVbXxV8SpocxaUspNvpm4fLJeOpyDj5hCp3sMZKKwY/Ez4o/FL9oL4gS+MPiBr+o+J/EurSrDCCpYjLYSCCJRhFy2FjQAZPTJOf2Jv+CS73K2HxQ/as05kjIWaw8FhyrHurXzKcr6+Qpz03kfNHWm6ZpujadbaRo9hbWNjZQrDbWttEsUUEagBURFACqAAAAMACqqqqqqqqqqqqqqqqqqtV8T+G9DUnWNesLMj+Ga4VGP0BOTWq/Hn4b6buWHVLi/deq2tsx/IttB/Or79pnR1t5f7M8NXjzjiHz5VRT7tjJH0HX1Faj+0V49vbeS3tYNKsi/3ZYbdmdB7b2Zf0q9+IHjjUGLXXi/V2z/AArdui/98ggfpVzres3ilbvV72cN94S3DNn8zWh+EvEviUOdB0O7vVj++8URKr7FumfatC+A3xB1pXe4srfS1UfKb2XaXPoAgYj6kCtO/Zl1iQqdX8UWUA7i2gaX8i22tI/Z28B2IVtSkv8AUnHUSzeWh+gTBH51afDP4fWSBIPB2lMAMfvrZZT+b5q28L+GbIq1l4d0yAqMKYrNEx9MCsADA7VVVVVVVVVVVVVVVVVX7Qv/ACQH4mf9ifq3/pHLX/BEP/klXxK/7GC1/wDSc1VVVV4t/wCRU1n/ALB8/wD6Lav+CGP/ADWz/uAf+5Gqqqqqqqr9sj9uz4XfskaCbG8ePxD47voS2meHLebDAHpNcuM+TF6ZG5zwoIDMvx4/aE+K37SvjiXxz8U/EcupXZLJZWceUttPiJ4it4s4ReBnqzEZYsea/Yl/4JR+IPiF/Z/xP/aZsrzQvDDbZrLwvloL7UV6g3BGGt4j/d4lYZ/1fDN4a8M+HfBmgWHhXwloljo+j6ZCIbOxsoFhhgjHRURcACqqqqqqqqqqqrWfGHhXw9J5Gt+ILCzl2bhFLOA+PXb1/SvGv7RlrAWsfA1oty44N7dIVT/gCcE/VsfQ14j+InjzxTahdb166ktGZlEcaiGNiMEghAA2AR1z1FeHvBfinxXIE0DQ7m7XODKE2xqfdzhR+deHP2ar+cJP4q12O3Xq1vZrvfHpvbgH8GFeGPhl4J8JbJdJ0SI3K9Lm4/ey59QT93/gOKqqqqqqqqqqqqqqqqqqqqqqqqqqqqr/AIJSf8npfGz/ALB+of8Apzjqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr9o39n/wb+018KtS+E/ji71C0sL6SOeK6sJAktvPGcxuNwKsAeqkYIJ6HBHx//wCCVn7TfwcuLnUvBuiD4keHYiTHeaDETdqvbzLMkybuOkfmDpz2rw/8TPjh8Gb+TSPC/j3xt4Ju7SU+bZ2ep3OntG/cPGrL17gjnvWlf8FCv20dGt/s1n+0H4jkXAGbtYLpuP8AaljY/rz3rUP+CiP7amp2/wBluf2gtfRM5zbw29u3Qj70cat39fT0rxb8a/jp8V5V03xp8VPGviprlhHHZ32sXN2rk9FWJmI6k8Ad/evgL/wTP/al+ON1bXd34Mk8DeHpcGTVvEsbWp2/9M7Y/vpCR0+VUPdxnNfslfsqeDf2Rfhm/wAPvCetahrM9/d/btU1G8AQ3NyUVCyRrkRJtRQFyxHdmPNVVVVV+w//AMpYfjh/2EPFf/p1Wqqqqqqq/b1/4Kkad4FGo/Bz9mjWbbUPEYDQar4phIlg0w8horY8rJMO8nKJnA3NnZ8OPhr8T/2g/iLb+DfAukaj4m8Ua3O0sjM5duW/eTzysflQE5aRzjnk5NfsUf8ABO74c/ssWNt4v8SLa+KviTLF+91eSLMOm7hho7NG+7xwZSN7DP3VJWqqqqqqqqqqqqqqqqtQ1jSNJTzNV1Wzs1xkNcTrGP8Ax4itV+Nnw20oMh8QC7kX+C1haTP0bG39a1X9pnSowV0PwzdTns11MsWPwXdn8xWrftDePr8lbA2GnLjC+TBvYfi5YfoK1Xx54z1sMup+KNRmRusf2hlT/vlcD9KstN1LVJfJ02wubuX+5BE0jfkAa0r4NfEjV1V4vDU1uh/iu3WHH/AWO79Kh/Zr8av5Zm1fRYwfvgSyMV+nyYP51Zfsz+HEgiGo+IdRlmA/emFUjU+uAQxH5mtJ+E/w80i3jgh8LWNwUHMl3EJ2Y+p3Z/wqDwh4TtcfZfC+kQ7c7fLsY1x69BSIkaCONVVQOFAwBVVVVVVVVVVVVVVVVVVVVVftC/8AJAfiZ/2J+rf+kctf8EQ/+SVfEr/sYLX/ANJzVVVVXi3/AJFTWf8AsHz/APotq/4IY/8ANbP+4B/7kaqqqqqqr9vH/gpj4b+AsWofCr4J3tlr3xGw0N5e4E1poDYwd38MtwM8R8qpHz9NjWdl8T/j58Sha2cOs+M/G3iu9JIG64ubyZuSxJ7AAkk4VVUkkKOP2G/+CZng39n6PTviZ8XoLLxN8RlVZreEr5lnob9QIgeJJh3lIwpHyAY3NVVVVVVVVVVRwBknpXij4yeA/C7Pbzar9uuk4NvZDzSD6FshR9Cc14k+PvjnVb+Z9DvV0myOVjgSJJHx6l2UnP0xj9aJ1jxFqZOLvUtQu3zgBpZZWx+JJwK8KfAfxt4gkjm1S2GjWZ5aS6H7zHfEfXP+9trQ/hP4K0XRrfRH0tdQiguTdBrwCQtKVCkkcDGAPlxjvjPNRQwwRJBBEkcaDCIi7QoHYAdKqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/4JSf8npfGz/sH6h/6c46qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqvGHw5+HvxCtlsvH3gTw74lt0BCxaxpcN4qg9cCVWFaz+wF+xprrO17+zz4UjLkE/Y4HtMY9BCy4/CtK/4J8fsX6O4ks/2e/DUhDhgLvzroZHtK7DHt0rwT8IfhR8NUCfDv4Z+FfDOF250jR4LRiO+TGoJz3z171VVVVVV+w//wApYfjh/wBhDxX/AOnVaqqqqqtW1bStB0u71vW9StdP0+whaa6urqURRQRqMs7sxAVQASSeBX7e/wDwU/1b4pnUvg7+ztqlzpfgxt0Gp+IEBhudZXkNHDn5orc8ZPDuODtUlW/Zl/ZW+K37VfjdfCXw50kpZWrI2r63coVtNMiY/ekYdXIB2xj5mwccBiP2Xv2TvhV+yh4IXwr8PtPNxqV2qtrGu3aA3WpSjuxH3IwSdsa/Ko9WLMaqqqqqqqqqqqqo4AyT0rXfHfg/w2Y01rxDaW7yjKJu3sR67Vyce9an+0Z4Es5GisbbU7/HR44QiH/vtg36VqH7S3iqS4ZtL0LSoIP4UnEkrj6sGUfpWs/FDx9rsjteeKL2NHP+qt5DCgHphMZH1zSpe6jc7Y1muriQ9AC7sf5mtK+FPxD1nabTwpeordGuVEAx6/ORWlfs2eLLrDatq+nWKn+FN0zj8AAP1rSv2bPCVqA2ravqN84/hTbCh/DBP61pXwp+HejhTaeFLKRl6NcqZzn1+cnFQW9vaxLBawRxRqMKiKFA+gFVVVVVVVVVVVVVVVVVVVVVVVVV+0L/AMkB+Jn/AGJ+rf8ApHLX/BEP/klXxK/7GC1/9JzVVVVXi3/kVNZ/7B8//otq/wCCGP8AzWz/ALgH/uRqqqqqqlkjhjaWWRURFLMzHAUDqSewr9vf/gqa90NS+DX7LmvMkWWt9X8ZWz4MgxhorFuoHUGcc/8APPHDn4G/AX4pftK/EC38BfDPQ5dT1K5bzLu7mJW3soifmnuJedijn1LHhQzEA/se/sR/DD9kfwsE0eKPW/GeoQqur+JLiECSX1igXnyYQf4QctwWJwMVVVVVVVVVVanq2l6LaNfavqNvZ26dZJ5Ag+nPf2rxV+0X4c03fbeGLKXVZhwJpMwwg+oyNzfTA+teK/il418Y74dS1dobVv8Al0tf3UePQgct/wACJrw14H8U+MJfL8P6NPcoDhpiNkafVzgfh1rwl+zdDE6XfjLVVnAHNnaZC/8AApDg/gAPrXh3wZ4W8KReX4f0W2tGK4aRV3SMPdzlj+dVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVf8EpP+T0vjZ/2D9Q/9OcdVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVfsP/8AKWH44f8AYQ8V/wDp1Wqqqqq8b+OPCPw28Kal448d+IbPRNC0iEzXl9dybI4l6D3JJIAUAliQACSBX7d3/BQnxf8AtTatP4H8FyXegfDGynPk2BfbNq7K2VnusduAVh5CnklmAI/Yo/YL+IP7W+vJrNyZ/D3w70+42alrzR/NOy43W9qp4eXBGWI2J1OThG+Enwg+HfwM8DWHw7+GHhq20bRbBfljjGXmkP3pZXPzSSNgZZiT0HQACqqqqqqqqqqqOAMk9K1v4q/D7w/K1tqHia2MyHBjgDTkH0OwHB+uK179pLw7ap5fh3R7q/kx9+ciBB/Mn6YH1rUf2ifH14xFlHptiv8ACI4C5/EuSP0rxB8QvGnimH7NrniG5uYc5MIxGhPuqAA/iKtbW5vriO0sreSeeVtscUaFmY+gA5NaP8BPiLqyb5rG101T0N5Pgn8EDEfiBWh/szQeQW8SeI3Mx6R2KAKv/AnGT+Qrw58EPh/4fTMulnVZs/6y/Ikx7BAAuPwz71Y6ZpumReRpun21pH/cgiWMfkBVVVVVVVVVVVVVVVVVVVVVVVVVVVVX7Qv/ACQH4mf9ifq3/pHLX/BEP/klXxK/7GC1/wDSc1VVVV4t/wCRU1n/ALB8/wD6Lav+CGP/ADWz/uAf+5Gqqqqq1bVtK0HS7vW9b1K10/T7CFprq6upRFFBGoyzuzEBVABJJ4Ff8FAv+Ck2s/HefUfg98E7+50r4dIzQahqC5jn8Q4PfgNHb8cJwXBy+Adg/ZG/Y1+J/wC1z4x/svwtb/2X4Y06ZRrfiK5iJgs1OCUQf8tZyv3Ywe4LFV+av2f/ANnb4W/sz+Arf4f/AAt0IWlsNr317MQ91qM4GDNPJgbmPOAMKoOFCjiqqqqqqqqqrxJ4w8NeEbYXPiDV4LQMMpGx3SP/ALqDJP5V4v8A2j7+5D2fgzThaIcj7XdKHk+qp91fx3fQVqus634kvRdavqN1f3LnCmVy557KOw9hXhb4H+OvEoSeexXSrRufNvcoxHtH978wB714W+AfgrQQlxqsb61dL1NyNsQPtGOPwYtUMEFtClvbQpFHGMIiKFCj2A6VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVf8ABKT/AJPS+Nn/AGD9Q/8ATnHVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVXxg/b9/ZO+COpT6D4v+LFjd6zbOY5tN0aJ9RmhcdVkMQKRsO6uyn2r4M/8ABTL9mj46/EjSfhV4LPiuLXNbmeKx+36UsUUrLG7n5lkbaNqE8gdqvL2z061kvdQu4ba3hXdJNNIERB6kngCtA8S+G/FlgdV8K+INN1myErRG50+7S4jDqcMu5CRuB4I6iqqqqqr9h/8A5Sw/HD/sIeK//TqtVVVVeOPG/hT4beEdV8d+ONbt9I0LRLZrm+vZyQsUa+wySScAKASxIABJAr9uj9ujxd+1v4uOlaU11o3w50a4Y6Po7Ph7hxkfarkA4aUgnC8rGCQMkszfsBf8E6/EH7SmoWfxO+KFrdaT8L7WY7QGMVxrrocGKE9VhzkPKPQqnzZZPDHhjw74L8Paf4T8JaLZ6To+lW629lY2kQjigjUYCqo6D/8AXVVVVVVVVVVXiDxLoPhay/tDxBqkNlATtVpCSWPoqjJY+wFeIP2jPCOnExaDZXWrvjhyDbx/mw3f+O1rH7Rnja+yml2un6cnYrGZXH4scf8Ajta14/8AGniKNodX8TX08TjDQiXZG31RcKfyrw/4L8VeKm2+H9CurxQcGRU2xg+hdsKD7ZrR/wBnfx5fyKNTax0yP+IyTeaw+gTIJ+pFaX+zX4TtlVtW1nUr2QdRHthQ/hhj+taZ8GfhtpTLJF4ainkXHzXUjzZx/ssSv6VHFFDGsMMaoiDCqowFA7AVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX7Qv8AyQH4mf8AYn6t/wCkctf8EQ/+SVfEr/sYLX/0nNVVVVeLf+RU1n/sHz/+i2r/AIIY/wDNbP8AuAf+5GqqqqtV1XTND0y71rWtRtrDT7CB57q6uZRFFBEgLM7sxAVQASSeABX/AAUL/wCCgetftLa7c/DD4a3txp/ww0q5IBUlJNflRuJ5R1EIIzHEfZ2+baqfsOfsJeMv2t/FY1TVRe6F8OdJmA1XW1j2vcMP+Xa13AhpT3bBWMHLAkqrfDf4a+B/hF4L0z4ffDrw7a6LoWkwiO2tbdcfV3bq7seWdiWYkkkmqqqqqqqqq8V/EDwl4MiJ1zV4knC5W1jO+ZvTCDkZ9Tge9eMP2iNf1UyWfhS2XSrYggTyAPOw9R/Cn4ZPvQGs+ItSwovNTv7lumGmlkP6k14Q/Z28Qar5d34qu10q2Iz5CYedh7/wp+OT6ivC3w78IeDo1/sTR4luAMG6lHmTN6/Men0GB7VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVf8EpP+T0vjZ/2D9Q/9OcdVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX/BW/wDa+8T/AAp0XSv2fvhprU2l614psWvdd1C2kKTW9gWaNIY2HKmVkk3MCCFjx0kNeC/hX8S/iPFfT+APh/4h8RQ6XC019Npumy3EdpGqli0ropWMYB5Yiv8Agm9/ye58K/8AsIXP/pFPX/Bb/wAJNL4T+GHjuGSVVttRvdNuEDnY5kjSSIlfVfKlwf8AbNf8ERfHF5qfw0+JPw6nnd4NA1mz1G3Vmzs+1xSIwX0GbQHHqxPfmqqqqq/Yf/5Sw/HD/sIeK/8A06rVVVVq2raXoOlXet63qNvY6fp8Dz3V1cSCOOCJFLO7seFUAEknoBX/AAUJ/bt1b9qbxe3gvwPeXNl8MdAuW+ww5aNtXmHH2uZTjjr5aEZUEk4ZiB/wTq/YCv8A9pXX4fih8T9OuLX4X6ROQqbjE+u3CHmCMjkQqQRJIMcjYp3bmTStK0zQ9MtNF0XT7aw0+wgSC1tbaIRRQRIAqoirgKoAAAHAAqqqqqqqqqWSKGNpZpFREGWZjgAepNa18W/h5oW5LrxNbTyrx5drmc59MpkA/Uitb/aY0yINH4d8OXFweglvJBEAfXau4kfiK8SfGXx/4lzHJrJsID/ywsAYR/31ksfoTipZbm9mDSyyzyucAsxZiT+vWvDPwP8AHfiMRTy2CaZayKGE1420kH0QZbP1A+taR+zT4ftwr654gvbxh1W3jWBfpzuP8q0n4RfDnRsNbeF7WZx/FdZnJPrhyQPwFRQwwRJBBEkcaDCIi7QoHYAdKqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv2hf8AkgPxM/7E/Vv/AEjlr/giH/ySr4lf9jBa/wDpOaqqqq8W/wDIqaz/ANg+f/0W1f8ABDH/AJrZ/wBwD/3I1VVVO6RoZJHVVUEszHAAHU1/wUv/AOCgEnxp1a8+A3wd1of8IBpk+3VtSt2GNduEbOEYdbZGAxjh2G7lQlfsK/sQ+J/2uvG7XOovdaR8PtBmX+29XSMhpm4P2W3JG0zMMEk5EakMQSVVvBvg3wv8PfCml+CPBWi2uk6HotsttY2Vsm1IY1HAHqTySTySSSSSTVVVVVVVVXi/4geFvBFv5muakonK5jtYvnlk+i9h7nA968Z/H3xT4g32egD+xrM5G6Nt07j3f+H/AIDgj1NafpmteJL/AOzabZ3Wo3kpywjQyMc9ST6e5rwh+zhqF0Eu/GeoC0Tr9ktSHkP+8/Kr+G76ivDnhDw14Stvs3h/SILQEYeRVzI/+855P4mqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/wCCUn/J6Xxs/wCwfqH/AKc46qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv+CuUWpp+2hrbX5PkSaJpzWWRj915ODj/toJK/Zc0bwJo3/BOvQf8AhX8NmtpfeAJ7rUJLcLmXUXtW+1lyPvOJhIhzyAgXooA/4Jvf8nufCv8A7CFz/wCkU9f8Fu/HuixeC/hx8L0uEk1e51S41iSJW+aGCOIwqzDsHaVwPXym9K/4IcaddPrHxg1YJi3ittIgLEEZdmu2wPXAU59Mj1qqqqqq/Yf/AOUsPxw/7CHiv/06rVVVV/wVE/byPxR1i9/Zy+EOtq/g7SbjZ4h1K2f5dXuo2z5KMD81vGwHPR3UEZVVLfsF/sTa/wDtb+P/ALZrMd1p/wAO/D06nXdSjBRrh8BhaQNggysCCx/gQ7jyUDeGfDPh7wZ4e0/wp4U0e00rR9Jt0t7KytYhHFBEowqqo6DFVVVVVVU7IiF3cKqjJJOAB61r/wAX/h94eLRXPiCK6nT/AJY2YMxz6ZX5QfqRXif9pO+uEa28I6MtqDwLm8Idx9EHAP1LD2rXfFfiTxPL52va3dXpzkJJIdin/ZQfKv4AVovgLxn4jVH0bw3fXEb/AHZTFsjP/A2wv61ov7N/i6+KvrOp2GmxnqqkzyL+Awv/AI9Xh/8AZ98DaQyzakLnV5l7XD7I8+yLj8iSKsNH0jSo1i0vS7S0VRgLBAsYA/AVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV+0L/AMkB+Jn/AGJ+rf8ApHLX/BEP/klXxK/7GC1/9JzVVVVXi3/kVNZ/7B8//otq/wCCGP8AzWz/ALgH/uRqqqq/4Ko/t4tpMWo/sufB7WmW9lUxeMdWtZceShHNhGw53N/y1Ixgfu+dzgfshfsoeNf2tvijB4K8P+bYaFYBZ/EGtmLcmn25OOASA0rkFUTqTkn5VYj4XfDDwR8GfAWkfDX4daJFpWg6JB5VrboSSSSSzux5Z2YlmY8kkmqqqqqqqrVdW0zQ7GTUtXv4bS2iHzSyttA9vc+3U149/aFu7zzdM8DRm2hOVa/mX943+4p+6Pc8+wNW9trXibVfKt4rvUtQu3JIAaWSQ9yTyfqTXgn9nOSQR3/jm72DGRYWz8/8DkHT6L+daLoGieHbMWGh6Zb2UI6rEm3cR3J6sfc81VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV/wSk/5PS+Nn/YP1D/ANOcdVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX/AAV+/ZS8Q/Efw5pH7Q/gDR5NQ1HwjZPZeIbW3jLyvpwZpEuFA5IhdpNwAJ2y7uFQ18Hf2tfj58CfDmreDPh547mg8Oa5BLFe6Pdwpc2zeYhR2RXBMTkHloypOBnOK+FPxQ8XfBf4g6P8TvAd3b22v6FJJJZTTwLMiM8bRklG4PyuevGcV8RviT8QfjP41vPHXxG8SXviLxHqrqJrqcDc+BhUREAVFAwAiAKOgAr/AIJn/s2av+zp+zpbL4w057LxX4zuv7X1S2lTbJZoyBYLdx1DLGNzKeVeR1PSqqqqqr9h/wD5Sw/HD/sIeK//AE6rVVVf8FTf233+D3heX9n34X6z5fjbxJaZ1m9t5MPo1hIMbVYfcnmHTuiZbgvG1fsv/s4eNf2pPizpvwy8HxmGJ/8ASNX1Jk3R6bZqQJJm9TyFVf4mZRwMkfB/4R+B/gX8OdF+F/w80pbHRtFgEcYODJO55eaVgBukdssx9TwAAAKqqqqqvEnjbwp4SiL6/rdtbPjKwlt0rfRBliPfGK8TftKMQ9v4R0MDqBc3x/URqf5t+FeJPHfi3xax/t3Xbi4jJyIA2yIfRFwPxxmvD/gPxh4p2tofh+7uI26TFNkf/fbYX9a8Pfs161clJvE2t29nH1MNsplc+xY4Cn/vqvDPwt8D+E1RtN0SKa5T/l5ugJZc+oJ4X/gIFVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV+0L/AMkB+Jn/AGJ+rf8ApHLX/BEP/klXxK/7GC1/9JzVVVVXi3/kVNZ/7B8//otq/wCCGP8AzWz/ALgH/uRqqqv+CjH7aNt+y18NP+EZ8G38TfEnxbA6aRHw502DO1711PpgrGDwz84IRhXwp+FvxB/aF+KOl/DzwXazar4i8R3h3z3Dswjyd0txO/JCKNzuxyeD1JAP7Mv7OHgT9lz4Vad8M/BMCzSRgTarqjxBJtTuyAHmfGcDjCrk7VCrk4JNVVVVVVTzwW0L3FzMkUUSlneRgqqB1JJ6CvHf7REdrPJpnge3iuCmVa/nUlM/9M14z9Tx7Ec14g8V+I/FU4uPEGs3N6yklFkbCJnrtQYVfwArwD8DfEPisRajrQfSdMbBDSJ++lH+yh6D/ab1yAa8LeDPDngyx+w6BpyQbgPMlYbpJSO7N1P06DsBVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV/wAEpP8Ak9L42f8AYP1D/wBOcdVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVWua9pHhvTZNW1u/jtLWLhpHyeT0AA5J9hXir9o+9acw+DdJiW3jbBub1Sxf6ICNvTuSfYV8NfEeoeLfBOm+INUEIurrzfMESlV+WV1GASeyiqqR0jjaR2wqAkk9gK0742fFvXWmOg+HYb9YSN4ttOlm8sHO3dtJxnB/I1/wALN+PX/Qiz/wDgln/xr4dax4m13wzHf+LdNax1AzOrQtbtAQoPB2tzVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVTKrKVYAgjBBFfH3/gk7+zb8Y9VufE/hI3/AMOtau5DJOdFjR7KVz1Y2rcIenETRr14JOah/wCCGuo/2hsn/aWtvsIPEieEm81v+AG7wO/O4/0r9mr/AIJmfs7fs56za+MzbX3jXxZZOJLXU9dCNHZSDo8FuoCIwIBDtvdTyrLVVVVVVX7D/wDylh+OH/YQ8V/+nVaqq/a8/aX8Ofsq/BbVfiRqohutWk/0PQNNdsG+vnB2Kcc7FALueyqQOSoN/e/ET47/ABPlvbg33ibxp401bO1V3S3l1O+AqgcAZIAAwqgADAFfsTfsmeHv2SvhDbeFYxb3nizWNl34l1SNc/aLnbxEhPPkxAlUHGSWfALkVVVVVUskUMbSzSKiICWZjgKB1JNeJfj74H0KaSz09rjV548gm1AEWfTeTz9VBFeMPjR408U3gkstSuNGtYxiO3sp2Q+5ZxgsfyHt1qzsNY8Q3/kWFpdaheTEsVjRpHY9yep/E14W/Z18T6oEufEl5DpMJGfKAEsx/AHav5kj0rw18GvAPhoLLFo639yv/Le+xMfwXG0fUDNBVVQqgAAYAAxiqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv2hf+SA/Ez/sT9W/9I5a/wCCIf8AySr4lf8AYwWv/pOaqqqq8W/8iprP/YPn/wDRbV/wQx/5rZ/3AP8A3I1VV8cvjN4N/Z/+FuvfFfxzd+XpuiW5dYVYCS7mPEUEYPV3chR2GSTgAkfGn4veOv2ifivrPxN8Zzvd6z4gu/3VtCCy28f3YbaFeuxF2qB1PU5Ykn/gnD+xdbfsw/DMeLvGenIfiP4ut0k1NnXLaZbnDJZqexBw0hHV8DkIpqqqqqqqviB8YvDfgcvp8R/tLVVH/HrE+BGf+mjc7fpyfYZzXjP4m+LfHDmPVb/yrPOVs7fKRD0yOrH3YnHbFfD74UeIfHs6zpG1lpSn95eSocN6iMfxH9B3PY+FvhT4I8JLHJYaQlxdx8/a7sebJkdxnhT/ALoFVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV/wSk/5PS+Nn/YP1D/05x1VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVeJvDWl+LtGm0HWEdrWdkZhG21vlYMMHt0x9Ca+PPh/RfDfw30/TdC02Czt11aP5I1xk+VLyT1Y+5ya+B3/JLdE/7eP/R8lVXji9Gm+Ddcvgfmh0+crn+9sOP1xX7NNh5XhXU9RIwbi/8ALGe4RFP83NVVV4r/AGhPDGg3smn6NYy6zLCxV5ElEUWR1CtglvqBj0Jrwl+0FpXiXV7TRLvw5d2dxezLFCYphOoYnA3cKQPfBxVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV/wUS/4KE/Fv9mP45+Hfh58IxoE8FroYv8AWYNUsjcJcSzSsI422sjpsSLd8rLnzuc4GP2NPjj44/aP+AejfF7x54Q03w7d6zcXC2tvYTySJNBFIY/O2uN0e50kAXLfKqtu+bA/b/8A+ClnxP1n4k618If2fvF914X8MeG7qSyvNZ0yby7vVLmM7ZWSZfmihVwVXyyC2CxYhgq/Df8Abb/ap+FviKHxH4f+OPiu+ZJd81jrWpy6jaXP94SQzMynI4LDDDswPNfsk/tIaF+1T8E9I+Kek2qWN8ztZ6zpyuWFlfRgeYgJ6qQyOp/uyLnnIqqqqqq/Yf8A+UsPxw/7CHiv/wBOq1VXNxb2dvLd3dxHDBAheWWRgqooGSxJ4AAGSTX/AAUB/asu/wBqf443mo6ReynwT4XaTT/DUBJCvGG/eXRX+9MyhuxCLGp5Wv8AgkP+x9FoujD9qn4gaXnUtTR7fwhbTR4NvbnKy3mD/FJyiHjCBzyJARVVVVq2s6ToVm1/rOpW9nbr1knkCDPoM9T7DmvFv7R2k2W+08H6e1/KOBc3IMcQ9wv3m/HbXifx94u8Xuf7c1qeaInIt0OyJfTCDj8Tz714Q+GHjDxswk0nTvKte93c5ji/A4Jb/gINeFP2ctE06WO78U6m+pyIc/ZoV8uLPoT95h/3zWlaJo2hW5tdF0u0sYicslvCIwT6nHU1VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV+0L/yQH4mf9ifq3/pHLX/BEP8A5JV8Sv8AsYLX/wBJzVVVVXi3/kVNZ/7B8/8A6Lav+CGP/NbP+4B/7kaqq/4KsftZv8a/i2fg74O1QS+C/h/cvFK0T5TUNTAKzS5BwyxDMSe/mEEhxX/BJH9j4fEHxh/w0t4+0vf4d8KXXl+G7eaP5b3UlwTPg9UgyCD3lIwcxMKqqqqqq1/xBo/hjTJdX1u+jtraIcs3Vj2VR1JPoK8ZftBeJtYnktvCwGk2XKq5UNO49STkL9F5HrWn+C/Fmt61Bo1vpFy99eRLcKJVK5jYAiQsf4eevrx14rwP8AfDmgLHe+JimsXwAPlsuIIz7L/H9W49hUcccUawwoqIihVVRgKB0AHYVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVf8EpP+T0vjZ/2D9Q/9OcdVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVftKf8iLY/8AYWj/APRUtfA7/kluif8Abx/6Pkqq+IGg6l4o8H6joGkTQRXV4iojzsVQDepbJUE/dBHTrirf4GfFHSrMQaR42t4UQllhhvZ4lz3IwuM8Cl8e/Fv4VatDY+LmnvrVzkR3biUTLxkpNyc/UnGeRXh3XtO8UaLaa7pUpe2u49y5GCp6FT7ggg/Sq+ILX6eB9dbTN32kafLs29R8pzj3xnHvX7OV54Vt7/UINRe2j1iYoLNpgAWTncqE984yByePSrjSNKuryDUbnTbaW7tiTDO8QLxnGOG6jg1VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVf8Fb7LULX9tHXp70v5N5oumy2m4nHlCAIcZ7eYknTjOe+a8C/t/fBX4U/8E6dD0/wh43sU+JGleGP7EstBjJ+1wajtMYuGTAxGCfP3/dP3c7jivh74G1/4o+PfD/w88MxCbV/EupwWFoHzgSSuFDMRkhRnJPYAmv8AgoV/wTx+E/7L3wM8N/Ej4a6zrtxqEWrQaVq41G4WRLzzIZG89VCjy23xY2g7cP6rk/8ABDrxLdvb/FvwdLIzW0T6Vf269kdhcxyH6kJF/wB81VVVVVfsP/8AKWH44f8AYQ8V/wDp1Wqr/grZ+1C3wk+D0PwV8Kaj5fif4iROl20b4e00pTtlb1Bmb90OxUS9xX7FP7NOoftTfHnRfh+8cyeHrM/2h4juo8jybGNhvUMOjyErEp7F92CFNaVpWm6HpdnomjWUNnYafbpb2ttCgRIYkUKiKB0UKAAOwFVVVr3iPQvDFidQ17U4LOAcKZG5Y+iqOWPsAa8ZftHXUxey8E2AgTp9sulDOfdU6D6tn6CtV1nW/El99r1fUbq/unOFMrlzyeijsPYV4T+BXjXxJsuL+3GjWh/5aXakSEe0fX/vrbXhT4KeB/C4SeSw/tS8Xnz70BwD/sp90fiCfegqqoVQAAMAAYxVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV+0L/yQH4mf9ifq3/pHLX/AARD/wCSVfEr/sYLX/0nNVVVVeLf+RU1n/sHz/8Aotq/4IY/81s/7gH/ALkaqv8Agon+1An7Mv7P1/daFqCw+M/FxfS/DyK+HiZl/fXQHXESHIPQO8QPDV8Bfg14o/aF+Lvhz4S+FMrfa/eBJblkLraQKC007gfwois2O5AHUivht8PPC3wm8A6D8NvBOnrZaJ4dsY7SzhA52qOWY/xOxyzMeWZiTyaqqqqqviN8YNB8DI2n2pTUdXxxbI/yw+8hHT/d6n2BzXirxh4g8aah/aGvX7TuMiKJfljiB7Ivbt7nuTXw0+BN94gWPWvF6zWOnnmK1A2zTj1P9xf1Ptwat7W3tYoobeJVWGMRp3IUdBnrVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVf8EpP+T0vjZ/2D9Q/9OcdVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVftKf8AIi2P/YWj/wDRUtfA7/kluif9vH/o+Sqqq+P1lZXXw3vLm5VfOtJ4XtyRyGLhTj/gLN+Vfs3SXL+CLxJcmJNScRE/9c0yB7Z/UmqIBGDzXjb9nvRtcuZtT8MXo0u4lJZrZ03QM3tjlPwyPQCtA8deOvhL4nj8MeMp5bnTwyiSOZzLtjPAkic8446dOCMA9AQwBUggjg1VVVVVVVVVVVVVVVVVVVVVVVVVVVVVMVAyWA7c1VVVVVVVVVVVVVVVVVVVVVVVVX/BVP8AYx8V/tAeGtG+Lvwp0h9V8X+EbZ7S80uFczalp5YuBEP4pInZ2CDlhK+MsFVr/T77Sr2fTdTsp7S7tnMc1vcRGOSJxwVZTggj0Nf8EnP2J/Fmm+LYv2nfir4dutJtbC2ePwnY3sJjluZJUKveFG5EYjZlTI+YvuHCqT/wW3+KkMfh/wCH/wAErTzGluLyTX747CFRY0eCAbu5JkuCR22qe4r/AIIh+BrvT/h/8TfiNcW7LBrmrWWm2zuuM/ZIpJJNpxnGbtAecZX2qqqqqq/Yf/5Sw/HD/sIeK/8A06rVeKvE2h+CvDOreMPE2oR2OkaHZTXt9cyfdhhiQu7n6KpNftPfHbXP2kvjd4m+LOtPIkWp3Rj0y1c/8edjH8sEOBxkIAWI6uWb+I1/wTL/AGYF/Z4/Z/tNd8Q6d5HjLx+sWp6t5iFZLaDaTa2xB6FEdmYEZDyuD0FVVXd3aWFtJeX11FbwRLmSWVwioPUk8CvHP7Q2l6esmn+C4Rf3I4N5KpEKf7o6ufyH1rWNa13xVqZvdWvri/vJmwu75jz0VVHAHsBivBX7P3iLXVjvvEsp0izbkRFd07j/AHeif8C59q8KfDrwh4MjX+xNJjFwBg3Uw8yZvX5j0+gwPaqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr9oX/AJID8TP+xP1b/wBI5a/4Ih/8kq+JX/YwWv8A6TmqqqqvFv8AyKms/wDYPn/9FtX/AAQx/wCa2f8AcA/9yNU7pGhkkdVVQSzMcAAdTX/BQP8AaZf9p39oTVdd0fUGn8IeHAdL8NoDhXgRvnuAPWWTc+Tzs8sH7tf8Ef8A9mBfAHw0u/2iPFWnbNe8cxfZ9GEqEPbaWj5LAHp58iBvdI4yDhjVVVVVTTQ28LzzypHHGpZ3dtoUDqST0FfE349yTGbQvAk5SPlZdSHDN6iIdh/tdfTHBrQvD+u+LtVXTtGs5b27mJZjnp6szHgD3NfDr4KaF4OWLU9XEep6uMESMv7uA/7Cnqf9o8+gFVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVf8ABKT/AJPS+Nn/AGD9Q/8ATnHVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX7Sn/Ii2P8A2Fo//RUtfA7/AJJbon/bx/6PkqtL+NXjm8+JEOlzCP7BPqItTp/kLmNS+z72N24dTk4z2Ar4pfEi5+HcWmTwaMb+O7lcTEsUCKoHAYAgMSeM9gai/aV8HNAGn0TWVmx9xUjZc/728H9K8UeL/GHxw1O38PeHdGeGwhff5QfIB6eZK/QADOB7nqcV4K8K2ngzw1Z+H7Rg/wBnTMsuMeZIeWb8+noMDtVeNPEqeEPDF/4ia2NwbSMFYgSN7EhQMjOBk8nsK0/9ozwPc2okv7TUrOcD5ovKEgz/ALLA8/iBXiS+v/jr4/tYfD+mTQ2kMSw+ZIvMcQYlpJMZA6nAz2A5JqONIokijGFRQqj2FVVVVVVVVVVVVVVVVVVVVVVVVVVVVVXiC1a90O/tY3KSPbP5bqcFHx8rA9iDgj3FfDTxzD410NWndV1K0AS6jHGT2cD0b9Dn2qqqqqqqqqqqqqqqqqqqqqqqqqvEtj8OLG8h8T+L7Pw3b3VuQYdQ1KOFHjK4wVkk5GOOh44r/hbHwr/6KZ4U/wDBzb//ABda147+B/iXTpNH8ReM/A2q2MwxJa3uo2s8T/VHYg/iK+GmnfCjRPD7aD8HrXwrZ6JaTu7Wfh1YEt4ZZCXY7IflUsST0qqqqqq/Yf8A+UsPxw/7CHiv/wBOq1X/AAWP/aR/4RD4d6T+zl4a1DZq3jLF/rnlthodOif93Ge482ZP++YHB4av+CbH7M3/AA0b+0RYXGu2XneEfA3l6vrW+Pck7K/+j2x/66OpJB6pHJVVXxE+Lfh/wHG1mrC/1Yr8lpG3+r9DIf4R7dT6Y5rxf498T+N7rz9c1FmiVsxWsfyxRfRf6nJ968FfB/xf408u6jtf7P09gD9rulKhh6ovVvr0968D/Cvwr4FRZrG2+1ahjDXtwAX99o6IPpz6k1VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX7Qv/JAfiZ/2J+rf+kctf8EQ/wDklXxK/wCxgtf/AEnNVVVVeLf+RU1n/sHz/wDotq/4IY/81s/7gH/uRqv+Cpf7SP8Awov9nW68IaBqHk+K/iN5ulWOxsPBabR9rnHcYjZYgRyGmUj7pr9kX9nzU/2nPj14c+FloJI9NmlN3rd0g/49dPiIMz57M2VjU9N8iZ4rSdJ03QdKstC0ayis7DTreO2tLaFdqQxIoVEUdgFAAHtVVVVWt63pXh3TJtX1m9jtrWAZd3P5ADqSewFfE34uar47nfT7EyWWio3yW4bDTYPDSY/9B6D3618O/hZrvj+5E0Sm00qNsTXki8HHVUH8TfoO56A+FfB+geDNMXTNBsliXgyStzJM395m7n9B2xVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVf8EpP+T0vjZ/2D9Q/9OcdVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX2m6dqkIt9SsLe7iVtypPEJAGxjODnnBP51aWdpYW6Wljaw28EedkUUYRVycnAHA5JNUnh3QI9VbXY9FsV1Fhg3Qt1Ep4wfmxnpxUsMU8TQzxpIjjDI65BHuKl+HngOeXzpPB2jF85J+xIMn3AHNWWn2GmW4tNNsbe1hX7sUEQjUfgOKqiAw2kAgjkEVd+A/BF9L5914S0eSQnJc2SZJ9zjn8asNN07S7cWml2FtaQjpFBEI1H4AAVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVrVylno99dyEbYLaR2zxwFJP8q8J+J7/wAI63BrOntkodssROBLGfvKf88EA9q0XWbDxBpVtrGmTeZb3KblJ6j1BHYg8Gqqqqqqqqqqqqqqqqqqqqqqqr/gp/8At0eJv2ddP034O/CS9Wy8a+JLE3l5qu0O2lWRdkXygQR50jJIAx+4qkgZZWXxN4r8UeNdYm8Q+MfEmp67qlwf3t7qV29zM/1dyWPU9+9VX/BEP/klXxK/7GC1/wDSc1VVVVVfsP8A/KWH44f9hDxX/wCnVa1vWdK8OaNf+IdcvorPTdLtZLq8uZThIIY1LO7ewUEn6V+0v8atV/aH+OPi34tam0qprV+32C3kbP2WzjAS3i9AVjVc4xltx7mv+Cb/AOzsv7Pf7M+hwavpwt/FPjADWtcLD50aVR5EJ7jy4dgK9naQ96q6urWytpLu8uYoIIVLSSyuFVAOpJPAFfEj4/KyS6L4DkOTlZNRZcfXygf/AEI/gOhrSdG1zxbqy2Gl209/fXLFm5LEk9WZj0HqTXw9+BmheGEi1LxCsWqaoACFZcwwH/ZU/eP+0fwArAAwO1VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX7Qv/JAfiZ/2J+rf+kctf8ABEP/AJJV8Sv+xgtf/Sc1VVVV4t/5FTWf+wfP/wCi2r/ghj/zWz/uAf8AuRqv+Ch/7QX/AA0P+034i1nS74z+G/DLHRNC2sSjwwMweYD/AKaSmRwcA7SgPSv+CQP7O/8Awrf4IXnxp1+xRNc+IsgNkXX54dMhJEY9vMk3yHHVREewxVVVV4z8b6F4F0o6lrNx8zZEFunMk7Dso9OmSeBXjv4ga74+1L7Zqcvl20RP2a0RvkhH9W9WPX2HFfCv4KXXifydf8UxSW2k/eig5WS69D6qh9ep7etWlnaafaxWNhbRW9vAoWOKNAqoB2AHSqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr/AIJSf8npfGz/ALB+of8Apzjqqqqqqqqqqqqqqqqqqqq1vXNJ8Oac+ra1era2kRAeVlJAJOBwAT1Irw/4l0LxXZPqHh/UUvLeOUxNIqsuHABI5APRh+dVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVTMiDLMAPUnFfaYCcLIGPtzSsHGR+tVVVVfGHWl0fwLexq+Jb8i2jH+9y3/joaq+Efj//AIRbVP7H1OfGl3zgMzHiCToH+h4B/A9q4IyDmqqqqqqqqqqqqqqqqqqqqqqq/wCCu+l6xYftl6reakjC21LQdOn08kHBhERjbH/bWOWv+CaH/BPv4N/Hb4ZXPxs+Ndpd69BNqk1jpmiRXklrAqRBd0srRFZGYuSAoYKAuTu3fL/w7d/Yj/6IFpP/AIML3/49X/Dt39iP/ogWk/8Agwvf/j1fB74AfB74AaZqGj/B/wAEWvhuz1WdZ7yKCeaUSyKu0MTI7EYHHGKqqqqqv2H/APlLD8cP+wh4r/8ATqtf8Fcfji/wu/ZmPgDSbzytZ+JN5/ZyhThlsYsSXTD1BBiiI9Jz6V+wB8BU/aG/ag8KeFNSshc6BoznWtdVk3KbS2Kny2B6rJK0MR9pCe1V4r8W6H4N0l9X1y7EUa8Rxry8rf3UHc/oOpwK+IPxQ8QeP7kx3EhtNNRsw2UbfKPQuf4m9+g7AV8P/hH4j8dul2ENhpWfmvJU+/6iNf4j79B69q8IeCfD3gjThp+hWgQsB5078yTEd2b+g4HpVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVftC/wDJAfiZ/wBifq3/AKRy1/wRD/5JV8Sv+xgtf/Sc1VVVV4t/5FTWf+wfP/6Lav8Aghj/AM1s/wC4B/7ka/b4+OL/AAA/Zb8Y+MNNvjba5qUA0fRHVyri7ucoHQjo0cfmyj/rlX7P3wj1T48fGjwf8JNJZ0k8R6nHBPMq5MFuPnnlx/sRLI3/AAGtA0LSfC+hab4Z0Gyjs9M0i0itLK2jGFhhiQIiL7BVA/Cqqqr4kfE7R/h/YbHZbrVZ0P2azVufZ3/upn8T0HcjxB4h1vxhrD6rrFy91dznaigcIM8Ii9hzwP6mvhV8DEtRD4i8b2qvN96305xkJ6NKO5/2eg788DAAwO1VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVf8ABKT/AJPS+Nn/AGD9Q/8ATnHVVVVVVVVVVVVVVVfEb4hab8PtGF7cp9ovLglbS1DYMjDqSeyjjJ9wO9abpPxd+MpbVLrWHsNJZjs3O0MJx2SNeXx03H35zmrj9nPxTpkZvNB8WwPdoMqux7cn6MCea8JfF3xb4G13/hFviQtxNbxPskknG6a39G3D/WJ37kjkE9DDNDcQpPbypJHIoZHU5DAjIIPcYqq+PX/JM9Q/67Qf+jFr9mz/AJEW+/7C0n/oqKqqqqqqv2jPFNzpum6X4d027lhuLuUzymJyrBFGFHHOCWP/AHzXwR1/WND8f6h4M8RXk7y3CPEEllL7Z4iTwT2K7/rxVVVVVVVVVVVVVa74h0nw1ax32tXJtrZ5REZihKox6biM7QcYyeMketWd9ZalbJeadeQ3UEg+SWGQOrfQjiqqqqqqqqqqqqq8WeLoPCeoaO1+4Wxv5XgmYj/VsQCr/QYIPsc9qBVlDKwII4IqqqqqqqqvdT03Tl33+oW9uMcebKE/nV18Q/DFudsF3JdMO0MZ/mcCpfiLJMdtjpqr6NK+f0GP50PEer3hw91sH92NduPx61bSSSndJIzt6scmrToKi+5VVVVXxk8ZR+JvEC6bYSh7HS9yKyniSQ43t9BgAfQnvVV8F/H/APa9kvhTVrjN7aJ/orueZox/D9VH6fQ1VVVVVVVVVVVVVVVVVVVVVVX/AAWEn/Zu17TtG8MeMPFk+hfFnRtN/tLQjHpM08d7YyySRmCaVF2qpkgcqckowPy4kJr9ij/goR45/Y/tNR8IyeF7fxb4M1W7+2SaZJdG1mtLgqqPLDKFYfMqpuRlIJRcFCWJ/wCH4vhT/o3bVf8AwpI//jFf8PxfCn/Ru2q/+FJH/wDGK/Yv/bG0z9sjwv4j8S6Z4DufC6+Hb+K0aGfUFujMXj37gQiYx0xzVVVVVV+w/wD8pYfjh/2EPFf/AKdVr/grb8XZPiN+1de+ELO9MulfD/TodLhRT8n2lwJrhwP726RYz/1xH1r/AIIu/BtPDfwe8T/GrUrILe+MtTFjp8jrk/YrTIZlPYNO8qkdzCvpXjTxjpHgfQ5dZ1WTOPlggVgHnfsq/wBT2HNeMfGWt+ONXfVdZuN2MiCBThIF/uqP5nqe9fCn4Gi8jh8SeNrYiFgHttPcYLjs0g7D/Z79/QxxxxRrDCioiKFVVGAoHQAdhVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV+0L/yQH4mf9ifq3/pHLX/AARD/wCSVfEr/sYLX/0nNVVVVeLf+RU1n/sHz/8Aotq/4IY/81s/7gH/ALka/wCC2Pxbk1Px34F+CVhd5ttD0+TWtQjU8NPOxihDf7SRxSEe03vx/wAEUfg0mq+MvGvx31O1VotCtU0TSmdMj7RPiSd1PZkjSNfpOaqqqvip8VLHwFYmxsWjudauE/cwk5EIP/LR/b0Hf6ZNTz634s1oyzPPqOp38oHTc8jngAAfljoB7V8Kvg3ZeD44tc15I7nW2XKr95LT2X1bHVvwHqaqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr/glJ/yel8bP+wfqH/pzjqqqqqqqqqqqqqqqvFbz/FD40jQjK32WO7+xoAfuwxZMhHudrt+Iq0s7WwtIbGygSGC3QJFGgwEUDAAH0qv2hvCVtqnhZfFEMSi80l1DuBy8LMFI98MQR6fN61+z94jl1rwSdMuZS82kTmFSevlEZT8vmH0UV468f6D4A01b3VneSabItrWIfPKR19gBkZJ/U8VB8cvibrjtdeGvAMdxZq2DstJ7kr7F0IGfwrxr8W7Txp4A1TQNU0yTStahkiJt3JKyASLu25AII7qe3c84/Zs/wCRFvv+wtJ/6KirxL4l0bwnpMus65diC3i4HGWdj0VR3J9Px6Amp/j54x125ki8C+BmuY4jyXhluXx2JWPG36ZP1rS/2hNX0zUV03x94SexJI3vDG8Txj1MUmSR+P51p+oWOrWMGpabdR3NtcIHilQ5DA/56dq+K3xW1P4eanY2VnpNrdpdwGQtLIylSGxjjt0r4ifHaHw1q0nh7wxp8Wo3tu2yeWUny43HVAF5YjvyMHjnnHgvWr3xH4V03W9RhiiubuHfKkalVVskEAEkjp3NVV45+Jfx6jgA8yx0+5Ceo8q3yW/BnB/77FfGW0uPBfxTsvF1iny3Jiu0AGAZIyFdfxAUn/fqzu7fULOC/tJA8NzEskTf3lYZB/Iiqqqqqqqqqqqqtb0ew8QaTdaLqUXmW15EY5F74PcehBwQexAq9k8XfC3xPeaTY6tc2Vzay43RMQky9VYr0YEYOCDXhb9pK+gEdt4v0hblBwbqz+R/qUPBP0K/SvDPj3wj4vQf2DrcE8u3Jt2OyVfX5GwfxHHvVVVVVVVVVVVV8evEdvqWt2mgWkocaYjGYjtI+Pl/AAfnXwW8fjU7RfCOrz5u7ZP9DkY/62MD7n1UdPb6VVVVVWt+PPBfhwlNb8U6baSL1ie4UyfggJb9K1X9o/wBaEx6St/qj/wtFD5Sfm+D+lah+0Xr99lNG0OyslPAaZ2mYfTG0foaufiL411okXniG5CsOUhIhH0wmM/jVs7yuZJHZ3Y5LMck1Z9qtO30qz/pVn0/CrToKi+5VVVV8Y/HLeGtHGi6bNt1HUkI3KcGGLoW9ieg/E9qqqsL670y9g1CwuGhuLdw8Ui9VIrwJ4xtPGugx6lFtS5j+S6hH/LOT2/2T1H+INVVVVVVVVVVVVVVVVVVVVVVf8FXf2bPjz8af2lPDmufCr4U+IfE2m2/gi2tZb2xtC0Mcy3t67RmQ4UMFkQ4znDD1r/hgL9sr/o3nxV/35T/AOKr/hgL9sr/AKN58Vf9+U/+Kr/hgL9sr/o3nxV/35T/AOKr/gkt8EPix8EPh1480r4seBNT8MXepa1bzWkN8gVpo1hKlhgngHiqqqqqr9jjVtP0D/gqT8fte1W4EFlps/i+5uZW6Rxx6mGZvwAJr4ieMdR+JHxB8S+PtTDG98TaxdajMpOSHnlaQj82xXwO8I6P+zt+zb4K8G6qEs08L+HraK+AGC92UDT4H955mkOPVq8eeN9U8e66+ragxjhTKWtsDlYI89Pcnue59gAPgx8HUCW/jLxXbZY4exs5F+76SOD37gfj6Yqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr9oX/kgPxM/7E/Vv/SOWv+CIf/JKviV/2MFr/wCk5qqqqrxb/wAiprP/AGD5/wD0W1f8EMf+a2f9wD/3I1+278Sj8Wv2rviZ4xjujPanXZbGyfdlTb2mLaJl9Aywhv8AgRPUmv8Agmh8MV+GH7G/gWGa3Ed94ohk1+7O3b5n2pt0Lf8AgOIBn2qqqvil8TLHwBpXlwFJ9XulItbcn7g6eY4/uj07nj1IJ1rxXreSZ9Q1PUZu/wAzyOf8/QD0Ar4WfCnT/Adit9fLFc63Ov72bGRCD/BH7ep7/Tiqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv+CUn/J6Xxs/7B+of+nOOqqqqqqqqqqqqqqq+C3z/ABcZrr/XYuiNwwd+Dnj6Zqq+J/lf8K88QebjH2CTGfXHH64r9mESeT4jJJ8vdbbfriXP9K15Jvij8a20aeZxareNaqAfuQQ537fTO1j9WqysbPTbOHT9Pto7e3gQJFFGu1UA7AV+0T4W0688Lr4pSBEvrCZEaUDBkjY7dp9cEgj059a/Zs/5EW+/7C0n/oqKvjXqWoeL/iXa+DLWbEVrJFbQqc7fNl2lnI/4Eo/4DXh7w/pfhfR7fRNHt1it7dMDA5du7Me5PUmvit4NsvGHhC9jkgU31lC89nLt+ZXUZKg+jAYI9wewr9mrxFcSpqnhWeQtFCourcH+DJ2uPpnYceufWv2mv+Q7ov8A16P/AOh18H/Blh4f8LWesTQLLq2rQi5ubqQbpPnG4Lk8gYIyO5yaqvHXiFPCvhHVNd3hZLa3byc95W+VP/HiK/Zzs9Msxq3ibU9QtoZpCLWDzpVVscM55OcE7OfY18fbXRvEHgxdQstTs5rvSZxIipMrM0bYVwAD/ut/wGvgH4hGt+AobCWTdPpErW7A9dn3kP0wdv8AwGqqqr43fEHxH4E/sX/hH5bdPt32jzvNiD52eXjHp941/wANBfEb/n6sP/AUf41/w0F8Rv8An6sP/AUf41/w0F8Rv+fqw/8AAUf41/w0F8Rv+fqw/wDAUf41/wANBfEb/n6sP/AUf41/w0F8Rv8An6sP/AUf41/w0F8Rv+fqw/8AAUf41/w0F8Rv+fqw/wDAUf41/wANBfEb/n6sP/AUf41/w0F8Rv8An6sP/AUf41/w0F8Rv+fqw/8AAUf41/w0F8Rv+fqw/wDAUf41/wANBfEb/n6sP/AUf414v8a6z44vIL/XRatcQR+WskMIQlc5AOOuCTj6mqR3idZInKupyrKcEH1FeFvjj468NhLe5vV1a1Xjyr3LMB7Sfe/PI9q8BfF3Q/HkosbfTr+0vQMtG0JkjH/bRRgD/e21VVVVVVVVVeP/ABVF4P8ADNzqu4faWHlWqkZ3SsDjj0GCT7CpppbiZ555WklkYs7sclmPJJNaEmpyaxZporst/wCaDblTg7x0xXgPxZF4y8Ow6oEEdyhMV1EP4JBjOPYggj61VapqNtpGm3OqXazNBaQtLIIYHmcqoydqICzHA6AEmvEX7cfgCyaS38L+GNY1aVCQHuCtpGT7E7m/NRWt/tkfE3WS0eh2Gk6LEfuMkJnlX/gTnaf++K1X4n/EPxUSNe8ZapdI4+aH7QY4z/wBcL+lWXQVZ9qs+1WXb6VZdqs+1Wnb6VZ/0qz6fhVp0FRfcqqqr+9ttNsp9QvJQkFtE0kjHsqjJ/QV4t8SXfizXrrW7v5TM2Io8/6uMcKv4D8zk96qq0u0jv8AU7Sxll8tLidI2fH3QzAE/rXgzxBffDfxm9vf5WFZjbX8fbaGxuHrg8j1H1pHSRFeNwysMqQcgj1FVVVVVVVVVVVVVVVVVVVVVVfHf/gof+yz+z7qtz4a8U+OJdb8Q2TFLjRvD1v9tngYcFZHysUbjujyBh6VY/8ABa39mye/EF98OfiPbWrNhZxaWchAzwWQXAwMcnBJ9M18CP2s/gB+0nbOfhL8QrPUtQt4/MuNJuFa1vYF7sYZAGZQSAXXcuSOeaqqqqqq+IfxJ/4Vn+0x+1jewXHl3uvy+KNDtRnBc3esJFKB/wBsDMf+A1+xf8OE+K/7U/w08GTwiW0k12G8vlYfKba1zczA+gMcLDPvXxj+JD+ONZ/s/TZj/Y+nsRABx5z9DIf5D2+pr4H/AAtXxFcr4t1+23aZav8A6LC68XMg6kjuin8zx2Iqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/aF/5ID8TP+xP1b/0jlr/AIIh/wDJKviV/wBjBa/+k5qqqqrxb/yKms/9g+f/ANFtX/BKz4iR/CP4J/tS/Et5VSTw3oGl3luG/jnSLU/KX6tJsX8a8GeF9W+IfjjQvBelu0up+JtWt9PtywLFpriVY1J7klnFeHtB03wt4f0zwxo0Pk2GkWcVnaR/3IokCIPwVQKqq8f+OdM8BaC+q3pElxJlLS2BwZpMfoo6k9h7kCtZ1jWfF2ty6nqEsl1fXsuAqgnknCog9OgAr4Q/CqDwVYrrGsQpJrd0nzZGfsqkfcU+vqfwHHJqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr/glJ/wAnpfGz/sH6h/6c46qqqqqqqqqqqqqqrWml+F/xqbU5omW1W9Nyu0fet5s7tvrgMy/Vatrm3vbaK7tJklhmQPHIhyGUjIIPoRVftC+KrfSvCQ8NxTKbzVnXKDqsKEEt7ZYKPfn0r9n/AMPS6N4GGoXMZWXVpzOoIwRGAFT88E/RhXgORPDvx0Npfnb/AMTC6tgx4+Zt6p+ZKj8ar4/XEMPw2u4pJArT3EKRg/xEOGwPwUn8K/Zs/wCRFvv+wtJ/6KirxhInhr48/wBoX52wJqdvcM5GAI22En8Mn8q4IyDmvE2pW2j+HdS1S7cLFa2sjtk4zhTgfUnA/Gv2aNPlk8TatqiqfLt7AQsf9p3Uj9IzX7TX/Id0X/r0f/0OvCP/ACKei/8AYOg/9FrVV+0t4hEVhpfhaGT5p3N1OB1Cr8qfgSW/75FaF+zhp+oaJY3+pa/eW91c26SSwpEpEbMAdvPpnFN+zLoZUhfE99nHGYVr4D6tceGPiDd+FNQby/tyvbyITwJ4iSP0Dj8aqqqvG/w58P8AxA+xf2692v2DzPK8iQJ9/bnOQf7gr/hnT4f/APPbVv8AwJX/AOJr/hnT4f8A/PbVv/Alf/ia/wCGdPh//wA9tW/8CV/+Jr/hnT4f/wDPbVv/AAJX/wCJr/hnT4f/APPbVv8AwJX/AOJr/hnT4f8A/PbVv/Alf/ia/wCGdPh//wA9tW/8CV/+Jr/hnT4f/wDPbVv/AAJX/wCJr/hnT4f/APPbVv8AwJX/AOJr/hnT4f8A/PbVv/Alf/ia/wCGdPh//wA9tW/8CV/+Jr/hnT4f/wDPbVv/AAJX/wCJr/hnT4f/APPbVv8AwJX/AOJr4paJ4T8NeJG0Dwq9zKLNNt3LPKH/AHp/hGAPujGfckdqrwt8IPHPispLbaS1nat/y83uYlx6gY3N9QCPevC37O/hTSQlx4iuZdYuBz5ZzFCP+Ag5P4nB9KsNO0/S7VLLTLKC0t4/uxQxhFH0Aqqqqqqqqqq/aJlkWHQbdXOx2uGZfUgRgH/x4/nVfBjTY9Q8e2kkhGLOKSfB7kDaP1YH8K+HNp/ZfjfxrpUC4tkuYpUUdEL72wPwOPwqqrxP8Lfhz4zdpvE3gvSL+d/vXElqqzf9/Fw/61qv7JHwbvwx0/TtS0pm6G0vmYKfpLvrWf2Nr23Jk8L+NIJv7sV/bGPH1dC2f++RWpfAH4qeHyWk8NNfRKeJLGVZs/RQd/8A47TabqGmTfZtSsLi0mHWOeJo2H4HmrPtVl2+lWXarPtVp2+lWf8ASrPp+FWnQVF9yqqq+OmtNp3g9NNhkIk1K4EbAf3F+Zv1Cj8aqqrRc/2xYbcZ+0x4z/vCvj3oC2PiG112FMJqUO2Q/wDTSPAz+KlfyNfCbWn1vwLp8k0m+a0BtpCf9jhf/HdtVVVVVVVVVVVVVVVVVVVVVX/BVn9r/wARfAbwJpXwl+GmsPp3i3xvBLJdX8D7ZtP05TsLRnqkkrbkVxyoSQjDbSLe2u9Quo7W0glubmdwsccaF3kY9AAOSSe1X/wV+Mml6Yda1P4S+M7TT1QObufQbmOIKRkNvKYxjnOa8H+MPE/w/wDE+meNPBWu3ej65o9wtxY31q+ySGRehB7gjIIOQQSCCCRX7E/7Sdv+1P8AALRviNcJDBr9q7ab4gtouFivogu5lHZZEaOUDsJNuTtJqqqqqq/ay/5Oo+Mv/ZQNd/8AS+av+Cb/AIA1q28R6/8AF2SIQ2Meny6PZSMpDPJKUMzIfQRqYye4lI7HHw18C3Pj7xJFpil47KAeZezKPuRjsP8AaY8D8T2NWNjZ6ZZQadp9ukFvbRiOKNBgIoGABVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV+0L/wAkB+Jn/Yn6t/6Ry1/wRD/5JV8Sv+xgtf8A0nNVVVVeLf8AkVNZ/wCwfP8A+i2r4Z/EU+D/ANkz43eEbW5C3PjfX/Ctg8YOGNvF/adzIw9t0ESn/fHbNf8ABLX4bf8ACxf2yvCVxPb+dZeEbe5126XbnHkpshb2xcTQH8Peqqtd1vTfDmk3OtatcCK1tIyzt3PoAO5JwAPU1478a6l471+XWb4lIh8ltb7siGMdAPfuT3NfA74VrpVvF4z8Q2w+2zLusYHXmBCPvkf3iOg7D3PFVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV/wSk/5PS+Nn/YP1D/05x1VVVVVVVVVVVVVVXxQ+Gll8Q9LQRypbapaA/Zbhhwc9UfHO0/oefUHRfGPxN+DYOi61ock+mox2R3CnYpPXypVyME9uRz0BNXX7SmsX8X2TQvBscd3IMRl7lrjn2RUUn868KfCXxd4917/hK/iS08FvIwd4pvkmnx0UKP8AVp2xwcdBzkRRRQRJBDGqRxqFRFGAoHAAHpXxs+FerajqX/CbeEraSa5IX7Xbw58zcuAsiAck4ABA54B55rSP2jtf0u0Gn+IfDUd/eQ/IZRObdjj+8uxufpj6V4otfH3xP0S+8X+I7JtK0nSbZpNPs1jIM0hIGcHk8fxHjoAOTX7OEckPgi+imjZH/tWQ7WUg4MUXP6GvjN8LZvHFpFrOiBP7XsoygjY7RcR5ztz2YEkjPHJB9R4c+NninwHZp4Z8ZeG7i6ktBsieZzBKqjoGyp3Dpg8ceteI/GXj341tH4f8N+HZbbTPMBkVWLKxB4MkpAAA67fX1OMfDnwJaeAPDqaTFKJ7qVvNu5wuN8hGMD/ZAGB+fev2lba4n1zRjBBJIBaPkqhOPn9q8JKV8KaKrAgjT4AQRjH7taqtVtbr4l/G0QSW8r6el4IgWQhfs8P3sE9m2sfq9VXxi0bUfCPxOh8UaNbyEXLR3sRRCQsqEBgcepUE/wC/WnX0Gp6fbalbE+VdQrKm4YOGAIz+Bqqqqqqqqqqqq8b67f6D4fnuNGsJr7U5h5VlBDGXJkI4YgfwqMsc8ce9eHP2ePE+tTHUPF+ppp4lYvJGpE07knJyc7QT65P0rwt8LPBHhEJJpujpNdJ/y9XX72XPqCeF/wCAgVVVVVVVVVVVVfGDwjd+KfDSy6bEZb3TpDLHEo5kUjDKPfofwx3plKkqwwRwQR0rwN4nfwh4ltdaETzRoGSWJOrqwxge+cH6ivhvo2o2tjf+Idbh8nUdfuTcyQngwp/AhHqAT+YzzmqqqqqvLGy1GA21/ZwXMR6xzRh1P4GtU+EHw91Qlz4fitJD0e0Yw4+ij5f0q++AdihLaLr80eBxHdRB8/8AAlxj8quPhT4u005jt4LxR/FBKP5Ng/lR0vUtOYJf2Fxbnt5sRX8s1advpVn/AEqz6fhVp0FRfcqqqv2iJWM2hQZ+VUnb8SUH9P1qqqvhtoM/iDxnptvFEWit5luJzjhUQgnP1IA+pr48WKXPguO62/PaXiMDjoCGUj9R+Qr9n2Rm8MajEScLfkjI4yY1z/IVVVVVVVVVVVVVVVVVVVVVVf8ABXLV77U/20Nbsrty0WlaJp1tagk/LGYfNIH/AAOV/wA6/wCCXfwG+F/gL9mbwl8TtC0Swu/FnjG1kutT1t41knH710Fuj8mNECBSikZYMTz0r/gsz8Afhh4b8L+F/jt4Z0Ww0fxRqmu/2Vqi2kQi/tON4JZVnkUcF4zCFL4yRKAxO1cf8EOfEF6X+LnhSSRns1GlXsSl+I5D9pRyB6sBHk/7A/Cqqqqq/aH8Mav40/bN+KPhLQbfz9Q1b4lazbW6dtz6jMMn0A6k9gCa+G/gPSfht4J0XwHoEWbbSrZYVYJhppOryED+J3LMfc18KfA8fgfwpBaTRKNRvAJr1schyOEz6KOPTOT3qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr9oX/kgPxM/7E/Vv/SOWv+CIf/JKviV/2MFr/wCk5qqqqrxb/wAiprP/AGD5/wD0W1CeZYGtllYRO6uyZ4LKCAceoDN+Zr/giB8PwZPif8U7m2GVWy0aymx675rhc/hbGqo4AyT0r41/Ek+MNXOh6TcZ0fTpCFZTxcSjgv8AQchfxPfj4HfDJfE1+PFGt227S7J/3Mbrxcyj27qvfsTgetVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVf8EpP+T0vjZ/2D9Q/wDTnHVVVVVVVVVVVVVVVUQGG0gEEcgiore3gz5EEceeu1QM/lVVTQQPIszQo0ij5WKjI+hqqqWGKdNk0aSLnoy5FKqooRFACjAAGMVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVr/wANfBniSdrvUdGQXL/emgYxMx9TtOCfcg1oPw08F+HJ0u9O0ZGuU5WadjKyn1G44B9wBVVVVVVVVTKrrtYAgjkEVc+HNDu8mTTYQx/ijXYf0xUng6xU5tbmWM9gwDD+lLoN3b/ceOQdsHBq3gmi4kjI+oqL7lVVV+0Lp7S6XpGqKvy288kLHH99QR/6LNVVV+zy9mbXWo1VftQeIsSOSmGxj2yD+Yr453kdt4Fe3ZhuurqONR64y3/stfAnT3tPBLXbr/x+3kkiH1VQE/mrVVVVVVVVVVVVVVVVVVVVVVf8FqPgxqmk/Evwn8drCykfR9e00aPfzKpKw3kDO8e49jJE5Cj/AKYNX7I//BRj4xfsoaMfA9ppun+LvBZnaaLRtQkaF7N2OZDbzqCYw5OSrK65yQASxN9/wXI006YTpv7N9z/aDR/Ks/ipfKR8dci2ywB7YXPtX7U37Xvxa/a38U2mvfEW6tLTTtKV00nRNPVktbINjewDEs8jbV3OxJ4AGAAB/wAEZ/g5qvgj4F+JPinrVpJbSfEHU4hYpIpBks7MSIkuD2aWa4A9QoPQiqqqqqr4aabYX/8AwUm+MM95axzSWHifxPPbM4z5Un9oOm4e+13H/AjXwM8Jr4l8bRXlzHutNHUXMmRkFwcRr+fP/ATVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVftC/8kB+Jn/Yn6t/6Ry1/wAEQ/8AklXxK/7GC1/9JzVVVVXi3/kVNZ/7B8//AKLaq/4JF+DR4X/Y00nWWhKP4r13UNTJOMsFkFqD64xajr9ehqq+PHxC/wCEc0b/AIRfSp9upanGfMZTgwwdCfYtgge2favAPgy98d+I7fRLUtHD9+6mAz5MQ6n69APcitL0yx0bTrfStMt1gtbWMRxRqOgH9ffuaqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/4JSf8npfGz/sH6h/6c46qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqsDtVVVeOvDg8VeFb7RkA85499uT2kXlfpkjH0JqSN4pGilQq6EqysMEEdRVVXws8TL4Y8YWs88oS1vB9nuCxwArEYY+mGCnPpmvid4pb4g+KLLw74ePn21vL5MDDpNKxALA/3eAAfTJ6GtA0eDQNFstFtjlLOFYw2MbiBy34nJ/Gqqqqqqqqqqqqqqqqqqqqqr4q/CvwL8a/AWrfDX4jaHFquhaxD5c8L/KyMOVkjYcpIrAMrDkECvj9/wSD/AGgvAGsXV98F/svxD8Nlma3RbiK01CBP7ssUjKjkdN0bEtjOxelW37Cf7Yd3qH9mRfs6+NFm3bd0unmOLP8A10YhMe+cV+y5/wAEePH2s67Y+Kf2nrm20HQbZ1lfw3YXaz3l6Qc+XLLGTHDGe5RmcjIGwkMNG0bSfDukWWgaDp1vYabptultZ2lvGI44IkUKiIo4ChQAAOgFVVVVVXwp/wCUkPxs/wCxh8Tf+nM1+z/4eTSPAianJHifV5mmYkYOxTtQfTgt/wACqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv2hf+SA/Ez/ALE/Vv8A0jlr/giH/wAkq+JX/YwWv/pOaqqqq8W/8iprP/YPn/8ARbVX7D/h6Pwz+yD8IdNjRVE3hOyvcDHW5jFwTx6mU1Wu6zY+HdHu9b1KTbb2URkfHU46Ae5OAPcivEmv6j4t1661y/YvcXkuQg52DoqL7AYAr4R+Ak8DeGES6iA1S/xLeNjlTj5Y/ooP5k1VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX/BKT/k9L42f9g/UP8A05x1VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVfGP4azNNN4w0G33q43X0CDlT3kUen978/XFVQBYhVGSegr4PfDaXREHijXbfZfSpi1gccwIRyx9GI4x2H1IFVVVVVVVVVVVVVVVVVVVVVXijxR4b8E+Hr/wAWeL9dsdH0bS4TNeX17MIYoEHVmZuB2HuSB3r4yf8ABan4b+HdSudG+Cfwz1DxcsLMg1fVbr+zrZyOjxxBHldD/t+U3tVh/wAFufjLHfeZqfwZ8F3FnniGC6uYZMZH8ZZx0yPu9efav2bP+CrnwF+OesWXg7xrYXXw58TX8qw2sWo3K3FjcyNwqJdBV2sTwBIiAkgAknFVVVVVVfChWf8A4KR/GtEUsx8Q+JgABkk/2ma0LTI9F0TT9Hi+7ZWscI/4CoGf0qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv2hf8AkgPxM/7E/Vv/AEjlr/giH/ySr4lf9jBa/wDpOaqqqq/aCvPGmn/Bfxbd/Dy3afxBHp7fY40iWQnLKH+VuD8hfrVfDbxj+39pHw58K6ToHhm9fTLLRLOCyb+x7ZswpCiockZPygcmv+Fg/wDBRT/oVr7/AMEtr/hVn8Qf2g9f0++0H41332d4p0Kad9jhgdSFzucxjkHcMAntnHSvgJ4H/wCEi8Sf8JDfQbrDR2DKGHEk/wDAPfb94++31qq8X+O/2+rbxZrVt4b8NXkmkxajcJYMNHtmDQCRhGQSMn5dvJr/AIWD/wAFFP8AoVr7/wAEtr/hX/Cwf+Cin/QrX3/gltf8K/4WD/wUU/6Fa+/8Etr/AIV8BdS+J2r/AAy06++MFpJbeKHmnFzFJbpCQokYR/KnA+TbVV+0r4p/at0Px1Y2nwL0a4vNBfSI3uHj06GcC6MsoYbnGR8gi46c+9f8LB/4KKf9Ctff+CW1/wAK/wCFg/8ABRT/AKFa+/8ABLa/4V/wsH/gop/0K19/4JbX/Cv2ZfEv7Teu67rUPx50i4s7CK0RrBpbCG33S7/mwYxk/L2NVX7RmrfF/Rfh9HefBGxlu/ER1GJGjjto5z5BV952vx1Cc1/wsH/gop/0K19/4JbX/Cv+Fg/8FFP+hWvv/BLa/wCFf8LB/wCCin/QrX3/AIJbX/CvAPjn9vK88deHLTxh4bvItBm1a1TVHbSLZAlqZVEpLKMgbN3I5FVWoNcpYXL2QzcLC5iGM5fBxx9cV/wsH/gop/0K19/4JbX/AAr/AIWD/wAFFP8AoVr7/wAEtr/hX/Cwf+Cin/QrX3/gltf8K/4WD/wUU/6Fa+/8Etr/AIV8NLrxXffD7w9eeOoWh8RTadC2pxtGsZWcqN4KrwOc8Diqr45eMf2z9K+KWt6f8JtAurnwpF9n/s+VNLt5g2beMyYZxuP7wyDmv+Fg/wDBRT/oVr7/AMEtr/hX/Cwf+Cin/QrX3/gltf8ACv8AhYP/AAUU/wChWvv/AAS2v+Ffsy658edd0HWpfjzps1nfxXaLYLLZx25aLZ8xAjGD83rVV+09r/7QOhWHh9/gHpk15cTTTjUhFZxXG1QE8vIkBxyX6V/wsH/gop/0K19/4JbX/Cv+Fg/8FFP+hWvv/BLa/wCFf8LB/wCCin/QrX3/AIJbX/Cvgb4x/bP1b4paJp/xZ0C6tvCkv2j7fLJpdvCFxbyGP5kG4fvBGOPWqr4l3Xiux+H3iG88CQtN4ih06ZtMjWNZC04U7AFbg844PFf8LB/4KKf9Ctff+CW1/wAK/wCFg/8ABRT/AKFa+/8ABLa/4V/wsH/gop/0K19/4JbX/Cv+Fg/8FFP+hWvv/BLa/wCFWDXL2Fs94MXBhUygjGHwM8fWqrx945/bytPHXiO08HeG7yXQYNWuk0t10i2cNaiVhEQxGSCgXk8mv+Fg/wDBRT/oVr7/AMEtr/hX/Cwf+Cin/QrX3/gltf8ACv8AhYP/AAUU/wChWvv/AAS2v+Ffs+ap8VtZ+HEF78ZrKS18Sm7mWSOS3SAiIEbDtTjpmqr9pjxP+1JofizS7b4EaPcXmkyaduvGj0+GcCfzGGMyDI+ULwOK/wCFg/8ABRT/AKFa+/8ABLa/4V/wsH/gop/0K19/4JbX/Cv+Fg/8FFP+hWvv/BLa/wCFfs1+Kf2rdc8dX1p8dNGuLTQU0iR7Z5dOhgBuhLEFG6MZPyGXjpx7VVfHvUvidpHwy1C++D9pJdeKEmgFrFHbpMSpkUSfK/B+TNf8LB/4KKf9Ctff+CW1/wAK/wCFg/8ABRT/AKFa+/8ABLa/4V/wsH/gop/0K19/4JbX/CvCPjv9vq58WaLbeJPDV4mky6jAl+zaPbKFgMiiQ5AyPlzyKqq/4WD/AMFFP+hWvv8AwS2v+FeEfHf7fVz4s0W28SeGrxNJl1GBL9m0e2ULAZFEhyBkfLnkV/wSk/5PS+Nn/YP1D/05x1VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV4v+Cvh7xDK99pEv9lXj5LCNN0Tn1KcYP0x9DV78CvG9s5W2+wXaj7pjn25/BgK0/4D+MrqRRezWFnH/EWlLsPoFBB/MV4O+EnhvwnJHfTZ1LUE5WeZQFjPqic4+pyfeqqqqqqqqqqqqqqqqqqqqqqq/wCCzP7Qeu6p8Q9G/Zx0TUpYNC0Oxi1TWoY3wLu8lyYkkA6rHEFYA95iSPlU18IPgX8W/j74jbwp8IfAuo+JNRiQPOLcKkVshOA0srlY4lJ4BdhntV//AMEmP21LLSm1GLwPod5MqbvsUGv2/nHjoNzBCf8AgVeN/AvjH4beJ73wX4+8M6joGuac4W5sL+BoZY8jIOD1UgghhwQQQSDmv+CT/wC0frvxx+Al54L8Z6k9/wCIPhxcw2BupXLS3FjKjG1aQnqw8uWPPcRKSSSTVVVVVV4C8WaZ4I/4KL/GrxFqu1o7XxL4lZImJXznGplhHkA43bcZwcV/w8v/AOqKf+XJ/wDc1f8ADy//AKop/wCXJ/8Ac1f8PL/+qKf+XJ/9zV+zh+1p/wANB+KdT8M/8IB/YP8AZun/AGvz/wC1ftXmfvFTbt8lMfeznJ6dKqvj58Xv+FHfDybx5/wj39t+TdRW/wBk+1/Zs+YSN2/Y/THTFf8ADy//AKop/wCXJ/8Ac1f8PL/+qKf+XJ/9zV/w8v8A+qKf+XJ/9zV4F/4KE/8ACa+N/D3g3/hUX2P+3tVtrD7T/wAJB5nk+dKse/b9nG7G7OMjOOo61VX919isbi92b/s8LSbc4ztGcZ7dK/4eX/8AVFP/AC5P/uav+Hl//VFP/Lk/+5q/4eX/APVFP/Lk/wDuav8Ah5f/ANUU/wDLk/8Auavhz4v/AOFg+AtA8b/2d9g/tzT4rv7L53m+TvUHbvwu7GeuB9KqvjZ+3H/wp34naz8OP+FX/wBr/wBkeR/pn9t/Z/N82COX7nkNtx5m37xzjPGcD/h5f/1RT/y5P/uav+Hl/wD1RT/y5P8A7mr/AIeX/wDVFP8Ay5P/ALmr9mv9o7/hoix169/4Q3/hH/7Elgj2/wBo/a/O8wOc58tNuNnvnPaqr9pL9oT/AIZ60PRtZ/4RD+3/AO1rt7fyv7Q+yeVtTduz5b59MYFf8PL/APqin/lyf/c1f8PL/wDqin/lyf8A3NX/AA8v/wCqKf8Alyf/AHNXwU/bj/4XF8TtF+HH/Cr/AOyP7X8//TP7b+0eV5UEkv3PIXOfL2/eGM57Yqq+I/i//hX/AIC1/wAb/wBnfb/7D0+W7+y+d5XnbFJ278Ntzjrg/Sv+Hl//AFRT/wAuT/7mr/h5f/1RT/y5P/uav+Hl/wD1RT/y5P8A7mr/AIeX/wDVFP8Ay5P/ALmqwuvt1jb3uzZ58Kybc527gDjP41VeO/8AgoT/AMIV438Q+Df+FRfbf7B1W6sPtP8AwkHl+d5MrR79v2c7c7c4ycZ6nrX/AA8v/wCqKf8Alyf/AHNX/Dy//qin/lyf/c1f8PL/APqin/lyf/c1fs9/Gj/hfPgJ/G//AAjX9h+XqEtp9l+2fac7FQ7t+xOu/pjt1qq/aS/aq/4Z61vRtG/4QP8At/8Ata0efzP7U+yeVtbbjHlPn65Ff8PL/wDqin/lyf8A3NX/AA8v/wCqKf8Alyf/AHNX/Dy//qin/lyf/c1fs5/te/8ADQHje98G/wDCvP7B+xaVJf8A2n+1/tW7ZLFHs2+SmM+bnOf4enORVfHT4q/8KW+G9/8AEH+wf7Z+wywx/Y/tX2ff5kipnfsfGN2fu81/w8v/AOqKf+XJ/wDc1f8ADy//AKop/wCXJ/8Ac1f8PL/+qKf+XJ/9zV4T/wCCiX/CUeKdG8M/8Ke+zf2tqEFp5/8AwkO/yvMkVN237OM43ZxkZ9RVVX/Dy/8A6op/5cn/ANzV/wAPL/8Aqin/AJcn/wBzV/w8v/6op/5cn/3NX/Dy/wD6op/5cn/3NXwg+IX/AAtf4b6H8Qf7I/sv+2IXk+x/aPP8rbIyY37V3fdz90daqvj5+2h/wo74hTeAv+Fbf235VpFcfa/7Z+zZ8wE42eS/THXNf8PL/wDqin/lyf8A3NX/AA8v/wCqKf8Alyf/AHNX/Dy//qin/lyf/c1fs0ftL/8ADRf/AAkf/FFf8I//AMI/9l/5iX2vz/O83/pkm3Hk++d3bHNV+0f8e/8Ahn3wrpniX/hFP7f/ALR1D7J5H2/7L5f7tn3bvLfP3MYwOvWvi1/wUS/4Sj4VeM/DP/Cnvs39reHr+08//hId/leZbum7b9nGcbs4yM+or/giH/ySr4lf9jBa/wDpOaqqqq8W/wDIqaz/ANg+f/0W1f8ABGv4TfCv4o/8Lf8A+FmfDPwp4u/sz+w/sP8Abui29/8AZfM+3+Z5fnI2zdsTOMZ2rnoKtLO0sLSGwsLaK2traNYoYYUCJEijCqqjgAAAADgV4v8AEtn4R8OXviC9IK2seUTODI54VR9SQPbrU0up+JdaaVg91qGpXOcDrJI7dB9Sa8CeE7bwX4XstBg2tJEm64kH/LSVuXb6Z4HsBVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV/wSk/5PS+Nn/YP1D/05x1VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVf8FjPh3q3hj9qa38dTQOdM8aaDby20+Dt862HkSxg/wB5VWFiPSVfWv8Agj3aeDIf2R0uvDwtTrNx4hvf7fZMGUTqVESv3AEHlEDp8xI6mq/4LgaZ4KSP4W6ui2qeLpWv4XKAebJYqIiPMxztWRjszxl5Md6/4Ie+DtYt9I+K3j6eKRNLvrjTdNtX/hlmhWeSb8VWaH/v4aqqqqqv2J7S1vP+Crvxwhu7aKdP7R8VnZIgYZ/tVecGv7B0P/oDWP8A4DL/AIV/YOh/9Aax/wDAZf8ACv7B0P8A6A1j/wCAy/4Vbabp1k5ks7C2gZhgtHEEJH4VVXFrbXcXk3VvFMmc7JEDDP0Nf2Dof/QGsf8AwGX/AAr+wdD/AOgNY/8AgMv+Ff2Dof8A0BrH/wABl/wqPRdGidZYtJskdCCrLbqCpHQg4qqIDDaQCCOhr+wdD/6A1j/4DL/hX9g6H/0BrH/wGX/Cv7B0P/oDWP8A4DL/AIV/YOh/9Aax/wDAZf8ACo4ooY1hhjVEQYVVXAA9hVVPpOlXUpnudMtJZG+88kCsTgY6kelf2Dof/QGsf/AZf8K/sHQ/+gNY/wDgMv8AhX9g6H/0BrH/AMBl/wAKtrGysgy2VnBAG+8IowmceuKqrqysr1VS8tIZwpyoljDYPtmv7B0P/oDWP/gMv+Ff2Dof/QGsf/AZf8K/sHQ/+gNY/wDgMv8AhUGk6VbSrPbaZaRSLna6QKpHGDggelVUsUU8bQzRrIjjDKy5BHuK/sHQ/wDoDWP/AIDL/hX9g6H/ANAax/8AAZf8K/sHQ/8AoDWP/gMv+Ff2Dof/AEBrH/wGX/CgAAFUAADgVVSaLo0ztLLpNk7uSWZrdSST1JOK/sHQ/wDoDWP/AIDL/hX9g6H/ANAax/8AAZf8K/sHQ/8AoDWP/gMv+FW9pa2cfk2ltFCmc7I0CjPrgVVXWnafesrXljbzsowpliDEfTNf2Dof/QGsf/AZf8K/sHQ/+gNY/wDgMv8AhX9g6H/0BrH/AMBl/wAKttM02zcy2en20DldpaOJVJHpkDpwKqri2t7uIwXUEc0Z6pIgYH8DX9g6H/0BrH/wGX/Cv7B0P/oDWP8A4DL/AIV/YOh/9Aax/wDAZf8ACk0TRo3WSPSLJWU5VhbqCCO44qqr+wdD/wCgNY/+Ay/4V/YOh/8AQGsf/AZf8K/sHQ/+gNY/+Ay/4V/YOh/9Aax/8Bl/wqGCG3iWC3iSKNB8qIu0D6AVVXGlaZdyeddadazSYxukhVj+ZFf2Dof/AEBrH/wGX/Cv7B0P/oDWP/gMv+Ff2Dof/QGsf/AZf8KtbCxst32Kzgg343eVGEzjpnH1qqubOzvUEd5awzqpyFljDAH1wa/aD0TRY/gH8SpI9IslZfCGqlWFuoIItJeRxX/BEP8A5JV8Sv8AsYLX/wBJzVVVVXi3/kVNZ/7B8/8A6Lav+CGP/NbP+4B/7kar9o3xeL3VLXwbaS5isAJ7rB6ysPlH4Kc/8Dr9nfwZ/amtzeLr2LNvpf7u23Dhp2HJ/wCAqfzYHtVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX/AASk/wCT0vjZ/wBg/UP/AE5x1VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVf8Fff2hPAukyWH7N/jD4NjxDc32iQ6/pXiSPW/sk2k3DzXEACR+Q+8YgO4FwGDYwCqsPgX+0p8af2bddn174P+OLrRXvVC3toyLPa3gXOPNhcFGIycNjcuTgjJp/8Agrv+2W6FF1/wqpIIDDQY8j3GTXxX+L/xK+OPjC48efFXxde+IdbuEEZuLjaoijBJEccaAJGgJJCIAoJJxya+FP8AwUj/AGjvgn4D0v4a/DVfCGk6BpCMtvbrogdmZmLO7uzFndmJJYnqfTAr/h7z+2V/0HPCn/ghT/4qvhJ4j1Pxl8KPBfi7WnjfUdc8PWF9dtGmxTLNbo7lV7Dcx47VVVVV+w//AMpYfjh/2EPFf/p1Wqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr9oX/kgPxM/wCxP1b/ANI5a/4Ih/8AJKviV/2MFr/6TmqqqqvFv/Iqaz/2D5//AEW1f8EMf+a2f9wD/wByNa5q1roOj3utXrfubKB5XGcE7RnA9z0+prUr6/8AEWs3GoXGZrzULguwUZLOx6AfjgCvAfhaHwd4U0/QY1XzYY91ww/jlblz+fA9gKqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/4JSf8npfGz/sH6h/6c46qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv2//APgnj8XP2uPjfovxF8EeL/CGj6Pp3haDSpk1We4FwZ47m6lJVI4WUoVnQZLg5B46E/8ADkv4/f8ARV/h/wD99Xn/AMZr/hyX8fv+ir/D/wD76vP/AIzX/Dkv4/f9FX+H/wD31ef/ABmv+HJfx+/6Kv8AD/8A76vP/jNf8OS/j9/0Vf4f/wDfV5/8Zr4YeFrzwN8NPCXgnULmG4uvD+hWWnzyw52SPBAkbMuQDtJUkZAOKqqqq/Yf/wCUsPxw/wCwh4r/APTqtVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX7Qv8AyQH4mf8AYn6t/wCkctf8EQ/+SVfEr/sYLX/0nNVVVVeLf+RU1n/sHz/+i2r/AIIY/wDNbP8AuAf+5Gv2jvEh0/wzaeHIJcSapNulAP8AyyjwcfixX8jXwK8MDxD46t7yePdbaQv2p8jguOIx9dxDf8Bqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv+CUn/J6Xxs/7B+of+nOOqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr9qT9v34Q/slePfDvgP4haJ4h1K413Tnv5ZNIjhlayh8wxxsySSJuDskvQ5Hlng5wPgZ8dvhx+0Z4Di+I/wt1O8vdGe5ktS91YS2rpMgUuu2RRuxuHzIWXORnIIH7a/wDwU48I/syeJJfhd4A8OQeMPG9sitqInuDFZ6UWAZUkKgtJIVIJRSoUMMtn5a+G/wDwW0+JMPiKFPi98JfDN5oUku2V/DjT2tzAh/iAnllWUj+7lM+or4c/EPwh8WfA2jfEbwFrEWqaDr1qLiyuY+NykkFWHVXVgysp5VlIPINVVVVVX7D/APylh+OH/YQ8V/8Ap1Wqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr9oX/kgPxM/7E/Vv/SOWv+CIf/JKviV/2MFr/wCk5qqqqrxb/wAiprP/AGD5/wD0W1f8EMf+a2f9wD/3I18edcOsfEK6tkfdFpkSWyc8ZA3N+O5iPwr9nTQBp/g2fW5IwJdVuSVbHWOP5R/495lVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV/wAEpP8Ak9L42f8AYP1D/wBOcdVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX/BXDUby+/bR121ugfL0/RdNgts5+4YBIcf8AA5H6f41+zT478E/s7f8ABNrwn8TGktzpmg+D31SRN4T7Teys7tDn++9zJ5Y9yBWu614n+I3jK+8QavPc6v4h8S6lJcXDhS8t1dTyFmwB1ZnbgD1xX7TP7Anxs/ZV8AeHviL8QrvQLzTtcuEs5o9MuZJZNPunjaRYZQyKDlUcbkLLlCM8qT/wRJ+Iuoax8OPiJ8Lr27L2/hvVLTUbGN2GUW7SRZFUddoa2DEdAZM9WNVVVVVX7D//AClh+OH/AGEPFf8A6dVqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/aF/wCSA/Ez/sT9W/8ASOWv+CIf/JKviV/2MFr/AOk5qqqqrxb/AMiprP8A2D5//RbV/wAEOriG0tvjjdTuFjhTQXdj2AGokn8hWsajNrOrXurT5829uHmYe7MT/WvB2jDw94V0nRSu17S0jSQf7eMuf++iaqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr/glJ/yel8bP+wfqH/pzjqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/wCC0PwA1uDxZ4b/AGjtD06WfSLyxTRtckiTcLWeN2a3kfHQSK5jz0BiUcFhmTx947l8HR/DyTxtrz+FYbk3MehNqUpsUmOcyi33eWH5Pzbc89a/4JYfs4an8Zv2jNM+IGp6U7+EvhzKupXlxJH+7lvVybSFSerCQCU+ixHOCy5/4LXePdO0r4F+C/hwJk/tHxB4l+3qmcn7PawSK5x2+e5h59j+H/BDnw5cLB8XfFsqOIZG0mxgb+FmUXTyDp1AaLv/ABHPaqqqqqr9h/8A5Sw/HD/sIeK//TqtVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX7Qv/JAfiZ/2J+rf+kctf8ABEP/AJJV8Sv+xgtf/Sc1VVVV4t/5FTWf+wfP/wCi2r/gkrrB0b4c/tCSo+JLqLw9bIPXzDqCt/46WP4V8N9F/t/x1oumFNyNdLJIMdUT52/RSKqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv+CUn/J6Xxs/7B+of+nOOqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqrX/AA/oXivRbzw54m0ey1bStRhaG7sr2BZoZ4z1V0YEMD6EVrn/AASQ/Y01nxCdct/DXiPS4Gl3vpljrcgtm5yR84aRVPorjHbFfCjwF8KfhX4Ug+Hvwh0XRtH0XSzxY6c4cq54LyNku8h2jLuSxxyTiv2lP2HvgN+1Zf2mufFDT9cTWdPs/sdpqOmarJA8EO5n2rE2+HO5mO4xkngEkAAfstfsw+Bv2TfhtN8N/A2oX+owXeqTahc32obPPnkcKq7tgAwscca8ADgnAyaqqqqqv2H/APlLD8cP+wh4r/8ATqtVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX7Qv/JAfiZ/2J+rf+kctf8EQ/wDklXxK/wCxgtf/AEnNVVVVeLf+RU1n/sHz/wDotq/4Jl6lJFo/xR0hfuXVxokzH3jW/A/9GGv2btHF54svtYdNy6fZ7VP915DgH/vlXqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/4JSf8npfGz/sH6h/6c46qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv26v2zNE/Y/+G9vqNnY2+r+NfETSQ+H9LmYiPKAeZcz4IPlR7kBUEM7MqggbmX4wftM/Hj486lPqHxS+J+uazFM5ZdPNy0VlDntHbJiNOOMhcnuTX/BN7/k9z4V/wDYQuf/AEinr/gqF+1N+0F+y/pvge9+Dup6Tp+neJTeQXt5caYt1PBPEImQKZCYwGV34KE5jPNf8Etv2m/iD+0h8I/FLfFbxE+ueJ/DmvbHvWgihL2s8QaIFIlVRh0mHAHGKqqqqqv2H/8AlLD8cP8AsIeK/wD06rVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV+0L/AMkB+Jn/AGJ+rf8ApHLX/BEP/klXxK/7GC1/9JzVVVVXi3/kVNZ/7B8//otq/wCCaH/NR/8AuFf+3dfs26WLbwjf6ow+e9vSo46qijH6s1VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX/BKT/k9L42f9g/UP/TnHVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV/wV+8Walr/AO2Bd6BdXEhtfDOgWNpaxFvlUSIbhmA9SZsE9TtHoK/Zf/4J9/ATQP2QYvi54+8G2fizxl4n8Gy65HdakTLDYJPaGWCOGHOwFUZCXYFt+cEDAH/BN7/k9z4V/wDYQuf/AEinr/gtxdaenwT+H1jJJH9tl8UySQoR8xjW1kEhB9AXjz9RX/BDe6uE134w2SN+4ltNHkcbf4le7C8/R2/yKqqqqqv2H/8AlLD8cP8AsIeK/wD06rVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV+0L/AMkB+Jn/AGJ+rf8ApHLX/BEP/klXxK/7GC1/9JzVVVVXi3/kVNZ/7B8//otq/wCCaH/NR/8AuFf+3dfBuw/s/wCGuiREfNLE8x997sw/Qiqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr/glJ/wAnpfGz/sH6h/6c46qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv+C0vwZ1jR/ip4X+OdlaSyaL4i0tdJvJgpKwXtuzsgY9vMhcbR38l6/Zn/wCCoX/CsvgFd/s/fFvwfqOtWFpo11p2haxpciGeGOSN1jhmikZVZULBQ6sCEAG1iMn9k74veHvgJ+0L4N+LvirT9RvtL8N3M01xbaeiNPJvt5Y1CB2VfvOuckcZ69K/bW/bD8Tfth/Ea08SX2kjRPDmgQPbaDpHm+a0COVMssj8BpZCqZwAAERedpY/8EYvhFqng34HeKPinq9rJbt4+1WJLFZFI820s1kRZQD2Ms06++yqqqqqr9gtjrP/AAVP+N+q2CF7fz/FFyWbClY21eMA4Pu6jHvVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV+0L/wAkB+Jn/Yn6t/6Ry1/wRD/5JV8Sv+xgtf8A0nNVVVVeO7yDT/A/iG/uSRFbaVdSyEDJ2rExOB9BX/BNK3lW3+Il0QPLkfTEU57qLon/ANCFeCbdbXwZoNso/wBXptup4xz5a5OKqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv8AglJ/yel8bP8AsH6h/wCnOOqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr4qfCvwJ8afAmqfDf4j6BBq+havFsngkGGRh92SNhykinBVhyCK+Pv/BHv49eBtVub/4IXNn8QfD7OTb273MVlqMCdcSJIVikwMDcjZY87F6VF+wj+2HPqH9mJ+zr4zEw43PYFI/+/hIT9a/Zm/4I7/E3xFrtl4h/aWurXwv4etpFkl0Kxu0ub++AOfLaSImOBCOrBmfqAFJDDw74d0LwjoGneFvDGlW2maTpNrHa2NnbR7I7eGNQqIoHQAACqqqqqr4V/HPw5+wt/wAFCfjT4j+Lmg67eWd3LrFgkWlW8b3DC5v4buCUJJIi7XiRW5YcOOtf8Pq/2V/+hC+Kv/gqsP8A5Nr/AIfV/sr/APQhfFX/AMFVh/8AJtf8Pq/2V/8AoQvir/4KrD/5Nr/h9X+yv/0IXxV/8FVh/wDJtf8AD6v9lf8A6EL4q/8AgqsP/k2v+H1f7K//AEIXxV/8FVh/8m1/w+r/AGV/+hC+Kv8A4KrD/wCTa/4fV/sr/wDQhfFX/wAFVh/8m1/w+r/ZX/6EL4q/+Cqw/wDk2v8Ah9X+yv8A9CF8Vf8AwVWH/wAm1/w+r/ZX/wChC+Kv/gqsP/k2v+H1f7K//QhfFX/wVWH/AMm1/wAPq/2V/wDoQvir/wCCqw/+Ta/4fV/sr/8AQhfFX/wVWH/ybX/D6v8AZX/6EL4q/wDgqsP/AJNr/h9X+yv/ANCF8Vf/AAVWH/ybX/D6v9lf/oQvir/4KrD/AOTa/wCH1f7K/wD0IXxV/wDBVYf/ACbX/D6v9lf/AKEL4q/+Cqw/+Ta/4fV/sr/9CF8Vf/BVYf8AybX/AA+r/ZX/AOhC+Kv/AIKrD/5Nr/h9X+yv/wBCF8Vf/BVYf/Jtf8Pq/wBlf/oQvir/AOCqw/8Ak2v+H1f7K/8A0IXxV/8ABVYf/Jtf8Pq/2V/+hC+Kv/gqsP8A5Nr/AIfV/sr/APQhfFX/AMFVh/8AJtf8Pq/2V/8AoQvir/4KrD/5Nr/h9X+yv/0IXxV/8FVh/wDJtf8AD6v9lf8A6EL4q/8AgqsP/k2v+H1f7K//AEIXxV/8FVh/8m1/w+r/AGV/+hC+Kv8A4KrD/wCTa/4fV/sr/wDQhfFX/wAFVh/8m1/w+r/ZX/6EL4q/+Cqw/wDk2v8Ah9X+yv8A9CF8Vf8AwVWH/wAm1/w+r/ZX/wChC+Kv/gqsP/k2v+H1f7K//QhfFX/wVWH/AMm1/wAPq/2V/wDoQvir/wCCqw/+Ta/4fV/sr/8AQhfFX/wVWH/ybX/D6v8AZX/6EL4q/wDgqsP/AJNr/h9X+yv/ANCF8Vf/AAVWH/ybX/D6v9lf/oQvir/4KrD/AOTa/wCH1f7K/wD0IXxV/wDBVYf/ACbX/D6v9lf/AKEL4q/+Cqw/+Ta/4fV/sr/9CF8Vf/BVYf8AybX/AA+r/ZX/AOhC+Kv/AIKrD/5Nr/h9X+yv/wBCF8Vf/BVYf/Jtf8Pq/wBlf/oQvir/AOCqw/8Ak2v+H1f7K/8A0IXxV/8ABVYf/Jtf8Pq/2V/+hC+Kv/gqsP8A5Nr/AIfV/sr/APQhfFX/AMFVh/8AJtf8Pq/2V/8AoQvir/4KrD/5Nr/h9X+yv/0IXxV/8FVh/wDJtf8AD6v9lf8A6EL4q/8AgqsP/k2v+H1f7K//AEIXxV/8FVh/8m1/w+r/AGV/+hC+Kv8A4KrD/wCTa/4fV/sr/wDQhfFX/wAFVh/8m1/w+r/ZX/6EL4q/+Cqw/wDk2v8Ah9X+yv8A9CF8Vf8AwVWH/wAm1/w+r/ZX/wChC+Kv/gqsP/k2v+H1f7K//QhfFX/wVWH/AMm1/wAPq/2V/wDoQvir/wCCqw/+Ta/4fV/sr/8AQhfFX/wVWH/ybX/D6v8AZX/6EL4q/wDgqsP/AJNr/h9X+yv/ANCF8Vf/AAVWH/ybX/D6v9lf/oQvir/4KrD/AOTa/wCH1f7K/wD0IXxV/wDBVYf/ACbX/D6v9lf/AKEL4q/+Cqw/+Ta/4fV/sr/9CF8Vf/BVYf8AybX/AA+r/ZX/AOhC+Kv/AIKrD/5Nr/h9X+yv/wBCF8Vf/BVYf/Jtf8Pq/wBlf/oQvir/AOCqw/8Ak2v+H1f7K/8A0IXxV/8ABVYf/Jtf8Pq/2V/+hC+Kv/gqsP8A5Nr/AIfV/sr/APQhfFX/AMFVh/8AJtf8Pq/2V/8AoQvir/4KrD/5Nr/h9X+yv/0IXxV/8FVh/wDJtf8AD6v9lf8A6EL4q/8AgqsP/k2v+H1f7K//AEIXxV/8FVh/8m1/w+r/AGV/+hC+Kv8A4KrD/wCTa/4fV/sr/wDQhfFX/wAFVh/8m18VP+Cwf7M/jj4YeL/BWk+B/ibFfa/oN9p9tJc6ZYrGks0DxqXK3hIUFhkgE47Gv+CIf/JKviV/2MFr/wCk5qqqqrxxotz4k8Fa/wCHbJ0W41TS7m1iZ+AHkiZAT7ZIr9mf9oDTP2Xrvxl4T+IPhDW31C4vYopbeBUWS1mgMqSRyK7LggsB9Qa03/gtV8BLLTbWyk+FXj9mt4EjYhbTBKqAcfvvav8Ah9p8Af8AolHxA/75tP8A49X/AA+0+AP/AESj4gf982n/AMer/h9p8Af+iUfED/vm0/8Aj1f8PtPgD/0Sj4gf982n/wAer/h9p8Af+iUfED/vm0/+PV/w+0+AP/RKPiB/3zaf/Hq/4fafAH/olHxA/wC+bT/49X/D7T4A/wDRKPiB/wB82n/x6v8Ah9p8Af8AolHxA/75tP8A49X/AA+0+AP/AESj4gf982n/AMer/h9p8Af+iUfED/vm0/8Aj1f8PtPgD/0Sj4gf982n/wAer/h9p8Af+iUfED/vm0/+PV/w+0+AP/RKPiB/3zaf/Hq/4fafAH/olHxA/wC+bT/49X/D7T4A/wDRKPiB/wB82n/x6v8Ah9p8Af8AolHxA/75tP8A49X/AA+0+AP/AESj4gf982n/AMer/h9p8Af+iUfED/vm0/8Aj1f8PtPgD/0Sj4gf982n/wAer/h9p8Af+iUfED/vm0/+PV/w+0+AP/RKPiB/3zaf/Hq/4fafAH/olHxA/wC+bT/49X/D7T4A/wDRKPiB/wB82n/x6v8Ah9p8Af8AolHxA/75tP8A49X/AA+0+AP/AESj4gf982n/AMer/h9p8Af+iUfED/vm0/8Aj1f8PtPgD/0Sj4gf982n/wAer/h9p8Af+iUfED/vm0/+PV/w+0+AP/RKPiB/3zaf/Hq/4fafAH/olHxA/wC+bT/49X/D7T4A/wDRKPiB/wB82n/x6v8Ah9p8Af8AolHxA/75tP8A49X/AA+0+AP/AESj4gf982n/AMer/h9p8Af+iUfED/vm0/8Aj1f8PtPgD/0Sj4gf982n/wAer/h9p8Af+iUfED/vm0/+PV/w+0+AP/RKPiB/3zaf/Hq/4fafAH/olHxA/wC+bT/49X/D7T4A/wDRKPiB/wB82n/x6v8Ah9p8Af8AolHxA/75tP8A49X/AA+0+AP/AESj4gf982n/AMer/h9p8Af+iUfED/vm0/8Aj1f8PtPgD/0Sj4gf982n/wAer/h9p8Af+iUfED/vm0/+PV/w+0+AP/RKPiB/3zaf/Hq/4fafAH/olHxA/wC+bT/49X/D7T4A/wDRKPiB/wB82n/x6v8Ah9p8Af8AolHxA/75tP8A49X/AA+0+AP/AESj4gf982n/AMer/h9p8Af+iUfED/vm0/8Aj1f8PtPgD/0Sj4gf982n/wAer/h9p8Af+iUfED/vm0/+PV/w+0+AP/RKPiB/3zaf/Hq/4fafAH/olHxA/wC+bT/49X/D7T4A/wDRKPiB/wB82n/x6v8Ah9p8Af8AolHxA/75tP8A49X/AA+0+AP/AESj4gf982n/AMer/h9p8Af+iUfED/vm0/8Aj1f8PtPgD/0Sj4gf982n/wAer/h9p8Af+iUfED/vm0/+PV/w+0+AP/RKPiB/3zaf/Hq/4fafAH/olHxA/wC+bT/49X/D7T4A/wDRKPiB/wB82n/x6v8Ah9p8Af8AolHxA/75tP8A49X/AA+0+AP/AESj4gf982n/AMer/h9p8Af+iUfED/vm0/8Aj1f8PtPgD/0Sj4gf982n/wAer/h9p8Af+iUfED/vm0/+PV/w+0+AP/RKPiB/3zaf/Hq/4fafAH/olHxA/wC+bT/49X/D7T4A/wDRKPiB/wB82n/x6v8Ah9p8Af8AolHxA/75tP8A49X/AA+0+AP/AESj4gf982n/AMer/h9p8Af+iUfED/vm0/8Aj1f8PtPgD/0Sj4gf982n/wAer/h9p8Af+iUfED/vm0/+PV/w+0+AP/RKPiB/3zaf/Hq/4fafAH/olHxA/wC+bT/49X/D7T4A/wDRKPiB/wB82n/x6v8AgkBrMPij9q34s+KLO3ljttU0O5uo1kHzIJdQidVbGRnB/Q1VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX7WH/AAUO+CP7KV//AMIlqq3vinxm0IlOhaWyqbVWGUNzM3yxbhyFAZ8EHaFYE6H/AMFxoJdaVPEv7Ob2+kuwDPY+JhNPCvchXt0WQ+2U+tfAP9of4VftLeBovHvwp8Qi+tNwjvLOdRFdafNjJiniydjehBKsOVZhzXxw/bI/Zu/Z3nbTPij8T9PstYVA40e0V7y9wRld0MQYxgjkGTaD618T/wDgtv4MshLZ/Bv4Natqr4IjvvEV6lmikd/Jh8xnX6uh9cdK/wCCcH7VnxR/az8M+PfF3xMXR7d9K1iC20+z0u0MMNtG0JYgFmZ2JOOWY+2KqqqqqviP+zP+z/8AF7X7fxV8TPhD4Y8R6xbRiJL2+sFeUov3UdurqMnCtkDJx1Nf8MUfsi/9G4fD/wD8EUP/AMTX/DFH7Iv/AEbh8P8A/wAEUP8A8TX/AAxR+yL/ANG4fD//AMEUP/xNf8MUfsi/9G4fD/8A8EUP/wATX/DFH7Iv/RuHw/8A/BFD/wDE1/wxR+yL/wBG4fD/AP8ABFD/APE1/wAMUfsi/wDRuHw//wDBFD/8TX/DFH7Iv/RuHw//APBFD/8AE1/wxR+yL/0bh8P/APwRQ/8AxNf8MUfsi/8ARuHw/wD/AARQ/wDxNf8ADFH7Iv8A0bh8P/8AwRQ//E1/wxR+yL/0bh8P/wDwRQ//ABNf8MUfsi/9G4fD/wD8EUP/AMTX/DFH7Iv/AEbh8P8A/wAEUP8A8TX/AAxR+yL/ANG4fD//AMEUP/xNf8MUfsi/9G4fD/8A8EUP/wATX/DFH7Iv/RuHw/8A/BFD/wDE1/wxR+yL/wBG4fD/AP8ABFD/APE1/wAMUfsi/wDRuHw//wDBFD/8TX/DFH7Iv/RuHw//APBFD/8AE1/wxR+yL/0bh8P/APwRQ/8AxNf8MUfsi/8ARuHw/wD/AARQ/wDxNf8ADFH7Iv8A0bh8P/8AwRQ//E1/wxR+yL/0bh8P/wDwRQ//ABNf8MUfsi/9G4fD/wD8EUP/AMTX/DFH7Iv/AEbh8P8A/wAEUP8A8TX/AAxR+yL/ANG4fD//AMEUP/xNf8MUfsi/9G4fD/8A8EUP/wATX/DFH7Iv/RuHw/8A/BFD/wDE1/wxR+yL/wBG4fD/AP8ABFD/APE1/wAMUfsi/wDRuHw//wDBFD/8TX/DFH7Iv/RuHw//APBFD/8AE1/wxR+yL/0bh8P/APwRQ/8AxNf8MUfsi/8ARuHw/wD/AARQ/wDxNf8ADFH7Iv8A0bh8P/8AwRQ//E1/wxR+yL/0bh8P/wDwRQ//ABNf8MUfsi/9G4fD/wD8EUP/AMTX/DFH7Iv/AEbh8P8A/wAEUP8A8TX/AAxR+yL/ANG4fD//AMEUP/xNf8MUfsi/9G4fD/8A8EUP/wATX/DFH7Iv/RuHw/8A/BFD/wDE1/wxR+yL/wBG4fD/AP8ABFD/APE1/wAMUfsi/wDRuHw//wDBFD/8TX/DFH7Iv/RuHw//APBFD/8AE1/wxR+yL/0bh8P/APwRQ/8AxNf8MUfsi/8ARuHw/wD/AARQ/wDxNf8ADFH7Iv8A0bh8P/8AwRQ//E1/wxR+yL/0bh8P/wDwRQ//ABNf8MUfsi/9G4fD/wD8EUP/AMTX/DFH7Iv/AEbh8P8A/wAEUP8A8TX/AAxR+yL/ANG4fD//AMEUP/xNf8MUfsi/9G4fD/8A8EUP/wATX/DFH7Iv/RuHw/8A/BFD/wDE1/wxR+yL/wBG4fD/AP8ABFD/APE1/wAMUfsi/wDRuHw//wDBFD/8TX/DFH7Iv/RuHw//APBFD/8AE1/wxR+yL/0bh8P/APwRQ/8AxNf8MUfsi/8ARuHw/wD/AARQ/wDxNf8ADFH7Iv8A0bh8P/8AwRQ//E1/wxR+yL/0bh8P/wDwRQ//ABNf8MUfsi/9G4fD/wD8EUP/AMTX/DFH7Iv/AEbh8P8A/wAEUP8A8TX/AAxR+yL/ANG4fD//AMEUP/xNf8MUfsi/9G4fD/8A8EUP/wATX/DFH7Iv/RuHw/8A/BFD/wDE1/wxR+yL/wBG4fD/AP8ABFD/APE1/wAMUfsi/wDRuHw//wDBFD/8TX/DFH7Iv/RuHw//APBFD/8AE1/wxR+yL/0bh8P/APwRQ/8AxNf8MUfsi/8ARuHw/wD/AARQ/wDxNf8ADFH7Iv8A0bh8P/8AwRQ//E1/wxR+yL/0bh8P/wDwRQ//ABNf8MUfsi/9G4fD/wD8EUP/AMTX/DFH7Iv/AEbh8P8A/wAEUP8A8TX/AAxR+yL/ANG4fD//AMEUP/xNf8MUfsi/9G4fD/8A8EUP/wATX/DFH7Iv/RuHw/8A/BFD/wDE1/wxR+yL/wBG4fD/AP8ABFD/APE1/wAMUfsi/wDRuHw//wDBFD/8TX/DFH7Iv/RuHw//APBFD/8AE1/wxR+yL/0bh8P/APwRQ/8AxNfD/wCGHw5+FGiyeHPhn4H0Twvpks7XEtrpVklskkpABdggG5sKoyecADoBVVVVVeKP2efgD441qfxL40+Bvw/8QavdY8/UNU8M2l1cS4GBukkjLNgADk1/wyb+yv8A9Gz/AAp/8Iyw/wDjVf8ADJv7K/8A0bP8Kf8AwjLD/wCNV/wyb+yv/wBGz/Cn/wAIyw/+NV/wyb+yv/0bP8Kf/CMsP/jVf8Mm/sr/APRs/wAKf/CMsP8A41X/AAyb+yv/ANGz/Cn/AMIyw/8AjVf8Mm/sr/8ARs/wp/8ACMsP/jVf8Mm/sr/9Gz/Cn/wjLD/41X/DJv7K/wD0bP8ACn/wjLD/AONV/wAMm/sr/wDRs/wp/wDCMsP/AI1X/DJv7K//AEbP8Kf/AAjLD/41X/DJv7K//Rs/wp/8Iyw/+NV/wyb+yv8A9Gz/AAp/8Iyw/wDjVf8ADJv7K/8A0bP8Kf8AwjLD/wCNV/wyb+yv/wBGz/Cn/wAIyw/+NV/wyb+yv/0bP8Kf/CMsP/jVf8Mm/sr/APRs/wAKf/CMsP8A41X/AAyb+yv/ANGz/Cn/AMIyw/8AjVf8Mm/sr/8ARs/wp/8ACMsP/jVf8Mm/sr/9Gz/Cn/wjLD/41X/DJv7K/wD0bP8ACn/wjLD/AONV/wAMm/sr/wDRs/wp/wDCMsP/AI1X/DJv7K//AEbP8Kf/AAjLD/41X/DJv7K//Rs/wp/8Iyw/+NV/wyb+yv8A9Gz/AAp/8Iyw/wDjVf8ADJv7K/8A0bP8Kf8AwjLD/wCNV/wyb+yv/wBGz/Cn/wAIyw/+NV/wyb+yv/0bP8Kf/CMsP/jVf8Mm/sr/APRs/wAKf/CMsP8A41X/AAyb+yv/ANGz/Cn/AMIyw/8AjVf8Mm/sr/8ARs/wp/8ACMsP/jVf8Mm/sr/9Gz/Cn/wjLD/41X/DJv7K/wD0bP8ACn/wjLD/AONV/wAMm/sr/wDRs/wp/wDCMsP/AI1X/DJv7K//AEbP8Kf/AAjLD/41X/DJv7K//Rs/wp/8Iyw/+NV/wyb+yv8A9Gz/AAp/8Iyw/wDjVf8ADJv7K/8A0bP8Kf8AwjLD/wCNV/wyb+yv/wBGz/Cn/wAIyw/+NV/wyb+yv/0bP8Kf/CMsP/jVf8Mm/sr/APRs/wAKf/CMsP8A41X/AAyb+yv/ANGz/Cn/AMIyw/8AjVf8Mm/sr/8ARs/wp/8ACMsP/jVf8Mm/sr/9Gz/Cn/wjLD/41X/DJv7K/wD0bP8ACn/wjLD/AONV/wAMm/sr/wDRs/wp/wDCMsP/AI1X/DJv7K//AEbP8Kf/AAjLD/41X/DJv7K//Rs/wp/8Iyw/+NV/wyb+yv8A9Gz/AAp/8Iyw/wDjVf8ADJv7K/8A0bP8Kf8AwjLD/wCNV/wyb+yv/wBGz/Cn/wAIyw/+NV/wyb+yv/0bP8Kf/CMsP/jVf8Mm/sr/APRs/wAKf/CMsP8A41X/AAyb+yv/ANGz/Cn/AMIyw/8AjVf8Mm/sr/8ARs/wp/8ACMsP/jVf8Mm/sr/9Gz/Cn/wjLD/41X/DJv7K/wD0bP8ACn/wjLD/AONV/wAMm/sr/wDRs/wp/wDCMsP/AI1X/DJv7K//AEbP8Kf/AAjLD/41X/DJv7K//Rs/wp/8Iyw/+NV/wyb+yv8A9Gz/AAp/8Iyw/wDjVf8ADJv7K/8A0bP8Kf8AwjLD/wCNV/wyb+yv/wBGz/Cn/wAIyw/+NV/wyb+yv/0bP8Kf/CMsP/jVf8Mm/sr/APRs/wAKf/CMsP8A41X/AAyb+yv/ANGz/Cn/AMIyw/8AjVf8Mm/sr/8ARs/wp/8ACMsP/jVf8Mm/sr/9Gz/Cn/wjLD/41X/DJv7K/wD0bP8ACn/wjLD/AONV/wAMm/sr/wDRs/wp/wDCMsP/AI1X/DJv7K//AEbP8Kf/AAjLD/41X/DJv7K//Rs/wp/8Iyw/+NV/wyb+yv8A9Gz/AAp/8Iyw/wDjVf8ADJv7K/8A0bP8Kf8AwjLD/wCNV/wyb+yv/wBGz/Cn/wAIyw/+NV/wyb+yv/0bP8Kf/CMsP/jVf8Mm/sr/APRs/wAKf/CMsP8A41X/AAyb+yv/ANGz/Cn/AMIyw/8AjVf8Mm/sr/8ARs/wp/8ACMsP/jVf8Mm/sr/9Gz/Cn/wjLD/41X/DJv7K/wD0bP8ACn/wjLD/AONV4H+FXwv+GEd3D8NPhv4W8JpfsrXaaHo8FgLgrnaXEKLuIycZ6ZPrVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVXxT8dWPwu+Gfiv4k6lGstt4W0W71OSIvt80QQtJsB9W27R7kV4x8W+JviT4y1Xxp4ov59T13xBfSXV3M2WaWaRsnA9MnAUdBgDgVJ/wTx/bRj8I/8Ju3wA146aIPO8pZrdrzbjP/AB5iT7Rux/D5efav2ev2lPil+y54m13XvhxqBs7vWtHudLuoZlyqM6kRTbGBHmQyYdcjsynhmB1XVdT13U7vWta1G5v9Qv53nurq5lMks8rkszuzElmJJJJ5JNfDX9kn9pf4vmFvh78E/FWpW85AivZbE2lq2f8Ap4n2Rf8Aj1f8E1P2Ufid+yr8NPEulfFSXSF1PxLqcV5HaWFybg2yJFs2yPtC7ic8KWGO/pVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX/AAUR1C40z9iv4rXNqV3vpCQHOfuyXEUbdD/dc1/wTI8I6J4y/bV+H1pr1ol1b6a13qMcUiBlM9vayyQtz/dkVHHugqv+CovgTRfAn7ZvjJNBtora216G11Z4I12qk88K+c31eVXkPu5r/glV+y98KvDX7PXhv45al4V07VfG3ix7i5Gp3sCzPp8MdxJFFFb7siPiPezKAxZyCSFUCqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr/AIKGacdV/Yt+K9qrsCmiifKrk/up4pOnp8nXt1r4V/Fbx/8ABLx1p3xK+F/iFtF8R6Usq2l4LaK42CWJonBjlV0YFHYfMpxkEYIBH/D0f9u3/ouv/ls6T/8AItfF/wCM3xK+PXjOT4g/FjxL/buvy28du939jgtcxxjCLsgRE4B67c+tfDf9v/8Aa5+EXgjSvhx8O/i3/ZPh3RI2jsLL+wdOn8lWdnYb5bdnbLOx+Zj19K/4ej/t2/8ARdf/AC2dJ/8AkWv+CWX7R/xo/aQ+H/jjW/jR4z/4SK90fWILeyl/s61tPKjaHcy4t44wcnnJBNVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV8Zfh5F8XPhJ4y+F0t8tl/wlehXmmJdNGXFs80LIsu0EFtrMG25GcY71/w458Vf9HFaT/4Tcn/AMfr/hxz4q/6OK0n/wAJuT/4/X/DjnxV/wBHFaT/AOE3J/8AH6/4cc+Kv+jitJ/8JuT/AOP1/wAOOfFX/RxWk/8AhNyf/H6/YT/Y41L9jjwl4n8M6l48tfFDeIdRiu0mg09rQQhI9m0gu+c9c8VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV4o8ZeD/AAPp8ereNfFej6BYyzCCO51S+jtI3lILBA0jAFiqscdcKT2NWfx4+BupXkGnad8Z/At1dXUqxQQQ+IrV3ldjhVVRJksSQABySaqqqqvL2zsIDdX93DbQqQDJNIEUZ6cniqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqvFHjLwf4H0+PVvGvivR9AsZZhBHc6pfR2kbykFggaRgCxVWOOuFJ7GrP48fA3UryDTtO+M/gW6urqVYoIIfEVq7yuxwqqokyWJIAA5JNVVVVXl7Z2EBur+7htoVIBkmkCKM9OTxVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX7efxT8dfBT9k7x18Tfhpra6T4k0YWAsb02sVx5Pm6hbQufLlVkbMcjj5lOM56itW/4KB/tna0XN5+0L4nj8xdp+yNFa4Ht5SLg+45qP9tr9ruKRJU/aO8fFkYEBtalYZHqCcEexr4ef8FTP2zPAd7DLffEe18WWMbZew1/TYZlk9cyxhJh+ElfsWftx+Av2w/Dl3FY6e3h7xnocSPq2hSzeYNh4FxA+B5kRbg5AZCQGGCrNVVVVVVVVVVVVVV/wWr/5NX8Lf9lAs/8A0gv6/Z6/5L98M/8AscNJ/wDSuKqqqqv+ConxC+JHir9rjxj4U8ZanqC6R4Zmht9D0uSRhBb27QRuJUTpul3Fy/U7gM4UAf8ABFX4i/EnxL4G8feBvEV7e6h4U8MT2J0SW5cstnJMJvOt4yedmEifYOEJJx+8qqqqqqqndI0MkjqqqCWZjgADqa/a+/a0+Jn7S37SGoz/AA28V67b6BHeLovhXT9MvZbcXEQkKJJtVgDJM7F8nkBlXoor9nT4TSfBD4L+Fvhxe6tc6tqmm2KnVdQuLh53u71/mnk3uSxXeWCg/dQKO1VVVX/Dfn7ZX/Rw3ir/AL/J/wDE1/wTh+I/jn4sfsmeGPG3xG8TXmv67eXuoJPfXbAySKl1IiA4A6KAPwqq/wCCkvhH4ofs3/tM31z4T+IXimx8NeL1/tzRFg1e4RLR2cieBMNhdkoLKo4VJIx2r9h39o5P2oP2edB+IF/LEfEVkzaZ4ijjXaFvoQu5wBwBIjRygDgeZjtVVVVVVVVVVVVVVVVVVVVX7efxT8dfBT9k7x18Tfhpra6T4k0YWAsb02sVx5Pm6hbQufLlVkbMcjj5lOM56itW/wCCgf7Z2tFzeftC+J4/MXafsjRWuB7eUi4PuOaj/ba/a7ikSVP2jvHxZGBAbWpWGR6gnBHsa+Hn/BUz9szwHewy33xHtfFljG2XsNf02GZZPXMsYSYfhJX7Fn7cfgL9sPw5dxWOnt4e8Z6HEj6toUs3mDYeBcQPgeZEW4OQGQkBhgqzVVVVVVVVVVVVVVf8Fq/+TV/C3/ZQLP8A9IL+v2ev+S/fDP8A7HDSf/SuKqqqqv8AgqJ8QviR4q/a48Y+FPGWp6gukeGZobfQ9LkkYQW9u0EbiVE6bpdxcv1O4DOFAH/BFX4i/EnxL4G8feBvEV7e6h4U8MT2J0SW5cstnJMJvOt4yedmEifYOEJJx+8qqqqqqqndI0MkjqqqCWZjgADqa/a+/a0+Jn7S37SGoz/DbxXrtvoEd4ui+FdP0y9ltxcRCQokm1WAMkzsXyeQGVeiiv2dPhNJ8EPgv4W+HF7q1zq2qabYqdV1C4uHne7vX+aeTe5LFd5YKD91Ao7VVVVf8N+ftlf9HDeKv+/yf/E1/wAE4fiP45+LH7Jnhjxt8RvE15r+u3l7qCT312wMkipdSIgOAOigD8Kqv+CkvhH4ofs3/tM31z4T+IXimx8NeL1/tzRFg1e4RLR2cieBMNhdkoLKo4VJIx2r9h39o5P2oP2edB+IF/LEfEVkzaZ4ijjXaFvoQu5wBwBIjRygDgeZjtVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV/wAFR/8AkxL4m/8AcJ/9O1nVeGP2b/2d7jwZpVnP8Bfh28MunQiSNvC9mVbMYzkeX3ya/wCCrX7JHw0/Z38W+E/HPwm0xdG0fxoLuO60aNiYbS5g8o74cklUkWXlOilDjAYAf8E0PF+seEP20vh2dKmk8rWbi40y9hTOJoJreQEMB2V1jk+sYPaq+NP7S3wL/Z5sIr34v/EjS9Ae4QvbWbs013cAcEx28YaRhnA3BcAnkiv+HwP7HX9p/YPt3jHyM4+2/wBh/uvrjf5n/jlfBv8AaC+DP7QWiSa/8IPiBpniK3t9v2mKFmjuLUt082BwskeecblAODjOK+Lvxj+HHwI8FzfEP4q+IW0Xw/bTxwS3i2NxdBHkO1AUgR3AJ4zjGSOeat/+Cn/7C13cRWsHx0UyTOEQN4b1VRknA5NrgfU8VXxL/b9/ZG+D/jjVPhv8RPi8ml+ItFdEvrNdD1C5ELPGsijzIYHQna65wxwTg4IIHwl+L/w4+Ovgm1+Ivwq8Sprvh68llihu1tprcl42KOpjmRHUgjuoyMEZBBr44/tLfBH9m6w0rUvjT44Xw7b65NJDYMdPubszPGAXAW3jkIADLyQByK+Gn7fn7JPxh8caV8Nvhv8AFh9Z8R607pY2K+H9ShMpSNpH+eS3VFARGYlmAwK+I/xb+F/wg0Ya/wDFDx9ofhixbIik1O9SEzEdVjUndI3I+VQT7V4n/wCCtn7F/h+eSDTvF3iHxAY227tM0KYKxzg4M/lZHv8AlXgX/gqv+xn43v4NMufHep+GZ7g7YzrulSQR5yeGlj3xoMDOWYDkc54rS9V0vXdNttY0TUrXULC8iEttdWsyyxTIRkMjqSGUjoQcV8Uvil4D+CvgPU/iZ8TNe/sbw3o3lfbb37LNceV5sqQx/u4Vd2zJIi8KcZycAEjwZ/wUW/Y0+IXi3R/AvhD4yrf65r97FZadanw/qUPnzyMFRN8luqLliBliBz1r9ob9t79nT9mO8j0T4l+M3fX5YxKuiaXbm7uwhGQzqMLED28xl3dRkZx8Bvjj4K/aM+GenfFf4fxalHoupTTxQLqMCwzBopGjfKqzADcpxz0r4qfGP4XfBHwy3i/4r+ONL8NaUG2JNeS4aZ8Z2RRqC8r4BO1FJwCcVc/8Ffv2OIdX/s2LUvF9xb7tv9oR6ERDj+9hnEmP+AZ9q/4Kk/Hb4SfH/wDYy8MeLfhD440/xFp8XxDskuPILJLaudP1AhZYXCyRMQCQGUZ6jIr9nr/kv3wz/wCxw0n/ANK4qqq+LP8AwUk/ZC+EGqXGgat8TV17VrRmSez8PWr3/lsuQVMq4hDZG0rvyD1Arw1/wVz/AGM9fu1tdR8ReJ/D6s20TanoUjIOnJ+zmUgfh2rwP458H/Evwpp3jjwD4jsde0HVY2ez1CylEkUwVijYI6FWVlIPKspBAIIr9rH4of8ABL7xV8QLvwP+1Jdabc+MPDBS3uSNG1eO5gDKsixm5sogXTDghd7KNx6Emv2Rfir+yR438K6n4I/ZHvtOOieEnhe/s7LR7uxWB7nzNjsbmJDKz+Q+Wyx+QZI4r4y/trfsx/s++Lk8B/F74m/2Drslml4tp/Y19dZhcsqtvggdOSjDGc8dORX/AA9H/YS/6Lr/AOWzq3/yLX/D0f8AYS/6Lr/5bOrf/ItfBv8AbW/Zj/aC8XP4D+EPxN/t/XY7OS8a0/sa+tcQoVVm3zwInBdRjOeenWq+NP7ef7K/wF1Wbw546+KNpLrts22bSdJhe/uIWxysgiBWJsY+V2U8jivDP/BXL9jXxDqSadf+IPE+gK7hFudT0R/K57kwtIQPcgY/OvB/jPwj8QvDln4u8DeJdN17RdQTfbX2n3CzxSDvhlJGQeCOoIIOCKqv+CiXxXuPg/8Ash+Pdc027MGqazaLotgyttYPdsInKnsywmZwRzlR061/wS9+GEPxN/bI8IPe24msfCMU+v3CkdGt1Agb2xcSW5/D8arxF/wUo/Yo8Ka/qfhbX/jV9l1TR7yWzvYP+Ec1R/KmicpIu5bYq2GUjIJBxwSK+Cnx7+E37RXhW68bfBvxZ/wkGi2OoPp89z9guLTZcpHHIybJ40Y4SWM5Ax82M5Bx8X/jN8NfgL4Mk+IPxY8S/wBh6BDcR273f2Oe6xJIcIuyBHfkjrtwO9Wn/BT39hm/u4LG0+OPmT3EixxJ/wAI1qo3MxwBk2uByR1qq/4JP/8AJkfg/wD7CGp/+lktVX/BZf4Yx+K/2bdH+I9vblrzwNr0ZeTbnZa3YEMgz2zKLX8vy/4It/FmXw58avFPwfvLkiy8Y6OLy1QnP+mWbE4Udt0MsxOOvlr6cVVVVVVVVVVVVVVVVVVVVf8ABUf/AJMS+Jv/AHCf/TtZ1Xhj9m/9ne48GaVZz/AX4dvDLp0IkjbwvZlWzGM5Hl98mv8Agq1+yR8NP2d/FvhPxz8JtMXRtH8aC7jutGjYmG0uYPKO+HJJVJFl5TopQ4wGAH/BNDxfrHhD9tL4dnSppPK1m4uNMvYUziaCa3kBDAdldY5PrGD2qvjT+0t8C/2ebCK9+L/xI0vQHuEL21m7NNd3AHBMdvGGkYZwNwXAJ5Ir/h8D+x1/af2D7d4x8jOPtv8AYf7r643+Z/45Xwb/AGgvgz+0Fokmv/CD4gaZ4it7fb9pihZo7i1LdPNgcLJHnnG5QDg4zivi78Y/hx8CPBc3xD+KviFtF8P208cEt4tjcXQR5DtQFIEdwCeM4xkjnmrf/gp/+wtd3EVrB8dFMkzhEDeG9VUZJwOTa4H1PFV8S/2/f2Rvg/441T4b/ET4vJpfiLRXRL6zXQ9QuRCzxrIo8yGB0J2uucMcE4OCCB8Jfi/8OPjr4JtfiL8KvEqa74evJZYobtbaa3JeNijqY5kR1II7qMjBGQQa+OP7S3wR/ZusNK1L40+OF8O2+uTSQ2DHT7m7MzxgFwFt45CAAy8kAcivhp+35+yT8YfHGlfDb4b/ABYfWfEetO6WNivh/UoTKUjaR/nkt1RQERmJZgMCviP8W/hf8INGGv8AxQ8faH4YsWyIpNTvUhMxHVY1J3SNyPlUE+1eJ/8AgrZ+xf4fnkg07xd4h8QGNtu7TNCmCsc4ODP5WR7/AJV4F/4Kr/sZ+N7+DTLnx3qfhme4O2M67pUkEecnhpY98aDAzlmA5HOeK0vVdL13TbbWNE1K11CwvIhLbXVrMssUyEZDI6khlI6EHFfFL4peA/gr4D1P4mfEzXv7G8N6N5X229+yzXHlebKkMf7uFXdsySIvCnGcnABI8Gf8FFv2NPiF4t0fwL4Q+Mq3+ua/exWWnWp8P6lD588jBUTfJbqi5YgZYgc9a/aG/be/Z0/ZjvI9E+JfjN31+WMSroml25u7sIRkM6jCxA9vMZd3UZGcfAb44+Cv2jPhnp3xX+H8WpR6LqU08UC6jAsMwaKRo3yqswA3Kcc9K+Knxj+F3wR8Mt4v+K/jjS/DWlBtiTXkuGmfGdkUagvK+ATtRScAnFXP/BX79jiHV/7Ni1LxfcW+7b/aEehEQ4/vYZxJj/gGfav+CpPx2+Enx/8A2MvDHi34Q+ONP8RafF8Q7JLjyCyS2rnT9QIWWFwskTEAkBlGeoyK/Z6/5L98M/8AscNJ/wDSuKqqviz/AMFJP2QvhBqlxoGrfE1de1a0Zkns/D1q9/5bLkFTKuIQ2RtK78g9QK8Nf8Fc/wBjPX7tbXUfEXifw+rNtE2p6FIyDpyfs5lIH4dq8D+OfB/xL8Kad448A+I7HXtB1WNns9QspRJFMFYo2COhVlZSDyrKQQCCK/ax+KH/AAS+8VfEC78D/tSXWm3PjDwwUt7kjRtXjuYAyrIsZubKIF0w4IXeyjcehJr9kX4q/skeN/Cup+CP2R77TjonhJ4Xv7Oy0e7sVge58zY7G5iQys/kPlssfkGSOK+Mv7a37Mf7Pvi5PAfxe+Jv9g67JZpeLaf2NfXWYXLKrb4IHTkowxnPHTkV/wAPR/2Ev+i6/wDls6t/8i1/w9H/AGEv+i6/+Wzq3/yLXwb/AG1v2Y/2gvFz+A/hD8Tf7f12OzkvGtP7GvrXEKFVZt88CJwXUYznnp1qvjT+3n+yv8BdVm8OeOvijaS67bNtm0nSYXv7iFscrIIgVibGPldlPI4rwz/wVy/Y18Q6kmnX/iDxPoCu4RbnU9Efyue5MLSED3IGPzrwf4z8I/ELw5Z+LvA3iXTde0XUE3219p9ws8Ug74ZSRkHgjqCCDgiqr/gol8V7j4P/ALIfj3XNNuzBqms2i6LYMrbWD3bCJyp7MsJmcEc5UdOtf8EvfhhD8Tf2yPCD3tuJrHwjFPr9wpHRrdQIG9sXElufw/Gq8Rf8FKP2KPCmv6n4W1/41fZdU0e8ls72D/hHNUfyponKSLuW2KthlIyCQccEivgp8e/hN+0V4VuvG3wb8Wf8JBotjqD6fPc/YLi02XKRxyMmyeNGOEljOQMfNjOQcfF/4zfDX4C+DJPiD8WPEv8AYegQ3Edu939jnusSSHCLsgR35I67cDvVp/wU9/YZv7uCxtPjj5k9xIscSf8ACNaqNzMcAZNrgckdaqv+CT//ACZH4P8A+whqf/pZLVV/wWX+GMfiv9m3R/iPb25a88Da9GXk252Wt2BDIM9syi1/L8v+CLfxZl8OfGrxT8H7y5IsvGOji8tUJz/plmxOFHbdDLMTjr5a+nFVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV/wAFR/8AkxL4m/8AcJ/9O1nVWP8AwV3/AGQdB8K2UAufGV/dWVlFG1ta6IA7sqAEKZJEXqO7AV+3R+2jrH7Y/jvS9Sg0F9A8K+GYJYdG02WbzZd0hUyzysON77EG0cKEAyTlj/wSL/ZB8VXvjm2/am8b6TNp+g6PbzReF47mIo+o3EsZje5UHrCkbyKGxhnbg/uzX7aX7Stn+yr8B9X+JKQQXeuXEi6d4fs5s7J76VWKFwMEoio8jDIyIyoILA1pGnfGH9rT43W+mLe3finx3431HBnupPvMQSzsekcUaKSQAFRE4AAArxV/wRM+KGkeA59b8NfGPQ9d8VW9sZf7C/sp7aGZwMmOK6aU5Y9FLxoCcZKjJHwk+LHxG/Z2+J+n+P8AwNqNxpGv6DclJoJAVWZA2JbadONyNjayn6jBAI+JFron7aH7EmryeG7YyR/EHweL3TbZsO0N6qCaGJvVo7mNVOO6nFMpUlWBBBwQRjFfAT4i23xA/Z88CfE7UL9Mar4Ws76+nduI5fs6mfcf9lw4P0NfGb4g3XxZ+LfjH4mXhbzPE+uXd+qN/wAs0llZkQeyoVUewFf8ESfiT/aXw6+IXwlurjMmh6tBq1ojHkx3MflyBfZWtlJ95Pev+C1vj4a38dPBfw7gnMkPhfw413IueI57uY7hj18u3hOfcelf8EVvhMdf+LfjD4xX1uWtvCekpp9kzJx9qu2JZlPqsULqfaYV/wAFQ/2O/iJ+05D8ONX+EWgW9/4g0q/uNPvmmuEt0is5kEgmkdyPkjeEjCgtmbgEmvCv/BDzxre6dHL43/aC0XSL4oC8Gl+H5dQjVu4Ekk0BI99or9sH9jH4hfsd+KNJ0jxZq+n65pHiGKWTSdWskaNZvKKiRHjbmORd6EjLAhxhjyB/wRy/aR8R6P8AEy8/Zs17VJbrw/4hs5r7Q4ZpS32K9hXfKkQPRJIhIzL03RAgDcxP/BSNVb9iL4phgD/xL7bgj/p9gr4X+OLj4Y/Ezwj8SrTT47+fwlrtlq0VrI5RZ2tp0lCFhkgMUAJxxmtK8D/Hv9pfxfrvizw74L8V+Pddv7t7vV7zT9NluiJZCWLSMilUz2BwOgA6Cv2B9P1n9nn9hiyuPjJoGp+FpfCi6vqOqWuo2zQTQW6TyzFtjYPMfI9civ2nP2j/AB1+1F8VdT+I/jO9lW3eRotH0vzS0Wl2gPyQoOmcYLtgbmJY9cD9m7/gnb+0b+0/4WPjnwZYaHoXhx3aO01TxDdyW0d6yHD+SsUckjKpGC+0LkEAkqwH7Q37N3xn/Za8Sr4C+K2km0j1JRdWV1Z3BmsdSEeV3xvxuZPMZSrAOu/kAOCf2ev+S/fDP/scNJ/9K4qqv+Cwf7RHij4WfCbw98JvBmpzafefESW6Gp3cEhSRbC3EYkhBGCBK0ygnuqOp4av2a/2ffFX7T3xb0v4SeENT0/Tby/ilnlvL9iI7eGJNzthQWY4AAUDkkZIGSPH/APwRT+N+haVJf/D74neFvFVzDGWNjcwSabLKf7sZJkQnp99kHvX7OHwwb4MfAXwF8Lpokju/D2g21vfBCCpu9ga4YEcYMzSH8ep61/wUi/5Pc+Kf/YQtv/SKCv8Aghj/AM1s/wC4B/7ka/4KXfsUftOftBftF2/jv4Q/DP8At/Q4/DdpZtdf2zY2uJkkmZl2Tzo/AdecY569auf+CYH7ctnby3d38Eo4YIELyyyeKdIVUUDJYk3WAABkk1fWcun3txp9w8Dy20rRO0E6TRllOCVkQlXXjhlJUjkEg5r/AIIx/Arx1N8Rtb/aEu7L7H4TtdKn0a0mmUhr+5kkiZ/KHdIxHhm6bmCjJDbf+Cq/7Z/iH4GeGtO+CPwt1Z9P8W+LrN7nUtShbE2nacS0Y8oj7ssrK4DjlFjYjDMrL+yd+yL8Sv2wfHN74b8G3drpun6TEtxrOt36s0VorkhFwvLyuQ21MjO1iSACa/ax/wCCWHxE/Zq+HFz8VfD3xAs/HOhaUU/thU0xrC4skZgolEZllEkYYgMQwYZB27QxX/gnN+1b4i/Z1+OejeHL7WJT4E8aahDYa3Yyv+6gklYRx3ij+F42K7iPvR7gckKRVf8ABbrxLNafB74deD0YiLVPEk964A6m2tig5/7eTx/hX/BEDw3HdfEn4n+LzDl9M0OysVk2/dFzO7kZ7Z+yjjvj2qv2hf8Akv3xM/7HDVv/AErlr/gip/yav4p/7KBef+kFhX/BXn/kzTUv+xg07/0Nq8Jf8jVov/YQg/8ARi1Vf8En/wDkyPwf/wBhDU//AEslqq/bz8Nx+K/2Ofi3pckPmiDw3PfBdu7BtStwDj2MIOe2M1/wT78SXHhX9s34UalbMwa410WLBRnK3UT27dxxiU/4HpVVVVVVVVVVVVVVVVVVVVX/AAVH/wCTEvib/wBwn/07WdVY/wDBXf8AZB0HwrZQC58ZX91ZWUUbW1rogDuyoAQpkkReo7sBX7dH7aOsftj+O9L1KDQX0Dwr4Zglh0bTZZvNl3SFTLPKw43vsQbRwoQDJOWP/BIv9kHxVe+Obb9qbxvpM2n6Do9vNF4XjuYij6jcSxmN7lQesKRvIobGGduD+7NftpftK2f7KvwH1f4kpBBd65cSLp3h+zmzsnvpVYoXAwSiKjyMMjIjKggsDWkad8Yf2tPjdb6Yt7d+KfHfjfUcGe6k+8xBLOx6RxRopJAAVETgAACvFX/BEz4oaR4Dn1vw18Y9D13xVb2xl/sL+yntoZnAyY4rppTlj0UvGgJxkqMkfCT4sfEb9nb4n6f4/wDA2o3Gka/oNyUmgkBVZkDYltp043I2NrKfqMEAj4kWuiftofsSavJ4btjJH8QfB4vdNtmw7Q3qoJoYm9WjuY1U47qcUylSVYEEHBBGMV8BPiLbfED9nzwJ8TtQv0xqvhazvr6d24jl+zqZ9x/2XDg/Q18ZviDdfFn4t+MfiZeFvM8T65d36o3/ACzSWVmRB7KhVR7AV/wRJ+JP9pfDr4hfCW6uMyaHq0GrWiMeTHcx+XIF9la2Un3k96/4LW+Phrfx08F/DuCcyQ+F/DjXci54jnu5juGPXy7eE59x6V/wRW+Ex1/4t+MPjFfW5a28J6Smn2TMnH2q7YlmU+qxQup9phX/AAVD/Y7+In7TkPw41f4RaBb3/iDSr+40++aa4S3SKzmQSCaR3I+SN4SMKC2ZuASa8K/8EPPGt7p0cvjf9oLRdIvigLwaX4fl1CNW7gSSTQEj32iv2wf2MfiF+x34o0nSPFmr6frmkeIYpZNJ1ayRo1m8oqJEeNuY5F3oSMsCHGGPIH/BHL9pHxHo/wATLz9mzXtUluvD/iGzmvtDhmlLfYr2Fd8qRA9EkiEjMvTdECANzE/8FI1Vv2IvimGAP/EvtuCP+n2Cvhf44uPhj8TPCPxKtNPjv5/CWu2WrRWsjlFna2nSUIWGSAxQAnHGa0rwP8e/2l/F+u+LPDvgvxX4912/u3u9XvNP02W6IlkJYtIyKVTPYHA6ADoK/YH0/Wf2ef2GLK4+Mmgan4Wl8KLq+o6pa6jbNBNBbpPLMW2Ng8x8j1yK/ac/aP8AHX7UXxV1P4j+M72Vbd5Gi0fS/NLRaXaA/JCg6Zxgu2BuYlj1wP2bv+Cdv7Rv7T/hY+OfBlhoeheHHdo7TVPEN3JbR3rIcP5KxRySMqkYL7QuQQCSrAftDfs3fGf9lrxKvgL4raSbSPUlF1ZXVncGax1IR5XfG/G5k8xlKsA67+QA4J/Z6/5L98M/+xw0n/0riqq/4LB/tEeKPhZ8JvD3wm8GanNp958RJboandwSFJFsLcRiSEEYIErTKCe6o6nhq/Zr/Z98VftPfFvS/hJ4Q1PT9NvL+KWeW8v2Ijt4Yk3O2FBZjgABQOSRkgZI8f8A/BFP436FpUl/8Pvid4W8VXMMZY2NzBJpssp/uxkmRCen32Qe9fs4fDBvgx8BfAXwumiSO78PaDbW98EIKm72BrhgRxgzNIfx6nrX/BSL/k9z4p/9hC2/9IoK/wCCGP8AzWz/ALgH/uRr/gpd+xR+05+0F+0Xb+O/hD8M/wC39Dj8N2lm11/bNja4mSSZmXZPOj8B15xjnr1q5/4Jgfty2dvLd3fwSjhggQvLLJ4p0hVRQMliTdYAAGSTV9Zy6fe3Gn3DwPLbStE7QTpNGWU4JWRCVdeOGUlSOQSDmv8AgjH8CvHU3xG1v9oS7svsfhO10qfRrSaZSGv7mSSJn8od0jEeGbpuYKMkNt/4Kr/tn+IfgZ4a074I/C3Vn0/xb4us3udS1KFsTadpxLRjyiPuyysrgOOUWNiMMysv7J37IvxK/bB8c3vhvwbd2um6fpMS3Gs63fqzRWiuSEXC8vK5DbUyM7WJIAJr9rH/AIJYfET9mr4cXPxV8PfECz8c6FpRT+2FTTGsLiyRmCiURmWUSRhiAxDBhkHbtDFf+Cc37VviL9nX456N4cvtYlPgTxpqENhrdjK/7qCSVhHHeKP4XjYruI+9HuByQpFV/wAFuvEs1p8Hvh14PRiItU8ST3rgDqba2KDn/t5PH+Ff8EQPDcd18Sfif4vMOX0zQ7KxWTb90XM7uRntn7KOO+Paq/aF/wCS/fEz/scNW/8ASuWv+CKn/Jq/in/soF5/6QWFf8Fef+TNNS/7GDTv/Q2rwl/yNWi/9hCD/wBGLVV/wSf/AOTI/B//AGENT/8ASyWqr9vPw3H4r/Y5+LelyQ+aIPDc98F27sG1K3AOPYwg57YzX/BPvxJceFf2zfhRqVszBrjXRYsFGcrdRPbt3HGJT/gelVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX/AAVH/wCTEvib/wBwn/07WdVqX/BGn9pNPCq+I/DfjPwJrVxJbLcRacl3cW8swZNwRGkiCb+QPmZV9WFRnxP8NPGuLrThp3iHwzqJWW11Gxjm+zXML4ZJIZlZGKspBV1IyORX7A37WVt+1l8GBrep2Vpp3i3wzMmn69ZWo2xb9mYriJf4Y5FDYX+FkdeQAT/wXD8UXkmvfCnwUjstpBZ6jqEijpJI7wxqTz/CI3xx/GeuePgP8ePHv7OPj5PiV8NX01Ncis5bWOS/sxcrGkmN5VT0bAxn0JHev+HvP7ZX/Qc8Kf8AghT/AOKrxl4q1Tx34v1zxvrgtxqXiHUrjUbwW8IijE08jSPsQcKu5jhR0GBX/BJnU77UP2KPC9teK3l6dqmpW9sWzzGbp5OPbfI4/Cv24PhUPgz+1X8RvBNvbCCw/th7/TkVcKtrdAXEar6hFl8v6ofSvgv+0GfDH/BHTxRfw3wXU9AOoeE7dw3z+beXAK4/2khvgw9BGDX7PHwTm+L3hb4x6stm0q+B/ANxrUMipkxzw3Nu+B6ZhS5/AGv+CSnxGfwP+2DpHh+W5Mdp410m80mUH7pdU+0RH677cKD/ALeO5r/goD45b4hftj/FLWhP5sVlrbaVDg8BbJFtuPbMJPuST3r/AIJafCUfC79kDw1f3dssWpeN55dfujjJKTYS359DbxQtjoC59c1+3X/wUL8Mfskx2/gnwtpFr4n+Ieo24uI7CeVlttNhOQstyV+ZixB2xKVYgFiyjbu8a/8ABQL9tX4u6uLWP4xeI7B7uTbbab4WX+ztpPRE+zgSt/wJmPv0r4waf+08dP03Wfj9ZfFH7Fczyf2dd+MIr/y5ZWAMnlPc8FiACdpycDNf8E2ZpYP23vhY8Erxsb67UlWIOGsbgMOOxBII9DX/AAUi/wCTI/in/wBg+2/9LIKr9lj4aeFvhL+z14C8F+ErGO3tYdDtZ53VArXNzLEsk0z+rO7MT6cAcACv+CoHiK88OfsRfEJ7B2SXURY2JdTjakt7CJPqCm5f+BVXgPwbo3w78EaB4C8PQJDpvh3TYLC0RU2gRwxhFOPUhcn3Nf8ABa/TbCX9mjwfq8lpE17beOoIIZyvzxxyWN40ig9gxijJH+wPSv2ev+S/fDP/ALHDSf8A0riqq/4Ksfso+L/2g/hfoXjn4a6VNqvifwFLcM2lwLulvrKcJ5oiH8ciNEjBBywLgZbAPhnxP4u+HPiq08TeE9a1Lw/4h0W4LW93aytb3FtIMqwBGCDglSD1BIPBIr9n7/gsn8WfCd3Z6J+0D4ds/Guigqkuq6dElnqUQzy5UYhmwP4dsRPUvXw7+IXg74r+CdI+IfgDXINX0DXLcT2V3FkBlzggg8qysCrKQCrAggEGv+CkX/J7nxT/AOwhbf8ApFBX/BDH/mtn/cA/9yNV/wAFTv285ddvdT/Ze+DusldLtXMHjDVbd/8Aj7lH3rGNh/yzU48wj7zDZ91W3/safsneKv2tvixbeD9NM1j4b0zZc+I9XVMizts/cTPBmkwVQeuWI2q1eBfA3hX4aeDtI8A+CNHg0vQ9DtVtrG0hXAjRfXuWJySx5ZiSckk1/wAFKfFF54r/AG1viVPdO2zTru30+3jPSNILaJMDk8FgzfVj06V+zr+2t8cP2WtA1bw58JrrQra21u8W6vHvdMW4kd1QIo3Eg7QM4HqxPevHv/BUT9qz4k+CNf8Ah74p1TwvPo/iTTbjTr6NNDRWMM0ZR9rZ+VsMSG7HB7UCVIZWIIPBHGK8Dale6z4J8PavqSMt3faVbT3CsCCJHiVmBz7k1X/BcyG5a0+C86K5t0k11ZCDwHIsNuffCt+tf8EMf+a2f9wD/wByNV+0L/yX74mf9jhq3/pXLX/BFT/k1fxT/wBlAvP/AEgsK/4K8/8AJmmpf9jBp3/obV4S/wCRq0X/ALCEH/oxaqv+CT//ACZH4P8A+whqf/pZLVV+1mVX9lb4ylmA/wCLf66Of+vCav2RLe4uv2rPg3FaoWdfHmiuQDj5VvYmY/8AfIJqqqqqqqqqqqqqqqqqqqqr/gqP/wAmJfE3/uE/+nazqtS/4I0/tJp4VXxH4b8Z+BNauJLZbiLTku7i3lmDJuCI0kQTfyB8zKvqwqM+J/hp41xdacNO8Q+GdRKy2uo2Mc32a5hfDJJDMrIxVlIKupGRyK/YG/aytv2svgwNb1OytNO8W+GZk0/XrK1G2LfszFcRL/DHIobC/wALI68gAn/guH4ovJNe+FPgpHZbSCz1HUJFHSSR3hjUnn+ERvjj+M9c8fAf48ePf2cfHyfEr4avpqa5FZy2scl/Zi5WNJMbyqno2BjPoSO9f8Pef2yv+g54U/8ABCn/AMVXjLxVqnjvxfrnjfXBbjUvEOpXGo3gt4RFGJp5GkfYg4VdzHCjoMCv+CTOp32ofsUeF7a8VvL07VNSt7YtnmM3Tyce2+Rx+FftwfCofBn9qv4jeCbe2EFh/bD3+nIq4VbW6AuI1X1CLL5f1Q+lfBf9oM+GP+COnii/hvgup6AdQ8J27hvn828uAVx/tJDfBh6CMGv2ePgnN8XvC3xj1ZbNpV8D+AbjWoZFTJjnhubd8D0zClz+ANf8ElPiM/gf9sHSPD8tyY7TxrpN5pMoP3S6p9oiP1324UH/AG8dzX/BQHxy3xC/bH+KWtCfzYrLW20qHB4C2SLbce2YSfcknvX/AAS0+Eo+F37IHhq/u7ZYtS8bzy6/dHGSUmwlvz6G3ihbHQFz65r9uv8A4KF+GP2SY7fwT4W0i18T/EPUbcXEdhPKy22mwnIWW5K/MxYg7YlKsQCxZRt3eNf+CgX7avxd1cWsfxi8R2D3cm2203wsv9nbSeiJ9nAlb/gTMffpXxg0/wDaeOn6brPx+svij9iuZ5P7Ou/GEV/5csrAGTynueCxABO05OBmv+CbM0sH7b3wseCV42N9dqSrEHDWNwGHHYgkEehr/gpF/wAmR/FP/sH23/pZBVfssfDTwt8Jf2evAXgvwlYx29rDodrPO6oFa5uZYlkmmf1Z3ZifTgDgAV/wVA8RXnhz9iL4hPYOyS6iLGxLqcbUlvYRJ9QU3L/wKq8B+DdG+HfgjQPAXh6BIdN8O6bBYWiKm0COGMIpx6kLk+5r/gtfpthL+zR4P1eS0ia9tvHUEEM5X5445LG8aRQewYxRkj/YHpX7PX/Jfvhn/wBjhpP/AKVxVVf8FWP2UfF/7Qfwv0Lxz8NdKm1XxP4CluGbS4F3S31lOE80RD+ORGiRgg5YFwMtgHwz4n8XfDnxVaeJvCetal4f8Q6LcFre7tZWt7i2kGVYAjBBwSpB6gkHgkV+z9/wWT+LPhO7s9E/aB8O2fjXRQVSXVdOiSz1KIZ5cqMQzYH8O2InqXr4d/ELwd8V/BOkfEPwBrkGr6BrluJ7K7iyAy5wQQeVZWBVlIBVgQQCDX/BSL/k9z4p/wDYQtv/AEigr/ghj/zWz/uAf+5Gq/4Knft5y67e6n+y98HdZK6XauYPGGq27/8AH3KPvWMbD/lmpx5hH3mGz7qtv/Y0/ZO8VftbfFi28H6aZrHw3pmy58R6uqZFnbZ+4meDNJgqg9csRtVq8C+BvCvw08HaR4B8EaPBpeh6HarbWNpCuBGi+vcsTkljyzEk5JJr/gpT4ovPFf7a3xKnunbZp13b6fbxnpGkFtEmByeCwZvqx6dK/Z1/bW+OH7LWgat4c+E11oVtba3eLdXj3umLcSO6oEUbiQdoGcD1YnvXj3/gqJ+1Z8SfBGv/AA98U6p4Xn0fxJptxp19GmhorGGaMo+1s/K2GJDdjg9qBKkMrEEHgjjFeBtSvdZ8E+HtX1JGW7vtKtp7hWBBEjxKzA59yar/AILmQ3LWnwXnRXNukmurIQeA5Fhtz74Vv1r/AIIY/wDNbP8AuAf+5Gq/aF/5L98TP+xw1b/0rlr/AIIqf8mr+Kf+ygXn/pBYV/wV5/5M01L/ALGDTv8A0Nq8Jf8AI1aL/wBhCD/0YtVX/BJ//kyPwf8A9hDU/wD0slqq/azKr+yt8ZSzAf8AFv8AXRz/ANeE1fsiW9xdftWfBuK1Qs6+PNFcgHHyrexMx/75BNVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX/BUf/kxL4m/9wn/07WdV4S/5FTRf+wfB/wCi1r/grp8MYvAf7W134msbQRWfjnR7bVMouF+0Lm3mH+8TCrn3kz3r/gjd8UD4R/aZ1H4dXV1ss/HmhSxRRZxvvLX9/GfTiEXQ/wCBfn/wW0+Fupav4F+H/wAX9NtHltvDt9c6XqToufLS5Ebwu3ooeF1z0zIo7iv2T/iD8Jfhr8bdF1745+BLDxZ4InSS01W0u7Fbv7OkgwtwkbdWjcKxA5K7gOSK8EfBf/gnH8SdGi8QeA/AHwY1yxlUESWltaOUyM4dfvI3qrAEdwK8S/Cf/gmn4OvLfTfE/g/4H6dfXc6QW9nMlkJ5ZHYKqrEDvJJIHArwR4C8FfDTw5b+EPh/4X03w9otq7vBYafbrBDGzsWchF4BLEk+5r/gtr8KvsPjD4f/ABpsbb5NVsZtEv3UYAkgbzoCfdllmGfSIegqw+MV3Z/s06x8Bo5XSLUPGlprxABw6paTQupPTG4wnB7qCOhr/gjx8EdNuf2dfiF4w8S2AeD4i376OVZRmWwghZGwewaS4nUj1jHtXgi+1P8AZr/af0a712Qx3fw28bRpqO1D8wtLsLOMDnDKjjA5wa8MaDrv7QXx10/w/Zll1X4g+KFjLnL+U93cfM7HqQu8sSewJrQND0vwxoWm+GtDtEtdO0m0itLOBBgRQxIERR7BVAr9uDXdd8RftefF2+8QvI11B4svrOLzDyLe3kMMGPbyY48e1f8ABE60+Ej+D/HF4RpbfEtNXCt5203SaV5EflmLPzBDMZw5XjIj3fw1/wAFofjb8PfFL+CPg74X1+01bXdBvLm/1hbSYSLYbkWOOJyuR5jfOSucqFBI+YV/wTe/5Pc+Ff8A2ELn/wBIp6/4KRf8mR/FP/sH23/pZBVfCf8A5JX4N/7F+x/9J0r9uj4Zaj8X/wBkv4leBtFtmuNSl0n7bZQouXlmtJUuVjUf3nMOwf71V+xx/wAFAPgh8Y/hJoNp46+I+g+FvHGj2MNprNhreox2ZuZo0Cm4gaVgJEk2l8AlkJIboGb/AIK2ftg/DD4xad4a+CHwl8UWXiaz0TVG1XWdUsJBLai4WJ4oYoZR8suEmmLMhKjKAEncF/Z6/wCS/fDP/scNJ/8ASuKqr4m/HL4SfBm40K2+KnjzS/DA8S3Mltps2oyGKKWRFDMGkxtjABHzOVXJAzkgH4vfsh/sqftUac/iHxP4K0TUr2+Q+V4m0GZYLpjxh/PhOJsAcCQOo9K/aH+HXhj4SfG3xj8NfBnitfEmi+H9Te1s9TBUmZQASGKfKWQkoxXALISAM4H/AARS1nXr79nzxjo980sml6b4sb7AztkI0ltE0sa+gBCNjpmQnua/4KRf8nufFP8A7CFt/wCkUFf8EMf+a2f9wD/3I1/wUx/blT9nrwc/wj+GeroPiN4mtT5lxC+W0OzfIMxx0mcZEY6ry/GF3eE9Cj8XeKtO0C88RabosepXSxzapqsxjt7VSfmllYAnaBknAJPQAkgV+zJ8a/8Agnx+y38K9P8Ahr4I+P8A4SmlUCbVtUdmWbVLsqA8z/KcDjCpkhFAGTyT8Pf2tf2bfix4ptvBHw5+MOga/rt4kjwWFpKxkkVFLOQCo6KCfwr/AIKyfC3UvAP7XeteKZLR10vx1Y22p2Mu35WdIlgnQH+8JIixHUCRT3Ff8E4/iN+yNpGta18OP2qfAfhK5j1qaOfRPEWu6fHNFaSBSskE0jA+UjAIyscKCH3EbhVh+zV+wPq2nx6tpfwp+D95Yypvjube1s5I3XH3g6kgjHetG+DX/BODxJ4yj+HnhvwJ8FdX8SyQyTDStPgs7mdUTG9mSPJXGR1xUEENtBHbW0axxRIEjRRgKoGAAPTFV/wWy8JS6l8C/A3jSGLeND8TtaSEA5RLm3c5+m63QfUiv+CI/ieCz+MvxC8HPIBJqvhqG9jUj732a4VDg/8AbyOP8Kr9oX/kv3xM/wCxw1b/ANK5a/4JBftU/Cr4beF/FXwP+JfjDS/DE+oayNY0i71S4S2t7lpIYoZYvNchVceREVViN2445Br/AIKxftifBrxr8KrX4C/DHxdpni3VrzVobzU7zS7hbm1sYYQxCechKNKzlRtUttCtuwSufD93BYa9pt/dOVhtryKSQgZwquCTj6CtA8Q6D4r0e18QeGNasdW0y9jElteWU6zRSqRkMrKSCMEVX/BPf9uv9lT4G/steG/hx8Uvip/YviKwvL+S4sv7D1C52LJcyOh3wwOhyrA8McZ5xX/D0f8AYS/6Lr/5bOrf/ItfAv8Aaj+BH7S39t/8KT8d/wDCR/8ACOfZ/wC0/wDiWXdn9n8/zPK/4+Io927yZPu5xt5xkZ/4KH+KIPCP7F3xU1GaUKbrR109BgEs11NHbgAf9tT9ACe1f8E4PCE3jP8AbS+GVlHEGi03UJdTmYjhFtreSVSfq6IB7sKqqqqqqqqqqqqqqqqqqqq/4Kj/APJiXxN/7hP/AKdrOq8Jf8ipov8A2D4P/Ra1/wAFdPhjF4D/AGtrvxNY2gis/HOj22qZRcL9oXNvMP8AeJhVz7yZ71/wRu+KB8I/tM6j8Orq62WfjzQpYoos433lr+/jPpxCLof8C/P/AILafC3UtX8C/D/4v6baPLbeHb650vUnRc+WlyI3hdvRQ8LrnpmRR3Ffsn/EH4S/DX426Lr3xz8CWHizwROklpqtpd2K3f2dJBhbhI26tG4ViByV3AckV4I+C/8AwTj+JOjReIPAfgD4Ma5YyqCJLS2tHKZGcOv3kb1VgCO4FeJfhP8A8E0/B15b6b4n8H/A/Tr67nSC3s5kshPLI7BVVYgd5JJA4FeCPAXgr4aeHLfwh8P/AAvpvh7RbV3eCw0+3WCGNnYs5CLwCWJJ9zX/AAW1+FX2Hxh8P/jTY23yarYzaJfuowBJA3nQE+7LLMM+kQ9BVh8Yruz/AGadY+A0crpFqHjS014gA4dUtJoXUnpjcYTg91BHQ1/wR4+COm3P7OvxC8YeJbAPB8Rb99HKsozLYQQsjYPYNJcTqR6xj2rwRfan+zX+0/o13rshju/ht42jTUdqH5haXYWcYHOGVHGBzg14Y0HXf2gvjrp/h+zLLqvxB8ULGXOX8p7u4+Z2PUhd5Yk9gTWgaHpfhjQtN8NaHaJa6dpNpFaWcCDAihiQIij2CqBX7cGu674i/a8+Lt94heRrqDxZfWcXmHkW9vIYYMe3kxx49q/4InWnwkfwf44vCNLb4lpq4VvO2m6TSvIj8sxZ+YIZjOHK8ZEe7+Gv+C0Pxt+Hvil/BHwd8L6/aatrug3lzf6wtpMJFsNyLHHE5XI8xvnJXOVCgkfMK/4Jvf8AJ7nwr/7CFz/6RT1/wUi/5Mj+Kf8A2D7b/wBLIKr4T/8AJK/Bv/Yv2P8A6TpX7dHwy1H4v/sl/ErwNots1xqUuk/bbKFFy8s1pKlysaj+85h2D/eqv2OP+CgHwQ+Mfwk0G08dfEfQfC3jjR7GG01mw1vUY7M3M0aBTcQNKwEiSbS+ASyEkN0DN/wVs/bB+GHxi07w18EPhL4osvE1nomqNqus6pYSCW1FwsTxQxQyj5ZcJNMWZCVGUAJO4L+z1/yX74Z/9jhpP/pXFVV8Tfjl8JPgzcaFbfFTx5pfhgeJbmS202bUZDFFLIihmDSY2xgAj5nKrkgZyQD8Xv2Q/wBlT9qjTn8Q+J/BWiale3yHyvE2gzLBdMeMP58JxNgDgSB1HpX7Q/w68MfCT42+Mfhr4M8Vr4k0Xw/qb2tnqYKkzKACQxT5SyElGK4BZCQBnA/4Ipazr19+z54x0e+aWTS9N8WN9gZ2yEaS2iaWNfQAhGx0zIT3Nf8ABSL/AJPc+Kf/AGELb/0igr/ghj/zWz/uAf8AuRr/AIKY/typ+z14Of4R/DPV0HxG8TWp8y4hfLaHZvkGY46TOMiMdV5fjC7vCehR+LvFWnaBeeItN0WPUrpY5tU1WYx29qpPzSysATtAyTgEnoASQK/Zk+Nf/BPj9lv4V6f8NfBHx/8ACU0qgTatqjsyzapdlQHmf5TgcYVMkIoAyeSfh7+1r+zb8WPFNt4I+HPxh0DX9dvEkeCwtJWMkiopZyAVHRQT+Ff8FZPhbqXgH9rvWvFMlo66X46sbbU7GXb8rOkSwToD/eEkRYjqBIp7iv8AgnH8Rv2RtI1rWvhx+1T4D8JXMetTRz6J4i13T45orSQKVkgmkYHykYBGVjhQQ+4jcKsP2av2B9W0+PVtL+FPwfvLGVN8dzb2tnJG64+8HUkEY71o3wa/4JweJPGUfw88N+BPgrq/iWSGSYaVp8FnczqiY3syR5K4yOuKgghtoI7a2jWOKJAkaKMBVAwAB6Yqv+C2XhKXUvgX4G8aQxbxofidrSQgHKJc27nP03W6D6kV/wAER/E8Fn8ZfiF4OeQCTVfDUN7GpH3vs1wqHB/7eRx/hVftC/8AJfviZ/2OGrf+lctf8Egv2qfhV8NvC/ir4H/EvxhpfhifUNZGsaRd6pcJbW9y0kMUMsXmuQquPIiKqxG7cccg1/wVi/bE+DXjX4VWvwF+GPi7TPFurXmrQ3mp3ml3C3NrYwwhiE85CUaVnKjapbaFbdglc+H7uCw17Tb+6crDbXkUkhAzhVcEnH0FaB4h0HxXo9r4g8Ma1Y6tpl7GJLa8sp1milUjIZWUkEYIqv8Agnv+3X+yp8Df2WvDfw4+KXxU/sXxFYXl/JcWX9h6hc7FkuZHQ74YHQ5VgeGOM84r/h6P+wl/0XX/AMtnVv8A5Fr4F/tR/Aj9pb+2/wDhSfjv/hI/+Ec+z/2n/wASy7s/s/n+Z5X/AB8RR7t3kyfdzjbzjIz/AMFD/FEHhH9i74qajNKFN1o66egwCWa6mjtwAP8AtqfoAT2r/gnB4Qm8Z/tpfDKyjiDRabqEupzMRwi21vJKpP1dEA92FVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX/AAUK+H/jP4pfsfeP/Anw+8PXWu6/qf8AZv2PT7VQZJvL1K1kfaCR0RHY+wNf8MBftlf9G8+Kv+/Kf/FV4bt5rTw5pVpcxmOaGyhSRG6qwQAj8xX/AAVh/Zb8dfH74f8Ag3xV8KvClz4g8TeFdSmt5bK0A82SyuUBdgCRu2SQxcdhIx9a+AX7J37aHwZ+Nfgj4pRfs9eLynhrXLa7uUjhTdJbq4E0Y+b+KIuv/Aq8f+A/CXxT8E6v8PvHOjRapoOvWjW17aSgrvRu4IwVYEBlYYKsAQQQK/aQ/wCCSvx8+GOs3mqfBmyb4ieFGkZ7Zbdkj1K2Q5ISWEkCUgYG6LO7rsXOBf8A7NP7Rml3LWeo/AL4i206dY5fC14p+v8Aq+nvXw+/YI/bK8canbJoPwG8V6TIsgZbnW7b+yFhIP3ibkoeMZ4BPoDWitqkmjWD65FFFqTWsZvI4m3Ik20bwp7gNnBr/gon8Cda/aC/Za8QeE/Cejyap4l0m6t9W0W0jALyzxPtdVyR8xgknUc9SK/4YC/bK/6N58Vf9+U/+Kr9lr4Ut8EP2dvh/wDC2e3WC80PRIV1CNcEC8kzLc4x1HnSSHNf8FCf2F/jz4n/AGqPFPjj4P8Awn1jxD4f8UR2+omewiUpFctGEnQ5YfMZI2kP/XQV/wAEz/2IPjV4H/aXtPiX8aPhfqvhvS/CulXNxp0moIgWe9lAgRQAxziOWZ89ii98VX/BSb/gnJ48+Jfjq8/aA+AWkR6vf6pCn/CReH43WKeSaNQoubfcQrlkVQ8eQ25dw3lyBB+y1+0xc6sNBh/Z7+I7X+/YYD4XvAVPqcx4A5B3HjHOcVcf8Epf2lNM+Bt38SNT8OSzeL3vrZLDwhYyRz3ItX3CWaZgdqsD5eI1JIBYtgjbX7En7IX7U3wt/ar+HHjvxh8DvEemaLpmqN9tvLiFfLgjkhkiLttYnA35/wDrV+3Z4H8WfEj9kz4ieCPAuhXOs67qllAlnYWygyTMt1CxCgkdFVj+Ff8ADAX7ZX/RvPir/vyn/wAVXw30+90n4d+FtK1G3e3u7LRbOGeFxho5EhRWU+4IIqv25v8AglT4ybxbqvxZ/Zh0eDVNL1SV7rUPCcbrDPZTMSXa0DYV4icnyshlJwoYEBbb9k/9p+81k+HoP2ePiOb9ThoW8MXabRkDcWMYAXkfMTjkc814V/4JNfGCw/Z98cfEDx9ojyePX06EeFfCenzpNPHJ9phaSWZ1bYXMKyqIgW4ckkMAB8Hv2IP2u/C3xc8EeJ9V/Z+8Ux2Wj+I9PvLl2iQBY4riN2P3vRTVV/wVh/Zl/aT+P+u+DNb+FHgweJfDXhfTbhZbW1vI1uo7qeQGVhC5XeuyGADYWbOflAGTc/s3ftS+G5pdJn+BXxPsWux5ckK+G75VuByMcR4cckdxz718HP8Agmt+1t8XdYtrW5+GV/4L0pmH2nVPFELWAgXuRA4852xnAVMZ6letfs3fADwf+zL8ItG+Evg2SS5g08NLe38yBJL+7fmWdwM4yQAFydqqq5O3Nft1/sc/tPfEn9rL4ieNvAvwW8Q6zoWqXsD2d9bRIY5lW1hQlcsDwysPwr/gkR8APjL8DP8AhbH/AAtz4ear4X/tv+xf7P8At6Kv2jyft3mbcE/d82PP+8K/bR/Y2/ao+In7U3xI8aeEfgt4k1nRtV1lpbG+hjVkmiEaKpUls4+XH4V/wwF+2V/0bz4q/wC/Kf8AxVf8MBftlf8ARvPir/vyn/xVf8E4f2Sf2kfhP+1l4Y8bfEb4P6/oOhWdlqCT313EojjZ7WREBIYnliB+NftZfsp+Af2tvho3gbxfI+n6lYu1xoetQxB5dNuCMFtuRvjYAB4yQGAByrKrL8Yv+Cav7XHwj1SeCD4Y33jTS0Yi31LwvG1+JlycEwIPOQ4AJBTHPBNQfs2ftF3d0LG1+AXxHmuSSBDH4VvWfI6/KI88AH8q/wCCeP7G/wC1r8Pv2k/Bvxd174T6j4c8O6NLOmpTazNHaSNDNbyQsFgZvNYjzAw+TGVHIqq/bd+D03x0/Zc8feAdPtftGqtppvtKRRlmu7ZhNGi+7mMx/wDAzX/BPD4rQfCD9rvwDruoXSQ6bq94dFv2c7VEd2piRmPYLK0TknjC8461X7Qv/JfviZ/2OGrf+lctaN+yF+0h4p+G/h74seC/hJr/AIo8M+JUmNlc6FatfurQ3ElvIskUW6RCHiY5K7SpBB64/ZM/4JOfFH4kXt34h/aG0m88D+HBYTpY2UrqL+5uXiKxSGMEmJI3YORJhmKBdu1iw+LH7EH7UPwg8U3XhrW/g34n1WKGUrb6poulzX9ndJ/CySwqwBIGdjYYdwK/4Jp/DrxP8Mf2PfB/h/xp4a1HQNcnutQurzTtRsntbmAteShPMjcBgTGsbDIHystf8MBftlf9G8+Kv+/Kf/FV/wAMBftlf9G8+Kv+/Kf/ABVf8MBftlf9G8+Kv+/Kf/FV/wAEiPgB8ZfgZ/wtj/hbnw81Xwv/AG3/AGL/AGf9vRV+0eT9u8zbgn7vmx5/3hX/AAWp+LFvoXwh8H/ByyucX3inV21G7RW5FpaJgBh2DSzIR/1xb0r/AIIn/B25vfFnjj476jZkWemWaaFpkrrw88rLNcFT6oiQg+031qqqqqqqqqqqqqqqqqqqqr/goV8P/GfxS/Y+8f8AgT4feHrrXdf1P+zfsen2qgyTeXqVrI+0EjoiOx9ga/4YC/bK/wCjefFX/flP/iq8N281p4c0q0uYzHNDZQpIjdVYIAR+Yr/grD+y346+P3w/8G+KvhV4UufEHibwrqU1vLZWgHmyWVygLsASN2ySGLjsJGPrXwC/ZO/bQ+DPxr8EfFKL9nrxeU8Na5bXdykcKbpLdXAmjHzfxRF1/wCBV4/8B+Evin4J1f4feOdGi1TQdetGtr20lBXejdwRgqwIDKwwVYAgggV+0h/wSV+Pnwx1m81T4M2TfETwo0jPbLbskepWyHJCSwkgSkDA3RZ3ddi5wL/9mn9ozS7lrPUfgF8RbadOscvha8U/X/V9Pevh9+wR+2V441O2TQfgN4r0mRZAy3Ot239kLCQfvE3JQ8YzwCfQGtFbVJNGsH1yKKLUmtYzeRxNuRJto3hT3AbODX/BRP4E61+0F+y14g8J+E9Hk1TxLpN1b6totpGAXlnifa6rkj5jBJOo56kV/wAMBftlf9G8+Kv+/Kf/ABVfstfClvgh+zt8P/hbPbrBeaHokK6hGuCBeSZlucY6jzpJDmv+ChP7C/x58T/tUeKfHHwf+E+seIfD/iiO31Ez2ESlIrlowk6HLD5jJG0h/wCugr/gmf8AsQfGrwP+0vafEv40fC/VfDel+FdKubjTpNQRAs97KBAigBjnEcsz57FF74qv+Ck3/BOTx58S/HV5+0B8AtIj1e/1SFP+Ei8PxusU8k0ahRc2+4hXLIqh48hty7hvLkCD9lr9pi51YaDD+z38R2v9+wwHwveAqfU5jwByDuPGOc4q4/4JS/tKaZ8Dbv4kan4clm8XvfWyWHhCxkjnuRavuEs0zA7VYHy8RqSQCxbBG2v2JP2Qv2pvhb+1X8OPHfjD4HeI9M0XTNUb7beXEK+XBHJDJEXbaxOBvz/9av27PA/iz4kfsmfETwR4F0K51nXdUsoEs7C2UGSZluoWIUEjoqsfwr/hgL9sr/o3nxV/35T/AOKr4b6fe6T8O/C2lajbvb3dlotnDPC4w0ciQorKfcEEVX7c3/BKnxk3i3Vfiz+zDo8GqaXqkr3WoeE43WGeymYku1oGwrxE5PlZDKThQwIC237J/wC0/eayfD0H7PHxHN+pw0LeGLtNoyBuLGMALyPmJxyOea8K/wDBJr4wWH7Pvjj4gePtEeTx6+nQjwr4T0+dJp45PtMLSSzOrbC5hWVREC3DkkhgAPg9+xB+134W+LngjxPqv7P3imOy0fxHp95cu0SALHFcRux+96Kaqv8AgrD+zL+0n8f9d8Ga38KPBg8S+GvC+m3Cy2treRrdR3U8gMrCFyu9dkMAGws2c/KAMm5/Zu/al8NzS6TP8CvifYtdjy5IV8N3yrcDkY4jw45I7jn3r4Of8E1v2tvi7rFta3Pwyv8AwXpTMPtOqeKIWsBAvciBx5ztjOAqYz1K9a/Zu+AHg/8AZl+EWjfCXwbJJcwaeGlvb+ZAkl/dvzLO4GcZIAC5O1VVcnbmv26/2Of2nviT+1l8RPG3gX4LeIdZ0LVL2B7O+tokMcyrawoSuWB4ZWH4V/wSI+AHxl+Bn/C2P+FufDzVfC/9t/2L/Z/29FX7R5P27zNuCfu+bHn/AHhX7aP7G37VHxE/am+JHjTwj8FvEms6NqustLY30Mask0QjRVKktnHy4/Cv+GAv2yv+jefFX/flP/iq/wCGAv2yv+jefFX/AH5T/wCKr/gnD+yT+0j8J/2svDHjb4jfB/X9B0KzstQSe+u4lEcbPayIgJDE8sQPxr9rL9lPwD+1t8NG8DeL5H0/UrF2uND1qGIPLptwRgttyN8bAAPGSAwAOVZVZfjF/wAE1f2uPhHqk8EHwxvvGmloxFvqXheNr8TLk4JgQechwASCmOeCag/Zs/aLu7oWNr8AviPNckkCGPwres+R1+UR54AP5V/wTx/Y3/a1+H37Sfg34u698J9R8OeHdGlnTUptZmjtJGhmt5IWCwM3msR5gYfJjKjkVVftu/B6b46fsuePvAOn2v2jVW0032lIoyzXdswmjRfdzGY/+Bmv+CeHxWg+EH7XfgHXdQukh03V7w6Lfs52qI7tTEjMewWVonJPGF5x1qv2hf8Akv3xM/7HDVv/AErlrRv2Qv2kPFPw38PfFjwX8JNf8UeGfEqTGyudCtWv3VobiS3kWSKLdIhDxMcldpUgg9cfsmf8EnPij8SL278Q/tDaTeeB/DgsJ0sbKV1F/c3LxFYpDGCTEkbsHIkwzFAu3axYfFj9iD9qH4QeKbrw1rfwb8T6rFDKVt9U0XS5r+zuk/hZJYVYAkDOxsMO4Ff8E0/h14n+GP7Hvg/w/wCNPDWo6Brk91qF1eadqNk9rcwFryUJ5kbgMCY1jYZA+Vlr/hgL9sr/AKN58Vf9+U/+Kr/hgL9sr/o3nxV/35T/AOKr/hgL9sr/AKN58Vf9+U/+Kr/gkR8APjL8DP8AhbH/AAtz4ear4X/tv+xf7P8At6Kv2jyft3mbcE/d82PP+8K/4LU/Fi30L4Q+D/g5ZXOL7xTq7ajdorci0tEwAw7BpZkI/wCuLelf8ET/AIO3N74s8cfHfUbMiz0yzTQtMldeHnlZZrgqfVESEH2m+tVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV+2h/wTP8Ajlb/AB+13xR+z18O5tf8JeJJjqkCWVzBAdMuJGJmtysjoQBJl02jAR1Xqpr9nHXfij4h+C3ha7+NXhW68P8AjiCyW21u1uHjcyTxfIZw0bMpEoAkwDwXI7V+0zYvpf7SHxX0ySRXa08b6zCzAYDFb2UEj8q/4JZ3MFx+wr8OIoZAz20mqxyjH3WOqXTY/wC+WU/jVVVVVftv/sy/tuftS/tH6344074HaqPDloy6T4dM1/aIEsYmbbKwMuV8x2klIIyPMx2r9mD4DaJ+zV8EPDXwk0d4p5dLtzJqV4i4+2Xsh3Ty884LEhc8hFQdqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv20P+CZ/xyt/j9rvij9nr4dza/4S8STHVIEsrmCA6ZcSMTNblZHQgCTLptGAjqvVTX7OOu/FHxD8FvC138avCt14f8cQWS22t2tw8bmSeL5DOGjZlIlAEmAeC5Hav2mbF9L/AGkPivpkkiu1p431mFmAwGK3soJH5V/wSzuYLj9hX4cRQyBntpNVjlGPusdUumx/3yyn8aqqqqr9t/8AZl/bc/al/aP1vxxp3wO1UeHLRl0nw6Zr+0QJYxM22VgZcr5jtJKQRkeZjtX7MHwG0T9mr4IeGvhJo7xTy6XbmTUrxFx9svZDunl55wWJC55CKg7VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV8af+CQHhP4wfFjxZ8Uh8bdR0ZvFWqz6jJYR6CkywPK25wHMylvmJOcDrX7KX7PEX7LnwesvhBbeMZ/EttYXtxcQXk1kLZlWZ95TYHccMWOc9+lVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV8af8AgkB4T+MHxY8WfFIfG3UdGbxVqs+oyWEegpMsDytucBzMpb5iTnA61+yl+zxF+y58HrL4QW3jGfxLbWF7cXEF5NZC2ZVmfeU2B3HDFjnPfpVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVfFH4//BP4KwLN8Vfij4d8NO6b47a9vlW4lX1SEZkccdVU1rX/AAVl/Yo0q5+z2Xj7WtXXJBlsvD90qjgf89UQ9yOnY+2fC3/BUv8AYm8TzR2snxWuNFnlOFTVdFu4VzkjmRY2RegOSwHI98eCfHvgf4l+H4fFfw98XaP4k0e4JWO+0u8S5iLDG5dyEgMMjKnkHggVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV+31/wVJ1ux1zU/gv8Aswa6lpHYs9trPi6ABpHlBw8NkTwqrgqZ8ZJ+4VADtq2rarr2pXOs65qd3qOoXkhkuLu7maaWZz1Z3YksT6k1Vf8ABOHwEfh9+xh8NdPmtxHc6vp76xM23Bk+1yvNGx/7YvEPoBVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV/wAFO/2h9Q+AX7NF9a+F9Sez8T+OLj+xtOmiYiS3iZC1zOpBBBWIFAw5V5UI6VX7J37Lfjb9rP4pwfD3wnOmn2NrF9q1rV5oy8enWoYAttyN8jEhUTI3HqQoZh8Lv+Caf7H3wy0SDTpfhVZeLL9YwLjU/EZN7LO2OW8s/uk+iIPx614v/wCCeX7GfjSwksb/AOAvh/Ty6bVn0gSadJGezAwMuTk9wQehBHFeGvD2l+EvDeleFNEhMWnaLYw2VpGf4IokCIOPRVAqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq8VeI9M8HeF9Y8X608i6fodhPfXbRpvYRQxl3Kr3O1TxWuf8Fov2WtPaSHSPB/xH1V1+5Imm2sMT84PL3IYcc/c9OnONO/4LXfs4z3Sxan8NfiLawtwZY7azlKn3X7QvH0yfY18Bv23/2av2jrpNH+HPxDgGuupYaJqkTWV4ccnYj8S4HJ8tnx3xVVVVVVVVVVX7I3/BSH9oX44/tb+Hfgh4zs/CKeHdUu9Tinay02SK4C29nczR7XMrAHdCmfl5Gemaqqqqr42fGr4f8A7Pnw61H4ofEvVZLLRdNKIRDEZZp5XO1IokH3nY9uAACSQASP2Xv2yPg1+1zper3vwvn1e1vdBeMahpesWyQXMKSbvLkwjujIxRhlWOCOQMjNVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVf8FvvEtzc/En4ZeDS7/Z9O0O7v1XGAXuJ1jJ68nFsO3GepzxX/AARX8GaVpf7O/irxxHCh1PX/ABU9tNKF58i2t4vKQnrw80x9Pn+tVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX7Qv/JAfiZ/2J+rf+kctf8ABJ/4R/DT4y/tHa74c+KXgzTfE2l2Hg25v7ez1CLzIkuEvbJFfbnBIWRxg5GGPFa3+wd+x14g06TTL79nbwbFDJ95rKy+xyj6SQlHHXsRX/BQb9iEfsZ+JfDvxW+Duv6rH4S1bUPLsnkuCLvRNQjHmoizLgsrBGaNvvL5bBiSAzfsHfH/AFT9pL9mjw14/wDEkgfxDaNJpetSKoUTXUBAMuBgAyIY5CAAAXIHAqqqqqqqqqq/4Jy/8pF/Bf8A2ENd/wDTbe1VVVH9pn4/+Mf+Clll8N9f+LfiObwrpHxem0y10WG8NtaC1h1F4443ii2rIAiKMuGJ6kkkmq/ar/Zv8O/tV/B3UPhN4g1u50YzXMV3Y6lBCJmtLmIna5jJAdSGZSuVyGOCDgj9hn9grRv2NofEWrXPjmXxX4i8SpFDNcrZfZILa3jJYRpHvcszM2WcnsoAGCWqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr/gt/4SuLfxx8L/AB0sLGC/0m901pAOFaCVJAD6Ei4bHrg+nFf8EUPjJpEvhPxt8BNQv1j1S11Ea/psMj8zwyRxwzhB6I0URI/6bZ9aqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv2hf+SA/Ez/ALE/Vv8A0jlr/gip/wAnUeKf+yf3n/pfYVX/AAWo8a6Bpv7PXhXwFcTxPrWueKY7y1gLDcsFtBMJZcdcBpok7ffPPGD/AMEcPDN/oP7Ic+q3kbrF4j8XX9/aFlwGjWK3tiR6jfbSD6g1+3n/AMFN7T9nvXbr4P8AwXsNP1zxzbIBqmo3mZLTSGZciMIpHmz4IJGQqZAO47lHh74Y/wDBX/8AaF02Lx4/xC8a+HbK/Xzrbz/Eo8Pearcgra27IyqQcjcigjGO1XH7VP8AwUd/YW8Z6dpPx6fVNe0m8O5LPxLKmoQahGpG/wAi/jLMsgBHG87Syl0PAr9mn9o/4f8A7UnwvsviX4BneNWb7PqemzsDNpt2qgvC+ODwwKsOGUg8cgf8FMfit8Ufgv8Aszt48+Eni2Xw7rNt4gs4ZbuKKORmgkEisgEisvLbD0HTr2PgH/gqh4+8L/swaNpL3cvxF+OXiLWru1tY7i0BWzg3IIXeKAKZWYttjjUAsQxJwAG1r4Tf8FmPidYf8J3f+JPHmnNcp5sen2Xi630OTaQOPssE0QQ4/hZQ3HTJ5+B3/BS39p/9m/4kH4eftPx6z4l0azuVt9WsdatRHq2mg9ZIpCFaQ4IbbKWDjG1lzurwr4o8P+N/DOleMfCmqQ6lo+tWcd5YXcJys0MihkYfUEcHkd6/4Kq/tWftBfAL4xeEvDfwh+Jd74c03UPDIu7m3gtYJRJN9pmTeTJGxB2qo4OOK/Z08Sa34z/Z8+GHjDxNqD3+sa54N0i+1C6dVVp7iazikkkIUAAszMcAAc8Yqv2Y/wBtD9p7x1+3bovwo8V/Fu/1Dwpc+JtTtJdMeztlRoYo7gxpuWMNgFE754qv+Ccv/KRfwX/2ENd/9Nt7TAlSFIBxwSM1r3ib/grz+0r4o1my8I6ZP8PfD+malc2Qay8rSLcPDK0bFJ5s3M4ypG9MpkHhelfEb4U/8Faf2bdEuviXdfGPxX4j0vS0a6v5rHxTLrKWkaglne2uuTGBkttjZQBk4AzX/BOr/goDdftUQX/w1+J1pZWPxB0S0+1pPZx+VBq1qGVWkVCTslQsu9RwQ25QAGVf+Cgn/Dbn/FBf8Mb/ANq/8xL/AISP7D9j/wCnX7Nu+0/9vGNvvntWn/8AC/P+Gmm/s77Z/wALl/4TGbdjyPO/tv7S3mdf3O7z93+xn2r4af8AD47/AIWP4U/4Tn/hJ/8AhG/7bs/7Y8z+yNv2Pzk8/Oz5seXuzt59Oa/4KAfEjxv8Iv2RfHnxD+HHiCXRfEWk/wBnfYr6KNHaHzNRtonwrhlOY5HXkHrX/BJ39oj4z/tBeHviPefGLx3deJZtFvdPjsHnt4YvIWRJy4HlIuclF656V8TPiR4N+EHgPWfiT4/1mPTNB0G2M95cOMnGQFRV6s7MVVVHLMwA5NeNv+ChP7aP7X3xDl+HX7JXhvUfDunMWNva6VDHJfvCCB5t1dyfJbjJHKmNVLBS79Tqn7O3/BYbwTYy+M7X4meN9XmhRppbC18eG+kUD5iFt5JSjnjhEDE9ADnFfsb/APBVnxbceNbT4N/tbRW8M9xdfYYPE5tRZS2l0G2eVfQqAqgt8pkVU2EfOpBZ1r9rn9sn4+fBn/goAngDTvjDcaT8O4NW0KW806W0tzBBaSRWxuQztGX2t+9YndkbzjGBXjn9sj9t79s7xjrPhb9h3wjqWj+CNIuWt21uFIYJ7odnlurnaluWGHWKMiQA8ls4r4iePP8AgqT+x7qln4q+Jvjjx1b2N1dYiutS1RNc02d8f6piWljjLBThDsbAYqBya/YO/bK079sD4aXepalp1tpPjPwzLHb6/p9uxMR3gmK4hySRHJscbSSVZGGSMMar9sj4lfGf4SfArU/GvwG8I2/iTxTBe2tvHZS2ct2wjnlEReOGIhpHDvHhemCSQQMVD8EP+Cx3xrgOveI/itqvgxblfMjt38SJo7AZGB5VgMp9HAPBz158QftA/wDBSf8AYL8Y6XD8ZfEGqa/pF+f3EXiC7Gr2WpIgXesd1uMkbgEZG9W5BKkHn9nf47eEP2k/hHofxb8GLLDaatGy3FnMwMtlcxnbLC+OpVgcHjcpVgAGFfto/tZ/tifst/ta6v4Vtfi1qVx4R+22+taPps9la+XPp0rbzb7jFvKK6ywbslsR5znmvCfifRvG/hXR/GXhy7W60rXbCC+sZhj95DMgdG4z1VhXiXxBpPhLw5qnirXrpbbTNFspr29mYcRQxIXdvwVSa/Y3/ay/a8/aq/a+sfDY+K2p2Xggahda3qelxWdsUttNjcutrv8AK37WZoYN27dh85yM1+0z+0h4B/Za+F158TPHcrzhXFvpmmwMBNqV0wJWGPPA4VmZjwqqTzwD4c+K/wDwU8/b8vr/AF/4Sa1ceCPBkE7QxzaXef2PZwN02C5Gbi4kCkbipYKcHCZArx/F/wAFV/2JbSP4jeJ/ibrnijw1byL9su59YbxDZR7iAEnS5zLErEhd4CjJADAkV+xH+2N4a/bB+G02uRWUWkeLtAeODxDpCOWSJ3BKTwk5Jhk2vtz8ylGU5wGb/go18TPiL8H/ANljXfiD8LfFs/h7XdM1GxVLuGCOVjHJOsbpiQMoyHBzg9PfNeHf+Co3xV8Gfsg6U97rCeLPjF4m8QX1nY3l7bIy2VlGIsTtFGqh33SmONMYJVmbIXY37AB/b+8QftFWHjj9oeb4nnwXqOjXe5Ncklt7LzWUNEfshKojZztPlj24Nft6/Fj9tTwh4j8H/Dr9k3wUNQbxZZXMk+rW2mC7uLOWF0DAtLmCCPbLH88o5JOCMVd/sr/8FgfEFr/wk+ofHbV7K/jPmLpcXjyS2djwcbIMW5HsWxwfXn4N/wDBRP8Aam/ZZ+LafCP9sCHU9Z0i0nSHVY9UgVtS01HAxPDOn/HwmCGwxcOv3WGc1YX9lqthbanpt3Fc2l3Cs1vPE25JY2AKspHUEEEH3r9tj9tLwd+x94Ft764s49b8Y66HXQtE83YJNuN88zDlIUyOnLsQo/iZfBvjP/gqb+3FdXPi34e+MfFGlaAkzRrd6TqY8O6dCQeY43RkeYqeDzIw/iNf8Lj/AOCmX7BV/p3iP48Qaj48+HrzJDePqF+mqREEgYF4u6a3lOcKZfkZj918ceA/29vjb8ZP2+PDnhXwL8Wb+L4X+IvE1qLXSHsIFH2MxK7wuShcMcMrfNwxODjFV+23/wAFLPFvg74hv+zt+yjpsWs+NI7v7BqGrrZ/bTBeE7Ra2kGCssytgMWDKGyu0nJWL9lv/gr94usU8Z6n8c9d0i/kHmrpD+O5rWTP3gDFbn7OM5+6SAOhx0r4Ef8ABRf9o79nD4up8Ev227S/u9NjmS3u72/tkGoaSHxsn8yL5bqDnJPzMVO5WbG1re4t7y3iu7S4jngmQPFLGwZXUjIZSOCCMHNVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV/wAFEv2b7/8AaV/Zu1XQPDNkLnxX4bnGs6FEB81xNErCS3X3kid1UdC+zOMZE0MtvK9vPE8csTFXR1KsrDggg9D7V4G8deL/AIZ+LdL8d+AvEF3ouv6NOJ7K+tmw8Tjg8HIZSCQVYFWBIIIJFfC3/gttq9jo0GnfGP4NR6pqMMYV9T0HUBbicgYy1vIrBSepKvjJOFA4rxR/wXC8PR2DJ4K+AGoz3jLhX1TXEijjPqVjiYsPbK/Wv2f/AIly/GX4IeBfindW8Fvd+J9Btb28ht1IjiuHjHnIgJY7BJvAyScAZ5qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/aF/5ID8TP+xP1b/0jlr9jz9qjWP2Qfihf/EvRvCFn4kk1HRJdJls7q6a3AjknglLq6hsMDbqOQRhjWv8A/Bbr4oX+mvaeEfgX4b0/UpPlhnvdUnvUUn/pkixFj/wIV4E/ZU/bL/4KFfFO3+JnxtOr6JoFwFWfX9Zs/sqQ2gYkQ2FqQpcctt2gJnJZ9x+bxO3hX9lb9mfWZ/BmlLb6N8NvCdzLp1pjcX+zwMyBiOrO4G5j1ZiT1Jr/AIJf/CGD9ov9rG++IPxKH9tw+FYZfEN6bsB/tupSTARNIO/7x3m9C0YByCRVftVfArQf2jPgV4p+GWsWEU13dWUk+jzugLWl/GpMEqHqPnwrY6ozL0Y1/wAEb/ihqPhL9pbUfhlJcuNN8caJMGtixA+12gM0b49RELlf+B+1f8Fef+TNNS/7GDTv/Q2r/gi18CfDuvar4v8Aj/4g0+C8vvD88ekaCZAG+yyvGXuZQD0fY0SK3YPIO9V/wWT+AmheJPg5pvx/06wih8QeEL6Gyv7hEANzYXD7FVz1YpO0e30EknrX/BG34m6j4w/Zr1bwJqt408ngjX5ILMM2THaXCLKi+uBL9ox2wQO1f8FtP+S/eBf+xPH/AKVz1+yd/wAmr/Br/sn+hf8ApBDVfsb/APKTDw9/2OGsf+irqq/4Jy/8pF/Bf/YQ13/023tV8RPi18L/AIR6SNb+J3xA0HwxZsD5cmqX6QGUjqI1Y7pG/wBlQTXib/gqp+xBpZlsE+It/rqkGOVbLw/dsjA5BGZY0DD6ZBz35r9g/wAR6V4e/wCCgngy88DzTDQr7xDf2diGQxl7OeKeOMMpJx8rIcEkgjrkZqvDX/KWGb/suF7/AOnWWq/4Kj/8mJfE3/uE/wDp2s6/4Idf8ip8W/8AsIaV/wCi7mv+C23xa1O3i+H/AMD9OupIrK7SbXdUjHAnKt5NsCe4Ui4JHTJU9hX/AATP+A2hfBj9lnwtrUWnxDxB4+sodd1a72/PKky77aPPUKkLJ8vZmc/xGq/4LP8AwJ0Lwl4+8J/HLw3p8Vq3jKOex1tYkCrJdwBDHMcdXeNipP8A0xB6kmv+Cf3xS1D4v/sifDvxVrN09xqlrYPpd7JI253ktJWgDse7OkaOT6tX/BWD/k9zxh/2D9M/9Ioq+B3wh8LfAj4VeHPhX4RsYILLQ7JIpJI4wpup8DzZ39XkfcxJ9fav2ivhxovxc+BXjr4d69axT22s6FdRxmQZ8mdYy8Mo/wBqOVUce6iv+CNvia+0f9q+90CGRja6/wCFbyGaPPy7o5IZUfHqNjAf759aqq+Kn7Yv7MHwVvZNL+JHxp8O6bqMBxNp8Er3t1Cf9uC3WSRP+BKK/wCChH7dn7J/7R/7O2rfDjwLruran4lt9Qs7/R3m0WWGISpKFl+eTaVPkSTDODnOO+R/wRD8Q3d18KviT4Uklc22m+ILW8iQn5Ve4gKOR7kWyZ+gr/gtB8EB4n+FHhr466RZb77wbe/2fqjovJsbkgIzH0ScIoH/AE8Ma/4JH/GYfEr9luDwPqF35mrfDq/fTHDNlmtJCZbZz6ABpIh7QV/wVh+Mx+F37KWo+FtOu/L1b4hXiaNCFbDLbY8y5b3UxoIj/wBdxX/BGb4Gr4S+Deu/HHVrILqPjm++yaa7Lyun2rFSV7jfOZQR0IhQ1/wVv+JN98Sf2tdK+DUmtRWGj+C7K0tN10+yC3ub1Y5pbhz2XyntwT2EdfDT9pX9h/4TeANA+G3g39oH4eWmjeHbGO0tI11uAEhRy7YPLs2WY92Yk9a8S/ta/sWeMPDup+E/Ef7QHw6vtK1izls722l1uErNDIpR1Iz0Kkiv+CWvjOT4c/tx2PgvS9ZS70jxRb6lo0s8cmYrhIo3nhlA6HL2yBT6SHsTX/BWD/kyPxh/2ENM/wDSyKv+CM3wF8OeOPiH4q+NfinS4r4+B0trbQ454t6R3s+9mnGeN8aRgL1wZt3BCmqurq1sraS8vbmKCCFC8ksrhFRR1JJ4Ax3NePf+Cif7Gfw6u5dO1n456Pf3kTbTDosE+qDOcEeZbo8Yx3yw6evFf8FN/wBpT4DftP8AjDwT4w+Dd3qNzf6Vp93YavLeaa1qWiEiPbbSx+YZe44wMZ9+P2BPEN34o/Y2+E2p30rySxeH0swznJ22zvAn4bYlH0r9rHUta/a1/wCCjF98OhfyJat4st/B1iQciyt4JxBM657eZ9olPB5Y4zxXg/wh4b8AeFdJ8E+ENKg03RdEtI7SxtIVwsUSDAHueMknkkknk14t8KeHfHfhjVPBni3SbfU9G1q0ktL6znXKTROuGU/geo5BwRgiv2SfC0fgb/gob4O8Ew3LXCeH/HVxp6St1kEDyxhjwOTtz0FftS/FG6+C37O3xB+J2nPt1DQ9DnfT2K7gt248uBiO4Erxkj0Br/glHrXwF8F/Erxb8avjt8TPDOia1plulroA1zUUjleW43m5uV3nJYIqpu7iaQV/w2v+yL/0cf8AD/8A8HkP/wAVX/BXf4hfs8fGHQ/AHjf4V/Ezwn4l8S6Vd3GnXqaTfxzzGzkTzELhSTsSRGxnoZj61/wTU+IN/wDEX9jL4f32q3LT3uiwT6PK7NuOy1meOEfhCIh+FVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX7cv8AwS40f466xf8Axb+Bt5YeHvG14TLqml3WUstYl7yhhnyJiOpAKOcFtpLOfir+zj8dfgjfS2PxS+FniHQRESPtc1mz2smO6XCbonH+6xqtE0DXfE2oR6R4c0W/1W+l/wBXa2Vs88rduEQEnqO3ev8Agm5oXj7wp+x/4M8I/Ejwlq3h3WNGlvYhZ6patbzmF7qSaJjG2GUbZQuCAfl6VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVftC/8kB+Jn/Yn6t/6Ry1/wAEaNH0jXf2nfFWn61pdnqFs3w/vN0F1AsqH/T7AfdYEdCR+NaR4E8EeH7j7XoHg3Q9Nn/562enRQt+aqD3P51X7b2hXviP9kX4t6Zp6F5x4VvLgKoyWEKeawA9SsZ/+v0r/giL4m06y+KvxI8HzyKt7q2gWt5bgtgstvOVcAdz/pKH8D+Fa5rOm+HNE1DxDrN0ltYaXay3V1M5wIoo1LOx9gqk1/wS10y98X/t4+G/EtrbOkel2+r6ncovzCOOS0mgGTxxvuUGcdSPWv8Agrz/AMmaal/2MGnf+htX/BEm5t3+BXj2ySZTPF4tEjx55VWtIQp+hKN+Rqv+CsHiPTtE/Yi8Y6Zeyqs3iDUNMsbMFsFpVvYrggep8u3kOPYntX/BELQLu2+FvxL8UOj/AGbUNftbONivBeC3LuAfXFymR7j1r/gtrDKvx48BzmJxG/hEqrlTgsLuYkA+oDLke49a/ZFuYLv9lP4Ny20gdV8BaLGSBjDJZRKw/BlI/Cq/Yzkjm/4KWeHJYZFdH8X6uyMpyGBiusEH0qv+Ccv/ACkX8F/9hDXf/Tbe1+1Z8dIP2bvgF4t+Lxs4r280e1VNOtZSQs93M6xQhsc7A7hmAIO1WwRX7If7Mfjn/gpD8T/FPxc+PnxF1qXQ9JuI01C7ikU3N5PJuZbW33ApBEijJwuFDIqrySvhj/gmv+xR4Vs1tbb4HafqD7cPPql9dXjyHjLHzJCqnj+EKOvAya/Zv0fTPD3/AAUw0Pw/ollHaafpnxHvrW0t4xhYYo5p1RF9goA/Cq8Nf8pYZv8AsuF7/wCnWWq/4Kj/APJiXxN/7hP/AKdrOv8Agh1/yKnxb/7CGlf+i7mv+C2WhXtv+0N4I8TSIfsl/wCDEs4mI4MkF5cu4z9LiP8AOv2TvE2neMf2YvhV4g0uRWhn8IabGwVt2yWO3SOVM99siOv4VX/BbzxRptv8MPht4KaVDqF/r1zqCIH+ZYoIPLYkeha4Xn2PXnH/AASi8O3uhfsU+E7m9jZDrOoajexK64IjNy8an6Hysj2Ir/grB/ye54w/7B+mf+kUVV4t/wCRU1n/ALB8/wD6Lav+CQ3/ACeVpv8A2L+o/wDoC1Vf8FbP2wfGvwisdF+Afwv1mfRtW8T6c2oa1qlrIY7iGyMjRxxQuOUMjRy7mGCFQAHDmv2N/wDgk98L/EHw30D4qftHS6pr2qeJbOLUIdAt7x7S3tYJl3xiZ4yJXlKsrHDoFJK4bGa/b+/Y3/Zi+EH7Gvjzxh8N/g7oui65pA04WWoxmWSeHzNSto3w8jsTlJHXnPDV/wAEOv8AkVPi3/2ENK/9F3NfF74b6N8Yfhd4p+F2vAfYvE+lT2LuVyYWdCElA/vI+1x7qK/4JZ+Ptc+An7ZGq/A/xlmw/wCEoW60HULWU7RDqdmztFn3yk8QHczCv+Co/jHXv2hv2yvC37OXgp/tbeHBbaRawrkr/aV+0bysSOMBDbK393y2yeOPhp4C0P4WfD3w38N/DUQTTPDOlwafbfLgssSBd7f7TEFie5JJPNf8FBvDemR/8FEPFul+Nbq4ttD1XWNIe8uoGCSJZy2tqJHQsCAVUuASCAV6EV/w5U/ZX/6H74q/+DWw/wDkKv8Ahyp+yv8A9D98Vf8Awa2H/wAhV8Dv+CWn7PPwB+KmgfF7wj4r+IGoav4dklktLfVdRtHt2aSF4iXWK2jc4EhYYYcgZyMg/wDBWD/kyPxh/wBhDTP/AEsir/giH/ySr4lf9jBa/wDpOar9vb9oj4sftV/tQN+yZ8NNVltPDOneIU8PW9hFO0UepagJBHNNdEfeSOUMFByqrHvxljXwf/4JG/sreA9Etk+Imkaj8Qdd8sfabu/vprS3EnfyoIHTC+0jSHvn0/4K8fAH4M/Az/hU/wDwqP4eaT4X/tv+2v7Q+woy/aPJ+w+XuyT93zZMf7xr/gm9/wAmR/Cv/sH3P/pZPXhe6j+E3/BUcS+KCIIbH4tXMUssp8tVinvXRJj1wu2VX+npVV+zxfWeqf8ABTrSdT0+4We1u/iXqE0Eq9HRp5yrD2IINf8ABS7SbzWv2HvijZ2IJkjtLK5bAz8kN/bSv/45G1f8E5f2Kfgj+2DpHjcfEfxR4v03VvCtzZ+RDol5bQI8E6y4ZhNBKSQ0LDIIAyPWv+HKn7K//Q/fFX/wa2H/AMhV/wAOVP2V/wDofvir/wCDWw/+Qq/Z1/Z88EfsxfDC0+FHw/vtYvdKtLqe5Fxq08ctxJJK25tzRoi4HAACjgDOTk1VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVUyqylWAIIwQRV38MfhrfztdXvw98M3E743SS6TA7NgYGSVz0AFafpWl6RB9l0nTbWyhz/q7eFYl4GOigDoAKqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/aF/5ID8TP8AsT9W/wDSOWv+CKn/ACdR4p/7J/ef+l9hVVe2VpqNnPp99bxz21zE0U0Ui5WRGGGUjuCCRXxg+GPxc/4Jn/tW2HjnwfbXEug29/Jc+GtRlVmt9TsHyJLOZhx5gjYxuODnEigAqa+GH/BVz9kLx14Yt9V8W+NLjwPrHlA3ekapYXExifHzCOaGNkkXPQ/KxGCVXpX7fn/BTrw38XPBd58DP2b31C50nXQYNb8QT2r2zXcBP/HvbRMBIFk4Ds6qSpK7fmJr/glT+x3r/wABPA2p/Fz4maRJp3jHxrBHDa6fcRlZtN05TvCyDqskrhXZDyojjBw25R/wV5/5M01L/sYNO/8AQ2r9hr9p7x1+x3r7/E+78M32sfDHxVe/2NrsUBA/0iFFlR4iSAJ40mJVWwHV5BkEbktP+Con7EVz4dHiCT4wtbt5W5tPm0W9+1K39zYsRBbPGQSvfOMmv20/2tfF37f/AMUfDXws+DXhDV38OafeNHoelmPddapdyfKbmVFJVAqAhRkhFLszDcQv7If7P1p+zJ8AfDPwoW4iutTtY2utZuohhbi+mO+Uj1VciNTxlY1JGc1/wWa+A+ueNfhp4W+NnhnT3u28DTXFrrSQxlnWyuNhWY4/gjkjwfTz89ASP+Ce3/BTL4Z/Cb4V6f8AA74/TX+l2vh9nXRdet7R7uI27yM5hnjjBkVkZjtZVYFcAhSo3ftIf8FVPB/ijQZPhR+x7Y634v8AHHionT7HVE06W2jtGl+XfDHIqyyTc/KCiqpIYk7dp/Yi8B6/8Lv+Ci3gn4deKkt11jw5r99Y3ywSeYizR2lwrBW7jI61X/BOX/lIv4L/AOwhrv8A6bb2v+Cknwp8RfF/9kPxloXhK0mvNX0k2+rW9pEuWuFt5Q0qAdS3kmRgByWVR3r/AIJYftqfC39nmDxP8K/jDqb6HpHiC9j1HT9ZMDyxQ3AjEckUwQFlDKsZVsFQVbOMg1+0L/wVS/Z98B+FbzTfgr4h/wCFheOb2PydJs9OtJTbRTOMJJLKyhWAJz5ce5mOFO0HcP2RNL8V+GP2/Ph/pfxHtriw8Rw+LwuqQ3xCypdOGLB/RyzdOuTjrVftPr4h/Zf/AOCimv8Aj260mSRtO8cp4tson+Vb63muBd4Vv7rbnjJHQhhwRx8Mv+Ckf7JHxZ1zwz4Q8K/EC9HibxRcx2tro93ot1DLFM+cJJKY/JzkY+WRgSRjNf8ABUf/AJMS+Jv/AHCf/TtZ1/wQ6/5FT4t/9hDSv/RdzX/BSX9lDUv2ofgjHJ4MthP428FTSX+iwEhTeo6gXFqCeAzqiMvq8SAkBiR+wB/wUJb9lSC7+BPx10bV/wDhEYb+VradIWN1oFwWPnRPAwDNEz5YqMMjbyFbcQPFv/BUz9inw14ek1vTvinP4iufK32+m6Zo139onOB8v72ONEPP/LRl7+mK1u7+Of8AwVP/AGp0n0rR3sNPUR26BVaW08NaUrk75X4DOcux6GSQ4UAYC/D/AMD+H/hn4G0H4eeFLX7PpHhzTobCyjJywjiQKCx7scZJ7kk96/4Kwf8AJ7njD/sH6Z/6RRVXi3/kVNZ/7B8//otq/wCCQ3/J5Wm/9i/qP/oC1Vf8FqfhR4i074ueEvjNDaTS6BrWhppElwFysF5BLK+xiPu745VKg9THJjpx+yz/AMFL/wBmbXvgp4a074l/EG18HeKfD+kwWWp2OoQShZXhjVPOhdFKur7dwQHcpJBHAJ/4KB/tv2v7V3hDUPgr+zTousa54T0OM634r182bwRyW9ucqFRwGSFXKMXkCszhFUf3v+CHM0LeGvi7brKhlS+0lmQN8ygpdAEj0O0/kfSq/wCCnvgDVf2c/wBsvw1+0N4QtRBB4lnttdtWX5EGp2MkYnXgfxbYJGPczNX/AAS48C6t+0L+1t47/ap8ZWrSxaHcXN9E0nzD+0tQeTaq56iOEzcD7pMfTiq/4K7fsh+JPiHp+m/tH/DjR5dS1Dw7Ymx8SWNtEXlks1YvHcqo5byyzh+p2Mp4VGNfsX/8FZ/BOh+BdJ+GH7Tkmo2V5okCWtn4ptrV7uO4t0UKguY03SiUAAb0V94GSAclr/8A4KY/sP6fZteP8d7OYKOI4NIv5HY9gFWDP54A74r9mX9sT4TftZ3HixPhWmriHwjNbR3EmpW6wNcCdZCkkaBmYJmJx82DkHjpn/grB/yZH4w/7CGmf+lkVf8ABEP/AJJV8Sv+xgtf/Sc1Xxqh8Tfsaf8ABQ3UfHGtaJPdQaX4zfxHYo2F/tDTridpRsfGNxjd4y3QOrccYq3/AOCmf7FM/hGPxdJ8ZreBXiDNpsmnXJvUfGfLMKxk7s8ZGUz/ABY5r9vf9ovxd+1z4qtPi9pfg7VdJ+F+gTvoXh2e8QL59wR5szsQSPNdVQsilgirGCcnLf8ABNmaGf8AYh+FjwSpIosbtSVbIDLfXAYcdwQQR2Ir/grZ+xv4obxXL+1X8NNInvrG5to08W29qpaWzlhVUjvAo58sxqiuR9woGPDMV/Zh/wCCxfhKw8E6d4R/aW0TXW1zTIEgHiLS4VuU1BVAAknjLKySkAbioYMcnC5xXxv/AOCo9x8abRfgX+xZ4G8S6t4y8XhrCHV7u3W3NorjDPbxhid4Uk+bIUWLG45xx+xn4Yv/AAT+3x4B8GarJDJfaB4yk0+5aFiUaSEyRuVJAJXKnGQOK8ZeE9E8e+Eda8D+JbX7TpPiDT57C+h6F4ZoyjgdcHax57Vo9/8AHD/gld+1TO1/pL6jpkge3YSK0Vr4k0pnBDxvyFkGFOeTHICpBBIbwH/wVS/Yx8Z6JDqOrfEa68KXzIDNpus6TcCWE9xvhSSNv+AuT7CvHv8AwVk/Y18H2zHQvGer+L7sEAW+kaPOgUnHJkuFiTHOSVLHrwTxVvcW95bxXdrMksMyB45EOQ6kZBB9CKqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq8RaBpHivw/qfhbX7T7VpmsWctneweYyebDKhSRdykMuVYjIIIzwRXwU/Yy/Zq/Z18VXXjb4N/Db/hH9avdPfT57n+2L67327yRyMmyeZ1GXhjOQAflxnBOaqvHHgLwV8S/Dd14P+IHhXS/EOi3gHnWOo2yzxMR0bDA4YdQwwQeQRXif/gkT+xt4g1Fr/TtC8U+H0diTa6ZrbGLJ9PPWVgPYHv9MfBX9hL9lr4BanD4h8A/C6zfXbdg0OrarM9/cwsOjRGUlYm/2o1U+9V/wV5/5M01L/sYNO/9Dav+CQHgbwd8SP2W/iP4M8eeGtP17RNR8WlLmwv4BLHIBaW5BwehBAIIwQQCCCM1rn/BIf8AY21jWDqllonirR4Wk3GwsdcYwe6jzVkkA/4H34xxXwN/ZS+AP7OFtInwk+HVhpN7cJsuNTlZrm8mXuDPKWcKTzsUhfaqurW1vrWWyvbeK4t7iNo5YZUDpIjDDKyngggkEGvHP/BKj9jXxvrkmvReB9T8OSTzCWa20PU3t7dz3AiYOsan+7GFA7Yr4E/sbfs5fs3znU/hX8ObSz1l4zG+sXkj3l4VIwwWWQkxhh1WPaD3FQ/sWfsy2/xiPx+g+GSp49OptqZ1ZdXvh/pTZ3SeT53k5OTxsxz0qvhx+wp+yp8JPiNZfFr4e/Cv+yvFenSTyW2of23qE/ltPE8Up8uWdozuSVxypxuyMEAivi//AMEz/wBkf4yeILvxXqvgS68P6xfuZLu58PXhshM5OWcw4aIMxzlggLEkkk818D/2Cv2XP2fdXh8TeA/hxFPr9u2YNX1e4e+uID2Me8lImH95FVueTV9+xN+zFqXxgX493fwzLePE1WLVV1Zdav0/0uMqUk8lZxF1Vcrs2nuCCar4/wD7LHwO/ab0q0034veDI9Tm03d9g1CCZ7a7tN2NwSVCDtOBlGyuecZANfCf/gl7+yv8H/HWkfETw/p/ifUNZ0G9S8059R1gskEyHKNtjVN2CAcNkHvmvil8LfAfxq8B6n8MviZoX9s+G9Z8r7bY/aprfzfKlSaP95C6OuJI0bhhnGDkEivgd+zL8D/2bbTV7H4K+CP+Edh12SKTUE/tK6u/OaIMEObiWQrgO33cZzz2qvjj+xb+zV+0TdPq3xN+GVjc606BP7YsZHsrzjgbpIivmYHAEgYD0rRP+CQf7G2lakt9f6T4t1mENk2V9rrJER6ZhWN8f8Cz718Ofhb8OfhF4cj8JfDHwXpPhrSY23fZtPtliDv0LuRy7HjLMST61XxY/YU/ZU+OXje8+I3xS+Ff9t+Ir+OKO4vP7b1C23rGgRBshnRBhVA4UZxzVXlpb39pNY3UfmQ3EbRyJkjcrDBGRyODXwh/Ye/Zc+AvjOL4g/Cf4X/2Hr8NvJbpd/21f3WI5AA67J53TkAc7ciqrxx4E8G/EvwvfeCvH/hnT9f0PUk2XVhfQCWOTByDg9GBAIYYKkAggjNXv/BIr9jO71o6rD4d8T2tuST/AGbDrsnkdemWDSY7ffrwt+yx+z94I+F+s/Bvwn8MdM0zwp4jtHtNXtLd5Ulv43TY3m3AbznbaSNxfcOxFfAz9lj4Dfs1yazL8E/AzeHG8QrAupD+1by7Fx5O/wAri4lkClfNk5XH3jmq+N37OnwY/aP0TTvD3xn8FR+IrHSbo3Vkhvbi0aGUqUJDwSI+Cp5UnacA4yAR8FvgH8I/2d/C9z4M+Dfg+Pw9pF5ete3EC3c9y0k7IqFzJO7ufljQY3YGOAMmqr4u/wDBOb9kb4zapceINe+GEWj6xdMzTX+gXD6e0jscl2jQ+UzEkksyEk9Sa0r/AII+fsd6fcCe7tfGepoCP3N3rm1eP+uUaNz9a+Dn7O3wT/Z90250v4O/DrS/DUd6EF3LAHlnuQmdgkmkZpJAu5sBmONx9TXxY+Efw8+OXgi8+HHxS8Pf234dv5IpLiy+1zW29o3Dod8Lo4wyg8MM454r4Ifs4fBf9m/SdS0P4L+DP+EestYuFuL2L+0bq782RV2q2biSQrxxgECq+OH7N3wV/aO0ODQvjB4EstdSz3GyuiWhubMtjd5U0ZDoDhcrna2BkHArw1/wSU/Yz8PawurXfhXxDriJL5iWep61IYFI6AiIIzL7MTnvkV8Rv2Uv2e/ix8P9E+Ffjb4ZafceE/Dc6z6VpVjPPp0VpIEZAV+yvGcbXYYJIJOSM818K/hT4C+CfgXTvhp8MdDbR/DektKbOxN5Nc+SZZWlkw8zu+C7u2C3G44xTokiGORVZWBDKwyCO/FeOf8Agnz+xp8RdVk1rxL8BNCW8mcvI+lz3Glh2OcsyWkkakkkkkjk8mvA3wW/Z4/Zg8M6vr/gTwF4d8GabY2UlxqmpRw/vRbxKXcy3DkyMihScMxA61/wT0sbn40/8FFbX4gW1myWkGoaz4lu0Vf9THIsoj65wBNcwj8eucGq+JXwo+G3xi8OP4S+KHgnSfEulO25bfULYSeU/TfG33o2xxuUg+9a7/wSA/Y41e+a70/TfF+ixMeLWx1wvGv0M6SP+bV4D/4JdfsX+BLuLUG+GM3iO6hIKPr2pTXSAjHWEMsTA4yQyHqe3FWtra2NrDZWVvFb29vGscMMSBEjRRgKqjgAAAADpVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX/BVbwr4n8Zfsj6jonhDw5qmuai2u2DraabZyXMpVXbcwSMFsDuccV/wR08EeM/A3wN8aad428I614fup/Fhlhg1XT5bR5E+yQDcqyKCVyCMjjINVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVf8FFvE37ZvhGDwL4l/ZR0PXbq00ae7udfk0m3jvXmJVFhiktCGaWPb5xJCMASpypANW3/AAWP/aY8GRLo/wASfgh4YbU4hsLS295psjEfeLRuzc8rnG0D05wPiX+0L/wUB/4KB2X/AArfwl8MNQs/Ct26m5sPD+mS21pcYO5Ptd5OxXaCM7S6ISAdpIBr/gn9+w/Z/sieCb7UvE97aap4/wDE6x/2tdW2Whs4UJKWsLEAkAks74G5scYRTVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVU/jrwQmsDw6/jLQ11XzRCLE6jEJ/MPRPL3bt3TjGaqn8deCE1geHX8ZaGuq+aIRYnUYhP5h6J5e7du6cYzVVVVVVVVVVVVVVVVVVVVVftM/tYfC79k7QtF8Q/FGDXJbXXrt7W1GlWiTsHRNx3BnTAx9a+DnxY8LfHP4Z6F8V/BUd8mi+IYXltFvoRFMFSRozuUMwB3Ie54xVVVVVVVVVVVVVVX7Rf/BQj4C/sv8AxAj+GvxJtPFUury6dFfq2madHPF5UjOq/M0qndmNsjHpVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV8QP2ivgL8KppLT4i/GLwhoF3F96zvdXhS46dod3mH/vmtR/4KafsNaXKILn482jsc4Nvomo3A446x27CvDv/AAUK/Yu8UyxwaZ+0H4chaUAqdRWfTwMju1xGgXp3xj8a8L+MvB/jjTV1nwV4r0fxBYNjbd6XfR3cRz0w8bEfrVVVVVVVVVVVVXxI+JfgP4Q+Dr/x/wDEnxPZ6DoGmKDc3tyThSxCqqqoLOxJACqCSegpv+Cr/wCxECVHxK1U4PBHh295/wDIdf8AD2D9iP8A6KRqv/hO3v8A8br/AIewfsR/9FI1X/wnb3/43X/D2D9iP/opGq/+E7e//G6/4ewfsR/9FI1X/wAJ29/+N1/w9g/Yj/6KRqv/AITt7/8AG6/4ewfsR/8ARSNV/wDCdvf/AI3X/D2D9iP/AKKRqv8A4Tt7/wDG6/4ewfsR/wDRSNV/8J29/wDjdf8AD2D9iP8A6KRqv/hO3v8A8br/AIewfsR/9FI1X/wnb3/43X/D2D9iP/opGq/+E7e//G6/4ewfsR/9FI1X/wAJ29/+N1/w9g/Yj/6KRqv/AITt7/8AG6/4ewfsR/8ARSNV/wDCdvf/AI3X/D2D9iP/AKKRqv8A4Tt7/wDG6/4ewfsR/wDRSNV/8J29/wDjdf8AD2D9iP8A6KRqv/hO3v8A8br/AIewfsR/9FI1X/wnb3/43X/D2D9iP/opGq/+E7e//G6/4ewfsR/9FI1X/wAJ29/+N1/w9g/Yj/6KRqv/AITt7/8AG6/4ewfsR/8ARSNV/wDCdvf/AI3X/D2D9iP/AKKRqv8A4Tt7/wDG6/4ewfsR/wDRSNV/8J29/wDjdf8AD2D9iP8A6KRqv/hO3v8A8br/AIewfsR/9FI1X/wnb3/43X/D2D9iP/opGq/+E7e//G6/4ewfsR/9FI1X/wAJ29/+N1/w9g/Yj/6KRqv/AITt7/8AG6/4ewfsR/8ARSNV/wDCdvf/AI3X/D2D9iP/AKKRqv8A4Tt7/wDG6/4ewfsR/wDRSNV/8J29/wDjdf8AD2D9iP8A6KRqv/hO3v8A8br/AIewfsR/9FI1X/wnb3/43X/D2D9iP/opGq/+E7e//G6/4ewfsR/9FI1X/wAJ29/+N1/w9g/Yj/6KRqv/AITt7/8AG6/4ewfsR/8ARSNV/wDCdvf/AI3X/D2D9iP/AKKRqv8A4Tt7/wDG6/4ewfsR/wDRSNV/8J29/wDjdf8AD2D9iP8A6KRqv/hO3v8A8br/AIewfsR/9FI1X/wnb3/43X/D2D9iP/opGq/+E7e//G6/4ewfsR/9FI1X/wAJ29/+N1/w9g/Yj/6KRqv/AITt7/8AG6/4ewfsR/8ARSNV/wDCdvf/AI3X/D2D9iP/AKKRqv8A4Tt7/wDG6/4ewfsR/wDRSNV/8J29/wDjdf8AD2D9iP8A6KRqv/hO3v8A8br/AIewfsR/9FI1X/wnb3/43X/D2D9iP/opGq/+E7e//G6/4ewfsR/9FI1X/wAJ29/+N1/w9g/Yj/6KRqv/AITt7/8AG6/4ewfsR/8ARSNV/wDCdvf/AI3X/D2D9iP/AKKRqv8A4Tt7/wDG6/4ewfsR/wDRSNV/8J29/wDjdf8AD2D9iP8A6KRqv/hO3v8A8br/AIewfsR/9FI1X/wnb3/43X/D2D9iP/opGq/+E7e//G6/4ewfsR/9FI1X/wAJ29/+N1/w9g/Yj/6KRqv/AITt7/8AG6/4ewfsR/8ARSNV/wDCdvf/AI3X/D2D9iP/AKKRqv8A4Tt7/wDG6/4ewfsR/wDRSNV/8J29/wDjdf8AD2D9iP8A6KRqv/hO3v8A8br/AIewfsR/9FI1X/wnb3/43X/D2D9iP/opGq/+E7e//G6/4ewfsR/9FI1X/wAJ29/+N1/w9g/Yj/6KRqv/AITt7/8AG6/4ewfsR/8ARSNV/wDCdvf/AI3X/D2D9iP/AKKRqv8A4Tt7/wDG6/4ewfsR/wDRSNV/8J29/wDjdf8AD2D9iP8A6KRqv/hO3v8A8br/AIewfsR/9FI1X/wnb3/43X/D2D9iP/opGq/+E7e//G6/4ewfsR/9FI1X/wAJ29/+N1/w9g/Yj/6KRqv/AITt7/8AG6/4ewfsR/8ARSNV/wDCdvf/AI3X/D2D9iP/AKKRqv8A4Tt7/wDG6/4ewfsR/wDRSNV/8J29/wDjdf8AD2D9iP8A6KRqv/hO3v8A8br/AIewfsR/9FI1X/wnb3/43X/D2D9iP/opGq/+E7e//G6/4ewfsR/9FI1X/wAJ29/+N1/w9g/Yj/6KRqv/AITt7/8AG6/4ewfsR/8ARSNV/wDCdvf/AI3X/D2D9iP/AKKRqv8A4Tt7/wDG6/4ewfsR/wDRSNV/8J29/wDjdf8AD2D9iP8A6KRqv/hO3v8A8br/AIewfsR/9FI1X/wnb3/43X/D2D9iP/opGq/+E7e//G6/4ewfsR/9FI1X/wAJ29/+N1/w9g/Yj/6KRqv/AITt7/8AG6/4ewfsR/8ARSNV/wDCdvf/AI3X/D2D9iP/AKKRqv8A4Tt7/wDG6/4ewfsR/wDRSNV/8J29/wDjdf8AD2D9iP8A6KRqv/hO3v8A8br/AIewfsR/9FI1X/wnb3/43X/D2D9iP/opGq/+E7e//G6/4ewfsR/9FI1X/wAJ29/+N1/w9g/Yj/6KRqv/AITt7/8AG6/4ewfsR/8ARSNV/wDCdvf/AI3X/D2D9iP/AKKRqv8A4Tt7/wDG6/4ewfsR/wDRSNV/8J29/wDjdf8AD2D9iP8A6KRqv/hO3v8A8br/AIewfsR/9FI1X/wnb3/43X/D2D9iP/opGq/+E7e//G6/4ewfsR/9FI1X/wAJ29/+N1/w9g/Yj/6KRqv/AITt7/8AG6/4ewfsR/8ARSNV/wDCdvf/AI3X/D2D9iP/AKKRqv8A4Tt7/wDG6/4ewfsR/wDRSNV/8J29/wDjdf8AD2D9iP8A6KRqv/hO3v8A8br/AIewfsR/9FI1X/wnb3/43X/D2D9iP/opGq/+E7e//G61D/grZ+xXZxq9t401++Y5ylv4fuFI/wC/gUfrVv8A8Fjf2Qp544ZLfx5ArsA0smixlUHqdsxOPoCa+H3xB8G/FbwXpPxC+H+uwaz4f1uDzrG9hDKJFyVOVYBlYMrKVYAqwIIBBqqqqqqqqqqqqqqqqqqqqqqqqrxN/wApg4f+yoWX/ocVVXib/lMHD/2VCy/9Diqqqqqqqqqqqqqqqq/ad/4KD/Fr4XftiaF+zL4K8N+G4dIuNY0W2vdUu4pZ7qWO7aEyCMb1SP5ZCuSrHvx2qqqqr/gt7/ySr4af9jBdf+k4r/gm9/yZH8K/+wfc/wDpZPVVVVVVVVVVVVVVX/BZL/k7m0/7E+x/9HXNVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVfFLxxB8MPhj4u+JVzp738PhLQb3VpLRJBG1wttA8pjDEEKWCYzg4zXxp/b9/a6/au8QP4S8JX2r6JpV67JaeF/Bsc3mzIf4ZZI8zXBxgEcITyEXOK8Ef8ABMn9tTx/GmoD4SyaJbz5Pn6/qEFm+evzRM5mB+qd/rVn/wAEXP2rbqASzeMPhhaNkjyptWvSw9/ktGH614g/4I5fte6NE0mm3HgPXmUcJp+tSIW4PT7RDEO2OT3Hua1z9mv9tv8AZZ1QeMz8PvHnhG4sQWbWtAlaaOFR133Fm7oqn0ZgCK/4J1/8FJPi/wDFv4q+H/2d/jHZWfiKfW4rkaf4jiUW1zE1vbSzlbhFGyUFISoZQjAnLb85FVVVVVVVVVVV/wAFmbYz/slaXItxLH9n8a2TlUbAk/0a6Xa3qPmzj1UV+yP/AME+P2PPir+zT8O/iH44+DaX+v65osc+oXS+INThE0uSpfYlyFUnbkhQBk8ADiv+HXH7CX/RC/8Ay5tW/wDkqv8Ah1x+wl/0Qv8A8ubVv/kqv+HXH7CX/RC//Lm1b/5Kr/h1x+wl/wBEL/8ALm1b/wCSq/4Jp/sy/A/9oL4vfEvwt8XfBH9vaZ4fs1k02D+0rq18hjcsmd0EqM3ygD5ia/4dcfsJf9EL/wDLm1b/AOSq/wCHXH7CX/RC/wDy5tW/+Sq/4dcfsJf9EL/8ubVv/kqv+HXH7CX/AEQv/wAubVv/AJKr/h1x+wl/0Qv/AMubVv8A5Kr/AIdcfsJf9EL/APLm1b/5Kr/h1x+wl/0Qv/y5tW/+Sq/4dcfsJf8ARC//AC5tW/8Akqv+HXH7CX/RC/8Ay5tW/wDkqv8Ah1x+wl/0Qv8A8ubVv/kqv+HXH7CX/RC//Lm1b/5Kr/h1x+wl/wBEL/8ALm1b/wCSq/4dcfsJf9EL/wDLm1b/AOSq/wCHXH7CX/RC/wDy5tW/+Sq/4dcfsJf9EL/8ubVv/kqv+HXH7CX/AEQv/wAubVv/AJKr/h1x+wl/0Qv/AMubVv8A5Kr/AIdcfsJf9EL/APLm1b/5Kr/h1x+wl/0Qv/y5tW/+Sq/4dcfsJf8ARC//AC5tW/8Akqv+HXH7CX/RC/8Ay5tW/wDkqv8Ah1x+wl/0Qv8A8ubVv/kqv+HXH7CX/RC//Lm1b/5Kr/h1x+wl/wBEL/8ALm1b/wCSq/4dcfsJf9EL/wDLm1b/AOSq/wCHXH7CX/RC/wDy5tW/+Sq/4dcfsJf9EL/8ubVv/kqv+HXH7CX/AEQv/wAubVv/AJKr/h1x+wl/0Qv/AMubVv8A5Kr/AIdcfsJf9EL/APLm1b/5Kr/h1x+wl/0Qv/y5tW/+Sq/4dcfsJf8ARC//AC5tW/8Akqv+HXH7CX/RC/8Ay5tW/wDkqv8Ah1x+wl/0Qv8A8ubVv/kqv+HXH7CX/RC//Lm1b/5Kr/h1x+wl/wBEL/8ALm1b/wCSq/4dcfsJf9EL/wDLm1b/AOSq/wCHXH7CX/RC/wDy5tW/+Sq/4dcfsJf9EL/8ubVv/kqv+HXH7CX/AEQv/wAubVv/AJKr/h1x+wl/0Qv/AMubVv8A5Kr/AIdcfsJf9EL/APLm1b/5Kr/h1x+wl/0Qv/y5tW/+Sq/4dcfsJf8ARC//AC5tW/8Akqv+HXH7CX/RC/8Ay5tW/wDkqv8Ah1x+wl/0Qv8A8ubVv/kqv+HXH7CX/RC//Lm1b/5Kr/h1x+wl/wBEL/8ALm1b/wCSq/4dcfsJf9EL/wDLm1b/AOSq/wCHXH7CX/RC/wDy5tW/+Sq/4dcfsJf9EL/8ubVv/kqv+HXH7CX/AEQv/wAubVv/AJKr/go3+w9+y58Bf2Z774g/Cf4X/wBh6/DrFlbpd/21f3WI5GIddk87pyAOduRX7E3/AAT/AP2Rfi9+y14A+I3xD+En9reItas55L68/t7UYPNZbmVFOyK4VF+VFHyqOlf8OuP2Ev8Aohf/AJc2rf8AyVX/AA64/YS/6IX/AOXNq3/yVX/Drj9hL/ohf/lzat/8lV/w64/YS/6IX/5c2rf/ACVX/Drj9hL/AKIX/wCXNq3/AMlV/wAOuP2Ev+iF/wDlzat/8lV/w64/YS/6IX/5c2rf/JVf8OuP2Ev+iF/+XNq3/wAlV/w64/YS/wCiF/8Alzat/wDJVf8ADrj9hL/ohf8A5c2rf/JVf8OuP2Ev+iF/+XNq3/yVX/Drj9hL/ohf/lzat/8AJVf8OuP2Ev8Aohf/AJc2rf8AyVX/AA64/YS/6IX/AOXNq3/yVX/Drj9hL/ohf/lzat/8lV/w64/YS/6IX/5c2rf/ACVX/Drj9hL/AKIX/wCXNq3/AMlV/wAOuP2Ev+iF/wDlzat/8lV/w64/YS/6IX/5c2rf/JVf8OuP2Ev+iF/+XNq3/wAlV/w64/YS/wCiF/8Alzat/wDJVf8ADrj9hL/ohf8A5c2rf/JVf8OuP2Ev+iF/+XNq3/yVX/Drj9hL/ohf/lzat/8AJVf8OuP2Ev8Aohf/AJc2rf8AyVX/AA64/YS/6IX/AOXNq3/yVX/Drj9hL/ohf/lzat/8lV/w64/YS/6IX/5c2rf/ACVX/Drj9hL/AKIX/wCXNq3/AMlV/wAOuP2Ev+iF/wDlzat/8lV/w64/YS/6IX/5c2rf/JVf8OuP2Ev+iF/+XNq3/wAlV/w64/YS/wCiF/8Alzat/wDJVf8ADrj9hL/ohf8A5c2rf/JVf8OuP2Ev+iF/+XNq3/yVX/Drj9hL/ohf/lzat/8AJVf8OuP2Ev8Aohf/AJc2rf8AyVX/AA64/YS/6IX/AOXNq3/yVX/Drj9hL/ohf/lzat/8lV/w64/YS/6IX/5c2rf/ACVX/Drj9hL/AKIX/wCXNq3/AMlV/wAOuP2Ev+iF/wDlzat/8lV/w64/YS/6IX/5c2rf/JVf8OuP2Ev+iF/+XNq3/wAlV/w64/YS/wCiF/8Alzat/wDJVf8ADrj9hL/ohf8A5c2rf/JVf8OuP2Ev+iF/+XNq3/yVX/Drj9hL/ohf/lzat/8AJVf8OuP2Ev8Aohf/AJc2rf8AyVX/AA64/YS/6IX/AOXNq3/yVX/Drj9hL/ohf/lzat/8lV/w64/YS/6IX/5c2rf/ACVX7bv7MvwP+EH7bnwn+EHw68Ef2T4S8S2ejSapp39pXU/2hrjVLiCU+ZLK0i7oo0X5WGMZGCSa/wCHXH7CX/RC/wDy5tW/+Sq/4dcfsJf9EL/8ubVv/kqv+HXH7CX/AEQv/wAubVv/AJKqx/4Jj/sM6fI0tv8AAe2dmXaRPrupTD8A9wQD71/wUN/ZH/Zh+E/7H/jnxr4B+D/h3QfENi2nJp99bo4lRn1C2RwpLHkxNID7E1/wShikT9iHwYzxsokv9UZCRjcPtswyPUZBH4GqqqqqqqqqqqqqqqqqqqqqqqqvE3/KYOH/ALKhZf8AocVVXib/AJTBw/8AZULL/wBDir4x+IdY8I/CLxx4r8O3EUGq6L4b1C9sZZYvMSOeK2keNmQ/eAZQSO/SvgR/wVx8XW3wu8b+IPjPDY+KPGcV/Y2fhDw/pVl9ka8eZZvMaRlDfu1KJkgFssqgfNkeOP2hf+Cxr6XL8T4vh1qXhjw5GhuG06w8K2k3kxdfnhmWW6VQAck4IGScV/wT+/4KSSftM69/wqD4taTp2k+OhbPPpl3YBkt9XSNd0iiNiTHMqAuQCVZQxAXbg18f/wDgph8RvFHxXl/Z8/Ye8Bw+MPEEVxJaza7JAbpJJE4kNrGGCeWhHNxKTGQCQu3DnxP8Rf8AgtB8G9Lm+JHjTStP1/Q7BTNe6dHp2k3iwRjk70sts5RRnLKxwBknvX7D/wC294S/bD8IXgbTo9B8baAqHWdGEpdCjEhLiBjy0TEYIPzI2Acgqzf8FCv2nP2wv2SvHmk+KvBN3oOpfDLX2RLb7XpCu9rdIMy2ssikHDqpdG4JBccmMk/Ab41eD/2hvhRoHxZ8Ez5sNatw0ts7hpLK4XiW3kx/Gjgj0Iww4YE/t5/8FBviP8LfjBoP7Pf7McFhq3i/zETWDJZfbD9pn2i3sokyP3mGDN1++g4IYV4Oi+K2hfBWKX4l+I9N1Dx9Do8s9/d2lmsdrFd7GYJHGPvIhwuTy20njOB+zZ/wVu8UQ+GviF4o/aTvtP1OTR7O0/4RrStJsUtJtQupGkDR7snC4VWLkYVQx5JVW1H48/8ABYT4yaa/xB+FvwnXwf4alUTWlrBptgs0kJ+6QmoM00uRg7kQA8EAA1+zp/wVf+I3hz4jR/CH9svwrDpLi6Fnca2LBtPudLmJAH2y3bC+XyMsoQqOdrCo5I5o0lhkV0dQyMpyGB6EH0r9s3/gpHJ8F/HC/Ab9n/whF44+JcsscE4eOS4t7GeTGy3WKIh7i4II+RSoUsMljlBe69/wW7stP/4TmTTbN7NI/PbQobLRJZNmPu+UoMxOMHYrl+2M5FfsKf8ABRKL9pPXbr4P/Frw9a+FfiZYLIYreBHig1JYh++VI5CXimj2sWiYngFgflYLrOsaT4d0i91/XdRt7DTdNt5Lm7u7iQRxwRIpZ3djwFCgkk9AK+Lf/BWP4nfEbx3/AMKu/Yp+FkmuTyytFBql7p8t5c3uODJBaIR5SDrvlLfKcsqYNL45/wCC3Wh2reKdQ8IHUbFsOLE6dokrqACcCGAi45x0PPT1q8+J37Xer/sLR/E/Sfh/a23xxvLaCWHw/BpciiPdqCRsGt53LK/2Tc7KxG1ieOK+N3i/9pjXP2rbHxV8V/C/2D4tx6hpb22m/YVi3TxiL7IPKBI+bEfGec+9fAH4y/8ABUnxB8ZfCWjfGT4Uf2b4Ku9QVNau/wCwoofJg2nLbw5K845xX7SX7Rvw8/Zd+GV38S/iFdSNEji30/T7cg3Go3LAlYYgSBnALFjwqqT7Hw7+09/wVQ/a2guPGv7OngTSvB3g5Z3SzuBbWZWUK2Nom1DP2hhgqzxRqoIIIU8Vof8AwUG/bJ/ZU+I2l+Bv25vh4l9oupsM6rb2UMNyIwdrTQSWp+zThcgtGAG5HK5APhvxFofjDw9pnivwxqkGo6RrFpFd2F5A2UnhkUMjqfQqQfxr9uz/AIKV6P8Asy6y/wAKfhfotj4n+IIiR7xrty1lpAcZVZVjYPJMV2nywVAV1Yt/CfDvxI/4LY+J7KH4haT4UlOizATx6ZeaTo1oWTAIHkTbLvBBHfcfWv23/wBsnxB+0j8KvDHw8+Knw/uPBnxM8Da/L/bWntbyQxyq8G3zFjk+eFgQA0bk43KQxBOP+Cb3/Jkfwr/7B9z/AOlk9V+1n8RPFfwk/Zx8efErwPd21vrvh3SzdWUlzbiaMMsiZDISMgqSPbOe1eBf+CsfjvRP2Wbrxr45sdK8TfFPWvF15pHhvTra0MFvHbw2tnIZ50jO4qJLkgKCGkJABAVmH/BP34s/tufFHxt4ru/2pPDmvaR4ffSop9ChvvCy6VCsxlwwRvKWRjsI4djxyPWv21/+Cjtv8AfFcfwS+CvhWLxr8TrkxRyQOjzW+nyS48uIxxESTzuCuIlK43qSSflNxr3/AAW7lsf+E3j0yyhtNnn/ANhx2eiGTYedvlnM2cH7u/fxjGciv2Gv+CjE/wAf/Fc/wO+OHhq38J/Eu18xbVIopIIdRaEEzRGKQl4bhArMUJIIViNu3bX/AAU0/am/aO/ZV1LwRr3wn1LSE8OeI4Lm2u0vdLW5MN5CysDvJGN8cnC/9MmNfAv4kQfGH4M+CvihCI1bxNodrfTRx9IpnjBljH+7JvX/AIDX/BS/9sLxp+yn4H8IQfC+502PxV4o1OYhr23FwqWcEf707DjkySwAE9g3Xt/wT8/aV1z9qT9nq18ceMZbN/FOl6rdaZrJtYfJjaRCJImVOdoMMsQPYsG+g/bO+Od7+zn+zf4v+Keiy2w1uxgit9IS4TerXc8qxISuRuC7i5Hohr/gm/8AHT46/tHfCPW/ih8aLzTZIZtZNloiWVgtsDHCg82Q4J3Au+wZ6GJvx+Kf7Sv/AAUp+IfxU8WfDf8AZx/Z8tdE0nwvq8+njW7yFJRdeU+PMW5u2jtyrrtcIqMyhwCSa8d/tF/8Fbf2VrWLx18btH03xB4XSZUnmn0zTp7VNxACyPp+ySHJIAZ8AsQBnpX7I/7UXhP9rX4SW3xH8PWLaXf21w1lrOkSTCRrC6VQxUNgb42VlZXwMg4IBDAf8FJ/2i/ij+zD8EtA+IPwo1DTrfUr7xVDpdwL6yFyjQyWtzKcAkYIaBfzNeP/APgqJ8V4fhX8N/Afwj8Mw+K/jR4t0OG+1qW10t7lLBpdzRxQ2seTJO0YWTByqqynDbiF/YG8ZftM+Nvg/rus/tXWGr2PiuLxRcR2UWqaLHpbrp/2W2aPbGkaBkEjzjeQTkEFjtwP2lf+CqnjnVPiS3wR/Yy8JQeIdS+1myGuPaNfvfXAJBWxt0OGQY/1r7gw3EIAA5tviz/wWh+F9mfiD44+Hi+KtBgUTT6bNpmlTuI+p/daey3K8dcg7ccjrX7c/wC0d4f/AGqPivofxW0HR7nSJJPC1rZ6jps77zZ3cU0+9A+AHXDKytgZDDIByBVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV478HaN8RfBHiH4feIhOdK8TaVdaXfCB9knkXETRSbWwdrbXODjg18KPgf8JPgboKeHPhP4A0fw3ZhAsjWlv++uMdDLM2ZJW93Ymqr4hfHX4LfCaeG0+JvxW8KeF7m5XdDbarq0NvLIv95Y2YMV4POMV4Y8WeFfG+jQ+IvBfibSdf0q4/1N9pl7HdQSf7skZKnt0NTfsu/Ao/GDQ/jzpvw+03SfHGhS3Ekep6ZELU3fn28kEguETCzEpKx3MN4IX5sZBqqqqpiqqWZgAByTxiv2lP+Cu3wp+Hsk3hP9n7S0+I3iPLRi/JePTLd+2GAD3PPaPapHIk7V/wTb/bi+NH7WPjfxj4e+KNt4bitdB0qG6tTpVi9uxd5dh3FpHyMduKr9uP9vHR/wBj230bQbPwFqPifxV4lt5JtNidjb2UaIwRjJNglnBI/doCcEZKblJuP2qP+CvXje1Hi/wt8FL/AEfSXJkitbTwYQHTjG1LnfM4wOCvXOfTH7Nn/BW/xW3xCt/hV+1v4Q0/w88lyLJ9ctrWWyfT584xe28hbaucAuuzZ1K4yQCrKGVgQRwRVV/wWS/5NFtP+xwsf/RNzX7Af/Jmnwl/7F+P/wBDeqqq/wCCN/8AyX74yf8AYPT/ANK2qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv+CvP/Jmmpf8AYwad/wChtX/BN7/kyP4V/wDYPuf/AEsnqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr9vH4rftnfDbVvB0H7KXgK78R2t/bXba00GgNqPkyK0QiBI+5kNJx3x7V+0H8Wf2qvHX7R/hHxR8Z/CU2m/E7QE0+DRtMbRTayPsunntgYOrlpZWx/eyBX7Inx0/4KLeOfjrovhv8AaG+Fl/ongie2umvbybwq1kqSLA7RDzT93MgUY75xVVX/AAVg/wCTI/GH/YQ0z/0sir/glx/yYl8Mf+4t/wCna8qqqqqqqqqqqqqqqqr/AIK/fEj4ifDP4I+DNW+G/j7xH4UvrrxUIJ7nRNVmsJZY/sszbGaJlLLlQcHjIBr/AIax/ao/6OY+Kv8A4Wd//wDHa/4ax/ao/wCjmPir/wCFnf8A/wAdr/hrH9qj/o5j4q/+Fnf/APx2v+Gsf2qP+jmPir/4Wd//APHa/wCGsf2qP+jmPir/AOFnf/8Ax2v+Gsf2qP8Ao5j4q/8AhZ3/AP8AHa/4ax/ao/6OY+Kv/hZ3/wD8drwx+1Z+1FceJdJt5/2k/inLHJfQq6P4xvyGBcAggy8gjtVVXib/AJTBw/8AZULL/wBDiqq8Tf8AKYOH/sqFl/6HFX7Qv/JAfiZ/2J+rf+kctf8ABG/4W+FvHH7Q+t+M/ElhFez+CdFF1pUU0YZYrqWUIs2D/EiB9voWDDBUVXxT0TT/AIC/8FYbew8D2qWdnafEHRrqO2gPloqXwtpp4lwPlUi5kQADABxjAr/goP8AFPUvhB+yH8QfFGhXjW2rXdlHpdjKj7XR7uVIGdT1DLG8jgjkFR061/wTM/ac/ZI/Ze8A+I9X+Kfie5svHfiLUfLdotFuLkw6fGi+VGJERgN0hlZgDzhMj5RTf8FXv2ImUq3xH1UgjBB8O3n/AMbr9kD4ieCvDn/BTq31P4M3so8CeL9d1Kys4Vt3tw1ldRySRReW3zKkcwiIBH/LJa+N/wAG/Bnx++F+u/Cnx3ZCbTNbtiglVQZLSYcxzxE9JEbDDscYOQSD8HP2hPi//wAExPiF8UvgZ4z0Z9UjuLSZtNtwcW41LYPsmoJu+9BIm0OBhiAgODGRX/BK79k/WNTvrr9tD41QzX/iHxHNPN4a+3jdK3nMfP1Fwed8pZ1Qn+Eu2CHQjxb/AMiprP8A2D5//RbV/wAErPg74a+MH7WWnjxZaQXlh4M0ifxALOdNyXE0UsMMII77ZLhJMHgmMA5BxVf8Fq/groU3gvwh8ftM0+CHWbTUxoWqTIu17q3ljklhL+vltE6g9f32OQBj9jj4/wB6v/BOCx+LWuTi81D4f+GdTim81v8AW/2cJRAhPUkwxwj1JPvX/BOr4/8AwA+Efxf8afHP9pbxXdnxTeRY0e4bTJr15J7mSR725JjU7JDhFB6kSyDpnP8Aw9g/Yj/6KRqv/hO3v/xuvj18efhNqP7f/h39ob9njV55NKn1fStRvpBZS2Za6WRUul2OASJI1BY4wxlfqSTX/BZn4uar4K+Avh34Y6Ne/Zn8fatIL4q+Gls7RUd4wBzgyyW5PsuD96v+CWHwB8N/Cj9mTQ/H40yI+KfiHD/aWo3rIDILYswtoVbqIxGFfH96RvbFV+23/wApWdD/AOxg8Lfytar/AIKyfEqy8c/tg+GvhP4r12fT/B/gu1sodQeFTKbU3jJNdXARQSW8hoPlAJPljHWvDv8AwU//AGCPCWgad4W8M+Nb7TtJ0m1jtbKzg8N3ipBFGoVEUeX0AAFf8FDf20f2Pf2mf2dL7wl4K8WXmpeMdK1G1vtBWbQ7mDDiVY5x5roAoMDykgnBKrxnGP8Aglh8WJW/YUvtW1+4ee2+HGp6rbAySYIt4okvNu49ABcEDPAAA6DFf8EzPAX/AA0z+2drXxc+JyLqsmgR3HiW4Ew3JNqUs6iHcD2VpHkUdjEnaq/4LcfDbw1bR/Dn4s2VlFBrl5NdaRfTImDdQoqyQ7j3KHzQD1xJjoBj/gm9/wAmR/Cv/sH3P/pZPVft+f8AJmnxZ/7F+T/0NK/4IwfBPw34y+JPjD4w+I9Ot72fwRb2ttoyTKGEN1c+aWnUdnSOHaD280kcjI+IHiy28BeAvEnjq8RXt/DmkXWoyqzbQywQtIRntwnWv2A/2gfgb8P/ANoHxf8AtC/tQ+KLk+IbqKSXSZjpk14XvbqVmubj92rbHVQVGe07Y6cf8PYP2I/+ikar/wCE7e//ABuv2uf2hPhBrf7avh/9o/8AZ01m4mtYW0zVNSnFjLZH+0LeYiQbHALBoYoSxxhi7Zyd2f8AgqN8Jx8Uv2PvFN3aW3naj4Kli8QWmEyQsGVuDnsBbyTN/wABH1H/AAR6+Jh8Z/sqv4IurgNdeBNduLKNCcsLafFxGx9t8s6j2SviwLP9rf8A4K1eHvhzcQpqXhT4ZxrDexN80TfZEa5n3AetzIsDeu0Cv+CVuuX/AMDP2qfi7+yj4infMkk/2VpTjfc6dO8ZKD/ppDKz5HURLX/BbT4numg/Dn4H6ZOZJ9RvJtcvoE+9iNfItuByQzS3HHrGPw/Ze+E6fA39nvwF8LGgjiutD0aFb9YxgG8kHm3LD6zPIfxr45/tbfs9/s4xrH8WPiRp+m6jLHvh0mBWur2Qdj5EQZ1U9A7hV96+Ov8AwVs/Zc+Ivw18X/C+y8A+P9Tg8SaNeaabi4sLWGIGaFkWRc3Bf5WYNnapBUEc1/wQ91y+h8Z/FXwyshNndaXp92yE9JIpZkBHpkTNn6D0GP8AgtX/AMmr+Fv+ygWf/pBf1/wRu+Cfhvw58Cr/AONtxp0E3iLxfqc9rDeMoZ4LG3YIIlPVd0qyM2MbsJn7or/goj8T9R+E37H3xB8Q6Jdm31TULSPSbORZNjKbuVIZGU9QyxPKwxzlR06j/gip8FNCj8I+MP2gNSsY5tYudSOg6XLIuTbQRxxyzsnoZGljUnriLHAJzX/BVX4beGvhx+1/rf8Awi1lFZ23ifTLbWp7aJNqJcS70lIH+28RkP8AtOaqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv+Cs3xm8c2H7YNvpXhPxZrOhP4N8O2lvby6bfSWrpLNuuHdWQggkSRKcH/AJZitf8AjBrX7LH7A2mfE7xl4juvFHirTfClmy3Wq3TTyX+qXSrsDucs6rLL9fLTqMZrUr74ifGfx3eavef2z4u8W+IrmS4nMUL3V1dykFmIRAScAdFGFUYAAFf8E8/2jPFP7O37SPh7RrrUrq38LeK9Ti0fxFpszssamVxEk7IeFkhkZWLYztDr/Eaqqqqqq0n9jz9n39nz4ffELxD8OvAdtHr2paPqk02sXv8ApF0okilYxxuRiGMA7QsYUEAbtxyT/wAEQ/8AkqfxL/7F+1/9KDVa34E8GeJde0TxR4h8LaXqWr+GnlfR7y7tVlksHlCiRoSwOxmCINw5461X/Bbn4d+F9M8RfDX4nadZwW2ua7Df6fqTogVruO38hoXbHJZBK67jk4KDgKK/Y18Q6x4q/ZS+FGua8ZWv5/ClikrysWaXZEEWQnuXVQ2f9r8aqv8Agsl/yaLaf9jhY/8Aom5r9gP/AJM0+Ev/AGL8f/ob1VVX/BG//kv3xk/7B6f+lbVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX/BXn/kzTUv+xg07/0Nq/4Jvf8AJkfwr/7B9z/6WT1VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX7ev/KUjwL/ANhDwz/6UJVVVf8ABWD/AJMj8Yf9hDTP/SyKv+CXH/JiXwx/7i3/AKdryqqqqqqqqqqqqqqqqviL8J/hn8XdLttE+J/gXRfFNhZ3H2i3ttVs1uEil2ld6hgcNtYjPoTX/DFH7Iv/AEbh8P8A/wAEUP8A8TX/AAxR+yL/ANG4fD//AMEUP/xNf8MUfsi/9G4fD/8A8EUP/wATX/DFH7Iv/RuHw/8A/BFD/wDE1/wxR+yL/wBG4fD/AP8ABFD/APE1/wAMUfsi/wDRuHw//wDBFD/8TX/DFH7Iv/RuHw//APBFD/8AE1B+xd+yXbTx3Nt+zr4BjlicPG66HCCrA5BHHXNVVft0i9/Z2/4KX23xaubOX+zn1fRfFFsqpzPDGIlnUdAcyW869e/Jzk1ous6T4k0ax8QaBqNvf6bqdtHc2d1buHjnidQyOrDqpUgg+9ajqFhpGn3OrarewWdlZQvNc3E8gjjhjQFmdmPCqACSTwAK/ZMaf9p7/gqJcfFHSYpG0aHxDqniVnZPmhso962u4c4JZ7ZD6biR0xX7Qv8AyQH4mf8AYn6t/wCkctf8EQ/+Sp/Ev/sX7X/0oNV+2F/ylqb/ALHDwp/6S6dX/BUjwZf+Mv2LPGx0uCSe40KWz1Ty0HJjiuEErH2WJpHPstf8Ervgn+yr8fPglrWn/ET4XeHfEHjTw1rbreSXiM07WkyK0D4DD5dwmQcdYzX/AAwH+xp/0bz4U/78P/8AFV4M/Y2/Zd+HnifT/Gngr4J+G9J1zSpTLZXsELeZA+0ruXLEZwTXxS+JfhD4OfD7XfiZ471RLDQ/D9o1xdSsfmbHCxoP4ndiqKvdmA71a/Cn4+f8FPPGfxb/AGhYVNpH4bsCNEsGUyJK6Hdb6VAeBu8kSMzgY8x1LAebkf8ABJX9r+XxboH/AAyt8SL908Q+GLdj4YluSQ9zZR58y0Oed8A5UdfLyMARc+Lf+RU1n/sHz/8Aotq/4I3eKNE8Pftb3umatfJBceI/B97p+nKxx51wJ7a4KD38q3lb/gNV/wAFrPiLpGlfBHwf8LluIX1fX/EQ1HyS2XjtraGRWfA6ZknjAJ4OGxnBx+yp8FNcuv8AglTf+BFsphrPjjwprd7aW4BDM90JjaY9Q6CBu3D/AI1/wSb+HP7O3xg8VeO/hx8avAWieIdc+y2t/oS6ip3eVGZEulTkcgvA2OuNx6Ka/wCGA/2NP+jefCn/AH4f/wCKqz/YO/Y90+8gv7L9n7wrHcW0qyxOIHO11OQcFvUCv+C3fgfUtU+G/wAM/iHbW8klp4e1e+0+5ZRkRm8iidC3oM2hGemSB3Ff8E1/iloXxO/Y/wDAsWl3URv/AAnaf2HqdsrZa3ltztTcP9uHypB/v+xqq/bedI/+Cq2iySOqquv+FyzMcAAC1yar/gpH4e0XwB/wUVsfHHxB0U3/AIU1+XQtXv7aVC8d3ZQCK3uIsDkgraupUc/N7itO/YT/AGJdX0+11bSvgN4NvLK9hSa2uIEZ45o3AZXVg+GUgggjgg1/wwH+xp/0bz4U/wC/D/8AxVQ/s/fDnwZ8GfF3wi+E3g/TvDOl+JNPvo2tbFfLRri4t/JMhJJ+YqEGT2UV/wAEd/HNj4A/ad8R/DfxQFsL7xRoktnbJcfI4vbaVZDDgkclBMcdcoBVf8Fv/FuhDw38MfAa3qPrLX15qT2ytlo7cIkauw6gM5YL67H9K/4Jvf8AJkfwr/7B9z/6WT1X7fn/ACZp8Wf+xfk/9DSv+CHX/IqfFv8A7CGlf+i7mvjb4RuviB8GPHvgOxUtc+JPDGpabCoOCXntpIlwe3Liv+CUnw9+APxR+Jnjb4W/HPwFo+vavdadBd6FDqqMGja3eRbqNBkfOVljYr1xCx6Ka/4YD/Y0/wCjefCn/fh//iqj/YG/Y3ikWWP9nrwnuQgrm3c8j23V4g0LS/FGgal4Z1u2W507VrOWzu4WGRJFKhR1PsVYj8a/4J0fEJv2SPj/APHn4V+PLgpY6BoWo3d0rME8+fR5HI29vnhecgjOeOtf8Edfh/q3ia7+KP7VPi5TLq3ivVH023nI27yzi6vHAPJDSPAAemY2HPb9uS0b9lv/AIKQfDb9pSz222jeKJrS71GRRtGYttnfqB0GbV42J7tISe5PjbS5v2r/APgsBbeHmButA+HV5brPuXcsdvpiCaVGH9171mj/AO2tftEfEuf4N/Arx38UbONJbzw3oN1d2ccille4WMiEMB/CZCgPtmv+CZ/7OHhb9sD4n+PfjT+0NPceMDoVxbyPZ30zFdRvbnzWMs5GCyIIuI/uksARtXafiT4A+Enw4+B/jWTR/A/hXw5pOm+Gb+RvsmlwWsUCJbyHOEUAAc1/wRD/AOSp/Ev/ALF+1/8ASg1/wWr/AOTV/C3/AGUCz/8ASC/r/gk//wAmR+D/APsIan/6WS1/wVI8GX/jL9izxsdLgknuNCls9U8tByY4rhBKx9liaRz7LX/BFT4j6NqvwS8X/CxriFNZ0DxCdS8ncA0lrcwxorgZycSQSAkDA3JnrzX/AAVy8W6F4p/bD1Cz0S9juW8PaDZabesjZCXC+ZKyZHdRMoI7HI7VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVf8FlPhZq/hj9o7TPigLRjo/jXRYkS5GSPtdqPKkjPoRH5Dds7j6GvjX+2f8bvj98LPB3wg8e6jpn9g+DEjEH2K2aGW/kji8qKW5Jcq7pGWUbVQfOxIJOa/wCCLfwDl0vw/wCK/wBonxBpBSXWGGjeHppYyCbeM7rqVM9VaQRIGHeGQetftKSWniv9vnxpD4LX5rv4ita25iXbuuRdLG7Lt5OZgxyOuc96qqqqqq+LP/JK/GX/AGL99/6TvX/BEP8A5Kn8S/8AsX7X/wBKDVUxVVLMwAA5J4xX7ZPj/Vf+ChX7Z/hr4I/Bqcah4f8AD8z6Tp9/GN8LEsGv7/PTylEYAI++sCkZ3gV4K8JaN4B8G6F4F8OwmLS/Dum2+nWUbHJWGCNY0BPrtUVVf8Fkv+TRbT/scLH/ANE3Nfspf8FMv2WvhD+zn4B+GnjLV/Ecet+HtJW1vUt9HeWMSBmJ2sDyMEc1/wAPef2NP+g54q/8EL//ABVf8Pef2NP+g54q/wDBC/8A8VXwJ/4KB/s5/tG+P4fhp8NNU12fW5rWW5RLzS2t4/LjALfMSecEcVX/AATT/ZR+P37P3xe+Jfij4u+Af7B0zxBZrHps/wDalndeewuWfG2CV2X5SD8wFVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV/wAFG/gx8Svj1+zPffD74T+G/wC3dfm1iyuEs/tkFrmONiXbfO6JwCOM5r9ib4b+NPhF+y14A+HHxD0b+yfEWiWc8d9ZfaIp/JZrmV1G+JmRsq6n5WPWqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq6urWxtZb29uYre3t42klllcIkaKMszE8AAAkk9K+PP/BTb43fGP4lP8E/2FPC91dMJZIU1uLTlvbzUNhw0sEbho4LccnzJATtwx8vkV4z/AGBf+CmHjbx/pvxG8WeHrrXPE+YJ11qbxZYGaweN8xgsZwwMZAYeWGUAjaSeB4b/AG5v25f2LPG9j4N/bC8Hal4l8O3T4Et7HF9paMYDPaXsP7q4KgglJCx5ALR5yNW/4Kz/ALGuk3gsz4q1+7zDFKJbTR3lTEkauFyD95d21h1VgwPIr/h7z+xp/wBBzxV/4IX/APiq/wCHvP7Gn/Qc8Vf+CF//AIqv28/+Ch/7Nv7Qn7NHiH4W/DnVdem13UbuylgS70poIysVykj5cnj5VNf8EuP+TEvhj/3Fv/TteVVVVVVVVVVVVVVVVVVVVVVVVV+3j+xXpP7YPw/s7fS9RtNG8b+G2kk0PUrlGMTq+PMtptoLeW5VSGAJRhkAgsD8OvH3/BUX9h3Tv+FbQfCDV/E/hqyZ1sba90KfW7G2BPWG4s3DIpPIjMgAyfkBJrx148/4Knftv6WPhy3wk1Lwh4Z1MeXfR2ujT6FZ3MYOG864vHLunBzGjkNjG1jgV+w1+xR4c/Y+8C3UFxqFvrfjfxAEbXNWjjIjATOy3t9wDCJckknBdvmIACqvxx0rUdd+CvxA0TRrKa8v9Q8Lalb2ttChZ5pXtZFRFA5LFiAB6mv+CRHwG+NHwh+I/j/Uvij8L/Enha11DRLeK1m1XT3t1mdZiSqlgMkDnFV+1J+zj8efFX/BTI/Ezw38IfFWpeE/+Ep8NXH9s22mSSWvlQW1isz+YBjahjcMexU+larpWm67pd5omsWUN5Yahbvb3VtMgZJonUq6MD1UqSCPQ18Sv2Ev2tv2M/i3c/F/9i+91LXvD7u/k2tiVmvLeBjk2tzavxdR8ABlDE7QxCMAasf+CjH/AAUbhhj0W6/Y5e71eNTEzf8ACGawjPIO5iD5z6gEc5xjoP2I/iD+3L8RvE/irxB+1R8PofC/hq5soP7BtRZR2RguFdvMVYWdrjDIwJMpwNgA6tX/AAUj0r9q79p/4naN8BPhb8H/ABfH8PtI1CJZ9Xm02WGz1C/c7DcPKRgW8KuQG6E735Gwj9nb4GeFP2cfhB4f+EvhGNXg0m3Bu7vZte/u25muH6nLvnAydqhVHCiv29/2L/jF8P8A9orRv2mv2TfCetahd6vqH9oXlrodq00umapGQzTbFzmKfksMEbvMDcOor4dePvE/xO+AcXjDxt4D1XwZ4iutHnXVdE1K1eCS1uVjZZNgfloyRuRu6sM8ggfsY/s/eNv2iPiNr2ifDHxc/hzxp4T8OP4i8PXazNAGvLe8tUVDKvzR5WZyrjowXPGavP23v+Cofwvs18D+Nv2WRrWuWymCPWB4VvbhbpgAN+6zk8iVs8nytqnI4Hf4W/sL/tWftn/GeH41/tkJqPh7QPNja4tdQT7Nd3UCHK2dvagg2sWCQWcKRuZgHYsa07TrDSNPtdJ0u0itbOyhSC2t4UCJFGgCqqgcAAAAD2r9rj/gnN8ZvBHxgm/aW/YuvJYr+W9bUJtFsblbW70+6fJlktSxCSQvli0RORuKhXU7V0n/AIKEf8FJ/Cdqnh7xh+yhc6vqdsdjXdx4L1O3lm7bmWJhGcnuiqp7Cv2UfjT/AMFFvjB8btM1T42/BuPwV8MltLlL+3bSP7MKS+UTA4S6drl23hVIXC/OSQMCvjD8JvBnxz+G2u/Cvx9YtdaLr1t5M3lkLJCwIZJYyQQsiOqspwRlRkEZFSfso/8ABQD9gT4i33i39ni31Dxj4fuTtafQ7X7amoQgtsS60/Jk3qCTlFYKWO2TJNW/7fP/AAU58W2o0Pw1+yF9n1Mnypb1fBeq7Ymwef3svlxnj+Mke1fsz33xm1L4G+Fbz9oPTTY/EF7eT+2YiYOX85/LbEB8tS0XlkqMbSSCOK/4KofsW/F74g/ETSf2ifgj4cv/ABDcRafFaaxp2mAvewSwMxhuYox80gKsqlUBZSinBBJX9kT9sD9v34m/Fzwf8Ovi38BHtfCZaWLXfEcvhC/sJlC28hR5JnfyEZpFTIEa56ADNftzfsY6D+2F8O7XTLfUbfRfGXh2R5tB1aaMtGN4Akt5goLeU+1TkAsrKrAEblb4efET/gqJ+w9p6fDa6+Duq+NPCmmsY7GK40ifWbW2jzgCC6tG3RoeNsbthegRTkUP23v+Co3xSh/sL4dfsqf2FcTkxjUm8KXqCF8DpLdyCBSMg/OD27V8LpfG8/w08KSfEy0jtfF50W0/t6FJEdUvvKXzwGj+QjzN33ePTiv27/8AgnJ8T2+KNz+0v+ylDJJqVzeDU9R0WwmFteWt8rb2u7RsgOWYbygIcOSV3bsL4V/bf/4Kq21lB8Pm/Zyuta1uFPJbUtS8BahHdk8DfIUeOAdR8xjC85PWvj9+wZ+1N4v+DV/8evjFHrvjr41+I9fskj0XSEF2dK00Qz+YjrCPLHzmP5Ivkj2jBJdgP2BvCnifwP8Ash/Dnwn4z8PajoetabZ3MV3p+oW7QTwN9snI3I2CMqQw9QQehqv21fC3iPxt+yp8S/CnhHRLzWNZ1PRHhs7GzhMss7l1+VFHJPB4r/gkD8Gviv8AB/w38Tbb4pfDzXvCsup32nPZJqti9ubhUScOUDAZALLn6iq/bN/4JyfFjTPi5P8AtN/sdXskGtT3p1G70SzuhaXVreHJkuLR2IVlcks0RIOWYLuVti6T/wAFA/8AgpZ4JtF8O+N/2VrnWtStRta9uvBmpW8s2ONzCErE2SDyiqvpX7MHxx/4KQfGP45aBqHxc+Ccfgr4Ywrcrq8D6OdLbmB/JbF47XDsJRHxGAOSSAOlf8FJv2SvjpeftR6j8TPgV8NvE+u6d400KNtTn0WwedIrgxtazwuFBxviSNjnqZCevNfsifB4fAb9m/wF8MZrZYdQ07Skm1RduD9tnJmuAT3xJIygnso6YwP+CqP7O/iL48/s9Wd/4D8M3mueLPB+sRXVlZ2NuZri4t5v3M8aKOT96KQ47Q1/wSe/Zs+KHw81L4kfGj44eFdc0bxT4huE0+1XWoGhuZkLfaLqdg/zMJJGh+buY269viX4C0X4p/DzxJ8NvEW8ab4n0q4065aP76JNGULL/tLnI9wK8B/Dz9u3/gmr8WNcu/AvwqvvHPhvV1WC5nsNKnv7DVYEdvJlb7PmS2mXc+A+CC7DDqQTc237f/8AwULW1+H/AMSPAB+CPwjlnSTXpGs5bW91GJWBESpcHzJMlcr8iRjhmL4VT/wSU+BXxt+C3xf8ex/E/wCFXiPw5Y6hoCR22oahYSQQyyxXK/IjMoDFlcsOQcKeOuP+CtPwu+Ivxb/Zx8OeG/hj4J1jxRqlt42tbuaz0u0a4ljgWyvUaQqoJChpEGfVh61/wTW8BeNPhp+yL4W8IfEDwvqXh7W7W+1B5rDUbdoJo1e6kZSUbkAqQR7GtZ0bSvEWj33h/XLCC+03U7aS1vLWdN8c8MilXRlPVWUkEehr4p/sNftc/sV/GGX4tfskJrfiHQhI/wBgm0aD7XewQOcm0urTDNOnygblV1barHY2ANJ/a8/4KwfG21HgPwX8A4PDl5dKIX19PCd3YGDIwXae9la3Q45+7n+6OlftT/8ABOL43fD1PBY8I+F/FfxN8V63Y3V/401rTLSa8hOoSTltqsV3fdPLP8znLkLu2iGRZ4UmVXUOoYK6lWGR3B5B9qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqtT1Gx0fTbrVtSuVgtLGB57iVgcRxopZmOOwAJr4f/ABb+F3xY0/8AtT4Z/EPw94ntwgZ20vUY7gxA/wB9VJZD7MARXxl+C3w2+P3gO9+HHxS8OQ6vo14Q6qWKS20q52zQyD5o5FycEdQSDlSQdH/4Isfs62HixNW1Px/431PQ4pd66RJLBEZACCEknSMMVIyDtCNzwwPNaF4U0Hwp4Xs/BvhHToND0nTbQWthbWESxraRquFEa4IGOvIPPXNeCv8AgkZ8MPAHx38L/GHRvin4j1Kw8Pawuqy6RrVrFcS3U8ZLxE3KGPGJgjnMbbgpHGc1VVVVVV8Wf+SV+Mv+xfvv/Sd6/wCCIf8AyVP4l/8AYv2v/pQaqv8AgoP+3drPxF1g/skfsnXdxr+o65I2n6/qejoZnu2f5TY2jLncCNwlkXjHyg431+wN+w9of7JPgd9W8QG21P4jeIoF/tm/jG5LOPhhZwH+4rDLN1dhnoqgVVf8Fkv+TRbT/scLH/0Tc1+x1+wn+yZ8SP2Yfhz458bfBnTdV13WdGSe+vJL26RppCzAsQkoUcAdAK/4du/sR/8ARAtJ/wDBhe//AB6v+Hbv7Ef/AEQLSf8AwYXv/wAer4W/scfs0fBPxZH45+FvwpsNA12KCSBLyG7uZGEbjDriSRl5AHaqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr4k/tOfs9/B/Uk0X4lfGLwtoOpOM/YbnUENwo7M0S5dVPYkAGvG//BTX9i7wTYi5Pxfh16dwTHaaJYz3cjY/2toRf+BMOtftf/8ABV/4UfF74C+K/hT8G9C8c6brniRFsZNR1OztoIBZlx9oCmO4d8yRho8FB8shzg1/wSl+BPhn4ZfswaN8RI7C3fxN8Qg9/f3uwGRbdZGS3gDf3Ai7yOPmkbOcCq/aa+BPhn9o74LeJfhb4isLeaW/s5H0q5kQFrG/RSYJ0PVSr4BwRuUsp4Y1/wAEnfgN8Av2hIPiN4f+MfwvsfEd7oDWF1p13NczxMkcwmWSM+XIowDEhGQT8zV/w7d/Yj/6IFpP/gwvf/j1f8O3f2I/+iBaT/4ML3/49X/BRP8AYv8A2Yfg1+yn4l8ffDP4S6fofiCxvbCO3vYbu5kaNZLqNHADyMvKsRyO9f8ABLj/AJMS+GP/AHFv/TteVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV4t/5FTWf+wfP/AOi2r/gip/ydR4p/7J/ef+l9hVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVXi/Qn8UeEtb8Mx3K27atp1xaLMy7hGZI2TdjIzjdnFfED/glV+2Z8Hr//AISL4dx2HixbImSC98Maqba8iHr5cvlvu/2Yy/XrXhr9vj9vz9mXVofDPj7Wdduvs4H/ABJ/H+kySSOAeplkCXBHbIkxzXwo/wCC2fgrUTFY/Gr4R6no0mAG1Dw9dLeRE9yYZdjIv0eQ18Kv21v2WfjQIIPAnxp8PSX9xgR6bqM50+7Zv7qw3ARnP+6GHvjmqqqqqqq+LP8AySvxl/2L99/6TvX/AARD/wCSp/Ev/sX7X/0oNV/wVW/bo8XeC9W1D9lr4XtdaNcTWUb+JdbV9kskM8Ydba3IOVVkYb34JztGBuJ/Yj/bC/Zx/ZE0u412/wDg74j8UeP9TQx3WuG4gjS1gJ/1FshyUU4BZidznGcABR/w+8+FX/RD/Ff/AIMbf/Cv2Wv2i9E/am+E1v8AFjw/4cvdDtLi+ntBaXkqySBoiAWyvGDmqr/gsl/yaLaf9jhY/wDom5r9gP8A5M0+Ev8A2L8f/ob1VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVeN/j98Dfhq08Xj/4weDdAntziS31DW4IZgfQRFt5PB4Azwa8bf8FS/wBizwjY3z2HxTk8RahaQyNFY6VpF25uHVSVRJmiWL5iAoJfAJBJA5rxl/wXA8LQwSRfD34C6reTMMRzazrMdsqcdTHEkhbntuX618Q/+CvH7XvjJpYfDGqeG/BVs+VVdI0lZpQvu90ZefdQvtitc/bO/a08QzGfUP2kPiHGx6rZ+ILizXoB92FkGOOmPX1Nar8afjJrtv8AZdb+LXjPUYcEeXd69cyrg9eGcjmvAvwy+JPxX1d9H+HPgfX/ABTqIBeWHS7CS6dR1LNsB2jg8nFeGv2Af2yfFmoDTdN/Z88U20hxmTU4E0+ID18y4ZF/AHNfE7/gl9+1L8JvhdrXxX8UWHhqXTvD9n9rv7Kx1Nri6jhDAOwUR7CEUl2IfhVY84xX/BKz41+Gfib+ytoHgm0v4R4i+H4fTdUstwDrGZHe3mC9djRsF3f30cdqr9on40+Gv2fPg34m+KfiW/ggXSbKT7DDIwDXl4ykQQID95nfAwOg3E8Akf8ABET4eavYeF/iX8Ub23lj0/WLuy0vT3YYWVrdZZJyO5A8+IAjjO4ckcVX/BWD/kyPxh/2ENM/9LIq/wCCXH/JiXwx/wC4t/6dryqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr2zg1CznsLpS0NzE0Uig4yrDB57cGvgP+xH+zr+zV4vu/HXwi8JXul6xfaa+mzzT6rcXQa3eSORl2yOwB3QxnOM8e5qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq8S+FPC3jTSZdB8YeGtK13TZhiWy1OyjuoX+qSAqfxFfFf/AIJU/sgfEwT3Wk+D7/wPqU2WFz4bvDDGG6j/AEeUSQhc9kReOhHWvir/AMEVPi5oXm3vwf8AiboHiq3XJFnqsL6Zc47KpBkjc+7NGP5ULv8A4KP/ALEzlZH+I/hXSbAYxIDqmjqB6E+bajgducV+xz8U/Fnxt/Zn8CfFHxzLaya7r1lLJevaweTGzJcSxghATjKoM9s56VVVVVVXNtb3lvLaXcEc8EyFJYpEDK6kYKkHggjgg14V+Gnw48C3E934I+H/AIb8PT3SBJ5dK0mG0aVQchWMagkZ5warxD8H/hJ4v1WTXfFfwt8I61qUyqsl5qOh29zM4UAKC7oWIAAAyeBX/DPXwB/6Ib8P/wDwmbT/AON1/wAM9fAH/ohvw/8A/CZtP/jdeHPCvhfwdpi6J4Q8N6Xoenq7OtpptnHbRBm+8wSMAZPc4qq/4LJf8mi2n/Y4WP8A6Jua/YD/AOTNPhL/ANi/H/6G9VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVUxVVLMwAA5J4xXjP9pz9nT4eF4vGnxx8D6VPGSGtptcgM/HXESsXOMY4WvG3/AAVj/Yv8IrKumeNtb8VzRA5h0TRJsk+ivcCFD9Q2Oetax/wW/wDhtBcbPD/wI8S3sO4/PeavBatjjB2qkgyecjPHHXPHj/8A4LN/tJ+INVaXwF4V8IeE9MVsxW8lu+oT4x0kldlVv+AxpXj7/gpz+2h8T9Pg8NwfEUaAspEZHhmwWzuLhzwMSrmVTzwI2XJPTpVxqX/BRLWrGXRbrUP2jL+01AFJbSSXWZY7jf1DISQ+7PQg5zXw6/4JU/tkfEWyg1S88GaX4Rt7ogo3iTUhbyYP8TQxCSVMdw6BvarH/giB8TJdDu7jU/jr4Yg1hLaRrS0g0qea3lnC/u0eYsjRozYDOI3KjkI3SvDH/BEb403c8a+NPjJ4J0qEn94+l291qDKPZZEgB7dxXw7/AOCL/wCzr4d8u4+IXjXxb4wuExuijkj022f1ykYaQfhL/jXh7/gn/wDsZ+GLcWum/s9eFpkC7QdRjkv27fxXDu2eOuf5mtM/Zg/Zp0WRZtH/AGevhrZyqoUSQeE7JHwOmWEWT0FaZpGlaJZpp2i6ZaWFrH9yC1gWJF+iqAB0FVfWNlqdjcabqNrFdWl3E0M8EyB0lRhhlZTwQQSCD618bP8AgnN+0b+zd8S7j41/sL+I9QlsyzyLpNpdrHf2KM25oAkn7u7t+FwjZY4AKORuNx/wVE/4KBeDNeg+FHir4R+Hz4zuGjigstT8JX0OpTvJgR7LdJkDljyu2Mhs8ZGK0j9kf9vv9vLxnp3iX9qTWtT8IeErOUMI9VhW1eBD95bXTkA2yEYBeVV45LPjafhX8L/BvwY+Huh/DHwBpgsdD0C1EFtGTl37tI7YG6R2LMzY5Ziaqv8AgrB/yZH4w/7CGmf+lkVf8EuP+TEvhj/3Fv8A07XlVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV+0x/wUF+C/wCyp8S9G+GnxF0rxFe3Oq6WNQluNJgimWzjaRo08xXkQncY5D8uSABxyK+H3xR8GfE34b6X8WPDOoyL4a1eyN7b3V/A1oVgGcu6yAFFwpOTxjkEgg18cf8AgtO2j+LrzQfgJ8NdN1jRrCZok1vXZ5VF9tOC8UEe1kQ4ypZtxBGVU8V+xT/wUk8EftYa4/w18S+GP+EQ8cC3ea2sxdfaLXU40XMnkuQrK6qCxiYH5QSGbDbbDTtP0q0Sw0uxt7O2jLFIbeIRopYlmIVcAZJJPqSTVVVVVVVVVVVV/wAFkv8Ak0W0/wCxwsf/AETc1+wH/wAmafCX/sX4/wD0N6qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqp3SNDJI6qqglmY4AA6mm+OfwSWa4tW+MfgcTWis1xGfENqGhCnDFh5mVAOAc9K8Y/wDBQj9jLwOJBq37QHhu8dAcJo5l1TcR2BtUkH45x745rxh/wWY/ZZ0JXi8L+H/HPiacBvLeDTorWEkdMtNKHAPshr4q/wDBa34r62/2T4O/C/QfDFsAQ11rE76lcN6FVXykj+hEn1rxZ/wVL/bW8W2/2GD4n22iRuMMukaPbQu30dkZx/wEio/Af7ev7S3l/a9E+MXje0uOY5tTe9ks8HniSciFR7ZArwX/AMEhv2xfFQjfXNH8K+Ekfk/2xrayMo+lqs3Pt69cV4L/AOCHuoSBJ/iJ8f7eEgDfa6LoZkye+JpZVx/3759q8P8A/BG39kvSYFTV9S8d63Nsw73OrRRAtxyFihTHToSevOetfC/9gv8AZJ+Eds0fhn4J6BfXMi7ZLzXYf7Vmb6G43hPogUV4V+C/wc8Cak+teB/hN4N8PahKzNJd6VoNtaSuW+8S8aBiT355qqqqqqqq/aj/AOUwfwy/7CHh7/0Oqqq/4Kwf8mR+MP8AsIaZ/wClkVf8EuP+TEvhj/3Fv/TteVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX/BW2XUJP20/EKXrMYYtH01bMMOBF9nUnHt5hk/HNftlftVfDPwb/wAE+vB/w3+EnjPSLvUvG/h7TtItrXS7tZHsbCKCMXO9VbMfCfZyrc5kYY+VsfsSfsc65+2P8QtT8MweIj4d0Hw/Yi61XVhafaTGztthhRNygu5DnJIAWNzyQAbLRfEX7JH7Y9joS60t3qHw78aQRG9t0MYu4UnXJ2ZO0SRHDJk4DlcnrVVVVVVVVVVVVX/BZL/k0W0/7HCx/wDRNzX7Af8AyZp8Jf8AsX4//Q3qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq1vxH4e8NWyXniPXtO0qCR9iS3t0kCs3oC5AJx2r4qftn/ALLvwZs1ufHXxo8OxzSLmOy0+4/tC5cdj5NuHcA/3mAXrzwa8Z/8FqP2edHd4PBXw98beIpEB2yzxwWELnHGGMjvjPqgx7141/4Lf+O7tGj+HPwI0HSmAIWXW9Xl1AN6EpEkGOMcbj9a17/grp+2brCTLp3iTwxohl+41hoMTmLjHy+f5n1+bPPtxWqfFH9uT9q+N9Bk8Q/E/wAfWV6+2XTtMguHs5Dn+OG3UQ4BHVlwvPTmrD/glD+2rqGk2OqD4faTbteqGa0uNdtkmtgenmLuwDjnAJI6EA5FeDf+CKXx+1ZI5vG3xL8E+H0dcmK1NxfzRn0ZdkaZ+jnt+HhL/giH8KbJkPjv43eK9ZA+8NK06DTc/wDfw3GK+GP/AATN/Y4+F97Bqtt8LU8Sahb/AHLjxJdPqC59TA2ICfcx8dsV4d+Ffww8H3Au/CXw48L6JODkS6do8FswPrlFB7Cqqqqqqqqqqq/bx/Yq+If7WmreDtR8D/FC08JJ4btruK4SdJj9oMzRFSPLI6eWevrU3/BFf4z3F4moz/tBeHpLuMjZO9ndGRcdMNnIxX7In/BN/wCLX7Onx10X4reK/jZYeI9N0y2uopNPijuA0hlgeNSN7FeCwPPpVVX/AAVg/wCTI/GH/YQ0z/0sir/glx/yYl8Mf+4t/wCna8qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq+LnxY8EfA34eat8UviNqUth4d0TyftlxFbPOyebMkKYRAWOZJUHA4znoK+An7R/wl/aY8N6h4r+EOvXOqabpd99iuZJ7GW1KTbFfAWRQSNrqcjjmqqqqqqqqqqqqqqv2gP2pfgx+zDZ6Lf8Axi8RXWlQ+IJZorAwafNdeY0QQvkRK23AdevXNfDj4heFfix4G0b4jeCL6S80LX7YXNjPJA0LSRkkAlGAZeQeCKqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr/AIKtfsU+Mfjba6V8dPhHokmr+I/Dlg1jrGk2yFri+s1cvHJAg+/JGzy5QfMysMZKBTo/w68feIfFieA9D8Fa5feJHl8oaTDp8jXQfIGDEBuHJAORx3r/AIJ+fsrT/skfAmWz8Y+SfGHiOb+0/EJgHmi22piK2VlzvEa7icZBeSTaSME3Fp4k/aN/b2jF54d1KyufG/xDWb7BfW7QTWtk90G/eIQCPLt1y3GcITVVVVVVVVVVVVX/AAWS/wCTRbT/ALHCx/8ARNzX7Af/ACZp8Jf+xfj/APQ3qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqrxN4+8CeC1DeMfG2g6EGXcp1PUorXIyBn94w4yQPxFeNf27v2PvAMbtr37QnhGdoyQ0ek3Z1RwR1BW1EhB9iK8f/APBZ39m3w8JLfwJ4R8Y+LbhM7JDbx2Fu/wDwORjIP+/VfEb/AILS/H/xCZbb4b/D/wAJeELaQHZJc+bql1H6YdjHGfxiP4Vd/td/t7fG3U5BonxZ+Jep3WSzW/hRZbTZjn7liqYGB6Vo37J37d37Suvf2prHw7+Iet3gjIOreMZprYbR2E98y7+RjCk89q8Bf8EeP2r/ABNqyW3jJvC/g/T1b99dXWpLeSbf+mcdvu3H2ZkHXn18Ff8ABE/4E6VHG/j34peM/ENwg+YWCW+mwufdCsz4+jivCP8AwS9/Yo8Izx3f/CpG1q4iPyyavq11cr17xeYIz26oen1rQ/2dv2f/AAwkSeHPgd4B03yB+7a18N2kTLxgnIjzk9z1NQww28KW9vEkccShURF2hQBgAAdBjtVVVVVVVVVVVVVVVVVVX/BWD/kyPxh/2ENM/wDSyKv+CXH/ACYl8Mf+4t/6dryqqqqqqqqqqqqqqqqqqqqqqqqv2mPhxP8AGH/gpD4t+Fltq0emS+KvG8enJeSQmVbcyiNd5QEFgM9Mj61/w458Vf8ARxWk/wDhNyf/AB+vgb/wSA8SfB/4xeDPilc/HTTdUi8K61bai9kmgPE1wInDFA5mO0nGM4NVVVVVVVVVVf8ABQL9u7Xv2OB4W0fwt8PtP8Q6n4rt7uWK6v7144bTyDGMNEi7pNxkHR0xt754+FnijUPHHww8IeNdWht4r7X9BstQuY7dSsaSzQJI4QMSQoLEAEk4xknrVVVVX/BUf/kxL4m/9wn/ANO1nX/BEv8A5ID46/7HA/8ApHBVVVVVVVVVVVVVVX/Bcb/kVPhH/wBhDVf/AEXbV+wH/wAmafCX/sX4/wD0N6qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqrV9Y0jw/pd1rmvarZ6bp1hC011eXk6wwwRqMs7uxCqoAySSAK+JX/BXn9krwJqU2keHp/FHjeaE7TcaJpyLbbh1HmXEkRYe6qwPY45r9kn9sv4e/th6R4i1PwH4a8Q6MfDE8EN5Hq8cK7zMHKGMxSPniNs5C4yMZofHv8AZo1r4j6L4Ltvil4E1bxtJNJDplnbahBd3cUnlsXVShYxMUDgglSQcc5waqqqqqqqqqqqv+CyX/Jotp/2OFj/AOibmv2A/wDkzT4S/wDYvx/+hvVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVfFv8Aap/Z3+BOpQaL8V/izoegalcIHSxkd5rgIejtFErOinBwzAA9jXxK/wCCxn7Lvg6/l0vwXpnirxxJE2PtdjZraWrY64edlkP1EeD614j/AOC4t46tF4Q/Z1hjOPln1LxKX/OOO3H/AKHXjv8A4KSftkeOdfvtaT4yaj4ft7pmEOm6JGlrBaoc4RMAucZ+8zM3+1XgrXf2h/Hnjy5v/hxrPxF8ReNdQtHW5m0S4vLvUbi3BVnDtEWlaPIUnORkAntXg/8A4Jwftu/FKZ9ZufhPqOnC5JeW98S38VlIzf7Ucr+cSfXZ9TXg7/gi5+0lrMscnjHxt4G8OWxPzhLme9nX6Isaof8Av4K8B/8ABEr4RaWY5viR8YPFPiGRTlotKs4dMjb2O/z2I+hB+lfDn/gnr+x18MRHLovwO0PVLpOTc6+H1Vmb+9tuC6Kf91V9evNaXpGlaHYx6ZommWmn2cIxHb2sCxRoPZVAAqqqqqqqqqqqqqqqqqqqqqqr/grB/wAmR+MP+whpn/pZFX/BLj/kxL4Y/wDcW/8ATteVVVVVVVVVVVVVVVVVVVVVVVVV4m/5TBw/9lQsv/Q4qqv2/wD9oL9pP9mLwVpnxT+Dfhfwbr3heGX7N4gTWNPupp7CR2AhmDQ3EY8lidhyuVYpyQ/y/sWftVaJ+1r8GrPxxGlnY+JtOcWfiTSrdiFtLoDO9FYlhDIvzpknHzKWYoxr9v79taH9kD4f6a/ha10nVvHniO426Tp2oB3higQgzXEyRuj7AMIoDLlnzyEYV+xt8Rvj98YPg5Y/E/4+6B4X0G78REXOi6botlcW7R2JHySz+dNLlpPvqBjCFSclsL4e/wCCl3jnS/20fFnwI+LkHgTRPh54a1LXYn1iCyu1u0t7KKeWIs5nZC5SFQwWP5mOFAJArXv+Ci/7Yvx41HUJf2Nf2bLu58J2UzRJreoaVLeyzEDruDrBE3IPl5kYcc84r4c/8Fbvjd8MfHi+Av2vvhAtnHG6pdzWmmzabqNkrf8ALVreVisq45wvl5ByCeAfC/ijw9428N6Z4v8ACer2+qaNrFrHdWN5btuSeJ1DKw+oI4PI6HFftd/tlfDT9kHwfbav4rjm1fxDq4caLoFpIEluyuNzu5yIolJUFyCcnCqxyBpn7XH/AAVZ+Mulx+PvhP8As+aXp3he5Uy2ONKH+kRHlWVrqYNMMDho1AOeB0r9l/8A4Kga74l+KEPwE/aw+H0PgLxdPcrZW1/HDNaxG6YgJBcW8xZ4WckYk3FSWHCg5qv2v/8AgqP8N/2d9fvvhp8PNDXxz44sHMN8guPKsdMlHWOWRQWkkB4MaYwchmVgRWn/ALX/APwVp8YwL4w8N/s4xw6O6GSG2HhC4RJUIJVlE0vmuMEEFTg8etfsjftJftIfGv4W/EPxD8XfgxB4U8S+Dne1060j0+7tzf3S27StG1rMTICCYRgOd3mYGMc/tx/HX9p3446r4Ruv2lfg3/wgFzpFvdJpEX/CO32lfa0dozK2LuRy+0qgyuAN3PUV8Pf24v8Agph4e8AeGdA8J/sff2nommaPaW2mXv8Awr7XJvtNtHCqxSb0nCvuQK25QAc5HBrxZ4q8OeBvDOp+MfF2sW+l6Lo1rJdX15cNhIIkGWY9+g6DknAAJNeIP+CmX7Snx+8a6j4M/Ye+A51ew0/Ik1bU7N7iYqchZWUOkNsGI+USMxOO3IF7/wAFDP28v2Z9b09/2sfgDZz+HdRlCi4iszZuTjlYbmJ5IC4HzeWy7jjqo5r4R/FnwL8cvh5o/wAT/hzq41DQ9ah3wuy7JInB2vFIv8MiMCrD1HBIIJ/a6/bF+G37IXguDXfFiSatr+rF00XQLWUJNeMuNzsxyI4lyu5yD1AAY8V4a/bx/wCCmfx28zxd8FPgTYP4ajmZYWs/D0s8EmDyhuJpAJGHRvL24z0HFftE/wDBQDxX8V/2X/if+zn+0b8LbnwD8T/J02WyjW0mt4L4R6layOhhmJkhfy0Z1OWR1ViCuAG/4Il/8kB8df8AY4H/ANI4KqcTtBItq8azbD5bSKWUNjgkAgkZ7ZFfs2f8FafE3inw/wDETxn+0RonhPTdH8H6Vbzadb+HLO4iudRvJpSiW6+dcSAlsE9BtAZicKa/ZP8A2+f2qv2lv2lvCejar8MLXQfhXrMt+s1xZ6NcSogjsp5YVe+f5C3mRoMqEDZxtGRX7Xf7ZXw0/ZB8H22r+K45tX8Q6uHGi6BaSBJbsrjc7uciKJSVBcgnJwqscgaZ+1x/wVZ+Mulx+PvhP+z5peneF7lTLY40of6REeVZWupg0wwOGjUA54HSv2X/APgqBrviX4oQ/AT9rD4fQ+AvF09ytlbX8cM1rEbpiAkFxbzFnhZyRiTcVJYcKDmv29/j78Xf2Zvggnxa+E2i+GtVew1eC31eHXLSeZI7aYMiyJ5M0ZDCbyl5JBEnqOf2Gv2k9U/aq/Z+074neJLHS7LxBFqN1p+r22mRulvFNE+5Niu7sAYZIWILHlj2xX7YPx7m/Zo/Z78U/FvTrewutX01IoNJtL4MYri6mlWNFZVZWZQGZ2AYHajciv8AgnP+3R4w/a+j8Z6N8StI8N6V4g8NtbT2kWi288Mc9rLvVmIlllJZHQAnIGJF46mvE/iLSvCHhrVvFmuXHk6boljNe3kv9yGJC7tz6KpNf8E9/wBub4//ALYXxL8U6X4y8I+CdJ8I+GtLFxJPpVldLci5llC28ReS4dMFEnYnYCTGMAZOP2v/ANtP45fB34qWPwN+Av7O17438T6hpMepxX0sM11B5Tu6cQW+HIVo3DM0iYx0wc14n/ar/wCCuHw30ub4geMvgJpX9gWkRmuYf7B8+OCJcFmkWC4MyKBkksQAMk4A4/YZ/by8LftiaRqOj3mhL4b8caBAk2oaWtx5sNxATt+0W7EBtgYgMrDKF0G5twNftSfE/wATfBb9n3xv8VPBttpdzrPhnTftdrDqcMktu5DqGDrG6MQVLYwwwcHnpXgr/grbqFj+yzqHxc+Kfhfw9dePr3xVd6L4c0DQ0mtYJ4obW0la4nMskrKiNckEg/NlFAHzOP8Agn7+1L+1B+0Z428Vn45+CYvD+gwaVFdaKINAnsYZHaXa2ySYsZAFI/iOODX7bv8AwUm8Gfss6k/w48G6LB4t+IPkrLPayTlLTSlcZT7Qy/MzsuGES4O0gllBXdoX7YX/AAVm1mzj+IWn/s6R3ugSr58dkPCFwiSRccxp5ouGUjkEE5zkZFft5/tnaD+1v8MPh5BceGLrwt408I6rqMHiDRZyWWJnSELJExAYqTG4KsAyMpByMM37Af8AyZp8Jf8AsX4//Q3qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv+CzP7RHiK88c6L+zdoOpyW2habYRarrkcRKm7upS3kxv6pHGquB0LS5OSq41/8AYj+O/hL9nY/tN+LtK03RfCji2e2tb25Zb+6juJFjikSFUIVTuDfOykqcgHIz/wAEOv8AkVPi3/2ENK/9F3NeLpk+Cn/BQXVLzRXS3t/CvxWee2VSQqQJqO5UOB08v5SAPWqqqqqqqqqqqqv+CyX/ACaLaf8AY4WP/om5r9gP/kzT4S/9i/H/AOhvVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV+0N8NNY+MnwT8X/C/w/rsejaj4j05rW3v5AxW3YsDuO35u3av2sP2B/jD+yd8Lovij4h+OEGvWsuqw6cLWyNzE4aRJGDZZsYHlnj3r4I/8Ewvjd8b/AITeGPixpX7QVlptp4nsVu4bS4W6eSEEkbWKtgnjtVVVVVVXjT48fBD4cTyWnj/4weC/DtzEPmt9T123tpfoI3cMTweAM18VP+Cnv7H3wx0x7mz+I3/CZ3+SseneGYDdOxHcysUhVc9y+ecgGvGH/BcKHyZIPAH7PzmU58u51jXsBeOMwxRc/wDfwdPfj4jf8FWf2yfH0rppnjjTPB1m/wDy6+H9Ljj+n72bzZQfo4+leIf2k/2iPFrM3iX47eP9SDH/AFdz4kunQd+EMm0DPYCvCvhHx18UvFNt4a8H6Dq/ibxBqThYrW0he5nlPAycZOAMZY8AckgV8Lv+CVX7YPxIuiureDNP8EWKNhrzxHfLFu9QsUIklJ47qqnI56keG/8Agh54uuDG3i/9oPR7HABdNN0CS7z0yA0k0WO4zj0OOcD4Qf8ABIP9mD4fSS3vj2TWviJdyxbFj1Ob7JbQnuyRQFW3dvmdhjoAea+GfwS+EPwa046X8LPhx4f8MwuP3jafYpHJN7ySY3yHgcsSeBVVVVVVVVVVVVVVVVVVVVVVVVVV/wAFYP8AkyPxh/2ENM/9LIq/4Jcf8mJfDH/uLf8Ap2vKqqqqqqqqqqqqqqqq+I/xAh+HmlW2pzaY98Lm48kIsvl7flJznB9K/wCGnLD/AKFCf/wNH/xFf8NOWH/QoT/+Bo/+Ir/hpyw/6FCf/wADR/8AEV/w05Yf9ChP/wCBo/8AiK/4acsP+hQn/wDA0f8AxFf8NOWH/QoT/wDgaP8A4iv+GnLD/oUJ/wDwNH/xFQftMWM88cA8IzgyOFBN4OMnH92qqv2mPBHiH4lf8FIfFnw/8Ja0mkaz4g8bx2VhfySPGttNII1VyyZYAEg5UZr/AIdSftpf9HR6V/4O9T/+Ir9lL/gn3+078D/j94W+KPxD+Pen+I/D+i/bPtmmRapfTNP5tnNCmElQIdryI3J/hyOcV4o8MeH/ABr4c1Pwj4q0q31PR9ZtJLS+s51yk8MilXQ+xBPTkVpUvjH/AIJRftqvZ6m99f8Awz8TDa0irvN/pDv8rgZANzbNwRwSQ2MLKDX7OXwy8Tf8FL/2ufEH7RvxX06dfhv4bvUEdhMcpIkfNppinpgKRJMR13NwDMCERI0WONVVVGFUDAAHoK+J3wxt/jP/AMFLvEvwsvLlre08SfEy4tbuVThkgNwTKV/2vLDY98V4W8K+HPBHhvTfCHhHRrXSdG0i2S2sbK1j2RwRKMBVH9epPJ5Nf8FUvgH4e+K37MWt+Pl0yH/hKPh5GNS0+9WMCQ2wdRcws3XyzGWkx/ejXpzX/BFv4n6j4o+Bfiz4Y6ldmceCdajlsgz5MNteKziMDsvmwzt7l29K+IfxT+F/xs/4KY6l4n/aL8TW9l8NvCevXFiqXsUk0DWunb0hh8tFLFJp03spHSV81H/wUd/YfhjSGH49aOiIoVEXTrwBQOgA8ngV/wAFV/jD+zN8ddQ8C/EL4IePdP1zxRYrcWGrtaWs8LtbDa9uzGRFB2OZQMHP7z0Ax4m/ac13w1/wTZtv2jhfGTxNN4Gsgly2c/2pOI7XzsEEnFxIZMdwOuOa/wCCO/7Oug/EDX/FP7Snj6wGs3Ph/UVsNDN6vnKt8yCa4uW3Z3SqskO1uxkZuu0iq/4Ljf8AI1fCP/sH6r/6Mtq/Z6/5ID8M/wDsT9J/9I4q/wCC1fxd1Tw78NPBPwb0i8khi8X31xf6qI2I8yC08sRxN6q0swfHrAK/Y8/an/YN/Zz/AGffCfw+tvjLodlrJsIrvxC66bd759RlQNOXYQndtY+WpyQFRQOBX7Sv7Zf7CPxv+BHjf4Zaj8atGvpNY0a4WwjbTrzMd4qF7aRSYeGWVYyD7c8E1/wRB+IOpTWfxO+Fd3du9javZaxYQknEUj+ZFcEdvmCW/p93v2+NkF9+2r/wVAl+Ger3k7aLB4nPh1I4WJMGnadvN0Iz23eTcyZ6BpCelaDoOi+F9EsPDfhzS7XTdL0y3S2s7O2iEcVvEgCqiKOAAABiv+CvHwi8KeMf2V9Q+KN5p8C+IPAN7aS2V6ExKYLi5jtpYC3dCZkfHYxgjvn/AIIl/wDJAfHX/Y4H/wBI4Kqv+CenwP0b9oL9p7w34H8WwG78M6ekms6vZM3yXUdsvyRsO6tLJGrd9juOM1Y2NjpllBpum2cFpaWsaxQW8EYjSJFGAqqOFAAAAHAr4h/FP4X/ABs/4KY6l4n/AGi/E1vZfDbwnr1xYql7FJNA1rp29IYfLRSxSadN7KR0lfNR/wDBR39h+GNIYfj1o6IihURdOvAFA6ADyeBX/BVf4w/szfHXUPAvxC+CHj3T9c8UWK3Fhq7WlrPC7Ww2vbsxkRQdjmUDBz+89AMaRplz+15/wT+sNL1uUXer+O/h9Esk0mcHU1gG2U554uow/wCHU9a/4Il/EWey1b4mfBHUy8U3lwa5a28gKtG0bfZ7nKnkHL2o9sc1/wAFgfGGq+PvGXwg/ZR8I3Jk1LxDqaahc2yjOZZpPsllnHu11kfQ18O9PsP2K/8AgrCPAWmp9h8I+N1j022UnAeG/ijaEAdOL2NU/wCAmv8AgqV8Uz8Mf2O/FNpa3Plah4znh0C1O7BKzEvOMDqDbxTr6fMPof8AgkX8Ij8PP2VYPGd/aGLUviDqk2pEuuHFrH+4gU/7J8uSQe01eOPHvgT4ZaDP4u+IPizR/DmlQALLfaldJbpnnaoZiMsecKMkk8CvFn/BWb9i3w7PNZ2PjHXfEZiO1m0vQptjHOCFacRgjrz0IHBPGf2GfHHhyL/gpfp2s/C+C4tvCXijXNaisLOWIW7R2E8NxJDEyKWA8siLgEj92Onb9vz/AJM0+LP/AGL8n/oaV/wR4+APhn4ofEzxL8UPG2mQ6pZfD6K3Gk2V0gkhW/ud/wC/2HILRpBxkcM6sOUBHjvxVaeBfBHiHxvfpvtvD2lXWozLnGUgiaRhntwpr/gmZ8MV/aj/AGuvEHxh+LES62PDiy+ILtbiPfHc6nPN+53g8bVJlkUesSDBGar/AILWfCLwp4c8ZeA/i/oWnwWeq+K4ryy1kxJt+1PbCEwzMB1fZKyFjyQkY7V+wH/yZp8Jf+xfj/8AQ3qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv+CwXgvVfDv7XM3ie7hb7F4r0CyurSXHykxKbd0z/AHgYgSPR1Pev2qv2ufgp+0P/AME4buz8KeMNJtfFcSaPFf8Ahia5WK8tpYriEShImw0sQwSJEBXbjODkD/glX+0T8H/2dPhZ8YPFHxW8aWGkqbrTpLKwMoa81BkiuMpbwg7pDkqMgbV3AsVHNeGbnXv2lf2tdP1JrFzqXxE8dpdS28blhALm88xwD12Rox57KvbFVVVVVVVVVVVVf8Fkv+TRbT/scLH/ANE3NfsB/wDJmnwl/wCxfj/9Deqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr9vn/AIKMXP7P+up8EfgfptrrvxJuVjF5NNEbiLSfNAMUYiXmW4cMrBD8qhkJDbttH9g3/go/+1dpsfiL46/E9dKtLxxdRaV4p1qYmNiDtZbG3R4oDhiNpEbLuI2jkVqvwC/4KafsIaQPF/gPx/ceJfCGhoZJ7PRdSl1OytoFJLGSwuEUqu3JZokO0ZO8YzX7CX7dHh39sDwteabqmnW+heP/AA9Cj6tpcTkxXERO0XNvu+byy2AynJjZlBJDKxqqv7+w0uzl1DU763tLWBd0s88gjRB6liQAPrXxK/4KG/sefCrWJvDviX4z6fc6nbttlttItZ9S8sjqGkgR41I7qWB9q+IH/BZ39m/QNNZ/AHhHxf4s1IjMcMsEenW/0eV2Zx26RtXxO/4LB/tV+NJJrfwOPDfgOyYkR/2fYC8uQp7NLc71J91jSvGv7WH7TPxESSHxh8efG9/bzH57Qa1NDbtxj/UxssfT/Z7n1rwv4D8ffEC6kg8FeDNf8SXKt+9TS9OmvHBPqI1Y5PvXgz9gf9sTx3dw2mk/s++LbLzWx5mtWf8AZSIMkEsbox4HB9+mAcjPgr/giL8T9Rgt5/iB8a/Dehu6hpoNL02bUWj45Xc7QgntkZH1r4af8Ec/2WvCPl3Pju+8UeOroD95FeX32G2J9Vjttsg+hlNeGv2NP2T/AAjHGmifs7+AQ0QGyW70OG8lXHcSTK7Z985rRfDfhTwfYta+HNA0nRLNVG6OytY7aMAdMhABgV/wm3gwy/Zx4u0XzR/B9vi3fluqKaG4iWa3lSSNhlXRsg/Qiqqq8aeONF8B2EGo62ty0VxN5SCCMMd2CeQSOMA14Y8R6f4t0K28QaUJRa3W/wAsTLtb5XKnIBPdTVVVVVVVVVVVHAGSelQzwXMYltp45UJIDIwYcdeRVVVVVVVVVVVf8FYP+TI/GH/YQ0z/ANLIq/4Jcf8AJiXwx/7i3/p2vKqqqqqqqqqqqqqqqq+KvgC8+IejWmmWWoQ2j21z5xaVCwI2kY4+tf8ADMuu/wDQz2H/AH5ev+GZdd/6Gew/78vX/DMuu/8AQz2H/fl6/wCGZdd/6Gew/wC/L1/wzLrv/Qz2H/fl6/4Zl13/AKGew/78vX/DMuu/9DPYf9+Xq2/Zq1yC4imPiaxIjcMR5L9jVVXib/lMHD/2VCy/9DiqqlkjhjaWWRURFLMzHAUDqSewr9pbxt4u/wCCnX7XGlfAj4NXdvF4L8IC6S11WWMtDtXH2rUHKjd5bFI441zz8h4Mhx/wTD/aP1z9nv4s6x+xf8a4G0mPUNZlh0s3J2mw1cEI1uSesc+1dhHG/bjIlyKtdc0rw5/wVzuNW1q9jtbRPircRPNI21UMk7xqSTwBudck8Cq/4KI+ONJ8B/sbfEy81SeNX1fSjpFpEzYMs10wiAX1IVmfHohPav8AgiB4OvbD4efFDx7LE62utaxY6dCx6M1pFLI+B/2+Lz/ga+EPwi+FN/8A8FH9e+CH7Q3h3+0tB1HxTrGnRQzXs9oHuGeVrR98LI5EhCBRkAmVciv+HXH7CX/RC/8Ay5tW/wDkqv8Ah1x+wl/0Qv8A8ubVv/kqv29vgtouif8ABPbxf8NfhhoZsdK8H6dZ3On2CSvKIrW1uopZcvIWdtsSysSxJJHJ61/wRJ+Imh3Xws8e/CU3USaxp3iAa2sLHDy29xbwwFlHcK1sAcdPMXP3hVV/wXG/5Gr4R/8AYP1X/wBGW1fs9f8AJAfhn/2J+k/+kcVf8FwvCl+mt/CrxxHE72U1pqGnSuB8sUiNDIgPHBYO+OefLPTHPwB/YA/YG+MPwS8D/Eyz+DSXh8Q6HbXNxKvibVRi4MYE6ELcgBllEikAAAqcV/w64/YS/wCiF/8Alzat/wDJVfA/9kr9nv8AZu1HVNW+C3w9Xw9ea1AkF9KdTu7wyxoxZVH2iWTaMkn5cZ4znAr4a39t8Av+Ctssvi5vstrH8QtVtllnOwLFqS3EVvKxOAAVu4nJ6AHPSq/4Kt+IdH0b9iDxtpmpXscNzr15plnp8bMAZ5lv4J2VR3Iiglbjspr/AIIl/wDJAfHX/Y4H/wBI4Kqv+CNv/J3N3/2J99/6Otqr4Q/CL4U3/wDwUf174IftDeHf7S0HUfFOsadFDNez2ge4Z5WtH3wsjkSEIFGQCZVyK/4dcfsJf9EL/wDLm1b/AOSq/wCHXH7CX/RC/wDy5tW/+Sq8B+BfCnwy8HaR8P8AwNpC6ZoGhWq2un2ayvL5MS9BvkZnb6sST3NeHtPk/ZT/AOCwX9jwL9l0HxxrLxwqBsSWDVo90aL6Kl46oB/0yxXwHsz+1f8A8FXPHHxbmkN14a+GEk32FusTNbr9itQvXhn825HTlCfav+C0Hw31HSv+FZ/tIeG2eC/0O9Oj3NzGvzRtk3NowI5AV0uOvGXXpnn/AIKSfF29/ak1j9nD4YeBPLe68Z6Jaa0LWMkhbrU3SCGNuTgxmKUEdQHJPavAvg/SPh94J0DwHoEezTfDmmW+nWi4xiKGNY1z74UVcz+JP+Cln7fj+CfGHiy9svBVhf3y2VtBLxZ6Xak8QqcqJptibnweXzyEVa8B/sV/so/DfTYtM8M/ALwY/lLtFzqWlx6jct65muA8nPpuxXwYs9C0/wD4K/Np3hq0sbXTLPx7rMFrb2USxwwqkNyuxFXCqBgjA4GK/b8/5M0+LP8A2L8n/oaV/wAEOv8AkVPi3/2ENK/9F3NfHjwre+O/gd8RPBGmRvJeeIfCmqadbog+ZpJ7WSNQODzlh2Nf8EUfHOkaH8afHXw/1GZIb7xLoUVxYiQ7TI9pKd8aju2yZnx6Rse1V/wXF8Q6O9v8JfCcd7G+qRvqd7JbKwLRwMLdEdh1AZlcA99jelfsB/8AJmnwl/7F+P8A9Deqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/bQ/ZA8I/tf/AA0Tw1qV4NJ8TaI0lx4d1jZuFrMwAeORRy0Mm1QwHIKqw+7g/GL9in9pv4HarcWHjT4S65PZwsQmr6TaPf2MqjowmiBC57K+1vVRXhD4KfGH4gajHpPgj4W+K9cu5G2iOx0eeXHTJYhcKBkZJwAOTX/BOH/gnNrvwH1uP45/HK3tE8ZC3aPRNFikWb+yBIpWSWWRSVMzISgVSQqs3JLYWqqqqqqqqqqqr/gsl/yaLaf9jhY/+ibmv2A/+TNPhL/2L8f/AKG9VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVqF7Dpthc6jcBjFawvK4UZO1QScD1wK/4JR+Drf9oL9p34l/tJ/ESCPU9X0F1vrdZvnWG/1GWZhMoPdEhlVey7wRgquKrxrp+gfsV/8ABV/RNQ8O3tl4d8H+JNQtp7iOSRba0tLPUlMNwrnhEhjm8yQZwqBE6Bc18WP+CjP7I/wq8OXusp8WtH8XX1sAsGleGbpNQnuXJwArI3lqOCSzOAAO5IB+In/Bby4k09IPhL8D44b1gd9z4i1EyxxnsBDAFLfXzF+nevFH/BW39tHxAZP7L8YeHvDgc8LpegQPs6cA3ImPY9f7x9sfFP49/Gf43Xq33xX+Jmv+JmjbdFDe3bGCE+scIxHH/wABUV8HP2e/jR+0FrUmgfB74e6l4juoELTSRbIYIABn95PKyxITkYDMCc8Zrwr/AMEg/wBsXxBdxwa1pPhXwzCxG+fUdbSUKO/FsJST2x0z3718J/8Agit8H9BihvfjF8R9f8WXqkM1npca6bae6sT5krj/AGg0f0FeDP2Dv2PfASxjQf2fPCU7xEFJNWtTqjgjvuujIc+9aPomjeHtPi0nw/pFlptjAMRWtnbrDGn+6igAflVVV3cwWVrNeXMgSKCNpJGP8KqMk/kKF14v+Pni6XT4r5rLSoAZBGcmO2jBwCVB+eQ/49AOG/Zm8OG32r4k1ITY++Y0K5/3cZ/WvhZ8NLj4dJqcVxq4vhdyp5JVSgVVB5KkkBiTzjPQc1VVX7S//Ip6X/2Ef/abV8Dv+SW6J/28f+j5KqtZv20vR77U0jEjWdtJMEJxuKqTj9K+EHxM134h6jq66rb2kENnHEYY4EIxuLZySST0HpVVVVVVVfGD4jap8PbDT5NIsrWea/eRd1xuITaF7AjP3vWvBmq3eueE9I1i+ZDcXlnHLKUXaNxGTgdq8e6HqXiXwfqeh6Rdrb3d3DtjdiQD8wJUkdAwBX8a+CPgHxP4HtNT/wCEhkjjF68ZitUlD7Cu7LkjgE5A49Oe1VVVVVVV8VPiB4m8E3GnReHtBj1FbtJDKXikfYVK4+6eM5PWpP2iPHMVwtnL4VsEnbG2JopQxz0wN2a+H/xW8aeKvFFtous+F4rK1lSQtMsEqlSqkjljjkjFVVf8FYP+TI/GH/YQ0z/0sir/AIJcf8mJfDH/ALi3/p2vKqqqqqqqqqqqqqqqqqqqqqqqq/aV8K/tAeBf2/fFvxo+HnwZ8T65Jofi9NT0uZvDt5c2dy0QRlyYgN6EjB2sPqK/4eTf8FJf+jUNJ/8ACE1v/wCSa0f/AIKO/wDBRy91axsr39lbSoree5jjlkHgbWl2IWAY5NzgYBPJr/gqF8a/i1ofw6HwL+B/gXxbq+r+MLZhrmqaRo1zcx2OnnKtCskaFRLNgqRklY92QN6Gv+CbH7I6/s0fBiPX/FmmCLx943jiu9YEi/PYQYzBZ/7JUMWcf32IJIRcf8FX/wBjTW/GYsP2mPg7oF9eeJ9PaG08QWGl2zSXF3ECFgu41jG9pIztRsZOzYeBGc/sK/HX4gfGv4MWsXxf8G6/4e8c+GRHY6sdW0uaz/tIBf3d3H5iqGLgHeF+64bgBlr4w/C68+Nf/BRXxz8KtN1dNLvPEnjvUrW1vJFJWGbdI0bMBzt3KuccgZxzVn+3h+3T+yhpqfDT9oX9ne48XXOkAW9n4hdp4vtsa8KzXMaSRXBwB8w2t/fy2TXjOy/br/4KieM9GtL/AOH83hHwJpspktnntZ7PSrPPDXDSS/NdT7GCgICQDwqBnJ+AvwV8Jfs8/Cbw98I/BaO2n6Fb7XuZVAkvJ2JaWd8fxO7McdACFHAFf8FG/wDgnx4s+NXiOH9oL4AeUnjqzhjXVNLWf7NJqXkgCG4glJAW4RVVcEqGVFwQy4fwl/wU2/bR+COnxeCPjx+zzd6/qOnx+THe6jYXWk3k23jdKfLaOU4/iRFzwSTnJ8H/APBRT9tz41+LvD+mfC/9kua10WXVLY6ndfYLq6zaiRTKouZBFBFuTcNzjjPGDzWp6bp+s6bdaPqtnDeWV9A8FzbzIHSaN1KujKeCpBIIPUGvjL+wl+1L+xj8X2+M/wCyINa13QIJnksG0qM3d9ZQty1rc22CbiLtuCuGCgsFbFaN/wAFcP2nLa2Hh7xB+ylFfeJI1w5tYr61GfU27JI4/wC+6/Yd+M/7Sfxt8M+KfEn7Rfwrn8EyLqEJ0CB9LlsEltWjO4Kk7GViGXJduG8wbeBx/wAFYP2WfHX7QHwy8M+MvhhoM+t+IfAlzcmTS7VN093Z3Cx+YYl6yOjQxkIOSGfGTwf2cP8Agop+1V8MvDPhT9n+7+AL+I5vD4g0q2lm069ivIrVCqJHJGoOTGmFBwvAXIJyT+1N+zn4V/al+Dmq/CrxNP8AY5pmW50rUlj3tp17GD5cwXI3DDMjLkZR2GQSCPAPiL/goN/wTT1DUPBk3w0n8V+BJLlptosp9Q0tj/FNb3EOGt2cDlJMZ6mPPNRf8FdP2iPFQOleAv2QpLrVn4iVJL2/5PT91FCjHn0YV+yN45+OfxF+C1h4m/aJ8DN4U8YzX1yJLFrQ2u638zdCwhZmeMBWCbXIb93k9cn/AIKYf8E/PEvx51CD46fBGxiufGdnarb6zo5kWJtVhjH7qWJmIHnovylWI3oFAIZAH+HX/BSv9tn4L6RbfC/4nfBO58UatpgW3t59a0q7s9RKr8oWXaP3pAAAYqGPVixOa+I/wf8A20P22fAniv46ftIeHr/wr4e8HeHr668G+C7HT5IJ7y+MfyFbVt0xB2gl5Pmb5Qg2scf8Ec/BvjDwP8EvG2keNfCOt+H72XxV58cGq6dLaPJGbWFQyiRRuGUYZHpVV/wST+E/xS8FftUXWseMvhp4q0GwPhO9iF1qejXFrEXMtuQu+RANxAPGc8Gq/wCCjf8AwT48WfGrxHD+0F8APKTx1ZwxrqmlrP8AZpNS8kAQ3EEpIC3CKqrglQyouCGXD+Ev+Cm37aPwR0+LwR8eP2ebvX9R0+PyY73UbC60m8m28bpT5bRynH8SIueCSc5Pg/8A4KKftufGvxd4f0z4X/slzWuiy6pbHU7r7BdXWbUSKZVFzIIoItybhuccZ4wear/grn8EviBqXib4W/H/AOEXhbV9V8Q6HcNp9y+k6c95PCYnFzZyFEVjtVxcckYyyjvX/BJj4Ea78Jf2ftT8Y+NdCv8AS/E3jzWXuLiDULdoLiO1t90UKyI4DAl/tDjd1EoPfJ/bV+D5+On7L/j/AOH9rZtc6nJpb3ukxxpudry2ImhVQOcu0Yj47OR3r/gmf+zz8W/E/wC1F4d8Z/FvwR4l0vRvhpocsunvrelTWqPJlo7eFDIo3FGuJJR6eWPYVXxv/Z5/aO/YN/arm/aG+Cvgy88ReE31O5v9OubSyku4IYLgt5tjeJH80YCuVDnAI2MrbgQtv/wUF/bS/aZ0oeAf2dP2Vrvw1rGpRfZ5/E1zcy3VvYbvlaZXkgiihI5I3tJjHAYivgF+zB8Wv2df+CjPg7Rtd8N+JPEWm6LqsQvPE8Ok3Bsp5LrT9zyCcqVKrLOVLM2flOcHIH7cujax4h/ZH+KOieH9KvNT1G70J0t7SzgaaWZt6/KiKCzH2Ar/AIIyeAPHfgTwx8U4fHHgrXvD0l5f6Y1smq6bLaGYLHcbigkUbgMjOOmR61X7Zv7Bfxx+Cfxrm/ac/ZJ0/UryxbUG1b7JosfmX2iXbEtLsgAPnW7Esdqq2FZkZdoyfDH/AAVW/bO1K0i8FWv7NNlrvjBU8kSQaPf75JOAGa1Q5zyMgMoyeABxX7Uv7JH7Vfir4Tr+058dbHXfEXxS8V+JrSxg8OaTZG6OkaT9luX+eKAMIsOkQCD7mfmJeRsfsLaXquifsi/C/R9c0q903ULLRFiuLS9tnt5oXWRwVZHAZT9R79DVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVfF39vnwp+z/+1nbfAn4sW0Nh4T1rQrS8s/ECZzYXUskqFbgdPIIjX5xyhyTlSSlneWepWcGoaddw3VrdRLLBPDIHSVGGVZWHBUgggjgg14++IPgj4W+FL7xx8Q/FGn6BoWmx77m9vZdiL6KO7OeiooLMcAAk4r4n/wDBVbxV8Yfjn4J+FvwDhuPDfgq98V6ba3+rXCbb/Vomu4g6qP8Al3iYEjAy7DqVBZKqqqqqqqqqqqq/4LJf8mi2n/Y4WP8A6Jua/YD/AOTNPhL/ANi/H/6G9VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVrMmlR6TdtrlzBBp5hZbmSeXykWMjDZbI2jB65Ff8Eu/i74V/Z2/aj8c/A7xT4p0uTS/F0/9l2GswXaSWlze2c0otysqnYUlSSYKwOCzIB96qurq1sbWW9vbmK3t7eNpJZZXCJGijLMxPAAAJJPSv26/jFB+0/+1d4m8TfD+C51fSYDFpOh/ZY3ma6gt12mVFAyVkkMsigAfK4yM5rwz+x1+1X4vdF0L9njx+6SHCTXOgz2sR69JJlVf4SDzweO4r4Qf8EYvjd4ttLHVvi1430XwNBOS0+nQx/2lexKDwCEZYQSM8iRtvGQeRXhP/gjh+yZoUMR8QX3jbxJOoHmm71VII3PGcLBGhUcdNxPPWvhf+xJ+yn8HLyDVPAXwS8P2+o2xDQ318j6jcRMDnckly0jI2e6kY6dOKqqqqqvEWmvrPh/U9HicI99ZzQKxOMF0K5/Wvg54wtPh34qvtM8TxvaRXaiCZ3Q5t5UY43Drjlgfw7VaXlnqFtHeWF1FcwSjMcsTh1YeoI4NV8avHOv+BtL0270CWBZLm4ZJPNi38Bc8V8Otcv/ABN4K0vXNUdGurqNmkKLtGQ7DgfQCvEF5Pp2g6lqFqVE1rZyyxlhkblQkcfUV8FviT4n8dapqVpr01u8drbq8YihCEEtg1+0v/yKel/9hH/2m1eDvifqWjeBNC8G+CtGbVtedJ2kXYWW3BnkIyB1OCDyQACCfStY8W/tD+GYzrGs2OLNBudRbQyxoPVjHllH1Ir4XfFex+IMD2N3Alnq9um6SBSSkq9C6Z5xk8g9Mjk18YdX+JVlPNZeFtL8/RJdLb7ZL5AbYSXD854wgU18MdU+IOmT6g3gDTvtckiRi5HkiTaATt6njvXwy1Px7qdheyePdP8Aslwkyi3XyRHuXHJwCe9W3xg1uw+JWraJr15bjRNNkui22AB9sYYqAc8ngD3Naf8AFH4qeMvEFpc+HdGuLbQjexpIYbHzR5W8Bt0jKRnGc7cY/WviX4s8U+F7Sxj8J+G/7Vu7+VowdrOIiACMovJyM85AG3mprn9pqZDfJbwwqBn7Oi2pJH0OT+Gc18NfjPqWta6PB3jWwjtdRZmjimVDHmReqOh+63B545wMVXxH+N9h4RvJNB0C0TUtVj+WUuxEUDf3Tjlm9QMY9c8Vb65+0prKDUbHTTbwEbhE1vbxZH+7L89fC/XPiLqzaja+PdJS0NnsEMhtzG0xOc9DtYDA5HrXxP1f4lapb6evj7S/skcbyfZT5Aj3Ehd3QnPQV4a8SfHa28P6db6DoHm6dHbItq/2RW3RgfKc554rxbreraF4AvNdhCJqNtZLIQ6ZCyYGcj6k18E/HviHx3a6tLr8sDtZyRLF5UWzAYNnP5CvF/i7RvBWiya1rMxEanbFEgy8znoqj14+g61B8S/jP8QJpZPAuhx2lrG2A6Ro2PYyTfKT7AD6VN8UfjD4Au4F8eaMl1bStgtJEibu5CSRfJux2IP0rwr4p0jxlosOt6LPvhk+V0YYaJx1Rh2I/wACODXxu+IHiXwG2jHw/PbqL4T+aJYQ/KeXjH/fZrV/jB4p16aLQfhlo39pXkdujXl75RZUcgZCjgAA8bm4zkAdzdfFP4z+B9QhbxjZebDKeIrm1SNXHcLJGBz+Jx3FeEPFWm+NNAttf0skRzAh42+9E4+8p+n6jB71VeI/FvhvwnbC68Q6vBZq2disSXfH91Rlj+ArxT4/8P6p8W9P8ZWTXD6dZyQFyYsOQh5wpP8APFeF/iN4O8Yt5GhazHJchcm2kUxyD1wrYz9RkVVV/wAFYP8AkyPxh/2ENM/9LIq/4Jcf8mJfDH/uLf8Ap2vKqqqqqqqqqqqqqqqqqqqqqqqqqqqqrw1/ymDm/wCyoXv/AKHLVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV/wWI/Z/wDizqXxWtfj3o/hK51LwVD4ft7C8v7MeabGWKSViZ0HzRoRIuJCNueCQcA/sd/8FK/iZ+y74eu/AOv6U/jfwitvI2kWFzemKXS7jBKrFIQ2IGbG6PHGdy4O4N+0X+1L8Y/2o/Ff/CTfFLxK08Fuzf2do9rmKx05T1EUWT8x7uxZ2wMsQAB+wx/wT5+PPxU8c+EPi/q+lf8ACH+DNE1W01SK/wBWhZZtRWGVZAtvBwzBtoHmNtTBJBYjaaqqqqqqqqqqqv8Agsl/yaLaf9jhY/8Aom5r9gP/AJM0+Ev/AGL8f/ob1VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVc3FvaW8l1d3EcMMSlpJJGCqgHUkngCvHn7WP7NPw10ObxB4v+OHg+3tYHdCltqkd5O7r95UhhLyOw4yFUkZGa+K3/AAWy8C6VeS6d8GfhFqfiBEJUalrl6LCMkfxJDGsjsp/2mjPqBXjv/gsH+1z4stprPw6/hDweki7Vl0rSTLMo9d1y8q57ZCj2wea+Ivxn+LXxdvTqHxO+JPiPxPLvLoup6jJPHEf+mcZOyMcnhQAM18M/2Qf2mvjFpraz8Ovgt4k1TTlGVvXtxawSdPuSTFFc8jhScVq/xj/4Kn/sReDbFPiHLfp4VVorWzm1xbPWYoXZCUiFwjPIpCoQEZ8DbwMVZ+G/+CoP/BQTw5YLresyaV8OdcUSLcTSQaTp08W7q0cI8+5XK5AZXGVB44Nfsc/sZ/D/APZA8ETaRoNwdZ8UawEbXNemi8t7plztjjTJ8uFSThckkklieMVVVVVVVVV4++EXhnx45vpS9hqYXAu4FB346b16N9eD05xxWq+Bfij8ImfWtC1OSWxjO6SazYlQPWWI9uOpBA9a+E3xUi+IFtLYahBHbavZoHkSP7kydN6g8jBwCO2Rzzx+01/yAtF/6+3/APQK+Df/ACTPQv8Ari//AKMavF3/ACKetf8AYOuP/RbV+zN/yHda/wCvRP8A0Ov2l/8AkU9L/wCwj/7Tav2fPDtlp3gaPXUjVrvVpZGeQj5giOUC/TKk/wDAqdEkQxyKrKwIZWGQR34q7tV+HHxvig04+VaxahGUVTwIJgMr9ArkfhXi7/kU9a/7B1x/6Lav2Y/+P7X/APrjB/N6rW9FTxH8aLzQ5GIjvNbaOQg4ITf82PfGatLO1sLWKxsoEhggQJFGi4CKOgArUNQsNKtJL/U72G1t4hl5ZnCKv4mtV/aD+HunymK1k1DUSpxutrfC/m5XivGHjLTvEvxEtfFug2k9ofNt3KzABjIhA3cEjoq/lXjXW38N+EtV1uEr5tpau0W7pvxhf/HiK/Z38MW+ta3qHizVE+0Saeyi3MnzfvnyWc+4A/Ns9RVV+07/AMeHh/8A67T/AMkr4a/8k/8AD3/YOh/9BFfFj/knOv8A/Xof5iv2Y/8Ajw1//rtB/J6+PWo3fiH4iWfhWCX5LNIoY0zwJZcEt+IKD8K0LRbDw7o9pommQrHbWcQRABjPqx9ycknuSa8W+HLLxZ4evdBvY1ZbmIhGI5jf+Fx7g4Nfs4azc2XijUPDkrkQ3dsZPLJ6SxsBx/wFmz9BX7UH/Ms/9vf/ALRr4O6HYaL8PtKa0iUS30IuZ5NuC7vzz9BgD6V8X9JtNX+HespcxqzWsBuImI5R05yPwyPoTX7Md5LJYa/pxJ8uCaCVRn+Jw4PH0jWq8Y+JLbwj4av/ABDcqHFpFlI843uSAq/ixArwD8PtV+L2p3PjTxpqM5smlKjYdrTMP4U7Ki9OPoO5EPwh+G0FuLZPCdoygY3OXZv++ic18TPgnb+HrJ/FvgKW4t2sP301qJWJjUcl42+8NvUgk8c54wfhB46l8c+FRPqDqdSsH8i6IGPM4yr4HTI6+4Paqr/grB/yZH4w/wCwhpn/AKWRV/wS4/5MS+GP/cW/9O15VVVVVVVVVVVVVVVVVVVVVVVVVVVVVWX/AAT3+Ath+0K37TMF34qPi9tck1kxtqEZtftEhYsPL8rds+Y8bvxqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr9oT48eD/ANmr4Wal8XPHWnaxf6Ppc0EU0Gkwxy3DNNKsa7VkkjXAZgTlhx69K/4fV/sr/wDQhfFX/wAFVh/8m1N/wWl/ZSuIXt5/h78U5Y5FKuj6Tp5DAjBBBveQR2r9q7xt+wz8Ur668a/s9+FfiN4E8Q3L759GutGshpNyxPLJ5d2z2p5JwiunAARck1+xR8cfgH8B/iVN4r+PHwZbxxaCFTptzCqXE2lzqSd6W0zrDLu+X5mIZCoKnqK8G/8ABYP9mfxx4v0LwVpPgf4mxX2v6lb6fbSXOmWKxpLNIsalyt4SFBYZIBOOxqqqqqqqqqqqqr/gsl/yaLaf9jhY/wDom5r9gP8A5M0+Ev8A2L8f/ob1VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV4u+IHgL4f2X9pePPG+geG7TBIn1fUorOPA6ndIyivi3/AMFX/wBkf4ZyS6foXiLVPHmox5Bi8O2m+FW97iUpGR7xl6+K/wDwWr+K+vwy6f8AB74Y6H4TjYlRfarcNqdwB2ZEAjjRvZhIP518W/2rP2i/jtbNp/xV+Luva5YNJ5jaeZVtrQsDkMbeEJFkdvl47Vo/wU+LXiVdEHhb4c+I9cfxDafa9Pj0zS5rlpYvOlh3YRT/ABwSD6DNfBD/AII+/tE/EeCHV/ifqml/DbTZQCIbtft9+VIyD9njYKv0eRWHda8G/wDBFn9nPR1il8Z+PvHHiK4QDekM0FjA57/IsbuP+/lfCj9jz9mT4JiKX4dfBnw7Y3sOCmo3Vub28UjuLicvIvPOFYDpxwMftDfCq4+OPwT8X/CS01qPSJvE+nNaJfSQGZYCWB3FAQW6dMiv+HHPir/o4rSf/Cbk/wDj9f8ADjnxV/0cVpP/AITcn/x+qqqqqqqqqqpEjljaOVFZGUhlYZBHcEelfB+JI/jIsejMWsY5LsAqcjyQrhfwzsr9piCRvDWkXKj5I74oT7shI/8AQTXwSu4Lv4Z6OIXBaASxSLn7rCRuD+BB/GvHV5bWHgvXLq6lCRrp8wye5KEAD3JIH1Nfszf8h3Wv+vRP/Q6/aX/5FPS/+wj/AO02r4DX1rd/DTTraCUNJZSzRTL3VjKzj/x11qvEcieNvjukGnESxHUoYdynOUiCiRgR1HyOfpXi7/kU9a/7B1x/6Lav2YyPt+vrkZ8mDA/F6qy/5OGb/sOSfzaq+Meq6h4w+J1p4GS8MVnBcQWsa/w+ZLt3OR3I3gfQcdTnw78O/B3he0ittL0K0MkagG4miEkrkdyxGevOBgDsBXx0NoPifbwWojHk21usioANrbicEDvgg/iK+Jmlzax4B1zT7ZC8rWjOiqOWKYfA+u3Ffs0a5axTav4cmkVZ5glxACcbwuVcD6ZU/n6VVftO/wDHh4f/AOu0/wDJK+GhDfD7w8VIP/Eui/8AQRXxY/5Jzr//AF6H+Yr9mP8A48Nf/wCu0H8nr41QzeHvi1Hr7xMyTfZruPjhvLCqQP8Av3+tWV5bajZwX9lMssFxGskTqeGUjIP5Vrer2egaPea1fyBYLKFpHJOM4HQe5OAB6kV+zrp1xqPja+1t0/d2lo25gOBJIwwPyD/lX7UH/Ms/9vf/ALRr4a/8k/8AD3/YOh/9BFfEv/kn/iH/ALB03/oJr9mD/mZf+3T/ANrVX7Q+qWLeCRp0Oo25uTfR74FlG/aA2crnOM4/SvhAlunw10FbYKFNuSdv94u278c5qrqO3ltpYroKYWjYSBuhUjnPtiv2Ymk+1eIUBOzy7ckds5kx/Wqr/grB/wAmR+MP+whpn/pZFX/BLj/kxL4Y/wDcW/8ATteVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVft+/Bvx98e/2YfEXwz+Gelw6h4g1C8sZLe3muUt1ZYrmN3O9yFGFUnk1/w6f/bc/wCib6T/AOFFZ/8Axyv+HT/7bn/RN9J/8KKz/wDjlf8ADp/9tz/om+k/+FFZ/wDxyv8Ah0/+25/0TfSf/Cis/wD45Xwf/wCCYH7Y/hH4teCPFeu/D7S4dN0XxHp97eSLr9o5SGK4R3YKJCSQqk4HJqqqqqqqqqqqqr/gsl/yaLaf9jhY/wDom5r9gP8A5M0+Ev8A2L8f/ob1VVVVVVVVVVVVVVVVVVVVVVVVVVVVVV8XP2w/2Z/gaLi3+I/xh0Cy1C2B36Xaz/bL0HsDbwh5FyRjLAD1I5r4q/8ABbXwZp8klj8F/g/qeskAhdQ8Q3i2UYYdxBF5jOp93Q+wr4rf8FU/2v8A4nW1xpmn+MdP8E2FwCrQ+GbP7NKF9riRpJkP+0jqfpU8/ivx3rpluJtW8Q61fNyztJd3E7fU7mY18KP+CeH7XnxeniOkfB/VNBsHI3ah4lQ6XEin+ILKBK4/65o1fDL/AIIhzl4Lv4y/G6NUBHnaf4Z08kn123NxjH4wmvhn/wAE6P2O/haLafSvg3putX9sQftviF31J3YdGKSkxKeB91FFQww28KW9vEkccShURF2hQBgAAdBjtVVVVVVVVVVVVVeKrXV73w3qVloMqR6hPbPHbu7lArEYzkZwfT3xVt4q/aA8GL9i1DQ7rU44hhWuLQ3QH/bSI5P4sa1LxR8d/H1s2j23hufT4JwUlMFm9srg9QZJScD1wRXwl+FcfgC0l1DUpo7jWLxAkjR8pCmc7FPfJAJPsMdMnx54Rt/G/hi78PzSCKSQB4JSM+XIvKn6dj7E1oQ+MPwlvLiw0/w/c3FvM+Wj+yPcQSMOAysnIOPQg9Mjji18OfFD4uXdv/wn0baLoMDh3tI4zC0rDsEYls+7cDPAzX7P3hbxJ4e1rVZNc0K9sUmtVEbzwlAxDcgE96/aA0LWvEHhzTbPQ9Kur6ZL3e6wRFyqhGGTjpyRVh4G+JXw9stP8T+BYnlN5ZQnUtLlXcyy7fmyhIJ59DuBJHStU8f/ABy8UwPomn+D7nTWlGyWSCwljcZ6/PIcJn14PvXwg+EZ8Eq2u680cusTIUREO5bZD1APdj3PQdB3JureK8tpbSdd0c0bI49QRg/pVt4f+J/we8Tz3eh6NNfQuGiWVLZp4biPORuCcqeBxkEc9uvwz8ReLvE2i3F94w0MaZcrclYYxbvDuj2rztck9c81aeDvFSfHFtcbw/fDT/7XklFyYD5ewk4bPTFV8Z/hb4gv9eXxz4Rhe5nIQ3EEX+tR0ACyIP4uAowOQRnnJxYfE3416lbrpVp8P2+342NdTWcsag9MkMQqn6nHt2rxj8KvHmn6lY6rLaXuvaheL9ov5LeMuqS7vuZA9Me3oMChhl6cEdCK8dfBTxFo+unxT8N3YjzTKttFII5bdz12ZwCvt15xgirT4q/G2yjGn3fgOS7uVXCyPpc4ZsdyFIB6dgK+GGrfEjWH1G68eaV9iify/sSeUItuN28bclv7vLV8ZfAt9448LpDpCq+oWE3nQxswXzRghkBPAJ4Iz3HvXhfxj8Z/B+mx+FbLwbcXKW5IhFzpkzvGCScAqQCMknnP5V8RbO/1b4f6vZ2VnJNd3FphII13MWOOAO9fs9+HNe8PWWtR65pF1YtPLCYhcRFN4AfOM/UV8TPh1Y/EPRktWlW3v7QlrO4IyFJ6qw/unAz3BAPsdE1H40/CmJ9DHhqXUtPRiYwYHuI0z3R4zlQT2PqeASa1aP4z/F9odNvNEbSdMDhmWSJreLj+Jt+WfHoMj2714b03wX8H/Dqabfa5aW8kh8y4nuJArzvjGQvXHGABnAH1NftD+GfEHiL/AIR/+wtGu7/7P9p83yIi+zd5WM46Zwfyq48TX3w0+EmjajcaV5l1bW9vA9tMxiKsRg54JyMdKsPFOqfE34S69fJpSpdyxXFtFb25LlyEGMd8nd0r9njwz4g8O/2//bujXdh9o+zeV9oiKb9vm5xnrjI/MV4g8ZeKtYup49Q8S6jcRCRgsbXLbAM9lBx+lW2n398CbKynuNvXyoi+Pyr4P/FaDwZE3g/xektvZq5aCZozm3ZuSrL12nOc44J9DxD428G3FuLqHxZpDRYzu+2xjH154/Gvit8atH/sm48NeDrsX13eIYZrmMHZEh4IU/xMRwCOBnOc18EfBF14P8KtPqcJi1DVHE0sbDBiQDCIffkk+m7Haqr/AIKwf8mR+MP+whpn/pZFX/BLj/kxL4Y/9xb/ANO15VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV448aeGvhz4P1jx54x1OPT9E0GykvL65cE+XFGuWIA5J4wFHJJAHJr4H/8FWfgB8b/AIt2XwntPD3ibw9Nrd0LXRNQ1SOLyryY8JG4jdjEznAUfMCSASCRn9pf/gox8EP2VfiLH8MfiF4X8cajqkumxags2jWNrLB5cjOqjdLcxtuzG2RtxyOfT/h9X+yv/wBCF8Vf/BVYf/Jtf8Pq/wBlf/oQvir/AOCqw/8Ak2vBv/BYP9mfxx4v0LwVpPgf4mxX2v6lb6fbSXOmWKxpLNIsalyt4SFBYZIBOOxqqqqqqqqqqqqr/gsl/wAmi2n/AGOFj/6Jua/YD/5M0+Ev/Yvx/wDob1VVVVVVVVVVVVVVVVVVVVVVVVVVVSyRwxtLLIqIilmZjgKB1JPYV8Wf+Ch37IfwfM1rrfxe03W9SiB/4l3hwHU5Sw4Kl4sxI3s7rXxX/wCC3FzIk1l8D/g0kRORFqPii73EfW1tyMH/ALbH6V8Xv25P2qPjc80PjT4w61Dp0pYf2XpEv9nWu0/wtHBt8wf9dCx96+HPwb+LHxfv/wCzPhf8OfEPiidXCSHTNPknSEnH+sdRtjHI5Ygc9a+En/BGr9onxl5F78UfEnh/wBZSDMkBf+071P8AtnEwi/8AI3GelfDj/gjb+y/4UkhvPHWs+LPG1whBkgubxbG1fH+xAFlGT/01Pb3z8OPhH8MPhBoq+Hvhf4D0TwzYKAGj06zWIykd5HA3SN/tMSfeqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq7vrGwj86+vYLdMH5ppAg468mtR+LXw40xtlz4usnI/5990//AKLDVrn7QvgPTos6S93q0pHCxQtEo+pkAP5A1P8AtOWxhYW3hCTzcfJvvRjPvhK1T9oL4h35YWlxY6ep6C3tgxA+r7q0v9o7xrZxLDqNjpuobRzI0bRu312nb+SitY/aI8dajCYNPi0/TQf+WkMRd/zckfpVyPEniO/33Eeoajetxko0shySfc9Sa8P6/wDtDadpVtYp4Qtr1I41WOS9QLJtA4DYkXnGByM+ta38OviR8TLGBPG/iLTtHiibelhZWxmAbGNzEvycEj7xAz7mvAvwv1fwAJYdJ8Z+dazsHltp9PBUtjG4YcEHAA6+noKms5buxksry6OZUKs9uDEQCOcckg9eQcj9a0v4Y/D/AEbBsPCen7l+600XnMPoXyaihhgjWGCJI0UYVEXAH0Arxh8OfCfjiNTrmnZuEXCXULeXKo9M9x7EEV4k+G+m6R8TrHwLaahcta3kkK+dIFLoH69AAcfSvCHwd8FeDp0vrSzkvb2PlLm8YOUPqqgBQffGR61VV/wVg/5Mj8Yf9hDTP/SyKv8Aglx/yYl8Mf8AuLf+na8qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/aR+ETfHr4FeNPhFDqaafceJNMeC2upASkU6sHiZwOdnmIm7HOM1+zH/wAErf2l9B/aC8L+I/ipoul6B4W8I63balNfxatDcnUBbyrIscCRMXG9kUEyCPapJ6gA/wDBSH9hj9pD9oz9oW3+IHwo8IWOp6LH4ctbJpp9Wt7ZhNHJMzDZI4bGHXnGOa/4dP8A7bn/AETfSf8AworP/wCOV/w6f/bc/wCib6T/AOFFZ/8Axyvg/wD8EwP2x/CPxa8EeK9d+H2lw6boviPT728kXX7RykMVwjuwUSEkhVJwOTVVVVVVVVVVVVX/AAWS/wCTRbT/ALHCx/8ARNzX7Af/ACZp8Jf+xfj/APQ3qqqqqqqqqqqqqqqqqqqqqqqqqqnngtYJLm5mjihhQvJJIwVUUDJJJ4AA718Xf+Cj/wCyF8HpJ7DU/ilb+I9Vt2Ktp3hmI6i+4cMDKhECsDwVaQHPbg4+Kn/BbjW7gS2PwU+DNpZjkR6j4mvTOxB/6doNoUj/AK6sPb1+Nv7XP7RH7Q00i/FH4n6rf6ezZXSbZxaWKen+jxbUYjH3mDN718JP2Wv2hfjs8Z+Ffwl1/W7WQ7RqAt/s9mD3BuZSsQPtuzXwq/4Ip/FnXDDe/GD4naD4Xt2IZ7LSYX1K5x3VmPlxo3uDIPr0r4P/APBLr9kP4TrBd33gWTxzqkQBN34plF2hPfFsAsBUnsyMQOMnnOkaNo/h/TYNH0DSrPTbC2XbBa2cCwxRD0VFAAHsBVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVe6hp+mxCfUb63tYycB55RGM/UmtY+IfgjQrVbzUfE9gI3zsEUvms2PRUyT+VXn7QHw5trZpre9vLuRR8sUVqys34tgfrVz+07CCVs/BrsMHa0t/t/QIf51qn7RXjq9BTT4NN09ezRwmRx+Lkj9K1P4l+P9XJ+2+LdRw3VYpjCp/4CmBTvc3s+6R5Z5nPViWZj/OtM+HfjnWMHT/CmpOpHDvbmNT9GbA/WtN+BfxJ1CZYpdFjskJwZbm4QKv4KS35Cl/Zl1043+J7AeuIXOK0b9m/whZxIdZ1G/wBRnH3trCGM/RRlv/HqsvhH8N7BAkPhOzfA6z7pT/4+TWm+E/C+jMJNJ8O6baOvR4bVEb8wM1VVVVXxU0z4m6hcacfh9eSQxokgugs6R5OV2/e696u/hd8bb7WYvEN2gl1KAqY7lr2LcpX7uOe1fD/RvjTZ+KLafxlqEsulKkglRrqNxnadvCnPXFVVf8FYP+TI/GH/AGENM/8ASyKv+CXH/JiXwx/7i3/p2vKqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr4lfE3wF8H/AAZqHxA+JXiez0HQNMQNcXlyTjJ4VVVQWd2PARQWJ4ANfE3/AILc+HbHUZrD4Q/BW71W0jYhNR17UhaF8HGRbxK52nqCZAfUCvB3/BcG+F9HD8QfgFbtZs37y40bWyJIl9RFLGQ59t61+z1+0z8IP2oPCDeL/hP4iN2tqypqGnXSeTeafIwJCzRZOM4OGUsjYOGODiqqqqqqqqqqqr/gsl/yaLaf9jhY/wDom5r9gP8A5M0+Ev8A2L8f/ob1VVVVVVVVVVVVVVVVVVVVVVVV8Sfjr8Gfg9bG5+KHxQ8NeGiE3LDqGoxxzSD/AGIs73PHRVJr47f8Fn/hn4cS40f9n7wXeeLr4ArHq+sK9jYqezLFxNKPUN5J96+O37Yf7RH7R1xIvxN+I9/PpTNuj0Sxb7JYR+n7hMByOzSbm/2q+Dn7JX7Rfx98ub4WfCjWtVsHbA1OWMWlkOcH/SJisbYwchWJ9q+F3/BE34kaq0F58Yfi1oegQHDSWWiWr6hOR3UySeWiNjuBIB718Ff+CZ/7JfwVmt9Uh8Bt4w1m3IKaj4pkF8VYYOVg2rApBGQ3l7h/eqGGG3hS3t4kjjiUKiIu0KAMAADoMdqqqqqqqqqqqqqqqqqqqqqqqqqqqqpJI4kMksioqjlmOAK1H4h+BdKyL7xZpaMucotwsjDH+yuT+lah8f8A4b2QIt7+8viO1vasP/Q9tX/7TWjx5GmeFryf08+4WH+QatR/aT8W3AK6bo+mWinoXDysPocgfpWq/Enx5rFz9qu/Feoq2cqkE7Qop9lTAFXl/q+u3iy397eajdPhVaaRpXb0AJyfwq08Pa9qFyLKx0W+nnJx5cduzMD9AOKg+BnxPnZA3hxYlb+KS8hAX6gOT+lWH7M2uTQI+o+JrK2lI+aOKBpQv4krWl/s1eF7fa2r67qF6w6iJVgU/h8x/WtM+Dvw30oAw+F7edx1a6Zps/gxI/SrHSdL0xPL0zTbW0XGNsEKxj8gKqqqqqqqqqqqqr/grB/yZH4w/wCwhpn/AKWRV/wS4/5MS+GP/cW/9O15VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVf8FkvjD4g8T/tAaf8HUvpE0DwZpcNx9kBIWS9uV3tK3ZiIjEq/wB358feNfsqf8E2/jd+1L4aj+IFlqOk+E/B80rxW2p6mHkkvChKuYIUGWVWBUszIMggE4OPi7/wRq+OngXwvdeJPh3430Tx5LYwtLNpcVpJZXcoHUQKxdZGxztLKT0AJwD+xh8bfEX7Pf7SXg/xZYX0trY3GpxaZr1q7lEnsppFjmWQHjKZ8xc9HjU9qqqqqqqqqqqqq/4LJf8AJotp/wBjhY/+ibmv2A/+TNPhL/2L8f8A6G9VVVVVVVVVVVVVVVVVVVVVVeNvH3gf4baDN4n+IHi7SPDuk24Je81O8S3jyBnALEZY9lGSewr47/8ABZj4R+EHuNF+BXhG98c3yZVdUvi1hYKc9VUjzpenQrGORhjXxg/4KOftdfGUzWup/FK58NaZL/zDfC6nTYwO48xCZmB6YeRhj6nOg+GPG/xF1xrDwv4e1zxPrFyxdoLC0lvbiVieWKoGYkk9a+Cv/BJv9qb4pSQX3jHSbH4daNIQWn1x990V7lLWMl93+zIY/rXwB/4Jc/swfBMW2ra7oDfELxFDhjfeIo1kt439Y7QfugOhG/zGBGQwqCCC1gjtbWGOKGFAkcaKFVFAwAAOAAOMVVVVVVVVVVVVVVVVVVVVVVVVVVUSAMkgADkmvEHxE8FeGI92r+IrRH7QxN5sh/4CuSPqeK1H9pHwZbqV07TNUvJB03IsSn8SxP6Vqn7S/iCcldG8O2FqOgNxI0xHvxtFap8ZfiRqwKSeJZbdD0W1jWHH/AlG79av9W1XVX8zU9Turx853XEzSHP4k1p3hTxPrAVtK8PaldqwyGhtXcfXIGK0/wCB/wAStQAY6ALZD/FcXCJ+mS36VY/s0+K5sHUdd0u3B7R75SPwwo/WtP8A2ZdGjwdW8U3lx6i3gWH9WL1ofwY+HehxsqaDHfO64aS+PnHHsD8o+oANaT4T8L6DKZ9F8P6fZSsMGSG3VG/MDOKqqqqqqqqqqqqqqqqr/grB/wAmR+MP+whpn/pZFX/BLj/kxL4Y/wDcW/8ATteVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX/AAWY+AviPR/irpH7QOl6XLP4d8QadDp2pXMUWVtb6HcE8wjoJIigUnvEwz0Ff8E1fjv8MPiX+zJ4K8DeGtcsIPEvg7S007VtEaRUuInjyPPEfBaOQYfeARlmBO4MB4++Ifgf4W+GL3xl8QvFOnaDo1hE0k11eziNcAdFB5Zj0CrkkkAAk18G/hxqP7Xn7bMn/CC+HLiLQNd8aXGu36rHhdM0prwzOXI4XEZCKCeXZVByRVVVVVVVVVVVVX/BZL/k0W0/7HCx/wDRNzX7Af8AyZp8Jf8AsX4//Q3qqqqqqqqqqqqqqqqqqqq1u+k0zRr/AFKFFeS0tZJUVuhKqSAfbiv+H2nx+/6JR8P/APvm8/8Aj1f8PtPj9/0Sj4f/APfN5/8AHq+JX/BZb9ozxZoFvpHgLwz4Z8FXhgC3mpwRNezNJ3aFZspEvsyyHvu7V46+JPxM+MXiJda+IPjLXfFmsTvsik1C6kuXBYgBI1JO0HAAVQB0AFfs/f8ABLv9pz44C21fW9BT4feHZsN9v8RRPFPIh7xWg/eN2IL+WpByGNfBj/gkf+y58NY7e+8c2mp/EbWIsM0urSm3tA+Oq20RAK9flkaTr9K8J+CvBvgLSI9A8DeE9H8PaZF9yz0uxjtIV+iRgD9Kqqqqqqqqqqqqqqqqqqqqqqqqq4uba0iM11cRQxjq8jhQPxNX3xK8AaeStz4v0vK9VjuBKR+C5rxN+0P4P0lVTw/DLrcxJ3bd1uiY9WZcn8AR71rf7SXie9ITQtHstNTHJlY3D59jhQB/wGtR+NHxK1KPyZPE0kKYxi3hSI/99KM/rV/rOsaq27U9WvLxieTcTtIf1JrSvDmv69IsOjaNeXrMcDyYGYD6kcAfWtO/Z5+IV6FN0mnWIOMie53EfggbmtM/ZkhAV9Z8WO3rHbWu3H0Zif8A0GtL+A3w303a02mXF+69Gurhj+YTaD+VaZ4T8L6NtOk+HtNtGXo8Vqit+YGaqqqqqqqqqqqqqqqtQ1bStKi87VNTtLNMfeuJljH5kil8d+B5GCJ4z0JmJwANRiJP/j1Wd9Y6lALnTr2C6hJIEkMgdcjryOKqqqv+CsH/ACZH4w/7CGmf+lkVf8EuP+TEvhj/ANxb/wBO15VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVf8ABYL4v/FPSPj2fhXpPxC1+y8H6j4UspbzQ7e/kjtblzNPkyRqQHzsTOc/dHoKqqr/AII2/wDJot3/ANjhff8Aom2qqqqqqqqqqqv+CyX/ACaLaf8AY4WP/om5r9gP/kzT4S/9i/H/AOhvVVVVVVVVVVVVVVVVVVVXt5Z6daTX+oXcNtbW0ZkmmmkCJGgGSzMeAAOSTX7ZH/BU74S/DXwzqHgv4C61pPjzxldhrZ7mINPpmnoQQ0jSrhbhscKsbFcnLNxtb4If8EpP2kfjlpUPjzxdPofw70nVsXEEWo25F1JG/IdLOIARLjortGemFwc18UP+CMPx88J6LPrHw58c+G/G8ttGXOneW+nXM2O0QkLRsfZpE/pX7Hf7PPwv+LX7RA+B37RHiPxJ4Lvy8lta2EFskEtzfRk7rSSSUHyHwpwDG24grlWK7vgx+yX+zv8As/QxN8Lfhbo+m38a7Tqs8Rur5uOf9Il3SAHuqkL7VVVVVVVVVVVVVVVWrarp+h6bcatqlx5Npapvlk2Fto9cKCT+Ar/hePwt/wChp/8AJK4/+Ir/AIXj8Lf+hp/8krj/AOIqqqqqqqq1/wAdeD/DMbNrXiGzgdf+WQffJ+CLlv0q8/aO8BwErbWer3JHQpAij/x5wf0rX/2ldauC0XhvQ7W0ToJbpjK/1AGAD9c1qvxY+ImsgrdeK7yNT/DbEW4x6fIB+tXF1eX83mXVzNcyscbpHLsfz5qw8EeMdUUPp/hbVZ0YcOto+3/vrGK034IfEnUpEQ6ALRGODLczogX3IBLfkK0j9mW0jnWTXfFEk0Q6xWtuIyf+BMT/ACqz+A3w1tZBI+kT3ODws10+B+CkfrWm+DvCekKo0zw3pluVGAyWqBvxbGT+NAAAKoAAHAxVVVVVVVVVVVUxVVLMwAA5J4xV7468F6fbPdXXivSljQc7bpHP4KpJJ9gKl/aA+G6OUS+vZADwy2jAH6Zwak+P3w2SDzV1G7dgM+Uto+76c4H61f8A7TOhRkjS/DN9cADgzzLD/LfWgap/bmhabrXkeT/aFpFceVu3bN6BtucDOM4zgVVfEHxX4/tPFmp6VqfinUt1ncuiLFMYV2Z+UhUwBlcHp3pvFnip1Kt4m1UgjBBvZP8AGgLzULjCia5nkPTBdmP8zUnhTxRGheTw3qiqOpazkAH6V8Aobi3+H6W11azQSR3koKyoUPODkA9uaqqqv+CsH/JkfjD/ALCGmf8ApZFX/BLj/kxL4Y/9xb/07XlVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV4u+HPwD8UeMba78eeBPAGreK7y08m2l1fS7Se+mt0bOxDIpkaNWbOBwCfev+GevgD/ANEN+H//AITNp/8AG6+LnxO/4JvfArW28M/E+w+FGl6xHjztPt/CcV/PASMgSx21vI0ZIwcOBwQe4r4XeHf2L/jZoB8T/CjwR8KfE2nRuEllsdAs2aByMhZEMYeNsc7WANf8M9fAH/ohvw//APCZtP8A43Xhnwj4T8FaadG8G+GNJ0GwMplNrpllHaxFyAC2yMAbiAATjPAqqqqqqqqqqqv+CyX/ACaLaf8AY4WP/om5r9gP/kzT4S/9i/H/AOhvVVVVVVVVVVVVVVVVVV4u+IPgLwBaLfeO/G+g+HbdlLLLq2pRWisB1IMjDOK+P3/BZv4c+FLq68P/AAA8Fz+MruFmQa1qrNZ2GR/FHGP30y54+byvYkYNfHz9sH9oT9pS7Y/E/wCIF3PpYbdDodj/AKJYQ85GIU4cjs8hdx/er/gl98KfD/xY/a88N23iizhvdO8M2dxrbWkybkmlgCrDkdwsskb4PB2YPBNVX/BY7wNB8NPjD8L/ANobwSi6b4g1TzUurqBQp+16e8EltOe7Ptk259IUHpXhjW4vE3hrSfEcERjj1Wxhu0RuqiRA4H61VVVVVVVVVVVVVVXxn+Ifh7w9o8/hi6tBqV7qMJDWokKLGh6M7DkcjgDk+o61pHwo8f8AiWIajpnhh4baX5ozLIIVwem0SNuI9Dz9a1XwB4s8EyLf+JvCTTWSnDkyFoiM93ib5Se2T+FfD/4haN8QdKN7p4MF1BgXVo7ZaInoQe6nBwf5dKqqqp54LaMy3E8cSL1Z2Cgfia8RfGDwB4bYw3OuJdzgf6myHnH8x8oPsSK1b9ppdzR6F4Wyv8Ml3cYP4oo/9mrVP2gPiLqAK2t3ZaeDxi2tgT+b7q1Xxt4v1wFdV8TajcI3WNrhgn/fIOP0rTND1nW5RBo+k3d6+cYghZ8H3wOK079n/wCIt/Eks1pZWW4ZC3NyMj6hA2PpWhfszTlw/ibxJGqDrFYxkk/8DcDH/fJrSfgh8N9KCsdCN7Iv8d3M0mfquQv6Vp+haJpChdK0exswOgt7dY//AEECqqqqqqqqqM8AmFsZ4/NKlhGWG4gdTjriqrxP4k0zwjos2vauZfstuVD+Um5ssQBx9SKvv2l/DEZI07w/qdxjoZmSLP5Fqvv2m9YkBGm+FbODjjz7hpcfkFq8/aD+It0+6C50+0H92G1BH/j5Y1ofx4+IE+q2Npe3tpLDLcxpLm1VWKlgCARjHFVV+C1jcKoyTE2Bj2qPw34hndYodB1GR2+6q2rkn6DFWXwQ+JV/bJdJ4f8AJVxkJPOkb/ipOR9DS/Az4oM4Q+G1Azjcb2DH/oea039m3xfcgNqWraZZj+6rNKw/AAD9a8PaW2iaBpmivMJm0+zitzIF27yiBc47Zx0qqvPAPg3UtYn1/UvD1ne3twirI9zH5oIUAD5WyoOAOcZr/hAfAv8A0Jeg/wDgti/+Jqy0zTdNTy9O0+2tVPVYYljH5AVVVVVX/BWD/kyPxh/2ENM/9LIq/wCCXH/JiXwx/wC4t/6dryqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/4K/eMNS139sGbQ5LiVYPCvh+xtLVA2ApkDXDOoHcmYAnr8oHQCvHvxQk/Y2/4J26L4k0/WHvPEVl4WsbPSri9czNcapdRqTJ83LBWeWXaf4Y8dq8JeCvih8dfHMmjeEND1vxj4r1d5budIVa5uJ25aWaRjz1OS7HqeuTX7E/xd8Z/sxftYeG01CO/0uK61lPD/AIp0q4Rom8qWUQyLLGcHfC5DgEZDR47kVVVVVVVVVe3lnp1nPqGoXcNra2sTSzzzSBEiRRlmZjwqgAkk8AV+0D/wVe8VeIvGp+EP7FPgZvFeqSzG3j1yWwlu2unHX7HaLgsoxnzJMgjPyYwx0T4B/wDBX/4p39l4n8a/F/U/C0L3Uc8ti/ildNYxhgSPJ08FVBXPyHHuBVVX/BZL/k0W0/7HCx/9E3NfsB/8mafCX/sX4/8A0N6qqqqqqqqqqqqqqqq8bePPBXw38O3Pizx/4q0vw/o9mpaa91G6WCNfYFiMsegUZJPABJr9rT/gsJd3hu/BH7KVs1tBho5vF+o2v7x/e0t3HyD/AG5RnnhFwGOua/8AED4s+LH1TxBq2u+LfEmqPgy3Est7dXDcnAzuY98AdO1aH/wRl+OGt6x4ZMvj3w/p3h/VtGt73VL65gkN1ply8aGW1+zAjzWVmYK29VIQlihwp+Af/BLb9l/4JzW+t63oc/xC1+EAi78SIktvE/cx2gHljsR5nmMD0YV4x1sfsG/8FTL7x54nspLbwT4xu57yWeGLCmw1EEyyKB2huskqASRDwMkVo+s6R4i0m013QNUtNS02/hWa0vLSZZYp42GVdHUkMpGCCOKr/goj8S7f9tH9q/4ffs3/AAYv4tbtdAuH04X9owlhe9uXQ3Tqy8NFDFAhL5wCkuOBk6Vplpo2l2ej6fGI7axt0ggQADaiKFUccdAKqqqqqqqqqqqqqqkdI42kdsKgJJPYCvhLpUPxJ+Iur+MNei+0R2j+esUg3DzHJEYPsqqcD/ZHpVTQw3EL288SSxyKVdHXIYHqCD1FabZ/8Kx+O8Ok6c5TTtRlWJIyTzFMPlXn+6+Mf7o9arV/GHhXQcrrHiLT7R16xyXCh/wXOT+Var+0B8O9PDLaXV5qLrwBb2xUZ+r7ePetc/aW1ifdH4e8P2tqvQSXUhlb67RtAP51q/xV+IWtEi78VXsan+C2byBj0+TGfxq4ubm8k826uJZ5DxukcsfzNaF4B8Y+JZlh0fw9eSg/8tXjMcY+rthf1rRf2adduUEmveILSyz/AMs4IjO34klQD9M1pn7OXge0AbULzUr9x1DSiNfyUZ/WtL+F/wAPtHwbHwnp5YdGnj88j6GQsRUUMMEawwRJGijCoi4A+gFVVVVVU88FtEZrieOJF6s7BQPxNXHjvwRa5Fx4w0VCoyVN9Hu/LOa8O+MfDXi1rpPDuqpe/YmVZyiMoUtnGCQM9D0zVV8WPiDe/D3Rra+sNMiu5buYxK0zkJGQuQSBy3Q8ZFTfH34lSyF01S1iB6IlomB9MgmtS+MfxJ1RPLm8UTwr2FsiQkfigB/WpvFviq4DCfxPq0u772+9kOfrzXwN1Ga2+Jmnl2dxdpLFK2STyhIJ/wCBBaqviJoFz4o8FaroVjGslxcwjyVZgoLqwYcngcqKsP2b/G9yA17qGlWg7qZXdh+AXH61ZfsxJgNqXi8n1SCzx/48W/pVl+zl4Dt1/wBKu9Vu2xzvnVAPoFUH9aX9nr4fpOtxE2poUYMqi5BAI+qk1VVVVVVVVVVVVVf8FYP+TI/GH/YQ0z/0sir/AIJcf8mJfDH/ALi3/p2vKqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr/gs18Ftc8PfGvRPjfaWDyaD4t0uOxuLlEJEN/bZGxyOBvhMZXPLeXJj7tfE39pj42/GPwL4U+G/xF8c3Gr+H/BcQj0m0eCOPZtQRq0jIoMrLGAgZySBnnLMT/wAEaP2fb/wd8OvEXx78S6Sbe88ZuljobTR4f+z4STJKvcJLNgc9fs6noQT+0He2nxS/b18WL4UKSR6z8RPsNm6EMszi6WAOpHBDMu4eoYVVVVVVVVV/wWG/aU1jwt4a0L9mbwNezx6n4yh+2699mz5jWO8xw24xyRNKr7gOSIgOQ5FfstfC39nD/gnn8KNLm+MHjrwl4e+I3iWxS41291S/iS5YkbvstuhO8wxn5TsGHdSx6qF+Gvx9+CfxkMkXwt+KnhnxNPAm+W20/UY5J4l4+Zos71XkDJUCqqv+CyX/ACaLaf8AY4WP/om5r9gP/kzT4S/9i/H/AOhvVVVVVVVVVVVVVVVeLPF3hXwJ4fvPFfjXxHpuhaNp6b7q/wBQuVt4YR0G52IAycADuSAOtftQf8FkNE0k3fhL9l3Q01e6GY28U6xAyW6HoTb2xw8nGCHk2gEfcYV8U/jP8Wfjr4k/4SX4q+OtX8UamxKwm7lykOeqwwqAkSk/woqjPav2YP8Aglp8e/jpPZ+IfiBYT/DrwdIVdrrVLci+u4+pEFqcMMjo8uxcEEb+lfs//st/BP8AZm8OpoPwp8H29pcPGFvNXugJr++PGTLPgEjIzsXagPRRVV+2D+y18Mv2ofhdd6L46tJLbU9EgmutF1m1VRc2EoTJAJ+9G+1Q8Z4YAEFWVWX4Q/tW/tFfAW1/s74T/FvXNCsN5cafvS5tFcnJYW8yvECSeSFye+a+Jf7dX7W/xe0aXw746+OOuXGmXCGOe0sI4NNjnQjBWRbWOPzFIPKtkGv+CW/7J3wy+G3we8O/tAxRS6t418baX5rXt0i7dNgZ2Bht1H3dwUbnJLNjA2jIqqqqqqqqqqqqqqqpolnheBycOpU468iv2f8AUF8N+Mdb8H6owhurkBFDcZlgZgVHvhmP/Aaqvix4vFx8VH1vRpo5To0kKW8hG5S8R3H6gPke+K134keOPEZYap4lvGjbrDE/lR/TamAfxrTtH1bWJPJ0nS7u9k/uW8DSH8lBrS/gh8SNUCt/YItI2/jupljx9VyW/StF/ZmuWKv4j8TRIB96KyiLZ+jvjH/fNaN8EfhzoyqTohv5V/5aXshkz9V4X9KsdE0bS1C6ZpNlaBRgCC3WPH5AVVVVX+saRpYzqeq2doMcGedY/wCZq8+Jvw9sU3TeMdKYDtDcCU/kmau/2gvhzbS+XDdX90v9+K1IH/j5U/pXhnxNpPi7R4tc0WV3tpWZRvTawKnBBH4VVfG34k+NPB3iC30jQL6G1triyWYSeQrvuLOpGWyP4R271ffETx3qTFrrxfqpz1VLpo1/75UgfpV5qOoaiwe/v7i5YdDNKzn9TVv4X8S3iLJaeHdTnVjhWjs3YH6YFfs++GPF/h3UtSl1nQrqxsb22Xa9woQmRW+UbCd3Rm7dqqvib4A/4WJotrpC6kti1vdrMZTD5nyhWUgDI/vA/hWn/s6+BLa1SK+m1C8nH35TN5eT7KBwPz+tWPwK+Glk4kbRJLpgcjz7lyB+AIB/EVB4A8C26osPg7RQU+6xsIyw/EjNWljY2EXkWFnBbxjokUYQfkKqqqqqqqqqqqqqqqqqq/4Kwf8AJkfjD/sIaZ/6WRV/wS4/5MS+GP8A3Fv/AE7XlVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVU88FrBJc3M0cUMKF5JJGCqigZJJPAAHeq8e/D/AMFfFLwlqHgX4heG7HXtB1SPy7qxvI9yOAcgjurAgEMpDKQCCCM1o/8AwSX/AGM9I8WJ4mPhPX763il8yPRrzWZJLNSCCARgSOoxja8jAjg5qLStOttKTQ7G1SzsYrcW8MFpm3EMQXaqx7MGMBcAbcYwMYxXhH/glr+zJ4A+Mnhv4yeDh4ms7nw3qIv4NIn1EXVm8yAmJiZFM2Uk2uD5hyUGe9S3FvA8MU1xHG9w5SFXYAyMFLFVHc7VY4HYE9qqqqqqqqv28fGfi7xD/wAFGddvvB1t/aetaFr2k2OgWTQ+aHuYI7fZFsONwa43fL0JcjODmvhv/wAElPBXiayk8c/tXfEDxT43+IWvAXGqvb6l5UFvKwBKBypklZfu7yyrjgIABX7cf7AepfsYxaT8ffgF42186DaaikUrzThL7RLhv9VIk0YXdGxBTOAVJUHduyP2Cv2kbz9qH9nXSPHOvvEfE2lTvpOvmNAiyXcKofNCjAHmRyRyEAABmYDgVVf8Fkv+TRbT/scLH/0Tc1+zz/wVw8OfA74J+D/hJdfBDUdXm8L6cto99HrqQrOQxO4IYWK9emTX/D8Xwp/0btqv/hSR/wDxiv8Ah+L4U/6N21X/AMKSP/4xX7J3/BTfQv2qfi/bfCTT/hDf+HZriwuLsX02spcqoiUErsESnnPXPFVVVVVVVVVVVU00NvC888qRxxKWd3baFAGSST0GK/aj/wCCs/wa+EAuvCvwZS3+I3iqItG1xBMV0q0Yd2nXmc/7MWVPOZFIxXx3/aY+NX7SWvjXvi143u9VWF2az05D5VlY54xDAvyqcYBbljgZY1+y9/wTN+Pv7RYs/EetWR8BeDJ8ONX1e2bz7mM/xW1tlWkB4wzFEIOQx6V+zt+wl+zl+zTBb3ngzwZHqviOIAv4i1oLdXu4d4yQFg57RKvbJPWqqqqqqqqqqqqqqqqqqq1LVtK0e3+1atqVrZxD+O4lEY/MmvEH7QfgbSS0Ol/atXlHeBNkef8AebB/IGtW/aV8S3IZNG0KwsgejTM07D6fdH6Vq/iTWNc1x/Ed7dBdQd1fzoUERDLjBG3GDwOetaF+0Z4u063S21ews9U2DAlbMUh+pXg/lXir49+MvEdpJp1ktvpNvKpVzbZMrA9RvPQfQA+9aH+zNI8ccviLxKEYj54LSHOPbex/9lrQfgx8PNACvHoa30y/8tb4+cT/AMBPy/8AjtQW9vawrb2sMcUaDCpGoUL9AKqtT1/QtGUtq+tWNkAP+Xi4WP8Amav/AI0/DSwYxv4mjmcHGIIJJP1C4/Wp/wBob4eRRl431KZh0RLXBP5kCrO7gv7OC+tm3RXESyRkjqrDI/Q1VfE3WPFeneN9a0q48R6o0cN2xija7fCxt8yADOAArCooLu+m8u3hluJm5Kopdj78c1a+BPG162y28I6w5zjP2KQAfUkYFWXwM+JV7CZjoSW425VZ7lFZvbGSR+OK+DHhfxH4Q8Lz6N4jtkhk+2NLCFlV/kZVHYnuD+dVXjD4Y+GPHOoWuo68t072kZjRIpdispOecDP6itP+E/w50xAlv4SsJMd7hDOf/HyasPDnh7S2D6XoWnWjDoYLVIyPyAqqqqqqqqqqqqqqqqqqqqqqr9oj/grF4e/Z9+M/if4O3nwU1HWpvDVxHC9/FriQLPvhSTIQwsVx5mOp6V/w/F8Kf9G7ar/4Ukf/AMYr/h+L4U/6N21X/wAKSP8A+MV+13/wVG0D9p74Faz8HbD4OX/h+bVbm1mW/m1pLhY/JnSQjYIlJzsx14zX/BLj/kxL4Y/9xb/07XlVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX7QnhXXPHXwC+JfgjwxZi71nxB4P1bTtOtzIsfm3E9pLHGm5iFXLsoySAM8kV8PP23P22f2Ntaf4aeI76/lh0g+W3hnxrZSTiBRkDy3JWZI8A7dknlkYIBFeBv8Agt/4amgih+JfwJ1O0lUYkuNC1ZLhXOOoimWMrz2Lt9as/wDgs1+yXdQCWbQPiPaNkjyptHtiw9/kuWH61r//AAWm/ZlsIWXQPAvxE1W4C5QPZWtvETg8FjcFhzj+A9fwr4of8FsfiXrFvNYfCL4SaJ4bDgql9rF4+ozAf3ljQRIrezbx9a/4J+6z+1l+01+2B4P+PfxKv/E/ifwr4UbUvterXmINPs3msLmFY7eMbYtxklQFIVyAQWAHNVVVVVVVX7U8cPwG/wCCrVj478RRfZ9Em8VaJr4nkBUNav5InkGT0WRLgdcZTt0AKsoZWBBHBFf8FPtY0fSf2IfiLFq8kedQWxtbSJwCZZzfQMoUHuoRn9QEJHSv+CJ2i6pafAXxxrtyJVsdQ8V+Vaq4IVmitYvMdRnv5iqTjqmMnHFV/wAFkv8Ak0W0/wCxwsf/AETc1+wn8N/h3q37IPwr1HVfAXhy9u7jQUaWe40qGSSRt7cszKST9a/4VN8K/wDomnhT/wAE1v8A/EV/wqb4V/8ARNPCn/gmt/8A4itI8BeBfD16NS0DwXoOm3aqVE9npsUMgB6jcqg4PpVVVVVVVVVVVftQ/t1/Af8AZXs5bHxbrp1nxYYt1t4Z0p1kumyMq0xztgQ8fM5yQcqrYxX7Un/BQH49/tRy3GjazrP/AAjXg12/d+GtIlZIXA6faJOHuD0OGwgIBCKa/Zt/Yu+PP7UuooPh34Va30BJdl14j1MNBYW+D8wD4JlcZHyRhmGRkAc1+y5/wTL+Af7O4svEviCwj8e+NbcK/wDaurW4NvayDBzbWxJVCCAQ7b3BGQy5xVVVVVVVVVVVVVVVVVVc3VrZwme7uYoY16vI4UD8TWvfFbwB4ftXnuPElpdOvAgs5VnkY+mFOB+JArUf2nEDsmk+Eiy/wyXN3g/iqqf51q3x88V65pWp6Ve21tAt7GFt5bNnie2YMDkHJJyBg9Ov4U8l7qVwvmST3U7/ACruJdm9h1NeHvgt8QfEWyRdHOnwN/y1vz5OP+A8v/47Wlfsy2ahH1zxTNIcfNHaQBMfRmJz/wB81YfAn4a2ULRS6NLeMwwZLi5fd+G0qB9QKvPhX8EdPJS/sLG2IOCJtWlTB/GT2NX3hv8AZs0+LzZ7nSnHpBqs0x/JJCaX48fDya8isbS9vLh5pVjRktWAyxwD82Dj8KqtWsm1LSrzTkmaJrq3eJZFYqULKRuBHIIznNTa54ihMtpNrOoLyVkjNy/UdiM1bWd7qM3k2drPdTMfuxRl2J+g5rT/AIT/ABG1RQ1r4RvlB6faFEH/AKMK1H8A/iXIyq2k20eRyWu4+PyJrwRpWqaH4S0rRtZkhe8srcQuYWLLheFwSAfuhe1VV14S8LX1/Jql94d025vJQN889qkjnAwOSCenFWtlZWMQhsbSG3jHRIowg/IVVVVVVVU/ij4m+K/H934U0XxbdwyPe3MduvnmFFVNxxlR/dSh8K/jqnzr8SNxXkA6xdHP4FMfnV/rvx6+GwF5rMjajp6H5pJVW4i/4Ey4dfxIr4bfFbSPiDA1q0YstVgXMlqz5Dju6HuPUdR+tazJqEOj30ukxLLfJbSNbRt0aUKdoP1OK+CeufEvVPEGoweKzqEtgsJLPeQlPLnDDCrkDtuyo4GO3eqqrXxB8W2+MJ0+SK+/s37eUaAwn7OLTdjeDjH3Od3Unj2qqr4n3vibT/BV/deEo5G1BAuDEm90TcN7KOcnH5DJ7V8ENT8a6p4dupPGAunCTgWkt0hWR1x83XkgHGCfUjPHFVVVVVVVVVWp/Dn4e61fS6nrHgTw7fXk5zLcXOlwyySEDALMykngAc1/wqb4V/8ARNPCn/gmt/8A4iv+FTfCv/omnhT/AME1v/8AEV/wVK+H/gPQP2MfFup6F4J0DTryO/00JcWmmxQyKDeRA4ZVBGRxX/BLj/kxL4Y/9xb/ANO15VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVXxT+Cfwl+N2ijw/8AFj4faL4ns4w3ki/tg0luT1MUow8TH1RlPvXxC/4Ixfs2+JJpbzwH4s8X+D5HzsthOmoW0fphZV80495T/WtT/wCCG2pJKDo37SdrLGSci58JtGyDsMrdNu46nj6V4f8A+CHGlRSRy+Kv2jbu5jx+8g0/wwsBB9pHuHz2/gr4Tf8ABKv9kL4XzQ6hqXhLUPHOoQ8ibxPdC4i3d/8AR41SFh6B0bHqetadpmm6PYQaVpFhbWNlaxiOC2tolijiQdFVVACgegFVVVVVVVX/AAUT/YfP7WXgmy8R+B3tbX4ieFYpBppnYJHqVux3NaO5OEO75kc8KxYHAcsPht/wUh/ah/ZB8NWvwh/aL+Aeo662gRrZ6ddapcy6TdCJBhEaYwypcKFACuoyQM7m61498Vftkf8ABVfxVoXhrQfhxJ4S+HGnXQnSV1lGnWrlcG4nuXVftMoRmCpGuQGOFG5mPwH+DHhT9nz4TeHfhH4MV30/QbbY1xIoEl3MxLSzvj+J3Zmx0GQBwBVV/wAFkv8Ak0W0/wCxwsf/AETc1+wH/wAmafCX/sX4/wD0N6qqqqqqqqqqqvij8Wvht8FvCdx43+KXjHTvDuj2wIM93JhpWxnZGgy8rnsiAsfSv2r/APgr1468dm88F/s12tz4Q0Jt0cniG5Uf2ldDpmFQStspGefmk6EGM5FeA/hz8Uvjt42Xw14B8Naz4u8S6nKZZEhUzSMWPzSzSMcIuT80kjBRnJNfspf8EgPBng0WnjP9pm9t/FWsqA8Xhuzdhp9s3X99Jw1ww4+UbYxyD5gINaXpWl6Hptto2iaba6fYWcQitrW1hWKKFFGAqIoAVQOAAMCqqqqqqqqqqqqqqqqtV17Q9CiEus6xZ2KkZBuJ1jz9MnmvEf7QngrSA0WjJcaxOOnlKYo8+7sM/kCK1z9onxtqStDpNvY6VGfuskfmyD/gTfL/AOOitV1vWtfuftOsapdX0xPDTSlyPYZ6fQVp/gfxjqzIuneF9UmD9HFq4X8WIwOnc1YfAee3tVufGfjLStAZhkwyOrso9yWVQfoSK8G+APgo2v2ulL4nk8Q6k5YxwEEQttUsT8oweAeCxz6GrHSNK0xSumaZaWgIwRBAsefyFV8TrrVrHwHq99od69reW0IljlTqoVgW/wDHQwrUfFHiXWARqniHUbsHqs107j8icYq2sb29O2ztJpyOojjLY/KrTwL40vnEdr4T1dz6/YnAH1JGBWn/AAO+JszR3C6JFbENlTPcxgjHIJAJNKSygsuDjkZ6VVTeD/CNzP8AabjwtpEs2SfMexjZsnqckZq3tbWziEFpbxQxjokaBQPwFVVVVVVVVVVVXw8/5L6P+wjf/wDouaqliiniaGaNZEdSrIwyGB6gj0r4jaJL8KPiRbat4czFbuVu7VAcBOSHi/3eCMf3WAoavBLoQ163UyQtafaUXONy7Nw5+lfDn4u23xE1S50uDQ5LI21v5xd5xJn5gMYCj1rxv8Z4PAvib/hH9R8OTSRlEkS5W4ADI3Vgu3sQwxntSOksayROGVhlWByCD3FeKvENr4U8PX3iC8QvHZRFwgbaXboqg9skgfjXw3+IzfEWC9u4dBksLezdUEjzh/McgkgYA6DGf94VD8Z7Wbx+fAY0CUSC9a1+0/aBjIz823b7dM1PNDbQvcXEqRxRKWd3OAqgZJJ7DFa9+0dpdvetY+GPD8+qKhwJ5JfKDY6lVCkke5x9K8CfHG38Ya7B4au/DVzZXlxuCNHKJFG1SxLZClRgH1rxr4nj8GeGLzxJLZtdLZ7Mwq+wtudU64OPvZ/Cvhz8QIfiJpVzqcGmPZC2uPJKPL5mflBzkAetanqen6NYTanql5HbWtuu6SWQ4Cj/ADxjua1j9pLTUuWtfDfhm51AA4Es0vlbsdwoDHH1x9K0H9pDRrq7Wz8S6DcaXltpmjk85U92XAYD6Zq1ura+toryzuI54JkDxyRtuV1I4II7V8SfipD8Obmxt59EkvhfRuyss4j27SARjB9RXjf42+G/BzrYRW0mpakYwz28UgVYcjIDvzg+wB98Vo37S+n3F2sGu+G5LSBjgz29x5pT6qVGR9D+FWF/Z6pZQajp9yk9tcxh4pEPDKRwaqqqqqqr/grB/wAmR+MP+whpn/pZFX/BLj/kxL4Y/wDcW/8ATteVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVf8ABZL/AJNFtP8AscLH/wBE3NfsB/8AJmnwl/7F+P8A9DeqqqqqqqqqqpZI4Y2llkVERSzMxwFA6knsK/bc/b78cfH3x1p3wI/Y0ufGqzaTqjFtd8LahcwXOtSKjIYoY7fDNbqSW3sSGKhgAoDNr/8AwTr/AOCjnxjs7XxN8TRfa1f28BW0h8UeM0vbqOM87VLyyBAeu0sMHqAa8YfAfxr+z/8AETR9B/ab+G3ivQNIlulN4tk0QlurcEeabS4O+CRwD2LDOASM5r9kzwn+zpoHwY0LV/2ZdF0228Ja3bLPHdwRk3F24+VjcyNmRplYMrBzlSCoAAAFVVVVVVVVVVVVVVV4n+I3gzwiGj1rXIFuFH/HtF+8l/75XJH1OBWtftM2aFo/D3hmWXrtlvJgn/ji5/8AQhWv/G34h68GjGsDToWH+rsU8rH/AALl/wBaVNS1i8wi3N7dSnoA0juf1JrQPgn8QteZWOjHToT1lv28rH/AOX/8dr4hfB4/D3w1a63Nqv8AaE8t0IZY1hKRplGIOc5PK4zx1Fab411rQ1A0BLLTmGf3sNojS/hI4Zx9Aa1Dxr4w1UFdQ8UapOp6o92+3/vnOKgtbu/m8q1t5riVj92NC7H8BzXwt8D+PbHxno2tr4Xv4baC4UyyzxeSFjb5XPz4z8pPTk9qqr2ytdRsp9PvYhLb3MTRSxnIDIwww49QTWm/D/wRpAH9n+FdMjZRgO1srv8A99Nk/rSRxxoI4kVVHRVGAKqqqqqqqqqqqqqqqq+Hn/JfR/2Eb/8A9FzVVftP+V5vhzbjzNt1u9cfusf1rwx5n/CpNP8ANzu/sFcZ9PJ4/TFfs0/8jXqn/YO/9qJX7R/hgX/h6z8T28eZdMl8qYgf8snIAJ+jYH/AjXwX8RnxJ8P9PaWXfc6eDaTZ6/Jjb/44V/HNftKeIxb6TpvhaCX95dym4nUdkThQfYsSf+AV8MPC/wDwiPgnTtKlj2XLx+fdAjnzX5IP0GF/4DVl/wAnDN/2HJP5tXxnW/f4aa0NOBL+Whk25z5YkXfj/gOc+2a/Zy1nwrbabeaVLPbQa3Nclh5uFeaLau0KT1wQ3yj609hYyXaag9nA11EpVJzGN6g9QG6gH0r44/8AJLdb/wC2H/o+Ov2af+RT1T/sI/8AtNK/aB8RX+t+K7PwLp7kxWvllolOPMuJMbc/RSuP9414J8E6N4H0WHTNNt4zPsH2i52jfO/ck9cZ6DsK+LHgHTfGXhq7nS0jGq2cLS2s6rhyVGdhPdT0x2JBr9m3xTczpqHg+5lLx26farUE/cBIVwPbJU/Un1r9p3/j+8P/APXGf+aV8G/AmlaH4XsfEFxbR3GrarCtzJcyDcyK43Kqk9OCCe5PXtj47eDdK1bwdd+II7SJNS0wLIsyoAzpuAZWPcYJI9CPc1+zdq8954Vv9ImkLrp92DFn+FZBnA9tysfxqqqqqqqv+CsH/JkfjD/sIaZ/6WRV/wAEuP8AkxL4Y/8AcW/9O15VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVXNxb2dvLd3dxHDBAheWWRgqooGSxJ4AAGSTXxJ/wCCof7G/wAN9Sm0Y/EW48UXlucSJ4csXvIwfac7YX/4C5r9mb9rj4S/tZ6Xrmq/Csa2ieHZoYr6PVLIW7q0ocpt2uwIIjbvxU3xQ+GcHim08CzfETwyniS+dktdHbVoBeTsql2CQ7t7EKrE4HABNVVVVVVVVVVVVf8ABZL/AJNFtP8AscLH/wBE3NfsB/8AJmnwl/7F+P8A9Deqqqqqqqqqq9vLPTrOfUNQu4bW1tYmlnnmkCJEijLMzHhVABJJ4Ar9v7/gqLp3ibSNb+BX7N14lzpuoQy2OueK8HbcRMCskNmP7rAlTMeoJ2DkPX/BJv8AZ68LfDf9nTS/i9NpcMni74gLLcT3zpukgsllZIYEP8KEIJWx1ZxnO1cV+0N8C/Bv7Rnwm134WeM7GCWLUrdzY3bxhn0+7CnyrmM9VZGPbqpZTkMQf+CK/wAQtdsLz4n/AAE12RxHpUsOrWlu7c20oY290MZ6Ei36cAqc/eqqqqqqqqqqqqqq1Tx74K0aaW11PxRp0E8I/eRGcF19ioyc+3WvEv7SOg2TNb+GNJn1JxkCec+TH9QMFj9CFrxN8afHviZGt21RdOt2GDDYgxZ+rZLfhnHtTEsSzMSSeSar4J+HvDnifxi2l+JLH7VF9keSGMyMoMileuCMjaW49q0rQ9G0OAWujaVaWUeMbYIQmfrjr+NV428H2HjnQX0DULiWGJ5UkEkQG5Sp7Z46ZH41Yfs+/DmzAFxa319j/n4uiuf+/e2rD4aeANMwbTwjpmR0aWASkfi+TVvaWtnEIbO2ihjHRI0Cj8hVVVVVVVVVVVVVVVVVVVVXw8/5L6P+wjf/APouaqr4taw3xF+JNr4f0CQTx25WygdeVaQt87jHYE4z0wuelX9jBpnhK4022H7q005oowf7qx4H6Cv2af8Aka9U/wCwd/7UStf0a18Q6Je6HeAeVewNExxnbkcMPcHB+or4Aapc+HfGWq+CNTHlyXIYBCfuzwk5A+q7v++RRiHxS+O7gjzdO0uX5geR5MBx+TSH8nqrL/k4Zv8AsOSfzanRJEMciqysCGVhkEd+K8Yfs66Pqk0t/wCE9Q/syZyW+zSruhz/ALJHzIP++vYV4E8e+MfAHjGLwJ4yuJZ7Rp1t2Sd95ty2Ajo/XZypx0x0wa+OP/JLdb/7Yf8Ao+Ov2af+RT1T/sI/+00r4gSDRvjwuo6g+II9Rsrgux48sCMk/QYI/Cq1C8t9OsLjULpwsNrC0shJxhVGT+gr9m60ml8bXl2mRHBpzhjjuzpgfoT+FftO/wDH94f/AOuM/wDNK8Bf8iL4c/7BNr/6KWvix/yTnX/+vQ/zFfsx/wDHhr//AF2g/k9VVVVVVV/wVg/5Mj8Yf9hDTP8A0sir/glx/wAmJfDH/uLf+na8qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv+CyP7Tfiiy1/SP2ZfCWrT2Olvpyap4kaByhvGkZlhtmI5MaqhkZejGRM/crUP2Svjxo/wKb9o/XvBbaV4GJg+zXl5cRxzXSzSCON44M+ZsJYEMygEHIJHNf8EOv+RU+Lf/YQ0r/0Xc1qsifAb/goJdy6EIrW08IfFVzbRhtqLapqJxGTxhTF8p9iaqqqqqqqqqqqq/4LJf8AJotp/wBjhY/+ibmv2A/+TNPhL/2L8f8A6G9VVVVVVVVV+0L+0t8I/wBmHwW3jP4qeIltVm3Lp+m24El5qUigZSCLI3YyMsSEXcNzDIr9rz/goR8Yv2qrq48PfaG8K+AllJt/D1jMf9IAOVa6l4MzcA7eEBAwufmPgz9m/wCM3j/4X+KfjL4a8E3k3g/whbefqGpyDy43AdVdYc/61kBLvtyFVSSQcA/8Eq/jJ4e+Jn7J+geELS+iOveAC+l6paZAdEMjvbyY/uNEQN3do3HY1Xxc+KHhb4LfDXxD8UPGd7Hb6V4dsZLmXfIEMzAfJCmeryOVRR3ZgK/4Iv8AhTXfEfjj4tfHTVrdlhvFi05J9pCTXE0puZ1Xt8gWEn081fWqqqqqqqqqqpZIoY2lmkVEQZZmOAB6k1qnxW+HekFlu/Flk7JwVt2M5z6fIDWq/tI+D7UFNK0vUb5wOCyrCh/Ekn/x2j+04xtJCvhEC5LnylN3lFXAxk7ck5z2HavGHxd8Z+MlW3ur1bG1Un/R7ItGrf7xyS34nHtWnaVqutXP2XSdOur642ljHbwtK2O5woJx71b/AAT+Jd1bRXKeG2XzWwqSTxowGPvEFuB9efatL/Zt8XXWG1XVtNslP8KlpnH4AAfrWmfs0+GYMHVtf1C7YDkQosCk/Q7jj8a034M/DbSwrR+GorhwOWuZHlz+DHb+lafo2j6Snl6VpVnZrjG23gWMfkoFVVVVVVVVVVVVVVVVVVVVVVVVab4mt/B3xau/El1bSXEVnqN5uijIDNu8xOM/72am/aY8MrGTb+HtTd+yuyKPzBP8q1b4nfE34mxtpHgzw7PZWk3ySPb5diD1DTEBUH5fWvhN8H4fA5/tvWpIrnWJE2oE5S2U9Qp7sehP4DjJOv8A/IC1H/r0l/8AQDX7NP8AyNeqf9g7/wBqJVfGmxvvA/xGs/GmikRNfL5qNjgTINrjj1UqT7sa/Z08Mf2f4buvE9yn7/VZdsRYc+UhIz+Lbv8AvkVVl/ycM3/Yck/m1fEXxjJ4G8NPr0Onm9kWaONYskDBPzEkA7RtB59cVbftJeC5LXzLrStWinC8xLGjjPs24cfUCtKttY+M/wAUI/EcWmPbaXbTRNK5+7HFHghS3d2x0HTPoM18cf8Aklut/wDbD/0fHX7NP/Ip6p/2Ef8A2mlfHD4Y3ni+1h8Q6BCJdTsYzHJAODPFnI2/7SknjuCe+BXgv4+yeHtOTw/440m/lnsR5SzxKPNIAwFkRyvzDpnPPcZyT46+LmrfEqEeDvA+iXqw3ZxLuUGaYD+HCkhV9Tk/gM5+Evw7/wCEA0B471kfVL9hJdMvITA+WMHuBk8+pPbFftO/8f3h/wD64z/zSvAX/Ii+HP8AsE2v/opa+LH/ACTnX/8Ar0P8xX7Mf/Hhr/8A12g/k9VVVVVVV/wVg/5Mj8Yf9hDTP/SyKv8Aglx/yYl8Mf8AuLf+na8qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv+CvnhTVNB/bDv9cvIXFr4k0Gwu7OQphWWOM27AHuQ0BJHBG4ccgn9rn9oj4RfHj/gmbdaj8O/E2lm7gj0SG80NZ0S602WO4gV4nhzuAUggMBtYAEEg1/wSP8AjF8NPgl8KvjN4z+J/jDTdC021u9NcG5nVZLgrFcnZDHndK56BVBJJq11XVf2kf2toNat7CWO8+Ivj5Z0tkbJtxdXoYJnB4RXxkg4C5NVVVVVVVVVVVVf8Fkv+TRbT/scLH/0Tc1+wH/yZp8Jf+xfj/8AQ3qqqqqqqqr9qr9t74K/so6NNH4s1hdW8WzW5k07wxYOGupyR8jSnkQRE4+d+SAdquRivjp8cfiN+0l8TNQ+I/xB1J7zU9Sl2WtnCW8myi6JbwISdqAY46sSWJLMSf2LP+CSiapaab8UP2p7eeKGZEuLHwZGxidlIBU3rjDJxz5K4YcbmHzJVr4V8M2PhpPBtj4e0230GO0NmmlxWqJbLbldpiEQG0JtJG3GMV8b/wBgr9pT9kj4sp8Yv2G9S1u/0m+nfy9L08Ga705SC7QTRuCl1bcYXdubO0MpIDtB/wAFOv2//Dlq2geJv2aLCbWbYGNpbvwnqltJuxgGSISD5s9QNgPTA61P8H/+Cif/AAUR8X6VYfG621XwN4Kh3XcbalpEmm2NqoYAmK1OJJ5iHOwyHlc/OFya+BfwT8Dfs8/DHR/hT8PbJ4dL0mM7ppiGmu5mOZJ5WAGXduTwABgAAAAVVVVVVVVVXxu+I3jTwd4gttK0DUIrW1ubJZQ4gV33b2DctnsB271fatqmpyyT6jqVzdSSnLtNKzlj75NVpOhazr9x9k0TS7q+mHVIIi+33OOg9zWhfs7eNdSCyavcWelRnqrv5sg/4CvH5sK0H9nbwXppSXWLm81aVequ3kxn/gK8/mxrTtH0nR4jBpOmWtmhAysEKx5x0zgc1VVVVVVVVVVVVVVVVVVVVVVVVVVVXGh6JeMzXWj2MzP94yW6sW+uRUPhnw3bNvtvD+mxN6paIp4+goKqqFUAADAAGMVTKrqUdQwIwQRnNW2mabZOZLLT7aB2GC0USoSPwFVdWNjfKq3tnBcBD8oljD4+maihht4lgt4kjjQYVEXaB9AKoaVpguPti6da+fu3eb5K7s+ucZzRAYbSAQRyCKm8HeELiUXE/hXR5ZQch3sYywP1IqGCC2hWC2hSKNBhURQoX6AdKuLe3uoWt7mCOaNvvJIoYHHqDVtZWdkhjsrSGBWOSsUYQE+uBVal4e8P6ywfV9D0++YDANzapKR/30DWn6RpWkReRpOmWllGeqW8Cxj8lAqrrTtPvipvbG3uCgO0yxB9vrjNRxxxRrDCioiKFVVGAoHQAdhU0MNxE0E8SSRsMMjrkH6irWwsbEMtlZwW4f7wijCZx64qqqqqqqv+CsH/ACZH4w/7CGmf+lkVf8EuP+TEvhj/ANxb/wBO15VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVft2fsY6L+2B8OLexsr6HSfGvhsyzeH9SlB8olwN9vPgE+VJsT5gCUZQwBG5W+L/AOzr8bPgNq82j/Fb4cazoRicql5LbF7SfnGYrhMxyDkfdY9cHB4rw94Z8R+L9Vh0Hwn4f1LWtTuDiGy060e5mk/3UQFj26Cv+CaH/BO3xf8AC7xRbftC/HnRxpmt2kDL4b8PysGmtGkQq1zcAZCvsYqkZJK7iWCsABVVVVVVVVVVVV/wWS/5NFtP+xwsf/RNzX7Af/Jmnwl/7F+P/wBDeqqqqqqqr/goB/wUo0j4DR33wi+CN/Z6r8RiGh1C/wBolt/D+R3BBWW454Q5VCPnBI2HR9F+KHx7+Iy6dpFprfjTxp4nu2dss1zc3crcs7uxzgDJZ2ICgEkgDNfsJ/8ABL7TPgVq1l8XPjpNp2veNrUCTS9Ktx5tno0nBEpYj99cL0BA2Icldx2utVVVVVVVVEgDJIAA5JrxF8XPAPhotDd67Hc3CdYLQec2fQkfKD7EitX/AGmrcEx6D4Xkf+7JeThf/HFB/wDQq1P9oL4iX4K2lxY6eD0+z2wY/nIWr4C+P9R8RW+raX4j1WS5u7ZhdRyzvz5RGGHsqkA/8DrxF8afh94d3RNrA1C4T/ljYr5vP+9wv614k/aQ8QXu6DwzpVvp0fQTTnzpPqBwo+hDVrniHXPEt39u17Vbi+mAwrTPnYPRR0UewwK8M/Cbx14q2SWOivbWzdLm8zCmPUZ+Zh/ug14V/Zz8O6dsufFF9Jqsw58iPMUI+uDub8x9K03StM0e0Sx0mwt7S3T7sUMYRR74Heqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv+CsH/ACZH4w/7CGmf+lkVf8EuP+TEvhj/ANxb/wBO15VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVfGX9sH9nn9n3xppHgL4vePk0DVdbsjeWoeynmjWLeY1Z2iRvLDMrgFsD5G5GK8J+LfCfxE8L2Xi3wbrthruhatEXtL20lEsNwmSpwRweQQR2IINfGP/AIK4fs8fBzxVfeCPhj4EvfGrafcNFeX2mzRWNi0i/K3lSbWM2CMbggU9VZhzX7KX7d/wT/a2+1aP4Nlv9E8U6fB591oOrKqTGIEAywshKyxgkAkEMMjcoBBNVVVVVVVVVVVX/BZL/k0W0/7HCx/9E3NfsB/8mafCX/sX4/8A0N6qqqqqqr/gpJ/wUKj+COm3HwS+CmtxSeP9RhxqmqQMH/sGBhwFI4+0uDx/zzX5jgla+DfwZ+Jv7SHxKs/h98PNLl1bXdVkaW4uJ3by7aPP7y5uJcHagzksckkgAMzAH9j39i/4a/sjeDFsdDgi1bxhqMCjXPEU0WJblhyY4gc+VAD0QcnALEkDFVVVVVVVVV+0h/bVpr1iV1O7/sy+tflt/ObyxIhwxC5x0ZPzrw94I8WeK3C6DoV1dKTjzQmyMfV2wo/Oo/gJd6ZaC+8a+MdI0OA9Mt5jfTkqM/QmtTg+DugqY9Pn1nxLdLkAlxa2+frt3n8OvrV7frczu9paRWMLAqIIC2Auc4JYlm6DqTXhv4e+MfFoV9D0K4mhY4+0OPLi9/nbAP0GTXh39miZ9k3irxAqDvBYpk/99sOP++TXhv4aeCPChSTSNBt/tCYxcTDzZMjuGbO38MVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV/wVg/5Mj8Yf9hDTP/SyKv8Aglx/yYl8Mf8AuLf+na8qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv+CuOqX+oftoa5aXjExaZomm29pkEYjMIlP/j8snSv2rvjNoH7N/8AwTi8KeHvh9q0VpqHi3wvpuh+Hmt5gZDDJbIbidSOT+535cYw8qHOSM/su/ss/Ef9rPx/P4C+Hkun2bWFk17f6jqTOlvaRAhRuKKzFmZgFUDJ5PAUkfC648cfsh/ti6Da6zPHaa34G8Wx2Gqm2lLRTwGTyrgK2ATHJC74JAOHBwDxVVVVVVVVVVVVX/BZL/k0W0/7HCx/9E3NfsB/8mafCX/sX4//AEN6qqqqqtX1XTdB0q81zWb6KzsNOt5Li6uZm2pDEilndj2AUEk+1fEf9on9qz/gpN8X9R+D/wCzZc33hz4e2LtvaO5eyja2DbRc6hOg3YcjKwKD6bXZS1eHP+CHmjjRgfF37QV42qugONO0BRBC3cZeUtIPf5PpXxD/AGTv2vf+Campn45/Br4gp4g8MW7pHql5p9s0amLdwt/ZMWUwnOA6s+wt95DtY/sb/tT+Hf2tvg9a+P8ATrWPTtbsZfsWv6UrlhZ3aqCShPJidSHQnsSpJKtVVVVVVrXivwz4dXOua7ZWbYyElmAc/Rep/AVrn7RXgnT1ZNHgvdVlH3Ssfkxn6s3zD/vmvEvx98c62Wh0yWHR7c9Ftl3SY93bn8VC1+zp4rvtTuta0XVdRuLudwl3G88pkY4+Rzk5PeOqrx1r/wAPtDjt7nxqbCWWHL2sM0AmkyepRcEjoOeB714q/aPnlgay8F6R9kGNq3V2AWUf7MYyo/Eke1arrGq67eNf6xqM95cP1kmcsfoPQew4rTdG1fWZDDo+lXl9IoyyW0DSkfgoNaTa6l4C1OPU/FXw+e7hQjbHqMEsSZ9Qfun/AIEGHtXgD4ieHPHlgW0cm2uLZQJrKQAPEOgIxwV7ZH44qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/wCCsH/JkfjD/sIaZ/6WRV/wS4/5MS+GP/cW/wDTteVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX/BYf8AZV8V+IdR0v8AaZ8CaJPqVrp+mDT/ABTDbJue2jjctDdlRyUxIyORwoSMnjcRLf6zq0VjpU19e3kdqDFY2zSNIItzZKxrk7csegHJr/gmB+y5rP7NnwP1DxV8RdPGl+LPHM0d9fW1wNkmn2USN9nhlz91xvlkYcbfMCnBU14u1mP9pb9uu61Lw2r3Fp44+IscWnNEuC1q94scTkdv3QViTwOSeBVVVVVVVVVVVVX/AAWS/wCTRbT/ALHCx/8ARNzX7Af/ACZp8Jf+xfj/APQ3qqqqqq/4Kc+K9T8JfsT/ABDuNIlaOfU47TTnkVtpWKe6iSUe4aMumP8Aa/A/8Ejfh9ovhT9jzRvF9jawjUfG2qX19fThfnfybiS1jQnrtVbckDoC7Hqxqtd0PSPE+iX/AIb8QafDf6ZqlrJa3lrMu5J4ZFKujD0Kkg/Wv+CR8978Pv2r/jL8ELW9efTLbTrp2O7IeTT9QS3jftyVuX5x37VVVVVX7Qmv+NNBnsI9J1ie00m+iZWEHyN5qnkFx82CpGBn1qz0/VNauvI0+yur65c5KRRtK7E98DJrRPgT8RNYCvNpsOnRN/HeShT/AN8rlvzArw5+zd4fsyk/ibVp9RcDmGEeTH9Cclj9QVrRPC/hzw3F5WhaLZ2QxgtFEAzfVup/E14m8X+HfCFkb3X9Tit1x+7jzmSQ+iqOT/Id68aftBeJNad7Twsp0iy5Ak4adx6k9E+i8j+9Wj6B4n8f3l9JYyyalqFtB58iTTFpZUBCkqWPzEZHGf8ACvDf7OHiO/WO48Sanb6ZGeTDGPOlHscYUfmaTwHo+t/Fg+BdDMsenWcpinmd90jCMZlYnpuLAqMAAccda0jRtL0DT4tL0axitbWEYSONcD6n1J7k8mri3t7uB7a6gjmikXa8cihlYehB6ivH2jD4PfEbTPEvh1Wi0+7YyrAp4ABAliH+yQwI9N3sKR0ljWSNsqwyp9RVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVf8FYP+TI/GH/YQ0z/0sir/AIJcf8mJfDH/ALi3/p2vKqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqplVlKsAQRggitQ0H9kP4a+MP+Eo1XRvg/4W8VeYX/ALRuLbTrG+3k5J81gsmcjOc9qf8AaC/Z+kQxyfHD4esrAhlbxLZkEd+PMrTrD9he6+IukePNCuvgwPGtpcEadqGnXthHdvNIpjwDGwMjkOQM5OTxziqqqqqqqqqqqqv+CyX/ACaLaf8AY4WP/om5r9gP/kzT4S/9i/H/AOhvVVVVVeKPE3h/wX4c1Lxb4r1e20vR9HtZLq+vbl9scESDLMx9ABX7e/7dPiD9rXxguh+HPtek/DjQZ2Olac7lXvpOn2u4UHG8jIRefLViMksxP/BIX9rbwXp/g2b9mPx7r1ppOq22oSXXheW7lEUd7HMd0lsrHjzVl3Oq5y4lIA+U5r9qz9qP4ffssfDHUvGPinVrSTXJLaRdB0QygT6ldYwiqvURhiC74wq56kgH/gjL8JvElw3xB/aV8UwzAeJJP7L0yeQbftZEpmvJQO6+YIVDDjcsg6qaqq8T+PvCPhCNv7c1qCKYDIt0O+U+nyDJ/E4HvXgH4taN8QdXv9KsLGa1NrGJITO43TJnDHaPu4JXufve1VXiLwvoXiyzi0/X7BbuCGZZkRmK4cZAOQQehIx71p2laZo9qtlpOn29nAnSOCIIv5Cq8V+PvCnguDzNd1WOOYrlLaP55X+ijnHucD3rxf8AtD+ItW32nhe3XSbYnAmbEk7D/wBBX8AT70zax4i1IFnvNSv7lsDJaaWQ/qTXgX9nnUdQ8vUfGszWUHVbKJgZW/3m5Cj2GT9K0Hwp4c8MW4t9A0a1swF2lo4/nYf7TH5m6DqarwjcJ4c/aF1e01FhGdQublIy3HMreYn5jA/EVVftK30d7qWgeH7QCW7RZZGRRlhvKqgH1Ktx9K061Nlp9rZFtxghSMn12qB/Sqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/wCCsH/JkfjD/sIaZ/6WRV/wS4/5MS+GP/cW/wDTteVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX/BWr9s3x/4A8Q2H7OXwp8Q3egtcaat/wCI9TsZjFcOspYRWqSLho12qXcrywdBkAOGlkkmkaWWRnkdizMxyWJ6kn1qv2ev+S/fDP8A7HDSf/SuKqqqqqqqqqqqqv8Agsl/yaLaf9jhY/8Aom5r9gP/AJM0+Ev/AGL8f/ob1VVX7V37Yfwr/ZG8J2ut+OZLjUtX1VmTSdCsHT7TdlQSXO4jZEDgNIc4LAAE8VD/AMFCf+CjH7Q15Pffs8fBP7DoYkKwzaX4dl1IRgN92W7nBhZ+o4VOh+UYNRf8FIf2+f2edVtU/aX+By3ujyTbJJdR0KbSJZsdRBcxjyCf+2b9vrX/AAUW/wCCgXhr9pH4U+B/BPwd1bULHR9WM194s025TyriOeJkEFtLtyrIDvk+UsrHyzwUwP8AgnZ+xX/w1h8RrnWfGPmQ/D3whJE+s+W5jk1CZ8mO0jYcqG2lnYcqvAIZ1YftVf8ABKL4Q/Gt08TfCCey+G/iSC3WJre1sgdNvAgCpvhQgxOAAN8fXqVY81afsV/8FX/BFp/wivhP4+Xcmk2wK25svHdykSr0xGJQrIPYAAE59a+E/wDwSB8e+L/GCeO/2vvjC+vMziS40/Tb+e8ur7HO2a9nAZB2IRWJBOHU814Z8M+HvBnh7T/CnhTR7TStH0m3S3srK1iEcUESjCqqjoMVXjT4t+D/AAWHt7i9+3X68fY7UhmU/wC2eifjz7GvF3x08aeJS9vp9yNHs2yBFasRIR/tSdfy2j2rTNI1rxHe/ZdKsLq/upDkrEhdvqT2+pr4U/Bjxb4d12z8U6xqEGnm3zm0T968isCCrEHaOD2Lc1VV4n8X+HfB9j9u1/U4rdSD5cecySH0VRyf5DvivG37QXiDWWksfCkZ0mzOR5xw07j1z0T6DJ96tbPWfEepeRZ291qN9cNkhVaWRz3J6n6k14N/Zyv7ry73xrffZI+v2O2YNIfZn5C/QZ+orw94R8NeFLb7N4f0e3tARhnVcu/+85yx/E1VV8ZPhXe+KXi8U+F/l1i0UCSIPsM6rypU9nHv1HfgVoX7QOv+Hl/sXxx4dluLm2G1pQfJm4/voRgn34rWf2lBPEbbwx4ZkNzJ8scl1JnB/wBxev5ivhl8NPEmt+JR8RPiEkonEnnQQXC4kkkGNrsv8CrxheDkDgAc1VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX/BWD/kyPxh/2ENM/9LIq/wCCXH/JiXwx/wC4t/6dryqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/4LF/DTW/C/7UEHxDnt5G0jxrolu9tcY+QT2yiGWLP95VWF/pKPoP8Agl98PvhHon7J/gzxl4H0TSJNe1mGZtd1ZIVa7kuhM6vFJIRuUIAFVOBtwwB3FjVVVVVVVVVVVVVf8Fkv+TRbT/scLH/0Tc1+wH/yZp8Jf+xfj/8AQ3qqqvhR4NH/AAUX/wCCiXi3xP8AEd5b3wN4TlnnNkXISSwtpvJs7MEfdEjN5jgY3fvsEFs1pGj6T4f0u00PQdMtdO06whWG1tLWFYooI1GFREUAKoAwABivE3hfw5400G98LeLdDsdZ0jUoWhu7G+gWaKdD1VkYEEV+2d+y1P8AAf8Aaovfgx4FtLi8sPEMttc+FreRx5jxXblI4dzHnbMJIgxPIQEnk1+wT+yrcfsm/A6Pwfr97b3ninXb1tT12W3O6KKZkVFgjbALJGiAZPVmcjggCqqvi3p2uaj4F1AeHtQuLa6t185lgba00a/fTI55GTgdSAO9eHfCniHxde/YfD+mTXco++yjCxj1ZjwPxrwf+zlplosd54yvjezDn7JbMUiHszfeb8Nv41pej6Vodotho+nW9nAvSOGMIM+px1Puaqrq6tbK3ku7y5ighiXc8krhVUepJ4FePf2hbO0EumeBo1uZhlWv5l/dqf8AYU8t9TgexFXt9rfifVPtF7cXWpX9y20FsyO57KoH6AV4G/Z41LUhHqHjSdrC3PIs4iDM4/2jyE+nJ+laB4Z0DwvZLYaBpcFnEBzsX5n92Y8sfcmqqqqtQ0fSdWQR6ppdpeKo4W4gWQD8wa0/w54e0h9+k6Fp1k2MZt7VIz/46BVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX/AAVg/wCTI/GH/YQ0z/0sir/glx/yYl8Mf+4t/wCna8qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv8AgsL+0P4utPHcf7OE/h3wxfeF5dGtNXiuLuwd721u2knQyQyhwEO1dvC8gsDkEivhd8f/AI1/BMXSfCf4n+IfDEV8wa6t7C9ZIZmAwGaI5QsBwGIyB3r/AIb8/bK/6OG8Vf8Af5P/AImvgn+3L+1v4h+M3gHw/rXx68T3mn6n4n021u7eSZCs0UlzGrofl6FSQfrVVVVVVVVVVVVX/BZL/k0W0/7HCx/9E3NfsB/8mafCX/sX4/8A0N6qqr/glLq9j8Kf2xPil8FfE8yW2rX1vd2Nr50gLS3NjdHfErN8zMY/NfryIiTnAqq/bP1e3+Mn/BVX4X+DPBrLd3fhe50LS75ozuCSRXsl7OT/ANc4p/mA6FGHUEVVeIPG/hbwxd2lhrWrwwXN7IqRRE5YZOAzAfdX/aOBVVVhpun6VbLZ6ZY29pApJEUMYRQT1OBVVXj/AOL3hrwMr2QcahqoHFnC/wBw/wDTRudv05Pt3rxl8Q/FHjm58zWb8i3U5itIcpFH9F7n3OTXgX4QeKfG5ju1h/s/TD1vLhCN4/6Zr1f68D3rwX8NvC3gWAf2TZCS8K4kvJwGlb1wf4R7DHvmqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/4Kwf8mR+MP8AsIaZ/wClkVf8EuP+TEvhj/3Fv/TteVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVXxx/YR/Zz/aM8fr8Sfiv4c1TVNXj06OwUQ6rNbRCKNmZTtjIO7Ltznv0qb/glJ+w9bwvPP8ADzU444lLO7+I7xQoAySSZOBirP8A4JW/sLajALnT/Ad/dREkCSHxNduuR15EteGv+CYH7HHhHxHpXivQvh9qcOpaLfQ3tnI2v3bhJonDoxUyEEBlBweDVVVVVVVVVVVVX/BZL/k0W0/7HCx/9E3NfsB/8mafCX/sX4//AEN6qndI0MkjqqqCWZjgADqa/aO/aQ+O/wDwUP8AjTN+zb+zBbsngfTLxnW7jbyReiE7Tf3NxgmK3BJ8tBgtuXIZ2VV+I/8AwSa/aE+BXhjR/i18FPiMfFvjXQLgXl1Y6TaNZ3Ns6EFJLNy5M7KQSVIRiMbVYnbXw4/4LM+O/BVqfB/7RXwSmvfEGmfubq906Y6bOXUf8trSVCFc9SVZRzwgHFfEz/gsF8WPinGPh/8Asx/Bi60zXNWHkwXszHVb7Lcf6PbRoFDjjDMZB/s96/4Jy/sF+MPhDr95+0d+0GzzfEPWY5Tp9jPP9om07z8me4uJMndcyBiDgnarvuJZyEdkRC7uFVRkknAA9a+I3x80/S0l0jwTJHe3vKvekZii/wBz++ff7v1oDW/FWsYH2rUtSvZPeSSRv8/gBXw/sfEum+ErCw8WSwyahBHsJjbcQg+6GPdgOCRxx1PWqqqvr6y0yzlv9Quore3gXdJLK21UHqSa+JHx8vNTMujeB5JLW05V78grLL/uDqg9/vf7taB4c17xdqY03RLGW8uZPmcjoo7szHgD3NeAfgNoPh1YtR8TCPVtRHIjZcwRH2U/fPu3HsOtAAAKoAAHAxVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV/wVg/5Mj8Yf9hDTP8A0sir/glx/wAmJfDH/uLf+na8qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv29/2zLD9kL4aW11o1pa6n448TPJBoNjcEmOMIB5lzKBgmNNyDbkFmYDIAYj4r/HX4vfHLW5fEHxV+IWs+IrmRy6RXVyfIg9ooVxHEvsigV4S8b+M/AGrR694F8W6x4e1KIgpd6XfSWsox0+aMg1/wTW/4KH+Ivjpqy/Aj433kFz4wjtml0TWlRYm1WONd0kMqKAvnKgLhlADqrZAZcvVVVVVVVVVVVV/wWS/5NFtP+xwsf8A0Tc1+wH/AMmafCX/ALF+P/0N6qv21fEuqeEf2TfitruizmG9i8MXcUUqnBj81fKLA9iA5IPqBX/BFjwJoWlfs/eKfiFFbRtrOv8AieSzmuNvzC2toIjHHnrjfNM3vuHpVeKPhz8PfHDRP418CeHfEDQ/6o6ppcN2U+nmKcdT+dfsUaDpXwc/4Kr+O/hn4btIV0ln13TrKONA4tbcOLiNAc/LtWJUz16g9TjUNQsdKspdQ1K7itraBd0ksrbVUfWviv8AGK68YyPofh+SW20ZCQ7ZKvdn1Ydk9F/E9gPA/wANvEnj2526VbiKzjbE15MCI09h/eb2H44614E+G/h3wDZ+XpsPn3si4nvZVHmSew/ur/sj2znrVVVeN/ix4T8D7rW7uTeaiBxZ25BZfTeeiDp159jUlz8QPj9rLW1uUstJtG3FCxEFv6bj1dyP6/dGat/2ZdCW3C3Xie+knx9+OFEXP+6cn9a17wB47+DM3/CUeF9aa6sFYCd40K4GeBLHkgrzjOT17cV8PPG9p4+8NxaxCixXCN5V1ADny5AOcf7JBBH19jVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX/AAVg/wCTI/GH/YQ0z/0sir/glx/yYl8Mf+4t/wCna8qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv8Agr/4r1DX/wBsG70K5lf7N4Z8P2FnbRk/KBIrXDMB6kz4J/2R6Cv2P/8AgnH+zb4I+E/hbxV478BaT468Wa7pdtf315rcQvLaJpY1k8qGBsxbF3Y3FSzcknBCj4j/ALAv7IfxM0G40LUPgX4W0NpYikV94c06LSbm3bs6tAqhmB5w4ZTjkEcUuian+yl+2dbaFYawbyb4d+OoY4ruP5TdQRXK43KDwXi4ZM8bmXNVVVVVVVVVVVVf8Fkv+TRbT/scLH/0Tc1+wH/yZp8Jf+xfj/8AQ3qq+Nvw3tvjD8IPGXwtuZ44B4o0S60+OeRdywSyRlY5SO+x9rf8Br/gmB+0vpP7NHjXxf8Aso/HqYeFXvdcaWyudQYRQ2epIohngmduEEixRbHOFyhGfnWkdJEWSN1ZWGVYHII9RX7Qv7RPwz/Zp+Ht94++I2u29uIon/s7TRKBc6pOB8sMKdWJOAWxhQdzEAE1/wAEnvAvjH4vftA/Er9sLxjbOlvdS3dvbyMDsn1C8mE0wjJ6rFGApHbzk9DXx18E6x4q8PR3+jXU8kmmbpHsVY7Z17sF7uO3qCR16/DL4Oar41ki1bVhJY6KDnzCMPcY6iMHt/tHj0zzjS9K07RNOg0rSrWO2tbZNkUSDAUf1Pck8k81VVXxW+Mun+GLWfQvDV2lzrbDY0iDclp6knoX9F7Hk9MG4nmuppLm5meWaVizyOxZmY8kknqTXwW0q10z4caSbeNQ92rXEzAcuzMeT9FCj8Kq9s7bULOewvIllguI2jlRhwysMEfka/ZzmlsPE3iHQRLvjEQYkHgtG5XI+oaqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqrWdX0vw/pN5rutX0VnYafA89zcSnCxRoMsx9gAa+HX7T3wc+K3iWPwj4J8Q3V5qckLzLFJYTQjYgyx3MoFV8QPiZ4D+FujDXvHviW00m0YlYvNJaSZh1WONQWc8jIUHHU1aft8/s+XOoLYy32vW0RbH2ubTCYh74Vi+P+A14b8TeHvGWiW3iPwrrNpqmmXi7oLm2kDo2OCMjoQQQQeQQQcGqqq/4Kwf8mR+MP+whpn/pZFX/AAS4/wCTEvhj/wBxb/07XlVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV/wWl+DesaP8V/C/xxsrKaTRfEWlLpV5OFJWG9t2dlDH+HfC42jv5LntX7MP/BWv4m/AzwPpXw08f+BrTx3oehwJa6bdDUGsb22t1GEiZ9kiyqigKoKqQAAWOBj4j/8ABbnxPqeg3GnfCv4JWeh6pPEVj1LV9XN6tuTxuWBIowzDqCz4z1Ujg/skfDzxl+09+114Wt9SnutWu73xANf8S38x3MYIphPcyyNxgufkB/vyqO9VVVVVVVVVVVVf8Fkv+TRbT/scLH/0Tc1+wH/yZp8Jf+xfj/8AQ3qqr9sP/gnb8Jv2sZ/+EujvZPCHjuOIRjXbO3EqXiqMIt1DlRJgcBwVcDAJKqFH7Qvwp/bK/wCCfmlaLpUn7Smo2+g+I3uLbSrXw14nvoRsiCF2MDBFiIDxjKEkFhg1+xT8BNW/bq/aFm0j4u+OPEuo6ZpOlyanrOozXzXF3OiyIiQiabeVLPIDkg/Kr4weR4A+H/gz4WeDtL8AfD7w/a6JoGjw+VZ2NspCxgksxyclmZizMxJLMSSSSTVAAAKoAAHAxVVV5d2lhayXt7cxwQQqWkkkYKqAdSSelfEz46ahr7TaJ4QlkstN5R7kfLLcDvjuin06kdcZIrwt4P8AEHjTURp2g2LTuMGWRvlSIHuzdv5nsDV78DLHSfhtqWlacPt2vSosxuNuC7Id3loOykZHuSCewHwF+ImmHR4/A+sXUdteWkjCz81tomVmJKAn+IMTx3BGOhqviT8Q9K8C6JPI13E2qyxEWdqDlmY8BiOyjqSeuMda/Zx8M3dnpeoeKr5GB1Jljt9/VkQnc/0LHH/ATVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVXl7Z6dbPeahdw21vEMvLNIEVe3JPA5xWnatpWswG50jU7S+hRyjSW0yyqGABIypIzgjj3FXXxU+F9jqn9h3vxJ8LW+ohtv2OXWIEmz6bC+7PB7dqVldQyMGBGQQetVVVVVVftZ/FLXPi74/wBN/Zc+Fk5me4vVj1ueM5V5Qd3lEj+CIAvJ7rjjYc/se6JbeG/2t9S8OWUkklvpS6paRPIQWZY3KAnHGcKM1qF/Z6XYXOp6hcLBa2cLzTyv0jRQSzH2ABNeENM139t79oq81HxPe3dv4Z05GnaFGP8AotkrhYrdOyvISNx9d7dgK8R/sh/s/wCv+GZPDdv8PrDSyItkF9ZBkuYWxw/mEkuR6PuB71+y14w8TfAD9oe/+BfiW9L6Xqmovp80ZJ2C6HEFxGvbzAEX3V1z90Yqqr/grB/yZH4w/wCwhpn/AKWRV/wS4/5MS+GP/cW/9O15VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVfFL4WeA/jR4F1P4cfEjw/b6xoWrRbJ7eUYKsPuyRsOUkU4KsMEEV8X/8Agin8S9O1i4vPgb8SdC1rRmYtDZ+IGks7uEdk3xo8cp/2j5f+7Xgz/gjP+0/ruoxReL/EXgvwzYB8TTG+kvJgvHKRxptY89Gdeh56Z/ZN/Y2+Ff7InhW40nwUs+qa9qqp/bGv3qgT3hXJVFUcRRAk4QZ9WLEZqqqqqqqqqqqqr/gsl/yaLaf9jhY/+ibmv2A/+TNPhL/2L8f/AKG9VVV/wWV+KkHjH9o/SPhtp9ystt4C0RUuFH8F5dkTSD/vyLX8c1/wRa+Gx8LeDPGvxA1wRW9/4vltYtKglbbK9nb+aXlVeu13kx/2zB6EE1VVWu67pHhvTJdX1u+jtbWEfM79z2AHUk+g5r4n/FrU/H050+0V7PRonzHbk/NKR0aTHf8A2eg9+tfC/wCFGo+PbsXt6JbTRYW/eXG3BmI/gjz1Pqeg+vFaHoGj+GtOj0nQ7CK1tohwiDknuSepJ9TzVfEH4GaH4wu5NY0q6/svUZSWlIj3RTN6suRgnuR9SCaT4R/GyyT7FY+NttuvCrFq06Lj/dxxXhj9nVEvV1PxxrY1B9257a3LbZD/ALUjYYj2AB96gt4LWCO2toUiiiQJHGi7VRQMAADoMVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX7VCJJ+zz46V1VgNLYgEZ5DqQfzAr9nVPi38U/Dc/7PPw81UaFo13qMura/q6FgywNHDEIzggkEx/cBBctgkKGNeKf+CcGiR+HJpPBvxA1GXXIYi0Ud/bxiC4cDhflwY89M5bHpX7C/wAZvFWg+PpPgR4uu7mWyuUmTT4LliW0+6hDM8S5+6hVJMr0DKMYyc1XxJ/bs8JfDbx1rPgW+8CaveT6LcmCSeK5jVZCADkA8gc1/wAPI/A//RNtc/8AAuKv+Hkfgf8A6Jtrn/gXFWjf8FEfBetaxYaNF8Otaje+uY4FdruIhS7BQT7c1VftZfH+D4JeAmtdGuoz4r11Gh0uPhjbr0e4Yeig4XPViOCA2P2MP2ep/h/oD/FHxvbvJ4t8SRb41uQTJY27ndg5582ThmJ5A2jg7s/sv/8AJ6/ib/r61n/0a1ftXa1JoH7O3jm+hl2NLpv2QnOOJ5EhI/ESEfjX/BNvRYYfCXjPxEEUy3eo29ru7hYoy2PYZm/ziq/bO8vwd+1VbeKLUrHK0OnakzKDkPEdgJx3xCKqqr/grB/yZH4w/wCwhpn/AKWRV/wS4/5MS+GP/cW/9O15VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVftWftYfDb9kn4fr4x8cPLfajfu0Oi6HauFuNSmUDcATwkaggvIQQoI4ZmVT8Tf+Ct37XnjjUZpPCPiHRvAumsxEVppOlxXD7M8b5rlZGLYxlk2A+g6V4N/wCCqf7a3hO+jub34mWPiW2R9zWWs6LbNG/sXhSOUD2Div2Iv+CgvgX9ri1m8K6rpsXhb4g6fB5s+kGfzIb6IcNNaucEgcFoz8y5HLAFqqqqqqqqqqqqq/4LJf8AJotp/wBjhY/+ibmv2A/+TNPhL/2L8f8A6G9VVTTQ28LzzypHHEpZ3dtoUAZJJPQYrRtH1b9sz9rPxD4kvjMdM1/XLrV9Sl5BtrDzcpED1B2eXCvcZBPQ14Q0O6W+0nw94Qt/sksRjt7CO2/diAKAF24+6qgdugFaVa3VlplrZ31897cQwqktw6gGVgOWwOmTVVXinxVovg7SJNZ1y68qFDtRFGXlbsqjuf8A9ZwK8f8AxA1nx/qxvb+QxWkRItbNWykK+vux7t/IYFfCv4JXPiZYfEHilJLbSjhobflZLoevqqe/U9sdatLO00+1isbC2it7eBQscUaBVQDsAOlVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV+1L/AMm9eOv+wU//AKEtf8E1YYhpHj2cRr5jXNipbHJAWbA+nJ/Oq8PqLX9v2UWxMefGt2TtOM7mk3fnk5+tVVVVV4o8Q2HhHwzq/ivVBIbLRbCe9uREu5/LijLttHc4U4FeH/jV4L8d/HyX4w/HxNQuLG0YSabpNlbidFKH9zEdzKPLTlj/AHm5I5Nf8PB/gL/z6eKv/BdH/wDHa+C3xr8G/D/9o3Wfirrseotot/PqLxLBArzYnkLJlSwA4PPPFfGX4keGfj9+yJ458TeCotQWygKIy3cKxSbreeCVjgMw24x39a/4JvXsEnw88WaarfvoNaSVhn+F4VA/WNqr9uGFfEv7Tdl4ftVZ5WsLCyZUI3F5HYgD0OJFqqqv+CsH/JkfjD/sIaZ/6WRV/wAEuP8AkxL4Y/8AcW/9O15VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVf8FgPFmua7+15c+HdQklGn+G9BsrfT4mY7MSoZpHUdMl5CpPfYOeMD9h//AIJV+EvjJ8L9H+M/xz8TazBZeIUNxpOhaSyQMbcEqss8zKx+fBYIgXC7TuO4qPi7/wAEZfgZrnhe6b4N+J/EHhnxJDCxsxqV0L2ynkHIWUFRIobhd6t8vXa2MH4NeJPFnwA/aQ8La2yz6drng7xTFBfwKw3Dy5vKuYCRwQyebG2DyGPNVVVVVVVVVVVVf8Fkv+TRbT/scLH/ANE3NfsB/wDJmnwl/wCxfj/9Deqqv2kPER8Ifs9fE3xQk3lyaV4Q1S4ibIB8xLWQoBnuWAA9yK/YO+FqeCvhH/wmd/b7dU8YSi4yy4ZLVMrCv0b55PcOvpX7OXgxRFc+N72IFmJt7HcOgH+scfXhR9G9aqq8c/FTwr4ExbahO9zfsuVs7cBnAPQtk4UfXn0Br4iePdV+Iusf2lLbyQWVqm23tQ5dYR3YnAGWPU49B2r4UjwKvieKbxzcMkMZBtkkjzA0mePNPZRxxjB7kAco0ckavE6sjDKlTkEeoqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/al/5N68df9gp/wD0Ja/4Jrf8gPx5/wBfdl/6BNVaN/yf9J/2Olx/6E9VXxJ/YS8JfEnx1rPjq+8eavZz6zcmeS3ito2WMkAYBPOOK/4dueB/+ila5/4CRV/w7c8D/wDRStc/8BIq0b/gnd4L0XWLDWIviLrUj2FzHOqNaRAMUYMAfY4qtT03T9Z0260fVbOK7sr6B4Lm3mTcksbqVZGB6ggkEe9f8M5fAX/okXhX/wAFkf8AhX/DOXwF/wCiReFf/BZH/hX7PHgXwb4j/a18QeEdd8M6df6Lb3OqrFYXFurwoI5WCAKeBtAwPSj8LfA9r4F1f4d6D4fsdH0bWra4guLeygWNf30ex3CjjdjHPsK/ZW8cP+zb8dtf+GvxJmTS7TU3Gn3k8p2Rw3ETEwSlj0jZXcBumJVbIGTXiXxb4a8H+HrnxX4l1q1sNJtIfNlupX+XbjI24+8T2AyScAZzXwVsNT/aZ/a7u/idLYyppGmaiNWnaRc+TFDhbOJiON58uMY7hHPOKqqr/grB/wAmR+MP+whpn/pZFX/BLj/kxL4Y/wDcW/8ATteVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX/AAWQ/Z7s9X1HSv2gfDWu6MupadpqWGv6PLfRxXUluJGMF1FEzBpACzowUE4VSAQrEfsFf8FLvgT4c+B/h74Q/HHxA/hHWvB9mLG0vpLOWe11C2TPlEGFGMciphWDgA7QwYliq+Pv+Cpf7Gvg7wxe61onxLbxXqUMTG00jS9OuRLdSY4XfJGscYz1ZmGBnAJ4PwJ17wD8UP2tYfi38ffHWl+EdBl8TS+KNZlmillFzKbk3H2WJI0ZjvkIUk4CpuOc4B/4eQ/sR/8ARftJ/wDBfe//ABmvhP8AGP4Z/HPws3jX4UeLLfxDosd29o13DDJGomQKWTEiq2QHXtjmqqqqqqq/aj/aX8Dfsq/Cq8+JXjMPdys/2bSdLhcLLqN2ykrEpOdq4UszkEKoJwThT4M+LX/BVX9thb3x98ItVj8I+D4rl47T7IYNOtSykgpHLKGnuCOjNkoGBHynivhx/wAFBP2sf2UPi5a/CH9uDRrnUtInZPNvZ7WIXlpAxKrcwTQYjuoQQ27O5vlIDAqVOmanp2t6Zaazo99Be2F/Ak9rcwOHjmidQyOrDgqVIII6g1Vf8Fkv+TRbT/scLH/0Tc1+wH/yZp8Jf+xfj/8AQ3qqr9u/TNT1z9k7x94e0adorzWobPTomUc/6RewQkfQiQj8a0HQ7XSNN07w3otvtt7KCK0tIR2RFCIo/AAV4X0ODw14d07QbcLtsrdYyQMbmA+ZvxbJ/Gqrxp4hTwn4V1LxAyqzWcBMat0aQkKgPsWKivg78OoPHNxdeO/GZa+R7hhFDL0nk6s7+qgnAHQnOeBg21ra2cC21nbxQRIMLHGgVV+gHSviT8JdB8Y6ZcXOnWEFprMal4J4kCea392QDgg9MnkfTIP7O/i271HS73wjqUjNJpWHtt/3hETgp9FYD/vrHaqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr4neB4fiV4A1vwJPqLWCa1am3a5SLzDFkg5C5GenqK/Z0/Zzsv2erLXLKy8Vza2NblhkZpbQQeV5YcYGGbOd/t0qrT9jzTbT47t8cV8d3LTtrMmpf2cdPULlyTs8zfnHPXH4VVVVVVVXw0/ZO074b/GXUfjBB42uL+bUZbyQ2D2IjVPtDliN4ck7c46c+1V8ev2XPAHx5WLUtTln0fX7WPy4dVtEDMydklQ8SKM5HII7EDINj/wAE4LqS6ig134yyz6ZAx2xQaSVcj23TMqHHs39a+Fvwn8E/B3wvH4U8D6Ybe2Db555W3zXUmMGSR8Dc34AAcAAcVVVX/BWD/kyPxh/2ENM/9LIq/wCCXH/JiXwx/wC4t/6dryqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/4Kv/Bf4xfEj9q221H4d/Cbxl4otU8I2SNPo2g3N6iss0+4FokYAjcuR23D1r/hk39qj/o2f4q/+EZf/wDxqv8Ahk39qj/o2f4q/wDhGX//AMar/hk39qj/AKNn+Kv/AIRl/wD/ABqv+GTf2qP+jZ/ir/4Rl/8A/Gq/4JQeAvHXw5/ZduvD3xC8F674X1RvFV7OtjrOmy2U5jaK3CuI5VVtpKsAcYOD6VVVVVVVX/BVfxfr/wAaf2y/C/7PWj3RWHQI7DS7WBiSov8AUWjkaTHuklsv/APevAPgfw78NPBGhfD7wlYpaaP4esIrKzhVQMRxqFBOOrHGWPUkknk1/wAFjvhNpvi79mq0+KMVin9reA9YgJuQg3CzumEEkZPXBla3b6r71/wSd+J198Rv2P8AR9M1S5ae68E6rc6F5jn5jEgSaEfRYrhIx7Rj3qq/4LJf8mi2n/Y4WP8A6Jua/YD/AOTNPhL/ANi/H/6G9VVfGHRm1/4caxpcWmzX8riGSK3hRmdpI5kdCAvJwyhse1fD/wABeK18baHLqfhTV7e1hvo5JZJ7CREUKd3JIwBx3qqr9oD4jQXW/wABaQ6SLE4bUJRz86nKxj6HBPvgdjXwFvLa6+GmnwQMpe0mnjmA7MZGcZ/4C61TsiIXdwqqMkk4AHrXwFYah8SPEer2i4tHt5ioHQb51Zf0U1VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX/AAVg/wCTI/GH/YQ0z/0sir/glx/yYl8Mf+4t/wCna8qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqvjL+2D+zz+z7400jwF8XvHyaBqut2RvLUPZTzRrFvMas7RI3lhmVwC2B8jcjFeEPGHhb4geGrDxj4J1+x1vRNTjMlnf2UwlimUEqSrDrggg+hBHavjj/wAFgvgV8L/F154N8B+E9W+IE2mTNDd6jaXcdpZF1OGEMpDtKAQRuCBD1VmHNfspft3/AAT/AGtvtWj+DZb/AETxTp8Hn3Wg6sqpMYgQDLCyErLGCQCQQwyNygEE1VVVVVVVeOy2qf8ABYe2OpQqxT4m6WFUrjiIweWcfRFOfxqv+Ck8MM/7EPxTjmiSRRY2jBXXIDLfW5B+oIBHoQK/4IjTXbfBz4i27lvsqeJoWiymBvNsofB78KnHbj1qq/4LJf8AJotp/wBjhY/+ibmv2av+CsXwY+CfwH8FfCjX/hz40v8AUfDOmLaXFzZrbeVKwZjlN0obHPcCv+H2nwB/6JR8QP8Avm0/+PV/w+0+AP8A0Sj4gf8AfNp/8er4Sf8ABW79nj4p+MYvCV34b8WeF45LeSX+0NStkmiUoM7Stu0j5Pb5cepFaTqlhrmlWWtaVcefZahbpcW0uwrvjdQytggEZBBwQDVV4q/au+AHgrxDfeFPE3j77HqumSmK6t/7KvJPLcDONyRFT1HQkVq37Vnwy8T+Hb9PhX4nbVtUQLHuOn3EC2+7Pz5ljUEgA4AzzjIxmtV1nTNKhfVfEOrJZ2vmDz7qcM+3c2CxCgs3XOACT6Gvhd8ctB8Na9fxeBPE1r4m0neBPF5UtqZQOkipIodDzjOCO3OARaftE+AJ7cS3CalbyY5ia3DEH2IJFeNfjVqnjmJvCPgPR7tBfgxPIy5mlU9VVVyFBHU5PHpXwl+Hx8A+HWgvGR9SvmEl2ychcDCoD3C5PPqT2r4mfGv4Y/B3+zf+Fj+J/wCyP7X837H/AKFPceb5Wzf/AKpG248xOuM546HH/Da37Mf/AEU3/wAo19/8Zr/htb9mP/opv/lGvv8A4zX/AA2t+zH/ANFN/wDKNff/ABmvAP7SfwU+KHiFPCngXxr/AGnqrwvKtv8A2ddQ5RB8x3SRqvHpmqrxh4v8O+AvDd94u8Waj9h0nTUD3Nx5Ly+WCwUHagZjywHAPWv+G1v2Y/8Aopv/AJRr7/4zX/Da37Mf/RTf/KNff/Ga/wCG1v2Y/wDopv8A5Rr7/wCM1B+2f+zRdTx20HxK3SSuERf7Hvhkk4H/ACxqqq//AGyf2bdMvrjTb74keXc2krRTJ/Y96drqcMMiEg8g9OK/4bW/Zj/6Kb/5Rr7/AOM1/wANrfsx/wDRTf8AyjX3/wAZr/htb9mP/opv/lGvv/jNeBfHnhP4l+G7fxd4J1b+0tJundIbjyJIdxRirfLIqsMEEciqr4jftDfB74S63B4b+IPjD+ytRubRbuKH+z7mfdEzMgbdFGyj5o3GCc8dORX/AA2t+zH/ANFN/wDKNff/ABmv+G1v2Y/+im/+Ua+/+M1/w2t+zH/0U3/yjX3/AMZr4bfHj4T/ABfvrzTfh34r/ta50+JZbhPsNxb7EJwDmWNQeewzVV8QviV4J+FWgL4n8e63/ZemNcLbrP8AZpZ/3jAlV2xKzchTzjHFf8Nrfsx/9FN/8o19/wDGa/4bW/Zj/wCim/8AlGvv/jNf8Nrfsx/9FN/8o19/8Zrw5+1r+z54u17T/DHh74g/a9T1S4S3tIP7KvI/MkY4VdzRBRk9yQKqr+9tdMsbjUr2Xy7e0iaWZ9pO1FGWOBkngHgV/wANrfsx/wDRTf8AyjX3/wAZr/htb9mP/opv/lGvv/jNf8Nrfsx/9FN/8o19/wDGa/4bW/Zj/wCim/8AlGvv/jNeHPEOjeLtA0/xP4evPtemapbpcWk/ltH5kbDKttYBhkdiAaqvG37UPwK+HXie98G+MvHX9n6xp+z7Tbf2Zdy7N8ayL80cTKco6ngnr65r/htb9mP/AKKb/wCUa+/+M1/w2t+zH/0U3/yjX3/xmv8Ahtb9mP8A6Kb/AOUa+/8AjNfDb4vfDv4v2N5qXw78Rf2tb6fKsVw/2Sa32ORkDEqKTx6ZFVXxL+Mnw2+D8Fhc/EXxJ/ZMepu6WjfY57jzCgBbiJGxgMvXHWv+G1v2Y/8Aopv/AJRr7/4zX/Da37Mf/RTf/KNff/Ga/wCG1v2Y/wDopv8A5Rr7/wCM14F/aa+B/wAS/Elv4Q8E+N/7S1a6R3ht/wCzbqHcEUsx3SRKowoJ5NVXirxRoPgrw7feKvE1/wDY9L0yEy3U/lPJ5aDjO1AWPUcAGv8Ahtb9mP8A6Kb/AOUa+/8AjNf8Nrfsx/8ARTf/ACjX3/xmv+G1v2Y/+im/+Ua+/wDjNf8ADa37Mf8A0U3/AMo19/8AGagniuoI7mB90cqB0bBGQRkHH0qq179rv9nfw1reoeG9b+If2bUdKu5bS7h/sm9fypo2KOu5YSpwykZBIPY1/wANrfsx/wDRTf8AyjX3/wAZr/htb9mP/opv/lGvv/jNf8Nrfsx/9FN/8o19/wDGa8A/EPwd8UPDqeK/Ausf2npckrxLP9nlhy6cMNsiq3H0xVV8SPj/APCL4R6ra6J8QvF39lXt5b/aIIvsFzPvj3Fd2Yo2A5UjBOa/4bW/Zj/6Kb/5Rr7/AOM1/wANrfsx/wDRTf8AyjX3/wAZr/htb9mP/opv/lGvv/jNfDj4/wDwi+Luq3OifDzxd/a17Z2/2ieL7BcwbI9wXdmWNQeWAwDnmqrx98Q/B3wv8Ov4r8c6x/ZulxypE0/2eWbDtwo2xqzc49MV/wANrfsx/wDRTf8AyjX3/wAZr/htb9mP/opv/lGvv/jNf8Nrfsx/9FN/8o19/wDGa0n9sL9nLXNVstE0r4jefe6hcJb20X9kXq75HYKq5MIAySBkkCqqv+G1v2Y/+im/+Ua+/wDjNf8ADa37Mf8A0U3/AMo19/8AGa/4bW/Zj/6Kb/5Rr7/4zX/Da37Mf/RTf/KNff8AxmvC3ijQfG3h2w8VeGL/AO26XqcIltZ/KePzEPGdrgMOnQgVVeOv2mvgf8NfElx4Q8beN/7O1a1RHlt/7NuptodQyndHEynKkHg1/wANrfsx/wDRTf8AyjX3/wAZr/htb9mP/opv/lGvv/jNf8Nrfsx/9FN/8o19/wDGa+Gvxk+G3xggv7n4c+JP7Xj0t0S6b7HPb+WXBK8SouchT0z0r47f8FVPg58Aviz4i+EPiT4d+MtR1Lw3PHDcXNiLbyZC8SSArvlDYw4HIHINf8PtPgD/ANEo+IH/AHzaf/Hq/wCH2nwB/wCiUfED/vm0/wDj1fto/wDBTn4SftMfs+a58IfCfgHxdpepapdWksVzqItxCghnSRgdkjNkhSBx1r/glx/yYl8Mf+4t/wCna8qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv8Agrjql/qH7aGuWl4xMWmaJptvaZBGIzCJT/4/LJ0r9q74zaB+zf8A8E4vCnh74fatFaah4t8L6bofh5reYGQwyWyG4nUjk/ud+XGMPKhzkjP7Lv7LPxH/AGs/H8/gL4eS6fZtYWTXt/qOpM6W9pECFG4orMWZmAVQMnk8BSR8Lrjxx+yH+2LoNrrM8dprfgbxbHYaqbaUtFPAZPKuArYBMckLvgkA4cHAPFVVVVVVVVft6Rz/AAD/AOCmelfFu6jePT7jU9B8TRY3fPFAYophnOSGe1lyB2bHSre4t7y3iu7S4jngmQPFLGwZXUjIZSOCCMHNf8FbvHdn4S/Y21vw7Jcol34x1aw022TcQ52TLdOQB222xBzx82O4r/gjv4EufCv7JT+JbyBkfxh4lvL+BmyN0Eax2y4Hpvt5Tnvn6VVf8Fkv+TRbT/scLH/0Tc1+x5+wj+y38Sf2Yvh1468ZfC6y1DW9Z0ZJ726eRgZZCzAk4PoBX/Dtv9jX/ojmn/8Af1/8a/4dt/sa/wDRHNP/AO/r/wCNaf8A8E7v2R9IuReaT8LIbK4ClRLb3UsbgHqMqQcVo+k2Og6PY6FpkZjs9Oto7a3RmLFY41CqMnk8Acmqrx7+yh8AvFOsax4+8X+HLuW8ui93fTrqU8YO1csQqsAOB0FWOgeF/DpuLLwho39mabJO0kVuZmlZQem52JLHAH/1q8N/sveBvH3hCyuvifpd5dTTP9ogto7yS3ESEYXcEIyxGTz03Y45q2/Yi/Z4sp0urLwxqcE0ZykkWs3KMp9iHyK0f4CfDDRrU2sehz3YLZD3l9NM446bi2ccVovhjw74cjMeg6LZ2O4YYwxBWb6t1P41XxW+Bfw3+NX9l/8ACwdInvv7G877H5V3JBs83ZvzsIzny069Me9f8MMfs3/9Chff+Da4/wDi6/4YY/Zv/wChQvv/AAbXH/xdf8MMfs3/APQoX3/g2uP/AIuvh1+zB8HPhT4lj8XeCfD91aanHC8KyyX80wCOMMNrMR2qq8beDPD/AMQvC1/4N8U2r3Ol6mipcxJK0ZYKwYYZSCOVHSv+GGP2b/8AoUL7/wAG1x/8XX/DDH7N/wD0KF9/4Nrj/wCLr/hhj9m//oUL7/wbXH/xdW/7D/7OdrcRXUHhG9EkLh0J1W4OCDkfx+1VVaj+xN+zxquoXOp3vhO9e4u5nmlYarcDLsSWOA/HJNf8MMfs3/8AQoX3/g2uP/i6/wCGGP2b/wDoUL7/AMG1x/8AF1/wwx+zf/0KF9/4Nrj/AOLr4ffD7wt8L/C1t4N8G2Ulppdo8jxRSTNKQXYs3zMSTyTVV8T/ANm74S/GLX7fxN480K5vdQtbNbOKSO+lhAiV3cDajAfekbnrzX/DDH7N/wD0KF9/4Nrj/wCLr/hhj9m//oUL7/wbXH/xdf8ADDH7N/8A0KF9/wCDa4/+Lr4W/s+/C74NX99qfgDRbiyuNRhWG4aW8ln3KpyAA7HHPpVV8S/hf4N+LnhxfCvjnT5bzTkuUuRHHcPCfMUMAdyEHox4r/hhj9m//oUL7/wbXH/xdf8ADDH7N/8A0KF9/wCDa4/+Lr/hhj9m/wD6FC+/8G1x/wDF14X/AGPPgL4O8Rab4q0DwveQ6lpNylzayNqc7hJEOVJUsQeR0NVWo2Ftqun3OmXiF7e7heGVQSCVYEEZHTg1/wAMMfs3/wDQoX3/AINrj/4uv+GGP2b/APoUL7/wbXH/AMXX/DDH7N//AEKF9/4Nrj/4uv8Ahhj9m/8A6FC+/wDBtcf/ABdeF/DWkeDvDmm+FdAgaHTtJtktrWNnLlI0GFBY5J4HU1VePf2Uvgn8S/Fl9428XeHLq61bUfL+0SpqM0Qby41jXCqwA+VFHSv+GGP2b/8AoUL7/wAG1x/8XX/DDH7N/wD0KF9/4Nrj/wCLr/hhj9m//oUL7/wbXH/xdfC34NeAPg1p99pfgDSprK31GZZrhZbqScs6jAILkkcelVXxU+Cfw7+NFvp1r8QNJmvo9KeR7URXUkGwuFDZ2EZ+6Otf8MMfs3/9Chff+Da4/wDi6/4YY/Zv/wChQvv/AAbXH/xdf8MMfs3/APQoX3/g2uP/AIuvh9+yz8F/hf4otvGXg3w7dWuq2iSJFLJqE0oAdSrfKzEHgmqrxf4T0Tx14Z1Dwj4jtmuNM1SEw3MSSNGWQkHAZSCOg6V/wwx+zf8A9Chff+Da4/8Ai6/4YY/Zv/6FC+/8G1x/8XX/AAwx+zf/ANChff8Ag2uP/i6/4YY/Zv8A+hQvv/Btcf8AxdW9vFa28VrAuI4UCICc4AGB/KqrxD+xr8AfFOv6l4m1nwteTahq95Ld3ci6nOgeWVy7kKGwAWY8DgV/wwx+zf8A9Chff+Da4/8Ai6/4YY/Zv/6FC+/8G1x/8XX/AAwx+zf/ANChff8Ag2uP/i6+HXw38JfCnwzH4R8E2ElppkczzLFJO8xDucsdzEntVV8Uf2dvhT8ZNXtNc8faHcXt5ZW32aF472WALHuLYwjAHljzX/DDH7N//QoX3/g2uP8A4uv+GGP2b/8AoUL7/wAG1x/8XX/DDH7N/wD0KF9/4Nrj/wCLr4X/ALO3wp+Dmr3eueAdDuLK8vbb7NM8t7LMGj3BsYdiByo5qq+I3w38JfFbwy/hHxrYSXmmSTJM0Uc7wnehyp3IQa/4YY/Zv/6FC+/8G1x/8XX/AAwx+zf/ANChff8Ag2uP/i6/4YY/Zv8A+hQvv/Btcf8AxdaN+xd+z5oOr2OuaZ4UvY7zTrmO5t3bVJ2CyRsGU4L4PIHBqqr/AIYY/Zv/AOhQvv8AwbXH/wAXX/DDH7N//QoX3/g2uP8A4uv+GGP2b/8AoUL7/wAG1x/8XX/DDH7N/wD0KF9/4Nrj/wCLrwj4T0TwN4Z07wj4btmg0zS4RDaxPI0hVAc4LMST171VfEH9ln4L/FHxRc+MvGXh26u9VukRJZY9QmiBCKFX5VYAcAV/wwx+zf8A9Chff+Da4/8Ai6/4YY/Zv/6FC+/8G1x/8XX/AAwx+zf/ANChff8Ag2uP/i6+FfwT+HfwYt9Rtfh9pM1jHqrxvdCW6kn3FAwXG8nH3j0rxL+zp+z54z1u68TeMPgT8PNd1i+YNdahqXhezuridgoUF5JIyzEKoHJ6ADtX/DJv7K//AEbP8Kf/AAjLD/41Xi79hv8AZI8Z21va3/7P3giwW2curaRokGnuxIxhmgRSw9jkV/wUQ/Yt/Zu+DX7KviXx/wDDn4cWmk67Y3lhHBdxuxKLJdRo4wT3ViPxr/glx/yYl8Mf+4t/6dryqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/4LD/ALKvivxDqOl/tM+BNEn1K10/TBp/imG2Tc9tHG5aG7KjkpiRkcjhQkZPG4iW/wBZ1aKx0qa+vbyO1BisbZpGkEW5slY1yduWPQDk1/wTA/Zc1n9mz4H6h4q+IunjS/FnjmaO+vra4GyTT7KJG+zwy5+643yyMONvmBTgqa8XazH+0t+3Xdal4bV7i08cfEWOLTmiXBa1e8WOJyO37oKxJ4HJPAqqqqqqqqr/AIKSfsa6h+1R8M7HXPAUEJ8e+DTLJpkMjCMajbuB5tqWPAYlFZC3AYEHAcsP2dv+Cpnjf9mrwVb/AAL/AGhfhJrmqX/hGMWVjcGY2d9BCnCQXEUy87F+VXBB2qoIONx8Uaj+0f8A8FbvjVosekeD5/Cvw38PyNElw2+az0mNipmlknKqJ7pwqhUUDoowoDufh74E8OfDDwLoPw78I2YtdH8O6fFY2cXfZGoUFj3Y4yx7kknrVV/wWS/5NFtP+xwsf/RNzX7Af/Jmnwl/7F+P/wBDeqqq+LXxek8ITr4a8NRJPrMqgu7LvW3DfdG3+Jz1A6DjrnFWvwm+LnjRBqfifxI9oZPmWK6uHZ1+ka/Kn04x6Vr/AMI/ip4X0y6bStbl1KxliZbi3tLiQM6Ec5jPDDHYZPtXwo8NHxP470yxdI2gtpBc3CyYwUjIJGO+TgY9/TNVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVf8FYP+TI/GH/YQ0z/ANLIq/4Jcf8AJiXwx/7i3/p2vKqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqUxrGzSsoQKdxbgAd8+1eN/2sv8AgmZ8FvHE2twx/D+78ZQSFpNQ8L+FYry5V8gk/bIYtm7PX95nI55r9nf9qr4NftV6VreofCfUr+9h0KWKHUIr+we2KGUMU4bhgQjdCcY561N8Kv2WdJ+LGieIj4M+HemfEWGWR9LligtrXUJZGifcVRdrynyy55DYGSMYyKqqqqqqqq1zwj4T8TmNvEnhjSdWMIxGb6yjn2fTeDira1tbK3jtLK3iggiXbHFEgRUHoAOAKqq/4LJf8mi2n/Y4WP8A6Jua/YD/AOTNPhL/ANi/H/6G9VVV8GbSHxj8TtZ8Wamona2L3EIIyFkd8KfwXOPw9KqvGtnH4L+POk3+kjyl1GaCeSNOB+9cxyD/AIFhj9Wqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr/grB/wAmR+MP+whpn/pZFX/BLj/kxL4Y/wDcW/8ATteVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX/AAWL/al8UaNqml/sx+CdYn0+yvNNGo+KJbaUo90srMsNoxGD5e1C7ryH3xg/dINz+y/8dLH4KP8AtDan4BurDwCGiEWqXc0UJuPNkEaNHCzea6FiMOF2kcgkV/wQ6/5FT4t/9hDSv/RdzV3cP8BP+ChFxPpEwt7fwl8VXEX7zaDaLqBBRj2DQkq3sTVVVVVVVVVVVVX/AAWS/wCTRbT/ALHCx/8ARNzX7Af/ACZp8Jf+xfj/APQ3qqrXvEeh+GLE6jr2pwWUAOA0h5Y+iqOWPsATXwl8Y6L4X+I16sl3jStUaSCK4kG0L8+Y2bPQEDB9N3PSgQwBUggjg1I8cUbSSOqIgJZmOAAOpJq9v0+KXxzsJNIJlsNNkjAlHQxQtvZvYMxIB/2h61VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV/wVg/5Mj8Yf9hDTP8A0sir/glx/wAmJfDH/uLf+na8qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv+CvPhvVNF/bH1LV76Jxba/oOn3dk5HDIkZgYA+zwv+fvk/tgfGz4XfG7/gmFc698NNc0+aK3i0OG60uGZfO0uRLiBWgljHKFSMDgAjBGQQa/4JA/E3wF8I/hR8afG/xG8U6foWj6feaY8lxdzBN5EVydiAnLuegRckkgAVJrV/8AtG/tbLr9jYzRXHxE8frNb2wwXgW6vQUTIyPkVwM/7Oaqqqqqqqqqqqq/4LJf8mi2n/Y4WP8A6Jua/YD/AOTNPhL/ANi/H/6G9VWoXttplhc6leSbILSF5ZW/uqoJJ/IV4e0PWvj14xvNa1y8lt9KsyAVQ8xoSdsUeeAcAktj3PUV4l+Afg3VNFjsdBg/sq8twTFcgtJ5h7iQE8g4/Dt6VbaB+0H4EUafo00t5ZRjEYikjuI8eirJ8yj2wKsL74ofF/VpvC1z4j2eUhe4glYW8aqrAHKoPmwSOCDz+dfDz4b6P8PdNaCzf7TfXAH2m7dNrPjoAOdqj0/M1VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV/wVg/5Mj8Yf9hDTP/SyKv8Aglx/yYl8Mf8AuLf+na8qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv2+v2LNP8A2vfh3af2Je2+meOvC4ll0K8mGIpw4HmWsxGSEcopDclGGcEFgfir8Eviz8ENfk8NfFbwDrHhu9RysZvLciK4x1aGYZjlX/aRmHvWl6VqmuahBpOi6bdahfXLbILW1haWWVvRUUEk+wFf8EyP+CenjXwH4us/2i/jtoM2iX2nwt/wjWgXa7bmOSRCjXNzGeYyEZgkbfMCxZgu1c1VVVVVVVVVVVf8Fkv+TRbT/scLH/0Tc1+wH/yZp8Jf+xfj/wDQ3qq+KYkPw718RE7vsT5x6d/0zX7N/wBn/wCEEuvK2+Z/aUnm+ufLjx+mP85qq8DbG/aF1g6ef3InvPN/M7un+3iqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/wCCsH/JkfjD/sIaZ/6WRV/wS4/5MS+GP/cW/wDTteVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV8eP2l/gt+zV4fg8QfF7xrb6QLwstjZIjT3d6yjkRQoCzAZALEBVLDcwyK8Of8Fif2Qte1xdI1CLxzoFu0gQalqWjRtbgE43EQTSyAd/uZ9q0bWfB3xH8K2uuaFqOleI/D2sQCW3uYHS5trqMnggjKsMj8CPUV4//aY/Y4/Zpuruw8S+PfBXhrVIcrc6bpNus14rcfK8FqjSKTgfeAHAq6/4LC/CXxH8RfDfw6+E/wAN/EOttr+t2mmNqmqyx2EMQnnWMypGvmPIAGyFYRkn071VVVVVVVVVVVf8Fkv+TRbT/scLH/0Tc1+wH/yZp8Jf+xfj/wDQ3qqvrK31KxuNOu03wXUTRSqe6sMEfka0PWNb+AnjK70nWLSW60i8IyyDHmoPuyx543AHBX8M8A1/wTZ/a++K/wAWPiz8R9H+N/xYbUtI0uyV9Khvore3ETm4ZTtKIpJ2gDkmvHfx88N6VYS2fhK5GpajIhVJVUiKAkfeJI+Yjrgcepr4CeA9Q0e2uvGOuxSJe6mmyBJfviIkMzt3yxAPPYZ71VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVf8FJ/i18Rfgn+zBfeOvhb4on0HXYtZsoEvIYo5GEbuQ64kVl5AHav2HfH/AIw+Kf7Kfw+8fePtcl1jxBq9lPJe3ssaI0zLdTICQgCj5VUcAdKqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr9ub9pr46/Cv9uX4SfCv4f/ABDu9H8K+IbLRZNT02K2gdLhp9VuIZSWdGYbo0ReCOnGDzVVVf8ABWD/AJMj8Yf9hDTP/SyKv+CXH/JiXwx/7i3/AKdryqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv2v8A40eIP2i/2kfF3jS4u7m9tX1STTtAthlvJsYpDHbxog6Fh85A6vIx6mta/wCCcn7ZegeApPiJqXwXvhp0Fsbme1jvbeS9iiCklmtlkMuQBygUuO6jBx8Mv2wvjZ8Ivgj4q+BngfxTd6fpPie7inW6hnKTacoDC4S3Ycx+d+6DEEYEZxy7GvB/w++IPxK1NtM8BeCtf8UagWy8GladLeSAnuRGrEZ55Nfsuf8ABMn9rG++J3g7x74t8HWPg3RdB1uy1KZ9bvkWeWOCdJGRIIt7hyFIAkCDPcVVVVVVVVVVVVV/wWS/5NFtP+xwsf8A0Tc1+wH/AMmafCX/ALF+P/0N6qq1zw/oniWxOna7pkF7bnkJKuSp9VPVT7jBr9gWNIvi98SIoxhUg2qPQC5avhP8LPClj4d0jxJfaSl1ql1bpceZOS4j3fMu1TwDgjnGc1VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV/wAFef8AkzTUv+xg07/0Nq/4Jvf8mR/Cv/sH3P8A6WT1VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX/AAUm/wCUkvwK/wCwf4e/9Pd3VVVf8FYP+TI/GH/YQ0z/ANLIq/4Jcf8AJiXwx/7i3/p2vKqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqtfvJtO0LUdQtiomtbSWWMsMjcqEjj6iv+CfelaRrP7Zvwostbjie2TXPPVZCAPOihkkh69/NRMe+Kr9qTwlpemftbfEvwdoMKWNj/AMJvfW9tGq/LAr3LYVQMfKu7AHoAK+GHwt8CfBvwVpvw/wDhz4dtNH0bS4VjihgjCtIQADJIwGXkbGWc8k8mqqqqqqqqqqqqq/4LJf8AJotp/wBjhY/+ibmv2A/+TNPhL/2L8f8A6G9VVXlylnaTXchG2CNnbJxwBk/yr/gn20lx8WPiExy8klsDwOSTcN2FadZpp+n2unx4220KRLj0VQP6VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV/wV5/5M01L/sYNO/9Dav+Cb3/ACZH8K/+wfc/+lk9VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV/wUm/5SS/Ar/sH+Hv/T3d1VVX/BWD/kyPxh/2ENM/9LIq/wCCXH/JiXwx/wC4t/6dryqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqeCG6gktriNZIpUKOjDIZSMEEehFXdvqHh7V57MzPb3un3DxM8TFWR0JU4PBHINf8Jb4q/wChn1X/AMDZP8amuLi4na6nuJJJnbc0jsSxb1J65r/hLfFX/Qz6r/4Gyf41+z94o8TTfHr4bQy+I9TdH8X6UrI15IQwN3FkEZ6VVVVVVVVVVVVV/wAFsLi7X9mzwdaJBm2k8cQvJJtPyutjdhRnoMhnOO+32Nfsnft//sb+Df2fvhz8ONd+MUGk634f8N2FnqNte6TeRLHcrCglAkERjYCQsNwbnGehzUv7f/7GcUbSt+0N4WIRSSFkkY8egCZJ9hV//wAFM/2G9OVGuPj1ZvvJA8jRNRnxj12W5x+NXH/BVv8AYghmeKL4o6jcKpwJI/Dl8Fb3G6IH8wK8Zf8ABVT9jXUfCmrado/xE1SS9urOSGFToF4vLKV6mPA61+xF8cfhb8Hfi1rviT4ma3PYaPqUkOyWKzlnLKtxvb5Y1JHy+1f8PYP2I/8AopGq/wDhO3v/AMbr/h7B+xH/ANFI1X/wnb3/AON1/wAPYP2I/wDopGq/+E7e/wDxuv8Ah7B+xH/0UjVf/Cdvf/jdf8PYP2I/+ikar/4Tt7/8br/h7B+xH/0UjVf/AAnb3/43X/D2D9iP/opGq/8AhO3v/wAbr/h7B+xH/wBFI1X/AMJ29/8Ajdf8PYP2I/8AopGq/wDhO3v/AMbr/h7B+xH/ANFI1X/wnb3/AON1/wAPYP2I/wDopGq/+E7e/wDxuv8Ah7B+xH/0UjVf/Cdvf/jdf8PYP2I/+ikar/4Tt7/8br/h7B+xH/0UjVf/AAnb3/43X/D2D9iP/opGq/8AhO3v/wAbr/h7B+xH/wBFI1X/AMJ29/8Ajdf8PYP2I/8AopGq/wDhO3v/AMbr/h7B+xH/ANFI1X/wnb3/AON1/wAPYP2I/wDopGq/+E7e/wDxuv8Ah7B+xH/0UjVf/Cdvf/jdf8PYP2I/+ikar/4Tt7/8br/h7B+xH/0UjVf/AAnb3/43X/D2D9iP/opGq/8AhO3v/wAbr/h7B+xH/wBFI1X/AMJ29/8Ajdf8PYP2I/8AopGq/wDhO3v/AMbr/h7B+xH/ANFI1X/wnb3/AON1/wAPYP2I/wDopGq/+E7e/wDxuv8Ah7B+xH/0UjVf/Cdvf/jdf8PYP2I/+ikar/4Tt7/8br/h7B+xH/0UjVf/AAnb3/43X/D2D9iP/opGq/8AhO3v/wAbr/h7B+xH/wBFI1X/AMJ29/8Ajdf8PYP2I/8AopGq/wDhO3v/AMbr/h7B+xH/ANFI1X/wnb3/AON1/wAPYP2I/wDopGq/+E7e/wDxuv8Ah7B+xH/0UjVf/Cdvf/jdf8PYP2I/+ikar/4Tt7/8br/h7B+xH/0UjVf/AAnb3/43X/D2D9iP/opGq/8AhO3v/wAbr/h7B+xH/wBFI1X/AMJ29/8Ajdf8PYP2I/8AopGq/wDhO3v/AMbr/h7B+xH/ANFI1X/wnb3/AON1/wAPYP2I/wDopGq/+E7e/wDxuv8Ah7B+xH/0UjVf/Cdvf/jdf8PYP2I/+ikar/4Tt7/8br/h7B+xH/0UjVf/AAnb3/43X/D2D9iP/opGq/8AhO3v/wAbr/h7B+xH/wBFI1X/AMJ29/8Ajdf8PYP2I/8AopGq/wDhO3v/AMbr/h7B+xH/ANFI1X/wnb3/AON1/wAPYP2I/wDopGq/+E7e/wDxuv8Ah7B+xH/0UjVf/Cdvf/jdf8FD/wBvL9mj9oT9m29+HPwt8ZX+pa7Nq9ncpBNpFzbKY42Jc75EC8AjjNfsX/8ABRP9lP4Nfsw+A/hn4+8c6jZeINCs54r23i0S6mWNmuZXGHRCp+V1PB71/wAPYP2I/wDopGq/+E7e/wDxuv8Ah7B+xH/0UjVf/Cdvf/jdf8PYP2I/+ikar/4Tt7/8br/h7B+xH/0UjVf/AAnb3/43X/D2D9iP/opGq/8AhO3v/wAbr/h7B+xH/wBFI1X/AMJ29/8Ajdf8PYP2I/8AopGq/wDhO3v/AMbr/h7B+xH/ANFI1X/wnb3/AON1/wAPYP2I/wDopGq/+E7e/wDxuv8Ah7B+xH/0UjVf/Cdvf/jdf8PYP2I/+ikar/4Tt7/8br/h7B+xH/0UjVf/AAnb3/43X/D2D9iP/opGq/8AhO3v/wAbr/h7B+xH/wBFI1X/AMJ29/8Ajdf8PYP2I/8AopGq/wDhO3v/AMbr/h7B+xH/ANFI1X/wnb3/AON1/wAPYP2I/wDopGq/+E7e/wDxuv8Ah7B+xH/0UjVf/Cdvf/jdf8PYP2I/+ikar/4Tt7/8br/h7B+xH/0UjVf/AAnb3/43X/D2D9iP/opGq/8AhO3v/wAbr/h7B+xH/wBFI1X/AMJ29/8Ajdf8PYP2I/8AopGq/wDhO3v/AMbr/h7B+xH/ANFI1X/wnb3/AON1/wAPYP2I/wDopGq/+E7e/wDxuv8Ah7B+xH/0UjVf/Cdvf/jdf8PYP2I/+ikar/4Tt7/8br/h7B+xH/0UjVf/AAnb3/43X/D2D9iP/opGq/8AhO3v/wAbr/h7B+xH/wBFI1X/AMJ29/8Ajdf8PYP2I/8AopGq/wDhO3v/AMbr/h7B+xH/ANFI1X/wnb3/AON1/wAPYP2I/wDopGq/+E7e/wDxuv8Ah7B+xH/0UjVf/Cdvf/jdf8PYP2I/+ikar/4Tt7/8br/h7B+xH/0UjVf/AAnb3/43X/D2D9iP/opGq/8AhO3v/wAbr/h7B+xH/wBFI1X/AMJ29/8Ajdf8PYP2I/8AopGq/wDhO3v/AMbr/h7B+xH/ANFI1X/wnb3/AON1/wAPYP2I/wDopGq/+E7e/wDxuv8Ah7B+xH/0UjVf/Cdvf/jdf8PYP2I/+ikar/4Tt7/8br/h7B+xH/0UjVf/AAnb3/43X/D2D9iP/opGq/8AhO3v/wAbr/h7B+xH/wBFI1X/AMJ29/8Ajdf8PYP2I/8AopGq/wDhO3v/AMbr/h7B+xH/ANFI1X/wnb3/AON1/wAPYP2I/wDopGq/+E7e/wDxuv8Ah7B+xH/0UjVf/Cdvf/jdf8PYP2I/+ikar/4Tt7/8br/h7B+xH/0UjVf/AAnb3/43X7an7UvwY+M/7afwr+Mvw/8AEV1feFfC1no8Wp3UunzQPE1vqdxcS4jdQzYjkQ8A5zgc1/w9g/Yj/wCikar/AOE7e/8Axuv+HsH7Ef8A0UjVf/Cdvf8A43X/AA9g/Yj/AOikar/4Tt7/APG6/wCHsH7Ef/RSNV/8J29/+N1+37+37+zD8e/2YfEXwz+GfjS/1DxBqF5YyW9vNo9zbqyxXMbud7oFGFUnk1/wS4/5MS+GP/cW/wDTteVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVefssfsw6hdzX9/+zj8L7m5uZGkmmm8H2LvI7HLMzGLJJJJJPUmv+GTf2V/+jZ/hT/4Rlh/8ar/hk39lf/o2f4U/+EZYf/Gq/wCGTf2V/wDo2f4U/wDhGWH/AMarTv2Xv2Z9I1C11bSf2dvhjZX1lMk9tc2/hGxilglQhldGWIFWBAII5BANVVVVVVVVVVVVfFf4Q/Df44+DLn4ffFXwpa+INBupEle1md4ysiHKukkbK8bDkblYHBIzgkHX/wDgkt+xTrLE6d4G13QsoVAsPEFy+CQBu/ftLyMZ9OeQRxQ/4I3fsiggnUPH5weh1mHn/wAgVZf8EkP2LbSRnn8I+Ir0FcBJ9fnAU+o2FTn8atv+CWX7CsECQy/BSW5dRzLL4l1QM3123IH5AV/w64/YS/6IX/5c2rf/ACVX/Drj9hL/AKIX/wCXNq3/AMlV/wAOuP2Ev+iF/wDlzat/8lV/w64/YS/6IX/5c2rf/JVf8OuP2Ev+iF/+XNq3/wAlV/w64/YS/wCiF/8Alzat/wDJVf8ADrj9hL/ohf8A5c2rf/JVf8OuP2Ev+iF/+XNq3/yVX/Drj9hL/ohf/lzat/8AJVf8OuP2Ev8Aohf/AJc2rf8AyVX/AA64/YS/6IX/AOXNq3/yVX/Drj9hL/ohf/lzat/8lV/w64/YS/6IX/5c2rf/ACVX/Drj9hL/AKIX/wCXNq3/AMlV/wAOuP2Ev+iF/wDlzat/8lV/w64/YS/6IX/5c2rf/JVf8OuP2Ev+iF/+XNq3/wAlV/w64/YS/wCiF/8Alzat/wDJVf8ADrj9hL/ohf8A5c2rf/JVf8OuP2Ev+iF/+XNq3/yVX/Drj9hL/ohf/lzat/8AJVf8OuP2Ev8Aohf/AJc2rf8AyVX/AA64/YS/6IX/AOXNq3/yVX/Drj9hL/ohf/lzat/8lV/w64/YS/6IX/5c2rf/ACVX/Drj9hL/AKIX/wCXNq3/AMlV/wAOuP2Ev+iF/wDlzat/8lV/w64/YS/6IX/5c2rf/JVf8OuP2Ev+iF/+XNq3/wAlV/w64/YS/wCiF/8Alzat/wDJVf8ADrj9hL/ohf8A5c2rf/JVf8OuP2Ev+iF/+XNq3/yVX/Drj9hL/ohf/lzat/8AJVf8OuP2Ev8Aohf/AJc2rf8AyVX/AA64/YS/6IX/AOXNq3/yVX/Drj9hL/ohf/lzat/8lV/w64/YS/6IX/5c2rf/ACVX/Drj9hL/AKIX/wCXNq3/AMlV/wAOuP2Ev+iF/wDlzat/8lV/w64/YS/6IX/5c2rf/JVf8OuP2Ev+iF/+XNq3/wAlV/w64/YS/wCiF/8Alzat/wDJVf8ADrj9hL/ohf8A5c2rf/JVf8OuP2Ev+iF/+XNq3/yVX/Drj9hL/ohf/lzat/8AJVf8OuP2Ev8Aohf/AJc2rf8AyVX/AA64/YS/6IX/AOXNq3/yVX/Drj9hL/ohf/lzat/8lV/w64/YS/6IX/5c2rf/ACVX/Drj9hL/AKIX/wCXNq3/AMlV/wAOuP2Ev+iF/wDlzat/8lV/w64/YS/6IX/5c2rf/JVf8OuP2Ev+iF/+XNq3/wAlV/w64/YS/wCiF/8Alzat/wDJVf8ADrj9hL/ohf8A5c2rf/JVf8OuP2Ev+iF/+XNq3/yVX/Drj9hL/ohf/lzat/8AJVf8OuP2Ev8Aohf/AJc2rf8AyVX/AA64/YS/6IX/AOXNq3/yVX/Drj9hL/ohf/lzat/8lV/w64/YS/6IX/5c2rf/ACVX/Drj9hL/AKIX/wCXNq3/AMlV/wAOuP2Ev+iF/wDlzat/8lV/w64/YS/6IX/5c2rf/JVf8OuP2Ev+iF/+XNq3/wAlV/w64/YS/wCiF/8Alzat/wDJVf8ADrj9hL/ohf8A5c2rf/JVf8OuP2Ev+iF/+XNq3/yVX/Drj9hL/ohf/lzat/8AJVf8OuP2Ev8Aohf/AJc2rf8AyVX/AA64/YS/6IX/AOXNq3/yVX/Drj9hL/ohf/lzat/8lV/w64/YS/6IX/5c2rf/ACVX/Drj9hL/AKIX/wCXNq3/AMlV/wAOuP2Ev+iF/wDlzat/8lV/w64/YS/6IX/5c2rf/JVf8OuP2Ev+iF/+XNq3/wAlV/w64/YS/wCiF/8Alzat/wDJVf8ADrj9hL/ohf8A5c2rf/JVf8OuP2Ev+iF/+XNq3/yVX/Drj9hL/ohf/lzat/8AJVf8OuP2Ev8Aohf/AJc2rf8AyVX/AA64/YS/6IX/AOXNq3/yVX/Drj9hL/ohf/lzat/8lV/w64/YS/6IX/5c2rf/ACVX/Drj9hL/AKIX/wCXNq3/AMlV/wAOuP2Ev+iF/wDlzat/8lV/w64/YS/6IX/5c2rf/JVf8OuP2Ev+iF/+XNq3/wAlV/w64/YS/wCiF/8Alzat/wDJVf8ADrj9hL/ohf8A5c2rf/JVf8OuP2Ev+iF/+XNq3/yVX/Drj9hL/ohf/lzat/8AJVf8OuP2Ev8Aohf/AJc2rf8AyVX/AA64/YS/6IX/AOXNq3/yVX/Drj9hL/ohf/lzat/8lV/w64/YS/6IX/5c2rf/ACVX/Drj9hL/AKIX/wCXNq3/AMlV/wAOuP2Ev+iF/wDlzat/8lV/w64/YS/6IX/5c2rf/JVf8OuP2Ev+iF/+XNq3/wAlV/w64/YS/wCiF/8Alzat/wDJVf8ADrj9hL/ohf8A5c2rf/JVf8OuP2Ev+iF/+XNq3/yVX/Drj9hL/ohf/lzat/8AJVf8OuP2Ev8Aohf/AJc2rf8AyVX/AA64/YS/6IX/AOXNq3/yVX/Drj9hL/ohf/lzat/8lV/w64/YS/6IX/5c2rf/ACVX/Drj9hL/AKIX/wCXNq3/AMlV/wAOuP2Ev+iF/wDlzat/8lV/w64/YS/6IX/5c2rf/JVf8OuP2Ev+iF/+XNq3/wAlV/w64/YS/wCiF/8Alzat/wDJVfD/AOH/AIO+FfgzSvh78P8AQodH8P6JB5NhYxMzrEmSx+ZyWYlmJLMSSSSSSaqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqviH8Tvh58JfDsniz4meNNI8NaRG2z7VqV0sKu+CQiAnLuQDhVBJxwK0L/gpB+xH4j1pNA0/4/aTFdPJsEl/YXljADnHM88KRAe5bHfNWd5Z6laQ3+nXcN1bXEYkhmhkDpIhGQysOCCOciqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq8T+KfDPgrQ7rxN4w8Q6bomkWKb7m/1G6S3hhX1Z3IA/E18LP2jvgb8b9QvtK+E3xM0fxPd6ZCs13DYyMzQxsdoY5A4J4qq+Jn7dn7I/wg12bwx48+N+i22q2shiuLSxguNSkt5BnKSC1jk8thggq2CO/WvhB+0d8DPj7aTXXwg+Jui+JGtlDT2sEpiuYFPAZ7eQLKik8AsoBNVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVWu6/oXhfSbnXvEut2Gk6ZZoXub2+uUt4YVHVndyFUe5Na5/wUj/Yi8P6mdIv/AI+6VLOH2b7HTr29izx/y1ghePHPXdj34r4XfG/4Q/GzTJNW+E/xG0LxRBbgeethdq8tvnp5sXDx5xwGUZqtf8QaD4V0a78ReJ9asdJ0rT4jLd319cLBDAg6s7sQqj3Jq6/4KUfsQWmsHQpfj5prXIfZvi0u+kgz/wBd1gMWOOu7HvXhHxl4R+IHh+18VeBvE2l6/o96ube+066S4hkx1w6EjI6EdQeDVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVXiXxDo/hHw5qnivxDepaaXotlNe31w/SGCJC8jn2CqT+FeANA+In/BWT9rTU9Y8b65f6R4C8OxtO1vCwP9lacZMQ2sIOV+0S4y0hByVdsEKqV8V/8AgkT+y14m8AXWkfC7QtR8IeKobZjYat/a1zdpNOF+UXEczupQkc+WEIzkdMV/wSk/aN8b/D34t6r+xl8UbucWxmvI9Gtrp9z6ZqVqWNxaqeyOqStt6B4+BmQ5qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqYFlKhiDjgjtXi79ir9tb9qKfxN8Sf2t/ifJovh/wtFfz6XpiqrGfyA+Gt7SMiK3jcIP3jkyFdpIcENX/AARD/wCSp/Ev/sX7X/0oNV/wVm/a68QfBbwTpXwT+G+sy6b4n8bW0k+pX1u+2ay0wEx4QjlWmcOocchYpMYJBH7If/BJ74Mr8KtJ8X/tJeHr3xJ4o8Q2kd22lf2jcWUGkxuoZIv9HdHeUKRvLMVBO0D5dzftt/sr6z/wT5+JXg/4+/s5eJNW0/Qru/Mdo08vmyaZeqpfyGYj97DLGHwr5JCSKxIwT+zf8aNL/aG+CHhH4v6XDHb/APCQWAe7tkYkW10jGO4iGecLKjgE9QAe9VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV+27+wz4y/a/+Ivge4i+LN14f8E6TaTprenMzzATBwYpreDIQzOrujO5G1UQgNkg6d/wSF/Y2stCGk3eieKtQuwmDqdxrrrOTgc7YwsWeP+eeOa/aI+EPxC/4JiftMeHvFXwm8Z3t1pt5Cb3RLy6AD3NurhZ7K7VMLIPuhiAAwdGAVh8vw78Z6d8SPh/4Z+IWkRtHY+J9HtdTtkY5KR3EKyqCfUBwDX7XXxS+I/7fv7Ydj+yl8MdbNt4N0TWJNPgAJMEs9uG+2ajMBjzBGFlWMZwVQbcNKc23/BIz9jiLwMvhe48N6/PrP2fY3iX+251uzJjmQRBvs45/h8oj69a/Z68deP8A/gnD+2ze/ALxjrz3XgfWdWgsdS3grBLBcBfsmpIvOx1V034zwJEOSqkVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVf8ABTzxdceEP2J/iDJZTrFc6utnpqEtjKzXUSygDPOYvMGPfPQGv+CKPg+3039n/wAZ+NmhVbvXfFZtC2Bl4ba2iKcg/wB+4mGCB696r9o8R/B7/grZb69pJFrbN420LUn2vt3JcpbNdAkDgOzzg8Hhqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr4s/wDJK/GX/Yv33/pO9f8ABEP/AJKn8S/+xftf/Sg1X7XC/wDC7P8AgqvZ+BdQKXOmDxToGheXuDBLYC3+0A84PzSTnbx1x1qv+Cp/hW08TfsS+OLqaBZLjQZ9P1C0LHGx1vIo3b6+VLKPxr/giz4on1X9mzxP4XubgyHQvF0rQKz58uGa2gYKB2G9ZW9yx/Gqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr/AILXeOtM8T/FP4afCnRW+2av4c068urqGBS7q988CxRYGfnItQ23riRT3FeFdO1D9nX9jfT7K8eNNS+Hfw6D3LOwCC4tLDdIck4C70bvwK/4IpeFItZ+Onjzx3eKZ5tF8NrbRySHcVkurhSWyTncVt3GcHhjyM81/wAFtfC9rp/xm+HvjS3iEdxq/hyazldTgv8AZrgspI9QLnGfQAdq+DXiebxv8H/AvjO4nM0uveGtPv3kLbt7TW0chOeM5LdcVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVf8FaNIm1L9ijxPeRE7dK1XTbmQBc/KbpIueeOZV55/XNf8EZb+2u/2TtVtIZMy2XjS9jlU8EE21qw/DDD9fSq/bdgj8Wf8FUrXw5YMzTS+IvDFgSqg/vHitPu88/6wDtzkVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVXxZ/wCSV+Mv+xfvv/Sd6/4Ih/8AJU/iX/2L9r/6UGq+J1unhP8A4LD2qanKVWT4k6NLuZduBci2dO/T96vPpzxVf8FML+2079h74ny3LECW1soUA6lnv7dR+pyfYGv+CJOkyw/A3x9rpLeXd+LFtlyvG6K0iY4P0mX9PWqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr9rz9rPwH+yV8NJvFviOWK+8QagkkXh7QlkxJqNwAOTjlYUypd+wIAyzKD/AME7v2ZPHf7S3xmuv23f2gVlu7GPVWvtIW6i2/2tqKkbZkU9Le3IUJjguiqDiNhX7SekTeIP2dPinoNsSJtS8FaxbRkLuIaSylUYGRnk9K/4Ie31tH46+KultJ/pE+k6dNGvqscswY/nIn51X/BcK/tpfHfwq0pGPn2+kahM49FklhVf1jav2btIl0D9nX4WaDOW83TfBWj2z7lwd0dlEpyOx4qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/aN+FSfHD4E+OPhQGjWfxHo09vZvIMrHdAb4HPssqxk/Sv8AgkB8b7T4V/FLxj+zX8Q5v7Hu/Elyr6ZFeN5Xl6pbFo5rUg8CR1xgHHMG0ZLAV4z8ZeF/h74U1Txv401u10nQ9Ftmub69uX2pDGo5PuTwAo5JIABJAr9inR9b/bA/4KMav8fbqwkTQtE1a58R3Pmr/qF+aPT4CRxvB8o+4gc1VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVXxZ/5JX4y/7F++/9J3r/AIIh/wDJU/iX/wBi/a/+lBqv+CwXwu8QfDn4+eDf2lPDMTRW+tQQQyXSJ/qNTsm3Rlj/ALUQj2j/AKYv6cfAL41eEf2hfhN4f+Kvg29hltdXtVNzbpJvaxugB51vJ3Do2RyBkYYcMCf+CzX7QegWHgDRP2ctC1aG413VtQj1TXIInDG0tIlJhSQfwtJIyuB12xZOAyk/8E8vgtqXwL/ZQ8HeF9ftDba5qySazqkLLtaKW5beiMOzJCIUYf3lNVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV+1r+1r8O/wBkr4dv4r8VzLf65fq8eg6DFKFm1GYD8dkS5BeQjCggDLMqn9nL9mP4w/8ABR74myftNftUale2/gWOQR2FnCGtxqSRtxa2i5zFaqch5B8zMWAJcu66VpWmaHplpoui6fbWGn2ECQWtrbRCKKCJAFVEVcBVAAAA4AFXNtb3lvLaXUKSwzIUkjdch1IwQR6EcV+zXryf8E/P+Chmr+AviFdGw8N3FxcaBdX9wxRFsbhkls7tuoCkpbsxJ+VWfJ4NfaLf7P8Aa/tEfkbN/m7ht24zuz0xjnNftN67H/wUB/4KGaP8P/hzeDUvDlrNb6Ba39ucxmyt2ea9ulI4KAvcFWB+dVTB5FQQQ20EdtbRrHFEgSNFGAqgYAA9MVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV+2b/AMExfCH7SHiiT4qfDjxND4J8cT4a+Z7Yvaak6gBZHCENFLwMyLuzgZUn5ql/4JYftqfElrPwz8cP2srXUfCtpIpihbXdU1owgcZjt7hYo1IAAHzcfhX7N37NXwz/AGW/h3D8PfhtYSbHfz9R1K6w11qM+MGSVgAOBwqgBVHQZJJqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqrxlocvifwfrnhq3nSGXVtNuLRJXBIRpI2QMQOw3Zr9gX9gXxf+x54v8VeJPEnxA0fxDH4h02G0iisbWSJomSTeWJfqCOKr4wfCDwF8dvh7qvwy+JGjLqOiatGA6g7ZIJByksT4OyRTyG/A5BIM/8AwSf/AGrfhRrt+/7Mn7UsOjaVft+8Z9X1DQrh0B+VZfsayLKRxz8oPXA6V+zJ/wAEoNJ+H3j63+Lv7RXj1PiH4ltLkXVvYJHI9p9pHImnlmPmXLA4IDKgyBnd0FVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV+2H/wAExvij+1B8fNe+Ldl8ZdI0/S7+C0g03Tb+2nmaxiit40dFwdqq0qyyYXAzIT1JqH/gjT8eLeFLe3/aS0eOOJQqIkF2oUAYAADcDHav+HN/x+/6OX0r/v1ef/FV+yJ8DvEf7OnwK0X4UeK/FcPiPUtMubqWTUIlcLIJZ3kUDeS3AYDn0r9s79hP4c/thaPZ3moag/hvxno8Ri03xBb2wmJiyT5E8eV82LJYgblKMxIOGYNF/wAEuP27BpC/DZ/2rdNTwAFEH9mr4k1YwiD+6LLyxDj/AGd2Pev2N/2Ffhn+x/o91d6Rey+JPGWrQiLUvEN1biJvKBB8iCME+TFuUMRuZmYAsxCqFqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv2g/+Ctv7SXgH4zeO/hr4S8L+AbbTfCfifUdKtp7jT7maeaO2uZIVaQ/aAuSEBO1Rz7U3/BX39sdpjKNV8Ihd2RENCXaBnp97OO3XPvXw7/4LX/GfS9Rij+KPwu8J+INN3ASNpDTadcqCeSC7yo2B0XaucdRnNfs/ftB/DX9pj4dWvxJ+GGqvcWMkhgu7W4QR3FhcKAWhmQE7WAZTwSCCCCQQaqqqqqqqqqqqq/4KIanqWjfsY/E7U9H1G5sbyCwtzFcW0zRSRk3kAJVlII4JHHrX7LPxO+JWoftO/CCwv8A4h+Jbm2ufHmiRTQzatO6SI19CGVlLYIIJBB4Oaqqqv8Agrf+1R8evhn8UfDvwl+G3jLW/B2hy6Cmpz3uk3DWlxfzSTSx4E6YdUQRD5VYZLktn5cf8Ejf2k/jH8cfCfjnwn8V9ev/ABKnhGayfTdavj5k5W4E2+CWU8yFTEGUtlgHYE4CgVVVVVVft8ftN/8ADLn7P2qeK9FuoV8W62/9l+HI3AbbcyKS0+05yIowz8gqWCKfvV/wSw+Kf7VH7RGseKPiV8X/AIt6prPg3w+o06zsZLS2iS7v5AHZi0cStiKIqSuRkzIexFVVV+2v8ZPF37Pv7MfjP4veA49Pk13QPsP2RdQhaWE+dfW8D7lVlJ+SVscjnB9q/Zy/4Kp/tOfFb48+APhr4nsfBK6T4l8QWlheta6VLHKIpJArbGMxAbB4JBqq/wCCpXxe/au/Zw8beHPG/wAKPjDquleC/Flu1s1glnbSpY30AG4BniLBZYyrAEk7kl6DAH7D/wC0lB+1H+z7ofj28mhHiSxJ03xFBHgBL2IDc4XssiFJQOg37f4TVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVftZf8nUfGX/soGu/+l81fsBfs1fs7+M/2PPh54j8YfAvwDrur6tZXL3uoal4dtbq5mYXUyAmWRC/CqoHPGOK/4KvfsmfCz9nvxL4O8cfCXSk0LTvGQvIb3RonYwwTweURJCDkqriUgrnaCowBuOP+CJPinXLb41+PvBELSnR9R8LLqFwMnYtxb3UUcXHTJS5l/Lv2nngtYJLm5mjihhQvJJIwVUUDJJJ4AA718VP+Cq/7IHww1WbRLXxRrHjW7tm2zf8ACLWK3MStnGBPLJFFJxzlHYe+eK+E3/BVL9kT4r61beHpPE+seDL68cR248U2S2sTuTgAzxSSxR+uXdRyOc8VqmoxaXpN3q7wT3MdpbvOYrdN8kgVS2EX+JjjAHckV/w95/Y0/wCg54q/8EL/APxVaFrWm+JdD07xFo1yLiw1S0iurSUDHmRSIHRse6kGvHv/AAVF/ZM+HHjbXvh/4h1/xC2q+HNRn0+++y6O8sazwuUkVXB+YBlIz7V4V8S6P408L6P4w8PXX2nS9dsIL+xmxjzIJo1kjbHbKsD+NfEX/gp9+yj8L/HWu/DrxPrviE6v4cv5bG+Fro7yxrNG211VgecMCPqK+Hfxx+HfxK+EVl8ctK1VtL8H3ttNdLfa0osRFBFI6PLJvOETMbEEkArg9DXj7/grH+xr4IupbHTvFuueLpoCyuNA0h3TcAeFknMSPnGAVYryOcZI8M/8Fj/2Q9fv0s9VtPHvhyJmAN3qeixSRqPUi2nmfA9lJrwF8QfBHxS8L2fjX4d+KdO8QaHfrmC9sZhIjEdVOOVYdCpwQeCBX7RH7THwu/Zd8Lab4w+K13qUGm6rqAsLdrGya5YzGNpMEAjA2xtzXwO/bp/Z1/aDh8T3XgfxTc2lt4OsVvtXudatvsMVvbndmQs7Y2rsO49BkZ618P8A/gpf+zZ8VfjdpHwK+Ho8Va1qWt3MlvaavHpscWns0cbyMS0kqzbcRnBERySO3Narqul6Hpl1rOt6la6fp9jC011d3UywxQRqMs7uxAVQASSSAK8ff8FeP2QPBOrS6TpV94t8YeRJse50DSUMOR1Ktcyw7gCMZUEHqCRzX7Tf7b/7Ov7T37F/xY0T4a+MZI9fh0y3kbRNVtzaXbRi8gJdFJKygd9jMV6kAYz+yd/ydR8Gv+ygaF/6Xw1VfGT48fCP9n/wyPFnxd8caf4esZWZbZZyzzXTqASsMKAvKwyMhVOMgnA5q8/4LO/sm22o/YYPDfxJu4Q+37ZDpFqIsZxuw90r47/dz7dq/Z2/a7+Bn7UtvqJ+EfiW7vLzR0R9QsLywltprZXJCE7hsYEqR8rN0Nft2/F/9hPR/EWjfCz9rnwdda7qI00apprW+nzGS3glkkiOy4gdJE3NbtlA2DtUntj9h74o/sp+O/BGs+Gf2TvDL6HoPhq6iN/bvpz2zSTTK22RndmeZysWC7EnCqM4Ar4t/wDBSb9mD4J/EXWvhb461bxDFrugSpFeJbaQ80YZo1kG1weflda/4e8/saf9BzxV/wCCF/8A4qv+HvP7Gn/Qc8Vf+CF//iq8EeL9F+IXgrw/4+8NySvpPiXS7bU7Bpo/LcwTxLLGWX+E7XXI7Gvj1+1r8AP2araM/Fn4gWmn6hcRmS20m2Rrq9nXsRDGCyqSCA77UyMbqsP+CzP7JV7qg0+40L4jWMBbb9uuNHtjCBnG7Edy0mO/3M+1fCf4yfC/46eEovG/wn8Z6f4j0eRvLaa2Yh4JMAmOWNgHifBB2OqnBBxgiqr/AILK/Fafxb+0dpXwutrstYeA9Fj82HdkLeXYE0jY7Zh+yj8Pev8Agnl8M4fhZ+x78ONINv5d3rOmDW7wkfM8l4fPXd7rG8aY6gIAelftE/twfAb9l3xPpvhD4r6jrNvqOq2H262Wx01rlTF5jR5JBGDuRuK/Zv8A2vPgz+1b/wAJF/wqO+1W4/4Rf7L/AGh9vsGttv2jzfL25J3f6iTPpx618dvjt8P/ANnP4fzfEv4l3N7DokF1FbO9nbGeTzJCQvyg9OOtfCX/AIKTfswfGz4i6L8LfAureIZdd16V4rNLnSHhjLLG0h3OTx8qNX/BUf8A5MS+Jv8A3Cf/AE7WdfsUf8nc/CD/ALHDT/8A0ctVX/BTH4YQ/E/9jfxzGtuJL7wrFHr9kxGfKa1bMzf+AzXC/wDAvwr/AIIu/Fe58OfHLxN8I7q5P9n+MdG+128bPwLy0bI2r7wyTZI5Oxc9OKqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/ay/5Oo+Mv8A2UDXf/S+av2c/wDgrRJ+z58DfC/wah+AS67J4atZYU1NvFP2YTl5pJAxh+yvtA8zGN5zjqM8ftC/tGfG/wDbv+LGjte+H2u7xA1l4b8MaHbyTCAOQzhF5eSRtoLueoQcKqgD/gmt+xVrH7LHgTVPFPxGjgHj3xgIhd20MolXTLWPJS33DhpCzFnKkrkIATtyf+Cw/wC1Z4h0S60/9l/wLrEtlBfWAv8AxXNbyFXnjkJENmSOQhCmRx/EGjHTcD+wF/wTb0z9qfwfe/FX4o+K9X0Pwol49lptro4iW6vpEA8yXzJFdY41J2gbGLENyoUbv+Cgv7DFr+x54g8P6l4Q8S3+ueD/ABSJ0s5NQVftVnPDsLRSsiqjhldWVgq5w4KjaCf+CPf7ROtfE34R618HPFupveaj8PHg/suaZiXbTZgwSPJJJETxso9EeNR92v2wfhUfgr+0z8RPh3DbGCysdbln05NuAtncYntwMcHEUqLkdwenQfshfGqwi/4J7eEfi1rNxHLD4O8G3K3gLc7dMEsOG75K26n33A9xWs6lqfiHVNQ8R6pI9xdX92891OR96aVmdiT6k7j+df8ABNf4wWHib9hvw5ret3w/4oGG80vUpSf9VHaEyR/TFs8NapdeI/jJ8VLu9t7c3Ov+OPEDyRxbuZLq7uCQuT6vJjPvVn8E/C1r8AI/2eQo/sBfCf8AwjjN5fLQm28hnIzyxGWJzkk5znmvAn/BF34b+HdAXWP2gvjtfrKEBuF0EQWFtbMQPlFxdLJvGc/MY0zxwK/bb/ZK/Zw+BPh/T/FXwB/aR0bxr5t+tpf+Hp9dsb3UbdXjZluF+zFcxgptbMYwXQ5OTj/gj/8AG/xD4K/aPT4NtfySeHviBZ3Aa0dz5cN7bQPPHMo6BjHFJGcfe3LnO1cf8Fq/+TV/C3/ZQLP/ANIL+vBGvePpdO1T4V+BIbu6fx5PZ2lzY2UTST35jlLRQKq/eBkKNtwclE6YOf2J/wDgmx+038Ivj58P/jR8QNN8O6ZpWjXMs99YDVRNeQq9tJGBtjVoyQzrkBz3r/gsr+0prdx4p0r9mPwzqklvpNlaRap4kWJtv2qeQk28D+qoiiXHQmVCeUGP2Lf2FvHf7Y+r6pcadr1v4Z8KaC6R6jrU9sbgmZxlYYYgy+Y+0bmyyhQQScsoP7Z//BMzxh+yp4JHxU8N+Ok8Y+FIbiK31J2sDZ3OntKdiOyh3V4mcqm4EENIo2kEtX7J3/J1Hwa/7KBoX/pfDVV/wUL+Lfin4s/tZ/EH+37+5ey8Ka1daDpNnI37u0gtJGhOxe290eQnqS/0A/ZB/wCCY/wN/aT+BWk/Em++OWrvr98sn26w0b7OU0mUOypDMjqz78KGOSmQeOMMf2Gf2Jrf9jPTfGtg3jKHxPceKdQt3ivlsDautrBGwjR03v8AMHlmPDEEEHjoP+C1f/J1Hhb/ALJ/Z/8Apff1/wAEOv8AkVPi3/2ENK/9F3NftR/8Eo/iD+0B8e/F/wAYNH+LHh3SrPxJcxSxWdzYzPJCEgjjIYqcHJjJ49a/a0/YIk/ZD8KWOteNPjd4c1bWNXm8vTNCsbKUXNyo/wBZKdxwkaDqx6sVA68fAj4IeOP2iPifo/wr+H9j52o6pJmWd1PlWUC48y4lI+6iA8+pIUZLAH4r+OtF/Yj/AGPzqsE/9qr8OvDFlo+kC7+Vr+4jjjtbbeF/vMFZ9vRQx4xXwu8C/FP9t/8AaRsvDeqeKZb3xP4zvZLjVNavh5n2eFELyylQQNqRphI1KjhEG0Yx8bv+CNHw+8MfCDWfEnwn+I3iu98X6Hp8l4LbVzbva6h5SFniRYoleJmAIUlnGcA8EsP2BP2ida/Z2/aN8NanHqbxeG/Et7DpPiK2Zj5UltK4RZWGfvRMwkB6gBh0Yg1X7dXiCTxP+2F8XNSlmWVofFF1ZAg9BbH7OB+AiA/CvCGhReF/CWieGYI0SPSNOt7RETooijVAB7fLX/BbT/kv3gX/ALE8f+lc9f8ABDH/AJrZ/wBwD/3I1/wV5/5M01L/ALGDTv8A0Nq/4Jvf8nufCv8A7CFz/wCkU9f8FR/+TEvib/3Cf/TtZ1+xR/ydz8IP+xw0/wD9HLVV8XNCi8UfCjxp4ZmjSSPVvD1/aOjnAYS27oQevHzV/wAE9defw3+2j8KNRimWMy60bPJB5FzBJARx6iUj8aqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr9rL/k6j4y/9lA13/wBL5q/ZL/4Jm/s1fHr9lnwZ8R/Fq+KbPxJ4jsZ3ubzT9UCBHW4ljVkjdGT7qLwQa/aP+CHiT9mL44eIPhTquovPcaDcpLp+pRKYjc27qJIJ1AJ2sVK5AJ2sGGTjNf8ABNX9ozWf2iv2bLG98X6i974p8I3baNqt1K2ZLsIivDO3qzROqsx5Z43J61/wU5k1ST9uP4mrqpbek1gsIJBAh/s+28vGOMbSD9Sc85r4S/A39srx74JtvEPwY8M+O7/wxJNLHBLpF+8cAkViJAFWQYO7OeO+e9a5+xp+314ohit/Evwk+IerRwMWiS+uDOEYjBKh5Dg4r/gk3+zV8ffgl8WvGmvfFX4c6z4X0q/8OLbQNfAKs8/2iNgAATkhVfk9M+5r/gtf8Kv7H+J/gf4xWNptg8SaVLpd86Lx9otX3IzH+80c4Ue0PtXw1+PT+G/+CVfxK+HK6h5eo3fjm30i02t8y213HHcOuPRltLsE9CHIr4W/AG48U/8ABP8A+MnxmayZ5dF8V6M1kxXB2WwkjuNp7jbqSk/9cvav2NvjnJ4D/Y1/am8EyXhjZ9BtrmwVW+YNev8A2dMwHb/XWwyOn5V/wSw+Eo+KP7XvhzUry1M2m+B7abXrnKfL5kWEt+egYTyxOB3EZ9Mj9t79p+D9k74F33xCs7K3v/EGoXSaboFpOT5b3cis2+QDkxoiO5AxkhVyNwIs5/2lP26fjHZ+HLjX9W8a+LNXkkeFL272W1lGBukZVOI7eJQMkKAOgAJIB/aP/wCCafxQ/Zh+DEnxe8dePvDF/wCTe29pJpulrPIQ0pwCJJEQHBBz8tf8E3v+T3PhX/2ELn/0inr/AILV/wDJq/hb/soFn/6QX9f8Eq9NsNR/bd8DtfWsc/2S11KeEOMhJBZTANj1GTjPQ4PUA1X/AAU0TUF/bj+KH9pA+abiwKZxzH/Z1t5fT/Y2/wBec1/wRxn0SX9kWaHSzH9ri8WXw1EKefOMcBUkf9cvKH4V/wAFKLi3tv2Ifii9xMkYaytEUscZZr63Cge5JAr9k7/k6j4Nf9lA0L/0vhqq/wCChH/BMfxT8V/Gmo/HT9nqO2utc1ciTXfDlxOsBupgApntpHIQMwALo5UEgsGydtX+l/Hn9mbxsi3tt4y+G3iiBd0Tgz6bOydMow2l0PTIJU+9f8Ey/wBv7xh+0LqN58EvjNNb3nizTNPN7petxosTanAjKskcyKAvnJuVgy43ruyAVLP/AMFq/wDk6jwt/wBk/s//AEvv6/4Idf8AIqfFv/sIaV/6Lua/aF+PXgb9mz4Wat8U/Ht0RaWCbLSzjYCbULpgfLt4gf4mI69FAZjwpr4s/FH4qftZ/Gq48Xa7Fc6x4m8T3iWmmaZaK0ggRm2w2lunJCruwB3JLHJYk/sD/sa6R+yX8LlXWYLa6+IHiSNJvEN+mHEOBlLSJv8AnnHk5I++5LdNoX/gs3Jqifsl6UunlvIfxtZLfAED9z9muyM+3mCL9K+EfhD4reOvGlv4c+C9hrN54nmglaCDSZzDcNGq5k2kMpwFGSM9BT/ssf8ABRyRDHJ4D+KjKwIZW1OQgjvx5tWf7Af7Z4vIDB8A/FVtIJV2TGNU8o54bIbIx1yKqv2x9Ml0n9rH4xWkxJZ/G+rTjKFeJbqSQcfRxz3696sbyDUbG31C2JMVzEssZYYO1gCOPoa/4Laf8l+8C/8AYnj/ANK56/4IZSRiT41wmRfMZdBZUJ5IH9oZIHoMj8xX/BXoqP2NdSBYZPiHTgAe/wA7V/wTe/5Pc+Ff/YQuf/SKev8AgqP/AMmJfE3/ALhP/p2s6/Yo/wCTufhB/wBjhp//AKOWqrx9fW+meBfEepXRPk2mk3UsmBztWJicfgK/YR0uXWP2xPhHaQFt0fia3uDtjLnEOZW4HbCHJ7DntVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVftZf8nUfGX/soGu/+l81f8E3v+TI/hX/ANg+5/8ASyev+C23wzfT/iF8Pvi7aWp8nWtKn0e8kVflWW2k82Pcf7zJcOB6iI+lf8EVPieNB+MvjP4UXdyEg8WaJHfWysc7rmzcjavoTFcSsfURj0Ff8Fgf2SvFGu6nZftP+ANFm1G3tdPWy8V29tEXkt0iyYrwqMkoEJjc/wAASM9NxX9kT9vX4t/shLe6F4bsdP8AEXhPVLn7Td6HqLMgWbaFMsMq8xOyqgOQykKPlyARqX/BcnWpbIR6P+zZY213j/W3Pitp0zj+4tqh6/7X+NfsLftK6z+1Z8Bofid4l0/S7DWodYvNPv7bTUdYYmjYPGFDszZ8mWEkk8kk8A4H/BVD4Vt8Tf2PfEl/Z23nah4JuoNetgBzsiJjnOfQQTStjvsHtiLXtWg0C68MRXrjTL28hvJ7b+FpoUlSN/qFnlH/AAL2Ffsufs22Nr/wTz0j4JavarFc+OPCVzPqBlBDJcajG8ilvRo1liX28od+aN3r/hSTXfDbtJZS3kR07VLduCVjuI5TG30lgjP1Wv8Agiv8JhoHwg8X/GG+g23Pi3VksLNmHP2W0U5ZfQNLNIp/65Cv+CxXwq8TePP2b9K8ZeG7WW7TwNrX23U4I1yUtJImjebA5Ox/Lz6KzMeFNfslftKaz+yj8ZbH4r6RoFvrkK2ktjqGnSzeSbm2l2lgkmG8twyIwbafu4IwTX7aH/BQ3xx+2R4Th+H/AIW+HEvhnwjo8q6pqcaXTX087J+7R5nVEWOFWlAxgguUJbOAP+Cb3/J7nwr/AOwhc/8ApFPX/Bav/k1fwt/2UCz/APSC/r/gk/8A8nueD/8AsH6n/wCkUtV/wWP/AGX9dk1/TP2n/CGkzXenS2aab4pEKbjaPGcW9ywHOxlPlMx4Uxxj+Ov2WP2xvi9+yN4gv9U+HU9hfaZrCqup6LqkbyWtyVzskGxlZJFBIDKR1wwYcV+0H+2x+0l+3fbR/DHTfC1lpug6dBPq11o2iK7eetrE0rzXMsjHKRorMF+Vc4yGbaR+yd/ydR8Gv+ygaF/6Xw1VeNf+Cw3jv4ZfHnx94Qn+Hmh+L/Bmj+ILmx0porp7C6WKFzEzeaBIjqxQuMxg/NycYA/bi/4KM6f+198PNH+HmmfB2Pw5HpuqrqLajd6mLyfcsbp5cYESbFIfLHJztAwMZr/gjj8FvFPib4+3fxqbT54fDXg3Tbi3F6yYSe+uE8tYFJ+8RE0jtj7vyZxvGf8AgtX/AMnUeFv+yf2f/pff1/wQ6/5FT4t/9hDSv/RdzX/BRL9p7xl+0J8e9c0HUXksfDHgTUrvSdG0oPlVaKQxy3D9jJIyZz/CoVR0JP7Nn7SPiD9l7xpL8QvB3gHwXr+v+QYbO88R2lxcHTgwIdoBFPEFd1O0sQzbcgEBmz/w+r/ao/6EL4U/+Cq//wDk2r7wzqv7fv7A1lb+Lo9J0vxF470GO/t5LWKRLazv45N8LKGZ3WPdGqtyx2s3XOKtZfjF+yp8Z4Lw22oeD/Hvgq/3ok8Q3ROAQeDlZInRiO6Oj91avC3/AAXD8XWWkQ2/jT9nrSdX1JEAlutN8RyafE7DqRE8ExXPpvOK/ZN/4KmeMP2k/wBpLw98Itb+Hfh/wxomu2t2IXiuZbm5NzFA0yjzDtQKRG4xsySVAPrVf8FW/h5P4F/bK8S6oIBHZ+MLCz1i0wuAd0QglP1M1vKT/vfjX7Inju3+Jf7MHwv8ZQzLI934Zs4blh0+0QRiGcD6SxOK/wCC2n/JfvAv/Ynj/wBK56/ZW/ar+Iv7JPxAm8c+AobG/h1G1+yappV+GMF5DuDDO0gq6kZVx0yQQQzA/tG/tiftE/t7y2nge28IW9tonh6C51k6FoUUkmfs8DvLdTu5Jby4vMCgbQN2ACzDPwE+L+q/AP4w+Fvi/ommW+pXfhm988Wdw5RLhGRkkQsMlSUdgGwcEg4OMV/wUa1ubxN/wTm8Z+I7mwFjNqunaFdyWokMnkNJqVk5TcVUttJxnaM4zgdK+HnjvxB8L/HWg/EXwpLBHrPhy/ivrFp4vMRZY2DKWU9RkDiv+HvP7ZX/AEHPCn/ghT/4qv2OP+Ck/wC098bP2l/Avwt8c6t4dl0LX7uaK8S20hYZCqW0sg2uDkfMi1+3P47g+HP7IvxU8RSTpFJN4duNOtyxwfNvALZNvuGmBH09K/4JDfDybxj+15ZeKWty1r4J0S91F3ZcqJJU+yov+9/pDMP9wntVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVeLf+CcP7F/jrxVrPjbxT8Gft2teINQn1DUbn/hIdTi8+5mkaSV9iXIVdzsxwoAGcAAV8N/hv4L+EXgjSvhx8O9G/snw7okbR2Fl9oln8lWdnYb5WZ2yzsfmY9fSvjT8BvhL+0P4Vt/BXxi8IR+IdHtL1L2C3a7ntjHOqsgdZIHRx8sjjG7BzyK+GX7Av7Jfwb8caX8SPhr8KpNF8R6MztZXqeINSlMe+No3BSS4ZGBR2UhlIwaZVZSrAEEYIIr4l/8E5v2OfinqU2t638HLHS9SuCTJc6Fcy6buY5yxihZYixJyWKEkjknJzoP/BKD9ifRbhLm6+Heq6wY33Kl/r11t9gRE6ZGecHr3yOK+HHws+HHwf8ADi+Efhf4K0jwzpAlMzWunWqwrJKQAZHI5dyFUbmJOFAzgCvEXh/R/Fnh/U/C3iGxS90rWLOWyvrZyQJoJUKSISCCAVYjgg81/wAOuP2Ev+iF/wDlzat/8lVBBBawR2trDHFDCgSONFCqigYAAHAAHGK8Uf8ABNz9irxn4l1bxf4i+Ckdzq2uX019fTJr+pQLJPK5eRhHHcKiAsxO1VCjOAAK+Gvw18EfB/wPpXw3+HGgx6N4c0WN0sbFJpJREHdpG+eRmdiXdmJYkkk1PBBdQSWtzDHNDKhSSORQyupGCCDwQRxivG3/AAS6/Yw8b+IJPEUvwyn0WeeQyT2+janNaW8jE5OIQxSMdtsYQDsBXhj9i/8AZi8HfDXW/hL4e+Eml2vhzxNbrBrMazTfab9FYMokut/nkBgGADgKemM14A/4J6/sffC7xnpPxB8CfCD+zNf0K4Fxp93/AG/qU3kyAEBtklwyNwTwykV8a/gJ8Jv2ivCtr4J+MfhP/hINFsdQTUILX7dcWmy5SOSNX328iMcJLIME4+bOMgEfCf8AYU/ZU+Bvjez+I3wt+Ff9i+IrCOWO3vP7b1C52LIhRxsmndDlWI5U4zxiqvLO01C0msL62iuba5jaKaGZA6SIwwysp4IIJBB4Oa8Z/wDBLj9i7xnrj683wwn0WWaTfNb6Nqc9pbuc5IEIYpGO2IwgHbFfDL9lX9n34OeENV8D/Dv4YaVpWm69ZvZ6qw3y3F/C6lWSW4djKykFvl3YGTgCvDn/AATY/Yp8JeIdL8VeHvgubTVdGvYb2xuB4j1R/JnicPG+1rkqcMoOCCDjkGqr4m/8E7f2PPivqlzr/iL4N2Fjqt2zPLd6LczaaXduWcxwusTMTyWZCSck9TXhn/glL+xV4cv01C4+Hep620bBki1PW7l4wR6ojIGHs2R7V4a8L+GvBehWfhjwfoGnaJpGnx+Xa2Gn2qW8EC+iIgCqM5PA6mvjX+xl+zV+0V4qtfG3xj+G3/CQa1Y6emnwXP8AbF9abLdJJJFTZbzIpw8shyRn5sZwBj4Hfsy/A/8AZttNXsfgr4I/4R2HXZIpNQT+0rq785ogwQ5uJZCuA7fdxnPPavEX/BNb9ijxXr+p+KNf+Cv2rVNYvJby9n/4SPVE82aVy8jbVuQq5ZicAADPAAr/AIdcfsJf9EL/APLm1b/5Kr/h1x+wl/0Qv/y5tW/+Sq8AeAfCXwu8GaR8PvAmk/2ZoGhW4t9Ps/Pkm8mMEkLvkZnbknliTXxf/Z2+CHx9sI9P+L3w00XxL5CFILm4iMdzApzlY7iMrLGCSThWAzzUv/BJL9iyS+F2ng3xBHEDn7KviC48v6ZJL/8Aj1fCL9ij9lz4F6ra+Ifht8H9Isdasgfs+qXby311ExUqzJJO7mNiCQSm3gkdDiqr/gr7+zpe/FD4Jaf8YvDFi1xrPw2eWS+jjUlpNLl2+c2AMnynSOTnAVPNNf8ABF749Wus+BfEv7O2tago1Hw/ctq+ixSNzJZzECdEHpHNtc/9fHscf8FtP+S/eBf+xPH/AKVz1/wTV/ZD+AP7Uv7O/iM/FnwW11qmh+MZls9WsryS1ulhe0tSYWZDh0DKSAwO0uxGNxz8Dv2YPgX+zjplzpvwg+H9jorXyhb28Z3uLq6A6CSaUs5XOSEyFBPAFeMf+CXP7GPjPxRN4qufhrc6ZLdTGa4s9K1Sa0tZGJycRK2Iwf7sewDPAFfEf4R/Dz4ufDm9+EvxC8Pf2r4U1COCO50/7XNBvWCRJYh5kTrINrxRnhhnGDkEg/8ADrj9hL/ohf8A5c2rf/JVf8OuP2Ev+iF/+XNq3/yVXw3/AOCf/wCyL8IvG+lfEb4d/CT+yfEWiSNJYXn9vajP5LMjIx2S3DI2Vdh8ynrX/Baf4921vo3hT9nHQ79GuruYa5rqRvzHEgZLWJscfMxlkKnkeXGehFf8EmP2dbv4O/s+yfETxLpzW3iH4lSx33lyxlZIdPjBFqpB/vh5JvdZUz0qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqurW1vrWWyvbeK4t7iNo5YZUDpIjDDKyngggkEGvDvwg+E3hDVI9c8JfC/wjoupRKyx3mnaJb20yBhhgHRAwBHBGea/4KafsS/Gb9qXxd4H8TfCO20e5GjabdWd+l9fC2KZkR4yuQdwOX+mPev8Agmf+zN8Yv2Xfh74w8H/Fuy0y3fVdZivrD7DerchgYRHJkgAg/InB/wAaqqqq8QfBz4ReLNWm17xT8K/B+s6nchRNe6hodvcTSbVCrud0LHCgAZPAAFQww28KW9vEkccShURF2hQBgAAdBjtVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVU00NvC888qRxxKWd3baFAGSST0GK8Jftgfs6+P/jAPgZ4G+Jen6/4p+yT3BTTw01t+5xvjFwB5bybSzbULYEbZxgA1VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVreuaJ4a0m617xHrFjpWmWMZkur29uFghgQdWd3IVR7kgV+0T/AMFe/gb8M/tfh/4NWEvxG1+IlBdxubbS4W6E+cQWmx6Rrtb/AJ6Cv2g/21/2iv2l5ZrX4ieO54tCkk3x+HtKBtLCP0BjU5lx2MrOR2Ir9ij4A/tTeMvit4T+KPwK8DXfl+HdViul13UWaz00BGAkjecg71ZSyOkQd9rn5aQuUUyKobHzAHIB74NVVVVVVVVVVVVVVVVVVVVVVVVVVf8ABUb9rv46/sw698PbD4O+J7PSofEFnfyX6z6bBdeY0TwhCDIrbcB26dc1/wAEs/2sPjd+1D/ws7/hcniS01b/AIRn+yP7N8jTobTyvtH2zzc+Uq7s+TH16Y46mqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqrxr488E/Dfw/ceKvH/AIs0nw9pFqP3t7qV2lvGDjoCxGWOOFHJPABr9of/AILM+AfDQuvD/wCzl4UfxZfhWVdd1hJLWxjbsyQcTTD/AHjF2618bv2m/jn+0Vqn9pfFr4h6lrMaNut9PDCCztj28u3jxGpxxuxuPcmv2ef+Ce/7S/7RhttU0Dwc3h3w1OVb+3vEAa0t3Q87oUIMk4I6FFK54LCv2eP+CUP7OPwbNrrfj21f4l+JIGDifWIBHYxOP7lmCVYf9djJ2IxVpZ2lhaw2Nhaw29vboEihiQIkagYCqo4AA7Cqqqqqqqqqqqqqqqqqqqqqqqqqqq/4Ljf8jV8I/wDsH6r/AOjLav8Aghj/AM1s/wC4B/7kaqqqqqqqqqqqqqqqqqqqqqqqqqqqq1LUtN0exm1PV9QtrKztkLzXFzKsUcSjuzMQAPc0jpIiyRurKwyrA5BHqKqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr4p/Gf4U/BHw+fE/xX8e6R4Z085Eb3s+HnIGSsUQy8rY/hRWPtX7Q/8AwWhJNz4e/Zn8Ejqyf8JF4ji/DdDaq34hpW+sfavif8Y/iv8AHHxCPEfxT8d6z4o1NmIhN7OWWHcclYYhhIlJ/hRVHtX7PH/BLn9pb45i11rxBo6/DzwzOocajr8LLcSoe8VoMSNxggyeWpByGNfs8/8ABNj9mT9n8W2rDwsPGviaEK39seI40ufKkHO6GDHlRYPIOC4/vmqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/4Ljf8jV8I/8AsH6r/wCjLav+CGP/ADWz/uAf+5Gqqqqqqqqqqqqqqqqqqqqqqqqqqqqv+CynxX8W+F/ghonww8OaPq40/wAXX/ma5q0Vs/2aK3gKsls8oG0PJKUbbnOISCMNX7Pf7d37SX7NrW9h4K8cy6n4egAUeHtcLXlkFH8Malg8A6/6pkz3zX7PP/BW/wDZ++KwttC+KiSfDTxBIFQyX8vn6bM54+W5AHlDufNVFUfxmtP1HT9XsLfVNJv7e9s7qMSW9xbyiSOVCMhlZSQwI6EcVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV8b/ANp74Ffs66WdQ+LXxE0zSLho99vpiv519cjoDHbpmRhnjdgKD1Ir9oj/AILMfEDxMbrw/wDs5eFo/CWnliqa7q6R3V/Iv95IPmhhPs3m/UHpr/iT4k/GXxl/aXiLWNf8ZeKNXmCI08kt7dXDn7qIOWPoEUYHQDFfs8f8Eivj38UvsmvfFu4i+G3h+X5jBdxefqcq+1uCBFnkfvWVl67DX7PX7DP7OH7NUdte+A/A8V94hhTDeItZIu75m7sjEBYc9MRKgPfNVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVf8Fxv+Rq+Ef/YP1X/0ZbV/wQx/5rZ/3AP/AHI1VVVVVVVVVVVVVVVVVVVVVVVVVVVVqOnafq9hPpeq2Nve2d1G0U9tcRCSOVCMFWVshgRwQeK/aH/4JI/s+/FcXWu/CxpPhp4gkVmWPT4fO02V+vzWxI8sdv3TIozna1ftC/sHftJfs2G41Dxp4IfVfDtuMnxDoe68slX+9IwUPCO371UGehNfAn9q/wCPn7N1+tx8KPiFfafYtL5lxpFwRc2Nwe++B8qCRwXXa/owr9nj/gsh8LPGhtfD/wC0B4ck8D6tIwT+1tPV7rTZCf4mXmaAZwMYkA6lwK8K+L/CnjvQ7fxN4J8TaXr+kXYzBf6bdpcwyY67XQkH8+Kqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr4t/FLwl8E/h1rXxR8cz3MWhaBEkt49tAZpArSLGNqDk/M61/w95/Y0/6Dnir/wAEL/8AxVfAn/goH+zn+0b4/h+Gnw01TXZ9bmtZblEvNLa3j8uMAt8xJ5wRxVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVftBfttfs5/s1Qy2vxD8eQT68iFo/D+lAXd+5xwGjU4hz2aVkU9jX7RH/AAV6+OfxN+06B8G7GL4caDKpT7TFILnU5Qep84gLDkdBGoZcn5zWlaL8SPjH4yNno+neIfGfinV5d7rEkt/d3LnALsfmduoyx6dzX7PH/BGv4leLvs3iD9ofxPH4L00lWOi6YUu9QlXuHkBMMHHQ/vT1BUV8Df2V/gN+zjposfhP8PdP0y6Zds+qTKbi+uPXfcPl8dfkBCjJwBVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV/wXG/5Gr4R/wDYP1X/ANGW1f8ABDH/AJrZ/wBwD/3I1VVVVVVVVVVVVVVVVVVVVVVVVVVVVVOiSIY5FVlYEMrDII78V+0T/wAEw/2afjwt3rOjaGfAHimcFl1TQIVSGR/Wa04jcEnJK+W5PV6/aI/4JnftM/AM3Wr2fhs+OvDEDErq3h6JpnjTsZrbmWPjkkB0Xu9fCP48fGL4Ca+PEHwm8f6v4cuw4M0NvLmC4x2mgbMco9nU4r9nb/gs/o9+bPw5+0v4NOmynCN4k0CNpIf96a1JLqMZJaNnyeiAV8Nvix8NPjF4dj8VfC/xvpHiXS5AMz2FyJDET0WRPvRtwflcBh6VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVf8FIv+TI/in/2D7b/ANLIKr/gkN/yeVpv/Yv6j/6AtVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX7RH/BRr9mb9nj7TpF94s/4S3xPAWQ6H4dZLqSJxxiaXcIocHAIZt47Ia/aI/4Kn/tJ/G37RovhLU0+HHhuXcv2PQpmF3MhyMS3Zw5ODj92I1PcGvhv8J/in8b/Ex8OfDPwXrPirWJT5kyWcDS+XuP+slkPyxqT/G5Az3r9nj/AIIvXtz9m8Q/tMeNfsqEK/8Awjnh2QNJnrtmumBUehWNWzniQV8Jvgb8I/gXoP8Awjfwl8AaR4as2CiY2kH725KjAaaVsyStju7E1VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX/Bcb/kavhH/wBg/Vf/AEZbV/wQx/5rZ/3AP/cjVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV+0P+wH+zV+0gtxqXinwUmh+JJst/wkGgBbO7dz/FLhSk/bmRWbAwCK/aI/4JM/tD/CL7Rrnw2jj+Jfh6Pc2dLhMWoQqMn57Ukl+MD90zknPyivBvjz4l/BnxYdc8D+J9d8I+IbB2hkls55LWZCDho5F4yMjBRgQehFfs8f8ABZ3xXo32bw9+0p4PTX7RQqf8JBoMawXY9WltyRFIfdDFgD7rGvgz+0T8Ff2g9GOs/CL4haXr6xIGuLWKQx3VrnH+tgfEkfJxkrgnOCaqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/4KRf8AJkfxT/7B9t/6WQVX/BIb/k8rTf8AsX9R/wDQFqqqqqqqqqqqqqqqq8VfEv4ceBbiC08b/EDw34enukLwRarqsNo0qg4LKJGBIzxkVoHiHw/4r0iDX/C2u6frGmXW7yL2wuUuIZdrFW2uhKthlZTg8EEdqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq5uLezt5bu7uI4YIELyyyMFVFAyWJPAAAySa/aJ/4Kq/s4fBZbvRPBOoN8SfE0AKra6LOBZRv/wBNLzBTHH/LISHPBA5x+0R/wUU/aX/aJ+06Rqfi4+FvDEzHGheH2a1jdP7s0oPmzcAZDNszyFFfBX9mz43/ALQ+rrpHwk+Hmqa2qyBLi/EXlWdqf+mtw+I0OOdpbcewPSv2d/8AgjH4M0A2viH9pHxc3ia8UBm8P6I8ltZqf7slx8sso9kEWD3IrwJ8O/Anww8O2/hP4d+EdJ8O6RbDEdnp1qsCZ7sQo+Zj3Y5JPJJqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr/guN/yNXwj/wCwfqv/AKMtq/4IY/8ANbP+4B/7kaqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr4+fshfs/ftK2jL8UvAFnc6oIvLg1uz/0XUIBjAxMnLgdkk3oP7tftEf8ABHX4u+A/tPiD4Da7D490aNS/9mXO201OIDsoJ8qfjJyCjHoENSRfEf4PeM/LlTxF4L8V6LN91hNp17ZyD/vl0P5V+zx/wWI+MfgH7NoHx20aH4g6OhVP7Sh2Wmpwp0ySoEU+B2YIxPJkNfAP9sP9nr9pSzQ/DH4gWcuqlcy6HqBFpqEWBk/uWOXAA5aMuo/vVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX7Rn7VPwZ/Zb8MJ4i+KniMw3F2G/s7SLNBNe6gw6iKPI4GeXcqg4BbJAPxK/4LX/F/VNSmi+E/wALvC+g6YHxFJrTTahcso7ny3iRSe64bGcZOM14Z/4LLftV6PdRtr+h+BNftgT5sc2mTW7sOPutFMAp4xkqRz0Pb4u/8FDvhV+1p+xV8VPCI06fwl47h0iCZ9Du5xMl0iXlvve2mAXzMDkoVVgMnBALVX/BIb/k8rTf+xf1H/0Baqqqq/bh/wCCgfgn9keyi8K6PpsPif4h6lb+da6SZikFjEchZ7pl5AJB2xrhnweUBDUv7X3/AAUr/aav7u5+G+p+Nbm0iYh7fwVoTQwWnfaZYkLj23yEnjk1f/Gb/gqn8GIv+Ei8R6n8Y9Os4f3j3Wt6NLeWq4/vNPE8Y6dD9a/4Jx/tF/GP9pz4K6h47+LunaPHJYau2nWF9YW7W7X4jjRpJJI8lQQ0gXKbQSGG0Y5/bd+PfiL9mr9nHxJ8VPCGkw3+tWr29pYi5jLwQSTyrH5soBB2qCSBnBbaDwa/4Ju/8FA/2gfjj8dZfhF8ZdVt/E1lq+nXF1ZXsOmQWkmnSQgPg+QiK0TLuX5gW3FPm5INVVVVVVV/wW9/5Kn8NP8AsX7r/wBKBX/BLj/kxL4Y/wDcW/8ATteVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVa3rmieGtJute8R6xY6VpljGZLq9vbhYIYEHVndyFUe5IFftD/8ABYD4IfDgXWgfBXTZviLriKyi9DNa6ZC/TmRhvmx1wihW7OM5H7QH7af7RX7Ss8sHxH8fXCaK7ZTw/pZa009MHIzCpPmkHkNKXYdjXwB/Yy/aI/aVuI3+GngG5OjlwkuvakfslhD6nzWGZMcZWIOwz92v2eP+CPnwU+HZtdf+NurTfEPW4mD/AGEK1ppkTDsYwfMmwe7sFbunWtC8P6B4W0i20Dwxomn6Rplmmy2srC2S3hhX0SNAFUewFVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX/Bcb/kavhH/ANg/Vf8A0ZbV/wAEMf8Amtn/AHAP/cjVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVXxl/Z5+DH7QOiHQvi38PtK1+NYykFzLFsurXPeGdMSR884VgD3Br9oj/gjD4l0t7rxD+zV4xTWrbJZPDuvSLDcr6rFdDEb9sCQR4A5djXjb4e/E/wCCviz+w/HXhfXfCPiCwl3xJdwPbSqyniSJ+Nwzgh0JB4INfs7/APBWH9or4PLa6F8Qp4/iZ4dhIUpq87JqESf7F4AWY9/3yyHsCK/Z3/4KB/s0/tHra6V4c8ZLoHiedQD4e17Frcl/SJiTHP3wI2LYGSq1VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVXx2+MPhj4A/CTxL8W/Frk2Hh6yaVYFbDXUzEJDAvo0kjIgPQbsngE18afjJ47+PvxH1f4n/ETVnvdW1WUkIGPlWkQJ2QQqSdkaA4C/UnJJJjikmkSGGNnkdgqIoyWJ6AD1rw1/wTy/bP8W+HF8VaN8A9bFg8QkQXtxbWU7KRwRBPKkpyOwT09a8ZeCvF3w88R3nhDx14a1LQNb099tzYajbNBNGSMglWAOCMEHoQQQSDmq/4JDf8nlab/wBi/qP/AKAtVVVVahLd/tZ/twi28YarPHF8QPHyWc0pYl7aze5ESRpnONkAVEB6bVFeDfBnhT4e+F9N8FeCNBs9F0PSIFgsrG0j2RwoPQdyepY5JJJJJJNVpGh6J4eszp+gaPY6bamV5TBZ26woZHYs77VAG5mJYnqSST1rxp4L8KfEXwrqXgjxxoNnrWhavAYL2wu498cyZB5HYggEEYIIBBBANfA79kP9nb9m/Ub/AFn4O/De20TUtTiMNxfSXdxeTmEuG8pXnkcomQvyrjOxS2SM1VTzwW0L3FzNHFFGpZ3kYKqgdSSegrRfF/hLxLLLb+HPFOkarLAMypZXsc7RjpkhCcfjVVWteIfD/hy2W78Q67p+lwMcLLe3KQKT7FiB3H51puq6XrVol/o+pWt9bSD5JraZZUbgHhlJB4IP41Vf8Fvf+Sp/DT/sX7r/ANKBX/BLj/kxL4Y/9xb/ANO15VVB8QPAV3qI0e18b6BNfltotY9SiaUt6bA2c+2KqqqqvPG3gvTtTXRNQ8X6Lbai5AW0mv4kmOeBhC27se1VVTTQ28LzzypHHEpZ3dtoUAZJJPQYrS/HXgjXL06ZonjLQ9QvBnNva6jFLIMdflViaqqqqqqsvFHhnU9Tn0TTfEel3eo2gPn2cF2jzRY4O5ASy9R1FVPPBbQvcXM0cUUalneRgqqB1JJ6CtL8b+Ctdu2sNE8YaJqF0hw0FrqEUrg89VViex/Kqqq1zxf4S8MvHH4k8U6RpTyjMa317HAWHqA5Geh/KoJ4LqFLi2mjlikXcjowZWHqCOoqq1PVdL0a0e/1jUrWxtowd81zMsSLgZOWYgDgE/hWi+JfDfiWJp/DniDTdVjQ4Z7K7ScL9ShOKqqqqqqqqqqqqqqqqqqqqqqrxv4+8D/DXw/P4q+IPi7SPDmkW/El7qd4lvGDjhQXIyxxwoySeADX7Q//AAWa8CeHDc+H/wBnHwk/iq+Qsg13Wo3trFT2aOAYmlH+95X418bP2lvjh+0Tqo1T4ufEPUtbSKQvbWBYQ2dqT/zyt0AjU4ON2NxAGSa/Z4/4J5/tL/tGC11bQ/CH/CN+GbhQ66/4hD2lvIh/ihTaZJsjOCilM8Fl61+zz/wSk/Zw+DQtta8dWb/ErxHEFYz61AFsYnHOY7MEqR/11Mntire2t7O3itLSCOGCBAkUUaBVRQMBVA4AAAGKqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr/guN/yNXwj/AOwfqv8A6Mtq/wCCGP8AzWz/ALgH/uRqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq+Ivwt+HPxd8PSeFPib4K0fxNpTkkW2o2qyiNsY3oTzG3oykEdjX7Q/wDwRf0HUftPiD9mnxo2kznc/wDwjviGVpbf2WG5UGRBwABIJMk8uBXxg+APxj+AetjQPi58PtV8O3DsywS3EW63ucdTFOhMco/3WNfs8f8ABSr9pn4AC10dvEw8beGbdQg0fxE7zmJB2huM+bHgcAZZBx8nFfs8/wDBUL9mf46C20bXdbb4e+JpQqnTvEMyJbyyHjEN2MRuMkAB/LYnotI6SIskbqysMqwOQR6iqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/4LcfEq703wL8OvhLZXJWLXNRutWvkXjK2yJHECfQtcSHHTKA9hVf8Ecv2avDfj7xd4h+P3jTSodQg8G3ENl4fgnQPGt+y+ZJcFT/ABxJ5ezOcGUtwyqRX/BXT4D+G/iB+zdd/F9bKKPxP8PJoZYLpFCvPZzTpFLAx7qDIsoz0KED77Zr/gkN/wAnlab/ANi/qP8A6AtVVVVftlfBnx1+yP8AtWalqOlRz6fay623iLwdqkaZTy/P86IKTkF4XwjKe6g42uuf2Vf+CoHwL+Oej6foPxI12w8AeOtix3FpqU3k2N5LgAvbXDHaAx6RyMHBOBvxuMM0NxCk9vKkscihkdGyGBGQQR1BFV+3rcXFp+x38Vrm0uJIZo9AkKSRsVZTvTkEciv+CMWua1qf7UXie31LWL27jXwDdsqT3DSAN9vsBnBJ5wT+dahZx6jYXOnyyOiXMLxMyHDKGBBIPrzX7Fn7To/Z3+Ng+J/jK/1LVtPsNC1GNLBriRzdXDQHyIweQm6UIpcghVLHBxUfwU/4KB/8FLJpPidr+o2+k+DruZn0iPWb+Sw0qNQTgWttGsjuBjb5xQ7iOXYg4+PX7C/7Uf7H9va/EXXbe3k0uzuECeJfC2oySJYzE4QsxWOaHJwA5ULkgbskCv8Agl1+3b4n+OiXfwJ+Merf2h4u0ay+1aPrEzYl1W1QgSRzdmmj3KQ3V0JJGUZmr/gpH/wUP1T9n+6PwR+Ct1a/8J1dWol1bVnVZf7DjcZjREOVNw6kN8wIRSpwS4K/Bj9gn9rj9tOMfF3xb4haw0rVTvh8QeL76aWfUEJ5aCPDyMnOQzbEI+6xxXir/gnp+25+x1HJ8Xfgd8QU1oaSnn3g8MXM0N2I05YyWjrtuIx3QGQkZymM1+1p+2L4o/ah1XwN4quRcaLrOjeF00/W1spnihnvFuZ2aWNQ3CtG0TYPRmZeQAT+zZbPZ/s6fCuzkuZLhoPBWjo0shy0hFlECxPqcZNf8Fvf+Sp/DT/sX7r/ANKBX/BLj/kxL4Y/9xb/ANO15Xxs+Mfgr4BfDHXPit4+vWg0nQ7feY4wDLdSk7Y4IlJGZHcqozgDOSQASL/4jfto/wDBUf4n6h4O8G38uj+E7bLy6XFeSWukaVbMSEN3IgJuJCMgblZmO/YiqCBrX/BEb4r2vh57zQvjV4W1DWkjyLCfT57eFmxyonBc+wJjHviv2ev2wv2i/wBg/wCMp+C/x9l1m98KaddJaazoOpTm6k0yNgCtzZSZbChWDhFJjkUnADFXXSdV03XtKstc0a+ivLDUbeO4tLmFtyTROoZHU9wVIIPvVV/wV91HWNE/a5C6Xrmo26XfhawmeKO5ZFVt8ycBSOyD8c18Tv2ov2qP27PElr8BP2SbTVrPwvpGl28esX2nzmxN6/lqsstxcMVMNvuDqkW7dIASQxIRfGn/AASA/a+8M+HZ/Edk3gzxRcxxmWTTdI1aVrtu7ACeGJGbrwHJOOMkjP7E/wC3h8Uf2W/iFYeAviNrGq6h8PHvBY6to2ptI0mifPteaBWy0TRnJaEDaw3DAbDCCeC6gjubWaOaGVA8ciMGV1IyCCOCCO9ftX/tN+Df2UfhLe/EnxRH9uvZHFroukxybJNRu2BKoDztQAFnfB2qpwCxVT4V8K/tt/8ABUzxdqerah4t+y+E9OnCzvdzy2miac33lhggQN5soXBzhnxsLuMqT43/AOCKvxq8O6A2s/D34r+HfEusWq+YunS2smmtIRyBFKWdd+em/YPVhX7Dn7f/AMWvgf8AFeH9nr9pvWdTvfDsmoHSXm1x2e98OXgfywGkbLNBuGxkcnZwylQrK1f8FGfEfiGy/bV+KNrZ69qMEMeoWwSOK6dFX/Q4OgBwK8KEt4W0ZmYknT4Mk/8AXNar/goz4j8Q2X7avxRtbPXtRghj1C2CRxXToq/6HB0AOBXhQlvC2jMzEk6fBkn/AK5rX/BaO+1jSP2i/B13pmuX9qtz4KiRoYZ2jXKXt0d2Aep34/AV+0H+178Y/jxoXw4/ZM/Zjk1q/tofCWkR6tJoZkN3rF2bGJpYmdcMkEQOHBwNwfedqiv+Ca/7D37Sf7PP7QD/ABF+LPga20fRrjw1dWYdNWtbl0mkkhZVZIpGPRG5GQMda/bq/bZ8O/sgeBbcWNnb6z478QxyDQtKlJ8pAuA1zcbSCIlJACghnb5QQAzL4G+GH7cX/BS3xBeeItS8V3l/oNrcFJtT1q8a00iyk4PlQQRqQXAIOI4yR8pcjIJ8Y/8ABGD9ojw3oja14L+IHhHxHqVqpkGnxvNZSuw5Aid12Fs9NzIOnNfszf8ABQ79oP8AZW+II+GHx7utd8Q+FtPvTZavpWtlpNR0YhsM8Ej/ADnZ18pyUIGF2Z3V4e8QaJ4s0HTvFHhvU4NR0rVrWO6sbuBtyTwyKGR1PoVIP41/wW6udQ07RPhBqWm6re2cq3WsRFbedow4ZbQ/NjqRsGPTJ9a8HftufHfX/wBnT4V/sj/swW2r3HxAvLW8j1vVLBWa8Tde3Jjggdv9WfI8uWS4yNgYYZNrEX3/AASE/bO8Q6dP4w13X/Bd1rV0plnsb3X55r6V8dGk8loi3bJlx718Kfjp+0x+wL8XZ/Ds41XSZdJuwuu+DtTmJs71DgnKglAWUgpPHngggspIPwl+Jvhn4z/DTw58UvB9wZdJ8SWCXduG+9ETw8b/AO2jhkYf3lNft0ftSR/snfAu78c6bawXvibVrldM8PWs4zGbp1ZjLIAcmONEZiP4jtXI3ZHwc/Zt/az/AOCkPiDVPiL4g8ePcaXYXZgudf8AEd1KYI5WAdoLSFFIyoZSUQIihl5GQD4t/wCCRv7W3wpUeMfhH470PX9RsF3xpo2ozaZqAYc/ui4Ve3/PUHOMCvgx4d8WeEvhL4P8NePPEV5r3iTTtGtotX1G8l8yW5uhGPNZm5LfPuAJJJAGSTzVVVVVVVVVVVVVVVVVVVVVfFf42/Cb4G+H28T/ABY8e6T4bsQpMf2ub97cEdVhiXMkrf7KKTX7RP8AwWgnkN34b/Zm8FiNQSi+JfECZY9i0NoDgdiGkY+8Yr4l/F34r/HLxN/wkXxN8baz4q1ed9sJu5i4jLH7kMS4SJSf4I1UZ7V+zt/wS1/aT+OP2TW/FGkj4deGJyGN9rsDLdSJ6xWfEh4wQZPLUg5DGv2eP+Cbv7Mv7PqWmqxeFF8Y+KIBltb8QotwUfuYYMeVFg9CFLjPLmqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/4Ljf8AI1fCP/sH6r/6Mtq/4IY/81s/7gH/ALkaqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqrxP4U8L+NtDufDPjHw7puuaTepsuLHUbVLiGUejI4IP5V+0P/wAEcPhP43F1r/wA8QSeBNWZWZdJvTJd6bM/oGJMsAJzkgyKOAEAr48fskfH/wDZuvWi+Kfw9vrPTtwWHWbVftNhMTwNs6ZVWP8Acfa3+zX7Pn7dX7SP7Nj29j4H8cy6h4fhYbvD2tA3lkVH8KKSHhH/AFyZPfNfs8f8FcvgB8Vfsug/FeGT4aeIJSIxJezfaNNlb1FyFHlDgk+aqquQN7da0zU9M1rT4NV0bUbW/srpA8FzbTLLHKp6MrKSCPcVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVf8ABbdr0/G74fxyB/sg8KuYsrx5hupN+D64Eefw9ar/AIIy/Y/+GS9U+y+X5n/Ca332jb13/ZrTG732bPwxVf8ABSL/AJMj+Kf/AGD7b/0sgqv+CQ3/ACeVpv8A2L+o/wDoC1VVVV8afgT8LP2hPBk3gT4r+FLbWdOcloHbKT2cuMCWGVfmjceoOD0IIJB+Pf8AwRi+JPhya71r9nvxjZ+LNNGWi0bWHWzv0HZFl4hlPu3lfSvhj+0n+11+wt4zPg97nW9GjsZA134P8SwSPZyoSeVicgoGwcSwld2PvEV+yP8AtY+BP2uvhsfGnhS3k0zVdNkW31zRZ5Q8unzlcjDDG+J8MUfA3bSCAVYD9vz/AJM0+LP/AGL8n/oaV/wRU/5Oo8U/9k/vP/S+wqv2Yfhtpfxg/aF+Hvw01xv+JZr2v20F+u4qXtw4aVARyCyKyg9iQasNPsdKsLbS9Ms4bWzs4Vht7eGMIkMagKqKo4CgAAAcACviB4J0L4leBtf+H3iW2WfS/EWmz2F2jLn5JUKEj0YZyD2IB7V+wdrGpeA/22fhh5MpW4HiT+y5/LPDLOr20g9xiQ1rerWWgaNf67qLMtrptrJczlRkhI1LNgeuAa+B3h++/bL/AG1dCsvHckk58e+J5tR1lBKcm2QSXM8St2HkxMi+gxgcYqxsbLTLG303TbWK2tLSJYYIIUCJEigBVVRwAAAAB0qv+Cifwz8O/Cj9sHx/4Z8KWsdrpV1cQalBbRrtWBrqCOaRFA4CiR32gcBSB2r9nr/kgPwz/wCxP0n/ANI4q/4Le/8AJU/hp/2L91/6UCv+CXH/ACYl8Mf+4t/6dryv+C2/xX1Fta+H3wOs7t47KOzl8Q6hCG4md3e3tyR/sCO5/wC/ntX/AAT2+Lv7I37P37MHhjw/qvxy8B6Z4n1xG1bxBHcaxEky3M33Y5ASCpjiEce09Cp9TX/Da/7Iv/Rx/wAP/wDweQ//ABVf8FefHfwG+LGv/Dvx78I/iL4Z8T6vHaXunawdJvY7h0hRo5LbfsJwA0lxjPr7V/wS08fXnjz9jHwhFqNy1xdeGbi70Z5HbJ2QyloV9gsMkSgeiiqr/gsl/wAnc2n/AGJ9j/6Oua/4JrfCfw78Lv2Q/BFzpNrD/aPi+zGuardqvzXEs/zRhieyReXGB0+UnqxJr/grl8PNI8Cftf3up6PbxwL4y0G01q4jjGAJmeW3kbHYsbbefUsT3r9i/wASXvi79k74T65qMjSXMnhWyhlkc5Z2hjEW4+58vP41/wAFjPivqPjD9pq2+GKXb/2X4A0eGMW+75Rd3aLcSyY9TE1sv/APev2bPjx+xf8AAb4GeDfhZpH7Qnw7hOjaXEL1l1uEGe8cb7iU85JaVnPPQYHQV/w2v+yL/wBHH/D/AP8AB5D/APFV/wAFUPEPwd8cftJW3xA+DfjPQfEdrr/h62fV7jSLpJ0F7E8kXzsnG4wpBweeK/ZA8fXnxP8A2Xvhj421K5a5vr7w5bR3kztuaW4hXyZXJ9WeNifc1/wUi/5Pc+Kf/YQtv/SKCvCX/IqaL/2D4P8A0WtV/wAFIv8Ak9z4p/8AYQtv/SKCvCX/ACKmi/8AYPg/9FrX/BbT/kv3gX/sTx/6Vz1/wR9+Beg+Cf2dv+Fz3GnRP4j8f3lxtu3T95DYW8rQpCueitLFJIcY3bkznapqv22fG/iH9pD9tzxVpthcNOR4iTwroUMjEJFHBL9mQL3CvL5kp95Wr4R/C/wt8Fvhp4d+FvgyzS30nw5Ypaw4QKZWHMkrY/jkcs7HuzE1X/Ba34H6PY/8IX+0Lo9jFb3moXLaBrciIFNy4jaW1dsdWCRToWOSVEY6KK/4I7fFC/8AG37MF54J1W6eafwLrstnaliSVtJlWeMZPpI86gdlCiv+C43/ACKnwj/7CGq/+i7av+CK3wn8Pad8JfFnxomtYZdf1nW30iKcrloLOCKKQoCem+SUlgOvlpk8YFf8Fvfh5pEUnwz+Ktpbxx6lcfbNHvZAPmmiTZNAD/ulp/8Avuv+COHiS9139kOfS7qR2j8O+Lr+wtgx4WNore5IHtvuXP1Jr/gtF8N/E3ij4GeE/H2iWtxdWHg7WpP7VSIZEENyios7j+6siImexmHYkj/gl1+3f8IPhB4Bl+Afxi1NfDQOrS3mk65MhNrIJgu6KdgD5TBlJEjfIQ2CV2jdoev6F4o0m217wzrdhq+mXib7a9sblJ4Zl/vI6Eqw9waqqqqqqqqqqqqqqqqqqqq+OP7UPwJ/Zz0z7d8WviHp2k3MkRkttMRjPe3Q7eXbplyCeNxAUHqwr9oj/gsr8RPFf2nw9+zr4Xj8H6Y6lBrerIlzqL/7UcQJhh7jB809wVNa74i+I/xj8Zf2l4h1fX/GXijV5RGj3Ekt9d3DH7qIPmY9cBRwOgFfs8f8Eh/jx8T/ALNr3xfu4vhvoLlW+z3EYuNTnTrxAp2w55GZWDA87CK/Z9/Ye/Zw/ZrhguvAPgSC816Lk+IdZC3l+WxjKSEAQ8cERKgPcHrVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX/Bcb/kavhH/2D9V/9GW1f8EMf+a2f9wD/wByNVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVajp2n6vYT6Xqtjb3tndRtFPbXEQkjlQjBVlbIYEcEHiv2iP8Agkr+z18Wlutc+GAf4Z+IpcsBpsAl02VvRrQkCMdB+6ZAOu1q/aH/AGCf2k/2bTc6l4u8FPq/huFyF8Q6Hm7tNv8AekwBJAOg/eqozwCa+BX7Vnx6/Zw1KO8+FPxD1DT7LzN8+jzv9osLkn72+3fKZIGN6hXHZhX7O3/BZH4ZeMPsnh39oXw5J4K1VwEbWtOV7rTpG9WjGZoMnt+8A6lgK8JeMfCXj7QLXxT4I8S6Zr2j3q7re+066S4hkHfDoSMjuOo71VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV/wW8+HF3d+Gvhp8WbODMGm3l3o184XJBnVJYPoB5E/4sPxr/gj/APtQeGvhj40134E+PNYi07TvG08N1od1cSBIo9RQbGhYngGZPLCkkDdEq9XFV/wV/wD2jvC3hL4JSfs96Vq9vceKvGlxbSX1nE+XstPhlWbzHx90ySRRoqn7y+Yegqv+CQ3/ACeVpv8A2L+o/wDoC1VVVVVf8FA/2fPBfx1/Zu8X3OtaVbf8JB4R0e71fQtS8sCa2lgiaVog/Xy5Qmxl6chsZUEf8EfPG2q+HP2to/ClrcyfYPFmgXltdQA/IzQqJ43I/vL5TKD1AkYdzX7fn/JmnxZ/7F+T/wBDSv8Agip/ydR4p/7J/ef+l9hVfsB/8nlfCb/sYE/9Aeqr9lX/AJPj+HP/AGUC2/8ASqvjRpN7r3wc8d6FpoJu9R8M6jbW4GeZHtpFXpz1Ir/gltrdjon7b3gAX8ixpfx6haRuxwBI9jNsH/AmAUe7Cqr/AIKuzQy/tveM44pUdorHS1kVWyUb7DCcH0OCDj0I9a/Z6/5ID8M/+xP0n/0jir/gt7/yVP4af9i/df8ApQK/4Jcf8mJfDH/uLf8Ap2vK/wCCztjeW37V2jXdwCYbvwVZtbtyQAtzdqR7HIJx/tA96+Bf/BIv4U/GP4M+CfimfjR4ktpfFOhWl/cW8FhAyQTSRKZY1JOSFfcvPPHNf8OQvhV/0XDxX/4Lrf8Axr/hyF8Kv+i4eK//AAXW/wDjX7Kf7NPh39lH4V/8Ks8NeIb7W7dtTnv5Ly8iWORpJQgI2rwABGoqq/4LJf8AJ3Np/wBifY/+jrmv2KP+TRfhB/2J+n/+iVqv+C1f/J1Hhb/sn9n/AOl9/X7Af/Jmnwl/7F+P/wBDev8AgpnY3lh+3D8To70EtNcWMsbHJDI9hblcE+gOPYgjtWmf8EU/g/rOmWmr6d8dvFUtrewJPBINOtyGR1DKRz3BFf8ADkL4Vf8ARcPFf/gut/8AGv8AhyF8Kv8AouHiv/wXW/8AjXwG+EGkfAP4Q+GfhBoWp3Oo2Xhq1aCK7uVCyTFpGkZmA4BLOeBX/BSL/k9z4p/9hC2/9IoK8Gyw3Hg/Qp7eVJI5NNt2R0bIYGNcEEdQRVf8FIv+T3Pin/2ELb/0igrwl/yKmi/9g+D/ANFrX/BbT/kv3gX/ALE8f+lc9f8ABN7/AJMj+Ff/AGD7n/0snqvD9wnw/wD29dNu/EsgiXw38Xon1BiSNgg1YGQ846bD1xVV/wAFoNcsLD9lzQdElkBu9U8ZW3kx552x21yzt9BlB9XFf8EQNFvIPht8T/ETofst7rllaxNjrJDA7OPynT86/wCC43/IqfCP/sIar/6Ltq/4I2/8mi3f/Y4X3/om2qv+C3v/ACSr4af9jBdf+k4r/gip/wAmr+Kf+ygXn/pBYVqulaZrumXei61p9tf6ffwPBdWtzEJIp4nBVkdWyGUgkEHgg1+0V/wRn8F+Kry78S/s6eMV8J3UxZ/+Ef1hXnsd57RTrmWFfZll68YAxVl4g/a5/wCCcvxbGjyXOpeGL9WWeTTpZTcaXrcAON+0Hy5kOCu9cOhyMowOP2ZPj74f/aa+C3h/4u+H7RrH+042ivrB33tZXcbbZYt2BuAYZVsDKspwM4FVVVVVVVVVVVVVVVVVX7QX7cH7OH7NkU1p4+8e293r0QIXw9o5F5flsZAeNTiHI6GVkB7E1+0R/wAFefjv8TvtGg/B2zi+G2hPuQ3EDi51KdTxkzsu2HIwcRKGU/8ALQ1ofh74kfGTxl/Zvh/SvEHjPxTq0hkZII5b67uG43Ox+ZjjjLHgDkkV+zx/wRq+I3iz7N4g/aI8UR+DtNcK/wDYulMl1qEg7q8nMMHHceaexAr4G/sufAn9nLTTY/CX4e6fpNzLGEudTkUz3t0BjPmTvlyCRnYCEBPCiqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr/AILjf8jV8I/+wfqv/oy2r/ghj/zWz/uAf+5GqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqplVlKsAQRggiv2iP+CZH7M/x6+06zp3h8+AvE8xZ/7V8PRLFHM57zW3+rfnJJUI5PV6/aH/4Jk/tM/AX7RrFh4eXx74Zi3N/anh2J5pIUHOZrbHmR8AklQ6Du9fCX45fF74DeID4j+Evj7VvDV8xAnW2lzFcAdFmhYGOUD0dSAea/Z4/4LQabdfZvDv7S/gs2UmFT/hIvD0ZeMnpumtWO5fUtGzZJ4jAr4afFv4ZfGTw8vin4W+OtH8TaYcB5tPuVkMLEZCSp96J8fwOFYelVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV+0T8E/D/7RHwZ8T/CLxEVii1yzK2t0U3GzukIeCYd/lkVSQMZXK9Ca+KXwx8afBrx9rPw0+IOkPpuu6FcmC5hblW7rIjfxRupDKw4IINV4f/bD/an8LeHx4W0D9oDxzaaYkYjigXWZWMKDosbMS0YHopFeCvBvxI+P3xJs/C3h6PUPEnivxFcMTLcTPNLIQpaSWWRsthVUszHJwO/Sq/4JFTwxftm6RHLKqtNoOopGCcFm8oNge+FJ/A1VVVMVVSzMAAOSeMV8Jv8Ago5+yV8Wby40eD4o6f4Z1S2upIBa+IpFsUnCsVWSKdj5MivjKgPu5GVBIqPxz4JmtBfReMdDe22FxMuoRFNo6nduxj3r/goj/wAFBPhF4U+EXif4M/Cnxjp/irxl4rsZdMuJdKuFuLbSraVSk7STISplMZdFRSWVm3Njbhv+CMPwJ1rV/iV4g/aC1Swli0PQNPk0nS5nUqLi9mKGQoe4jhBDe8y+hr9vz/kzT4s/9i/J/wChpX/BFT/k6jxT/wBk/vP/AEvsKr9gP/k8r4Tf9jAn/oD1Vfsq/wDJ8fw5/wCygW3/AKVVX7YfwN8b/sV/tTPrPhaGbTtKOsDX/BOpxxfu1RZhKkQPILwPiNlPJAViNrrn9mr/AIKR/s6fHTwnYSeKfHOi+BPF6wquo6Prl6tnH52AGNvNKQkqMc7Ru3gdVHf4+f8ABQj9mT4FeF7nVB8SNF8X675R+w6F4e1CO9mnkx8okaMssCdCWcjjoGOAfjV4x8dfEn4l618TPiNaSW2teMJv7WZGjMaiGYbofLU8+X5ewIT1QKcnOT+z1/yQH4Z/9ifpP/pHFX/Bb3/kqfw0/wCxfuv/AEoFf8EuP+TEvhj/ANxb/wBO15X/AAWH/Zz1j4kfCvQ/jZ4R017vUPh+00erwwoWd9Nm2ky4GSRDIgYjHCyyMTha/wCCXv7f3gXwD4Pt/wBm/wCN2vW+g2djPLJ4Z128fZbKkrl3tZ3PEWHZ3WRiFwxUldq7tO1LTdYsYNT0jULa+s7lN8NxbSrLHIvqrKSCPcV46+MHwp+GMaP8QviN4d8PNI6pFFqGoxwySsxAUJGTuYkkcAH1qqr/AILJf8nc2n/Yn2P/AKOua/Yo/wCTRfhB/wBifp//AKJWq/4LV/8AJ1Hhb/sn9n/6X39fsB/8mafCX/sX4/8A0N6/4LOfs56wdb0H9pjw3prz6a9mmj+IzEhP2aRGY2074z8rhzEW4AKRjq9f8E4P+ChXw58S/DXQfgT8ZfFNj4b8V+GLWPT9Jv8AUpxDbataRjZColbCpMihYyrEF8KyliWCwzQ3EKT28qSxyKGR0bIYEZBBHUEVr3xg+FPhjxHpng7X/iN4dstf1i8SzsNKl1GMXVxO5wqLDneSTxnGKr/gsH8MdR8H/tWN4/a1k/s7x7o9tcw3BB2tPbRrbSxg+qpHAxA7SL61/wAE+P20Phf8Xvgb4V8D+JfHGlaX4+8LadDpd9puo3awS3iwKI4riHeR5odFQttyVfcCACpb9on9tD4Dfs3+E73WvFPjXS9T1uOAtYeHdOvY5b29kI+VdikmNCSMyOAoGepwD8ZfFnjP4jfEbWvij4706Sz1LxtdS6wqNGyIYppG2eWG5MYC7VPcKOT1rwl/yKmi/wDYPg/9FrX/AAW0/wCS/eBf+xPH/pXPX/BN7/kyP4V/9g+5/wDSyeq/4Kz/ALNes/C749XXxm0nTHbwl8RJBObiNMpa6kEAnicjoZNvnLn7258fcOP2Bv2+PAP7QXw+0fwP498VWOk/E7R7ZLW8tL64WE6xsXAubctgSMwGXjX5lbccbcGvGPjbwd8PdBuPFHjrxTpegaTaKWmvdRukt4lwM43MQCeOAOT2r/goB+1dcftm/GrSdB+Gmn6jd+FfDzNp3hu1WBjPqdzM6iS4EQG4GUrGiJjdtRcgMxUfsPfs8yfsy/s5eHPh1qkMS+ILgvqWvmNgwN9PguuRwfLRY4sjg+VnvX/Bcb/kVPhH/wBhDVf/AEXbV/wRt/5NFu/+xwvv/RNtVf8ABb3/AJJV8NP+xguv/ScV/wAEVP8Ak1fxT/2UC8/9ILCviV8bvhN8HLrQbX4o+O9L8MnxNdPa6ZLqMvlRTSooZg0hG2MAEfM5VckDOSAdP1HT9XsYNT0q/t72zuUDwXFvKJI5FPRlZSQR7iv+C1HxW+Gfia58A/DTw/rOn6t4q8PXN7c6n9klWVtNikWNRDIy52u7Lu2dQIwSAGXP/BIDwdr/AIV/ZBj1HW45o4vE/ia91PTklBBFt5cNuCAeil7eRh67s981VVVVVVVVVVVVVVVVX7RP/BSD9mf9ntbvR5vFQ8YeKbcFRofh9luGR/SafPlRYP3gWLj+4a/aJ/4Kk/tJ/HH7XofhfVh8OvDE5Kix0KZlupE9JbziQ8ZBEYjUg4Kmvhr8Iviv8cvEw8OfDHwTrPirV533Si0hLiMsfvzSthIlJ/jkZRnvX7O3/BF+eQ2fiP8AaY8aiNQQ7eGvD75Y+izXZ4HoyxqfaQV8KPgj8Jvgb4fTwx8J/AWk+G7EKBJ9kg/e3BHRpZWzJK3+07E+9VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV/wAFxv8AkavhH/2D9V/9GW1f8EMf+a2f9wD/ANyNVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX7Q/wCwP+zX+0ktxqXizwVHo/iSZTjxBoW20uyx/ilwCk56cyqxxwCK/aI/4JLftC/CP7TrnwzEfxM8OwqXJ02DydRhUdQ1oWYyeg8pnJwSVWvCfjb4m/Bfxc2r+DvEev8Ag/xHp0hile1mktJ42U/NHIvBIyOUYYPQiv2eP+Czni/Q/s3h79pLwgniK0BVD4g0ONLe7Qd2ltziKU+6GLAH3WNfBj9o74I/tCaP/bHwi+Iml68I13T2aOYru2/6628gWRBnjJXB7E1VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV+1B+xt8Fv2stChsfiJpM1prVhGy6br+mlYry0B52biCJI89Y3BHJK7Sd1fEH/giv8ftFv5m+HPxC8H+J9OXJiN602m3LdcAx7ZIx2GfM6nsOa8D/wDBFv8AaM1u9hPjrx14J8M6e2POeCabULlOmdsQREbAz/y1HP1zX7Kf7FPwc/ZJ0WaPwRZz6p4j1GIR6l4i1EBrm4UYPloBxDFuGdi9SBuLFQRff8ERviVNe3Etj8afCsFs8rGGJtPuGKIT8q5zzgYGa/Y6/wCCX/j39mT4+aH8YNc+KHh/W7PSra7hks7SzmjkfzoHjBBbjgsD9KqqqtZ0q017R77Q74yi21C2kt5jDKY3COpVtrLgqcE4I5B5r4r/APBEfzb2e/8Agj8ZY4baRiYtM8TWZJiHYfaYM7h2/wBUD7mm/wCCMn7WYvPso8QfDcx79v2gavc7Mf3sfZt2P+A59q+C3/BFCzsdTtdX+P3xTi1K2gcNJovhuF40nwc7WupQHCnoQsatg8Mp5rwV4I8I/Djwrp3gjwJ4estE0LSYfKs7Czi2RxLnJwO5JJJY5JJJJJJNftHfCvUPjf8AA3xl8JtK1W30278T6a1pDd3CF44WLKdzBeSOO1fsH/8ABOnxp+yH8XtX+JPiP4j6J4gttS8Nz6SltZWksTo8lzbShyX4wBbkY9WFV+zj/wAElfiJ8EPjl4N+LGq/Fzw5qdp4Y1JbuW0t7GZJJgFIwpY4B571VfB//gkt8RPhp+0B4Y+Md98W/Dl7Z6D4ki1aSyisZlkkRJt5QMTgHHGTxVfGf4HfC/8AaC8Ez+APiv4Wt9a0qVvMiDExy2swBCywyKQ0bgE8g8gkHIJB8bf8EPtFuNUkuPh18fryx093Pl2ms6Et1LGvvNFLGGPQf6ta+BX/AARy+CXw51y08S/Fbxff/Ee5snEkWnSWK2Gnsw5HmxB5HlAIHylwrdGUg4r/AIKjX+kXn7avjey0QRLb6Xa6bZFIlCpG0djBlFA4AXO3HGCCO1fC/QJfCnwz8I+Fp4jHLo2hWVm6MSSrRQIhH/jtft9fsC+L/wBsTxf4V8SeGviBo/h6Pw9ps1pLFfWskrSs8gcMCnQY45r9kf4H6r+zf+zz4T+C+t67aaxe+Hftnm3trE0ccvn3k9wNqtyMCYKc9walijmjaGWNXR1KsjDIYHqCPSv2j/8Agjt8NPiLrN74v+BnitfAWoXjtLLotxam401pCST5W0h7cEnO0b1HRVUYAj/4I+ftkaLdvZaL458CJbu4zPba7eQo3+0V+zhuB14PtmvAH/BEb4gXd7FN8VvjX4f0+2DZmi0CzmvZJB3CyTCEKT6lWx6GtNs30/TbWwlvZ7x7aBImuLggyTFVALuQANxxk4AGTVV+3F/wTb8b/tY/GiH4o+HviVoeg2sWiW+nG1vbOWRy0TysWynGD5g49q+A3w5vfhD8FvBPwu1LUoNQuvCuiW2nTXUCFEmaJApZQeQDjoar9vD/AIJ0+NP2vPi9pHxJ8OfEfRNAttN8NwaS9tfWksru8dzcylwU4wROBj1U1+zj8K9Q+CHwN8G/CbVdVt9Tu/DGmraS3duhSOZgxO5Q3IHPetd0LRfE+i3vhzxFpdrqel6lbvb3lndRCSKeJxhkdTwVIJBBr47/APBFnw5rmqXWu/s9/EVPDsc7ll0HXopLi3hz2juUzIqj0dJDz97irX/gj9+2XbM2jW3jjwHb2TMys6a/eLCQRydott2D/u555r4A/wDBG7xb4G8d+HPiD8RfjfY2114b1O21GC08OWLyM00EiyJiefbtAZBz5R/Cq/aV/Zp+G37U3w5m+HnxEs5F8tzPpmp2wAudNuMYEkZIPBHDIeGHB7EeNP8Agix+0TpWryReBvH/AIH1/Six8m4vJp7CfH+3F5cir+EjfhX7Pv8AwRdstD1218Q/tHePrDXba0cOPD/h4SrBckEY825kCPs6goqKT/fHQ/tkf8Evdd/aO+LNp48+H3jbw14R0iz0G00uLS30+TEf2feq7BHhVQIUUKAMBa8L6ZeaL4Z0jRtRu1uruwsIbeedV2iWRIwrMB2BIJx71+3r/wAE9/GX7YHxH8PeN/DfxD0Xw/Bouif2dJBfWksrSN50km4FOAMSAY9q/Zc+DupfAD4B+EPg/rGs2urXnhu2lhlvLaNkjlLzySAqrcjhwOfSq+Inw68E/FnwZqfw++Inh211zQNXh8q7srgHDDqGVgQyOpAKupDKQCCCM18YP+CJerPqtxf/AAG+LWnixmcmLTPFMMkbW49PtMCvvHpmJT0yT1rRv+CNv7VPiK9gh8afEfwRp9jB8okbUbq9kRT18uPyQvYcF17V+yT/AME4/gx+yveweMftNx4x8dRxlF1zUIFiS03DDfZYAWEWRwWZnfBIDAEiq/b/AP2LvE/7ZGj+C9M8M+NdL8PN4XubyWZr63klEwmWIALs6Y8s5z6iv2Hf2Ztd/ZO+C03wu8Q+J7DXrqXW7jURdWULxIFkSJQuG5yPLP51X7fX7HniX9sTwf4V8NeGvGOmeHpfD2pTXcst9bvKJQ8ewKAnQ9+a/YP/AGWNf/ZD+EGr/DXxH4r0/wAQXOpeJJ9WS5soHiREktraEIQ/OQbcnPT5hX7dv7BrftlL4e1ax+KFz4a1TwtbzxWVrPZC5s5fNZS5IDK6OdiAuCwwg+XvUv8AwR9/bG0SaTTtB8eeBpLJ3LB7bXbuBWzxlkNuMNgDPX0ya+BP/BFZNM1u01v9oj4k2OqWVq4Z9C8NpKI7rHQPdShHVOxVYwxB4detaLoukeG9GsfD3h/TbbT9M0y2jtrOztoxHFbwxqFREUcKqqAAB0Aqqqqqqqqqqqqqqqrm4t7O3lu7u4jhggQvLLIwVUUDJYk8AADJJr9of/gqz+zf8GRdaJ4Gvn+JXiSJWCwaLOosY3HaS8OVI/65LL74r9oj/gob+0v+0Yt1pGteLz4a8MXKlG0Dw+WtYJE7iZ8mSYHjKuxTPIUV8FP2avjh+0Rq39lfCP4eanraRyBLi+CCGztSf+etw+I0OOdudxHQGv2eP+CMngbw6bXxB+0f4ufxRfowc6Dosj21iPVZJyBNKP8Ad8r8RXgf4f8Agb4Z+H4PCnw88JaT4c0i35jstNtEt4wx6sQoGWPdjknuaqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/4Ljf8AI1fCP/sH6r/6Mtq/4IY/81s/7gH/ALkaqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr48/sjfs//ALSVlJF8Uvh7Y3epmMJDrdov2bUIAB8u2dPmZR2R9yf7NftEf8Ecviz4IN1r/wAAdfi8d6QhLLpN4UtNTiX0BJEU+B1IMZPQIamg+I3wd8a+VPH4h8GeLNDnzhhLp97ZSDv/AAuhx9Otfs7f8FhPjN8PFs/Dvxw0iH4haJEAh1BCLXVIl6Alx+7nwB0dVZjyZK+AP7ZP7PP7StrCvwz8fWzaw0e+XQNRxa6hDgZb9yx/eAd2jLqP71VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV+0R+318Ff2YfizoXws+J8OrodZ0r+0JdSsIBcJYK0rRoJYwd+G8uQ5QMRgfLzkeHP22f2RPFViuoaX+0f4AhiZQwXUtbi06TB9Y7gxuD7EZFftDf8FRv2aPhB4bvR4C8X2PxD8VmJhYado0hmtvMx8rTXK/u1QHqELOegHOR+xZ8FPH37aX7WKeOvGMdzf6RZa1/wkPjDVWjxEzeaZVtx/CGmkAQIOQm8gYSqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqrXvEGgeFtIuNf8T65p+kaZZpvuL2/uUt4YV9XdyFUe5NftD/APBYP4KfDs3WgfBLSZviHrcTNH9uZmtNMiYdxIR5k2D2RQrdn6V8fv2z/wBon9pWeSL4k+Prj+x2ctHoOmA2lhHzkAxKf3hHZpS7D1r4AfsWftFftKzxTfDjwDcJorsQ3iDVQ1pp6YODiVgfNIPBWIOw7iv2ef8Agj98Efh0LbXvjXqk3xF1tAr/AGIq1ppsDjnHlq2+bB4y7BWHWPnFaJoOh+GdJttB8N6NY6VptlGI7aysbZYIYUHRURAFUewFVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX/Bcb/kavhH/wBg/Vf/AEZbV/wQx/5rZ/3AP/cjVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVXxh+AHwb+P2hjw/8XPh/pXiK3jUiCW4jKXFtnqYp0IkiP+6wz3r9of8A4Iv+INNNz4g/Zp8Zrq8ALOPD3iCRYbhR1Cw3IAjfngCQR4HVzXjr4cfE/wCC/igaD8QPCWu+EtctX3xR3tu9tIdp4kibgOuRw6Eg9jX7PH/BV39o34Ni10Px7dR/Evw5AoQQaxMY76JR/cvACzH/AK7CTtjFfs8/8FCP2aP2jBbaVoHjJfDviWcKv9geICtpcO542wuSY58noEYtjkqKqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv2l/2Bv2fP2p9THifx5pmrab4nS3W3XW9IvTFMY1B2IyOHiZQT3Td7itb/wCCG/h64uS/hz9o/UbG3ycR3vhZLp8cY+ZLmIev8POR0xz8Pv8Agil8EdCvYbz4i/FDxT4rSEgm0tLeLTIZuOQ+DLJtP+y6n3r4a/C34efB3wna+Bvhh4R0/wAO6JZ8x2lnHjcx6u7ElpHOBl3JY9yaqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq8d/ELwL8MPDlx4u+Ifi3SvDuj2o/eXmo3SwJnsoLH5mOOFGSTwAa/aJ/4LNeC/D5u/Dv7N3hJvE16hKL4g1qN7eyB/vR242yyjp98xc9iOvxr/aT+N/7Q+rnVvi38RNU1tVkL29g0nlWdqen7q3TEaHHG4LuPcnrX7O3/BO39pb9ov7Jq+leEj4X8LXBB/t/X1a2idP70MWPNmyM4KrsyMF1r9nb/glV+zh8Fls9b8a2DfEnxNAAzXWtQAWUb+sdnkpjj/lqZDnkEdre2t7O3itLSCOGCBAkUUaBVRQMBVA4AAAGKqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqvEC662gakvheSyj1k2co0571GeBbjYfLMiqQxTft3AEEjOCK/Zw/4KT/B/4xas3w4+JsQ+GfxGsblrK70jVrhRbT3KMUdILg4XdvBHlvtbJwu/Ga/4Ljf8jV8I/wDsH6r/AOjLav8Aghj/AM1s/wC4B/7ka/aQ/wCClHwf+D2sL8NvhlF/wsz4jXtwtlaaRpM4+ywXMjBESe5GV3byB5abmzw2zINeH111dA01fFEllJrIs4hqL2SMkDXGweaYlYlgm/dtBJIGMk1VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV8Q/hf8Ovi14el8KfEvwXo/iXSpQf9G1G0WYISMbkJGUb0ZSGHYiv2h/8AgjB4d1MXXiD9mnxk2j3G1nXw7r8jTW7H+7FdDMkYxwBIJMk8uor4x/s9/Gb4Aa2dB+Lnw+1Xw/Mz7YLiaLfbXP8A1ynQmOTgH7rEjviv2ef+Ck37TX7P5ttIHik+NfDMJVf7H8RyPc+VGONsM+fNiwOAMlB/cNfs7/8ABUb9mn45m10TxFrDfDvxPcMEGna9MBbyv6RXgAjPYASeWxJwFNRyRzRpLDIro6hkZTkMD0IPpVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVfFj44fCT4GaCfEnxZ+IGj+GrMqxhF5Pia5K9VhhXMkrf7KKxr9oj/gtBdz/afDv7M3gv7MhUoPEfiGMNID/ehtVJUY6hpGbOeYxXxJ+LPxU+OPigeJPib401nxVrEzbIWvJjJ5e4/wCrhjHyxqTj5EUDPav2eP8Aglj+0n8bvs2teK9LT4ceGpSpN5r0DC7lQ9TDZjDk4wf3hiUjoxr9nf8A4Jx/szfs8/ZdXs/Cv/CXeJ7dg41zxCq3MkTjoYYsCKHB6ELvHdjiqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv+CtPwHHwo/aXl8faRZ+VofxKtzqaFVwqXyEJdoPcsY5ifWc+leLfil8RPHug+H/AAz408Y6nrmneFY5YtGiv5jM1lHJs3xo7ZbZ+7TCElVx8oGTnwh8XfiX4A8K+JPBXgjxnqWh6R4vNuNbhsZfJa9WASiNHkX59mJ5coCFbd8wOBj/AIJK/Adfiv8AtLRePtXshNofw1thqbl1yrX7kpaL7EMJJgfWAetVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVeJ/Cvhnxrod14Y8YeHtN1zSL5NlzYajapcQzL6Mjgg/iK/aK/4I6fCPx59r8Q/AXXpfAetSEuNLut11pcrei9ZYMk9QXUcAIK+Pf7If7QH7Nl46fFL4f3ltpYl8uHW7MfadPnP8O2deFJ7K+1/9mv2e/wBuT9o79mqW2s/AnjmW+8PwN83h3WM3diy9wqEhoc+sTIfXPNfs7f8ABXX4C/FL7J4f+LlvL8N/EEgCm4upPP0uZ/8AZuAA0WeTiVVVeBvY1pWraVr2m22s6FqdpqNheRiS2u7SZZopkPRkdSQwPqDiqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv2uv2sPAf7JXwzl8YeJpVvdc1BZIfD2io37zULkKOv92JMqXfsCAMsyqf2N/HXjL4ofs0+CPiL8QNRS91/xLbXGoXcsabUXzbqV0RFH3USMoijnAUDJ6mvjn+1P8Bv2cdNN78WPiHp+mXTLmDS4mNxfXHpst0y+OnzEBRkZYV+0R/wWU+JXi77T4f/AGePDEfgvTSWUa1qapd6hKvYpHgwwcdQfNPQhhWraz8SPjH4yF5rGo+IfGfinV5diNK8t/d3LnJCKPmdupwo6dhX7O//AASE+OfxN+y698Y7+L4caDKof7NLGLnU5Qeg8gELDkdTIwZcj5DzX7P37Ef7OP7NcMNz8PfAVvca6iASeINWIvL9yOpWRhiHPGViVFOBkVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVf8FPvgOfjZ+yvrmoaVYC48QeA3/t3TtqZd44lIuoweuDAXfaPvNEg9MVX/BMH4Et8FP2VtCvtVsfI1/x2/wDb2oB0w6RyqBbRk9cCBUbacbWkcetVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV9Y2Op2c2nalZwXdrcIUmgnjEiSKeqspyCD6Gv2iP+CS/7PHxd+0658NEk+GfiKUs5bTIfO0+Zj/ftCwEfYDyWjA5JVq/aG/YF/aU/ZvM+peKvBT614ciLEeINBDXlqqj+KUAB4BjHMiqOcAmvgZ+1P8ef2cdSF78J/iHqGmWrNmfS5m+0WNx677d8pnr8wAYZOGFfs8f8FlPhr4u+zeH/ANobwxJ4L1MhVOtaYr3enyt3LxgGaDnoP3o6ksK8I+NPCHxA0G38UeBvFGl6/pF0Mw32m3aXET8A4DISM8jI6jvVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX7Sv7R3w+/Zd+GF98SvHt15nl/utM0uKRVuNTuT92GIH82bBCqCxzjB/aD+P3xB/aV+JmpfE34iakZbq7YpZWUbnyNNtgSUt4VP3UXPXqxJY5LE14T/bL/Zy/ZL/ZZ+Fvh3x744ivfElj4H0kv4d0dlvL5pWs42ZWUELEMk4MrIMY5Pf9or/grr8efil9r8P/AAit4vhv4fkJVbi1k8/VJk/2rgjbFng4iVWXpvYVoPhz4j/GTxmNL8O6Tr/jLxTrEpkaOCOW9urhifmkc8seuS7cDqSK/Z2/4I0fEDxQbPxD+0Z4pj8JacxDvoWkulzqDj+683zQwk+o809iAenwP/Zg+BX7OmlLp3wm+Hmm6TcNF5dxqbx+dfXQ7+ZcPmRgTztyFB6AVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVU8EF1BJa3MMc0MqFJI5FDK6kYIIPBBHGK/a9+B837O37RHjL4XrC66bZXxuNHdlI8yxnHmQYP8W1GCE/3kav2QvgfN+0T+0R4N+F7Qu2m3l8LjWHVT+7sYB5k+T/DuRSgP9519agggtYI7W1hjihhQJHGihVRQMAADgADjFVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVUyqylWAIIwQRX7Q/8AwTM/Zl+PYutYsfDY8CeJ5lYjVfDsSwRySHndNbf6qTnJJAR2zy9ftE/8Exf2lvgN9r1rStBXx94WtwX/ALV0CJnliQd5rXmVOASSvmIB1cV8Kvjb8XvgR4h/4SD4U+PdZ8MX4YeelrMRFcbTws0LZjlUH+F1I9q/Z3/4LQWdx9l8O/tMeC/szlgh8R+Hoy0YH96a1YlhjqWjZs54jFfDH4vfDD40+HE8WfCvxzpPibS2IDzWM4ZoWPRZYzh4mxztcKfaqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqvjh8bvh5+z18OtS+JnxK1pLHS7BdsUSkGa9nIOyCFP45GwcDoACxIVSR+1N+1B8QP2r/ibP8QPG0q2tpbq1voujwyFoNMtt2Qi9Nznq8hALH0UKq18Lvg58Ufjb4jXwn8KPA2q+JtUwGkisYNywKTgPLIcJEmeNzlV96/Z4/4IvOxtfEP7S/jYKAwc+HPDsmSR/dmumHHoVjX6Sdx8K/gt8Kfgh4fHhf4T+A9J8NaeSDIllDiSdh0aWVsvK3bc7Mfeqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/4LS/AhdZ8G+Ff2h9Gs9134fm/sXWnRMk2kzFrd2PZUmLp9bgV/wRa+BK6P4M8VftDazY4u/EE39jaLI6YItIWDXDqe6vMET2Nuaqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv2hv2Df2bf2k1utR8ZeCY9K8SXAz/wAJFom20vS396QgFJj2/eqxx0I4r9oj/gkl+0F8Jzda78LTF8S/D0bEqmnwmLU4l/27Uk+Zjgfumcnk7VFeFfGPxJ+DPjEa14Q1/XfCHiXS5TE8ltLJaXELA/NHIvBxkYZGGD0Ir9nb/gs1408PrZ+Hf2kPCS+JrJAEbxBosaW96B/ektztilPX7hi47E9fgr+0n8EP2h9HXV/hJ8Q9L1tljD3FgJPKvLUdP3tu+JEGeNxXaexI5qq8DftV+DPEn7R3jz9l/wAQ+TovjDwpNBLpcck4K61aS2kVxvi9JYxJ80fXbhhkBwlVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVXxP+Jngr4OeA9X+JHxC1uLS9B0S3M11O4yT2VEUcs7MQqqOSSBX7Zn7YXjf9rr4jSa7qUlxpvhHSpHj8O6CZMraRHgyyY4ad8As3OOFB2qKqvht8T/H/AMHvF9l48+Gfiu/8P67YH91d2km0lTjKOpyrocDKMCp7g1+xr/wVT8AfGr7F8P8A45tp/gvxs5WK3vzJ5em6qx4G1nP+jyk8bHJUnG1ssEFVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV8Zvhhonxp+FPir4VeIABZeJ9Lms2kK7jA7L+7lA/vRyBHHuor4NfDDRPgv8ACnwp8KvDwBsvDGlw2ayBdpndV/eSkdmkkLufdjVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVfHf8AZL+AH7SNi0HxU+HtjfagE2w6xbD7NfwcYG2dMMQMD5H3Jxypr9of/gjf8VvBX2nX/gB4ji8daSu5xpN9ss9ShUcgKxIinOB1HlkkgBDV5Y/Ej4O+MfIvbbxH4K8VaRJkB1m069tH6ZH3XQ9eRiv2eP8AgsH8avh2LXQPjbpMPxD0SJRH9uDLaanEo7mQDZNgdnUMx6v1r4Bftnfs6/tKW8UXw1+IFr/bLJuk0HUx9k1CI4yR5TH95gdWiLqP71f8FUZdb+H37dt3428NahcaZq0mm6VqlleW7BJIpYohEsikc5BgHJ9PSv2AP27tB/as8IJ4U8X3Vpp/xP0O3zqVkoESalEuB9rgX0ORvQfcY8AKy1VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVfH/8AZW+FH7Ti6RafFxdd1DTNFZ5LfS7XVpbS1aVhjzXSMgu4GQGJ+UEgfebNl/wSv/YYtYfKn+DU9424nzJ/Euphvp8lwox+FWX/AATr/Yp0+UzW/wCz9oLsV2kTz3Mwxx/C8pAPHXGfzqw/Yh/ZC01I47b9nLwG4ibcv2jRo5yTnPJkDEj2ORVj+zD+zTpbtJpn7PHwztGcYZoPCVjGWHviKtP+Efwo0lYV0r4Y+E7MW5zCLfRbePyznOV2oMc+lBVVQqgAAYAAxiqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqvi/wDAL4OfHvQ/+Ef+Lfw+0nxHbojLBLcw7bi2z1MM6YkiPujCv2iP+CL+s2C3XiH9mjxmNUiVS6+HPEEixT5/uw3QARs9lkVMAcua8f8Awz+J/wAFPFX/AAjvxD8I614U120cSRR3kLQP8p4kiccOuRw6Ej0NfED4n/EH4rajYaz8R/FuoeI9R0zT49Pt73UJPNn+zozsiPIfmkIMjfM5ZsEDOAAPAnjvxd8MvF2l+PPAev3Wja9o1wJ7K9tmw8bjrx0ZSCVZSCrAkEEEiv2H/wBtHwl+138P/OYQaV460KJE1/RvMHXoLmAdWhc/ijHac/KzVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVa1rWjeHNJu9e8QatZ6ZpthC011eXkywxQRqMs7uxAVQOpJxX/BQf8A4KWeBvi54f1P4H/BnwppPiDQ5SUvPE+tacswLYwTYwyrmNhyBOwDDkoFwHNV8JPiz45+B/xB0j4m/DnWX07XNGm3xPglJVPDxSLkb43XKsvcHscEfsg/ta+BP2tvhpD4r8PywWHiLT1WLxBoJm3S6fOc4IzgtE+CUfGDyD8ysBVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVfHr9mr4ZftKaNbeHPip/b13pVqS6WFlrM9nA0naR442CyOv8ACXDbecYyc3//AAR6/Y8vFRbeDxrZFScmDWwd313xt+mKv/8Agiv+y1cmZ7Hxz8TrR3B8tRqVk6Rn6G03Ee278avv+CI3wXkKf2Z8ZfGsAAO8T21tNn0xhFx+tX//AAQ48MSrMNL/AGjNTt2LHyTceGUm2DPG7Fwm444yMevtXwR/4JYfF79mv4l6b8S/hF+1TZLeWq+Xd2954TeOG+gJBe3mjW7bfG2AeoIIDDDKCBkqNwAOOQDmqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq+KHxe+GPwV8MyeL/ip430rw1pUZIWa+m2tMwGdkUYy8r452IrN7V8Y/+C1vgXR5p9L+Bnwvv/EUiMVXVden+xW5x0ZIE3SOp/2miPtXi3/grr+2Z4jld9F8ReGPCyt91NK0GKUL9Ddecam/4KV/twT3Ju3+PmoBy27CaTYIuf8AcEAUD2xivDP/AAVo/bW0G4WbVfHOheJEVsmLU/D9tGrex+zLCcfj3r4Uf8FuIZZrew+OHwa8lGIE2p+GLwtt+lrOc4/7bfga+CP7S3wP/aL0htV+EPxB07XHgjD3Vjkw3loOB+9t5AJFGTjdjaT0Jqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv2xvgP+0d+1L+3P8RPDfw68Ma94ns9Cu7a0gubiYpYaVEbWFzH50pEcQLOz7AckkkAk18Kv+CI91NBDffG34zLBIygyab4Zs9+09wLmfAzjj/U9e57+GP+CSn7FWgQLFqvgrX/ABIyggyap4guY2b3P2ZoR+Qr/h27+xH/ANEC0n/wYXv/AMerxJ/wSk/Yl1+KVbH4aaloUsob99puv3mUJOcqs0kiD2G3HtXxQ/4IjeFrm3muvgv8ZtTsbhQTFY+JbRLlHPoZ4AhQe/lOauv2SP2qv2QfjR4L8S+I/Dmp6dZQ+IbOOHxP4euWmtCrzqhDTR4aIOCV2ShCwJGCM1VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVSRRxgiKNV3MWIUYyT1P1p3SNDJI6qqglmY4AA6mvh/8d/gp8VwE+GvxX8KeJJec2+natDNMuP70Ybev4gVc3FvZ28t3d3EcMECF5ZZGCqigZLEngAAZJNfEL/grj+yF4E8QTeH9P1HxT4vNtMYpb3w9psclsCOpWSeWLzFB43IGB6gkc1+z5+1b8Dv2n9IuNS+EnjBL66sVDX+lXURt72zBOAXibqp6b1LJnjdnIqWKOaNoZY1dHUqyMMhgeoI9Kqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv2w/Fd54I/ZY+KviXTZ3hvLbwpfR20qfeilliMSOPdWcH8K/4JofDnTfiV+2V4Es9ZiSay0GSbWpI2H3pLWJnh/Kbym+imv8Agsd+0TfeAvhdonwI8L6mbfUvHbSXGsGJyrppsJAEZx0E0pxnPKwupGGrwD8MfiL8VdWm0H4aeBtd8U6jb27XE1rpFhJdSRxAgF2VASFyQMnuQOpFfs7/ABh8Vfs0/Hbw58RtPe8sZ9B1JYdYsmDRtPaFwl1bSIR3TcMEZVgrYyoxDNDcQpPbypLHIoZHRshgRkEEdQRVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVTukaGSR1VVBLMxwAB1NeL/+C2vws0rxBead4O+DPiDXtLt5Wjh1C41SOyNwAcb1i2SEKcZGSDgjIB4H/D8Xwp/0btqv/hSR/wDxiv8Ah+L4U/6N21X/AMKSP/4xX/D8Xwp/0btqv/hSR/8Axiv+H4vhT/o3bVf/AApI/wD4xX/D8Xwp/wBG7ar/AOFJH/8AGK/4fi+FP+jdtV/8KSP/AOMV/wAPxfCn/Ru2q/8AhSR//GK/4fi+FP8Ao3bVf/Ckj/8AjFf8PxfCn/Ru2q/+FJH/APGK/wCH4vhT/o3bVf8AwpI//jFf8PxfCn/Ru2q/+FJH/wDGK/4fi+FP+jdtV/8ACkj/APjFf8PxfCn/AEbtqv8A4Ukf/wAYr/h+L4U/6N21X/wpI/8A4xX/AA/F8Kf9G7ar/wCFJH/8Yr/h+L4U/wCjdtV/8KSP/wCMV/w/F8Kf9G7ar/4Ukf8A8Yr/AIfi+FP+jdtV/wDCkj/+MV/w/F8Kf9G7ar/4Ukf/AMYr/h+L4U/6N21X/wAKSP8A+MV/w/F8Kf8ARu2q/wDhSR//ABiv+H4vhT/o3bVf/Ckj/wDjFf8AD8Xwp/0btqv/AIUkf/xiv+H4vhT/AKN21X/wpI//AIxX/D8Xwp/0btqv/hSR/wDxiv8Ah+L4U/6N21X/AMKSP/4xX/D8Xwp/0btqv/hSR/8Axiv+H4vhT/o3bVf/AApI/wD4xX/D8Xwp/wBG7ar/AOFJH/8AGK/4fi+FP+jdtV/8KSP/AOMV/wAPxfCn/Ru2q/8AhSR//GK/4fi+FP8Ao3bVf/Ckj/8AjFf8PxfCn/Ru2q/+FJH/APGK/wCH4vhT/o3bVf8AwpI//jFf8PxfCn/Ru2q/+FJH/wDGK/4fi+FP+jdtV/8ACkj/APjFf8PxfCn/AEbtqv8A4Ukf/wAYr/h+L4U/6N21X/wpI/8A4xX/AA/F8Kf9G7ar/wCFJH/8Yr/h+L4U/wCjdtV/8KSP/wCMV/w/F8Kf9G7ar/4Ukf8A8Yr/AIfi+FP+jdtV/wDCkj/+MV/w/F8Kf9G7ar/4Ukf/AMYr/h+L4U/6N21X/wAKSP8A+MV/w/F8Kf8ARu2q/wDhSR//ABiv+H4vhT/o3bVf/Ckj/wDjFf8AD8Xwp/0btqv/AIUkf/xiv+H4vhT/AKN21X/wpI//AIxX/D8Xwp/0btqv/hSR/wDxiv8Ah+L4U/6N21X/AMKSP/4xX/D8Xwp/0btqv/hSR/8Axiv+H4vhT/o3bVf/AApI/wD4xX/D8Xwp/wBG7ar/AOFJH/8AGK/4fi+FP+jdtV/8KSP/AOMV/wAPxfCn/Ru2q/8AhSR//GK/4fi+FP8Ao3bVf/Ckj/8AjFf8PxfCn/Ru2q/+FJH/APGK/wCH4vhT/o3bVf8AwpI//jFf8PxfCn/Ru2q/+FJH/wDGK/4fi+FP+jdtV/8ACkj/APjFf8PxfCn/AEbtqv8A4Ukf/wAYr/h+L4U/6N21X/wpI/8A4xX/AA/F8Kf9G7ar/wCFJH/8Yr/h+L4U/wCjdtV/8KSP/wCMV/w/F8Kf9G7ar/4Ukf8A8Yr/AIfi+FP+jdtV/wDCkj/+MV/w/F8Kf9G7ar/4Ukf/AMYr/h+L4U/6N21X/wAKSP8A+MV/w/F8Kf8ARu2q/wDhSR//ABiv+H4vhT/o3bVf/Ckj/wDjFf8AD8Xwp/0btqv/AIUkf/xiv+H4vhT/AKN21X/wpI//AIxX/D8Xwp/0btqv/hSR/wDxiv8Ah+L4U/6N21X/AMKSP/4xX/D8Xwp/0btqv/hSR/8Axiv+H4vhT/o3bVf/AApI/wD4xX/D8Xwp/wBG7ar/AOFJH/8AGK/4fi+FP+jdtV/8KSP/AOMV/wAPxfCn/Ru2q/8AhSR//GK/4fi+FP8Ao3bVf/Ckj/8AjFf8PxfCn/Ru2q/+FJH/APGK/wCH4vhT/o3bVf8AwpI//jFf8PxfCn/Ru2q/+FJH/wDGK/4fi+FP+jdtV/8ACkj/APjFf8PxfCn/AEbtqv8A4Ukf/wAYqf8A4Lj+G1hdrb9nLUpJQp2JJ4nRFJ9yLckD3wa8Cf8ABbvwRrPiWz0z4gfAnUvDej3EqpNqVj4gXUmtwTje0Jt4SVHU4YnGcAng2F/ZarYW2p6bdxXNpdwrNbzxNuSWNgCrKR1BBBB96qqqqqqqv2jfhzdfFz4C+P8A4aacEN94h8PXlpYhzhftLRN5OT2HmBM18FfjH8Qv2afivp3xL8DrBa+IdAklie11G2Z43DK0csM0eVbBBIIBVgQCCCAR+0D8fviH+0v8Sr34pfEu7tX1S6hS3htrONo7aygTOyGFGZiqAlm5YkszEkkk1/wSH+AzfDL9nWX4na1pvka58R7sXcTOmHXTospbD1AZjNKOxWRD2r9tTU9G1j9rX4t3+grGLRvFt+gMf3XkSUpK49d0iu2e+c968A2d9p3gTw3p+qSM97a6RaxXDMSSZFiUMSTznIPXmqqqqq+I37RnwF+Ed0dO+JXxg8J+Hr4Lu+w3uqRJc7T0Pk5MmPfbivh1+0r+z98W7xNM+G/xk8Ja/qDglLC11SM3LAZyRCSJMcHnbiqr4k/tLfs//B+/Gk/Ev4w+FfD+okA/YbvUU+0gHoxhUlwD6kYr4a/Gz4Q/GS0lvPhZ8SfDvihLcAzppt+k0kGenmRg7kz/ALQFVVf8FZ/jV8TPg1+z9ob/AAv8W3/hu+8ReI0sbvUNPlMNysAglkKRyrhoyWRMspDYGM4Jz8P/ANhP/gor8UvAfhv4maR+1VbxWHi3SbXV7VL7xzrInWK5iWVBIFgYB9rjIDMM55PWv+HbP/BSX/o6/Sf/AAu9b/8Akav+HbP/AAUl/wCjr9J/8LvW/wD5Gr9qf4J/tufsh+H9D8R/En9pXUNSttfvJLS2TQ/GWqTOjom8lxKkQAx6E89q8G/sA/8ABRbxx4P0LxrpP7VVjFY6/ptvqFtHc+OdaWVI5o1kUOFtyAwVgCASM9z1r/h2z/wUl/6Ov0n/AMLvW/8A5Gr/AIds/wDBSX/o6/Sf/C71v/5Gr/h2z/wUl/6Ov0n/AMLvW/8A5Gr/AIds/wDBSX/o6/Sf/C71v/5Gr/h2z/wUl/6Ov0n/AMLvW/8A5Gr/AIds/wDBSX/o6/Sf/C71v/5Gr/h2z/wUl/6Ov0n/AMLvW/8A5Gr/AIds/wDBSX/o6/Sf/C71v/5Gr/h2z/wUl/6Ov0n/AMLvW/8A5Gr/AIds/wDBSX/o6/Sf/C71v/5Gr/h2z/wUl/6Ov0n/AMLvW/8A5Gr/AIds/wDBSX/o6/Sf/C71v/5Gr/h2z/wUl/6Ov0n/AMLvW/8A5Gr/AIds/wDBSX/o6/Sf/C71v/5Gr/h2z/wUl/6Ov0n/AMLvW/8A5Gr/AIds/wDBSX/o6/Sf/C71v/5Gr/h2z/wUl/6Ov0n/AMLvW/8A5Gr/AIds/wDBSX/o6/Sf/C71v/5Gr/h2z/wUl/6Ov0n/AMLvW/8A5Gr/AIds/wDBSX/o6/Sf/C71v/5Gr/h2z/wUl/6Ov0n/AMLvW/8A5Gr/AIds/wDBSX/o6/Sf/C71v/5Gr/h2z/wUl/6Ov0n/AMLvW/8A5Gr/AIds/wDBSX/o6/Sf/C71v/5Gr/h2z/wUl/6Ov0n/AMLvW/8A5Gr/AIds/wDBSX/o6/Sf/C71v/5Gr/h2z/wUl/6Ov0n/AMLvW/8A5Gr/AIds/wDBSX/o6/Sf/C71v/5Gr/h2z/wUl/6Ov0n/AMLvW/8A5Gr/AIds/wDBSX/o6/Sf/C71v/5Gr/h2z/wUl/6Ov0n/AMLvW/8A5Gr/AIds/wDBSX/o6/Sf/C71v/5Gr/h2z/wUl/6Ov0n/AMLvW/8A5Gr/AIds/wDBSX/o6/Sf/C71v/5Gr/h2z/wUl/6Ov0n/AMLvW/8A5Gr/AIds/wDBSX/o6/Sf/C71v/5Gr/h2z/wUl/6Ov0n/AMLvW/8A5Gr/AIds/wDBSX/o6/Sf/C71v/5Gr9pb4d/tmfsreL/DPgr4hftH6xqN94qhMtlJo3jDU5o4wJBH+8MqxkHJHQHiv+HbP/BSX/o6/Sf/AAu9b/8Akav+HbP/AAUl/wCjr9J/8LvW/wD5Gr/h2z/wUl/6Ov0n/wALvW//AJGr/h2z/wAFJf8Ao6/Sf/C71v8A+Rq/4ds/8FJf+jr9J/8AC71v/wCRq/4ds/8ABSX/AKOv0n/wu9b/APkav+HbP/BSX/o6/Sf/AAu9b/8Akav+HbP/AAUl/wCjr9J/8LvW/wD5Gr/h2z/wUl/6Ov0n/wALvW//AJGr/h2z/wAFJf8Ao6/Sf/C71v8A+Rq/4ds/8FJf+jr9J/8AC71v/wCRq/4ds/8ABSX/AKOv0n/wu9b/APkav+HbP/BSX/o6/Sf/AAu9b/8Akav+HbP/AAUl/wCjr9J/8LvW/wD5Gr/h2z/wUl/6Ov0n/wALvW//AJGr/h2z/wAFJf8Ao6/Sf/C71v8A+Rq/4ds/8FJf+jr9J/8AC71v/wCRq/4ds/8ABSX/AKOv0n/wu9b/APkav+HbP/BSX/o6/Sf/AAu9b/8Akav+HbP/AAUl/wCjr9J/8LvW/wD5Gr/h2z/wUl/6Ov0n/wALvW//AJGr/h2z/wAFJf8Ao6/Sf/C71v8A+Rq/4ds/8FJf+jr9J/8AC71v/wCRq/4ds/8ABSX/AKOv0n/wu9b/APkav+HbP/BSX/o6/Sf/AAu9b/8Akav+HbP/AAUl/wCjr9J/8LvW/wD5Gr/h2z/wUl/6Ov0n/wALvW//AJGr/h2z/wAFJf8Ao6/Sf/C71v8A+Rq/4ds/8FJf+jr9J/8AC71v/wCRq/4ds/8ABSX/AKOv0n/wu9b/APkav+HbP/BSX/o6/Sf/AAu9b/8Akav+HbP/AAUl/wCjr9J/8LvW/wD5Gr/h2z/wUl/6Ov0n/wALvW//AJGr/h2z/wAFJf8Ao6/Sf/C71v8A+Rq/4ds/8FJf+jr9J/8AC71v/wCRq/4ds/8ABSX/AKOv0n/wu9b/APkav+HbP/BSX/o6/Sf/AAu9b/8Akav+HbP/AAUl/wCjr9J/8LvW/wD5Gr/h2z/wUl/6Ov0n/wALvW//AJGrx78O/wBsz4dftI6F+y5rf7R+sT+K/EE1nFa3lr4w1NrFGujiPe7KsgA/ixGcds1/w7Z/4KS/9HX6T/4Xet//ACNQ/wCCTv7ZXiMKfG/7T2jymRi83/E41O++YZwR5iJuOMcnGMnrS/8ABETx5fD7XrH7RekG8kJMpGhTTZOeDvaYE8Y6j/GvjX+xR+1p+xJ8PL/43eGP2l0XSdBlt1nj0rV72xuWaWZIUCRYKSDLruUt90HggV+wR8YfG/x3/ZW8GfEb4izrdeILsXVtd3ixCIXnkXMkKy7VAUMVQbtoxuDEYzgVVV8QPi78K/hRaRXvxM+I/hvwtFP/AKk6vqcVqZv9xXYF/wAAa8IftifssePNSj0Xwt8ffBV3fynbDbSarHbySt6IspUufZcnr6VVfEr43fB/4OW0Vz8U/iZ4c8Li4UtBHqeoRwyzgdTHGTvfp/CDXw1/aT+AXxjv20n4Y/F7wv4i1FFLGxtNQQ3O0dWEJIcqO5AwKqq/ag8aeIfh1+zn8SvHXhK9W01rQ/DF9dafclA/kTLCxSQKeCVOCAQRkDII4r9mH9lj9rH9s7wPq3xZ8O/tH3Nklrrkul3I1vXtQeeWZIIZS+UDgrtnUcnOVPtX/DqT9tL/AKOj0r/wd6n/APEV/wAOpP20v+jo9K/8Hep//EV/w6k/bS/6Oj0r/wAHep//ABFftXfsj/tR/sj/AA6074keNf2g31mx1LWotJjg0vWr8yrLJDNKGPmBRt2wMOucke9fCj/gnR+198Xfhn4Y+J+iftLWlnYeKNLg1C2t7rW9S82JJUDBX2qRuAPOCRX/AA6k/bS/6Oj0r/wd6n/8RX/DqT9tL/o6PSv/AAd6n/8AEV/w6k/bS/6Oj0r/AMHep/8AxFf8OpP20v8Ao6PSv/B3qf8A8RX/AA6k/bS/6Oj0r/wd6n/8RX/DqT9tL/o6PSv/AAd6n/8AEV/w6k/bS/6Oj0r/AMHep/8AxFf8OpP20v8Ao6PSv/B3qf8A8RX/AA6k/bS/6Oj0r/wd6n/8RX/DqT9tL/o6PSv/AAd6n/8AEV/w6k/bS/6Oj0r/AMHep/8AxFf8OpP20v8Ao6PSv/B3qf8A8RX/AA6k/bS/6Oj0r/wd6n/8RX/DqT9tL/o6PSv/AAd6n/8AEV/w6k/bS/6Oj0r/AMHep/8AxFf8OpP20v8Ao6PSv/B3qf8A8RX/AA6k/bS/6Oj0r/wd6n/8RX/DqT9tL/o6PSv/AAd6n/8AEV/w6k/bS/6Oj0r/AMHep/8AxFf8OpP20v8Ao6PSv/B3qf8A8RX/AA6k/bS/6Oj0r/wd6n/8RX/DqT9tL/o6PSv/AAd6n/8AEV/w6k/bS/6Oj0r/AMHep/8AxFf8OpP20v8Ao6PSv/B3qf8A8RX/AA6k/bS/6Oj0r/wd6n/8RX/DqT9tL/o6PSv/AAd6n/8AEV/w6k/bS/6Oj0r/AMHep/8AxFf8OpP20v8Ao6PSv/B3qf8A8RX/AA6k/bS/6Oj0r/wd6n/8RX/DqT9tL/o6PSv/AAd6n/8AEV/w6k/bS/6Oj0r/AMHep/8AxFf8OpP20v8Ao6PSv/B3qf8A8RX/AA6k/bS/6Oj0r/wd6n/8RX/DqT9tL/o6PSv/AAd6n/8AEV/w6k/bS/6Oj0r/AMHep/8AxFf8OpP20v8Ao6PSv/B3qf8A8RX/AA6k/bS/6Oj0r/wd6n/8RX/DqT9tL/o6PSv/AAd6n/8AEV/w6k/bS/6Oj0r/AMHep/8AxFf8OpP20v8Ao6PSv/B3qf8A8RX7I/wL/aP/AGxNR8S6b4H+Ot7osnheC3luG1XWr0CUTNIFCeXu5HlnOcdRX/DqT9tL/o6PSv8Awd6n/wDEV/w6k/bS/wCjo9K/8Hep/wDxFf8ADqT9tL/o6PSv/B3qf/xFf8OpP20v+jo9K/8AB3qf/wARX/DqT9tL/o6PSv8Awd6n/wDEV/w6k/bS/wCjo9K/8Hep/wDxFf8ADqT9tL/o6PSv/B3qf/xFf8OpP20v+jo9K/8AB3qf/wARX/DqT9tL/o6PSv8Awd6n/wDEV/w6k/bS/wCjo9K/8Hep/wDxFf8ADqT9tL/o6PSv/B3qf/xFf8OpP20v+jo9K/8AB3qf/wARX/DqT9tL/o6PSv8Awd6n/wDEV/w6k/bS/wCjo9K/8Hep/wDxFf8ADqT9tL/o6PSv/B3qf/xFf8OpP20v+jo9K/8AB3qf/wARX/DqT9tL/o6PSv8Awd6n/wDEV/w6k/bS/wCjo9K/8Hep/wDxFf8ADqT9tL/o6PSv/B3qf/xFf8OpP20v+jo9K/8AB3qf/wARX/DqT9tL/o6PSv8Awd6n/wDEV/w6k/bS/wCjo9K/8Hep/wDxFf8ADqT9tL/o6PSv/B3qf/xFf8OpP20v+jo9K/8AB3qf/wARX/DqT9tL/o6PSv8Awd6n/wDEV/w6k/bS/wCjo9K/8Hep/wDxFf8ADqT9tL/o6PSv/B3qf/xFf8OpP20v+jo9K/8AB3qf/wARX/DqT9tL/o6PSv8Awd6n/wDEV/w6k/bS/wCjo9K/8Hep/wDxFf8ADqT9tL/o6PSv/B3qf/xFf8OpP20v+jo9K/8AB3qf/wARX/DqT9tL/o6PSv8Awd6n/wDEV/w6k/bS/wCjo9K/8Hep/wDxFf8ADqT9tL/o6PSv/B3qf/xFf8OpP20v+jo9K/8AB3qf/wARX/DqT9tL/o6PSv8Awd6n/wDEV/w6k/bS/wCjo9K/8Hep/wDxFf8ADqT9tL/o6PSv/B3qf/xFftZfsq/tP/sh+C9H8beNv2gJdZtdZ1T+z4YdL1q/Lo/lPJubzAo24jI4OckV8N/+CbX7YXxN+HXhb4kaT+0xZWtj4r0Wz1a2gudb1LzYo7mFJVV9qEbgrgHBIz3r/h1J+2l/0dHpX/g71P8A+Ir/AIdSftpf9HR6V/4O9T/+Ir9p39kH9q39kj4dW3xY8U/tFyalbf2tBYxR6TrmoCdJXV3VwXCgAeUec5ziv2RPHniL4nfsyfDXx34uvWvNa1bw/bvfXTgBriVRsaRsADcxXccDGSaqqr41f8FNf2TPgnqs/h698YX3i3WLRzHc2Xhe1W88lh1DTO6Q5HQqJCQQQQK8If8ABZr9lvX9Sj0/xJ4c8deGYpGx9uutPhuYIx6sIJXk/wC+UavBfjXwl8RvC+n+NfAviGx1zQ9Vi82zvrOUSRyrnBwR0IIIIOCCCCAQRVftE/8ABSH9mj9nHXrnwbrutan4m8T2ZKXWk+HrdLhrR/7s0jukaNngruLr3Xpn4Uf8Fc/2VviT4gg8Na8fEfgae5kCQ3evW0Qs2Y9A00Mj+Xz1LqqjjLdcQzQ3EKT28qSxyKGR0bIYEZBBHUEVVXlpb39pNY3UfmQ3EbRyJkjcrDBGRyODX/BFXw14b8T+IvixaeJPDul6tFFZaU8aX1lHcCNt9yMrvBxwe3Xj0r/hU3wr/wCiaeFP/BNb/wDxFf8ACpvhX/0TTwp/4Jrf/wCIr/hU3wr/AOiaeFP/AATW/wD8RX/BZnwV4N8M/AnwTd+G/CWjaVPL4tCSS2NhHAzL9knO0lFBIyAce1fss/DH4a6h+zD8IL6++Hvhq5ubnwHokk002kwO8jtYwlmZiuSSSSSepNf8Km+Ff/RNPCn/AIJrf/4iv+FTfCv/AKJp4U/8E1v/APEV/wAKm+Ff/RNPCn/gmt//AIiv+FTfCv8A6Jp4U/8ABNb/APxFf8Km+Ff/AETTwp/4Jrf/AOIr/hU3wr/6Jp4U/wDBNb//ABFf8Km+Ff8A0TTwp/4Jrf8A+Ir/AIVN8K/+iaeFP/BNb/8AxFf8Km+Ff/RNPCn/AIJrf/4iv+FTfCv/AKJp4U/8E1v/APEV/wAKm+Ff/RNPCn/gmt//AIiv+FTfCv8A6Jp4U/8ABNb/APxFf8Km+Ff/AETTwp/4Jrf/AOIr/hU3wr/6Jp4U/wDBNb//ABFf8Km+Ff8A0TTwp/4Jrf8A+Ir/AIVN8K/+iaeFP/BNb/8AxFf8Km+Ff/RNPCn/AIJrf/4iv+FTfCv/AKJp4U/8E1v/APEV/wAKm+Ff/RNPCn/gmt//AIiv+FTfCv8A6Jp4U/8ABNb/APxFf8Km+Ff/AETTwp/4Jrf/AOIr/hU3wr/6Jp4U/wDBNb//ABFf8Km+Ff8A0TTwp/4Jrf8A+Ir/AIVN8K/+iaeFP/BNb/8AxFf8Km+Ff/RNPCn/AIJrf/4iv+FTfCv/AKJp4U/8E1v/APEV/wAKm+Ff/RNPCn/gmt//AIiv+FTfCv8A6Jp4U/8ABNb/APxFf8Km+Ff/AETTwp/4Jrf/AOIr/hU3wr/6Jp4U/wDBNb//ABFf8Km+Ff8A0TTwp/4Jrf8A+Ir/AIVN8K/+iaeFP/BNb/8AxFf8Km+Ff/RNPCn/AIJrf/4iv+FTfCv/AKJp4U/8E1v/APEV/wAKm+Ff/RNPCn/gmt//AIiv+FTfCv8A6Jp4U/8ABNb/APxFf8Km+Ff/AETTwp/4Jrf/AOIr/hU3wr/6Jp4U/wDBNb//ABFf8FqfCvhfwx4n+FMfhnw5pekrcWGpmZbGzjtxIRJb4LBAM4yevrX/AAqb4V/9E08Kf+Ca3/8AiK/4VN8K/wDomnhT/wAE1v8A/EV/wqb4V/8ARNPCn/gmt/8A4iv+FTfCv/omnhT/AME1v/8AEV/wqb4V/wDRNPCn/gmt/wD4iv8AhU3wr/6Jp4U/8E1v/wDEV/wqb4V/9E08Kf8Agmt//iK/4VN8K/8AomnhT/wTW/8A8RX/AAqb4V/9E08Kf+Ca3/8AiK/4VN8K/wDomnhT/wAE1v8A/EV/wqb4V/8ARNPCn/gmt/8A4iv+FTfCv/omnhT/AME1v/8AEV/wqb4V/wDRNPCn/gmt/wD4iv8AhU3wr/6Jp4U/8E1v/wDEV/wqb4V/9E08Kf8Agmt//iK/4VN8K/8AomnhT/wTW/8A8RX/AAqb4V/9E08Kf+Ca3/8AiK/4VN8K/wDomnhT/wAE1v8A/EV/wqb4V/8ARNPCn/gmt/8A4iv+FTfCv/omnhT/AME1v/8AEV/wqb4V/wDRNPCn/gmt/wD4iv8AhU3wr/6Jp4U/8E1v/wDEV/wqb4V/9E08Kf8Agmt//iK/4VN8K/8AomnhT/wTW/8A8RX/AAqb4V/9E08Kf+Ca3/8AiK/4VN8K/wDomnhT/wAE1v8A/EV/wqb4V/8ARNPCn/gmt/8A4iv+FTfCv/omnhT/AME1v/8AEV/wqb4V/wDRNPCn/gmt/wD4iv8AhU3wr/6Jp4U/8E1v/wDEV/wqb4V/9E08Kf8Agmt//iK/4VN8K/8AomnhT/wTW/8A8RX/AAqb4V/9E08Kf+Ca3/8AiK/4VN8K/wDomnhT/wAE1v8A/EV/wqb4V/8ARNPCn/gmt/8A4iv+FTfCv/omnhT/AME1v/8AEV/wqb4V/wDRNPCn/gmt/wD4iv8AhU3wr/6Jp4U/8E1v/wDEV/wqb4V/9E08Kf8Agmt//iK/bT8K+F9L/wCCpfwW0DTPDml2mmXV/wCFxPZQWcccMofVGV90YG07hwcjkda/4VN8K/8AomnhT/wTW/8A8RUXws+GFvKk9v8ADjwvHLGwZHTR4FZWHQgheDX/AAWy0zw5pPgH4YQ6dolla3lxrF8wlgtUjJjSGPcpYAHGXU46ce1fAWwl0v4F/DnTJ/8AW2nhPS4X+Yt8y2kYPJ69OtVVVVVVUro5ZUdSUOGAPQ4zz+BH51X7Xn/BKjwH+0B4tvvid8M/FKeCPFOqSGXVLeSz8+x1CY/emKqQ0MjdWZdwY8lQxZj8C/8AgizDoviu0174/fEjT9b0mxlEn9h6DBKi3uCCFluJNrKhxhlRNxB4detXulTw+G59E8KXFros0di1vpsq2YkhsmCbYmEIKhkQ7TsBUEDGRVn/AMEa/i/pnxe8PavrHxJ8M+KvCLa5Dca7cuJre9ltxKHmJhYOrM4BH+sPLZORmqqqqq/ag8H/ABT8efAfxd4X+Cfi278OeNbqyzpV5bTrA7OrBmhEp5i8xAyBwVKlgdwwa+FX/BFv4fJpaar8fPil4h13xFdZluodBkS2to5G5YGWaN5Jjkk7yI8nqtftu/8ABMGy/Zx8BXHx1+A/jPXr7S/DtxFNqWn6k6Nd2SNIqrcQTRKmQjsmVK7lHzbjtNf8EyP2ovEv7SnwJuLbx/fNfeLfBN6unX98+N99AyboJ3x/y0IDox/iMe4nLHH/AAUt/a71T9l/4Q2mj+A71IPHfjaSW20u4Khjp9vGF8+6APG8b0RM8bn3c7CD+xl/wS58OfGT4dWnxz/ad13xFe3/AIyj+36bpdvemOQ28nzJc3MzBneSUEOqggBWBYsWKp+2V+yD4p/4J7eMPC37Q37OXjfW4dDbUvs8U1xIrXGl3e0usUjKAs0EqLIMMuMKVfduGf2WPjzpn7SvwK8L/FywgitbnVLdotSs433C0vImKTR+u3cu5c8lGU96qv8Agtp/yQHwL/2OA/8ASOev2Tv+TV/g1/2T/Qv/AEghqq/4Le/8kq+Gn/YwXX/pOK/Z6/5ID8M/+xP0n/0jiqqqqqqqqqqqqqqqqqqqqqqqqqq/4LIf8l++Df8A2D3/APStaqqqqqqqqqqqqqqqqqqqqqqqqqq/aj/5TB/DL/sIeHv/AEOqqv8AgrB/yZH4w/7CGmf+lkVf8EuP+TEvhj/3Fv8A07XlVVa5ZX2p6JqGm6XrEulXt1ayxW1/DEkj2kjKQsqo4KsVJDAMCCRyCK+GH/BIPxP8RNSuviD+2F8ZNb1HX9SmMk9no92J52xwDNeTq+SRj5FTCgYD+n7RX/BGvwJp3gDVPE37PXi3xK3iLSrV7iPRdZliuotRCLkxROkaNHKwB2ltylsA7QSw/wCCPn7Vvi/xs2r/ALNnj7WJ9VGhaX/aXhu7uX3SQ2ySJHLaljyyqZY2QHlVDj7oUL+2V+0dY/stfAXXPicYYbnWWK2Gg2c2dtxfSg+XuAxlEVXkYZGVjIByRX7GH7EGvft0Xms/tM/tP+NNfvNH1K/kjtEhnCXOrTI37xi5BEVuhzGqIByGC7Ag3ftxf8E1dG/Zz8D/APDQv7NniPxDax+FbiGfU7C4u989km8Bbu2nUK4KOYyynJAJcMApFf8ABOj9qm//AGpPgSt74tuY5PGnhK4XTdcdcKbrK7obraOF8xcgjpvjkIwMAVX7a/8AyaL8X/8AsT9Q/wDRLV/wRU/5NX8U/wDZQLz/ANILCqqv+C1f/Jq/hb/soFn/AOkF/X7FH/Jovwg/7E/T/wD0StVVVVVVVVVVVVVVVVVVVVVVVVVVV/wQ6/5Gr4t/9g/Sv/RlzVVVVVVVVVVVVVVVVVVVVVVVVVVX/BbT/kgPgX/scB/6Rz1+yd/yav8ABr/sn+hf+kENVX/BZL/k0W0/7HCx/wDRNzX7Af8AyZp8Jf8AsX4//Q3qqrXNF0zxJomoeHdZtludP1S1ltbuFiQJIpFKOuRyMqSPxr4Df8EtP2Xfg9YrceKvDCfEbXSxZr3xFEJYEHZUtf8AVYxjlw7E85AOB+0V/wAE4/2bPjH4A1TSvCPwv8N+CfFKWrto+q6Dp8enrFcBfkEscIVJIyQAwZScElSpwa/4IyfGHxLoPxc8UfAHVryc6LrGly6la2krEi1v7d0V9qn7u+J23Y6mFPrX/BRP9o+//Zr/AGbtV17wxf8A2XxX4knXR9ClU/PbyyqzSTqPWOJHYHoHKZ64P/BLX9iHwF408CN+0x8bvDVr4qvdevJl8P2Grwi5t0ijcpJdyRvkSSPKsgG8EKE3DJYFf+Chf7Bfwi8cfBDxF8SPhh8PdF8M+NfB9jJqUb6NYpaJqNtCpaaGWOPajt5akq2C+UVQcMRX/BHj9ovWPiT8Kdb+Cni3U3vNR+HzwvpMszlnfTZtwWLJySIZEKg9lkjUcLVVX/BDr/kavi3/ANg/Sv8A0Zc1VV/wW0/5ID4F/wCxwH/pHPX7J3/Jq/wa/wCyf6F/6QQ1VVVVVVVVVVVVVVVVVVVVVVVVVf8ABcb/AJGr4R/9g/Vf/RltVVVVVVVVVVVVVVVVVVVVVVVVVfGX4jwfB/4UeLPilc6U+qReFdIuNReySYRNcCJCxQOQdpOMZwa/4fi+FP8Ao3bVf/Ckj/8AjFeJP2nLD9rX/goV8FPilp3hCfw3FF4n8N6d9invBcsTFqasX3hF4PmYxjt1qq/4Ljf8ip8I/wDsIar/AOi7avhP/wAkr8G/9i/Y/wDpOlVVVVVVV/wUTP7TPwe/bI8ffF74et8QfCvh3VjpjW3iDSftNvZXXl6bbRurSx4jbDxupRj/AAnIxXwq/wCCwP7UvgYQWXjqPw94/sY8B21G0+x3ZUdlmt9q57ZeNz+PNfCn/gsl+zd4yMFl8StA8R+AbyQ4eaWL+0rNP+2sI80/jCB718Nvjj8HfjHZi9+F3xN8OeJlC7ni0/UEkliHH34s70PI4ZQeaqqqqqqqv+Ch/jTQ/BH7G3xNu9bljH9q6SdKs4mI3S3FywiQKD1K7i5x0CMe1f8ABETwRqml/Df4l/EK5ilSy8QavZWFqXGFc2cUruyjuM3arnplSOoNf8FS7+++Kv7eOjfC1biXytPstI0SCNSPle6fzWZRz8x+0qMkfwjsBWm6bY6PptrpGl2qW1nYwJBbwoMLFGihVUewAAr/AIKL+ELXxp+xf8T7GeASPp2lrqcLcZje1mSYsMn+6jA+xI5zX/BEXxZcX/wq+JHgiS53xaNr1rfRRl8mP7VAyHjsD9l+mc+9VX/BbT/kgPgX/scB/wCkc9fsnf8AJq/wa/7J/oX/AKQQ1Vf8Fvf+SVfDT/sYLr/0nFfs9f8AJAfhn/2J+k/+kcVVVVVVVVVVVVVVVVVVVVVVVVVVX/BZD/kv3wb/AOwe/wD6VrVVVVVVVVVVVVVVVVVVVVVVVVVVX7Uf/KYP4Zf9hDw9/wCh1VV/wVg/5Mj8Yf8AYQ0z/wBLIq/4Jcf8mJfDH/uLf+na8qqqq8XeKdE8D+FdY8Z+JLxbXStCsJr69nYgCOGJC7tyR/Cpr/gjv4T1Lxb+1V4k+IkFm8GmaB4fuXnaP7iTXUyLFCTjuomYdP8AVV/wXB8ZXT6r8LPh5DOy20Vvf6pcRg8SOzRRREjPVQkw6fxn8P2TPBVp8PP2ZPhd4RtLcQmz8K2L3CAY/wBIlhWWdse8skh/Gvif4QtfiD8NfFfgO8gE0PiLRLzTnjOPmE0LR9yB/F6iv+CJviy4sPjx458E/adlrrPhQXrRl8CSW1uokTA7kLcyn6Zqq/bX/wCTRfi//wBifqH/AKJav+CKn/Jq/in/ALKBef8ApBYVVV/wWr/5NX8Lf9lAs/8A0gv6/Yo/5NF+EH/Yn6f/AOiVqqqqqqqqqqqqqqqqqqqqqqqqqqqv+CHX/I1fFv8A7B+lf+jLmqqqqqqqqqqqqqqqqqqqqqqqqqqv+C2n/JAfAv8A2OA/9I56/ZO/5NX+DX/ZP9C/9IIaqv8Agsl/yaLaf9jhY/8Aom5r9gP/AJM0+Ev/AGL8f/ob1VVVfHH4s+HfgX8JfFHxX8TzxpZeHdPkuFjd9v2ib7sMK/7UkhRB7sK/4IxfC3XPE/xj8Z/HjVIGbTtF0x9OiuGQKJr+6kSR9vb5YkbcB085PWv+C4niSeTWfhN4PjmIhgtdTvpYwD8zO1vGhPbgRuB/vH1FfsmeHLbwn+y/8J9BtVQCDwdpjybBw0slskkjDp1d2P41qFhZ6rYXOmahAs9rdwvDPE3R0YEMp9iCRX/BIHUJ/C37Y+s+F/tDSJf+GNQsXwCFdop4JA2M8H902DzjcR3zVVX/AAQ6/wCRq+Lf/YP0r/0Zc1VV/wAFtP8AkgPgX/scB/6Rz1+yd/yav8Gv+yf6F/6QQ1VVVVVVVVVVVVVVVVVVVVVVVVVf8Fxv+Rq+Ef8A2D9V/wDRltVVVVVVVVVVVVVVVVVVVVVVVVVX1jY6nZzadqVnBd2twhSaCeMSJIp6qynIIPoa/wCC1PhXwv4Y8T/CmPwz4c0vSVuLDUzMtjZx24kIkt8FggGcZPX1r9sjQNC8Of8ABVX4H6d4e0Ww0u1Op+FXMFlbJAm46q2W2oAMnA59qqv+C43/ACKnwj/7CGq/+i7avhP/AMkr8G/9i/Y/+k6VVVVVVVTokiGORVZWBDKwyCO/FfFb9hT9k/4yma58YfBjQodRn5fUdIiOm3LP/fZ7cp5jf74bt6Cvit/wRI8PXQnvvgn8ZL2wfJMWm+JbNbhCT2+0QBWUDn/lk56c9z8S/wDgnN+2b8FLv+14/hrf67b2bb4dV8JXH28qV53LHHi4THXcY1HvwcfDr/goZ+2h8D74aHJ8TdV1WLT28ubSPFtv9uKleNjNL+/THTasi9h2r4U/8FuNJn8mx+N3wauLVsgSal4YvBKpz/07TkFQP+uzdenHPwW+M3gP9oD4dab8Ufhtf3N3oWqNIkL3Ns0EivG5R1ZG6EMpGRkHsTVVVVVarqumaHpl3rWtajbWGn2EDz3V1cyiKKCJAWZ3ZiAqgAkk8ACv2nfjD4+/4KbftIaH8APgNFI3gbQrp2tbuVGSKTHyz6pcZAKxqh2xqfmw2AN8u2vgt8I/CfwI+Fvh34TeCYGTSvDtmII3cDfcSElpZnxxvkkZ3bHGWOMDAr9tu1Og/wDBVa21jUHMdtdeI/C94HMZ4jSGzjYj+9zE3T6dRVftxXcFl+yD8XZrhyqt4TvYwQM/M8ZRf1YV/wAENtLni0b4xa027ybq60a3TKEDdEt4zYPQnEy8duPWqr/gtp/yQHwL/wBjgP8A0jnr4WeHv+Cx8/ww8HzfC+41MeDZNBsW8OhNU0FANPMCfZsLK4kA8rZw43Dvzmv+EZ/4Lkf8/Oq/+Dbw7/8AHK/4Rn/guR/z86r/AODbw7/8cr9tHSf+CgOneF/Dj/tjy3j6M9/KNGE97pk4+0eX8+BZsWHyf3uK/Z6/5ID8M/8AsT9J/wDSOKqqqqqqqqqqqqqqqqqqqqqqqqqr/gsh/wAl++Df/YPf/wBK1qqqqqqqqqqqqqqqqqqqqqqqqqqr9qP/AJTB/DL/ALCHh7/0Ov2mtB/4Ky3fx18WXH7Ps+or8PnuYv7DEWo6JGoj8iPfhbhxKP3nmfeGfwxX/CM/8FyP+fnVf/Bt4d/+OV/wjP8AwXI/5+dV/wDBt4d/+OV+1Lon/BUiy+DGrXH7T09+3w+W4thqAm1DRpV8wzL5Py2rmU/vNnQY9eM1/wAEuP8AkxL4Y/8AcW/9O15VVVV/wUr/AGxNR+MviGD9jD9nF5Nfl1HUI7XxBc6afMN/dCQbNPhI4Kq4VpHBxuULkBXz+w7+yrpv7JvwTtPBs7wXXinV5Bf+JL6LlZLoqAIkPXy4lARfU72wN5Ff8FudLuIvjN8PNaYHyLrwxLboSpA3RXLs3PQ8Srx2/Gvg3dW9/wDCDwNfWknmQ3HhvT5ImwRuVraMg4PI4I61e3cFhZz3905WG2iaSQgZwqjJOPoK/wCCMGlz3v7Vmt36bhFp/gm8d22Eglrq0QLnsTuJ/wCAmqr9tf8A5NF+L/8A2J+of+iWr9hz/go3on7IHwl1b4aal8Kb3xPLqXiKbV1u4NXW1CCS3t4fL2mJ8kfZyc5/i6cV/wAPxfCn/Ru2q/8AhSR//GK/4fi+FP8Ao3bVf/Ckj/8AjFfCz/gsb4a+KHxP8H/DS3+A+p6fL4t16x0hLt/ECSC3a5nSESFRCNwXfnGRnHUV/wAFq/8Ak1fwt/2UCz/9IL+v2KP+TRfhB/2J+n/+iVqqqqqqqqqqqqqqqqqqqqqqqqqqqv8Agh1/yNXxb/7B+lf+jLmqqqqqqqqqqqqqqqqqqqqqqqqqqv8Agtp/yQHwL/2OA/8ASOevhl/wV+8N/B/4XeBvhZc/AvUtUl8K+FdK057xNfSJbgxWUKlwhhJUHHTJ+tf8PxfCn/Ru2q/+FJH/APGK/wCH4vhT/o3bVf8AwpI//jFftrf8FKdD/a1+DkPws074S33huWLWrfUftk+sLcqREkqlNgiU5Pmdc9q/YD/5M0+Ev/Yvx/8Aob1VVWq6rpmh6Zd61rWo21hp9hA891dXMoiigiQFmd2YgKoAJJPAAr9pH43/ABN/4Kd/HfSf2fv2fbG8j+H2jXRmFxMjRxzbTsk1O6HGyJFYrHGfm+bpvkCL+z98C/Bf7OPwo0T4T+BYCbLSoy1xdyIBLfXLcy3EmOrM3boqhVHCgD/guLoEsXiX4S+KFjcxXVjqdmzDJCtE9u4B44JEp784PpX7LetWniL9mj4U6zYspjufBmlEgHOxhaRh1z6qwKn3FXt5a6dZz6hezrDb2sTSzSN0RFGWJ9gAa/4JA6dL4q/bH1vxStqY4rDwzqF6w3kiNpbiCMLnHJxK3p0J7VVV+xX+xVqn7ZmqeK9M0z4g2vhY+Fre1md59Oa78/zmkAAAkTbjy/fOa/4cc+Kv+jitJ/8ACbk/+P1/w458Vf8ARxWk/wDhNyf/AB+v+HHPir/o4rSf/Cbk/wDj9ftvfseeI/2rPhF4R+Gvh3xlpuh3HhrUo7uS6vbd5EmVLZ4cBUOQSXB/Cvg34Huvhj8IPA3w1vr+K9ufCfhvT9JmuYVKpO9tbRws6g8gMUJAPrVVVVVVVVVVVVVVVVVVVVVVVVVV+3/+wl4s/bI1jwXqXhnx5pHh5fC9teQzJfWskpmMzREFdnTHlnr6iqqqqqqqqqqqqqqqqqqqqqqqqqr/AILjf8jV8I/+wfqv/oy2rX/+CJnj/UvEN1qFp8ePD/2S8uXlcyaNMskYdySAokIbAP8AeGfav+HHPir/AKOK0n/wm5P/AI/X/DjnxV/0cVpP/hNyf/H6/bV/YO1b9jTS/Cmp6n8SbPxSPFNxdQokGmNaeR5KxkkkyPuz5ntjFfCf/klfg3/sX7H/ANJ0qqqqqqqqqq+I3wX+EnxfsTp3xP8Ahv4d8TxbNqNqWnRzSRD/AKZyEb4zyeVIPNfFf/gjl+zP40868+HOr+I/AF85JSO3n/tCzXPXMU58z6ASqBk8dMfsgfADUf2YvgXpPwc1PxJba9NpN5eSrfwW7QLKk07yLlCSVIDYIyenWqqqqtV1XTND0y71rWtRtrDT7CB57q6uZRFFBEgLM7sxAVQASSeABX7TX7Vnxa/4KD/E9f2Uv2VLGceCZrjF9qBDwnU442G+5uX/AOWNkh2kIRuc7SQWZI1/ZD/ZG8A/sjfDr/hE/DTrqmu6iwm1zXpbdY5r6UDhQBkpEmSETJxkkkliTX/Baf4Rapo3xA8D/tA6PbslnqVl/Y19cRAjybuB3lgZj/eeN3A/69z04z+zV8bvD/7RPwT8L/FXQbuKR9UskXUoEbJs75FAuIWHYq+ccDKlWHDCv+Cv3xy0TwL+zkfg9bajE3iL4g3cK/ZVf95DYW8qzSTEDkK0kccYz97c+M7Wx/wSg+Duo/Cv9k/T9a1yya31Lx5qMmubJFw62zokdvn2aOISj2mqq/4Laf8AJAfAv/Y4D/0jnr9k7/k1f4Nf9k/0L/0ghqq8cfDH4a/E60trD4k/D3wz4strKQyW0OuaTBfpA5GCyLKrBSRxkVp2nafpGn2uk6TY29lY2UKQW1rbxCKOCNAFVERcBVCgAADAAAqqqqqqqqqqqqqqqqqqqqqqqqqq8ZfCD4S/EfULLVviF8L/AAj4ovtNXbZXOs6Jb3stsM7sRtKjFBkA4GOeaqqqqqqqqqqqqqqqqqqqqqqqqqq1X4QfCXXfGFr8Q9b+F/hHUPFdi0bWuu3WiW8t9A0f+rKXDIZFK9sMMdqqq/4Kwf8AJkfjD/sIaZ/6WRV/wS4/5MS+GP8A3Fv/AE7XlVVV+3j/AMFCfEmq+Jpf2Vf2R5J9X8UapOdM1XXNLBllSZztNpYlT/reqvKPuchcMCyfsAf8E/tF/ZY0YePvHbWur/EzV7ULNKqK8WixsPmt7du7nOJJR1xtX5cl6/4LNfBu/wDGnwO8OfFrR7Lz5vAGpyJf7V+ZLK7CIznHULNHAPYOT61/wTF+Oui/GP8AZW8M+H01BG8QfD+2j0LVLQv88ccQ22sgHXY0AQA9NyOP4TX7fHxz0T4Efsw+M9ZvNRhi1nX9Om0fQrYviSe6uEMe5B1PlozSn2T3Gf8Agij8HdR0fwb44+OGrWTRReIriHSdHd1wXityzXDr6qZHjXP96FhVV+2v/wAmi/F//sT9Q/8ARLV/wRv8D+CvE37MPia+8R+ENE1W5j8eXcSTX2nxTuqCxsSFDOpIGSTjpkn1r/hU3wr/AOiaeFP/AATW/wD8RX/CpvhX/wBE08Kf+Ca3/wDiKtPhj8NbC6hvrD4e+Gra5tpFkhmh0mBHidTlWVguQQQCCOhrxR4N8H+ONPj0nxr4U0fxBYwzCeO11Sxju40lAKhwkgIDBWYZxnDEdzWl6Vpeh6bbaNomm2un2FnEIra1tYViihRRgKiKAFUDgADAqqqqqqqqqqqqqqqqqqqqqqqqqqqvCvw4+HngSS4l8D+A/Dvh6S8VVuG0rS4bQzBc7Q5jUbgMnGemT61VVVVVVVVVVVVVVVVVVVVVVVVVV4p8EeC/HVnDp3jbwjoviC1t5fNhg1XT4rtI3wRuVZFIDYJGRzgmk+Efwoiijhi+GPhNY4kVI0XRbcBFUAKoGzgAAADsBX/CpvhX/wBE08Kf+Ca3/wDiK/4VN8K/+iaeFP8AwTW//wARX/BXnwJ4I8Ofso2moeHvBuh6XdHxbZIZ7LTooH2mG4yu5FBwcDj2r9gP/kzT4S/9i/H/AOhvVVWu65ovhjRb3xF4i1W103S9Mt3uLy8upRFFbxIMs7seFUAEkmv2uv21PEv7bnxMtvgH8KvGuk+CfhabvyptT1/VY9Jg1TYwJuruSVl2wLjckHLNgMVLlVT9lZv2C/2TfACeEvA37RvwputVvljfXNdn8YaeLjVJ1B+Y/vjsjXLBIxwoJ6sWZvD/AO0h+zt4t1m08N+Ffj58OdZ1a/k8q0sNP8U2dxPcP/dSNJCzHjoAa/4Ke/s+6p8ev2ZL+Xwrp8l54k8E3a61YW8KFpLmNEZLiFQOSTE5cKBlmiUDrX/BJz9sLwNqPwptf2cviB4osdH8R+GJpBoJv7hYV1KzlcyeWjMQDLG7uNnUoUK52tj/AIKL/tkfD/4NfA3xF4D8M+LtO1Hx74x0+XTbGwsbpZZbKGZdkt1LsJ8oLGzbM4LOVxkKxX/gjj+z5q3w/wDhPr3xr8T2D2178QJYY9JiljKuunQbiJRnkCWR2I45WJGBw1VVf8EOv+Rq+Lf/AGD9K/8ARlzVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX7Vn7X/AMK/2R/CFvr/AI7nn1DV9ULJpGhWTL9pvWUfM3JxHEuRukPAyAAxIB/ag+Of7Qf/AAUG1fR/Feifs/auNF8Jw3MVmvh3S7zUiiyshkM86qVYjy16ImOc5zx+z/8A8FgrLWPF1v8AD39qD4eweCL6WZbd9asfNS2tZTxi4t5syQJ0y+98ZyQACwhmhuIUnt5UljkUMjo2QwIyCCOoIqv+C43/ACKnwj/7CGq/+i7avhP/AMkr8G/9i/Y/+k6VVVVVVVVfGjx8vwq+EHjX4lmJZW8L6Be6lHEw/wBa8MLOif8AAmUD8a/Yc/b3/bO+KHx98GfBvU/HGmeKNN1y/Ivm1jSYjJb2caNLcMkkHlNvEUb7S5YbiowRxX7d37VUX7JvwQuPF+lR2t14s1q4/s/w7aXA3IZypZ5nUHJjiQFj2LFFJG7NfEL4tfE34seIJvFPxI8d614i1OeYyme+u2k8tu3lrnbGo6BUACjgACv+Cdv7fPxE+FnxR8P/AAm+J3i+/wBd+H3iS7j06MalcmZtEnkYLFLE7nKw7yA6Z2hWLgArhqqqqq/b9/Z3+Mn7T3wfsPhn8H/GWj6Cz6ulxrKarez20N7apG+2ImGKQviUxvtIC5QHqBXhX/gkr+3L4ENw3gf46+CfDxvAouDpXifVbTztudu/y7QbsZOM9Mn1r/h2z/wUl/6Ov0n/AMLvW/8A5Gr9hz9kf9rf4B/FnVvGHx7+N1j4x8P3nh2awtrCDxLqOoNHdtcW8iy+XcwogASKVdwO4b8YwSR8X/hH4F+Onw71f4YfEbSBqGiazDslVTtkhcHKSxNzskRgGU+o5BBIMf8AwTc/bo/Z08R6nJ+yt8dLVtC1OTkR6pJps0ijO0zwMrQsyjgOrE8nAUHFfBv/AIJUfErxt8Sofi5+2z8T4/Fs6SLLJo9vfTXst6V+7HcXMgXZEMY8uMEEHAZat7a3s7eK0tII4YIECRRRoFVFAwFUDgAAAYqq/wCC2n/JAfAv/Y4D/wBI56/ZO/5NX+DX/ZP9C/8ASCGqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr/grB/yZH4w/7CGmf+lkVf8ABLj/AJMS+GP/AHFv/TteVVV+0n4H+IvxL+Bni/wD8JvE1v4f8Va5Yi2sNSuLmS3SDdInmZkiR3TMQkXKqTlh06jw5/wR8/bO8H6mmt+EfjB8PdE1GNGRLvTvEGp20yqwwwDpZhgCOCM81/w7Z/4KS/8AR1+k/wDhd63/API1fs8/sIft3fDb42eD/HfxJ/aO03XfDGi6is+p6bH4w1a6a5iCkFRFLAqPyRwxArxD4f0TxZoOoeGPEml2+paVqttJa3tncJvjnidSrow7ggkV42/4JaftFfBX4jXfxA/Yq+MK6ZZ3DMIbK51OWxvbaNjkwGQK0dxEMDmQqemQSN1aF/wTB/ar+P3jbTvFX7ZfxyFzpunnYLW31GTUbwxZy0UW5Vht1bAO5d3PJUmvBPgnwr8OPCOk+BPBGi2+k6FodqttY2cAO2KNenJySTySxJJJJJJJNVX7a/8AyaL8X/8AsT9Q/wDRLV/wRU/5NX8U/wDZQLz/ANILCqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr/gsl/yaLaf9jhY/+ibmv2A/+TNPhL/2L8f/AKG9VVfHb4P6H8ffhL4i+EPiTU77T9N8RwRw3FzY7fOjCSpICu8Mucxgcg8E1/w5L+AP/RV/iB/31af/ABmv+HJfwB/6Kv8AED/vq0/+M18GP+CTvwZ+CXxS8N/Ffw/8R/Gl/qPhm9F3b214bbypWAIw+2INjnsRVftC/wDBJz9n/wCNvim98c+GNW1XwBrepymW9TS4Y5rKeU8tJ9nbGxieTsdVJyduSTXwe/4I5fs/eANftvEXxD8Ua38QHs3DxafdQpZWTkHI82NCzyAYHy+YFPOQQcVb21vZ28VpaQRwwQIEiijQKqKBgKoHAAAAxVVX/BDr/kavi3/2D9K/9GXNVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVeL/CsP7Z3/AAVl1PwH4/kkn8K+Fb2W0ksWcgGy06IkwLznbLcbi2DkCZyOgrR9G0jw9pVroWgaXaadp1jEsNraWkKwxQIowFRFACgDsBX/AAVY/Z18F/Ef9nLXvi3HotrB4y8CxxXdvqUcSpLcWvmKk0EjdWQI5dQckMgAxubP/BLL4kax8Rv2OfDCa7dPc3XhW7udDWaQks0MLBoR9EiljjHsg96r/guN/wAip8I/+whqv/ou2r4T/wDJK/Bv/Yv2P/pOlVVVVVVVX7b+i3/iD9kT4t6bpiM9wPCt5OFUZLLEnmsAO5KoeK/4J/8Axz8D/s7/ALTfh/4ifEeWaDw8LS6sry7ht2na0E0RVZdiAswDBQQoJ2scAng/8FKf2tvDv7U/xh01fh3qFzdeB/CFi1rpc80Lw/bJ5WDXFwsbgMgbbEgDAHEQJAzgf8EtP2HPhp8evDXin4t/G/wqdd0KC6Gk6JYSXMtujzKoeecmJ1Y7Q8aLzjJfuox+018O9H+DX7RHj34ceE7qY6Z4b1+eDTneUtJHEG3RqX6llBCk9SVzXgjWZ/Efgrw/4guhifU9LtrqQZHDSRKx6ADqfQVVVVVVVVVVVX/BbT/kgPgX/scB/wCkc9fsnf8AJq/wa/7J/oX/AKQQ1VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVf8FYP+TI/GH/YQ0z/0sir/AIJcf8mJfDH/ALi3/p2vKqqqqqqqqr9tf/k0X4v/APYn6h/6Jav+CKn/ACav4p/7KBef+kFhVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV/wAFkv8Ak0W0/wCxwsf/AETc1+wH/wAmafCX/sX4/wD0N6qqqqqqqqqv+CHX/I1fFv8A7B+lf+jLmqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqYqqlmYAAck8Yr4l/tifsvfCFXTx78cPC1ncxA77K1vPt10uPWC3DyDPbK8/ga8QftZ+Afh1/wULvv2pfg8L7WvCd5qZuLu1ntPsktxFcW4ivFVWP3tzSSIW2/Nt3Drn4e/tn/ss/E7w7D4j8N/HTwhBFIgaS11TVItPurc9xJDOyuuDkZxtOOCRzX/AAUv/b8+Fni/4W3/AOzr8C/Etv4u1PxNPFDrGp6bmW1trdJFk8qKUfLNJIyouY9yhdwzuIA/4J//AAN1j9n79lrwn4L8UWbWviG+Euq6vbum1oJ7htwiYf3kjESNnPzIe2AK/wCC43/IqfCP/sIar/6Ltq+E/wDySvwb/wBi/Y/+k6VVVVVVVVXlpa6haTWF9bx3FvcRtHNFIu5ZEYYZSD1BBIIr9sb/AIJ3fF79nzxtqeqeBfCGr+Kfh3dzvLpeo6dbvdSWUTHIgulQFkZMhfMI2uMHIJKr8C/2Svjz+0N4rtPDPgL4f6qLeSUJd6xfWklvY2CZG55ZmG3IBzsGXbHyqTXwh+F3hX9mX4G6T8O/Ctpf3+m+D9LkZ/s1qZbrUJQGlmkWJMlpZZC5CLnlgo7V4t8LfFL4zftLXGmeKvCWsaJ4t+JHixnNhqFlLDLBLeXJ42OFIVN+OgAC9gKsrOCws4LC1UrDbRLFGpOcKowOfoKqqqqqqqqqqr/gtp/yQHwL/wBjgP8A0jnr9k7/AJNX+DX/AGT/AEL/ANIIaqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv+CsH/ACZH4w/7CGmf+lkVf8EuP+TEvhj/ANxb/wBO15VVVVVVVVVftr/8mi/F/wD7E/UP/RLV/wAEVP8Ak1fxT/2UC8/9ILCqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr/AILJf8mi2n/Y4WP/AKJua/YD/wCTNPhL/wBi/H/6G9VVVVVVVVVX/BDr/kavi3/2D9K/9GXNVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVarrGkaFZNqOt6rZ6faocNPdTrCi/VmIArxT+2F+yr4MtJrvX/2h/ACfZ13PBba9Bdz4xniGFnkPHYKc18V/wDgtb8LdAvpNN+D/wALNZ8XLG206jql4NLgb/ajQJJIw7fOIz7cc/EX/gst+0v4psJNO8C+HfCXgvzFI+2QWz31yh/2TMxiH4xmviN+0J8c/i67/wDCy/i14p8QxSHJtb3U5Gt1/wB2EERr+Civh98D/jH8WZUi+Gnwu8UeJQ77PO03SpZ4kPT55FXYg92IAr4ff8Eg/wBrvxlFHdeJbLwv4KhbBKaxqomm2+oS1WUZ9mZT64r9s39izXP2Nr3wjp2u+PLHxLP4qtrqb/RLF7dbbyWjBGWYl8+YOcDGK/Zz/wCCa/7Nn7OetW3jDTtK1DxZ4os2D2uq+IJEm+yP/ehhRVjRumGIZ1PRhVV/wXG/5FT4R/8AYQ1X/wBF21fCf/klfg3/ALF+x/8ASdKqqqqqqqq+LXxt+E/wJ8ODxX8XPHel+GtNdykL3bkyXDgZKxRIGklYDkqisQOeleJ/+Cz/AOy7pF49n4f8J+P9eVGx9qi0+3t4XHPK+ZOH9OCg6/hX7N/7Q3g39qD4YW3xW8CaVrOnaXcXc1oINWiijnWSIgMcRySLtyeDuz7CtX+M3wFi8U6R4S1j4n+B38SXd8kGl6ZLq1tJdtct8qiOLcXDEnbkDqQO9VVVVVVVVVVVX/BbT/kgPgX/ALHAf+kc9fsnf8mr/Br/ALJ/oX/pBDVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV/wAFYP8AkyPxh/2ENM/9LIq/4Jcf8mJfDH/uLf8Ap2vKqqqqqqqqr9tf/k0X4v8A/Yn6h/6Jav8Agip/yav4p/7KBef+kFhVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV/wWS/5NFtP+xwsf8A0Tc1+wH/AMmafCX/ALF+P/0N6qqqqqqqqqv+CHX/ACNXxb/7B+lf+jLmqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq8vbPTrWS91C7htreFd0k00gREHqSeAK+I/7dH7JHwrWVPFXx38MSXMWQ1ppNwdUnVh/C0dqJCh/3sY6nA5rxj/wWY/Zc0OB18K+H/G/iW5BwgjsIrSI+5eWQMP8Avg14j/4LY+Idbm1fTdK+CsOjaVd6Ne29nOmtNNewXrREW8+/y0QIr/eQKW5BDfLhvF3xB+IfxI1BL/x5448ReKb7IVJtX1Oa+l9AA0jMfQV4e/Z9+PPi4xjwt8FPHerCU4RrLw7dTKeAc7lQgDBBz0wc18D/APgkV+0t8Tmh1H4itp/w10aTBL6li7vmX1W1iYY9xLJGfY14N/4Ij/CTTrqCbx58Z/FOuxR4MkOm2EGmiQgDjLmYhSQcgc4PBB5r4YfsPfspfCGGD/hD/gj4be8twNuoarbf2lc7u7CW43shP+ztHYADioYYbeFLe3iSOOJQqIi7QoAwAAOgx2qv+C43/I1fCP8A7B+q/wDoy2qqr/guN/yKnwj/AOwhqv8A6Ltq+E//ACSvwb/2L9j/AOk6VVVVVVVVeNPFmjeAvB+ueOPEVwYdK8PabcahfSAZKQwxtI5A7nap4r4+fHL4kftT/F2+8e+LJrq+1DVbn7PpOlQbpVsoC+IbWBBzgbgOBl2JY5ZjXxP+BXxe+C0GizfFfwBqvhY+IYZJdOi1KMRSzJGVDkx53oQXXhwp5r/gkXFHN+xhp8Msauj69qSsjDIYF1yCPSvHNlefA39oTxBp3h6Vorv4f+M7mKxcNgpJZXjCM5BPIMQ5rStStdZ0uz1eyYtb31uk8JJByjqGHQkdCOhqqqqqqqqqqqv+C2n/ACQHwL/2OA/9I56/ZO/5NX+DX/ZP9C/9IIaqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv+CsH/JkfjD/ALCGmf8ApZFX/BLj/kxL4Y/9xb/07XlVVVVVVVVV+2v/AMmi/F//ALE/UP8A0S1f8EVP+TV/FP8A2UC8/wDSCwqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/4LJf8mi2n/Y4WP/om5r9gP/kzT4S/9i/H/wChvVVVVVVVVVV/wQ6/5Gr4t/8AYP0r/wBGXNVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVXxM/aJ+BXwct5pvid8WfDHh+SBSzWt1qCfaWA/uW6kyufZVJr4mf8Fm/2c/C8ktn8N/CXinxvOhOy4Ma6ZayfR5cyj8YRXxf/AOCwv7TPj2GbTfh3YaD8PLCTIWWyh+3Xu09QZpwUHHdIlYcnPTGlfB79uP8Aa0uI9dHhj4lePIrht0Op6zPN9kOe0c90ywgAY4VsAY6DFfDz/gjR+034meKfx34g8I+DbUgeYkt21/cpn0jhUxnH/XUduteE/wDgiJ8KrGSN/HHxu8U6yq43ppmnQadu6d3M+B1/Mc8c/D3/AIJnfsZfDxRJD8IrfxDd7NrXPiG7lv8AcP8Arkx8oH3CA14E/Z8+BPwvvTqfw6+Dvg3w7fHreadosEE+PTzVXfj2ziqqqrxt4z8M/DrwhrHjvxlqf9n6HoFnJeahdeS8vkwxrudtkas7YA6KCfav+Cqn7TPwP/aT8QfDq++Cvjf/AISKHQbPUI9Qb+zbqz8lpXgKDFxFGWyEb7ucY5xxXwL/AGo/gR+0t/bf/Ck/Hf8Awkf/AAjn2f8AtP8A4ll3Z/Z/P8zyv+PiKPdu8mT7ucbecZGar/guN/yKnwj/AOwhqv8A6Ltq+E//ACSvwb/2L9j/AOk6VVVVVVVVftxWWo3/AOyD8XYNKYiZfCd7K2M/6qOMvL0z/wAs1f8A+t1r/glJN4Hh/bM8NDxmbYTvp16uhG4A2jUDH8mM8BjH5wXvuKgckV/wXG/5Gr4R/wDYP1X/ANGW1f8ABJ/WNK8PfsOw6/rmoQWOnabrGqXF3dTvsSCJCGd2J6AKCSfavjJ40tviT8XvHPxEs4TFB4p8SahqsUbDBRbi5klAxzjAcCvh/pV1ofgLw3ol8uLnT9ItbeYYIw6QqrcHnqDVVVVVVVVVVVf8FtP+SA+Bf+xwH/pHPX7J3/Jq/wAGv+yf6F/6QQ1VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVf8FYP+TI/GH/AGENM/8ASyKv+CXH/JiXwx/7i3/p2vKqqqqqqqqr9tf/AJNF+L//AGJ+of8Aolq/4Iqf8mr+Kf8AsoF5/wCkFhVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV/wWS/5NFtP+xwsf/RNzX7Af/Jmnwl/7F+P/wBDeqqqqqqqqqr/AIIdf8jV8W/+wfpX/oy5qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqp3SNDJI6qqglmY4AA6mviR+2n+yp8JxLH40+OnhaK5gyJLOxu/7QuUI7GG2Ejqc+oFfGH/AILW+AdHmk0z4H/C7UfEbrlRqeu3AsIM9ikKB5HX/eMZ9q8df8FZ/wBszxizrpHi/QvCMEhOYdD0WI8Y6B7nznH1DA+9HxD+39+0Kvl2+o/GzxnaXSnMdr9vltNpOCSqfulXPBOAOgr4ff8ABKL9svx5JHLq3gzSfCFrNhvtPiDVo1OD1Jig82UH2ZRXwt/4InfDLSooLz4w/FfXfEF0AGez0O3j0+AN3UvJ5juvuPLP0r4U/sN/sp/BmSC88FfBjQm1GAfJqOqRtqNyrf3le4LlG/3NtVVVVVVV498D+HPiX4K1v4f+LrR7rRfENjLZX8CStE0kMi7WAZSGUkHqDmtV/Yq/YJ0vU7vTJPgp4jkezneFmHie6AYqxBI/ee1fsmeAv2e/gh4k1Pw38Efh9q2gy+MfJ/tCW71WS8D/AGVJnjwJGbbgSSDjGdwz0qq/4Ljf8ip8I/8AsIar/wCi7avhP/ySvwb/ANi/Y/8ApOlVVVVVVVWsaRpviDSL3QdZtEu7DUbaS2ureT7ssUilXU47FSR+NftYfsx+Pv2RfjBc+F9SjvV0mS5e68La6mVF7bK+UdXX7syZUOvBVsEfKyk/FX4/fGP44WmgWfxa8e3/AIoHhiCWDTJb9UaaJJCpfdKFDyklF5kZjx15NJ8d/i9F8Jh8C7bx5qNv4E+0vcyaLb7IoppGYMTKygPKNyqQrsVBAIAxX/BPH9lrX/2kPjzot9daTMfBPhC9h1DxBfPGfKby23x2oPGXlZQpAOQm9u3NVVVVVVVVVVV/wW0/5ID4F/7HAf8ApHPX7J3/ACav8Gv+yf6F/wCkENVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX/BWD/kyPxh/2ENM/9LIq/wCCXH/JiXwx/wC4t/6dryqqqqqqqqq/bX/5NF+L/wD2J+of+iWr/gip/wAmr+Kf+ygXn/pBYVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVf8ABZL/AJNFtP8AscLH/wBE3NfsB/8AJmnwl/7F+P8A9Deqqqqqqqqqr/gh1/yNXxb/AOwfpX/oy5qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqrxZ458E+A9OOr+OfGOh+HbFQc3WrahFZxDHX55GUfrXxZ/4Kq/shfDOKe30bxhe+OtTiyBaeHLRpUz2zcSbISvqUZiPQ9K+KH/AAWt+Lutmez+Enwu8O+GLdiVjutWnk1K5A/vKF8qNWPoVcDOOeta78Vv21f21Nbbw9Lrnjnx80jjdpOk27pZQ54DPBAqwoADje4GB1NfDf8A4I6/tUeL1guvGt54V8EWz8yRX1/9suUHtHbh0J9jIK+D/wDwRn+AHg2aHUfiv4r134gXUYGbRR/Zdkx90iZpT2/5agdcjnj4f/s7fAf4VRxp8OvhB4S0GSIDFxaaTEs5x0LTEGRj7liaqqqqqqqqqq+OehaHa/D7UdQtdGsYrozxEzx26q5JkGTuAzznmvgdpGk/8K80TVf7LtPt3+kf6T5C+Z/rpF+9jPTj6cVVf8Fxv+RU+Ef/AGENV/8ARdtXwn/5JX4N/wCxfsf/AEnSqqqqqqqrVdU03Q9LvNa1i+hs7DT7d7i6uZnCJDEilndieihQST2Ar4i/DD4YfHLwa/hT4jeFdJ8VeH79VlSK4XevI+WWKRSGRsHh0YHB4PNeO/8Agip8BtcvprzwD8SvF3hhJTlbS5SHUYYeeiZCSbcf3nY55z2rwF/wRX+AWg30N74++Ini3xUkRBNpCItOhl55D7Q8mMY+66n3rxj8Rv2Zv2G/hvpnh+RNI8JaUz+Vo/h/TIgbrUJmIUmOPO6RiSu+VzjkFm5qqqqqqqqqqqq/4Laf8kB8C/8AY4D/ANI56/ZO/wCTV/g1/wBk/wBC/wDSCGqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr/grB/wAmR+MP+whpn/pZFX/BLj/kxL4Y/wDcW/8ATteVVVVVVVVVX7a//Jovxf8A+xP1D/0S1f8ABFT/AJNX8U/9lAvP/SCwqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/wCCyX/Jotp/2OFj/wCibmv2A/8AkzT4S/8AYvx/+hvVVVVVVVVVV/wQ6/5Gr4t/9g/Sv/RlzVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVqPjnwTpDXaat4x0OybT4/MuxcahFGbdOm59zDYMg8nFfFz/gqX+yF8K3lsbDxtdeONSiBH2bwvbi6jz2/0h2SEjPdXYjrjpn4m/8ABbT4laoZbT4R/CHQdBhIKrd65dyahMf9oJH5SofYlx9a8S/txftz/HXUG0Sx+K3jCeW44TTfCdv9ibb02gWaLIwOcfMTnODXhj9gD9uX4y6h/bF/8JvEkUtwf32oeK7tbGQe7C5cTHr2UnrXwv8A+CJHjO9eG7+Mvxk0nS4hgyWPhyze8kYf3fOmEao3v5bjPr1r4W/8E0P2OfhYsE0Pwsh8U38OM3viic6iXI7mE4g/KIVo2h6J4c02HR/D2j2Ol2FuMQ2tnbrBFGP9lEAA/AVVVVVVVVVVVVeJfEmkeEtHm1vWrnyreEYAAy0jHoqjuT/9c4GTXiz4keNvixaXPh7w74UkbTS6sywQvNL8pBG5x8q8gcY/E14W+Jvjb4V2Vl4b8SeE3XTIS3liaB4ZcMxY7XPytyx4x7ZFeHPEek+K9Hg1vRbnzracHGRhkYdVYdiP88HNV/wXG/5FT4R/9hDVf/RdtXwn/wCSV+Df+xfsf/SdKqqqqqqqv2hf+SA/Ez/sT9W/9I5a/YF/4KR+IP2erix+FPxfu7vWvhtK4jtbg5luNALEfNH1L245LRDleqc5VvDnibw94w8PWHivwtrVnqujanbrcWd9azCSKeJhkOrDgjFftlf8FWvAnwi+3fD34ASWHjHxigeG41bd5mm6U/Q4I/4+ZB/dU7AerEgpWs/Erx78XPitY+OfiV4s1DxDrt9qNv515ey7mwJBhFA+VEGeEUBVHAAFVVVVVVVVVVVX/BbT/kgPgX/scB/6Rz1+yd/yav8ABr/sn+hf+kENVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX/BWD/kyPxh/wBhDTP/AEsir/glx/yYl8Mf+4t/6dryqqqqqqqqq/bX/wCTRfi//wBifqH/AKJav+CKn/Jq/in/ALKBef8ApBYVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVf8Fkv+TRbT/scLH/0Tc1+wH/yZp8Jf+xfj/8AQ3qqqqqqqqqq/wCCHX/I1fFv/sH6V/6MuaqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqWSOGNpZZFREUszMcBQOpJ7Cvi5+37+yX8GI5YfEnxg0nVNSiJX+zNAf+07jeOqsIcpG3H/LRk/Wvif/AMFub1NWubL4NfBe1fTonxBqHiO+bzJxx8xt4cCPvx5rdjx0r43/APBTX9q/45aO/hy98WWHhDSZlK3Np4UgksftAxgh5Xkkm2nnKhwpycg8Y+Df7Lfxv/aI0bVtd+EvhGbxG+k6paWN1DHMiOHuUncSlnIVY18jDMxGDInXJx8Hv+CKXxC1kQal8cPifpfh2BhubTNChN9c4/utK+2OM+6iUdPw+F//AAS//Y5+GUcMsvw3fxffxAZvPE9015vx6wjbB/5Drw14R8J+DNOXSPB3hjSdCsUAC2um2UdrEMdMJGAP0qqqqqqqqqqqqqqr9oy8u7/XvD/heOXZDIplx2Lu+wEj2AP/AH0a0HQtM8NaRbaLpNusNvbIFUAYLHux9SepNeIdA0zxPo9zomrW6y29yhU5GSh7MvoQeQa/Zwv7q01bXvC80u6ONRMq5yFdW2MR9QV/75FV/wAFxv8AkVPhH/2ENV/9F21fCf8A5JX4N/7F+x/9J0qqqqqqqq8ZeF7Dxx4P1zwXqs08Vlr+m3Gn3LwELIkc0bRsVJBAYBjjIIz2r9r3/gnN8Yv2X5rvxRpcMvjH4fq5ZNcsrc+ZZJ2F3CMmL08wZjPHKk7a0D9oP41eFfhhqnwZ8OfErW9P8GazP515pMFwUjc4IZQfvIj5+dFIV8DcDivhv8LviH8YPFNt4K+GPhDUvEetXR+S1sodxVe7uxwsaDu7kKO5Ffsm/wDBILw14MnsPHf7S2qQ+IdZgZZrfw3p0zrZWrjBUzTDa8zA4+VdqAjkyA1VVVVVVVVVVVf8FtP+SA+Bf+xwH/pHPX7J3/Jq/wAGv+yf6F/6QQ1VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVf8FYP+TI/GH/AGENM/8ASyKv+CXH/JiXwx/7i3/p2vKqqqqqqqqr9tf/AJNF+L//AGJ+of8Aolq/4Iqf8mr+Kf8AsoF5/wCkFhVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVWv6zZ+HNC1HxDqAkNrpdpLdTiNctsjQs20cZOAeK/4fJfsi/8AQO+IH/gmh/8Aj9fs0/t3fBH9q7xXqng34X2viaK/0jTvt9wdVsI4E8rzEj+UrI5LbnXjA4zVV/wWS/5NFtP+xwsf/RNzX7Af/Jmnwl/7F+P/ANDeqqqqqqqqqr/gh1/yNXxb/wCwfpX/AKMuaqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqrWtc0Tw3pdxrfiLWLHS9Os0L3F5e3CwQwqOrM7kKo9ya+M3/BWD9lL4Wyz6X4Z1rUfiFqsJK+X4fhBtVYH+K5kKoV4+9H5g5H4fEf8A4LWfGzXUmtfhj8MvC/hWKTIS4v5pdUuIx2Kn91GD/vIw/nXxd/aZ+Pfx4u5Ln4q/FPXddic5Fi9x5NpHzn5baPbEvQdFzwK+Fn7M/wAfvjbLEnwt+EniTXoJTgXsVkYrRT0+a5k2wr36uOh9DXwY/wCCMXxk8R6o83xu8X6T4P0mKMFYtMkGo3c7nHygDEaKOcsWY5HCkHNeAP8Agjh+yv4Vu4NQ8W6p4x8ZSREF7W+1BLW2cj/Zt0SQAnPHmdPzPhHwZ4S8AeH7Twp4H8NaboOj2Kbbex0+2WCKMeyqAMnuepPJqqqqqqqqqqqqqqqqr9onwnfX1hYeL9Mid30vdHc7BkrGSCr/AEUg5/3s9jXgr44eEde0uFde1OHS9TRAJ0n+RHYDllbpg+h5HTnrXjj44eEtC0qePw/qcWqanJGRAkGWjjYjhmbpgdcA5PTjqP2dvCd7p2lX3irUUdG1Uqlsr9WjXJL/AEYnj/dz3qv+C43/ACKnwj/7CGq/+i7avhP/AMkr8G/9i/Y/+k6VVVVVVVXjD/gtT4n8L+Ldb8Mx/s/aXcLpGo3FoszeIpFMgikZN2PIOM7c4r/h+L4q/wCjdtJ/8KST/wCMVP8A8FvvEl1BJa3P7OGjTQyoUkjk8ROyupGCCDb4II4xXx7+J3wr+LHiI+Kfh78DbX4a3VzIz3tnpusNc2UpPeOBol8k57IwT0Uda/Y2/b/8cfsfadrHhzTfA+ieJ9C1iT7Q1tMRZ3EdwABv+0IjM67RjY4IHBUryG0f/gtv4p1TVrHTG/Z60qMXdzHCXHiOQ7dzAZx5HvVVVVVVVVVVVV/wW0/5ID4F/wCxwH/pHPX7J3/Jq/wa/wCyf6F/6QQ1VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVf8ABWD/AJMj8Yf9hDTP/SyKv+CXH/JiXwx/7i3/AKdryqqqqqqqqq/bX/5NF+L/AP2J+of+iWr/AIIqf8mr+Kf+ygXn/pBYVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVqOn2Wrafc6VqVulxaXkLwzwuOJI2BVlPsQSK/4K8fAH4M/Az/hU/wDwqP4eaT4X/tv+2v7Q+woy/aPJ+w+XuyT93zZMf7xr4E/s4/A34PWlr4q+GPw00fw7q2qaTDDeXdnGyvNGQjlWyTwWUH6iqr/gsl/yaLaf9jhY/wDom5r9gP8A5M0+Ev8A2L8f/ob1VVVVVVVVVf8ABDr/AJGr4t/9g/Sv/RlzVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVa1rmieG9LuNb8RaxY6Xp1mhe4vL24WCGFR1ZnchVHuTXxk/4Kt/snfCxrjTvDviK/+IGrQ5UQeHYA9uHHTddSFYyp/vRmT6V8Wv8Agsv+0P4va4svhX4Z8O+ArF8iKdo/7TvU9/MlAh/DyePWvid8b/i/8aNQGpfFX4k6/wCJ5UbdEmoXryRQHn/VxZ2Rjk8KoHJ9a+Cf7Av7U/x5MF34T+GF7pejzEY1nXwdOtQp/iUuN8q/9ckevht/wRD0eOKC6+MHxvvJ5Djz7Hw1p6whPULcT7t31MI+lfB7/gnz+yZ8E5odR8NfCmx1bV4QNup+IGOpTAj+JVkzHG3vGinmkRI0WONVVVGFUDAAHoKqqqqqqqqqqqqqqqqqqdEkQxuoZWGCpGQRXxq+H3w+0TRbrW9JEVhqvmJts4rgKr7mAYiI8gYOflwBXwa+Fng3VPCumeLtW09r69uDITHO+YkKSsgwgwDwo+9nmgqqoVQAAMAAYxVf8Fxv+RU+Ef8A2ENV/wDRdtXwn/5JX4N/7F+x/wDSdKqqqqqqr4h/8EzP23td8f8AibW9K+CXn2Woaxd3FtL/AMJJpa743mZlbDXIIyCDggGv+HXH7dv/AEQv/wAubSf/AJKr/h1x+3b/ANEL/wDLm0n/AOSq/wCHXH7dv/RC/wDy5tJ/+Sq/4dcft2/9EL/8ubSf/kqvD3/BML9uWx8QaZfXXwO2Q295FJI3/CS6Udqq4JOBdZ6Cqqqqqqqqqqqr/gtp/wAkB8C/9jgP/SOev2Tv+TV/g1/2T/Qv/SCGqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr/grB/yZH4w/wCwhpn/AKWRV/wS4/5MS+GP/cW/9O15VVVVVVVVVftr/wDJovxf/wCxP1D/ANEtX/BFT/k1fxT/ANlAvP8A0gsKqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr/guf/zRL/uYP/cdXhL/AJFTRf8AsHwf+i1qq/4LJf8AJotp/wBjhY/+ibmv2A/+TNPhL/2L8f8A6G9VVVVVVVVVX/BDr/kavi3/ANg/Sv8A0Zc1VVVVVVVVVVVVVVVVVVVVVVVVVVVVWua/oXhjS59b8S63YaTp1qu6e8vrlLeGIerO5CqPqa+Jn/BT39jb4arLCvxNbxZfRE/6J4ZtGvd3+7MdsB/7+V8af+C03xL18T6X8Cvh7pvhW2bKpqmssL+7x/eWIYijPThvNH518Uvjj8X/AI26p/bHxX+I+u+J7hWLRJfXbNDAT18qEYjiHXhFUc18F/2Bv2qfjt9nu/Cfwsv9N0i4GV1jXgdOtducb1Mg3yr/ANc1fofSvhD/AMETPC1kkOofHX4tX2qTgAvpnhmAW0SsOxuJgzOp9o4z718I/wBi79l/4HPBd/Dz4O6Fb6lBzHqd9Eb+7Vu7LNOXdCfRCo9qqqqqqqqqqqqqqqqqqqOAMk9K8RfFbwF4ZYw3/iCGacHBgtf3zg++3IX8SK1H9pfw1CCNK8P6jdEdDM6Qg/kWOPwrU/2lvEs4ZdJ0DTrQHODM7zMPpjaM/hWt/Erx14gctqPie92k/wCqhk8lP++UwD+NMzOxd2LMTkknJJr4HfEvSf7P0T4cfYLv7d/pH7/C+X1kl9c9OOnWqr/guN/yKnwj/wCwhqv/AKLtq+E//JK/Bv8A2L9j/wCk6VVVVVVVXxu/bW/Zn/Z38RW/hH4rfEy303W541lOn21lcXs0EbdHlWBH8sEcgNhiDkAitL+Kvw7134bN8X9D8WWWo+D102XUTqtmWmj+zxKzSMAgLEqEYFMbgVK4yMV/w9H/AGEv+i6/+Wzq3/yLX/D0f9hL/ouv/ls6t/8AItf8PR/2Ev8Aouv/AJbOrf8AyLVp/wAFPf2Gb+7gsbT44+ZPcSLHEn/CNaqNzMcAZNrgckdaqqqqqqqqqqqv+C2n/JAfAv8A2OA/9I56/ZO/5NX+DX/ZP9C/9IIaqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv8AgrB/yZH4w/7CGmf+lkVf8EuP+TEvhj/3Fv8A07XlVVVVVVVVV+2v/wAmi/F//sT9Q/8ARLV/wRU/5NX8U/8AZQLz/wBILCqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/4Ln/APNEv+5g/wDcdXhL/kVNF/7B8H/otaqv+CyX/Jotp/2OFj/6Jua/YD/5M0+Ev/Yvx/8Aob1VVVVVVVVVf8EOv+Rq+Lf/AGD9K/8ARlzVVVVVVVVVVVVVVVVVVVVVVVVVVVV8cf27f2Xv2fxcWXjX4m2V9rVuGB0PRP8AT7zeOqMsZ2wtx/y1ZB718bv+C0vxH8QfaNJ+AvgCw8K2jZWPVtaYX14Rzh1hGIYm6cN5w49+Pib8Zfix8ata/t34pfEDXPFF7uPlfb7ppEhz2ii+5EP9lFA9q+F37C37WPxgjt7vwd8E9fSwucGPUNVjXTbdk/vq9wU3r/ubvbNfB7/gib4mvZINQ+PHxYstNt+DJpfhiE3EzAjobmZVSNh3xHIPevgn+xB+zF8AVguvAXwt02TV4ACNZ1Zft96WH8SyS58on0iCD2qqqqqqqqqqqqqqqqqq1PWtG0aNZdY1eysUY4VrmdYgT7FiK1v4zfDrQ5BBJ4hivJCM4sh54A/3l+X8M5rUP2m7MJMmleFZmbBEL3FyFH1ZVB/IH8a1r49fETWE8qC+tdMTGCLKDaT/AMCcsR+BFaj4r8UaujRap4j1O7RuqTXbuv5E4rw/4O8T+KpBFoGiXV4M4MipiNT7ucKPxNad+zj44ugrX17pdkvdWlaRh+Crj9a079mSwTDat4snl9Ut7UR4/Fi38q0D4MfDzw+qsmiLfzD/AJa3584n/gP3R+C03gPwMxLN4M0IknknTov/AImrHwj4T0u6jvtM8MaTaXMWdk0FlHG65GDhgMjIJH41Vf8ABcb/AJFT4R/9hDVf/RdtXwn/AOSV+Df+xfsf/SdKqqqqqqr/AIKAeCvHXgz9rr4lnx1bXQfWteudS0y5mQ7bixmctb+W3RlSMpHx0MZB5Br9gTwX478I/wDBNf4p3njC2urSx8Q2Ou6hodtcxlW+yHThH5oB5CPJG5UdCBuHD5NVXhL/AJGrRf8AsIQf+jFqqqqqqqqqqqq/4Laf8kB8C/8AY4D/ANI56/ZO/wCTV/g1/wBk/wBC/wDSCGqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr/grB/wAmR+MP+whpn/pZFX/BLj/kxL4Y/wDcW/8ATteVVVVVVVVVX7a//Jovxf8A+xP1D/0S1f8ABFT/AJNX8U/9lAvP/SCwqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr4xfGr4ZfAPwVc+P/ir4qtdE0i3OxDJl5bmU9IoY1y0jnB+VQcAEnABI/wCCiv7bfgr9sXVfBlv4F8H6zo+neDG1EJc6pJGJbwXX2bB8qMsI9v2c/wAbZ39sc/su/wDBU39n/wCL99o3w18T2mo+A/EEscVpZNqkiS2V3KAFVFuFxsdscCRUUkgBiSAar/gsl/yaLaf9jhY/+ibmv2A/+TNPhL/2L8f/AKG9VVVVVVVVVX/BDr/kavi3/wBg/Sv/AEZc1VVVVVVVVVVVVVVVVVVVVVVVVVfFb49/Bj4Hab/anxZ+JWheGo2QtFDeXQ+0TgdfKgXMsn/AFNfGn/gtT4A0UT6X8Bvhxf8AiO6XKpqmvP8AYrUH+8sKZlkHThjEa+Nf7en7U3x58+z8X/FG/wBO0ebIOjaETp1rtOfkZYzvlXnpKz9vQU0E6QpcNC4ikZlRypCsRjIB7kZGfqK/Zm/4JBeOvi14L0z4h/Fzx3/whFhrECXNjpNvp/2m+eBwCjylmVYCykELh2AI3BTkD48f8Et/jD+y/pzfHj9nv4p3PiKTwex1F4o7M2WpWKR8mWIq7LOFUEsPkJUHCtyK/wCCd/7X11+1l8IbibxcbdPHPhGaO013yEEa3aupMN2qDhPMCOCowA8bkAKQBVVVVVVVVVVVVVVVVfajp2mQ+fqV/bWsf9+eURj8ya1r41fDnRtyHX1vZV/5Z2SGXP0YfL+tax+01AN0egeF3f8AuyXk4X80UH/0KtZ+PHxE1ZWjg1G306NhgraQBT/3025h+BFTT6rrt8HnmutQvJjgFmaWRz+pNaX8O/HOs3ItLDwrqJkxnMsBhUfVnwo/E1pX7OXjS+tRPqN7YadIWA8iRzIwHckpkfhn8q0H9nDwnZLFLr2o3mpyqPnjU+TET9B83t96rD4eeBdMCmz8JaUrJ913tVdh/wACYE0kccSLFEiqqjCqowAPYVVVVVX/AAXG/wCRU+Ef/YQ1X/0XbV8J/wDklfg3/sX7H/0nSqqqqqqq1jw54e8RLAuv6Dp2pi1fzIBeWqTeU/8AeXcDtPuKeGGSE28kSNGylWRlyCMYIx6Y7V/wiXhT/oWdJ/8AAKP/AArxb8b/ANkjwJevpnjD4ofC3Sb6J9klnc6nZLOh5+9Hu3Dp1IxXgj4i/s2fEyZbX4d+Nfhx4luGXP2fSr+zupRxnmNCWBx2I4oeE/CykMvhrSgQeCLKMY/Sqqqqqqqqqqqr/gtp/wAkB8C/9jgP/SOev2Tv+TV/g1/2T/Qv/SCGqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr/grB/yZH4w/wCwhpn/AKWRV/wS4/5MS+GP/cW/9O15VVVVVVVVVftr/wDJovxf/wCxP1D/ANEtX/BFT/k1fxT/ANlAvP8A0gsKqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/azi8Qfttf8ABSbSf2ZX127s/Cfhm4GnjyjkQJHbfab+cDBAlba0Sscj5I8+lfDT9nT4HfB/w5b+Fvh58L/D2l2cEQjZxYpJPPwMtLM4MkrHAyXYmv8Agpx+w38KNS+CuufHX4YeCtL8NeK/Careagul2y20Op2e4CbzIkATzEDeYJAAxCsDnI2/8Ey/jVrvxu/ZO0HUfFN+19rPhe8m0G8upH3PP5Co0TOepbyJYQSeSQWPWq/4LJf8mi2n/Y4WP/om5r9gP/kzT4S/9i/H/wChvVVVVVVVVVV/wQ6/5Gr4t/8AYP0r/wBGXNVVVVVVVVVVVVVVVVVVVVVVVTFVUszAADknjFfH3/gpH+y58BRcaZP4zXxh4igJT+x/DRW7ZHHaWbIijwcAgsXH904NfH3/AIKz/tJ/FhrnSPh5c2/w00GUlVj0l/Nv3Ttuu2AZT05iWM8dea07SvH/AMVvFf2XSdP1/wAX+JdUfcY4Ipr+7uW4GSBudz0Gee1fBX/gkN+0x8SUt9T+IUulfDjSpQCf7Sb7XfFSBgrbRNgH1WSSNh6V8Fv+CUf7KfwpMGo+JtBvfiHrER3GfxDIGtlbvttY8RlfaXzPrX/BWzQ/DvhT9oj4MWk+hWen+C7TRokNnb2yxWqRpfMZ0WNAAB5bJkAdCKt5re4t4p7SWOWCRA0TxsCrKRkFSOCCMVqNxYWmn3N3qs0EVlBC73Mk5AjSMAlixPAUAHOeMV/wRdjkuPjT8XL/AEGNo/Dw0uBVQgnaWunNuCeBny1l6jP05qqqqqqqqqqqqqqvL2y0+A3N/eQ20K9ZJpAij8TxWtfGv4c6MrKdeF9KvSOyjMufo33f1rxB+0tqMxaLwxoENunIE145kY++1cAH8TWr/Fn4ia1uFz4pu4kP8FqRAAPT5MH86kkvNRuN0sk1zO5wCxLs39TWkfC74ga4V+w+Fr1VbpJcJ5C49cvjP4Vo/wCzT4iuQr63r9jZKedkCNOw9jnaAfoTWkfs6eBrEq+pz6hqTj7yyS+Uh/BAGH/fVaH4V8OeGovJ0HRbSyBGGaKMBm+rdT+Jqqqqqqqqr/guN/yKnwj/AOwhqv8A6Ltq+E//ACSvwb/2L9j/AOk6VVVVVVVVXt5aadZz6hf3UVvbWsTSzTSsFSNFGWZieAAASSa/be/4KIfEr9ovxVqvhDwB4jv/AA98MbWd4bOytHa3l1VB8pmumU7mV+SIvuqCuQWG6qt7i4s7iK7tLiSGeBw8UsbFWRgchlI5BBAOa/4Jvf8ABR3xrceNdH/Z+/aA8Tza5Ya3Ktp4e8QX8m+6t7psCO2nlJzKkh+VXbLh2UElSNlVVVVVVVVVVV/wW0/5ID4F/wCxwH/pHPX7J3/Jq/wa/wCyf6F/6QQ1VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVf8ABWD/AJMj8Yf9hDTP/SyKv+CXH/JiXwx/7i3/AKdryqqqqqqqqq/bX/5NF+L/AP2J+of+iWr/AIIqf8mr+Kf+ygXn/pBYVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVXd3aWNu11fXUNvDHjdJK4RVycDJPA5IFav8AtGfs9aAxTXvjx8O9NZXCEXfiizhIYjIHzSDnHOK+K/8AwUn/AGQPhVpEt8PipZeL75SRDpvhYjUJZiPSRSIVHu0gz2zU/wDwXD8FrcSJa/s961JCCfLeTxBEjMOxKiEgfTJx714y/wCC2Px31G9nHgL4V+B9DsXDrGupG51GePP3T5iSQoSPeMgntXjT/gon+2d46klbUfjxrmnRSZCw6KkWmiMHHCtAqP26liffmvE/xV+KHjYyN4y+JPinXjKT5h1PWJ7rfnOc+Y5znJ/M1+xr8b9N/Z1/aT8F/FbXY5ZNH026kg1RYk3strcQvDI6qOSUEnmADqUA714N8beD/iJ4ds/FvgTxNpuvaNfIHt72wuFmjcEA4yp4IyMqcEHggV/wVH/am8AfDT4BeJPg3p3iKyvvHPjW2GnrpdvMHlsrR2/fTzKPuKUVkUHBZnBAIVsf8Ep/hVrHwu/ZC0efXrJ7S88Z6pca+IJAQyxSpFFCxB6b4oI3Hs475qv+CyX/ACaLaf8AY4WP/om5r9gP/kzT4S/9i/H/AOhvVVVVVVVVVV/wQ6/5Gr4t/wDYP0r/ANGXNVVVVVVVVVVVVVVVVVVVVVVfEj4vfC74PaMde+KHj/Q/DFlg+W+pXiQtMR1EaE7pG/2VBPtX7Tn/AAWUtrOe48K/st6BDe7cq/ijW7dghPrb2pwx9nlx/wBcz1r4pftUftF/Gozx/Ez4yeJtZtbknzbA3ht7M56/6NDshHHHCV8Ff2W/j5+0LepbfCf4aatrFqX2Sam0Yt7KA999xIVjBGD8oJY44BPFfs//APBF3wrpBt9d/aP8cPr1yuGOg6A729qPVZLlgJZB1GEWMjH3jXw0+D3wt+DeiL4e+FvgHRPDNjgB00+0WNpiOhkf78jf7Tkn3qq/b1/ZAtv2vPhNBomkXttp3jLw1O934evLkERFnUCW3lIBKxyhUywGVZEPIBB+Gn/BQ39qv9iOeb9nX4w+BtP8RHweEto7DVrox3thGY1eKJLmIujw7HUplXwrKFYKAB8R/wBvv9rH9uy7i/Z1+EPgjTtAh8VBoJ9P0mYvdXkIUmRJrqUqqQhQS21U4BDMVJFfsL/sj6d+yL8If+EZu7y31Hxbr0y3niK/gB8tpQuEgjzyYogWAJALFnbA3bRVVVVVVVVVVUkkcSGSWRUVRyzHAFa58VPh/oAZb3xNaSSL/wAsrZvPbPoQmcfjitb/AGl9IiR4/Dvh66uJMYSS7cRKD67V3Ej2yPwrVvjf8SNVkZl10WUZ6RWkKoF+hILfma1PWdZ164E+r6pd383RWnlaQj2GScfStG+F/j7X9raf4XvQjdJJ08hSPUF8ZH0rw/8As06nPtl8T69DbL3htEMjH6s2AD+BrR/gd8ONICs2jNfyr0kvJS+fqowv6Vp2i6Po8flaRpVnZLjBW3gWMf8AjoFVVVVVVVVVVV/wXG/5FT4R/wDYQ1X/ANF21fCf/klfg3/sX7H/ANJ0qqqqqqqq/bfu9Xsv2Q/i5caG7rdDwpeKWQ4IiZNsp/79l6+DHwy1L40fFjwn8KdIu47S68UatBYrcyLuW3V2AeUrkbgqbmxnnGO9fCr/AIJ6fsjfCrw/baNB8GfD/ie7jjC3GpeJrKPVJ7l8cuRMGRM+kaqOnHevj7/wTU/Zf+MvhW/svDfw70bwH4lMDf2bq/h+zFmkEoHy+Zbx7YpEJxuBUNjoynmtY0zXPAfi690a6ka01nw7qUlvI0TEGG4gkKkqeuQ68H2rwhqt5rvhLRNb1C2Nvdahp1vcTwsMGN3jVmXGB0JI6Cqqqqqqqqqqq/4Laf8AJAfAv/Y4D/0jnr9k7/k1f4Nf9k/0L/0ghqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/4Kwf8mR+MP8AsIaZ/wClkVf8EuP+TEvhj/3Fv/TteVVVVVVVVVX7a/8AyaL8X/8AsT9Q/wDRLV/wRU/5NX8U/wDZQLz/ANILCqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq1XVdL0PTbnWNa1K10+ws4mlubq6mWKKFFGSzuxAVQOpJwK8Tftmfsn+EEdtb/aJ8Ah4874rTW4byRSOxjhZ2B9sVr3/BVz9iXRpDFZ/EnU9YZW2k2Hh+8AzznBljQEcdR6jGa8T/APBan9m/ToCvhb4f+P8AWrnblRPbW1pFn0L+c7D/AL4rxl/wW/8AH935ifD74E+H9LxuEcms6tNf5/ukpEsGPcbj6Z714x/4Kv8A7afip3GnePtI8MwueYdH0O3AA9A06yuPwbP614r/AGrP2mfHBYeKPj749vYnHNv/AG/cRw/9+kYJnnritR1XVNXn+06tqV1ezAYElxM0jdc9WJPU1ofwq+KHieKObw18N/FOrJKu6NrHR55ww9QUQ5FeDf2Jv2tfHtxFb+H/ANnvxsomI2T6jpT6dCR6+bc+WmPfNaH/AMET/jpf6Hb3ut/FPwZpepyoGksFjuJ1hJ/hMoUAkDrgEZ6EjmvAP/BELwrDaNJ8Uvjhqt5ct9yHQNNjtkj+skxkL8f7C496+H3/AASu/Yw8CW6LefDy+8WXaDH2zxDqkszH6xxGOHt/zzrwv+zX+zx4JVP+ES+BngPS5E6TW3h62WU855k2bj17mv2jP2a/g9+0P8Mp/A/xJ8OBrLTle4067sAsFzpsoU5eBsEKSOCpBVuMg4GPjH+yP8Ufglq1qfgd4l8X+KdO1lZlnbTrCSzmtlTZhJhDK6sG3nByM7DwK/Y0/wCCavwjuvF0Pij4ya9qXjLUtPRL9NGks1trDfuHE+Xd7j5iDtOxTghgwJBjijhjSGGNURFCoijAUDoAOwqv+CyX/Jotp/2OFj/6Jua/YD/5M0+Ev/Yvx/8Aob1VVVVVVVVVf8EOv+Rq+Lf/AGD9K/8ARlzVVVVVVVVVVVVVVVVVVVVVftAftvfs3/s2xT2nxA+IFtc69Cp2+HtIxeX7NjIVo1OIcjoZmQHsa+P3/BYn42+PWutE+CWiWfw+0dyVS+k23upSLyM72HlRZHOFRmU9H71qmr/EH4s+LPtmr6lr/jDxNqj7Q88st/d3LcnAzudu/Ar9nr/gk3+0Z8W7q21X4lWS/DTw2xDSSarHvv5V7iO0BDIe374x46gN0r4O/wDBO/8AZK+DFna/2X8KdN8R6rbqu/VvEsY1KeR1/wCWgSQGKNs8/u0SrW1tbK2is7K3igghQJFFEgRUUcAADgDHaqqqrxT+zz8AfHWvXXinxt8Dfh/4g1q92fadR1XwzaXdzPsRUTfLJGWbCKqjJ4CgdBXhP9n/AOA3gHW4fE3gX4JeAfDmsW6ssOoaT4atLO4jDKVYLJHGrAFSQQDyDiqqqqqqqqqrWPEegaBCZta1mzslAyBNMFJ+g6n6CvE37R/h6xLW/hjTZtTkGQJpiYYvqARuP0IX61qf7QfxCv1dLWawsAwwDb2+SPxctWra9revS+frOr3d84OQbiZnx9ATx+FaN4J8XeIWUaN4dv7lTwJFhKoPq5wo/OtH/Zw8Y3qrJq2oafpynqm4zSL+C/L/AOPV4f8A2ffAulQg6vHcaxPjl5pGiQH2VCP1JrRfBXhLw64l0Tw7Y2sqjAlSEGT/AL6OW/Wqqqqqqqqqqqqqr/guN/yKnwj/AOwhqv8A6Ltq+E//ACSvwb/2L9j/AOk6VVVVVVVVeKtG0LxH4X1jw94oihl0bU7Ce11GOZtqNbyRlZQx7AoWya+IHhLxJ+xB+07p+q+EvE2i+JE8LaxHqWgalZ30V1Ff26OCqTCJiY3K/JIh2kZJHBVj4A/4K1/sceKvDdrqni/xjqngvVXQfadKv9Fu7tonx822W1ikR1z0Y7SRglV5A+L3/BXP9lbwj4Nvb/4WeIr3x34kaF10/T4tJu7KETY+UzyXEceI88nZubjGBnNfCEfDv4i/HKz139on4hQ+HvDV7qj6n4i1CSxubmS8zJ5kkUaW0bsHlYkbiAqgk5yApT/gqJ+wfGixx/HNVVRhVHhjVgAB6D7LXwU/bM/Zq/aK8VXXgn4N/En/AISDWrHT31Ce1/se+tNlukkcbPvnhRTh5YxgHPzZxgEiqqqqqqqqqr/gtp/yQHwL/wBjgP8A0jnr9k7/AJNX+DX/AGT/AEL/ANIIaqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv+CsH/ACZH4w/7CGmf+lkVf8EuP+TEvhj/ANxb/wBO15VVVVVVVVVftr/8mi/F/wD7E/UP/RLV/wAEVP8Ak1fxT/2UC8/9ILCqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq+vrHTbSW/1G8gtbaBd0s08gjSMepY8AfWvHP7cv7Inw6kkt/E/wC0B4T86LPmQ6bdHU5EI6gpaiRgeOhGa8a/8FgP2P8AwyjDw7e+LfF8mDs/svRWgQntk3TQkD8D16dq+In/AAW78NRabHF8JvgpqdzfuT5k3iK+SCKIdsRwFzJn/eTHvWpf8Frf2lZ1C6V8Ofhra/KQxms72Y57EYuVA/EGvFn/AAUt/a4+MeleI/h1rMmh6xpXjLSLjSZNFstDwYxKuDJAYz5pkAzjczrjOVPWvD/7NX7RPitkHhv4EfEDUlfpJb+G7t0HOMlxHtAyCMk9a8O/8Eyf23fEiLNB8EbixhOcvqOrWVqR9UeYP2/u14c/4I4ftb6zJENXvPAugxsR5hvNXklZBxnAgicE9RjIGR1xzXg7/gh5M3lz/EH9oFF4/eWujaCW79ppZR2/6Z9/bnwd/wAEeP2Q/Dnlv4gHjLxW4ILrqWsCBGPcAWyRMB/wIn3rwl+wv+yB4IEf9hfs8+DZWiA2Pqdh/aTKR0O65Mhz75z09K0P4eeAPDGz/hGvA3h/SfL27PsOmQwbdv3cbFGMZOPTNVVVVSRxzRtDKiujqVZWGQQeoI9K/aM0DQtD/wCEe/sTRbDT/O+1eb9ltki348rGdoGcZPX1NeH/AA54e0e3iutI0LTrKaWBVkkt7VImYYBwSoBIzzVV/wAFkv8Ak0W0/wCxwsf/AETc1+wH/wAmafCX/sX4/wD0N6qqqqqqqqqv+CHX/I1fFv8A7B+lf+jLmqqqqqqqqqqqqqqqqqqqd0jQySOqqoJZmOAAOpr9oj/gqN+zT8DTdaJ4c1hviJ4nt2KHTtBmBt4nHaW8IMa9CCI/MYEYKivj7/wU5/aj+OIudKsPFK+BPD025f7N8Ns1vI6HtLc581uMghSinJytfDH4OfFf44eIh4b+FngXWfFOpuwMosrcukO48PNKcJEpP8UjKPev2ff+CLeqXqW2vftKeOxp6Eq//CP+HHWSXGM7Zrp1Kqc8FY1cEdHFfB79nT4I/AHS/wCy/hH8N9H8PBk2TXUMJkupx6SXEhaWQZGcMxA7Yqqqqqqqqqqqqqqurq1soGuby5igiQfNJK4RV+pPSvEXxz+H+gO1vFqEmqTpwUsUEij/AIGSFP4E1r37S2qTqIvDXh+3tuDmW7cyn8FXAH4k1r3xW8f+IgY77xJcxREY8m1PkLj0OzBI+uatrS+1O4EFnbT3dw54SNDIzH6DJNeGfgP4618pLfWsekWx6vdnD49oxzn/AHttaZ+zd4Mto0Op6nqd7KPvFXWJD9FAJH/fVaB4B8HeGEUaL4es4ZE6TNH5kn/fbZb9aqqqqqqqqqqqqqqqq/4Ljf8AIqfCP/sIar/6Ltq+E/8AySvwb/2L9j/6TpVVVVVVVV+0L/yQH4mf9ifq3/pHLVVVV/wRU/5Oo8U/9k/vP/S+wqqqqqqqqqqr/gtp/wAkB8C/9jgP/SOev2Tv+TV/g1/2T/Qv/SCGqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr/grB/yZH4w/wCwhpn/AKWRV/wS4/5MS+GP/cW/9O15VVVVVVVVVftr/wDJovxf/wCxP1D/ANEtX/BFT/k1fxT/ANlAvP8A0gsKqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq+IHx9+CHwqEi/Ef4teE/Ds0YybbUNWhinP8AuxFt7H2CmvGn/BWP9i/wkXj0zxtrfiqWMHdHouiTdR2D3AiQ/UMR718Z/wDgtlo6afaW/wCzv8LbyW+csbu68aRLHHEB0VIbWdi+e7GRMYxg5yPiP/wVX/bK+JUsFtpHi3SfBsWAhtfDGliMzP0BLztNKDzjCuB04zS/Gr/gof41Il03x/8AHa937ph/ZdxqMYIJ5IEGBtyQMdBxUP7LH7e/xruYU1j4VfFPWZN48qTxOJ7ZQdvBD3rIoGDjOcds14D/AOCMv7TXiNI7nxr4m8G+EoWHzwyXcl9cpx/diTyz6f6yvB//AAQ98LW8yTfED4/6rqERHzW+j6FHZsOe0sssoPH+wK8H/wDBHb9kPwvrEWq6tP458VQxEH7BrGsRLAxHr9lhhc/TditP/YW/Y90ws1t+zp4JcuRn7Rpqz4x6eZux17de9eDfhT8LfhypX4e/Dbwr4YDAhho2jQWWQTk/6pF7mqqqqqqqqq1jWdK0DT5dU1m/itLWEfPJIcAew7kn0HJr41/EbQfH8+lxaDFdbNN8/fJPGEEm/ZjaMk4+Q9cda8EfGLwX4oNvpEV1LY3xVY44btQnmtjGFYEgn2JBPpVV/wAFkv8Ak0W0/wCxwsf/AETc1+wH/wAmafCX/sX4/wD0N6qqqqqqqqqv+CHX/I1fFv8A7B+lf+jLmqqqqqqqqqqqqqqqqqqaaG3heeeVI44lLO7ttCgDJJJ6DFftH/8ABVb9nf4Jvc6B4FuD8SvE0JKNbaPchLGBh1El5hlP0iWTng7a/aP/AG+P2jP2mHudL8VeLjonheYkL4c0MtbWjL6THJeftnzGK5GQq18Bf2Rvj/8AtJ30UPwt+H97daYZCk2uXim20+Ag4bdOw2sR3RNz/wCya/Z//wCCN3wi8GLa658efEdz461ZDubS7JnstNjPYEgiabBHUtGDnBQ14O8EeDfh5oFv4V8B+FdJ8PaPa58mx0yzS2hQnqdiADJ7nqe9VVVVVVV8Y9O+I7S/254R1maz03T9PeS7WO7MRJTczEKOp2gV8OG+LvjG6t9XsvFF7Np1nfRpdLNfEZUFWYbe42mqqqqqvL2zsIGur67htoU+9JNIEUfUnivEXxw+H+goyQaodUnHSKxXeP8AvvhcfQmvEf7R3ijUN0Ph3T7bS4iMCR/38v1yQFH02n61rXiLXvEc4uNc1e6vpAfl86UsF/3R0X6CvDfw28Z+K7jyNL0OdEAy09whijX/AIEev0GTXg39n2ystK1Ky8ZS2t5NfLGIJLXIa1K7slXYdTkdsccg1o3wM+HOjusraTJqEiDhr2UyD8VGFP4irOwsdOgFtp9nBbRKOI4Ywij8BVVVVVVVVVVVVVVVVVVX/Bcb/kVPhH/2ENV/9F21fCf/AJJX4N/7F+x/9J0qqqqqqqquba3vLeW0u4I54JkKSxSIGV1IwVIPBBHBBq28EeC7KBLWz8I6LBDGMJHHp8SKv0AXAr/hEvCn/Qs6T/4BR/4V49/aA/ZC+GGtv4a8efEn4daRq8L+XNYTTwNPA3pJGoLR/wDAgK8IXPwk+IWgweKPAcnhPxFpFySIr7SxBcwuR1AdMjI7jqO9f8Il4U/6FnSf/AKP/CrLQ9E0yUz6bo9jaysu0vBbrGxHBxkDpwOPaqqqqqqqqqqv+C2n/JAfAv8A2OA/9I56/ZO/5NX+DX/ZP9C/9IIaqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv8AgrB/yZH4w/7CGmf+lkVf8EuP+TEvhj/3Fv8A07XlVVVVVVVVV+2v/wAmi/F//sT9Q/8ARLV/wRU/5NX8U/8AZQLz/wBILCqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqvFfxv+C3gPV18P+OPi/wCCfDuqMu5bHVvEFraTkYzkRyOGxj2rW/8AgqB+xDoWtNok/wAZhdNE22W5stFvbmBD7SJCQ4903CvjZ/wWd+DXhXzNM+B/g3VPHV4MgajfbtMsl9GVXUzSdPulI+o+avE3/BZX9q/WiyaHo3gLQI8YRrXSpppB9TNM6k/8BFa5/wAFLP24vFcq2p+Nt7b+a22KDTNIsrZsnHCmOEOSSB1JPJx1prn/AIKCfHVRB53x08X2s6n5AdSmtQpUZ4H7tQQV54ByPWvAX/BKT9s7x00c2peCNJ8J282CLjxBq8Sde7RwebKv0KA14J/4IfahIqz/ABH+PtvC2PmtdE0Qyc+00si/+i/yrwN/wRr/AGZPCviWz17xD4k8ZeKrWzkEn9l39zBFbzsDwJfKiV2X/ZDLnvxkV4C/Z0+Anwt1U678Ofg54P8ADupldv23T9HhhnC4wQJAu4A55AOD3qqqqqqqqqqqqqq+JU2o/En4s2nw+trtorGzcRnB4DbN8smO5C5UZ9Pc1ovwx8B6FZrZ2nhmwmwuGluYFmkf1yzA9fQYHtXxm+Eug2mgz+LfC9kmn3FiQ88EI2xyJnBIXopGQeMDAPFfB7xTdeLfAtpe38plu7V2tp5Ccl2TGGPuVK598mq/4LJf8mi2n/Y4WP8A6Jua/YD/AOTNPhL/ANi/H/6G9VVVVVVVVVX/AAQ6/wCRq+Lf/YP0r/0Zc1VVVVVVVVVVVVVVXiTxd4T8G2I1Pxf4o0nQ7MttFxqV7Hax59N0hAzXhj4l/DjxxNJbeC/iB4b1+aEZkj0vVYbpkHqRGxIq5uLezt5bu7uI4YIELyyyMFVFAyWJPAAAySa/aY/4Kx/Ar4OPdeGfhUE+JXiaIFS9hchNMtn6fPcgHzSODiIMD0Lqa/aF/ba/aK/aZlktPiJ44lg0Fn3R+HtJBtLBMHI3RgkykHo0rOR2Ir4A/sbftDftK3ULfDPwDcto7ybJde1HNrp8OOGPnMP3hHdYw7D+7X7N3/BIv4H/AAs+yeIvjLc/8LI8RR4f7LPEYdLt39BBkmfHIzKSrDnyxWn6dp+kWFvpek2NvZWdpGIre2t4hHHEijAVVXAUAcADiqqqqqqqqvif4o8O6R4T1nS9R1i1hvLzTbiOC3Z8yOzRsFwo5wSQM9K/Z58U+HNJ0W+0fVNatbS8uL3fFHPJs3jYo4J46gjGc1wRkHNVVeIPF/hjwtF5uv63a2eRlUd8u30QZY/gK1r9pPwzaM0eh6Ne6gw6PKwgQ/T7x/MCtf8A2gfHergxaa9rpMR/59497ke7Pn9AK1HVtX1u4E+qaldX0xOA08rSN+GSa0P4XePfERVtP8NXaxN0muF8hMeoL4yPpmvDn7NKApN4r1/dj71vYrj/AMiMP/Zfxrw98OvBXhYI+jeH7VJk6TyL5sufUM2SPwwKqqqqqqqqqqqqqqqqqqqqr/guN/yKnwj/AOwhqv8A6Ltq+E//ACSvwb/2L9j/AOk6VVVVVVVVftLePbv4X/s9/Eb4gabcGC/0PwzfXFjIGxsuRCwhOf8AroUr/gmv4k+M/if9qTwP8O/DfxW8W6V4cN1Jf6vY22qS/Zpra3jaZo5ISxQiRkWMkjI8zgiv+Con7U+tfs5/BG08OeA9Wk0/xn49nls7G7hOJLK0jVTczof4X+eONT1BlLAgoKmmluJXnnleWWRizu7EszHkkk9Sa/YY/ai8S/sw/HLRNYi1mdPB+uXkNl4n05pD5M1q7BfP2dPNh3F1bg8MuQrsDVVVVVVVVVVVf8FtP+SA+Bf+xwH/AKRz1+yd/wAmr/Br/sn+hf8ApBDVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV/wVg/5Mj8Yf9hDTP/SyKv8Aglx/yYl8Mf8AuLf+na8qqqqqqqqqv21/+TRfi/8A9ifqH/olq/4Iqf8AJq/in/soF5/6QWFVVVVVVVVVVVVVVVVVVVVVVVVVVVVV+3Rf/tc6f8PNAl/Y7juX8TNrW3UxBbWMx+x+TJ1F4pQDzPL5X5vwzWk/tf8A/BVTXfi5c/AfSfHV1cePbN5Y59FXQNDEiNFH5jjeYNnCAn71fsb3n7R9/wDBO1n/AGqUnXx5/aNyJhNBaQt9nDDyvltQI+me2fWqqeeC2he4uZo4oo1LO8jBVUDqST0FeMv2s/2Y/AHmR+LPj34Gsp4h89qutwzzjnH+pjZn6/7PY+lfEb/gr3+yN4NWWDwrfeJfG90hKoNK0treEsPWS5MR29sqre2RzXjj/gt946upHj+GvwM0HTUXcI5tc1SW+LehKQiHb7jcfrXiv/grj+2b4jSSPSvE3hrw0HBA/srQomK/Q3JmPt/9fmn/AG1f2rNUk8RLefG7xjd3niq0hs5ZY9VmhkgEc6yL9nETL5ROHQqgAZZXBBzx8O/+CaH7Znxa0y68Uv8ADuTQVkbzFPim6+wXN27N8xETgyA87i0iqDzgk8V4N/4IifEvU/D7Xnjv43+HtC1g4Mdhp2ky6jEB/tzNJCVb2VGHv3r4Sf8ABFX4V+HNRi1P4xfE7VvGSRtuGmadZ/2Vbv8A7MjiSSVh/uNGffjnwp+xr+yj4Khih8P/ALPPgNWhUKk13okN5MAAR/rZw75IJBOcnvmtE8K+F/DUfk+G/Del6UhGCtlZxwDH0QD0FVVVVVVVVVVVVVVVVV4hvF+Hvx8GvaorrYXb+Z5hXP7uSPYzf8BbPT+771bXNte28d1Z3Ec8Mqho5I2DKwPQgjqK+OHi3TtA8FXmktcI1/qsfkQwBhu2n77kdgBkZ9SK+AWjXGkfD2Ga5Qo2o3Ml0qkYO0hVU/iEB+hFV/wWS/5NFtP+xwsf/RNzX7Af/Jmnwl/7F+P/ANDeqqqqqqqqqr/gh1/yNXxb/wCwfpX/AKMuaqqqqqqqqqqqqqq/bc/aftf2T/gZf/EG2tbe98Q3866d4fs5zlJLt1Zt8gBBMaIjucdSFXI3ZHwJ/YC+PX7eCL+0b+0R8Wr/AErT9fy1jPc232q+voFYgGKMskdtBndsABGOVTaQx+Mf/BGLxT4Q8PS+LfgF8V7nXtb0pfPi0nUbRbSadk+YGC4jfCycDarKAT/GK+Mn7Yf7Tvxm8O2fw++KfxO1m60vSYhbT6cFFoLl0P3rpUCmaQEAfvM4K5wCST+zn+xV+0D+1BeI/wAOvB72+hB9s/iHVd1tYRc4OJMEysO6xK7DuAOa/Zu/4JPfs/8Awb+yeIfiXH/wsvxPFh86nbhNOt3/ANi0yRJjpmYuDwQqmrSztLC1hsbC1ht7e3QJFDEgRI1AwFVRwAB2FVVVVVVVVV8V/GkvgfwhNqVky/brhxb2m4Z2uwJLY9lBPpnHrXw7+Cn/AAmNini/xvqV4/8AaBMsUKPh5Qf43c5PPUAdsHPOK8R/s6eFL2xkPhu4udPvFXMQklMkTHHRgfmGfUHj0PSvgX4w1m11e8+G3iSR2lsg/wBl81stE0Zw8We47j0we2MX+o6fpds13qV9b2kCfelnkEaj8TxXiX4/+B9FDQ6U8+s3C5AFuuyMH3dv5qGrxR8d/HPiAPBYXSaPbHolpkSY95Dzn/d21bWmra9feVaW93qF5MclURpZHPrxkmtB/Z88d6sizaitppUZHS4k3SY/3Uz+RIrQ/wBm3wzZssuvaxeaiy8mOJRAh9j1b8iK0LwX4U8Moq6FoFnasowJFj3SH6ucsfxNVVVVVVVVVVVVVVVVVVVVVVV/wXG/5FT4R/8AYQ1X/wBF21fCf/klfg3/ALF+x/8ASdKqqqqqqqv2vPBeo/EL9l/4oeENGgkn1C+8MXhtIY/vTTRxmREHB5ZkC/j261+yL+0RN+y38dNF+Lg0E63Z2cM9rfWCyiJ5oJoyrbHIIVlO1hkc7ccZyP20/wBq3Vv2vPi+fH82jPoui6bYpp+i6W8/mtBArMzO7AAGR3dmOBwNq5bYGP8AwR1/Zr0C3+F+u/Hzxl4as76/8UXb6dohvbVZRFYwHbM6bgceZNuQ+0A7E1+2J4e8KeFP2pfil4c8E2cFpo1h4nvIre1gTZHbkOd8aKAAqq+9QoGAAAOBXw/nv7rwF4budVBF7LpFq9yGHIkMKlv1zVVVVVVVVVVVf8FtP+SA+Bf+xwH/AKRz1+yd/wAmr/Br/sn+hf8ApBDVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV/wVg/5Mj8Yf9hDTP/SyKv8Aglx/yYl8Mf8AuLf+na8qqqqqqqqqv21/+TRfi/8A9ifqH/olq/4Iqf8AJq/in/soF5/6QWFVVVVVVVVVVVVVVVVVVVVVVVVVVVVV+2F/wVHuPhz47n+B37M3he18Y+Mba5NnfajLFJcwW91kqba3hiIaeZW4JztVhtw5zt0nwB/wUj8FfGC6/bAtPgd4j/4Sy8lmubmU6EkpbzYtkgNkv7xQUbGAgI56EGvg1/wWf8GHw1fWn7Rvw91bSvE+mttT/hGbUSxXpBwy+VPKrQSA/wALOwPPIPy18Q/+C31ywltfhP8AAuJDg+Ve+ItUL/TNvAo/SWviF/wVN/bO8fGSG1+I1n4UtJM5tvD2mRW+PpLIJJhj2kHvmvGvxU+J3xJuDd/EL4ieJfE0pbO7VtVmu8H2EjHA9h0rwX8D/jN8SAjfD/4TeL/EaOARLpmiXFygB6EuiFQPcnFeAf8Aglx+2f48eJ5vhlb+GLSQj/Stf1KG2C/WJC8w/wC/deAP+CIGoyLHcfFL48W0JA/eWegaQ0ufXE8zrjv/AMsvy6V4a/4I1/sm6K0cms6v498QMMb0vNWhhjY98CCFGAP+8T718Jv2L/2Xfgjdwan8OfgzoNnqdsd0OpXiPf3UTf3kmuGd4z/uEflVVVVVVVVVVVVVVVVVVVVea5omngm/1mxtgOvnXCpj8zXxO8S/BrxVYjS9b8SRvdQEm3ubGNpmhJ64ZQVKnjIzz7EZE+r3/hmdrPwl43vpbNuQ9q81rn6pkc/nXhH9n6Se9i1zx1rY1AtiQ20TM3mHqN8jYJHqAPxqOKKGJIYY1RI1CoijAUDoAOwqv+CyX/Jotp/2OFj/AOibmv2A/wDkzT4S/wDYvx/+hvVVVVVVVVVV/wAEOv8Akavi3/2D9K/9GXNVVVVVVVVVVVVVVf8ABbP4h+D9YT4d/DzRvFthe65ot3fXOq6Xb3IkksxJHCImmVSdjEB8BsHBJxg5P7PuoaBqvwH+HV/4Vlt30iTwrpv2LyMbFjFtGAoA6bcbcdiMdqr9nP4Y/Cv4+/8ABUX4j2Gs+FNO8U+D7TWde1VbZ132jlLnakrKDtkjMkgwpyrbhkEVYafYaVY2+maXZW9nZ2kSxW9vbxCOOFFGFVVXAVQAAAOBVVVVVVVVVVX7S1nPL4V0y9jUmOC+2vgdNyHB+ny4/EV8OdUsdX8C6HdWDqY0sYoWVf4HRQrL+BB/SjgDJPSvGfiueL4k6v4k8L37W7faZFhuIT1G3YzA+jDJz75q/wBT1bW7kT6lf3V9cMcBppWkYk9hnJrw78HfH/iTZJBoj2cDf8t70+SB77T8xH0Brwz+zh4fsdlx4o1KbU5RyYIcwxfQkHcfrla0fQdF8P2os9E0u2soe6wxhdx9SRyT7mqqqqqqqqqqqqqqqqqqqqqqqqq/4Ljf8ip8I/8AsIar/wCi7avhP/ySvwb/ANi/Y/8ApOlVVVVVVVVftjf8EkfHlx421P4g/svWen6npGsTvczeFpbmOzmsJXOXW3eQrE0OSSFLKUGFG4AGvgX/AMElP2l/iB4rtIvixokXw/8AC8MwN9d3N5BcXUsYI3JbxRM/zkcBpNqjk/NjadA8IaZ8Lvhva+C/hl4ct0tPDelfZtF0wzeWjmOPEcbSHJG5gNznJyxY5Oa1D/gnr+2N4i+POnW/xT+Fep+V4x8TCXWdespI7y1iSeffcXDyQM/lKAzt8+09BjJFRxRwxpDDGqIihURRgKB0AHYVVVVVVVVVVVX/AAW0/wCSA+Bf+xwH/pHPX7J3/Jq/wa/7J/oX/pBDVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVWt65onhrR7zxB4j1ix0rS9Ohae8vr64WCC3jUZZ5JHIVFA5JJAFf8ADWP7K/8A0cx8Kf8Aws7D/wCO14J+JHw6+Junz6t8N/H3hzxXY2s3kT3OiarDfRxS4DbGaJmCttIODzgg96qv+CsH/JkfjD/sIaZ/6WRV/wAEuP8AkxL4Y/8AcW/9O15VVVVVVVVVftr/APJovxf/AOxP1D/0S1f8EVP+TV/FP/ZQLz/0gsKqqqqqqqqqqqqqqqqqqqqqqqqqqYqqlmYAAck8Yr4k/to/srfCSSW28c/HPwvbXcORJZ2VydQuIyOzQ2wkdT7ECvHv/BZr9mXw9bzR+CPDPjLxbeqG8nbaR2Ns5HTdJK/mKD7RH3FfFz/gsF+0/wCPGuLL4e2+g/D3TpQVQ2VuL68CnqDPOCucfxJGhHUc4I/4Iq/CfwxqWneO/jvrFql/4kg1JdHsLi4Xe9mhiEs7oT0eXzUUt1whGcM2a/4LT/CLwj4P+JPgX4p+HtNt7LUvHFrfQ6wsKBBcTWht9k7AdXZLgKzY6Rrnk18Jv+CLXxB8V6TpviD4i/GXQdDtNQto7lLbSdOmvpgjqGVWMvkhGwcHhgD618Pv+CPv7JHhERzeK4fFPja4GC41TVTbQ7v9lLURMB3wzt9ccV4C/Zk/Z4+F6ofAPwV8HaPOhGLqHSImuDjpmZgZD+LevrVVVVVVVVVVVVVVVVVVJJFEm+WRUUdSxwKvPFvhTTxm+8TaVBjtLeIp/In2q6+Lfw3swfN8XWbbf+eQaX/0EGtX/aL8D2Mhi0y21DUsdJEiESH/AL7Ib/x2tR/aZ1Z5saR4YtIoh0+0zNIx/wC+duP1rXf2g/HeqoIdNa00pMcm3i3ufX5nzj8AD71feN/GWpEm+8VarMD/AAtePtH0XOB0qe9vbrP2m8mmycnzJC2T+NW9pdXjeXaW0s7D+GNCx/StK+GPxA1o4sPCd/js08fkKfoZNoNeKfAvijwWto3iXTRaG9L+SvnJIW2bd33CcfeFWH/Hhbf9cV/kKqv+CyX/ACaLaf8AY4WP/om5r9gP/kzT4S/9i/H/AOhvVVVVVVVVVV/wQ6/5Gr4t/wDYP0r/ANGXNVVVVVVVVVVVVV4v8X+GfAXhnUvGPjDWINL0bR7WS6vbubO2GJBl2IAJOAOgBNftef8ABXHxr46mv/Af7MzXPhbw4cxSeJZV2alejoWgH/Lqh7HmXGDmM5Wvhr8K/ir8fvHC+Ffh14Z1XxV4h1CRpp/Ly5XJy008rHai5PLuwGT1ya8BfGT9vj/gm2Z/hT4o+Gv/AAkXg62b7Raw3dtLeWUAkdiXtbyAjyw7BiY3zgknYrMSfHn/AAUo/bP/AGl9BuPAvwP+Ekvh631dPs8994fs7m+vFV22ELckBIFOSpfaGXqHUjNf8E6v2H7j9k3wbqXiPx3PaXfxA8VpGt8LZ/Mi022T5ltkf+Jtx3SMPlJCgZCBmqqqqqqqqqq8T/FPwP4T3xalrcUtymQbW1/eyZHYgcKf94iviF8dp/F2mXXh7TdAgg0+5G13uj5kpwQQQBgIQQP71eFvHXinwZI7+HtWkt0lOZIWUPG59SrZGfcc+9WviL40/Fmzk0/TJT9gLeVcPAqW6cjkM33iMHlR1HY14a/Zv8P2aRzeKNTn1CbALQwHyoge4z94j3yv0rRPCHhfw2gXQ9BsrNgMb44hvP1c/MfxNVVVVVVVVVVVVVVVVVVVVVVVVVVV/wAFxv8AkVPhH/2ENV/9F21fCf8A5JX4N/7F+x/9J0qqqqqqqvE//BTj4RfC39onxf8AAP4zaPqHh+HQL6KCy8RWqG7tpUkgjk/fxqPMiIL7coJAep214D+J3w5+KekLr3w28daF4n08gZn0q/juVQns2wkq3H3WwR6VVV4m8WeFfBelSa54x8TaToWmw/6y81O9jtYU4zy8hCjgHvXxW/4Kv/AXw54q0n4f/B6Of4g61qupQWT3sG620608yVULGV13TEbsgRrtb/noKqqqqqqqqqqr/gtp/wAkB8C/9jgP/SOev2Tv+TV/g1/2T/Qv/SCGqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqvix8OtL+Lvwz8T/C/Wr66s7DxRpc+n3Nxa7fNiSVCpZNwI3AHjIIr9q/8AYy8CfAD9q74dfAHw14p17UdH8ZWuly3d5fGL7RCbrUJrZwmxAuAkSkZB5JzxxX7J37Jvgv8AZD8F6v4J8E+JNa1m11nVP7Qll1UxF0fykj2r5aqNuIweRnJNVX/BWD/kyPxh/wBhDTP/AEsir/glx/yYl8Mf+4t/6dryqqqqqqqqq/bX/wCTRfi//wBifqH/AKJav+CKn/Jq/in/ALKBef8ApBYVVVVVVVVVVVVVVVVVVVVVVVVV47+P/wAE/hl4c/4S3x18UfDulaR9rezW6kvldWuEXc0KhMlpAoJKAFvavir/AMFn/gF4WklsfhX4J8R+OriPO25mI0mzk9CrSK834GFe3vjx3/wWi/aO16OS18D+CfBfhaJydszwTX9wnphndY/zjPPp0r4r/taftI/G+OW2+Jvxj8RatZTAiTTkuBaWbZ9baAJEfxWvht8BfjT8YpPL+F3wt8S+JUVirz6fp0kkEZHUPLjYnb7zCvBX/BIv9sjxWYW1vQPDPhKOQZLaxrSOVHutqJjnHOPzxXws/wCCJvw80u3N18Y/izq+vXbxMFtNDtVsYIXIwCZJDI8oHXpHzwRjr+yv8e/Ev/BMz49eMfgV8evD9+/hbVbtPtF1awEvEULLDqECn/WwSIfmAO7AGMshRrv/AIKP/sUWmgt4gHx60maARl1gis7prhvYQ+V5gJx0Kj3wOa+KPjfxV/wVa/a68LeEfh/4b1Ox+HvhgCJpp0Ae0sGkVru8mIJWOSQKiIm48pGOpY1BBDbQR21tGscUSBI0UYCqBgAD0xVVVVVVVVVVVVVVVT3FvaxGa5njijUcvIwUD8TWv/G34e6BcmyfVZL6ZDhxZR+aF/4FkKfwJq7/AGkfBESYtdM1ed8cAxIg/Mvn9K1b9pq9dSmheFoImB4ku5zJn/gKhcf99Vq/x3+IuqxCGLUrewAOSbSAKW/FixH4Yq58deNbw/6T4u1iT/ZN9JgfhnFXF1c3j+bdXMsz/wB6Ryx/M1aaNq+oECw0q8uSRwIoGf8AkParb4Z/EC7wIvB2qjPTzbZo/wD0LFaR8AfiJqbgXdla6bH/AH7q4Bz9Am4/nitL/ZkkKlta8VKrY4S1t88/7zEfyrSf2a/C9rMJdX1q+v1B4iRRCp+uMn8iK0/4Z/D/AExFS08IaYdv3WmgEzD8Xyag0DQrXb9l0Wwi252+XbIuPXGBSoiKERQqqMAAYxWowTXOn3NtbSBJpYXSNiSNrEEA5HI5q/8AgF8UNV8v+1PFOmXnlZ8v7RfTybM4zjMZxnA/IV8LvAPjvwjq11d+KfEcWoW0tr5cUSXksuxtwOcOoA4BHFVX/BZL/k0W0/7HCx/9E3NfsB/8mafCX/sX4/8A0N6qqqqqqqqqv+CHX/I1fFv/ALB+lf8Aoy5qqqqqqqqqqqqnngtYJLm5mjihhQvJJIwVUUDJJJ4AA71+1h/wV98K+A7+98C/s2aVY+LNWg3RT+JL1mOnwOOCII1Ia4I5+fcqZAx5gNfET9p/4+/Fnxm/jrxz8TNX1PVntLqxiXeEghtrmJop4I4FAjSOSN2RlC/MDzk81+yZ/wAEqvi98aL2x8V/Ga0vfAHgk7ZfLnjC6nqCHB2xQt/qQR/y0lGRkEI3b4O/Az4UfAHwpH4M+Evgyx0DTgQ0xhUvNdOM/PNKxLytyeWJwDgYHFVVVVVVVVVVeKviL4P8G/u9c1iNLjGVtogZJT/wEdPqcCvEv7SsKgweEtCLnHE9+cAH2RTz/wB9D6V4h+KHjvxOrRal4iuFgbOYLc+SmPQhcbh/vZrQfCfiTxRP9n0DRrq9YHDNGnyIf9pz8q/iRXhj9my/uAlx4t1lLZOptrMb3+hc8A/QNXiDwf8ADjwL4N1uzsLbTrbUJdJuRE9zKrXEhMTAbSxzyeMLgZr9nLV9JsfDd/Z3uqWlvPNqP7uKWdUZ8ooGATk88VVVVVVVVVVVVVVVVVVVVVVVVVVVVV/wXG/5FT4R/wDYQ1X/ANF21fCf/klfg3/sX7H/ANJ0qqqqqqqv26v+CW+r/HPxvrXxv+DPi6KPxRrGyXUtD1htkF08caxgwTKP3bFUUbHBUnneo4rx78HP2hf2afESSeNvBnivwPqUDlLfUUV4Y3b/AKY3URMcn1RzXhj9uv8AbB8IwLbaP+0P4xljQEKNRvv7QwPrcCQ/4V/w9H/bt/6Lr/5bOk//ACLXiT/goN+2f4rjki1T9oTxLCJc7jpph04jPXBtkjK/hjFaZo/x2/aL8TrbabaeNfiNrzHBbNxqk67uSWY7io4JJJA4JJ4r9kj/AIJD+PE8RaJ8SP2i9cj8O2+l3UV5b+HNMmSe7lkjYOgnmGY4l3AZVN7EZGUPIqqqqqqqqqqr/gtp/wAkB8C/9jgP/SOev2Tv+TV/g1/2T/Qv/SCGqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqtU1Ow0bT59U1O5WC1tk3SyMCQo9cDmv+CiHi/w3r3/AAUP+Ceu6TqsdxY2dhoAmmVGAQrrN0zcEZ4Ug14d8X+G/Fsc8nh3VY71bYqJSqMu0nOPvAehqq/4Kwf8mR+MP+whpn/pZFX/AAS4/wCTEvhj/wBxb/07XlVVVVVVVVV+2v8A8mi/F/8A7E/UP/RLV/wRU/5NX8U/9lAvP/SCwqqqqqqqqqqqqqqqqqqqqqq8d/E/4b/C7TDrPxH8eaB4ZssErNq2oR2ofHZd7DcegwMnJr4gf8Faf2N/BJeHRvFGveMriMkGPQtIfaD/ANdLkwoR7qTXxa/4LZeN9SR7D4JfCXS9EQgj+0PENy17KQRwVhi2IjD/AGmkB9K+Ov7TPxu/aS1i21j4weO7rWjYpss7NI0trW2HOSkMQVAxyQXILEYBJAGPBfgf4/8AxO8GHwb8O/AHjXxX4ds9SbUDbaNotzewwXTxiJpD5SMFZkRFJP8AcFfCH/gk5+1l8TY4r/xHomleANOZhmTxBckXDL3K28IdwfaTy+nWvA//AARF+F1gYZfiP8a/E2tlRmSLR9Ph01SfTMhnOOg7E+2ePhh/wTr/AGPfhTJBeaP8HNN1nUICCL3xC76m5YdGCTExKQRnKotWtra2VtFZ2VvFBBCgSKKJAioo4AAHAGO1VX7cNr+ymvgTSb39qXwppOp6bcXps9OuriVra4t5WRnIinjIkTIQkqDg4GQcV4S0P9lST9rK+0nxNdXzfBqK6uTat/aRSR4xCTErTgZIMmASACR3HWv2Prf9nc/CGHUf2afC+k6R4Xmu5InayzI9xNHwWllbLysM4y5JA46VVVVVVVVVVVVahrei6Su7VNXsrMAdbi4WP+ZFat8aPhvpIKt4ijupB0S0jaXP/AgNv61qH7THh6Jiul+G7+5A6GeVIc/luq4/advXBFr4Pgj448y9L4/JBWt/H34harJmxvbbS4h0jtoAxP1Z9x/LFah8VPiJqa7bnxfqCgjBEDiD/wBAAq7vb2/k829vJrh/70shc/ma8J/D7xP4r1W3sLTR7pYXYGaeSIokadzuPHToOpq3/Z2+IM7bZG0uAeslyT/6CprTf2ZL1mVtY8VQRqPvJbWxcn6FiMflVj+zl4CtZ45rm61a8CfeilnVUf67EDY/GrX4Z/D6zULD4O0psDjzbZZf/Qs1Z6Lo2nADTtJsrbHTybdU/kKqqqqqqqqr/gsl/wAmi2n/AGOFj/6Jua/YD/5M0+Ev/Yvx/wDob1VVVVVVVVVf8EOv+Rq+Lf8A2D9K/wDRlzVVVVVVVVVVVfHT9oX4Sfs4eD5PGnxY8WW+lWpDLaWq/vLq/kA/1cEI+Z26ZPCrnLFRzX7Y3/BTL4oftL2V54A8G2kngrwBMzLNZwzlrzVI+gFzKuAEI5MKfLz8xfAI/ZL/AGMvir+1z4rbTfCNuuleGtPkUax4jvIibezB52IBgzTEdI1I7FioO6v2df8Agnr+zX+zg1lrOheEh4j8VWoVh4g10LczxyActCmPLg5zgou8A4LGqqqqqqqqqq7u7Sxga6vrqG3hQfNJK4RV+pPFa98cfh5oYaOLVm1KZf8AlnZR+YP++zhfyNeNfjz4q8SBrPQydFsjwRDJmZ/rJxj6Lj6mtJ0TXPE1/wDY9HsLm/upDlhGpY892PYe54qy/Ztv7u10qS81aKwkMDf2kmPNYSbyVCYwPukA88EZ5zXh/wCBvw90LZJLpj6nOo/1l8+8Z/3BhcfUGre2trSFLa0gjhiQYSONAqr9AOlV8QvhHpvxD1O21O81e5tHtrfyQkUasCNxOefrWl/s6aJpep2epx+JL2R7OdJlRoVAYqwOP0qqqqqqqqqqqqqqqqqqqqqqqqqqqqr/AILjf8ip8I/+whqv/ou2r4T/APJK/Bv/AGL9j/6TpVVVVVVVVd2dpf20llfWsNzbzKVkhmQOrg9QVPBHsa8XfsVfsmeOZXuPEP7Pnglp5c+ZNZ6WljI5PUs0Gwk89Sc1cf8ABMH9ha6uGupPgRCHY5Ij8Q6pGvHoq3IUflXhv9gv9jnwpcC50r9nnwjLIGyP7RtW1AA8fw3DOOw7evqa0Lw74f8AC2mx6N4Y0PT9IsIf9XaWFqlvEn0RAAOg7VVVVVVVVVVVVf8ABbT/AJID4F/7HAf+kc9fsnf8mr/Br/sn+hf+kENVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVftb/tQ/A79nDwI3/C3dVupp9djeOw0PTAHvb7bjcUBICIDjMjkKM4ySQD8fv2mfC/xe+Pnhj4u+HfhvdaJpXhiOziGlXOsfaJL0W91JOWMoiXyi4k2YCttxnJziv2Jv2+P2bvi7rH/Cv9O07UvAfjDWGTydM1i8W4gvnUHCW9yAoZ+T8jKjNxtDcgVX/BWD/kyPxh/2ENM/9LIq/wCCXH/JiXwx/wC4t/6dryqqqqqqqqq/bX/5NF+L/wD2J+of+iWr/gip/wAmr+Kf+ygXn/pBYVVVVVVVVVVVVVVVVVVVVfFr9pH4EfAq0kufiv8AFPQPD8sabxZTXQku5BjOUto90r8f3UPUetfGb/gtX4C0hJ9M+BHw01HxBdKSqanr7/Y7UHH3lhQtJIvThjEetfE//gph+2P8UGlhl+K0/haykzi08MQDTvLz/dmXM/5yHFT3HjL4h+IRJc3Gs+Jtd1B8BpGlvbq4b8dzsfzr4d/8E1P2zPiPDDe2nwfutBsplBFx4huotOK56Zhkbzh/37r4Yf8ABEfxZdyQXfxk+M2l6fEMGWx8OWb3TsO6iebywhx38txn1r4X/wDBMf8AY5+F1xaalD8NX8T6lZsrR3niS8e9yy85aD5YCc4/5Z1b21vaQJbWkEcMUYwkcaBVUewHSqqqqv2lv2W/hn+1d4T0vwb8T59aisNI1H7fbnS7pYH83y2j+Ysj5Xa54wOcVpH/AAS2/ZnvvjBc+BZr7xp/Z0MkqqV1SISYWPcOfJx19q/Z++APgX9mn4cwfC/4dTapLo8F3NdI2pXCzS75SC2WVVGOOBiqqqqqqqpiqqWZgAByTxitS8ceDdIyuo+KNMhYHBRrpS3/AHyDn9K1H49fDaxBEGqXN8wH3be1f+b7R+tal+01piZXSPC1zN6Nc3Cxfoob+dar+0T47vgU06LTtOX+ExwmRx+Lkj9K1b4geNtbyNS8U6jKp6xrOY0P/AVwP0qC2vL+by7aCa5mbJKohdj6nA5rS/hb8QtYIFn4Tv1B6NcR+QPrmTFab+zf4zulD6jqWmWQP8O9pGH4AY/WoP2YXODc+NFGDyqafnI+pk4/KtE/Zy8G2Egl1i9vdUwP9UzeSh+oX5v/AB6rD4L/AA009g8fhiKZgetxNJL+jMR+laf4e0DSQBpWiWFntHHkWyx4/IVVVVVVVVVVVVVX/BZL/k0W0/7HCx/9E3NfsB/8mafCX/sX4/8A0N6qqqqqqqqqv+CHX/I1fFv/ALB+lf8Aoy5qqqqqqqqqqr9tD/gov8Mf2XLK78I+GZLTxb8SShWLR4pcwacxHD3jqfl4OREp3txnYGD18Vvi78U/2hvH0vjL4jeIb/xHr+oyCKCPaSsQLfJBBEvCLk4CKOSe5JJ/ZA/4JEa14ttrL4gftRyXuhaZKqy23hO2fyr2dSMg3UnWAdP3a/vOeTGRg+CvA3g/4b+GLDwX4D8OWGhaJpkfl2tjZQiOOMdzgdSTyWOSSSSSTmqqqqqqqqq8ReLfDfhS2+0+INYt7MEZRHbLv/uoMsfwFeL/ANpC4mWSz8F6b5A6C8uwGb6rGMgfUk/Sta8Q674luvtet6rc30xPy+a5YL7KvRR7ACtB+Fvj3xGEfTvDlysL9JrgeSmPUFsZH0zXhX9m2GKRLrxjq6zhefslnkKfrIcHHsAPrWkaJpGgWSadounQWdunSOJNoJ9T3J9zzVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX/Bcb/kVPhH/wBhDVf/AEXbV8J/+SV+Df8AsX7H/wBJ0qqqqqqqqqqqqqqqqqqqqqqr/gtp/wAkB8C/9jgP/SOev2Tv+TV/g1/2T/Qv/SCGqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq8W+OvBHgHThq3jvxlofhyxOcXOr6jFZxcdfnkZRwKuP2yP2Tba6Wyk/aT+GzSMQAY/EtrInPq6uVH4njvWrfti/snaNYS6jeftJ/DWSKFcstp4ntbqQ/7scTs7fQA141/4LHfsmeGryWx8O2PjfxZsJCXWnaVHBA+COc3MscgBGcfu+3aoP8Agtl+z00yLc/Cz4hpEW+dkis3ZR3IBnGT7ZFfAPwzp/8AwUt/b38UePviQL2fwRpUUupJpVzIY3NhFIsNnZHy2IjzvV5NhwSJcHL7q8O+CfBvhDQI/CnhTwpo+j6NDH5aafY2McECrjGPLUBcY9q/4Kw/sjeAfhx4b0n9pv4P6Pb+E9Tt9YhtNZtdKQWsTs4ZobuNUwI5VdArFMbt6twVJb9lH4paj8a/2cPh98TtaZW1PW9FibUHVdokuoyYpnA7BpI3YDsDVf8ABWD/AJMj8Yf9hDTP/SyKv+CXH/JiXwx/7i3/AKdryqqqqqqqqq/bX/5NF+L/AP2J+of+iWr/AIIqf8mr+Kf+ygXn/pBYVVVVVVVVVVVVVVVU88NtBJc3EqxxRIXd2OAqgZJPtiv+G1/2Rf8Ao4/4f/8Ag8h/+KrwR+01+z18S/EVv4Q+H/xn8I+Idbukd4LDT9UjmmkVFLOQikkgKCT7Cq+Nn7YP7OP7PYltvif8UtKstUiXP9kWrG7vjkZGYIgzoD2Zwq+9fGT/AILZ30sk+m/AL4TRQxjIj1XxVMXc+4tYGAUjnBMrdsr2r4m/t/ftefFgT2/iL4263YWUxINlobLpcQX+4fs4RnX/AH2b3NeFvBfxC+KniBtK8GeFtf8AFmtXLF3h0+zlvZ3JPLsEDN16sfxNfBv/AII/ftL/ABCWDUfiLeaL8OtNk5KX0n269x2IghbaOOzyoR6enwn/AOCRf7KPgCK3ufGdjrXj/U4xl5NWvGt7bf6rBAU+X/ZdpK8B/Cj4Y/C2w/s34a/D3w74Yt2XDppOmRWvme7FFBY8Dk5Jqqqqqqqtf8TaB4XtBe6/q1vZRE4UyNy59FUcsfYCtE8b+GrX413PjC4v2TSppZSs5hYnDR7QdoG7r7Vo+uaP4hsl1DRNSt723Y43wvuwfQ9wfY81VVVVcXNtax+ddXEcKD+KRgo/M1P498D25KS+MNGDDqovoyRxnoDnpXiD49eANGVo7G7m1acA4S1j+UH3dsDH0zWr/tLeIrglNE0Cxs1PRp3adh9MbR+hrUvjL8SdTJEnieaBSeEto0ix9Coz+ZrUNb1nVju1TV728PrcXDSfzJrTvD2v6xgaTol/e5OAbe2eT+QNad8FviTqe0p4be3Q/wAVzKkWPwJ3fpWnfs0+KZyp1TXdNtVPURB5mH4YUfrWlfs1eF7ba2r65qF6w6rEqwKfw+Y/rWlfCX4daPta18LWkrr/ABXIM5z6/OSKtrS0sohBZW0UEY6JEgQD8BVVVVVVVVVVVVVVVVVf8Fkv+TRbT/scLH/0Tc1+wH/yZp8Jf+xfj/8AQ3qqqqqqqqqq/wCCHX/I1fFv/sH6V/6Muaqqqqqqqq+Nf7Zn7NX7Oviq18E/GP4k/wDCP61e6emoQWv9j313vt3kkjV99vC6jLwyDBIPy5xggn/h6P8AsJf9F1/8tnVv/kWv2yf+CtXinx2178PP2Yrm88N+HjuiufE7qYr++Xp/o4PNsh5+b/Wng/u8EH4SfBj4r/tD+OI/B3wy8L3/AIi1q7fzLmQZ8u3Un5priZvljTJ5ZiMk4GSQD+xV/wAE4vhz+zBa2njTxibPxd8SCgZtTeHNtpZI5S0RuQRyDMwDnsEBK1VVVVVVVVXiz4p+CvB4eHUdWWe7Qf8AHpa/vJM+hxwv/AiK8W/tC+KNZ32vhyCPR7YjAkB8yYj/AHjwv4DI9aA1jxDqOFF3qV/cN6NNJIf1Jrwl+zx4l1fZdeJblNItzz5QAkmYfQHC/iSR6V4V+Gvg3wcito+kRtcr1urgeZMT7Mfu/RcCqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv+C43/ACKnwj/7CGq/+i7avhP/AMkr8G/9i/Y/+k6VVVVVVVVV4w8a+Dvh9oM/ijx34q0nw9pFsQJb7VLxLWFCegLuQMnsOp7V4n/4Kr/sU+HLx7G3+JGo628TbXfTNEuXQHnOHdEVhx1UkcjFfBL43/Dv9obwHB8SPhfqlxf6HPcSW6y3Fo9u4kjIDqUcA8E9elX+taNpdxa2mp6vZWk99KIrWKe4WNp3PRUBOWb2HNVVVVVVVVVVVX/BbT/kgPgX/scB/wCkc9fsnf8AJq/wa/7J/oX/AKQQ1VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV47+N/wY+F84tPiP8WfB/hi5Me9LfV9bt7SV19VjdwzfgDWmftSfsza0qnSf2iPhrdMYw5SPxXZF1U9yvmZXqOCBivjt/wAFBf2XfgDYWc+v+PovEl9qMRltdN8LPFqU8kYON5YOIkXOQN7ruwcZwcWn/Ba/9mySWRb/AOGvxLhjH+raG0spGb6g3S449zWvf8Fp/wBmWxiddA8CfEXVZwuU32VpbxMcdCxuCw5x/Aa8W/8ABcLxNOzx+BPgBpdmoGEl1bXZLktx1KRxR47cbj9eeF/4LY/tGC8d3+GPw3NqUwkQt70SBuOS/wBpwR142jqOeOfjJ/wV2/aV+JvhVPC/hK10f4fNKuL3UdEMjXUvHKxySEmFSecp844+frmH9qX9py2lmntv2jfifFLcNumdPF98rSEd2Il5P1pf2t/2qlheBf2l/ikVdgST4vviwIzjDebkDnkA88Z6CtY17x18S/EK3niDW9d8Va7esEWW8uZb66nbsoLFnY+1Q/sx/tJ3Nm+oW/7PXxLltYs7508JXzIuOuWEWBVt+zh+0Pezpa2fwF+Is8znCRxeFrxmb6AR5NfDX/glR+2L8RrNNQvPBuleDbaQAxt4l1AW7sD3MMSyypj0dFNXH/BFj9quGB5YvGvwtuGUZEUeq3wZvYbrMD8yK/Y1+Isn/BPz9sDUvCXxwaO10jVdPbR9Vv7UNNDb73jkhu0OAzxh49jEDIDOcZXFaB4r8L+K9Ch8T+F/Eml6vo9xH5kWoWN2k8DqBksJEJUjHvX/AAVu/a0+HvjXwjpf7Nfwv1+18SatJrEd5rs+nOJ4rbygyxWwdch5WkcEhSSvlgHlsD9jHwNJ8Nv2Wfhr4LuXzdafocZu1JzsuJGaSZAe4WR3X8Kr/grB/wAmR+MP+whpn/pZFX/BLj/kxL4Y/wDcW/8ATteVVVVVVVVVX7a//Jovxf8A+xP1D/0S1f8ABFT/AJNX8U/9lAvP/SCwqqqqqqqqqqqqqqqv2gPjR8LPgV8M9T8ZfFvxINK0d4ntkSIb7m7ldDiG3j6vIRnA6AAliFBI8fW/wl1/VnsvgH4I+IKWtuzMZdf1a31CaaID7xhtrSIQnPX55BjvX7Mn7RXwg/Zg8T2fxS074VeNNU+IuiW8ttAl34ptk0uZpYWimeSEWImj27iVQSN6FhjJ+Mn/AAU6/a4+LzXFnb+Px4K0iYkLYeF4zZMF7ZuMtOTjg4kAPoK+H/wu+Knxt8SyaJ8OfBmu+LtZmbzbhbK2e4ZNx5klfogJ6u5Az3r4Kf8ABGL4w+Khbar8bvGml+CbJsNJptgBqN9jjKsykQxntuDyYx0r4Zf8Esv2OPhwkM174AuvGN/Fj/S/El+9wCe+YI9kJB94z9a8L+DvCPgjS10TwV4W0fQNOQ/LZ6XYx2kS9uEjAA/KqqqqqqqqtQvYNNsLnUbpsQ2sLyyEDoqgk/oK8JeG9U+OfizUPEPiS+mh061YBkjPKg52Qx54AAGScfq2ak+Bnwyks/si6A6NjAmW6l3g+uS2Pwxj2q9stU+AnxAtJ7W+luNGveTu4MsOcMrAcb1zkEeo6ZIpWV1DowYEZBBzmri5trWMy3NxHEgHLSMFA/E1f/Fr4cadI0Vx4ts2ZM58kNMPwKAg1rv7R3hGwJi0PTrzVXHRyPIjP0LAt/47WoftL+JZgw0zw/ptsD0MzPKR+RX+Vat8aviRq6tG3iFrSM/wWkSxY+jAbv1q91HUtVn8/Ub+5vJifvzytIx/EkmrXwl4qvlDWPhnVrgEZBispHz+QrRfg58RNckVY/Ds9mhPMl7+4C/g3zfkDWi/sywhVk8ReJ2Zv4orKHGPo7Zz/wB81pnwI+G2nBTLpE1869Gurhj+ikD9K07wZ4R0kA6b4Z0y3ZejpaoG/wC+sZrAAwO1VVVVVVVVVVVVVVVVVVVVf8Fkv+TRbT/scLH/ANE3NfsB/wDJmnwl/wCxfj/9Deqqqqqqqqqr/gh1/wAjV8W/+wfpX/oy5qqqqqqqq/aw/aN8P/ssfBTWPitrdoL+6hZbTSNO8zZ9uvpAfLj3dlAV3Y9QkbEAnAPw8/ZR/ay/4KZa7L+0D8UvGdhoeiXCmz0/Vb61JDwxyOfJsrWPGYY3aQbmZQWLcu24j40f8EZvjJ4C8NXfiP4W/EHTPiE1hEZZdMOmtpt5MoGSIE8yVJG6/KXUntkkKf2Gf2RYP2ufi1e+B9a8Zx+HNO8P2f2/VIliLXtxCJFjZIFI2qQzIrO5+Tep2v0r4NfA/wCF3wA8F2/gP4U+E7TRNMhw0pjXdNdyYwZZ5T80rn1Y8DAGAABVVVVVVVO6RoZJHVVUEszHAAHU14u+Ovgzw3vttOmOs3i8eXat+7U+8nT/AL53V4t+M3jfxXvtzqH9nWbcfZ7MmPI/2m+8fcZx7VoPhfxD4puvsmgaTcXsmfmKL8qe7MeF/EivCf7NrN5d34z1bHGTZ2Z5+jSH9QB9DXh7wn4c8K2v2Tw/pFvZrjDMi5d/95j8zfiaqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv+C43/IqfCP8A7CGq/wDou2r4T/8AJK/Bv/Yv2P8A6TpVVVVVVVV4i17SfCvh/U/FGvXiWumaPZy3l7cP0hhiQu7n2CqT+FftZ/tV/ED9q74mXvi7xNqNzb6BbTOnh/Q/M/c6bbZ+X5QcGVhgvJ1Y8cKFUeJfAXjrwZa6bfeMPBeu6HbaxG0mnTalpstql4i4DNE0igSAblyVyBketf8ABIuNJv2MNPicEq+vakpAJHBde4rxRqvin4LftJajfjWL281z4eeMpVhurqdpJWlsbwhdzMSTzFzmrK7g1Czgv7Vy0NzEskZIxlWGQcduDVVVVVVVVVVVf8FtP+SA+Bf+xwH/AKRz1+yd/wAmr/Br/sn+hf8ApBDVVVVVVVVVVVVVVVVVVVVVVVVVVVVq+raVoGlXmu67qdpp2m6bbyXN5eXcyww20MalnkkdiFRFUEliQAASa/4ax/ZX/wCjmPhT/wCFnYf/AB2vA3xT+GHxQhu5/hp8SPC/i2PT2Rbt9D1iC/FuzAlQ5hdthIVsA4zg+lVVVVVVVVVeX1jp0H2nULyC2iBwZJpAi5Puat7i3vIEubS4jmikGUkjYMrD1BHWviz/AMFEv2Rvg1r954S8UfFOO91vT3aO6sdHspr4wuvBRpI1MauDkFS+QRggVpP/AAVd/Yh1JVN58TtT0sspJF34cvm2kHofKifk9eOPev2jP+Cxfwo8GWlnp/7OWmx/EDUbuPzJtQv4biws7MZI2GOREmkfjOAEUAg7jyBaf8Frf2nEnVr/AOHfwvmhAOUhsL6Jj6YY3bAflXiL/gtR8ftW8K3mk6J8OvB+ia1cRFIdWh86b7KSfvpDIzKWA6byy55II+WtW/bF/ax1q/l1G8/aT+JUcsxyy2nie6tYwf8AZjidUUewArTv23f2vNLaJrX9o7x65hTav2nWZbjIxjkSFtx9zk96sdL+Ifxb8Wzrpem+IfGXifVZHuJlt4JtQvLpycs7BQzucnk89a1L4G/GvR3WPVvg943smYsFW58PXURJHXAZB0yM/WvBHwB+NvxJ8QxeFfBHwq8T6tqckxiMUWmSKsTA4PmOwCRAZGWcqB3Iqf8A4I0ftbRWUF1HrXw7mllj3PbJrFwJITgfKxNuFzzj5WI4PPQ1o/8AwRt/a41NkF9qPgDSQxIY3eszPtx3/dQP1/8A14rwf/wQ916ZUm+IHx/0+0IPzW+j6E9xu+kssseP++DT/wDBEX4Pm1jSP40+MRcAnzJGs7Yow7YXAI/M/hXh/wD4IifCi11FZvFHxt8ValZCTJt7LToLNyvpvYyjPTnb/wDW0P8AYT/ZB0HQtP0C3/Z78F3cWnQCJLi+0tLm4lxyWlmcF5GJJ5Yn0GAABJ+w5+yBLdRXjfs5+BfMhBCqukoqHPqg+VvxBxXgH4K/B/4VBz8NPhd4V8LvIpWSXSdIhtZJAf7zooZug6k9BVVVfFH9lb4NfHjxhqfgD4wW9zpt9FJNDpGu2TrFc2cu/dH8zAq8bgn5HBB3AjaSGH7S/wCxnefs6/tI+FP2c9P+Jb6zbePIdPmi1BtPNt5KXV7LbIssQkYSFGiLZ3DOei14X/4J5/BD9le3sNXvfEN145+ITzB4b27hFtb6fGAcvFbKzbXJwA0jueCV2kGvhhplzo/w/wBDsLtSsy2odlIwV3kvgj1G7FV/wVg/5Mj8Yf8AYQ0z/wBLIq/4Jcf8mJfDH/uLf+na8qqqqqqqqqv21/8Ak0X4v/8AYn6h/wCiWr/gip/yav4p/wCygXn/AKQWFVVVVVVVVVVVVVXiHxJ4c8I6PceIfFev6boul2a7ri+1G6S2ghHq8jkKo+pr4wf8FVv2Svhlp97H4Z8W3Hj7W7dWEOn6FbuYpH6DddOoiCZ6shcgchTwDrnxj+LH/BSv9qDwD4D+IWqRaVot/q/2Ww0vTUxFpVo2HuHTdkyTGKIku/UqBhV4Hww+FHw7+DHhCy8DfDLwnp+g6PZRhUhtYgrSkDBeR/vSSHqXYliepr/gox+yf8PPjf8AAjxb49Tw9Z2vjvwfpE+qadrEEISedLdDJJbSkcyI6K4UN91ipBA3A/8ABLT9jn9nf9orwd4j8e/FjQtU13V/DmuLaR6a980NiYTCjxuyxhXZ95kBBcrhV+XrnwR8PfAfw00OPw18O/Bui+GtKjORZ6VYx2sZbAG4qgG5iAMsck9zVVVVVVVVVVXi2xn1TwrrOmWi7p7vT54Yl9WaNgP1Ir9mrW7FLPVfDcsqpeeeLiNGODIu0K2B7FRn/eFV+0trVjeaxpGh28qvcafFLJOAc7PM2bVPocJnHoRWoftC+MptPTT9JtbLT9iBBMFMsmAMdW+X/wAdrXfEWueJrsX2vapPezgbVaVs7R6KOgHsK0nwB4112BbnSvDN/PC33ZfJKq30ZsA1ovwA+IOqOPt1ra6XH3a5nDEj2VNx/PFWH7MenoAdT8WXE3qsFqI8fiWb+VaZ8Afhxp5DT2V5qDA8G6uT/JNorSfDHhzQVC6LoVhZEDG6G3VGP1IGT+NVVVVVVVVVVVVVVVVVVVVVVVVf8Fkv+TRbT/scLH/0Tc1+wH/yZp8Jf+xfj/8AQ3qqqqqqqqqq/wCCHX/I1fFv/sH6V/6Muaqqqqqqqv8AguB4jgXwl8L/AAjDrFr50mo3t9caeJx5u1Y0SKUx5zty0qhiMZyAetfszw+Gof2dfhjF4PMR0YeEdM+xmMAZT7NHycfxE5Ld9xOec1X7L6Wdt/wWC8eweBmH9kHUvEAvRABsA2sZR7KLnaOO4HaqqqqqqqlkihjaWaRURASzMcBQOpJrxn8ffCugb7PQB/bV6uRujbbAh93/AIv+A5HuK8W/Erxf40dk1fVXW1Y5FnBmOIenyj731bJrwp8OPF/jR1Oi6S/2fOGupv3cS/8AAj1+i5PtXhL9nbw5pYS68UXT6tcDnyVzHCp/D5m/Ege1WOn2OmWqWWm2cFrBGMJFDGEVfoBVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVf8Fxv+RU+Ef/AGENV/8ARdtXwn/5JX4N/wCxfsf/AEnSqqqqqqqr9uP+0/8AhkD4u/2Rnz/+EUvN+P8Anls/e/8AkPfX/BK7wr4D8W/ti+HLTx1a2d39h0+7vdHtrpQyS38SBoztPBZE8yRfRowRyAa/4Ljf8jV8I/8AsH6r/wCjLav+CR9xb2f7Fljd3dxHDBBrupPLLIwVUUOCWJPAAAySa+Ovi7TfiD8bviF490cn7B4k8VanqVruGD5U91JKn/jrivhvYXOl/Dvwtpl4my4s9Fs4ZV/uusKAj8xVVVVVVVVVVVf8FtP+SA+Bf+xwH/pHPX7J3/Jq/wAGv+yf6F/6QQ1VVVVVVVVVVVVVVVVVVVVVVVVVVVftIv4Km+Avj7RPiB4903wZomv+Hb7SZ9b1AqY7P7VA8IfYWXzGBfIjDAuRgHJr4reCfgz4R8b2OgfDH43XPjzQJAov9dPheXSxbsZCG8uCSVmlAQBs5TJOPev+CVPhf9n74f6J4z0r4R/tJWPxGvvEMtrc3GnzaE+i3lisCyKSbeWV3kUmUZdflBAGTmqqqq6vbOyWNry7hgEsgjjMsgTe56KM9ScdKvLu0sLSa+vrqK2traNpJppnCJEijLMzHgAAEkngV4k/4KS/sReFNYn0LVfj3pstzbNtdtO0y+1CHPT5ZreB426fwsa8S/8ABWH9iPQtOa90v4j6t4imUHFnpvh28jlb6G5jiT82rxL/AMFxPBtrdsng79nrWdUtg2Fk1PxDFYOR6lI4ZgD043fjUn/Bcx3ikSL9mNYpDG3lO3jLzAr7Ttyv2JcjOM8jjNa//wAFcv2zNX1Sa+0zxP4b0S3kY+XZWWgwyRxDJ4DTeY54wOW7Vcf8FYf22LjTJtPX4haPFLKTi8j8P2oljBGML8mzjrkqTnvjivH3xK+IPxT1yTxL8R/Gms+JdTkZj9p1O8edkz1CbiQi9PlXAGBgcVbXVzZTpdWdzLBNGfkkicoy/QjkV8Ff2dvjR+0Z4gk8PfCHwLf6/cwjddXAZYbe2HrLPIVjQnspbc3YE8Vqv/BMP9uPSWPmfA6W5TeVV7XXNOm3e+1bjcB9QK8E/wDBML9s7xh4ig0S8+E7+HbZpdlxqer39vHBbqMZchHZ3HI/1atnn0OIf+CI3wgOkWkU/wAZfGA1RYkF1MltbGF5NvzlI9u5VLZIBYkDgk9ak/4IeeCWut8X7Qetrbb+I20CIvt7jd5wGffb+FfCX/gm7+yT8K/CcXhy5+F2m+Mrwv5lzq3ia2jvbmZyO3yhY0HZUUD1yck6j/wT3/Yv1RPLuf2fPDaDfv8A9HM1uc8943U456dOnpXwn/Z9+CnwLtp7f4SfDTQ/DJu1CXE9nbfv51HRXmbMjgHnBYjPNVVVVVVVVVXxN+EOl+PyupW1yLDVok2CbZuSVR0Vx7dmHI9+Mft2eFvGnh79uv4P6BrWqfaNSNjogsJlumcIp1a5CBWOCoDBjjjHWvBXwFng1ZPEXj7VE1G5VxILZWaQO45BkduW/wB3GD6kcVVf8FYP+TI/GH/YQ0z/ANLIq/4Jcf8AJiXwx/7i3/p2vKqqqqqqqqr9tf8A5NF+L/8A2J+of+iWr/gip/yav4p/7KBef+kFhVVVVVVVVWta9oXhyzGoeIdbsNLtS4QT3tykCbj0XcxAycHj2r/hbHwr/wCimeFP/Bzb/wDxdabqema1Yxano+o2t9ZzgmK4tplljkAOCVZSQeQRxVVXxs/bG/Zu/Z7klsPif8UtLstWjQN/Y9puvL3kZXMMQZkB7F9q+9ftIf8ABXzw74Q8NeGLz9nzwpYeJZvFekPe/bdZumjOkSrK8Rt57SP5jICm4/vVUgqVLKwavjZ+0b8af2iNc/t34ueP9R1xo3LWtmz+VaWme0NumI044JA3HuT1r4UfsB/tbfGSC21Dwp8HNVs9LuQGTUtaKabAUPR188q8i+8atXjj9n74vf8ABND40/Cb4teK3sPEEC3KXsk2lGQW/mRuVubHzHCncYGBDFVB3nAIRq+EX7S/wN+OXhS08W/Dv4j6LewXEQea0mu44buzYgZjnhZt0bAnHIweoJBBP/BRT9t/4UfDP4KeKvhX4P8AGOm6/wCO/F2mz6VHZaZdLP8A2bDOpjmmndCREwjZ9i53lipxtyw/4JHfBjXPhd+zC3inxNZS2l98QNVbVraGRSrLYiJI7diP9vbJID3SRDVVVVVVVVVXuq6Xpq79R1K1tRjrPMsf8zXiT4/+B9Dna009rjV5UOGa1AEY/wCBk8/gCPevEn7Rnim+uHTw1aW2m2w+40iCaU+5J+UfTH4mr/4rfEXUgRceL79c9RA4g/8AQAK02bWDqcVzpEl22ob90b25Yy7/AFBHOa0/xt8bNd1JfBdprF6L8gqYJI4oJVwuTudgGBA9TmrT9nDxfqDtda34isIZZmLOQXnck8ksSBk59zWn/szaFEVOqeJr65A6iCFYc/nurSfg98OdFuoL6y8NxtcW5BR5ppJfmH8W1mK5/Cqqqqqqqqqqqqqqqqqqqqqqqqqqqr/gsl/yaLaf9jhY/wDom5r9gP8A5M0+Ev8A2L8f/ob1VVVVVVVVVf8ABDr/AJGr4t/9g/Sv/RlzVVVVVVVftu/tueCv2RPBWxDbaz4+1mBv7D0IycAcj7TcYOUgUg8cGQgquMMyeMfGPxL+PnxIufE3ia+1PxV4v8TXgGEjaWaeRjhIoo1HCjhVjUYAAAAFfBf9tf8Aae/4J3On7P3xt+FL6ppFlGLqw0nU7r7Lc2kMrE7ra5QSRyQFxJxtcBwwDLhlr40/8Fo/iF4v8L3Xhz4OfDG28F3l5EY31q91P7fcQAjBMMYjREcdmbeP9kHBH/BMb9ijxD8AdF1L40/FcIfHXjK0WOC0MgmbTrJysrCRxnM0rhGYAkKEUZyWAqqqqrxt8QdB8AxWk2uJdMt4zrF5EYflQM5yR6iv+Gi/h/8A88dW/wDAZf8A4utf/aA8G6bpUV1pIm1G9uI9yWoGzys9PMbkL9Bk/SvGXxM8W+OJGTVdQMVnn5bK3ykQx0yOrH3bP4V4O+GXi3xvIraVp5isyfmvLgFIh64PVj7KD+FeD/gL4P8ADvl3WrodavU5zcLiJT7R9D/wLP4UiJGixxqqqowqgYAA9BVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX/Bcb/kVPhH/ANhDVf8A0XbV8J/+SV+Df+xfsf8A0nSqqqqqqqrxT4b0jxl4Z1fwh4gthc6XrlhPY3sJ48yGaMxyL+KsR+NfG/4P/FP9jz453XhW9vNS0nV9AvftWga5aFrc3cAc+TdwOp4yByAcqwZTypr4/wD7VXxh/acg8Lj4v6tYapd+FLeeC0vYLJLaWYSlC5lCYQn92vKqvetJ/ax+N3h74CS/s3eGvE8ekeDLueeW+js4AlxeCUgvHJNywjOACq7dwyGyCRX7Df7N+s/tL/tA+HvC0emSy+GtIuotR8S3RT91DZxtuMbNgjdKV8tRzksTjCsRVVVVVVVVVVVf8FtP+SA+Bf8AscB/6Rz1+yd/yav8Gv8Asn+hf+kENVVVVVVVVVVVVVVVVVVVVVVVVVVVX7QFx48/4KL/ALfVx+zzoniabTvA3gi9ubMNH+8itIrQ7Ly82ZAklklHloemGiGcZJ8I/wDBMn9i3wp4cTw/N8H4ddlMQS41HVr+4muZ2HV9yuqxk+kaoPav28P2J0/Yr1Dw/wDtM/sx6/q+i6XZ6vHFLaG5aWTR7k5aJ45WJZ4H2lGWTcckAlg+F+E//BUf9kzxh4J8PXnjr4qWPhvxXdadA+saZNpl8sVpdlQJUWbyTGyh84IY/LgnvXxw/wCCtH7MXwthhtPAl5efEnVZl3eVo58m2hH/AE0uJBjJ7BFf328VH/wXMtWgmMv7MkqTBR5Kr4yDqx77j9jBUfQHPtXxA/4LHftT+Jr0t4KsPCfg6yU/u47fT/tsxH+3JOWUn/dRa1f/AIKn/ttaxpdzpbfFW1tFuojG01podnDMgPBKOI8q2P4hyOoIPNeI/E/iXxjq02v+LfEWp63qlyczXuo3b3M0h9Wkclj+JqPXdci05tHi1m+SwcYa1W4cRNyTymcdST0718J/2Sv2j/jnok3iT4V/CPW9d0mCTyzfKqW8Dv3VJJmRZCMchSduRnGRWlf8EzP23tWvY7JPgbdWu4/NNd6vYxRoO5JM36DJ9q8F/wDBE7426tZLc+O/iv4R8PSuuVt7KCfUXT2ckRKD/ulh71ef8EPPG6QbtP8A2gtDmmzwk2gSxLjvyJWP6V8Jv+CMXwM8O6JLH8X/ABjrnjHWJnBWXTydMt7dR/CqAuzn1Zm9MKOct/wR8/Y7a9W6Fp4zEarg2w1z92T6k+Xuz/wLHFfBf9lP9nv9nyHHwm+F+k6PdsuH1GRWurx/XNxMWkAOT8oYL6AVqPgvwdq6yJq3hPRr0TPvkFxYRyb2znJ3KcnPOTWnaXpmj2cenaRp1rZWsQxHBbQrEiD2VQAKqqqqqqqqrWNb0jw/ZNqOtalBZ26nHmTPtBPoPU8dBzSfGz4XyP5a+KkyP71rMo/Mpiv+FyfDP/obLf8A78yf/E1rn7RXgvT2MWj217qrDo6p5Mf5v83/AI7TftPrkhfBBIzwTqWP/aVS/tPFoCIfBe2bkLv1Dco9/wDVgn6cfWtS+P8A8R72QtaX9nYLnhILVWx/383V/wALx+KX/Q0/+SUH/wARXxS/aT8XeFvh5rPiPxXrt5LpelWxnulsLeNJ5VH8K42DnIGMgepr4u/tJeB/iB+0p8P/AIyaRpeuxaN4Vk01r2C6giW5k+zXzzyeWqyMpyjgDLLk5zgc1+yv+2H8Lf2v9H1/WPhnpfiTTl8N3MMF5BrlpFBITKrMjJ5UsqlTsYdQQV6YIJqv+CsH/JkfjD/sIaZ/6WRV/wAEuP8AkxL4Y/8AcW/9O15VVVVVVVVVftr/APJovxf/AOxP1D/0S1f8EVP+TV/FP/ZQLz/0gsKqqqqqqqq/a9/ZjsP2tfhRF8LNR8YT+G4otXg1EXkNkLliYkkXZsLrwfM657V8CP2KtL+Mv7Xfjv8AZiufiDdaXa+DbnV4Y9Yj05ZXufsN2LcExGQBd4O4/McdOetfs7/Bu1/Z9+DHhj4O2evS61D4at5IUv5bcQNPvmeTJQMwXHmY6npVfH7/AIKA/sxfs7i503xN48i1zxFb7lOg+H9t7dK44KyEERwkHqJHU+gNftEf8Fb/AI//ABYjvPD3wthh+G3h6dWj32Mvn6lKh4ybkgeUcc/ulRlyRvPWvAfw2+KXxv8AFraD8P8Awprfi7X72QzTLawtO+WOWllc8KCTku5AyeTXwh/4Iu/GvxZZrqPxf8e6L4DVmGLC1gGr3QHffskSJewG2R+pJHGD+zr/AME4f2aP2dbq18Qad4bl8WeKLVg8WteIdtw9u45DQRBRHEQejBS4/vVXxm+G3w6+K/w41jwh8U/Cdr4i0CS3eaaznBVgyKSHjdSGjkHOHUgjJ55NfE+T4J3WofbPg3beM7C0eQ7rHxE9vcNEpH8M8O3dg8YMfQ/e45/4JxfD79lb4nfHPwz4J8eeDvFXinxFcW8t1HFqEsEWkRSwQtKweBd0k4ymAGYKf4kIOBHFHDGkMMaoiKFRFGAoHQAdhVVVVVVWu+LPDXhqMPr2uWdkSMqksg3sPZR8x/AVrv7Q/gfToCdGS61af+FVjMKfizjI/BTWo/tH+NblyNO0/S7OP+EGNpHH1JbH6VrfxK8d+IGb+0vE96UbrFDJ5Kf98pgH8ajiubybZFHJPK56KCzMf514Y+EXjvxTIDbaNJZW/e4vlMKfgCNzfgDXhn9mu5W7WbxdrVu1un/LCwLEv9XZRgfgfwqw+C/w008KY/DMUzr/ABXE0kmfqC2P0rTNE0bRIfI0bSrOxjPVbeBYwfrgc14d/wCTkr7/AK7XH/ok1VVVVVVVVVVVVVVVVVVVVVVVVVVVVVf8Fkv+TRbT/scLH/0Tc1+wH/yZp8Jf+xfj/wDQ3qqqqqqqqqq/4Idf8jV8W/8AsH6V/wCjLmqqqqqqv21f2yvBf7Inw8OpXZh1TxnrUUieHdE3f66QDBnmxysCEjJ4LH5V5JK+K/FXxO/aC+J0/iDxBd6n4r8Z+K75UVUjMs1zM5CxxRRqOAPlVUUAKAAAAK/4J8/8E+NF/Zl0WD4lfEq0tdT+KGpW/J4ki0GJxzBCehlIOJJR7op27mf48/sR/s6/tK+L7Px18XPCV7qmsWGmppsE0Gq3FqFt0kkkVdsbgE7ppDkjPPsK/wCHT/7Ef/RN9V/8KK9/+OVZWcFhZwWFqpWG2iWKNSc4VRgc/QVVVVVXxP8AHHwrtSun+J7WLXL2zLbLSFd5iY4zubIC9BkE59qDEIUAXBIPKjPHv1rwt4K8S+M7v7J4f0yScKcSTN8scX+8x4H06nsK8EfAHw3oAS98SMms3w5EbLiCM+y/x/8AAuPYUkccUaxRIqIgwqqMAAdABVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV/wXG/5FT4R/9hDVf/RdtXwn/wCSV+Df+xfsf/SdKqqqqqqqq+PH7OPwe/aU8KL4S+LfhGDVYbcs1jeIxhu7B2xl4Zl+ZM4XK8q20bgwGK8d/wDBD+B76a5+GPx7eKzZv3Nnr2jCSRBn+KeF1DHHpEvI9+PAX/BECyivorn4ofHiW5s1I82y0LRxC7884nmdgvGP+WR6+3PwS+Avwq/Z38GR+BfhN4Vt9H04P5lxICZJ7yXGDLNK3zSNgY54A4AAAAqqqqqqqqqqqv8Agtp/yQHwL/2OA/8ASOev2Tv+TV/g1/2T/Qv/AEghqqqqqqqqqqqqqqqqqqqqqqqqrx98SPAHws8PyeKviP4y0fw3pMR2m71O7S3QseiqWI3MeyjJPYVH/wAFOP2GZb3+z1+PNt5oYrubQtSWPIHP7w2+3HHXPNfHj/grh+zv8K7qw034dW1x8Tbq7h86eXS7oW1tarkgK0rqSZOM7QmAMZIzirj/AILi+Gjazm1/Z31MXIib7OJPEaFDJj5Q2IMhc9SMn2rxt/wVr/bP8T6xJf8AhzxpovhCzLHy7DS9CtZ0Ve2XukmcnHU5A9hX/BMH486N4R/bOvfFHxS12CG5+I+m3unS6lcbII/t9zcw3IZtoCp5kkJQAAKGkUcCq/4LD/Frwj4Z/ZsHwnuNTt5PEvjHU7WS2sFfMqWtvL5slwy9k3RrGCepY4ztbFh+wr+1zqvhfTPGWlfAfxJfaTq9ml5ZTWyRytLC6B0by1cuMqwIBUHnpXgD/gnP+2P8Qtag0i2+Cur6FFJKEmv/ABABp8Fup6u3mfOyjuI1dvQGpP8Agh98QAbYRfHzw8wYD7SW0WYeX0zs/eHf367c8dM8fDT/AIIpfBnQlW4+KnxQ8S+K7gD/AFGm28elW/0IJlkbHqHXPp2pP+CQ/wCxmrqx0DxS4ByVbXpMH24Ga+E/7OvwQ+B2gHw38LfhrouiWkibbh0t/NnuRjH76aTdJLx/eY9auvhL8Kr67XUL34ZeE7i6VmZZ5dFt3cFvvEMUzk9/WoYYbeFLe3iSOOJQqIi7QoAwAAOgx2qqqqqqqquLi3tYHubqeOGKJSzySMFVQOpJPAFXPxn+GNpKYZfFcLMDjMcEsg/NVIqL4vfDWYlU8W2gwP41df5itb/aV0Gzu3t9D0C41KJGIE0k/wBnD+6jaxx9QD7VF+09bMv77wZIpzwFvw3/ALTFeKf2j9YvY44PCempp3GZJrjEr59FH3QPqD+FJ8c/igrBm8Sq4H8LWUGD+SZq5+OXxNuU8seIliHfy7SJSfx25H4Vf+N/GOqNm/8AFOqTc/da7cKPooOBS6lqKMHTULkMDkESsCD+dXWoahfhVvL64uNh+USys+M+mTUHwr+IV1aQ3sHhS9aKdd0Z2gEj1Kk5H4iv+FT/ABG/6FG//wC+R/jWifAb4h6vIoutPg0yE9ZLuYZ/BVy2fqBS/sx6iVBfxfbBscgWbHH/AI9S/sx6mZdreLbUR4+8LRi35bv61ov7OXgyxjB1i9vdTlxz83kJ+Crz/wCPGv8AhQvwz/6A0/8A4GSf414r/Zj+Gnibw9qOgyae7R6hA0LRXTmeEq3BDIfvLjjBNf8ADtX4Kf8AQo+Df/BAn/xVfs//ALPnhT9nzStX0fwjp+lWdtq1xHPJFp9iLZS6qV3EAnJIwM+1VX/BWD/kyPxh/wBhDTP/AEsir/glx/yYl8Mf+4t/6dryqqqqqqqqq/bX/wCTRfi//wBifqH/AKJav+CKn/Jq/in/ALKBef8ApBYVVVVVVVVajqFhpGn3Oq6peRWtnZQvNcTzOESKNAWZ2J4AABJPtXxE/an/AGvP+Cg/xc1P4T/sl3Gp+GvA+nM2bizuW095LYNtF1fXQw8YfnECHkZG2QqTT/8ABIb9rLwgsnjnwP8AHbw+fGOWmlaz1G9sppXLbm23WwMzFgDlgoJ5JHWv2f8A/gpn8WP2cfEHiD4LftwaL4g1O88PRMLe7S3jk1SGZVDLBKdypOkikFJi2eQSzqwK/taf8FR/jD+0BHdeDvhwLn4feCpNySQWl0ft+oIRgiedcbUIzmKPA5IZnGK+D/7Pvxn+PurnRfhF8O9X8RyxyKk9xbw7ba1LdPOnfEcWevzMM4Nfs+/8EXLSEW2v/tKeOjcPgMfD3h1iqjjpLdMMn0KxoOnDnNfC/wCD/wAMPgr4bj8I/CrwPpPhrS0wWhsYNrTMBgPLIcvK+ONzsze9VVSxRzRtDLGro6lWRhkMD1BHpX/DPXwB/wCiG/D/AP8ACZtP/jdeHvg/8JPCOqxa74T+FvhHRdShVlivNP0O3tpkDDDAOiBgCCQeeQaqqqqdkRC7uFVRkknAA9a1z4r/AA+0Dcl54ltpZVB/dWpM7Z9PkyAfqRWvftMXJlaPwv4diWMH5Zb9yxb/AIAhGP8Avo1rPx0+IusQm3TVYrBGGG+xwiNj9GOWH4EVBbav4gvzHbQ3eo3s5yQitLI59T1JrwT8CvE2qXjjxVpEun2Uls4jlMyb0lx8h2A5xnqCBWifs0aPFEH8ReIbu4l67LNFiUe2WDEj8BWi/Bn4daGwkj8Px3ko/jvWM3/jp+X9KtbKysYhDY2kNvGOiRRhB+Qqqr4oeMdd8F6PaX+gaQmoTT3PlPG8bvtXaTnCnPUCrHxR4xsPHc3j6LwvI15MzsYGtpfLG5dpx36e9fDrxLq/i3wzHrGt6atjdNM6GFUZQAp4OG5qqqqqqqqqqqqqqqqqqqqqqqqqqqqqv+CyX/Jotp/2OFj/AOibmv2A/wDkzT4S/wDYvx/+hvVVVVVVVVVV/wAEOv8Akavi3/2D9K/9GXNVVVVVqHxI+Hek3sum6r4+8OWd3bttlt7jVYY5I29GVmBB9jX7RX7ZPwX/AGfPhfqXj+78XaPr99GPJ0vR9N1GKae+uWB2IArEqgxl3PCqD1OFPxg+LvxF/aJ+J2o/EXx5fy6pr2tzhIoIVYpAmcR28EeSVRc4VRkkkk5Ykn/gnL/wT4sP2f8ARbT4wfF3R4bj4majCTaWsuJF8PwuMbF7faGUkO4ztB2KfvlqqqqqqvFPjTw14NtPtWv6nHAWBMcI+aSX/dUcn69B3NePfjr4i8UCTTtB36RpzZB2P++lH+0w+6PZfxJr4ffCzX/iG8lxZyxWlhA+2W6mBI3ddqqPvHBB7D35rVf2Zb+GzMujeKYrq5VeIp7Uwhz7MGbH4j8a+CXjrUrDVG+GHia1W3nt962n7sIyOvLRtjg8AkN7dTkVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX/Bcb/kVPhH/ANhDVf8A0XbV8J/+SV+Df+xfsf8A0nSqqqqqqqr4neONP+GXw38U/EbVF32nhfRrvU5kzgusELSFR7nbge5Ffsj/APBVP9oT4w/GHwn8G/Ffwz8J69N4l1Fbb7bp7zafLbQgF5pmBMquI4ld9oVchMZGc1+1t+0p4d/ZT+DGpfFHWrRdQvvNWz0bTDL5Zvr2QEom7nChVd2PZUbHOBXxC/b6/a9+I/iCbX7/AOO3inRfMmMkVj4e1CTS7aAdkVIGXcoHHzliepJPNf8ABPf/AIKY/EfU/iRovwP/AGh/ER8Qad4jnSy0XX7lFF1aXbnEUUzqB5scjEIGYFlZlyxXO2qqqqqqqqqqq/4Laf8AJAfAv/Y4D/0jnr9k7/k1f4Nf9k/0L/0ghqqqqqqqqqqqqqqqqqqqqqvF/wC0p+zz4Bu5NN8afHLwJo17C22SzvPEFtHOh94i+8flxXxs/wCCxPwO+G3i+fwn8PPCOo/EOKz+W41az1CO1s2fGcQuVcygdC21Vz0LDmtY/wCC4Xh86BcN4f8A2f8AUBrZ4t0vNdQ2y/7TssQY4/ugDP8AeHWta/4K6ftmaprD6lYeI/DOkWzOCun2mgxPCgHYNNvkwfd8/Svi7+1H8fvjnrsmvfEj4pa5qDlsw2cNyba0th2EUEe2NOOM43HuSeamvdc8QXNva3F3falcM4jt43keZyzEAKoOTknAwOvFWf7Av7ZF/oNv4jtv2e/FJs7lA8avCkc+CMjMDMJV+hUfrXhr9gX9sTxVrFvotl+z94rs5J3C+fqdp9hgjHdmkmKqAOvXPoDTf8ERvjCYrdk+M/g7zGZfPVrO5AjGPm2kA7iDxg4z6ivg3/wRq+AfhPSHb4yeIdX8eaxOoB+zSPpdpb9/kSNjIx7bmfBH8K18Zv8AgjT8EPEPhSVPgj4g1Xwp4khcvA+qXT31pOMf6uQY3oOmHUkjurdviN46/wCCjn7IPijRfgHr/wAaNXin1uCMaKkWqw6jHLE8rQR+VNOpkhXcpAB2bRjgY4+An/BKn4leNPiDB8Xv22vHreIblZUmk0P+0ZNQuL5lxtS7umOBGMYMcZbI4DqODDDDbwpb28SRxxKFREXaFAGAAB0GO1VVVVVVVdeI/D1jMba917ToJV6xy3SIw/AnNS+LPCsNs13J4l0sQoDmT7WmBj3zWuftH+FbC5a20bS7zU1Q4M24Qo3uucsfxAqH9prQmC/aPDF8n97ZMjY+mcZrxB+0L421C7kbQ2ttLtckRKIVlfHqxcEZ+gAqD49/EuEASavbT4GP3lnGM+/ygV4n+K3jjxbAlrqWsNFAgIMVqPJWQ+rY+99DxSXVzGzPHcyqzn5iHIJ+tT6vq11EYLnVLuaNuqSTswP4E1ofw98aeJLE6jonh66ubYEgSgBQxHXbuI3fhmpfhd8Q4cb/AAfqRz02wlv5Zrwp8B/GniGRm1SD+xbZRxJdJl2PoEBz+Jx+NTfsyayDiDxVZOMcb7dl5/AmtM/Zm1VrpP7a8SWiWwI3i1jZ3YdwNwAH15+lN8Afhs0QjGm3YYAfOLt8n9cfpUHwC+GsL7pNMupx/dku3A/8dINad8Mvh/pYH2PwjppKjhpoRMR+L5NN4b8OvH5LaDpxTGNptUx+WKtvDfh2ymFxZaDp0Eq9HitURh+IGaqqqqqqqq/4Kwf8mR+MP+whpn/pZFX/AAS4/wCTEvhj/wBxb/07XlVVVVVVVVV+2v8A8mi/F/8A7E/UP/RLV/wRU/5NX8U/9lAvP/SCwqqqqqqqq/bnvNTsP2P/AIuXGkSSpcHwvdRsYgd3luuyXp28tnz7Zr/gizpHhy2/Zq8T61p6RNrF94wmh1GT+PZFbW5hQ/7IEjsPeRuewr/gtJ4c0W6+Pvw7bRIYm8R6z4eMF1EihWkQXLLbsxzkks0q5I6IBk4wP2ev+CLskd3aeIf2lvHEMkCFXbw54ddv3hznZNdMAQOAGWNcnJxIMAnwH8P/AAT8L/Ctj4I+HnhjT9A0PTk2W1jYwiNF9WPdmJ5ZmyzHJJJOaqqqqqqqta8WeGfDq/8AE716xs2AyElmAc/Rep/AVqv7QXw808MtncXuouOALe3Kgn6vt4rxd8efGOu3udCuW0ayThIoiGdvdnI/QYA9+ta14r8TeIj/AMTvXr29XsksxKj6L0H5Vongfxd4k2nRPD17dI3SURFY/wDvs4X9a8Nfs3a7elZ/FGqwafHjmCD99L9Cfur9RurT/wBnb4f2bq90+p3+OqTXAVT/AN8Kpx+NaN4e0Lw7bfZND0q1sou4hjClvcnqT7mqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr/AILJf8mi2n/Y4WP/AKJua/YD/wCTNPhL/wBi/H/6G9VVVVVVVVVX/BDr/kavi3/2D9K/9GXNVVVVXl3aWFpNfX11FbW1tG0k00zhEiRRlmZjwAACSTwK/wCCgkf7M+vfFvxb8Q/hR8c7zxn4m8Qa1591YWejbdOtUKYfZemTExBVcFFKnJ+biq/4Jc/sCL4TtNN/aa+M+iH+3LqMT+EtHuo/+PCJl4vZFP8Ay1dT+7Uj5B833iuyqqqqqkkjijaWWRURFJZmOAoHUk9hXiz4/wDh6wvZtH0BzcnyJVGoqN0cU207Nq/8tF3YyeB6Zq8vNb8Vaubi6mudS1G8fA4Lu7HoAB+QA4Hata+D3iPw14Mm8Wa88du8bxqtmvzuoZgNzkHC9RwM9ecV8EDZn4ZaQLQqSPN83HUP5rZz74x+GKrxkIn/AGidKGlkeeLm0+049QBu6f8ATPFVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV/wAFxv8AkVPhH/2ENV/9F21fCf8A5JX4N/7F+x/9J0qqqqqqqq/botdQu/2Pvi5Dpm/zl8LXcjbSQfLRd0n4eWrZ9q/4JqfEr4ffCn9rfwz4o+JWt2OjaS9ne2ianfSCOCzmlgZUZ3PCKeU3HAG/JIGSP+CsP7Ufhf47/FnQ/Afw28TWmueE/A9pJuv7GZZbe7v5yDK0cikrIiRpEgYEjd5mODk/8E5P2BfDH7WVl4p8cfFHUdcsPCmiypp9iNKlSGS6vSoeT53RxtjRoyQBkmVecAg/HP4dXPwA+PPiz4c6XrMtxJ4N12WCxvyAsjrG+6GQgcB9uwkDgHNeE9b/AOEm8K6N4j8vZ/aunwXezbjb5kavjGTjr6n61VVVVVVVVVVX/BbT/kgPgX/scB/6Rz1+yd/yav8ABr/sn+hf+kENVVVVVVVVVVVVVVftr/8ABSHwD+ypft8PvDWjp4v+ILQiSXTxceVbaWrgFGuXGW3spDCJRkrglkDKWh/bl/4KpeIbJvHOhfs/3P8AYJHmolr4BvZLdo/VCzNI6+rBiBg8jBr9kD/gqnoHxq8Z2nwi+OPhW18EeML6f7NYXdtI4sby5zj7OySZe3lLcKrMwY/LuDFQ1apqul6Hp1xq+tala6fY2kZkuLq6mWKKFB1ZnYgKPcmtY/4KL/sUaHqx0S9/aA0OW4DMu+ztbq7hyvX99DE0ePQ7ue2a+Mf/AAWR+BXgHxO3h34b+D9V+INvAo87Vbe7WwtWYgHERdGd8ZwSVUZ6Z618av8AgtH8SPFGhW+l/A/4fW3gi+Z2N3qepXCapKF42iFDGsanrkur9sAdauf+Co/7cF5o9zpFz8ZIyblGQ3Ueg2EM8YIwdjRwrtPoQMjPB6Y8Q/ELx94t1aXXvFPjjX9Y1OcsZLy/1KWeZyww2XdiTkEg89DXgz4VfFD4jsyfDz4b+KfFDRsVcaNo896VIGSD5SNg45r4S/8ABP79q/4seL7PwwPg14o8K2ksqi71bxLpM+m21pGfvSEzKpkwP4UDMTjjvX/Dkb40nUFi/wCFyeCfsO4Bpvs915oGOSI9mCc5438+orwH/wAEYf2ctF8Nmx8f+LvFvifWpQPMv7adNOijI/55QhXxnvvd+gxivgr/AME7v2UPgXf/ANteHPhxHrmr7Sqah4jk/tGSIHrsRx5SHr8yoGwSM44rR/h38PvD19/anh/wL4e0y9JObiz0uGGToAfmVQegA+gFVVV8dvClz48+CHxD8DWU5huPEXhTU9OikERkKPPayRg7Ry2Cw4HWpf8Agmj4ghdopfivZo6HDK2jOCD7jza/4Jk/s06t+zp4c8dw6l4iXV4/EN3ZSQSrYtbqoiSYHGWbd98dOlVVX2q6Xpiq2pala2gb7pnmWPP0yaufG3g60t3uZ/FWlCOJSzEXaMcD0AOT9BWoftG+BLWcw2dpql6qnHmxwqikeo3MG/MCpP2j/Av2ZpYrDVzKFO2J4EXJ7AkOQKvf2lvFslwzadoWkQwZ+RJlklYfVg6g/kKvf2iviBdQmKCLSbNu0kNsxYf99sw/Sr34mfEG/fzJ/GOqqc9IbloR+SYFN488cMpRvGeuMrKVKtqEpBBGDxuq2sL/AFAsbOzuLkr97y4y+M+uKOkaqJfIOmXfmf3PIbP5Yrwl8EfG3igGe4s/7ItgOJL1GRn/AN1PvficD3qb9mjxaob7Pr2kOQfl3mRc/XCnFeFP2bbe3n+0+MtVS6VRxbWZZVJ93IBx7AD69qn/AGevh3McxxajDznCXWfw+YGtP/Z9+HlhdrdSxX96FbIhuLgFPxCgEjjoT3o6DoTRJAdGsTHGu1ENsmFHoBjgUPCXhQEFfDOkgjofsUf+FIiRoscaqqqMKoGAAPQVVVVVVVVVVVVVVV/wVg/5Mj8Yf9hDTP8A0sir/glx/wAmJfDH/uLf+na8qqqqqqqqqv21/wDk0X4v/wDYn6h/6Jav+CKn/Jq/in/soF5/6QWFVVVVVVVqep6ZounXOr6zqNrYWFlE0tzdXUyxRQxqMs7uxAVQBkknAr9uD/gqp8OZ/C3iP4K/AbS7Txi+t2Nxpuqa/exk6fHDKjRyrAnDTttY4c4jHBHmA8fse/to/EX9jzxXfan4Z0+217w7raousaDdzNElwUzskjkAJilG5hu2sCGIKnCldb/4Le/D1PDjS+G/gd4hl19oyFt77UoY7WN+xMqBnYDrjYuemR1r9lD4EfHn9uP9pix/a9/aK0uew8JaPdw3tgk1u1vFfNA2+1tbSNsk2qNhmc5D4YZZndlqqqqqqq/1HT9Ltmu9Svre0gT70s8gjUfieK8V/tD+FtJL2vhy2k1i4XjzMmKEH6kZb8Bg+teJfjJ4+8TbopdZNhbt/wAsLEGEfiwO4/QnFWWnarrV15GnWN1fXDclIYmlc++Bk1pfwP8AiRqiq50NbNG6NdTKmP8AgOSw/KvCv7NtrbTx3XjDV0uwhybS0DKjfVzhiPoAfetG+GfgLw+wk0vwvZLIDlZJVMzqfZnLEfhWABgdqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv+CyX/ACaLaf8AY4WP/om5r9gP/kzT4S/9i/H/AOhvVVVVVVVVVV/wQ6/5Gr4t/wDYP0r/ANGXNVXxu+NXgb9nz4dX/wAUfiNcXkOhabLDFO9pbGeQNLIsaYQcn5mFf8Pef2NP+g54q/8ABC//AMVX7PX7Rvwz/af8FXvj74VXeoXGk6fqkmmTNfWhtnE6RRSsApJyNsyc+ufSq/4K/fHLxtPrHgr9kv4eXVxFN4vjjvdXit5Cr3wlnMFpanH8BkjkZlPUiPsOf2ff+CWv7M/wq8J6evxC8G2nj7xY0KNqOoasWktxKQNyQ2+RGIwc4LKznqT2H7ZP/BLb4KeJfhrrvjf4C+FR4S8Y6JZS3kNhYyubTVBGpZoTExIjkZQQjJtG7AYEHI/4JHftGeJfjL8E9Y+HfjXUptS1f4cXNvb297O5aSWwnV/s6OTyzIYZUz/dCDtk1VVVV45+JvhjwHAV1G5+0XzLmKyhIMjehbso9z+ANeOvir4p8du1vd3AtNOzlLKAkL7bz1c/Xj0ArwJ8L/EvjydWsYDa6eGxJfTKQgx1Cj+M+w/EivBXwz8K+BYg+lWXm3pTbJeT/NI3rjsoPoMe+a8QaJY+JNFvNC1FS1vexGN8dV9GHuDgj3FWlx8SPgPf3Fs1gL/RppN24oxgkPQMGH+rcgAEH06HANXv7R+v6pD9h8O+Eo4L2UbUczNcnPqqBV5+ufxr4QfDPXbfWpPiD42Ei6jMXaCCUfvAzj5pHH8JwSAvbJzjiqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/4Ljf8ip8I/wDsIar/AOi7avhP/wAkr8G/9i/Y/wDpOlVVVVVVVWvaJpXibQ9Q8N65Zpd6bqtpLaXlvIPlmhkQo6H2KsR+NftZfsl/Ef8AZT+It94a8SaTeXPhue4c6Dr4hJg1C3zlMuBtWYLgPGcEHkZUqx+Gfww8e/GLxlp/w/8Ahr4Zvdd13UnCw21tGW2rkAyO3SONcgs7EKo5JFfs4/Brwv8Asjfs6aR4CutVtvI8NWE1/r+q7SqTTkGW5nPfYOQueQiKO1fELxDrP7Rf7Qmu+IdKtHOo/EPxZK9hbMPmQ3VziGLjP3Q6L36d60fS7XRNIsdFslC29hbR28KhQuERQo4HA4A4FVVVVVVVVVVV/wAFtP8AkgPgX/scB/6Rz1+yd/yav8Gv+yf6F/6QQ1VVVVVVVVVVVVVX15Bp1jcahckiK1iaWQgZO1Rk4H0Ff8Et/h7pf7TX7TnxG/aI+K9jBrmoeH5U1KG3uwJY11C+lmZJdjfe8pIZQgxhSVIwVWq/4LQfA/wno+geD/2i/D1hDpniGfW10XVJ7ZRG17vglmglfHWSP7O67+uGAOQq48R/8Fdv2wdc8OReH9M1PwroMsdskDanp+kF7uXaMGRjO8iB26kqijPKha8d/Fv4pfFG8kv/AIjfEbxH4lmkfcTqmpy3IBHTarsQoHYAADtXgT9kD9p34neHP+Eu8DfBDxTqmjscRXa2RiSb3i8wqZB7pke9fCP/AIJrftbfFPxPbaLf/C/UvBumNJi81fxFD9kitkAyWEbfvJT0ACKck8kAEjSv+CH/AI/k1G3TXPjx4fgsC/7+S00eaWVV77VZ1BPbkj+lWf8AwSE/Y5t7W0hutH8U3U0EMaTStrbobh1UBnYKMAsQSQuAM8ACvBv7E/7JXgPTrbTtB/Z68Dyi0A8u41PR4tRuMjncZrgPITnuWqw0+w0uyh07S7K3tLW3QJDBBEI0jUdAqjAA9hVVVVVVqHirwvpM/wBl1TxJpdnMP+WdxeRxt+RINfEfxvrvxE8VR/DvwHeo1kzbJJoJfluTjLEuv/LNRngdcHrxVh+zNoy2ijVPEt7Jc4+Y28aogPoAck/Xj8K13w740+A2q2+t6Jqwu9NuZCmWUhJDjOyWPPXGcEHscEVH+0r40AIl0bRGOPl2xSrj/wAiGtS+LnxF1Odp5fFV3Dk8JbEQqvsAuP1qT4j+PpUMbeMtXwRzi7dT+YNMb/VbosxuLy5lOSTmR2P6k0PD+vMQq6JfknoPsz/4VoHwL+IOvWhvGsINNTGUW/cxO/0UAkf8CApv2e/iKGKrBp7AHAYXXB/TNeH/ANmiJrQyeKdfdblj8sViBtQe7MOT+Ax71b/s1+DI5A9xrGrzKP4A8a5/HZWm/B/4caZEI4vC9tOcYL3JaYn3+YkD8MVJ8Mfh7KmxvB+lgf7NuFP5jmtK0XSNDtfsWjaZbWUGcmOCIICfU46n3qqqqqqqqqqqqqqqqqqqqr/grB/yZH4w/wCwhpn/AKWRV/wS4/5MS+GP/cW/9O15VVVVVVVVVftr/wDJovxf/wCxP1D/ANEtX/BFT/k1fxT/ANlAvP8A0gsKqqqqqq/aX/at+EX7Kvg//hJfiRrW6+ukb+ytEtCHvNRcdo0J+VAcbpGwq565IB/au/bo+NX7WGqS2niXVDofg6KXfZeF9OlZbZNp+V5m6zyjj5m4BztVMkV+yz+wL8dv2p7qDVNB0j/hHfBvmbZ/E2qxMkBA+8LePhrhuCMLhQRhnWtK+DP7Hn/BNH4R/wDCaeK9An1Z9Tnj0rUvEF5pwv7y8eVXbyxH92KEiNsogAOF3FiM1+zf8ff2b/h1+214++MPjvQIpPh3qt5rMugWq6Ek4thPeiS122+MRbYgQMY29BXwj+KPhH41fDnRPif4DluZNB12F5LJrmAwyFUkaM5Q/d+ZDVVVVVa1q+n6Jp8t7qGoW1oiI2155AgLY4HJ5+lf8NBfEb/n6sP/AAFH+Nat8bfiPq0QgOu/ZExz9kiWIn/gX3h+BFXl/qWr3Imv725vbhzgNNI0jkn3OTXhb4MeOvFBWX+zDptqes98DHx7J94/lj3rwz+z54M0YJPrTzazcAciU+XED7Ipz+ZI9q0/S9N0m2Wz0qwtrOBekUEQjUfgKqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/4LJf8AJotp/wBjhY/+ibmv2A/+TNPhL/2L8f8A6G9VVVVVVVVVX/BDr/kavi3/ANg/Sv8A0Zc1VfE34W+AfjL4PuvAHxM8OQ67oF7JHJcWM0kkau0bh0OY2VuGUHr2r9ub9nf4MfCv9uX4SfCv4f8AgS10fwr4hstFk1PTYp5nS4afVbiGUlndmG6NEXgjpxg818Ivgl8LPgL4aufB/wAIvCFv4c0e8vnvp7WCaWUPcMiRtJmRmOSkUYxnHy/Wq/4KTfFXwj4S/wCCiHgj4gaBqaaxN8PotEuNatYefIurW9kuDBnoW8oxEjJwWIJBBA8G+MvC/wAQ/Cml+N/BWt2ur6HrVstzY3ts+5JY2HBHoRyCpwQQQQCCK/aD+M3hT4BfCHxJ8TfFup2ttFpdjKbOGZwGvboofJgRf4md8DHYZJwASP8AgiD4C1iz0D4n/Eu8tpI9N1S5sNLsZCMCWSBZpJ8eu0TQjI7k+lVVVUskUMTTTSKiRqWZmOAoHUk9hXxJ+PyQmXRfAcqu4ysmosuQPURA9f8AePHoD1qysdb8U6sLWyhudR1G7ckjJd3PdiT+pPTvXgL9nzTdNEWpeNXS/uhgrZIT5MZ/2j1c+3A+tQQQW0KW1tCkUUShUjRQqqB0AA6CqrxH4u8N+E44H8RapHZrclliLozbiMZ+6D6ivgL4u8N+ErzWJfEWqR2S3MUSxF0ZtxBbP3QfUVpmp2Gs6fBqmmXKz2tym6KQAgMPXB5qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv+C43/IqfCP/ALCGq/8Aou2r4T/8kr8G/wDYv2P/AKTpVVVVVVVVWq6RpWu2Eula3plpqFnOMS211Assbj/aVgQfxFaFofwr+Gwm0zwzo/hTwt55DzW9jb29jvPZmVAufqa/4S3wp/0M+k/+Bsf+Nah8Nf2btY8X6d49vvB3w/m8T6XdC5stXFrbLdxTDowlGHJB5GSeeetVVVVVVVVVVVX/AAW0/wCSA+Bf+xwH/pHPX7J3/Jq/wa/7J/oX/pBDVVVVVVVVVVVV48+IvgL4XeH5PFXxG8Y6P4a0iJgjXmp3aW8Zc9EBYjcxwcKMk9hXjr/grH+xl4NLxaZ4y1vxbPEDui0LRpTz6B7jyUb6hiPevjj/AMFpPH2s3UNj+z74DsvDtggzNfeIYxd3UpzwFjRvLjXGM5Lk54Ixz8X/ANv79rH41w/YPFHxc1PTdNKbG07QT/ZkMgxz5nk4aQHrh2YegFf8E+P2urH9kn4wXOr+K7O6u/B3iizWx1pLVA81uVbdDcIpI3FGLArnlZHIywUHTf24P2QdV8Pf8JNa/tG+BEswm/yrnV47e5x/17SFZs+2zNft2/tPn9vn4neCf2b/ANmrTb3WtJtdUZ4buSB4Rqd6ylBKEYbo4IYjIS7gHDuSAFBPg/8A4Ir/ALO2lpbT+M/H/jnXbmNVM0VvcW9nbyN/F8oiZwp4xhwR688fCn9kj9m34KWUVr8O/g94csZ4sf6fcWgu7xiO5uJt8nXnG7APQCqqqqqq48ReHrS4Npd69p0M6nBikukVgfoTmvEHxs+H3h+6NjJqj30yHDrZR+aEPu2Qv4AnFa3+0n4agtM6Bo97eXJOALkLCi+5ILE/TH4il/aV8Y72LaLoxX+EBJQR9Tv5/StU+NXxH1S4Mw8QNaLn5YrWNY1X9CT+JNT/ABR+IVzA9vL4u1ApIpVgsm04+o5FWemarq8rrp2n3V7Io3OIYmkIz3OAa/Z7eLT/AIizWeoIYbiTT5Yoo5V2sJAyMRg8g7Vaq/aCuLOH4dTQXDL5s91EsAPUsDk4/wCAhq8DfAbSvFfhHTfEF7rF9aXF4js8SxqygB2Ckd+VAP414e+BHgHRIWF7ZSatM45kvG4X/dVcAfjk+9L8Kfh0jBl8IaeSDwDGT+ma0zQ9G0WLyNG0mzsUPVbeBYwfrgc1VVVVVVVVVVVVVVVVVVVVVVVVVf8ABWD/AJMj8Yf9hDTP/SyKv+CXH/JiXwx/7i3/AKdryqqqqqqqqq/bX/5NF+L/AP2J+of+iWr/AIIqf8mr+Kf+ygXn/pBYVVVVVX7ePxz/AG5vhR8TLG1/Zs8AXOseC4/DEd7qV+vhw38cF2J7kShpRwu2GOFivYHPev8Ah7z+2V/0HPCn/ghT/wCKrx54/wDij+0D8RpfFPjLVdU8V+LNfuEhiVYzLLKzHbHBDEg4GSFWNBjnAHNfsS/8EmtM0SKw+J/7VOmwajqLAS2Pg0sHgtvRr0g4lfv5IJQfxF8lVtLO0sLSGwsLaK2traNYoYYUCJEijCqqjgAAAADgV8WPg38M/jn4WTwV8V/Clv4h0WO7S7W0nlkjUTIGCvmNlbIDsOuOa/4du/sR/wDRAtJ/8GF7/wDHq8AfD/wf8LPB+m+APAGiRaRoGjxtHZWMTu6wqzs5ALkscszHknrVVVTSwwRNNPKkcaAlndtoUDuT2rxD8cPh7oLNDHqr6nOnBjsU8wf99khPyJrxP+0Z4k1JWtvDVhDpURGPNc+dL9RkbR+R+tXF5rvirVYhd3d1qV/dSCOPzZC7MzHAUZPHJ6dK8Pfs16LHZI/ijWbua7YZaOzZY40PpllJb68fSviZ8DH8IaXJ4h8O3817Y2+PPhnUGWIE43ZUAMM9eBjryM4+Dd54c1vwhbavpWgafp95Exgu/s1uqHzFAyc9SCpU8njOO1VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVf8Fkv+TRbT/scLH/ANE3NfsB/wDJmnwl/wCxfj/9Deqqqqqqqqqr/gh1/wAjV8W/+wfpX/oy5qqr/gpN/wApJfgV/wBg/wAPf+nu7qv2tPjpa/s4/s/eLviszxHUdPszBo8MoyJr+Y+XbqVz8yh2DsB/AjHtXhrw/wCOvjj8T7Hw7pj3Gt+LfGmriNZbiQs9xdTyfNJI/OBlizMegyT0r4p+A/j7/wAE/rvUNP8Ahj+1/wCGrq3F8tte6N4Z8VBL6GZkJ3XOlsxKEBcb8NjK5I3Yr4RfALxt+234j8MeIP2iv21vB6w6pIqWWi3vjKG/1xt7Y8iCxL7bd3xjB5BIPlt0r4VfCvwP8FfAGj/DP4daNHpmhaJB5VvCDlnJOXkdurSOxLMx6kmqqq8Q+I9F8LaZJq2uX0dtbx8At1duyqOpJ9BXxK+L2s+O5X0+zL2GjK3y2wb5psdDIR1/3eg9yM14A+F/iDx/ch7RPsunRtia9lU7R6hR/E3t0HcivB3gXw74G0/7Dodph3A865k+aWYjux9PYYAqqqvGPgLw947itYdfjndbNmaLypSmC2M5/IV/wz58Of8An1v/APwKNaJo1j4f0m10XTVcW1nHsiDtuIHuaqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr/guN/yKnwj/wCwhqv/AKLtq+E//JK/Bv8A2L9j/wCk6VVVVVVVVeINd0jwtoGpeJ9fvY7PTNIs5by9uJPuwwxIXkc+wVSfwr9rL/gob8bf2jfFeo2vh/xVq/hHwEsrR6doWnXbW5mhHAe6eMgzOw5KElF6Acbi7vI5kkcszElmY5JPc1XhL/katF/7CEH/AKMWqqqqqqqqqqqr/gtp/wAkB8C/9jgP/SOev2Tv+TV/g1/2T/Qv/SCGqqqqqqqqqrx/+3J+yX8NrbUZPEXx38KzXOlkpPY6Zei/uTIP+WaxQb2LZ4PZT94jBx49/wCC2nwg0l5IPhv8IPFPiNkbCy6peQ6XG/uNonbH1UH6V8YP+Cv37TvxDs59H8CW2g/D2xmyPO0yE3N9tPBXz5sqvH8SRowPQ+ni/wCIPj34g3g1Dx54317xJdK7Os2ralLduGbG4gyMSCcDP0FfCv8A4J2ftefGDRrHxL4Z+FE1nouowJPa6hq99BYpLG4yjqkjiVlYcghCCCD3Gfhb/wAES/iBe39rd/Gb4t6Fpdgrq09l4eglvJ5E7oJZVjWNj03bZAPQ18PP+CVH7G/gDVl1m48Fap4rmiYNDF4i1FriGMj1ijEaOPaQMOelfEn9kf8AZu+K/hD/AIQnxd8HvDX9nIB9mbT7FLCa0YDAaKWAKyY44B2nGCCOKvf+CKn7NNxqhubX4hfEe2sWbJtVvLNmUeiu1uePqCfc1+z5+yJ8BP2YbSdPhP4LjtdSvYhFeaxeSm5vbhAQdplb7qEgEogVSQCRkZqqqr/WNI0squp6rZ2hcfKJ51jz9Mmtb+IXgrw/ai71PxJZKrD5Fik813+ipkntz0q6/aM8BQxube21W4dR8oWBVDH6luBWoftNaq8hGleF7SJAePtE7SE/98hcV4g+M3j/AF69NzHrcumxAYS3smMar+PU/iaufiP48u7RrG48W6m8L/eU3BBPtkc49qgsb67R5bWznmWP77JGWC/UjpXhr4b+M/FkuzSNDn8sD5p518qMf8CbqfYZNWn7O/j+edYrhtNtoyfmka43Y/BQSaj/AGZdEFvGs3ie9M4H7xlhUKT7DqPzNaH8APh/pSE39vc6rKR965mKhfoqbf1zS/Bz4ZqwZfCduSDkZlkP6bq0vR9K0S1Fjo+nW1lADkRwRBBn1wO/vXxc+FeqXmqf8J94Ln8nUYcSXMQkERJQcSoxwAwAGQeuM9c5g/aG8faVD/Z+qaVp09zEMGSaF43J9WCsB+QArSfDfj74361Dqnim7+y6TbHkKAgjU9Vij5OWx95vzOAKsbK102yg06xhWK3toljijXoqqMAfkKqqqqqqqqqqqqqqqqqqqqqqqqqqqv8AgrB/yZH4w/7CGmf+lkVf8EuP+TEvhj/3Fv8A07XlVVVVVVVVV+2v/wAmi/F//sT9Q/8ARLV/wRU/5NX8U/8AZQLz/wBILCqqv2tf2vvBH7IHhzQfEnjfwzrmtQeIL2S0gj0oRFo2RN5LeY6jGPSv+H2nwB/6JR8QP++bT/49Xw38baf8Tfh14W+JGk2lxa2PivRbPVraC5x5sUdzCkqq+0kbgrgHBIz3r4q/Ezwn8Gvh1r/xQ8c3xtdE8OWbXN06rl2xgLGg7u7lUUcZZgO9XPx3/wCCg3/BRrxJrOlfApLzwd4DtHNvPFp999gt7dHB+S5veJJ5GUjdGnGCD5YByfip/wAEnv2uPhh4bn8T2+k+HPGVvZxGW5g8NX8k9xEgBJIimiiaQjH3Yw554B5x/wAEYrL9n7UtW8Sx33h0v8YNKU3Ntd37LKq6ccRs1ouB5TqzBZCcsRIuG2llFVVVVXd1a2NtJeXtzFBBCu6SWVwioB3JPAFeNv2idPsHew8F2iX0qkg3lwCIgf8AZXgt9TgfWvFPjrxp40g8/XdSnms4nCiONPLhVjkgELwWwDjOTwa8NfDXxr4sVJdH0KZrdulxN+6jx6hmxn8M14e/ZobKTeKvEAwDzBYp1/4Gw/8AZfxq70TRfAPx202yit/s2lpLA0AkcnG6MLvLH/ppk+g9qr4iXtlp/gXXbi/ZPKNhNGAxxuZlKqv4sQPxr9mm2uI/C2qXLgiGW+2xg9yqLuI/MD8Kqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/4LJf8mi2n/Y4WP/om5r9gP/kzT4S/9i/H/wChvVVVVVVVVVV/wQ6/5Gr4t/8AYP0r/wBGXNVX7ev/AAUY1r4K+LU+AP7PWkw658RpzHHfXT2pul06SUAxQQwr/rrlgynBBVdyghyxC+IP2Jf+Cmnx48Q6Z8cfiDB5niuwjhbTJNV1i1tby2WKQzRBIo8JDtkdmCHaQxOQK17/AIKJf8FD/wBmnU3+F/xhsdMfXbNRIreJdEVp3iPCsktu6JMmQR5g35II3Eg4/bO/bq8dftVeGvBPgnWbeKwtfDtot1rUUELQx3mrsGDuEYkqkSN5agnO4yHJDDH/AARq/Zxvdc8d6t+0p4i00rpXhyGTTfD7ypjzr2Vds8qZ7RwsyE9zOR/CcfGf/gk78Gfjb8UvEnxX1/4j+NLHUfE16bu4trM23lRMQBhN0RbHHcmvhR/wSP8Agr8I/iX4Y+J+ifErxteX/hbVINQtre6Nr5UrxMGCvtiB2kjnBBqqqviF8UNA8AWpS4cXepyLmGyjfDezOf4V9+p7A14t8Za/421M6nrt55jDIihQbY4V9EXt9ep7k18MfgVd60Ytc8ZwyWthw0VmcrJP7t3RfbqfbrVpZ2mn2sVjYW0VvbwKFjijQKqAdgB0qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr/AILjf8ip8I/+whqv/ou2r4T/APJK/Bv/AGL9j/6TpVVVVVVVV+27ZavqH7InxctdDDG6/wCETvXIUcmJYy0oH1jVxXwk+Hl/8W/ij4T+F+l3kVpdeKtZttNjuZRlYPOkVDIR1IUEnA5OK8Hf8Emf2LPDmiQab4g8Bar4rvY0Al1HUtevIJJG7nZbSRIoJ7beOOe9f8OuP2Ev+iF/+XNq3/yVVp/wTC/YZsbuC+tPgdsnt5Fkib/hJdVO1lOQcG6x1A61VVVVVVVVVVVf8FtP+SA+Bf8AscB/6Rz1+yd/yav8Gv8Asn+hf+kENVVVVVVVX7bv/BSrwn+y9qz/AAy8BaDb+LviD5Ie4hmmKWek7gCnnlfmkkYEHylKnaQSy5AMf7VP/BXzXrL/AITnSvhBq0Wj48xbOHwN8rp1+WOQG4ZSOhBORjB71+0B/wAFBP2uvjBd33hrxh4xufCNiLdrC/8AD2iW76bE2V2TLMGJlYtzuR3KjJAUV4G+GfxF+J+otpHw48Ca/wCKL2Mr5kGkabLdtFuJClxGp2g4PJwOD6V8Lv8Agjn+0342sbXVfHOseGPAttcKGa1vbh7u9jB6ZihBjBxjgygjOCAcgfDj/gid8ItFlguvij8WfEnidoyGa20y0j0uFz3Vixmcr2yrIe/HSvBH7Hn7LPw7s9PtfCvwA8DQy6WQ1re3Wiw3l4jDo32mdXmLe5cmqqqqtX1zRtBtvtmtapa2UPZ55QmT6DPU+wrxF+0F4G0mIrpDz6xcY4SFDEgPu7gfoDR/aduSkoHg6IOVPlH7cSFOOMjZyOncVP8AHX4nzTPJH4iSFWJIjSyhKr7DchOPqTWp6nq/iTVJNQ1K5mvb66f5mIyzE9AAOg7AAYHarLQNc1K4W00/R725mb7scUDMfyAq0+APxHukR5NPtLfeAcTXS5XPqFzWkfsyzNGH17xQiPjmO0gLAf8AAmI/9BrQ/wBnTwfpl8l5qV9eamsZysEoCIT/ALWOSPbIr/hUPw284Tjwlabgcgbn2/8AfOcfpVjp9hplsllptlBawRj5IoYwir9AOKqqqq13S11zQ9Q0V5jEL+0ltzIFyU3oVzjvjPSv+GYrD/ocJ/8AwCH/AMXXw2+GsHw5gv4INXe++3OjEvCI9m0Edic9aqqqqqqqqqqqqqqqqqqqqqqqqqqqq/4Kwf8AJkfjD/sIaZ/6WRV/wS4/5MS+GP8A3Fv/AE7XlVVVVVVVVV+2v/yaL8X/APsT9Q/9EtX/AARU/wCTV/FP/ZQLz/0gsKqq/as+K37H/wAPNF09P2p08G6oqs0+l6TrOjR6vcMx+VnhtykjKD90vgL2LV+zx8Wv2T/DP7Zvjn4gfFbwVpF18KNau9YOh6deeGY723so5rwPaYtNjLGEhBUBFOwfKBivg78QPhH8RvAOl6v8Edf0LUvClrAlpYpo6rHDZpGoVYPJABg2KFAjKqVGOAMV/wAFf/2uPDet2Uf7K3ga9W+ubHUIr3xVeQyZjgkjUmKzBHDOGZZH7KVRfvbgv/BPvwv4d8K/scfC218N28Mcd/oceoXTx4JlupyZJmYjqwdivPICheigCtD0yw+HX/BbKfRfBCR2ljfatI9xDAoCMbvRftFwMDj/AF0jtjsR7VVVVa1r+ieHbM32uapb2UIzhpn27iOyjqx9hzXiz9pG2iLWvg3SfPIyPtd6Cq/VYwcn6kj6V4m8ceKvF8u/X9anuUBykIOyNPogwPxxn3rwL8H/ABV42Md0If7O009bu4Q/MP8AYXq36D3rwp8LPCfhXSJdIitPt6XEkcs7XgEm90+6duMDBzge/egAAFUAADgYqvi58MIfH2nxXtlcRW2q2CkRSS8JInUox7c8g9ufXI0z40fEPwdENGnvdM1VbcbUadxc7QPSSJxu/EmoL7x58c9Xt9Jv9c0+1tVfcIBKkapgcssW7fIQM4znHqAc14Y8Oad4T0K10DS0IgtUxuYfM7HlmPuTzVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX/BZL/k0W0/7HCx/9E3NfsB/8mafCX/sX4/8A0N6qqqqqqqqqv+CHX/I1fFv/ALB+lf8Aoy5qq/4JraRY/Fb9v34p/E3x1Cl1rukpqmpWcNyCz291PerE0i9QPLjkeMDPHmDA44r/AILK+BvDOtfszaZ45vrWFdc8OeIreKwudoEhjnV1lhz12naj49YwfWvgV8Kr344fGDwl8JtP1S106bxPqcdobu5cKsCHl2GSNzBA21OrNhRyRXw0+HPhH4R+AtD+G3gTTFsND8P2aWtnCOWIXq7n+J2OWZjyzMSetVVVXxU+Ntt4baXw/wCFJIrnVBlZrj7yWp9B2Z/boD1zyKvb271G6lvr+5kuLidi0ksrFmYnuSa+Bnw48Ky6Xb+Mbq6g1TUCflh4KWTA8Ar/AH+hyemRgd6qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv+C43/ACKnwj/7CGq/+i7avhP/AMkr8G/9i/Y/+k6VVVVVVVVeK9T8P6L4W1jWPFkkSaJYafPPqTSxGVFtkjZpSyAEsNgbIAJPTBr40zfCv4XfHgeMv2Uvig+t6BY6nHqugXX9n3NrPpUySCRImW5jXf5bgBWwdwUbgD18Ef8ABbzRE8N2kPxG+CF++vRRhbqfRtTQW07AcuiSLujBOfkLPj+8a/4fefCr/oh/iv8A8GNv/hWmf8FrvhbqmpWmmR/BPxSj3c6QqzajBhSzAZ6e9VVVVVVVVVVVX/BbT/kgPgX/ALHAf+kc9fsnf8mr/Br/ALJ/oX/pBDVVVVVVXxx/as+AP7OdoJfiz8R9O0q9kj3waXFuub2YdisEYZwpPG9gF9WFftP/APBWf42fEvxLPpnwF1S68AeD7WTFtIsUbajfYP8ArJpDuEQPGI4zxzuZ88f8Eifhlpfxr+Pnjj43/E6R/EmteEYbe5tpdTk89m1C8eX/AEttxJeRFgkwTnazhuGCkV/wWr+C3hG38H+D/j7punW1pr760NB1KaKPa19FJBLNE0mPvNH9ndQx5xJjkKMfsMeC/h74Y/Zh+H2t+A/BGl+HZfFHh2w1HVvscO1ru7a3TzJHc5ZstuxknAOBiqqqrWvid4B8PzNbap4otEmTho4t0zKfQhAxB9jXxJ8f3/xS1m08FeAoJLm1SYSJMqtG80gUgn5sbUAY9QPX0qL9nXxnJbiefxVZJdBeEDSMB7bsZ9O1a/Z+Pvh1qMujX2p39jJOobfb3ThJ0GcEMCMjrwencVJNqGq3K+bLcXlxIdq7maR2PoOpNaN8FviLrRjKaA1nE4/1t64iC/VT83/jtP8Asx6kIEaPxdbNMV+dGtGCg98NuJI/AV4c/Z08K6fbuPEd3Pqtw54KEwJGPYA5J9yfwFeGfhX4H8JXo1PR9HAu1BCTTSNIyZ67cnA44yBnHeqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv+CsH/JkfjD/ALCGmf8ApZFX/BLj/kxL4Y/9xb/07XlVVVVVVVVV+2v/AMmi/F//ALE/UP8A0S1f8EVP+TV/FP8A2UC8/wDSCwqq+M/xJ0/4O/Cbxd8UtTh8+Dwvo9zf+RnBneNCUjHu77V/4FX7Ev7KOof8FAvHvi79pX9pbX9S1TRI9W8h7aCdoW1O72K7Qhwd0VvFG8ShUIOGVVKhTnXP+Cbv7Feu6A/h9vgbpdihj2x3VjdXENzEQMBhKJNxI6/NuB7g1+0X4J+Nf/BNz42a54F+EPxV1/S9C8YaUlxY6hayiKW7sy7AJLgYWeKRHXemGAbI2iQrXwe+DPxR/aO+Ilt4F+HOi3Ot65qUjS3E8rkR265y89xKchEGcljySQBliAf2Wv28/iP+webr9mr9pb4Za7caPoVy5s4oQiX+lh3LOsayER3EDsWdCHUfMSGZSAvxS/4LU/B3T/DdxH8Hvh54o1jxDLCRbPrcMVnZwOQQGfy5XkfacEoAoI43jqP+CY/7NPxU8W/FvWf22/j5ZXcOpaqLiTQft0XlT3lxdAia8EeBsiETNHGMAMJDgBVUmq8WeLdH8F6WNY1ySVbcyiIGOPedxBI4/A1/w0F8Of8An6v/APwFP+NeM/2jkMZs/A9idxHN5dp93/dT+rfka1TV9b8TaibzVb251C8mO0FyWY+iqOw9AOK8IfALxb4gEd3rZXRbNucTLumYe0fb/gRB9jXhf4NeBPC+yaPSxqF2n/Lxe4kIPqF+6Pyz71gAYHaqqp4lnheBiQJFKkjtkYr/AIZo8J/9B7Vvzj/+Jrwd8EvD/gvX4PEFhq2oTzQI6qk2zadylT0UHvVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVf8ABZL/AJNFtP8AscLH/wBE3NfsB/8AJmnwl/7F+P8A9Deqqqqqqqqqr/gh1/yNXxb/AOwfpX/oy5qq/aR8JfFr/gnL+2XdftO/Dfw22p+AvF15PO4KsLZxdNvutOmdQfJbzAZIjgjCoRv2Oo8Of8Fh/wBkjVvDg1XXP+Ev0TUkjBk0qXSfPkZ+hEckbFCPQsUyOwPFftHftG/FD/gqF8TfDfwF+AngbULDwlp16btRfkB3faUN7elCyQRxIzhVBY5duXZlUft1fCzw3+zl+1D+z34e+Fjhda0Lw/ocZdAIpLqe1vWjhuXOTh5DGQT0HlgdBVVVV8Y/jM1u03hPwde4lUlLy+ib7h7pGR39W7dBz0+Cnwq03xlHP4j8Rq8thby+VDbhivnOACxYjnaARwOpPXjm7+FHw6vLU2cnhKwRSMBok8tx7hlwahs734JfFe0sbW+kl0nUigbf/HA7bfmA43IQTn27BiKqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv+C43/ACKnwj/7CGq/+i7avhP/AMkr8G/9i/Y/+k6VVVVVVVVfHXTtQ1j4I/ELSdJsbi9vr3wrqcFta28Rklnle1kVURFyWZiQAAMkkCrb9jD9rW7gS4i/Zu+Iqq44Evh24jYfVWUEfiK/4Yo/a5/6Nw+IH/gim/8Aia/4Yo/a5/6Nw+IH/gim/wDia8M/sXftZ23iTSbm4/Z18fRxRX0Lu7aHMAqhwSTx0xVVVVVVVVVVVV/wW0/5ID4F/wCxwH/pHPX7J3/Jq/wa/wCyf6F/6QQ1VVVV8c/2qvgH+zhaRS/Fz4i2Gk3lwm+20yJWub2deQGWCIM4XII3sAue9eI/+Cy/7LWnaTqNx4c0HxvrGo2yH7Havp8dtHdvnAHmGRii9ySuQP4SeK+Nv/BVf9qz4sS3Vj4Y8R2/w80SbKpaeHk23Oztuu3zLv8A9qMxj2FeGPCPxN+NfjP+yPCuia/4y8T6rIZHSCOS8uJmJ+aSRuTj1djgdSRX7M3/AARz8f6xrtj4l/abv7PQtBgZZJfDum3gnvLvHPlyzR5jhQ9CUZ2IyBtOGHw68Za//wAErP2z/E/hnxxoWoXvw+8Rq0SSW6Za505pS9rdwgkB5IvmR0z1MoznBrTP+CgX7GereHh4lt/2g/DEVrtyYbqSSC5HTj7O6iXPPZa/bW/abvv+Ch3xZ8Efs7fs46NfX2h2upM1rc3UJhOo3bLtNyyH5oreGLecsA2GkLAYAr4a+B9N+GXw68L/AA40Zy9l4X0e10y3dhgukESxhj7nbk+5NVrXiTQPDsKz67rFpYq33fOlClvoOp/CvFH7RXhrSbl7PQNOl1hk4M3meTET7HBJ/IfWtU/aO8bXgZNNstNsFP3WEbSOPxY4/wDHa1TxHr+t3cl7qus3dzNKTuaSU/kB0A9hxWg+EvEviiQx6Bot1e7fvNGnyr9WPA/E1+zLBZnV9dmlA+2R28SxAjkIWbf+qpVftLw2beFtKuHC/a0v9sXrsMbF/wBVSvhlplha+CNCu4dNtoLibToWkkjgVGfKDkkDJJGDk9aqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr/grB/wAmR+MP+whpn/pZFX/BLj/kxL4Y/wDcW/8ATteVVVVVVVVVX7a//Jovxf8A+xP1D/0S1f8ABFT/AJNX8U/9lAvP/SCwqq/bH8G614//AGWfij4S8O27XGp3vhq6a1gVctM8a+YI1H95tm0e5Ff8EWvi54Sv/hF4n+CkuowweJtJ1yTVorR3Ctc2c0USeZGCctskiYPj7u+PP3qr/gql4ph/aH/a08JfBb4Qwf8ACR69oNkujyx2bqwfUZ5izQBs7R5a7N5JAUlw2Nhr9in9kfw1+yN8KI/C1u9vqHivWSlz4k1eNDi5nAIWKMkA+TGCyoDjJLMQC5A/4KeftJSeAvGtl8GtQ/Zt8K/EXT9T8Ki/g1TV7BribTZ5priE+SQjbGUQI2QQcnniv2XPjnqP7NN5qWrXH7MXh/x7qt1NDLYX+v6W8k2lmMNnyG2HYW3AkjB+Uc9a0e9k1LSLHUZYhHJdW0crIP4SygkfrVEgDJIAA5Jr4k/ELUvidqDeBfBejfbrWObcJlQu8rLxvXsicnk+oORV18CfiVbWn2r+xopSBkwxXKM4/DOD+BNeC/B48U+K4fC2o6iNJkcsrefEd+5eqBeMN14OOnrwfB/w28JeCIl/sfTle6xhrycB5m9fmx8o9lAFVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV/wWS/5NFtP+xwsf/RNzX7Af/Jmnwl/7F+P/ANDeqqqqqqqqqr/gh1/yNXxb/wCwfpX/AKMuaqviR4B8PfFTwB4h+G/iu3M2keJNNmsLtRjcqSIV3KT0ZchlPZgD2rxD8MdL/Yj/AGirvwL+0x8FNP8AiPoqR+dZxNqtxY/a7JpHWK9t3gkAyfLcGOVWGVZflI3Dwz/wVe/ZL+C/gBtB+AP7M2t6NeOu99OENnp9tJL0DSXEbyySn/aZM44+n7H3wU+OP7bf7UNr+2V8e9NlsvC2jXsV/pvm25hivZYDm1trRG5NvEwVmkOQxUjLM7kVVV8afi+LRZvB/hO9/wBI5S+u42/1Y7xof73qe3TrnHh/wtrvimW4h0OwkuTaQNNMVHCqoJ6+pxgDua/Zy8T6dP4fuPCkk6JfWs7TRxk4MkbAZI9cEHP1FV8UtStvHvxW0bw7oUy3K2rx28ksZyN5fdIQe4VcZPsaqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv+C43/IqfCP8A7CGq/wDou2r4T/8AJK/Bv/Yv2P8A6TpVVVVVVVVXjLxt4P8Ah34cu/F3jvxPpmgaLYruuL7UblYIo/QbmIGSeABySQACTXjP/gsD+x94WvnstIuPGfixUfabjRtGVIz6kG6lhJHbp9OOa+G//BV79jr4h6hDpd74r1rwdcXBCx/8JJpvkR59GlhaWNB7uyj3rTNT0zW9OttX0bUbW/sLyJZba6tpllimjYZV0dSQykYIIODVVVVVVVVVVVV/wW0/5ID4F/7HAf8ApHPX7J3/ACav8Gv+yf6F/wCkENVXjXxz4O+HHhu68YePfE+m6BoliUFzqGoXCwQxF3CJuZjgZZlUe5FXn/BSv9h+y1T+xpvj5p7XG7bvh0m/lhz0/wBckBjxx13Yr4a/Hz4K/GS5ubL4V/FDw74pubKBZ7mDTL5ZngjY4DOoOVBPHPev2lv2gvBv7OHwn8QeP/EWt6TFqdnps8mi6XeXaxSapdhf3UMaFgzguV3bQSq5Pav2Gf2Nbj9ubxD4l/ac/ac8QarrGkXGrPClqlw0Mmr3QVTJvkHzR28asiKse3kbQVEeG8U/8Ezv2K/E/h86AnwattHZYysF/pd/cQ3MJP8AFvLsHP8A10Dj2rXP2XvBX7Mv7eXhX4IfG7Tv+Eu8A61qVr9nllnkszc2N47QwzO0TKymKYEOAcN5L4wGGPh18Jvhj8IdG/4R/wCF/gLQ/DFgceZFplkkBmIGA0jKN0jY/iYk+9Oyohd2AVRkk9qPwT+H/wC1t4j1g/GPw5Frej27GZYZGaOSJnJWNY5UIePCqeVYE7QDkE1ef8EY/wBk251Nr+DxH8SbWBn3Cxh1e1MSj+6C9q0mPq+fetR/ZO+EP7LFhpev/BHw1/YsskqwXd5JO9xcyyr86OZZCW52sSgwmQMKK1X9pjUGtoE0Pw5bpcbB50l25dd3cKqkHHuT+FeKvi1438XCKK91Q2kMQ/1NkWhVj6tyST06nA/OlXUtZvFjRbm+u5iFVQGlkc9gByTXh34NePNf1CO0n0S402DIMtzeRmNUX1APLH2H6da0z9mfw7CAdX8RX92w7QRrAD+e4/rXh74XeBPDKt/Zvh+3eRwA0tyPOY492zjoOmKiiihjWGCNURBhVVcAfQV470PxD8HfGh8c+GTH/Zt/MwCt90M+WaF1znadpII9B3FRftN6ebUGfwncfaQvKpdDYT9duQPwq2bxP+0B4xiF5stNM08AyJG3y20RPOM8s7bcZ9uwFW9vBaW8VpbRhIoUCRoOiqBgAfgKqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr/grB/yZH4w/7CGmf+lkVf8ABLj/AJMS+GP/AHFv/TteVVVVVVVVVX7a/wDyaL8X/wDsT9Q/9EtX/BFT/k1fxT/2UC8/9ILCqr9pr442n7NvwP8AEnxqvvDsuvQ+Hfsu7T4rkW7TefdQ24w5VguDMG6HO3HfNfGv4ueAvEvxdk+Lv7P3hbxB8L7y6uHvJLS21gMLO6YktJaSxLG0KnJ+QZ2knaQpCjXf25f2u/Enh8+F9W/aD8XvYNG0brDe+RLIp6h5YwsjAgYIZjwSO5r/AIJK/sfaL4U8DWX7UfjjTjceKvEaTL4dSbkWFgTsM4B6SylX+btGRj77VVVXizxv4b8FWX2zXtRWJmBMUCfNLL/ur1P14A7kV4/+NXiPxj5mnaczaXpTZHkxN+8lH+23p/sjA55z1r9m3SrKHwpf6wiq13c3pid8cqiKpC/mxP4iq+PltDoXj7Q/EOloEvZUSVxGMMzxuNrfUjA/4DVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV/wAFkv8Ak0W0/wCxwsf/AETc1+wH/wAmafCX/sX4/wD0N6qqqqqqqqqv+CHX/I1fFv8A7B+lf+jLmqqv+C1f/J1Hhb/sn9n/AOl9/XwK0n4m3XxF0zxB8KfhXefEDWfDUqal/ZMOgzatFhGAV5oIQSYw5TrgZKg9cV+zt4n8f+Nfgh4O8U/FPw6+g+LNR01ZNV017CSyNrNkgp5Mnzx8AfK3NVVfGj4vrpUc3hHwpef6ecpeXcbf8e47op/v+p/h+vTwh4R1fxtrcWi6PGDI/wA0sr/dhQdXY+n8zXgrwVo3gXRk0jSIsscNcXDD553/ALx9vQdB+Zr4p/BvUYtZg8QfDuwuWmupHe4hhkVBA/BDISQRkk8dj0wOKu/C37QN9amxvDrs0DDDI+oghh6H5+fxr4Q/Cyz8HWEWuanbudcuYsSCXH+jA9VXBIycDLZ/LnNVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV/wXG/5FT4R/wDYQ1X/ANF21fCf/klfg3/sX7H/ANJ0qqqqqqqqvby006zn1C+uI4La1iaWaWQ4WNFGWYnsAATX7bP7X3i/9rH4pXupzajcW/gnR7mWLwzpAYqkUAOBO695pAAzE5K5CA4WtE8MeJfE0jweG/D2p6rJEMullaPOVHqQgOK1fRNZ0C8Ona9pF7pt2qhjBd27QyAHodrAHFf8EvP2zPEPwe+KWk/A3xnrklx4A8Y3gtbWO4fI0nUJSBFJGT92OR8I68KCwfja26qqqqqqqqqqq/4Laf8AJAfAv/Y4D/0jnr9k7/k1f4Nf9k/0L/0ghqq8cXnxX/4Kqfte6l8OfDviiTSPhn4Pnma2Y5eCysYpPKN35YIEtzOTlQTkBtudqE1pv/BH79j6z8ODRr6z8XX9+YtratJrRjnDd2VEURD6FCOnWvjp8HfjD/wSm+Neg/FD4M+M5tX8K+IBLDavqEZ23ATBksb6OMqsnBV1ddmSCVClDj4kfEz4jfG/xxeeN/iF4hv/ABF4h1WX5pZSWI/uxxoOEQdFRQAB0Ff8Eb/i34V8R/s83vwjjvbeHxH4R1aeeWzLBZJ7W4bek6jqwDl0J527VzjctV+3B4q039o7/gpF8Nfhr8M7mLVJfDN1puj3lzbESolwt49xckEHBWGNvnx0aOQHpVeIPFPhjw9aTPr+s2lsiRkvG8g3sDxwg+Y/gK+CXjbR/Bvim9sNRvBHpupqI1uX+VVZCdjN6AhiM9sjtmori3ngW5gnjkiZdyyKwKkeoPpXx68a6f4nl07wT4amXUZI7rzJmtzvBlwURFI+8fmbOPUDrnHh79m7xFeNDP4j1W0sIHUM8MOZZl/2TwFB9wTVl+zn4BtZ0mnudWu1XrFNcIFb67EB/WtL8P6DoildG0Wxsdww32e3WPd9do5qqqr7TdO1SEW+pWFvdxK25UniEgDYxnBzzgn86/4RHwn/ANCxpP8A4BR/4VYaNo+lF20vSrO0MgAcwQLHux0ztAzVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV/wVg/5Mj8Yf9hDTP/SyKv8Aglx/yYl8Mf8AuLf+na8qqqqqqqqqv21/+TRfi/8A9ifqH/olq/4Iqf8AJq/in/soF5/6QWFVX7cH7fPwd/ZntZPh1qfhmDx94wvoEmbw45UW1tGTujkunZWC5IDKgVmOAflBDV8Wm/aP/aL8ZS/tAaT+zdq2i6e1pHFDL4V8IXY06KNFKq4fa67yG5fI5wQBxXhDwZ4k8e+LNN8DeFdOF3rmr3ItbK0knjgM0x4WMNIyqGJ4AJGSQByQK+Auk+I9B+Bfw50Lxjp6WGv6d4T0u21W0SOOMW91HaRrNGFi/dqFcMMJ8oxxxiqpmVFLuwAUZJJxiviN8frXTzLo3gdo7q5GVe/Ybo4z/sD+M+5+X61cXGt+J9V82eS61LUbt8DOZJJD6AdfwHSvBP7Ot1dwfbvG101pvQ+XZ27AupI4LtyB/ujP1HSvCfinXfgZ4ivfDfiXTZZ9OuH3Hy+CccCaLPDAgYIOOg5BGKuf2hfh5DZ/aIJb+ebHFutsVbPuSQv6mvDttr/xt+IsPifUbMwaPpsibhglERDuWIH+JmJ+Y+hJ44FVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX/BZL/k0W0/7HCx/9E3NfsB/8mafCX/sX4//AEN6qqqqqqqqqv8Agh1/yNXxb/7B+lf+jLmqr9rvQ/jv4j+BWs6T+zbqU1j49kubU2E8V3HbMsYnQzYeQhRmMMOetftQ6p8eLn4u6h4d/aP8UXGueNPC8Eem3Es13Hc+RGMzLEJI/lIBnYnHQsR2r/gix8Iz4e+D/i34x6hZMl14u1ZbCwkcfetLRTlk9mmllU+phHpVVXxn+LI8MQP4Y8OXIOrzL+/mXn7KhHb/AGyOnoOfSvDPhnV/GOtQ6LpEJluJ2y7tnbEv8TsewH/1upFeBfAujeAtGXTNMTzJnw1zcsuHnf1PoB2Xt9SSaqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv+C43/ACKnwj/7CGq/+i7avhP/AMkr8G/9i/Y/+k6VVVVVVVVftwa3e+Hv2RPi3qWnSlJz4Wu7cMpwVWZPKYg9jtc1+wl8G/CHx7/al8FfDXx5mTQbuS4ub22WQobtbe3km8kEEEBjGA2CDt3YIPNeFvCPhbwNoVr4Y8GeHNN0PSbJAlvY6fapbwxADHyogAHQV8TfhN8N/jN4WuvBfxP8Hab4i0i6jZGgvIctHn+KNxh4n4BDoVYEAgjFfGnwVB8HPjh40+H/AIe1qS6i8HeJbywsr4MBIRbzsiMSvSQbRnHRga8Ga1L4l8HaF4inQJLqmmW906D+FpI1Yj9aqqqqqqqqqqr/AILaf8kB8C/9jgP/AEjnr9k7/k1f4Nf9k/0L/wBIIar4n+P/AAR8MvA+qeMPiH4xs/C+i2kJWbUrmXYImYYXYOrOT91VBYnAAJr/AIJ6/tm+AP2ONZ8dXfjbwnq/iBPE8FlFazaSI98XkNMWz5rp8r+ap6Z+UdOa/Z3/AOCjH7M/7SOtW/hHw54gv/Dnia6wtto3iKBLaW6bpthkR3ikb0QPvI5C8HH/AAW18d+Grf4R+A/hi9xC/iC/8R/2vHEGBeK1gt5oWZh1Ad7hAOgJjbrtOP8AgnD+wZ4X+BHgrSvjJ49soNV+InibTY54hNFlNCtp4w3kRg/8tirYkf3KL8u4v+1V/wAEyPiv4F+Jtp8XP2Fzq0FxqF3LLPpOn6vDpcmiORkvb3Es8ZMTkkeWMlOmSpAXxR8IP+C1HjHw8/hbX7nxTLp0kRiljg8W6PbPKhGCryRXCu4I4O5jnnPU1+xB+xV8OP2NtHtvin8Wtdt7n4n6ppiCa1lMbjQlkQGS2gWNn8yTOUeYEggYXCli/jn9oPWdW8zT/B8baZaHKm5fBnce3ZPwyfcU5vdSnkncz3U7ndI5y7E+pPWvDvwW8feJLeG8i0tbK2lbAkvH8sgf3tn3sfhzR/Zq1xM28PjC2+zsfmBgcZ9Ttzj9a8A/Bnw34HnTU3lfU9TUYS4mQKsXqUTnB9ySfpzVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX/BWD/kyPxh/2ENM/9LIq/wCCXH/JiXwx/wC4t/6dryqqqqqqqqq/bX/5NF+L/wD2J+of+iWr/gip/wAmr+Kf+ygXn/pBYVVf8E4fAOifte/tcfEP4+fGCxi1x9Gk/teDT7tfMj+23M7eQWU5DJDHG4VCMAiM/wAOKCqqhVAAAwABjFf8FlvgL4O8M6P4U/aT8JafFo/iS815dJ1WezHkm9doZZ4Z224/ep9ncb/vEMoJO1cfs4eOdV+J3wA+HXxB15w+qeIPDFheXzhQoe4eBDKwA4AL7iB2zWo6hZaVYT6lqFwsNtaxtJLI3RVAyTWpfGL4ieN9Wm034a6VLDboMKywLLLjONzlsqgPHHb1rxknx7h0Oe38VDUG01xmfyvKcBR/eMWSF9c8eteAvAeqfEDWDpemzQwJCm+4mkP+rTOMhc5Y+w/EivBPw58M+BLQR6TaCS7ZMTXkozLJ68/wj2HHTOetV8YPCGq+NPCsel6JBbyXaXSODMQu1ADnBPTtX/DPnxG/59bD/wAChXw/0bUvD/g7TNG1gRi8tY2WURkEfeJGCPYiqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr/gsl/yaLaf9jhY/wDom5r9gP8A5M0+Ev8A2L8f/ob1VVVVVVVVVf8ABDr/AJGr4t/9g/Sv/RlzVVXxt8av8SfjH44+IDzmUeIvEV9fxsWzhJZ3ZAPYKQB7AV+yP8Pofhb+zH8MvA8dusEtj4btJLuMDpdTIJrjp6yySH8aqvir8WLHwPaTaTYEya5LCGgRozsjVsjzCehxg8euO1XFxPfXUl1d3DSzTuXllkJJZicliep5r4dfD7Tfh/oi2VvtmvpwGvLnHMj+g9FHOB+PU1VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX/Bcb/kVPhH/ANhDVf8A0XbV8J/+SV+Df+xfsf8A0nSqqqqqqqr4w/D+D4r/AAn8Y/DOedYV8U6FeaaszZxE00LIr8c/KzA/hWjaz8RvgF8U4tW0m4uvDfjTwVqrKCVHmWl1CxR0ZTkMMhlZSCrAkHINfDv/AILeSQ6PDZ/Fb4Hm51OKMCW+0HVBFHO2OoglUmP/AL+NXxm/4LVeLPEHh+70L4JfCyPwze3UTRprer3y3ktvkY3RwKgQOOcF2dc4ypxg/CL4deKv2g/jR4d+Huny3d9rHjDWUjubpiZZFV33XFy5OSQieZIxOeFJqys7bT7ODT7KIRwW0SxRICTtVRgDnnoBVVVVVVVftP8A7R3gv9ln4Saj8UvGKtdNE622l6ZFIEl1K8cEpCpP3RhWZmwdqIxwSAD4A1X/AIKd/wDBQWTUPiB4G+KL/D3wjDcvFZy2uq3Gh2O9fvRQm2V7ifb0LvuAORuBBAi/ab/bv/4J6/FLSPCn7S+rX3jvwhqbbs3t7/aH26BSFkks7yTEolTI/dykDkblG4NXgXxt4Z+JPg3RvHvg3U01DRNeso7yxuUGN8brkZHVWHQqeQQQeRVV/wAFtP8AkgPgX/scB/6Rz1+yd/yav8Gv+yf6F/6QQ1X7ZN741/bd/wCCguj/ALJ+k61Pp3hbwxdiy4HyxFYPPv7wocBpAgaNATg+WuMb2z8Pf2Df2Rfhv4eh8Pab8BvCOteWiiS98Q6VDqtzMwHLtJOrYJ6kIFX0AAFf8FIf+CfPwv8AAfwxv/2i/gFoh8J6l4Xmhn1bSrCRlt5oGkVPPhTJ8mSNmRiEIXaGOAwyf2BP2VfGv7ZXxOP7SH7Rms6v4m8LaNcqI7nUb+O7fWL2Api1lDSGRIUUoxUoFZSFBAJq4ngtbeS6uZVjihQvI7HAVQMkn2wK1b4jfEX4o65Pofw5jns7CEn95C3luVzgO8h+5nsox+OKuvhR8cLC3a7tfGD3MoU/uYNWmDnI5HzBR+tNpmsXWrNpTWV1LqTSmNoCjNKZO4I65rwV+zxrepul54wn/s214P2aJg0zj3PIT9T7CvDvhbw/4UsRp/h/TIbSPA3lVy0hHdmPLH61VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV/wVg/5Mj8Yf9hDTP8A0sir/glx/wAmJfDH/uLf+na8qqqqqpZI4Y2llkVERSzMxwFA6knsK+PP/BX7wR4R8YzfD74AfDyf4j6hBO1u2pm7aG0lmBIxbpGjvcLkfeGwHqpYYNfDT/gsvbReL4fCn7RHwTvfCEEsirLqOnzyStZ7ujS2sqK5TBBLKxbHRG6VoOvaL4p0Sw8SeHNVtdS0vU7dLizvLaQSRTxOAVdGHBBBBzVV+2v/AMmi/F//ALE/UP8A0S1f8EVP+TV/FP8A2UC8/wDSCwqq+C/j+5/4JgftreMPAnxL0q9PgTxBm3+1QxMzfYWlL2d8i9ZAi7kdRkjdIBllAPg79pj9nn4g6fFqPgz41+DNVSWPeIYNZh89Rxw0JYSI3I+VlB5HHNf8FPPHvxL/AGldQ8MfBr4R/DnXr3wlo+qLd3etzWjRQ3d86mKMqG+ZIYklk3SMACXJwFUM3wU8E6b8N/g74I+H+kagt/aeHfD9lYxXajAuRFCimXHbeQWx71+0XfXFp8P44IJCq3moRQygHqoV3x/30i18HtDsNF+H2ktZwqst9ALm4kAwZHfnn6DAH0ogMNpAII5BFaLZw+Ev2jH0jR1ENpNKymJRtUJJB5hUD0DEYH+yKqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr/gsl/yaLaf9jhY/wDom5r9gP8A5M0+Ev8A2L8f/ob1VVVVVfFb9oD4KfA62huPiz8TtA8MtcIXt7e9uwLidRnLRwrmRxkEZVSM8dTXww/a8/Zn+M2sL4d+G3xn8O6vq0hxFp5na2uJsDJ8uKYI0mAMnaDiqqq/4Idf8jV8W/8AsH6V/wCjLmqr49+NYfhv8EPH3j2WQr/YHhu/vI9pwzSJA5RRnuWCgZ7kV4F8MXHjfxv4e8F2au0+varbafEE4YtNKsYx15yw7VBBDbQR21tGscUSBI0UYCqBgAD0xVeJPEGneFtEu9d1SXbBaRliB1c9Ao9ycAfWvFXiXUvF+u3OvapJma4b5UB4jQfdRfYD/HvXwQ8HeA/FlpqEfim3gub1JkW2ha8eJ9u05IVWBPPfnpVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV/wXG/5FT4R/wDYQ1X/ANF21fCf/klfg3/sX7H/ANJ0qqqqqqqqr9u7/gmpof7TOozfFP4XalY+G/iF5IS8S5QrZ6yFGFMpQFo5goCiQBgQFDDgMPG/7BP7YfgG+ksNW/Z98W35RiFm0SyOrRuM8MGtfM4PXnBHcCvh7/wT2/bE+I9/BZad8DPEOjRSH57rxDB/ZUcK5wWYT7XIHoqs3oDX7DH/AAT+8I/sj6dL4q17UbfxJ8RdUt/Ju9UjjK29jEcFoLUNhsEgbpGAZsDhBlTVVVVVVVf8FkPiDrXjn9obwb8ENHlea28P6VHKtsmfnv76THIHX91HBj03tjqa+FHw60L4RfDPwx8MfDcEcWn+GtLgsYtq7fMKIA0hx1Z23Ox7sxJ61/wVV+F2nfET9j3xLrb2sb6n4JubfWLCUjDKBIsU67uuDDLIcdCUX0BH/BGb4kXvin9m/W/AGoXJlbwV4gdLMGTPlWtyglVQOoHnfaD6Hd7Gqr/gtp/yQHwL/wBjgP8A0jnr9k7/AJNX+DX/AGT/AEL/ANIIarxj4jsv2Pf+Ctt549+IUZsvCviW+kvGvirFRaajbsjXA6nbHcFw+O0T4HStP1HT9XsLbVdJv7e9sryJZbe5t5RJHNGwyroykhlIIII4INf8FT/jz4L+Gn7MfiL4c3erWsvivx5CljpumLIGlEJlVprh1BysaorKGPBdlHrj/glT4F1X4efsZ6HqHiRDZ/8ACS6hd63Gk/yeXbSbUjc5/hZIRID/AHXBr4s/G6z1Wxu/CfhJRNb3CmK5vmyAy91jHoehY9s4HQ1+zxZWUHgD7VbhTPc3khnbHOVwAPpgA/8AAj61Xxlhi0z4v+H9Q00BLuZbaWQKOWdZiqsfqFA/Cqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr/grB/yZH4w/7CGmf+lkVf8ABLj/AJMS+GP/AHFv/TteVVVVVX/BW74+al8JP2dIPAXhu/e01n4k3cmntKjbXTT4lDXW0/7W+KI/7MzdOK/4JRfsx+GPhj8BdL+NOq6LBL408fRNci9mjDSWen7iIYYifuq6qJWxjdvUHIRcf8FHf2bfDPx4/Zx8T63/AGLbt4v8FadNq2i6gkQE4ECmSW33AbmSSMSDZnG/Y3Va/wCCLvxv1PxT8OfFnwN12/knPg64iv8ARhK5YpaXJcSxL6KkqbvrcH0qq/bX/wCTRfi//wBifqH/AKJav+CKn/Jq/in/ALKBef8ApBYVVft7/Cn4V/Eb9nvxFfeP/BFprWsabbC38M3RV0ns9Su5EtrUrLGQ6xm4lhLqTsKjLAgcfso/sm+HPDXh1PDnheCOys7NEGq6uYQZ7+fHJJ7nqQM7UBGPe6/Zs8GSWnlWmratFcAfLK7o4z7rtHH0IrwhrniD4NeOl8GeJr4No90w+Zm/dqrHCzJn7oyMMPY+gNfHT4ixeKLm10LQ761udHiVbgSRcs8vzL82eVwM8cdcnqK+Evxr0vw9o0PhfxYJkgtiRbXcaFwqE52uo54JOCM8cY4rXfj38P8AS7NpdNv5dVudv7uCGF0ye25nAAH5n2r4M6HrPjDxvefE/XISsSvIYGKkCSVgVwmeqopI+uPQ1VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX/BZL/k0W0/7HCx/wDRNzX7Af8AyZp8Jf8AsX4//Q3qqqqqv2wf2h7D9l/4CeIfinJFDc6pEq2Wh2krYW5v5siIH1VQGkYDBKxtgg1+xF+xVqX7cura/wDtM/tPeKda1TSL3UpIoY47nyp9XuVx5jM4H7u3jBCKse3kFQVVMH9un/gmD4C+FHwvv/jh+zfNrOmz+ElW71TRprx7hfsyEb7iCRv3iPH99gWIKhiNpXDf8Ex/2rtZ/aW+Ct1onjvUTe+NfAk0VnqN3I2Xv7aRSbe5fvvISRGPcx7icvgVVf8ABDr/AJGr4t/9g/Sv/RlzVV/wU78U2/hb9ib4hmSYpPqyWenWyh9pdpbuLcP+/YkJHcAj3r9hS10+4/a5+GFxqibrTTdaGoTHYzbBbRvPuwvPBjB+oFf8L5+Gf/QZn/8AAOT/AArwp418O+Nrae68O3bzx2zhJC8TR4JGe4Havjx4/PiTXf8AhGdNnzp2kuQ5U8TTjhj9F5Ue+49xXw1+Cl/44sRrmp3zafprMRCVj3ST4OCRk4UZGMnPIPHetc/Zqt47Rp/DHiKf7XGNyR3aDDkdgy42/XBr4J/EDWrvULrwB4ulle/sg32d5zmT5Dh42PcjqCewPPAqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv+C43/ACKnwj/7CGq/+i7avhP/AMkr8G/9i/Y/+k6VVVVVVVVV+0r+2X8Cv2VtOik+JXiOSbWbuMvZ6BpaC4vrhefm2FgsaZBG+RlUngEnivFn/BcLWHvXi8C/s/2cVoj4SbVtdaSSReeTHHEoQ9ONzdOvPHgD/gt9o9xfQWnxR+BN1ZWjMBLfaFrAuHQdyIJUTPc/62vgt8d/hT+0L4PTxv8ACTxfa65pofyrhUVo5rSXGTHNEwDxtjsRgjkEgg1VVVVVVVftoi4vP+Cr8NrqMPmW3/CUeFY4UliG14jBYlh0+Ybi45z3HbFV+20iSfsifF9XVWA8IagQCM8iFiD+YFf8ENDeGw+Myvv+yibQ/KyPl8zF9vx742Z/Cqr/AILaf8kB8C/9jgP/AEjnr4Q/8FjfDXww+Evgn4aT/AfU9Ql8JeHbDSHu08QJGLhra3SEyBTCdobZnGTjPU1/w/F8Kf8ARu2q/wDhSR//ABiv2s/+CgHwE/a88I22ieNf2dtd0zW9J3to2vWXiKIz2TNjcpUwYkiYqu5DjpkFTzXwi+Iv7TFrcL4D+BfxF+Ill5+5xpnh/Wrq2QDPMjJE4VRzyxwBnrzX7Nv7D2j+IPEY+Kv7ZHinXNTvo7sTP4bR/tk2pFcENdXhkOVJ4KKcnH31HX4i/FK48VInh7w/bjS/DdkqxW1nCgj3ooAXcq8BQAMIOBx1xXw9+Fuv/EC4823X7JpkbYmvZEyM91QfxN+g7kcZ8PeJ/FHwI1668O67pputOuX8wIG2iTHAlibkcgAEH0AOCKvv2lfCkdmX03Q9UnutvyxzBI0B92DMf0rTvjH8O7r4p/8ACTfFjx1p2mSWTLOtrskk+Zf9WihFbaFIDHPJ99xNf8Ne/s3/APRU7H/wEuP/AI3XgX4+fCD4ma0fDngbxtbatqSwNObeOCVCI1IDNl0A4LDv3qq8X+L/AA34C8O3fizxdqsenaTY7PtFzIjMI97qi8KCeWZR071/w17+zf8A9FTsf/AS4/8Ajdf8Ne/s3/8ARU7H/wABLj/43X/DXv7N/wD0VOx/8BLj/wCN1ZftZfs8ajeQafZfE2yluLmVYoYxa3ALOxAA5j7kiqqp/wBrj9nO3me3m+KFiskTFXU2txwRwR/q6/4a9/Zv/wCip2P/AICXH/xuv+Gvf2b/APoqdj/4CXH/AMbr/hr39m//AKKnY/8AgJcf/G68IeL/AA34+8O2nizwhqsepaTfb/s9yiMok2OyNgMAeGRhyO1VXjr4+fCD4aa0vhzxz42ttK1JoFnFvJBK58tiQGyiEclT37V/w17+zf8A9FTsf/AS4/8Ajdf8Ne/s3/8ARU7H/wABLj/43X/DXv7N/wD0VOx/8BLj/wCN18PvjL8MvirNe2/w+8WW+sSaciNdLFDInlB8hSd6jrtPT0qq8ffEfwT8L9Hh1/x5r8WkafcXK2sU8sbuGlZWYLhAT91GPTHFf8Ne/s3/APRU7H/wEuP/AI3X/DXv7N//AEVOx/8AAS4/+N1/w17+zf8A9FTsf/AS4/8AjdeG/wBpr4FeMNdsvDPhv4h2d9qeoS+Va26W06mR8dAWQAdO5qq1TUrHRtNu9X1O4EFnYwPPcSsCRHGilmbA54AJ4r/hr39m/wD6KnY/+Alx/wDG6/4a9/Zv/wCip2P/AICXH/xuv+Gvf2b/APoqdj/4CXH/AMbr/hr39m//AKKnY/8AgJcf/G60bWNN8RaPYa/o12t1p+p20dzaTqpAlikUMjAHBAKkHkZ5qq8VftKfA7wTr954W8U/EG0sNV09gtzbPbzMYyVDAEqhHQg8HvX/AA17+zf/ANFTsf8AwEuP/jdf8Ne/s3/9FTsf/AS4/wDjdf8ADXv7N/8A0VOx/wDAS4/+N14B+JvgT4pabcav4B8RQ6vZ2k/kTSxxugSTaG24dQejA/jVV8Q/i/8ADf4Uf2f/AMLB8VQaP/anm/Y/NikfzfL278bFbGN6dcda/wCGvf2b/wDoqdj/AOAlx/8AG6/4a9/Zv/6KnY/+Alx/8br/AIa9/Zv/AOip2P8A4CXH/wAbrwT+0F8HPiPrqeGfBPjm11TU3iaVbeOCZCUUfMcsgHH1qq8VeKfD/gnw/eeKfFOpJYaXp6Brm5dGYRgsFBIUE9SBwO9f8Ne/s3/9FTsf/AS4/wDjdf8ADXv7N/8A0VOx/wDAS4/+N1/w17+zf/0VOx/8BLj/AON1/wANe/s3/wDRU7H/AMBLj/43VVWqftWfs+aNqV3o+p/EuygvLGd4LiJrWcmORGKsuRHjggjiv+Gvf2b/APoqdj/4CXH/AMbr/hr39m//AKKnY/8AgJcf/G6/4a9/Zv8A+ip2P/gJcf8AxuvBvjXwt8QvD8HinwbrEep6Xcs6xXMaMgYoxVhhgDwQR0qq8ffHb4S/C/WIfD/jzxpbaRqFxbLcxQSwSuWiZmUNlFIxuRh68V/w17+zf/0VOx/8BLj/AON1/wANe/s3/wDRU7H/AMBLj/43X/DXv7N//RU7H/wEuP8A43Xxf/4LGfD74afEfW/A2hfCa+8VWOkyokOsW+tC2S7DRq5YRvAWXBYryf4c1/w/F8Kf9G7ar/4Ukf8A8Yr/AIfi+FP+jdtV/wDCkj/+MV+13/wVG0D9p74Faz8HbD4OX/h+bVbm1mW/m1pLhY/JnSQjYIlJzsx14zX/AASt1Kxv/wBhn4fWtpcCSXTp9VguVAI8uQ6lcyBT/wAAkQ8f3vrVVVVVf8FvNbnuPiz8NvDbO5hsfDtzdopxgNNcbGI75xbrn6D3r4F6TbaB8Efh7oVkP9H07wrpltEMY+WO1jUfoBWq6ba6zpd5o96pa3vrd4JQADlHUq3XI6E9RX/BF/VprH9qjXdMUFotS8FXaMu7AVkurVw2O/AYY4+9VV+2v/yaL8X/APsT9Q/9EtX/AASX/aG+Bfwk/Zx8ReG/id8WvC/hfVLnxtdXcNnqmpR28rwNZWSCQKxBKlo3GfVT6V/w2v8Asi/9HH/D/wD8HkP/AMVX/Da/7Iv/AEcf8P8A/wAHkP8A8VXxK/ah/ZR+I3gy98G6b8c/A+s6hqEtv9gsLbWo2lnuknjkgVFVss3momF7ng5BNfs3X1pN4MvdPikX7Rb37PKnfDIu1vodpH/Aar9pS9s7jxXp1lA6tPa2X77HVdzEqD745x6MPWvBngy11eS31nxXq1noXhwTiOW/v7lLZJmwT5UTOQGchW6dACe2K1CP9mnUrOCyn8SeDFS2jCRNDrUMTgD1ZZAW6nrnrWnaH+zJp0wnPjDwxdMpyoufEETqP+A7wD+INQfFD4TWsEdtbfEXwlFFEoVI01e3UKB0AAfgVZfEn4c6ndw6fpvj/wAN3d1cOEhgg1WF3kY9AqhiST6CqqaWG3heeeVI441LO7ttCgDkk9hiv+Fr/Cz/AKKX4V/8HMH/AMXX/C1/hZ/0Uvwr/wCDmD/4uv8Aha/ws/6KX4V/8HMH/wAXX/C1/hZ/0Uvwr/4OYP8A4urS7tNQtIb6wuorm2uI1khmicOkiMMqysOCCCCCODmqrUviJ8PtGvZdM1fx34esbyA4lt7nU4YpIyRkblZgRwQefWv+Fr/Cz/opfhX/AMHMH/xdf8LX+Fn/AEUvwr/4OYP/AIuv+Fr/AAs/6KX4V/8ABzB/8XWieI/Dvia3e78N6/p2qwRPseWyuknVGxnBKEgHBBx71Va74s8K+F/I/wCEl8TaVpP2nd5H268jt/N243bd5Gcblzjpketf8LX+Fn/RS/Cv/g5g/wDi6/4Wv8LP+il+Ff8Awcwf/F1/wtf4Wf8ARS/Cv/g5g/8Ai60nx/4D1+9TTNC8baDqN5ICUt7TUoppGAGSQqsScAZqq1DUNP0myl1HVL63s7S3XdLPcSiNIx6sxIAH1r/ha/ws/wCil+Ff/BzB/wDF1/wtf4Wf9FL8K/8Ag5g/+Lr/AIWv8LP+il+Ff/BzB/8AF1/wtf4Wf9FL8K/+DmD/AOLqqq5+J/w0tLiS0u/iJ4ZhngcpLFJq8CsjA4IILZBBGCDX/C1/hZ/0Uvwr/wCDmD/4uv8Aha/ws/6KX4V/8HMH/wAXX/C1/hZ/0Uvwr/4OYP8A4utJ1jR9fsU1PQtVs9Rs5CQlxaTrNGxBwcMpIOCMVVa3428F+GrpLDxF4v0XSrl4xIkN7qEUDshJAYK7AkZUjPTIPpX/AAtf4Wf9FL8K/wDg5g/+Lr/ha/ws/wCil+Ff/BzB/wDF1/wtf4Wf9FL8K/8Ag5g/+LrRPGng7xNcPaeG/FujarPEm94rK/jnZFzjcQjEgZIGfeqrWNb0Xw/ZHUde1iy020VgpnvLhYUDHoNzEDJ9K/4Wv8LP+il+Ff8Awcwf/F1/wtf4Wf8ARS/Cv/g5g/8Ai6/4Wv8ACz/opfhX/wAHMH/xdWnxM+G+oXUNhYfELw1c3NxIscMMWrQO8jscKqqGySSQAB1Jqqd0jQySOFVQSWY4AA71/wALX+Fn/RS/Cv8A4OYP/i6/4Wv8LP8AopfhX/wcwf8Axdf8LX+Fn/RS/Cv/AIOYP/i6/wCFr/Cz/opfhX/wcwf/ABdWV9ZanZw6hpt5Bd2twgeGeCQOkinoysMgg+oqq1X4g+AdDv5dK1rxx4f0+9gx5ttdanDFImQCMqzAjIIPI6EGv+Fr/Cz/AKKX4V/8HMH/AMXX/C1/hZ/0Uvwr/wCDmD/4uv8Aha/ws/6KX4V/8HMH/wAXWh+KfDHidJZPDXiPS9WW3IEzWN4k4jJ6bthOM4PX0qq1zxN4b8MwxT+JPEOmaVHMxWJ767SAOwHIBcjJx2r/AIWv8LP+il+Ff/BzB/8AF1/wtf4Wf9FL8K/+DmD/AOLr/ha/ws/6KX4V/wDBzB/8XWl/EHwDrl/FpWieOPD+oXs2fKtrXU4ZZHwCTtVWJOACTx0BNVXiL9rX9mLwlrt/4Y8T/HnwTper6XcPb3tldaxFHLbyqcMjqTkMCMEGv+G1/wBkX/o4/wCH/wD4PIf/AIqv+G1/2Rf+jj/h/wD+DyH/AOKr/gqn+0b8Bfit+zFa+GPhr8XvCvibV08U2dw1lpmpxzyiJYpwz7VJO0FlBPuK/YD/AOTNPhL/ANi/H/6G9VVVVV/wXD8Y3Udj8Kfh9b3BFtPLqOp3UW7hnQQxQtj2Ek4z/tfWv2K/Btt4D/ZL+E3h22gER/4RWzvJkx92a6jFzL07+ZM9eKfD2neL/DOr+E9XjEljrVhPZXSEZ3RSxlHH/fLGv+CNmv3/AIZ/ao8TeCbubZHq3ha6jlg8zg3Fvcwsp9GIXzh9GJqqr/giFdWtn4m+Lct3cxQp9g0obpHCjPmXPc1/buh/9Bmx/wDAhf8AGv7d0P8A6DNj/wCBC/414v0b4Y/EHQ5fDPjvTfDPiLSZ2DSWOqxQ3ULMOhKSZGRk4OMivB37Df7H3gj4kL8R/Angqy0rV5LWW1NrZ6tKbdvNwGZYi52NgbR5e0YJ45NXXgrwxH8dl8GR6ZjR/MRfs3nSdDbBz827d9456/pXjibw18GPBd3D4QsxY32sP5cCid5DuxhpBvYkbVPbuV9aJLHJOSepNfDO5s7v4feH5LFlMa6fFG2P76qFf/x4NVWN7Yy/tITXtncww20VzKJJGcBQVtyr8njlgR9TX9u6H/0GbH/wIX/Go5I5o1likV0dQVZTkEHoQfSqqbV9Jt5WguNUtIpE+8jzqpH1BNf27of/AEGbH/wIX/Gv7d0P/oM2P/gQv+Nf27of/QZsf/Ahf8atryzvUMlndwzqpwWjcMAfwqqur+xstv229gg352+bIEzjrjP1Ff27of8A0GbH/wACF/xr+3dD/wCgzY/+BC/41/buh/8AQZsf/Ahf8ag1XS7uUQWupWs0hHCRzKxP4A1VTSwwRNNPKkaIMszttA+pr+3dD/6DNj/4EL/jX9u6H/0GbH/wIX/Gv7d0P/oM2P8A4EL/AI1/buh/9Bmx/wDAhf8AGqqn1rRo3Mcmr2SspIZWuFBBHXjNf27of/QZsf8AwIX/ABr+3dD/AOgzY/8AgQv+Nf27of8A0GbH/wACF/xqC4t7uITWs8c0Z6PGwYH8RVVc6lptm4ivNQtoHK5CyyqpI9cE9OK/t3Q/+gzY/wDgQv8AjX9u6H/0GbH/AMCF/wAa/t3Q/wDoM2P/AIEL/jVtqOnXrmOzv7adlGSscoYgevBqquLm2tIvOuriKFAcbpHCjP1Nf27of/QZsf8AwIX/ABr+3dD/AOgzY/8AgQv+Nf27of8A0GbH/wACF/xqPWtGmdYotWsndyFVVnUkk9ABmqokAZJAAHev7d0P/oM2P/gQv+Nf27of/QZsf/Ahf8a/t3Q/+gzY/wDgQv8AjX9u6H/0GbH/AMCF/wAajlinjWWGRZEcZVlOQR6g1VT6tpVtKYLnU7SKRfvJJMqkcehPpX9u6H/0GbH/AMCF/wAa/t3Q/wDoM2P/AIEL/jX9u6H/ANBmx/8AAhf8atr6yvgxsryC4CfeMUgbH1xVVdXtlZKrXl3DAGOFMsgTJ9s1/buh/wDQZsf/AAIX/Gv7d0P/AKDNj/4EL/jX9u6H/wBBmx/8CF/xqDVtKupVgttTtJZGztRJlYnA5wAfSqqWWKGNpZpFREGWZjgAepNf27of/QZsf/Ahf8a/t3Q/+gzY/wDgQv8AjX9u6H/0GbH/AMCF/wAa/t3Q/wDoM2P/AIEL/jQIYAqQQRxVf8Fxv+RU+Ef/AGENV/8ARdtXwl1TTH+Gfg20TUrUzjQbFfKEy7twgTIxnOfaqqqqqqqr4v8AxF034Q/CvxZ8UNVi8628LaPc6g0G7aZ2ijLLGDzguwVQfVhXxK+Ivi/4ueOta+I/jzV5NS13Xrpri7ncnGTwEUfwoqhVVeiqoA4FfCz/AIJ+/tc/GPQbfxV4N+D98mjXcYktr3VLqDTlnQ/dZFndXdSBkMqlSMHPIr4z/sT/ALT3wB0l/EXxM+FGo2eixEeZqlnNFfW0WTgGR4GcRAnj95tySPUZ/Y7/AGkvEP7L3xw0Px7pt7KNEuZ0s/EVkGJS7sHYCTKjq6D50PZlHYkGOSOaNJYZFdHUMjKchgehB9Kqqqqqqr/grh4a1b4Zftf+FfjDp9sxh1rSrO+t5WUqrXdjLsdM+yLbk+m8V4Q8U6N458KaN408OXaXWl67YQ31lMhBEkMqB0OR/ssK/wCCoPxBsPAP7GPjaCedVvfFBttGsI2P+sklmVpB+EEczf8AARX/AARd+H9z4f8A2ePE3j68gaNvFviNktiy4ElvaxKgcf8AbV51/wCA1Vf8FtP+SA+Bf+xwH/pHPX7Pn7CX7Onjj4AfDDxjr/he6l1PWvBWj3d26XAUPLJZQs5A28ZJJr/h3T+y7/0KV7/4F/8A2Nf8O6f2Xf8AoUr3/wAC/wD7Gv2ff2V/hX4c8Tal4a+FeiHStFSaSW41CZ/tF3LDuAVTK3PzbVwv3RgnBOcw/A74ZQ2gtG8PGU4wZXuZd5PrkMMfgAK+K3wxg+HWp2Wo2Be90i7k+WOc/MjLgmNiuMgjoRg9fTJ8OS6ZceH9NuNGt0gsZrWOS3iRdoRGUEDA9jXxQ+Bfgf4u6hpeqeKpNWiutHjljtZdPv3tmUSFS2SvJ+6Kuf2R/hhdwfZp9Y8XNHjBA1uRcj3I5P4037BX7PjsWbTtdZicknVGyT+Vf8MEfs9f9A3XP/Bm3+FfC39l34T/AAd8TN4t8FWepR6g9q9sWuL0yrsYqT8pHXKjmqr4h+AfDvxQ8Hah4F8VxTyaXqXl+esEpjc+XIsi4YdPmRa/4YI/Z6/6Buuf+DNv8K/4YI/Z6/6Buuf+DNv8K/4YI/Z6/wCgbrn/AIM2/wAK039hf4BaTqNpqlnp2tCeznSaItqTEBkYEZGOmQKqquv2EP2fru5luptN1syTOztjU2HJOT2r/hgj9nr/AKBuuf8Agzb/AAr/AIYI/Z6/6Buuf+DNv8K/4YI/Z6/6Buuf+DNv8K+HngHw78L/AAdp/gXwpFPHpemeb9nWeUyOPMkaRssevzO1VXxT/Zd+E/xi8TL4t8aWepS6glqlsGt70xLsQsR8oHXLHmv+GCP2ev8AoG65/wCDNv8ACv8Ahgj9nr/oG65/4M2/wr/hgj9nr/oG65/4M2/wr4Rfs9/Dj4IXOp3XgO1v4pNWjjS5+03RmyIyxXGen3jVV8WPhF4N+NPhy28LeOILqWxtb1byNbacwsJVR0ByO22RuPpX/DBH7PX/AEDdc/8ABm3+Ff8ADBH7PX/QN1z/AMGbf4V/wwR+z1/0Ddc/8Gbf4V4L/Y2+CfgLxTpvjHw9YaumpaTOJrZpdQZ1DAEcrjkc1Va5o9l4h0W/0DUVZrTUrWW2nCNtYpIpVsHscE81/wAMEfs9f9A3XP8AwZt/hX/DBH7PX/QN1z/wZt/hX/DBH7PX/QN1z/wZt/hX/DBH7PX/AEDdc/8ABm3+FeG9A07wp4c0rwto6utjo1lDZ2qyNuYRRIETJ7nao5qq8efsffBf4j+LdR8a+JrHVpNT1R1e4aG/aNSVRVGFA44UV/wwR+z1/wBA3XP/AAZt/hX/AAwR+z1/0Ddc/wDBm3+Ff8MEfs9f9A3XP/Bm3+FfCb4NeCfgrot5oHgaC7itL66+0zC5uDMxk2qvBPQYUcVVfF/4B/Dz44/2T/wnltfTf2J5/wBk+zXRhx5vl7846/6pfpzX/DBH7PX/AEDdc/8ABm3+Ff8ADBH7PX/QN1z/AMGbf4V/wwR+z1/0Ddc/8Gbf4V8Mv2VfhH8I/FMfjHwbZanHqUcDwq1xetKu1xhvlI9qqvHngnQviP4S1HwT4mjmfTNUjVLhYZDG5CurDDDpyor/AIYI/Z6/6Buuf+DNv8K/4YI/Z6/6Buuf+DNv8K/4YI/Z6/6Buuf+DNv8K/4YI/Z6/wCgbrn/AIM2/wAKqq1z9h/4DeIdav8AXtR07WWu9SupbmcpqLKDJIxZsDHAyTxX/DBH7PX/AEDdc/8ABm3+Ff8ADBH7PX/QN1z/AMGbf4V/wwR+z1/0Ddc/8Gbf4V8N/hz4Z+FPhK18E+EYriPTLN5HiWeUyOC7Fmyx9yaqvix+zJ8LPjT4itvFPjiz1GW/tbJbOM214YVESu7jgDrukbn6V/wwR+z1/wBA3XP/AAZt/hX/AAwR+z1/0Ddc/wDBm3+Ff8MEfs9f9A3XP/Bm3+FfDz9kP4C/DXUr7VtF8F299c6hEscx1VEvBhTkEB1OD71/wqb4V/8ARNPCn/gmt/8A4iv+FTfCv/omnhT/AME1v/8AEV/wVT+Gfw+0r9jbxHrOi+C9E02+sNU054biy0+KBwWuVjILKoOCrtx9K/4JU6PZab+w54DvbVWEuq3WqXNzubIMg1C4hGPQbIU49cnvVVVVVf8ABcDwrcQeM/hb42WAmC90u+055QvRoZY5FUnPcTsQMdj17fs3eKLTxt+z38NfFVlIjR6n4U02UhRgI5tkDrjJwVYMpGTgjqa8d+JrbwV4I8Q+MryRUt9B0q6v5WYZCrDE0hOMjsvqK/4In+Fbi/8A2gPGvjEwh7bRvCJtCxTOya5uoWQg9jst5R7gmqr9tf8A5NF+L/8A2J+of+iWr9gb/gnb8OP2uvg7rHxI8X+O/Emi3um+Jp9Jjt9NSExtHHbW0oc+YhO4mdh1xgCv+HJHwN/6K/46/wC/dr/8br/hyR8Df+iv+Ov+/dr/APG6uf8AglX+zt8Fde0XxncfGfxq+p6TfQ31jZmC1kaaSGQOuUCD5NygEkr6ZzWi+I9S8I6sNS8KarcwuihfMkRQZBxkOgypBIzjntV3+0P8Qbmz+yxf2ZbSYx58Vsd/1+Ziv6V4b0PWfiH4sh04Tyz3V9KXubiQlii9Xdj7D9cDvXxo+AHh74yfD7S/h1Pq1zothpF9FdQPaxK7fu4pIwpDeolJJ65HvX/DtzwP/wBFK1z/AMBIq/4dueB/+ila5/4CRV/w7c8D/wDRStc/8BIq8BfsE+EPAPjXRPGtn4+1i6n0S+iu44JbWNVkZGBAJHIBxVVrWmR61o99o8srRpfW0kDOoyVDqVJH51/w7c8D/wDRStc/8BIq/wCHbngf/opWuf8AgJFX/DtzwP8A9FK1z/wEir/h254H/wCila5/4CRV4M8NQeDPB+heD7W6e5h0LTLewjmkUBpFhjWMMQOASFzVV8U/2G/CfxU8f6x4/wBQ8datY3GsSpI9vDbRskZWNU4J56KD+Nf8O3PA/wD0UrXP/ASKv+Hbngf/AKKVrn/gJFX/AA7c8D/9FK1z/wABIq+AXwI0j4BeG9Q8N6Pr13qseo332tpLmJUZG2KmAF7YUH8aqv2hf2bdD/aF/sD+2fEt9pP9gfafK+ywq/m+d5Wc7umPKH5mv+Hbngf/AKKVrn/gJFX/AA7c8D/9FK1z/wABIq/4dueB/wDopWuf+AkVfBr9i3wv8GvH9j4/0vxrqmo3FjFNGtvcW8aI3mRlCSRzwGzVV8SfBFp8SfAus+Bb6+ls4NZtjBJPEoZowSDkA8dq/wCHbngf/opWuf8AgJFX/DtzwP8A9FK1z/wEir/h254H/wCila5/4CRV/wAO3PA//RStc/8AASKqqvE//BPfwZ4n8Sar4kn+Ieswy6rfTXbxJaxEI0jlyAfQbsV/w7c8D/8ARStc/wDASKv+Hbngf/opWuf+AkVf8O3PA/8A0UrXP/ASKvg18LdP+DXgCx8AaXqtxqNvYyzSLcToEdjJIXOQOOC2Kqvj3+yT4c+PfjCz8Yav4u1HS5rPTEsFhtoEdWVZJJNxLc5zKR+Ar/h254H/AOila5/4CRV/w7c8D/8ARStc/wDASKv+Hbngf/opWuf+AkVfAL9lLw98AvEmoeJNH8Wajqsuo2P2RormBECDer7gV7/KB+NVXxs+Emm/G3wLL4F1TV7nTYJbmKcz28auwMZyBg8YOa/4dueB/wDopWuf+AkVf8O3PA//AEUrXP8AwEir/h254H/6KVrn/gJFXgz9gDwd4M8YaF4wtfiBrNzNoWpW99HDJaxhZGhkWQKSOQCVxVVe2q3tlPZM5UTxNGWA5GRjP61/w7c8D/8ARStc/wDASKv+Hbngf/opWuf+AkVf8O3PA/8A0UrXP/ASKv8Ah254H/6KVrn/AICRV4B8I23gHwVongqzvJLuHRLGK0jnlUK0gRQAxA4BOKqvi9+xJ4V+L/xE1b4ial431XT7nVvJ320FtGyJ5UKRDBPPIjB+pr/h254H/wCila5/4CRV/wAO3PA//RStc/8AASKv+Hbngf8A6KVrn/gJFX7Pv7O2i/s+2etWWjeI73VhrUsMkhuoVQxmMMBjb67/ANKqv2gP2f8ARv2gNG0rRtY8QXmlJpNy86PbRK5csu3B3dq/4dueB/8AopWuf+AkVf8ADtzwP/0UrXP/AAEir/h254H/AOila5/4CRV8If2JPCvwh+Imk/ETTfG+q6hc6T52y2nto0R/MheI5I54EhP1FVXxa/4JK/s9fF3x94n+JeseOfiJZa34p1Ce/uEtr2zNtFNKxYhEa1LbAT0L5960z/gid+zlF5n9s/E74kXWceV9muLK329c53Wz57emMHrnj/hyp+yv/wBD98Vf/BrYf/IVft+f8E5vgh+yr8C4Pid8PPFPjjUdUl16208w6zfWssHlyRysxCxW8bbsxrg7scnj0/YD/wCTNPhL/wBi/H/6G9VVVVV/wXC0qeHx18KtbbPk3ek6jbplcDdFLCzc9+Jl47fjX7Nl5bal+zr8LL+zk3w3HgvSJI2xjKmziI47UxVVLMwAA5J4xX/BJS1HiL9tzUtbsWcwWvh/VL3/AFfVHmijGf7vMy/jx3qqr9lz4L/tMfG+PxT4c/Z3Hmx2i2k+twHUYLRX5lEBPmkbsEy9Omeetf8ADvb/AIKQ/wDQFg/8KSx/+Lr/AId7f8FIf+gLB/4Ulj/8XX/Dvb/gpD/0BYP/AApLH/4utI/Zwg+DPxC0DUrD4gSeINc0OaG4lto9JEKR3S4Ij3CV/MweoAHpnOQH8PfGu48R/wDCwj4euxqeRIJPIjBGECAeUefujGMZ/HmvHnjDxL4w1WOXxPGsNzYxeR5CxGLYQSSSpOQxPX6D0r4SfCay1jwLqV/rsIWbXoTFaOVy0EanKyAepcA47hR2NQQ/Fv4Jz3EFjZnUNHLlywhM0DD++dvzRnA5yR+OAam+MPxU8dQPpXhbQlhMg2STWMDu6ZHPzkkJ354I9e9eL/2K/EXxK0qxi1j4lRaF5bGWWzj0r7ZmQ8AtJ5yAkAngDqTya/4dof8AVa//AC2//umvAvhn/hCvBHh7wb9u+2/2DpVtYfafK8vzvJiWPftyduducZOM9T1qq+L/AOwl/wALX+JGt/EH/han9l/2xKkn2P8AsLz/ACtsapjf567vu5+6Otf8O0P+q1/+W3/901/w7Q/6rX/5bf8A901/w7Q/6rX/AOW3/wDdNfs4fAT/AIZ98K6n4Z/4Sz+3/wC0dQ+1+f8AYPsvl/u1Tbt8x8/cznI69Kqv2mP2Z/8Ahov/AIRv/itv+Ef/AOEf+1f8w37X5/neT/01TbjyffO7tjn/AIdof9Vr/wDLb/8Aumv+HaH/AFWv/wAtv/7pr/h2h/1Wv/y2/wD7pr4GfsS/8KW+JGn/ABB/4Wb/AGz9hhmj+x/2L9n3+ZGyZ3+e+Mbs/d5qq+Kfgb/hZnw813wF/an9nf21aG3+1+R53k5IO7ZuXd06ZFf8O0P+q1/+W3/901/w7Q/6rX/5bf8A901/w7Q/6rX/AOW3/wDdNf8ADtD/AKrX/wCW3/8AdNVVeLP+Cdn/AAlHinWfEv8AwuH7N/a2oT3fkf8ACPb/ACvMkZ9u77SM43YzgZ9BX/DtD/qtf/lt/wD3TX/DtD/qtf8A5bf/AN01/wAO0P8Aqtf/AJbf/wB018DPhT/wpb4b6f8AD3+3v7Z+wyzSfa/sv2ff5kjPjZvfGN2OvNVX7Rn7IP8Aw0B43sfGX/Cw/wCwfsWlR2H2b+yPtW7ZLLJv3ecmM+bjGP4c554/4dof9Vr/APLb/wDumv8Ah2h/1Wv/AMtv/wC6a/4dof8AVa//AC2//umv2cP2S/8Ahn3xTqfiX/hYH9v/ANo6f9k8j+yfsvl/vFfdu858/cxjA69aqvj58IP+F4/DybwF/wAJF/Yvm3UU/wBr+x/aceWScbN6dc9c8V/w7Q/6rX/5bf8A901/w7Q/6rX/AOW3/wDdNf8ADtD/AKrX/wCW3/8AdNeBf+Ce3/CFeN/D3jL/AIW99t/sHVba/wDs3/CP+X53kyrJs3faDtztxnBxnODVVf2v26xuLLzNnnwtHuxnbuGM4/Gv+HaH/Va//Lb/APumv+HaH/Va/wDy2/8A7pr/AIdof9Vr/wDLb/8Aumv+HaH/AFWv/wAtv/7pr4c+D/8AhX/gLQPA/wDaP2/+w9PitPtXk+V52xQN2zc23OOmT9aqvjZ+w5/wuL4naz8R/wDhaH9kf2v5H+hf2J9o8ryoI4vv+euc+Xn7oxnHbNf8O0P+q1/+W3/901/w7Q/6rX/5bf8A901/w7Q/6rX/AOW3/wDdNfs1/s4/8M72OvWX/CZf8JB/bcsEm7+zvsnk+WHGMeY+7O/2xiqr9pL9nr/hoXQ9G0X/AIS/+wP7Ju3uPM/s/wC1ebuTbjHmJj1zk1/w7Q/6rX/5bf8A901/w7Q/6rX/AOW3/wDdNf8ADtD/AKrX/wCW3/8AdNfBT9hz/hTvxO0X4j/8LQ/tf+yPP/0L+xPs/m+bBJF9/wA9tuPMz905xjjORVfEfwf/AMLB8Ba/4H/tH7B/bmny2n2ryfN8nepG7ZuXdjPTI+tf8O0P+q1/+W3/APdNf8O0P+q1/wDlt/8A3TX/AA7Q/wCq1/8Alt//AHTV/wD8E9P+CjC31wtlpUEluJWEL/8ACRWS7kzwceZxxjiovgt/wWighSCHx5qipGoVFHijT+ABgD71f8Kb/wCC03/Q/wCq/wDhUaf/APFV+2r4K/bg8JaX4Uk/a88Q3ep2lzcXQ0NZ9Vt7zZIFj87AhJ25Bj69a+B37F/9keJvCHxu/wCFlebvEOr/ANl/2Ntx5se/y/N849N+N2znHQVVVVVVVVftX+AtU+KH7NXxK8B6FC82p6r4bu0sYU6zTqheOMdfvOir+NfskaN4P179p/4YaJ8QUt20K68UWcd3FcgeXKfMGyOQEEFGk2KwPBDEHHWgqqoVQAAMAAYxWsaVpWu6TeaLrthbX2nX9u8F3bXMYeKaJ1KujqeCpUkEHjBrxh4f024+KOt+Ffh4ralYSa/cWWhiE72uYjOyQbfUsuzHrmvCOkXHh/wnoug3lz9ouNN06C2llJz5jRxqpb8SM1VVVVVVX7dH7Jmn/tcfBt/CtndQWHizQpmvvDl9MvyLPtw8Eh6iKVcAkfdZUbB27T8FP24v2mv+Cf2gv8C/jl8DdQ1rSdImdNIi1O6k06W1BYlkhuvKljngzuZdoOMnDFcBfFEf7XX/AAVf+JugK3ge48FfDbSHJhuZIZDp+no5HmzmVwn2y4IAUKgGOOI1Lufhd8N/C/wf+Hfh74YeC7VoNG8N2EdnaK5Bdwo+aRyAAXdizscDLMT3qq/4Laf8kB8C/wDY4D/0jnr9k7/k1f4Nf9k/0L/0ghqpYxLE0RYruUjKnBH0r9n7ULfw74v1rwrqzJBeXAEcYc4zJEzBkHudxOP9mq/aW1yxGjaZ4bWVWvHuvtTIDyiKjKCfTJc4/wB014GsLjS/Bmh6fdoVng0+FZEIwVbYMj8DxVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV/wVg/5Mj8Yf8AYQ0z/wBLIq/4Jcf8mJfDH/uLf+na8qqqqqv+CiH7NN9+01+zrqOgeGLNbjxb4buF1bQY84M8salZIAf+mkTOAOm8Jnpmv+Cbf/BQDwJ8IvA0n7OX7RWsT+GT4evJhoep3tvIY4kZ2aW0uMAtEySFypYBcMVO3aN3/BRP/goz8LfFXwm1D4F/s+eJv+Eo1PxcgtdW1WyikWC0tCQXhRmUebJKMIQoKhWbJ3YFf8Ewv2X9Y/Zz+A0mq+NtMksPGHjy4j1DUrSVNslnbopW2gcdQ6qzuwPKtKVPKmqr9tf/AJNF+L//AGJ+of8Aolq/4Iqf8mr+Kf8AsoF5/wCkFhVV4K0dPjN8SNT13xC7yadbHzTDuIymcRRcdFwCTjHQ+uag8NeHbWz/ALPt9C0+O2xjyVtkCke4xzXx5+G+j+GBaeJvD9strb3cxgntkGESTaWVlHYEK2R04GK+A3gm38P+Fk8Qzqr3+soJNw58uH+BR9fvH6gdqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv+CyX/Jotp/2OFj/AOibmv2A/wDkzT4S/wDYvx/+hvVVVVVf8Fgvgxf/ABF/ZusviFolkbi++HWp/bLhVXLCxnURzkDr8riBz6KjE9M1/wAEpPj7ovxV/Zm0r4dXGpxHxP8ADoHTru1aT949mXZrWYL12bCIv96I9Miv24fj3oX7Pf7OHi7xRe6rDBreqafNpnh62L4kuL6ZCiFFyCRGGMrdMKh5yRX/AARP+DWpado3jn48arZmK31cxaJo8jDBlSNvMuWHqu/yFB6bkcdqqq/4Idf8jV8W/wDsH6V/6MuaqtRuTZafc3qqGNvC8gU99qk/0r9nPR7bV9Z1rxVqQFxe2xRYnkGSryFi7/U7cZ9z61X7S2gWEcWleJIYkS7lka3mIGDKu3Kk+uMEZ9x7V4PuFu/COh3SRJGJtNt3CIuAuY1OAOwr4r/FbTvC0l74QuNJuJ5r3Tm2zI4Cr5gZRkHnjFfCn4rad8PNOvrK90m4vGu5lkVonC7cLjvVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV/wXG/5FT4R/8AYQ1X/wBF21fCf/klfg3/ALF+x/8ASdKqqqqqqqvHviqHwJ4F8R+OJ7N7uPw9pN1qL26PsaYQRNIUBOcEhcZ7Zr9pD4n/AA2+LvxV1L4j/C/4ZXfgKHWJjc3unNqy3kRumOXlhCwx+SGOWKZcbiSCoIUeAP8AgrB+2D4D8N2vhqbxJoPidLNBHDea9ppnutgGAGlR0Mh/2n3Me5NfFz/gqD+1l8YPB974G1HxLo3h/TNThaC//sDTzbTXMTDDRmV3d1Ujg7CpIJBJBIr4LfFvWPgZ8Q9M+Jvhvw9oGrazozGTTxrVtJcQ203aZUR0BdedpbIUncBuAI/4fJftc/8AQO+H/wD4Jpv/AI/X/BOf9vv46ftVfG7W/h58TrXwvFpeneFbjVITpenyQS+el1axDLNK4K7Z34x1xzxVVVVVVVVVVX/BbT/kgPgX/scB/wCkc9fsnf8AJq/wa/7J/oX/AKQQ1XijxZoPg7TDquvXogiztjUDc8jf3VUdT+g74rXtfv8AxBr934iuWEV1dTmX918vlnttx6ADnrxnrWlfFH4oXXk6FY+LZiZyI0M7RKc9syyDI+pavAPwW14+KD4i+IxF0YsSxq1z5xlmzx5h5JA647nHbg1VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVf8FYP+TI/GH/AGENM/8ASyKv+CXH/JiXwx/7i3/p2vKqqqqqvjf+wZ+y5+0Frcvirx98Nok8QTkefq2l3MljPPjvJ5ZCSHGBudWbAAzivgz/AME/P2UvgVrlv4q8GfDKG7160cPbanrNzJfy27A5DRrISkbDs6qG96qq/bX/AOTRfi//ANifqH/olq/4Iqf8mr+Kf+ygXn/pBYVVfC3WLX4XfEPVvC/iWUW0Nw32f7RJwqsrZjYnsrKx56cjPGcI8cqLJE6urDKspyCPUV+0F4y03XBYeCNDmW+uYrsSzmE7wsm0okYI6t85yO3FeEtJl0LwtpOjTsDNZWUUUhByN4UBse2c1VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVf8Fkv+TRbT/scLH/ANE3NfsB/wDJmnwl/wCxfj/9Deqqqqq1HTrDV9PudJ1WygvLK9heC5tp4w8c0bgqyMp4ZSCQQeCDXxT/AOCTXxQ8D/Eef4l/sY/GFPCfmOzwadeahc2M9gG5aKK6gDmSMngK4X5eGZup8P8A/BKr9pL4y+MbDxJ+2F+0MdW0+x+Vbe01O51S8aPIzFG86rHbhsA5UP7rk5HgfwR4U+G3hHSfAfgfRLfSNC0S2W2sbKAELFGvuckknJLEksSSSSSaqq/4Idf8jV8W/wDsH6V/6Muaqrjyfs8nn4Mew7wR2xz+leA/Htj8M/GF+bGdtT0K6fy3eNCjvGCSkgVgDuAJGDgcn2NL8bPhk9oLo+JlXjPlm3l3g+mNv/1q8YeI9S+OXi7T/Dvhqzli0+1Y7HkXkA43zPjoAAAB/VsVZWcGn2UFhbKRFbRLFGCeiqAB+gq+0DQtTmFxqOi2F1KF2iSe2R2wO2SCcV/wiPhP/oWNJ/8AAKP/AAqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/4Ljf8ip8I/wDsIar/AOi7avhP/wAkr8G/9i/Y/wDpOlVVVVVVVXxY8M6j41+FnjLwbozQi/17w/fWFqZmKoJZrd403EAkLuYZODxVt/wRZ/aruIEll8afC63dhzFLq16WX67bMj8ia/4cqftUf9D98Kf/AAa3/wD8hV/w5U/ao/6H74U/+DW//wDkKv8Ahyp+1R/0P3wp/wDBrf8A/wAhV/w5U/ao/wCh++FP/g1v/wD5Cr/gnp/wT0+NP7Jfxp1n4jfEbxP4J1LTdR8Lz6VFFol7dTTLNJdWsoZhLbxqE2wOCQxOSvHUiqqqqqqqqqr/AILaf8kB8C/9jgP/AEjnr9k7/k1f4Nf9k/0L/wBIIarxfBN8TvjcnhS5unXT7FzDhW+4iJulx/tMwIz9PStH8PaH4fs1sNF0u2tIVXbtjjALfU9Sfc18cvhvod14buvFel2ENpqFhh5jEgQToSA24DgsM5z14x6Y+C/iK78SeALK4v5WluLR2tpJGPL7Puk++0qCfUVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV/wVg/5Mj8Yf9hDTP/SyKv8Aglx/yYl8Mf8AuLf+na8qqqqqqqqqv21/+TRfi/8A9ifqH/olq/4Iqf8AJq/in/soF5/6QWFVXxc8D+Cdc0eXxF4lu30uSxRV/tCGIyFQzBVV0UEuNzD3GeoGavbqSxmudL0vXJ7nT0kZY3XfEky5+9sJ4z1wea/Z98FeFbzTR4ulb7bqtvO0YR1Oy0I6YBABYgg5yQMjGCDVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX/BZL/k0W0/7HCx/9E3NfsB/8mafCX/sX4/8A0N6qqqqqqqqqv+CHX/I1fFv/ALB+lf8Aoy5qqljSaJonGVdSpHTg18cfh94W8E2mkS+HbKSBruSVZd8zSZChcfeJx1NKjSOEjUszHCqBkk+1eA/CGn+DfDlnpttZwx3XkJ9slRAGllx8xJ6nknHoKqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv+C43/ACKnwj/7CGq/+i7avhP/AMkr8G/9i/Y/+k6VVVVVVVVazq+m+H9Hvte1i6S2sNNtpLm6mfpFFGpZ2PsFBNfBj/gqT+yx8Z/EeleC7S/8S+Hdf1q8jstPsdW0hn+03EjBERHtzKo3MQAXKjnnFfET4heDvhR4J1f4h+P9cg0jQNDtzPeXc2SEXIAAA5ZmYhVUAlmIABJr4hf8FufE7eIJofhT8F9KTRIpiIZ/EF5I9xcRj+IxwlViY+m6QD1NfsXf8FJPh9+1Zq//AArzXtBPg7x4IGlt9Pe68+31JEGXNvIQp3qoLGJhkKCQWCsRVVVVVVVVVVVf8FtP+SA+Bf8AscB/6Rz1+yd/yav8Gv8Asn+hf+kENV4pupPhl8cR4mvIJG0+9czEov345E2yY9SrZOO+B61pWsaVrlkmoaPqEF5byDKyROGH4+h9jzXx0+IGi6f4YuvCljfRXGp6gBG8cbBvITILFsdCQMAdec9q+CegXXh/4fWUV7G0c9673TRsMFQ+Nv8A46FP41VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV/wAFYP8AkyPxh/2ENM/9LIq/4Jcf8mJfDH/uLf8Ap2vKqqqqqqqqr9tf/k0X4v8A/Yn6h/6Jav8Agip/yav4p/7KBef+kFhVV4u0nStc8N3+m61CZrNot8qBypOwhxyOeqikRpHWOMZZjhR6mvCPgzQvBOnvp2hQPHHM/mS75Gbc+0AnknHQcVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV/wWS/5NFtP+xwsf/RNzX7Af/Jmnwl/7F+P/wBDeqqqqqqqqqr/AIIdf8jV8W/+wfpX/oy5qq+MfxUu/Bxg8O+G1V9Yu03mQpv8hCcLhehYkHAPTHTkVB8EfiV4wiTVfFfiFYZpBuSO8meaRM+o6LxjgHj0FeKfh74t+GF/Z6rqFvbXEEc6vBcxZkiLqchWBAIPHQjB5wTzXwb8WeKvGWh3Wq+JY0C+cFtZEh8tXUZDY9cMCKqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv+C43/IqfCP8A7CGq/wDou2r4T/8AJK/Bv/Yv2P8A6TpVVVVVVVV+3Hq15ov7IPxcvbEkSv4VvLckHBCyp5T/APjrtX/BKnQdF139tbwc+sNGW0yz1C8s4pAMSTrauFwPVQzOPQpntX/Ban48uZvCP7OWhamQiodd1+KNupJMdpG2PTEzlT6xNjoa/Ze/Y++L/wC1zrGtaV8L00e1h8PWyTX+oaxcSQW8bSEiOINGkjGR9jkDbjCEkjjNnN4+/Zq+OEM1xC2neLvh14iBliWThbi1m+Zdw4ZGKkdwyt3BrTNRtdY0y01axk3297Ak8LeqOoZTx7EVVVVVVVVVVVX/AAW0/wCSA+Bf+xwH/pHPX7J3/Jq/wa/7J/oX/pBDVeLfBugeNtM/svXrTzEU7opUO2SFvVW7fToe4qP9m3xhFkxeI9LQkYO1pBkf9814D/Z+Gia2NS8XTWGpwQpmCCMuR5mRgsCACMZ49aqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr/grB/yZH4w/7CGmf+lkVf8ABLj/AJMS+GP/AHFv/TteVVVVVVVVVX7a/wDyaL8X/wDsT9Q/9EtX/BFT/k1fxT/2UC8/9ILCqr4hT31v4I1ttMs5rq6eykjjihTc2XG3IHfAOfwrwVpUmr+MdG0oRkma+iV1I6KGBY49gDVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVf8Fkv+TRbT/scLH/0Tc1+wH/yZp8Jf+xfj/8AQ3qqqqqqqqqq/wCCHX/I1fFv/sH6V/6MuaqpUhvP2mVj1IBkS5BQOMYZbXMeP+BBSKr4rwWlx8OteS8VSi2jMu7s6kFPx3AV+zfJK/gK6WRiVTVJVjBHQeXGePxJqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/4Ljf8ip8I/wDsIar/AOi7avhP/wAkr8G/9i/Y/wDpOlVVVVVVVXxX8A2HxV+GPiz4Z6lOYbbxTot3pskwXJh86JkDgeqlgw9xWrab8TP2f/ijd6TNc6p4T8a+D9QeFprS4aCe1mTILRyIQcEHIYHDKwPINeKfFXijx54ivfFfjHX9R13W9Tl33d/f3DTzzvgAFnYknAAAHYAAcCv+Cb37O2o/s7fsz6TpnijTGsvFXiq4bWdZglTElu0iqsMDZ5BSFI9yn7rtIPU1+1Z4ztPil+078SvF+gEXVnq/ii8GntB8/wBohWUxxMuM53qqnj+9Xg/RW8OeEdD8Ou25tL023tSSc5Mcap1H+7VVVVVVVVVVVf8ABbT/AJID4F/7HAf+kc9fsnf8mr/Br/sn+hf+kENVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX/BWD/kyPxh/2ENM/wDSyKv+CXH/ACYl8Mf+4t/6dryqqqqqqqqq/bX/AOTRfi//ANifqH/olq/4Iqf8mr+Kf+ygXn/pBYVVUPB/hpfES+K49IgTVVjZPtCDaTu4JIHBbGRnrgmqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/wCCyX/Jotp/2OFj/wCibmv2A/8AkzT4S/8AYvx/+hvVVVVVVVVVV/wQ6/5Gr4t/9g/Sv/RlzVV8a/h7rdxqlt8QfCEcj31oFNxHCMyApykqj+IjABHoBx1rRP2lY4rQW/ibw5MbuMbWktXADsPVGxt/M14o+IHi/wCM8yeE/C+hPBZM4aWNX3l8HhpHwAqgjOPXueK8D+FLbwX4YsvD9vIJGgUmaUDHmSMcs30zwPYCqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr/guN/yKnwj/wCwhqv/AKLtq+E//JK/Bv8A2L9j/wCk6VVVVVVVVV+0V+xP+z1+1DJFqXxN8IOuu28Qih1zS5/st6sY6IzgFZFHOBIrBcnGMmvgZ/wTC/ZY+BHiq18baZpGueK9a0+UTWFx4mvI7lbOQYKukUUUUZYEZDMrFTyCCARrOk2mvaPfaHftOLbUbaS2mMEzQyBHUq210IZGwThlIIPIINab/wAEhf2c/DnxP8N/ETwp4j8VWtroOrQahLod7NFd21z5Lh1j3FFkVSyjOWfIyOM5qqqqqqqqqqqq/wCC2n/JAfAv/Y4D/wBI56/ZO/5NX+DX/ZP9C/8ASCGqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr/grB/yZH4w/7CGmf+lkVf8ABLj/AJMS+GP/AHFv/TteVVVVVVVVVX7a/wDyaL8X/wDsT9Q/9EtX/BFT/k1fxT/2UC8/9ILCqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr/gsl/wAmi2n/AGOFj/6Jua/YD/5M0+Ev/Yvx/wDob1VVVVVVVVVf8EOv+Rq+Lf8A2D9K/wDRlzVUzKil3YAKMkk4xXiC8Go69qWoK4cXN3LKGHfc5Of1r4bXfh1vCml6XompadcTWljALqO1mR2V9gBLhScEkHr3zVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX/Bcb/kVPhH/2ENV/9F21fCf/AJJX4N/7F+x/9J0qqqqqqqqr9uL/AIKpePNY8X6p8L/2ZfEI0Lw7pU72134mtgr3WpyoSHNu5yIoMj5XX53xuDKp2nxP428Z+Nrx9R8ZeL9a167kfe8+p38t1Izc8lpGJJ5PPvX/AASUur6+/Y006Sa8eWca7qKRyTkybcOoXvkgemR+FSf8FW/2tLH41aVpPj7U/DWi6JoniWO28R6TpWkIqSwRThLiLzZzJIvyh8MGBBA7ZBqqqqqqqqqqqv8Agtp/yQHwL/2OA/8ASOev2Tv+TV/g1/2T/Qv/AEghqq/aL/al+E37LOiaRr/xYvdTt7TXLp7W0NjZG5YyIu45AIwMd68K+I9M8ZeF9H8X6I8jafrlhBfWjSJsYxTRq6Er2O1hkdqqqqqqqqqqqqqqqqqqqqqqqqqqvj7+2R8E/wBmrxPoHhD4oX+rwaj4liMtgtnp7XCsokEfzEH5fmIqqqqqqqqqqqqqqqqqqqqqqqqqqrxn+2R8E/AXx50f9m/xBf6unjPXJbWKzii09ngLXJxFmXOB7+lVVf8ABWD/AJMj8Yf9hDTP/SyKv+CXH/JiXwx/7i3/AKdryqqqqqqqqq/bX/5NF+L/AP2J+of+iWr/AIIqf8mr+Kf+ygXn/pBYVVV+0n+0v8Of2VfAtj8QvidDrEul6hq0elwrpdqs8vnvFLKCVZ1AXbA/OeuOOePhr4/0L4rfD/w98SfDCXS6T4l06G/sluoxHKIpFDLvUEgNg8jJqqqqqqqqqqqqqqqqqqqqqqqqqqqv2Y/21fg5+1rfa/p3wstvEUUvhuGCW8/tWxS3BWYuE2bZHycxtnp2qqqqqqqqqqqqqqqqqqqqqqqqqqr9pb9qP4Z/speE9L8ZfE+DWpbDV9R+wW40u1WdxL5bSfMGdMLtQ8564rwF4y0f4j+BfDnxC8PLcLpfijSbXVLFblAkoguIllj3qCQG2uMgE4Pc1Vf8Fkv+TRbT/scLH/0Tc1+wH/yZp8Jf+xfj/wDQ3qqqqqqqqqq/4Idf8jV8W/8AsH6V/wCjLmqrUY55dPuYrRUad4XWIOcKWIOMn0zivjZq2kfs5adp+t/Ge/Ph7StTvRZ216LeW8R5dpbGIFdh8qseQOlfBPwv4KtfDVl488Ba/eavo3i7TLW+sLm4iMfm20ieZE4RlVl3JIDhgCM4IBzVVVVVVVVVVVVVVVVVVVVVVVVVV8cv2rvgD+zbd6RY/Grx9/wjs+uxyyaen9lXl35yxFQ5zbxSBcF1+9jOeO9VVVVVVVVVVVVVVVVVVVVVVVVVVeOf2rvgD8NvixofwN8aePv7O8b+JJLSPS9L/sq8m+0NdS+TAPNjiaJd0gK/M4x1OBzVV/wXG/5FT4R/9hDVf/RdtXwn/wCSV+Df+xfsf/SdKqqqqqqqv2wvGGpeAv2Wvil4q0W4eDULPwveLazR/ehlkjMayL7qXDD3HfpX7Dv7OGnftTftB6R8M9e1Oex0KG1m1LV5LYgTNbQ7cxxk8Bnd403YO0MWwcYP/BVz9nf4M/s76n8LtD+Dnge18O22oWGom8Mc0s0ly0bwBGkeVmZiNzd+9f8ABIb/AJM00z/sYNR/9DWv2oJrC5/aX+LdxpTRNZS+Otaa2MS4Qxm+mK7R2GMYr4bXtxqfw58K6jduGnutEs5ZCBjLNChJ/M1VVVVVVVVVVX/BbT/kgPgX/scB/wCkc9fsnf8AJq/wa/7J/oX/AKQQ1Vf8Fvf+SVfDT/sYLr/0nFfs9f8AJAfhn/2J+k/+kcVVVVVVVVVVVVVVVVVVVVVVVVVVX/BZD/kv3wb/AOwe/wD6VrVVVVVVVVVVVVVVVVVVVVVVVVVVX7Uf/KYP4Zf9hDw9/wCh1VV/wVg/5Mj8Yf8AYQ0z/wBLIq/4Jcf8mJfDH/uLf+na8qqqqqqqqqv21/8Ak0X4v/8AYn6h/wCiWr/gip/yav4p/wCygXn/AKQWFVVf8Fq/+TV/C3/ZQLP/ANIL+v2KP+TRfhB/2J+n/wDolaqqqqqqqqqqqqqqqqqqqqqqqqqqr/gh1/yNXxb/AOwfpX/oy5qqqqqqqqqqqqqqqqqqqqqqqqqqr/gtp/yQHwL/ANjgP/SOev2Tv+TV/g1/2T/Qv/SCGqr/AILJf8mi2n/Y4WP/AKJua/YD/wCTNPhL/wBi/H/6G9VVVVVVVVVX/BDr/kavi3/2D9K/9GXNVVf8FtP+SA+Bf+xwH/pHPX7J3/Jq/wAGv+yf6F/6QQ1VVVVVVVVVVVVVVVVVVVVVVVVVf8Fxv+Rq+Ef/AGD9V/8ARltVVVVVVVVVVVVVVVVVVVVVVVVVVX7cP/KWH4H/APYQ8Kf+nVqqv+C43/IqfCP/ALCGq/8Aou2r4T/8kr8G/wDYv2P/AKTpVVVVVVVV8a/h6nxa+D/jX4YtOkLeKdBvNOimfOIZJYWRJDj+6xVse1fA/wCLvxE/Y++Pdt41stGEeveFby4sNW0a+JQTLzFcW0hGSpyDgjO1lU4OMH/gob+2R8Pv2w5/h3rXgnw/rmjXXh2yvYtUtNTSPCSTNCV8p0Y+Yv7tuSFPTiv2fP8Ago7pn7NP7Hsnwc8DeGtRvPiJPfXstvqNyiLY2CzMNsv3i8sijJCbQucEkgEH4RfDjxF8cfi34Z+GujGe41TxXq0Vs0xy7IrtmWZieSEQPIx9FJq0tLextILG0j2Q28axxJknaqjAGTz0A61VVVVVVVVVVX/BbT/kgPgX/scB/wCkc9fsnf8AJq/wa/7J/oX/AKQQ1Vf8Fvf+SVfDT/sYLr/0nFfs9f8AJAfhn/2J+k/+kcVVVVVVVVVVVVVVVVVVVVVVVVVVX/BZD/kv3wb/AOwe/wD6VrVVVVVVVVVVVVVVVVVVVVVVVVVVX7Uf/KYP4Zf9hDw9/wCh1VV/wVg/5Mj8Yf8AYQ0z/wBLIq/4Jcf8mJfDH/uLf+na8qqqqqqqqqv26L+HTf2Pvi5cTAlX8LXcIA/vSL5Y/VhX/BF2ymtf2UdenlK7bzx3eyx4PO0WdknPvlD+lVVf8Fq/+TV/C3/ZQLP/ANIL+v2KP+TRfhB/2J+n/wDolaqqqqqqqqqqqqqqqqqqqqqqqqqqr/gh1/yNXxb/AOwfpX/oy5qqqqqqqqqqqqqqqqqqqqqqqqqqr/gtp/yQHwL/ANjgP/SOev2Tv+TV/g1/2T/Qv/SCGqr/AILJf8mi2n/Y4WP/AKJua/YD/wCTNPhL/wBi/H/6G9VVVVVVVVVX/BDr/kavi3/2D9K/9GXNVVf8FtP+SA+Bf+xwH/pHPX7J3/Jq/wAGv+yf6F/6QQ1VVVVVVVVVVVVVVVVVVVVVVVVVf8Fxv+Rq+Ef/AGD9V/8ARltVVVVVVVVVVVVVVVVVVVVVVVVVVX7cP/KWH4H/APYQ8Kf+nVqqv+C43/IqfCP/ALCGq/8Aou2r4T/8kr8G/wDYv2P/AKTpVVVVVVVVX7dH/BNLw1+03qEvxN+GWp2Hhb4hGPbdm4iIs9Z2gBTOUBaOUAAeaqtkABlPDL47/YA/bE+H19NZap8A/E+prE2Fn0K1/tWOQZwGU2xc4OQcEAjuBg14C/4J9/ti/ES+hstM+A/iTSkkI33Gv2/9lRxDOCzfaCjHGOign0Br9hL/AIJ5eGv2TYZPHHi7U7XxJ8RtQtzA97AjC20yFsb4rYNhmLY+aVgCRhQqjduqqqqqqqqqqqv+C2skY+AvgOEyL5jeL9yoTyQLSbJA9BkfmK/ZPUr+yx8G1YEEfD/QgQRjH+gQ1Vf8Fvf+SVfDT/sYLr/0nFfs9f8AJAfhn/2J+k/+kcVVVVVVVVVVVVVVVVVVVVVVVVVVX/BZD/kv3wb/AOwe/wD6VrVVVVVVVVVVVVVVVVVVVVVVVVVVX7Uf/KYP4Zf9hDw9/wCh1VV/wVg/5Mj8Yf8AYQ0z/wBLIq/4Jcf8mJfDH/uLf+na8qqqqqqqqqv+Cm18dO/Ya+KFwsYkLW+nw4JxjzNRtUz+G7P4V/wSS09bP9izw9crIGN9rOpTEbcbcXBjx7/6vOffHaqqv+C1f/Jq/hb/ALKBZ/8ApBf1+xR/yaL8IP8AsT9P/wDRK1VVVVVVVVVVVVVVVVVVVVVVVVVVX/BDr/kavi3/ANg/Sv8A0Zc1VVVVVVVVVVVVVVVVVVVVVVVVVV/wW0/5ID4F/wCxwH/pHPX7J3/Jq/wa/wCyf6F/6QQ1Vf8ABZL/AJNFtP8AscLH/wBE3NfsB/8AJmnwl/7F+P8A9Deqqqqqqqqq1jUoNG0i91i5I8mxtpJ5MsF+VFLHk8DgV/wQ306eTVvjDqwIEMNto8ByD8zO143HbgJz9RVVX/BbT/kgPgX/ALHAf+kc9fsnf8mr/Br/ALJ/oX/pBDVVVVVVVVVVVVVVVVVVVVVVVVVV/wAFxv8AkavhH/2D9V/9GW1VVVVVVVVVVVVVVVVVVVVVVVVVVftw/wDKWH4H/wDYQ8Kf+nVqqv8AguN/yKnwj/7CGq/+i7avhP8A8kr8G/8AYv2P/pOlVVVVVVV4m8TeHvBnh7UPFfivWbTStH0m3e4vb27lEcUESjLMzHoMV8T/APgtZ8IvDWs3Gl/C74Va540trdyg1C81BdJhnwfvRgxyybcdN6IfYda/Z0/4KyfAb43eIrLwT4y0a/8Ah3ruoyCO0bUbqO4sJpDgLGLkBCrE8DfGqngbskCtW1bStB0251nXNTtNOsLOMyXF3dzLDFCg6s7sQFA9ScV8Uv8Agqt+x78NpJrHTfGWo+Nr+AlWg8NWJnTcOmJ5THCw90dq8bf8FpfiB4o1m10D4S/CbR/D1teXccH2/Wbt7+fYzAFljQRojc8BjIPrVVVVVVVVVVVV/wAFP/2Xvid+0z8IfD1t8JbG31PXPC+rtdtpctwlu13DJEUby3kZU3qdp2sQCN2DkAN4fj/4LZaH4f0zwToGnatZ6Zp1nDp9lCLLw+nkwxoI4181l3DCgDczZ4yT3ofCn/gtdqY+3XXjLVrSWU/NCfEmlpsxx92JygyBnj19c1/wpv8A4LTf9D/qv/hUaf8A/FV8VP2Lf+Cpvxv0+x0r4sSN4ntNMmaa0hvvEtiwhkZdpYYcckcVo3wD/wCCyfh7R7Hw/onjLUbTT9Mto7W0t4/E9gFhijUKiL83QKAPwr/hTf8AwWm/6H/Vf/Co0/8A+Kr/AIU3/wAFpv8Aof8AVf8AwqNP/wDiq/4U3/wWm/6H/Vf/AAqNP/8Aiq/4U3/wWm/6H/Vf/Co0/wD+Kr/hTf8AwWm/6H/Vf/Co0/8A+Kr/AIU3/wAFpv8Aof8AVf8AwqNP/wDiq/4U3/wWm/6H/Vf/AAqNP/8Aiq/4U3/wWm/6H/Vf/Co0/wD+Kr/hTf8AwWm/6H/Vf/Co0/8A+Kr/AIU3/wAFpv8Aof8AVf8AwqNP/wDiq/4U3/wWm/6H/Vf/AAqNP/8Aiq/4U3/wWm/6H/Vf/Co0/wD+Kr/hTf8AwWm/6H/Vf/Co0/8A+Kr/AIU3/wAFpv8Aof8AVf8AwqNP/wDiq/4U3/wWm/6H/Vf/AAqNP/8Aiq/4U3/wWm/6H/Vf/Co0/wD+Kr/hTf8AwWm/6H/Vf/Co0/8A+Kr/AIU3/wAFpv8Aof8AVf8AwqNP/wDiq/4U3/wWm/6H/Vf/AAqNP/8Aiq/4U3/wWm/6H/Vf/Co0/wD+Kr/hTf8AwWm/6H/Vf/Co0/8A+Kr/AIU3/wAFpv8Aof8AVf8AwqNP/wDiq/4U3/wWm/6H/Vf/AAqNP/8Aiq/4U3/wWm/6H/Vf/Co0/wD+Kr/hTf8AwWm/6H/Vf/Co0/8A+Kr/AIU3/wAFpv8Aof8AVf8AwqNP/wDiq/4U3/wWm/6H/Vf/AAqNP/8Aiq/4U3/wWm/6H/Vf/Co0/wD+Kr/hTf8AwWm/6H/Vf/Co0/8A+Kr/AIU3/wAFpv8Aof8AVf8AwqNP/wDiq/4U3/wWm/6H/Vf/AAqNP/8Aiq/4U3/wWm/6H/Vf/Co0/wD+Kr/hTf8AwWm/6H/Vf/Co0/8A+Kr/AIU3/wAFpv8Aof8AVf8AwqNP/wDiq/4U3/wWm/6H/Vf/AAqNP/8Aiq/4U3/wWm/6H/Vf/Co0/wD+Kr/hTf8AwWm/6H/Vf/Co0/8A+Kr/AIU3/wAFpv8Aof8AVf8AwqNP/wDiq+Jn7Cv/AAU6+MmsaZr/AMULZfEeo6MhSwuL3xLYlrdd27C4cfxAGv8AhTf/AAWm/wCh/wBV/wDCo0//AOKr/hTf/Bab/of9V/8ACo0//wCKr/hTf/Bab/of9V/8KjT/AP4qv+FN/wDBab/of9V/8KjT/wD4qv8AhTf/AAWm/wCh/wBV/wDCo0//AOKr/hTf/Bab/of9V/8ACo0//wCKr/hTf/Bab/of9V/8KjT/AP4qv+FN/wDBab/of9V/8KjT/wD4qv8AhTf/AAWm/wCh/wBV/wDCo0//AOKr/hTf/Bab/of9V/8ACo0//wCKr/hTf/Bab/of9V/8KjT/AP4qv+FN/wDBab/of9V/8KjT/wD4qv8AhTf/AAWm/wCh/wBV/wDCo0//AOKr/hTf/Bab/of9V/8ACo0//wCKr/hTf/Bab/of9V/8KjT/AP4qv+FN/wDBab/of9V/8KjT/wD4qv8AhTf/AAWm/wCh/wBV/wDCo0//AOKr/hTf/Bab/of9V/8ACo0//wCKr/hTf/Bab/of9V/8KjT/AP4qv+FN/wDBab/of9V/8KjT/wD4qv8AhTf/AAWm/wCh/wBV/wDCo0//AOKr/hTf/Bab/of9V/8ACo0//wCKr/hTf/Bab/of9V/8KjT/AP4qv+FN/wDBab/of9V/8KjT/wD4qv8AhTf/AAWm/wCh/wBV/wDCo0//AOKr/hTf/Bab/of9V/8ACo0//wCKr/hTf/Bab/of9V/8KjT/AP4qv+FN/wDBab/of9V/8KjT/wD4qv8AhTf/AAWm/wCh/wBV/wDCo0//AOKr/hTf/Bab/of9V/8ACo0//wCKr/hTf/Bab/of9V/8KjT/AP4qv+FN/wDBab/of9V/8KjT/wD4qv8AhTf/AAWm/wCh/wBV/wDCo0//AOKr/hTf/Bab/of9V/8ACo0//wCKr/hTf/Bab/of9V/8KjT/AP4qv+FN/wDBab/of9V/8KjT/wD4qv8AhTf/AAWm/wCh/wBV/wDCo0//AOKr/hTf/Bab/of9V/8ACo0//wCKr/hTf/Bab/of9V/8KjT/AP4qtf8A2Ff+CnXin4k2Hxg8QWy33jPTHhez1mXxLYmeFoeYip34+XtxX/Cm/wDgtN/0P+q/+FRp/wD8VX/Cm/8AgtN/0P8Aqv8A4VGn/wDxVf8ACm/+C03/AEP+q/8AhUaf/wDFV8Q/2VP+CtvxY8K3Pgj4jazc6/oV48bz2N34msDHIyMGQkBweGAP4V+xF8HvF/wD/Zc8DfCfx6lomv6LFePex2s3mxxtPez3AQOOGKrMqkjjIOCRzVVVVVVVVVftO/B2X4//AAC8a/CC11GGxuvEWneXaXEykxx3EbrLCXxk7fMjQEgEgEkc18OvhF/wV/8A2adAb4a/CfRrqz8PQ3Mk8MdmdE1CFmZvmdGn3uoY84O0+wofC7/gtf4wUQav4t1nR42CqJD4j0u14Jzn/RXLDGBnjPOOea/4Zc/4LB/9F01X/wALp6/4Zc/4LB/9F01X/wALp6+If7Bn/BT74t6JB4b+J3j5PFGl210t3DZ6p4vNxHHOqsgkCsCAwWRxn0Y+teHf2O/+Cs3hLQrDwx4Y+L13pek6Xbrb2Vla+NWjit4lGFRFHAUAYAr/AIZc/wCCwf8A0XTVf/C6ev8Ahlz/AILB/wDRdNV/8Lp6/wCGXP8AgsH/ANF01X/wunr/AIZc/wCCwf8A0XTVf/C6ev8Ahlz/AILB/wDRdNV/8Lp6/wCGXP8AgsH/ANF01X/wunr/AIZc/wCCwf8A0XTVf/C6ev8Ahlz/AILB/wDRdNV/8Lp6/wCGXP8AgsH/ANF01X/wunr/AIZc/wCCwf8A0XTVf/C6ev8Ahlz/AILB/wDRdNV/8Lp6/wCGXP8AgsH/ANF01X/wunr/AIZc/wCCwf8A0XTVf/C6ev8Ahlz/AILB/wDRdNV/8Lp6/wCGXP8AgsH/ANF01X/wunr/AIZc/wCCwf8A0XTVf/C6ev8Ahlz/AILB/wDRdNV/8Lp6/wCGXP8AgsH/ANF01X/wunr/AIZc/wCCwf8A0XTVf/C6ev8Ahlz/AILB/wDRdNV/8Lp6/wCGXP8AgsH/ANF01X/wunr/AIZc/wCCwf8A0XTVf/C6ev8Ahlz/AILB/wDRdNV/8Lp6/wCGXP8AgsH/ANF01X/wunr/AIZc/wCCwf8A0XTVf/C6ev8Ahlz/AILB/wDRdNV/8Lp6/wCGXP8AgsH/ANF01X/wunr/AIZc/wCCwf8A0XTVf/C6ev8Ahlz/AILB/wDRdNV/8Lp6/wCGXP8AgsH/ANF01X/wunr/AIZc/wCCwf8A0XTVf/C6ev8Ahlz/AILB/wDRdNV/8Lp6/wCGXP8AgsH/ANF01X/wunr/AIZc/wCCwf8A0XTVf/C6ev8Ahlz/AILB/wDRdNV/8Lp6/wCGXP8AgsH/ANF01X/wunr/AIZc/wCCwf8A0XTVf/C6ev8Ahlz/AILB/wDRdNV/8Lp6/wCGXP8AgsH/ANF01X/wunr/AIZc/wCCwf8A0XTVf/C6evhx/wAE6v8AgpD8H5765+Fniiy8Ky6miJePpXiv7ObhUJKB9oGQCzY+pr/hlz/gsH/0XTVf/C6ev+GXP+Cwf/RdNV/8Lp6/4Zc/4LB/9F01X/wunr/hlz/gsH/0XTVf/C6ev+GXP+Cwf/RdNV/8Lp6/4Zc/4LB/9F01X/wunr/hlz/gsH/0XTVf/C6ev+GXP+Cwf/RdNV/8Lp6/4Zc/4LB/9F01X/wunr/hlz/gsH/0XTVf/C6ev+GXP+Cwf/RdNV/8Lp6/4Zc/4LB/9F01X/wunr/hlz/gsH/0XTVf/C6ev+GXP+Cwf/RdNV/8Lp6/4Zc/4LB/9F01X/wunr/hlz/gsH/0XTVf/C6ev+GXP+Cwf/RdNV/8Lp6/4Zc/4LB/9F01X/wunr/hlz/gsH/0XTVf/C6ev+GXP+Cwf/RdNV/8Lp6/4Zc/4LB/9F01X/wunr/hlz/gsH/0XTVf/C6ev+GXP+Cwf/RdNV/8Lp6/4Zc/4LB/9F01X/wunr/hlz/gsH/0XTVf/C6ev+GXP+Cwf/RdNV/8Lp6/4Zc/4LB/9F01X/wunr/hlz/gsH/0XTVf/C6ev+GXP+Cwf/RdNV/8Lp6/4Zc/4LB/9F01X/wunr/hlz/gsH/0XTVf/C6ev+GXP+Cwf/RdNV/8Lp6/4Zc/4LB/9F01X/wunr/hlz/gsH/0XTVf/C6ev+GXP+Cwf/RdNV/8Lp6/4Zc/4LB/9F01X/wunr/hlz/gsH/0XTVf/C6ev+GXP+Cwf/RdNV/8Lp6/4Zc/4LB/9F01X/wunr4i/sC/8FOPi7pdtonxP8dReKbCzuPtFvbar4u+0JFLtK71DA4baxGfQmtD/ZC/4K2eGtE0/wAN+HvjFe6fpelWsdpY2dv42ZI7eCNQkcaKOFVVUAAdABX/AAy5/wAFg/8Aoumq/wDhdPX/AAy5/wAFg/8Aoumq/wDhdPXj/wDYT/4KifFbQl8MfEn4hjxNpKXC3C2Wp+MDPEJVBCvtYEbgGYZ9zX7M/wALtX+CvwC8C/CrxBf2t5qfhvR4rW8mtd3lNLyzhCwBKgsQCQM4zgZwKqqqqqqqq8V6GvijwtrHhprjyBq2nz2hlC7vL8yNk3Y743ZxXws+Df8AwVA/Y21XxN4S+Dnw6uXttVuYmu72xsbTUre98oOI5I3kyVBDsdpCtz8wBGAvjP8A4Lg6mPt1rpmr20UpJWE6JoMezBxjbJHvHTvX/CS/8FyP+fbVf/BR4d/+N1/wkv8AwXI/59tV/wDBR4d/+N18bfg//wAFaP2jfD9h4W+MvgfVfEOmaZefbLWDyNGtPLm2Mm7db+Wx+V2GCSOeleE4/wDgtX4G8K6N4J8K6Rqtlovh/T4NP021/s3w/J5FvBGscSb3Qs2EVRliScZJJ5r/AISX/guR/wA+2q/+Cjw7/wDG6/4SX/guR/z7ar/4KPDv/wAbr/hJf+C5H/Ptqv8A4KPDv/xuv+El/wCC5H/Ptqv/AIKPDv8A8br/AISX/guR/wA+2q/+Cjw7/wDG6/4SX/guR/z7ar/4KPDv/wAbr/hJf+C5H/Ptqv8A4KPDv/xuv+El/wCC5H/Ptqv/AIKPDv8A8br/AISX/guR/wA+2q/+Cjw7/wDG6/4SX/guR/z7ar/4KPDv/wAbr/hJf+C5H/Ptqv8A4KPDv/xuv+El/wCC5H/Ptqv/AIKPDv8A8br/AISX/guR/wA+2q/+Cjw7/wDG6/4SX/guR/z7ar/4KPDv/wAbr/hJf+C5H/Ptqv8A4KPDv/xuv+El/wCC5H/Ptqv/AIKPDv8A8br/AISX/guR/wA+2q/+Cjw7/wDG6/4SX/guR/z7ar/4KPDv/wAbr/hJf+C5H/Ptqv8A4KPDv/xuv+El/wCC5H/Ptqv/AIKPDv8A8br/AISX/guR/wA+2q/+Cjw7/wDG6/4SX/guR/z7ar/4KPDv/wAbr/hJf+C5H/Ptqv8A4KPDv/xuv+El/wCC5H/Ptqv/AIKPDv8A8br/AISX/guR/wA+2q/+Cjw7/wDG6/4SX/guR/z7ar/4KPDv/wAbr/hJf+C5H/Ptqv8A4KPDv/xuv+El/wCC5H/Ptqv/AIKPDv8A8br/AISX/guR/wA+2q/+Cjw7/wDG6/4SX/guR/z7ar/4KPDv/wAbr/hJf+C5H/Ptqv8A4KPDv/xuv+El/wCC5H/Ptqv/AIKPDv8A8br/AISX/guR/wA+2q/+Cjw7/wDG6/4SX/guR/z7ar/4KPDv/wAbr/hJf+C5H/Ptqv8A4KPDv/xuv+El/wCC5H/Ptqv/AIKPDv8A8br/AISX/guR/wA+2q/+Cjw7/wDG6/4SX/guR/z7ar/4KPDv/wAbr45fs7f8FS/2k7vSL741fDjVfEU+gxyx6e23SLPyVlKlxi3MYbJRfvZxjjHNf8JL/wAFyP8An21X/wAFHh3/AON1/wAJL/wXI/59tV/8FHh3/wCN1/wkv/Bcj/n21X/wUeHf/jdf8JL/AMFyP+fbVf8AwUeHf/jdf8JL/wAFyP8An21X/wAFHh3/AON1/wAJL/wXI/59tV/8FHh3/wCN1/wkv/Bcj/n21X/wUeHf/jdf8JL/AMFyP+fbVf8AwUeHf/jdf8JL/wAFyP8An21X/wAFHh3/AON1/wAJL/wXI/59tV/8FHh3/wCN1/wkv/Bcj/n21X/wUeHf/jdf8JL/AMFyP+fbVf8AwUeHf/jdf8JL/wAFyP8An21X/wAFHh3/AON1/wAJL/wXI/59tV/8FHh3/wCN1/wkv/Bcj/n21X/wUeHf/jdf8JL/AMFyP+fbVf8AwUeHf/jdf8JL/wAFyP8An21X/wAFHh3/AON1/wAJL/wXI/59tV/8FHh3/wCN1/wkv/Bcj/n21X/wUeHf/jdf8JL/AMFyP+fbVf8AwUeHf/jdf8JL/wAFyP8An21X/wAFHh3/AON1/wAJL/wXI/59tV/8FHh3/wCN1/wkv/Bcj/n21X/wUeHf/jdf8JL/AMFyP+fbVf8AwUeHf/jdf8JL/wAFyP8An21X/wAFHh3/AON1/wAJL/wXI/59tV/8FHh3/wCN1/wkv/Bcj/n21X/wUeHf/jdf8JL/AMFyP+fbVf8AwUeHf/jdf8JL/wAFyP8An21X/wAFHh3/AON1/wAJL/wXI/59tV/8FHh3/wCN1/wkv/Bcj/n21X/wUeHf/jdf8JL/AMFyP+fbVf8AwUeHf/jdf8JL/wAFyP8An21X/wAFHh3/AON1/wAJL/wXI/59tV/8FHh3/wCN1/wkv/Bcj/n21X/wUeHf/jdf8JL/AMFyP+fbVf8AwUeHf/jdf8JL/wAFyP8An21X/wAFHh3/AON1/wAJL/wXI/59tV/8FHh3/wCN1/wkv/Bcj/n21X/wUeHf/jdeOf2dv+CpfxJ+LGh/HHxp8ONV1Hxv4bktJNL1TbpEP2drWXzoD5UZWJtshLfMhz0ORxX/AAkv/Bcj/n21X/wUeHf/AI3X/CS/8FyP+fbVf/BR4d/+N18bPgP/AMFW/wBpmHRdL+NHgLU/EEWizSNpwdNGshA8oUOS0BjyCEX72QMdua8C6LeeHPBHh7w9qDRm60vSra1nMbZXfHEqNtPGRkGqqqqqqq/4LV/HHVdM03wb+z3ot88FtrETa5rkaNjz4kkMdqhx1TzEmcg8bo0P8NfCn4I/Fn4561N4e+EngHVvE99axiS5WyhylupOA0kjEJGCQQNxGcHHSvi38A/jL8CNRttL+Lvw61jwzNeqWtXu4QYbjGNwjlQmNyMjIViRkZxkV8ZP2xfjR8b/AIVeBvhB4v8AEdy+h+DbAW8yicltXmR38qe4PBdkh8qMBi3KM5O5zjSdG1jxBfxaToOlXmpX05xFbWkDTSOf9lVBJ/AV8Cv+Ccv7XXxI8Q6Rqn/CqrzwtpMV3FLLf+Jm/s4IqupJ8l/37cZPyxke9VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX/AAWG1K4vv2wntZjlNO8L2EEPJOFJlkP0+aRulf8ABJjwToPhr9jDwz4l0yzjTUPF2oahe6lMEAeR4ruW2QE9SBHbrgdAS3qc/wDBRn4e6H8Q/wBjr4iw6vaQyT+H9NOs6fM65a3ntiH3IexZBJGfaQjvX7H3wa0j9oL9pTwJ8I/EE80Wla3fSPfmFtjvb28ElxKit/CWSFlz23Zr4d/Cn4a/CTQofDXwy8DaL4a06FAgh0+0WIvju7AbpGPdmJJPJJNVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVf8ABYfTzZ/tgtcmQt9u8LWEwBXG3DSx49/9XnPvjtXwf/4KBftR/An4f6d8MPhv45srHw9pLzNaW0+kW9yYjLK0r4eRC2C7scZ7mvHv/BSn9rn4l+Ctb+H/AIu8e6bdaL4hsZbK/gTQrWJpIZF2sAyxhlJB6g5r4R/Fjxv8DfiHpPxS+HGpQ2HiLRPO+xXEtsk6p5sLwvlHBU5jlccjjOe1f8PYP23P+ikaT/4Ttn/8brw//wAFVf21dQ17TbC6+I2lNDc3kUUijw9ZjKs4B58vjg1VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVft5/8E6/FH7XvxQ0L4jeGPiFo/h9tL0BNKngvrSWUybJ5pVZShwB+/YYx2/L/hyF8U/+i4+FP/Bbcf41/wAOQvin/wBFx8Kf+C24/wAa/wCHIXxT/wCi4+FP/Bbcf41/w5C+Kf8A0XHwp/4Lbj/GtG/4IofFHS9YsdSk+NvhV1tLmOUoNOnBYKwOBz7VVVVVVVVVVVVVVfGv4z+Af2f/AIb6r8UfiRqps9H0pB8sa75bmVuI4Yk/ikc8AcAckkKCRaft9f8ABQz9qTW9Suv2VPg/b6b4d02XaHg0+O8ZOOFmuroiEyEc7UVSAeh6n4Rf8FSfjd8KPihB8JP24fh6NJRpEjuNWj017G7sA3CzyQjKTwEjO6IKcEsu/AU2l3aahaQ31hdRXNtcxrJDNC4dJEYZVlYcEEEEEcHNVVVVVVVVVVVVVVVVVVVVVVV4/wDiB4L+FnhDUfHvxC8R2eheH9JjD3l/dMQkQZgijjJLMzKoUAklgACTX7Qn/BZu7jv59F/Zj8C291ZWjjzvEPiKCQrMuQP3VsjKUUnADyNk5xsU81+wR8ffHX7S37PVn8UPiJFpcesz6rd2rLptu0MWyJgFwrMxzg881X7bP7c3gf8AY98N2ttLp48Q+N9biZ9J0JZvKCxglTczuASkQYEAAbnYEDADMtj+1J/wV2+IPh8fFnwb8KjD4Wnj+0W9tZeGIis0PUNFFMzXMqkEYK53DkHvX7DH/BSuz/aN8Sj4O/GDw/ZeFviDsf7C9qXS11RowTJGschLQzKqk7CzBgrEFSApqqqqqqqqqqqqqqqqqqqqqq/bX/4KM3H7NnjmD4K/Df4W33irx7eW0M0bXsciWaibPlCNI/3l0xIwVQoAeN5YMo1P9p3/AILCy2n/AAmafB3VbHSwnmmyh8EA4Xk8xOGuBx+OAPfP7F3/AAVTHxl8c2Xwd+P3h3S/DPibU5hbaVqmniSK1u7nO0W8sUjM0UrHhTuKsx24UkA1+3R/wUZ8O/sr3kfw38CaJa+KviPcwrK9pcSMLXSkcAxtcBMM7uCCsSsp2ncWUFQ2pftPf8FfdB8Pn4tav8LJI/DEcYupLSTwtCVihxkl4VP2pEABJZiMDkkCv2Fv2/8Awt+15p934X1zSbfw18QdHtxPdaZFMXgvoAQrXFsW+YAMQGjbJXcvzMCSKqqqqqqqqqqqq+LvxZ8C/A34eax8T/iNq40/Q9Fh3zOF3ySuSFSKNf4pHYhVHqeSBkiP/goL+31+1J4k1OP9kj4PQ6foGlyYMkdhHeyoD90T3NwRAHYDIjVQQM8tjNfDP/gqH+0F8FvidbfC39uD4bmxt5ZEW41OLTGsryyRjgXBiXMdzDkHJiAONxUuQEOmanp2t6Zaazo99Be2F/Ak9rcwOHjmidQyOrDgqVIII6g1VVVVVVVVVVVVVVVVVVVVVVVV+1R/wUA+BH7Kz3Hh3X9Qn8Q+NY4Vkj8NaYMyqHAKGeUjZApBB5y5BBCMK+C//BVn9oj4vftKeCvh7d+E/CGh+GfFXiGzs3s0s5pbmG2mdRxM0gDMVYHdsAOQQuKqeeC1gkubmaOKGFC8kkjBVRQMkkngADvXxn/4KkfFr4j/ABPk+Df7Dfw9TxJMsrxR61LYSXk19t+9LBACqxQjn95LuBXkhK1f9vP/AIKOfsu6zp2o/tN/Cm11Hw/fy7MXmmR2iy9ykV3aZjSTAJw4c4ydvp8Bvjn4B/aN+GOlfFT4c37zaZqKlJbecBZ7G4XHmW8ygnbIpIzgkEEMCVYE1VVVVVVVVVVVVVVVVVVVVVa5q9r4f0TUNevY7iS3021luZkt4WmlZI1LMERQWdsA4UDJOAK8S/8ABUf9p746+Jrvw1+xr+z9dXNhayAG+utMl1O72H7ryLERDbA4Iwxf2atf/by/4Ka/s8T22ufHL4aRS6NPKI92t+GPs1u5P8C3FsUVXxnAJP0OMV+yH+114B/a9+HknivwtA+l63pTpDruhTSiSSwlYEoQwA3xOFYq+BnawIBUgeINf0XwroWoeJvEep2+naVpNrJdXt3cPtjghjUs7sewCgmvHH/BTb9pb4/fEi7+G/7DnwrN1Z2pfZqVxpxvLueMNtFwyuRDaxHIwJQxyVyyk7ah/wCCi37c/wCzD4007Sf2vPhNHfaNqJyxfTksZ3TPzNbXEBMEjKDkoQc8Asmc18MfiV4N+MXgHRfiX8P9XTUtB162E9pOBtOMkMjL1V1YMrKeQykdqqqqqqqqqqqqq+LvxZ8C/A34eax8T/iNq40/Q9Fh3zOF3ySuSFSKNf4pHYhVHqeSBkiP/goL+31+1J4k1OP9kj4PQ6foGlyYMkdhHeyoD90T3NwRAHYDIjVQQM8tjNfDP/gqH+0F8FvidbfC39uD4bmxt5ZEW41OLTGsryyRjgXBiXMdzDkHJiAONxUuQEOmanp2t6Zaazo99Be2F/Ak9rcwOHjmidQyOrDgqVIII6g1VVVVVVVVVVVVVVVVVVVVVVXj/wCIHgv4WeENR8e/ELxHZ6F4f0mMPeX90xCRBmCKOMkszMqhQCSWAAJNftCf8Fm7uO/n0X9mPwLb3VlaOPO8Q+IoJCsy5A/dWyMpRScAPI2TnGxTzX7BHx98dftLfs9WfxQ+IkWlx6zPqt3asum27QxbImAXCszHODzzVftoftseA/2PfCNrc6lZHX/F2tq39jaDHN5RlVThp5nwfLiUnGcEs3AHDMul/tVf8Fb/AIqaH/wtX4ffCpYfC0q+fbQWHhqIx3EQ5zClwzXEykcZQnPav2H/APgpkPjx4xT4JfHPw7Z+FvHrlotPubZXhttRmjB3wPFIS0E42khSSrkMBtYKrVVVVVVVVVVVVVVVVVVVVVVftu/8FEB+y74rsvhL4H+GV/4p8darZx3NubtXiso0lZkj2hAZLliyMCibR235BA1D9p7/AILB6jaf8JnafBrVNP0tV802Vv4JB+Tr/qpd1xjH44r9jr/gq1dfE3x9Z/Bz9o/w1pnhrXtRuBZ6drFikkED3e7atvcQyMxidm+UMG27jgquc1X7c/8AwUN8LfsmiDwN4U0e38U/EXUIBNHp80rLbabE2dktwV+ZixHyxKVYjJLICu66/ac/4K/ReHj8Wn+FcsXhjy/tRs18KwkLDjO4wZN0EA+bcegGScc1+wl/wUM8N/tbJc+CPFej2vhn4h6bbm4ayglZrXU4BgNLblssrKSN0TFiAQwZhu2VVVVVVVVVVVVVVVVVVVVVVVVV/wAFkPiprfjj42+Df2d/D8ry2uh2cd3NaocefqN4xWMEdCViCbT289/WvgT8HvDPwF+Evhr4U+FLWCO10Kxjimmjj2G7uMAzXDerSPuY59cdAK/4LEfBbSfGn7OcHxet9PQa54A1GHddKg3tY3MiwyRnuQJXhcf3cN/eNf8ABJ34sX/xN/ZH0zRtXunnvfAmpzaEHkOWa3RUlg/4Cscyxj0EWO1VVVVVVVVVVVVVVVVVVVVVVV8XfhZ4U+Nvw2174V+N47l9E8RWwgu/s0gSVQHV1ZGIIDBlUgkHkV/wUc+B/wAJ/gP+wdqHhH4S+B9N8O6cPEGmmUW6Fpbhg7fPNKxMkre7sT+Ff8Ehv+TNNM/7GDUf/Q1qvhdbQft8f8FMbnXPFqrqXhSLVbm/NrKu+M6VYDbawlc42uVgVx0Pmue/McUcMaQwxqiIoVEUYCgdAB2Ff8FWPh9L8Af2sfC3x0+HSDSLrxPFHrKSwrsC6tZzL5koAxywNu7erMxP3q+HfjCy+Inw+8MeP9OTZa+JtGtNTgUn7qXEKyqPycVVVVVVVVVVVVVVVVVVVVVVS+BPBlx4zh+Is/hbS5fFFvYf2fDq8lqrXMVtuZjEkhG5ULOxIGM55qv+CvXgnQ/hv+1jpPjDwWiaXqHiXQbfV737MBGRfJcTR/aBtxhmEUZJ6llZiSSan8ew6B8IpPij4qt2totN8NnWNRiHBiWO286VfwAYV/wTU8EXH7VH7Znif47fFKCLVX8PiXX5kmTfG2o3Eu23G09FjHmOg/hMKY6cV+0Zp1v+w7/wUosvGng2H+y9AOqWevQ20I2Ktjd5S9hUDACE/akUdAuB2qqqqqqqqqqqqqv+C03xf1TW/iF4J/Z80aeR7LS7IazfQRZPn3k7vFArDuyRo5H/AF8Hr2/Zt+Cegfs9fBTwr8KtBsreJ9JsI/7RniQKby+ZQbidj1JaTOMk4UKo4UV/wV1+C2k/EH9mC4+JUOnxtr/w8vYbuC4VB5jWk0iQzxZ/uZeOU+hh9yD/AMEhvixf/EL9lceENXunnu/AOsS6ZC0hyxtHVZoefRTJIgHZY1HaqqqqqqqqqqqqqqqqqqqqqqqqvFv7H/7P/j/41p8e/HXgaDxB4nhsLe0gj1A+baRGFnKzeRja8uHC5fcAI02hSCT8SESP/gslZRxqqqvj/RQqgYAAgtugqv8AgrB8Zb/4U/spX+gaHdtBqfj/AFCPQw6HDpasjyXJHs0cflH2m/Ef8Ef/AIG6N4G/Z0/4XDdadE3iH4g3kzLdPGPMisIJWhjhB6hWkjlkOMbtyZztU18e/hF4f+PHwf8AFXwo8R2sMtvr+nSwwSSLk21yBmCdfRo5Ajj/AHfTIr/gjH8TNY8KfGnxr8C9XkljtNc0xr6O2kOfJvrSQI4AzwWjkfcR18pfQVVVVVVVVVVVVVVVVVVVVVVVXgvwJ4M+HOgQ+FvAXhbS/D+kwOzpZadarBGHY5ZiqgZYnqx5Pc1498D+GfiZ4L1rwB4y0yLUNF16yktL23kUHcjjGRkHDA4ZW6qwBHIr/gj7q2reHP2vda8J2Fw9zYah4bvoLwocIfJmiaOUjnuu0enmHnnn/gs38Zb/AMF/BHw38ItHu2hm8f6lJJflDy1lZ+W7IcdA00sB9xGw9a/4JjfA3Rvg/wDsqeF9dTTok1/x9bJrmqXflgSSxzDdapnrsWBkIHTc7nA3Gv24/gtpPx2/Zh8c+Er3T47jUtP0ybVdEkKAvDfW0bSRFD/DvAaIn+7Kw71/wRJ+LF/daf8AEH4I6hdPJbWLQa5pcbHPlbz5NyBnoCRbkAcZLHvVVVVVVVVVVVVX/Bab4v6prfxC8E/s+aNPI9lpdkNZvoIsnz7yd3igVh3ZI0cj/r4PXt+zb8E9A/Z6+CnhX4VaDZW8T6TYR/2jPEgU3l8yg3E7HqS0mcZJwoVRwor/AIK6/BbSfiD+zBcfEqHT421/4eXsN3BcKg8xrSaRIZ4s/wBzLxyn0MPuQf8AgkN8WL/4hfsrjwhq908934B1iXTIWkOWNo6rNDz6KZJEA7LGo7VVVVVVVVVVVVVVVVVVVVVVVXxd+FnhT42/DbXvhX43juX0TxFbCC7+zSBJVAdXVkYggMGVSCQeRX/BRz4H/Cf4D/sHah4R+EvgfTfDunDxBpplFuhaW4YO3zzSsTJK3u7E/hX/AASG/wCTNNM/7GDUf/Q1qrKJP29P+CoD2niUm+8J2+uzoIH+aM6RpocpFjPCzGMbgP4rhjVvbW9nbxWlpBHDBAgSKKNAqooGAqgcAAADFf8ABXj4Yj4PftGeDPjz4DU6ReeLITdvPbDYU1SwkjPnjH3WKSW592Rm5JNfB/x7B8VPhP4N+JVvGsa+KNCs9SMYGPLaaFXZP+AliPwqqqqqqqqqqqqqqqqqqqqqqu/Angy/8YWfxAvfC2l3HiXTrN7Oz1WW1Vri3gdtzRo5GVUnrjGar/gsn4B8O+C/2jfDfjbwzBHYaj4q0IXOpC3AjL3MMzItwcc72TYpPfyweua8G+N2/wCFJ6F8SPGJe2b/AIRa31bVNwy0R+yrLN6cj5vSv2AfC037Y37duv8Axp+KFnHqMGkG48S3FtOvmRG5aVY7SDB/hi3hlB4xbqOlV+2Zow/Ys/4KI6b8TvAdqdP066urPxTbWtsu1fLmd47yFQMALI0dyNowAsuOmKt7iC8t4rq2lWSKZA8bjoykZBH4VVVVVVVVVVVVVVVVVVVVVVVVV+0hKPEv/BXuws9TTfA/xD8LWbRhjjylFjGQPTIBJx3Jqv8AgoLawXf7GPxYiuIw6roRkAPGGSWNlP4MoP4V/wAEPb6eTwL8VdMYr5Nvq2nSoMc7nimDfpGtVVVVVVVVVVVVVVVVVVVVVVVX/BXn/kzTUv8AsYNO/wDQ2r/gkN/yZppn/Ywaj/6GtfEPUrjRvAHibV7Q4nsdHu54jkjDJCzDkcjkCv8AgifZW0v7RXjS/dMzweC5I42J6K95bFuP+AL/AJNV/wAFyLWB/D3wgvGjBmivdXjR/RWS0LD8Si/lX7CN9PqP7HfwjuLggunhm3hGB/DHlF/8dUVVVVVVVVVVVVVVVVVVVVVVVrOsaT4d0i91/XdRt7DTdNt5Lm7u7iQRxwRIpZ3djwFCgkk9AKlTUv8Agp3/AMFBxfaZY3J8A6XLEskkiFfs+hWj5y2eVe4dmwvVWuO4Umv26dQn0j9jz4t3FnhWbwtdW+ASMJKojbp/sueK/wCCHFlbJoXxg1BU/fy3ekRMxP8ACqXZA/N2/wAiq/4LZWsCftD+CbxYwJpfBaRu/qq3lyVH4F2/OvhvfT6p8O/C2pXRBmu9Fs5ZMDA3NChOPxNVVVVVVVVVVVVftuzDxB/wVVi0XUU8y1g8R+F7MJuPMbwWTsPbmVulV+3BbQXf7IXxeiuIw6r4SvnAJxhkjLKfwKg/hX/BDS+nk0/4zaYxXybebRJUGOdzrfBv0jWqqqqqqqqqqqqqqqqqqqqqqqqq+Jf/ACmVs/8AsoGjf+iLaq/4LlalcRaT8G9HQ/uLm41qdxk/ejWyVeOnSVv85r9hyytrD9kD4QwWqbUbwpZykZz8zpvY/izE/jVfsVS/2P8A8FW5NL09Fjt5vE/im2KcnbGtvfMAOfWNetVVVVVVVVVVVVVVVVVVVVVVVX7fP7UWkfswfAbVtUt9RRfGPiWCXTvDFqG/eG4ZcPcY6hIVbeT03bFyC4r/AIIz/s46l4Y8Na/+0j4nsHt5fE8H9leHlkXBeySQNPNj+68scar0/wBSx6MDX/BbnUriX40/D3R2P7i18LyzoMn70l06tx06RLXwZsrbTvg94F06zTZBa+GtOiiXOdqrbRgD8gKurWC9tZbO6jEkM8bRyIeNykYI/I1/wRkvp7X9rLVraIrsvPBV7FKCOwubRxj3yg/Wqqqqqqqqqqqqv23Zh4g/4KqxaLqKeZaweI/C9mE3HmN4LJ2HtzK3Sq/bgtoLv9kL4vRXEYdV8JXzgE4wyRllP4FQfwr/AIIaX08mn/GbTGK+TbzaJKgxzudb4N+ka1VVVVVVVVVVVVVVVVVVVVVVVf8ABXn/AJM01L/sYNO/9Dav+CQ3/Jmmmf8AYwaj/wChrWt30mmaNf6lCivJaWskqK3QlVJAPtxX/BGC2i1D9rHxFeXhaWe38C3s8bsxJ8w3tkhY+pKu3X1qv+C39rA/w0+GN40YM0Wu3kaP6K0CFh+JRfyr/gnZfT6j+xT8Kbi4Kl00mSEYGPljuZUX/wAdUVVVVVVVVVVVVVVVVVVVVVVVqOoWGkafc6tqt7BZ2VlC81zcTyCOOGNAWZ2Y8KoAJJPAArx3eal/wU8/b/ttJ8KRXLeBdLMdmt0FKeRolrIWmuDnBVpnkfYCAQZo1PQ1+15dHw9+yT8Wm0qJIRB4H1OCFU+URK1q8Y246YB49MCv+CGllbSXvxn1BkzPBFocSNnorm+LD8TGv5VX/Bbu1gT4u/Di8WMCaXw3cRu/qq3BKj8C7fnXwLvp9U+CPw91K6Kma78K6ZLJgYG5rWMnH4mqqqqqqqqqqqqqqqqqqqqqqqqv+Cicc3wX/wCCkukfFq9SRLW4ufD/AIkiZFySlqYoW2jHXdZNxz196gnguoI7m1mjmhlQPHIjBldSMggjggjvX/BUHxnaeDv2KvHcc1wI7rXzaaVZruCmR5bmNnUf9sY5jgf3fxH/AARb8G3ejfs5eJ/F92rIviPxTItspHDQ28ESbx9ZGlX/AIBVVVVVVVVVVVVVVVVVVVVVVVV/wV5/5M01L/sYNO/9Dav+CQ3/ACZppn/Ywaj/AOhrWo2Nvqmn3OmXQJhu4XhkA4O1lIOPwNf8EoNRn+E37b3iD4WeJZBDf6ho+qaE0R4zeWs6SsOechbWcY96r/guD4ztLvxb8LPh5b3A+06Zp2oandRBh924khiiJHUc202Pqa/Zu8G3fw9/Z8+G3gfUEdb3RPCunWt0rjBWZbdBIMdvn3cVVVVVVVVVVVVVVVVVVVVVVX19Y6ZY3GpaleQWlpaRNLcXE8gjjhjUZZ2Y8KoAJJPAAr9sz9rHx/8Atw/E20/ZG/ZQS51Pwrc3Sx6je26Mi6xKjgtK787LGIhW3EAMRuOQEr9j/wDZO8FfskfC+LwZoDpqOvagVn8Qa00Wx7+4A4AHJWJASqJ2GScszE/tKeDLv4ifs9fEnwPp0Rkvda8K6jbWaD+Kdrd/KH/fe2v+CH/ja0tPFvxS+HNxcD7TqenWGqWsRIHy28ksUxHc83MP5VX/AAVfv5viz+3B4e+F3huXzr6w0fS9CESndi8up5JVGB3K3UPHsKsLKHTbC2062LGK1hWKPccnaoAGffAqqqqqqqqqqqqr/gqXY33wq/bw0H4qeRI0N7ZaRrUDJj5ntJPKZRnjcPsynB/vD1rTdRsdY0211fS7pLmzvYEnt5kOVkjdQysPYgg/jX/BSHxnZ+CP2LviVd3FwEm1Wwi0q2jDANK9zMkRUZ64jaRiP7qtX/BE3wbd6Z8FPHnji4R0i17xHHaQBhgMtrACWHtuuGX6qaqqqqqqqqqqqqqqqqqqqqqqqqr4l/8AKZWz/wCygaN/6Itqr/gtn4Mu9U+C/gLx1bxF4tA8RS2c+P4FuoCQx9t1soz6sPWv+Cb/AI2s/HP7F/w2u7e4DzaRYSaTcxkjMT20zxKpx6xrGw9mFajqFjpOn3Oq6ndR21nZQvNcTSHasUaAszE9gACTX/BLKxvfin+3dr/xSEUyQ2Nhq2szM/Z7qURKjH+8ftDH/gBqqqqqqqqqqqqqqqqqqqqqqqv2jP2kfhj+zB8Pbnx/8SNXWP5WTTdMhYG61ScDiKFD16jcx+VAcsQK+AvwS+L/APwVC+O91+0B8fJbmx+HWkTrEsMKvFFPEjll02zJ6IMnzZQdwLHne+RpGkaX4f0my0HQ9Pt7HTtOt0t7S1t4wkcESKFRFUcBQAAB7V/wXB8GXY1D4WfEKGMtbPDf6XcP2RwYpYh/wINL/wB8V+yf42tPiL+zN8MPF9ncCb7b4WsUnYEHFxFCsU68eksbj8O1fFzxnZ/Dn4V+MPH19cCGHw7oV5fs+4KcxQs4Az/ESAAO5IFf8ERvBt3e/GH4h/EFUf7NpHhqLS2bHBe6uUlA+uLNvzqqqqqqqqqqqqr/AIKl2N98Kv28NB+KnkSNDe2Wka1AyY+Z7STymUZ43D7Mpwf7w9a03UbHWNNtdX0u6S5s72BJ7eZDlZI3UMrD2IIP41/wUh8Z2fgj9i74lXdxcBJtVsItKtowwDSvczJEVGeuI2kYj+6rV/wRN8G3emfBTx544uEdIte8Rx2kAYYDLawAlh7brhl+qmqqqqqqqqqqqqqqqqqqqqqqqr/grz/yZpqX/Ywad/6G1f8ABIb/AJM00z/sYNR/9DWmVWUqwBBGCCK/4JqSn4Hf8FCtW+FetyG3ubmHWfDB8wgAywSCYAk+ps8D1JAHWq/4LheM7RpPhV8O7e4DXUa6hql1FuHyI3kxQtjryVnGf9k9ecfsjeDbvwB+y/8AC3wlqCsl5ZeFrFrqNxzHNJEskif8BZ2X8Kqqqqqqqqqqqqqqqqqqqqqrm4t7O3lu7u4jhggQvLLIwVUUDJYk8AADJJr9t79sPxp+174+t/2Qv2SftOtaHqFytvquoWKnGsyqwJVZB92yjwGaQ4D7Sc7AC/7GP7IHhD9kT4Zjw3p0sWqeKdX2TeItaEZU3UwB2xxg8rDHkhQeTkscFiB8bPB9x8Q/gz488A2a7rjxJ4Z1HTYRnHzz20ka/TlhX/BEnxnbaT8V/iR8OrqYxXOu6Ha38Ub/AC7jZzMjDnncBd5x6Bj24r/gsLrsnj39rHwn8NPDpN3faV4etLMwKwJF5dXEjqmB0JRoDz/eHHc+GNEi8NeGtJ8NwSmWPSrGG0R2GCwjQID+O2qqqqqqqqqqqqqqqqqqqqqqqqv+Cn/7H2tftKfC/T/Gfw6077X448DedJb2aD95qdk4Bmt09ZAUV0HfDqOXFfsi/wDBV/wz8KPhlYfB/wDaQ8L+Jm1PwhCNPsdU062SWSWCL5EhuYpHRkljACbhncFG4BgS37Rvx9+Kf/BUb4ueG/gv8B/BOp2XhHSLg3EMd8ACHYbXv75kLJCkaFlVQWPzMAWaQKPgZ8IvDvwG+Efhb4ReFzvsfDdgtuZimw3MxJeadh2aSVncjsWqqqqqqqqqqqqqqqqqqqqqqqq/4K8/8maal/2MGnf+htX/AASG/wCTNNM/7GDUf/Q1qv8Ago/+zd8SfgD8e7X9tn4IWcp06TUIdS1N7aEyf2TqMZUNLKo6wT4BYnjezq2Ay58Pf8FqfgNP4Gi1LxT8PPGVr4rS3xPpdlFBLbPPgf6udpVIjJPVkDAZ4bjP7OHwl+K3/BR79q+X9pf4taAbPwBpeoRT3CujfZpo7c/uNNt92PMHyjzWHGC5OGkANVVVVVVVVVVVVVVVVVVVVVX/AAVE/bo8XeO/GniD9mXwI11onhPw3fPZ6/Lv2zazcxNhkbB+W3Vhwv8AGRub+ED9kH9vn9mf9kTwH/YHhv4H+KdW8SakqtrviGa6t0mvnHRFHPlwrk7YwT3JJJJr/h958Kv+iH+K/wDwY2/+FfBH4p6f8b/hN4Y+LGlaVcabaeJ7FbuG0uHDyQgkjDFeCeO1ftLfCb4o/wDBOb9rO3/aW+FWhm88B6pqctza7Ub7PEtxn7Rpk5GfLBBby2P8OwjLIwGv/wDBan4DReBZNS8M/Dzxlc+LHtj5GlXkUEVtHPg/6y4WUkxgjqqbiCPlXnH/AATq/Zt+J37QXx+uv23fjtYSrp66hLqelG6hMf8AauoPnZLCh5FvACCjfd3JGq5CNiqqqqqqqqqqqqv+Cl/7Imq/tP8Awhs9a8B2aT+OvA8ktzplv906hbSKPPtQf752RumeNybeN5I/Y7/4Km6J8EPhxa/A/wDaS8J+J/tfg5fsGm6hYWqvMkMZ2rbXMMrxsjxAbAwzlQAVBUs37UP7THxL/wCCmfxH8NfAb9nnwLq0Hhexu/tSQ3oCyTzYKG8vChZIIYkZgoyfvtyzMqj9nf4KaB+zv8GfC/wh8OyLPFoNmEubvy9hvLpyXnnI5I3yMxAJO0ELnAqqqqqqqqqqqqqqqqqqqqqqqqqviX/ymVs/+ygaN/6Itqr9oj4K6D+0P8GPFPwg8QSiCHX7Ipb3WzcbS5Qh4JgMjOyRUYjI3AEZGa/Zc/aX+I3/AATN+Jvif4E/tA+BtVn8NX12LmeCywZIJgNi3toXKpPDKiqCNy52KchlZT+19/wVP0j43fDu6+B/7MvhLxSdQ8ZJ/Z9/qF7aKk7QSfK9tawxPIzvKDsLHGFZgASwZf8Agml+yDqn7L/wjvNa8e2UcHjvxtJHcalACGOnW0YPkWpYZBcb3d8cbnC87Axqqqqqqqqqqqqqqqqqqqqqq+O/xe0X4CfCHxR8Xtf0661Cy8M2XntaWxAkndnVI0BPCgu6gtzgEnBxiviH+0nJ+0f8e4Pir+02dZ1Xw9DN8ugaHMsIgtVOVtIGc4iQ/wAUnLtyfvHcPDP/AAWX+BXgzw9p/hTwp+zv4g0rR9Jt0t7Kytb22jigiUYVVUDgYr4I/wDBWn4dfG/4seGPhNpXwk8R6bd+J74WkN3cX0LxwsQTuYLyRx2r9sv9nCz/AGpvgJrfwwE8FrrKsl/oN5MDtt76LOzdjorqzxMcHCyEgEgV+xt+3H4j/YRutb/Zu/aU8B+IU0fTdQklt0tolN3pMz/6xRG7KssEhw4ZW4LMy7w/H7Zn/BRG+/a80C0/Zu/Zh8D+J5LbxPcxx6lLcWyi81LawZLaGKJn2x7lDOxIJC4wF3bv2Cv2WT+yl8CLPwlrYt5fF2uTnUvEU0JDKs7KFSBW7rEgVcjgtvYcNVVVVVVVVVVVVX/BS/8AZE1X9p/4Q2eteA7NJ/HXgeSW50y3+6dQtpFHn2oP987I3TPG5NvG8kfsd/8ABU3RPgh8OLX4H/tJeE/E/wBr8HL9g03ULC1V5khjO1ba5hleNkeIDYGGcqACoKlm/ah/aY+Jf/BTP4j+GvgN+zz4F1aDwvY3f2pIb0BZJ5sFDeXhQskEMSMwUZP325ZmVR+zv8FNA/Z3+DPhf4Q+HZFni0GzCXN35ew3l05Lzzkckb5GYgEnaCFzgVVVVVVVVVVVVVVVVVVVVVVVVf8ABXn/AJM01L/sYNO/9Dav+CQ3/Jmmmf8AYwaj/wChrVf8FOP2YfiJ8K/jHZ/ttfBG0uPIiura+1prOIu+k6hb7dl2yDI8mQIm44wHDbuJBXg//gtT8DbnwJBfeO/h94vs/FsVvi50/TIIZ7WWYDrFM8qsqMefmXK8j58ZPwY+GvxW/wCCn37WL/Hf4l+HXsPhvo93CLsOGNstpA26LTIGOPNZySZWGNvmSOdpZFIVVUKoAAGAAMYqqqqqqqqqqqqqqqqqqqqqq/4Km/t0eLvEHi7Xv2Wfh611onh/RZvs3iW8D7JtWl2hjApB+W3GQCOsh64UYb9jb9uz9mn9kHwW+n6R8EvE+t+LtWQf234hlureOS4wciGJeTHCpx8uSWPzMTwF/wCH3nwq/wCiH+K//Bjb/wCFfAD4w6Z+0B8HvDXxg0fR7nSrPxJBJLFZ3MivJCEleMhivByYyePWv2vvgx8Uf2Cf2rbX9rT4P6QbjwbqWrvfxMsbNBZzT5F1YXO37kcu+TYeBtcBfmjzV/8A8FqvgEngZtU0z4c+NJPFbW3yaRNHAtstxj7rXIkJ8vP8Yj3EY+Qdv2Bv2e/il+1X+0lcfts/HTTXTRLXVW1Sxa4hMaapqC4Futurc/Z7bahD5IzEiAth9tVVVVVVVVVVVVVVVVVVVVVVVVV47/Z0+AfxP1I618Qvgz4M8Q6kcbr7UNFgluGAGADKV3kY7E4rwZ8PvAXw40n+wvh74K0Lwzp2dxtNI06KzjLepWNQCfc81VVVVVVVVVVVVVVVVVVVVVVVV4k8K+F/GWmNoni/w3peuaezh2tNSs47mIsvRijgjI7HHFeHPCvhfwdpi6J4Q8N6Xoenq7OtpptnHbRBm+8wSMAZPc4qpoYbiF7eeJJY5FKujrkMCMEEHqCO1XX7JH7Ld7rB1+6/Z2+HMt+z72lbw1a4Z/7zLs2k98kZzzVhp9hpVlBpul2VvaWltGI4LeCIRxxIBgKqrgAAdhVVVVVVVVVVVVVVVVVVVVVVWq/A74K67qNzrGtfB/wTqF/eStLc3V14ftZZZnJyWd2QlmJ6knNf8M9fAH/ohvw//wDCZtP/AI3X/DPXwB/6Ib8P/wDwmbT/AON1o+jaP4e0y20Tw/pVnpmn2abLe0s4FhihX+6iKAFHsBWp6XpmtafPpOsada39ldIUntrqFZY5VPVWVgQR7EVYfsk/suaXrA17Tv2d/h1Bfo+9JU8NWo8tv7yrswp46gAikRI0WONVVVGFUDAAHoKqqqqqqqqqqqqqvHv7PfwJ+Kd+NW+I3we8HeI9QUAC91HRYJp8AYA80rv2+2cdPSvBHw3+Hvwz0s6J8OfA2geGLBiC9tpGmxWaOR/EwjUbjyeTzyfWqqqqqqqqqqqqqqqqqqqqqqqqqqf4V/DC68TDxrc/DjwvN4hWZZxq0mjwNdiVQAr+cV37gAADnIwKqvHfww+G/wAUdNXSPiR4B8PeKbOPJjg1jTYrtYiRgsgkU7W9xg+9eAPgD8DvhVeNqXw2+EXhDw1fOCGu9M0aGCcgjBBkVd+ME8ZxyfWqqqqqqqqqqqqqqqqqqqqqqq1zQdD8TaVcaF4k0ax1bTbtQtxZ31ss8MoBBAZHBVhkA8jqBX/DPXwB/wCiG/D/AP8ACZtP/jdf8M9fAH/ohvw//wDCZtP/AI3Wj/BL4MeHtTtta8P/AAj8F6bqNm++3u7PQLaGWFv7yOqBlPuDVfED4OfCb4sQxQfE34aeGPFQtxiBtX0qG6aH/cZ1JTqehHU+tfD/AOCvwg+E6yj4ZfC/wt4Wedds0mk6TDayTDOcO6KGYfUnHFVVVVVVVVVVVVVXj39nv4E/FO/GrfEb4PeDvEeoKABe6josE0+AMAeaV37fbOOnpXgj4b/D34Z6WdE+HPgbQPDFgxBe20jTYrNHI/iYRqNx5PJ55PrVVVVVVVVVVVVVVVVVVVVVVVVXiTwr4X8ZaY2ieL/Del65p7OHa01KzjuYiy9GKOCMjsccV4c8K+F/B2mLonhDw3peh6ers62mm2cdtEGb7zBIwBk9ziqdEkQxyKrKwIZWGQR34rUP2Sf2XNV1g69qP7O/w6uL9n3vK/hq1PmN/eZdmGPuQTWmaXpmi6fBpOjada2NlaoEgtraFYo4lHRVVQAo9gKqqqqqqqqqqqqqqqqqqqqqq1j4JfBjxDqdxrWv/CPwXqeo3b77i7vNAtppZm/vO7IWY+5Nf8M9fAH/AKIb8P8A/wAJm0/+N1/wz18Af+iG/D//AMJm0/8AjdaJoOh+GdKt9C8N6NY6VptopW3s7G2WCGEEkkKiAKoJJPA6k1f6fYarZT6bqdlb3lpcxmOa3uIhJHKhGCrKcggjsatv2SP2W7LWBr9p+zt8OYr9X3rKvhq1AR/7yrs2g8ZyBnPNQww28KW9vEkccShURF2hQBgAAdBjtVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVfET49fBL4R3cOm/E/4teE/C99cQieGz1TV4beeSIllEixM28puRhuAxlSM5q+/4KI/sV6c6R3H7QWgOXGR5ENxOPxKRnH41pX/AAUK/Yu1mf7NZ/tB+HI3yBm7We1Xn/aljUfrx3rwd4+8CfETS/7a+H/jTQvEung7TdaRqMV5ED6Fo2YA+1VVVVVVVVVVVVVVV+1L/wAFRtA/Zh+M+rfB2/8Ag5f+IJtKt7aY38OtJbrJ50KyABDExGN2OvOK/Yv/AG/9H/bI8T+I/DOmfDK88Lt4dsIrtpp9UW6EweTZtAEaYx1zzVVVVruuaP4Y0S/8SeIdSt9P0vSrWS6vbu4fZHbwxqWd2PZQoJJ9qtf+Cxn7KF145Tws1h40h0h5/JXxFLpkYtRzjzTH5vnCLHOfL3/7FWF/Y6rYW2qaZeQ3dneQrNb3ELh0ljYAq6sOCpBBBHBBqqqqqqqtQv7HSrC51TU7yG1s7OF5ri4mcIkMaglnZjwFABJJ4AFeEP8AgqF+yv4/8a6V8PvBl/4r1bWtbv0sdPhg0KQCeV22qQWI2r3LNgAZJwAaqqqviZ8V/hz8G/Di+Lvih4vsPDmjvcpareXrFUMrBiqcA8kKx/CtO/bs/ZB1bULXStN+P3he4u7yZIbeFJn3SSMQqqPl6kkCqr4y/Fzwr8Cvh5qfxQ8bW+pyaFo5jN8+n2huZIUdwgcoCCVDMuSM4ByeASPgD+3t+zj+0r40l+H3w18R6iddjs3u47bUrBrXz40ID+WSSGYBgSvXAJ6KSKqqqqqqqqqqqqqqqqqqqqqqqqqqqqqpZI4Y2llkVERSzMxwFA6knsK1r9tz9kTQZjb3/wC0Z4DkcHafsmsR3YBxnrCWH/1+OtN/wUg/YkUlT8ftJyDjiwvD/wC0a8I/tofsn+OriKy8N/tBeCZbmYAxQXWqJZyOTjAVJthLc/dAz144NI6SIskbqysMqwOQR6iqqqqqqqqqqqqqqvip45h+GHww8X/EufTn1CPwloN9q72iS+W1wLaB5jGGIO0sExnBxnODX/D8Xwp/0btqv/hSR/8Axivg18R4PjD8KPCXxSttKfTIvFWkW+opZSTCVrcSoG2FwBuIzjOBVVVV+09+178HP2S/D1jrPxOv7+e91dnXTNH0yATXd5s272UMyoqLuXLOyjnAyeK/ZY/b6+Bv7Wmp3vhjwV/bGieJLCA3DaRrUMcck8IIDSQtG7q4UlcjIYZzjHNVVVVVVVX7RP7Unwd/Zb0HStf+Leu3FnHrd01tYW1pbG4nnZF3OwQc7FBXc3QF1HVhX7Of7Vvwq/ansdZ1T4TJrs9hoMscN3d3+nNaxmVwSI0LH52CjJA+6GXONwzVVVePv2vf2Z/hd4svvAnxB+Mvh/Q9f0zy/ten3crCSHzI1kTICnqjo30YV8Kv2hvgn8cbjUbT4SfEjSPFE2kJG96li7MYFkLBC2QOpVvyqq+Pn7evwD/Zp8cr8PfitJ4lsdUlso723a30hpobiByQHRweQGR1PoVP1r4QfF3wH8dvh7pXxO+G2sf2joWrK3kyMhjkjdGKvHIh5R1YEEH2PIIJqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq+Ifxe+FXwktbW8+KHxI8NeFIr4uLQ6xqkVobkpt3iJXYGQrvTO0HG4Z61qf/BQn9i7Sjtuv2hPDch3Ff8ARlmuOR/1zjbj36GrH/gon+xTqM4trf8AaB0FXxnM9vcwL/308QH614D+LPwt+Kdq958NPiN4a8UxQqDKdI1SG7MWem8RsSh9jiqqqqqqqqqqqqqqq/bJ/wCCjeifsgfE/TPhpqXwpvfE8upaDFq63cGrrahBJPPD5e0xPkj7OTnP8XTiv2Q/+ClOh/ta/FeX4Wad8Jb7w3LFpE+o/bJ9YW5UiJ41KbBEpyfM657VVVVMVVSzMAAOSeMV41/4LA/sq+EfHU3g+ys/F/iKytLhoLnXNKsYWtNwOC0XmSo8qA5ywUAjld4Iz4J8a+FfiR4R0nx14I1q31bQtbtVubG8gJ2yxt0ODggjkFSAVIIIBBFVVVVVVVV//wAFT/2R7XxfP4I07X/Ees6lHqJ0+H+y9FkuI7uYSeWBCwP7wM2ApH3sgjrUbNJGjtG0bMoJRsZU+hwSM/Q1VVXj7x/4M+F3hK+8d/EHxFaaHoGmeX9rv7tiI4fMkWNMkAnl3Rfqwr/hvz9jT/o4fwp/3+f/AOJq3uILy3iuraVZIpkDxuOjKRkEfhVfETxzpHwy8Da38Qdftb+40zw9ZPe3qWFv58ywRjMjqgILBVBY+ymvgx/wUZ/Ze+PHxB0/4YeCPE2qxa7qqyfYo9S01raOd0UsY1ckjeVDEA9cYHOAaqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/4LV/8nUeFv8Asn9n/wCl9/XwT+BPxO/aJ8a/8K++Evh+PV9aW0kvHhkvIbVUgQqrOWlZRwXUYGTz0618R/8AgnJ+2J8LvDdz4t8Q/CGe60uxiaW6l0q/t794EUZZmiidpNoAJLBSAASSK+GnxT+Ifwd8V2njf4Y+LtR8Pa1Zt8lzZy7d655SRTlZEOOUcFT3Br9i39pGD9qn4BaN8TZ7a3tNbhmk07XrW3z5cN9CFL7M5IV0eOQAklRIBk4yaqqqqqqqqqqqqqv+CsH/ACe54w/7B+mf+kUVf8EQ/wDkqfxL/wCxftf/AEoNVVVX7Rvwz1H4y/Ajx18LdG1FbHUPEmh3FpaTuSEWZl+QORyELAK3+yTVp+wJ+2Fd+OV8Af8ACgvFUN6Z/Ka+msyunr82C5vP9QVHXIc8dAa+Cfw/ufhR8HfBHwxvdUOpXHhXw/ZaZLd5OJXghWNmXPITKnaD0XA7VVVVVVVX/BX34/3Hwy+Adl8J9Av/ACdZ+JNy8FyUbDJpsG1rjpyN7tDHzwytIO1f8EX/AIGWvif4l+J/jzrVmJIfBtsunaMzrwL25VvNkX/aSAFfpcflVVVf8Fkv+TRbT/scLH/0Tc18Jv8Akqngz/sYLH/0oSqrxV4Y0Pxr4Z1bwd4m0+O+0jW7KayvrWQfLNDKhR1P1ViK1WLxt+xv+1Fdw6Pef8T74Z+J3FtMcqt3HFIdpYDny5oSMjushFfD7xvofxM8B+HviH4amMmleJdMt9Qsy2NwjmjDgNjOGAbBHYgjtVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV4t/5FTWf+wfP/AOi2qvhB/wAE+/2p/jr8P9N+J/w68BWV54c1dphY3c+tWkBn8qV4pCEaQOoEkbr8wGduRkEE/Gr9nP41/s76tbaN8Yvh/qHh2W9DGzmkKTW91txu8uaJmjcjIyobI3DIGRX/AAT4/bk8e/AD4l+H/h54r8S3Oo/DPXb2Oyu7C8lMi6S0rBVuYCcmMKxBdB8rLu43YYVVVVVVVVVVVVVVftZf8mr/ABl/7J/rv/pBNVfsUf8AJovwg/7E/T//AEStVVVX/BWz9kv4xfGjU/CXxX+E/hy+8UJoWmy6dqWkWK+ZcRKZfMSaKIfNJkuysFBYbVOCMkf8Euv2Lfj14Y+P2nfHH4j+Ctb8FaF4YtLpYYNYtns7jUZ54JIBGIHAcIokZyzADKpjOTiqqqqqqq/4KOfH+4+Pv7UHiS4sr8zeHPCEraHoiK3yFIGImlHY+ZN5jBu6bB2FfsU/Ay1/Z5/Zr8G/D82Yg1eSyXUddJXDPqFwoeYN67MrEDx8sS1VVVf8FR/+T7fib/3Cf/TTZ1/wQ6/5Gr4t/wDYP0r/ANGXNVX/AAWC+Blr8QP2d7f4t6dZoda+HN6skkoX55LC4dYpk467ZDDJz90K+Opr/gjP+0BceGfidrX7POs3xOl+MIH1HR42biPUIEzKFGf+WlupJ6/6hfU1VVVVVVVVVVVVVVVVVVVVVVVVVVVVVV/wXP8A+aJf9zB/7jq+Gnw18Z/GDx1pHw2+HukLqfiHXJWisbRriOASMqM7fPIyouFRjyR09cCvFH/BMD9tjwto82tT/CD+0obdS0kOl6ta3c+AOqxJIXf6ICfavD3iPxf8O/E1vr3hjWdU8Pa/pE5MN1aTPbXFtIpwRkYZTkEEfUGv+CcH7XOrftV/B28XxvLA/jfwZcRWesywoIxeRyKTBdbBwjOEkVgON0TEABgoqqqqqqqqqqqqqq/4LV/8nUeFv+yf2f8A6X39f8Ebf+Tubv8A7E++/wDR1tVVVV4j0dfEXh7VPD7XclqNSsprUzxffi8xCu5fcZyK8af8E/v2uvB3jubwIvwO8T604naO21PSbB7mxuEB+WQXCjy41YYOJCpGcEA8V+xT8EvEf7PH7M/gz4U+L75LnW9Mhnnvlil8yK3luJ5J2hQ9CE8zaSOCwYjg1VVVVVVX/BTj4/3HwJ/Zf1e20K/Nv4j8cS/2HprI2HiSRSbmUdxthDqGH3XkQ1/wSM+Blr8U/wBpN/H+t2Yn0j4a2S6ioZcqdQlYpag/7u2aUH+9CvvVVVV/wVH/AOTEvib/ANwn/wBO1nVeEv8AkVNF/wCwfB/6LWquba3vLeW0u4I54JkKSxSIGV1IwVIPBBHBBr9p/wCGmr/sn/tX+JPDHhK7l05vDGtx6p4buomJaGBytxalWPVkDIpP95DX7Ovxf074+/BDwd8XNNVIx4i0xJbmFDkQXSEx3EY5PCzJIozzgCqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv+C1f/J1Hhb/ALJ/Z/8Apff1/wAEbf8Ak7m7/wCxPvv/AEdbVX7TEnhCb9oj4mS+ATbHw6/izUjpv2YAQ+Sbh9vlgceX/dxxtxX/AARR0PWbL4AeNNcvEdNO1PxWUslZcb2itohI6+oJZVz6oR2NfHj49/DX9nD4eXnxJ+J+tfY9Otj5dvbxAPcX05B2wQRkje5we4AALMQoJHxX/wCCz3x48RarcQ/CTwf4c8I6OGYW8l7C2oXrDoGZmKxKcYO0IcHjc3WvA3/BYf8Aax8OalFP4tXwr4usd3763utLFpIV9EkgKhT7lWHsa/ZM/a/+GX7XfgqbxF4LMuma1pZRNa0C7kVriwds7WBHEkTYbbIAM4IIUgqP+Cgn7e/7RX7Ivxe0vwj4L8J+A9S8Na5osd9ZXWrafdyz+aJHjmjZo7mNTtKo3C9JF5JzX/BP3/go58Vf2p/jRqPwv+KXh7wXpUC6BNf6dJotpcwySzxSxAxsZbiUEeW7tgAH5OtftSfGST9n39n7xt8XrSCznvfD2ml7CG8DGGS7kdYoFkCsrFDLImQGBIzgjqPDH/Baf9ombxLpMPivwN8NI9Ee+hXUntNNvlmW2LjzTGWu2AfZuwSpGcZB6VHJHNGksMiujqGRlOQwPQg+lfG3/gsL8efBPxg8aeDPAPg74c3nh7QNdu9P0651DT72SeeKCVoxIzJdop3bdwwq8EcV+wT8fPi5+0z8Dm+LfxZ0Tw1pcl/q9xb6RDodrPDHJawhUaR/OllJYzCZeCBhB61+1B/wV8+IPw6+Inij4WfDD4Pabp154Y1S50241DxLO9w0kkUhRnSCFkVQSpKkyOCCpxjir7/grd+2ld3ourfxf4ds4gcm2g8PwGM85xlwz9OPvdB6818CP+C0Xjez1u10j9ofwRpWpaNNIqSavoETW91ag9XaFmZJQO4UxkDOMnivDHibw/418OaZ4u8Katb6no+s2kd3YXkDZSeGRQyOvsQR15Hev+Ch37f3xv8A2R/i14f8E/Drwz4H1LSNY8OR6g8ut2d1NOtwbieN1BiuIlCbI4iBgnJbnGAP2b/+CrOqeJ/hB8U/ir+0LoPh2xXwPNpdvo9l4ct5oJNUuL0XW2ACeaXLf6KWyMBUDsQdteFv+Crv7UnxR+PHhDQ7C48O+GvDWueJbGxk0qz01Jz9mluY0ZWmm3OX2MQWXYMnIA4A8R+IdC8I6BqPinxNqttpmk6TayXV9eXL7I7eGNSzux7AAE1+0H/wWd8Y3WvXWh/s3eEtLsdFgcxx65r1u89zd4/5aRwBlWFT2D7yRgkKeB4I/wCCwX7W3hzVorrxVP4X8W2G8edZ3elLasU7hJLfYVbHQsGA9D0r9tH47eHf2lfjxqHxh8Mabd6dba1plgstjdEM9rPFbJHLHuHDgOhwwxlSDhSSB/wRD/5Kn8S/+xftf/Sg1Vftrf8ABQH4f/shw23hmHSG8VeO9Tt/tFto0c/kxW0JJUTXEuCUBIbaigs20/dHzV4m/wCCvX7Y2uX73Wj6x4V8PQFiUtbDQ0lVR2Ba4MjH65Ffssf8FV/2mviB8YvB3wq8YeFPCfieDxTrFtp7zw20lhc26yOFebcjMhVFLOR5eSF4Ir9of4h618JPgX47+J3hu1sbnVPC+g3WoWcN8jPA8sUZZRIqMrFcjkBgfcV/w+r/AGqP+hC+FP8A4Kr/AP8Ak2vhZ8Q9a8cfs9+EPixq1rYxavr/AINstbuYLdGWBLiazSd1RWYsEDMQAWJxjJJ5r/h9X+1R/wBCF8Kf/BVf/wDybX/D6v8Aao/6EL4U/wDgqv8A/wCTa/4fV/tUf9CF8Kf/AAVX/wD8m1+xH8ePF/7Sv7Oug/Fzx1p2j2OsapdXkU0GkwyRW6iG4eNdqySSMCVUE5Y8+nSv2xP26/hb+yFpMFlrEEniPxlqcLS6d4dtJ1jfZyBNcSEHyYiwwDtZmIO1TtYr4z/4LC/tb+Ir+Wbww/hPwraF8wwWeki5ZV7BnuGfcfUgL9BXwa/4LPfGXQNZtrX43eD9F8V6G7qtxc6VB9hvol6F1G4xSEddhVM9Ny9vhb8UPA/xn8B6T8SfhzrsWraDrMPmW06cFSDhkdTyjqwKsp5BBFVX/BXL4iTeNP2wdT8NLOWtPBOj2WlxKGyu94/tUjAeubgKf9wDtX/BKbwJD4L/AGLvCt+LfyrrxXf32r3QIwWZpjBG34w28Jr9s341+Kv2df2avGHxj8E2Gk32teH/ALD9lt9Vikkt386+ggfesboxwkrEYYcgZyMg/wDD6v8Aao/6EL4U/wDgqv8A/wCTa0O9l1PRNP1KdUWW6tY5XVAQAzKCcZzxzX7fn/BRj43fsq/HSD4Y/Dzwv4H1HS5dBttQM2s2N1LP5skkqsA0VzGu3Ea4G3PJ59P+Cen/AAUL+NP7Wvxp1n4cfEbwz4J07TdN8Lz6rFLolldQzGaO6tYgrGW4lXZtnckBQcheeoP/AAWS/wCTRbT/ALHCx/8ARNzXwm/5Kp4M/wCxgsf/AEoSqqv+CxngKHwt+1fB4rtINsfjHw3aXkzhcBriFntmHuRHDCc/7Qr/AIJD/ES58bfsiWvh6+uDJN4K1270qPect5LbLlPwH2hlHoEx0FVVVVVVVVVVVVVVVVVVVVVVVVVVVVVXi3/kVNZ/7B8//otqr/glx/yYl8Mf+4t/6dryv+CtsnhFP2LvEK+Izbf2k+r6cugeaAX+2faFL7PRvswuc47ZrR9J1HX9WsdC0e1e6v8AUbmO2tYEGWlldgqKB6liB+NadBc2+n2tveXHn3EUKJLLj/WOFAZvxPNftnf8FMfh5+zDqs/w68HaOnjTx7FGDc2q3Pl2mllh8ouJFyxkxg+UoBweWTIzr3/BXj9snV9Q+2abrvhXRId5YWlloUbx44wuZzI+Bj+9nk8+n7O3/BZzxH/b1n4d/aV8I6bLpNzKI28Q6DC8UtoDgb5bcswkUHljGVIHRWIwbrXLjxD4Em8R/DTUNK1O41HSWutAupG860uXeLdbuSjDdExKE7WGVPBHWrn/AILR/tX2lxLaXXw8+FkU0DlJI30nUAyMDggj7bwQRiv2RPjdqP7Rv7O3g/4v61b6dbarrcE66hb6ejpDDPDPJC6ors7KMx5AZicEHPNf8FDP+CiXxI/ZR+KOgfDX4WeH/COqzXWiDUdUfW7W4nMRkldIkTyZ4tpxE7EHPDL07/8ABOj9v74g/tceLvGHgr4o6F4V0zUdG02DUNLGhW08HmxeYY7jzBNNLnDPb4xtxuOc5GP22/2h9V/Ze/Z41z4q+GrPTL3X4Lq0tNJtdTjd7eWaWZQwcRujECESsMMOVHavAX/BXr9r74i+OPD3gDQPh/8AChtS8SapbabaBtK1DHmzyLGpP+m9MsM+1ftQfHuD9mX4Laz8Yr3whf8AiaHR3gjks7OZYOZZFjV3ds7E3uoJCsQWHB5I8b/8Fm/2mtevJP8AhDPC/gvwvZZPlJ9jlvZwP9qSRwjfhGtaF/wV1/bN0i7W51DxH4X1uMMCbe/0GJEOM8ZgMbYOR/F2Hvn9iX/gpP4L/ao1aP4beLdBTwl4/MDy29rHOZbTVFQbn8hm+ZXCgsYmydoJDNg4vTdiznOniI3QibyBLnYXx8u7HOM4zX/D6v8Aao/6EL4U/wDgqv8A/wCTa/av/wCCq8/wn8NeGPCnwr0HStT8f614esNU1m5uwz2ejG6t0mWFY1bdJLtkDYLbUBXO8kgf8Etv2svjb+1G/wAT2+MXiKz1MeGjpJ01bbTobUQ/aPtnmg+WoLZ8mPG7OMe5r9rz9sv4afsg+D4NW8Uxyax4j1ZXGi+H7WUJLdlfvO7kERQgkAuQTk4VWOQPGn/BYL9rnxHqj3Xhi58K+FLIMfKtLPSFuSF7b3uC5Y+pAUewrS/+CtXjvx98JfiF8I/jz4X0q6/4Snwfq2mabrmjQNBJFdTWcscQmhLMrIzsqlk2bc5wwziv2KP+TRfhB/2J+n/+iVqr6+stMsbjUtRu4ra0tImlnnmcIkSKMszE8AAAkk9K/aP/AOCzWpWev33hf9mjwhptxp9q7RL4k16N3N0QSC8FsrLtTgFWkJJB5RelWn/BW79tK2vzeTeL/Dt3EWJFpN4fgEYGc4ygV8dvvZx3zzX/AATu/a9+JP7XngfxPr3xD8F6LpEnhu+gs4b7SXlWK+kdGeRfKkLGMoBEc72Deb0Xbz/wUW/bw+L37IfjTwh4c+Gvh3wfqVtr+lzXdy+uWdzM6OkoQBDDPEAMeoPPev2I/wDgpn8eP2lf2idB+EXjrwn4BsdH1S1vJZp9JsLuK4Vobd5F2tJcyKAWUA5U8enWv+Ci37XfxJ/ZD8FeEPEfw10TwzqVzr+qTWlymuW08yIiRbwUEM0RBz6kj2r/AIfV/tUf9CF8Kf8AwVX/AP8AJtf8Pq/2qP8AoQvhT/4Kr/8A+Ta+CH/BXX9pP4l/GnwB8ONd8E/DSDTfFXijTdKvJbTTb5Jo4bi6jidoy12yhwrkglSAcZB6Vc3FvZ28t3d3EcMECF5ZZGCqigZLEngAAZJNftOf8FkNO8La7feDf2a/Cum+IDZSPDL4l1hna0kcEgm3hjZWkX0kZ1BxwrLgmP8A4K4/toJqf29vFXhuSDI/0JtAhEXbjIxJ2/v96/Yx/wCCqvhr48+JtP8AhX8ZtBsfCPi7U3WHS7+zlb+z9SnOAIcSEvBKx4RSzqx43BiqtVftEfEGT4U/Ajx/8RreTbc+HvDt7dWhzjNwsLeSPxkKD8a/ZS8Dx/FD9pr4Z+DL6Jrm21TxRZm+Q8mS3SUSTg/WNH5qv2/P+CjHxu/ZV+OkHwx+HnhfwPqOly6DbagZtZsbqWfzZJJVYBormNduI1wNueTz6f8ABPT/AIKF/Gn9rX406z8OPiN4Z8E6dpum+F59Vil0SyuoZjNHdWsQVjLcSrs2zuSAoOQvPUHxxrd14a8Fa/4jsI4pLnStLubuFJlJRnjiZ1DAEEjKjOCPqK/4fV/tUf8AQhfCn/wVX/8A8m1+yV8XfEvx7/Z18F/FzxhZaZZ6x4jtZpbqDTYnjt0ZLiSMbFkd2A2oDyx5z9K/4Kj/APJ9vxN/7hP/AKabOv8Agh1/yNXxb/7B+lf+jLmqr4ueBrX4nfCvxh8ObuNWi8TaFeacdwB2maFkDDPQgsCD2IBr4FfEC9+D3xu8E/EWOR7d/DPiC1urlWBUmJJV86Nu4DR70PfDGqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv+C5//NEv+5g/9x1f8E3v+T3PhX/2ELn/ANIp6r/gpfJ4Ql/bX+JDeDTbGAXNst79nACfbRaxC56fxeaG3/7e/POa/wCCHuh6y3iP4reJVR10lLHTrRmK4WScvM4APcqqtn03j1FfEf4j+CvhJ4K1X4hfELxBbaNoOjQGW6upzwB0CqBy7sSFVACWJAAJNfGv/gtL8SNV1e40/wCAngLSNC0aNysOoa9G13eTqOjeWjiOLP8AdPmduR0rwz/wV8/bE0TUku9Z1bwr4htg4L2l9oiRKV7gNAY2H1ye1fsXft8fDr9r7TrjRI7A+GfHWlwCa+0KecSrNEMAz20mAZIwSAwIDISAcghj/wAFGv2wvjr+yD/whWtfDPw14N1XQvEn2q3vJNbsrmaS3uYvLZADFcRAK6O2AQTmNuRxX7Hv/BVH41fHb9orwl8JPiV4W8A6boniR7iBrrSrG7hnSZbeSSIK0tzIuGkRUxtP3u3WvH/i/Tvh94E8RePdXI+w+G9JutSucnH7uCJpG5+imv8Ah9X+1R/0IXwp/wDBVf8A/wAm18M/G1j8Svhz4W+IembfsvibRrTUoVU52rPEsm38N2Pwr9sj/gqf8ZPgP+0R4o+EXww8LeBNS0bw39nha61exu5ZnnaBJJRuiuY1wrOUxtzlTyetf8E6P2sfjj+11oHjHxf8UPDvg/StG0O8t7DTG0OzuYXnuCjSThzNPKMKjQYAA/1h9K/bK/4KreMPgH8VPEHwW+H/AMILNtX8PSJHPqviG5Z4pfMiSRGjt4SpKFXUhjICQeVFat/wVw/bQ1K486z8V+G9LTJ/dWmgQsv/AJF3n9e9fBz/AILRfF3Rdat7T43eCNC8SaHJIBPdaPE1lewqTyygs0UmBjCYTOPvd6+GXxK8GfGHwFovxK+H2sJqega9bCezuFBU4yVZWU8q6sGVlPIZSD0r/go9+2T8Wv2QLDwDqPwx0HwlqcXiebUItQGvWtxMY2hFuYvKEM0WARJLu3Z6LjHOf2RP+Cq/jL4o6/46uP2hNI8G6D4W8GeD59ee70KyuIpmeO6toFiAmuJAzSNcqqqACXKjPNfFj/gsV+0p4p8TXE/wrttD8F6BHIRZ2z2CX1y6djNJKCpY9cIqgdOep0nVUbwvZa3q13DEpsI7i5nkYRog8sM7E9FA5J7Cv2nP+CyV3pPiK98Ifsx+GtL1Cys3aJ/E+tRvIlywOC1tbqy4Tjh5Cd2fuDgnw1/wV7/bE0XVlvtY1fwt4gtd4L2N7oiRJtzyA0BjccdyT261+3X+1BoX7XHxG8I/E/R9BuNFurXwdb6ZqunyyeYtvdx3l3IwjkwN6FJo2DYB+YgjINf8Ebf+Tubv/sT77/0dbVVftjftr/Dj9j7wta3Wv2c2veKtZRzo2gW0ojaYLw0sshB8qEHA3YZiThVOGK+NP+Cwf7XHiLUZLjwxN4U8K2e/MVtZ6SLllUZwGe4L7jyMkBeRwB0r4Pf8FeP2r5vFui+GPEHhvwj41/ta+gs47d7NrC4mkkdUVUlhOxCWI5MTDnpWuXsumaJqGpW6o0tpayyorgkFlUkZx24r/h9X+1R/0IXwp/8ABVf/APybX7Gfxr8VftFfs1eDvjH42sNJsta8QfbvtNvpcUkVunkX08CbFkd2GUhUnLHknoOB8VP+Cwf7THgf4neL/BWk+B/hjNY+H9evtPtpLnTL5pXjhneNS5W8ALFVGSABnsK/4fV/tUf9CF8Kf/BVf/8AybX/AA+r/ao/6EL4U/8Agqv/AP5Nr/gnf+1b8RP2uPhh4k8a/EjRvDmm32ja8dPgj0S2mhjaPyIpMsJZZSWy5GQQMY471+1J+1l8K/2TfBUfin4g3kt1f37NHpGiWRU3WoSD720EgLGuRukbhcgcsVU/EP8A4LJftPeJtQmbwHo3hTwbpxP7iJLM39wo/wBuWY7GPuI1HtXwx/4LKftJeGdVt/8AhZWheGfGmk7gLhFtf7OutueTHJF8gOP70bDp0r9nr9of4bftN/Di0+JPwz1N5bSRzDeWVwAlzp1wAC0MyAkKwBBBBIYEEEg1Vf8ABar4hza58dPB/wANYZy1p4W8PG7dN3AuLuU7uP8ArnBCf+BV/wAEY/AkOgfs0a542lt9t34r8TzbZCMb7e2ijjjH4SG4/Ov2h/iHrXwk+Bfjv4neG7WxudU8L6DdahZw3yM8DyxRllEioysVyOQGB9xX/D6v9qj/AKEL4U/+Cq//APk2vgh411X4l/BbwB8Rtdt7SDUvFXhfTdVvIrRGSGOa4tY5XWMMzMEDOQAWJAxknrX/AAUW/bw+L37IfjTwh4c+Gvh3wfqVtr+lzXdy+uWdzM6OkoQBDDPEAMeoPPev2I/+Cmfx4/aV/aJ0H4ReOvCfgGx0fVLW8lmn0mwu4rhWht3kXa0lzIoBZQDlTx6da/4Kj/8AJiXxN/7hP/p2s6rwl/yKmi/9g+D/ANFrVV/wW58BQ6b8UPhz8S4INp17RLnTJ2VcAtaTLIpbH8RW7xk9kA7cf8EUfiJc678FPG/w0urgy/8ACJ69Fd24Y/6qG9iOEHt5ltM31c1VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV/wWr/5Oo8Lf9k/s/wD0vv6+AP7QXxF/Zp8dP8RfhfdWEGsyWEtizXtqLhPKkZGYbSRzmNea+Kf/AAUl/a++Lfh+58Ka18TV0nSb2Joru20OxisWuEYYZWlUebtIJBUOFIPINfsffsl65+1v8RP+EO0zxr4f8O2diqzahJeXaG8aAH5zbWud8zAYyflRcjLDIB+FHwt8F/BX4eaJ8L/h9pYsNC0G28m2iJ3O5JLPI7cbpHcs7N3ZieK/4Kh/HzVfjL+1Fr3haK+kPh34czyaJptqWwq3EZAvJSOm9plZM91ij9K/4Jhfsvfs1+IvAV18bPj/AHfhHWdRvL+S10XRNcvofItoYsK80kDsBIzvuUB1ICoCB82R/wAFNv2Zv2WLX4OS/GX4HDwT4f8AEXh28t0vtN8O3dvFDqFrNIsRxbRHaJEkkjYMgHy792cAr/wT2+MGpfBv9rTwFqdtdyJp3iLUY9B1WENhZYLthEC3skrRS/WMfSv+Cz3wqPin4AeHvinZW2+68Da2Irlwv3LO8Ajck/8AXZLYf8CNf8E8vHJ+H/7Znwu1UyERajrH9kSLnAf7bG9soP0eVD9QK/4LVfEpdB+CHg/4XWt0EufFmvNeToG5a2s4+QR6GWeE/VKvNL1CwtrG8vLV4odSgae0dhxLGJHjLD23xuv1Wv2dfj/aaj+wJ4f+Ol/dRzy+GvA88moM7ZL3GnQvFJuH953tycd94x1FMdR13VCx869v9QuOwLPNK7fmWLH8zX7Pnwut/gr8EPBHwrgSMP4b0W3trlowAJLnaGncY/vStI3/AAKvid+wn+x9f/FbxR+0n8cobeb+2JYJriHWtXFjpVm8cKRlyAY9zPs3N5jspJPy14/+Kv8AwRt03SLnwnq2ifDG5UoyFtA8ISyTHsdl5awZB54IlHTg8V8VLPwBY/EfxJa/CrVrnUvB6alN/YdzdRNHK9oWJjDq4DbgpCkkDJGcDNf8EafiDqHin9mXV/BepXbzHwd4kmgsgxJ8q1njSZUGe3mtOcD+90Hf/gtp/wAl+8C/9ieP/Suevhb4D8d/F7xno/wg8ART3mpeKNQijgsvNKQvKiviaQdAI43mYuQdqlz3OfhB/wAEZV8DeIvCnjvxL8ejPrOgapaalLZWOgZty8MqSeUsjzBmUlMbyqnn7or/AILTfG7VPDvgTwf8CtCv5IF8VzS6lrQjbaZLa3KiGJvVWlZnI9YFr9nj4I+Iv2jPjJ4a+D/hm4S1udeuSs15JGXSzt41Mk0xAxnbGjELkbm2rkbs1b/8Ej/2MovBY8Mz+Fdfn1X7P5Z8RNrtwt5vxjzBGG+zg55x5JHsa/aT+B2s/s4fGzxP8Hdavlv5NBuV+zXqpsF1bSIskMu3J2kxuuVydrZGTiv+CIf/ACVP4l/9i/a/+lBqq/bk1zxD4h/a++Ll94maY3cPiu8tIhLnItoHMNtjP8PkRxY9sV/wTDsP2EfEHhnVvDXx+03wrP8AEW61U/Yh4r2i2mszGgjS3Mh8oPvEmVOHJZcZHA8H/sEfst+Afi34f+N/w78Bf8I/rug+c9tFYXshs5jLC8W5oXLKCFkYrs2YODziv21/+TRfi/8A9ifqH/olqr9l6xh1P9kf4R6bOzrHdfDjRYnKHDANp0IOM55wa/4cqfsr/wDQ/fFX/wAGth/8hV+3x+zp+yT+y5qFt8NvhX4u8eeJPiA+ybUI9Q1WzltNJgIBVZlitUdpnUgqgZdqkM3BUN+zz8BPHH7SnxU0j4V+BLb/AEq/ffeXjoTFp9qpHm3EmP4VB6dWYqo5YV4I8EfDn9kL9nk6DoEM8fhf4faHc391K7BprgRI89xM54G9yHY9ACcDAAA+LfxP8VfGn4k+Ifij40vWudX8RXz3MxLErEp4SJM9EjQKijsqgV+yd+yL+xX8Mfg/oK/Ea3+Gfi/xlq2nxXOt3mvXNpfCGaRQzQwpKzJGsedgZQGbbuJ5wP8Agpt+z/8ABj4M/E/QPEvwJ1LSP+Ed8ZWlxJNpWm6glzHp93AyCTYFYmON1ljKqeAwfbhQFX/gif8AGDUrXxp43+BN9dySadqOnLr2nxO3ywTwukM4UesiSxE/9cR+NV+2vrDa7+1z8X71pS5i8Yaha5K4x5EzQ4/Dy8fhX7G2lW+jfsmfB2ztsbZPBOlXBwu35prWOVuP95zz361/wVH/AOTEvib/ANwn/wBO1nVeEv8AkVNF/wCwfB/6LWv+CyX/ACdzaf8AYn2P/o65r/gip/ydR4p/7J/ef+l9hX/BZL/k0W0/7HCx/wDRNzXwm/5Kp4M/7GCx/wDShKqq/wCC5OlW8OtfB3W1x513a6zbv8uDtiazZee/Mzcdvxr/AIIc6u0/hj4u6B5jEWd/pNyFKgAGWO6XIPUk+SOO2B6mqqqqqqqqqqqqqqqqqqqqqqqqqqqqqrxb/wAiprP/AGD5/wD0W1V8HP8Agoz+0z8B/hlpHwl+HmsaBb6DognFmLnSEnlTzp5JnJcnn55XIyOmBXxs/aO+Nf7ResW+tfGLx/f+IJbIMLO3dUgt7UNjd5UESrGhOFBYLubaMk4Ff8Eyv+CfOj6fPoP7UPxK8Q+H/EUkY+0eGdL0m8S+gtJ1JHnzyoShmjIO2NSdjfMTvUBf2u/jTJ+z3+zl43+K1ltOpaVp/laWrKGH2yd1hgJU9VWSRWI/uqa8P6ZrPxP+Iem6Nea0rat4s1qK3l1HUJ+DPczBTNLIx6bn3MxPqSa+HX7KX/BPf4f+ArbwPNoXwt8RyC3WO81fWri0u726kx80nnOxaIscnbGVC9ABiv23/gv4E+BX7Q2ueDvhfrltqfhK7hh1DSGgvVu/s8UoO6AyKTnZIsijJ3bQpJJOT/wRn+MGpeNvgP4j+Fur3clzL8P9Vj+xM7Z8uyvFd44/XiWK5P0YDtX/AAUK+FX/AAqH9rz4g6Fb23lafq+of21p+F2qYrwecwUdlWVpYx/uV/wRZ8cnW/2ePFPgWeTdN4Y8TtLFz92C5hRlGP8ArpFMfxr9uLxtffHL9s7x/eaCG1BpdfGh6TFEdwlFtttYxH7O0ZYepfPev+CZ3xKX4aftleBJbm5EVl4mll0G6BbG/wC1IVhX8bgQce1f8FvPiALPwN8NfhZb3GW1XVLrV7mNT90W8SxRFh6MbmXH+4fSv+CR3wlPxE/avtPF97aiTTfh/pc+qOXXKG5ceRAv+8DK8o94fz+MHww0D40/C/xN8KfE8ksem+J9Ols5pYQDJAWHyypuBG5GCsMjGVFSfs7/APBMP9kbT4tN+I1j4DOpxxhpD4wuk1W/ucKPn+yvu6jBxHEq89Oa/bm8af8ABMj4i/DPULj9n0aTpPxD09oX00+H/DF1pdrdr5qrLDKhhji/1e5w+3OVX5uSK+HXjfWPhp4+8OfELw/O8Wo+HNUt7+3ZHKktFIH2k+hxgjoQSD1q0ure/tIL60k8yG4jWSJsEblYZBweRwR1qv2Ov2Dfib+2pPqfi+68Xp4e8K6XcLZ3WuXsD3k1xOsanyYY9y7yiGPcWdQoZcZ6D9ib9hzS/wBjA+M/7M+I114qHjAWG8T6Ytp9m+y/aMYxI+7d9o9sbe+eP2xfjdqn7QX7RfjP4hXl/JcWB1CSy0WNmysGnwOyQKo6DKjeQP4pGPcmv+CbX/BPbwn+0lot/wDGL4ztfP4Qsr1rHTNJtZmt21KZADLJJIuGWFdwUBCGZ93zKEIb9uj/AIJdfBjwp8F9e+LP7PekXvhvU/B1lJqF9pL6hNeW99ZxjdOwad3kjkRA0gIYqQpXbkhlr9ij/k0X4Qf9ifp//olar/gptrniHQf2JfiPceHGmSW6hs7S5kizlLaa8hjm6fwsjMh9nNaO2lJq1i2ux3L6atzGbxLYgStDuG8JnjdtzjPGcV8JfgB/wTM/aE8CW9p8K/h94A1+xjt1MsVrI8Op24x/y3O9bpG56uRk9zxX7Pv7O3w1/Zk8E3fw/wDhXaXtvpF7qs2pyJeXHnuJpVRSN+ASoSKNRnJwoyT1r/gt7/yVP4af9i/df+lAr/gk/wD8nueD/wDsH6n/AOkUtftT/sifDX9rzw/ofhv4k634l0228P3kl3bPodzBC7u6bCHMsMoIx6Ac968S/wDBHj9jjwd4f1DxV4p+KvxL0vSNJtnub29utZ0+OO3iQZZ2Y2XAAFfFVfhYnjzVYPguniM+EIJjHps3iG5imvLhV481/KijVA3UJtJUEAkmv+CUv7EM/jnxFp/7UHxN06SPw/4fu1l8KWbgr/aF9E3F0e/lQuo2/wB6QdcRkP8A8Fifj5qvw3+Cei/CLw1fSWt/8R7mdNQlibBGnW4Qyx5HI8x5YlPYosingmv2H/gb4J/aA+P2l+EPiX4htNI8Jaday6nrDz3q2huIYiiiBJGYYLySRqcHcF3EcjI8d/sq/wDBPTxv4AuPAUPh74W6AGtmitdW0i5tLa/tXx8souFbfIynBxIWB6EEGvEWkXvgzxbqegjUI5LvQtRmthdWkuVaSGQr5kbqemVyCD6Gv2Q/izd/HL9mj4e/E/U5fN1HV9HVNQkPHmXcDNb3D47BpYXOO2ar/gqHrDaP+w58RvKlKS3v9nWqELnO/ULfeD6ZQOK/4JY6Vb6p+3D4AkuNpFhDqdwqMuQzCwnVfpgsGz6qKr/gsl/ydzaf9ifY/wDo65r/AIIqf8nUeKf+yf3n/pfYV8Wf+SV+Mv8AsX77/wBJ3qv+Cb3/ACZH8K/+wfc/+lk9f8FR/wDk+34m/wDcJ/8ATTZ1/wAEOv8Akavi3/2D9K/9GXNVVfGrSrfQvjJ480O12+Tp3ibUbePC7RtS5kUYHbgdK+EWrt4g+E/grXmkZzqXh2wuSzKFJMlujZIHAPPQVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVf8Fz/+aJf9zB/7jq+EvxS8WfBP4i6L8UfAs1rFrugSvLZvcwCaMM0bRnch6/K7V41/4KmftoeNNGl0RfiTaaDDOpWWbRdLhtp2B/uykM8Z90Kn3r9n74GeKf2nfirZ/DvQ/Feg6XqeqyNJJe69qSw+Yc5fy1JMlxKRuOxAScEnAyR+zR+zp4G/Zd+FGnfC7wMjTLCxuNS1GWMLLqV4ygSTuATjO1VVcnaqquTjJ/4LJ/HzVfFXxk039n/TL6SPQ/BNpFe6jbhsCfUbiPerMO4S3eMLnkGaT1r/AIJg/s1fAv4r6x4h+Jv7QGqaDPpHh2aK00rQ9U1FLeO8uWXe8sqF1Loi7AFOVYuc524P7d37LX7HHiv4A+KPF3wrtfhz4W8Y+EdOl1Owbw7cWlmLxIF3ywSQxELKWjVtvG8OFwcFlb4DfFjWPgb8YvCPxW0OeVJvDuqRXE0cbYNxb5xPCfaSIuh9mr/gp58Mofi5+xp4m1PTIhdXfhIweI7FlXOUhyJ2z1A+zSzN+Ar4IeOT8MvjL4G+IYkKp4b8RWN/LzjMcU6M6n2KhgfY1/wVZ+JS/D79jnxHpcF0Ir7xlfWui2+GwxV382bj0MMEin/eqTS9Qi0uDWpLVxY3NxJbxTkfK0saozqPcLLGf+BCv+CUnxOh8dfsb6Hpl7eo114Hv7vR7lnfBWNW8+In0URTooPT5D6GvjZ48k+KXxh8bfEaSRnHiXxBe38e7+GOWZmRR6AIVAHYAV/wTu+Ep+D37IngLRbq1EOp65ZnXNRBXaxluz5qBh/eWEwoc8/JXxw/Ya/Zh+Knxauf2hvjfBLOttpVva3Npdan9h07MLNi4mZSjltrImC4TCAEHNeMPiX/AMEdPBmnXHhbVNI+EV6vlshbR/Ch1KVsdlu7aByCccN5g69ea/aItvgtb/GLxEf2edTu73wBNMkujm7hljkhV41MkREoDkJIXUFuSoBJJya/4IifEHUNR8F/Ev4X3t272uiahZanYRsSQn2lJY5gvZRm3iOO5cnHU1/wXG/5FT4R/wDYQ1X/ANF21eHbPxFrOqQ+FvC6XtxfeIZYtPjsrVjuvmeVDHCVH38yrGQp43Kp6gV4A/4Ih6vf+G7W++Jnxzi0nW50DTafpOjfa4rYkcqZnlTeexwgAIOCw5r/AIK0fF/VPg/+y9ovwq8P6pJDqHju4GmXM8f7tn0+3iU3AHUjezQIRnlXcHg8/DH4eeIvi18Q/Dvwz8JQpJq/iXUYbG18zIRGkYAu5GSEUZZjjgKTXgv/AIJGfsg6D4Jt/D/izw5rHifXfIC3OvTavc2splI+Zo4YpFiRc52qyuQMAljyf2z/ANma6/ZP+Oeo/C9dVl1TSJ7WPUtDvZ1Cyz2UpZV8wLxvV45IyQAGKbgFDYH/AARt/wCTubv/ALE++/8AR1tVV/wVL1zxDrP7bXjq211phFpUNhaabFJnEdsLOKQbP9lnkkf6ua/4JlL+xTea54l0j9qC08Pv4iungHh+TxOcad5WGEqgsfKWUsU5l6jbsOd1W3/BPv8AZDbx74W+L3gz4d22h6t4d1OHVbCXRLx0tbiSNg8RaEloigYKwKBTkDnHFeLf+RU1n/sHz/8Aotqr/glx/wAmJfDH/uLf+na8rxl/wR8/Zn8ceL9c8a6t44+JsV9r+pXGoXMdvqdisaSzSNI4QNZkhQWIAJJxjJPWv27/ANj/APY0/ZD8KwaXoXjX4j678RdbjL6VpFxrFiYraLJBurkJaBhHkFVUFS7AgEBWYfDr4eeLvix430b4deA9Hk1PXtdult7O2TjLHqzHoqKAWZjwqgk8Cv2R/wBmPwv+yj8HdP8AhtoNx9u1GV/tmuamQR9uvWVQ7qD91AFVEXsqjOWLE/thfHzVf2kv2gfFPxHu76SbSvtb2WgQs3y2+nQsywKo6DcMyMBxvkc96/4J5fsp/sl6L8EtE+J3xqPgHxP4w8VxG7+yeILu2uIdMtmJEUQt5WKbyoDszLuBfaMBef8Agqd+zl+zv4A07w38X/gDc+GdOGp6idM1rRNEvoXg3tE0kM8cCMfKGIpFfaAhPl8Aklv+CQ3xg1LwH+1JD8OXu5P7H+IWnT2k0Bb5FureNriCUj+8BHLGP+ux+oqv+CoesNrH7cXxGCyl4rH+zrWIFcbdmn2+4f8AfZf86/4JkaVb6T+w98M4oNpa5hv7iRgu0s0l/cNz64BC59FFftr/APJovxf/AOxP1D/0S1V+yd/yav8ABr/sn+hf+kENf8Fvf+Sp/DT/ALF+6/8ASgV/wSf/AOT3PB//AGD9T/8ASKWv+Co//JiXxN/7hP8A6drOq8Jf8ipov/YPg/8ARa1Vf8FutKt5vg38O9bbb51p4mmt0yvO2W2Zm57cwrx3/Cv+CIertD8WPiToIkbF54dtrkrtGCYrjbnPUH99075PoKqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/wCC1f8AydR4W/7J/Z/+l9/X/BMX4N/DP45/tI3Pgr4r+FLfxDosfhm7u1tJ5ZI1EySwBXzGytkB2HXHNftz/wDBMX4H6X8E/EXxR+AXhWfwx4i8JWb6jcWEF5PcW2o2sS7p1KSu5SRY1LqUIBKkFSW3L4K8aeJ/h14t0nxz4K1m40rXNEukubG8gba0Uinj6gjIKnhgSCCCRXwL+KFj8a/g54O+K1hEkKeJ9HgvJYEJIgmZQJYgT12SB0z/ALNfHSHULf42/EG31Yk30firU1ucjH70XUgf9c18MP2M/wBpr40eELfx58MPhTe6/oNzLJFFeQXtsgLxttddryqwII7j36Gv+Hbv7bn/AEQLVf8AwYWf/wAer4Wf8E7f2y9J+J/g/VdX+CmqadY2WvWM11eNfWjC2iSdGeTCzZO1QTgc8V+0V8L4fjT8CvHXwtkiR5fEWh3FvaFxwl0ELW7n/dmWNvwrw3rep+CvFuleJLON4tQ0HUYbuJHBRklhkDgHuCGUfSv+CuPxXtviR+01YaNpN35umeGPDFjHEoOR5l0n2sv9THPAOP7or9sP4AXHwy/ZD/Zg8XmxMU93ol5DqbsmGWS7kF/Are+2ecYPTZivgj8c5NI/4JdfHH4cfa/9OsvEVhbWg/uwak8e9OOxW0uz9WP0r/gnR8JD8Yf2u/AulXFn5+meHro69qOeix2mJI9w7q0/kIR/t18TvH+hfCn4d+JPiV4ldhpnhnS59QuVQ4Z1iQtsXP8AExAUDuSK/aN/aZ+Kn7T/AI7u/GnxG12d4DKx03R4pWFnpkX8McMecA4wC5G5jySa/Z//AOCQvxm+L/g7S/HfjnxvpPgLTtat0ubK1lsnv70wuAyu8QaNE3KQQpk3c/MFNftI/Bn/AIZ7+N3ir4N/8JJ/b/8AwjNxHD/aX2P7J9o3wxy58re+3HmYxuPTPfFf8EQ/+SVfEr/sYLX/ANJzX/BbT/kv3gX/ALE8f+lc9f8ABImGGX9szSXkiR2i0HUWjLLko3lgZHocEj6E+tV/wWwt79f2kfBt1IzfYpPA8KQgnjzFvbsyYHrho/0r/gj9qOlWX7Ytta6g8YuNQ8NahBYh1yTMPLkIX0PlRy/gD61X/BVu4t5/23vGqQTI7QWWmJIFOdjfYYTg+hwQfxr/AIIh/wDJU/iX/wBi/a/+lBqq/wCCgP8AwTUf9ozXJvjJ8G9QsdM8dSQJHqem3p8q21gRqFRxIAfLnCgJlhtYBclMFj8WfgF8ZvgVqa6T8W/hvrfhqWRykMt3b5t52HURTpmKXp/Axr9jb9vL4q/sv+LtJ0u/8Q6hrnw4knWLU/D91MZUt4WIDTWu7JikUZbauFfow5DL+2Ve2mp/scfFfUdPuEntrrwVfSwSoch0aAlWHsQQar9k7/k1f4Nf9k/0L/0ghr9vj9srSP2S/hcx0ae2uviB4kjeHw9YPhxDgYe7lX/nnHkYB++5C9NxW1tPHfxf8ex2lrHqfifxd4r1HCgkzXN9dTNySTyWZjkk/U4FfsL/ALHvh/8AZK+FUWm3MNveeONfSOfxJqaDdmQD5baJv+eUeSB/eYs3cAftzQ6hP+x/8XU0wnzh4Vu2bA/5ZquZP/IYaqsf+CdX7aOp2NvqOn/AnU57a6iWWCVNRsyHRgCrD990IINf8O3f23P+iBar/wCDCz/+PV/wTR/Y1/aU+Cv7TMXjv4qfDO/8N6HBoN5Cbma6t5FklfywseI5GPPLZxj5fpVV+2bpT6N+1p8YbOQODJ411S4G9cHE1y8o/DDjHtiv2PdQt9U/ZP8Ag5c2pOxPA2kQHOPvRWkcbdCf4kP/ANav+Co//JiXxN/7hP8A6drOq8HSRzeENDlhkV0fTbdkZTkMDGuCD6V/wWS/5O5tP+xPsf8A0dc1/wAEVP8Ak6jxT/2T+8/9L7Cv+CyX/Jotp/2OFj/6Jua+E3/JVPBn/YwWP/pQlVVf8FytSt5dV+Dejof39rb61O4yPuyNZKvHXrE3+c1/wQ40lofD3xf1wh9t3e6RbAn7uYku2OPf98M/hVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV4t/5FTWf+wfP/AOi2qv2Bv2IP2V/jN+yB4F8efEr4RWWteINZXUlvr9r+7gkl8vUbmJP9VKoUiONFyoHT15r/AIKJfsgaX+yV8WtPs/BlxeT+C/Fto93owvH8yW1kjYLPbF+N4QtGwYjO2VQckFj/AMEh/wBoTWvh/wDH5PgpqOqSN4Y+IEMqxWskhMdvqMUZkjlUdFLojxNjG4mPP3BX/BYOHUJf2O5nsj+5i8T6e13x/wAs8SAf+PmOvDnh/V/FviHS/Cnh6zN3qus3sNlY24dU86eVwkabmIUZZgMkgDPJFf8ADt39tz/ogWq/+DCz/wDj1f8ADt39tz/ogWq/+DCz/wDj1f8ABJr9mL45fs/ap8Tb/wCMfge78MJrNvpkWnxT3EMv2oxtcmRh5TtjZuQc4z5nHev+C3PwqCv8OvjfZWw+YTeHtRm29xuuLYZ/8C6/4JX/ABwT4N3nxzuJ50SO3+Hs/iCNHPEs+nMfLTHcn7UwA+tf8E7/AANefFH9tP4dRXSvcrpmqvrt7O/zbTaI06u3fJmSJc+rivivoGqfAD9pHxPoWkp9nu/Avi+f+ziVwNtvclrdwPQqsbD2Ir/grL8U7L4oftG6DNo85bTdO8Faa0CE97oPd7vqY7iIccfKPev+CNXwkPg79nbWfijfWZjvfH+sn7PIf47Kz3RR/T98119Riv8AgqX+2R4o/Z18EaP8NfhfqR0/xj41imkl1OM/vdMsUwrPF/dlkYlVf+EI5GG2sPhB8IPil+098U7fwH4Hgk1nxLrLSXNzdX9yQsaDmS4nlbJCjIyeSSQACSAfi/8A8EjvEfwV+BPin4weKfjfpt3f+GNNN5JpFhoTvFOQQCguHmQgc9fK/Cq8Jf8AIqaL/wBg+D/0WtV/wSchhj/Yl8JvFEitLqWptIVXBZvtcgyfU4AH0ArWIby40i9g06QpdyW0iwMG27ZCpCnPbnHNOjxuY5FKspIZWGCCOvFf8Ep9R0q+/Ye8DWunPG1xp93qkF8FXBWY388gDep8qSL8CK/a5uLe1/ZU+MklxMkanwFraBnOAWaxmVR9SxAHuRVfsUf8mi/CD/sT9P8A/RK1XjXwb4b+IfhHWPAvi/S4tR0XXbKWzvrWQcSRSKVYZ6g85BHIIBGCK/aZ/wCCT3x5+FGsX2s/BzTJ/iL4Q3M9uLML/adqh6JLb8GVh03Qht2MlU6CCfxd8PvEyz202r+G/EWjT/K6NJZ3dlMvoRteNh+BFf8ABLf9uTxh+0Db6p8Fvi9qP9p+LfDtgL7TtYdQsmo2ausbrNgAGWNpI/n6urZPKszf8Fvf+Sp/DT/sX7r/ANKBX/BJ/wD5Pc8H/wDYP1P/ANIpar/gqF+3O/xo8UT/AAF+FetlvAfh26xqt7bSfJrl6h6Bgfmt4iML/C7gv8wWMj9gj9jLWv2s/iakms2t1a/Dzw7KkniDUEynnHqtpE3/AD0cDkj7iEscEqG0LQtG8MaLYeG/DumW2naXpdsltZWdtGI47eJFCoiKOAoAAAr/AILcw6gvxl+Hc8h/0F/DEywjH/LUXLeZ/wCOtHXwk+C3xO+O/iabwb8JvCk3iHWreye9ktIZ4omECMiM4MjKDhpEGAc85xgHH/Dt39tz/ogWq/8Agws//j1f8O3f23P+iBar/wCDCz/+PV/wT0+Ffjv4M/sneEPAPxK0abSPENpcahJdafNIjtbCS9mZBuRmU7kKvwf4/XNV/wAFTNKfVP2HPiDJEHZ7GXTLgKi5yBqFurZ9grMfwr/glXqFvY/tw+A4ZyQb221OCM8YDfYJ25ye4Qj6kVX/AAWS/wCTubT/ALE+x/8AR1zX/BFeSNP2qfEyPIqtJ4AvFQE4LH7dYHA9TgE/ga+LP/JK/GX/AGL99/6TvVf8E3v+TI/hX/2D7n/0snr/AIKj/wDJ9vxN/wC4T/6abOv+CHX/ACNXxb/7B+lf+jLmqqvjfqVvrPxp8f6vaHMF94o1KeI5B+V7qRhyOOhFfB3SW0H4ReB9CcOG07w3p9sRJ94FLaNecd+Kqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv8Aguf/AM0S/wC5g/8AcdX7Dvw/8H/FP9qz4feAPH2iRaxoGr3s8d7Yyu6LMq2szgEoQw+ZVPBHSv2iv+CUv7N/jL4baw3wY8Et4P8AGtnZvLpM9pqFxJDdTICywTRyyOu1z8u9QrKSDkhdptbrUdG1GG9srm5sb+xnWSGWJ2ilglRshlIwVZWAIIwQRX7DPx2vf2i/2ZfCHxC1y4WbX44X07W2AwXu7dvLaQgcAyKElIHA8zAr/gonDqEP7avxWTUiTMdXjZcj/lm1tEY//IZSvg/+yv8AH74/aPfa98H/AIcXfiSx0y5FtdywXdvF5MpUMFKySKeVIOcY9+DX/Dt39tz/AKIFqv8A4MLP/wCPV/w7d/bc/wCiBar/AODCz/8Aj1aT4Mtr34V2Xw98VQ/a7efw/HpepRv/AMtVNuIpQevUFh3618SfA+p/DP4heJvh1rWTfeGdXutNnYrt3vDK0ZYD0O3I9iK/4KQ/HiT4nfCT9mvw4l5vlm8Cw6/qiBs7rmdEt+f917a5A7/Mc14q+AFxpn/BLLwh8W/sJF5P8QbnU5XZPmSynj+xAk9QDLaQkeu/Pfn/AIJ4fHOT4a/BT9pvw0bvZMPAkms6YP8AnncRq9ruGPWS7ts/7o6V+zt8LLj42/HPwP8ACq3heSPxFrUEF3sJBS1Db7hwR/dhWRv+A1qN/o3hTw/c6pfSxWOlaNZvNM+NqQQRISxwOgVVP5V+15+2F8Sf2r/iBe6vrmrXdl4RtblxoPh1JisFnCCQjugOHnZeWkPOSQMKAo/Zg/4JYfGb9onwXp3xK1jxRpPgjwxq8fmadLdwPdXd1FnAlWBSoEZxwWdSRggYIJ/a9/Zu/wCGU/jLc/CP/hM/+Ep+z6fb3f8AaH9m/Yd3mqTt8vzZMYx13c+gr/gh1/yNXxb/AOwfpX/oy5r/AILjf8ip8I/+whqv/ou2r9geGGf9sj4SxzRJIo8RRMFdcgMqsQfqCAR6ECq/4Lk29+NU+Dl07MbI2+spGM8LIGsy3HYkFPrj2r/gmlqOlaX+3D8LbnWHjWB7u9gQyLkedLp9zHCPqZHQD0OKr/gtPcW8/wC1X4cihmR3t/AVmkqqclG+3XzYPodrKfoRX/BG3/k7m7/7E++/9HW1VX7fv/BPbTv2sYbbx/4F1Wz0L4iaTafZkkulItdVgUkrFMygsjqS22QA8HawI2lPjN+zB8ev2frs2/xZ+GWsaJb+Z5cWoGIT2czdglxGWjYkY+XdnnkCv2W/2zfjN+yv4osrzwl4iu7/AMLfaA2p+F7u4ZrO7jJ+fYpyIZSORIgByBu3LlTY+MtD+I3weh8f+GZ2m0nxJ4cGo2TuMMYprfzE3Ds2GGR2ORVf8EuP+TEvhj/3Fv8A07XlftY/tN+Dv2U/hHf/ABE8SNHdalLm20LSfM2vqN4R8qDuEX7zt/CoPUlQfiP8RPHfxu+ImqePvG2pXOteJPEV5vlYKWLMxwkUSDO1FG1EQcAAAV/wTY/Yeh/Zt8Dr8SfiJpKf8LL8T2w86OVQzaLaNgi2X0kbCtIfUBeikt4/h1C48CeI7fSSft0mkXS22B/y1MTBf1xVeDP2D/2tfiH4U0vxv4M+DOoapoetWy3NheRX9oFmibowDSgj6EAjvX/Dt39tz/ogWq/+DCz/APj1fsXfsI/tX/Dr9qP4d+OPHXwk1HQtB0XVDcX1/JeWzrEghfgqkpY7jheAfvelVX/BTvSn0n9uP4lxkOUuZdPuI2dcbhJp9sxx7Biy/hX/AATO1C31P9h74XzWxOIra+gcNjIaO/uEPQnuuR7EV+2v/wAmi/F//sT9Q/8ARLVX7JUkcv7KvwaaKRXA8AaGpKnPIsIQR9QQRX/Bb3/kqfw0/wCxfuv/AEoFf8En/wDk9zwf/wBg/U//AEilr/gqP/yYl8Tf+4T/AOnazqvCX/IqaL/2D4P/AEWtVX/BbnUreL4LfD3R2P7+58USzoMj7sdq6tx16yrX/BETSWm+LvxH1wB9tp4bgtiR90GW4DDPv+5OPxqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr/gtX/ydR4W/7J/Z/wDpff1/wRt/5O5u/wDsT77/ANHW1arpdjrel3mi6nbrPZ39u9vcRN0kjdSrKfYgkV8TfBF/8M/iN4o+HWqB/tfhnWLrTZS64LGGVo930O3IPQgg1/wRq+J48W/s06p8Obq5DXXgTXpUhiz9y0u/38Z/Gb7V+Vf8FWf2Ztd+Enx91L4uaXpksng/4i3BvEu0jJS11Erm4gkI4DOwaZc43B2AzsbH7F3/AAUA+IX7H5vvDkWhQeK/BWq3H2m40We5Ns8FwVCmaCYK2wlVUMrKyttH3Tk03/Bbf4H/AGPevwg8cm72f6oy2oTd6b/Mzj32/hX7F3/BRi1/bA+LHiD4dx/DZfCkOl6IdSsXk1T7XLchJo45A2I0C481CAM9+ar9v74VD4P/ALXHxE8NW1t5Wn6hqR1fT8D5TDeKJ8L7I7vH/wAAPXrXw48PeI/2kfjx4O8GalcNPf8AinU9O0qedeCkCLHA0h6/chj3HH904Hav+CmvwltPHf7Ffimz0jToxL4HFtrWnQouFhS2OyXA7BbWSerTxPq9j4Y1Pwja3GzTtXu7a6u4xnLvbrMsffGB57nGOuOlf8ESvhKLTw54/wDjjf2g8zULqLQNMlZcMI4gJrnHqrNJbDPTMRHY4/bb8Da78SP2T/id4P8ADMEs+qXWhSTW0EIy87QMs3lqO5cRFQO5bFV4K/4LTfC/R/hRpdj4m+FXiu58cafpscEsFo1uun3M6IF3iZpPMjViM48piucZbGa+LvxP8S/Gr4meJPip4veE6t4lv3u7hIQRHFnhY0BJOxECouSThRknrX/BEP8A5JV8Sv8AsYLX/wBJzX/BbT/kv3gX/sTx/wClc9f8Ehv+TytN/wCxf1H/ANAWq/4LBfs66v8AE/4PaP8AGLwnpsl5qfw6knOpQwpl302YL5kmBy3lPGjY7I8rHgGvAPjzxX8L/GmjfEHwNrEmma9oN2t1Y3UYBMbr6g5DKRlWUghgSCCCai/4Lg+JF8FC1l+AOmt4uEOz7YNccWJkx/rfI8rzAuf+Wfm5x/HXxK8W+OPiV4u1P4p+Ppbi51TxbeTXst5JEUSdy2G8sdNikbAF4UKF7Yr/AIIh/wDJU/iX/wBi/a/+lBqq8a/8FgdO+F/x/wDHnw08V/DD+2vCvh3W5tOsdT0a88u6HkERy745PklzKspBDRjG0YPWv2of+CrH7Nnxc+APi74a+Fvh/wCLNU1bxVpM1lbxa1p1tFBYySLhZ2ZZpDviYh02g/Mg5HWvDXhvW/GPiHTPCnhnTZtQ1bWLuO0srWFcvNNIwVFA9yQK/aS8Mz+C/wBgTxx4Nurr7TNoPw2lsJJs58xobMRls+5XNV8NPihpHwV/YC8BfFbXbG5vbLwv8LdGvJLa3wJJyunwbUUngFmIGT0zmvjv8bvHH7RHxP1j4qfEC+87UdUkxFAjHyrKBc+XbxA/dRAcD1JLHJYk/slftU2X7J/iO+8b6X8IdC8V+J54zDZanql3IjadEy4dYUUYVnBIZ+u35RgFt3/D7z4p/wDRD/Cn/gxuP8K/ZF+O1x+3F+zjrviPx94SsdIt9Wvb/QLuwsp2kSS3MCBzufnJWZh6cCv2h/gT4y/Zw+LGt/CvxrZyLPps7GyvDGVj1C0JPlXEZ7q6jt91gynlSB+yh/wVu8U/BfwPpXwv+L3gmbxjo+iQJa6bqlleLBe21sgwkTowKTBQAqnchAAyW61rX/Bbr4PwWTP4d+C3jG+uwDtivby2tUP1dDKR/wB81+xP+1MP2vPhFefEubwzB4fu7HXLjTJ9OiujcCLYkciHeVUtlJV52gZBqq/4Ku/D+bwR+2X4l1RbYxWni7T7LV7b5cBswiCQj6y28hP1r/glt43i8afsWeCoDMr3Xhua80m5x/CY7h3jH/fmWH/9Vf8ABUf/AJMS+Jv/AHCf/TtZ1XwN/wCCxXjH4W/B/S/hz4v+E1v4s1bw9YR2OmauNZNoJYo12R/aI/KcsyKFBZWBfHOCS1fHL4ufEX9oX4g6z8afiEDLeaxcrCXt4GS1tQiAR28XUAKgGASWPLEkkk/8E+P2jrb9mX9oW28VX3hi51618Sac3h+a3tZVSZPPngdHTdhSQ8KDBI4Y81/wWS/5NFtP+xwsf/RNzXh7W7vw14g0zxHYRwyXOlXkV3CkykozxuHUMAQSMqM4I+or/h9X+1R/0IXwp/8ABVf/APybX/D6v9qj/oQvhT/4Kr//AOTa/Z5+IetfFv4F+A/id4jtbG21TxRoNrqF5DZIyQRyyxhmEauzMFyeAWJ9zX/BZbxzF4j/AGo9M8IWs6tH4S8MW0E6A8pcTySTt+cTwcf41/wR7+H0vhH9kv8A4Sm7tyk3jPxDd38TsCC0EQS2QY9N8EpHru9MVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVeLf+RU1n/sHz/+i2qv+CXH/JiXwx/7i3/p2vK/4LLfDU+Kv2Z9J+IVpbF7nwP4gieaTbny7W6HkP8ATMxta+F/jm/+GPxJ8K/EfSgTd+F9ZtNSiUHG8wSrJtPsduCOhBIr9oL4W6L+1H+zj4k+HljqMP2fxjoyTaTesDsSYbJ7WU8Z2eYkZOOduR3rxR4Y8Y/C3xrfeFvE2m3ug+JfDt6Yri3lBjltp4zkEEfQMrDggggkEGvgd/wWnvNF8M2Xh/49fDO81zUbKFY313Q7mOOS82jG6S3kCqrnAJZXCkk4VcYrxT/wW9+G9tZMfBXwN8S6jdlflXVNTgskB55JjExI6cYGeelfs4fF+L4+/A3wd8X0sIrF/EunCee1ikLpBOrNHKiseSFkRwCeeK/4KIfCr/hbn7IXxA0W2tvN1DRLH+27DA+YSWZ81wo7lollQD/brQfE+seGotVi0e58kazp76fdnHLQOyMyj6lFH0zX/BEf4Vm41z4h/Gq9thts7aDQdOkIzlpCJ7jHoQI7b8HNf8Fgvhj/AMIV+1Z/wmtpa7LPx3oltfM6jCm5hH2aVQPXZFCx9TJnqTWqar4l+I3iLT1uFe/1Wa10/SbONASzpb28VpbRjJ6+XFEv4dq+Cfw2sPg78IfB3wu05Y/L8MaNbWTvGMCWVIwJZPq8m9z7sa/4LbeBddtvi54B+JbQSto2o+HG0hJAMpHcW9xLMwPoWS5UjPXYcfdOP2G/2o7b9kf44J8SdV8PT61o+o6VNpOq21q6rOtvJJFJvi3EKXV4IztJAIyMjOR+25/wVS8OfHv4TX/wd+C/g3X9I0/xBsTWdT11YopzAjhzDFHDJIBvZVDOW+7uXb82RXhL/kVNF/7B8H/otar/AIJP/wDJkfg//sIan/6WS1X/AAUB/Z11f9nf9pDxJpv9myReGvE93Nq/h25CYjkt5X3PEpHGYXYxleuAjdGBP7GH7ePxD/Y7vtR03TtFg8UeD9alE99oNxdG223AUKJ4ZQreW5VVVsqwYKoIyqkftY/8FOfiH+1d4QHwS+HXw3bwto2vzww3sUd61/f6o3mKY4EKogRGkCgooZnwBuALKa/Yo/5NF+EH/Yn6f/6JWq/bB+PF1+zV+z14o+LulWtjd6rpQt4tNtL0MYri4mnSNVYKysQAzMQCDhTzXw7/AOC2Hwc1Owhi+KXws8V6DqG3Ej6O8OoW5YDk5d4nUHHA2tjPXvX/AAUS/ax+Gn7WnxK0HxJ8NPBl/pVpoWnSWk+o6nBFDd6iWfcu5Y2fEaAHZlifnbIXpX/BGT4beItd/aG1z4mQ200eheF9Altri5wQklzcsgjh9zsSVz6bFz1Ff8Fvf+Sp/DT/ALF+6/8ASgV/wSf/AOT3PB//AGD9T/8ASKWv+Csv7X3iL4NeELL4DeADc2Ou+OdOefUdXRtptdPLtG0cRBz5khV1LcbUBxksCtfB/wD4Kvar8Cfh9pfwz+Gv7OfhHTNF0qPCL/aU7STyH780r4y8jnksfYDAAA8G/wDBaH4neKPF+heGp/gt4Xhj1bUre0eVNQnLIJJFQsBjqA2a/wCCr37M2u/HX4H2HjjwRpkuoeJfh1PNdrZQRl5bqxlVRcpGo5Z1McUgHcI4AJIFfBb4yeOvgD8SNH+KXw51JLTWdIkJVZU3xXEbDEkMq/xI6kgjgjOQQQCPB3/Bbv4dT6NEfiD8FPEdlqqqBKNGvobqB27splMbKDwdp3YzjJxk3P8AwWv8Jap4q0XQ/C3wU1CHTb7UreG91LV9XSNre3aULI6wxowZghJGZAAfWqr9qj4fzfFL9m/4k+ArS2Nxear4avFsogud9ykZeAY/66olfsdeN4vh1+1N8LfFtxMIre28TWkNzI3ASGdxDK34JIxqv+CyX/J3Np/2J9j/AOjrmv2dPj34x/Zp+LWjfFrwSlvcXeml4riyuSRFe20g2yQvjkAjkEfdYK2DjFftKf8ABXjxx8Z/hhqPwz+H/wANIfBMfiCya01jUZdV+3TtDIu2WKECKMIGUspc7jtY4CnBq+sb3TLyfTdSs57S7tZGint54zHJE6nDKynlSCMEHpX/AASJ/aOtviP8Gl+A0nhi5tb/AOGlqTJqYlVoLuG4uZXjG37yuMspHIITORnA/wCCo/8Ayfb8Tf8AuE/+mmzr9lX9sj4n/sgah4i1L4aaF4X1OXxNDbxXa67azzBBCXK+X5M0WCfMbOc9ulf8Pq/2qP8AoQvhT/4Kr/8A+Ta/Yz/4KgfH79or9pTwd8G/G3hH4f2Wi+IPt32mfS9PvIrlPIsZ502NJdOoy8Kg5U8E9DyPiX4xtPh38OfFPj++dFt/DejXeoyF+mIIWkP/AKDXwg8E33xf+MvhDwDtkuJ/FfiC1s52Gc7ZplEjkjoApZiewBNIiRoscaqqqMKoGAAPQVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVf8Fz/APmiX/cwf+46v+Cb3/J7nwr/AOwhc/8ApFPVftu/DQ/CX9q74meDY7YwWo12W+sk24Vbe6xcRBfUKsoX/gJ7g1/wRE+J4ez+JHwXu7kAxy2+v2EOeu4C3uW/DbaD8a/4LIfsza7YeNbD9p3wzpkt1ourWsOneI3ijLfY7qIbIJnx0SSPZHk8BowCcuor9kP9sf4jfsf+MrzXfCVpbazoetJGmtaFdyGOO8CZ2OsgBMUq7mAbBGGIKtxiw/4LcfBaXT45NU+DnjW3vimZILe4tZo1bHQSF0JHvsH0r9mr/gqhpX7Sf7Q+hfBjTvhQ/hzTdbt7s2+oXerC4mkmhheYL5axqqArG4+83OKr/grr8Kh4A/awufF1nbbLDx7pNvqaso+UXEY+zzKPf90kh/66/UC51XxP49uvDnh1t1/dWNtHpGlQqMMyNM7RxjnGd8zAdOo+tfFf9nCwvP2FtZ/Zt0m2jun0vwOljYKi8T3tpEskL9uWuIUYnHUk1onifV/D1nrVjpVx5Uev6f8A2fe4zloPOimKjB7vAmc54zX/AARc+Eo8TfG7xR8Xr+0D2vgrRxa2bsuNt5eEqGU9yIIp1I7eavqM/G7wlqXj/wCC/j7wJozldQ8R+F9S020YNtxLPayRoc9vmYc1e2V3p15Pp1/bS291aytFNDKhV43U4ZWB5BBBBBr9nH/gr38L/hl8A/C3w4+Ifw38V3XiLwho8GlWsmkLbva3kNvGI4Wd5JUeJiiqGAR+QSOuB+0p8evEv7S/xk174veJ7SKxm1Z0S1sIXLpZW0ahIogx+8QoBZsDcxY4GcD/AIIdf8jV8W/+wfpX/oy5r/guN/yKnwj/AOwhqv8A6Ltq/YD/AOTyvhN/2MCf+gPVf8FRf2ddX+Pf7N82o+ENNkvvE3gS7/teytoU3SXVvsKXMSDqTsIkCjljCAMkgVoet6t4a1rT/Efh/UZ7DVNKuo7uyu4G2yW80bB0kU9mVlBB9RXg/wD4LdeKtN8Dw6b40+B9nrfiq2txGdTtdbNnb3UgGPNeDyXKHuVVsE5xsGMfHL4rfEb4++PtU+NvxGUveeI7toklggaO1i8iOMfZ4AScLHG8Py5LfOGYktk/8Ebf+Tubv/sT77/0dbVVftAf8FWE/Z7/AGnfFHwg1T4bweJfCugLawte2F4YLyK4eBJJuG3JIqlwm35CCrfMeMfE/wD4LA/sv+LPhXr+g2Hw38Z6xqGs6bPaLpGr6dapbOzoy4ncTuPL6E7Qx5HA5xb21xeXEVpaQSTzzOEiijQszsTgKoHJJOBivgL8Ptb+Ff7I/hL4e+JVZdX0TwakN/GzbjDOYC0kee4RmK/8Bqv+CausWXh3/gnt4C8QaiX+y6Zaa3dT7F3Nsj1O9ZsDucA8V+17+1N4u/ay+LN3471zzbLRbPdbeH9HMm5dPtc8ZxwZXwGdu5wB8qqB+zT8cdH/AGd/iNb/ABOuvhfo/jXVdMUNpEeq3Lxw2E4OftCoo+aQfwk8KfmA3BSv/D7z4p/9EP8ACn/gxuP8K/YA/bQ8T/tkaP4z1LxL4K0vw83he5s4YUsbiSUTCZZSS2/pjyx09a/bu/Zm139mb4+a7ojaZKnhXX7qbUfDN6Iz5Utq7bjCG6b4S3lsOvCtgB1z+xj/AMFN/HH7L3hWH4X+LvCg8Z+C7aVn0+Jbv7Nd6YHYs6xuVZZIyxZtjAEFjhgOKv8A/gtv8E47Nn0v4O+N7i6A+WKee1hQn3cO5H/fNfsL/twJ+2dD43mk8Dw+FZfClzaCK0XUDdtJBOsm12fYnO6FwQFwMiqr/gs/8P5vD/7SHh/x7DbFbPxb4ajVpduN9zayPHIPwie2/Ov+CNnjeLxF+yxqHhF5gbnwp4muYRH3EM6RzI34u8w/4DX7a/8AyaL8X/8AsT9Q/wDRLVX7Kn/BWDxV+z18JLL4S+LvhlH4ztNBjePRLyPV/sMsURbcsEuYpA6qSwDDBC4XBxmv2mP2hviZ+1d8Qr34u+OrNIYLWOLT7O1som+y6ZATI0UAY5yzESvljljvIwAAv7Ivx8t/2Zfj94a+MN9oMmtWWlC4hvLKGURySRTQPExRjxuXeGAPB24yM5H/AAUt1Jda/wCCfPj7WFtpLcX1ros4hlxvj36pZNtbBIyM4ODiq07/AILO/tRaZp9rptv4E+FjR2kKRIX0u/LEKoAzi964Ff8AD6v9qj/oQvhT/wCCq/8A/k2v2M/jX4q/aK/Zq8HfGPxtYaTZa14g+3fabfS4pIrdPIvp4E2LI7sMpCpOWPJPQcD/AILe+OYr3x78M/htBOpfSNJvNUnjB/5+ZUijJ+n2V8f71f8ABEv4fS6R8JPH/wAS7i2KHxJrkGnwOwPzx2cRYsvtvumGR1KH0qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr9rv/AIJ0+Cv2vPiTpvxJ8R/EbW9AudN0OLSUtrK0ilR0jnnlDkvzkmcjHT5RX7J//BNvwP8AsnfFGX4o+HfiVrmvXUulTacbW9s4okCyPGxbKc5Hljj3qv2jP+CVXws/aF+L2t/F64+Ieu+HL3X/ACnvLKytIpIjKkaxmRS3ILBFJH97J71+x9+wToP7HfijXtf8KfFHW9dtfEdglteaffWcUcZeN90coKHO5Q0i46YkNeOvAfgz4m+Fb/wR8QPDWn6/oWpx7LqwvoRJHIAcg4PRgQCGGCpAIIIBr4n/APBFD4S+IdSm1D4VfFfXfB8MpLCw1CwXVYY/9lG8yKQL/vM5561pv/BDfVnuQNX/AGkbOK3AyTbeFWkduRx81yoHGeecccGv2T/+CcHwn/ZR8XD4i6J4x8TeIfE/2OS0M928cFsIpMbgsCLnJKg/M7dBjFV+19/wTx+Hv7XvjXRvHuveNdX8OanpWmf2c5sLaKUXMQkaRN2/kFTJJ+De1fsz/wDBLj4Zfs1/F7SvjDp/xD13xFqGjQXCWVre2kUUccksTRGTK8khHcAerZ7V4p8OaX4w8M6v4S1uETadrdhPY3cZGd8UsZRx+KsRX/DkL4Vf9Fw8V/8Agut/8a/Zz+BPhr9mz4PaF8HvCt9cX9novnM17coqy3Uksryu7heM5faPRVUdqr9pL/gkV8IPjJ4ov/HXw08W3Pw81jU5WnvLOKwW80+WU8syQ742hLMcnaxUZ4Qd/hv/AMERvCGma1DqHxW+Nl/r2nRPltN0fSRYGXHZp3kkIU9CFQHHRh2+OX/BJf4HfF3xPpmt+G/Emo+A9P0rRrfS4NJ0mzikhCwl8SZc7i7BhuLEliCxJLE1+xz+xj4c/Y40zxRpPhvxzqniKDxPPbTSC+tkiMDQrIvy7Dg7hIM8fwj8P2v/APgnv4N/bA8b6N438SfEPWfD8+i6V/Z0cFjaxSrIvmvJuJfkHMhGPav2Wv8Agmb4F/ZZ+LFv8WPD/wATdd1y7t7Ge0FpeWcUcZEoALZXnIxVSxRzRtDLGro6lWRhkMD1BHpXx9/4I9fBH4n67c+KPhV4pvPhte3jmSewhsRfafuPJMcJeNosnPCuUGflUAYr4Z/8ESfA2kavb6j8WPjPqfiOyicM+maRpY04S4P3WmaSVtp4yFVTjOGB5r9o3/gmb8Hfj3beC9L0fW7zwHpXgfTJNO0/T9Gso3iMTyb8nfzu3biWJJYsSSSST+x//wAE/wDwn+x74v1zxZ4Z+I+t+IDr2mrYzWt9axRIm2RXDgpySMEf8CNVXxh/4I0fCDx94i1HxP4D+KfiXwnd6pcyXNxBe2yatCsjsWYplopMEnPzSMevNaf/AMENr43xGq/tJW4s1bg2/hQmRxn0a6wpx3+bB9a/Ze/4J6/AH9lrUE8U+HLG98R+LljKLr2tMskluGGGFvGoCQ5GRuAL4JG/BIr4vfDmx+L/AMLvFPwu1LUp9PtfFOlT6dNdQIGeFZUKllB4JGehr/hyF8Kv+i4eK/8AwXW/+NeIf2dNE8Q/suxfsuzeI72LS4vCtp4eGqLCpnMdvFHGJdn3dxEYJHTmv+HIXwq/6Lh4r/8ABdb/AONf8OQvhV/0XDxX/wCC63/xr/hyF8Kv+i4eK/8AwXW/+Nfsn/szaF+yd8Lpfhd4d8T3+vWsuqzaibq9hSJw0iRqVwnGB5Y/Ovj7+zN8GP2mPDcXhv4t+EIdT+yFjY38TGG8sWbG4wzL8yg4GVOVbAyDgV4w/wCCHmhz3kk/gD9oO+s7Ut+7tdY0BbmQD3mimjBP/bMV4e/4IcQrMsviv9o13iDYMGn+GNjMOORI9wcHrxsPY57V+y3+yf8ADj9knwdf+Dvh1qeu38WrXS3d9catcpK8kwQJuVURFQYAGAOwyTjNVX/BZT4CXXjf4Q6F8cNAsTNfeArlrfVAi/MdPuWUbz3IjmCcdhK5PAr/AIIqfG220nxX4y+AGr3axjXol1rRld8BriFfLuUA7s0XlPx/DA5Pt/wVH/5MS+Jv/cJ/9O1nXwi8OaX4x+LHgrwjrcbyadrfiKwsbtEcozRS3CI4DDkHax57VP8A8EQPhy/ik3lv8ePEcfhwvkac2jwtdhc9PtO8JnGRnyffHavEP/BPn9nDWv2dYv2bLLw5caZoVrdfb7bU4JA1+mobdpu2lZSHkZflYEbdmFAUKu3wH/wRbbwb8TdC8X3P7QEOoaRoWrW9+to3hgrLcrDKsnlMftJVQdpUtz67ecD9rD9mbQv2sfhdF8LvEPie/wBBtYtVh1EXVlCkjlokkULhuMHzD+Vf8OQvhV/0XDxX/wCC63/xr/hyF8Kv+i4eK/8AwXW/+Nf8OQvhV/0XDxX/AOC63/xr4b+DvD/wD+DOheCp9fMmi+BdCS3l1O9Aj/cW8XzSyY4XCqSewr4keIvFn7XH7UGsa1oNlNdax8RfE/k6Vav96OOSQRW0bHsI4hGpboAhJ4FfCb4c6N8Ifhj4W+F/h/5rDwxpUFhFIVCmYxoFaRgP4nYFz7saqqqqqqqqqqqqqqqqqqqqqqqqqqqqqtUsU1TTLvTJJCi3cDwswHKhlIJH51/w5C+FX/RcPFf/AILrf/Gv2cPgfpP7N/wX8O/BfQ9cu9YsvDv2ryr27iWOSXz7qW4OVXgYMxX6AV8Xvhl4e+M/wx8TfCvxSZF0zxNp0tnNJEBvhLD5ZUzxuRgrjPGVFf8ADkL4Vf8ARcPFf/gut/8AGvg98PZfhN8LvDHwyk8SXWvL4X02LT4NQuoljlmhiG2IMF4ysYVc99ue9ftLfsRfAL9qiFbz4h+HJbPxDBF5Vv4h0lxb3qIAdqMxDLKgJyFkVsc4xk14h/4IcFrxpPCn7RgW1Zjsh1DwzmSNe2XS4Ac/8BWvCn/BDvw9BNHN45/aE1G9iyN9vpPh9LVhyc4lkmkByMc7OPevgV8E/Bv7PHww0n4SeAZtTl0TRjKbdtRuBPMWlkaRyWCqOXdjgKBz0q6tba+tZbK8gjmgnjaOWKRdyurDBBHcEHGKuv8AgiN8JJbmWW1+NPiuCF3YxxGwgfy1J4XPfA4zX7LP7NnhT9lT4TwfCrwnq13qsQv5766v7uNUkuZpSBuKrwMIkafRM96/bG/Yo8E/tkab4YtPFHifUfD914WnuHtruxt0ld0mVA8bB+2Y0P4e9fCL/gkF8KPhT8T/AAv8Sz8UfEOtv4X1OHUYbC6sYUjmlibfHuIycBwre+Kr4z/BX4c/tAeAL74a/FHQI9V0a9YSKN2yW2mXOyaFxzHIuThh2JByrEHxT/wQ7tJdZeXwT+0LLbaS7ny4NV8PCeeFc8AyRzIshx32J9K+An/BKD4BfB6x1C88Vanf+OvEl/p89kmo31utvBYrNE0byW9sCwWTDthnZyvBXaRk/wDDkL4Vf9Fw8V/+C63/AMa8MaL/AMI34a0nw79ra6/suxhtfPddrS+WgTcRk4J25xnvX/DkL4Vf9Fw8V/8Agut/8a/Zl+AOj/syfB/Svg/oPiC81qz0qe5lS8u4ljkczStIQVXjgtj8Kr45fAH4VftG+CZPAfxY8MRatp+8y20qsYriymxgSwyjlGGfoRwwYEivE/8AwQ602bWDL4M/aHubTSnl4g1Tw4txPCmf+ekc8ayNj/YQcds8fss/8EyvgZ+zTrtr46u7y98ceMbPm01LVIEigsX/AL8FuuQj/wC27Ow/hK81q/8AwRQ+FOqareaknxp8UwC7uJJRENPgIj3MTtB74zivgx8M7X4NfCnwv8KrHWrnVrbwvp0djBeXKBJJUThSwXgYGBx6VX7Rv7Onw+/ai+HD/DH4kS6tDpv2xLyGbTLv7PNDOiuquCQysAJG+V1Zc4OMgEeJ/wDgh1pstxJN4M/aJubeAn93bap4bWZgPeWOdAeP9gfhXgL/AIIh+B9Pv4Lv4l/HPV9btkYNJZaPo6aeWx/D5skkxIPqEU+mOtfCj4RfDn4H+CrP4e/C3wtaaDodkSy28ALNLIcbpZHYlpJDgZdiSQAM4AA/bD/YF8H/ALYniXw94k8S+P8AWPD0vh6xktIorG1jlEod95Yl+h7V+zL/AMEv/AX7Mnxg0r4waD8Udf1q80qC5hSzu7KKONxNC0ZJK88Bs/hX7X//AAT38G/tgeN9G8b+JPiHrPh+fRdK/s6OCxtYpVkXzXk3EvyDmQjHtX/DkL4Vf9Fw8V/+C63/AMa/4chfCr/ouHiv/wAF1v8A414Y/wCCL3ww8MeJdJ8S2/xp8UTy6TfQ3aRPp8ADtE4cAkHoduKr9on/AIJZfs4/HfWLvxdosV94B8SXrtJcXOiIhtrmU5JkktmG3cSckxmMk8nJJNXn/BDfXFuith+0jYy2+Rtebwq6P7/KLlh+v5V4C/4Ik/CbSLuK4+I/xk8S+JY42y1vpunxaUknPAYs07YxwcEH0Iq1t0tLWG0jd3WGNUVpGLMQBjJPc8daqv27vgPe/s6/tOeK/CttaNbaLqd02s+H3QEKbO4dmVUP/TNxJF9Ys9wa/ZM+NVn+0H+z14K+J8Nykl9f6ckOqoGyYr6H93cKR1GZFZhnnaynvX/BZL/k7m0/7E+x/wDR1zX/AAT5/Yp8CftlaD8TrHxR4m1bw/qvhaTSG0q/slSVUE/2wSrLE331PkxkYZSCvXBIPwC/4I/fBX4U+KbHxj8RvGWofEW90yZZrWyn09LGw8xSCrSwh5GlwQDtL7D/ABKw4r9rr/glr8O/2lPGt18UPCfjS48D+KtSwdTI08XlpfuqhRIYw8bRyEABnDEHGduck/sIfsKH9jGLxfdX3xFh8W3/AIuFmrNHpP2NbNLcz8KTK5ff5wJ4XGwdetftH/8ABLH4f/tIfGjxF8aNb+KniDR73xD9m82ytbKGSOLyLWK3G1m5ORCGOe5Nf8OQvhV/0XDxX/4Lrf8Axr/hyF8Kv+i4eK//AAXW/wDjX7OH/BLH4f8A7N/xo8O/GjQ/ip4g1i98O/avKsruyhjjl8+1ltzll5GBMW+oFf8ABXH41w/DT9mCbwBp98setfEa9XT40VsOLOIrLcuB/dIEcR9p6/4I5/AWfxx8cdR+NurWTHR/h/aNHZSOnyyajcoyKBnrshMrHHKs0Z4yKqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/bL/Yg8Kftmf8If8A8JN441Xw7/wh/wBu8j7DbRy+f9q8jdu39Nv2dcY/vGvgD/wSj+HvwA+MHhr4waP8WPEOrXnhueSWKzubGFI5S8LxkMVORxITx6VX7W//AATh+G/7WnxCsfiVrPjXWPDWq22mJp9wLC2jlS6VHZkdg/RgHK57gKO1fsu/8E0fC37K3xZtfit4R+L/AIh1OaOynsrrT7qxhSK7hlUZVipyMOsbjHdBWtaJo3iTSLzw/wCIdKs9T0zUIGgu7K8gWaG4iYYZHRgQykHBBGDXxi/4I0fAjxxqk+s/C3xprXw+luWLNZC3Gp2UZJJPlxu6SKOenmkDAwBUH/BDfXXugl1+0lYR22TmSPwq7uB2+U3QHp/Fx71+zb/wSn+Ef7P3j7Rvijc/EPxR4l8SaBP51g22KxtUcqVJaJQ7twxGDJjBOQar9sb9ijwR+2Rpnhm08UeJ9R8P3fhae4e1vLGCORnjmVBJGwftmOM+2PevhH/wSA+FHwq+J/hf4ln4o+ItbfwvqkOow2FzZQxxzSwsHj3FecB1Vsd8Y71Xib/gix8Itf8AEeq65Z/FzxLpkGo301xFYw6fAUtVdywjUnkqoIUewr9kj9lLwh+yH8O7/wAAeFNcvdbOqaq+o3eoXsKRyu7RxxqmE4CqsYwPVmPeq/aq/wCCWvwc/aO8UXvxE8N+ILvwD4t1JzJqFzZ2a3VpfSH70sluWQiU93R13EksGYk14J/4IfaPbaxDc/Eb4+3V/pkbjzLPR9CFrLKOM4mklkCdx/q2618aP+CT3wC+J9l4S0rwlquo+A7HwnpsljHBptuk5vA0hk82Z5DueTcz5YkkggcAAV+x7+wP4U/Y78S+IPEXhj4h6x4g/wCEisY7WeC+tY4hH5b7lZSh68sMEd6/bH/Yu8L/ALZGmeF9M8S+NdU8PL4XnuZYWsbeOUzGZYwQ2/pjyx09a+CP/BJb4dfBD4seGPixpXxb8R6nd+GL4XcNpcWMKRzMARhipyBz2qq/aV/4JQ/Av47eIbvxt4L1i7+HPiLUJWlvn0+zW6srqRjlpWtiybXJ5JjdFPJKkktXgX/giB4cs9WjuviT8er/AFTTo5PnstG0RbOSVR/02kllC55GPLP19Pjh/wAE1fgj8Wvhn4J+FPhe7u/AeieB57ma0XS7dJnuXnWNZHmaTLO58pCXJJOMdAMfsrf8E1fBf7KfxVT4qeGvijr+tXI06exezu7OKON0l2kkleeCin8Kqv2iP+CSnwg+OHjjXPiVoPxE8S+FPEHiK8lvL8SImo2rTyMWZ1jco65J+6JNoGAAoFQf8ENdSN/suf2lbX7EDxInhJvMYf7pusD/AL6OPev2Zf8AgmX+z7+zhrtp44db/wAa+LrFxJaanrKoIrKQdHgt1G1XHBDOXZTypWtUsU1TTLvTJJCi3cDwswHKhlIJH51/w5C+FX/RcPFf/gut/wDGvgv+zponwW/Zys/2c9L8R3upadZ2OoWi6lcQqkzC7mnlZiq/LlTOQP8AdFf8OQvhV/0XDxX/AOC63/xr/hyF8Kv+i4eK/wDwXW/+Nf8ADkL4Vf8ARcPFf/gut/8AGv2Of2LvC/7HGmeKNM8M+NdU8RL4ontpZmvreOIwmFZAAuzrnzD19K+L3wX+GPx58HT+BPit4Rstf0iY70SdSsltJggSwyLh4pACRuUg4JHQkH4gf8ERPA2pX8958Mfjjq+h2zsWjsdY0hNQ25P3RLHJCQo6cqx45J61o3/BDe7a43eIf2kIVgUj5LPwsWZxzn5muQF7dm79K/ZJ/YY+F37H51a/8EeIfEes6tr1vHBqN1qc8flusbFl2RRooXBJ6ljyeeaqv+Cr3wEuvjF+zLceLdBsTca58OLltXjVFy72WzbdoPYIElPtB74r/gjt8bLb4fftBan8LNYuxDp/xG08RWxd8KL+23SQjnj5o2uFHcsUHev21/8Ak0X4v/8AYn6h/wCiWqvE/wDwRG+GWo+K21Hwr8bNf0bw/JMXOl3Gkx3kyKT9xLjzEAA5ALRscYySQSfDX/BPr9mzwx8AdY/Z3tfDN3caL4haObVNUnmU6jc3Mf8Aq7jztuFdDnaqqEG5ht+dt17/AMEOFfXidN/aO8vRWclVn8Mb7lE5wuRchWPQbsL1J28YP7RHwJ0T9of4Ia98DtT1i50PTdcS0Q3VnErvAtvcxTqFVuMEwhfYGv8AhyF8Kv8AouHiv/wXW/8AjX/DkL4Vf9Fw8V/+C63/AMa/4chfCr/ouHiv/wAF1v8A41+zl8EdI/Zt+C3h74M6Lr11q9j4cF15d9dxrHJJ59zLcNuC8DBmK/QCv20vizdftJ/tYeL/ABP4bMmp2dzqi6R4eitx5nn28GIITGByfNYGQD1lr9lb4K2/7Pf7P3gv4TIsZvNH04NqUiYPmXspMtw2R1Hmu4B/uhR2qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqvEnhzQ/GHh7UvCnibTINR0jWLSWzvrOdcpPDIpV0YehUkfjXw1/4JLfCT4R/ErQvil4F+Lnj2y1bw7qKXtkrvasnynmJ/3QLIykowyMqxGea/4KN6Q+t/sTfFSyjgWYx6ZBc7T2EN3DKW/AR5/Cvg7qqaD8XfA+uO0YXTvEmn3BMmdoCXEbc47cVVVVVXx4+Edv8d/hXrnwnvvFereHrDxDEsF9eaXsE7QBgzxKXVgFcDa3HKlh3r9mT/gmh8Ef2YfiSnxT0HX/ABD4k1m2tJLew/tgwlLJpBh5UEaL85TcmTkAO3GTkVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX7Wf7F3wr/AGwNI0Oz8d3ep6TqPh6d3stV0oxrcCJxiSFt6sGQkI2CMgrwRls/sl/sieHP2QtC13wv4M8feJNd0fXLqO7Nnq/klbWdV2NJF5aLguoQNnOfLTGMHP8AwWq0h7X9pvwrrCQKkV/4It0Lj+OSO8u92forRiv+CHGqpD4h+L+hFk3XdlpFwAc7sRPdqce374Z/Cqqqqq/am/4J5eBP2tfH9r488f8AxO8YWB06wSxsNO042629tGCWYgPGxLMzElieyjoor9nn4A+BP2aPhbpvwp+H0U7WFk8k093dFTPe3Ehy80pUAFiAq8AAKqgcAVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVzbW95by2l3BHPBMhSWKRAyupGCpB4II4INL/wR9+Bul+Pl+IHgv4m+O/DV3aasNS0uCxltimmyLL5kaxF4mbahCgbiTgDJNftSaPNrn7L/wAWNFEa3Nxc+BtXSJSuA0v2KXYcdvmANV4c1VNf8PaXrkbIV1GyhuFKZ2kOgbj25qqqqr4l+DJPiN8P9f8AAUXiPUNBHiCwlsZNR0/b9ot45BtcxlgQGKlgGxxnI5FfAb/glP8AAX4EfFLRfitZ+J/E/iS/0BzNYWmrNAYI7jGEmIjjUlkzuXJwGweoFVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVqulaXrum3Oja1ptrqFheRNFc2t1CssUyEYKujAhlI4IIxQ/Z7+ASkMvwP+H4IPBHhm0GP/IdVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV4q+Gnw48d3EF343+H/hvxDPaoUgl1XSYbtolJyVUyKxUE84FeFvhj8NfA11NfeCPh74Z8P3NxH5U02laTBaPImc7WaNQSMgHB4zVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVzbW95by2l3BHPBMhSWKRAyupGCpB4II4INf8M9fAH/ohvw//wDCZtP/AI3VpZ2lhaQ2FhbRW1tbRrFDDCgRIkUYVVUcAAAAAcCqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv8AhdPxU/6HjUv++x/hX7O2o+Ltf8G3HiPxZrd1qDX10VtBMc7Y4/lJH1fcP+AivGNrq174V1W20G8mttSa0kNpLEcMJQMrj6kAfQ0fjR8VQcHxxqWR2LD/AAr/AIXT8VP+h41L/vsf4V/wun4qf9DxqX/fY/wr/hdPxU/6HjUv++x/hX/C6fip/wBDxqX/AH2P8K/4XT8VP+h41L/vsf4V/wALp+Kn/Q8al/32P8K/4XT8VP8AoeNS/wC+x/hX/C6fip/0PGpf99j/AAqvjv4x8WeBPCdt4h8KzQKy3iw3ImhEg2Mpwfb5gB/wKvht4nl8Z+BNG8SXDI1xd2w88oMDzVJV8Dt8ynivi14yn8B+AtS8Q2LR/bIwkdqJF3AyOwA474BJ/Cv2f/itr3xGXWbPxNLbtd2BieHyovLyjbg3HsVH/fQqR0ijaSRwqopLMTgADqa+CvxT8ZfEvxhra3slsuhWEbPCiW4VwXfESluv3Vb8qqq8RfE/wh4Z8Sad4Q1G9mbVtTlhjgt4oGbHmvsVmbhQM++eOlVVV401/Xbf9pjTdIt9bv47FryyDWqXLrGQVTIKA4578VVVVVXia/uNL8N6rqdoyiezsZpoiwyAyoSMj6gV+z98SfE/xHstan8SzW7tYywrD5MIjwGDZzjr90VX7QXxT8W/Dm80WDwzPaot9FM03nQCTJUoBj0+8as5Xns4JpCN0kSs2Bjkjmqr9oXUL/TPhZqV5pl9PaXCTQBZYJDG4zKoOCDnpX7PeoX+qfCzTbzUr6e7uGmnDSzyGRziVgMkknpVVp3xY+IF38b5/Ac3h6NdIjuJItogbzEhUHbOXzjDYB6Ywcdaqv2hdQv9M+FmpXmmX09pcJNAFlgkMbjMqg4IOelfs96hf6p8LNNvNSvp7u4aacNLPIZHOJWAySSelVVV8bvGur+AfAz67oUsKXhuooUMse8YbJPH0Br4BfF3XPiFcaro/iqS3+32scdxbeVD5e6I8Nx7Epz/ALdV8cviNffDvwtbz6G0R1bUbpYbVXTfgDl229+ML9XFfs+/E3xL8SLXW28Sz2skunyQiIQwiPAcPnIH+7/OqqqqvG3xP8IeAJrW08QXswur0Zt7eGBnZxnBOfugZI6kVVVVftHajqGlfDKe70u/uLOcXkIEsErRsAScjKkHFfAS+vdS+EuhXuo3k91cSfaN8s8hd2xcSAZJyTwAKqvE/iTS/COg3fiPWZJFs7JN0hjjLtyQAAB6kgenPJA5qb9orx7r7NP4A+FV7e2SNgTvBNcFvqIhhT7ZNeBv2lLbVdcTwx460BtCvZJBEkwZtgkPRXVgGTPryOecDmq+I/x207whrK+EvDejzeINfYhWt4SQsTHopwCWbH8IH1Io/tC+NPCt/bxfEz4bT6bZ3LYSeEOpUewbIcgdRuBrSdV07XdMttY0m7S5s7uISQyp0ZT/AC9weQeDXjb4jeEPh/arceJdWSGSRSYbaMb5pcf3UHbtk4HvXgT9ow+PPH9l4RtfC32S0vWl8u4kut0gVImcEqFxk7OmeM9TVfEr40eJPA3id9A0vwFNq0CQJILlXcAlhyOEI4+tf8NMeNP+iTXP/f2X/wCN14e/aX1zWfFmleFrvwCLSTUL2G3ctcvviV2AL7Sg6A5/CqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqoopJ5UghQvJIwVFAySTwAK8F+HYfCfhPSfDcIX/QLVI3IGNz4y7fixJ/GgVYZVgR2xXxo8LDwj8SNZ02KPZbzTfabYAceXJ82B7Akr/wGqqqqqqvi5oH/AAkvw28QaUqb5DZtNEuOS8f7xQPqVA/Gv2Udd+3eCNQ0KR8yaXfblHpHKuR/48r1+1Hfz6pJ4W8AafJm41O981kHrkRx/mXf8q8GxQ/D79prUfDsQ8uz1WJ4os8DDxrMv47l2/UmvjN4gPhr4Za/qMcm2WS1NvF67pSIwR7jcT+FfsueHRpXw6fWZI8S6zdvICRyY0+RR/30HP4143+NuoaD4nufBnhX4f6rr2p2gXzigKou5QwICKxYYI5O3v6V/wANHeJvDd9Bb/EX4ZXulW1w2FnTepA7lVdQHx6BhV34kt28I3Hi3QoTqsS6e93axQkg3OELKo4JBbAHQkE9O1eOPiBrOv8AxX0PxfeeCbzTrywktGi0uV2MlwY5SygExg/MflHyn8elfDv4s+IvG+vto2q/DTUdBhW3aUXVxK7KWBUBcNEoycnv2r4gfGe68J+Ij4Q8P+BtU1/VhCspWEEJtYcEbVZm6YPAHvU37RXjLwzcwnx98K73TrKdsCZS6EeuN67WPHTIrQNd0vxPo1pr2i3QuLK9jDxOBjjoQR2IIII7EGviLd2th+09Y317cRwW9vc2UkssjbVRVjUkk9gAKv8A9oDxHq7TzfDj4YatrenwMVF+8UgRyDztVVP5ZzyMgdK+HX7QuieMdYXwvr+ky6Dq8j+XFHK++OWQfwZIBVv9kjrxnOBUjxxRtLK6oiKSzMcAAdST6VrX7RMl7rM2g/DHwZeeJ5rfPmXMe7y+OMqqqSV/2jtHpnOa0T9ouez1yHw/8TfBt14alnxsuH3BBk4BZWAIX/aBI+lKyuoZGDAjII71Xjf/AJEvX/8AsF3P/opq/ZD/AOQb4m/672//AKDJVfte/wDIR8M/9cLn/wBCjrTf+Qba/wDXBP8A0EV44/aK8N+GtVbw74b0u48Saoj+W0ds+2NX6bQwDFmHooI7ZzT/AB4+Julp9t174KanFZE5LqJkMa/7RMZGfrjPtX7RjvL8HdQkliMbtJbFkJyVPmrkZFfDX4waT4B+FeiaLZ6Zca3rt1JcNFp9r1VfNbBcgEjPYAEn6c1pv7Ud9YawmneO/A0+lQyEZdGcSRKf4jG6gsPoR7A9Ks7y11Gzgv7G4Se3uI1khlQ5V1YZBB9CCK0/42fb/i23ws/4RnZtnli+3/bc/ciaTPl7O+3H3u+a1jV9M0DS7nWdYvI7WztIy80rnhQP5noAOpJAqT9pHXPEGpS2Xw5+G1/rMMDfNM28kr6lEU7AexLfhXxQ+Mtr4x+Her+E9e8O3vh3xBG8DizugcSKJVyVJCkHHOCOnQnmv2bf+SR6X/13uP8A0a1eN/jbqGg+J7nwZ4V+H+q69qdoF84oCqLuUMCAisWGCOTt7+lf8NHeJvDd9Bb/ABF+GV7pVtcNhZ03qQO5VXUB8egYVpep2GtabbatpdylxaXcSywyr0ZSMg//AFqr9rC5MXw80+2VsGbV48jHVRFKT+u2oXX4W/FbwV4hQ+VputaNYpOxPyhWhWF8/Qqrmq8VTN8S/wBo7SPDcR8zTvC+HmGMruTEkhI928uM/Sv2UJI7fW/F+nRFVQiAqpPOEaUf+zfyqqr4l/Gjw78OZotJFrLq2tTgGLT7dsEZ+7vbB257AAk+mOauf2gPiJoSLqnin4QXtnpRI3SnzIygPTLMmM89DjPtXhXxtofjXw2vibw5MbiAq26JhtkjkUZMbDnDdPXqCMgivjH8QNZ8ba/o2o6r4JvNBksYyscFw7MZ/nByN0a/Toa8EfGbxP4t8UWXh/UfhTqmj2915m+9nlkKRbY2YZBhUclQvUdfwqvGviyy8D+GrzxPqFpc3MFmF3R26hnJZgo6kDGSMntT/HX4oX0X9o6L8FdSawHIeRZnLr6ghB+gNfCz42aH8TJptKawl0rWLZC72kr7w6g4JRsDODjIIBGe/Jr9p3/klU//AF+wfzNfs7/8kc8P/wDbz/6Uy1Vahp9jq1jPpupWsVza3MZjlhlXcrqeoIqC3t7WCO2tYY4YolCxxxqFVAOgAHAFftc6PpqRaBrkcSJfSPLBIwGDKgCkZ9dpzj/eqDxJc6L8KIPFmpLuurXQEupVc5LSiANg49W4/Gv2WPD66rca78Q9WJub97g28U8nJDsN8rfU7k59M+tfEzwtaeMfA2r6JcxK7vbPJbsw/wBXMoJRh6cgD6EjvX7JviSe/wDDWreGLiUuNLuEltwx+6kobKj2DIT9XrxH8O/CHi3WLDW/EekR38+mxskCS8x/MQfmXo2McA8cnj00yOOL9rvyokVESWUKqjAAFg2AB6VVa5rFh4e0e81zU5vLtbGBppW74UZwPc9APU1+z7o19458ba58YPEMW5/OaOz3DIWRh8232SPag9m9qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq+Afhg+J/ifpUbx77fTWN7PkZwI8Ff/Hyg/GvFeuweGPDWqeIZyu3T7SSYKTjcyqSF/E4H41+zn4sn8U/DqOK+uDLeaZdSQSs33mBO9WP4Pj/gNftbeFxJZaL4xgj+aF2srhgP4Wy8f4AiT/voVVVVVVUyq6lGAYEYII618ClbwV8a/E3gVzthmWaOFT/EYn3Rn/v2WP41sHj39qQ5Hm2fhWAY9jEv9Jpf0r9ouNvCfxL8IfEC3BUAqJCo6mCQMc/VZMfQV+1brby6L4f8K2DGWTU7prjYh5YIAqD6Eyce614V0KHwz4Z0vw9Dt26faRwkj+JlUAt+JyfxrxZ8QPBHgNfN8Ta5bWUsw3LEFLyyAcAhFBYjtnGPevi38d/hv458F6h4XsrDVZrqXa9rPJaoqRSKwIbJfcMjIyB0Y+tfsxX1xefCyGCZyy2d9PDFnsvD4/NzXxe/5OS8If8AXfTf/Sk1Xivxv4O8Dwi+8T6za2LTLhFYFpZQP7qKCzAEntgZ96+Jn7QHwx8WeENW8LwWWrXcl5butvKbRAiSgZjY7nDAbscgZ68V+yhfXFz8P7+zlcslpqjiLP8ACGjQkD2zk/ia+LuiQeJP2i4NAumIgv7ixhlIODsZEDY98ZqysbPTbOHT9Pto7e2t4xHFFGu1UUDAAHpX7Vvhe1sLjRvHWnKLe8llNtcPH8pdlG6N+P4gAwz6BfSvjF49vT8BdL1SCYpdeJ4baGZk4Kh4i8oHsdpU+zV8I/ix8IPh/wCCLHR5dVlj1CRfOv2WykYtM3UFgvIUYUduK+OXxS+FfxB8EvY6TqUk+r2cyS2W6zkQ8sFcbiBgFCTjuVHtXw/+Jln4e+A+k+MPEcV5cx2CfZJBbqrSNtlMSEbmAPyhc5PrX/DWPw5/6A3iP/wGh/8AjtS+LNN8dfCbU/FOkQ3MVpe6XeeWlyqrINgkQ5Ckjqp79MV+yH/yDfE3/Xe3/wDQZKr9r3/kI+Gf+uFz/wChR18WvE914S+Ed7qmnymO7ktYreBwcFGk2qWHuFLEe4FfsveCdOsPCB8az26SahqssixTMuWihRimFPbLBifXj0qv2k/+SR6p/wBd7f8A9GrX7L3hnSrDwCviSK3RtQ1SeQSTMvzKiNtCA+nyk/U+wr9pnQNP1P4YXesTwobrSJoZIJNvzDfIsbLn0IfOPUD0r9nDUJ9Q+E+mJO5Y2k00Cljk7Q5I/INj8K0D/k7eX/r9u/8A0lkr9rbxFc2+n6H4Wt5SsV28lzcKDjdswqA+2WY49QPSvh/4QsPA3hLTvD1jbxxvDCpuXVcGWYgb3PqSfyGB0FftReHdN1D4enX5YE+3aVcx+TMF+bY7bWTPocg/VRX7Nv8AySPS/wDrvcf+jWrxZ8QPBHgNfN8Ta5bWUsw3LEFLyyAcAhFBYjtnGPevi38d/hv458F6h4XsrDVZrqXa9rPJaoqRSKwIbJfcMjIyB0Y+tfsxX1xefCyGCZyy2d9PDFnsvD4/NzVftdXBXQPD1oC37y8lfGePlQD/ANm/nX7Q/hA33wo0TWoolNx4fSFZCo6RSKqNj/gQj/DNeDfiLb3Pwat/Hd/MJZLDTXN1uOC80QKkfVmUY/3hX7LmgXNzaa78RNV/eXmsXRijkZcFgDvkYezOwH1Sv2cmNp8V/GGmMEUiOYlB22XAHHt838qqrq4is7WW7mbEcEbO59ABk/oK+EnxC8G2fjrW/iD8Rr511K5bdZf6O82xnJ3kbQdu1Qqj2JFXn7Q/wZ1C0msL3V5Z7e4jaOWKTT5SrqRggjb0I4r9lvWIbfxr4l8M6ZcSTaZNA1zbM4xkRyhFbB5BKyDP0HpX7U3/ACOvhX/rgf8A0aKqrqS1itpJr2SJIIl3yPKQFUDnJJ4AGM5rXP2j/hXos7W0Wr3GpyIcN9hty6g+zNtU/UEitL8ZaL4g/aJ0nxX4RtbmyttSvokkjnjWNizp5cpIVmHzZJ68kk1+07/ySqf/AK/YP5mv2d/+SOeH/wDt5/8ASmWqqrm5t7O3ku7ueOGGFC8kkjBVRQMkknoAK1KS6/aP+LEFrYJIvhbQuHmIIzFnLN7NIVAA7AA44NfG6NovhH4iis0CBLRQFUAAKHXIA9Nua/ZXCf8ACs5tm3P9qzbseuyPr+GKYqFJYjAHOa/ZFEp1zxEVJ8oWkO7njdvbH6ZqrD/k75v+u8v/AKQtVV+014tu76TS/hVoBMt7qk0clzGh5bLYij/Fvm/4CvrXgbwpaeCPCWm+GLQqwsoQJJAMeZIeXb8WJP0xVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX7JnhgW2iav4unj/eXs4tYCR/BGNzEexZgPqlftS+JP7J+H8OhQyYm1q7VGXOMxR/Ox/768sfjX7KXiUad4yv/AA1NJiPV7TfECessWSBj/cMh/AV8VPCw8ZfD/WdBjj3zyWxkthjnzU+dMfUqB9Caqqqqqqr4qPF4B/aH8P8AjORvKtL8QvcSdNowYJPyj2n8a/Zg0+41QeKPiDfpm41i/KKx6g5MkmPYtIv/AHzX7UGgjVfho2pomZNIvI5sgc7G/dsPpl1P/Aa8J3zfFL4s+B4ncyw+H9HtTcnqN8KbyT9ZCgNSuY4mkVC5VSQqjk47CvgRpOl/FT4ha54k8dJHqd1DGLiO2uPmRmZ8ZKngqgAUKePmHHSvjfc6N4e+Emt2wSC0S5hFvbwxIEDSMwAAA9gT9Aa/ZY/5JjJ/2FJv/QI6+MzpaftF+Erm5YRxK+nOXbgBRctk/Tg/lVfDOw0/4sfGzWr/AMcoLz7Mk00NnOflOyRURCvdUU/d7kc55z8T5dC8L/CzxGiQW1jbyaZPbQxwxrGC8iFEUKMd2H61+yV/yJWsf9hQ/wDopK+K+sWvh79pC21u+J+zWVzYyzEdQgRNx/AZOKtrm3vLeK7tJ45oZkDxyRsGV1IyCCOoIr9qvxJb6lc6H4E0t/tN8kxnnhjG5lZgFiXj+I5Y49Cp7ivjl4JvLX4F6Lp9vFvl8Mi1Nxt5+VYjG54/2mBz6A18FtJ+GPjX4f6Zcnwf4cudQsYFtr8SadC0gkQY3OSuSXADZPXJ7g1/wrX4c/8AQgeG/wDwVQ//ABNWWjaPptgmlabpVna2UZJS2ggWONSTk4UDA5JPTrX9mab/AM+Ft/36X/CrzTra70240vYscNxC8TKq4ADAg4H41+zdra+CfHeueAPEMgtbi9YRIJDgfaIWYbQf9oMceuB6iq+M19D8VfjHovgjQZhcw2ZW1lkQ5VXZt0zAjsqAZ91NfHvw9c+IvhZq9rYRGSe0VLmNFHJEbAtgf7m7iv2XvGOn6p4HHhB7iNL/AEaWQrCThnhdi4ceo3MwPpxnqKr9pP8A5JHqn/Xe3/8ARq1+zb/ySPS/+u9x/wCjWr9oj/kjniD/ALdv/SmKv2Yv+SVW/wD1+z/zFaB/ydvL/wBft3/6SyV+1xoNxJBoHiaGMtDCZbWdsfdLYZPzw/6V4G8V6f428K6f4i0+4SQXMK+cqnmKUAb0I7EHP6Hoa/ai8R6bp/w//wCEeknQ32q3EflQhvmCI25nI9MgD6mv2cnMfwf02RULlZbkhVHJxK3Ar4EaTpfxU+IWueJPHSR6ndQxi4jtrj5kZmfGSp4KoAFCnj5hx0r433OjeHvhJrdsEgtEuYRb28MSBA0jMAAAPYE/QGv2WP8AkmMn/YUm/wDQI6r9rhmutQ8JadGV3FbkjJ7sYgM/9814h0Gz8Q+HL/w3cKFgvrR7c4H3Ay4BH04I+lWnifVtH8Aa78I3hk+33etxL5AGT8uRIo9/MiiGPc14J8NQeD/CWleGbfaRYWyo7AcNJ1dvxYsfxr4PY0/9o7xdZhQglbUI1Vuu37QrDH4KDXijxn4X8GW0V34n1mGwinYrEZASXI6gAAk9RWl6nY61ptrq+mT+daXkSywSFGXejDIOGAIyPUVf2iX9jcWMjYW4iaNjjoGGD/Ov2fdP8K2fi/XPAXj7w/pNxqJcJai/tUlIliLCSNS4PJBBGOu2v+Fa/Dn/AKEDw3/4Kof/AImtJ8JeFNAuGu9B8M6Tps7pseW0so4WZcg4JUAkZAOPYV+1WRD4u8LXMvyxLA+WI44kBP6EUrJIgdGDKwyCDkEVX7WviDU7LSdD8O2srx2movNLc44Evl7Nq+4Bckj1218OfAHg7wp4c01tD0qzeaS2jke+MQaWdioO/eecHqAOBnivGmqWOqftQ6R9gnWUWl9Z28jLyBIpG4D6E4PuDX7Tv/JKp/8Ar9g/ma/Z3/5I54f/AO3n/wBKZasvGXhTUtbn8N2HiGwuNUts+baxzAupX7wx6juO3equbm3s7eS7u544YYULySSMFVFAySSegArxj4w8T/H/AMQnwB8P43g8OW8oN7fsrKJAD99/ROPlTqxGT0+XwN4H0L4feH4fD+hQkIh3TTP9+eQgAux9TgcdAMCvF+hjxN4V1fw9lQ2oWUsCFuisykKfwOD+FfsqeIY7A674A1I/Z76K5+0xQycMWACSrj1XYnH19DXxQ8U2vg/wJrGtXE6pIts8VspOC8zgqgHrycn2BPav2T/DU+n+FdU8S3ERX+1rlY4Cw+9HECNw9izsP+A1Vh/yd83/AF3l/wDSFqrWNVsdC0q71rUphFa2MDTTOeyqMnHqeOnc18DNMvfiZ8StZ+LOvwkx2spFqrcqsrDCqPURx4H1ZTVa/wCJvD/hayGoeItYtdPt2bYrzvt3N6AdSfYVZ3lnqNpDf6fdRXNtOgeKaJwyup6EEcEVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVXw48Mjwf4F0Xw80YSW1tF88D/nq3zSf+PM1ftD+BfiP478XWp8P+Gri70zTrQJFIssahpGO5yAWB6bR0/hrwT8J/i/4T8XaR4jTwZdYsLtJJAs8WWjzhx9/upYfjVfEb4FeO7fxlq8vhzwvc3mlT3TS2skJUja/zbcZyApJXkdq1vwl4o8NMF8QeHtR0/dwrXNs0at9CRg/hVVVVVV8ZPhGvxVs9Mjh1WPT7nTZJCsrwmTcjgblwCO6qfwr4c+DIvh/4OsPC0dyty9rvMs4j2ea7MWJxk+oH0Arxb4fh8V+GNU8NzyBF1G1khDlc+WxHytjvg4P4V8Hfgh/wq3UtQ1W61uPUp7uBYIiluYvLXdubqxzkhPyqvEv7O2qW/iiXxZ8MfFx0K4uHZ2gbcgjLHLBXTnaT/ARj8OBF+z/quvW11d/Efxzc69qTWc0NirhjBZyOhAkCkjcQcHHyjjnPGPhJ8Pbr4Z+GJvDt1q8eo77x7hJUhMW0MqDbjJ7qTn39q+MHwYsfijHa31vqX9narYoUimMe9JEJztYZBGDyCOmTwc8fC34e/ETwjq0t54v+IEutWf2NoILQzSyKjF1If5z1AUjp/F1rxt+z1eX3iqTxp8PPFDaFqM8hlkjO5VEjfeZHQ5UHnK4IOT0HFWPwG1zXZPtnxR8d3PiCSGNxa2gLeRE5UgOQSN2M5wAucck9K+D3wxu/hdpF/pNxrUWord3InRkgMWw7QpHU56Cte+B1t4l+KL+O9Z1C2udNmgMMumvbklgbcxZ37uCCdwIHGB35qX4FfEbw+H0/4f8AxXvLPSnY7LW5d1MKnspXIz7gLmvhx+z/AKV4P1geKvEery6/rauXjllQqkTnq+CSWf8A2ifwzzVzb295byWl1Ck0MyFJI5F3K6kYIIPUEcYrV/2bLrTNXk1v4YeNrvQJJM/6OzPhc/wiRCG2+xDfWj8GfjDrSfZfEvxmuUtmGJEtPMbcPQjKA59/1qwtWsrC2snuHnaCFYzK4+ZyoA3H3OM1VfFH4D+HfiNdDWre8fSNZCgNcxRB1mAHG9MjJHADAg465wMH4F/GS4g/sm7+MVwdOxt2i5uGJXpgqSOMdt2OlfDH4MeGPhkj3VnI+oapMmyS+nQKQv8AdRRnYpxzySe56YIBGCM+xrxd+zTp15rB8ReAfEM/hy8LmTykViiMepjZSGj+gyPTAr/hS/xj1iFbHxF8ZbhbQHDLbGVyw468pn8Sa+JHge48feCZ/CS6qttLOYibl4t/KMGJ2gjrj1718MfBUvw+8HWnhabUUvXtnkYzLGYw29y3TJ9cV8SfB8nj3wVqPhOG/Wze+8rE7R7wmyVH6ZGc7Mde9fCzwJL8OfCUfhmbUkvmSeSXzliMYO49MZNaf8FLqx+MLfFE+IYniaeaX7F9mIb54Wjxv3dt2eleIvD2keKtGutA1y0W4s7tNsiHg+xB7EHBB7Gl/Zw8beGdRll+HnxKm0+0nb5kkeSJwPRvL+VyPXA/Crb9nK2k0PVzr/ii51jxJqsHlLqd0pcW3IPyqWJJOMEk9OBjJz8MfBU3w+8HWvhafUUvWtpJG85IjGG3uW6ZPrivEv7O2qW/iiXxZ8MfFx0K4uHZ2gbcgjLHLBXTnaT/AAEY/DgRfs/6rr1tdXfxH8c3Ovak1nNDYq4YwWcjoQJApI3EHBx8o45zxj4SfD26+Gfhibw7davHqO+8e4SVITFtDKg24ye6k59/aq+KPwdufiP4l0bXF1yC0h0pQGge2Mhl+fcedwwMADpVXPwF0q5+Ky/EttY/dC5W5Om/ZMgzKmA3mbv74D/d61WgfBq50L4sX3xJTX4pIb2SdjZm3IZfMH9/d2PtX7QdvceO/if4T+HFg7FhGXlKf8sxK3zsf91It30NWtrb2VrDZWsYjht41jjQdFVRgD8hVfE74E+HviJdjXLW8k0jW1ABu4Y9yy7em9cjJGAAwIPTrgVB8Ivjrbr9hj+Mri0UYVzJK0mOPXkf99V8NvA+p+BdKurLVvFl5r9zeXPnyXFyCCGKhSBlmJ+6Opr4tfCnTvinpFvazXrWV/YMzWlyE3hd2NysuRlTtXvkEA+oPwz+FnxI8Ha5aXGvfEWTUdIsonjj09ZpWVgVwvDHAA6gc4xxVfEz4a6N8TtBXR9Tle2ngfzLS6jXc0L4weP4lI6jjtyCAa0j4G/FzTrZdAT4vS2uioNipbtKXVPRVJG0ewbFR/s36fpPi3w74i8M60LWHRTC1xFPCZJLyRJS7uz5ABYEL0wMCvip4El+I/hKTwzBqSWLPPHL5zxGQDaemARXw38HyeAvBWneE5b9b17HzczrHsD75Xfpk4xux17V4K/Z5uPCfxLPjSTxEk9lbSyy2sIQiVjIGXDnpgBjyOuOgzVfFfwJqPxG8LL4b07XjpQa6SWd/LLiWNVb5CARxuKt/wABFWn7K/iXT0aPT/iYbZWOWWG2dAT6kCSv+GZ/Gn/RWbn/AL9S/wDxyvA/h278KeFNP8PX+qNqNxZIyvdMCDISxOeST0IHXtXxF+Atr4p19fGPhPXpfD+uhg8ksanbI44DjaQUbHUjr6ZyTF+z34u8UX1tcfFH4jXGq2tqcrawFzn1wzYC5A5IXJ9e9adp1jpGn2+laZbJb2tpEsUMSDARVGABVW/wbuYPjKfiqdfiaIu7fYvs5DfNbmL7+73z0qvit4H1f4h+Gf8AhGdM16PS4Zple6doDIZVXkJgEYG7BP8Auivh94Ms/AHhKx8MWkiym2UmaYJt86Vjlmxz34HoAB2qvjZ8Jbr4qadpqadq0dld6ZJIUEykxyK4XdnHII2DB+vrXw/8Jr4G8HaZ4UW9a7NhGwaYrt3szF2wOwyxAHpiqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq1zXdH8N6bJq+u6hFZWcRAeaU4VSTgfmSKsfix8NNSkENp440cuTgLJdLHk+27Gaubax1Sza2u4ILu1uEw0ciCRJFPqDkEV+0F8IrXwFqEHiHw7AU0XUpChizkW02Cdo/2WAJHpgj0qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqtfAPhm08X3XjtLN5Nauo/LaeSVmCLtVcKvReFAyBnr6mqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/aE8UN4Z+GOoLBMY7nVGWyhIOD8/L/+OK/5iv7T1L/oI3P/AH9b/Gv2Sbq5udR8Si4uZZdsFvgO5bHMnrX7Sf8AySPVP+u9v/6NWq/ZO8WaldHVvB95dSTWttCtzaI7E+T821wvoCSpx0zk9zXx80yDVPhPryTKN1tEk8bEcqyOp4+oyPxqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv2svEv2zxFpPhSCUlNPt2uJgDx5khwAfcKuf+B1X7If/ACEfE3/XC2/9Ckr9pP8A5JHqn/Xe3/8ARq1aWN7qEggsLOe5kPASKMufyFfsz/DTX/CsWo+KPElhJYzahEkNrbTKVlCA5ZmU8rkhcA88H2r9o7XrfRfhZqNo0yrcarJHbQKTy3zBn49Nit+Y9aqqqqqvEfj3wb4RuIrTxJ4is9PmnTfHHM2Cy5xnA7ZFWtzb3trDeWkyywzxrJE6nhlIyCPYgiqqqqqqqqq8RePvBfhOdbTxH4lsLCd0DrDLL85U5AbaOcZBGcdqsvjV8KtQmW3t/G+nq7HA84tCP++nAFQzQ3MKXFtMksUihkdG3KwPQgjqKqqqqqtX1bTNC06bVtYvY7WztlzLNIcKgJA5/EgV4d8V+HPF1rJe+GtYttQhhk8uR4WztbAOD+BFVP408K23iGPwnNrlsusSgbLPJMhyMjj6DP0qqqqqqqqqqqq0fxd4X8Q3l1p+h6/Y31zYnFxFBMHMfbnHbPGemaqqqO7tJpPJiuoXcfwq4J/KqvL2y0+A3V/eQW0K9ZJpAij8TxVhqem6rD5+l6jbXkYOC8EqyAH6gmqp2RELuwVVGSScACrfxL4bvLkWNp4g02a4JwIY7tGfP+6DmqmurW3IWe5ijJHAdwufzr+0tN/6CFt/39X/ABr+0tN/6CFt/wB/V/xqtQ1/QdJYR6rrdhZsei3FykZ/8eIq3uba8hW4tLiOaJvuvGwYH6EVVVUkkcMbSyyKiIMszHAA9Sasdd0PU5Wg03WrG7lQkMkFwsjKR1yATiqqWWKFPMmlRFHVnOAKimhuE3wTJIucZRsjP4VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV8RfEp8YeONZ8RCTfHd3TeQT/zyX5Y/wDxxVqv2Q/+Qj4m/wCuFt/6FJVxdWtr5f2q5ih86QRx+Y4Xe56KM9ScdKrxZ8cvht4QNxb3murd3tuzI1nZoZZN6nBUn7qnPHJFfFP4oav8UNbS/vIha2VqpSztFbcIlPUk92OBk8dB6VVVVVV7eWun2c1/ezpDb20bSSyOcBFUZJPsAK8Xxan8TbDxd8X7zzEsrK6gtbCNj91C4AX/AICpXPYmQmvhBqI1X4YeGboNkrp0cJPvGPLP/oFUzKqlmYAKMknjFWuoaff7vsN9b3HlnD+VKH2n3weKqpNS06K6Wxk1C2W5f7sLSgOfouc1VVVVWu6ZY/Ej9p59E1SEXWmWS+VLGxOCsUG4rx0HmEj8a8Qfs+/C/XNNls7bw9HplwVPlXVozK0bdjjOGHsR+XWv2XPEGsWmv658Pby4ee0s4nniBJKwukio4X0Dbwce2e5qm1PTFuhYtqNqLk8CEzKHP/Ac5qqqqr9qHVhp/wAMGsFb5tTvoYSM9lzIT+cY/OvgNqV18OvibL4F1aUiHXbOBlLcATGISxn8Q7p7kj0qvBIHi79qLXtaLeZHo6z7GxwDGq24x+ZP61U+qaZbTra3Oo2sUzfdjkmVWP0BOaqqqrm6tbOEz3dzFBGvV5XCgfiaBBAIOfSrvUtOsWRL7ULa3Z/uCWUIW+mTSskiB0YMrDIIOQRVXWqaZZSJDeajawSP9xJZlQt9ATzQKsoKkEEcEVTatpK3QsW1S0FznAhMy78/7uc1JGssbROMqwIIBxXwv+B+kfDDWtQ1qz1m5v5buIwQrLGE8mIsGIOCdzZVeeOnTmqqvjl8TJfBGhR6HoEhk8R61+6s44xueJScGTHrk4X1b1wa+AujXPh744S6JeyiS5sra4jnI/56BRuHvg5Ge+M14m1+x8LeH7/xFqLH7Pp9u0rgdWwOFHuTgD3NeAvBmtftDatfeNviBqt2mkW0xitbSB9o3YBKJnIVFBXJxlieuQTXxM+GV/8ABCa0+IXw21i9ht45liuYZX3eXnpnGN8bEYKt3xyc8eB/FVr438J6Z4ntFCLfQBnjzny5Adrr+DAiq+J3wfvPiZr1lNeeLryy0WC32zWEWW8yQMTvUE7QSCBkg9BxXib9lLwuNFmk8KatqkepwxFoRdSJIkzAcA4VSCcYyOBnpX7L3j7WfEWnal4W1y9lu20tY5bSaViziNsgoSeoBAxnn5iOgFfFH4L6X8Ur+xv7/W7qyaxhaJVhjVgwJzk5r/hkbw5/0N+pf9+ErVvhRpr/ABZtfhl4W1a5vwGRb26kRR5Jxukxjj5Ux1/i4rxno2p+IfC2o6No2ry6ZfXUBW3u43KNG4II5XkA4wcc4JrT/wBlHwYLUnXfEOs317IP3k8TpEu7uQpVj19Sant9e/Z2+Klhpmn61NdaNftFJJG3CzwsxRgyjjeuDgj29SKqqnmht4XuJ5FSOJCzs3AUAZJ/KrBvEf7TPja+gudXudO8I6UwPkRHBKkkJx0MjbSctkLzj0PjX9mbRNN0OXWfh/qOpWusabGZoUefd5xUZIBABV+OCDjPYZyP2fPiZe/EDwzPYa5P5uraOypLKRgzxsDsc/7XysD9Ae9V8cvGd98R9V1Hwp4auD/YPha3kutQuEGVmmQYAyOoDEIPcs3IAr9lj/kmMn/YUm/9Ajqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/ZD/AOQj4m/64W3/AKFJX7UUssHw2t54JWjkj1aBkdCVKkK+CCOhr4H/ABUh+I3hwW2oTKNd01Al4nTzR0EoHoe/ofYiv2lPhH9vgk+Ivhy1zcQJ/wATSGNf9YgHEwHqo4b2APY5qqqqqv2k/F90unWHwz8PlptV8QyoJYoz83lbsKv/AANwB9Fb1rxZ4AtPC/7Pep+ELVVdrLTvPndR/rJUYSu//fSnHoAB2r9mTURffCm1tQwP9n3k8GPTLeZ/7Ur9qnVBZfDm305X+fUNRjQrnGUVWYn8wv514E8BeJvi74R0mPxHrF5onhGws47e0sLMhZL1kUB5nJBGC4OMg+wH3m+IHhO4/Z88d6JrPhPWbuW2nXzVEzDewRgJI32gBlII7DqfTNajew6bp9zqNwwEVrC8rknGFUEn9BXwWk8ceIrvW/Dfg+5/s+41t45dT1jktaQKXyE772Zz3zx25I+J/wCzno3hfwbfeLNI8Q6pdajYKJrj7Y6Msw3AMRhQVPJPJPTHvX7PnijU/FXw1tLjV7h7i5sp5LUzSHLSKuCpY9ztYDPfH414qx4v/ap0rSwC8ekPAOegEMZuD+G4kfWq8M48X/tVanqPLxaQ8+Qeg8mMQD/x8g/WqpmVVLMwAUZJPGK+DPjvwnp3xM8TeN/FmuR2Qv1m+zmSN23mWYOcBQcYCgfjXjH9ojQHtX0b4aR3XiDXbpSlsILV9kTHjcQQCxGcgAEHuRXwB+FOoeAdMu9b8SKBrerYDx7t5gjBztJHVmJyevQe9fEDxx4t+I3jlvhL8Nb9rKC23LqupRsQV2nDgMOQq/d4wWY46dfEf7L/AIO0zwnf6hb+INU/tKytZLg3M7p5bMiljuULkKceuR6nv8DdX1n4g/CLX/DWtzS3jQpNZQTTMWZkki4Uk8kqT17Aj0r9kbUvM0TxDorNzbXUU4Un/nopU8f9sxXx/wDH48EeBprWyuNmqayGtrXacMi4/eSD0wpwD2LLX7Pvw8HgjwVFf30G3VdaC3FxuHzRx4/dx+2AckerEdqqv2opZNb8ReDvBVsx8y6lZmUdSZXSNP5P+dftO+G7jQtS8PfEbQl8mS0ZLZ3Uf6t4zvhb9GGf9lRWg+L9O13wXa+NIWAtZrE3LgHOzapLrn1Uhh+FfspWc1/N4q8W3Y3S3c8cQbH8R3PJ+rJXxa+I3ifVvFdv8IvhnP5eq3PF9eo2DACMlQw+7hfmZhyOAOc0P2U/CculSf2l4m1efV5ELPelkCeZjrsIJK56gtn3Ffsr+Kda1TTda8M6leSXltpDxNays5fy1feCgJ/h+QED614Zx4v/AGqtT1Hl4tIefIPQeTGIB/4+QfrVfA3Hiv44+K/FxBeKIXDxE/w+ZKFT/wAcDCv2ttRh83wtpErN5e+eecL12/u1GPw31p3wl8UfGeZPHXj/AFubTLG4YPpmkxpv8q3/AIepAXKgdsnOTjpXxy+K+r+Hbuz+H3gQF/EOqbVMiAM0Ac4RVB43t6noOe4I0f8AZb0W9svtvjrxNq2oazcrunlhnAVGI7F1ZnI9T19BXhS/8Q/Ar4uW3w8vtWmvvD+rSRrCsnACykqkij+Eh8q2ODg+2Pj7481LwF4EN1okvlahqNytrDMBkwgqzM498LgehYHtXgj9nHw34r8KWPifxbr+r3epazbJdNJFOo8sOoZRllYs2CMknr2rwV4U074eeE7fw9b3zy21gJHa4uCASCxYk9gADj0wK1PxZ48/aA8V3fhXwLqT6R4YsTi4u13IZEzgM5HJ3YO2Pjjr0JB/ZK8Fmy2HxLrZvNv+tJi2Z9dmzOPbd+NfF+S38GfBLUtNtJW2xWEWnwljlnDFYzk+u0sT+Nfsz6P/AGZ8K7S6K4bU7ua5II9G8sfpGPzqq8W+KNK8GeHrzxJrM2y2s49xUfekY8Kij1JwB9a+DXhbVfiJ4qufjV42gzvlI0i2bJVNvAdQf4UHyr6tluoBPw8/5Oh8Qf8AXe+/nX7UWovZfC5rVHIGoahDAw/vABpMfnGD+FfAXT4tP+Evh+OMDM0LzOR3LyMf5ED8K+NFlFqHwr8S28oBCWLSgEZ5jIcfqor9ma/1K4+FGr2mnbHvbO+nFmsnC7miRkB9t5OfrXwBu/jDP4q1ePxyNZbTRAxc6mrgLcbl2iLd227shflxj2rxV4k07wf4fvPEmrLM1pYoGlEKBnwWC8AkdyK8WftRabqNi2jeANMuhqV4PKju9QaO3jgLcbhliMj/AGioB5ORxXwF+Fj/AA70Ce+1K6guNT1jY8pgffHFGoOxFbox+YkkcHIAzjJr4geL7TwL4Q1HxLclWa1iIgjY/wCslPCL+JIz6DJ7V+zJ4TuZbXU/idrhaW/1qZ44JJByU3Zkf/gTjH/APeq8W/tGeBvCGr3+gXdjq1xfWEhjdIoU2lsdmLjj3x+FeFNG1n9oD4g2/wAQ9eays9E0lkENjHcrLKVRiyoyjkZbJZmC5HAGORVV4zsbvU/B+uabYZNzdabcRQgDJ3tGwXH4kV+yPq+npDr+gPIqXzvFcIh4LoAVOP8AdOM/71Xl5aafaS319cxwW9uheWWRtqooGSST0GK/Zguo5Pif4hj08H7DNYTSR8EYAnj2f+OsevP618efiVc+GNLh8HeGC8viPXsRQJDy8MbHbuGOdzH5V98ntzcfDS1+GvwC8RWUixvqt7Y+bqEy85fIwgP91ckD1JJ71+yx/wAkxk/7Ck3/AKBHVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV+yH/yEfE3/AFwtv/QpK/an/wCSYxf9hSH/ANAevBvi3VvA/iK08R6LLtntm+ZCfllQ/eRvYj+h7V4R8VaN468N2viDSJBJbXceHjflo26MjD1B49+vQivj38Jm+H+u/wBsaNbn+wdTkJhAHFtL1MR9upX2yP4Saqqqq1TUrLRtNutW1GdYbWzhaWaRv4VUZJ/IV4csvin8UPHGo/FfwdbWqS292Y7d7xlKwjZhUUMCCVQrk46nPU1q2j/tSalpV5p2oXuizWt1byRTxhYcujKQw4XuCRX7I2omXw/4g0jdn7NeRT7fTzEK/wDtKv2pZ5db8VeEvBls4EkoZwP9qaRY1J/74b8zWm6faaTp1rpVhEI7azhSGFB/CigAD8gK+OZPij44eEfCKEvHF9nWUD+EyTEucf7iqa+Nur/2L8K/Ed2rANLaG2UHv5rCM4/Bya/Ze8Pw6X8NxrPljz9Zu5JGfHJSM+Wo+gKuf+BGv2jdWGl/CfVIlfbJfyQ2yHOOrhmH/fKtX7P+knSfhNoaOpEl0sly+RjO92Kn/vnbXwNB8U/HHxX4tfLxwi4eIkfdMkoVB/3wGFanqFrpOm3WqXsgS3s4XmlY9lUEk/kK/ZUtJtV13xV4vu1Jml2Rhz3aR2d/1VPzqq+JWr/2D4A8Qaqr7Xg06YRk9nZSq/8AjxFfs6fDDwj4k8EXOt+KPD9rqE01+6QNMpO2NVUcc/3i35V8afB9v8GvEuh+Nvh68umpcSMr26SMUWRNpxySSjgkFTxwfXA1nxCtp4JvvFdqOINKkvYweekRcfyr4KWvxfMOqa78ObPT5/tMyw3lxeMhbcBuwNxzj58nHXj0q++F/wAfPH8X2Dx149srHTHOJbe1GSw91jVQw74Zq8EeCtE8AeHoPDmhRv5MRLySSHLzSHG52I7nAHsAB2r4FofDfxs8aeFn/dxbbgoDwCI5xsOP9xya0qJ/j38bZtYmXzfDHhkgRDHyyqrHYP8Ato4LH/ZGO1VVa8B4t/as02wI8yHREjyey+VEZh/5EcD618RvCcXjfwVqvhplUy3MBNuTxtmX5oznt8wAPsTXw38cT6b8FfHnhS9lZJ9OgbyEfgqtwfKdRnphzn6vX7N2mpovwkttQkBH2+4nu3GOQAfL/lEDXwqm+KmveKdf8YeALKxutSkbN7JdsnyCZ2bC7iOpQ9PT3q+8A/tG+NoW03xR4307S9Olys0drgMynqMRqCwxxhnArwH4C8PfCfwvNZae8k21TPe3Ui/PMyrycDoABwvb3JJP7KlpNquveKvF92MzS7I957tI7O/6qv514512Pwz4O1nXnl8s2VlK8Zzj95tIQD3LFR+NfskaP5HhrXNdZMG8vUgUkdREmePbMp/L2r4t2cfjz9ojRPCEh3wWyW8M6f7HM0n4lG/lSoiIERQqqMBQMACvhiieLf2lfEWs3/7x9Na8lgDjpsdYE/JGH5VXxrdfFPx88M+G9OIkltPssM+08oWlMjZ+iFWr4jeAtM+I/hafw3qMzQEuJba4VdxglXIDY7jBII7gnkda0Txd8RP2d9atPCfjSIah4bmYmB0O8KmfmaFuCME5Mbevbdk/HrxCdO+EWqXumzg/2lHFBFIp4KSsNxH1TcPxr9m7QrXSPhXp95Ei+fqsstxO2OSQ5RRn02ov4k1X7WmsC28IaRoaOA9/fmUjuViQg/hmRf0rwBo/9g+B9B0cj57XToVk4/j2Asf++iaqv2h18feJfF9poVn4N1vUvDul+XKVs7SVkupGALEuqkAgHaOuPm9as/jP8TNOs4NPsP2f9Xgt7aMRwxJBcBUVRgADyumBXhfxt4p034wan4ssfA95e6tcyXBl0hEkMkJc/MCApb5e+V/KvjZeaz4z+A1n4k1nw9Po95BqKTT2U4ZXgAeSEEhgDyGU8j+KvgPfRah8JfD0kTDMUDwsB2KSMv8AQH8a+NV9Fp3wq8SzysAHsjCM+shCD9WFfsqae9r8OLq8kTH23VJHQ+qqiL/6EGqri2t7uFre6gjmicYaORAyn6g14q+FPgPxfp0tjqPhyxikdCI7q3gWKaI9irAZ49Dkeor9nXxBrHhzx3rfwp1C8a5tLV7gQAkkRTQybW2+isMkj1A9TVfHrVb74ifETRfhH4fn3LBKrXRAyFmYZJPqI48n/gTDtWj6TY6FpVnoumQiK1sYFhhT0VRgZ9Txye5qr/wz4b1QSDUvD+m3fnf6zz7RH3/XI5r4zeF4vgt4x0Txv4AkbT0vXfdaqx2KybSy4z9xw3K9Bg9OMeJX8Y+JvBVre/DvVrXS9TvUhnimu0DKsbAMQQUfnBH8Nf8ACK/tUf8ARSvDf/gOn/yLX/CK/tUf9FK8N/8AgOn/AMi18OtM+JWm2N3H8SfEOn6tcvKDbPZoFCJjkHEac59jXjn9naPVfEL+L/AXiOTw/qkkhldFDBPMPVkZSGjzySORz2rV/hX4og0W61z4x/FC81PRNLiM8unW0jolwV+6pY45JwPuk88Eda/Zb0e6t7LxP45WxLgp9ntYY1x5jKDI6r/5DAxXh3U/idpnj29+Iuv/AAj1/W9UuMmASWM8aWpPHyjyz0XCj0Gep5rx/wDF34j674N1bSNV+DuraXaXVuUmvJYpwsIyPmO6MDH1Ir4C/EDxjoOjWvhvSPh3f6rp11qn73Uoo5SkO/YrZKoV+UDJyR+FVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX7If/IR8Tf9cLb/ANCkr9qf/kmMX/YUh/8AQHqvgb8VZfh14iFnqMzNoWpOFu0PPkt0EwHt0PqPUgV4h0DRfGnh650PVI0ubDUIcblIPXlXU+oOCD9K8f8AgfVfh94mufDmqLu8s7recLhZ4j91x+WCOxBHaqqqr9pjxlcywaf8LtA3S6hrMkb3EcfUoWxHH9WcA/RR618P/CFp4F8Iad4ZtdrNaxZnkA/1kp5dvxJOPQYHaq/ZrP8AYnxN8XeFshQkcg2YxzDPs/8AZzWtY8X/ALVtlZgK8OkSR4J5A8mLzT/5EJH1qvDGPF/7VWqal8zx6Q8/HYeTGLcfhuIP1r9q/VxaeBdP0dGw+oagGYeqRoxP/jzJXwz0j+wvh74e0sqFeHTojIAOjsoZv/Hia/a41VhpXh3w7FuZrm5luCijPKKEXj38xv1rw7pSaH4e0zREHGn2cVuP+AIF/pXw48Xn4BeO9b0PxZpV29vPiF2iUFxsY7JFBIDKQT37j0xXxD+JGu/Ffwnq0Pg/SrzTPC9havNqGp3abGuSgysCAEjBbAPJOOuOjfss6Utl8NZL8qN+o6jLIG/2VCoB+at+daZ8c/GWo/Go+BxpFsNLGoyWXkCI+cqoWHmls9eNxGMY/Oq/ae1cab8LpbFZAG1S9ht8dyATIf8A0WPz96+CWkf2L8KvDloVAaW0+0NxgnzWMnP4OBX7WdoZfAmlXirkwaqqE7eQGik/IZUfpXgqK18U/CbRrK5bdDqGgxW8+B6whH/XNfD3xnqf7PXiHVPB/jrR7ptPu5BJHNAufmHAkTJAdWGM85GB3BFal+1F4JCrb+G9H1jWL6XiKBIBGCfQkkt+Smo3EsayBWG5QQGXBH1HavjZNqnhD4yazc6IrCbXdPEaBAS2JohE+3vklWx7mvhF4Bi+Hfgqz0d0T7fMPPv3A+9Mw5XPcKMKPpnvXx9+J/iH4baTpbeHLWBp9SmkVrieMusQQKcAZHzNu79lP4fDbxJqPjDwNpHiTVrNbW7voC0kaqQuQxUMoOSAwAYezCq+A4/4Sj4x+NPGhBeJDKsRP8PmzfJ/45GRVftAeHrrwV4/1Z9PJj0/xTbiZgOjZkV5F/7+xhvowrwJoy6H4H0LRHjGbbToY5VI6tsG/j3Oa8P6pqv7NnxC1Ow1zSLm40DVTtjmiGTJGrExupPBZQzAqSDz16Zv/wBqT4exQKNJsdY1G6k4jgS3Cc+hJP8AIGnE3i7wSwe2eyl1nSyDDKCGgaWL7rZAOQWweO1fBz4n2/wZv9a8L+NdGvo/NmUv5KAyQyoCCCpIypBGCD27g5Hxa8ceJPij4LvtY0rSrrR/B+mNG3m3S7ZNSmZ1VVUA42ru3cEj5eucAfs9aWumfCXRSVAku/NuHx33SNt/8dC18LVHiz9pDxJ4iIDR6ebpon69GECfmhP61XjOy174HfGCb4jWelS3ug6tLI0zIMAeacyRk/wsH+Zc8HAHrjW/2pfCR0sr4T0nUr/Vp1229vNAEVHPTdhiT9Fzn1HWvgh8LfEFrrd18UPiEjjW78u1tBKMPDv+9Iw/hYg7Qv8ACCQR2Hif9oDwj4O8X3vhPxBp+pRfZFQrdRRB0fcoJ4JBwM4yMgkGviX4ul/aD1vSPCPw90a7lhspGkmu7iPYq7gBuOM7EAB5PJJwB0z8Q/h8fEvwtuPBGmspntrSJbJn4y8O3aPbcF257bq+EnxutfhrpLeAPiDpOoWrabK4gdYctGGJZkdCQR8xJBGevbGa8H/HXwr488VReF/DlhqMhaF5HupogiLtHQDJPPviv2gv+Kq+MHhHwUrB4gIVkHXaZpsPn/gCKa+MXjbVPh/4GufEOjWcc92JUhQyqWSLccb2AI47D3Ir4G+Pde+Ing6TWPENrFHcwXjwLLEhRZlCqdwGeoLEHHHH1qqr4ef8nQ+IP+u99/OvFXh2y8W+HNQ8N6gcQahbtEzBclCfusB6ggEe4r4eePNV+AOqX/gP4h6Vd/2dJMZrW4gTcAeAXTONyMAvQ5UjpkkD4j/ETUvjtcWnw/8Ahto97JZ+est3cTJsDY4UtgnZGMk5PJOMDjnQNCtvh34Eg0bS4Huxo9i7BEGGuJACzEDsWbJx718DPjT4s+IvijUdD8QWFr5Mds1xDJbRFPJIZV2NknIIbg9cj8vFn7QsHgbxnfeHfEXhDURp8DKLa8iGGmG0biFfAYZJwQ3avEH7VnhhbJoPCWh6le6jKNsAuY1jjVjwM4Ys3OOAOfUV+zv8NPEWm6nf/EbxjBLBfakjLbwzLtkPmMGeVl/hJIAAPPJ46V478W2fgfwnqPia82sLOEmKMnHmynhE/FiPoMntX7MnhS81K51b4q68zTXeoSyQ2zv1Ylt00n4thR9G9ar4p+O7/wCHnh6HXbDw9Nq+bpY5o4wwEceCWdmAO3oACRjJFW37V/w+ktBLdaTrkM4HMKwxuM+zbxkfXFaxN4u/aZ8Y2C6do02neHdNJTz5OVhRiN7lujSMFACDpgdsmrS1gsbSGytU2Q28axxr6KowB+Qqqrxt+0R4M8E61e+HLvTdWutQsmCusMSCPJUMPmLg4wR2rUbn4p/tH3sFjbaU+geFIpA7SSAlGx/EWIBlb0VQAOM4614X8M6V4P0Cz8OaLCY7Syj2pu5Zj1ZmPckkk+5qvjT/AMkq8S/9eLfzFfssf8kxk/7Ck3/oEdVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVf8KG+Ef/Ql23/f6X/4uv8AhQ3wj/6Eu2/7/S//ABdeFvh/4O8EyXEvhbQ4rB7tVWYo7tvC5x94n1P514l8K+H/ABhpw0nxJpqX1osgkETsyjeAQD8pB7mv+FDfCP8A6Eu2/wC/0v8A8XX/AAob4R/9CXbf9/pf/i60jSNO0HTLfR9JgMNpaJshiLs+xewBYk4HYdhx0rxT4E8I+Nlt18U6HBqH2Qt5JcspTOM4KkHBwOPav+FDfCP/AKEu2/7/AEv/AMXX/ChvhH/0Jdt/3+l/+Lr/AIUN8I/+hLtv+/0v/wAXX/ChvhH/ANCXbf8Af6X/AOLqmtLV5RO1tE0o6OUG4Y96qktbWOUzR28SyNnLhACc9eaW1tUlM6W8QkOcuEAJ/GqjtbWKQyxW8SO3VlQAn8amtba52/aLeKXb93egbH0zQAAAA6dBU1ra3DBp7eKRl6F0BIqtR0TRtY2f2tpFle+X9z7TbrJt+m4HFC1tltxarbxCFRgRhBtAHbHSooYoUEcMaIo6Kq4A/Ck0zTY719Tj0+2W8kXa9wIVEjAdi2Mke1VNbwXChJ4Y5QDkB1BwfxpVVVCIAAowABjFTQQ3CeXPEki5ztdcjNRxRxII4kVFXoqjAH4Vfabp2qQ/ZtSsLe7i/wCec8QkX8jmrDRdG0kY0rSbKzyMEW9usf8A6CBVSWttLIs0lvE8ifddkBIx6GqvtO0/U4PsupWNvdw7g3lzxCRcjocHIzSoiIERQqqMBQMACqhtba3z9nt4o93XYgXP5VU1rbXO37RbxS7fu70DY+maq7srO/gNrfWsNzC3WOaMOp/A8VYaBoOlNv0vRbCzbnm3tkjPv90Cqv8AQdC1SZLjU9GsLuWP7jz2ySMv0JBIqS3gliEMkMbxjojKCBj2pI440EcSKqqMBVGAPwqK1tYGLQW8UbN1KIFJ/KqlhiniaGeNJI3GGR1yCPcVY+HtA0uY3Gl6Hp9pKw5eC1SNj+IANVqGk6Vq0Yh1TTbW8QdEuIVkA/Ag1Z2Fhp0AtdOsoLWFekcMQRR+A4qtR0LRNYKnVtHsb4p937TbrJt+m4HFWen2GnQ/Z9Osre1i/uQxBB+QprW2eUTtbxGUdHKDcPxq5tra8ge1u4I54ZF2vHIgZWHoQeCKtrW1srdLSyt4oIYhhIokCKo9gOBVVS2tqkpnS3iEhzlwgBP41V9p2n6nB9m1Kxt7uL/nnPEJF/I5qy07T9NhFtptjb2sQ6RwxCNfyFVaaZpunvNJYafbWzXDb5mhiVDI3q2Byfc1c2lrewm3vLaKeM9UlQMD+Bqw8OeHtLk87S9C060k/vQWqRn8wBVTQQ3CeXPEki5ztdcjNRxRQxiKGNUVeiquAPwqquPC3hi7uPtd14d0uafOfNks0Zs+uSM1HFFDGsMMaoiDCqq4AHsKqqp/D2gSXr6lJoentdyHLztaoZGI9Wxk9BVVUkccqGORFdSMFWGQfwqKCGBPLgiSNc52ou0Z/Cqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr/9oACAECEQE/AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAACUWtwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADEUL4KjWXmn97a/UAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAwuF9vlCp9Wm163bQAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAWADBU/Z4anDskZthvYYuSWz1Xz/AOCAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAGYYZ0K1ujSf3/8ABuABa5ZAwMDh/b4iFPu/w6hxFhuejGqt46fJ/wDB/MAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAw9F1q0aa6uxxDhlKhGrH7Onyf/BAOVsUUMGMDhygnOdV9NF89wxNGNajKm+qJxcZOL3QAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHD9FTxTm/sr/gGNjGdF05fa0+/YatoCSAYMYGS0VTwce71/r5BiKns4c3mvxaX6mdUVTxkrddf6+YAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHDdueovJGatpUbfzxKtnOVu7AUi6YwYwMLDkoQj2S/IM5ly4Kb9PzRxBaUqVRdV/X5gAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAFScKcXKbsl1ehlvEmVZliJ4fCVlOcVd2T22unaz+TYAAAAAAAAAAAAABk2KjQxS5no9DM61Olh3OW629egAFzcYwIfAg4grKGFUOsn+Wv+Rm7vhcN/h/SIAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAZnnmXZbTc8VWUbdL6/KK1f3GaeLDjVlDA0E4raUm9fPlVrfff0Mzz7MsznJ4mtKSbvy3fKvSN7I4W4Rz547D4mFN0o3T5nZNLrpvqtLWs766AAAAAAAAAAAAAAA5Skld7AABcAws1OhCXdL8g4lg3Tpy7Nr77f5Gce7ChT6qK/r8AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAM943ynKZSpTnzVEr8sddezey+eqXTa+deJeaY33cL+5hbprL/wKyt8kn5lDC43Ma9qcZVJyfm235v/ADMm8K8TXpc+Pqezf8qSk7d7p2XpqZLwjlOUwj7GmpTX2pJOX320+VvzAAAAAAAAAAAAAAAAAAAySsqmDiusdP6+QY/BrFUlB90/8/wM5rKrjJW2Wn3f8EAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABtJXZn3HuU5XHljL2s3sotNfOWy/F+RnnHucZneCn7Om/sx0+97vz2XkZPw1mmbythKTaW7eiXze/orvyMk8LcHh5OeYT9q+iV4r563f4L1MLhMPhaSpUIKMV0SsvwAAAAAAAAAAAAAAAAAAADIcYqNZ05PSX5/wDBDHYqOGoSqPfp6km5Nt7gAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADaSM58RsnwKlCjL2tRdI7X85bW9Lmc8b5zmilCpU5ab+zHRW7N7vzu7GRcIZpnFS1GHLDrKV1H5d/RfOy1Mk8N8pwCU8QvbTX83w/KP8AncjCMYqMVZIAAAAAAAAAAAAAAAAAAAAMPxBXpw5ZpSt16mNx9bFzvUei2XRAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHEXG+WZRGUObnrL7C7+bs0vz8jPOLs1zdKNepaKv7sdFr3XX5mTcC5zmkY1IU+Sm/tS0Vu6W78tLeZk/h1k2BjGVWHtZrrLa/lHa3rf1EklZAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAGfcX5Vk6ca871P5Y6y+fRfNryucR8fY/NaSpU37OGt0nv2u93putFfoZJwtmmcSTw1P3L2cnpFfPrbsrsyDw9yzK5wrVP3lVLd/Dfuo/hq33AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAMyz3LctssXWUG9k9/uWtvPYwHGWWZhmSwGFbm7N8yXu6euv4WAAAAAAAAAAzLNcFl1H2uLqKEfPr6Ld/JHEniZicWp4fL1yU3pza8781Z+7f5vzRlHD+aZ3Wf1eDlrrJ7J+bfXy1fkcP+G2X4Dkq4t+1qLX+4vl1t5/ciMIxioxVkgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADMczweXUfbYqooR2u+/ZdWZl4k5JhLKlJ1W/5VovVu34XMd4s4ud1hcPGPnJuT9bK36mP4rznHU5U6+Ik4y3Wy+5W08tjAZXjcfNwwtJza3sr2PDjhbMcFjZ43F0+SPK4pPR3bWtu1k97b6AAAAAAAAADaSuzibxMhhqksPlsVOS0c38Kf91dfW9vVGIxWa59i4qcpVaj2X+SWiXeyS6nD/hap01VzSTT/AJItaf4nr9y+8wuEoYSjGjQgoxWySsgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAMy43yLATdOpXTkukU5fK60v6tGaeLNNRtl9Bt956L7ovX70Y/wARM/xcXFVVBP8AlVvxd2vkynTxGMrqEU51JO3dtsy3w+z3GyadL2aXWen3JJv8LeZgvCSFk8ViflGP6t/oYLgfIcI4Shh05R6ybevdpu34ehGEIK0VYAAAAAAAAA4g4qy3Jad8RK83tFayfy6LzdvK5xRxjjc7qtX5KS2gn+Mu7/BdOrfCnh9iM1gsTim6dLpp70vS+y82nforamT8O5blMOXCUlF9XvJ+revy28gAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABtJXZmfiJkWDj7lR1JXtaKv8Ai7K3o2Zn4qZjVqNYOnGEel/el69vlZ+rM14izTNEli6zkl00S+5JL8DLOE85zKCqYag3F9XZL5NtX+RlnhRjKk08bWjGPaN2/wAUkvxMF4Z5Dh2nOMqj/vPT7o8v4mHwWGw8FCjTUUtkklb7gAAAAAAAAAKtSFKDnNpJatvRJHE3idBRlh8rV3tzvp/hXX1f3PcyvJ8zz3FONFOcnrKTvZecpf030TOFOAcLlD9viGqlbo7aR9E+vm/klrcAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADOPEHJsvvGM/azTs1Hp31en3N6mc+KOYYlOGBgqUe71l/kvub8zFZvmGKv7evOV+8m/wuZRwTnOZqMqVLlhJXUpaK34v7k++xlfhPhowvj6zlLtDRfe02/wADK+Ecly2XPh6C5u7vJ/K7dvlYAAAAAAAAAADOM7wOU4d18XOy6Lq32S6/kurRxTxtjc7k6Ufco9Irr5yfX02XrqcNeGuKxyjiMc/Z03ry/aa/9tv56+RgcBhsDQjQw0FGC2S/rV929QAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAzji/J8qvGvWTmvsx1l9y2+bRm/itiKi5cvpKKad3LV36NJO2nnfXp3xWd5lim/bYicr95O33XsZNwdm+a8kqNK0JX96WkdPxfbRMyfwpw9NqeYVed/yxul83u/komF4dyjCpexw0E115Vf72r/AIgAAAAAAAAAABxZxvg8poSp0JKdfZR35X3lba3bdv5tVqub8QY9Kd6tWWy7L00SXXourOFPDzDZa44nGWqVlql9mL8u783oui6gAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAFevSoU3UqyUYrdt2S+bM58Tcqwl4YVOtPy0j973+Sa8zOeOc5zO8ZVOSD+zHRfN7v5u3kZTw5mmay/wBlouS77R+96fLcyvwnqOTeYVkl0UN/m5L9H6mD4XybCQjGlhoadXFN/NtXEklZAAAAAAAAAAAAcZ+ISoP6plVT30/enZNLyje6b7u1l0d9si4czHP8V+7T5W/em9l1evWWu2+vRamU5HgsspKGHhZ2Sb6u213/AEgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAMfmOEwFF1sTUUIrq/6u/kZ34q04PkyynzO+spp2+STT++3oZjm+Y5rW5sTUc23oui9FsvkjJvDrOcwtKrH2UO8t/lHf77epk/h1k2AtKpH2s11lt8o7fff1IxjGKjFWSAAAAAAAAAAAAK1alRpyqVZKMUrtvRJHGPiDUxylhMvbjS2ctnL07R/F9bK6OEOBMRm0o4nE+7Q/GXkuy7v7vLA4DDYHDxw+Ggowjsl/Wr7t6gAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABmuc4DK6PtcXUUV07v0Su39xnnipWlN08sglH+aSu36LZfO/ojH5hjs1xPtMRN1Kj0X6JJbeiRknhdjsQ41MfJU4PotZ/5L72+ljJOGctyiko4amuZbyaTk/V/orIAAAAAAAAAAAADHY7D4LDyxGImowju3/W/ZdTi3jLE55W9lTvGinpHq/OXd9lsvXU4P8OXPlxeaRst1B9fOX/yP39iMYxioxVkgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADF4uhhKEq9eXLCKu2+hn/inLmlSyuGn88l+Uf/AJL7iU81z7GqLcqtV7en5JfckZD4VzmvaZpPl/uxav8AOWq+Sv6oyjIMuymm4YOko33erb9W7v5bAAAAAAAAAAAAAGZ5nhcuw0sTiZ8sV+Pkl1b7HE3FGM4hxahFNU07Qgtddru28n+Gy8+CuA6eXRjjMdFSrbpbqH+cvPp07gAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADaSu2Z/4j5ZgE6eF/fVF2+Fesuvyv6oznP8AMM4rupiZ37RXwryS/pvqzhvwyrYqnGvmMnCL2ivia83tH0s36GVZFl2VU+TB0lG+73b9W9X+QAAAAAAAAAAAAAZpmeGy3CzxOJlaMfvfZLu30M/z/H8SY5JJ2vaEFrb/ADb6v9DgvgellEFicUlKu/moeS8+7+S0u2AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABnGe4DKaPtcXUUey3b9Fu/yXVo4r44xedSdKleFD+Xq/OVvy2Xm9TIeAs1zXlqOPs6b+1LqvKO7/AAXmcP8ACWW5LTtRjzT6yer+XZeS+dwAAAAAAAAAAAAAK9anQpSq1JWjFXbfRLqcWcS4jiLHxpUE/Zp2hHq29Ltd30XRad2+C+DaWTUFWrpOvJav+Vfyr9X19AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADi3jzDZQnh8Padft0j/i8/7u/e3V/wBq8QY++tWrL8F+UUvkkcJ+HdDLmsTj7Tq9FvGP+b89l07gAAAAAAAAAAAAAEpRjFtuyRxpxrXzatLDYeVsOn/4Hbq/Lsvm9dvD7g5YGkswxcf3sl7qf2U+v+Jr7lpu2AAAAAAAAAAAAAAAAAAHGnGcckgqFBKVeSvZ7RXd/dovm9N8nxVTF5dh8RU+KcIydtruKbsNpK5Q434vxNNVaGF5oPZqnNp9N07eRlFfEYjAUauJjy1JRTkrNWdtVZ6r0YAAAAAAAAAAAAAAAAAADaSu2cZeIarQlg8rk1rrNO17dI21s++nlo7nDPCWOz6tzL3aafvTf5Lu/wAurMlyHA5Ph/Y4WFu7e7fdv9Nl0SAAAAAAAAAAAAAADxC40lUnPK8FL3VpOS6vrFeS2l322vfw94MpOjHMsbC7esIvZL+Zru+nlr1VgAAAAAAAAAAAAAAAAAA4zyvC/wBnYvHON6vs+W/ZJ309b6v0OG/+RLg/+vcP/SUZjU9ng6s+0ZP7kzgin7Ph7Cr+7f723+ocUcT4bIcLGrUjzSk7RitL2316JdXZ7rQ4W4mpZ9hZVoU3Bxdmm7q9r6PS/wByAAAAAAAAAAAAAAAAAAr16VClKrVkoxirtvRJHGfHlXM3LC4JuNDq9nP17R8uvXsuD/D6rmChjMd7tLdR+1L/ACi/va2smmUMPRw9KNKjFRitklZL5AAAAAAAAAAAAAABx/xhPKaccJhJfvpq7f8ALHv6vp237HAvBcsfiZYrHx/d03blf2pefkuvfba4kkrIAAAAAAAAAAAAAAAAAADjT/kQYr/D/kcN/wDIlwf/AF7h/wCkozTDVMTga1Ck0pTjKKvtdppX307lfL+JuEowxftvaUY2TipNxS2s01onsmtn2MLiaeKw8K9N3jNJr0aujiCpRhxbl7xNuTlla+3M72/Hlt52KdClTcnCKTk7uy3drXfd2SAAAAAAAAAAAAAAAAADF4qhhaEq9aSjCKu2+hxfxjiM8reypXjQT0j1fnLz7LZeupwX4eqPLjczhrvGD6ecv/kfv7KysAAAAAAAAAAAAAAHEWeUMmwE8VU1a0iu8ui/V9lcyDKsVxTnMqmIbcb805eXRLtfZLotehCEYRUYrRAAAAAAAAAAAAAAAAAAAHGn/IgxX+H/ACOG/wDkS4P/AK9w/wDSUHFVOFTI8Wp7ezk/mk2vxSODHJ5Bhebfl/4b8Dizhijn2DVJy5akdYy7Pqn5Pr8n5HAOY42pQr5fjXerh5ct3q7O9tetrOz7WAAAAAAAAAAAAAAAAAKk4Qg5ydktW30Xc454vlnNf6rhW/Yxf/gb7+nZfN+XA/AsMDGOOx0b1nrGL+x5/wCL8vUAAAAAAAAAAAAAACUlFNt6I4rzuvxJm8aGFV4J8sF3b3l8/wAElfqcNZBQyXARw0NZbyfeXX5LZeXncAAAAAAAAAAAAAAAAAAAONP+RBiv8P8AkcN/8iXB/wDXuH/pKMo40x0spxuOxEOZ0pqytbRtLl0X2V1d/O5nnHSz/CxyzLaMlUqtJ3tt1Ss3o+rdrK/yy7BwwWDpYaG0IqP3K34me8aYLJsfHC4qLtKPNda9WrNfL+tzgSc8diMdmvLywrTSjftG/wDmr+aYAAAAAAAQoe0wk5reLX3P/hkAAAAAAAAAHiPxbKtUlleFl7sfja6v+X0XXz06a+H3BTg4ZnjY67wi+n95/wDtq+fYAAAAAAAAAAAAAAA8TOI/qeEWX0Ze/UXveUf/ALbb0ueGfDHsKP8AalePvS0h5R6y+ey8vUAAAAAAAAAAAAAAAAAAACrSp1YOFSKcXumrr7iFOFOChBWS0SWyXkUcHh6MZRpwSUm20kldvdvu31MJk+XYOo6mHoRhJ9VFJ/gGNy3B46KjiaUZpbcyTt6X2KVKnSgqdOKUVoklZL0SAAAAAAADIaEatCtCWzsvzKlOVObhLdaAAAAAAAAAcb8UQybAuFJ/vp6RXbvL5dO78rnh7wt/aeKeOxSvSg9L/al+qW77uy7gAAAAAAAAAAAAAAGZZhQy/CVMVWdowV3+iXm3ovMyfBYnizP3Ur/C3zT8oraK/CK+8hThTgoQVktEuyQAAAAAAAAAAAAAAABicRSw9GVaq7Rim2+yW7MJi6GLoRr0Jc0JK6fcAAAAAAAAAAAADh6ny4Vy7v8AyM/w3s8TzraX5rf9AAAAAAAAAx+No4LDTxNZ2jBNv5fq9l5kKeP4uzxvZy+6EF/kvvb8zLsvoZfhYYWgrRgrL/N+b3fmAAAAAAAAAAAAAAAeJ3ELxGJjllB3jDWVusui+S/F+RwTw6smyyMZr97PWX6R/wC3V+NwAAAAAAAAAAAAAAA4uxuY5rmcciy6XLZXqS2suza1ta2nVtIjgcz4IxlKvKr7TDTdpWvb7tbO2qa3s0xNSSad0cUf8iTF/wDXuf8A6Szgz/kQYX/D/mAGY5rXjxvBqb9nCUKbV3a84vp6t/cAAAYrFUsLRlWqu0Y6t9l39F1KdSFSCnBpp6prZryAAAKtBwpQqdJX+9P/AIYAAsWGAZdQ9jhYQe9vxepnmG9rhHJbx1/zAAAAAAAADxUz1+5llJ/3p/8Atq/V/JnAfDiynLVOov3tSzl5LpH5LfzbAAAAAAAAAAAAAAAOI85hlGW1cVLdK0V3k9Ev1flc8Pcknm2aSzDE+9Gm+a7+1N6r7vifnbuAAAAAAAAAAAAAAABwLBYnF5jmD1c6jivJR1t9zX3I4zwUcXkWJg1tFyXrH3v0sZPntLDcLUcfiHpCCv3bXu29W1b5mJ49x2LwNenjcN7OlVhNQklK1+V2V3pK+11bXocMYujhOGKGIrO0YQu/RXIeJeYKssRUwlsK3ZStK/8A4F8Lflbyv1OJM+eWZQ8fQSmvdt2ak1rp6mTcVSzTOsRg6UV7KknaWt3JNJ+Vt7aed+gY6Mq2V4zNo7rEqS9I6L/0ohONSClF6PUAADEUIYijOjNXjJNP0aszIOLMbw1jJ4KvedKMmnHqmnZuP+Wz8tzLsywuY4eOIws1KD/qzXR+TAAArYFyyiGmsfe+/V/gwC1zlGlYGMDLcP7fFQg9r3foglFSi4vZmKoOhWlTfRgAAAAAAAYrE08NQnWqO0Ypt+iV2cL4OpxLxJLF4hXinzy+/wB2P5L0TAAAAAAAAAAAAAAADxMzaeOzKnllDVQ3S6zlsvkrL1bOHMmp5RltLCR3SvJ95Pd/ovJIAAAAAAAAAAAAAAADwz/5FFTv7SV/W0TOLf2diL7ckv8A0lk1LEcK5VgL/wAaq0/Tnkv1T+Rxjl9Gtw9iKXKkoRuvLl1Vu2it6Ga1ZrgjBYeD1quEfxb/ADSMzyehVyWpgIx91Q5V5WXuv5NJmIxUsR4eRcnrFqP3TsvwscOYCGCyqhRiteVN+cmrtv1ZmWKWFwVav/JGT+5NmAo4V8Czw8qkeeUZStdXupOS0ve+iOFcV9ZyTC1L/YS+aVn+KA4u45zfLM5rYTDyjyR5bXjd6xi3+LP+Im5//NH/AMBP+Im5/wDzR/8AAT/iJuf/AM0f/AT/AIibn/8ANH/wE/4ibn/80f8AwE/4ibn/APNH/wABMyx9bMMVPE1rc8tXZWV7W2Mg4ixuS4j2uGlo94vaS8/Ps916XMgzujnOBjiqUXFPRprZrez2a81+DugADDUfbVoU+7J8ipu+1vwMVR9jWlT7MFEsDBjA4coXnOq+mn+YTdoto4gpL2sK0dpL8v8AgWAAAAAAAA8T82+q5VHCxfvVXb/t2Nm/xsvmzw5ydYHJo1pL36vvP0+yvu1+YAAAAAAAAAAAAAABm2Y08uwNXF1NoJv1fRfN2R4e5bUzbOqmY4jVQfM/Ocr2+7V+TSAAAAAAAAAAAAAAAAOAJrD18wy97wqtr0el/wDyVfejjHGRwmRYmbe8XFesvd/UxmGeCwOQynoozi35czjL/M4qlGOR4tt/8u5fimjNGqfCuVVntCcG/wDyb/IxdWNHD1Kktopv7lcVJw8PG39qV/8AydL9DBf7rT/wr8kce4r6vw9iGnrJKP3tJ/hcw/hPhalCEp4iSk0m1ZaO2p4a1pSyT2Et6U5R/wDbv1AzfgHKc0xk8XXcueVr2aS0SXZ9Ef8AELci7z/8CX/yJ/xC3Iu8/wDwJf8AyJ/xC3Iu8/8AwJf/ACJ/xC3Iu8//AAJf/In/ABC3Iu8//Al/8if8QtyLvP8A8CX/AMiZ7h8Lh8yrUMLf2cJOKu7vTRu/m7nB/h3PFcuLzKLjT3UNnLzfVLy3fkt6VKnSgqdNJRWiS0SXkgADJUnjqfz/ACZmEuXCVH5P8jMXzV+buov/AMlQXaFJMGDGBkFNRwd+7f8Al+gT+BmNftcpoz6p2/NfogAAAAAAAOLnLPOLIYCD92LjD9Zv5Xf3FOnCnBQgrJKyXkgAAAAAAAAAAAAAADxXzfkoUsvg9Ze9L0Wi+93fyOCcmWV5NSpyVpy96Xq+nyVl8gAAAAAAAAAAAAAAADi3D47Is3We4GHNCStUXTotbbJpLXpJa72P7RzDjjF0sOqPs8NB3nrf/wAmstWtEktLtu/TjjJamZZNKnQXvwalFLutLL5N287FXifNeJcDLLlR5eWLdSavqoK9rWXLdpK13d9lcoZLLNeC6WEivf5E43/mTul89vmVOLs3zDALIoYd/WGuST62WjuraNrSTbta766cW5WsBwf9Tp68nIvV8yu/m7swS/2an/hX5HiBCWJpYPBRV/aVY39Fo/lqg4QhPC5zmeEasudTX/b12/waAAAzLETw+Dq1aceaSTslq2+i+bsjhHw/p4JrGZilOtuo7qL7vvL8F0vuAABl9ZUcVCb2T/4BiaSrUZU+6aMbOM68nHbZei0/QAu0c1wYwMmVsDT+f5sMXUVOhOT6JkXzZM12l+qAAAAAAACvWhRpSqzekU2/RK7PDbDyzDOsRmNRXcU3/wBvTb/Tm+8AAAAAAAAAAAAAAAKS/wBZ+MHJ601K/lyQ2/8AArL5yAAAAAAAAAAAAAAAADcjGMVaKsgUYq9kHLG97agAAAAAAAH9qYr2PsefTbzt6gABcYGRyvgYLtf8wztP6jO3l+aPhyf1l/X5AAAAAAABxbOUcixbi9eSX4qx4W4JUcmlWe9STfyWn5pgAAAAAAAAAAAAAAHGGZ/2dktesnaTXKvWWn4b/I8KMqVPDVsdJayfKvRav73b7gAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA4cqp0Z0+zv8Af/wwY2l7XDzh3TM0h7DCUMO99W/6+bAAAAAAADjD/kQ4v/AzhPCrDZHhaf8AcT+cvef4sAAAAAAAAAAAAAAA8WcwfLh8DF73m/yj/wC3HD2WrLsroYW2sYq/q9Zfi2AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAGTYpUMUr7S0/r5gZriliMVKSei0XyAAAAAAADi9XyLFJfyMw9FUaMKa+ykvuVgAAAAAAAAAAAAAALIx3/C3xzGktYQkl8qavL72n94WRZDFYsiyDSxZDsFkWQKxZDtYNLFkOwWRZArFkO1gsiyHYLIsgVh2AAAAAAAAAAAAAAAAAAMBn0I01DEbrr/mZlnkalN06HXd/5AAAAAAAAZ7hamKy6tRpq8pKyRYsFrlhoLFgtcsNBa5YaCxYLXLDQWLDRuWLBa5YaCxYZuNABj8XDCYWpiJ7Qi5fcrnhXhJYjHYnH1NWla/nJ3f5fiAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB4jY76tkFSKetRqK++7/AATPDbAfVshhNrWo3L9F+Cv8wAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPFrFuUsLhI7+9Jr7kv1MswaweCo4ZfYil9ysAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABxEv7R45oYbdQcF8l77/NgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHDS+u8c4ivvyOo/ufIvwYAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAE5KMXJ7I8K4uvj8ZinvZf+TNv9AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAOIK/sMpxNTqoS+/ldjwmw/Ll+IrfzTS+5J/+3AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAceVXS4dxMl2S++UV+p4aUPZ8Pwl/NKT/Hl/QAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADxD/5JvE/9uf+lxOCqPsuH8LH+7f723+oAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHiH/yTeJ/7c/8AS4mQU/Z5ThYdqcF/5KgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPERpcN4j/tz/wBLiZfFRwlKPaMfyQAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHFOT1c3yqpg6UlGUrWb20kn09ClBwpxi+isAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAGdZlHLcBVxbjzKCvba+tj/iLlD/oFf8A4Ev/AJE/4i5Q/wCgV/8AgS/+RP8AiLlD/oFf/gS/+RI+LdCUkvqr/wDAl/8AIgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABisxweEcViK0YN7c0kr+l2RkpJNO6DM8wo5fhJ4qtflgru2/YweKp4vDU8RT+GcVJX3s1dAAARlGUU4u6YAAAAAAGf5bPMstrYSEuVzVrvpqj/iEmN/6CY/cz/iEmN/6CY/cz/iEmN/6CY/cyPhNjYyT+sx08mAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAATnGEXJ7IynhWfFiq5rjazjzyaglbRL16La2mzd9Th3KZZVltLBynzOF9e9238tHsHG/8AyT+K/wAP6o4b/wCRLg/+vcP/AElFep7OlKa1sm/uRwpm1bNspp4yskpSctFsrSaXfogA4mzSpleVVsZSScoJWT21aXT1Kka2Pydxi7TqU9+zlHf72cJ5bictyahhcS/finfra8m7X8k7ABxLmeIy3LKmJw0OecbWVm73aT212Zk/iDnGOx9LDzw0VFyUZNKWl3Z9dLeYAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAYqk61CdNfaTX3qx4aZlSnlbwEnarScrrrZu9/vbT7fNHFOY18vyiviqDXPFK19eqRg8ZVq5TTxMvjdNSfq43/MrZric04HrYrENObT2Vtp2IcScRSy6jLKaf7ihCClKyd2oLm31stny6rdsxXEGOzLhlY3LoXqPSSte3SX+a8mjw5xeeNUaDp/7J73vW66vf/FoS40ziu1hMNHmre1mm1G9qcWltrvd3b2t5nEWP4rq4+ph8ppWp00ve933m0nZc2nlZK/VvVHCGfVM4y72laPLVg3GS21XW3S6f33M34pz3MMxqYPIYXVLSUrR1a31l7qWll1dm0cW4/iCrw3ReIpW5uZVdLW5ZxUOul/LR+hwxmWfrLcRPG0+WNOmnTdrXtF/fokZJn8qnDsMzxr1UZOVlbaTWi87Jeo+LuLFBZs6K+q3+Gyta9t/j/wC3vhv06HEGfzo8OyzPAyV7Rcbq/wAUorVej+8zPjXiBzePwdP/AGSDtflVpW0bb3Sb0TVktE9TAYynjcLTxNP4ZpNfNXMbjcPgsPLEYiajCO7f9fh1ODOJ8tqZli6XPaVao5QurXT6eT8n+YAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHGHCeKwuJlnWVScZx96SW/nJd7/ai99d72M2zlZvwVVxdrScUmu0lJJ/LqvJoy3/kn6P8A16j/AOkIy7/k3tT0l/6WcI4Snh8hwsIrRwT+clzP8zguH1ahmmCXw05zt9zX/tp4df8AJOUPWf8A6XI8O8JBYjMMTb3nUcfkm3+N19xxBxXluSKKxMm5PaMVeVu+6SXq15HBmbwqrNsbRVo350n0upvX7jw5wcaOQ06v2qjlJ/e0vwSOP/8AknMT6R/9LiVf+Scf/Xn/ANsMXVmuAcLRi/4kuX/yeT/Qq5bRnl7wKXucnJbytYo15VfDutCX2JKP/lSL/UjgKMeFfqyXu+x/Hkvf1vr6nAE3PhzDN9pL7pyX6HF2HnnOcYTJ+ZqnZ1J27LRfk0vN3OJeBMt/s2VTAU/Z1aa5otN621s9dW+j3vbXc4ezF5jlVDFS3lFX9Vo/xTAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABpNWZlcL8EY9Q+Hnlb0XJ/wTLa8P8AVqlUb0VFP7oamXf8m9qekv8A0s4XqRqZJhHF/wDLuC+aik/xRwg1Wjm2Kj8M6k7fK7/Jo8Ov+Scoes//AEuR4e/w8d/1+l+hw3gaGZ57mGOxMVOUJ8kbq6SjdXS72S9Ne5k1FVsRntKn1bSt3tNfmeHmIjW4doJPWPMn68zf5NHH/wDyTmJ9I/8ApcSrrw4/+vP/ALYY2LXAuCqpfBUUv/Jpr9SpiqUMM8Q37qXNfytf8jD0pR8Pa839uV//ACpFfof80/8A9wv/AGw8Pf8Akm8N/wBv/wDpcjB1aVXjKu4yTaopaO9veV1/mTUXFqWx4d3fDeH/AO3/AP0uQAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABjYVZ4apGi/fcWl620/ElnHG7ovK5UG5NcvPyvmtt8d+T/t753vqZbwzDDcPf2XJ6yjJSf8Aele7+TenkkZTheJ8RTWQVqbhQTtKdmnyXu0pbNPpZXfe1zA5XjocD1MJKlJVfe92zvrO+3oTlxVkmH/szB0XOnNXhJRbcOZXkrrRNO/xbbnC+QrKsohhJ6yd3L1e/wB23yOCf7cyzHPKK9H9zFyfNyv5Wl8Nnva19fkcD4DE4SGMVem481WTV1a601XkYtZ/kOdYn+z8P7WGIfMnZtKTu9WtFZt3u0mrM4GybG5Zi8bDFJtycHzPaTtJys+tmyrR4g4Ux9aOX0XVoVHeK5XJJ/8Abuqa27SSXynhMzzfhedHGQ5a84y0215m4ry0SWuq66nDGLzrF5PisHjaDj7OnyQ91py92Stru1ZLRLz3MmyCpiuEYZbiYuEnGSs1qnzNxbXrZiqcXVcIsgdBpfC52fw/4vh5baXWrWm5n+X01wxWyjAfvKlONNOK1fxp3aWzdm7Es3rqqslVLX6ve/W/LblsZ9UzHLOGcvwrcqbfteaOqb9+6uu1nf5nB2ayybM6eLrRapSvFuztZ9fOzSduyZxRxfl+Eyubw9aM6k01FRabu9L6bJb6+hwxl8svyfD4aatKMVddm9WvvYAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABUqQpwc5tJLq9EZn4hZFg4SUKvtJrpFN3/7ety/iZt4q42vBQwNNU+7fvP5aJL5pk/FLOZYX2SjBTtbms7+tr2v8reRwlieJqWJq4nL6LqOWkuZaN97tx1WvXrqYHhPNZ5lHOMViVGu1rFRvFK1uW/Nqrfjrd7mKyXBY2rTrYumqkoKyutFfdqLbWum99kZnlOEzLBywmIjeD+VrbNdmv60ODuDcv8A7SxNSq3P2FRxina2mzfd/h5AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABVq06UHOckkt29EjNOOcjy+bpzrc0kr2iub0V1pd+b9TM/FfGVYOODoqHm3zP5KyS+dzMM3zDMp82Kqym+ib0+S2XyRl/BOe45RcMO4xfWXur1s9beifkZX4T04VebHVuaK6R0v6t9PT70YHhfJsE4yoYaKa2drv73dgHEtHMauWVIZc7VtLapdVfV6bXMhyDi/B5jCpUlaEpqVT3o66q7evYAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAG0ldmZ8c5FgLxnXU5LpD3n960XzaMz8Wask44Ggl5zd/wDyVf5sx2dZlmMmsRWlPmd7Xdr9LR2XlZGV8J5xmacsNQbSdm3aK/8AJmr262vYy3wmgnGWNr37xira/wCJ30+S+RlfDmV5Yl9VoqL77y+93f6AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABmWaYPLqDrYqooxXfd+i3b8kZv4r0opwy+jd/zT0X/gKd382vQzbiTNc0b+tVm122j9y0+/UyzhHOcySdCg+V9X7q+92v8rmXeEtR2ljcQl5QV/8AyZ//ACLMs4UyfLlD2FBc0dpNXlfvfv8Al0sAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAE5RhFyk7JGc8e5Nl9GTp1VVqLaMXe7/xJNL+tDNfErOcXKSoSVKD6Kzf/AIE1e/mkiEMZmOIUY81SpL1k3+bMp8KsbWtLHVVTXZe9L79l+JlHB2TZWk6NFOS+1L3pfjovkkAAABWrUqNN1KslGK3bdkvmzC5rgMXLlw9eE32jJN/g2AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAV61OjSlVqO0Ypt+i1Z/xELhv/AKCf/JJ//IgAZhmOEwFB1sTUUIrq/wBOrfktTOfFbDqm4ZdTbl/NNWS80k7v52Mzz3Ms0nfFVnPy6L0itF9xl3B2d49x9lh2k+svdXrra69LmU+F2WYeCeMk6sr+cY+lk7v7/kYTAYXB01Tw9NQj2SS/IAAB2sZpxvkmXyjGpWUm/wCX3vvtovzMw8UsoowX1aMqkvTlS9W9fuTMNW9tQhVtbmSdvVXPFDG47C4GjLDVXCLk1Kzs3pdarXo9OpXxuMxdoVakp9k23917nhtgMXhs8brUpRThLeLXVd/QAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA4qzShl2U1q1ZcyacUtruWlv8/K5LhPOKOFWYVMN+63av8AZ80nzJW+aWr7mUZnRzPA0sXS2mr27PZr5O6AznPsvymi6uKqJdlvJ+i3/TuzN/FPH124YGCpx7v3pf5L0s/UrYjMM2xK55Sq1Htu38l+i0Mp8LMxrzjLGzVOHVJ3l6dl63duzMo4WynKkvq1Fcy+09Zfe9vlZAAEpRjuw4m43wWR1lQqQlOo1dJWStdrVt+T6MzDxUzStphqcaa/8Cf3uy/8lPD/AIjx2c0a7xck3BxtZW3T7ehU+B+hlnCmcZlJqhQenV+6vvdtfIwHhlnledq8Y0l3bT+5Rb/GxhqPsaEKV78qS+5WMfluEx8IwxNNTjF8yT1V0mtVs93o9DC5Pl2Fnz4fDwhLvGMU/wAEAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHFtKOMzTLMDPWMpyk135En+N2iUVJNNXRwJT+rU8ZgVtSqyS/wu1v68zOeNMnyq8KtXmmvsx1fz6L5tGfeI2aZg3Twz9jT8n7z9Zb/AHW+ZgsvxuZ4hU8PBzm/n82+nqzJvCmjGMZ5jVcn/LHRfOW7+SXqZbk2X5bDkwlJQXktX6vd/NgBjMzwODX+01ow/wAUkvzZgcdh8dh44jDy5oSvZ662bT380cQZnPLMsrYuEeZwV0ntul+t/wDIxXiFxBiIOHt+VP8AlST+TtdfffzMizPF1s6wkq9WU7VI7tveSvu2HGvBmOzzMqVag4xioWbb7NvRJN9fQy/wnwNOzxdaU32SUV+r/IyDhjA5Iqiwl/fte7vte1tF3AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADiWaoZ9lWIltzTj85xSQZ1xHjI43GUsLVcadSpJu2jaTstd7W6depk/C+a5s08LSbj/ADPSP3vf0V2ZB4Y4HCWqY9+1n22iv1l87LyKGHo0IKnSioxXRJJfcgMdn2V4FtYnERi10bV/u3/AybjHK83xcsLhZNySbu1ZNKy069eqQca8U5ws0r4SFdwhB2Sj7unm1q/vsU6GMx1RuEZVJdbJyfz3ZwJg8VhMipUcTBwknLR72bb26b7PUzvLf7Sy+rhObl51a9r2+WhhPDTIaPK5xlNre7dm/RW+4weUZfg3fDUIwfdRSf3pXAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADiHi3LslhatLmqdILf59l5v5XOJuN8bnajTlFQhF8yS1d1s+bfr0SK/idmtXL/AKtyxU2rOave3dLZPz+5I4e8NMDgnGtjn7Wfa3up+n2vnp5EYxjFRirJGIxNDD03UrTUYrq2kvvZi/Efh/DycVVc2v5Yv83Zfp5kJxqQUo7PU8Uq+JoZdRnRqOK5rOzavdN6232MJleOxr/2ejKfpFv8UcCcI5xlmZwxWJpqMHFp6q6vton5f5hU4OyerjqmNrUuec3f3tUtEtI7dOqZh8LQw0PZ0IKMeySS+5AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAGa55l+WUnPFVVGyva65n6Ldma+KWJxKlSw9FQg1JXbfNqmk01ZJpu/XbcyfIcxzvEcmHi5d5PZebf6at9EZR4X5ZhoqWMk6su2sY/cnf53+RjeHOBsGn7dQi109pO/3c9/wMk4jwGcqo8HJtQsm2rb32vr0K9P2lKUE7XTX3lSpi8TNU5ylOS0SbbfyWpg+AeIMTBTVDlT/mai/ubv8AgZdRqUcHRpVfijGKdtrpJO3zMVgsNi4qGIpqaTulJJq/ez66kKcKcVCCSS6LQAPEDGZjSWEw+BquEqs+XR21dktd0rsxGX8a5DTeMeI9tGOslzSnp1upJO3fl1W+m5k+dRzbKo4zDr3mnp2kun3/AINM4JxGY4jKITx9/aXl8Ss7X6/ivQOLsTmlCWD+oJ61EpWXTs/J63DOqmIp5dXnhv4ihLl73s7W8+xwtVxlbJ8PPGX9o1rffd2b82rAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABiMTRw1J1a01GK3bdkvmcSeJ/LKVDKlf++1/wCkr9X93UpYbMc2xDdOMqtR7uzb9W+nz0Mr4JynBYijHNsUnVk0lSj3b0Umru3R6JedijQpUYKFKKjFdErL7keKmPxtHHUqNOrJU5Qu4ptJu7ve2+ltzCZVjsZb6vRlO/aLf5Hhzw9mOUwxDxkOXn5bK6b05r3te26DD5dg8PNzo0oxk92opN+rSAAA43/5GGVf9fo/nEaUlZnBaWXY/MsvXw05c0V5O/6cpLjmmskhmvsHyuXK1fVb63t5bW6mMxUMNhamIlqoRcvklc/1zisBg8W6DX1ifKlfbVq+2u17HFfFdHIqUPc56k/hje227b10+Wv3mUcWwzDJq2Y+ycXSUrxvu4x5tHbr5rRmV5xSxuWQzCS5IuLk7vZK99fkR8VcM8Woug1Rvbnvr68tvwvf8jPs7p5Xlkseo88VbRO17tK99e5mnibg8HmX1WNJyhF2lK+z62Vtbequ/vKdSFWEZwd01dPyYAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHE3iFgcqm8PQXtaq3Sfuxfm+/kvm0Zxn+Z55XTxE3LXSK2Xou/3vzIcB4+llVbMMX7nIrqL+J+v8q/H0JZ/mH1dYalU5Kf8sfdXztrL1k2zhrLMwxWYUauGpSkozi20tFZp6vb72GKyjA4qvGvXpRnKKsm1e3yegkkrIAAADjf/AJGGVf8AX6P5xDL/APkrM0ttyR/9JiZZhPrXh/Vh1XNL/wABlzfkjOc15uCPrF9Z04x+btGX6nEuD+p4HJ8P1hUpr5pK/wCJmtejR41wssQ0oum1Fvbmbl+L2+aOIqcIZNjeVJXp1G/N8j1Zj8VOjwHh6dPepyw++Tf4pWMbw7hpZBLLIxVlCy/xJaS9b6srYyWK8PLyesbR/wDAaiS/CxV4fw0OD5YblV1T579efl5r39dPTTY4JryrcP4WTe0bf+Atx/QAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAx+ZYPAU/aYqqoR7t2+7ucUeJVfFKWGy33Ibc/2n6fyp/f6bD1PC/LcF/Zn1tU17Xmacnq7K2i7aPpv1MywMMdg6uFm7Kaadt1ddDLeA8hwNnGjzy7z978Ph/AhCMIqMVZLoAAAABxv/yMMq/6/R/OIcKzWMzHNcfHWMpcsX3UU/05Tw+oRxHDCoy2k5p/NtGDqSxWR4DK5b/WHGXpFuT+7mRx98eX/wDX6JxbwtRz7CqDly1IXcZeu6fk9PTfyeS5li8RwxmGExjvVoRqQd97crtd9bNNX7JGZPk4Qyyo9o1It/8Ak5VqRp03OWyV/uKEHHw9qt9Z3/8AJ4r9DE/8k/P/AK9P/wBIOAP+Scw3pL/0uQAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAZpnWX5ZT58XVUO1936Jav5Ix3izhIVHHC0HKKvq3y3fTTXT8fIzXNcwz7He0qXlOWkYxTaXlFav/N6mXeGmd4qS9slSj3bu/uV3f1t6mU+GuTYO0q6daX97Rf8AgK/Vso0KVCmqdKKjFbJKyXyQAAAAAHiPi44Orl+JkrqFTmt35eV/oZl4iYnNqf1HKKEvaT0u90utkr2/xN6b+Zw/kaybJVhb3lZuT7ya1+7RLyR4a/8AJPU/8UvzMmy+ceNatB/BSlUqJedRRX6q3ocffHl//X6JnvHOEybMXhcTTk1yppq27b0s2tNFrfcySNWvkObZlUjy+3VSSXkoy/VtfIw+UTzTganh6avPlvH1Um7fPb5lbxArYrJ1lkKUvrUl7Nvp2bWt+ZrpZWbvfQ4kyv8Aszgh4R7xUL+rmm/xbMT/AMk/P/r0/wD0g4A/5JzDekv/AEuQAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABxxmuMyzJ5YjCStNNK9k7J6dStXq15udSTlJ9W7s4d4OzHO5c1JctPrN7fLu/TTu0cOcIZfkkL0lzVHvJ7/L+VenzbAAAAAAArYejXjy1YKS7NJ/mYfB4fDq1GmorySX5DSasU6NOlHlpxSXZKwqNNTdRRXM+ttfvKlGnUtzxTtqrq9n3RisBhMWksRSjNLbmSdvvuezg4clly2tbpbtbsQpwpxUYKyXRaEcvwca7xEaUVUf2uVc332uVKVOrHknFNdmrjhFx5babWKdOFOKhBJJdFoAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAZxneBymh7bFz5V0XVvsl1/JdbGceKmNrNwwFNU493rL7vhXpr6mTcY4TD8O0cZmFW83dW3lJptaL0trolfVo4o40xudydP4KK2iuvnJ9X5bL11OEPDurjXHFZinGluo7Sl69o/i/Lco0adGnGnTilFKyS0SXkAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHFeX4/HcT18LSUqk76LeyaTW+ySfkkYzhnLMjpc2aVues1dUof+3Sey+S8rmDwGLzbFqjhKd5PZLZL1eyXdv1d2cLeHuEyzlxGLtUrLb+WL8l1fm/kluAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAcQca5Xk6cJz56q+zHf5vaPz18mcP8AGWYZrxNQlXlam+ZKC2V4u3q721fystA4t4zwWTTqUcHFSxMrXdlppZOT6tLZffbZv61j8Q3Z1Kk35tt/mzw5zGnl+JllWKoezrS1TaacrdGn5X5baPXrqwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAOO8TjMNkVarhZuMla7W9m0nbtvvuZJwvmmdT/2aHu9ZPSK+fV+SuzhjgjAZLFVGvaVv5mtv8K6eu7720OPONP7Mh9Swcv30t3/ACL/AOSfTstexw9wZmGfTjiJTtTldym3d3vZq27l1101vfoeHmTYWhUxeKpracqcb7qMf87q/oeIOFjTwdLM4L95QnF362bSa9L2/piaaugAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADMsywuXYaWIxM1GK/qyXVvsZBnVHOMDHF0o2TbVnurNrX1Vn5XMZg6GMoSw9ePNCSs0UaNOjTjTpxUYrRJaJeiOOOOo5apYLBO9Z7vpD/7b8uvY4U4YxPEONc6rfs07zk92+yfWT/Dd9E8Hg8Pg6EaGHgowjsl/X3vr1OA5JYXE0X8UK1RP70eIdRR4erQ6ycUvXmT/JMpQ5KcY9kAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABnmd4TKMJLE4h6LZdW+y/rTdmdZ3mPEeOXMm23aEFql5JdX3fX024HyHFZNlnscTJOUnzWX2bpK1+u2vS/fcOPONFlcHgsHL9/Jav+RP8A9ufTtv2OEuEsTn+JdSo2qKfvS6t9l3b6vpu+ieCwOHwWHjh8PBRhHZL+t+7er6hnONxXCmeVMbGnz4fEWbXaS316Pd+aflpRzbEcY5tQp06ThhqMlOV+rW1+muyXZt37AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAeJWXY7F53h6VG8+eNors03zemlm39+iOEuDsNkdFTlaVdrWXbyj2X4vr0SOMuKqWR4T3GnWn8K7f3n5L8Xp3a4X4bxXEmPlUrSfInecurb6Lzf4b9k8JhKGEoRoUIqMIqySArUKVem6dWKlF9Grr7mYfC0MNTVOhBQj2SSX3IAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAMwzDDYCg6+ImoxVld927IHSpympuK5ldJ9Una9vWyv6IM/zzD5PgZYqt00S6yfRL9ey1MNh8y4rzh3d5S1b6Rj/ktkur83cynKcNleDhhcOrRj97fVvzf9aAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAANpI454knnmPjhMJ71KDtG32pPS/6R+/qZXTxFPA0YYl3qKKUvVLUKlSFODnN2SV230S3ZxNnWJ4mzaNLDJuF+WnH/wBufm932Vr7XOFuG6GR4FUY6zespd3/AJLZffu2AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAeJHESwGX/U6Uv3lXTzUer+ey+fY8NeFHpmuJX+BP7nL9F832YHF+e1M2xceH8td5Sdpy6K2rXorXl6W7mZ+H9fJMOsxy6u5VaXvNNWulu1by3TvdX16PKMwhmGAo4qP24p+j6r5PQAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA4o4SxWZcVQjdunUSk3/Ko2Ukvwt5yXmUaNOjSjSpq0YpJLslog4sz6nk+WTr399q0F3k9vkt36eZ4V01VzmrVnq1B/e5R1/P7ypTjUg4SV09Dw8b/ALApQbvyua/8mf8AmAAAAAAAAAAAAAADRa4AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAFScYQc5OyWrfYz/ADTF8U5yqeHV1flpx8u77X3b6LyRhsDQ4Yz7BU27U6lJ03LvPm5m/m2l5XM8zOllmX1cVUduVO3m+i+bOBsJLDZBhoz3acv/AAJtr8GgAAAAAAAAAAAAABDGMAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAMLjKGKjKVGakk3F26OLs1/XrsB4mcRrCYRZdRfv1F73lH/AO229L90eGfDkcLg/wC0a0f3lT4b9I9/+3t/S3c4n4do55gXh5vlkneL7P8AyezX+RgvDvN8TiIRzXE89GGy5pSv5K9uVPq9/LtGMYxUYqyQAAAAAAAAAAAAtjdBYsHQe4x7AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAGcY9YDL62Kf2Itr1tovm7I8J6WJksVXlJ8jaVu8tW362t9+vQMdjKOCw08RWdowTb+X9aeZlGCxHFnEEqlb4W+aXlFbRX4RX39CFOFOChBWS0S7JAAAAAAAAAAAAAHQQANXGhj2AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPE/GewyP2SetSSXyXvfovvOAMAsJkFDTWd5P/t7b/yWweJHFVKtCWUYa7aa530015fOzs32tbvbwr+o/wBm1FSf73m9/vb7NvK1/ncAAAAAAAAAAAAAOgguXC4xjAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPFus5TwmHj/ef/AKSl+pg8PHD4anRjtFJfcrGYYpYXB1cQ/sRlL7k3+hwBlFPDZXHGTV61a8pSe9m9F6W19WVsFTyni3D1aC5YYmMoyS25oq97eenzu+rAAAAAAAAAAAAADoAAPUuAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHHWV43GZ/g3SoylBKN2otpe+73aVlpq/IPEHNI4HI6sV8VT3F89//Jb/ADscLzjPJMI4/wDPuC+6KT/FHEzUs7yqmt+eb+SirgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAFrniPiquY55Ry2jq42Vv707fpy+mpkPFUeGvaZPmkX+6b5ZJXum77X2d7p672drHDmMq8R5/LNeRxo0YuML73e77Xs3e214rzLFgLFgtcsWCxYCxYLXLFgsWAsWC1yxYLFg3LFgsWLBYsG5YsAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABcKlSNODnLZK/wBxwNTnnPE1XMaq0jzT9HLSK+Sbt6GaZDlmaOLxdFTa2b0f3qzt5bGEweHwlGNGhBRgtklZFy49QuXAuN3C5ceoXLgXG7hcubhcuFy49QuXAuXC5ceoXLgXL3AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAOLM8eT5XPEQV5v3Yr+89vuV3braxX4K4nlhpZhKu3VabceZ81mtV2vb7O3TyPCjCezyytXe852+UUv1bAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADiuEa+bZXhpapzlK3+BJoODqaoYvMsNH4Y1W0u3Mv8AgIAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAOMc4weFzrLuaa5qcnzLtGaSu+3fvbUbSV7nA9RYqeOx0fhqVXy+aS0f4gAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAFWpGnBzlslf7jIuEqPEGX4jMsa/wB7XcnF/wAiT0dvVW/wqytcxVLi+i6WSV6toVHyxelmu3MlzWt0ettLdDJcqo5XgKWDpaqC37vdv5sAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAzq/9m4i2/JL/wBJZwby/wBgYXl/lX/B/E4vt9dyy2/tl93UAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAKlONSDhLZqxwTmSy/CYnLsa+X6rK3M9Fyybt+N7eTRSzOnxFxPR+rPmoYZSk5dHJ6afO1u9n0AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA44wlCGSYyvGKU5Rim+6Ula/pdnCuT4XLcspQoxs5Ri5Pq5NK//AAF0AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA46nfIsTBfyr/0pFKmoU4wXRWAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAz7LMTjqdalBXUowS16qbcvwsAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHFHFNHIaVOdSm587a0aVrepwzxFSz3CSxNODilJxs/JJ309QAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAOLcNRq5LiZVIKTjCbV0nZ8r1XZnC1ClSyXC+zilenBuytduKu33b6sAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADj3iCvTisowVPnq1ou6Sbai7rRLq9fRa9TgPiGvWi8pxtPkq0YpJNNNxSS1T6rT1TuAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABkNKNfiXMsTP4o8kF5Ll1++yM9pRocTZbiY7z54PzXLp9zbAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAWaUMj4qxMMXLlp4iMZKT2Tira9r6/hcWaUM84qw0cJLmp4eM5OS2bkrad7aa+oAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHHmV4DF5RVq4mylTjJwd7Pmtt53tt16anBWTYPAZRRqUI+9VjGUm923FP7lfRfrdgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAGbZzgcqoe2xdRRXTu32S3f6ddDNPFlWccBQ/wC3pv8A9tX/AMl8jMs2xuZVXVxVVzfnsvRbJeiOBs2pZjktL2cWvZJU3fq4xjdry1AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADN+JcqyuL+s1kpL7Kd5a/3Vr89vMzvxSxmJi6eAh7NP7T1l8ui09fJohHMs5xcYJyq1Xtdtv73svwRlfhPialpY6sortHV/e7JfczA8H5JhKCoxw0ZW6yipN+bbX5WXZGEwOFwkHTw1OMIt3tFJK/eytrogAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAM44kyzKYXxVVKX8q1k/ktfm7LzM78UcdiU6eAh7KPd6y/yX4+phMtzHNKt6FOVSUnq7N6vvLZebb82ZF4Vy5o1cznpa/JHe/Zy7d7fJmAyrA4CHJhaUYLySV/V7v5gAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAGc8R5blEObFVUn0itZP0X6uy8zP/ABHzPHydPCv2NPy+J+sunorerMvyjMc2rOOGpucur6fNvRfNmSeFVGFqmZVOZ/yx0Xzlu/kl6mEweHwlGNDDwUYLZLQAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAzXOsBldL2mLqqK6Lq/Rbs4i8TcXin7LLb04fzO3M/wA1Ffe/NGEy/M87xT9jGVWb3e/zcnovmzh3wyweGiquYv2k/wCVfCv1l+C8nuUMPSoU1TpRUYrZJWS+SAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAqTUIOT6GX8d5bjp4anST56zkraXjb+ZX0vugAAAAAAAAAAAAMViqGFpOrXmoxW7bsjiHxRrzqSpZWkoL7bV2/NJ6Jeqb9NilQzDN8U1BSq1Jb7t+rfReb0Rw74XxhJVs1al/cTdvm1b7lp5tGCy/C4GiqOGpqEV0St8/N+bAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABpNWZkOWYGnxNmKhRivZ+y5bRS5bw15dNL9bWuAAAAAAAAAAAATnGEXKTsl1ZxD4nYXCuVDL4+0mtOZ/Cn5dZfgut2jMs2zHOcQpYibnJ7LovKMVt8lqcNeGMq9NV80bjfaC0f8A289bei17tbGX5bhMvoKhhaahFdF+be7fm9QAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADEV6VClKrVlaMU22+iW7OH+MstqcSYuo24xr8ii2usY8uuul3t+NgAAAAAAAAADi2lmFTKqn1CfLUVndO2i1evoeHv9q1cvliMdVc1Ozhd3dtU/TUAzziTLsnpOeJqe90itZP0X6uy8ziXjTMM7k6bfJS6QX/ALc+r/Dsjg3gPE46axWNhy0bSVnpJ3i0mlbSzd031WhknDOWZPC2Fp+895PWT+f6Ky8gAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAOP5VKmX0sFTdnXqQh8m7/mkcScHZX/YdSGHoqM6UXKLS968VfV7u9rO/qcP4yWMyrD15u8pQi3621/EAAAAAAAAADPcZh8Ll1apXmox5Wte7VkvV9jgHGYevkVCnTmnKCtJdU7vdBjcdhsFQlXxE1GC3b/r8NziDxRq1VKjlkOVfzy3+S2Xq7+iZgstzPOsU1RjKpN7t6/OUnt82cL+H2CytRr4m1St/wCSx9F1fm/kkAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHHk/q9DCYt/DSrQk/TW/9eZxLmFHDZLiK7krODS83JWVvVtHCuHlQyTC05b8kfxV/1AAAAAAAAAA4iy98QcRU8tnJqjShzyt1bdrfdb0VyOSU+GOIcLUwjfsa94NN3s+n42t10eupxFxVl+SUr15Xm9oLd/5Lzfyu9DPOIMy4gxS9pd62jCN7L0XVvvu/TQ4f8LatRRrZnPlX8i3+b2Xor+qMDgMLgaCoYaChFdF/Wr83qAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABxTmeR/UKmFx9dJTVrLWSfRpK70dnqrdzhXIKme4t4eeJc8NQadry1Tvbli/hvbXt0uJJJJAAAAAAAAAAYCUafF+LhLeVODXorJ/ieKeNlh8JhXTlaop8yfVcq3+9oyzJ8zz3FuNFOcn8Um9F5yk/+HfQ4X4MwWSQU/jrPeT6eUV0X4vr2QAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAGKx2FwseavVjBf3ml+ZmfiJkeDg/Z1Pay7RX6uyt836GY+KmZV4Thhqcad9nvJL56X+Wn4mLzTHYtt4itKbfeTf6mXZJmGYzUMJRlP0Wnzb0XzZ4Z5dicuxmOwuJjyziqd1dO11JrVNrZoAAAAAAAAADN84wWVYZ4jFT5Y9O7fZLq/wCnoZ3xpi8bm8cxwy9m4K0ertr8XR3u9NvzMl4ezDiutHMc4m/ZJe6lZc3pbZd3u+ncweBw2Coqjh4KEV0St/XqAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAGd8RZfk9JVMVOzeyWsn6Lt5uyMy8WMLGDWBouUu89F9ybb+9GZ8aZ3mE7zruK7Qbivwd382yFOviqloJzk+123+pl3Aee42aj7BwXefu2+T1+5GB8JsPHklisQ5W3UVZPyTevrp9xg+H8qwcVGhh4Rt/dV/ver+bKkG6bjHR20OH+CM/wABmsMXXxMZRv71pzbkkmle8Ve3S70AAAAAAAAADinxAweVOeHw69pWWlvsxfm+rXZfNoxOLzXiDHLnbq1JaJLp6LZLv06t9TKeBcHk2AqYvMJRlV5Xa/wxdtLX3l599u5wLXpVMhw8YSTcVqk9Vq9+wAAAAAAAAAAAAAAAAAAAAAAAAAAVqsKNOVSbtGKbb7JblHO8vrTowp1U3VTcNHqlu9vzsAAAAAAAAAAFfMcRTrVaccPKShHmTVrSf8q8/UwladahCpODg2ruL3XkwAAM24mynK4v6zWSkvsrWX3LX77LzMZ4t0lJrDYZtd5St+CT/MzHj3PsZKVq7hF9I6W9H8X4mX4DF5rjI0qac5Sau9Xv1b/VmX+FOY1ZJ4qrGEfK8n+i/H5GVeH+R4BO9P2rfWdpfcrJfhfzKOHo0IKFKKiuyVl9yAAAAAAAAAACc4Qi5Tdkt2+hxh4i0VSlhMrneT0c1sl/d7vz2XS+64V4MxOexqVW+WCslJ9XdXt3tG/le12ZFw3l+TUuTCw1e8nrJ+r/AEVl5HEOSU85wMsJUm4ptO630dzhfhajkNKpTpVHPnaeqStb0AAAAAAAAAAAAAAAAAAAAAAAAAADE0IYijOjP4ZJp+jVmYbhvAYephqlNO9BOMdej0d+4AAAAAAAAAAcQZrXwFXBxpWtVqxg7ro97eYABjcxwmCp+0xNWMF5tL7u5mnijlWHvHCRdWXf4Y/e9f8AyX5mc8eZzmd4up7OH8sNPve7++3kZdkmY5nPlwtGU/O2nzb0XzZgfCbGVIJ4nERg+yTl+N4/qZb4b5HhIR9rB1ZrrJu3/gKdret/PpbD4Whh6ap0YKMV0SSX3IAAAAAAAAAAAxuNw+CoSr4iajCO7f8AX3LqcX8aYjO6rpUrxoLaPWXnL9FsvXU4X8PMZmE418anTo72ekpei6J938l1MPh6WHpRpUoqMYqyS2SAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADjnO8BQxuBpTqe9TqxnJau0V1dvy38jD4iliKUatGSlGSumtmgJyjCLlJ2SM142yXAUXN11OS2jBqTb+Wi+bM38Ss4xjccO1Rh5ay+cn+iQo4zMK+nNVqP1lJ/mzK/DTOcXaVZKlH+9q/8AwFfq0ZN4c5PgLTqx9tNdZbfKO333IQhCKjBWS6IAAAAAAAAAAADOM4weVYV4jFStFbLq32S6v/h3ocScT47iHFKNmoXtGC1/4eT/AOGOCuAYYBRxmPinV6R3UPN95fgumuoAAAAAAAAAAAAAAAAAAAAAAAAAAABPEU41o0W/ekm18rX/ADQ2krsw9eGIowrU3eMkmvRq6AAAAAAAAAAq1I04Octkr/ccL8JYLOsJPM8yi5zruTWrXKrtaWe91pfS1tDgWFTAzxmUzldUJrlv/LNXX5X9WZnnuW5ZFPF1lC+y6/crv8DOPFaMJunl1LmS+1K+vpFWf3tehnHEmaZtL/aqra6RWkV8lp83d+Zl/Cmc4+pGNLDys+rTUfW70/XsZN4X5dhoqeNk6s+20fw1fzevYweAwuDp+zw1NQj2SS/IAAAAAAAAAAAAOIuI8HkmF9tXd5P4YreT/Rd309bIzHMs04lzBcycpy0jFbJdkunm36tnB/A9DJoKvXtOu+vSPlHz7v7rLcAAAAAAAAAAAAAAAAAAAAAAAAAAAA4txH1TMctxUpWipuLf+NJa+WhxpjY4TIsTPms3HlXrLTT7zJ6MqOW4elLeMIr7opAAAAAAAAAAY2nKrhakI7uLX3o4BxEK3D2H5XrG6fk03+ln8zPOL8RlefZg8Gk3Pljd62cI8rstm7330XZlLDZlnWLk6cZVaktX1fq+iX3LoZL4WYvEQVTH1PZ/3VZy+bvZfj52Mm4OyjKWpUKV5r7UtZfLovkkAAAAAAAAAAAAAcTcTYXI8L7SprN/DHq3+iXV/qN5rxPmf89SX3RX6RX9Xb14W4SwmR0Pd96q/il+i7L8935AAAAAAAAAAAAAAAAAAAAAAAAAAAABxHkdLOcunhJuzeqfaS2f6PybKnB+Nxyy+njal4UV76u3zST0+VtG3rbQAAAAAAAAADiDivLslpc1aXNPpFW5v+AvNmdeIecY6rJUKnsqfRR0dvOW9+9ml5GRZjnuHXssvnJKrJR6Wcntq9FJ9007dTLvCipN8+PxGr3UVd/+BP8AyZlGRZflNJ0sHT5U9Xq22/Ntt/oAAAAAAAAAAAAAZtmdDLMHPF178kbXtq9Wl+pPxOyV0Kk6alzRWiaS5n0S1fz7L7j/AIVOKs2/mqS/8BjH9Ir8X3b14c4awmR4X2VFXk/ik95P9F2XT1uwAAAAAAAAAAAAAAAAAAAAAAAAAAAMRiKWHpSq1ZKMYq7b2SI8e8OSV1il/wCAyX/tpmHixg6NZwwtB1Irq5ct/RcrdvW3oZr4qZhiIKOCpqk+rbUn8rpJfNP5FPjbiTHSWE+sfxPd+GK+LTdRTXqjh/L6uX5ZRwtVpygrO17fK6T/AAAAAAAAAAAM/wCNcqyjmpznz1UvgW9/N7R+ettbMzrxAznMoumpqnB9I3T+bvf7rJ9jJ8ix+b4hUsNBu+7d7Lzb6fn21OHvDXAYSkpZilVqer5V8tL/ADXy6nFWHo4eeWUqMVGKrwskrJb7JAAAAAAAAAAAAAAcUZhlmDy6cswXNB6cvWT3SS+V+yMBgMRmGJhhsPHmnLRL9X5Lq3sjhfhnDZHhFShrUfxS7v8AyXRfPdgAAAAAAAAAAAAAAAAAAAAAAAAABKUYxcpOyQuOuHZTcFildeUrffaz+8xHi1hoV5QpYZygtnzWb87crt95mviZm9fEueDl7On0jaLfzbT/AAt+pmPGWeY9ctXENLtH3V8+W1/nceKryg4ObafS7t9wuDM+lFNYWVmr9OvzMr8LcyxNH2mJmqT6Jrmfzs7L7/WxlnhPQp1ebG1ueK+zFct/V3b+S+8q+G+SKN6EXCa2fM3Z9Hv0epk3DWc4TGwr4rMHVhG/utPW6a6ye17gAAAAAAABm2dYDK6PtcXUUV0XV+i3f9XM68T8xxTnTwSVOD0T3n632XyWnfqcN8NY3O8UpcjdO/vS7X31e762V33Ml8NMqwXLUxN6s13+G/8Ah6/Nu/YhThTioQSSXRaBx3nWYwzSlRo0OaNGUaidpO7S2dtLehlGLq4vAUcRVjyylFNrXRtba6gAAAAAAAAAAAAGYY/D4DDTxOIlywitX/l5vZLuYvFY7jPO40ovli78qe0YrVt+b693ZbWMvyOvwjnmGfOqlKv7jdrNN2st3bWz31V77AAAAAAAAAAAAAAAAAAAAAAAAABiMVQw8eatNRXdtL8ziDjfLMppRkpKrOW0YtPTu3rZfn99s38Va1bDqOAp+zn1btKy8vP1X56Yzj7PcXhZYapVVpaNpJP71bfqRxVeMXGM2k91d6lLhTOq1CNenhpOEldNLdd7b/h57GW+F2Y4rCRrVaipyf2WndLzs9H5WMl8LcHSpS/tGXPJ7craSX6t+asjAcCZBg5c0aCk/wC9eX4PT8BZVgFJSVCF1s+VXXpoAAAAAAAAAGMxuGwdF1sRNQiurdv69DivxKpzo/V8pk+a+s7W07RT117tKy23uo08zzvGac1WrL5v/gJfJIyLwsw8aUamZzbn/LF2S8m9352a8m9zDYWjhaMaNCKjGOiS0SAAAAAAAAAAAAAABtJXZx3xTHOq8MLhG3CMmkl9p6JSX3tJfPrZcDZHVyfP/YYlr2kqPNbteaVvNq2vz6anHtvqmFS+L21O3rr+gAAAAAAAAAAAAAAAAAAAAAABXqOnSlNK7Sbt3t0OH+P45xmEcHHDOLd7u97WV9VZenqwrVqVGDnUkoxXVuy+9md8c5PlcY3n7ST6QtL5t3svvv5GceK0FGCy2ld9XNbeSUZfjf7zNOP89x9l7X2a7QvH8buX42MRjcwzGUYVqk6rV7Jtyfna9+xknDGY5tilQpQcV1k00o+vn2W5l/hRiI4uLxdaLpLdRvzPy1Stfvcj4Y5CqqnaVk9ubR+T0vb5k8py+duahB27xX+QkkrIAAAAAAAAAAOJ+LcFkdH33zVX8ME9fV9l59elzNc4zLPsWpVm5yekYpOy8oxX/Dvq2cPeGeOxFWNXMV7OmtXG/vPy02v1u7+XbLcpwOW0fY4SmoR8t36t6v5sAAAAAAAAAAAAAAA8RuMHHmyrCS12m1/6Sv8A277u54ccIpRWaYuOv/LtP/0r/wCR+/szjijjcDicPneCjzOleM13i+/lq79rp9DK80xvF+b0KsqXs8PQfM9b3kttbK7vbS2iuAAAAAAAAAAAAAAAAAAAAAAAcbZ5F4P+z8DVi69WSp2UlzRvvdJ3XbXuZpwRW4cwsM1wVdyqUmnJNWTWztbW3dPdX1Mz8R87xtKNOMlStu4XTfzbbXyfqYzNcxzFxhiK0qltk23r5LuZPwnmuaVvZUaTjbdyTSXzt17JP7rmVeFmOqV7Y+ahTX8ru2/LSy+f3dsr8N8kwUuecXVf9+1l8kkvvuYLIMrwVX22GoRhK1rpJOwAAAAAAAAAAAYivToUZVKklFJXbeiRiPEfP61J0/aKN+qik/k+hlGS5jn2L5KKcm/ik72Xm3/TZw5wngMkpJUlzVHvN7v07LyXzuAAAAAAAAAAAAAAAHG3E0clwH7t/vp3UV27y+X4u3S5wNwpLOsU8VitaMHrf7Ut7enWX3dbqMYxioxVkjHY7D4LDyxGIlywju+13bp5s8NszwlTKKeDhO9WHM3HXROTs+3VAAAAAAAAAAAAAAAAAAAAAAGbcVZTllJ1K1ZNp2tFpyv6J6ed7GbeK81WSy+inBbud7t+ST0+d7+RiuN8+xE5SeJav0WiXkv6v5nCVKpTznB1qkWoSmkm07N7aPa9ziyUI5Fi3Lbkl97Vl+Jw54cZa6NDGYhufNCMuR2tdxTe2rSvovvuYDh7KsBU9phsPGMu6Wv37oAAAAAAAAAAAAM94iy/JqPtMVPV7RWsn6L9XZeZxLxbjuIK6p25ad/dgu/Rvu/wXTrfNvDKlhMnlXpVHKtCPM9uV2V2krXVltq7/PThueHq5Th6lCChGUU7JWSdtfxAAAAAAAAAAAAAAADF4qjhaE69aVoxTbfkivUxvF+fWhpzOy7Qgv8AJavu35mW5dh8uwkMLQVoxVl+rfm3q/MM0y2hmWEnha9+SVr20ejT/NGR8I5Zk1aVbCJqUlZ3d9L3/QAAAAAAAAAAAAAAAAAAAAAx2dZdgoTlXrRjybq6uvlvr0VjNvFfDQSjgKTk+8tF9ybb+9GN4zzvFVak3iJRUr+6m0kn0S6fn5mXZNjsxrQo4em25baaebvtZdzKPCnEVIyeYVeTso6/N9Pl/Twfh7kGHoxhOjztdZN3f3NL5WOLOH/rmT+wwaUZ0mpQS0Scei7aXt52bIZ7nfFc6eUVEoxuudpWbS3vd26Xslq/Ip0404KEFZJWXyAAAAAAAAAAAADi/iyjkWF920q0vhj/AO3PyX4vTu17LN+JMxvZ1KsvkktvRRX5+bMXwXRyDC4XF1p81T21Pma+FR1ul31tdv5Ja3zWpCngK057KMm/SzOCISjw9hVL+X822vwAAAAAAAAAAAAAAADxN4m9tU/svDy92Os33fSPy3fnbseH/DP9lYD29aNq1TV94rpH9X56dAAAAAAAAAAAAAAAAAAAAAAOIeLstyWDVafNUtpBbv17LzfyuZr4o5piYwWDiqNr32lft8UdLenzPZ4vH13NRc5zfRNtt+hlPhZmNeaeNmqcfL3pf5L7/kZZ4d5JhKPJVh7WV73l+VlbQp0oU4KEEkloktEl5IDE0VWozpXtzJr71Y4f8O6WUZhDGRxDk4305bbpre77gAAAAAAAAAAABxfxph8kp+yp2nXe0ei85fot35LUwODzPijNbSk5TlrKT2iu/klskvQyjJsLleGhQoL4Va73dm3q/Vt+VziXKFm2V1cJeza0fZrVfK6s/IhxHnmcQp5HKrBKTUXK6u15yvaX/busttbmDwtPCYanh6fwwSS9ErAAAAAAAAAAAAAAAHF3EEcly2VZP95LSC8319EtX8l1OAcuoZpnilipXcU52f2pJr797vvbtcAAAAAAAAAAAAAAAAAAAAAOM+KcLXwssry+oqlepJQsr6Xeuu176b6X8jPOE854fw8cb7e62bi5JxbVvLR7X+RlORZnndZrDQcn1k9l6t/8F+RlnhRRpYhTxlbngl8KTjd+t9l+PkZRk2CyrDfV8JDlje/dt922AAAAAAAAAAAAAAY/G0cFhp4mt8MFd+hm/ihl8MJL6gnKq9FdWS833t27+RkWQ5hxJj5Pmdr3nN9L/m30X5IyTIcFk+GVDCxt3b3k+7f6bLoEkpJoy/wzy7BYuliYVptwkpJPltdO/YAAAAAAAAAAAAAAAMZVp8V8WxwsnehSuvVR+J/9vSsr9rHFWRYLJ6mFzXAwVN05xUktE4vR6d+nmnqAAAAAAAAAAAAAAAAAAAA2h2DibjPAZLBxb56v8ieuvVvWy/HVaHEPGmZ5zaFRqEFtGN197vd/l5HBGX1qed4KtWg1Cblyt9XGL2+bRxxGEuHsUpbcv43VvxMmo06WXUIwio+7HRK2tlfYAAAAAAAAAAAAAADj/NKlDL44GhDnq4h8qXl1frqkvW/Qr8HZhg8bh8NjUoKq0lLdK7S6dfL8bamU5VhcrwkcLho2ivvb6t92/wCtAAAAAAAAAAAAAAAAA45zz+ysonKDtUn7se93u/kvxseHang83oVqytCspxi31at+unqeIVWMstp4RP36tSEUuu9/8vvQAAAAAAAAAAAAAAAAAAAHFE6tXNsswlObjzTlJ2/uRv8Aim0ceU6sshrTpTcZQ5ZaeUl/w/qkcQeI+YZjSlh8PFU6bS1V+bz1von2S23ZlGQZjnNdKhByTdnJ3sr7tv8AHv8AeZF4d5Vls1VqXq1F1l8K9I/5tnHuFrU8DRzDC/HhpKS/w7Nem112uZhxdV4rdDKMPS5PaSXO276LV202Vr93ZEYxjFRjsgAAAAAAAAAAAAAAzHMMPl+FnicRK0Iq7/yXm9ku5wxnNXPuLY4qvooxlyR/lVrffrdvv5Wt4iwX9iOr9qE4SXre2nybAAAAAAAAAAAAAAAAA4yx9XiDiGOCwzvGL5I9r396Xpfr2VzijhKNXIaeGwUW6lC3JbRt3Sl23+J+aOFOG86rZvDF5ypNUlePNJS97p1e2/qkAAAAAAAAAAAAAAAAAAABxbnmBwmf5dN1Fem5KX91TSjd9rau29jj/MqVHI50ou861oxS3d2rteVvxa7mR+FdFUlUzKbc9HyxeiXZtq787Wt0b3MPhqGGpKlRgoxWySSX3IKtKnVg4VIpxe6auvuKOVYCjNVKVCEZLZqKT+9IAAAAAAAAAAAAAAHZI454nqZ3jlhcM70oO0UvtS2v59o+WvUhkWI4Rhg82mnKV2qiXRSVkl5rXXbmstjOOJMPxNicLleATcZSUptq1lHVr5at9LpJN3AAAAAAAAAAAAAAAADjDOv7JyipXi7Tfux/xPr8ld/I8K8j56lTM6q+H3Y+r+J/dp82AAAAAAAAAAAAAAAAAAAAcc8bU8tpPCYOadd6Nr7C/wDkuy6bvpfJsgzHO8Q4YeN3u5PZa7t/02cP+GscDjoYnF1vaKGsUlbVbN3b23suvXuAAAAAAAAAAAAAAAFWpClTlObskm36LczLPOKs5oV8VhIyjhdVZKPw7Pf3m7buOi8jwx4Zpzj/AGrXSbu1BdraOXr0XbV9jiHA1cdlVfDUknKcWlfa5wFwjmmTY6pVxSSjKNtHfW6f5XAAAAAAAAAAAAAAAADxMzyONzGODpv3aV0/8T3+5JW+Zk2WU8swFLCU9oK3q+r+buwAAAAAAAAAAAAAAAAAAAlKMYuTeiJ0uKOJp1MwwdV06SbUI8zjdLtbRvu3ZX0vZacL8JYvO8Y4zTjTi/fk979Uv7z/AA3fng8HQweHhh6EVGEVZJf19/d6gAAAAAAAAAAAAAAAY/PcswFRU8VXjCT6N6+vocWV1/q/ialN6ODt6NW/JmQYanh8pw1KC0UI/krv5vU4EpqjRxuHj8NOvUivJKwAAAAAAAAAAAAAAAAGLxNPC4edeo/dgm36JXZw1Twuc8Qr+0UnGo5Nptq7abSumnvsrgAAAAAAAAAAAAAAAAAAAcecRU8qy2VKOtWqnFLsmrOXyvp526XOEKcIZDhFDbkT+b1f4nC0VDNs0hHb2ifzabf4gAAAAAAAAAAAAAAAVqsaVKVSW0U2/kZLwXDiPD1M1zCpJTrNuNraK9le6d9rJaaLz0xee1sDw5iMnrzTrQm4a7uG91+S8nptpkPiJllDJYQxMn7WnFR5bP3rKys9tVa97WdzgHD1oZR9YrK0605VH/29t96V/mAAAAAAAAAAAAAAAAGc8V5VlFaNHF1HGTV1o3pdrovI474ioVeG4zwsrqu7Lo7J3lo/S3zMn8M5Y3KY4qdVxqzXNFW0S6X66rttfZnBeZ1sfk9OVd/vINwlfe8e/na1/MAAAAAAAAAAAAAAAAAAAzrizKsoqRpYqpaT1sk20u7tsYieL4v4gtT0UnZf3YLr92r7ydupk/G/+r2FnlePpSdSi2o2tZq91e70WujSelvnwFhcQsBUxuJVqmIm5/J7fLdryaAsWC1yxYLFgLFgtcsWCxYCxYLXLFgsWDcsWAAADOYuWW4hLfkl/wCks4LqQqZBhXH+W3zTaf4ozXKpZ1xjWw1HVOXvPsopKT/T1sitwFkFavGs6FmraJtJ22ur/wDD9bkYKMVGKskBYCxYCxYCwFiwFiwWLAWLAWLBYsAAAAWPEDKMFiJ4KtVheUqtOm3d/A+ZtaO2/XfzPEWWHw+Mo5bhVanRjort2cnd7tva25lEoTy7Dyhs4Rt6cqOBWpUsdOPwuvUa9PdAsWCxYCxYCwFiwFiwFgLFgLFgLAAAAAAAXOG+HMHxA6+a5jHn9pKSirtJRTsrWa9F2t3OCsqo5TnWPwSV3HlcW9+V3dvxV+7XoYzJsuxlWNXEUIzlHZtJv/hvJmysh6hcuBcbuFy49QuXC5cbuFy5uFy4XLj1C5cC5cAAA0M7x2EwWX1a2KdoJNO27vpZeb6GV8Z5hlFKrhsC17KTbjzK7jfqrWV7b3TV9bHhTgpyp4nMKmspvlTe+msn82180XLhcuN3C5ceoXLhcuPULlx6hcuFy49QuXAuN3C5ceoAAAFzifNs4zzMJYfK6TlTw01K6S+ON9bvTuklvvr04YwdXiLiGrUxq15ZOStaza5ErPa19F5Ec74nyWDyKNLmnqoSs27d49Guzfw7PbThbJ3lOVUsLLWau5f4nq/u2+QFxu4XLj1C5cC43cLlx6hcuFy43cLlx6hcuFy49QeoAAAAADV1Y8OqkXkUKP2qcpxfrzN/kzKqka/FmOnDVQhCL9d/818gAAAAAAAAAAAAAAAA41oLH4zL8tl8FSbcvNQSbX3NnH/D+CqZHOrTpqMqVnGyS0uk1p0t07pHCWXxwOSYait+VN+svef52AAAAAAAAAAAAAAAAAqz5Kcpdkzw7oRhkFKp9qblJvu+Zr8kjJaFOhxXmEYL4owl82tfverOIOM6GG4lp1XSb9hzxeq1urXQAAAAAAAAAAAAAAAAAABjsbQwWGniK8rQgrt/1+HmYHiHNaGLqVMJV5JVm77Wu356LffS3dHBWQV8pwM/rWtapJyk739Nfx9WwAAAAAAAAAAAAAAAA4wrxwWZZbjqjtCE5Rb7c6Sv8kmzj/H0qGRVKd7yq2jFd7tXt8v07mCouhhadJ7xil9ysAAAAAAAAAAAAAAAABK3K77HBPGWV5fgp4HE1bKnKXI7O0ot37Xve7s0t15nBVeWZ43HZs1aNSUYxv2grf5fO5VynL6s3OdCDk924pt/gAAAAAAAAAAAAAAAAAAAcfReIwuGwKdlWqwi/TW/42ZxTkGDr5DVoQppKnFuFls4q+nraz73OHcVLFZRhq03eUoRv62V/wAQAAAAAAAAAAAAAAAA4ly7B47K61LFvlgk5N2u48qvzJbu3bqtOpwLgsNjc/hTrzc4003C99XF6aa2X2rabW8gAAAAAAAAAAAAAAAABpNWOMcrw2XZzVwuFjaC5bK7e8U3vruzK8DTwOCpYaCSUIpad+r+b1fmAAAAAAAAAAAAAAAAAAAHH03h8Nhcba6o1YSfprf9F8zinOsLh8hrV1NNVItR83JWVu+935I4bw0sNk+GpS3UI39bK4AAAAAAAAAAAAAAAAY6hSr4WpSqq8ZRafo1Z/geE+X0auJr4qS96moqP/b3Nf8AJAAAAAAAAAAAAAAAAAHFmcyynKamIp/Hoo/4novu3+Rm/BedYPCf2tiJqc1aUk23Jbb33t17ea1OGsbicblNDE4n45q70t1dtPNWAAAAAAAAAAAAAAAAAAADMMBQx+Fnhq6vCas/+B5rdeZlfhdi6OMpVMVVhOnF3cVza+W3fcAAAAAAAAAAAAAAAADNJTjgazpq8lGVkt27OyXqeEtFxweJqd5Jfcv+CAAAAAAAAAAAAAAAAAccJSjgIy+F4inf8TO4wllmIUtuSd//AAFnBrbyDCX/AJUAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAYXA4fC8/sIKPPJydusna7+dgAAAAAAAAAAAAAAAADjbK62PyeccPrUg1ONt7x7edr28zHcf4rOsAssw1BqvV91u+mu9l59b/AAq++5luCjgsFSw0doRUfuVr/MAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHZI8Mq+Hlm2JlOSUpL3btXd227dXotbdNwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAKqk4NR3scC8DYvB454zHRcJU37qumpXUk27XdtVbYAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAACSViKTRZFkWRZCV2W1sNakkkFgQANajWiDoDSFuPcErsasyysAW0A+yCVxqxylhqxyprcasAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAEtiIALct7w9Whq7GhpWOVNCHHQSuNWCW49kcoldDVho5S12NWFuSHsCVxx0GlY3HFWPsgbpD3HoyXQJbgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABLVm0gAHub2N2NWQ9kJMW5HctpoNaWHGxLc6IlufZPsnVC3Grsashbm6H1BLQtoPZAkfZAe6Q9yW4+hHceoAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPWQ9wAHsLYWx9keyHvYW4lqdAJbnYktT7J9k6oS1O59kjuR1FrctoLYux7IjuLVn2RoSOo1qS3H0FsAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAArXJNMumAF9LF9LCdhu6sXVrHMX1LoTsNphdWG7l0dLF9LDdzmRcburCdhOwmPZCdhtNF1awtDmRfSw3cTshu6ObQHqX0sLRj1AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAtYWqAAAAAAAAAeoAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABFaD+IlsR2AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAFsP4iWxHYAtcAALMAAAAAsAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAS3ErltACIwABu1iW4AtyW5IAABu4JaC0Vx6q4tC90c2hJ7Cd9AL20GtRr3QAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfxGzADoj7RdXsLYeqJdCQNXZo2Xuhq8h6sjuPVIerG7okdTd2L30Foha6l7j3LXD7J0HZvQasi1yW5ayuBLcaux6oAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAH8RJC1QA7H2hbn2T7JLcluF9bCtYbVrIe4IWqNNRtWsh6s6i2I7is1Y+yLcZG1xJdx7H2QWqI7hHsaDtbQvc0LKwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAP4hq6I9gErsep1EP4R7Ia1uWTbFudRa3Q1YeyLKwtmR3FrdDVi2tzuRLWQlbUWqErMsrAtmLVWHtY5VexayOglcSsAlbUep9kAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAuwuwuy7AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA//9oACAEDEQE/AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAACMoyWgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABg8YlmuJwr/uyX/gKT/T8QAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADMMxWF4lda+iaT9OVJm4AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAGbVlXx1aousn919PwOG8d9by2nJv3o+6/Vf5qzAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADJccsXhb31i3F/J6ferMADQbADN8YsHgata+qWnq9F+IcEY72eJnhm9JK69V/mvyAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAMdio4XDTrS+ym/69TgnHuGLnh5P41deq/zV/uAG0NgIA46xbjRpYZPdtv5aL8/wDA4qWFxNOvHeLT/AM/vWhTnGpBTi7pq6AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA41xLpZcqa+20vktfzSMqlOniVWhvD3vkt/wABNNXQMAEAcWYp180mr6RtFfLf8WwwWH+sVXT8pP5qLa/I4UxX1jK6d3rG8fu2/CwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHHifsaD83+SOHYQk8TzdKU/0KCaowT7L8gGhoBAGY1Pa4urPvJv8Q4Xgp5tSi/73/pLOC706eIoP7Mv+B+gAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABGEpyUYq7Zjckx+CoxrYinyxlotvxV7r5gAAAAAAAAAAAAAHFWXTxuXtU1eUXzJd+6+4yDC18TjFSp7NWl5R6/fsABZDQgCt/El6sOCsK6mYOr0gn970/K5w1HlzDHpbc/6zAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADA5VjMdNRw9Nyv16ffsYHw9UqaliqrT6qKWnzd/yMDlOBwMEqFNJpWvZXfq9zPuIsoWErUJTU5Wa5dXr0120eu+nqAAAAAAAAAAAAAAEacItuKtcAALAGY0vZYyrDtKS/EOA6qVWvT6tJ/df/ADOGLVKuMrraVR/hd/qAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABlXC2YZjGM4R5YN7vT5pbv8ArzMs4Hy/DLmr/vZeei+67v8ANtFWvhcHSvOShFL008kZnx/QpT5MHDnXd3S+61/yM04izDMJP2k7Rf2VdR+6+vzAAAAAAAAAAAAAAAAAAA4twroZpN9JpNfk/wAUwyfM5ZfXlVju4tfNrT8UjhbCuhldO+8ry+/b8LAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAJNuyMp4RzDHu8o+ziusk19y3f4LzMr4Sy3A2ly8811lr9y2X5+ZmWd4DLo/7RUs+i3f3L9bLzM049xNaKjg48nduzfy6IxGIrYibnVk5SfVu7AAAAAAAAAAAAAAAAAAADjHK3isIq9NXlT/Lr92/3hlGXTzDFwoR26vsur/rqQhGEVGKskAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAWvojLeC8yxbjKovZwfV728lv99jLOFstwDjKEOaa+1LV38ui+SM24iwGXQvUneX8q1f/A+ZmvGuYYu8aL9nHy3+/wDysSk5Ntu7AAAAAAAAAAAAAAAAAAAAGrmO4Lwdeq6lKThfdJXXy7fkZVk+Fy2lyUVq9293/XYAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAybhbHZk4y5eSm/tP9Fu/y8zK+Hcvy67owvLu9X8u3yMz4qyzAScJT5prpHX8dl99zMuM8yxcmqcvZx7Lf79/usNtsAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAynhvH5laVKNofzPRfLq/kZLwjg8vqOpNc8tLNrbvZf0/MzTPsBlsbVp+9bSK1b/AMvnZGccY47HxlSh7kH0W9vN/wDAQAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABgspxuOT+r0nJL7vv2+Ri+GcdhMD9bxCUVdKz31/D8QAAAAAAAAAMFl+KxtT2eHg5Py6er2XzMl4HoYfkrYt801ry/Z+ff8F6mY5vgMrpr20lHTSK3+SXT7kZxxtjMXzU8N+7g/8AwL7+ny+8lJyd27sAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAwWAxONqeyw8HKW+hguCM0xF3NKml/M9fklf8bGF8PMPGzr1nLySt/n+hhOH8sws4zo0UpLZ7v8AG5i8dhcJFSr1FFPu7HGufYLFYWOFw8+Z3TbW1kn173aAAAAAAAAAEm3ZGR8DSrwVbGtxT2it/m+npv6FLD5flOGfIlTgt3/m3q2Zxx7yz9ngIpr+Zp/gtPx+4xGIq4io6lWTlJ9XuAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABguFc1xcVOFK0X1dl+D1/AwHh7Nu+Lq6do/wCbX6MwnBmUYdpum5tfzO/4aL70TnRw1Fyk1GEV6JIx3GGU4VK1TnfaOv6pfiYrxEk3bD0PnJ/ov8zFcU5tiFJSrNJ9FZfLTX8SUpSd27gAAAAAAAAGUcP43NJ2oxtFbyeiX+b8l8zIeG8LldNO3NUe8rfguy/Pr5cQcYUcvk6GHSnU69l623fl97MyznG5jK+IqNrotkvRf0wAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAC19EYHgzNcS/ehyLvL/JXf3pGB4AwVOCeJm5y8tF/n+P3GAyXAYC7w1JRb66t/e7sx3EGWYKThXqpSXRXb+5Xt8zHeIGFjG2FpuUv72i/Btv8DFccZtWTUHGHov8AO5WxNevJyqTcm97tsAAAAAAAAACEJVJKEFdvZGR8CybjWx70/kX6v9F95j8ywOVYdOq1GK2it35Jf0l1OIOLq+Yr2VC8Kf4v1t08vvv0AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAy3g7MsZZyjyRaveX+W/32Ms4DwdC0sVL2j7bL/N/evQoZdg6FvZUoxt2SRmPE+WYHmjOpeS05Vq7/l97Mf4g15SthKaS7y1f3J2X4mP4izPHR5K1V8vZWS/C1/mAAAAAAAAAABluVYrMa3ssPG76vovNv8AryMh4WwuVxVR+9V79vRdPXf8jPONsPhXKjhFzzWl/sp/r+XmYrF18VVdWtJyk+r/AK28gAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAy3hzMsws6VO0e70X/B+SZl3h/Rh72MqczvtHRfO6v8Adb/KhleBoJeyoxVuyX5mZ8SZdl/NGpO8l9lav/L72jMvECtNOODp8vnLV/dsvncr5zmNe/tK8nfpd2+7YAAAAAAAAAAAOH+FsRmNWM6sXGlvfa/kvXvsinTy7J8I+W0Ka39fxbf49DiDjGtjk6GF9yn1fWX+S8vvfQAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAApUqlWahTi5N9Erv7jLeBcfibSxDVOPnrL7lt82vQy3hTLMDZxhzS7y1+5bL7r+ZmGdYDAL/aKiT7bv7lr+hmHiDBRSwdK76uX6JP8AUxWe5liZOU68teibS+5aD1AAAAAAAAAAAA4Z4N9qvrGPh7rWkdU/V229Pv8APNs6wWUUPfaul7sVu+i06Lz2/IzHNMVj6jlWlpdtLor9kAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABhMFiMZUVKhByk+xlnh/OS5sdO3lHf5tpr7r+pg8uweApctCCilu+vze7+ZmXGeWYS8YS9pLtHb79vuuZnxnmWMvGnL2ce0d/v3+6xKTk7t3YAAAAAAAAAAABSpTqzUKavJ6JLc4b4OhhOXEYxXqdF0j/m/wX4nEfFlHL1KhQ96r+EfXz8vvMXi6+LqurWlzSfV/1+AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAYDLMXj6ns8NDmf4L1b0RlXANJRU8dK7/ljt83u/lYwuDwuAoezoxUILX/gtv8ANmaceYSinDCR55d3pH/N/cvUzTPMbmU3KtPT+VfCvl+r1AAAAAAAAAAAAAwuErYqtGjRjzSeyOHuGqGVU+edpVXu+3kuy7vr+BxJxmo82GwEtdnP/wCR/wA/u7km5O7d2AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAYfD1cRVjSpRvJ6JGUcAx5VUx8tf5V+r/AMvvFDL8pwraSp01/Xq397M24+hF8mAjf+9JO3yW/wB9vQzLN8bmMlLEz5rbLZL0SAAAAAAAAAAAAAMDga+NrxoUI3k/6u+yMjyHDZPh3JtOdvek/wBOyX/BflxRxbPGt4bCO1Pq+sv8l+fXsAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAJNuyMp4Jx2MtOv8Au4efxP0XT529GZblGDy2koUI27t7v1f9Izvjmlh5OjgkpyX2nsvRdfwXqZhmuMzCfNiKjl2XReiWiAAAAAAAAAAAAADA4GvjsRGhQV5P+rvyRlGUYTJcI9Ve15Sen/DJdF+pxPxTPMZOhQdqS++Xm/Lsvm/IAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADLcpxeY1fZ4eF+76L1Zw/wrh8siqk7Tq9+i/w/57+mxm/FuX5feClzzXRfq9l+L8jOOIsbmc/3krR6RW3z7v1/AAAAAAAAAAAAAAClSnVmqcFeT0S8zh/I6OTYRzqtc7V5S7Lt6L8Xr2txPxLUzOq6VJ2pLZfzeb/RdPUAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA4e4Sr5jatW92l+MvTy8/uuf8J+T4S2lOmvz/Nt/NnEPGVXGp0MJeFPq+r/AMl+L69gAAAAAAAAAAAAABRcmktWcMcMUsvpRr1les1/4D5Lz7v7tDjHiT63N4PDy/dx3f8AM1+i/F69gAAAAAAAAAAAAAAAAAA4Z4ZlmknVqu1JaebfZfq/l6Zlh4YfGVqMNoykl6JtCV3ZFbhbhyjN06uI5ZLdOcU/uaMyo0KWLqU6ErwTaTve69VuAAAAAAAAAAAAAAAAAAAk27I4a4NdOSxOOinppBq+/WXT5fqZ5xDhMppcr1nbSK/Xsv6RmmbYrMq3tMRK/ZdF6L+n3AAAAAAAAAAAAAADg7hiMIxx2JXvPWK7f3n59u2++3GXE1RVHgcLKyXxNd/5U/z89O4AAAAAAAAAAAAAAAAAAHDOPr/XMPhFL3Ofmt3dra/cZ3/yM8T/AI5/+lMwcOfE0492l+JxTPnziu/O33JIMiyKtm1d04PljHd727adWzP8inlNeNKU1JNXT2+9dPvYAAAAAAAAAAAAAAAAAFKjUrTVOmryeiS3OGuEqeBSxGJSlV6LpH/N+fTp3fEnGFPBuWGwvvVNm+kf82vuXXqitWqVpupUk5Se7erAAAAAAAAAAAAAADhHhuGYTeIxC/dx0S/mf+S6/wDDnFfE8cJQVDCS9+a3X2V5eb6dt+w227sAAAAAAAAAAAAAAAAAADhn/kb4f/EZ3/yM8T/jn/6UzAVoUMXSqz+GMk3beyabsUsZkfETlh/ZclV3abSTv3TW/dp7mIoToVpUp7xbT9VoZPCpPh3GKh8d1e2/Lpf8L/K5OrOaipSbSVl5LyAAAAAAAAAAAAAAAAADD4eriKsaVKN5PRI4c4ao5VS9pOzqtavt5Ly7vr6HE/GPNfC4GWmzkvyj/n93ceoAAAAAAAAAAAAAAZNlVXM8XGhDbdvsur/y8zN8fh8gyxQopJ2tBefd+m77v1JScndvUAAAAAAAAAAAAAAAAAAA4Z/5G+H/AMRnf/IzxP8Ajn/6UwyCcoZrh3Hfnivvdn+BxMorN8Rb+Y4fz2plOJc0rwlpJfqvNHF+CwsKtLGYVWhWV7eatfTputO9wAAAAAAAAAAAAAAAACMJTkoxV29kcK8ORy2j7euv3sl/4Cu3r3fy9eKuK5YuUsLhJWprRv8Am/8Atfz9AAAAAAAAAAAAAAAEnJpLc4fyujkmXOrXdpNc0n28vl+LM8zermmLlWlotorsv8+/mAAAAAAAAAAAAAAAAAAAHDP/ACN8P/iM7/5GeJ/xz/8ASmZjwxhFmOFwlGXKqkXd76pPXXv2+4yrhT+yK7x2NqRcKabVr793dL5JXu7GNxMsVialeW8m397Mq4YxWZ4R16EldStZ6dE73+ZxbCOFoYTL+a8qcXf1dv8AL7rAAAAAAABWxSo5jTpN6VIu3rHX8mwAAAAAAAAA4K4dVOmsfXXvP4V2Xf1fTy16nGPE6mpYHCy02k1/6Sv1+7uAAAAAAAAAAAAAAAcD5L9ZxDxlRe7Dbzl/wN/Wxxznntan1Ci/dj8Xm+3y6+foAAAAAAAAAAAAAAAAAAABCpOnJSg7NdVoycpTk5Sd2yriK1Vxc5tuKSV3slsl2sYnMsZiYKFarKSXRtsMNjcThZN0Kji32bX5FSpOpJzm7t7t7gAAAAAABxli5YfF4WpDeN3+K/yKNWFalGpB6SSa+YAAAAAAAAHC2QyzPFKU1+6hv5/3fn17L5HGOffUaCwmHdpyXT7Mf0b2Xb7gAAAAAAAAAAAAAADA4OrjMRDD0leUnb/g+iWrMyxNDh7KFClulaPnJ7t/m/uJylOTlJ3bAAAAAAAAAAAAAAAAKNGpXqRpU1eUnZLzZicPVw9WVKqrSjo0AAAAAAAAAAAABxvW58wjBfZivvbb/Kxwbjvb4D2LetN2+T1X6r5AAAAAAAABhMLUxVeFCkryk7L+vzJywnDuVJdI/fKT/wA/wXoY3GVcZiJV6rvKTv8A15LYAAAAAAAAAAAAAAA4FyZUaDx1Vay0j5R6v5/kvM4pzl5njm4P93DSP6v5/lYAAAAAAAAAAAAAAAOHcLgsvwDzXGx5tbQXn3Xne/ok2SxWA4pw1SlGnyV4q8b7/f26NdL3GmnZ7mRf8jTDf44/mjib/kb4j/EAGCy+jLhaScVzyjKe2toyXX0S+8AAAw9CpiKsaVNXk9Eu7JwlCTjJWa6MAAAoYqNTEVaPWDX3NJ/ncAAbNwAzzFrF5jVqp6XsvRaL8jhHHfVsxjBvSfu/Pp+OnzAAAAAAAADgHKU1LHTX92P6v9PvOLc6eY41xg/3cLpefd/Pp5WAAAAAAAAAAAAAAAMlyyWY46nh1s9/JLf+u9jjHNIZfgI4OhpKat6RWj+/b7wAAAAAAAAAAAAAAAOLJeww2Dwa2jBN+r0v+D+84ZxTw+bUJJ7tR/8AAtP1MyympXz6pg6K1lJ28k/ev6JFHhLCYfFUp4Wvz1KcouUW1tdXdlqu9ncz3D1MTntWjTV5SlZfgT4HwbpujDEXxCV7XVv/AAH4kvP52MlyhY7MVg6rcXrfuml/wDM+H44HK6OJnJ+0m9uiTTa879/uDCuNPH4bL3s6DT9Xq/8A0knBwk4vdAAAUasqVSNSD1i016ozfh7C55ho4qj7tSSTT73W0v8APdfgY3A4jB1nRrx5ZL+rrugAAMLm0Y8TVNfdm+T5pJL8Vb5gA2bggDPcZ9Uy6rVT1tZer0/DcITlCSlF2a1RgMXHF4WnXj9pX+fX7noAAAAAAABQoTr1Y0oK8pNJerM+xMMkySOHov3muVf+3S/P5tAAAAAAAAAAAAAAABwPl0cLgp46ro5fhFdfm/yRneZyzHGzxD2eiXZLb/g+YAAAAAAAAAAAAAAAHHP/ACMYduSP5sy2/wBdo235o/miPLSz/H4v/n3Tv8+WL/RnDeMq084o1L3cpWfnzaO/339TAU4PinE1pbU1KX5L8mzA5lVhmcMXJ+9zXfzev3rQo4eNHjFqOzTf3x1/G5neLliswq1G+rS8ktEvuMFQ+sYqnR/mkl97sYyrXXFcayg+VSjG9na1lF/LVnEFD2GaV4f3m/v1/UDh3hTLcdllLEVk+aV72dtpNfkj/UbKP5Zfef6jZR/LL7z/AFGyj+WX3n+o2Ufyy+8/1Gyj+WX3n+o2Ufyy+8wWDp4TDxoUr8sdFfUzfJcLmlH2dZarZrden6rYzfKquW4p4epJNrqu3p09H+WoABmGJWGwtSs/spv59Cl7SVaPI/ebVvW5gMUsVhadZfaSfz6/iDYAIA47xdqVLDrq238tF+b+4KcFOcYt7s4Lry+r1cLPenL8/wDgpgAAAAAAAcCZd9YzB15LSmvxei/C5xrmX1vMnTi/dp+6vX7X46fIAAAAAAAAAAAAAAAy/BTxuKp4eG8nb/N/JanGWNhl+VwwVHRy930it/v0XzYAAAAAAAAAAAAAAABxfD21HB4xbTgl81r+r+44awzxGbUIJbST/wDAdf0MNWWKxWbRjq3Fpf8AbqaMgi5Zrh0v54/mYBc+f4+kt5Qlb8DD03UrQhHdtL72Oalxikukbf8Akt/1MV/Hn6v8zhGh7bOKK6K7+5P9bFbxBrwqyjGimk3bV7HG9KKzT2sdpxi/0/QDLuLcwwGGjhqKjyxva6d9W338z/X3Ne0fuf8Amf6+5r2j9z/zP9fc17R+5/5n+vua9o/c/wDM/wBfc17R+5/5n+vua9o/c/8AMyqtXrYKlVr255JN2031/I4k4yhh74fAu8+sui9O7/BefSpUnUk5zd2929wADittZPWt5f8ApSMlpqpmNCL/AJl+DMjhyYNQ7SmvunILJjQAgDjStz5ny/yxS/X9Qo/xI+qMqX1fiPE0ukk383Z/qwAAAAAAAOHVHKuHpYuS1acv0ivnp95OcpycpO7eoAAAAAAAAAAAAAAB4f5bz1amMkvh91er3+5W+84pzL6/mVSafux91ei/zd2AAAAAAAAAAAAAAAAcPVsJm2WvKcVK0ou8H/l5q706p6bH1LB8LYapW9pz15K0dLfhd6dW79LHC2ZwwWZRnVfuzTi2/Pq/mlfyKeRZfkmKjjfac3M0oR7OTte99bJ9l95WzOOA4nniG/d5mn6NW/Df5EOHMuweLeayrL2S95LpfpZ3112SV9l68O4/63xH9Znpzcz9NHZfdoYrWvP1f5nB8o0Z4nFN/BTdvX+kHEco4jLMDiE9eVxfyt/kwAAMFRhWxNOnN2i2rt9F1fyRxHxfPFJ4bBPlp7N7N+nZfi/LYAADOsLLFZfVox3a09VqvyMDiHhsVTrW+Fp/czKqM6WDpqatJ6v1k7v8WANXLAgDip3zet8v/SUGW0HXxlKmusl+ZOPLxRFrrD9Gv0AAAAAAAClSlVqRpx3bsvmcb1o4TK6OCg97L5RS/WwAAAAAAAAAAAAAAAVH/YXDaS0m1/5NL/L9AAAAAAAAAAAAAAAAAJNyd27sG20rsLtq19AAAAAAAAD/AFey7619Z9n7179bX722AACwBxdT5c2qPuk/wS/QOE+X+16V/wC9/wCksa9pxMrfZp/r/wAEAAAAAAAOHYqWa4dP+ZHHuJ9rmapLaEV971/KwAAAAAAAAAAAAAABw3gPruZ0qTWid36LX8dvmeIOP569PCRekVzP1ei+5fmAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHHWG5cTSrL7St9z/wCCGVYj6vjqVW+ikr+nX8Dh+osZmWLxq+HSK9F/wyfzAAAAAAADhz/kbYf/ABI4ir+3zXET/vNfdp+gAAAAAAAAAAAAAAB4e4JXrYp9LRX5v9DOcb9dx9WvfSTdvRaL8EgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA4py54zLpcq96HvL5br7vxsBw7l7wWXwpyXvPV+r/wAlZAAAAAAABw5b+1sP/iRXqOrVlN9W394AAAAAAAAAAAAAABhf+EvhRz2lKLfznovuTX3AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAGdcG1J1XVwVrPeL0s/Ly8uhkXCEqFZV8Y02torXXu/8v8AhgAAAAAAAMprwoY2nUm7JMAAAAAAAAAAAAAAAwmHlicRCjHeTS+92OPsRGjhKGEhs3f5RVl+f4AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAcF4X2+bwfSCcv0X4tHG2L9vm0oJ6QSj+r/F2AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA8PMOlGviH5L82/0MfiXicVUrP7Tb+9gAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAZM/qXCtWts5KT+b91fkgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADPP8AZeFaNL+ZQX3+8/yAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAjFyaS3Zx/JUsHhqC7v/wAlSX6gAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAZPS9rmFCHeUfzR4h1b4yjT7Rv97/4AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHCVNVM5oJ92/ui3+hxxV584lH+VRX4X/UAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADg7/kd0P+3v/SJHFFT2mcV352+5JfoAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABwd/wAjuh/29/6RIzifPmNeXecvzYAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHBqvndH/ALe/9JZjJc2IqPzf5gAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAZDmVPLsfDEzTajfRb6poqSUpuS6gAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABlmBeOxdPDKVnJ2uf8Q6q/8AQQv/AAF/5n/EOqv/AEEL/wABf+Z/xDqr/wBBC/8AAX/mS8O6qV/rC/8AAX/mAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAFDBYnEJujTlK29k3b7hpp2e4YHB1MZiIYelbmlormJw88PXnRnvFtP1TswAAJRcXZrUAAAAAADKMbHBY2niJK6i72+R/xEPC/8+Zfej/iIeF/58y+9H/EQ8L/z5l96JeIWFcWvYy+9AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAEYuUlFbszHP48Pezy/C01LlScm+rfp1e9/NGdZhHMMdPEqPKpW09El+gcLf8jjD+v6Mzv/AJGeJ/xz/wDSmUoKpUjDu0jiDLqWX5hPDUm2opb76pNgBkeX08fmFLDTbSle9t9E3+hB0sJmSbV4wnt5KX+SOIcbQxuZVa9D4Xa3TZJX+dgAyTA0MdjoUK0+WLvrotk31My4Oy3C4SpWjXbai2k3HWy9PyAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAChUVOrCb6NP7jjjBVI45YtawqJWfS6VrfckzIcFSxuY0sPV+GTd/ubMThqdPMZ0I/CpuPyUrFPL6GA4ppUKPwprfXeNyWSZNHGVI5hP97VnJxV2rJydttLvz06WKGT4XBZ48LjJWgtU9r/AMv+T8zjTDZV+9qqf+0e7pf06eguGMspL6xXdqfs4tXdrzkn102008zJsHw9DBwrZhUvOd/d10s7X93Xzu9PLQ4kyiGW4zkpO8JJSi/J/wBfdYy7IMpweChic2nZ1NlrovSOrfV9F1OHsJk9POqio1L8vL7PXe8XzetvPYz3A5Q8bRhhZ80pztNX2u193UzXJ4wzmWAwq6pK/mk9fvP9XeHnL+z1U/2i293e9u3w/Le3UyfKIVc5jgcUusk7abJv9DA8L5OorCYmf+0SV7Xd49Uktm7bp3vvsYzDTw2InQnvFtfcYXC1sVWjRox5pPZHE+RY2GCw1TluqUFGVunn6eYAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHDfENDEUI5XmEbxekW/wT7eT6aepl+WPLuJ6eHvdJuz8nFtf5PzMd/wAjip/18f8A6UY3/ksYfL/0k4ixE62bV5N6qTX/AIDovyOJ5e3q4DEvecI3+9P9TjT/AJHVX/t3/wBJRxniJOjg6N9FBP5tJfp+JlHD2NzRt0ElFbt6L06t/JepxNl0oPLsLUd5W5G15OK/U41xDq5tOn0gkl91/wA2cIf8jqh8/wD0llP/AJHS/wCvn/txh6cXxdXqS+xG/wD5LFfqU8bUjjFir+9zc3zvcq0Yw4ypyX2k3/5JJfoSxdSWf+2vr7T/ANutb7tDi+CjnVdLy/GKZw7WjluW4jMrXndQjfvu/wBH8jI+LMb9djDFz56c3ZppaX0uvLutrGc4JYPH1aC2i3b03X4WAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABXT0MfL/AIynCOW/Ir+vvGOpSed1IJauo/xkY3/ksYfL/wBJM+hKGaYhP+eT+9t/kcRp0nl1B7xhG/4L9DjT/kdVf+3f/SUcZfFhf+vcTO8VVwOU4PCUJcqlHmlbS7dn+r/DsZnUdOjlNSfTX8YM4yoyp5zVb2lZr7kvzTOEP+R1Q+f/AKSyH/I6X/X3/wBuMM0+K8VTb+KFv/JYv9CGHnOsqKXvN2+d7FaopcY0or7Mbf8Akkn+p/zOP+4n/txxl/yO6/8A27/6RExNOcOGaSkmr1G//JXZkW01bc4zS/tqtb+7/wCkoAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAw0qca8JVPhTV/S+v4Cy3hZVFjo1VyrXl5la/wDhtzfL8LGOzyVfOPr8VpGSaXlH/Nb+pmNfIqU3m9OalVa92N18VrJuO6a630+ZisfhZcUwxCqL2envX0+G24lkGa1vr2JqqM4P3ldJStonbd3VtvQz7NnmGYyxEfhWkfRbffv8zilZVjsIsxpVP3rUVy3Xzut7rbscVYuhiJYb2U1LlppO3R9jDvKM3yyh9crckqKs1dJtK3fe6S263RxXmWFx2HwssO1pzLl6xV0o37XSIVcn4gwlN4yr7OrBWbuk39+jT38n+McRgcuz2FTDSvSi1rvpZKT8+r/Iz3D5Xh8yw+Jw1VPnnzS1TS1Tvpst9zM83hQ4iljaElKKa26rlSf4XQ4cOU8Q83VW735br4v8O9/J6J67GU4ybz2nmOL9yE3NpvRfC1o/K6Qstoum8z9pp7a3la973MphgsdnmMxCUZpez5Xo7e7Z2+44ky+OZ4GeGpyTqR95K/Vf5p2+ZkPDmMxGPgq1NxhF3k2mtumvfb8TPsZHGZlWrRd03p6LRfggAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAACEJTkoxV2+hgeDs2xMo81Pki+srK3y3/Ay/gDC0pc2Km59kvdXz1v+KI8BZYq/tOaTj/LfT0vvb8fM4ioZHUoU6GMqcij8PLuvlZ6fIxfEGXxwLy3D0HKl/M5Wk3e99tP8uiKGZ4rDU50sPNwjLe277aqz/LcwOYYjBYhYijK0l+Pe/e5xLxLjPqNCFO0fawvJrfXdLsvxAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAACFOdSSjBXb6LcwHCmaYyKlGnyxva8tPw3/AwPh9hqclLE1XPyS5V992/yMHl2DwULUKaivJa/N7v5mM4oynC3UqybXRa/LTT736mP8QZzp8uEpcsu8tfuS/X7jF57mWKTVWvJp9L2X3KyAMkqYKnjoSxqvT1vo30dtF52M2zfhzE4KUIK84xah7r000t8wAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAASbdkYHhTNcZZxpcq7y0/Df7kYHw9pqzxVa/lFW/F/wCSMLlmBwcU6NKMbK17K9vXf1uzH8QZbgWlWqpN7JXb/C9vnYx3iFJpxwtK3Zyf6L/Mx+dY/Hv9/VbXbZfctAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAwWAxONqqlh4OTf9avZfMy7w+m2pYyrbyj/AJv/ACfqZfkmX4BL6vSSffd/e9TH8RZZgrqrVV+y1f3Lb52Mb4hQSawtG/nJ2/Bf5ox/EGZY1y9rVfLLeKdl6W/rzAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAACMXJqMVdmW8I5ljKkVODpwe7ktvlo2ZfwRlmHjF1V7SS6vRfcnt63JSw2Do3doQXokjMOP8LTTjhYOb7vRf5v8AAzHiTMsfdVKlo9lovw3+dwAAAKdKdWahTi230SuzEZfi8PHmrUpRXnFr80AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAUqU6tSNOCu27L1Z/qbnf/Pj/AMmj/wDJAAYTBYjGVVSoQcpPt/Wi9TLfD+q5qWNmlHtHf72rL5XMDlOCwMbYemo+fX5t6mN4kyvCJ+0rJtdFq/TT9bGY8eY6tK2FSpx+9/jp+BicXXxM+etNyfm7gAAWuYHhbNMZFyhSsl/N7v3X1MJwDmNWX76SgvvfyS/Vor0vZVZU73s2vuOBMLhMRiqsa0FJpJq+ttbPTbqilhcNh05U4Rj3skjjbF4evlVqVSMrSWzT6PsAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAZBgKuNzGlTpOzTvfslrf+uouIMtq13g4V/3myduvk7crf4GZYCpgcVPDVN4v7+z+a1Ay3KMZmNVU8PC/n0Xq/6Zl3AOEpJSxU3N9lov83969CnRweX0HyKNOC9Evn/wTMePcFSjKOFi5y6N6R9e/wCC9TMc+zDMG/bVHbstF9y/W4AAot7IMj4WxOa0nVhNRgnbW976dPn3RhOAMvpq9ecpv/wFfcrv8TjDJMJllSksOmlJO93fa3+ZD4kY/iDLcFFOrVWvRav7lcxfHOVUoXpNzfZJr727fqV6iq1ZT7tv7zCY3EYSUpUJuLatdaO2j36bFfMcZiI8tatKS7OTf5sAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAOHajw2X47FR0lGKin/idv8AITaaaepxZL288NiutSnFv16mW8MZlmCUqdO0X1lovl1fyTMp4LwGDSlWXtJ+ey9F/nf5GKxeFwNHnqyUYr+tF+iMz8QKkm4YKnZfzS1fyWy+dzHZnjMdLmxFRyfnsvRbL5IAMNgMVif4FOUvRN/kYvCVsLWdGtG0la69Vf8AJmT4GGOx1PDSlZSe/wArlDg7J6UlL2V2u7b/AAvb9DNsDh6eV4iNKCjeEtklsvIOGOJsJlWBnTqpuTldJLyS3b8jGeIOKnph6Sj5vV/ovzM3z3FZpyfWLe7e1lbe1+vkAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAGRwdbKcfRW9oy+UW2wyzJcM8Lhp14JzhBJX6aXene5mWe5fl6tXqJS7LV/ctvnZGb8dYrEXhg17OPf7T/RfLXzKtapWlz1JOT7t3YGFynHYpJ0KMpJ9bO337GZ8NY/LsPGvXSSbto7tPz+7oHDGQZa8BSxEqSlOSu3LXXyT0X3E6uGwsEpSjCPS9kji3EYfEZrUq0JqSaWq2ukl+hleN+o4yniLX5Xe21zEccZtV5lCSin2Wq+buYnMcZiVatVlJebb/AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADJuHcbmkv3StDrJ7fLu/T52Mj4WwuV3kpOU5Kzb2t6f53KXAuX08X7fmbindR6ffu15fe2ZzxxisUnTwq9nHv9p/Pp8tfMlJyd27so0KteahSi5SfRK7/AAMPwVm9aKbgo+rX5K7JwcJOL3RwFSoVcZUhVgpPlurpO1mtr+piMdhMKv31SMfVpHFnEWW47Ayw9CfNJNNaO2m+oR4kzKnhIYWlU5Yx7aP79/usVq9WvLnqzcn3bbf4gAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAGAyrGY+ahh6bfS9tF6vZGA4CoUHGpWq80k07W00abTWt77dN9jMs2wWV0eatK3ZLd+i/pGZceY6u+XDJU197/HT8PmYbOuKsTb2Lk79eSNvv5bGa5Ji8s5FiEk5XtZ32t/mUpqE4y3syFPD0IucUordvRfeYni7J6EnF1rvyTf4rT8TG1KdXE1J0/hcm1fs3oUMVXw8nKjNxbVtG07dtCc5Tk5Sd2+4AcH4bBVHiK2KpqcacebVX2u3p6Io4zhfNpLDKj7OT0T5VHXycW9fXRmZ5W8vzB4as/dTWveL6/d+JxRRwVHMZQwluSy21V7dP63Dh2hgKscT9ba0g+W/fuvPawZZChPG0Y1/gclf0vqZ/Tw1PMq0MNbkT0ttsr2+dwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAo0Kteap0ouUnskrsyTgS6VXHv8A7dX6v9F95UrYLL6KUpRpwW2yXyMfxRmGKo1Hl9BqEU3zvslq0nZemrfkVas6snKcm2+r1ZwDhMLUws6sqac1Ldq7Sstu3UxGYYTDL99VjH1aRxpnGCzGVFYaXNyc19Gt7Wtf0CtjMTWioVakpJbJtu33gAAcLf7nmH/Xp/lITad0cT/7bg8DjH8U48rfmrfrcXCknmksv9quZR5k7b7aWuYbDyr14UVvJpfe7H+rLeLxOHVVP2MeZu2+l7b6djh/h6pm1SXvcsI7vf5Lb/gGZcOyweZ0sF7RP2nLZ22UnbVf8Ex+W1MNjpYOL5pJpKy3bt0JeH9dYa6rJ1bX5bael7/ja35mU5VLH46OE5uVu+62sm9vkYHgbEYnBe3dRRk1eMbdOl3fS/o7FSEoScJKzWjAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAyPg7FZhFVar5Kb201fou3m/lcy3J8DldJqjG3eT3fq/+GRPi3CVMfTweGfNzOzl0Xp3/AC9RZRg3V9tOHPPvLV/K+i+SSM8x2DoYOrTr1FFyi0lfV3VtFuFDMcXh6UqVGo4xluk7X+7UbbYAAAHC3+55h/16f5SDGf8AJPYC+/PL/wBKkY7Eew4vpy78q/8AAlb9TLcvtxT7G2kZyfyV2v0MkxP1nF5jW/mhN/K7t+BgKVSrwviI0dWp3lbflSj+HX7zJpylmeFUne04JeS5kYTDwq8W1pz2heX3JL9TC5zXWbxx0pauWvo3qvS2hTw0aHGVo7Su/vg7/jcp5xXlxIq3Npz8tv7t+W33a+upxRSVLOK8V3v96T/UAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAwmBxOLnyUIOT8kZFwRRw/LWxvvT/AJfsr17v8PUOO8bivr31Zzfs7JpdPn3+ZgsVLCYmnXgruLT+4x3FubYu6dXlXaOn47/iSlKTvJ3YAAAAHC3+55h/16f5SDiCLw2Dy/CP4kuZrzbX63OMKsqOeupHePK/uMTTjh81xePjt7HmXq1Zf+ks4Q+DGf8AXuRw9n9TKa7klzQl8S/Vea/ruZpgcPRz3B4jDK0KsoSXrzK9vXR+pglzcR46C3cJJf8AkpCDnNRW70K0lLjGml0j/wC2tlD/AJHEf+vi/wDSji//AJHVf5f+koAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAMBleMx8+XDU3L8l6vZGF8Pa84J16yi+yV9PXTX8PMwGX4PKcJyQtGK1bdtfNvQxvG+V0Iv2bc5dkrfi+npczHjfMsTeNJqnHy3+9/okVatSrNzqScm+rd2AAAAABwVhniaeMoRdnKFvvujBcGUMvn9azGtHkhrbo/Vv8ktTOM1eZ5n7dfDdKK8l/nv8zjf/AJHE/SP5GZ4uL4Xp1V8U1CD/AO3G/wDJnCHwYz/r3IyrhTEZngliKE1fmaafbTXS/wBxmsqdLNsvwUJX9i4Jvz5l+iT+ZWzGOA4qnWm/d5rP0aSv8t/kU+D6VDMnjpVI+wi+dd+68rLvfVGS4/6/xSsQtpOVvRRaX4JFD/kcR/6+L/0o4v8A+R1X+X/pKAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAOFsvw2OzGNHEK8bN2vbVehTpU6UVGCSS6LQzriTBZWuWb5p9Irf59l6/JMzriPGZpK03ywW0Vt8+79fkkAAAAAABSrVKTvTk0/J2K2IrVnepNy9W3+YVKk6j5pu78x1JuCg5Oy6dCFScL8smrlDF4jDtujUcW+za/I55c3NfXe/W/cnOU3zSd2SxmJlRVF1G4Lpd2+7YhOcJc0HZ+RzS5ua+pOcpy5pO78wAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADLcqxWY1fZYeN31fRerMt4AwtNKWLm5vstF/m/wMz4axFbOauGwdO0VZ9kk0uv3+ZkPDGFyuKn8VTrJ9PRdPz/I4j4yp4ZPD4JqVTrLpH07v8F57FWpOrNzm7ye7e4AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAZBjMHhcio15tQjbV7XabT9W7erMNnmOzWpy4Cly0+tSX/ALbHq/m/OxicXh8vw/tcROyXV7t+i3b7JfgZ/wAY4jHJ0cNeFP8A8mfr2XkvmwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADKOF8fmVpRjyw/me3yW7/LzM44aweAyKqqMbzVm5Pd2av6K3RfiHD3DOJzOEKmJk1Qjsr7662XRN7v7vL9xhKKWkIRXokjjXBTxlCOPw9Xnpx0srNLzVvxvqvTYAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADhOhhq+a06dePMneye10rq/3bGaZ7gMsh++nr0itX93T52RnvFGLzNuCfJT/lXX1fX02/M4S4YWOl9ZxK/dLZfzP/Jde707mc8TYPKYujGN5qyUVorW0fkun6HGeZV6sMNh5v7CnLtzP/LW3qcH13PE1MDJ+5VjJW87b/dcas7AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAGCwOIxtZUaEeaT/q77IzfK6mW4p4eo7tJO/qv89DDYmrhqsatJ2lHZlWpOrNzm7t7t7nC3CjxrWJxStT6L+b/AIH5nEGe0MnwqjTS52rRj282uiX47d7YnE1sTVdWtJyk+rOLY3xFCotpU4NHBsHLOaUukVJv/wABa/UqS5puXcAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADK8rxGZYhUKK16vol3ZlmVYLJsI+VpWV5SfX17Lsv1OKs2w+Z472lBe7Fct31s3rbpv/wAMHCfDH1+SxOIX7pbL+Z/5Lr327nEXENDKKChCzqNe7Ht5vy7Lr97WKxVbFVZVa0uaT3b/AK/AMtwuH4hyqGFc+WtR0XnH07bLya89auX0eGsuqznNSr1Vyq3RPe3Xzb72QAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABwRjcLh8rrVKlo8kryflZW/VJf5nEXElfNanLH3aS2Xfzfn+C/EOGuH55riPe0pR+J/ovN/gtexn2dYfJcGoU0ue1ox7W6vyX47eaxOIq4mrKrVleT3YFOrUpSU6cmmuq0ZXr1a8+erNyfdtt/iAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAGEwdfF1VSox5pa/gCqTUHBPR7r02+67+8MoyqtmWKjQpfN9l1f9dSvWwPD+WqytGOiXWT/AM31fQzHMK+PxMq9Z3b/AAXZeSAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAtfRHCuSRyrBvEYjScld3+zHe36v8A4Bj50Z4qpKh8Dk7el9AhCU5KEVdvRLzMjyuhkeXOdZpSteb/AE+W3m/Uz/O6ua4p1JaQWkV2X+b6/wCSAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAOCsmeLxf1movcp/jLp92/3dzjjiBWeX0H/jf/tv+f3dwOHMpp5dhnm+NWkVeK667P57L1v2MDxfSzWs8FjKSUKmi177X+ezVrOxmODlg8XUw8vstr/J/NAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAGQ8Q4fBcPydlzwbSXdvVP87+SKtSdWbnN3bd36sOH8pnmWOhSt7q1l6L/AD2Rx/PkyynTjonJfckyEnCSlF6o4yS/tecl9pRf/kqAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAjCU5KMVdsyjAYfIcsc6zs7Xm/PsvTZd36lfFVc+ynEzSvOE1NLtG1rfJJv5GV4CeOxlPDwV7vXyXV/JHFeIjXzetKOydv8AwFJP8UAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABiMNVoSUasbNpNejV0wOB8leIxH1yovdht5y/4H52OOc6dfE/Uqb9yG/nL/gfncyLOauVYpVoK6ejXdf59jFcZZbRoyeX0OWrLryxVvN2vd/gSbk227sAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAMtwjxeMpUF9ppfLr+B4hToRdClGK5km79lsl6Xv9wYXDVMVXhRpK8pOyMxxNHh7J4wpfElyx85dW/xb+4nKU5OUndsAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAOBcL7XNfaP7EW/m9P1Zxfi/rOb1ddI+6vlv+Nw4K4fnTlHMa+iafKuuunN92i73v68frFPGw9ov3dvd9ftfPb5WAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA8PKSUcRWf91fm3+hiazr1p1XvJt/e7mDw/1jE06K+00vvdji/MZ18c8NB2p0vdS6XW7/T0RTxU8x4drU6rvKg4tN72elvlr+AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHCmOwuGyfEqpVjGbcrJtJv3VaybvuHB+XvF5rTfSHvP5bfjYz6DjmmIT/nl+LbMjVsrx83tyxXzbYAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAcFYengsqqY2ppzXf/AG7G/wCtzN+H5Z5yZjgGvfS5k3azWn4bP0v1M6w1PJcojl/MnVqvmlbsv0ulbvqwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAACEHOSjHd6HFc4ZbkUMHTertH5LVv5219TA5vjsCmsNVcU+nT7noYjEVsRUdSrJyk+rAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA4eypZlj40ZP3VrL0X+ei+ZS4nyJV44ONK1NOylZWv0fe3nv18zxBxHPjqdJfZjf73/wEAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHD8nSy7H1o7qCj/4E2g4lk62HwVaXxOmk/kwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA4ay3E4jK8ZaPuzirPu43dkWbdkcVQ+rxwmFfxQpq/q9/wAgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAACEHOSjHd6GbcQ1MnxdHBYb+HRS5l/Nda/g7+urKE+HKqqZpRheUFzNa3T78u179dr636maY+pj8XPET3k9uy6L5IAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAyy316jf8Amj+aOJr/ANr4i/8AMzhz/dcdfb2b+/oAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAEJuElKO6OKMD9cxFDG4Vc3t1stXeKV/w/FMqYGeTZDV9vpVrtK3VJa/le/qgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAOFcRVlmmGpOV4xcml2bi7/fY4gzKvjsdUlUldRbUV0ST/AKuAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABwnC2a0Jeb/APSWVJc83LuAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABlGOoYWdKcnrFzb07xsvxAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADIcgqZvOcITUeVJ6+ZnmTVMqxEaE5qTavp6tfoAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABw9WqU8zoKEmk5RTs7X1W/cz+rUqZnX55N2nJK/RKT0Xl5AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAcJZPSm/7RxM+WnTat0u1rq30Wnqzi3JqNNrMcLPnp1G763tJ3ejXR6+j0AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAzao6WR4GhHaXNJ+bvp+bMqm6uRY2hLaPLJeTvr+SAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAeAq5rkFCWHXNOi5JrrZu/+X4jwFXKuH6zxC5Z1nFJPdJO/wDn+AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHCeOxeHzGFOhrGbSkrX0vv5W79PQ4ozLEYzMKkKsvdpylGK6JJ2+921YAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABl2WYrMKvssPC7/AAXq+hgPD3aWLq/KP+b/AMjBZdhcFTVPDwUV+L9Xu/mcVZdUweZ1Odp87c1bopSej89AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAy7I8fj2vYU20+rVo/ft8tzK+AsNRkp4uXO10Wi+fV/h5om8FluHcrRpwXZJL8N3+Jj/ABBoQvHC0nJ95aL7ldv8DF8SZpiarqOvKPlFuKXyT/4JiMVXxMlOtNye1223b5gAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAZbkmOzGVsPTbXd6JfP9NzK+A8LRtPFy55dlpH/N/h6GIxuCwFO1WcYJLbbTyW/wByM24+jyungY6/zP8ANL/P5oxePxWLlzV6jk/N7ei6fIAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAyzJMbmMrYeF11b0S+f6bmUcFYHCRUq69pPz+Fei6/O/yMbmOCy+levNQXRf5Jav5IzTj+pK8MFCy/mlv8lsvnf0MTia2JqurWk5SfVgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAGAyvF4+pyYeDk/wXq9kZNwLh6C58bacuyvyr8m/wAvIxOMwGV0F7SShBbL/JLf5IznjnEV26eC9yPd/E/8vz80VatStNzqScpPq3dgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAQjzSUV1MZwnjcLCvOo1y00nfW0r/y6dOoAAAAAAAAAAAAFDD1cRUVOlFyk+i3Mn4DoxgqmOd5fyp6L1e7+Vl6lSrg8uw65mqcF8l8kZ1x45xdLAK395/ov1f3GKxdfF1HUrzcpd2AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAACbTujN8dip5Fg3KrJ8/tObV62lpfXW3S4AAAAAAAAAAABGMptRirtmTcC166jVxj5Iv7K+L59F+L9DBZdgstouNGKjFbv9W3+pnnHUaU/ZYBKVt5Pb5Lr67eT3MZjcRjKrq15uUn3/TsvJAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAFKjOtUjTpq8m7JeZnHDWNjkmHgrOVHnckv7zvp3st/wuAAAAAAAAAAcPVMHDMIfW480Hpa19XotPU4xWXwxcaOEgouF1Kytrpb1AMqyTGZlUUaENOsnsvn+i1Mj4YweVxUkuap/M/0XT8/M4l4soYWDoYaV6ujutlZp2bv1Ss156maZ5jsylevPTolol8v1d2AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABwhGEMZUxM1dUoSl81p+TZknEmPeawlWqOUZtJpvTXTRdLeRnGGjhswrUY7Rk7el9PwAAAAAAAAAAyrDVcRjKcKUXJ3T07J6s4uw1almtWc4tKTun0ei2DC4WtiqqpUYuUn0RlHAVODVTHS5n/Ktvm938rerMVjcDllBOpJQitl+iS/Qz7jDE4+9Kh7lP8X6vovJfNsAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAOEoe2q4jDLepSkl66GSYOpXzOjRS1UlfySd3+RxBVjVzTESjtzP8NAAAAAAAAAAMmxiyfJp42EU6tSXLG/RJb/n87Es0nnuTV4YhL2tK0k0rXXX8L36aoybh/GZpUtSVoLeT2X+b8l87GV5Pgsnw75NNPek936vovLb8zOOPKcL08DHmf8AM9vkuvzt6MxWLr4qq6teblJ9X/WnoAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABkOBzX63CvhKTbjrd6LzV3ZarTucQZvDKsMq0aCjXqpq+mjVr3a3tfTv1sNtu7AAAAAAAAAAxkXPhvDSW0ZyT9XqjgLCqtiK6mrwcOVr1f+SZjsxwOU4dOo1GK2S3fkl/S7mfcTYrNZcvw01tH9X3f4L8QAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAChhMRiHajByfkm/wAjA8GZriZLnhyR7y/yWv4GC4AwNKUZVpudt1sm/wA7fMw+AwmHSVGnGNuySMbmeDwUebEVVH1evyW7+SOOcbQxuGwtehK8Xz2dmtrLrrugAAAAAAAAAMuy3E5hWVHDxu/wXm30RlnDGHw2WywVZ86k7vtfTbqtl5/kZnnGD4fpPBZbH959pvW3r3fZbLqYrFV8VUdStNyk+rAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADK8lxmZVOTDx23b0S+f6bmB8Pa8pJ4qqku0dX97sl9zMDwxleEjaNJSfeSTf47fJInOjQheTUYr5IxvFmU4aDftVN9o63+e33sxfiFWkpRw9FLs27v1tt+P3mJzfH4lt1a0n83b7tiEkppvXUzjinKMXl8sNRotO3u3jFJa3drN2v5IAAAAAAAAAMh4PxOYKNas+Sk/vfovPu/uZRw+X5PhXypQgt339Xu3/wyMx4rxOZ4uGHwaap3V7fFJX1vbZeX39jiylUhm1aUotJvTTfRbAAAAAAAAAAAAAAAAAAAAAAAAAABTpyqzjCCu27L1ZVyrGUo1JThZU2lLbRvbqAAAAAAAAAAFLBUZ0qc3WScpWa1vFfzPyMRSjSqyhGSkk7XWz80AABl+RZhj2vYUm13ei+96fcYbw7m4p166T7JX/FtfkYPhHKcNGP7rmkustb/Lb8DGYvDYDDOc2oxinZbbdEjGeIGChG2HpuT89F+r/AzDi/NcW1afIl0jdfjdv8SrWqVZc1STk+7dwAAAAAAAAAAjCU5KMVds4b4MqOpHEY+Nktovd/4vLy+/zz/iWhlThTS5pdYrorO3prb5XM2zvGZnU5q8tFslsvRfq9TJ80nluKWIhFSaTVn5mfZ9UzecJTgo8qa08wAAAAAAAAAAAAAAAAAAAAAAAAAAo1ZUasakN4tNeqK+d4utCtCTVqrTlp1W1uwAAAAAAAAAAZPl9HF08TKpe9Om5K3dd/IAAwuCxGKnyUIOT8lcwHAWPrWeIkqa+9/ctPxMt4TyzA2koc8u8tfuWy+6/mY3NMFgY3xFRR8uvyW7+SMV4hYaEmqFFyXdvl/R/oY7jXNMRJ+zkoRfRJfm9fyK1erXnz1ZOT7t3YAAAAAAAAAABhcLWxVWNGjHmk9kcO8MUMrpqpUtKq9328l/nu/Qz7jLDYOMqWFfPU/Ber6vyXzK1apWqOpUleT1bYAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAcK5Vi62FxVSENJ05Ri9rvy/wA9itRqUajp1FaS0afQCMXJpRV2YDhfM8ZUUVScF3kmkvv1fyMu4Iy3DJOsvaS89F9y/W43hsHS1cYQXokv0MfxxlmHvGk3Ul5aL73+iZmfGmZYu8ab9nHy3+/f7rE5SnJyk7tgAAAAAAAAAAAZdlmJzCuqNCN3+CXd+RkuRYTJ8O2neVvek9P+GX9M4o4uli74bCO1Pq+sv8l+foAAAAAAAAAAAAAAAAAAAAAAAAAAAARozlSlUS0TSfzvb8hJt2RWoyo1JU57xbT9VoAAAAAAAAABCDnJRW70M+4hxOWYiGAwMlGNJJPRO7tfr/V76nFkoYuGGzCKs6sXf1jo/wA7fIwOU43HNrDU3K33fe9DLfD5yipYypZ/yx/z1X3L5mXZJgMvj/s9NJ93q/vf5bGMz/LcJByqVo3XRNN/ctTM+PMbXbjhUqce+8v8l93zMTi8RiZ89abk/NtgAAAAAAAAAAABk2SYnNK/s6Sslu+iX+fZfpqYPA4DJMG7NRitXJ7vzf6L7jiTimrmcnRo+7SXTrLzf6L7/IAAAAAAAAAAAAAAAAAAAAAAAAAAAA4eo/WcFjcOleTgpJf4XfT7zhnCvE5tQildJ3fotTM6kauNrTjs5Sf3tgAAAAAAAAAYWahXhJ7Jp/icXUZU84rX62a9Gl/wxlfDlHH5Tg1iW7Q5nZaX5ndfK1tipWwWWYeKnJU4LRdPu7macfYelLkwcOfzd0vkt3+BmfEmY5gnGrUtH+VaL/N/NsAAAAAAAAAAAAAyPIq+a1+SGkV8Uu3/AAX0X6CWX5FgP5YR+9v9W/6sjP8AiHEZrV192mto/q+7/LoAAAAAAAAAAAAAAAAAAAAAAAAAAAABkuaTyzGRxEVdLRrunv8A5rzIcSYXCvFzwsLTqv3XZLlVtfx6LS+oAAAAAAAAAGT8PYzM6lqUbR6yd7f8F+SMs4Ny3C04+1h7SfVva/ktreqZmuCymt+8xkU3TTl1uorfRateVmjG+IMIrkwdHbZy0/Bf5ozHNcXmNRTxM+ZrRaJW+SAAAAAAAAAAAAAMvwNXHYmOHpW5pXtfbRX/AEI8CZn7WEZuNm9Wney77L/hz/YMhy/tCP3yf6t/1ZIzvO8Rmtf2lR2ito9Ev8+7/QAAAAAAAAAAAAAAAAAAAAAAAAAAACjRqVqkadNXk9EkS4RzqLt7B/fH/Mwfh7ialJSr1VCXZLmt6u6X3XMv4AwdGTeJm6nZfCvnZt/iifC+SYWLxHsfg97eT212bszOMZTxmOq16aajJ3V9/wALgAAAAAAAAGUcL4/MuWcI8tN/af6Ld/LTzMs4PyzBSU+XnkustfuW3527mZZrhMuoupXklbZdX6L+kZzxvi8RU5cE3Th6Lmf52+X3mQVqlaGOnUk5SdKV29WAAAAAAAAAAAAABkODx2JxsVg3aa6/yrZsxmLo4ShKvWlaK3/4Hr27mfZ5WzXEc8tIL4V2X+b6gAAAAAAAAAAAAAAAAAAAAAAAAABGLk0krsfCmcqKl7B/er/de5R8PK0qSlOulLty3t87/oYDgbLaVBRxMeefV3a+5Jr8bmD4ayrCO9Oir937z/G9vkLD0VJSUVddbD4lyhNp4iN0Y/j3A0KnJQg6nnsvldXf3GP8Qas6fLhaXLLu3e3ov8/uKfGuaOVqslKL3Vlquq+aMzzvLcThZUqGEVOTt7ytpZp9lvYAAAAAAAAMvyvF4+p7PDwcn+C9XsjLOBMFQUZ4lucluto/du/m9exned4XK8O486U7e7H8tFsvWyM043x+K5oUP3cX2+L7+nyS9Sc5Tk5Sd2+4cJ5Zgp4CpUqVbSqRlBq6Vl3XmZlh6eHxdSjTleMW0n3t6AAAAAAAAAAAAAGDwdbF140KMbylt/XbuYfD4ThrK5Tlq1u+spdF6duy17mMzWjxFlVZcvJUpe+le6aW/bpfpo7AAAAAAAAAAAAAAAAAAAAAAAAABRw9WtLlpRcn5Jv8jJ+FsdmNRprkit3JP7kur/Iy7gClTrOWLnzx6JXWvn/wGYbhHKcPXjWhT1WybbX3O5KhSlJScU2vIqcQZXTqyozrxUo6NPv+RjePMFQxDp04OcV9pPr5f5mZ8e4mpOP1JckVvdJt/wCSMZxZm+Jjyuryr+7p+K1/EePxbi4+1lZ9Lv8AzAAAAAAAAADDYWtiaip0YuUn0SOH+CJxqe2zBK3SN76+dtNOy/4eUsDlmG15acF8v+Hf4szbj2s5uGBilH+ZrX1S2Xzv8ivXq16jqVZOUnu2AAAAAAAAAAAAAACTbsjhTIHldKVfEJKckv8At1a3X5Nv5dDivNaeZZR7Wh8EavLfvaL19Hf8jhK/1iv29lO/poAAAAAAAAAAAAAAAAAAAAAAAUoKdSMW7Xdr9jOOD3l2DeJddStayta9301+YU6U6slCCbfZasyvhTMse3aPIl1ldfdpd/kZb4fSbl9dqWXTkf43a/CxgOEMqwn/AC7533nZ/hZL8ClhcHg4uVKEYJ72Sj99rGaZ7gsvw7q1Jpvok1d+n+exjPEGjLDyWGptVHte1l576+lh8c5s6bheN31tr+dvwI5hjIr3asl/28/8x6gAAAAAAAAABkXDuJzWr7vuwW8v0Xd/0zAZbgcpw7jSSilvJ7vzb/pIznjjCUacqeCfPPa/2V567/l5mOzDFY6p7TETcn59PRbL5AAAAAAAAAAAAAAAHBfDaly4/EL/AAL/ANu/y+/sca8RNt4DDy0+2/8A23/P7u5wtUwuLoVsrxMrKpZxf95dvPb1s11MfgMLw5ltamqnPWqrlXT3eul3pbr1dgAAAAAAAAAAAAAAAAAAAAAADhfKpfWfrmKptUqcXO7Ts7bWvo+/yMBxTSzqvLAYqklCpdLuu1/Ps1s7GB4KyvCzcmnU7KVnb7kvxMNl+CwacqNOMO7SS/EzLiDL8BS9pUqJ9lGzb+V/zMw4+wkKP+yRcp+asl+Jj+Nc0xUeWElTX93f722/usYrNsdiqfs61aUo72bAAAAAAAAAAACjSnVqRpwV2+i3KPBWUU5qfI3bo22vuMxzPBZThuao0ktord+SX9JGd8Q4vNKn7x2gtorZevd+b+VgAAAAAAAAAAAAAAA4XyJ5pi/fX7uGsvPsvn+RxXxBHK8OsPh9Kklp/dW1/wBF/wAAk3J3buzC4Wtiq0aNGN5PZficb4HEQzGeJlH3JWSfdqK+fRgAAAAAAAAAAAAAAAAAAAAAGX5BmGOqKFKm1pe7TSt6/wCRl/h9F0m8ZUal0Udl6trX8ChwtlFKMYqinbq9X8/6scRThPLMTShJOSi21fVL09Dh6MpZrh1H+Zfnr+BnfGmNVSrhqKUeWTXMt7J2+VzF5xj8XDkr1pSXa+n3AAAAAAAAAAAABlWS4zM6nJh46LdvZer/AEWpknDuEyik5/FO2sn+nZfn1Mv45qYjMo0ZwSpSdlvdX0Tetteumn553CtDMK0Ks3Jxk1du7t0/AAAAAAAAAAAAAAAAMPh6mIqxpU1eUnZIpQwvDuU+9ry7/wB6T/z28l6GOxtXG4ideq7yk/6S8lsGBxtXBYiOIo/FHa/mrfqZrxFjszpRp4hppO+itrsAAAAAAAAAAAAAAAAAAAAAYXK8ZipRjRpN822mn37GX+H1abvi6iiu0dX970X4mF4ZyqhThFUU3Hq1dt93/VjG5lhMHSlUrTSUfv8AS29zMeP6MHFYOHN3ctPu/r/gYrjHN61RyjU5U+iSsvvuzh7N/q2Ze1xD5o1LxlfW6fXz1t8ieU5Vw/CeYwbk7PkT1V32t913svMnOU5OUndsAAAAAAAAAAAAOHOHqmbV9dKcfif6Lzf4b9r8+W5LgrXUIR+9v823/Whh+J6mb16+HpR5Yeznbu3pZv5X0X3swEJTxdKMd3JW+9HFUoyziu13/RAAAAAAAAAAAAAAABwNkXs6f1+qveekfJdX89l5epxhnn9oYv2VJ/u4aLzfV/ovv6gAAAAAAAAAAAAAAAAAAAAAZPw5jczknTjaHWT2+Xf5fOxl/AeAoOX1l+0vtvG3fZ6nNhsJSUW1GMV1drJGY8e4KlG2Fi5y89F/n+HzMdxnmmIqc1Ofs1a1l+evUnOVSTlJ3b3b3Ao1PZVIz3s0/uM44yqZlg54Z0VHmtre+zT7eQAAAAAAAAAAAAcOcMVs0n7Sfu0lu+r8l+r6GLxOByHL7xjaMdElu3/m+r+ZmWZV8fXlVqvd3t0WiWnyS+4yTMXl+Pp4i10nqvJ6P/geZPJcqy2U81jCTaTko2dk/JWuvnpHysYnETxFadae8m2/mAAAAAAAAAAAAAAAcO5O8zx0aT+Bayfl2+e349Di/G1cDlTjh1a9o3X2Vb8Oy9QAAAAAAAAAAAAAAAAAAAAA4ZyDEUq8cdi4clKCcru2tl23t126GVcQZZnFaWG9lZ7pSSs0v162MwzXA5XSTrSUeyW79Ev+GMd4g1KlFxw1Lkn3bTsvS27/AAMxzLE5hW9tiJXla3ay8kAAAAAAAAAAAAAAYTC1MVXhQp/FJ2Rl3AeMliI/W2lTW9ndvyXr3M2zbB5JhIq3S0Yrrb8l3f6maZtisyre1ryv2XRei/p9wTs0zGcc4zFYedCVOKUk116q3cAAAAAAAAAAAAAAAMNTnw/w9KulatO3yb2+5a273MgzXE5lCvl+Km5qcG1fdNf1fyaAAAAAAAAAAAAAAAAAAAAtctcMj4ZxeaSTS5af8zX5d/y8zJ+GMDll5QXNN9X+nb+tTinF055ViqVKSc4qN12vJb/JM4VclnFDl3v+jv8AgZnUlUxlVyk37z316gAAAAAAAAAAAAAAcIYCFbGPFVZcsKK5m/Pp+T+7zKXEmDxOFrVsK+d003bZ6evQzDMK+PxEq9d3k/uS7LyAAAAAAAAAAAAAAAADhXKv7QzGCkrwh70vlsvm/wALnGXLictq06bvKk4ya7J3/TX0ODqbWNniPs04Sbfyt/XoAAAAAAAAAAAAAAAAAAABkMacMvx2InFO0VFX/vP9GkzhOVNZtSjUimpXWvmn/wAN8zKOCsHgpxrVXzzV97cvlp39Xv0MxzbBZbSbqySaWker8kvw7GbcZ4/HRdOn+7g+2/3/AOSRwjXpTxVTB1/hrRcfn0/X5mD4cp8PqrmNapz8ifKrW1eiv5u9u2pKTk23uwAAAAAAAAAAAAAAweDrYyvGhRV5S/q/oupnuWU8p4ddClvJx5n3e/3aWS7HBkm80VPpKMk/S1/0AAAAAAAAAAAAAAAAA4awlPJ8mliays5Lmfe1tF934syLiFwzadfFSXJVvzX1S7fdt6M4gzvK6eXSw+WOKc3aVk1p16Lfb0YAAAAAAAAAAAAAAAAAAAHD2VYrEZRjIqDtNR5fNxu9PXTXY4QwNSrmkajVo07uT7WTt+P6ma8fVHNwwMVy95LX1SvZfO/mkVq9WvNzqycpPq3dhCpOnJSg7NdVoypj8XVg4TqyafRttfmAAAAAAAAAAAAAAFrnCuQwyvC+3r6VJK7v9ldv1fn6Es1ocRyxOXwdlZODfVp6v56ab2uzLckrZHQr4/FtKSi4xSd9Xpf/AC8r3AAAAAAAAAAAAAAAADhzK/7RzGFJr3VrL0X+ei+Zx9mvLThgab31l6dF9+vyQAAAAAAAAAAAAAAAAAAABwrwvPHVFiMTG1JbJ/a/4Hd9dl1tmWb4LLKKlWlZbJLf5L+kZxxu8XhJUMPT5HLd36dUrd9r9gAAAAAAAAAAAAAAAKdOVSahFXb0Rgsq4fyyrSw+IkniNHq3v08l5J7nHWeTi/7PpO3WT/JenV/d3MmxVPC4+lXqP3Yu7t2OLeIsBmeEhTw7bkpX1VtLNf5AAAAAAAAAAAAAAAABwNlTw2CeJmveqWt/hW33u/4GZ46eOxdTET3k/uXRfJaAAAAAAAAAAAAAAAAAAABGLk0luRnkORxhg8TTU6jS5nyqVm+99l2Svpr1M+4hw2V4ZOLTnJe6v1fl+ey8sTiauJrSrVZXlLdgAAAAAAAAAAAAAAAYTKcdi4OdCk5JdUjh+j/wsUITWqkvvX/BRnFadbMK05PVyl+en3I4sm6tTC1pfFOlBv11AAAAAAAAAAAAAAAAAw9CeIrQpQ3k0l6vQzuVfLMmf1LRwUUtL6JpN2afTcAAAAAAAAAAAAAAAAAAADhLJZ5hjo1HpCm02/Povn18vkcRzlPNsQ5b8zX3aL8DP25ZfgJPfka+SasAAAAAAAAAAAAAAABSpyqTjCO7dvvM04mlktaGX4OCcaaSd76u13az+966mHyqlis6o5lSi/Zyjzf9v9n+b899zN+DMdWzOUqCXs5u97r3b6u639LHF9anLMfY0n7tKKgvl/le3yAAAAAAAAAAAAAAAADLeHsfmNJ1cNBNJ23S10fX1OE8lq086ca6s6Su+urVl+d/kZlxwsLmLw8aalTi7SfXzt00/HyOKMDSwmZTVL4JJSXo/wBL3sAAAAAAAAAAAAAAAAAAAZZw9mGZQdTDw91dW7K/ZFGOH4cye89XFa/3pP8Aq3kkZlwt/bNeOPwlRKFRJu97ro7W/FO2v4cXV6LxcMLQd4UYqPzW/wCifmmAAAAAAAAAAAAAAABljSxtFv8Amj+aOJ4Shm+IUv5vzV1+Bl+PjlnDdOvU3UdF3bbsv66alPi3N6dJ01V36tJtX7P+rdLEm5Ntu7AAAAAAAAAAAAAAAADhDMcTRhiaUJWjGnOaVl8Ssk9vw28jgyNathqmNru86st7JXUVZbab3MyjKOMrRlupS/NnFacamFjL4lRhf11AAAAAAAAAAAAAAAAAAAM7zrE5OqOX4J8vJFczsrtvXrf1fqcUZhUzHK8Jim7J8ya6cysr/g7dkzDZnjcNTdOjVlGL6JtD1AAAAAAAAAAAAAAAAyvC18Vi6dLDq876dtNbvyXUx/DODzGpTrYpe+kr8rsn69bfc/M8QMVFToYOGiir2/Bfck/vAAAAAAAAAAAAAAAAAMjy7LcqwarY+padeLVm38Ltppr2u+m2hnuJp5Nk1OGFfWKj9/N+NtfUlleRZnJZrKdo6OSukr9pdU+669N9c/zJZjmFSvH4dl6LRffv8wAAAAAAAAAAAAAAAAAAE7M40ptZrKr0nGLXpZL9DMIOlw9hYy3lKUl6bf5AAAAAAAAAAAAAAAABwxV+qYbGY2PxQikvJye/3pHCOcYmGaxpzm5RqXTu7620frf8DiLGPF5pWqPu0vRaL8gAAAAAAAAAAAAAAAAKceeaj3OM6rnm9SHSCil5Kyf5szSrOtw/g5SfwuS+7b7loZPwzVr5HOmqiXtuWS0elu4AAAAAAAAAAAAAAAAAABhcLVxVaNGkryk7IxeTZfWw8IYinzRprTe9kvLV7bdTijN6WYYqPsP4UFaKtb10/D0SAAAAAAAAAAAAAAAADhui8VgcdhIaylGLS78rb/VHCGDnWzWE7e7Tu2+1l/mYqoqtedRdW397AAAAAAAAAAAAAAAAAje6tucUcNY/GYqGKoU7uaXMrq6klbva1u3Y4opRwOFwuXJ3lBOUvWT/AOH+ViGYYyEVGFWSS6KT/wAwAAAAAAAAAAAAAAAAAADhFqjXr4pq7pU5SXr/AMNdGQ5viaWbU6spt88kpeabtr6bozrDxoZjXpR2Unb0voAAAAAAAAAAAAAAAAGR4zE4XH054ZXm3a3e+lvn+D1OK8TWwuUSnSjyynZS20utder6X87gAAAAAAAAAAAAAAAACdndHDeOrY3LKeIru8ne/TZtfkjH4qeKxVStJ3cm3/l9y0AAAAAAAAAAAAAAAAAAADhGKrV6+FvrUpyS9dP+CZBlmIrZtTouLThJOXkou7v+XqZ3XjXzKvUjs5O33gAAAAAAAAAAAAAAABhas6VenUpv3k016p6HiFjKlOhRoRfuzu3/ANu8tvzAAAAAAAAAAAAAAAAAOHssjmGYQoz+Hd+i/wA9vmZdxNleJxH9n0IuMdVF2ST9O1+nf1M8wtDC5hVoUPhi7Lr01/G4AAAAAAAAAAAAAAAAAAAYPF1cJXhXpO0ou6/rs+pj+PMPVw1SGHhKM2rJ6afiAAAAAAAAAAAAAAAABgIwli6Sm7LmV2+iutTxEqJ4ihDtFv73/wAAAAAAAAAAAAAAAAAAA4WbTxbjuqM7fgZW5LHUXHfmj+aOJUlm+It/MwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADEYqtiOX2sr8qUV6LZfiAAAAAAAAAAAAAAAAAcL46lhMxi6r9yScX6P9L2uYXhChlmLeOrVU6VP3l38r+nS277bGOxLxWJqV3vJt/ewAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAtc46pVll1CMItxjvo7LRJX7fMAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAACDipq+xxXxVhsThFhsJJSU172jTVmmu3Z33AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAQwAex0OhugBgB0A6gPY6AzodQADqDNy4blzcAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAEMADodDobhcZcZuC2C49zcLnQ3HsHUGAXOoB0N0B0AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABbHQAAYbsHYew9jqF7i2DodTqdB7HQ3Y9jqAzRgM6gdDoLYHsAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB0FsAAMZ1DodB7HUBbG50Op1Og9jsdR7DAZYHsM6gw6C2BgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPYQ0AFiw1csWLFiw1csDQWAsWGtCw9QBq5YsFgsNXLFgB6oAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAGgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABnQQwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABnQQwAAANAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAWwPUAYAAWuLYAewtgAAAHuPUCw1cSLWAtc6HUAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAOhugAOgMAL6HQsdDoPYOhYOgWGrjLWOgHU6nQ3DobgLY6AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAdBDANToM6nUFsFtBlgGMLC2OgxjOo9gewzqAPYGM1LHQ6gAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB0BgPYOgwPI6D2OgzcOoPYZudAZuPUZugGM3OhuBuBuHUAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAsgsgsiyAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA//Z)
Figure 2. Subtractive computation in dopamine neurons. (a) In one task condition (no odor, black), different amounts of reward were presented without any predictive cue. In another condition (odor A, orange), the timing of reward was predicted by an odor. (b) Prediction. Division should change the slope of the curve, whereas subtraction should cause a downward shift. (c) Average response of 40 optogenetically identified dopamine neurons. Prediction caused a subtractive shift. Data from Eshel et al. (2015). (d) Three example neurons. Although individual neurons exhibited diversity with respect to response magnitudes, their response functions were scaled versions of one another. Data from Eshel et al. (2016).
DA network
![[[@Watabe-Uchida2017-gi]](https://doi.org/10.1146/annurev-neuro-072116-031109)](data:image/jpeg; charset=UTF-8;base64,/9j/4AAQSkZJRgABAQECWAJYAAD//gAuSGFuZG1hZGUgU29mdHdhcmUsIEluYy4gSW1hZ2UgQWxjaGVteSB2MS4xNAr/2wCEAAMCAgICAgMCAgIDAwMDBAcFBAQEBAkGBwUHCgkLCwoJCgoLDRAOCwwPDAoKDhMODxEREhISCw4UFhQSFRASEhIBAwMDBAQECAUFCBIMCgwSEhISEhISEhISEhISEhISEhISEhISEhISEhISEhISEhISEhISEhISEhISEhISEhISEv/CABEIEaEKjAMBIQACEQEDEQH/xAGiAAABBQEBAQEBAQAAAAAAAAAAAQIDBAUGBwgJCgsBAAMBAQEBAQEBAQEAAAAAAAABAgMEBQYHCAkKCxAAAgEDAwIEAwUFBAQAAAF9AQIDAAQRBRIhMUEGE1FhByJxFDKBkaEII0KxwRVS0fAkM2JyggkKFhcYGRolJicoKSo0NTY3ODk6Q0RFRkdISUpTVFVWV1hZWmNkZWZnaGlqc3R1dnd4eXqDhIWGh4iJipKTlJWWl5iZmqKjpKWmp6ipqrKztLW2t7i5usLDxMXGx8jJytLT1NXW19jZ2uHi4+Tl5ufo6erx8vP09fb3+Pn6EQACAQIEBAMEBwUEBAABAncAAQIDEQQFITEGEkFRB2FxEyIygQgUQpGhscEJIzNS8BVictEKFiQ04SXxFxgZGiYnKCkqNTY3ODk6Q0RFRkdISUpTVFVWV1hZWmNkZWZnaGlqc3R1dnd4eXqCg4SFhoeIiYqSk5SVlpeYmZqio6Slpqeoqaqys7S1tre4ubrCw8TFxsfIycrS09TV1tfY2dri4+Tl5ufo6ery8/T19vf4+fr/2gAMAwEAAhEDEQAAAPy9J3EaAFQAAVNBoABMGlARQQAUERpUTFBEaVQQBFAAVUwTVAAFQBABpFBUAVNFaEAAVAVAVAAFQAaBNBpRNyJgAAigDSACjBAAg0ACpoVIJiggANIoAmI0KDiaBMAVARQQaFTAEGgBUAAAAaQAAVAVAQYAlEwBVmkGgBAHICAKgKgKjQqYAgAoINKgA0AAAJooCNKqYiYNAComDQAAKgAANJIqYjkQBQAFaCqCAACACocqpABAUBFAAEBUAAAEdUoJgAAACNKJoNKgCpiNDk0AVAABQQBUTEaUBFaBMQFAAEUABBoE3IAAI0qAqJqoINIqYoIAAgGI0qAqAAAAoNUBQQBAFcmgmqADSg0BACiaDQoIADQJiNKJgAAIAoA0ICOTciaoCtBQBAUAAEBVTQaAEGlAEAAVoKAICgAAAomoMQQAqAIxUQ4BAAAAFBqgDQJoNAAoIADSKmAOE0UEAABRtBKAIArQAgCoCoAoDQBpyJiggANIAqpgmAA0qJorQJoNKgAAAqAitCJqNCAKAgKgCAKCKm4TAEUBGlRNQEUEUFQBAAFGhAVAFAAQaAAAAABU0AUBAAAUEAVAFBAAaEBQATRWhE1GhE1VNFYIIAAAAAAFRMGgaBMAEaVE1BIMUBABQQAAUmhQEBQEAFAAAEAFBBoAAAAARQEBUaVAABUxQUmkGkUABFaRUwAARQRWkAVABMGlQAAABoABMGldNIgwQo0IAqaDQoIAAAAjSiaAA0AAAJqNIAAACioVDVaQaUTEBUBUBUAUEVpAAAUBAFAExGgAUEAUTAEGlE0AABQQAGlRMRpQAAABoQFQAAQFAEBRNwmKmhUhNIrSAAAoIoINAAAqJgANAAjSomo0CYjSqmCaDQqaK0gAJitCAICo0qJuQABAABUaFAJpVBo0igACAKANAmACAKCoAgKADQgKgACoAAAK6aRAUAAQaUTEaABQEAAAAARpRMGgTAEGlQAAVNRNFBUBFBFAAAEUEVpAFRNRpAAAGgTAAaRQABAcs0CYjQqYjSiYgA0ACporTVAAAEUABFBBpUTBoARwAmqAAAmDSomKCDQAACAKAAAAAgKjQAACggCqmiggAgA0omIAADQAqAACgiggAAg0KCAAAACb0TFBEAAUaRQQTVAVrTkAAVAAAAAAABWgqNKJiNKqYiYNACgIgKgCgigigg0qJqjSiYNIoIgDgBMUEUEAAAaBMAEaFTAEUEGgBUBUAAVAVAABAFBVmkUAAAEBUaABAUTBpQEABoVNBoEwaETFaQBUTFBRNFAQFQFAEAUAAQBQQAGgAAABGlABMABoAVZpWpgArEEAwEKgA0AAmitACKAAg0AACKAAACgE0ACsQEAAComqADQDAQAAAwEANRMGhGhUCMcs20coIBoQFRoUEAAEUABBpRMGhExWkAAEAUBAFUB00igiADQqaKAAgxRCoCKCDQAAACoANAmI0qAADppFBAAaAATBpUAGgTBoE0VoABMAEaRQVE3CYgA0KmI0KDVAUEARQRQAEUABBoUEAFAQBQQBRMVNFaEAAVAUTQaVAAFE0GgAABoEwBBoAAFaCqmqJqgCtAmgCgAAAI0oCKCKCKCAAAAAAAAAIrQqpRCtGoJAFQFQBQRQRRoCAFQEVoAAYIEaFAQBGhyak0isBAAI0KMEINKJoNAAAAANHhZ2MvRe3Pqje05ZHA5CdFRpsTiKvpzV4L5ayXzKA1WkUFQAAABoRNRpABQQAAAEUAAUbiaQEigICgIoA0IAqaKCFSKqEBUEAAAAitCAomg0OmlQAAQBWCCVU0RoAAaoIAEaUARNUaABU1QATVABoAAEUABUAAAEGhQEBRNFAGkUEAAAAAFTABQQAAAFE0RpyADQAAAmAAwQSoComK01QRQFTVE3ImDQgKAigigIAAACgIANAAAIDkAGkUBAVAV00ImKAgCtCAKNEEomI0oAAAACKAjQAqJorQAI0qpuSaRQAAAAVARQFBXlMaJASoAAKBsz17zOT6CqmnAIAA1wCAPBAFE3uHFjac1KLbzEVwgAADQAAA0AAmitCAICoAAqAOTUTAAAAAAAQFQFAAEGgAAVEwaAAAQFRpUAExRioEYCAAVoQFQAAEwaAAAARQBoRNRoEwAAAABUARoVMAAABAAaAFEwaQAUEUEUEAAHImKmAIrQJorQiYrSKmI0qJqjSoAAAIoIAoCDQqEViOTVFSgCCAAAARwIoACI0qAomDQAgAgKAAK0FABodNAMAEEACggAgKoNGlExAFBFBBoAFTEaRWlE0AAHk0gwEINKg3vVz2Y6bTs+pwaIgqFHbzoyuZAB49Ld5vZmhy7hlYtr1k1alqgqIc8aAKgOE5XTaZt45dbo8gSswAAAAaQBQEAAABUAQFQFQBQcTQAqAAAxFQitIoAmI0AAAoIAAAAjSoAAKmg0qpoqaK0CYAg0oNAQAKCIA4EQBQRWgTEaFByTaK0Ig65r6j7lLyT5UbjVpBMAVEwaEByADQs0g0iiRWIAoAAgDhNAFRNQEGgAAFBAFE0GgAGgTABGnImDTVAAVAHJqioBAAACYNAmDSAAAMVEKAitIJg0qJg0DQAAKs0rQVAAAAAEBQBAUAAAAAQFAQaEBQAAABoVNyTQ5tGpKBa1c+7Qdj6aIpZG5RGmtaXMvBjNOSOJ4L0uXdK3LundPQ1SI61ZRF5qggCWSbYNCA4AkTc9W2hWeC3p8RAQNAAAAACK0AAAAAAmiNKJvRUqAAgAAAAAQAaVAAAAEwaABrTgATEaVAVAHTSomDQAigAIAoCK0gmoCDQAAAAoIAAKJgCDR++RP0dQefT/AAWP8WxaqJiADSiaDQgKAigI0oAiBWCJitIgOWaAaAlRoVNFAQBGlAAAAAQBQFQBAVAVGgAAABybmzQoINI4GqCAKAKCAAAgCgAIrSAKgAAoIADQAOmkAVE1RoUEUEUEAAAAEwaBoRNQAAARQQaHjldO6SDtS59k8sblQiaytbKR0iGaMIQEjGNbUy1QibWTCsn343kI1WRaSsnVleueNbhqiRRySzUatOiTcjTkQOG+RVUxNuKszbzRBA0AINKJoNAAAAIoORUiOVE3KmgNASgAAAIAAKAjQAACggAAqADSACA5AAByoRCgIoAAgKAAA0AIAAAComqNAANIqaiYgAC+uC4X10P3G9PrH+Z7i1somgCgCAoCDQNAAqaAK0FAAAaRQFmgBzBqCAAAQFAEBVBEYAgBUAABgiBQEaAFQFcmqTQjFRANKJoNAAAACAK0gAAAAComDQgKAI0oCrNIoIoIgKAAAAgKgCgigg0omigCaFSomSq7ssd6wilRW6RXI+bmfGj3JiDcFNUlRMBoxyxtQiCa1j1Q+SaVpSgPVWjHMSXk1QRAe+aVAexCq21yTRgoLCyGC8MZu/kA0KCDQJg0ipiAI0qAOBoCqDkmmq0qJgAoIAoCAAwUSAAAjQJqArQBpQBAAABVmgBUBFBHAgAgKgKAg0qJqjQJqNIAACNKApNKA1QP1IF+muvWLJg/nP8AGFsgAqYAACNKAgAAKDRpRMQFGhAAHE0igIDkAAAAAE0WpQBQEUEAAABFaQAEwaRQVAUTek0gwQSgIoACKAAACAKwEINKgAAAgCjQgKiYrTlmkBgCohUYCABFaAYgAJUAAUAAJC7djPusyZ9TohtictHBMqlfNudND01cComrQcFAAwTGVCFS4mgakSWKNXK3BcbnmyJw0qWgOUayS2OByAOQA41aSQb1Wubz/T47TTiAEBAxFAQFAVExGgBQEVN4m1QEYCFRoUEUEBqiFQFQEVoABpUmkVpFARoExWhAV00gAAKNqiFBABABioIAAABFaRU0UEABpxNIAoz6G/oRM/lH8OSv2p+y3n/Of4xOoDQaUBFBAAAAVAAEGlE0VpFTRQEacTQAigICoAoCptGgaAAAGlRMAVoKAg0Cao0KCAKqYs0g0ACNKJiAqNCpoAKCDSoAAAIoACK0gmo0IAArppAABFABo4EBCgrUxWhEwaVoK4dy3n3z2M+pJJtzRwRVCI5RQJE7BNDlQBK6aAYIY+kqKkQhjKT2gIBKCKqHjFVoAiOEYSiNIlS9qpXIRQVU1UFUpgSjWPeqjzLwyI+nxmjgUYjSOBqgACAKCAAA5NRUAkUEVoEwAUBEwaVAQaUAAABAFBAAaBMRpQEfNIAAAmDQAICoANAAoIAjgEAABpBMAcqpABBfUX77Vl4N8AJ/qr0bj+c/xhbIqaKAgA0AAAAAAAADQJoNCggCiaiaDSoCoAAKCKCAKgAmDSDSomoCDQAACggAAom4mkGgAAAAAAABARWhUxGKggBUAAABAcgAAjTlmlRMUEGKiAYqIa5oAEAAUGoD9rPtkky7hQHJsa1ElJUUqDJU7CxoqgwE8KcKhjkUBQVExjQoljBwCvGINVKQQI5a0kY0qoWOiYTQEiA5w3ImigjRNexyJ4eZ1eQ0vlBoQABUQDABFBAFQByHIqABU0GhAVAFAQFARQRWkVNFaQAABoE0GlAAEAVUwTVARWkAUTBpAFRNUBUaRQBMGgBFaEAUFJoQFAf+wzn7r8hefgX0un+HniRoImAKgA0KNAQAAACK0CaK0CYjQAqzQAIAoINKiYNKgKJoNAAAqADFRAAACNKgKgKACaiYgKgKgADkAAQaVEwaAAAAAABBpRNBpUBQEUFdNIDEQqggAAACgiAqAKy9bz9CaTPpspGka1LQEAGgqMcIHArk5mzauQigijAYAitKioVtAQInrrVUrBQaoO1X0SJlSKQ0JY1po1KhUjE3gAMQSDQqFQZGIc1UytuWgbeUitIqAGI0gCgACKAgKA8i0GgBFYCAAAAAAAEaFTQaABQQAABpFTEaUBHKhEK0HI0KCAwEqAAAAADSAKiao0CAYoCpNCgIANAmK0igAAm1WgBFBFBFAQFGkQHAIAqAACA4AEwAAAFQBAFAGgBFBFBABQEBHAICoAAAAAqbkTEaFBFAE0GlRNQBABpFAQBWgTABGgAAVAAFAVZp0i0VytEBiOJZZ1VhTYXiweYBJsz22lx7gBzBoCUAcnIxN7AaoKAkgSE2igAwEAwAkZez7aLFIJkq1Y1yIZiuQG5HaIswUkbTHkxqxrwkE1AFKEBqkuClRMjaY5w6o1j0unxwrEGkAVExGhwCAICgI5NUTVAUAQFABNFaBMGgAAAEGlARQQAGhAAAAUBUwTFBFBEBUBQEGgBAFAABGlAAATEaUByTYokUEAAABqtAMUSKCAKgAAAIArQAImKCKCDSoCqmgCg0BAComDQAACg0BQAAAEAGgBQABAAAEOFQAAIAK0CaDSiYNIAqAAxRIAAA0igigImqNKgSzxvavZek182WVTHqpnLRBiUyKNXbzURxp28u96TqjBPaMUTnzTkTVjHIJsgAhEZIi0UGgIBigAFmls89vkhz7gNZZ6YkfOI5UBUGIQipNCRrTUaEBlkAVKAoMBiKRWwGjCSNyOCjlb8NU185FaQBUAAVAFBAFQEcm4mkVgggGKAglQATUYgIEwaBNFqUABpFBFTQaVAAHE0qAICgAwRCo0oMQQo0BCjQEqAigiiAaK0pNACDSiYAigigigigDSKCAK0FVNBpFaBNBoUBARQVAVZpFAARQEAUBGgBUTAAaAAaVEwARpUTVAFaAEAFTUTRQEBQABFBBpRNFBAAqQTAAaBMARQABAUAcO5Yz77M+fXK+NJSbVXaAKo0QlHpsgeciEoAqjFQKMVAFDWnAIJKgxGnotBWhEKDB0qDAkBijRRCq0RwqCABBAQxCWqCoDUaRWnTKmPTEBQYDFQisQG1xMbCEViVKjFh6fGFcIAg0omNacAiAoAgPJpFBAAaFTUG9o0UVieOoVaNgeccdc6JWYNKiYADSAIoKgIoKs0IAAKAAigI0AKJgCDSK0CagNVgIAEUFVNqsBAjQqaDSgCArQHAgAAKAgAANACIA4BAVAAFWaVBgIGKiYCAEVoEwAAQaUAExGhAVQaADSoCoAqaiYgCtCAqAKCAKACYjSomDQADSKmACgAIoPuR0OYtrs8dkj40WVN6zqqFuU0FKYGaKQjSI2vN6jcKhBKowQijEYKIRCoIRjlWqoUDkBqKlRgggBRWISoAggcIJAJGOFYDSpQBBoQHzTbVBVTUGAxRohKg0QhqCjKQ1qhnbcFdNeBRMGhoKAKCKCIAqHCoAFAQFUc8s9M0sdDlnWYWitCRU1gEyO8qVTXzkSsUUAaRQEAAUTUBUTAAYqIEAVpFTRWgTQaAFAQAAFAaCjSkWitCAKAADQiao0AAmK0gAADQJqjQgKJoNKAigICqqEQoCACgICoCo0omAA0jkxoKgKNIAg0qJg0KAgCpqJiAqAoAAAINKqpBANCNKiYNKiYNAmqADS6EdHRrze/Wo3yyzIVU5JJ0V5spOyKwQSI4QIEIaEPQoAAFQATFKQJFAQAJEHI01UKFBFBAJo6pooFTQaQIAQiAcggaS0aa1pqOQYMCRgSSzQDcJuQFQagANoQoONGmo5RGqkOmGfDr5iDgQFRoUEAVAVARwKsWg0AKJvuronsZd86To1lJJAETVyBwOnmzHvLKj6PGQciAK0AAmNac+aRAAFQEUBQEAUbVEAIAoAAIArQiYNADkTEBQEGlRNUaAFE0AVGgBUAAVAQaUAE0VoQAAAVAVZpAFRpQYIGtKoIoIAigIA4BAEaUAABMGkAFBFBFTck0DQAqDRXKKmAACiYgKjQAAIrQJrZWlx+ffRq6cNvfy7xmfSxlKw6KcDeqpXGzhbIAqA1zgE2q4aECEqDRRAUjgVpSBAo0CQQAPQ3EcqgOE0BTw20epjFKKCCQCEJVolY0NBHNMY01tJRDmg2QHPTHJuE3A1QYoCCEBASNBqVLUarQXjFDpyVY741VpAAAAAAAE3k0KxqiLi2nnz77L89wAUJXTsgJzUNHLZBIwK2fry0o9vNEcAAAAA0pNK0HAIAICgAA1iqIE0GI4SAKgKgACtBQFE0UBAAFRoVNBoUEAABQarQAAIqYgKAg0AIAoCKCk0K0CaK0ICoCoCoAAAA0ipg0ICiatByAI0qAACgjkwmgBQEVgiABUAGgTVGlQABUTFAvrbTtc/qoxOGrpkaSbBW5qb5VThW4NEBK40HLUVNAYjhoSg8mqIBAA40aEqKkQzVWCFIqQrHPnpYhIOTR1nOXO2ikRyBIgkVQiOVYCIJzQQY0aaNNAEBEacickqaCbnpijUKABhIgkVjWiGtDWqdfTngkvlZUeaJWAgA0omIANDk1FSogGWLsdlqbLue+LR45nT1RteCqJFActGNeaMJka1QobclaHTzhHKo0gCo0rpoQBU0GhGlRMGlRMGlRMUEGgAAEBQBAUaUVAhFAAQaVAVAVAAEVoExAVAVGhQBNFaRQQBUAQFAUTkYrQcCjQEAAAAKCKDXAgAAigICowUSCYrQiagI4FFQAiiEGoJABQEAUEUBAFGsiu1cz7J7uXa5k6RsrOGteUjxzTRo6NOWRWKaOQ1aGTnLdUVq8tqoajhBwiEA0IJRoVNAHIraPJXLS7RndEvkFHIk9CI4ByCYjSBAhI0lUJRBCgxGlaAjSOBoCI00YgCI0r03rLkaNVQ9yoChBEYmK5ABAalSkTVetpzVmXwxFZIoAJFGI0CFRiibyaRQJSrmhl6cz8+hwhz1q4WzB4ClK0YECjYwzajRE1Sp7cVRuvngIGhAcTQjFEACDQJqgA0oMQQAICgAAAIANCgpNI4EAAEViKJrgETVGlQBQQBQEAAAaETFBFaAEAFAdNIArQFAAAEGgBUAEwaAAaEBRMRpQBAUAQBQ/QX7WU/iN6utP1m/MFx85/eSrS/OkEFSKCKCSDklneWVbSzR1SunYlTlWdWhDIyGpUxtctkYlqaek05xo5VuICK0qrRwtFVW1Hm0eYIEaEJVrHI5BN7S3OjRkgr+b08/Ucrxeyib+Ms8dDJZ2hTTnRHmACEopCBLWEiiajABqNCtIgDk0GmoDRpGg5QY4JXKnulqib0bcqtAkjJZHUjmkaCoEUdTWg05KrL4UHKK0AAAJgANKs0qqlaC2jTRuYeqssaOam5SxVoiOFBuRaIPJFENKiUxYNMrXnQpbeYxNOYUBAVZoQABQABABQQaBMRyoMAVAAFRMKlFBBN4mgAAg0omDSAAAAqAqAAKAgAoIgKAitIAqAqAKAqYiagIrSKmDSKCAKgKgAAMBAAAA0ipg0igipoNfsj9sGf40/Ri0/Qj8myfL/wBKStj8E1Q1aDxyXY6J7OfZZkjpVV0grGuFaS0MQUticqxpQT3JvQ0FBQtyrVASqWqK1QHIqQeaKxRNASI0qCcy80dFSIbJcy6ay1lqZE9PT8t1eW1chBp+rh6mWzXhRKwAlEIGiEJjVitARoUGjTVAQBGKgKwERpABwNclJIrdJNNkTEGoUqA2MyiZcqjFaDo0mQ3FetpwwJfKIJUByprIXNPn0258+9zDRXJOFUGfpxxWrzuy49ksk6qipFEjgVpaqA9asR5iEjykYTAysFBV6WmNGHby0WsxoOdNIAAACK0igigipoNAAoIoNVoRNRoEwBFaFmgBAFRoUBEwAAFaQAAEaVE1GgAEwAQAaAABQATVABMGgTBoAAAAARWhAUAE0GlRNRoQBQQAVP8Abqd58J9Gp+NfPA/zg+0Z6P0g/CiOi4mPpMgrN9ybETcK1UtFGINUBAzBACABCRGlE1V25y6GoJ4UCoUEUaqqYjzHMbLNxOcjQRKzejB0Xo5+qdZ5PQjhrLpeej0d3mtvNRt8AESNnoVrho8UCEBCAxWmoDSpc1NUBEaRwNRjXCcgwBjRI0T0YORNJNOcrRwKKlBgDGERR3AjQ1iNEyCsa1fXgRVm+FssDuWMvQt2se6/b5/eei6KbL4WFYIx5sq3xZ82vLM+NZZI0ULEIYCkctUQFHQoxCQAAiieDVqVgCpQ24KxpwgCiYAACAKAgKAAAAgKjQqaDSoAADQJgDkVAIUGq0ipgAgKAIAAoCDSoCgIAAAAjQJqjSoAoCzTVaVAFTEaFBAAAAAAaBMARQBpAFABMQFAAP3l+FA/VLxtPxvmo6vzZ+web6D9Bvw308+NM+mFtZK8TnTYGiqWorUGAIhKoqc0aDzQMQTFYA3OW4DVjcjVQonC0c1jUUItQ5k6DxI1oHLnvRiolVbt6Xk9XBg2850VZPhebE04kQzcFoEoCYpKoDEaEaQBo0rQVgAxjmORJFGrQcwGjlUQjm3iJnTQDRwSLNooDQZG4a2pImmxuXMarx6ckdWuWw5W5pT9COnSt83sOuZei5I6nPKrtrngSuZ7AhbpxtgrmrJeMs029825CmsIHhIq6WjlUGoA0lUEITXbeIgFRxSpb+Y1KwVU1E0GhQQAQFAAEAVGlRMGgAAAFQAaAHJFqjQAACA5oKAACNCpiAK0IAAAKgAANIoAAgKJipoNAAAKAgAAAqAAA0AAAACaoAjSgAA9VZ08PS/fn8rVp+svwHWPpF8r8s/rzH1P0K/DXTiamXUxpm4ac5aPWetVVgNUQICDlBwImrXKDzFbRFAEueuhApUae1WPBVnWzUsa1YkhToppZBxucoCdPekbJEtWQvx2ZA8FR5xl8rVIYjiQVohAgRK05oNRoUEGmowQFjErWDgUTSMkcx4mxrTmIVQlQqSSaAQoPdNgDUEwajKlsbUDKzSKsYm3ztiMrbVurh3NPD0rkmHrzOz6Z0XU1xlXjrBGVk9FUcV8jGXx1XVi55Ujp0EBrlDFYSFysnpalYKDEBUEgs1r0msqEUIKmmFCHbyx6SAIAqNKgCggAoAmitIAAAAAI0qAoCKCk0igACggCOARMGgARpUBQATQaVAAAaEBRMABoEwAGgTBpWpqjSqmg0AAComDSKComAAAAqJvvTvtafF9MV1r6/5rpxaWKtOsuOPPOinXsvKahEQivNQpXLVXTqiqgAUbUJVoCkgAgDHKOcoOECFcagqei0c0EmdSmXXPT0ljVyyyHR0vM83qj+jzdDLneWDbjX6K9P5352PM+gd7F6Xz3iq+X7pC8xj5xlYtUSoOVFSsEgJjalWgqAK0ghEYMYIIARrAE0AUJ41QjQ0QgSA5Hy3/AKlXzfW3zTrwfkp9Qc/q/pN3nR4/5ffIHP64TsxGkiaZXrKJmnMx7mN4hoRXBdDZ4vpx8dCvnR71pGjUbKxQecjJ0ZHfKxunHHIQBQAqJg01xDHMVy1cxaIryVCwM0aSOCKvWbXNDGqdDXhgdfE0GrQByqAKmKCCaDSo0qAAACiaK0igiA5AcJgCAAAAiggCgAAADSAAAgKAAA0ICgACACgCagCAomg0ACg1WgAEwBFAAFBAAAcnobfN7mk/k96uzXkYlcwoORbuE3oU4ViCRHmDSKxzo2VjT0Vq4aMUqxyI81CnNVI2sRQGkoPJZFsjlq9FoiuXzrp39Lv5U5Dg9mOCfOLppCwS3Vetg5+jGbeUDW1hdvAxnLq1pirSUYc41pqNA1MK1iEjXIgDgarSAIARMRWmIJBjhD64Oa5dIqcwbVErQc4Y5CyJ+4/u/t5f8/D8/Qf/AEC3w/jj84Z9lLlp2kkWgiGxOa8OmTkY0JjRwROGbs66MnJ9Ax41dOrlVRpUo1wqEzRTokV87GacY4lqgAIpKoWIZgpY5j1XS1HioUIIBJGZuQdZLzVQbSrOpW281AyQBEaVU0RoUAaBNBpUAAAFAQAAAABRUAgAQFARWkAFTEAUABFaQTBpUTBoABoAEBVTRQBMQAaAABUBUBUAAUBABQQAUATJitvZ4vppIsu1G3g1L5wkc9bvkz6muKBtr2KyXE2+RXMarHrGwlSKmjxkQCGaDkUHMVojyR4IjBCVmnWO2tZ4464G680ttdE0q6/T8L1fmfOTyvYjtR0Qx68ZLGkEjmFlc7gGFZjTNqksR840lGNI1oGSpLVoDRKrGgKgINDAaykjROaIQHRgqAg05yciFCgrkKwHoDnJ6f7hacPqf5hXzn6FD/Ap2HqvdOj1WgpSsHAr5nINrHL2DSo837E9MrsfUc+dAG5FaR1CI81QU8E6Kysom6cio5VZpVLRrho8UCUaZo5iyTsNdgQICtJVpDWiGtRvpMa1BSvmopr5ogkUAAAQaAAAARQEBRoE0BCsRU0Rp5NAAAICgIrQJgAjQAKDVAAVAVAFBAAARpHA8iwBqtAmitACKmg0KMBIAAAAqYjSic2nHZpXOT35446mJWAoMdUq6dHLOytGoEotY7AQKQ6uCO052PFI+OqPVjry565oXNPjTY+pjVZslZJFn0tNOZsqaKUOnSvYvBdHUqxY9jmvOX0nXbT43z/tNDjvQ+PiZyX0vQnr+fUun5ppOZC83wkK2sxwo2vmASNIGuRBKDajT2oljKcqFRiqMjcta5RomKxHAIDUYrBIrBqT3lPSREY4Q9oKgOBoC+uv2u38n8X/ANeh/wA9XM83rveuhxPQ9JtWtNKyVVaKJrXDGvnt2svVa6dpWrR7laIS1jhEaaEoxwvRYepyh2fOKpTnRsA2srJqVgiGQwlHCVxT1XU1Hk9Fqg80CEjMx4IrccLzKlRl1+jyBHCoJFAQAAGgTUYgIAFARMGhAFAQHk0A0UQgKjQAqJitCAACggAAqAqJqjQADQAgm9ZoQBAHA0aRUwaAAAAEBQATVAUC7q5eno2OX3WtBq1miIfNHQ1rFa5fJOqINFE6RaxjlyxrJTeaO05y0t4HVLxXBci4HifEE9VolkrvvOwmHfWm05JEz6WlwyR5s7bm93j9qsqda4mWq6zN0mKXi+g1U6vMq8tx9tmx6XzNGhXGxcxkTwI6wLuuVJIaIYoEjHLWko5ocm1RuQJGjco2BLGWkjUukGohUTVw42VICjZQjhNQSg3uTRwK1A4FARQ9r/ZjXzqnG3y/iF+yeXofS/jW3nfn18h8v0blz7HCtRUAIohBjXnGVz2bWPpqk7yCoUlrgSJpYnLlBgSnU4evxjuz5lHluZOgrQwliXzNHgAowlJGEi2BajgGMRTFEAQlj3LIiRlLOq7efDFfII4BgIEBzAcAAIrSKCCajSKCACgigpNKgACK0igipoNAAAAAAAqAqAKAgIrQAAAm9FSohFaEAAVAAAAAAAAAFAE7evl6Wq/l95SdI23gkipyzsOVMKgWkqxpOT0RK8lUpIbznbnpMydar9+EknWNwnSXUNrLV8RrK1Xq4vRyPj5t2yOOgm8/7Lnm9nzKuz7puiz9PBo9PgO6TD1uc0GoHa8m1zno+HE7j3BVr9dr0edpnjezdML+bpgkeFMj05GpWAoCEohmISkblGuUVpypq5N4WoxIxQNrNZU3rNOFSoxUABI0TRpqCcwGMpOAkahUAAVQVo1QS+ludDycHLGnoXUufKaM9rlnVyzqKWAkcNEENcNc85L2Xe5J6kWaaVgqFIx5tVJWse1DRzVXXjSzG8LXmj2kAjZfMiPEQhASsYsiqRF1IGbmlsR8quLRoo2PKWMayja1lKtpy0q2vnIlYqAipg0IAAg0qJqNACKCKAgCA9ZtAQAICoANACiYgxXKCYNAAAqACYjSgINCg4ixAVAVGlRMGhRoCAAAVNFBHBb18vS0Zuf24hyOilknocTq1XLHOUcDJ9ZSPPS6j6GGVOgczxaZtnzbm4+tVXo8Nw6aqb1KmjL6fn8va383t8RNnyPreXTq+eHvLavZ+/qcXxfQ3eiz0mwOjn1uN9P45FzzSxne1jx3s3PZ+M5OTzvUeU4ZSYWqZoan1LmPU+b5Bvm+9Ghi1KxEIAhEJaOWo1Grkcm56pVG4NYYL5x8kj1aioUEAUbESULEcKgkBKJsHCAKo42OWjSOB7U0GhQJE3PVjlqrlsotHOnQUtJFYiaDlrHkDiS9Hdr5L1RcgaUyV5pGCI5kRN0Y2hNV+nIlid6zb5kHKPAYDSsGDwGiEcjkLIukQpwrY2uZFIe01he8HIqVxccLzVriOveeZV28sR5A0AIAqgg0CYAAIoA0iAKDlmkAEaFBAAAAUTVAQaVAEBQBAFaRyaACADSgPSLVAQBUaUBEBQAAAEBQEUJtXPt15Ob3GjlBDpJ3mlx72xiQqFBvdOjG6ZrMqe/P0GOTdA4lNMK88w6zy/QR52/kW49OaCyDHClhs9Q4H0OWqnleyuny+zk9EipnvK2/TbWhnpXe836jM5Tp8TawerxEb0+Er8+n1nhfH/TuZd9D+PxplsIYDWkY82uqOih9DyueOHvjHgIJAhUctaS1GhBCg14SCp0hrFWeZIRI5aAgaIUAAQEBKo1G1QagkE0QlUAUI4qkHKPBFAaCtCZZuVy39f8AMnGc6etRUqrRXLR0kaPSba5kUV4MKwlu5+grsvQjdXExzTJBEQ2K83qqa4IUcV5b50etKz65o1qBGldOjI650HijTMa0jwfIuhzZ3a55o1yxTFymrHOBVcbHiqgkTl4EOZpzZ7NvLEcgAAig1WgAAAAAUGqgRjyaABAVABoAAABRNBoAEaUTBoE1QAaRQQAUHEWqAAIAAA0ACpoMBAoJMVt6XN7gTsA1knaQz6llnWWCdGuvJJFVewqe1atkuZlz9Ilz7IbaKcvR5taQwn6DzfsauB0eNu42uLJL4bLY7PX/ABv6D4/p5uX1eUb5H0G3gYekb1YbPOcvv1eg044ueqOkpZehhO7PmL+dfIrtOdzahYsepkO/kuY4a5yxHA0lCskGmhCI8lRCBmNE1rTo2kUlrm1lluc7jicxSkTk6iIVolQQowBoJBggnOKcFACAhBACqqjirJqtKDAEQTlBZZ1eq3+ke904/jR+PrI5NAqaXPpJFbhaCmqIYRw1lYt5dlrOjvRmvluCWuEjGmMqJGIkcqiivOGw8WPVV0vmEaVHKuVjBRlcaROEkE16bnm70npjWuZWq0a+ZzzcQYExOMlaDYnEsy1rx1jRydvOjTTjFBBoQFGgTAABABWkVMGlJpFBUAAAEGhQBMQFRpUAUBAABQQAAAERpwDyLRQarSoCCaqDRoAFBFABroz0brub2BVY5azGfWqJky1fJn2shrCwxN6z0Dp6Y5dvOt6WX0Eedzd896dceXq8fTzNIg9A836ePjHFfX9n46lpcHvRZ23L0XQeh5XnfT8+0NPyvqp8J4QL1/OIqlijUnoCudFeUT9OVdjS+k8/6fLdV5OyaJyNeYxwqg1lZtHgAI15KhIglYkDlGMRwmqDHA246SBuF6zmdOiom1zlHJtcDEE1WPExQaCRWnKqcPREIAAQDI43CDSSIQYiA980rzRXz0O/VzTj/KQy9NXrQknSeSOp17L3H0dsJmZdkDq4L+ZI8eDAQx8yjzRGLASjnLVYqIInMbq51ergS+ZHAjXKjTidGMrmGmUY5WQqVy60E4x4PVbxI+QcNzTUHmOWjUM0a5bOqrw1k6vUYsO/joVmigDSKAJiAK0gAAAADybQQDQNAmKCICoCgAANIAICgKgKgA0igAIJyJNK0ABUAGlQAAAAEUFE1156dSXn9ZRavdHS4nVbE715Kh8sdMkeXqsmvz2qiRufpPS42Dl9/p+d5vatYGvmenc1Xdir6Xyzeo4vZ7PlfG/QuC9W+x/EeMzPF+yxpejwfSuufteFW15mYleeEbNXTyRZ3Y14Kjzax5rOadbzfH9RRueh8pA3bGuzGZYVTDTmRrl7U3RtCPERw0MwEiiY0lHMRURzNJXKHOFkZVkCWWLGjFQKONHDEaUBWg5AVRvE1ARQlDpAlGmYCFTa1wxtSgKQmmNpSSzSi0c9dB+kl83w3wmffYnjuYpb7efrWLufrLBvz6mVy+pUfpxdDyGGiG/zytbry351eRyrUxrRHUjhDQWMlkjxY8cJWCOYjSUHm5y2IiEHzta4c5VKw2AckKp7m42nOqg5y6Ih86qUPWsDnikThFIWMK+Ptw1jXz0UEUEVpAFQEUAAARWhAck0oCDQAqAKmIAADSKAAAAAAitCAqJqjSoAA8iwBABGlVNBoAHJoAATa8dtp+XdI+dVknocsdI5Urlq9Y3erck/P9BXS/Me2Ox1zPsYxb3vRub6foMPyfu8Gft+R5C76PyvSMj3eh4evO9N1Pl/3DyFn0P47Nye3mQ2ejx4uojtrZ2e1Ot2fMO2J62ZENvcZdfO5W/j0XdHjooWCOl0SpfQ+b6Hhuv9j43z056Y15KjhUGIJWtAQiECPJUBqEjRDgVo2OcthanmNKsLzszRYJoohgMVpoxrhKgIoIMEQTDSRNHKnrWqrOzWmaDxRQEQ1jzR4IwBzUizq5TdXrU9OH9kfnq4suy96S7j68F5dvW5nn/U6EnpeJZ57k9anPfm18Q4qpv82qVzxteDH1CtVRDzalQDASEajUxHFMSskHCKDB4CrSVs9EaVxohACc41QEoajgIyRSJZF21h8aPKdJPVFHXPDK8IkIHA+oGDX6/EB5gAgCgDQgKgAAAKDkmlRNQEGkUAAQFAAABAAaAATBoUEGkUEAFTcTQAoCAAAgKAitKJlxa70/P7bUlOU0kfHYKrJZ1c6ehkxTXiHZd08GfdqbXN9DyOpt5W1ocP1WXp7YdTu+d9py+rBR5bp8fs/PQZF1/Odnz3N7WNU9L5GTNrz1nZDdHVi38cUzSSO19edUdfDp4U6RL0eQPVK1W6Wejr+Nx9dncer8XwLMOp6Q2GmLQhFEANcQ0aQIAGoS1WiRVJVVI2sicp9Q0ZO8Zkm2oJzUNZUqAIDkAcNqCFBEYAnPmpGjfNWqLG6ICIQDgQQ0gaSjWlkTcq1HG6uWkksdX6u/mBvzwzcn2XYcpj7a7HT5e7h8H02pT25uk5Xn7NmErc8k9X4Wq3m86RDBs2+NWOG5qI26c6oAoIoCAxTJko2w1kqPNQaIZig9Z6UjrIQ52DhylKFOFsIJ0YCmUtufQrwPBHuJJI6qzb4nQinhAQxVCqmLvwVU281UEACMFQitCAqAoAgOJoUEAAEAUBUTBpABQRGlAAAAAAFQAaAABxFgCKCKCI0omAAxwjS3sPackdCClXroc5dIS1VORx0DErNl7xpye3rnJ7tfd0nOWNNj0ny/vs/N934h3oPJ7fiPYce+nw+foS871/MJBt5zs7Tlib1+DITqwaa2vPe15MmoYZenuVsPVx29nyKxhE+8WOIarJev4/o+Th6/DbHfK6N5vRONaxRSRBqgkUhEEg4c1UiOBonKNjHKTFRvaqjm2+LagKwHQicjQAqDAQoJKOJBINKJo4HuTk6jpnnG8voopADECRitra5ECGoJ6qhxqrjZ0kdE+lj7H2j5X0cnlF7k+37jz7P0Z9Ht8Oe7h61TLw7eqyjS3ja8OPz3R8xWrYebNXG6DXgY2sRGkaQ9rQqYjQoMYYkhRCZotYuQ2VpAEK9bjQaj5kaZq4pAUiLqaryVpSBg6wu10a2SdJsTSR1zvYkNebXGaoWVnng0+rw0W8QBFTRQRQQaVAAABxNCNKiagIAqNCpiADQAqAAAAAACggCK0AIoOIsAEBUAGgTVAVAdpz0bi83tgSrlq5Fs5yt7zZjZJJY3VFo4V6W5x/W3ul837Xm63V5XXYcd2f0Gc18G8HbvTzVbvF7HY4PB9fwGj635re5gvMZ2/NPh0852rG+R1x0cgb/POgYOeRNh7Sddl38O3u+QsGXdVNfPjSsgcKKlY0NrNWuVdNMSsBRIg1GmuUogkHI4psbzciFBwlRJK3FVafaliTSKJIgUaAaAlAAFcmgxgIa0KCoD3qpvSe7i4eh5v0SJXEK2CqVs7RtvkEM0cgUtXG6vW0lyO2W9h9Ju/pDt53wIzzv0eNvb5Ffe5fXiv+v81nWPO9uXTN+M3+Ts5nmPQ+MwUr5Lcwce6wys6ht5qI5GkA0oCtVOQEY80e0RGaOrJ6Tug5RpCyK3ItokrkRSEQlVKVTVEIVBorzWWeqZk9KxOHgEbgaS0fMgS563jV5swdOPMTp8VFAABiKhAAaVAUBSaQBUAAUARoAVAABQRQQaBNBpRMQAaBMGgTkJpAEUEUBGhU1aAoau7h7Ktz6h4OWekC3OVuebTQzbXyr2lxfT5UW/j2pcvf63b837jouWnv+qPmrq8/nuy8/wBXzGz3fH9NR9f5zqud+b/V7May4uH0vh6Bv5lc1821WndOrjrTOx7sev3/ABskVZMdWep3flfonncXd8fBF0ePY0cPRxE6PIRRNVyg5fEMVwI0OQ1KyAkRUK0ghAka5eitiViNGrmiVawRvKC01KkWxROiGg5EBVGgJUAeNRCKDUY0coAAK4ep0Pfy8vH53qoEAJRUqFNHiISKNXLdyrZ8k73bWPv296fdv/pNp5/5yR5fQdvka9Ldry/q7OV9B8BkWuL0oKuO2lzN8WFXjz8Wnt8iF4wTogLwQaRCBQUKFTcwbWPFHtSV0IVzqFuCxFKK09Fs1r50QyByKmKaK0BWIED1rIs9b1Vsc02MzY55Na8lQQCAHRiy8To8ZpfKitINKAiAoAAiNPJoAVABoRNRpFTEaVABNRpAAaBMABoAQTVGlQHkWqAg0qAAAAqa2Vr0Nnl99gZveutyGionNPHSiTpNKdddkLQ7Dz/seXmvnzLFZdxynVPWe/8Aj/c+C6GfR0PluPZck6vn4cPr8vczurzKDctNHO25bWbhVY28zuMLz/r8TT7flsyPbx4V05GFQPVOljq9F898/wCypw+p+fKjgY8wcIMQIVy0aOWq4R42tMxHm5FqDlFJaEognsHG2s3W56CMtazzhKwmnmgVNQTGuVBiAqoFYOGr0wVIjTGPNWgiNKAoF2TfSJObUQAECHIWjTEVgJucdCyTu+3PZp2ub6/bl5voew983np/kHPPsW+jpuV8vY16fT40du/Po+S35JX8z1uXH4VfMryFdfHGtZhSasjRw4KUVIjiRitEecTqxlgVg8UcNWjcwStIe5bDBNSuURSo2OViFgSI4R47D49InW0CkRwvmErNEM1eqUNmseDQwM2scSLq8VUcipoNKiYNACKCiaKGlnLZo8QBUAVpBNQEGgBUBUTFaQAAAAaQAUHEWACAK0ipoNORN+xHXsHN7jVeUpPYo2oqehTruPo9XzvF9W1nX4sa5mh7L4/6f49P3fJuyNefQ6fm9fPyqxvUie65vi+kscv6Hze37L6HzXz6/m9LUxNcGV8skXTyxrl92NqFPbzUXTmYtRIydHCc8+HpVId/LiW+YY4VGnorI3mr02DhQAAQQOEVCK0iCVqGjl8dCDmzpR62RDfl11cOnmpidGhCxDaOVAQBUTkcU5VQFIoCCRjzRBRpUqAKy87eok5dQEigCpVLajxRwDls4W8lmeqeefS27fJ9l6Fk+f8AaaF7q5/0G+COv4zoMfo9/wBlwu35uHy3zfsev5z3/wAqz+W8f6qfmMsueSfCzqteSj3lGlQI4AYEijAJI1atJdA4RawninoYVzClACoA0hXKlaW1K5hFIORxTkW7R4gDVIlknuddnsrqFaOuRUIYPFyrUQJIhtHyxKRSwejzmJpwitIAACAqNKApNKAiAjmAgAFAQFQAARpRMRioJUAUEGkUAAE3E0IArSKCKCKAJz7ufo3WY9zpVs5J6ZGlKssSm+zn09HU4fq7ObvElVZS7/L72NU38a71XH9Fxad3zm3iZeg31nk93zDT38ijf6OHYxujDWocPt4a6+TTS/MlDKBu3nz1biCRZwF84jSgKs6LtY+i3CrJh0eOrRopmKWISKqGtOaJzSXNGI8wQDTQTHuY5WnK9RlZ269dGeyOQlRLPGgg2vAgGDzaA4GSjc6dVU2EEBSBCtJa14JG05glVlmXWoUwoUAKESUKjV5OFcgdL3zvY1MvXbKvX0Nfn9/ftcf1DZOng/Qv5A7PmW8Xh9x9a+c+9+deWc98/wDo+y73vzXjrXj/AFHPUvR+Hw60ceXBz4sJyRjkbWStEDSuKAbmAgSRPNFecxPU1hkhWQgKAIJqmciLpa55NYZK0zVW1FsimatdMUwdIb2LefqwgQRPla55xo8BxUpHUr1UEd8jGvmkz6jn2dfhAJFaRAcJg0igqzSACptGlAAEUATBoVNAEUAaABAVAUAAQaFByTSKDVaAAAAHDs9Bj6c6Y9w8pZDocyNpGjVRvtPd7c9ppl6en13L9RxPdXz+cVX4K6WfZ1nB8/sNsdnz/c8LwfVss9nz/a8fxfRR6Pb8/luvJYb5ep47j9b0nL4vpeQzPa/L1W+WCy4gjrJFrIBgm0rNyTaNrEUTVJRRqA1ROaWqEjmI+dYi+VrkNlLiHk+NWJpjKTvYoK67NOSJ6JZ5qR82rSx6b4FcSXzqoMlB752VxuCt7E5GFIlZIENQyAlo4R7HlW1sSqArUKMFTaEukOhHGz7EdN3p+P7Otox60XRbx7N5z6HnZd3h9/036S2+e+Wuay+19W8rH37fofyvku06PK8s4nwPs63Uej8T5tR8z0I7GGz8iZRu3mgJFcooKpSAAAgLG4YrwlI6rEK1ZK8oWXgISqEojzlJ6mFc6oqajwBJXGyNcPaEamTnGiuXQKW9J0aOEaQ+ZbQqK2T01FrmgbXC9oZ3PdHkNXbhBNFaBMABpSaVAFBAEGgBRMAAEUBAVGlExGgTVGgAQFRpRN5NCAAAxAQoCJ2Oly9KaPLtW5VRPz7RqcpG7msfo1fb8Lz++1z4PVOf69+m8ny6Uh5OdVvmFNQIfHPYO04+z3fC/WOV5j0/hln6PIqPOfcwsfUYuvl1jXz4VrnGUhHmiOFAVgKjlUJc1Nr3DUJchoKJEJcq0RQRHmiiQqVWdHNoHBPUekDtOCCGaWIkUSIJ15VK6NkEPknrpuvjASqNXrRHmyqtRoDXkOBRW0eaOVPRW1KyQcq81jacw0kaS5xTg0VibnFvdPVPZjuubeX1m9nz6btBa9G6Pb009HwLP6NY8/57ZGP1nuPnXbzYtDf5Pj8OPPbsVwcTS5dVgrzm6PF62ZQ28dDXhEAQSgPaUIJQTmqkY82lYyE7K9aRyCbE4G1gK00UK9bNGmucOhUg8letURtEMYZHA9WqrUHbnTuKmKUgJRVapDQrndEpYVyrjXjjHT4jXNIoIAAACiYAigAIrQJgmDQo1E3vWk6Ld6Ko21kyJ4tSsgAAQaFTeTQgACAA0AKJu156dVcPVW/oqTst3vXW6XPoWbPsJVvZj6Odph3O0L173zLs8iNPL9dg+SJ18zmgSrqYGFixPqQU3wIacksZqm1tljR5REx5MSudzGlRy1XCIQoNrnCCYqpqvJzC1UatGBChQ+dY1rNFJRWIo5W22za4Nzs6rhztARohjmGrfHb2I65TLpcq0dEaV7FcoTT1NhCnPNUYQgYICAAYOVoCe1bLIrjLyFGNTEeQqTVamSeyIrle5dMlvPsmJ7tPfy+t6a95H3ehoe787QveP9nB2PrfN2fobp+V855Ppw1vLPK+ywsKOaty3o/FUbuecbuP0cqHbwq1qMaMGnC0vBASKIBqrpFQgS4WiRPJhfO9Z1eLYVMQTI75kV5iJkipGsEeSom0MBz1c0tEMYXPJZFoBaqtHOXQKnO2egY5sxq31nm+Jy15nBFXCDjmqvX4YPMGgTBpBNwmigIArQqaqUry5ZI2knneZ8buWdFaCuBUQyvecFLTjij05Gq4arQA8iwAQAAUEAUAtGm/Ly+y2RWoaPdPTcdtpLe5fd6XH8r7iu31vjJs3bgkjlXqs9vYcF1eL23E+J9ZG7t+aYNJKqVpUuvrEuBObBVNAqSWNp8t4yQ3zI2+aRZ2iLwQeSAhQRCRWlQYKgEKNAStYqhZMfTrt185rnm5pbo6lHopF4NYCPSbC5bHWUNe+SErnv72PquXLpJFq+qrZLXO5VauOlUTVrlqHMKNFEIwBCIUbBCKArpQpHjBbACzxu1hMLr5Hyz2TXJ75bOH0PS9/j9hQh5fT9a839n5ydPM+v8AQvQvo/juK6/g+g9T+JL+NtZfD9ZJznR4N3g158Gnt5Gf0PL6/L7uDwcvo+aidr5yCEbUAACUBUG5rFVVG0yalc49aK9agrarhrXm0rBWjHKnItBHmxHzoqkeaIgxHnAjzdIm4Wiq7e+Ot6rZyrRjXnYiWkjQgdWEbXiRGCFY0ed38tDXiRQRQQaUBSaBMGhQmlnoJDR1iN7M2XS9025Rky2YjAJRpmsQlcU3K0xpJt5rUJalZujQISigAIoIrQJlpabtrn9lFmxVs+Seh6Gs2iu7p+25f0HlW9Hicx2Xp/I40V8nNy+P9Br4k7RVn5iSVhGXLokTTTrFqXl2XnHT50tfj9VFvJ7JrRyc+gl24YmPNk4oUrAHIDapmIDkaQQ5pY4SKMQHOWrErGzVz6WrpxQzVEiG9dtc8QhsbToiYYb5iN5xRVztHmoF3oMfVDPpfLPRHWeUjyXKtHKdKtE1DmABWNUQCBMQFAGtKoAMVAOnPnpc9dKujWGK+FHVM0s9lzTz97R0eL6/a7/T0sylh6rqfT52l12Hv2rm0bflPT8/DoZb8NW18GpHPNRudXic5czI6mfTVs6eNQpZ8ka1zXKUdCGnGA2qSqlClIhLxaxpWDG1yjk3C3cipUBEcNK50Bo4SunojSuVWiVCVctEAGOKy1i6dWqTTnm8lmO+N06PUtK7hz0mWSoRxWjvkRr5muMoMDbjppv5SoCKAjQoOSaACRVJKbOlnont5dUyxo961RQAaqbIEopKISNIVyBrbInnG2phZeLoxVq+nJEPmGgDQAALenfcl5vXVVSuesksd1gjtfLO1j0RfS+ncF7Xy3nVTxPf9n8G9P5CuvB2b2wvSx+aXE19+eotHIm0cWZdKpJEysjVrNMZ2To2xN03acSsGjaylhVKVjbq6ZtTNgJRpqClSd4krnsNnpHjHLWuXxqhLY2mNvOxAVC0l0BMMV80MN8jEeSoCgCNKidrocvSlTLrdLO9aOsXSg9Z2VTcaZIGQDATEcq4FFSKIBogka5AHPKRyZej0I3TtDDpxvAJTR1pdfR9HP007vP+t7bnj1uhueJ99gb3035rndRwfV43Tb8HCTxhDaVXuC9v8zqUvP8AZ0Mnp8ymzLWnSvy06zt8LiqPGmOMEaDkEIIAcFKDFB742rM15Go+YBgqUKUTBjWvnc0FcrVFowrnVqFQkc2NTWCs45Hm6SbFKnlj0moN9iOh7VqNcwteKqCoyulcghMJXIyhWOCnV4YgKADQApNKDmmneR89Nyzl1yyRqBSuNGqCtVPC0CQG4VCEtSpEWYOUQhGiVjI4Kzo09vOjSuYARQRQLq16Jeb2kcm509Mr47Lrub2kftF/SXuamyvZyuP08KKDf5qCuuUliUgTRzlsqaMBzBP7jpx4hOfpZZlwxVir20WakqmblcLC4cNMV46dDP0YzTgRHm5Bo1w9Y2IHFx09sElYLGrGXytY80jCNKlwiOJzXqacMKVyorEBIoCNKAgnZ38/RsGHa562ihrFZQeq0FWzQwAECUhpNcJsgKqblKGia1zGxpJGSyxbhWt/P0akdc7XVhKxdTpFpYtT3aurz/YbXZZfS2c3g+mi0KJNXeM93P6D17fLyui5uznM6ON/PfQfm96Py/pcQ7PCdmysiB+MqvnTNOKJdPNUKRpCg0BPaU9BjhvSNYW68kY+cBoKRWKK1QpEeA4piqVU2ajxVpIKVU0GtMjeDZgc5WKXbjn0XtTJpt8k9LlKrI8IH1gkZKOebGPKNXzR07zwY+nwwcgCDQA4mnWFrPJHZNZz6Z589QKFGqrcVW5VogJBoBKqoQGtrMa8lRQAgBR2gpbQvLIg6fHapk1QAB4+kt8/uiZ6vculzlqsj1ns83uaepHv+mYWX1K+Z93yLfafS/NvBl5vSz245K1JHOFFbmqhwMdcWa2fQ1L5p259MJtxyJGyvjWpNpwscXFI8kaGpQz7YXa8CDGhmNcyujsgirktPnqhkrJYhxpfMqp1o6kAGE1INOOs2uMAQAGgAAAACXdz7rpj6BKtI67zdIh71oNKEIUBIXMMdw9RNcNVQ5BigxjmNGlspzujRQsE4pXDxdaqqfZjqmtL0rm9zfW9Tfy+juV+H6PSpdfD2lh9jLC7us806vAp4i5K2A/MqWd/JZY15L0F4YHX8vseZXY8XBTTxq8F8CI8gEAkARQeFACoVKTrVL4WFZAAggEOVPFtElcygxE3OWiMeQjzBIcaI1xElZLLNOU1Rw3ItnSTrHKrusjvZMoqsvBQTkbIyIm1xta+dlWs8ap1eMi1iIAoIA98aTyrqtW8e2WfPYC0UT1OhxOqiYgKCGNKNAAghHKBmiggQ1xKi2Boapyczp8uFL4QAQFA0+hx9pqxsONFWeidHo60b3eq09PrYfB/VKHBex8HUt5eXl178yJpyxlc7nLRFKcxUrSVHJcnavG4SQGMrFZCo3ImqJzRkqysnorRalqkCDehcK1ySpHVXbfJM5aRSsIGmteUsTUUbGuJWsKNl80deudivFABAUaQAAUTXYjq0nYemsitK4klcvfOghQEo1zDDUOlHqUXbCFCOYypAFYAoppZuRZtWtoCVxRIuh7J2mt59k81ejf7Lg+26q9zfaGX0+azb2xsejcP2tPkuvy9LnvO9nKXn4cvA9H5PP0ejyMh2nLNn3zy58KGjPBXl7fmM9s4tQzQciCUABuCgAB3ay3qpfmoEj1bUcIogY9s6tSuUUFRUONBoDXgAhxaRGY0l8po+NbD3LXlEaFcEs0d7pc94UqEBMR5tHnG2+ZqHPFRvKrlb+WJfMAA0IE88dFixn3P0MuiRZ2RRj1sKtAGCQjBHINgkAlaNUEiGUbaxYjlXJzLNiugV18Pbzoa+3miOFRNRqXrcfUVM+1HrWSTPtmctpFXVpeh8/1/GfQf1X5np+NdfF5tnfKfVxMXCjKxEIRXCuWrRyCbR5uaFuGOtpWMscbRrryTMjoUSrWqwiQGvctkWjGvJ7SkRwDyRVpAlc9gWkM1JrkV1ecVOorWbwdJG8EF4Ix5olZ1WnMDSDQJg0CYoGtPTst5/UJFSxlMkcAm8VI1pK7hklK0/PeevcoQ0cDEcpA5rlZvnVSumnKtblSOxqXwq8tO16s6XO8/bp7Xn/U5Nne5u9PV6XJ877ddeevabvoqcvpsqusqXPlz89+Xla5w8x0PZ51bn3FrN6/Gu1MddflcXSZ2/HQMvjGktR5OBoglerQbARKr0sZ7RJXligijVBoOBypGGSo0BmobIqSqatDJB5uatWQ1zveh7zcRsEOetoB8gDes9D7EdaNWqoIY4aysEY+cR5Mq1lSyN/MaacaK01QEDRky9Ca1n13ZMupBNymqisSkpNIOQBFEgJUExDJriFaCMaalTG1yihOKnTzSuXS3KvlxmdPiNVwgCgGzHVuph7zUIc6dpHVvO02venev46c/h61n2vzvvvnHAw+YdVPNSPTkRKxVw2Kk5C0USIJAzfPPUxiZMDYnKStMljR9UFbeCo4tx497K+3Akjyq2HI8uRufVSm24FREbnELhOgHSr68dqadkaqjZWb4gbHWEEF8YEgAjSomqNABqx0bC8/rJIqcq1he8muBVKISYlqbDo1UKEJRwIDjrVjCVE8ipzlThOR89cKPmUcvQ2c+drEJvdXD0dS3PvN6mfb6HuF7vJX/ADvo9Onn7WxT7vDy4uPqzV7fm5s/h76T1x5N7o4d3np7cqr6Pyl3XFy1bu+QwIseRY3mskXC28AJa4BRqpbQl887uz9MEFgrlbUaRXCKCCGPeSI0gZKhahYIFGijajxgV4zquhZVuxjyVQljW0SPiFTV5sr46nKtmuctQSNeTW1gBKRmVPO24s9u/lCg0aEDUnw9W3cy65Cd1RU9TZEEBAjFQBDOMM3KqGOBokRpGNNRpiOWI0qjsEU9ydh09Q1lDI28uo3XzQAGlcq6TS5vpEjKR8uSa+lZTXWsdePr3ZfQfM+P938j+x+GYi+TjdHH3HJ8P0+Uej8cjxDGIpmoaNVSNJFanXPsY9kkcbRt24nrOpbx7a7bwe0TY6xdC8RawSQ0fHO0Vi8qMj5pIGSqEdcTHVMVesZ70a5pUNieb3lpE4rV9OFqGSo0AAmAKgLrT06T8PTcwc6zojVL2je0oa4HAKmAwGAmVqiFazkmVvSW4G5ytULjR4jwcLZ7l0PJ6Jbkdd27h9Pv9X5v3a+s9XpZfF9nhdLzvl/ZW2Z3CyuXJx+r5rXv+b7nMrr4d2vHbkQ+v8r2PnG/m0YjztOqow5L8KJtRtY2HfCdHlI8TEJVRuVaoNTTx0Va98rVrB6To0eY0TlKY14iuRqkCBW1CwVCggCQvFs5MqLskfPQyKuYGnNTYHM1WnKt1fOz2LUUSStldw0fMqAIQynWWVS6fIaVzg0Aa83P7F+fLqHLZyLVQpGvICAGjDNrXD1RG5prWmDTYxOQGvaYAIJFZOk256dh0buDeOpXJgwdXhiOAEonN1ufrTMy9UcnJJHUrW0kJ1zfok9NXT5rjZ402zwXIM+ioa+YjmlVaox5KqoAFGjVKvHYWe2N8UkbRNUwzy5Fx9JiVhnu6fEkgm3K5WOiGYzarUdgTm1b5FvY0Ozg51nPTasSitCp0q+vJIgkRzHUfMDzQaAABUAUDTjp1nYeoIEiKnvnRABBCNKIFGAxjmGCs0VqZ804G9E1VD3nVFE+YcJXlK5dDhbSOnd93L0umk4vujrs/WubFexf6ne8mryexq4Ovn5OVny14q83C2r4Ktvk9LmbPq/MU5pqrbWfIrt8zark0Y9fJEcPjkYViICqNqimYbAK6uXoVauvmNWsHNVNc4QcoJNDJR0jDJUJFBwWKUKUIEcb57D57JBbijhSuN0Y3NIaGSoMcMeuh7Z2VGiZaTUQHPAY0BDYnhSzNuKk3byxGlQN67y+5dXL0WqIVMawQyRTNjHKgka0waY1yoEbmhAVqFRiANVpHBO2be+alfNvDraPny8Xfyo004QBQF0Ft1Def6ZHS36F6dRzG8RWfK96hN0ebemXr7HK5aNvdnhYCRwtDNHCApHMRFI+m1FLidmPeazz1Ds93yZdlZ+3GkLiYnZFRWi28qzWnaWC8HxuQakrEvkvN9h9U8b69jH6fOi6fzPcx6Jx6/LRpUh7PCjbWESXkqCSuZRq8hABoUEAVAW7O+0/n9YQAHO+NALfGUBmACCGNQRXmj0pHKpXKkEIMdPO5XdNR8qSDd1vTngwcnqPI6CVXNfj0+x5vh+2ubHXzei9afY8pq8Xpa/G3y9RyM9FejjzP5/0vlMHoMCCDPeh13b4/OPvGTXx7vO6z8Mhvx2xacjXEJA81Rw+JJUcq4pwrLJrdz8u6qzfxGSEI0pFcICATVeatG1hgqgPG0EINQHIUyULMUegoQqNsbXKoNGmKISAOU1eq6B820aa8HRq5GAiPJrXi1hg2leeVU6fHFrJoHSX+P6CRY6xSlRUNcIwxRjyEaRrTUaRjSOE6MaqJqgig1QYrQIEZOTbnpummnOnrRjxr52vHkpv5AjQ4aAdFtYfTRplu4Ha1r6MJuUOsRvuaPL9bDz3Z4NaHXyYSeVqvNHKhBtc81Q0JayhJbkWitJmF0OkjosHN7BWvjgb0eTNEqa55pFWLoSVtLatDpy2oU7S1vI3o5tRz76bnPVVe3zui86yzSv43t0am/kxlS2NwxlYrG4ZWfOiPIAEaUTAAAuLbZm5/WaAgC2FbknVyDahm8pMbJFXvN8gSE09AHDGg5RvWeiBK5nKhzjV8h1uJ3fNnvG9l22du0zL6a/PHoewPy+0xVjXtPPdPPu4ePauVr48+P0eLh9NwdtWrOmpoaY4kfsfKX+Zz0ya/K4Id/nWtrnhWs3REtaYvjTcjQ5kxWq9Pze72nj/AAfQQt9b84SZXGyskUzRQVBgMa8Ygya8Q4aoMaKKRpzlTpJ6VFuKCtbYlcqoqRHgjSRwDi5Gz0Dih60jUhRWriox5sSsGRvnYxxRyd/OiNeFHB1F3i+kUXQOmmgIlZJGYIxyxHMY0xzSIDmgAKoIA1RCAigisRQmbNOnG101JJOpA1E2scap0eSivJAAal7bH2xuHqxpXMSzq9psjpJ7u2up0Xo8XG4/n9kTY4UFkqvRUKc2W115OSdWpWDhW9c+pErN809Ng5/UgTThrG3nrEKRqbqtZSI8Yri1KTV+d9j9Hoz2Mb0fJ263F7+PTnHrs71fA2OB4dok8/qiiecSuUrCjjvBHOYa9c8QYCNA0AKJgBeW2vLh6rUTACVOYnUQQ0StabA5a9qd82CGjSg1QHtKhHDnJyOWzpF0o5aTMW7pCrKvRL3P6cu70drdzl+i6Lc5Ptc3pqqhlc/f2nnXf85Wsc2kWIubEua+PoxcfqR6yvMzvoPhKWXjVfQg1uSzzmPR+Vqyc/XTXTnarhjIQhLp6q5FpXRYZ1MzsISKvOSZVEysRDJwaKFICa187GPMcD3KlBoglUYFSKupyToqjWKoar5xBo4TEMVaIUbhbyOjoFLFVNR5jiiJ5jHijHzMiqIc3XmzzbykeHXv4vpAKcq2ajgaYxxPJY2mI0gJzQcgPYAoIoKo2oIYxzGSMQiNACypzWI2cRoyC8YGXnRpbec0fOitIDk7fL2Fjy9aMnncO1v59UanRL2ndzHd8H1nP+b9nyeO3DlaGKIQ5KBysVWisRSRFcjo3RGrUcdU65d8bL5I2a8SIRLCmJeIwznnjueVqy1vDndGUfX9/h38CerAref6DI1ysTKkbObUefQc/vztoc/RBHpzNcQlOsom1yqAACAoAoSb8d2oT7DRSISxk4vYqaqTybI2qz7iw+NAGAgGoN0k9MTHzMc4mJ7JVXQsi1bYWjoyn2DRQ2Fx9W5uL0+xvV7WtHyfTVbmPTuN7/Mxsry/Zrbfo/OcLpc8Y6b+Vo0o6Clpy597t8DOpTEeth1YMevjynR5jG49qM14FRBBNNaZOnjeu+8nOy681/V4clTKlmHGx5CuUUBy1Rhmg8BBDhqGjlWwoxBjhvfO4FtR5tY8VkKhHzoo0VSkLyVylVNrCT2jppFGyQToyhCI475Ua+ZsTiOtpnkx7+Q5iXZJyfQji3ItRrzGmccbyIalqOVQBw0chqsRBPE0RiAntG1RNcDWtLGJ7yr02PUA0heUcNZsp6YZi6+WitIqZ0a7dqLm+jjcudGCWenq049KFVlJvbdHWcJ6Pz+K3y+1EMUaZCgObc5dLUErimoQ2UUsEdN9cvQryvOWtGkKdPmI6XGtZNkBqOU363m6bfp5Sfr8vc5WsU1JrpuK7PNo1fL9MblYkt8VZiZmlnVEtDLWnFryRNrnmrFUDTzxBKgKjFRCgWuly9WyL02ooAlWLNAlETV00yEl8oxShEKMRRrZnqpSVygWjkLIup756GuKc9aS6GXfSk1yFNXvx9axo5+j6jy/fHoWb5H6T0fKXm/Ww7cnEry4NauPCo7eZczq5u6571Pn8Gjy7pSgbFXJAwwYj5RHziLR7duMbNRQrNOo6+HAn4Paz7tcsNSFmW+nw1ijYeCxqgrEVMc7YhkgswGqmivXQ9y3UVDXLUM1UbgtEJjbXOOcilNQkmWkDHyKCVy1c6ep5Oskk7xvBkdZNSsUY8ljeLGGMbLzy6W/mCPDr15Pohxo5q0RHijTKFKzZE5c0BGDwEBVTa4TFYIDVEqAKDASKyFWnWFV1+PWCtGvCGO86y3nnVNfNaVgoMkT7GTm+rRkiq1K2NmrpzsUZ2Xdy8yzk9FvYed73Bnb8i1zhUWiKJXvZyLVFSVoDXnKq6nW8e6hNfDOmXdRsb8ETHErFSRvnlu3v0faenPq3nf0fxVDl/N9TN6nNcpW87262XzdoiwjlUxLWMcymxSz0asogUxwVhWq6czZKzoR3wiCFAVNAFkHuauXuDjpRCQUiEohmIgWbQmhQFbEGoqnjnqQrEQHKtERzKsdD3GrlWkjVVhTQdPSO0wfocv00c5v0dy+/wCnOR+n+S46p8n+p0Y8qjr6cR2m/Nw+HzLd5/o8uk0xLGnH1vEdvkx1vL+ioxdPhxo+OaO4lrxoqGD0nStJr573O6unlvm6rWBIuWs3bhvZ2XQw082Rk7tWsEUbyd4kfGiOFeU566pVnseq1QAaSjXmNcNVwrEmpWIGauNFFs1DIaIHkrls9Z6FljeZZ3iVxGVDGvIa4ajwbG8II7wp5m3nqlc/WnJ76vWyOVsSshiyjZWcbKlUEAkUABrhoCFBBj4gc5NrWkGgRGlJ1iauSZdSorQMoW3nHDUV6GvHWXTgQTBrR6rH3hMvSajxY1c7pFtE8STaT0OB0j+w8r6zhG9PyTUWbJKQ4octJHrrRE2zAwcoiI7q0SGaco0Y4mp787pM3HcL6mzr7PaWvrfz58Xi/a8Hna/NwM5u+nFy9lo158yPBtM1Y9a935vyR/k/Qdo7j9rgRcDIr5CB89WlpzQt05KrK5UHKiYoNUH7Mdew+PYEKVBCCVFIrTnR2Trh3V23yojzVVQKhrlQFFQNADXiV87SOOg1Mu6BjhZqcbp6EKmeTL2dJ8d1nuM/ar71ehdoef8AT48PR4EkebsVuvii6mK4qSebn4N/H1bvf4mTDy+ps8/zejSo9fzjzfzLEFEFlZQMSHzVSfThraC0jqKNHMw761i/Nid0c97Fy3iWvIek9Ea1yqFKGkCnMKDXBLLPbbbHog0ICI5VqSMrJjXzojwRDMUFcbq80gDmRWh02jzYdOrp46JFnoidUEDhqPB0blqPnYPGulY08nbhjdfH1xyfRK5auatEbWKIYxsebIqlUEigKCuHGomghRtcKNWlBooIxpzAfEBKF6fHqEWqoZpFWUTNIhjeeXFt5QhIoGyujoocPo0YuYQzCW5G3SVrsYmuIY6vu5duUzTy0RoeEr10g6kdPUjpuSAEV85C4ljTlQ0lgTketJpXvccel2Fn3fnr3f8A035t5bY8n6ilzeG0dfzPcr6aI3XjmZ3F2G7vx5kmHdqc/wAfoWIezzIW5ZCpRo+elR05Kqa8NVlcqDkAFTSYrf0MPccGoNiEqgAJZFuNx6HomKNBocm1rlHEoqpUGAhRscS6Q3kkncmWjmz0v7nj+o5Sn1ePM3Tmlmn0p+ix9et6FfazGfRqbXmfX806/Nrxzk2Dq8+9R5+pssHLu9H5eSsqZpdnh86/HZahNq26hpb+c2/fPTrwFh53MvfirWeXqr1oys1ptr3jCdHNbyYlEOCRZ64nPnarkUqNqwEaJSp7efsCrVzRtRyAmKJjHixtYNR8qNIATlLVTRBJGuHoaKqpXLaaSOpY5uRRrCxjawRHkxK5mhnXKxgx9eSNNOLrU5Pfeq6HNTRrzGmLG1lHFUiEqg2uEKNqiaoOaNrmkQEGkAAFYAxpzk79jLpUW6BCQvFkdZ1TTMx7443XxgmODf2cfo44ZSJOatBz20UaOVq4qxo4exgs38BGtK5DlKcpq6Q2dLPWiTdUvz7NWN3M053PnSaGOlNldTel7+e/S7/Nuel/Q/FcH02PV52zyfovSOLenHX+Ppt6HTxHJed7VJuAy4T6Zz/bnzC8XbRZpxQtyySbPSDRjbFzejxaibcEEL5kSs1EwHb38/StrHogwAASKA9aIuW6PVSNWyoDVIAbRwxXCrNuaqUBVKjuLWGQHbOXp14KkVtZDps2josU9I1s/m9S3sc/sWNzbo38jv8APu2vO+hx6PF6KRdvj6270Y8dS4OmpIYUjbgbl3zQP6/Gilz0sSx12Y898A6/nd/B2lj1zO6Hp8042cpIuX0Iqi5ZnqmrUsbrklfLKIfE6SeqNrwBwKVEGAILBtbdHsCA8TRrQBEPFoIYQxtc0avmYEK5WqukQSqMQHOW6uW7lnSWOdVcxoJjXmxXixKwEJgirmZla81dunB1E3J7hIul7Y2al5I14JG8mxuWNcjmCIVAEB7AEYrBOENVjJAjRoQBrFULdzLZSd1QaNMYmXnFWuJFU5NffywIAHdFPbpx5etATgKMAc5WjSXWta6vkXrEZ4thUJKDlVte0spu+yu1I5uqlcTnK0BMfUMkTta1eh7Yn2HwPK8d5X09S/y9PUT+hyY2lt5OZa8f7DkKq4Z7XL6+dAueiHLLew77mj6vgPxs+jFi4OgSsQimI8K1O+SpFrxsqVzCPFRNNJbb8uXtC1aKmgkikI962q+HcweKvVK5aCFIEoEoCY5yrp0QYAmqJrwmnjss9Lt1cynPYOZB9lurn1uludN2PrJ6Hu8Ts56eX1cfXsM836nHs15nQWO7PO1O75rjdTl7ucN/FyIsbjqPCxudvj5MfN6mnn59durpzUB8va8P6fgR5/n9kgZwW75c5mURyON7F6cYbGOsVd5kayhRcw9XJFOsaXxKFNac6qNb897kXa6dUPVxwOFcKArMQlEJYhixtczB4NcJAFAFG4W8zV0uVU5Z2c1UI5aPJWA1Hg1KxFCtHXNHn6YV6m3m9ivH7quOiRM90beTUeKxrNsVZsjqVUHNAUFBtUQ0Gq014RqJBjUBUY6NJVbsW89JVjrapIhEUN4pG5rFw3NvjYlcgArh9Bo4+42ORrSRVUjVQ9yhSdIaI1BAoZK0qg5VSOG+xfVeq4evWbflsnFGhKSMlkOrrYfa8ebr+7yV5zh+nyY8MtD0Hsw4BpyYvouPX57l8Po9P6j2x5Vzfl9zapx2ZZ2rt0w7vh9uCBvJ2aR1cmYcnQjCKteuSnDpyMq3ytR4KDv9Hl6rxdqoMBAIFK2I7oCQEJIU4WygI0hEJUBRUjXCjHtm3CbHVM1jPtJDRwbq5bpYOmezHdmdfh3061uPo566+5e0Sb82pf8An/0BsNxv5nt/M9b58pqbfO/PN+vPzK+W2W/p8eb070vn/OKnJ6/Tc9j6XodPPfjK/V40tN5Q0tPMZAcUsEqpKuZk9LXpb11nP7KGhzukyOZY5hjlmr1qwJWJ5xLXEoqkvz6ak9T3FNQTY3M8U1AXyqpTYzIR5iOWxvmapm0EKCOG6Q2RVtIs7uFoiEojhqiGuBrzGkNQyhZeUdO8s2Pbyetl4/fV66XpO4xwJWSNMmBnElRG5jUEqNuRAghrFAQE0E9g4lpIA5EPYEs6qzJn0iKxCIYrwGNNgqUhrKjFp5oEgD9yezSZh6TGEukHGCV1KMQ+QtUQsQRyEq4bhtHDllntncdtWO+Fr4mesaOeVLrduWmzv8/X771PC86r+P8AUU9JKzn1GZJx+hWdnp0OG95e+e3B4mEPp1yN6Xl9GDQ9HycHZ8z3uWZv4aOcsRS+F5V4zjoVteRtSuVg8DQW3QS5e4DaoNUaFQkk21M9xUCgoaSLOogK0Y0zVRojSxqRHDgpHCUVzSz1vnba6ek2MfXzdM6qK7crerjqzs3TB1/Lt6Hd8j7urc2q1d836LGtdvkT4nR5/U870+bVo6+bkdKRe5Dz/ZrX/Q8IjjavHXNX1ujg9u8b34mZnm/RQRd/ztOicJFOTaenDFJOM089Nq1vrv4WPsQM6fn8injxo+c0FSKhXK0rVztV8iqrfpR6bZF1K4pjBRxvElBrhsjeYx5PCkYQsLwaj5Y5GkEI4BSnKtBTWRZ6GqS0EiiaVKyzddK5GI84FqKdTTnpRa+Z1MvL7k5HY586o1iJWQJIwxYlQwckbQgK0Fa0IDhDWsVBI4TVbjY5c5NEZO6alsRbyehAlsbzjbeaAVWVmlO+asmnCqIJSurOf3I2KXxgK0+MABVCWQojVsDN6Jo4GS0Soum85+hBDXIiRNiAubqN89XL7b6WXt4KlSIyN/i9Xm7OnF6Jy/sfNchD8p+hJZz21cldcVvTz2594S+i9fFxuR5vrTprm3Z4/V5yv0+IWJqs6sHVXzwxPhr5187KunEwrCx02XqTLPoiNKCEGKDjLoastRUqlNeDnrZg5UTGCkjAGmKZiAAnKrEG9TS30eyxWc/pE2fpT6u9YkuG3RHF9Nianr/H2c28r9vy/sZ9XY1NrH2eXs8/Uy1j2RZfV5l2OuXEmxnGs9Plx6vTysl4Peox3yUbXd4aMm7lOdaVbXzooL4KsK8yuXyRzy9fe4PtMTvPe+Po8rG8Ui5sKhhyteueZmfXGGQ8cDK4RCHBUluPRnSex7BsRxGx4CiQac1U4b7PkezhhOPsSuYRpXA0BzQVQAHKUorWQ6UFStJRHAEqg2NOeNXMcNxRg14oYL49+5ze1Zdn3Ek6o0ESoRCBCBjxRBMKlGNOYm1HKDTkGRtSxoAGjTEaQEr03vVrInafn0CFgRHFWKI1BDebXCq1dOKNl8qiNfax9eukatVoUEcNWAPQ/T0fRQ+V9tyi9/xpG5a8lFB7q0ke+ueu9FjeMzMN7nqnueFzmZHbLHl1s1leLDlpFHnpc0uvgpZfN0juP0ZP1ivz/q7L6/jfAvyjn6D6M/V2vOf+OGXR89t5/pW9dxe5xsHd8+jpSWqVKOcYI682nn7cqVL5I5Hh0V/D6BCmCQNiiFjZFjRrxq6NFa6FJehqKhBqRk6K1w1HmgSrRqghpMjlqgPQfPcLPVLrc/0FA1xVlZTJn12O59P5/gncnp7F++jdq8/vyWvP9+Lay9DN08tqWV1eXasep8vl2uTu5l23n9VubzmUvF+zysvt+ZKvX4q7/B7eTT6fMpydHnQ0ny1oNvnoyFY63Z0Dk+ll9f8Af+A8gOX1eozI35JVx5UWDSxj0Q1JwGVzNHzI5N0h0yTT3Kpbo5axOGo8nKVE4yVxo5jrvOL9Hy6y+d6LIHywq+JXDFWgraCAGqAOKe1ao15ikoo0CGRmUbLhlO8oItOOOF827e5/andHQ+SOxEVMSshKgQgQzIlKPcRJSajTFaRABAjBBNeCICwtNHKuTfKrcwJLUaOWdwAYY126ZrEEcV5qIza+3lKEzdLl6Do8+xqNCA5AFAcEiTavAcWMJa4lFG99Wr62WRdMcl5xzTeruelx0d7fC5j8fuc1X587Wv2+bn5uW8qc3Z0Gd18Oaeb6tz9gL4vFfz50v9q/NO34vqOBt/lPV8/7KfNzu2nN6MPfeh4nBO4fTi3sfSx6d+bXW/Nx6XT47K98kW0q2lz9xByoCK0KqVuPQCHKUj1bmlMkcOSdSNy5yHtNFYS6MlHEjBqOEAcDkdO9uVdoi65XZ9zkNxazknz7LlTr86q/HXS2n22kXbdPO+j26C67GiXk5enny1llIZ6rBtz7Rz+vLV05sar6PycMKVfemsKhXHbb1ebHc18zAzM+OTe5fWt799vNVe7wbfTPfldJ4bnd79nm1pedxufyuq/KaEc8DA5kEKrb5J7LEkdzBjpFqys8Wq8ABBpyqhGCtMkkrRX56BioMeatcgVbvFq1XLB5CCQMmqSAxCYkcRQXENXTljW+eGF4dHc5fce+OoknZ6Ts1KzaOGjzQeYiBjyWMTW1IxpqNOagAYNCgxwmOYxrSAppJscNROzJGqi2EcMjeSQ1KVaze4DFvjYXxruz16UWHpQq5aA5rByElG5iavB6lSMNGos1QSqyVa6Xi0sw3qtrXPqfUPY8fz/F8v6en15z8tiZQSRvE1S6ai9NtnhyY079hY4/Cfjf0eT719znyvzs/U7Lp0/x2VeJU+f6IsY9K2jbQk4fosyh6HyjbHN2ZVbfwc/P38tta+OTpI67BPoitjGleNCNBMtBw5GmrnKmoIeN7Vq0eTFJsRzu1HmjXmOBBMHLQTkKsJO8+lptHbx9iBue6uXQj7zQKfb0ips8/oZ+r18OtW5vcurl29TT4voaWnS1fM/T+Q6WhhrN1/g/oXJyej89ov5fbtdz9B8px/mPZ8Xbh5PTu6PH7fG3d/Fo53Z4Nns+r5zzhnL1yaeXq9Jz99pZ6fHghM9Houziw/f+nn8zw/D+8qZd+TXxH5eTHl58SLmAgcqkfHY950NkG2QtkDwZMQjAkQpo5c0bVeawkMR8bQzGtOVUCayG4q2UKGEo14iGKKIQEY5YNV4Lyq19eGGOuQGugu8vuyz59aOnoeTsweTVqGtcCECOVFMY4aCjSpY5qEaVghQRwMGkQBgKjUr5pQaoOSaXM6OlFGjDOBmmDYWkc06JTipv5BMV0jef2UAYxiuBrgsNmgErk2zFPU2iHg1HKvmnOvQG32a6JtXpXoXmvb50mtD5t3F6r9Pz/e5tvR4L7RtPd0uLo+T3+L7f1vh+N/YPBfP3w/h6H7f/ADF6Pyv50cn5X3H7H+Vb/L/mfmY/SyTTU7J7gJjnUyaRryUc+Tj5/V4Da+nJrbGXqoPVVYgqVRyTY9tZJyFGri1QcrVo1wpIy2oZoEOcasCGK4VBIoANqkvFo4Kl2ddIoc/UtVef0Ju07eDHg8n63Mn6PKZb2yguxvX0+jkh1M+/crY+xty8X0GP0WfRq8f1+Lei8z6aYz1aysdEfeuL1eXz2h6fw+fnrK7HG9S7XPF2Hs/B+ZVeH15WXz9DJ181Veb0dLHvFmjrw9lnbdfd/QEehc8P+s/J+dwfK93By/P9fkqHBVOvy4Ij53OWkliO1spqx6bYnEba5p2mzYzGZFqSWRsmBpDIa4noQNErXMhOyKSKaqFK5aIo2I8kQxEJapLUZFXuIqWnLDDpwPaDEefQzcvuWZY6JSO6RJ2QM2JWSJUoOEFIOEas1ayMcxNqRggaaCAGqxqBNAMa5knVKktQGqyW1Gqk7Kg44a50jqIlcskDIrbeUFZ7V/n9dEWjGsarCdMSQGnqmk6u370ej8/NvgayYHic57q25lRb6Or111ud6ng8tu8fpc5Fy9rW5bSwrFbEdSR6Z3idW9P18XJ/rljx/V/NngfLH5sx9Z+m31lXgcl+PeXreNQ4e6korfo/fw8NS8v6ek7Xx1rKa1evOz6GvlRppzbtjL1gYDBRq6dppce6OAyRTN6Fig5FSOG5xq1hm2cBk21tZNHmAKpQgkBDylFpPce9mB+g1uT1TamP4fdqs14ppapky0t9/wCz8151t+R9b23JnT0PVcv0PIycnpdPy/Rx7OXyevpwef7GlDn6a2dJj5T0Pmsf1X3vzjieYirVXDefJrHWwK5JKl82saT2uN18sUPL6O/zHofJdDgb4+pct08Gx9Z8f1O58u/V/kPR53L7ePw3L6fFcr8/7uZS4sHzZ6V5bykdPRHIJ7VpAlc6qnGriNTOxGdAEI1yyKuSBHxiiVxTxUoWgkqFqEq82UWrB4oOUUlERVh05qUOvHXirjRzkBsRx0knL7klidnSx2uI3RHKIQiVCI4ByCYjlEMUYSkVS2OpRBOQEQTBj3IYxgjT5VThMUBU0cE0kayC6ECBhEMN4tlc52btwMW+Rb09GpJh6Q0bHMdEAMcsiShIpVh2m/e+d9vixy+b689eNXLrk9LtHJWd7bt2ey9n53heS4fQfJy92/z22fb4PX5tTV4/S5/UjeRc9tbnIrazPN+lztJ8OVYBtitNTL3b62cVzXytGndWX5+nRbHCMedJj4KWdr5NbQvHTdn6QEiugbkny7iSOhkJkxK51erEKc9XE5xYZPQ1pCShG1wgSiOBwxE0HLVErimPCxaz9CSXTthZkOlerNDPuqJeck5o2zHS5/VwR6mPoaUB1ppY9txfP+hl1cvTSRayPx7r9bi96d631OP974erDPK/mb87cy+jiv4PT5EyYb5Paen8/e9A27eJx+n5zNh5OjpMH1Pm/VPNaXb+l+r4HKbHhfpvpHlfv/l/ufme3B4p7Lx9/mHinle5QoeH70VjOKrng5VckZaOIga8nwia14rMaNlVwjzbDXPFDXnsRzI5ORg1eNUWgqSoagZjjVQtEM2x1hVjvCtVvkjhvmaGaI0KCAKjWvPh7Fm5n0OfHXIufWgwQhqPNCpEEKJEaQMkaQyNwxlSxtS4QiMVARBOjBzQncqQY5qEcDkG4QSpySLdRaDTNqkQ42vJXi28+MMrGjPZorh6BGDY6QjHqh6oZO0986rvdnGznpmU5e/0vzX0/Jc7g9lqi6OLp6OtsdWGTzkxnTcXoK3Pc9D7/Jz4O/yuYn8X6ebX4Ppq+AuTW2ddaOfxd8XQ53zR1+JPsR7Gu7qnh0r5yRFSXjo1sbk9anH0eHBoYa8tS7fk6uvUXGx3CmSo7cptM/H0HNVMR5sQxUGr1bmFKoTRzrK1UsDzQrNUTa15qoxrSBmAxQUVutrouT16ldmeqOWa2K6WasdlB146Ul2+hl1aUe/H1+x046XCcnracvifdakvH9DG3Xntz83szqdmrTjq+pfkf6T8b584Pclp3LKfT4+zibYUrF+ZV2jbtbfo8GZ5z6/513+DNRS8voZfTVrp+6advkvOnPr+6ez8H4X0fzn69ynmWvyHTeXYwyhw9lR+OV6PH0xNeSBVlK01ZDWQGaMM5Yhq1zRdp5z4VETaxaMeqLDp1VFoISOKInL0FDBWLYKxgbeDfbemfF5eXYRUADSYK180KXzAIRrbdh7Ni3nvI6emUz6XC0QctRwiVAglQEVwjSAIia8mJUxsacwTVaEBAABECWRUqDRwhRooCoRwIoSSzblWogEbiDO05a0evCyN429CO203PskSQViNBpcks1YdNojTWuXui3pdKrpUb7St6OFHXz7NHnunhyoPM9WeN09XPSZjup2XVwq7m93jDzyK5HTbr5enXi6vn3w3HW7Xl/oMEb6OPi9H8+e6N9Gl0YXsmLWouWxXwrNpdHg1b9420jrBwqvRZ467iT2pBKY4zha8FUFcaKToKKZFqKhInI1wk42ROBCEa83NAGnIqe5azSx1WZr9J8M7RquVH01tT2mvq68y2Wjz9bdno5rK3TbvnfSbG1HrN0+X16MnB7s6rtr6tq+6fW6Xy7p+VivdvgUn4ehQTu8CSBzm7b56j439bue58f4pFfjM47p+d6bZ00sOnvo9vp58nLb54HpPPttecfOfrdXz72fyXFrclsh5d3Lnu9bgSaCdZKlYshrjaryknXRWY8YsjTzGy3zW2xrCPNzQVQndOoDBCqaEDmRyaoaFZ89J2nMKqBtGqUcxEBWhHDWNaDTgRKy1LeHqulne5Jn1PSemRZ2QQiOGpWao0IJB5oCEJawyZDcrG00JEaaoNAFakgTsCp4hBtVAQFQBwDkWHxuAxCUHKsCOtWVaprxpDWDrKu1bz7ZnzsyNzExp0g53xYA1lQrpb3Fqt6OSn6vk+34VDK5fX6fmApv4/Rr69Z5ew1ku4+2pb6ObQq0zR8n6XLTfzWI8p73RjkPzfXSP3ci7y92JFr4ljYw9Wrla8KR1yRRnMlZYx1782lNfNcdPQDShpYmx9R0prHGiBa5HQoR7getFetIXVFuPPpEcsHLoULKyNrhGvJEIUGMcPFbnzpZkjulu36HrHknVw1jj6dDPjsfLrM9hdMektct9ZX447Fmy7bWxh6ljrF62Ra4vX0rvn/Yxvx7WzdXHrp1Tg5K8aK36HzML1o9+ucGZE0O56OGs7h+myMv6H8mz6k8/dZPqfKU9TP1PSb3s/OeOr5XtUOhjWpP5n2Gv5Vy9tfC6/iKxngQxRIQybLrdr8/tYJ0+PDDfmRjye6dGvc14K5clu3kWCaeDiRpXoAJJZpRWA2yFyOnsdPPQQpxRaefHFXIxtQjaFEkQFUEaEaVKV3lSh183RM+26+Oq3NlvKi7HvjYFTVctRwI5ASI4QcCENaZxsqWNpDCVQERpzRoCHATKkcNEB7kIo0BI4HCcjU0ViOQgxyIRQcA0Zn5uvBVk047mnl6F6PLsjbcV5WrLosQTRqOYHOVvWt0dRvx90373VwZsXB7dWXLRu1VZ9adaFxYaef3ea5nme7Pn8/Tbm00mj6PN6bGO5OfvxuqtX3Ylfm6ZVjpZq6YY8/L3U2XxVh8TITOGA8+Bl8lpy1HFk0dMyz3SRja8ljCZIHET3zjp1ehbXCtxT0I1w1rzAFmWkLXiiOBBI8csKsRw9VbpVvdmPT1M87K7J4pow6Cj006zRYjz6aMeXRPNHZbXH002Vt11efcmueX9Nq2uP6+Gzh11trfnvY3o+Pj1l51+L0vnbNHD0HXOry8rnedXr+vL0vI7xzmZ1fFZtl8k3e9fLnz8n0NTrvW+S8/jwK/W8P0WXzPT4vX/ALAT8pL87nkfll+0WPvdr5Zr89+ZX675+ySV5Xg/5ac/2WLHl0VkfKsypJVpXr3zNytPNpyaeekqpYwie5llm3vm5HTsiExxVnes5euxS5Em2scRQ350cd8rWtK4JJJsEK0GjTEagiuI61YKPN9idp7+fXIkdaSqlJ0EENaEcg00cIECENaZsZUjGmNaHAjGkcJGA9gTyq1RA0AY8Q9E0AciataHAgMQQMAT1VSKqKosKp1eKtsN+Tl9lrKSRtySKVVNi0o3okcre57NVztu9Gdr1D2PB8RPnfr9hnJ9BXz+vw9PV2WnzenH03GdHHJleT7CzYdHtXLe98zndd5P6BymXwejk6Pb8xS6Xo5trE7/AC8TT8r6PGj5+m30WPp8W/bw30b5a8mU1YzkigvgmBOctrDo9BZDWWFWOahHi6Cs2CyHqtChoBncxO6MENeY8b0WsbK53IIBNeEjJ2aVlIxWTFS259Czfz+hizb4Q08zWq63JPn6ELMuhZ6l2obVrnN7NjVz9Lu8XXqnsfPfcWky9qzZXoN0TWTH7/Irs38Ik9D52Wpn1X7Pd4XJM4e2/nLbb5br8jIzuj5O3Jphn9Jphu7uvp6vN9Pm1/PI8fVwfP8AfwdDo+Y779w5+a/nz/dGO786/wBTsd/xA/RLXLgvhCOn9f8A4v18b4+k5fqfuPy3XweY+UMvWlijqmUurDfHUxNvDvTSUX3CxhUg049TQx9Cdufa8NGteKyrSdM+4nKha4ZDXLHDfno1w1WnzTai0RSQBABBFdqozTGot8jnFvtxvfdl2D1qSTs1wxFTSskRoBIjzVCUaZxxuESpY1pUBrhNAYrTQLKzYA+MHACgr0xUxoKgKANaAEGkAFBz01jHQxt/Nat8ezew9hRXWmalcqWUpjCUcmSJvedSLc+m4Pv+LBe4Pdxq2Zd7bwv0DgovV+Nm7+trnHdnm40Hm+jdq8/o6OZJf9K975Pj/SV6WH53432OdHXzOhf4/qD0f0fC8y7DP0sjEw3Wr1+DH1fm/Q87nb+XYyt/GSPOGV3xxy3zKqu47P1nyzq2QtrjWil+cITGPJzlojy0BTxzsiEqjRKm+IuFK5kcJAFRNyMEEqA9VrNZz7ptGfdSjSRtcDn2r0GHrDaiy+qS3n1WLePps0Vtvdd5/wBxjXOD2Xyr0rr8PYlLNTN9HxqS9PhHT+l87zb+b0jn1hn7eFU9j0/mmcNwezT1fd/N4/Xu3wOKi5fpczptMqnc+r8T41m/P/c7HGYdOU3p+Z+kv2P7/hvhf7N5o/Ef9v8Am9r44+p9/E/NnzH1sv0q+D/F9L43/bmO/wAW9Q6PE4X8jef2JFw9gaORB0qenlVkrgRGhoQ2nNqfL0Xxz0I2sCUqUjrLC0hnVRRvKtHr5bWvBEEycHC0VBuU2ESQeaNIY9y2o1mm/A5o1equ2seuWWOtsq1EVioaOUSpAlEcAQ1pnE2oGtNGIJNGIgmuaYBNLNCjaoACoCgPkio1BFY5Bogmo0DSAADHMUQ3MvHOTbyberl6Q5dD0RMTTngKxjgZLNOleqPvTsr/ANB4OLmeZ7z7SqrpxrnbPRxY/QF7x18nP2fj/wBa5qt3fHJKi51Pdhs53d5UnE+V7ObJzXZReiyUW5Ux921nTVN/X4DHvmsQYerqcz2/LVkwaQrLLk28m5ay9F757bUc9LHkQq84xzAj5hHCyLVHmiipzExCWK8xRzRTrGXzICRQUdIihbi3qI8pHraSWemSxHpbNrl+tyaHX89rZqtVvFFna21b2bS6pLfP6rblzvaXH9Zct+f9Vafl7Fx2XpDrhbvXwtodXjZ+l0+I9trOx88q0+DydP6T8wv8Tw+5I3s+Z0/SK9Hd8a9X5DTh4vaoZ8czeSwrLkPLbpLr9t/aOPG/Kv5zj1eI/oK6vmfze+ZMO/xuPD0/22+EOn5T5K/oK5/d/FH6p2871H8kloLj60ujh62e3bz4K9csJXGxjxVpCqORs9ANKoDlb3TuSq40cwNvjjHzLItUYAOBRq6dQKRQYPJEeSV3ksgsNN+CVi0VHL7UbWLmXWouhXTaKUNBB5olQBKBLWGTGVANRDTXijUEUEQBjU8s2OTGBIwHNBwCSJgDhMUGDBBIgDKkARARtCMJy6u3km1PRIufcXm1ZnTxijTVE5Z0kV6uW76St6PLp2o9DZ5HzfpUzez5276PPfj8j1eVr1/K+r1r0+jylOvE9br9PRxFjp+e5tfM9SpazcGz0+fBHl6MsWXpR39eLQXk+mrZ/R5Tn4dEu5z+ryVb0PjIEnmjp1wQ1ujyP2e9618mn+X+fo9D+pQfiZ6Dj7H27+Wlc37K+pdHz/53/QMdv0Saeav87vP7gmPtKqqWJaNHkiuUQTmqhrgRw4VojlyJo4SA1kWks0dUkkd+l1PF9xzeb3fNtleDUqVlpNdHVcmnssXub1o7LrptTy/ub1zk+qNDL0ZGLrss6cJNju8qozfz6E2vnXMZrDoR4tGf1fjrmfh6HRcF0+X1/OdHg9lav06GTh1fQ/F+78t5RleZtg5cec6rFOt4ep7H+zL+W/nndj9JB/Q7v8p+HXj/ADe25k1+3Hwl1fK/G/6WY+v9p5Pb8f8APn5L8n1cKZenY6Ll97Cq7+ZXZp5jEeTWPIHKqqa8SIU+SdGtCZ66BE68enEBADcKwBrawcqscrRwxSox5tR5DDKGOsMdvR5s8kdMSOJHK57uXXI6bVTZXTo5qtFctSoVpIhmNJZG80SpRjGjSKkijRolRqWSbFAVNAFUGuBHJrImqAomqMVo2BLWtCOUGMRkY5jyr44b9YaCZenJYVPSWOYr6TGhtUOb1Yn9Hnyx6a9lZ93w8vd5/T9h8L7POxrPjfQu6rp58nFmFn4/U6qOPa4yy/Ftei5/Q4vm/T86q8O9luXdE/p8+05dN6udrW53tdhxfXVuI6fMf3/P3wedd/zK1s8YEz4aDNfLz6u/kfu/8HPD2T0Qr5q/UGs/F/gaOv7v+RDL7k/DKd5quXo/oP8AQPR4f5AZfN9EiiVxb41oiGSoJGuRRIoIjSqhEYKJEE5VaPKfZnqmXL1LHS8/0+bF0clLRnDNj282029LSO0v5egaHP6kj56tTp/O+tu6Hm/ct0o77tLpdvvvR+d4TZ0b6nb4tre14OK1+bo8m670/wA2zeH9P5V1PxvrqN/s8Crp83qbXovrefzcbjb4JGPS4lx+zPD7B5X6XFk3fN9N1zg9PE52MjtunxeOilIwXU5ROP7vXL3f6AdHg/Mf544fQ04YaXFRVKZI8I0ciA1Hi1HkoUqCkatpVnWNHmNcOVWIIUaoU1HiIJyJqpQgOaNBwiOEQxa0z5Q6/BksT0SItlQJXzrZs57yPjoUNnCoQSI5EcASISkZDG1DUaRglQBGhBOaORyqRE0VpHJgCqm5o5GolFSDSomoDEcjWmgMWpQBkbTqTyy7W3najsfWLEtypucmvSdvJ6D4v1eXfg8b6mSPp5pUOnu9r6HwM3B8z6DtuZ7PKpVvK+g1ZbnSrd3jcpD5PvPdphXnjXasV3ZOVhirM1bfHalW+bSfq03uf3PQ/KN+HuOr8j9D5nAzM+zv42vxPo+BvX+f0uFq18qlPThzjXx/3W9E38TtvyantvfU8dXtHzHfJ6x+QuXofsp63v4v5N/HXL9H+hP0Pt5n44O5vfeLRoJ7S0a8h6bGVk9qascOVUiMVrlVltGlUbnTs+Weiy7L1rF/H3tfOj0qtPp8WNdvJnbpMskdmlPHqNmw77yvWx1Hm/Vadzg+31g9iVqqaXo4uj9Y+k+Dx+Z189Nue3zjcjzOf5Po+P4xZM2nOSXN/Oz7WfR0HovXc3ce58L534p5Xuttxy0djDv3eG5vTkystYqPOqqa+PfyoZNryRiOh5ro5Ivp/PL638018X4Oxef6ekleZLJPRZrroajxAShUaPBg8xQGvNZlskY2hkoqkFuIJQGhADUGigqIQaFBrXmK5aEIhHHJ2fNvsraV09QxCuCWaOjQnx7WjShahYAgQiEASjSYUqBjTUaGicg0QSKKV6tEB6pjwjUHOTAFEx4ANQoEINOZUtRDWOURpGNNBYy7eZrPy9MnVOsuYHRc7+iaS4N6t9Did2D9rDpZ6J3LN2NdOXg873qlTXyrCc3pP7Hu8fD5jzfoHQvjs9X1Yd/yXn/Y8xgbfMIc2li70RE5beq3tvY8gk5cOji5PpLneeR9tzXDd3zbG+n8g92RHV1wzoJ8tKV+fXdfm/uf8D6+do/o83+dH1VHT8Q/sJXP8/8A4uZ+hm/oDO33Z+CF8f6FfQtY/jkvN76KNJE3Rq2lZo4SNcqJujE9owcDQBpAkUFknVZp6rU2Xqz2uf6Sd73gbtw566ealuk3b6MYtTl93Obn0aq8/q3+h836Pb0z6u23D3knmmX+nm9E9H+t/MXcp2eJ0HD8nRneFcx3vlnsfm3K3+Lo0M64SONa2hn0bns3R6Jj+58T5Hj+W8nrMunmtny/ptTPfbc4XXxMjofO7szG08xztvPGodADgmOB5KopK4o1fPJLO6RjeDYjzWMlAzatQrU0l1x7Lj+zmjTz+9BJUVPC1VWgJBw2RNjgaNKgIohAaPNEcCgILi17PlyVXbWexqoVxSqVbmy6bZnux7ShbkNAJQJEJEJZG4SGpGtOjE4Brga1qw6bR6BRiJo5pRN7k2oKVo1BijFLRQEIECNrhqNI1qNzVHO18/Zkz722BybWuWTJGrq8WwUveN5opg7Oj6rfvq9j73yG9wOmWJz/AI30PQ2/L+wwb1c8GZp5728vo2esrq9K8d9H52nkcpv1MPSypK5b2fc+h1ul8s3zvcNOejah836ulsS+X676T8287Tg6p4G5aK486LXxGK+b9ze26PC1/wA9J7cD6njp/HT9uJ6IPie+P9QbOvl/mn8Ccn036F/Qe3m/jk/m+ga0RMqYgI4Go4c0aOJVCkQkBMWoBNByoA5VLJHXasY+3Yuc30k8C1bBvyQJpwOmuXvtSanN7GhmcvqS7VaxbOPpdlq19Nqw4+/FY5+qjtaHd/RH1H5H4PzcfU1uf49uI53s/Oeo869f4Ola8314KwtePXCHqp3sHH9X7v479X+U8PS8zsyYuD2alyH3nN67Z/Mc2dUz5IIK41sCWKLVlS+NETaxHJsR5oCfLOzYyFRhKqqufOgDHtMa8hzlR6CQIOEerFVyvnqYCY2sFBKKkGkCURpUE0eQhIo0cVxJ1/LKBbmnsc1aI8GgtGvn13J4ssRvCrycpsBbR5AIRSQVJE5axyiuQTRKByJJFTZEMcDXMRyYqciJuUEcNFTAYNqg1QQCaEsY5RGmNqW5F81+zOyTLR7kPeVPQjpcy+QkZM6qkW99y9v1VOl7vH+zfnz7r8a8txPzv96Nbz/do78d2NDv5epc5/Zk4w5nx151a/pybONxfSwy9/zuhnRss3L6sU9rqw9LnpcOjIk28lNEtlnk9Ki7v+c5+CfOrNvyRrixuk4kseoscbIjzQFp1SKahILVjmlYA5DmjdESkjSIACVFSI4EcgJUGAIjzFTtux9SSwu+xNz/AEN1i6dPPdQ09eCxXrG7HvzaEnN7VmLm9Be73vj+h4/R6Td4/wBB9BqdmPKzcPu6+v38PrnWfTflXn/oOufk/jfm/RcLk38Hiw7/ADzp8e2zDvx6jM/Sm9d6OPxp78/0byrXKSPn68fVnHO53LTXxces1+Tvx6WvjR1tPNksFLGraJOiaEJkjm43ViSFOJ06Hluf00d1eK6ETo3mNM1RoaSoS5DRoQKCqtFFcsk9EBWKtQitK1JpUKiSJUoohCQB6GiOFxB2fKAEthbzLHVNErYjzHhpWcPQiCVByPTUWwOQAEmxkNjuEa1GORQRoCNTSzbVBwmigrU3KCuTeqbXMAoVNAQNqDQEIJWCjRw1tStdzWuVCzRrKibpVr6L5rw/WxL6HxjHMlv6y+borpt7fvxsquH2/X9fzfEaXg/S4kvNoy4t5m9HF0GNvi3F4+ls3D3tt9HOvS11czocfrVLnT4uTb5vUdDtj02UuqTsOX3PPbOnlUWPCXet471liUX4UZfmjHm1RWZ8/XYibB5PiVI55pIrY8Ix5ijVVciTsjHmDSKSA0cCIIY05ElAHq5qQlfWRZnpsT5+imly/SWLWHryVioYurhbFpyuu6ZblOfQrSZbD89tXouP6DY2+T7zQcvZTa1jT6X1fA+ksr6f8T471/s8H5t+Y/F+3z6GvymDW5biv6cz5dM2pG2zY6eaehj6Gvhxd/svQ8PiOD8z2JMPkjb5xadQ7TNOP4uyOPt+XdfdRMiyVU2GpcilkTyks6EtG4a/n1mpptWueeutFa82teSFYiqhjzVSUClU0BUoqcK2K83CsAEBEcoOECFaCgCjUWgouJO35NBNZB2nT2WIZ2S6nRfWdiWOici2PZIs1KToK7QEAhhDGOWlTGgmo0okiMnnnSIaRyHipitCg9005VQCB0CYoIjkUFCgTQIRjlrGmrU1lqLT40c5UIN0AwrJoEu314Oft1dje9TgwO3y26/zSu3Cl8X6KDT7PEwb3j/Vt0fd+J4+HzPV087j9liX5iTTc1jTSS1pd/lnzWJcOy9d6M7GJHRoV8fSzXvzbtng+jxLfo/N5bcuas2uWBNPOawwjkFbdHrek6JzeQTLohY8QE506MKhRUqJq4qQndiPNEeagNcJFGqJg0IJARKtLFCbUvmLa3url7zp8fWdu4+pRtZ7PdVRUtOWSPp5UttGnx+vay43s9Fl3X+q8z7/AG4b+iudb6Pjdb6L9R+dbHlvR4fr3ykcOV51fzvQ8/5/0nDUD5yayqimrPQb0cvTdt6XjUudw9vb888/1ex4jfzOSrcIy3yehRr35ao84GTg2O8tBDrRiiZi0ga+dWiatQ6RtizaOlxx1hHIQiJrG4RHmg80DJjHk9wxy1ELVy0EVOFSPG0BEeahQgA5GkKghRIAoWKPiTs+UVExWp7s9gydWuJBslmrDo6FkTRw1sTThaijQBUJSIhjalI3LRpEAcKSRWigICqmKAgK5Nypq5MBgMUaBKDSiEUYAIJrWolqWQVNmSamSbaNMjqHsBzmP9m9n57A6PH6Pj71b5aRXQdL6ngeeM8L67povS8ru/PdeDJb5fv4ely9lpdFm1sCbS5fUyLF80kfd52nGbdZzfi/oNrn+n52TW9L5/RkXo8ta4+2nUXE598zKk5uc5os08xra445EXPa8/S++vyu08/HTL0GDycK1etGgAhEacqqRDVqkNRw9ipUcijQErQVolQQ1yOJaCklnpvmPvSyZerMq6rDzTo2cntZTOrzehwse6vo9PnrPx+nDT055Ooy7eh6jy/0rpMXX2NXb7uD0rufrPy6rJ18t358y5eD80y+cp43JGBYjj0LMelZbcUN3q8j1JevDzWvx+mulpy4uHw+pkpPDTjjlvUduWN+GzYFDoLxtyvWFyCMSxiRFJv9GG3w3Txvh830mq4HDRhCI4ajzRRKjI2HG9g3qMUpVWopYg1FTlVoCYOFQAcohCIZKFKjaiEUFU04k7PlRAVQdOtZY1sMebZQW5PRIufTPGaPjFYlipHToKWoaCCGGUaPONtSRtK0TlbVyaNBwJyJorEchVTAHrLcDcpQpQFCEiEI0TgEAAI1aVQa8EcBEDSpfEDZKT+n7Oefc7s9WPojGfx+t1/Hdvm9Zi+Z9L23I8H1enhez8B2vBcH0/MP4+SLrvQ8yDJy6K/qHzX6j5k3v+TSTs8ONZ6H9Jl6WNbz9LmtP0PjbWYttC75H3GCdvzRW28tSLmcujMgfmxsvzUkVfTX11eP5dQc/srEJo8BytVWiISOTEbkarbG8ZIwcAgJzlbUaAlABBCNAkhUjk5Hz06Fjm+oJp752Z9N5Y3u6EehibETQrdfJYmKqLmzO05t/t/N+u2ui4/0iHb6Kl6P0/J9H2vpfzz2PE6/zvx/yTD63xfneP57Pzc+WrokXLmXpVN/P0Kuh6Pz09/v8bo/JHwauJ530dDIxxr9EYZmXKpx44SavP6WC7fykivOezHTNUWzkeLWpPiB9miOF5RJKQJAbkCIrAQStcta+aBTKVVaKCqWqFiqwYPVo0BRIhKKSAKA0JULEJVSwGAcS7s+VaoAA4Y5OJXLgctiemWWOqVY1dG3JInNNGgqtXGiBYhCMMYkco2pUBisRQFQAKgOQaOEqJqAr01cgcUKWAxSkQQpLWkDhMRprgV6aACMSJy6NoVpjwn7/wBbzJui9HP3TzT6H4azQ5/dqZ/yH636x8+et8HpwfH/ALFNQ9X5Wrn8PftZXP6NVerxdmpn00tuOuDCfPdmeVLXq7z/ACfs7PJ6ctdnqfDTw3M+hl6Og/m+hZznf8jYtLoir5dFfMrya4/Kb9i1z+1/mzPSyTPvaIY0yeKxW2ooFG4VqFMa8VQTXg9q0QcK5UjRCiarEQkQUsSpFrNbUdFuTH6Ga1j7TQpjhSddyew3W5Pdwl6/KdV1yhlWZX0jq+p8f73X0ef7mfX9DkOp7vL6z1L6X827vE7vz7w3z/n935wb5UZmTt408aL/AFS7Y7la36vqfOYnY83bQf1+PzNLzPd5jM5+RO95+/i87TzY43xviQ1XmsTi7Yns7nh+f6H1vxv2/gmSeZ7Sir0nzrzvs6aen8L1WLxfR5kfb8uKQKU1XMamaIZiPKMM3A43g4LUWg4aA1BgUjkhG0CEQhHCQEqFKgKpYpaiOIO35MEwaQBRNFaAH3p6J4o6o52nRq3zq7suPS0FIpoqmog0QyRCGwuFGhAEaY5iiFaDpE2I0IDxUqIFAkQ8NRQAaKNUGgQqCQJY5pRMUGo00ajRyiAOpW+m7cO+yvUj6n85+g+R5vkfI+m9Awvl/wBg5nne/wCIm6rTHoPNbzzdHwPtKFbXzHRxs7dnagnV5lbP5NXxOLXcba3eY8r7fqeL5eyhF7v5xK2ndqY9la/pw9XzXRt1M28czX4PQi5+OCu3TwvvG/r5nwQzD3lRJ7JprXk9VaISgSPLVqsc0jUkVyigqJqrbVEoqYrkQkAQEqAg5Wadr9jD6aZ+PsIysoppL0+Xdpw5ejfmnqv42e+jVSkms6Hb8/7nQvx9Prr6nnal3q4vbPWfqfw7jua6V4n4X5Xs85nZfOUa9ebC6sOl9R9XzOPp8Pu+sO9/4Xyfb870KOfz+jk43D69DJ5ZYPhqIYO7XWuDTPFYyVYRfmXbNY0rZ5e+dq8nbA94OjbjirCevKjc8EAVoIVKIZCtMQyaGauVwTOFDRFGjhoLSRhQohBqhKg2hmISKAABYo3C0BPiDu+RAEUEVoRNQABXpzzT11nVk4Vzzx1W5sul8ap0qHBqKWA0CGoZDRMKTVExWmoDkBVQ5Bjga4JGqmIIkBZFSoWOGBQgK0FUGhAEiAIJBpEaVgmo02Wlqdp6vnZux137t7V9T8Z8m3/if2r638+y7/lDXfiYVec9lJ2wdjHuhg5vR2+O9X5Wx6vy+lzvnvV5FeLx/bgt68mntV6k+Py+2zOyztx1jFc6/L6LkOvz6zOOtn171eX2rxn0a8m9X8fu8bxfN6an6OdHy3lnxytmNz6pEnQcDI3k8T1IdueguHQkitGgKJqORUANVGkqtsVTI1CKQ1WkAAEUSNcvsxvcsY/Tatu+jnVw1axxY6Ln9W3Xz9GTVvSJ/N19PzfL7Nixvx7nScP31nc29T3/AMk9/wDPPQuP7dPcem9b8yweZ16fJvmnn8+hn83kurPlqdv1+Pu+fm+ld8766hU6fEr9bh35WVh3afEdvzVWnwdFZpwMBCA0UjQRHlb09Omzn6DV4+1sBnFIYo5uJSGsExr5kcCI5VrlFJRrloYNUhyFoqQrYFKFiI6bnejnjdz9SKNAQoNRwomgSiiFGoWBKi14k7fkwTRWkGhQETBpRNZlaPLAc0k9Fuzl2WIY1c4lz1YpahYDASIQqCaERx1ICFaaA8TVqBWTOnRGiY1y543KqFLEKcDQE9Fo1XmgSgSikogNHLUaVrTpKWj6h6mXMG62/bfR5o+J9DxaXe+b7vj1D5/7DsNd+lkcVnisPN0+j5Pm/c8/Y9780lyHD+jxvz+bzfXrz0t7K6+SzR873Ojh4vocet63xl6vz+o/J7vmG6evLL9Q9Xs/UPxV18vjel8v9dQwb8T9L/lDv/P/AJjdn6wsWCbhUxDIVuc0zrGVrJOo1iISISrQcDAEcAgxQETQcCgiMEIVoLcnazJj7z560p2M+ty59jrsdGjfy9S7Pj6cmry+rtYnJ72g3Q1NLD6bd3/W4uq7b6D4Sbmu3m7+Hs8LZ87WPn/zwfKc5V4ua3Ht5+XPBJd0mSjG80GfVP2HoeBw8vlfS8/c28jEo8rjdGcaOX11MjUwctmQ6zes6XUb8OisxZRzOXot4G1yuiUtR5ASg5SKsFUlEcoqljxK0sUQAgCqUNVdtyXXywnJ2IDcwQrhqoVGK0kEhWArRQkateKb2/JqoIAg0qAomigAAA8aip0y1t2su9Zp1lSaSVU4ViulU0EG5E0a8QJa0zRGmK01AUae1NVGWFaotWsMmDhzxuFoKMUaKNRW1XKKA0lUcI0hFE0acgyS4vfQnrnHZfoeTb9KXdpcn1Z7S7+VxvM/Hfq3S9T28HP1vM+qwcfb5vsbFzyF7xvs5Mjp8eMI05eH6CGjlqZ/d4HQUcfTtt5+zm9v0/laWbjtE/XyUt9fn6f2hf23qXyxr63h+P5nz139OOj474R8FSR06JJOkjY0VyddlYIrgmnWEM3SLVqjYjlriBoIOUBKJuClQaAhw41eaipo5GidfjpnbHqWXZezI86K1ic5iezT0MvS215vavbfm/U6OfzfQWjSrWxfd0PRe38/t7/vfLeg+Z9vhfVvn+n5x4v435/6rwnH7fnuWteNz0WWb5o2v3N8swkgWav0teaGjmbVDfjwoPN62qQyAzfGk5okc5WRaySXUtu9c4wUM8uFHBK22JSsTh71TITJjdONAzRSEGIIRwkAFGgNQABRavbTaqkVUIIQFQBRNVw5q0AkARwOWd+HO75EBgIQFAEBQABBpRMUCRXavZ+gySdbRGr7E6tQlzjVFKVZ0VChHKI8kDNWjQcJGS0aRWkVDpFcqroRBNiMUc5kVaAMUAGAwGKNUVCPNoZKwGlQ9o5Fsudr63LsdLfq5WXfP6D6F9J8nb5nv+e4jkvgf2ut6F1+BzVjg+r4TM5efU9Dw+gpR492Jh9fy/QaGm3Lx+d7Lbab7nF723ja0zlTymWfQ+Ymr8/s1INfBdPph0f2z6Ppel/Pu3v/ADRP43n/AKY/nFt8T5dTXZpZ2HWti+djYpsqdcrKJXk5FYiQ4qRVqNFGx4zQpqOUVpUVKo0ARCXtGgSAKwSjLNrH03q/QldPa+bDupyacthhtq7vF7+jq8P09jS4fpZos/TvS6elP0O0XO7935vsdb6X4OBjn6Kydvybwv5q5v0zhee5/nM3J0+fWis/s39CD5On8mOfz7/b7Tq+afzBw9D9S/c9vm/zo/THD0kS/G/NX4z5/qmNz6o2mb2pqiQo7MS6JJalmpW2Qi5ySNYphOYmjCZIWP6Xm9flYer5+Jt8DkGAIEqDRXCCAAAc0pwCBbmuXE6tUkQQFKgkVw5qtAgUaKJFKAOLXs+YQAAQaAFE0VoRNRpFARNQJLS6J7OXbG9VPZjd80btRgDAT1W6KAgNR5AQAkBIhLYnINJYG509CoqRHlCryeqahQDVBgNQocUgAhKBmjHDkAVy2zdaHvX0vzuf0Xs+JxXR4el2XpHv/LW/BfN93gPaOfez5j531+NzXkejRt+d7N7ovL+37zyT3/i+bdycbtPi+kz9XTk9Y8j6VLe8P6/pjx/uOAqfR/lSQ9vg0bUZdHxvN6k1jq8f0D3Lf7HM3PP/AEDzT9Aun8T/ADG4YqtFp51mtj1I4xfUmi5OleGsI0fPI1WqzTZRucaLGD4FLCsGq0CbgpEE6+HQ8c4VioVoBK0ayq7UkdznL0luXqyLDuc+NJrhvsdP5P2MOpz+reu8n1Mz+f29E6tOku+r4299QfS/mUWB6Xy3U8ULN9F0z5X5b5/Z8gweP57OpbeBm1s8v0O9Yfzf54/u3Wf87n9B8dXLfg4n+5Xqmnzn883UZ/dfvz+BtfMeb++8/ufUnmOnk/PHimPrObOwCFb6jmOf1Vl6POmYta6Vzo6WWFrZgx7oY9vPfp3Ojyu3HHFjCF84s0oWiEoPNriVCxElRtzZajYA4WqIIVwCsAEBB5ACoWohVLYGIpXFnZ8yAIoCAo0ImoAAg0omitIA56uaSeiRZ3ksxvoMy6rcE7TV3KueKhqoWoUom0cIEgSrRiGaMEg5fItFDdEMmoZRuac9NApQYAAx5oIWqEoGaI80QTmtLNouq97+gx5B/o+H7fofX/lS28/a8wl8D6zkree/W4vp+FyvN/CfssvJcHsVNTG9Ch2eXo5PX85R1uD6LYpep4kvaef9UcJ8r+o9rl+f7vn+x9J+T83Y6/CiracNaWJfbrbs++5vv+jn8f8AWPXvin2/56y6PV5/oPl/V4Q7z/XgM3Iuesc446ZmNvncyWOE5Vo4dSSTo2AmJK51lJ/Vv6k7PlfljzSOv7XNfPX4Hz7f0K/AKOpW4+y5qSo0rRvuR0SrPYr59QeavhnWSWdZtSO7f6LyfttSTzvsNFvP9I6e7u3+vm0fpP6L8+671P6T8VzfEL9PRsr1vRvNd/ls/wCUuH7ny/msPkc6J+XDWU/oJ9138VyXz6L85f3Vj0vH/m2+Lv8A7pOP+eHOz+0/ov8A53a+Zb/QPK/J79PHz/lv8YYe6iT0KiFQFlKY4qdt02CZmjkkiTkcVE5qw+dDL24RixRXzimiIhUaQUIEKhSq2IqUJQActAbABBCpoDVBoGaowBKK0CFQbkd8Ydfy6ACAAKAigI0qAAAmqADSqm8VzvXQludZn59S2p0lnz6687i5QjraunEKMctBChryAQpQ0lQSBKAnBo5pQ0zax5K5MUFUpASOQrnsqLQGkQzRHmiEOa299HT+qfRef9Bdj9d+bevfLml9DvZ/UbnmPPXmvbcfT6D5ptzebeb/ABP651O/l3+QX+bmsUujho7PT48vQ6bdr5ud6Uc+XtuR4Pf7Tl/nf02vznufn+ja286pmXxZ7L82Tonfou10+72LPl/3XyLju78szZO/4qnUOJrYta+N2FndqyFUUbb5EaoUG6SdZUWkrp1KrUSXyN+g65v2v/APbyLGXj7H1N+nXR4P4hes4ex+sPwB0eL538fc/vMWOhzAVwT2c+1yr1HOnrsE9j41aWRTamHpdW7g+t29PyfuNFM/obk2/Pa9j9L5TzP2P3eH1/O+i/I9vnZXhWN4P7F9P8j9D+N7Py/l3+E8txeFThryalTJ/oX7rXy/h/044/KX9np6vjD7y38n8jv2Gnb8XvDOr0P6KP57sPN9m/Xnfj/nc/czzfY/P34vjsVI63NB7BjgQamYaDpaSKrFbPthfr5zFFJEVAl8LmqkV5IBNEtER5KhKKJHNIDEJVBigorApBJWtKpQglQEFANqJgwAEQDnjU6/nBU0VpABQQBFAAAEUAAAVAWRUjymSlTST0vdG01mdrUuPbHI5ge8gKRRKpoqFI4EQkAUGgQ0M2uaVy2EKGGTEeYoSIqY8TBkjp0cp0IoDHCDxEUg0r6eh697/n/Ucf6F+S6Xqnn+9y3nOXvUej9D4H5pg+G/dukqbc9XkFx6HX+J9v5mvN06tXk9fV5H3fhs5vB261/k9/ndrr8W/V3ja3vE+8wOG6/DOjnOvjdfjz6npfOc72Pl/VepfV30fheWcj8L/T3iXAdv45BodvyOdWzIq+cWos9p452ja+exXnSKOudgYoCfLOzbK1YBJXBjaxZqvH9q/Xu34/8AMn4X5fpvpb9Z9fJ/AL6Xw9r9pvzT6vnP1A/nTjr5xvN9CoJ0k7WXz1ulPSs6XP71WV02moltF37/AC+r19/zPv8AWl837WfS1div0c1rU6uX070H7D8tfzfZxexeaY+J4Tm/N/tO97r7HwWn8ld3yviHNcPk1KmviCxp+g/rGvx/5ofv3Ov40/pqz8mf2iR/P7/QSV+H3nU+x/Rh/PDt83zn7XT0dP3FeH+Uvxly/UuZHUogcOSN22MwtxPdHTLkpzwxrHM4iZWUaPlcipwXIxWwePadM/X8mYeKBmigIJUYqIBpHJgNUKBMG0USogG0CEUkQFQoFIMEFxydfzgDBANCggAJqjQAAINCpiAPG9iaoIeVPIul8sdF3Qw74JBRPcOljeNKgUBTRAQo0BCEA5aJJG5JRuet0BpG8kQhwgQSIN8hQ42VCxpIhmNeYEuVuz03onuH0H938BFyXV5/imz4H2Xaer/Tfl/zQ34r9t4b1z0Pl+8+YObom9V5fc8oxvK96a55f1rqnb4elS8v6izz/q/Jdhz2/Hqej73rcX6PzudyPy/6PBSnivdRzezzDvT+S6bpPT4PoD3ft9PyvjvmP2f5ArL8lb1eXXQ57v8An6kvH0yQx0MWUxijVxtuZkcZRhgKrJp0VzsaCvTijrNFeUX1vXJ+zf8ANxr5n0b+s1Z/gF9MYe1+sv4C6ef/AEjfgJXN5kc30UiLSSxGr70+s2yenPJHddr49i1Fm63Sua3F7G9t+d93ck877fR3O/hrQObfY+l5XU9T9T8JmYPJ1afGT04eD5vp9h2fo8HDeMZ+ZxWH2/llGkeY9ydqxM5PofRzcXaw0552XTH0BzYrbe1y088H2FXFa/VzfwvxB8T5/fR87MUTXoFKEoe0aIS+NJRg9U1pMaPmHOnqaoxSiPN4rcUQiwQeQpQhSoSCEUFGArUWiAgEA0UBpCgIOQEAhVaBPGp1/OKAiVKiYAKDXAICKxFQI0KAiaox4miCRWnTTq6Zbvt59T1naeSNHzxvFM7WTPphZfOLUIglQGMeLBwjWopGnLNEo3unZWjjR4ioe0EAJBqGjnLdqsa0xVpmDlVZJo9OvoH2L99+WeRxdWfm3sHL63SZXsfFZPkfyv63F0fH18/0OfV576byd/nGvL5yx4f21Dp/N+hrWvU8LquFw9HQ633fi+F3ozl670eLH848n38Svj5tnew9Wr1fX5vOega6/Q/fe32eY+QfLfpPi9PX8pk1evyLOLU5XQ+L9dFhyRpp5ixxu9jxqtrgbG8Vcm987OcUgCoVGPNvu9837BM7PjfF/wAbOf6D6W/Wa+H8A/pbH2v1n/AW/P8A6RfwCvm8zdzfQrNOs08+leta9zJ8u+1WndY895q0XflN9HqvL+p0tDyf0C1Jl7+5rev4tFud72x7fiZt4SYWNVKMdHS8h0+J2+FW+B5RPzufkex+VZdOONHKUmqI9nTHEZlcb8OiOJ8yyXFeuZR/eNeb6z8y3xfJ6YeyAI5IRseiRlaA0jrxgIlXzq1CRjzQMx9v0zzj0fPhXzfSQlq0FcUxKxaGDkWqBKo0qDAQqQGiKgGwTBgqaAkR5I4BAFEgIFXHN7PnBUAxAQAAAmICo0KAgCggCoCgINKgK5MkVzvXTI6d3vjSWedJJZ22KXL7j47545KyhbpzIGcbXFWPTGNlZRuediaOgcrWROV86tiIVwNcIRCyFKpsqFORWg5aPnEEK059ve9H+h8braH0Py+Jm+P9Hq9v1R1HmXVx+pYWnJ4/1nyP6Vm7XB9LxWXt5ED8OitqcntdHS8v7D0Txn0fm+l7Pp4+Jy3jvdXW2PjXhmY3V8rndrkc5692ebxvoHh/rvrHtv1HzPFeNLH54oeFjF6f63x/Et4/b5zoPG+q6rn/AB/uuIf735KrsvRjVTHG+KCOuZ7BSSzsKN6KlZSYOXxh21iuTz90dVjob5OPvrTouPDt+RCu6bnnns1J9PXfFPVJPO1unzem+Os57uHbFqR1bfR8P101vxvv793atG/6vlVkT0/YfW+T8tgXqaHGY2zro56HHWtLSx7uH4Pu/Ps3l/S+Azo587cp9fm+p+S9Hm1F8v6HqeS24Wx8vQ6tWIOWwqWIYOCkRSAKJgCATNrR7Bk+mNVuaGuAUoNDk3o9JIqRLGkNrTGBkUaWfTms04kV5gACEAQhXK0BoAoqBoUpUVg4apKBKKIQFQFQQo+LOv5sAAEVoWaarQAICo0omI0AAAAqAAAAAASzzvKT0Olm3MV2LMbWGZ9dgWkdpaRxVi1HlSr7csDL5YmVgKJ9md5JZ6bDI2lmlysnSIrN7psYJZG1ctVQtyFoqlo8kR5itOkNUsdBo3+/lmt75dJp+jxew8L7nyepieP9NxSeF9rb7almcJ4f6FZn7Pl8B2PTo3HffeP5exJ0evjHVr1brtPP5Wp0eV6F577nwvK1PK9eZkbb30Fwfp3uftf6B/OvDfMWXR8/4nwv67jdF2/LZNLDrpTc/RpVMPZzx+O589iMnSKN+e1ryibXO6xOqunWJzU8atHtNa0rRRMeKNeb0LVAlI1bMVsoe502fvbmc/ebWfZochw+xLY34Jtzl9aru8/pT6Ee1u3PG/Q7mh38FnT9Lzs+DLaz2Pd5fI+lb+ZyPA8vdo8xj0dDe3w9B8LXn8/w+nyVjk/V+JpIvMur08cmZmI3LV0IpGky5+nIxYFhSY0xUBEIcLSSNUjiGuZP1PZzco3m7WkysUisrJWkOa5VVSoxRWiuUlZEOUfF2qc7It87B4iiVq0EM3IWAhU0ARzEUY5W1HIqSNeaghBASIMUONTq+eRWkUEcEkatAlCpAFQABUAUEE1QFQFRoAAAB6zb5DRAbhUsy1RyuaxGzEHGjlGuYI75YmPBqkgA4ayF27eXoWn59E7JuNtQrk3omyUHKaKGqqNAEQzEcikLI9mLcOm1q3Y6ot7/AGcmpznRyaOt531EdOo2NfbHQpeH+j5+v1fPctmY3r3e/wAz0DzvzPtaVrGNfF2y6PnvT+T9U8W876DQzunwbVL1flsqv5/qdH9w9/TtfTfvfnPyj8q8v1vN4fgeriw8fU6q84HY9MYs4kxpyroRM+xlZ8Ea3yRsMJLE7ujWizlK1W9jlQcU6dFt8jHOEAVSpHxtYu5+10GZr2TPNZEjfSh5fXZA8J0nZbLH6PN6W5v+P95p3uP7C90ns+PBt+j52RncnY207n5/fzsv06eTleXx6b5e1XlX5eDjd3wCcn2fM1YzxnJURvljHm+N5kcOenpzxKlGihiLJHA0cqJuaqRzQo3d73cnCR8napI+ukrKwUQi3CPi2K0qq2DzUGKNwtNTHz6Ht040R5COHST0taSiuWqQ5q1B5ilAmisBMRygZq0AIAEBAHHJ1/PCggA9NGsFBAQAigigitIAomg0oCKCAAAAihIs6PaNg4UTFG55aNEqjbGQrXKAlE0GgBUCe9HZffl6E5Ftjcigig6ZUKrcGjkWorGhmDlFJJTSNayRacul2ZxWdFBVz1kjw6owkmm+qfzfV+00dPc4+5k+myPY+b6/G9f4itS+V/XcevPHt9b6fkc3yvn+nzNbo+T3fRvb+Zh83i73qnl/ffSml6nF5F84z5mdnedctjzPpaVTv+dSXi9mTHnBjtOMWdFjy9BGrlhgrz2DzLKtEVPercjFaUKhEcxOJ+kLW3lfLhl3SC0mlnqt6MfS29mPSz4hrWvnltGlWxz9L1nZ2jPTJp8Xq7PTcP3tyxn7Wntev4/Vz+x8vyOL430mdHz+t1/n+/mMm5+x1nr8qujz6HgNfJ4nF2+Kv8P2fP1U0+aJXpYg1xrpjEtelGTlGqUUlTXSudpEo5iNErk0BqAKOR72Y0kRpqrNInmrTNyMcTaDlRWI5FBUG+7HZUiMGl8wjkE1ctEAEcoCASoCqtEcDBw5FQjkaQqEKjaChWjRhHJp1+CICoCoCgACACgigitIAqAAACoCDSgAmAAD1mgBABQQAAVoCgigiNKJooCAoCKxb076cmHpyOm2NaBpHJulVOVaAUrlsittUhAQjyJVbUqBab9Ts58x+W6wpoZijY5aks7b3sOvodBzXB9v6C/o9LyfX+i/HtXOw9LibHh/Xb2h28mRQrz+JF5Hq/Q/U/F+ZU/nfr+ln8v7r0XgOvTIf1fF8zl8PbDPfG5sdENpODLWKSZsfPQMy7iM5mQzxwJpzSWo3aporZTh2OJVHUY80al+p35f6+XRXL1FeaPln1dTQvu6j0Pv5vMZfM+hxaGedmy3Ziy7ejz+H36lfo8jY2uD6LT2/H/Qtux3lzq/W8breT9Tws2Hy/d5jq8O3nMTG6zVnLq9fl5OZmYXWRx+VV+/4W3xHd89WZfzQ8I5y4XGTY6ygFksYRteBHWaOmUUFRUjgSQpY0waertJptIkmF4Na4RxAOlJoBIIRHLmhIxu9Y688vQ4O/MNOFFEKmPLjlTjRwiOFAUKHJtAQcgIQlUABIGaISAK1PkE7Pn1AQAAABAUAQFARQEBUaVAFAQAAARWgAAek2oMQSiYCAABFAAaNKgKg1BAmIxb87XZcfTkmmyIEa09QLE04WooDjQUtEEoNB5oGcjTRS5FsFGKJqIAauoaS3zLo1bm/Xb9Xz9Te476b8/qt+a/UOh7SPovMNnr+YwNP3PzryxPD96Ta6uCg7D0O/xvQ45svy/pOe38KxJwp9J1eRzZyem92vN7d88ej83Evj/QSkdTosu10UVHXXmsZfPJbnSFzapLkWe1i1zPHsIQdW8/v381b855j7KvOu7e067Gtq/TsPp6aOZ4f1eZBt5M25x+xlS9HHr53lfQVJN/Nv6fF7fRdtz/AHFPZ6bveje38tyt7ZZXGeT7fT9H2eR5vlef9C/AnBd3v+Wx8zz/AKGtV5uPz2D3vy/W57v8OlSr5lxl2C1mxFCDKjHyzKriirFzbhpEADnrRIiFGgGqjcq0bMNkSlErIapBwKWiOVVCISqlNVyoXK26JMtIErnUVI15yE7K0Yhm1Hk5TUcraOGuEICDkCUCEBKghBNaS1WuTXp8Jo0AAComitCgiMVECgDQg1RCoCoArSAA0AIqb0mgBABQVAAAAAAEGlE0GgAAUTW0aT2cvQlsx0SEuJlSOBLE09VoBQpTkWgjSoDmgNeL7F6SRb2OSayRgsRyoQbpNFC3NK4uVbrTt8/vd9J3RPr+P+jZvf8Af5nqngXV4fJY/Z8ZSq8919bLrkoaZdcX1NxpVO5z7o3o4Yen5vZ5mgcrXypK23HDSwuxJl1viz7kjiYWnFA2sJLEaIjbZU3ItXMLejQ5wff+nH8p+WR2udPdbs306/f9eHJ3N+T1Dg775+q8z6rkYfP9PMdz6SxRvp5vJ3Mg287R6Hg+k39Lk+80dP1PN1r3seL1fEdHBzdjwfqIK86dF59rxVafM+h4P1viL2Nw+g7ltfDpU+z51mZ2eBSK8CUXQOZHC+VsyIa9ZOENeQxlywWQoDihSolMQYqq5WrZVTbGZubUIrSOuI1zFQaioaTPCUiVi4KVXTVSVgAkrWlE1cqbZWtNK5ZEWiK0gIBIOBAEJVE0R4qAgIYANcivV4LRiohQAAAAAAARWgTRWkUEUBAVGgTAEVoE3oqAEUQgAAAAKgACKAAACAoAMBE2jHbYky7plmpKwx7kmVK9W5q0cFKFCCUYoNEMzR2qeDffQt6KJuXTi7m3lc6nJoqIFKclS0kHFO1F6O5cr0r3RL1Ny7qYdLyPsYsH3/y/IsZ44l8wldvz2/RtNMviebr6PSz7uOy5yb0hry9aMXQVzI+8HVo1svjpSPPYdh3wwTzxxVwS2I3IrmKYzc2akWekWmoaL+qO/i/lhJHs6Mr9CUtWLO/F13Eb8/d4GHq62r4f6JjLzdK3+fty9xXSzr5861p5d/p/J+16HWr6izb9Dzp9bu8/Bl5evBj831btHpzq5fNdWvv4dHntfm7GbXmUoe7xa3PdHg59vf5yqyuOZY6UAHVmiDghfMxXm0cNSoaqzEG4VCjaily079DWVkx6ESXE95FhalJSpeCvtRkMQQEq620kUKliVjayFJs1LhTPRAgfOrFctHA9aNCQGqNNUzQEINUIGuFRCIJYwa+o5Jenw0AVBqAiiQAAQBQEUBGgBUTVAFaRAc0FGlRMGnpFqA1WgGCEUBGAIEwaEAGhUwBAFALq20HY+pNLNPbNRFS6aWr1oqmqKCOKQE5CnBaJWWvH3ZbN7pLPX752uMy9DndCOPMoZVTZzaNJh7WDnbCVLpX1aHTdWXO3suijtY+ju0d67Pmfb+PwU8b3q3bdnB7j8udvk4Ot43sbVCPS6lOf6Hn8/p+chjx6o7OXSYT5XzdHnRpnvaSekjimJla0lMleMpJo6UWso1ctYks6tVNH216v0N6l1/PfGK8/0+lW1wk6fo5easJWq8daXMuu5c5voLd7yfsIM2Mn3LVKGeTVTHv0dnzvtNrQ29Lc0vc8GrTjVKvP0R1ufpgz+btgz889nlfT+Vw+bfyWi1xiVe3wedze74Fl+sYWOXyK5hdIqccVcr4RNR5wurFGtNQxBJQtyNjJTpbe9j9fNAcXaIiN1Yut59cuWmqacaq2xSFRMQHtGKDXCex21TIbRLE8hsUoqcKmlZqK0VoBiAKQrE1HCIQqARvNWpCsa9NGVPKJ0+GqCAEAUBRMQAaEBRNUaRQRQEBUaFTQqVRMGkVNwqBAAI0qAACgigIAAAAAAIxUEomjx6VnH1ZJ42mbNIxp0qoemBoqlqg1UaKaOQBHlJ3nqcvSWPa8Xu/M89Oy4jD2dGa+R/R68HA5fJ63L5/ldsSYt+tl6GXHpxP1N3Y6Dpw5l3P1Mny6e6Ti+2od/wDUflvn647a/r3teL5F478p9fEuPidZ6j6GnBWef3qhfNtc5pk3S8z6KTma5q3p/s/Gee2PF+xhtdPBXkjeiRmsGdsXKopZauKG3LWTD0FJJV6Wr2Zfpn+a9+fEvN9dFdVb/M9PnPpeX9Ntno+DJg8/dob/AJv2Cb3D9Hh19/G3Mfm9c2OfVHY92tp832G7Z7I6nofpfleXzPN9bpOZnLP2uP0MLK5vRrtjHAwdfkMHm/c/OtKpeWJl9PxddunjveaTxrYkbsk7ujaVqJapTG1yMdeaNrOInNWoUGpQ10iqaOlumEU1qzSWhGIdFIqOBbkSKASDQEjlYMGiEcqNIjhUkBUK2iEqDFQoWig1RKgIPNBQ0eaNJaoK1gA1rXLr0+GgCKhBoUaIJQaogGkUABBpUTVGlRMUEGlQAaAHJFqjSomDQAjgEGAgAAFBEBQBGKiBQRwS3o7J7WfdaXPVIwbKJyqlUsUtVLApUTcjFG5ut9DPY7r0/FxOg5vsOE5ePI3fVTflvRPX8rudf1/nfEPOfmPt+aPG9OhRrirtMLHRabPk0qi2UybPazFn36sno+F3ut1dm3wvf834tl/M9iLlm/ut+u1a6O3Iny1Ssr9J8x4/Yg6/1PlOB2eD1V28PY5zW7/BZI7yafPTWzDEx0R+ezQiRa2ZEc7J9M5Ox7ObrJPQ8v7j+Aebfm5fN+0S/eOMcftX7K3lr68iPw779jl93ayOb1qGht5WnV5PV0drzPraF+Oi3t37nUp7PidTD6njUMDzPX082bo1+D0tXiOe0z+eMDnfY/McTL9n4afGrzM+Db5ceZqwq3Cbve3NNn0MbadGiSuKCV88TazVWmIoVAABGlE1c7HKkVUrh1nVyhOiOQiNKgxHCoDXoRRuYm5KTRwqAg4EUKGgiAJBiqrQByltQkUTXANEiPEBJEZuY01rSqgYzmTo8MAAGq0AAmDQAqAAACKAjQAACpg0gAAig9JpBgokAAAAUaAgAUAGg5AATVAABGiwrvT4+pZsxvKkaMc0omoWijVSwKcKxRjqJun7lqb3qedaoO8XmPN7+o6nzPud/wCjvs/yvXd9T+efNvBfF/rEuP5vqw7FYbvnOFVO5ntfsx6fm91+Xl2K5qudFaVzs8v6N9c9H1flXyLDwudj8RBWZ1d67z9/XodLryXuW8r7LQ5uuSvn93ykKcnZqw9nHY18+7Rl2MjL59K8GcImGsiz1ROmXPOlsN8SPcP6Tpwp/cpz/NnmU+lXdz+ywx7DocPS1Ocx9K/D0eeTF9jFPt5jsXDY4eyTdy9R0nB7967HtX9Ho6e3571fA9l8t9HwuS2/A+wyVz1rPjbPyOPZsxzYPCev+b8vB7n5fWzL8JqX5zB5SxrWdpo9S5Zp3VRxvNI4TCJ9YRrUiJtR4iA9o1jMnPNVfWiKmLNtrHM2Z4OF1K6bSOsxWmuQgCOG1SQBBoGhEDXm5AABCQEA3NKUTUVisRBIpKoNqmaIQjXk1HKtCNGkRnNrv4iDSAAAoIAAAAoCAAACKCDQAAAAgKAigqpiAKDVARgIUBFYCEUBGKIQaAAAUEAAJ9DPttT5d9tZ2FnVFEiCUKUKFBXrZqgqundD0Vqd/wC74HD9XFdT5NyfQ43TcXoe+4HrR6nm+j4HlXDeT7PRJwfRYzvQ+S3bO/FxN/zfoOk5TbHO2YmtFfNY5mKzeo2836Sp+xz+D+W+MVGeZSBL+jrt6TvPQ7sbijk1m49tbPgmZ0ePk5/B6k83Rx3+w6Kn0t9MrG5eilQ5dYm4dCSrWSNaTXc+6nF0eTGyJerbv1Mrzvy3sZ+8rl0RT59fR3c/T5pyieqnPscXt9RVr1cm3gS6PF7Gmzl+jJJNDTn19W53YbGx63h6nneTpS+V71PTa55eH0LeFpz1Nd8Xm/Cez+S4TvU/PKlTTySN8rVecjDRzhk86NmNpIVc8Sp8YoH1hAtYgDmjalZPSbQciprLdWqbuVMda8F+eltNA6URS5o1RyCBAEEo0IhFqUcNEJRXKKMExpCoxVTRAkFqjhoCRCRrlQGDxGmaBDBpoxiiRA5henw2jFEINKJgAgCtIACYNAAjTkTAAaAABFBAHpNKgIoAmDSK0ImACgIAADQoIAAIoAAASqtC7j6li1n1K+O1UHGPnRSXBqKWSK0UpHsbo7T0fovq8vke/hr6bl92/m1nztO8u+hhr6se1wlXze6x6Bze1w1rv8Kr0bjWwOzzqHObeHPreT9lDj9HlW63ofO8l7tXkW/F9ax+Q8P3K5xZRuc6XqE/T51j0fOdj5dexDvwv52+WvHkuh4zKoZFWx6B6XDa0t+vp+Wvl53I8b6WhT5t3aGPoLNtiUs9dvL5+6DI1820jj331/XwvmL2D0vM+etnk+n5mTz/AHO/qdnDzjPO9+J+XRsR8vuWm7Y7t7zvsJ72HtJR5OyRcS/1/R3Nv9nJ21D1vm8nnvP998XH11jLfJr8HfC2uKa76fxvkHLel+e85a9L87pMvnZUfnrI8xTZrhOmjpbI7WSdRVojHEbnDYjFRzcijuY/TCzSnRiXySbOvPlRnQ9MdtGiuilHXirKVKyesUSom5G0QkbUqJujScgKjkGCCAkRiomg8lFoKNivOdkdKtaEcJIJrQRDJiPBzRMcJkdSqAj6ULk+ZXo8NABGhU0UEUEGkcCAAAAAAAAAAxASgIAAOFSogARQAEcCAACDSohUGI04TQaABQQAAAHWp2u6OPp25cfTci6GNfO1XkKWKU5ztEQ5xomp1YrqdOeN3Ws7NXrx5u5ydPT2uuYb/N6VLT4Pa2u++o+L8p575/7CL0o3x8n2/hPbPEO356lH5X0uw99bM3t+eyofN9epzvH16eEZUavG0vqvVc31JesbZ9Tk9R17Tno7HT4nMZHP1VJOXrXW6eJvS9OXQTd2U+Vx+x6t5F0+ZnRfP/anR8/rrW0WxzfV49DS5PQ2+QNul0fT+e+8Pzj4OeG7PvN1t+bk4vP9bbsYeq3QrozNOsNTZw+jtLx/TWE5vVSsZhh0Q2eHpt6E+rdvd/DoJ6HB1PE58uOeP9G2PWjO59H0sJqUfW+C8ryPc/KM2br+RjirODntfGv3FKotkewmnoJZ6UcVIk7NHnE55o1pFJsMjpl7jl+jp8VfOrez5u7u9PPyyc+znTt1fH4erWOn5MmW0hPa1rzehTkTGCkCmNM1RoeDUEK0rU0HAjkJYxxKw0HEtUBShytAUbqwQQ1HiQOHqhkdSsYOQIypc0fPJt4qgINAAAAAAAACAKCKAgCtCJqjQADQJgDiaEaRQQaVAUTRQEaABQEBUAARQEaUAEwaAEVN9ud9jQ5vckfn6DEJRryByKUrqdupo3tx1nXddbp+34mf7P18vE53N1clN5/rdrqepwa1DxPv/o7rfsfx/sOD5fqovlDEpQ/L/oO9z3sfC9D699B8Rx69F/aHjHnel83Z9RylXzPRymZF1sdMmnl3VrlDqqqOuR0N86s7nMdW2unw+18q14m0uL02dOXU2+jHdk7+Xr+Ry9aKLz/RtVOL6Cxp5929k5ezHi9fy0PpxWf6T6XzvmnM+b73uvy0/H1+q8L9P4Ppe3xUp9nz3PdLwfR7KY/Q6nVV6NvOy9mzByepCzk9BxUlqOhubjT9jHtN3p4M3WrKvXsza/nelDFsmN5tI8J+Hkc/6n55yuF7H5y/2rp+c/QD8j9vM3/1T18rxv3SuVFF+QvrOPr/AKRfBN83zS/m9/7S8h05PpH6j18z54+OM/R/TP8AN6ubkf0cV/lI7H1dfNz755XtmmvktcJ5VjEnyRoUbwjV+ctmetzZ6Wjh7RtcJqE269thMiA2UlAtFqWkgNI0hyFKghyGjFVUiNORVJCISuYBNaZLCJXsSISsaYjGo0qCwDbxkGgBUAUFQEAEAGlExGgAUEAFBABGlQAAVNxNCNAAjQqYAAIoINCggAAigACAqgiCVBqNIqYBNr592pPze490dLWuUKga5cqdmLd6NHk9Ji78U17ddHv9j2uI5/as6XqfIfR8v3f5B33z54P3fa+wYex7N86ep8Z1fh3gfrHk+H879Jb6Tt8m92n2n5Rzv6IcPJH8oely+feG+R9FHo8HsZFwz6Kp2+Yy3xfRakPVyZ+T4n1eU2/Kl2Lzo08eugycrWMKSCCK2T1Nzrrteb9r47bMve5OLwfpuirYe7f9q7PP4HO8D9LZpfSfkXE9b8/+mUa/rfG8T0WDs835/wBRo08ui52Hd53LaXD7VDo7ezfy+ifWy9FE5PUscznn6NU17asPH6sUXL0RRZbRv5dbG/6XndLy3b5dWLzvX6Llb580ndtTLKHj9Pl8jmfR+Dx831Pg5NfXyv6Jfwp6/l/avuu+T8lPudafT35fkfMX6e59n1d4ptwfj1NxfW/tL+de3kfs/wDirfPWsY+v+4Pzz1/Lfk9+2GPq/wA9/ouPs/Sfz48eAzK6hmdPah5Tl9D9LzPPzzPaZQfG1z5HdRr1YrOTuhL5UBIhKOavUtBG5DVcI4FC3NGjSEWso3GSi3EJVBuFQrEQSibSskaYIjgaCNEjxvFSV3CNpRNqRBYia+OK0iAomFSCaKD2DQaFEgDmpqjSoAAAAANCAom4TEAAEBQEUAAQCRONalAAAAAAAAAAaAEUATWVVa3MfVtrz+u8WoNMbWTlbVW5HnRp5m2cL8i51enoQ6vXz9f6P9p+Y8fuZd/o+Trh5XrfNfo7YfR8JO48D7zlec8r69nU/Ufjr/UftPyuf7B5OSh8gd/ieO8R8/8ApOI3xPfZ33Th6/Q+g82LpujzudyebqwOb8L6WtTyUexw+4zKJuX/AFfluVo8Hb0fExvnbXPt6JB7fhVOo4vfhh6efIbhdmrh6HQ7senj7/N6+Psd/wAx0/OvbEg8/wBvoeA5fWll4fasdBHfy2r1+LL0Gfs2Npepciz9bQrFWMfh9mzncnpJ0+uGbXy6m7Bhn3M3y2R5nrRbd5vdGvWYvqeRV7Qz86uaYZd2fM835/L57GxvR+ToZfb8TMt8n7LfO/Z8p738sXyfnj94T1fcX4gS3/0EE/lh+xVZfzo7HL9L+wP5F6cH7w8J0eJ+ZWPy/R/q7N1fPfE/6BR0fiF+75fxN8Svu8Ady+qrSnIA8bum6M8WHLorV1zIhzy72m+XBzdLErneOxhA4BgCpI9uNSHtKcwGuJGOBBKqbRwoNUG4LVE0R5ojxG1kIkjgREOFaoOJXm1HNeG4YVGKa+QAINKiagIoNcxASgIoCAqAAIoACAIrQAAKJuFQCAEAVAQYohFBFAAQBUYoJEBVBEBQBQQAUEAV6olV6Ozh7Exj6b2ttjvlepo4NH38+6/pdvNxzuVX+l27u02+6+26v7z8mk9aifIPOeL1s2787+i8z0G/lb/aL1/JKPhfVVLPq/EfS3ZfoP8AOW76nyz8x/P0/XRTc/p0MTi9DU9M309o4/1vF6/I9H4zjMTzvseEq/P/AFPOt870et4PTh1szyvsNfmu/wCd6PjtvFuyZd+d1LnI0npFeNegzt1NhVx3c7Hq627vjkwc3q9N3laeZ9JO3D3fH+q6Zef6eGHXhvWuH6HM7Ln92nY6+DS0neLa4/fswLevT4/Su2Y07DnO3wue6/j9fjNTLsyKPH3ZGHxqtqHA7Z5/a1MX0Oegzfw5cxG1wWnzubzK8DJh9H5fOo9fxY6sPtj7h6/mvcv5574/P/vmej7e/D6b9f8A6D6w8Q9p05PxG6bm+g7z4Cjp2PvFX+nX4lb+P+r35dFfrNLr5n5q/d51/iJk8/0Vmlh3C1gTFNjp9O/s4b3PVeXHwdLWvGWSelqJDaiaKhCBASKk9lA1JFJarQIRHIoCqxrhFJAHC0alZqqbR5IjzapKKkgUAkQHRiWNp7Qjjqa7KzxzbxwAAEBQEUBGlQFE0VoQFQAAAABAVAFaETFaUmkGlExQRQQAGkAUTBoRNRpAABUAAABQQAUGqA5NAHXZ32rfN7j0XUwfI5z2HmutrdnHzvaeb9XwknR4lnsNezc7z3vI9/8AYfoPi4thTw3I+h8v4Qz5X9nrbdzs9pj6HnOX5/fzd/0fnvqH6Q+q/B7dLg5fjPw/m/W+e7LTj3b3seV0vW93jexpj87zHFet8/z3L+d+j85jeD9Zh0/F+giXl9CpFz9Tzs8bI5vnmzt64Lee3dX/AKv4ePnefsu1uT0uq5HTCGHg9iWxUOvbY8T2HnexodV38+B32PoRdFxfQz42m1FnmfXaWsvTzeky7NjavLhc3y/qsVOTr7Db387F9V18/l8fTfhNrxvp+f6bNO5G65XjfJ2zdW/np9vz/rzW6itx3p/KScrPm7PD34NGjp5NWv2fPVafR8ui3y9J/RV0eL86fgjpx2vvnPt+2Pw/F+jv0U8fxh/XqOrK2Lw/Nbouf3P03t9Hg9p+UM9X60fgrO37YewbeT+JH7wZ93xv8Qrp+azD1UeNH0OjLSYU1exGwxCZEWiOkZe0y6Tiu3gVnB2OE0GOYS5gK0hRgIAaDgAEQoIUp8ZQhCo5apk0JQqVam9VqgNK7xJghBJOOqlTFC4x038ZUAUEAUAQFAEGqCFBAAAAAAAGhAFBABQETVGlEwBUAAFBFBqgAAAgKAI0qJg0qAqAKCKCACggCic+xn36rsPZY0we41BuSffRujyenlSacmn6Tv6jvdvf8H6T6X6P8892Z87fF7nU/kTwnr+j5zo/P9vmvUObu8i6nD01zPpfyv3b1j6H8pueCef0fPU3z37P2/T+14XZaHtfGeo5nV+d4Wf2TgVOT7Gk/Ps1vNOH2MTF8X6zN1OL0c7mePsze3wnm+TUwyYbM1NMupv+z4aW70ust+1cJ1+JwGnw+713OdnkVtLL08HP831+v3fR49zmTq7jO8f7TvvNvR+WytTwP0uTcn0ot57WKHn/AFO3w/m+9m6XJ39n1nq/NV73Vx+h+RX53JWPH+7rUObpz8zg9Tj/ADXn+cxZOz4PW6/zfsYaq3y+c9v4qzPv5PN08ojivhrUt/mmR6eZCtcVf3Hbi4jzK8L3c59vZeJuPaM1HmXo8bdt9EXl8B6WXpfRNm+X5z0o9H1bwSuftfTlHzz6Ia+5+B1ycKkdr2toqSo3rQZepVTTgCk1UlBzQRqjrxRokas3IqAQMaGYOVRWDhRNBgglAVExonIxFSBw1CVa5hsOUSacLRQbEebVFC2oRzIZBJEGCdHiCAqArQUBFAARQEBUBQBAUAAEaFAQAAUGq0AAmomqAAAAAAACoAgCgIAoKgCNCgigigIAAAKgCgr1fQaPL9CiKh1QKtdP233fm/FqHk/VRu5ttv0D0Nu69q+y+K+g/cPO26dcPi7HJ5+15N5T7nRw9nm9j5Z9e8L6/wAk6bPtwl7PE1vU/q/yLwbiPnvtOh6zZex+ge95mroe3+Wlzr+WoZc+jDXOqDOz9XJ4HzvqM1nkfS18/n36Lz/zfos2DDmbHj0ZT74M/Qmuhv8Afyvi0z7zm+rjv+k4+95DtvwYeg6OHz7b5Pd1uo9r5uChj6K+wc3s+veL5x2HkF+hq9bh6XVcLw/ScrU4Pc9SweT0ur848n7uL1txx5XX0x3eL6l5Xp40lxerxVPy/o+O5/5/6fgeHf51zr+75B23h1wahtFX9fwunxe7xcKDh7KFPPmpVtPnkhrkSCuTLZvxXJ8+gidI8JHzRMh6rVXrcRaOVjru0Z6YaIrnbq9vHnHF2Elpi52rW5LGjrLnT0WhNpDGpxrCaqQo4WVySotRrQEqDQaEUqroRAPMAFKQBUGgZDQcwhVYwIQE9Vqija0hqOWI1NEm1tTGNJGLENvGEaBMGhGlExQRQRQQBUAAQYqCFTEaFTarQNAmitCJuVNABU0GhU0GgAUEAABABpRNFAGkUEUGqAAI0omKCrNTdHn6V2PD2BHzuClk0eonXdCPh30/Zu3q+j8v9B/M/tb0z5P3Z+gz+ZdkY99Xzvr6/OfBu/7Dxev5P0OXgeB9lR0fZ/PfPm7/ADGvc9HyOwtd+HtHq/qY0Oy9j8gp4/V4tXp+b3ud5nUz9LP2uN5Tj+hy+T+c+1Njfy6k7rAw+Lo1cHPKFkXLoaHR6PpedYl6/N28nk96q7p8fnYvO9f0ap7fzHIWPF+rzNV59X1Xs+Fj2+L6Pt6eq9L4yOHsLi28g1fD/T8LzXlr2nv8/S4X1Hy/s/Ie65Potjkt+X1FnR8xxfXV6vT7+nxXjvnvD+ncDwfyH3nMctHwXNZ3qfERWHhqry+v38303x+DUOPiDzuyOnPHVi28dlC/PoVd/NlsLTQMuwjYwczOmpHTUgtFU1cGgoIoCKnKxC5asCAnC1JCu14v0PPuUuTvFytr7Hxsa3ORUlaNo8xAFY1RADVIGtOVWiiQJEaUTABARzhqGataWNy1wmvQSFoiYwUauGq0rU45WNiajRzgrt4yDSKCAOVNqtIAICqDVAAEBRoEwaRQBNBoABoARU1WaEaFGioQaABQRRoIBpUAAAAaRQETBoAVAAAaFTBUthX00/L9KiEI5wqvSW1t0bGNvzv+uJ97s/UvtPz36X7jw+abqeXzLuFjvBw3oZ+IeZ9f6v5V5Fx3y8WHl0l5e7AZPH0ur9L8R6F2Prc3Weu+h4PXUb+B57I9HnodLPRRzteLHz+X7PI5Lh+jd5r8x+kZEx4cPrfd4vjUPm+567w/tfF4OD8/95saPrfM9z0X0fy+snd40kcdWanP3Y+dwez3Op2/LcFjef8AWU8nzfXt9TWm/wBF7Pz1Czze/Ynjp7H2HPyOJm5vc0eKx78303i+w8+4vwP0zRfx/T71Dbl7TG6fJ0Paa+b7G0vzbivm6f07yvzv4n9l4Xk9fy/Cj9D4oRaSMJOhoytHs4Iskxz2ZefXi083Lyujxopr5pbkdl12XW0AYxXokdNSC0WVdLWuR49rCy7XppyutLoqFcwKkRw5RyI7mboW6IOn5+2GbbngZmDSBybQlGPNRgqaDSAkcJyLRHhGPMGhCXIrAlUGjHk5CCbWIFzGxBI5NJDRYURMrFzgSQGPHG0SRUqyVlkJt5CibXNIAqAA1QFBIgKAqAICo0KmAACDQAAKgACKCiaOAE0ViKhBoRoVMAAAAAEBQBGlEwBBoGhAVU0UHJNAy/0vP7Spn6At4CrSSXTf0Pq/q/jPK+g8P7L0T6O+o+c+w/bvG8e3F5u00byxs70PJ888y91/LPinz37hSo7fLV48qltdnk6Pp3vfM7/cdz7zX0+W0pb8TAyezk53sejy+dovsyMnm+qt1PK+wp+UcunIXPE9+u3DuY+uaL2v3Ph8nxHzvVt9MehsdH73zTMnXmj3I0xtEWZBj1dhT08vNZHreieUxh69x6587Sz+p0vZF6UnO7+R1MMdt3t/I+t4H3vPxeV6HHbd+buD7zUzcvtL87fNZvJ7vrfd9H5p6T1U/mFT50v6b5C8g+M/pXz/AJjt/I6zej5QUyG59UHWeh42cr7MuLjVOHXy6+Nt5dRm3mDwk0o7bJl1IMSMTng6ebkSdXOWqtKc0aDzSUtrldiutkKwEBUB3Zcn0OPR9b5mucvWqlNCRYtEcKgEZIhmoU5qpQQAiOQaEEDgE1a0KS5q0EJRrwdGJstSRCEQ1GnOVRDklVPgKepCRBCy4hHmg1kpr5QoNUEUEcDXAgCoAAIwUQJgANCAKAJiNAAoIAACpigAI5No0KmIxUEAKgACoCoAoNVpFTRWkAAAABCjUmhQNaenomcv0si9fLA7Cpo62m0eiZdDePRPQfe4vpL6s9D5D1i58ls7MM8Hn/U+fr4Xp8PzZ86+j9fwfMfJ/oR2nq/Kyr6nh3HVyemekejHpdzk8/P0Ov59eSq1f2fPc7p66VvOPP8AvemqaZbPhvN6PC7vg/X0L61t9B7nxNbP05vZfF9fGn6aPfv9F2eDgclze3W7vNcDT5vUdDNy0JW103M/NeG8v3Ol3evO1u9Vau2vR+guJ2+MoWPL/X27Wnm1un5urkM/g+69J4Pn7tfIz9X1LlV4fOdS/W7zcPgfSd3L8/zvlq/svl7xD5T934HH6vyuKO/BRTKuj57Lzogi05WNnKCpr5mBB1eGjiFcWmnPRdMe5VBUTY1olCV86OFaqtkViIQ5S78+Hr0q2vApWIgnsKcqtRt46clUjs6RohZEa4aOHxqnoga5VBg5RHCBA4NjGy71TbiW9pjWhjRUmxjxGGTm1D2qmBMUVJygjkOE1eNrhNaFZlxGjhquUY5zDTzAAAABQRAFAGgTAAAAABABoEwaVARWgAQFE1VMRNQAAAQaAATEaciaoCo0ICoCo0AAAJg0AAAqaomAK4N3Y5vorNfemLlQq0V+j6Fe/m6L1L2fK9Q+yPoviPauy+M9CXJ59VwOvgyuF9K/JuK+s+a8k5HzvueCzfE9XP287+hKn03hemQd3ynU0+b19/n+742zx76+rwdvMzd6qVmHfsYcaMr8f1fhNXg9Wn3S52eW+X9Xep9HiWYEauz1Z99ne98PydXzPo39Zvzcfn8XozdfXPmT9JLgcnZW5/y/W6Cku7oNXs5fW4ejnubPT5tb3TzPT+a7/m/qGtv5PlJOP7jr+c5dYOhfT0q3851WceVqa783oduflOU+e9ftvnXyL5n9I4nn6+AgReQyJ8LGVk5jmNK54Kenn087fy4CuNFBAHaS6dFcPScDlbLjbUuUJpJ1BaKpTXCRRoNPFbXDa5oJ0AVqJdLJCNbofT8/joOPuRcbCkiQKgNHCtQ5AVBijQJaOEHCBNypHUGnMa15UYZ2RstUeTUeSPTa1xHG1ElSrho9E0SaK0+aLq17zRHMaOGsrNo889L4VARwIoIAigqAAACDSiYAAA0gAjSiaoAADQAgDlmkAUGIJUGKJAAAAABQQBFAAAEVpAFQYCVAEAcComqJqNTdTh6thM/YXT7OPMXl6rlbdd103ueBy/pXTl9xeta+NX9l28HZt+D7GXndPDy/hns7/PPe+0rXkfp/A8Tm/NfrWNR831PaPRfq/hfROax1bU1dCl0+XpWCNTL6PAwvp7zPW8l2Ory5Fx6PNuAr6TL5Py/awvQOH24PLeTtm2ezxduDu8afpPL/AED03hvqfxbz7F8X7ne6J8HjZ4v1Fra7OAm6eaKhzd7dLPf6T8F9P57u6i+i3dfp4U18+q/p59OW/n9672nn+h47t+B+xxbcvc7/AF+Vobdefw/QnoU5Kz3OfrhwPKJ+h858q+b9zz/n9PzqKR+XZyNOGFs87YTOlS18qjFt50Q8BAEEALeW2jPj6hJNPVVHBUuUU0s7CqmuGBShQNOQYJq1t1nLprJpyy7/AE486c3Vd6L0+HmG+b6isllrXOqmTa0lwmrCXNARoUByGK01RIjhQEVwDQ+dmNeLkco1QrGNjIjSk1lJrhD0CjcqbUE2JqOKsWtrFUFG6s6Y+dqiFTQAGgBUGAhAUAAAAAAEaVAAABFaBMQFGldNNVNFBFAQAaAAAAVEwaABAVAUBFAGkAVABMGhQRUwTVAdoLfp05PqEn35q701C7vpfq+dtfpjfmfK3JfVfP8A037pv8ps9v4HdUysenzzyf6Lg+dvVuv2Kfi3rfnnkvHfAfuHU931P2X0L6X4/ocnNc3hbb8fanXf0uz5/oco56X1/wCV0/OW363ze3R5unH4w9vyl3m/dc0vNePg+f3Oiy29Ow/b+C6j0KvXyeY9f5Sh5/5fvu6fz/b4/NwvT6zfm5eCNMyDh9Nt4PQ+g+g+YzYuf2+3p9nlN6HD3I+m5fb2LmPRZ0ub28rkPJ+y9J03wcFPz/Wy7GWEMLVa11azJr53H+fYdPCec+RycXhr4+m88q7zmvDn0F5NNl8UEdcyBCAgBABQbOVo3cfWfLGj3K213LW1Nixns0BVLRRilI6ptVqtpmlQoCUcwu2UzPV0+jbct0m4aOBjZEQlVQxrzVQVBgxFUoNooka4FJaoAQqO0UxcyxqTEbHA0I2UmgmK5GjWRNEBrXKwPOJb51VWiExyOaQ+VAEGgAAUTAEGKAiiRRiAAhRoCAEUEGlE0GlQABwqRUAwBAmI0Kmg0AAComDQoNUEVpFTAEVpABQGgoDkTAABVT6DW5vogXUnfet4/G1+D1LX6AvPrvzx2wudF7HH6t9V/a/k3d91j4VG7yPjOV9XPz7o+rfzD548L7nmeF+S+21Oh1v07276n5tKvZnz8VcsGTLkzste2h6eD2rrfMvz1n0HzXeVo8XleWn6jhZ+L6Tir3D13uLx3fzvm+36Df8AoPg7mpXZ0vAd3zmhxPJ7GHreJ9bgx46VZDPtMru8Pjr3m+/SuK/SMP0vLw73N2+i9F6/y9/p+/kii8n9E9G4bm5vTOFz9DUya713fN+vreq8/NQ5jl+iyI8vfw2R61ubbyuF81z4uE5Dm+X5Shj5efQnyc6gvHrQ6clWG+EUhFBAEQBWhUxR3b2fo2n59Lnzboh1pLze+alSdR5oBSsJVWACo0qTSDkUTn1qizqqKnK6V0aQueT2rRjlDFJdGNq1kICoAo0cJEEiOFBojxVVoiOBDNAUaOWxOSKkjRCCGND0NRisFHBfNXdXLM5btcDBykidFXzOQbRyAxRIoIo0BACoNUQI0oAADQJgCKCDQJo5pBNVAExAAAAUEAVAAAaFTQaAAAQFQABUaAABFAVNUTUAUEAk6/H17jJ9uLe6vOyvXcNv0P8AhG+D5+v161q/04dJ6T9F8r0fZfWfC99qep8Fr8rXleR8V4H6jw2B8p+h4lfx/d2Nf0fL3NX0eGddSfuO/wAfFxEaeUtrpn0fc/zDjz9Kn0/zPQ34fH5K6aGLzd+VzHl+xB2PPo6Tv+f7flfR86v0PB9Nz51eBzy+d9Jx8/ndzXXk3Nla2as4Ou05MWB9XpXGa8lTs6dG3rX0F13tfm+XxHJ9h0uX5v6FpVMfV7LM6vmH9th04/o/H05UPn/V8ztcv2mPjvui1Kx8x4Pk7/OMzH5XmOIx8XnMzP5qnVrzIq2nJXSuNXJogAAgKgKgKBLpT16Bj6CSpyrOlZt5LKqlWdVQsBOHTZE2jSISoAAKNzzoaqJHx0Nmpkc3GrxRWkahAh0Y1QhFaEaUTQaAFjUjqzQBAhFSSNwjHLI6liUMFKoDUaVAax5wR3yMY8moQ6QpGgrymgkRpqkoAAkAVExWCCAAAAVAAABBoVNFaEAAVAAAAAUTc0YokBqiBWkUaAgAAABAFARpUAUEABoARQABU1FQIRWgGdDn29C3L6WvcWP6GXtvH/OrOz9Zz3rZu9M9NL2Zr6L7vyHeYf0/wvc+YxXm2L8T+m5DuD0Lti60l9DyLsmsSOubml0YVeg356tm88h3J6H6JfCMc290H0vj2Lm/JT5rDtWDOnZPJ15nSY6rzuvH6u/2/iOLv8H1eVhwug4Dk6MaPyPo1vZ9OK9ZSLXPczZvocnfN1jLt1bHZxe5+V+n8zq/Q/Rz+bbfS/SsLyvc6nfXP5+3f0neo8PQz0fzPR8tuYfaZvM8v2+NTy+gdjc+ebzOWvK+VYfPcby+nwGBmR80yK+WmXxwteLlGIAglBgCKIQbrS1058PUcKnTTcUblHNTvnZQtVGA2qpFGiNASoCOWjXAqmgqtwtXLNMKhdK3oc3rgic9vYMHCtGqADQCRRDSQeRe0UdMza7Oo43DGVMTXLRoaJXDbXeULq5q8Vc4OEAEZITTQSoxiuQBxNIMBANCpoADSKAgKAIAoCAqAqAqADFRANAAAqAPTETVAAABFAQBQQBQEAAAaAABAFGkABNUBUBUBVTEAGlVNJyuuk4/rPdv0B6fK/PvwqO5HToj3S2NjS0evnS30cs/QdnB3PDev4XGx/M/XR1IcspWta9HyY7jHyaRJubZ0dLSM9mWulu1P2h8WTl0Ot9B5OLsKr7NoxukfLmQc/XxkPm+rJavm12d/nxdA54q9zXl83y+hmUvL9aFcOqMTsw7ct2SjosTq4blbPqsaNrW2Nuib1P3Pjuzpd/znovO+d7vm1i/WX2Vedzelze/6pP5scC7m/Rus47n7Oa5fm+q4Wv4/wBNXwK8rmfIdPhuUxuf5nPYefDWvkhlrCqPFRMAAABFAAFCQNNC9l6TljVXA6NNWtPmm5Hz1MHKKIUpByiiRyEaSoWAKpQ40ck6K4bVBVdkbzCRUEjKhwmqjajkBCEK0AMx9JK14tdAkbUMNSDQCWIUUd81aOuRFJQAGlRMRpUAGgTBoGnkWgCgIAACsEQICgA0CaDQJqAg0ICgCgiAqAK0CYowBFQICgAAAI0qpojSg0BAAAIArQiYrSAAAqaK0CpUAchAFA6P9PI7s/8APqfXowx0W7/Xx4juXpc7V2NLatLU7uB/W+j4e1xXRyZXN/O/bXa/T5NSxh1bDe7zblvo5mzaY9JkV0dPW6eWhNnV36NzjmvHp7tep6Pn6cHTy6mCqbqkdLV7fn+fh4vYzcrk7+8zurg9J4Hfi4ff8/0/ZvmG+HO9V4vW8XZ5H0To6yJ7yhlVFhvUu9vG5FpNZXRe+hfZ+H0vO+rH13i95vvw9nn/AEDHp4nqp7+p6Cc93ief17fPc/0+p8+eT9Bz55nr1edzy4ngz43Dp4/PVa98lRb4VkLrteKqmgwBAmitAMkTmrF2NSOycz61EOUJYVZOJz52AaDAIcLUGKipEMhXQJgNXrV6Ts9CkCWOcIMRUkQBWkUGqIa0qCapmg4QUEVJkVZsRohEg5EFFBfNBHXIgSitIoDQVAUBFBFARoVNBoGnpFo4BAVAVAAABAFAABGhAcgAAgCtIqYNIAigAACuVIIAAAAEAVpABAAFAQAUEAVAFaQBRNBpUTBo9ADf8jVNBCp/aHjjnxFxX1jxk78RVz+ojZgWHdXPUXGh1D7Ouut0nocGdc6OOzS35asfN2Us7n2kuldLU7uHqMvp5LOHFXL7L3QdvNmOCv8Aof523wLb9F6jezjgr4dGTfytz9+XqMjr4OfzPD+pk0ejyu31fY+T7Pyvg9rzmv5/txZmcVK/H6EEeWm1g7YNtZ2xWSKVpW+jDRTq400p6tr1r2/jfL9bn93va3tfN+seabeT2uNyejyfoHl/YY2Dfpa/N+T9Z0Lu/wCes8lw+35aeJ7GDw+Xmc3kc3hZkJ59SO+JJBPDSBHiKmqAAIrAQAPmNIEILq1t2cvTc5UOQipuUHip7lsgAomuEDpBACVS1aUqjFVjlbhWrRsKwVGkQQhKqUgIBqg0DNAgRpUJhr1jEy4RwI0Go86te+FrXiACADQACaDSgINKgA0CYADTiLAABFaQGKAIAAAQFQFQFQBWkVNBoBoohABpUTBpxNK1jkQAAAAAIoIrQiagCNAAAICgCAoAjQAqAf0FE+5fzfhwpOitD93fhus/ggnRHj6bQ5PpxZ6WJeCLSUKHWuiqd2Oou2FquyCOWQGcNibl1ujmitaRHNpjet7Rdtalmz1cVH9DvO1/O5OvTUdtlnVufq0LOuaVU93Z9LxedwfJ+hp5nLrZ9e9Dw+G5rn9Ch6FfPx3JcW7IeP068kXDDNj1NpmubLIWHGyW7yv3ern7bnvQ85/U1Of0Pl/b9zyX0/5pNT5fUXA876G7eNd77I08D5owdfo8Th+PM5Lze3jeW4MObzc/nWJXOIBInI0uJHkjwRU0UEAABR2FW9YfOAXr+fpyOjoVUKoIAsqqRJtwU5hQPMVUICjb2rZVGIA5U5Z0RWLYneulYsKxRwNEINOagG1BICRRNQxbHWVSK8431nGwmKO+aFr5mjSOQNAVpFTEAGgBFEINQEGgAABNwNABQQEomigitAmigisEEqAigAAAAitIoCAAAAAqqlRAAqAjgQBFAGCCUTABGlQAAAEaVAUTRWhARwegf0GkfN/yMP8APz9E1X6H/Erj6U3muw/NpP8APl06zdRj6tl2HuFrbHX5vo5A5usWput2du899HP7PPqxZaST6c1QRLJStyb83o2B7Hh3ulvZbWyzIMq9A+l+K/jjJ7FrMtVIubstR64aHZ2odrq5OM8+8P6je5jp8HouZ8X7f0zy32/z6tr+Z9NxUeExVs9CxOlNkjnjlmvN7qi22qaTT79mxp+j52h2/bGT6r6vzlzhu3wsCv5H17+9gtZenbp+w+h83758BfI/R8Jws+xgZXnb87hYePSYcICQEOHNEW1pCqIEwAAUAnLfWKaGQoOurouyZekrk3IgAWUcizY0HIhyDVAUY4WqhogAolctRVQpRPOsbKhrK53IAAIICUUGhCoCI8mRuII7iOCsWRVzQtrnYjhRAjAQDQAAAAAIANKgKgKgA0AADyLABrAEomAAwQFBIJqMEECYNCAoCKAAICoANCgpNCggCKAAAI4BAVAQBUBUAGgTBoAEAGlRMGgD9Cf0AI8Y7EPx2/o9H/OV5vOn7qfLFZeC/tMH8z5Oq2lr0drl+mdGao4zarAqUex9itr8fXhVml9xyXo+JSTj9Gz0/ZwYVpPb3fT8vYOpSWCo65P2V4vz5+c4fYT1+bupJhs/a6eanFj160fV5uJl+V72f2O/k81sXhu+V8+vWcFh1UqvB6qLNQxxcaRmszsm1zllNJWMc4a2am5r9nHq9j6/zvpns3s/I8X1vVz8d47we7Izx/0Onk7+Ne0e7xq/3B4Xb85fGHn+vxlHgjNrTxNVwgIRocOeBWIQoMECgIAoTvN465mqkIoDh3bEehM6OhzU3A3KD3yyRWqCRAJCgAVU5HuKgUY5WCpyDEB7loxryaXi+NoSZRXCCYAjXmMJqR6ZNqVjXbpxMjeIolRNWgqAoAAg0IAAKCKAAACAK0gAoIA4mgAABiohFAQFABoQagCCAAAAAEUEUAAQFRoAVZoUEARWgTAEVoRNUaUTQBUaAAAARQAEVpFBAAD91ebM/KP0pD+ef+iIf8/HkC0/bf53efiP6vh/N4k6gF/pMPbsRY+sivMGBUjqUr985q7pWZauW5kl6OZ2/wBPJV198NDf9Lze25r0OKuzy/oE0NuX9CPz/wAeYwejeOTm6a9aG7esizbymwsNYnYb0JIbuv7vH4zH833NDEyKPRTWFFnrTh58mEQ6R2+yd8rToc3TKQuJJdMLez082t0HseH3/R+l47va/a+Iq0e75HleY4f0HK8y8X7Dn+j8/o4y95H0/wBRdhxeL+WFLHFgoQYgIGkcOeCdFZWaqgE0UAYlkpVWtZpgDQo0BSTraeSOuwR0uRtXySOTmZNSNGjWiYBVSKm4NRTQcMWdVaDwaAh5TGkRrfNZr6ZtTMa4QgIhLIaiOOphq3zV49OFqPMRpzpoRMAAABRtoSAAAAitCAqAomg0AACKD0mgAAAEAUBAFAEAAAAAABoAAABFAAEaVAAFWaFBBgIAEUABBoE1AQaVE1ARWkEwaAEUEVpACwH7q/jsT5b+4Sr8yvokX3t8Uue06gPb/wAuB/IgtFQHakdHRu5PqWqSisa6olt9mDGXlTOToYrwePYntWE6ItX+nk0/SPb+drxa60pOL1NCrrH1L8vmMDmZuvGnPN5uqeBX6z5x6PzuEeH9hVvXlkVs5RJe7xWvNVb53p6Gh08vMw8RCyMGKD5ByPN7JPohakfTe/bKTR257Wj3+V6JudWtH076P4T0qH2vhOO4zi+n6LO8z6X5z5/5b6jDocN5XtmHH+gv5RLk4tFQgKjQAiharLUa8lUAGioHDleakYoFeKKmg0omAOkLdOt55s+xwrkUqV8UjgawTng9FQ4HC1Fdqq0HK1RUrikaJrnAwmFtcTZHMbGQyuXLLZFShrXhFXvkiSuZBoARWldNDU1QAaBMAAAaRQRQAEUEUBABoAAAByqmqJABQAaAlQFQFQFQAAUEUEUaAgBFBFaRAUTc1oAVU1EwBAAAGCoQaBNUBWgqNACiYjQAigigAINK0FcCk0isQBUAEKAIArNTo+b6B7cvTSS8mTa53F7eWvS5deoxu7z8xfL7Fa092rc+6mm6Mpuk7fOs2Nq26uyt1o6OgwunjlqYa6eMyuvL1tBX4+jkzsrh9Clu8nqZNTXzrWBy+lKk40hZNgjGJi5okEiA6YHSFvebOVdAtp82yXStzRdPn9FH1Ydt0/0XyUXqfo+Hy3m3j/c4WZyc3KZPj+lh5nAqrVz9f+rxn+fHzuUIAAACNW687I2sXCoVCqVI80Rw1qksHiACgICiaKAoSWl02Js+0lWiqm5ETIm5oOaN6BI6dRXSKm5VuqqxRogoyudI3lBDeCRuK0F8bkpSE3HFWUA+ZG1IgKADQAqzQiaoCgACACgitIAI0omI0KAgAAAAIoPJpFBAEUEGlEwAQAYACgggGAIExGlQBQEBQBGlRNwComqAAIoAAgCggDmgAKMRUACDQJg0ICgCNCporSk0AACoAAqAAABprfopuP6lLO6qooUrO9naRPVjJTOxqiSbchRJz7ck+2d7b6smbWr28DUZcw644az0auuNN2G1d4EFYx2ovnrXP05leJihjO5r3jn4HO4W5DATUaRQHBJIOVyt4tVWqlNRZtJdUrLVj6uPc0ezk7DiO3wsznPK9vFpcLayCqyIQaSwHvfz4IEwaQAAleta5WKuVOVUA3NGwcOYhByAAAJqjQDFAVElhbyST1Tyxu9FaPAchXpqwHSqlFQpTlN1RUqDGPFEeUUNZNr1FWlp57HVhIhaNJRpKI5BoRpRNBpWg5ZpUTVABpUAVNBpUAQAAUAaEAARQFTQaBp5FgCKAgAxRAgACoAArQVGlE0GlE0UEAQqQAVNFaETcqYAgxUQoDVYCBQGgoCAOaCgACK0ig1QRQQaAFQBQck0DQqaAIoAxUQjmIqH9Fn36Duf3kQSFQiPJEM3v0bBDVUIEitqaHZS3XMbt9HbYmk97lhwT62fb2581uV03xT4byvc9WMJy9LZL5qDsJdRz0iUxGJsRoQSKAAso5Hy5XDcrtzr1SbbGvjRgtRc1pdIgrPGs2IQSAEViKhFBqsRRIgA0CaylIwHKNUTVQY0hBpRMRpUARiiAAABgIAFAdMtbtjL0HItVeNZJaAPVORVQ9UBQpSAETiMeSQtR52nNUbp5qPbeipYhIjhQABBoAABGnOmmiagINCgiggAJg0AKJoVIgKgKgKJoNAEiK0VJAFQBQQAUAAAEBUAUEAQBUBQEGgGiiEAVpBNXDBCAACowEINAComoAAg0jgQAAAFQAAQAaVExWnNmgBVAQEUAAAEaVE1ePT3sPblTH1GpWSJXPBMZoNKqoRgomhIOf0F+19/M/Cn9GZ9P6N8f04PszDOTxL8kX7szteqGOaqOISFS2CeaIyhzAWCTMR4PetSVNqA0BEAUBQe9OVRySjc27q0q5G1jKGJ8w0kaSqMBJUGIIRg5CAIAI0DTkTEac1NVAVMUbUEKAgA0CaK0AIqYNIoA0ipgAASKrk0d0ro6AKlE7L4uGRpzU5ELcCRgiNqOKoSGoz62vlorzABRujAHIADQgAAoIArpoRNWgqg1WgAABMGkAkaUgSgCoAoINAAA9JpUAGKIRQQAAAFAGgqNCgCYjQqaKwRCK0AIoCNKgOWaBiKhFARggCiRUwYCBABoUEQFAVAEaUTQaFBAFE1BghQEAABGlBghFAVhKq1dLD2p35d6McV7FczWiHIBtQoRSI16J/Qzv5n86v7Rro+CvaKnX+G30fuP/OQZytPZVryWN5RQZ88EKxc8c0pVfn1zrLeMjFcauB6cjlsKMYJzilcrc6aatTAjxjhrJkbxRhAhIjSgAAIQYAk/VYXzp8agn7xNfi5x6r94vwtc8+qoAEBVGjgRUINAmDQgKgCgIAAAKgKgCAoAoNUFeqlsLpe6Oh8q1fNOjlltVokVK0cLagYJsTipT185qViAhAUAAQaUTQaEaUBBNQFcmiACYrSKNAQJqNIAqA9ioHIgKAI0AA0APSLFAQFQBQQAARQEBUAAUBABQEAAAAaRQAEUABRNQBARQUBAABFAQFQBQQaBMQFGgTEaFARoEwaFTUTAAAAQAGhU0AFAAFAkL1dLD1rEcdrJ3mjUgJAAABiOV/eDXi+HP1L05P52f0ly9H7uyt/E+APzjx+hFXSi1CMWVekYVI75WNeSgQMMR4CAKNQJHmsbSCUpXrR751a1qOKsBRKwIx5IJCCQYKgQFaAMUQn7kOPkX88Rn9QTn+c/gVf9Qf8yLz5YWiK0AKTQAqAigACAoAoNGAgGlamo0AAAAigACArxvlW0skdMks9EoreiaxsVBIgMrVhHXvkia+cGkUEABpRNBpUAAAAAGhAV80gmgAAi1IJgAADSKmACAKCK0gAADQA4iwAAEBUByAigg0qDBANACKAAAAAAitAmDSAADiaUBWgKCAAAoIjBRCJqgKgKgKAg0AACK0ImqADQqYqaK0gCoAAAKAipoNKJooAwlVXLOffJoR2TGfYiPJEWajABEJd+jd4foB4Pv5P4vfqLj6ux+UjgSPQYlZkJMFesKEGnnta8gEIhQEAFBXliA2wNsAgIVSnBSKJXlqg3CpoSDI2mTWuRQABBCgA0/chx8ifniC/1BC4yu49Q/mRK5ZFaKAAqzQjFEAxASomKAAAA0gAAAIoINKs0g0IANKgKJueXKs7SyTvIi0GtDCWNeTYqyjR4oCVARQABAFaQBUAARWgAEAxXTSCYAg0qJigg0Kmg0DQJg0igIAAqAAAD0mkUEAVAFAAEaAAAAAEAUAAAAAARWkGhU0AVAUTFGqIQAUEAFBBoAFARMAEaUAAAAARoAVAABU1RNQBAc0FAEBUBFaUGghFaAAAUck8727OfbctZeiNVoPERBG83erNfsX+bG3m/JH35HZL+d1ZRwq4WXgoIhcVkfKACMFQI0jgVqaDQoLOWkSECQBZzSZFujXAoNaSr1SMaa0zRpICAEUEAVAX9x3HyJ+eQz+oJz+A3Dlf0jfzIkcshoiiAHE0gANKgKgAmKAAgANKiYAigDQAICjEEKAg0AKJuE1BqAiAIJ6DewbQkAQBUARpQABAABUaRWgTEaUBVmkVNBoAEBQAAE0WpRU0GkUAAGgTBoAEB6TYokUaIJVBAAAARQAABFBFAQFAQAGhQQAAAAByqkEIrFENGlQFVNABGgBUTVGlRMSpBNQBABoAAFRoVNxNIgKjSiYAIAADQAAAmDQJg0AK5U+dbXruXpTNjobXeLEvOWzGsjlSV3NeKs2V6xYt4o4S1SYkfMAAAAI0qAqJg0qAsytsYkHL3K3BaoEaPMVCAkVgIQYCFa0qJg0ACjT9yHHvPCPP8q/3RVfzn8AX/UH/ADIkcqFgAAqpoqaDQAoAAgmqNAAoIAgCoAMFQI0AACiaFSAAm9JoUEARQFBEYKgGkUEAEBUBUaFTQaAABFARpRMAHJjQUBFARpQATQaAAAAAAaBMGgTBpFBSLFaQAAFTEAGgAEwAGgAAAABABWkAFBAAAQHk0KCAAAAjga5jVAECAAKgKjQAICo0AKAgAAACg5JpBpUTBoVMRpUGAgAAEBQBAUTBpFAAJBuVaOaqdMXbt5d1t2fQkLVWrpzwxacjZXLGiY8msGKDBUACDSoADRRAA8bUErk3BbQlyDaEooIoIoIMVEAwBACKCAAB6OHQAvO/QWeUVA9l8eFAoxAFTFBAFE0GgGAhQQAAABGlE0UBGgAAAEUBGlAABU3JNCgIAAigrQcgINACKCDSgIAKmi1KCaK0CYNACCaqAJiAoAAACggAAICgIrSAAAADQJg04i0UBGAIAEBQAAARQEaVEwaAATBoAAAAGhE1AVU1E0ARwIjFQAECYoCNKgwQqNIoCAAKgCgAIrSACg5JpUAAABAUAAAFQEUEGhU0GgTBpWgoAAigigsy0szR2OctGROImVgxrxe80eiuNjyjQyEBUaVEwaEAUEAAFRoAes2qjYOVE0BNVoEwAAAAYioQAVpAABAFQBWhAVAVGgTBpSaAFQBAUAQFAEAABpRNBpUAAABAVQQBUBFAARwKs0gAAAIoCgiNAAJg0AACKANIoIArQVGlAABAcs0rU1QFQAaUGgIAEAVpBMGgBUTBoABoRNwmigomg0AIoAAAAAAgKAI0omIAADSiaDQAAA04mgTFBAFQEcCIAoCAqA5oKgAwQSgIrQJorSACggAoKTSoAAqAgAoAAAAIoCAqADQgKAigIArSAAAA500INQEBDhzBsrRNiIAhABABgglAVEwaAFQBwORWoDHCVjAEKhAFARQQAAAEGkUABFaQTUaQAAAAAGgAdNCDAQAAAAAACggAAigg0omitIjSiYNCAAAA5NxNIoIAAIAK0gCKAAICoArSKCAKAgANAmAA0igrppqpoNCNCpooAAAitIAAKgAAADQJgAA4mgBBoAVARQAFRMUEGlQAAAAATEaFaQAAAAAAVU1RMQBQRQEBUAAVGlExoKNAAAICjQKhBA0oCDQs04TABAVAAABQEAAEUBAAAaBMGgTQaVABoAFBVVCoAaDRIqmaapXeaooABGlRNUaRU0GlAAAAQBwOVU0EOGiA1wlQBAAFaCoAoCAAKgA0AAAgKgKNINAmAAA4EQFEwaBNABQEaAFQBQQAAAAARoUEAVAFBABU3E0I0Imo0AIoCAoAgCtCJorQAAK0FGgTAEGgBFBVTRUwBFBFBFaBNAFARQQaAFQFQAAAAABVTQAVNBoQFAABAUAAEaVAABQQAAFAQFaDkAAFAWaVAVABoQYohAAFQABWgqCFYIAAoCACggCiYjSiaiagIoIMEQ5ABoQABUAAQaUTAAAAEaFABgIAFVOahQEAWwaDRtheaiQNIoCAigICo0qAACgACpqDQEAxBCgACAoCKCACgICKAoIAigitIAigKAJg0AAComAAAAACNKAgAAAA0CaoAADQJgCAnIMAEBVBUmhWkAFGIJUAAAAAGgTQaFBqtAmqADQJo4EQABypiJqgKjQoIAAAAAAA0gCoAAqAAgYCaiaoAgKgKJg0igACAoAgKAitCAKmg0ACggAAqAAACoComDQAqAAAAgKAAINIoK0FUEARwI5MEwTBpFBAFAAAAQFQFQFQFAEBWtKAAACOBBoQHLNAm5oAAAqAACgAIAigAIoCgiAKCI0KAAg0AKJipoNAAo2qJFAAAFQAAQBWgTAAaQAAAEVpFTQBVTUTQAGgTFBBoAVEwAGgARgqEGhQRQQAGhAAFE3E0jgaoAAjQJqoIoIAgAjQ4BEwAABpFBBoAFTQaVUxU0QAYCABQEGhU0GKgAACABQQBFaQAAEBQAD9TAhQYAACIFYIIAVBgIAFGgIABgIAAYCAAAAAYCAAAAAAABgAAAAgKCEGoIBgAACAAAAAAAAAAGAAAACIUYICgCCVBqglQagDk0AABUIADBAVUCADBUIMBCKwEIrBAFBAAEqAAKDEBUAUEUAAQFQSg0UBBKgwEAAAADAAEAAAAKDAABASg0BACoMBKDQEAAAAAKgwEAAwABCgwAAUQgwABAMBCDSgwAAEEAwABAAAAACgwQDAAVDVYCAYICgICBNUYKAgCgIACAAAABn6mBAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAFblw5jmg5vngwMUMTIDLywo1gAAAARQ09ANXYDf3g6Xpg6vrg7G0AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAc35+Hn/AAQcJwgZCAOBALPUB1HSh0nQhv7QbWyGpqBesgAAAAAZeeGVjhz+GHMcyHKcgHHVQVgHSB759JB34AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfhkGf+7oAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABD54HmXmQeacGEAD2h0voAegd6Hdd4GwAAAAAAAAAAAAAAAAAAAAAAAAAAABzIfP/zWHnmgH1V9ZBogAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfy+hk/wBTAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAFPzEPKPJw8tyQALHoIem+nB6V3wSgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAGYHyl8kAv2kH1GoAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB/L6GT/UwHLawaYAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAcsHkPj4eRckDXBc9OD1f1gPUtYAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADL+Kg+WPdg+/9sAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAD+X0MX7ID4p2g/YH9DwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAKfh4eG+OBzwCdUHr/rwevdcAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB4CH5+elh+ivQgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB/L6HL/fQe5flKFf+lUPSwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADj/lwPm7LC97iHufsodCAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHjYfm/7UH6IWgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAD+X0MH+pQLn4lB8FfuCH3SAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAGH8sh8u4wdj9SB9JawAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfNwfnf+gIfUAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB/L6GL/AFNgv4vh+ev7ih9ygAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAJ4SHx95uG79Th9R7IAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABF+doeV/qcG6AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB/L6HL/rIHuX4qhzv9LAekAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABjB8l/LQUvdQ+wvSAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADyj8sw/R/6EAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAD+XEPXPNA5KwH7AfpEAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABwHxkHhG2H1n9TBfAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAKwflx6cH6BAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB4uHS6gfPPtgekgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAeQh8T+Yh6B9nB7sAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfEAeAfqyEoAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAeI/EYcL6+H2v6cAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHzd8DB+vl4AAAAAAAAAAAAAAAAAAAAAAAIgz9QFAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADxEPiLhQ9y+1g70AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPDQ/OT9fwvAAAAAAAAAAAAAAAAAAAgfP/AMlB5/KCzhpZQMaEfYh9M/TAaYAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHkPw0Hnnt4fb/cAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAeH/nGH7CzgAAAAAAAAAAAAAAAAcEH57efh9DfXYeigAAAAHGh8wfMYZP0oH2ptgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAcAHwt48Hr/3MHogAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHzkHxZ+sAAAAAAAAAAAAAAAAcaH5nzB+iPpYAAAAAAAAADfmgPiWcP0C9qAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAwvicPmzvw+6PYQAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAD4j5gP0DAAAAAAAAAAAAAAPgb54D9PfQwAAAAAAAAAAAAOYD8+vIA+/8A6MAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAIw+VfjQLv2wH0moAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAH5Vh9se+AAAAAAAAAAAABk/kqH1t9lAAAAAAAAAAAAAAABnB+ffhIfpD7UAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHlQfBXAh9VfZoaIAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAVA/GX9kQ1AAAAAAAAAAA5oPyD/UcPWgAAAAAAAAAAAAAAAADFD82eaD9O+qAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADM+KA+XvVA+9/QAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPE/h4P1NAAAAAAAAAAMT8eQ/WTvQAAAAAAAAAAAAAAAAAAA82D8wPpUPvJQAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPFvgcMX7YD6iUAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPy1D7l9lAAAAAAAAAYH46/p6HqAAAAAAAAAAAAAAAAAAAAAB8eB8U/qSHqAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAUQ+J/lYPafvkOtAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAMQPyP/YsAAAAAAAAD80g+j/p4AAAAAAAAAAAAAAAAAAAAAAA5z8sQ9t/QMFAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA87D89+ID7Z+qQUAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPzq+iw+igAAAAAAA8F+Nw/UIAAAAAAAAAAAAAAAAAAAAAAAAD4/+LQ/VztgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADwIPz/64P0K9FAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAzfyMD9hQAAAAAAiD8Y/2UC6AAAAAAAAAAAAAAAAAAAAAAAAAcUH5Q/bgfW4AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHzIHwX7oH6BbIAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfl8H3j6gAAAAAAfAYen/VoAAAAAAAAAAAAAAAAAAAAAAAAAAAn52h5x+pYXwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAD5pD4A+lA+9bgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHngfn1+pAAAAAAGd+PwfskAAAAAAAAAAAAAAAAAAAAAAAAAAAAB4b8bB+nYAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHhIfnP8AQYfoBbAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPx6/YUAAAAAD89feQ+kQAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADzkPzN9JD9GtUAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPzw+mw90AAAAaH42/soAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABjh+avOh+mnYAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAebfCQfpsAAAB8oBxf3GAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAACB8R/I4fpX7UAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAH5CB+vYAAAH5AB+uF8AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA8b/ADKD7F+1AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPyHD9d1AAAMH8sA/W0AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAzPzMCr+m4bIAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB+a4fbfpoAAB8NehB9RgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHy2HwF+mIe4AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB8ign12AAAfkB+vITAAAAAAAAAAAAAAAAAAAAAAHmHwUH2d7sB8Yh5CB9n+ugAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABzQflr6yH6IygAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB5AHyZ+iQABEH5D/r6AAAAAAAAAAAAAAAAAAAHAB5Z9JAAfiJ+yAfjF+4IT8mGX+Vwfqj6AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfInxCH6memgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAZAfl9+rAAB88h4Z98gAABGHBgAAAAAAAAAAB12kAAIHyD87h9i+sh+Sv7Jh+Tn6FB6yBwf5jB+uoAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABzn5Wh7p+gAKAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfkj+twAB+Z4fcnpgAAAZgfnMAAAAAAAAAAAH2x6oAAAHEh+P36qh+Zf7LB+SX6SB6SB+T/wChYeqgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHyuHwV+qAemgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB+SX62gAH5Bh+vgAAAAAAAAAAAAAAAAAAAGCH529EH3tsB+J36ah+X37Dh8x/UIflD+x4AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAGaH5b94H6NvAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAD8kf1uAAPyGD9eQAAAAAAAAAAAAAAAAAAAOdB++ABxvyCH1ruh4r34TehgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB4f8AmQH6de2gAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfkl+toAHCfn6H6egAAAAAAAAAAAAAAAAAAAAAAAeaB03SgAfNsYez9eAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAn52B51+pQXwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAD8kv1tAA+R4A+vwAAAAAAAAAAAAAAAAAAAAPLvUQAPF/yPD9LfqsDmg/Hv7TD6T9DAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA5APyd+2Q+uAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA/JH9bgAPzV+3w9LAAAAA+JuXAAAAAAAAAAAPefogAAA+EfLQ+7/UQPz+D1b6rA+fQ/MD77D7GAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA+Zfz8D9Yu7AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADng/NL9TgAPyN/XAHAAAAAfze+LgAAAAAAAAAAH6ffrYAAAFQPyg+rw+u/z+D1b6rAqBl/mEH039ggAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADPzTCD9MQlAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA8bD5X/QwAD8jv1xAAAAAA8TywAAAAAAAAAADvPRAAAD58+PA+i/rsD8/g9W+qw+efaQ3PzgD137DAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAOSD8lvvEPqUAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA+RwZ9dgAfkd+uIAAAAAAAAAAAAAAAAAAAAAHyP9bgoB87h1vrYfmv8ASgfE/tYfoAoAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHz1+cYfrL3gAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB+cofYPrIDQ/JT9bQAAAAAAAAAAAAAAAAAAAAAAAAMINtwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAH5+h5b+pgWAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA/I4P1slA4oPz8/TgAAAAAAAAAAAAAAAAAAAAACpbAA+PPnIPBP3LAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADPD8oPpYPtkAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAD8hg/XkA8MD55++QAAAAAPgLkAAAAAAAAAAAD6N+nwAAD8y+7D7utgnHB+Ov7eAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHn/AOTIfpx7cAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHOfmQH6tAHymGX9gAAAAAAfze+LgAAAAAAAAAAH6ffrYAAAHlwfAX1wH0F+Xgcf+wITAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHzqH54frgHUgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfEvaB9TAHxIHpf0gAAAAAB+QnnAAAAAAAAAAAB9p/d4AABz3wSEP3wHSgfkJ+lIepAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfC3iIfqhOAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB+Qf66BYAPz6D6f9mAAAAAAAAAAAAAAAAAAAAAA/OT7rDsAPzw8hDp/1GAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABv5eh6R9+AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHMB+YH6zAAH5tB9v+jAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAGYH5G/cYfUQAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfm+H1d7kAAfmAH6LdSAAAAAAAAAAAAAAAAAAAAAAAAEf5whx36pgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHJfkQH6geyAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAFf8dg/ZEAAPypD9QdUAAAAAD8YPMgAAAAAAAAAAD7m/QYAAA/Mb7RD2gD82/WA+rd8AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA85D8nf1vDuQAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA+BPWw+oAAA/KAP1VtAAAAAAfze+LgAAAAAAAAAAH6ffrYAAAQ/EYeW/dIfi59mB8u/q0HtQAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHl/wCUwfsP0QAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAZQfkD+yYAAAfkmH62AAAAAAH83vi4AAAAAAAAAAB+n362AAAB80h8qfaIflr+3QfIfm4foIAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB5eH5X/sKG2AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB+YAfZ3uIAAAfkmH62AAAAAAHwpygAAAAAAAAAAB9BfSgAAB+Yf0WH1iB8I+Ch51+tgeggAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB51+UQfsJvAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB438Gh+qwAAAH5MB+sUoAAAAAAAAAAAAAAAAAAAAAAAAGUF6cAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAOLD8jP2CDpQAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAK4fjJ+xYbAAAAH5UB+oWqAAAAABzcAAAAAAAAAAABqa4AAAAAGB+cQfa3swfnp4iH2p9SgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABzv49h+uvYgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfl4H1z9FAAAAAflQH6haoAAAAAH83vi4AAAAAAAAAAB+n362AAAH51/d4dKB5t8BB9N/VYcd1Qfhr+6IAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAZwfjj+rIejAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfJHiwfpAAAAAAfliH6X74AAAAAH83vi4AAAAAAAAAAB+n362AAAHKfAoaX3uG5+fwerfVYHKB+R/7NgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAEf49B+kfswAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHkIfm1+wQKAAAAAfl8H6LdWAAAAAB/N74uAAAAAAAAAAAfp9+tgAAAeeB+eH1mH0/+fwerfVYZX47h+rXoYAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB+UX2UH0mAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABx35Eh+yWqAAAAAAfmaH3v3YAAAAAH83vi4AAAAAAAAAAB+n362AAAH5qdOH3nogfn8Hq31MH4MfTYeofoCAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfnIHpP2kAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABgh+O365B2oAAAAAAfm4H216SAAAAABxNUAAAAAAAAAAA3uiAAAM7RAAOXC7sB4uAe1gAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfGflofo8AAAAAAAAAAAAAAAAAAAAAAAAAAABkB8Tb4faKB5L62AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAGP8AjsH6p+nAAAAAAAB+d4fWnrgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB4EHwj+tYAAAAAAAAAAAAAAAAAAAAAAAAAAAAH45h98bAe+fnoHxJ+8wAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAY4fjn+pIetgAAAAAAAfAgfRPuQAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABn3QeAAABmBpgAAAAAAAAAAAAAAAAAAAAAAAAAABwX5Oh+ytoAAAAAAAAAAAAAAAAAAAAAAAAAAAAD8pw7j9CQ1wPxT/awAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA5n8gA/VH1QAAAAAAAAD4cD1r6JAAAAAAAAAAAAAAAAAAAAA8eD+fP+iAPTQAAAAAAAAD48D0v3gAAAOGD8Cf6Fg/IHgw/Xv0gPyw/V8D8Iw/UH6bAAAAAAAAAAAAAAAAAAAAAAAAAAM4Pxq/XsOrAAAAAAAAAAAAAAAAAAAAAAAAAAAAAED5J/NQP3L1w/FP9rAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADzgPyc/WkPQQAAAAAAAAD40DqfqIAAAAAAAAAAAAAAAAAAAADxQP5wP6VA8W9AD5o+pA9y+Iw9i+Xg+tfZg+KvfQn+cw99/nqD7h/VIOO+Ug+kfo8D82Q86+lg+JfpsPG/Sg+iPqYD4SD5D/AGpAAAAAAAAAAAAAAAAAAAAAAAAAAPyJD9DPagAAAAAAAAAAAAAAAAAAAAAAAAAAAAADmA6f8ag/VL0MPxT/AGsAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAyw4big5Hlg57nwys0KlQH6YW9YNzpg63sw73ugnAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPmb4DD9c+iAAAAAAAAAAPlAKP16AAAAAAAAAAAAAAAAAAAAB4oH84H9Kgfh9wYeneGB/Tb/ADOh3PqAfMf9Kofze/rYGt+NAfu7+DQfRf6yh+BH6SB9Lfa4H4pB9degh+Xf0OEvBB9L44fo34yH4hf0YAAAAAAAAAAAAAAAAAAAAAAAAAAfnd6MH2WAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAH5/h4j9HB9wAfB33iAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAGUHiPigePc8CKBqh3Pah1/VB0O+GvpBauAzMCpkhg80HI8YHA8IFcBvYB7l74HqigAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA383Qxv05CQAAAAAAAAAAPnEPLvt8AAAAAAAAAAAAAAAAAAAADxQP5wP6VA/D768D7V/nbD+mv+dEP1l+9A/lp/fcPxF/WwNb8aA/qC/mnD9V/u0P53MsP18+zwPwuD75+1w+BeMDyztw8L2Q+l/rYPwG/pRAAAAAAAAAAAAAAAAAAAAAAAAA+aQ+Uf1EAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAhmVA5AAAAAAAAAAAAAAAAAAAAAAAAAAAATxYPlzxcG64e3e3B7D0YAAAAAAAAAAAAAAAAABxweG+BB5SwOu+pQ+ldEAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADiA/KD7qD6sAAAAAAAAAAAA8PD5x+/gAAAAAAAAAAAAAAAAAAAAPFA/nA/pUD8PvsAPtf+dUP6av50g+0frEPxB/pND8EfqoNP4KD+oL+bgPsb9TA8s/OwPnD+ioD8cA98/RMDx8Pzu9UDzDZDqPpcPyH/AKDAAAAAAAAAAAAAAAAAAAAAAAAA4f8AJwP2WlAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADmvh7H7Xw79cPB/Q5z6T8HAAAAAAAAAAAAAAAAAAAAAAAAADkw+QPnII/aw+pPageAAAAAAAAAAAAAAAAAAAAAAAABx/yyHzNmh7h9pB34AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB8dfGgfq72YAAAAAAAAAAAAHmgfFH6RAAAAAAAAAAAAAAAAAAAAAeZB+MH7Xh+Xf0WH01+N4ft9/OAHt2KH6C/ooHwj+aQfV3i4fur+dwfnj+sAfmBQD9OvtMD4iD4s/bAD8xQ/QmwH43yh+xP57BzP6tAAAAAAAAAAAAAAAAAAAAAAABCH4z/raHYgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAeP8Axl6vo/Pf6Pfmn9S/SZ9t/JIAAAAAAAAAAAAAAAAAAAAAAAHEfEYeH9YH2F9HhMAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAeG/FYefe2B9zdqAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAc1+V4e6/f4KAAAAAAAAAAAAAc0H5ufqQAAAAAAAAAAAAAAAAAAAAAAAAAAfzJB+tn6AgAAAAAAAAAFQPx1/YcLwAAAH5JB+l3fgAAAAAAAAAAAAAAAAAAAAAAB+XwfV/wBJAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAeV+XfR+F8Odd+L/wCgv6sr+i/wuAAAAAAAAAAAAAAAAAAAAAGSHxV8xh2/3CHt4AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAeJh8L8aH1n9kBbAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA+Pg+Jv1OD0wAAAAAAAAAAAAAAIw/Jn9agAAAAAAAAAAAAAAAAAAAAAAAAAD4WD6J9gAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA+WPnwP0qAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPLfNvpPnuW+EvyD+5v0o+ifrv5lDt+UAAAAAAAAAAAAAAAAAAA8JD4LpB9r/TwOAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAIg+SvjYO1/QcPRwAAAAAAAAAAAAAAAAAAAAAAAAAAAADhA/Lj6ID75UAAAAAAAAAAAAAAAAPyR/W4AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADjQ/Jj9lgeAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAedeOfSfPfS/5G/mv9Qaf6zXz2j6P8IAAAAAAAAAAAAAAAAAKnxOHyj7uH3t0gAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHnf58BxH3oH0cAAAAAAAAAAAAAAAAAAAAAAAAAAEAfnj40H6i9qAAAAAAAAAAAAAAAAAB+SH63gAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAn43h+rHcgAAAAAAAAAAAAAAAAfk+H0X9qh4t+T4fuT8xB8NfsEAAAAAAAAAAAAHF/PX0nz31z8xfGfqX51/b3yX9M/aZ95/GoAAAAAAAAAAAAAAAHM/nqHkn22H1eoAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB5IHnnvgX/ggPnT7YD68UAAAAAAAAAAAAAAAAAAAAAAAAPlz8/w/Qj6bAAAAAAAAAAAAAAAAAAAPyR/W4AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPz49GD7BAAAAAAAAAAAAAAAAAPx/DO/ZEPyt8dD9tPKA/Kn9rQAAAAAAAAAAAA535V+k+d+zE+b+i/MDxr86/uj9V/Tvvv4vDfxwAAAAAAAAAAAADgvzmDC/QsPaAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA/JrtQ9h9lD2oCL4nD4/+0g+ylAAAAAAAAAAAAAAAAAAAAAA8Y/NsPoP9AgmAAAAAAAAAAAAAAAAAAAD8kf1uAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADyMPzw/WsAAAAAAAAAAAAAAAAAA/HANP9SQ/KzID9nQPxT/awAAAAAAAAAAAAK3xH9J879ynzf0XE/k78p/SPX/qv0eLpH0f4QAAAAAAAAAAAB5r+cQWv0aD0MAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADxYPgf9YQAPzp4sOs/R4GfFgfHH3aH1gAAAAAAAAAAAAAAAAAAAHk/5wh3X6PhugAAAAAAAAAAAAAAAAAAAAfkj+twAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABF+MIfsRtgAAAAAAAAAAAAAAAAAH40h93/CAfXfxyH7PuD8U/2sDw7wr5/9sDk+kRCp+128r0X072fyrZOz5cAAAA+EPpPnfu8+b+iPmrh+v/OT6X+K/rH9GF/Q/wCHwAAAAAAAAAA88/NcNP8ASIO2AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPkoPJf0PA5YPws+5g+Cv6CwAIfhAPmD9Hg92AAAAAAAAAAAAAAAAA+efgcO7/RgOrAAAAAAAAAAAAAAAAA47gQb6cHRgAH5IfreAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfm39FB9NAAAAAAAAAAAAAAAAAAB+MQfsx/OyH9DX5HB+rvSh+Kf7WAAAABBxGHreceY+N+rYpx/U+i+4ev+Ydwer+cgB8IfSfO/d5839EAfBfyF8B/an279qfV/zcHqfngAAAAAAAHNfmkFH9Kg7YAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAOQD8X/ANwwucsH5C/rwDupAAAMz83Q4f8ATsOyAAAAAAAAAAAAAPIg+MvHg9++9Q2AAAAAAAAAAAAAAAAh+Og+Tnh3HoYO8YDuP0rAA/Lb9Ig6cAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA8eD4B/V8AAAAAAAAAAAAAAAAAAAMQNuqFrIDTkDE2wAAAAAAAaHnHh/j/qPDXPL/Q/b/bfoPxO18H/AGP5r94HzX0QBD+aXhfsXgf6CeJ+t/VZ9x/IQAAAAAARfn0Hgn6Xh6sAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB8w/CAdJ9xB4H48Htf3yAAAAHn35fh75+gQPAAAAAAAAAh8UD5o8QCL1oPsr1gAAAAAAAAAAAAAAAAD49D4l+1w+vbAAAB+aX14HuIH58fT4e0AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAM/GIP2H2QAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAApLTw/xH579ti84+q+N/Sq7zfCA8zMnb8wPJfzn+6v0M+oPuf5BDv8AjQAAAD53D85vvwPqUAA+TfMQ9H+rQueTB8nfSge7AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAUfAfX8n1vsfP7ww3AAAAAAAA+TvhMP1E9ZAAAAAyA8O8EDxaoE3tQfSvtwM8YDxDygOKYA0BwIoRND6x+wQUAAAAAAGh+VvqoffygAAAB5x8KB+mQHx5qh9XAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfnt6wH1iAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAARfnX9D5fXxfmX9Ue9e4fQfiMp2fM4f5h/Ofuvlf315P6T9ZH3P8AIIAAGEH5e9iH6R2wAAD8LP17DxP8/wAP2x/EEP1ttB60AAAAAAAAAAAAAAAAAAAAAARQhb+Vw+dPpIPqH5PD5A+gA+7/ABcPz67oP0z50Py1A/VXXD8v+SD9EfXA+CPCA+m+c+s+W+UsH9Y/Lvqb7r/DP6Gn8287j9E64AAAAAAAMQPyw9xD755kPF/FA8ayATaD2/3sPaLQFIPiT5kC77aHunrAdzIAAAABWD4y+Tg/X6yAAAAAAn5Oh9j/AE4AAAAAAfkkH62gfP8A4gH3eAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABwofl7+vgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfE/0PgfX2j4PveDeHfM/u2t9J7+P6Vh+z+V/md45+d/3P9pfbn2X8quPV/OQD5Y+BQ/Uf1oAAAAPw+/cED8vQ+s/yrD9P/qQAAAAAAAAAAAAAAAAAAATxQPAPscOk/MgOJ/RAPXfMg0e2CYAAAAAAAAAAAKXxd9J898X537F+SdxyP5B/VMn2h+Qf0v9RfIH3H8d1v0HDmfy2Bv60hY+UQ+jugDA8iDx7x4OK8PD2zqg9o9rD2XVAAAAD4v+PQ+6/qUHAAAAAAAAAHzuHyz+lwAAAAAH5Z/TofV4AAAAAAfkmH62AeffCYfpaAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB+QQfqN24AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfMHseT6Z6nydQcXZifNXmfoHmvo/lfo/0N8A9HifPP0j85+7fobp/oP8R5n5a1j6F+jAPAAAAA/D79wQPzDD609pD89fPw/WkAAAAAAAAAAAAAAAKPx+Hzp+qYS/AAe7fQAWgAAAAAAAAAAAAAAAPKu3j/N7zT9x/F/d/WPzL9q+UF/AP9D/ALE+vftv5J3z2PzAAi84DybyUPKPkwPdOkDd9eD2D2EO1AAAAAAAA/MwO6++QAAAAAAAAAAAA/ID9aQ1gAAAPzPD1z7UAAAAAAAIvybD9aQKX5Uh+sYAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfL/hofokAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAARJ8hxnD9fyXL+d9zgU+X6RRWcZ6fh5/ofV5xN4X1nQ9P1/N9fn9fzfi+V5H6b415T4X7D6V9hfQ/iP5+/p59R/PHtYAAAAAB+H37gh5R+RAfuFkBc/FQP3CAAAAAAAAAAAAApfLIfOP6ghn/MQfSc4fgL9Qh9E8QHhtUP0I+jgAAAAAAAAAAAAAPlL8+f138pv/oBnpm/A382/wCiXpX256XwOV5V9P8Az75VyoKBJ6OHrPrQesbYAAAAAAAAAAAAfKvkAfoSAAAAAAAAAAAAB88/NYfo0AADA/LT24PuAAAAAAAAOJ/PsP06APyRD9bgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAgD8ZP2cCYAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAwfI+H7DyrC8P9bdLO/Ydp3/AB3W9R6Pw3QXOv5sHmYGufyT9q+34pX+e+g5/l+X6Tj+I837vj6XB9p8/cH4H7QVs+377+vvv/4rU9D4kAAAAA/M/mw6778C7+XYV/0CD2cAAAAAAAAAAOOD5f8AtEOW+Ww+rt8AAyg/Gj9rAPy+D7B9BD8uv2EAAAAAAAAAAAADP+G/pvnPjLl/1X8w9a7H8A/pTy/v/wA8/syX0T9F/hj1P0esNXLDiePDlOZDnqgOQFAAABGge3h962wAAAA/In9bwnAAAAAAAAAAAAAPyRD7k+igAPMfy0D70+pQAAAAAAAA8F8ED71APyTD9bAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA/PEPb/p8AAAAAAAAAAAAAAAAAAAAAAAAAAAAMzw/h+v8cq+D+y7/rvd8j6tve3+RBfMAAAAAAHwX96fS/Oh819EAAABnfjxj6n1Z4f8r/R/lXvnlfo36Cdj+gfxKGvAAAAAAAAAAAAAAAAAAAGHuBwX50h9n/QgAAAAGUH87/2SH1X8/hwvOh+gf00AAAAAAAVfLQ8h8mDy+iBG18+9t+3fjP0n9lfA/daf5e/kn9c8X7Vr5+S76X8DboB6F3wdt2AdX0odFbAAAAAAAE+bA/PT9gQ3gAAA/I79cQAAAAAAAAAAAAACsH5seYBpVw679Hg6kAAAAAAAAD443Q+qgD8kw/WwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA5z8mg/YgAAAAAAAAAAAAAAAAAAAAAAAAAAMid/nbyv5v8AeNP3PfyfZLXvfjIOAAAAAAAAAD4J+9vpvnA+Z+jAAA4QPyU/TgPbqyv4r+M/hP7Hv/bunD9bzfb/AMig5AAAAAOF+JvA/aPpn3r1Pz0Ov5kAAAAAAOM/KsPSf0+AAAAAAADKD8aP2sA/L4PqT6ZCQAAADhg+cfn0OTcFn1UPXPWg9QaHknyZ9Z8v8u85+g/Be2S/if7J5Tu/nn9vfod7f9z/AB9150+GAAAAAAAAAAAAAAHmv5VB+ydwAAA/JP8AWwAAAAAAAAAAAAAAAAAAAAAAAAAAAAPzt+tA9eAPyTD9bAAAAAAAAAAAAAABOdDl+cDBxwyM8KgABPrBu9SHZdwFkAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPyy+8w9aAAAAAAAAAAAAAAAAAAAAAAAAAPFuX6H54vfO/uP0Z6t9J+EB0eIAAAAAAAAAAAB8J/cf0vzlk+a+jAADwv8ANMP106wAA8v5/Z/PryP8/wD7a9N+8/c/H/cT6z+bAAAAOU+LPD/XflWt8X/V/dfrB9X/ADhrn0v4EAAAAcOHwL+lwO+cw+FrAfafLB8n2A/Rz1EAAAMoP53/ALJD1vjw+wfoAAA8uD5M8MBvdh9G/QodZzweHeGB47RBdEPG5v2z8c+uf0H/ADv75fO/kfuvyW+pvhP7E/Qc/Qf4mAAAAAAAAAAAAAAAAPCQ+Df1oAAAPyS/W0AAAAAAAAAAAAAAAAAAAAAAAAAAAAPyp/TwNoA/JEP1uAAAAAAAAqB5N4+Hk/mwVFAAOkDqeiDf2Q1tALgABXyQwuVDjeFCq0Heyh9VexAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB5+H5t/rIAAAAAAAAAAAAAAAAAAAAAAAAFf5m8/7XyH2vyf0n6KtfS/ggOAAAAAAAAAAAAAD5f9K+m+c9WPmfowA+Tg+OP1sC4AAAEfyr533Hw/znwX9me/fcf1X86eoH0n4MAAHy9+d/xH9dB4v6qfRvb8n+kTv0T+HQEAAFT8tw/QX04PNvyfD9n7QH5ph9Leph+Wn7HAAAAAU1oXAKzz+XfkYMT1MPrT3UPP/nIPn7mwOgD3H3IPY74Hxt7/AIfwP9P/AKl+Z/pA78O/Zz4D8f8AUPk/9cvK/Se6Prf5nAAAAAAAAAAAAAAAAAPh7ND7zAAPyT/WwAAAAAAAAAAAAAAAAAAAAAAAAAAAAPyS/W0ABn5LB+tgAAAAGf8APAfNHmwOuh6x7GHrHpQWwAAAAAAAAAAAAAAAi8VD5Q8QD10D5387D9C/vEAAAAAAAAAAAAAAAAAAAAAAAAAAAPyWD9MO5AAAAAAAAAAAAAAAAAAAAAAAIPlvzvuPLvpvz/tfYT6H8SAAAAAAAAAAAAAAAA898M+m+c+tT5n6MA+G/Hg/URQAAAAAMT4x8f8AUfkan8R/XP0Z9q/W/wA0emn0X4YAN/LvwP2fxQ+N/qgA+t/vn7r+P1PX/MDkg/NT9Ew9FAAPz99DD6/+FA9Y+Zg2+MD0T9AAAAAAPmDg+y/O37C+e/cPnyt9h/Lv1J9bB5L8qh5U4PRPosPoTpQAAAA8C/MD9X/L/bf1F+d9+0fFfYwfjp8z/QHrn6bdPgB7v48AAAAAAAAAAAAAAAAAH5NB97e3gUA/K39YQAAAAAAAAAAAAAAAAAAAAAAAAAAAA/JP9bAAPPfhIP0vAAIfnoPkrgg0fooPpn0kAAAAAAAAAAAAAAAAAAAAAAA8dwQLwXvZAAAAAAAAAAAAAAAAAAAAAAAAAAADy78/w/VEAAAAAAAAAAAAAAADzcPFvrED4tD6sQPmH34PR/n7j+p+fPqfyv0b10+h/EAAAAAAAAAAAAAAAAABPgn6b5z73Pmfoz8+wyP0bAAAAAAAADkPi3w/1/5arfF/1Z9AfY/1X86e0H038/nFZej+UWB+bf3qEdJ9Xej8T9+zff8A8UH59Ofrj00AAA+KuWD7+8UD8vvoAJfsEPZgAAAAPK/z04frvLfrT5v95+wPk77L+WPnNQ95+rA9UAAAAAADiujn/JTa/c/xX9Zeh/Cf20MtfE/y6+N/qr9C/qb6L8JD0PiwAAAAAAAAAAAAAAAAAzw/Gj9hQ6Lx4Pk/9EQAAAAAAAAAAAAAAAAAAAAAAAAAAAA/JL9awcB8kVQ+weCD4o8XB/0OH1r3oAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfkf+oAdgAAAAAAAAAAAAAAAB8WB+Xv9CQc7/PCH7j+aBufmMH7p+ec/teqHR4oAAAAAAAAAAAAAAAAAAHwJ9N8599nzP0f57Th+gQAAAAAAAAABxvx14f678r5vxn9W+sfXP0H4p9JXvrv5k8N/Mn47+pax4H7MAe3+7/Y/y59ln0H4mAAABm/iEH2x3QfCH0AH0t9LAAAAwPkf4xD036M5fofz+6T5P+lO0+x/tv5K+jZQAAAAAAAAi/Kz7X47yP8AV32/G9XPy/8ASgD4m+J/gv7M/YHsPuP5CDXgAAAAAAAAAAAAAAAAAADzX8sw/Yz88w+oPdgAAA5wPzFxg/UfZD82uND3n70AAAAAAAAAAAAAAAAAAAPzl+tQ9lA/LL9Ag/PesH3l7SCgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHnAfnp+qQAAAAAAAAAAAAAAAAH59h89fc4eL+NB9kfU4H4uB+xG2AAAAAAAAAAAAAAAAAAAAB8c+94f0j8WeT6lv795+gAAAAAAAAAAADn/k/wAj9L+S+S+J/rbofp70fh/qnzb6f+e/zng+E/sgFR2OvB+iPt/6F/EQdXzgAAEAfOWMHvXhXL9Fy32jh6oej8QUg+EPmgPrr2UPgmvn28B9u/D/ANe/ah95/GoAAAAAAAAB8W/CH7L+Rff/ANhfnH6EHg+2Afmd5d+f/wBs/sEfoH8TAAAAAAAAAAAAAAAAAAAAHhAfmT24frsAAAB+bge6/VoJ+OQfo17qCKAAAAAAAAAAAAAAAAAAAcJ+VgfsLXD8IPXg/TP18AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPyhD9JO6AAAAAAAAAAAAAAAAA/M0PqP4TCr9bAz7SD5S+cg/ToAAAAAAAAAAAAAAAAAAAAA8v7OT81fZ+7i/RM8X2AAAAAAAAAAAAACv87cP2Hyf4l8P/XjPWY6NLxSsQ5foQB/1Z6PxH3Lu/e/xgF8wAAfJ35+/rv5Sq/jf9vS/rv8AM9/a0vrf5o+AvnoPvOuHwt7MH6BdKH46+v8A59/bX6Vn6D/EoAAAAAAAB531cv5H+z/tf47+pbvwr9sAAD8iPQPz/wDtf9Nz9A/igAAAAAAAAAAAAAAAAAAAAA+PvcQ9QAAAA8dD8j/vMPt/8Pg/cEAAAAAAAAAAAAAAAAAAAAA+fw+DbAfZf0+AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHLfl2H62gAAAAAAAAAAAAAAAAH5NB+l35Mh7f9DB5V6oHx9+qgaMoAAAAAAAAAAAAAAAAAAAAHyQHyd+tH03zgfM/RgAAAAAAAAAAAAAAcJj6fy78w/G/1TxJ436mAAAB0X2z7f5N9XWPtf5LByAfJn54/sH5PLb/ABr+3T6v+E/UtP5Y/QP4o/QSmHwJ9RB92WwAPzW8Y/Pv7b/Y1/6D/EoCAAAAAAA/Lb7L5Hxf9dPa8fvT8z/RgAaP8Wfqn4D+1Pv8+/8A4qAAAAAAAAAAAAAAAAAAAAAAAAAAAAg/LAPov8+A/dIAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAD8z/s0PYwAAAAAAAAAAAAAAAAD47D65+dw9Wrh5r8+B50B+hnpYAAAAAAAAAAAAAAAAAAAAeFfn8H0B92fT/N2j5j6QAAAAAAAAAAAAAAAAaPxr5p+e/b/nLmPk/6TDPsAADu/tL3/wAZ+nLH2P8ALAOflf8AOb9e/KLFj8f/ALWGfmP9HfXX0h+kfwZ+bX0T0eL9+2QAAD5G+B/hf7C/Vj1n7L+WQ6PDAAAAAADwj8tf1f8AL/t77j+L+wD5z6AAMKOn8a/tn4X+wvtg+9/jQAAAAAAAAAAAAAAAAAAAAA4foQ1wAAAPnYKnxCH2X84Bl/YAWfoEAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAgD8fP2KAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA4sPyS/ZUPCLH0/zfvZ8x9IAAAAAAAAAAAAAAB8JhnffoAEXjXL9H86/K/xn9V5R5f6CAAHZ/Xvt/kv1PtfZ/yl80fmr+vfkfc8N+af07oH49/Y2z6L9d/M36dbP0f4SAAABwn5FfI/019o/b/s/lYer+cgAAAAAB+XXj/7l+LfsbsfiP7KGegAcbl6H5Afd/xX9a/YR91/HQAAAAAAAAAAAAAAAAAAAAB+f30sHtYAAAHAh8wevh78B4N4qHu/tgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB8ZBb+wgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAg/GcP1s7MKnwv9P8AN/eZ8x9IAAAAAAAAAAAAAAHmny+H3WAAABxuXofCHzh8D/aSHD9aAAGr9Nel8F13wl/SH+enRffHzX1f57J+Bf6On0d3fJ/o/J+hfw6DQAAH5QfPft2b+uE7KfR/hYAAAAAcF0834/fYn7D+TffR+K/sQAAHE5ej+Qn3p8V/WX16fdfx4AAAAAAAAAAAAAAAAAAAAAfD30GHrwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAH48h+wMoAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAflz9eB9FAHwR9N85922vmvoQVAAAAAAAAAAAAck4OrAAAAAADxjl+g+GPHPgf7QDm98AAGUv6G/grqbq+I9J8Q/DP7wDzfvPqz9A/uP5Dcez+WAAAfHPm/ffCn6oeB+0ewn2P8ALIAAAAAfEX0XgfDf66fXfKemn5f+lAAAcvn2fjt9yfD/ANefZp95/G4AAAAAAAAAAAAAAAAAAAAAAAAAAAHk/nH03zn0Vq/N/QhNAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB518AB+ogAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHy6Hz3+kwAB4V6XnY30dpAeT6gAAAAAAAAAAHy6GT5QHlqB+ndcPyK/aQDyYPzmlD7I+pQPBuP6b4u8Y+E/skOX6AAbR/f/wCGFv8Aj/Wel+d/kH9RQHP7J9M93yH6GXP0L+IweYAcvn2fkB9K/Bf2f+iR+g/xIAAAAAH5JfcfF1/11aD4b7UAACFV+LX1l8D/AGl98n3/APFYAAAAAAAAAAAAAAAAAAAABi6QWAAAAAPzq+U/2/8AGb/cfg392fVv2bzfnIen+fgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfFnzCEv6Sh8c+WBJ+oIbYAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfmT9yB6kAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAGB+P4fs48AAYL4A/QL6f5wPmPpAAAAAAAAAAAPhQNX4+D9bPnwPCtwPlL9MA94/D8P2t3wAAA8f5ve+I/Bvhv7BkOD7BKH7t/FJoY9Ifh/9mgAB6/8Ao/8AW/zV2Z9J+CgAB+c3zn+e/wBufrz1n338YhpwgAAAcd+OH6Z+c/af3b8v9KHzf0IAAAH5Eeg/n/8Aa/6bH6B/FAAAAAAAAAAAAAAAAAAAAAB+dP2AHqwAAAFVz+M+B/Qf4P3vJfyZ/qL9nfcftfz2Hq/nAAAAAAAAAAAAB+LYemebh+uXhYfmB+8IHwyHy9tB9cfWoAAAAAAAAAAB+X32CHUfm8GD+tgfGHWh9mgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB+PX7CgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB+Tn6Gh6sAAAHxb9OfUfNdsfL/SgAAAAAAAAB8bB9RfHYavx8H2r8dB9zfAofqX+cwfc/yEH6fgAAAeAh+a36VY+p+clL4n+tadD9W/Akvej4Lj8D/uEAADcvn+8PqP77+LHHofFgHlv5Q/H/ANQ/Xn3p7P5WHr/mQAAAHy1+cf69+U/qh7j+Zfowed6AAAAH5n+W/n/9s/sEfoH8TAAAAAAAAAAAAAAAAAAAAAH50/YAerAAAAeCen535c9L+4fi/a+a/wAaf63fpN9F/d/xsHb8mAAAAAAAAAAAAfh8H7g/CgaXy+Fr9Dgt/mcH7DgAAAAAAAAAAAAfl96AHl3s4fKfTB80/vEHSgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB4kHyl+jQAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfPofKX6XgAAABzHyJ9T819xHy30oAAAAAAAAB8Nedh+lPwoGr8fB9gfRgaf4YB9Y/KAftV+bwfr4AfB3kfpfy2z4r+tKlSOphOwAUv1/+U2XfqfzN5/O/98AAAAHpO/kfSX0d9f8AzD6Qe3+Sn5r+H+u+BfrX533PdH1v80gAAAfnh8s/tn43+01/8V/YgmgAAAPiP4q+C/s39iOp+5/j8NOIAAAAAAAAAAAAAAAAAAAAAAAAAPz8+n+a+QfqX9H+A+dqf8g/64frt3f6V/AwbeYAAAAAAAAAAAAfhSH054QH66/k2H6QfGYe1uD7f/L4O7/QYDxsPJuuD6InD594UPdvRgAAAA/L7qg+t/YA/Gj9bA8C+cQ/TYAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPy8D9Be+AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAE/GEP2SugAAAAfBP03zn3BrfOfQBFgAAAAAAAc90IHwoGr8fB+tnVh+dHt4fVH5hh9TfBAe0e3B6NyHB9l4X8bfOfu/2j833y9573fHdu6+fd+SP0T8W8z+q/t/zDc+cP5v8A7/5c8r9FAAAAAPRvo32vyj6Nv/U/zr+Tft/xP9cfpsv6J/DIAAAH5JW/278a/WI/Ef2UAAAAA8B/Mz4v+r/0l+jPp/58Dt+TAAAAAAAAAAAAAAAAAAAAAAAAAD8e/u/iOC+8fZ5/h6//AC5/qV+z7/0//PUBAAAAAAAAAAAAH4eB+0m+HyT8DB7d8vB+sPw2H6ygfin+1gH4eh+k3nofCf7Xh+H36gB+a366h6gAAAH5ffWa0+jh5/jR9kh8t/UIfaIAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfj+H7AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB8ObwfYoAAAAB5z18vz79kep5oeB7gAAAAAAfn390BuAHGBJyoeoWQ8j9WCbmgwPRBeCcOP6G1A/If75D5T6UP0eUAA/PP6n5n5K+1fv8A4X7si/Fv2Hxb5x+e/cfnXlPlP6QDPtAAAAO928yhyM6/fP1x95/HAej8KAFdr8Ufqr9r/Hf0MPxP9jAAAAAMb8bflf6O+svvn0/z8PX/ADEAAAAAAAAAAAAAAAAAAAD88frsPUwAAA4P8ev0783sfpH5v135yelfzz/ol+s5+lfwOAAAAAAAAAAAAAH4fB+4IH43frEHX/lCH3V8OByHcB51+yYH4eh+4QHwwB8S/t8H5FfoyHr4AAAeUfmP83+9fTv1B7359V8z2+G9b9NAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAOC/PsP1BAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAACmH45fsyAAAAAAAfAH3b9V8zpHyv0wAAAAAfCAU/vsAAAAAAA4z4N+0+P82/WzTNx8P9pxv5Eh+kP51h+hf0gAAAHx173h/Af0Z+rfmH6an4X+1gEfjPL9F83/O3xv8AU3Inlfo4AAAABZ+s/T+C+jfZPtP5PkOr50896ub8gPv/APWPy37CPx39bAAAAAD8rfn/ANq5/wDXKOoPo/wsAAAAAAAAAAAAAAAAAAAD83vtYPSwAAA+UPzt/X/yf2X6s+F/Xvz++gfxX+4f0wP0X+GAAAAAAAAAAAAAAAAAAAAKdANsA/D0P3CA+OeID4F+ig8//a0AAAAPz1+WPz7+2+v4f+nv87v1r9P/AC76sOPrAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAD4K9tD6IAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAD89A91+lgAAAAAADzn54+q+Y+zT5X6cAAAAAMgNcAAAAAAAMb8bP0X8/yv1K9Lz/AHfzf8o/UPyd/QIPgT9aw7gAAADwj0vO/LXu/wBu/Gv2GP5//dgAAGD8d+cvn/2v5y435P8ApQMu8AAAA6nTj+h/oz6r+cfbvC/tf57/AC5/Tz9H/PPoU/Jv1IAAAAAPi3y/0L4g/WvxP1z0Y+v/AJhAAAAAAAAAAAAAAAAAAAA5rdCwAAAH5z/K37f+Mfaen+Qf0V8U/V35d/W36Bn6D/EwAAHwGHsP02B+A4fUXEB9wfPIfN1gPpT9KAAAAwI6vHNSPo8Pvfp/zT00+e94AAD8PQ/cLKD8Wf15D8gP3BD8dv1ODuwAA+PPN+9+Eet8T+i+F+kv6f8A8zv0sPw39pAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA/Ij9cglAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAKn47B+ygAAAAAAAHwf9L879l9L4HuhjqAAB8iB7534AAAAAAAAH57fUfNfI/0B+m/nH3T+Wn4V+2fXvzqH6yXAAAAAOd1y/GFf6E/Bv2g3/57/eQy1AAAATyTm935x+cvj/6i4U8f9QAAAAAN7tvtPxDxv9Y/27+FfXj8u/SgAAAAA84w9b8lftr4r+tftc+8/jQAAAAAAAAAAAAAAAAAAAAAAAAA/IXzj9+/Cv058K/n7+sfkv7R+F/pL7gPvf4xAAA/OkPa/qsD8Pg/cGIPwp/dMPhzVD7W+CA8t9PD76APD+T6P8vvWOb9w8X+qv6D/gX9Fz8Q/ZwAAPzcDjGB9y+6B+bH6Th8u7QfRAAB8ofAHw/9fdVy/wCxfzN0n60+v+Pb5+ffeAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfkN+vIAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB8EerB9QgAAAAAAAcztj8Zfefv+GHzX0QAAfPfw4H6zgAAAAAAAAHnvVzfkLH+/fhXZ+y/gv7i79JuHtAAAAAAD8euD/oD8J/UL3/8AEv2QPM9IAAAAAAPLPnPwv2D5y86+S/poMPVAAAh+v/KqvR/p/wDNvv8A9FflP7D79r/T/wA+hWAAAAH5P5f55/cf65O/Q/4dAQAAAAAAAAAAAAAAAAAAfAX2OHbAABkaR+LDP6E/BP2Y+E/5j/s/5j+7PmP3T7FPu/46AAA/OkPa/qsD8Pg/cED8dv1RD5M1Q9A+UQ/TX8CQ/oFAPnji+q/NX0h/rvk/2f8Auv8AC/3kfi37CAAB5fz+z86euL6n0jt/R/NQ5+gAAD4z8z9A+HO0+R/qrAzf67/yq/Vj1T8S/Xw4uwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADiA/Pb9QAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABA/Gv9lQAAAAAAAAAPkb1P6z5f2Y+T+oAADjuqCcAAAAAAAAAA/Nf5m/cfxa/9n/n/AN59xnx31oAAAAAAfm/8vfuH4v8AbP3R+Zfo4fOfQgAAAAAAAHiXL9F8Z+EfCf2Qhx/SgBB9n+RVr/6J+BqfhH9s3fcu35T6L+hPsf5b6M9P8/AAD5F+CPhf7C/Tz3X6/wDmIOv5oAAAAAAAAAAAAAAAAAAPzT+7A9AAADxf0OD8put/cvxb9lPzc/mL+0/nH778H9m+tj7n+PgAAPzpD2v6rA/D4P3BiD8MP3RD4X1Qy7wfbn4Zh+5gB8k+d9z8Ceh+h915Z9+fsf8AGP2Kfjv62AAB+cXifrPzq/6jXD9a/bv4v+/fafxT9jDz+8qK/gL5W/P/AO2PSOd/Yv5S5P8ATn7X8B9+Pyn9QAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPj4LX1uAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfKgcH9zgAAAAAAAAAR/nn9V8x+gd35f6UJrG8bD30AAAAAAAAAAODD8nOd/Tfzc/Ufq5/ej8o/UAAAAAAA+TPzx/YPyf2f9WvlPpw+K+xAAAAMqdvMPLfH/U+LPN+6Ok05PQO99X84n+esfS+dPI/kP6eZD9N8PVvfpv8AOjj8D/uEAJWvZvon6D8T+h+2+r/m4NfOxvx/+W/oj1/9Ouz5oPd/HAAAAAAAAAAAAAAAAAAA807sNAAAPkz2/G/PH3z9b/K/1F/Nb+bf7K+d/wBAfC/YvrA+5/kAAAD86Q+WfUA+7/xWD7d85D7r+pA+FNUPq78aw9z+Mw/oVAPgjyP075L736Tq8s/TL9N/lT6PPyX9TAADyf8AKn4/+ouw88/cP5cH/pP4F+v/AH/4B+6hy9PF/m/81+9+Rey+d+q+d83/AFp/l7+k30d+L/rweT6gAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB+Y4ffvdgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAH4+B+u10AAAAAAAAAA896ub5d+5PZ8gT5z6D8gPscMX2MPcPDQ9P6MPH/Tgw/lEPauaDlQNj6pDnPxzC/6B9R81496T+rfmH63WvwL9yBUAAAAAcH+PX6d+bz/s9jtuH5v+hADfNsPY8c4DwP2k1c+31H1H2Py3tV9L4MAAAA5DLv8Anb5cx/UPGbv3/wCePP56/vMAAABfUejxfoX6C+s/mr1b4c38j4z/AFE8L9h9tPs/5VAAAAAAAAAAAAAAAAAAAAAAA+CPpvnPjX7D/Rfz/wC/vzh/nn+vfmz718D9p+vD7r+PAAAAAAiCK0BUqZpUzgU7X5Ch+1wB+W3gfs/O8P8Ae/H4P64fZ/z76ifl36WAEKr8rvOvzr+6bvG/19/lefQkYfp6fhX7YfN3D9b+fud8B/avuPhv9I/5zR/ph3/E+8n5Z+mgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB+RwfriAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAcN+cgfqoAAAByodUAAAAAAfKHY/XfK+/Y/yP1XgX5+h94/EIfdfwcGv+uYflR+i4fjr+m4aUQfJPHh9rfRQfjZQD9FPe9cvyB5v9+/C/ub7b/Hf1oPnvfAAAAAD8gfuvifPP0n+i8D6a86/HP2vwvh/nv3CX0iuf2Tv/oPxVTbywAAAAAAADx/v4fye+mv0P5+p4Z/NP+iVM4vqwAAAADrvZfS+D+bfSOT6X9W7v6T/AAWDzAAAAAAAAAAAAAAAAAPhj7IDfAAD8y/r/k/nL9Afs/kfsD89vwT+rflf7f8AnP3f7SPvP41AAAAAAAAPKO3j8a+vPQ4Yvzo8L2sP75DvwK340/L/ANC+x/PX61+FS/tZ6n5doH5z+ggB8V+X+h/EXpHN+yeTn9Tf5tL+r/xP2PsPPflP6v8ABHzN8B/avc9p99+FeC9z/Qf8Qfpt6f8Ahn7UHH1gAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB+RwfriAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfmr9lBD+SgfuiAAH4fB+4IAAAAAAfAX0/zf136F4HuH40c/R9vfHwfot+ageh/VYfJP6Lh+NP6fh9LgfH3n4foD+RgeX/ooH1eB8sfnL+vflFz9Z/P7vTj8x/SQAAAAA+C/pfnfjL1D9G+P9/8ARv5t/tT3TuPovw4NfOAAAAAAAAAAAyLj8WPsb9n/ACD74x/xP9p8C+dPmP6C8Eyfmf6ACdgAAAAD3P6q+l/Avfdf6r+dArAAAAAAAAAAAAAAM4NHxIPyQ/bEOrAAD8lvt/jPJ/05+i8D6I+PPxT+j/m/6S+f/avsE+y/lgAAAAAAAAA+A/u36n5q+fLfSgAB41+Wfxv9UfRfzJ+9fx36H+v3D4ofC/bAHhX5k/Gf1X3vB/r/APOeQfrv8xfTf6S/kf6r8x/CH49/a3O+v17nIeef1b/mT9G/ot+b/fbx8Z9eAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB+SIfrcAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfjv8AsQH5Z8OH6He7B4f8Wh9cfmQH3V5yH6H3gAAAAOYuPxJ/cj6r5jV/Gj5L6r61+bQ+wfhsP2D/AA9D3b9IA3vzvDzT9fQ+PvPww/j0P0G+2QAE/L77H5LwD1T9L/Of1hufgv7iE0AAGd8/cH2fj2Nz/oPyVp/07/B/7M6v8+fvIRYAAAAAAAAAAAAB+Q+/+3fjX6sn4j+ygFLwvi+s+d/nz47+pOaPM+/AAAAALvunb8n9F/Qv2X8tdAel8CAAAAAAAAePh82ehh9YfNAfEf0AH3nx4eG/SQaAAAfjR998Nyv60ep5nrPx1+W/pnyfYD9HPlEPGQPtz4cDzn0YPtj6UAAAADyTy76j5r6sPl/pQAA+HfjH4L+zvqb5I/p7/OP6Z/Sj4v6QPkvqjzPD2Py11Pz7+3bHnH9ff5Sm33+T+kXx3/LX97+Ldb8x+99L5L/U3+ZGj98R5X1gfkf6qAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfkgH63gAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHjwfHv6Rh+MH6jh+fH6UB+QP7NAfh0H6zeXh4X+nwfMPt/v+F154HugB8BB7R4T9h8n+gf44fIfV/rZ4gn8q+WB+y/ykH4y/0Dh1ugH4afucHx94EHx/9uh+kIAAchvh+R3Pfvv4Z9g/oB+RfqwfMfSEafh3gfz/AO32voKuf2J3v/jH4+cF/QH4R+jP1R+M/rweL7AAAAAAAAAAAAAB+ePyv+2fjf7R6f4t+whFgBGn4589+B+0/Pvnfyv9IBh6oAAABYc+1/Rv0X4b9FdP9V/OAacZih476cHXfn6Hh326H0t4gGf7EG6AAAAAAADPxH/RPgIf2Q6MOx/NP8z/AEb0Dt/M+/8Aia76fwH6g+Th8v8A6cB+Hv7hBX+MQ9Q2A9uAAD8/fp/m/wBAj5j6QAAPyTu/nn9wdJ88/wBb/wCWn3v9lflX6cHge755+XXyn9IU/U/pPC+fz+kP4OOh/Pf3Whe/mr/RHpua/pL/ADq576N+w/If0B7P8T/ZA59wAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAD8kg/W0AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA/MMPvXkA/JP6MD5h/VkPnD9KgPw6D9xeYD8jP1j6Of8d1/oD8J9f+i/jfr/or0r80/RA5Os816+X5Mi9zxfJNX5j6T9J/iIPv4D8w/wBDw/MGqH2n9QB4T4YHzr+uoAAAAfOX5m/rX5Yn6UtfS/I/kH7F8ucz8x/QPufvnvfjlg7fkwD4i+G/2b8h9n/Vr4760PivsQAAAAAAAAAAAAD5n/Nb9c/Kv08+hfzf9CDy/TAAAjT8h+QPnP3n5w0PlP6QpxLQEwAAAJPZ+r53c+2vsf5a9Q/OH1vzfp/sMO0AAAAAAAAA+QfqUNkAAwvxb/Sfzw/bGXofmn+cfoUfJZej678/a+d9YfKofZH1mH4e/uEH5PfUQeQ8KH6rgAB8Z+/4X0z2fkesHPueU8/t/lJ9w/Gf1Z8W4H9f/wCX36u+yfhX7YeU+R9T+ZdD88/uH6R+R/6g/wAykPtPzqbe/m7/AEG6Dn/vPwjm/Rv0r+efvr6B/Ff2MPL9IAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPySD9bQAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAD8fA/YP8oA+/vYQ/Lb6tD8yfs0Pc/zSD6J8nD7j5/3/AAvgk/avx4AO75uj6E+gvzr9A9w0PhvtPKfl77H5L7xm+N+uBMAAAD4y8S/T/wA28tw/zH9I/ZjQTAAAAPgT6b5z451f2D8n+quz/mH+6fqPrPU/Hw04QAA478d/0v8AOoP1p3x9XPy39LAAAAAAAAAAAAAML8ZP0j89+qv0O8D3A+W+mAK3hnF9V84/Pvx39TbP0Brwe++0/W/zPjfPfP7Hy34t8f8A1HMcX1YAAAA8XsX0b9B+K/R3Y/WfzUGvAAAAAAAAB+X36LB1YABxf45fpn5zZ/bjm6D80/zz77J9W4Ps/pj80t/G+mvpfo8X1AD8Pf3CD8P/ANwA8T+FA/VcA/NHw/1/6/7D2/zr5/8Ar/3vig8L2z8/vH/UflX9Icft/wAyNT+pP8+/2ks/z1+9/PH51fF/1bc+pvV8T4yq/wBS/wCeJ2nH6PoXFfAfsXn3Wfo34j9ufWP41+u2DwvbAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA/JMP1sAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA4b88Q/UflQ6oDMhDR8rD0vMCCEOx/Jz7b43yE/U/zMAAAC/wC3eV6fvnVfBfc+K/fXF13j5X6cAAAD4h+HP2b8h5v9ififsfXD4/60AAABvzdx/V/nlz377/Pvc/rF8h7XTH5796AAAAfmr9b8r80fT36J8D+kZ+HftAAAAAAAAAAAAAAfl19l8j5R+yndxWD8/wDu/Ivlz579u+bu0+a/ffo/2v6z+a+r7j2vyj5t+Hg9t/QYPOfAp36v51+Y/oPwPM+Z/fwnYAAAB/rnT4P0d9H/AGP8udqev+YgAAAAAB8r/TIXwADzn8hP0/8ANtv9puDtPzT/AD/7zc+Xvi/6u+rvAfc/Iv1B9A9j8uAPw9/cIPy+3Q8+vB+q4EX5D/N/vPN/qhy/R/Kv33+qfyAHzP0fP/kD8r/RnunqX1/5l8LfQf7h/Kn6cfEX86f1r8aM+E/sWxkfv/8AC6H63/LZuZ6YfUS/s765/Hv1rSPnffAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPyTD9bAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPg32YPowAAAAAAAK/53fWfLfO2B+sflwa5gAAAAFn1jz+76691/J/1LsT573gAA8O/Hj6T57t/eP1P8y/S7b/DP2oM9ABnzbw/YeIfRfk/o/qf5Pfsv8mcL7F9z8V+p2j+EftwRYAAHkXdxfk5L+9fh367+ifgP7oHL1AAAAAAAAAAAAHyr7Xj/AJ0fpj978Tk/I388f3h4lwfN736U/b/2f8qfG3znt5X159Mgx4AAAAZ3g3B9j84/PHxv9U5p5/2oAAAAoer/AEh73439Gd79b/M4beYNH5b6pz+yHR4oAAAHln5I/qP5p1H7M+Z6J598D915v+a/xv8AVG9+mHX816jpfTfz6DXjvsQJ4cHkfnYfokAcPl6P5Qei/n/9s+v/AHV+1/55WT5n6P4e+L/gv7O/U34E/af4f8d+xvsPi/K/Hf5b/wBNxvXjST+vv8pA6OE7Ln2+yfrT8f8A1rSPnveAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAD8kg/W0AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAD8iP1yCUAAAAAAAACv4h6fm/Onzx+ofmvDn03zoAAAAAKHofufzP0fuft/wCc/oHUQ+F7f4V+jB9i/CH7J+R+h/pl4Huekn51+gHgHH9R88/SXkfp3uZ9H+EnmnXyflLh/vX4h9Dfpr+V/psx8d9aAAAB+XvgH7t+JfTf6Tfm36IHyX1IAAAAAAAAAAeS+OB9QQUfjSv7B8P7l9ffzl/ZW91H1X83+hK5AAAAAAAAb8+cX1XyL4h8N/YLO6y9LCwo6QjpAAAAX1Do8b6N+jPr/wCYvRz2vyb8ufpv5H+nPqs+u/mP5f8ApYLQAB5X+SX6j+adV+y/mekHwP3MX5DfN/vXGfVnhfsH0t7f9v8AyQHV84AfN3xwDv08DaAAPjP4a+E/sj6r+p/1f+NvX+Q+f9D8kPYvz/8Atb9Cfx0/pD/NRl/5z94Xf/CP7U7/AMT/AKA/gtp+r/zeencfX9l/T347+tWjxPZAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA/JIP1tAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA/IT9ewAADK5Tt4+X5v2PJwML1PNxaXdwqlIUDkA8AsfS/OZuT63lxGuYAAAAAOD0ja+Y+k+SP2v/PPvvZfm/m6Pzmi/bfxv769l/EP3f5L92+G/f/oxfpfwEAAPD/y7/VPzKl9Se54v6Nu/D/2YBgAB5L+Tf6l+Zp+rrXsR+WfpoAAAA0OB40PcuTD8uZw/WpQ/PH1gNb4l+P8A6j5/A/pH+If2L678l+0DDcAAAAAAAA+f+P6j4b8y+B/s/f8AqHu+S+oPUftP5OreGZej84fPXx39T88eZ98AAAAAei/RXtflPnHzvxfWfVn3z9V/OH5afqF7X5TqgAHlf5JfqP5p1X7L+Z6QfA/cnx75v3vwgfAf2l9l/c33n8cB6v5uAAAAAAGD+O/yv9Hex/Qf2/5F9gfmx8t9Z89/q15n6F86/DP9I/wTJ7X+Vf1b9gb3H8Z+Z2H/AEJ/MJPcfox9Q/iH7Op43rgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAH5IB+t4AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAVvFuzk/Pf6v+o+a5dfW8tUDW6zh7On6Xx/W6Dd8v09q7w9oTQAfjmH6/3QMbTPwrwP7r4nwTg/0L4MOnnAAAAABQ772v5v6Dp/lBVzvSdfT+qXpf4z+mhzbAAAfN/reX+asH7l+K/W/6E/kH6wp8t9MAAB+cP1ny/wAuesfqH5r+sc34J+4xY6e787B8Pe5B98eLh8be5B9ftBVAA5j8+vn/ANr82+9eD7Lp/wAl/wCgf4W+2fuj5D6sPnPoQAAPJA/O4D5n/ZgPkr9G1X5r/Pn53/cvVfZ3f8b9T6X2n8nhWQAQKvDvj35j+hvCD5b+hwAAAAAAPXenwvT/AL4/Q/4cumvAAeb/AJDfp/5tvftHwdwfn/3eb8KeN+q/JZ8P/XXqv6ufYfy6H0P4kAAAAAAB+Y3hfsXmf0j73L6X+e/w37L73ZV/NzP6v/zj+0/uv8h/Ufn781PvvzvNPs/kLX6QfIfV/S5+S/qYAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfkeH64AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAcptj5R5V9f8pgr63l2vE+Ls+k/pv5f6XqDyfUAAAAAAAAADODRAAAA4Hp5vB/CP0H4Tw3nfuviQ1zAAAAAAAAu/Yfzvv/AGpu/jn62GegAAfK/wCdP69+Us+v+7i/QFfxP9jAAA+Ud8fzht/v34V95VPxr9e+bPsf5/3fqbGDQsgAAAAAHiXL9F+cn0p8d/U33Jofc/x4flV9t8V5r+xfdxah+e/egAAAHxSHCfUwfCf6rh8HXvk/6T+0NP6r+cgrEAAOG+ZPG/Vfm7zP47+ozQy9L03i+v5vlji+pAAAAAOm/SD6L8N9rPr/AOYgDhPx5/Tfzi/+2XJ1B+e/emRO/wAWfIfwn9j1/wBifR+D6U+s/m0AAAAAAD4n8v8ARPib2fp+98lpfnX9FDPZ+VpSf17/AJS/rZ4/+Kftf57Qftv4gXUfpT9F/hv7SHleoAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAH5Hh+uAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAFXyT0fP8Y5T7T5BerD1n1T5D6vePJ9Q/J0P1P0wAAAAAAAAAADjNYOGA7vbDmZw3wABnlndxfN/yt+pfmfGn1PzQAAAAAAAF31nzvQ9i9j+E+29i7T4r7AOfoPk32/G/PNn7d+NfW33/wDj36zc/L35f6Xj/wBGA9a+cw8n+Iv2D8mu/rB53oeqH5h+kgAAAAAAAfI/xN8R/XH6GfQv1X84h2/Knz16nmfmH97fqH5t9ln43+uAAAAcN+RgftX4qHwn+q4AAAAEafz78kfLf0Z4cz5j+gfS/oj2/wAn969b+s/muj88Yev8odF8r/RvlfG+d90GXeAABYc/df1999/Fwel8Hzf4xfpX52ft1npOfnP6AAHn+PqfB30x81++fTJ9X/N4AAAAAAUPinyP034zjf6tIfCfthV/Sf57hm/av49dX4/WD7P8il/TL4X776KPzD9AAAAAOTD45uh9gdKAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAH5HB+uIAAAAAAAAAAAAAAAAAAAAAAAAAAAAABD5H6HB4Xzn2/xtz1nj6/aOs+L+wDDYAAD8iQ/XYAAAAAAAAAAPCA+WOzD5V9/D6/9XD8gLgfrnGLxjwT7P5Hx/wAt+9+H5Y9nyAAAmhQDAAAAAAAOuw29f9R+M+v8r/Rz4b7T5o/PP9J/PGfV/fw/odJ+JfsoJgCfmD9j8l8++h/p35v+sm1+CfuIZ6AAAAAEHwL436n4d+lvn/b+in1P86gA0PyW5H92/Ev2H0vwz9qCLAAD8XP1dD0bxQPhP9VwAAArqvmD42+O/qbz7e8P9e+oPqX7D+XvTj3vxrz35X8P9em+iZ29Zd7342nkvN7vyl8sfD/1608/7UAAD6Z7vkP0G0v0H+JKv4kfpH5yfs9pHRn5r+iAAAc5HX0ZfIAAAAAFCdfzS8w/P/7a5mD6f476S63zOPh/nD9m/mFH/oX4uw7/ABR3J6du/wDyP/qh7v8AdnsfhvoR9F+GFYLNYLPzSHxt9ph+Z/7eB+P37AgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAH5UB+nu0AAAAAAAAAAAAAAAAAAAAAAAAAAAHM7ZfPvlv3XxMvr0X7p0fw/2YZagAAAAB+RQfrqAAAAAAAAAAH5mh+ifxmFf7IDoqgfkZzAfpz+dP6b+ccSfefEdz6x4Huekd58n9T2vxh8/7vs/2h5PrfH0/jfsFvxX9b/mDxXkPuPjA3xAAAAAAAeHoHP8An93OHocP07+jf5J+qWT5H6oAOZ2x/JXjv3z8N+gf06/L/wBKmPjfrgAAADJnf83Knw39g/pLv/cfyCF8oAAfO35j/rP5Z91fa/w/2ofPe8AHw0H50/XQH1wHxj+q4AA0fzT8RfHf1Jwfd+L+sfYn1D9z/H+iel8FhfGvj/qPQfWN8eser+dAHJfFfh/r/wAu0fjP6sCdDWrDr+u7/jeu6/0fhfdfbPpfwU/Fb9L/ACHH/Xzu4vRT8w/SQAAAAAAAAAAofml4H7R4VJ8d/U32a79T/ir6n7fX8+/Nn5k/afx4PY8oA9N9m/I/6G8S4n8G/vf0L9WvqP561fkD6T8F+FYA6yYPRvqIPlH7aD81Pt8PyI/XcPrYAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA+CA9798AAAAAAAAAAAAAAAAAAAAAAAAAA4z5s+m+c4fpvovA+hPU/hPtlOLsAAAAAAAA87+Ew/TAAAAAAAAAAAAD4UD439yD77+cA+e+ND6/wDnn7n4r6Y+h/K9Tsz5v6EAPxUD9ifz54P0nvtL738h884H7L874jj/AKLwEN8QAAAAAAAAAAO55uj171X43671P1P4z6/rzxPZ8e/K79R/NKH1d7vifokv4j+zAAAAcv8AmT83+9ekfoZ0/P2j2/yUAAAA/Kf7X47zD9gPS8/pz84/QQAygyQDoApbAgH5B8D/AC39FeS9z85+6fa30395/G056HxR4hy/R/N32r5v3fVnufkIAeeY+r+anAfnf9yfRnv30H4p13Xex+Wdfseh8WFYAAAfj3938RwX6q+14/th+Vfp55gHzH4YHl3GhUkDpPRQ9n+iw+hbAAHmXjnj/p9mXD2PYfGPs/xX2LA28r8pW/t/4yAH0D5fpfd/E/lf6b5/4R7eGz+kvz/tnx94fXxf2IH5UfsQH4m/o4H5g9mH2x9kh+Kf7WAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHzwHi/3eAAAAAAAAAAAAAAAAAAAAAAAHN65fM/nf3/w3bfSXhe13B8x9IAAAAAAAAAAHyZSD7AAAAAAAAAAAPJfVQl+FA1ftYD8Xw+78cPjv9jQAADP8Y9XzPEvh37/4TocP7D5MNc9jvfO7+57L533+s6XxPZ36XF2eTfMP2vx3Jn03zoAAAAAAAAAAdH6f5Hqenef+L7Pz+fY/JfXv6Bfj36y4+X+lADicvR/MH6I+L/q77qd9z/IICAAAAPH+/h/KH6s/X/yj9Ej8R/ZQAAADwL0/N+FP1I+p+af8K/lH9J/LG18Z/VX2r9a/d/xxaPT+AAPk6r87+5eqfBv0X4b9AY4eXgb2CH1f33l/ofqh6n54AAHlHq4AAAflP9p8d4t+lH1Hzf0N+eP5H+q/nj5oH1xygfI32eHzv5oH2Z8ehVA7L7uD9NfVQAAA+Vva8f1f83/q/l/MD9E+BAF3stMA1zAPYP1h/LP0w+PvifsuV+wA/Kn9gQ/FP9Fg+af0iD8T/wBug/En9tgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADKD8sv1iAAAAAAAAAAAAAAAAAAAAAAh+f/V8vwboftPkPpn0X4D7kOXqAAAAAAAAAAAA/OX7DD1cAAAAAAAAAA/Hb9iQPkcPhXtg+xfk8P1XA/JT9agXz3q5vnj5+/R/z3xWl9t8adrhv7J6z8Z9f6p6X8Z9f16+L7B5z18vhHiX3/wnjnF/b/Gh0YAAAAAAAAAAAAAAAAHrvD2+r+hfGfXegd58x9I78z/zj+pvq77P9P8AOQ9P8+AAAAAAD86Plj9v/Gf1X9n/ACD9XDz+8AAD549XzPzK9X/YPyrnOY/kD/Wb6i+6frv5h6Q9z8fAD458377uvozXzfwx/bXt+TyNAPlTxoP0J/HoP17rh3IAAAfkn+tgAABzf5+fZ/IfMv1j938T8HfK35H+qSf0meb6PyfhB+R39S4fJf4FB/Ur/MkHLzBCBrftCH3mAeU9vH6t8a+l5/yb51+v/kwdfMAdT7N4fs+vepfDfa+jdh8t9NadydR4mGP7YHx99sh8HfToeV/VAfn/APoAH5n/AEIH1YAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfkN+vIAAAAAAAAAAAAAAAAAAAAVPjX6L5/lPoL2PJ96k+G+1BMAAAAAAAAAAAAA/JT9ZAnAAAAAAAAAMwPzy/SAAAAAADz75j+s+X+aPMv1H8zOx0z98sfm3678v/tR+d/q0h2fCDQ8b+aftvjfnXhP0r87Dp5x4d56X4Huehd38v9L2XV+B7vSb3k+pd82qfEPjz9L/ADrmIvqvmKZrmAAAAAAAAAABY5+71Hr/ABfR63qfE9nqer8X2Ot6/wAT2erseT6gJgHN/kL+i/AdJ+tfN0Xz4P7cAADMnb8s/Hf1f47U/T38+/U/cDy/lwAA+Vtnwv2L6QPd/Hfx89WD9Dt8PlTxQP0W/FMPbfnYP1Q94AAAPzM/TMAAA+ffzR/VvzDOPs/kPjrm/wAB/dLn9OHJ1fnT+awcp/UEHgX5bB+4n8zAfr39YB+b35lAWw/pS9NDxD5B+/8Ahfob8/fb8bHPr/lD0jk6vrT3z8q/TvSF+W+mAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA/JH9YguAAAAAAAAAAAAAfG31+FoD5R+TA+k/s4ABOS3w64w3AAAAAAAAAAAAAAAA/I39cgAAAAAAAAAAAAAAAy7j5k+U/wBN/OPGk+5+L9N+lPkfqvoz1P8AMf0jwv4b+T/dof1f9/8AFTkN8PlH5X/VvzDgz6z5cuS/Zvcvjfr/AGj1v4D7raPJ9QAADNuPnHyH7z4jV925en2hfjfrvNPKfpPnvKvK/tPj/LsP6r5gNIAAAAAAAAAAAAAJOr59+y7Twfb7bt/n/e7bx7l6vij6/wDs/kP0CPxP9jAADzb83flv6J869F9/Hyn7Q/c/4b+8T8W/YQA8d5vf8b+xuX3w9P4E+Yw/Mf8AdQPlTxQP0W/FMP2V+Ug4X7+AAAAAAAD8w/sPkvnrK/U/zP4rP58/eT24P1J+wg/nR/pfD8D/ANcg9K/DkP3K/mVDPAAP6LfXNcvyR5n97/DqZ084B9WfoF+R/qmmfJfVAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHw/6EH08AAAAAAAAAAAAB+Kf7Lhq/GgeDfpuH5sd6H3uAAAAAAAAAAAAAAAAAAAH5H/AK4AAAAAAAAAAHlQeqgABwHTzfH/AMr/AK/+U4vYe1431J9Q/lv6Z6YfH/WHjHL9B4R8+cn03231X0f5Z8Z/N37F+SVz2fJ1PonxPZ+kvfPyv9N1jxfYAAAAPPurm+UWfoHwn0h7D8J9spwdwAAACcT0c/z58VfqP5ruSex5PndH1vKCkAAAAAAAAAAAAAAAoet95819F3Xd/N/Q93y/yX6D8BzfkX97/oV9E/e/xb+avzZ/QH81fb33H+X/AKUHzn0GV8HeL+s/oI72vycBeJd6H4ufumHyp4oH6LfimH7L/KYef/oCAAAcl1oAAAefdXN+Reh+zfkPwXnfh/7OfoGHCfuOH88n9Gwfyq/vWH1z/O6H9Cv4gh8/eTgfoOH7RfmX9f8AJ/OZ+tfloBufoV8d9b9Nn5P+ogAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAByX5sh+qYAAAAAAAAAAAAH4p/suGr+JQftPeCD8Qg/cQAAAAAAAAAAAAAAAAAAAPyO/XEAAAAAAAAAA/IMP18APJO7i+KvnL9k/I099uPrr6B/H/1iY8r1ADzP5c8L9j+7vwR+s/Dvqfwf9n/ACBD0/O9k+tPhftfpXU/M/0YIsAAAA4Xp5/j7rfvfh/rPb/PfvAz0AAAAGi+d/BfvfiO2+lPK9Puj5f6WLhOrl8586+p+Z81+WvfW3g/W/n8Z1cQAAAAAAAAAAAAAAAC9Dx+j6h3Pgev1vyJtjhfXvRz/fNv8V/YviH6p8P9Y7U9r8pAPgDzoPuP2EPNPH98Mil7XkfY3mnz3vYfuAAAB+Sf62AAAAB8I9p+g/B+hfkz+ffefJv6wB2nxeHxX+xQfjX7aH6T/kCH2B9uh8YfP4fqV99B8j/nx+w/kwfUfNHsfB3fpT2v4b+0Bz9AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAH4+B+wYAAAAAAAAAAAAH4p/suGr+IwftVfCr+Jwft2AAAAAAAAAAAAAAAAAAFIPyp/WMAAAAAAAAADi/iYP0N8k7uL4Y8B/Z/yC99UcvT9i+i/j36yHL1ABy2fb8R/W/L978LfOv8AQX8lE3qeZ9LfZX5l+ketnwn2wAAAAAQNfHGT+g/B/Zez8D9yGegAAAAB498o/cfF/Rv0B839C48v0wAAAPgry34r+tP09T7b+TOJ4n2/E43kPf8AD5Tzf3/C8MZ9d8oDAAAAAAAAAAAAAAAAJes594vafnPrev67x+nrOp8T2eo6fx/W6Kz5vomc1+QGT++fhv1f9gfl/wCk9ufI/VgAB+T/AOsAAAAAZXP9fJ2pydfm/wAch8yeFh5fxwU3h0vooez/AEYH119HA7zDs5Pydz/3v8OCpNjO/wBAPqn8U/Yw8X2AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPznD6o9wAAAAAAAAAAAAD8U/1YDU+Yg8B/TgPza9uD7aAAAAAAAAAAAAAAAAAADzkPhv8ASoAAAAAAAAAA896ub4T+bv2j8e0frnk6/svqvx79ZDHYAADJnb4C+Wf2n8frzfQ/N/UH25+WfpvfnxX2IAAAAAAcr8LfY/JfXfrHzf0AcXaAAAAAGFpn8r/YPt+PIeB7gAAAB818P13wl+rnjfq3Sn0/89gFf539fyvlPw79b/K/avoT5v6H3nqPgPuuP829TzPh31X9D+Cl4jHXh+R+h8GI6MAAAAAAAAAAAAAAAAF2s73t3zfR5TlO/hJ9sfo/7G/NP0b2U+C+4AAAAAAAxw+cPAA8d84Dk8QF2w630gPYvoEPo3VAAz/yg+7+Jo+l/XfKel+m/I/V+l+h/JfU3Dh7QAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADjw/NP8AVwAAAAAAAAAAAAPkTw4D9Bw+dvmEPqT6oAAAAAAAAAAAAAAAAAAADwAPD/u8AAAAAAAADE0z+IPkL9l/Ij61a+2ep/G/1w8bx2536GDE+Tg4fywPoT7TD84vlv8AcPxjI/Yv4z6/00+C+4AAAAAAA4Tp5vin73+l+e2j5H6oAAPyg/V8D87A8izQ/TPXD8vJw6n9SAeAAAHE/lJ8r/R/6W+yfQ/h4dPgZvyR9F4Hx1036l+Z/WP0x+Wfpu+fPe8AfP3zv958R9nd38Z9eHN0FVzxXCe34/C/Enq+jydn6v5qgfW/l4AAAAAAAAAAAAAAAAAAexfY/wAH9v8ASM/5n+jAmAADPjUPzu8gDuvlgOg+sQ809NAA+RPeg6P5KDW+vQ/RP7DCg/TO6Z6AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAH5AfrYGmAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB8nBS+vgAAAAAAAQPl74J/V/y/l/o33PF+9O//Fv2EOXpOQD8j/2fD4akD4e/cEPyU+5g9d8g9rx/zk/aPxfYAAAAAAAA5v4S+v8AlP0FufJfUhNAAB+Hv7hAfkmH6LYQfDvmwfq/34fnj1IfcoAAND8v/Z/jf6q+2z7L+VYvk33fE+H/AFn9S/NPtT2r8g/Vw8/vAD4y9/wt76vz0DxPZAAAA+OPhX4T+xpqP73/AB7XP0n+fTRivde0+R+r4zh/Q4OIx/d8QNIAAAAAAAAAAAAAPSP02/Ovv/zW/Wz87+/ADhg/Db5XA+iw/oA/mFD7k/SMPzw+8Q94+Ng/Iv8AohDp/wCbQONAPrYP3D7EAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA+a/EQ/QIAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAD4hD076QAAAAAAAOQ/Ob7n4rwb0b9F/P637Cfif7IHm+iAAH4h/tgH4y/siH4mftEH5HfpmHsnggfMn6LAAAAAAAAB+fP3R9b8tvHyX1IAAAH4e/uEB+SYep+DB92/nGH7ggeafmkH68AAB8sef9p8hfq35v39nyz6/wDAPzsf+wfk/wB9+0fkX6qHn94AAfFfrH2fyHvh8Z9eAAAAeH8v0f5h1vmf6wqJ/W/+WBaa+zPs78f/AFjdPl/pQAwtM/gL6W6vpeO+av0D8W8IPu/iehtcfVzcHXzA0AAAAOD2n6q+E+2+ldf80/RfyT/WyLAD8Lw+JQAD+lr8XQ/Qv5uD8zftMP1B/nrD6H/ZQPOvigPgAAA+7v28AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAD8aA/ZGYAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPz5D6e9oAA538tQ8A9yD6t+zAAMP8AJX9A+E+qPtPw/asP+b+hAAAA/PPug+Rv2LD8GPrMPqz6IA/OkPq/2cAAAAAAAPnX1fLk+hkw8v0wAAAPw9/cID8kw+rfpgOn/DEP3HnD5a+eg/SoAAxvyN+Y/oT9JvZPe/Gvjr4i/Z/wn7j+vPz/AO7kPB9wAAD5k9jyT6bAPH9YAAADI/Jb5n+gOMpfpX5wh+y/yR0X6c/C/a+zH5h+kgAAcFj6fgv1fy/S/Cnxr+8/zNv/AG9x9n1nd/JP1NmFcc3z3q+Zg4fped8H8b+q/mJ2wfUv1P8AlX6d358n9SAc70QAB/PaHzOAAfsA0Pvz+dAPNPtMP1B/nrD9WvsIOY54PwrAAPq79/AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA+Uw8y++QAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA/NkPt/0cAAA/Gj9bA5DBD3j4sD7TpVN0mgAAAADh/wAD9ifq4Pw9/cIAA/HsP2CcAAAAAAAH5+fUfN/oGfL/SAAAAAH8736Fh6L8yB+i3qwHx6HxH9Oh8hfssHUAAHw7wnx/wDUH6I/nZ+vfxr5R+l/reV6Ofnf34AAAHPfFX1fzH3sfKfTgAAAAHwt8b/A/wBoU4/6r/zkNb2fj/1V9W/Bv28OLsAAA+SfO+59j+Cf0H8n8H+kv0P80/Qzo/xH9lDLUAAD5z9Xy/zc+kP1n8u+qPdPyT9UeeZ6IAAAAAfNofgryIAH1J+7IeJ/z3AfaYfqD/PWH9Fvw8HDfr4H8w+QB2wfvd9DAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB+N366huAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB+XIfpB0wAAB+NH62B80+fh+gP4fB+4IB5oHxz+iIAAAAHw4H29KHyR9bgAH4zB+zIAAAAAAAReOejwe0Hnd4AAAAB5SBraYdFbAA5TzoPaZwADBjp/If9RPmP6J+C3/0b/CX6Wa/4/wDrARYAAABA1OJgAAAGFHT+P2R8r/YdM/rb/LU/SL5P6j6fPyL9WAAAAPgnyv0ryjw3+jP5H+7ftL8o/UA+f94AAAA8E9PzvY9jg7Qz0AAAA/MH9PgAADz/APLkPgLmAl/YMPkH41A98D6//NoP18/IsMH9fA/OTig/QL9Qw7kAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADyD4CD9XAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA/KAP1RvAAAH40frYHzb5sH6Dfh2H7iAHgIeG/eAAAAAAAAAAGd+P4fsiAAAAAAAAAAAAAAAAAAAAAAAAAfGXmfoHk2H9X5Wh+lPsflFk+P+sADwQPy29rDj/1EDwD4cDe+6A+oQAAAObz6/kfwrt+z8Rb/AEB/Ifv/AOoX5P8AqIfHfWgAAAGeP4p3f0L4H7JPz370AAAAAAAAAAAAA/JP9bAAAAAzPlgPlr5+Dx7zoObAAOl9GD2P6CD6m+pA0AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA/NkPoP6jAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPySD9bQAAA/Gj9bAxfxeD6m8AD9vwDDCn1AAAAAAAAAAHg/wAoh+kwAAAAH89v7rh3f54hyP2AH5a+ph7398B+Hn6Oh9TAAAAAAAAAAAAfEXlfoviLP2P8I/UK5+Z/YBNAB80h80/pYHAfkkH2Dqh77+d4frSAAAAAAec/In2XyP1R6z839CHD2gAAAAAAAAAAAAAAAAAAAAHmHp4AAAAAAAAVswF0gsgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAwPxs/VMPQwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAD8kw/WwAAAMbWB/NBZeGyAAAAAAAAAAAAB8OdiH1kAAAAB+CH2YH6R/kMGT98h80/pYH4efruH5sZIfsSAAAAAAAAAAfDHzaDf0hDtZOvl6E5OoAAD5pD5p/SwD8Pg+1vikOS/UcPqcAAPiwPtMDxG0HsnjYVfbgAAD5o+RA9I++Q5P8APIO49UD58APTA+hfcAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADmPx7D9hulAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA/JMP1sAAAAAAAAAPlcO/9pAAAAAAAAAA/L775D0gAAADGD8oeLD9LfnEPm39CA/Mf3EO0gD7C/MIP2fuAAAAAAAAAAfl99gh2f5nB9veEB9x9oAAAHzSHzT+lgcX+RofXeqH2f+IIfuKAAH4eh+4QHwpqh9rfFIZX3WAAAflR90h5d8/h4d+vQS7AeY/mkH60/n4H0b6uHpQAAAAAAAAAB+d36IgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABwQfkt+uIdqAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB+SYfrYAAAAAAAAAfCQe2/QAAAAAAAAAAH5C/ruDwAAAPNg/PqgHG/rMH5C/oaHyV+mIbX4Yh7v4sH379hgAAAAAAAAAfl9uh5l9PB9DcwH5wfsmAAAHzSH5efSgeVfqUHzJ8ih23rIfosAAH4eh+4QHwp8jB7x4OH1z91gAAH5UfcgeJ+dBS4wP0r7kPP/y+D9gfzTDxywH0/wDdAAAAAAAAAAH5J/rYAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHHfkeH6NfQ4AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfkoH61gAAAAAAAAHj4dx1AAAAAAAAAAH5GfrmAAAAB4KHyr9XB+W/7fB+LH6Ih80/pYHkv55B+s/GB+Sf7PgAAAAAAAAAfl96aH1v3Yfjl9DB8m/t2AAACBgXw0QIecDetgAAB+HofuEB8KcyH138iB033WAAAflRyIfTX3KE3iIfkJ+6Acx+XwfsD+aYfSnqofmH+v4AAAAAAAAAHgXvoAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAFYPzQ58P0t6sAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA/JMP1sAAAAAAAAAAAAAAAAAAAAA/I/9cAAAAAIgjsgAUrAR2QgAnAoXwAAAAAAAAAPy++wQ+gM4Pwv+8w+MP29AAAAAAAAAAAAA/DUP2KcHylqh9rfFIZX3WAAAflR92B7WB4z6GH4yfssGP8Al8H7A/mmH0t6sH5ffsCAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB5aH5yPD7G+lgvAAAAAAAAAAAAAAAAAAAAAAAAAAAAfkmH62AAAAAAAAAHzuHp/cgAAAAAAAAAfkl+toAAAAAAAAAAAAAAAAAAAAAAAAcbvBqAeeVA0e9AAAAAAAAAAAAA+FQ89tB9d2g9d8iCr7OAAAfNPs4deB8ZeAh9ffRoYfzmH1R8+B6F0ofOf1QAAAAAAAAAB8OfcYAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAYIfIXzkESgAAAAAAAAAAAAAAAAAAAAAAAAAABwwdyAAAAAAAAAceG3qgAAAAAAAgKB5J62AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAeSetgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHswfoMAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHxt8Yhg/bofbAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB+SAfmJ0Qep/oWH6VAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAeb/AM0Id9/RgHcAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHyN+Bwfb37mAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHiX84Qekf0Zh14AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfhCHxn1oewfdYfpkAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAZgfnH8rAfeYfcoAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABxYanQAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB+TDA4r9RQ9aAAAAAAAAAAAAAAAAAAAA/OkPa/qsAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPxo/WwPFfmYO/wDu8PmvuQ8K8jD9CfgYOe/SAN3jg+O5g+i/h0PcO+D0bIDkPeQ+WoA+e/tUOz+Uw2PUQ+LPOw+9vqsAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPxo9JDyD9Sw/J/wDcIPzp9rD83v1WD5j7EOm+ew/T/wApD8dP3UD8VP1bD8ufu8PD8AOZ+mQs/VweBfmoH6g/mIH7X/huH7mfk0H2F9VgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfKnrcvkp9d5gAaEv71Pz33AD8aP1ND0lQ/Dr9xQ/OP3EPzp/cEPyT9RDsPaQ+gvKQ/On9bA/AT9EAPscPyd68KuUH6y/i0H6IfnOH6ffHwfpR+NgftZ+cAe6fVYAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAH5nfUed8vn6j86AG7m/20Pwr7IA/Gj9bA6sD8u+JDzH9Gg/Oz9wQ8H/MsPdvs4PaPKQ/On9bA/LrMDtP0kD8ivpwOp+Dg/aH8Kw+qPl0P1O+Pg/UH8dg6zxMP0b+rgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPzO+o875fP1H50AN3N/tofhX2QAAABkBrgABXDK3QAAAxgvWwAAAgCvfAAAMbZAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPzO+o875fP1H50AN3N/tofhX2QAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAH5nfUed8vn6j86AG7m/20Pwr7IAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAD8zvqPO+Xz9R+dADdzf7aH4V9kAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB+Z31HnfL5+o/OgBu5v9tD8K+yAPy7eHhH65h5p8Jhnfp2HqQH5i84H1v8AHAfrb8GB7z48HhkIfqoB+anEh99/BAM+qwwPmoPt35DDR8fD9Ae3D824g/QbwoPHvKQ/SjkA+QvtAOl+GQyP1XDowAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA/Gj9Uw8D81D5K/bYPO/zND9f/Cw+Hf1WA/D0P3C/NMPpb8mw+5Pi8P2V9RD8k/ugPN/iwPon5PD6C64P0H/HcP1H70PxW3g/XXUD8XPowPffWg/N/wBcDA/SEPxe+nQ8N+pg+5wAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA/Gj9bA+f8AxQPlX9qw8u/PwP1p8OD4E/WwD8NA/cb8sg+vPzPD9mwOmlD8k/0WDwvzwPuK4G/8iB8scUH6idmH466YfqzrB+QHvYfS3qwfl9+wIfNPwcGB+soa24F8AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPzb/QUPI/Mg9R/PwLf6eh2QH5ocaH17yIfJvTh93+WB8n+hB+l4H58/ZIdr+VgSfWIefeFB9efBAehcqH6MbYfmhOH6O/P4e198Hwx6EHz99MB6n+dQdZ+lYdOAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB+NAfrZ1YAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAcAHcTgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAMwQ5vBDCxAx8sM/LCCgAAAGkFjSDW2Q298Oj6kOgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADM0wAAAAAAAAAAAAAAAAAAAAAPN9IO2AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAACgHB8EHCcEHE8cFdQRQRwP3w6PeDd3A1tUNDVCe+AAAGaFbNDJxgxeeDnOXDngBARQ9G92D6I6sAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAD8hw0vNw/Sz6CAAAAAAAAAAAAAAAAAAAAD8/vQA+wQAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAOM8yDzLzQPOc4EcCXg7zvg7zuw7fsgnAAAAAAAAAAAAAAAAAAAAAAAAYHm/g4fOvHhr/AFmH1PcAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPx+D9QfPg+PupD9Gfl8Oo8CDyz9EQ/P7nw/R/uQwfzqDoPpUPi/0APfNENrxIPsH4bDqfnkPqj2YPhfvw9E+RA8y+8g+wQAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAYHAeTh5R5UHKACh2Xpwenekh6PpAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAGL8kh8pXg+3PpMAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAD8fgs86H6i/kmH7g/n8HoHwCH6v/JoL6AHzX+socp+HIfvd+HQfph+dwfon8dBv8CH2HzIfXnyuH56frSH5bftmH4XfugH5j+8h9ggAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAGb5OHkHkAedRAkgegesB6v6qHVgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABiB8O/NQfSX3eGiAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHh3q+VwZ9V8oAGjF/Th8H96AH4/B+oPoAH4dB+4n55h6Z+fwfuF+WAeq+nh6P7IHKfkmH7L/g2H6cgfRv5Mh2GoHnH61h+LX6rB+V/6oB8J/qcH4j/uCH52+oh9ggAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHL/OweC+cggGj6yHr/rwejyAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfNPwQHuP6FhogAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHyl9B8/5KfoP58AGzlr93H5H+uAB+UQfpH3QH58h82RB92/DAfsx5eH5VepB9w+5BzH5ah+uP55h4t3Yfp1+aQem+4h+Z37QB+LXtQeGfqaHxV+mwflnyIc592B9YgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB5z8qB8+wh1P0SHv3pAKAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHzz+dwfW33ACgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfKX0Hz/kp+g/nwAbOWv3cfkf64AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABznyiHzFQD0f6rD6DsAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB8ZfGQfp568AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHyl9B8/5KfoP58AGzlr93H5H+uAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAACeDh8e+dh0n1eH03ogAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABiB+XXowfow4AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA+UvoPn/JT9B/PgA2ctfu4/I/1wAAAAAxeMD00DH+Mg+5QAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADKD5P+VQzveA+xfQwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA+Ug+GP1nDrQAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAD5S+g+f8lP0H8+ADZy1+7j8j/XAAAAADz7yAKuaH0j+SYfsuAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHI/GYfPGmH1j9VhpAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHCfkqH6a+4AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfMfu+F5afd/CABrZ6fbB+V/qgAfjoH7F/hCH7jflkH2v8ANQeLcSH6m/i2H1jWD9cQPAfzqDv/AKzDyT78D8Z/1yD8qNAPrL67Bn4KB9f+qh4HxQfrN+QIe1/oaH4nfQ4eV/s4H5c0w5H9hQ/Df3APK/2xD8a/Qw/U38og1O8D9FAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPPA+KPFg637MD6IcAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABRD8kPtQPqkAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAD8kg+7fzdD7T+QQ/Q34+D2sD6r/IcP2j/DoP3FA/E/8AZoNb5UDxT9Fg/D36+D5n9aDxn9qAPweD94fwTD6r+dQ/Rv8AN8P0t94D8fP2uD82e7D5s/X4PzX+hw/ND90A/HL9UQ/HH9Bw7/8AKMPp/wCWw/eUAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA83+Hw8k9AD7Z9qAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABv5Wh9G/Y4AAAAAAAAAAAAAAAAAAAmMG0AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHyyH5eftaH48/XofQvx8Ht9APrb8kw/Zf8Pg/cED8aP1ED1P5xD5F/SkPw/8AvwJft4AA/D0P3C/E4P2ivglEPxK/XUPx4/dYPyd+xQ/Ov9sQ/JP9AA/Kn9wg/H79QQ7j8dA+0/mIP1dAAAAAAAAAAAAAAAAAAAAAAAAAAAAAODD4d8YD0b7eD2EAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPyu+nA+tQAAAAAAAAAAAAAAAAPnP4hDK+3w+oAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAArB+YX6jB+cf3SGF4MH0l+WAfpf8sB9/fnMH6MgcJ+Z4bP6jh+UfVh2338H5j5QfYf0kB+cwfoz4YHwL0Ifol+dIb/6DB+GH1KHov6IB8V/OQe1fe4fnN+jIfB31oH503A/T34GDyb6LD7RAAAAAAAAAAAAAAAAAAAAAAAAAAOZ+JA+ee8D7i9pAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAT8kw+6PoUAAAAAAAAAAAAAAA+SPh8PpP7oDcAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA5QPyT/AGXAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAACiHx78nBv/AG2H0OAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABAH41/q4HfgAAAAAAAAAAAAeaflyH1B92hIAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAMDJ2QAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAD52D4fzA+w/rELAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAByv5EB+zU4AAAAAAAAAAAfG4fJX6tB0YAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHC/BYeRfRQfcHQgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfOIfK36bgAAAAAAAAAAH56UA/RoAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAACAPkH4/DtfvMPXQAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA/LH7iD2wAAAAAAAAA/P4JvvsAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAD4+8j7rxs+Q/eAA29Ob9Cj9C/lYAAPE/awXmAwu3DnuuAAAAAAy/lUPr4AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA82D4A4APr/7ACwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABlfj0H7NAAAAAAAAAfJnhAfpUAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfnn8/wDqPzmfn39TAB02vJ+uR+ufwmAAH84n7VB9I/loHG/peHhv2AAAAAAByn5Jh+y4AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAR/JAfGfoIff8A6OAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHw+G39igAAAAAAB56H5dfsMCgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAH55/P/AKj85n59/UwAdNryfrkfrn8JgAB+J/dh+t35FhifpcHzF5yH6H/m4H6efmKDPqsO8/OcOr44P2XAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAOa/PwPGfrkPsyyAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAwPxm/ZsFAAAAAAD8bw/WfqQAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAD88/n/1H5zPz7+pgA6bXk/XI/XP4TADMD8k+YD7rxg+Mv0hD4+/QQPw8/TMF+LA+ifk8Nv8AY4Oa/KoP2XAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA85/OkMr9BA9tAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAD4a7YPq8AAAAAA+JnB9rgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB+efz/6j85n59/UwAdNryfrkfrn8JgB54H51+rB8NfuuH40/pKHx99ph+UX1WHq3ngfcVwPx1/X0OT/MAP2XAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADy784A3f0XDvAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAKv5CB+woAAAABSD8c/2ZAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAD88/n/wBR+cz8+/qYAOm15P1yP1z+EwA8xD5w+sw/P/8ASoPzO+1Q+dPKQ/Sn80g/Rn8zAk+sQ7/80Q9pvh+joAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB57+awbX6Qh2AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAflUH6RdmAAAAB+ege6/SwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfDvifonhB8R/RgAdDpzfpwfp38cAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAcz+aoZv6Vh24AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHg4fNv6FAAAABF+OIfsmAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABW/PIPEv0tD1YAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA/ID9fwAAAA+NQPsoAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAD5RD4O/QkPpYAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPyL/AFrCyAAAH47fsEE4AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAebh+YP0KH37IAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHwN7wH0CAAHEB+cX6qAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAYfzXRRPzTvACTVdMftfkgBwAeV+nh6IAAAAAAAAAAAAAAAAAAAAAAAAAcKHcqAAAAAAAAAAAFYPzi86D9QOrAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA8Q+Zg/QgAAPztD6c92AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA5j4zryz8W9UAJd12x/SvgAB+PwfSHzaH2L9fAAAAAAAAAAAAAAAAAAAB+YIfp8AAH5gh+n3wMH3JqgAAAAAAAAAAHzz+cQfop9HgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAFT8oA/WoAAPx6D9hQAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADmPjOvLPxb1QAl3XbH9K+AAH4/B+oPkofM/CB+tvxcHa/Kgcb+kQfnXVD9B/bgj/LMIfoQPHvt8PgP9Aw/KT1MOS/XwPzl5QO5+9A/KXoA+k/yYD9NvTg+XtYPp38aA/Vf55D9S/wA3g5N4fq1+LIeiZIfryAAAABk/l8HRfpUFsAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAD8jg/XEAAxvywD9aAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADmPjOvLPxb1QAl3XbH9K+AAH4/BZ0A/T/8AGcP3B/P4PQPz+D9k/wA9w5fqw82/X0Pl786A+ovj4P2u/D4PvT7LD8bv28D8yvYg/N77eD4T+4g91+qwPxTD9rPNg+JviAP6AfxTD9rPxTD9R/zsD9ePyxD7Q/KIP3b/ABUD9jN8AAAPmMPz4/T0PZgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAD8kA/W8AA+RKwfYgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABzHxnXln4t6oAS7rtj+lfAAD8fg/UH0AD8Pg/bP8AOYPZvz+D9wvzKD6M9+DT2A+YPmgP0LtBx35Hh77+jQfjV+4Aflv9GB8T/rSD/gsPW/sED8UA/a/8QA/ab8Vg/cL8Tw/bD8Uw/U780Q/Yz8mA+/vynD9wfxoD9bOrAAMkPy47gP0glAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPyQD9bwAD8pf04DogAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADmPjOvLPxb1QAl3XbH9K+AAH51B9z9cB8lB8UdwH2P8AJIfqHzIflh0QfcPuQN/LcM/6fDwz9Cw/PP7SD8TPrsNT9QQ/O/y0Pc/tQPym3Q/Qj4cDvuSDA4kP12/KgPojxoP0l/PoPM/Qg/R78vA/UP8ANsP0F3wAPnf84g/T/wBmAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAD8kg/W0AA/IT9ewAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADnvlOnOPyb0gAl3XWn774oAAAAAAAAAAAAAAABygfkn+y4AAAAAAAAAAAAAAAAAABX/MsG/pqFkAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAD8kg/W0AhD8kv1zAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAgDh+/AAAAAAAAAAAAAAAAAAPKw/K/8ARgPowAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA/JIP1tAPFQ+W/0OAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAD4Y+bg/VnfAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPySD9bQD4bD1P6RAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAMX8og+kfuYAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPyRD9bgD8rQ/TDogAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADBwrPODQAHtdWerkAAAAAAFLkw7kAAAAAAAAAAAAAAAAAAAAAAAAA8RD8xv1OD1kAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAD8kA/W8A/I0P1yAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADleW8k8vUAJGd4e7zgBxAflVuBg/rmHmmoEf50h+tgAAAAAAAAAAAAAAAAAAAAAAAAHwR4mH6p6AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAH5HB+uIB+SAfreAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHK8t5J5eoASM7w93nAD8dA/UD0sPlT5rDwe2H6Q/m8HqP2eH0Z8cB4B94hw3lgev/QQAAAAAAAAAAAAAAAAAGX+UAfSv28AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABgfmWH6pgH5IB+t4AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAcry3knl6gBIzvD3ecAPw+D9wQK34nB9c+rB6r+Qwftx+IAfrl+e4fo3+VIfSPOB+mFkAAAAAAAAAAAAAAAADysPyr/UQPZAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAD538cD7rAp/lSH6xAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAByvLeSeXqAEjO8Pd5wA/EwP2L6gPA/iQPVvVg9V/OkP1s/D0Pv/4rD6W7IPOPaw+qwAAAAAAAAAAAAAAAAPkj4wD9atwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA/PH6jD2sDzj4hD9JQAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA5XlvJPL1ACRneHu84AeGh+WH0OHhH7AB8/fA4fpf8HB+tn4eh+0f4yB9M+1h4/7WH1WAAAAAAAAAAAAAAACfmmEv6TgoAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfkN+uwPA+fvFQ+6wAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA5vnrNPO1AB7XZnsYgABByQdoAcoHTQhbohejDiu0COQJQAAAAAAAAAAAAAAyA/JH7TD6yAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAiD8h/19AA+TYA+uQAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPOvydD9S/XAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPmIPNPugAD4i9PD6OAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA+eg/Ov9dQ6QAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA/I0P1Y2gAPgP6PD20AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPjD5pD9XpwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAOY/LoP1sAAPzn+xw9RAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAD86wqfo+AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfmX9oB7IAAfmn92B6AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAIH5dezh9vgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABzwfk/wDsCAAAfmR+gwdiAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAn5RB9SfXQAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAflmH3b6+AAAfl9+iwdWAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB+VP06H1yAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHkPwOH6rAAAB+X36LB1YAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAH5eB9CfX4AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAMD8Zf2BDcAAAD8zPvoO5AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAD82fTQ+2QAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPzdD336lAAAAD80/uwPQAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA+BAT78AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAD51+QA/UkAAAAD80/uwPQAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAD428cD9LAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAOHD8mf2WCwAAAAB+c32KHqgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB83h8XfrCAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAGV+Nofrx14AAAAAH52/W4etgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAeR/maH7FSAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABAH47fpiHr4AAAAAB+f30sHtYAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHIh+Q37IhrAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABH+RYfef0KAAAAAAAfCXtwe/gAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABS/GoP1q7gAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAIw/Ir7fD6aAAAAAAAD4m9GD6TAAAAAAAAAAAA8oD1cDzTigAOk9cDxT2sAAAAAAAAAAAAK3Lh2IAAAAABy3g4ei+uAeQh6jdDz3y8PVu8A5UMj0EAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAD8i/0ED3AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPnHxAOu+sg3eCD429wD6WAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAKn5DB96fSAAAAAAAAAfHmyH1UAAAAAAAAAAAB+Hoftjoh+HP3MHxf7EH1L1gfhX/Q+HZAAAAAAHyzeD6YAAPij3AOl+dQ+zAAAAAD4yD5D+vQqfa4Vf54g/Vj7PD8aPrQPj/9IA+g/wAow83/AGkAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA/OMPVvsYAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPw9/WYPPfzJD9q/xeD9X3h7QAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAc8H5DfpEHvAAAAAAAAAHyxgB9jAAAAAAAAAAAAfnwHo3sofnz+sQfmn9LB9LflAHvXLB+hwH5UaIeSfpmHxp+goec+DB8o5IfrR8Gh5j+pYfjD6YH6P/MAeRfreHxl24fN3PB6N+gwH4ah+5KgAfIfAB8v/ALNh+NH6NB8Kfdwe8/jKGt+qgdiAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHyj89h+mQAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAH4e/uEB8UBh/GgfeP2SEgAAAAAAAAAAAAAAAAAAAAAAAV+GDguKDjuWDmMQMeEFAECfaDoOqDr+9D0H0YL4AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB47+XQfq96SAAAAAAAAAB85+VB9wgAAAAAAAAAAAcYH5R+lh9re3B+af0sH0r+CIfon+fwfu6B+Gf7YhjfkyGz+oIeU/NIeg9UH1/8SB8j/TwcV9WBq/CYeg+5B8Hfp4H43fagfDf79Afh2H7ifGgfnR+6Afjj9Xh+eH70B+QHvIeG/foS/nEHpHq4foYAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHl4fmH+xQOAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA/D39wgPi4Od+5A+EvlYP2lAAAAAAAAAAAAAAAAAADNDwvwgPHM4BQsd0He9qHZdUHT7gbEoAABBihzvLBx3Ch515yFBAT0kPfvoQOoAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPh0Pm79Xg1QAAAAAAAAADxH5uD7/AAAAAAAAAAAAAAPxU1A/ZcD80/pYLXxMH3L+ZYfrv3ofhn+1wct+YYa36Fh5L4eHofRhBwAel+fhy31IG38Jh+lv5EB6V+hYfmR+rASdOB+HYfr16UH40frkH4xfqEHyd6gHyT+tgeTfGgc39kh1H5eh+4gAAAAAAAAAAAAAAAAAAAAAAAAAAAAABmfjUH7FbwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAH4e/uEHGfjSH7MWg3vw4D90AAAAAAAAAAAAAAAAOKD5T+cwp6we6e8h7HpgAAAAAAAAAAAAAAAAAJ5oHgHz0HKzh9C/WwdwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAYYflR9DB92gAAAAAAAAAAB5n8VB+kIAAAAAAAAAAAAHxFlB95AfFHtYfOv1SHpvzSHO/XAfhx9BBwv6cBy35pB7H7AH0J+XAfo18AB3/0EHoX50h+m3zQH3t+VIffHrwfnH54HvX3KB58H5g6Yc9+g4ee/ZAc/wDngFjhQr/poH54/qGB+Y4fpDrgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfkb+hgezAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHwN5EG394B6V+WIYH28H0gAYGk+RcF9JwXfpDxOqW35XTbXokAAAAAGh88/G4ch2wfV30aF4AAAAAAAAAAAAAAAAAAAAAAACuHzv8kBwnoofa/swAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB82fneH6m+rAAAAAAAAAAAABy35xh+ooAAAAAAAAAAAAAAAAAAAAH4ffuCAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfBgaX3CAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAFH5W9rk+UfGP1D52I78fWP05/I/pA+E9d+ptO1f9TINEAHz0HxHzQfRf2UHYgAAAAAAAAAAAAAAAAAAAAAAAAAAAAHnQfEnjIepfeAeiAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABl/l6HTfpSE4AAAAAAAAAAAAEP5Oh+tAAAAAAAAAAAAAAAAAAAAAHnPowAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHi3wAH60AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfNcen+efHfu354G0AHrH6c/kf0gfCeuAGlouls+xhwXwRR5T9DB9u9OAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB5T8Hh559aB9mWQAAAAAAAAAAAAAAAAAAAAAAAAAAAAPl8Pz9/TYPbAAAAAAAAAAAAAAA/JIP1tAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADLD8a/2VDTAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADOnX4C+dfzH+xubyv6v/hoPa8YA9Y/Tn8j+kD4T1wAAd8udUeH/fPqZesAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAARfJIfGfpAfoV2oAAAAAAAAAAAAAAAAAAAAAAAAABx4fl364H6JygAAAAAAAAAAAAAAB+SIfrcAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAH5Mh9/wDs4AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABkfmv8v8As/kx8N/SEfG/v38ksP1z8LAPWP05/I/pA+E9cAAADo9318Q3kAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPKw/POgH6H+wAAAAAAAAAAAAAAAAAAAAAAABCH58+EB+oPoQAAAAAAAAAAAAAAAAH5JB+toAAAAAAAAAAAAAAAAAAAAAAAN4Ds4/MeH+z+M5pnqeT9ZS/iP65av9M6lvvzC0AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfHnngfoMAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAECr82fFPyz+1Q4voDH+8/JeeP6e/i8A9Y/Tn8j+kD4T1wAAADqOqNg9TIAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADlw/OzzgP0R9xAAAAAAAAAAAAAAAAAAAAAA+T/g8Pv76iAAAAAAAAAAAAAAAAAAPyRD9bgAAAAAAAAAAAAAAAAAAAAACL5l8v7L543/0H4IPtPyk1MNvrM/B/wBjAC9ouh0fWxDRAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAcUH5RfsiCgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB8X+N9/wDG5+b/ANcAAmvLyVT+yP8AOsO3j9Y/Tn8j+kD4T1wAAABzOzt+1gFoAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADDD86/LA/R32YAAAAAAAAAAAAAAAAAAA+c/wA+A+nPvMJAAAAAAAAAAAAAAAAAAA/JAP1vAAAAAAAAAAAAAAAAAAAAAPP+f0/z68s/MP7LsfQP6v8AzZYP2H+UzUw2+sz8H/YwAANXVdPJ7GAMAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAD8cg/VXuAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADgPyu+I/oyufK/tYAAUuT/o/+NA/S/yD1j9OfyP6QPhPXAAAAAL/AGPp5B1wAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAYAfm/wAaH6Zd+AAAAAAAAAAAAAAACB8s/DAe8/oIGiAAAAAAAAAAAAAAAAAAAB+SAfreAAAAAAAAAAAAAAAAAAAAB4b+d3wn9MZR8z+vnqP2/wCPegH9F/wQamG31mfg/wCxgAAAWOv787J35gAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHwpqh9rAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB+e/zf8Al/8AZged9WAABzn6b+I5J/Rv8h+sfpz+R/SB8J64AAAAAdf1xonp5AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABxAfmX2YfpNpAAAAAAAAAAAAAeZB8SeMB9RfcQWwAAAAAAAAAAAAACP5SD5FqhM8KfXB+o7wPyoD9PtkAAAAAAAAAAAAAAAAAAAPEeL6H84Kn5Z/aYKzT7fI98d/Wn+YBqY6fWZ+D/ALGAAAATUuyse1gFAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHDB+Wf7CgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABz2fV+SFf8j/uoFYAAEXd5PHx/2R/nf6x+nP5P9AHwnrgAAAABqarrD2cAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPCQ/Nb7mD65AAAAAAAAAPLQ+aPnoKHpQfafsAAAAAAAAAAAAAAAAfGIfG314H2PqAAHyOHC/eQH51h9c+tgAAAAAAAAAAAAAAAAAAHEflt8X/QuOfJfuQAAeydb/Un+bhqet8h9Zn4R+xgAAAAFvs/RycdsAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfjwH6pdqAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHzB5n2HwMfmP9jgAAAAGPz39Q/wAP+sfpz858yHwnrgAAAAA5ndv9znBgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAN+Aw+fv1QDrwAAAK4eP+CB4Rggeoh9L/QQUfCA8b80DlgGoCPBIQ1/uIPegAAAAAAgD8m/oAPupQAAAA/Ib9eQPi4Oz+ngAAAAAAAAAAAAAAAAABA/Mvxz8q/tkOP3gAA6T2395/ig2f0H8F+rj8I/YgAAAAA3d56M9fEAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPjbmA+8AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPz98D9O+aT89/qkAAAAATkv17+fPTf05+g/nwPhPXAAAAAADr9H2sA0QAAAAAAAAAAAAAActVDs/mkPjT6ED7d+eA+GPZw/QfyMPzv7gP005IPzAsh+sYH5LRB+o3UB+YvFh+jHqwfD3Ph9Zenhz2wFkAAAAAA8//KkPu/6UDx/xQPE+WAmD2D3sPd9sDjA+APLw6P34PZ/TQ6xQAAAA5z88g0v0eAAAAAAj/IkPvn6BAAAAAA/JH9bgPnEPLPuEAAAAAAAAAAAAAAAAAA+evP8AqPzsPy7+zwAAAD3b9P8A5y29n9r/AI7+rj8I/YgAAAAAVnb2Pb5woAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAMMPyB/ZkFAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA/Lj5j9j8uPhf6UAAAAACl7fzHWfpz+x/xAHwnrgAAAAAB0u37GIbSAAAAAAAAAAB4CHyz9Gh9L/EgeI/YofQmGF2+AAAAAAAAAAAAAAAcj5+Hd9mHx58tB9e/VYfIXPB9Hbg+h8oF5B48HnDQ819dD2H2oPbOpAAACMPzY88D9FPWgAAAAAAAAAPzS+pg+hwAAAED8l/tgPpEAAAAAA/JP9bAPLg+Mv0eAAAAAAAAAAAAAAAAAAPy8+Z/YvKj4T+lQAAADt/pfz71rZ/pX/O/6uPwj9iAAAAAADb6X1cQ6ZAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPyqD729fAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA/JP5T9u5M+J/okAAAAAAO0/ST9W/i8PM8AAAAAAAN/ofWxDokAAAAAAAA+Ow+UPusPojwgOl9RBQAAAAAAAAAAAAAAAAAAAAABnn4eT+Th5NzIFUNv1oPXfYw9EeAAAAAAAJ+QIfcv00AAAAAAAAAAAZ35Fh+woAAAJ+UQfVf1mAAAAAAAfkl+toGMH5ifqoAAAAAAAAAAAAAAAAAB59+VHw/9Hh8t+0AAAAEl5fQHTf1f/mF9XH4h+pgAAAAABJ3foZqd2YAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHi/w8H6kgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAH5G/J/uHMnxX9DmnOO1tc/g7W1zfP7ezz/AD+3tc3z/Velfpf5iHueEAAAAAAG90XrYh0SAAAAAHywHxv91B9C+fB3U4AAAAAAAAAAAAAAAAAAAARedh5Z5WHlfJAIEnoAer+sh6x0wAAAAAAAAAAAAfE2gH2SAAAAAAAAAAAB+ZQfX/uwABQD8l/sIPrUAAAAAAAPyP8A1wAA/JH9bgAAAAAAAAAAAAAAAAAPkr4c/Of6yDxP0UAAAAA9J9h/or+Avq4/OO4AAAAAADseyL56WQAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAH40/r6GyAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfP/wAF9La2vyj19mxh4oKQAAE74jP27ygYAAAAAAHQ7/r4hvIAAHGB8Xe4B9KeQh6bogAAAAAAAAAAAAAAAAAZ/mAeV+WB5fiAATehh6nthU50POuGCsAKAAKAAAChHGHZ/pqFsAAAPyI/XUHAAAAAAAAAAAAZofjj+ggfTUQfJvwoH6Ye6AAAAAAAAwPyU/W4AD8k/wBbAAAAAAAAAAAAAAAAAAPz3+b/AMv/ALLDzvrAAAAAC99MfsH8m/WR4XwQAAAAAAHSdM7h6uIAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHx0HO/c4AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAFcPyw+xfhuz1c/HPUAAAAAEj/QOMPvuMAAAAAAADqdf2cA1QBynKh6r8hB33vwKAAAAAAAAAAAAAAGMHiPiYeO4YIoaHp4ep+phuebB435EGOgO7cPYfUA9G7gLYAAAAAAAAAB4H+dofsHbAAAPyQ/W8AAAAAAAAAAAAACn8Jh4NZD6X+ywmAAAAAAAA4MPgf8ATQAD8k/1sAAAAAAAAAAAAAAAAAA/MLyT8p/tsOT3AAAAAA91+2/GfuM7f52AAAAAAA3N56Q9fEAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAK341B+zqgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABzH5Fh+ivs/y3Rhn4Z7AAAAAURp+3+UHbAAAAAAAAHbWvcwCkfGofNH6Ah7AAAAAAAAAAAAAAAVvn8PmTywF2g9u9uD2OyHjfigeLcuAgdv7IHsXsAbAAAAAAAAAAAAAAAAHg4fA364gAAH5JfraAAAAAAAAAAAAAAAAAAAAAAAAAAfPIeN/dQAH5J/rYAAAAAAAAAAAAAAAAAB+VPnv5N/cYc/qAAAAAB2nb4P6Sn0H8ZIdch1SDAAANjaeoPYxAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAD88/cQ+mwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPLQ/Kj9aA9Bo+ZpyR/O3uAAAADPp8Gn6v5wAAAAAAABYpdxD7mHxJgh9/gAAAAAAAAAAAAAB44Hx35SFv6BD6d9NDxvwIPDMMDXD2P2sPaepAAAAAAAAAAAAAAAAAAAAAA+NPPA/Q4AA/JL9bQAAAAAAAAAAAAAAAAAAAAAAAAAD4+DV+rAED8kv1uAAAAAAb5KHz94cHNOA/RMPXgAAAAAD8svNPyf+4A5vXAAAAAAD759k+N/kcObwAYJ1yHXInXJsfQ4dKfrPmAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABkB+RP7GgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB4L+cAfr9tgCOMr/AM3e8HLQAANZ+tecH0mAAAAAAAAB0HRPQeI+tj5p9cAoAAAAAAAAAAACB8+fFQcv7GH2PbD5i+fgqTB7H78Hum0AAAAAAAAAAAAAAAAAAAAAAAAAAfliH2R9DAAfkj+twAAAAAAAAAAAAAAAAAAAAAAAAAB8Mh7B9DAcaH55/p8AAAQfPQfKPnwS+uh9A+5h0oAAAAAAAAH5seHfln9qhw/RAAAAAAB3X6PfO/gcp435IAAAAAG79Nz7x+7eOAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAH5rB9Ze7gAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB8zB8NfroFoAAxud/EPXD5LpABO+Iz9u8oGAAAAAAAAYn5rd2f7Av8AQzAAAAAAAAAAAAPNA+CfOA+h/qwPm/5vCD04PqD38L4AAAAAAAAAAAAAAAAAAAAAAAAAAAABXD8Zv12Dq6gflL+soAAAAAAAAAAAAAAAAAAAAAAAAAB8OB6z9FAfNIebfb4HkAfJXioT/Q4fV/oIAAAAAAAAAAAAfDfyX+af2AHk/bgAAAAAAfZvnfAfSB8r/M4AAAAAdP8AZ8uoftPkgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAGJ+SofsKAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHyt8dh+tMgAAAnH/AJ53VT8m9IASP9B4w+94wAAAAAAAACp6H6+Ng3kAAAAAAAAAAI/kEPjj0gPsj5nDw7rA+vfpELAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHBflCH7JfPgeM/eQAAAAAAAAAAAAAAAAAAAAAAAAAB8zh5z9vAfm0H2f88B8j+7h9ae0goAAAAAAAAAAAAAB8/wD5z/Af1EHzv6qAAAAAABuzzfpZqfEfxuE4gAAAHa/o3DMfq3mgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAH5s/WQe6AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB8sh8c/rUDgAAAKfn3yTf5190AjT9w8kOyQAAAAAAAC9ouxX2sAAAAAAAAAAAy/gIPn/7ADY+Nw6f7kD3YAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADwUPzkzA/YTpAAAAAAAAAAAAAAAAAAAAAAAAAACgH46/sqEn4mBtfYofagAAAAAAAAAAAAAAAAGd+Rnx/73lnx374AAAAAAAB9Q/Xnyn8xB5/woAAAF3rv1/yw+94wAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAACgH5C/sSAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfOfw0H63uAAAAAAyuZ/FvWGcltP1bzgAAAAAAAACx2XoZzHdmAAAAAAAAAY/54h4796B4h83h9afagWwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADzrkR+5ggAAAAAAAAAAKznzf5z+u+f8An/j/ALLD66+nPyH6Wxc8np0dHvzU0QAAAAAAfN3wSF31QP0P1wAAAAAAAAAAAAAAAAAD4S8P9I+VD86/q8AAAAAAAC1+k3i/j/Wnz34QAAAB02r/AEB4oe5iAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfn+HtX0sAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAeNh+bH7CBOAAAAAAGV4e3MR/OdAfQ4gAAAAAAAFq111j2cAoAAAAAAAAqfnsHhH6Gh84/PgffH0uAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABmXn4L4t+ifnmd5f73neCnqfS/cX1f+E/XB892k1G3uepi86JAAAAAAAAAAAAAAAAAAAAAAAAAAA5PLt/KPP8AyX+5QnUAAAAAAD3Xl+b+6D47+TAAALXbPYL/AEd4IMAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAT8bQ/ZMAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA4r8kQ/ZHSAAAAAAAACjy3n6Vjh0AAAAAAAA1+m9TJ51QAAAAAAAAfGwfFf6Fh518dB+g30sAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABw+OHqAAAAAAAAAAB5F80fffA5h9H84c7Wnyuet9V9xfV/wCE/XB892gBNR1Gl7GAaIAAAAAAAAAAAAAAAAAAAAAAAAAAPkn4d/Of6zDxP0QAAAAAAAD9DPUfi/5BDDxgBes+847x+weWAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfKnCB9zAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAFIPxp/XEOyAAAAAAAAAGoxMLzNYTlsAAAAAn6nvz0DvzAAAAAAAAPIw/Mf6vD3L81g+yPtMFAAAAAAAAAAAAAAAAAAAAAAAAAAEwQ3wAA8BD34AAAAAAAAAA+afPw+1gAAAAAAAAG/NfueH4cfov50ACB8i0PX+s+4vq/8ACvsA+e7QAAOh6J3z1sQAAAAAAAAAAAAAAAAAAAAAAAAAAb+c/wA7+reAnwH9QgAAAAAAAelfol85/Pqnj/lQBv7n7v44fS84AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB+Pn67hcAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPycD7u91AAAAAAAAAAABAzcziulW4tI1h+e+U6z8s/HHdn+lf6M9UgAAAAAAAFQPzX5cP0h/OENj9LQ0wAAAAAAAAAAAAAAAAAAAAAAAAAAxg/GP2cPqL4/D5U+gw+sfiEP3BAAAAAAAAAAPGOTD6TAAAAAAAAA+XvGf078xD0/MAAD5x4P1Pp/uL6v/DftQ+e7QAAAN/ofWxDokAAAAAAAAAAAAAAAAAAAAAAAAAKE6/m/4t+W/wBpBxfQAAAAAAAfcPmfnXvh8v8AzabP0WHRH714wAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAcIH50/qiAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB8SYofeoAAAAAAAAAAAAAAAHPh8bfNAfmqB9A/wBFIAAAAAAAAeHfmSH6QcWHw7+n4evAAAAAAAAAAAAAAAAAAAAAAAAAAAAeUB+Wn7Pg8D8k/wBFg9W/CYPpSYP0/AAAAAAAAAAAAAAAAAADwf1vJ+cD9L/NAAAAPIPFPR+i+4vq/wDEPug+e7QAAADqeqNc9TIAAAAAAAAAAAAAAAAAAAAAAAAAAKfwX8/+o/M5+ff1MAAAAAAB1Mcf6UXfif4439v928IPpMAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA/M8Ptb1oAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAKvjvZj+WH7D7T0lnx+mc2QAAAAAAAAAAABjB5L7mB4WH85IB/QeH02AAAAAAN/OEOL/RsPy/93D9AVAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA+HPh8P03+jA/JP8ARYPVvwOD94/yED9PPTQAAAAAAAA5XNDvAAAAAAAA4b41+w+WjPsfhwAAAOP+Ze/3PuL6v/Ev0APnu0AAAAeztp/b5woAAAAAAAAAAAAAAAAAAAAAAAAAAD55+HfhP6W48+Z/YQAAAAAA+3/q33/5DuH6F8IAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB+O4fsQAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAGJHR4Xw31Pgee/HX6N8c39IPlvqvcT8e+ml0t519L08g0QAAAAAAAABxv5Fh+p/q4AH8/nywB9vh+5YAAAABxwfkx92hm/Eofqn6IAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAGMH4v/ALch+Sf6LB6t+HwfuD+VAfdntYAAAAAAAAfNPn4fawAAAAAAAfmt5f8AAf1j6ru/0Z/CAep84AAEPyB2ev8Abv1f+K/oofPdoAAAABsdR6uIdMgAAAAAAAAAAAAAAAAAAAAAAAAAAVFfzb8x/Ef0X5JH8v8AtAJgBcrP3H6k+w/Bvdl+v/BQAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPDPl4P0UAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAKU6fIPyR+d/1cvvX6r/Pp8p/rn4Fg/pB8v8AX+4n499MAFy10ej6+IaIAAAAAAADm6QdiAAfE34XASh/SF7GAAAAHzn+dgfqb+cwezfoICgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB5SH5sZIfaX2CH5a/foem/jQH7L/AJkh9q+wgAAAAAAAB8744fUAAAAAAAeX83sflufk/wDb6939T+bel2v6S/z8DfjAD5j6fS+tPq/8Y/Sg+e7QAAAABe47s5z0MwAAAAAAAAAAAAAAAAAAAAAAAAAAAADC8s876vhsXxf0Neicegeo+7+bXzt+dAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPyx/RkO1AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA87/O74/8AfPPD4797Pcf0z+eOg+eP37+PfPP0g+Z+09xPx76YAADb3npF9bEGAAAAAHjnDB9OAAABD/N8HjgH39+1gAAAfAoeUfZYfnF+pIetAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHwn4f6P8pn51/WABc9O/RPwbtl/ff4hBo8S27PoX6v/ABv9QD57tAAAAADoeid89bEAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA/H79gQAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPHuT3fzfyPyz+0wjc7r6f859X8d/qH/Pjxf9IPm/uPcT8e+mAAAA0es9LJ52QAAAAeYh+XH7BBpgAAAfn0H4tAWP6Pg9kAIw/K72sOT4cP1DtgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAc/GHSAAAAAAB+T3yv7XxB8R/RgAAdD6r+y/yZtn67/LRwN7+rfV/wCPfqofPdoAAAAAX9F2J7WAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB5cHxH+loAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAea/mP8R/ReSfK/tgAWduT6D4f+vP8v8A5y/SDwftvcT8e+mAAAANLVdavs4AAAAFUGXQAAAAKQfze+SAfcwfuIBmfkMH2v8AFYfZ312AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAFap848/wDp/mOTyPoPnvbe0/LP0Sxdx6tO13QFIAAAAAAPlTJD7BAAAAAAMD8hvi/6FD5L9yAAAc57z0r+hP4Ws5/334p1H1f+QfrQfPdoAAAAAOZ3q+7zn4zB3frofbP5GB230sHmXiIVfVg/T4AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAD87Q+qPagAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAy52/LPh/wAn/uAMPRAAD1/Z/qL/ADl+Tv0grX3E/HvpgAAAADoOg9bEOiQA8qDwn7LAAAAAAAPgj8TAAP6Fvbw/IH7kD4V/VsPTgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAxtMvBfGv0T88oHs+MAfSHb/iP62Hndxdtbut62IaoAAAAA+ZMsPq8AAAAAA8q/Lz4X+lQ+Y/YgAAAC70+f6f2n9K/579z9H/B+yHgegAAAAAAHcz+5zn4fUfuD+KofqH+cofsD+Fwfuj8fB5/+gIUPi8PW+jD2EAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA/IMP17UAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPh7xP0T5IPzn+swAAAOo9o/fP4j+Qv0U+n+F9xPx/6cAAAAAXtO2LZ6OQBz348B+yGwAAAAAAAQ/wA7IeAgfRvuYfXPzYH636oAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAgeI/On6F+e0j3/AAAD6Q7f8R/Ww87uAC/ouos+xgFAAAAAAAAAAAAHh/wCa/wAD/TwfOfrIAAAAAdH7Hyv13719d/IAceYbyFAAAAHa2/bwPwJtfp/+dIfsN+Pwffnw+H7WfHwef/oCH5A/a4fMmsH6WAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAJ+Q4fryAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHJfk98X/AENVPkv3EAAABfoD9Y/mP51+8P0n8B9xPyH6sAAAAANTVdYezgAGYGmAAAAAAAB8nB+AwB9uB9Ifp6CgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAEHyx9D895MfoH5+AAAB9Idv+I/rYed3AAA/q+yNI9LIAAACmoWwAAAAADxX80fgv6cD5v8AWwAAAAAA7L9Gvn/wrRPE/HQANUJug6JDddla/d/GPwG6p/VD6TDlfxuD9NPpMLXx8Hn/AOgIfh/+4AfNHzUH6WAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAeR/H4fo4AAAAAAAAAAAAAAAAAAAAAAAAAAHjYePfYoAAB+V36ogAAHxZ8dfm39dh4/wB4AAAAem/afknHfZf7X/HfuJ+R/YAAAAAAB3Fj3OeD4bo+6wAAAAAAAAAD8MfiMDqQ/pK7cAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAia+RvNv1L8tDu4QAAAPpDt/xH9bDzu4AABWdZp+xgGqAAD4+1g+qwAAAAADzj8rvhv6RD5f8AZgAAAAAAPoDk+Y+3T4/+TwAAAAAA7eT+lvAPw+2X7ggef/l8H7AgHx8Hn/6Ah+Tfq4eEesB+lgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHwt62H0cAAAAAAAAAAAAAAAAAAAAAAAAAAH5Jh5V+4ALAHlnpYfib+ygHWgAflFwn5L/coYekAAAAa3f4nVfU/9G/57+4n5N9oAAAAAAHR7vr4/lX9kbz9GAAAAAAAAAGAH43/AAgFMD7y/bgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA+Y/d8LxQ/RPzsAAAAA+ku1/Ef1sPO7gAAED58994/0H0c+q/NgAA+PtYPqsAAAAAApfj38d++wHx/70AAAAAAAB9we+/H/yeHJ8wAAAAS7Ltj+lvAPHQ9iAo+ch6oAcqFHuAZ4UHg1oPucAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAD8pf1CDYAAAAAAAAAAAAAAAAAAAAAAAAAAIvxrD3H6ZD2z8aA+uvooPxr+7A+Yf08D6A5DHu/JY/JP7pAAAAAAD0X6i/oX/P73E/PfSAAAAAADS6708uA7/rgAAAAAAAAAo/jcGc8PgIA/dsPs8AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADzH5G+/wDgA+i+dAAAAAPpXsvxD9cDz+0AAIPj7xP0n5v9J8D+kP1In/V/4gByAAAAAAAAAH5g/M/sHkp8J/S4AAAAAAAFj9JfG/GOwPnfw8AAAA0/Zy6g/oPxAAAAAAAAPlj5PC3+noXwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPyM/XMAAAAAAAAAAAAAAAAAAAAAAAAAAA+Wfy8D17UD6gyw+6wPw3/cgPA/jMP1I8I4fpPzePyz+0wAAAAAA7L63/X/469vPD+VAAAAAACr0vo5dKdsAAAAAAAADQ/IPzMP0h+oQ/nr+cQO9D+jbtwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAia+Iua/WfyYN8AAAAAD6V7L8Q/XA8/tADDy6vhnyn4X+yAz937u+qv03+Ow9T4wAAAAAAAAAPkPyfufiM/NP6+AAAAAAAic/NP3J6n5N+hs/wCYfgoKQAAOm+y5dU/avJAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA/I/wDXAAAAAAAAAAAAAAAAAAAAAAAAAAAA/Hr9TQ7j8Mg/VX83w/YSuH4L/uCHxD6cH2n8ueZ9j8FH5j/YwAAAAAAWfvf7j+d/WDr/ABgAAAAAAIe0789I78wAAAAAAAD8xA+Uv0nD7BA8O/nYDNA+0v3XBQAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA8i9HzvlU/UPy8AAAAAAPpXsvxD9cDz+0A4D4S+W/f+TPF/cADsP1j+2/nap5D9X+I877+H51fQA/qYEAAAAAHN59f5L0/wAk/ugJ0AAAAAAD5o+lfb/O/qP68/OfyYPP+FAACXtP1HznH6XwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfkh+t4AAAAAAAAAAAAAAAAAAAAAAAAAAB8dfYoHy+Hu3gYfLv3AHmvjYdh9zAfJvlfbfDJ+Z/wBggAAAAAB1P6Aeh+F9efZ/ggAAAAAAHfd+bjvzAAAAAAAA+KvzJD9GvvEAA/M0PyHAP2QD9GgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA+Pvo/m/OT774IAAAAAAPpXsvxD9cDz+08N4fqPiih8V/YQGoAfpP9H+TWvmP77+YfefooPjreD6nAAAAAAAD4e+SPzT+wA8n7cAAAAAAPCvVPa/Pej+/PkOL18+Z/ncAAOg2/3jxg+kwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAMqiHRgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfkj+twAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfMXmfYfAh+Y/wBjgAAAAC/RvH8n9e6Hm/yow/YfHAAAAAABe/78w78wAAAAAAPFw/Ez7dD9PgAAiD8DfkwDTD+hb6BAAAAAAAAAAAAAAAAAAAAAAAAAAAAAApfBP2XxsZ9f8gAAAAAAH0r2X4h+uDPP7vlH5Z+K/rRDh+6AAA+rvV+K+6j9M/j0AxLAaYAAAAAAAZH5W/I/unIHxn9BgAAAAAB5J0Xtfnvc9D4X3H6Pb3xX8dhHMW+6OvX+jPCCgAAAAAAAAAAAAAAAA/HUP2KA/IrVDxbrg+lvs0Pwr/oBC2AAAHz760Hz54oH6LAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAMD8l/1rAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADw/i+i/Nc/K/7VAAAAOtz4ftf1n5D+UhK8GM/dPJAAAAAACp3no5XztgAAAAAAMb8Ig9X/Z0HAAAB53/ADohw4Hrv9EYdUAAAAAAAAAAAAAAAAAAAAAAAAAAAAB598c/efBh9L80AAAAAAH0r2X4h+uZvxH8p+zeKny/9LAAAAHq/X4v6gn6t/EQAAAAAAAAAAHkPJ7n5oVPyr+2AVgAAAAHmFv2vz70Q8X9B9j++Pmf52ceV+Yv6/8ARuGyfq3mgAAAAAAAAAAAAAAAAB+VgfbfqwfkD+0Afmn9LB9LfnSHkX1sH16BwX59h6V92B+Cf0QH118jBt/fger/AAwHmX6Hh82YgfR/ogfI/LB5x+kIcv8An6HqP3AHxZ9ph8bfX4fIHBB91fneFz9Eg+X8MPDf0nDg/gEPtf3gAAAAAAAAAAAAAAAADKD8vf1XAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADnM+r8i0/Iv7tAAAcL6T+uPl/5s0DzPzwAiP3fyA3QAAAAB8cfkN6mX2X9p9UfXvSgAAAAAfkH88B+8m+AAAAB8iB+CTQPswP3akAAAAAAAAAAAAAAAAAAAAAAAAAAAADxn5e/Q/zwPe8EAAAAAAPpVfwv9t+EeG/Nv7XDL1AAAADe05v16P13+EQD5E7gPoMAAAAAAAAD5+/PD4D+oK587+rgAAAAHluh7X596EeL+gn2F5/wf02/wCX/mPqr39DeIHp5gAAAAAAAAAAAAAAAAB4SHxJ64HZfYYfmn9LB9LfjCH67/kqH7IgfiL+xofDvXB8/wD3yHnfxEH6mfkIH3z5iH1B+f4Zf0+H2zZD8Vv0bD5q9UD5E/TgPz5+5g/LD9wQ/FP9lw/n2/c8PzH/AEnDzv5YDnvqEMfkw8G/S4PRtQAAAAAAAAAAAAAAAADKD8vf1XAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAD8svNPyf8AuAOb1wDsvtfxfx71Q+e/CQAAY39l8wPZyAAAAAPyQ+BPYxDabv1GH2L9gB7ooAB8ah+Pn7rB7uAAAAAAflR+VgAH6nfq4AAAAAAAAAAAAAAAAAAAAAAAABUD5J8aD9HbIfmR2H6N+ch7fiAAAAAAifbfKH8zf3dkHy/7SAAAAAD2v2VP2D+CQD4p9AD6WAAAAAAAAAPEeL6H888H8v8A7NDPpAAAA8i6b2vz3tzxf0IsKP0X9t9L+Mrh+ofMAAAAAAAAAAAAAAAAAAAAfiTuB+yFoPzT+lg9T/EwPp/5RD99NEPw7/cQPn75ADI/QEPKvFA/Rb8PQ+k+KD072EPjP9QQ9AA/FP8AZcPlPz8Phv7PA+yg/Ij9ww/FD9mg/FL9wg/Dv9xAyfxpD0D9Fg5X4+D9Avy3D6g+5wAAAAAAAAAAAAAAAAMoPy9/VcAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPkL4k/OP61Dxf0J/0zxfHfWd75L+XQWYAANZ+qeeH1XOAAAAZzP51+O9znBgAAej/XQfXX1kGl/P2H6E/pMAAAAAAARfg+Hx0AfuMH3IAAAAAAAAAAAAAAAAAADA874EPoXiQ/MLZD9WqwfK3swelqHhXzb+jfnIe34gAAEca4PO/n37pznPfkn9Q1j5z74AAAAAACZz+yR+wfwUAcheDoQAAAAAAAAADlPhX5n9h+fD4T+lgAAAPD/TPa/PekPF/Qn/THp/H/AHBu/pX8ghpzAAAAAAAAAAAAAAAAAAAAflHSD9aQPzT+lg+b/o0Ppn8/Q7L7QD8sdIPDP1RD5O8kD6/8UD9Fvw9D9Y/ysD6o+hw+Af1BD0AD8U/2XD5T8/Dm+WDrP0ZD8QfpQPhf+gQPxU/cIPgbw8Ob+5g+Ov0WDlfj4PbfKgT9KgAAAAAAAAAAAAAAAAGB+TH61AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABhfkz8d+/ZXbfF/tH2x6f8AH/yiGHjgAAAEX6Fxh95xgAAAHyXtP4lt9jEAAAAAAO3YH7P/AFWHWAAAAAAByofzyeNAbAf0CfRwAAAAAAAAAAAAAFPng63xcPgP08P0V8nD4s94D6zAAAAAAPH/AJY/Qvz0Pf8AAKuHZz3O/m39Bc7g/k/9ORni/XAAAAAAAAAB0+vH+uJ+ufwoAAAAAAAAAAAAAAHlHyh8t+1fP+b8T/RATqAfO3vvtfnvS/RMdP1h6N+hfysHR5QAAAAAAAAAAAAAAAAAAAABQQNADMtBnbAOrhS1QOEDqtEE89DuIQ0cMNzMDh/Rwy9QJAMTaCpGGhxgSdeFPlg3tQMbcA5EL++GRpAymC8sHeAAAAAAAAAAAAAAAAAAH5J/rYAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB8heR9zw/1f+F/SWzzvggAAAAAj+x5kP03gAAAAd2/dnwHxb6Gfxf8nhmgAAAAABIHv/1WH1P9Rh2QAAAAHi3884csB3/9DIepAAAAAAAAVAt/M4fIH0gH2P8ALgfHv1EH2HlBYugAAAAAAAAAcR8O/X/M87zvyv7RzuP+V/0sh5vvgAAAAAAAAAAAHs3Z4H6ZH6r/ABOAfJ/qAewAAAAAAAAAAAAAU/J+L6Lzbkvnv1WnoYd/wj+tP0H5d6pP6nxgOQAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAD8kP1vAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAASTkaf8AOvuh59gAAAAAz38Wn6/5oAAAHTbXsYhtJhB8m/HYfH/lAAAAAAAAKHuf1GH1D9Qh6iAAHyQH4LVQPcA/oP6sAAAPCw8B+iA9q+EA+fftkPpjjg19oAAAAAAAAAAAAAAArfkF8h+85p8b+/AAAAAAAAAAAAAAAfYnr/Cfah+lfyGAfCntYfQAAAAAAAAAAAAAAAAAH5GfrmAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAH5IfreAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB+e/wBo/mHoRn5l6AAAAAAJ3TGfuHkgAAbO09OexiAAAAeNh8ifIwfKfMgAAAAAAAB6T9PB9OfTQfQF0Pzx/GMAD6o/ecNv8zA8/wD0WD135jDc9qDWAAAAAAAAAAAAAAAAAAAA/O75/wDU/ng/Pv6kAAAAAAAAAAAAAAA/UP6f8d9VPuv5qAPNuhDqAAAAAAAAAAAAAAAAAPyK/XUAAAAAAATzzpz8t5H6Tg9c9O+T9Ge5hpObSAAAAAAAAAAAAAAAAAAAAAAAAIGNR6+Pel4O2U0QAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB+R364gAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfkR+t3HXKQfzx7gc1AAAAAyI/efIDRAGx1Hq4h0yAAAAAAHw98eB698nh8x44AAAAAAAAa30SHuvzUHgAB9wh+4U4AAAAAAAAAAAAAAAAAAAAAAAAHiXF9D+ah+V/wBrAAAAAAAAAAAAAAB6d+pX3X82B9P+NgAAAAAAAAAAAAAAAAAAcsH5ufqYAAAAAB5/8f8A1Xm/MXF/pHghvH3d9R/g/wBeHgdlrX6o2pfTyBgAAAAAAAAAAAAAAAAAAAHM+Tdh51xH6N+W073Ry/XJ+D/skt/adTV9LN5tIAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfkj+tYPAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA/Ij9dwi5n8/7c8/I/TAAAAAI0/afLD1c16PpncPVxAAAAAAAA/En9ew7sDP8Am8PlP5WD5lxAAAAAAAAAAAD9E/1hDtgAAAAAAAAAAAAAAAAAAAAAAAAPzF8h/Kf7bDk9wAAAAAAAAAAAAA/Rn3/9S/i4O/5kA+ZfRg9UAAAAAAAAAAAAAAAA+WQ5b7PAAAADC+B/qPO+SoP1D50GAH3d9R/g/wBeHgdgA/e6Z3l9XEAAAAAAAAAAAAAAAAAAOc+MPnv1T5j9H5/v/QD+iv4LNPDb60Pwf9jACRm9ueri46JAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPzZ+2Q9NAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA5cPzd/UoADKwfx/1K58H2AAAANZ+secan1fP0Vr18QpAAAAAAAc6H5KfsaBCHK9gHkv51h2n1OHy38vB8wcYAAAAAAAAAB6j9Ch9DfQYfQXpIAAAAAAAAAAAAAAAAAAAAAHmf5d/D/0hXPlv2cAAAAAAAAAAAA947/m/0gP1L+LAAA+FPaw+gAAAAAAAAAAAAAAAAPzND7t9FAAADh+rl+RPj39c+Dwz0/TAAD7u+o/wf68PA7AAAu9X6Gdg7swAAAAAAAAAAAAAAAD5/wCD6f4FwfzH+xzc7/H91P6v/wAyDUw2+sz8H/YwAALVrq7fsYhaAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA+SqgfYAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHzQHnP26AAIjPy/wA476Nf817w5aHUWr/655n5s/oz95xdaUHB9EeJea/XeXy32b5m/V2fjfUu3eyFLR8Chn/oUP53+GhfSX22FWYJAOa+ew8C+KQ+U2gAAAAAAAAAAB2Ye+++B7x7wHuG8AAAAAAAAAAAAAAAAAAfJ/lfa/C5+Z/2EAAAAAAAAAAAB136j/ZfgHRn1/4OAAHjHXh3AAAAAAAAAAAAAAAAH5CB+vYAAAecfJP3nwdXj/qPA+Z2+r9QAAH3d9R/g/14eB2AAATUuxs+1gFAAAAAAAAAAAAAAAfInxJ+c/1m08T9EAPd9r+sP8xjU6/G+sz8H/YwAAAH9X2RpHpZAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAcV+eYfqCAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB8Ah9Ee7AAAAANzcKeZo+XuhTpn8e/wBhAo/KPtcnyH5N+p/OB2ZH6pfn/s9+flH0RLRsbnpZT/if1R+1rwAADn/koPkn5VDw1gAAAASdGG/hhyoAAAAAAAAAAvqYe3e1h7T7KHr/AKWEoAAAAAAAAAAAAAB8Q/Iv5p/X4eT9wAAAAAAAAAHRacv6Zeifqv8AEwdHlgAAAAAAAAAAAAAAAAAAAN/IgP15AADi+nl+Nqv6t+VBU+I+R+l9GG3WAfd31H+D/Xh4HYAAABP2foZzHdmAAAAAAAAAAAAAHyj5f2vwqfmX9hAAB6D6h/Rv8DGp9d+TfWZ+D/sYAAAA5nX3/ZwDRAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfkMH68gAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAflOH6f7IAAAAAAAAAGV4b6XP8Anjwf6/8ALh0QAH6pfn/s9+flH0QAOZX/ADU9XH9cTokwQ+JvhUPkfOAAPSA+gvcQ9j9XD0jvw6qQBoeP/OAfOPzkHz7hgAAAAAAAAAABfD1L1MPS/RQ7/vg9B9ED0fdAAAAAAAAA+QvJ+5+KI/zX+vQTAAAAAAAA7/8ARz7H8D70+w/BAAAAPm/1QO8AAAAAAAAAAAAAAADwv5qD9CQACn8T/WfJ84fa/FAEPy71epzp3e0Afd31H+D/AF4eB2AAAAGjouvPawAAAAAAAAAAAAAPHfzO+E/piI+Z/XwAAu/QX7T/ACK7U/Uv5n+sz8H/AGMAAAAAl7XvzmO/MAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAD8hg/XZ4AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfkcH64gAAAAAAAAFMPg74t/XfmGnucYAAH6pfn/s9+flH0QAAFi12XhPs4fnB8IUc+Aeqh9gfWIfUHogAAAAAAAAAEHiofO3z0Hz58/hy4AAAAAAAAAAAAAAdSHpnpweoepB6p6sHq3oISAAAB5D8E/Gf0H52fI/ugAAAAAATfVHrfEfa+n+k/yKFYgAAAfn99LB7WAAAAAAAAAAAAAAAH5k/dgelAAHzj4N+lfmoev5AActWvzFH631AD+7vqP8H+vDwOwAAAAA6nX9nANUAAAAAAAAAABUV/lXw35N/cYYeiAAB7N+i/gvV7H7r/ABR9Xn4R+xAAAAABq6rqz2cAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPz9D6G98AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAD8jg/XEAAAAAAAABn4/fTef5kfqfzgAAAB+qX5/7Pfn5R9EAAAByP4D9+fnZ35+q/eQfcvvoKAAAAAAAAAAAAAADQ8q8CDwTwUPCfHArgAAAAAAAAAAAAAAAaQeqesB6z6yHrHrIerdcq+cflX4n+ivJU+V/awAAANj6O9f4b615z9D/lPg/pzfzQAAAAD539PDuQAAAAAAAAAAAAAAA/Hr9hQADI0z+GK363+SBUgAB41416X0Ybdf3d9R/g/14eB2AAAAAFnt/RyDtgAAAAAAAAAAD5P8r7X4XPzP+wgAAADsfofhfYNn+mf86Pq4/CP2IAAAAAAOyve3gFoAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPPg+AP1AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAD8kA/W8AAAAAAAAD4J+JP1z5gPd4wAAAD9Uvz/wBnvz8o+iAAAAPE7X5xfop7WH1jIwAAAAAAAAAAAAAAAAAAAAzQ8Y8RDxLxUPG/HQ5kA27z6fotccfzzl6qxQAAAAAAAAAAB0Qep+qh1OZ5P3Fnp/n/ANS9C9cz6fRvSfZ+B/Mn9gPoPy3o67j+c/oQ/TL7cCyAAAAAAAAAAAAAAAAAAAB8MB5p+mQAB4J63kfOZ+l/moAAAEfzD0+ny53+393fUf4P9eHgdgAAAAAdd1xpHp5AAAAAAAAAAAz8n/lf2zij4j+iwAAACS8voPo/6x/y++rj8O/VwAAAAAA1uq9TEOqQAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA/Hj9hwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA/JEP1uAAAAAAAA8D9DD8rG/tHyYMAAAAP1S/P/Z78/KPogAAAADZ6f1cQ6ZAAAAAAAAAAAAAAAAAAAAAAAAIWvnb5U+v+Qt9R9Z8mfRfy30PeU/y/9D8M+T+qPh7yX0cgoAAAAAAAAAAAAAADr/QALYfUXZh4P8UAeph+lP6Eh0oAB4T6aHUgAAAcWHz944HmvIBiAXOpD0D1oPdfZQeAfy2h+l/6wg4A+LvpvmeJPu/hAAAADna0+XYfV+p+7vqP8I+wDwOwAAAAANnaenPYxAAAAAAAAAADyP8AMP4T+lw+Z/YAAAAAD1b1T+k/89Pq4/L/AHAAAAAAAd3nfm478wAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA/OD6zD2wAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAxPzGD9VAAAAAAAAi/Ov635z5gxv0rzAqwAAAD9Uvz/ANnvz8o+iAAAAADsuyLx6WQAAAAAAAAAAAAAAAAAAAAAADBeK/P/AOgfAYp9D88AH0X8j9N3h+XfoYAz571XDec+vh5d5ZR5V5WGQAAAAAAAAAAAAAAAAAAHSh+hv6UB6gB+dP2AHqwAB49+fIfFvhwXvWQueUBhgO9mDheRA7EPrn72D6b/ACJD9ldAACl8EfZfGsPr/kAAAAADyPxH0fofu76j/DvvA8DsAAAAACzS7c9zAAAAAAAAAAAPjX4x/N/65DxvvgAAAADoPor9v/jP6uPk/nAAAAAAAOu640j08gAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA5D8zw/V8AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA8u+Mg/R4AAAAAAAPIPlf8AQvz2DyP6XzPIa3o/Qg2AAB+qX5/7Pfn5R9EAAAAAGnqutPZwAAAAAAAAAAAAAAAAAAAAAADnvlP6v5PhT7b4oAAD6L+R+m7w/Lv0MAAF6PpncPVxGh+ef43hTAAAAAAAAAAAAAAAAAACwH3F+mwc37yHagYv4/h+ekQAH1R+8IfIv08H5U/nQH0t+/ofEP4lAAAHuf7Zh9CAAHE/F33XwofT/MAAAAADfmfp9L6++o/xb9LDwOwAAAAAAO+d7vODAAAAAAAAAD86Pn78u/tAPP8ApwAAAAA+lP0v+c/sQ5/xwAAAAAADouid49bEAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAD8iA/VrdAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA+f8AxEPu4AAAAAAAPjbgf1X8qDs48jxPp9LzlO/3AAAP1S/P/Z78/KPogAAAAAA7mf3OcKAAAAAAAAAAAAAAAAAAAADi+nl+Qsn9T/LA1zAAA+j/AJD6fuD8v/QgAAANnp/VxZ+T/TP5cAAHoX6HB9se3h5/5UHlflgeW+YB5l5oHOgAAAAAAAAHuYff33qHogAB+HHw4AAAfVH7Lh/OB9WB+8/8vYfqztB99fzGhVAADpQ/o89JAA8P+APC/ZLdvp6rdv3fi7dv3fjLVv3fi7c3tfJGHrn6V9M/jv6yHg9oYNTzNBfM0F8yw8zTtp/2ryA3QAAAAAAAAB+XXln5R/bwcvsgAAAAB699P+Z/fR7n8uAAAAAAAbe89KeviAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB4SHyj+kgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHyvgh9jAAAAAABk6Z/CDf1z8iBgByPhvZ7HKnZ6wAfql+f+z35+UfRAAAAAAB0u37GIbSAAAAAAAAAAAAAAAAAAABy/xn9p8XmH1vyQAAAB9IfIfT9ufl/6EAAAAbO0/kR+efsYgGv+qIfpbqgAAAAAAHOedB555+HCcMHFceHz940AB2n3eH3/APRAAHj/AKGG6B/L9ywAAB9UfsuH84HqAf0tfy/h/Qh8LB+j/wCIIfJoAAAfuh9tAAfP/wCc/wCf/wBRB89+qgAAAEumNu37PyXce0eb8b6h6P8AG/k9s8vxQAAAADtJ/wCkvBDqkAAAAAAAAA/KbgPyb+5A5/TAAAAAD28+X+6k+m/j4PdxAAAAADY2XUHsYAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfjx+tgbYAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHxn1gfUAAAAAAAeP638g6v6Z+cOPQ8IAGh5r4p3+7mHT6J+qX5/7Pfn5R9EAAAAABD1+p7sg9PMAAAAAAAAAAAAAAAAAACrU/FnK/q/5QHRzgAAAH0h8h9P25+X/oQAAAAB+P8A8K+xge3bL9z/AG0AAAAAAAAAAAADMD+dDyMPsj7/AA+yL4AAB+dP2AHqwH5O/luAAAfVH7PB/Pl+qofQf4mh/RR+WwfYXg4fjuAAHsof0RdGAB4n+afwX9OB83+tgAAAAAAAFhR6N6h5n5z6f6h4n490Bx/JAAB20v8AS3gBsgAAAAAAAAD8tvMPyj+3w5fYAAAAOmjj+vfefkv5aU5PmRO6ET6rnRPq+cPbyADa2npj2MQAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPFviQP1EAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA+FvZQ+hAAAAAAA+Q/J+5+I5fzf+uNjZ+1/ItrZ/RPwbavfefiZU6uDx7yf0voof1S+F+k78/KPogAEKvz+ynyfSq/LdFzr/1XzQ+/4gAAAAAAAAAAAAAAAAAAPnL2vF8FP0f84AAAAAD6Q+Q+n7c/L/0IAAAAA5v+fLvz9e/ervz6kAAAAAAAAAAAAA+LA8t+7w7YAAAAD4u+kQ9AAT88g/KTz4AD379pAp/OQfll9KB9R/kYH1d+gYficAWA++/10DswADh/yf8AiP6ND5X9qAAAAAAAAAAAA6/1Dg+J9Q9P8T8d7ybi+A7g/pj58AAAAAAAAAAPzi8E/Lf7SDg+lAAAtKPp36m+W/mjQPO+CAAAAABPQhE+r506D7Xk3z6zlAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPynD729iAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAD8+/pwPaAAAAAAA+IvF/Q/kQ/OP60AAAt9HDtbX3f4tst/Rvwnyn9Gen5vv1/F/p1X5LoVflOkOKgAAn7T9X80P0XhAAAAAAAAAAAAAAAAAADD+G/tPi4D635IAAAAAPpD5D6ftz8v/AEIAAAAAPk7afsjqfYxAAAAAAAAAAAAAAAAAAAAAAAAAM/42D4r+Ww8mAAAAAAOl+lQ+v/ukPSAAAAb+QvyX7phnxf8AQYAAAAAAAAAAAAAAan03p8H90n67/IgAAAAAAAAAAfFXx5+a/wBeB5H3YAAe1/YXzv4D2B435MAAAAAAAAAHU/b8mkfsvlAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAGMH45/s0FsAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPzn+wg9WAAAAAAD4f8T9E+Rz85/rMAAAAAAF/Sby/yHtj5j8GAAAAAACbtf1bzQ/RuEAAAAAAAAAAAAAAAAAAPnbwH9J/NQ9jyAAAAAA+lPj/AKfsz8w/QgAAAAAczupPc5wYAAAAAAAAAAAAAAAAAAAB5p3YaAAAAAHnPzd3Y+bc/wCT9HpzbeNr9NWfoXrIemPAAAAAAPz0+f8A1H5yPz7+pgAAAAAAAAAAAAAAPtf2vz77BP0f+SQAAAAAAAAAA8V/NH4L+nA+b/WwDtvsbxfyH2E+e/BgAAAAAAAAAAXt/wBL4Hn6j5wAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB4H8Lh+sgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfmn92B6AAAAAAAHx15H3nxYfmv9dgAAAAAAfpN435L2587+BAAAAAABa7H9c8wP0DiAAAAAAAAAAAAAAAAAAD8qY/l/6O7LoP3P8AjdT1vmAAAAD6V+P+n7I/MP0IAAAAAAOo2PYwDZAAAAAAAAAAAAAAAAAAAAfmn92B6AAAR/O3pYfMfhP6X8/xp7nH3sf8o/3W/wBB+c+p9w+kvtP589SPpPyEAAAAAADwnh+k/N0/LP7TAAAAAAAAAAAAAACT9ZPrPxHsD7X+dgAAAAAAAAAAg/JX5L9y5g+L/oXT+rfN/Pvpaf5f+awUgAAAAAAAAABpdT+y+UH2/IAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHxUHmH6SAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAH5k/f4duAAAAAAHzn531X55n5f/ZoAAAAAAH6TeN+S9ufO/gQAAAAAAafUfs3lB9tyAAAAAAAAAAAAAAAAABw+Ho/k+fkv9zGl2eT2XZ/uX8bbR+ifgwAAH0r8f9P2R+YfoQAAAAAAa/U+piHVIAAAAAAAAAAAAAAAAAAAHwF9jh2wCfOHnfWfGHlPo/e4Z+8/zGAd7H/KP91h8n9+Aezfbv2f8/eqn134SAAAAADR/ll5t+T/ANwBzesAAAAAAAAAAAAAfSH6EfoX8sB7/wCXgAAAAAAAAAAfG/j/AHnxj9Bflf8ASX1z0HyX8thHKAAAAAAAAAAAB1eh/Q/hh6uYAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfC/lYfp2oAAAAAAAAAAAAAAAAAAAAAAAAAAAAAH5ffosHVgAAAAABxWPofk4fkn9zgAAAAAAfpN435L2587+BAAAAAABv7n7v44fS84AAAAAAAAAAAAAAAAAfPnn/UfnUfl39ngABsdn+h/hPZan7j/Ggd3kH0r8f9P2R+YfoQAAAAAAW+19LIOyAAAAAAAAAAAAAAAAAAAAAA57Pq/Pzwv8w/soh93xeMP6v/AISAO7b/ACf/AHWHyv34ASNfXv2l+jfyZKe1+eAAAAAHiX5qfBf04Hzf62AAAAAAAAAAAAaNZfqp2f6z/DgbcAAAAAAAAAAAGVG3inov86fXhw+UAAAAAAAAAAAAaHq59Wf0P4YAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB8oh8O/q2HagAAAAAAAAAAAAAAAAAAAAAAAAAAAAfl9+iwdWAAAAAAAH5V+d/k39xBz+qAAAAAH6TeN+S9ufO/gQAAAAAAdja/o7wg7oAAAAAAAAAAAAAAAAAD5N8r7b4ZPzP+wQAAADf9X5rsux/ef4ov/SunxXZH5h+hAAAAAABJ3noZh3ZgAAAAAAAAAAAAAAAAAAAc1uhyP5p/H/vfEnx374P0x4+n/Z3+codHL3bf5P8A7rD5X78AAD3ju+b/AESvfqP8YBWYAAAB8I+H+kfKp+df1eAAAAAAAAAAAAfenu/mv1Ifon8pAAAAAAAAAAAAAGby34z6ofE9YAAAAAAAAAAAD+w/R+GwfqvmgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHDfl8HpP6GB0oAAAAAAAAAAAAAAAAAAAAAAAAAAAfl9+iwdWAAAAAAAHyN8QfnP9Zh4n6IAAAAAH6TeN+S9ufO/gQAAAAAXPQjrz+jPCAAAAAAAAAAAAAAAAAAA+QPI+6+Jj81/r0AAAAAFc9J9YfY/z79Bmv4uHZAAAAAAvf8AfmHfmAAAAAAAAAAAAAAAAAAABw82XZ+ZfG/lP9uBj3gGVzf9T/wyH2f533Sfyf8A3YHyv3wAAAHvf6M/f/zBIfRfk4AAABWV/m74l+Wf2mHF9CAAAAAAAAAAH1b91/ov8nh7n5wAAAAAAAAAAAAAGB8t0YZ+GewAAAAAAAAAAArOn0/6C8QPZyAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADxX4EDG9dD1yQAAAAAAAAAAAAAAAAAAAAAAAAAD4Z+yA3gAAAAC4FoDI+I/kP3mY+N/fgAAAAA/SbxvyXtz538CAAAAADyL7Xkcfs/lAAAAAAAAAAAAAAAAAAAeD+V9v86n5n/YAAAAAAAAHo/1l43457EbfgiJ9nyofRYAwAq+Z+hmHdmAAAAAAAAAAAAAAAAAAAB4txPzX7B44fB/0uAADOK/eP5aQ/Yv567Ob+Tv7vD5b7sAAAA+qvY+B9YP0n+RgAAAADO+HPm/12ifBf00AAAAAAAAAHX+t8T9ou/S/4/AQAAAAAfRoWQAAAAAAA53G/BPZD57cAAAAAAAAAXpPsuXWP2ryQAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABPKg8tYAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABTCqAct5n2PyifmP9jAAAAAB+k3jfkvbnzv4EAAAAASfMP7H5aH3PGAAAAAAAAAAAAAAAAAABxHn/VfOh+Xf2aAAAAAAAAAenfWXh/jHrp4H4eJ6EIn2XMifY8tH539zAO2AAAAAAAAAAAAAAAAAAAAPAPPPzr+sA8P9HAAA5r9J/G8s/pH+Oezm/k7+7w+W+7AAAAInP0D6N+r/AMQB0+SAAAAPTh+SPzn+ta54v6CAAAAAABs3z/XmZ+mfx6HpfJAAAAAAAfZIXQAAAAAAAMb57fn2/g3shIAAAAAABLuum0P6A8QPVzAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAQPzo+f/AMt/tAOD6cAAAA/SbxvyXtz538CAAAAJdl2E39G+EHTIAAAAAAAAAAAAAAAAAAHAc/p/lMfk39yAAAAAAAAAAB6r9Y+F+Kerng/iYAFmn1H7f5AfVcwAAAAAAAAAAAAAAAAAAGdOv5Mc/wDkn90Bn1AAAUfa+c5Q/sD/AD47Ob+Tv7vD5b7sAAAAA96/R77/APl8PovygAAAAAPPuf1PjD58/N/64iPO+rAAAANCsvp37G+//l/pT6D8tAAAAAAAAAAAAAAAAAACtz3533Zx+T+kAAAAAC0a+/8As3lSH2nIAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAZn5n/L/ALP5UfDf0gAAAAfpN435L2587+BAAAAP6v8AQOK6frnmAAAAAAAAAAAAAAAAAAAACB+SnJ/kf91Bl2gAAAAAAAAAB679Y+D+H+nnhfjAB0ex+++KHv4gAAAAAAAAAAAAAAAAAAfLPl/ZfBp+Zf2KAAAAAcjU/sT/ADy7Ob+ZP7SD5b7sAAAAAd+rf1X4r3Z9v/OQAAAAAAHJ5dvz34t8d+++bcr85+s09LHv6/0/1/hfbvoH7P8An3ZPV+JAAAAAAAAAAAAAAAAAAAAAKmT8J2Ztf8p9IOSgAt+hGnrfrvmTn1/KAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABjxv8Anb4j+Xf2eHF9CAAB+k3jfkvbnzv4EAABLuupu/0J4YejAAAAAAAAAAAAAAAAAAAAAB8O/JP5p/YAeT9uAAAAAAAAAAAB7F9Z+B+F+kL4v452sv8ASfgBugAAAAAAAAAAAAAAAAAAPzM+b/XfGj4L+mgAAAADK5v+lf4z7Ob8T/pEPlvuwAAAAA+z/Z+A+yT9I/kcAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAACLF12fPbktqw/6DENEAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAARfHvj/ffHVP84/rUJ0AD9JvG/Je3PnfwIAANHpP13zJj7/iAAAAAAAAAAAAAAAAAAAAAADiPyk+J/oqE+U/bQAAAAAAAAAAAAD2r7Xy/nz1c/ZvwsAAAAAAAAAAAAAAAAAAADP/AB/+O/fq58f+8gAAAADeN/Zf506a98D+qB8x9uAAAAAHsfZ4P6an6r/EwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHGfH3zn61815/wAH/TATqfpN435L2587+BAF/c/TODQP1DzgAAAAAAAAAAAAAAAAAAAAAAAD4Q8P9I+Vj86/q8AAAAAAAAAAAAAvfqv9Z+H9mfa/zwAAAAAAAAAAAAAAAAAAAB5X+Xfwv9Kh8x+xAAAAABh/bfms2txekHy33AAAAAAa14fsEfr38HgAAAAAANDkLQdPGGJvgZAW8AAC/uAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABl+G+b9f495/8AJ/uHO/dfzXz/AGNjm/Frml+g8U59XzAAAAAAAAAAAAAAAAAAAAAAAAABjR0fljx35P8A3AGPeAAAAAAAAAAAB9xfW36Z/H4er8QAAAAAAAAAAAAAAAAAAAB8+/nT+f8A9Rh89+qAAAAABF1cVHT9r5sPm/sQAAAAAA/YHX/Xv4PC8AAAAAA+aQ/Mn6BDI/QoPwA/pOCz+EofoD4uHyT76H1P9pgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHlv5lfD/wBH5x8t+0AAAAAAAAAAAfRno/K/oWv6h/GQAAAAAAAAAAAAAAAAAAAAB85fnp+ff1OHz/6gAAAAAADH6YhnsAAAAAAH65dN+ufwoGvGAAAAAHzSHzT+kofiT+mwflZ+kAe2fkaH15+ggflR92B7X8Wh82zB96e1AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAeMcfv8A5xZf5b/aQRsAAAAAAAAAfQP6G/f/AMv2j6L8oAAAAAAAAAAAAAAAAAAAAAPnv86/z/8AqQPnv1MAAAAAAAAAAAAAAAD9ftr9e/hAL5wAAAAA+aQ/KmwH6TyBwPzQHqXvYeJfpcH5Ufdgew/iQH7g/lUH1/8ASwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAeG+CB928gHwJwAaNIP1WvgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHnH53fHfvvn58f+9AAAAAAAASfX/sfB/aMn6R/IoNAAAAAAAAAAAAAAAAAAAAAHk/5f/Cf0sHzP7CAAAAAAAAAAAAAABqXj+wh+vfwcAAAAAAB80h80/oCH5F/Wwdx8OB0f6TB+eP6sh+VH3YHrH4rB+3n5bh9ZfSwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAVfxHD9wvz1D1H89g/cL88Q9R+vQAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAApTp8gfJH53/V+UeJ+iAAAAAB671+H9x+sfpn8fh1eIAAAAAAAAAAAAAAAAAAAAAAGZ+QHxv9AQnyH7uAAAAAAAAAAAAAAB671+H+nZ+rfxIAAAAAAB4CHgH36H5AfSwe6+VhifYgfmx+owfm99jh61+cgedeEB+pX0aAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB+NQfrJ+P4ftT+GIfoN8Hh+vnogAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAGLHR82fPPxP9E+S1Pmf2MJsA6/3X2vz/AOlfX/vP5lDr8IAAAAAAAAAAAAAAAAAAAAAAAA/MP5r9g8jPg/6XAAAAAAAAAAAAAAA+x/Y+D+0T9J/kQAAAAAAAAAAAAAAAA+dw0Py5D9m+jAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA+QA8O8+D9cfwxD9RPbw2wAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAArcPzetzVXzvq9jtevw+jO/5oAAAAAAAAAAAAAAAAAAAAAAAAAAD5O8r7X4YPzP+wgAAAAAAAAAAAAAAP1T+p/GPRT7j+bwAAAAAAAAAAAAAAADxIPC/psPRgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAACgH84P7kB9C/h6H7hAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAGRG/5L435J/dIRuAAAAAAAAAAAAAe2/pX97/ADEH0n5KAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAeX+oAech6MAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABWkCUCjeAoheAAAAAAAAAAAAAAAAAAAAAAAAAAAD5C+JPzj+tQ8X9CAAAAAAAAAAAAHi/UD1X9X/iMOrxQAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAD80/pYPpYD8Pf3CA/D4P3BAAAAAAAAAAAAAAAAAAAAAAAAAAACv+YfzH7L5WfC/0kAAAAAAAAAAAAfW/rfEfcJ+l/x+AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAH5p/SwfSwH4e/uEB+HwfuCAAAAAAAAAAAAAAAAAAAAAAAAAAABxuPf+X/M/lP9uBl2AAAAAAAAAAB7P+lH3n8z2T6X8gAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAD80/pYPpYD+bn7FA8zD9zQAAAAAAAAAAAAAAAAAAAAAAAAAAAAPO/wA0vi/6E5g+S/cwAAAAAAAAA9i7PB/SDW/Uv4tC8AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA/NP6WD6WA/D39wgPw+D9wQAAAAAAAAAAAAAAAAAAAAAAAAAAAADkce788vIvy/8As0OT3AAAAAAAAPqX7n/Q/wCVbh735kAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB+fPzMHZ/fQfnR+xIH47B+xIAAAAAAAAAAAAAAAAAAAAAAAAAAAAABF8r+V9t8cc3+b/wBbBn1gAAAAHpf2/wDW/h3tx9p/PIAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAUfnby/svnbxv4P8ApvLPO+qADqNeP3D6S+x/A/bV+r/EAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAjT4zmPO+sr38uzsem9L5INeMAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA8d7dra8uAGAAAAexYkvTYGfthzvRAAAAAecs9GEAAAQhkboc9vg4DnAXRg8LdA5wDowADMCvthnUw3QAAAAAAA5iw1viZlhZthFRDTArWQpwg++AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB+cX27c91+fieWXIAAAH6O+X52uY1x3v4eHfQCKt5PwD3pp1NPQ+X6X0rPLkrAloDFDxT6Fa+aPo4J6CfANa9EvJ+Xeosp+cCf6UFe8meOteffUbPBM9n0WQxAztIPn/3pqYTzdAPKNKl6OS8Pytrzr6nZ82bTPR6sni31mzzDmw80+vgCWAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB+f/wB+UsL5vDxMuQAAAP0do5X4DYtU+0R0mMjz709nzz9BhDwYeh+Gh6h5uz6E8eRT99DbxZfin0K180e9h88d/Ro+XIg+gA8K9XDz7Ba9Yjl+e+0M67wlHK/STPA67PpIhnzc10kjPH/eWeoYUPzniqXf6QejksA+b/T6XK+yI+cPpYPmj6XA88HmerCBMAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAD5Y9opecdiHxuXIAAAH6OmV/NmnakwA+mvn2St1TOK+l0eBxMvciz6h8GkxLFFvs0c16EjyX6LD5l9DZZ9OR5eBZzWecTNfSnmstOKZU7gPWfnVHpvj7PVfMg+lSXT+RaXqegHF/S4HAp8HwNT6/pJ+jks86Zy/etcH7Ej53+kg+aPpcD5kD6aVMAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA8y84pebfawfn1nXIAAAH6OmV/OnZUug8OD6o+Sw63pQ5L0sPE+1ZPw4d9xIe9+Ho7H2NE4n8u9baxPekfNXoDWw+XZzWecerh5nTa9K8hH03aI9b+bEfSfxwz7F+ag7H1dF/wCZGdl3SPLOtZ6r4YiLz2j2HSRw/v6OI+V6PWvZpPn3So9b7SH80fS7XA82z2LyBHf9AmAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABFzAZ/eC4jh2fNmJpIAAfo6ZWAABCD3gkIOUJAIJgVgRWAACoFsBK4WQAAAr2AIAlcAAAAHj/sDQJ1LYFcLAFayABFKAAAJCE4AAAEQSgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAYXgtL0fUR85fQ7Oe+RaQNAH6OmVgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAeCfLuk/Q3o8v4y97pez/DqApAH6OmVgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfFnmus/Q3umb8r9ZDwj53tA0Afo6ZWfOLXuuymAAAAAAAB4TXpdx6MmCAAAOJDjfaGvGvZQBMAAAAAAAAiDyX19oE8zyZr2zxEIfcwXzAOI9dZzvChR97DxnVD2klgHk/NUvZdiX5votduJgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB+c8WsfQ3sEV0/zcHgZcgAB+jplfHeH0vXNVHIYjPevFkYvs4cb6cPzH04QJ/Nv0O186+xs819AB2+ngd0g8Ga5L6lZ86+wh5d6WGD6cjzG0HM+nB5H0zO88bR61zYcL6oyHsJYB8651r1OiiPQC33qeD4019Csl/N/0i075aZ9Sksyw8I9+a+eveWfNvpLPYiGAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAH5xGsfQ3axXkn04Hwzk3IAAfo6ZX8seg0ocsPS/Hg948HD1jFDg/pRHzX9KAUk/kx+k/VHzHL9C0A5RGe+3oZ86NdpnMh88Z6fCjk/o0Pnmiz3vwVH0t80h36B3njwfQPjaO07BM8ea2/JmdX6MivxLOy3UfPfq7PSvGUdj2qfjPCWvojiJDpg+fPQ6OIuB6ZziPbCWAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAH5xGsfQ3axXyJ9U0ui+NkNKQB+jvC53xHoYvHo6PpP5tk77VDlfQQ8P9cZwXrSN/z4efybXpfiYfQuzL+V+8teqaMOf50a+h/i5n0p4qH0hYl+GdhSrcUH0p8yI+lfmln058wI9B0Wem/N6PoBqel5yLD8cte0PTr9KHQbEuP5ppfTvzMH03FLl+ZmvoFqZfCx401rZ7Ocy2fTJDAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAD5ELn1jo5fglql9SeIy/PykAfXfmc16U5HmitelebDtdoHBeji8L2GdlwqPZeKT6Hn2bvloq/t4eR+tB5v2YbPnQei+Uh6f5IGf7QHhe+z0LkkelebByHrwef1Weu+GI6qYNyuHbCZ5o16X4qE3rYeWVmYvsKEzA7Lkk/LuoperEsA8c2KXpZLxMlrsRMAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA//2gAIAQEAAT8AODRIAxXXqax6nNYHaiQO/Whg96IHAB/SiAawOgPSjjHWuMfyo4x1rHvWAOlEA1jgAHpVYz3+lN8wxms5GKBxxigQQAO1YyOKxxjNDgdawCelMwNcE4FEjsaIz0NZAOCaAB4r2A6USSOBRHaiQeAaHPBokEYxRUAUCKwpFAgAYrKnnvSkdhROaLZ6UWrIAwOaBAGAKIGODRII20ME4AogE1lfTNMA1HHTNcHiiFxWAOlAZ4zRAIxmtq4yDzRHajg8URxgHmiBjg0RkYodOaogY4NcHkHpWAe/Shg4INcYxQwRgGiM8g1nqBWAR1pcEYz09qIz0NdeCK4PGaokcYNE8YBocjrmuCOKByOTRA6A1nAABo80Rk4Jo4PSuvFEAjIogY4NZGMCuoqq4o4PSjgjAPSjgjAogY4NVR6YBodOaOAM5rjGKI44/CsA9RRweKAGOBXUYr6GuM4B5rp90VketYB5B+lDk4B4okDiiAaz2B/CjjHWhggAN+lAk96wTxmlGBkmiAwyOtEkd80SDyDzTHtWR0B/DFE4GAaIBPX9KIBA5rjpn8MVgZ61nHejgng0SCMZpiCMCmIK4B5+ldsZongAGsA8ZrA/vfpQAJyDTcng1k4wDWQTmjz3onIxmiflxRA6A/pR6YFUcA7RVZ9DXBHBonI5NHpxRwTg1wooDtXGcA1xn3o44rIPQ1gE+4oY6GgMjGfpWMjiguR1onH3fyr6GqpgAMCivGc1j3+gqsZxzVYz3o80eaIyODVYI71gEdazjoaqqOTxmicDg0QCMUR6GqyOma68Vj3rrxXTivbFdsZqsZ4IoAY4rHGM0SDxmgQRmgcd6OCetY96xxjNYP8Ae/SqyQMA5rAPGaB+XGaJIHFEDqTWeRx9KbBGTzRGBmiAaIGODXXtXAwBVY9/wogGjgnGKZU28DJ7VwRkUTgdKIGOmaJB6iqHPSiAR70SBwKJGcCs5ORRbIwKKg0QMcUAp4o96+XHrQx3rA/vfpRx0B/ClANEDHBoYxkGjyAKOD0r6Gq4PFYHTNHpxWM8A0Dii3b8q69TWB2oEDoarp0rr1qqAHY1jPAqiAeKqs5GSaPoKIA6GhyKonpk/hQ5FEgCv4qBB6UTjkUSAcUSD2okDvQOaOBgigeeK4xitoPU0QCOaJDYIogEYogEdfwrAxgUCCoArAPGawBwKwCME1gDjNEZxRANMc8A9OlEA0QCc96IA5FD09KIyMA0AccmscYzTAEZBrAPIoDHSscYzQAA680Dj5c0BxigBjANdORROMYP4UQB0NFc9/0rBxgH9KIXGAelEZGM0eOtZBGA3P0rA7UQCcelY7CsDOAfwonAwKAPXNYz0HNbSBjFEZ4okY60SD07VkHoa4ajgjAokjpRIxzRPYVyeMUAMcV17USOlE5GAM+lcnjFHpjFEgd6AAFMRjFDHTFcAZFEYORWc9PyrAPIokE0T78jtXB/wokDg0wB5BraD0P6Vxj+VAA8ZoY6CiBjg11H8qOOgOaK+9Yz0o4x1rqMjmh0znmuMcmuD0NYGeDTAAgZrB61gHjNEADk81gngVQAxjNHnvQ57VkHoa7DA/CuowD+lEZGM1XXkUAOhNdelAdhXJ4xXA6fjWCRjPNHAGaoAEZA5FFSaA7DmsAc0RniiBjFELjANbQeAaUY4BzRwVyKxnpQGBkiu2AaIJGMVRGKOCOBRGOM0fYfhRJPBornAFAZ4rGRxR5PFbSOpoAGiFHGKYKRgGiBjg5oAEcHmiMjiiARih8wwDzQGehojHGaOMDFEZGAaqtoxz1oDJ+U/pWM9KIOOBmsAcA0dp4Jqse9Fciguegoj0NZB6GlAI+Y/higARwf0oAZ4P6UwB70QNuM9KYA8g1gY6/hRAJ4NBRxzxWBnGaIBO0GmHGKIxxmjjHWsKRwaIG3g0MEdKwOKAyODQxj5jXGeKwTwKI6ACmAAwBWeOaABGSKIB4ArigMjg0R2rqOOaOemK5Nbe9HGOaGDQAPOKOOma78DPtRJNDjjFEZwM/hXArFEjGBzRIGADWM8CguOpogdjXHpRAJwDRAHANEZGc0QCo559MUASOtUASOlBfWmAHeq4I4NEYOAaIHQGjjAGeao9OPyrGeBR6cVjtRAHFEE/Litp6flRBIxRyRjPSlGen4UenSgAa9sUccfpR5o4IBBrORkUCCMA1QAIwDmguO9DnpWM8Z5rafStvQZojtRHGQc0BnpWOOtAZGCKA4yRVEDoDX0NEdqwSMgVRBxkChyOtHt+lZB6GsdjXXtRHFHngUCAAM0QCMZrGRg0FGMZrIPQ0eSMdqIz0NHAxmgCTxR7AVtHSiMng9KIBHWhg8d6Iyf5CiMcZraMdaIB4BojjINKBjrRGOM1gHgH9KPIAFELjAPNYxxmgMjj8K2k8UV3Dg0Bzg0cHgVgYzn8K25wRRHbNEcAiiCfu/lRU9BQBzRGTjvRHQAUQccDNEego5JwRW0npRHamwDihz0rGODR5wK68UR2rGRxRAxyaOegFexP4V1OAK244okY4NEgd6o9OK69q2jtWOelA9jRHHX6cUSOgPNHGOOtHB6UckcCgQMbjXQBc/SiePpWAe9AqTgmiRnAoYI5OKOOxphnpWPemAHQ/hRGRjNYBHBrBJ4NAgjiiM9DR54H5UAOxrORgc0PQmse9EHr+lDJPP5UUJOaIJGBWOMGiuSBTAgdKOBwG/SqAwARVAZHBrGeBW307UAB0NUQDVUSOmaYDFHHTNVRGOM0enFYGMCiAa4Jx6VgDpQGR06UQBzRANEZ4FEDqazk4BogHg1kDpXAFHB4NAKaAB4IrAxgVwRwaJBGBzTYxgVkEZ9KIBohemOlYyeDRBPGaxgc/lQ5GaAA7VQ4o4IwKPTiiMjOabBANVgE4z0qgcYo896wCOtfQ0cHoaqsDGBRHasEnrRAIxXU4zVEZ6GqIz0NE4HBpueTRPGQa2jGCc0R6URkYBogDv+FbRQGRgnNA54ogdBQUDGDRAIwOorHQV25ojPShgUT2xWVJ5FEY4zRIPSjknk0BngGiMcZoECq4xjNUcFcUeRjNZ4xmhjvRHbP0qjycg0QCMVjI61Q6c1jI4qsDtRAIxXB/CqPPeqxngGqwO1f5FUcZwKIBoAAYBogGhnvWB2rAxgUqgjGa2rj+VfKBwayTxmjkDGaKgDrQORyaYAnA60BgcnpWRnIrdQAYAZ5racYplyAB1og4zmv4aPHFEZAArGeBQxjmgM8ZrGBRI7VRHFYyOKwB0qj6CiAepo9OKAwMZrA7VRANEAjFEcVgEdaGCMA0RkYBogEYrgcCsAdK6igAeCeaC5GQaIBOAa+UjANYGMCj6A0RnoawMYFEDHBrGRgdqIBUc0y571jPTtRGRXGAB2quSBmqxjiiKx71tJFVj3qiAPlFY7A0QQME1jI61jA4quMYoAEisZ4FY7VgdaAx3rA60VBogYwPyrp/SsAdKCjGAefpRXHQ1jPAojjBFf5FEZ6GsHHFEcDmqogHpWAOlVxiiARisDtRwenUUQCORXbFYB4JrAxgUQMcdaIB4NYA6VgYxWARgUVB5o4PFDnjPSuD+FYz1P4UCCMCsDtQAI96I4wT9K25HWioxgdaAPUVjHJNEEnINHGDiiBnFMMnANAds/hQUDjPSgMjAo5I5NADHBo896GM9aHOBRAB+lVxjGaroMCiARijzzVHnvVEDAxWB1qiCOBVEZGM0AMcUeRjNVRIHesDtVYwOKqiRnFHHH6UBwOelUQDVDaBnNcEcGgMnIFDqaPPejtPAojHGaxgZzVEDqDzQGeh5orkDNe4o5xgc0RkYBqiAaojiiO1Hnmq2j/CiM9DQ54NN0rbznNfQ0RkYBpunBqiCeM1t9D+lAEcZrknANEZ6Gs5OAa+9wK2iuooAnAH5VgEdaC5HBo9OKwe4o5PArBIwOTXI4IrqP6UQccUORnFHd0zXXpRGRgGunU/pXoM0egzRIJwDR4ogkHB6Vgkdf0okjjP6V25NEYOM0eeBWARjOaGGIANNgcZo4HNcEYBoD3rIPOelZzwDn2xXXj8qO3oe1EADgUw4AzW0dKK4HFHkYHNfLjANEEDAP6Vz/wDWoDjg1gMMZ5o7em79KPTitpxzRGRnNEYoAHjPPpiscYzR45ArHvRA4wc0BkcGsHuKwSOKIwetcngCsZHFAAjANUeeFPPpQBAANY4xmiOxrnpjNEZwBWCBzW3PANAcYNEDGM0QCME5oggcmj2xRGe/0rHHFEcdaBBGCa57UMkYNd8CjknANFQRjNEYxg0Dk8GskjINYyOK4I68+lcEfSiVPegAeKPoOaOMDFYGMCsDHXmjgrwelYGMk0QDwpo4IA9KodMnrWR0JogdAaPTijwKwCMg1QXPAojtRAxk1XHaqxjgmiMjANUATxiqIJHA6VgDpXXtROO1HGOtde1AAcA1kEcc0DnjOaAz3rBIwaIzgYrGBgc0SCuCeRWNwzjmtvHX8KIJ5B59KxlcA0BkYArAHehw3FEE8CiMd6IJHJokEYz9KGSOO1BTkUOeaK5HXmj020OOtHpxW0jpTA5wOaK8UTnvXSuTzXWiBjg0ATwDzR9BRHGRXWiei5rnGD+VVgAcUPmHFY96Kjgg0x4GDQI6ZrGBkGgDgYNFc1jJwOtYI6VtNAAjAHNEYGM/SiDitprbx/Sip6CiDn5RW3PH5UQRQ5PWiPWsE80ASMA5pgcUM9qxnvVDJ+71FEdqI4HNY5xmiBjg1gEdaOB0bn6UCAeTVYz0NZHQGmA4APP0o/WsVkHoa6igM8Zo4PFdeKAJOAfpWCeCaCmiuTwaOcYP4Vg9MURiiOOKIY8gVg+uawD3oqego8kKTWCODRGRgGs5GB+FEdx1roOTXBAINcdM0QOgNHp1r2zR7YokdM1jI61weAaOMZzXHQGsDtRGRx1rA6A9KJycdaweg/OiOOTmicdqODyDn0og7cAUQCMZ5FYz1FBe1FcmsHrjJpV/+tTDvmgDjituRxS0VJ6CsE9OaI7UQSOPwrBI4GaK8UfQUVyKPAzXUVtNf5FAEgY5o5xgVg4611FVRHbNHpxXB4BoAniiCOAKIyMZognof0oA465rB/vfpXbr0oDsTR+tEDHBzRz6/hR5GM80SSOKP1rAHAP0rHGBQ4ABP4UADyDQwRjNYyOtMfeiR2NHGKJz05oA45NEggAGsZHFL0GDRwR16UOOAKJ7gUxBGBTDigARgGiDuwD0pskYFH2FA5oAgYAoruwKw3aj9aBwOv0FEZHFHp1oAkUfSunA5onHf8KYE8UCKOe5ocjrXBHBojGMVgDpXbANUee9VwOBVVkYzmuOgoA45NDkZFHBO3NEA9BzWM456UcgcGgM4OaJBGc17UcY60vIxmuv+FAE8A80AT1NAc4FcngUOnNEEjANHpxQOeCaAHUGh0x6UQDWCetYA6UVBGAa2jgUQcYzXbP5UBkcGtpPFEHGAaH61k+tVnjGaOTxmsHHWvoaOD0ogY4NEAjFAZ4Br2zWBjArr0NEds0Q3QGqHTmscYzVfQ0RxWP/AK1HpxRGeM0eBXbGa+hrGeQaOMZ9KHSjx/ShnvXB4Bo9OKHPQ1jJzmh05ok4x3ogngdqxkdeaJA4FDPQ0owetHngURij2/Sh9a4YcV06URwD+VVj1rHGaOQMZoknvVfSutHpj8qxxmiM9DRGehrisZ4BqqxxjNEURkYBojsarHvRGcYND0FYOOvNAce9HGOtUQDgUcY60fehgjigBjiuT/hQ5HWlGe9AZPJogEYzRIIwRmscZ/KjxgmsngmsgnjrQPHHah04rBPAoEDtWccUQOuKIyBk1gE4FcYxR570cEYFA4460QAcYojtVAZOAaJyMGs8YzR7YpRyBmu2Aa+ho46ZoZHeh05qiM9DVYOeKrHYGiAaIBGKogGgpI4PSiOxNckYFfQ1tz3rjGDRBHArGRxW3jr+FY9e3atvFDIGRWMnGaC470Ru6UQSMdxWCeBRBxigCRgGsDbmiQRgCvoaxnpTAYGBR44rrR5GM19DR9KPPeiMcZrOOQelYzwK2OeAK8t/SvLf0oow4xWxvStp6YrY2MAVgjjFbGI4FFG9KEUhHSmgkAyRRVutDJIBNN/+qicjGaY9s0R6UCegNDONtYzwKzjnNE570ee9MRjBokDvR6cGj04oA4Ao9OKAJA5ogGiCRkHmgD1zRAPU0emKHTmj04ojPQ0eenbpRBJ4NEZ4BraM5BoAkjNHOcZorkYFYyM5oLkda28fyocjBo8cVtzxWBnAocHNYFE8cUSCMAUOF4oNwADWe2axxjNV14rsM1XJGAaIBGMUCBjFVwRwaJJ4zR9KY5AxR5H8q4I4NE5GBRGehqgQDjNHjrQPcGiARiqOe1YyOK56CuCODRHPFEZ6GuKIBoA4xn6UQD3ogkdfwrGABmgBj3pQelbSeKIOcUeeaIBGKwMYFVRGehrHvWBjAqiARisdgaIA4BqiM9DX+RRGehoYIwKAx3oAdBQ569RQHGM0AAQB1rHvRweKIBGKxxjNVgDpR4FHsBRHp1okkdaAJHB/SiMjGaJ4wDyKIxxmgSOBz7Vgk8CiG9Kx71j3ojPQ0yntQUkVnA61kHgGiO1E4GM80CAcA81g4HNEk8V+P4UTjv8AhRz1H5UORXfG79KIz0NEc9KPPAo5PQUAfSuTxito6jtRyRjFbfx9KGOmKIweBzRGeSKIB7c1jjgfhXbAFEZGMfhRHGAKx6CgOMYodBgUQTxmu3Jrbk5BrGBxRA6VweAc/hXQdOKxkcUAOgGf6VweAaAyODmhxRIJ96I4yDWMjkYrHGM1jHJNEZXGKIBGAaAAAANdRQ7YFEA8E0Txg9qxxjNGuCK7Yr6114okDvWQeAaBBOCK69q68igSfegR0BrPvRI6jmiQRweaz23fTisjHWjg8CiR0zzSnPWs5PA6USAOmaPIwB0o+gokE4FAZ4zRGOKJ4waII4AogkcDpRHasZA4raa24HNcYxiiCOMVjPQVjHFAEjpRBxRxx+lAZGBWM96AzxXXqa+ho4J4qqo8jArB9aI7Zo4PGa5zWPf6VwRWQRwaJ46UT2B5rnH8q6jg0AzDgVtNYyNufpRU5wBRBIxiiCaC8Yog4wBRBIAxWCflU5rkDGKAJ5rBHANEE8da2msHG3NHjgGjkrgUFJ6HNDkfL+VAE9KIJOQKI9TWDnGaIJGMUVxROeSKxngVxjFYzwKIOK4onJ4FYyMAURxjFV1GRW0kZAojtQBI+lEehrb6GipGCDQzjgURntXPTGaAB4xQ6YAzQGetEEcA0ckYxQBPANMvqaBOME5ojPQ5rBAwKGQMYo5K5FAEjGaIz0OaIx3/AEoHPB61jHAPNY7E0QOgNZ3cZoAYwTRA7GsAjrX0NAAngUxzxQPcGgjudqjPtUWl3U/Gzr7j/GrPRvJiDzDP+fY1LbwlsxjI/HimjiYAjr6c0Y1Jxj6UtsmNrR/rQt4T1X9TQtYM8J+poWsKHLR5/E01tZt1XB+pqOG1jQL5fI9zUq20gGYf/HjWEcYRMY980GCDDLn8aMFq4yUz7ZNS6ZaykFRj0HJ/rTaHE3Sb/wAd/wDr1JoQLbVuOf8Ac/8Ar02i3IGEO4+mAP61Npl7CuXiwOx3D/GipHynrX0NA85Jo+grJPIojuKIJ4xRAIxRIIxij7CuvIH0rk4H6URgehrBJAFdeKIyMV14ojPANYGMCsehogk8UcgZzmiCTjtWD60Vycg0R2HPtRXI4/Km6YxQBHaiOMA0RngflQPq34Yrtgdaxg8VkkdaHI5NEYAUmhyMYraemc1gjkDPtQGeAc/hWM9KwemeaIIHJoHHSiMjOaBxgZ5rHGRWAeM19DXBPJoEY+U1g5xn8KI7E1juTRwRgHpQweh6URnoa4I4o8jGabaRgCjwMCiPSiPQ0QMnBzQ5A5quvFUORnNAZOaxjAxRGeMUQCPU0R2NMOOfwrqAA36UeRx1pcDknpWe9EbuQMUQBxijjoB0rHGaOCMAV14xXBHBoAEcDNEDoDW3PIo8HGea6nIFMK4PANYzwDRGOpojpxWfQ0ee9cHgGuvSuvb8KyOhNYI5BzWAOlEgd64OMH6VnJxisgjjmgQeKPHQ0ecDH0psEYzQHGKPNVVdDXB5Bo+gojODmqJ6D8qr7w6USAMCuCODVUeRjNcEcGgc1kHoaogEYrIHFMPQ1kdjRIxx2rA7VnJwDVEA96OAMZoDj3rAJrg8A0xGMVgZ60wyBg03PQ1kYwPyogbeKIyo5rAI4NDg4oEHgUtYGMCsDGBRUGsVgHjNAYGM1WB1rvj8qrA61jPANEgjA7VQwRjFE4XFDpg9aPYVwRwaPYUQCMUcdM0enFDpzRx0zXAGM1kZwaPAzmgQRxQORWciqOTwK6DmuR1okHiicUTnpQP40QAcUTxjvRyeM1wRwaJGMAVQHoORRHatoJxmjgHGawD3qjjoKwCAB1qiOBzRIxjPSu2KxngGiAaqqrA60R2rGR1qse9UCCM0cdM1gdqIBojPQ0wOODR570enFYA6V16GiM8A1gYwKoYo5I460T2A6UGwMgUCSODRzgc80TkYzR5GM0M96OR0NA4GBRPYdRWCOlEkdaPNEnsa3E4ArJxwaY8daycdfpWTjg0PaqzjoaBxxmm5Ao8nivoa7YBrjH8q7YBqs46GiRt69O1Vj3rAxgUTkYFVnnFHHTNVweAawO1E44qgcURkYzQHaiMDBrOBgdq3e1Ebjmmzjg1ketZ3DGOlE4xkfhRwQQKJxQOBig3HuKAGMisEjOKEbuAB2owP3FPG6gCiCRnFYz0oYH9KU4GAfpRHasdqOAcCgfU0celKm84WtK0/zXDPyB2/P3oRBM47dqllAXaOlBflyO1Ku7gHpRAyAaJHY1kE4J5FNIDyaEgI5P0pXVqLjvQcHgdqVwTlTRfcMUrYG2s+/SgxBHNNISCKEg6mmCSY3DpVxZQ3CbSOfXmrnR5EZijZA6cf/XqS3kiHzCiMcHrVcEDBo4xXB4BogEYo4x1oYxxRx0zXXiqIBogdc9KwO1YGMCjwMg1weRQGBjNHHTNYG7Arg8A0cMMA0FyOtYA75xR44rdxis8YrkjANAZwM0ACMVnHQ0DjpQ+bgVnjGazxjNE5GKwccUOBgGgcUee9dRjP0rGeBRy3SsZHFdeM1xWSOBQxjBNUc561n0NHn+lHngUOTgUcY61juDVVRIIwKIyMZquCKOMdaJGMiuAKIDY5rgiiATjFHHWlo4JxnmhyMVhSMCgQa4z/ACqqPNHBGBVYGMCq46A1QVfWiB0BrOOhoEHgUee9EAnIqiM85onj+VA9iKwO1EgChjHFHpxXBHBrI6ZrHNcEcGhk8E1RHGBROKOSOOtEn06VweAarBI5PP0rPG3NdelE4xz/APWqsZ7/AEr2FE9ADVHGMBf1ogEYosAMA0RkYIoAEDNEgDANZBFbR2P6UQDxmioxgGioHSgMAAH8MVnnGM1gYxRwOB2oqMYAzRAHatoPOc1gE8H6UMY4oqCMA/SsAjiscYzXI6miARyc18vQUfQUDnjd+lMARgHNc4GTRPGQaxngnmgB0Ark9DXHRTQCkcH9KxnjP6VtAGAc023HBpsEYz+FADG011H9KJyAoNAjGM0FBPXmiAeM0QMcGicDGaABAIrAPeiB2b8MURnHNKMjH6VgHvXXgGuCBhuntVEA/wAX0GKyPTijggYo9QM1wOBWcEUeeMURxgUSMY7+lDBP9KYZ6V8uODWQRgUMAYAzQwRx2rAzigQRigBkgUcdQelMBivlIzS44waJUjFD0BoEE4o4Pf8ADFHaaIBxzRCsM45rgcdawoGM5ogAcnmsD+9+lZ7Z/wDrUVHc8+lAADrRIByKbBHB6UCDzW4E4BotnpXtWF6j8qpT70QDznNfSsAjBGaIBPFcVwO34UcZAodMdcdKPPBFZyOBQORisDtWQD1o4PSuSMg/hXBHFcYyDRwRxzQA7UMNwD+FYPY/pWAcDdn8MUyrgYNEADAb9KOCMbv0rH8Of0o4xgHNBQRweaz75o4IzmgFAGKIGAAelNzwDQwRnNEA8V8o4AoEYxn9KwKI9K4/+tRAxgHp0rGRxRye9Y75oqMYFYA4AoqpHJqscYzVdRXXt+FHBHFEDPBo+go8GiAawBxnNcY/lVEA96oDJxn6UfYVkdM1gEZJzRAA5OTQIxjFE4puRjFZHpRzjk0SD2oKWGFHPpS2s78LHn8RQ0y48vcU59Mj/GoNDZxmRsfh/wDXpNDtkOXfd7YI/rUmnQINqDj05/xpLeGM/LHj8TTEs3Iz6dqmggc58nn03Gls7WQhTb4z33mrzTY4RuHPoP8AJprcg8CmXbwV/WgFIHFZHYUdp6daGMYPNLE8zBIlyfTNafp0izKLiPA+uex9DULmJg0JxiiZp1ZtufU5pgW749qZcqQDg9jQiCoABuP5UYwwznGPxqQ7e+aMmcDHNNg4wc/hRAyTn6cUBgYDY/ClIKglv0rocA8fSlPZV/HNZGORQZcc0zAgL+VA4HX9KB6KaZi2CetBh3FK46YpX3DBFNErjb/Ce1XekQsMpw34/wCNTWM0PVc/jTDHGKwMYArIIwKJ4wBXUYo4I4ogHjFYJPWjjoTWBjAo84FEjGRR5GBzRBPQ/pRAPevZaHt+VEd8/pRAPfpWAOc0CMAZrIPQ02COKzheDWSBwazngGid3Ar5Tx+VdOho5ByaBAOQaJyozQOKJJPBoHA4Nc9z+ldOKPPNDBwK7YBrIxjFE8YAzQGRyaYDHByaJBAwarKkcCjzzimxjOaByuKyfSiAeQKHuPwrkDCn8K9yOlHkcGuCMEUcHgDpRwTgdqIB7VjAwK4IweaOAOBmuvQ1wRzR9j+FHg8n6VwTwM0QG6GjkDg0AO1EDufwo8jaBSgdO4o+gNZI4FHBPBogcAdqIHY0TmgBwQ36UQvUHmuCNprrxQIPymsDOM1xngYrHGM0cCqJBOM/hR5PX8MUCSB60enFAZ6fhXDe9ZAHNbl9Kzu7UMYwDz6USB3o8YP6UOQCDRHoKyMgUTgYHOKyCADRAPfmsZ6H9K/hojjIP4UF9K4xjFcH/Cqr4d/CT4n/ABd1X+xfhj4B1zxNeKQJE02yeZYc9DI4G2McdWIHvXg//gkj+2B4ot459W0rwp4V8wZ2axrQZlGMjItUmx2468845w//AARa/aBFuWj+KHw9acISqNLeBS2Om7yCQM98fhXxR/4Jh/tefDCwudXHgWz8W2FoCZZvDN59sfaO6wMEnf6LGT64q5tri0uJLS7gkhnhcpJFIhVkYHBBB5BBGCDRx0zWAeM0QD0rAJ461wOBXUUQOxogEYo4HNFQa4IxQGDgCjjsaODisDoKwMYFdeKJGOaIBGK68UDnvQGBjNEYGFFYGeDRAJApu2KIyMUenFAg8ZrqKAwOTXy44rjGKAB4zWAT1o460QCODzRx0zWM8CsjoaPTigc11FVRIxkUcY60cEYFVR571xRo9MCuoo8CuoqsZP8AKj1xmqwOtdORTZ4ANdsCqOMdaJBHFEZOKIwMA80uP8K5JwDRIHemOOhrkjigAaIXaMGsA8Z/SsAj734YrA6ZrAzwa2jpn9KIA4BogAZzRUEZz+lEKRjOawB0P6UQCMjqKI4oLnisDpmiBwAarA7VVEjgCq4x/KqGO9EDoDXBHBojAxjmqodOarAPGarrxR9BWPeqyOmaAwMZrgjg114BoHHSsE9TRx0zRJIGa/hyKODwK+hrJ9a4OAaGMYJoY70cE9aIBGRVYGMA/hRIIwBWQOgoYIyKHFcelHB6UQOgNEDGc1gYBz+lEAcA1gdM1gEZBrAxgVxjFEDsaOOmao44GaKgnOa68VVEDqDzQwfrRGRjNUeoqhntWCTgVtbtXluRwKaNjgAUUP8AEKwT0ogkYzSoz4VRUdhNKQNvB9xVvpKKoMnX0/yaWGONdsa/rWB2rvisnOCalYbdoPIocjBNMCRx1oqCNoPWmjK8qaUsrYFXSgx4J57cUIzu9akiyM4oxCniJ6GnTawUikQyHagzWj2q2UfmHlj2/P8AxqcuZt6jP/6qTkhlPNLIwYIBwetEFk5pQ5TFAbePSpeMKehqZVIC0Ao+UHpR5G0CmQFdpPIrGOnJ9KbptHaiAwHNIxYbcUyArhetMqg80UXgg1sG3OaBOMEURjgc0QMDmg2e9IyjIJpXUng81uB4zUsayrjvV5pKyxlo/ven+TUkTRnDCjgjAptoxzVDGOKIBGKOBzRAIxWB2qiMiiOMCiARisDGBWMnNAcYojJHNdcYojJzmj6Cj04ojPANHgYH4UOlDryPwpgc4WhwOaxk/LR3Y60DkYogAbSeaJyBzRAIyK4AyOtAZ70eRjNEjpmhjpmgARgmiB2NAjGCawCOtYBGCawDxmsKRgmsAdDWB1rgjg1wRXB4BohTx6dq46ntRAIxQAxgGsg8A0QDQ560QCMA1RxjFHpwawCOD+lYUjOaA7D8KwOtAY71tHaiQOlEA0AccUBxigD0BoAdM0VyBiioyOfoKIANEds9KwDxmiMnA/CiBwM0BgYzQXIzWM8CiO1VRx2rjH8qPOBmiB0BqgOMVgEdaIHQGsDGBVEAjFEAjFdeKoj0FcEcGqI7UeOBzRANcHgGqIBHArgjnrRxmiQRgCsAdKwD/SsDtX/BM7xto/jf9jrwW+m2FnZ3WhefpOox20Kxhp4HwrttA3O8JhdmPJLHJPU+JPGfg7wdbrd+L/FmjaHA33ZdSv47VT9C7AV4b+KXwx8Z3p03wf8AEbwvrt2q7jb6bq8F1IBg87Y3JxgH8qr/AIKZ/sS+G/ir8PdY+O/w90OGz8e+GLZrvUltYQv9t2ca5kEir96eNBuV+WZVKHOU249/wrHvQC96IA4zmscdKxkYJogHqao4PArkcCgOMVgdq68UwBGfSuCKGOg7Uxx0NYHascYzRAHQUQAKIBGAPwrAHSiR2NDGOtY4xmmxjkfSsLnGaIA5FYB+tEYFAD60BxigM8ZrpwaIU9D+lEE9D+ldxR5rHOQaVSRjNYGPeqPJwDxTAEYBogHoeaC9hzQAxjvWRkChjGBRBK4BrgcH8KOcYFVwRxRweKOAcA8elZzwDQGBjNHacAiuPSiF6AUMdB2rAJyaOewrIBwKJA71gHBxR2g4FYBOCOawAME0QAAM0cAcCsZHB/CiATwaADEc0QCuR+FYyMAc0AMc/hRC44psEYBogj7p6Ueen5UwAAGaogEZ71X+cUMd6Ixx1ohT2rAzj8qIU9BRA64ogHoKODwKCgHGKrgdayPSvl6AUcHpQA44yaOD0FHk8fhWABgCuPSgAO1EA8D8qIAXGKIBGKGCcGiARgDpWQRgUQCME5ogHAx9Kx2FUASOtELjigfejyOB0ogAYHWgAeAOaxgAAc0QMYNAAgA9a2g8Yrj0/CtoYdKbvigAegojAxmjgLR24wK49KK8UQR3oYPGKO3GBQx6/Sic9+lfSiQRgVxt4rAPUZoY6Zo47UccFe1AZogHgHisAjrX0r2qsAcA1SxsxwKS3B6nNNbpn5V4o2wI+VfpzUOmySZOen+fWhpcQGT1/wA+9JbwAbFTkdTk0ttCV+5+ppLW1PDRf+PGptNtJMFYuf8AeP8AjTaLCx+Vfwyf8aj0W1AzIvPYZP8AjS6dYxnKQYI/2j/jS7AQAvHpmiQcc0cHpRAIxTAHjP0FMrH7orbnG4ZPc15RIwDijGBxnJoqB36dq27uW5x2ps/dz9BXkoyZemiQAADr1poMnANfZskkt+lNbkjAHNRWQlXDr9OaNkiDMQw3YVaNKqqsn40yh2IL4H0ouxcZfIHXijJuk2O2VPQYxSu5QJIefpULIpCSDcncdKdVZsqOPSnQsAFbGO2KkjOeD+lGMgYU9e1NGQAAaZSGKg/SljbGd/T2pgD90/hRVTjaenWlBHTilPzjJ6dakAzigoYgE9KeNAuM5PagmOAOK28Yz+lMmSDnNeX6flWGyBnP4UMfw/lWcdDSkjjf+GKBGcZyfpV7aRypllyO4z9KlsCHwnSpIJI+WHHrWAR1rjHH40V/Os5HXNEZHHWqogHgHpRCjvWBjFDpjH0ocnNcYxn6VkAcflWAR0oYzgCuB2ohSMA0cYwDQxjmvlpBGfvD8KbaThRWATisZ6CuDyRQ254rAzmgFPFEDGM81hSOTR54BrAxzya4xjFADHTpWM8CsAjkUcHoK4xjFDaRg1jnBNVjHBNYHb8qO0j5TRwRgURkYBogEbRRGRjNHnGKIyMZrA7UAvpRGeAf0o+go88A1weK4xiiMjGao+v5VjnNEAjkdKznj9Kbn/CiMjGaAyOD+lFfxoBegPNYycA80fSmGQApqgV/KiB0BrBI4NcAcnNcYx3rj0ojtR6YHWuK4I6dO1EAjgUSDwO1HB6USMcUSOtcUSDwDzRANEc8VgDpWASD6Vj3rjHvVZyMZo4AxmsDoKz6Vj3o4x1o7TWBjAoEEevpQwSMCiRtxW5SAM9KGD3/AErIxgGsg9Pyr4YftXfHX4MfDrWPhh8LfHNx4c0nXr8Xt3NZIEuvM8sRny5vvRAqiZKbW+Uc4yKsdJ+InxV8QSf2ZpviPxhrlyd0vkQzahcyE9zgM5/GvCX7Ev7aF1eWes+GPgT430y8hkWW1uJoDpssDqAwYGVkZCOMHg5468V+zBdfFy8+Ang5vjxpk1h48hsnt9YinkR5HeKV40ldoyVLSRJHIcHq56dKngguoJLa5hSWKVCkkcihldSMEEHqCOMV8bfB0Hw7+MvjvwBaoVg8OeJdQ06EEHmOG4eNCM4OCqgg9waJHSsZ4BoYPGa4PANEgDrWAR1ogY4NYB5FUQMDFAAjijjHWgQenNDBHFYHaqPoKojiiARiuOgoc8UAM8fhWQTimByADTAd6OMZ9K4IxR9BQyawAMZokE4FDrgGiO1DHemVQvHWuOlEHFYz0rpWOM/lWM8CiCKxnvRIJwKIwOawCuRRBxkCscdaxnpWAWwKJXoO1Egd6PUAGuCODRwRXGOKwSM4onHasg8A0cE4zzWB1o+grgjg0VGOBW0YxXA4FEAjBOaK5/pW0dCawD0P4UwGKbpgGiM4oAHgn6VgYwKAwOaAJxmsDpn6UQR92jk8URxRIIor6fhWDjFAEjFbTWM9KwcYFbTWMdO1A4GBXbGaIIGKOAOTROBkURxRGRQORR4HH4UKI7VR9BVDGMZo8Drms54Brgjg0SOOaIJ4xTDIwDQAxjPSsjpnmjyQKIz3rAxgUpBPBohfWiRjGKYD1okcAH6UcEZHWgcDAqjgL1rAPeuPT8K4B4Oab0FHGMCuCME9OlYB4JoYPGaJAHFEgd6IGODVEcZFZzzmuD0NHAwO9AZHJrg8A0ACMDr2oAHiiATgGvbFHGOtAZ7UcHgURnC1FaSy4Cio9PRW2kc/596fTQfur+v/ANeptICxhgOe/wDnNJag/KB0owBRjNCME4HNFccAfhUTkZxRJbvSqq8N+FADbgVgBAQeaQ7hnPSsgAA0GViAp5+lK2VBNdRRxjrW9ScA/pTDPegM9TWB2ogGqIA+apEVl3A/Smj28NThevpQYEbc/pTRA4IPTtXlZPFCMZyDwOtGJQ2T0pkCsPLHFLsBw1eWAy4OaCqDtx1rAMQYDmkRVXc55FRncfm6jpTuFGAeewqtwLbc/SpgDgZ/CioIwD9K2AnIb9KKEjOKU5UgdqjQbBu60CM7QOvSgpC8GlGWGOvakCu53HNOBn5TnHSgEC7s0ME4BpQpOGNLGCCSfpW0kbs8UsRbk9KdccA0ACOTz2ojAyTQYlcqaDb/AJGPFXNqGXKCobISnZKOD2q/0bYu+AZx2/L3ooQdpFHHTNHnoKOMdax2zyKxngV1FYDCqPsKBzR4/pQAJ4FHpxWOMgc0wB4A5phxgUwHQUACOlY7VggcUM96yT0NEdiaHbFDHaioIwDRAHANZB4zQIPTmuCKrg8A0cdM1wRwayMYqsjpmu2AaIAGSfwrII61weAa46VgYwKo896OPWqyD0NUcY60xx0NZXsaY4HBrgjg0MbcijgjAoHNZyKJ5wPwo9OK9j+FE9geaO080x7A1hsYFDJ70o7/AJUnJzXvWcjIPShz92mA64rtgGmIBxWM8CjjpmsADJrGRxQAIyDRIPSsgjAonPBNE45FEDAxRHaiARgc+1ZB6GgueldeK69aGSACabAHBzRI6ZrAIznpWAR1okDvR5wBXHavpVNjoD+FYBGMVRwOMVjKkA0dpGM0oBoqCcbv0oY71gA4zX7Ef/BK7RdZ0HS/i1+03ZzzjUIludN8Iq7wbYyAUe8YYfcevkqRgY3k5ZB4R8EeDfAGjReHfAvhTSPD+mQj93Z6ZZR20S++1ABn361PPBbQvcXEyRRRqWd3YKqgdSSegq0vbLUYBc6feQXMRJAkhkDrkdeRVftng/8ADWnxcOeP+Ev1D/0c1AA85ojHU9KpgNvWuMDB5o46AURzgflTAdB2oKCMGjgjAojIwDRHauKOSc5o9OKKgjjtR7YquCMY5rIIxnmgM5BNAAdqAU9BxWBjFEDsKIyMY/CuvFEZ7ZokEZ9KIz2o88EUCCMY5rgmiAcUeDxRBK4JzRHTB6VtO3Io8gCsUBkY646VjPShjAHWiAeMfhRwRwK4ogEcCsd6yCM5+lfhX4UcEYxRAPQUCDgY5ogdAaqse9YHaiARjNEg8A80Bxw36VjjGaY9ga68CsADGawCMA0Qp6GiA3AP6UADwTWAeAenahgjANHb3rt8v5UBjgGsAdK4btWM8URjjNcHtzR6cUcEYFHoCDzXUUMY5FAZ4oYPGK/ChjoRXFcen0FEBuAKYDoKIGBxWBjAo4xwOa4xiuMYxQAAxiiARg9qwMYFdsA1gDpRAPAqgM8E/hRC9OtFh2/KuCMCiARijg/LRANDB4Wsd81x1zmsj+9+GK68Uo4xmuO1FRwKIUcGiONoraMYrAI4oBT0oKDxRBPyk0RzgcisDGMVjnA7VgDoK28ZHWsDgUVBGBRUYwDzQXJxTAnjPSiueAaA/hzRXjAojgAdaIxjmgARjNFRjjrWMjOM1t5xW33rYWOF5qOznk4SP9RUdgFB8xcnt2xUVmHGAn619lRSAi4A7UbLPKrz9aVVRdgHIpnccZoyMmNr4/CpDvORz70Vx/D+tKMcgc0FyOFz+NRqBytKMDJXPtmgoB4H0FYAIAFAjHI5rccYBpSH4c5plBO1aBJHSiPQ0Qx6HFEZwR2plOPl5NAg8Z6VQ5HI/CqyDwD0p9wGd3T2oLlc5oqT3/SnBPCnntSqQfmbPoMUCG6GvlPGaYZGAa2gjhuR7VHg/LQEaAMOSKQrJxn6URIpA8zP4USzOFPSkg3fKX6e1KGHyI3HcYoqCduaxhwqn8KYqRzzTqo43dOgxRUE8imUDgLnHfNFTjk0FAA3DPoKGQduPpSKc/KKYhBljUchIyrfhiuHAKtz9Kx70qkjH5UyNnCNx9KBBzlskdOKCODlW59MUF/hIyB0FOCeAefSnXPVv0ooTjAz6UVbbjbyPeghPIFI2Bg1WSBhT+lanp8FwvnKMSnqeeeg9fSpI3jbYx5o4x1ojPArA7H6V7YognjNbRjGM1tB4xWARgitpPFADpisBhmiARwKx6CsZPPNYGMUBnrRHTI6URkZArAI4FFT0FYzyB9KwBwT9KIAOKVQxxmtoB4NbfeiAQBigMnrQC4rA7UB2JojtmiBjAP6Vj0NYP8Ae/SiAcY60enFbcjIraT0P6VjPeiFJwTXX/CjtxxRGRjNMuAADVbeMZraO1EZOc0RkYrAIxisAdKxwOaYZGO9YGAOtFRgYraMdc1gHt9KIBGKIB6DNAe1EAjArHGMVgdBRAPQ80AD2+lADsOe1YBOMZo8HCisDODRwTx+VEe1cYwBQAxkjNYGc4r5SOlYBGAOaKgDnmjjPFAAngUcZ4pgD0FVtx161gEdaIDHBNBSehzTKOgrHYGtoFYGAAKxxgGgBng1gY4NADHBrAxgUAD0/CiMjANbcnk/pTAdjRAIGDWCOAaZRjGaOMDFAADGaGCduaOD0oLnnP0r9gH4VaZ8Yf2s/AXhfXbNLrSbO7k1S+hkj3pIlrG0yo46FWkSNCDwQxHOcGv+Cjf7UPir9mL4J2mofD5oovFXinUf7PsLyaESrYoI2eWYK2VZwAqqGBGXyQduD43+I3xA+JOpya18QPG+t+I76Vy5m1O+kuCCc/d3k7RycAYA6DFf8El7eeH9jfR5JYmRZ9c1F4iwwHUS7cj1G5WH1Bqv20Qf+Gs/i4R/0N+of+jmoDPetoI4OaI7A0RxnPSjk80BkcURjjNDBBx2rGelVzjHWqIHXPSuvFUc44/Cjx0/KiCeAaIxzQAA60SCODQ54BzQXHGawSQM/SsZ5z+FFc964I4NADGc1gAdaIyRiiOevP0oDnGaCknAojFEdqrBI4ongAGiCRwOlAZwM1VEjGPSsZ4FEDuaC4+WqIyMA1jsDRI6VkdM0QOuelURk/SiMjGaCgKOaJA61jvmgMd6wDxmiAelAYGM0RkYzW0EcGiAeBW0YxQAOCDRUHjvWMnOK4JyKIzkUAM4702OlHHHNDJ4A57VtJ4FEHPBrPfNEc4FbTjFbTWMnA60enFEEdTWMjrRBxVfQ0fYVWMjOar2qsAjINH6/hiqPPeqIzgDvRIPAPNEciqK571RPvWPesDOMURxgUo96K5qto6jtQHvRUHpRAA6UemBQ6USCMZoHp7USCME0QOxoAEAZo46A0MDnP4UegrA9ayDxmgMHANHrwaLAjGa7cGiflzmiR0zR55rPOaGGyKABPB6URk8VtPpRHatvHvVEECo4zIdq1FYM+AW59MVFpkZGS+fbH/16WzgUcHp9aWJFG1BRi3DGKSIoMAfSmC+vPpRnKjAGaeXkqOoouMgGiueSaCgg7TRGe/4USpwAfwoYPCmoiCzADNcfdzRGDgUCTxQ2gHPWlRmGAM4oqQcd6BG3LDGKOAMZ6USAQvr0oEEcGqJA4zSqS7DPSq+hokDvQAwMHNEblKqaIK8EdKY5YYH4VgOgZRzSqGBJb7vtQj4+UUV2gEigoccGhGo5zmkTccg0VUuVNCPZyozisZGCKZQwxROBkGlO4bs80CCMimIB3N270+FGTRAkAOOfSmYH5QKwGTIPI60VIABFFc4zzRUZA60ozyDU65XAOc9KSIhATSxMDxW3PehkHGOR0oszLgCgoB2kcnpR5GB1rYScBv0oL6mmCngHPpWMnrTDj736UykqCDwKBBUAHJ9KLEgAd6qWMNgg9O1X2neYd6HB9Py96uIDA+xjyKx2o/L8vpRyRiiCeM0eMDrXJGKKcdax/dNcZxQ44z+FEZwBRw3ANZzxjmjjGC36V0HBquCNoNEcYx0rAI6/pWAeM/pROe9EYHX8KTHrRUAdfwxWRjGaAB4zQwBnPSqIGcA1wRwaABIGaGO9DDcrRA6ZogYyDQGeM0VyAM0uCMCjj7tEAnbmiMcflTLgZFEYrGeAelUOTgdqx2ojA5raSARRGDgGiveiMUVPX0o9OK2k4IogkcGsE5waJHQdaI6YrGeAaGSMGs54FAY4zR9fSh0ApQM7c0V4wKJ6DFEY4zWAQCDWAR1ojA6/hQBxwaIyorGRnNA46H8KOGOBRGOM1kEcGjg9Pyo9v0okdM1kHgGjjHNZB6GjgDg5rHaiARjNcEcGsjqDRHA5rGR9KIyMZogDknrQUE0Vz8o7USMcdqGCeD9KK4GCaJAHFFc96Y54BoEAAA/hRA7Gs46GgSOhrOflzX/AATV+IOl/Dz9sXwPdazcJBZa4bjR3ldgAslxEywjn+9MIl/4F+Br9sn9lTRP2uPhSvgW81xtE1fS7wX+jakIfNWGcIyFJEyC0bqxBwQQQrc7dp8C/wDBFj4sXHiWFfib8V/CVj4fSQGV9C+0XV3KgPKqs0USRkjjcS4BOcNjB+HXw+8KfCnwNovw58DaYthoWgWi21lbhixCjkszHlmYkszHksxJ61X7aOT+1n8XMH/mb9Q/9HNXBGPyroMCuKzjoaJz3rIxg81Xb1rgjg0MY4rGelUSM4FYHase9HpxROeB2ojIwDRAIxRAIxRAJyDWc8YqiPSse9Ed89O1EAjIHNYIHX8KoHI60VGODQwehok9RWcgA0Sw4FEk4BojGCBWPeunT8BQOOM1n+HNMueM02MYzQI7Gic96IyMA0SSMZo56AVjjGawDxmiAeh5rOMDNVWR0zXDcelAdgax2BoKMYzWOwNEZ6GiMnAPNFc9KK56GsZ4zQXIyDW3jANbcdD+lbR0FbewNBQRgGiABgDNFQRjNYzjnpSqOldB/SgAe1Z9D+FEYHWjk8E0BnpXGAB2o5I5NE5GKGABgVn3/Ci3HWskjFAk/KK6jGfpR5onPGc1TcYrtjNe4o8967YBojIwDWTxg9KIyMA0RkYBqic8Z6UQCMVj3qsDrRGRgGiM8ZoYGFBrAI61jjGaIz3rGRkGsc5zRGSATmlHGAaBA6nmicjrTAHvRyRwc0BwOeaYcYBrDYwKGCMg9KJxwDXOcA0Tk4FEgChgnOaIBonK5FYBPXP4URwFBojjAoE8AUTnkUCQAAKIJ75oqW6VhgORRR26Co7aaQhVFQ6fHEg5z/n60LdSwApoFDbRRGOAKI7VuYDArzC42g0Tk8CiwPHesLyQKKBunWiuRjFCMMMUVwMjmlQtgflSxZ4A5qJGDlc815QxkHpUiAtuLcn2rylIwG/SlUKcA89uKIBJDHpTZTgUVUx8Gh8uFonjrXHQUwBGCaaJWHy0qjbgHk15aHqKKhlFFFxgjPtQUAbRRUEYFBFxjH4U0YJG0U+8DHagqqh4z7VswMKaAHQ/hW0lMDmgNwTB9aMYPGawCMYpl3LgGsBlwRRHHqaIAOOtEZ4U0o2jaRnFH60y7ximVUGRTnLDJpl3szA/Slba2FogZz3pRu+TPWohhdg6DtUqs8gVTW3PynnFYVhjHSgAw2gUIgTnOaEe8ZBrYM8nOKWNW+Yj8KADJgDmh0x6UAAoAHTtQ+cYI4rGBg80SHXBHHpTKWOc8/SlUHg/hRcg7Qc0CD1NSlguUNXsZYkYqRSjY/KjgcE0AR3/AEon1o9OtY96zkcGhjHA/CuCMLQBxyaJ561uzxRwO/NYBGSaPoT+lYyM5oEYxmgARjNMAOhrAHAb9KAz3oAHjNKozjrQG4ZJ6UVJ7URwBmgCTwaI4ofNnJrAI60QelYBHWsDGKxkda7bRVEDHBoY9aAwMZornGPwraB1og5xn8KwCOtFeaxkcVWM8CqxngUQDVUQCMVXX/CsDrWPQ1gEZA5ojGADRGRgGiM9fwo4A4rrxRBIyOtDBGM5pTg4zQOAMUWJ6UTnvTE9AelZJ4NZyMjrRHFYzznrR5oZAwD+lYHUHkdKwDxmiMYwf0qiAeh5rAI612wO3SunQ0MEZFAkDANMR1PWiAcYogEYrAxgUQD3rAPFYGMCh0xRGRjNYzkZr0GawO1YxgA0RngnNDkUOeAaxkcHp0ojjg1gkcChk9OtAA5PWre4uLS4iurSeSGeFw8UkbFWRgchgRyCCAc1+xr/AMFTvh94x8M6f4D/AGkvEEPhrxbYxrAmvXCFbLVVAwryOMiCXH3t2IyeQwzsGlfEn4c67p41XQ/H/hvUbIrkXNrqsMsZGM53KxGMc18Z/wBu79l/4IaZcXPiD4o6VrWpxKfL0bQLhNQu5HH8JWNisWfWVkHvX7I//BTDwv8AFzxb8Sb745eLvDXgDSIJ7JvCdhf3qRbbciZZVMrY82QFYmY8cyYAAAFH9s/9kzH/ACcV4B/8HkP/AMVX7a+s+FfEf7VPxH8Q+Cte0zWtF1XV/tdpf6bKssE4liR2KsuQTuZgf9oHPNYBOAc0BkcGqIBo46Zr6GjjHWj6CiARigARtHUUQMcGjntRGcc1jAwOaAzxVHngH9KHI60VyBQIIrI6Z5o8Ciew5o8jBokdM0QCMZokdM1wwwPwonHFDB6CsgjmgAehog4wBR5AOawAcE1jjkUTjkVweAaOOp6USOmayR0HPaj15FcEYo8cUeDiiO1HpxRyRgc0FwOR+FcNwKx2NDngdaohs8ituT05phjpW0k8GjyOBXJ4xWO1YzjiuTwBXUfyo5xwKxjoKA4xXTijnoBWcnjmgc9KB7AUBnijyMA80c44FAmjg8A9Kz70eMYNdRWMnbmiM9DWCAMnmqqmBAwBWCRnFYz0ojFe2Kxn+lE4XBrOcAfhRAIwKo+grk8Yo+wrk8Yo9OKxnisYGKrnoRRz2FE9OPpQPGetZPpXUUTzgCs84xXUcGhkDGKoDAxWMnNHGOtIeMUxByaxkAgUTxgiu2AKJ3cCieMisnPFEEnINYyelEc5HNFSTx+FENnAH4VjJBxTDPSiMjFYxzijk8CiM8qaIOMUFJ6UiM5wo59Kg0yeT5mX9R/jSWe19u3GO2aFpGRgr+tC0iDc9qUxR/d/KpH3DYF/WivHB+nFZyME1uB5BrHvRYjgHNBSTxzR3A4A5og55FGBh16V5TD7ozRgfGVGaCEdePamGcEDJ9KUdyKzngClAdxg80oC4UGpFBKgn1pl+XaoojJoqG6isZHNFQy7QaOMYJrAxjNYHWjg8UMY4oIByKxkdK2gDA7URkV2xWAe9UfQUQDQUE5B5qWKBI8rLlh0G0itvTcOewojPQ0FCjGelEbhwaPXFbgflFEjgZ+lDTrprVrtYv3UeNzbhxk4HGc9aIJXIHFAoRjdz24okE4B59KJG3JPSsg8A0qZAJPSiUY4ByfSgy8Yb8MUQp6N9OKLBjkNz9KLqpA3c9uKiwqnn8MUgDuZA3TtitvOQefpTIx6daYqOCee1YwODSgAYBzW0FduelHgbc/SincHmgTj5eaJ2jBbn6UACAA3J6cUAcbSaxzgN+lEAcE0yK/Q9KZGXmg4J61lm+UVc25Kl1Gc96ubcn7o5phg4Jo46Zo4OOaIyMUck8dqI3DAokAAAVxWQeO9EZ5FAc4Nbc8Vg4ximBzjFYyuB1rAI460Bjt0rBznFEdyK+ho4bGK6HAFEkD3oHPAFEMRgDNDcRwKI7A5NcjpXt+VYz35rGelFTjitpoqR/SuvAoA44rBPSsE8CutH0oL60Rk4pjjr+ArOehrGABn6VjPAojnBP4USPWiR0zRAJHNcEcGsZHFc49678V7Yqj04o8CiMjrR4GcUenFDIGMUenFDAHWhgnANcelEknAFEEcAU2cD1rJPAFEEdTWTjmuwGKwSOBmh0wRWc9BR9hQBOABRGBjFYbGcURxxXJ61k4wBXJ4xXJHSj7CuvFZz0okHoOlE4GMUQcZArJI4FEHHAzWQRkCug6UAQMAUTjjHNE+orII45rqKABXAFEDGc9K4I4ND1Hau2AarHvVUckYBo4x1rJIxR7CsZ4FHjiuPrX0NNg9KBz0og9j+FA5FEZGAaBJ4qgc96HI60QcYBonOBmjjHWsA9DzXBHBqiAB1zQI7npRxjIrgcelMQMUWz0okYyKGD9aPH9KyCOtEjHFZGMAUMEYFAEHIP6VRANYHaj6A0QAM0QDgg1wRwaJBIGelHB6UORRI9a2igARg9qx2Bogdc9KwO1H0FYU9aIAHHauDyKIHc/hQAAwDRH8IojgVgEc9awMYFDBGRRAJAoEAe9Z3DAPSiMj1rJAwaGCOKJA+XFA5IAo8cAfSiOMkURnIFAAY9aJINFs9KXOeayMYAokEYFHkAVQ46VnvmgcjHpRORiicjAoHAwKHIrAJ4617VgHv9KOD0rgnmiQelUcY60cY5rI6CuCODRA61hT0o4P4VtB4AoYPTtRANEccdaxnmqIAHSvlIpiOmKIA6UDk/yrAxgUTjgfhQIIwK3A1gYwKIHWlOeKYjoKVWbBUZPavs0pH3fwzX2WQ8CvscuMU1pIF6c01rKvVf1oxOOCv615Ljov61FZTzfdX6cil0i5PQfhx/jTaPdE7QOn0/xp9LuFAyv6j/GnsbheNnT3FNE6naV/WiOcEUDms84rd7VDA07gKKtbJIEwwyaVW25ArAJwBW3IytHI4o7cc9e1ZPY0/IyDRHoaxnAAraaCZ4A5oR5wKMewAg5NNgrjvQ+7hjTA44HSiCPumim8YArbj5aaNVwQeaCAnLc1Co5xTKGGDQUAADtTKGGCarAxgGgQQMUAB0o57VRANEAjBoqpGCKwMYFQW8l1MsMK7nbOBnHatZ0DU9AvZNP1W18maPG5d6tjIB6qSOhFHBOGrAPC0V6AGgAR1rjHB5rGegqGB7g7V7UyBDgjp2rqeDRwTzR55o5/hNEljknmiMjgVgbRiixVding0CCMGmx2ognoaJAGCabGODz2oEgAd6CFvut+lFjkjNHa4wDzQGCCD0rcQQRRACnn6VCXVBnpRGRnOaHUAdqzg89aBzzmguOBRBJGDWB2rBx15rYc5Br5h1P0oLjoa+ho896AOME80RnoaZccrXIOR+FB2x1oFZBhuaurckZQVdW5DcCiDkUwyMA1WB2rA60FGMZonBoYI4okHgdaHXmicnJrI9KOO9YBHFZBPA5rB7UeTR+boK46EUwGBihgcimHJIrGefSiOBzWBjAojihjPNEAjAFYGMCgM8A0VxjBo4IwBRHaiMjANHkYzQIAwBR57104riic1nHQ0TnvWeMUDgYrtgGqo8jGa7YBrHGM0c4xmsDGBRGehquvFVxkAGiQOBQOBimwRkVkEUCuOKGDwDWecA1nHQ03J57VxRY5wDWcjrW7PB5oHAAxWeMVwVwBQAAx+VUCM8dqyoOcUcNyKPUAGsDGBXBHBokEYNAcYI6VgYwKqs9sUQDR4AFYGMCicUDk4FV1NEZGM1gAdelE5bFE4rHGM0SMcGiqlfvfhitoHGf0oEYwD0obc8N09qwB3ogE4BrAI4P6VgEda+Xuf0okEAA5xRAA4P6USBwD+FAZHJrGe9deKHuf0rtgGuvGfwr73A60cjAx+FdRRxtwKPoDn2qvpRCk8VwfwrIPQ1twODzRI9eawDyKzziiq9BWQehoqCMCio6AUQDx1xWFIwe3asA8imA9aIUd64xwaBGME1gAcUBnihggEGsZ4FHjjFAgck0PcfhRAOOawMYFYz0/GmwelEA0wHrXB4Bo4HTrRwB1/DFHngUeMc1njANcYxigBjAOaPIwprA4A/8A1UcY60AMA55okdB1qsgjANEgcZo9CQaJHQigMiiQeBzXXgn8KJA4zRx0HWsjpmsg96JB4o5A4NZyQBROeAaLZHFAnOO9MAF461xjFc9PSuO5ojPShjvRBxwM0AR159K68UV4waIAPSiBjA6+lUfYVwaYDtWM8CiBjGefSiAeKxkcihjvQAHFEAjrk+lBRRwK7YBoAHkGjgDAPPpR6cUcdM0SBxWc9DXU9Kx7VgE5BrrwDRwRxzRG7jPSs+tEA89a69qOCMZqGMsKSAE4IqKFA3HFSBHXav50lsoAO7PtivJUjBGaSAF9zH8KWBCfmXP409pGxyq5/GltLfpt/U0EVQAg49KwpGAefpW05xmiisMEVgEdKnsIXOV4P41PpEjE+UuR25/+vT6TfJh/J4HfcP8AGv7LuByU/Uf402nzA5I/z+dWFgYY9zDmghxyfrQ+7gN+GKbGSAaCttOOlMS3JNIochR0oLwCaZAcAUsZYYApVKncq5xSrn5WX9aKgEYXP40RlhxRAC4FHHRVz+NAlx/SmBCkgfSghKjC5/GguFwODRG5RkZ9qKIBg0FJ4K8fWo+pAFcVXBGO9cYqsZ71VUee9EDGD1ojHUVx1H5Vpt7Jp15HeRDLR5wPqCP61418bav461241/VpMz3O3cMKMbUVR91QOijtTNuGKxgbaIOMUTkYrGfu0BjjP0qKWSD542wT7UzliWPJNHHauvU1XWjyaIO3Bo4xgVnjGaLKOCaJyQByKVlJ45osgPJ6dKBVzgHNMzDIA6dDQLDkDimZfTPvW7AwBk0cEAYoEAgAZ9qUeaeDj0pVUKEzn8KChSAG/St3J+bOO2KIU85ojA4NAgjisZGDRIAoZ7mjnIANbcjH5UAR2rAxgVjoBQ9x+FNuONhrDkZBr5mXhMjtzTZIxj8KwWOAPwqQsF2kVdQZyQv61cwEKWx0o+wo56AVwOgoYIwKyCMGjjoarjqB0ojcOlHkgZogYwTRweRQ244NDkUTgYB5rIxgnmsg8A0RzjOaxkcn6UpHAB/SgFHBP6UUHZv0rA/vfpXUcmiPlxmjgKcHNAKRy36Vgf3v0oYIxmiMgZPSiAeQf0ogY6/pQAAzmiCOCKwP736UQOmaI5B3dPamIIwDQA7HPtTYPQ1xjGaK8VjjINDHc1getcH/AArGD0zQAPAPNHA71R4FDBGRXQdKwDyKznjHPajx0FEgjPpRIHAFYz1FEgdOtAAjgVjAzSkHk0ADxmsAnkUSCeKO04xWeelEA/dNEErwOlAdjQIJxXBAA5o+uKyB3oAHgniiB0Bo4xxzRIPNAcYogHvmgFIwT9OKOMc0cDoayCOtEKRgHNEZGM0QCODR2leDVDPeqLZPHNHPam46c1kEcc1z6VkdCaAGMg80TkYzXGMYrIxiqrigRnFEdqCjbxVEgjAFEZGAa49KJwMCjg9K4HWvpR570cHpRAOOa4xjFcEcGsDtWAR1o4JGDQIPTmjg9Ko4x1oKB3rA7VWR0zQCkYFEgcAdKGD9awO1HArIxgCiQe1Eg9qOCOtEgjGKHP8ASsZHWqI9KIGODXH/ANagAOKJBHSuPSsZ6GjjHWjgjArAI60BRIHeq49PwrqMiiM9DVVRPYUORVMT619DQAHFAgngVjPSqI9fwrg8A0QSaPTigfU0SD2oDjFH0z9KHIrABwKGGGD2o4xweRQ5FYGOT9K4xjFEA1x060cdqxngGiDnk0cEYFcY/lWeMVXGMVwRwaPUAGgATim61RIPSqzkcGjx0rAxgUe2KrHv9K4PANYAHWgQRxRAxtBqjjpmiRjjtQz0NHOMChyKPUCj04oCuooHIHNEjoT0qNQx5qFRtwTSxAjioYs44opngUiDGSajRDyRzQUCq68AfhRxwSKPI/lRORg0ORxQBAANEDbjPSiMjNFQajQO22pV7ChCCfrQGBtBpjnJB69KBAXg818pOSOe9Fiw2ijGo6CtinnFMuMc06owz6UQGA2ml2sPej6CgQVGKIBqmUM4oHIBBogEYo4QYHSsBuT+FUFAPv2pjjmkwzE55ogEZFUQCMVjI4rAI61R571xjFbc9KYDoKbpjNZ4wDWT60BngGic9KOemaHHWsk9KGO9KecCiSeh+lEnHB+lEnrR5okngURnC1xtxQx3oknoaBBGRRIGBmmIUZJpnBAYdR0plUHimOG4PTpSswOM0zhlAIzTKijgc9qY55pSc5B5rIzkHpTsNu4VCAowD9KwCOK3f3aBBGM80NvQGgMjrQCqMChzyBVVRwelEAjFdsA0RnoaIBGKZcL8pokEdaIJTAPTrTdcZr2B6dqBB680ygE5q4t1dSAOO4qW3aMZxRAIxmiARgGjgjArIHWjgnAPSsZOQfpQIIwKJHTNEA8UcH5ayCeawMYFHAHFABu1Hb0Ar739KwucDrTAUBk4BoAkAZzQGRj1ojtms8YqmGeAaxxjNY96rlhgUEJGaEbNwooQsBz1oxsAARzWxsYAqOFpOFNJYSOMml0pm7f5/OjpZxgj/P502mhu3+fzptMwvHUf59aawlHSmt5E4xTKV4I6UVI61WPeqIGODRAPQ81RAIxmjwKAGOKIA4ArAIojB4rAPNZBOKyMYxRwRgCgARQABwBQxnrTHPQ1RGRgGgM9+e1YGMCgcDFHnkVxRx0zXGMYrisA8iqIGeDXFHpxRweldKIDdTVN2I7Vx0BojofyrgisZHFHkYzRIAwKODxQxgAVTZHA60e2KbpiuemM0CQeD+lMQwwTzXbFcEAZoY70BwcVgdM0SCcLRximIIwDTHjrXbp+FYJHA57CscYPX0rgjIH0rI6ZrqMAdOlDIGMVgk8ijg9BWc8AZ9KHuPwoHIx3FcHg/lQB6E1xWOMEc1xjA7VRHpVHAXA7UDkVgE9KxjgGgQBxR5FcH8KbGMZokEYDfpQOeM/SsjO0V0XGK6f4UvbmiQ3esjAAP6UfmOAaKgDrQKgZJogY4NcY/lWQBjvXHY0Qp6H9KIBHB6e1EL0B/SiBjhv0rHGM0CCo55o9OD+FfQ0AMYJogdAawAcg5piDjHWiQR8pzWD6/pRIPArAPGaPNAZ4o4PSicnC/lWfUUSD0HShk8AZokAYrcDRxkUTnkdqJyOlA5AxXBORzWcjj8qJwcGgQcAHNBh25NHBGMYxWBjrzQUDgj6UcAgdaOD0oAA4BogDgVgAcGiBjg/pRx0zQHHJ/CiBjgVx0FHA5JogE8Gm60QOgNEKAADRwBwaByOtNgjA/Kh05o4x1rOaI6YbP4VjjGao9OKxx15oYx61kHoaBB9669KyAORXHp+FEDHBo9OtZHTFJbTyfcjz7ZAqPSLpkDsmPxB/rT6YFwgPP+feorN0OGFRW/y8CkiUrgNn8KVCvAbP4UAQOTSAngH6UoBAAOfwrAPBNAZ4zQXIzQUnofpRUg8dqwB1NYweOcUzBhgUxUj1NNkisepoEYAzR5ogsM4otg4Vf1oZYEkc0qAjGaCnJYjntS4PbpW0etMMjApVwMEUFwcg59qUYAB5rAzkdaxQUDgUaIBpsEYAyfSgqjoOaPIwKOCMGjkYAFcHgGmUsMA/SgOODSKQSd2c+1UBnjNKeQKwOmaxngGiO1YJGRWAR1/SiCRwc1ntjJoAAYojJ6ZoYJxniiB0z+lZwBRA6AUc5yaJ9vwpsEcH6USCeKABwQKyAeRRIPBrI6D8qyAeBR571QyRyMH0rg8A1nI4GaBGM4xii28ABf1pm2jaRkelbwV2qKPTg0zkgAnNFgOrdegxW4Hoa3E8A0owclv0o4wAtMV7ngdaQq33jisqQFBz7UFCng/pQdS3K4P1oFCcA0ASo4zRAIwawM8GjjpmsDPBoglcBufpQAPBNMBkAc+9EZ7Vk9MfSs55zWCwwTzXQ4AzTLliC36V5a4wR071IMn5R+FEHbgChg5JohSPmNTwEqOOKnt2A3L/APqrHGM0Vz1OaIz0NZGOv0ognof0o4OMH6VjnOaPHANZ4BJ/Cjg8bv0rp0NZA4JonPIPSsj1rr0P4Yo4JrAPegQON36UXJwP0rJAwBmgQVAzQ44FcYxu/SgQBjd+lEA9/pxRIOBu/Sj8wwG/SlUuMDmkt3YYB/CliYYXGcV9klxuVc/jR06QqARn29P1o2DKuQufxpLXyzhmxj2zQcIMLz70khHAOfQU5ViCDz6YoA4xmlkBzkdKDoRgihHEy8np0FTWCSDIP4ULTA8vyc4/2qk0gzL8iYI9/wD69XGk3UAzsyO3I/xplKnlenvRwegojPI7UCAME0AMDBzRIHej9f0oegb9KI55oEHjNE4PJokdjRPYGgeMk11HBzS4PB/KsEjiiABwMkUQvZv0o4BwB0okE9aZRjGfwrAP8X4Yr5du0GhisA9627hjP6VgAYBzWFOMH9KI9DWARVNg9DWeMUQOoFZz1FdOPyrA4BNYA5rIIGBRwelAgjANV15H5VkHjvXA4FcUwJPBo88EURkYoDjFEnpmsDbmge4NE5GCazzmjz0qjg8Cs8YzWAR1rjGMUcHpWcYGfpWOMZrj0/Cjg8CuDwDVEAng0cDvRxivav8AIrHvXXiqo9sUTgcVQx3odKIBGKx8uCaxxjNVgnrRBNEZGM0VGMCiPlwDVVknpS89a4IxmiOKrkjANEA8ZrqP6VVZx0NDHeqwCOtYz0NEdq4I4NHBGBQGRjPNYzwKI7UQOgNEZ6HNcHjNBC3AFMuOlY96OMDFA+nWiSeDX0NUe2KY4GBRwOe9Upz15rCnpXfiiQ3ANf5FN0o4AxXG3igOMA0cFcZ5oAHvRAxwaojHGao80V4zmiBjg0FBXIPSiB0ArIIAFVROTkUxBAA7VgkcVt4/lW0kUQDwPwojtmqIGMA/SqqjgjAqiARiqwKIBrGRxXUVVAZ4FWGmtM4L9B/9f3qKJEG1KkRS2QelXEqcAHkUzAnI6ioCzggClUYwK25HWgAOCaRRuyDWAOlYzwKIHTNDoP0onPCmgO2aI3AUAOxrAPArr0pgD0H0oj0rGe9EE9+a2+h4FBADnFFFXp3oAYwBRGe9YGMCiMjANH2FAdhRHamPHWqrA7UeBgVVYHaiARjP4URjjNEAjFEZ4zRVR0OaoAAnBogEda6np+Fbe1BQRjPNbcDIP6Vj3ojjGaIwPvfpWCFwK68/lQhdoGmTomM/iaIJGQawOgNDnv0ocDA7UenFbs8miPlxmic1zgYP0o8gCjyAK+hrg8A1WBjArPOKJAGaBBGc0W24HX0p5AeBSOQu0dq3guCadlJJWgwJ4piMZrIYDB6VkHgGioPIoAA4JosBgVIysduaQgnAoyBSVHWtynqaKhjgHrTMvTP0osAQoNEjGRWQwwKIBGKzk7fSsAdKJx1P0ociuDwDQALdaJA5FMAcDNHbjiicHd+VO27oa4zyelONh3ZokHkU3Az6VMpJ46DpUyCQYBqeAqcrRHbNUTgVkHgGiue9U3selMCcYo4xkV8xHBojIGTWOc5o8cChnvXzVgkcUOnNEYAoqABmivFA4GM/SuMYoAHvTDJwBRHGSKiiL8E9KSFVGBSJztFRrsbnmjjHNFgOKZtxwKdAevNBc8KaKgjAox5ApkzwDSjI4rbniguAB6UCRgClIJGaVgw4qRVZcNV9ZoVJX/PSpYdmMGiAOAaIBGKAwMZqsYI5rAAzRGe9YHaiM96IwPlogEYzVc9AaCgd/wBKAwMZokDC5o4xxR55oLjrTDvmuDxVYBGCaIBHFADGTXU5FYzwDRx/DWRjrzVY4zR5GM1njGfpVVjPArIIwK4JrGRxTADoarg8A1gHjNAA96Zc8ZrrxRxjrRxx+lUVJ7Zo8DAFHBHAogHgjpRCkZ2/rRCkYArgDGK4IGDRAI6UVA+WgFAwBQBPFEA4BojPQ0MdDXB4AohT2qiABjNUBkdentWAeDXB4qiAeKAGBxRGOM5ogHAJo8cAUV4wKwO1Y4xmtvfP6UQCMUVBGBWPeto7UVzwBW0EYBo57VtzyTWOeKx2z+FBaAyMAVjPGax2zn04ojI4NEDHBokAYNYGcZ/SguQRnn1rHGc0MEYJogY4Oa4AGDn0ojPQ1t4zn8KI44ojgEGvlAxisZ/pROMEj8KPPT8qwCOnNEdxRJPAoMwHWj04OTWABXUUAAOlEA9qPYUVHQURnjFYHrzRIAwaIBrAxjNBQO9DBAGaABGAc+1EBug+lHAximXPQ/pXGMA/pQAHBNbRQHOAaYZHB/SjjoKznqKAz3ph6HNcEAGhkjBojBxij83agFxmsDH8qxngURjjNEAe9YHQCidxAzRwOgo7egFBMjkfSuMYoAAY/KupwBRIPAFAA8UQQ2AaIGOlAA8Ac0RgcjNBdwwFzUNg8vApNLixgj/P50+lwbMAcj6/41NYsDle1OhQ7SasLcSOCelLsjX92KMrno36UzseM0yKTyaCKTwfwqAoqkYrAYfKv60q4HK/TmlTuDmggyDnkUc4wKBIPFA54ofdGBXDcAUx5xjNDkYApgOmOawQM46UQDzQQ8EijyMEdKC54HGKIBPTPtQUAcGmA6CiAeAK49KxkVgHvXbANEZGAaIB4AoLnAFFT0xRGOKAB4J+lBVIABzmiOcCqqiBj+lY460RxwaI44NEEjCnntQBAAJyaVQDxSqSORRUdBQXHetvt+FDAABraelKARgHmsYGBzQ4bJFMADkHn6Ul1OkD2ySYjlxvXaOcHIo4xk8e1A5IyPpUgjzlBijyMk0MY6VWccA1jI5FVRAIxVK24kZ6VkHoaLLuGD9a3g/d5osCAQ2D34rkkESfjiiwA2kc9qZgBgHk9/So1RuSfwphlvlPHYUVxzmnYbcgdO1McjG7cfpikAA5OfQdKIHUmivygq3T2oHPVskdOKwpbBOcdKAjbgD8ahOB8rfpTAZwDmlUHkilAOBu/DFMUPBGfSiFVckc0CAMAUCCQAOlHIHAyfSuT0pgTjFEDsazzjP0oEHgmsAcA/QYolW4xzTEfdY80XA4IzTbWPA+tALuxninGCcNn8KYgjIPSiMjBPSiqjipnJwo4pwAhYPyOgx1osj/AClfxzT2atyjfhinjZDgisA9B9BXBGaxkYJor6H8KK56GtuRj8qIGOetbQe1EDGcUACMkUQD0GaI7kUFHSiM9B9KVTkEHmhhj0raKJBAJFbdw6c9qAB4AoAAdKGWPFW0CEZYUsajgDio4Q4BC5/GtgA+Rf1pUGBgYoqCcY5raDwKIxwBSxl/lC9Peo7YZ3McUYohxisRkFRHn3zQWLgFcH0yaESHgDmnhyMIfwoxOeAKePC/e59MURkjFF9oBPNb2I+Y5/SmijcbQMnsKn05CPlPHpj/AOvVxYIvKDB7GpIGXpzXFAZ4BqiCeh/SucY61jPNH0FEHoDn8KIyMUAAM0OR1oAZ5NAgVtyOtEZ6GjyBzWOMKf0o8rweaxkcUB75/CgMnANEdwaABOKxzjNEAY2mipHINE84FEZ4zRAxtPNEA8E1gEAqK9sVjbwBmjg/MKOD0oEk4J4FYHajyAKoYPBNH0FZyMelUQOgNYGMA5oHIGKIBrkjHpR5FdRQGQM1jPAoj0NVjuDRHFY96PTisAjrRHHBqsDGAaAGMZojBxn6UcY617Yrgjg1jPINEcUBigc8VQJxg1R6gA1gk5AojPQ0BgYzWCec80OnNYyev0ognjNAds9KOTxRBwMUcjmsEnjrRJA5P0rBA4P4Vg9qGe9cHgGiv/1qojtWOM1gHjNUQMDFYyOKIyc5rGRxWPf6URxWc8Z6VxjFY96IzyD0oAkY70y54zRGRjNHsa4AoAY4NEZGAa4AxnpRwRnNY96YZ4B6VwTj0puRgVgHjNEEnkVjI4rAHSuCODWABgmgBjijytAZGM0B2zTDuDVFQT1pcE4Bogkcn6UMkYFHkdelHJHJokHkH9KGBwTXWmGBitoIABrAPU/pW0Yzn9KIwOv4URkAk0MkYH4UQRwaJz0NY46/SjxwKzhQBRAxwf0onK4zRyeM1jI4rAJwKwcDHWiM9TWOdoNZAGAaCM5wo57CrbS3kALD/P51DpqqBkfh/k0IEQYWpI8KCtE7V968xdvz1dx28qARtkjtg1bIsRBzx602wcqf0o4J4NBT1HfpWzisAjrSJlcr1pVyOar2oAEYNFT2FEDsaJzjmixIwKC5Gc0Bx6+1cAcHpWA5yG+nFBfl2mgoIznmiARgHpW0gUcdjzQx3okAYzXA5BogHGDVEcUQMcGiO2awCKwDxn6cUB3HNE+ppgB0NYyODSgg4ApgScVt4qsZHFKMjFMuOho8DBPSqxngUD09qAycgdKAyMZ+lIoxtNEMBhTWDjOKAOOD0ofNRHGCa1ZNLWXGmybo/ow7D1/GmBIwRQA70wweT9KJDd6bg4B5FHJ4zQGeBRAxwa6dDRxjrQdTwD0ogkcGiTnGKZjhQp60rZGTRJBCiiNpwxp25wp6Uh2rk0cFAQeTQxkAmu/NM2DjFE5HB5rleF5plLAHNHg4Xr6UFBPvTKAeD9KYbu9EZY5PSgFB4NCNSAD+FOpXjP0qNcfLmgEzgHp04o5xgmlGTgHkUuAMnrRUEZ70WOcZrnj9aOcYAr6GqBDAHNUGDHg5oKpOBRVgeDRBcjJ60UDD5e1BQflB57UwPQUApPNN6D8KKk04GNpPPapeWxmps4xmiTnANQyruAY9KmmjZsZ6dKkKs5KnjtXFUewo9cZrGBxRBIwOaxxjNADHFEgd6A5wDRHBGelYA7/SsZGAaCjoD9K2jGM/pW0EdeaYAgYNAY4z9KC0RnvUMXFQwknANLbgjnrSoF4WgoHApkwMA8isZ4FLECASfwpYVIwK2Khw5pyrfKDn8KWNRQULyDRAPFBAn3aK4GB1odOawMYzzTIGHNPGegp0AGR1oZ7Vu9DxTQI4xUtoRkYyO1TWg5B7VLbspAUUFJOKIxVHkYzWcnAogkYzmioPQ0R0ANHOBilGTgmgCKIHTPSiOODR6AA0eme9EDsa28dfwo8gc17YrIK4qqOe1YB4zQAzyaPoKB9OtHnmh0ABoketYB4zRxng9KJA71RxjrVV+FbR6/hWQehonFUSDyD0rIPQ1kDjNZPrRLY4NEkjGa6jbVckYFYPTPNHd0Bo9OK+hokketAYGM0SQODVAEkYNY4xmsEd81gYwKIyeD9OKwMYoAYwprHvQxjANYJHXPtXXiioJzmsDGBRAHAPNFRjgUBxijnGBWB2ojAwBQGe1YxwBRHoM0BgjmmAxwK6jIFYB7Vjjj8qHBwDmiCeAeKKgjAoAYxnNVFB5pO1sY9qdQDtzmgDjJo46ZrqMiiOAQee3FYPY/hRCngCiMis54J+gogGiBjFdBxXbFAkcZr6GvcUee9EkrjOawDxmmAPAFEZGM0RkYBoAYAz0oDjhv0rIHejz0P6VgYAB57VweAaJUjGazjoa6DAP6UBxw36VgdQa288UcHiiAe9Yzwp/ShyOtEZ70eRjNAA962ZGTzjpW3tmgD0BrbnjNBc8GioPQfStvGD1ogDgjNFRjAFFV60VAHFFcjBoqccUAQODWCepoqMYHatpNFSeCaK4HPNFQRxW0EY60MDnNEYGB+VKhIG0UFc8Ac1HazSdB/KodJmmyCeR7f/AF6GgqWwz5/D/wCvUWjwRAEHn6H/ABpI1jXYp+lMwAAJ+lOwOArfpRc4wW/SnYAZzUqqwwTTxFT8rfpUaoybc89qW6kUYdskdBikuISuCvP1NKzMBtGAelPgJgtk/Sgqk4BqMArjNFSBgCsZ4FKM8g9KOSvTNEkDAP04rBxkUOePyFbT0A/CgDtxmlJC4B+lAdiM0F6YNAZHX6VhiME/SsYGc5PpRfIwTRORj0rOT1ogEYJr2zRXgEHn09KAB5oqCMAUQMcURnoPpRUbMg1yBWPT8KwG61tx1rAU8H9KUBmBNFTuwDmiCeSeaK5FYAXpSJkdOaTRdRuLNbyGDdEejbgO+Omc9aGMbTWR0BrjGM0RxkGgRnAor6n8KwAOBzQBBxmiQygdTROBwfwrIJBBo9euTROTgHn6UGJIGc/hW7HQ0WJ70SSeDROeSaokA4Bovno36UWJGQaLLtJzk9hTHByrdfamYnADZ/Cg5bjd+GKZQVCk9OlFyBgN+lbgAMGiQ4yz/pRcHqcmm5GCePTFMV3YUdKz0Oc0XL89aJVvlIo4I4PNADjNBCcAt+lFQDgHntT9QQ2fwokZDLRyB8xz7YooGX5TRTAwBTMD07dKI3Jkn6cVGduRu/SgARwaYsTtDZ/CirIcgc0AODnNAMQRnpTAYxihyOtHpxQFH2FBeACelFQRgHmmULxmj7CmIP3RQVcgBvrx0rcQdqmiDj6Vkg4U/pRA6UQTwBRDAdeakbOATnFSlQflHSpj2FOQTgUc4wG/SmG7vWATx1rPOM1jOMmioNHOMCu3J/SiM9DRHHB5oDIAB/SiARx1rGep/CiM8URkcn9Kx6GgcHrmgSRwawemf0okgZU0SxGB1oDjH6VEgcY9Khi3Yx2qGHYQcUFA70oIIIP0oj/a+nFEMcEH6Uigc5yaH16dOKWQKME81Jh+Qc0FAOR3rjtRAIxQx2HNH5zgDmqKAnJ/CqOTwKPzAYooSMBv0oLtI28miCODR7ACiEZcEU9vG3QfhU1vtOCM1NaqTlTTxshwaPoKII4AogGiOeKxxgGug5NDJAAoAk4zQGaxngUB2FZ4xmjyAKIyMg1nHFHkYrjpWB2ojPQ1jPAogEYPaiBjAH4VjIwTRIx1o896yCOD9K7YzR6daYbu9AADrzRXPf8ASs5GM9KbkYzROeAelEcYBo5IwDRBPQ/pRGRgGgAO/wBKzjgmuMcGmGeh/CsE9/wo5ziipxgGuDyKHA/lR5PFEY6USeo60Rnk9qPoKYc8VRGKIJHBojFAEjArrwKxngVWCRxRGRgGjx1oDI4rAHAoL6VtPYVgnoaA4xRXkD8qYYFYPXNURkYBoEkdaK7jgGhnoaIOcVjnk0eOCaIHXPSiPQ18x4Boc8k0TwMGj6ClAOKwM4/KgWQYBojnGabHIzWc/KDWBjAoqccUQOgNAAkZrBzgdaIAx61RxjrXBGAa4IoAY4PSqJHT0quvAonFEAng9KBB4zRI6ZrIxnNYwOKYZ6URRI6ZokDjNDaRgUcAYzRG4ZBqhxwfwoDAxmsc5/Ksc8UARRGTjP0orng1/wAEx/2PPgn+0t4S8c6/8YPDlxqp0fUbW10/yNSntTHujdpMiNlBB/d4znoeneT/AIJT/sVvE0afD3VoyykB18QXeVPqMyEfmK+IPgzVPh1468ReANaH/Ew8N6pc6dcnaVDSQyNGxAPYlcj2Ir9iX4DWH7Rv7R/hj4c69bzS+H8y32tiJ2Q/ZIULMu5SCu9/LjyMEeZmv+HVn7FeMD4c6p/4UN5/8cr/AIKJ/Ab4dfs7ftBp4D+F+nT2Ghz6Ba3y281zJcMkjvKjfPIxYg+WD7Zrwz4W8S+M9btPDfhDQNR1vV719ltYafavcTTN6KiAk/lXwE/4I+/F3xxBFrnxv8R23gLT3UldNt0W91B+ONwVvKiHQ8s7dQVHWv2s/wDgl54C/Z1/Z+8VfGTRfilr+sXvh/7H5VldWUMccvnXkNucleRgSk/UCiAB8p5ojOcUR8uAaKk0QR92sA9D+FMCTgUck4pIXbkc0LKUjGKTS7hui59OR/jS6NdMwGz9R/jX9gzlckfy/wAaXw7IwwT/AJ/Oh4dhAAJ59cH/ABpNCtFGXXdj3I/rX9kWYONn6n/Gl0uBB8q4/E/41HaQqflXn6miirkA80SCOOvaiR0zRIAqRgVBFHnn8qYAjBNMBjkUEzwKKksAByKEeXylfZTMCMcjpSWTEcjJFRgZKsOV7UwB5oKrHb6Ug4wD0oDPGc0ACQc0CMAZouAMA0dpFY44Oa2gHANcjgCgT0oAMMg0MZwKYBevFZOMA1uIOAciiSSCO1EEjmmB55okHANEE9q2mguOCaIOMZrAwD6VtyOBQHOKAJQgUBxg1njigMYGc0FDYJ79KwCeOtIDvAHWp4vLYA1sJOAORUFs9w/lp1rVdLhsYI2X77ZyOfUe/vUbhTkGrfxNJYaT/ZscW5Ox3Y/iz6e9FQRkHn6UcHAFFTjpQ255NT+QxBhOcdetFCR1/DFG3YxJKD1zQAb5Qa254o4IwayD0NE7jkdqYA96IBXANAjpWRjrXGMVkHoaIzxiiMnArKD7xoOvQdqIHDA0yhcKTz6UdnQHntxTkH5lPNEtIvXhabBGM802ccmgMcCiARmie5NZyc5raSMA0AAMA0eOaxnpRVueaZSBgHJqRssCTQwRmipXCmshuN36URt4J/CiwcBVPNAKDjPSge4PT2pFBHFYwMCgp/KgrZDHmmUClQkEE80F4wazgqgPrRGehrbxgUenFMCcYphkYzTABcZpgAgIPJ6Vkk9aJJXCmlIBosQOtBveiMr1onYAc0zs7Eg8dqlAKZB6VI2OlOSe9SEk5BrtRxjrRAHJPWiOcg0SPXpXBFNwMZ5ogEYzRA6GguBWBjAo9M0cY611GRRANYz0oIT2o5J4/CgpJwfypomAB9KKnHFKu4YByfpUUeBgVbRLgMaAXHynn0oAHgGhHjHPTtW0USpbaDzW0V0NFVPehjGRTYPIrHGM0ql3VVGT6URtOPSlYAE0V3dDRHv0rGTjNAD16dKz6GiueBTRg8g4NMjDrXl45JoAdAayMdaaNZByamtd3Ciri2zxjmpI2jbH5UenFYGNuaJ7A0BgACm6UCBwTQwcAGgAehrbx1/CqG09+lURxWAR1rGeAa28dfwrGOCaqgM8ZquvTtXBHFcEcGuvShwMGjgjrVdTgfhRIHesZ6VWQR1o+wojij6CiSBxWM8A02CMihjFE8e9bc9TWeOua5IxmsEis5x7Uee9ZGMCj04qjgjAokHtROO9E54zRx0zWOMZqsDGBRAIxjJogGiAegr2xRIxgVRAIwBWBjArr1riuoogHoOax70MAYFAY71gDgflVDJ61gkYP5V1OAelEZ4J+lEg9aPPANEnpQyOAKIyOv6VzjOaJ7Chzxn8KHFE56da46V9DWT61R5GM0RkYBrA7jNE5ORR5719DROTxXXiiNx5rjGMVj3oEdKGMcGiQQADRAIxihjAArI9awpGAKYAgDvRwB1o4PFEiiARWB1oYPSiF6E0QOAKPsKHTGPpR5+WiM0AM8fhR56CiQTgURnjGa/4It6YIv2efGesbYgbrxnJb5C/OfLs7ZuT6fvePqfWtC8eR6v8UPFnw5ZI1l8N6bpl8CD8zJdm5UZ+jWzCv+Csfwr/AOFf/tWXviqytfL0/wAd6XBqiFUwguEHkTqP9omJZG95vev+CLXwiFpoXjr4539tiS/nj0DTHIwRHGFmuSO5DM1uM9Mxnr28A/EKz8c+IPH2jWbxsvgzxIujlkOdzf2fZ3LE/R7l0+qGv+C0GnvH+0l4R1JWTbceCIIgo67kvbskn8HH5V/wTR+OPhb4D/tJDXPHviODRPDWs6Be2OoXlwT5UW1VuIyQMksXtwigAkl9o+9XwU/4KL+Af2if2iLb4K/C7wlqMmjmxurqXxBqLiAzGFQQIrcAttYn7zspwPuDPH/BTsFv2GviWF6/8SrH/g1tKW2c8A0bdyMZ/SjBKMALRjcjgUtrK4HFJp8h4J/DH/16t9H8wEN/n9aj0dFOS2Pw/wDr1DYJEeHz6cUqADGaypH0rdxzTPjo36UZOME0JCTjPHpiieOlE4HAonAG089qcg8ZzXBHBquDwDTHA4/KiidcU2CeDmmUMMqOKMRxz2pYyBkDNGIdWNCLAyG/SgArjJ+tBlPQ5oqpbcBzRUE5oKByKC5GM0ATyO1AEjg5onIxmmAwMDntWGPAb8MUFJxk8dhWPelIxg/gKxnqKBGM5xQDE4pl8wZB+lEnGFP4YoDPAFE84AoqM4BoqpOAOlMozgCmAxjHPal+Xk0yjdnFFTtzRILfSlbHbPpRxjgc0pypUGio7CtpoLnpQIUdOlPsZsqMYoqW6dqnAMRIbKjtiooWYZU8CtzRtlT0rXfDOqaMRNd2u2GT7j71OcYB4BJ6kUsTuNoXOO2aufBs9p4UsPFEl3+7vvM2R+WONkmw87v6CnG45I+lEE96B7E5xThc4Wl6EZoHbxn6UniHUV0tNJW5/wBHTO1Ni8ZbcecZ60jbRjOfSmYbuD+GKyp4NAgDiiQRgCiSR60SAOTTEDjNOQAFPJNElhgH9KDBuC36VuB43fpTFn+63J9qYkrtPNOq9uR2rccYDcemKGD0okHgj8KAOOmfSgQuHI+lE7jlRRJI4NAgtnOSKLKMCmAYZFEYXKjkUHyBxR+U/KK28YzQfdgbufpTYAwDk0ZBjFA5f5eh7U4BOFNAnqTk0wZ+c/pQBxyKwcbienSsngZ/DFA84Bxmky2T+VDGzAHPasMOKQ4GKIDtkc+1AAYAFYx0P4UBknIqjyAK4rAxgURwQwzmpAobC04wOD+FA7T7UQSMEdK3MBtbmgfkGTQIHvTkEDZ2pgWHB57cUSwG3d9OKdiOd36VI+R1qWUgYzmm5OaySMGghC7gM0ASMGuMcGgcgCvoaIJ7UIpG+6v60LSRuFFR6ZPJ8o/p/jSaHct98fQcf41D4bZ/mxkD/PrR8Oxo2Rx7f5NHRYvug9P8+tf2KgHL8/T/AOvX9kRhcf5/nSaVEvQ5/wA/Wv7Mt+CF/U/40dNtiPmT6cn/ABqbTkDFlHHYen61PblTgD8KWNM4IqKFTjioxkYHalXJwvWo0wcE1WB68UQG4NBFAwKIBGKZgBkjpQA9KAB4P4VsJ6nOKGAwYdR0o7SMgc0uAMDv0o7SPlODRUE5BoLu4BrGTwaIyMZoD5iQKZlYYrbnv+FPkJlTz2ojIwKZarAYbWOanthtyo5qWAPwRyKlt2TODnFde1EAciuT/hRGRnHNdRgVgEYFAHgZokkYz+lfQ03PFEgnmiAeQOlHkAZqiSBgH8KAIGM0DjofwrA7UORg0cAcCiRjAFYGMCm5OaBB964I4og54NHBHFcZwT+FEDGAKA56VRwR1ogMMd66jj8KxkYNE7uB0oADoKO0dBXHT8qIz0HNDpzQ9x+FZwMCs8YzRBAxVV9DRGRgGqI7VgHjNYHaiB2NAYGM0RnoaIBGKHPf6Uee9Y56/hXUcGug5NHGOtAYGM0AQMDn0rI9fpXGc5/SsE9/wrHvROKIyMA1kHgGuM4FZB4BokkZBrtzRIBxWQSAKJ7AdKJHBFEjgitwIrI6Gs5OBQOaAB5Bqs55NZB4BrORwa6iqx711Fe1ZB6GiQcYP0okDjNHHTNdePyrqKoEEcUfQVRIziqGD07UdpPWgAABmvbFcEcGsAd6yCOaGD07URkYoDAxiiPSj04NDGMA0cDnNZBHWmGRgGiMjGeaAAGc1gDkUcY61/wSB00WX7IzXQEf/Ew8VX8/yjB4SGP5vU/u/wAsVpXxQfw//wAFW9d+H15eq1t4r+H1taW0OQCLi3DXSflH9pOOvzZ6Cv8Ags98ORrfwa8GfE62g3z+F9ceynZRytvdx5LH2ElvEPq/ua/Y+8IaT+z5+xl4LTX2FlFp3hpte1mRl+aJpla7m3epQOU/4AK/4JReNdR+JPgb4u/EHVyftniT4i3WozAnO1poInKj2G7AHYAV/wAFrtOMXxM+GureSgFzoV3AJB95vLnVsHvgeb/48ahRzxjkV/wSZhZf2wNOcj/mAahz/wAAWv8AgphGZf2I/iRGoyT/AGXgf9xS0qGzc5BX9aSyhUYK5/E09nCw+Vf1NPbW6jKjn05p4M5CDio4AuMHmlZR8tM2ehocE8V1FY96BxXBODWO2efSu2FFHJ6Cs44rOOAKI7VVVRGRjNbVxgCgABgGio70BgYoENx6USGAOfpQUEZzn07VtB6iiPmGD+lURxSDIIzWFHBpjlQFrGB1rGQKGOgorxknNZXsaAG3JPSgwIxmsjHX6UAOvpQJUkZoBcgZyKY44FABuh5HtRO3jP0rgDI611xzz9KaVUTZjJP6UPQGmC7V5ye9EEpwaYD1/CuDwDWB0JqMDcFocHmiueh6dqVdwwKsNKa806+uw3Np5fGOu4kevtTqFOCcVEm/KkVGgkTyz26UZUjHlxHOfwog5wRWv+MbnXraKKe08tYt2B5gPUj0A9KWXByh61PBrNz4atGMu6xG/wAldqjHz/N79fWiC3IHSiQVwDz9KBx3onNKc9OfSgwK4I5rpyOSKBJTFEcZPWuB1o+g/Cmzt6UWAO2iSMhh+FFgTyKI3sMHpTMAOvPpTfMQAOaZlHAGaB2sMiiwZuTxRJIxmgM8ZpnBAwv603zDFBsgCiScLnPpSYQ4HNAhTjPNHhsrz7USDjH4URtG7r7VuJJBPSiQOM0G2fKeaVu+elDDNnFOMEsT9BTBG+VT+lFSuCKZcDBrAUdeD7Ur4OA2fbFbtw4Oadty4U9KYK2CD9KHPymkGBtBoqAuA3PpilbJxnNMCFzmkxnOaAIGc59KXpjP6URuwQ34YocjrRxjrWQelE9s9KYhlznp7U+M5pgAMk9KOMAA05DDAb68VuIOc5NFiRtB+lEgcZ5onB4P4VvO4ACpGJOB2qRu2fwqUnGQadt/GOlbSTtHNGM46c0hVEwaJxxQIzg1Dbb+SP8A61R2iHjqajso3favaorJSu3H0pbKNOi9feo7EGPPfsKWJFQIB0oKAAF7dqkRXODUiiM4U7v0plIXJHFMBk46UiEkHOR600eBwaKg8Z+lFQRg1PZiQfKea+yMTyPrUcRQbSaVcAAHmkJBwaBGM7ufpSEMuAelAg8A1gDgGj060TgDJ57URuO0mjJj7pzSncBk/Ssg8A1tJFV1wBXTigAQBmiAOhoAlcA9KA3cA0fl5NZBGBQPAwaIBGDTxEn5TTKehOMe1A570eANvNHk4qW3EhG0/pVxAV4PNT25ByOtMD93Fde1HHTNEAjANKQDtokHpXTtRwelEZAIoDPGaIGcCq6j+lcEY70Rg4xVceldeB+VHmgARzX0rjGMUADgCgMDFEcV17URnHNEg9KPsK4PANMvGM0cCgABjrRwO9e1HGOtEZI5ojIxmiARjNY4xmmPYGicjFAgjpQIHbNAZOKJB4FYGMCuoAz0rHGM1njFHGBii2Ris8YqvoaJyeRX0NVWOOKPIxmsZFYA6VkjpRznrzRxjmjz3rOOAaY5GD+VY7A1XU4zVDB6GjwM5rAPfNZzwDRIziiRnAogE4x0okY47UWGOKxkZ/IVQwOAelZ75o4B5NZxzmhgDAotgYrIPQ0DjjNdsA1j3rOec1160SKLAnP5USB3rtzQxjiiAawBwK4PANcfdFVVHArgihyAKwO1AAcUcZAoAEZArhhR6cUSB3qsDGBWPes84ojPGaAA6Gse/wCFEA8EV/wSz077F+xJ4GudyH7fdapNhVwRi/nj59T+7/LFfth/Ei5+GP8AwVIf4kWyukfhLU9Dku8HmSD7FbtMo5/iikda/aI+D2lftDfBzWPhZqE8Qs9cks3M2ThViuYptykc8iMjjrn0Nf8ABTD4jL8PP2T9d8PaZdrZ33jOWLQ7VY15EL/NcAKP4TBHIh7DzB6iv+CMlvJZfC/4iWLXJdF1+3kWMnhS1vgnGeM7QM99vtX/AAWe0ZJp/hHqyQtu26zBJJyQMfY2QegPL/5FW8MUPDDcR+Ff8EtWV/2s9P2/9AK/4/4Atf8ABSIZ/Yt+IoB/6Bn/AKc7WiRjA60ckdelNjb1ohduRRCjpRA3ZxQIJBz+FAL0oDA61QBx1zQAIyTQUY5orkZJzQGDj9KKAL0oqAOK2kmjGB05rZkcDNbATgGioB4oKMDIrapGVHNKCQQD0og4xmiARkHP4UV3DbQXA2k0ABxVVgHjNcYxWM8CkIHFEY5zQOTgVweOtUVBPBo/doEA5A5okE9M+1Ab24FDAUjHSlCk5FMnOB2oKAcE5o4IxRwU4FEZ6HNAYGM0EGM4olSeBQA60OEJBz6UTlMjtRUE4A6UV4oDBwppF+YEflRUA0w7ClBB5NREIxHUGiiswC9qkGH25zjrXhPV7XQLkXE0HmMv+0R2Yeh9aWBUjCA8p0NOVZsk5IouB97kmkZ1fcG5HtU04lGAKkcjAB6UevP4UcdqOMdaUDkA0TmjzVHGOtEqBjNEqe9SOrDaDW/5Mmo+Rms5OQOaZBjcD+GKZgF2jmgVIBHUUTk4ogdCKbnisDtROOM0SSoUnOOlEAj5RRIK7SM0xIOCeB2xS5zgGmGRz26UCx5z+lFj2bkdOKdmK5B5oqzfKT+FNGFG4N09qJU9D+FGQYCk5oNjgGty4HGfSiT1B5oOX4bt0oMCOuaLkKFBrGTgGsgnAP4Up52qeaC4UKG/Ss84qJVc4NSIvIUYxW0BQQefSmAIDYoDAyKUoQAoyaZcjaDWBjAoqCc5ogE5z06UQGGDTDuppmVsZFEAdTmjjFSKCMZojCdOlEgd+aD4yAaZiTtB4+lblb+L9KkcAYpnz15okHrU+AvzfhRPGCahjG3PeiQDkn9KCK3Wjabm4PHpilsCrjccj6Va26SOEI4HaoLOFGDJHg9uTQTLZzk/SgilsCo1jC7M9KSHYDsbn1xSgMAQOaUKhIzXBHBq4B3CpJVKBAv60wGDzkGkUFunSmCsDgcelMq7flHPaiD0HOKZccUQFHAqcF2DHt0FZC9DQbkYNK2+lVMAAc1g+nNIAFAxRwelUGJO3P0ooD938qUruGKQjaAOlBgCB6UVBor2FE54JrIxyayCc0QCM5ogN0P6USQNpPTtR9BRAxwa2gdKzkYBp13jbn9KdCnBNE5woPNAFeAf0oOu4cc06hxkmriAEkg1Nb5U4p4yo5oEgcChgjANAjsaxk8GgVHQ0QCcZo8gCiB0BpsE5FAgDFEgjFbQRmsAcg81RAAwDWc8k9KOD3rtiqwO1YB71gYwKxjgGiq9jWMDisZ6isDGBVVWOMZqgMnGaIBGK68VgZzmj7flWCe/4UcY25rAHQ/pRwBgGhjrmiFPQ/pX0NcE0R6Gsgjg0DkYBrIK4zR68fhQAPGa6jH5UcYGD09qJB4Haic8E1R9Pyo4A4NHHQGiMjANHHTNE88UQOgNDkVR9BWc8E80fQUearr2rr0o9OKAA4Ao5PaiM0R3ArORkUAT0FHB6igAV6117V05H4VjPAonsOaPIxXUVweAaA4xRx0zVYJ4ArjGKJAxg8iqOOmaJBGB1okDvWAR1oDjFVgdq4OMH6USB3o4zgH6URkYBokZwKIz0NduaJBHFHGOtAg4GaIHQGiRjINdRwaOMYJokDvXAGM9KzkZoAYwDWfWo4XcYQZqLSL6fBSDjt8w/wAat/Dco+e5G0dh1/ka/YH0tNH/AGPvhjYpIHH9lPKCFx/rLiWTH/j2K/4KAail7+2L8SZ5ZllZby1i3L2EdnAgHHoFA/Dnmv2LfiOPip+zD4A8Ty3ImvINLXTr0k5bzrUmBmb0LCMP/wADFf8ABXP4qf8ACQfF7w58KLC93W3hHSzdXka8bbu6IbDepEMcJHp5h9TX/BGy7il0L4qWQkzNDd6U7LjGFZLoA/iUb8q/4LHWav4F+G2oeYQ0GrXsQXHBDwxkn/yGPzrPvX/BLE5/a008f9QK/wD/AEBa/wCCkRx+xZ8RSP8AqGf+nO1pjngUxJ4x0ogEYNOMjAHNDngDn0rAPbmlTAyBzQHtQGetBVY4U0yAEAUFXoG/SiFxwfwrIK8Uzqeh5r7wGOgonnAGTRU96JwNtBcHr+FMRtI79hWBjaOcdqODwaBAGM/QUAw4PeivGCc0wydoP4V0o881gE4HNEZHWiQelVRx0zSgY680wUHBP6USCNqmiPlwDRTjOaAzwDQwE2g1gA4HX0rHGQ3J6cVGrMcKOaUFEPFRqG4ByfpUawbGDSc8Y4NMpOMHNMAvBPPpRAJyDn2rgHGaMTAhlGQelTHcAqmlTJ459qAbcBUcDMCiHn0opj5CefSmXgep7elbAQBTLx8tBQw5NNCVRZSOKfMjkKOvQUylTjHNZAYYPWnjkgIDL83YZpgQu5h97+lTS7k2xnPtUk7NGATyKG5+AKZ8zEsf84ouuSQ304okDgUWwMk5xSszclf1psEYByaLnGMc1EMk4o9sUQc8CiM9qxnqa2qvU05yMAc0w29BmlGR8xreFXavNCTC7aIIGc9Kb1odRzQbjGKOW4ArAPGayxOB09aIb7oGc0jMy5I59KIYHcOlEkpk9aYsy/yrcQcLTBgfm6etMpAxQO0AN2okHkN+lNJ82c1M4K4ApSw9/SnJbG00ANvzGoTsIYnIHbpW4lssMUFV8rn6cUqqvVvpxRXIBBo4LZB6UFJbcfyocngc0CMcHNDdgBVzn3qJiFK45qMc4kPHenOThTkUpzwTxRUdC2B9KQDgqc+1blByrZ9sUxJI29O9bgeAefSssTx0piT05x1phkZB/SiCBzUgUn5TQBAAIosMYYUWyCAeO1OD02/rTfLwBxRYE46+1FscGpSSOtMSvBFBix24q4JYBQKWMsQpFEALTKGGDRBA6dKjlCgBhSlrr5Y15+tWdmYuXHJ6UoAOFFQ7S+CadccxjNQgKSxPA70CCMg1s+bcD+GKcAAsRz2FMNy4qUyMmQMgUAZDgCpY0Q7A3I9qTg4NJwpAH/1qKgjr+GKdQOgpyrAHd+lEDORUyAjIPT2oxBjkGlhyMGkh4wtGJs/KMilDBcHrQJKgHrVHA5NOgIBXrSoFGM5p0UHr9BW35cKc0o3HJ6r2oNgdayhGQeework4A59KPBwD0quowPwojA60ckZJoiu2AaPoKYt0Vc/jS7ifmXp05ohX680ybRkj6U3HIrAZtw/KiCxwBTAHgGpIMjg1LGWjxj6VJCy8AUQFGMc+lDgZApSCMCiD0Apj6DmsjOMUenFEADGea7dPwrHGc0TgYrg8A0uMc1XfgUQCMUR2zRA7GiBjg5rORgdfSsDHX8KokdjXUcc4qgQSBmqyOmaJA70Dn+lDOOTROP6UATzms55Jogdc9KbBGM0OR1rA9abHY1gYwDWQRigcd6IB6GgccZonB4rAPGaOMYBqs5GM81njrzQIAoA43Z6VnvmgSOhogYwDRwelUeRjNUBxkHpR571jI4otniqJB79KJGKIyMZqifesAdKIGOTRIxkVkDjNEjpmqJHTNEAjgc1jjANEA0VBrgjg0SCcZ5okAYHWsDGBRANYyME0AB0rAPUUQCMA1xjFd+KokZxmgBjiguVJB6VXB4BrrwKHP9KOOtY5ocVRGRjNUcnla4P4Vk4wDWB2rOec0ewBrHvXXj8qPArGepzR5GM0ORSwO/Crn8atdFurjBKYXtyP8atdBgt1LSfMfXkf1rbFEAkY4H1p7t3TZn9KMhbgnNeGv2vf2l/B3h6w8K+F/jJr+naTpVutvZ2kMiBII1GFVRt6AV4p8V+IvHPiK/8AFni3V59U1jU5TNeXk5y8zkY3HHfgVoHxR+JfhHTf7I8J/ETxNotj5hk+zafq89tHvOMtsRgMnA5x2rW9c1rxNqc2teI9YvtV1G5x513e3DzzSbVCjc7EscKABk8AAV8Mfjj8WvgwupD4W+PNT8N/2x5X277E4Xz/ACt/l7sg/d8x8f7xr4k/tDfGv4waVa6H8TfiPq3iKwsrj7Rb2946lY5dpXeMAc7WI/GiR0zXgH4jeOPhZ4jTxb8PPEt5oWsRwvCl5aMA4Rxhl5B4IFeOP2qf2h/iT4XvfBXjn4s65rOiaj5f2qxuZVMcuyRZEyAo6Oit9QKwOtY3Dmtue9bQa25/CihIyB+FBcgAdaKqOBSgBcClAJyKIJOc/SjyQCKY5GM80QVxk0AAowa6cZ60fmGRRGBjNAgDAFEEAE96wOtAA5NAYHB6UVBHXpQO3AB5plzwe1EgnAPPatmTk8mm5PP5UeRkCiCAF7UQQuDQ9Sf0o81xS8dDRAJ5NA4PBoKCMEVtwucUFz1rIIwD0rqKIGMAZqGLJwoqBGdXTGSMYqNAr7TQiM29c/dxigpYBc1IhVtp61wVwfyo4dc1a5VHOemMVpGhafd6Q13JLtmH+yT/ABEevoKikVMgDr2p1Vlyh59KsrWW5ukiiG53zgdOgrUNHv7ECW7t9iDvvB9PQ+4pBh92OKeJHUuh/Ciqngc00JVck14b0P4Z6x8GRLqtx/xNU+6myfvcnPKkL90CpkMaKT3zSoWOTzWnTRQ3qSSj5Fzn8jXxz+Jnh/xxa20elR7ZI9+fmc9TH/eUdlNO25euMVlGjwRk1JsEYA6isZGSafDtvxzTMFpTxzTkH5SOKATOMcViMngUFRhwKhOSRRAzwKJJOF7UTimAUHFHYEyvUU4UjKnkVgMcZoAYzQbgqTRXAwKClhuzTKR3oICMg9OvFFQRkGjhhg0cqM1ghMA80SwXIPNYO3aTyKA5BzTM5UAHikK7ivftTy4XAoOdvJreSuFPFfMduT0zROTigQ2MGm3dM/hQUsAWP6U5yMA0GBHWrceYNoP0pmKNgGl2lgSeR0pthOFNKxI2kdKGW4P5UwwQRQAIOaBJG0GiikZHUdKQEqBmiM4UmoYpriVLe3jaSSRgqIi5LE9AB3Nfsu/8Eo/Cf/CN2HjH9pZr691a+iWUeGLS5a2is1PIS4ljIkeTGMhGQKcjLdRZfsU/snWFl/Z8HwD8IPHtxunsfOk6Af6xyX6Drnrz1Oa/bg/Yn/ZO+HvwG8WfFjRPBVx4Y1jRrVfsJ0rUZEjnuZJEjiR4pC6Fd7gkIFbaDyMZBLN1Nfsn/sGfFD9qFU8TPcr4V8ExSFH1u7tzI10ynDJbRZHmEEEFiVQHIySCtfD3/gmn+yZ4FsoY9Q8C3Hiy/RMPe65fSSFzjk+VGUiH/fGRxz3rUf2Kv2T9Utfsd18A/CCpgjNvYC3fkY+/GVb9fev+CkvwN+CPwA+Jvhrwp8HtDudJm1HSZNQ1O0e/kuYkVpSkOzzCzqcxy5BYj7uAO/3upqRQAAKdQeFNFfQ/hTANx1raANuPpRODgjip0UoCo6UwwKYZHFMhI4HNMrdMVGpUbj26CmQ9+aihCtnHSmGe9FgBzTn5s5q1gNycKM4q0txEoVe1AfNwaC4b5TSspyuM0AWIUH6VIeAp7U0asm70ppMNtzUjhgVH8VGRcbSelM3y8nOaK+lYUg+opQGwCaUKEYEc0QGwqdaYA4BqRVGDW33pgT0PNRKRyfwojtiqqiQMZoKWGAarHcVTJuPIrAAoqN2R36UAPvHrXB6HpQbAwDW0kZBzRBzwK/CsnGAaZtyhc9OlbecZojBxmjgjgUAB0NOobrzimbbhVFEj1qRFIyKfCsB2rKryOtEAgURt5AqWIOM1JGwfkVNAQcg80R/CaAAGAaBA6CjgHinwQMVWBjg9OlUeQBVVxjFcVjcMg9O1HGAAaOMYBogGiMng9OlAcZB6UQDQAJAzRBziicY5rHGM1jHQ/hVdsA1nHQ0STwDWcjANAHoTX0NEE9/0rnOM0TkdaoqMcVtGOaCgjANbfetvfFEE9q+ho896IboBRB9K68UwxWM8CqIJojtWM8CiprGRxXFEEnAFYzwK6iuvBFEEcdaI6YFYxxnmiATjFHOOBRBI4HSgcisA8GuCMA0QDijjHWiMkZ/KiAaxjgmioNdelHHQmtpA4NAZGCfwr2oAHkGjjpmiOKODwBTY6KK69q+hojPIokDvWQeho46ZplHasDGBRAxwf0o49fpRAHes54FcHgGiM8GiR0zXtXBHBrqMGqCkDrWOeldeBWM8CqOOP0rGe/0o4x1ogEYoIW4Ayewq00S8ugMQ8eu4f41Y+HIEyJxkr9R/I1Hp1nDgBOPqf8aZ4o8rCuB2Oadwx2hv0pm28VnPeqpfStmcZraeteXleOcUQCOvShHvGQOlbMnjk0ISxxjn0rygTjb+tCPHAFNGCMnt0oqucCiFxkCkODjNFQc889hisbu9BSF689qUfLkD8KJBGF6imTA4PNEYOFpcHkjn0rac4IrGenFJHvO3PSio27QKZQowBRGRtY9OlOARhTn0FFSOM80c/dI+tYOMAUoBU7/wqJWclSv61JHsYrjpRIxmgpdsflRGMe1MMjIb9KK7lwOtY4AzWO1EcUozxmpFxyRn0pQCc5ooWfAHNCNywiHNBTnaetMBuxmlXJAzXl4XeRwelWYLScDIFeB/D/8AwlXiWDR422m43Y4z91Gb1H931rx94HfwXq4sLsYLdD9FU9mP94Vqdho6arewafc74l8vYNjD+EZ6nPXNSp5EpjePJ7c+1XIAYFTmiBvAqMbzgnj6VDAHjYDhRjBpZivy4yvpSyIGKlevbNKrB9oXJ9OlWN1JpV1FeQJl48459Rj39a1/xpeeIIPs1zD5SdxuDZ5B7KPSpUVEBSonXftY8GnHlLtZM56HOKjm3x+WVzn3qOFmcFTx64rWL/SJNHjtbZcXUec8tzlge/HSluAq42bj9cUxBUBTzUrcYSsll5PSt5xgflTEE4x+FZBGKJyMA9KYsTt25/Gi7ZxjmmJDfNxntTg425ohQoIOT9K8w9qic78AcUS54UUzYIAPWvvcdqYEkgc4pSM8igzBcCm+760DnoPpRGSCTShSMM2B24re23aD07Uw96bk5HSidvJ/Kgdy5U0p3cYp2QMVVs49qZ2c4Bpiw+8fwrOV+UZP8qLHbz+VZywOOe1M/YLn8aDEr057UVKDCjNJKUflcj0zU0iuQyrjHvTHABAredud34YpT2I/CghLADv0oxEx5JqFsfdfntxUzB+RQP3ec5zUaM3AFFgo65PaizEZWlAYDPNAKrBWFMFIBpWUnjr2qBXI+YcdqJHYZr9iHRtF1/8Aay+GOm68sbWn9upMEkGVaWJGkiBB65kRBj3r4hT+LLXwD4luvAVtFceJodHu30WGXGyS9ELGBWzxgyBAc8V41+Mv7RMvjC/uPHnxJ8d23iO2uWF1Heapc281tIDynl7h5WOygALwAAK8VftK/Hjx38Pj8L/G/wAUdc8QeHDdxXf2bVLj7VJ5kYYJ++fMm0bz8u7bkKccCv2aPg5c/tAfHHwp8Ko3lhtdXvN2oTxEBobSJTJOyk8BvLRguf4ivrWm6Z4R+Gvg2HTNLtLLQvDfhuwxHFGvlw2dtEmSfYBVJJ+pNftRf8FIfjH8XvE2oaR8LPE+p+CvBMMjRWUWnSm3vLxBx5s8y/OpYZPloQoBAO4jcdM+Nnxl0nUf7U0r4t+M7O8yT9og165jkJJyfmD56iviL8TfHfxY8QDxV8RvE95r2rrbR2wvLsgv5UYwikgDOMnk8kkkkk0CMDJqMOSN3SiqgcCtoPVf1ryl6mhF1Vhn0p0ycGmkKkKw4qQB2wgzQtiFztyewzSIB94ZI7VLEDytGHI+Y/hTwhTwOe1KuflBzmmhycqPwqSMDkimhy3K/hmtNtwI8qv+eajZY3+YZqRkBYR8g9KXcw4OfSg6hSpHJ70hHc//AFqlYyc7cDsM1konK8fWmZMEs3zfSgCq7s596wzjAGaAPKnr29qYk8A0EYgn0pCM4PFZR4+eCKHXK0VyM1IuRiiu4YFOCOFGTSKdoGOaA3DrVYJ5xmguaKZ4/KiMcUORgUQduDXGMVjI4odOaPOARXBHBokghMZ9KY4GcUjlflBzmmC7cgfhWMDkUfYUcHgCiOBRGBgiqojIphgYAzRJXIJzmhwmGqSFG5J6VJGyngcdqwS2e1Myn5SaBAGKZUY7SOauIdpxj6VJbhjkDjtTKVO0jmjgDGaOCKwOoNYUjiiuTgUeOKOCoAPPpQUEZzWB0Jo4x1qsDHX9K4xjFMAOh/CiMUR2onnFdeK+hrOeM/hRxjBNUSM4zXHQVwRj8qxkcVxjiuoo+go8cd69iKIoYxxRz0Ao47VgEdaB4/lQ56HkUSCeOtHnvQ4rA7GiO1YyOtHnpXFHGOtVR55FH0BrOec1RA7GiB2NYBHWqI7VRAxjNUQDxRA6A1gdqOMdaIBqqwOtVzmqIyOKxnnPNEE4AoDPGaC+lYzwKGBRANYwMg0RnoaxjgmgMDGaqiAaIBrGeBRBzg1jPAoA4GaK5GM1tJ5BojPFYz0NYzwKx2rHGM1WABxRU44NUc9qraT0ojAxWO1bS33TVtaSXEgjXmtP0BWIaZc+2cevvUemWluMLHz25P8AjUkm0AR0WcjJPWizMME0SQcA/SgfU1n3rGeBQUkcUI88YpYxkAn6UEBPt2pYUZcjrUkcYOF/KmZcbVGKKEMVBzSjBwDWCDnNFSY8mlAMZ55pY96EHr2ogsME/hR6YJ/SlABwDRjBORQXnBNBeMAdKRS5wKCkgkdqTcVIJ+goo2OKUZyMfSmBC4B5oKepOT6UkbMMqa2FlyByOtEMw5NBDgjv2ooSM0Fbjn9KcYIA6ihngDrSgk4A/Co1Z3wh5qZWDlD1pQTwD0oKeuelcsf5V5Zxtx9KVCWwDRQqME9KWPzDhRzTptOB2pBnPrUigIFalAb5cfSoIVVN5qK6exuEvIB+8TOD9Rj+tXd097J50h+Y/wCf6UAGIpE3HGelKASCatIGeUMp6f4VFNcR6m9xZna4xtPBx8uO9X91dajMJLqTe49gOw9PoK+Avwxb4ka5eC4OLay8vzj/AL6SbejKeqdq+NXhjw94d8QpbeG02xNnecueiRkffJPUmrhDvG4/pXkxlN+KWMsm4c0QBbspHXGPzopGX2sKXashLjOOgqRy/OailjVQuefSnO44JqUBmEcZ6dKlQjjPSmcuOTQJAwgoMqoFHUUWLJgnJoEABB1pwAoANBqxkbs9K2DbkHpWc85pgMdeaPUkGpMhck/TilVlXcDzTOXHBohjyDmmLEYz06UFyOtICrkg0Qw5Joc9efSgSq4A5osc5HWjk8A03PFKC3ejEw5oqx4x0oqx+UGhGeATTEhselGOTsa8tyMZoxPjBpgyc56UzFScGtxY8GuCmGPIpQW4B4p41B2nrQDNgelMMjcDSkKfmpwcbs8elNkkHNMH27QciiqgYxzRX0P4VtIUECgjEZzyOlAOTtU0kbNhFpkycKOaKH7oPPagDnFEByVTtSRdj+VGJSMZpo8nBPSlQbf5CnXbGdp5pEI4B+lEEDGa8L+Jta8F+JNK8X+G75rPVtEvYr2xuFAJimicOjYPBwyjg8Gv2U/27vhN+0jolhpN9q9n4b8eLEqXmhXkwj+0SAcvasxxKh5O0fOvORgbj8XP2dfgn8drMWvxT+HWk65Kkflw3rxGK7gX0S4jKyKM87Q2M9Qa/bM/4Jp6t8EdCv8A4qfB3VLzxB4PsFMupadeANe6ZH3kDKAJoV7nAZBydwDMP+CPuiWt9+0Z4k1i5VXk0zwhP5AK/deS5t1Lg54+UMuP9v2r9vDV9Q0T9kL4oXmmS7JpNG+zMQD/AKuaWOKQcescjD8a7YBoAEcHpUaBjk0sQIAPWtoHArAIwOtFewoLk4oLgUQcYHWmRX+UjOO1LAo4ApkAICimhJwBzRty3OKa2cHIORX2VMYPNPZoFJUc1PAFGF6VNEQBgdaisXkGQat4GSPaD0oKW61s+Uq3JHSkiZhknIpoj/CfpQgYcUbeQ4KmniZoyp5oRMV2jrTQuOCfu9K5xjOfSiCe1HJPArB6A0oIYYNDBJJpiAOKZjgZ6U+09enaj04/KmXnjmgBjg0TxkGtgI4FCMGgooKAMiioJ5raByD9KOD0ogYphxx+FU2OM0SFGQaDhjhu1FgeKAbuf0oYJAzmpBj5QelEYGMUcYGKGTwDW0kUenFEDA5oA7jk0QccGjHuxk/hTxk9DRcgAZ6U2XHSjF1HfvSxAtxUkYHDDntQi4yaeMseKaNVOSOO9XCRE5ApgpOFHTtX+zQBJ29hWRjAFcHpWAR1ojivoawMdfwogdAaogngGqOMdao46Zqv8imJHQ1wTgUe2KLDtR46URkYBon36UMkYNEkAVnPANZHQ9RTHA4Nbs0QDRJAHNZHpRKkYBoqSODzQAIyDQwRgUMA9aIB6H9KOCODmiMcg5zWAep/SiB2NUcdBW0Yzu/SuMYoYOB3ojtQUkgEfSuvFdeK6j+lEArnNYJHArv0ojPSsdiaIBojBxmto60VyMDmiMjGfpRBHAFADGPSj2xRHbNe2fpWAOAaIzgA80wBHHNEkjBrgjg0ORgt+lEgHhv0oAEdf0ojA+9+lYB4Bornv+lAgADNMGPBH0raaxxjFbfeivHAoDI461jcOtEYOAa6Dg1jB60U460xJ4Jz6VhsYFEE0ATwK2k8VjtRB6URiivoaIOM5o56g5/CiCeQaI7g0F4wOtQ2ss/EYq30OR8B22n0xn+tQ6JCgAY598Ef1qLT7ezcOgz6jkf1p7g42RjA9KaUlQFbP4UWBOCeaZsYAP4YoMMgg/pQww3A0EJBGeleXhRg5ooMciigXp+AoEsQAOR3oxsw3KK2AkAHJ9OlcjgCiq4BU8+npW0HrSopG4dunvRTc2Dwewpk2jA5PpSorKN56dqxtPI/ClYrKHxgdvyqeJQMKOKZQp55/SgoPU/hUgBA56UCQOaAxzio32ndjPtSlS5AXg9s0ow2McelafaadcwSjUdU+yMm3y18hpPMyTnp0xx+dIqZ4k+nFFA8IYckdKjZCf3hpGyNoP6VuERU569alTLBUOacBGCk59KVVMRJf6jFKBtAJ/SpE24HWgQoAAoL5TBpB16UYgkgIbg9sVPGofdu69sUI+wOR9KUDawU+nFKNuM/lTIpwxbAPQYpCE5FTRlXye/SoyUBYLn8amhBzIp57jHSioIyDV1GzgMg+orTpYba6SeaHzljzmPcVzkEdRWoXdnqMnnW1l9mQ9I/ML46Dqfp+tMI5YQqHHvikj2ts3dPahbNNGNvy7ep65qVI0AUckUscbbWTkc8elfAjwroXi3xzaaV4p1z+xtNl3+Zd/ZXuNmIpCPkQhjllUfjntXjKy8GfDn4k69oen6l/a+nW/2f7LdeTLb+ZugVn+Q7mGGcjk84z3pI9MfVzcapL9itZOj7Wk6LjovPXH51c+LdV8OTXWk+GNR2Wlxs8xvJU+ZtG4cOCRglvrV15kbiPfnd149Ka1kmk2KnzDqM1LbPDuhfg8ZHpXw78KT+LNbi0e1XLNn0H8DN3I/u+tfGnwHJ4D1ex0CaDE8Xmed83TKxsvRmHRuxq/hjEihX557fSpFy26U4PpimEcrYHAoxDdsJ4PQ+lJGC3lg59D0pX2nC9RRZ5F3bunQYosrDOOaDqFxjJ7UwAGQefpQK7cY57GmdFOFfPvincuc55o4+tHngUSSMCjjHWiAFzn9KyDkKc07qF2Gix28HOaGE2gj1okI3y0NrZAPX9K20xXd14pRkcNj8KDKABu5HQYpTuXg0EDHCnpSowOd3I9qEYGB1olRznpRbcuV59qAY4AH/ANaioxjPNbVAyTRljXhRn0ozbuox6U0xH3evpQldhgjkdqcZ74oxFjkN+lGFc43fpQjBGBXl8cGmhJUNmii4IpggGN34YoImB83X2plGNu7PtimwxyBRIXqelBQSCadV6KaJJ4VenbNBtyhS36UpJHBpWYLhTj8KBLcEUwA53fpQIY+opVGcj8aDc9a+RV4PIrI4wM1k4wDQCsQCentRb+6aLKxBLfjiiN5Cqck8DApSQdyNgjoRxiv2P/25vjz8MfiH4X8Dal4l1Lxh4T1fUrfT5dH1KZrmWFZXWNTbSNl42XcCIwdjdMAncNQsLLVbC40vUrWK6tLuF4Z4JUDJLGwwysD1BBII96/4J4+NdD+EP7bQ8NyX6f2Z4iF/4bguWIIYtKrwc/7clvEgx1Ljsa+Lfw8074tfDDxT8M9UnMFv4l0qexM4XcYGkQhZAO5VtrAeor4l/DHxn8IPG+qfD34gaLLpus6TMY5onB2yL/DJG38cbDDKw4IINBcHaH+oxQBwMCgMdDQTnBP0rYAMZ+lBQVIohScdaKgchuB2xTKrcUY1BwD+lBADyKMak43fhiggIGKKjOAelPEuMhsn0xRQYwDTJkYzk+lOgYYJpIoy+4nO3oMVaxqzYB6URsyA2fUYoKWUhTXl7o95/OkUhAFfI+mKADNmMcDpWFY7SefSmIA5NAMWztx+NOXA3A/hUjEx7j26e9OqLwRz60qs7DacgVMoVvkNI+zjNFSeR07GlCbc7ufTFckYFBRINhOPwrRfBOqeIrGefTovMaHbhdyjOSR1LD0NEgjAPSgQe34UFGOtBMkAURt4H5UcMMYxiiqlcK3PYYopkYLcj2pk5wtMMADNEZ710HB/DFY4xmiQRjPNBB0xWxc8GgvUKuT65pUA78+lZIyM/SsgEAHmgd2AKK5UZNMvp2oDPANEhhgCthJx60QMc9aKHbkdB1rigMjpmicDI5pzGxzuz7YoAkfIKMbMuN2T64pIwgJzn8KZVbGRUpyoAbj6UCAeaZVkGMZq4toyMjj2qa3KnKmmU54FfN6UcjjFAEjIFNkckUx4GDz6USSMiicjbmugwPwrBznrRzgYojPBOawMDnI+lE+hzTDI611HBo8jGeaOeADRDZwKByAAefSsbu9c9CfpWCR1o4x1raSOTWBgHPSsZPBrI7GiT0BonI61gEYB/SiAw4NEkcA0V4zmumP0rkDIo896ojtWAeM0BwBnpRAI5NY5qsd80eRjNURkYBrGeBRHaiM9DVVRHbNcHgGqo4HNEYr/ACKPIxmsZ70R2o+5qjzwO1Y4xmv8ij2FHtimHGQelDJoZIHNHpxQ6c1j5sZogYwDRAzjNYAAwaPGDWMjOaIA4BqiO2elfQ0p5xXFYrkjANYyOtdsA0TkYzQPbNUWPTNH5uBQycAGiex5o9RRJI60eBwKCsVyDVtpstwAQf8AP51b6EijJbJ+mP61a2MUakIMetHaqYA5HSt+V460XbGKOSMihyc56U6kjilQkDB+lKmTg08QQ4JyaJEfXnNKMpkmmyVxnPpRYkhSaIK9T+lBSAATn0poypwTW1RwfwphtJB7VjPQUhKttNSKSu5TyKmYO+4UrgMAKuFEajA/CkKyqFI5XpUq7lKjtTgseOooMQARXLjGefSkGTgnNWtmbqKWTdjytvGPWhgHZ6USm7cg6VIVADgcipFO3APPenwMAde1QfNCFHJFSnZN0/zigQzZX8qlj3x5Bp/3ihx27U1nJcoRGM+V1/GjGVO09aePY+30oIJV+U4IrGQGz9KLbwAe3SpopHAKDOOtPayTxGaM4Cdfx4ooyjYTzUMIAYE/SjGCNpOaKK4AI5pLYAZJqZGdkH1qWJkXk1GwmjK4xRtiRhT0rQfB9rqugi/ml2s33flJ6OQehHpU9h9mlaEvkjHOK8LeH5Ndne0R8bMYOPUE+o9K1Kxk02/k02Q7mjxk4x1APv61GApxmhM0cLwBf9ZjPPpVukMMm+fkH6/0rWvFmjXvg3RfD8Ee2403z/MO5jnfIGHbHQeprQvF91oDrJaR5YdOQPX1B9avPE9xqGuNrtxH+9fGVyOybew9B6V4i8TR6zEqLHjrnnOOnsPSr3wpNa6DaX3mbpDv3HGP4wB3rxZ8NtY8PaHD4iun3QTbsDCjGGVOzE9T6V4i8VxamI1hhwyZ/i9ce3tU8jTytK3BbH6CtL1S60i7W8tWwy57D0I7g+tat4ivNYvJL28fc8mOwGMADsB6Cr54pnGzr/8Aqpo3IPOcVFE0zYU9O9Ooi2ljnr2pZ8PlRQCk7ifwp5ABwaVtpx6UxzytaqNPjuj/AGaf3Xbr6D1565pyGOQPwrIPP6URzxWM8Cv8isDGBR5GAeaLLjA60p2nj8KkIY5I6UfmGAaz8u0d6Y9CaU7zx27VJ1pl+6Cc0o+TANCPGCKC980FCnIoEkZH4UVJ4zR2g4JoyqvQ9KN2pOAKedmPBpnbbgnIpgzDIqOJn79KMZU4pUDc5posrkHmvKI4BzijHkdfwooAdw5pjuXbnrRU7c5oqB0NGMN1FFBjAp488g0F6York5ArBwKYluDRTnNbgo5oAE9aUsxEY6Uzbev40wBIP5Uqiiue9E4GRSOHTBoDAxmmBCg4+lUSB3rPpX7Fvg/TfHP7U/w20PWLq2gso9bjvZjcyKiOLYGcRnJ53tEqAdywHeviP+xZ+y58V76bVvGXwc0WTULglpbywMmnzSOf43a3ZN7e7Zr4U/sO/swfBnxDB4u8EfDG3XW7V99rfahdz3r2zYwDGJXZUYc/MoDc9emP22v21fA37PfgPV/DPhvxHaah8SNTtpLbTdOtZg8mnM67ftM+3IjCZ3KrYZ2AAGNzLaXt7YX0GpWN3LBd20qywzxuVeORTlWVhyCCAc+tfsa/8FE/h/8AGPw/pvgb4u69ZeGvH9pEkDz3siwWuskDAlic4VJWx80Rxlj8mQdq/FT4F/B/446bDpvxT8A6T4iigB+zyzxlJoAeojmQrIgPcKwB49KsP+Caf7GtjffbT8Kp7kAkpBPrt80anORwJgTjphiR65r/AIKZ/Bbwj8Gvjlo9v8P/AAzZaHoGteHIJorSzhEcaTxySRSYA6kqsTEnkliTX0NfM3FEk4Ao4IwTRGR1oHIyKxu4oKW5z06VjJxmivSsHtTr3B59KPr6UFGMZorkHHWmUHoaCkjirYYNFA/BpIRGMBsk+1eUSWUt+GKAATAPSmYIBtGc0pVzkdaVt6hgKdyCADwK4b/CgoycfhTsSNpFbwqqPSnJk5A6UvIBJzTqCwBoJgg5yKVFZs9hQAV+TXhrxlfeG4Jbe0j3CbGeQMYJPcH1pY9x206BGKg1jPAqOMu4UGnUbgMdKHJ20y85zQJAx6UBxkcmtgZcg0RkYzTc8UTnjPNYHWic96KgjAplYN9abPTNMG3YzQAPAPPalbmgS4xXJ/wplzgA0pwMA033dy0QCODTAjjNfQ0Dk4okYwDWAVxiigK4FO+1cA0JSRgigQ6gg8UwXZtA5p0CtzzQYcCpYRIMKauIDu2gcijZS55qS3ZOhziiCKGSMUenFYyf5UeOlAYUAdfSsZwaphnAzRwelN1rJOBRBxgdaGSMGjwKzn+lZz0ongEUOnNE4/pRJx0oHPQ1/kUTu4FDJGDQB7npRGTjP0onjis9BR3Y60OlHB7c0cdxk0QMdKIBGMcDtQAxgCjkZBFY44H4UMAYFUcHoKPoKJBGAKogHvRx0HWqPYAUQCPpR9hVHGKoAEDnNcemfSs45HNE56D8KPPevpRyf4v0occg0ee9EZGCelHA6n6VkHgGunU0ORRwflo4Hej8w4rg8igCABj8KJNAHHynp7URjhRR5A5rZngH9KPIwDzWCeM9KPI5P0rrznntQZiMA0c4wTRBHQ1yRwf0rr0PNZJA55FZPQmiTjg1kn+L9KPNZyeaKkjCmsAd8n6UCScjrX0NYLDk0eeA3P0pVZuFFQadPcNhf6f41a6Epx5jZ9sY/rS26RoFXtS7VABNK6qxJGfamfzW+b8qdRuwO1MARjPSthbAPahEepP0FGMDnOfahCrKNoz7UCqjAGSKkYv869e4piHAAGCtDhQAelEr1xwO1FCQCq/rTLlcEUkhTCnp2qSMKDIjZ/DpQiMkYDHmmQk5PaowQMk8VIhYbxznpUbtGdp6U8IxkvlexxUg2/dOfeiftIGTyOlRDbKAe3+FKCuEzkdjToASgGM9D60c5255qMAnrTx+W4yfwpz5TlImxn2zigrL8zHNI5U5I3e3SmCyAEv+GKild02DnFCMnjFWuQQp71eWbI2SOvQ+tRqQoYjNIhwxDZz046UYowNpXj61o2sjSobmM23mfaNmfnxjbn2PrUrq8xdV257Zz2qRI5FJB+93qAqG2gcjvTwqgyfm9ulJCjgYNaLq2l2VjNbz6X50j7cSeey9CT0A9DivDtqNWgn0iM7GuNu0dfukt7elXNq1rLsmh2uO27Pb2pYwhLA5z0HTFYbbgHJ9KVDIwjI+Y9BVtYXM5KW8W7H+0B/Oh4Te58KW+upJsK7t4253fvNo78flTaTeCzN+8P7lerbhxzjpnPWlCqdoXaB15zRRCc7cn64qS+0+Xw5a2QgzLFv53Hu+f5UQxXJb9K0bWb3R5HlsJtrvjnaD0z6g+pq5uZ726ae5ffK/3jgDOBjt7CrG1N3dJbq2C2e2cYGf6V4x8PS+Gb6K0mfzC+ccYxgKfU/3qkLq3LcemKbyQ7Mx5GPWhLzgrj8aaTegKPgn2pJA/wAvftS+KtcGnx6UL3/Ros7I/LTjJ3HnGetat8WPFmv+CpvB+r6z51idu2L7PGuP3okPKoD1APWmLKPmbc30xSkn/WH8MdK1TTLnTpDFOu78QPT0PvRQSuEJwVqGw+1WpeCLfIO27HejMwUMh+ZutQTNDJujb5vpUuS+JR83cU7KWGxcY96Y7mLEdazkYx9KI3HIND0NBgTyf0on5sA0CSQAadmIwT+lHB5JrIPANEjoT+FB8YBPNFsHIb9KMqEbVTOO2aByMg1wRwaABAAPPYUFYigqAhc5z1GKKL07UqKV6c0V7fpWCRgfhW09jRCEcdaZ1QAluewxRnBGCM/pUkgY7VT9aKlxgL+tJbsOSKFuSMk5owqRg0IlAwBSLg4HWgmTyefpQUAYFBfU/hRAH3aIxjAoqCMUEVRwORTKSMA0YyT1rYTxTI5Awa8sng/lQjJxjtRhA6HJprdmznqvapIwyhVGMdaELEYLfjijAeBu/ShEAOuTQDL91v0oIc/M2fwrHGMZx0obiOKA5IJzTAgYBpCo6jPtRUMMg4oEhQuc+lAMDnOR6YrAPf6UAcZB/Sic8ZzXg/8AaY/aF8A20Wn+EPjX4y02zgAEVnHrEzQJj0iZig7dq8Sfta/tOeLrV7HXPjv41lt5F2yQw6vLbo454ZYyoI56Hrx6CndpXaSWRpJHJLMxyST1JNAADdj8KZF2gquT9a8EftBfHT4dWsen+B/i94u0WzhULHZ2usTLAoByMRbtnb09R3Nap+2V+1VrMIguvj94zQAEZtdTe1bnH8UW09vXj8a1nXdb8SahJq3iLWr3VL+b/WXV7cNPK/1diSfxNDpyKBzjPFbQx3A/hiiAcDFFNwyBQUAZBzRXuF/WiABgDjtQUlc4oDJ4ojK8DPtRGehojAwvNMM8g0V/u1tBOAaaN403ngHpULHANBS65Jye3HSvLJXLHOPwohgwZWxjtimBZSAcn6UQSP6UYypJR8e2M0R5mCr8emKyC3lk5oK55Z/wxTuY1BI4FeYVOFXP40VLDb+tKkZOCn606KOF+961JGBzv/SkRDGBjJp4yF2jvW1CgJPJ9qkkaNRjqOlEYPFOvQnrW0etWyDduHJFSgu2GOcVvB+UCt2e/wBKLKY1APzd6RgnCHJoKCuKdd3QdO1EKOAKIxVcE/TtRUDn9K27h0+gpUQ9qZc9DROO+cdqC5wSeaVihyD0oMp5BzTEMMAYoKDyDmsnG0n6UcnoaK7uAeRRB6A0eO9YGMCgARyfpRAzweO1Om8cGiseCp6ikQYypyaDscKRz2pgZD70YWxj8qSIjgHNfZIg5kZeT056VcxpKArNjHtT2iE5DZ9sVPZbuduPfNPZSBdyNnHtTh14PWs/LjNZOMVlj3/CsnqDR3Efe/SiQQFz9KJPRW/SmGACDmsnGAaY5A+b9Kz2zWCeAf0okEYAoAk57ijWSegrGevasn0ogYwBVAAjgUSB3rGQCDmiAcA0QCORTc/KKYZ6CiAOMZoj1ojgAGuSMA1jI4rp0NE9s0ee9URwOaHI61VE4xzQXjArGRxRA9fwoA8YquM4B5onPegOM1gYwKrIIwDVHGOtEYPWuortgGu3NZB6UCexo56AUB6nmj6CuCODRznrxROKbJ4FDoATzRI2gAUOwDfpR5AIoDIxmgM9awetYJPJ5o5Axmjux1o5I69K5xQzjg0ScZH4UDgcd6JI71nvmiTjrXuKPNZPY1knjNDpgHNZ5zRIzkH8KJ4BzShjxVtps07cL+o/xq30lUYM5z7f5NLEEXHp0pZdq7c0WyME0xyOtBgep+lKT1U0VLfMOaeHKAgU0aquQeaZgygDtW0n5hQyBlecUUbO5eaiYq+G/GpAUOR0oplNyjjtWABg9KDMibWGBS7Gby15z0p03Hj8atHhysczYBz2P9KnUs25egp1BHmRjIPXtQUvGVJqL/V+UfvL1o/K+HPStnlggnj+GjC8gyKgYI+AaZUEuSeW/wAK6AA0V3MD6dKkTa5A/CtmPmzx2qZi4DEVLtZUYHnmkBZgop4yj7QelIQw2nr2FRqEcAtz9KeRY22k/SkbbhSfmFaveWt/otnHCcyW+/cMHuwx+go46ZrwLqmnaL4ht7vVX22o3eY2GOPkYDhQT1IrWJrW4vWls23IcY4I7D1rRrV7zUYbWPlm3YH0Un+leK/A1/o9mb7ysQL945HHKj+8T1NBAqhc9O2KYELhRzTFmxtGfWgABxxWp+FrvTNKt9RZf3c+7aeOzAep9a+HAZPFliCOP3n/AKLeviVbNBqck6J8nGOf9lBRYSIWI4XrWlaDd6m262TcfqB6+p9qm+HWrpbx3Jh5XO/5l45wP4quLKfwTH9oSPJk68gdOPf+9Q8T3g006Tt/dH3H97d6Z61FN4bHhV7W5ucXxxtTY/8Az0yeRx92njUnp06Vj3+goMANpNEehrTWZb2NlGcZ4/A1dafdPrsMflc3e7YNw/hTnvUqTWc+SuHXtkelXeo3etagklw2SM44H932x6V8QvBw8MXVvFG2TJu4x6Bf9o/3qWB3PzU5Zm24q1tHlRkA+YY/CvL8ldveklZzg9aYlIySee3FTElVzweaDKcZb9KfUJLhi8zZz7Y/lSuY3Lg/f6/hXwD8AzfEPXZtGRNx+XaM4/gkY/xL/c9a+J3gS68AeJptGuo9gG3YMg/8s0Y9Gb++O9RMFbJNXDK8m5TWQB15qQ7uVpsbeDzRYdqZsjaDR5AGa9qYg80Tx1rcDTFcnB5pnDDC8mixK5oliNoHNFQORQUdCaCsR8o6UFzgCgWJGKdAQOelALjAPSsDGBWwkZzW9V6t+lPMAcA/pTTMzZU9KCO/+FLAWHNCBRznmtigYFAYGAarr0NHmuM4B5qtwHFEkjGa4Ixnmsjpmj0FHkcDpWAR1rg8ZrGeBRUGmUNRXPSgOMGsUBhulBFIwtKihs5oqCMA8isjPNGNAOtMFAOD0rGV4oAcEGjyMYqMFUCk80DgcV5hAwKJ3URRwRwele1Hk8fhSEg8UGwxU9aRAOprGf6VR4OaKlhgGkT0P6VjOABQ44FEHgE810GKYbl4NHDLgGmBOFzQXgAHmnQoMnr3pVJ+WmA4CnpTgngdqYZGMc0ykc0DxgHkU7hvlB/Cl4GBUR3KBmte0yPTJESNshs8Y9Me59aYAEGmBK8igR369xTEYCk9aXAjGT+lRjcxZetEjO0HmpFDDbnntQQsMilOw/L0pVV22k/pSLuOD26UIsSZHI9KEYV9wNTRluQfpTxEsQTUse6BsH6D8aA7E1IgCg0FJ6VbkKrHPTHamJJzTDJ+Xp2oKAMdfSuSPl59aBIOAaU45U8dq+/0FMmenNE1I4c/KKogEYonHFK4PBotnjtRYdBRLZwD06UWOMDrSkEZBzQx1zQXAwDRB3YJoZI4/KhzTAdqYkDijkDGaB3cA1yRwKYDpSoTkCtnOQaIA6GgNvFcEcGllVFAB5qZyxDA9KGWGQKMbbAQPrRA2+9MiOME/hU1iJBwOlTWDoMqOPWjxwaOMYBonPeqI4461ROBjP0oZPemAIoDAxmsHHBqsAcCmGRxVGjgc0CCOKIyMZoDnGelA5oe34VwfwpjzwaIJ4NHIGM1kAYHNZB680TgYxWOOlH7uMZ9q4xisAdKIyMA1gYxnmqONuDQA4GaAwMZokAZ60D3BrJI681wTyaJxjnNHmgRwD+VEZGAaJ49ayMZNBgAB2ohcYocEAH8K4HWiQTjPNEAHNZBHWjj16UCCMUCCMAZokdM0SM5IokY4NHaDwKBB/pRIIwDQAHFf5xQwRxQLdAaH51kjnNE56H8KyfWiTnANA+poEkct+lEkd80SSMA17ZrtjP0GKycYBrknINEg9Kz3zRORjNdAMV160OKPPeiM8CgvHAqGBp2CKOas9JAO5z/AJ/OoYY40wKJHbrRJPHWgCVwPwojIyK6/LXA+6KRGI3jqKVEmTC/eqMBl8lhyKlJAwBTEFFDLx25pSwGFb8MUynGUHTt6UhY/KgxUilHIxjPagsc8e3G0jp3qCRlURP+AqWHCEEc9jSs0i+XnOKjPlyAkZIpgI23KMj06UYFGGU8duK2iRSqnPtikTYCitkDpxTwFeM49KVAo3H5s/hmrmJB+8Q0n7yNUboueKeN1AZXwB2xRURPhGyfpT4+9nBpQHYJs+nNWsUWZYZo8k4wcnitQ0+SJgCu1exzn096YeYFUHO3OBTRLNEEHGOlGFRFsAyR0HSlG1gAOR2qSISHao696aNomDFsge1GOMqHjTP40YkeMlo8sOnOKQM7YArw98K9W1fwVfeLIH3w2Pl74sKN2+UoOd4I5GelX9g1pO0THgdsewpioXJOPwo4HKigXi2TA4znFX/i3xFqtobC/wBT822PWPyUXuD1AB6gUAZCAnJPStR8G3djpI1Tzt6/3NoH8QXru9/SrXwrq99p0WpWVv5qSZ43KuMNjufajZ3IZl8nlOvzDjNLqN5qEUWnz3OYkzsXYOM8npz1FWl1c+H9YW9tpdskOdh2g9VweuexrxL4on8RIXeDYD/DvBx074HpVjdaJF4a1azl0ffqE/k/Zbn7Qw8na5L/AC9GyvHPTrXha71eGTZpw37eq/KPX1/Gh8QdbW8aS5GIBjMOV449QuevNeKPHUmuwi2W32IOg359D/dHpUz+Y+4phu/Oa1Xw9f6dZpfsfMgOcngY5A9c9TTFiOGwfXFBiRgncR+FMxc+WY9x7DOKVSF2k9O1RBncGI89uK8Oaj4UgsrC81CXN/aeZsbbJ8u4kHgcH5cV45vbDVNaku7QfIcYHP8AdUd/pUhSJtkJ2nvxmnuVZgQ2D64ppXdyittA6cZpJmf5ScH+9/8AWoStHIwI3Hv2qSUsmwjFRuoG5xk/lTTM52uePSp5C2AH3+nGKC5XeD07UWGNofI9MUmxxgnGPavCnxD8V+A7r7f4P1P7BcHrJ5McueCOkisOjMPxrxj458R+Pr5dU8T3/wBruxnMnlJH2UdEVR0RR+FEk8Zpm4BonJyDmicHAb9KY5GMUQOx/Siegz9KwMYFEjbgdR0rII60XU/Kx5HtQZAcKKBHJHJHSgc8hv0pnyNvX3pstigxAxn8KCgruPOKClhwaCgp16e1AgDgZrbnpQX0rbhae4iUZByR0HSncyPux/8AWoIzjAFC3AHXp2pY0UcDmuCODR6cGucY60TgdKJJ4o5JBHbtQJPBFZOdu79KBGOTRJxwf0rPHLfpTZUZArccZJ/GtxPGelEgjJP4UCCMA5pgTgBsfhWT2NHJ6Gsn1rJIwR9K9gaPTgU2QM5/SgWPApuRgGgCeCK4XgDpTBm4BxUilQAG69eKY5GA36Uqg8CmXjBOajjO0D0ooSOO1BecA8isjGCKK4GOvtQAHAOCKYKVxt5+tBAcc9KjxnPpSq5bcR0oDJwOtMFVcdT6elfs3fss/Ev9qXxBqGh/DuTRrZNGSKTUbvU7zyUt0kLBSEUNI+djD5VIBxkjIz4l8Oat4T8Rap4T123NvqWjXs1neQkcxzROUdfwZTWzsopiwXOcg0EyMKce1bQeg+lEEncD0obvXNEEAArjPvQXI4qKIyMSp5HSo0YPg8+1MpDkK2PTijC5baZM/hTRN3OfQ0kLKcsOe1Oj7vmXHtmpEY8Yx6UyAgc/Slj5Bz+FIhPQUhJXg9KySQAaEblC4GQKcuCNp/DFFiDnOAKO5mDryB0FSAlcA0iMhOOPU9aGCMjvTEKuc5xRXc2wLgDpzTD5trDPoKKqBhD+OKWMFfkbp2xSuueF5HvTMcghuT1GKLMoBK/Q5qQEADdnPfFMSigI2B6YphjgH9KYYReOtA9gaBAgkIPTH86yAPlX9awdmfSmYk5BonCgA8emKIyME0RgjaaDA9DQUAYI/CmQMcDgntRA7GiAcVRBzwf0oFCeaJU/KDRwV2A5rZgDaeRXzFeKAA4HNKTjIbp7U0ZA4rgoQfwrJ6E9O1AqR97n6UQCMZojtTIG5z+FAALhTR5GM0fmHApFKrj0rAIyBQAPGaKZHWpG2kAN+lFyx4NCJtuA3PpigPLTGM08h25WlBf5c01sFHLfpRiAGc9KkgRxuXgnrVxpyj5g238M/wBaeNozg1xjFcVwO/0o8jgc0cEYxyKwQODn0o7ScbenvRGRgCuD0okEYAojI4FYAXBGfajuPSj9K2ggAGiD2rGT0ojPAFDHXHSiMkgDNAegrGenNHJxxW3Peip6CiB0Bz+FHAGM80ACME80BxkHpQ4PNY96GO9UcdM1jIogGuDwDR6ZHaq68VnnFcnAFYxwaGOCKPTihyOtDB4Bo49eldR0onFHpwaxn+lYzWB2ogZ4NHB6UBjpWB1qiOMA0eOaABwQKIAxTAnGKOMdaGCBg9KIB4BoDjBNYGev4UQAOKUe9HkA0R2oEYxWfT8qI4o9NtAdzQ4oYJ4ogBevNcf/AFqOCMCs5G0UQCcA1gYznNAKRmjjtUEfmEDNabZCNt7fh+tbQq4H5Vzisn1rBJ460vtQU4yDSJk4JzSxKBgn6DFKSp25yKhYRNkmmdd/nLz60oSVMg5p0ZVyOR2oKpGM89hSOMeWRwetMBGRg/Sm8tlJJ5bpWGjOVNOvmL5inmifNhwDzTL5ZDY6U8TSHeoyKjB2BT1FR7CRAx+nFK2GwtTo8TbW7dRSnzRsPWmBX5D2qz0m41BMwpn8QP6+1KoDEgfhTHI2+lTx4YFT1oKRGM96s4WuHEajkdKh8L6oZLYxw5+0b9vzLzt/GvElnc2spjnXBHbI9B6Ukao2SOatV01tHO9v9J9MN/e/LpTwlHYN3xiiiu2ep70FJ+72rT1t1mEl4cRr14PofT8KMUD3Ijtmynbgjt7146+HPhjw74Mh1y0k3XJ3bhiQY/eqo6sR0PpU9u0MwUdv8K0zx/r2laFc+H7afFtc7d42ofuuWHVSep9av/CerXXhKHxn5WbW43bWyv8ADIIzxnPX2/xpnMqpGq9M96hhaV/KTr2FXmgXS6dHG0fz2mfM5H8Tcd6dShwRSnZhl5I6Vf8Ai2/v9NGmuPkH0/vA+ntXg7xtbaNpn9mXz7Ui+5wTnLMT0U+orUNTinuZJYG+V8cY9APatHlhg1CKa4P7td2ev90+leNb7SL65tJtM5Ub9/3vRcdfoan0tIfCMOpf8tG3Z/CTHr6VBMEkVnbKnOeK8Fa7Y6LevNeyYRsY4J6Bh2B9RXiHUbW/u5pbRvkO3sfQev0oEYxn9KYnPJq71u/u7P7HLJmMfw7R659KZtpww47VEADvY5rdt5xnFEhvmA69aBKcoelS3MghJHWo5RJyW59MVPhWyBmhtLZWkDyg7RkimcKu0GiS37wnrTDdjac0Thjk0WUng5ohRwKd8DapogAYzTMQoAPWshuAa4Ax+VEknpWcjbQAHAPNEAcZotuXA/CiQelUenFHHc0GHX8ql244PJoJu4BplROT1rcpXHeghYZFOqn7pojPNfLtwTSrtJUmjtAAoKCMY5rbgjJqSZI+VPPpUly7jCnGKMQPJFRRBjwOKUKBtFcEcGjgc1uB6HNECsg9Dz9KIz1NAZHFAjdtBognjOaJAyTRA28UpJXBFDDDArODt9KZsgKKVtw5oMScUGwQO1MygjJonI60eaIrGRxXUVRAIxRwMYrbjkGiDjA/KgMgHNArjBqQAkZPSmUAcGgcYx2o5xzUbExijkD1okdM0VJGewon0NHaSKK7hwaCEDFIoByT9KV9wIAoMD2p8MxbrX/BPr4wD4O/tP8Ahm5vbvydJ8UsdC1HJwu24ZREx9As6wknsA3IBNf8FTPg7/wr79oYePdOtfL0vx/ZC8BVcKt5FiO4Ue5HlSE9zKaUZ6mpWEgwB06VGCefTpTBQAAefSiAV2g80MA5JosrEHsKfy2UCOlTBAQ8+lIpByTz6VsBYsTmlXb3zRUEYzRA7H6UVU9RRjQjBGfapoA4+UdKEaqvzUq4UKtadYT6lexafaLullztGQOgJPX2Fa3oN9oE6298m1jnHIPYHsT6iizAbQeO9A570Nzkrj5aYhNqjpzTbtoHenYFSpHNLwNpPI7USGDLnpTMrIAp5+lEEruz92ssRkVu+VU785oMFOCacKFVhzRcS4VTk0wJGF5qUMCMmicgc0rAxhRyR0oNgdPwoqfK4PXtRIHBP4UzEfLn60WBGAKALDNFlC4HJonjbRK5wpzQKnOTRxt3A9OlOvGAKPoKpjlgtFVBwT+lDIPHUdKIBGTQZiOlDcw5FAd80CMAAUWMigelAHGF6imbJx+lAZGMUBuHBph71Te9KcDigfQ0QCciuAMilAJ5PSmPGBUgzgDrSphRkVkEU5VgFBqVBtGO1DKnIPSo2Ln5j06UUDLkc00ZK5BpkRxhhkVd6eJFBUcj/PrU0BiO1hj0ogdAayOmaIBHJokAYo4JxnmiQO9UAOoNEDqayDwDRGcc0SMdaOOmaJGOtEgcZrI9aGMYzzXy4wDQwO9ZB6GhjoKyD0NVgYwKyTwDWSehojIwDQOBjOaBGOTQOK7YBrGRxR570Tkda6jg1gdhVA+honPegcjGaBAqgcDAqicjFUeTgCgRjisg89aLLniuD0rrxVHB6DpWAOlAAjaaxxjNHHTNcdK4ogA5pgCBg0DngHNDHagAAADR6daBOcGiARgGiOxNALjg9KIB49KYADg0wA6UDgYz9KAzyB+FHHTNADrnpWARwKwDxmiARxWF9f0rC4yBz6UwHQUQCuQKG3GCawBwaUA9K06Nd2e//wCuo2UpgHmse9D3H4UQMcGtue9BeMgVtUDkUMH5cUygkAn6CjtUgZo4J55HpSFEfCH5T1FACNtyDI9KG1ueuaaIKQYx+FMu07s8+lKFdAAaMarwRx2oKrLsPPoKiLREKD8h6CpGGSIV5PvW3tmhGythRwaWEYDAc96ezN1arLHHkw9eeuTWrabpSuh0o8c7l+b2x1P1qawR7GO6Y/Mc8fjikSFrUhR+87dfX/CooY5UguZ1zE27dz6cduetawul2E8Muhvs+9uOGPYY+99TV5bi3uniAwRjP5VFD5hBbgVr/huTQyq/afODdDs29Me59aEAmgYjll6D15qGNPPCyLkfX2rS9R+yWANkf3sf3Rj1Jz19s1f2l7rpLTLuP97IHp2GPSr/AE2fT5zDLHtA+7yD2Hv70gUOCe1XGj393ZJqVvbboOdz7wO+OhOeophHLBhjlx0/OvCUWivfFdaGYew+Ydm/u89cV4kttOtrzGmXO+Fug2MNuAPXnrmoArSZP4CjpV74ekg15IsKm7ncOM/L6n19K8c2viHxw7eL/sO6GTHz+ag6bY+nB6r6UY1XlhkirzU/FVnoEVlLd409M7IvLjOPnyecbvvc14HfQ4dRZ9ZHycbD8/y/K2fu/hXiO30u0vBJpT/Keow3oPX8a8NeJpNHluiTk3OzPb7oPsfWrq485nJHJxigD0xzQB4KmscdfpTBCoGc/pQYrhQeDXGcA8+lNkABaDshDKeady7ZY8/SgAON30GKABOVNNwCAPwoh1PmM2SPapCzKChwTSmRTl3yPpWsaHqGnWsV7dW22GbOxt4OcEA8A56mvCmieCb/AMD6xeatdeVq0PkfZf3crZzKwf7p2/dA6/hRd5DuRfwzTEyQ/LyaRgOAOanXMQYtkp0GKyJDkt+lBZcbwOB3rapPTNEAqEY8CnfacKelOfObcxyaaWQJsByPSiVZcDrRVQM5oqzcE/pWSRt646USAME1kYwe1MQTlaPPWge2KxkDB/SjjHWhjoPyogdTTZYYz+lBVK4JzQVCMZplBGCeOwok42g8fSiAON2R9KKsRtU0QWPJzmgqk7SayAcAU0iqoVTx3FAbjhT+lRIoG0mpGjHAPHpipp1ZdiH6ituT/SljZuFFJEq8k8+lEEkYNHpwaBIXAP0o7iOW6e1BSRgmigC4J47iiABwaJyvDfpR5HBpnGNqvz9K3A8M2cdOKLODnd+lGUjgN+GKLgYUGmYKOG/DFE5OQawSeH/DFAcfKePSgFJBB57UUYn5m49MV/sj8KGAOPyoHODS8nAphk7cc0RzgGirE4DfpQUnoc0VGMA0wJGBWOMZoAccdKp03cAc9qKgDkURntTgMuCOaQbQFB6UQZB8zfTigh2570wBGWNMC33W6e1AEcA5NRoOpOcdsUAXJCtgemKRFTgHJokngmiATgGhtKg9c1GzxSrNDIyOhBVlOCpHQg9q/aNhi/bD/wCCd+hfGSyiS48S+FbVNUuti/MJYAYNQT/ZUgPLj0jSgSRwefpQI4wfpW0k8nntxXJGCcGgoPAOc00ZCjDfpQUlQqtyepxXl5G3NBMrkGtP0W61Aho1xH3bjjr2z7Vc6PcI8r2o86GPH7zhevsTnrxQiMhwtfY5ZELImdvXnFGJhHtVP1ooe4pUz1GcdqvdCubS1F1J9w9Bx649aeMMQT27VHDJM4SJdx9M4qWJ4n2SLhl6jNE5ODTKAc45ojIxn9KfdvJByB2x0pcgbieKLEnCn6UFRVyg+ambd8pOaVl2lXOfQUyYUHvTMSwOcYpmEo5bB7cUgVjtY5HrTBQ2xW4+lOqLwOcVuDEEtjHtRcAEqevtTAfdJ+lNkrx+VRn5cYrKnjOTTMrRbQvI6HPSmyTyfwxQ45IyTRwOBRII2g5A6VkAcnPtSkkZoNgcGg3ZTSsc7s9KDZ/wp0C84/CqIBGK2gfKorYOp/KgFPb8KVVx81Iq9AKCCQ7EqWNY1245qMoy4ccijyOD9KKjHSh6+lKFzzThCAAPwogEYpgCMZ/Ck6YBz6VnHQ1Hg8Y+lE57/hXFDHemGOlRru4/KipBxmggY4FeWCmB2oI6cFcj60hUkFuorzQq4FNKwG1TwKKkjIFBCRnr6VcQh0KFcg9Oaubcwvwvy/WmCnGKrAxyeR0ojIxnJrAxijgDGaIAH0o4HSuorODwfpQHHBojOMUB2B/SiQRjd+lEYHBojIBBoqAMZoEE8UeTgHn0phnoelEg45ojJ461gkdf0raCMd6YYHFY9RQGe1HB4ArGRxWM8CiO1EDoBWOM0ACMAc0V4wKK8DFHjitvH8qogHG2iD1NYyOKJGMd6IGBijgjAqscYBrAxgUQOgFADpmq4xiiPQ1jI4rA7UfYVjnP5UVB6GgMDGaIBGKwCMCioI4oY6ChgjrQORyawCRg1gdM0cZ6VgHgUQAMZ5o4wB1riiQRgCgQBgmiQwyO1ZBGBXbFYB4BogZwKIHQH9KO3OBXB4rIIwO1AKRkmtvvQPIA61plszAMT/nmkCg4U/ShyKwDxmgMjAoL0BoqFrAUZJz6CkAbNKeQhprfcBg8jtW0ZxmgMoSB061DIFODRRgd8fIPUUBjoc+lPH5UhVhknoKQHzMA0wUgAnntQUA4I6d6V1cZBqKIyOIk6t3oAEgE8CmjwNyn5aBHTNeHHkl1OOyRd4lzkZx0UmrzS7uwu/Ldcbu2R2H1968KaO2r/arBV3La7Pmzj7249Mj0rU/Dlxpuoi3kOCenT+6D6n1pdPjnuLrToWyE2bOMdRk96ayKTtbytjb1OPbNePdB0640+G5t12H5sn5jnlR3NeBfA+oeONSXSNMbdM2cDAHRWbuw7Ka1Ke7lkENzJux04A9PStFtTdatDZRDc77sDp0UmtV0W60e9NreLh17ZHoD2J9RXh7TpLo274wo3Z/X3q3tIbGb7MTu29+R2z/WvH2gTXMVvfQJhF3bjkf7IHepbZYcFm5+leHPiD4ftPhnc+FbjT9178nly+a4/wCW5c8BcdOOteCtNt9T1xLacYVs+vZGPY+1eI9GTRNZudMSTcsG3a23GcqD0yfWm+Y4pMjBA5XtV7r2kaj4PltPM/eLjs3H7wH09BWhfFbRYvhuPCzabuuF/j85h/y3L9NmOnvV2wMpA/zxUVxe30Itmk3qv8OAPf8ApTgHnsaZi/Bb6cUOnymiMdBzQyF5OT60VyMD86AyMHtQicxmQDgdaJAHXpSspUtnrQkBUEHIPTipVBiIU89qbdNHlV5+tLulTg4K9KiYodrHmmJ6qOa8BQWurXaaTcwYL5+fcTjAZugx6Vq9pDp2pT6ckm4RbcNgjOVB6fjUOGJBbIHtitS1S+vbOOGW43wpnYNgGMkZ7Z6itNkDTrC7fK2cjHsalUxAtjH9KVjEQ4OR6dKmXY3FPIpt8E8noPxpCsfUZ9qMrBQu78MUZMjIoEnJJ6UApXGc0MEYFBmPBNHavyg0QBwDml5GCOlY9KBBHSgB1o4HeiAOc0cY61xjimIIwDk0CCMimIIJLfpTgMuVP4UhGASfpxQXJyD9OKkGQATQCjvQUgkZzigoK7SeaLRKgjB5FMc/xfpRZRwTQQMp5+lQYY8dqkl2YGKdnfjP4UkIClg3I7YpIc8sfwrgcCiMjANPknANEHGcfhWMAZPSioPINZyMipHBXGf0ppC4AAptwGzNOXJ+WiMkUM96IIO0Uy0FYgcUxI4NBiOAelIwLAE0zpwAf0oOpcKpzmhlgDigxzjHIrIY4z+lKRkqDmgMHigASBmmAJ4NbSpxmmGTgCjxxRI9aokDvTBeimmIGDWVPU06g/dNNgLg9aRcjOc0cnIHagAQCTyfanXpzRB6A0BkZz0pUBXjr6Uq4HPWqAyOD9BQUg4BoA46URzkH8K/4JJ/FCx1S08cfs6+JjHc2WpQHVbG1mOVkVlEF3Hz1DKYTtHYOfWvjv8AC2++DHxg8W/DC+3k6DqckNvJIMGW3J3QSH/eiZG/GkRmHTmoYt7AjoKkgUHAFNbqo3LyR2owk8Kc+1RxsoAY5JqODd82eBTQJLGFUYIqwv5bCLy0Xcv1x/nrUF5JHavbKPkbGT9DmlgAT92ufxrSL+0so7mOaPd5mzbyRjGc9PrUuxWCgcmhbYxntToFxtHHrWqaBcxaKl7Jd71OcL5YH8QHXNOoGSDnFJI8ZDRn9Kd2c75Gye5xSnackZogNjmriVkbarY/CpAFRmBpX3HGfpSAtyOlMhRCxPToKc7Tgcgd6DJtIXknpTlg2WHNb9nJXPtmmw3I/Kj8uFbtSLuIxTPvJbHWkYE4A6UvzLtA5o5AI9KYfLwKhKiFlP3u1NkDAoAFNuetMDuz1o9Pp0rIz1yaBGNp5NA5GKOCMCscYxmhwcGh1AzQAUAnrT4YDH4UwA471VMoZgPSnQk8GlUDjNDA4HaoZFTJJ+lSMXPJzRwPumgQwyDRU7eRR9AaXAAOfwoqGHFMB0BojIyOtADHFYBXjrSYA4qRVPIP4UcDvWQTimP+1+lK+OAaALNmmUKuc/hUshTOB17UkyNHhhzTRKT1qQLjAb8MUke8delIqk7SfpTlCoVT0ogdPyq4tUnTg1c2nknhuPpWATgGiABjNUQM4zRANYBGCaIB4Iog4ojPAPNYGMCiAOQaornmiMjFAccdu1EZ4zWMcYrGelFcng1tFHngUTjis5HPWiMjg1gADBrGR1qsdxRANHnoKxkcUQCMVjjijz3rtgGsDrVcEcGs5HBogEYoDJwT0o9ODQHU5rA61getEA9TRANY9DXXoaOMdaYZ6V1GAeaIBFEYGc9KzkZFfe6GiM9DWAe+fwrAHQ0cgfe/SjyAc0Bnkmh0Ao4JwRQUHv8ApQA6AURk4FHBGAK7UQD0oqB3/SsAnpRA6A0QCo5oDPegM4GawCcA0Rg4BoqCc5onFAA8ito9agtzKeBxVtpaOn7wY/z9aSwggHC5I6cmgCBjH4VGuO9YGMCgM8A0qjgU42sVHIrZg5zW0sMChyelBDIwXOTTxyQMMn6cUVWZQW+93NRYVwCMMKEUcq7l4NWxHIUYosTwKhaNl2Scj0qWJRIdo6dBUfIGDSwGRhtHI6U0ewk4waUHcNp/SriKJ4/OAy3rUaxTx/OuSvQ5pokbBQY96sbuWwuVurd9skGdpxnqMd/rWu6NNexRXtxcbxzhdgHoOx9q8NWkFnpSSg4Zs7uvZjXirRX1CNr1xuK9O390evtWlTx2dxBNEnyQbs89cg141e1l23qxZJz/ABH/AGRVto9xfTsLdeD05Hp7n2rT7fUvDepI4+V+dp+U5+U59fWvDmhQ+IrVl1Bf3iYwcnjJPoR6CpFXw3qcMmmnF1bbtr9cbl9DkdCa+M2v+J/F+oS6z4g1Lz3lxu/cxrnCxr/CB/dHavBtxNc+IbXSVbCTb93HojMK0iB7NzBL8xHfp61cwx3kH2d+VbqPof8A61eIfDg0q98m4m80N0G3b0A9CfWvCWt2uh308Zt8xvtx85GMBvY+tXOp2P8AwkAu3hwh7bj/AHMelSztPKzM+Tx29qc9iaF3D9lSAJ8wzk5PrRndgIwfpxXhGys7y6DX/wDq+3X0b0+grx1p+nWGoI2nnIlzu4bsq+p9zWieIb/Q5DLZS4J/2R7+oPrWoXk+o3b3c7Zkkxk4A6ADt7CiADnNIxZc5wTXhbRYde1RNPlbCtnJweyk+o9K8TaLFomoS2EUm8RY5246qD6n1rAPGKZcsTu+gxXgDRdD1W3uF1Y7mG3aPnHds/dI9BXiPS7PStRe2spvMRcYO0j+EHuT60qhTknk05HQCg+AAv40jD+JfwzRfcNgXPoM1JGqqCx5+lJKHBAHIrcAwUHntQKpJuJ5FSxBnAAzntXg7+xDobQ333hjj5+fnb0/CryK2jv2jtBiPjaOeOPerrwJqN94Eg8YQxeaZN2TuC9JfL/ve3pT/ON36Uyqyg5yfpWdyfyosM7SaIBXGMkUFBX3oFWGD2ogE43fpW4EbSc4ogZx2FFtygg0eVJoFSeB+FFucgUCOmKbrRx2/KjzwDXBFEAAkDmiSBkGgoXgGnJxnPSmz97P6UFLHIODSRgHGc+2KkQ9M4JrLE53fpTN6Ci2V+9z6YqT5VwWz+GKJJ+71qKEv1P0qWFIUGDz9KuJwMJEeBSIZWwT+lRQDGc0qBelEg9B0okHgGscZB6dsVtJwGOaIGcA/pUjADANeYcYBokhcCuT/Su3rQJJ4FdRTAYG3n1FISDxRJIyK5Y9KIAGRQAxkCjg8Y6UAAOn0rcemKWQBeBSyc4xzRI4B/CtgPU1wOBQBPGc0oz36U20DmmXocUykcGgAeCc1gE4BojIzmicDrQAJxTbTjb2o46ZplUnpTrleRUaApknJoKuNuaKAfKG6e1MSuMHkVkcCljBOM8Vt5zn9KzjnNBdwzmtqngGiqjoKIyAoP1odM+nSv2bPirN8Efjh4Q+JsUzrbaRqKC/RQcyWkmY7hcdz5Tvj3we1f8ABW74S26a74P+Pmgoklnrlr/ZWoTxDKNKgMlvJkdS8RkXPpCtRLkBVGSaEQXgGiuWwKMaqoCnJ7VpemR3krJPJsC4xxnsfQ+1G3RGK9+xq0t43kHmcitYsNI07Rrd7SP97cbsnLcbWHr7GvDGmafql4trfj72eefQnsfYVaadb6X4iNpbyZEX3Xwe6ZPGT614ruGuJ4o3l81/mwdu3HC1oPgiTVw/2WX/AFON52+ucdWHpV5p6DUDZW4wR1P/AAHPc1NpNxAczQ+X6fMD/WoikaKrpv64GcVD4avZy0gTZGMY5B/rVxaskxTP449qZRnC/lQcN0okFcg5ojnIP1qSISNkngdqCqHKjqtO4U/KORVs2I/mP0FSQgEODnHUVnIyx5pmwpHc0i7kxnn6USSMelDimfdwT0rHpQGDxWAeM0TkZNH5uKYA9D9KRCQWA4FEgjGKQ7RgDPpUg5yeaADAgdqbBHFZAGOuaJAFcEZ3fpTAAYHX0o4PQURxtBpR2xzQUqpOfpTKNvPWtvHX8KwOtNxiio6iiwxtzQOCBmqJAGAaPIAoEkgUGDcenSicjIP0oAY460pOMZp1Xbkdu1OMj736VwBgmgMjAqByDtFSQ7Vz+VEcYIojgVQXfwKVCM5PT2pFDDJOKcbgMHpTKGXGaRGifAPy/SmRZABitjelbcjGeR2oKDxmh8pwG6+1B1UAVfxJcR8Dkf8A1quYDGaJzx1ojIxmse9YGMCuCKJHTNYB4rABwD+FHGOtYHasDtR6cGgO1deKIPWhkjGeaIP1puQCKOCMkURkDH5VjjGaYZ4AorxRGR06UFyOtEZ5JraCOD+lYX1/CsZ4HNHHQGs8YonIwKwMdaIGOD+lcHjNAA9TWB0zXHY/pQx3oAHqaYKBw2fwrr07VR9BXB4Bo46A114oqAcA1tGOtVxjNAADANcEYB59KIBGAawO1FcnArAHANBRnBNFeMD8KI+XFUFyBk/QYosO1BRwQaGO1EjpmsjpmiVI61kHjNFQRjNBcd6ONvFbQRnNAjGM0o4ojnANHPGBVUFA4Bz+FW1g7gEjrVvZJGvTJpsA4FNzQHegPagpJwKSMHvRPGMfjRHyAZ5FHBXBb9KyFGSaSHzF4bp2xQDJJnbg+malZp5AhHIp1eJvlFF45VEanDd6CtEc4/GlKsd0bbvUYxUkgVwpFZyMZ5qOMSx4Y/MOnFGMxsUIwa0u0FyjMjfOuMDH1qbwzDfeH7bV7WffJJu3JsI24faOSfQelMCvykYI7VpyRTEwF+B04PvXhbTIU163W5GYm3ccjOEPoa8ReFINNuC0Mm5ew2kdh7n1poLMoGDfKn3zg/hR8SyMUtzyoz/npV94gs3UWdvL88P+sXaeM4I6ipm1HSnSw1ux+zPLnaPNV845P3c+o/OtX0+1srwwzSeVG/T5S3QD396sNI06XTjFPL5ino21hjk9ga8L6NLPvlUbTxgdcdfevEj38N95txHuSLo2QOoHYVqGj3Gm2r3VnJknGRgDHIHcn1qO7QyyXlzcbpOMrsx2x1HtV9q19qn7uZsj0wPb0HtWm3E+jXa6hHFuMWcHcBjII9/WrTxQL2EXUkmzd0GM+3p7Ve+LL6GfbDLlU/2R3A9qvdbnu5PNmO/8h6eg9qiu8yklM59/arp4prJpHf8AeDG1ce9AkDA6isknkVaWj3QYIM7MVtZHwRgihq0i2y2iR4xnDZ989MVLK853SNkjpxRKgZJpSr8qc0QDgYroMelW13cWUgntpdrjodoPt3+tPdG933FzNmRsY+XHt2+lEgclv0p5Y1+Rm5HtVvePA+6KTBPQ4ptJiGhW2qPc/vZt+U2HjD7eufSvNdH2uOfrXB4zSr8hOOnShhzkUgy25Pmx17YqaZWwFOQOlfMBvAotvIKnn0ojcgZl/Wr7wjFbeG31lLjLR4+XZjq4Xrn+lSmVGOHz+FRSKjeYOSO1aP8AFvVNJ8Hr4NSz8y3XOH8xR/y0MnTYT1PrUg5xnIoZDAiiNw2HrTKhXrz6UCQaPopz6USBwK3r1PalGDjNZOMZ6Vkfdz+GKKkDGetDrgGmXHAPWmHfP4URxmqyMZzQKnjNHB+WmUdzR46nNO4K4xRKKMA5osoXHeopAjhgu7HbOKvbgXM5lVNuei5zjii2/gD8KZWCAEUUJOFp0A4Y8+mKjhQgE9aaVI+AeRU1yZWwB+GaVOOTmoIwoyBRwBxRII+U03C5zSKDkg8dqPAximcb+DwKZy5wp5o/MeRRBC5A4FMSehokYx6VkdM0y/KMHOa68UByMt+lMVAyp/SjzjAoE560SABzQKnjrRPOM0QDgZ/SsA8Z/Ss4OAc0HXoDQZQAM9PbpTMqjBPP0o4bDA/SkYE0ACASaIVhjNbieM0SWGAfwoqAMA0cMMA1gYAJrYCcA1gDAz9KKk4xRHasZ4FFQ3BpUAXaBnFFSOQM0SewpgCcAZNBCxz+VAALnHNcHpQAbjPSgOMA/SiOOv4UVJHWgMcn8KSMvhV5b0pYSoBYcetfCdE/bL/4JzXvw8lYXfinwnaHT7bcfn+1WQElmQTyPMh8uIseu561bQ5tEvmsbobJF6g844B7E+tCBHOA+fwqzsXmlVCMZz/KtT8L6Zp+ipdC833DZ/d+WwxhgOucdDRBQkE9ParGL7VexQ44bOPyqzW3dzHGN2O/Ip9Gs20eO8e7y5ztj8s/3sHnNQ2Uu/z4Gww7Y/CrKFFt5Fu22ynG04znnnp7YqO2Zp8Rne3bt2q08QXWmRztbx+UJNuRkN0+oPrRuDLI1zCcyHGO3t/Kr/S9fu5JPt1p5fk4yPMQ4zj0PsKfRnsNKGtKPMiHXt/Ft9c9T6V4vsrew8Lpqmman5pfPyeSV6SKvU/U9qnneQEsfmPXinOOgzUY2L159K4A5o5xx+FZbb16VZWqXMdxJI+1otuBjOc0UEgKdCOlFiRlR0pZm2k4zWFYbi/6U5BztPTpSOawOuaLbjx1rBYADk0QxXPatoA68dhXBFHBIwKYkCpCCMZ6VGwCld/4YrgcYqDYHXc2AM9quSgbCNn8K+7+NBwBgiivQVtXGAf0ogkZBoqCOtEAHGawOuaG0HmgQQefpxRAHfinjG0MDW0E4zRUkYxRGRgU6AjPWkVRzn8KyC2M8+lYzzjpRHasEdRRCn7pyfStjMcLyfSmB6A0OOMUVGMA5NMMDkVgAccmh6+lRuuMZ57UtwkkYRl6d81cW5C+Yp+X1pgetEAgEUgzjmldXOc4A6e9JGHTJbA7cVtZhtUZxTKynBFBVZNvemBiO5TkdxihETGHA47GiozxQAAwBn0oxg80YQRxTRBRkDJ9KubRZxwOR2q7s3iO5RxRUnoKIJGB1oqegFAEjgVtBrBzjFEY6j6UQSMAUoOcAUc44FMM4FHPau3T8KGTyDRBroMZ+lADGAelEZGKogk8UDgYFAdxWSRwaBPGTXfCmu2BWOxNdeO9UcE9ea+hrPfNE+hrtgGiMjANZ4xmiMVgYwKrAx1/CiARg9qIAxzWB2qs9s0V6UQO9YGMCiARzQOTjNYGMCsDtWMjiiM8ZqiCRwaJ4zVH61Wfl4/Kgcjk0DngVkk4H5UxBAxQOeKJzWQBgc0TkYqicjGaPI4HShgcn8KyB0onIxRAyAT0rIxnNBcng1ZaYJvmx0/z60kUcaBFHC9KY9gaJz3ojPeiqkYxQUsdoraFHIokZwtKwIOD9KIOeBRKlcDt2oDI4PPpW4qP5CmcvGoY5xSZIyOfancn5fyFFMDcelQN5nB/Oru1i08RFXyJd2eOmP8A9dFDJgR84qKCSdwsI3Me3SlM0AEcq4P1q60W5uk+1Qx5P8XI+nrVnvtZMMeR2q9S2mtY1gX5TnAyeOal0GdwpU7uuOg/rUukXugXG+5XC9+R6exPrVpMWeO5tj84ztPp2PWtHnXV9NWdzktnPGP4iPb0rwjpvhmLxJLpviuHda3WNvzSDG1GJ+4c9dtW3hG2l1aS2W58qNcbDsLfw5Peo/EFpey2ukN88y78NyO27pjHQetfETx1c+I9GOl3cm+f+E4A/iUnooHQCtQuLu5nDXhye3T0Hp+FeEFS7RY5ORzj/wAe/wAKXRptPmkFscK+No47DnqfevEMd8bjybjgHvx6D0q88S6hDZraiX5XzkbR2OfSrR7Y36CQeuevpWo27Wl2Y5D0/wABU7tOVjibhc44pL54IhCrfKO2KuJY2O5D160zBuc0CMkg1AQz4/SpoXt5THIuGXqM9KLZOCfpWGxjHSgoUEkU6kgBaUADA61IQY/lGagZFJVRzW4LyT0oyK5OG/Sjg8V4RSxk1iOK+HyNnHXj5W9PwrxBbxRanMtufk+Xb/3yPWmSNclh83elkEQ4GPSpUVgGjNEFvlY/dpHLd6DsV2nkVgAgLTMqjaTk9qVynINOxY4bnHSomA5B57VLp+gjwhaXq/8AH6+/d9/jEmB3x0r7VKqeWD19qEjR5UHP4UxIGQeKRiORTndg5oMc4B/SgSAGPamwfu0DkYJ6UGyMA03SlUsAuKLkfKD0rPy5ogMNxPNHpwaYDPFMOAc5xXWqOcciigzkGhjPWpF2uQD09qcBgOORRJ24zT425bnFcKcHrW4hcg80oYAZPSmAbkjIFSFD9ztSo0hwD0qRYVGIVw3bk0ImLZb8KMxjXaKdtwx1qKAsMY4oRqqbSOlKBtAFOoPegQcgfhQPH8qLKjYFeah4zWwMcnrW4oMAUF3HAphgYBoHIyKdtw57UQCMUclcfkKViOlE570BnoM17EdKkGF60eRg0CQcrRJHeiSe9Z+XNbuODzR570rsCMHp0rfx70W5C1Gu7jP4U3gXW08NWviZov8ARLvf5Z3L/C+w/wAWevtRBI20V4+nSiDtzmlORwaABHWtoxmiQTjNFVPzA0sakgqMmmXHBrbnj0phge4peUGaYAcAUoyowOe9NGDnIrhQAKqguMUFz3oAscYrbnAFKhKgAdKiQrhxzirew86B7gPjbjIxnviv+CWPxcXwV8br74ZahdbNP8dWO2FWbAF7bhpI/pujMy+5KV+2x8Hn+Ef7SfijQbS28rS9Xm/tfSgBgeRcZYqoz0SQSxj/AK51Yabcu4CLwOoyP8a0bw/dazrEOk/clm3YPBxhS3qOwrU/A1t4JuJdP1G4825GOdhXHAPZiOjCtVtLcSkKOPx9BXg/VZfDHie01+0O17TftOM43Rsp6g/3vSkR7m/actw+M8egxUttpttr228j3WA+8uWH8HqOfvYqXQtOvXNxoKbIf7uWPt/Ec9QaitcWUksrYcYwuPeo0lkcCBufpVtpHnaMpkl552rt/wBr61/YMenBJL/5kbPHTp9D7itR+163dm31S68xx/FsA7A9Fx6CtfW4tbc6ZuzDF0OAM5IP161HPbmzaG8G5R93qMc89PwoWk19N5NuuT6Zx29/pSmCIMs6ZP1P9KSRX4UdKmjmgXEiY/Go5MlsmpVJT92OfSjJhyxPWhKN2QeaJJHWtoKFs9PagQRmiRjaDQGOFoEg8d6Ld6wuCp6mmb5dgNA4GM/SgcDAGaAyOeTQAYcUy9waBBOaY9qj+VgVPSrmIIAScms4G1TxWMDrz2rr8pNEYArjGB+FMARgUykjJNYAIANdeAfpxRIPHemxjCmgN3APSinPWioI256VxRUEYoxqvKigABgnmiQRxXbmjjHWlUMvNKxUnaaI7UrY4oKD0psHBFEA1gYwKT1qIAck8jpUjs0ZAPXrS7Am3NJD5rhB1qeGOCMBGy3fiktSsYkB/DFMVP3aEZ2kA9O1c5GaKAoGzTDI5FRnbH5eelFRnrW09BQBIx3rbxgflQTPAogA4zVzZrLGVBq5sJIyTjgUwOMUAcAA0QCORWAOhrAxgUVGOKPFHd0zQGBjNEE8CiARiucCgc85oZI4611P0oAdqJ5xmm5OBRBJ4okNwKxkYFYAGAarrwKokGjkYH5VRAzgGvoaIxRAPSjjtQGRjv6USOgH4V17UOetHHWuvaiO1MAeg/CsDtRAIxWM1jj3o80R0xWMjBojNYzwaAJ6Gq4oAdjX+RR+v4YrgjmjjGM/Ss56UeQABQGBzQA7VjHAFMBjgUPQdaIGcAfSiM4A/KiCRitpojP3aIJHJo4Iz1ojIGBXzUQe9IpchVFWOlvK4ZhhfX0/WooEhTagokdM0TxQGBjNAbjgD8K8sDgmkDKCQaznn07UAGXJNAbVyKXDjFLAQQcZ9ulSKUQBhzQ2kfe+gxSko20jPqKYEAmIcjtQaKYZB+b0pQq8THj+Ef8A6qZtq4jHHaiAwVZRyc4FaLdwWlzvu4N0Z6/MR2Pp+FTX2hw+JJL3T2xZnHljD8/Jg9eeua1fVNDv50aBuOc8P6D29q8NXOkiY2MzZ837nDfNgEnp0rxl4ZXTL5J4DlJM5HpgKPU+taEyq4huU3JL0OcYxn0p/CesWcBvILLzbYdZPMVcc46Zz14rxfNe3qZngwnZtw/2ew+leHXjEi2Lt88v3Bj0yTVudY0U+SG2Rr0TCn9efWp/Ef8Aad2lwY/KlizsGd2cjB7ccCtT16e9l3Sj5l6dPQe3tVk0qxxajanBizvOPu54HXrmovFNzql7GZbfHX+Men09q8bWFxp94ZVH7l/udOwXPv1NeBJLo3aRk5Bz6Ds1WdlqsUzrdy70XGxtqjPHPA6dq8dWU0kC3sgwsWc9O5UVMsks6F2yDnAxjtWs+Gn0yxXW1OU5z7chfU+vpVimg+LtP/f3XkXw+8mx2xzxyMD7q1d2v2K6aGNt4jxk4x1H/wBemkB+Vnzj2oRgnAOCegxUcC7ts7bPTjP8qZFD5DZ/ClYqwZTzVzbS6nZf2gke6Vf9dzjHOF/QdqJ5Geo6Vp/hifWtHOqW9psT+E7wc/MV9R6VLFIGMbnaR1HXFCMqc5z+FfL2NGT59sbfL2GKMsSyYB+tB2cbQKhhZW2kY9KaFo1yy4980HKPvjfJHQ4rxBqnhO58OeRp8mdTHTiQfxj1G37uadklYnNM5YbTz6VZwTTv5MS5bsOBWpadeWEwW7h8vf8AdG4HoB6H3FSKsT7Qc1gBSxbp04oSZ5FShWbKtn8KxwCOcU+4YP5Ujgjk0zRmTgVK3ACN+lO4blTSuxXyyetE44zzWQw25+lHgdaBDDbnrSHgg9KIAGKwOoNEDOTSNjgGmxn1ogEZArfngmiRjg1kEcHPpRYFMelD6fhRIzj8qzgc8UpYjk5NAc4AqVcOcjJ70SFHIoyAsRnPpTAsu3FMOdxPFAKRlTnFFi3Crn8aZmYbVOc0oYjC0sTLxnr0FABACx5FPOCMKMkdKd2JwTSQ5AYmhGNu3NOxAxmgSQBmmIxhm/DFAALnPT2oyhTtIpiGPJraO1fxZJ5ojdwTROOM/Ss7urfpWMcZ/SmwRisg8A11IANADHBoLuyc5I6UTtG0HFBhknrmmk3Dk8VuGcE1kYwDRK85NcEZBoZ6hs+nFAgnrRIPahjOR2pCpHJ5+lcuMKPxqHO8EDmtT+LuqXXw+0T4eGy8u30b7Rtk81Tv82YSnjZkYPH3j+HSr22toYg0E+9vTaR6U5YcZxWMjBo4AwDigcjANZx3onPeugAI4qLLNhevapQwba45ojPGfpRwO9BgVyDxRI7c1uIwBQGRyeaxxjNKATn0ojnIoDKgDqO1YyRxSgHoKwCQAf0qGEKCG5pYgcKgoQOsfkxjk9T614H13VvAXi3RfGuhS+VqWhX8N7asTwJInDLnnkEjBHcZr/gon4P0b4ufA3wP+0n4XTdHYRxee4GSLO8ClN5z1SbauMdZW9KutY0+1s1+ycy856+vuPSoNWmXURqFn+7kX7rcHHy4PBHpWtalfapqBvNRu/Mlb7z+WF6KAOAPQCpNHFyP9FfzR642/wAzUOlGMssnylsY75x+NaZaWscwW+fyl7HBbsfT8K8UxWK3UZ0ubzYlzvbaVxwuODz1zWkWU9uixNdeX5nQeWG6ZryJ7oSK0X7zjPzCl8MapbaNPqi23yJt/eb14y23pmtRvLQmE6Tced5W7emwrjOMct+NRW1r4lsXklm8uWHG2PaTuyeeeMcLQwrYlkyw9qA07UdBljlH72LGD83d/wDAVryaHPcyJp1zmFcbTsf0Hrz1zVq95bzu+nn5xjnj09/xq6nmZz5i4PfmoCuMr17itR1C0vtIdGOLoY2pz/eGeenQUzIke1R83cVY3sFvIr3EPm4/h3Ff1FXdpMUW7EW2J/uncD04Pv1pgoACHNHnkVgkY60CV4IzjvUhVuAcUpwBzWO+fpQKt/Fz9KA7k8UeeAcUchQoPPrWV6A5oEYyOvelIwADmiVI4Oa4AwB9KJ7EVEOeBU8glOM9KI447VnJwaABOcU5BACjpR6YA/CuG4plBP3eB70y55C9PegMEKoogjBND1HagpZdyD7vWmGOtMBjNYz0oilUuQooqAShHI6igABw36UCCOK28ZJoMx4Bo/dJ7etNjGK5xwelDPQH9KIAG0UQCMUQcUD0ANIpYEg9O1RqXOwcn0pgrtiM89hTxywsrMNvXHenXc2Sf0p5nEaqreueKReMjn0oRyO5VBkntRVlbYy/N9ac9umO1Z4xmkVFcnd+lEgng/SidwwPzoHC8UuC3JqRU2jyzn14rbgZIoKWXI7U1vG6/OvFahpbxEvEMr/+r3oqw4xzWCOCaGSODn0o5BxijzwKIAGCaJGMis4H3v0okEUQSODWQOCf0rAPANAgAUDk8nntQHb8qOMYBrI6E1wRwaAyMZogdj+FYGOO1Eehr2U0QDVURxWAeM0cHpRySQTVZB6GscdfpVEA1RUHmiOKIyc5rGelAYGM0enFcdBROBxWecZogkYH4VweAax7/SsDGBQ6fyoHPGfpXJHBon2oEGjx0pabpih0x6V1xg0AB0NHPasEnOKK9Bmif4RR5oA5GaAz1okngVjI4NEArkVgAYzRGcYFYx1FbdxwDWm2AkYMw4H/ANekVUUBenamJIAqiRnAoICMDmggXGBTNxgGgwBwaAVjxRVlGO9KN3A5pVQLgjmkkIG1jwKWJH5I4PapIXQFVHFK5zuY5NIxeUEj8Pwq1tGuv3cf3h2qw0OKS2uxfxYdNnl/N6k56H6VLAYHKDoKjJkJYjOOlRRLNkHqOlJEvliJjlh7VO2lX2kKsEWLpejbmP8AF6dOgrwTpkd9CGk5eL7vtnd7+1NoravdCAHO3rx04+o9K1PT38P620NwvI7Z/wBgehPrSx2/hX4U3NkW8z+1tu0YI2+VcZPr/e9q1PTWupzYzjdjp27A9jVvYro2rgyxcw9G3dMr6c+tTarYajaYKfN9T6/T2q9uHt58QnkdPyFT6bHPaC7jXPqM++PWvhFrWk6DqUz+IYvNsLjbvTcy/dV8coCfvEVrcGkS6p9p0NNkPplj/CB/Fz1zUN3pXirTxb3qZ8vqcsOpz2x/dFLrdroMzLanHl429e49wfWpviI7RRhX+c5zx/8AY14g8WLcWK2sr7vMz2x0IPpV1JHO4aEcDpV5ez/ZzbSPmP0wB3zSXLxnMRxU900yBAee9EhTgCmbIBBqO1ubmHzUXdjtkCnyp2MOayMcda8E2dveTyWt8cw3GMjkfdDHtz1Arxx4f07RJYTpi4R92Rlj0C+pPqa07xbq2l2JsLWXER7bVPcnuCepp5TcuZC2SevGKDOFwhwO3FAl49jHn6UXYcqeKO0Pxz7UjYIKVpdtHfahFaMeGznj0Un+leKvC+j22kNPZD9+uMfe/vKO5x0qVDE/lsPmHWiysxUjkVKzMNmcY6UGI4qyu3s5lmjbDDvj2rXvEM+tLB57bmg3dgOuPQD0pjn5sUHONprBHHbtRK4JoFiMA0DuTaT0oEgYPasBhkflSIJF2CjyMZoDnI59KYdwaJBAyee1EjFKMkHOfwornv8ApXUZHXvQBIwaIIGM9KJ9DTDIzQ5GBTLg4zTDJGPyp2TooodMDvQojJzn6UQOtEqORURV3xTMXJ/SnYgY71uVjknntRkycZppN3y0occigjE5BpIgxKnpTKsQwBTzlXwevpTSnbgmg5zgUsYcBqChRgUTj7potnrQYFcA80WUHAp5c/L61kkc0eo/Ssk/KKzuOe9HGOtAbjgHmiAeKB7g0FLHg1kEZoYJyOtMMAL2pRjIooD3qQZwB2oICuDzTqN20dO1MAeAaK56UyjHFY4zmsZ6VQwcCtue9BgMBa0CaCC/jluFyi5yMkdj6V4/1bRtZ1o3OirthPbLH+FR/EAeoNSNkYJofMMCs/Lijz3oHHSiMCgcD+VZ7ZpCxcbTz2pyWOGP6UDkDBqhjGBRIPAoqD0NAZ4zWCeQaAzx1oDsDQGe9AAniljyQAOtIqEAY5qKDeFZhkc8VBbiNshf1preyeyiaNcSc7uT6/4VoWn290jpJHlhjHJHr71+xRdWfxo/Zk8Y/s3eLrkGfS4prWFnO5o7W6DPFIBnJMcwc+g+QVqfhjW9F1nUvDOr2xg1LSLuW0uoSRmOWNyjr+BUitEt42voor0ZTncOnY46fhXiPR/DlrJZXGkw7f8AWeaNzn0A+8fr0r+zYILjfGPkPbn0+tXdukl95UC/In3Rn1HNaRBa3brbXK7m57kep7fSvGXgeWHU3TS03R8ZGcY+VfVvrXiLTrvRrOK3eT5nzjgcYIPqfWvM/s6wWZTljnJx7/8A160/Ub2S1e3ST93LjcMDsTirawvNGxevDiGX7p3Dtx7nqah8VzWc8ogfCvjjA7D6V4t1LRdTtYbvSU2H5sjLHuo/iHsaubO6k0432/5B2wP72Ks5bSW6+y3I3A9OSO2e1Xdu9pqjW1nxuxtH0XJ61rWnTWNwqTHJbPb0A9D70qtECFbBPtUMhxtz+lSuspAXtTLg49Ke4keIRMcgdKKcYBplI/CiCRigOMA/SnAPFIBjjrRHIWgoP4UwxxXGNq1t4BzzQUOMEUYysSvu657Uw4wDRHGBR5yR1rn0qN9rhsdKmiRk3KKJzwDnFFO2aBK8A9aGSMGgM8ZrAAznNEgCj2FEEHAP6U2MdKDD7pFE54U0QAcE5NMuDwKwB0FNySKojJya4I4oADiic0DxgUW+UA80UJHWiM9KLZ6UBgcmm57dKJz3rGR1pW96PyqGB6UQUIBrzHI8s856Cprdo03A0w81FBFLG0J6/SlZo23g81KS7bs1jPOaRVcEZ6dKXAU55oDccCmUZwDRAHGaAJGBRBUZzXJGAaXk7VNOwxtFOqOuxhkHtV3pAc7oh/n86kjKHaeooDIo4I5PNHnpR571gdqIBGM0QR3rGe9YyMGiPTrQ6c1j3rA7VjPf6U3HQ0SemaZuwGayCOlcHgCiOg2/rRwDgCjgjAFZBGB1r5ccc1xjHWiQTgVuUdKOCciiAR9KxnoKxngVkdM0AOtVkHjPNEccGjjAGPpQOOM1jvXA4FD0/SqrHvR+v6VkYzmse9HOMDmuorI6ZonI+U1jPQ/pXPSj14NcHr2rIJ4P4UDlQSKzjkVndgAUQcYFFTjigCO1bTng0VPY0QccnpW0kZFEEChkcisbh1/CjkDAFKCeQas4Glfg1awbEADZP0qRCOorB/vfpQz3pIiwyRmiFPRsfhSxOVL9QKKcDn9KO3GAcigEA659sVHLsQoy5z05xSvaphlbH4GmiWRt6Nn2xRUO/lMu0/XNNA8BwRlx17UHQJmQY9anhDNvRfwzSqFBkI2gdutaZfrp19HfCPzBHn5c46gj+teMPG2meItIjiit/LukztG9jtyy/wCyAeBVray6uggt4/3npn/HHYVqfhDxDosIm1DT/KRuh81G6YHYn1FYWUZSTLDtirWFXRcf6w5wPWtWvPCUnhgi0vPL1NOkXlyHOXHc/L93mtP1+/05mazutmcbvkBz19R714M8Q3YvDczyb4/41wBnhgOgrVdN+FPjr4V6TqV34t/szxXaed9pg+wXE32jdcAL8wIjXbEoPHXPrU/ijVNPieN5NpXGOFOP0968M6+msah5MjZI6HHsfYeleJ9V0yz06S2vpMM+Nq4bnBB6gfSpdZhhvMW8uR/un09xWu+G794f+EgtIt9o33m3AbeijgnJ59q8GwTyzPBcN+6OMHA44b0rxBo194buDp7zeZA3+rl2gbsAE8ZJHJxzUNzcWLEq24+mAP8APWvD8U2ro1kF3IcZOcbep9vStX0q50O88i8g4b7g3DngZ6E+oqZ4CFKyc/Q0AJDlrjJ7Lsq103U7lHOnx+aiY3HKrjPTqfrWoTR+Ubd12yL15z6V4c1DSrGWQ6pD5iNjaNzDoD/d+orUZLaW6kks49sZxtGSccDPWl4AFcf4V4G8TaHo0DQ6lJtzjBw57t6A+orxhe6Vf6vJNo4/cHGG+bn5VHRueoNAZ4BrT9IvNUs7iSAblttuRwPvE+/tTrIpMe3DD3oyIycH8KeRolBV+vtTyZIcDIHQU7mV8Ada3KvG/A+lAYYFj8p6UrgfcX9aFyDgngrVldsLlXkbIGe3tV5f+A73RjKZsXh7bZj/ABfl0FTtB55WJsjtwfSrgOZCwGRWdp5ouqnJPPpTOrHKim4G080X3HAFSSMwC0N2MkUcDr+VEAjKjpR4GQOKDDdxz7UrYYNnPoKlKlsKMe1AgnAXP40WUjaB+FDG7GKGGAXFYGORSrkZLfpQxkqD19qPB4okHOD9KOMdaLZUADkVnPIondzmjyvB5FEgjK8+1I5ByBwKJxwOT6V2HHTtRY9AfoKByMDp2pNpG5TRc9B19Kdwxzmio6hv0pAS3ynJ9Kh0+eQhlTj6j/Gp4hCSrH5h1FRMJDkmpZ4FOyN95HXgiprrzD8o6UxDkbW/SlUE8Pn8Kjgyck0qjoDmmYfdpQCQM1LtBwBmgQigg59BTOpGCMmlCgE96Izxn8KJBGR+Ve2KIB7c0FLcA0Mk8Hmjg8HrRKn+H9aCMw+WhFkDLYPpihCB0b9KAwMZqug5NMhKgKcYorn5Sc0U25UDOKKDHSjGyrmiM84/CiuBjOawGxiiO1HB6UtHcMAHmrCJ3YBRuPp0qRTkOD97t6UwBOPSiM8Uf7vWic9/woKAN2cmiSRjNYBXgUFyMCgdp+U8053HmiB1FEgd+aQjYFBrGBnNBSeh/SsZ5JzjpQxjAOayAMA0AWHFIgJwTWxlOCfwq3hYgYbPtjFTaXd2bqtxDtJ6fMD/ACNJAThFHzCrTSbi62pEnzc8ZH+NTabLbTNbzPynVcdMjPXNeGdFlsZUuZm2b87BjOcAg9D71+x98XR4F/aP0nUNW1jbYeJcaRdKybV/fbRGxPAGJViJJ6Lu9TX/AAUG+GMvgb4wy+OdKtvKsPFlqt2XA4FyhEcygH/tm5PrLVnaySyLJNwOcmtG8MxeIr2Gztrjy92759m7HBPQkelXGimCb7GY95HRs4zxnpmp/AWoIlm9v999/wAnHb3LUvhzS44/Nj8Q/vx/yy+yN/POOnNQv4i8O27XVhf7YJcbn8pDuwcdDkjBJrwppuneN9eGm63qfkm46zeQzY2ox+6pH90CviT8PNP8HeNL7wUutfaILLy8Xn2Ypu3RJJ9zcSOWx1968LfCq48QxSWXhu6+1s2MyeWI9vU9Hceh/Ktf+JZGmf8ACJT6V9ohs/8AU3Pn7N25g7fIFyOeOT71BBO7+VFHuf1yBWqaLq1iUt7yLyiM/JuVsdO4PuKudXng02TTWlwrY42js2fSnlaKXzI25HTj2r7ffJcrepL+9TOH2jjjHT6Vq93NesJprnz2XvsC46Dt9KZ2kAyM4qFyXHy9O2aKs5LoMY7elEEnmicnAolgMA8UzsRhqYbhgGivfNEEjg5qPaVwW57cUOmD1NKMng/hTKDj+VP0wFx+NAAjmtwHJH4V5gCBCM46URgbR+FHgdM0sig4Zc/jTLnIpMEgEdKckjaDxRAUECirE56+1bWOAT9KxngnOKCkYCrz6ZorkZog+lAgD7v603HRvwxRGRgGsjvzSsCevI6UxOBmjg89+9EZOQ36URngN+lE9geaYMOM0eO9EY6mqPAzQG7hm+hxRJPG79KK55HShx3o8cVjPSmIx8pqMsFwaDBRk1FudeRx6UE81d+a3MCHjPI6UZ2aPZImW+tKHSQlRn0FMWZtzmpQZHCIv4ZqUENtJ/SgCE47Ur4OAKI4+VvwxQOeM59qBJ4zRiyMhv0pFQDBf8cU4jIwHz+FORgKpzRUrinJIAFHAGM8imYBc1faUJ3MkTYJ/hx9PeriJ7dtjrz9az6DNAA8fpVZH5UMY5FEjoDzRGRjFYyAcVnIxiuvGKODxRwTj8qIBGKIBAwM1gYwKOCMCuDxVcYxVutjNaBX4kH19f8ACnAVtoP0olT0NDBGRVEAjFcYxiqOMc0QSMg1VHBGBRx2qgcjrVVnjb+VUcEYFZGM1nnFHB6UTgcU2MjFUORVAZOCa9vyr/Io9R+lEjOKJ7CgMDGaABA5o8deaPAABoAjjFd8VjHNE570RwFpiDwK3cYokkYzWKGCMA0AOgNRqWbINaXGC2Sf880CqjjmixIwTRz2oAscA0WwuFP1ojCggUrEjFE4PFDaTlRRJbtRXcgUHmtvy7c5qBtrAg8UpzhiK0/wVqWuWEeowTYV84G1exI/vD0q+s5rGZoJfvL3/Af401vIkauyYBzg5qa1aaEqpoRmMBDzimTgc9elaJMba+Rk9/5GvEqpqOj/AGaMZ877p9MMD/StU0ybQZxDKdxb6DsPc+taRpjayZDBJtaDGeM5zn3HpU9hKWKI2SnUYqSILJsz16VY35tQY16GrLxjdm3TToV4OcHI9c+leI9Nvra3NzcHK/QeoHY1ourNpk4mUZI7fgfY+tXniCbVb1Xn+6c8censB6VdRnftVMD61pPjbTNM0uDTLtN2N3dh/ET2U+or7fHZXX2mH7p7fhj+teO9fg8QpY3FuMCLzM9e+0eg9KaB54jOhyB2/SvBmurok0kkwyJMYHTGA3sfWvjl4k8JeJPCkep6bHsvkzhcyHrJGO4A6KaNq5gW4TlWz+HOKhhMOHk/KtK8U3GkRTQxRbhLtz8wHTPsfWrxvPfzz/F1/Dirjys4jojuOtEY5BoHcBg03A64qKXzQVxjFbSTnP6VZazPpdtdQRLxc7N3I/hPHY+tGdXBkz068UxwM0zGQbSa3kx7QaJPUHmgMjn8qDb1CDt0pm2/LTOpUZ/EUj7SCvXtTTFVyT9BUFpJeNtt/mbt2/n9KlimhfyJ12uvUZB/lTkZ2ntRIIoDgYrGRtBrABzQIIxnmiTtxnpSqHTryKDHbgClORtNYAAANE5GKX514ocHGaHFOD94UCMYzTA5wDWc8CmIDcmmGc80qgjGaI7Vj3o8gCsdgaJ4C5onA5FAgnAFEj8RRYEYB+lbstij87Go0PT9K2FjwetMmOM/SghIxmoYwMKvJ7VczNDB5CdR+nNTlpOS36VJcKvyIelPKXJUDrSRgAgmlTBqOIZzmgABgGse9OoJ4NEEdK3AjNSPnimIJyaIHXPStwIwKDBaB3cCsegrB6g0QCc96RM8nrSJgYNEY4ArGeBRHYHpRGO9EH1/CiATgVjPAqiobnNGIMeadCTimjCjIP0oqQKIHQHpWOM0VA5PWjz3pcZ5q1kEZ3g00hYBcZx0piDxRwQADXUfSsZHFdsV2wDXBXFEDoDScHJ5xT8kCjyMZokEcGk5UZonPGaC5Gc0AcbRWPlxjmgueFPNKpJx6VFbk/N37U8QyNoyajUFgB+FSTB0BJyRVrGu8SkdO1eGLZCEuM5xnj8xTPb3mrTPIP7uBz/d/wDrVeXEc9qYg2HT7vGep5rRIG1K4a4tJGjkgZWR1YgqecEHIweK+PlvB+0v+xTo/wAVLZUm1vw1brqN0EHIeIGK9XGflXAaXHXCL61BpK2urRWBPmAZz2/hz61rekyWEsk0a+VtxsGd3YZ7+9WcdublYS+c98H0rUvgd4i0LwW3jf7d5lhF94eUi9ZRGP8AloT94+n+Nah8G9UtbNNdE++E5yNqjuF/v56+1eM9N0q78LpJpMv+uzxtbjEi/wB4/WtInm8OXsV+DuaLPoOoI9/WvH3xBHxa1H7bZw7bqf7/AM2eiqB1VR0jrQ9Zvfh9rEV5qB+Tnjj+6R2Df3xXiufTNY1m7u9KX/R32eWMt2VQfvYPUGrCJTepCpw3PP4V4rv5b+4dSf8AV45+oX/CrXU7KK2uEul/eNt28nseegqa2lnzNEmV+v4VDqAtrC5tGTPn7cHPTac1NH5qjYachDsU/Wgu0bh2rzdylR1brWwlStDkYFNmtuRwacdMdu1MSRgGiuMKPwoAE9KC9B6dqCnHBo7h3oAsOT+FBkMAUDkf40EU4DUORj8qbsPyFEDYMGmGRkUwBGAaUZOAaJ3LtU1ICDj1rGeBWM4XPSmGOBQJPA/CiCBisZAwaYZ4FH5jx+FMDjA7UQcc9KLHHy0OwNKAegplOaOD8o/CiuP6Ux3d6I6GhnvROTtqge1KA3FOMMVzRwelLjGTR54BzTrxgUuCMCqicggZ4pgynaDx9KXJzg1JEBGDnkUy+fjy+SKYYTYaZtrE0w3HcKONu0H9KaMhQVPWiBgAGhyAoNRqGOCacfMeaKgjb+VBMk57UsY9f0ooQcGiAeDRUE9f0qQY4Bp8huKvtO+0rlfvf/q96mhMLbGHNYB4zVcHgGiB0BrjtVZ5xWBjArgcCiAOnaqbnigAABmqCg/0oqDxu/SiABQORgH6UQM4zWM8A0QM9KYAAAGgM8ZogEcN+lYz0P4UQMAjrWOMZ/SjwMFv0oAEYB/SgM4GaCgAAGmHYH9KIHTNY5wDTAE8H8MVjOOK4IAzQGTgGiOcE0QQMA80y96PTiiOBzWegrHGM1nHQ1gdqIHUDmgARwKHTmiMkUQDwaIyMCsDGAaBG3GelH0LfpWMjbmunGf0rA/vc/SjnGQf0ogDgHmtvfP6VgHv+lErjAo+gNADHJrC4xmlx3qFA7YUVp0GxMj8vzojHAFM+DjNAHAwaQDHBzQQH73FMAOA3TtigAB1ogEYWkxg5NMV+6gxSspxuGcdqJbOB09KUqSNw6Usm5SGGa8N+ONT8PKIon3wj7sWFGOvfaT1Oa1rWJdauTcSx7P7oznHAHoPSrrxC19o9vpdxbbnt9373fjdubP3QOOmKjBUqSME5xV1DJMfNB3H06UmA21xx3FANFJ+7+YevSm8TeK4tMFtFc/6KvbZHx82fTPWtQ1S/wBWYSajceay9PkC46egHoKjmkhfdG+GHQ4rTNVktbgSyHd69ux9qufB9rfaOniC1vMQy5wnln5MNt6lsnJHpUytBIUV8gdDjrUczxsro3zDocdKn8YarqGnPpepS+fG+MHaq7cHPYc9BTtHjKr+tCVkXKNgjpxV23hTXNNV5Z8zDPO2Qfxfh2FXN28kewNkfTpWmpHfWpjnTJHTnHc+n0q4DwkwynaU6d+teF7JtS1aKxjm/wBbn+H0Un+lfEf4b6v8MfFt14d1JvMittm2bCjfujRj8oZsY3gdan0HUpbb7XHBuh9dyjvj1z1rwBJp00l3aaqu6E7Ni5YY+8T93nrir6JtV1VtL06D93H9xd3qoJ649DU8FzZuYLldjemQf5VbrvR7djw+McenNOCOCOaLdFqS1dYxMg3Rno3T9KtbC9ut5tovNVMbvmC4z9akyzGMNgjrxmki+cEnjucVMkMRCwvuB68EVLsYZLdegx0pX25G7I+lAo8Z8wZPas8cDmnIUfKaLbR978MUx5wD+FFhtBB5FMTJyeCKJBwD2ocNUzB3IQfSvB/iBPDGprqbW32krn93v2fwsOuD/erxpqsGr+Kb7UrdMJN5e3k8YjUHqB6UwAwxOTTFRwKBwuM/Sg2RgCmAxjOfwoYHFMAQcD6VuAGFNA4GAfpQCkZI5o4JyB9KyCMYpOeBTHA4FNg8Yrgx4HasDPBrIbk9e1bieKAVkCkciiT0zQOeCKI7GgoHAFH5hg0duMZoAFgTW0vwx/CmO3hPwoozt0pU2DluR2xQOeQPwpVVDnv2FM4xhV5+tImFBxzQjLEADOOlFQX2qahlitiHU5I7c1fagxkJVt34Y9KknkfILde2K35ONufxraAMAfSk3NwDzSRENlh+FAAdKJJOP1piegFYAOCeaAx3pogeMVIqhsBeR70QpHA6dqHPyk59qB54oncc9TSgD5s8jtRJA4NKu7jNCJQMHmgoAwO1be1AE9+a25wc0yg8A/hRUkdaAB4BoqMAg0VBAINADOMfhRVScjj0FFRjGc0UVhyOaKY+UdKkAYZUVjJ606rnCUFJ4zQXJxmkTJGT+lIATgt+lPtChUOcdDRHYD8KZuaGCMCgRjFYB4JrIx1/CvlYUSCODSnBBBqViRkHJ7DFBieD1ocHAX9aA7AVQXcME/pQUDGKKqAM8/pSLxx1q3gydxXNWenXd5n7PFvK9RuA/mfajbMowEwR71FbhQdp3H06Vb2QkbDN+lQWbmNkd8kYxx0rQbLWoWWXTm3Y74Uevr+NaRBs1sw3sGc993+wfT8K16WCK7Eapx9T6CtCutYlulsNFkyZM/LhewJ/i/Gv2BdZuLS08U/A/wAawI8Gpwvf2lvLIrLIhVYrmPhjwVaE4H+2fWvGPhDVvgf8Zdd8LJI32nw/fzRW8jqMyQsp8p8HI+aJ1b/gVfEPxvr/AI71L+19eu/NnX7h8tF/hVT91QOijtVnbzT3HkM3yj2HpWv+MNFk/Z//AOEKtb3z5ZPvw+Wy9LwSD5iPTnrV98Z9V07w3/YGnWvkxHqfMVv4w3dPc96trPUNeMgsU3OmN4yB16dcehrU4b+wlaCZ8SDtgeg/xr4azjSdXWZLryJ3z5cmzdtwr545HQ4r4r6Z4/1JkvvFa+ZbJn7PLmEZzsDcIc9Qo5rwprsGnyj7fF56n7y7imOGxyB7ivFM3h4wy3WjxeXK+Nq7nOMYHVvbNabplxb6ibG/0/zZj92PzQv8JJ5Bx0xXirwhrNpNdXR0z7NGmzK+cr7MhR685/rVlrF1YQ/ZCfk7jA+vp71r0elfZbZtOjxId/mjLeox1/HpUChkwzZPpiriDyzujb68dKkDAbiOfSoYTIDIpxt68ZqUqWBV8/hTGMqAgwR1NAkAjPFFsnC9qQgDJOfwrCkYPX1plyMLSncMEcii2DwKIzwR+FAAqAPyoEAcClCnORn0FKAGwDiiFzgHmiAe+MUwwAAM1zuwKPoKBHTNLtKDaPxpjvBUDJHSnjBHynmivzYNeXu5J4FEkrkc1nK4FZJGAKJPTNKwxgDNHA4FMSOPSjgrgdq3EH736UVYrkHOPaiSy9c0CAcD8KlkVgAqYx71kAbh1oAtwRTEsMZ/Stnof0rB9aABHSsgcqaJ3HFMAOAKBBG3GaxgZUVJgjANRsNvWgc96yRwK8zuRmreZIyABnPb0p7iNhsJ49at3SJioP6Vcvvb5DminmKXC9O2aYJ6YPpSqWGVOfalXK8nHoKYBgCoogjg1kg8HmgSxxmkjDcFsY6cVIAOCeRSycggZ9KmIkfc0ePxqUKR8oz6Vg4xTxqDk/lW4K20DpXBHBrVLD7UgeNfmHXn6VNE8LbGXB+tEbuQKwB0HTpTAEAAUeO+axxgD8K6ke1HryPwrHPSj1xmsA9/pRGeKJAGBzRz0Arbx1/CiAByOaJJ4FHI4z0oj35ojAxn6VX0rBPIo47UDiicjGaPTAo5PGaIxxms+hoHPU106GsnOQeaZs8iuozQJHQ1WSBxR5PFEYrPfND2ok+tVxR56CuvWuDXGMUcE4BrAxgUcdM1gdqOCMCiCFwDWMcZ4ogcc1jJ5FFcUDjvRGKOCOvNYBHBrjHvQOO9MQeBWM8CkIHBNWwBkwDVqmVyPwqiAwxmsdgaAYc1nJwW/SkYE5NMA33TQXDdaAVhj07UUAOR1rG5QM0AFxg8ilwQVBzVvas4CIMnsKngaGRty4b0zSMAMMeaBAHymmA8rOfpWkOyMokHC5x+tPoFr4tsybL5rq1/h5H3j7kDotXlg+l3bW9wMMMcfhntn1r4X+O9H8L6drdhrBzHqH2fyj838Bcn7qn+8OuKvpo5rlniPyHGPyFcE5x16V6Zo61crYpp0Z/dJnHTuc+metay2lS+F/OifM0fbDd5AP5U7M53etbdwxmrMxpOjSn5BnP5Vr+iaPP4ROs2suZovuLtYdZAp6nHT2qOeUfIpqCQwyK/pUF89u+7satvDN14h0uDVbKPc77uMgdG29yPT0rULa80u4ww2OOnQ9h9fWrnUrzUWD3k/mMO+0D+Q9hUHiq8tdNbTIxlD34H8WfSpJivKnDd+K07UJdPuftKHJH+BH9a1XUX1Kfz2GD6fgB/Sml3RDnBTp+NXXhW81jS/wC2reLJ7jcP723uR6elMoHBOMda+HIsXsliuW9ex9W9Kg1638I6/qdvD80E3lc8jon0J6tWs6quo3JnjGPT8gPT2obic54pZAHwDTxgAjP0oDmpWR3BByB3pyofK9KYg42npTEfdJ/CicUAMY70FDfKprGDtJokFcZ4pTnoaf0FKFAyTRbIxnrRAIxmiAOKABGM0ckc1wRwayDxmqZdwAPXtQBOK6rtFCicfdplOAQeaJBGBWMxnBojFHLDA5xRPGaAAHX9K4ZdpPNA7RgGmYAZzmlYsMDr6UwJAxWxmIFMoVdp6jtQG7nOa2mjwOBzQfD4J60qjflTR3FiAfpSLhetSyj7qnk9qlKw2+0Hlvb0NTXCRDahz6inuWfqeKKkruzTZOB+VKTjB6+lInykjrUUbAZFKS5wR0ojtRxjk06llwDzQHORWMrwfpUgKoF70xGck1tJGRWM0Qo4U1hdvXmgOxFBAy4z0oIByB0oAEYzQXAwawAME0F44NYIwKwOx/CtuTyadVCgKeTRQA8mivpTJzgGiuD1plB6GtueM0BxgHpRANFQaMYxwaCAYp493KmgM4Hp2ogkcVg7QAOaY5Gax3zmuCODQIPU1kHgGsqTgGiwHA6dqOG4BoLg9awOtFScEUKU9CTTqMAA0B2J/Stp4wfpQyRgfnVsmXGOaVOw7Vo2ojTrZ1gGWlxu7YwTj+dW8L3U+0Dk/wCFaloT6YqiQcvn9Me59atF3MClWmrRxabNaOPnfbn8Gz6Va6N4c0zwF/aWlPu1AdsOP+W2P4iR90mvFHi2bxN4km1lYtjSbcLuzjCBfQf3fStUtp7xUnMf3c55HsP6V4bvE0hWukPznG39QfX1r4IfFq++HnxY8OfESW4c2+m3qC+VRy1tJ+7mGBjJ8ssR7gV/wUc+G6W3iTwx8Y9FjR7XV7c6ffSx4KtKgLwvnuXjLD6RCtY1u21nS7S1h/1lvv3jnuwI7D0qSQLLtRufp7VZ+OBf+Dv+EWdMTfwnPX95vP8ADjoPWtWguTJviOR+Ax0r4LeNLP4f3mo6lqLYFx5XljB52iQHorf3xXx0m0GXXYNRs5vnu92Btf8AhSMd68P30OkajZXcnSPzMde6ke/rX7Q/jm08QaLpNvYyZVPO3jB4y0RHVR6GvCGneGJvDt0+qXO28GzYuyQ5+ds/dOOmKvAYbNoWGGfGPwNaV4ivNE1C31Nfnkh3Y6DqCPQ+teNfjTrPi1THc2uBL94+Yvbb6IP7tXg+0SGQHrTEMTGD8woBo8AHk9KtIBcyLCDyenFa94evdFYLdJgt0GR7ehPrRyoKA9evFAD7oNMcfJnpRACkgc0RjlTTYHAFEAgYNNgKMdqRVK4J/DFbSRkdutY4wDRIOMGgB2NAHHPWjgDAplyeDmmAKZJoDK4pgMYBpuelK2D/AEoEkcGgNvJP1pCjrljjFMuSQaB4wTkUvzRrGBUi87QaIB4X8qKg4AogLwDWdwzn6UwIxQXOQDRAzjNZBQLRXcOD0onIzmiMjGayNuKQFSMdfSmAHAo46Zo9OKOMdaBI5BrDEAAUy4HAqlIxyefpRIPNKuRxQAPU1gYwKUAjk0MEnaf0osTwD+FbugzXmdwfwpGLHaDxTKM0h2HAP4UAGO4UVx8p5PYUDg4JpVVjkmoxGiEk8joMUWJNDBTgZx1q1dVUnHNLcxyoyuee1PCE6mmEbxkK2W+lMhAwaRFA4PNADGSaGDwDWs2omg8xRll/xFEYOM0QCayD0NcHoelHt+lYBAB60wyOPwo4PFYHaiAOc0RkYzXB5FVnI61gbc5ogYyKIzglv0o8isBhgdq2miAOMZqiM9DXXih83QUqj1/CuOmM0ADxWBnGKwCQMVj26VgdxmiATgCjjHAqgMgADmuopgM8Vk4xmgATkCs4PB+lYHWioHFEDHBrggYNUee9AZHJ/SiM8Z6e1ADIyc+ldejfpRAIxmhjGAa2/LitoPQUVA4zmiD2NfeGcfhWN3SsYHFEdxTDHSgpyABmhz2zSgHoKI7jrQOe3NWzBXzVo4khBH5UwwMA80CxGD1pWJODVrY3d6jPbRbxHjd8wGM/X6UNh4K4P1oRxtwrdPamjKnjnHQ1lh0OD3pHjAwy59DnpWxs5Q7h6dKCLKC0abSO2c0UOcMvP1oAq4QjJ9K0y9utPuVuIWyR2wPQj+taxeeEfEIm1xNL/s+W424tvPkm2bcL97jOQM9O9ECUFQcGgpA2g0CR0NSweB9Q+FFnKsfl+Jhv8+XMp3f6R8vy/cH7sY4/nXgLWzp+rLaXEu2OfO47c42qxHQe9fEn4W6zY2g8RLFvtpfuvuUZwUQ8b89T6UrOowhpkV13Kef7tEZbA6dqHIwDxRIIPOcdKKKeQaz0FEED5TQYjkD6U0sjxlN2QeoxQORgijyBgUsigiN169DmvAfifVNPvI9E8/Nsc7U2r8vDMecZPJ9a1PRtOm8SQ2mtv9ptZN2Rhk6ID/Cc9cV470LTND1aSDS18uIYxF8x2/Kvckk5JJrcMZY9KBHY1atbLIDcweav93cV/lREoICHjuKwu7BGc9qHibW9PtzYWWo4tuy+SvrnqRnqaeQ3DMz/ADHjjpQAc4ZuR7UxAIXdz9KUpuw/I9KlZCP3fGOoosCAOppnPVGzj2qRhJHuX7woOQcZyaVsP8w3D06UwAbaWz+FEAkgnisAHbnp2xXAbGenbFK2D8rfpTsHOc80ewz+lEgYIb9KUk8Fv0okEcN09qByOtBd3Jbp7U4xyWyfpWD2P6UmCcMf0ojHGaAwSQaJGMg0TxhfyoEZGe1cA8GiR65ojI4NDJGB2p4xsDDp6UB8pGaZAybgM0uAOtZx0GaPI44pSQME0V3jGaKjGO9RxgnJNOAWAA49KDKAAB8xpsltvc0yjoopUBGB1qW18pRvHXpUkSE8nBHtRlyMRvj8KNzHGAqJkjvmnu5WbIbA9MZozknOcHt7U0zyffk3Y6cYouCMFf1osQMEfSt7H7p/ChIQM45FNKQ2Q34YpLli20N+nSonJGCfwoEkfe+nFAgrnNA5PWmHGAaUe9K0afM6bvQZxUk7ucjj2oMNoXOa2KBzz6CgFJBAoRqeAee1NHkAZzjpSoCPU1syAc9enFADrnpSkAYJogHjOfaim4Dn9KO1OAaZ+MA0ZT2P0rdxwfwrfnjNCQDt9K8xmXZngdBimZWGCv60E3Hhv0pxJEMkZ9KSRSuSnX36UhjYYZsfhUkUYfCPu/DFSQhQNpzn2p4wvQVhSMAVtBOAKK84H4UqseAf0pgmMY5pgAeaxgYAraMcdaIA4BojI60wY8KKIYLgnP4VjIwTQxjAogA5zQQEcGtoJzQXIGD9KO0YBpVBUHdn2xSoSQoPNLGDgOeKgjjBwhx71FGgOAdx+mKsLSORQ7np0GK0/SoWjLvcYPYbCfX3qeGWGUw3Um5x1GMdvarlYFjxAm1u3JNaXYi9Rlbhx1H5/wCFW+gi1mjdrnYef4M9vrWoaMsFwyWzb9uMPjGeB2JrwxNtL6fcHKS4yfTGT2rxd4TtvCWojT7bVft8Q/5a+QYv4VPQknq2PwrQ7fRPtMbXUeUGdwy3PB9Pwrw8bH9qD9j2+8N2FsV1bQ4jbWsTSGV0uLUB4MM3J3x7ULH+83pWgWtnc6utrfzbPNz+82k7cKT0HXsKurFp78lH/djpx7Vp3wwvrnw3Bqvh8efK27nhfL+cr/E3ORntxXjDwxZaJqENtFceaW3b02FeiqRzk+teMdDsH8T3dnpaeTBa7PKjyWxujUnknPXPWvGg0QaWLi1k3X8PThx1ZR34+7QurhrWLzx0zg8evtXiPV76SNFmfcq5wMAentUV1a3KRzRrgHOYsn5fx/WviH4j8Panp1pBovhn+zZ4N/mS/bXm8zJXHysMDABH41dMUjG0fSkdnkw7Z9sYxUyqnfJ/KhhHDkZJ6U7OzZzitP0K41Cxur6NsJa7MjA53Ej19q1W+u7zabmTzCM44A9PT6UAQASPpTkA5Aorg8CugwRmjyMBfxzRK7cAc02B/D+tMQVANIOMZpZCmUQ9a2+honJI3dOnFAEc0yHpuz7YoAnjdjHtR27Rjr6Vg8At+lDGDg1kE8n8KdehDfpQAOAOfalJYYDfpTcgAtk/SlXPIOMV3G7kmihZsZ/Sgu9hgZX0op8/pjoKdRnpmgqkUwyOBX0rbkZFHGDxWN3B7VnK4NAIBwN36UyrjC/lXGMCgVAyOvag54w304q5KE4jNYHrzRIJwTQIIzjpWcdDRICgA896JPdvwxWQe/0onsTR6YPNI+On5UGYjlenvWcjA/KgOOBRAxkdaBUDFYB7/SuvakPIAH1p8EdPoKCgjOOaKOgyRRUk8ihGDwi5PbmljDZVRTIqjDL+tBVI2benvQQKwCr+GadNqBAmR9aZGjOCOaYOR8oraqjOOe1K2/Cyru/HFOhRtp6inBIwDSlQMAfhTbXyMcVq2nRonnQ8D09Og9aYDhc/pRA7GiecA0QMcH9K27uM/SsD+9+lHB4U0TjHNEZ70AOmaIz0NbQetEYrB7VgkcVtNURjgVjJxmiMcZo4x1ogkdaxngUD0GaK5+7QHfNYyOtYBOBRXsKKk8DmhEzcAZoRknANG3ZjwaSzZhyaXTiFyf8AP619iU9/pUdkjrkDOKktogOOtC2BHy09qDyBzTW/QflTRsuVIojHGayDwDWcc5rjtVAAcA0SD0PPaiNvGaOD36UAOgoA4A9KBIHJrgjg1jnNEDOc0MDgNz9KPvRIPAPSioA61kAYHWhj1o4JxmiA3Q1t96U471pF05DITnGMfrSyK2BnmiFJxn6VgKM5qx1WewieKIffxnp2/D3rcG70GxxmlcEjJzj2onJ4FJEJeE5NPHsO0HmosogJ4zQCygEHIoRIrBscihgjINWzwSJ5dw2COnBP8qubZ7bBA4bofpRUNhfSgYwhU9e1JOyHKN+labKJ5t2PnX7v5HNav8Z/EGoaJH4ZuYMwQ52neg6sH7JnqPWpotvK9KK4GVNJD5rBQct2FMjIxUjpQAOcc5/CpJPLOzNFwoAY9alYrjB4piNmS2QPajMrLlTwKKqY8jr6UB/GW/DFMPmHGcVa3DRzLIj4kGdpx7Vd6tf3MwlluMuOh2AdvpV3e3N9KZbubzHPU7QO3t9BVwxEe0GkkXYATyKW4YtnPHpTjK5Dcj2pmCkMWyewxSSknynPPavDdhpl3qAi1FsRnrwx/hPp9BV4Iobt1gOVGMce1CQM+Sc+nFSgRPjrTLnOD161kBsA8ikxuw1SBEI8s5/CnAVsE0FXbkmiPfms7uhr5QeTRwRx2oHsDzQAwQTWQ3AP0rGBjNAgrwax6mgewrI6A/hRAb5QeaYgDrX0NMysoB60AM8n9KHI20wwRkda2qw60wwKxuXAP0ojAAzzS4xgGgzEeWTnFMuOAaRSV61tzwDz6U6belEcDJ+lBc85piB0PNIm4cmgyk7R2ohRyTU0axKF/iqFQxyT0pAZDtXtUDxRBmkPIxtFXdyxXfI3A9qluTJ8w7VvOdoNblxuJ+lM24ZqNAzBSefSnCA4zzQ+tE8YJpjt+UH9KluAowDzUlwXOAf0qzw5GTUQU9DTAAAHqaBUDCmhj7uaGO9KoAyTVy5C8ClTsDzSrhcEZrA2+9BRnAoAHgGtgAxRUZx+lE84z9BQVSM46UkYbjPIpiBwDzTORwPyondzmjyMZo9QAaqiR0zQIIBBrOOhoNjkUzluCc0Np4Jo88CmUBRtNB26g9KV1JBY8VOYG2+Ueec8Ggw9aIJOSc/hWcdDTbTg5rKkbSeaK55zRUYBH5UQCOtFQBg1hSvXn6UeB1zQAPANQrGGBc8fSnCj5hW8Mxwfwoc8ClUYAJ/Cgm8Ag5IpIwWx+VLajIOa8qN1AA4HSlQDKgfSrGFJMMOpqxt3E8MTDh92PwFWdkgIjJyfSvE+nxzSWzCPBbfuOfTHvXhPwPY6/qkdlIeDnPX+6x/vD0qfStH0Y3Gno2bmHbkYbvz6kdDU1pNcMrFuBnHFRkXLSTAfI+Nv4cVHbBJDIhxjvWh6Np2papFZay++Js/Phh0Unopz1Ar4w/DIeA4re705f9Fn385/u+WO7MerGv2B/iG3hX4nXXw+vpdtn4rtT9n3NwLqAM6/TchlHuQo9K/ad+E83w5+M3iO0sbYR2F7N/aGnAAALDOSSqjPRX3oP9yvE+v+FbzwrFZ2q7NQXduGZD/y0BHUY+6K+FXxv1jwlpl1pRi81bnZ5fzKuNrOx/gP96r7xHNrGsPf3h54wP8AgIHYD0Fa9rt6NZkvYR/rMbunZQPStSu0dwxOS3UYx0xU1+Zovsjpgx+/rzWpTNNOsbHIGf5CrWbyrjaDgH29qv2S6jKA8j7v6VdW8C2ySBsl89j2Nad4W1HVVaaxj3KnXkDrkdyPQ1dRNE5jk4YdRSW8s4YxrnZjv0zUkhB2E4x0GKg1e6s7aW3WTak+NwwDnac+lTPDKcl8/gakYZ2qcjtRwP6VkFuKY54BpsAbR+VA4OM0cdCefSmGeB1pBkc9awDyDRHoaPPFKNzFRRBKgg05AACtz9KCg9TRQEFgc4pkXYCDRBcYXrROeM0VJPHalIAwDmgcUCCODR+Z+DwOlbSRvDcfSh8i8USzfMTwKYIeEOR2plB4HWieMZpgDzmsg8Z6VtOMg8UQBjmgSODz7UFwMk5x0o42gnrRAIxmgMjGKZduAawpABNOoB+U8UMY4rj7tVgA4BptrfWiACADTYz1pwdvFRgHpRdSPlNK4bgGiQRgUwGMg1jK/wAqHPNALnk0EwcD8K2gnAP4U2V+U9aeQkAA59KEqBAjVGYzyW57cVKgjb5T+FSIW+YVFbK0Ycnk1OgR1UHrmoiyruJz6VzKdxP4VINgwetKoKZ3fpRwBjNGTLkvz6VJgdKfOMCuVXJ/CnjW4hKPzmr60NtL5YH0/IUBk8GsdwaqgPesADrQ54FFeetEHPBrbx1/CiG6A1gkcV9DRAPTrQGTgmsAnGKIz0NEdjWOP5VjjNKpbgV5eeTRhLDihbkLnNLAFOMdKW1LjIX9aXT5MbgvHbn/AOvSaa7kEdf8+9LpCL985/z9aTToGyFTp7n/ABo2dtj5IsH/AHjSxRbSBFn/AIFXljHB/CmtdwGBQgKnCHDduKSyZRgH9KFnEcAjkd+aa1j27QfwoW0K8KuM9eTT2iNgAcin05GXh8Y6DH/16k0lduRz/n609g6sQBTWjqdpp0KnBqjz3ojJwD9KIyMA0Rx1/SuPuigCBx2ogngUQxwAawScA/hWB1o5HvTbuuaJI4zRPoaIJOBz7USSMiiCQMGskDAPND3NA84DfhirJ3jkwp61BuKhmPNAHHXPtRy3yk49KA3DIOfQYog4waxxmoyF5ArBxkH8MVpgikuVinbCHrx7H0qeREkKpyBUJSWMIDyeoqw09bydYN2wt0OM9Aff2rXfh5NpFstwbveDn/lmBjkD+8fWtMSwtLhZdX0/7Zbr1XzTHng/3eeuPyrWE0w6lMNJg8q0+Xy03M2PlGeW565qOaW3j8svlG6rjril0lruIyWabSv3kznH4k+1NCkbmGVMHuM0bUlN0J3D8qhLxnBGGHWnvjcIIpzu9+n8qZHidVc7omzz06V4i8E6dpenJfWl9uJzlfKYfxAdSx9akidn3I2D9KsW8G32ikala4ul6HfL/e9uOgFbVCbIjg9vag+MhxlvyqNyUw3UdKeRmUjdx9KbDLtWnTC/fyewxUU6hQGbB+lbY5jvQ4YdeK82RDhzkUzI64J4rw9ok3ii9XSrVtsz52tgHoC3QkDoteLfA/iXwTqD6X4jsvJdMf8ALRG/hU/wMf7wpVWP5SeD046VJD5bBkPXtipXC4ZW59MUkqsm4N+GOlSpFKu6Nvm9MU82/Ck8j2pXWQbS3T2rUtA1DTkE9xHtjPTkH0HY+9BypwDyOhostxH8vDClkLDaTzTKSOTzR+YYNRMCQjnj0xTxkja55HTigcHAbn6V3yKcgkc0RkZz+FLjOCeKKgfxfpROTkVgYyKIyMg80TxyaIPbt2rimII4NA4GB+FNhuf0ooQuRQIzgGnRY2APU9KBx900BuGCa6GsA9T+lEBeFNHB4Jo5Hyimjwfm61BGjDc5+Udq3KSSzcduK357/QUWGME59KYDIYn6U2GHAplRUVvvE9umKciRfLI49KNusADHv2oSsJNxPTpU8xU8nOaaZFt9yHB7/nTzl12LxSzqEwTkjpx0qSR5Dh3z6DGKCkgjd+GKA3HDHNLtDFA36U2wHANNJzlD09qaYBck5P0r7RGgyH/SpLtNpCHkVJcyPyHwPpRZnPJ5+lHjv+GKsT8wUH6VAiheKkKngjpRAZcD8qXnjP0pUHBK59s0xXaFz06VIomIG7p04oRlSA1DsB2rHGM0qgnFYA4BzisA8AVt5zmioOCKBRThhuH5U0hJwD9KJLdTRAIxRANdeBRGOKI4rGRxWAOlUQexo5IwBTAFcHmgB0FEY6DmsZGDQOMbaAB5IogGj7Cs8YBoMF+6aJDdTRPPBrd2zR+YcdqCA8kZ9qKgdKZCMDOaERznP0qK2Mp+UUZBazDI6f4VcyxzEMlHB4z+NBQTwa2ggc81GTGcoefWood52g9KWP5QOuOlLb7l3E8HpUFn5jAImWPQ5xivC0UdpcrLPzjoOnY+laFcL4n8RW+nSRZRd20bsZ+Qn2/u1rlhdaPqADrkduR/dH19a1i5gvXinhi8t0zv+YnrgD+VfDrw/wCBdc8LyDW23akcbDiUY/eNn7pC/dAqdF0lnsZjmXjB9O/v2NRX4KBIm+b1xTxm1ixGmG/iGeleHIr/AMO6cmqTL8j5xyOxK+/r6Vpep/D7XvB50+50TOsr96f7TNz+8JHygBfuDH/16i8D+INSsWh1K7zZxY3/ALtBjJ46NnqBTWXh74V6lpviHR9d+2azot5DdWz/AGeWHDpIrgEZIII4I7j61+1n+0b8MPG0PhjXfhtrVxceILGNhdgWZVEhlQHy2d15dGyMKCPnbnIq9v5LudrycbpG6t06DHQfStP1qSN/PmG9U6L0x1HpWpXsk7h4zgD/AOtUGqaffWcq6i+JFx5XDc889B6Adau1jWTyi2VPTjFaxdWUVtBa2g+Ybt7c88gjg9Kvi6XBKN+nsKG+ZldW554x0oQAsixtuY57Yqa3dGztx7ZrQPFN3o9jNb252l9uTwehPqD61dSvPKzu3J9vatI1m68L2hfTL797fffHlD5NhOOoOchj6VrElpe3ImtofLJ+98xOeB6/Sp2diFPOKKtnGfwoNuIReoplJO3blh2zTRlDsJ5Haj2WiS3U/pRYgZHatu75gfwxRJAxjOO9KOM45ojPAHStuTyc1t+XAoDBwBxRUgZNcAbgOaUkgBaYFTjGackYxzQOAeM+lOuTg1kYwBmgR6c0PWgodsg9KQ7vkB+hxRCgYfnFcNwwzjpR5XB/KiAy4PQdKC7sADinAxgHntRJPBojPQ0zZHFELigSBgmlPqc0SG7/AE9qEbEDB/SlITBVufpTuzHLHNMSe/6USuKHPamA7GsDGBSAdFOKKgcZo88/pRHFEKF60jdj0piTyRzSEHoKXPQ0enFdRRO7gUMnB61kkcflSRbucfhUiOgAIqONXIUnmsRldpXkdeaKIVATr6Ud3fn2pyCeRUcjMu1pMfhTq6tknIPQ1AI2f5xTlWYgLgn3osQdpP6UYiSrA9c4oqFbGcmiAFAIp1aQZJzjpxRRSckcjpRjQnJFCJAMYrVbJbi3yq/OnTn3FOm04NHJOPTpRUHkVggYBrHfNbQeMUAB2ojPAo9PWiARzQHGKIOM5oj0NYyMA5ojFAZ4zRXA4NYJHFBD60sIYAlv0oIijjmjHngUsBKjBoW4Ixmls84JOPakQRQsAMk4x2qNswhSKVdrKQPoKCbxkmgVBCCjGpGBSqANopTD5JUn5u3WgmV4/CgihsZogDjOaIHQGiPSiUwSTk96JUmnQN846UF8wZUVDbKOSKlsIpmJA5q40xi2AOn+fWnsHU4HUV9gcnaBn0r7DJ0z+lNZuB1prY4+9n2xX2dyODQhcDAonBwB+FdsE/pQYHgCjzwaGMYzzRwflPbpTEdK4xgjpW70onIxQJOBmiw6CtwNYH8NWSlpCSelW75AGKAyuAaKA8ZoKFXbnpVDA5P4VjIwDSoSoYc0sbhlKjrRbnDVauokAU/Tiobl4pRIh5HTj2qw8RNqTrZX9z8rZ52fj2HsKsfhFqmpRC40w+er9Fwq9OO7+xrVPAMkOuz6VaT+YsW3DbcZygPQtV/8M/ENq+1oflXr8ye3+1VtqE9jNvU59V454/8Ar14e8Mad44SWQT+TeceXFsZt3XPOQBwua8NXljoOq7NZGIx9f7p/u59RXjfUdGv9SN1ozZh+jf3VH8XPUGg5ZRxg0lwTEUcZA7+lWNprWuwssUnmRr22qv8Ah6Uy7W2t1HapGI4Uc0ZSkhPelkydxGfat4cYUV5mBsxSkJgg5HpTAS4K9u1Egt1qGfbzjOaYLcrhTyO1GRIT5UnUVoGuzaJqUWo2334s8fVSO4PrXxe+M1z8VtQOo3ej/Y5G+8ftAkzgIOyL/cH50ro8ZjZuPXFSyBECgZHrTJ8ocnIouFXA70oITzQOF60bdSm+Ntw+mKWUoc4/CrnUrrUogs0mVHQbR/h7U6DkqKtplibLD8KnjRv38RyO/GPaiytHjFMAOcUcnABppFmUc4Ip054onAwBQXdhR19K6HbRX0ogN0PSsYHFAijx8ufpQGeRRGeCeaJCnrWRjGelcEcGjgjOM0WIG3tQII4reJcAHkURjjNYwuQaADHIFYGcGihYfKaCqoyx+goMB8xHTtWd3U/pTSj7MVUcDv6c0zHGBzivmyGJ47CiAOGPPapHLkDGMdqDArhKjUyEIBye1TMsHDfe7CnuzIdx5FPMgXcDn0FSSByAOR2p5dy+WPyp1VRkH8KDqpwaMqM2PSpbhVyDT30ajHejfgHIHNNqAZeU/WmvF7Cnui3Rf1osxHNYyMAUWBHFAA4xRIB6ZrT2DS4z/nBqI4QCmO7kdqBZjgCkwRjPSgwAqaRR8oGTQ+U8Nz9K8wHnH0pGDD+lZB470Bz1okHpRAI4FZzwDTMDhcdKYg9/pRHpVdetVkDmiynAB+lAgjg1ketH1zWR0zXB4BokdM0wDLjNZB6GiR0zRxjrW4AcHIoFTwprjOM0CD05rcBxmgQRwa+hoso4xWc8UpwcEUmX5x0oIGOR19KMYAGe3alCgeYei1FeiJtyR7s9s4qcicBieT2rbs4IoAEdKUAkDNKoAyKiUk5A6VBDj7pzmrOyaVthXP41Hpbxx+WydPf/AOvS6fFbQRyxvuPORgjv9a8MQxW98j3Ue4LnK5x2PpXh7xXpHhHxlbeJF0/7Stvvzb+ayZ3RFPvYOPvZ6Vfyz+I7l7podnTA3A9gPb0rS/DbX0N1cSzbdmzC7c5zkevtVndS6a+LeT5h04/xz61aaLq3ifWkS3+Znz/dHRfcj+7TafLoGrrYXw+YZz/3znsT6ivEeh2EoF5plz5kT/d+QjOMA9TnrmpfEQn0pdNmTCpnnOf4s9hXw9019V8URQCXZHLnc23OMRtjjI9K+MVxa+HLxtH8Oax9rMuPMH2cx4wI2H38+p79qn1O6LbLp8uOvA/oKjvYbktAj5Y47EVrvh6x06xDW135zjqPLK9x6n3qLVMuImXAPv1/SpdVCt5Trhu3P/1qLxvGshP+qz+tauQGSe3feBnjGPQd6uJ3kto5mOC2eMdMHFaANBkugdcvPIjH8Xlu3Y9l/CoEMEzQye2PyqCdbS5R25xnjp2q6nDpvA/CkuUC4J60fmOB+FJp9w9tJdKPlhxnp3OPWshiRnBFSvghc9K3K3O7n6V0O8fhVtJaMgMsu1vTaTQJLcnNZJGBz7UcdqU7hgDNMMLzRwRjPX2oEY6803JyDRI4BNAEAYORWPf8KyGwpP4UQWOFNIAcBT+FPkgqDyKfHQHp2oMBwT0pxz0oAjgCsZOQaDHGM0GUHOf0rfzkcUknOSM0rjPSmYMMHpRICgZ6UDlcCmwTyaYDPSuD+FEDGM1jK8UwJ5oHKgHmlK9M807HG2jnGKCgjg1tUjg1gZCisZHFEZ4BojtSKWBI7UxB75rbkdaK4G2nGFwayCMA0WGOKC/Lgc0oyAM0RiqK5HHWkjL8A0F2HB6U0SrH5iv+lSMcBW7V0QFB0poAR5oOfwpECIXY1JblRkHPtTKNpI60mCducmpEdHGR9BUkckbbm4BqNEcCVu1TWjL+9PQVPJtTymXke9Eb1z3o5wCRTNngGiO4NOeM+lMeMA80TuHJzWo2phkLA5/D6VjI60OO9AZ4Bo9ODQORise/4VjAwOaI54P0FEA8k1QHaiM/hTdaJBGB+FZJ4FLGW79KjiOfelQnn8qEW48HmhbgHrk9qjg3HApLBcAs36ULZQwIbp7UIR0DdPahCh4IziigC4HSqxngUBnjNHB6UQem79K9s0BxiunJ5ogE4FY7ZraCOtFQMYPTpRQEdKwwO0DiggX7prAPGazjoacZHWpYEcDPans0YYJo2UYXKnkU9qrdetSWAIxu/HFfYABzzTWgCjDdO2KkskZcg801iCeDUkLocBuPpRzuxmuCdoPNEjsaYg9DRyR16UTkYzWM8A0Dihgf0ogYGKI3dDWmpuBOev8A9eoowF3E8ClYFQSK3DpW/J25oxqxGT06UEBHWgNowH/SkBA3B/wxVpZTX9wsEIyzZwOB0H/1qubGW3kEcq8/WoooweB8wqMmIYXrUO1UDMmc9814Y+KHiLQVVbC52oM8bEPr6qfU18T/AI9WPivXF1yx077PPzv/AHxfPyIo6xgdFPbvWsfF7UNRk+0XA3FenIHoOye1XNtZSWkc0I2SNncOT3xSyyWrq6vuxnHGKi+PWn638L4/h74o0T7a9vnyZ/tJj2bp/Nb5UjGc4Ucnt+FTQLC7KrZHpjFPKc7AelCfIweorwj4jk8OXT3ESbt2M846Bh6H1qKxj+IGr3urG8+ysfLwPL35+Xb/ALP9z0714ivpYmfTpZfM8rGG24znB6UQDyDzRIByetByh3DmizTuMnrQJU9ee1NKCCD1NZSVsE/NUh+YRN2pHKtwfpTFbsBH/h6f5/CtT0i90llW9i2bs4+YHpj0J9RTuCApGR2ogxHcrcD2qJ0kXy2/CpB5Z2tz6Uw+X5T07UJiF2Co2UfPGce1OnmMXzzUc3knaRn1qRFnUOnX0p1AOAfwq3kG4xuflbr+FTRKj5Q8dqCK7AE9fapI/LIBOfSlYoeDRDFDIp+oojA4pWxwKILdTVg2niOUXsPmN8vl/MRjrnp+FMV35UYHpTZIyPyrqKyDxSsynNN0yD+lEFlwBz2oA7cE0XAGcUuGXaTRYkAZ6dOKL5GCOaKuWDAdOlNHhck0BlcVtBG3NKBsAIpEDdT0q5kVsIgwB2qQKE45osBhRTElfLzw1OQDjNL+8Yt2HQUZgrYFO6hiAenSklCDgc1HOAuCentVxNvbAP4UzFuKx71LIX+Y085A5PSnuVIyDk01+QNv5Ubxj0NPK8vLN+lDBODWPejjoK6jFEDHBogkZrA4IPNEA8GsZGBQAA96sDiQkGrYBk5NKgJIz1oLgYB/SlBzjNKeOKnGW3A/hRAIyOoodKRQV4oAYwDQ64BoADjNHpgH8KLAfdonPeiAe9Ace9EAng9KPNZ460SB3pnBxg0XJHShIG4IoEEYAonigQRgUCCQMUeBQwRxR4FZGMg5oEkgA5HpTOAevNNIKEwJ6U0pzhaVsjIoZIwabnjHShxQ6YB5qO3ym4H9KmmVk2IvPbmrm6YjCrx65p7l3j2dqyM5FDB4z9KYHpnP4Ui5OAawNvX6Ug3Djt2qJP4cda8LeHm16doFufJ8vHOzd1B9x6Vo1rbaFqzsW3z2+PLfBH3lOeOR0NT3d14hujcXL8v14HGBjtj0oaVBDcrarJknPOCO2fWovh9JY+Hl8Spe7VkzhPLz0fZ13e/pWkxG6mkjPEvG1vTg9vpUZisZAbfmQ9T/APr+tPaW1tFLKBl3x69qZQH3oOnatE8OX1/pc+v2c20Wm3zF2g/eYqOp9vSohcXd2Jd21v72Ae3p+FS3Q0jfp8t1v8vGBsx159/WpLtLmTKjaPzrQ9a0fTtGKLb/AOld5N7c/Me2MdDip72Y3LXLH5j3/DFDSpHb7Up3A9e3t61Olrbp5kC4J68mvDniMeGL1rhRu349ugI9D/er4p/D7TNM8Haf4usLzdPe+b5kXlsNuyREHJYg8EnpWieHbXUZibybGOnyk9j6H2FNp86l7a3i81OP4guO/c00PlkrIvA6rmrCXRYhc22p2XnF9nkN5jLsxktwOueOtPbqtxtiGF7flT2TzZcrkr15p4EeMOD93pVxNsXanKipZA+DjkVICh25rQY5z4V1q8gv/La38j915QbzN0hHXtjr71K7lyW71Io3FRQCgYJoZxjNADOQ3T2oEAEkZ9KZQFyD+FbsfKDWT1BrORyc02AcYzSgD7opzuwc/SuB0o5xgUTgYzXbGf0oAk4BqEDIGMmmIVWXGSccU4BxRGWwp+gpyehPIo4x1rqcA17VgEdaLZOM80rfwg0Tn7poYAAJpjgYBzRAOCe1FV25B/SiFxnP4UQM4JrJIxnOaYAjigcHANLgHOaAAGQc02SeTXB7fQUeQBRG4ZBzivcUW3DrXXiiARigTjAotxgUAMYzTNgcGpSWGTSAOMEc+lCMc8/pSLgYBoDAxmiAaxngVDA8uWHQVGNg2qamiZGIPNYIXJGKDg4B5x0FEgLleajZQPJB5XpTZhiLAcig6FNrjNNEPKyrYHpipI9rbVGaZvLO7+70FGZpcBhx2oSmORVAyFzipJS77h+VSASjfIfm9MVIFwABxRZSAMU6BwCp6UxBFSYY4IzRiQjihGC21ua1S2R0yDk//qphjGOaxxjNADHAoAg8UeepojPeuoo+wrFYP979KzkdaB6DNHaaADcDrSQFzyOaWPbxSoBwBUcY65z6UlsTyOaFrkZI5qOEJ2+lVWBjAodMA8+lHnnqaCknAorjgcmgCeRRXnAoAYBPaguaYEjkUF4GaCEDituTjv2opx1raCQBRQgdaKAcg0ygcAZornp+VEHFUQCMUVBG2igK7QaZAw5OTTRgqMnpQjJ4owhhgn9Ke36bea+zFxjGCOop9Od/ujOP8+tXOnOo3Kv6/wD16e3dT0ogHgjpRUEYFFccAUQCPlo4B4ogEZzQXNEAjC9qC1pkoilIPQ9PyNIQ8YYD8KIPIAoAngr+tFVzkVtYgHGaVQDgmkSNict9OKEIbhGz+FWck1hMlxC2ZEzjgcZGP61PNLO4kd8sPbFbmGGK8Hoc0JAx5HAq1BnAjgG5l6L0q1t2PylNvrzmtM02LUQtvM2JGz2PbJ9fam+E2t3t9BpulQfaJJt2F3KnQZ7v6A18Q/ht4i+HWqPo/iSy+yTpjKeYkmMqrdUZh0de/enhUwb1bIHTj3pd0OWxxUxRl5bG/wBvSnhUxgocn1qRWjOCv60sxToKtL65tDutpNp+gP8AOpLpp3LTPlz1OMfyqQBThTnHSg6n5WFDAXKc0p3DC00qSgLt/WpE289fSnAcg4waZlYgO2PfFFccA5qNinA6jtXi/wCIg8XWIs5NK8iQdG8/d3U9No/u1IvlrsYcDoaDIw2OeKIMfB/A0rJOojc89q2OBgjpTDjAGKHy4JH4UH3LkGpIix3qKikMR4PHcU6pKN0Zz606BcEHIqOUbdhP408Tx4DDAPSi+6Py25HY0wK/KRyKinCHHUelE8YzTYHGazu4BzWi6BqniGVodJtfPkTG5d6rjIP94jsDVxby20pgmXa69VznHFAjpTqVGCa68U54wDQKMm3PIrnAoOCdualIC7c9ajYEblPWgQGyB07U4Tbwc+1JKd2C2R6YpmDJtU596EiiDywnI6nPvSAM+AayCuScelEhwADwKcg9O1KEdCoOTXl4O4c0GJbkYqZiW4qORVYgnr7VKyFsk/SvkblDn1HpXyg80jM3anQsM0ZIwvDZx7U9wpwBU15GPlDZPYYqS5L8LzRZtoUURkdOaxnoKA4xisjPFAZHFMvGKwCOtYJHFdeKxngVxjFcHgGjzVmQsgBq3YFMKc11IIPSmYDjNKcjH5VnAxn8KkjDDfn6UACvWlXeetYx8oHSgdwz+lYz0H4URjoaY9gaJ7k0RzxR6daJAGM0CCeDRJxwaaRAOv4YoyblwDzQOSDmi2F4P6UJATtB/Ckc96MmSBn8K8xV6n8KDKeh+gosvrQYFeDTMpGAcmmcEY/SjMApANNJgdaMm5gQM47UZATkUrbsEUnJ4NAhgCB07VjPArHalwDzQuVCAhf1ppELhg3T2qSYSHapz7UM4oIW4AoQkHAplI4P4UBk8DHpSqScUIcDJOB61HHiPOa0m6mspN0Um09+M56/41dXct/KsyRbSmcndnt/9avLZYkkjbHXt1r4eWGi6jqKx6leeXI2dq+Wx/hbPT6CvEE+py3J0mI/6JF90YXuA3161nTbeFltbvdOuN6+Ww2+nJ46Vay3McpkMW4f7wHapNQeWNIZY9pXPOc0kltKfKjbOe+CKS/udEsLqxtbn5L3Z5ybBxsOV5OfXtUdwXg8qNOexz71d2/2q8F4W3MfvLjGOMCr2KEKkTt5ZfOBgnpipNJhs7FDFc+YxzxsK9/rWmwG5u/JuRtHYdex9Kvde0+2BsIm+YdeD9fSrnWdLW5OWzHH9/hhjI47VqMmnzzb7abcvb5WHYetf27DqthPZT3P+r27U2HucnkD2q/nKxZgmx/wH6V4V15rItHKm4yYzzjGM+3vWrPHcXRliXg9s+wq6YwnDdE/rRu1mQYHSpGZImWI5L4yPpVujp8r/hSy4DBvwolQMBv0p5d42nrSXMkSMEbCtjIx6Vq95HdFWji2Yzn5s+lEBVyBnNMCego8YAFDAOAeaOD9aZcjGc1hgNooD+FRTAE7RRXHBPWlPAwM+lDnnGcdKwAetAgHIoncc4oAfwnNKcY29aiLKwCjNSMC+AOamIUAA0v3C2elHBGDWPelYZxn6U2DgijzwOorHTPWgO1DgdPwpjkcCvYc0BkYNfKRjPT2or2P5UQB0HSuoyB9KJUcAZojpk0BngHituADjOKJDLg02Pr6VtNZwNuKIyMA1tDciqAzxmsEKDj6UwAHBrGBxTnup/CmB2AE9KQEnI6iiu4UkZHGentW3HyisZ6UFJNRXCxRbCM0rqT979Kkk3kbWz6DGKMjMNhXj60BuIycCoptg2ou4/XFRsWkLhP1p2WcbA36UyAoAw+goRlYwoG7HbpUqOZtqjk9B+FXS+UiqfvHNKfl60pC/eGc9KdmZsIvT3ptzPkj6GtxLYJz7V9nZgGC8duaKgDbu5PbFMMjI6VkL1PFNIoPWpT0IP0q5OV25qdCrbcdKHI60B055FZB6GqJHTNEnOM0Sc4FZ9qySOBW4kVxjGaAZgMUiYAPpQUsPpSw5GAOlLA7EKKht9vBpVRWBoEEcGiRjg0CCMg1gdqxkcUoHU0fmOBRGSMmto9aYDtRwRgUQCMZpQByKK5NHHc0cAcVjA45NHk4rI7CjtPemwRzROeAaOCMg03PANUcEYFVQGBjNZB6Ghk8E0SDjJriqx708SydRmryzA+ZBkVLCB0HSmKgYHWmAHBFMQAABRwRnFEfJQI6ChjoD+FEAnAFIdrBhWnTpLEqg8jt+dEHvRxgYrqMigRjAFZz0HSg7EYJpHw2Qf0rcXIIPShK4GN36VFckkKT06VCYFkBc59uaihtZXCwrtH1JqztL9RiF/l7DAqx8O2On2iXMkOdmdw3H1x6+9eK7+11Ge3uNMfY0e7sT1Cjv9DXim28U+MLUeINam87d/HtjXuF6Lj+6O1XICfcPWrG+msLyO8hPzpnHA7jH9a+Jvxo0Px38INM8HeVjU9P87L7n533KSdNgUfKvqajZ7Ygqenar67W7YMh5Xr+n+FOG6KeaLKeCK3BhhuaPHA6dqJycfpQYIuM/pQKuMr1HSid65PWhKVXYTTrjnNcOArfhQAVdrDimGAB27UjgdDyKBS4jwTzUkbRDnpSttTb27UAcZXpQlDrtfr2qaIAB1PPeg2Bg0rFehqJwOh59KkjLEsg4q2kSJiGPXvUsQZTLGcqe1Mp9foKWTfH5LHp0ooyjD/hUhMoyx5pgoOCeaV1KbSee1YwuMZ9KIIrRdeu9DmaazbBfGeB2B9QfWruZ7iQzOclv/1UAMAZ6UG+XaTmhkDaW/SsbuBS/K2R170xPUGiAGyTUmGXaOvalVRGAp/CiioAzjg9KZweFNYKybM5NMV2YBoOEi2j+Pt9KjITjNA71VBzjpUuEAU9aaRVyCaBUnKmlkQ4wanO7AHanCuoINSYXhaJDdTQ2t1rABxn9KknCMdh5p71zHtBp5ixwDUpYLgUQSOaIBXANMQVHrWQVx+VAY5otnpXHTrRGOM1gEdaJyMUSCMCjz3oHtmu2Aa5FYzwKtiEcZNWTqy5zSHjP5UTuGBSKR3oEAYqRSwwo6Uy4OD1FYIOM5pPu4/KlHGaJA4FE91onJwKoqDyDRGRjNZwOTRYDoakkwMKadyw2g80w3HINKGIAzwKO7bgnmgz5wTzUbMeKLfNtzRdcY3fpQl4+U0ZBjrTzJuPzfpQkQKeeT0rzN3Q5xTncCBTEAYJoLg8flSqHO01GoAA9KVFB4oDI/lRxjrQYY963Ajk0XBGCaMgY4A+lBQOlKAoyDzRGea68KaKkfLmkXcoIPSra2e5bZEMkdqUBl8tO1RjcuzOaRDIwKjr0rT9KuLwiGJNxPuB6+/tVnpTC0CyH507fU/WtD0CSa5+0SNmM/eGPYgd61GWR75wvTjA/AUluszMko+lSavcpALZnyo7YHrn0q7u1u73cOncfh/9avBXhbTvEmox2877VOezH+FvQj0rxjocFl4luNP0lf8ARRt8oZP9xSfvHPUnrX9mTQyxxSDyxzk8H+teE9P0zULy3sbv/VLu3/e54Yjoc9QK+LXgTw/o0WlXWky5WTzt52vxjYB95j6mtNtC0n2mI/KnUfXIrxbqFrfXVvNarjyt24ZPcKB1+lX90+ouWdssfb6f4Vp2nJdXH9nyNsE38WM4wCemau5hp7lTyR3/AM/WrW8DalG8Z+Vs5H0WpoVVSAOtaLLaQX8SXnzRc7hyOxx0564rxhNpovEOlx7Yud3LHsvrz1zWkQaLfWEsd6uWbHl8txgnPT6CvFvgtPDFrbyR3vnfa92F8vbt2lf9o/3qsbMytI7TbdmMDbnrWhQeDm0K91HUk/0638vyBmX+JyG6cdMda1JrV7otarhO3J9B61fW+jR6MHiT/TG75bsw/DpRKjrUgIjLEVc479aJBG30ohW4BzW3IwKJwuMUrcfyFMB2PNAAHKjjtShcEgUBk8GioDYBoKSBnk11GCax2AoA9CawSMZpCTwB9KICNg9ewpHBkDGp5GzgHr2qXnANMVZFUdaIGeKz0FYB5FdeKIGMAUMH6muvIFEjoTRAHINDHekJDqU69qmcyvuNMARQx1oYzgVxnAP4USDwTmgAeDSgHFMoJANMoxwKK4HvWMDGKqsDsfpXBP0peeBRHamYA0SWPy0xJBUdulRLuUgUFBGaUdhTEjiiNwC0qnHFMUIAFFQPumlBJBPNStGEyhpnD4JOcVEQT8rdOnFRyoYwA3PpiplWNCV60wG4HNMzquV/CiWVs5oqcFyc1EqHg0DGThhyOlRbSfmFSyI/yoeB0oRRhPl5NNKQCF6elOQRwfwo4C4NFc85ryl7U2N3P5VdlSMD8Kvoyrcfh+lUB2quBwKJBGAaHPAokZwKAHY0TjisZHIpY9wAI/CljA6CljLHAHNRxKBtIqCEbeV+nNJD3UVtUdBRRSMBcn0zQj25UjFFMjaBSrtULnOKCg9aCqQBmghIx+VBQAMHmsD1rAJwDRAHG7P4UxzwDzWMZz+FZBHFMwcYzzRwe/Ppim5OM0CScKelEjqD+lE7hgVwOd36UWPQ0SMdaPXr06cUSegNEY6Gj6CiOKOB1ojPQ0BkkA9OlAEdTn8KI4BBr2zXHp+FEZ6VxjAP4VIu9cAZPap7YoMsKuIwD8tEHo3WsgnJNcAYz+lHngt09qJxwetADAJNZBbIPP0ok9j0rRrgC4VC3rxj2NMFzgHIojjp9KG0DANBgeooOTyDSknnNAMTgdKtLS4u5BFBHub0yB/npTRsjFGXBHbNIDkEdqMRYh0bOO2KsZHd0EjYBz2rTbpI08vftPrjPrVv4yL24tbiLJHU7uvOewrxLY6TLpBuNOX9/wCmW/vAd+Omaj8Y3ttoC6Iz7UH8OAf493p/WrkFA0YHXGKIZT8w59K0xNOmlYajc+SBjHyM3r6fhV20IlIjkyB04I7VI6MoCjkdqY9iv60RGeCeaJGcAZotwAawGxsFfKVwxxW0L+FKykYz0obXwGb5fXFbwDgmtqtwD06cUZCPlzmt4HB5U1tyMIcj1qOQLgZzj2pZ4ZI9jc+3NTQmJsDlT0PSgVUjnjvWGyCD9OKjcPhXP6UVyMDr6V0bBFHHVT9KWUKuwNwevFPGByORVtIqMVkPyt1qa2Urvj5X19KIIOM80riaPYx57UrY5BplRjwaIBHHakIb5WP04pk2HBFAEUpXGGH0p1KjgZ9KJBXI6ih8wH8qJwflNMqhjx+FbQeTRLs+GHTtTqSCVHPaoNmOv04q7lYworj7uajjG0NnJ9KxmdZCeP8A61ElhgflRcAAMfoKd0L5HIoEodyn6VIxugOeRU0bhASOfWojxtJ5oIAMgVvjZdpPP0oyxqflb9KeWNmBDZ9sUJEJwTzW/DZI4PeoUMj7mGQKdUOWc8noKZo1GCOfSgM9qYg96ZAR0+lSRkDij6A1wwAzzQ64Br5ehP0ogdj+FEY7/hWAeM0QOxojIwDRIHfOK4IwDVDr1o4J4NZzjB5rS2Z1Izk+n50qsRgN+lAMo5b9KDEnaDmhkd6YhRnNOAxyT+lDPTd+lKzDAUZx2okhsAZoEdB1oEYwaJx2oHIo4x1onsKLjjnpUkoJIBrLkDJoI7MFUZpLaRwAy/rV94c1HTbWC6vLTy4rjd5beYrbtpAPAJI5IpoVbkCnhwQFHNJDjJYU0TKB8uPxpohj5aMbfdUZz2oxuRyv60Y2HVf1oR7zkdBShYxgN+lM6l8K2R9MVIqHgHPtikUHg8+lRwgfdH4UkakZJ6UygrwaA4xQXdwKKYPH5U6AgFTzWAygjmljUdqOCMCskDGaVSRn8qKgoNp578VjnaeagGXVT75q0kntJg8R2k/Q9qigdEy1K21doOT2q0immZI4Bludo4FaHNqGhTh7yHbIfuLuBzwc8jPqKnsbi3aK4ki3CXdn5gMY4pZbeOc2ltL5by99pPQZ71q9gNEtUXU5Nmoc7o8ZxyMcjI+6RUgeQiRn+U/dOOtWwuiSrpu9OQK0/wALa1rFrJLY6d5skWN/75VxknHUj0NaZdizfyZV2Sr75x1/pWhaVaXOiS61JcbTBt2JsJzlip5z/SvEl3Yapo7Ns8px0OS38Q/wrw5cW0Wohr+Xy7Q9X2lv4T2HPXFanqdle3Bszd77f+A+WR2BPbPUUl3/AGbMUeTEfc49vx9a1aeCSVJ7Y7o+d3UdgO9GLS0szJHNmQf7LetLeWM1nKjSfv8AjYuDzzz7dK1y1mkj8wR8fUe1aFYzT6hFagfvXzsT1wpJ5+lX8M8UvlgZx9PQVqkTWbI4bDHOOPpSzrdW4WQZ/wD11bLvtgYTlR0/Ori4leZFeTO3OOMY4pbOW/u/3Yyvr+H/ANatVbyJjEr52e2OoFXOjahbBHmt9obODuB6fQ1dKY3KMOR1qRWIAWpruL7K9r5WWbGG3dMHPSpZA5AJ5obQeefaiFU8ChwOfyo4PAolQuAf0oAHqaJ7Z6UoA4zj8KJJOQaG1uM5/CkOVxjpTDkY5pFXPzHHoMUQw6jrRI6DrQ3gA4+hoDcc1Z6bcXBZ4otypjJ3AdfxqZlZ8r26CgS5JAzTfeBz0onPIojgH8qAA4qTytgKNk9xjpXU8GsAdKweAPwoAHgGiuOBRw3GfwpWKAOpwR7VIwYe9bcc5/SgSTxRwD159MVkYwDWNwGDzW3HQ0pNYBI+b9KJzxmiM8A0AcYB5HtR9BR7Cj04rGeBQOAB+lMcjgVIScYHFRqO9MhzkGok+X5W/SiABgVFGZDsVcnsM0RjjP0oDPehjOKx3H5UoyMYzTOQcE0ctwKOegpGwMg186YfselGYOu0mivmRhkOSOtRtlArfjTMrvlvwp0CxhgevWoxEB8x/CgihCWH0pDg4dsfhQ2qm0LluxzQZlxjrT7cerUwU8k03XBFEjGRTZI6VIcDJHSp23c9fap1V4txHSmOTwK9s/Sie2KBBxjvTcjAo4HQVkcGs47U3WlBPINKnAANJEBzVugEnIoKRJmolyvI+lHAGM1gHjNEHIIPSuvHWtue9FARkflWB0Ir5SMA0Mk4zWOcg0SB3oAdqJBHBoMcYzRJIxmjgc56UOOlHpxQ4GAabgfKfwxXXoaHFFRjIP6U3BxQ+anODgUWBGKGDU7uoxGcGvMcjr+lOz44f8MUEAHyt+lEHGA3P0pCQQSc0CrDANUCCBQGeM1jsOtFewq8iV4ye4qdwgwOtF1ZcHrRXPQ0FogHoawAOtYz0oA5okdutWr+XIGBqNg6BlPWmAIxRHaqBIGKVSSFPWkiBOCf0q3m+yyCRD8wo3aFixGTUb+Ym5FyR1GcVbz72CN1p4Ftp9mcjt27VaWmjz6KHEOXHX5m/vU0lzFcMA+DxxgelfAbw14d8aWt1b6+u4ps28uO8hP3SP7orwX4E8F6xa6hp2txfvbXyvL5l/iLE/dYdgK8V+Cbezvr0acv7m38vaMn+ILnq2euavFwSE6imllVNoP0ogjqeaZSDkUSMYNDCNyPpTbW6Uy7iT6UM5wD9KGFGCeBRLEcHiiBj3onjA7UuDyabDAleooqQcEdaKgHGfpQyDkH9KYbhuFAsBjNQSKybG79Knt2iPXIPSky3INOCrgAUyBf8KYZ7cU2R1plL/MO1QkZ2k1NEyNtzn04q2kKSBWORVxDhzjoOlHcGwDTOtxGsg+93pSUcMpqQBW+XtR55rlhjPTpQViDUKhjhhUikcE9OlA5OM/Si2xdpHNJhuTzinlVRk9TSsGTiiAeTQxt4/CoUKjk1dAskak+tJtaPGeac7UX15xSqAvFNHvQMDxQRQeaDhk3LW0k71bB7jFTYYcUse7gH6Uis4+9wPaiFYbfWpnAbAOaMjSNsU1FCccHJpIhs68ii4SMgDmrkMFAQ9KC5wTUjZYqK5PGaxngU6KVyw6UyFflrGRwaHtRHqaPHWsg9DRGRgGsDGBRAIxTAHgGtoHAoe/NUR2oYABB5rTnOcKf880BleDTKScA0GXOB1FAEjg/pXBHBqZMgYNFCFDKetRorLk8+lAY6GkYkkGqoAgYJyaL4fYKkcEcGiSehoqX6Goomc8mlVVGAKAIXIP6Vc6hcXUUcMsmVjztGAMZ60eRjNHGMn8K4pgCQM0VAXivLQjAFBFPGPwpjHHlTTXIb5FPAosCPftSrntyKSEMQQOaCAAAfhQRQMAUAOlYBGBW3A60Iojb7o+GH+Nbw2WHes5GAaUArhTQz3oqgjDZ5peBkDmvhj4PtfGmsPp1y20LjHBP8LnsR/dr4geHIfCvi290GA7o7fZtOCPvRqx7n+9600YVsCvDl7Fp2qwXkoyIt36qR/WvFOs2+tXiTwLgDPc+ij0HpSksMZ6dKELSRFwOlQTOrKVPPOK8D6zPoniGy1e5bfHb+ZuGAOqMvYH19K1C8mS8a4Zvv9sDsAKnuJWvVlRuedvHtRluL2P/AEl9z/QD+X0rQrMT74JVyBjaM/WrDUDpT+bEmD9f/wBfrV/4iu7vUJLu3faz4zwD0UDuPatC06DXLkKq/Mfc+h9x6VJqN7pjvo8x2NHjcuAevzDpn19ajuzI22RuPTFXsgiDRp93sKkvjEMSHOeleILd4bSGSBuRuzx7ijeG4gGzr3/OrlsW/lg4P096t5nlm+yE9fu/lmoZf7RsBGRkjt+P/wBalJgkjnDfMM84/CrwtclJIRwM5rxDaCS2tpAmc78c+4pcp8gGcVbXCxWRRTye340VZ5ckfT8qS2mSbYDj8vSrjTJJopp2f/VbcjHrx61qnjAX2mQQkfvl3bj/AMCGO2Ogq8vGulVXGdnSiy79gq4YBNoHNSKueKVkU9eT0p2BXJHNEdwKIVe30pgOVB/CiwKY6mioHOaYAMNorGRg0BkYzRVVG1R0onBGDR56D8KIDNjP0oKAcHtShm+9UiBEGKS4ntLTahwG68DsalbjGa+7wKJ/Gh0wKABGAKKhulIoJxTBQ3y0QDyKJwuKPI4NEZHAzWwDp+VOVIAo80DnGDR5OQeaxnpQJPANAE4ANBccVjAwTzStjp27UOevWiox61jIBBorxwaIwcCiMjg0uFGTR+bmpCduBXJ4z06cUEJGForkYzUYAXANIBRYg5Q0RnqfwrBJwKGegNMo4ANTgR4CGid3U0CB2okE5onP9KDkrtB6dKJI6UrEDaDUM4QEGhnAK0SxBB5xRIKg9xRkbacHr1otghqkKs+9OnanUsOK6DpTDcvWh7URiiMjGadAEOKeJWGMc1eBUUxjvTpto9QAaGD0NDnGfwrv/KjyAa3e1ZGAB+VHB6UFXHAqNAe1AZYBTmoETOQcn0pYVHU5oAEYBoru4BoDsTRUnjNbUHOaJAxznFEY6H8KIyM5ogHjNA9s9Kz2zTEY681jjrRI6ZrIPANbjjANArt65NEAcZ47UxHY80y84WmBBwT0ouh6Nn8KMoGRigZG5B4rO/gNjHUYptxGFbp7UQoH7w9egxSgEcJkfWpHXOAeB0osMcGmYkkDtRkAHByacMnJX9a3sTuzwO9Fyx4OKAwgzJ83pii5YbQ/6V5rAY3fpQJI5oDPQ0uCBk06hsqKvtMym5Gz7Y+nvTwOpxn6UyHOM9KOeMmiCV5rGeBWD2pulAZXAFAHOcc1pF+dy27njt+tEAjgZzRUEZzRCk8t+lYB4B+lA9DQVmIwfwpEB7ZNAqvBGfao7ueAEW77A33uAc4oO2/fu5+lW13M0iqi5Iz3Ar4ceKvCGna1BL4ttN1qm7ePMkGcq+PuDPUrXxmt/C03xM1aXwTPnSn8n7P8snaCPd/rPm+9u6/yq1k1GzQiC4x7bFq11m3uZne8iwRj5txOePQD6V8GPCug/EiU6DD4s/swzfeH2B5ugkYd1/uevevFmlTeH9Qe0mfzGTHOAOyn1PrTThhuZd3pzjFSxrOgkQ9afeh2suPxpUaQ4Q5I7UysvGOfSicjGKxgYH5UF3HAPPpXlhhw2D6YphggA5onAwTz60yfKCDn3pipAANAKOopyx++fpRYbQrD8aZSOAaGScqefSgQ52scH1psoMEVFcZIEj/KPapbWWPJUZA71G4b5GpleJtjjBqICQFCPvdDRCo211yPrTrtwQM1jn5aQ7l2uOvT2qSB0bGeOxqGdVPlztwOhx0/Kri3jYBlPHc46UUeA4ZfwzWCuHAyveuUPA+lS5Kb1XjvzQBzuU1IXFvuY5X0x71HMqR4KZJ96lVSM9xUrlB1z7Vw43KPwqMAptqcMJSS3Hpj2q3OUwKjy6jnmiuGKgVGY0Yb+RWqbH8vyzwM/wBKgcK+GX9avCsTKwXI5wM0siONqmkIQCIN168UVw5AHFMGCAoOKmkLAKg5rzXiUB2z+FRyp95m+gxUzSBdyNx9KcyyHJXP406N90n9Ks7cMjtjJGMUkRRs/dH500sSHaxyaldDwg+lHcck/lW5SuB1p0jfGODUkAP3TRhI4XinUjAapl4xn9KIyAAelYz0o8jA7VjPAogdMUw7A0VA5o4wMUfYfhWOM1ggcmiD0zWOMZoqcitJRgc/570CFbhsgd8UCCeTTKdxwM47UvyjGc0rKwwG6e1MFIwppwAcdfSgAUAHegpXA2/rROcleM0oIHJzXB4BonI45p2AbCnP4UwY9fyqK3ckFuBSRog4bP4V1HBrAPGabDHjimABwDn8KyCMA0QTyD+FEg9KUY6nOOlHnhTipJ40XO/ntxUl0rptPB7Gicjg59KVCOQaEQIGTzSRgDIP4UI+MA/hTFQuM/jitwIwW/SgARlTn8KQ7T60rRiMhuXPT2ppMjAb9KUgjINEg/xdPahjGAcnsKAB7UcEcflWflwppHKYwOfSri6kuggkfO3O0Yxj1pz3HNaXbQXtwIpp/KTu20tjg9vwpoliuAEbcD0OMdq0w+aTZx2nnyy/d/ebcYyT+lOLyyiaxmh2CTqNwPTnt9agiVzkDgdqsne2RHZcdcDPWpLDUrm5S5uZ95kzgbFHQY7VpHw4S+0e61yXU/s0lrs2R+Rv3bmKnndxwPTvW1XnIeTcw9sdq0/UZLe7jJbCR5zx0yDUt2t7yi5Ppn/PpUEcT3Yt3Ozb1HJxxmpW/wCEbvY7qM+aFz8v3e2OvPrXiLVZPFOoya7MnlyTY3pnOMKFHOB/d9Kuoy0Q8s81cXp2qkkeQc4bP9K1h3nEbqcBc4GPpVlfxSw7LmXaE7bc/wAqhuNPmVmVMAYwMmtftAkp2jB7j8qikt0gFs7fvG+62Dxg56Vp14un3DK8e4H3x2P+NanZxwTqitw+ccegFW1wLdNjJt/HNafe2MunlLuHcsf3fmI6k56fhWpLpcd2GtJN27+HDDsO5rUbKWwl8tkyfr7D/GrvUBPEsK2mxxn5vMz+laXNc6lqSre3OQc/NsA/hPYfQV44l8LWmnzW2nS77s7dvyyDPKk9eOma8PaFc+K7tdPspcTc8bR6E9yB0WvFPhy98KavPouojE9vt3dP4lDdiR0Yd6lJD7C2fTjFSZzsLc+mKR1CbXry2YZUZx3pVDLw34YqSEqCqv8ApW3bwecdqHLYDfpQIbjd+lODnGc49qIOcflWABj9K3LjGOvelOR1okHAB570QMdefSgOwPSgScAH9KAJ4HUUhBXMgzj8Kvbt5sRScBeg/KpMnisEcEfQUSCev4UFyRk/pRGcYPStysQM0TgEZr6Hp7UQpAGazk4J4oBScZoAdM05YKFB/Csbvl70QuMA/pRHYHP4USRjNbSelAHNE5HHah0wD+GK4PIrOeMdK5Y5JoDJwDn0ogEYAzRBxgigARnHSiQRhTk0wOKYnjFSPu7fhSIAAOhNKueAKdMHApVJGAKQHbjFMoPQc0eeKwMYzRGeg/GoQoVmJ5GMD1qcnOCKLfLwv60GJ42/rQ55xTAjGBQx60pDEAGiACcUnJ4HNM7AgU0m4ADtSsQ2CKkVWbcgxSsfu4oqQuFP4YrDqu4jig4J+Zc/jUjAnKjArp2o/N0FZGMYqpFIOVqeJXGQORU1vnoM47U67TtIwaIyMCsDtRxngURk8V8p4Bo7e35VkelIuRwaQA4GaijBPNRxIACKIAPBocUwz0oAE4xQBBHFEYHBo8jr0onHU0cZArqODQ56H6VnPAo4yBRI6A0eev4imYdj9KWTI5HSvNXpQkUDg9elHO3jrWD2P6U+7hQaZVUZoyDGBRlxwtF8HOaaUA9f0rzQVBY00/GFrc0gytO+Bgmt7j7orewbBFM754P0ouxXbikZjwaUs2MHp1oOelADHFJIXOCaHFAgDjnFADpnmioIwKubVcZxV1AVBIqjzjNAjPNZOMZrr1NCicHionMbhwa0y+EybGPI/wDr0EBXGaZRj3oKOhrOBil+YYFJuPyjr6VtUHB5p0UHANLBuTJ6+leG9ck0K9W68vftzxux2I9D61d3kuq6g10o2+ZjjrjAx7elfCj4fWXj4XEup3vlGLbt/dls53+jL/cFWeg+BP8AhHdXthcfv7TycDZLzufPrjpWgNYatMEX5XHXqex+npXiLw9qUOgK2kR/PHnLZXu47MfrWrrJNKTMcsfb2HpRQK21ugosVPyn8Ka4YEFhXg7wXpviKwa6d/nGMcN6sP7w9K12ybTb+TT5my8WMnHqAff1pgDwp4HalXcOKYY4BotuUA0xyNuaKgrwayw79KyDwKJyOKOe9EE9DW4ryKKq2MGm60CTwTTDH3TUVyyjaec1NbjG9D16Uj5wjdaaJo2A7HoafDJ5idfSnGFVwM7utMuBlelZOOD0qNt3yMaki2HY3ao5mhODyPSrhBJgKKMLQkEnK/lUsOH+Xp2FKojhZ27YwKSAlPMJzTzeYBGB0oAt8oP0p0KMqk805G4Lmhwv0oDJxUihxnNQy+T8wHIqzkjKFiM+lTcygD+KvPBkEfr0qaMRbAzffz26YqVRvwp5ousjGE9RSIIxgGpFZ5Nyj6UoDJgdaVcLtz9KkKnCg8jrTiUNjHSnD/ePbtRLAcGmuJEXYB1oxlzjd+lW58p9rn5fpTW4IyrfpRUl9rdaZNuAxp3whVaQkAgUq55DfTigdp4NGbIwTTMm3rmnAb7pqRdpxmh7VX0rryarHvRXcMCqquo+lUMluTWnEpxj/PNZGBjtS8nINMc9KLcYzSqAowaJyMg80yhjg0FKjH5UcHj8qYADFMoCZU1nI4oDAzmiQRkmmG5cCiD1odODQIIyKIzxXBHBo881RweDWR0zUkyxnaTzUs5dgVGMUPnOfWvKLDg0sBPU0IlUgZyKCgjBHSiQWCA0QQMAZrbnOelbc8ZoJkZFYBGBWQMYoru4U0TtAyaDAnApTuHNYyOKxkYFAErgfhSKCeD+lBDvwTkjpWCBtzn0q1cQsGHeti7t3etLuXtLyO6iGWTOB06gir66ku51kk+8c/yH+FaHpcMka6gGzjouD7j1qRbW/lhaZdoG7IyT29vpUL/6UMngdOPavAttpWp3tpp+qXPl28u/zPkY9AxH3eeoFfEvwdpXh121DTpN0bYwNrD+4O7H1NWMIluVlXqO34Vd2I0tFkcZZs/57+tXs7tdCRByen5VNcTSKC/OKt2URbWPJ7UHG44PI6Cp7qG5sxcMv05PripJhPGxA6dKn822dlU9e1eGnWVXvJThbfGV9c5Fa4YZ4zMDz9D7U+x4gEGSOlXCLON6nmtFntZbyOK6H3s4PPYH0/CvFd5E7hrYYUfX/Z9a0fU97NE56Yx+vtWpSsbraBx/9YVoFnb6x4TvZZZMS2Xl9jzukP07Cr8+Q+4HI/8A1UuoSLyp+lPI9zMBI2c+3tXh+2uZ7g/Y32svU4B7H1rxEs41GVLiTfIMbjgD+EelO5R9p6iikcicnmpECnAOaRij5U1ba2bbTzZvH97vn3z6VOWJLAcdq37h70QAuM8j2oYXkjmhEXTfu/SjnAAP0FMSR1zQ44FDrRXJGK6GuCcA80QMcCoULkADpUsZkIVTVyAZ2Uj/ADimPzYFA/LjvQAY4z+FdBgc4okHoelMcdDTE9jWcjaBRGQFBoKcA+nalwBg/lRGeFNE5xzR5GM0BkfyonjIog5waqhzzn8K6jFDpgUctwaAyOv4U2MbgaIAGM80QdvWjlePyrGDjOaODwDUjZ5FRoSelKm45B5oxkLgUqZXOeaVBggGmABABqXb2PIrGBxyaHHegR25rzCrAipXMhDVRAPGaGc8H6UGDDA7VgEZzStjFK2Md6LHOVHPejnHH4U2ScCsluM0zZPFEknIqNgSARU0e3oc0cY60xB4FADHNHpj9KOD0oDPGawMYFPEr1cwKOnOKngVm+lSLtfArgjg02MYzRx0zR4wBRAIxQGBjNKowCDSR5wQeRUULEA56e1BCO+cVnJxigo7msgdBQAzkisg8E0SDxRNEgniic965xWOORmsHpXXimlYHG3n60z7hgDGetKvcCiq7ioX9elLGp75oHB6UZTjkUZSO/4YrcW+Y9KL+tAFiBu4PtW4qdu39adiWxmgqlcg/hRIOBjpW5TxigSB8oohj83p0oNu6jHtWQRjFB03EfkKIzzjp0oHaelKWCjB/Chkn5hmgCVG00FJ6tz9KUcAZz+FA9s0OQBnmiMHipI94x0q5slaM45q4gZDRGe9FCRkURjHNAZ4Bor2BrbnrRXPere5kt3DLzVpMLiIOoxntTgkbQaxn+L9KC5+UGtm3oeahKmQBuBUgRXyhz+GKlZdwkRcE9s008gGd2T9Kd3fG58/hUQlYghse+M1Zarf2ke21udgPbYD/MUZJpZfOZ/m78e2Ktkd38yNsY6cZrwL8e/F3w5RIrdPtNumf3OY0z97v5bHq+a1q+fU7lr3ydgboN2egA/pU8DyR7mb5h90YplYc7efrRCt1PIrTdZ1LR2Mmm3Xksep2K3r6g+pq9ubnUJTd3U3mTP95toGcYHQcdBWRmiSRkH9KJ7k9KAUjAP0piGwMUR3BraGGepoqccc1uCjkc+tDnqeBRwDlaA3HBOKZcNjPIoBSeTj8KZCvGc0xKDAGc/hQUFQRUc7qNpPHpipIRON+cn1xikUxxsJTvQYx2p0kjO6M5WoiZVYhfu/w5/rTRhl+U8eleSCVy3B9qkQxEFWz+FF/NiLM+SOgxiiWKEHj0q2KJHhpM+gx0qGUsSJD9KlmSNQxOSOgo7buIgnbnp3x/nFRElfKXtQY+aynquOKDFuSakf7rE7iOg6VtDEOPypgVHI+lQMhYbhn2pWUfIecVMSFyh5FJdNEM569qL8LMTwuamiDPlGzj2xSyySKY5m4X7vHT16UwQnBfJ9cUIFKeYp570Qrx9cn6UkYHAPNeST90VKqsMKuCO+a0jR7XU7xYbm9+zg5+fyi/Y9gfb9a8YaVpmmak1lpV/9tjT/AJa+U0WflU/dJPqR+FSwDdnOa+zLjPQ0bcNgE5A709mwbhv0o2B25LfpRllh4U1G8bnLrz65qS2GNytux7YqQBxtJwRQJQ7c5oTDGMU02RgCjID/AA/rQdV/h/Wp5O6p+tY3ck1wRitgJ4P6UR2zXUVj3r2zRGeho88A0ADgA/hiiD0B/DFHgVgHjNYB4BqyYAfKc0hDDAP6UFDD5T+GKySMg0FBPLfpSzZUKTk+tBlVSSc+2KQA87s/hQUnkdqIyMHrRAxweaIBHBocdD0ogOuM0qAcA0cYwpx74okngHNBAK6imIHesgjNHsDzTyovAOakuDtyo5HamlIGd36VkPyW/ShHu+6/6VHAuNwPNCPBy1eWCcbsenFZIO3OaUDbwMGgo3biaoA9COtBd3f9KeIIqlZMls8Y6VIxHAGa3luMfhQDE9elMjEgA1swM5oYIwaCZHNBRjFBDjINKrZxn8KUAMCetBS3CmoF3tjP4UoDECtM0KzktbOdr/Ek2/enlH5MEgc55zWv6VbaVLD5d39o8zd/yzKbcY9z61pM6wzLKs20R5wu3PUGrLxNbzOfttj5hP3W80rj14A+lWS3k5L29r5ip1beFxn2NRW017FvLY9OOn+cVqcD3VosU1znGcLs9x3r4f8Aw6g8V6Zq2py679h/sjycJ9lMvmeazDruGMbffr2rxBHcQxfZ5JvNC9G2hfTtVrpOnvpn2v8AtDE69Y/JPPzY65x0rwFaaHqt79j1u98hT0by3bsx6Lj0FXenWxuXMEmQuMHB54+tXcYwUBw3Y9akna1DRsPMjOMDpUcyo+Ubg/w4qe2nnRpIPn24yOBRvjDkq2cfeXH9akS5SD7RvzEegwPXH160JVVBMhyF6rjHtWjWcWol2ebYeONufX/CnunRQFfp7VLJ50eJZsgf7NBI4o47iM/Mc5H6Vco0tt9qY/d7fjireO4vZBFbr8z9FyOcf/qrxNob6fbQGaXl93y7emCPQ+9FGVsjoelABcEirHVbuyRjbNsMmM8A9PqPepZ3nkLyNlm6nGKuFzIcCkUsSuaZSq8HPvWCcYHNTHDAA1JI8hwxzjp2ogHBxSjBxnpTYxkUN2MKKIY8EUW7DrQJbj1raScZ6UigkKW/Ss5O0dKIx0NBS7bQc+nFKvl4RTn3q3BlkKs+APagQ+5mGafaVJB+nFYyMAc0oAPXp0rKt/F+lEHcBmuvAoj5QSM1k4wDWATwKJDcYrBz8vNE560RzgCgMY5/Cm6cUTkYokjk9qBJ6CgST939aJ6HFBs9RRbPSgcnrWeMZoEHgjNZHXH0oYK4FEkjntTMCApHNSOFGM/hSqZep/SkiBXCmlVRxigwPBP6UDgYHPtSNk9Kkb0rd7VwRwaVdwx+VMMniuD+FHDDBP4UxAwF7USQMntQIIAAwfSnz0U5pW3cCsccml9KABGM0eVAJz6cURzwaDZGBRGeAaweo/KoipcE9qlljZgNmfxputYIODzRUE4BogHgGtoPAP4USp71gHgDNMQCADTruOdv61cQocsp/DFXFtuG4N+lMu04BpunFHPask9DROO+aIU9D+lImTioIAcE0FAGBQHGQaZecZoHsDWSTwKwAOTRXFHJ4FNnHBrGRxR7YokY5PSpGKruU9KMrHgClUsRk9adRkAUzKTlaJJHJppARgGi2F4PNM5AGD0oln5PWixDZFOcnGaKlu9ZxhQfpWwscKaBY5UimCgfKc/hRBIOelbVK5B57U6YGFNEZGCeaYKGKg8UGCkGgUbqaJxyOaVw3A5pVJG0U2Qf5UgOMn8K5zwaWQkgEVWAwAAogDjNNk9a7YBq8tRjkZPpU8Xlylc/SlOe9DGcVgdj+lEDsc1jAyDWM/MKyD0NaXftbuFLcHqMfWlJdBimTjCmhuAwBW4HlqQxu4UmtOjtPtsbTP8AuxnPB9DXxGvvDupS2MHhq485bfzPM+R1xuCY+/8AQ1LprxwGVjwO/wCP1qezCWqTqfvZ/Q4rSLaxuCq3TbSfYn19PwpFfywwPSknkMW4HOKso5rkFrYbgvXoP50l28U20jleo/CtU8UR6rpa2Mse2QZyM5/iB9Pap4QcAN+lWSRzTrDKfl55/CvFOlWukXEUdm25XzzgjoF9SfWm5OVo8cA/pTE8URnlTRXFEEDgUVBGQee1EEjg0CCMA/hX3gATzWMHaRxWCOAaGc08TR4DCiOxor8uBSnPFOgzwaIHQGgEddoPzdhQLodrH8K3pLHsB59MUDLGuVGVpRHIcofoKikUnDcelNGrrtPaigVtrHio4dsLtng4xUaqIGz2x/OhIF+VTmrUZJYnp0FX7q5Cjt/9ard0jTLHH4U22I7geO1SogbeD9/+lK2w8fhUyh4Ek7nOfzpl2LkihJmDy2HTp+dQoWO0dRUsRSMS9j0qCQSttY9Ke3DqEbqKgfyyYXPB600exAKJUSgMeDTW5lO7PFLkLgHOe2KVSRxSqrqMdajkMabQvP1ogsmAeasfC+r3mmzahbRfuYdu47l4y2B1Oeop4HT94ejdDTKQQF+96V4M8MR680sVwcE48sY/3s9CPSvEXhnUfDU8dvqCYaXO3kdgM9CfUVYvFHco05+QZz+XtWvXeh3b389tPuP7v7N8rjPQN1+nepmhlHynJ+holB8y9qZmzjP6U6knituTTKQcA/SiMjANEdqbkUwBGCakIBwKoDPegAeM0eOKoYxgGq4PFVjjGaOD0qgM8ZqyIzgnpUJBTAPSiNwyDRChdpPPpiiRQAK4zWQR1pCpyFNZwODR55oAE4pkyODW3PU59K2jGQeRSqB1NEDsaAwcg0ee9fLjGacgLkmmmUr8pppWVgFpizHOKRWwQD1ryeOTSxErwM4oL6UFwo5rBJonbxmj93A61kEAk8+lZGdueaC+p/CoLdpsgdutS7A20GmYliMdOlNlv6U4CgAGl55Nbcnr0ojJABrwP4Th8S/bVm+9b+Xt6/xbs9x/drUbP7DeSWv9zH6gH+tFc8Z5oKWAQfhTRleCeawcjaKUqTkmrZBICF/h6VbwRzTrEp5Of5Ve6c/hG8YyD9zJjYfoOe5PVqvLk3c7OjZz7Y7f/Wqa1liIV1xnpzWkoVkUyn1x+taTbzWNxLYgcvjI47An+tPoNpp+gSSyP+84wMH+/wDX3q71KKaQwocgf59K0KW4ty628mFuMbhtHO3PrXj3wxrmgJHLqNvsjfODvQ9NvoT6irZ90ZmB+YdRXiFdBfSTNp0+bgdtrj+IDvx0zRvWSISK3H0pXecmaPkflV3JEzOin0xxVxp0eneTJdHCy7sd+n0PvWq3strObeM/KP8AAGmUrOADnd/hVvK0kQhU5QfhVwr28gK9KBmt5Sr8H8D2qaNpHKDr2rw/4cuPEzNZ20W5kx/EB1ye5H92r7Sp9Mu3s7pdskeMjIOMgHsT2NI7NH5IGRS2tvC1tIj4f59/B49K8XwWsujxyRybpVzgYI/iX+lXUTIdrDnvRX5gDU8aqdob9KcbpEwfWriAqcg8+lRAElCetYI+Wo1G8ZPSpYyw3elBACOelFc8A/SiRzg80i5jLAdOlAjAyaC7lyp/SjxxSNtOAK7c1GhdiAelbccmo0Mh2ihaNGm5evpVvps7WTTqmdmNxyO5x61JPtj2qfmpmbhVqUgkYNA+/NLkkD1pwpHBonOMHmvbNZAGAaGccUDuHHaiQfu0CVXiiMjOawD900cY60cgYosCK3EmgwK4z0rgjg1uHAB+lYBHWvcGgRnANE45NAgDg/hRdRwT0rzFIwG/Ss55BokDvTrvOCKiQA8UxCx7VogdAelYAxk0VA5BoKGXjr2oIXHJzTAgYJognABpgegNbDuwO1E+n4UwAGAfpRGcetH0piB0NKAeQaA4yKC5HHalQkAA067TjNEHoDzSR71Jz92nTHANKpI60OflzQxtyDRCyLjPNMgUnNRLvbaKZVU4FEAnANMM4GaZRxtNU4PY0eD8x+lBNxNXUDHgCruEqdw7UVz0NEnOBR7GtpPFFT0FRW6vwDio4digBs7enFEYGAeaVugJrGenWioAwDW0DpRwTgmmB2/KaZjjFYJHFM5Bxt/WpSSMDpRkOcg4rzdw2sM57dKLAYAFGZs5zyKMpbgmnkK9Dmgxbo36UGXGc5xRkJOBWSx3Z5pmK4Kmt5BwG/SmDg5Jzn8KRiQNr8fSjIVYYGTQlfdwcUZDghufSlVS3J/SjIdpQHgdsVuGODzWC3Jb9KZgcbj+lEkjhs49qcKQAh57UrEqSTQIP3m/SgApwppC+Rls57Yp1xgk5/ClYNxjp0oAkYNISxDHv3rcS2BzQPQA9KYZHWmBPU5rJ3bcfjUzK/JXke9apZ5i8+NPrz9BR4HNdelYB4J5rb2FEnpmg74KhuvXijkcZrO37rdO2K0fV5JCsEgznPP5+1GMlcqfwxRJBxSoXPAoIN4G79KMhB2qas7xrZ8A5I6cVea7G+lvYTadvkbGJvOxjDZ+7j8KMkrJ5Ybj0xQC/wAY/CoWZxtXiom2KQ74x04zXhfxLcaHHKLdd5m255AxjPqD61oWiDxn4gSzF79nuLzOG8rfjahPTIHRa8e+DbvwRrsmh3k+6WLGTsA6ordmPZh3qYCRAhfJPfGKkh8ptuentUzFgAx6dBQJHQ0RkZLZ/CuvBFFVPAPTtiskDgZNIx28Nj8M0QAuT2oqwGQazzWCwyp/ClYg9ckdKHXg9Paio3bWNXF1PcENPJuI6cAfyrcp5BrIAyDnHtTbTz6d6LEcbs+nFMFxhDn8KB8twRzUk4nOX608ZRQ8bbvUYxilunA2KeBUKcloTtI6rjNRxxqdxXafrmgROPlG0enWpY4wgjVuT7V5rvEULdO2KjmCgxn8qfAPyHpTLhA6N9RipnMj7t2T9KYgjbnp0pZC6CJm+UdOKnURork8HNMiyLlT9KDsyFDyB+FPLMIip6elW05cCNj9KhiRZAyyYb/dq4llA8thuQdB0xXklMSQHPtUReeMS4571IgdyW4PrUCpIBG77vfGKe0UNiM7/wBKSwldMEfQVHbFmCGP9aHhy/hhEvk/Ieh3D/Gr3Qr3T4IZruHy0n3eU24HdgjPAPHUdaWHzE3E5x+FKkIOA2zHUYJrTtW8YR6XLomj3ubS62+bH5cfO1iw5YZ656GtI0a98SSJpanywmdvRsdSe4/u18QvhPH8PbWzvZda+0yz78R/ZtmNpUdd7dnq3vrqyu0vLSbbLHna20HGRg8H2Navruo6zci41Sfz5O3yqvYD+EDsBVxPbzHdbWnkD08wt/Opxk4zkdjipE9Wz7YpUBIUn8KaNQeDTwDGQ2fwpo1A+U4qRI+pbJ9MURGOFPNGJWH3vpxTrtOM5plyOv6U+RyDQywxms44FEYOAaB3Dlv0rJ9a69DRwBkGsADANA8Yz+ldsA0AD0PP0o4I4PNAgDr+FWQzzmrcoy5HBoqMfKenbFHB5LfpW4jgHNEA8D8KJwME1GxHIP4UpDLkVjPSgecDtW9vWjyOv4YrdxgCg27GF/WlAJAzUwSOUqjbgO+MVvJHyrn8aBUfKDnFSHAJxwKwznOM0sbdCce1CFG6rk/WjEoHyjmjGBxmgMDC/lXUbSKG4/KF/WiSBt60wIxgUGy20L+tFcjg59qVFABxyKUADkZ9qe5YLsQfhRdmJG39aLbiMDntQznDL096Khzz+VCMAfKKChuR+VBQBnFR46d+1NHlsk0wHDD8qCKwUL1qONXOHOSOlSRbGGW+nFCFg20HFafdfYS0bQ+YHxg7sYxn/GtLtZL69VIZPLbnBxnsf8KlW/vNRifXrjejZ3tsA6Lxwv0FatBYRyqmkJtHO5sk56dm/GtV1afWCsc8eJFzlsg56dgB6VZRSnCM2ce3StF8O6PdeGIPEx8TbL+bdvtfsTHGHKD5s4+7z09qv9LN1o41C61HDt0Tyc5+YDqDV/4bey09NVhuNyNnHyY6EL6+/pWlXz20f2yFtxh6DGNuciviB8Z/FnxAsIdN1KT9xDuwMR9yp7Ip6oK+0EzeTIcbuh+grVd9jakRNn2xjuP8alnRovs7L8rdOemOajuJILYx2cmG7HGe/vU6vJGJom2k/wAOM+1T6hqWpwJbXEu8R52/Koxk57fSvEEw1q4NwI/KcdRnd2A9vSrOCNbpftPKHO4fh7VZw2d08n2VvLEeOxOc/Wr+SO5tVI/i/TBFQfa7uXYDvA+g7VZ6Hp9yZXuZdpTG0bSevXoa8JeOdf8AA9xJeeH77ypWxubykbOAw6Mp7Ma1XxBdeMNduta1WPzp5dmeQvRAv8IA6KO1K1tIDHDp3luP4vOJ/StZk8Np4WsJ7az8rUn8zzR5jt0kAHX5fu+lf8JLILR7SW28yE43Jv255yOcZ61qmotqt7JdxxeSrYxHu3bcADrx6UYQ5VQfqfSpUKttzzRV1cYbdjtjFRku+cfhUqCIhiN2fwxW0kZK/rSoVIDDr0qR3UbEfI+lEKRtA5oADnGTQOT0+lLM8ZwpxWwEYHWhuAwD+FEB2yT+lNxgZpY/MwoHPpRge34dMH60d7rtPJ/KlH2faSnPPerK40S7jS1Oi7Jmz+++0sff7vToMV4kk8M+H9FuNI0+PE7bdwzJx8wYdcjoTVw8e4qq5/GgzhyUbB+lMrE5brSpkdaWNTHyee3FMgBxnNMAfumhnvRXBGDmgN3OeKycYBpQRgqelEZX2raegoggetMe9Nz0NOBkKeT2onPyjmjgHg5oN2DfpWQCBmiSeAfpQGAOa25OOlMp/v8A6UQSMBv0oksfvfpSNlcZzRYAc0CCMigoJwD+lIFQfL1pm3dTTEYwO/SiQen5UMMME0hyMd6ck0Tu6mhx07UQnBzk+mK3NnKmiFPAGaIHBBplB4Jz7VgAYAoICuP0rACjmgxIwKBLYUCkk8sZByad97bx17UCScZpSyggNnPtRVm6n6cUAF4Bye/FY5zmmBxkHNBWA5ODTID3z+FAlTgCpIlzgNu/DFFVU4aicgYGfSiSAA35U6gAFaKq3JFSZHA4oFt2c8+tOu84IyavLdQTg9auIfLPynj0o56LWMjk0R6msn0q3iJbJ6UuOmaPJzmiABnNIBnBb9KKjJA7URjAzRPYnpRI7HNEgjINHsK8wFsjpUjbuFNEZIpnAbAOTTOexzTuG6UWJXOayxGCaDEjA61k42gZooNuRRKr9aJLDAFNk8issRz2rGTgUVAI3H6VtXseaUE5HXFZJGAKACAHHNbsjGKIwcKaYF1wBmgGJwBTBg2FHSmUpgsKd1kG3v2pjyMdqzt2tn1rKtwW/SkZAAM81tJXJ5FKCBtJ6dKDhDwajbIx6UeazxjNO2WIBokAc0XEiNEx4PQVeWbRuSKAxxnmiD60ckDFbfWjkjNHpj8q28dfwq3eSOVSh6ZqzmEsOc9adQDWdq4U1uOc55rcByKUsxyDUkzsMMfwpSGTOeaZQx4P6UrNjC0WYjBNQzG1OVGattXurW7ivbSTy5UztbAOMjHQj0rX9UvPEl815ezeZM/U7QOgA7AdlFaF8IfFOvW0d9YW2+EZz88Y7kd3B6g1qdo9rK0Eo+ZMZH1AqRdy5NMBjANEHGAa4xiuSeDzW7BwawpXANFSVyDQkUnYDyKKKRgHmnRlGBW8ZwTQOenWshhtPbpTAAYNEDbkV14FEYGCaJBGBzSwME3oOKBjYbSeaaAqu8DjtUUrRPjOR6U9uSvmKKtpVWN2J544pJA6ZIq2lUxlTww6CppcyBd304poSr7V/CpYvLbJqVdoyKEny7c5oegNOmQSD0pzt4IqZTNCYyenSmcx4UUpAOVPXpRZX+QH8KiVo5Rkf5xTwSMgkQ/jU0Aa1SQHk5z+dBZIsBe/UVpfmNGUC5/H609ixXIHPektbZsiNskexrQrC0kaaa9fEMG3ccHjOQOnPXFaVq2jaTeuyfvYjjH3l7H2PrV9Lb3WoyTW/wB1sbRz2UZ61P4kl+wramLhe+73z6VeS3+uW1pM4xEm/wAocdzg+h6jvVxA9u3k45PagS77SOleGtfn0lt8cO/23AevsfWrLxDJpuofaLZ8Z9s4+XHcH1r4h+J7DXIrE2mpfaZF8zzV8kpt+7jqB6Uzg9Dz2p13EHOcdKJwcZqRCenaniVjimgwcCjGG+XvUqFV2iicd6kVTwTijGgH8hTlkIBOawkuOefSnQr8p61MhHI79qAPRTXuK5J4PNEdqPsKI44puRgV14rGeBW0gc0AT0raaAJOAKskwME1bxsU4FbHHAFMDnpR6cUFbANMCVwDQ+XkUj5UAH60rZUEGsjpmiQD1ornHPTpW33oEE8Hp1pnUcZpirNx19KLEAgGmlI4BpWZztJz7UkeO/TtQjUAYOfWsqq4U0MDkmsA8ZoEZwGz+FHaW69KQkjrzTHAyTQBbqaIUuADWBjjtUjhV4PPYUJ2PGf0rcDznmkGTgGljIOc1jPSgmehzQXjaKWMAYxWMdT+FKoPU05Uxk56UDkcmgDwVPIrayAVvRGRnHrTusrZAxitpL8jIFaUlw90Psp+cdOnofWnu7i54kfOOgwKtbzTbF4prU+Yw3b1+YfTr9atIjc3pnYYHp+GKZfPkBgHCdfxrTruS0RY2OFGcD861LVvtenbN+AvQY6cj29qj1+5ubNNMZ8xrnHA9c+lWegLb6FdapLxENm7/vvaO+etLcWipsHJ7daaJXnV5OMZ2/lXjHWdD1y0WHTh+8XPGX9V9QOwNXFsyXZVjwvT8qlvYYJfLDfhg+lWmnTTwkxDI7dv60NShtS1s4weOefr6VJayec6IPu4z+VXURD4U/WtGv2sbgEnh+v4A/41dxxGfzE/1R/Tj8+taXfRWErO/Q4x+R/xpbqW7nMMDZLdOPb/AOtUFukdqbaVfn+vvmg3hy00KC5tZ92o/N567HGPnwvJ4+76Ulw4i+0g4J//AFVdGW63SxnOMcdKeDdGwJyxxxTEK21e3WtLhF5fRWa/x5/QE/0rxx4Xj0CZWhHyPnB+gX3PrU0uxdo69q03TpdRu1tbcZd84H0BPr7Vq2mzaXqMmnXI/eRYyOO6g9ifWljYqF7V5S4AIzipACcUVCkEUVBbOKktniG5xjPSmiO3cT9KVSBzXU8HpR/WsZGBUD/Z5Q5Gfary4S7zKwwfT0/zitPliW5V2OVGf5GvFF9pGqLbtZfeTduHzd8ev0qOR43Dq3TpxV7eSXqNczSbnbGTjHTjt9KWPzR8vavJcR+cBx/kU8m8gjjHagc/KDSoXjIB6U5KjJND72RR65NMQGwDz2osAMUB8v8AKgCRgdK3cAVg46/SmOeM/Sic45osO1M2QCOvaiT2NBd3Q0qkdqAVwCRyO1DBGAaIxWM8Cio71tzwDSxjGa2/3aKZ7UEycAUBiguRwelFARkUV55pR3HNDgdeTW4KhA6mi5IxR5o9MYoqCtEdOaAB5AplAHX8KYDPWiuRgGgoVcZ5p+VwKHOOOlA9wa3AjB/Cjz909KRC4IFNGQPegCAABmghzgGjGAMd6K/LgdaJzwT09qIcjg0oLDAHIo5OArc/SpFyQneiuDtNP8xAA6VkjjNY7A/hTKAck00R7H6UykYqZTIcAc1eQkjaOtHjrR5wRW3ceK2N6UiBFxnmvajg9OtAZ4zQGOhokEYFE4HB+lYyOv6UcHp+VEEDIOKJbozfpWCDgGmAHzelFyTgUwAPyjntTDjrS4DZzRJPOa3cAbv0ojcev4VIEI+X8qPzdK3ZULjpRO0ZHFNk4weaKluWP6USxGAaySAM8jpSqpGWPNSKrDOenbFN5RHyrz9aULuz+lOQT8vHtRAxnFE7lwBTLu4PalZ9oIbkdBimVmYEnOKcszYUdOhpY2Y5PNAGMkqOaKL3FKnUgcdqVkK4K/rSrvTKjFRIAoIFY460eRgGpRkDFNwwI/AVKcDApnkTawPT2q4t2lbeh5NOuw7TWTnHWicgEVk+tNz8orPGKOSMConMTh1PIqzkW4iBU5HapBhiuaZMHgU46AGlXPasbjz27UGReAKRskEjOO1ane2t0I1t4dmzOfmJ9PWlsvMgEitz6Y96t4CceYvH1qG2gX7zbgenBFaDqPhuLw5fWF5aZvJPL8iXzHGzDktwODkYHNaYthJYxrJe4uTnJ8s+p/DpWt63441HwVb+HL/xbt0W33eVD9giO3dKHPIG4/OAeT+laF8GtU8WKw0bWvtTLjj7Mqev95x/dNeK/D954Y1y40LUBie227xx/EisOhI6MO9Fd3ApgR2+lYB69a9qIz0NHIANByVwTmmjRhuUYPahvC5JyfyoFcYHWnUMoAH1pQxGOtEHsf0pgCP5GtrEYzmmUYyDzTkEABcH60CBxnNCViNjNkDoMVsUkDOCe9eY4HlyHIHbpREb8L07ClZVjCzLuUdDnFXEaGJGQ+uawZothOSO9RHzY2idsMmM8etQxASEMeRUv7uQqv3fSp7fYquD1/SmbzDuY5NMBjOelbGJ3CkmUDyyeG/TFOhU9aRmAKA5DdqaEIM7cfjWdw4pX2nDc1B5DsyvxnoeeKWOe4UwIN4HQ8CoDHOgimXJXvnH8qaNICQq717jOMUC0p3KMEVC85Uln6ewqNHgG5G49MVMzkJI455xTl1fLflUfgTXrvQpfEIt9tqmMvvQ9X2dN2eo9KPws1DVtM/tDQ7T7QifebeExlsDhnHoat9d1+CwTw5bXubSPOYvLTud/UjPXnrV4zq5ikOAO1eYqoFjXkd80JnPLtkjpxTZJyzfpQ9cURxR9MZpyT8ufwoqFGVpmzwRWm+CNY1jQbjxHbQ5tbbbvbcv8TlBwWB6j0ogbfLI/CpoiybQOe1TQkDLU8YC7QeRTIpHTpRBI96iC7sEc1JEyrjb+tTAhsGsDGc5rJ6CiO1HnnP0FZ4wDRyeCa4orgYogAfKM0ORigoHPcUQScgVbJukAA5rToGHzKf880FyvJraMZAphnv07UM44NHBHXmiB0Bpl3DIocnaeabnhaZQwxnpRRR92kJJwzfpWM8A0cDoKKgc/pUhIHA/CmUyHAPSkgJGQOnU0sYC4JzQAAwDRGehrAxisDGM1gZyeaIHY/hWM9KxnoPwoKSOK2ljgnGKiSNpFWTkc5phk4A+gog4wD+lbCxyTQQ9KXbjDLn8aVcnaBn0oxBF5pVymMc0oBwAakZeiNkDtigNxAzn8KkwSNv5UUhDAg5P0NMhU8njsMVG7A/KMj0pQOoFA44J+lAnPBo5I649qVpPuk/hUEyqgXHNaTp9pexM0935W3H/ACzLevofarEWmm6Kkflb3fPzbiOjenPrVqmjXZtra3h8uQ790m5jjuOD7DFX+laNbX3mfbMAfej8tjjgd81fJp126JGmYec8t7fj1FRWptpDLaR8L05/xq313VLwrb303mRxZ2ptVduevIHPQVoSW6XQnu4cxr1XcR2I7fhXjS4sNa1B77S4PIhbGxNxbHyqDycHqDSvBYSiTdlh7H/Per3VHa53KmQ3TnpgD2qL7HLOJJxx3Xn09q1DVbVLCbQrQb449u1uR1IY8Ef1p2ljl8tG2/hntUeqy6dO9yPvTYz+Ax6e9SWS3moq8lx5ay5y2zOML6fhT2dvFP8AZo5t+P4tpHbPSo7CSdUtY23sc4XGPetY06+0m6MF1H5bDtkN2HoT61YxX9kBqMUmFTvgfT+tXV3PcN+/k+XsMD+lbUkXaoxt+8aj1a0i0xtPNvukOMPvI/iz0xR1OaGMwxzYR/vDaOcdO1SmJh04pYgyqCcdcd60zXJNKjeO1OGOOfTr6g+tSht2S278MU8YyHI5HSoLiW1bdA+0n2zVxc+YQzjLHqelMxcbQeKIfbuHIHWmQBdyDB7UEDSeZjntTgj5gc4oKzymRm6+3tU0SOnynn6UwRjjb+tPGsakbPoc0Yzu4Xr0Ga2MjAKOfSntnnBMaZce+KnRo0VCORnIqF1iXBGTQuAXDxjaR+OKubnzIirSZ9sUZlICYyaeRSgCUzRtaoueRnj8axjgHNEdqUqYyuPpUm4jkUAuMD8qA28DmmOBg9a4YYB5o4+6O3auvArI6ZpiFHAonPJP4VxkZoqOmKIB4P4UyhjyfpQTnOc+2KOOgFYGMY6UF6UUyKCkjFGMnjNBAoz1ooSuAK2kd627uO9RnbwPwrORgHmuegojPI/KiwHJFbQx6/hRbBwTwKlIIAU0MGuOg60VjRMjr6Uwx0oAHgmtq9CelSNv43Zx7UE3Ngn6U8ZzwacAdD+FNyRSrxyfwrAxgVnHT8qGAM/kKhlCLhfvetSNuYEmgc9aPHQ0BnnPP0oLuHI+lbGPAP4UIsr0zUIRVJZN2O+cVvwQycZqVllHI+b1pnVkACYb61I5IGOoorzuHWiKMZbknpUiljyeKdcd/pxToG6c1cwErnb+tTQsXJz+FCMAg5zTDJytMMnOKI4ps4+X8q6igMdvwo/N0FEj1ogEYoggYrAx1/CiwzgmpctxjpWQOKPzDgVIcNlaeQHtitwz70e5702QuBWCBtPU15ZUZx0ogE5FAgDAHNEEcA9KEe4Z3fpTKcYBogkbR+FDapwaZc/cOa3AD5q5cYB6UqlfemA6A0FA+YDPtT43Y6UwJ+UHmjtUYJ59K3BWBBzRYMRk/pTLh8Icg+2KIcNjPFONy4ByaA3Dk9PakfCbQOlRgNypzUCjbg1njFEdqPIAogEYp1ySQOlOiBMgZNOQVCjk+lIqg4rULdMho/x/SsYPHaiQQDWM8ChtxjNEA8LQGBzWQwxn8K0G/BTymbkdBj61Id7Z9aY5OKI5GaJUDg0W6AHmsDsc115FQxCQ4B/Sr+zn0i4ewuPvpjK8cZAPbPrRuY16vz6YoXW0bUGfevNlZvvfpUc8quCJcH/dqXxBqEunfZHbei9sAd8+lfD228VatI9t4fufKkTGfkjbruP8WPQ14rkv5tbuX1dt12dvmHAH8C4+7x0xRGDwabrgGtoJ5b9KZRnANYOMAU2elKM8A1tHSgCMAN07YpQpwM80Rxwc0rqByKKAHg9KDAcCiQpwBn2oYcjaPwqQEtgCsADA60Sc8VEd3ygVJGASjcGmUpxjr0qF1kjEbJwO+aljjaHaPwNW0ux9jtj8KYGG43E8SdsdMCmQhgwHA61cLney9sYqOUONkjfTiijBc04UIB39KXrycCpNhYhWyB0OKxuHJoKQ23vRfPB59ulFcNnHBqSHzUwpqK1cjIOa0XTbm6xb267pD05A9fU1b+H50iWcrw2cHI7cetQ/Di+uNGm1iIboYdu88DGW2j+LPX2q90cwy+Wj5I7Y9vrWkaV9q1CLT7iTasmecZxhSexr4i+H9J8Oa3Jpmjah9st0xtl8lo8/Ih6MSepI/CjCzRmMtwfunHT1p9rLlzzUfi3VotOOki8zbN1Ty1Hfd1xnrXh34vX3hnRn0i2sfNV8fN5oXoxboUPrVjqkdpM80nzM2Pbtj0q/uVupWnA64wPwApj8vBpHC5BGfSiQwrAIxihjoOaMa+XkHkdBimGelOgYAA9KCgLtaoPEGr2umtpNtd7LWTG+Py1OcNuHJGetQ28lxKIYhlj0rUbC4067eyuB+8ixkcdwD2+tThWbkdKkjGcK36U8Z4AoqB3okLwD0oTLIuO9XUO4DiioBxmunQ18o5z+lUQOgNcHgGsjGKJBPBogEcHmiO2aAwM5ogetWMLPKNtWkHlLyetEg/dNEAHGaIHUn8KI44P0rqMflWAe9cHiio6g0DhsZoqC2QfwobScVtBOQc0vIxmgARzTH+EU248GkhJ5JrAA4NVkAYxWM9KrGRxWAeM1gdqC5oqenpUZgijzu3N6YIpQwdnJ69qDZIApumKz69qIBNAYyBQQk4FBVBzTEEZBz+FMVdcKKC5GQaCBhtQZpIyOnWnGSCp6UkRAyRzV1pE8Voty4+U5x09cetLHtyB+FcDkflX3uM0pAHHWljMuSvQVJbyLgHmo7dUhSXd1zxitHtDdylC+E78ex/wr4ieJtBv/ACoNCi+Vc5O5/wDZ/vD2NaVL5coJb5h04+ta2mkJPKWkyWxg7W9BVnbJsSXP7kZwf889aF9pkelJalcOM/Nlv72elXaNHdKYBndnj6CkvrouIzx7cGrxIlRBG3yjPar6a2cbUfL9hg+1TkrEFYfMlW0QeYb5OPp7URAsELxn5/m3Dn14q8tt9xlTyen5VOyk7JDnbQ0ye60+TUI1zFb43Hj+I49c9a0i2lutRSMr8z5wM+imrfTBot41lct/ps2PITHXAy3IJHQ96+Imoz6lrsk88Pktx8m4Nj5FHUfSra+lnVbQ8RnPPX3q+uFZwgH4/lSWkrZdR8rf0q5gZZRsOR3H4VJbgxkZzjpSwsYyCvP1qSRnIIH0qRigyDyaZ95yO1EksFx07VIMNg0xBUAUrAnmmlVBtPfpRAZME1gAhQelEIFyW6e1AB8MrfpTyqG8v9aD7AFAyad8YA5H5VY+HDr9s15px3SR43LjGMnA5JHoadowzFmwR7UmoxpAIo15HfPvTEyktnPrV/pVzZsokX72ccj296v7CWztoLth8txu2/8AASAaltpVtTPjgf44qFnaTG3jtzSruG3NMjFAAOlZIHIo4IBpCAcE0xVmwTTYJwB9KBGPegcjJ7UGHX06UACNwNYJ6GnKjkdaJBxg0RgYokDvRORxzRANUFBOc0AMe9KhIBxSpkYBpUAGAa8sDgUFAXNFQMAGgFHFMu44BopjkUwyMg0DkYB5rdkcHJosBwDyOlFsqDSkDjNMR2NMeMiuOlE980XyMYrrgipdpbKiixJzito7UWwSKglVfvDNMOMGmGelD09KxngUR6GgOOtKwXmiAwyB+FABhgdRQBHGKEW6PcKjtwzE7uB04qSPYxBpIyI9xPJ7Ur+X0G726VIhUYNRKruFL4Hc4pz83y9O1MpJJNYx0OfwoqrAkHp7UBgYzQXexDHpUqjIANA5+XNELnBbj6Vfxqr7UOf8iiM8A1tx0P6UQBxjNUcEYFDrWM8A0BjoaJJ4zRGeM0RjqK68UYlxgDmiuD1ogcVIxAxnPtS4Jyafaw47Uw9BRBIwe3SgobrQAPWjGB8wP6UjgDAGakCucg/hRCtyTXCrxQG/BzRbA2g0pwaJ25INA5GM0oIzg5op2Bphg8H8KJ3DANdD0oqWGc/pUaKF5/Knw/FbQrZNSMw+ZTxToQvDc/SiS3U0VXGQfwpATyDUJ3EAVEPlAz9KAzxn6cUBnAzRGOM/SqlBKdelMMjimQjpUiMW4PFY2n37VdxAkMBTEYwPwFcen4VnjFZG3APNDBAANBQO9QTGJwwFWE63Furgdc/zpowD1plUDaDx9KZAzBV5oQEjlvwxRRfuqaWKPy9xPI6cUZwFwO3Sp7h7mUzSHLN7egxUFvLcNtiXJ9M4onb8q0xJwSaVQDgHNeGPD+m6npV/Pc3GyWHy9g2Mc5Yg9D6CpY/s85VHzt6HHtTgzMWZ+fpSWaswUTYz/s1deD9Ebwla6nDc7bxd/mnY53/vMD+LAwPSiMcA0SSPWvmosScE0yIOFP6UQSM17UCSODRYgYFLzwK2sAMHj0pvmOR+VEA9R06UoxyDmmIbgn6UcEZ61IRJ8xPIoHPIpX3cnrTbWXGetQSmM7AeRSIssPznkVLw1K6SW5VhynT8TUbEWwcH/OaL+cGiYYHGBTRlHwTmjtkBVeD6VKcpy3T2pSDjn9KuERo1ZB65pVCjAk/SpEUSDYfwxUqFW4qBmxs/hqzsLm9mFvEMlug47D/61XPhnVNCn+xX8G2VOq7lOMjPYnsRWh+HNY0qNNUePyV5w2Vb1Xpk+taX4b0bTNKg1/Xpvt0Nxu8u32tFs2sVPzKSTkkHp2rxXd6fLfpPYWX2KLnanmGTPCjqeeufzq98Qa54l0+28P2MPnQ2G7au5FxvIY9QD1U961HSLqF51uRtkXbvj4OOmOQfSrfS2uYPMV/px15+tfDjwt4d8QeJrPTfFzeXYtv3Nh2/5ZuRxGQfvBavrCzg1lrO3uP3HGG2H+6D0PPWvEmmxaVqsthaT+dEmPn27c/KD0JJ6mrXQby+iMkLcDtgf4+1NGynGeaU7TtzSRqyEM+Me1AbTnPT2qZo5H3RrgHtmgMMADTQsBuA4o5zgGorcyICrfMegxV9pGoabHE99b+Ws+fL+cHO3GehPqKZCKAwBg0hZHDo3I6cVPM88hmc5Zuv4U8YYYz+lGIkAiniBJyfpxUqEnpUijOBWOnPSpZi0YXP3elS5J4NZx0NHnvXB4Bqh2Ao896OD3/DFNyMA1k+tE84B4qKPzGwDn8K022AO5u3T9aBA71wR1o4xkn9KJBGM1x2/Cs8Yz+FE8DB/CgQ3bNEZOc0VAGDWDwQelMMdOaI/hNADoBRUnpRVu3NbDnNEZ5HaiuDyKyOmKABHWiOeDz6VjI4P4VtYnIGaCZOM1DDC/DHkfWmfGVU0FyMZoqBxmmUFcg5rac4P4UB0waGMjmpACcisdMGlGOM/SsZYDP0pVBYDNFQVLA/So3K8Y5plLHaTyKiRgcsPpTlsgFs/hSjavA/Chz2okZpULkKoye1SBo8qw5HasKUxnmrc7vkzj0p1Bj2q34Yq2tpZxtiXJX3xSK0aFCMH0qez82Jb65fJfOBjHTjtVlaSXAWMjbuzg9a8G6d4G8O6O2la7b/AG6XjMu+WL+Jj0UnsQPwrX9L8NXF0Y7M+Xar/q0w528DPJOetatdiC+a3txuhTG09OoH49auLGK70y51Vp9ssGzYmwndltp57cD0qG/zGUI6dqsHWVXEvVscfTNXLO84DcgdvwrVLkXUihV+WPOOfXFQOEjyvT0rw1YeGdR8OSS6ku28hxtfMhzlzngcdAK14pbOGhOcd/ypomlGZOo6CtH8WX2j6Tc6RAf3Nzs3DjnaxYdiepo6dLFB5gXDfX3+tTSyIwR+SKupy6hA2QPakeW+xb2kO6Vui7se/f2FX2g6lpUijVrfynOdq71bPT+6T6ivAvhS08UW+o6hd6h9lXTfK8pfJL7/ADCwPIIxjaOueta3AseqSxRPvRMYOMdVFW1nc6guLWPdt7ZA/n9KujJaSCORee/NOdrbNvC9OamYFgR0p4xHhQaIY8AZzUgLLwaG4EADNLDGyblbB9MVPGHlyx5H+FPGFG0GiDjAPP0qC3DsE3498Veae+n3klu8m/bjnbjtn196kxFGQvU9KEIRdoOfSkRJAFc81aNskaMDiriF1ldFORxUOgNFoiau1xgvn93s6Ybb1z/SraV445FXocVrHiO+1qFbe7l3Bc4+UD0PYD0FSBVYrnke1RmSWPYTkU8RRtvcdqUFmwBn2qzgEiGUnr2qfaJSB2/wonK4zQOOvajkcg4qTG0EH9KIGMg1/s5rBBwDWP4SfpSjAwDzRI9ac/3TTZAwD+lEZ5zWSBgmlII2g9KABIzXB6HpSx54zRjBGM0qkDGaU4IUHrQARgc5pwN2AaYADOaQKR834USc9azxgUU3cCioIxmiSvArI7Clb+Hd09qds8CiTnBNE8ZokEYzRII469qGMAA1u529c9aUMRwKGwcDrTDd900+OFzmsjp19qYg8VgHnPSioIBzQPGKAOBg/pWwAcGggzgnpTBcYB6UowMZpCwBVO/WkjYLknj0xR2x7QO2any74BzTSlIxFt5HvQTIwp/SvLUruzgDpxQEbDNFVzjP6UQU5B/SmUMuAORS5bIPatoxwc/hRJHyg1KBjAHJpkz0P4Yp0G3PU1dx/KRu+nFEAcZr+HGfpVVQBPSgAehoHGBTDI4og4wtE5GKPIxmmXndnPtTEN0FMASADRGOM1gAYByaChl5P0FEoDgt+lFlY4J+lKo65+nFOABkH9KClztU0yhGKhunTis45K/rQIY5zRG8fIaQbjgGnwOhogE4Vv0orsYFxxRPHFeaNmAv60zRsowcGmIDcHNEKBkGhgtknINOpwT2pmGeDmlBJwB1piDxnkU+HTbnmiymPlsn6UVBQtnntSkDqfwpMg4X8RUR4BzUPK56g9Kxu4H4CmHygg8URxVMARjr7VIVzkGsgjI5pkB57dqmK4BpirDBFXMAB3LRC9j+FEKOKI44NEelcEAE0BnitL1E20gQvwe2PrUMq3MQdDuz+FMsZUEHp2xQCr8wHPagc4y34YouPWnfPOePpXXkGgQAAT+FQXckBzE2M+1Eg8jrQbcBuPTpRcjgVBcSxkiKTG7qMelP4J8QR6JHr0tjttpM7ZPMQ5w23puz19qYxDDI2T3GMVaywpLukP6Gp/EPh+fwrb2CNtuo924Yc9ZM+mOlOBnAH0ojFZI5x9KPzZApD/Cpokg4xxRYFhntSAHp+FOdpw3SgehU0HUnDcH0pQCMEc9qIZCcrkD3plUjcjZHfjFYDDApbfaAxP0q4QDDjoaWASQs6nle2K3xum0nB9KbAUAnpTkK2Cat5o2G0v06DFTRh5dp/wA8VEqqWRjyenFRSlT5W79KduFVzyKkQxgZbOfalRWYAtgjtipkj2YzyaIBXGaidoxtYZU09tuAMR3Dt2pUbOMc08KSRCNDl/yrwx4f1ZLE3w0vzref/VP5yrnaSDxnPX1rRLCbS9TW5uV2Kue+cfKR2z61qniCPXr43B/eyt1b7ucADpgdhXiCHxPZ2SLqbbLVs4GIz0I9OeuK8MXlzqd9Fppk87du2R4C9ASefwr4jRyWN/HbXsHlmPO1d2eoQnp9RVxquo6PcNPps/kF+p2Bs4GO4Pqam1G/v3kuJbnfJJjedgGccD+VW95NENpfAHbFahffa0iDvlVzuOPp/hUzQyAMh/nUrbzuRvl78dKtLnUogIrafardtqn+daxpFxpMka3C4Muccjtj0J9alQg8dKbDLwOlIw+6wogk7QeajgJwFXJrUvEvgOTwFDpVjL5mpru3jbMv/LUMOo2/dzTIioT0I7Vazm1uEuRHuC5wucdsV8QfiL/wnmm6NanQ/sJ0bz8H7V5vneayn+6NuNvvnPavDngK01/SW1CS68tlxhfLJ/iI/vD0p4yhAdfmHbNSOrj5RRACAYpsHisccVLEScgZqdMjcDUyArmmKkYFOQy/MadRuyDRAxwc0OetYA4BpgCMA0cAURxRC4wOtYz3oAYz1qKMyttqw07cAT2/z60kaoMA9KJB4BrrnBrGTg18vY0DjvRAHGaPIxRwOOtHB6flRBbjGaA52j8qEeeMUkIc/N0p4obdwSM+3NXMkJVWU88+tbwRTLk9aIFMN33TzSxDaSTyO1FQF5rapT73I7YrC4965OMfhQUvyKUEHANYHYUqjOSaYjPBpVAX+lMpZTgcjtQHGM15eB1z6VgEDmgADgGlABoY4I5ox5bAOTRQD5R0HSosbua0kWJlla9l2Zxt+UnPBz0/CrnyhIyxvlexwaAj6g5PagSTx+NK6kBdvP1qQAMRSsyHKtg/SpGZ33Fs+pxQ5GOvtUJRSJCMelM6n5kP4VpcxhG5G+/7emalKSuR1NQTzTTNG75RcbRgelWEovLEwpHgp0Off/61Rahf3Ds0j5A6cAVpV9DNYzRTn94NuBz6n0qzgt7uAW8cmX78H1z/AErUrdEU2MLbmX2x6GljO7y2bDfSpIpYboKG4Pt04qaYNMoLYHc49qkgYbkTnGMULfbagudp9OveroPDEkjL8pzzmrxZp0T5ODnHNRWHh8eFIbk3mb9t2+Hy3+XEmBz0+7zU0IDqQeBnPtWqam+oyMlrFhf976eo9quIJ0lDSL+OR6VPHJCcFefTNeE9TXQ9btdSaDzDFv8Ak3bc5Rh1wfWvHHja58X3SXFzB5Rizgbg3UKOyj+7WnatJZthmxGOq469far+4juZfMgTCn3z2FaTrUPh7T0kjTfLLnK5Ixgnvg9jV7dfbp2nZfmfHGfQY/pUux4xsOW7ipAwIXHNCPcck5qNVaTap57cVcIkQ2KM0+0RgAc+tYzwrfjilt2lUlBnb+FOpDbSOacIpVc5PPGK2lTkmp53mbzJWyx6nGKZQq5AzRdWACnJq7LMyqnUZ5qytJLk524A681qOxHiSB955zxj0qa8unT7OH+QdFwOO9aboV/qNvLcW0G+K1x5rbgNu4kDgnJ5HartCGwnOKMZwCB97pVqSjeWV+nNTyCVzIVx7ZpLoRwNCsXLdG3dMH0qwCpbZP8Ank1cFXlYqeKIHTND5vwpMHjFOoBCk4oEHgHpRAxgHJ7CjgDINE5kK9h0NUwAABPNTEHAFAsw+YYrAznGaZs8KM4pQQSQKGc8CkXOMdqRwTgDP6U4UAbBkr1qKPzW5OBTIQ+Sv0GaILZJFMVZRjr3psEYU5onK7QvT3ogAZJo4Y8Uy7WwD9KwB1NPhuFNEA9D+FMoBxmiMHAokgYz9KJyOTWVIyGz+FZXpn9KGMYzQIC81u4xnFKhOATTlRhVNPwQBRAA6/pRIJ4NF1B2saAyODn0rY3ULx9aAJ4HbtQ+YZBzimJJ4/KlwTnOTWADgGo3CcgZpDvG0N+lKm6Qhv4e1MQH3Kfwph5qCQjJ7io2iRCGbBPQYpypGCeO1FRjafwpl2gDOakbMW0jB7UJCABiicAg9+grlu1df6UV3rgU64bA/CnUkZIxVyjlOlSqcg4xiip7VgkcUADRAzQA6elAYIGeaIyMA0cdAfwxRxTdaoElMnrTRjZ60DkDmn2jvRVTzmgR1B5HSiSxyT+lEEDIP0GK2kd6+UHaDTbSuAeaCgjLH6VgDhTSjcME0qiNhmmUg4BoqV4BoEt3/ShkDcG5HTit2Tgmn2N0HAr5c4pmI+UfjSFNmD1ph/DmnCgDb3pSneiq4yTQQCiAq4BzmnAwAKIjRAQOTWMjK/lRJByevakJH3TVq2UGaBGMVtJGR+FUQOxqmRW5Jp1APymnTIwKcAYA/GpFVsE/lU9uJwwByRjtUsHlNg9qOBWRjjtRHAwaAzwDQ5HNYweK0W9Eb+WzfTj60VDx71PNMCDjNHkc0xJHBoHcMii2aLDGc/hQyR/KiB0BrP8AdNBQwzioyY3DKentU/irUZ9LXSHkzCmcDA7tu9M9aJH/ANasZ/pXI6UkTSYCjNXWhX0FhFqRjzDJnByOxx6560WJ+X8qC4AIFSAYG2kbI20AQwqNlxgVnHQ0pVhjvW5hxmiARhT+FI23GeMU22TletFgx2t2oF4upzUixytle3UU+FO5RgDtUqANjNQSDd5Ln5T1qWNWTHcUCVwD+FFjLCEJ6VECUIJyV6cUGA5B5o7nG0nkVHKrxmGT8KYbTgH9KDMyZQ8jtilxN8rnNRW0QYo/I7CrazZJMJz6Uts+xwR84xgVa2mZNsifrVreNYaZHbQt+7Gdox0+bJ/nU8K3uCgy3bt/npWhG5j1iC1n4C7sjj+6T2r40WTSWNvNHb4Rd/O/3joahPDsu3b94M84H0/lVl4skjvFvLttxXO3jHbHYfSvEU0+tTjUCmY26HI7AD29KkizJ5EI+90qRFL7JWyPpWoRxoIvK5U5yPyo2UzqGSL5T0+an8P6pBZiS4ttqN9071PQ89DWjaFqt1L59na+YE6neq4yCO5rVdQN9fy3R5MmOPoAP6VPIjHaDzRAzj1oJgHAqL+ym0eFlj/0sbt7Zb+9xx06UUaQqXfp7UI45Bk/e7irnw5awaKdWWXLD+Daf723rn+lSSu7cmkUGIgnr0ppWVNiHis7htJonBxS5YYNSQuqbyvH1rp0FBQ3ergAZXNSJlSc1tz1ojHANTqC/JojjGelFQOBVUQCMVRIPSqgt2mIVRVhp0apiRee3P8A9eljSMYSsY5oDPAojnANFfQ0FyKwO1EYGM0TkYBogdKIA780BngHmgme9M+0YFR7mbaoogDgGiAcKBQUdCM1tBXrQQHpTBSAFplwuKK5GAfwphxkHpRBHSjnO00AAvFAbhwaByMMaUKRjHArbg8VjnGOKOD0618pPJogjoeBRVd3PJraD0P4UwToOtAfLgjmkB2/KfwqJQ6EZ+ntT/KuAeaWMBcHrRwOlBSWBJ49MUSMZBrwtpPh6TQ11C8/4+uf7/8AeI7HHTFassbXskidGxj8AKSNpWCLyau7G4siBPHt3dOQen0+tRRBzgDNXEKrtLLye2atofOTYgyfSoY2jfy5DyOtaRptjeZFwOe3X39D7Va27LOFiPHf8qE15pen/Zlf5Ln7wwP4Tn39as7Bp0Fmy43dBnPvWvTzWV0mjzNmOPOBgdwG7fX1rw9az3U4jVvkXoMD0Naxp2pKI4pDmGTPlj5e2M989an0iSyuQSnX39vr71cyRrIuPu81dSb0yDxTTNJEpBywzQLSR/OOlM7sPLccLVyj2kCGc8Pnb+B56VI7pEJT0bofpUN4IsTqcMnT8eKS0utU0035fIPsP72P6VIrEFSOe1S28rRgoOKtwZ4nJb5kxt49etXUhB25ya05RNcBHPB9vY14st9LtbiBNJGAd3mD5vRcfe/GrhzwCee9QgsNufwpkjkgAI5H+NSRgrtU81FaOyhUTLfWotNEa7rhdp7DOf5GrshJjHngdPyqV0cGPPJohV+VTzWmWsN15jTtgJjHB759KuVRidp6dKGVGUPNH5xz19KsHRLjMh+VOv5GtYu7W5jRLUcjOevt61Mqom0HLetCMylSW6dsVcShYvLQ15fl8g5zUjjfhD06VBezW9vLEDw+MjHoahLOCR+NTQtGAuc/hUI2w7j1FQ7SctzQjaV8ryBVzdbYEjjbpnt70x7nrXDcZ+lEEck81Ge279Km2nBzmgMEUVXt+FYycE81nnGaIz0OTRBxjFMOeaByBzRIUYFAblz3rBGADQAPANRx4PT6VFGpHNINrgL260SobPesCMBgOtOQq71PNA5jABpQpQgjntWwLGN3U00SmPeBSqpGG7U42qCDnNEAjg80xOcA0V4ye1Eg8DtR6cUeaIyMCgoIwKK5OB1rPoaDZG3NAgCgSegokDisZwc5raDwKwMcGsHPWm24wOtDOODX3ulDAGDzQC46fhQLZwDx6VDGHB3H8KEPlKXY8/w8VJsK4H3qwFcgH6VIVVNsR+agwEKlevOajKN8zHpUyxgfu+lMAqYz9KAUjDUWAwqmih6Dp2rAAxmmwT8tHkYNAZ4qQANha3kjaauEDJlRUqZyAOlAEcEUwUng/pWcdDVAYGM1gHgGjjHWio9aIHQGiM80VGMg05wvFFXIwvNGIhf6U67uCKZATgH9KdVVcA80CnTd+GKYKVA3fTiunBP6UQp5U59OKCk8EUUxjHalZcYJolRwDRYEdc+lFhnJHNMGYgA5P0rbjjNAAkKKYDoBmgilc7+nbFUuwAl2wR7UdpI2Vg4AJpwMjac+tEg8Z5pmHQmshh8prAIAz9KfYT8pz7YrKldpP4UGI4WiwIII+gqEKeCatiB8g7UTk5FZBxk0wHABo4PQUQMcGiMjANPgEg9aYjoDUoUfdb6cU7EHBP4Vbu2/GM5q9tI3UlT8/YflUkZRtpOT6UQTwBmtpxgH8K4xjPPYUVyMA0e2e1I7KdyN+laJqrzSm2mbOenHTgn0qRARkD6VIoUYplJ4NBcnAPFMVXgt+lKqkfMMU8mBhKQhhjOaVQvU15jYwfyrLMAQPpSkk4zyKZ0GAGyfTFBgRgrn8aDYXGM1FM+MRR5x71deKry50iHRWGIYd2Ondt3pnr70doH0o4J+WpVBwAetAIThW5+lEjoW6e1RqGGfypWBO3PIosAcZ5oMAME1lc5HHpQZScs3H0ptpwVbI9cdKdQVDKcjsacKYCxOcf408TRjc4qWOKWAvGeR1H40CGUAt+lMAFzmoGiZcSN+lbctt6+lKwXOOtEBfmU/UUrKy8Hmg4QiTGfUUEgmyYT83pg1KqAAqc+1Q8uAwxVvZWez95LsbsNpNGOJpkRhwc5P4VpFuNJuFubqw+0R+nm7OxHbPrWv32n6zqzXOm6V9iRuieeZeigdTj0P51rek3ejBWnXBbODke3oT614MtrbW9Xh0y/uvLjk3fNsJxhWPQY9BU1x4J0LS5Vgk+03y42JiVP4ueeR901f6xcXmqtdQQ+Ux/2g2PlA9Pam+LV3cWi2Otab/aMUWcJ5wh6nPVUz1x+VNPJccyvtI7YzUckUsoiVs/h7V4B8UeEIYBoHiex3Qj7tz5sg28sx+VBnrtHX3rxzFo9n4luo/DV559hHs8uXy2TOUXPD8/eyKnuB5uwDNPYeXbLcSnAbO388V/wkEB0uOzj035kz+98485bPTH4Vqnj+61fToNPFh5Sw7ufNDZyQf7o9K8P+Mb3QozBavtV/vHAPTPqD61rM1vb6pcRadcedB8ux9hXPyjPB565oSBhiQc+tSxZjEgOc1EGkBAOTVgYFUxSS4LdBtParX4TWmi3emzePLz+ydMvvNxc+WbjGwf3InLfeKjt1z2Nazf8Ah+7T7PY2Plf9NPNdvQ9D9CK1BraLMVrJvA/2SPT1qw0LUdXBXTbXzivUb1XH5kehqVSgZO4xxUYhwd7c9uDTAA5UVwTgiiCGwDk087PF5RFMARgN+laNDpk0wTUrnyY/72xmxwew/CniDru/IVMhbOO1MMH5aII5NToGIwa2kDg0eaAJ6GiD6V15z+lEZ4zzQUkDIq3snmIFWenQQKCvJ/H/ABoArwD+lcZyTXJ6VjPT8qI7CgQB1rIHAFDgdaPQfpRAxwc1jHFBc96baAADQchetbtwwBUQflkHSjGQuWGKVARnvW3PGaZTgHseho5HCnn0rORx1oqDgE5NE4OCaA5C5zRYk4BxR+Y5B+lFjnArgAEHOe1Km7g15TIPmH4UwBXIPTtRU45oqAMqeRQCHG5se+Kfyw4WNsj6Yp1DENmnitWtS0T5kHbB9aSOI8u+D6YrYxOETP40oUttU49q0LSP7XvUtIzjOc9+xPqPStRsFsrySzIzsxz9QD/Wskk45HavYn8KGDwT0rAKkg8joKWR1QRrJ8o6DFWlrc6nMtraxeY5zgbgvbPf6VpEFit/5Woz+QF77C3Y+n4V4pmjudQZIrzzreLHlN5e3qozx16+taRKltMJGg80f3d23sa8VRwXcqalFbfZUuM7Yd5fbtCj73fpn8a0xQ0wEMmD64z2NPbF9Qa1HzuMYPTPy5qPSp7aSMXh2Kc7eh+vQ/SrW3AGIl3yt0XOOn/1qee9ltvs04zIPurwMc5PSvD2tQ+X9ovrLcD91/MI9R0A+leJ7ez1LUReWz+YG7YK4+UDv9K8MalBoupq92MqOq5I/hPoD6ivHfia4vfMvtLfEMWPJTA4ztDckZ6g9aOtzTwBpU3v9QO/0oqZoXJXkYwM00btbn5c+gzjvWn74z+/j6dOfr6VYxJLcBnh3R9xux2rxH4QuGjk1mzsNlm+Nv70HphT1OevtWoXEskI+0dI/ur9cZ6V9okuLdImX5Fzjn3q+GLdxGPTA9eRWm6lNa6b9meTbGvTgH+In0qN/t9szwx5EeMtn1Pp+FSiWKIRflT2IsI/MlfAPt/n1q5aPdgPz24qKUxnCr8w6c0pLtnGWp4txyBkireFicIuT6ZxX2BkkMNwNjj7w64/Kk023eJ2XouOef8AGvD1xDbO8t1DvPG0biPXPSvEV8b9/tS2/kiPou/dnOB/SpJRId0j8n2r7OpXca1vwde6Lp6X9zHsR845B6EDsx9aZgY1RX6dRipQo4U0s9rHwwz+dM0DpujXH4mvBqeF7XxBA3iu68ixO7zpNkj7fkbbwnP3to4rx3e+D5r+NfCM/wBogGdz7ZE7Ljh+eu6owDJ5Rfcfpip4RbqSnLenSpiWO1TmrpwsWzdknpxSbmGFNSOTEAO9QkIny/ercZZCrnB7Cpl3YijGf0qaMxLhkwfrUckkY+Q0kgciNkyfrU8exytAetYyc0AMYA6UV8xMZyaZGAwRR54NDk5FMBwAK6AUeBmnJJGOaKggYFYBGCc4ohcYWlUtwaijBOSOlKilhtPTviiHjGe1O3GYz1o73Gdv61jzMAHOO1JCXO1hgU8a5KL1FKv7tvaiQyAM3ToMUHygjZuPpWDnGaUFm2g5NEZ+XFbQ3fp0pkAHBpowCAPxFMoAplQYAOfatvoawMjFMGzkVtyOaAAxg1QIBwDXynq34YpnUjArd/Dn8KO0fKOtADgd6YAjIoDBFPtxhRj0pFDjaOvalVQADQVA2EO78MVk9FOKcs+Axzj2pYHCZIoAgZz+lRnLBeh9aRjuITketOwBKjpQdQNhXJ9c0SDwOvamJYZJogEFTz/SiSoCZp8HkGmAxxRTkKDn8KJC89cVL97APPpTHI4PNMzsuD+FXAKk881gt0pigON36VlwwGa7AZzWcdDSgEcnpQA6iiMim5wBXtn6UQMZPNEHsaBcHANOzZ2k02AMg1tLNgnj0xTJ82R0o7R1rjHt6VtLDOaCjnJ6UzMByaEiuu0jmjEzcrWQRtFEHGM1hCucVyeB+FdRwaAAOVPIpuOp5rJYAA8dxTHHf8KAVtxYfSg4+6KLE/LTgFcLREaIAeTQAJANKpAwDTnAwKATIXPIoquelHb0BrClhg5zTBl5A4q2cAYzSkMoIoAkYFHnv9KZRjijgjgVUiblwDzTqVOAa4x/KplRh0p02kbagfByTU1tb3KcL+89cmrq0e3fBFckZFEAjmmAxxQ5GKJxwKjdomDKea0nUmnJjlfJ7cfX2qUDPy9OwogHoeaZiDjNDOQW/CmYAlfTpUeGpWDD5aY8YU8ijgnANKCACxpnz8q07Yxk1GVJ+WoBCiq7jJ59am+IF6IY47Y8x5x07/Vau7l7ydriT7z4z+Ax/SmGFziiVXk0wU4zTIc8nNYyoGaVRjCdqBwuD1ocjrRBJyDR6YFfd6VEQe+a3DOUNQrHMNw6jrSNsPksc+lTI8A2qflbr+FMpABBog5HNKAW25qZSCp781JtDAqKCgx5646CgOgFOhBwO1AsCCp5qJmY5z07VBMQ2GGc1aac9xLvUfT8vrVnqPgTS/hw+lW+n/8AFQXOMTebLxtm3fdOU+4cf/Xrwnd6Fpdw0+v2PmqcbR5rr2Yfw59RXia6hu3860XbF6ZJ9B3rVZp7plV3yBnHA9qhaSOTejbWXocZqDVZo3HmvkfQD+leG9Jn8WNJG3VcbTx757j+7WpxS21rJp+eUxzj3BqwNxY6yjaedr87eAf4eev1NXbxz6sZtXG6b16fw/7PtitUv0vYkhQf6vP64/wqxsr/AFi2hhaPfHFu2DIHU8+npWqrHazEwpjHTn2FPqLTMoc/Kuc8VGAAAo47U+j3cqm5VMKnU5Hfj1q8chgYjg0wLIZYxy3X8KYFmyetRXDqNucimjYJ5qj8K0SC3uboRTD5fTn0PpWq6tcfbDayzboofuDaBjIBPavCGhweKdXj092wjZzweysfUf3a8V+Gx4b8V3OmWxzFa7cHp96NT3J9afXbzT52aB8McZ4Hp9Penk3lmJyTRK5oDIOKwDxmgoDY/KmUGv8AIpiwXINSPgbCaljUqzAdKwCu0jmiBjAHNOFYYxTEA4AobScYogA4xn2ohSMgVtHQmuMYxVhZLI3zL+tR28UY2qv60CCMjtWQR8tAg9/oKJ7A0QAMij8xxmjzwaz6GhzgVjI4pVB4qFLaNN0wye3WpWDZ29B0FHGMgZoAAgCsAnANRDLYU8VvyduaPABHWsgjOc1kEY9KHU0T2oHPyg/SjyME5oAHBBogCuoyO1KATwaPykqKWTYcg8jpQYuMscmjktgVnIyDzTHOABQxjr+lErjrz2rPOaU9gc5pWATAPNQgFQQeatkgaZTMMoM8cjt7V4hXw7b21nL4eh8qb95553O2eRt+97Z6U7F2LscmieM1weh6UOR1pDnoaEbA49Ktb2exYGBsE+wP+etSuzyb93J9qjZnfLHOKEgUgMatrW51EiGI529Bxx/nFC0a3QqRhhUTgOsin5h1NeE10C+sZZdbh8ySPb5fzOuMls/d+gpEltSmxvnXPavDPxF1fSraXRYW/d3WN3C/wksP4T6+tf2jc2cX2VGwi9BgfWtI1G2aE/aF3E9OSPX0p4UuGN3AMAdB+lTBNWsZMjLxY2j6nn09Ks41kn8h+g7fhW6O31B4kHyDGP8AvmjoWoLb/bQ37sew9cetWvhkavpdzrcL7fsezcMZzubaO49PSpdSaeNUccLnv/8AWpPiXr+o+H18Lib/AEbsu1P7+/8Au56j1qa2McwSc5z/AIe1ST2jXUU1kmETdkZPce9eKdQgvr1RCPXI59F/wopawaX5DJ8w75P96keSG1IDfhj3qOZ5lCMeBWq3LMNgbIHXj6VgSgEVNFC1vE6r83OeT61oXg+/1i0+2W0WUHU7gO5Hcj0q9t4LV9gHzDr1q3uBE29DyKu7e51O6+3MMJL06dhj+lXKNDOIEPXrx7VZweQWOc4xU6yXsjAHjsP8/Sr61hR/lHI+vtVvA23ePujtWn+IrWezNvqQ3gdByO5PYfSriJJLmR1+4MYH4VK0ElqysuZe3J9f8KJ2YFeaxXYpqed5Sd5/SkcsdsZ5FTxwRDMQ+bueauHBQgHJ7VC/mgluSKYecXYdsVbEH5fyFaX4Ym1VJir7PI29s9c+49KuYRbyGInOKlR4ypH3qt0kOG3c/Sr6AibAP+cCnGBgH8KQqqHA+Y08ZK4JyTTJg8D8KIx3okDhaUgYJNBRN2polIJXtWCRjNDk1yeDTcjAFEA8A89qIBO0VjIxmh0pQGOAaiiUrjHIqIAkgH6UzZG0GgmMbuc9KEQ8vA60iBVBf71HhfloLtj3E/NSjd8pPXrRjIXIPSoyrj5vvDpUhUHC0TtAJNMvyZzS7QuT36VnByDTMCeTzTEEdc0wUrxTLjjNMuKYZ4z+FMCRgGghznNBhnBNEnPBrJ9aJOMg0Wz0oEk4zWQTilAx0oKSeaKLjao5plK98mlXeRnp6U+1W+TtRBAyPwp95QAnjtTOT8uf0oOGAJ6CmbPNI4AyfwoE55ocHOaJUEHv2p2BGO1ZDYGOR0r73JNbuwFHAOBQAbgHmsADBNSthuaLLjrTHK5zzUyhlwOtXDLGoVetHnvXyluDzRPQZoHtmgq5xmj82BmiM85zRAboaIB4A/CmAHQ/hVEDHBp1VsgHkUyADIH1plHSioK4AxTIx6Lz9a3ZUgr+tHAAJNFAwG0Z9qbsAKCEDJX6c0AQODRjcndmm+bgmt5IBJosh5B6dBTFWX5Wx7YosNmAefpW5icMc0AoBJbp0GKYjOF5owsqBmPXoMUDjq2fbFbssdq4HYZoBhyV6e9fKzYDfpTFQcbv0pn4G00rbucdOlY3HKt+lMgC5dvpx1rA7N+lKpKkqPpRcg4bn2qMgdD06CoZVcbQeRWc8E02MYqiBjg/TisZHFYHamAdsDimZQeadVI65qUKcBW6dsUwAbg0GKn5W6e1Thphh2z6cYqVcAArj8awCMCiM9/pW33rGOA36UewqORom3K36Vp2qi4QrK3zDpx9fagAwyr/AIYqRVZeTzRKkYzWFPUUXCdBQfC7VGaTk4HWiNo45P5U7sRkdB2oupPytn2xXDjcTjFCXPCGgwB3FuntSyAnIf8ADFJG0z7FbGehxUqtExQnJFAgnIb9KZVYcHpRcAdd36UAWGAfoKjO5evPpRwo5X9aZiDgflRAIziiuR8rfhigMKFI5pcnjrQcjpSuifPGcn0prX7WvmKNhH4/56VK7mPyCee4pSG/d/lTqAdgHPas9gPxpMTYjPU06bTgc0V7A9ayRxnp2oSFlEZHPrUSxj5XXr/Fnp+FRWMY+ZJt3ttxXhHStEEvn69feQo6Dymbsw/hP0rT9Va1/deXx2Ofr7Uly6XX2gjcfTp2xXiH4iS+JoE0yHRfsjDP737T5meQem0f3cfjVxot5pNisN3b+bJqOfJ+cL/qz83Qn1HXFa3o15pUim7iyGzgbhxjHoT6iprYkfaUm3KvVduNvbrQsrm9i8y1jyn1A/n9K0PxnqOhqLa1k2Y/hwpx1PdT614Ltrrx3cLaxptklz82QegY9OP7tWl81vf+W03lyR/dk25zkc8V42tdUtjDdXE/mmbdhtqr02joKsr2RWDs27HUYxVp8aI9H0QaXpegeTeL/q7v7Xux82T8hQjoSP1q8vZr0bpDnPfA/wA9qZVBIJyO4xivD/h2XVIBNBebf9ny89z3z7VpnhvxCNI0+w1K5+yWOteb5c+xJPL8piT8oOTlsDtjPevHXh6Xw3eGxe4889n2beynpk+tQTrFGSWye4xUwcp5ifdplAPBz+FW90y/L1Hp0o26l/kXKnpz0qaIAkHmleaMB42x6cVofxB0vS/CGpaHqfg/+0bu88ryL/8AtBofs22Qs37sKQ+4ELyeMZq9CmQyKmM/w56dKK7hgN+lFRt65/CvDa6U2tW41m8+zWXzeZL5bPt+U44Xk84H41rtvpaXROkX32mM/wAXlMnYdm/GiGHOazuHWm6YzTEKOTmpUXIIOanQEct17Ypl29/0pxjkH9KYZGM1ICGwD+lAZOCefpWMHAb9KIyOT+lKp3bQc+1WOnPcPgt+n/16it44I9qdq6jaKwP736UAMYHNEZ6HNbegzWB0Jo8cA9KwPWmUEZzQPGAelKvoadoY4tiDcx78jFMS33jn8KK9CBW3OOaI7E1twcA0gYHGKYAcgc+lFc4xW3PFcAZ71gHkUQD04ogEYByfpXVeayCcA5pwCeDn8MUFB5I6VLGIzyc59qICtkHNFgTkGiRjjrSuMct+GKePy/lbkjtQALYqIx7trDOelSIEkKH8DSlWOAaXk4XmoShYK6/rXlpEhZTn2qGRXG3OCOgpCwGWOD6USpGM0ME4AojFKgbI3fd9qA3EDNKBjcGyPpT26rGk6tuXnK4x7V/ZVtHpSagbzMj5zD5ZGMNjrn05p1wFMZzu9q1bwe1pYpeQ3W8nPybMdwOu6rLUJvD6GeGb96/8O304689jUl7PdxmSU5P4f57VADsDA9entX9s2NzZC0j0/wAmUf8ALXzS3fP3cenFW13JOd0ce5/TOKMEEUH2l7jEqdE2HucdfpU+qCdRDJa7X/veZn9MVbtJGdpXjsM1ZXshIt0GQ3/66ttEvoJ2lifCHGTgccfWhpEtxN80G3H8W7Pb0zUWlyrc/Zc72P3R07Z9aZNfswLXUn+zWzdsI/Tntz1x+daTq8T3qaRJP9ltJM+dNt34wCy/LjPXjj1qe1s9YsJNA8O6X9tumx5dz55jzg7j8jYHQEde2avLOfR7rySNrxdRwcZH4+tCIalLuurrygO/l7u3t9Ks7KwstAW7/tDM0mf3XlEbcPjr0PBzV0J5LgeW25j0GAM8Vpuiz63K9vDN9zGBtHcH3HpWpaTcWd19kkbdu6HAGOAfWri1kihAU4/Cp7JXKyCTOM4GMVBZTbtu3OPeo0QziN+UHT8q8H/GbVPBuiSeH9LtP3MuNx8xezM3dCerHvV8Uu5jcOnzP1GfQAf0qS2ATIXgds1qPixr7SbWwig8vyd+Pmz1YH09qhidiWaT8NtaB8JvFmpeGf8AhJrW13Wc33G3xjo5Q8F89R6Vp3huH+xHluTtuDjAwTj5j6HHSriyia4aJTkrjHHtUoaIBWbgfdGPzq1WBkKu3zdhg0UZmIA6dRUsSxHexwBVzNG7/IMEUnB5HNTTheozjtTlIYDOW+YdBj3xTyAJndn0OKQndndj3xStGsWVOG9MZqJcxl2fAXrxVtkkSKcsOlNrNxNHhpMyfxDA4/ShA8rYYZY9D0p/C+g2/h+G/uPE2NSuN2LH7E/y7Xx/rAdv3fm/ShYyIo3rz6ZqO2EgIUfd6mpbQvIVU7R69akjKHbnOKDAMCOfapFiDbfNz/wGpEUHCNkduKwAARzWcgVuwOG/DFeYuMDrQCtwDzTRIpGGyfTGMUYgy5Jx+FOR68U+SAQeR2pCfSmBPSgBj3qJcsCDQY4wr5/CiikAq+fwoQqMYfj6UhDMAGyB0OKkYpghs+gxQYOmTRdQu3NdT+9bntxTYc7V5z0qVkbCoefSiPLfJHTtUjc/KKYAjPU0Mldob9KSNpDtUdKbgcUxBxgcim55LZ9OKZGABx9K25HrTp2JplyKbPTNBc8A/hXlBj1pVIbAFMSCOOa2ljg0VA4AzQBYYAonA24qlbGDjPtTOTniicjBoMSMZoAEAYyaaR24Jye3ai2w4Iz7UzL0BolVXG78MUG3DBOaZhwCeRR55B/Sug4f9KJIXOelfKeN2PwonHTtTAD60VJB74psKMk/pRkQZAb9KkfjGM5phn5if0ooHGA+fbFTnIwGxj2q42qmQ2fwxVwctktn8KwSOKjIJzmqBA4oUgzgA0wJOM9PasDGBRAB4rAI4ogEZBogY4NFQSSOpp0YLxzRAP41IpVtpPSmXcM5ogEilXeME0EAPB6Vkk4BphkcGuSeDzQ3Bcj8KzxgnpQVTwO1SYIwB0rkcA0D1NMCy80fmABHSpMZABp8sQmfu9qPy8YpTkBhQkKjApsMS1OpYgAc1gAcc05GNgFKcEKDTDedoHSggHfNbtvy+vSg247SKiBPIpHAIUGo2D9Dz6UW7CgRjGaI5xmjyAKqRSoyDQUZz6UcYwaYBmGOtNEGbb3qSLaeDTSFlCmpoVkGFqa3aIYBrnH8qOePWicHAo/MBivu8VDK8Tbgas9ULkIw/wA/lTNkZFOfl4pXyoOOlFix3UiFxx2p2Ea7epoSAd80JTkUq5AanmyoVRRJUYFAnAGaBA6HpS3I2hfSt5cZBpRxgUT2FSNjgUCQMA1Eo25p2KgEDNAlgMUsg6AUwHUGuMYI6VjHNQqJPmzwK/dtgRmoZ3gIQnjtU8aMWnjbpjIx+FSQsjbgaYl23g8imGec1C4RtxGSOlXoRXBXp/8AqonI4oAOvJpIiwJXqtRh3QqV/WrO23pnb+tWqeYRCBz608CLLtb7wrTY7aeN1mPzDGOvv6Umm6X4Hma9uz5itjAww6DHYn+8K1jxTdX98sqDAhzs6dwM9vatT1q61Ir5y8jOOR7e3tTn7OhVmyz9Rj0rTJp9QK6bargydDkdsnv9K8SeFpfD1++l3pzPFjcceoB7EjoRWneILzw9J/o3JXp09/UH1qS8KkRsPoautWeeNIp+QM49vyFQTxmQMenersQ3Ev7v7o6daPhVrZMTS5I/2cf1rXNOWzdXRsnnjH0960nVBBIoJwo6j8/ar/xRHqdvaadpz52b9gx0yQT1Hsa1qN5SWmbLj2+lJEEUNmt6oPLIzntTR4YqegpVHQCvDeiprbSQPJt8vH8OeufcelTaLHBfm13Z2+2O2fWpLHQbeyZb05lGMff9fb2xSCM2wU/55rVbfQFtVfTx+/5z9/1Hrx0zXHPr2o5PBNFARgUCQQBzRUgnFBcGkieUEoM7etNgjipGUHNSkOmRVwhD5xT5zij04/Kpm5IoHsaI54NEMTgVYWJuZAScD/8AXUFstuMKaHIwKIAGPyo9OetAZFYwcCiCeBQwSMCsE8E0V4wDRyTgHNQwtMdoNPgHANMc9D07UMngH8KHPAPNUowAeuKYAcAde1BQGxn6UoB4JpxjBBre3l7QKJycGjkKVHeuoyBRAzg9aCgjg0Vwu7rihyOKb5sqT0oHccGlJAINVjI4rGOM0TkZzzQwACOtA4xjrRJbBJooVOQelA/NkdfSonVH3NTTkHAFPHuIZTzUdvPcK7xpkR43HIGM0hyMHqKA28A80RkbQa1a20y3jQ2ByTnP3vb1/Go4ixAXr2q+islh/wBEP8/b1pLoxoEIyKMvnRBAfwq2iaSXyga0rUrGdhb6u37kex9z/CM9cV4oj0htUdtG/wCPUY2D5v7oz97nrmlkKpkdKhnxtUDpnFWl3HGxEgyTVk9l9q861+6vQc+nvVpqMD3QaZOB2yfT2FNYwX19vjbCHpwfT/61XthawxyIrfOuNvB/GtDgSRfsx+/6/ma1OS70+08huknQ8diP8ahvbiCXdnJPbiryKeJU1QN1zgYH+7/nip/Ft3qSKkycrnHI/wAPaoWUTMp9sflWh+KJ/BN/Fq1onmMuflyF7Fe4P970rWNZuNcvnvZl2tLjIyD0AHoPStd0ubTLeGZnyJd2Bj0I9z61c33m2KWoPK5/nmo1ZJlkbquf5VomvW+mTMwXLHryfQ+x9aNzp2qSyXbHDR42j5j1GD/KhdmKZ0IyvGPyq3hjIIJyDUMKucqatbFLhiY/4On401v5bsT2qW38tBIDx2FXJZ4woryy7kZ+nFWtv9kUO5znr2qf4sWvhT4d2XhLSItwTzN53EdZt4+8h/vHvS3c8s4vkfhP4cDuMUoE18sc5wGzk/Qe1a/CkGoSRQnKrjb/AN8jNeW0K7lOaivS+4AcmpmYHYTnNNGA2T1NBthDL1ppU8zawq9uFuFWOPtnNMWc4PWjkPtzSgFgc9KMZlUuD9BUKle/XtSxApkHk9OKSIwrkHPtWieG59X0661R5cR2ezjaP4iV9R6VeuzykK3C+3sKY+VGRnJaomEiMpHNSwuw3E/hinJPGfpW4nqaJyKCAnC81jJ21jsDWOMZoA44oZLZP4U7grgc05ycCiCQRmiM9DQBPOfpWOMZqM87QajQhcE1wGz6VGyyKVQ0n7tcA5JrZkYJ4phhti9qnIXAJ69KZywC56dKkkPrSktgZ5ppGb91mmjw+1jUyKgBU5pSEPXNbstxTjcOtAZ5zWwMvFEZHvQOBjPNHn5R1FMpVaK4ojA4NZAGB1rbyQpoBWGCaPzDAoY29aL4HApTx1rdngGsg45rGRxQYjjFA5pW5xmlG44B5+lSE9+fSgMncD0oneciuce9deR+VM+Bk1v+XANEljx0okHgda3ADaTxUyxBPkPNZIGAaMiqcE067sMD0p1JXdnpW4k7aZti5qUnccGrkgYx+NTrjArdjjNRSKBkL+tK5kO0D8KC54xWPQ0rY4zXB4H4VRAJwKI5xW33og7cYrBA6V14FCM92yfpUiMDknP4UV3nCinjwMEc9qAwMA5oBQckfhTIGbcrfhimBxhTnFEbOvNeYCo7+1Mwk4zilAAAByPpTqAMhs/hWO279KYjPA5qR8jGPpTyqV24oIActWVBAHJ7mm+b7vFbQqcHOPavvdqYoQGU8nqKLAEYP0FNHswuc05CscL+tcUp43A8jqKCkjO/PtilY7ulAgtuzz6VC2RjH4USByo6UjqpABzSuG4NHpgV1GMfSqp49xzu/SmhYtweKMbdAc/hTIO56UFYNwePpUigL8o61KpUYLZz04o4PTj2poRKv3unbFXFnJHyoyKbK8Vt5zms+vagSeAawCcg1bymJw2OlQTJcxB0PWioPGaZVUAF/wBK2qeA36UXVV2pWAMUxBGBS4ONw4FGTKYA47UGI4BzTFj1b9K44welZ/hpRggk59q6DA//AFVBFLOwihj3t9cVOjQOVlG0r1HWpYyfmWijY4FRhyMEcVhcYAz/AEotgcDmhtbBNHJ4U/hSnc3XFKTjIOf0pHKAhBjPUUBg4B6UJ/MQrKOR0NQOjoVZuB14qV5FTypE5HfNbcd/xplYDGeKAVeozmhGZYf3Y3EfhQQOcBufpUFjPcTLbwruZ845A6Cr7wF4s8O3KRarpXlk548+M54H91j6ipTosluo8ry5os7o9zHdk+vQcUJZAggK4Ze2a03TbGW1IuNS2buv7knHJ9DWkaFpWpRCXUNb+wMP4vszS+vYH2H514jj07TriKDT7z7Uq7t03lsm/gfwnpjJFaRZ674gkNppkX2or/DuRMdT1OPQ1q9lNpOpyWV7FtmjxuTdnGVBHIz2IrwzbaKiPc6jqO48bYvJYeo6j8DXiq7tzr0w045iXbt6/wBxc9eeua8Maumm6it3NHv25wM4/hI7D3rSvEumeIrHUdQ8YS/adUufK2T7WTG0kH5YwF+6FFeJfDdnZW32201j7Rn/AJZ/ZyncDqT70/mbNqjJHbpSSBsIxye4xT2+x9yttPYYzUKtdzJaxjMhzgevGf5CtUvNX0+Q2upN8w6jC+x7fUVJKLpiVO4np2pEKcscn0rTtZuNHv4bq2XEibtpyO4IPUH1rWdRn1OcXNwmWHfI9AO30qOb7OS3k71f/axjFNH5yedG+30GM1HJovhvWNX8Pa34Na61CdrdYFW+cGE4zgBMhywZfpV74W1SW/eOPSZ7YyAFIplKHAHUb8Z6GikquFcYx2qK4ZHDRnOO1XG5xkNuP0xRQGAs7bSvTjPetQ8NzWWmLqQut6NnHyY6MB6+9NEVjd2XkYxzTrkYPPpSjHbJ9KltoxCZRLkjoNuKG1xwea2KUyX59MUlzPbBhC+3f1GAelMvpUihVyeTRUFck9ParpVKZ3fTipB3B+nFFSeQM1LwQRQYsfWs5IBFQW7ysFUYq0thbxhQcn8qLZ6VkkZFEj8RRIIz6UBjit3oKBzgYogdd36UQCOD09qAJwAMmgu3mixPANY7AVt560V4wDQAxk0EIOc0CSc9QOtHLcg/SlUkkselYHQHPtRCnIA+lEAcA0V3HIb9KhfY+7qfSsqBxWAec8+lIDgjrSkKpJ5A7UM4yD+FAAgnvWQOPTpWQOh/CiNwyDQGBjrRRepoqSMrzTMGGAmMd80oRzgnH61KUVtgbd+GKRtx+Y5orhsjqKbnGRQPrzUQ8xMA4PrXhbxVH4civ7aTTfta3vl7v33l7dmT6HPX9Kv5luJ3ngh8sNjCbs4wAOtEjhQKII6ilIEYIXr2z0qN0KghenvRvbD+xVt/sWZxnMnmH+9np06cUpBUBuTSMufkjyPTNeD7TSHv/tWrX3lRw/8ALPymbzMhh1Xpjj614ufSDfKdHP7vnJw3ov8Ae/Go1eYBVXn0z1rTdEGp3aWs159mVs5fy9+OCemfb9a1/wAL2Wk+JpdBsNd+328OMXn2UxbsxhvuEkjBJHXtmpEjgleBH37cYbGM8elafdRRorOM5z/WrrVo412rHhj91s9PwxRFyi+ZC3A9hWi3ltIwa/kwR7Hnr6fhWi6hbpefaphwOg554Iq71CPxNdCCJPKWHoM7s5H4f3avE2y7C+D2GPart59kcEjYjXOBgc0IyjhYjuHp0prMQxpJPF0zt+b73r06Vd2j37KAuxewzn/PSreEidWM21TnJ257VfkXLxxS3f2jbnaPL2Y6VqOmxaJetBfQ+ZIOo3bccD0J9RV44u5fKt48Fui59B/9aptDe0Qz3dx5Uo/5Z7N3t1B9MUt88l0jvHu254zjPFGxeW1Fys3J/g2++OtabDJPLsEXH+914NPpV3agtNFj0G4f41ptrqErqtumVXO7kcdcdauoGmcx42sO3Wrm3kJEdwNmOh6/yq4iit4wWXdnpzirSyMjmYSZB6cVJOYpVBbIGeMe1Xc8hjVBLvxnnbiop5I4TgZXuKis47uZUt7zD84Hl+3v9K1rT57Z2EjZkHsPb3qGZkO1Ex+NGJjLleAe1T25PzBvocVKURdud2PwpyWPTinZZ1ZIxkjv0qKMrJkjOKnKuw2DFJDvkCFsE+1T264BVv0rw7or63qEGlpP5TS7vm27uik9Mj0rxF4Gk8NQ+e+ofaB6eVs9B/ePrSlGUKq4P1rwX8Lda8YXRsYT5TL0OFbsx/vj+7XjCx1Dwjcz+EzecQ7fOHlj5shXX1x97saTSb2e3NzDFmJerbhxzjpnPWrWya4lRg/Jz8uPatTsVjaNbSTe3O4bcY6Y61LDIybt2Se2KdCDyc+1bc8LzSgD5M5zT27RJycGpIdq780VYj5RRGBzSk9BxTsqHBOT27U7EnOawDzn8KYYGAaUAgDOcVwcZ60BgAZqJSDnPSkLNgk0wAwqr17ZoMicBOfrW5Y0Dhck+9NKD1Tn61vKHnk08nm4BHSnZCcKvT3pgWODSgjvW7bz1oysfvUSGya2s3IX8c0Bk5FMA5wDSqM7evpXhT4V+K/F9t9q0TT/ALQP7vmxp3YfxOP7pq8tJ7Kdra5TZImNy5BxkZ7fWmAK4zQ2leD9KJAXaRmmOeAelMwBGD07UxyMg1hjwG/SipJznNYY5w2MdsVhycE8fSmDYwGz+FckYVv0pRngHPpRDYxu59cVgjq36UeB1oEgYBrPOaDEHINFwykMeaJ7EUCABg0wLcE4FMwHQUWyuMZosMcVjI2k9fahgHAOa4UfWi2RgGshvut+lF0I5H4UXIG0Nx9K3gLtK5/GiQAGqefaMKM08pboanb5QKmXdGrA9K4xjFRnKDB571AAMA9fSlII4NEAriguDmgMjbituOh/SmX5hiiD1NHnrRHHX9KJJ4zmiDjGaogHvUikHK1tL8selMgyApp1wcCsADgUAM4FBSDkc0u1VGe1bl3ZzTqVOMfhWCOo/CtpakDq+GFFgWw1HnoPpQJK4brQQkjHasDbxRIIxmlJRgQMipCCcgdaYLwFPSgWHGeO1MCWyOaOCMk8/Ss8YBrOOhoIWGRUm1hhTUYBTI60V+XilUqc5pJeeaDKw+U5/CgQBihyMURkZArtgGsdqkX5eDRAUgmiB94GnkDR7FPIqRSy7WPPapEIA4rBUgg0jnG0jOau7beTtGTTW7xnJqQjIANdTgH9KI+ajnPXitFuQreWW6+31piCM1KhZiRQIxjNEE/donIwO1ckfSiwIAx06UDgYoAAYBoBc5Jo4J4NEHpS4cBc80q/wk1pGoHS5zKi7s9s47H6+teF/EulWHie31PUZNkHzeYcMcfuyB0GepFfEHVNM1vV3v8ASpN0Jxg4YfwqO4B6g0wDDFAZGBRXHINMm7jPNMBkLnpROBkUxVjgHmgCq4AoEsvSgT0pWI4JzSHDgqakdbhMD7wogkFT17UBkCnQk8CrRzG5UnrVyCJ2CdsfyrT4bh5VlhHK5x0968GfELWdOvGuXg86QYyN6rjhh/d9DWteRqN9c6l92eXbleTjAA+nQVdpPu3KvPpkVFqPkqA5+tT3skvysflFX+pxXEcEUTZEe7PB749q8O+O7rwtdNc2Ee8vjI3AdAR3U/3jWt39zr+sy6rMuJJ8ZGR2UL2x6elW6l28s9aubIpdM4HI7fhUkb7OBUNzlAh7UzzWbpJMcqc44H9KvGe/v5rsLhZNuOfQY/pVzAUfdH17CllWQYkOD6UzmFfMiPToakmec7pn3E98Yr/glv4H8Gal+zvc67qXhLRrvU18TXSLe3FhHJMFWOAqokILYBJIGeCa8WfE74QfCUwWPjLxx4X8KvegyQW15ew2rzDoXWMkFhwAWAx0Ga0vWv2dfjjDNa6VqHw98dKylp4Ea01FgOM70+Yjt94elfHz/gmf8FPiRpd1qPwtsYvAfibaXgNruawnfsskGT5anpui27c52t0PxD+Gni/4X+MNV+H3jbSX07WdIm8qeFiCD3Dqw4ZGUhgw4IINfsGfsZw/tH65feLPHzXEHgXw9OsM8UDmOTU7kgN5CuOURVKl2B3YZQMFiy6Z4d+C/wACPDxn0/TvCfgbSY1Ecl0/k2KPxx5krY3tx1YknFaP8Tfgv8TA2ieHviF4K8V+dlWtLPVra+38cgorNnj2r9vk+A7H9pzxP4e+HXhjStD0zRVgs5YNNtlgikuBGHmfYoAB3uUOAPuU0MuBcKv7uP7xz0zwKdmkbavWo43jyDXhe40JtKEeo3eLkdU8t+PmPccdMVc7yhAHApkKjB60AxUAHpWwmPBpIjnYB9Kk0q5ZlijTeWzjkDp+NSq6MEdcEdRTncAKYEjrUiFwFXk+lSsWi5HNOcNgdajYd6n4YHNEg8ireIyyBV7VYWYjUFhz/wDrpx6Vg4yKySODRyeM1jnANdegojHGaJLDAoAY96wx6HmgABtJrO45z0oAkUR6t+lAcZz9KX7vFDaUKg/hTAgYXqKQDdhulEDPWkIIPrXH3fyrpjNEDPNFgVCk9KBArII5NBiMEc4ogMMg0z5TZmgVIxmiMmiBjINYJIJPJ7V0GBR6Fc80Sdu0muAMUFAGK4K/LRUDBJpYmIDLyKyDgKeaGe5pQXbAP0qNQicnpSAEZU0VyMZrGDnPNQYaTDfhWr6Dc6QVSX7pzjp7e59aijbaQtDYQEBz7VcW6pGkgPXOOKt12tkmlJ80jr6flTBmOP0qxkj8+N5mwoznj29q+IFp4asZIpfDd35indu/duv93+/9TS3TlCpOSepxVpIhkGW/Sv7P8q0guWPyybsfgcetal4burWOKeQcSbtnTtgHvWpyaZZTfYrOXci9DtYdge/1qzitTcBpmwn0PpUscQthLbtlj0GMd/etBnls7gyIf3h7fga1O3lWdZZh6/yHpV5eWWpi3iV+V3ZGD/ntUjPE25TwK1VP7QjtBAd3kb9/brjH8qnvI4IjAzYYdOKmcCQxIc56VpNpay3Kx3L4XnsfQ+leK9Fjg197Kyk8wcc7dv8AAp7mp7VLPcqn98mMD6/p0rxHq1v4gm3wvnf14PYD2HpVza29peCNGyR2wfStjkhAa8NQ22lXsGoXB/dpu3HnuCO2fWvHXiS18SPDJbDHl7vXvt9QP7taFeJaRyxsPmfGR9M1eFGZ3J5OKuZAICCck/41cpPcKvH3c46U6bYFgVuRnPHvV1H5agKfm+lWaoLhFmOFGc/l7V4luLBtKEFi+XPTg/3h6/jVtYStZC+Q4b/6+Knv5WVWkPrk1JKznEYpcrFxyy9qlQFclv0qcorBIzn1q4jduR0FRyQwblRuO/BolWO5TQgLqCKWIAAqOlXmgweHt2n62PKvExleW689VJHQitOln0ucX0S9fucj0IPr61rmrzXxMRbKj2+nt7V4TNiNdtG1N9tqN/mHBOPkbHTnriviv430641qG/8ACc2IV3bjtb+6gH31z13Vq1zNrdy92X3NxuOMdgB6elaefK/dTviP6fX0r4cXeneHtXj1TVWxEc44b+66/wAOT1Irxdplxc6ld6zaJm3k2beR2VVPU56+1Jp08gVNvrjkf41f+G720UNPDs3/AHfmBzjHoaexdAFI+ahG0L7iOR2ppGZ+efSphvtwT1H+NSNAseEb5vTBqRwx2iiVHBPNMSTnNUTj+lBfWlA6flSAE4zQjAGAaSPPAPNSMSoVegqJwGGOSOtOVJwDzSSBBsc8DpTMDwv4UItvDmn2k4U0CCoANY2jINFwCMU6iTDKeaKlcA0FA70G4wD+lMhVQwNFcnJpiHOR1rwd8UtS8H6Z9isl3EdBlR/Ex7qf71X1w1xKZGbJPXj2FBScAmpAA2E6Vx3pmytOpduKAPTFBTwB2rA7UVOc4oqQORW044FFTjigpIrBPAraT0FEcZFEMeDWCMEigentS8/KaOOmaLAHaBRbP3aPP9KGO9bgBgUSCP5UQAMA/pRIPWiBjg04wQQafaT8pzTBSODTMDxmpHAGzuKmZSAM8/SsAjOM1cDcOtAqU2k0RjjNIu9shunbFRD5hihkd6ZiwxmhyAA2fbFKcrwea2k856USWGSenSjkjkYpueR+FHJGAenagoxjNEEjGaxxmiAQMdaI7GiAwwaZAWwgp1C8Bs/hSqpXbnP4UFwMVt5oqTxinQEYZsY9qfcT8rfpRwRgnNLyNhPT2rJUgKc560QHO7bz9aAABGfpRDdEOKVmDDBo4zyM0V3HI6elMSTgtnHTigg2gZxilVQ2WOQO1AqWAIyPSpQuNyDj0ooSMA9K6HBFEDHBz+FBsDAFF1fgr+tRqVOVP4YoBmHDY9sUGXGAeaJO7BHFK4U8GkcMBg8/SqBzgFunTisc4BoiiMVIm9cd+1MrD5S3H0raR8obn6Uq7xh/zpzuA45p41ZSufwpSR93rRDSDO7J9MUnljKyRb/QbsYq6t/KIAPFYycZojd0PSjgnFW0nlSqVPr/ACqOTzIg3Wnyy4K/hmmAxhRQwBgGiBjpmiQRgGtuBjOT9KDADNKcjGa69D+lZyMUNueRmgivkqcY7YpIscg/hipJPMznp2FYUjAODW/KgkUwJIANJnHPP6UwBGT1ojHA5pmYPwcj0phuG3PFBABkHNKpByG/SuGOAaBGMGvlxjdn8KDApyentTI4IbP44ouSNobOenHSjmghkQgDpTRlflA57VHFOoKjj+8OKtHnihAjb8MCtL1nUPDjm9sJMSSffXA5xkDqD6mhcJf37308f7x8ZGfQY7ewoabcvmeY/IOnT6etWPhjRtYt3uJZtjR4wdrHqceo9KufB2ovcGK2Xeh6N8o7ehatYsp9KuJLCZcOmM8jjIB9/WoZlK+VL8w9OlWiqTslb/dOKluNN0uZZrRPMznIyw7e+fWtW1kahem5ig8pf7m7d2A649qaV3O4j6UpAkKv0bt9KVPJl3KuV+vtXhy50me9+zaxDm3X7p3MOxP8PPXFa2bEarKtm/8Aoox5YwePlGevPWpoow2EGR6UjAkQk/Kf0pGUn5+R2r/glnHGn7Mlx5bZDeJ7w9On7qAf0r/grNa+X4u8D6nBd7Z10m4QReXnIEy856fxH8q8PeJNY0HVrXWNH1O703UtPkElrd2k7Qywv2ZHTBU+4NfsP/tEX/7Q/wAHxqfiWWN/E3h66+wao6Lt+0fKGin2jgF1OCBxuRiAAQB/wUe+D3w01nwjYfFfX7JLfXbaSPSor5ElYurMZFVhGQCAqzYLDjdjOcV/wTWOhx/syWthpEySXFnrd6moMBgmYlXGRk4/dNEPwr/gon8Bfil8Rtc0Px94Z8P33izw7o+lm2n0WylczWcwkkd7hIl+aTzFaNTsBYeSOMGtYhmstduI4LWaye2kA8l9yvC6gZB3YYEEHr0rULy91S+nv9Ru5rq6uZGknnnkMjyOxyzMx5JJ5JNQ6pdJp02krLiG427xtH8J3DtnrW+Qnex+YdDUdyJI9sh+Yd6RWRcbs49qDtMQgbaO/Ga8Y/D/AMO+HbGO50rxT/aMj5zH9heHGCo6lj6n8qaICIyrx6CoyFOGb8MU6ByXU4/2a8J64PD17HqDWn2jbn5PM2dmHXB9a1a4W+vZLpE2b8fJnOMADr+FfZXOCRn3qWIocZziotkJE2clei9M9qmDKm0CpM9R+FBQeCc1cYJC/lSqSQFP4Vo1ojbnfquMfrQAHQ/hRJPINA9gaZs8Z+lBVA5b9KYDsaA6c0RnjNFRjAb9Kx2DfpQYqdytz24ogscsayTkA/pQyeM/hQUsMYz7UQccN+GKYEKBjPpUQL/Kx696ZQp+Vs+hxTHA5OfTihljuYUqgEgHP4VjJ55ogk5J49MVtU8kdOlICyhifwrBIwBSgFNpXOPfFR8ggPg9jilRm5DZ/Ciu3Knv0NFR1FSYCja24ntjFHB4AplDMMGo1DHDNgfSpFw5CHOO9FWAOT+ldT0rGOd3P0orxhaKgjkc0juF2K/H0oqc8Gvs5K7jz6ClJjQBX69eK2o6bQePpUUTNHkduvtW3jOelA9P5UkLuN4TKr1OagcoTuGc00hj4U5HpioEd5Nu3J+uKnhKuEDY9RjpUJZmwWyfXFRMRLvLZI74x2qFlyTu5+lJZwXFo1wJ8OuMjYT3xUsbKm3OfeirA7UOMd8VBBIyNIDkDGasIY5VAebC9vlzWmW0F1cCF58Beh2n0NT6bNCDJnLfl/WkEf2VpGbbKMY4z3/wqMQLpylBiU5y3Pr6fSoHMZRY5t7jO47cVLEl3aZafcw/2cd//rVp4guZ4kuz+7TORzzwfT8K1W0tFnC2Ayvpz6D1/GrVby0tlUTeSLjO/wCUNnaeK1awnWbfd8jt054Hp+FQXIaXY8ecdDmpdryAH7vcVNZusivF17mpoJHjNuWwH68enNWuiyXEgtSMZ7/r61qWgWelLJJMN8sWNg5Gc4z3PY1a2Tlg+eOwxVxb3sOmyBXxG2Ow4w1GyieIAHcfyqDT5ox9rz+6bp7449alha5ZgIcnt82K1exmsCIZE2HuM59P8aVgYSqLz35p4yg8zbkn3xVrFeXk4igg3ueg3Adv/rUwM8u3dux3xjtU2klNIbUZD8sWML65bHrUUzQZSF9ufbNXJMZGT1q1neBMCHJ9d2KEzKXB+YtjPaiqOu3zP0pfBetvpaa7NZeXYyZ8qbzFO/DbT8obIw3HIq/eMsIwdy/l6UyKnyA1bxswJJ47itStdKWKKLTUzIc7mywz07H2zUmlW+n6exuY/MlfGxcleh56H0NfFb4keIPip4vufFfiaf7Re3O3zH2omdsaIOEVR0Re1Xd9dXVtBa7sRwbtvA4ycmmhAG4vn2xirItayLMqbm5xzjHatU1SPUdMW9umzdLnC4/2gOoGOgFXEMZmMiSb2HtjHFWNvPeR7IoP+Bbh/L8K8H6dpV1MI74+XIen3jjhvQ+mK1f4dT2miSarJbZVcbZd4G35gp43c+nSpJbtZlnsodxjzj5gOox3rxj4213xlDa22unAs9/lD5DjdtJ+6o/ujrWp6fHp90Y7W585OzbNvYdifemtzLJuZ+vbFC3ZpsRjn/61ajHJCIxIOucfpTAHoaZQe1EZOCaIBAwelUVB5zXH/wBahgHApQSQFGSelKFDAn5vbpSNGnzKMsentTbmQEtn2xilGF+U4x1oMG53YP0oR785X6c9Kt7XDBmbn6VdFQ+0HPqPSn45oMpwBzTgscCmSQAHGaErKOvI6CslzknNFSRgHgUwUcK2fwoAsuM5PbimXtnJ7UoywBb6nFSIo+6+fwpnyME5x0pcg5BxU7AHar7vfGKkPvTKScZraBgA/hXTjd+lHjHP6VndwG6e1F2BwD0pnLDlvwxQckbQaVzjBP4UpQrgilwBgNz9KjAzwfwpl3EkUV3Hg5FMBwDRUkfLQXIyTyO2KKgjg9KYKANp5rrxmsnAANMGONrY/Chk8EURkYBogGjgjAP0picYB5+lM+7APUVJgfdpiAcj8Klc4469qkkOME07ZOAadwTsBzii0WMl/wBDTtFjCHn0onPJNWqLsJPWo0UAEdarYCOPyrHYUCSMA0QAOKOCMCsA8Y5ogHpRAPSgpY9acY+XNAcYNEDAxRIJzR55rGOlEA0M4BXtTJkYAz60AW+UUfb8qcqQAaKALj07UwQsADQwhpmUjI/KsblwooqAACcmiR0Jp9hOAaC7zwa2MigZzTBSuSaLHYFJ+lFcrtz19qEIRsMakUjJVuPTFYU8A0yc4z0plA6GiBnAqojhODRGEznmgGbAUdelBWB2kZpwD90UpbAUdulJISduc1WcDg10PNH5uAfoKp4y5yaWMLJtY0/BIXnFSYLbV6imTC5xyKdcHrUUZb5hRwzBavbZJIgF+8P/AK1SRlDtNAEHANYAOAeaJA71pdyDGEY9Pb61IARjNNtX5aIB5BoEADmiEYcGgecHt0pyg6mt24/L2pmJGFojHTvRIA69KhTzAG7VK4A2qaO0KADzQIIyDz2plJbbnjtSkDCk/pSYJ4/CgmQT6dKIGcA0wIOcUQCcGtoPAP0puEwOtKcjJNAkDOaCg8GsYOPzqOQAbX6dqUEYA5PamRDGHx1oEgYXr2pstg96t1aUA7sluvFeErO2udWgtLw/I27PXsrHt+FfGrwp4U8M63Lp+gzeYY8bvlkHVIz/ABE/3jU9jp6aXJKh/eDGOv8Ae/wr+1Ly4hwJvlHbaP8ACoNTvIHKRzYVu20f4V4Q+JF14fv2v1Xzc4wuQvZh/dPrXi3WX8S6/c65NHsM+3K5zjair6D+76VcNDFLtJ4/GpJScorZz04qCdUGJDkU0kbMSDwenFEll5HApT5gVQ3P0pgJE8sPz2GK023guHEROfz968ReHpdO8tyuFfOPwx7+9RsXcAHP4UwjWYq3T/61RuVXbI3H0r/glVn/AIZguATnHii8x/36gr/grnK6/EDwFGrcHRbg4x/02FaddeBrrwrcW11c7dWTb5KbJTnMhLcj5fu+v86/4JD6fe22n/FK6lD/AGWWbSY4iTxvVbsvgeuHTP4V/wAFEfD134o/Z5/sXToFlvJtdtDbg7BtZVkJILEY+UMOOefrX7OH7U/jD9k3x5eo1p/a2haj5a6zozS7N+BlJYmGQkqhj6hgcHsV+DX7XHwC+O1vax+CfH1lHq1wozoupOLW9Rj/AAiNj+8I45jLjkc18Z/2avgx8fNMlsviN4Ls7q8aPbDq1uggvrbjAKTgbsDj5W3IcDKmv2uP2Z9W/Zi+IR8L3V0b/SNRhNzo+o7Nv2iHIBDDJxIrcMPoejCo9Mujp0upKv7qDG48cZOB3z1pkEjjaakQxYXFQSb1wx5oW6q4Ze/apr2R1CyS7j2G3Fapo0Nr4c0rUB9+787eOf4XAHetBbSGsfst0379ugw3Yk9uOmK1rSH0qdoJBhl7fgPc+tRuQduc0xDybQOB3ppxHbtER97HP0NSKWG5jTklTtNXC4Gc1KuByaUAjHpVwAWAHUdKs4RJIOKsoBFH05NYXpRBzgGgoIxjmsAdRRUkYFdetEZPWgBjNMPemQg8/hXB4z09qOScA0AOoOaYAAAGkxtw1N83Q9K4ahhOR+Aotnp+Ap1yOuaG0DANAKTwaLApigOSBzTkDhaKrgYNMc9OaDEDOaEeyPfjNEgKCDz9KY7uAeRQII4ogt0FHBHXiiSDx2rHGCentSgkZz16UqkjAPTpxRAB4o5AHtRzilQkc0yqBiicjaOtQzhRg96KlxnHTpSfPDx1HStLvTa2ssZXd5uO+MYJqYqx4HWtS0W602NJZ1wHzjkdsehPrVmssrfZ4xnd2+nNQ4Ykv+FGJZCVU9elW92IHyBk1PI807MB/nFWqKrcnmnUxrkfxdKiRm5Dc/SoWdCybuDjjFFyUww4qVRv2g9aOhXVvprXzL+6HXkf3setRSgcJ07UsqWkaOrYY57VbeIZJM2zrkDvn8fSmSe6u28jlF7celSb4rVIyvPPf3qC3nVw6Dr24rSLRQN8n3e/61q4sY9slk+QM7uDx09atJWVhKxyBWoX73qQANjy92Bj1x/hXxAggXTIbi0fLDdng/3lHeob5y29jwOtePbvRrKHSxpcnMnnebw3bbjr9T0qNwyAluvTirLzIpwJhkHp27e1C1hhhIi++Oh5rVVnlfdKd2zr0HXFacr3MQYDj/8AXTQXEsCxOfl5449agiEB2MeT0FXFvOtrDbIeE3ccdzmvA1l4esLyGfWOUG7I+f0Yfw/UV8aNb0LWPEN1c6Mm2BtmwZc4wkYP3gD1BpA78IaM6yQhAOaS5ms3EkPysOh614Y0JdVdlAztxnt6+49K8QWlvYRvYvNiNcZXafY/1qYKz4jHAqbRZprMXir+7T7xyO5x61dMkACqeaeZiu0GrSUQuGcZArU/jJqd14QtvBIssW1nv2P5q8bpBIeNmevvUpDsDimgEx4HTrRQKm1RXh94YL1Zrhcouc8kdj6Vqt1a313HMgyq5wOfQD+lXChnMjDGKiRZ2Ck/KelCyjWMHrV2XjRA3B5xRlLRgbv0qKxFxGjxLk855qCyt7PTjIZNsg7YJ71YtcQaqjk4HPYf3a8HfFezl09tG8Qr/o/Hc88s38CZ6471Haw6jeS6pYp+4XGBk+m3vz1Bq/ni1a7LGDbs6fMT1A+npVl4Vm1aFrpEyi45yPp6j0q70qOzkeILlhjHP/16WwgWPzAOe3WtZt5RsZunO39KZOORTqMciimea2jtW0dPyoqOgNbRjANADGTUTANu9K8lljEg/Co1BYY60xK/LUjk4UUhVhtHUUXYrgGoXcsEzkVdcSdaZwEKd+1INowTzSgoAxGaaZmAyOnaiAx+U81GhK8is4XGfwrGeBQLA4HWsEjBP0p1LDaDRBI2imUFQBQAbg075703PfNFeM5o+oopuNFWLcdKZtgwB+FcgZI5ocituDgH8KIyOlYJAxUfzHAFMw6d63E8fnSjC80MOwAPSmQKcZ5o8de/am5GBWMNiiMcZo4x1oMDxmsg9DWQeAayuevNF13cnpTnnANEA0ShHLfpUzLjIPIqR0xjPNSMHO0GppBt2Kf0piCODR5wBR9BRI6ZqAg9TzUUin5Sf0rI3bc0hUgYrDE8HilJIxW0nk0cZ2itoJoqCOBzW0HrRAJyBTDcvA/CiN3Q/StvvTADpRAA5PNEZ6GsVjI60ykHcG/SlVHGSMmmQFioOMdKdAo47UwDAYGawAOn0FJwfm5qQRvwBRULwBmiSB1zT7nUbW/StxA4OcVu2kEt+lM25sigRkKW57cUqryvU9qcKoyDTIhIK8mnjGRk/hRIK5zmlCHkDp0pQGIUmgEJwDj0NDb0IzSAY+Q0zkgDPSlJ6mgGIJBo7FOMUQWIKHHtikG0c/lUcoxhj9OKBB5BoD0oD05ogA7T2ogYyDUiIRk9aZQPumpDtXAHNBlYbaSNNvK5Pbmim1SwPNIpBxHwRUm5cO7fpV1apLHlDyPaiAOK2+h/SsDseRUErwsCGx+FGXzEDIf0phnqf0ojnGaJAGSelZYHG7P4Ux6gnPtSgkdfpxSDjG7PtihgDaTk+tKu1eT09qjjXaHJ/CmkLNhWwB04oscj5s49qwTglsnvximUA4DfpRAY4zTBVbCn9KU5Hy10IxUoXOAMUxxwx6dBTEEcDNDA5H4VuOfvfpSMH4A6dKYAjjqOlAkcZyaHTGM+lEDtzQYE4xxUUjLgK34YpDGWHOAetKuH+YcVo01ohLNFwnbce+avPBc8vhZ/F1vb+XDDjI3A9ZNnds9fap5ndtznOOtWUu6TY75Ru2K8SeAPEHheJJ7+x8uOTOxvMQ5xgHgMf7wppZGj2B+fpSzFBjd+lLLGy5Z+e3FW3i7V9Msha6bfeSf+uSt3z3B9TUlzOHJZ/mPXgV5wI2k8/SkY7wSaSGJDuJprJkHnEfIelWU2kRJi6h3N67mH8qtZYIDvKfN65NXmu6pr+gy2cg32645+UY+fPoD1FXETxP8AJ8o7d6+H914AisLpfFmifbbltnlSfaZotvLZ4Tg5G0c+lQoJUCFssfbFf8Eq4Gt/2YrqJjz/AMJTeH/yFBX/AAV6kK/Ej4foO+h3P/o4V4I8J+K/H/ie08L+C/Dt/rer3TbYbOygaV2zxkgD5VGRljgAckgV+yV8Bh+z78IrPwtfpCde1GY32sPG29RO4AEat3CIqrnoSGI61/wUr/aA1LSdV0b4TeCtVVLjT0N9rLKiSbJH2iCMhgcMELufaRKuvAPxH8T6Ff8AxFtfCuralotncC2vdTtrJpIbeUIpCyMgwnyleuByKtWa3cSIxGDlSDggjuDX/BNDxv8AFrxx8B7q6+Jl3qN/ZWOpm30DUNRLNNcWwRSw3t80iI5Kqxz3XOEAH/BYCTRv+EZ+GkU7R/2oL7UGtx/EIfLh8z8C3lflUt3ItsbYS/u5PvDb6HNPG6NgNkduKO1rYhR83f8AOjNNNgSPuHbjFWrhnaOX8KmtyrgscjsKclGKqeO1REhwwPI6VfPLcNunl3Oep249PStO02a9O2Fdx7cgevv7VpFxZWt2Lu4h81fTcV7EdqXXPC8zTtcaPvZ9uw/aJBjHXoKkMbR+WoyfWo0gELlzlxjHWrrJ+8eRToOvWlGep5qVCzgZrSbVCQSM/wCTWMDFZzwTRAPAFEAY56UwHSuBx6UQD3/CgQRWAeQfpUUZlcIDzVykK4VF5+popyBmuc4z+lHKnBrCjkmgMnBPPpSqpG08UwB4ArBPANJD5gO04x7UqFmC+lMvYCkOG2LxT7FTYBz60QCMVtA6CiRnb3pwSBg0i5ABNYIH3s+2KyCMdaIz36dKcBcBTWTjBPH0pQvAzyKlfzJiw6Ht+FYGeaOAMA0DxjNEAjGentQUdBWCRgnn6URlgM5HpQXcAAOaRG37ByTUkLRthOtEYAIPPfikAKZzz9K0jTft8UiK+3y8cYz1z7+1XNu1vKYmO7H4UcEYzWm6ne6RcrdWM3lyLnB2g9iO4PqaBAUAmrdMyBs9O1fELwppPhTXH0zSpvNVcYbay/wKejE/3jXh7ws+vXflifYF/wBnPY+49K1LSH0u8a3Mm8LjB247A+p9annkJ2MchPu8Y61G8f2XaF/e+uff/CkkJIRv4akKtDwOahaRSHU9KvNQuru3EM0+9F/h2gd/b6VEEOFHXvUUsSgGT59v4Usiud3erTU9RtUC2FzsRvvDYpzjp1H1qG183SxeSndnt0/ixSoZY1eOLkZz81aaL1MQSNsU9sA+tX9jPN9qezh822t9mTuC43Y9eetRrJHwItg7/NmtA02HVpYYbY7Zfm3dTng46kdhXxG+HmteCnj+3HcsucH5RjGz0Y/3hTXazW0NrOm4R7sHOOpz2rS7CzusiWHcB0+Yj1q9toTIwiixGMYG41aae8xURnaPz/rVzbRufMjGG9KVJDGUMe7f1GcdKgtZom/1e0fXNLCjpvVeV9+lDSpi/myrhfr/APXq8WQl3iG4nHHSri8kfiY8ir6/Y3jWyNuC4wMY7A1pulT3gIgg3ueg3Aevv7Ve2M9s5SZNrDqMg/yq7UwbXjPPNWd5dxRmW3fBPfA/rV0LmWI3E8m9n+9wB04HSo1aNPlXg+9W7STRNbH5kONy9PetEtLeXVooL9f9G53jn+6cdOeuK+MenfDey1a3i+HFl5VsN3mnzJ2z8seP9ac9S/T/AAq6iDfOgxjtRRpeSeR3pLYSuEUU8JZ9iDg9BXhvwhqHia9j0+1TcxzxkDsT3I/u1qVktjM0EI+71P4D/GjE/l+exyF7fpRMc3zOKivn07TZkt5sCfbldv8Adb/69T3izRBAfm9cUYJpxEVOetWNnBBcKZ4fMUZyu4r29RXijUNC8QR2ttoGj/YVtt+R9oeXduwf4sY6H86t7m2htfsUkG6bs24jHOenTpXhCG1jubPT9XtvMt7rf8u8rjaGP8PPXFeOP2ffCOk+CYPFOnaf9iifd5n72STfiVUHWQkYJPbvWmfC3wX4d8GxeH59C8zVDnzW+0yj/loWHAcr90+taj4L8G+A9aWbxJYfbouf9H82WLPyj+JGb+8p/CvAvw5uPG95JbQPtXjBwDjhj/eH92vHHgy58E+ILjQpX8wR7ecBc5RW/vH+961b6Rd204unTZGOhyD2x614ru7zWpYYZpfNS33eWNoXG4Lnpj0q7tEVwqtz6YqcmJmijXCt2zUsewYY0QM4BrGeAaZecUoHTPI6Uqlh1qKIsQqjPtXlnGHP4UyKmCp69vSgzSR7T1oRoCEI59aLLGMIc1IEXBH3qLHOSakfzG5NLhvlU5pQF4I+b0ph2JoqCuCf0ogcACkUYApwMYUc9qVQSM9KkRATtbI7DFMcjrzW5lIwaYlup5ouo4ppBjAP6U0gzlT0qN0IywzQGe/6UQAC2/p7UEZuC2R2GKC4GCM+gpg55BpORyM0wGRtPHpXBHBogY4NDA7/AIYrgdqJUdB+FN8v3T+GKGCcg8UpBIGabI69e1FiwAz06UwIGO9Oc85rAIyDWO9Y96IwODROcAn8KK9jQXn5m4HQYplj6rUsigZDcfSjM2cE5/CnkU9etSyKzcH9KlfcMA0EB5FLCjDBbP4UYoxwDn0plC8A1jjGagjBGSa2EMCKDblDKOaRhwCKHzHAP0ox45NJtIwT+lFcDGaI4Bz+FFQBwabGPlontmjzyKxk47imGR1psHjNHBBNEcYz9KJBPrXbANEA8E0UHr+lHYT7ipMEcHpXBpgjDCnpTpnoeaYBVBzXlgqCDk+mKO3pmtwJxu/SmUHOD0okBeRQOUyB17UyLgEc0Su8hG6e1OCyk0qjoG6e1HliM5x04qPDYQmp43jjC7OvfNAheOpoKpXJNYBHAqIE9KRMnpn2rytw4PT2oRnGc05U8GkAAwTTDaQM0QSABzUUrKoXP4UGDcZ5Has46Gsg9RWARgGmUdM5polboaaPcoJoIqnI5PamQ9F/CnQggZ+tYw3XFTEFcbue3FIiiLO/n0xV5EqvuU/Sto9fwrAA4NY3d6sLxXhWMnkf/XrKOMlsfhTYOCKblduK2bu/04ooM8Ghwuc0VIxiuCcKc0iKI9znk9KZgRtB/DFAhcAmnAI60oz3pgpXAOT9KRiT060ykEZqONlUMTxUVxGMI6cDvmtfsfB+g/DwazZ2/n3LdRvkT/lsF75HQ+lEIGIzn04ohSODRQDAFSLhcg80MLjAoHOBihjoKwCuQelFR0FBQOAaKr5AZTz6fjStwBnmrR1kGHP0rRkVTLBtznbznHrWmeLbzSVFlNJ9os+8OAmep+9gnqc14m1rTPEl5cahZ2v2dX27Y97PjAA6kD0pI1LhFbB+laz401bXLaO1vDuWLOD8o6kHso9BUhEikIuCPelIZcDr6UhUghuCOlGMMOTVpZPfT/ZkHJ7+nGf6VeabJauFJzn2x/WrYGF2ikGemDUsfkjeTkenSkmkkXyg24duMU0IdgrNz9KF2sIZRDuzj+LFW+pTJE9tE3yvjK4Hbn0qQLNhs5H0rT9F06fQp7tZ/wB8m3am1ucuQec46VPG0DiRD8vbiv8Aglj4k0K3/ZuutOvdd0+K8XxNdubeW5RJApigAbaTnBIIz7V4w+FPwu+Idzb3vj74beFvE1xaRmO3l1jRoLx4UJyVUyoxUE84Ff8ACU/s2fA7TZEg1z4deBrNl3GG3ltNPEgHoibSx9AATX7RP/BUnwB4c0u78OfAGGTxJrsqNGmtXVu0NlZkj76I4DzMOcAqqZwcsMqdZ8f6/wCKr+/13xHfzalqepTGe6u53y8sjNlmY46mv+CcPxf+AvhX4E6f4Guvif4f07xde6ldXup6df3Is3MzuEjCGXaJT5MUP3CccjtVz8Pvg3r7DxLd+B/BmosnzjUJNMtpiMc7vMKnp1zmvin+1d+z38FdLkl8WfEbSDc26lY9J0yZbu7cgcIIYySnTAL7V9SK/ar/AGi9d/ad+J83jXUrNtO0qyh+yaNppk3fZbcEnLEcGRySzH6Doop4mZNrc+lSRMMLuz74piVAix170zrGoCjNXN+LmCCBbfyzBu+ffndk+navhynhK4sJW1+98mT5do8uRu7Z+7+FWmm295qi2ss+yLn59pP8Oemc9RV1BHBMY4m3Ad8Y7VIwmbKNkD2pb6a3I+zvjHsP61DJA8YVG57jBq4RIkAB5PSpSq/KOTTICDtbP4VdKoGc1MhKlh2oAHvUcPnPgdqsYVjXrnNMDnINEfjR6Y9KxkAZpiOmKJBGBXUcGgAOKVRnk0h2ZKnntUMIdssc/hV1GgbKH8KKgLkGhhuCfpRIIwTSjAzjmmYHgcmiQOC3T2pTu5/Kt5AwpollBIPP0oSAnINRDc5ZRx3ouCcUyKqo2c5zUiEkKB17VIm0mMjkUFzwTmhEI22k59OKkTaA2KEZ2hia2ljhT+lEZ4xRA6Chg/Wicck9OlcHn8qwMYB5NLgrkUVyoFE5OwCsgjGelHBIFK+TxRJyGzW4jkn9KJBGQeaiQOuM1DdSWmPJfbu68Zp/3xBdsk+1MvYimXjigcDAGTWi3ttBJ5NwmVfvk8Yz6VqmpXWoXRubiXe577QOwHYe1Wmp3VspNrNsY/7IP8xTXE87FpW3se+AKdQVZmHpxQbPRv0qI+bgBefrQZVXaW5+lFzvCDp/KkjXBJbI+laZcQ20qvKm7rkZI7GrrwLZyeDNU8VJdYltfJ2Q+Wfm3S7Dzu4456VGG83yiMGre3kjKQn+POD9K8IeHYta0+4imvfLEG3aPLznJb0I9KgvJIJQoOQvbp2rTNX8+XMqcdufY+1XPiDTtK8P3Flbx+a95t3Nll27HyOCDnOatWbViLZF2k9859/b0qY3vh7UlZON+dvTsv4+ta/40v8AxlHFHqDcx5xwO+PRR/dFC2jjCvM2M54xVlBHKBHbt8x6cVcqWfAfAPtWnyiIeSp3Fvwx1prdkfDt8w7YqxVJIiWTBTpz61KFVdpqC1ZY2cHI4zxTFigVnzj2p7cSQllb6cVrAeGNyqbjx3x6VoejSTOLqZss3bHTGR614D8TaZ4KuRf31h9q29E81kzww6gH+8K8baxaa7qc2oW1v5CS7cJvLYwqjqQPShFFKuwvn8KEaxWq7F9e/vTM0mAw4pIka32qu49ucd6dpoZSiNg9+K06zbUYDcRDDL1H4kf0qe/NzIWWPaT23ZzU9gbeHfMcfh/hS2hkeMxHIOc9qs9JeQblPTv/AJNJorC4Qk8NnH5fWvBsV1oesw3dsd5G7jgfwsO+fWr/AEyR55GT5gcc9Ow96t7GOGLypmyrdeOmPpWpaW3h7Uvstwu9o/4c46qD2J9a8L6rolx4hs9R1+3+zWNlv3Dez7t6ED7oyMHH51rr22t6q91p8PlwtjaNxP8ACB3weoNWiWmm6PJHPBuabG1txG3Dc8DPrURMs21e3f8ACvCPxJ0zT9FXw8/hXz5LTObj7cybtzM33dvGAcdfepdKkv8AVJL9Y/LV8YTOcYXHXPtXh/xFFZ6laancJvSHftTJHVSDyBXgn9ozwf4n8Py+GvF8H2eNMeQu+R8/OztzHGP7q9TWheNPDGpW32nUp9t5F/rhtkOMkgdFx0A6V8QtOvPFevT3lm3mQR7ecBcZRR3IPVa8IeEvFsMaNpNttjuc4ffGcbd3Zj9a+Nfgi4+H17pd5qt39rurvzt6+WI8bRGB91mHRx+Va3eaX4r8Mx3ttb/ZZzn91vZ/4wOpwOi5/GtN0rTtN1CCfWW2253bjhjjggfdJPUivEem2X2rzLN8ofYjsPU1dQBHEbDrV3blG4bcPpimhA5zUMIkIXP0qbTXWMTA8Hpx/wDXoRDoKEQCgg8VGyowI7d6clhwaIAYHOfwxSxuxwp4qYAMqL15ohAdqnJqXLyfKOTQU52k800IByTzSBU56n0ppMnjk0HDck0W6KTSgFqYoFBXmhhs/wAqIB5JpTgjmnIBosABgfhTsG4NO20ZFGQuMDr2FRncSCKYFTkHgUAxwqnj6VvKsI+uOvat6qcFvwxWMnJPT2ogdDQGRtFEY4zQx0HahycVj1PSjjgZo81kECgARjNHAwo60+3aAG/SgoZcn8KYg845oqMDnNbQePyokD5cUBkgCgpYYB6Uy44NPtVck8VNdrtwF/WhfRLxIMVJeAjBH0p7wdM4prkuTgdKLl2yTQYg8/lRcHgCiwU5HPtTSbuAKY9iaq3iiaANu+b6e9JgrjP6VEVJweaA7A0DtOBTOCAAc0FA6GuOoP6UACetcHgGmX0psN2oDI6/hQAIwDzRx0BraKIGMA1gAYNAA9DTDvmiMng/SsDHBz6URnoaZQw2k0UGPWmQYJB+goqMZ3fpTAsu0H9KK/Lgn9KIJPXp7UCOm79Kb5jjd+GKDqFwR9KViBkHk0RuBJNNxjuaKFvlB/CipQgHt0rf/ADk9qGB8pqQ7lxn6UAX4J/Siu0EGmAH3ahyCRRBPApnLDAoIpwCazjgGsjbgDJFMUABU80Mlck/StmeRRbDYzj2oTnoeaSRX4piP4awDwT0ogZxmuduKABwAaaJSBtOKktmYkqaeAgcnkV88kigjIGeKijEYyo5q9j8xxkY9P0qSLb3oA+tDBO3PNKxQA5q11ArwxzUMokTIPNDJHvQUEdfwxTDgHNFcYKn8K2MQCzcemKRUXoKd2ZgM8GnwOlZB6igABwea4I57UpwCQKVS52g/pTMI12L1qNi2AxyaDcbQePSkm2nei8/Wo/iRbzeBLfwpdWu+S13eW28jG6XeeAv070yqeAKJwdobge1QL5zhQ2PfFahaCyunthJvCYw2MZ4B6fjW0EYFAY7/hQI6ZoAFeDk0RgfKaGPWhz93tVvIwPXn6Vocc2oTG3iG6Q9BwOx/wAKktp7C7W3uE2u2cDIPbPb614kudIh8C6Xpa6ZtuYPO8y485jvzKCPl6DA4qLXp7TSrjSonxHPtyMD+Fs+lSAo4QmiWRcxPz9KtyZACOvrTxaemktNs/fjGOW/vfl0qJI5J/nPH/1q8EpoUPiKAatH51v825cuv8Df3eeuK16W2k1AtZrtQdOSf4R60lq8zAtyBQ8H6neaY+p2kGYUxk7l7tjuc9RTRoI8j73pWgQWk84F4Mr6c+h9PwrxVYWUOuXK2Ee2BdmwZJ/gXPXnrmvDeg22rX621w+1T3wT2J7EelfETwlbeDPEUuiQS70ixg7SOqK3qf73rSzuI2hjb5T7V4U0HRtT1F4tZGYxjaPmH8Lf3T7CvEFjYWWrz2mnD/R127Bz/dBPXnqTRijJyo5q5utFOnRxRWW26Gd8nmMc88cdOnFRSGTCscmjEycMufQ5rjOSelWdw1tKs8Jw6ZwcevFNrN1cQskzZBx2H+FOsZyEXn61Dbo1tJID864wPxqSIPznmny0gVxg9jTAb+DkflQsybV7ktwuOMe+KaKJeR1qE4AQnI7DpTRmNStSwBhhWx+FLBH5ewUSIzsRcAe9TvGRwMntTsGfDU0ShCyt06DFXCZXAPFTJlTu/ClUNxVnCW+UHr1qJAowoo7QcVx061hcDmjtIwKyu3bjJrK9jWQRgGo1B60vy8kZpkwOuaUqzDcalVBKVHQdqlVc5QYrgrjGTRHGM/hQI3AZyKbaR8o49Kb5eCOawBznn6VgE4xUiYXIP4UCByKiO5ipFRqoUgtj8KlYMAB1HWiCFyvNOCCSTW3AyT19qi27GDHkdBRYFSKyWPXpTgAfKaUZPBokA4B6URu6DkdqIbdhTQUsQoOT9KwCNueawo4zQGQFzRCggA0BnjP0oDHOaKqFzjNBflLZ6e1AljknpQIIB/SklUDApWDDIOaGWXFMAEIJpgB0NEFhkUDgYFDbIo3DmslePTpWnWS3pOZ/Lx/s59f8KZyQFB+vFBC4y3Jq3lNuwZTz3qZfMXOeaSNyQS3HfipERYso3P0qDiMLnjtS3r27ho35X2qCZioJ5PpVpcSMRIeqfd/Gg6SPsVcA9OaktnjlCDnPQ/hRjlVvLQVZI8M6Oz+vb2oM6y+bE/I7Yq+mlkki3vuxuwMYx0qCEXDBG4zWm2RWMylskdOOnWpY9zbS+Pwq3S2hdRcHep69R/KpRa7ybVcL6ZP9aeADkVBAoiB6Gpt6XRXGR2H4UkfmHpRiDj5R0pbaaPwx/aEKf7xz/wBNMV9nknha4kPPH+FJcxQyFQMmru+eYiGJfrzQtrS7tD56fd+7yfXnp9KWwAukXqhzkfhWo2iwxgoeD0/Smty8O8nJ9MVbaXLJbfaDwn/18etXUCm4KZzu6dscVp+oyaLc+ZaHBPUY9j659a1XUpJ7hrorjfjPPoAPSnhWUgnoeteG7yFvDjaQ8W5+PKbcRt+cseO+fetH8PahdL9jZvkPfA9z6140K+E/D1rpVmnmqd+Xztz86noc/wB6rBr+8haC0i3SPjA3AdM+v0rRNKjjnLX7YUdFx7H0P0rU1ZJ5Zre5wqY2rs9hnrVol5r1zsC7z35A7fh6VongmbVLTUhG2JLHysrgHO8n/a9qhjt/ta2t5HuiXORkjtntz1xXjXXIJNA07w94aby7OLzfMTBbq6sOXGeue9abA6zFYm+Y9Bj2NeBbbV/DXimO+DfZJ+fmwsn/ACzYdOR0P614g8deIfHtz/YOpy+dat947UXPAYdFB6qO9Wvhqz8Lv/asB/ewf6vg/Ln5T3PZvSvhVceGZfGiX3imP7TA+d65kTOInA5TnrtrxL8aPDng34n+IYdMs/tejt9m+zL5jx4/cDfyULffY9fT0rxRr8PiyeXWBZ5t4cfu/MPcBeuAeq+leI/jbNrdrF4a0iL+zkXO6XcJt/IcfKyDGMEde9fGTwdpmm+F4Z9L1b7Zqt3u8xPs7R79skYHJYqMKT+VLo19od5DLeQ+VMu7Me4Njj1BI6EUuueBvGWgwaB4i0vZcW+7ZN58p3ZfcflQADhQOteLfg3qGjExx332nPRfKCY+733n1rUI4xMVQ/c9vUCpIl6h8/hU1u0jZB+lKgUbvTtTyeZCI93A6cUpKfKv51tcjawyaW3CggnJ+lLAzqA7Yx2xSrHGMIaeQO21TUseX2R9+tJCI1JLfpTxqoBBye9IQGwelMEkcgngdOKZMMAT9KKA8n8KJJPBpUDGgoC9cilQPxmmt2C5J+oqQoBgDn1osrKFPXvTEAYqRgG2g8UzqvU00pYcUDtHPOabDHGeaQB18st9OKjZQQhH0pgoGVPStm/gmljCsMGge4NElhg1gYwKHI61jI4punFAZ/wrGR1rAI60V4pSAMnmnbPQ1kBcGiR60zZ4zRYDqentUl1Eo5bJHbFPqCjhe1S6vs6H/P5VJrcuMIef8+1TalNMcM34Y/8ArU9yzDbmmkYjrWcjg0VBrHaiAeKIGMUMCqPPQUQBwDRANQBiMKaJYHhvwxVuCCCarAY8UrBeCaUqQPm/SiARlRnHasAjg1tAHXPtWARgnpTqo70QBxmsAcD8qK9/TpWB0J/CiBjg1kHgGsc0RjjNcc80eORRA6A0R2qiFwcnipEUJ1rhu1NnHAoqSAqinjBOAc0Qo4A6VGpYY20V28d6ZDtwrfpSxNI2QevSnDI2D1FEgkFjQCmTKn6UzDfkjin2qVZTx3pASflosEYknJNbhnCil+UFSOaVQQOaJAXCmm9cVtV/umiQhwpzRUE88U2dpwfpTOUGAK2bsEnmgBjOaUhTkHge1GXawIbntxSXGRj8qW4RjyenSg6sAF5r5WIBNEgjANEjbgUQDSooOVoqp4Iq6hEiYRcsOnOKuIHjH7xMfjQiXAOc+lNGQdoH0oAYx+lAHGDUd08I+U1Z6gJsKx5pGUjGKZA2ADTRoBgHn0xTNuGCOnamYHgH8KwTwPy9KIPXHAoKScgVlQQM8+lRjC7SeaZ0UYPNCTHQ/hTSBmwD+lRlQOvNLIG+XPT2pCCMimyuAK8G6Lpmt6qlnq159mgOcyeWz/wseikHqB+deP8AQ9K8O+LL7R9G1D7ZZ2/l+VN5TR78xqx+ViSMEkcntSDZ0OKkmMrb3bJPfFK2flbj0rx/o3gW18M6ffeFPEX9ozS+b5yfY5YfLw6heXODkbunpWMjIPPajjGAawrLjNE5GKAAOFPNWHhu5l0L+2VjzGejZH9/b0z/AErStRl0q5S5hX5xnAz7Ee/rXh3xJZ3Wrpe642FGecH+6R/CPpXh3xRPpuoQyX9r9uist2YvMEWdwPcDPUg/hXi/X7PxDfLcQ6f9jVc/L5pk7KOuB6frV5bmOMMBnP4UCoGM1bzeVjI+lTXZdwCOBRQO3y0kvlHD9T2r4d2nhjXLeWy1y/8AsYTbtk8qSTuxPC49APxpLM+TdNZjzILbZmT7uN3seevFWnju/sNLfSoj+6bGenZt393PU1E7OchOfrUEjMcEYpzNIoPWvDj3sFx9qt7D7QE6r5oTqCO9eKvEJ8Q3JumtvKY9t+7so9B6VmSABwOB1FWN3GDunGPb/wDVWgeHbXxPqyW6TbF5ydpOPlJ9R/drxPoLaBqL2ZbcFxzjH8IPqfWprS2a2SVJdz87htIxzWmxWJmK3l15Q7HYW7H0/CrDXdMghezli3o2Nsu5h056Y/CtQngFwwgO5T35HYUpABKt9Bioi0gwp/DFWfhzUL+xa+t4N0adTuA6nHc+1X/hRNN0pNVim80nOV27cfMF9T6+lLoekRaGLy5u9kzfweWx/ix1Bx0qUrHhX+92FGJWTfHJ+lJcSSW5iLYU9Rj3p0QrkNn8KeSRQDGufXnFRzpdRhCce9eGJ9Fs7qYazP5Snbs+Vm7HP3fwrVJIGv5Hsh5kXG08jsM9eetTsuemM9BTMJQIim0j3zT2oZPlO49uMV4XbTLLVoJtdm+z2a7vMfaz7flIHC89SKZJFXEh6VOoZjz06cUFUNgHmrKP5c/pQG35fyogdjRPYUADwDz9K29BnNFcD+VBFI4NIm47QPpSps+UjmmOF2saKER5HQ9KEZIwTRIPJpiFGSaK5GQaYqeAK29waBAGAa3Z4NKY2+Unr3qMA/MTyO1MxdixGB6VM0chzHSow5IwKkBK7s9KYAnIPFIV4KNnHtTxB4w4PI7UANgLjp0ogbcA0ULKSO1HAGSK2g43cA0QoI2n9KZc9Dz3pQV5H5UQcgqPwoHByDQXIA70y5O00FJHWm54AoAE5Bz+FOMZ5oHisBuCaKjGM0SCQQaBXG0VAR5YQn6VbkAgE1KY2TCNnHbFOmBkj6UBkYAo5OT6UoJHWhlzgHmrcFzsVsH6VhVbaTnHSl0ppNN/tBThf/ssetCJSATwR1FBwW2K36Vp1iLl/JB+dvujHpnNa54f1jQGhGqWfkrcbvKbzFbO3GeFJ9R1rTtHudYR009fMeLG4ZC4znHUj0NS2rxTGGdcMvUZz29qv7C3sjtt5vMB6fKR6ev1ppFSFI425Oc8dK+H2hJ4h12Ownl2RtnL7c4+Rj0BHpXiiDTfA/jvyrJv7Qjs+qYaLO6Edzn+8fyq9h8Map4XOqy3/wBm1Hta+VI//LTb977v3Rnp3xVyDHJG5bg5zxUSxJHlpOfpQEc0aTINx5z2xVpc2FtEBO+1+wwT/KizJCsoP7t+h9MUyJKBJGc4/ChBG5+ZuR2xSRIkf7tenvUTB1G7jFRTh8oeB271aRiacBm6+3tWr2K2U6Roeuf5D/GtOszd3SWythnz29AT/Sm+ANjonwSj8V3epfPBnfB5J5zdbB8wkPqD0rWtSW6ujDBBtTt8+ew9qe0aOQgDJb8MYo24gG6Rtvpxmrd2kt/s/lcL33e9TrFEwZPmI/Cr4TCMNKOO36UhJ2hVyOe9SSSR2wt0OV+nvV1amN1lUZHOT0xQg3KZBHn3zUdo0tnu27iO2cd6j0u4uZRFDFnPbcB2+taPYLY3UazybFGcnbnHB9PrWqX0NvbrBpF15+7OW2FMcjs3418QPBvjrwb4dsJ/F+m/ZkufM8o+dE+drqD/AKtj/eXr614T0jWLuxl1XTxxBtyfl4yWXufY9qXWbia7V3G/Gfl4GePpXjz4X6vongxvGU1rste53qcfvVj/ALxPU+n+NeF9V0Sy0pLg3e29bOYvLY4wxHXGOhzVz4h1GzlM9nJte6/1gwp+6MDqKvodEXwfDcJqG/Vju8y38phj94APm+793mtF1CW3iYqMK+MH6ZrwXptpf3sgv7vyFTG1vLLdm7A+wqbxS6eKIdQlH2uODdhP9XuzHjrjj/61WfiVPGug7dE8KfZ2X70n27f/AB+jAf3SPxrS/FMOm6r52qL5lov3hkj+E/3QT1IqbXkt/EJ1GytN8R+4vmY/gweSM9a+z33xA1uVNF0jDvjEX2gcYT1bb/cJr4L/AA/1DW1vNJ1TTfsyW2zzj5wfG7zGX7rD0HT1q58U3t9dgaZp/wBotW+7J5wTGB6EZ6gitN1fV4NZ0u4sdL+3Xlt52yHz1i3blIPzEYHGT+FfG+48d3HiObV/Emh/2dM+3Mf2mKbGEjXqnHTb271earcwGCeSfzJV3YXaF29vT0rTPinq9kHu7qb7Q7YwNqpjGR2T3q4jaL5wc7+nbpVuIZv9Y+D9CauXdHKKeF6GmBPU1GIioJfn6Va6ddXR/wBGh34/2gMfmaMRxhuo7UFVSWJqRgFyvPpQj3oSrc9hikVUIQ9fStq+YCG/Spl3gKD0oQhI978GvlLdfxxQRC+0t9DipAo+Xd+leSWyUOQKEYY7RyfSnhKcA80wZflNKwJAU8+lTXDRr8zYH0qXUYwduc0l5DIDh849jRbI4NSvtOAfwosGOScn0plD8KefShGy5Gc0qkD1ryy4yPwoqSMjp2pFBXavOO1BAeB1HakRcZJ+lHHY0QMYBrAxgVQUHGTTYHyg156BsFv0oMrDg0zovVuntT39sgxvz+BqTVIsYiG4/l/Sm1GYjKw/+PCpNVuAMCH6fMP8KOr3A4xz+H+FPqVzI2Mc/h/hXmXMnJX9RR8DfB9PhPo2vP8AEXPiO68/7VpX9kXI+z7bjan73dsbdH83A46HmlsG25d/0o6fCRgyc/Q/419it9uA36GhZw9/60bWEgjHT60LOEnBH86+wwE53Y9sGn06E/dl/wDHT/jT6cAfkbJ78Y/rT20iHAX9aKEcUQMDJogdAa60RjgGqRgnb6UAzkuBmrWdActyaWVWxtodcA0RxSAHqOaDAHaKPAwKIzwDRPGBzROPlzROeBRHamPGKIwOTQG4jJojHGaJBGM8iic96xnv+FEgCvlH1qiobrTruXFOpJAJ5p0I+UHJpgBRAbBB6dKZdx4oKARkUr7eAeKYByCaJwMGoySuQa3f/WFDpj8qXBbBHNH5mwabbna3OOlblC8CsMxy34U+Q3HWohuXJ5oANwxogINqnmgcjYO9RqoGD+FMigEKKBBAyeaYBuQeRTZ6E0oUKAT9KO4DAP6UCMAA9aZQWyDzRAJzTEEjBoAjjNKpbjd+lBS33j+FHpxRA2+9EcDmsDGBRAwB+VSQRyDDD8KubISDK9RU9vKvBXp709urMcL+tNCo4xTQ44BokqeOoqC/eLhjk1DqkbYBP+fyoSLKoZaEKu4z+FGHHGKaIsMYrywi4ryiOopoiPmI6UACSMdKBVW3GmcudxreCNuKjHGaBBAANZOMA0WDJtLcjrxWCw60+D0PSsgClxnbnpRx0zSsQuAetAdCBWMDbmj7Hp0ogk5z+lMABgV4DtNU1aR7TT7fzmGPl3quPvHvj0Najo02l30ltdxeXPHjcm4HbkAjkEjoRQkaCTLHirCa3a8UO2YnzuGD2H59a8Q2mlQwRNpiYLZ38t6jHX8aVV5LGpYzLIQ3Slcq2VraHjLsf0oBc5Q1IWcAseR0FRMF5df1q18RzppH9j2x2xSfe6Ho24dR/WtY0SXRmSOeXfvzj5cdMehPrWlPGL2MSfc5z+Rq70bTbq8gi084D7t3DHoOOp+ta7o1rZXz6WiZmixuOT3APr6GtH1TUvCStFFPhJcZG1e2fY/3qSJUUCLipW2Akn6VAAgywq2aETD7QMoOo59ParjUFeEW0Jwi9B+tWlt9ruVtw+3fnnGegz/Sp18lipOSParSWSK4wp5P+Faiw80EDOev5Co3aIBW6dq8NaC+qgqyfuz15+vuPStUeHSWa3sZt0LdtpHTHrk9Sa0DU7PSdQivZV+VM8ZPdSPQ+teKNHmt1+37dsbdFyD6D1qfLNkHJFbMHCGlQY3L1HaiqvgLzjtWkalJpl0JwcAf4Eeh9akvWF0Zg3B9vbFXF3Z3MbTun75sYOT24+nSppdsZfPIqaBTGjk/Mc1KCGw5qxW0jnRrtMw87xk88cdOeuK1htOe/lbS0223GxcsccDPXnrmplDc08IIIA606APj0q3bYMCgQBjNHpxVDjpWT61nI+9+lBS3C0iFG5/KoIxK3mE8jpTRiWYgdB/hUodAESnZ88nJpVAHyD6053HJPT2o/nRI6EZrGTwfwpgCAAaPOBQXC7h+NbsHg0QQAB36UVUfK9Qc5APHpSEk4ajICdpGR2rOUKLTbSojPai24eXngUE3jKnpTqdoHrW07eOlEAEEGnCgbQM4plIw2foKIydpplUdBRYAYH4UW7jrQck5HWgcjmie4puACDmtyk4z9KzgBSKJBbb3oEYwTXpmjBld6mrdi2WY8mo2JPy9ajVwx2nimjyDtGKEMjZKLnHXmm9COlIATgnH4UxIbAPSlkZSMH8KZiH3jrSX8pt/s5b5fTHvTkkJGrevatAj04y41Fdw+rD19PwprqRHwh+grUNTlvY40nbJjzt4x1x6D2rQ9dutEkkazk2tJjdwD0z6g+tXlzdXVw9wWyzYzwOwxVom18scmgI2t2kDfvF6DHvWj65cadOtzDLhlz/CPQj0961nVp9X1L+0Jm3O/XgDooHoPSlLKflP4VBFLNwzZC+1WlsLsAl8ke3+fSvDA0iO6xq8W+CTtuYdAf7vPXFaiLOW8kj075YONo5PYZ689c1aX88eirppf5OeMD+9mpYwI9qjmorVXX9/y3+fSkjIIQHjvxRi2nigWkuBHjp/hULFAFRuntQiN03Lc/SoLV7ORblGy69OPwrX/il4k17w7BoOoT5t7fdtG1B95wx6KD1A71crB52AuSOnJHap4wzElunUYrWPDL2VjHNPJuEudo24xgjPQ+9LJGifZgflToPTPNTW6iDKfd7j8antGPzN1Hao7V3QkjNLasykd6udPkiiRSfv54+n41JDsj6V4f8AC9xrtlPdxJ+7tdu45H8RIHcelSo1pEUjHyirPT5tS/49hnZ1P1+p9q+EHiqx+G9+mo6nD5pXOF3FezjsG/vivHnjrxX8edUJ1u93rb/6seXGMZVc/dVP+eQ/z11+O6huZNHZ9wXHYDsG/wA81f8AhX7BGksj4L5xx6Y9/etUuPEWqad/ZkEPnr2G5E7g/wBK0+C6trpJZeAueOPQ1qF++sTWaRD/AFG/n0yB9PSrrTtNkt0s7GL/AEvnJ3N657nHTNXXhG40iZYb2LDyZ2jcOw56E+oq80280mIPKm1T05B/qfWlYTzLI5xjOPyrwn8X/EnhKxfRdMvNkEmMjy0PQs3dCerHvXg3xL4QggkXxHbedI2NnzyLjls/cH0rWiLm9kutNXEIxtGenAB689c1pHivVPCuotPp0myZe+FP8JHcEdGNS64I/D17rYu92oTeXj93jOH2+mPu+1WXhC+0TTVJn/eJnjYOMt9T2NeJvG8tpcXNnNJmZNvOMdQD/dx0rxx8S9c8bWoudRn3xnvtUd1HZR/dFRalFbubkH5k+7178elahqkssmc5/wAj2qa9llTYT937vFR2xb5weaaMs7BRik0/zDln/T/69LDHEgOf0pp3kwi9BRwq4FON4AJpl3LgUQCu0HNQwx78sKuDAZMWy4A7ZP8AWvNJ+XNTSZARTjHWhkc96JLHOc0FBIJFQyGMlVNeTgb1P4Yp5GfABpjtyM9Kmm8lC27p7VcXbXTjaelCDPI60luCoBHNQLkbSanBVtoPSnOBnNAkjJNRvtPBp4RG2FOKG4DIbp7VGuBhjyKSQZGD0osOoPNE5HBqsDtVYz0oEY5NMmUznmltFD7zyaSEdulNZAgEr+tCwjVNwX9aFuFyQOlEYOM1LBGWyBzTWqs2Qv4ZpbMYzjmmtwCNooQhSABzT8HGOaYAd6DBl4FMOMkc0vbHWsk8A0cLworKqMgUQTkgUGJOwnNMqOuGGaayjcjA+lSWbrkjpRDdKJz3qg5PerS7ihBEgzUTKycL+tRyqBhWyfTFRybhkdfSg4PX8KVwOQc0WUsCKHqBTHI4oKNoAFEHvW0YyBRGfujNFSRg9aYE0O2B0pjnjFEDOAaIIHSgc9/wo4PQ0VPaj6GiO1HkYpl3HGK5D4U5NMM9TTxsD1+nFINq5Jp8Ho2T9KOQuQaLDgA89q4YYLfpSgBcK34Ypk28hv0okY5PSirMdynkdqLADnrUigqCD07YpvkAH5UoWaPrgr0FMSW65P0pA0ceHGDSFXHAosrNgr+tExq2Fbn6UxCgACi24bVP4YpAhG08mpAGwMZxTEBcMM0CMBic0Sp4BpcrxjOOlEjOAOaYkHIFHaDuasEgHvQEfANHGMDrWQOKPNHB6VnHOaMikZzW7jkUCGGAeaKRv8p59qFpCDgDB/GpdNWRsK/4Y/8Ar1Jp0q52R59Of/r1JYTKwV0xn3Ff2e5XcvP+frUltLBhiPpSX08Y2hsj6Co9YdTyOn+fSo9Xjc4L/p/9ao7tJFyJOO3FI0bYIOTTq7DhfpzTFtuM80yADO7k+1Oh6luPpQJ6A8CjxjBqJxtxnNIRtyvUdaGOrcGtozwKyOgpiuNoOTRCt8pNOACNppDggE/pRIA5FA7hwf0o849uldRkdulKVI5bn0xRBxnuK+Afi7U/Bmtzanp+nfbGO3K+csePkkHUg/3j+VePtcl8RePdW1l4PJe48nMW7dt2wqvXAz0z0qz0a41l/ItIN7+m4D+ZHpUcYVmWI8Doa8PabpN5ZTyX+peVOu3y4/JZt3JzyDgcAVdhonxsyfr0oOWbD8D1o2sMwwh5+hp4pYiYmX9a/wCEcuUsF1JkxG2cHI7HHrWwO4UHlelX9n4fi0vzLXVfMvP+ePkOP4h3PHTmnm8iIKsvz9htqafVNTHnXkvmBeh2quO3b6CkZUIOf0q01KeyYNDJ+g/w96ufEV5cXBu533Sv1OAOgx2HpV5q91eKBI+SOnA4/T2rTI3vLhbdfvNnj8Cf6VJpdtBqLWWoXHlKOrbC2Plz0B+lSWzluTgelMskDKZeFfOD9KbEjb4WyR2xihdbyM/KR0709vJOgCD8Ki0C8MYvpF2p2bIPt0zV2+ZShk3Y6HGO1I4lAR15HvWneJZraxNrBb7fVt4PcnpipvD06Rg2fIHXoP5mpnmhkEU3ykfQ1qPijUNVsRp9627HfAHcHsB6CmjKDDCgpLjnAphEJQyNle/B9K1W40q7RRpFtgjO4729v730NTahb/2dHYtZYnTO6TzD82WyOOg44p4lkB4qBg0XlM2WX2q7GYioFN5joHzjPtQyyl25C/hUsrsNpGCO1MCzbcZ9qkGPlAp48DI/CniZ3zUbFRtPJ70pyADRAI4PNe35UAe5zRAxxWTnA/KkjcASL0FMpk+YCkdl+RRzSyIpLdTUkjTMPLHIp1aMgOvXpzTyMhIUZz1oEEcHkU2CoGeaYY4PWiccg5oEt0/CmIztFIHcbEOc9qxsbDDkdqeZSgwMFelbgRnr7UhIOSenapB8mVFBUDkqvPpnpTOijCNk+mKd4zjJ6dsUXU8YzSsEBANNKrQlScE9KBITBPHYYoFVAPXHalZMfM/zemKLDOCee3FZB70xGOtMA4AFBSQB1xTFQcCiMLx1oqp4B59MUFyPXFEZYAGnO4DnmlHGdufTmlVmICjNBlU/OPpUMiiPBPApFC8xjr1pTjlTVnLG6+UxwexrxL8HbfQfBS+K5L7IOfl8oj/lqE/vn19KM0kK4iOPXiiSTnNM2eM1a6JqF5p8+oW9vvhg2723gYycDgnPUUV2na3aiqsBgfWreJG+V2x+FPGI5CEbIXrxjFSvbscW0u/HX5SP50sc884MSbs/d5A7UlhE8DLO+2XjauM5556e1MhB60GZcED8KS3gkkKWJ84L/Fgr+h/Gp1MUm1+GHUdaiiTA3H9KP7t8KM4pLlSwXbx9atonlDCHpxVgzRThc49vwNWMFlJYtFJH88PQZPc1KqpIfL4PYVZXJeJWkbAGccUFjKhmOQe9Q2guV3ocgd68oBto4xQiLjaRk9qt7dYLkPLFuPpux2pImlfEa49s1Y6d9qvobMybPM3ds5wM+tQ201wzpAm8JjPOOtXRabhJOD0G3pV4EtASzYcdOKt4Z70mQjj8P89q1C9upkiivZsLHnYNo74z0/CnsJLi4aa1O6F8YPToPfnrS2rxOq+dvReo24xVwtvKocPgn2NeUiWm5Bgjofxpontoy83B/P8AlV1fb49yNuK9BjGKtRLdExIu4+mcV4Y1R7bTLvSbRM+Zs8059GLDqP5VqejNDYNcyxfuxjJ3e4HrWnXulaNaq0bcS538N2Jx6+tat4szLutkyo6HOOw9RVp4r1WxneeyuNu7HOxT0GO496n1d3vPtlzJhz1OOvGOwrUvFMmryAyXPyp9xdnrjPYelfDH4kWvhK6Euq6P9tQ/xfaDHt4fsqnOdw/KvE0eg63Obnw/H5ef+WWXbHAHVsehNWVpNpM0ckttvSTO35wM4HPr61eTs96LzTYPK9Bu3dsd/wAa8JeINOub8X/i+1+3sv8Aqo97RdmB5jH+719K+NelzwtC0ulfYYG3YXzxL/zz989f51JLA0EY3ffzhsHsfSpNOa0tTcv09fxx61ok+nLqsUWqXXk28md8mxm24U44Xk5OBWo3+mQ3CxeH7v7XDzvPltHjgY+9z1z+VeNPDckOrzXlxY/Y/N24h83zNuFUfeB/H8astD0G5jjiXV9krZyv2dz098+laz4u1a3uHjW+81uMnylXHA9q1eG61PV5ZQdzSYzwB0Uf4VeXE8FqLFjiJO3Hrn69avZY3CJF75pkDD5lzjtmnhBOUXn69KLlVKqKlbADDnPQUrSycFPpzRRNoXr7UwIGFGT6VbWdzd4WCPcT0GQP89Ku7Wa1fyZ12uOoyD/KtK8L31/pzakIP9H4w+4epHTOeooKS2F5NTFVTCtkj2ouQu7OD2rYCNzNg/SmOW3E9aALDiltXYYxj3qSJYgCOf0qKNgTuXOPehOBxjOO1Hp8tTyKuXY8DrVzevMCq8CreJCCe4pzGnLNj8KaVHACnFRyALlG/SnbcPnbn6VIVIwTRcEYWl9SadiwB3Z/ClOTgis5GCf0pFx060FyOtAADGaGOn6V1FMQoyT0qKVZf9Ucj8qETsMYz6V5een5UI1C89fSljDEKKMW9QhGNtSMoXYOQKVQykfpUiqDkflTIhORUiLjcDR3MML2pv7pFOCfun6UELA881tRhgt9OKMQxhTRt227ic+lJbs5+WhbuvAGTXkuSMc4pbcHgn8KMahsM36U0SbtxoxktgHge1FHPAFMMDbt5+tSWqSfd61PaSRjpmuorkVn3qGZfL2ZqMBmAQ80jlV2k1GpA60AuMGiACCKBz1NZBXANZGMUxDHGK6gZNLtPaiOetMPWiMkDNMM0Tz0pj2okd+1HBHBrPGBR6cUTnvRIAzRAJ3E/hRAL8HI+lFRnmnGeB2p0yODSoCT7UcCgpHIPHpigMrkihGpAIHI6VyG56U6Bm+U80MKwXPNOhJGB0pvmIQd+tMehLdO2KWTJwKOyTkDmoy8rbScmjGEQt/EKGCOetGML8w69qYhl5pRkggciiCeQK47miocYFCJXU8dKCKDhakjD4ANLGAMDmgvzYHSmjBHSiuDgii6kYoSMBtB6U7uQMnpQZj3/CmYngGgyimO4AA81uOMZo8Jg9aRtvNbw3IPNK3OSayGHBpoldemSOlR28ajaB07VLawSrhl6e5q50lWOY1/z+dTadJH90Z9P85o7lPNRTOnAPSoNRkj4DdParLUBOAHbJ+lNgjOaIDDK04JG0HI9KaEBA4/KnGeEPT2qJQQCDzSyKF5NbkNAk9DTAngCmIB25pjGcAjmiwLZ7elL83T8qXHTNA471jIx+VEbRgihjvW5Tkd+1eGtSfTy5RsE47fX296uZme9eZzy+OcegrTJbxHAsm+c9Bge/r+NN4Z1WwV4bm32uuMjepx+R96dpbFi6Phl7Y/z61qF4mpyAlsMfb6f4VNbEAKvWrSye5bZGvzfWr21e1lEUqfP6Z9hUN5dTacoab93zhNo459aj0gzM3PI6DH/wBerqB7ZtzJx9akhadvMTtRkmmj+zwjcx6Dp71p/hu4n3JPFtYYz8wOOvoa1WwFqxhUcr3/ACqbQ7qDTYNQlT93Nu2nI7Ng96t1RgEHSr61htJf3bZb1wfQU1xKSxZ9zeuMUhlx8y8fWo7db1jbhsenH4/0qy8MXEw8yKTdjttA/rR0W6ZTL5XA6ncP8a0KSys9PZbiLL8Y+YjufSpBdzr5KfdXtxXibQ7HS7VQr753zzgjGCPcjoaS2KyKD+NXVnpkGnpJbf6znP3vX39qT+07TTotQC5t5s7TlexwffrV/cm8YMTnb/8AW/wpI5WwW7USZVCE5PrSWgxuY4qCFJ5FiJwWzzjNanpMejFUW48wtn+DHTHufWiVZ8noai0a+uU8+GHei99wHt61IyFyyVKglTZiiswjCuOB06VNH+7BU5xRRZSWVsAe1FdrZB+73qU5Gcc0VZRkUYlLEyjPpSKX4z9KXgYNZHrQORigcDAoAn7wpUBAz1oMFiKgUMjkGmDbNy96zg4Qc0WfqT16cVOplYd8VhVwpHNEDOFoqgh5+9RG4EUOVwOtKwC5zzSYIzmicqSD9KUsBkGiQxwxzntTgDAX8qiIJOD0rzE27c/pTSOsewH9KEgVMDrRKNHyPmpULDpQOBkmi24ZB5peVGDRGV9T2qRgFwopgByOSKPHANAAkA9+1MCOCa6YBpskZFKSOAaxnnPTtTQlYwwXj60VZ2AXr2ojng80cE8HpRO3GaX5hkjpSsMYNIXfaB2oMMAA1auEmVseuPyq51rU7mD7LLcbof7uxR3z1xnrU675MqePSnUKdoPHpRAJpbiZImhjfCt1GOuKxjvXh7wxd67p+o39uuY7DyvMOQMbyQO49K1KxNoAFbOO2PpWlpLMz2sS5aXGOfTJq/thbuLVVwT2z9DWgao+gaxb30R+aDdj8VI9D61q+qSapqMl8zctjt6KB6e1F1OcdD0osNuM1bu0YKBvwxQCyHa/NIysmFP4VLCfK3A8npVtbYGQfwqwil24Q4PfirPSrie3+1rDujXq24DvjpmrlJt4Fuef4v6daifGBIOR1q2tkRFAX5RnvSrblBx/Oo7uNIzCPwqFUc5B5HtUihYQyD5qRGkYOWzj2rRFVZysi8N0/I1dWEOkTAq++T+FsEY49OfWp70MSLZtvrxn+dXEIabYDwnT8RVxAJLhWcZUZz+VXJiJJtRgH/PetF0iHXb4W1y21e5wT2J7EelABP3ML5VOhx61eSqoIQ/NVlbwSERRjd6dRUttDZ3KSTxfKudw3H09q8R3Qu3aSFcIcYGfp/hVvatK3l4yzfh0rSLZrCYtcdD/AIH0+tRXEdtdPJGcLJj9BVhp+oeMQNMs33Rt14A9+5H92vFbw6dctYk529Ovop/rVzKjQYTn2/Go3kghLk/NU/2m4BYt09hVvYLboGK8HtmoZCZgiDj/AOtVgdOXSI4LOLdeLnPzMP4vfjpU9xePdR2t7Fg87RuHpk9Pwq4ikljzAvzfWo5tX89BAm7y855UdRWuanr/AIu09ZJG3JF7IMZI9h/dqeOSzullLZC54x7VqGqzagRGr8dhge3t7VZWeZhJcn5O35e1W7aTexiGI7WXr94/56V4v8VnUb2SW6m3u2MnbjsvoPYUvilLNMIfm7H/ACK/Z+i0v4hfHXwJ4R8T2v27Sta161tL233tF5sTuFZdybWXI7gg+9af+x5+zFprK1v8FvDkjKCAbmFrgnPr5jNn8elX37In7MWowG3ufgb4Q2t1MWnLE3/fSYP618ev+CWvwo8UaJear8DZJ/CPiGNGeCwnupLmxumAzsbzC0kRJ6MrFVz9zGMa54e1jwhr2oeGPEmnS2Oq6VcyWt5azDDwyxsVdTjuCCPSvhh+xV+yyngbw7qcvwZ0a6urvTLW4mkvJJrkvI0asx/eO3Uk8dO2MVJ+yL+zBLG0bfAbwVhlIO3SI1P4EDI+tfFD/gmn+zN4902ZfC/h+58E6sVPk3ulXEjxhu2+CRihX1C7Cf7wrxt8BPFPwS+P2l/Cj4iWEM2dVtNkseTBqFpJMFEiE87WAYEHkEMDyKP7FH7KZGP+FG+Gv+/L/wDxVf8ADFH7KWMD4G+Gv+/L/wDxVR/sX/ssQ4MXwR8OLjpiJ/8A4qv21/2XP2e/BvwOvvEnhj4baPompR6haoLy2icPtZsFeG6EVL4iu4rEadBLiIdBtHrn0z1ouAw2jrUz4Xg0z56Hg0FeXoKS2dzknArZHGu0nmlMzYIb9KKlk2vyaSRIyQDn0pzg7v0qVwFq/uS48pDnPX9KCMR8q5/Go2lhPAoytK+CenasKJQT2/woqGbA6VdsVYFDx2pLhCvzUHVjhTmlXjmlUlcChGwHHXtQiz1pIyPuj6UUY9BRHaiRjIppGzhRRjuLg7G4X8Kt7VUAVF/WnUKoBOcVjPANeW7HgdKeLnKdO1bCcFj9KYBcg1IwCgA896kcMMimb1NPJnheaLk5wetPkrwfpQQjpQVjgZpYCOMc9qWFiOT+FbMjk80FHAB/CthznOaK7htz0rbuHNGFWOR2prdGPBo26A4xz2poQBgdTTwnGKMZU5NSKG4bp2prQMcqOO1fKetEArkCoyF6moG2tlaWeMv8x/Co3R1yp/SiQRyaJBGFPSsjoTQIYDB5rbgYBogfwnn6UQDRAPINdRyf0okHgmhggEUxDDAbp7UeeQentXBHLZ/CscYBoDpg5ogZ+U/pRA7GjyMZpgWGAefpTBTwW/DFOoRflPPpjrTEk4NbWJ65okNwT+FMApOD1pgScg5pgSPlbA9MUucYPP6UVKvgDp2pmUgnGKIIOd36UQGOSeRWW6KPxpVQHJbn6VIQzbRxii0bYzxjpRQqCBzVvtVwS2B64qQBzkHPv0oopAdW/Sjg/MT09qfaThWz+FAgHCmmY52qOaIfGAc+2KDgjANKrt8qmgVPAXn60UYgE/hQUjoc+1EEDGOaySNpb9KOSc7v0oqCRh/0oAjnd+lDkcnP4UpH9/n6UQeDmgMcZ+lHk4BoAdQentTMxwC36VnAwDmivQqf0rPHXmiQBhTSsQB82fbFBlYZBocjrRAIxVzGCMDv2q9i2H3pgQABzQbsasphG4WopRJH8poYwF/KmXaPlOaVlD/P0q5jjZd0XaoiCuNvTtmsblwD0pTxjP0FKcEkNk+mK3KwwOKKrjcT9KIBGaKEck0pAIIb9KJB5U0ATgg5H0oAjv8ApTcjJrBPAPTtS4zwee1aRZyTxl0Gf8mpLaSeTbEd2PwrQWlsL2KZYPM2Z+XdtzkHv+Naz4tsfE8+ua1cWf2An7P5MHmGXPAVvmAH93PI71dXbz3sjgcHH8qs0F0RGkeWP3eavNJvdJWNdTtvKMufLO8NnHXoT6itLvILLFxJJ+8HQbT7j+tarePqcwuk6n/AD+lQ3BifErYI9qtNWtxMJJDx6YPp9Kums9VQRJJ5RHbBb/D0q90PwNbeDbWS18W+frJ3/arP7BKvlfvRs+cnacpzx06HmvC1vbR3qTXM3Azldp9CO1X2u2+o3d5qMH+hRTbNkPMmMAA/Nj2z+Nak0s8xZpdynvtx2FLrGpXNhHp13cZt4c+UmxRjJyeQM9cdaZjG/wC5fOO2KWcj5Su/0GcYpE3NyvB6jPSvgpB8OdTmlt/HF39jVsYfy5pNmBIekfXOF/P61468GHStZu7nSj5+kPs+zXONu/CqG+QsWGGJHP16V4f1CHTZipi84t/DuK9j9fWrbRLy90+4Wa2+ytJt8r5w+7B56Hjp39av9E1/RYTJd2+Y+zb09uwJ9RUesNZtIssHmbsYO7HT8KmkhupWYrgntk+lX2m20UQe2u/Nz1HllcdPU0jKBsDfXioNemudLt9Bm+SC23bOh+8249s9fevCnhbwvcagg8S+If7NtTnMn2SSbHyt2Q567R+NeI7K0tXMFo249jgj09aW3ljZVdPl5zzWm+GNf1qPfptj9ojH/TVE9fUj0NNGUPzjAFNI7EK3P6VLYFo1kVtoOcHGa0fxpHpVg2mxaZ9pLY+bztmOSem0+tGBoxsc5ZevFQhgMuvA961FJJCIym3HXnPpRjcLtHzD16Ux2DC/jREfkgLyx7UyErtB/SmVmA44qSMuOlSgKyrH+NAFiNgzTAklQOlZZR0z+lIVJwD07UY3GGYfSgFbq2PbFBcjG7P4Vg7ApHNK+Y9g4IrcA3AoL5hWPOGGcD1qRJIfmLfpTklvmPPegAx+XrUrDywrDkUcYzmipPC80TxjP0ohgMA0F2gbhnNadfWdsZPtNr5wbG0byuMZz0ogFsrTnoKiPUfrTFcYVefXNKA/yu/A6DFMqnhD06DFZwOTW98YA60VIAUc0RkbQaRQCABnFFd7BVGM9BTEY2lefrRI7HPtRO4cmuAQVbn6UGOcZzRAPyk8/SsgHg0QTyfyocjGfwpriR02FuO3FA4YFeookE4B5oFQcE9alCbQRJk/SgQRwa3ds81DKyDaDyaSTPyM2PwqMkHjqOgqMtICpNaZa208wS5m8tP720nsewrUbS0huHEdxvQYwdhGeBR2qNoOSKS3mZDMoyi9egokNx+laJ4l1jRrW8sNPu/Ktr7Z9pTy1bfsJK8kEjBJ6Yq5ufOfJbOfarSafTniuFO1udpwD7H+dPdy6hdKztl+xxjt/wDWq9jmt7gxyjDr16egokuhG3HvmtmQBGcAdq+xFGBm+Xd0HXP5VZWUs8+1jjPt04rVdFutFkhkuR8lxu2Nx/DjPAJ9aALtiIdehqa0u4YYy8fD52nI7da0uCFiY2b5vofeoIRE5tzLlx0+XFeG/Ej6XocmizWW9pMYk8zGMMW6AH1x1pFuolVzbYVs/NvHOParkLKoEUWD/vUo2AIzYI9qDgx7QufxpmUcN1q2MecbuT0GKlleRNg49qijUHZnJPQ0shjQKDgjtjNPdC4i8uGDLnod1SqikhGyfTFSxS+X8q7nPvirdSnFyM+3T+VOHll2QJ9RnpUMBKNAvyu2OeuMVFHbiERQyfOOvB5q4t4kidXbDnGFwea8K+Tpt+k1/DuiTPy7iOx9PqK8Ya5puo6472kflQNjCZZuiL3Iz1FaldeVECy5Xtz9K0i8iiuPtDQb0Xr82OxFas8KzB3j+gyfQUsctzDlIMle27pWh3utaLYOlh+5mOMfdbufXI6E1q0EzXkcuoNtxntnsPT8KleAXGIn3j0wR2ozSSTbFXI/vZ9vSrSwF02xRgHv1qVBcx5zsx0XrU0UluQQ2GP3RivCurmxu0ujH5oTPGdueCPT3rUdYOv66mpi18kR52R+ZuzlADzgeleCtAl1TUyb648uHu2zdj5W7A56gV4m0dvBlwLyI+fa33/Hu+Nu7YAG4ySOW74oeKTY+Hns7V/3smMrj0fPcehq08HR6r4In8ST6h5ci7dkPk7s/vdh5DDsM9KtraeScI44/D0q8kgtFTzW8zZnK4Iq51gI7NCdnoOv9K1DUGuJScYx2/AV4j/sS4sYBo975lyu7dH5br3GOW46Zr9kqyuo/wBpv4Xs0Xyr4oscncOP3q18SWZPh14peNyrLol4VIOCD5L8itE17xL4X1eLX/Dmv6hpWpwtujvbK5eCZDnOQ6EMOfev2SfiP4n+Lf7Ofgnx/wCMyG1rUbOVLyXy9nntDPJCJSowAXEQc4AGW44r/gov4a8FaJ+1Dq+ozXsdvPrGkWV5cQpCx/e7WiJJXoSsSE565z3zXw1A/wCFceFQD/zBLP8A9EpX7WV5eJ+1H8TLiG7mWaHxTeeXIrkMm2U4weoxjj0r/gmB8Y/iB8TvhX4h8PePNWu9Y/4RO/hh0/ULyRpJWhlRj5LOeX2FMgkkgOB0Cgf8FcdLs7LTfhl41srj7JrNrfXltFPEdkpTbFIpDDn5HXIx0Lnpmh+0r+0gx4/aB+JP/hV3v/x2v2D/ABL4j8Yfso+BvEfi3xBqWt6td/2h9ov9Ru3uZ5tuoXKLukclmwqqoyeAAOgr/gpV8ZPi74C/aLttE8D/ABU8X+HtNPhu1mNnpWuXNpCZGkmBfZG4XcQACcZ4FeLvjX8YPHuknQPGXxX8Ya9phcSNZanrlzdQs6/dYpI5Ukdjjivgz8D/AIj/AB98YR+Cfhpon2692ebcTSt5cFnEDgyTSHhV5x3JPABPFaF/wR8nk0lX8S/HaODU3j5jsdAMsMT4/vPMrSDp/ClX3/BLj4uWXxc074fX3iixHh3V7a4ltPFNvZPNEjxLu8maHcDFIw5GWKkZwxIYD9qn9gbUv2WfhrafEDU/ihZ6+t/rEOmRWkOkvbne8Usm7cZGGAsLcY79aimRE2k89uKDmRAV4J7U0bu4V257cUoKgKR071IWCZUc+lMiouO9NGduCM1dOUiJI6UkweUbjx/9aonhVRmXp/smomSU5UZA71OvlS7lGR2ry/MPmiT8MU4ZhmIUPMXCyjIHUdKvcFf3S/QZq3knUgBcj6io2ckADdnt0pELHlcfjRiUpx1oKQMfpQUkZJoDH8X04qRB0DfpTIVHJoRqQDn8MVHHztVv0qMBRuJ5HaiPMG4H8MUsTAbyf0qEFRvMnHcYotJsDbuPpRZsjLdOnFSMxbcadyRjNc496kYOcKelMQRgD6UV9KIyMCgpPGaKEDA6ikXC8tk/SiOhB/CgcimXIIXrWOMZojOOaPJxTAhgQPrQIAxmiVIGT9K2gkYb9KaJWGM1NbkrlTx3qS3dQCRRQdN36VRHFHmhIVHrigxcZNW0xAIzSMHXINAA4FDrTcDAPSlySFzWAB1pgdvXigMrgGiABiiuenbpRGc4raD1OaABHNEY4zWOKIBoLiiB0FDgDArr1NEZxzUhwoYdqLkjFHHamJwMdqZgRgHmmUNwDzXlFRtB6VgBcE1uIOaJBYVwOn4UxDnOeaYgDapoAEtRRevekQMCCaVAD979KgVDHz1HSgC64zRUE7c0VCsOfpRIIyDTKG5JrBHANFM45/StwUbAOlBecqaBx3otlQoPSgu7gUODtFFQRxRGK25G6iARjNBASFrHamTBBzWDmhzW0dM/SgM8A0BnjNHJHH5VgEYP5Uc9qzgYzQfB4oSAijICMCnIYbhV5BvGcfSpFKnaawCOtKQO9abMGQq34frTNuOM0rhTyakKnp1p2J5BqE5UYrAPygdaZiOCaGCBjtQjV14NeXhipNSAn5QaAJOzHSmjzgg1GNwyT9Kx2BrGBjNY4J9OlFegpRzkV4Js7m38PnUI/mRu3A6OwqTT7jSH8mZfv/d6dvz9at71rO53YyP/AK1Nq6XNkysMMMfz+lRQQzI75+Y4xwagikj+cHDCpbma52GVt23OOAK03XNOj8OtpyW3mTLjnew/jJ9PSoZPJVVfqM5oq07FGT9atbAszKX478V8KND03Wdc+yalNsiPU7WP8DnsQeoFfGvwvofhjxpcab4cn8+1TbsfY65/dRk8OSerGp7CeztTI6YHrkHvUm/cSrcemKkLsMk/LUF7BJKzSLwe2T6VLDE0xdDx6fhVh4eF3ZvciTAXHGPfHr7UpWIkjkDpWnzTqQIHwT7D3rxHo9/pvhPRby7ufMj1D7RtXYBt2SAHoT6j0oQQmTCnDfjUWqeH7fwnJZB830mMHD8Ykz6Y+7Wqy2NlpKo8u9jn+Ej+If41fhJpi6ngf4Cp41YblPIpZ5GlJzken4VbKYCJZF47c1f39veXBljGPz9B/hSXcjRmAPkN7elfEP4xXvjrR49Mu9P8kpnnzg3VlPZB/dFMZUYbRkGvAfi+48PgwouQ3Q5H+17H1rxBo09jsvJTxPnAwONuB6+9QeWr5/Sr+7JKxKMKucfpUUrQNvQZqQmRt2eteFNIs9WtfIuJNufYnu3oR6VqNncW8rLIeeOePQVEpzsIrVNEvtOIW6h2Z6fMD6eh96GYznrimjwgAP4UVDNtzwKaPLbQau4igXHbOaWYqAoGaj4O4Hk0xJGc81bx7WxnpROV6/hSgnAzSJhsg9Kh2kc1IQ42KOaigCkAnPpUkIhuUJbrnt7Vf7WUKD/nigSScdqUEHJOayM4z0pxkYJojLZpvmXC0AByDk0ynZ1/SlAxtBpvmUKB0ojPFDaPlH4UiBV3Hn2rBd/c0VA+XvRY7ApHSuAu0Uj4XaOaVwuGPbtUbKQF7ip85Ei/w1Iwc5FHngUxK/4UpLEAdqBwvFKQRn9KOSM0yA8g/hQA65zW7K8mh0znNfKeQaKknJOaY8bc/SsjrQKgUo5BHehhV3jnFRtuUNioWG7k/pUb+YvyGpUeRCh615TFq3silPWlGTgUuQcg/WiwLbwautSN3ZWtnsx5G/Bz1yc+ntSyCIAZz6VcyF3Lk5J9qRd68mrYIhKsfpTyh+C33enFWzE/vAeaeaa5UB3zt6cYqFVWMMo5pHmZVZjnPQVbNI74PFK7pLyc5/wrTyGjyrfN6Y+tWksd1ZpHKeYs7fxNXEuJmUdun5UqgfLSKTyT0ohmk2mkiVVyvJFRwlSMGmAYAenSjFlgwbk+1aZqL6JqUF5FHvaPd8ucdVI9/WpLRrmVpycZ7Y9sVKFjcq3WpbRHh8wHntxVhGgiLP8AeTp+Oa8HaVZ6zdMl2+xR0OCezehHoK0zyzLsI6dfyNX1qk9yEjPynv8AgKv4gGCRtwvXj6VeMoucr+f4VPH5m0ZzntWjaXCsq3UhwIugwe4I9a8QTo84dGyB7ewrwhrMdhp13Fcx5MuzHPozeg961LXYI4/OiHI7c+3tWr37X1zJI4xnGPyH+FJbiKPIO5j07VNoscFvGs0nzLnPHv7GnhjgjxAM18K/hboHinwdcavqFxsuYtuxNjnOZHU8hwOijtWr+CL21eJ7iLCHO3ke3o1eKfBp8GztZzS73XH8OOwPZj/eq3mBWONBw2c1b+I7rRbQWsceQO+R659D61e6tf8AjW80Xw3G21bTz9rYB+8A54wP7vrV/pWkfCzVDYagfP8AO6H5k6KD2Lf3xXii7aF1tLWXEQzxt69D3561odu812js+AM9vY14uhtUuCiydenB9Fq4iiRwN2R9DVyLeGTy88j608ILKwOB24r9mDxpHqPx0+G+nJbYMniSzBO/p+/U/wB2vEOkJ4g8P6noMk5hXUrOW2MgXcUEiFc474z0rQ/+CTPg6LWobzxV8X9S1HTUYGSzsNHWzeQDt5rzS4Bx2XPXkdRLcfDv4IfDqIXV5p/hjwj4WsEhR55dkVtDGoVRkklmPAHVmY9ya/aW+KzfH34v+JviqI5ILO/uBFptvIMNFaRARxBhk4YqoZhyNzNivhoP+Lb+FBn/AJgdn/6ISvid/wAEurD4nfFnxJ8Sr7413Fla+I9Vmv5dOh8PBnhEjFiizG4wcE/eMf4V8Efgh8Pf2c/h9H4F8CQSw2EUjXV5eXkoaa7mKgPNKwAGdqKOAFAUYAr9v342+G/2hPizpnhPwjqsN/4Z8FwzQw3sDb47y7lwZpEZThowIo0U9CVYgkMDXxT8f+BPCuhH4b+G7bzZrH702+Vd290lHyupA4Yj73+Ff8E75fP/AGOfh9Jjr/aXH/cSuq/4Kr/8nO2v/YrWf/o2elJDAgV+wR8GtL+EX7OPhq5SxjTW/F9rHrOq3BXDyGZd0MZzyBHCyLt6Bt54LGv20P8AgoZrvwg8caj8JvhJaaf/AGxoqxf2jqV9A0yiZwrmKJOF+VGG5mJ5YgAbcn9hD9rvVv2ofCuuWHjPS7Ky8VeFpIRdPZIyQXcEwby5VViSrZjdWXJH3SMbsD/grqAf2bPDY/6ni1/9Ib2ooMnJHFSAxy7wcg9vwpkZ33KaZtsYjJy1ZJTaTQAcg+nSp4g6fL1qaAAAVLbqwxinhGKVChCqKEbkbcZooUJXHWmRsYU09uxQuD07YoWZYgk1b2SqgBpIEQ4FOio20HNMoVQVoeoP0p3/AIQPrQcYxQVmOR2pVzyT+FNtcgrQ6YHUUAScE9aKMigntRJESgn1xSgiHKnntSuYowFOR2oHJMgPTtUjbunamJC/KadWbgGvKOMZrY23ANNGwwTQjYtgGhGAoGfpXXiqIyMA80RnoaC5AFbQTg0U9qCgd6ZcHHpW3PAplBIz2plI5XtSschc1vVjs71LCripYEDkA0UIGCaIGMDn0FAZ70UOeTWWHy5qMk1bTMBgNz9KLuw4b9KjJbALc/SiQR15oDtmsf3T+FMAQATTZ6A1jAwxrG7mj04oqT1b9KIBGe9HnvWCR979KI4wKA4wD+lEA9KxxmjzyaZSRwakyBx361kkYNAluCuPxrBAornn1ogfwc0BmMHFKATg8VuyeF/WihY9cUVwoPX2ogtyBTDK8Nz9KAKnk0UGOGz+FKADhfyoDzGypzjtUY3kLjp2phjhRRj3AfN+lGNWAVTk08RBwT+FIAFwaAxx+tYzwDzW3njk1jPQ89xWDjkUDjvSnoc020/dNMWYgHmj85JzTDJAUUFxgqc+1M23DAZPpSrnDA05Ldax2JrGeCc+lbT6VjI4ojPAFYGMCiOmKAJA5ogHjP0oudvC/rSsSM4rOV61uHUdu1MQVxV9brklV5+v0pkKnBoYAxitPnEb4Iz/AJNbsNgHOPai4I6/Tio2K4VjmpCrLgioEYgY/CgpC5LfpRdmGM5pTnKg/SlJC8HmgN3DH9KIRjtB5+lMrFid/wClOmFwD9TigQThW5+lKCR15+lDgZB/CkwRtzWOOG5+lBlEZUD5jXhPWnsbT7Owyo6DOO7H096ka/8AHWtQ2DXmw/N5f7sHb8uT6Z+7Wt6PNot6+nTyb2GPmxjsD0BPrUtpcRx7ohuH4CrVtzAFcAdRmpCWXcyfrSyOylAvB9+laX4r1DQ42i06bYsmNw2g5xnHUH1NXJilc3Dr97tn04rUPFFld3RnsfDv2ZD/AAfay/YDqR9fzpkErmWP5d/8PXGPevBkNhLqBhvrzyE/veWX/hbsPwrx5pelaP4jmj0nUft9m+3yp/JaLOEXPysSepI/CtZ8XnWtAl0RbPLSYw/mdMOG6YHp61F4cube6S31OL7MJM87g+MDPY/T861jwNf2O17GX7VC2cttEeMY7Fs9Sfyqe1ezne3lG11xlevao0BkLFunbFMlw7hIV356DIFCCa3fZImGHVc9K02C9urkQ2abnb7vIHQHPWrfTX062uINZttlw23y235xzzwpx0IrQdO8PXc8a3+q+V1yfIdux9D7CviPFYaD4ovLDRrr7RYnZ5Muwpn92hb5WyR8xI5r4c6p4QsJp/8AhJ9P+2BtuwebJHjhs/cHuv5VHYajq5dtOt96xY3/ADqNuenXGehp4p0mMMnysvXoe1bWjYsTuJ7YxTXtzdxhLiTIT7vyjjPXp9KWOKRcNJtx0OM0A6Ajdx2OKS5BYAcj8q0vUordgzxecg6x7tuevfFS74596/uj2H3u1arqd9qUcAnl3rHu/hA649B7U8cJXB+U/iaHB2uenSlXC7d2R9KK7WIAq6aMBTCOec1c3FxeW8RnvNxOfl8sDHPqPpXllOc9O9eK/GviLxkQ2tX/AJ/l/dHlIuM7c/dUf3RTInk7ivT3pCCuG4x2ooSQ36UFXdkir0JuJjPHbirhnCYU4ParTMceG5Y/hSqpOWPPpioFQqxz9KDJt27eexzUBj3YYUYkK4zippF27V5qN9ox0pZixyD+FZZmyDk9jjFXDHIBOT60oUuMniplUHCGtrZ57U5DYAplHTrTDIFKhrGBnGfamU4wBz6UpUEZ59qcKDkHNQqHYqx/+tQDAEKcUOeAOaLKq42ZPrmiQTwfwpsZ6/pW0A4LfpTLg/KaBOMFs/hRJWMKWzntjpRbIxt/HNEkgjFLkjGelFQTnfz9KJ4xmgFwMH6CsnG0iiccbue3FAADIbPtiiDjGcfhRIOOaALkKpqKCOFCZX5+nSmGW4P4UeMDPNFQRhTSgN8hP44qLAPlsfxqIyZCsaBUMFPeoZAh4b8MVZQHUbiKFDhznaPXj/61N4W1CS5NrCu6VeqZAxxnrn0rVNNudNnMN3Fsb03A9h6E+opkP979Ki0m1fS5r1b/ADLFtxF5R5y2OufTmmiZDjdn8KUFXBIqdRjKnP4UibhweV7UhVeCfwpIhKm9Bn17YoBd2zbyenNWZRI9rNlvTFLJGRkfLjt1pJSGG3hatrgSzBSeO59OKId5ORgetLN5DFQ2c+2KsF8wjbwPX86sQjMp8vGc4OaeFWbay5/HFPHgZ6+gq2tHnhMgOAvXioIFMmAc46Uyxqc+lIykZBz6U4BAPWreZWckjOO1GQtcAHn0H4U1sJk2Ec+tQ6RBOoRZM7evB/xrUbY2knlQnLdhjHpSx3UDiGRM7fvDIFaNam6laO4OBxgYz6+lNGbVctyx69quliNvi3bDn2Pr71OskUZR35PtU8scaNHK27OMHGKtJkSQPLDuA/2sVq+si7jWztYtgHVt2c9D0I9qs/LivAJ5d6d/lx29q169tbm6CWkWIrfp8x/ebgPXkYNaf4aLr/aF8fKiX/lljdnqOoOeuDWumG7vnkthstxjYvJxwM8nnqKjgvHthJaDEQ6vxxz6HnrTRboBObndIf4dmO+P5VpdlNIASdwPbAHrXgK807Q9XS91X5o4s4X5hnKsP4c+or4v/FPTvFE8Taav2ZIt2Eyz5yE7lB/dP51qet3mp3G+5k81/oF7D0HtVpa3tlCt9cWvlrJnyxvB3Y4PTp1q81G8uk2eThR0+YVb3VxpI84j5h9w8fj6+tX3iS91STfdt5jjocAY6eg9hVq1s1ncz3cu9oNuF2kZycdR+FX+sPK7CCPYO3zZ9PapbmdhlpPwwKt3HU80luz8HrV5JatFGkaZIznk1+x+9jH+0L4AimGHPiKz8vr18z2/CvGGpXWjeEtb1ewZRc2OnXE8JZcgOkbMuR3GQOKl/wCCmP7Smp2ckY1Pwnpcg+7Jb6IzuMgj+ORl9D0/rXxI+PHxY+NmsQ3fxW8fanrsUDForaRhFbxHb1SGMLGp9woJrWrmyuZALO08hV6/vC2enrXw05+HHhTn/mCWf/ohK+Pn/BRv9of4dfGHxx8PPDkfhUadoOtXVlZzT6Y7zLGkhVST5m0kADnbXxV/a/8A2hfjVaS6P48+JV7LpEvD6XYxpZW7j+66xBfMHfEhb9BUjCRsA8dhTAA1/wAE6P8AkzT4ef8AcT/9Od1X/BVgFv2n7Uf9SrZ4/wC/s9eH9Ki1jUIrKa7+zo+cyeXv24UnoCPTFfCS907UvhT4Mv8ASGU2Vx4fsXt9q7RsMCFeO3GOO1ftw+F9a8J/tXfEWz1yCRDqOrPqFrIw4kgnAkjZT3ADbfYqRwQQP+CO+lX41H4oa2bd/sRh023WYrhTLm4YqD3IGCfTcPUV/wAFdSP+GbPDgP8A0PFr/wCkV7SMyIAF3fjipCSwDNj8KMiswQLn8agtmmk2x/MfTpTeHNYMXmizyvr5i/40VYDkUxB+62PwqVFKhl6+tSRoUyDz6U0Q6A0I0HJGT6UVRei0QpOAKKKwwDTwoSOM0sfoaCg4GaBB4P5UwGOT9KHQLS4HI59qVFY7j+VMgJAXmlXbweM0owcZz+FbdpwvftS70yCPoKQjfuJ6VJKFHXOe3SlCAkE/Q4qZkGDE2T9KMmeFP6U7nGAf0o5aiOgxSqx6UVBHSjHhcUUJOBQTOOawM4xRA7fhQBGFFGNTwDzTIAOvNFQFxnmtuByc+1YDmipAyPyogE5ArOeMVtBUEHmiPamGRgD8KZTjGaDNnlv0ounQCic0ABwTW0YxmmXsKrdnApTtGQeagmLDBPSo3G4EGsEgHHFAjoaBxgZosCcGmYDiicgHNbsiiD2PNMM9TROelMMd6xxgGiMnGelNgEEVnJ4putfQ0wyOtNtc7T2plXPBrAwcGqYKR1oqegqRh91TTElcUpCxg55NHnvSBGHLfpTYBwD0o4J60xDfKDUZCnFMwB3E9KZGK/0q2/1YxTE7NtA4wRW8KwK81Ihcbz3rAXqeaK4GfyrknIoDjFEcUzEgBqPKBR2peDkGkwx3ZplHY/QUrHG0UUUJgmiFj6UDkgCmJ2DFFODjtUZGcUeXyPwpgCcA5otk5okDjNPgknv6VjPIPSsj7tcYwxpVAXGaIBOQelZAGAc0SucA0XBOM1MFZOBkirqI5zigM8Z+lZ28g1A6Sxgg8mk25xmmKgDmgQSFznNW6gJ8ppiQM+lAAnIFKuGYZrOBgGhlyFFBSrAd6IPc0wZl29qCBQADQHIGaGCMA1jGMDmipB4P6UAD3rRrSUWH2xh8p7/iRVjdvFfebG+CehxntUlwJyXkfc30xWjOlrercy/6tc/yIrUvC41Szk1PT13bcZ5x3A7n2NTDylVCfXmhNAgAjOT6YNOrOeasHMUg8wfIK0S005/DMF5MfmbdkfN/fI7V8Q7fRbU2cmkybt3mb+GHTbj7341bSpE24Gpdfkn022010wlvvw2eu5s9MUqr95G5+lafIL2Z7nUnxtxt4z2I7fQVNrUv2ZreI5TsenfPpU4S5dpd2WPtTxxg7QeT7V8NvDXh6TQZNVvJc3i42Jtf++6nkHHTHarrwTq3icS3ttb/ADLjI3r9O7D+7UEM2lSJeMuHXORn8Pf1rXPEY1u3CSHEi9PzHsPSpGYS7Fbhfb2q4unkI84/d6VMRIcg9OhrTL2exm86A5DdRgdgf8avZ2mmZm6tjP5CvC/ha68U3j2ll8zJjHQdie5H92tR0ybS7iWymHzx4yOO4B9T61aWjXMixgcNnH4VLbnT7jaBnH4dv/r1cmNnLQ8ira4kgdZFHIzxV1qTahc+fIMH0/DHp7VdyW0tpCsLZb5t/B454oWMjgF48D1zV5ZoJ22jO3H8qffG20dKiRp28tBuY9B0r+x7wvjyvoNw/wAav9IksZ2t3XDRYyM+oB9fetQ1vTLjQDYxNmb0w398H0x0pUwpVutT8qYe56UsCmQIDk1cRFXBIwO1PHGz4DcD2q4dWbC9O1R2T3UnkxGluIgflOT2r/gnb8Dvgv4i/ZN8C+M/EPwl8H6tr+o/2j9q1O/0S3uLiXZqFzGoMjoWwERVxnoBXie+/ZQ+G15D4e8Z3nwm8LXZhEkNjqkmn2MnlkkBljk2naSCMgY4Nat+zt+yh8Z/D7XX/CsPAGuafeBlXUtHtYI3b12XNttcHJ7N1r9uz9iuP9mDWNO8WeC7+61DwP4huHgt/tXzTadcgFxA7AAOrIGKN1wjA8rub/gln4X8KeMv2gdf0rxX4c0rXLSLwbczJb6jZJcxpILyzAcK4IDYZhnrgn1qX4DfA6eJ4ZPg14HZHUqw/wCEetRkHg/wV+15+z/e/s4/GrV/AwSRtEuD9u0C4f5vOspCdgJ7shDRtnqUz0YV/wAE1fDnh3xb+1BYaR4p0DTdYsW0W9c2uoWqXERYIMHY4IyPXFf8FFPhb8MPCv7KXibWfDPw48L6RfxXtgsd1Y6PBbyoGuowQHRAQCCQee9fBD4OeKfj18TNG+GXg+OMXuqSHzLiUHy7SFRukmfH8KqCcdScAckV8Fv2E/2c/gxpVrHD4D07xPrkSKZ9Z161S7lkkA5aNHBSEZ6BACB1ZjklLX4NatL/AMIvFb+C7ySP5f7OVLaQrjAx5XOOoHTvX/BVr4bfAr4a6F4OfwR8N9B0DxZ4i1GeWa50u3+yhrSCMBw0UZEeWkmiO4ruOw4P3q/4J8/Cb4V+Jv2QfAOt+I/hp4U1bUbn+0vOvL7RreeaTbqNyq7ndCxwqgDJ4AA7V/won4H/APRG/A3/AITtr/8AG6PwJ+B5GP8AhTXgb/wnbX/43Q+BXwQHT4N+Bv8AwnbX/wCN04LAsO1FgRyaPHI59qzu71Eu9jjtWShxTtuwidulEZGM/pUgIb5TQwVwRRXI4b9KcjIIPSl4Oe4qRiwzmgQRjP0oE4xWWPTmsbTmm6dK4PIo7scUy85pgxOc/SiQR70ygDjrSnbyDRLHJJ/SgpzwaKknP5UAQeTS8qCDzSLgg5+lIWMgcfxf0oZYkEfSkUMetWjlHXDY6846Vpusw2aG9kk/eHoMH3Hp6VrF2+q3TXMp+eTt9AB7elSIV4JzQJX5VPFRI7LyM1MojO0HOaByu3P0pVIPPbrXyqMqOtRO6ZCnjtQfyxyOagaMJndz9KRd54P0pE3LgnpSPG1thDll/wAaUmMZY80yb5Ny84rTVY2hK9f/AK5rTV/0ZZSeTnH5miHYjA6USrW4A6j/ABqGVy2zPHpUBaNBxk1KweMhO1BcLgmlVRblc9en51LGtuAo5IpGWe6jBHyjPP4VDCzQl0XK/WlJQJGDxzV3G8Dh3X6c1Ivmzbz/AB/0FWVtKk4Ur97oc+1apFHK22I8dv0pke2GXHSpY5Zi0kpxR0271LdDaRbyMZO4DH5n2qSCUzhCfu9vwq/KW1jIqn55MbR9CM1pXhu0uLY3V5JtYdOCe5HY/StKgjv9SaFW4bG3j0U5/lXjPXluNummTA57Z/un0pzHPcBIzwOn5Vq3j9LHwdJ4Usotxlxk7sYxIH7r7+teD3lub2OW6T91HnPPqG9OetNcDxFqCaZpy888Z9s98ehrxBpH9m3b2UR3KuOenYH1961XQLOPTRPDJl+4wR/EB61/Z+xtyrlj7/8A16+HHw/HjbwlFcas3lRaPnyuN27zZHB+6wI5Udc/hXxE8I23hrVpbK0O6FMYbkfwqehJPU1P4a1HWp55bBN8Vpt3cgY3DjqQeoNX1paxWn2ln/eDqMH1A/lWpXMV5bCKJunXj3H+FaZpUk8wjC5//UferrTGjYxleV9//r0Ld4zgJz6ZpdsYyOpq3sLiV1Ij+9nHIr9nS9s7r9qL4ZtaNlP+EnsscH/nonrXxLOPhx4qIPTRLz/0S9LI5HNM4iQMD1oyhmzmvhqP+LceFR/1BLP/ANEpX7Xcqj9pn4oLjp4ovv8A0aabAG9TmsDsaK5ZWHWv+CdP/Jmvw9/7if8A6c7qv+Cq5H/DT1qM/wDMrWf/AKNnoSvGN0R+YdK/4JyftreDbLwZZ/AP4t+JbbR73S5GXw7qV9II4LiBmLfZnkbhHVi2wsQGUqowVAb4t/s7/BH4+QWh+KngDTvED2abbW6MklvPGhOdqzQskmwnnbuxz05r4La/8APDmuan8A/gVaaXCnhW1W81SLSAJLe2klbYqyzZJedthzkswCDcRwK/4K6vGf2cPDMJkXe3je2IUnkgWV7kgenI/MVGgVMA5oRF0yzdOgxQjB5J6VDKYpBtbkdOKuPGd7Pp/wDZ5jwo75Hrn+7TzGRNjdulSP5YOKaQGMKp+nFRguQoNMrBsCmOCMHOe1bCeaYZ5xW3B4NHrx+VHgYz+lBtqgGiRnINAbsY/CmBBxnpXBOQeaC7hg9ugqNkXqaGXfaaUZbGayFPJ57UpBXcTk1KqBsq2T9KR40XJPPai5AwtbucimXPPcdq+prp0oDnJNMox/SmAAwaJHQH6U23bk1247dKAB6GuCAM0ABwDWD2PStuRkGgAeAaO3GM0MEZFAoRgHke1MuOM/ShggCmUdBW0etMFJyO1N0460UyOnSjntTOQMg5rzSTtBqJ1dcE81gbeG59MUQQcg0SByOtRBnO0n9KJCNgHmkeRQGByfpQvJTGI5G4HQYpXDKNoyfSqJB5HWifM5JokgYBpTnnOfSiw44+lEDrTAEcUQCKYYAAPNA84IoEH3NY3d6YDoKIxWCRgdaZCF+U/N2oqGGe9EkjCn8MVTAEYAzRAC4A59KKk8DgjtTLkZpm3cbs/hWccA0R2DZx7UMdhk0UTbjdz9KUkDAPNH5jgtz9KYBVA3dfajvc4DZ9sVG2FC9aHJyG/SmUtgBunbFCMsdq9aDuVCMc49qRC7BQf0rIYZDfpQJzgUduMEc+tAoRjGfT2onPGKI28Kc4rJzigf4QaUAnBP6UhUr8o/CiOMda2krgjJHQUc7c4yfSgg/+tRGRxSqBzisD8aIU8Kea+UnaT+NKqngjPvThCAQKIPQDpTAYAJo4AxnNcEcGmAI60wUHKjmmjBXJ/OtvH+sx7YoAEDFTQBlzip49rEA1getWb5jC5yB0FbQF4HPYUUUDGKWJdu4cn+VQggDDfhimOOA2c9OK8vcOtKm/lT+leXx8prhDk9ulBVfG0fSnXbz19qR8dsCtD8PXmupO1rybfbu6fxZ9SPSru3ltJ3tpVw6YyM+ozWc8GhtPXijEzL5gHHag2ADjNeFUa9thDM26I/w4xjk9xz1FeM/D1ho11byWbbo5d3y4IxgL3JPrVhAskgjAxnt+dadpE9ydhfAPQ4Hv718HPh9qviG5k0YL5SS4/wBI+VtmBI33dwznGOvvV3bxycyja3p1p0ktm3gZFW9u9w4SNPNc9BnbU7XdhIYZ7fy2H3l3g/qPrWh6ppZtwlzab5e58xh3PoPStW0e4ubgHS7DzEX7370DHAx1P1ozEP5Tx4I7ZoSplt54HQVBOiOrSJuA6jOKn13w3qUFnpVtoH2eUb9832t33c7h8pHHAxXij+ztJ8qCwXbuzkZY+nrn1NNcoZ2Vk5GMnPtVlp8WqXAja9+z4/i8vf29M+1aVcR2d4ouG3gZzxjPB9Kk8a3VitvLpDeRFHu3LgNnPT7y56k14413w/eTMlhovlqMYP2lz2X1HsadwZQI2wv0z2p5Jo22mTP4U9wTgjkdxUFkt2f3cuMdtuf89KdWSVvMHC/dHpxUksYJKvz9KguikoNudjDoev8AOhO7ZZzuY9TjFLICm1TwOlaZNZxo3nW/mPxtO8jHWnhgZsRLz9TUkEeShGH7GoIPNbBPFWGgazd2l1qdlaeZZ2Ozz38xRs3kheCcnJB6VL4jg1SBbCS02beh8wnvn09qsfCwuEnF3c+Sibdg2bt2evQ8dq1KFY5WjQ4x0OPpWi3jabfRXgbc8ecHGMZBHv603i22kuY9Vjt/NkTOU3lc5G3rj09qPhnUvG1pN4rjX7NDLtxDlXxg7OuQeq56d6kh2kgtx6YpwrgEmrpFPEfX19KWJUk3KMFehq6lV0y7bmbvjFSqQABSRhweeR0q/DquFfHtj6VaQgPgnmv+CbQx+xV8ORn/AKCn/p0u6/4KwgH9qS1J/wChUsv/AEbcV/wSK8FfFTR5/GfirVdP1Gw8D6paQR2i3SGOO9vFbIliVuoWMspccHeoySvH/BWfx14b0n4CaT4CubiCXXNe1yGe0tiQZI4YFcyTY6gZZEz38w+hr/gkQSf2k/EnP/MjXX/pbZV8S/ijo/wx1HwRDrskcVp4w8TR6As7nAhnmtriSE5/2pIEj+sgPGK/4KBfs3D9oD4Kz6hoFh5/i/waJL/SAiEyXMe0efbDHJLqoKgdXjQcAmv+CXiov7Wenqn/AEA7/wD9AWv+CmuP+GPvFQJ/5ftO/wDSuOv+CQbaKfjb4xFxsGqHwvi0DHkxfaYvNx+PlV+3R4F+LfxG/Zy17wv8GZ7n+2p5oWurS1m8ma+swT50CNkcsNpK5G5VZed2Dr2g614Y1S50HxHo19pWpWb7Liyvrd4JoW9GRwGU+xFa/wCMPFniqGwtvE3ifVNXi0qEwWKX1484tYyclI95O1c84GBmrbXdcsoEtrTWr6CFM7Y4rh0VcnJwAeOTX/BILVtV1L4kfECPUNTu7pU0O3KrNMzhT555GTX/AAVYvb2w/ZgtZ9PvJraX/hKbNd8MhRseVPxkdqbxP4mIAPiPU/8AwLf/ABoBsYAwaByMdxQGThevamjIA3c1GFbd82Me1OqBTg5P0oZJ+VvpxQG48mlGW2jjHeioPAOTSo7jI5pVDHBpiR0NDgYJzW0EYJzWSxwDkfSkwBkN9OKADd6IA4A/ClPGMc0cHg1j1P0oHPFHIORW3HWsZOTWR93NcAZPNEgjkVux7+1DHpRAHRs/hSMHII49qUEdXz+FKNx2g80i7cMDuI7YxUk2iXGlqsVr5Vzz829m/i9OnSoIDJIC0W8r/tYqe1UNsjGD6VcQSQt5b9R+laZqUFrC0E9p5ynp+8K46+gqY4IDDJ7GgADjrUJRkMbHr0pwqgrjIH4VpNxaW7sLvT/tIONo80pjr6fhU4jkmLJHtU9FznHHrUKdweKt4g0eC+PwzShg+0Hr7UI0t0SWHqM7h6+lTRLKAyGs7YwUTLL0OcVpLXKyBJGwp9h71YBBbxjr156dzRmlVwgGfevISc4Rtv4Zq2tysrAvux04x2p4ljOwNn8MU8diti+yLMxxg7j6/wCFOgdwpPWkh2dF3EdFzivGvhXwVYeF4dW0mf8A4mD7t0WyXjEiqOWOPuknpUS+XEGZOfXNWt1dfZfIhlwp7bQe9aL4dudWvI7WN9oTPYHqCfUeleI3jOpCyvJvOMXT5dvVQe34Vo3hddc1SDRNJh/fSbtvzeilv4iOwPevHPgG+8A/ZYr1syXG/bwBjbt9GbswqWUXLhJl2MO+c/yrWvhp4sOjnWrbw/myh/1s/wBqjG3LBR8pbPXjgVf6nPcJsjG5R0HAx09q8J/EiHwd4N1TSE8PfaLu/wDJ8q6+1lPJ2Ssx+TaQ2Q2OcY61qOm+HbfQVvxe+fqL53DynTGHAHfH3at0lv7ryo48sn8WfUen4Vfy3wXAuMkdtg9qsb2aCOUeZ8wxzgVFO7v5p7dqW1zO0kb4DdeM4wKu/IR9rtk+uDVjqCWUO61m+Yf7P+P1rSLuWScSxy7JT/Ftz2Pb6V4bcS2/9irHteT70uc9CWHy/p1rV9At9Fknjebz2Tbj5Suc49z61pNzZWqLeXsPmouf3e4rnqOo/A1qfj+e6tYRpD/YYod37rAlzkjuy/U/jXifxf4gJNnd3PyjoNiex7D6VoHjTVvDZuVtLzCXGzzI/LX5tucclTjrXiyKKKNbuCbO7OV2+m0VPcSOwMYxj8a8Px3hnRhLjOcfKPQ1f/C14vC0Pip7nesu7dHsxtxIEHO/+lahcojbU+cD+HpjpTBCwJTOPep/E2tPLCIZ9nk7to2qeo57VY2Xiaw1NdQtL6403UdPkSSGe3l2SRP1VldCCpGAcg1e/GT4zSiSxu/i14ymikQpJHJr1yyupHII34IIOMVo8Ogy2cjaoN0oxt++O5z932xW4KMsMDt7VkHlah+NXxks7eK3tfi74zjjiQJHGmu3KqigYAAD4AxxitS1PUdbv7jVNU1K4vr26kMlxcXMjSySueSzMxJYnuTRAA9cUCCMqc0QGb5Thh1rQvir8UvDmmRaJ4d+JfinStPts+RaWWsTwRRbmLNtRHCjLEk4HJJPevEPibxL4rvV1Xxb4j1PWr8RiMXOoXb3EgQE4Xc5JwMnjpyalZhIArfpX7Mf7HPjn9qbTfF154K8Q6Xpc/hdbURjUo3WK7km8z5PMQMUIWMn7rfeGcZzUv8AwT7/AG7bEHwpa6BNLo8h2sbfxVbJaEHqTG0ytjgfwelfsL/sjXH7LHgjVW8Tapaaj4t8USQyam9pkw20cQbyoI2YAtgyOWbAyWAxhQT/AMFc/itpeqa34O+C+l3azXOkCXVtWRWyIXkUJbocdG2eaxB52uh6NW3C5Haprhg52P8AL2GKkunddqnA9Kjckg96hct8pbP4VcKWGFb8MUySthc/Sgh6MeR2pZQiFVNB8tyM+9A/P149KKAcCjGAMk5ojcMgUQMcGiMDn8KOD1H4UQMYznNHBGAKI2nIpXwMDmoznJUZx3pi2fl6elRJGzgMaZcNgHNM4AyDnFecDxt6e9eac5B/Ct5J5NGNWAbNMUGPLXn60QDwfyplULhfyoLkbiufSsAEHb+GaKIwzn8KIBTnn2o8rnb+tEZGB+FbcAEHn6VhWAIogMoOcUFGOT9KAwMA89qJBGBR4GM/Tih6A10GCfpxQXuGz+FNyoyeaCjOM0ygjA/CipHRvpxRAJ4P4UwUcYplDDBoxqei/rQkfHJoOW61FJtwFNLIW7/pXGNpoKCpIPNJlVyDzRYMcg80spxsB/Smldjkn9KgkbAKnn0pZTtwB0pWDjg/hRAABH4UpBPJzWQDkdqYAigQwxRx0NZyaJDcg0cZ+lYAGBzXGcijgnIOaPPevoabnpTBQOtMiqOTyelOFx8porkYzRHG0daXAPWiNx25okKSD0HSgUY5zxW0NwpogA9aK4oDqV7VhT0olNpDHp0pSMfKai27cE1vKjFK3GFFKxLYzzRIBxmtzbhsOCKBVV2seRQGRkD6UQCMiiAo4FZB6GsZHFMqFcqaABOCaCIF5pSpbJP0piAM0RnoaZugHU9KJwuT1o8jFdOCfpR9BW0A5FOFAyKDKRgGiqgYJpwAAQawGxzUiADj8KYgHAogEYIohTwaPPAGaKqw46isheppCr8Zq+t0OSO3/wBamUZwD0qyOOp+lIQV4r7wxilznAqIkDrRYkhQaLMeAaU4+VTz3FE7eCeaXBHzDmvAmn6dqGrrDqLYi57N/db0OeoFfHyb4epcx2Xg+XdJFnzBtmGMiIj7/wDwKmBcDFabrF3p4kFnLt8zG75QemcdR71PcPczNLM2Xbvj0FTWktv/AKxf1rC4+lJIVAXt2oIGBIP0rwM8CWpWQ8/j6tXiKC1urMTs+SnTgjGSBVtoTnQX1iIcJjB+r7fX+lQa1e2zArJ06cD/AArwv8V9e0NilnNsDdTtQ+vqh9amkjJ2SnLd61RxOiIU24zznPpWmX8+m3iXKJuKZ4yB1BH9anvpvFusRxrDtkkzhd2ei/h2WtTtE0e4+x3A2yL179ge2fWvD/iaXT5pNkHm+ZjA3begPt714o0LS7SP7Zat+8fqMN2wO5p4FZ1QnntQdkba7fhirFjJcbkb94Pu8e1ahdSyTgzPuK+2OwoCNjuz8x61aR72JR8Ee1adpZmd3cZ24wfz96nZY18pX69BirmUTIUxzQhDsUzjHSnjkI+YfjUsfCHsc4q3doVBQ4qW4Fy+4nkdaa3STkL+tPauDlD06VpEDO5M/Qf/AF/Sri12/vIuUPSrYiORS44rQdJi1S9EAHH/ANY+/tWuDwDpngXVtDmmxrz+T9kTbNziUM/PKfcPc/TmmQx4AH0rTr3V9B8H6narHm21XyvMO5ePLkJHqep9qsvD2nXUCzKvzc85b1+tatfy2HiCax/5d124Pp8gP16mvFek2tjfpHC3yvnsewX3960vSYtRml2DKrj27H39q0a2tZL5rSZsBe2D6E9qvJdb0DRIoIp9lvLnau1T0b8T1NELIuQc1p2n6bpc4l1lsRn7vDHsc/dJ9RV7B4e8T+K9K0vSmzBP53mcP2jBHXB6qanR4JjDIOR/hUjCSRVC8LnvRiEo3HpU7CMZQ9anIlOCfpUMah9y1/wTb/5Mr+HP/cU/9Od1XxL+Pf7Ofwz8Tx+H/il478NaPrpt0njh1BP3oiYkIwO04GVbv2NeGPGfg/4k+Hm1v4f+NNM1nT51aNNR0i7iulifb2I3KHXIO1gccZHav+Chf7K/xm+H3iJ/jR4n+Id/8Q9B1WcW76peRCO40xzkxwyRJ+7SI8hWjCruyCqkru/4JFDH7SfiQDt4Huv/AEtsq/4KuGe3/Zu0XVrO6mt7rS/Gdlc28kTbWWQW9yoIPUEbsgjuBX7I3x7sv2jPgdofjwyp/bMC/YNdhXA8q+iVfMOB0VwVkUdlkA6g14a+BQ+BH/BTfRtQ8PWkdt4Y+IOl6pf2MaIAkUnkM1zAo7bZQrgDgLIo7Gv+CnAz+x54rAP/AC/ad/6VxV8HPi34v+BnxE0j4l+BrtItT0mQkxS5MVzEwxJDIoI3IykgjqOCCCAR+z9+3r8BPjzYWlm3iW18J+KZQFl0LWbhYWMnpBK2EmB5wBh8dVWvid8FPhR8Z9KOj/E/wFpHiCHYUjkurcefADn/AFUy4kjPJ5Vh1r9ub9gyf9mxI/iN8Pr+61XwJf3XkSR3PzXGkyt9xHYY3xtyFfAIICtkkFiM8E1/wR0z/wALK+IQJ/5gVt/6PNf8FYhn9lu2APP/AAlVlj/v1PTHdzmlYEVGquck1JtU/KfpW5m4B6VEc5AoqpX3HQUUYqdp6daGG4J+lMSDgiiwxx19KjYgYzTbTn26UTkYI4pgNoxRzjmtpZcHt2pjyBSrkYorgZzQAC8mjhhgGiDt5PSjyPpRIxkUAGPJ5HSshlAJoqMZzWCeCaBOcZrAPINAADINR4b5ahUCUqev/wBauuAD9KU/pSkgjI4qGAkBlHSrfUJBp8tqkPXGX3dOc9KmDeYWPWrnMjhnHLdPwqSPYcKfwognGTmgoJyDnHbFKYVt1w37zuMH1p+VHPP0qMBED55NKDnJOatbESISopYAuMHp2pIkXlzzThnT5Dkd6tbZ7mPKjgVEituXP1q3E5cFDkD2FWBfylA6HO39aLFSuR0pkuYCA3XuOKtbWMKrSHG7P6VKoV9hP0pyZCi49eKjTEpHXH+FSSkcr1qNp74AyPlewwP89q0Xw+dQl8gDJf7v4A57+1eHPgfe3rrcxw5h7/MPcf389RXjDwVqnh3xLrGk6UmG07yN3KnG9Fbux9T61r3w+VZoboDMkm7eenQAD+KvBMc+m6yjWh/0nnYeP7rZ65HQ14/Or3YiOtP5knPl8KMfdz936Cl0z7NbLcT8zPnj6HHY+laj8WvEEnh6Xw4fltZceYfkPRw4/gz196+Evg3QfGGr3Fpdn93bbd3D/wASuezD+6K+Kej6Zo3i++0rTXzbW3l7eGH3o0Y9ST1JpZDnDSZz2xT6hFYMpg6nO4/y/nU06SvlHz7YqeQW5LDo3T8KikjERkI4Fec7QhlHyirm7hKlQMt+IrSlMrAFPl78/WtIbzbvy7YZI7fgfWtB0HV9WuZk0sZkt9u4/L/EDjqR6GviF4UfQrKKWV909xuzxj7pX3I6GvhNP4afw/8AY9Xs/McfdPmOv8bk/d/CviC/g2ytLqe3n2X0mzyotsp6FQeTx9055rUdWGoXBkds/hjsPb2qxks53KXTY3fd4Pb6UdM/tC3MZfci9OMd/r7Umj20MAnk+736+uPWrm50BLVF0qbdcDPG1x39+Omaj8aXc/hldHuNS5XOYvJH9/d1C/j1rUEWEB85z/8AWqO4SyUkR7gehzj/AD1q8mMrmYDbj8av/Ed/eXD3Pm7WfG4bQegA9PaovMmDTNyRimIJ+brRUH5RWkNpySMdQfA428MfXPT8KLkT7M8f/WqFc5IPStoIAJpisbEk8H2pVAO5uvpXiO18PWelwvps265bduG1x0YevHTNTMeCDx2oHe4JNfsh/t0a5+yro+peE1+Hel+JNH1bUPttw/2prS7V/LVMCXDqUAQEKUzlm+bniz/4LA/CSS03aj8J/F0FzjiOGe3lToP4iynrkfd9/avil/wV38X6zps+l/CH4aWvh+eVSq6rq12L2SPP8SQqqoGHYsXHqteIPEGt+KtavvE3iXVbnU9V1Kdp7u7uZC8k0jHLMzHvU9w5G0HA9KdicAHpQOeM80mABzyaSQg4Vv0p2PlhieaaVuq9e1MSQOaeMEAKc0yADANNg4A59ayAME0AGGM05A4U0MDvR9RTAk5A4pV3YBP0rbg4A6UxBBxRHGBSAj5QevWgUHGeaHLcGmkZiDmgcHGfwrGeBTAAcUFGOtAKFAz09qwoGVOaBA+WmwMYosMAE02cHJoOD0NDk4Bo8HaDRTCgmmwy5FEKqDnn0oLuUBfwojB96ZcDNDkY60fmGfSiARgmgynAAoKM0QQMjpTAk5ArGBjNKAxxRGDjNMo6LTAcEVRyRkGjzyKUkHGf0pTkZBpW3Cgfm5GaYnOFpjzwaHPNKO+elI7DhTSTkPgtkduKWRWAO79KUxNCCp+bv1ojtXHQGix24BrcCOTzR4HBzQbIGTz9K68UQCOBRxjINbmHANHB5z+FVTDIwDRVic7/ANKddw5OfSiDj736U2D/AIVtyN3pQ5wVP04pgc5zzRBIBIpmZuD+VBeMg0X+XBGT2FOjA43fpQYr70RntTEYwx/Cgqjtk1EoPIHNZXowpWWPBzx2ossh4FEdFFICpypxiiC53MfxphgBfMzn2okE4zQ9M/SgVPRc/jSqSu5T+lAMT97n0xTBgMDoKQhjgjJ7UFwBtbr7UxYDGc4oMrHAPSiuTkGjntRANY96IDDFYA4BoqGFFARgmn2sBtXB7804GACePTFOu7AB/CjERxngUyjoDn8KY5QKT06UAOxojIwopowx69KZQM5HSlAzipIQ8ZxV1FsfB5q0wH/lUR3JjFHKjaRTEbQFPToKgAcZNBSSMCp7C7hi8yaPC+uRSumSB1oEMMCoo127n6dqS9e2B8g7f1pmz0NJIB35pQxHyn8MUGIbDdq1rxXqeuQeTe3PmA/7CjuPQD0FKwJK5zjtQJ6itTvPDsumWcWmaV9nu03/AGmXz3fzMsNvB4GBkcde9aNeS2i7424+n1rwj4c1TxtpeoT2p806Z5W5cKv+sZgOpH933qW316WGTRo22wjG6LCH/aHPXr71YfDZR4Ug8XXN1vjuN22DZjbtkKfeDfj0o2li8u8rtUdsk9qvLkySKWHy88VdTb2Vccc05WR9yjHvWm/aY7xJ7aTbKudr7QccEdD7VeSTXFwZLiTzH7nG3t7VYzPb3ImU/Kv8P4YrxPr3h/X4kGl6N9kkTOT9oeTPT1A9D+dRau9hp13ZxPhbrZuXH905Hb3prVWO5T+FJHPCcoMHseKaOWVzvOCe9Sok5EUEO0jr82a8NeCbnXZ1toLHdK2do8wDOAT/AHh2Far8KtW8OwQ2Os6X9jvl3bj5yyZ5BH3XI+6R+da/DBBP/Z9/p/myJ0fzSucgHoPwrxXplrpt4UsNL+xoOieeZMcL3JPqfzp4xIRjhu4pJVChCeaWOOSVUVcY6808rwjy0GRWm22l3EbGdNr9jlj6+lahZzWEwCJlW+6cgdh/jUF7GsixycgZyOladZ6TrGiGx0aDyZpPvtuZs4YkcNgdAa8c/CQ/DG6k0fWbn7XOMY+Ty8cK38LsOjjv2q6t2ExAXaPTOe1WN5dafOlxby4K5wdo9Md/rUPi21uplOrweeDneN5XPHH3R9Ktrd7mQvBFiPsN1Ikgh3KmxV6DOc14R1u0uIGhlTkYwMn1b2rxHcQ6hfvYg/MuMtzzwD0/CrizvbCQhhhR9P8APeo5Q2TGfvdql025VftATaPXIPt61N4g1GWWMXF15ix5wNijqPYVrmtrqjQMibTFuzznOce3tV7q97qsKLdz7xHnaNoHX6Aegqy1GfTr2K9tH2Sw52tgHGRg8H2NSlpF+Q59sUqGPczDDL0prgocRnFXBMz5JzRjUnpzUSKhIP51/wAE3MH9iz4dY/6if/pzuq/4KzYP7UtqB/0Kll/6NuK/4JJXnjOL9oXV7DRZLk+H5/D0z60g/wBSCroIGbtv3sQvfDPjjdX7YWn6Jqf7LXxStvEAjNqnhe8mjDjI+0RxmSD8fOSPHviv+CRBz+0l4kwf+ZHuv/S2yr/grC239ly1I/6Gqz/9FT1/wTn/AGj1+Bnxri8M+Ib8Q+E/HJjsL8yNhLW5BP2e4PoAzFGPTbISfuivFvgLTPFXiLwf4rlEaan4N1SW9spWjyds1rNbTR56gMk5PpuRM9K/4KZLu/ZA8UqD/wAv2nf+lcdaR4e1/wATXcmn+HNFv9UuoYHneCytnndYkGXcqoJ2qOSegHJ4poyMqwwRwQRiv+CWPxv+M178WR8HJtW1DXPBJ0me4mgu5GmXR/KA8t4mJJjVnZYyg+UlwcZGa/bet9Iuv2TPifFrbKLddBkdN3/PdWVofx80R/jRx0PWv+CO2P8AhZXxCA7aFbf+jzX/AAVj/wCTW7XHP/FVWX/oqevvDNBTtyF4HXmsgjGOazvOG5xRI9MVCFLFSMk1IoXvzRBDFSv60y8ZPaipJ3HkU3XFKmeScY6DFOQTt6mijYyTQGRkD8aCHjI5pkBAOc+lMoJwooAbcA8mgpIwefaiSQB29KAA4xTksAB2o7wuM/pWSOCawSMg8/SiDjg0SSMjmgORg0Tj5QaL8AAVG5AKk9aQ5wSc1CwB5bke1ICxzn6VGvAOKDbSADn1pPMIADZA7YqGeXBVnyD2xTskiB0OG+maLZAIOWFQRQyuRd9O3X+n4VPH5chVenamcuuC34YoElsZqOFdnJz7UjuDsPAHSv7ItksWu5bv5hjCeWeecdc1a6jdMhCvjPbArTbuCIZuYfNA6jcV9fSrl97Eo2B2GKtvmkCu/H0q3NtFHtZMk9OTRjWXdgc9qjs5YwHxg1p7KI0ULzz3+tW0cchG5d3tnFanC9vdMkhx6j8BVraySxhlOQPamje7bcpyfT/P0p0ljcAHGz+tWlq0iGdlx+OfamiSRgiJl26c+lWgkbbaq3T2/GvAGvyeFr28khh/0ltnlybvuYDZ4wQchsVF8WfGKae1smo4gP8AD5MXrnrsz1rwn4+0yGMLren/AGuM/wDHw3mtHu+9t4VeMcdPSvH+t6NdXDTeG4fs0BxmPcz9lHVxnrn86+Bt3omjrceKNXfdLHt+zLhxjPmI3K57EdRXxJ8Y3XiXXTLPPuiH3BtAx8iZ6AHqteMfiXptx4atdD0+08mO034HmM33nVu6/XvU1/d6hcq3mYSPpwO4/wDrV4EtPEEWgaleeGLnyYm8r7Q+xG34dgvD8jB3dK1LUpCjxtJlhjnGM9PapLi/uZ/Ijbcx6cAdqSxOnRtLqAxKMbB6evT2Irwf4WXxVfSQvqf2QJjDeR5mchu2R/d/WtTjWx1ObTZpPNWHbh8beqg9PxplldPKj5Q9qa5xGLNjhR0GOnerm1hhi8zOR+IqwnMqrHI2UHRcY/WtNnl0e8jurBvKfnBxu7Ed8+pr4fXlv4YvZr+51HM0u3yx5J7BgemezVqmma5qhvfEd1p/nWsHl+a/mouM4QcA56gdqufB+ladbrc69rHBzmL7O3rjqre4NeN4dPjvpI9MO+DjYeRjhc9eeuam8N38mhR6tAv388ZHZ9vr/StOs57tyT/B9OM5/wAKE93Don7l9sHfgH+L8+tXMscJEKj73etH8Ky3hWdbzy19fLzjqPX2rTvC2nXl61pHP9olHQ7WTsT6+grUdOvbW4aKVPLji6HIPUD3rxD4DvLDS7LWZbzEF75mweWONjBT/FnqfQVr9nFZNGkV15u7ORs246f41E6BgCuQO2cVqHieCTRV0qztvJ67jvLZ+YMOorXda/tefz5F5+vsB6D0okiRmJ3bcYHSvMjJ+U5P0phGp3bvmPtW4jAJ/SssJMD8Klcqm7PI9qyJH3EZPegplOGbIHtUu0/L2HSpFBjJA6dKwTznmiQTnvQCbcbcke9LdNHGU6+lOCw4NbQeAf0oJgdfwxUa7mCg/SngwQEGT3qSSQnypWzjtiggzxTLsbbTIByp/SmORgGiBt+Uc1uyMA0SwIBNBVY4J6UyHICmicHB60RgDn8KYsRjORTMAMAUAMdajaFWAli3j03EVfWPgu40C3k023+y3w3ea++V9/zjHB4GFz09aniSOUqkm8DocY7UwPRT0p1HBLZ9sUCF6Ci244zRIAAB5ocjrQG5QR0oEAYBrbkcN+lFQeSelcryBTMznBOaKjoa+UcA0SpOM0SrKFA5HeiQvBNMeMA8dqOQM5o88rz6UCMYB6Uc9QaBYnJ5IofMMGjx0o5HAOfalIxgH6UTzjOabDHgdKY4GFNEZ4xXTtW0AZB/StuR1ogdMYogBcA0G56Ujk/LmoWjxljTMDwO3SieeDRAI460rADGKUEJwc0jYORQYEgA0hZI9xPAqOZARg0oV+GOB60cDvRcgAkVwRwaPHFcYxil5U+1EYGVNZxwTTDvmgM9BmiO1HB6Vw1AYGM0wBGM9KMWRkH9KPPH5UTtTb1ojYeDTnIBJ60X2gc0rBh15HSsluAelbWb7tMSOCa2FgSOgoDK5J/CioJznNZ4xUJUJgHJNMwLYp0XaBu5HtR2bMIcnvSSKFAIya3g4bHAold21Tmjg8Z6VtB5Bpfmwoo8cA0D23dPaldSRmsBhlT9KEbZzmg4ChfSsBgDQQAfLWCeDQYYGKHA60SB3qsZ4FYGMCiB0BojtRAIwaKgjGaYZG0U6Bhx19KMTMuFFFOw60y7ScmsAdDUikjGaEdBSgxnNXtvuGVH0/Sovkbr+lQ/MuFPIpju5JoLv4U9elROF+UdqOWHB+lXXiFb2w+w/Zdp/v78989Me1GIKuQOaRIlG5n5HbFTShxv24HYZp5ARjP4VgKASc57UzKVwBSSALgHpQdWXBFZH8JoLuJIqMjAFNjoDWli3FmsT/e9efU1p2tN4OtXXS7rd9tx5w2YxsJ29Qf7x6YqHxHLNcGdEyzdefQY9K8DaPq/iyeLwxb3WIhnCeWvHDP1JB6r614t+Cuq+AmS41VMQS55yvbaP4XY9WFf8I8bi48hRknp+X1rXvD93pLkzjhenT29CfWo1aNsMeKEmmLo5WKfFweg2t/e/LpVj4Rm1HThewS7ie20D+LHr7VEp3hSOtW+mXV7KEtevpx/U+1eIIX0u8/sy8Ty5I/vc56gHtnsRUpUjMZ6VFJ50I3HDDpQXc+CKQpboGA3E/hUPxBltmtjp9v5ckO/J35zn6r6ZrXvE/jn4zTSeJrHTfKSHG4edE3UBO4T/nme1an4iul1B2vIcumONw7qPQfSvEniz/hIbgzva+Tntv3dgPQelBAzeYD+FXXgLVrfw9F4qkhxZzZ2tuXs+w8bs9farfTNGGnzXMt/tnXbtTymOecHnOOlAupxjPoelLauW3hsDsMUbx5UEcy5A6HOKIiWIShs++MV8LfG9h4VvTcanHvhPVtxGOHHQKT1YV448US+INRa5t9X+2oekn2fyv4VHQgen6VFJFIdtzJs98Z/lUMTSYUHk9Biru1e3YAjpVlqNzYIAI9y9uQP89aTxApgiV7bCxbtx39cn6Va+JLLTkJjky7dsH/D3qPxBHLMZyvzydeemBj0qK+0m9011vZdknGPlY/xe30FWkQmQBBjPevFWug+H101dM2Zz+887P8AGp6Y/CpwpbCmkQkYNCJogVbj0plAbAOfwrRZra2YvMu70GSPWr+4E0rME2+2c9hUkme9MR1BraT3oAA4z0r/AIJtkH9iv4ckf9RT/wBOd1XxZ/ZB/Z1+OfipPG3xT+HY1vWo7RLRbk6te22IkLFV2QzIvBducZ561oPh39nz9l7wpLZ6RD4S+HuiO3mzyT3Mdr57DgM8kjbpWxwNxJ7Cv2//ANvTw18XfDs/wR+Ct1LeeHpp0fW9baNolvvLYMsEKthvLDqrM5A3FQB8uS3/AASKQr+0j4kIOf8Aih7r/wBLbKv+CsWP+GXLXP8A0NVl/wCip6CgjIr/AIJ8ftIj9oD4I22n69fibxd4MEenatvfMlxGF/cXJ7/OilWJ6vG54BFf8FNiR+x74qK9ft2nY/8AAuOv+CVWt/DHwr8ZPEfiXx/420PQtQGirY6NFql2lt9peaVWkMbOQpZREq4zkiTgHnHib4FfAf4jyDWfFXwn8Fa/PcAOL650a3mkkB7iXbuIPHfmrDR/gv8AALwxPNp2neEPAGhL89xJHHBpsDFR952+UMcZ5OTzX/BQ39uzwv8AGPRR8Evg3ey3nhtbpZtb1ho2jTUHjIaOGFWwTErgOWYDcyJt+UZYupO5Rk1/wR4XHxJ+IJ/6gVt/6PNf8FYzj9lu1wf+Zqsv/RU9HHQGlRnXYpokY2Z5+lMcg4HSuD1FW7KcheSelOS7cmsljxyaC8cHn6UAWG0Gn4O0Hmtpf5jTgcYNHptJ/SiOMA/pSuc4zmirNlgeBRIPQ/hQPy8cn0rtgGgQBgU7hhjFAnHX8KJ34+b6DFEjr+lEZHH4VkE4NE5PSgQOBQweD+dAgDgVuHl1ABIuBSEAgHt3pJFflT0qFlAwT+FLGpGC3PbirZ0GB1J7VIVM8a7duM8Zz2osgbIOB24qZcgSIODTI+fvfpUi7X+dvpxWYz8oPNCAo4BP0FDzFxgcUkDum7NPLMyrGWyozjiraJ8kqa0zyGuhDPNtB/2Sex9Ku9Bdg08D5jGOcY9vWorQMdyNnHtRIWPGc1aaBqE+myauqfuIsZ5Hdtvrnr7UL2V4xIRwO1aRf+dMiqnAz39jWl31vZzq80e7rxkjsa1m+ttdvmaAbSuMjk44H09K0a/gtbSS3KbmOMckdya0e4hsZN067s9Bkj19PrWqyx3E5EI27ug69AKklaOzW2VcAZ+bPvnpVpIi3aPjIXP8qsLMtOLjPB7fgRWhaPLqeppZQL81xnBz02qT6+1eNPhnoXwt8LRaze3n2jz93yeW6Y2yKvUM398dqnuIDHNJDHhJduBk9qms5njZymE47iob2dLMQ28m1E6DAOefetSvUY4ZssPb6V4j1VJZAluMjv29ParadXulj3bVGdxxnHFH4hQaXa3GnaK+6CXbngjoc/xKT1Jq6vFun37cbevNRSLaXSSryVzx07VqetLfvtkiwe3zfT29qtb64sQXtJtjH/ZB/n9akuGvZDPK3zn269v6VoesQWcoE8O/0G4jsfQVdxxSRRKF+Zs85NG2dhtdePrXhfwDqGqQXOpW43Q2uzzOgxuJA6tnqKvNEtoYDci6yyfw+WR1OOua0a8n+1RmabfjOBtA7Gtf+Klvofgqfw42k5k1Dbz5552Sh/7h9fUVpVlZ+JLyWPXta8iOPbj/AEYt1B/u4/uivFNxotvefZdIu/tSr95vLZMcKRw340t1fGyCediDsu0evr160ZFiRikm0/7uc1FqF1JCbQPlPTA+tXMcsUuZjz/L8qtNYu48RR3OF9Ng/wAK+HmryaFfrqlxZ+akvQ+ZtxgMvYH1rxt4qOq6jKIYvLHHG7d/CvsPStT8RarqtjFZvPmK33bF2qMbiCecA9RU87TAq5yR+FEqMBT0ppABk1LMoGMfShuUZB4bpVscvxzWwht5PFOwABAzipHJZSRjrToX4DZz2xWCj4xz2FRkjOOpppQV2kY9KdjjaDkUDkciiOcYpAAclufpToCcg/hW3HFbeMk0wBAOelRgnBUfSkmEZ+c81KgcCYHluox+FKwQhic4rdu5JoHf8oNbMcjrTQboPMVunt70xxjBokEAlv0rYAM7v0osFAweaOG6UwU8L2ph0yaYjHA6V3rGTn9K3EkD0onPSi65wDRw3emKk5FFgRgChksMigcECjnp6dKUAYIP4UiF+AM4pgoUoRz6UdpAUDmsqowetEjPFFMndmioYURhhg9PamVWGcUSpGB1og7eD+FY4wDRGeQc1t4wDnHtRGCDmicHbmiSAAKPHStoGATVHgYzk0VHrQBIxmvagOMZrC4wD+lEkd6JI71nAwprIyMH9KiO5etBQed304pkI4BpRwcHNEc5B5oMQm3OaXgYzmgwA561DqCC0e2Zclsd/fNRuBgE1GwYYHaiSDkUz4Izzirm0SOPzFl3HsNuKGQACea3DH9KH1rIIwTSjjrTYxiqqjjHWsDGBWAoyOtNwMdaAGRg9OlMu8c00a5KjnHSiCD0pkyMA9O1FQOAfwrapPNADOFP0oqD8u79KZZANqmixxgCgSuRnIPQUhGOTUWWGFrgjGPxoFQMA0gTksOe1ZBxg8imkcnaT1oheoFOyFcqvPbmmIIGKC5wAf0powvAbP4UoBPJpcDgfhQkJ780RuIodMk/pTqG6/hWPejgjjtRAIwKACjHpRxx+lVQOOlHGOtADFMAR1quAKZAeR1raN/SnjwMk1tz0plUnisDJYdaY/Nim3HjOaeMyJwOlXMBjfdjNWTggLjntTHcMClHf0qFSVABoDAK55pAynAP6UGc/KDUaE8CrsMlqkT9ef51yBgmnwzYDdPam55JzQG7gHFKOgLfjitxzilJxjNKwXAJzSr5zAKcUtsbNAobdntjFT2c8dol48WI5s7TuHODg1DJhcjt0rwh4outG1Vb+Bsvzngf3WHofWvhz41m8d6BJb6paebBBjcfMC4y7/3VB6qK+Lfh9fBevTWcUnmsmOdu3qiH1P8Aeq41UajaSy3R5jxtX6nnoPagERgqt+lXLmLB3Zz7V4X8UDSGd5I94bGOcevsfWrS4XzxKTx/9arPxT/Z9z59uvP19senvXiHU5te1R76bq2Mj/gIHoPSrDRhezrbxjaWz79Bn19q1nwtc+HL37Jex/J2bI9AegJ9RWkDT/Dl/Heu3njn5cFexHv614r8NW+mTGMv5co6rgnsvfJ7GpbcsdpPI6GjeXEcX2aOTCr2wPrUkbxRB5RkN0/CisYIB+bHTtVtp6T2V3fG62m22bY9md244POeK0OxuNei+wC78qOP/pmG65Pt6Vf2atdtHE+RH0OPUChAF+Vhz6VLM7ERdk6fjU9xZC1Ebpufsckd6tNNk1G6+yRj51/wz6+1eRJZytA/VcZ/Kntp/KS7CfI+dpyO3FSOxwHORWnwLcXiQibyw2fm25xgHtS2drbX0Wm3Mv2qGXO75TH0GR056/yrxvaaTZ6q0GjQeTAMYXczfwr3bnrmlR5Izl/oMVdaUyRb5H/T/wCvUqFH2K2ce1RSODhW5HQYoH95wcMOtRyLLa+bLJ8y9Bj3rTbK31S5aMzeR052lux/wrXfDM/h4xzifz43z82wJjGB0yfWtOtf7RneKSfyhx82zd2Pb8KvdIgtoroC83m32bf3ZG7d178YqJ2iQqT+FSMCcmjgng0FBGM81tGMUwBHXNWGr6rpTl9K1O7s2Ygk287RkkdCdpHTJ/OpfiT8RbmJ7efx/wCI5YnXa6PqszBh6EFuRU09zdztcXVxJPM/3pJGLMfqTzUZ2jA71IxVAing9ax8vT8aYgrxShc9c0BycflRUEbV7VpHivxP4fVk8P8AiTVNNDZ3CzvJIc+udpHoPyrUtV1TWZzd6tqV1eztnMtzM0rHJyeWJPUk/jQwOAaVVHTmiU2AY570se5CQelKwK461vVVwOtDBP3v0pmCjbjmsLjNRDJLKeBTDzPu0FJ4IzRKlduORTAev4UCzDg0OBzTEMvBot/DnpRIJye1KcjaTRcthSeBXtmuAMKaIJ4U0RgYU0VwcKawegPA6UemAeaAVl57UxGcL+VYGOuaYADAHSgFxjP4UcE4B6UDgjFKDJhAMk9KhnWFBGw5FOQ48wHJParVgQVJ/ChkHNK4cAFsY7YqNGOGBz6VHNPEp2Dn14q3toriT9636Vem1jhigig5XduO4/N0x9KWQxHdEmM+9TkTYc/ePWjAA+A36VPDlCyyZHbjFCZkAXFQJHJCHZufTFK5DAKOlWkEjw7gfrUceOU7VFdNHEY0b5fTFQSbJCU6ntV0C2ARg+lWnxMubTwLL4PEX7qbHO4dpfM/u/1oD7TKrY6Z4rTYZWcFenf9ahYuQueRRkjUgJHhx9456/hXwguNGtPECz63B5tuM5Xcy/wOP4eeuK8eXekXmsyz6FbeTBx8u9m/hUfxc9Qa83jzDzt6CoxJeRiRhg/5/wAKs43NsA5yB1H41YWzCNTEcn0/OvhVbyLr1tqdyMRWm7j13I47c9a+Oeq+IJREdQn3Q/NtG1B/zz9BnrivAXh+Xx7ImjmfyDDnc+3djO5hxkf3fWvFvw1XwhZCaYeZDH1b7uclR03E9WqxsNKm0eeSebEg2+Wu1v73PT2rxFcR6XLI5k3HjHGOw+vrWo6wLpVCcKM8fl7VeGdY/Oj4X04rduy78n8qZguCFwD70lxHINhHPrUUNr5Dy5w6YxwfWobKNbbzWl3N2XbjvSWkizBZOr9PwFSWUNtBuZcP9T61NLHeRoh/5Z54+v8A+qrG4W0lAUcf/WrwXfXWu+B9Z0ixv/sqx+R5h8oSb8ysw64xjB7961uCTTJPLEvnMOh27fT6+taLqTyavayzW+Nm/J3+qn2r4ovpevW9o2jWm2Rd/mt5jHPKY4bHoatbZ9cvliefYWz823Pb049K8Q6Lb6S7Wqzec4xh9pX0PTJ9aluLm50mPTT91c46f3s1qmkzWc0cLtgtnjHoB71BLp2l6LkQbr0/x7mH8XpyOhqSDzdLW7lTfM+dz5xjDYHH0oDy5twfLDpx7VdeIL2azjtZZ8iPOBtA6nPpUsoc7icmo3RiFI59aPha7e1a6jkyBjI2gd8etQ2F3cozwx7lTG45AxmpztAUnrTgNEGzRIIJJ6VFgdDzQLOoRhkevSiqx/IooRhhuZuewxQORtI4p1y2R+dNGzKMHJpkGeuKZGIABzQAYAE08QWIkvkjpxii24YNEluBQQvwBQXIPP3e2KiSNz+8GauDGcJGMBelMBNGAp5FNBokfh60kEGbs7953MP4+PbpROQSTn2pSCMAUgBIBP6UXwNo6UzENgDpRICbetKN3Bb9KZsjA5osd2Ac0TnoeaYBeAcke1FsjJPSieOtDcQDRPG2klITYO9M+RgGi3GCawKIxSYHU1vBPy05HBFMSxyaKgAHNKwONv4VI6yLgjkd6yM7c0zAjHpW4FcA0W3cDtQJPSmHzZzQIxiiADjFcL0rOOQetCQAjFE+lHaTx0pskYz9KIJ6GiCRgmiMKO5rHGCaAVu9EAc5rGRgCioB61g9z+Fc4wBRJI5/Cj0xjpXBGMc0ASMAUku0Y2/rSuSM0rlhgc0mADx9KW1mZPOCZX1zTlQuAaHOMCmA64ocYKilPvVgjPhh2q4liJ2K3Ppis5+UfhSEoc4p5lkIweRWfelZdoBPJoEHjNfQ1xj+VHBGBR5rgjFHHasZ4FEdqZlXgn9KIQsct9OKJA4B/SuxAPNM4GBn8Kwp6n6cUyZ+6aYIQArfpQAHBPIosvTP6UHxzn6cUcHvQQH5i/TpxQGeAagOFwozQwev5VsDdDyO2KBfGCKI9ByO1OzMRhenvTJu4zTbVAYHOOoximC7sqeKBBwBTgHhTmgATgmgRjAOT2oxk4IHPpQIIwDk1W0k9elHJOBWAOlYGMCjzwD07VxVe2KqgB3P0rGRxVV160VyCM9fasZGCaKA52n8MUQQcE1woIAz6UVwOuaZQ3Ab9KQ7flz1q6gLjaBn0q2RoZQpPXoPwoqQaAG0ru/Srds8U64JIGcUAOvf0oAk5UUjbWDDtWnabpet+BVkR8XltnemG/ilOOcgdBVxG0UrRuOV6j8KlVVbg0uGGAOaBxwKDErgdqUqRya0vS73V7uOx06HzZ3ztTcFzgEnkkDoDV1p91Yzm2vIvLkHVcg9s9j7irSBc53cjoMVo2hahrY8uwg82Req7gvr6kdgavLXVGit/Cb2X+mRbvLj8xec/OeenT3rX/B+s+GWWLXbP7LK33U8xXzjGeVJ7MPzrTIzbzqJTjrn8jXwz+IjeEL5JjcZgbO+PZjOA+OdpPVq+J2v6D441m61uO78sybMJ5bnoqL1wP7vpV3baddu0EE25+w2sP8APSvFnhy50DVW0+ZcOmOOP7qn1PrUkco+Vmzjtio7Scp86YHbkVGyiMgN06cVYrDKCpkwR/smpbC7t1E7x4ifo2R246detWWpXNhcLOg3FM4HA6jHp714u+IT+LiovbfyHTOfn3f3fRR/dFRm1njCpN/46a8RvqV/qk008nmH5fm2qM/KB0H0o3U0ZKn5j6cCopVAJkbBPQYpLlpxtd8qn3Rj86tZYYJklHz4z8vI7VqM0d27SQR7CcYXOfT/AAqymuLchbd9pPsDV54Mv7PSU1Zk+STODkdm2/3v6VvEjDDZ/Cp9Hkmso57Zt7c7+MY5wOpqXT7mBRJNFhR/tD/GvGWo3yeLL3UIrP8As+aTy8weYJdmI1H3u+Rz+NWslxdOzytvdu+AOlSyXrItpC26Nc4XAGO9SabeRW7XEsOI06ncOMnHrSW7g71FeD9F1fxFqkVrplp50x3bU8xVz8rE8kjsDWtS38t65uYdjLjI3A44HpXh+4t7fUYrm6fKJnK4I6qR2q3xf6e8rQZXjB3e/wD9apY3jmKPxt/wq5trOKPzIbrc3psIpyJBkH5u1K/GxzxUZDMVjOT6dK0+LWNcX+yo185v4Uyq+pPPHpUzxD5EfOO2DU+j35sH1M2+LePGW3jucdM561dCMcK3zDtipBnjOSO1YB4zTA9BRGehogEY3fpRCZxu6e1IRnAOaiIXgn8KChQCTj8KJBILH6cU7EgAnj6U5DLwf0pBzRIQb8ZosGOQaCkcGnfK4FAErt70ACmM/pTYI+U5xSMFQndz6YonbwBWSTn0oDJ5P4URkjJ6UyhhjNIWyVX8RTvhdqrz9aj2qQSacgv8pyPpRyXyRQKKMseewosScEdaYZAAFMQTtU8iiAx5PI7UGBG0DJpjkAA0CMcCh1wKOcZUVgFcDqK2+9MDgYFbc9BRBACgcUVBGAOaABGAKRcD5h+FXmmXunkC7g8vd0+YH+R9xRA65ooM4H5UpeJlcDGM4Nbtz7ieahCk8Hp2xSKm4MD+FZ4pTkgAVZusYxI2B64pG8xysT5X1xigsaAZXp3zVzdQXCx7DyM8YNOjE5HTtQiZ5NrpjPvmmgDtgnGKWIxYQDOO1SxFug+tRqAgU8kdqhjRRk9O9afqttHatb+VnOOdxHcn0qSdERo4xw3f0pcscKM+9KPKdHHOM1JIZBlj83pTiQqhePpnAzWlvE0isE6e/wBagn2wKIBlh1H41byhY18zhvSre4Dx7WHNQTiOBsHH/wCupJdg2IelQQtMrBU5471FC5jyY8H0zWmQSyISY/lHU5+teHtB1TVLrytOtPNc9BvVex9T7GvAfw+vbe5sfD/iKD7BLc+Zv+YS42hmH3G9Md+9fEDU7fxFqS6frC/ZrePOHyXzlVPRQD1A/OtEnsfB3juWCzm8yyONku0rv/cnPByRgtivE/xD8G+O7AeFxq2xX++/kSnbghxxtGfu+teN/BqaFqWp6paT+do0XlfZLnZt83Kqr/IWLLhzjnr1HFeK9Q/tG9aKN927GOMdl/wq98N6xol09lq1n5LxY3DzFbqAf4SfUVc3DlhGx2xdz1/+v1qa4jdyIW3fhipmaQDafu9qtEjN4iTPtQ5ycZxxWsRW5nWGwl8xRncdpXsPX8atrj7PHtbkj8KSS5vj5sIyYu3A6/8A6qvLu8UA3KbX7DIP8qsZXYGQj6irmyJsvtG7Ddhj3xVrfXdnYCz0+bBl/wBYNo5wcjr+NXlhBaWYnln3znomwjvjr06VBcwxxnc3zdhg814bXW47K6vo9O8y3j2eY/nKNuSQOOvWrPRoob0xXFx5CL/Fs3dvQGn0S3ke4mhufPhg2/PsK9eOmc9a0TwDrHjS7EfhPTvtrp95fOWPGQe7sP7rflXxMNrBry2b/JNbZ8xeTjciEfpWoakbhgI0xjvn6e1Qa00WiNYEZJxg5xj5s+lQqzSAEVJ4cvjpKasYP3D5w+4dm29M56+1SxJvG2Tr2xWgWvhWTTpH1bWPs9zxtj+zyPnk55XjpirHxS8dnLpDnEM2P3vXbg7vu4yeeK8QWiaAvkWF95m/7w8rHTGOufU1dMGYDPSnO6IKD0oAY2g9ajTLBVOaYKmMHp2xU7AYK9akcEhVHzDtQRR99se2KUKcoRyO1TDGNp/Cp3R2yp/So2y6gnpmpCkcny8gVcOjnKmgMnNKc/KByaW3ZRuYYq6e0SyhhifdIN28YIxzx+lRuo4NFgFwailVDyaZlEWGbGenFMQFwOaXA4zRKkAKeRUxRowU5ooAN3fsKByQAKfCnbj8KdgwwDz6Vxniic9DmiTjAohcbQabaBjPNKw6A/TimJJwaXkdKIyMZogYBJ5on0okYwDWCRxS47mjkii3GByK5K4BzSqRiiSByaJBPBoZJytAY5JoAYypzSlTwawueG/SiARhTTc8iseg6UxU4waK7iMfgKcrjANAY4BoAE8N+lZBGc0ScdazxyaBGeTRAzkH9KwKBAGM11Hy9u1EgEetZHrTYIHNMcDjrQxigAOQazjg1G5BxngUjA8CiCGwDnFR3DiPyQeKYcmhhuAajaPyCpPP/wBeiAG2iiMYXNfaHEXkg8fSoQGYAinAX5fypCGGAaUIDkN9OKXBI5onymKg0JQeCM0CCAc1wRwaoDPGaqqogEYzUilyAD0pzkbT1FMAy8HmgpUYJ+lMgxwentTNjjNFFAGD+Fbs8elN60Cw6H8KPNMxYBc1jA4P6VbnGDnOKkkBG0CgQ2TW1SQAaDnIJOaDhsYFZBHFMQyhfSmUsOD+lbQeetZxxStzkDpRfLbgKWU42kUrBuB2okAUQCMUBzjOaxgcVVYHrWBjAriqJzwv5194cGj6CjgjAogY4NAZ4o896oD3+lOoODjOKBIAKjJbrTEhs9DRBxnNNkjAraCMk1dorISp5HSrZxIuG60XPl7c8CohuXaTRQHGfwFEfwg0qgEgUzAHjnFRzmNSinOetSEFs460yY4H50oU8Dr2oKG6HpSqo4NA5wAK8N6tPoeoJf2rYdc9h6EdwfWtU1OfU7k3Exyx+noB6e1afB5jbsZx26eteFPFF34cDNYfK5xluD/e7EH1Na/rz6otrqe79/Jv3nHTGAO2Ogq4u7ie6Se7GVXPoO2O34VqR8Jarp1odOtPs92N/nyeZI+/kbeDgDABHFeF4tK0/S5TKvmt8u1csv8AEf8AGvEPhrxFo6D7TpuxH6N5yHOMdgT61o93HYXay3C525yucdj6fWpvJeTzY2wp7YPFXNtblw8Zy/bg09pe22nq8L5g5wMD1/PrUVpDG5YDCjqOa1/TtPgdDY++evt6n61c6ldy6fFp8zfu487Rgdzk9qv/AAwbXS2vLN/PC98bMfMB3NSRGYknnH4US0DYWrG3leDJHT/Gr1ssoByVz/Smd3OSeaALHBFRRhl2nrSlolBk59BUkytEEQ8j2qC9nuY1tCc4ztHH1NS27RIFkGG7CtLnlhJYnAHTj614gjlt0W4W68wnOBsx6ChHJLJ5oGG7moomlOB1FSLJA211we3NW2u3C27WEpzHLjPQdDn0rws6waxEsc21RnPy5/hNePVhl1uZ4Jd4bb/CR0Ratb2aB9u7j6Cjp11NbSXdvDuiixvbcBtycDjNT3csMXkq3HpincsRGTkDpXkrLyz8/StPv7G2t54Lu28x327TvIxgnPT8K1XTdLt7EXFpL85/h2t6gdzQIAHNQ3bRIVjPNRIYl+Y/hjpRuCH2g8elSDacA/pRjJTJok5+XrRGT1phhflpvmGQelSMxXGahwe/Sohkhc0zl+WNfAL4AfEb9ozxzB4J+H2mGQgq+oahMpFvp0JODLK3bocKPmYjAB5r9qL9nLxF+zD8Tpvh7rms2WrQTWy3mnX1uQpuLdiVVniyWibKsCp44yCRg0uSQFPNO4VQoHSpZGcBc8DpWMnGaBAGK4JxigATyawAOlbc8A9PajycDtTKrDivurx+FKAe9NgcA9OlHBXgVCoJOTSBTwxoqoUEHJPtTKpTcW5HTiic8g/pRUHBBokY68+lMC3ANMCFGTzSg54b9KBD9TRA7nNHB4/SllZFZQc7qxkgg0MZ3UzAcnnFA54zmmORhaBI6Gsc4HWieeBmojGG3OM+lXuqy3iBZTkjp/nHtWB1U/hXCjDL196jbEewN17YrGBgc1bY8wAUyjbjOaVc8A/hUS7X3DqO1Fwx2g5NDccjFaG2kCMi+h3nt8zD19PwphEx/wBGXYB2zn+dQwxyLiRv0oxM37teQO9R2ckUwYHP6dqdPLvmmuBu29R07Y7VNIrzMUXAboM+1NackKMgUCVxErde+Ks2xHhjnHSnjV1BHBNJGWfaB0rSNIOrXn2RZNp7HGccE+o9K1HQvsDiO4m3Ht8uPT0PvXkQD52bOOgwaUqAHbn26Vb3aRaZJK/O3GB0/irQwLnbcbcdcDOfUVFKzsC653ds1aQJIgVxRAlnKjoP8KtiBdRoo4Gf5VoWmW+r6pFBIcB85OCcYU+49K8SaPb2M40jT23FO+COoDdz7mvBPilvh9drqMsP2qVs/Ju2YwGHXDdnr4keMLjxLKLprfZLLneu8HGAgHYdlrxFc+KPGF3ALqPKpuwcx8ZA9Mf3RXi+10yw8AWWhPaedeW3mbX8xl+9MrdOnQ+ta94dkgvmDS7Qeo257D3r4i+ItQufDVro0826C03+Uu0DG50Y8gZ6+pr4f6PY6zrX2XUJvKhbqdpborHsQeoFWmr6R4b1ic2cG6I7cDcw/h9wT1NfEzxDBr0sMluu1U3fLknqE9QPSp5LWCPYeT2601zG52Z57cVGDcgR4yaa3MMfyyc+mKI2ptLfpVqklk6XUL8844/D+teIdfv9d2z3bbnTOOAOuB2A9BWhadNq88djFN5RmzltobGAT0yPSn8JQ6SQZ7vzyO3llP5E+teNPi1deKbWy0r7D9nisvMx+9D/AHyp/uA9VrWdP8NX+iecsu2buNrn+Ie+OgqWysYLC3ZYsE7sncfWrvX5NH0R9L0eXcbzG9duPuNkfeB9TXiddQ/tNhfcuMen90en4V4K8RaV4Pszd3sP2pr/AKDcyeXsLDsDnO4emK1PxNdLcvNDJgtjsOwHtU902/Ibr7U5MrZJqRhGSM5zWnaxPpwzAefw9/b3qWdpmJI69KZijYNI6soGealmVAOM1Gzu3y84qUHzCT1orlM5rrxmrMoE45YVduZQrg9M1HGCA2efpQVy3mBuvbFRM0OCrZP0p3MSBF5qRmYfMajjMhxuxj2p2MTFQc1I5kO4mmUcAHNFSrZx+FM7ScE5IpJ5ANoPAqRzJJ5mM57UshU7VOKVThtp6e1QBV+Ynp2qaZ5eB0rbtXOfpQUsaAz8pNO+PlApDhsZp5DuypouSxJ5JrA6A4o9cA80QCcg1it3GPyrKnnvTNk5A6VvJ6ikJJBHQVk44Ncen4VwBRPGM116H9KwCME1gFQAc/hQKgYIyaUgfKKUDrmsA9Pyrblsd+1K7IGQH73UfSmIK4I6UANu00Of6UozwTRGPumgMd6wpGAfwxQ4FZBOCaIBOAKkPTBoEE9aIB4BoKTgUcfdzRAI4NMcnINA45zzXU4B/Cm44BrHvW0EcH9KyMYoj1NEgjAphgAZoHPINDOMmoXHADfpSOmxgRknGD6UTgZFFiRuJo4H3T+GKL5PWgQTy36UzID979KaXPKt09qjmJAG7H4Ukoc4Z8ntxSkM2C3H0ooMYQ5xTNEq4Rsn6U0m5uW/SmxgYb9KjZ3G38qQr90Nz9KDAj5TR9h+FUee9VRwRgUSAKKo3HemXDbV5I7VIrYx+lKAF5OTUiIRhevanHYH8KCsDmm5+6MmiScKOtDeDgjIokY461tJGOtQDAwetED6jsKIOOD+FHGMDrQAAwTmgCxAUfhQbjAoEFSSeR0FcDo36VuBG0DNOFBypoEHnNYJPWjgqCzc+mKATAIbp7UWUnJb6DFMCRndx9KAwMZocjGeaJA70Ae5+lV/nFcVWMdD+FdBjP0osCMBuntRIIwDWDjGefpQIb7rfpVH0FV26fhRXByD07YplLHI5/SpVBGVP4UAQRxRC9M00YI6Vbq6tgigOcEfSoOpApulHOeK+YsARTjBwvXsKIUkkGjycA1IWPyg5FYIINLtxy3TtisIfunNIMng18IPhVonjXRNbvtW1j7JLYfZ/Ij+zvJ5u9nDcq424Cjr1zXjjwvoeiWLy6dqHnyJjC+U69SvqT6mvhp4b8Jaj4c1y61jV/s97D9n+yRfZ5H83LuH5U4GAAeevahC6SskfI4qbQtSOmjUreDdaL1fcox82OhOetNM1zEEaXK+m2rUeVITG/PbitJ129s5Ns1xtj7jYD6+grVfjFreoysupL/a0HHlJlYPL4GeQmTkgdemPermWLULt5RD5W/Hy7i2MD1/Co4LiUiCNvm/u4FaP4atX027vb678ue22bY/LJzuYjqDjpin1dvsy6VFyi5wOnfPpXgT4e+CPEd0mk+J/Ev9lPPnY32KWfOAzHhGHov5+1eL/DF34avvsxTcn8LZAzwpPGT60plulAdc46DPSk1nWrK2bTra68u3bquxT0OepGepoTSK+1egqzktA4FyuR3OT/SoYboL5IbbnoMA1qfgfxFpbRS6lp/krc7vKPmo27bjPRjjqOtXOmzQ8hee/I/xpGC5R+GFWmyKcSmPzF/u5xnj1rWLWN4lvRb+Qr52rv3dMA80lrNMMRr07ZrTlmSQzpFkxdDuHGciry+e8nVmbJX29v8A61XNzPCxQjIHQcVZr9vVUd89eMY/z0o2M6qLmOPdbno2QM9unXrVzp2hy6J52ly+ZdDqm1x/F6k46ZrUopZMSSxYJ7bvpSafpreG726Z8XUfl+UuG5y+DznHT1qKSe3lEkTc/QU14002+WTLdztx2qfR5vs4miXep6HOO+PWvD+vS6dZ3WlTruivNm8Zxt2kkdBnqa12TQxZBLRd8x75cY5Hrx0po/mKY+YdvSpre3WPFvJvP0I/nUiDdhxzUocNszx64pJRI2zoR0NSSbV2o3P0qZlA2jkHtRRdwAb8MVFGsrYzV4u2NVTpzkU6gHg/hRGeM0SG96wp6HJFHDHp+FRqVAxUahULHr6V+y/+yr8QP2o/F40Xw1AdP0HT5EOs65NGTDZoT91Rx5kpGdsYPPUlVyR8Z/j38Ff+Cd/w1j+BvwF0iy1Hx3LCrzGYCQwuw/4+791wXlI5WLjAxwqBQfF/i7xN4+8Rah4v8Y65d6vrGpzGW7vbqQvJKx/kAAAFGAAAAAABQXcMCnOTgH9KPHAP0GKGSQAc1hSCO9KQOCeaIBxgZx0o7SMA1gDtxTheAgogfdzRG47VNMOcGl2k4NfLtwe1RAHOTTBew6UoUN8wokBsfkaRS7lccU+M7VOcUgOcGpNpGEbP4Vnd3plOMgUox1op0yeO1FQeVGaIBGTRBPAFIAOCelY3Hp+FABTg0QCQAfpSrgdPoKOeopiAAMfQ07bhgGgrEgD8qA8p9si8jtmnkMh5HPYUAPX8KULt4PPpirY7XUZ57CicjGaBwOKhBdwM8d6BhDAK3P0NEkthTwO+KiSNjl3/AEqCKZzsg+Zj24FXMF7byCORME9BkVpb26aTKt5Fsmk2+W24noxzwPbFRtMITcLHlV9/wqedrnMjDO7qemcVLCqqpjTJ5zzWg3/h+LTwmpnEp/3z3Pp7YrVY7SGYPaSblbpwRjAHrUKKMlE3enOKi06UJHLcR7EbO3nPT6Gi9tE2yNNw9ckVa38tnKslkcOc/wAvervU7u8bN0/mH1wF/kParkR8tE34Y6UjOrLGaS0WVYrVDmM539vcVZRCFBHEnC9Oa8PWy3d7HFcnahzk9ccH0+la+NOtXhi0u48xvm3jYy46Y6/jUQKPmStNtlnuo2J+UZzx7GtOultJjNby4YdDt9j618PtKl8SXBuYpftV/wD8s49vl7uGB5yAPlX9K8aeHrbQXVll8y7XOY9pGPu98kdDVlqX2i/luNXtdjNt2JvznjB5UfStPi8Qam5tNEsM7ereagx1PRvoa1fxRBok81lri+fcx7drZK5yAeigjoRXiy6fVb+aa2bAbbgYzjhR3x6Vr8NtdWci6ldbHjxtHlk7skZ6dO1XM7WKyG1f5jjacfn1+tXVyI4AWkzIevGMc1NNJ5DZbnjHHvUr3M7gu2T6YFCMqu4r846c0blLeMLG37z6f57Ukczwi4mPHpj8O1SPa3ZDGPywO2Sf89K1bULSextrbTh+8i3+Yee5BHX8atXubg+WP6VpMk+mkXCS7JF+78oPqPf1r4UeCp/iT4iis9T1D7JaPnfP5QkxhHI+UMp6oB+NfFzw1p/hHxffeHLa9+1Q2fl7Z/KMe/dGjn5SSRgtjr2q5iLIzImUGMnNS+IXa2ELT5CfdG31P0rSJtRLf2htxFD1bK8ZyOnXrXiiLULJ3k1VvtN0cdgnp/d46YrTtN1LWIJIoE3NFjKZUYyT3yPStSiMBIIyw6jp6VO0eQVPHpTxTC2FxsxH65HripJkCjacke1IXZ8qOKg068uQXijyifeOR3/Gpw5l8sLnHv7UiKEy/wCFR2VxKjTrFmNMZOQMZ4o7EyqfepmZnOTzTI4XKjIo4BxUDqic9aPltEwU9KiII2lufpWXULkcDPNByG3MOO1SsS+B17ClRyu4r096YmNv3ZpwzncxyTRRgMMKAA+6a8LaRa69e/ZLmXyx2O0nsT2I9K8e+A/+EKultidwlzsOMZwFJ/iP96g23g0skSAuOSe3pTuGbIP4UHZQAG/DFM4x8pz7UpB+836VM6EBVP6UhXBG7p7UwYEUw4yDS4xyf0onPC81+HNH3rAzgHmvbdz9KYkDBpznAU1lmYAHIogYxRwRzS7cbRzijjtWBgHNEZGFoqSME0FyOTRUbsHtX3hgH8KIGOKVcnAFZXAAPIoYbj0oKobk0wG4HOaZgRhTmtuV255+lD0BoqBweo7UQAeDQ244b9KUhQTnmmO7vTAEZJogDo36Ueeaogg8GiT0zRAPU80RTBegNFewNEEHk0TwADmun3vyon0NEgjAo8gCuMYo46ntRHalIJ5pHCtzSOCBg0ZNy4zzTvjAJrzF29fwxTyjGQelLM3HPFEhjgn9KOAMZqweEvtl6fj71csgn/cn5e35Ukm/60rBlwTzQ4PNABiTn6VKz42qahkIXKmvM3A880oJ6mqHvzRHFDkdayeMHNDkdaxu5ojimXqQeakU4yKIAUEDmnUlTg89qZenNZbofwpmJwMUUO0MtEMRgGgvUAfSjwOayQoweaB4yTT5IBBoq+ME0xcHANJIUGBRII+Wh83BNOGx8pzQLBeRzWCeQaYEBQpoAgZNAAj3o44UVz9aUnIANYLKNp6UNwHXntWGzyaAIJGfpWPegMDGa49KIyOD9KKkjg1t3cVtGOBW30NbSQAT+FbCBhT9KVWwT3HSjuA680VHY80AV+XP6UA2Thv0ps8AUQSoIPSj70iqV5FMqAgEcelGJCMAc0iqG2sMmlUL92izbiuayGOaX1PWpGDNhTQUt8oHNFccE0SoO1aYZHvRBBAB6UqqDknk1DuL57eleHp9YvLhdM0Y7pZc4T5RnAJ6t7A1f6XqkF19i1FNsg6rlT2B6g+mKvtI1Tw5Hau52i937R8p+5jPc+teE/A2reL7p7XTmxIuMnC+jHuw/umtTs7zSrufRbpsi325XA/iAbtn19auNOkT5yMDsP8AJqx0eSXUFgAzuzjt0X61q3hUQaXuljwx6c/7Q9601RNlHHzdv1qL4f628CzJY7kfPzeaoxg+m6vF2mQeHtWextJNypjDYI6qp6En1rwhDYzaZG+ryf6PLneMHnDNj7vPXFeJlsNN1ZzZ8xjG3qP4R659a0OWDWNM1S/vZNs9p5P2b5SfvMQ3TA6AdaIub27EQm+bsdo9P/rVpmn6HplrJDqx3zrja3zjPJzwpI6EVrEtsJmKDKdhz6CpUjDAqMnvUVrNMwWGPcfTIFRyXCsAW5HsKtL5NRhEF0+5l+7xjHr0+grxBbJYIkUffP8AT/GpNBv2sX1URf6PHje24cZO0cZz19qsla1ZSG+7nHFaZfaR4o0J9IuB+8TG773OXLdsf3avrdre5aMdsY/IUolgZo1OM9eKtClvOJhyR/hXiO9/tbVpdQgPEmOPooHt6VbOVTBbn6VJcO0Oxj8q9BVpcsq4R+PTFS6Sy+GLHVTPkz+Z+724xiTb1z/Ssbx5ZOFPUYrxJodrZRb7Z8+gwfb1PvQVgvltz71bavd28a20b/IM8YH19K8Qaha3FvbLZja8e/eeT1Ix1pbqTIYNyPat0c5F1K+Wf73HpxU1mVszcKenQY98VIDuPPPajbyTxbQOantI7a1TAy/OfzowZXIPPpQhfbgH5fSlt8qGxz2pY93K1JbF4XJXIGMnNLAZG2gcVLGN7Ih4p1A4HNFfSto9efSo1BIFfsgfsWeNf2oddXU7oXGieA9PmA1HWWiwbggjdBbZ4aQjq3Kp1OSQrftOftgfDn9kfwSn7Nv7K1jp8OuabG0FzewYmh0Zs/vNxOfOuyc7t2dpOWyflGpanqGsajc6tq9/Pe317M01zc3EhkkmkY5Z2Y5LMSSSTySaWJQeakCqdo59KKAngUSD8o7Uoz3oDIBxQVdvNKBgBTWB1okD5SfpWQB15FE7jkGkYqd2enSiQ3NEAYOOacIygKOaYbeM074GCaLlmyDQYEYzzQODkHmsheSazu5H4USSMD8aAPWipAzn9KWMOODzTjA4P1FKM8/pQJJOfwr733etAEDAocdaOW6c+lbSeR09abAGKIxwBTdODQw2B3ohemaIGAQeaUEnA7UQD0NIExyc4qAMx3BuntShSME0Y40G3vSIN64P6UIhgDHSpLVBEGDcntirZYI3/ejP5iluLaMs0Y+Y9OtG8jZt8xzjpUeui4gitXORHnaPqcntTacY7RpiuB6fjj1q1tVbYGXK896s9MhvG8sPjHTg/wCPtV5p/wBlna3LZK9ePb609v8AMN3NQxEuc9BRkDzqHb93FnAx6ip5HZsR/lUFvI7Ak89quYwnEZz7Yp7h7WQButabEbqVXK569/Y1p8EKAELgt7ntmkgBXA496s5Y7e3Lp1P3fz5piz7nLZY47Vpmii7tBdlvoMe5Hr7VbMqMIVXBPfNaNpz3UyxwxbnOcLux2PvWgeJLvwXcxWvleVcndgbg3Yn0I6NXhbU59Z1mT7WN6tjHQdFPoB6Cl+Fmj6D4Q1TxBrUeZk8n7OcsNv7za33XI6EdRT+K/I0sWOnNsk78Z/iz3H1rx0mnajrs8cj/ALqDbgYbui/j1Fa1pN1bQz6lGuyzTbh8g9cDpnPWvEWuSXk7bJMg44x7D29qnk3AMTz61LMFkYOeB0q8ulc7EPFRqGwpWobeDyGLDnt19asXW2mWQjgdvwrUdSN2V38oucfp7e1Wuly3ZJUcDt/k+1aTo2mR2E080eZl27OWGOSD0OOladawrcgleT05Poa1vTG0y9hMjblfdxjGMAe59a0+8mhlBsZtrdvlB9fX8avdE1uN/t+sJ8kvQ5Xtgfwn6V4z0TwCng0TaOmNSP3jmb/nquPvHb93NT+VA+Ej+vNXXijXNftE0CzGUfPy5QZwd3UgenrWpLe+dsuWzj2HoPSrm+a1QrBJtB68Zo3jnhnz+FErM67m45zxXh3wbfeJ2aLT3+VMZGB3z6sP7prVNITRNSk0+8G54cZPTqoPYn1FSbC5MRwB0prh0lZUfg44x7VJMXA5qAEFJCfWtU8U3l7ZCx35j78D1B9Pahll3Bv0qMkKWJ5HandmX5TWckE9e1RqTzSDzQY6+wzxR7pIsAd9wqTcZjk8dvyoRGQgZ/ClcgBgOlPM78KfwxRhZRgj6UEHkMCf85qJS5+Y15WXwo+lI00GHhPzVfald37hribeV6fKB6eg9qjhLDcTxQB6gcdqdGIyo+lBc9+aIUDIohSSAOaK8jJpIwX2k9Km2q+1TxTMT8oOfwpUI5I+lEY4zTDPSqOD938qJ4xTgYwKBYKTn6VEWzkGsknb3ojIxmgQoAJocjOaGAPeiR0BosCeDW7tQOabIHSicjIo5JwK5xkUpBHPPtRwRgc1u9T0ocjGfwoYYUc9AKYn1/SsEnpmscY60ACuAaJU8D8qJwMCuCKK9gaIAwDW0HpQUE4o4AIFcH6+lHGetDB70SD0FABvl/KsA8ZokdM1QxjrRzjbW3K9f0rAPAOawCOD+lHnqaVsDg9KMhJyKeQlcA802SMiiSRjGKyCvNF8jioRvXk0i54ApRhtrdKZQoDB8+2K87ocYoEnk80ScbgOPSnfjpUb4XaD0pWJ4HUUXY9DzQ4UO1MSOgzQOR0rg9KycYxXXtkisY6D8KIyOBzRHGDRAIxRTcMDimUdKIBPNEdiM0QhHyrk/WlXoB+VSKRwDQIJxjGKIBGAefSmUr8uM/pQAYYI5rBIwO1AZOCaJGOaK78YP4UXVRt28jvmmwxBByaMbOMKcUzGMdc0pGM5pSOhPSmO7jP6UAQcA/pR56Nk/SsEjhv0o9ODQwFyD9BSsoXBb9KJD42t+lHj5d/04quvGaxnvRA6A0PrR9AaAGBgZpiCeKrrxXbGaYE8A/SghOTuyfpRXJwT+Fd8gc+lMGPOfwoliAVH0pgx6nmo2UrhjmmXdgA0MEYB57UwB/i5HtQyFALZ/CjhTlm69OKZl6KK80chhRYA5AoOCMg8ipX4z1onKbs4Pp6UoBXJbPtisBOWbPpxQwSDnioxjgNnHbFfDXU28Pa5a+Ibey+0tZb90fmbM70dRyQfX0rxTrt14k1Z9bMHk7sYTcGxhQvXA/u+lXNrqmt22lzahf8AmKnneWvlKu3JwemM5wK8J/AnxboGjv4itvEH9n9M/wCiRy5+Zk7yHHX071YeFNY+I1/dXei232y4+Tcd6x7uCP4ioHCH8q1LwPLaEWet2n2N0+8fM8zGcEfdb6fnXhbwK+qeJrmwTUvsr2uzafJ353Rkn+L2/WvFutSGaTQpJMyw4w2PvZCt0xxx71o/g5LrTU1Zrzy3Gcx+Xu/iK9d349K8A/G7S/D1jNp/iDwt/aCPt2N9uaLy8FieFQ5zkflWtzLcXDTB+vbHsK8Na/YRItnrEv7gZwdp46n+EZ64qbTBqFr9ut5so3QbffHc+1W8ANuttK+12zhsZxg56U10Ipf3Jwy9/wAK1XWLzUCjCTiPODgc5x7e1QrPdSrCHyx6DA9K1jwfeaQscl021pc4XAPTHoT61El7aS5t5NjDodoPb3pIt7g5yT0HSriaW1cBTg1r2uWuqWcKR23lSpu3HeWzkj29BS65rJ0iXR1ud1lLjzI9ijOG3DnGevvRQbQwPPpitA0rUr6yvbzTZtj2Xl7l2qd+8kdSeMYNarZPCCZ2/e9xj6entT3IkYoz8jpxWi29o7lNQuPK/uHYWz1z0/ClsorGSS31R/JkONgxu+v3fYipI0DkRvuHY4xWoQaWzfZrK43f7Wxh6Hoa0bwHq17E86RZCY2jK/N1H97jpUNnNY350jVovJmP3U3bu248rx0IrW/BOp2Fu1zC3mqOowq45A/ve9TTTsNkx4/CpomeMtEc47UiuACrc+mKmmjuEWMptYdTnNNC2Mq/44qC3meMlF37evIFRamJY/s8gwPXrj9K1HSNNh0m0uor/wA2eXfvXymXbhgBznHStEjF3EdNtLDzrpujebt9T0PHQV4w8FeJfB2rzaN4n0z7FewbfMi85JNuVVhyjEHhlPXvTQsSCrdO2Kt0SXMYPzUunSRHDrnHvj+tSQAp8nB9KaRwjQFsBsZGPSpbG5iiMqxZHruAoIRGSepogE4HbtRAPQZor3xX7Ef7BPiD9oa8tviF8QYrrR/h1by/I6/u59ZZGw0cPdYsghpfUFVyclP2vv27/Dnw60Fv2cf2TGs9LtdLhNle65pYEcViqnDQWRXgvnO6YdCTtJY7xNLJPI0sjs7uSWZjkknqSaBzzjJFE5GWPNEljg8+lMAPlU/pWwjJI57Uo3fdFKpzgjkdKJx2oAbflHTtQZTwaDAHAGaYFjwKIyMDj2oHI6/hQBxRAAwRRYAbVNOVCgB8n6V8rDmmVFA2nJ7jFFHZcgcUQvaiPl5NNtwMNn2xUqhU4bOe2K2gLkNj8KJ3jhv0oZAwGz+FMAflU/pQUE4DY/CkCBySc0SScg0SOoPFAKep4+lbQxznHoMUDuBUN9OKwcBT1oHsTRQgYJ4ogY3A0hg8nBb5/TBpR6msA8A5oHKYzk0FyMBv0qBlJCp19KGDxmmYt8wFRICd7vgD2pllCjadwHbpSQvIQu7GfapIDGdhf8cUSyZ2rkdjmvOWVgCuMds5oQiGRZoxuAz7Vplw2txfZDdbc9tmfU+3pUlvb2eLUHJTvz35qWfw7JYiKzfF13OH9ffjpUli07G4ebBb/Z9OKmtoViU784zkYNW2l3Uib44tyDqdwH9aniVJMxvkH+HGKVXiYkw/+PUkUpTcflLfd7/WordWk2qct37VdWK3LEovzL3z1rRNNNvGJJVxj3+vvWnxRzxK6jjnHt1oxJMohjf5vTFSIBGqochc81FZPLgq/Xpx0rcbaDy4ZMr2GPerESSuHKYI7Zrw14iGg6xHemPzzHn5M7c5Ujrg+tajrx8deJYNTW2+yn5sR79/8AHXA/u/rQ8AX/g+1g1a4v8Adu3fuvKA6Hb1DH+9npWrfE+DxLJDok2jbbNd32kfaCc9GXooPVex+tfE3SNBa5hv/DN59lh+bcPLeTsgH3znrn86+IukzWwjvIpPN+0Z5wFxt2D1r4jfE7SJPh/ceCIbTZcS7f3/AJjHGJkk+7tx0461fkrOzI/A9vYU8wki5XJHQZq/nhlxInHtz7Uw8xtsa89hmpbSSOEMX5PfFRzuqBC/TvirYvdzLGjevb2/+tWvDSYNHs7awXzLz955vLDb8wI68HjNWBNtDiQ/O3Sr2KTTLaBWT55d2efQj6+taVpsFzZGSIf6T/COfU5746V4m0fxDp4RdWgwzZ2NuTtjPCn3FWM8lmdxG9/yrXfitqniPTl0q70zesOfn85RjLA9Ag9BWjXOgS6bP9qhzd/L5abn5+Y55HHSviD8N/FfgO5EHibTvsTN0HnRy9l/uM395fzrRPENnoF2uoxndNFnYvIzkEHnBHQ1qmv215pywtbf6SM7n3nnkdsY6CrsFo97HrUgLDCvj8KhVpBtjXOPfFeGby60DRH1K11/+zXbGf8ARRNnDEd8+v61qetzavcveXknmTNjfJjbnAAHAAHQAVNOzL8jfL64ryFKhzJgfSlVGbBfj6V5gVMBs+2OlZBO7ORQCnhWz+FbgvBb9KkCufkbj6UACwG7P4VaKAMs304pYwkm4Nkt046V4s1jw3PojWWlNuuDjDYcY+ZT0Ix0zVlYT6hfJp9su55M7eg6DJ6/SviR8HIPBPhOHXBq/mzPuzD9nK4xIi/e3ns2elMu75EkyPpQRVYBuaaXzGGBx2qWJGTl8e+KtY43cK8u0dztziotE8MJ4PnvWnzKm3MuyQbcyY6Z5yOKuiVY+U+V7cVHGJGwp5PSpHIbYD+leBdCsNduJ4by78sx7di+WW3ZDZ5BGOgrX7ZdO1SWziO5Exz06qD/AFpmzwvX0ouSMYpQWOSKZuMd6DAHO7J+lN+85AxROOQM4ouCBiiwzgGmYgYXrRYgY70oGOTWQTgUwGOKCq+QBQGBgjFAjGQM/pXJ4xUr4ZcDOK5PI/KuAMEUWBGM80AAeTRIJGB9KJA6HIokEDBzRPGBz6UpB5zWTjg0QRyp61gZ5pjuXArBxgnmgQVwBzW1QMBsn0xRwT1zROB1piM4BzXTqfpRA6g/hQ6YNMRtxmhjjBonPQ5rBIAWip6YraBzn9KODyo6UcEjFBQ2ADRBAAB6UOMGi2aMannP6Vt7A03HfNHnmsAjk0AD3/CsDHX8KIBGKwBwDRBAxRGBwarAIwOtYyMAUR2BqN9nAq3kTcARxU6oxHlmnyOQaM7HANK+5cZoSHGzNNKScUhJGc0jfLkGll82JUI6Ui4PJ49MUzFx8p4qNxnaTStnkVjI61nHBocjNZB471tzyaI4yDRGRjNbF4x2ooCeDUiKq4HetgHNBcc0FJAJNEAYyaIUjGaxxnNMcDNBiSD+VMB60QFGAc0MtwDRAPGa2gHOaB44NKQvQZpl2nFAhuc9KI4yKByMZrAAHP0rjoKzxtzQJxjNHJOc0WJ70rEnJPFIxZqqq7YBojHGa7YBqmIIGKKjHFEY71VYzwKK5xzyKK57/pWMADv2p1UjOKiA28U4yMUASOtA5OBRIAwKB3HbnpW4g4FMSTgD6Uy7BnrigpbkUcBfeiAeAaBIG00WyoyKQb0wp+lRoFUDOa8M+IrnQorqGCHf9r2Z+YDG3PsfWvD/AI3trbRTpk9tlm/i3nsxPQLVrcs85g3cr9wY9ua+BdrHrvgK/i1W6/fx+VsGzrmWTP3cDoBWmaZq/hieTXNFGIo8fL8vOQV/iJPVj2rVfiLqmqXEtreQ4L4ydy9gD2X2FR3l1b6qWWTGepwP7teHfDVxO39qanBi0H3n3D3UcA564rVNLns2Wa3GQ2eOPb3q8tppZgtwdobp0PQe1X1qVPHbp+lWunIbSS6kODHjA+px61pWl6S2lRyXD/Oc5GG/vH0Nah4GjsPCtz4hguN0KbMrsx1kCf3s9T6VfxiGLcgyP/1VFIzyhCOP/rV9sgtUwi/N25NXN1d3Ch5HyF6cAULmQtgDJqMAKDjkVKTIcNUUSSLitJ1WXTLSW1ih3iXGfmxjBJ9D61DpV3c2vnRJkD3Hr9a0rXbnQ5Wmtxu39RwM4B9QfWtR1CbUZzPJ95u34Af0ooMD1FElV3FunfFPLPOVnc8joOPpVorSsAOprw74BN9Nb3moy/6LLuyNvoCOzZ6gVca5c6HcT2Gnx7oE24OQO2e4J6k0JZLm/WaZueece1a5dxaJZJd2svmSc4XbjuB3z614gELXpZDx6c+gqcRqFEXQ5zTNk4UcelLBlN/YUqgnbWmJAb6OG6OImzu4PYHHTnrV9Gkc5NqNyfl2HrSXcjDYT93oK0+5eCUSI2CPb2Na9qVzr7fb705aTvx2wOwHoKaxLnC9e1eHfC5lgNy45/8ArketapY/ZlZmXjjBz9KlWIEqevarfT5LqVgozsx/nrWqXrzL5O3hff6VKCAqkcnNOozxRABwfwr9iT/gn0fHlpbfG79oO3Ok+BrWM3VnpV2xt31KNBu8+cnHlWuATnILgZ4TBb9tn/goDH4ts7v4F/s73CaZ4NtojZajq9mgh/tCNfl8i2C48u2wNpIwZBwMJ984HOelFsnBNMVA4rbu+bPPpSKCQCakXnAqQkKDjNKvPJooQcqevSiMDk0Du5og9zn8KxgYFbSec0B6mtoPANBc8A9KIyMZpc49fSlI5UnjvRIIyKAAIz17UykgjNBcL1ojC5HbpQClcH8qyc5B4FYLd6CknA/CjgUVzxn6cVgHgUELKTnpSxlhgGjgDFBRjOf0orgAZ+gxRwBz1HSiAwGTzTEHhTRGcLW0E4H5UBgdaOAeaC8dawQ2FNKpIwTUS/vNoNAE9DUMSyRgg8ilZ4yABn1p2LDcT1pVBILHge1SNtb5elGYl8EcVCIydxNRtB8ynkr0HNWF4Ypgycf/AKqZzLPuXn/9VCZkdiwp9RWbaqD5RnFLH55DLwBV1ql29mLeNeD1OR659Kt7Z5J40c8LnJ/CodPViQTkj/PrV7FtkiiXqc0YcMFxVvbxtKSB06flQtn8rArSrNkgaTPAxxj3NWrEyblHPao4PPkKg89hVrCscR8zqO1X8bi4WIDkdfyFSWh8uOMHk57VZQxpPtAyf/rV8E9K0681y2tbxfll3ZOW4wkh7H2FeL7ay8Q3AsLGTlfY+invj0NeM4pPAaJp0X7xr7OT0+5tP+1/e9q+Ixl0mCNIWyXzkY9Cn19a13WL+7g8phuMH0HUj29q8Y38lzqBRx+H/AVpYfOJwenWr4sq7FH+eKKAysvX0/KrgNGwAHWjIpjEZPzCrS2mupxGOn/1q0eGGJ/KPUfX3rw74Bs9StoNU1G5xAm7cuw+pXs2eoFfEG307SdUW30l98fOThh/Cv8Aez6mtJ1GSEvJIN3TA6Y61oUs/wAQLZZrsfZ4Y+vR+pI7bT1QVd6r8PL+3S91OT/U5+XE3c47D2FavNo7X0kunH90cbPvegz1565r4UaDYeK/EiabrUnlWxzlsFv4HPRSD1UV46/s/QtdfT9MO6GPHOCOqKe+T1JrxR4pvPFEgmu5Mt6YHsOwH90VBoF/fwtcWyZSLG7kDrx3PtWkW8V7deXdH939PY+n0FeIIoItQktbQ5jjxt691B71hU5NLcmQYUY20zTyKI1k+UdsCihZ8E9OtTNhdpqHD7VJ4Gc14xl8M22l6emkr/pDeb5vL9mXH3uOmaf5mODxUi7VBB/ClTIwpqRSDgHOKPAwtIMtyagComSefTFRwgsNpyT0po3lXOasJRZul5GfnTOB9eK8U+L9T8UWscV02FizhcL3K+gH92o4fLXJbP4UkZL5FNGGPIzipOZPLHQUHEcYA/CheTiNoN+UbGRgdqmkLHZTEoAqnkUeTg1FdS20m6Btv4Zq5naRy7Nlj14pQTjJplA6flRwBgVtDDk9O1FFA4o4A4HPatwFFcDPrTEZ47UxGBzQAJwTUm0nC0RxnvT8jANIu1cZor2BojAwDXylcbufpUg+fINHkYzRXJwKI29DWfeto7Uc96xxxWOMA1jceD+lEAfKKbg4ArGWAB/SiMHg9KJzyD9KZRkYPSmBIwaVcnk0QCMg1gbc5o8r/Sgu4YHaio7GiOBzQHPWsA9D+FMecA9KBJ6c0CSAK2Z6GtuBnNDngdaXkYojJGKx70QT0P6URjjNYGOv4VjPSgM/hWPeiBiu+MUQOgo4IwK4I4NEAjGaIGcCiBjOaPNAgjbSuUOAfwqGYhhlc496lcsuQMU6Hqp5pS4GM0XJX39KV2fjFIxXgHNRsCMd+1RMcYzmly3BPH0pI3ZAwPHY0SqEoRk0jnOAelBgelYB4zWewP0rHqefpWQRtFdeDXy4xRAPAOaIHY0VDdRTxhuhrbgYBzinjJ4U/pXkbjgt9OKMaj+LP4UyAHAbP4UybVHzZ9sUFzgA/pTADgH9KbDDAb9KVSoyGz+FEAY+b9KdgVyrZI9qUBuDWwA4DZ9OKCbuSf0pl52g4NZIG3OQKRFkOAelMozgHpRBQYJ60YwVwD9KMW0fLzTp0yaHPIP4UPm+UCkO3oM+1btx+7n0GaHYdaJJ4rr2qgN3U4ph70cEECiMdD+FZx0NAgjBNUQTRXjg5rB6ZzVEAjGaAwMURkYoqM8UQCcAUxI4A4o9eB9KZMnOaDDIAGfaiQxwaGFGBx6iigbpRAA+Xt3p0JXcR0oRgAKPyqOP5sZ+laXYS6nfRWNv/rJM4/AE9/pXiv4a6z4Kt9KvdRbKan53lcL/AMsyoPRj/eHXFJ+4AwufbNaRAt5eeZJN5Rh+6Nu7OQf8K+HvirS/B1n9rlXzrnsuWXPLD0I6NXifV7T4gfCgePYfDH9jRL0T7abjzf8ASBF1wpGCnpzn2zWq6jaRaqLm3iyD1Xcf7oHetG0q31DWJo3bER27FwePlOec+1eIfAlqPh7DZax8QPO075t9t/ZRXZ++BHzK245bBr4o694LuC134Ti8m3H3E3Stn7gPLjPXdXiHWbzU71VlXaozjkHHA9vatRsbK10ePUhqPnyvnNt5JTGGA+99Oan826wsUmxH6x4z096ube4tyBKMg9DwKV/EtvpD2twuzTrnHOUO7a2e3zD5qvdctToQ0YaX+9/5+fPP98N9zHpx196hjVUyW3+2MYrUtFsHtDc21/vcdI/KIzyB1Jq10ia2iZLmP5ZMeW2R268A1BaR2c+ye28w9vnx29q13SbbTFHn2Owt0Hmk46ehPrVzGI5gV5U/4VY2Fxeq0tumVTGTkf1+lfArwboPirxRa2viPUvsVp8/mTeS8u393IR8qMCeVA/GtZ1e48KXMul6JqP2q2fHPkhOgB/iBPVjUylpNuMAdfaiVPAOTTx+bhlfHtind3IUnpUYZnCtzmltoQoO/afXBNP42ur7Q7fw6LDIg3bZfNHzZfd028dMdaXULrRNONtc2PFz90+aP4Tk9M+tTO8kQZZtxPVduMU+myzaXcXhlwsO35dvXLY9aS3UyKjvtBzk4zir7TrW3khEF75wk3Z/dlduMe9Q2kcpKtcbQOh2ZpJnZRbqa0qziu7gRPN5Wf4tu7se34V4m8HQ6Q5jh1D7Wg/j8kx+nbcfX9K+0SW7YSTj6UYkb543x7YoeYHyeAPxqHWr2a1jtJjmOPOxeBjJye1fC7wJZeL7pdPkuvJd84fYWxgOem4f3a+IHwhs/AklvZ2Wp/bg+7J8gxYxtPd2/vH8q+Jktsqx6XFZbNmdz+YTnOwjipkSQllGMdutfDnV/BuleB/Ekmq6N9p1V/sv2D/SJU6Sv5n3QV+6R978K1ZFN28gj2BsYTOccDvV5EVYODn2ooWcKoyTwAK/Y7/YF0LwhoS/tEftYwWum6Xplub6y0DVMJFBEoDC5vQTxxyID3xv5+Sv23P2+tb+Pdzd/DX4XXNzpHw6t5CksgzFPrm0jDyDgpDkZWLqeGfnCoGJQKB0okleT+FDJOAaYADGeaB2gYbP4VyTwaxnktz9KJ4+nQUMtxSjHANPgDCnNeF9Bn8T6zBoltJsluN204B+6pY9SOy+ta1pLaPqc2mTSbpINuTtx1UH1PrWCTtB6Vt3DB4oIB0NRqpAVm6d8UxAb5OR2ohTkE8+lD7vH5UCBwOtE8YVv0o/MNpOdvTil5ODUmCxAOf0ohCMA8/SiAF47d6C7+E5PYUoDAAH6UOHAYdKYJIoAO32xmnjTYCH3eoxjFDcegx7UDsyu7j6Ux7AV15NEYUnFEcDn6VgFcAUQByDmgFYYxz2pl6Zpgp4Iog9QenSmBYAZ/SmBAwDmsE96PAwOat1LNuJ6UDnjNBGRQe1aHp7a9qlto0EuyS43YbbnG1S3TI7D1rxb4W1DwfqDaXqkn75MZXaO6qeqkjowqW5JAUflSEMo2t+lF16hcEe9IzE4Dc+mKgidpMZ471aWtlGMmff7bCKeZLaMiBckdRnpUU0cuTJDnH+1UccbTsI04Hv04okrHlZMgdsVcLcSxAxybPUbQasLSSaLymf5j7fWorCC1tTcM36H1xU6Ga489lwqfdOfUYq0jZx5kp57VbxtK+8jZt+73zVmJ3ASQfjxVsQLeO3z97OTj0NLD5OFU/jVnpyFEvFkwecrt/D1r4TeC/Bl34T1LxH4ovPKS08ranlytu3SOh5RuOi9q0XR28Q3qxD5ZDnjr2PuOwrUvhtFpUccl1f4YZwnlHnp3DH1FQC3Nx5cZwF+p7V8Htb8O2drM14P3ybdjfP6vngDHTFaDaxX++80y58mYY+XZu9R39ga1u+WfSpdXvrf7TfDH2Vd+zHzBX6cfdx1FeMdXvfHgiLWnkTJnA8wP12+wHRa+JFjqvw+lNnPDm5l/1g3L8uAhHTcDkPWv3El3qEkzj52x+gA/pTM1tCAj5J68VcMJ2UN1oKrxpbxWWZlz83mfe/DtxWq2NxCEW5TymbOFyG9PSorSPo4z/StP8AFfh7TPC9zo6eF/tGoTbPKu/trp5eJCx+TBBypxyeOtan8QfB0/h5NIsPh95F4ud15/a0rbvnDD5CuBxlf1pdZ1OSQWzTfuh0XavHfrjPWtPhglQvO25vTBHr6VJFb21sty827f8Aw7SMYOKi17VBAbe1k2wnqMKe+e4z1o3z3kLLGeuMj/P0oo9sOW/SvD1re+LrE6NpOj+dPD95/tAXOSWHDYHRT3r+y7a0vHsdSufLkixvTYTjIyOQfTFX0Lq5aSPym9M7vStL8e6x4d0m5061uMx3Ozcu1f4WJ7qfWmuUEexlz+OKMxIHljbn8auSFO0Nn3xigzDIVsH0xSTvCMoPxogGPeWyT7UTlNpOaC4IkAwKeV3ILt06DFMccA/jWwleegraccN+lMu4bSeaYFeAf0pFDOMVDhWwV+nNWRW3lWRjkDPGK1ExNODAdqd+p7Ci6NwpzjoelSZxgH9KEZK5UcDr7UxCDJoTFFwFyT2zQyG3EfhWRINqpj05qNDG4+Xcewzip4gTvZ/wxUjFlBYfSnXKgsPwoQiRwqvwenFTRiFuTu/SgRjdmnkLAYFbtowBmlVmUtnj6U3zLgGiccZzQQdd36Uz7hgGt5xgCo0DkKpqRQpxnNcEcUxJ4B4pgB0NKOMZz+FZ7ZohQM0TkdaOD8tAA8Z/Sh0AB5rGOM/pRKkcc0MAfe/DFY4xmjyMZohSMbv0oEEcnpXhzwHr3iiwuL/S7bzIrXbvO9BjcSB1YHqpqeCS3lMMy4deoznHFA5Ix2rAHHX0rI6ZpoyFyR1rjGKIHY0UwOufbFMCvAOayp70wHQUPu8de1FeODWAQOaK4HB/Csg8g81njANDjiiccg/hRII5GaC4IKmsDtW49B+FHH8Jr5q2kjH5VjPesZ6V24qgCRgVRANUeOK25FUQD3rgjg0PasHOB1pSVYEmjNuXANAgcZpXBGCawv8AhUeAcig4J60rY5zSTKB8xpHVuFOaE7LEIOoHSnCuck89qRcgA0QEHy/jSOWOCa4I4NVgHjNcDkGsA8GiAOh+lEZHWivHBoqSOTTBTjFFAORQGRwaMYxgUFDLg9adFYDnpRjIUc0UyeOTQQgYFHAXFYwcA0wz0oAYwD0oqT0PSgRjAPSmIc9aCNjFMpU4BoqCMCmUgBhyO1LgYXPPbitooopGKaLJ+U1sKgMO3Ss55zSkYAz06UJB0BrcpGQf0rIPQ1XGP5UTnpRxjrRJxx+FAErgfhWMnmiMniskDANZyMZ6VyBj8qCgjBPIojHGaAJwpNEA11FEKTyeaIBGKAA+UGjjtXyj5V61nbzRAduDmnj7g0UIXGelMwYYzTKCcnrQXBHPWtJuJ7G/iuoRl0zgcdwR/WtVu/EnizT7F7y2xDa+Z5Z3p/ERnpg9VFXVrI9wVQcjt+FfaZITsIwxqy1R5oA07dOnHv7CvBnjrXfEng2y+Glqu+2g8zyhlB1kaY9VB6r3b/CrHwL4e0ad4tfm8q+XG1Nrt255ViPukV40nWC9aa0bDvjIx6BR3qx1C9mUC4ucg9tg/pXjj4X+JdKjj1WaD91cZ2fMn8O1T/ET1PpU0Z+1NbyJ8/GRn2zVrA9/frAD8pzj8v8A61DS47GF4iOeMf5z714f0XwpeMyeIrjy0GNvySH1z9w+wpvESw3semxputbbOw5xncMntnrWru08heNePr9K0OawGm3y3b4kk8vyBg84Y7untjrVrE7OFQ/LWlTz6lZw6RHHukO7aMgdy38h61rdo0Nz5Vwm14+oznqB6Vr/AIhuNc1N57g9cZHH90DsB6Vb2WiahLGXm+VM7vlbj0/lV/cabo0kiaPJvglxzhh0x/eyepNW2sXNiZPszfLJjsO31HvWhxLcXcI1Btts+7c2M9AccDnrivHOl/D7TbBI/D155l3LnK+XMMYK/wB4kdCaSylmcJGuc9Oa1Pw7faOI/tEWN+dvzA9MehPrQ02WYqoXk5xz/wDXrVNMu9E1F9PvE2yxYyMg9VB7E9iKsI7WWxzOcN6YPqfStKuYbO7illPyxZ9e4NeI9Yk1N4Xx8se7b+OPb2qwRr65SzH32ztH0BP9KPh6ax1G30zWx5dtc7s9D91c/wAJz1Iq/wDC1lHqDx2z5txjacH+6PU561d6ZBHKYLd8uOgwR29zVpaT3BITqO3FeF9Qs9L1uG7vmxGm7dwe6Eds+orVtSW6u/OhOB2/ID0p9fnksVs3Ocd+B3z6VJiTg1EVDCLNS6Qy2BvmH7tep/HHrWg6e9/eJEgyDnH5H39q+EPgK8stVhvrtNkQ3YbIP8LjoGz1IrXPhjpl9okviCe5z5eONjDqwXs3t6V8RLKCTXLiWQ4tTtw2D/cXt161NuUFiOTXhjU7HTdRgvtQfbHBu7E9VI7A+orxTc2Wq3wl05tyDrwR2X1+hq3sb3UryDS9Ns57u7u5Fit7eCMySSuxACqq5JJJAAHJr9lz9jP4f/sveEW/aU/aqvdOt9W0yEXNnp90wkg0jK/KWXB867J4VFyFbG3c2Cv7ZH7bfjH9p7W30HSTc6H8P7CctYaSX2yXZGMTXW0kM/GQgyqZwCTliADxQHHvR+tAAHGfpTKzEbe1EBhgCgxVCBTAKcE06k8KaCkiiQTtzmhGNuQa0++m026jvLdvnjzjj1BHfPrV3LNeyPeSnLNjJ+nH9KORyOaY5HWivGCeRSLuwAee1FdrYz0ojccg800ew4J/CmwThTRAboaZS+AOoraCNpNY6ZNBQcKtKCeKVCDtDcn2oEqNo7UcFsMaRiBgUQCOtMcDg0oMnyqcntRUk4z+FBcsRninbd8graSAAelMAVwp6VgEYJoKDGSvOKP3cCtwBxWR61kHimUqQucimwDgUFyMD8KtgXO0DIFYxzmjLlcE8CrW5ktZluIGw6ZwcdMjFajqN5qsxu7p9znvgDsB2x6Use7kmgp8sIRUceBlhwaliRQNnJoSMoBPfrVmXkcIvWoFWOxBdvnPUY96tnVoMAZ//XVtp6pGhJ5Oc/5zTx4kAiGcdqWzYp8vIHStM01GQS55/wD1+9NYSXdl5bjAH+P/ANan05UAt3Hyn/PrUej4jyoyP8+9GxjWaPf74/KraOPzQM/Sre2czqR/DnA/CrayOQZB9K0XTmurxLRh8j5z+AJ9a8SW0enWcemRT5SbO5dvoQR/OvhBe+H9G1ae+1WTaBt2fK5/hcH7ufUV8VPFcWt6mktq37nnnH+ynqAeoqPS7O2vZLQHJ4x19M+tfDnwQt/dRW1yuFkzz9Ax7N7VBrz6HqMkduflGP8A0H3B9a0X4geHLeW50/U58Sz7PKG1z90Et0X0x1q88cWtnqPnwPlF6HBH8OP7tfHH4ijxrfDVpX+afOeP7ojX+6P7vpWqRRpKbgn9PYCpZmAJY9acvORsXge9WOoPp11He9SmcD6jH9a8Qa8NbuPtEvyt6dewHoPShK+Si8mrO4S3mVpT93P8qjnSWXCnk9B+FW1m9xdrBGMuc4H4VNF9nfDnH4Vd322YKrZRPb1FW+pWqQFI3y56DB9fpUM0UKk55HTio7yPb5kzYA9v8KsvHNj4a0Vo9Ck3yzY8zgjGG4+8p7E19qfVLqW8kb94+M8egx/StW1t9Wm8+QbfbOewHoPSp5GkckdBUHhe/ubc3caZQd8j1x61NkExjmnjcjJFIu5sg9KkKtgmpF24x0pn3AKRQDND8vIFE4ODQIxtJ6U2D8pNZxzmgCx4NMDI+0V9naPGfxpWZm+lJKrDCnJrdvGCK2RxpkHlv6UVDHd6UDgZFTMJMIp+tCKSQblFKCxABoSeUdg5qM+Y5kB5j/rUyM2WJyK4Axn9KJJwCadwVUL/AA5qUmU8n9KaMqNpOcUY8cA0UxwKLFRjNKcfKTTlByDzTybgAKYe/wCFZIPFBivSid3OayMAVt45NNjGM0vI5H0rgjNN90iiBjg1gHjNDHQGj169KYZwAaIHQHmgOaJyB836V1HBojisEdK8EfEnU/BenX2nWUe5L3y9/Kj7hYjqp/vGru4a6ne4IwWxn8Biic9DTMWGAaGcVyRjPA6UVwOaOB3o/MMDnFcmsZHFEYxmgAQMURkYFAYGM0OeBWOM0QAPetpPGfworxwaGV5xQYcYNAAnGelEZIwaIHGDXzEUSSMgV7CsZHFDgZxRGOM0DgYFH0FVgHjNH161yBkURkZFe2aPHANZx0Nck8HmiMjBNZOMA0WJ6n6UsjAY60JCQAe3ag+e9BiOQajmPXFI6sMEZxURB+RFyT0GaMrRHLH8KE/mAEt+GKErdCPxp5M8A9KU5GCM0okK4CdPeiQRgDkVXB4BoEYyOaBFcelY7A0wGM4oKuOBTDsBRA6Dr6Vgk4oKo6GmQMMUUBHJz7UQgOAPpRjzyOKaPC5K/jmmjX1/ChFnAH4UUCknb+tAYGAaA7E0AOhNFhjapzWCDtPWicURuQKG+gxSnacbuR04rIIwKAGM4/GsZHB+lOu5doP6UYfRv0plI4J+lcUpDLgUGUDr+lYGeDVVwRis46GixIwDWTjANE5wB1oEggCtwzkHmiDng0wznPUUF3dDRG4ZzRXjg0QDxmigK5BojIwDQHbrQQg5H5UUD8k4xRBYbQMe9DgAAfjTlguAKXGMdfSjCGHB5oKcYLfpWBgD06VYyMkqMo5GcflXwt1DTPE9r/Yeu+KP7Gt7f7rfYmuPM3F26LgjBUd+/tWu6TZ6ddSPDN9pbj+Epnge/vVtpFgLCbUbjWf38e3Ft9mPOTj7wOOnNXEulXGjhIbXZL/e3sf4vT6V8DPiEvw81231tY/tkltu2pu8vG5JF67W7P6dq+KPxNPj3xHNrTad5F1Jjb++3YwiL/dA6L6Vc6HfSRRXcz/ujnPA47euetan8PNIsvDz69Z+LfNnTH+i/YGX+ML94tjoc9PavBfj3xT4YsZdLV/7QE23YMRw7cFif4TnO79KvryXR9UOo6rYedI38Hm7f4QOq59RS3Fm04njTYw6DJPbFabfR6xJFbzy+Soz823djqegx6Ve+EtU8OWf22aLdbt905UdwPUnqa020gt7ySeey8xGxj94RtwCO1X2kx3cBTT+T3/Mep9jUNzp2n2i2txZ5lGfn8xuec9B7Gnu3LZhGRXgHx1qXhfxVZ+Jlj8z7L5mItyru3RsnXacfez0rXPH1vrWtXOqT6d80+zMfnHjCheoUenpQsjcyid/3SP0/i6cUNDNvdIsmofZ4nz+88rf0HpnPXj8a03w/BPp/wBtvpswy/6ltpGcEhuAc9cdan0tEkkiiucEYwuz/wCvWpwX1on2SaXcg6DaB6GtP8PXGpsojbCDO98D5OuOM85xWueFLjw5Al2NR87rgeSF7gep9au9dfULZUvF+0smeM7MZPsPYUdR097dUbTfLkXO1vOJ788Y9K1O9utd16W/1q43yTY8xtgHRABwuOwFXMEkbeRY3O6M/wCxjH5/jV1p95bHE0OCe24f0NeE/DFnr0UgvdS+zFMYXyS+c7vQj0H51qekS6BqANtefvI/uv5fqB2JPrWna1cazr1n/wAJTefaLWPzN37sJ1Q4+4AeoWvFuuwS3z2umf8AHoMbV59FJ6jPXNCTe+5m/DFaTpY1DaLa48uTuNmfX1PoK8QW01nqk0Eq8rtzyP7oplLLyKllWPGRnPQUELjK8CkijMhZW5HbFPqNzPCbR5d0XddoHv8AXrXhoPHeefG2COnHsRXwv1fWvE+nxWUMnzRZwMLzkufQf3a8VWWtabZGwvpM5/hwoxyp6jPqK+KemP8AZbuIptVdmOc9096mjOChNXQdpPL35A68V4b0DxJ4y1+x8KeEtIutU1fU5hDaWdrF5kkrnsAPbJJ6AAk4Ar4Jfs8/B39gf4eH49ftB6rZ3njUxbbZUUSmykZD/olmmf3s7DIaXgAZwVQMzftVftX/ABB/ae8Ufb9edtL8N2EjHSNBim3xWoPG9zgeZKR1cjjJChRxRG85FCPDcn9KC5XbjmsYO3NMhbDEZFJEojYocHjFJEHGFbn6UYCDtB5prZWHLY/CmA5I/ho5PBNYwcZ5NYx900FJUbe3WtwMHlhuR7e9BMrt6gURgYPSo9h5YZ9BWYxGQo2+g60ygDGMmnComNvzHvnpTpwMLg/WioQYZeT0ogEjIoDaNyvz9KEUkhwFyfXNMq7RkZrbtXcRTBiAaAUKcHFBVI60Dnnd9BimUDABxWPl3EdO9AFjgDIoHachv0o5Jzn8KOCcZ57ClHOS344og7tob9KKAcE49qIGMAUGwu1W69sUFTFHJHPWmGeAaARhhm5+lEkD5TR+bkn9KRWbjOat49i8Hk0emCa2lsL+VLGkcf7yPJPTnGKUs4CBcfjUcTbwFf8ADFSRqRyKe5ZWC53AdBjFW9yrfM0O723YqW4ilUJFFsA/2s5q2d1kBifn6VbxCdlLzc85O2obq5hkCAeZH3HC0zXEq4C4Ufd5H41aGQRuFGDxVs5cCGNs/hVrbIke2R847Y6VPJG0G2JsHsMe9AbpFO7ewzxjGKgm8yIxIOaSyMhBZMlOnOOtW0UQfhee3NWFmZGDs3I6DH1qFIFjG4bsdByK8I+GbjxTqUelW821p85O0HbhWPcj+7Wq+EV0y9GnNfea0f3v3W3GQD6n1rw7oEelW6ahqI+1LLnEf+r6EjqCfUflVz4U8M6p4I1TU0sPLu4fJ8l/NkPlZlw3GcHIHfpWl3M/25dQW23k5+TeBngjrXwNk0e7gYXmmeTdT48s+czYwZM9OOgFeMPg7p+n5In/AH79RsPGNv8At46GviD4PufBV6BNP5zSfdOwL0C56Mf71ahrRnlMcN1sYdtmew9q8Q3crXrrcJlVxsOcdhnpWo3cl0rRlMKPf6VBbCTEjSZUdsYq7v7KzsZIorfMjYw288YPp9KkmuHQBVwv1FbCVy4/CrWXBLkZpiWbIPXp7VDBHGQySfN9KF69k63ET/vec8fh/I1d3tzdPvjPP4U1tdpEBIuN3fI5xTeTb/MRn86guRMyqr5HYYxWqWrLZ+YsHl477s9xVjFc6grRRjzBHjK8DGf/ANVAMHaBI/mHbNPezMu1flHpwavdV0J9BmsYNF23ku3bcfaXOzD5Py4wcjj9at/E2rWll9htbnbD/d2Ke+e4z1qa4JYHGcdaM4ZcAYPpUbRwjBTdu7ZxQRJRgjaPzq4WBUVYjk8+tBgOM0S6oCBwe1Y3nO78MUVBHB5+lAk8ntRzjOaXkfe57DFRkq2QelNIWySc560pV0wi9PeokVQGUZNRXaFwGGPSp2SS3wFyR05xjmklYDDD8aldvL2k596VmQZAzUMkiR7FTPqc06knapyaaNlOC304pfMRtobr1GKWVujPu/DFMvmDYZOR0+WiWJxu/SpUbbj73oelKOyGijKdrdauGXaqq/r2p1/chlXr3zTAjktz9KYc5JonIwBRGRjrWegogDqM0QCMUAOxpsZAJo9MCjkjJNDjpWMDrR54z+lE570ecHvW7t1rjGBQOB/KicjI7USMYPNEZGAaJAHXpQPHrQOBgjNDKp161kgYB/CsZPA5oc+9BgDjH0rIxg0WLDBFMATgfhRG3oKAPc/hQGBwc03zDgUGA4/KnII4NZJ4zn2raM5BpmBUKBg0CNuCufxrOehobuma69DWCOhzRPpW0kcH9KYbBjP6UACOv6UeaJ4wByK4IokEdeaIHY1nHQ0ee9E570R2ogbQBRAxwa68UeRjNdsVkAdaOeuaJ7k1wRwaPPeiecg1yRzWQBgc1vbpmgwI+WoZBnGaidov3wHTpUshlOSatupAokqOTRkUjaDyKUqepqK5ESMvc02WBIHWkJcYYdKokDpWRnIo4PINAjPBonNAAqABRA7GhgDI5oqDzRGBxWM9K/yKKgjBorgcU2CMUY1NMmcbT0po9wyOtMM4NFeODRA2+9DGetZyck0Tngmi27kH6UUIAzSAgbQelAjGM9KPPAojd0o896eMMowfpTDBxigSBilAcbe9AkjpXBHBoHI4o8jAqjz3o4PT8qGDxij83AP0FcgcGs54WicjFA8YzQ6UQccCsA96I4OD+FEDoDWDk8/SiD2NbQBk11HBoBguCaBGcE0eRgGlBHOf0rarjBPNKueD1qzGZl4x/wDqrQoJ9OnW/m4jOcdPQjt9a8W6xa6hrLz6a+YTjHB/uqO4z1BqEeF5vCd2Jxm+Ozy/9Zx+856cfdrU9PtU0UR2oy578/3h6mtKs7uwEs0vU49Pcf1rwP4YfxRIGVcv25x/e9x/drwr8P77xGn2C0TzUuf9XyF+7knqw9KHwd1OzuTpWqJsifryp6DP8L564pvDll4avxLOPmj6DnuPYn1rx0U1DUYrqeLbEM/xZ/hUdueorWLCxku2n047oexwR2A789c0v2iCEyW45X6f1rUfGfiPxR4di014/lXd/En98H+6P7tXeq/ZAsQb1zx/9ap7yBrX7RavtPcYJ7471qUsVw4Ytkj2PtUErK20nira7jEJQNz24rwt4N0bWtO+0NL83c7W9SP7w9K1jwRqmk+G7HVZYsR3HmbTuU/dcKf4j6+lXk9vrehfZbUZkTqeRj5898dhX/CR3Fx4asNJt4vlsfMw24c733dMVeX1xcz+ay7WfoMg9BV5pEijfeNuI9sfyP0qy12Kx8La1otouZb7yNrZPy7JNx4I54PqK1O7vZ0ENxPvA6DaB6elWWiajcwvcxD5Fx/d+nrRtLgQfaJBwPp64rwzoFz4jnuGTnytueg6g+49Ke1n025e1K/MuO49M/1rW9ahvbwGKP5F6cn0Ht7Vo2nxXMscmmHI58wdMcHHU/WvFuox6pfCbbtB6DOey/4Volzpdlpd7FfLmV/L8vlh0Y56exHWriJVOB2qOFpF3EcCrS8msnDxybce2f8APWry8OoXUl1I27djJxjoMf0rYgcDORV/FDHIGRsp/DwfbNC5KptUVGRIA44LdaK8YU814TgZrry5B1/wav2ePE2g+GophqBw/wAu37//AE1z0B9RXi3x1pHjnWpI9Fk37sY4Yfwr/eUf3TXxg0ea3gvrWRf3qeXuGR3KEd/SrpkwXbj2rwD8OfGHxY8Z2PgbwDos2qaxqcgWKGMYCKMbpHboiKOSx4Arwb4D+BH/AATW+Ff/AAnXj+8h174haxD5YeJQZ7mXbk29oCMxQLkb5TyeC3VIx+0J+0B8Q/2jPHDeMfHepfJCGTTtMgZhbafEcfLGpJ5OAWY/Mx6nAAF2Q6YzzUIxJg1FpjXQ3Rrn8f8A69MCzkJz6UVQgjPzVIiKoDHHoKDggqp5ogIuQf0oT5cDdkD2pDkbic/hXDHBP1FSqq8D8KJBIOaU4bk/SgCmcnn0o8jIPNBc9+tNkDA7dqBJXiuWUDNIpbgj6Uqh1Kt1HSj8y4zyKba7YLdOnFMF24B57USQQT2qGUBgSeKxn3oZbhu3SgRnaD+FMTnaOlAKelHPQCmweM/pQA4HamKrwprZvGBRAPGaPYitwJyDSYA5HNEsx5PSmAIxnmhwcDqaJyMAUcY4P4UI3dcL2rIHAodeD09qIYtgVbRlMFqCkjA5oggYzSEBwf0qSUswAFAlPvH9KSRt+U69qMw6kVJGWmPHXp+VQsU4A49Kn2BUaMevFWu5pRk/5xVmzJIITyRVvEFIOck9qt5SI0ic9M54qa0Frb7x/H0/A1ZaekMfmKfxx/8AXq0VXTBOakjOMioIlEmQeT1q3iVTtUc0Q6vsHTvVlbxmTLHJPTirdY4v3Y+8OtWg8yTy1NeCNK1a126rYNtIz82FOPvL0J+vaovDOo67I00Xzt/G3yj6cZHpWgFdF0Qwar80x+4OmPmOfu57EVLNqMeh3MRk2rNs+Xap6NXhTwRrWvzpdwxYtRnJyvow/vA9RQ8T6F4J8J2cVmP+JnHv8v7/ADmTnqCv3WrX/Hi6hZf2ndJ8jfdbPoQDwF9q1yK+8cWdzeXXzrYbN54GN5AHTH92p9PS0vZHuTgDGPy9jWvXRmvnkJyjY2j8BmrlVwSOallZU8tT06VeyRudjHp/9apTGkSlvfirm4jK7Iz1r/gml8GPhb8XfEHjqL4keDrLxAmkWlk1ml0X2wtI0wc4DAHIVeucYqf9nb9l3wxave6l8G/hxZ25IDTX+i2pQHnADSqQO/Tr+FN+zh+yl44tJJNO+FHgC8gxseTSLKCHb1/it9uD175r9r//AIJ5+HvDnge++JHwKS7hGhQyXOo6FPM9x5luMFngdiWyigkoxbcM4IICtoklr54N02EHXg+h9K0b9mj4A3vhzSU1H4R+FL147OLMk2lxksxRdzcg8kjJr/goX+zB8MPDnwAl8efDT4e6J4fvvDmp28t7JptkkJmtZT5LK20DOJJIm9gD9as2EBLucenFfsD/ALNvw48X/AK38efFDwBo2v3fiTUJ5rE6nZpMYbWNvKUKGHG545G9wVPoa8cfst/s9af4E8Sz6L8F/ClpeDSLpoZbXSo1kSQQttZSBkMDyPev+Ccng3wh8Q/2hLvR/HPhvTtbsh4euZ0tr63WVFkV4lDAEYyAxGfc037Jv7MzHLfAnwUT76PF/hR/ZM/ZkIwfgR4J/wDBPF/hR/ZK/ZjIx/wofwT/AOCeL/Cv+Cn3wz+H/wAMPi34V0f4deDdI8OWV34cE89vptqsCSSfaJV3kL1OFAz7CsI78ngVI6o3ynOelFiI85psHGKxzRYsuzPTtQHYVIpU4NA4wAaIDdDSrk7RVvCSSrDp2qWIxtjqG6VBKqHbnrQYxgY5FeQQdxPPYVYX8UNtNBKmWk27TkjoTUjhpSCePT8KL7jsFFyq+WBkVG+2MYGW7inLFsng9qc+YgXHPaokYHBHIplEbZoBSN7dRTqM8Vgt+6bqOtFCj4PWlJdtzCvCenaHqO9dUl24xj5XP97+6fYVK24BVPPpT4Axmic9TQPqaIHY80eBweaO3GPSiue/6UwHRT+lKAepoqDyDTbcYU/SgTtwaJA4zWMjrRAHGaxWM8A1gEdaBzzRI6Zo4PSsD15ogEYBrAA61nPTmicjGa4bgGtq4wT+FNtwAprAz15olexrHvRA6E8/St2VwawMcmhgjCmsgHINHmiBgDNMqhVIPPNE9s0ST3o8DjrSg4yT+FEgnrR6D9K2sVAAoEE5JosByDRywwaBAGCaJB7V8pwT1rqOTk0BkHFYUjGeaxnpWOMZo4x1qiATkH8KPIxmjgjANVjI4r6GsDOcUOK4qjzRxjArC4xiiQeAKBCjAFRuQODSykptJz+FQyIFyTk+lG4jeLaF5HTmvPA6/hUcqk8/lSOrLleaiKupUdR1oAp8pqIKWwy7vbOKYAHgVjI4ojHemwQADR44ongYNAjHWiuTRIPANZBOBRI6ZrbxyaKkjOeaxjH8qPIxmj0wD+FHnvRBGOaIyMA1jHBNMu5duaKADA7dKCMeSelMuQG3fhiguepohnO7dz9KKsxAzUfKjn8MVz0BoEnjPSjjgKeaJY96JJGAaPTighPLdu1LlvlJ/CsIo4PNAZHX8KOMdaBDDg5oDIGetEelYBHWj6CqIBAArI24xQIxg1kYxiiwxgH8KJ7VkYwpok4wR0oriiAeKY4GBTDIxW11OAOKJYcA/SlVSeBz6UBgcn6UBgciiS3JOaiAYhc9atNPeaRVjPTPb/69SeAdI0r4Yr4kk1PdeNnbbeQwzifYfm3EdDnpWkeAdNvPD8er3WoeVM2cR+SzdHK9Q3oPSvDHirxPp/hi98OWGoZtr/y/Ph8qP5tkhdfmIyOSTwRVrot7GSk0GSenzD/Gk+G/iC0gtri6tf8AR77f5Z3p/AcHo2ev0rRPhtaRWomZ8OO2Ce5H96vg5C3h7Uzq938kn/LN+uPlkU8DPYjtXxS8c6lc/wDEu1/Xv7Ssbj/ll9lWHbt2Hqq5POD+HvUFnquoxzyWI89E27h8q7c9OvXoakXUY7/y5V3Rd/ujtXgC68IXEQbUYNrr0O6Q/wB70/CvF954dvbG7jsLPLts2nzH9Vz1+hrXn8RWNiunTfurFc7E+Rscgnkc9TV7BbDTY/M+bGdo5GPmFGGaCQraPtU9eM/zqaG5Zt5HHrxVtBZtbFnb94OgwfX/AAqXVvtFhbaePuw7+fqc+laZcWkdoAsm6TscEdzXiHxPo0ukRWUcHneVnHzMuMsD6VF4p1nSmmWxuPKV9vGxW6fUH1NJNI7bI2+Y+1S5EgSU5ZaEzSvuZsn0xitAm06LV4ZdTj3w/NuXLD+E4+7z1xV7cpczsIzhR0H4Vp2pa1awGzsr3y4JPvJ5anOCSOSMjmtH8dbfCB0Cz07eP4m87H/LQt0K/wBalsbWOVQvGc4HPpVjbCQ7RFufsN2K0WCyuJ44tTTAGcrk+h9PwrVPCmjtbpNpttgc7hvbnkep+tbtDuYBYa/DtWH7h3Oc5OT938K1X7CJ5E09MRrjacn0Hr+NQQXeoyGO3G8L1HAx+f0qcRwoYo1/ed+akEQjGThqQuJigbI+mO1faXBDFuntVl4zFr4am8Pyx7422+Ud2Nnz7j/Dzn3NJGoySfwqJUZRJ69Kj3EgL+FaaLZrWGO2bEq7txwfU+vtXgnx3b+E9PZYrja743DYTnBbH8J/vVoU17aXK3llLsHc7QexHf8AGvGeqTa3pM6yXGXfbuOzGcMuOw9K+GnwV8d/HPxrbeA/AWltczyENdXMilYLGHI3SzOM7VH5k4ABJAq9vvgP/wAE2PhU1hpMUPiD4g6zbq0gZtlzqTgkCR/veRbKc4UdcY+ZtzV8Wvir47+M/je88d/ETW31LU7v5V42R20Q+7FEg4RFycAdySSSSTdKd+VGT2po3cbW5P5U9uqAoTtYdRjNR3DLB5ajJ/LvUkEaMcDj0oohHPApEQyHYOKaJQp2cH1pyzHLHLemKMbBd36UAD71NGqJnOT61uZRw3P0pl53DrQ5NEbuQfpxR/2etYCgEN83fikC53ON3t0oYIAFRjawZT070xbzOvH0pQiuXDZ29BjFNEuciiSWAU5A7YqVNy7geacAoqHgjOTXzbgN3H0oFifvfQYplIAwc4oEFeK4IyDkUrEEcU7gnOeKGQQQ3PbimDsSxOa43YHagCeQc/hWiaLr/iXU4NC8M6RfarqNzu8mzsrZp5pNqlm2ogLHCqScDgAntWo/B34v6Np0+q6x8K/F9jZWkbSXFzc6FcRRwoOSzMyAKAOpPFMMnaT1o8gAVg7gc/pR4GaLZG3NEFuhrGDgn9KY/Lwf0ocjrXyvwDUESuSR2qJOdxP4YpckcnJoY71jPINKQrhgelT/ALw7yufxqN4gp+XBHTnNM6gZJz7Us7ldhP04qNMnJowqW+arMqkgCpkemait/Pk8xRye1aNo7XakMPlHUfn7+1GzeCcRyDhf8KKNNcxS7spHnjGOopoHmwynIH4VGhRcAcjpQLEGPOfaraApNhh16flQsliO9Dkn2qOFpHcE8DGOK0dYIgDNHvH1I9fSotJZpXdU+U4xz1/WtN8Gz3Uq/Zk2sc85B7H1avAfwOXxL4Yggu28m5G7cuN2PnbHIcDoorxZo9r4As/7LtB5ksnXqOhU9y3Zq+HnjfS9G086b4ig86Sfq25l+6WPRFPYivHttoXjlLDw14dj3iXzN0eXHTa45bH90968QWPiTwLoJ8mf7Lbr91NqPj5xnn5j1b9a8L+IdS8UQWsN7L56jfu+VVxycdAP7orxx4btLq/WPT49tuM8ZJ7L6nPXNeNL9dPubi203iD5cj8FPfnqTXifT7jzHdT8vHp7e9a8ZDKfKGMdT+VSTZXG/wCf6VcyOVw5waKhVLK2fbGKuY5HYgDgdKCEMEB+tf8ABImExeIPiYD0ay07j6PcV/wVgwf2dPDw/wCp0tf/AEjvK/Zn8aeNvBHxy8G6h4BvruLULvWbW0ktoGbF7FJKqtA6j7ysDjB6HBGCARffY/sVx/aHlfZfKbz/ADfu7MfNuz2xnNQ2lpqniCPS9Pd1tbq9EULFcsEZ8KcdzgivG+oXekeC9f1bT5vLurLS7maB9oO10iZlODweQK1XT9L/AGgPgHLZSGJbPx94WBjccrF9qtwyOOv3S6kdelaf4U1zU/Ftt4Et7Fhrd3qSaals/UXDSCMIcZ53nFeNL/Rf2ZP2Zbn+yp44LTwV4dhsbGVxtHnBUghdvUmVkJ7kk9zWpWY1HTrrTzJsFzC8RYDO3cpGcfjWg+JfEnhC6bVPDXiLUtHvTGYzcafdPbSFCRldyEHBwOPYV4e+NPxpl1/TI5vi/wCNJEe8hVlbXrnBBcZBG/pVfEj41/Gez+IXie0tfi34zhih1m7SOOPXrlVRRMwAAD4AA4xXijxj4t8X3cd94w8U6trtzBH5cM2pXsly8aZztDOSQMknHTJr9lz9hP4q/tN2H/CTwXdt4X8ILK0Q1m/haRrllOGFvCCDLtPBYsq5BG4kEVq3/BHbw42klNC+OGorqaL8r3eio0Ln0KrIGUH1yceh6V8L/wDglF8QfGM/iXT/AIi/EFPB17oGpi0txDoh1CDUoTEki3MMpmiyh37cbcgqQ2CCB+1r+zlH+y58TbX4bw+Nz4pabR4dQkuzpn2Hy2kklTywnmyZwIwd2R97GOMnOeppyOdoxQJXkN+GKLMp2q36UQSM5pSNvXmgxHIbn1xUfikxeFYtHgXEy5y+ev7zd0xjp70BIDsIyfStrJww5pHKptDZ/CjIGUlHzj2ou7ptDfTiiWRuDyKDArgHntR4HrSncNpbp7URjgnIFSSq8XXn0rSY7Wa4jiuW2oc7jgnHBx0q6aMOVjbOPamd2Qqxz+lKFU/MM0AY2ABzntitI0pNY1COyE3ll887c4wCfUelfEDwiPBOv3Xh5r/7W1rs/e+V5e7cit0ycfex17UzADAHNMwA5FE5HJojIoDA4GawfWiAehraCeKBBGCPpRC5yDTKrHIH0FFQBxTBRyOtKQRQXC9aPK88+1EA8kVkEcdqI7VtyBitoHOKwMYogE4FHk4Hatqnv06CiSwxQJIxnNfKFx6UcE5zTKM8UOeK5BwG+nFBQBgGipYZJ/SgoYcnn0oHBwD0onLYJoADocmsejfpR6YBzRX/AGvpxQYjvmgM8A0AAME5oY9fpWcgY/KiCOc/hQZei/hTAEAKaYDPAojtmguR1rDYxmmA6CjyMZrIBwetE56UcDgmiOBzRwen5UcHgUMd6Ydgazxigc8UeQBXFHrzzRwelYPrmqz6Gjg9KHPWj6CiD1zRxjrQbbwOaSTI+U0soxjNK4GMUql+RRxmkLAZDc/SoJmDZDfhilPnpuUUl0qgYOTVtNbs2ZG/Q1kt908VDC87BEGTS7zjmmYqwBPH0rIPANduD0rdW7kAHpQI6A1ycU2DwKOGOBRUHgHNFQOpornoKHyjbRIPajg9KBGcnmmA6/pRBxwKwMYpUXaBTJlcCvKYjpQBB6dO1EFTjNIxI6dKHPH5UACRgUwDcA9Kxt69aoLyWPftRILYUZpjk5pjhcCgeBgZFAY6CsjGSaBBzg02OOa68UfrVcEcGgQBkGqBBUY60AGIHSiuOAa7YIorzzQADAk9KOCeDRGRjNEnGO9OM8A8/SgQBgdRTcgECs5GaBqJhvANaascc4KnI7cexr4Q+BpviHdf2de6p5Fnb/ePkBtu4ORwGU9UFeObu20rU5NK0y982OPGP3ZXqqnvn1NeFZ7gu7IMnj04618GfCMevapKuo2+fu7Ru/2ZM9CPQUuj3kGpXnhR5vOurPZtj2hdm5d55zg5HvXh34V+JLi9bUb268u0jxv+RGxkEDo+euK8U2Fzbo3h7R5fMmXG1toH91jw3tnvXizw/wCJfDixtrcW7zs7DlBjG3P3SfUV8QPEWj3uuzXXhIeRZT7fl+ZuiKOrjPUNWpaTpyWDyC6/enGBsb1Hv6V4K8N6dqGpFNZ1b7BAOj/Z2l/hbspz1A/OrTw5ZwSXsIu/NtB5flT+WV3evy5yOeOa+I8JW9NlbJ5qL0OdvZD3rQZm0vVftt5p3npF9yHztu7KkHkZx1Bq6sk1B3u4x5ecfJ1x26/hS3mnvoZtDB+97tuPHz56fStQt1gGYG3DucY9KDgQhS3zemK0K+NrcrI67guflzjsa1a5vp2GpRR7IZvujIOMYB9+tXwVYVVuGkz+mK8OpaaFcxarJB9pjXOV3FOxXrz6+leLdatvEfiK51Wzs/s0Mu3EfmF8YRV6kA9R6d6tLKS6kEMBy1aZ4buri7a3k6rjPT0PvWq+CJdNhE79fT8h/ePrRzGkMSx/Md2OetWWvPpDmOCLzd38O7bjH4H1rQPAGn+J9CF1Hfbb5f8All5RP8ZHXcB91c1ZeE47BWhvH2Spjtnr9D6EV9kYXDGR9vTHGe1eCTNcRzR3Byq7dvGP73pXjwwJOqKOTn19Fqe3dDlTkDtVjqtjZaN9lLbLnucE5+bPpjoaYu3zA89zioYxduIxwT0FPplxaX0lrMPnXHHHpnsaeRQCpNOC6KEP3eopHGPu5PpmrYBV2k8+mKjHzDaMmvDNlN5hYn+XvWpaQtrp/nLPudui7Mdx71oWYLXY0m7PTjHc1rEhXTHzwTjH/fQr4N/tseD/AICfs/33hbwn8ObdfH/2nal5s/0e/DElbi4bO8tGDs8scHjBUFgvjfxx4q+Ifiy/8XeNtfutX1jUZTJc3Vy25mOOAOyqBgBRgKAAAAMUFj8rzM7iO2MVJfIJWUxdcc7uleMvhvZeFvDb6yt95syYxH5RXOXVeu49m9KuwZZBt6jrUUewcNk+mKl+bJI+b0pxtXaRknoKjnUcbevvQkwcZ4oQoFweT61MArIM/Kc5oFSPlOR2q4I2EgfSm5ORzRUHknH4UMY4/CgQByfpRYDisgcE/jQyRgVI6A4ApUyoO7j6UCrDBP6UAqIxz6YoDeAS2PbFSRglQB65p2DttxWEdiWbAHfFMvOVbjtxSqCeDTYPA7VYWF5qd7b6bp1rLc3V1KsUEESF3ldjhVVRySSQAB1Jr9mv/gln8OfD3h6w8SftCwS+I/ElzGJZNFhu2isrAkZEbNEQ00i/xHdszkAMBua1/ZP/AGYbOxOnw/s+/D5oipXdL4dtpZOmP9YyF8++c55r9vb9k/8AZY+HPwF8RfFLRPh5D4e8Q2bQw6YdJuXt45p5ZVQK0OTGVCl3ICqcIcGv+CYHwD+Dfxj8BeNdS+J3w80rxFdadq8ENrLeozGJGhyVGCOCea/4Yh/ZNxgfAnwz/wB+X/8AiqP7EP7JpGP+FE+Gf+/L/wDxVf8ADEX7J23aPgV4Zx6eS/8A8VXwU8K+HfBH/BWCPwl4T0mDTNI0zV9Whs7OAEJCg0m4O1Qe3Nftff8AJrnxUxz/AMUpff8AopqBB4PUdqIGODW3jIOcURt4Jo8njmlB7/lWVzz3o4xtPNADjP5UiKfumoI1QBCc56UEA4Borjv+lZUjg/hSFQOTUY3uMH8KIYjG7P4UYiUHepEIbGahQNnJ/Sk653Z/ClJKg5yfStOjUyBWOAf/AK9WkBSb92Nyjv07Va3clohMSbie2cVd6fHLM0qvkHGBj2+tWunq5ClsDvx/9eoY4ok2Mee3WiqHoc+nFbAi5LfpVpCXl3k/KtOHk+aM4UVDpk0Ful1MMLJnZ0OcHB6GrFfLYK/Sv2eX8C2UWoz+LG3G68r7OmJRjb5oblPqvWvHU3hqHxjbajoEH+hpvym5+f3Sjq3PXNWOo313rP23QoPkuvuRbhxtXB5b8TXiDQrXUZWttQf/AEt8bRg8YAJ6HHQCr3wRLJrrhmxHb4zxn7yD/aqPxHY+BtctNTs3+0Ou/K4KY+THUg/3q1n+zPE+g2+lPrHnXUm75Ps7L0cHrwOi1c+ENN8B/CqeVrXN5Bt53sOtx6ZYdGq71G+Nz9jM2XXr8o9M14u0+5MP9oou6GL/AFhyB12gd89a8eanNFeGJJMA+w9F9q1gSwljIeGxj8MVP8v74t09qnlMjbg+R9KClyNp4HWrhNmPm47HFZRf3nXHav8AgkTIZPEfxMBOcWWndv8Abnr43fArwB+0H4UtPBnxGtr2bTbLUUv4ltLkwOJkjkjBLDttlfj6elfCD9jP9nz4Ia8vivwT4MZ9ciDLBqGo3Ul1LACMHywx2oSCRuChsEjODX/BRX9rTxT4D0i9+BXhLwlrOlXPiK0eO88Q3kXlRSWpwJI7QgnzCwO1342BiMbmBX4N2I1P4ueCdKDENd+IrCEFVyRuuEHA79elfHG6Wx+CnxAvXn8kW/hbUpDIDjZttZDnPbGK/wCCd/jz/hOf2WfDVtPOJbzwxNPo9zznHlPviGO2IZIR+FeEf2dwn/BULWFax3aPpF5J4t4TA/fRiSMj0C3coHT+DjHBr/gq58Rxovwv8L/DGzuCtz4m1Rry5CnP+j2qj5WHbdLLGR/1zNV49tjp/jDXNOkZS9rqVxESvQlZCDj24rww4bxDpIU5P26HjH+2Kr4oKT8SvFbn/oN3mB/22avhJ8P5/ix8WPCfw3t5mh/4SPV7ayeZRkwxu4Ej4/2U3N+FWVj4U+GfgmOxsLa30jw54Y03Ecca4S1toI+w64Cr9a8f/wDBVz4/an4+n1X4e2+iaN4Xt7giz0q6sFuHuIQeDPITu3MOojKAZwM43H9n/wCLdl8d/g34W+LFjYmyXX7MvNbE5EM8cjRTID3USxuAe4APev8AgrAcftRWuf8AoVLP/wBG3FNhvu/lRA6Z5puRgCgTjkc0xI4Bo8cUw44oE9O4qSYFE4+YZqSQtyTSsMEflRi2LkN83pih8kg3flUyDfkHiiCrYqJFccn9KkdS2IxUamRCoH40QQMA1A2wEkZFTFGbcj8/SlOeCawpPLc/SolSQAg8r1FRTzWt0JLd8OvTj2961jWbrVrl7u+l824fG5toXOAAOAAOgFFy/GfpTYYYU0ycYFcFQM1jjGaIAGc0VBAANEZGAaC54zXGMVtyMZojBwDRAPBNYBGAawcYB/SgM8E1tJPXp2oDBFE44obcdeaI7Zr+HIoDPOaIxxmgAeAea29s1gDGT0rAHQ1gEc/lTHd161gAcD6URkYBrGRlR0oemfwrPaiCT0zWFHfmhgjk4orzwaK9hXBGCaUZ6H8KPJo+nXNEZHAokY4P0rG4cH6UVxgKa6cURk9K+U9qyoGKyOhNHB5xWRjANY45NDGQfSs85o46ZrqMVROeRROe9VkEACsgc1jjGazxiq68VRANV7HmsA9/pVGvoa3cgjtR4FBycKKjl2nbnmjIOx/Smk3EAn9Kb904WM7v0qKR8bQ36VIsZiAZuV9qjZSMg5xSXZSPYDVtemNCqN+lQTLFEYwM5oorNnFJKCojcZArTbS1vPO85sbNu3gnrnP8qkUK5VTkDpXXpQAI60OKLcYFBu2aU470QCM5o896Ybu9EY4zQUEZBo896BxxmifamYsMZrAxgVjPArGRxTIGBBogGgAvegR60CAMVgHnFEArjNMAOhrYCuMVtUcgc0U+XANbQGAIzQRc5HbpQIyB60QuNqiugwOcdKyCMGgABgUUUnp0pW3LnFbUI3GgVIyDmiMjikCgda46gVweK2gHB5oqCM4oqCOKI7UQMZHJrGTmmUE1tAGAKzuGD2oopOQKHIpAFYEHmvD91FHKkcgyOf614S0W/wBXknFlZ+bAm3cfMC7c7sdSD1FfB/4F+GfFc0V3qNjhznc/mSHHDjoJB/dFSfCKyn8d3Ftbz+ZbQbcNsK4zDnoXz1r40eItEsre3szH5wXdgZZccxn096m8O3l3qOpT6ZDmKbytvzAYwuD1Oeua8I6Nc21xGVmxMmfl2juG9/StNv7/AMOeK4nup9hXOF2g4/dn0z618V7PXPiNp0V00+57Xd5Z2oMbigP93+5V74Y1Twvqcery2/n+Tncd6rnKhR3P970q50bTr7xM+ssNq3GNyfMcbY9vXPt6V4V8S+DItIFnd2PmXA6t5so/iJ6AY6YrWIYNe+GK6dpd55UU+fNg8stnbOCPmOD1BNaB4L1nRp5LtD5kfH90diP7x9a+I3iLUdRa38q38uNd20b1P9z29quSyuARzXgi1FxexG9GIZ8/N64Ddhz1xXxa8JaLocTR6Neeeh6ny2X+5/eJ9TQt2upAqDJNRaPqNiBdSw7Yx33Kfb196uL67kgW2DYjXOBgeuajtry+mEVqm9x0GQO3v9KZLi1bEq4f0yK8O2dje3QN7Hu9RkjsfT6CvDTpbXwmB2jtxnHBqHxVJpuqytJN5iXW3Py4xtX6H1rxp4y0rV9LSOFcuM92/vL7D0q0ggS5xO2304J7e1a3oUGmyLNHcednOPkK46e59at9Tmtblipz0/l9K8BHwp4u8T28fjmPzbNN2VzIvWNv+eeD1Vf85prS3bUGS1G5eNo5GPl561Fq7aVp4jkjw5+7z789j61rjx3shkHUdB+X+FaPayxW0zW8O6Q7cHdjHJ9alRDHhRgCtM8OG90+XUGu9ixY+Ty85ySOufapPCxhlVEbzBzk4x/WtTtv7PvXS3H7uPG38QM9frU+lfaU83dyPb/69S6fcBAyJ09x/jRgaNNqjNRxqsYUivBllHd6kiSrlOe+P4W/wp7BbW7ka2G5eMDp296ttG1W42zfZMKM4bzF+nTNaRpWoapOYEttzRdRvA6g+/tWvW82nGS2vIvu4yNw46Ht9RV+0i2bifnGP5j0pLO2ljaZzj06mhEjMQR06GtPFpFI88se9lxs5I9Qavrhrl/MlO4joOnpRYqS6jPtVw4ZQw/KkcEZI+7196aXOAFx+NSwqjbaSNWj2sOD2qREfqKMSEgbeB70AoAAPSnCsNpGQe1TKAdoHFM2V2joOlFQRkmhkAKDWwE8mgADxTjcOtKAxyaEYY4DcfStueQfpxQDNIoY8DPaoxt4U1ktgq36U7ADNAEBsnJ4xRGRyaYHAUHNH5QBn9K/YF03RdX/AGwPhra68sZto9Qnnj3jI8+K1mkg/HzUjx74r4rN44T4Y+K2+Gaxt4tGi3X9iCTGPtflN5X3vlzvxjd8ucZ4zXjLxl8YbTxbeSeO/E/i+DxNbzMLv+1Ly4juopMkkMHO5TknjjrXiT48/GTxr4Lh+HnjH4k69rvh+3vUvYbPUrtrny5kR0Uq75cALI427tvOcZAI8I/Fj4p/Dm1uLP4ffEvxV4Zt7yQSXEOjazcWSTOBgMyxOoYgcZNfsB/Hb43+M/2t/Anhrxf8ZfHGu6Rd/wBo/aNP1LxDdXME23Trl13xvIVbDKrDI4IB6iv2ptY1fw9+zh8Sdc8P6reaZqVh4avZrW8s52hmt5FiJV0dSGVgeQQQRX/DTv7SYIA/aG+JfP8A1Nt9/wDHa/4J96/rXij9ujwLr3iTWb7VtUvpNUkub2+uHnmnf+y7r5ndyWY8Dkmv2q9M1HWv2bPiVpGkWE97fXvhq8htra3jMkk0jRkKiKMlmJIAA5JNfCr/AIJF+O/EmgQaz8V/iRaeE7u4QMuk2Nh9vlhBHSWTzERWHdV3j/ar9p//AIJv/En4A+Frn4geGfENv418M6eu/UJYbNrW6skz/rHh3OGjHALqxI6lQASMKUwRz2ojPTmnOQBnpQX5dxPPpRwwyR0oqCOKAK/KD9agiDckZxSgDgCgAO1DYQMjmvLzyv5VDAzjg1HYkjKvyO2KFqOx/CgqoMAc1ITnC0qF8KD07UkIU4LfTiktUBQlsk54xVlGpnUKMY/wpABCrAcnOatoy8o2jJHarUuxClenfNIGZ1QHjvUkKyLuY0IAq7R+FQQ7hsxnFTqqQbVPTr+deH7WKfVrdJj+7+bI/wCAmv2iNf0rVH8OaNoiYj037VvOW58zymH3h7HuasIJrq4WGNcnnuPQ14ZvSPDthpLxc2fmc7v7zlq8HaR/aNktxFPs2/7Ge7D19q+F2svFrTR6U/lXfGDjdn5X/vDHTNeG9I8SeIfG72d/c+W0mMPsQ9IiegI9BXxB0ZvDOoy6Zr199ocY2t5Wz+FT0Un+8K124huPMtIHwjY5wfY1pAeK8Eu/I9MY7GvAPia01FNH8P6tNs0k+f8AahtJ/vMn3Ru+8B0P14qbTrjVXeXSI9tsMYbIP6MQeoNX3jOS4+HGpaAq4hTytr565mDHjbnr714g8CPqJfVYpNq8cbc+i/3vb0rxfbiOREc4Zc9v92rgCKPcGye3FXMrX0flO3T2/wA+lPKlsgiU5z3qS4aQYc1O24YPOOlf8Egx/wAVH8Tcf8+Onf8Aoc9f8FOfG/jXwD8A9C1jwH4w1vw5qE3i+2gkutJ1CWzleI2l2xQvGykqWRTtzjKg9hX/AATm/a7+JXij4n/8KW+KXjDUPElrrVnLJo93qU5nuILmFC7IZW+Z0aJZD8xJDIMYya/b++Elh8Uv2bvEV8tisuseD4W1nTpgvzIIRmdc9drQ+Zx/eVDztFfsr2B1P9pj4XW6h/k8W6dN8oyf3c6Sfl8nPtX7TV8NO/Z1+Jd0Yw4/4RTUUKk44e3dT+jV/wAEqPidCNW8VfCGaYYksk1WyVmxzHJsmwMckiWLv0Q+9Wnw/wBItPibqfxQQj+0dR0O10llCY2xwzTyls9yxmUfSMde3/BRL4kf8LI/aa17T7S5aSw8Iwx6NbqTwHiy0/HY+c8i57hB9B4Wlin8MaRNC4eOSwgZGHcGNcGv2pPh94H8M+OPFM2lMv28a/qAuYlSVQjC6xtGWK4GWHHHFeH7NYfEukhpN3+mw4+XH8a1XxB0kXXxL8U4lwDrd7/D/wBNn96/ZE8P6L4Q/aV+F3iS+1NZBNrP2ZY2hI/eTK0EfOT/AByL2/xr40eGNS8b/B3x34L0Yj+0Nf8ADOo6faZ/56zW0kaf+PMKu7a6srqWzvbeWC5t5GjlhlQo8bqcMrKeQQRgg9K/4J76Lqmg/sdfDiw1izktriS1vLlY5FKkxT31xNE2PRo5EYezCv8AgrESf2orUA/8ypZ9v+ms9HOMCgOOTzRxkGmJ6Dmh6Z+lEDYDn6VklSAeaHzcYpvulcc9qXHQ0GxyKjZJDgjntTSF8A0CNw56VhlAcD8KGQwYGlEbnc35UrIQCp5FXQUtxUZBcAmpY0VsKenSsgjBNBc/L+VRN5YIDfpUrA8g05z160ACcE1tBUADp0rb70Fz3/SmAA61gHoaHHyg1BGJAV/KsDoTRUECgcjGeaAzxXbBNbV6g0ygjpzRHOBXB5FFe4NDPQmiSBwcntQ55Jongc0cE5rBYdelbTjIPNDkdc0Du4JrBK5HNDqBniiPQ0oLMOelHGeBRyBjNLjHvROBweRRO7gGmGe9HgVuwOKOO1YyODR5AyaORj9KzkcflWc9TROAMflQbHQUCTxRI7jmmIIFFTiiM80eaIG3ArGcACiMDGKAB7fSscfyoADr+FYJOcUcZo4xgGjtHHeiMUQAcVjPFDHesDODRKlqbB6UcEAA1RA7H8KwP7304rAPAP6UF7GgoYYzRUY6/pQAJ4FBiOKAyODTLjkmjgDg/hWSOgqK4eM5HOOlJcs3OPwpZsj+lJJk4Wgxboc/hUUgQjB/CluFkTanarYRtCSzYYdBilkQR7SuT9aAqK4+zZJi35/2sUjD7p69qY7etZyOPyqgSeo4osGwAaBxWc8CiMjGaIJ75o+gNfKOBQAPGKPzdPypsdBWMjigBkYNFuMChxRAA5P6URkgUFBOM9KLDJUGkYEbQc/hSsCcA9KBDEAGio5wePpRAJwDRXPy0yA85oqc4U0Qudu7p7USAMK2ce1IcnO76DFAAjNOyqApPP0ra27CmlUgbSOBT7cYVvwxSEMAu79KBGMA5NKexFHngGgCBkim5PBokEcGgBkDOfSmojjiiMnOaOCcDnFHkZxTEEY70p7ZrgDBNJgjg9KssvKCp/zivAHj/UPC1rPY2Ceb9t2+YMhduwsR1U/3jXwV+IDnQ47S0+eZs/L06NIe6+lXXibxN4c1TWITb8t5HmHenyYXjsc5zXhzxZa+KrI6dfJvvP4RkjuSegA6KKuLKz8F6XZ+Gnj26pdeZsjyTv2tvPPKjCt614pTTvB91HHqM32eeXO47WfbgL6ZHRhWseI9M8Za5c2Wi3v2yRNnlyeW0ecoCeGAxwpH4V4S+LUmga0dLuPmCff7YyrEdEPrXjb4sXmtac1tp+n7opMfP5oHQqehQHqDXiO6vLI4kGM9+Pb/ABoaLfaDBFqt9DiMZx8w5529ifUdq1L4uPeWtpZ6Xa+QYd+795uzkg909jUvxO1aysvIW7yD/wBM19f933qHxW17p1vBqGn+WI93lv5ud+W54A4xgVqghvJA8MeAPc+3+FW8mp3OnpY2tr/x652tvXnccng1r/iy01Tw4+gHSPKuZcYk+0FsYcN0xjoMda1rSDoF/LbQyZMWOduMZAPqfWrTW7u4Ihnfen93AH8hWk+D59SnmF9J9n27dowH7H0PsKgivPB+r8w7yfufMBn5efX+9U+mWuqh5mGyXj5eT+uR2FOG0bUD5sW4joN2P4fx9ahea6bEa8+manTClZmwV9qsLa2t4I795sk5yu0+uK8a6hYarLanSmz5W/dwR1C4+9j0NQ3F7OcM24D2Aq7v7K50u0tQNs0W/ceTuy2fw4rSNEjvQpY+v9ff2oxSWczXMC/KOhz7Yp57jVBDd3MPlwwbtzbgevHQc9RWt+C9Q8QaOvibw5bebYNn95vC9GCdHYH7wI6VpwB8OXt9DdbJbfy8DZnOXI/lV4ki/LjntVtdXJtzAr4z1GB61oXiqbT7XyJrbzm9d4Xufb3qeaS+YTSx7UPbOf8APSrxntZNgbg+1adrkdjG0NxD5m/HO7GMZ9B71aW1pPaMWXnjHJ9avrfZK20cHp+QrwNNZWtqWnfEnbgnu3p7V4EgafVHSSDzEnxtO7GMK2a8eaVYaF8OobeKy2SPu/eeYTjEy9snscVpMrWDNeW8uWkxkbfTI7/WvGV+upyXJc5c7cn0+7/hXj2eO3VraBt2Mc4x/dNNI5QKTwO1SuU+RWz+FaPqNraW8ySQ+YX24O4jbgmponKtJGufbOK2HYM/lTqoj55NFXV9g5J7U3KDAyamjYyKy89eKKgrkGiCGyDkemKuHCpgNhuwxShnUBOtFQV4PNSxtI2ByKkjZX+716DNFc8DmmBBwOayScHrTDsPyoAfdJpRkEgZP5VEzmMhU/HNRFhwT+FAlWVSvXOOaBaN8kZFBcAE8e1OV2ZI6VjzHO2nAHyZ5oDYckZ9qJBPIrwl4q13wR4n0nxj4Wv3stW0S8iu7K4TkxyxsGU4PBGRyDwRweDX7L37ePwi/aG0iy0nVdXsvC3jnYqXWi304iW4l6FrR2OJVJ5CZ3r0IIwx+KfwH+D3xssP7P8Aij8PtI18KhSK4nh2XEKnPEc6ESIOf4WHNftm/wDBN+8+C2g33xU+DuoXms+EbEeZqemXfz3emITzKrKAJYRn5iQGQYJ3DcykBhgHNf8ABN8Efto/Dr/uJ/8Apsuq/a+/5Nc+KnP/ADKl9/6KagN75JwB7V/wTgC/8No/DvB5/wCJn/6bLqvH3jPRvhz4I17x94haX+zPDunT392IV3O0cSF2CjIyxAwBkckV4a/4K++PLn4i2o8U/DXw1beCJ7wJNFavO1/awMwG/wA4v5cjKOSPKXdjGVzmr6y03XdLuNOv7eC9sNQt2imicB45onXDKR0KlSR7g18UPCaeA/iZ4s8CxsxXw7rt7pw3HJ/cTvHyRn+5SxjbyeR2oqXYADpRAIxnNbB1DfpUUZc4U0qlWAI5NBVHA4pcYGecUoyetLHFgEyfpUYhJADZ9ODVxFBcvttWyfTB/rSI1u5VjyOoryCWCHrUkMYk8pWy30qSI55P6Vb7MhgcnsMVHGsp+Ybf1qK0iXJjfcT04xWnRBJ1Vj+GPY1a2Zmt/OcYz0H44rTbAo+8r9Bn61NalJWKjjt+VRRBoTtX5u3NK5ChcVKsVzbQqvDLuz371Hbqi4J5HQVJEXbIPB61YxgSZjOSPwq2ha7usu2ZOy4xnivC3hfVfBetRT+ItM8oPnaPOVs4Uj+An+8K+H+h2kdjqtxcSb5Y/J2JtIzksDyD6VG+o6fOxaf7OjYwuxX7ev4/rQ8K/EDQLt/FVhpeLWHGZfPh4yNnQknqSOlfB/4t6BpkVxqvjO++yOm3Y3lPJ13qeI0Pqvbv9a+O/jGy8Z6kNcsz/o8n3JOecLGp4IBHK46U1ysz7VOPfFW+ozRyM0bZ24xwK8PajrevXMGiac/mS3W7EeFGdoLdTjsD3rUvElx4A8KQeHp4fLuU3bhuB6yBh0DDo/rXj3xb4NtvhDo+mac+69u/P8wYlGNtypHUY6ZrxP4x1CKI2ts/yemB7HuK8TX8l1csjHJTr7ZAqSRidrc+gqSTaCENO+5uvNOSWx6V+7Rcsc/hX/BIQA+IviZgf8uOnf8Aoc9f8FZzj9nHw7z/AMzta/8ApFe18C/Hp+F3xk8GfENpWSDQtZt7i62jJa33gTKPrGXH41fWWmeItGuNOu1ivNP1O1aKVQ2VmikXB5HUFT+tfsufD+98Fft4eG/hxqglN34d8S3lvIVXljbRzEN/ukIGz6HNftr3q2H7KfxKnNw0O7RWiDAkZLuibeOx3Y/Gv2MPiQnwv/aW8C+Iri7ENjc6iNOvmdtqCG5BhLN7KXV/+AA18QPin4S8A+BfEHje81mwni0HTLi9aGO6QtL5UZYIoB5LEAD3IrV9d1TXdZv9d1e6a41DUrmS5upn5MksjFnY+5JJ/GvhRex6n8LPBupRBgl14fsZVDjBAa3Q84781+1hZNp37TXxRgEboG8V6hLtcYP7yZnz9DuyPYitAd/+El0hCOt9D/6GtV8Tr6W3+IfipI25Gt3mOP8Apu9Wmvavp95aarZajJb3ljOk9tNGArRSIwZWUjuCAfwr9k/9tf4aftE+FNN0/U/EGn6N4+igWPUtGuZRCZ5QMNLbBj+8Rsbtoyy5wR0J+J/7Pf7JkusXHxf+LHw98GW91AxmvNW1TbbwyMBktMCyxytgdXDE4r4QfE7wj8YPAlp468ApL/wjtxcXNtp0rw+UJ47eZ4DIidVQtG20EA4AyB0H/BWCWKT9qWBI5EdovC1krgHJU+ZOcH0OCD9CKDAtgDpTMBwfwojdxmicY5oqH5U/SiwJwDRIYALUXlkbieR2pyCcigABnP0rdjqatljK7i3PpiiFY4zz6YrgnANK25QC34YqV1lUDPK9q3KTtHNW5QAqTz2FTyKz8Nx64obTyrZx7UzqTnP6U5A61uOeKLc5zzQbIwDzUi4ODSqCcA0/zHap4pUBIDN+lPDtAOKcIQADRC4xmgARkV8NdLtNT1KSO5bCjGOD/df0PtXxC8D6W8T3mkXHmfZMeaNjDbu2gfeb69KIBGAaAwAM11FMMjAFKMjkUMY4ojtQyBjFAcYplHrWM8A104HNYzwaI4wtHjla69qxnqKAGOetHgYxRHasjp3oAEEDrQH40VHpzQAGMdawM5AojIziscYFY3cY+lNgjFbRjFNtHA5oAE4JzTc8UCcZrjOc/pR2nvRbIwDz6USCcgUeRXB6mm6bQaJGMA0SOmaIAGMdKbnkUADxRGOM1x9a+XPSgAeP0o81kdhRAIzmiBjijg9K6Dg1xiscYJrj60OSBmjg9KJzxmsAjrXXtVKAeAKHtRGKqkYn5SaOSMZpjg8Gm+YAg1kEcGlYjkH9KVz3oOSOucUGOAc0hLNgGo5Cv+FW84eRdx/SpDJZZ2t+n+fWlfzXDHt0FTEzMC5ziiitgZ47DFbFUetBeAaxF5BO35+xz70V+UHNMhbG0UBwKAzxmsAjAHNZJ4BojHHWsA8A0OTzTYJwOgogdAKIAGM80QBxnmsg8A0T2FEgcZ5rdlgD2rcPukUwQ4Apju7/AEpHCtg0JFJwKDBu+cUcEYzRwBjNBfU/hRAHANSBh8wPT2oMGGCKHyjIbP4Uj5XbnmmJJ2g5/ChkjB/KiMEEUWLfJ0zQK4+YZ9KwEGQPwokEDBrrRwBwcnvRIIwBROBwKB5wKIz0ogY6VWBjGKYhVximIx8p/CgAoBBodOaBB71A5BABrSbpchCfr+tfDfx/c6HHGIvuDPHH+3/sn1rwZZ6d8TG1a6nT7Oi+TtXLPn7wPTb/AHP1q7ig+G2lvod3dfaJZMbTsKYw27tu7OO9eJ9In8QeDk8T2kmZUz5o2jvIEHUjsOwp/Guq6fFJpsUmEbG5cKfcfw+9fDjxJqGmGaG0k8n7Vt8w4Dfd346g+teAvCd1P4kbU72z+0QnGT5gT+Bh2OeuK+Lx0vwra2ljo6b3k3+YMsNuChH3s/3jWuWkcmqxyta8c5G/r8orxZ4/1W+tm0qyby7FfuxfKccqx5K56jNaY8cF0jFM9c847GofHWjW88TWdj5bpnJ81j1HutfGXxx4Z8aeE7HVp/nvbPzNqfOPvSRqeQoHRfSr+9aa43pHtHYZz2FWF8LY/d+Y9efrXw60Ww1x/Pv7jbj+HYT/AHh2I9BWqaydVd4mTOMc5+nsPSi0tu2IRnHbpU17PdJ+8PPrxX2CSG3W58neq53HdjvirPXJo3cW5xuxkf8A6xWpyNcIzzpjOMc/T0rwhb2C3plvl+UdOT6N6fhXiddOudUdbJMdOctz8o9atPBt1JpyahDHuRs4OQO+P71XUUiXByefT8KsfDVlY2RuNdt/M3fd+Yjvj+E+4rRNEl1O5ihik8vzM4OM4wD7+1axZXmgzrbNcbxzzsA7A+/rWg+GI7jT4rfWodkD52LuznBOfunPXFSXHgyz8O3+j22kYuW8vY/2iX+/uPB46e9XGmakdPFsJMxp0G1e5z61pUh0E+eY9yD7wzj1Hv61qCW330XA7DJ9qa+EcG6JMMvQ596lDI28Nz9K07UMqYJBkD/69Wvgk65am9s77ygv8Plbu+O7D0qTQLqRiIo9xj+9yBjP41a3U2nSB4zyvbj/AD3qK/h16xd7m33TjG1t5Hf0GB0FfDbwrL4g1JdPt5djnPO3OPlc+o9K8N/CO5SS1nvbzMUu/A8scYyOz564rRvhf4c1u0mSaXLWu3YNrj7xOf4h6VoOoWItrmK4j5+TbyfU+lfEPxjEvhG80W3t9udmW35/5aK3Qj+tahqE2oT+ZI34Y9h/hU15KVEYPC9KQeYcEZrTbPQpdEupJFxOuzacvz8xz7dK1fw7PplubmQbox1PA7gep9auEUgLHzntWwL8oXn61Y23mMI2NXkUMK4Ucj60FBQEdfSnjGAcc9qCgjr+FSxK4wy/rSZjOzP0ppoJdLlTGHTG0892qVnj5zTzM/I7dBRz3p8kbc89qUAgDNMOwNFdwwKUleAeaXleRknpQWNl3BfwpX+YOTnHQelSoSdxokGUBTmkARxg/hUoOxdh6ZqQADOMmtq9Suc9Oa8sEbj27VHbyTyJBBG0kjsFRFUksTwAB3NZIGMc1+xT+2H8cvAPxU8H/DdvEeo+KPC2v6vbabLo1/K1wYFmkWMPbu2WjKbt2wEIcEEc7hq2lafrulXmiatax3NjqFvJb3MEgyssbqVdSPQgkfjWoxQWl/c21ncCeGKZ0jlH8agkBvxHNf8ABODn9tH4dgH/AKCf/psuq/a9Gf2XPioB/wBCpff+imrBBwG6dsV/wTg5/bQ+Hf8A3E//AE2XVftejP7LnxTA/wChUvv/AEU1BAeCOa8Mj/imtJGf+XGH/wBAFftNbT+0t8WAT/zPGtf+ls1Ft/JPNZyQoNA7Btb8KVA2XzwtZ3cr1oDIBJyaC44Bqys45oZZZWwV27Rjr61FEFfOOBUYjdsuPwoRxngnj0qOTLBUOFPQVdi3aERwJ8575NAl12K/44oQIsTsW+ZcY4qOKOY7dv60lqYm2jkt0qNto2kZHpUcQkjUR/LjOR1qztYxhkXn0qzk8oiBhjHaogq4AHWly6gAcDpQUqQc/SrO1jl5Y5xUUMcSgoOexq4Tam4Hn0qEKhJA59KsDlySK8LW0dzqto0r7Ui354z1U14x1n+1b6OaRvMMecHGOoX2HpSXz634YtreJ9hi34XGer+vHpWnv4l+H10utsfLZc7fuHsVP97+/wClaj8Qdc1y3NtfXe+DsuxR3B7KD1ArTLzT7jSZLueTatvj5cE53MR1FePPiFrPjy9U3p3gZ4+UY4X0Uf3RWr+FtZ0LT/7Tul/0f6r6gdiT1NSqIv3atkj2rwBcGy1CK9ifE652HHT5WB9uhrx14ouNUut18+Zu7YA7L2Ax0ArVNVDWNzKJMiPZgY9SPatZ1JyxjjXJ+v09q1ZjOVQNyc9vpV3EkGC5zUm0ZUDp0qVsnCioFQ8OalZAMJ1rRfEviDw9LK3h/XtR0xpwBKbO6eEuB0ztIzjJ6+ta34v8Xa9aLY674o1bUoEcSJFd3skyq4BG4BiQDgkZ9zW3YeTz9K07UvijN4Xvdbs/H3iGKz0pYl8pNSkCgM+xQB5gwBjoB+VDXvEEGsHxCuu6guq5JN8LpxPkrtJ8zO7lTjr04rU/HnjjVrKXTtT8Za5eWs4AkguNRlkRwCCMqzEHkA0o28ZzippARuA/CiFJAJ/Crf4kfEC2tY7W18eeIYYoECRxpqcyqijgAANgAAYxV1qF9q9zNe6lez3V1M26SaeQyO59Sx5J+tRu6qGDlXQggg4IPqKk+KPxJC7D8QvEpz0/4m04/wDZqmllu5HmnlaSR2LM7nJYnqSe5p+AQT+FfDv/AIJp/Ff4q/BHwx8XPBXinQ/tfiC2kuH0XUxJbNHGJXWMpKodX3oqvhggG7qa8Pf8Evf2svFmuW2m+MX0XRNMiYK9/e6wt2Io8/N5ccRZicDhTtBOMkdR8Jvhr4b+Cnww0D4aeHJX/svw3YiFZ5yA0pyWklfsC7s7nHALHGBX7Z/xb0742/tI+MfHGhXQuNG+0rY6XKrblkt7dFiEi/7MhRpB/v0uCODROeaRfm+WiA3b6Vtxxn8MUTg4zmgFIyKyAOaUhhgDJ7Uy7xt7jtRjGMYpVUDGc+1OuDuUcCiQRxSspOc005YbCeO1KQBxRPOM0CWGcc0CAABRZWTGOaKrjbmmIAwKBUdDWABkmiQeM/So1DfLmnCk7VPSljOQalbnbnpRAODnPtTLkY702DxnpSuVJCmgRgjPXtQRRytTCIou0c85ogY4FEZHWl5/DpQGRjt6UADxnNZGM5oBQpOeRRJBokEcUQD2rAOBmjgD5aySME0fu8CqJwOKxkcVgYwKPB96ZyTtBokEbawM5PNABuCKOMcih82ADRIJwOtEBgMGsZXOKK5GPSsbuM1zjJNOQ33fyoAYwBTADpRHOKHFEcVx6fhRxgYoFQRzRYYwTWewNEcYzRIJxmuQMUAMcHntRwelEHjIo4IwKIGODRHbNYG3rWTjANMMAAGqwCOKJ3cCiBnAojpRHA5rqK2miuKPTiuor6UcngmiR0HWiG6AUQQOn4UrZGDTjIyRQ3AfKaIOKIyMA0WYAAUsjAfLzQmYKMn6VHOd3yjJ9KafJwDz6Us/IANXGozzDaZtw/3QKS4VUUk5PPapL5pMALtHfnNOwRU3P1z2qHZLICXxj2zU0iqNoOce1BIFUM03/jppW3ruP5VbyJG+WHFXS24kxbybl+hH86aC1dokhl3sd28bSMelGBmciEbgvfpTctgj8KOD1bP4UFLjcooEAdaHA4P0piCODRYKMGmkQjg8/SgSFBA4ppTj5TRYuB836UZSpwW5PtTyju34Yoz7TgH8MUkqMMKefSgSSMdaJHY0km3oaE65wT9KEiEAA8/SgfU0RuxRAIwaY7DjFLtdguOlMi7AQaRWDcDj1oFSMjtQzgFTn2ogDkjBoqSNg7Ud7JtxmhgjOfwr6UMkYHNABjjPNBVzgmioOADRA6Zp+cAHNVjPApgCKKjb8o+lKRjAOTRII61gEZzSMM4U/pVjMQ4WM8mtC1MxWsaKvTPf3PtXwNtrLW/CtythqPl3ibN6eSTjMkmOTgdAa+Lmoa3bakJb64+0Y6PsVMfKnYfhXw88cebo+p6HqWsfZYJfK8pfs/mbsMzHouRzjqe9XNxZL4gkaRPNt1xl8lc/J6detaTZ+EvEWmz6npEHkCHbiPdI+csR1bHoa8PfFNtAj+wyxeTZn/WPu3Y6kcbCepr4i+NdM8UeJpdQ8KXP2yzkxsbY0fSNAeHUHqrflWi+FNW8YTyJYW3mSxY43qMZB9SOympmuYbnzHs9zf8APPzAM8etXOmajdTecbD7GrfdHmrJ0HPf/Oa8B6BpOpXTprt55CLj5vLZuzdlPsK8TXOnx3RtdJbzUi6HBXOQD/F+NQ+A7DWvDL6va32LmPGYvKPd9vUsB0GelWsm5JJLmLbtxtbdnd68DpXg/WpNE0KV77TdsnG2bzs4+dv4QD6gUtm1tIRLBtU992c14a0vRr/UFh1S9+ywnOX8tnxweynPUD86uY7ZZSY5sjsNppPEk0Vu2nSJujk6nIGMHPpVgI4ZdxOFrUoNPvLCAH5PvYPJzyP8KtdPs3xFcyZz/sn+hrSvClpd6ssXm7Yudp2k/wAJz3z1FaTq8mnyP4ce4228WNp2Z65c9s9T61rWmwW2oujvmJsbXweyjPGc9aa71bxgy2Cr5mzOBlV68+392rfRofD0aaHqcH2HULXO5txlzu+bopI+6R3714olee88ua683b0bZt7DsK03xL4m8TQrbPfbxZ52L5SDG7JPYf3a0i+iOq41CPLp15PPyn0/Cpbt7SRY2g3RHO75sfSvHIsFsUGnx4DZ3Lk84K46/jUmkNNCRCNzDqOn9actb7rd15PbP41NKDJtRc/jircwROsh+Y8/LyM0niy8hixpyfYoh1TIkz+JGeufzrQ/El4++4D/AH8bhgc4yPSj8MvFep6HL4qstG8zS4dvm3H2iNduW2D5S245YEcD9KZNX8PSPZXVt5anG071PuemfWvh7q93o7rqOn3G2cdDsB/vDuCOhNeDNX8Sa5cxWBvfO2Z2r5aLtyGJ7D0rT9e1bTNZbR7eXa748x9qnGFLDgg+vrWrv/ZyM8abguNxzj0/xrxh4qm1AyadapvE2OM4xjae49qu7IQSYC8fWp4UQYjbPqMYrEcEO4nn0rxL4dn0TRtE1VuBqf2jA458tgvqfX0FXGv6hrtj/ZLrkP7jsd3oPStb0yfQ79rO5TaV6cg9gexPrUud4ZTnPSvDdlZh2u7ptyrjC4I9R2/CtcsYZpQ9rH8p7ZPoPWjG8SbXHNMSw4OfQUCV7/QVuZuC36UxUHceooHA2huvXinRZPlNSqiMFVsnvxTAZxmpAucBs+hximOBgV1HFJyNpOBQVD0PSojuYL60FI6DihGQpKLk9hmn3yjG3B9M15aoMlee3NI8brtY8+lMwCblbkdsUxBXLHn0oICACc47Y6U2Puk5Br9hvw94f8Q/tWfD+PxHq1hY6fp2onUWkvJ0hVpbeNpYUXeRuYypGAo56ntXxC/ZK/Zt+Kl7Nqfjf4O+Hry+uCxnvLeBrKeZj1Z5YCjsfckkV8Lv2Rf2cPgvrK+Ivhx8KtM03VY8+VfTzTXs0ORg+W9w8jR5BI+Ujgn1r9uj9t3wJ8JfAOufDjwD4ktNX8f6xbS2Pl2UwlXR1cFJJpWXIWRQSFj+9uwSMDk4AyTX/BN8qf20Ph2Qf+gnjj/qGXVfte8fsufFMg/8ypff+imps/eAr/gm/j/htD4dc/8AQT/9Nl1X7Xhx+y78UyO3hS+/9FNSMWcAmvDGP+Ea0nH/AD4w/wDoAr9pwf8AGSnxYOenjjWv/S2aiQVzSEKetbzIcAZ9O1JFgYDcnqMUkQVeBQhJxxVlpzTyBFXOe/8Ak0dNcbmBzGn9fxrygh2E/SrWG2Fwolb92epwfStQWzlKLYjcBnceRjpjr+NW0WUyV+gzSSMr/IPm7e1AOvCNx9KMNy6Axpu9eQKQzRynYnP1qKOaSMHHzevHFQW8inaV3H0zitOilMrRzR4Axjn2NWdjbxgM3H500EUsqt5GHOcnd7VDEgxuTkdOaSI8gDmordpDkjOKtoY4oeevYVa2Uk8Su3CjqfSmQPIRjIFS23O5BnPatD0aKRybkYHb9fQ1eJFp8dvHbnLndv6/h/OoJr6PNxMuYx3yBjt2q08Tajp0xaCTMafc4XuOe3vXi/4o+IfFnl219c7Y484XYhznaeyj+6KijuLawju5eQ2dq8c4ODzU2p3fktC77YnxlMA9PfFQXc7kmzkwT/FgfyNa/rnjCbSorDxFc5sX3cbIx0YH+AZ64qS9jkutick9vwpINK0zSo723vv9OOcReU397HU5HQ5qXVL/AFy7LSNub6AdvoPSvFN9NYWU8I4Py7x6crj+dahqU0kxKNj8Aew9qngMzmXfgL1GOlaoscvVskdOMelXCkNsDc9xinQxuAO/SlIByTmjg4A/KkiAbeDyaMan77dO2KZdzZz0qDxhr9pol3oFrqHl2V9s8+LykO/Y25eSMjB54IqVnnOd2T9KIO0A8460Tk8dqJiypL8tnPB4qZYwMg9PaiscJILZz2xQcLwoxmpnaNgoGc9auyigbBk1IQvCNn8KcZ+Ynivgd/wVY8V/DPwXovgHxb8I9H1vT9AsILCxn0y+bT5FhiQIm9WSVXbaozjYD1q8/wCCxfghLMPYfBHXJrvb/qptYiiTOP74jY4z/s/4V+0X/wAFHfjT8etDu/BWmWll4L8L3yGO7s9OlaW4u4z1jluGwSh6FUVAwyGyDiiF6Z6UePu1ntmlfY2UPNM2DgH6UdrL15+lEFRwM0HzxRII2k5pCFIIPNIFfJD8/SixbIZufpTJt56UHwhWlVCuCahsri5jd4E3qmNxyBjNMGUbehoegrGeQOfSkIX5mGfapXVmyoxjtRkIGFX9aL8c0WLDFYPQUAM4IogHgVEMNg14jX4fW/hDTIdKu/P1VfN+1J5cy7f3g2ct8p+XPT8aZ2YjIwBTZPJ5NAE/d7V160oK4J/KmUkZUYoliMAUr7eAc00bv84X5frTDBwDzWPl4ocjAonjkUqkjANEdqb7vFZPQ126fhQ55FMAFznNZB4zRIz1oEng0Tg5FDlaz2zQwQcNz24pmz0NAcAAUSD3/Cs5GB+FAn1/CuByaDdgax70QMYBoEhcA5oc9T+GKIQDAPNEA4waPzHAonPejg8k/SiuevPpRx60SDyDQ+bgijjPBorkYxRB9KAJHSiRjrRG45FNzxWRxWcdDzR5b1rg1jPSj15oYPGelZ4xmiAeSaPQfpXbGfpX0NUxBAxVUcjmsds1WOOtYBHWiPQ1R571X0NEk8Zqq4NYx8ooZxwaBXaARQZVI2ihIpfrUciKwYHNK2eDW5jjmgxGMGvMcnaW4+leZIjABunbFG7cHY3GKilD4Ut+lC5Kv5Z/Ctyk4FW01s1oI40+dOvJ9agLFWYN93HarTUprFWji5DY9P8AD3qNrZnUkcnO7r+FYXOQOO1ReZJG0US5z15xisEcEdKI5wDVMoK5BpYi7bQakkEZ2E9OlPMoOPyo3ZJwtPOzHOfpTTgcMeaMwIwDk1HcMp4OKF2rY29qjlV1wOtCUD5T1FFgRkDpW84HNJMcfe/SllUjFAhhlTToGGe4ogjoOaDEDbQY7dpPI6Uu7+GgmTgfhTFSQD1FHLNjd+lIoB5pQNoAoDPGaxn5VNEFTtPWiwPenGQCvaqIGMflTKetURkYFDnjPSgtEArkGuvGfpSjio5GRtwPStM1yS2QDfyPb6+1eDfi1rPhkSyadeeWZMZ/dqem71Q/3jXhvxjp3iy4aLxDLvzjB2sOx/uAf3RXxGXw7od2LHRm3SJnLYcdQh6Nn1NeE5PD92ceIY/M/Fx/e/ufQVYapfDbDBLtxnjaD/SlutRvF8qRt+O2FH+eleDbe4bSH1KxvvLLYzH5QPRmHU/jXgq08SR3D3+mT+VOcYbYjdmHQ8dM1q9ndaXqQW/XEidsg/wj0z6ivhTrvh7UtSWDxGmRHnYcvxlXz9wey18T9H0vR7uVtLk/dnGBtb0T1J9TU1uzTqoHJ6/lU+rWFn4WMOnHF4ep5/56e4x0zXhjwzJ4itLvVr0b7LS9nnr0/wBYSq9CD1A6A1rWszXentHYQ7LPudwPceoz1BrW5LkQqCflGcjjjpTak8iCOFuR04/+tXSXao57/lWoWYgCs4yWzj8MVHK4jDK3B7Yoajvt4oF++M/zrTr1rWVzAuAMcZ9jUd3Pqck8sj/6vbgYHf8A/VUNr4KTwgtxLBjWTnL7pf8Anpjp937n+c1qV1M4Rd2d2eMemK8H+LL/AMJ3vnWTbJG9gegYdwezGvEvxAOvpJPfv5l3cY3NjH3cAcBQOgFTWazRm7Z+n8OPwq1tr2zlN1att3dRgHtjvWr2mr6atrd6hDt87f5Z3Kc4wD0z6ivDUus6jDNHp1t57R7d3zquM5x1+hrxFqt7qRXTbgbZBnjg46HsPahp91asYnT5h7j/ABqTQp9Uv0ijT52zgZHYfX2rUtFmtZ2imTDL7+w96QMzBQeBXg8+B5IJIvFdp5zjHl/vJV7tn7n/AAGrWyS2kKyDKr0NeBviHYW/g248Las/yzbcjB7Ss/8ACv071E8ur6TZ6pevuebfkYx0bb2x6CrDw7appST2iYY5zyf72O5rwfousWaR6jbjCyZ2t8vbcDwTXhH4e+ItTvFvGODJ0Pydgw/ve1eJfhnqWkXctvdw5EmM/MvYD0Y+or4keBtO0DS5r24ttjfL5Tb2OfmQHgMfXvV1C8ilm5/SmtmQZ9egqQjftbt0rwr4X1f4yaVDodvcYOgbvJGxePOYs3Ur/wA8+5P4VZ2L6ZqhtpX/AHkft6rn39a8XIb29ku5pNzHHOMdlH9KCqBgirLW7dbWGz7pu3Dnuc+lR3ejRWoEKfvW7Zbsf8K8U6NZQwLPbv1zxg+qj1qRIypUdTQB2gE0AM4NBN3AoAZwKwCMCnjRmJPU0YEAyO1FOdh60y87aMfyBlOfamJA2g1HGzHC/lQRiTtHzCgzHAAzXzoNwPPalJxn0oOsx2j8KdNpDL1FOWIGD9aZmYYWghLD3pkREJbqOlJGXXCnmvCnxy+NngWGOx8GfFzxjottEMLbWOt3EMQ4/wCeavtx+FeJf2hvj14ytZLDxV8aPG2qWky7ZLW5164aFx7x79p/KvNYOGI5FBhIMOM1oeva74Z1SDW/Des32k6jbbvIvLK5eCaPcpVtroQwyrEHB5BI71qXxj+Lus2E+k6v8VfF9/ZXcZjuLa51y5ljmQjBVlZyGBHUEYrknGelaHruueGtUh1vw5rN9pOo227yLuxuHgmi3KVba6EMMqSDg8gkd61D4x/FzWrC40nV/ir4wvrK7jMdxbXOuXMscqEYKsrOQwI6gjFHI6Covjh8bIo1ji+MXjdERQqqviC6AUDsB5nArUb2/wBTvLjVNTvZ7y8vJXmuLi4kMkk0jHLOzEksxJJJPJJplxxS7DgsM+lBNwG0fhUVvn5j/DUWi3a232mSLCdjuHrj1qz02a4R3jTcseNxzjGenesmNf3fUUJZGAjU8elPakgFqtrZZz5WMlajtSqkY+VetSOqACIVEN5APJpFKDLioblgTHHyG7fSm08MMRr+v/168uWEhUGAat7GXh3XB9c1BDHNJlV+devPWpbO0ht8ocy9uD6/4VAilNxH0qCJnPH4Ulm0hJB6VaWIJIIqKMSNtQ/KvSirtAwBwo6DGe9GFEGAMikjBbBGQO1G5FrEHVufpVj8JnufAY8YC43j+7sxj975fXf/AEq9099QgYGbCDGV2+/19q0PTZ9WuxolkPMfsOB2LdyPT1rxR4C1TwxOkF+m13ztGVPQLnox/vCp9bnmbyJZchfujaBj17UbiW9kWItuIzgYxj/OK8K+N9G8C2t35kf+k3GzB3N/CT/skdGrxJ411Lx3aRQyNuSPdkYUYyR/sj+7U1rLbQi5hGSvT+VaXd3F2kElw23du4wDjGfT6V4itm8J6ubMS7m9duP4Qff+9XjXUHuIp7p2yH2549Noq4i0aSy86Bf9I9ct64+nStSdv7O+0Acyd/oQKeYyD5jVw/71iG/T2piZcCtA8MtrSTssu0wbeNuc5z7j0o2W2fyE5cdB+FeMPhHo/gfSYZtSuPMvH3ZGxhjDL6OR0YVcSIwMiHjsKLEDgUpJwGPFap4xa/8ADY0LHA/+LDent60xJbbu/SiB0qVNxBJ6e1BA6bVp12xhietCUlMMc46U8yuQU6inBKE+tABhk1JyAM9O1KflyDRbsRQwOCaDHgE0x54NDOMmgM8A/hQPGKYlmHOPSgxFMCQWx0rPGAM0o9B07Uint+VAc9elY38qKcZXilJKbAahvGtoWhjOC2N34UfmOcc0cA4FMCelZzwDTKc8UAAcZ5oMQCBQGf8ACiBnkVGc/KKlg8nndnPTigACOakuGkYsTn0osTgE8UACdoNcRjIFEAnIqzsHvCUV8Y9v8+lSLsfyz1FBQOp+grA6ZoavMumx6en3I84HHds+lHA5Uc0RkYBpRgcmjg9K5xjNHgUMEEH8qAJGAc1jPOKPHAFEE8Ch7Vt3HApkwBkURxxWCRgdaA4HFZz060oGME1k9D2piTjFHJ6mse9bcnANNyMZoHHGaJ5yDWSR1ojIwKMeRwc/hRAHGKBGaIzz6UTk4Bog9B2rbxjv2rAI4FDB68mhgdaJwOKzkda3NjAP6UQcYx+FEErhRROR0ogFcVxRHYCvYDNYOeBWN3QU67Tg9aJ4wBWQRwOO1YzyBQBxt70MY4HSjzwBWCBzzWM8A0FJwMc0RwBTAAYHJ7UcEYrHGM1QxgbTWccE1gDgVz0ojiuoo+wrAAxmhz0o8cVnPWgQehrqeaZgeM1HyB834YoSY4xQmQAFuKEu45zgdqadDjD8/So5tzA46dKmuPNkL7cZ7ZpJwnNJdqXBK/hmnvEYqNv61b6sIgyRrnOO/wD9avt25WQttJx2zUM9s0XmBsj6EVJLHNGoiODzg9c0LhR8obOParO7SBi4XdnpzipGDuQpyTTyRwSGKVsMvUYqWVVXNJNHGqpjJ54qe7cS7ozj079qkvA4AB5FPPkcUZkAznmmuBnj8qacE4A59KEwA6ZPpX2j/Z/WlnRRjHNR3JQcDFG4RiGA5qK4U/ffH4UBG6gqcj6UCMglvpxQ4HBqJuMbvwxXBHBpyS21eo6U25W+Y8/Six5OaEuBgJ+tAswBAoRtnJb9KZQ3KnPtiijADa36UhJQc5/CieOtAnGRRORuJya2k9TRztwTzQJHAGaOCeDjFEjbgjNYz0/KiOxNEY4BrjGA3T2oAkcGgM9+aAPSo4y5wp+tBiDgVFdOg+Vv0rTdae0OQ+M+319qOrrdztI77mbvjHb6e1LqHltuA4rTNVjS6WVl4GcjPXg+1aV4ogtbzzVg3j+7uI7H2q3nhtZAsF1sPceWTWm+N/DOnaEbeTW/KeT70v2aRvLw5I4285zj2r4peJ/D/iXxXNqWi3Hm2bbdjbHXOI0B4YA9VNeHfFPgjSNEuE1DTftV5Pt8tvOlj8rDHPABByCPpivF3inQtS8Oxx28Xk6hzu+Z2/jXHUY+6KtYJ4pBIzecG6HAXpUIM+6Mt5S+mN1aPqes6bpN7penzbItQ8vzflU79jEjqCRgk9MV4f1s6x4Yl8GR+E911cY2332/Gza5kP7vAByBt6+9eJvCd34cu5dE1aDzLlscbgNvAb+EkdCO9Xvh17KYnOD2XGe3rn3rSdT0rT4JrfUNI8+aTbtf7Qy7cE54Awcgita0CO0sIruW+3sc7I/KxnkA85p7WCKFXSTrnK4PH41cQtapDdpFuX5v4se1WNz/AGjG9wLz7IOMDy/Mz2/p+teH1eyP2q/i/cTfxbvTI6DnqRV9Y2t/oSNaWOc5w3mn++Ox+lXEElpqEUU8e8jdlc47eoq5WeScSsNuOg4PatFHnzBmst4PU+ZjHBpbIvqJjjXK/wB30+X1q80HTLa8eC6vPIIxhPLZu3qD9K8UeO9G1q2s9Kvl842G/wAp8suN5BPAUeg65r4dfGSP4VXN9PF4d/tVNQ8v5PtnkeXsDDrsbOd/t0968R3z63dyawYvsjTY2RbvMxgBTzx6Z6d6h8UWstyY759pbq2CcYHoB9K+H/w8vfFrz+JdBn82z0rb577Au3zNyLwzA9VPQH8K8Z3On3GoSQxriVcZ5P8AdX+lW/h64uXW3tfnlfOE4Gcc9SfSmtZ7WQrINxHQcCra/jaMQyRbin+1jrVy8DHc77T6YJrwuss19BbyNvt13bUxjsT169asNBuTp0bR2mxecfvAe5968E3+lW/h+0s9StfLjbf5cm9m6OxPA/Ac14a8Ux29uttBFujTpJuxjJJ6Y/Cvi548l0m8EF7pfnNJ0l8/bnAT+EKfUCvjP8QLHxLpcWlS2OxrXOT5pOdzRn+6P7tS3FlLpqiO3wf7+8/3vStQIKxBVx97nP0qaXL4T5sfhXgrxdrPhG7nutJu/I8zbv8A3atnAYD7wP8AeNXd3PNdNdyPudsc4A6DFXN7JMoQnLHpTTlDtfr6VEDHdFmbA7DHTiotTNuwmK7tvTnHt6VdXs1yu2VunTgV5JyGLZx2xTMcfKN36UpyeTz2FKNvJf8ADFBd/TtWMHrzUh2jeTnH6Ucn5yePTFSMHXGNrL0HWj8oGUz75rJU4U0SGyM9KSQ7s5pHwNyfNnqOlKWVsgcU0yEEodxXt0oM7nCvn2xWF4KDBXtSneNxP4Vj96ygZx2/CiQPkBwTS7RiM/KR0HWnQSLsHWgFjOScA9KYxggjp608BU5B4qUKG+9u/DFCTC5FY5yRwOlKQMjOaIzwo69BWM8A0AegHPrWCe+ajjUkozYx0GKcs52AcjpSxF/lU81HbuGAB6dBWm6DqGqhms4fM2Y3DcBjOcdSPSktzEWRx6ZFXHiKS40saathtUfx+bn+LPTFWd5Jp0E1tBLuE+3cduMbTkfzoRF0wYse+f6UEt4IwY5PNI9iuKMEkxDO2AegxWmaQ0yxyJdbPPzg7M4xn3rxHpy6ZYCGM+Y8n8eMYwR2yfWhA7HekmR/u1FG0Z3M2fTimTzEIz+FWVjHGBM03A/h2/hTz6c+npClz5Nzzn5Gbv8Al0rbGXji3eYDnLY24/CrzUrCZVSAbQM5OSf6VpccbkCSXaOzbc+varW2PnZZ846HGO1JbK0KsH/DFWyCHhhyaQgnKj8KtwrLnO0jt1zVvGY24/EVHatewMifKBjB69//AK1ajINPPlkZ9B+X+NC6VYDIY8HsM+9TgyxhgcD061oXxSv/AA/pNzo87/aoJtmxMBNmGLHkKSck+tNqyl2l8varY+XOenvivhd4x8B/DnQLbW9Ti/tLW7jfstt00Oza7qfmUMpyjg9O3rXj74uX/jnXP7UuLTywv3YvMB2/IqnkIP7ua0+zm1O4Sy021+03U2dqbwmcAk8njoDWtaBr3gmWB9c077O8+7y181HztxnlSf7wq5ee5ska4O6TnI4GOfavCCGC1llF1h124h2ff5b+LtjrXhHUNO0zWIZtYt/tFqu7dFvZP4WA5XJ6kGvin4m+Fut6D5vhKD7HfRf8sN1xJ1dP4nAH3Qx/GtX8S3epP9qnl85/720L6DoB7V4h1VL3R5Y0Hz8Z/wC+h7e1WrouTIdv61qFw82lxW8afLHnnPXLCpJEQ4K8/WpsZKk8ioyS2E6irDVLyy3fZpdu/G75Qen1HvQ1Fo5vOA+cdD+GK1/xPrPiSf7Rq9558g6Hy1XsB/CB/dFIwI559BUh5wprcWITGcdaZMnAP04p4iWyH6e1EqWCk00YwyEYzjFaZp19qE4tbKLzZOw3BexPc+1XELx5jf7y9qQOUwB+NYUnC/lUkpA2dRRAX5QelPj7o5pRgYI+lEgjPeiQRk/lTDAOOtBgRjrSEEgmmcbsr09KjTcBhc/jThS+McDtTAYwD+lKxZMAcVgDgGlHfP0qGQRnlcn61D8LPG0+ht4lXSM6fHjdL9oi4y2wfLv3fe46UXMZKAfjSqpBBkxj2rG04A6UwyOvWkgkk+5zRBXgN9OKOdvXmnweRWCR1zQUjGaCkDGevavlByDRxjO78MUATyDn8KkZnwC2fwxQBIxnOK2jOA3H0pRk7Q36UPl4bmi+4YzRJAxmorqWLDRPgj2zTNvbJbJ+lMdy7Qc0Rno34YpRjg9qudLu7aH7RJHiP+9kfTpmiQBQUAgk4FSRlT1zRXniiQTxRANZGOOtcsvAphtIBFEZGM0Vx0OfwojAyOfSi27vRPABPShxTDjBoAnkdqLAcd6Jz0NcZxmh06USBTYJ4onNAKFBzzRwR70TgYrIbgflQIP3Vz+NHJ4NHBwAentWOAMZoAjjFHrgCs45HNA5+7+VAH0pgSeDRUngn9KwMYFYBHX9KPTHpQIA4quKrJ65qlRmAry1XBzk05DYwaxx1+lY4zVVVHkYzVEc8mqJzRGcc1WMjiqOOmaPPej04rJI61/kV2wDX+RVDg5FHpxRPOKOT0odOaRgOK805yRmg5Y5B+lDOOtADNBypyBXn5GCf0rzl9aWb0NGdNvB5pLkDvRnVzuJrzliYFW6dsUdR8yRSq4C5zzUlwGf5RT3jfKoONtG9cNuVufpTzu7+ax5anu9w2k5o3QCYB5p7gE5J5ozLjrTSFhgGgxPGaPPQ0QBzR5IpmJoPjoKEhJxQcjoaW5IAUmoLgkYBpHLDAFLwOetJg9+tAgj5TTDcDg8iiM8GjgYxQcgbSKVmCBRSNleTWQ65JrJ6A1GAEAqgcDAo8/KOtZ5wayDwD9KGKZQfmJojIyaIUjA60TnvW3I4ojsaCgDAoAEVnOADUbYBI/CjyMZoEgCg+D1qKfY3BzRvfMXZ+VW0/kFgeasb9hcYUf5xX9tj7WgI45z+X0rWfElrJa+TCfm/H1HtUGvsoEfXH+fSl19ZF8phwOv+cU3i3SIYBBMmce7ev0qLxrpK2+IkzIvTlvX/dqfX9OmlDxDHqPm9PpWteO7K50jT9Mt0w9v5uTkn7zA91rw9f3Wl6b/AG1HNkRfw7R3Yr159fSvGHjafxz4lk8SumHkxuXI4wip/dH930pb63vLhXmHTORz6e30q28OXepyulunK47j09z7V4W0fTn1xLLxIf3C53DDf3CR9w567a8SWGlx3jJpfMXGB83oPXnrmrkXV3aJaxrhYc5OR3Of6V4f8G3+sWhW3fG3qMD1Pqw9KSynj0+bT7pvki29h3Oe1eH/AA/eXWgR3dsMxHOOg/jI7nPWvF/hW6lulvF4Jzxx6KPWhpWy2zj5x/j9a8N3M2lyG4cbon69BjGR7nqa1iaG7jNzb/Kv4n0H9KjuL68uNsp+ZvYen/1q13wJd6Rp0N7PJuFxux8oH3SB2Y+taVLvULImSOnP1rxDoGj3nwi1jxFbz4vNM8jbHtb5vMuAh5zgcD0NHwzcvMGeTBP+yPT61pXi3U/Beq+RbvuEf3uFGcqcdQf71a3qFjPd+fB1br17ACre/jtW+12y5eP7ozjrwetXE7XfzlcMff8Az6VDZTQxMWHJx6VGdp2tya8Gu41aKWFclc4Gf9lq8OeLvD2p6HZx38OJYd+Rufu59B6AVoOh3H9s2kqNug+fHAH8J989a0TUtJ0+7EssOVXtuYdj7e9fEjxbdazbfYr5NskXQ5B6lD2A9BXjjVo43mticyptwfrtPp6Vo2pW+sRW+geX8vzbmyfdumB6eteMrCOw1p9JtWykGNpx6qrd/rTQm4HkQplh74pYJoVlikXHTuKitGljyTUsCjCE5prYs2c9O1OExhetEjGwmpFJO0j6VIcjaprGBiigIGTRGehogkbR17UEJqRFk61IAThO1bSQSD09qGCuKAU8MaYYHH5VEQvUUvyqXPbtW5XTA7Um4MyofSlVg+BTKHGR1oKaVcMWB69KJHQnmnXftK9RW0kgj8qZAwB9KYea4UCo/LZT5ox6Cp1Il2g8f/WpwSelHJ+Un6Vx061yDwfwxR570ARwDmlXd8opdsfHU96CM4wKtYfMIjU4NfZBAeWyw6cVoutXWjQPFax7vMxnkDoT6g+tNFNFkyL97pyKtGLfKwpYFjk3seewpnfGWPParZ1jypGc1p9mdRulhD7c57Z7H/CrhWtLkwbshehx7Vc3s8kW2U5A6dKiQyYCngUxV2CqOnWpFKp5a9aTzIoQVOc9ulLIhhCuPmqJJY1Zyc+lQR71ANaWjKMt0PT9agj3EFRVvKSgjAqNCQCe1RW0kCB5B16VpUMMUf2pjkp06+4qRfNlIQf5xWlgQwmNuVPU/nXiaKN5AI/89KlQLBHH354/GlPnfKTwK8OW8Gpar9ilbCP1OD2Un+lJq8Wm6qbiCLfFH1+bHVcehPU14p1Fb26N5CcK33R6cKD/ACq3RrqPIbD+uK+EfxB0f4eD7VdwebcjoN7L/fHZWHRxXxF8ZTeOdY+3zx7I1+4M5/hQHoB/dq5VrhY7WIZ8rOfx5qGc27xqRwM5o6gkkjAL16c1etI1lKIl5OMc+4poQI9jcH0rVTteS0z/AK7GPwwaRI1PzHPtVywa2O3p/wDXq4y8pWp2+YoeopW29KSQBMCiSOc5oMHyKKkAEGpG3DBNArtAB6Vv2kAjmp5NjAY5PSmkVT1p5sEFahu5rZhLCeR0qTcCxc5LY/SlYg5zS5CkEc1uJIxR+bhT0oqSeD+lHAXANZBxzSgYwTR7DFKuSBWCDgdqKgjJOTSuQuKJ4zWMjINAYxg/hTqWPWiFYBQaABPWtM+J+o2fhq78NiLMdzs53D+Fy/8Ad/rUo3HJ/KgeMA0OelEevaorgxA8URk8UwJGM8iivYGjg/LmiecE02SQB+FIm/gGipHWgWCYrrxQ+Xv9KZSCDn6VjPQ10NA8daJA6mm5GBRAIwTmh0wPwp0YDB60ThdxPWrnW5LjTTYleD3z/tZ9KIyM/lSgNGQTz2p2ZgATSnkUR2ogZwD9KIPY/hSZIIJpYw3BNAKRg/hRAJwKCljwaZQeKxlcDrWABjNKFZeT9KPoDRBPANZzx6VjsDTdawCMA5NEEHk5pgCMKaU8YJo8jJ/Cjg8E1wDwc0MAYIphnAzRAPOaIJAwaxkcURjvQ4rA7H9KI7UPUVXUVjjGaJOcA1xQGeM596B7VnPQc0cDjrmsbuaD4+UD6U5LUVx0NEHvRBIwBR4GMfShx0ogYwTzWKPPSjyMZogdAeaIz1NcdqIxQ5HFUOKJya4PANAg8fpXbANEgDFUTxgCqIA6Gj6A1weBVHA/pRPGDxWPeg2QABQkfGCM0JQDwM00m7jFM2TgVkZxigeMY/Cjgjg80XI4xxTS56fhSyMRjNLMu3AHNGY9VFGVmORQnOPepJ2cDI6dKLt0BosTwDTI5ABP0oKQOv6VsJwAc0AR0P4UBkYzWCOhrb0GaxgYJojHGawSM5onnrisMOQ36VDJsG2oJWxkGl/eDd+VDjAqMnpu6e1FTklW/SnPbdn8KKk9+nSgzA4YUWYrtU8DtigW6A89qJwPWgisclsH6UmdvPWgO2KotkYFEg8EUQCMDv29K+nIosCAB+VAEr05rZzgmiMDr09qxkcH9KIPUGsdhzRAxjPT2oDA4Oah4yc/hXbANDBFcUAQRxUGZJFRRyelXlld2gEsyYDdDkGobsRuMHP6VLeJI25Rj0HWmnLDDck9O1CcZwDzSXxjbO3J9M4qe4EzbiMegovk8CmkOOKLMR16Vb3UkDbkP4V4b8bro8U8Ull5/n7efN27cZ/2T61JrhnuDOj7Qei4zjjHXFeE/Ftzf6Uuo2F79hd/vfuxL0Zh3HsfzrQNSsZtde68RzeaDj97tK5+Qjog+gq4u7SO8aa0fcjdRgjGBjvVvcW6Qz3CSbvM2/JtI6cda8HeGdfk3tpt1iNsZ/dp/tep+tan4emfUTYahefZ1frL5YfGFB6A/QV4M+JU+lWCeFra/wDs8EGf3vlB85LP0Kk9Tjr3rxH4ludQ11niT7VFa9HyI925B2xkcj9KuNWi1xNtlbeU46rvLfzA9KBtbNZ7O7ssTxbdx8w/LnkdOOmK1m/gaDyIYcZ7bj6irFjFqDLc3OVGOdnT5fatQ8Q6jqqx2y3nmxQ5wfLVcZwfQHqK0DTNJmiJWHzAvRtzD17ZrxzoSafE14t7v9F8vGPuj196vbnWWsc38Hlx9juU9/b8KljVmwJfmPQbaWModoHPpWjtJDKkc8X7uTOTu6YB9K8OeAH8b6kun6Bc7JBnPyZ/hJ/iYf3TXxD8DT+Ctffw3eXXnvb4/e+WF3bkV+gY4+8B1rVdNNk+UG4/l6e/vXg/WZNEvU1IQ+b1yu7b2YdcH1rTfFl1rzhiPKz24b19h6V8GPHFjo9lFZa9J51vb52NgrjcZCeFUnqRVz+0BpGl63Nb28X2i0l25fcybMJnoYyTkmvi9JYKsWqxeL/tv2jdmL+zzF5e3YvXvnr07Vq7vczzzRt5kSbctjHUAdOvWtJ1ObSx5tq+GfqMemfUH1q7nc3z3bybnfGeMdBitM1ybSrnz4o8k9RnrwR6H1rVtefXbiSYW3lg4yu/OOAPQelWSSSSiHGR+XatStns714JGyVxxj2FIBIODx9K0TRzrGorp/2jyt+fm2bsYUnpkeleJPDv9hTxwrefaN+efL2YwB7n1qSYTIFUbSOp60p2Lkr+OaChjkn6cUoJ+QCiuD1/SiUYgpxipCWOTXAOMdKdAy4I+lOgCeWDhaU4bAbj1xTKSQ35UybhgcgUBg4xmoQWOCvB9+lXC45UUgk243Yx04osYwAV/HNOCSCjdO2KbgZH5Uka53Z+g9Kd4wNrHmvL2kMH6e1FsMFAzmgpztLZ/CmQmRcHjvxQiKrtJ3fpUkaq65G39aEb9d2fbFf8E/fgqPjD+0toA1C1E2ieEx/bWohlyreSV8lD2O6Zo8g9VD1/wUd1z4f6n+0xrGgfDrwto2k2/h23js9Sm060SAX1/wAvNI+wAFlLrGSRnMbcnNBW69QO1eWx5xxSxkg/J16c9KjicP8AMf0pYju3M+fQYry3bhXx+FWtt5mRvwR04qOFAozJuP0xWnW1013GbeHzCM4TcB2Pc1rq3krotzaeSecDzA2OnpV1pEsLqvm7s/7OP61JbyMoCPjb1460tqIBulbP4UWUoQsmT3G3pVgywyCWYblH4VdStLiURbd3+1nGKmWWcYYZA6UirDGGUb89+lPb7gJVbaR7ZoKHXOeTUUckEZEbc9uKhhhmXBi+bv8AMas445bhWf5gucdu1QWkhfO7C/SrVQ7qsbZRenGKjvICAIbPy8dvMJrT7GZ4fOxwenTjrQQ/cU5P0pHe4iVZnztzjitKtl8s7+fQfnXiL4e6v4SnEeoDiTo3y9gOwY/3hXgnw3L4p1yHwvB8k1zu2y9cbUZzxkdlx1rxxoFz4c1uTTbs/OmOw7op7E+oqMo7rvXcBnBzjFPqarNtMWR6bsdvpUVvI8hkY7FXt1rSPEmm2ehvosWg+fM+MT/amXGGLfdxjpx1q4tGSRhMmNnbOc5qJoZX8vGxR9TUt0LdGeJd3+1nH6VaQya3OlvE20nPOM9s+3pVzbroF60Fncfax/f2eX29Dn1/Srqc3NvJCf3bSY+brjB9KsZrGYLFLHgc/Nk/yrXvG+gaONNtrXw95qQ+b5jfa3XzM4I4KnGM/jXiyV9Vvmvbez+zRN0j8zfjAUdTg9R+ta60bO+D8y44/KkkCNlxz2qdQNJSRFyz5x26NVyz+cRt5+vtTseveicDJNE570gOQDzS/Zohnb5hPuRU+WJlCbQO2c1I/GTUQQDcO/ag+fu8VNMjOBIMehphj52+YflRG6DBH4/jW5gmFNSyCSMMe3amdWwAMUX3R7iM579KyvQn9KfPQClfaMd65I+9+ldTjvShicA5plJOA36VtI+UHPpWMjGaxjAJorgcN9OKIO3Of0oI2ME1tJ6N+lchsbv0pBk9aiTe2N+Pwp4337c5A79KfceDQBwBnn0rGO9EBhgHmnXA21EMPtZ+PTFThTIQhz+FEYHB/SgB2PSjz3o5IGKHBxTcDOeaJJHDfhilHOAaIx1bOPat6gbSKVh0zRB6GgAP8KLZ4UVkHjrijlhwKG0cYye1SytJgbf1qw063vY5mnvvIaLbtXyi27Oc9DxjFSoIZDHu3BehxipOBj9aDfLgHIo565rHGQaPPegB3NHnoKC5GM8iuex/SiQRhTk15ZHOfpWNvQ5x7U2SMk0wAGQc195cAUq//qrI6EfSiMjgVtBGBSjI5/KmB7DmtpIyDTHIyTz6Vz0xRHalOOo4okHgflQwMEGiQRxRAIye1HJxiiox1oDoM5/CiuehogA460wzyDj0oDOBn9KPoDTMAOOaMgxRKnnd+lEDhQawB3/ChgjAojHFYB70RyDisA8ZqiBjg0ACp5rqNortgGmGRgGuCODVE5ORR5IrrwKIGciic1wBgHpXBHBrtijjpmjzwaPoKFHBPHaic8CiAaIBPX8KAwOBzRx1rg8UcEYNAepojkYNEZIxTHA4NcEcGgPlFHJ4JoEdjRwTwaJHQGsZ4FHrgUxBAxRGRjNYHAJ5oBSMBv0pgCuM1kEbQaxk5zRBJxmgcgVgE5BrgnAokngGjkcA1njOc0ScUDQYjjNdT9O1FewNdT9KOM9aI+bg1kY680D6GrUkrg1EyhMA/SlIIBJpGAPNE5UgHn6UQccUeBjFM3FblIz2oEcYNFiQKQqCCT07UgUqCpo+gogdjWM8A0ee9AgHmlI9aI9DQAK4zSru60Rjgc1yVyfwrPqKPzY/SiOwPSs+hq1KiQBjxTg7tpFUMd6wMYz+lQuI5A4PTpV7rAurVYSeVzj8x7UHAOaEo65p5AV+U/pQxmiQe9Egjg80SCMDtQxjmg3GAayDwDTMB8ooXG1cKfpVnrt3ocs1m7c/LxgccZ9D61+z18WPC3gfxdBqnjIbtOXdvGZB/wAs5VH+rQt95l/zmvjN4y+GPjHxo+u+BrjNg2Pl2T8YijX/AJaAH7yt2/pVx4h0O/U2djLub+H5XHueo9jXw98bRQyrpVo24N14x/ePda+MHhu40yw0+6iTfJc+bsXIH3TGD3PrX9jXl1EGSDDHvuH+NeHPDz6LrUX9rfLE2c9/4T/dJPUitC1208J6/JeY+R8Y69kI9D/erx5c6XrmpXPiG0m/e3W3cu1uNqqg5OOw9Kk0u7ZPN2598j/GrbSJ7pZDGNzDGeg/rXh2z0m2SSO+k2ucYGGPr6fhWsXd7aWkb2afK2ecjsR60kNx4gu4rBmy0u7jA7DPt6V4uj1mxX+ydRt/LCdfnU9drdvqKu0RPmVvnHTiraJZHBZunbFaf/wiV34LvUe7xqcfl+Qnlyc5lO7n7v3R3rxJ41sNNt45/Cr75xncMEdwB99SOhatZ8XT6vfzXWpviV9uBgdgB2A7AV5v2qQPnIFaDqWkWOoXj3D/ALp9mw4bspz0GeprRJopNZNxC/7o9OP9kj69av8AxlYRacLWCXJX/ZPqD/dq7muPsst07Yj45wPXFarrv2iwS1STcy57Y7g+leDoX1vd4aT/AFt3jH/Adz+3YetXsNmHaS1fMY+6cH+tXDB4d4PPepFMqhVPIqFTDCxb+LGPwqxnMb8nFXKSyzvIeQcc9O1Rh1OxhxSNK7DyvvjpWoQ3TpvuO3Tp7elNGWbap4FSyKyGNjz2FBVVAinkUEckE/jWDuxnpWCQdvXtRG4/MakweF5xQIwRmpE8xcAUFRBhjXAORT5PC1bsoYAmiVAwT1pZUK8nBrKPwKZkA+9+lAqv8X3unFBg5Az0og7jk8dqcISD3oAHBB+lYz0phkYFMhQ5Q1GvloA55qUKG3E/SmjYKAK/Ya0XSv2Yv2NPF37SXia0UX2uwy38KSAq0lvBuitIeenmTM5B7iVDWv6vqHiHWr/xBrN491qOp3Ul1dzyHLSyyMWdz7liT+NFF4JryzjdHyfSljd2+YdKaLcuzNKgAxUMTs4Aq2tkMYVzzVpYKD5jn5VrSUkjCtax7iuc849fWvFHiXRvEfhG00+0bN/b7/NXDDG6RSOoA+6KuLdkm2qM/wD6qh0ua8t5TAuTFjcOO547+1X0bLGUzlv/ANVR2ckgCIuT35qKOW1OHTp71HbXVyuwp19xUlrHBGFY8jtipmSZ1WM1Y+H5by1dQvHHf3+vtU2nrvMsAyP8+9XMRZM9xVtJLGwUdfSrV085CxxnP8qVUYjDfL9K04AklTx2/WoLck7jzVlcOsKwgcc1LbkEMoqO3MyiOPt0q306806GNbyHYJM7TuBzjr0PuKvvG97rtsYr1cunTkdyPRR6CrPxBdaBqQ1C1P72P7vTuuD1B7GtYvLnV7n+170ZRurcdgF6DHoO1eLdU0m70qxh0uTdJD5nnDDDGWXb1HoD0qxtG1CYQQLlz2zjt/8AWqfwfcWNzGbwYRc+YOOOOOh9xWuXNjpeqxvpsm4DORgj+EeufU1qUtxdzyX83HmY447AD+lJbvdkCMZH5VdJtcwg8R/1qK8cxbbY/Ovf/wDXWkwyX0LzRpv2YzzjHUf0oMwmaVj8oxn8q1HUGS2DKPmH+Iq9u7rVGBYcJ06d8f4VDcRTQeVIflHQ/jWrPHBfzjflfl28ewq5uLS4cGN/0NTSZ0iFEOdm79Wq6BE7Hr/+qpCRwRRb0rOeh5oSsMKKDnbgHkUsxaEoTWAW2g5qRGTb6HNM6qMA5qVRIhwOe1c+VtbrSShVCseKmkQptjPFFQYwSetO29dpNBuxP4UygrkHpROV6/hR+7gHn0pdpTGcmm9AaBGOKIzg5pkz0NAcYJoAEcGiR0zQYFCuKiZQhDfhTYIIFBQVGOtKhJzSZ9aLM3Q0CeuagjRyWY8D2p40bJRufpTAdj9KfpwaJIGBTAAZzXsetEEAgUFAHB5oYK8npQAbgGiRt2j86TA4FAAnINOABjPFLjHJ5pj6UfugDtQG7gGioHU81tBOAayo4BohRzmgkDWhfOWHQYPrV/JYtpOnRW0mblPN89cHjLDb146elMN3U1aaTLewO8XPlY3e2T9fagARgc03/wCquT+FYyMmgNwAB5FMoJwnNEZbmpBGThDn8KwTwDmmUGLBPPpiiBtOD9KYkjA7UB2J/SiO+awCM1gYwDXJGBWCO9cZ4NMSxzWccA5o880RnoaxxjNYzwDWOM0aAAGAelA/Lgc0SeAPyoAAYzQIJoEYHtRIPA/CjnHA5oghcD8qAJ4zTKDgCiFxwaOO1HB6DNFeM/pRbIwKKg8A9PaiMnbmjk8Zz6URxwayQMA5/CjjAxTHtmsZGAKxj5c9OlEjgYoY6EUQPX6UB360TkdazxgGiAeQKONoFHGMA1gYxiiQR1/CiAR1rj0/CiAaJycAUQCM55rGeBXPp9BRGe2axnmvbFEA8YroPpRAxwOldRRPHSsgjAPNAd6GNvJ6e1Eg9DQKngGhtHAPNZGcZo8dW/SuCdoNMDnG79Kxn+L9K2gnNHA4LZ/CiRnANEgYyfwoYzjPWsgHIb9KJB6N09qLDgg80cY60eQOabkcN+GKBGMZ/DFZJ4z+lAjHXP4UOT1rP+1+lZ4xu/Sic9DRJJwDTZbgUQCOKJJ6H8KgBKdahII2k9OlBsnYH49cUjcYDZP0qIBhgnPtTAgbQfpRB4xW38aZnHyjoKBBI4onjA6UOflpRhQN2R9KAG3C/hR6cV/nFHg4z9KyTxmgADxR54BoDjP5VkA/Si2eSa3ggAnp0ogleTz2pgAuO4oHPGfpRGRkCsDsf0pkLqHU59qB7fpQAJwBRGenX0rbx1/CgoYUAAOtAg8YzQx0xWFIwRRG3pRBPau2DQAJxmlwaKkjAGMUcZ69Pasj1oFQoAPNRTjYYmGQe3StLLtEzwLwOoz060NSgQBWhyw77jx+laB4t1LSJ/tdtJsZuvCnsR3B9a8D/HrWtF8T6d4l1yH+2ItK83y7Tctvu8yNkPzrGSOSD0PTHGaf9ocz+G08OJ4X3bM4n+3Y6vv+75f4dag1ia9eOW8uNzJnZ8mPr0HsK1WN9SmBuxuA6N07DsPoKs/C2o6isrwy/ulxsXavPr3z1FaFo9teRixuZ/LZevyk+p7H2o+HFhS5n0Gx84LswfN27u38R471q+h3D3Ek01r5chxlN4PYdwal1C71HS001JtyQZ2naBjLZP8AKvDEc9t4ntYXvNu/flvLBxiM9q+LHiWTxPfyXU9150z4ydm3oEHYAdFqaF4myzfUYqXY0uYPl9uvaotQmhLKh+X+Icc1pOsT2zloJ8g9V2j39q1y0NzfSXi8xS4wOm3AA+vWtOfynVjJnH8OPrVrZWniCxjFhpmbxc+a3nHnnjg4HQHpV0TpNwYG+WRevfHH49jVus15bCWPgDoOPX/61azqFytkbdn+Vu2B2INWSzCTYj7iPYCrOwkulkukk8uWDGTjOc8fyqSOSQlA2QParC3tHyLld3oOR/KhbGSUsnygfj2rxTYaNFomjPo77pz5/wBqGGGPmXZ97jpnp+NR2jKm+Rs47YqBmcbQeW6mhEXYLKuV9c1pl/DbXgknXzVXouSueD6V4q0H4a678N2uvDcGdeXGF3TjH79R1chPuA//AK6EBHzFOOxzTKRnA4NR4dASefSgygcdKBJbOaAVQSfwFSOCc7f1p9jRgjr6U4AICimdRwTzTohfBf8AHFCJGXaXwfpRVQ20dulWxBY4b9KU5GN36UIX+8D+FBSikhst2GKVQ6880qkLtzgCiCo+UZxUaFiFL89zinSRGwr5HpiiCFGx/wAMUBxgn9KGcYzWAeM0ckDJyRRyRgV8Kvh7rHxZ+JPhr4baGD9s8Q6jFaCQLnyUY/vJSP7qIGc+ymv+Co3xC0jwF8OvAn7MHgt1tLSO3iu7q2jI/dWVsvk2sRHoWV27cwqaKZ7c1tGMHtWCRgHmlViRzk+mKSCWTgCotNeRvu5x2z/9el08oMEbR+f9ajicYFunI9/8asdMlmAu5ZM7eq7evb1rwv4y0Lw1pbtJoH2yeTHP2p48YY/7JHQ/pWjaffeItVaa1Xybdv8AWJw2MKccnB6ivEFkXnEcIyF6/kKm06GO2ieIbHfPmHk9Dx3qa3lCED5h69KQmFUUR/Mc55qQlnVWj3Zz8ucUlzapp8QL/v8AnccHnnjjp0qW3kvcxhvve3+fSvsFtZIAr737nBFW2qXNvZOkJyxxjoMc/Srm8neAWp+UL/Fwe+elRhPLaNo95OO+MVFZwvMWaTAPRdp44qLT4mlCqOF7+vH1qz08yp5itjH8OP8A69aNbRtcBJhgdz+BrxHdaLcyxroulfY0Gdw89pM8DH3vx/OrWFRsD9ea023e8uBAX2J3OM9j/hUE8VtIjIm9RnjOM14s+KEnjvT7DSbjRfsosfM2N9p37t5VjwFGPu/rTOiqViGM9a0bwrDqGpWen6tdfYYr7zNk/lmXGxST8qnPUAduteMryTSYW8NaZqPn2UX3W8nb12ueDk9Se9WE8EkFxb3YzI23yzyNvJz0/CvhlodjrerywXOqf2bsxtl8gzZyr54BHpj8a8d6RB4Pjhu5/Ef9sXWobsv9jNvs2bR0BIOQ3t0961fw20MovbYeZnqnT0HUmry0RAsNzebMZ48vOfyqxv7fTR5cEXmj+9uK/pz61fywI5UJuc+5HpVnqlzp8Za0fDntgf1HvXhfx9rPh7SZrWzvfIW52+Z+7Vt21iR1U461MUktGljutx7DZjvWtrLpmIYxuB6Dp6fX1qa2ltrb7ZK+5u6YxjnHWofEkaK1k9vtjGMPvz79MVftHCpSI7tn3eMdcUZSgVgvJ75p5Fjt/KBxn296ie0W+33ke9B2yR29vwrVrq3u7+Wa2t/IhONke8tjgA8nnqKRy2QV6e9FO+evtWOMHtXBX0pSCpXNDC96EjAkMMAdKkZRgnnHSjKXiIUfhRMgXBNEAjGckUFVYvlOT9KETSQcPk+mPegmByelEDOAK5xt/KiAFzmiAQRmkUkct+lAZ4A6dKOQchv0peRgnn6VEqs+Hfj6VPHiQhW4HTiiCOAaByMA0Bjo36UwJGFb6cV1GM89qYEjOf0ohtv3ufpWOMg0wBbANMGBwTSxqYdxb5h7Uyts3KenbFYJGAenSmJK4Jog9c/QUxG3J/8A1UVJGc0FNFc9qODxTqMgqc0eRjPNAEjINYJ4A5orhcdaJPVTROeMUADwTmgVVsLR5bGPwo4HAoc8VhQcgZqPaRjPXtWcHAHFMQw5PSku54UKwPjd94YBqMgckUVJGQKVRxmpFBPynNIMgijzSBguRSAgcN+GKdlBAHFEl+MZFBFI5PSipI69OlEZwBQAzgmsEDjrXI6miqkjmgoLAA4zRUA4BziixxgN+GKHJwT+lAKTy36U57Bs/hXGPegc8k05zyDRxjJNEAjGaIAGAf0roOBnFAgjBrCkdeaIHTOTXGP5UwBGBRBIxnNAkcBf1p2ZRlTW5+o5rzMj7v60QQOO1EAjANEDOKJIGRXJPB5odDQIFYxwaOT1psZxWe2aPoawCOOtbcjGOaxu6VWOOv0qiR0zQGeAaOM4FdsA0ASOlHGMGiBnAokHAB+lEDOBRAOKJyMCqrGRxVEHODRAIxWOaODxR9BRGORQwTjNAKVxnPpRC4oqCAQaCgd6bg5FD5uCaYAdD+FEZHvR4FFhjiioIxmsADNKu6gO5rbzRAyAKKjHFEHbituRxRxnGOnStoPSgARRAzgUcA4FAA45/CmGABRGKOD0/KiuelADOM8+lA4+UGoX44OaimD9G/So2GAueaUgd6gKnkNz24raG5Y0yxnC557CnQrwaJQHaT9KIUOADnHamChjg8UCoIANIBsGTWBjbmiO1YyOKwOma6HFFs84rAbkGgc8Z4orhuDRXsDRXIzms5A5/CgoAG40QO5piMYzXXikkKcA0luZVaQDj0pLYSLuHSnjCng0YgU3A9KW3YruA69Kkh28Hqa2DGBRBzkUoyM0SQcZzXBHFFQMYNbcng80Fxznmi6su0D8KcdqI7UMYrA7VDczQKUSTGevAoSNyC2fwpHgTTlk3fvPTB9f8K8OWOoa/dLplmcyv0HHYE98dhXh/wAB32p65LoAX97DjIyO6Fv73oPWtK1ix0e41C1vBlovK8vr3BJ6A+orR76K7Z/MbPTHH1/wrRdQgj0mWGK4/fxYyNh4yx/DpVtdX2m6uszN97PGBzhce/rXhL4oQ6PHLY3Df3ex/wBo9lPrXgDwt4J8aQsfEE3ly3GPKO2U/d3Z+6R2UdaiaXw5e3NvdpgnbgZ9s9s+oq5vt175tu2JD9046ce9JI9yd0jbiPbFXgEzBRz60p8gYIyaKsyGQDLdxWhaNe6ozLbL0xnke/qfahLLAJbK5HMWM/jz2qfdF80R+leCvF114b3SlN5lxnkDpu9j61eCXxFPJqZHXH4cY9vStXuZtJc2sRwnbp7H39avSbiMANk/T6VaiRQ0iD0xVtqEsELxGTBlxnj0qNTFLmQcf/WrUJ4VkjW2PPOeD7etQyLPiNTz9KXwrNHAHmHDdPwPsalVZXMSPnHbFRShZWjDdMdvajJJI5QP+GKvzCkoMZ5PQc+gqDWrvTuIJtpPT5Qf6e9SzII8DoOlSxGWAyL/AA/41Gocbi2Me1ALIm3PXpQQMcZq5j8tV55OcVhcYNMoA4NOARjPNGAZznOamWIAc0EKqGPQ9KI3LnNWoPJIpQXJJ6UyOTkUHDMeeaaQjAQ8V94ZNAg8ZoH5yAenSsZOM0F2gAijyMrSnKg05OPkGT6UArKCp/SgpJwK/wCCUHwhg1Dxb4n+O+uQotl4atzpumzS4CLcyrunkBPQpDtU9sTn8P2n/i5N8c/jn4r+IizM9jeXrQaWrHGyzi/dw8diUUMR/eZvWiAO/wClIu8hccUsZkbbGKis2iUFx9aCAKPL5rShtBlccjqPzr4Z3Pg/UPFFnbeJJfLgbfu+WQ9I3I+5z1C1qulrpM5gi/D8h7n1rRr0Wa+bOMgdvz9KutA0HUfA0Wo23/Hzzu+//wA9cDqcdBVnd3elRPNDJtjOOwPt7+tWURvyZXbOPb/PpXi21g1HRNHazg27fP3HcTn5lx1+leHLK0ub4QaiML6c+h9PwrVtNt2lYWyfKPu8n0GeppLaKP5X60bVJ5Cr8A9KkEcIEcY+pqMBocsMk9K0az8yUPKPkHUfgaKLcvsI4qSGK2kyp4X61CBI2M8ioZmdggPLdBWmx8YH+etWFurAyHr2pUYyqCeKVVeQbelRXJtQCg6V4Ysra/mzdthe3B9D6fQVexrBM5gHyJjn6irGz0caDvuG/wBIHThv7/tx0q+1S7l8spNk22dnyjjdjParm6eV2eV8nvxip9F+yWMGsBcq27d+e0d/6VDqX2SH/RX2yH2z/MelatfajPaq+pTbymdnyqMZIz0/CvA0umXJP9t3P7od9jf7X93nritUma4ulGcqM4OMdhUMcxfIPyipnDSgnlv/AK1eH9AsbvS7rVr5sJBs5weMsV7H2HaortiPLduB0GKtboOCC3A9q8QancMwBfPpwPb2q3ju7nTGkeXcox/CB/FVxeyITE/bpQzcr5Crln9/SpVkVtmOnSkvXkyWb6cVfHdLx/ngVO5TBHU0jgnb3NOcDGenasfLwaIHQUihu9OwBwamid2JXoKkGWAZske1OyqmV6DrUjIYjgcilUjI9aVyDsFbiAAG/StzA5JzQQxdR16Uwyc0zArjNEgkYNKCPvHj0puDxVYGMCmJA4pXJADmpFBHFNlcEVncMj8aYkHArk9D0ojPGaAB4z+FKIwuM80wKsBjpRUu2AelEszEg8DrUhyMRnP4UGJGM9KjTzW2g04AO3OcUQDgCnKgbR2oNwQBRMZXAPPpWADwfpRKscA0wXotEqQMUoUEAH6cU+CMbv0p2UgUpUkcVK8DIAo5H1oAEcGiF70QMcGlAOAKKjueaAA+bP6URkkg8UcAcHNcbcA80pUgAmgeymmwRgHp0ogKODzQbKEA8miBgZNZYjANF8jAPIrKueTiiQsewUgDEZo2yRxlpDj0oDAyKIDcA0i5HXNSAZ2g0VA7/pQByBnNE+hoLnpRXHOawQcA1jPfpR9AaAGaJx2rAYVkgYpRlqZccVRYAcGhhhkGmYDqeaZhjKmskjrRZR3pirDCnkVEQUx6VtB6GmwelYBXBFUATyfyoY6GsZ/pRxjA60Md+aJwOK4rI9Kxnn8hRx0xRUDkUcEcCgAeKO0AYo4PQUABwPyo4PSmA4xWARgij1AzR9jQ54FDGQCaJGcCsUwBHBrAxgnNEDoDXBGM5raO1EZ5BrHvXtn6UQTxmse9Y96x7/hQVSo5zQUEdf0ornGDQUYwT+FHrgHp7VgHvmiFPAPNMPeiuRgHmiuFznmutFQMHriiAMY/KgM8A0VycCiMDAP1rBIwO1FQB70ABzRGQMUADwaCjHA+lAgDrRHFEAgAdRRAPBrjGMUSO1EehrAPBrAI4NAA8VAwHGfpULgryc10HyjNRybuM8ig27rWSRyakjjkGQMmmUg4B6daUr1J5oAtyDnHWtuOtRqHQHP0pwxGF7VwRwaCjbmqxx8ooAHgDNE+nbpRHoaIGeD0oHPU0RnoaAJGMc1klcVjjpRHY0wXoDzQUk1p9xJsa2V8K+O3pk1JHcxx+SG+XsMCjbsCqr1qGx3pt2YJ96OnMkJVE5HTn/69QaZLMpLx5YdOf/r1qNnmFQieuefpRi4xt/HNQeELmbR11XdhTnjA/vbfX+lFTnAFFT0B6UCSefwrj0ogZwKC8YzQgd/mUcCimOCaZCehoAdzTYPSsjHWgvTJrQryWwv47uJ9rJnBxnqCP61FM93dG4L73PU4x2x/Shd+aRBINz/wnpj1rSwj3Kx253vzgdOxrxEk1jc2ljqMeLiDfvTPqARyOOhFeMT4Y0+1tU0Kbez796bZBjBXHLfU01zKbk6Hq9z51nYfcGzb98bj93nrjvXiNdNuSnlyblTO0YYYzjNaPZ2ur3JGpNvXsMEdj6Y9BXiLStO0+9hi0ZMk7srlh2H94+5q5QRjKcZqchXJxgCpfDkpsl1Py/3T5wc+hx6560TKVI3/AC9hitK1HUrJibK68onr8it6+v1qKa/tLpbsv865w2B6Y6VJcyajcCZ0xjrz7f8A1qa2jmna4EWFOMDd04xXwj8H23jHWYtHur/7LHJnJ8ovjCu3Yj+6O/eviz4Kk+HPiu88KXtx9ouLLZul2bN2+NHHAZgOHA61LOrQcD5h0/OodB0mbRku432zc54Y5+bHrjpWi/C9tX8E6x43mvvLg0nyN0XlZ3eZKY+u4EcjPQ/h1qxjk1S0YrFvdevzY7//AFquEWKHMifMferZSw+U8ipvGOv2GnHSLXUPLhuf9avlId21tw5IyOT2qVMjfnPvVq7H5mOc9B6UwKtuJz7VCTIfmOSnWmkjd8Fcge+KZ2L4AzTyPGAjPwfahEFXcW/DFW8ZZCWH0oKobGc/hU6owAU/hQwVAUdKJBcgmisJjypy3406gHAHSriQouU61LEioGC5P1ohSuV6+lQBAMAc0ACuKITbsP5U6+SAVPTtQeRTkvn2xSyE4HX9KYZTINK+0bZZMn6UBwMGjz15oAYxmsZ4xTIVxkUyNgc0qPIwjRCzMcAAZJPtXxVcfsZ/8E77D4exMtr4s8WWn9nzgcOLq8DSXhOOf3cO+INxgiOkhduAPxqOCLI38+1RRWRcWsdtmRunzn61Na3Vm+yRMHsMg1AcACdcMPf/AAreRKAGyPp7VDbzXcyW0C7mbOB0zxn+laro17ozxrfwbN+cLuBzjHoT6itU8UwXHhez01Ids8fmbpNxOcyAjjGOnFQzkTBHG9ux6dqt/HmoxaCuhJdYiGfk2Kf493Xbnr71PfxLYsjH5h1P41pV1O6CJJOPXA96m1uwtojEx8zH3RyMevari8zbtHHHiU9Gz059PpUFtJJF5afO57dMVLb2yW5ivIMn/eIxz7fhVpodrc2kzXqbRHt8s5Jzk89D9Ksle6ujaomEXtn2zV8LXUdqrHgJnByec4/wq5meBNls20D2z/Opru2kHlsmJPqanaQDCjI9ahs5mjAC89+RVvZoZ44tvXOOfarK2SJcIv4ZrSXEbK6pwue/1qaY3VwJUHI/wqMkMFmbco7Yx/KrOze6ZhC2fbFRxbpBA68r15ptTGnw/Z4ZuH+8Nvp+HvSajOs+923e2AO1HURHcNcRjbJ/CeuOMVdt5khdz8xqOJzgxtgn2q6li8oDy9rDvnNXV+81ugkbcVz2x3oRhBiEYz1rUXhLKsS4Jzk5PPSlaNpAJOVH4VHHFOT5w5PQc1dW8ccg84fL2FXt9LPOwtm2rx2z296sppkwVf5h7VrRNy+5Hx+GfSo5LmOFnknyI+nyjvRhjkkJYbh2PSr9RZxhmP3v/rVcX0hYlXwB7UbzL8mid67s5xUhJbJOQvahOCwwcAdqJxxnJpGJTDdaVi6gnrSo5IJbOO2KESKfkFMSAFB69aGBxRZN2wnk9BVygAwDx6V5blM9u1DCY5pym0Feo6mmJI4b9KySNqn9KCjO0UcDoaJyOKBO0AHpQ4HXNAY6mjzgEVjjANAA/hStzgimAbGB1oqBiiARjHNIm47QM1caNdW9kL10wjdOR649aOMZA+opcDkCm7BT+lEgcY5oOgyCvJ6HNFjn5TSsCABRQnoKs9Pku5xbxrlz2z7Z9fatWsJdNuTazJh16jOewPv60RxgGvajg8CgMnANSGPIKL+tEhjle9dOD1rg9aIUDg/WiCw560MY2mimeTTOhjCgcivlPI60dpH9KCqBkNz9KG0nFQQ+eNqdaBKqFz07UxXHWiMjANKKwMYFLp0rWTXoHypjP4nHrROelcdq3DpijxwKwCMA80QQASfpSomzdnn6VgE5z9OKb0BrG4ZWhkcg9KxkbRTBQMqc0y8ZBoAHqaK9gaOOgpgDwDzRAHAoc9/pVbSRxyBTHjAPSshhgCseopsEZB6VnI60QCOKKDORTdQtMFHOKVgeAenapHGMqelARlcKaYgEc5FFgTk/hQYkfKOO1Yx8qmiMjrWARkGiPlyD+FBd3I/KsZ6UFz1ogHoee1EYHWiPlrAK5B6URxkGh6jtWAeM0RxnPSsZ4BoDJwDXGaIHY0QDwDmiMYquvFUAMYzRUHgGiMcZqhyOtEdMGqC981gEdaI3Dg1kHjNVjPSq9qoAFRgVtBGAaAwMZojIorgdaGM4FEDuaZc9KYYGQaIyODmiMHiguO9FQRgGsep6UWyODRGO9fQ0w561gEYHWiQRgdqwNua68CmHOSelHGOtEgnGeaOD0o4JzXA6HNVWAQCDQAIxnntQGehq2lGQpNK6lck4xUZAwwPWkkUnAFEg4xQKjgGpAoAAPNMBnIFBgvAOfalO4jmhhYxg0p3DpVYPQUCDwK6Hg0VwMZrBxjP0FEjGRX0HPpQG4YFMF+6DmuvApUIXNLgjAonPGa2qT1rSLCwuo91zPtb02E+voatLZQzMG4GMcVaabcX5xEN2PoP6+1JorR3DxXQ2Hj37exqSyQEQxnB7V8LvBtl4y8RxaPf332ZJc/P5RfGEdugYf3R371rvglPDmr3NjFL58ce3a23ZnKg9Nx9a1aziUbM/Tg+1S6YAcj8P85pLi7isxp5fEY6DA9c02irJna3Ttj/69TaY6E459P8AOaMDEE4zimjwODRXHQ0GxzSyuoxmi244FBd54p4igBI+lFQehoLnio1ycH8qgtxHskY8HNQ3XkABFyfrXh/QL3xJFeXVnHj7Ds3nI435A6kf3atdTns7pljX51xzken0q48QXepXRubs7pO7cDtjoB7CrjU7gybi+QO+B/hWma1dX0/lXlxtWT77bAegOOAPYUSiwtGjeYD0bGP0pZ5oijQt97Pb0rwUNMm1SKXVpcKuc/K391v7v4V4vjjjvDDbrj8fZfWrnTl8gzF+V6rj3+tSX88sAsxJiNegwPXNJYwTXJVZPl7cH0rQvAsN7F5plwR/sn3/ANr2rQfgp4Uvvhvd+KbrWtskezMX2aQ7czlOof2z0rXtb06wneCyXeBjByw7D1HuaXxO+eY8L9f/AK1ab42vbFxNp03lSDodob19V9Cav9ZvtcnfVNUm+0Ty43NtCZwAo4UAdAO1SI9whdY8Ads5rwXHa3dpOlxPgJtwNh7lvT6VqeuXdpouoaFZ3P8Ao115W9dg52sGHJGevvWi3bWSmNV+/wBTnpjP+NX7KJnt9mQuMNnHYHpUSIPutyOgxVyplAdm5+nSrYBhgnJ9MVYaXNe3gtLdcuvv7E9z7VrWkz6fOILgbWPToew9D700yiRjEeKVw5CAYpV2rtByB36VcxAwmQtnb0GKUIy72bB9MVCcIQRjNFCxyo59KaJs4FbQp2mnwzFFPK+1KAvDGpEJGfXpTkqctzijCxGCeB04poSvLcAVCqgnac+lF41woPPaiwdcD/8AVQjwDk5B7VJCMZB/Cimwj9BTOXbaTg1KxL7SvI6c0JCo2qMn0pN7cMuPxoEHgHpQBwMmg2SN3aiEmXaowa/YQ+Cq/F/9o3w9baha+do/hpv7Z1LcuVKwMpiQ9iGmMQI7ru9DX/BTzx5f+O/jPZ+EbSQnRvBVmbdAGBD3c2152HPZfJjwRwY29am09oZ2hLfdx29vrSWyxupc5FXpgluhcWowsfQ8nORjvR1Brlw0nLflV1Oix7CvznpzVh4am1fTleRPKB/iyG7+mR6Va3ht5knjf5lzjj2xWo61carcxvdvkJnHAHUD0A9BSIZYVVugzg1dvbwOnlnLLnPUVDdLI+4jFSyW8SqC24yZyMEdK025eZ/KiOP8ml0o3d/9kL4eLrx6jPr7VeOrKU8raR33ZrSZIookdBvZM7+3XOK1XULdpPNWPd6rkj0r+1J54wxGE9OD/Sre3eBvtZl2j/dz7U93HDbqXOCc+/erouIw0fzZ/CtLsory4Rrh9qtnJxnHB9PpWoWFjbXLfZ5fMijxztK5yB+PWoYxM25Dj2q0t1ADMOR0NWEQdsA/SrdAjlQeO1Wqb5gF/L8KsrA3rpAg+Z849sf/AKqg019NDKDl+MDp/U+tSzCY7Il+buc1cwPIQqnJHXirqOOKYRk4Ydfyp3jjukDjIOePwrVZ4L28LrH5YPbJPYf4UsJjXzWPCdPfNakXYhOh7fpUiOoAYcCobnPyk1dLJkgD6VdeFhp+gnW3n3OOkezH8e3rn39KmYRo0hbmPqMetarfgoJXf5R3x9Ka9F3ct5PyquPft/8AWq3eRNPuLxeWi24XpnJxWsQJZLsnbD9xjp09Kecbzht0Y6nGKvNajVNka9Pf/wCtWpazJcsqE5xnA9OntU1w7EAn8KcgYOc5q2n3xbAKc7ZSjD/OKdQvHrSzZA56Ujl3BPXtRXHArgjisDGBTgMuM0CCck9KeN5T5iGhFu+82fwqXKDyx0WhCpi80t+GKaEhMMMelBTnaRW3aoVRmsAjjkiggYYzz6UV/hB57UvAwaYZOAelE571njAPSgdwwDXUcGhwOOa3jG3Ga4b8KBAHJqEoCGY0NW/tK2GlSptA+6c5zzn09qvtPlspPLkPXp+nv70V/hU0TyPalAZgCf0pgp4J5HSjhjgc0hAPWmJXof0rTr97K5W4RdzDPGcdiP61rl6+pXJvJE2s/UZz0AH9KypAAPNMqYBDZP0oAHjNBQRkN09qC5HB/Svu/h0rIY4FbQOCfwojIpdq9a4PStwJyfyp8Mc0FXAAPNMoHQ9O1AjPXPtQUHkH9KjkeNsoaAyCxFHJ4FEgDigAVznn0o5IAr7TOIGtxJ8hxkYHrmsYOAa68iiB1NEjriulSAnGD9OKBAXArAPOea6jiivyjH4VwRwf0rgcg197t+FH5jxRGBigMjA59qK9BRA6A0eeaCcZNEE8AUDxtHejkjj8qPzDAFZyMZog4wDRXHAo4xkU0mOlZU9+aPzHbmkTJwTRiAO0n8KIUgAH9KMSkYPNFUQYNF0PQ0RnkHmmDFcV/DwaJJGM0AeooHAwKrAxgVgZxmiD60SSM56UQvrzR4XAPNHGOKzxjNY4xn9KJIGBRyTnNdePyphgAZoEHqaIBPByaPIxQAJAqsnoDXTgUST1P0rHYmiOwNEZ6Gs8YzVEA1jI4qqIBogAYxVAAgDHHpW3Ixu/ShgDrQHHBo56A5oqCcE5oqD0NEBTkGjwME5oHtmjg9PyoH/61fQ1weBQ5PFE54BoDPJNNnGM1kgcGjyMZojiskjg0cHrzRweBQGDmjz3oDnjrRGehokkYzVZJ6GvcUee9RuUYEN09qtZg6jNOwxtFIT2bHtikORwaK7u/wCGKK5bGabI+TNEAHAFIT0HUUiKyAkc0GC/KBQII+U1j3rA7URlRzTAEgA0ea5xhT+FA56GjnHNZDcgdKCg9TTDaMA0T8uCee1FQec0AD1NQrkgE1eaNpdvpNhcQjfJN5nmD5h0bA7+leFfFV54Y/f6OfKkP8XDeo6MD2Y1/wAJLceJfFSav44/4mCrnzukW/8Ad7V/1YGMbV6Dt9a8SeN/Dmu6G3g/wvpX9mW0uMjz3mzhg/8AGoPUN371Z6FfvBDe24xG+7B47HHc1reoadpOgARR7rk98sP4x9R0NaNFb6tqsFpqs37ht247TxhSf4cHqBWraTpdnqr2dmN8C4w3zD+EHoTnrUR8J3ejrENI23/OZvtEhz8393p90Yrwj4FTxP4p07w/Ld/Y1v8Azf33l+Zt2Rs33dw/u46jrXxM+Clx4EnNl9p+1MvRtgjzwh6b2/vfpVn4FDuyXEe0P1Oc9Po1eJvCFnpjrHD85OfUenufWr3RXt/uL+v096ktZF6ijHjqK29s1G4ByRmtOn0on/TrPzl/u+Yy+vp+FSiPd+7GB6daAA6GkXcQBQjdYkIOSc9qK7QEU8ivD/i/XfDenanY6ZdeTHf+T5w2I27YSV6g46npiog8krSOdzHr27U6MuQG/SkkcPtJyKtUiWRWI+UZ4rQYhe3sFnBcbVk3bvkzjAJ/pWvra2V60NpJv/2sEdh2P1rSvEGo6BqEep2U+yaHO1toOMqQeCCOhrx1431b4hxya/4gvPPuJcZPlqvTav8ACqjog7Vo6zy3SOTuVc8YA7GvEFk13ckw/IPTr2H+FfDnwjdeJtb+wQxeasX3huC9UcjuP7tQaP4es8mO32unUb3PX8a+LXxEe/nt9CsB5UNlu4zuzvCN3XPUHvUzkvuY5NM5bjNLK6jIPNeDPEcOn6tBLqsH2q2Xdvi3FM/K2OVGRyQa+J+oeD9bls7vwR4f/siD959oi+1yXHmcIF5kwRghjx6+1LGAoSM1FOyQtalvv4zx6c1dErlVOcd6ghimiDtyw71PqK6RZfYWXeD0Ocd8+/rU0TGNdjcPnjHpWm2rF8htuPbPrWgatPba2l0x3Bc8cDPyEeleNdcOr3SzRxbGXP8AFnsvsPSry3miK+cuQc88CmcBdsQwe3etG8R+Bo/C/wBhvfDvnX4/5b/a5V/5aE/dAx93Apoo3k5HPal8OajLbi+RMoPceuPWkWTgMevXipoXddwfJ+lWkYZ2LDhe1T25LcHPtUkSjLgdeopNp+ZfyqV2ZdpbPoMUiZO5l/WlRy5JbI7DFOVYbM8/SoAiIA3BNNGjEEjOKUgqCKAGOTQUP94ZqWMMNuPpRijYYxRjUna67vxxivKQrsU8elJESo3846UoDcrzihGW4x9BV34c1Swijmu7Xyo5M7W3q2ccHgH3FeGvCV14hvF0+2bEh6HA9Ce5HpX7HXgWw/ZM/Zi8c/HjxgkcmoajDJeIHAUvb26lLeEYJwZJmcded6elazreteL9Yv8AxV4gvmudQ1O6kubqZlGZJZGLOxx6kk1oFyn9oJJOf3QzuGP9k46c9a1rU7e9ug6w/KvQbj6D29qv5rD7Lbra2+yb5vObeTu5GODwOPSvD0WixX0cmtWv2m3Gd0e9kzwe689cV47u/BU2vPceE9E/s+zOPLh+0yzbfkUHl+Tk7j+NauY78qZX3bc4GMY6f4VfanLNYW+l2w8uODdtHXqQe4pNNufLM8pwq9+Pp61o2kXtzaTTWyZRNu45Hcn1NeFddGgeIbTVJF82ODflc7c5Qr1wfWvE17b6rqz3dnD5SnGF3Fv4QO+PSvLkEuVbDdjjParKOYDKHLeuBUlw+nWyMr4klzuOPQ/4GoNInv79I5bjcGz/AAAdvr7Vcw6fZW72unReSRje25m3c5HB6dTUf2WeTyxPtb12k1YpJeyeQBvZPw6//qpdMwVkuXyUzgY9foau71JJMhMj1zVss0ziNT8rdeKOmsIwyfKvp1/rT26iEhGz6nFWEaEB1T9ahUN8pqwjOc7vwx9atrVZIFLHn6e9QJ5UysRwP8K0u9clJrZcSxZwc+ufX2rTpVuLpXueCep/A+laxM1nfyW8RzEMYX8Afr1qW424liTaV6DOa8I6DpXiCC51O+uPLng2YOxjuyWHYgDgCvEc0Ut0s1tHt25yc57Ad6SaKGdZZU3gdRnHavE2uaNqlnplvoWl/ZJ4PO+0v57SeZkgrwwwMAHp1zV9aXlioN5xnoOD6en1FTMVIaNuvXika3DiSRdwHUZIrXtXgu5glnF5MKfcTcWxwM8kZ6irrU3tRkfP7dP6VqOpXBncGX0z8o54HtWp6oETZcDfGeq9M9PQVbX0gmY27bd3XjPQe9JcyxHf5mGHQ4q/1N3zNNJvf6Y9ParzV5pEMQf5R/Dj/wCtT3cp4D/himbBznmmyx+U/higS6YzmraUxuADV0AX3gc0XLgKT0pQG+XPTtikYxuK3qSAGz7YpwW4DfhiiQBnNOznAL5DdsVIm4dc4pWVixYcDpWA4+U/SrsqgXA9ajdWTYOD60W3yZc5PepFAOFP6UsrouEPJ68UpLAtu/DFXERgbYDnFAbhkGsA9aKkjg9KB3dTQ5GRQPOSaGckA9Pas9weRRO7jPTpQGFyTyOlbSwGDyKDkDb1p2xgYrzpNoVW49MUzkHANAAjAHPatpY/LUhYYDdRSk+tRNGMF0yPrip5VkICLjHbNBioBDfTipZWYKSabkfL+VAZUnNYHTNE8bQetAlhwf0o4IyTk0CegNAA4FEBRgnp7UcNz+lEAjg8UTxkGgM85or2rIA2g1tBAC0CBxTkE4B5+lZwMZoDIyBQO6kBPANE5HBokEY60TxyaI4AzTYJ20R2J+lHK9Gz7Yrgjis8YBpgD0H4UCCMZ5ojKgZoAE7QaJGcCsnGAenSiOMCkAc4B5FEgjArr25FNjGQK25Gc0WZRgGmUAYJrAAzmiSBxXU7TRUYyDTHjg1nPANEZGCaUg8fpTIGPXp0FAnO3PSi4XgUZGP8X04oPxgDk1vk6k/pTuSMk0GGOBWccGsgjg1kFcLzQxn5TRGeg+lEZHT8KIxX1o47UenIr2zXXj8qHOMDpXUbRQ5GB1o8YB/Ks+lHBGBWBjANEk8Zoj0ocYNYxwBRJHSiMdRVYJ6UQSMDrRGKJA4qscZokDvXBHBo4PSuCODRx0zRbj+VYyMGqAyoANAZrb261jBojJ4NAA8ZpgMAA9KJBPJoYK+uKypGAaogEcGgAeAM0CAOv4UcZ4NEZHB6UG44onPUfSjg4FEggAHpQAJxWCB0rAI61XGBmiQeBR9AaGDRzjAqgMAYOTWM8CsZ4FYOP5VBPsO0/hUUu7jOaRl7n8KiYZAPfpQAHyg1kZwDyKaPcRipEwcCgCOQajPyjBzTgMPlPP0pQAMA9KIz0NY4zQ5G3P0pVz/SjyML1oAD5awoPB5HtTEEAk0UKoCKOdvH5UB2aiuTwKYZABFEZ4BqFMtz2rRLN7zerN8qY7dM5/wqO3SC4ELzbAP9nPatb8WeEb/wFpegWEG/U7Xzvtkm6QZ3TBk4I2/dB6H61pHh65m0z7Uh2BercHHzEdM0fFFp4Zhj0VU8/wCzZ4yU+983of73rXifUxqT/aI02e2c+g9PahdrI27zOvQYqy1p1CxrBk84G7/61eK9A8BXmmxz+BPEf9qzc5X7HNBj5lHWQ46Bvy9xVpPeLdxGMfNbbtrccbh6ULjWvFLJl/O69lX/AA/u14Z8DXOr34s7vT9kB+/J5oO3gkcBgeoArxN+znpuuSBtIvPM29R5bL6f3pB6GvH3wwXw3qH2S5fHl/e49VU9mPrWpeFomObd96+uMenqam8PNIxWMc/596fwtH9lLh8zDomPf1zjpV94Z1SzjR5LbCvnad684x71Jpt3EoaSLHvuH+NNGwwDTIfu1bRMzjnirazFyYzGc7M7h6ZqeJY5Nm35vrUto8dg+8fM+MD6GoYJDHwnT3qaBwAzClhIGCPoahjDK8YXkY4zVrcy2CtJA+HHQ4/z61BO7yYJya3osfzrluy561aXbXSiOM719OleEPCd9q8Yu4bfFvD/AKx94+XO4DjIPUV4i+FniP7YBp1h50LfdfzUXOAM8F89c14Cu5vhHe3tzNFuuZfLyu4DGAw6/MOkla14806IyiCbfK+MjawxjH+z6VrF4bq6eXOWOM/kKc7jtFFSAMVjI4pW2HOa8MawL22axf75xj8yfT2qRoY2/wBZgjoMGraWyntZI3kw7YwcH1ovPIm0x9O+RWl2st1cfZoV3M/QZx0BNapFdOMSLyvuPaoopplVI1ztz3rVvA5074Y3PjKRdoh2cdes4j/ve/p/jVnOUcyZwVq5uWaQsw+lQeMdEk8L3Fhd2+24fb5bb3PSTJ4Ax0xRFvkMs2T6bTXh+z0iS4kXXdQ+xx8eWfJaTdwc/d6dvzrWbBraVns1823XpJnbngdjz14q38bvZ6X/AGclnvU9H8zH8Wem2orlJX5P4UsAmjCQNux7Yq109pCQo+b+f60ts7Ph0x7ZrUrWaCQoY8E+/wBKKEL8gzW0hQG60ZWT5VG79Kkco20igoZwwGR+VMD5mNufTmmwikk/WuWUBBQYltpHA71uAGBX3sDNfKTlealkCAAnr04pAQQzHilkc8Bc/jUbKVKxj8K8DeDvEPiuaSXQdK+2Czx5o89I8bg2PvMP7p6elfEjx7L4i0uLw7eaV9jks92T5/mfeZG7KPQd+9fA6y1XxR8TfDfg3wwfM1HXNQjtImK8IHyrSHg/KiszH2U1/wAFOfiBZeF/h74R/Z58O3Jhju1S9v41blbS3+S3RgMZDyBm+sIPerPRje2gDLsUdDnPf6+1XKJDI9qhyeM/zqO4tNvl4yfx4qzuLOyvoZLl8qm7cMHuPb614n1bTtbvoptNGxEzkfMeoHqB6GncK2GHHajIrLjPzdhVndbhkxbgOnzYq91Z54xbum1fTOc9D6Vp3iPULK2e0shmOXGTwOhJ7j3pEjWMo/ysPxp7iNmBR+R2xVwoKK8J3u/UdMYrTLeUMCDg+n51qkEtzcrDnDHO0fgM1GX0hBJcHgd//wBWfWrl544DcM+TdfdGB/CcGtOtUuJd077B9M9j6VY3n2K78yybzVHVsbcceh+tXGpG6GYjuP8AEemPTtSql7FtgXIHfP8An0q3CQwhVHzL1qWeKSH92cqaNuWcIRtX86it44owsZ+lWtuSrDGcdK0jTp7uQQwpuJ9wPX39qn8K61aCQSWmPIx5n7xeM9O9RwQNa+UrbpOwwR3/AMK8I6fHCLqW6O0Js98Z3ehq7ugZGSEbgcYPSjqegiKY313tuW2/J5bn9QMdMV59vJCZJZNpX7owTmvDl3ZQW9x9obCtt2nB9TnpU00EvmRxnKnG04Iq3shdMw34K47UscsD+ZKNkS/xdf069a8c6xo+oSQxaRqH2tF3bm8po9v3ccMPY/lTu6nAPTtTMGTAX9au8CElvwNPNOGKxx7z2GQKZ2ku3Fw+0cY4z29q1+4TAUHIHT9K024RJizjgf4GtSvY5GyjfTj6VcXAxkn9Kdi7EqfwpjROVBA6UFZxgCgu1fpQbBz1xRJkgXAzjP8AOs46cmlyGBxUzhiMDpUagKCDkVkA4FKuBtP5VsLKy45GMU5kIwBimjZhnHNRAqmG7VMwYsMZ9PalXsDTjEakL696YYAIoE52mirqcAU7M6/McmlYLkEZpjn/AApW7Ggu44HHpTYXjPT2oDvWVPINMVGMGl4G8fhWM4zxQ46UoAIJpiG6GhlRyM1kEcCiQpBBpGKN93P41JPEecc/jUjBj8v5UmNoOc+1Agn5RUgIQZHFAFxjHNFABg/hQUFOD09qZcDpQwV47URt4rg8msjORQ4FMVPC/lSjAwBRQAYJorkYU0VYDGKYZTrRxjAOT6UhBGBTADoaAJOKxk8H8KJyMfpXAHHNBiBgGiOMAURzgVQGfl79qwOhFYHcUSMYz0raCOBR6cUFAGAeaAA6cUMAZoKOpNEg96JDLkHpSpuGBQKhsjn2piucqaKjbnPNAAtipF2naDW7PGc0RnFHBGCaYDGAc0MBuTRGTgUVxwTRHoaIB4z0piA3TpSuGJGPoKYkHBPNNzwOawOhNBsHGc4oygA7hxTSA/KBReMHB60RnoOaIGQcc1u9qAHY0Tg4zRIIwKIBGKPIxmgAeponNZ420SDzXHaiQeRRxjAFdDRIJ47USD0oAHnNYHQ0MY5rGRxQCnGaOO1YyOKIxxWB1ogGjwM0RnFdeKIGODRwRgVxQORWB2qiAR9KYjGKrHAGaByPpRBb8KJyME0oJ7/pQJzg1k+tZPrW7jFZPrRznrzWTjGaLEjGayR3/CixPesn1oHAwKycYzRJIwDW6i3pRYkYzRyRgGvoaGAcijz3ocgAmic1njrR570COorIxgCjzWAe9VBJg4J5pJVfvUZOMgUsoIwTUcgYAE81xTLng0IxmgvGCc1jsDXB4BojHGa4IojtmmAHSt527c0GwOuaBDHJpgDgdaIDDih0wK9qGCuc81tJOc0pz8oFRlSOnNaChRJJV4Axx+dWc6yy785PY/hVj4fhm+0XLXW/ydvmDyyOvA71b+I4NEha0EW+I9RuI9/Qnqa8J6dp2vPdR3a8RbNh+bvuz0I9BWtaebS6aONtyJjbxjqB70kKI3AwB0rw7pmlzQSXV3LlkxhdrdyR1B+lW/iz7Mhs7K3wh/2+nfuPrXg7wnNfXQ1Bp/kPVdvsw9favg9eWPh+1/szUoflX+PcfVz0GfUV4A8OXfiC88TyaPJ5VrZ/ZN/AbduDY+8QRyDWpXVj4VsHnX95I2Pm5XGCB05/vV8QvGF9Jus7g+Yo6HhcfdPYVp5W9kFu/Ct1PXGMn+leI9HtdNvZLTT5/PC4wdpXPAPcn1NQ2t0X8t4efTcKuxdyMLNW+WLpwO/NWNxYzQ+RdxeYV6fMR/L8K8XfA2fRroRXN55Oeh8sNnhfSQ+ta54FGm3LQx3nmFf4vL29h23H1qPQXjk8x5d3/Acf1r4b6/beD9YGq3Fn9oVOieYU6q69QD/e9Kv7eLVR56Daz9+uMYH9K1f4X2dh4QXW73Wd0s2fJi+zkYxIFbkN6EHkVAhX5QM1qFvaXVpbokex03bjknPIxX2RU4HJqw0X7bKsQl25zk7c/wBfatT0w2zmNWzjvjHp70YmVQC+fTira/d08gnJ7VptuouBIh4Hb8DXwx8RX9j4I1TQ44sWupeTvO4f8s5XYds9T6iotP8AE2peDIo7m82Qx52/u0PWXnoc9RXjrXo4kFtEdzt1PTptPpWoSJE5ZG5Pt9KckuTnmlHOQaZQ54NGMADac0o5ABrwDLBHq6rcLuXnjJH8LelQhbhfnGSKl0y4tVF4Y/3P1H09c9ajiJYszfhivh14qs/BPie316e189rfdhd5X70bL1AP970q28VWt9fSXF3b8PjA3njAx2HsKvzaxak08EeEbGFyeygVrfiS41rTG0y4X5Wx3HZgewHpSxKmVC43deaurdLhCqjBHerps/u1HHYU1u5wR27VG6253Pye3bFXms3eo2JtpG+QdsD1z6e1GVHj8scDtVlGqEMV6e9WcUjPvTgdqt42Em5Dg9hX2OaMEsvPbkVLavIdsjZPc4xVxbqHyvHtTW6jhucdDRVnbGeBTHccKOKkjV4yEHNIgXI71hWOccjpTwiQbTSRiIYFKrYyB9KeJ2IcjjtSruprdXXJPXoahhMPIOcdO1Rx/wAQ60AXON2Me1aD4r1Lw6ksenPtWXbu4U9M+oPqau9Xkvd5uBlnxk/T6D2r/glN8IItX8c+Ifjfq8Ciy8L2x0/TZZBhRdTLmVwfVIflOe04r9pz41T/ABr+O3irx3b3Jk02W7NtpIPRbOH5IiAem5V3kf3nav7ReRt0j5HpihcZkLqcGg43fKufxqd2llJ9O34VseQbQf0qKMlQoO4j8K0Xwpd60AFm8pT32hvX3HpV5caboyvpEI8ww4w/zDOfm6c+vrUrxFcnlu1WZjitik0e526HOMc+1KDChLtnPXioY1C7nOcdKspJZrsRvzEPvL07fn1rWLK2mnjGjrsTndyT2H978aa1ihcSXLcHoMdfyq+mdz50g+Qfw/lVnCdWgyz7I4+2M4yfw9KvLBIV8iF9y+uMVFYw21ssUUfr3PrVrYSTSBC+1e4xmjpUdodiy7gOny4/rQTevzDAHakiYyeazfhirXTbm6RrmNMpHjJyOM8etW1lHIAG/EUbVY12KOvStIYWz7gMkdP1rRvGOp6fpOo2URzHf+V5v3f4GJHb37Yq1tZru9SOFMs2cDIHb/61W3wnvIfCkuv6jd+TMmMxeWGxmTb1D46YPStYvpFOxHyU6HHripZFmbax59amitbawXym3M2c8EdDRnMc5RnyT7e1XLMnKnrSkP8AMG/SmuPMmgt5W/dru7e2a1Dar5Rs+gx9KkBYg5ppCvyihpl9qNkwtotyjHO4DHPufahfppSNDImWXGef/wBfrV0sExNwj4Y9sHjtWpy+Ydmc/h9KtZdm4kZ6Vd3Cr82PwpmDdKIyOD+lMEaiMjIoYPfmmYFAB1o9OtRHIC9Qv4U+Cfl/KlO7gfhTEr8jDmrdwpwTmnIJBBoKMgg0AEcYPWp2YkKO1RoSm8SY9sVKyhvlHNSgbypoDHQ0wdowB0oNnAz0pSQMEUHIIwaZwTgD8KIz0PShnoaYkHg0CTyD0pyWAPpWc8A/pTZU8HmnJkOAaXhRQOeAaVtvQ0CCck5PpTAtzinOVVc880gzxTKSQCOadwowT9Kds4IP4VnHQ0pBQKe1fQ0wBO3tWABgGnlWVcuvzeuaDMI2Vec4oNxg81gkDFUSD0onI20EyNpPSsALx1oKCMk9PaiRsxmuCeTTH5cLRJAwDQ3LyPyokHtRBODmsKRtApWwM0Tk7sZxQYsuAaUZGa60QvTPSiuTgUTg5zR+boKbONo5xRAIxWM9D9KIz0NEAjFcAUMbeaYkjOelbcjihxz3FMxcgGiSeAaAxwaI2NgHNELiskjbn6URxSkt0PNMNo65ogY4P04on07UrL3FM2TkGi2RgmiQRgH6UykjrRxnB/CiijnFZB+UGpTjGDyOlFlZjxzR6YzTKcfKa8pSOetZIHIoHIontRII460ee9DgUDk+po56H8KJAOD+VM248DHtXGOOtYAAwaIIHSscfyrAPPpR5rII4NA9qOMcj8K+XHJoZxwaJzgUBgAA80cHpXIOMUQCcCjxxR4GaOCME0TgdaJ461jjGayD0NcdBWQehokDgVWQO9HBFUckDB5oE49aJOM4oMMYFZHQVnPArgjg0TmjjoBXB5FE9yawG4FEgdKyMYJyfSqJz3o4rj0rI6ZrPGM0WwuBRJxgUOmKxkcVjIwa69qOOmaPPAFbTjmqx2FYz0onPej04og4xjn0pT3BqGfaQG5qG4BHByPSkBkGVFIGUAdxSlWXrzRHOM0BngGthPTmtrAYoDnAPSmYNyK69K4IwT07USCOlKobv9BRyRgCgCAMCgTjiiBjBNADG0CsFhjNCMgZB/DFEtjAFBWUZZeKV9oyDVhflLXyn+QSfdPXODzWmWdzLxbp5nryF9fWvh/Hbo+o3WpRZsIfK+2NuPyZ3BOByctjp+NeItT0+fVZ306TNp8vl8MP4RnqM9c18G7Rb/XE0uOz+1zXufKj8zy/uJITz9B3x0r4i/DTXPCupxw67pn2NrrPkp5yyZ2qm7lWP94dfWtU0eKAYJ2nsME+nvVlbGGdWkG3Hfr2qwvY7d1kZcoM55rwpN4Wb4fX+s2us51CDy9ln9mk+bMzKfnPA+Xnp7Vd/Eic2qmUbyP4sgd/TbX7Pnxxexm1XQ44/wB3qfk7zu6+WJWH8Hv6ivE3jCPxi7afpsflhcc53eh7gf3TXivTrzSrSeHWbLElxt8h/MHO0ru4UnsR1q1nNrOJAm5R1Gcdq0o6KDFDE3mXfOVwwx1/DpXiDS9dt0F8lr5Ea9G3o3oOmaNpcR23nlNzP/FkDocdK03REnQv52yQ9F25z196vPHXiTxHPHB4j1L7VImdp8lE6j/YUegrxN4D1Oz02DXHhzFd7th3KPusFP8AFnr7ULWVm2SJgntmrPT9PR1hv7zyFfPz+WWxj2B+leF/Dl94tu20rw/D50iYx8wXqC38RH9014kuNbkuV0a/ud5t87U2KNu4BjyOueO9Wtp5X+tl4/3astCm1WJr6CDfbwY81twG3PA4yCeR2rxPY6RaPG2lXvnLzvHlsuOmOv41p6RwOmTtznB6+tahpjSgSMvB9/p71qWm+Uu+LkDt09PehG6YljPIrwrbJqc8QV8hs7jj0B/wrwNq2gRC38PQxee8e7cm50zncw5x/Wta/ac1G4ibwxpGjboxjaftKj0f+KL696+KGgar4Og0+61eHy5dT83yV3Kc+WUDfdJ7MOuKuZ9/zFsk+1Oc96Bz0P6UWH3Q36UHLcKc0gI5IrwrCq3YmkHA6fkas3jOQO1a14z8PXHgRPDdvpub0ZzN5zj/AJah/ulcdOOtaX8PNXvvDX/CRRwb4+53KP4yn9729KS3kcmVPm/TFano9/4enRNSt/KZ87RvDdAM9CfUVYaRfa3crHY2/nOc4G8L29yPQ1rFncWkvlS/I47cHsKkvFMIUrnPTmtF0nUPEF0LOxi3ynou4DsT3I7A14j8Kax4a8R3HhnU7XydQs9vmxb1bbuQOOVJBypB4NTxzxPsK4I9xTRu7bmGcVFcgjykXOe2amhdG2od2Pwq1yyYf5f1rSowbVV8rcOx3Y7mreFBLuIwW7+mKCXJG2SHPp8wq50yaORiybS2Nozn+tX2kvA+JU2n659PeriyURkwndnocYxQt2A2kfWnhYEheMda2BBgCoQ7Egr+tPCVU4GaW0kZN6rkfWkgZ5AGXJ9M1GhZhGi7vbOKud10kUUVtt25535zUtpJGNoj59c1LaypEFZcY6c0ITs3EdO9EBmwPyoRlhuA4HU0jO3yx8/pSJJJIEMe5ugANfEUxfsa/wDBO608GQsLLxb4utBZzDpJ9tvVL3RODn91AHjDDoUj9atraWeQCFdzdhnFJp88oxCm5vqB/WvsckJxIMFvuj+dMrqv7tfxzQDKCzfePSoAzkKg5rwp4O1XxFdfZtLtfOn/AIU3qvZieSwHQGrh9f8ACl41ld/unTrF8jds9Rn+8DTQy3s7SsvX39v/AK1Naxq3L5x2xVxeOw3Rpx25pftF0+Wb5e/AqaRY8JG+cdsYrRZoQWcxbh/F8xHrijqlnBGI9PiwT1+Y/wBR9a1wW18tuLS681k3b12FducY5P0q/jupoPKSPr2yPUVbRXVrY+SY8G46jI/hNW9jtGJpPm/u4/wq2sbosMp8vbkVDpyKpcjOP8+teXgYBzjoKILkFzwvUUJA/JPPpVnqN7awvbW0+1JMbl2g5xyOTWm2s8igLye/T3q0jsmiuo7ybZMNnlDaTn16e2OteHNJfUZnSI52Y/UH39qsLeS5kOlQRZL9DuxnHNX/AIf1LwpexmRfLm52nKt2Ge57NV78UvFOrWklhq15vWTG4+XGOhB/hUegrULtXdismfTj6VaaNqF5Cbm3h3qvU7gMdu59qjOIctziikZmEhOSen5VqdpHDpDSySYnfHlpjrhhnnp0oyhYwobBHUYqWeJo1kK7iucDJFeKL/Rb8xPpEewjO5csc9P730NS2N0IRcMnyeuR9Kcg/dfP4Vp3iifRtIuIYJdrNt2/KDnDHPY+ta9qccsrPP8AKWxk9egHoKllcSMyvyMZ4q7lDOQ74z3xSuQMKfrV2Sxxn6V5eBkGmxtweKIB4zzRbjGc0QCcdaIUqCDW3nIqNnVto/GnZWGQc0jlDuU5IqfLgPjpUZBPBo/cyKWSJk2q/P0rMhOAc7enaiSo3uPwp3CkMOVPSmZJvlUfj6VcbXYBOtOhTG4c0ZSYtinpWFJ4PP0oNngnp0oFM8Hn6URzkn8MUFB5z1ojjINOuRgGgCOOvtQG5dm3n61sC9eoqQE8ZoBSMAc+tE9j+VKdvOKwMcioow7BWOB607w/Z/LRcn+9k+vpRXg4HSoFBPC5P1pnZm3EZ9KlKt0OfamwTjPPYV7YqWKNAAjZPcYxQGBjH0plJGQelFfly3FEcAYoNuXaOp6igoAwTz9KUZycUwJPWiq7c7ufTFKMjrTggYBo5HG79KKjGSf0oqSMCghIyTQYf/Wo5HNHJ5zmm+bjP6V14FBeOvSiGK8UAxGAM0eBgGgTtxmicf0oc8iiQOM80wBAwaAx35pmBG0GlAPejkcA0QSM9TRAIzRXPf8ASgRtAxk0ScYBpkLDKUI3ZdwGQOtKQBz+VAqBkHnsKK4XJP6U20ruD5PpigFIxRXJ+X8BRQEYY81nAxTEkAA0wC8E89qcgjI/CiABgiiM8LzTHjmtqsAAabdjANMCFw3avMUrtBz+FY6BTk0yHoT9KCk9BRGflZsfhTDspzXOMYrOBgGt3GKogEYrj0o47VXfBHSjg9KwMACsDGc1kEAelHngVRAIxRA6A1gdqqsZ4Bqj04rFVTAnGKPPejzj26UQDVVweRRxjrQ6miOOOtDkZzTen5V14BpjjoaJ4yOaByME0T2NLyMZoHFUTt4JogYGKJzRAI4rAIojIr6GuMYxRA7GqrA7UMEZo4PSqIyMYrAxgVVdeKwDxmj6A/QUQDXUcGiMjAGa6DPeuorA7U3Sqx2BqopSjcVDdAchv0pLwOgBPI6VDKTgA59qOGGRSjPGaAyMUyADIoqCOKKjPHaiPSiQTigMnANAgAA0CDxmhjqKJBGAKAycEUFCgHPSgFJGe1ZXcCO1AkjINJtOQ/IpQAeKutTe5gtIH6W+/H/Aj9K8Na89ldpNGfu54/A+3vXjX4kXWqT63c2h8qLWPs+8cNnygoHVc9R7fjSTxxoVY/NXhnVJbK6S6ik2umcfLnGQR6e9XniuXVoIvtVx5jpu2/JjGTz0A9BVnp2j30FzNfpv27fL5YY5weh9hV8F86REPyjG38hUFs7oNg6dq8Ef2zqDroVg+0XPXhT93c3f6HvU/hO9tZ202V90qYycAds+vpXwwm0/w/4kWTVXwi9ODx8j/wB3PqK8e+INMM/2nQX2J34Y9lH8Q+tJdPrsub+4zjp8n+GPQVf2Mdmvlxy7z2O3HpWlaPetLHfW67jJnnIHTI7mvF/hvVYtGhFzJ5aybudqnGGX0NC2YWqsbnaR/Dsz3p4tPi043Qb9+OnB9cfTpVhosN3iVLra/f5Cf6+1aRNqMd8LJ286C36HCr94E/XrWr+HfD9jp0WsJd+ZcvncnluMYIUc5x0PpXiLSdMutJkvbeTMwxtXaw/iAPU46V4NuPEGhym7tOAe/wAh7MO+fWtakd9SaVTl5cZ7dFFaNFZxXJbUFyvYZPofT8Ku9Yt7eO4i0yXENzt3LtJ+706jPXNeEzaQ6iJr/kduo/hb0/ClguBdmWUZj/D0ozNONgHA6CpoY4rd0jXO7Hf0NT6dK+YlGCen+c14q+JXh3xJp9tovhix2XY34Pmue6t/GoHRW714s+Jv/CG+HbXR7aL7Nqx3+Y27fn51YcFSv3W9e/rUV/DdXH2sPiU9Tg+mP5V46v7jV5YpZ5N4izs4AxkLnpj0oIWG3HNSRsrbB2pR2x9KZTnAHTpQU9Mc1pOlyajOkKDjnJ9OD7+1XPh7TNJsENlHmQ53NlvXjgk+tW9tLI5hgi3O/wB0bsdOtPpc+nTKupx+W4z8uQe3qD7io/Empw2X2KOfEX93avrn0z1pYoo5PNI4avEniW48Vuk9/JuaHO04A64B6Af3RXh2+urOVp7cYKY9O4I70lxa3t5u1A5B+vp7fQVfpE1xKtsn7sY28n0GetQ3Mtn+8t22uOhxn+dTatezao2p3Mm6dsbmwBnC4HAGOleCLrwnqXiOF/G1r9osF3b03yJ/A2OY+fvBf85qKCaZS5j3AdecUpMMgZBzU0kyuFfr3rTpN6/PyB2/OtH0wy6al4kuFbOF29PmI65q3jjih81V+f1zVmiXq7GHzjp/n8K+HmuaZ4Y1mSXXIvNh4yu5l/hb+6CerCvGV7puo3LvZR4RsYGWPQL6/Q0vhvULi2Nza2+YezbxxzjoTnrU9siZiZP3g96uIHckyDg9KewJQ4GRVnZOBsx07U+lSoFMyYD/AHec5x171JZXCWQuhH+7PQ5HrivDkInuhHKu70GcdjV/EZnKomMe/wBKjs3LpuHAzmtZ03SIId9gfm9Pm9vU/WvIe68uJUyTnIzV1pl1bHynj256DIP9aEDBuB0qWylCBUG4t+HSltpbM5kTBPvX7DfwgHxl/aO8O6beWfnaNoDf2xqoYZUxQFSiMD1DymJCPRm9K/4KM/E6w+Jn7Q2nfDA3Qk0PwNaPDOFYlWvZkEkp+Uj7qiFMdmV/UiviB4f8L+FtUtv+EW/1Z37v9Z/dXH3yf7zV47+Kvw/8R+DLfS7TTPLnj3Zbz5jnMqnoUA6CtOtTqd0Iifnbv+B/wrWfAOq2igRDzB/wEen+1VsgiiCscnvQuFQbU618KPiJc+Cb5riOTazYwcA9nH90/wB6vG/iCXxNrLajdNuDe2OiqOwH92mV5XyDhOwqW3mkcRxL+tNZyRNiTkdhRt5JGCxj5fStP0aykjL3Sc9uT7+hqSGSA4tjtXuOv86+03kMQPl5PY5FWd7eu7qIuWxj5hVpa3yOJJh8o+lSXMFw3lumfL+7yeM1CibN4GAKK24tQ0Y5Hfn1qBh5LDGRxTI5CgfjRaMKFx+FJGqjcBzVnAzsN1eGdSsNPDLcJnpjk+/oPeraG2vrqJGOOu7g+nH8qtYktLny42wD7e1aWreHJdP8Q2lzll83C7OnBXvn19K8Q+L7vxLqyXd/JuZc4OAP4QOwH90VfzLNeTpAmXbb39hVxCqsdx5HUVDqE+nwtawyfKevA+v9aWZBEUDfN24pZHVMg8j2q7vJ5Y0V3yBnAwKeN2ySOKkbZG0atycYGKDSNk56dKe6m8vyQ3HpitLfTVuY11JN0PO4ZYdjjpz1xXxat/CFlqKReDYvJsucrukb+FP+enPXdV9rduqvbyjJGMHn2PpV5cwMjSRjrjHWpB9omIPIHbp2pucAVIjSNhR9KYE8A1IFCZYcimlUnGa3Buhr7wwDyKJAwF70zArtBoNjgdqBBFcAfN+VNIrx7U+9Sh/X9KV2Qgqf0pCFOT3pHQNuHJ9KMjOu0mgkcsewcFentS4jGFPNSsBJmmyTknNR4DAY/Cp1AIKUQDypplGAKBA4NWFk10WJOFXqcU8OHKK2ce1PEcjaaW2kmOyJct6ZxV5pF/phVLuDyy+cDcD0+hPqKKgjHUimRjyKZdo4qOBpmEaDLHoK8SeDNW8MRWkupJtF5v8AL5U/cxnox/vCoo8qc/hRUlgq9R0FEEDBNIxXmrfTZ1g+1snyHvkeuPWr0oJfk6f/AFhTuCcCqAzUZDEAmmUFetEHaDmiTtAJ5FFsnApSQrYPXpTYJyKGBwKI5wx+lYBGCfpXB4BpwTjB6UVJX5RRUkZBoHI25pmO0KD0ogcEjijkHaDmiBjGOaPzcDtVMSRwaDdFDfpRJH3TQG4fSgQVwOtMGPB7VkjHNEk9D9KIOM+lAk9DRwTgn8KAPQmjggZNHIGBxStjjNMDxg0Oe9HgZFRzLGpU9TQfIIBpSFPX9KZtzZWnYsMVgjAB/SslTkGpZt7ZApn3ClAIyaZlPK0xUnjmpASnH4UrAjBPNEgdDzTEEYzRABG00WA70z7zgHiiVLYzRYEAr1osGOCaZwp4NOylAVOaYlQBn9KDoRRIPasY5x+FHb0AoFRxnmjg964I4NZBHHNEcdKOMUcHgVkdM0QOgbPtiieMZ+grBI60ORijz3rOfrVZB6GuDwDRwRhW/Shg8Z/SiQOM1kHoaoEHpzWQeAc0DnihyKIyMCsg8A1wOM/Sgc96PBokYyK4HH5UecEc0cEZBo4OCDRIAx+lKR1/KuvU/pQxjrQIxis9wfwoYY1gYwKyM5Fcf/WoY65qgRgc0cEcHNYBHJokHmiQw4ojnANUTngmsH+9+lE/LgH9K4PANEgcGjjH3v0oYOOaI4wD+ldeh/SiR0B/SvQZo+35VgkdaHIAJolegNDB711OAfwoc8Gjk96PI4NLLt4BqG6AGCefpUF4q4wfwxVvciQAZ59MUrdMUpGAM0yAgDPSmAxhTRAIxTA/w1tA5IrOe/0rORW1Rzu/Sgeev4Vgfw18p6DBoDKYAp02nFDcRimwDhRx2qPao3MKAL8j8qsYLaOFpb1tvTyxgnPr06dqs0ukYtFFkdvmArVb+6urO1tIrTY8G/LeYDuyQenaktbiWQIyYY+4rwXoD3VyBNB8qdfm65DehqfS7qTUTa6fBuPpuA/hz3P1q6huLCz8iU8ntgev/wBepHsWXEi4cdOTVuzK4S2bch/ixj9DWk6zdaFJHJp0u2UZ/ebQdvXsQexIq28SahPqLXNxdZZsZOwf3cdhWuxJODNbyb1Xq+3Hp2rTZtEvPDx015P9L/v4b++T06dBWm+BPEOsX1nYWFj5015v+zp5qL5mwEtyWGMAHrWufC/xn4G1VNJ8aaV/Z0z5wnnxTdFDdY2PZl/P61pWhajNcGy8O6l58EHQ+SF6gn+M565rxPpfiPxV4aEEmn+ZND0bzUHVx2yB0Wr3wvNNaJdyx7ZFzlMg98dc0NSt4S0NxY/MfujzT/QVYQI6s6rg8YXPTrXw+8T6X4dsLu41qx3xDZl/NYbeWHRQSeSKtda8JXHhuJR8+pHO4fvBj5+O237tWKQfacyvtj7tgnHHpXg9PDN18OYZYYPMufm3S7nX/ls3bp04rUdMs0MV1b3PnHnd8hXHQDqasbNL2bddSeWP93djj2+grXfCy6bfkaXffbYP+enleXt4HYknqT+VWHhPWL9Y206DzZudo3Kvr6nHQGtR0y+t4Ps09tslX7y7wccg9j6VoPhe7vbO5u5J/LeLZsTYDnJIPOfantnBYum0DHfNX0EYQTRN932+grTtWbQdRTV7RPOkjzhM7eqlepB9fSvHfjG98fat/ampxeTMPuruDY+VV/hCjog7UkEysCG47HAqazN7bPGT8y4wfTn/AOtU2kTxMXUbvbgf1qXT5XG/HXt/k01pcKclOPXIpbWV+AvJ96ttAvZ8FU/Uf418NdH0jRNO1P8A4SSz3vP5X2dvMYbdrPu4QnPVetXMgjt/s8Fxuz1XZjvWl381peRagg+eDPy+uQR1rX9cm8Vau2oTvmX+CHA4+UA/MAOy5q50XU4FLTQbF7fMp/rU1pdPBtZMKnQ5HGTVvo91dLhflA78H+tW2oJYxy24Tcz7fmzjpz0/GnsW1WYW8DYkfoMegz7eleN/hn4z8AWmnnXtN8gan5vkfvon3eWV3fdZsffHXFXOl31tCGu4dgPT5gf5H6VqEUiPuJ3FugxjHSoDIWxI2F78VBfXERManAb2FWnlG9UzNhOc8e3tXit7K81BrnTE2Q9uSeyjvz1BrRiACrcg9/zq0v7m2URI/wAg6DAqO58uEPLyo6D1qxvZJ382A7fbrWp3BgfzpnwX7Y9MelXPmFg8R3r37V4X+JHhHSvBEOiXWiebdDdvk+0yLn96WHAUjoQOta1LHPqMsttDtV8bRuzjCjPWriImXyyu9R93nFTW5C7BFgd/m6VZ2c7fvol3Ae4HtRkW+toTcwf8e+7B3f3j7fSpws1mtgjYQZwmOnOetWGjXdhO0zrs/uNkHPHPGfetK8F6trLvFDHs2Yy2VPr23D0q8soWhxbXPmbPvHYV6/WoLAbvMXkjtSQS2zpeRpkDOBnp2rxvBazzWcum3Xmk+ZvXYVxwuOv41daUbZgkqfMv3ufy714J/wCETFz/AMT29+zxjq3lSP2bsv4V8QdN8KXN+ZfDeofaIT0PlSJ2X++c9c1+w14Y0n9nP9l/xl+0Z4qtgLjVoZLqFGO1ntbYMkMQPZpZy4HY5jNa1q+q+LPEOp+J/EN0ZtS1i8lu7uZuTJLI5d2/FiaTRJowZM5T1wP8avriN7dYEGCM5H4ihFLpkaSCHBkztfd6deOfWoPFWtGMqb3fnoPLUf0q40a8SNJUOd+do47fjUMaIwV1y3rmtLeyt7qSSeLzV4x8xXHB9KuZbaY5D/L2XB4q3NmWG+TCj/ZNTS2SpugXJ7Nk8fhSwFnLXU/J/wBn/D8KtbJJHQwHevPbFW/hzVrxgxh2xf3tyn9M+1WvhmETxi6izGM5fcfT0Bo2Vkw8qKLeB/tEVJpMMkkclomzy87xknr06mpLRG4efcP7uzFLpFvGpcDDSduT0/GpbSJI9sr4QdsE1uWWTyo22Qr2xn/6/WricbPKt0wPXOf50yidQpfaOxxmpbdo+V+dl6ds0YpRGGSLK+u6rRpXKqg4Gc1uRQE/iFQKqIrofmGcVI8jKDGnTvmhdKLdYojuYZz2xzTSpJHsiixJ2O6vAeteGrW11ldbg86+n8j7Iu51+6W3/dGPu46/hV7GJb9zP+6D4wPvYwParqVnmxEPl7c+1CQuQQ2T9Ke7MalDwaRmlU7TnH4VcStIvlxR5b0zTiNZVVzgjOR1xxTvGiYQZzUrRh9zdq1B1vIfJsPmkPQdPT19ga8beGPEHhzTbC6121+zx3/meR86Pu2Mob7pOOo6461qB81id/A6cfSr9lihAU9Pb6VaszkkcjvUjMgAByaU4HD9O+OlSKCflq/lS3tWdjluMDp3FJdyM+c8dhUDZ+YtyO2KRsHinIGApziie279KBBGCaUA4C9qOSM5zURRRgGkPUA0MyYVRzRJI2g1ECrgA8io1Dg5bB+lRyokfJw1Bip4bI+mKkAdAwHNEKBlWyfTGKLI+WUbSO3WkYDOT+NPGydRQ2sMMeaKEnOajuZfKa2hP3uv4c0FAGWfLemKijwdsg+gr4d6bpayPf6rJ8gxhcNx94dVP0r4keKtO8QahHBpq7orbO2bLDzNwTPykDGMEe9YKjDH8aZ0KgAYNAqeG6VYzwW8sc7vhkzxg9xivG/jzUfGyWEF/wDLHYeZ5f3T9/bnoo/uj1rzGYY3YA9qDAMCD09qZvMJOaQLIwXd074q/wDEllcaP/ZMFp5Y/v8AmE/xBumKuTjA3Z/Cj70QOx/SsDHDfpUKFmAFSxkYAP6VgMMbuntTIeBnmimRgNz9K6Lgt074oAqOTTY4OfwpsE8t+lBdwABo8Njdn8KVCw479BWCp24z7UwXHy/lWM8Cjwvr7UACmD+ApSCOOtFc4LH8KXcO1EA84rOe34UU2gEDr0oAY561gr3/AAoDHBogkYB5ogseTROD93P40zbhxQC7QSeawCKAPbmiAVyR+FFsr0rgDGelAZGB1FYHQUBuxk1tJGSOe1L8vJFE5PHSioUjLZz7UQMYFOhxkDNHk4Bon5cAUGUDGM0WwNoFZzWQAMUzEjgU6MhGDk0hLDDHkU6EncD+FBlDZJ/CmJbofwpiCgC9e9OCCAG5+lKqgBS/PfimAByG/SnUls54pdrfKr5P0pgDwTRj9DRUHoaOCMZ6UoBGQaIUDg59qAB4z+FYBGAelYB717UdpHHWiMD3o4IwK6UQCcZrGRwaIB4H4URXBxk0cdqwDxmqojtVEDHBo47UcY61RxgYrGRxVdsA0ORVYGMA1gYwKwMYFV14FEA1weAaAGM02CcZokFaABHWhyoFBQe/NHHArAxgUQMcGiMjGaxngVgEdawMYFcHiiO1EDsao8c1WM8A0R2rgjg1gHjNEAj6VVHk4HaiARgGsdgaI54NDkUB71gYwKIzjmjgDis8ZocijgDIraTznmhnGM1HKUOAc1Bc981Z3iyDaxpT8uAaVu5NHDcA06gjAP0plyMA0oGCD1ojI4oDJxTDPFAZ4zTZAAHasE9/pSnt+lXluI5CsYyB07VgseKWPPzDrW0k9enaoX2cEUshdwSenavDssay5bkf/WNan9ne9mOP3Y246+gqxCy3yvBHuC9s47V4O17R72xvo5Yvv+Xt+ZuxbPb2qz8X3nhTxK+o2dvu2Y2jeB1TB6g/3q8Y66dfuPtDrsf0znso9B6VKNybkGStW07IqIBzzkVY6NeXdtFKsO5Wzg7gOh+vtUNncSECKPJPbIrX4rjTtBk0m6gxMcbDuBz84Y9OOhFeFdIMYj1Jm+UZyuPqPWviR4i8La5b+ENT8NRbDp327zl3SH75QLy4HoemasDJ478LzFrf9/8ALt+f/poc+g6LXwM+B7+DIbq61Jd89xs3r0xt8wDo5HRhXj3SZdP1S4urIYRduV/4Co7n3NfFzRtI067h0zTI8yru80Zb0QjqfQnpXxC0VrDVAxTb7Zz/AAr7+9ahJjRrWeNcMd+7n/aAp9W+22AgHHl/e75yfp7V4J8U6Po+mz2ssO6Q7dp3MP4mJ7H1pNauJIntUT95JjByO3Pp6Vof2ywthHJNwf4do9T/AI18EovD1xY63b6vwbX7P5f3+dxkz938K1O8tbx5JGj2FcYGSfT/AArw1oa+JLSOGCHIGcnd7n3H92vgbpHhuS0s7e8t9r22/J3uc7jJ6H6V+038KtMsPiVe32gy77efy9g2sMbYIQfvPnqTWo6fqlpqBtnXYD05U5+UH1rxzILUqYj1zn/x2rKcXUbQufvYxU9qyOYmH41dWpjkDDkChAVPBqOdxG0Ma9cZ5po5M7MZp7CNwi7fm5zzS+Ey1mbovkDtj3x61FpcEM5Q9uvX0+tWQijm2qvH19qnvpb4l9uF7DP+fSnBeMgHntTxs0IQNx9KYxwKNnVavvFNxq9uIJYtvvuB7j2HpXhrwLe+I9L1PU7B+NP8nzFwOd7EDqw9D61o2vPokkttJFuPH8WPU+h9aW3aR5JSeFxgYqa6MUizIOV6fyqTxLdz3kF7cN5jw7tq4C4yMdQK8d/FS38VeFE0KK02MM5bzCf+Wit02D+761qLm6kCgdf/AK1TaZi2aRD93r+f1qG0lkhyhyRUC7nMbjla1KxGmxfYpW3svfGPQ+/rVlEplCp2qSFi4B5FBJDY4lXgdOferN1CAA8itK8NjxbGYoGw8XbHrn3H92r6Jre4NqR07/gDREUI8uQ/SgsMLeYT+hqDRmv0DRjkd/8AJ9qksZoLgRSjI7/lQtTHctGBwf8ACvsMbweWh+YdPzrR/BsV7pYupJcEdtv+0R/e9qvLdYNUsrdGyv7zJxj+HNX4u7WPJfaT7A+lSp9kgKLzv7/Q0sUSgCMfNXgX4Sah410iGWwudrLuz8gP8Terj+6at725+HXiLfcw/aJLf/aCfeT23f368STeBvHukz+IjF9nvLzbld0r42ME9h0X0rxb4L8Px+F7rUNMm/fwbMDa/OZFHdsdM1pFrFb6WJJG3M/bGOjGvjd+1zqXj/8AZ10H4IWPgmDw9BpjW0V3La3ZeK5t7ZAIkCEbk+fYxBLcoDnk1bgSYdxitDmZL6NAcoM5HTsa8SRafYRfbkOZH/h+YdMD+Rpo5bqdRKcAdvwq70p7W0BYdf8AH614D1lPDfieHUp4vNiXdld23/lmw7Anqa8X3ttrWqfarRNq9xkn+FR3x6UYJFunCjOccfhXh7TxDItzPyPT8x2NJp0TpujPyr0H+TUOhtqEoTOB/n39q0bws/mZm7f/AF/erTw/FGSw/D/OamhgtoyFOSO2CKa6Sc+WF6dOaNorxhXGw/nXkCEth8g47YowOfliGWq2knLYmH3Og471cQiR978D0p4Zbk+ZGmFHbNJBHghhz6VZ2cV+xeTgL0/z+FapoD6evns2Ufpx6YHr70sDyqIielANAwCHmra0juIBM74Ppj3xUEhlZYh71rc0dzCiQD7uf6VAzo2wduprw3qstlqazpFvH+9jHykenvWoJ9kYQxnLDqcfSrWcGYGU5x+Hap2BcleprL+YGTrWouI5DH1P/wCqrabaNpPSgGlmJBx6flUqtBIrScg54q7mjeD92eR/jU1+gtyWHzf/AF6F1OhwG4+grX9bnadYZJPMWLOOAMZA9qv5mkcsp6//AFq1CcvKUU8D/AVboRHkd6/ZG+HPwz+L/wAdtF+GXxUn1KDS9fhngtprC5WB0ulTfGCzKwIbYyYxyzrXjb/glR8BY/BuuSeC9Q8X/wBvx6bcNpQuNRjkjN0I2MQdRECVL7QQCDjuKdHjZkdSrqcFSMEEV8Kfh5d/F74peE/hlZFxJ4l1i2sndBzFE8iiWToeEj3MeOi1cf8ABI79kmxtpby61fxzDBBGzyySazCqoqjJJPkcAAV+yD+wJ+zP+0X8Lb34mXtx4ygt5/Eeo2umRw6lGmLKKbEG8GEkyeWV3HgE54HSv+HS/wCyz/0EPHP/AINov/jNH/gkv+yyTn+0PHX/AINov/jFf8Ol/wBln/oIeOf/AAbRf/Ga8a6PaaD4x13QbBnNtpup3NtCZDltkcrKuT3OAK/Yz+Atl+0f8eNH+HWuSXcWhLbXF7q8to4SWO3iTjaxBALStEmSON1f8Omv2Wh01Dxz/wCDaL/4zX7en7MHh/8AZf8AilpWh+CptRuPDWu6St1ZyX8qySrMjsk0ZZVUHGEbp0kA7UORgUgwwIPSv+Cc/wCyh4F/aH1XxhrXxT0e6v8AQNCtoLe1iiupLbfdSszFt0ZBOxIyCOn7wH0r9s/wf8LPhn+0Jr/w++D+kSWOi+HYoLWYSXklyZbryw8x3SEkbS4jI6ZjPrmv2a/+CWGu+PfD1j42+O/iPUPC9nqEYlt9C0+JRfeWwyrTSSBlhYjny9jMARu2nKi1/wCCYH7I9vYmzl8K67cyFcfaZdbnEgOMZwpC57/dx+HFfte/8E6vgp8GvhB4h+Lngnxt4i0s6IkbDTb/AMu8iuXklSNI0YBHTLOPmJfABOK/ZX+Gnhz4y/Hzwf8ADHxe94uka5czR3LWcojlAWCSQbWIIHzIOx4pf+CUH7LqnK3/AI4/8G0X/wAZr9r39nW9/Zo+Mup+B4muZ9BulF7oF5PgtPaOSArMAAXRgyNwMlQ2AGFfALwRonxM+Nfgj4feInuV0zxBrltZXZtnCSCORwrbWIIBweuDXxn/AOCZX7OHgH4PeOvHWhXvjFtS8OeGtR1KzW41OJ4zNBbSSJuUQgldyDIyOO4plww5oHjg14d0J9ZmkMcuzysZ+XPUH3HpVzGYrhoic4x29qswVJLHr0q4uZIYtsT4H0pjnknmgQeM0q5zu60hyenSpCGOcUEJxzTR5J29KKYIINBMgZpYlaRVDdenFSoY2wG/Spm3HIokk0BngGiOwP0FWsJkHy9qYEHaTUoCnYozim5AzQXA559KCKUJJ5XpXOPehgnnrRALYzRIB4p1DMDmhYTfZxcgYjP3Txzzig3UjqKUk9/wpQOoPNbSOO/asEEZNDIbaDQIHyigRjkVjIyfwoHAxTtuRV9M1jPANMe2aABHWicjAougXA60STxTKFGFoLu4zRIAwBR5GB1pDnCk020jANEEjINBWPyk1jNFQU4onjB/CmORgGkUkYzQIDZz9KdFVd36UGJFHGOtA5OBRXnk5o4K8GiARgGgoIxmiMDANSfNwKjLEYFBSy9aKIDnvTKN2M04JUAjpTAkZHWhk8GiN/U9OlH5SV/KkIWvLIHJ+nFZUcGmxjArGRzQBIwBW0kdf0oqOgPNKCec1tJOBWMDbmiBjGefSjgjOaCj+E5/CivHBrAPJP4YrGeBRGRwaoc9aI7Bv0rOOhojtRHAINH60fQV17UeegogdM1jAwDWOOtUQBwDQGBjNY4xmgc1gEYB49KIBGKwCciiATgnNEKRwabGOa68VhSoHeiVIxmiox1rAPAb9KYDoKwD3rAI6/pRwO9EcYBrA9awAODk/SsAjrWAR1rAx1/CicmiB2P6UenWsADg/hRHTB/CiOBzRGRgGiM9DR6daKkjr+lEADHWj7CiM8UenFAd81gE5BrqOlcV17fhRGeAOlcHgGsADpVdRRAA4FZGMflR5GMV7VFOyEDNWeobUCnkVDMkgDIfwxQkzwDQIPFBQwxnHvQGF4HSig6k1nnCjpWGPAogKcAdKAzxQxjihzyKgvJYo2RWwD0GKDbsYqxltooysse8/UjFSFHmZlXjjAz7VtyMiiM45qK5ljx5bfpX9oTXSqkzcDOTj/CvCmtHR7tWhi80c8btvZvY+taE91pV9c3MC7kj2ZXIGMgjrz61b32n+J43lvpvszQY2jaXznr0xj7o/Orj99hs4PpirC0EoaIJuLe+MYzWmaRBLtaQ7uvGCPX3r4R+IIdD8OXdpejzIDs8scjZ87k9ASckivh/4Hs/Fes/brC98lOy+WW/hYdSw/umvjle+H5tY8Oy6jBuS2+1eeu5xncse3p+HSpbKPVNQaTTV8mJMfLndnK+/PUGvBGo6fcappKb/I2eduXDNjKtXww+KF14h8WR2kejfZ7dM5l+0B+sbnptB6jH41o3xM0W7spJLtvOeLHGGXGSfRfYV8RfiLZ3epT29jbfM23B3n+6h7r7GtIsrDx9rX9raa27Uk/4+EwRnKlV5O1fuoegr47ast/rb6Ze2X75cc+Z935Iz2GOgFeIwbJY9Ohk3Qx5xxjrg/XqafSmuYGuUbCR/eOPU49asLBngWK3GSM5b8fQ1a6fox8N6vcB9moWnkeQuGPmbnw3fAwvrXhXTrfWLsST3G0d12E9j6Eelad4R0nWNWvobafyo4PLx8rNnKn1I9Kk0O4m1VLO6iynODuHPy57Gvg18GNDXwFp/wBii+yXU/m/aBueTbtmk29Xx09PWr3wHoPhyK4uNXvvMlTbtHlOOvB+6x7EVrvjO8tdWeBLPaLfG9vMBzlRjjbXxt8aXPiKPT7tNL+zra+blvPD53eWOm0Y6U2pXdxcGHUJPNA6HAXHHsPpV1DBYuJ7V+B2wfp3+tJf2V3CJfL6/wAGTx+P4VcmIz7oB8g+8vPpT2MM4zCcZ6cE14a+Fupav4e1fxA9x5SaR5GE2K3m+a5XrvGMY9Dn2qfRRG4Jlzjtt/8Ar1caa8wEsLbC3UYzj9a0vR7oxlJEyfXI9/etG8O6beX8kd1L8gx821vQ9gfar7QbMXTW0bYjGMNg+gPTNappSaW5trW781R0/d7fQ9yfWjYSIMXEG0N907s9PpUsizSGNFwo6c1LF5Q3Ffl9c0MRny0GWrR/EHiDw5DNLpV35Im2+afLRs4zj7wOOprm7iM8j7pD14x7f0qQ28CEFclugye1S2he3LA5bsPxqzKxEFkyB1GamhS5jJRMD1zmpo2SQqI8gdDmvPcwmEvhW7Y9KS7azjJgkwfpmnKyqZz949f5VpEJ1HUorW4fYsu75sZxhSeg+lav4ZPh7UIo4Ln7VFLuydmzbhR7k9TUcCwoGZfwzXw88C33xGe70621T7I8GzZ+4Em7O4nqy4+5+tCCKCd7eNt7JjLYI6jPStD1W60nUFNqcFc+nPB9QfWtXI1KXci9evPsP8KuofJnMbzZA6DbjtX2cE/O+fQYrR9cuNIDIibvXkD19j61fahdTTC4jfdn+HAHYDrWh6lHdTGS7g3Ieg3YxwfQfSpPhDoknw4vPHsOvc2/l/6L9lb+Kbyvv7/x6e3vTDS4IREl7n/Z8phmk0Y+L9d0/wAN6DBm8k8zZ82M4TefvEDop71420fUvDdzHpGrtiRs44U9lb+En1HetU+C3iBfCcPiu1n8yybdldiLs/eBO75OT7VZraGLbPNgj/ZP9K0651C102KbR5PKjOcPgNj5j2b8avZfEehXq3+o232hTnjeiZ4A7Z9RXjK+/tq/kv7Cx8i0bHlxebu2YCg8nBOSCaurq8tyUZ8xntgV4X1exjt5muLDzm+XA80rjk+gq8Meq2EzvB5e3btXduzz68elW8K3jrErYBzjj/PpUmjzaK6zLHuZ87RkDGOvc+tS27yMJHOW7mpra2lRLiE5Zc5HI9q1DxBouo2a29xb8rnHztxyD2HtV5ZxJKJrN/3TdFx93j3561beDhfWZuobraw7eXnvj+97VbaVLG7ySPk8YGMf1rSPCou4xLcz4X02/X0PtVl4b06NMNyw6n5v8as0sTcLBFZbsd/MI7VeThW2xfM3Zen60nmMf9Ibyx6Yz/KjLawMYiN5/EVbxSS3O6F8Beq49q8Zzy+Jpo5orH7P5eePN35yFHt/d/WtMSazu1tbpvmk+5wOwJPSobMeCb1JdV0/7ZuzhPN8voPVc/3h+VS2drJK19GmEGMR5PpjrWoJHcx+ZFB5WO27d6VYm6hBjEnEn3uBzjpV1ZRyONhy3pQvFiQIiZbvzirjULu7QQTTZUdF2j+Y+lNAsjHyxwOgox/vAuc/0qVRCNpPNaNdQRXRluU3ov8ADkjPB9K0eFda1JbaBvs6c5ON/wDCT7en61r9lonh/Utd0KST7Q9v9n8mXDJu3KGb5ecdccmrDUrqyjMMDcH2Hv7e9NJNFceW7bh6Yx2q5jlLZjjyO/NeCvAsXjW1vy2tfYWsfLwPs3m+ZvLf7Qxjb+tXMMkDNAJvmjxzt65p5TNKSTz2/KjayPcKAnIzk59qkhnhuNgO/wDADtV/5ryGMtnb1GOnSoLd5o2eabaox/Dn+VX8qSyJFBLuLZ424xjFarK9sPJSTcT7Y9Ku5lWPywdzHvjGKkY55bP4VdEGQ81bykcV4N8U6t4H8X6J410WTZqGgajBfWpzj95FIrrn2yorwf4o0nxx4S0XxnoU3m6drunwX1o/rFLGHXPvhhX7aHw5X4T/ALS/j3wnbW3k2UuptqFgoGFEF0BOir/sp5hT/gBr/gkv8MD4s/aB1P4jXdsXtPA2juYZCpwl3dZij9v9SLmv+Ci3xcHwh/ZP8X3dpd+TqnieMaBp2H2sXuQVlKnqCtus7Ajuo6da+DX7cH7RnwE8DxfD74Z+LrHT9EhuJLhIJtJt7hhJIcsdzoW5IHGa/wCCcv7Znx9/aK+OGt+CPin4ps9S0mx8KXGoQxQ6ZBbFZ0urWNW3RqCRtmcY6c+1ftz/ABb8cfA/9m/X/iN8OtThsddsLuyjgnltknVVkuER/kcFTlWI6Un/AAU+/bIZgD8QtLx/2ALT/wCN1rGqXut6rea1qMoku9QuJLidwoUNI7FmIA4HJPFf8EePhgLTw144+Md7b/vNRuotF0+RhgiOICWfHfDNJCPTMZ98Xfx3it/2x7H9n1rpfs1z4Gk1QAEc3v2niPHXPkRu/wBCPU1/wVi+F/8Awl/7Pdj8QbSHdeeBdVSV2CZItbkrDKB3H7z7OfTCn8Ovf8KXa52A1+wJ4V0/4G/sY2fjfxGv2carb3nijU3IwVg2ZjOTjj7PDG3plj9a/ZRtIvjx+254W1TxykVydf8AEd3rV9HJyjzRxzXYQg9VMkYGD1HFfFbxPrvgn4Y+K/GHhjRP7Y1fRNFuryxsNpb7TNHEzIhC/MQSBkDk9BzitV/b8/a41fVm1pvjPqVrIzkrb2ltBFBGMnCiMR7SBnHzZJ4yTXxa/bb+PHxx+F0fwq+Jes6bqdnHqUV81/HYJbXMpjVwsb+VtjKZcNwgOVHNfsBHP7YPw1P/AE/XH/pJNXiXxLpHhLShrWuXIgs/tVtbNKcAI886QoSTwF3yLk9hk9q/b9/Zu/4aF+CN1JoFiZvGHhESahomxcvcAKPPtgO/mIowP76R8gZr9kRGX9qP4VhhgjxXYggjH/LVa/acx/wzZ8WP+xH1n/0hmplI+6f0r4Rfs7/Gn49Xslp8KvAOoa1FA4Se8AWC1tz6PPIVQHHO3O4joDWnf8Enf2o5dOe4l13wDZyFc/ZZtUnMjY5wCluy+33q+O/7OHxQ/Zx8RWPhz4oWFhDNqcDT2UtlepcJPGp2swAwy88fMq57Zwcarf2s9na29tD5bw797bid2SCOD0xUbsBjd+lSR4GQeO1Bcd/wxWzgMenpTqQNw5HpUYVxgvx64pz2ByB0PSoHTOx249MVN8NJ4fA1x45juN9tabfMTYBjdL5Y53Z6+3+NXhVpcIuPx9qijVpI0QfNzV2eQgPSnXBwDXbHesYANRQtKcJyfSpfDWqaJbQz6la+Ws27A3qehx2J9RV2gEo2Dr/hSwhyQrZx7VIpB2Z6U7DHIrIAyBn2oEkChknIFE96ByAQPwoDPGPwqTVb+Sxi0xpt1vBu8uPaBjJyecZ60AFYbh+FPJuYkDjtQBC4JokkbcUAe4o5PAFHgciuc8UQDTZY7VHPapU2AKwoEEYI/CmBPIXGfeh7CiBjApgCeRRAzxQyegzQbIx0oDI55pxjBFKQeM0xx3oYxgHmjhOQ2fbFNgjOc07FgMCmUKMHrRAAwB9KIIwVP4YrAILA9OlE570SMc0xBHBrHOc/hRbsBmmBJ3K3HpilUkcn6UScYDc/SsMBktn8KYqy5zzSKWOxX69eKaIqMl8+2KMRUZHOe1FPfmiwAAIpyWOQv4Zpc+n4VJkDIogkjBpsA43/AKU7sTgHpWQ3JXP40QSBiicjGaIyM5o8rjPNYz0rHvQA7N+lbT0ojtRAAwDRB28GjzzRGehrpyKJQx4z83pXJ4H4VweM1txyDXB4BojFZzxVYyowaojHGaIBGM0cHpRGRgUB70fQVRGcc0VBrHNHkYoAYwDWBjANEAkCsDGBVYBAGfpRAPAPSiAQOaIHY0BkY70BnjNADGetUQOxrAI61wRwaIBGM0FBHBoAHjP0ogZxmgB2NUpQRsG6npVEcUeOKwB0orkYrHGM1TcdqAwMZojOOaqqOegFcEcGj04oYPQ11FYGMCiAOc0RjoKHIoqDXB4BoAdc9KiZgoxVrdtHIDn8MVbXiTAHvW4AAA5okt0P04rHYUQM8Ggobj8qIwcUQeortiuD0bn6UhyOtA5HHNKc8E0ME7e9QxM0m1BkmoNKWWDKff7j8frV1bSWrBZBjNAEHGaXcOhq1n8twUPzfSrDWLm2s5YGGRc4yeP4SfapBIQBGevtVppUrS7VGSv+H1rwrpM0upxRpFuc7sDcB/CfevDOr/2bon9nXFr83c7/APaJ7A+teJJHjiJs3+Ruox6Y9a8B3euWTpeWEeYue6f7Q789Sa1bxpomv20lp4gn2RTY+baxxgg9FUHqBXiLwPpZ1EDwjc+eg+8djL/CP77f71SRXc0E11YHIt9u48cbjgdfxrQPH2uaNF9jMeyJ+rZQ9MnptPrT+PNVsLjdbv8AOnbC9x7r71rPj241kiS7+Vx9D6eij0FeFvFV/pt0bi0m5k6naOwI7g+tavbaclza6k8u+4u9+4bWH3QAPbpXjTw3b3Wok2jbwvTgj+FfU1FaQhX02Y7S2OME+/art00WNhH9/jA/zn1rSdR1D4ieErqxgg/faFs2ruHPnSEn+72T3/CvDnhbUNN1P7NqcflOOoyG/hJ7E+oqLTbbw7eRqW4us7zg8bRx3PrVjNps/iOOZH3QDOThh/Bj69a0TxVLoGnGe3fMM3Q49CR3BPU1rd3aeJrZpWm+Zu20+o+n92vHtlBaTXEySZd9vGD2Cirm6jRpJJ3zGMZ4x/KpbOSXcVXLHpzVzHPat5Lr97oMjtTW5t0Lxt17Yq1WQTqrDOc5/KtO0wyMksS5698evvV34O1c6DBflNtpc7snKnO18f3s9an0ODywducd8n/GtO0F5EISLMfru6dfetS8P6ZDZYsG3Xn8KYYdxnknHTNJdWumaikki/Mudwyf7vHr614jvItQvfPiTA7DPsB/Sk1u+htzYpDuC99wHfPpV/LJInly/wAPT9Kht7OCNkZvmPsautOmkgwi5U9Pz+tWNsImEUw+b/PpUF29hFLbFMpLt3HPTHP9au7O4luw7Jhe3IPar62RXj3fd5/pWnafbtdI275RnPB9D718QP7Dvp4X0aTeY93mDa46hcfe+hqy0jVdTbyLODex6Dco/mfarPbdXQglPB6/lXi/wva6XbWZgfJn8zPB/hK+59av9KeMBcf5496EM4/dHp+FWltdX0QMUfK+4/z2r4bSaNa6pDceIbnEabs/I3dXH8H4V4tu7aTUJH0s74eNpwR/CuevPXNabfXmnYntZdpPX5QfUd/rXhTw4msTsLg4PGPyb0I9K1Gyl0rVXs7gYEWPfqoPbPrR81W3ofkq/s5ZL0SyJhexz7CntxLKDu4Ht7V9lJdl7nFW1yI5Wic8rjH5V4YuLSKWSMHJbG0YI6A5rUtU1JbZ9OD4ifGUwvYg9cVpUkRuRBdttX1xnsfT8KaW38Ja1ZeINKl3TJ5m35SOq7e+R0Y9q1ue78UTidvnlHToOwHt2WtL8ReJPDcEcVwmIWz8mU7E9wCepq/vdD+KGoRJKPKuecJ87fwjv8o6JXi+S++GWsX3h62TIi8vIyB1VW77v7/rVz461XULBoJYsq/U7lHQ/wC77VZxz3NwsUC5354yB0FeIvh5bRaQNVtG3vH98YIxlgo6t/Sjaps3Nw3pWiwyQ3fnXB+Tt+R9KliliucRx/ux0OfarWee91CG3ZeBuxyP7v8A9avFHh/Fsstku5znIzjuPU1p+mNHKGkGHX7o/A+9anoU67Gto89e4Hp6mrWyuIo40aP5hnIyK0qBFhKq3J7Y+tWei2kBSR3zjOeD/jUFqk6Ew9BUTrZTuCvHHf2/+vSW1zdXaC1j+Vs85HYe9Xtp9vIQptC9DnP+elQaBZGQK8vPYbT/AI1q/h60icxW3zfmPT1NQaebG5K45PT8vr71I2xdqjmr3S5b99wT7v3efz7+1W15M94kGoSZPOOB6e34Va6Qk9rMFbldvb3+tXljLbR7JBg9qKRGQJE2W9MYqSH7LIGk+8a8L+BbrxLa3kyDDWnl56fxE/7Q/u1feHYtL8ne2fO3dvTHufWvG3hL/hEdYfS3OAMdv9lT6n+9VwkcDkRtwe+Kkld33DkVHKBLtB4PtVpcNEd6vj8Kn1QyTPI77nfGTjHQfSoJ5A4AGM1p/gjVNX0eXX4l/wBGgxzle7Ff7wPUelR6oIYJLQj5Wxz9Dn0rTVvbSxF3FJtWT2BzgkV4L8GTeK9MuL+cYhh27j9WYdmB6rSSC3UvIcE1qOs6e3h6W2gkzeHGwYP98E9sdK8OahY219bXGpSYjXf5hwT2IHT8K8aaroOtal9o0WbdGn3jtcfwqP4gPQ1rd7NA21Pu/h7VLf8AmhZGGMZq4uRKSN304q7IzjNOAO9Slmbmrcbhszk9qLEZUHkV/wAEvvioPH/7NVt4TvbjfqXgW/k05wx+Y27nzYG+mHeMf9cvxr/gsP8ADdLLxP4E+Ldpb4XUrKbR76RVwA8LebDn1LLLMPXEY9Bj/glT8Mh4L/ZmXxldW4S+8c6tNfBmXDfZoj5ESn2zHK49pa/4LIfGA698TPCvwU0y5DWvhWwbUdRVW63dzgRqw7FIUDD2uDUbEoa/4I3nd+054m/7EO7x/wCB1jX/AAVAx/wxv4r/AOv/AE7/ANK4qDt90CgWGP5V+x/8Lh8Hv2bPAfgia28m+j0pLzUUZcMLq4JmlVu+VaQpz2UeleMf2lDbf8FEp/jouoD+y9I8XrZCVWIX+zIv9DkZfQNbh2+rnPU18UPAun/FH4a+J/h5qLott4k0i4sTKV3eUZYyqyD3UkMPcCtX0q+0PVLzQ9Vt2t73T7iS3uYXHMciMVZT7ggivh74Qv8A4g+OvDvgPSf+PzxDqlvp8B252vNIqAn2G7J+lf8ABQ/xfpfwU/Y5n8CeHG+xjWxZ+G9MiU8pbKoaQYH8PkQMh7fOPUV8LPiLrvwl+Inh74k+GjGdR8O38d3DHJnZLtPzRtjna6llOOcMa+AX7Rnww/aO8IQ+KPh9rkUlwkSnUdJmcLd6dIeqSp1xkEBxlWxwa+Nv7A/7OPxvnudX1Hwk3hzX7nLPq2gMLWSRzk7pI8GKQknJYpuP94V+1b+wj8TP2Y428TrdJ4o8FPKI01q1hMbWrMcKlzFk+WSSAGBZCSBkEha/4J/kj9sL4bA/8/1x/wCkk1ft4W1xdfsjfEmO1cLJHpscoJOMCO4ic4PrhT+NfsX/AB7tv2hfgNofii6vUl8Q6XGNN1+MvlxdxKAZCOuJV2yf8DI7Gvjt8BIfgN/wUJ+FnjPw9pmzwv4/8Y2F1bJGoVLa8e7jS4iGBgANIsgHAxJtH3a/ad/5Nr+LP/Yja1/6QzV+y/8ABO8/aJ+Nnh34YQzyW9leStPqlzGPmt7OIbpWHoxACL23OueM1omhfD/4LfD+PStDsLHw54V8NWTNsjXbHBEi5Z2PJZjgkscsxJJyTXxY/wCCrVv4G8THSfDfwVnv7CNyBcajrK20s4G05CRxyBBg55YnkZAwRX7XX7TMX7UnxHsPHkOgz6Jb2WjQ2Eeny3In8pleR3IcKu4FpDztBwBxwKftmo0DjA6jpTMAMA9Olfe5FeYWQITwKCMBlelAkjC9B0pQDjcaWEP82eO1aX4ovLbS5NKd82z43DA7MW9M9aMzzykMuM+/tUzJEQYjyvWg2eT1qRfQ0FJNN6Zq3KhhuPFat4gutUEYnP3c4HHt6D2q6JbYV560cjvTZI2nv0oDPGaAGME/pRUZyDTqAeDx9KIB4/KgAVGD+FE4XCiiwzgnpTjdwKCgjBFKQTgdq2hQP0r5QMg9KK8UcngHNLtz8xp8E4z0oMAOvSnfec1jjgYoMrAKT0oYByvXtQK9QfpRII460QCflrJ6qelKVCZJ5pjlQc1tyODyKUHPJoAMcE0VAPFMDjA/CkAIwTTlVyAaUb+TTHa+e4pgCuQaBUA5PPYUxBOAaJB4zW0YzmmXB4NAYHWlyQMHiiSW4rgUxGAc0q7+MUYwpwaODxmiDjBrO7of0qXCuGHPtRmXbxyaYoUJ7noKjVWBBP0p0JY4PA6U64AINE4xzWR61jHANbQe+a4xjNY6AHrXXigobjPSsdqyT3rBx1rqOD0o9OtE88GiATXUYFdBgCiAeKwAAR+FdsA0AMAZ6VjjNEcZFAHufwrJ6A0MgYDfpRAPWsAd6244DfpQGeM8msdhWB6802ccGipxgGgSSBmsE9DRHYn9KwCaA5Az+lZAGO9H6/SgM/xfpRIIHzdPalAPO78MUcYGG/DFYGcBv0rAJxu/SsAcZ5+lEDGAaAXrmsDGQeR7UFHBzzWB2punFDHrWAe9UcEYB/CqJJ4zX0NE5GCOaJ7CgMjOKJJGBzRGRgGs+ho8967dPwok46VkngnNDkgCi2ABWWJ60BjoaPHehkHrWcdDRJboaPbnpXtiiSeprIHQ1kk4JzQ54NKoxnvRBB+9+lRXEsRG09Kg1iQkK5z7f5FQzJLGHHfpQYkYJzmgRnGOlYYjAPFNxyRTzEqEA6UCT1Nbs8ilPTAoqMYFBAMYpSWOCK0y9gsbxJpbX7QozlN5TPBHX8a1DUoLycvZ2P2Zey+aX7D1Ht+ta/4L1HTtD07Wbm686O983YuxV2bGCno3Oc+lFSeBSAKwVzkVawIZFaNs+2MVbW8zbQ0vAztG0VHa+FRYbTBm7PRt0nHPp06V4Q1Pwlp8ly2teEv7RafZ5Lfb5IfK27t3CjnOR16YrwJqV34f1+HVdNfYE3YXAOMoy9wfU9qm0vVbgG9a23Rt1Xeo9vWm8AaTrmoaNa2CYe88/wC0nLfutq5Xqw3ZwenTvWm+FvCXw30tYL9d1qc4TMg/iPcFj1cUum/DzWNeubJ9G8jShs8qT7RM2PkyeMhvvDHP8q1LToNHdo7AeZEcYHI9PUk9Sasrp4lljt22JPt3rjOdvSvEesvqlkS1v5W3od+7uPb2o384uWgnTejdZMgdB6D8q07StG1C6jYaXgDOR57fNwffjpXgrwl4f1/VT4cbUPsQ7S+U8mflZ+mRjpjr3qL4ReEdHurqx1HW/tUsGzym+zSR9Rk8Bz2I6+lPc6rDIDpwyB95/l/Dg/jX9nXN5dS3011mXjjYPTHY+grVyivvLbn78Y9K8JeIdZ8K+ILXUvDj+TdtvwcK2fkYfxgjoxr4jeAtQu7WLxlqGo/2he3ed6eSIsbSkY5DY6Adu3414D+E/h+fw5Fq14vlS3WdyZdtm12Uch+cjHatH/Z/0J70XEuo70HRPJcdiOvmVq2h6Zptrb2lhp/k2kW7915rNuyQepJI5JNeO/ivrGn67JpmjN9gjXGGwsuPkU9GT3P514i1/Ub6/kklu8dMjyx6D29q8zTWiUQty2d4w3OOlRQ2Gh3XnW1r9oU/w7ynbHU59a8Q6fpOvA3Mdl9jkHRPMaTPQdeOw/Wj4URX8uNsH1x/9erDwSQ/nyLuCdDnGc5H96tH0GY2ccUNpgrnjfnufev+EG8QS+FLW5kvd1s+/Nv5SDGJMfe3Z681ZeBdX1GQwi0wR33r/wDFe1WPw/t/DVgF1KDzXuPuruK7dp56MfUVb29tpl8L1NK2Sr92XzyccY6c9jitd022vPEMutxw+T0wu4t/AF9fb0rxDd22oERPbbJl+7LvJx07dOgxV+t4jthNpGO4PpTQWFxZSPPY/vxja/mnjnngcdKeziJZ/L2kY7k1GZlZTDcdOibP614d0rwhqFpcXHiK28m7j2+XLvkbOSQeEOOgAp7qa4mlt07Y+b8M9KttSu75MNPmQdBsH+HtV+lwAkF2nXO3kcdM9PwptOf+y2mtrDy1GPm83d/F6E1c3l5HPJbmLBXG35hzxk14H+J3izwZeJdaPP5LpnHyxt1DD+JT/eNWFvpYZVg+Rv73zH9Ku9J+3bWN7uaPO0eXjGevf2ptM1AQnzLjzAO2wD+tf2Laj95JHtx7k/1rw3fxaNaPEYfOL4x823GCfr61PZXTSNLEmRxtGR+NWVtNPaAyJh/qPWnglitger/ljmtN1vUracGCXyz67VPY+1fZr/X7iXz5PtE67cNgJ29BgdBU/wBpsH8sxbsdRuA/z1rVdQk1aCKGOy2KM4Hm5xyPp6VKWt0MYTdjtnFWmoSCP5I9uehzmirSzvtO52xk4x0FeD7620WSK8t7bzZ493zbyvXcOhyOhrxVrt14k1ptUvJtrHGV2g4+VV7Af3RUlpDIu5Gznvg1F8kcgLeYkWPkxjOfevDHiWxstRSWxtvsrJnI3l85U+o9z+dfGj4rn4lw6co8Kf2dJYebl/t/nb9/l9ti4wE9+teG9FWS+Sa31L7PMM4/c7+xHc46V8RND1PUobfxBrdv5k9/uwdyrnZsX+E46AdhSaNPMVO35V6DI4/WrOS8Vwtk2116PgH9D+NeH3mtb1H1GL7UXzg7vLxgH0/D8q1fRLG7YTS2f2eRuo8wv0x7+1WPgGG+njji1Dy3Odv7knHH+97Vr3ge/wBNlKXh3KOjYA9Owb3FeFvhnrniEMNJuvL24wfLU46/3mHoakg1DQJGtr/9+yfxcJ79Bn1FW9ncvqCXUkfnqM4TIXPy461p9ncXkBSKw8lh280N3P8AhSeGo0d1v38t+MHGf5H6VZ+GvD8EflJYeaW/j811/TNTeEdD3KLGPa653Llzj8z9a1nwzfaDaK2pJ9njOcch+49CfUVNawzv5kV3k/3fLNWlzqAnS3WP5Wzk5HYZq4je3i2XV35meg8vH8vwrS7PR5M3F/bbgOjb2GOo6D8KmOJc2Mu4dhtx29/xq7iuZn3DiU9Dx/npX9lX/CXcO3PQ7gf5H6ULZ7OTy0fzG7HGMV4F+G/hTxPJ/aPirUfJ2f8ALLyZGzncOqMP7qmtRgjjb/RdP8uNeqebnHTua1K+uruURS/Nt+6vAx0z/KofhzdyeGIfFen3fmL829PLC7Pn2DktznntWpT3PmI8z79ucHAGOnpSWzvD5sE+5n+8m3GMe9PDDIxQ8yDp1GK1B7m9Yi5n3N67QPT0+lXDPDGQr/d9qa5dyRt47c0t4VmBKbx6Zx2qS6LRjyk2/jmreZppcO3X2qxvFinDFcqPf2qbxFqCQf2RFd4tf7vlj13emetahfYVUhg3AZyd2PSn1/VtOt0jkhxbSZ2ncvODz2z1NPr+pXEJtbRNwPQZUe/cUpS7tw1zPgjoNvv7fSvt1vbXipO3mwjO6PBXPHqOeteMLaxWRryx8J/2PEekP283H90fePPXJ/H2pdXuYpQ8KbVTouQeo+lav4mkvIxFfdug/L0HsK1TUvNVVgX17/T2p5HWQMHz+FTzK2WAyT3qdmPylcg++KlGw7VPTvUUjRnIb9K38ZxX/BKL4qN4Q/aAvfhzd3BWx8d6W8caFsD7Xahpo2Pb/VfaF+rD6H/gpH8NpviR+yj4iTT7P7Tqnh68tNUsIwuSXWURSY9P3M0v+ea+G3hHSvhH8KPDngsTww2PhPQ7e1lmJwuIIQHkJ9yrMT7k1+0F8T7r40fGrxl8ULt5CPEOrTT2yyD5orYHbBGf92JY1/4DSHc4UHiv+COKAftNeJsD/mQ7v/0usa/4KhkD9jXxYc/8v+m/+lcVKzDlTnFfsmfDFvjP+0V4D+H9xbGazvNWjn1FR0+yQAzTgntmONlB9WHrVxPb2sLXFzPHFFGMs8jBVUepJ6Uf2aP2NWJJ+EfwzJPUmxtq01tOawgXSZIHs40CQmFgyBV+UAEcYGMfhX/BSH4Xj4aftV+JLi1gKWHi+OPXbX5cAtNkT89yZ45j9GH1P/BLT4aHxz+03b+Krq132PgjS59QZ2Hy/aJB5EK/X947j/rkfx/4K8/Eoa58VPCfwrs7jdB4X0t726VW4+0XTDCsPVYoUI9pT60wIXINeFvE/iDwhq8HiHwn4h1HRNVtTmC80+5e3mjP+y6EEV+yL/wUq+Kx8d+Hfhj8bJovFOk67fwafBq4gWK9s5JXEcbPswssYZl3ZXfgk7mI2nxt4P0L4g+D9Z8D+JbNLrS9dsZbO7icZykilSR6EZyD1BAIwRX7CFo9h+2f8PtPmdWktdUu4mK9CVtZwSPbiv23v+TTPifg/wDMCf8A9DWv+CeX7R4+A3xvt9I8Qaj5PhLxsYtP1Qu2Et5tx+z3J9AjMysegSRyegr9oL4URfFLwtos9nAX1zwb4j07xFozoQGE9pOrsgzx88XmIAcDcyk9K/adGf2a/iyAf+ZG1r/0hmr/AIJCrpo/aG8Ueey/bh4Om+zhs/d+12u/HbP3PfGfev2/NQ8V6T+yj411XwdK6XlmLSWYxoGb7OLqLzTggjAXJP8Asg14s8Xa74y1BtV1+9+1XT4y/lonRVXooA6KO1QSLGwZ03qOq5x+tandWN5IpstP+xoufk80yZ6dz9P1ogk8HFBu3U9qJyMAUGLDbioEd2CKcZ74qRDH8inOOlFSepzXghfDRvx/wk9h9sgPRPNePHDd0567fyrxeNB/4SS6Hhiy+x6cuzyYfMeTHyLu5f5vvbjz614K07Qrm6ml11/kXbtTD88NnlT9K8RGwbV510uDybc7dibi2PlGeTz1zSJk80687GHI6UFyeDTLkjFRJgZx+FcsOBTO+MCmJB4rJPPeguTgn6UYCIi7rwOnvTZ6Zz7YpjnAPNZwMg/hWDswKydvApiSME5pMngNz9KnieE7XXBHbNAhRkHNZJHPat204Bo5+7QcKOtH5m4FZHTNZBGAaIAOAefpRwcjdx9KZh2P0oEgYBrAI4bn6UwQgKvWig28N+GKAzwx4o7RxijgjIojK9KPQEdaVSwLE0FLc5zUhXGAaDHb8prO4deaXjgn9KYhhgHOOlHkYB+ldRgHmsAYGaY5xg/hTN2FHjkHNFQ3vRRcYx+FBVH3RTRq3SiCV2lc+2aICjco5oMzjcy8jpzQky20L+Oa2kYLNn8KZnVsBce2a2M33m57DFOu1uKCq4+VenvRibGAvPY5pdzDB4NSxhFLA5IoxnkE9O1HB6UQSMUVOOKxngVt4/lQX1rbg8UTjkmgD0Jr6GiCBgmqIwMZoqSODVFSRwaojgc0RxWR0zWMjijxxWQO9EdqGDwDRIHeh6GqIGM+lEZ6GiR60RgZJoZGBRxjrQOep+lBe1Y5ogEYornpRI6ZrHGAaxkcVnA4NA5wfTpWQehofN3rGeBRwDjOaKj1rGRwaK8delbSeKI7D8KxkcUAT0NVx2rAPGaK4oAnpWMDBNbcjINDGOK68CiO1Dnp+FcHgGiMcVWAeM1nHOaIBog44oc/KKI3Ec11FYJ6H6URjiuoyKUHp+VFTW0k8Vgg4JoZ7mskDIPI9qLE45rJPU/pSsQdw7VDqMqgKW6dBirfUIZQF3c9+DQkyMjmg5wADmmY7duaPAxmg2OVoDByDRPYHmtx/wABWSvXtXhnTotb12202X7k2/P4IT6j0rx94Zi8Ja7Jp0C/IMbef9hSe5/vVHP5Z5P6VaaN4m1vRzI83mWlt/qV2oNu5vm5yCeR3qQNHM0TjDDHH4UIGdQVGc1Y70lEY6//AFqsIrlwHIyqfT3qz0uWZvMZPpz/APXrw/4N1HxAqwWNvuaPr86jrn1YehrR/goukeFLjW9RfbHBt8w4ztzIVHRz6jtVt4Z0T+xFaFN8Z6HLj+L614U+Cuu6zqceqeGrLzUXOT5iLj5WX+Nx6GvE/wAN5LnOl65F80fUbumcH+FvpVx8EdEtbaZYeRPt2j5+Np5/jrxT8ALmytTPaQb4u53AY5Ud5M9TVz4by8qRJxDjbz6/jR8KaprluY7CDcnf5lHf3I9DVn4f0640qS4nbk42cN/ewehr4f8Aga91m6BtzlR06ejerD0rQ/Cdto2sw3ssux5N2DtJ6KR6n1rxd4B1aGyn8SQ/Oi7ct8o7qnTd/SviH4l0pI1tdGgwozn5m/2T/EPrWjW9nF4SvtcvxsaLy+OT1kK9vw7VrENpqt2Rpwyo+o7D1x6GvBNnEfGOnNdRZjg83jcRnMTeleHtC17XbxLyRfMtTnC/IOxHXIPUVp3h3SDp9vY2w+Ybt6/N6kjqa0dtP0aRv7UG1RjHU+v93PqK8d+LtDGkm6VsLF7OerKPSvjT4s8NLp88umR7rj5fmy47x9iMdM0thLe2TX5GVbGD9Dj+lRyyCcxoeF/wrRrW7uFy8e7PTkD1o+GVuHRgnzjPGf8A69Josc822WPHvnPb61pugyTmK0gg3Rndzux6n1qz8P3aMu2PAXPcf414c8Tte2C6ZPDmKDOPm9ST2GeoqS+gsJTLbx/jk/5714k8XWGpMCkm516DBGOnt7Vqmr6dbQNJezbR6bWPceg+lX4tbu1RbZ8sc8YPqPX6VPo4huPNm+4P8PrV7a/2gDdwcxv0/DA/pU+kTqn7lcg+4/xr7DK67ZByaazt7eYLO+3HTgnt7Ub24I8ll+X6itK8NX+vXMqaXHuaHG8ZA6g46kehpI30m5KSD5h/h+PrV5dtfOZSOR93+v8AKor26DraiX5Gzldo+tanOJpbdVi/fx7tw3eoH4dKa9NjMZLhNp9M57e31qIrGCFfDjpxUGoTwSK1w+1PXAP8qh8RXBBfb8n1H+FQ+I7OeZI7g4Q5z19PYVf6ppdve/ZIJOO3DegPpWjrpN5pcMdoPMmO7ePmGPmOOvHQVHolgIxJKdpPbk/1q78LQi3M0a5U+59cetXmnJbTAqv6+31q11WWwmkliOGOOevb6e9SajHeTgtz/wDq/wDrV4c0U31pNqMEeYLXbvOcY3Egd89RWsfY0u3ZBwcevoKAs3jxE2T34NWWmNdSMtqMnv8A5JrTLS8sGkVxyuPT3/xqKOe5DzgZ3Y447cVbTFboeYMAdvwrWJbaOwku4RgRY38nuQBVvqi6hcbLGT5x7e3uPY1ZXV/JG0dwc46ngfyqxkms5hPGv3ff2xWv+L5Nd0+ytLg4Fn5m0f7xBPQD0q3DK3mqMp3rQY/Ao0m5jtpt2oPswu2UYwx7nj7teAPDtnr2uvb6g+2FMZOCcZRj2IPUCvjNqfhW2vYtP0Qfu4t2D+87iM/xD618Mvhdd+ObeW+0pt89vt2LgD7xcH7zgdFNeK7TVppDYXKfOPdf9k9q8O6ze+DLpUWXaTnjaDng+x/vVcWl5r8pkhTzM+4Hb8PStJhj86OJov3gzxu9jUs9yx22sWP+BD+tTaM89639pybPbGe3sfpVh4e06OIJ5nPb5W9/eoPClmhN5E+XXqMEe3rXjC/g8XabHZ38e1I885J6sp7Y/uirzw7pmksWL/KfZv8AE+tK0UB+0253Benb271q8V09wFvG2+gwD2Hp+FXOqwrbLpq9DnLc+uemK8C/DfVvEUclzp8e9Y8Z5UYzuHdh6GvDXw81HxLfXFrZQ7ns9vm/MoxuBI6sP7tXPh/WLnVl0srh+eMr/dz614R8K6Z4X1e98Oa7b7r288v7Ou9h91Wdvukjow6mvFXgnXfDetrZXkflF87VyjdEU9mPqKvfDzSaZNq1qd0S7ecY/iC9znr7VOiTP5oXDDvmtM8Tapp+jSaLBLmCfGV2rxhi3pnqfWruzkjwLpflbpz/AIV8NtetNF1SRdQh8yO4x/ERjar+gPqK8cxaC+tLNpR+V85Hz9lUd/xrV7ewt7p/IbK8Y4PoKvI/NEpxwMVq1+tuSu7Hpx9KOpYARG57cf8A1qh1COSPbLJgj2rStTtZ38tJc4/2T7+1SXUccZEbc/SoLp7mdYyeuf5Vpd3otyJbaWT96uMDDe5/lV9fJf6DZ27w4WHf827rlwen4VceJbfTpkVGwq5ycH0+nvUfiOzlsAu/98OoweOfpjpWoawzMz7/AJlx2+ntWu/FqXWdOWGdsTDqv4j0QDoKbxPPO626L655H+FSTC4y8zdOnFTTBAqk9M4qSfccqallCrgGpbkMODk0zljnP6UX54ppyBzXw28e6n8MfiF4a+ImjnN34b1S3vokzgSeXIGKH2YAqfYmtOvtC8Z+G7TVbNoNR0jWrKO4gZlDJPBKgZTg8EFSD+Nf8FH/AIuL8I/2TvFklrdCLVPFgXQLAZwSbkETYxyMW6zkHscVNKSc55q3tjJ8xPSv+CO6Bf2mfEoH/QiXf/pdY1/wVEwP2NfFnP8Ay/6b/wClkVbwBkV/wR0+GB1Lxl42+MN9bZi0exi0iwdlyDLO3mTFT2KpFGD7TfWv+Cq3xKHgn9l+Xwpa3BW98batb6eFVsMIIz9olb6ZijQ/9dPTNI2Vx6V/wSN+Jo8TfA7Xvhnd3Je68Gav5luhP3LS7BdQB/12S4J/3hX/AAV8+F51v4Y+Evi1Y2+6fwzqT6feFRz9nulBVm9lliVR7zV/wSS+Ga+GPgVrnxMvIQlz4z1cpBIR960tAY1Of+uzXIP0FftK/Es/GD48eOfiKk/nW+r6xL9ifdnNrHiK35/64xxiv2L/AIa2vxc/aa8BeD9U06G90waj9u1CCeMSRSQWytO6SKcgo/lhCDwd+O9fEH/gl3+yz43vZtS0fTNe8HzylmKaHfgQ7j38qdJAq5/hTaPTFfBP/gmx+z/8F/GWn+Po7rxB4n1rSLhbjTm1e4j8m1mXlJFjiRNzqeQWLAEAgAgGv2xv2nfC/wCzb8KtTvn1iD/hMNWtJIPDumrIDM87KVE5TORFGfmZjxkBc5YV/wAE/wDP/DYfw1H/AE/XH/pJNX7cH/JpfxPx/wBAJ/8A0NaZtwyRxX/BOz9pBfjv8Erfw/4g1HzvF/gdY7DUvMb57mDBFvcep3IpRieS8bE/eGf2nv8Ak2r4s4P/ADI2tf8ApDNX7K3xzm/Z2+OHh74lvDLcabbO1tq1vF96azlG2QKOMsvDqCQCyLk4rw/4h8C/FzwPFrWgahp3iTwx4is2UOmJobmF1KujqfYlWRhkHIIByK8af8Ejvgbr2uT6p4T8beJ/DdnO7ONOUR3ccJJ+7GzgOFHTDFj71+3F+zD4c/Ze+JeheDvBupatqOk6poEd79q1N0aRrjz5kkUbFUAALGQMcbup607Nn+VK/rWcfdpG3/KeoreBwO1QXTxPvUZNTTGVt4FCQngUJSeQfpSTuOFPNfbpInIRuPpUk5kk8wnk9KLkc55pmJJJpSDS54WgSo4ppCFC46UjK5wTQkC5xyaLEHNaQltcahDb3RxEd248/wB0kdK8fJ4ftoLG30STcy+Z5o2uO6kfe/HpTuc4J5pnzwo/Ci7YwOtbj0zRYkYBrkjGc0CRyDU0rSHc7ZJ9qDk8Ci3HXmsEjimcldo60M4x1oHHSmOeKDYG0USScmiQ3FEADAFDGMCicDr0o5Izis9BmiwxgdqIyBtrcwHFKSeB1pmzgUG2jaBSuAMjtTMW6H6UrDpRGWyBQxjkUzccGg+eM1nAzmsknANFwOAaZWAzmlORyelF1xgGlYOcelNhRxSjHy5p3VQFolHGAeKOEBPapHUnK9q8xmKlm6dOKLgrmmlcHrSXH8OM0swDbVPPpRZ8jinLKBmmkLDIHTrUoY8CjGw4xRUMORzRUAcCiBtxmgTjg0c8Z5rthjRHFZwMCsKeBWFHGKyAABRIPaiRjAr5exoZJ60eDnFEk4H6Uxz0NEYOAfpRIIwaJyMA8iiSRgGgR3onA+UUcA5Bo4I5PNcen4USCMenahjoaJ4wDWTjANEjGPyrjGMUeTxQAHQUcEYAo+gogHtWBjFHBGBRCnrz6VXGMVgdqABPy0RnjPNAHgHmmBPAP0okkZFD2o5I6/SgSBgGh6A0AfWsk8Zo9cCtpPGa5Pyk1k9AaJzxmuSMA1jPAo5PINdepo4wMUeRjNHmqJz060RkcmiM9DR9BRIJ4ogEYo81gVgHkV2wKo4xgUAB2o4PAo4x1risCiAe1Fhtxil9qiuDGMD8KsdRDAK/PoKRtwyprkjNOCeAfpWSON36UCSOtAjGKJyMUCCME0o5BHNKzgHYevtTEsMGofFPiCGxk0uDUdtrJjfH5KHODkc4z1pSHOWPNKBkBTn0rSbR7i4VWbnnt7GrXQWt9NV5W3LJ0GMYw31rQ4bQXXlu+fbB9DXwn8NeMNdhuL7RLzylg2+avlxt13gfeI9D0rwtqjz+Hriw1HWvNB25h+zbc/OT94D6GtD8C6wL1Lm4n/dnORsX0I7NXhKI6XCqwHKxZwfru+vrWvaDDr2ptPbN5OcY4LfwgdyPSviR4Oml02M2KYuos4myDjLJ/CWx0yK0ayvbXSSLqL7VIep3BMfMfT8K+L3gGfV/H19p2kD55fL3JxxiFCOWYehqaLxX8PLxrB5PIzj5cRt2z/tf36s4LyYNDprYDYyMA9Pr+NN4a8c+E9J/ts232Wy7y74n/i2/dyT1bHTvUPia5jgU3M2V52/KB356Cte+Od5beBhoM0e+KfoNwHSUN2TPX3rXvFtt4h1J7y3h8qLjCbi2PlA6kA9RXhXxbJZeGtU06W086xufJ8xfM2/dckds9TXw50vwprsbHWLnyIxjB2SN/f8A7pHoK1W10rTNakGht9oiXG04ZP4Rn72T1Jr4ZfFy20W2jtbi03zDOD5hHdz2Q9jSeJGbUINZ8S6h5EUO7yx5W7OV2n7gz129qn+L/gGNzFHrO6QdP9HmH/snpXxb/aWtry5bw3pzfu7f/XHB/ebgjr1i4xg9DzXibxWdbdtr7QcYGM+nsPSvCltPqNvHA5+c5449W/wq/wDhfd6VZpe39rhJs7TvHYgdmPqK8A6DHGq2M0f7tvf/AHj614u+GsGjCBtMh5l3c7jxjb6sfU14e0O40W9WW8g3E5yNwHY+hPqK8Ltp1i9w6J80+3HLcYz/AI0LpLPVJAjfKcY4/wBmriHWdJnupJRsWXZsk+U4wOeOfXFM1rIPNudT2v6eST/KtSiW6vPJkXdH2OcY4Fap4XXULM/2fe4n7Dys9x6nHQGtM8O6ppdwNK1k/wCq6H5e4Lfwk+opNOsku2N82FGMDB9PY/SviLP4ZvNUEOn2u4xffbfIM5VMcHp0NeJobCR43tE2Iudy5J9PWp0Jt1aP74zh/wAfSrrS765kFxqEnmR/3cBfbsfpUml2tvciRY+B1GT6fWrq8shZ4tD5ZP3up78daOgXsNmb+MZj/D1x61dNa3KtGU4OMcnik0sxtlbrHodn/wBevBHhOx8VWepWup+Jv7OlsPK+zH7EZvM3li3QjGAo69c1q/hiOCd4otY+1GPGB9n8vdkD34/+tVxpjK/mF9p7jGcfrRtllO2K62kd9mauZjJiJZfkHbbSw2uNpXg+5rRPC8etXqQrJt35+bGcYB7ZHpT6PdeE7qXTEvvNAxhvKC9s9Mn+9VvcyFsSybj9MVLdypGZ7c/Mn9eO9W2vyIc3Kbx9cfyFRXmnX6+ZDb7NvX52Of8AOKSwtID5jDb78mvgl4rs9FxpcY8+Ob7kXK9PMJ5wfWvGXhG5vNXuryws9wbZhfMAxhVHc+xq2lsPOZJJdx452kdqi0qB7WWW2vPLYYwPLJ7+5r4gX3hK6vRd+ErXyIW+8m+Ruyjq/PXdSataW1o0Bk2tPjPBONp+lNBouofuxFuk7ncw/wA8CrfwyBEpgmwhzuXb1/WtT8CanEfPsfmP/AR6Du31rSW8SWDssz5j43DCe+On1rQ3t9UuVgnbk5+XB9D6fSp2ttHvZdOmn8vGMHaTnjPbPrVppVhfQrbwnzd/fBXp+PtWqeHL6wuGa3PmBMY4AxkD3968FeDfE2oQnUNPs/O8r7x8xFxncO7exq40O8m1+PUbmz3rFnA8wDOUx2NeJPGuua21pJoI/sxLLfvHyzZ34x95R6H868W6PqGtXrXYvef+uQ9FHqPSrH+3rZjYPN50DfeXai9OevXrWkWd9pd411aXHlo2MJsB7EdTn1oxySTs5ucStjA2deP8K04X93MLa3m2N67QexPf6VqcOtQ6x9jlvPtDw9T5ap1UGtRvdUgVYoZd2c8bVHpVp4k8TXrG2W55X7w2J9fT2qaa4WZm1W747jyx6f7P4VrGhahp2i2uo31r5cF1v8p94OdrAHgHPUjrU9hexOPJudwbtsA6VIuqSESSvk9+Fqx0u8v5YYnh3ht2BuA7fX2rRdf1jRYmttNufsxbG8bFfPUjqD6n868KfEFtF8O65pqPtnv/ALP8+M/cdj02kdD7V4Kk8P6jeSPrVxsHGG2Oezdl+grWrHRItViv7S+xCm75fKbn5QO/PWvCniiW01iK+YeZGufm6fwsOmK8cw2ja7MLOXfbPt2DaRjCLnrz1q8srdVzNDlP94ihoDqEvDHiHnAz+HrnrWqW1jOUWG22MucneT6VJY6nCUumi8nrtO5W9jWpavPeyqt1Jvdc4OAOw9B7Vqqq0ZMUu4Dvtx6VqWpC3BBk6dBj6e1eK7x7rJibj6f7tRas6QIgm9eNvTn6U+qIx4+Y/l/StH1y3h2SOeUzkc98+1HXrWWDeq59sn1+lR69YqUR22qc54Jx+lX2r2bHdbycDvg+3qKuPEktuTFHccDoNnT9K1HV5LmNljP7w45/L2rRNP17xg0sGj2vnNFjcu9FxnP94j+6a0vwTrE+rS6Zdv8AZ54MeYuFfGVJHRsdMVqNobO5eETeZtxzt29hSsATsOCOtIzg5DfpU0jyf6w81LKqDGeamkLvkN+lEs3Ct+lFsDA7U5JHynn6UATwTV3MsC5z1rwd/wAFBf2vPh94S0rwT4R+MD2Wi6HaJaafbPoenzmGFBhE3yW7OwAAA3E8ACvjT+1L8fP2j7bSdP8AjF4/fxBbaJJLLYxDTrWzWJ5AodiLeKPecIAN2cc4xk5js1yCwzSrGiYUfSvhB8c/ij8APE9z4w+Efir+wtXvLB7Ca5+wwXW+3Z45GTbOjqMtFGcgZ+XrgnPxS/bZ/ad+NHg27+HvxM+KH9s6BfSRyXFn/Ytjb72jcOh3wwI4wyg8Ee9IemT+lfCH9sD9on4DeFpfBXwm+IKaDo09497Jbro1jcF53VFZy80LuSVjQdcAKK+M/wC0t8bf2hhpCfGLxy/iBdC846en2C2tFhMuzzDiCNAxPlpy2cY4xk5QqDgcY6V8HPj98XPgBq1/rnwi8YvoN7qlsLa7cWUF0ssYYMAUnR1BBHBABGSM4Jr4i/twftQfFnwZqPw9+IPxNTV/D+rqi3tm+hafEJAkiyJ80cCupDorAqQcgVoX/BSXwP4I/Zjg+BHgT4aa3ZaraeFW0i31SS7iEa3TwlZLnaOeZHeTHqa2k4xWgeIvEPhXVYNc8La7qGkapanMF7YXT280R9VdCGXj0NeEv+Cjf7X3hOCOzHxRGsW8Qwqatptvct07y7BI3bqx6fWvE3/BSr9rzxLaS2UXxHtdHimXa39maTbxPjviRkZ1PupB961vXfEvjnWrnxB4p1/UNZ1W7bdcXt/ctPNIfVnckn86+H/j3xZ8K/GGn+PPA2rf2X4g0WQvY3fkRz+UzIyMdkish+VmHzA9a8c/t1/tU/Ejwjqngbxr8VP7R0PWYDBfWv8AYenw+bGSCRvjgV15A5BBqSMp1FfCP41/FH4EeJJvFnwn8WTaDqlzatazSpbxTrLExDFWjmV0PKqQSuQRxivE/wC35+1v408M6v4Q8R/Fv7bpGuWM1hf239g6dH50EyGORNyW4ZcqzDKkEZyCDQAJ4618K/2gfjL8DbuS5+FHxD1XQBO26a2idZbeZgMBngkDROQOAWUkUf8AgqP+1z9k+zf8JPoHmBcfaP7Dh359cY25/wCA4r4ufHP4rfHnW7bxD8V/GE2v3tlE0Vs728UCwoTkqqRKqgZ9q+8NvXFEEHpmgBsyevajG6rnt2oEkBCaCgcg0ThQo6igVHXrWiyaVDvk1K287ptG9lx1z0/CrqW2lu3ltotkZxtXcTjjnk0wG7IHNOGZhls+2K3EnLc46UWJGPTtWB1oFSBgUxB96cY5BzUX2cREum5uxyRW1QAG60FycfkKQ7enNNhju700ZC5Io8jApSoPIzWAOD+VbgOMfhQJBwO3aic8HqOlNjH8qYgEACmDMuaxQAxjH4UFHWsZ+6KGCOR9KAwcg0TnjPNY4yDR3Ock5NEZGAaJOMA11GAf0oEkdKPOPasEjOKKqByMmiMjANMFIAAyaBwciiQTgCieME5oqT0FA5HJ6USCMGgQARRIPAPNFcnrmmUjpUi/d2nnmgWB2k0wXZkHmtvHSlwTkGvvcA/pQAOAeaIydoHPpQUgbM/hTSDeFJ6e1NJF0IyakeNz8poyRlcg8/SnG4BuuelKVztFBQWzind48FGx+GaaZicuc1mNhgmnUbc4/CgABgCgVPNFgeAaYAjrWcjA7USSOKDAnPWmbPIFHHArg9DijnvRweBRAIyBROcACiARwKznkGup6fhRyMfpVHPeuCODXJGaB7CsEjiiO1A9hRz+NcUef6UemPyo+wrr1odAQaAJ6URj/CjjpmjknrRGCOelEdq6rxVEZGM1gYwKBB4FY75/ChgdKPJ4okgdeaLA8YrORgGs9hR5GM19DR5rOOc1xXQ81nJzmi2OaO3GAaIxxmvoaxkUemaHzYHemGOlcEcGuDwDWB0NbePesdqI4ojIwDVY4zQBPGKIxRIxgmiOKxkcVgAAE0CD0NEYPWiM8ZrqcA1jPAqiAe9KMcA0BgcHNRylGypqwvkkQKxwe1YBGQc0y54ornjNK4XKEZNZHTNUADgDn2rw9pi6vqkGnM+BLu5xnGFJ9R6VrH7LPi6HQrfxBo0Hn2j7vMfdGm3DhRw0ueue1XenyxTNbyrh0xkfUZp4mi5POelYBIGM0AFb5T06V4flBuFUn15/A1ptxcSsEeXMcfbaO+a8B/D6fxXqWNNbcR14A/hb1Yf3TXg66n0+7MGgvujk/wBZwB0DY+99TVn8G9e0zQ5PGbputIcbhlBnLeWP4yep9P8AGvCEdhcaTGZpck5x8p/vNUdm1pHiFN8cnfOOlLa3LyYWPb+INahpy3tm9up3Pxx07j39qOi6ilm0TzbAcYO1T3+teLPA11oupReKmk8xp85O0D7qqn94+vpXiD4K6j4o1EahqT+XF/ewp/hA6Bx/dFeLfh5b+FvENva20eLdd25snn5FPQsT1avjH490jU/An/CG2Iy39/5v+escnQr7HvV8jxu2nuMND1/HBr4baT4P8W+HL3T/ABLqv2KWPy/JbyJJM5dy3CEdlXr60bWCzl+zRLvLdOSOgzV3qqaD4XltZrT/AF+Mt5nTbJnoAfWvDczyn+zLI5YdT09T3qeSfRdReOZckY4yP7vtn1q08QXNqI9RtoPMU5wNwHt3FfFrxNrkXgnStd1KD7Otz53lLuR922VFPIHHbqO9af4/ZnecLvY4wM49R/dq8urm61J76Z/mbG7gf3cVDZmWBZkP+c14Vcwz275wV3e/Y1Nr114ltYLS6HlxW+7ByDncc9gD1WvD0en6VYq00WZf4RuI7n69jU2s3XimeHT4bXa67sfOD2z6D+7WkWtzdYutTTy4e7ZDY6joOeuKn12x068NvYP58bfdfBTPGTwR7mk1WGZwJLb5/wDfP+FaxfX+pWkyRW++G12+a+8DbuIxx16ioNCa9YJDHu9TnH9fatU0NEYWkcn71evH0Pr6Ve6JLpUIDyYb12/T3PrRtbrVv9NVt7n2A9vb0rV9DYWrX91LsVOo256kDsavvDrw23255txuOg24xtIHrWsyCKRbZznOefyNRa3JagRS2eVHQ+Zj+lXmpWWpyRq03k9fl2lv6e1XFsoG0rkDvnFJZ20rtHI23f0GCelaNqUiWUlitt5qHH8e3HJPpWrwQPeOwj8vp8uSewq4skkXesuMf7NK8j7o4jsYY5xnNRWF3E/miTOfYUQY4DA8W9m/i3Yxg+lPYJv2xn5j2xV94eltIlmkP3s4GPp70YlICk89qglmtiGtpdrDp8uf51Y3N1dNG1yPMds5PA/lSxTMfMUfhU0ga3yq4z94Z96jdHbEp2+3WvD3hi9n8I3urW1pvtYfL3S7wMZkK9Cc9eOlT3bPAYmX8c+9eDda+z6xYypJ5ajzOcZ/hb2rV/Fcmi3YsbW3+0ibq+/Z0APQg+talpQ065lsiMvHjJ+oB9fevhv4m0jw/rsdx4gg8y0XO8bmX+BwPugnqRWoappms6l9m0Vcwt9w5bnCgn7wB6g1r3w2gh+H2reKft+LnSfJ3Q+UefMmCfe3Y6c9D+FaVIsUnmFufTH1q48ST+QIIlxs75HOT9K0XXbm0O+ceYD2yF9fQVDr3h2/t5LZoNjy4yd7nbg59Parjwc1kv2qCXGOh2/h/e96uI7fUJlNzJ+8TOTg9x7fStI0XUbbUVngvdy9k8tR/CR1zXw3+F2ofEeW6MVx5ckezcmwHGQ/fev9yvDWg3fw18Ha1Jpo/tKceRlOIdn7w9yWByHP5V8ObLU9a1lPD9zDl+cNuUfws3QY9PWviD4fl+Ht6+iXNr5k9zjyxvxnaFY9Cw6P61JI2qReQj+XJ3GM/wCeBWl+GXtLR9Qmm3CPHG3Gckjsa0P4eyazZprMMmI2zldvuV/vD09K8dP8P9T8JQWGijy9Vt92f9cd+6RT/F8owoNJ4ccuJo5/LP8Au59vWte069tdQnmiud80O3cNgHVRjv6V4a8QT39yLeRt0npgDsT6e1eAfh/qfjC+nSzk2sNuRtU9m9WH92viV8NNc0DUP7Mvpdsjfd+VD0CHsx9RXiHxvHr3gzS/DFxaeXJpvnZfzM58yUP0CjsPU1NFOrqsPzdfQVZTw3KBVO5h16ivhj4D1fxvFJFoFr5tzBjA3qvXf/eYDoprxVZS6TqElvdjy5VxlfvY+VfTPYirDRFubF5BN97GTt6YP1qx8BXlhp5ubgfum6Nx2OOgbPU1D4bt102Rbm8/fRY2p5Z5y3PIOOlJJa3Hhh9Fjg8m6kxtk3Fukm48dOnvV14OuLPWtQ0y9m+ex8vd8o/jXI6N9Kv/AAjZrZ7Eut7t0HlkdCPetV8PWNno8Wnw3/mXsW7z4vKK7MsCvOcHI9K1TSJ7JVlcZXnJ49vf3q5vVmhW2u28tUztOM5zyen4VrCJ5pEJzjofyrV9Rjtsxo2cdsfStYvDeTtHGMbsfyH+Fa2khYoGwOxxn0qE+VlSdxH4VJOAflqK8WOMgDJq08QfaIPs3lbW7fNnvn0q5u3JVWGCM5rR5W1RG0y1i3TSY2/Njpk9+OgpvCesTGIiP5pd2OV7fjWoeGNQ029jtZ1w75wMjsAexPrXhm313wLNci3tvMkn2bhvRcbQf97s1XHiGRNflu9Vu/spm2+YfL34wmB90fT861O40UXDGwvvtCH+LymTsOx/GnfSoLdporjfM2MR7GHf16dKku0I4jx+NSODz6VKS7YIqQgHk4xSvge9O4A560ZFIxmp7pIl4OannNw20dKZPNwinmoLQRADOTQjGMZ5FMgIGD9OKZA3CnNbSRnb096Vcjg0CCcUpB4JpTtINbgxoEHgUVz0OayR359KikCPvxk+lOwY5B5ork5B61FK8bZjfBHtTMSTu60ArJgnmr/Uo7uGOJINhTPO7Oc49vahhunboKJKcChgncpogE4B5rwf4WvPF+twaNZLmSXdxkdkZu5H90961vTBpF61nu3FMdsdgfU+tKx6g0RgYYc0MkYz9KM5ZdmPpW0MePypSOlYyPp2ooDyDmoYXncIgyTVxayWgCyDk9KZSTkGlOcg/hXyhcAZoH5SCOlJHuOAa+wyFQ6j8KktXiXLinUFAB2oEH5T2oKCME8/St2QMHpW4HgdafgBetM+6MKB0phyMUASeTQAIzmjtPemJBwDRYEjFH2FdRwelFgBgVx0AzTEdc114oNjmj1+lAg8UYzkAGiQT0occCiQ3Q0Rjg0OOB2rAxgUCCMYoHAxRAIwKyOmea2kDgdKBU8U3B6UV9K3AfLQKk7Qc+2KKj1p2wRSkE4B5pyBzmmkB4B5rIK57+lMExk9qymQA2cdsUWXpmiuPl79q5/hFMoAAJ59Kc4Ge9K7M2M1Ku1xkUyJjG7n6UiFxlecUVDH5fypUYKFK/d6c9aeMq3SiQG3VIxkYKB0okBiGH0ryyxyOgozErtAz6USc4I5okgdaBJOM1wBwayBwDQ4onIxmhgjBP0onPBPIrGRxTdMg80CMYBpiCBisn1rCgYAoHHf8KyCME0cdjRJHANZz3+lEg9qBXGKwuOB06VjJz1oYxzWB2FbV9KPPej04FYBPWiQRgUR2oBT/Sq+vNFR2NDkZojIxQ5FEccGiMjANDI+UGsAdKB6ihnOM5rPOc1u9q6jGfpR9AaJGcis57/hTcgc5onIxXB4oYHFHBGBXHpXB6da6jB61xjGKJG0ACuMYxRAxwf0rjGMUcYwOtcelHnp+VYHYUQD2o4PSgR6VwRxQCngCjg9KI9KPPT8qOCMCqwCOnPaiBjOK4Ixj6VgYxWOc0QCMU3ajjt17UBxiiAaHAwDWc4GaAPQmrecx4A/CrC7WZQm7ke31okY4NEZGM0Vz3ojHQ1kHoa69aBOMA1HNJHjY36VpuqfZ+W5Y9+nr7VdRLOAwGcVPA8TbWGKxgYB61ojYvEQn17exrRPCWp3MdtNbL9pWbdtGVTpkHqa+E+mSeF7WW703UvKuht3ReTu7uBySR0YmvD15Lc6lDpkqbQN3Gc/wk14d0J9L8OeVIfMY/dTp/Gc85PrUtvI2nRxPL5ac4G3d3qyZE2Wztu252nGMdzVvbtGgeHp2FLaMJhOB87/AKYGKitmWc+Y+V9Me1alb2lusphh4fG8bjzjGKOnWWpW0kd2mF47n19j7CvGejeF9M1601BrHfI+/cfMkGMIo9T2NeK9Q0Z9almh8O+YRjB+1sM/KorU9EsdS1CWS1ssebjH7w9lHqfarFbbw8JIZkxIcYGT/TPrTapFB5s0kPmFsbTu24x+Fa/rvmWaq9vuDZ434xgj2rwR4jtoL9Zrm33Kc8byOzegrxG8GteIBcWsO6M/eTcR/AAOTj0q4vby0laKCTy0XoMA44rzhqMZkvG8yQdWxj9B9BWkeH0vVIikwfXGfX39qh0GC2vEtr1/NxnJwV7Z7H6Vp/hmaW28+3ts2q9W3j1I6E561Z6BLMY0iOFOcL/k14DsrWzQC7GQvQc/7Xp+FFn1fSIYrKLDRbtx3dctx1x6Gh4fmgspbxrfMkeNp3jucHv6Vp91NHZiO7g8jPRt27HJ7D8KOlafA73cj+buxlsFenHTNRXcMExm8jB/3j6YrUFtdTmS4jg2u2cncTngD+lX2radpCo9hY+S5zk+azenqD6mn06+8QzTX8UuXbbtO0fLgYPcelSaTrGpXX2S8ut6/wDXNR2z2PsKsLW4s7ye2M3Me3Z8o7gk1eWkY0OV7yfzRxmPbt/j9R+Fa/FbtGkyS4D5wu08YxUoeMM/m7yMYG3FeItAGmQx3DDO7Py+nQep9anCNIs0Z29ffHaoVknuRI918h/6Z+1a3aRaVbW93bXW7zt2fkxtwQO+fWo5I5ot6LsPrnNR24SKRXfcsmMcY6VrGn2jf6hfwyfb3q60hIn/ANFTYn8S5zn8zXhzwdqmrRme2jwg/iyvHUdCw9Kt7afUBcXEce5U27zkDGeB/KkSGJTIsedvfJqUyzgbl3jsM4q30USkStDu2/w7sf1pPCzyphItq9huz/WtH8EXcluFC8H7vI45P+1UHgy4Q7VX5h3yP/iqm+HlxHALkScHqMD1x/eqLwJcSOu1to+gP/s1aj8PZdO0+1nt5/ll3+YmzrggDnd714b+H8l1qYWe42xH/Yz/AAn0bPWrv4evZ6g1pFdbvJxz5eM5XP8Aeo+FplOx5ef90f41qHhCUXUkcMmAMdvYe9W+jrb/ACTS7sdtuP61HqO6KygtJNi2XmbOM439eteIbe98R3t3d38vmrcbNzbQuNoA6DHoKOim0k8kQ4x0+b/69XKQW9oYJYtyyfeG4joeKhthAxljfavpjNaddXN7Lstn2lenAPY+v0rRfGGo2jC2c7l7j5R6n0oR6FrE7ytF5U5xltztnj04HQVf6WkEpTPnL2/h7D3rwz4g8S+HbuT+ztS+zyHGP3KP2PqD6mrXxfrlvbSQnUt/2nHm/uUGdp47VpWsPFcCSJcXI6S59j/DjHTivAl74e/tUXviyP7Xej/Uy5ePZ8rBvlTg5G0c+laDrGnTaqltq1n56nO4+Yyfwk/wj6V4nj0e3VYtJg8u3bPybmPp3bnrmhaa7omiKun3+yB+ieUhxhvU5PUmrvwv4us5hdW9/uC9B5UYxxjuasNZ8V2kwW8k4HtH6H0H0rxfDNrd8+otBkXGMncP4Qo9vSrXS9Ts9XQ2SbRz3U/w+5+tfD698YeHfEEUenXu2Y5wfLjOPkb1yOhNapo3if4l62i6xrHkXC5w32eNv4B2UqOiCtOso9N11LSeXzYVzvO3bn5CR056mvGfh/4bXmim40Kx8i6Tq3mztnLKOjHHQGtI8NvFHJJ5uAMYG36+9fDDx3rPw81FrvRrvazY3Dy1OcK4H3lb++a+J/iCz8f6wdbNj5V5N1fzS3RUXphR0T0q80QadGp1FfNWb7oztxjr0PuK+EuoWNhMf7ZvN8A6ReWRjh+689SDXxb1/RfEOqWMeiQeS8XmbjuZs5VMfeA9DVt4Xa8PM/ly9m25/TPoK+G+hSalDqMWn/N5vlefJ06b9vBPsRxQ8FRrL9jgucue+z2z/erxZbafqPhq1tTaeVcW2/L+YWzudT06dBXiXwpcadafbDLuDdtoHcD1961m3srzTInitcSnO5955+YY4+gq8umt4HiHUY/nWpkMxlkG70HT0rUUZlaRZtoGONufSriaKG4ORk9+o7VqN0EvHaI4DY/DAFGRixBbn6UjAP8AMc/hVlMYJgVb9PahcI5wHwPXFR6zd6TdR3WmXPlyrnDbA2OMdCD2Jrwv8Zr/AEWPToLyTz49O83y1wExvznoh9R6141+Jtj4qtr/AFSayzeSeX5T+YfkwVU8bQDkL3rxV8X/ABV4r1p9c1HU/Mu3xuk8mMZwqr0CAdFA6Vc2Ufi3SLzXdQ1L/S7fZhfJ+/ubb1GAMBR2qSEBtg6Uy+UMr1pmDdDTS5DKOc4xTEgbSefpTkdAeaLKo5NTXaAHB5FTXu7GD09qkmMhwOgpAAm7HSrSPdL834UVGcVgdqKqeAPwoopX5RzTREn2HQUka56VtGelKq7hkfhWxOCo/ClKE7QKIBFYFFQKJBChR0ohcAAVgZwDz9KAJPymmVkOxhz6UuM8imIY4Xj2o8DAP4UBkZB/CirFcAVuQRhdvzeuaMhVcKeKWVl5U81tBPBpQM4NEA8KaAB4NK5BxTMATtpWGc9fSvlAyBmvBnjaw8M6Td2rWe+a52fN5hH3WY9Np7NVzcSXU7TTNlm68Y6DFQaddNZi8CYjPRsj1x0zSEAnApgMZrcD8qjpQkOeK3A8E/hTSjy/KU8elPwAtOxYAelBscA4p+Fz1IoBiMg/pRJbA3Z9eK28dfwojIxmslflBonK8daBBGCa68D8KCjqTRAJ4OB9KUZ6mmBBxn6cUc560x6AGsMecUo9D9BTAngUBkYB57UAxGM9K3ep6U2CMk5FMwwCornGM0enApvY1xgAdqKg47+1MMYANZAIwKY5OAfpRwflxz2oLkYzS/LjB6UwBOQaIAGQKLADAHNMCV5P4UvTg0zE/LmggztzW8KSoNICzY3fhisbX6cdqcZO4N+GKYA8Ec0TuwEOKKkAAtn8KKKSMn6UHSM4U0XWRuakk38Coxj5FNAgfL1osUPI/CpGMg57V5YJ55qO3Tqw+gqX5ySp6+1Om3BDfpQjQfNu6dOK80H+HP40T2PNZHHPTtRJI57VgEDmqbB6GsnGR1o5PasHGMUQCOmaI7USCODQyeCOBRx2og7eBXUUACcAUSDxjNUCMYNHHaiBjg1jsOaIAGaOCcA0QCODz9Kwvr+lEA9DRx2piGGAeRRAIwKAAGCawP736USCcg0RnoazkcGj9a4PQ0DkYJ6UenFYwODk1XUDnmsg9DS5zjP4VhehFELjg1wTgUdoGM5NDDcE0QOgNDBHPaiFI4NMOcCiB2P4VwR0zisAjr+lHG3ANEKRwayCOTWVxzRx2/KiAe/0o47UMcAGjzWAe9EDoDRAwADWAR1/SsDaBnpWAe9HA7/pRAIwDz9KJXoDRyTyaJA70OR1zWQehrg8ZrAPeiR0B6VwRwaOByD+lAg8Cs4HJonsBQIxkfhRJA4oEDvR5+lWlz5TjLY/CrO4jubcFWy3cY96PYHtRGeSaIDDj8KI5wDWCeCPpRwOMVgEYBpSpHB59KgmIwhPTpV5Cl/GGVMFe2f8+lT2MkJyoyPWrJXinWQjgZ5/CvhZ4xt/DHgvRNYtYPtU0f2jzI9xj25ldRyQc5DenavEOsaH4h0s654PvN+or/roPLcbfmCr80gCn5Qx4FfDr4V+J9c1ltQv9F8iJceXJ9pjbPysDwHB6gCvhr4O8SabqM8usRboDt2HKDHyvn7rE9SKuhcXmLVYt23oNwHv/StL0m8hvBJew+Xs6LuDdQfQ/Srd1jUKpz7UZEZMx8sO1RaXPJJh+PTp/jXiLRzqFhJbwybJmxxjPcH1x0FR6R8htJ0x75/Hsa8b6XdN4qcTRfu4scZHeNfem8U6Lo3iE2moW25O53sMfJnsD6ipY5tP1KWTTLL7VHBjYfMCZyvPX6mvFtzqd3qj3lzZ+WeMJ5in+FR1FXN/fRMweHBPbcOKg1O0ukNtdTbGHbaT79h9Ki052nxYneB+Hb3P1qO18UR34Gm2PmSf9dYxnj3/ABrU7DVLSQi/h8uTuu5T6en4Vb2mLcsjZ9Rj3rwlqNnCzx3tx5I4wdhbP3vT8KsLWXU5GjK7m4yc49f8K0vVNTsfD0nhsRYSXHzbl4w5fpj39a0yzjMUZiTL84GfrWmWV1PGUitd5HX5wPWvB1vPpWnM1kv2nzMb+Qm3BbHXOc5NXt9HJoUiwQ7nGON2P461xdU8QWyLa2O+IZ+fzVHcdjg9RWpSWUEX9lTyeU56DaWz0btV7AhbzLeXzUPQ7dtRW9/cXji3hzHbY3fMBjcPernQL7WYFkjjxsznkev1HpS66fDF5PbWq+Yh24Ocds9wf71aXrEerymONtrN3xnHB9h6VfXEFreyovzyrjjkdh/StFeEahnWZfLjbttJ/hP938KvfDR8QXNxHpjbwm3yxjGcjnqR6GrTTNTt5ngvbLYI8bj5inqPY/SviHa6vbiMNp2Y2zhvOUdNvar+PFoV27WHQZz3FMtysflrBx67hWoXyW2lpaOdzS52jGMYYE1pOpSXyfZ1XIHv9fb2rW9IsNb8FaVceHz5t1pvnf2gnK7N8oEfLEA5AP3c+9TXojyJm2EdsZpNQkjmXzBlDnn8KT4kaLpPhYWWmS75D95drD/lpkdVPqa0SaOy0jUdPMWIdV8ry5d2f9WxJ469TjtWqeHF0mCNRPvWTP8ADjoR7n1q0uHik8tPmz0HSodXWEmHOHPb0/SrTX3cqhfAHtn+laTryi1QGfB5wNvufarLVFcbC2GPt/8AWqO9iWCOJJfMVc7ztIxzxU/iLT7WURCTcvrgjt9KbWYL2JLe1G92zheRn8x7U9+bUGOaDyx67s/yqzvYLtnkU/O2OMH/AD2rVLmPT5Fj1FfJBzhs7vTsPqKgn0+4jIe4w3+4ak0SxmLP9qz6fuz/AI18L/Alvqd6wluNyjHGwjs/+17V8QfC/wDZGpy2VmcpJj5cY6Kp7k+ta7dfZrhke3zj+Lf7DtVwbHVLN4oJMz8YXBHf16dBU+iXdjHslh3Z6fMB/X3qKLU9Nl+1W1vkN0+dfTHf61a6xIr5nTa3pnP8hVrMBPHczHakmeeuMDFfabCSVBp955x5z+7K9vf8a06bTNStwt+32d+74Z+57D6Crq3AZjaz7guMHbjP51aubCA3Mh/d/wALevODx+Naf4m0YXcct7c7cZx8jHsfQfSo9OuhJ9otV809hkL7dzXhjQdV1qeKGWDY3OF3Kex9x6VrM+i2+iRWK3W+RM7hsYYywI/SvC3inRbXxhYf2lJnTI/M+0SYb5cxnbwBu+9gcV4zsY9Q1ZZ9EP2q0Ocv9zHyqOjc9QabWPhonhVfDd5feVqgz8nlTtt/eb+oG37uO9T/AAp1SGZbmC2zBz829R2x03561e2t5pniCVYJP9IgxgbR3T8R0Nadp97reib7L97qZ6JwufmPckL90V4n8C+KNKtVj1DSPJkGd58+Nu4x0Y+oq3kntEaPZ1x3FSpqAhLqmE+orRoZ97XKxZVOp3DvkVL5012EgG5z90dO3Na5Dr0UqpqkO3yc4XchxkD+7+FTaNpr3Ctd3HkpzhtjN29AfpWm21mk8l3DP5jQY2DaRuyCD9Ke6vJb9Jb6L7LFzk7g/b2564/Ovhbq3h7wzY6tp9tc71vPJ8h9jjfsLluCDjGe9af4ZSC1fWHb5jjaMe5X1/pWleBtU1WKaW9s8RybdjeYvbOeAw9BXxG8GvbWQVm/dr146cp/tVrFrHp9ubWE78e2Mcg/1q4tWv7h7eL5pDjC9O2f6VqjGDKucEdvyrVr3NwV3/L9PYVqV3G0p8l8/hjsKuZBIwwcnvSyBMHqaZlds4xRKqcq34YoyBowpPIoMe5r5c/ez6cV4ftrS/jeGeXaxxj5SfX0+lRWV28PnpDuQ9DuA9q0Lw/rOsXCLp9l5pOf+Wir2PqR6GtUgOnym2lXEi9Rn2B/kaurjKhgvA680kzK3XIpiqEhDkfSnZSeD+lTSpGNzmr27DthOlTOWO0nmgMcA1Evqf0pgCu0HrVmpJBNcEAg1gDkGiR2P6VgDpWwnvwPamQk8H9KVSABn9KUKSATgfShtAwDxQUYyDQHbNUQegP0pGVSARmiwLYBpsA8GgSOh6VI7SvknJNajYaRBppe2v8AzJ/7nlMP4h3PHSgWBAP5UMMOaYZOAKDMF25qKzWaMyFuB04qTap2jt0NY3KAp59KwynBFMrbtpogAYB5FEcZFd+lIhJ6VPazW7ASx7T6ZBpVL5VRmre2mupNsEe4jtnFXNrJbSmKZNrL1Gc0urzLpy2AP7sdPzz6UGUDr9Kj2kjngVdPC0cYhbJ5yMEYqMRkbSf0qSG2WJHSbc5zldpGKKgcFsHtxTDLZPIrg/dopgbQa2kkDriipCYFAnPWhweaPK8dqZixx6UBkYBoYIwaAB4zRGVwe9bewNDIIA7U2SOBRyRkmlGRx2peOaYq3AokHAA5oLu4FBwoZcZNEbuKJJGMUVGeOtKqkcnPpW0sOBxQ54BzQyPlBrdjgmjkgEj6UQSAcUMkDA5FFTnOaZeM5piRytEg9qB3cAVHIqSKzDIHap3SWQvEMA9BR3HhRTY4BFEoV4OaQMTkDNCPHQU4VaYbhlVz+NBgV4qOSONtzjNSTrnIqWYvHhV/WlAYBRyfSvLZBnb096dEOAjfhiljctlRTKIwSBmpGEhGD+FRxqV55qUAthefQU7blwW6d8UHQD734YqQxuMFv0p33AAmqIGMnrRwRx+VcDgUccc1gZzisgjmvl6UQGOM0QpGM1tGMg0SCuO9YXHB5rI24B5oBR0NZA6Udp70VU8ZrAPIoYHQ0QCetEgjANZx0NYBHXpWO4ogDgGiAaPPeu2KOMAA1gYIBojOADmgRjBNEA0enFAY70OnNYPr+lEZ6GiSRgGsDrRPRc0RnoaIHQHmhg8ZpVB4okEfyogHHNMARXDDANbRjANYXGAaGOgNAgd6wOmaKg8E0QMcGjjtWARkUB2FEAjGa2rjCnn0xQxjFYBHWiB0FYBOAaYDpmiBxzRA6A0VUjg9K4AodACc0MHg/hR571jjGaPPesAN1rj0/Cic1WARxRI6Zrgjg0RkYzWFzgdaIz0NYBHWiR09KXsfyrcAMgc1kUQCAc9KBHQGtLvvs77D0PX9aglSZAymmIJx+lE5wAKC55zyKYE8k/WsLjrmgSOR+FKpIyBxUIUvkmvhjcaIuqGPWo98fblh/C/938K+L58Paj4hjsPDi+XbWudvLnO5EP8AHz1Bq6zHOY25K9PyrwtMiqyA9cYHp1rw7fy2Mu/OV/8ArH/GvBvxYv1VbWLjrt6cfeJ/gr4WePpGiit7+Te0mcNjGMb+wX6VELfaJw/XpwfpSl2gLJ989D+NSW3nTb88+uPaolaIBCM1ewmZBGx4HatTm+ywJ5UuWXPO33FRXjXMZG/n1xXinwsNTvTcedtl/vbc9lHTPoK1zwBdJK1xLN5mMcbQOwH96tfivo5p7AwYaPbxuHcA1q8N1Y3TPKNp4wOD2H+NTarE0m5jz/n2q4s9P1SbJl2Me+0nt/8AWq60MMo8iXJHbbj+tJeavoU8c8UW4DOPmUdse/rR8QTXb5uIefTcP6CrW8hn3xzR7VOO5P8AKtP8P6XJJ5sV183p5be/vXhKygs7G5lS6zc3WzcfLP8ACWx7dKsfB0xtmv0XzJOO+O+P71QaUJbGNZxsePPHXGT7GvC0Wl2mk3D3Sb5ht2csP4jnpx0xV54aj0HTIYLWfzZm3eadm3OG46kjoa0a1cW3lIPmbt6cmjqLwwGzni3E/wC1jvntQ8LpqFx5kr7U7jGe319q1LRrnQWYWg+TsOPb1J9a0hZ9QvJPMTy0bGeQ3Y/4VZ6prSIdPt49sfrlT79xV/oWn3k4ttRHlPF1blt2QD0B46CltrrS23QDMPrx/wDXPU14guTaSqCfMdM8/d64qDXXluRNIMbf4fwx6V4YS61W9kjVcx8YGRxwfp6V4r8ES28pe3ut0bfeXy8Yxtx1avEPg290/So7trnzBLn5dgGMMB/e961jTY54jCUx68/T3pLa7hfG3K+mRVxaTPEEXgn+GrK2u9OfCLknvkf571d2N1cXKvK3HPYelXXh2a9XZbvu9sAf19qs/DNxp5luWtc+XjPzjjPHr71b2lxPNlkwD05B7V8WDJYeDNF8KW485tO+0Zk+7u3yo/Q9OuOprxj4ul8RfDT/AIROHTvJfu/nbs/v1fpgenrVtoWqNH5jRYx05X/GodFYgl3+Y9sf/XrTQbcgpDk/731qxvL+a+WIJheccj0rTJbvUrg2szYZe+AexP8AStQvZ7JTBPDhV6HcOenpWp6/cyKJAm0D3B9ParHxPdl/PU4MX3eh68elaj4qv7mxJkbK+uAO49qm8Xz3dqISMqvTp6/T2qbxfBqenC1uG5XpwfXPYe1W+qxJbrDGmNucHPv9Knu5b51EKZK57/59K8A+P7nwbaz2zR/vTt2/MOOWP909mr4g/Em88QGxvLnmWDzMnjnO0dlHYU/i86pxIMHsM/8A1varO6F7M2n7c+b3z6DP9K8c+CbHRfDsF0dR8wvu+XySMYdR/ePrU9lHbX/2vzt5j6DbjqMV4V+EPwp17TWv7lsX4xt4uP7xB6OB90VLpj6DbS2Jm8wQ42HbjqcnufWksYr47JhkN35/z2qaN7ef/R3xC3QY9vfnrWj28IZmVck+596stVjnYWc0OfK7bvXnsKvvBtn4hgkvAfKaHG0YLdSAf4h6V4E0iwukkm1A7DHjZ1PXcD0P0q1kkjig1C2uPLnTdtbZnGeOh46Uz61qLGSJd6DocqP89K0JrfU4pYLsYxjJ556+n0FWPiHUbOL7JpcmQv8ADhfr3H1rxlcWfxF1J5tdtPs19puNzeYXz5iqOi7R0QevWk1qbw94N+xWeqZJ6J5GP+WmepB9an8HaH4ktZtWuLjyrm424Gx2xtO3swHQelfAvxDFo+qnTru6xHF0Oz1WQ9gfUV408f2+oapqVnepvtofK+znJHVVLdFz1A61rmqWMlyZXgyq/wC0fQe1ah4e0K40qG4SHyHO7J3O+fmA9a0aw0+G6jt5LnzonzkbCvQE/XrWt6XLahYEjyFzgZ6dPete8LXUfh621WO93h926PygMYcKOc/0ppblIPJhlx6jaPWl0HUdVExhtvMRdu5t4G304z7V8OvhZb24UXtxukOcDYR/e9H+lHSdJ1HSZbK8h2SQbdh3MerZPQ+wrwvBFpenwW1mm9fm747k98+tadLPBapJNzHJn5eOME+leJNCtdbijQ8oM56+3uPSvin4L0XT7lbuxlwsmdw2txgIO7e5rxs/k6hOYThRtwf+ArWubCxRTkf/AKq8Q3ISUJH/AAf1Aq6kLtgGmG4bajQIcE59OKZA757isArhWwe3FE9OfwppVU4z0oKCNnUDpXg7xRHoDGCWHfHJ975sdN3sfWrO5MMflF8j6U2sz25IhfA+g/wp5gzk4yT3qVgxAI+tSAcYNPIWABP0qW6RV4PNXFwXfrUrFjjNNGGXIbp7UEBXGfoMUh9KADMBmrZB3OfaiAeB+Vc5p4w469KaJSaYsi4UUZGHzH8qWbJwy/Tmt6n7vNBgCQB0pXyAAc+ldsA0QCQAc0CAuAeaaYtGseemcU0YMe8NkntilIGATWdpyDmnGMBT+FByBjNEqRjNc44NIQQQTTcjg/pQAUYB6VHY3EtrLdouUhxvORxk4FFfL6HJpzuxzQZux5rcVNMQTlaKhlyOtLtAxn8KkJc5BrT9L0VvDF/dyHF5F5fk8N3chu+OnrWgeJbzw8kosTgzY3dO2cdQfU1qF9NqV495O2Xkxnj0AHb6UxV+BWMKRn6UEK9DTDaBu/ChtxjNIMDg0w3Agn6cUrfKBimwBwa2nGSeaI4wD9K5HU8074HBrrwT9OKUAnANFc85oAAgE/Su4GfwpuTwaJAFdB0rG45Bz6Vnf1FMApyD9KY54zzQYDgdaJBOTRbIxRwSOaxuIOaBAGBQIPJpiPr6UWJGDQIAGKIGOtE44oYPeicfKDkVtGMCgoHANHBOKznnPNbgOP1pTtzijjJxW0tz3qMELyelOyqucUylhwM0yjZt/KsIvGaMjEYBpm3HINJLtGDSzDoR0piHO7H4UkYfgmjGEO1TUBCNkjPtU8qSuAopZCnGK37+MfQUSGIBHSpSAwCN93tipplU5UcijPuyw608rFeDWT60STxRGehornvTdMYrBI4FYJ6isE9aoAjtVfMO1UVOOK4PFMABwa4IwBz2ojB55ogHjOTWO2KIB4U8/SicYXNYzwKPsPwqiMURwOazjoaPPIokGiozgdaIPSgMjAogkcGqxlc0BnoM0QM4BrGelY4xmjzwK6iiM9DXFdeK4ArHeiMnpQ+aiOwPNEgjANEjqDWBnANMRjIPStvHJrAPIpiOoNAAjg0QAcdaOD3plHZv0rCjgN+lEAj5aKgDrQUHvRA7Gioxyf0rAPGaIBwAa284J5oqAuAelYBHWiFI4NYB4zW3IyDQAIwDzRGcc1gE4B/SgD93FEHFHOOBRHAFYGMCsYGBRAIxRBHb6UQTwBmuDzVH2FYz2oDjJFY3D+VADOD1odcDt0rb3FAAdK0i+EbiOR8L9PrTIGQMOQehpgD0HPpUIRevarho2ACDpQIBGTQAxkUZVCBBzig+D8pqOZ0OVP04qw1GZJhIZsY/2enBq/cNKJFbOepx7CvD90kc4+bj6exrR7kzPh5Mr6bfrXha5+zXGPtHzj7o2dODXhjxrc6daxz3Vzs25w2wHuR0C+9fD7xPqer6fDLOu9Tuxyo7t6AelWd1OkaO8eIznccikYNKWQZTsfwoMkkoUPkemPatVuJ1kCof5e1XenXM8NzOZMqNm0YH0PerS1veWEuxR/sg1NbTXFzM0UnmKm3BxjqKu9V07T7jfqz7Yh14Y9v9ke4r4jweH5vEs2safa74Jtu353HREU9eeue1fEjTEvtRSW1ssRjOR5n+ynqfaofC+n35kjlfyXTGBgtn9fatV8O3Vm8qwDO3G1uPbtmrKDVLfaWb17LXhvTptWkaK7+dFx6DHX0I9BUejeGjqD2t3+724w3zntnoD9Kk8LWKxzhoN0T7fLbcR068Zz19a+Gvg3RNR1CWLUbvyEGMHy3bs/o3sKv2g0zxPdaVY3mUh2bH8s85jBPBz6mvBfiO/tVWKa93xjr+7A/ve1aJexzai667b/u2xht/op7L+FWPhbSbm1+1QDci9B8w749auJ9OtJPOkk3bunDDpS6hOrNPanYo78H271YjTtQuFtXuPMl552MO2f5Ctahg053jFzgJjI2Hvj/GvFHiRJ9MUMcSc8/8CHtWmaVBdeDpJ1OLl8bG57SEHjOOlT+KbjTEWKax+UZ+bzR/IA+teNru21p7S6tT853+cuD8v3QvXHp2rTL/AE6exOl3L5LdBhuxJ7VZaN4VSdo9dudsg/h2SHt/sn6Ve6bYyeIXjtm/c8bTg/3B6nPWvB0yaO95GYdoXy9h3Zz97Pr61DenVr4wmbaG+8m3PQevHpWp6taxae9lf2PmGTHlDzSMYIJ6D6da8U6YbeVg0fl9OM5xwPenS9V8i33KOh3gVb6e1zpzT+RmfjaN3vg98dKuNK1GVN0afjkf41q2l240uzkDYkXf5hwefmGO9eG0sbS5QmHewz8u4jsa8TapZ3dzPY2WnbRc7fMXzidu0AjqOelav4U03S9HW8Aw/PHzf3gPU+teGdcPibVILO/m8zbuwNuM/KT2A/uivHfhfwdF4UihtI/I1CXO05kbOJFz1OPu1F4a1BUeKcbWGPl4P6g1YeCLJkNxcJ9zqMnvkdmrS/D3hlG8ucYPY/vPf0NabpPh59WS3SPJ5wcuP4SfWvEHhDRtHvN1smGPXlvQepPrXifRLC5tN1ydoHRsE9x2Bq/0PTmCoBvTnn5h6e9P4e0tdsUUHyc87m/xqLwn4dm8MShkxLxjl/7/ANfSm8D20cJS3j+Xscn+rVF4GsWnCFsHvwfT/eqbwuliGRo8oehzj+vvVhpUcCsYJ8vxgbfr71PoNzrl79ojixnr8w44x6j0rxH4YWO4hjc/LHu38eoGO9aV8LHm0a0165l+zwT78tt342sV6B89favjJ4M8IfD7QPCuo+EfEn9oXupfbPtH+hyReX5bxhfvswOQ7dMYxWs3HjbV9DXULo+ZZLnDYiGPmA6DB6ioda1Oa5cNJlRjIwoxx9K0XxDqVpOv2e42Y77FPY+opPFEup2axyvvl5y2Md/THoKsJDND5f2ra/Zdmai0m6vggtpd0fPy7QP5n2q1t7iyjZVi+Yf7Q/z3rSNavo74h5/PkP3jtC9jjt6V4T13wtrscGkWqb7p929cyDGNzDqAOgqw8D37aJ9uWPzEXqcgfxY/vU6yw6S06wfKmON3TLYrQfF+rwaeLe3g246ruU9ye4qC8hl+zi3XZJNu3LknOOlazez+DdTjlsbjz2fOV2Beij1z/eNav4l07WdZbVNQ0nyPtH+uHns27aoVegGOg6CvET6XeTxPp2teXEN2V+zMccD156g14gu9VsZSdO1fz4x9wfZ1XsM9fqa0S+8QWl+t0G35zlcIOxFa34c8U3vhnT9Wey4m83cPMj4w4X19vSrjQprlGhlhxux/EP8AGteml1DSIrB7TyjFu58zd1YH+laR4UmtbZNQQZ25/Hkj1qTwj4iS+t7zV7PFrdbvm8xONox0U564rxNa6pa+J7zwrZXXm2ybPKXy1XOYw5689c9TU+mQRz/vbnYR1Gwnt9a8E3cLC+kt7jZBD5fnRbCd+d23nqMHnivBvi3TtU8XW9w6eVbJu3Llm6xsOuM9RWv2ltqXiy4bTJ82kW3cNh+XMYx15PINWX2OTQhHo1z/AKUvQ7Dx8/8AtcdM1f21xaaUIWfzGX2Az8wq21Ge2tyZE+X0yOOa+JGtGe6mt4pt8Z27RtxjhD6V4peAtLE8memeD/s1rV55RKl+B2x9K1SZJndw3p2+lSIWbIGaIPGD+FPEWOS3TtionLEqxwR0FSkIhYdR0qJ5W+VTnHSghYkE5PpSxoAMr+tM6quScegpLkMSQ2PbFSzbjy2ce1CYhgQeKkl3jAb8MU7k8Z6VdXAjjzmprt5DgNgfSjIex/Sshh1oOAMUsiE4JpinZv0pJBvA3fpVtJghSefpRGSBmhz3+lDB4BonP3TRQkZHSmRWGMUUVgFC8jvmggBwDQRwRhsnvxQUg0nmkYJ6dKUMMHPSiCBgmhxRDEcUpGMs304onJwtFQDx+IpgDyBRyRgjmiABg/lRYnAArRrCPVNThsCcCTdzj0Un+la9p0Ok3xtYpN69jgjsD6n1pNRvIbWeyhnxDcbfNXaPm2nI56jn0pzkgDk0DnjNEADAoqxUccdqCk8EfhTgJ905/CsZGSaQlRnH0oSMCdpx7USCuQKKscbaELxgF0x+NIrSyqiLnrxmnODjHSnJcAsPpS7GOB1FFeducgUUB5BzRDbcKOnas7l6UGHQjP6U2QePwplI4PXtTDLAZrGONv60E3K2wUqlucZrKlADy3pQHHIrOMD0rIIAxzTKSa254HbpRI6ZorkZzxROehrgDgdK3Z6msADINEgjg81HgEAnj6VqnhfVtMsxf3dp5cXZvMU9wOgJPUigRjGc1wRzQIIxnJo4IxnpWmaJqetM8emW3nNHjcN6rjOfUj0NXNlPaXDWlzHslT7y5BxkZ7e1EheCKKkqNozWAeM03AwD+FZJPSiSRhR9KJweBmmJC+9Keck0yAcDrRbjA4p3OQpFTShFBU8005bvTSAjANIRjAbP4VjcMZ/SnBIxn9K8tm6D60CoXaBmlEjDjtRYhvm5PpUhQj5T+lKrMMryfSn2lcbcH60hQfX0qSQN14pwS+d+R9Kdh93P4UcYIB5NYJ+X8qbk4FDPembaOTRl9BRORgURxQ6AHmgARjNY96GAetZHYUee9YyOKOCME817iqIyODXB6mj6Vn0NEEng10oHHejjPBrjH8qwCOtAE9DXUfSiO1YzwKxngUQc4rHvRGRgGqA45P6VQzng/SsZHFYGMCvY81WPejnt+VUPQmqIBGKXtmsKRxR2jgCsDGfyFYyM5z7UAGokEdKBHXNHBGR1pcdKYDoKBA4Bo46g0QDyD0roODR6AUMVWVJ5NEUOwo5FE5AGa+hroODX0o9BR6YBrtgGj6A17YongA0RkYBogEcHmqqq6ij1wKPoKwMYFHGOtDGcV1+U0ME8V2yO1ZJ4BoHOAKVyrZU81o2oSSkwytnb04+tNg1ZtAoJcc9utXBVpSVPHb8qIBOAaKNs3dqAwMZpVB6Vt496jJyAp5qxs/t0ARV3N25x3/8ArVDGttMNnBH+FaZqDDlHww6cfWvBuoX15ffZ4Blj1HA7N/hT+LNXjuI9OkmwsecrtXjIz6V8I9X8a6vbLb+HtMlukiVmdlChUGW6kr1PYdTXhDxFLrdmJ5yQMkYK4IILA9h6VaXSrECzcfSmR2YSZ47VcSRNJ5T9fWpp7aGDy4Hz5n3xgjoeKkltw/lKOT0HNagjadNNfMML8vH4Af1rxX4ssrqMxhuO4wf9n2rWLnR7q5xDFujXp8zDsK8Z6ZYXWoWi6fbZHz7hvP8AdXHU/Wtf0jT3kAmtvJPcby3p6GrzwHaXl2hhb92+c8HsP972rVtEvtIYoUwn1Ht7n1qfU7uOJINNOJBneeOfTqPrT3GoQvvuG+b6D+leFtVnvtct7OU7lk3ccDohPp7VbW0Hh91hnj8sPnuT0+mfWr/wvow1qS4a3y5xzvYfwgeteE/hub3SGu7aHYnH8Wf4mHdvatFj1O9iV7MYWPOT8vfPr9Kt7+9tLY75cMP9ketXmv3cKxxPN8z5/hHb8Kn05JdGMlzL+9b7ny+jc9D6VZSeRfLaCTk55x7ZrxA02kzl/N3KnX5cdQPr61rfiW61m7+zw8RjoOD2HsPSrrV7yw0yLTHfCfNkYH97Pp71f6jIhCTHd6dvT0rV7+aKXzIG4P3uB6CrG3gmkQxjk59asY20vXZLN1zHxzn/AGM+/rWv2MUhe8tUyVxgZx6DvXhHR7u50iPULiLcsmcjIHRmHY1cWr2eoebZRbSe27P8Pv8AjWsw3mg3Lw6sPnbG0cdgP7ufUVrVr4Un8CX99dx79Qj8vyTmQdZQG6cfdx1qO0BceYcqO2MVfXDwzOIztj42jr2GakuppMGA4UfSvht4Rg8W3V7HqUm6K28vAwR94P6EHqor4jx2/h5mbRUwF6HJ/wBj+9n1NTa/qNxqLXFzJ+9OMnC/3cdh6Va61cXNwBeTb4x22gdvYfSpbvUbMvJAMMMYPy0fE/iK6REluPkXP8Cf4UPF2qqgjM/P+6v+FWGv6zqLrZQvmR87eFHTJPb2rSo5XkCzt+9HbH19PavBOg3Gq6qIAMTN93p2Vs9wOgrxhPq3h/WJLa4myUxxtX+6p7Z9a1DxVcTxlbmTOztgd8egp9chdR57ZHbg/wBBVpr0caPGr5VcbePz7VL4hht7TEUuR3+U+v0p/FMX2hgr/KcYGPb6Vc+IVE+1H49Me30qXU2uFB8zI+n/ANatB+IS2WmTaVIm/ftxzjoxb+771p8UGmuA/wB0dOvv9fWp5ft5mdE+X5cc/wCfSjql6mjppTP+6jztGB3bJ7VqWom8WCK/PmBN23tjOM9PoK/tm9hsjp1pLiE/w7R657jPWorS+aQps+vIq00YjTnuPs/Ixzu/2setWthbmMRLH8w6cmh4ZnliV7deOc8j+pqysb+G1iEDYHzcYHrXw28P6DrtvP8A2ycTQ7dnDnqXz90gdAKjtrJLKz05bb/SIt+87zzk5Ht0q4tDp2q201knlH58jO7+H3+tHX76zt20uN8Jx2Hrn0qwtriRNsqbwfu8geuatPE3h7SNLEphyR/tP/e+h9a1vxHfav4kurmBsJNs8tcDsgB7D0rTdVnkjMd2u5h0GQPX0H0rRLvT9Z1SDSb2DBn3ZG4/wqW7Y9BXj7TY/DWox2scOIufm3dPlU9Mk9TUl3b6yuTJmVfvcH/63YVb2i+cotm45zx7e9aZ8ZfEuk2MGiCXEcG7A2pxklv7h9fWtY1C/vnk1ETcjGRtHsP6VaT+IY2CSPvDdBhBWg2uoXGoLYXx2Qz5z0ONqk9ueuK1Hwr4jt/DcWoSXOVGcfIn98D1rwNaouqRXOsncs2cNjGMKw6L+FeNbHwi/iKMJBuj543SD+BavvCc93qmlx6ecLded5a8cbVGeSfY1No2qaTqSW6xbGGf4lP8OfU+taRqdvNHeXl6+J7vy8jB/hyOwx0xWm2k0EAubWbB7HaPUjvV/wCOLU2k0Eku6U7cDaR3H+z6UfEMuo2sttby7JuNo2575PUY6CvEy2t9E8qSfMuP4T7f4V4ps5PNeaNsgYycf7tatBHO5YDIP/1q1mFIpCg69v0qcgvtHSm6YByawCOtSblAKnp1pptw2A00bIMAfN3pd4BbNJLIqAhv0p2Y9TQAxxRJAwanuI0GSf0qbUFQcNz24q41N8YV/wBP/rVcXcs3O76cUSDx3oAEZBoEEYFDkdaBAXA5onjCnpSbdwOahbJGDmlwVGDXXihg8VjmgF20cd+lHnmg2Ttz0peMZrgml4wO9Y4zQ54zTDHGKDEDANMAVznJoDNOrIduf0ogbeD+FEdieRRwR1rAPQ81FLJC4kifDDocVJK8reY75P0xW4EY70oDHB/AVnHANH5uooMSMHt0o5xgUGz1NE4GM81Z2YuuGPTtVygilMcZ6UhG3moZNsisecZ4pmvfEt4Ax5P09Pw/u14s+GWo+DdA0zXbyXfHqnneUNoGPLdVPRj/AHh1AqB4VJMoz6daJDNgdqKsPut+lbgBgHn6UMk4Bp1KqDnAonjGaAx90c0xwNuaJU8UD83JoZPGaB6gn8MVjOADRyOtBs8bs/hW4E4o4PGPpW4NRZCeOtHBHPWmA25Ao4PU/SnAU4AoDsKG1jg9qxu5NAggAdulXGpXN3D5Mz5A7YA/kKJ29BxW7IwDRxiuCOK8P+I77w88kti2DJjdwO2fUH1NahfTaleSXk5zJJjJx6ADt7CiRjB60GxwKZs8Zo42hs8mmAIyDQP8Pp0oKcgA1kMcA8jrQxjGaDFzwaYKXGeTUxwcA1IQ23b75piNuVpW3Ac8UGGetDkYzz9KYMD979KBLLkN+lRuFGCaaUoeDTOXYDNFSw+U1CcZCnp7VJtK5J6Ufm+6entRDlsbv0piAu0Hmg6BMd6IxxmiVx1/Si644NNISMCsk9+fpWT60xBAwc1nIANZyMA0BkYBrBzimUY4oDPGaIBGAeRRHGM02M4qiMjgUFGAc/hWM9BW3PWiAOBWAeAaZcYwaojIrGRkHpW3I4oA4BBo9MelEY4zQHQ46UR2og+ldRQBI5HSqx2B/CiMjANdRQGeAa69qIz0NEgd6rtgHJoc8VjnOa4AAzWcjihjvW33ogcfN+lEA/xfpWFIwG6e1AAjGf0ogHkH9KABGSa2gjOcfhQXIojPegoI65op6Nn2xWMHrn2rBIzmiBjg1gkZzRA6YxRQEdaK4oAEYz9KYHGAc0QcciipxxWDwM8+lFTng0QSBg1tNbTRGfu80M962kDk0fYURnisZGc1g9c/hQB6E0BjjP0oDd0NVjPeiCTgGsE8Bv0pRnv0osDwG/SicjArAx1ogdVNEg8g80AD0PP0q3na3kDq3T2qzuo7qPKHn6dKOVHWiCRuBz6cVtBIG79KSztxYySzz7CMbBtJzzz0psDoc0gAPJ5pxElrsaP5+zZPr6UpVSDmvhdq+nWniKGLVbbz4X3fLvZeiP8A3RnrivjJ8Nb3wF4kuBbv9q01tvkXGAm/5I93ylmYYZsc14bjS+uxH5m09jjPY/4VJY+KPAutx4jYXMjBUjRUdnYqAAAN3Pzjj3r4BfsfeJvF9svjb42WEvhrTXQSrYPLsu5lA6yDAEC465+fg8L1rxV+1D4A8Gw/8K++B+n2d+1ruQ3sX/HpC3OSh6zt8py2dp4OW5FeC/iFqqWhTUQZr65keSRyVG9i7sTwuB1rS/EbLYRi5g5Oed3ufQVb64L2PyYjtYdG6/pj2qKwVx5ly27HbGP5Ve28C3IaCTZGevBPatdm02zgNzatvkXthh3A7+1axrE+sq1tGnlpL/HkHGMHpgeleINJvBrT2c2o+XGcfN5IP8IPQGtd8cahpeqTWlleed5e3I8sL1UHup9a+HniWO6uE1HUl8wQ5yudvUMOoH0rWrTw54w1Ca71GH7BHHtwdzy5yoHbH90fnXjKDTtKvMaTdedbp0by2XqF7Nk9Sa8SazbWzCDWLHag/i80n07KPpUFj4e1XL6Tc7ZB22Of/QsehrVfDN7MDII9w9cgenvWhaf9juhdxTYmj+78vqCD3x0pPE0mvLGNSiw0ecHd6/QD0FeHobHU5o/Mtt0j553kYwD/AEFXZv7HRitrqXkQL1Hkhv4vfnqatorzw6TZQ3HnRr95dgXHcevrVnfDVJ3jeXGMfLtznj1/CtbigeRUjm3lc/wkY6VrXjMRWaQSXPzRZwNnXJH+zVjqF1Pc5W7yB0byx6elS64/njTrweas/WTO3bgZ6AfQVcW+jx28ckNxvkGf4GHepv8ATXju9Q/dwLnMnXPboOeoArUHspZMWUv2lB/FtKenY1Hf6eH2XEn+s6Jg/Lj3HWrH4YeD9E/Z1g8YaxpAk1+5g862uBK6NmWTEQwDggIQ2CPWmltjqccd5a4353Sbyei8cD8BVxYabHAb4ReXEv8ABuY7ucde1aVNcWFgy2s+Ymxti2j5cE55PJ5OaNpp8Vr9vnfMi/w4b1x2r4g3+r65qPmXFtlx/tr/AHV9Melas2qQW7RGHcpxlNyjPI71oc9rp909zdaH9tiONo+0mLHBHbnqR+VeJWe61F5Es/s8UmNq+ZvxhRnnr1/nU8GqRIBAu5T2+UV4J8UweHbCWM2++eTbvG8jGC2OxHQ14t8Wx+IfDkmmWvh/7JcNj/SftfmfxqfuEAdBj8c1NG7yshfzJe/GP88UttLAA/l8jtmrS9W7iiiEfPO5s/0/CrHwfqPiWFzo0HmCLG/5lXr0+8R/dNeJfBUXhGUWGtReXcjqN2cdD/CxHRhXhrUdP0qZLqG18x0ztk3kYyCDxz64q28GapNF/b1tcb1/u7FH+z1Lf0r4dXWnSS2TQzf8TRvM/d7W4wG7/d+7XxStprTxROmqXXI28bB/zzT+7+FatYo8TXQT5e3PXoKvYzDHhxxWi/D6/wBS8L6d4kW+8qPUvN8uPyg2PLkKHnd7egrx94JHgfSDZHVftU8nRvI2YwynpuI6NV0lzBMQLnd6DYB2qSUou8y7m9NuKOpGG3SFRllzlumefTFS6xawWSNYDzJTnceRt546j0zR1PVL1QGbay9RhT/T2q01XWLSJ1kPycZHy/4VL4kaQGM3nzenl/8A1qvdTyFIvOUzkeX/APWqF3dt0V3n28utLa9e7D/bN23qvlgZ4Perf4f+Hn8DT3LeJMP8v/Lm5x+9x/epD5M6ytFtDZ+TdnbgetaNqEAgVXbavPGCe5qN7mC6CGXKL935R6c0tzcWMYubKfzXP3htC47d/rV74nit54bqyud7DdkbMZ4A7itZ8a3WsukqwfZ5I84+cPnOP9kelQXU2oQfappNm7ouAehx2+lW3iKTSY44pTgLnB/yD61FrVpqsIggvsE/w+Uf8B6Vc+G5bCZNSRM+XnAzjqMevvV3PFbXKBrTyyc8+ZntWlWdo/iWRxdbbiPG1fLJxlD+HSvGGqXmuaY9lLo3mSnGJPtAX+JT0wOwxVokOlSrHNLiV+o2nsPbPY1b+fJMv9nzevG0env+NagmpgmV7f5l6nevtUnhHxFZ+H5dbubbFsNufnT+/t7HPX2rRNWvtItba+u/C326Kbd5cn24RdCQeACepA/CvFEeg6l4EvNWtH+y6qdnl2uHfH70KfnPy/dGenfFaF4nfxDZjRby9+zJH/D5e/OSW7Aeg796nv8AwFMlhoK6z9muofM85vs8z9fmXjp09D3q78R6TZ6r9ovLrzYR919jLj5cdAM9cV4bl8U6xfS6rokX2mCDH2aTdGmMgq3DYPUEcir34q6y99u1G28x06jeo6j2T6VY+KrC81UXN9beTDP91d5bGFwegz1ArX/iDBbRC00Kbzou52lcdD/EueuatdYuBdPc3F1vPHGwDHGOwrWvGd9a3i3On3GWjzxsHcAd1+tazrU8K7JW2B/YHpj2rXv38TiK744/5Z/StXUISIhkev5VrS75iwfn0x7CplIbB60wBXjrVqkFw2wzbffaTUmi3p8x4Yt8MWMvuA6+2c9auYhGNynkULjafn5p50YhQevanvII1AkfGOgwafUrcpgn6Hn/AAqfVIkGEO79P6VNrDscIcD/AD7VLevKfvZ/CjMzcZ6Uxz3zQKkYDdPaiuOc59qwMYB6dKCrkY5oDjANdOKwM4B/ChgEY/CoHyBxVu5KhTVEZHAoJngn8KUEjAH0raGGCc+1SIM4U9O1YwcDmjg8A/hSsDxmlZX7YoZ25AzXJPB5pjntWPf8KALDg9KOQcCj83OfwpQzHaDRyTisAjINFTjCn8KIIAyKAUHBNOowCpz+FAkciiPQ5oMF4I/CggZdxbp2xTyWwtyqjL9uvrQRSmSeO1Hngdajnlh4ibGevFTeBvEEOhXPiO7tfLtLbbvfeh+8+wcBs9cdqYAgAfhS8nGasdQuNPcPbtyOnA/r9a1XxF4n8U6DjUr3zrTSf9XH5ca7fMcZ5ABOSo9fwpxvXITaPrmmRVAIOT9KMZCZHQUyheMUpJ+7wauJY5rWONIdpXOW3Zzz6UQMYJ6UQQMg0cEZI5o5xxS4OOM0pU8A05XOAcn8qY7SOeaPzAHP6UQQcmmIPQ/pWAed36UTtAGayOp5o4YZXtQJbgiic8VkYznPtXDcg49qQIVxnJ9KJA4B/CiucEGhtBwTx6UzBgB+VA5GAKIyMZoYAxnpQIIwDmiCR8p5oFSMsv605zyD06URimUkAE0R8vBoqMZBzW4A4DZP0ogdqMiE7RzUjFWwDii7k7lGTTsScsfwxRIYAKckdBTbcYDZPpitxA24ohj1pWJYFDRYng8+9CQKAFXP40Zg3CjpRHmcgUiAAZNEhRtBovtUEDk1HMzrtY8dq3EAlV/Wi7E4xj2pmUHDH8KabPyijIWGAKLFhgUVfaDijgDANM2RheaBOMZogg4HamXPStuKAyOKHXmm681jjNHk8VgkYHasAjrRHArBPAoqQmc1gkcUVxRHaiMVRHYCsEdKI9KxkZzXUYzRHFYzwKIBGAaI7U2cD1o4IwK4PANYzwKPHFZHTNVQ6c0cZHNDpzVUcY69KLDoKPOCK4I4NKR1z9KrpwfyognoOazxx2okngURjBBpuRwaA7flRJHet1dQMfhRGByaxnpRXiiMUAT0NEHGDQB6Vjtnn0o8jGaHtRBNYz0ojFVgkYFMCBxRz0zX0NDkAd6xngVkEYFYyOKrvj8qxngVVAYOQarAxgVweAaGccGsZ6URih0wDzWMnGaCgnGaPHANEcDmtLvDbyAHof8A69BcqOfpWw5wDxWBnAprt2hMBPFZ5zSwvJylNvxhj0oAkjJqzneznS4j+8mcD6jFa38Vr3xV4ZtdCvYubffsO4d3DHog/ujvX7PX7Pfxb+O/igaT8N/Drz20Dhb3VrnMVnZDH/LSTB+bBBCKCx6gYyRfaP8As1fsS6Xa+OviprcWv+OpYS1iGgSS7lbADfZLfIES/LgysR3BcZ218Z/2ufiZ+0DPNYCc+HvCgYiLRrSU/vlycG4k4Mp4HGAgwMLnk/DbxPFY3cV3MMgZ4/Bx6e9eCvGtn4g1u2lkXao37Rkn+Bs/wj0qPxNauALboOnX+orw9rji42gZ2+/sfarPxQDaF5ExjoM+/wBKsdesJUdmHL4zyeMfhXiu6sp9PeS05cY45HdfWtM1gWOuRH7yx5yOnVD7e9fGbWYvEN/LbWJ2txz17RnuB6GoNFk0+7a+C+YG6DOOgx6+9fC3R7C7mnuZDh22/LyegcevtXiLVYvDuqXemSLlTs5zj+EH0PrXiX4j6fa3U+lmLKrt53Edgf7teN/Ez6/ulUbQenOf7vsPStO1u70y5E0T59sD0PsfWrPx6ciOYc/5/wBmvD8OnarcRySPgc54b0P+FWPgexmRRpM2Rzn5SP8A0JvrWj6VPoenwyyjLtu9Bjk+59a0/UbYWLTXh9OOfU+n4Vq+nw65dTmzfGdvYnsPUj0oWsfhsu6yb5Djtj/H1rU9WW9czxN846jH0Hp7VrTy36+TLxu69/T/AArRr23hjMBf5z7H3rSoIra7gvLqTKJuwMY7Edql1nQ7CXNz8wHb5x29h71qPie113TIp9Pi2rbZz8xP3mA7gehoXv24HHb/AD/SrLwrL4k1iw0excfaL26jtoQVJ+eRgo7+pFftNX9h4e8HeHvBdkwSCDDiMD7scSiNAcdvmP8A3zXhiOz1q5a7WTaq44wT2I9vSraOO6ma2kk+Xjt7ZrQbGCa38qZ/kHTg+pq5tVS5MVs2fTjHb3rUtJj1C0igueJI889epHofQV4j8D2k2mz3ay4e325G085YD+9TSqX8oCtd0xv7HN4h+aLt9WA9ae7Vow8i9O2a1G4tjaxPEuGOd/J9Rirprs2ZIX5PqPWkllkvQiL87e/tV9bzxKTKcH049q8OXR026WRWztzgYx2P19a+HPj2/wBCmu4LRMrdbN3I42h8dVPrXxO+I91461RJbmLaVzj5gf4U9FH92vhv8Ob7xNIvlvxzxgf7f+0P7teH/G2m2PhJ9CltP374w3mN2kLdNuOh9a8O6rYaE3mumbofdOTx1z2I6GvGFx/bF+90zZLY4x6BR7elQ+G7zW0h0u34eTdjp257keleIEls5/sjD5v/AKwNWOr3tnbxWhb5Yt20YHGTk9qlu9U8X3Kh25TOOF9Pw/u1PE75yOT0ou8cnzLkD3rS54yGW6TrjHP19Pwqz1nwoujT2F3D/pb7fKbdJxhsngDHTHWl1DexTHNXV4baPLDdn8KuP9I/eJUrOy4JzijevbuFIyK0y4nF35yN09h6GrrxpqFzZizibCH2Hrn+7WjaDpFz4c+06hJi7HQYb++R2OOmKmsniG4SfKOgxUnihDE0bJ+v/wBak8PX1toH9qzTYR/urtHZ9p7/ANKbU1sZTABuK9T07fT3rUkv7u2NxbDAHXp6gd/pVvqVyNMjV++fT+99K1KG/ggjv3+aLnHQdwPr1rwe1tZILrUep6Dn/aHb6itO8WfZ729NzHvgXy8DOOx9BnrQWLxZfCGwi/er23H098dlNNYf2HeSxXy/vExu59R7Z9RWgxSxzqYmyp749jXxL0uxvr9ry2XEnfk+iDufavDmnXt7qi2dsdrc9gf4Se59qXWH8HJdeD9Vh8y6tNm5t2PvfvOigjow71q/xIXWvBsnhmW32rJjnfnpKH/uj09a1jxGy6da6RGvyW+/Bz/eIb0q1vtRvp7HT4m+WfzM8LxtGf6VbWN9e69bWtu20vuycA9Ez3+lR6onh3xxqml3yeYlv5OXzj70WegB9RXinwVGdEOtQXGd3Rdn+0F/vf0rwJ8Q7jwtb/ZyvyP05HYsf7p/vVqmrwyXf2iHkN1HPYAelSPOBbzKcrNu2jAGMVrDTx24kcdPp6irm5kFq0g5HHH41rupiG33xct6fiPatb1iGeIMwyfqfb2rVNWSVSq/56e1axdBlO0/p9K1GFZZNxPLdP0q7tgHIBp4XLgA9O1CIRtgdaXxJPZadNpypuW425OQMbTn096luSybc1PenGQOlTX0uRg4xT3Du24nNSXJxtJozFxxTAnjNfe/ChkdaPArgDNUeBXUcUPu8UOetAkjFL6VA5BCjtUT5wQfwpJA4wKABOAaQHNIeMelcEcGpE3DGalQgZB6Uw7g0Bk7RUakKGHekIIwaxxgGsjGM9KwDxmiDiivGc03IBFKN3yg0chuDWc9PwqM7CD1xV5eG6CfLjZnHPrTfOc0CVGQc0rZ4H5USVPWsAjIPSgx4xTNuOM81ltu0UVwOtEDOAal1W9mtTZPL+6b7y7Rzzn0z1rGOM0RzkH8KyTyKF5IkRhRsK3UY64qHY7gOeK1FLONwtmeB97r7etFztwBx6USGbGfpWCTgGvbNbQ3epOBtofMAtYBO3NEAHCmmOORRXJJBrHqKXnABz7UWBOAOaUK2A5+lOFIwpps8HNMfkIFZyMijjGAeaK570FJO0UUOcE0oJO31p0K8CidxFEgdTSqSv8AKhx05oAEbfSjyNvWiAOgoZJAB6UST+FEk96GCQQadgCBTEHvRJPQ1Jg4FMxxXyE8mnbec0G2sMGpWBO0HpW4DHPSmbB5NMoKjFEYGM0nWm+Y4zQ6c0CM4zW9TwD+lGQjgc0ZFzyaZw+AO3SieBg0zgISx+lC7RuFFFnY8GgAODRIPyigQozmjKCcE/SnYFcA0DxgCtoo4x1rAOCD+GK2g9DQGO9YB6H8KAz3rHOM0QOgogk4zRAzgGsA9TnHtRGTgCtuf8KCnGQaZcDigMjkUBnj06UVGMAUeRgHpQGB/SicHg0MngGsDGQawW4zRGTjNYwQKAHZv0ogA8mjzRIPSj6ihgj6VgUBkDn8KoDOBmvl6VgEYz+lY4xmqxxwfwqiOmD+FUFHTOa29lNEDoOtEcYz+lFRjis7htzRXjANEZXg/hQJxgc0QT1NFQehogDis4PPWiOAwFEEckZoAlRxWSMD0rJAwDWSRjNYx0PPpWMkZNFcDg1jPANZI4JokEYxW3Izms+ho896JyOOaJzgE1z0rAPGa4o4wMUAccmiBjg1RGOM1RA6A1xRBxgGiM8ZqsAnINdTgGivYGsA8Zo8nitpPFFc96OVHWtK1IyoY5jyOn6+1BwV+UUFJGc4/CtoXHelXLAZ/GtA8JLfabNdx3PEG3cNnqxA7+1akm2cqo6f4ClYjBH/AOqrO2vdRuYdP0+0murq4kEcMMMZd5HJwFVRySTgACv2Z/8AgmVqOpafD8SP2oNQbwzoNrGbk6CJxBO8QG4tdTZxbpjqoO/GcmMivj3/AMFGfh58JvDh+EH7HuhaWI9OjNuutRWgSxs+cE20RH7985PmuNhPP7zOa8QeNfE3jrX7vxT4w1++1jWL199xe3s7TSOfcnsBwB0AwBgCtL8W6npnEUu722qPX2PrWieMp4nBjXB+o9/9mvAnxRvtJv4bkpvKbsLkDOQw/un1rwp43uYtAi1m7+YPnC8Do5XqF+navBfxrs7O4W8a287d0i3legYddn41dfFa21eTzUt/JH9zeW/XYPSrXxU1zNcNbR4jO3aM5zx7itL8XRabL5lxHuj7pkjse4B9a17WdPmnmutPT7Or7cHJfoAO4+v50+g2bSfb7a4yx7bD9O5rxHpFxo3imfTrS/3Ww27F8oDP7sE9ST1NDxJpHhSMC7st8r/dHmsOn0B7MK+I/jnTNatLXW4rbyPtG/zI97Nt2lFHO0enpXix7IXP9pW589W+8nK9Ao6mviZ4z0PxJeyP4f8AC/8AY8DY2xfbnuNvCZ5ZQTypP4+1RSpO3ls2PwqOxiZBKlxnPbZWiT3GnuGikyfTA9/8a8KfEvUrNseR/wCPr/tf7NeGvivHqNslvOmWGctnpyewT2rxD4xs55xbRr5gbvkjoAfSovF9xa2AlSbn02jnn6VB4wiv7plvX8v0OM9vYewq2mm1e4a80ux2oMbh5uccY749DWo62t4PsSQbpO3z49D6e1apeNpM5We23OvbfjsPTPrQ8UzX1j5sMXlyRfdXcDnJ57e1P4rnmuMTw7l/3gO3sKtPF1zYaTd2NuuLe62buQfutnuM9TVt43i05CsUOWPfcf8A4n3r9kDSW8a/GLSL14w9vpcE2oTgjvH8kZ6dRI6H8K/aK+JNnrvxl1Tw+LD7TBo0cdkkonKgMql3G0Ds7MvXtVza2um6hJZQyZhGMPg+menJ6mhrItb5I1jzCM5fOO3pjPWrrXtFXTs6bbfN2O9v73uPrVlqltBpMt3PF5kvG0biP4sHoPSta8XSajq72kC+QBjBzv8A4QfT2rxfq2ppA1mt754GPl8tVxyp9KsppILgPcWu1B1Tfnt6ivEOrR3UaR2UfkQyZ387tuMY6jPWpLQO+Y7jzF9Nm2tRvdN1ifR9Hc/Y0h8/zJ/mkzkBh8vHpjr3rxHqGmaFEfDtlN/aaS/8vG1oduMN9059cde3vWi6S9zq0KCfDvux8vTCn3rXbKaW4kWa73NHj5fLx1A9KaweJygj3Z6c4r4deGLC7nLahc7MdBsJ7N6H2FfEH4Zf8IPLCsupfaJH3Z/c7MYC/wC0f71eBPGOo+GNRg1E3eYId26PyxzlWA5wSOWrVtWi1fUG1CyufKRui7C2MADqQPStP1bSfDPhq6vNQj/teW42bbfLW/3XIPzAHswP4e9XN5bLKWI2+nU44qw8U3llqdpqdnLloN+1No4yuDyR714iWbULw3V+20nvjPYDt9BUs1rGmwy5Y/7JFadqJt7Rr2GXy2XGPl3dyKu74yMgZMA5wc9KuLpJovIRfn7HNLKzsI1bp3xQj8wFAuPfNfDj4dy+PrqaGC48ryNuRs3dQ/8AtL/crx7osXhHV5NMW781ocZHl7cZVT6n+9QvmMhCwZXsd2K0nSb/AF66MVnHtKe4PUH1I9KMMwnMbjle3HpWnus8nkiLbjvuz614N8Ba14gsnv7W33W8WNz7lGMlh0LA9RVx4hmEZQDfn7w6Y/StS8dz6euWj3gdBkD0/wBn3q3vL+xnRr+ywWz8vmA9vb6ivB/ijSLzUof+Emf7VbRbsQ4aPGVb+JBnqFP4V4s0vQ49dfVfDl79qsWx+78t0z8ir1c5657UFt9fjjXTNP8Asac74/NMm705OMdD+dafovh82jQ3U/l4xhtjnuewP0q6msDaxaDp9150cm7fJsK7cHcOD+I61ceRcQgWkeHHU5P9aW28JWPwtlkMHl6rLjfLukOcT8cfd+6cV4P8a6hoWq/a9J/dzHq3DfwsOjKR0JrXdKm8VvLqt5r3lOccfZQ2Oi9iP7o7V4Z1kS6qujyz8N/y02+ilumPw614k0nVdJnFjJL9z7vyr82QCe5x1q3e50p/tpXbMvQ5B9vfsa0+31LWr6bWtYvM3N1t3yeWoztBUcLgdAB0rUbvSYNLlWKPdPHjD5YZyw7dOla5o1it6kFo3nqud/BXbwMdTXgrwTpi6Yt29tuL58qTew2YZgeN3OenNXVtpeg6rIGg80HG35mXb8vPr61d6Ldaxc6jHpqfvLjysnI+baPcjGADVp4a1e5k+wCDgdRuX6+teKPA9zbpGmnJ5wXPmjIXZ93HVuc89Kh8MTW1sZ5eIx1b05x0zTzy6WQtvcbh/CdmP559av8AV9RvbHDz7l7jYB3Ht7VBqZi0+4huY8Odux93XnngCvEouIYxIp3Z6ngY6VqZuDBv83K9htHqK1W6dMoE3Htzj0rV0dSQRx2/Sr6acuAiZx05A9KlBcbnGKuLk28ilW3e2MVdX8kpyBt/WpLhm6dRUl3IRjH4U8mVGDyO1fO3VsfhUku07U7d6LbuM8/ShwMA0RmsDua2k8A9O1H2FAe9MMjANAZGAaIOOBRXJyDTEjoaPt+VZJxk89qU4YHNQPuxg9OlW7knANd+aU/3TQfGAD17UuTyDRAIwalDEAE49KbltoHIodgaEj7dobgdsUhcruz+FZDDPesZ4Br8KYAgAHmsZGQ36UyAEBTmgp247+lMMHaKJIHNMCp25/StueRWPeiVA5OayCODRZWGMc+tEEnCmsg8A0T8vBrOSMCgCRjvW0kYHbtSRtI21Tz2FEYGFb9KT5eCaYYGM0qAsADzUiqjAK2fwpiRnDdevFRr5hEYGfapkCOUU9KJAOAaLYIAFIyjk8+3SmAJ4b6cUCScA9KZccAc0ApXIbkdsUGycE0YyoLAZ9+lEAcY5oFSMCsgcZpmAwtDGME1nKjJpyMDBoAf4UQoHDZPbisE9/woAZ680QTxigcnHU0SSc9KK5x7dKYDAJNFc4A603HHeifQdO1Egjr+FBQV3FufTFE46Gs85rAPQ9KJx2zTtu5AraCvB5ouQuSKjkViQT+lO+5QAOakwRgmg6kbQOnSpDjoPpRIHeggYjHNY2jGM0AoOAckUFU8Ac0QQ2AcmiCx60zBCUUZxQAXAJyO1HIPBoEoAQfwpmCryaa4RRyfpUt3jlF/Wmld+rfhiolAGc0GUgc0WVRyelMyZyBn0pZFAxjNMQeBRCtwW/SlKgfKc0WXpn9KwSODWPlwTSsAODXJ5o9eK57UQTWMnBPNEY4zRBPAoqAOtYOABTD3peRjNAbjjNFcUOuAa5HU0Tg4FNnpR6DNE9waI7g1We+eayfWvpRwTmjg9KHTg12wDXOODRORg0Seo60ST2rOeQaokDvVHB4FA+/Wu2AaqgRnGaByOTQODg1u9qGMZzWCelAcZz06VnIznpQx1zQ64BoccGgPf6VknjNEcDmsgDA5ogHnNZx0/KiSRgGjyOKBAHX8KzkgntRyeM1nHFZ5zWeMZrtiuBwKJz3rGeBVDiqGD3rjpmh6CsDGBXB4Br/Io9OBWB2ps8AGqrAPGaI+XANEYPFEk9D9KIyME80CAMUecEnp7V1PB6UM96gcpIDmrZg8KspzmooC/Cj6VIpQmNhyKBAbIPPpWj+IpbG3mty3Em3PHoT7e9XU5uX3Cv2df2Ufi9+0trYsfAmifZ9Ht5Ql/rt6pjs7TuRuxmR8Y/dpk8jOByNL8Lfsg/8ABNDwymt+JNRXxR8Srm1JhkaNJNRuCRgiCLJWzhJyN5OSMgs5AWv2l/22fjB+0veTadrWo/2F4RWXdbeHbCQiHAPytO/DTuMA5b5QeVVc0pVDkGrMQocTHHp1pwobK9BVoisQYjkjoMVY6mLcIrNhhnt/9atL+MV+NLi0Zn/cx5wcDuxb+5nr71Z/E+N5ongmyY93G31H+7Xhz4gXN2/2dpfm7fKPc/3faoPGms+H4UlI+WTO0/L26/wn1rSfiNb6u4W9n2juNpPr6KPQVqGr2C6cJHm+YdBtP94e1eI/ippOkW4hW5/eDtsb1H+wfWp/i5HZ3TX7TZPG35cdsH+CvEXxln1+5FzcScr04HoB2Qela146N+23zeB2x9P9n2qTX3kG0H5R1P8AkUI4Spdm5+hq3hMjEAfN2qOza2iV36c1GqTSKUbhc44pNNurQCVxgfh/jWl65awLIs02JDjYNp989BVt4luln80t8o6cD0+lS+IZ7ksI3yhxjgf4VZajJayrK545x+VeHfGj+HIZo7UbjLt3Dp0zjqp9a8QeM7O8eOS3b96M5GD7eq+gpru4uW3t8zH6CtPtxiLzmwXzgY9M0Wjnc4fgdOKg1uOOWOz35xnAx+PpVzqsEieSrc/Q/wCFf8E+NKtvDvgDxv8AFXXMR2Vu/wBmSZlGUht4zNOR3wfMT8U9jUnjG78T+IdR8RX7j7Vqt5NdTd8PI7O3b1Naf40s4pZVupPv4wcHtn0X6VZ6ml9dERtujPTjHb6e1eHbULK6zPjOMjH19K1a9j0fCyyfK/3Rj0x9fWl8TwLN9pkbBHTg+mPStW1+yvrszCXPmdRtPYAentXiS50+fUGfTZd6cY+Uj+Eev41FLBPqKWkjYEucjB7Lmp7aOOYqjfL9PatWjhEoLPynTg+gq1nSPDFs49q8BXMNvq0PiCyHmC03ed1GNyso6/0FfEfVbHxPq51O1GFH3uv91AOoH92vBXhfTNcvZbO9l2n5fL+VjnhiehHoKutD8O6NF5GmXG+Qf6z5HGPTqT6mvEt9d+LL4S48zd06DHA+n92hpenx6fcWkkv735cDa3rn19KsvCkMHh5L2J8kZyMH+/j1rRdHHifxLb6JLLtjm3ZO3OMIW9R/d9a8VQwQX72Fk29Y8c4I6hT3ptRS1nUI3HPb2rxd4gk1aNYyMAZ5z/u+w9KkZYiATnHXihfJJD5W7A+lXjwRBImflc9jVvfwAbYjlh9RWoyJpmqS24PAxjj/AGQff1qfUZCzhOnFeHfF2o+H7g31m2HPUYHoR3B9a8VeJ38QapLqF837ybGePRVHYDsBXh+0sdR1eHT55sCTd/Ceyk9vpXj7QbTwZdwWul3G55N275COgUjqT/eNackVijT3bcNjBx9R2+oqzUpIsp+6+cfhXhf4yav4T0mXQrH/AFM2N33ezFh1Qnqx71d3cM1w0kByhxk8+lXz2c8BjY5J7c+orxH4tj8UadZwzLgwb8HPqw9AP7tQeDtZurc3tlbbol6tvUd8dC2eua8O6xcWp+yS8L+HufSvBF14GSa2tzqGLqbf8vlS8YDH6dKfwKNL1srcrizP3W9fk54DE9TXivxt4Y0mG68Lwt/pL7NhxJ22uf4SOh9aXXJ45wiL8h6HI9PpWs+G9dstO+2zw/6Fd/cbcn8JAPAOepq28S6LZXKQxyfOc9m9Pp71441TUItcbS0GDHjC8Hqin0q00a/EI1COP7nU5Hfj1rxD4um1uWGTVPkX5sHr2H90D0FarLp3li4jlyo/2WHoKuZpLOVLOYYPOB+tR+E9Gh0aS7mk/wBKbG1cN/ewec46VHZTPJOIU3Ftu/nGMdK8KuLXwhPZxNmQbdgxj/loSf0rXNN+3o90/OMf0Hr7VpVlPHbQtCPlbdg8dian1K50LVTJbDdI3bgfw+4PrWj+Lr1U1BWhy9z5WBuH8OfatQ1m5Gjy6PdRbUlxzuB6MG7D+tRWulwaXcvcyf6R8vlLtbn5vm6cdPWtOjWaHyoU3Meozj1rXdJ8ODSpILa4/fcbRsf+8M9TjpWoW0yx+RdDCdj+XpWvaPNBEUC5Hbke3vV7pjPkgcj/AD614gs9MgX7PDLux0O1h6GvEeiJp9rbTueLjft/4CQPX3rUJVRPLB+Uf/Wq4kUE5bp7VcXKt8kRye9TSNBJtJ5707l2zmpHVFyTTysw45AoknkGuSeDzS5P9K6celHmmOBgUxwMCiQRigTnBNHnmgecA1mgR603QZonocfSgRgZNRSbTgGreYHkH8KMm8YJr6Giwz15pZOgoMCME80wJX5RTqQcGlYdT2pTk470j9EFYGMCsZOAKwCOuaJOcZrOeprODkHNbieQKIB49KznqfpTjOGPWt2OM0Tk5pgGPB4or6UPQH9KyBxQBwCDRGcEGgQeaBAGSKLZGAc0km0YzQGRwc1ktyPwpZA5wxoEDMSnO7+lGMKcZ5qGWOPcjdT0oROTlam3Fst1PWmOeBWA/fJHSipJ4NEh+hpkIUkDp1rIPJNNgtkHPtRwD8pzTtkAA/WmOeCaUFV5HWiqkZJ5rG4YHWioHGeaA42k/hRA6ZoYK4oqD3/DFcHjvRIIAWicH5TSjnOfpRBIzj6UTkDnmiQwwTX+RRBPGaKkjP6UBkcfhW3A5NUF3dDR2g4BqQgkHNO4wMHrQXI3K3I9qOT8v5USI/lz+lEEjIP6UwLYANEFRj1oZznOQKYDqKChsbTRUls55ojbgZpVbGRTAFuDTMBxmjNGON36VLdbflHSmu2HQ0blyOD0oyueCaJLd+R0o9ge1KAOT+FK+BgGmc8dyK3lu/4Vnjr+FKcYOad8cE/pXmKF25+nFGYnoK85qIAGAKwCMChwOBWc9BRXngVkHoKwetY9aJBGCKY54rtyPpRIIwBQwRgmhxzn8KJAGQKz3zQ46UTmic1nPeiQRjNZAHAyaHHAOfwrisgnIH4UMDmqx71gdqPTANDjvRJIxmiBjg0QDRGRgGgAOKIUDijhh1odMYo4HNHBx+lYz1OazxgCuSMUPagMnGPwoggY6mipPeuSvJoAjAzW44wDRJwOax/CD+lEY6GiSRjNA4HB+lYLck0w4xn9KxgAg1k4+9n8KJJGAee1dsA5FZxwKOTxmsnGAaz3zRyR978MVXXoaBIwBWcjANYGMCjz3ogE5BrHcVxWB0FHHTNckYrnGOuOldRRAxkcmtvpR9hRAIxXUUenAoAkcCgQeAKAB60y5wB27UFOeB+FY3HisKelBST8o6USR/hSnuBzWi3wdFt2HI6fqatrgQyBwuce9XUy3EzSkY3Y4/CipPAHNaNo2q67qtromh6ZdajqF9KsVtaWsLSyzSHgKiLksSewFfs4f8EyLDSNMX4rftb6va6Lo1jELpvD5vBCqRgZLX1xkCJQOqI2eRl1IK1+0V/wUu8N+CNI/wCFR/sgaJp9hYafGbVdfWyWK3t1Axiyt8AHH/PRxjg4Vshq1/xFrvizW7zxJ4n1i81XVdQlMt1e3k7TSzOerM7Ekn61k9RWQwGDzSyEDaOlRXMgXCt07YqO63YYNtb0xmtOudLm0uUXcGbgY2Sbm4+Y54HHTFWV9bWq5a23L2+cik8RFJAGi2oe27P9K0HxhpzW4a1h2zjqdzHufUY6VH8bVhsRYX0PmTQfcG7GMnJ6J6Yrw18QJJdVVUOVOcjp/Cf9mm+K2kDxJJBcxboRjd8zDPycdEz1rx1rVl/aj3Nhc74mx8uwjGFUdxnrmr/xGLyEQyRfju6dPb2qWONYtqjJ7GnDK5XdnHfFJMwYAHI71ptjBeu1rPc+SF6NsLep6D6Vb2FvaXkkIutyjG2TyyM8c8fpWnRWCyN9rbzhxt4K4656fhVoGYkzHdjp2/lVzqV/dgLPc71HRdgH8h7UssaWSvCu6Rs55x0P+FWd47DyJFyD1XNPeS2s0bfZvLibO19+c4HPHXrQ8QFsRyruA6c4x+lJdAx+TcS+W3Y7c/yotAild25uxwRQ8RTwWKwtFgDPzbh6+mKXVmEovYOJR91vTseMelPBo1vZidtQ33J6x+Uwxzjr06UL9o7mJ44fmTdxu65FQGe4cyFOnbIrxQy/BD/gnrYaQwFvqXimwihYdGd75jLIre4ty6/8BrS724juljsxnr8vHofWrfUZXncyJy2MHPt9K8Pa9caZOLkJ5rL0XIXsR6H1q48W6je6nJK91tDYz8in+H6e1Q2dxrd0ot9X+Yd/s44498elXGkW1pZrJDLvkP3htIxzx39Kvp7iKaJIE45zyPQUt80LebHyfWorabVdUjFpzPLnB+i++B0FXemXMdyLdr35m6Dyx6Z9a8V+E73QpEg1FMPLnHI5xt9CfUVbXS2I23NhvHr5uP5fWvDHxCh0HQZ/DtnY+YLzb5svmkZ2OWHBU+uOta7rEtp4XmEmj+Wjbf3v2jPSQdsfhXg3W7vT9Sg1Nh5kSbty5AzlWA5xnqah+KejaxA+it4T8gTYzP8Ab2bod33dg9Mdaj17/hV9/d2lhZ/bpJNn7/zPK24GfukN/fI/CrK4Ou64rW9rvup88b8Z2p74HQVe2HibTLYieH/Rl/5abk459Bk9TV3bTNpY1Oyu/OVf9YfLC7PmwOvXNafqDRSqZrjapzkbM+tfEbQ/DGgWtjq2keKP7Ra68zNt9ikh8vaUX7zE5zknpxjFXd1bXsW9X2j0wT/npTvbOkixnjjA54qO7tUTYZP0NT3dg7Fwd2evUYo6np9lAr2Ufltzzknv7j3o3XnHy8ZC9G/+tVvqbgtEsfAxzu/+tWoXMZhR4Zsu2crt6Yx3rwi3h9/FVl/wl+nfb9JHmfaLbzni3/u22/NH8ww208enpXjZPDUfiGSXwVY/Y7AY8uLzXk2/IueZOfvbj+NNNezDMzbnb7hwBtx1q1g06fQjJPqHlXHaPyWb+P1HHStMuXtC0Qh3AYx82PWorqW5nzLa7V7/AD57VJOQo8qXaD1G3NaQmmS3CNqke6MZyMsOx/u/hVrqUCWqItl83OD5h9fpWj/HpvBfhI6Jp1p532z/AFj+Zt2bZCw4MZznce9aPfqXLeR9oH97ds9e1PfWl1KqRWGGGct5p449Ks/jpH4r00WGoaPtU/dn+0Z2/Nn7ojH90CvE9jZi7muLO8+3ebt3HyzFtwAB165/pXgnwpfa7p639uPLAzhuDnlh0JHpVprOtXnhh9F1fV/s1pJjzB9nV9uJNw+6MnkDv3rVfgv8O7a6TxBbeJft8Mef3X2KaLqAvUvnrz07V4W1X4ffEfxVrXjbxSn9nfa/I+z2eZptu2Mxt86Bf7inkd8e9ST+ALuJ9JgPkBsbR++bp839KMNnq6xaRIv267g3YOTF1+b2HQfpWoeHrvTbNp9Qi2wJjI3A45A7EnqRWiRLrGqR+KbDWvLtkzsT7NnflSh5OCMEHtV/8Y/7e064tbfwv+8G39/9u+7yD90oM5xivDWuGe4ml0jQ8Lebd032n7u0HHysOe4q01jSfBmjviH7fdNjcu5ot2G47EDhv0q313UNGhlhuI/Ojkx82Qu3HsAc5zXgrxCt5cy2F8v9oJFj7Mc+V5eQxboOc4HXpir+xuU1Jnijxntkf3RXiHxLfaZJFbwJsfnPIO7p7HGM1deJWvFxN+9P937v9ParnxhbxTtElnjOOPMPp9K0r4nax4a1BL7SLbEqZwu9e6kdWU9iau/E2seLta8myi23lx9xNyn7q88kAdFNeJNU1vTEGl69B5Tt0+dWz0P8IPqKAll8uNY/P3ZwudteMtN1fREa4u9L8hex89W/ujsT6itW14yQmKODbjvuz6e1arr5UhJjvA6Dp6egrUtUhmJCR4P+8fb2q4kMgKq+PwqQspwrfjimLP8AMz5PbipW2Dg1LK0hwW/SlU7Mhv0o5I2g9K+lKXIwDnFAnqTRZs4B/SskgAHp2piegP4YrJIwTRYkDmicDANAkY5o4IxmgAeDzWCOSc0OR1pt3Y0uT0NRSlMA9RVvOZiFC7ce+a6cE0MHv+lAAkUrshx+lB3KkYzTDeSQOlHBPArAUkgUMDjFKwAAJyaJDDGfpRUYwa4IwaBIGB1oLuHrRGOoo4IwOaxu4UUCMACvlPWsbhnH0rBxkCgcAAVwR9KKnoKxuGGFZ7Uee/04ouCMUTnjNBgBx+VB+wPNI7A4B57CiSx2saztwc9ayp4BzW9UOKMpY4ppZCRg4A6DFP8AMdxFFwMALkjpzQbJwa0WOwk1KBL45g+beOf7px0564rxpDoltqCJoZ/cvncPn4wq/wB7nrmj8wwBXAXjrW0uQAMU6qx2jtRUdhQJKjHWiQODzTYzkHj0oA8cdKJJ5xTBhgdaIAGQM0CT8w7UwBGMVhein8KKEd/0oAngHp0ogEdc0eeB1oAgZNYOOvNHd61YWMeoP5Ul15WPu/u92ev+FX1n9juXt1k8wJjDbcZyB2p2B4BoNklSf0rIAyDThuoOaZi/QU6qRgj8KJIXAP4UJCANrdO2Kdgw+bg9qB4onI5PSmAHVv0rODgmsbh1/CkTJwrfhimkC8EVNdIownU0t8VGCn61JqPy7Ej/AFp7l24zj2pnZuC2fwrJwDmmGRkGu2AaOSOvSvoaAzxmgoIGDRKgdeaLADpSlQaZgelF26A0dp4HWjjrXBHBrgjg1gHoaxjoaOMYBo9MCgB6/hR570ARjNHmuMYBokFaAyOKK8VwODRAxjPNEZwDRx2ojFUeRjNE5GKIBGQPpWcjiuorAwOKIA4A+lUcZ4o89BXJGBVceldR0qqIHQGqIyKGMcVRweKAA6Hp0qsjGBVKT1NDHUVnPOaJyPlNEAjFEHGAax8uKz0Ga4zjvQORk1gHoaPUUQc5FEE4BNAcEUwBGRS4/wAKAyOKAyCCetZHXOTR4waxngUQM4FHjihjoaPHI/CjyBjtWKwCKHqO1VRwelYz0qqqsDpmqIHQGsDtRwRgVWAeAa2g8Z/SiAR16e1H680TkYzQGBnNAnORRGRjNZz061ZTmCdJN33c9vaop0kUBWz+FE5OK/Zt/Y4+MP7TGpJL4T0oaX4ail2XniHUEZLaLB+ZYx1mkHPyJwDjcVBzRX9kH/gmj4aVIox4o+JdzbdSUk1K6JXqTytlbn8yP+epBNftL/tefGL9pbVSPGGrDTvDsEu+x8PWDlLWHHRn5zNJj+N+hJ2hQcUQCOOooEEYArIzxRPTFZAHNK5HQ0Zgz5TpVrcI6eUzYJ9qEoaMRA5xUaxy8MeR04q1F1by7rbj8vT3q4mnkuzJO2Hb2Hp7V4JvW07WormY7oxnI6fwsO31rxPNoVx4nbWIG220+Nww5xtjC/Xr7V4llWS7ZrI7oe3GOw9eeuaMF4HeZo/3bY2nI/GorxCShbP4VcRJKQyjrTW7bsFuB04oTiOTKHI+lW0pecGRunTj2qKEbt6nj0xRnQHaD9KaR2bCHPtVjugQynkDr2xU14HJMB+arq4ku7SK0kGDHnb+Jyf5VbWE8MRlnTgdOR/StW1mLW7gTRR7SO24nsB6D0q5MmNyjJHQVPqRu7QQMmMdDn3+lW/miXaW4HbHtQzNcHzG57ce1TWbiJZ0Pr/hXwt8J6h4++JHhrwNbhv+J5qtvaOVx8iO4Dt9FXcfwr/gpd43htL3wN8LdNKRxWlvLqdxAgwFB/cwYHYAJOPxFabq0ljeLcRrnbnjOOxHp71our2k8+b3jb9fQ+g+lapqmmrAf7OkzIOgww7j1HpmjLecuo+vSvC+sahFOYo+N3fg9AfaovFarEiXS4Jz3/wH0q5utIupkuYZc7c7htbjjAqfULZiYOn514RSx0i5g1W+bEa7sHBPZl7Z9R2rVjHLfC9jPyL9049gDXizWNb8QyxzXkO4pnZ8yjqFz0A9BWqxXkTgypgDryPaoLR5YRNbR52decdeO9a3Pfy6d9nu3/d/3cD1Hp+FeC77SYtUis798QPu3cN2Vj2564rX9NMV8sujjdFzg9McD+8c9c1fanc3kfmXMm5u5wB6eg9qsLq90/8A060Xb5fR8g4zx0P1rxJrWow6P9mmkzGe+FH8QPYVLrMsGkvYrJthkxuGM9Gz6Z61eeFJf+EXg1G2+e4bduHTpJjucdKjt3vtKluphu+x7eOmNzY/pWoqqjyrc/549aiMcETxyNycY4p4iVMqcqOpqORVyg5zRnUjYWz7YqSScvvh+726U0zopIbk9eKE0GwYbJ+hpo/MkBY8dq09UWTa5+UdP1p53+0ED7q/d/KmTw02mCT7T/pf93Y/9716dKhkkMzEDjj+VbwRhjg1dwSJOVBwo6flW6ZoxHFyewq2v4AqxynD84HNNqumtGqF9xHUYYf0rQtT0f7E1u0m2Ttwx7k+npVt9gL7jPjb975DXhIW11ZRaVp0e9hnJyR3Y9/xq68DJp2nnU7lNsS/fOc98Do2eprwt4p13TrCOy0mLNvzzuQdyf4hnqTSa74eHi3w7Y3a5th9q+1jL/N+7ynQZHOOn414ltvDek6r9ng1D/R2/g8l+yg9eT1NeKvElnb68sXh5v3MOcjB7ov94Z65q48VXIjBZtr+uAf6Vaa1PZ62ms2c/wAyZyNo5ym3uP6VpPiaPxbpP2KYeYz9R93oxPoP7tWCXM1w/hjT4tkdpjA3A43Av3/HvXgmzj0fxhapdybo1370xjP7tscjPqKmudNguXg0o+XjHy/Me3qfxpo2u7FoZebqTHlD1wefboO9eIvDfiHT7ny5k/cP0OU7Ae+epqHQ9X8O3Fvcou1H3beVPQY9T61ZanpyxiW7f5x14b+g+lfELUvDOq30c1o3MOcDEncL649Kn8D30VgfEKw4sI/vvuHGTsHG7P3vavFMnh/S9sYm+cZ2na/P3fr61ceLbVpvJjkyw6HB9PpVt4m1DRruLU9NOZ7fODwPvDHcEdCa8Q+I7vxOwur2bM69BtHsOwA6KKsvHF74bvEuWG5VzjkDtj+6fWviN8Z9Q8X5in6L0HH+x6IP7tX+psAW3c/T6e1andCdsbsnsMfSrmRz8qGnkOQA3P0psHqad9uDmjJ5jbSf0og4qFQynmiSOM0BnAzSgEdaz2zXHWiATxRUHgUAG49KIB4FY4CmgATigRgeorAAyKIHTNHgYFMMjANQxngkUzEtkVaSKDwaHzmuA2AelcEcGu/NB+NuelIyqDk1jJwtSOHA4xihtPAND5sHvSFh1NDbtwT9KXy9mCee1DJPXpROOAaIyM5oHIwD9KBA6GnVMAKcmtvHvQOV20BngVggcGsHGAaAB68U2M/Kax712wDXQYHWip6CsfLwKxuxmjkHKnp0o9cisbmGT9KKYHy0VJ680VJXgc0zAqqntnFHLd6K4xg0pUjntQGGyD0qZt+MnJFY2jGawDitvanGMkmmIA4FDgYP5UV5rAxgHNEHG0H8KGBxn9KYBmXB6Vt4xmu1c4xmgcdDz2o5PQ190j07VkHvWMEUQM4FYzwKPPAOajlaNgytgj2q4uWkkLyNk9zimdSvB57UGJOR2p2BXAbn0xRlGwAmmlTpu/SpXU52mi7Ada6gY/CpGDAA9qALD5eaUZOMZpimBzzTyx9c8+mKa6jTvT3xxhDT3DPzn9KYknJNE54zRAAxnmgARzQ2kYNEgjAoY6GiAOhocjrXBPSqLHoD+FHGOtdR/Sq4PSqo4xzWBjAqjjGCawCMA0cdCfwxQUDJJ/SsjpmsDoDWewOaPPANMM96JUjGaJXbj9KJBGRRK4wv5VgAZxWAefyrIznNEj8RR+bgGjjpiiqntR54xQCjgUTxgUT3B5pjnoa4Jo8HNZDcYpgAeD+FHGOBRAxwaoY702DwtDijtJ5NEDoDVHBGCK4IrgcCvoaIBHA6UBxgt+lKCMYNcnlj+lAZ4z+lNuPQ/pR5HX8MUPTP4V25P6USx4B5oEjgt+lZB4Bpj/tfpQ3DAz+lH5jkHNEgjg116N+lMTnaDx6URgcGsKB1oH1rGRxWQxAolcYAzRIPShg8UdpHHWiO1EDHA5r5ce9EgniuegogEYoEADHajz3o9MCqwMACq2k8A0cdT2ogHvRweKJz3o88A0ADxnmlyeM1getAYGM1urAJyBWQRmuSMZ5ontRIx1o4PFeCtF8SeLfENl4V8KaLfaxq2oSiK1sbK3aaWZznhVUEmv2dv+CafhXwFof/AAt79sXWtPsrPT4hctoDXyxW1oB3vLgHDH/pnGducZZsla/aM/4Kb21lpbfC39k3R4ND0Wzi+yp4gNosO2IDG2yt8AQqB0dxu54RSA1alrOpazqNxq2tajc399dyNLPdXUzSyyu3LM7tksSepJzUzmQ81tAO0Vp9g2oXCwx/54P+Fa5pMOmRrErbpBnPBHp7n1ogk8DpQ5HNKRjg9KRlAGfwqJkbgDmoJlU4P4Vpds9/cpb2/wA0jZwOnQE9/pV/Yajpsqpdw+We3zKew9M+opjbz24luTukH3TyO/PSvhkdEn12GDxAP9EO7d9/+4+Pu89cV8ZF8PJ4xvNP8GHGkrs+zp8/H7qMtzJ833t3X+VLPawQmG6XB9cn+n4VeapqItRYxXH7hfupsXjnJ5xnrUECwOflz7ZqD7Gspdk4H3Rk+nNagIpZGmhfcxxtXGPSrcBlxMPm/wA+lWsY81ecr6VID5YBbp2xTXZBWP8AjGcVZw3G8bkwvcZFarJBa3pS1Gy3f7vU5wB689afSbuCE3yL8h75H09aFxa3cEKFcEbvnyfX0/Cra7k1Wz/sua/8uL18rPfPseoq12WWYmO5vXp/nrXg61Goa5axXyeZC2/Izt6I3pz1ArU7DTkJMU+VH3U2Nx071aoA+zZ1+62enrxUmnSr+8jg3D13Y/rVxrFhc6etq9viaLO1t57nJ4x6V/wTl8Ct4p/aCPiu5tw1t4T0ua5VmGQJ5QIUH12vIw/3a/a58Uj4lftK+LZ0uwbTTLkaXbEfMFW2TY+OehlWQ/8AAqh05zdCBBuT16dvrUmgz6fOwR954wMAdvr71Ak8N1i4jz7ZHp7VHdERRtEOTnctXZulfYz/AOs6Lgdver/UYtXhy9xl16fJj0+npToYQQ75Q/eXGM1byrcXmwjao7de1XM0zD7Ij7o26DGMY5rVNdOraTHpFnb5ZM5O/HVg3cD0rw78Q7/wNrf2iFN+/wC+uQM4Qgc7T/erxj4svdcuGkmmwjfw7R2C9wB6V4T8UQWE12ssPmLJs2DcVxgNnt714j18X90WUYXsvXsPb2q9WxgugllJ5hHU4I7D1rwD4qGjadeWd1Bv8/y9rbsY2lieAD6itAtdIutPll1C7/ejGF8tvU+ntivB2keHfFHh28sbpvsyrsy+Hf8AjY9AR/d/WvFfja417SY9LNx5uzOPkC9WU/3R6VqMU0EKm7j2Bs7ec56Z6fhXhHx3f+GSRZvkyew7bvVT61qE3wn8LfAHUw6/afFGqeVkZuEz5d4PrH/q29vzq61GBpDJAnX3PFJcbb5Yy+1D1OM9q8aJ8KNJ8DSQ+GpvN1VsYO24XGJV/v5X7pNPcOXDI2GHXijNuOSfm9ailEKglvmHXip75HJwc56jpUd8u3DjJHbPSv7Sby90cu0D/Zz/AEr7UrHBb9KS7XB28Y7VLdyM2Uf9Ks7hpmYGTB44xSCQEKy5HpnFFoHfcybNvbJOaMkTuJkk2qvbBNSyxLIJ40yewz+FKE3rKZsZzkbaUGFNtvPk/wC7/jWj6ZqGsXiabaxedNNnbHkLnAJ6kjsK+C3gnXNL1pbzVrP7HGucnzEk/hkH8LH1H518dviHrml3K2Oma7/ocud0H2VOcCMj5ipPUk1Z+I720s4ZrW62D5uNgPfHcV4e8Tah438Z6JGJvIVvtHlSbQ3SI54wP7uOa1t9d8S+LlivYt7rnJ3IP+WY9Mf3RXxJ+HWpWXxBubbRk/4lzbPLbK9oUJ4Zt33ia8d6TpOkOkKjdJz/AHh/d9z61ZaRqWtRi4sNM3Qp/rP3yjrwOpB6g14D8QP4K0qWbStM8zbjevnY/iYDkg/3jXg/xA8PizUNeurn7Is3lYXZ5nSNl7D+nem8S6pLftqekPhx91sLzxg8MPrVl47m122iurO6+yy22fMGwP5m44HVRjAH61pvxNGp6pbG8n8h49+1tu7qp7BfYV4i8bR6zpa2ouOOc/J/tA/3R6VDry2s720r7oGxtXGO2TzjPWtRgmaNZ4ZPLT1259BWsa14UjtYUs7jbfru80bJDnkY6jH3a8S/EvVIvA0+g6m215du18KcYlDHgL9O9axf6ddMLpm3yt97hh0wB+lPqSQSZE+1fTbn+lTXv2rMaNjPUYqRordSz3GCP9itRv4LhwWmyB0G0+1azdQIxeI5YdOvtWp37uxjEmR9PpU90qtnP6UbiQHMTYH0ppUUjeenQYqe7VhhetSSF+rdOnFQcsOadSGxUAycE/hTccZrJ9aUEjANEc4J5o8/dPP0pQQME0QccH9Kyemee1ckYzzQJI255psjp1pQuAB1oDHem45FMcjANIhY8GpWVI9gPJotgDFW8oDYB/SoJFX5cfStoYdck0ThduKB3AYPPpQUn7xzWOxolSOBg1PDGh2o2fwrbt5B6UASfl60AAm4tnHbFBsDFA8YA6USQQc/SgQTzQIFEdhR571kkYJoNggqeR0oNgcHn6USME5oYHK0cEdeaIBORRGRjHSic96AGcY4oAHhTzRPAAPApgAOKIBGF7UAACc9K4Ixu5HtQO4bQaUoqY28+uazlOvIoklNucimRXQMG5HbFBML1o8jrzRBPOaZy7ZzmiARz1plBHApBk1kDgCiMg85Paj9f0rdk4okE4DZ9OK3A8Z6dKYhiAvUVkk4DfpXHTPNZynWj04P4USGGBRxwM1k9M0ST0/CiABjNHJ+UGmyRgGgMdDTMFGQcmmk3EHd07YqZxtBB+orzQRndn8KExBKA59qMjZwG/Sg6n7/ADjoKdgTkVx3P6VlScA5oFAOBk9qaUHGDjFSXRU/I3H0pr0scg59RTzsxxu/SiSVwDROBjrRbpj8qIHY0cAYBojoMU2MYrIx15qgM0een5Vx6VgY4HSgAMYo896IyMZoAEYJo4HWiAeAee1EA1xjGKx71R9BVHGKI3LweaAB61wRWB2rHYCsDtRGTwaOMVgEcDpRAArAI4H4UVBHAo4AoDPFEAHAFDDHGKAB6CjhjgdqYjGBTDPSsZ4FZBHFAgjANEA8A0SCOOtHmgQMYrGe9EZ6GqwBwRVEYOM1xjFEdqIz0NEgd6OGGAawMYBogHgHntWQehrHGM1SkE4BogHjFYOOOtEZwf0oqAKAGM0RxgDNBQKC+p/CioPAogZzmieAc10ogAYAo4I4ojIzmscZBrqKJHTNZyODQwADRxjOa4I4NVweBQAPGfpVcdKwD3o44FUQMdc1XXkVkHqKAyOKrGThTXFUQDXBHBqvh78APjb8WtJn134Z/CvxN4n060uDbT3WmadJcRxzBVYoWUEBtrKcejD1rV/2RP2oNA0m913WvgH44stP062kubu5m0aVUgijUs7sccKFBJPoKABPIojAxQAJxRODgUFB6iv2a/8AgmV47/aW+EWl/F3QvifoOjWmqT3EKWd3ZyySIYZWjJJXjkrn8a/a2/Zf1/8AZJ+I+m/DjxF4p0/X7nUtEi1VLmyheJER5poghD85BgJz0+YVkYxQPy4FfsJ/tneF/wBkafxldeIvhpD4juNZsUOl3VskcV3FcIQPIedgWS3cHc2A2GjUhTk4+PH7Xnxj/ah8RnUPiHr5i0qCQtYaDYlorKyHOCqZJd8dZHLNzjIGALCeARhGHI+tTMHlLAYx0/Kt3PSiuBkVZ3LW0gkRsEe1X17JduWkfcfpj0/wokdjRIA5NKAcDNAY4z9KRsMGJ6U1yH4BqxnlgxJA+HHQ4rTdWSdCdTm3uOny49fQfSlZrq4ZYeU42/lzVi81pdBc89unpUmrbbl5pTluMdu30qe4a6YswzmmZnBYcg9KubfdbF4TnHXtjmhswVDfpVyzLMArdPb2oSq8WSfxrR5VjulkkG5OeOnY1qOqafcWyrCvzemT6j2pJYr+WK1SLa4zg7s9s/0q31qwgthplzBhz/HuPrnoBV3oUmpeKhommyedE3+qfbtz+73HgnPUHqavUvdHuTpU43lOq8DqM9s+tRTLbsbcckdB+tafcyIzBj6YH51JIwcq/wB5e9aRqV1HOskPBTp07g+1a1HYu4Fq+4ntgj09atoJJr9LNDy2cflmhqF3bymHy9yjvuApIm1C5EMEfJ6c+3/1q/YQsbX4Ufs5ePfjXrECJ5zzzoWH+st7KElQCOeZXlXHqBWoXt9Pq93fahcF7yeVpZZGGSzv8zE/UmtNjubhsRnn1496077Ub8xTSbmXr8oHY+lX14iSspXJGOc+wqbXIpExBB+O7/61ahrsksccph2mLODuznOPaoNUe3BG3O7pz/8AWqzuFnjcNJknGBipZkt7jIPPp+FLqIdsI3I6cVN4ktrudZorbyAucjeW7fT2rWbuy1WXfbNjyO+DzkD1xjpUU0LwMs7/ADcY4Pr7VaSLZTSurb/MxtGMYwP/AK9GJL9trSbSfbP+eleBo9Nk8T2cGoSYhO/ecN/zzbHTnriviJY6Jp0sJ0K987fuyPLdcYCf3vqa8M/ArxZrltHfwQYtWzl90Z7kdPMB6ivHvgyD4Y+FZNVjbzo1x5jYK7f3iqONzf3/AErxL428FN4Tgs9Lgzfru3ndL/z0BHVcfdzWo6q2qxxMTwm7j649vamnLfLAenev7Tvpyq3Mm+NM44Ax+Qq6AcYjOD6VLIghUl/nXPalu0+6wzUl4JCNkeB25pJCx5PNafo41G3e4EmNuP4fcj19qdNjsAeBii6ZGTj2pTHJF5aHO7v9KQhR8o3enapJxhVK4Jz3pldowy8e1WSyFmaNumM8dKW6kK7Q/wA3YYoanseNHXJ5zzj+lLq9k0fl+X+p/wAKNo88QaCTIbpxjpVvI9qjxyruPGDnFWey7jJDbWHQYz/npXhvT9cttThksRmU7trfJx8pzwT6Zr4g/GnVLuBdKhtPs0iZ3N5iv1KkcbPb9a1jxbLqs6Pcnzdmefu9QPQD0qy8Z2bQJA1rwM87z6/7tR+OoZNPtLa3i+yyWe/ZNuL53HJ+Ur+H41ZeMLY3RmS83SH+LyyO3pivh/YXF54duL+91TzpjtwPIC7fnYduDwBSXuo6zqJgvIMqej7l4+X0GPQU1hqvwotJLO9Hm291jy5PlXO05PA3Hq4FeC/iRYWfiFRfW26xOd/zkfwNjoueuK8a6tYap411HUtFXbp9x5XkpknG2JQ3LDd94HrXgbx7pVq0ej6lp+yOTOZvNY7cbm+6F57DrVteW/hhpWuZsq+MDae30z/erWfiAkxDWVtjHRt/07Fa8KfFjS9NsXsdXf8AeHG04b1Yn7qHsRWs/E26lmMdnBuWLq24DqB2K14X8eapFCJLhfNjHUZVfX0Wo7u313xNea0ibITs2rknH7vb7HqPSvFfja31S3MAt8qO+8juv+z7VH4eTWfDus+JI22R6Z5GRjO7zH2eox09DRuZL2QiMZB/z/StRlnSZ2ilwVxxtHHAq61S8dGVzkcc8f4VqPiO6yVQdPp7e1XOq3dw2TJ+g/wq6uTsyDzUtyG+UmpLsBsKalnaU5xzRIJxnmiBjJHNW43OABUyjbkVCTninHzVgEcUpB71nI4ogdqYZwAaY+hoAdQaYDoKGMfSiRjmgBjI60AMe9BR0zSxgkAnp2xQGxMHr6VIS7ZBogYwDS/IcjrUMwIGDyKSViMg/QUo4yTQAJDKeO1Egjg0CB9405QkbWz7YpQScA0Vw200gwc56UDvJGcZ6CkGRjPSiwT5Q2T2GKRgwyeawpBwelDaeM0QpGB+FEDoDWwdjWB2oEDjH4UQCM4onK5U11HB6Vyeh6UTnoaAB4IopgDBojAwBRUEUxBGAaOMdaABGDRUKcKaHJyDS/McUQCM5pdpPB59MUWwxFZBGCPwpgpbIb8MU5BIyaIAGQKXLHGaJJOAa25GAefSgSBgGsg8A0RnoelKBjrRYdSOlDBAIPJrHHWiQBxWM8A0wBGBQRTgZrGBgHmsc4PNMO+aJB5FEdqKnGM9KyAORRPGakbjAp34wDTyAgZNK2OKLEsCOtJ1xnn0rGQSDnNHGME0xAGB2pp1WPaBzUt0pTAozM3INMxbqaHQkDpW0EfKM0Ac4FEHPSiD1Ipueh+lE57104FEkjGawcYFYGKzjgGuvFE5ojtVUcHpR5rIx1rIPQ1VA5FHpxVHPQCiFxQAIGKz8uawpHFYGcZ/CmAxgflQAzisLjOKI7g0w5xQIA6c9qJA4ArAxlaABGSfpQAIwDRUZwDRA6A0QAMCsAgAGiMGiQOM0AD3/CgA3WvavlIwKI9D+FEA8LWB90Gj0H6VkHjNHngHNHBxzQPYGiATnP0qsDoDn3xQx3qqrjH8qwB0rA7VjPANAZ79KK8YFEAjFdO1NjH9K4OMiiQRjFAg9KJXGAKIGMY6VwTjFEhsACtoo4646UeTxTY9KBA7fSgQw4o4Py0QBx1oEZwKIA6CiQRiuMYoAdc9KHFcE8msDGQentTAHkGse9HnvQyMEnNcZo4PArGeBRHAwaIHQGiB0rHHFEAjFADpVEA1/wAE9vhQfhD+yV4E0a6tRDqWuWh1vUMrtYy3Z8xAw/vLCYUOefkrw9r3hj4k+DrTXtHmi1LQ9es9yFlyssTghlYfmpH1r46fDS4+Dnxk8Z/C+5DH/hG9ZuLSF3BBlhVz5UnP96Mo3/AqyT0oDPUUQD1rIHQV/wAErOf2K/CI/wCn/U//AEslr/gs8Af2ofDB/wCpBtP/AEvv66jg1j3raKXMZ3IeR7Vpd200IBbJHXj60DxjNHrjP4UwUgnP0oqcdaYg980oBOfSjg/LQPTmmBPINfQ0jYPWkhtn0qSRflmGNrcnHzc8dOlQXLxNgnJ9elQ37BdwPTpX2q6LCVp+nbaKFtcXSh1+dF6DgVPBGBhRhvSlkwRETyO+KmnuLVwUkz+AFG7LJ8o5NSwRyKGK5J681AhD7F4X060zqjLsPK1YTNFJujfGevFaXDFc3Ts0vllcbTtJ7HNTX10SVkm3Y6fKBVnNHbrBdYyU3ZH14rUb7zpvOCYPYZ9hWnwW09u73Ay/GDyO59Ks7CBLeRlXc/G3kjHNQ6fNcrhRyOvT/GrfS71cIse5/wCHkD696C28soYR8n3PpVzBHbOj245Ofl//AF1farpd5arDDp/lzDOX85j3HbGOgpbeCJN4+9268V+0PEPgZ+wd4X+F0RW31DWYbOxuI14IkfN1dED0Lqyk/wC370UeeXJPzHqfwpZbiKQKHwR0OBVlBqBnEdncfO/X5B2HvV7p9/ZTgTS4Y99o9B6H3q6hltn2hsY68U0jyfI75PYYrSngEpilXKt7kYxmrm2iiuylucBen5e9XTRyneq4HpmluYFV1ROeMHJrT4HvEDoee59Ov+Fab4Jttc1eeylvcfZ9uH8o91J6bh6Vq+mSaXI0dymGTGDn1x6E+tXGqSWl6LyCX5l6jb7Y9KunfU7rzbiX73U7fQe30qd45pDEhwP4R6VYXckUiwibnnB21pHxf8Y6pbyaE+v7LWPGE+yxnqS3XZnqPWrhvEfie+bT4LnzQuP4EXtn2/u1p2ofZy0BX5eMDPTrV0oMwKrgDqM+1eFrHVrPQk1q1uP3RzgbF4+cr356+1a/4/8AEvi+2/s/WdV8+2T/AFKeQi7MkFuVUE5KjrTDyXx5uT67auZQ4VjyTnNOsaJkdaM0RyxXP40ZkeLCJg/WvtA+6pxRky20UJGZirH6cV9odW8sGo7lS2Hq4OGyDn09qhIU5U4qC4mDMqtwOnFG7dhlmzj2xUk5lbg5x+FNMq8FfwzVvcDfuL8fSl1mWAbPM3g9sY/pST2btmWPnscmvC2v6haaXd2Flf7Vn2ZXygc4YnqRTx3UTeZJPv8AbYBVu80swaKba3Y7c44q08aeKLLTBpMep5tl6L5Mf97d1256mrV2mmjhlk4mz/D6Crr4fP5kckFzgtnA2en/AAKtB0XWfB7R3r2/2qOLO471j3ZyB3OPvVqfiGYzGSxTyCe2d3Yeo+tah4n1XxLbxaRqF5kWu7yf3a/LuIZugH90dasNHjkm+y3N95QPVvK3ds9Aa1fwu2mRItveecpzj93t9PUn1qPW9f0qQ29reYDfw+Wnb6g+teIb2e6uttkmyFfurkHHAz1561oPhy31CAzT3Xlzt0Hlk9z6HHQVBpHhNWSO8hkjjDqJXj3OwUnkqpYAkDoCRn1Fav8Asq/CD4t/DXTPFH7Omtw2t9Y2iwkSyFlvWVfmW4U8xT55zgA55GCGD/Ci48K6rc6X4ysJrPULVtskEyMrKef7rYIIxgjg9RWqajHZFkhnzEn8O36fj1NaV42u7Z2ntosoMfLuA9R3WtV1spawyTx7iu7vjPI9BWp/EC4vdEOmEbYz7g/xA/3fatS1XEZiU8Dv+XtWq6mVUlG6e309q1DUdwypx7flV1fSPkh+ntUkrucls/hRbJAzzWBuwKIUj1rgnAFcN90VARvAAqcbRxzUBBbgcimGTQA9KByP6UMdAK+UnBGaO0/KPyoFcYAxXAFAjoBRIIwRQAPavlIzSgdc/hQVW61EikdKuWRV2qMkUu3HAokAYxRIxkUWZfun9KhnbAO7n6VFJuXJOTS4IAJ5FAEd/pxRAJwaIzwayMcigQSABxRAPArIAwOtEjHA5oLuKgj1qmx0FAALgH8KGD1psEBRQUkcH9KPTANDkUenBrjGMVyTkVRJPOaAz1PNAAjPehgcUAGxg0AGOAeaZQDgHNAfwg0ASMZ/SlOO/Sm2nkUMggE5obhzmgdwznNEDOAelJaPJF5iN07Ypw6HBPPfipGRlAHUd6AxwDQGRnPNKcnAp0CjIP0rAJAB/SsYHB/Sic96xkYHWo4wwIJ5FHA4phu5HaiAAADXGMUSCMCsZG0D6UcDpWCOhzRxgYptp+7THJAzTvtqSU4wtSSFxz1pm9DXmDOKyh5x9KMjA4Bp5QOh5p7g8AHP4U1y38LdPammkbq36UzZGCaOMYHaiBjrn0oEDtWAOlHBHWiVHahgkCiBTY6AUQMZFADGc0cA8UQpGKIGBiuPShg9TRwBxRx2riqODwKwCOtHt+lV14zWB2ogEYoDGMGiAT8prHGM0QcHNAEgAUBlRntWMDINEgnBNAZ49KIz1NEADBP0ojI/pRUbck81gEZzRwV4OaIDDKn9KKg9DQA6g9Pasg9DRAA5NHHUc0cHAz9KIXPX9KG0jIPSsAj5aIAHWgA3Q0Svr+lHGcCiVJwKKgcZogY4P6UV4zmguaOAcZzQ5OBRHOAawBwKx2rHagMDFHpxWM85/Cj0z6VyR0rb3rIPQ0FzQHIGf0pgD900cEYB5oqAOW+nFEBuhrIPQ0FAI5rbnvXB6H9KJ5xmiBgDP0r5ehNAADOaIB5zW0etFcjg9KIUjg1gFcA1gHAJ/CiuTgVg4x+VADgE1jPSiBjaDzWO1Yz0rAIwDmiKAJxtFYPTPNbT061tNYHatpoqevpVYJHFEHGM0BxgGgCetEEDpX7P3wwuPjR8bPBPwvgiZk8Q6zBb3JXOUtt26d+OfliWRvwr9oX4gWfwW/Z/8cePoJEtP+Ec8PXDWCr8gW48vZbIMdMytGo9M1/wSQ+JyeOP2VYfBtxPuvvAmr3GnlWbLG3lb7RE30zLIg/65V/wWQ+FB8LfHbw98VrKDFr440fyblgnW7s9sbEn3he3AB5+Q/gSSMiv2H/+CXSfF/wvp3xf+P15qGmeG9UiWfRtCsn8m5v4Tys80hBMcTDlVUb2BDZUY3aH+xh+x34C00RwfALwCLaJdvm6zpsd+RnHWS68xieByTnr6mv+CrPwS/Zr+E/we8Ma58N/hZ4d0HxP4h15Y7e70mI2yGzSCR5SI42EbZYwAZU8NkEV/wAEqjn9irwhn/n/ANT/APSyWvjh+wv8Mv2if2gbH4y/GC4m1HQdD8MW2l2uhQzPbrNNHc3MzyzyKQ3lhZkAVSMkMScDBj/Y4/ZD1Lw+ulWv7Pvw8msJEIWeDRoPNYYxkXCjzM++7Oeetf8ABQ7/AIJ1aN8BtCf41/BJbxvByTrFrGjzu0zaSZGCxyxyMSzQliqENllYr8zBvlIx3oL6Vb3T25BUVZXa3Me5WyR7UeRgCguRkH6URldprSbW1u9Rit7qXy4nzubaTjCk9B+FeLtI0LTVtv7F1L7SX3+YPJdNuNuPvE56miMijjoK68CiAeKzt4FPqBe2Fvsxjvn3zUUx6Dn9KhmAwHbn0xSzxOMA5xWm60tiGiU7s/w4x6+3vV3eebK0oPJxhauZpRyiZ/GkuZBgSr+tbo5APKOSPwpEKr5kjYA9qkvjGwZYf/HqNwLmVcLtJzkZzio4WPCnke1QyMWGGyfpQVnPyjcR26VKwWBG3/Mc5GOnNeEfB2meKdPS7v7z7Ltzz5bP3YdmH90fnVxaWZgUpLhjn5dpPetJtVRSZWwPpn1rSrOBrWSdJ/nTHy7T6kVaX9zBKl0Y87M9x3GK1KPR54BPazfP/d2t6gd6uUje3d1fBGMce9EREDdx718AvBUHxM+NPg3waIvPhvtWia6jwebeI+bN/wCQo3r/AIKU+JLzXfHnhnwPZHfbaDpr3c+GAxNcPjaRnqEhQ/8AbT61MluujW6xvm4XdvGDx83Ht0p2LLuY5IqxE1tapqLN5a87GwGzzg8VNrz3R8xx5pXv93+lS3DSR79u7PfOKtbSOX55HwR0GKuhpiTMbe73IMbW8thnjng06gwm6EvA/wBn8KsdR07+xfJlbM3phv735dK8K/CPS9Y8I3nie51zyri28vyrf7KzeZukKH5g4AwBnkc14c0y3tNV3z3G20X+LYT/AAnsDnrXjCS68SXcN14W0ryrS33b5vPDb9wUD5XwRgqwrVIbmyzNrq7J/wCGPIOemeV46EVqtwJ7tpRB5e7qN27GAK0e90C70Z9Je4/05sbV2P2Ysfb7orWND1Dw7rMunatB9nkixuG4PjKg/wAJPYir/wAOaRYaE2qxah5jjqvksMfOF9ff0pL57XZJjBbOe9eDvAvxZ1TTbrUPCXhb7VGNmZvt1um35mH3Xb2YUsyuuU5b0qXUSsTw7Ms+Oc9MVp3jO7sdJ/skv+7HQYH94t/d96IkQAIM461I4PU0JHdCCenSpJ3f5RR2+WWB/SriKS2tw5HXp+dFtoyw59KZyxyD07UXAPytn8KRPM5Dc+mKTapwz5I7YqNtzbW6etCNUblv0qEASfKcj6UFRVwPyoOqnBOCe1SOsjhFOTQL7tgX9aW48shCcnsOlG9R0CHkDoa0rWP7FUzrH5jD7q5xnqPQ+tWepfbQsLpiRv4M+nvj0FNbvZyKksexueM5q71PwZLpKQW+pZuxncnky/3vUjHStPjtrmeMEYCZ9e4NaB4Z1LV7YtZXPnFei7FXHJ9SPQ1LJ40W2bQdPsvtNwuPtMXmRJ5PO5OTw2Rk8HjvWrzX6XQ07XNG/s8n+L7QJuwPRfw/Orx7a31F7cTcJja+085XPSr+4imhMSyb2PT5celaVqd7BdRSFPNiiz8uQvUH2z1rxD408F6xDHbqv2WfnPMr+n+yB0H61q3iHwGdDtbCxvN8qb/Mk8uYdWBHBHpkVp9t4RZDMPEHzL0P2ST3FPN4Y80bLrzAvQ+XIM8V8K/i3H8MbW48SeHtQNvJF5YkgfzHiuFLkFXXGPx6jPBFWevfA39uHwk+jy3A0Txnp8BZAAPtVoeMvHkATwZwCOMZ/gJVq/aD+EfxK+BPiMaD4207/Qrhm+wapAu62vVGOVbHDDjKHDDPoQTpfjP+yrMwRWnm5/i8zb3PbafWtX1kzrs8zIX2+ntU+u4jMbPkD2/+tV14gkEZRl/X/wCtV9qvmZBf8MfT2q6vmk+VmyfTFSNv4zXXofwxQAJ6/pWR60CCcKaOPWiB2NREBwAauACmM/pUDKGyTUpBcjPNZB4zzS4JGDRyTtBraMYzQXFMcDAoAY60dpHXpWAeQaYDGM/pWARxSLng0kYx1okIm7PTpTMDyTz6YrAI4oBSMZzivlIxmtowBnNRuynH6VDLlelRSbuo6VweFOaIbggc0GYjkUeRxS88UpJGDRPy5UZPahk8Y5pJMDLdugreSm4CmZSAM80CGHWgXGRj6UwyMlv0rawPXpTjK4zW7PAHPpRPGaBJwCK6ihk4wKwQpFdBjFZA7/SioC9a4HGfpQ5GBRK54P6VgE4B5HagAAMmsJ0B/CguQOeaIB5zRYNwKGEUYGc0MEdaF08UflqeKdi5yRyaKLng8Ucdq79K2jpn6UPbnFDnA/KuowKI7Cs8YzSsQeD1okZxmieODQODyKLE8A/Si3QVu44NEg8Y5osCKLDGRzRlG3OefSnmLAYqWYjgHmmk3chv0qU4GQf0ppkYY3Y/Cgy55f8ASpbj+FTRn3HaDTO5HDfTisknIogH+L9KI4ABrGQB6dqyAMZ5HagM8A0V7CiBjGefSgARjNHBGAaAB6t+lEAnAP0o896ODwDRA7GmxnAPNEcUQM4zQCngN+lcHgGsA8E1RC9N36UfQc1gEZz0qsZ6dqogDgV25rgADNYB4JoDsD0qmXPANAYGM0AAMA0cY60RxnNFQT16VwTjrRPJANAkjANEgjrROAADR9Cf0rA24J+nFcY4NHnpRA4JNEg9D0o7T3psEAA0QBxnNcYxn9KzgcHNDB5zX0NHB71knjNEAnk80AD3o4IxnpRAIxR5GM0ee9FQcCj6Ec1tB5z09qI7UQOgNEA0ee9YHaj04qqOD0rPOBRHcGgTgY60x4zmu3JrqOKJOOtDJOM4ok4ABoA9AaBJOM0Cc4JoknoazkcGug5NZwOTVYBGBWMd+aOTxmgSMYNEAcA0QexoDtiiOgJorimAPINDp8tEY4zR7EUeaAzwDWAR1rA7VWB2rGeBWB2qqPTiq/4I1/Cg+Jvjd4k+LN9as1r4L0j7NaSFeBeXZKAg+0Mc4IHTzBnrz+3v8FPi3+0N8CG+FHwjuNGgudU1a2l1R9Uu2gja0h3SBVKo5LGZYT24U1/wTo/Y5/aG/ZP8b+LJfiLeeF5vDPifS4ldNM1CSaUXkEmYWKtGo2+XLcAnOclePT/gqt8KP+Fj/snar4hs7cyaj4EvodYhCLlmhyYZx7KI5TIf+uQr9nD4e2fxY+Pfw/8Ah3qUZew13xDaW98gOC1sZAZgD2Plh61O/wBL8KeHrvU5ohb6do9k8zxwR4EcMSEkKo9FXgCv2g/2h/iV+0h4+1Dxt8QPEF3cRzXDtp2mGdjbaZCT8kUMf3VAXALAZY5ZiSSabUNQexj0t7+4azhkaSO3aUmNHIALBc4BIABPsK/4JVj/AIwq8IDP/L/qf/pZLX/BZf40eObPxz4V+BOl6zc2PhqbQF1nULe3lKC/lluJokWXH3kjFuSFPGZCSCQpH/BM/wCP3jT4V/tJeFvAdrrd0/hXxxfDTdS0ppC0JlkBEMyryFkWTZ8wGSpYHg8fGzwfYfED4O+N/BGpRRyW+t+H720YSDhS8LBW6HBVsMDjIIB7VyQBn8KA75/CiAOAasb97VyB0q0u1uUzjpQbHC8+1MCy4rgck/hRAbkGgCR1okevSiMcZofWiDnGfwrOWyDWT6/SkndFwDSXbEbWOR6VChulJt49xXrzjFAASbGfA9MUs7hhHnIHSpiZMZPToK8L+LtG0rRZLOfTvMkfGG85lxhiegB7GryW3u3ZwmD25PH+cVNFtbP3h+VIUVgQmD65q1jDyBAcE961bwReaJp9vqgufMS53cbAu3aQv94+vpRTd/qxVraLM4BPJ6Go4JbdRHb/AJ//AK6s7I3TqFkw4zj5atrGfcY7x8AfdOB/T8K1myFvN5kMm5T0OMdhVpPNcWzQyS4TjPyj1qXTjZSBklyP93H9fepLoRPgrkHqc4q2mjjkEcqZ2984r/gnL4RsvEHxY8Q+Oo7ELD4d0tYY2LE7Z7g4UjP/AEzimH/Aq/aF+JEvxI+NvjHVIrkS2bapLb2rDBDQw/uoyOOhWNT+Na7pr6Y4WKTIboNvpj3PrUdzF5ZMy5Ye9faw+nQQmTCfNgY6c1MIbldyy7Nv+yTXnSlfJDYC+1SQxRp5m/JHQYxUFlGyFRNgL/s1puky6hCYVm2o3X5c+vv7VHbxWxHnpv29s4/lUF55Gofa0XYg6DOf4cVrepanp7RiRcA5wMr7e3vWs/ETVL22giaTYse7Awp6kf7PtWr60+tuJ76TfIvRtuMdB0AHoKsUlvIbu6km5t9m0beu7g/ypJ4rd/OP3/X9K09V+JS3+qaveeXcR+Xn93ndnK/w7QMBBWq6Pb6JdRC4HmRtn1XHA9CfWtAs/Csel3La3Y+fL8nlt5siY+Y54X2x1rQPj5440vTToGjX/k2cn34/KibOGLDloyepPepp4YbsgR5x05I7VpHw3vfFXhXVfFlrqPlx6V5O+HyQd3mSFByWGPu56H8KNw0EzRgfMuOfwq4uzIG3DpRm2jJX9aSdpeFHFLblTuJ/SpiCxOefpTOAM9aMgYc00iJzii47VFcMv3D+FCZg29TzUN25JTsaa5BPynNWe64Dgt93HGKkkVlyo6V5+7jPTpSEScBsenFGRVQhTjPbFW1yhJRz9KhmSaIBh92rO5jLqirxz3q3byJAqfnQv50CyFvmTpwO9aDdeFLiNE8T6D9uJzz9qki9f7n/AAH8quPhz4a1HV/Cvh/wxJ9mOvfbPOO132eSu5fvtznDdCOverb4i+NPhro3mSzeYnYbYh/F/ut/fqD4t6p4z1rT2hb7DcQeb5z/ACy+ZleONgAwF7eteN/Dtp4s8Qw3trcbI13bhsJ/gUDqR/dr4kfBqzm0C48WLefMu3H7s/30T+//AEoPNa+E7jTIm8zft7AdJM1oOkubd5JT5ZTGV69Se+avNBvrCM61Hb74YuvzAdfl9c9T6Vdalq0VmkgixE2cDcvr9Kk8SXC7InTcDnjIH9KXVrpFV5F+T0yP8K037H4pvI7O8m2RQ552k4yCe2D1UVe6vd/DjxNZar4a1e4t7uyk823ubd2hkicAEFWGSPvf5zXwf/a4+FX7S2hS/Bn9ojQ9OimvwsVvd3RC2t6+ML8/ym3uMk7WGAT0KkhT+1r+xj43+Asd34v8FxXXiPwJuLNchd1zpYznFwqjlO3mjC/3gpIzq9+03Q4C9B+VXurNDMyKOR/gParjVnnjAC4P1/8ArVNOzcCiSwzmgxHet2ADW7/9VHmgQMYokE4zR560n3gMZq4c4wDUbEMMGpMFzisnGAaUZ6GiCehrgcA5oHIAosOgonOMGg2RigTjBPShgjAPNIN/GaVFAwTQBxyankONgPNEknnmsgjr+FcEUcAYzQAPNAgjANI5BwpqKUFeKSbaciln3HBH0oyZOAfwotgA4reTwKY4GQaDgjkdKDArgDNFiTtI+lI+3nFYL5bHNIoZtpNPGFIKtn04ogMOTWBtzjmmUyLkdR0FMoQcdaEvGCKU5AOPwpmGQCa4Io4YAClJzXB4onI+U1nPANEDHBrHHX6VgEUADxSr27VgKcA1tJOM0QGFEAjIPTpQC460SOBj6UACOTTDceD+lEDIxT7VAzzTNu6Vt5GalgkiVXZeGzjn0rj8a3YGM5r+Hisn1onIwBRAIwD0rJxgGtxxgGixPeqZgMEHpQkUjinlCgAVLdFvlFeaWI3HgVK6AZBp7zaSqjmmumb7xzTNuGN34YppR0UfjRYnvSjJwDzW3HQ/pXXiicUQD1NZJHBrgjBNZIAwa6jk0CMYz+lZz3+lNyODRUEgZrPoawDxmiO2aonPeiAawDxmiM9DR570TwBnpR5719DROe9Z4xmqAwMZogdAaIBojIxmse9fQ0Rnv9Kojjk0MHgH9KOCvBoAEYogNwD+lbRt4P6UBxwf0oqCeDmse9NgDbWAR1/SioNFRjAOa25oAA4zRIIxmioPAor2J6VtzwDWB0JraCvB5orjqce1EKRgdaxkYFMB0FYyBiioA60cHGKPoKIIHAo4xgGgAaPoKIxxmiOOK2kisZ4BogDgGsAjrRxjrWM114ogkcGsgjAPSgMnANEjpnmiR0zRUcEGiNoxnmhgnOeaB9TRPGQa4IwD+FDJ4JpjngVgDqa5IwDROOBRBIGKKtjBFHOM5rkjAognpXXtRXoM0wwAM0VI4BogntQBHGM0c9APwrrwKAz3rlqOQcCunSgCRkmiOODWCRxWOMZraSOKIz0NHkYzVdeKPoK56EV/wSs+FC/Df9kzR9eu7cR6l46vZtZnJHzCIkRQDP8AdMUSuB/01PrX7Zn/AAUql/Za+Lsfwp0H4aWfimWLSYLy+uZ9Va1MEsrPiIKsb5/drG2cj7/Tivh3/wAFl73xZ4+8N+F/EnwV07R9K1jVbazvNRTXXlNpFLIqNLsMI3bQ27GR0rxb4Z0rxp4V1nwdrkRl03XdPnsLyMfxQzRtG4/FWNfCi+n/AGZ/2u/Dsni+byP+EB8bJa6xIY8bYobnyrhgDn+DeR+HPer6z03xBpE+n3Sx3VhqNs0UihsrLFIuDgjqCp/Wv2if2I/jr8AvHOo6DP4D17XfD4uXGk69p2nyXMF3Bn5CxjBEcm3G5GwQc4yME+IPhr8RfCWjWviLxV4D8QaNpd9O0Fre6hpsttFPIq7mRGdQGIHJAzjiv+CVeD+xV4Qwf+X/AFP/ANLJa/4K7fsyfFPx34v8NfHPwD4Z1HxHpthoQ0fVbXTrZp5rLyp5ZkmMa5Zkb7Q4JAIXy+cAg1/wTb/ZE+Kvir9oPw18U/FXgjWND8I+DbhtQe91Kze2F3cxqRDFCHALkSFWYgEAIQSCQD+1D8R9O+En7PPxA8e6heJAdO0G5W0LNt33UqGK3Qc5y0rxrx60doHBocgYNAqR1/Ssg960+/8Asp2nlT2/OrKZLiPegz605wuB2o/MMj8qUAdTR9PyFYzwBn2ojsBQAPGaIA4zRwRwfpQRn4HaobV5X2KOR1q5ihjcLC+fwIrT9Vm01JViGfNxk8cYz7e9PcPOxdzkmmnTpt5HfNG4kbkNyOnFJJIqYEmB6Yrz2C7c/pUNwSuwtwPag0coAJxtpZGRsxnp0OKfxlrWo2UWk3d5vgjzsXy1GMnceQueoFRS24OC/wBODVnbRND5sT/OO2KtJJ2XIPI+laXcB4lt4LfdMucPvx69unSrXQbbUdPe51XU/I8rHHkl+px/CfYVOiCARNccr/Ds9/WoLm2jtZbeaXaz4x8pPQ57VbarcyqUlXefqB/SkuZGjCTHAXr/AJFTXdj5Chbbe3PzbyM8+lfsroPg3+xp4s+LckAtNQ1gXl7aszZJMa/Z7ZT7GZSR/v59KlspdIulWU/O2cZ5zxz6+ta5qd55+29GHToODjIHoPpUxlYec8GAO+78KSSR3WKVfkGcc1qdtthBhPB/+tUtwkioElwecnbU1wA21W3fhira8eGQF03L6Zxml8REQlbNvKYdFxu/mKk+0vCJ2bPvgfSr+92ph+F7fpWteL7nVmRbhtxTODgDrj0A9K1Cd5iJAu4+mcelJdKybpjtA7dae4uA84sz+5Xbv6fh1561JG1ycI2Se2MVpd9dWYP2ebYT1G0H19avdVvNT/dXMvm7eg2hcfkPavEXheKx8KafrVveeZ9r835PLK42yBeuff0qCKa0tXug+0JjsD1OKkuTIS6DJ/KtH8beJ9HsLrS7DVPs9pebPPi8mN92wkryVJGCSeKvr+3OWil3SHr8pHp/SmlEn3jj14rXPAviPw/brca3YfZo2zg+aj9CB/Cx9R+dB4ogSpyT0HNTXbFdoP6VJOzE4bP4YppmJ2lvwxXmggYP0rzgRgjpSzZGc0sh9aWQlutWQiadUkbCnPb2q9tDbsCg4NRXEkT7UbBPtQut64J59KWQsu5Tk9xihKeBvx+GaWYhsuM/pS7JuVXn0zQk8pduenbFW8qL84b6DFWFyyOA77sdsYqXVbW1YSzJ5iemSP6V4W1mOScPcQ+ZH2XdjHB7gVe+PI7RbdtItPs89pu8m88zf5O7G75GXDZGV56da13xN8VfEOhxXerabjTUziXzrfnLAdFAb7wAqbW9GOmabquk2Hlsnm+dL5rHOW2r8rD6jgV4W0nSNX0VtWtvE+2GPG//AEJjjLFR1IPUelaV4t0jXrOLw2J/t+nT58ptrRb9pLnjAYYYdz2q/wDhr4G0eVodT1H7IZOi+TLJjGO4Y+o/OvEvheDRLaLUWTcku7IyRuwQPU460/xHie0k0EaVi0uMbv3/AE2ncP4c9fetPTzNOeG5b7THHjbxsxknPStdMkV401tDiFcYG72A789auPFV55ggnm3RjttA/kKufGWkwaEtvp1rsu5M75PMY9HyOCuOmRV/rs90+6SXnt8o9varPxhaaXYtCwy74yOex+h9a/ZT/wCCkF98OBb/AA5+OT3Ov+C5AIINSZTPc6YhG3aykZnhA6r95RnbuACV+1j/AME+ND+JXhr/AIXv+x1eadqNpqEJvJfD1jKrW94h5aSxccKwIOYDgZyFKkBDq1vqOn6hcadqlpPaXlrI0U9vPEY5InU4ZWUgFSCMEHkVuI4U80Seu79KI4wDk/SlIPBb9K6cbv0oDd3rA/vfpQGeM0Rjgc0cY60n3hV2AOlICDnPFSgbuKIz0NAnoDRIBwTWc8bufpQUAjmmPbNYx36VketE9cHn0pVLnCilj2jAFEE8AZpjtTPf0pzvPJorituRkGgSOP0rBPUfShnHI/CsHPArBHSo5WXg0k+e9CXBxu47cUs2TwaWZHXG76DFBweAefSiSRgmiDnFK5X5c0jiQfe6e1bsjFK5BIB470pVwMHkUWkDBWNBQ/GadipwDmjKAuFOD2oncAc5NBugC/hmgwIojcARWBjAroxA6CsjGQaALDPagARgUFAocjOa5Jzn6UoBGD+dEDgK34YpfTNEZO4c0cnFY7LzW0j6dqIBOCfwplBXdSvkYpsgYNMCRkU5yvB+lZI4JwK3kjFPftNAsMq7tudpzjHPNEgDBNHB5BzW4A4zSyKxwD0rI6g1vHAzTOByT9KMijjNNKAuBTTELkN+lSXCggFv0oz5GC3PpijKzNy3H0qeWGPpJk/Q1JfqMhD9OKa6eTqf0p2JPXn6UT6GmJOADXAFZyMZ5pRg8Hmuo4Ne1ZwcE1z0xn0rI9a2+9YBGAelY+XINBQRSmPGGGfSiVYbQPpRHOBQC45NfSjg9KI7UdpHymsdq25HFBfSiMcZrAIximGOBzWAcAHmuooYIPtWM8CiMcCtuelMvUVjPAogGsdgaPoBVA5A5ojPGelZHQmiSRhT0onPTrTNnoazz1onPANdDR6EE80DxgHms8cHmiSeBRbPSiMjrn0oHJApvT8qJI71g9aOTxmivvRAJwDWMjrQGRgUcYwKJOAc0ST3rGeBR46Hmic9/oKzjoaAx0NcYxWM9+aYEDijz3roOPwr6GutVnjGar/Iqh1wDRw3ANAdzTc9aYE8A0CQMGsgDj8qbpRyRkUQQQo60Bzijk4wetbfQ/pTDvmscZo+hrBI4raaAPAzRUHoaK9gawD05ojng0F9awRwK2+9BQeRRUHoaAzwDW3HWgCRgHNEHgYojFEccGqIJHBqsZ4BqsHpmse/4Vptva3moWtpfagljbzTIk11JGzrAhYBnKqCxCjJwASccV4T/wCCi/7Angrwpo3g7QvjZ5Wm6Dp8FhZxnw1qvyQwxrGg/wCPXsqgV+1D8VYvjf8AtC+PfilaXLz2Ot6zKdOkkQozWcWIrbKnBU+THHwenSlwVIJr4Q/8FQv2Urj4WeEn+Jvxck03xcuj2ya5bSaFqM5S8WMLKd8VuyMGcFgQx4Ir9v7xp8F/iZ+0lrHxJ+BvixNe0TxLZ291eSJYXFp5F6qeVKmyeNGOREkhYAgmQ85BA/Yq/wCCpr/Cbwvpvwl+P2n6lrOgaVGtvpWvWIEt1ZQjhYZo2I82NBwrKdyqAu1uMaN/wUM/Yy12yW+tPj1okKN/BeQXFrIp90ljVv0xX/BT79rf9mj4+fCDR/Avwt8dN4i8R6T4ihvw9vp1xFDHD5E0cimSVFVs+YhwueV6jBFfsDftvfsp/A39lrwp8PfiV8Wf7K8R2lxfy3tj/YWoT/Z/Mu5WQeZDA6NmMo3DcbsHBBrwH/wVv+DEnxZ8XeEvH99cJ4MGpZ8L+KbPT5mRrXYoKXEGwTKd4chghOGwQNu5vEP/AAUc/Yx8O6P/AGxJ8bdP1DMe6K10+zubieQ4yF2LH8pP+3tA7kV+3T+31r37WV5beEfDGlXPh/4faRc+fbWVxIDc6hMAQs1xtJVdoLBY1JC7iSzEjac45rGQAO1YBXANAHHFDPrWj6mLVykrYVuhx0xn2qOaO4QMjZz7YoIDwfxFFVPy96wCMEUSoGR+FYXAXGTRADZFCMufkGTUFlLO5RByOtC3ayTMn4f5/GodREFwJAc+v5VfXKT/ADKf88UNx70zMuOaDknKn9KDHrmi5K9a38fLQc96jmAOCaSZSuB1qJwW3E4I6U8jHBDc/SrTUpFHD/hj/wCtVvrAjXlufTH/ANarVrCza21XQF2uN+Tkn/Z/i/HtV54mvrq4K3M2c/7I9PYVeXsc8mA36VMY43UvyPSrGVIrpZIzgjPH4VrF5bX08Yj4kGdx59B/QVHd+Q6xs/TOBivG37Wdx4g/Z2sPg/D4YtNPWyS2ia6t7h8SxwsOGjKn5mYK5O7GQeOeJdTgvZ/MlTOOnJ9Pp7VcXV1fDc0mW7jAH+elT6vdvZGykfKnqMD1z6Uiu52B8Y7YrTtYurRBFC3I9h7+1TRxKoJbn6VLbv5PnKfl78fhSyKRgnOOlSqUdZw2MZ4x+FeA/C/hzXtKkfUZtsq42ja5/ib0I7AV4hht1v5tNhO7yduzqOoBNFI0bBHIqzMdzDMHfBXbt4qVo4oSC+T9KsJbO4sGAT99xubJ9eOOnSroOjFVb9KM8axlVFRG4liaVE/dpjLZHH4UdeuxGts0uY1ztXaOO/pVvqlpf2Lw3bZAxgYPr7D2FNfEng4qWdmcuTxRCsc4/CtMit7q6jiuB+7bOevYH0rxPrlzdFbaW43pz/AB6H0qSf8AhB/SnlyMZppCMkGmk3DrRkAOAORRYYyTzSuBwDSyZI5pX96Dtxhv0qK8bcDIcge2KkmWSYupz6D8KE5A2qetLOyHhufpQuFZMHrQZRGGU5JpJVWM4PNJMFOX5PpSSxlCVHParW+eQYB5q6u/9FKqen+NeHL53TaG5Ht9a+HHiOx0HVX1PUo/MK42DJH8Lg9AfUV4j+Mut67ENGsbrFiP+WexfZupQHqD3pNQkTSQivhz95cf7XFfDH4n3eh2f9itcbIZvvfID0Lt/dJ6n1q08F6Ff+J5XF95aLjb+6c4/d8/xe1fF5IdMRLGK480DOH2Ff7h6c+teIPiDf6qhtL2ffFb/cG0DrgnooPUCtD/AOEavp5472H/AEiXb5Lbn+XAO7gcdAOtWuo+E9T1qO2VPLiTOeZD1XPpnqK+Nn/CGI62PhxN3l53NmUdfLPR/wAa1mS3V2SA8fj7VPqrW8flb8+nH/1qk1SZ8hX/AE/+tU2plT+9k+nH/wBarnViRhD/AJ/Kv2Vf22vip+yt4kEmg3Taz4SvZg+q+HLqUiGboDJEcHyZsfxgYOAGDADHxJ+Bv7Mv/BT34dy/Fv4J69Z+H/iNaRKlzLJGI5UlwdtvqMC5JBwQs65OBwXVSlfFz4PfEj4GeNLvwB8T/DFzour2vzKkg3R3EeSFlicfLJGcHDKSOCDgggMSR1oE45NHB5Bo4PQUCOAKOD0oe1Y7UaGAeanG63WRT1z/ADpQCQM04+Yij04pRnvRAxk80Np6daJJPvTcnGaxhcGsbufyFKmcClRTgAVgEAZ+lKAanbHy1k4wDTDI4Nc4x6URk0R8pANHkYzQGRwaWjg8GgMcg0JWHShOQBnrS3W3BzSXYPANJMCeDQdSM96AHUGucCkPOBQOTnuajkCnBPFSOATg9aSQ+tLKFBApVBPPNfQ0oLY/Si2w7SeO1K2c46DpRZX4BrBJ4OaBB+UGgSw2g0MZ61jijgDAocDrzQbI4oHnIo4yOM0MEbfyoAqcEZpTzhRQI65z6VjPQfhTcEACiwAouGAzzTPnAByPSiwHC1JKPuj8K87AwOa88DBJoyqRkGvtAUYFPOGHX9KMxRsA8UJyy4Vv0ppmIABrzcjFPIMBQelSTbejfpUl2qj5m/Sn1CNTtUU2oseFP6VLeTSH736UzsThjn8KPPeqJz3qiAaJzjFcEcGhjOKOScCiMn6U3/6qLDHFHnvQOKAyMCu3Tj0rAI60RxQxjg89qIx0NAcgGjg9Kxzgdaoiq6jGfpWAeM0QOgNURxRA7GiOKPHFY7ihyKx3BrA7VWB1o4J5PNE9hWOMZo8cYoHPatueKAIPAzQOTnrR25wRXB4Arbjt07UcE4IrAbkfnWMjAFEDsK4xkCgB0AzXUYFAYwAcms5IwKJ3cClHGM0SB901kMCQaAyARzW04A70AOxo47flQYEcVnJyTR9hzR5o881jPAogdM1TEZ5rIPANZHVTRGeho/736VR6cUfu4FAY70RkcGsg9DQIJwPyrAHUUASOBTAHjFHpgCgMjGOKK8YHNAZ61zjAFAHPHJoA5wBmtvcflRBPSsEjIFHLDAojPaiNw4P4VzjpRz0ArJ9K6iunaiC2K2kDk1yBW3I4rBxgV81YOMA1973x2pcEjBzRGTgHNUckAkVRbAwOtfQ1jPArg8A0SM4ArjH8qyOhogHjFKcjArGTgCh7imIzyKA4wF5+tYA4AoYPQUckY2/rWO2KPPGKwMYA/CjzxjNHnkChjHFdsAUCT71g8ADmtK1RbRxHKvy/Xp19qinjuFDQnOfathPUc04OOBQXIwDUdtNO22FdzDtnFSRvG5jkXDDqM1byiHnbk/WrO6Mcm9Bjd+mKvrmOeLy9+W+n0oaVcvbG6UfIO/Hrj1rPGK8MeHbbWo2Se8+z+jeWX9ewI9K12yt9PvHtLe583Zj5thXqAen40o+XrXHTGaJ3Dg0GA43fpS7idqjntTAr94YNLIF6GvNUgEDpST+YNuainjXOT+Nfa42YAfnWneJLiwt/s8D4TuMD1J9PeptctriLIl/eDqNp/wAKttRhuGEzScemD9K1HW9NvI4o7Wx8uRd2T5rHPT1HHShdIjjHzfpXmRyYkEWdvX5qFzAtz580efbJ9MVqXie11XTlhtdP2BM4Pmk4yR6gelIJkfKxZB/2hVjrEFhqLGWx+0dPk80p/D6496XUYDqKySRYj5zHuP8Ad9a1ZF13VLm90jT/ALNG23bB5u/b8oB+ZsZ6E/jUk1zbMYXXDjtkVNFKwEjjr2qe5dbfyWXCN7+horPCAF5H4CpIp5gMnKntxWj6jeRuFSbYjdflB9amubaw1c3DN5q9xgrn5cf1rVryGSbdCm0HqMk9hU1zDb7RjJOcjkU10smVTn2rS9SsYdDu4XX9/wDJtOTx8xz2x0pbppYwrr831qS6idvlX/gOaHiiOLSp9Kj07mbb8/ndMNu6YpbhifvZI9qupZTBuMWB/e3e/pVvax3EPmG52HsuzNAgDJPXtRlGeuae6IGAenb0pp17U0wPQ9OlGQYxu/SjKp4DZ/CvMToTTOqngZNF06Hn0pZFz1reCcZ4pXHRT+lK4ZuT9K3Z60jA5Gc4reB25rIYYJ5ouMbQKWQjAB6dqEygYrerc55+lQvltqjJ7CtNe2aQh5fLx/sk+tXc8IV1ifevHOCK0vVWtnwWwOwx9fao9dWMhA24P7Yxj8KtNUkhu90ZyW7fh9K07WUkk8m6X5x1OenX0FeJ/Dt/olzGEu8q2cfux2A9z60/inxBG3mnUP3h+9+6Tn9K0HxhomteEZLTxFL9nuuNs21n/wCWhJ+VAB0AFfFDRbnwBKr3epeYs2cJ5IXGAncFv7wq48cWsQWWGHlf9o//ABNH4gCJ900mPfGf5LWq/EuGWOS3t0yZMZbJ4xjsVq48SQMpIG4n6j+lXWqxSJuaPZ6nOf6VPqaKxCNn8P8A61TXu9uDmnmLHGKYgjA7V8Jvi78Q/gf42sviB8MvEtzo2s2RwJIjlJ4yRuilQ/LJG2BlWBHAPUAj4YfHr9mf/gpz8P4/g58ctAtNA+IltCzWkccgjl83HzXGmztk9AC0DZOByJFUsP2tf2J/it+yfr5bXbVtb8HXkxTTPEtpCRDIeoimXnyJcfwscNglS2DgkEYxQAIxiiMcZrIPAFYxwaxnpXSiQ3FVD+8tjGp5Xt+NKpB5HSpOWJB+lY96HTAGaxk9enaiCeRQUnqKMYPQ/pQVe5/CgitwrfpWwAAA9KUrnrzWU/vdPagF7Gpjvb5aVcHBNEYraepP6UTnvWAR16UORtAzWCM4PNcnoelEZNHtmicnA6iiCepok4oHjNJcFTjGRUd2jYCn8KjuBnjn2oTK4J6YpZUHQ80pUjCnJocna1FwBwcigy4yRkUpXGQc56UsgUYK5B96DRkYPWn+7j8jSlehOaDlQVx9KOYzjOaDbVL9qWRSRgUpUjCnNAjbjGabB5FAqAAD+lHBAFAdgentQIPQc19TRIBznP4UCDznJNZ2DANblx1oNkYDfpTjA3E/hRYE5FMe35UzqMAN19qkmTblXyewxTMDgmvNJO0N+lPLGQOfpT30aDDCrjUgxxEmMe//ANavtzlsEfrUlyx4HWlu5F5xQv3UAlevvTamVHCZPbn/AOtUt/Ix+RP1p7h2HLY9BimZ3+8c0MH3xRYHoKIBogEYonr7VyeAKK46CqOccCu3T8KAxxQPYc1k+lE+34UeD1zTAUcY6VnHagfQUQSMAVt46VgDqM1weAKJHcUT0OPpRG7kCjk9qI9qPJojtXHSsY4NEdqIOKojPQ0RxRII6812xWOKOTxXX/CiAaJxwazgg9/SmPQA0c9AK4IrKgAYrGT16dqYZAGetYG7ArqcflQIFA5PFFcDrS4HApsYxmjj7tEYXGaBwvFDg4or0GaIHQGtoIzmvcdqJB7USCMCgQBiuccUODjNEDrimAHSvpROTmiSehoDIwKJPTNHsAKHsKOc80RkYBrjGM1xVNnpROe9ZJGK69qzjoaoDPOabBHBogdvwonIwBTFCMUORWccdaHGaPGCaPI5phkdaznGKPPArIxisZHFY4xmiBwAaIOMZpFzX3qJOee1AE9KAJNE5GMUMHAoEk8UCAMUWyMCiSRjNDijg9KABwBRwRgVnjGa7YBrg8CuK+ldetAggUSOAKIGODVKM96zk4xQIAwaZgcd8VkEdaBz/SlonA4o8jgV94YocrgHmiR0zW72ogfeBokEdaBB4FZzyO1BsmtN1NrdwjnK/wD6/arfUbaZCitlu3BFPKpwRTMCBikkQR7wfm+lS/M249aLccClkKng1vOc5r+0iLT7OB19/fPpRYHp+FW99PajETYz7Cpp3nkJJ5PfFDgbc1nHQ1TZxgGgwBzijKsyhCeaY4OKLcDFCQjpRmDAJ3FLKCMZ5qO8ZFCZ6VZahAjZk/rWrXmji7L6I37k9OG9B/e565q21EC4Qk+v8q0PUtPGrJHqBzDzn7390+gz1xWu3ulx6gx0g5g7D5v7o/vc9c0knnyM6tgHtikma2iO05HpihqzKgQDn/PtVxewiBUx+8GcHn1qG9iRhKRyK0LxG2lXi3qrkjtnHYj0PrWuXsmpXbXpXbu7Zz0AH9KguhnEh6Vf3pfCIMr/APqp9QZSFbpSXmctnnsKgmkQEg81JcNJuBOScYppyQA1WUOkvp93Jdn96mzyuG7nnp7Y61HPF5m1TyelX8KWce1HyJfbHTFLfCPAA6dKljl2eYOh6UJcLgimlUKABmrrVZb22Fs6YA7598+ntXgrQLfXtQFpdy7Yh1+Un+Fj2I9BXjOxs9F1+60zT23Qw7NpwR1RSeuT1JppiTwKeRj0HNF2YdaEjFcflRkOMgU0hPNFi3ymvMI4IpmyMGlbbyK8wk5P5UrgHIoOOoNb8jFLICeDzQYYwDWcdDQHQZrd2pSCmDRckcGkkwwGPpSSbeh5qS5YptNJc4kyak1SJIxn/P6VFrcaHLdun+cVpniJC7SE8nGB+ftV94yu5fvSZA9gPT/ZrSfjFpFj4ATw7LFuvhnc25h/y2LDjZj7p9a1D4hu8OA2Mduvp/s1rvxOudVVRO+4DOOAPT0UelXHjFQMId36f0q78RSTKFA6d8//AFqGqSbslufp/wDWo6mSck5/z9Kn1B5F2jp6U0hY5NE5OKYZHFck0TzkGrG+vdNvLfUtNvJrS7tJVlt54JDHJFIpBV1YcqwIBBHIIr4yfty/H747fCDQfg14/wDEcd1pukTmW8vIlMdxrG3b5P2og4fyyCRgDcSGbcyhqxnpQGeM0cHkGsA8ZogdAaqqOOmat3CPgjipkKfN1zVV1GAKCccGggzgmiuMVgHvRAzkGhgcg5qSX+EGt/tQJxyaDcYzRGe9AYAIFEZPFEEgDFfQ0cHpQ+tZwMA0OeBRGFxms8YonPIoHHSiexFMQRgGiBt4NDjgHpST4ODS3Q9c0J1PekuV3dea+0jpSSI4wKBGODRUr0ouThh2oMCMk0ZsAAHp0oMHXg0WJ70pCjJNAh1OfwoYwdvWoyU4IpWDDIrr0NH2FUD/AAg0GxwaJBAUdaLADBNKxzjv2oP2JzQbPU0W96LE9668UzADBNSMWPFEkdvwp3AwQKaYKeOamnAGQcmpJixyTRbceah5bk0z/McGiQP8KJLDApmAIBPSi3HAovlcYonIxmunIqj2FUMHGBVUSB0oHAxRIxwaBxxmhySKIAPBzXyk8mjzTcnAo9BWMjigcAAUMEYBrdRzjGaAAGaOCeDQ5G3NZ4wO1ZOMA02elVVHgV9DR5rtgGuKqqGTgVX+RXXgUck5zROTiqOegFHGME18uMjt0rgjg0VG3OaG3AAOTXH+FYA6VgE5zTAEdaIA4U1jIwTRA6A0eRtz9KHIxWMjG79KwCeufwoAdBRAYcGuRznOOnFH19O1EAjI6jpRG44zQ5GAaKgcbufpWB/Cf0rjGc1gf3v0rPfNZyBg0QAMZ/SjjHDdPasgjk19DRGecV/nFZB4H5VRJxgc1tB/i/DFEDoDR55rAI60CABjrVKBjGentXB6HpRG7BB/SjjHWiVIwD+lAAc7vpxW35eDRUYwTQUAcN+GKPoDXJ6npRXnIb6cVkDgn9KK85BoLjoaJA70QOMGs8cce1Bh61t9DQHPzc0cHoKyCODR+bgUQB0obcA0RnBFEAdBmsA9KAB6CuAeDmgMkHNEds0duMCqGMDjmifQVgdqIz0NH0FEjGAM17YojJ6UCB2rAI5HNEAnGaJA4FbR60SoGRQII4okEYHNEgjHWiFPQ/pWBjANMAeM/pXBG3P6UAMYzRUHoaA54OaJGcCuMcGiBng/pUUpjOQas7yKdcF8H0waSRSeDk0wJ6U2emfpRUdhWc4zTegocUxHQU7A5CnpSgYzWQeRTbABz0rOen4VgEYJot6D8KDZ4x9KbnGabpgdR0oMemKLKcjH0pWA7/hW8sa8FfCP4t/ELT5tV+H/AMLvFvieyt5vImudH0S4vI4pAA2xmiRgGwynB5wQe9eMPhH8Wvh5p0Or+P8A4XeLvDVjPMIIbnWNDuLKKSUqWCK8qKC21WOBzhSe1QXpUgFcj0zivCvhvxX451iPw/4J8M6tr+qSozx2Wl2Ul3OyqMsRHGpYgDknHFXXwK/aF8PWN1rOq/A34g2On2MDz3V1c+GLuKKCJFLO7u0WFVQCSTgAAk14a8J+NfHdw9t4I8H65r00ZUPHpemy3bITwMiNSRk8CtO/Zh/aWuYi6fAfx2IycfvdBnibj/ZZAa8QfAf47eFIJL7xZ8I/Gel2cK7nubjQZ1hUY7ybNo/OrliZWAk3KMYOMdqhuIIxhhv9+RSXcRKyE+uR61eaxHd7ESPy1Ge+f6e1ebGWLM24HoMYxTToBw2B6Yp53GB5mT24o3LRoCDlu1NeSryJPwxRutzLtkwOc8ZqW7sTpvlsMzdjz/e/LpUzuQpY/ShIoOd3TtikuAjkkZHcdKYmd/lGB6dan1Ii2NnGuQO+ffPpW5scHp2rIKgY5ouqrkt09qmvMNtTmjOWG7PTtijIScjmiy8AmnfJwposAeB+FA54Ar7wxWe2c0fQ9aJyOBR44FA9ya3dlNIwIOWz+FA45U0HBOAM0rgcE0rA85+lEkjO79KB5yDn0rcSMCixPIbB+lPkYbr6UzQpHkPz9DUlwAuAc1JdFXyDk1Nfb4woGDTXEmc0t/lQFGMU2pyL8oOR6dP6VcaiZCOOnapLx24H5U8jOcg/hROOM1ROe9USAMZrK4wDROec0DmiaJU96JAHX8KY5AIrGRgN+lYB4B/SsADk80ORtz9KrAI5PP0qiOmD+FVgE4BpZzt8t+R+VNDnlDkduKEbdCaUKBjd+lF1A25olSODRwQMN+lbc8A/pWNoBJ/Si2RgGiQRwaX2oKRwG/SuAck81kZyD+lEgjGaY54BoHIwDzWR1xTEEYqj020D2zRYE4xWR6UcEcCiPlyD+FE55okkY60DgYoHHGKPPOKyD1FbvQUXIOQaExUYqO8JbkfhUN4B1HHpUd2pGCM0JgMALnPvQct2oFHUqRio3AH3c496MhAAI+tNcIy7FX9a3j+EZxRf5gMc0zEjJPTtSSMPlDfpSt8oA5rcxHy84pWDDJP6VgHjND2rJHfkdKYk980CT06iid3U0TgYBonPAqRmxgCgzKMg0zqenamkDLtpnXGFPNSzAAH06VNcKMlDmnYMOvNN0wawCAB1qIhTkHNMVOMdfSu2KBAGQKYAndmunQ0QDwOawCOtcHisD+9+lYBGM/pRIHGax70QD3okdM0RkjmgO+c0QDznpRGeN36UhUcdaOMYAokEe9E5GK6cAZxRIZcAc1nnIo4IGKyAMDrQ5GK7YoEYxmhjPWgAO/6U2CcCgN3eic8E1gdM1gHjNDHGDSruplAOAaY56GvoaPPese/4VkYwKo4IxnpVAZ4zQwD1rqP6VXBxg/SqYBhwa2nbxRXIAzRI9a6n+lA56GgDnINFWzxTHIwTRGRwaOQuaP1+lAcZBodOayBzRBBxmj61RHajk8CmIK4FDIGKHSsBuBzRGB8tc96AycU3WvmojPXr2o89RQwByaxj8K3ZHrXFHHauOlEcCiOOTzXB4Boj0ogY4NHsBWBjApRzkUQw5xxTDceKZeMA0VBGAaKgjGa2ggAGgOMGiCVxmguRg1nnFEZ4BrBzgdaycZzRJHeiMd6I4znmipI4raMc0CCME1weh6Ucg9aHFHk8URnoaAz3o56HtRx0FdsA1nHQ1jPege2a3cYo4PShxRIJzXXqaOOgNVR571wRwapcHoaI4znpSjPesADg80STwKA569KIyMZoqMYFEACgAR1orxj8qbkcGl+Xk0M96IJOAelcnoawSeOtc4xnntRUmgCRgmkdkIKnpVnqflkKTn/P0qG5SRco2fTitwAwBmmXPGc0QSRg0wI4zRA7H9KIBGRTNh9oPSiQFwBRBI4PSi2TtzXAGFNHBHJo5I6Vn15o4I6803Tis9gaTHc89qc84BoAnvX/AASF+KjaJ8TfFXwhvbg/ZfE2nLqFkrNwtzbHDKo9XikYk/8ATEV+3/8ADT/hZ/7KfjWytrfzb/QLddbs+OVa1O+TA7kwecoHq1egAr/gj78MhfeKPG/xfvrbKaXaRaPp7suQZJm82cj0ZVjhH0lPrX/BTr4rD4d/sz3vhqyuQmpeOL2PS4gDhhbj97cMPYqgjP8A12H1H/BHJt3hn4oc/wDL9pv/AKBcV8df2ivhj+znouna78TdQvbe31W5a3tBaWbXDO6ruPA6AD1r4J/tg/AH9oDVH8PfDvxp5mtxxtL/AGXf2slrO6L1ZA42yADkhCSByQBX7fv7HXg7xx8Pdc+MngHQrbSvGHh23kv79bSERpq1ug3TeYqjmVUDOrgZbbtOcgqlwXOAee1I5CA/lU0yPblQ3z9hj3p2VQfLbOPbFGRZFBLdPallcJwM5pnkkCqallY85oS9s8/Sprgs4JNS3BYFQeKLZ4zSsAQSaL45B6V160QSKmuVRM7ufTFS3LSn736UGJ7/AIUGyvFBxnGaYg8ZpUBXBP6UVUNgn6UVUnCmioB255+lYBHB/SiB0zTAEda4zgHmiMjrQUDjNAgHAOaLBcbTQl+bAPFIQTkmgQQAT9K3YTANIQVAzRdQuN3P0oTKf4v0q5vF24jbJ+lNckkAP+lPcls5PH0qYhuQac8YHWt+O9Fy3INFyeQaY+/NYzwKPXg0wBGAaIBGAaAAGM0SAMA0VBGM9O1UVz0NMoBwDRwRnNYHTP0o4PCmiox1/SiMDg0cEdeaOCAAaA4yD0qqoLkZP4VtyQDRGOM1WTt4PTpVKQeBTAHAoDjGaIGQAaBIIANSPuGPyoDAxQGRgVnjANdFyKyCOTXy5xWBnBrGRwa4xweaJJGKPOBWM8CsAcEVjPAqjjGB1rPAANA44zQ46USCOvNHkAUSAMV1GRXGP5VxjFA4HSsgCiQRgUCR0NRzFeCaS4UgAHNJKCAAaWXIC55oSbfummcsOvNKNjDJqSRW4U5xUbhiBnpS/OMDtRKLyT0rLE7RQYocd/SidgCjrRkZTgiopMjJNSSljgDpSZK5NBwTjHSt6gcmmnAOF59KWTH3j+lNJuOSakcBc5ovvHBp3+UDPTpU06qvJ/Cp7kOuAaY7u9UeQBWeMUDg5Fbsmg3IwaDLg8/pVUcdM0AOmawDjBquCODRx0zQAJ4PSioPQ0VyeDW3jGaPTg1gEAZ+lEfLgGkGePSgQBgmjgnBNfQ0RnGDVbQRnNEDABNVgHvWM9KAB4B5raaIGODXXgURjjNfQ0cE8GqpAQM/lT5J5ogY4NEHFVQAIwTVf5FcVVDJ60e2KIyMZrAxgVjAwKIBGO9YBGM5psAc0ADwBQAPGOlYJPFMxxgGsD+7+GaYIpworHHBxRAx1oAY4NEAnAGTQweKwM4xRGeM1gdqbg4HNZHQflQORgCjxjAoDPHpW30ojPatvGM0w75oAMQAKIA4IzQweg6UeTgmiB6Zrbnt+FcHiqwCMZ/SgM8ZpiD0NcdSfwrAPevoarAPeqH+yfwxRJPG78MVgjkGmHfd+lMBgDNY4xmsEDg/hWMjBoqMcUARwDmtvNOOhFcYxiiABwK9MH6URntmlGeAfwrbgetbcjGfpQXPBrjOAMVg9qbrRA64rI9KBGOBWQeoqjgcYocVjAwTmuDwBRHagQMA80BkcGiBgYqsDtVHkAUenFEZ4B/SiBjg/pSj3rhh/StoPAraKAA75o+wqiAelYUrwenWtoA5NHBGR1oAEdKOCMY+lAD15rgnBXJ+tYGcYogHgnmjtPAo4HUZohSPlFELjkV8pGAKjup4iCj4I7YFWeszFcS8n8v6VFexSLuJ5/Gkk3cj8qABGQfwxWAwxinC4yRUWGfBHPasDHBpjxwKHsf0otjhRnFIdwz0NMwyB1oDORnp7Vkdc5rORgmiBjg/pRJHAP6VkDkVkY65r9nr4mT/AAc+Nfgz4lwylI9C1aKS6xnLWzHZcKMf3oXkX8auINO1vTJLadYbyxv4CjrkOk0Trg9OCpU/ka+LXgO7+F3xQ8V/Dq8DmTw5rFzYq7jmRI5CqP8ARlCsPZhX/BPb4Zf8Kw/ZT8H29xAI7/xLG2u3ny43G5w0RPv5CwA57iv+Cr3xXHjP4/2Xw6sbrzLDwHpixSKGyBeXIWWUjt/q/s6n0Kn6D/gjaP8AimfiiM5/07Tf/QLiv+CwrhPht8PSRn/ieXPHT/lgK/Yn0DxZ4o/am+G8Hg+G5a4sNdt769khBAhsoXD3Bc9lMQZOepcL1YA/EnUtN0b4deKdY1kr9gsdFvJ7rf08pIXZ8+20GvMJOSaSUsgXdj8KaUk8HmjOQMKcn6UTuTIoTYG3HTpQnZeQee1FyVyW+nFSEgZPNSXQHyg005chVbGfbpTO4IBbd6cYpZNzMoGfSkSMgZGT+VHAGWFXFwoj+WXYR/s5qa7eQ43fpSuAuQ36UrA4G7n6UsgC9aa62nCmmvXI460LuRl4bn6ULuXPJzihdMwwTSS7upz+FF8H5W/SvOzwT9KEy9M1JMoGVNCZiMlv0rzVPGaDAcA0GXoBWVPQ/hQkVRgChPhQSM0Z3JwrYH0qS5ZY8Kfxo3cg5Vv0oXDkblf9KklLDJNOxI5bP4Uzqq5JoyMehpnJHJo4PRufpRz0Bo896wDwT+lYA6Uxyc5/SiML7UwHZv0oqCOG/DFMARQAxwaA5zRyOC36Vjd/F+lcEbQawPWv+Cd37En7MHx2/ZssvH/xV+GP9ua9Lq95bvd/2zf22Y42ARdkM6JwD1xn1r/gp7+z/wDCL9nb4teFfC/wc8Jf8I/pmpeHPtl1B9vubvzJvtEqbt08jsPlRRgEDjpRPGAc0QAM5zXh/wD4JmfsQX+g6be3XwS3z3FnFJI3/CSaqNzMgJOBc4HJr9oHwtoXgj48/EnwV4XsvsWjaB4v1XT9OtfMeTyLeG7ljiTc5LNhFUZYknGSSea8P2lvfa/ptjdJvhuLyKORckblZwCMjkcGv+HYX7DP/RDv/Ll1X/5Kr4g6XYaH498S6JpUHk2en6vd29tFuLbI0mZVGSSTgADJJNduv4UODnNEgjgUOKyMAdayR0NAZHFAAjr0ojnHWiM4ANbfQ0cAcVyRxQBI6/pRAPAX9axuHWiABgjNAAgcVjJwKIxXGMgVkelZGMAVj/Z/Wjg4wM0QAcVx6Vx6Ufm6CiRjHeiFA6VkelYBGcVkelZHpRGehogdjmjgjAP4UACa4JwKPHQ/pRA6ZobRgg0JSBjP6VFcEjBPT2pZ2OBn9KWcj7po3DMcnnFK+88HAFKVxhDzSN0yPwoFXGM9OlMfQ0uwKWPOO1Z75ognnvQIA60pj24U81JIBwp/SlZE3d/SpHzyBn2ppDwFP4YrzQR1xSuTyTRkLHAPTvUkyp97r2qe7Yjap/GncuOW/SsLjGaIwMZokAYBoDPGawMdfpRUGgM8ZoqMY3fpRxjANBQRwaI4wDRHpWOM5rAOOf0oKOhNcdBRGRgGgQR1rgiiOMA1twcA5rHGBQAwBnpRHGBSnsaIBogZwBmiAOM5rOQAPyo4I4FDnr2rAPAFYXHHWsA8YrI9KwBwRRHfH4UACQAK2jpiiQeCKwpPA5rAHBoAHgCtobihtUYPWiBnGa4rjGK4xjFVxjGKIA4zn2o4PQVRA7fhWAOAaIJHBo4PBoj0NEjpmiCRgdaIJApRx1raCaCkDFAAHGaIz0NEZAHftQBAx+VEHnFYJxk1jK4zRXIAzW3jrRUev6VhSMA0RgAZrb2zTDJ4NEY4zWM9+fSgoI4NEZGQfwojIxnkUMEYrH8Oa4J5rg8A9OlFQOpo89625GQaC5HWivYGsDpmsAjrRGOM1xkDNEZ4A5rAPGaKgDrRGRgGsA96IGODXbANKARnPSiM8A0eRjNHGKIBHNbRnINAHAINCmyQADQA24B57UFx3ogZwO1bR60cD/CsZHWmGR1rG0Ag1jI4P4UBgZz+FEY4zQ5NEZOM1tBOM1jtRAAxnn0raD0P6UBkYFUemPSgM8ZrHOM0QOgNEDHBqjjHWiOMg0BnjNUVyM5rvgGjjpmgMDGaIGMjk0oyeKC8UuRyKBHc0ApGBQGDgGhk96IwME80BleDRGBjNYyOv0rJxwa4PU1gE4Bog54NMOetYwcA0RgCiCeM0R8vXpWPlyDQIAxmj04NIcDg80lxImAG/SrTVyQEc8j/AD6VDexyDAb9KRldCQePpU2DGeattpyQaIA6GivpTLRUYwBzQAx70cDmjjpmhz0P0ojOOawMYFEA8Zo8DgU2CuR1qMkr0r9gf4pf8LZ/ZZ8Gapc3nn6loVudF1DJyyyWuETce5aHyXJ/26/4KF/AC71/9tzwlYaPC6R/FldPiMqDGydXW1lI7fLGsTk/7RJrWtV8O/DXwNfa1dhLHQvC2lSTyBeBBbW8RYgZ7BE/SvH/AIy1X4i+ONf8ea4+6/8AEOpXF/cDduCvLIXKj2GcAdgBX/BGzB8M/FEA/wDL9pv/AKBcV8Svg/8ADH4xadZ6T8TvBem+I7XT5jNaxXsZYRSFdpZcEEZHFaH4S+AX7OOgXFxo2k+Dfh9pco3XNy3kWCy7R1klYgvgf3icV+31/wAFBfCPxB8H3/wO+BeoS6hYamwTXPEARoopYVbJt7cMAzBmC7pCApUFRuDkgN2BrcRxnpSkng0ScY9KLN0zRfjOaY8Zz9OKecKM7v0qa9yCFP0p7jJ+U01yB/Fz9Ka9c9G/SlvJB0b9KOoTgfKensKfUbmQgF8/gKkmlfgv+GKI9DRJA6/pQlb1oyMw5P6UcHkmiR0oFjxXJGAaUnsaR+2elMzkYBpnYDA715rAgE5rzCRjNCUKMbufpQkBOQc/hSuSev6UGx1P0oSEg5PTpTTg9D0o3HPJ6dKkunIwpozueCc+1CX1NGVRwG/ShJuOAeBUjY4BzTNuXGfwoEYAHNMdw5P0oDNHB/pXX/CiuaJA4FZB4zTAkYBpgMZ60TxkGiMjGa6DIPPpR570ADxmgB0zRAGKwDxmv+CSeD+x3puD/wAx/UP/AENa/wCC0wH/AAvnwPzz/wAIgP8A0rno4xgVx/8AWrwpz4W0b/sHwf8Aota/as/5Oi+MQB/5n7XP/S+avCoz4p0fb/z/AMH/AKMWq+K//JUvGP8A2H77/wBHvRIJzR5qs46GjgjArtgGgRjHpWMnOfwo4Py5rOAADRIAoAY4NY5xmgO5NFgBwaOOoNbuMVnIxmgcHANMM9K2gnAP4U2Acg0cEZz0odaAzxmsZOM0TmgcdKbBAOao80CBjFZ7ZzRAIyDWMjg17UQOxrAPGaPTijg9KIBHB5o4PT8KIGODVZwODSynGAfpQm9TSTK33W/Slkx0P4Us2DkH9KWU8HP6Us64+Y9KEisgJbnvxRcAgA/pRY9zQc7uTxTOpGVP6UZNoznn6UWY85re3r+lHkdf0piqjBP0ppkUZJp7rJ+U5p7psYDfpUkpfgmnJY4zTAAAA0VwOtH0Bo46ZrAx1/CuDyKqqwOmaOCMCq7cmjyMZqqwKx71gDpRAIwDVH0FVW3jGaVcnAodOawTkE80QSeKxnpWOxNEehoDPANAZ4rrwKxngUcHpRIHIrOeQaJz3orxWD0FH3qgxAwDROe9EHGDRHFUeCMGjz3rg8A0ee9EDrnpXBHBoAYwD0ojIxmgMDGaOO5qiR0zWDjr+FHOOtEAciiAVwKC471hTwKI5zR+bgU3+RW7tWcjIofMMZo5HANHJ4FcZwDxWF6g03Ix6dKBxxmsFuM0eeCa4PU0T6N+lEk9DXAPBzQxjg0SAMg80cE8GiCDwaznqfwrII4rJxgGs8AA1gdj+lEgng1RwTwa69aPPevoaJz3rg8A0BjvXtmsAdKXGeaY44FZJUd6PTihkjANDOcZrJwNponI5NDJ5zRHoawxHWhzj17V3xn9KrGRyaOcYJoE9AaJJ4FEHGO9EE8GicDrzRBPBpgB3ya68g1ksuPSs8YBoEbQM0CBwDRJIwDRY9M0TkcGiAQOelHj5c0DgYojIwDWeMZogY4NEds0Md6IBGKxgYBqqAB4zRGcY/Csc4HSiMdvpW04yDRBPGOaAx8oojJwBk1yOAaIzzQyBkVsx0ojjINAAjpRGPu0VJogke9HJ4JrBOATQXI57UFJGM0VI5xRGehrrwprBz1oZJAJqO5ljbAf9Ki1WRVCl/0/+tQ1Dzl2s2fwqK6EWADxUd3HIACc0CjrkN+GKZCcc5pkJGAefSiuMj0oqCuAfpQX1OcfhQAAwKAOOuaPYVTcDC0NpHDfpSAY+U1/wR8+Ka2Pirxr8Gr+6YJqtpFrGnRs2FEsJ8ucAf3mSSE/SE+lfEP4O2Hjv4rfDD4mXIhaX4e3eoSrG68uLm1MYx/uusbfUA9q/wCCo/xX/wCFffsz3PhOxuvL1Lx3fx6agU4YWyfvZ2+hCJGf+u34gkkcCv8AgjWP+KZ+KPP/AC/ab/6BcV/wWE1G+0z4bfD640+9ntpDrdypeGQoSPJHGR24FS3V1ezNd3tzLPM+N0krl2bAwMk89ABWcDANFl/hNPPGvBbn6U08S9W/SmuY8ct+lSX8S/KpqS/J4B/Spb+VgV38fSpbtyMK36UZmIxu59cUZWfgt+GKBLHGf0rnuf0o9QAaJwMZr5ick80Scct+lEkD5TWSR1/Ss8fe/SgcjFDOOD+lDAwCc/hReNeo5oyRkcL+tFwBlaDEng/pQOQADTAnjNMADjOfwo5PANdsBuR7VkkfK3P0pZGTgt+lPO2Bg/pQnc5+b9K3Pn736UA7ng8UIM8lv0oRJjGK2KeCM0YkIxQhATCnHrTWwcYD/pTWTgZVs/hS22OtNAAev0rykxjFNHk/KfwpoyON36Uyjpnn6Vgg4Bph70QSvJ/DFFQec0QAMZzWCRgmsAdDk0QQME0QDwKC9iKxkdfpWMjAr/gkmP8AjDvTR/1H9Q/9DWv+C0wH/C+vA+P+hQH/AKVz1tyvSgAFzXhT/kVtG/7B8H/ota1b9mj9nHXtUvNc1z9n/wCG2o6lqNxJc3l5d+FLKaa5mdizySO0RZ3ZiSWJJJJJqL9lj9mGCVJ4P2cPhdHLGwZHTwfYqysOhBEXBqviuP8Ai6XjHn/mP33H/bd6PPeqPHJ5x0qjkgDNYA4Apc9h+FA5og9j+FZJGKIK8Zoglcd6+bGAfwrDAYHasFh1+lZPrQB9aPNZI4JpueKPA60RuA5rtzQBHAP6Uck/KaIY9DWCOe9DPaic8E0TkcNz9KPPNZ4xmqGccGqPOMV14FEgjAoLng0Ru4ojHGaK4GKKgYx+FHnp+VYBFV2rJU4BpXYZ+b9KS4bOCaNwoOM/pQuAVwG/Shc8jn6UtyV5B57V9sbqH6e1G63HJb9KF2gTaD9Ka7QjG79KN8uMBs/hTagQMZ6UdQdh8pr7VIwwX69sUWLjBOakbBwDRbdwG/Sj0waJIGAaOTxmm6YzW0YABojtWO1DoADzQGeAaYY4BrGRgnNY4xmiMHGKoDAwDWOM1WAecc0QDWACMGgQehojIwKo9OtYJHWiARjNDkUvXisA8Zo8dKIJGM0AcYH4VgAcijjPFc44NcEA55oc8KOaxngUACuB1ojPAHNAY4FEUQduDWeMA116mvoaPIxmvoaOeuaJyME0SSME5rpyKPPeu2AaqqPPeqPPej9aJAGCa4I4NYHamxnAOaYA8g0BkDBokZyK2571z0pgB0NEDHBo7ScGjjAAojIyD+FEALQGRgGjgd80QOgNDBIFcdKYe9EAHGaYAmu2AaAGMZqhjvTAHhTWB0NcYxWAeM1gHvRx2ojtXFYzwKrGeBVEZ6GiMjFEcDmsZ6VWMjiqXkEg0cY60ST0NZBGe4ogYyTRYdqK4HWjgAEmjgjGelMAcDNYB4z+lAYGayCc0SPu0cYABokHmlAPINE5GBRORigAQAD0ogAcnpWM8A0QAME1wOOtFeM5o4x1ogEDmgMjg0eRjNEZOO9Edq2miD1NYyOKI4qqxngUQQOlEHHAojjiqPTAoAkYoDjg0ck8daBIGD2r73AoAEYrgdaKg8A0RjGDRBA4NHBGa2+9AcZB6UNvBBoAEdaoAE9ayM0doHBz6UQMcGjyBjtXB4BogYyDQUEcGmIPArjGKO0cg0cEDAoOw6GlumDc8mkvSBgGor+VVwjcfSotWKDDjJH4f0qDUYpuGGCOlKySHKn8MU0RAxiipAwBRGBjFEZGRWMjOaIBogkYB/CimCeajAC4Br9lz4rJ8Ev2gPBPxJuZzHY6Zqax6iwBOLSYGGc4HUiKR2A9QK/4eBfsd9P+F4aZ/wCAN3/8ar/gpB+0f4X/AGgvi9pNt8PddXVfCfhXSxDaXSRuiT3MxDzuocBgMCGPkdYj1BFEDbnP4V/wTE/aO+CfwK0L4gWnxZ8e2nh2XV7uweyWe3mk85Y0mDkeWjYwXXrjrX/BUH9pj4G/HXwF4J0j4S/EO08Q3emavPPdxQW00ZijaEKGPmIoOTxxX2sqMDoKa8kA+99OKa5kP8X6VLIzdG/Si8h/iz+FeZIw5b9KBYYy2fwrOBuJz6VI6nODz6U/TGc0B3zRA65rHoaLc4NAAjOa4ogAYzRXjg0xBFAg8ZraDyDRIAzRIPGargjg0D3BogE4zSfeFFtoximbPBFNhjgURj5R1oAgcHmt2DimBbGOaEbMcYqK1yeWz+FeUgGc80AoHyjmjg9KzwCD1rcAME80XC4HpTTjGAfoKEhxjNCZgeDSXHYipJlxkflRmBGQK8wMMgdKLk1wwzRAJ4PNMoxgfhRGf8KKgdTRUGtvHJoLnoaPNBd3AorxwaYe9EZOAa/4JJjH7HemjP8AzH9Q/wDQ1r/gtMM/HnwOR/0KA/8ASuetuByf0oDPFeFMf8Ito2P+gfB/6LWv2nfjh8atJ/aT+LGk6T8YfG1nZWXjjWYLa2t/EF1HHBGl7MqoiK4CqAAAAMAAV4Z+Pnx2m8S6TDL8a/HkiPfQqyt4kuyGBcZBHmdKr4rDPxS8Ygf9B++/9HvRGOM1R+5j0ocjrQGBjNdeKGMdaGDyKxn5c/SiMdDWQec0TnpWNvf8KJzwDQB7VwTRwR15rIPescZ/KiBgDNEYGM5r5QBg59K4HHWjg85o8gCvoaAHSjhe9cHn8qwDwBmic96IyBgVxjk1j5etAA4ArGR1rBI4rHbPIojtRHavwrgjg1j5eDWAR1ojjiuoojJFcEcGmGO9DGetEAjrWAOhoHjG79KLMcfN+lB2Awp/ShIwGM0zMwxmgCeM/pWOM1gAYJpSoHFFicZ/Kg+BwKYE4z+FdByPpWNw47dqZegFFeMCiMcZ+lEehorxgfhRGO/P0rAKgiqx2ojtRHajyMZoAYwDVVWMjiqI5yDWKPAokY60RkYzRXPf9KxnHPSgMDGaUZ71ye1EE1gk80QSOT0oc8CiMdDQBPSsDHX6VnHOaBx/hXbiicjArGeBRXiiCRVdsA1RHGQayD0NUwBwM1VEjHWqIyMA0SB3rgjg1gZ4NH6114FHB4ogEYoqD0NFcj3or3FdRgHilA7UFzWQelZPrR571k+tMAy5o8gDNEHselEcZBoDPSiCKI7VRGKAycZo88A5o896HtWB2rqPpWM8+lYzwKbB6GscYzR6D9KABHJqscdaIyMZrA7URnoaPPeiAaojPQ1WPeqxxgGsAdKU8bc/Sse9HBHJ+lYJ4Jok9M0SQeDRBPQ/pTE4GO1EnsaJJGAabPTNAk8A1g/3v0o8CucZz+lEEryaHTGfpRAHINYBHTNDB6UDkgURgYzWAcA9RWe+aIyOTQz0BrtgGiM9DRGehqjz3rHAx1rj/wCtR6YFEZ6GsZABPSiAaxxjNEA0fQV9DRANEA9TSjnrRB4INEZ4z+lEHoprJPANBjnrRXIokgcn6UCCOtH5uAaIwMZ+lEsBgN+lZJ6Gic96yOg4onng0ckcmsZ70RxgH6UM44NEkjGaGccGsA9+fpTDPAP6V9DRIJwTmunQ114ojtnpRGefStuRkGhgjOeaWRwMK/6Us7L1OT2qK8kBB38/SodalBwzZ/DH9Kh1QTMEK/jn/wCtRuYz8pb9KW9jIyDUl/gbsZ9qbUxnA/z+lHUnYYU/5/Kvt7sBmre7Zv4v0oTswznPpQYlcZzQZiMbufpThivDc/SnB9c1LHhcA5oIO7fhipNoOAelSls/K36UXYjANFQybh1pXwcA9PavOHX9K2pKMZx6cVKhjbbmskfxfhigRj6VkE4DfpXA4Fcf3ufpQ4zmgMjk0QDgE1j1PSscZzQAxkmjgjj8qAyOT+FEAnAogEcHOKwOhNEds/SmGOAaQDOB+FEjqecUXLDg0ST0NE56daAJ4FCPHU80pC8qOaMrY60srEHLc9uKEjdA36UZG7H9KLFmwG/SskgYb9KBJ4LfpQyR97p7UeBwee1FQR15+lHOPvfpRzjAb8MUQOgNYGMUoycA0VB6Gu9EHGAf0oqCMjqaKgnFMMjg10onPes8cHp2rI6D8qGegNZOMZongc0c5681/wAEkv8AkzrTf+w/qH/oa18Xf2WPgB8edas/EXxb+G9n4j1HT7X7JbTz3M8Rjh3F9oEcijG5mPIzzQ/4Jz/sVAYHwG0r/wAD7z/49X/BTH4P/Db4IftG23gz4V+FYNA0Z/DVpdNaQyySKZnkmDPmRmPIRR1xxXhTnwto/wD2D4P/AEWtftWEj9qL4xYP/M/a5/6XzV4VyfFOjEH/AJf4P/Ri1XxX/wCSpeMf+w/ff+j3ok5xnrRANdvWhkd/wxWcjg1yBgHNAE9DQJ6ZotnpWSR1+lEkd6PI4/Kju9aO7HWgc9TQJAyDWSRgHmsZPBrH900eeCaJz0NEcUwAAweR0rJAxmiTgAGtvGQaOeoP0r6GsDtVHJHJojA21gYwKIyMZ+lY9/wrGRxRGOM0RwAD0rBxjP6Vj3ogHjNDpQ6GiAegrGec9KIJ6H9KGGGAKxxjNHGABQ9+aOcct09qI3Dg1gEYzmgCRtB/SicjINDB5xWMng0RnoeRQG7qelHGODWM9T+FEEjg8UCcAA0TnofpQG7qfwo5A681jpg0TwMGsn1pgM8GsA8ZquMYrHGM0QM4FHJ5B/SiM9DQySOar8KOSOOtc4AzmiM9DXU4zQBHANZJGeponAyKJweTmicV1HBo8DrmhyKxngGgMjGaAxwDVHBAGaPIxmsA8GiBjg0eRjNV2wDX0NZ75okkYzWD6/pTEkYBocd6Ybu9dBRGRgGscYzVZwQBWM9D+lVRANEE9D+lYGPeqOMdaIyMA1gdqOOgP6URnjNYP979KJPTNdBnPTpWAR1ogYwtD3rjGM0cEYFZx0NHB6CicjBoAEVkdc80TkYFD1HajyOv4YrBI4rHrROR/Kiuen5UBg4oj0FHB6CscZxXXgCsA96zk5FEA8LWAR1o8jGa6j6VRPGAfwqiO1de1HmuvaiO1EZGBVEEc1jjGaoEngnmgB680ME5zRz2okHgflR6YNHp8tbgeF5onP3TWcjGeaAGOOtHJ4zRx1okHqeKJLDA/Cs8YzQBI24ziiD2FZIGDWMiivFc8Y/CiPU1gDv9KIJHSiDtwBRz0A/CtuRxWDjAog4oDPFEDHBo8DNYJojIwD0rGeBW04xRHasdjWMnr+lHOOBXUVnPQV1FKM96bkYFbu3WjyBzWe9DGOaI5wBzTc8iuMYog4o4YfSiTjBo4I47VhSOuT6Uee9cYxWO2KIxxR9K7YBoggdKyCuKrGelcelYBo8nAFEYraaIwOaBJHy0Pfms46GjnGCaIJFBj1zSzup4Ffan7ULtwAAM496+2SN1FGdieRSs78gUuScA81BGzcCowCo5qMoBw3T2rKnkHinkReSf0oTI/QZqWZPu56U8wLYz9KeQn7p4rzRnaTRkUcZpHC9s1ImTuTvTAYAJ5oEr0NOwkjyaPPGKwMYNYA5BzQIPBogHgfhWQBjNAdzR6ZPUUSDjmiB0BrAK4BzWMjg1gjpRwelHgZoAHAFMB6/hR96GRg4otkcCmwRwaPA5ogt905oKoOAeaJBOM8isDHX8KZhjGaQjPHNN8w4FEqRgGiBng1gYwKAwOBWRiiQwwOvasqR159KYZxgZorkZxmiCRgc1k+lALjINE8dKHPNHaOS3PpimweM0VGOv6UCMYJojdyDRwRjPSgRt47VgHvWCRwazzmuM4Jo8814G/aP+PXwy0BPCvw8+MHivw7o8crypY6dqckEQdvvMFU4ycc0f20P2s+37Rfj7/wAHc3/xVD9tD9rMjB/aL8fZ/wCw5N/8VXj34kePfilra+JfiP4w1XxJqy26263mp3TTyiJSSqbmJO0FmOPc1D+2T+1bbQR21v8AtD+PI4okCRoutzAKoGAAN3TFa3rWreJdZv8AxFr2o3Goanql1JdXt3cSF5LiaRi8kjseSzMSST1JqCeW2njuLeVo5YmDI6nBVhyCKP7aH7WfGP2i/H3/AIPJv/iqvr271K9uNS1C6kuLq6laWaaRtzSOxyzE9ySSTR5Ix2rA6Zo8DGa4I4NDBAANYBGAefSlz2H4UcEYzXH1oEHkn6UxBxjmgRjIo880TwF7+lcDgisEjg0RgYJputEEEYNMeMUQccCsZ+XNEADn8KwSOKK+hoAnpRBAwTQBPb6UeuCeaIOKI7VjPeiB0BoqTgg1j3o9sUfrQBIDZojIwDXtmiCRjFHHTNMMjigD0rkjAquQcgUSM4FEZxzQIzjNEZ4oemKzjtXXt+FAYGByaIIHXmhyMCiO4rp7Vhu1E4HSjzj9KVRxk0eDxROOBRGOM0Tz15ojHGaGCOD0rGelHkcdqr6GuvAFH0rPqKo8jGazkcGu2AaJHTNAgjAOazkYHNYzg1jjGaIOcYo4x1qj7CuCODQGOlAEAACguen5UQcYrgYxTADvXGP5UCD3o47VjPSuMdKAHXNEY6igARzWM8CsIR1ojisAjjrRHaiADisZ4FUcHpRHasjGc0cEYFEA1WMiq4I4NZ5xRGeKPoKOBWB2rGRgmgMDGaY44FZ7Zo4PQ5pQCQAf0plCng0ST0NdetdRgH8KzxjNAAjBNcEAZrAxgUTkYFEcVgHvRAHHWhxXXqaI7ZojPSh0NYyMA1njGaIB4NH0FUeTzVHkYFVRGcUORWOMZrB6ZzXXiqOSetH0FHOMDmjyNua5IxWc8Z6VjjGaH50xwODQ5FMSRwaJ44NEbhjNHA6Gs4GSPwrgDFDHGfyrgdKIycZokEYxRwe9A9BmmHPWgAMEUCMcCtoIyDW0HjPNADPBrIPBOaOCc0cEZFEDbg9axkcUQDWOM5rAI6114qq4xiqrGeBRwRgUQMcGqOCciqxxjNUvpQIIwDRIPAP6VjjGayDxmtwx/Sgc0DkZNFsjiiRjAPNcZ4NEg8EUMdqYggd63UcE4zQIJwD9KLA8E1nPAo896yCOtNg9DWR1xRwayMYArPpRIJzROazzmjgnNfQ0CAABVfQ0OKLbhxRORgVRORjNHpxWT2NByO/4Ulzg9PpzUV+MYAoXoP8AShqAPQ0NRAAB5xUl+rZAHHpT3gJOB9Ka5DDAFG43dOcU0wPApTk5JpVDHIrpxSkA4IqUKBnPFMA/IpmwOKJzyKOCOD0ojIwDXGMdaIDDArGPlBoEAYxRw3BNYyOKwCOBQwSAKxxjNdBgVijzj26V1HSiM9DVZ4xmqwO1AZ4zSoccmtuDwawOpontmj9aB5xjNbe2aPJIHajwOuaIyMA10NEZGCKOcdaxjgc0cdR2pjjoaJBGBQOaLcD+VEDbxQBxxW0Yxmm5Arkjiic1uOMA0TkYrOAKHFEkjGa+lE5qsDGBQ4rtiq7YBrOOhqhyNuao896qqwCOKocUOtN1onIxmmGQAKAB6mic1gdqOCMCsDpmiBjrmiBxg0MZ60SD0rg8UcHisDGBVVgE8daPNfQ1gYwKOOmaPPej04NEZ4zWMDIrA60QDXUUDnpVHnvR6dawMYFZ4xmuvFZwcg197nNA9RmiR0zRIIyaGB0oHNHgU3ahjtRIxwaJxxTDuKBzQOKJxW4Gsg9DRyBwaI7V9DWT2NdOKJzWc8E1nHQ0TnvWSeAaycYzWT60ee9ZPrR5719DQ5FEk8ZqiewrOOho5ztBo88Cs5xz+FH0FE5OM0RkYBr2qgSeKGBQ5HFZyPlPNZPrRPHB+lZwMCs5rPOaBwMZ+lYBGQaY8DBrdxigcDg0TkYzR5GM0TkYzRx0Aqq4okAcVVY96qjyAKyOmawO1EjOBRGcUfQUwGKPAo57VvOMCskjFbcjmqABHHXtQOeD1ogdTWO1Yz1NYJ7UQSMA5rBPAFE9jzR2kcdaIx3ojjJrituRxTHPFEcZBo4PT8qIPf8ACunSjnGM0BnjNfQ1kHoawP736VgEdaJyMCq7YzWR0zXXiscYzXtmiMcZo57UR6GqPTih05rkjFAgjihgjANKR0ohjyB0rJA5rOeM/hRAPGaA7E0RxgGlHqaJGOtcdT+FE+hz+FAcYrPG0msLjGaBAOAaIGMbv0rqNpPNA8Yz+lZHrQIJzmiQec0OuAawDwD0rA7Hn6Uwz1PP0onjANYBHX9KpjnoaxkcVWAeM0QMDBrqPpVEcdelVwRwaojtRGehrgjg0enFD86HA5FcDtz6UORjPNEZ7UQAeB9KBB4ArgCiARhRXB60QMdPpXB4AocdR9KJ4FMSBkc0Npxgc0Rz8orgnIPPpRwOcUcEYAoEEYNfLjANEqRxzRUdc0SM5FYz3o4PAogmuOlNg9DQ2kYNHHQGhjrmiRnIrI6ZoH0ok54ojIwDWQRgHpWQeho46Zr2FUnrTE4wBWRwRTMAMAUSCOlZA6CiRiiQO9cEYB5oYIwKVcDHpTZPAOfSi3HH4UzgjGaMm1cE0xDcKf0rcOxoYwADWc8CunWhgcZonB4NZ4xmicDCnNE54JqhgHrX0NEAjINA470dp6H9KH1puelE570SCODXXjNfQ0pyPeicdT9KLA8A0MnqaJA4FEZ70gB5DfXiiRjAPNA4HJ60SDwTiieMZ5pRxRPcmiRxhvwxRPoaJDcZrIPQ0CMYB/SgRng80TgdaDDGM/hRbHAonI4HSm56dqYkduKwBwDWSTVA4Bx+FYBHWjk4qiOBzVdRR570eao4x1oHIHNUMHvROfrRAAwPyqiDjg17VwBWQehoHuDQBJyTRwTgnimA6KazxjNEDHBzXUY/KgFIABogHgHNYH979KIB/i/SiBjg/pQAOADTYPQ/hiiFI4NYB70QNuAf0oEYwTRUDv8ApRA7GsDGSaGOCD+lEA8ZrAIwDk0TkZNEZ4XmjjGc0xGBk0SB3rAI61ketAYIANZBGAelEDPBrhuKx70enFEnHSsA8A/hQ9ulHpgiio7CgO35UDzzQxgAGsqRgHNMMjg0AMcCiQOKJBGDRxtwDQ9vxrG7vRAxwf0rORyaIGME80CBxn6UQBxmuvasdqwDxmsKRw36UxyODmiQeAeaxkcGuvFDpzWAR1qsA8ZqqIxWAR1qiAayDwDRI9ayDwDR9BRGRgGtvf8AKj1ABrrxXOQM0AMYBrGeBRIxzRyDwKYjGR1ogkYA6UeRjNHOMZr2FYOOTWPyojHFc4rj0rGRxWO2OaPQAVRHaqHIo4x1rA7Vwa4PQ0RkYBo56AUcAZBoEEcUQCMVkHoa69KIOeDXB4BokDij0zmuCODRYN1okAYFbvagcn+VE5r73GaJAHFEgjgVu4xRORgVXHpRIIwBWRjFDG3FHHasYwaJB4rtgGgQKOCOD06UcEYFcYxR570QOgNHHY0TzkGiO1HGBiiARweaxngUQDjBqsZ4FHpxVE9yao9OKqiMjANUcY60COooHJAFHaRjPNMBjA/CiQRjNAgcUTkYoEbeO1ZyMGl2k4IpguetEZxzRUAetEcDmvlPaiO2elFRwBWBjrXyEY/KiB6UQM4qh9aJB7UQPWiB0BobSOtYB4zWAR1quPSsDdgnpR5GM1nAwKOCMAUORVUee9HnvR7CqIz0NL6UMBeaBXjJrIzkUCM4Jo7S2aBBOM0Tk4xRAAwKOCM02COD0rCkcUdrDg0BxiiR602OM0cD/ChjGRWB0zR244o4IwtcEZNDGetEnoTRA6A1jPesZ4FHnvXbANMB0FY7VjI4rPGM1VEcDmqqqxxjNUcEYFKQBg1kYzmid2DnOKIAOCaYqTgV8p7VkFfegQTg0CCv8qjUgg0QDR56UwULgmmbPSiwPeiQRgGiRjbmsAYBNZUdDXTnOfSgSSKIycE0SCMUdvQURkYBrAI461VY7Zo4/ho47VgZxms46GiB0Bo4IwKrA9aU8YBpiD0NAjGM0QOgNEg9T0oEdRQAJyKyCME1gEgDr2pgfWsDtQXI60QSODRUkDBzRGBwaIGPl69qJIOQaHCk96zx978MVkY680eTxQPBOaBG3BNNzyOaOCNoogY4NNg9KIGME81gY/lQx3oAHg0cHpR6CjgjArAPGaOCOO1UAMYzRGRXFHB6UcdM1XBHBqsDrVcdBRGB8tAcYrcDxXHpROflBo4IwKJBGBWM8CsZHFAgYxQx3rGe9cEcGqOD0rII4o+wrg8ZqjyMZqjjHWvoaqqPHOfwo46Zqq68UenFEgnFEgcCjjpmiQOlbhj+lEjGRWQR1rKk0SOmayM4zWVHAo4yOaBB4okYwD9KB7E0dpGAayfWiQR15qs8YzXI4BrJ9aPPeqPPeic964PAonPesnsa+hrqOKr6Gs8deaokkDFVVcEcGq4ximOBgUQCMVwBQIPArrxR7Yo4PGaLdhRPOBTYIxnmgQRgmsk9DQIPFDDHANZ4xmq+ho8gc1xjGKogYGKokYwBVYB71xWAeM1RGeho8gCicnIqiOKqj2o46ZogHgHNUQOgNAAdDVVgE9eldRVE8Zrr0NUxz0NfeGO4rjH8qHzDCmjzzRGRk12xXHTrX0NEdOawD3pjxisAjr+lEAjBNYyOKoAHABohem79KIwOfwo4IwKIHQflRIHGax71g4xn9K/yKyOKwQcGiOnNEZ6GsAdKoEEAZojIwDRA6A0BnpVYBGM0Qe1EHFdeKA7CiMjGaIBGCKxkYFYA4xSgEfe/DFfKR979KOG4Dcj2oquMA0dmAN2fwoAHkN+lfLjANAKeS3PpisDs36USp43fpRwMDdz9KJH979KwMYDfpRAIAz+lEjPBokEY3fpRKk8N+lfKRjd09qz2BrI4G79KyB3rKHofpWQeho46ZokcDr6UpXg5/Csg9Pyo46CgRnHf0o4PQ0APSiynpQwff0rHvWM9KAJHSgM8VgngCmGOMUBngDNHpxWDjkUoHQCvvDpW4H8KwO1DB6GmweM0QDRYEfL+FE8YNHAGKGMdf0ojjANFRwKBGOTQwO/6U2DxmtpIrHYnpRx0HWihHFEDs34YrAI68+mKIHY/hRIAwaJwOtHFDGetHaeAaOO1Eg9DWBjGf0rAC4zzQGe2aOD0qsEnAP4UVPAAoqwGQKxxmiMVjIx3ogZwTRUAdaUADJ5xXyjo36UCoIAP6U3agFIHPShjHFHgZFZBGK4IoSRFMlsEdBg15sech/0pnQ8K36UzR7eHz+FYBHBzRUY4FbQRjFFeODRA6k1gkcUAMc0SDwKKkd6I4zmsZHAogisZFYzwK68VjPA5ojHFAA9DVdRRGK4xjFAZoDIxilolcYB+lAqR1pmXOAabaRx1oFcYJ/StxYEA81lc5osMAg5o4xjdj8KBBwf1oFRxu/SiQRjNAqOAeayG9jRK447USucDtWcjOM46VkE9KJBAPWgVAyTXHQDNcH5cUSAMCjgjgUSvr+lfSiR1NBgefyrGTjNEDsayOlEDucUenFFccbv0og9KokY4/CjjHWhgjIP0o8jGaC5rGeAaKkAAc0FIGKKmsEjgUEyMhv0rafWsHGKAzxn6VgDof0rHrW0Y4P6UQMcH6UQMcGtoxnP6VhfX9KIU9D+lYB4zWBjArAPGaPHFHGOtcEcGiO4FZBGMc11HTpWc8CiAeMV7Yo+wrIxwM0SD0o+woj0rqKyfSjzwKzk4FHsSKJGcAV17fSsgjGKOGOAM0BkdPwoqSMbf1rYT2/CtmeooqT/D+tbc8YoqeBjOKCknkUVHXNHGOlFT2OaC9iK28cmgM9RRHOBRUcAGtvvRBx0oqMYAzRAx06UBnjNYBAA61jPArHOMfhQBHasAHGc0QDyB0o4IwKIAHvXGMUR0P5VRAIxQx0IrHHymsDtXB4Bo88A1xjFbeK9sUQc8GscYzQXPIrZ26+1EEdTQUnvmtuR1oLjoaK56GlAHGfpRIIx1rAHQVjP3a2+/6UcHgUSSOKAB4zWBjpmiQRk0RzgGgAeAeaxngUAOMmiB0BrAIyDRHFEAnGf0rG7kHNEDGSf0oAHo36UQM4z0rgnaDk1kdc1nAzmjjpmiCRxQ5HXNbSelVgDgV16VRAPFYHaqAB4zQGe9YG2snsaPPeqPPAqgMc1x6UeRnFdOhojIwDRBPU80BkcGsZ4FYI6VR5AFYyOKxnpRANAEjgVjtQGBjNEdqI9DRHTBqiBjg1RBxkUOnNYwOtADGM1nHANEZ4ogGj0wDzWO9EdqwSetcHgGqrHvWM8CqJB780RkYzQ5FYB6UQCMUMdB2o4PSj6CiORWBjAqiM8flWOMZrbznNHHc1weAao4Axmiue9Ac9elEA8g0eeCMmh05o4PAoHHAP0rPHNDrgGgOo61tJ4rAIxmgD0NKQRzRwBkH6Upz0NZA6CuG4HWu2AaIGODRAIxROACDXBHBquOBmhzxQAJx1piDxmgcDFYBHWiQwyDWQeAaIGOD+lHtg9KIGODRGcc0fQGgcjg1jsTWccA0ee9AYGM0R2zQOcDFcHgGqPJwDXHQVweAaIBoYxjPNHAHNY5yDR46USMZFAep6UOtNwcj8qK570VBAwa254JoYJojjmhyOaIyMA0TjgfhRGSMUwz0phkdazzijyMCqxkcmiDjA60OeBWDjANH7oFHgYFEYGQeaByOTRyeO9EZxmgMnOa68UVBGBXaiMqOa6jbRGKrGSOaI4FdsUBnjNEj1qj2FUBnjNKM96C5HWiBjGf0o4PSto+6DRUkAVgdjmiMcZoqAMZqsAgAGjwK25GQaAz3raCOD0ogY4NYB4zRXjg0RzgGtoxkGtuRitobvVVjPSiB2NEZGKIwOtYBHWiB0BoqMZzzVEZ4zRAIxR4OKIBrjtRB7GuvAoqAKA9TRHHB6VtB6nNKMd+nSsZ6CiOAAaYCivoaC5HWsZHFDrRGBgGsAnANEAcA0VGOv6UR2o4PSjg9BWBgc0V44NYB78/SsZGO9bRjANAA4AojjINFcVjjNHB6flRANADpmsZHFdeKrAxgUR2qiuV61RANURnvQAAwDQAIGD0oqD0NAAgZogGgOMA1hscUc9q6e+KIBGAawSeOtAE96xjgHNZzwBW0HgH9KIwOtA981jIyTWMjaD0oLkbc0R3B5rkgCm4xiiMDINYBOAaIyMisgjFVWQBgc0TzjNHB5/Sjgjj8qYDAxXBAGaYY6VjjNAZ71gdqIGODWAOAaOMVRGcc1WM9KogGgM8ZoAA4B6dKYDrmmXPGaIAXGa6Dg0ACcUee9EZ6msEjiiMnbmiMHANNg8ZoL3BrgnA/CiOKxke9AZAWtpIBFFSTg9RTAZ2+lEY4zRAIyK25GQaIx3o884+lBQRwawCMZojHGa+9xRwDx+VEY71RAHANYyME03HTmqIBGK68UQpoAbRg0QM+9YHaq/4cf8A/Vz3/lk//d1H/gh/np+09+H/AAhP/wB3Uf8AgiBkYH7T3/lk/wD3dR/4IgZ4/wCGnv8Ayyf/ALur/hyBxgftPf8Alk//AHdQ/wCCIGP+bnv/ACyf/u6j/wAEQMnI/ae/8sn/AO7qP/BEAH/m53/yyf8A7uo/8EQARgftO/8Alk//AHdX/Dj/AP6ue/8ALJ/+7qP/AAQ/yMf8NPf+WT/93Uf+CH/YftPf+WT/APd1D/giB0/4ye6f9ST/APd1f8OP/wDq57/yyf8A7ur/AIcf/wDVz3/lk/8A3dR/4IfgjA/ae/8ALJ/+7qH/AARAx/zc9/5ZP/3dX/Dj8f8ARz3/AJZP/wB3V/w4/wD+rnv/ACyf/u6v+HH/ABgftPf+WT/93Uf+CH4IwP2nv/LJ/wDu6v8Ahx+P+jnv/LJ/+7qP/BD8EYH7T3/lk/8A3dQ/4IgAYx+09/5ZP/3dX/Dj8f8ARz3/AJZP/wB3V/w4/BGD+07n/uSf/u6v+HH/AP1c9/5ZP/3dX/Dj/wD6ue/8sn/7ur/hx/8A9XPf+WT/APd1f8OPx2/ae/8ALJ/+7q/4cf8A/Vz3/lk//d1f8OP+Mf8ADT3/AJZP/wB3V/w4/Hb9p7/yyf8A7ur/AIcf/wDVz3/lk/8A3dX/AA4//wCrnv8Ayyf/ALur/hx+O37T3/lk/wD3dX/Dj/8A6ue/8sn/AO7q/wCHH/8A1c9/5ZP/AN3Uf+CH4P8Azc9/5ZP/AN3V/wAOP/8Aq57/AMsn/wC7qP8AwQ/zx/w09/5ZP/3dX/Dj/jH/AA09/wCWT/8Ad1f8OP8A/q57/wAsn/7ur/hx+f8Ao5//AMsn/wC7q/4cf/8AVz3/AJZP/wB3V/w4/wD+rnv/ACyf/u6v+HH/AP1c9/5ZP/3dX/Dj/wD6uf8A/LJ/+7qP/BD8H/m57/yyf/u6j/wQ/wA9P2nv/LJ/+7q/4cf/APVz3/lk/wD3dX/Dj/8A6ue/8sn/AO7q/wCHH/GB+09/5ZP/AN3V/wAOP/8Aq57/AMsn/wC7qP8AwQ/z0/af/wDLJ/8Au6v+HH//AFc9/wCWT/8Ad1f8OP8A/q57/wAsn/7ur/hx/wD9XPf+WT/93V/w4/8A+rnv/LJ/+7q/4cfcYH7T/wD5ZP8A93Uf+CH+en7T3/lk/wD3dX/Dj/8A6ue/8sn/AO7qP/BD8H/m57/yyf8A7ur/AIcf/wDVz/4f8IT/APd1H/gh/np+0/8A+WT/APd1f8OPsjB/aez/ANyT/wDd1H/gh/np+0//AOWT/wDd1H/gh9np+0//AOWT/wDd1f8ADj7/AKuf/wDLJ/8Au6j/AMEPgf8Am57/AMsn/wC7q/4cf8Y/4af/APLJ/wDu6v8Ahx8MY/4ae/8ALJ/+7qP/AAQ+z/zc9/5ZP/3dX/Dj/wD6uf8A/LJ/+7q/4cfD/o57n/sSf/u6v+HH3f8A4ae/8sn/AO7q/wCHH/GP+Gnv/LJ/+7qP/BD7Ix/w0/8A+WT/APd1f8OPj/0c/wD+WT/93V/w4/4x/wANPf8Alk//AHdQ/wCCH/r+09n/ALkn/wC7q/4cf85H7T34f8IT/wDd1H/gh/np+09/5ZP/AN3V/wAOPx2/ae/8sn/7ur/hx+MY/wCGnv8Ayyf/ALuo/wDBD/PT9p7/AMsn/wC7q/4cf5GD+09/5ZP/AN3V/wAOP/8Aq57/AMsn/wC7q/4cf/8AVz3/AJZP/wB3V/w4/wD+rnv/ACyf/u6v+HH/AP1c/wDh/wAIT/8Ad1f8OP8A/q57/wAsn/7ur/hx/wAY/wCGnv8Ayyf/ALuo/wDBD/OP+Mnv/LJ/+7qP/BD8EYH7T3/lk/8A3dX/AA4/H/Rz3/lk/wD3dX/Dj/8A6ue/8sn/AO7qP/BD/PT9p7/yyf8A7ur/AIcf/wDVz3/lk/8A3dX/AA4//wCrnv8Ayyf/ALur/hx+MYH7T3/lk/8A3dX/AA4//wCrnv8Ayyf/ALur/hx//wBXPf8Alk//AHdX/Dj/AP6ue/8ALJ/+7q/4cf8A/Vz3/lk//d1f8OPuc/8ADT//AJZP/wB3Uf8Agh+CMf8ADT3/AJZP/wB3Uf8Agh/np+09/wCWT/8Ad1f8OP8AjH/DT3/lk/8A3dR/4IfZ4H7T/wD5ZP8A93UP+CHwH/Nz3/lk/wD3dR/4IfZ5H7T3/lk//d1f8OPhjA/ae/8ALJ/+7qH/AAQ+x/zc/wD+WT/93Uf+CH2Rj/hp7/yyf/u6v+HH3GP+Gn//ACyf/u6j/wAEPs8f8NPfh/whP/3dX/Dj49v2n/8Ayyf/ALuo/wDBD7Ix/wANPf8Alk//AHdX/Dj7HT9p/wD8sn/7ur/hx+O37T3/AJZP/wB3V/w4+P8A0c//AOWT/wDd1D/gh+QMD9p//wAsn/7ur/hyCcYH7T3/AJZP/wB3V/w5AyMH9p7P/ck//d1H/giBn/m57/yyf/u6v+HIJPX9p7/yyf8A7uo/8EPwen7Tv/lk/wD3dR/4IgEjH/DT3/lk/wD3dX/DkHAwP2nf/LJ/+7q/4cg8Y/4ae/8ALJ/+7qP/AARByMD9p7/yyf8A7ur/AIcg/wDVz3/lk/8A3dX/AA5AOMD9p7/yyf8A7uo/8EQM9f2ns/8Ack//AHdX/Dj/AP6ue/8ALJ/+7q/4cgDt+07/AOWV/wDd1f8ADkH/AKud/wDLJ/8Au6v+HIP/AFc7/wCWT/8Ad1H/AIIgZGB+09/5ZP8A93Uf+CIAIwP2nf8Ayyf/ALuof8EQMDH/AA09/wCWT/8Ad1f8OQBjA/ad/wDLK/8Au6v+HIP/AFc7/wCWT/8Ad1f8OQDjA/ae/wDLJ/8Au6v+HIHOf+Gnv/LJ/wDu6v8AhyD/ANXPfh/whP8A93Uf+CIGcf8AGT3/AJZP/wB3V/w4/wDT9p7/AMsn/wC7qP8AwRAzyP2nv/LJ/wDu6j/wRByMD9p7/wAsn/7ur/hyBwAP2nun/Uk//d1H/gh/n/m57/yyf/u6j/wRAzyf2nv/ACyf/u6h/wAEQBjH/DT3/lk//d1f8OP/APq57/yyf/u6v+HH/wD1c9/5ZP8A93UP+CIBwB/w090/6kn/AO7qP/BD/P8Azc9/5ZP/AN3V/wAOP/8Aq57/AMsn/wC7q/4cf/8AVz3/AJZP/wB3V/w4/wD+rnv/ACyf/u6j/wAEP8/83Pf+WT/93V/w4/8A+rnv/LJ/+7q/4cf/APVz3/lk/wD3dX/Dj/8A6uf/AA/4Qn/7ur/hyB6ftPf+WT/93Uf+CIGf+bnv/LJ/+7qP/BEDP/Nz3/lk/wD3dX/Dj8Yx/wANPf8Alk//AHdR/wCCIGf+bnv/ACyf/u6j/wAEPwRgftPf+WT/APd1H/gh/n/m57/yyf8A7uof8EQBjH/DT3/lk/8A3dR/4Ifg9P2nv/LJ/wDu6h/wRAwMD9p7/wAsn/7uo/8ABEDP/Nz3/lk//d1f8OQOMD9p7/yyf/u6h/wQ/wAf83Pf+WT/APd1f8OPx/0c9/5ZP/3dX/Dj8f8ARzv/AJZP/wB3V/w4/wD+rnv/ACyf/u6v+HIHGP8Ahp7/AMsn/wC7qP8AwRAz/wA3Pf8Alk//AHdR/wCCH+RgftPf+WT/APd1f8OP+MD9p7/yyf8A7uo/8EPwen7Tv/lk/wD3dX/Dj/8A6ue/8sn/AO7qP/BD/P8Azc9/5ZP/AN3Uf+CH+Rj/AIae/wDLJ/8Au6j/AMEP89P2nv8Ayyf/ALur/hx//wBXP/8Alk//AHdX/Dj/AP6ue/8ALJ/+7q/4cf8A/Vz34f8ACE//AHdX/Dj/AP6ue/8ALJ/+7q/4cf8A/Vz3/lk//d1H/gh/np+09/5ZP/3dR/4If56ftP8A/lk//d1f8OPzjH/DT/8A5ZP/AN3Uf+CH+RgftPf+WT/93V/w4/OMH9p/P/ck/wD3dX/Dj/8A6ue/8sn/AO7q/wCHH5/6Of8A/LJ/+7qP/BD/ACMD9p7/AMsn/wC7q/4cf/8AVz//AJZP/wB3V/w4/wD+rn/w/wCEJ/8Au6v+HH//AFc9/wCWT/8Ad1f8OP8A/q5/8P8AhCf/ALuof8EP/X9p7P8A3JP/AN3Uf+CH+Rj/AIae/wDLJ/8Au6v+HIHAA/ae6f8AUk//AHdR/wCCIGRj/hp7/wAsn/7uof8ABEDAx/w09/5ZP/3dX/DkDv8A8NPf+WT/APd1D/giBjp+09/5ZP8A93UP+CIGBj/hp7/yyf8A7ur/AIcf/wDVz3/lk/8A3dX/AA4/9P2nv/LJ/wDu6v8AhyAf+jnv/LJ/+7qP/BEDIx/w09/5ZP8A93V/w4//AOrnv/LJ/wDu6v8Ahx//ANXPf+WT/wDd1H/gh+CMD9p7/wAsn/7ur/hx8O37T3/lk/8A3dQ/4If4/wCbnv8Ayyf/ALuo/wDBD8Hp+09/5ZP/AN3Uf+CH+f8Am57/AMsn/wC7q/4cf/8AVz3/AJZP/wB3Uf8AgiBn/m57/wAsn/7uo/8ABEAEYH7T3/lk/wD3dR/4If5GB+09/wCWT/8Ad1f8OP8A/q57/wAsn/7ur/hx/wD9XPf+WT/93V/w4/8A+rnv/LJ/+7qP/BD8ZyP2nv8Ayyf/ALur/hx//wBXPf8Alk//AHdR/wCCH4PT9p7/AMsn/wC7qP8AwQ/yMD9p7/yyf/u6v+HH/wD1c9/5ZP8A93Uf+CH+Tn/hp7/yyf8A7ur/AIcf8Y/4ae/8sn/7ur/hx/8A9XPf+WT/APd1f8OP/wDq57/yyf8A7ur/AIcf/wDVz3/lk/8A3dX/AA4//wCrnv8Ayyf/ALuo/wDBD/IwP2nv/LJ/+7q/4cfn/o5//wAsn/7ur/hx/wAY/wCGn/8Ayyf/ALur/hx/xj/hp7/yyf8A7ur/AIcf/wDVz3/lk/8A3dR/4If56ftPf+WT/wDd1f8ADj/jH/DT3/lk/wD3dQ/4Ig4GB+09/wCWT/8Ad1H/AIIg56ftPf8Alk//AHdR/wCCIAP/ADc7/wCWT/8Ad1f8OQOc/wDDT3/lk/8A3dX/AA5AHb9p3/yyv/u6v+HIAzkftPf+WT/93Uf+CIOcf8ZO9On/ABRP/wB3V/w5B/6ud/8ALJ/+7q/4cgDt+07/AOWV/wDd1H/giBn/AJue/wDLJ/8Au6v+HIAxgftO/wDllf8A3dR/4IgEjH/DT3/lk/8A3dX/AA5A4wP2nv8Ayyf/ALuof8EQABgftO/+WT/93Uf+CIAJyP2nf/LJ/wDu6h/wRA/6ue/8sn/7ur/hyB/1c9/5ZP8A93V/w5A/6ue/8sn/AO7q/wCHH/8A1c/+H/CE/wD3dX/Dj/jA/ae/8sn/AO7q/wCHIGRg/tPZ/wC5J/8Au6j/AMEQMjA/ae/8sn/7ur/hyBxj/hp7/wAsn/7uo/8ABEAdv2nv/LJ/+7q/4cf56/tPf+WT/wDd1f8ADj8f9HO/+WT/APd1H/giAD/zc7/5ZX/3dR/4If5GB+09/wCWT/8Ad1H/AIIfgjA/ae/8sn/7ur/hx+P+jnv/ACyf/u6j/wAEPwen7T3/AJZP/wB3V/w4/wCMD9p7/wAsn/7ur/hx/wD9XPf+WT/93V/w4/8A+rnv/LJ/+7q/4cf/APVz3/lk/wD3dR/4If5GB+09/wCWT/8Ad1f8OP8A/q57/wAsn/7ur/hx/wD9XPf+WT/93V/w4/8A+rnv/LJ/+7qP/BD8H/m57/yyf/u6v+HH3Of+Gnv/ACyf/u6v+HH/AP1c9/5ZP/3dQ/4If4/5ue/8sn/7ur/hyAOv/DTv/llf/d1f8OQOc/8ADT3/AJZP/wB3Uf8Agh/kAf8ADT3/AJZP/wB3Uf8AgiBkY/4ae/8ALJ/+7q/4cfj/AKOd/D/hCf8A7ur/AIcgDr/w07/5ZX/3dR/4IgAjA/ae/wDLJ/8Au6v+HIA7ftO/+WV/93V/w5B/6ud/8sn/AO7q/wCHH4xj/hp7/wAsn/7uo/8ABD8Hp+07/wCWT/8Ad1f8OPxjH/DTv/llf/d1D/giBj/m57/yyf8A7ur/AIcf+n7T3/lk/wD3dX/Dj/PX9p7/AMsn/wC7qP8AwRA9P2nv/LJ/+7qP/BD/AD/zc9/5ZP8A93V/w5A4wP2nv/LJ/wDu6h/wRAx0/ae/8sn/AO7qP/BD8EYH7T3/AJZP/wB3Uf8Agh/n/m57/wAsn/7ur/hx/wD9XPf+WT/93V/w4/8AT9p7/wAsn/7uo/8ABD/P/Nz3/lk//d1f8OPx/wBHPf8Alk//AHdR/wCCH+Rj/hp7/wAsn/7uo/8ABEDIx/w09/5ZP/3dX/DkDjH/AA09/wCWT/8Ad1f8OP8A/q57/wAsn/7ur/hx/wCn7T3/AJZP/wB3Uf8AgiBn/m57/wAsn/7uo/8ABEDIx/w09/5ZP/3dR/4If5GP+Gnv/LJ/+7qP/BD/ANP2nv8Ayyf/ALuof8EPyP8Am5//AMsn/wC7q/4cf/8AVz3/AJZP/wB3V/w4/wD+rnv/ACyf/u6v+HH/AP1c9/5ZP/3dX/Dj/wD6ue/8sn/7ur/hx/xj/hp7/wAsn/7uo/8ABD8H/m57/wAsn/7uo/8ABD7jA/ae/wDLJ/8Au6j/AMEPyen7T/8A5ZP/AN3VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX2pabpkay6lqFtaIzbVaeVYwT6Ak9a1L4y/B/R1Lav8V/B1iFfyybnXbaLD8/L8zjng8e1al+1r+yzpKyfb/wBpD4Zo8LbZIl8VWbyKc4xsWQt+nFaj+3t+xvpjSrc/tEeEXMKbm+z3TXGRjPy+Wrbj7DJ7Vf8A/BTX9hrTii3Hx5tXLg48jQ9Snxj12W5x+NX3/BWH9iS0WY2/xG1e9MRIQQeHbwebg9V3xr16/Nj8Kv8A/gsL+x7aeX9nm8a32/OfI0QLsxjrvkXr7Z6dqvP+C0n7KVtOYYPB3xPulAGJYdJsgp+m+7U/pUv/AAWz+AALCH4U/EFgCdhZbNc+mf3xx+tP/wAFvfhaEJj+B3ilmAO0NqUABP1wcV/w/F8Kf9G7ar/4Ukf/AMYp/wDguagciP8AZeLKCdpbxtgkfT7CcVff8FydZkkU6b+zXZQIF+ZZ/FjSkn1BFquB7Yr/AIfi+Kv+jdtJ/wDCkk/+MUP+Cz/xn1PN5of7NukyWbnEZ+23U/Tg/OqAHnPYYr/h8h8fv+jadK/7+3n/AMTX/D5D4/f9G06V/wB/bz/4mv8Ah8h8fv8Ao2nSv+/t5/8AE1/w+Q+P3/RtOlf9/bz/AOJr/h8h8fv+jadK/wC/t5/8TQ/4LafEbSybTxB+znpKXYOdv9tTW+FPT5WhY/jmv+H4vir/AKN20n/wpJP/AIxUP/Bc2VYUWf8AZgR5QoDsnjUqC2OSAbI4Htk/WrL/AILkaJJCTqX7Nl7BKGwqweK1lBGBzk2q89eMfjVl/wAFwPh1JCTqXwG8RwShsKsGsQygjA5yUXnrxj8atf8Agtt8C3t0a9+EXjuKfnckT2kijnjDGVSeMdh/WrP/AILTfsrXEkcVz4K+J9ruHzyPpVkyIcf7N2WI7dKsf+Cwn7Hl4jNcXPjSyKnAWfQwS3uNkjfritP/AOCrH7D16+y5+K1/YDZu3XHhy/Izx8v7uFjnn6cda0z/AIKPfsS6uYxafH/R081SV+1WN5bYx6+ZCu0+xxWlftp/sj6yENn+0h8PY96kr9r1+C1xj181lx9DWl/Hr4F66ofRPjT4E1AFN4Nr4jtJcr/e+WQ8dOasNV0vVozLpWpWt4inDNbzLIBn3UmqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqrXviX8OPC28eJ/iB4b0gxEhxf6rDb7SDtOd7DHPH1rXP2y/wBk3w8GGpftHfDwtHnclt4gt7plIOCCsTMc57YzWuf8FOP2H9CDJL8cILyVc4jsdFv7jdg4OGWAp+bDPatc/wCCwv7Hukhv7Pm8a61t3Y+w6IE3Y6Y8+SPr2zjpzita/wCC23wLg3Dw78IvHd9g/J9te0tcjHfZLJjnjv6+1ax/wXI6x+H/ANmz0xLeeK/zGxbX9d34V/w94/ao8UD/AIoD9mfR5vMXMX+h3+oZ3/6s/uimR9Mbu2K/4bh/4KteMG3eHf2abqxjkb5Xs/h7qPlrtHI3zu4568nOenpR+I//AAWy8WRg6Z4Y1nSkZMlf+Ef0m1JD9Bm4TcCPbBGeaPwk/wCC1fisFNX8a6to4kPlsf8AhJtMt9oHO7/RXOPTI549KH7DH/BVfxaceIP2m7qySZsvFe/EPUjGhXodkKOozjjA7845of8ABJb9r/xCQ3jT9pzR5GYmSQjVNSvf3p6n94iZzk/NwfatP/4IeeJbxzN4g/aTsIJJFLSNb+GHuSZCeeWuUyOvzdfbmtM/4Id+DYtn9s/tC6zdYTD/AGXw9Fb5b1G6Z8D25+tab/wRL/Z+iMX9sfFb4g3QUHzRbPZwbjjjbugfbzjrmtP/AOCM/wCyTZIi3Ot/EW/KvktcaxbqWHofLt14+mD71Y/8Ekf2LLQubjwd4hvdwGBPr9wNv02Ff1zVl/wS6/YXsvJb/hSH2iSHHzz+ItTfeR3ZftO0/TGParH/AIJ3fsV6c7Pb/s/aA5cYInmuJx+AeQ4/CrP9hz9kCxgFvB+zn4FZQSczaSkrf99Pk/rUP7JH7KsEKQR/s0/CwrGoVS/g+xdsAY5JiJJ9zzQ/Z7+ASkMvwP8Ah+CDwR4ZtBj/AMh1/wAKm+Ff/RNPCn/gmt//AIio/B3hCGNIYfCujoiKFRFsYwFA6ADHAqx0vTNLRo9M061tFc5ZYIVjDH1IAqqqqqqp/CvheRzJJ4b0tmYkszWcZJPc9KvPhp8ONRm+03/w/wDDdzMRgyTaTC7YHuVzV78CfgfqUwuNR+Dfga6lC7Q8/h21dsDtkxk4q6/ZX/ZgvZ2urz9nD4XTzOfnkl8H2LM31Jiyau/2Hf2Qb2A2837OfgVVYjJi0lIm49GTBH51ff8ABOz9irUZFln/AGftBVlXaBBPcwD8QkoBPuavv+CV37DN3Gsdv8HbmyZWyXg8SakSfY77hhj8K1T/AIJA/sb6gGFppni/Tdz7h9l11m2j+6PNR+Prz71qn/BFX9mO53PpXxA+Jdk7PkK9/ZTIq+gH2UN6cljWqf8ABD/4dTAjRfj14jtfmOPtWjQ3HHYfK6c+/f0Fah/wQ/8AFWnyLc+GP2lLCeeIBo3uPDUloVfPYpcyEcYOfXt3p/8Agld+3J4YBXwH+1BpKRorIir4i1SwJTrgCOJwMkdM475ofsu/8FhPB7EaJ8cdX1vDcE+O3uc5HJ/0sjgYx/L1pvEP/BbvwKpF7pmtalEhy0YsdE1Iklf70Id+3QHr9eW/4KIf8FJvhlk/E79mtJ7WPJkn1LwZqNkSBkkrKjrHjHfaRhfqa8J/8FvrRJBZfEX9nu5t3jbbNPpOuhyCOCPJliXBBHQyfyrwL/wVw/Y38XmOLWvEPiPwfK4xt1rRnZQ3putTMAPc4Hrivh/+0D8DfisI1+HHxd8JeIZnHFtY6tDJOv8AvQ7t6n2Kiqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr4gf8FSv2ztA8eeJNB0v4iaXHZ6bq93bW6N4fs2KxxysqjJjyeAOTXh//gqr+2rqGvabYXXxG0pobm8iikUeHrMZVnAPPl8cGqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq8vbOwgN1f3cNtCpAMk0gRRnpyeK8Q/tLfs6eEwy+Jfjz8PtNdBzFc+JbRJDwDwnmbicEHAHcV4k/wCCkH7E3hcMt/8AHrSbp1yAum2N3fbjzwDDE4/hPJIHI55FeIf+CxH7H2i7hpreN9e2nj+z9EVN3IHH2iWL1zz6H2z4g/4Le/C+2Qnwr8DfFOosB8o1DU4LIHg90E2OcfmfTm7/AOC03xZ8TXBs/h1+zbpf2gnCRTancakwyRj5Yooie498j05/4bz/AOCpHxAIbwV+zZJZwSD93Np/gLUZUA5IJkmd0yRj0BwMDnlta/4LdePwY7XT9a0uFgfkFpoul7eME7pAj989evTpwf2RP+Cufj3C+Jvj9qGipMcSRz+PLiBACdp3JZhgRgZxg8H1JFf8Ohv2qPGf/JT/ANpfRbjzD+9/0y/1TOTlv9cI89AecZP0zWhf8EONIjKP4m/aOu7jIXdFYeGFh29dwDvcvntg7R9OeNC/4Ip/s32ZR/EHxG+IepuoXKwXNpbIx5zkeQ7YPHRhj1NaH/wSS/Ys0nZ9v8G+INa27c/bvEFwm7HXPkGPr3xjpxitF/4J1fsU6Bs+wfs/6FLsAA+23FzedDnnzpXz+PWtH/ZT/Zi8PrjR/wBnj4b27bSpkHhe0aQgnJBcxliMgcE9hWkeDfB/h5g+geFNH01lYsDZ2McJDEYJ+UDkjj6VVVVVVVVVVVVVVVVVVVVVVVVVVVVVXi74YfDT4gRmHx58PPDPiRCMFNX0iC8B/CRWrx5/wTJ/Yu8eJI7/AAhi0C7fOLnQb6eyKfSIMYfzQ/zr4hf8ERfBV0Jbr4T/ABv1rSpFO6K217To71SfTzYTEVHTnY3Tpzmj+zP/AMFZf2YSJ/hZ8SL/AMW6TaH93baVro1GBE9rO/Uc44IjRjnucZrw9/wVp/ac+DmpxeGv2nf2fUmmX5SzWlxoN6RnlykqvG/0VUB9e9fCT/gqt+yJ8T/IstY8WX/gTU5cD7N4ktfKi3d8XEZeIL7uyfQV4f8AEnhzxdpMGveFNf03WtMuRmC90+6S5hlHqroSp/A1VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVXxa/5Kp4z/7GC/8A/Sh68Jf8jVov/YQg/wDRi1VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVqus6Podqb3W9Ws9Pt1zma6nWFBgZPzMQOgrxN+1v+y74PLx+If2g/h/bzREh7dPEFvNMuPWONmcflXib/gqT+xH4cDRxfFubWJ0PMOmaJeS/k7RKh/76rxJ/wWp/Zu0/dF4Z+H3xA1iRejzW1raxN06Ezs3r1UdK13/guBql3MLPwV+zhEJJG2xPf+I2lZ2OAB5UduOc8YDHPHSh/wAFGv8Ago78ScR/DL9miJIJceXNp3g3Ur5lBxgmRnMePmXkqAMgng0fEf8AwWz+JwI0/SNb0O2lB/d/YdI0jYD1O6fbKMBumc8eopv2Jv8Agq58Sc/8Jp+0Lc6VFPnzLe/8eXgiGck/urRXTuRwO4HTpaf8EW/i54pnW7+JP7Sumefk5kh0241NhnOcGWWI9l/M+nPh3/giH8JLby/+Es+Nni7UsEb/AOzrC3st3Izjf52OM+vUfj4e/wCCQH7HGioi6lpvi/Xyo5bUdcKFuO/2dIv0x1rw/wD8E7P2KvDO3+zf2ftBm2Hj+0J7m/7k8/aJXz17+3oK0D9nP9n3wqF/4Rn4GfD/AEtlHD2nhq0iY8AZLLGCTgDk81a2dpY26Wljaw28MedkUSBFXJycAcDkk1VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV4i8MeGvGGlS6F4s8PaZremzf62z1G0S5hfjHKOCp4J6jvXxd/wCCVv7InxSW4vNI8HXfgXVZssLrw1cmCMN2/wBGcPCF9QiIfcda8Q/8Euv2wv2etVn8V/sqfGxtV2ksILK/k0O+lA5CMhcwSjsd8gB/u88eHP8Agp1+2N+zjrEHhH9rL4MzaugbaLm9sG0a9lUcF0kVDBMo45VADj73OR8D/wDgpN+yh8cGttNtfHq+EdcnwBpfihVsWLeiTbjC5J4AEm48fKM4pHSRFkjdWVhlWByCPUVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVXxa/wCSqeM/+xgv/wD0oevCX/I1aL/2EIP/AEYtV4j+K3wt8H3YsPFvxK8K6JddoNS1mC2f/vl3BrQPFHhnxZZDUvCviPS9ZtCBi40+7S4j5GR8yEjkVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVeKvit8LfAoc+N/iV4V8PCP7/APauswWm3r18xxjofyNeKf8AgoR+xf4PLLq37Qnhu4KHB/ssTan3xx9lSTPXt9a8Uf8ABYv9kTQgy6NF438RsB8h0/RkiUnHc3EsRA7dD9DXij/guH4agMkfgr9nzU73kiOXVNfS1wOeSkcMme3G7v145H/BU79uD4kAJ8IP2ZtPninwYmtPD+patJjgja0bqpyD12nhh061/wALA/4LV/FFRHovhXXNAhlBPlf2NpmkEcf37sK69D/EOv0pv2Jv+CrPxPynjv8AaCuNIgkz5lvqHjm78vnIP7q0V0zgkdOhxnFaX/wRR+IniK7/ALS+JX7SdgLpzmZ7XR5tRd+ckeZNNEectyQeecHNeGP+CJvwFsRG/i/4q+OtXdMFlsRa2KORjgho5WwcN0YHkcjHPhj/AIJQ/sUeHjG998O9V1+SMDD6pr11yRjkrC8anoeCMfMeOmPDX7Fv7JnhLYdF/Z28B748bJLzRYrx1Ixghpw7A8DnOfzNaD4R8J+FYfs/hfwxpOjxbdvl2FlHbrjjjCAccD8hVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVXiXwr4Y8aaNP4d8YeHdM1zS7kYmsdRtEuYZP8AeRwVP4ivjl/wSH/Zt+Ja3OqfDWXUfhvrMoJUWBN3YM/PLW0jZUdOI5EUelSfCD/gpr+wBM958NtYvfGvgezYsbbTC+rWHlgZ+eycebb8cs0QAGPvkCvgD/wWS+F/ix7fw9+0H4VufA+q5CSarp6vd6ez8AlkwZocnPGJAMcsK8HeN/BvxE0C38U+A/FWk+IdHus+TfaZdpcwsR1G9CRkdx1HfFVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVXxa/5Kp4z/wCxgv8A/wBKHrw5cwWXiHS7y6kCQwXsMkjkfdVXBJ/IV+2b/wAFMPir8ePEeo+FPhP4i1Pwd8OYWaGCGykNveaqg4MtxIvzqrc4hUhQpw24807vI5kkcszElmY5JPc14L8eeNvhxrsHibwB4t1fw7q1ucx3mmXj20o9tyEEg9weD0Nf8E7/APgpNdfHPUrX4I/He6tYfHDxn+xtajjWCPWtoy0UiLhUuMAkbQFcAgBWAD1VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVatrOj6DYvqeu6tZ6dZxffuLudYY1+rMQBXjf9vP9jz4fGSPxB+0D4Vmkhba8Wk3Dao6t6FbVZCDxg+nfFeK/+CyP7JuhFotA03xz4lf+B7PSo4Izz3M8qMOOfunqPfHif/guDC5Nr4F/Z3llkfiKfU/EOPmI4/dRwHPOP4x+vB/4KK/8FH/ikNnwr/ZtSC2k/wBXcab4O1C+ZRwctK7mPGCOSoHze4r+y/8Agtf8WgPtN9rfh6zk43efpWi+UOR0j2zevYnp7Uf+CZn/AAUB+JmJvix+0taGGYfvItQ8Valqcq52nGwpsxwOj4+UdsGvC/8AwQ70SJUl8a/tC3tyT96DS/Dywbee0kkz54x/AMe9eFf+COP7JGhBG1y88ceJHHLi+1dIUJ46C3ijIHH94nnrXhf/AIJ3fsW+EQg0z4AaBdMn8WqSz6jk88kXEjjueMY6egx4X+D/AMJPA7I/gr4W+EdAaIjYdL0O3tCuORjy0GOQKqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv2gP2Gf2bf2kY7i78deAoLLXpuR4g0bFnfBv7zOoKy9TxKrj6da8bf8E+f2z/2Odbn+I/7JHxJ1TxNpsTeZNa6UfIvii5ws1kxaK8UA4wu8k5PlivgH/wWCt7TUl8Aftb+BLvwxrFpJ9nuNb06zkCRuOD9ps2/exEY5Kb+TwiivAvxC8C/FDw5b+Lvh34t0rxFo90P3V5p10s6Z7qSp+VhnlTgg8ECqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqvi1/yVTxn/2MF/8A+lD1X7F3/BK7Xv2g/CFj8V/i54nvfCXhLVB5mlWVjCrX+ow/89g0gKQRt/ASrlhztClWb4v/APBE/wAEjwzcXnwI+KPiGPXbaFnisfE5guIbxwDhPNgjiMOeBuKuPUdx4q8LeIfBHiXU/B3izSZ9M1rRbuS0v7OdcPBNGxVlOOOCOo4PUEg1ous6t4c1ix8QaBqM9hqemXMdzZ3Vu5SSCaNgyOrDkMGAIPqK/Zr+Ltv8evgP4I+LkKRpL4i0pJbyOM5WK6QmK4RfZZo5FHsOcVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVXj74xfCb4V2xu/iT8SvDPhlNu5RquqRWzuP9lGYMx9gCa+I3/BW/9j/wQssHh7Xtf8bXcZKiPRNKdI93vJcmJSvuu72zXib/AILLfFzxxqDaB8Af2dbZryQbYTfzT6tO+eNwgt1j2nnGNzDOPpRm/wCCzX7RGQkfiHwRpVwc42W3hv7OD7nbd9/9oitH/wCCOvx8+Il5DrHx7/aRsftROXaBbrW5wDglfMuGhwx6EjcAefmrwV/wRl/Zf0ARzeLvEnjbxTOpG+OW+is4Gx6JFGJBn/rofwrwl/wT5/Yy8FrENJ/Z+8NXZiOQ2rrLqZY5zz9peQHnt07dOK8MfDn4e+CEEfgvwJ4d0BVGAul6XDagDntGo9T+Zqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/aD/ZG+Av7TmlGz+KXgqCbUo022uuWOLbULXjA2TAEsoz9yQOmeducV49/YL/bH/Yk8S3PxS/ZH8e6v4n0WIb7iDTY9t95anPl3Fid0d4g4+4GJOT5aYzX7Mn/BXvwR4vuIPBH7TGix+CPECP5B1q1Rzp8sgOCJYzmS2bPBzvTOSSg4rS9V0vXdNttY0TUrXULC8iEttdWsyyxTIRkMjqSGUjoQcVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVXxa/5Kp4z/7GC/8A/Sh60Wxj1TWbDTZXZUu7qOFmXqAzAEj35rRtH03w/o9joGjWkdrYaZbR21pbxrhYoo1CoqjsAoA/Cq/4LK/DvTvCv7TeleNdMtli/wCEz8OQz3hAA8y6gd4Wbjn/AFSwDkduvpX/AARw8QT61+yJcabLI7LoPi+/sogwwFVore4wOembhj25J+tVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVeL/HXgj4f6Wdb8eeMdE8Oaeuc3erahFaRcdfnkYD0r4nf8FWf2O/h351tpfjLU/Gt9CSpt/DmnNKuf+u0xjiYe6u3414r/wCCxnxk+IWqN4Y/Zx/Z5ia+lJWBr0z6xdSr/eW3t1j2t043SD69KPw9/wCCwn7URA8S+Itb8DaLcjG27vo/DsSKf4HhtgLlh/vo31rwD/wRN+23I1b43fHq7vLiZt1zbaBY/Ozdz9quCxJPvDXw6/4JkfsZ/DpYZU+FEfiW9iIzdeI7uS+34/vQkiH/AMh14a8I+E/BmnLpHg7wxpOhWKABbXTbKO1iGOmEjAH6VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV+0/+wP8A/2o7a41PxFoQ0Dxc0ZEPiXSY1juC3bz0+7cL04f5scKy1feGf27f+CWOtvqnh++Pi74Wvc7pWRHudKmBIyZYc77GY8DeCATgb5AMV+yh/wUG+B/7U9vb6HZX6+FfHBT974b1O4XfK3c2suAtwMAnAAcAElAOaqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq+LX/ACVTxn/2MF//AOlD14cuYLLxDpd5dSBIYL2GSRyPuqrgk/kKR0kRZI3VlYZVgcgj1FV/wWp8X6brH7QnhLwhZTJLP4e8Lq93t/5ZyXE8jBD77ERunRxz6V/wR08OTaJ+yC+pyxlV8QeLL++iJJO5Vjgt8+3Nuw/Cqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqp3SNDJI6qqglmY4AA6mvip+3L+yj8HDLb+MvjVoEl/DkNp+lSnUrkMP4Wjtw5jP+/tr4m/8FsvB9tLJpnwV+DOr63M52Q3uv3a2a7vUQQ+Yzg9hvQ89ulD4tf8ABXf9q9f+KI8L6x4J0K75WXTtPTQINvHzJdXTeewH/TOQ9+OOPCP/AARx+L/xA1L/AISb9o/9oWNr6Y7pksBPq9zKDyQ1zcMm1uf7rjP518MP+CV37HXw3MNzfeBr7xnfw8i58S37XC575gjEcJHsyGvC3gvwd4F0tNE8EeE9G8P6dGAEs9KsI7SJccDCRqAPyqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqvLO01C0msL62iuba5jaKaGZA6SIwwysp4IIJBB4Oa/ax/4JL+E/GVxcfEj9l67h8F+KopPtJ0IyGKwuZQdwaBxzaSZ6AZjyFAEfLV+z7/wUo+NH7Nvi1fgT+294Z1u4h091h/tm5gP9p6enQPKOl5DxkSKS5GSGl4FeCfHPg74leF7Hxp4B8S6fr2h6lHvtb6xmEkcg6EZHRgQQVOCCCCARVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVfFr/kqnjP8A7GC//wDSh6r9iX/gq38PNH+H2i/Cn9pS7vtJ1Dw/ax2Vl4mS3ku4byBBtjE6xgyJIqhVLgMGxuJU5z8YP+Csv7KHgHwzc3fgHxNdePtfMTfY9N06xnt49+PlMs8yKqJnrt3sP7tfFz4p+L/jb8SNf+Kfju9S51vxDdm4uDGpVIhgKkUYJJCIiqigkkKoySea0PRNW8S61p/hzQbCW+1PVbqO0srWFcvPNIwREUdyzEAe5r9nH4R23wH+BXgr4RwSRySeHNKjhupI/uy3TEyXDr7NM8jD2NVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVXizxr4N8BaRJr/jjxZo/h7TIvv3mq30dpCv1eQgfrXxX/4Kx/sjfDkTWfh7xFqvjvUYwQIdAsj5IbsDPMY0I90L/wBK13/gqx+1l8c9Tm8L/svfARbSSRtqSW9jPr17EOznaqxRjGCd8bAeuOaT9ib/AIKY/tVEXP7QPxUn8O6PcEO9lres7kIP8SWFnmFWwej+We3FfCz/AIIwfADwuIbv4p+NPEnji7TG+3hI0u0b1BSMtL+IlH0r4Yfs3fAX4LxRp8L/AIS+GdAmjGBeW9grXTD/AGrh90rfixqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/aG/Zh+Dv7T3hM+Ffip4ZS6eFW/s/VbbEV7pzn+KGXBx2JRgyNgblOBXij4b/tf/8ABKTxvN44+HurP4q+F+oXQFzKYmexuhnCpewA5tpsYCyqcE4AY5ZK/ZO/bZ+D/wC1r4f8zwnfHSPFdnCH1Pw1fSr9pt8YBkjI4mhyfvr0yNwQkCqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqviDfPqnjzxJqUkao13q93KyA8KWlYkD86/Yy/Zug/aq+KGr/CttWGl3X/CM3t9YXjZKw3MRjERcDJZCzhWHUKxI5Ar4sfCP4hfBDxtf/D34m+GbrRda09yGimQ7J0yQJYn6SRtg4dcg/nVaVpGq69qVto2haZd6jqF5IIra0tIGmlmc9FRFBLE+gFf8E3P+Ccmq/CPU7T4+/HrS4o/FaRE+H9AkAc6TuGDcT9vtBUkKgyIwST85Ajqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq+J/x++CnwXtmufin8UfDnhtgm9be9vkW4kHqkAJkfp/Cpr4s/8ABZ74C+FjLY/CbwZ4h8c3SZCXVxjSrNvQhpA0x+hiX6+n/DWP/BUb9rdRB8EPh5eeF9Bu8eXe6JpQtYXT31C8JXPIJMbIfbGc+E/+CQnx7+KurR+Lv2nv2gFW8m+aZIZp9avmHdGnnZVQ9eR5g+uePhT/AMEuv2PPhcIbq5+H8/jTUYSCLvxRdG8BPobdQkBH+9GfrWheH9A8L6XDonhnRNP0nTrZdsNnY2yW8UY9FRAABwOgqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqrUtM03WdOudI1ewtr6xvYWhubW5iWWKaNhhkdGBDKQSCCMEGv2tv8AgmX4u+GHiL/hoL9iS81PTr7TJjeTeGrCdkubNxktJYODuZcdYDk9QpYERr+w5/wU50L4vz2Pwf8A2g5bbw38QlkFpaalIvkWusyg7QjA4EFyTxsOFdvu7SQlVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVU/wCxd+yXdTyXNx+zr4Bkllcu7tocJLMTkk8dSa+Hn7PHwL+EmtTeI/hj8JvC/hfVLm1a0mvNL02O3leBmV2jLKASpaNDj1UelfEn4Q/C74xaMugfFLwDofiixjJMMepWSTGBj1aNiN0bdsqQferr/gld+w5c6h9uHwhuYkLbmtovEWoLE3rx5+4A+gIx2xXwn/Zt+A3wMQ/8Kn+FXh/w7cMmx7y3td906/3WuH3SsPYsRVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVfFz9rj9m34GGa3+Jnxg8P6ZfwA79MhnN3eAjsbeEPIvPGSoHXng4+K/wDwWt8HWjvpXwK+Eeqa7dMxjiv/ABBOLSLd0BWCIu8gJ7F4zT6z/wAFdP2xAy6fZ634C8NXq/8ALGMeGrVUbPKu5+1SoRxgNIMfXn4Y/wDBFW1u7r+3P2g/jbfapdztvubPw5DtLP3Ju7kMzg/9clPvzx8Jv2G/2VPgt5Nx4K+DOhPqMOCuparEdSug4/jWS4LmMn/pntHtVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX7dH/BNjwd+0hb33xI+F6WXhr4lqpklYrstNcIH3ZwPuS8DEwHPRwQQyfsd/8ABRDx18CvFg/Zi/bTh1LT/wCyZVsrTXtUDfadKbjZFeHkywEFSs+SVBBJZCGS0u7TULSG+sLqK5trmNZIZoXDpIjDKsrDgggggjg5qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr4w/tqfsw/Aoz2vxA+L2ix6nBkNpOnyG/vA391oYAzRk+r7R718T/wDgtLcanf8A/COfs5fBK61K8uH8u0vPETlnlc8ALZ2xLNnt+9B9qb4c/wDBWr9seRX8XazrHgPwze9Yb65/4R61RD/C1tCPtMo6n94j9uelfCT/AIIrfC7RHg1L40/E3WfFVyAGk0/SYRp1tu7q0hLyyL7gxn6dK+E/7Mn7P/wNhSP4VfCbw9oM6KF+2x2vnXbAdmuZS0zD6uevvVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVftlfsRfDj9rvwpjUFi0TxtpkDLo3iKKHc0fOfJnUY82EnPB5QklcZYN+zH+1x8YP8Agn58R5P2Yv2rtK1NvBVtLss5vLMzaUjOdtzavgGezc5JUZK4JUBg8baHrmi+J9FsfEXhzVbXU9L1K3S4s7y1lEkU8TgFXRhwVIIIIqqqqqqqqqqqqqqqqqqqqqqqqq6urWyga6vLmKCGMfPJK4RV+pPAr/hLfCn/AEM+k/8AgbH/AI0jpIiyRurKwyrA5BHqKqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqvjD+2p+zD8CjPa/ED4vaLHqcGQ2k6fIb+8Df3WhgDNGT6vtHvXxQ/wCCz2oa3e/8Iv8As0fBO81HULpvLtL3xBukkkb0SytiSxxyP3v1XtUXwX/4Kvftkrv+I/inVvBPhi9Hz2+rXX9iW5jPVTZWy+c+B0Eqcnq3Uj4Pf8EX/gj4XEGofGPxtrfji8UAvZWY/suyPqp2M0zY6bhImeTgdvhd8BPgv8E7EWHwp+GXh/w0CgR5rKyUXEwH/PSY5kk+rsaqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv2tP2Svh1+1t8PH8J+LYlsNbsA8mha9DCGn02YjnjI3xNgB4yQGABBVgrD9nX9or4v/wDBNT4v3H7NX7Slnd3Pw9ubjzLa5iDTJYpIxxfWRxmS3c5MkQGQdxAEgdH0PXNF8T6LY+IvDmq2up6XqVulxZ3lrKJIp4nAKujDgqQQQRVVVVVVVVVVVVVVVVVVVU7pGhkkdVVQSzMcAAdTXxV/b2/ZK+DrTWnin4z6LeajCSradorNqc4ccFGEAZY29pCtfEj/AILceAbAyWvwl+C+t6y2SqXevX8dggI/iEUQmLD2LIecnHSh+3B/wVI+N53fCP4HzaJZTj/R7zTPB80iAHoTcXpeEn3wB7UvwH/4LL/FIrN4m+J+t+Fll+YE+LrfTcAgHldOJK9BwQCOcgZNJ/wSh/bQ8VhZPiF+05pEjE7iG1vUtQIIGF/1iJzgke3YnNWn/BDzxVcSRPqv7R2mRGTmdovDUkxVj1xuuF3c9zjPtX/DjD/q6P8A8sj/AO7qf/giL4808/a9D/aN0kXcR/csdBmg9j8yzMV4z0B9Pem/4Jbft1eDl834d/tQacnkZ8pYfEmqae+AcjaEjYA7u2QM85pvgr/wWd+GJ8zw58Rdb8TJbnIceKbTUsgZ52353N0HGCTuHHXB/ba/4KpfBwFvid8Cp9btYVAmu9T8FzrHx1InsjHEGI+o9q8Ef8Fu9DM62XxT+AmpWBjbbNcaHqy3DZBwcQTJHgjngyH8K+HH/BTr9jT4jmK2HxTHhi9kPFt4ks5LHb9ZsNCP+/leHfFHhnxhpcet+EvEel63p0w/d3enXaXMT/R0JB/Oqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr4x/ta/s5/AOOWP4ofFjRNMv4gf+JXDKbu9JxwPs8IaQZ6ZZQvqRXxa/wCC0trcXh8Pfs4/B671e7mby7bUPETFQ8nQbLO3JdwT0zKp/wBnniT4V/8ABV39tdVl8c61qXgfwpeqf9G1K4/sG08s9Va0hBuJRjkGVGHP3q+DX/BF/wCDPhcw6j8afHGr+N7pQC2n2KnTLPtlWKs0z+m4PH16V8Mvgd8Hvgxp/wDZvwr+G3h/wzGy7ZHsLFI5Zh/00lxvkPA5ZieBVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX7Wv7KHgD9rb4bP4M8Vg2Gr2BebQdbij3S6dcEYzjI3xNgB4yeQAQQwVh+yZ+018Sf+Ce/xivv2VP2mYJ4/Bj3wEN2zvImkmQ/LeWxI+ezl+8ygAg5YAMHRrG+sdUsbfUtMvILu0u4llt7iCQSRyxsAVdWGQykEEEcEGqqqqqqqqqqqqqqqqqr4u/Hn4O/AfQz4h+LfxC0jw3alS0MdzNunuMdRDAmZJT7Ipr4w/8ABZS91jUz4N/ZY+El1rGoXT+Ta6lrkLyPM2P+WNlAd7HupaQHjlO1W/7K/wDwU2/bSkXUfjp451Hwh4ZvTuay1y6NlGE7BdMtwMMOB++VG7lj3+FH/BGn9nXwgsF58UPEviLx9eoQZIfM/suzf/tnCTKP+/1fDj9nP4DfCFIh8NfhF4V0CaH7t3a6ZH9p/GdgZG6d2NVVVVVVePPgx8IvijG0fxH+GHhbxMWXbv1TSYbl1/3XdSyngcgiviT/AMEk/wBj/wAd+bceH9A13wTeSZIk0PU2aPd6mK4Eqgf7KbfwrxJ/wSP/AGkvhBqknin9mP8AaAjmuo+VX7RPoV6QOQiyRM6Oc/3mQUP2yv8Agpn+yM8dl+0B8N7jxJoVs+x7zW9MDpj+6mo2Z8st7uZD7dMfBj/gsF+zZ8QRbab8SrHWPh1qsvDteIb6xDdgs8Q3jnu8SAcc9ceEPG/gz4haJD4k8B+LNI8Q6VP/AKu90y9juYmPpuQkZ9R1FVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVXxV+P3wV+B+nnUfiv8TdB8Nrs3RwXd0DcTD/pnAuZZP+AKa+Mv/BaT4b6M82j/AAH+HGqeK7wMY4tS1hvsNqTnhkiXdLIDxw3lHmgf+Csf7b4yran4D8H6iMcZ8O2AjYfjd3EbD/rqDXwd/wCCLHw60gx6r8d/iXqvim8Lb307RU+w22e6vK26WQdeV8o8j05+FH7O/wADvgZZLZfCf4X6B4cKpsa6trUNdSr6SXD7pZP+BMaqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr9tn9jfwj+138N20qb7PpvjTRI5JPDmtMpHkyHBME2AS0D4AIwSpwwzghv+Cen7YHi/4BfECT9iv9p03OlLZX32DQrrUWJfSrokbbSRySDbyZUxOPlUuMEo4KVVVVVVVVVVVVVVVVfFX4xfDD4IeFpfGfxW8a6b4c0mLIWW7k+edv7kUa5eV8c7UVjjnGK+Mn/BVf4yfGrxIfhX+xT8ONVjnvSY4dUl0/7bqc46F4bdd0cCD+++8gcnYenwh/4JK/FP4r6+Pib+2X8VNRN5f4ludMs743uoyn+7Pdybkjx02oJODgMuK+DX7NvwO/Z+0saX8JPhxpGhMyBJr1IvNvLgf9NLh90rjPOC2Bk4AqqqqqqqqqqqWKOaNoZY1dHUqyMMhgeoI9K+NH/BOb9kz43LcXeq/DS38NaxOD/wATXwwRp0wbuxjUGF2PcvGx968Zf8Et/wBqv9njW5vHX7I3xjudVaIbhbQXh0fUWAziMjd5E64JzuZQc42c18Pv+Cq/7RnwH16H4e/th/B+9vpYAokvBYnStUCZwZTEwEM44ONoiB/vGvgN+2N+zt+0hbRJ8MviJZS6sybpND1A/ZNQix1/cvy4GRloy6/7VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVeMvHfgn4d6LJ4j8feL9G8OaXFw15qt9HaxA+m5yAT7dTXxq/4LDfs5eABPpvwt0vV/iNqkYISW3U6fYBh1BmlXzD9UiZTj73epP2jP+CoX7bTGy+DnhPUPBvha7+UXmiW50yAxnPzHUbht7MB18l1J4+XJFfCv/gjFf6zqI8VftNfGi71O+uX8y7sNALSPK3/AE0vbgFm9CBFn0bvXwa/ZE/Zx+ASQTfDH4UaLYalAuF1a4h+13xOOSLiXdIuc8hSB7VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVf8FJf2GLb9pLwY/wATvhzpaj4l+GbQiKOMbTrdouWNs3rKuWMbdySh4YFf+CYP7b9z8XdDX9nj4u6pIPiB4agK6XdXjES6xaRD5kfdybiED5s8sg3clXNVVVVVVVVVVVVVVV+2F/wVU8AfBq4vPh38CobLxz41jZoJ70SF9N0yXpgsnNxIDxsQhQer5BWvhN/wT9/ag/bS8VRfGr9sbxxrmg6VeAPBbXSr/aVxCeRHBbkeXYxckgMgIPPlndur4Lfs+/B/9nrw0PC3wj8D6foVs6r9puI033N4y9GmmbLyHk43HAyQABxVVVVVVVVVVVVVV8Qvhj8O/iz4fk8LfEvwVo/iXSpMn7NqVosyoxGNyEjKN6MpBHY18e/+CNXhHVLiTxT+zN44n8JanE4lg0bWJpJ7VXXkeVcrmaHBAILCU57jtoP7Y37fn7CWrQeD/wBpXwRqPjDw0soht7jW5C7Sgf8APtqce8SErztk8wjjheRX7OP7fH7OX7TCW2l+FPFy6J4omADeHNcK212z9xCclJ+/+rYtgZKrVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVXNxb2dvLd3dxHDBAheWWRgqooGSxJ4AAGSTXxu/4KafsnfBb7Rp0fjg+Ndcg3L/ZvhdReAOMjD3GRAuCMEbyw/umvEX/BSj9tX9p/Vrjwh+yb8HrjQ7ct5bXOnWR1a8hB4BluJEFvACO5QbSeH714N/4JR/tLfHLW4/G/7W/xtms55fme1+2vrWoKDyY/MZvJhGcY2NIo6YHFfBX/AIJ4fsofA37Pe6H8NLbxBrNuQV1bxIRqE4YYwyowEUZ4zlEU89aREjRY41VVUYVQMAAegqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/4KbfsoeIfhT4ytf22/2f2uNKvLK/jufEcdgu1rK6DDZqCAcbXbCyjpuIY5Dvj9i79qnw7+1n8G7LxpaNBa+JdL2WfiXTEbm1uwv31Xr5UoBdD/vLklGqqqqqqqqqqqqqvFnizwz4E8N6h4v8Za7ZaNoukwGe9vryURRQIO7MffAA6kkAZJr9ob9tv46/t0ePH/Zu/Y90PV7Twzelo7q8gzBdanADh5biTgWtpyAVJBbIDH5xGP2NP+CbHwu/Zpt7Lxp40jtPGXxFCh/7Rnh3Wulv3FpGw4YdPOYbzjjYCQaqqqqqqqqqqqqqqqvEPhzw/wCLdGuvDninQ9P1jSr6Mx3Njf2yXEMy+jo4KsPYiv2j/wDgj58NvGD3Pi39nHXn8Ca+pM0ekXTvNp0sg5Gx+ZbY55yN6jgBFrwh+2f+2/8AsGeJLX4c/tSeDdT8W+GshLWTVZ985jBxutNQXcswA52SFyOBmOv2dv2wPgP+1BpYufhj4wjOqxR77rQdQxb6hbDuWiyd6jj54yycjnNVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVfGb9qT4Afs/WsknxW+KGjaPdpHvTTBL599KMZG22j3SEHj5ioXkZIr4s/8FltW17Uz4P/AGXPg5d6nf3TmK01DXY2mlmb/plZW5LMcAkEyemU6ioP2Sv+ClX7a9xHqnx/8b3vhLw1O4kFnr1wbZFGTjy9MtwAHHT98I2x/Ea+Cf8AwSQ/Zg+GPkal47g1H4kaxFgl9XbyLJW9VtYjgjH8Mryj6VoHhzw94T0i30DwroOnaNplou23stPtUt4Yh6LGgCqPoKqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqtY0jS/EGk3mg63p9vf6dqNu9vd2txGHjnidSroyngqQSCD614p0vxn/AMEn/wBsy18RaBFe6h8LfFu8xwF2YXmmM4823JJwbm2YqVYnJBQkgSsK8LeJ9A8beGtL8X+FdUg1LR9ZtI7uxu4Gyk0MihlYfUEfSqqqqqqqqqqqr4j/ABH8FfCTwVqvxC+IXiC20bQdGgMt1dTngDoFUDl3YkKqAEsSAASa8a+Pv2iv+Cs/xmX4ffD21uvDHwo0G7EjtKjG3sYjkC6vCpxNcsFby4QcLyAQBJKf2bv2Y/hX+y34Ci8EfDXR1WWUK2qatcIGu9UmA/1krgdBk7UGFQE4HJJqqqqqqqqqqqqqqqqqq8Y+CvCHxD8O3fhHx14a03X9Gvk23FjqFss8UnoSrAjI6g9QeQQa/aL/AOCRF5o+pn4mfsceL7vQ9YsJftNv4fvL94WikXJBsr3IaNgQAqynqc+aoGK+CH/BUP4z/APxOPg1+234H1i4k09hE+rtZ/Z9TtV6K0sWAlzGQARIuGIy2ZCRXw1+Kfw6+MfhW28a/DDxjpviPRbofJdWUu7Y2ASkinDRuARlHCsO4FVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV4j8T+GvB2jz+IfF3iLTNE0u1GZ77UrtLaCIerSOQq/ia+OX/BXn9m74ai40n4aQ6j8SdZiBCtYD7JYK47NcyDLduY43U+tN8d/wDgp7+3XIbT4S+H9Q8FeDrxiovNHjbSrQJwCW1CU+ZIQDysLc5+50A+DP8AwRe8PQ3EfiL9o/4o3viHUJW8y40vQS0MLSHrvupQZZQSeyRn3r4T/s//AAW+Bem/2Z8JfhrofhtCmySe1ts3EyjtJO+6WToPvsaqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/a//Zp8PftU/BPV/hxqYht9YiBvPD+oOObK+RT5bE9djAmNx/dckchSP+CU/wC0h4g+H/i/WP2JPjIJdN1HTry5Ph6K8O17W6jZjdWB+pDyp7iQZO5QKqqqqqqqqqq8beNPC3w58Jat468ba1b6ToWh2r3V9eTnCxRqOTgckngBQCWJAAJIFeMPF3xs/wCCtn7QyeAvBUl34c+E/hiczq00Z8qwt8lftdwFOJLuUbhHFnCjIBAEkh+CPwR+Hf7PXw7034ZfDLRVsNKsF3SSPhpryYgb55nwN8jYGT0AAAAUACqqqqqqqqqqqqqqqqqqqr45fs6fB39o7wufCvxa8GWmrwxg/ZLsDyruxY/xQzL86HpkZ2tgBgRxXxO/Yg/a2/YP8U3Hxh/ZL8a6z4j8Nw/NdRWUW+8jhGSUvLMAx3UQH8aqcctsjwDX7Jf/AAVb+Fvxma08E/G1LH4f+MnIjjuZJiul37/7EjnNuxPGyUkdMOxO0AqyhlYEEcEVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV8cf2xv2cf2eIpofiX8TdNg1WFeNFsW+2X7HGQDBHlkz2aTavvXxJ/4K6fGL4ra23gL9kP4MXovLoMtveXlm2qajIOBvjtIcxxkEj7xlHPIFeG/+CcX7a/7VWs23jT9rT4tXOgWjN5i22o3X9o3kSk5IitYmEFuCO25SuRlOor4Gf8ABNz9lT4GC31C18CJ4u12EAnVfE+2+YMO6QkCGPnOCE3Dj5jjNIiRoscaqqqMKoGAAPQVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV/wAFZv2cdX8F+ItD/bU+Eol07VdMvbaPxDPaLh7e4jZRZ344xkMqRMT38ng5Y1+yP+0Ro37UPwL8P/FHTzDFqUkf2TXbKM8WeoRACZAMkhTlZEyc7JEzzmqqqqqqqqqqaaG3heeeVI44lLO7ttCgDJJJ6DFftWfHb4if8FHP2gNM/Zd/Z1lkl8CaXeFpLxdwgvWjOJdRuGHS2jBxGv8AEWB5aRFX9nL9nnwB+zJ8L9O+GPgC0Pk2483UL+VcTaldFQJJ5OvLYACjhVAUcCqqqqqqqqqqqqqqqqqqqqqqv2t/+CaPwX/aTF54t8MwxeB/HsoZzqtjbj7Pfyc/8fUAwGJJ5lXD9yXAC14C/aP/AGx/+CZ/imz+Ffx88LXfijwAXMdhFNdGWLyhgF9OvMHAAwfIcYGcFYy26vgF+0p8Hv2mPCa+LPhP4rh1BYwBe6dPiK909z/BPCSSvQgMMo2DtZhzVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVrmu6H4Z0m513xJrNjpWm2UZkuby+uFghhQdWd3IVR7k18d/+CuX7N/wvNzo/w3W9+JOtw5Vf7Nb7Pp6tj+K6cHcOnMSSA88in+L/APwU7/b6b7L8NtHvfBXgi9fb9q0oNpFjs6EteyEzT8feWJmBz9zoB8D/APgjD8O9DeDXf2hfHt94w1At5k2k6Qz2VnuP3leY/v5RnJ3L5J9vX4b/AAj+GHwe0QeHfhd4D0TwxYHG+LTrNYTMRwGkYDdI3+05J96qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq8ceC/DfxG8Haz4C8X6bHf6Lr9jLZX1s/G+KRSrYPVWGchhyCARgiv2NPF3iT9gj9t/xD+y78RtRI8L+LL6Owju5spG0rZbT7xc8KJVcRvjgGTBP7viqqqqqqqqq/wCCp/7XusRyxfsc/BSa4vfE3iUxQeJG08F5lSbb5WnxheTJNuUuB/Ayrz5jAfsFfsdaR+yX8Kkt9Vht7nx74kjjn8R3yYcRsBlLWJh/yzjyRkffYs3QqFqqqqqqqqqqqqqqqqqqqqqqqviD8OfAvxW8KXngj4jeFtP8QaHfrieyvYd6k44ZT1Rxnh1IZTyCDXx9/wCCb/xx/Zc8WH47/sS+KtdvLbTi0p0u2kzqlgh5ZEGNt7DwAYyCxGAVk5av2Pf+CrHgf4rS2nw5/aFWx8E+MhiGHVWbytN1GQcYYsf9FlP91zsJBwykqlAqyhlYEEcEVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVXxg/aF+CnwD0n+1/i38R9H8PIyF4baebfdXAHXyrdN0sn/AVOM84r4w/wDBYzX/ABPqh8D/ALJ/wkvdR1C7fybTU9Yt2uJ5nJx+4sYSSTjlSznrynGK0X9hH9vj9szVbfxb+1J8Sb3wtozyGWG21mTzpoQ3UwabCVig9CHMTcDg8V8B/wDgmp+yv8DBb6kPBa+NNfhw39q+JlW82uO8cGBCmDyDsLDj5jjNIiRoscaqqqMKoGAAPQVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV/wWH/AGcH8a/DLS/2h/C9kTrfgQi21Zol+eXTZH+V+OT5MzA+yyyE/dr9gn9os/tL/s3+H/F+qXgm8TaMP7I8Q5bLNdwqv74/9dY2jlPbc7Afdqqqqqqqqr9tP9qDRv2UvglqXjuVobjxFfk2Phuwk5+0XrKcMw/55xgF36ZAC5BYV/wSr/ZW1fxTrN9+2p8aUuNR1rWbq4k8Nfbly8ssjN9o1FgerMxdI+g++2OUIqqqqqqqqqqqqqqqqqqqqqqqqqv2xv8Agm98Jv2nYLzxf4ZitvBvxFZWcavbQYg1J8cLeRr97PTzV+cZ53gBa+E/7Wn7Uv8AwTm8a2/wP/ac8L6n4h8Ex5SxEkplkhgUgebp1y3yzQgEfuWI28LmI5FfCH4z/DL48+DLbx78KvFlnrukXHys8JKyW0mATFNGcNFIARlWAOCDyCCaqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/aE/bk/Zw/ZqjuLPx746ivfEEC8eHdGAu79j2DICFhyOcysg9+leL/APgoh+2j+1zr1x4B/ZF+GOoeHrAnZJcabELu9VSThpruQCG1BA4wFIOf3h4r4O/8EevFXjDVh49/a4+LN9f6jeOJbrTNKu2urmZv+m99NnnjBCK3s4r4P/s7/BP4BaX/AGV8I/hxo/h4OgSa5hh33VwB/wA9bhy0snTozECqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqvE3hzRvGHhzVPCfiKxjvdK1qymsr62kHyzQyoUdT7FWI/Gv2Hdf1r9ib9vXxT+yv4yv5ToXiu8/su2nmO1ZJxmXTbnHTMsb+XgfxTrk/LVVVVVVVVdXVrY2st7e3MVvb28bSSyyuESNFGWZieAAASSeleKL7Xf+CqH7ddt4c0q4vYvhZ4RLKkqZUQaVE486ccYWa6k2qpIyA0YIIjNaHomkeGtFsPDugafBYaZpdtHa2drAm1IIY1CoijsAoAA9qqqqqqqqqqqqqqqqqqqqqqqqqqq+Lvwa+Gnx28GXXgL4p+FLPXNIuQSqTLiS3kxgSxSD5o5BnhlIPboSK+Lv7Jn7UH/BOPxtc/HT9mXxTqWveB4juvWWLzZYLcHPk6jbLhZoQP8AlsoAHLYiODX7HP8AwUW+E/7UtvbeFNYkg8IfEMJh9Eupx5V+QOWs5D/rPXyziQc8Mqlqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr9or/gof+zV+zn9s0fV/Fq+J/FVrlT4f0FluZo5P7s0mfLgwcZDNvAOQprVf2ov+Cg//AAUD1S88J/ADwte+D/BruYrmXSJTbRxJ3W51N9pLbScxxbCw42NX7PX/AARx+G3hWS28T/tFeJ5vHGsZ8yTR7B5LbT0kPJ3ycTT885zGDyCpFeEfBfhD4f6Db+F/A3hjS9A0i0GIbHTbRLeJOAMhUAGTgZPU96qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/4LKfBa+07/hCP2pvBsclrqWiXKaVq1zbrh48MZbKfI6FJBIhY55eIcYFfsy/Gax/aD+A/g34tWbRLNrmmob+GPpBeRkx3MYHXAlRwM9Vwe9VVVVVVV/wVv/agb4WfCOH4F+EtRMfij4iQst8YmIe10kHbIfYzMDEPVRL3Ar/gnH+y2v7NHwBsn8Qad5HjbxoI9T18yJiS3yv7i0PAI8pGO4dpJJeSMVVVVVVVVVVVVVVVVVVVVVVVVVVVVMqspVgCCMEEV+2N/wAEqPDvj26uvip+zC1t4R8ZRyfapdCR/s9lfSg7t8DDH2SbPTGIyQOE5Y/swf8ABTj4gfBvxN/woT9t/SNXt59MmFqPEV3bOL6xIwALyPG6ePHImTLkYJEm7cNA1/QvFei2fiPwxrNlq2lajCJrS9sp1mhnjPRkdSQwPqKqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/aQ/wCCg37OP7Na3Oka74pHiPxVACo8PaEy3FxG/pM+fLg5xkO2/ByFatZ+P3/BQf8A4KM6pdeGPgv4evPB3gGWVoZ202ZrO1SPut1qDANM2Dho4sBh/wAsjX7N3/BIP4M/DX7J4j+OGof8LE1+PD/YNjQaXA/p5ed8+D3kIUjrHWj6No/h7TLbRPD+lWemafZpst7SzgWGKFf7qIoAUewFVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX7Qnwl0347fBPxl8JtSWMDxHpUsFtJIMiC5A328uP8AYmWN/wDgNf8ABGb4talo998Qf2YPFhe2vtKuG1jT7WY4aJ1YW97F17MICFHcyGqqqqqqvEfiDRvCXh/U/FPiLUIrHStHs5by+upThYIYkLu59gqk1+yz4d1r/god+3lr37QHjyymfwb4Ouo9RjtJQCkaRsV02yI5H8BlcYw3lyZ5eqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr9qH9jz4N/tX+Gv7L8f6P9k1y0iK6X4isUVLyyPJC7iP3kWTkxtwckja2GH7HX7OP7c37Jn7TEHwktNTh1T4QX3m32o6lKjTabNAuBviXIe2vWLIvl555Y+ake6qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/aO/bU/Z//AGX7GRPiH4wjudeMe638O6Xi5v5uMjMYIESns8pRT2JPFeJ/2uP26/8AgoHrt14A/Zs8I3/g/wAJq5jvH0m5MTIjf8/eott2fLk+XFsLAkbZOK/Zs/4JAfCnwCbTxV+0Dq5+IHiFSsraXFuh0uCTqQw4kucHu+xTyDGa0rSNK0HTbbRtC0y006ws4xFbWlpAsMUKDoqIoAUD0AxVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV+0REf2MP+Cpnh74vwObXwx40vodVvHAwqwXm621Dd2Yh/Nmx23J35qqqqqqr/AILD/tEn4f8Awc074FeH71V1n4guZNSCt88OmQMCRjqPNlCqD0KxyjvX/BP79nZf2b/2a/D3hvVNOFt4o15f7X8Q7h863MyjbC3p5UQjjI6blcj7xqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr42ftDfB39nfw0fE/xb8cWOiQOrfZbZmMl1eMP4YYFy8hzgEgYXILEDmvin/wUe/af/ay8Uy/CL9i/wAAa3olpdBoze2yLJqk0Z48x5s+VYp1+YNkHB80dK/Zx/4JAaTb36eP/wBrPxVJ4s1q5lM8ug2N3IYC55Jubo4kmYknKptGR95wa8LeE/DHgfQLPwr4N8P6fomj6fGI7WxsLZYIYVHZUUAD+tVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV/wWg+Eq+KPgT4b+LdjbBrzwPrP2e6cDGLO8ARiT3xPHbgA/32/H9h/wCK5+NP7Kvw78bXE/m340lNP1Elssbm1Jt5Gb3cxb/o4qqqqqpZI4Y2llkVERSzMxwFA6knsK+FkMv/AAUO/wCClt98QdQie98A+DLj7bBHKpMf9nWThLOLB4/fzlZWjPUSTdQKqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqvF3jHwn4B8PXfizxv4k03QtGsE33N9qFysEUQ92YgZPQDqTwK/aL/4K4ap4h1Y/C79jTwlea1q9/IbaDxBc2DzSSyHp9js8FnPo0o/7ZnrXwR/4Ja/GP47+JF+MH7bPjzWIJr9llk0gXn2jU7lQchZpjuS3TGR5aZYA4/dkV8LfhB8Mvgn4Wh8F/CrwXpnhzSYcEw2cOGmcDHmSyHLyvgAF3LMcdaqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/aS+GEfxn+Anj34YGASz6/oVzDZqe10q77dvwmSM/hX/AARM+Jkl54J+IfwZ1CYrJompQaxZxufm2XCGKYAdQFaCIkesvuaqqqqr/gpp8dD8Ef2VPEUWmX3ka942P9gaZtbDqJ1b7RIMcjbAsoDDo7J6iv8Agkf8CR8Lv2a1+IurWKxa58Sbr7eWZMOlhFlLVCfQ/vZh7TD0qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqpiqqWZgAByTxiv2rv+Cp3wX+BK3nhL4ZS23xD8axFo2is5/+JdYSDj9/cLnewPWOLJ4IZkNeEv2aP23/APgpF4hs/iN8ffFN54S8BmTzbL7dbtDEsZH/AC4afkZypA86QjcOd8hGD+zn+x/8Cv2XdHWy+GfhOM6tJEEvNf1DE+oXfHO6XA2Kf7kYVPbvVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX7NSj9nD/grf4w+GPNnpfiu81OxhjIwqQ3MY1C1XH/AIkB/2u2TVVVVVf8FFNa1X9q39unwJ+yp4TvpGsdAlg064aL5hDdXRWa8n9xFbLFkc4MTj1FaBoWk+F9C03wzoNlHZ6ZpFpFaWVtGMLDDEgREX2CqB+FVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX7Rn7WnwR/Zd0D+1fih4qjj1GeIvYaHZYmv77nH7uLIwucje5VBjG7OBXjH9or9tj/gpd4ju/hv8C/C914U8ACTyr1LW5eG3WMj/mIX2BvyDnyEABH8Dld1fso/8Eu/gp8ABZeLPH8Vv8QfG8JEi3d9bD7DYuOR9nt2yCynGJJMtkAqE6VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVf8FNUb4Ift5fCr4+2cbQ29xDpt/cyKv+tlsbvbKPfMHkqR70CrKGVgQRwRVVVV4+8Z6P8OfA3iDx/wCIJNmmeG9LuNRuyDg+XDG0jAe5C4A9SK/4JLeCta+M/wC0X8Sv2sPGsCTXNq84gkZcj+0dQkaSZoyckbIQ6Y/u3AFVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV4m8UeG/Beg3nijxdr2n6NpGnRGW7vr+4WCGBB3Z2IAH1r9pX/grFrvivWz8Iv2LPDl7rGq6hKbWHxG+ntNNPIeMWNoVLMe4eVfX930av2cv+CUnjH4i69/wuL9tnxRqV7qGpTC6m8PLftNd3TH/n9ugxKjGP3cRzgj50IK14P8F+Evh94bsvCHgbw5p+h6Lp0YjtbGxt1hiiA9FXuepJ5J5JJNVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVf8FufBX9o/CX4dfEJISzaH4guNNZh/Ct3B5nI9M2YGexPvX7MXjJviF+zn8M/Gcs5muNV8K6fJcuzbiZxAiy89/wB4HFVVVX/BXz4uf8IB+y3/AMIJYXgj1L4g6rFYbAcP9khInnYe25IYz7S471/wTV+ESfCL9kHwZb3Fr5Op+K428QagSm0s11gw5HXIt1gXnuD06Cqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr9rP/AIKFfBL9lm3udAmvF8WeO1T9z4b06cAwsRkG6mwVt1xg4IZzkEIQcjQvht+2/wD8FTfEkHivx7rMnhL4Yw3Be1llgeHTYF5/49LbIa7kGSvmsxA5UyLwtfs0/sc/A/8AZX0QWnw58NrPrc8QS+8Q6gFmvrv1G/AEaEgfu0CrwCQTzVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV/wVV8Jr4n/Yn8ZXaQrLP4fu9P1GEEDI23UcTkE9MRzSH6ZHev8AglV4qbxP+xR4NtJZRJNoF5qGnSHjPy3UkiA49I5UH4A981VVVf8ABRm8u/2mf2/Ph9+zPo9wxs9FNlplyUOfJmvHWe6lHstt5JI6gxn6CysrTTrODT7C3jgtrWJYoYkXCxoowqgdgAAKqqqqqqqqqqqqqqqqqqqqqqr9oH9o/wCE37Mngr/hOPiv4hNlbzO0VjZW8fnXeoSgZ8uGPI3HGMsxVFyNzLkVqH/Bcfw3HrBg0r9nLUrjSt2Bc3HidIZ9vr5K27rnGOPM/Gv2W/2yvg1+1rodzefDvULuz1nTEVtS0HU0WO7tQTgOApZZIyeA6k9gwUnFVX7Uv/BTHwL+yz8WLj4Ta/8ADLXdcu7exguzd2d5FHGRKCQMNzkYr/h958Kv+iH+K/8AwY2/+Ff8PvPhV/0Q/wAV/wDgxt/8K/Za/aL0T9qb4TW/xY8P+HL3Q7S4vp7QWl5KskgaIgFsrxg5qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr4ifEnwF8JfCl344+JPizTvD2h2IHnXt9LsXceiKOruccIoLE8AGvjz/AMFJPjn+094ub4FfsReFNcs4NR3Q/wBqW0WNUvU6NIhztsoRnmQkMBhi0fIr9kv/AIJLeD/A09r8Rf2m7m28aeKTJ56aErGXTrZyc5nLc3UmeobEeSQVfhqtbW1sbWGysreK3t7eNY4YYkCJGijAVVHAAAAAHSqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv2wfDo8Vfsq/FvRAm+STwdqUsK5xulit3kQdRjLIvJ4r/gih4ha9/Z78aeGJJCx0rxe1wgOflSe1gAA9t0LH6k+tVVVeXdtYWk19eTpDb20bSSyOcBFUZZifQAZr/gmxZXP7Rn7efxG/aV1iKSS30gX2pWpkHME99K0NvH7Bbbz1HpsFVVVVVVVVVVVVVVVVVVVVfH34oXPwV+Dfiz4r2egf25J4X09r3+zxMYvtCqw3LvCtt+Uk52nGOlfs1ft/fs7/tOLb6R4b8Snw/4rkUBvDmtstvcu3cQNkpcDrwhLYGSq1VV4ouNS/4KSf8ABRVfCN5qtz/wg2m309rbCF8C30ey3GR0/uvcMud3JDTr2UAaP8KPhloHgRPhfpHgLQbfwklv9nOjCwja2ePABDxkEOSAMlsknkknmv2h/Ddz/wAE5P2+NK8ZfDJJ7XwxcPDrVjYRudr6dO7R3diSeqgxzKoOSoMTdQGqwv7LVbC21PTrlLi1u4VmgmQ5WRGAKsD6EEGq8SfCP4UeMtTbWvF3wx8J65qLIqNd6lotvcylV6KXdC2B2GeK/wCGevgD/wBEN+H/AP4TNp/8br/gqNceCPE/xx8Ffso/AX4ceFdP1iG5gOpHR9It7WWfUbwhLa2Z40BAWNw5HT9+pPK8fAL4P6J8A/g54U+EegFJLfw5p6Qyzqm37TcHLzzEdi8rO/8AwKqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr9r7/gpB8Hv2Yo7zwlocsXjT4gxhkGi2VwBDp744N3MMiPHXylzIeMhAQ1eAP2aP2wv+Clni+z+LX7QPia98MeAj+8sZZrcxR+SedmnWZONrDA89+GGCWkIxXwH/Zw+D/7NnhNPCXwn8JW+mxuq/bL6QCW8v3A+/PMRuc9Tt4Vcnaqjiqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqrxjow8R+Edb8PbC/8Aaem3FrtBAz5kbLjJ4HXvX/BDfWDv+MXh+RiQRo1zEAowP+PxXyev/PPj2NVVV/wUC+JQ+Ff7IHxJ1+G4MV5qGktpFntOGMl4wtyVPYqkjvntsr/gjZ8Mv+ET/Zm1L4h3MOLnx1r00kMm3G61tf3CD3xMLn86qqqqubi3s7eW7u7iOGCBC8ssjBVRQMliTwAAMkmv+GhfgD/0XL4f/wDhTWn/AMcr/hoX4A/9Fy+H/wD4U1p/8cr/AIaF+AP/AEXL4f8A/hTWn/xyv+GhfgD/ANFy+H//AIU1p/8AHK/4aF+AP/Rcvh//AOFNaf8Axyv+GhfgD/0XL4f/APhTWn/xyv8AhoX4A/8ARcvh/wD+FNaf/HK/4aF+AP8A0XL4f/8AhTWn/wAcr/hoX4A/9Fy+H/8A4U1p/wDHK/4aF+AP/Rcvh/8A+FNaf/HK/wCGhfgD/wBFy+H/AP4U1p/8cr/hoX4A/wDRcvh//wCFNaf/AByv+GhfgD/0XL4f/wDhTWn/AMcr/hoX4A/9Fy+H/wD4U1p/8cr/AIaF+AP/AEXL4f8A/hTWn/xyv+GhfgD/ANFy+H//AIU1p/8AHK/4aF+AP/Rcvh//AOFNaf8Axyv+GhfgD/0XL4f/APhTWn/xyvD3iXw34v0qLXfCfiDTda02ZmWK80+7S5hcqcMA6EqSCCDzwaqqplVlKsAQRggiv2j/APgmJ+zr8eHufEPhzTT8PPF0hLrqmhQKkEsnXdNa5CNzklkMbk8ljVv8Rf8Agoh/wT3k+zfFTRJfjX8KLMbV1SGd7iWziH8RuNpmgwMDE6vGPuqw61+zj+3P+zv+07FBp/gbxeuneJZEy/hzWALa9BAy3lrkrMAATmNmwOSBXxW1+bwp8LfGPii2kkSbR9AvryNo/vK0Vu7gryOcr6iv+CJeh2l18dPHniKQKbjT/Ci20QK9BNdRMxHPH+pUdO/Uc5r/AILk6VbQ6z8HNbQDz7u11m3c7f4Yms2Xn6zN/k1+yVrU3iH9lv4R6vcszTzeCtKErMBl3W1jVm445Kk/jVV8dvi94d+Avwi8UfFvxOytZeHLB51gL7TcznCwwKezSSMiD0LZr/glR8KPEHx4+P3jP9sL4nxtqEmlXs5s7iVPkn1e6y0rqDkYhhcgL/D50ePu1VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVXxB+IvgX4U+FLzxv8RvFWn+H9DsFzPe3suxQccKo6u5xwigsx4AJr48/8FFvjz+1j4tb4DfsVeFNd0+x1EtC+pWybdTvo84aQODtsYOeXLBgMEumStfsff8Ep/AXwpNl8RP2gxZ+N/GpInj0xx5um6bIeclW/4+ZR3ZxsBPCkqHIVVUKoAAGAAMYqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv+CQIOg/tOfGXwiEXEWmycxHag8i+2YC+n7zj0A96qqr/gtv8AEc6d8Ofh58J7Wch9c1e41a6VTgiO1jEaBvZmuWOPWP2FfszfDhfhH+z58PPhyYPKuNE8PWkV4u3b/pTRh7g47ZlaQ496qqqq8T6HD4n8Nat4annaGPVrGa0eVBlkEiFCwHqA2a/4chfCr/ouHiv/AMF1v/jX/DkL4Vf9Fw8V/wDgut/8a/4chfCr/ouHiv8A8F1v/jX/AA5C+FX/AEXDxX/4Lrf/ABr/AIchfCr/AKLh4r/8F1v/AI1/w5C+FX/RcPFf/gut/wDGv+HIXwq/6Lh4r/8ABdb/AONf8OQvhV/0XDxX/wCC63/xr/hyF8Kv+i4eK/8AwXW/+Nf8OQvhV/0XDxX/AOC63/xr/hyF8Kv+i4eK/wDwXW/+Nf8ADkL4Vf8ARcPFf/gut/8AGv8AhyF8Kv8AouHiv/wXW/8AjX/DkL4Vf9Fw8V/+C63/AMa/4chfCr/ouHiv/wAF1v8A41/w5C+FX/RcPFf/AILrf/Gv+HIXwq/6Lh4r/wDBdb/41/w5C+FX/RcPFf8A4Lrf/Gv2ZfgDo/7Mnwf0r4P6D4gvNas9KnuZUvLuJY5HM0rSEFV44LY/Cqqqqviv8avhT8DfDjeKvix470rw3p4B8trub95cMBysMS5eVv8AZRSfavjqnhX9vH4pxx/sP/sq6jp2p2F6JNS8aRN/ZkMp6h5YkIt4Dn5xIzec/wDdyMV4H+F/j7T/ANlS2+EPxM8QDxH4sfwjPpWqX5vJbj7VPJC6E+bJtd/vAbmwTjNf8EU9eg0z9oDxz4TvAIbnUvCpljWQbWLQXUQZAPXEpOOvyn0NV/wXF8QW1z4t+EnhRHQz6dp2qXrqPvBbiS3RSfbNs35Gv2aPDM/g39nT4X+FLu38m60rwfpdvcx+ky2sYk/8f3VVf8FgPjvqfjfxt4U/ZD+HxkvrqO6gvdYtrc5ee+n+SztfqFk3kHgmaI9Vr9lz4G6b+zl8CfCfwlsWiludJsw+p3Ma4F1eyHfcSZ6kF2YLnkIqjtVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX7Xv/BQn4Pfsq2lx4eWdPFfj5o8weHrKcD7OSMq91KARCvQ7cF2yMLg7h4D+AP7X/wDwVA8Y2/xU+NviS48L/DyGTdYyvbNFbrETzHptqT8+QADO5IOOXkK7a+Av7OHwh/Zr8Ip4P+E/hWHTonVftt/LiS81B1GPMnmxlz1IAwq5O1VHFVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVf8ABNBFs/8Agox8ctNtCYrVNO18LCpIUbNatQvHsCQPqfWqqq/bXb/hoP8A4KkfD/4Mx/6Rp/h6fR9MvIguV8st9uum75Pky4Pb5MHoaqqqqqqqqqqqqqqqqqqqqrx5458K/DLwdq/j/wAb6sum6DoVq1zf3bRPJ5Ua9TtQFm7DABJzXjT/AIKR/Hn9o3xDc/DL9gj4Napeyg+XN4n1S1Vzbg8CQIx8i3XI4ed2znGwGvhF/wAEsJ/Fuvp8Vf23PiZqnxH8V3BDyaTFfym1iHURyTnEkgGfuR+Wi9BuWvCXg3wl4B0C18K+CPDWmaDo9ku23sdOtUt4Yx3wiADJ7nqe9V8efD/i3/gnR+35a/GnRdBuZ/BWt6pNqdgIhiO7s7nIvbJWOQJIzI4UHkDynPWtO/by/Y/1LwOnj5f2gPCFvZNb+c1lc6gkeoJgcobMnzy46bQhz2yOat4df/4Ki/t7Jr9loV9B4A0p7dbpp1/48dEtmJCSEEhZbhzJhAThpmxlUZq+NHxV8O/Av4UeJfix4nt55tM8MWDXMlvbAeZMchUjTPALOyqCeBnJr9iL/go/pn7XvjvWfhxqnw2PhHWLHT31GyMeqfbYrmBJER1JMUZV1MiHoQwyeMYPxh+KHhz4K/C7xN8VPFc4TTPDWnSXcq5wZnAxHEv+1JIURf8AacV/wTF+F3iD9pv9qbxT+1X8T42v4/Dl6+oCWRP3c2r3JYxqoP8ADCm5wo+4RD0GKqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqvEniXw74O0G98UeLNcsdH0jTYTNeX19OsENug6s7sQFH1r9pn/AIKe/EP4yeJl+BP7D+i6vNcapKbUeIbe1Y318SMEWcRGYEAyTM+HA5Aj27j+yF/wSg0HwpdQfFH9qp7fxb4qnk+0p4deT7RZ2srHcXunyftUueqnMec58zIIhhht4Ut7eJI44lCoiLtCgDAAA6DHaqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/YQjXS/wDgqn8btNsC0Nut14pgEYYnMa6tGQpzyQCqnn0FVVV+wiJPjz/wU6+JHxpk/e2WiPrOo2cp5CrLL9jt0z6/Z5W59EPrVVVVVVVVVVVVVVVVVVVVVfEP4f8AhX4qeCtW+HvjfTmv9B1yDyL+1WZ4jNHuBK70IZQcAZBB9xXgbwD4J+GXhq08HfD3wtpnh7RbIEQWOn2ywxqT1YherHqWOSTySTVVXxY+Dvw0+OXg+48B/FXwjZeINGuDv8i4BVoZACBJFIpDxuASAykHkjOCak/4Iz/skvrP9prrfxFS23lv7OXWLfyAP7uTbmXH/A8+9fB34G/Cn4A+FF8F/CPwZY+H9ML+ZMsIZ5bmTGPMllcl5GwMZYnA4GBxXjzwN4W+JvgzWfh/410pNS0LXrN7S+tXJXzI3GDgjBVhwQwwQQCCCK/Zk/YT+A/7J+uax4o+Gtvrd7rOswG2e/1m8S4kt7YuHMEWxECoWVCSQWOxctxXx9+AHgL9pPwMnw6+JMmrnQxex3kkGnXptTNJGGCB2AJZQW3bem4KeoFfAn4DfDb9nD4fw/DX4W6TLZaRHdS3UhnmM0080hG55HPLHaqKPRUUdqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr9p/8AbC+DP7KPhr+0/iDrf2rW7qItpnh2xZXvb09AwUnEceQcyPheCBubCmG3/bJ/4KyeOfOuJD4R+FWm3fBw66XYY7KOGvbrb36KW58pWFfsyfsg/Bn9lPwyNI+HWhCfWLmILqfiC+USXt+epBfH7uPIGI0wowCctljVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV/wT/wA6/wD8FRfjjrUYEKq3ie7MbHJw+sQrtz6jzB+VVVfHnxsPhr8EfH3xAWZ45PD3hu/vYWQ4bzY4HaML7lwoHuRX/BEDwOLbwb8T/iTLbk/2jqdlpNvKRwv2eJ5ZVH1+0xE/Reneqqqqqqqqqqqqqqqqqqqqqqqqvi7+0n8CPgN9lT4ufFHRPDk97zb2k8pkuJF/viGMNJs4xu27c8Z5r4cfEjwR8XvBenfEP4ceIIdb8O6t5v2K+ijeNZfLleJ8K4VhiSN15A6elVVXP/BRr9im0uJbS4+PekrLC5R1+wXhwwOCOIfUV/w8h/Yj/wCi/aT/AOC+9/8AjNfC34tfDr42eEo/HPwt8UQa/oUs8kCXkMUkamRDh1xIqtwSO1VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVMVVSzMAAOSeMV+2R/wAFVfD/AIEubr4U/svm28W+MpJDay67HH9osrGUnaFt1GRdTZxjGYwSPv8AKj9l/wD4Jj/ED4x+J/8Ahff7cGravcz6nKLoeHLu5f7dfE4wbyQHdAgHAhTDAYBMYXadA8P6F4V0Wz8OeGNHstK0rToRDaWVlAsMMEY6KiKAFHsBVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVf8Ew9+r/8FA/jh4kiVUgk07WsxsfmBl1i3dfbgIc/hVVX/BVHxmfCH7FfjG2hmMdx4iurHS4WDYPz3CSSD3zFFIMe9f8ABKXwUPCH7FvhS+eHyrjxPf3+qzKRycztCh/GKCM/Qiqqqqqqqqqqqqqqqqqqqqqr4R/tM/A/45XmoaP8OvH2n32taRcSwX+jzN5F7bvGxVyYXwzICPvruT3qqr9r+0+Pt78Bddt/2Z9QktvHrz2sdkIVhEkkTzpHMEab5IyI3Z95wVCHBBxX7YH7CHjj9nj4Gaf8dvjL8UJvFHxB8U+K4LC+t45HuIoUktbmVmkuZf3k8uYIxnCqPmHz8MP+CXH/ACYl8Mf+4t/6dryqr4neJx4J+GvizxmZVjGgaHe3+9ui+TA8mf8Ax2v+CZf7EPwg/aj8H+N/FnxesdXuIdI1K2sdM+w3zWwVvLZ5t2B83Dw9+Px5/wCHQ37Gn/QD8Vf+D1/8K+BPwJ+H/wCzn8P4fhp8NLa9h0SC6luUS8uTPJ5khBb5j246VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVfFX4tfDr4JeC73x/8AFDxVZ6DoliMPPcMS0r4OI4kGWkkODhFBJweODXxj/a2/aZ/4KMeNrj4C/sweF9U0XwTNxeqHEUtzBnBm1C4XKwQn/nipIbofMJUD9jX/AIJ1fCv9lm1tfFmtLb+LviKY/wB5rdxB+6sCRhks4z/qxjgyn942TyqsUFVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX/BGoDxF8ePjD412vN5umx/6Q7Yb/SLtpOVz1bys9ONvbPNVX/Bbzxa1l8Kfht4EWTA1jxBdaiV9fsluI//AG8r9mfwb/wr79nf4aeC2h8ubSfCunQ3C7Nn7/7OhlJHYmQsce9VVVVVXxR/4K0fs4/CT4i+JPhj4j8F/Ei51TwvqU2n3k1lptk8DyxMVYxs92rFcjglQfYV/wAPq/2V/wDoQvir/wCCqw/+Ta/4fV/sr/8AQhfFX/wVWH/ybX/D6v8AZX/6EL4q/wDgqsP/AJNr/h9X+yv/ANCF8Vf/AAVWH/ybX/D6v9lf/oQvir/4KrD/AOTa/wCH1f7K/wD0IXxV/wDBVYf/ACbX/D6v9lf/AKEL4q/+Cqw/+Ta/4fV/sr/9CF8Vf/BVYf8AybX/AA+r/ZX/AOhC+Kv/AIKrD/5Nr/h9X+yv/wBCF8Vf/BVYf/Jtf8Pq/wBlf/oQvir/AOCqw/8Ak2v+H1f7K/8A0IXxV/8ABVYf/Jtf8Pq/2V/+hC+Kv/gqsP8A5Nr/AIfV/sr/APQhfFX/AMFVh/8AJtf8Pq/2V/8AoQvir/4KrD/5Nr/h9X+yv/0IXxV/8FVh/wDJtf8AD6v9lf8A6EL4q/8AgqsP/k2v2Vf24/hL+2BqHiLTfhp4e8W6bL4Yht5bs67aW8IcTFwvl+TPLkjy2znHUdaqqqv2hf8AglV8PfHPiGf4o/s++MNR+GHjz7S17HJbzytZyXJJYyAq3m2zljnfGxA7JWj/ALZ/7af7FOqWvg/9tL4YXfjLwiJVgt/F+mhTKV6ArOoEU5wOI5RFKerNXwO/aZ+B/wC0bo39r/CTx9p+ryRxh7nTmbyb20/66274dRnjdgqT0Jqq/wCC1f8Ayav4W/7KBZ/+kF/X/BLj/kxL4Y/9xb/07XlVX7f/AInPhH9jP4s6qJdnn6A+n53bf+PuRLbHUdfOxjvnoelfsRf8FGvCH7IPwiuvhtdfCbUvEd3qWvT6pd30OqJbBd8UMSoqlHLYWEHqoyfqa+An/BU79mP4363ZeEtQvdU8Da/fyCK2t9fjRbe4lONqJcRsyAnOB5nl5PAySM1VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX7X/7enwj/ZM0uXSr+4XxH47uIN9j4as5gHXI+WS5fnyIzwRkFm/hUjJHwz/Z5/aq/wCCn/jqH4x/HfxJd+G/h3FIfscywGOLyt3zQaZbMSMcBTO+RkZJlZStfBn4HfC/9n/wVbeAvhT4UtdG0yABpWRd015LjBlnlPzSyH1Y8DAGFAAqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq+Imtjw18PvE3iMymP+ytGu7veJPL2+XCz53fw/d69q/4Ib6MU0/4w+IXiUiabR7WJynK7Fu2cA+++PI9h7VVV/wAFai/xK/a6+DnwVt3Mnn6faxbVGSsl/ftCRwc52woccdRzzUcUcMaQwxqiIoVEUYCgdAB2FVVVVVX7a/8Aydz8X/8AscNQ/wDRzVVVVVVVVVVVVf8ABDr/AJGr4t/9g/Sv/RlzVVVVWraRpWvabc6NrmmWmo2F5GYri0u4FmimQ9VdGBDA+hGK/b4+Cf7EXwH1KXxn8Jfixf8Awy+LOnkz2Wg+EZmut03UbolkU2OeAGEiKBkiNzxX/BNX4v8A7Rnxk+C99rv7Qmn3ZktryOPQdWu9LNlLqlqY8mQ4CrKA2AJFUZ5yWOTVf8Fq/wDk1fwt/wBlAs//AEgv6/4Jcf8AJiXwx/7i3/p2vKqtW0fSNesJNL1zS7PUbOUjzLa7gWaN8EEZVgQcEA/UVqHw98A6vZPpuq+B/D97aSLte3uNMhkjYYxgqykEYr/gqX+wh8LvAHw3b9oj4L+F7Xwz/Zd7DB4h0mwj8uzeGZxHHPHGPlhZZWjQqoCsJAcAg7v+CWfxx8Q/Gz9lmyj8X6jLf6z4K1OXQpbudt0tzDHHHJA7nOSRHKseTyfKyckkmqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqmmht4XnnlSOOJSzu7bQoAySSegxX7Y3/AAVKmj1ab4IfseL/AMJF4kvZfscvia0g+1IkrHb5dhGAfPkJ483BT+6HyGX9kD/gljcXepx/Gz9siSXX/EF/N9tj8M3dwbgGVjuMmoSEnznJ5MWSv98tkqLW1tbG1hsrK3it7e3jWOGGJAiRoowFVRwAAAAB0qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv2yvEDeGP2T/i5q0chSQeD9Rgjdcgq80DRKRjuDIDX/BFPw99h/Zx8XeJJE2yar4xlhXI+9HDaW+DnP96SQYwOnfIxVV8VYz8Uf+C03hvw8v7+Lw5qmmGLIBA+x6et82ARxhw/4jII4Iqqqqqq8U/sVfsqeNvEepeLvFfwO8N6nrOr3L3N9eTwuXnlc5Z2w3Uk5r/hgP8AY0/6N58Kf9+H/wDiq/4YD/Y0/wCjefCn/fh//iq/4YD/AGNP+jefCn/fh/8A4qv+GA/2NP8Ao3nwp/34f/4qv+GA/wBjT/o3nwp/34f/AOKr/hgP9jT/AKN58Kf9+H/+Kr/hgP8AY0/6N58Kf9+H/wDiq/4YD/Y0/wCjefCn/fh//iq/4YD/AGNP+jefCn/fh/8A4qv+GA/2NP8Ao3nwp/34f/4qv+GA/wBjT/o3nwp/34f/AOKr/hgP9jT/AKN58Kf9+H/+Kr/hgP8AY0/6N58Kf9+H/wDiq/4YD/Y0/wCjefCn/fh//iq/4YD/AGNP+jefCn/fh/8A4qv+GA/2NP8Ao3nwp/34f/4qv+GA/wBjT/o3nwp/34f/AOKr4Vfs8/BP4HXGo3Xwk+HGkeGJtXSNL17GNlM6xligbJPQs351VVVftGft1fs6/szQz2PjXximp+JI1Ozw5o226vS2OBIAQsA95WXI6A1H8Wv+Chf/AAUIkez+D2hn4L/C26OG1uWWSGW7hPXbdbRLMSM8W6omflZ+9fs3f8Ezf2fPgJcW/inxBZP8Q/GaN5jaxr0KvFDL1LwWx3IhzyGcyODyGFVX/Bav/k1fwt/2UCz/APSC/r/glx/yYl8Mf+4t/wCna8qvCv7e/wCy741+MzfAjw98Q/tHiY3cllbubORbS6uUzuiinxsZsqQD91iMKWyM+KPHngjwTLpkHjLxfo+hvrNybXThqN7Hbfaptpby495G5toJwOaBVlDKwII4Ir/grn+1X8P9P+EN5+zV4Y12z1fxP4ku7d9ZhtZllGl21vMk4WUgnZK8kcWE67QxOAV3f8Erfgrrfwd/ZS0678TWT2mq+ONRk15reRcPDBJHHHbqw7bo4lkx283BwcgVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV8U/ix8PPgr4LvvH/xO8U2ehaJYLmSe4b5pGxxHGgy0kjY4RQWPYV8Yf2o/2l/+ClPj6X4D/s2eHdR0PwExxextJ5Rngzjz9SuFyIoeOIFJBPH71toH7HP7AHwo/ZP0yHXDFF4n+IE8O288RXUGPIyMNHaRnPkp1Bb775OTjCrVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV/wVT8TL4d/Yl8bWqyBZtcutPsISfU3kUjjBBzmOJx2xnOeK/wCCVPhk+Hv2JvBl28YSXXLzUb9x35u5IlJ+qRIfxFVVfsiIfiP/AMFcfih4tCiVNAvPEVxE4H/LOOcWKN0GMrKvbPPOetVVVVVVVVVVVVVVVVVVVVVVfBj/AIJn/s9/DTxfqnxI8Z6b/wALC8VanqU18k2tW6m0smkkZ8RWuSpI3D55C5yoK7ORSIkaLHGqqqjCqBgAD0FVVftx/sq6h+2B8JdJ+Gmm+NLfwzLpniKHVzdz2LXQcR29xD5e0OmCftAOc/w9Oa/ZR+Bt3+zb8AfC3wVvvEcOvT+Hftm7UIrY26zefeTXAwhZiuBMF6nO3PfFV8Iv+CQ0vwz/AGl9I+Kk/wAUrW88GeGNbj1jSdPS1kW+kkik8yCGVidoVGCbnBJcKRtXdx+2n+xN4c/bL0LQbDVvH+s+F9Q8MvcNp8ttAlzbEzBA5mgJUuR5S7SJFwCw/io/8Ehf2odJT+wvDf7TekpoYBQRG5v7YbDwR5Kbk+723Y7e9fs2/wDBIT4SfCjXbPxl8XvEr/ETVrFxLbaa1kLbTYpB0LxlnafBAI3FUPdDQVVUKoAAGAAMYqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr9rb9tj4S/sk+GzN4muhrXiy8hLaV4Zs5gLi57B5WwRBDnq7Ak4O1WIIr4e/BT9qn/gqf8Qovit8Y9buPDfw2s52S2mjhMVvFGG+eDToGJ8xuNrTvuAI+ZmKhK+C/wO+GH7Pvge1+H3wq8L2+j6Xb/NKyjdPeS4w008h+aSQ+p6DAACgAVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV/wWz8Wf2f8C/AngmOUK+teKWvGAJBZLa2kUj0xuuUP1Ar9kvwo3gn9mH4VeGZYzHPaeEdONwhz8szwI8o5/23aqqWSOGNpZZFREUszMcBQOpJ7Cv+CNkLeL/j38YvidNH+9k05FLMMMpvLtpiOh72/r279qqqqqqqqqqqqqqqqqqqqqqqqqrUvHngbR78aTq3jPQrK+JAFtc6jFFJycD5GYHk8dKjkjmjSWGRXR1DIynIYHoQfSqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr9t//AIKgeHfhBJe/Cb9nuaz8T+P2Y21zqiAXFno0h4KqBkXFwDxs+4rfe3EGM/slf8EzvGfxV8SL+0L+21danfXeqTC9i8N387NdXrk5V79uqJjGIBg4wG2AFDp+nafpFhbaVpNjb2VlZxLDb21vEI44Y1GFRFXAVQAAABgCqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr/grve3PxP/ah+D3wG0yV5JvsUSoqEEpPqV4IQAP72LeM/itWlpbWFpBY2cSxQW8axxRjoqqMAD6ACqr43a7/AMIv8F/H3iYSBDpHhfUrsMX2bfKtZHznnH3evav+CHug/Z/AfxU8T+WR/aGr6fZ79p58iKV8Z6cfaOnv71VVVVVVVVVVVVVVVVVVVVVVperaVrlkmpaJqdpqFpIWCXFrMssbFSQcMpIOCCD6EVVV+3p8Af2q/wBoDVfBnhH4EfFB/DHhK5huk8UI1+bOJGUxmJ3MQM0wcNIvl8oCik4yWo/8EObRtHJf9pCdtXMZJb/hFx5HmdcY+07sds5z3x2r4NfGb4+f8E4P2nY/gl8SPEs954Pt9Rgg1rSxcvNYyWk4BW9tN4BjYK4f5QpYoUfpxVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV4g8Q6D4T0S98SeJ9Zs9K0rTYWmu728mWGKCNerO7EAD61+1D/wUG+K37VPi9v2bP2K9H1ltO1V2tZ9XtEaK91ZOj+WTj7LbYzudirFfvGMblP7EH/BNfwL+zbDY/EL4lLY+KviVtDxz+XvtNGb0tgwy0g7zMAeyhRktVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVWjEftGf8Flrm+R/tGk+CdYkYkZYQjSrby1I9jeIpz0y5PtVVX7cutHQP2P8A4u3yybDL4VvLTO8L/r08nGT6+ZjHfOO9f8EYdEXTv2Vdb1Rk/ear40u5AxQglEtrWMDPcBlfn3I7VVVVVVV8Xv8Agrv8OPhD8UfFPwu1L4Q+JNQuvCuqz6dNdQX8KJM0TlSyg8gHHQ1/w+8+FX/RD/Ff/gxt/wDCv+H3nwq/6If4r/8ABjb/AOFf8PvPhV/0Q/xX/wCDG3/wr/h958Kv+iH+K/8AwY2/+Ff8PvPhV/0Q/wAV/wDgxt/8K/4fefCr/oh/iv8A8GNv/hX/AA+8+FX/AEQ/xX/4Mbf/AAr/AIfefCr/AKIf4r/8GNv/AIV/w+8+FX/RD/Ff/gxt/wDCv+H3nwq/6If4r/8ABjb/AOFf8PvPhV/0Q/xX/wCDG3/wr/h958Kv+iH+K/8AwY2/+Ff8PvPhV/0Q/wAV/wDgxt/8K/4fefCr/oh/iv8A8GNv/hX/AA+8+FX/AEQ/xX/4Mbf/AAr/AIfefCr/AKIf4r/8GNv/AIV/w+8+FX/RD/Ff/gxt/wDCv2Of27PCX7ZGpeKNM8M+BNX8PN4XgtpZmvrmOUTCZpAAuzpjyznPqKqqqvHv7Ff7ZH7LfjrW/jB+xl8U7nxHpeqahNqGoeErxljeUu5ZlMDnyLkAEgMPLlAICgnmvgX/AMFXvAus62vw2/aj8G33wn8Z20ot7me5glFh5vT94rjzbTJI4kDKByZAK0rVtK17TbbWdC1O01GwvIxJbXdpMs0UyHoyOpIYH1BxVO6RoZJHVVUEszHAAHU14y+M3wj+Huiy+IfG/wATPDOiafCpLT3uqRRhsdlBbLN6KoJJIAHNfE9bj/gpr+3tAnwq0S+fwXp6Wljd6rJCUEWl27lprqQH7m8vII0bDNlAQCSBVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVfG/47/DD9nfwNc/ED4qeJYdL06HKQRD57i9lxkQwR5zI5x06DqSACR4t8f/tT/wDBWP4nHwP4B06bwv8AC3SLpXlikkYWdigPE95IuPtFyR9yJfu/wgDfIf2XP2RfhL+yd4P/AOEf8A6b9q1i8Rf7X8QXcYN3qDjsSPuRg/diX5R1OWJY1VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV8SPGum/Db4e+JviFq7L9i8M6RdalOCcZSCJpCPqduMepr/gi74L1HxN41+K/wAftfVprydYtLiumX/WzXEhubrn1zHbn/gdVVf8FRdXOk/sOfEYRyFZb06dbR/LnO7ULfcD6fIH5+lf8EoNIGm/sReDrwIB/auoanc53Z3YvZYs47f6rGPbPeqqqqqqv21/+Tufi/8A9jhqH/o5qqqqqqqqqqqqv+CHX/I1fFv/ALB+lf8Aoy5qqqqr44/sy/A/9o3Rv7I+LXgHT9XkjjKWuoqvk3tp/wBcp0w6jPO3JUnqDXxL/Zi/aI/4J16ZffFr9mX9oyzn8CWrmW98L+LryG3WTvsVZCsE7kA8x+TMeFQMa/YX/bD/AOGxfh3qviW98F/8I5q3h28jstQhiuvPgmdk3CSIkBlB5+Vs46bm614q0JPFHhfV/DMlybddWsJ7QzBNxjEkZTcBkZxuzivE/wDwQ7KaTLJ4M/aGE2popMUOp+HfLhlPYNJHMzIPcI30r4EftU/GH/gnR8TLf9mr9pHwDocPhIyRu+oaRpkUE6QuSqX0csKJ9tjyDuMgMvysNwZSlWd5aalZwahp91Fc21zEssE0TBkkRhlWUjggggg+9VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVftfftt/Cz9kfwx5muzrrnjG/hZtI8NWswWabsJJmwfJhz/GQSeQqsQcfBz9nL9o//AIKc/EaP46/tDa/e6L8PIpXW0eKMxLJEH+a102FiQkeRtadt3K8mRwcfDP4YeA/g74L074ffDbw1Z6HoWmR7YLW3TG4/xSO3V5GPLOxLMeSaqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv8Agrl8WF+Hv7J914Ps7nZqXj/VINLRVbDrbxnz53H+ziJIz/12/Ef8Ez/hQ/wo/Y88FwXdt5Oo+Klk8QXg24LG6wYSff7OtuPwqqr/AIK/6p/Z/wCxxeWgeUf2l4k0+3wnQ4Ly4bnp+6/PFf8ABOjS10j9ij4VWirEN+lTXGI1wP3t1NLn6/Pz75qqqqqqr9o7/glZ+058Vvjz4++JPhi/8ErpPiXxBd39kt1qsscoikkLLvUQkBsHkZNf8Obf2uf+gj8P/wDwczf/ABiv+HNv7XP/AEEfh/8A+Dmb/wCMV/w5t/a5/wCgj8P/APwczf8Axiv+HNv7XP8A0Efh/wD+Dmb/AOMV/wAObf2uf+gj8P8A/wAHM3/xiv8Ahzb+1z/0Efh//wCDmb/4xX/Dm39rn/oI/D//AMHM3/xiv+HNv7XP/QR+H/8A4OZv/jFf8Obf2uf+gj8P/wDwczf/ABiv+HNv7XP/AEEfh/8A+Dmb/wCMV/w5t/a5/wCgj8P/APwczf8Axiv+HNv7XP8A0Efh/wD+Dmb/AOMV/wAObf2uf+gj8P8A/wAHM3/xiv8Ahzb+1z/0Efh//wCDmb/4xX/Dm39rn/oI/D//AMHM3/xiv+HNv7XP/QR+H/8A4OZv/jFf8Obf2uf+gj8P/wDwczf/ABiv+Ca37FPxi/ZL1zx5qPxSufDssXiS0sYrIaVevcENC0xfeGjTAxIuOveqqqrx38QvAvww8OXHi74h+LdK8O6Paj95eajdLAmeygsfmY44UZJPABr4k/8ABUXxV8TvEsvwp/YU+Eeq+O9flBUa5e2TiCFc481IPlITkfvJ2jVT1Uivh/8A8E0vil8bvE1t8U/2+vi/qfinUPvxeF9OvSIoAeTG8ybUiX1jtlUZ5ElfD74afD74T+G4PCHw18HaT4b0e35S0062WFWbGC7Ecu5xyzEse5NVX/BbzxN4T1D4h/DLwpp01vJ4h0fSr+fVNhBeOCeSEWyPjocwzsAecPnoRX7K+k67oX7M/wAKdG8TB11Sy8G6XFcxyfeiYWsY2N/tKMKfcd+tVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX7dv/BSrwx+z1HffC34Py2XiL4lMpiuJT+9tdCY8Ey44knHaHop5fpsb9j7/gnB40+MfiYftJ/trS6nf3Gq3Avrbw7qbt9p1BzgrLfA8xxYxtt+CQAGCqNjWNjY6ZY2+maZZwWlpaRLDb28EYjjhjUAKiqOFUAAADgAVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV/wUM1W7/aq/b4+H/7Lnh66lfT/D0ttpt20Z3CGe6KT3sy44/d2yxZ9DE34adp9jpOn2ulaZax21nZQpDbwRrtWKNAFVVHYAACqqv+C1eorb/sxeFNMSaRZLvxzbMVUkB0SyvMg/8AAmQ49RntX7GOnNpf7JXwetWSNS/gvTJwIxx+9t0kB+p35PvmqqqqqqqqqqqqqqqqqqqqqqtK/wCCaXxO+P8A8TNT+KH7aXxf1bVbAajO2j+GtOvvMaK1MhKRvLjy7dNoXMcCnOc71avhf8H/AIYfBXw3H4R+FXgfSfDWlpgtDYwbWmYDAeWQ5eV8cbnZm96qv2wP2dP+CgEnx+1T4+/sw/ExprK9sba0g0Oy1RbKW3ihjx5ckNwfs86mRpZAWOcyEbRxmb4g/wDBbPVY28Nx+GdXt7h18prtNE0iEnAwWEpURqT/AHhjrxjiv2Z/+CV3xF174lR/G39tDxHDrF6Ltb59BN8dQn1C4HIN7PypQEL+7QuGAwWCgqaqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqlkjhjaWWRURFLMzHAUDqSewr9tH/gpX4g8Y6+/wCzj+xi93q2sapcCwufE2lgySzysdpg04LySehuB77O0lfsJ/8ABMzQvgo1j8XfjxBa+IfiIxFxaae5E1robnkNnkTXI6mT7qH7uSBIaqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq+J/xA0L4UfDrxJ8SvEsu3TPDOmT39wAcF1jQsEX/aYgKB3LAV/wAElPAGu/GH48/Ej9rnxxEZ7iK4nhtp2X5X1G+cy3DJxx5cR244wLgD6VVV/wAFuryZPg78OtPUjypvEs8rAjnclswH6O1fs3WcFh+zr8LLC1UrDbeCtHijUnOFWyiA5+gqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqvE3ibw94M8Pah4r8V6zaaVo+k273F7e3cojigiUZZmY9Biv2mP2yvjN+3p4/P7Mf7JGi6mnhK8kaO7uYwYJtXiVgGmuHP/AB7WY4+U4LZG/lhGP2LP2Dvh1+yRoC6pJ9n8Q/EHUINmp+IJIcCJT1gtVPMcXTJ+85GWwNqrVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVf8FmPj03hr4a+Hv2edAu2/tPxncLqGrRxk7hYQP8AukI9JLgAj/r3Yd6/Yq+BC/s6fs2eD/hzd2yxa19l+366QOWv7j55VJ7+XlYge6xLVVVf8FxZJB4Q+EsKyNsbUtUZkzwSI7fBx6jJ/M18KUeP4XeDo5FKsugWIKsMEEQJVVVVVVVVVVVVVVVVVVVVVVVVVXNxb2dvLd3dxHDBAheWWRgqooGSxJ4AAGSTX7Rn/BW+10vxa/wv/ZM8ExePtbM32ZNZnilntp5uhS0t4SJLn2fcqkjgOpDHwbB/wWV+J/iTQ/EviSbUvD/hz+0raa5s3bS9Kb7OJAZEMIxPjYWyJOT7kcVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVXxU+KvgH4K+BtS+IvxK8R22jaHpce6WeY5aRv4Y41HMkjEYVFBJNfEH4n/tIf8ABWD4uj4Y/C7Tbjw78L9FuVmlSdsW9pEGIW7vWH+snZSdkKk45C9Hkr9mL9lb4W/sp+A4/B/w+00TX9yqvq+t3MY+1anMB952H3UBztjHyqCepLMaqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqrUdQsNI0+51XVLyK1s7KF5rieZwiRRoCzOxPAAAJJ9q/ZxsNS/4KD/APBRTVvjlr2nyv4K8GXSanFDMPligtzs022I5G53QSuvRtk3rVVVV/wXOJ2/BNQxwTr+R/4Lq8Jf8ipov/YPg/8ARa1VVVVVV+03/wAFKf2ufhp+0L8RPh/4Q8e6ba6L4e8R3llYQSaFaytHDHIVUFmjLMcAck5r/h7B+25/0UjSf/Cds/8A43X/AA9g/bc/6KRpP/hO2f8A8br/AIewftuf9FI0n/wnbP8A+N1/w9g/bc/6KRpP/hO2f/xuv+HsH7bn/RSNJ/8ACds//jdf8PYP23P+ikaT/wCE7Z//ABuv+HsH7bn/AEUjSf8AwnbP/wCN1/w9g/bc/wCikaT/AOE7Z/8Axuv+HsH7bn/RSNJ/8J2z/wDjdf8AD2D9tz/opGk/+E7Z/wDxuv8Ah7B+25/0UjSf/Cds/wD43X/D2D9tz/opGk/+E7Z//G6/4ewftuf9FI0n/wAJ2z/+N1/w9g/bc/6KRpP/AITtn/8AG6/4ewftuf8ARSNJ/wDCds//AI3X/D2D9tz/AKKRpP8A4Ttn/wDG6/4ewftuf9FI0n/wnbP/AON1/wAEuf2u/jr+09r3xCsPjF4ns9Vh8P2dhJYLBpsFr5bSvMHJMSruyEXr0xVVVVrv7Vn7cn7EfjjUrX9oz4d3Hj/4W3GqTHT9dtyJJLa3eQmNVu0GMhTgR3KhzjAYDmv2fv2wf2f/ANpqwST4X+OreXVhHvn0HUMWuoW+Bk5hY/OB3eMug/vVVf8ABX79p3XPCXh3Rv2Yvh9fXEer+NLf7Vr5tQfNawZzHFbKV5/fyLIGA5KxhTlZCD+x/wDs2/A39hL4X6b4l+LHibwnonxC8QWivrOt67qVva/ZywDGzt3lZQsacBip/eMu48BVXwh8QfAPxDsm1LwB448P+JrSPAe40fU4byNcjIy0TMBkVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVXx8+Pnw4/Zu+HF/8AEv4l6uLaxtR5drax4a41C4IJSCBCRudsH0CgFmIUEjTNE/aQ/wCCuPxpfWdYuLjwr8KfDt06xuEMlrpKED91EPl+03jrtLMfu5ydq7UPwZ+Cvw4+APgGw+G/wu8PRaVpFkNzc7pbqYgB5pn6ySNgZY+gAAUACqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr/grp+0x/wAKt+DMPwQ8NX2zxF8Ro3S9aN8Pa6WjASkj/ps37oZ4K+b6V/wTr/Zt/wCGb/2b9H0/WtP8jxZ4r26xr+9MSRSSKPKtz3HlRbVK9A5kI+9VVVV/wXLvo5NR+DOmqjeZBBrcrMehDtYgAe/7s/mK0qwXS9Ls9MWUyC0t0hDkY3bVAzjt0qqqqqqr9tf/AJO5+L//AGOGof8Ao5qqqqqqqqqqqqv+CHX/ACNXxb/7B+lf+jLmqqqq9srPUrSbT9QtIbq2uIzHNBNGHSRCMFWU8EEcEGvj/wD8EqPgz8QdRPjn4Hapc/Cfxnby/aLabSQwsTMpyrCFSrQNkDDQsoXrsY1Z/tQ/t3/sOSw6N+1p8OJfiX4ChcRR+L9JkEksSZwC1wFAY+i3KRyMf46+A37RXwk/aV8IN4z+Efib+1LO3kEN5BLA8E9lMVz5csbAEHHcZU9iRzX7ZfxD8V6z/wAFHfEPiPwppkWt65oHiqwsdA06SIzRy3VosMcMXl8bw06ZKdGLEdDXhP8A4JJy/FaF/iL+2B8cPGWuePdbQTXsWjXUCrZMefJ82aOUSBc4wixovRQVAJ/aq/Y/+LP/AATq8TaF8bPgn8T9Wn8P3F8La31SJRb3VhcYLrb3KqSk0cio3O0I21lZB8u79j39oS2/ae+AHhz4qNBFbarOr2etWsXCwX0J2yhRk4VvlkUZJCyKDzmqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr9ov9or4b/sx/De8+I3xG1PZFHmLT9PhYG51K5xlYYVPUnGSTwoySQBXww+F/wAdv+Cr3x2uPip8VL260L4aaFceSzW5Ihs4chhYWQYYeZhtMkpHGQzf8s4z8Pvh54K+FXg/TfAPw88O2eh6DpMXl2llaphUBOSxJyWZiSWZiSxJJJJqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqrxZ4p0DwP4Y1Xxl4p1OHT9H0SzlvL66lOFhhjUs7H8AeO9fsy+GvEH/BRf9u3Wvj94906R/A3hC5jvFs7gFoo4oyV0+xA5XJK+bIPuttlJxvGaqqqv+C43/I1fCP/ALB+q/8Aoy2qqqqqqqv21/8Ak7n4v/8AY4ah/wCjmqqqqqqqqqqqq/4Idf8AI1fFv/sH6V/6Muaqqqq/aQ/4KDfs4/s1rc6RrvikeI/FUAKjw9oTLcXEb+kz58uDnGQ7b8HIVqbUv+Ch/wDwURjks7CzX4GfB/U0KSzSK6T39swwRk7Z7oMpIwohhYEgk9/2T/2Svh9+yJ4GvPB/gfVNV1WfVrhLrVNQ1GRd1xMqbQURQFjQDOF5Pqzda+I8Np8Gf+CvMWqeJ4hDp0/j+yv/ADpThFivhGwlyf4Uack+mwjtVf8ABVu70e3/AGH/ABtDqbRfabq90uPTg/XzxfwMdvv5STfhmv8Agivp2p2v7Mnii9uiy2d742uGtUYYzts7VXcexIC/VDVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVftF/tFfDf9mP4b3nxG+I2p7Io8xafp8LA3OpXOMrDCp6k4ySeFGSSAK+EXww+NP8AwVb+PV18WvjFd3mjfDHw9MYhHakrDBHkMNPsyeDKww0s2MjgnkxpXgvwT4S+HPhbTvBPgXw9Y6JoWkwiGzsLOIRxxKOTgDqSSSWOSxJJJJJqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv+Ctf7Sura3f6P8AsZ/CiSe+1rW7m3k8Rw2eWkkaRlNpYADqXYpKw9PK7Mwr9jb9mzSf2WfgVonw4hW3m1yZftviG9iX/j5v5FHmYPUogCxqeMqgOASaqqqq/wCC43/I1fCP/sH6r/6Mtqqqqqqqr4sf8Ej/AIK/F34l+J/ifrXxK8bWd/4o1SfULm3tTa+VE8rFiqboidoJ4ySa/wCHJfwB/wCir/ED/vq0/wDjNf8ADkv4A/8ARV/iB/31af8Axmv+HJfwB/6Kv8QP++rT/wCM1/w5L+AP/RV/iB/31af/ABmv+HJfwB/6Kv8AED/vq0/+M1/w5L+AP/RV/iB/31af/Ga/4cl/AH/oq/xA/wC+rT/4zX/Dkv4A/wDRV/iB/wB9Wn/xmv8AhyX8Af8Aoq/xA/76tP8A4zX/AA5L+AP/AEVf4gf99Wn/AMZr/hyX8Af+ir/ED/vq0/8AjNf8OS/gD/0Vf4gf99Wn/wAZr/hyX8Af+ir/ABA/76tP/jNf8OS/gD/0Vf4gf99Wn/xmv+HJfwB/6Kv8QP8Avq0/+M1/w5L+AP8A0Vf4gf8AfVp/8Zr/AIcl/AH/AKKv8QP++rT/AOM1+yP+wv8ADz9jzUfEupeB/F/iLWpPE8FvDcLqphIiELSFSnlovJ8w5znoKqqqvE2rf8FFv26/FWt+FPBar8HvhRaahPZtqmZbRtQhSRk3CXHn3O4K3EQjhOdrHvX7NH/BNj9nj9nV7bxBdaUfHXjCLDHW9dgV1gkH8Vvb8pDzyGO9x2fHFVX/AAU1/YY1r9o/RLD4tfCi1Sbx94XtDby6eCEbV7IMXCIx4E0bM5UHG4Oy5yFFfCL/AIK6+Pfg94ei+F/7THwc1vUvEvh2JbV9QE5sr2YIAoF1BMmfNwOZARuPJXOSfiz8XP2mP+CrfjjRfh38L/hrPoPgTRbwStvlaW1tZSCpur262qu5UL7IlG7DMFDk5r4BfBfwx+z38IfDXwh8JFpLLQLTy3uHXD3c7MXmnYc4LyM7Y6AEAcAVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV8fPjz8O/2b/htqHxO+JWqm20+0PlW1vEA09/csCUt4VJG6Rtp9AAGYkKpI+Hfw5+Nn/BWj4+XHxS+JE0/h34X+G5/suLZsx2cIIcWNoWGJLhwVaSYj5dwYjHlxnwJ4E8IfDLwhpfgPwHoNro2g6NbiCysrZMLGo6n1ZiSSzEksxJJJJNVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV+1n+0d4b/Zb+CutfE3Wnhn1FUNroWnSNg39+6nyo/XaMF3PZEYjnAP/AASw/Zy8R/FX4ga1+298aEl1G7u9QuG8PyXaA/bL2RmFze7SMbUJaOPHAbfgAxrVVVVVf8FyLWdPEPwgvGjIhlstXjR/Vle0LD8A6/nVpdW9/aQX1pJ5kNxGskTYI3KwyDg8jgjrVVVVVVVVVVVVVVVVVVVVVVVVVVrnhTwt4nWNfEnhrStWEP8AqxfWUc+z6bwcVY6fYaXZxafpllb2lrAu2KCCIRog9FUYAFVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVfGH4wfD/4EfD/U/iX8S9dj0zRdMT5mPzSXEh+5DCnV5HIwFHuTgAkeHNE+OH/BW/8AaIfX/E8t/wCHfhR4XnZP3AzDpdueRBFn5ZLyUBd74O3hiNqoh+HHw48FfCTwVpXw8+HmgW2jaDo0AhtLSBeAOpZieXdiSzM2SxJJJJqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqtR1Cw0jT7nVtVvYLOysoXmubieQRxwxoCzOzHhVABJJ4AFfEHxD4y/wCCrP7ZVh4E8IXd5Z/C3wm0ghuRGVFrp6sPPvWVv+W9wwRUUjgeWCBtc14O8IeHPAHhTSPBHhHS4dO0XQ7OOzsLSIfLFFGoVR6k4HJPJOSeTVVVVVf8Fz/+aJf9zB/7jq8Jf8ipov8A2D4P/Ra1VVVVVVffEv4caZdzafqXxA8N2t1buUmgn1WGN42HVWUtkEehr/hbHwr/AOimeFP/AAc2/wD8XX/C2PhX/wBFM8Kf+Dm3/wDi6/4Wx8K/+imeFP8Awc2//wAXX/C2PhX/ANFM8Kf+Dm3/APi6/wCFsfCv/opnhT/wc2//AMXX/C2PhX/0Uzwp/wCDm3/+Lr/hbHwr/wCimeFP/Bzb/wDxdf8AC2PhX/0Uzwp/4Obf/wCLr/hbHwr/AOimeFP/AAc2/wD8XX/C2PhX/wBFM8Kf+Dm3/wDi6/4Wx8K/+imeFP8Awc2//wAXX/C2PhX/ANFM8Kf+Dm3/APi6/wCFsfCv/opnhT/wc2//AMXX/C2PhX/0Uzwp/wCDm3/+Lr/hbHwr/wCimeFP/Bzb/wDxdf8AC2PhX/0Uzwp/4Obf/wCLr/hbHwr/AOimeFP/AAc2/wD8XWh+L/CXih5o/DPinSNWa3AMy2N7HOYwc4LBCcZwevpVVVVVVXjnx94J+GXhq68Y/EHxVpnh7RbIfv77ULhYY1J6KCTyx6BRkk8AE18e/wDgtH4e0LWW0P8AZ28AxeI7e3lIl1zXzJbwTgf88bdCshU8fPIUPBGznNfse/G3X/2jf2cvCPxl8UaTp+map4h+2+fa2AcQx+Rez267d7M3KwqTk9Se1V+0n/wVi03wB4+1H4PfAT4W33jTxVpl7JYXNxfxywwpdxsVeKO3RfOnKlSDzHyDjcOa1r/gpf8A8FEfh6i+K/iR+zbYaT4fkxltS8Gapp8G04+5NJLweRgksPm6Hiv2LP29/h5+2Bp91o0emHwv440mHzr3QprkTCaEEKZ7eTC+YmSoYEBkLAHIIY1VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV8Q/iD4P8AhV4K1f4hePdbh0nQdDtmuL26lyQijgAAZLMxIVVAJYkAAk1rWsfGz/grh+0bHoOiC/8ADvwm8LXG8M8YMWlWxyPOlAO2W8mCkKmTtGQPlV2Pwj+EvgX4HfD3SPhj8ONHXTtD0aHZEhO6SVzy8sjfxyO2WZu5PGBgCqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/4Kk/tbax4q1mH9iv4GNPqes6vdQ2/iZ9PJeSWVyPK02PHVmJVpcdPlQn/AFgH7Dv7Jmifsl/B228MyLb3Xi/W9l34m1GMZEtxg7YUPXyogSq+pLvgbyBVVVVVf8Fz/wDmiX/cwf8AuOrwl/yKmi/9g+D/ANFrVVVVVVX7a/8Aydz8X/8AscNQ/wDRzVVVVVVVVVVVVf8ABDr/AJGr4t/9g/Sv/RlzVVVV4f8A+CoPin4PfFHU/hL+2R8JtT8Mxw6jPHp/iKysZEElqJGEcslucmRCuCZYC2eyV8PviX8Pvix4bg8X/DXxjpPiTR7jhLvTrlZVVscowHKOO6MAw7gVVftDfAfwh+0p8KtS+Enje5vbbTNTuLaZ7myKrPC0M6S5jLBgCwQoSQcBzX/BUL9nn4Ofs8/sa+FPDPwj8C6focDePrIXNyiGS6vCLC/+aadsySHPQE4HQADiv+CXH/JiXwx/7i3/AKdryq8M/B34X+DfGfiL4ieGfA+k2HibxXOJtX1WOAG4uWCKmN5yVUhASq4UsSxBZiTqGn2Gq2NxpmqWVveWd1E0U9vcRCSOVGGGVlOQykEgg8GvDegQ/s0/8FXdM8GfDpJbfTLf4gW2n21tESwistR2K0PqUSK5K854QEkkZqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqrxJ4j0Hwh4f1DxT4o1e10zSNJtnub29upAkdvEi7mdiegABr4z/ABV+L3/BVT9oOz+C3waSfS/hroE7TJNOrrEsKna2pXa8ZYg7YouCN+3gs7V8AvgJ8O/2bvhtp/wy+G2lC3sbQeZdXUgBn1C5IAe4mYfedsD2UAKAFUAVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX/BQ79tLTv2Vfhr/AGF4WvYJ/iR4qgePRbbIY2EPKvfSL2VTlUB++/YhHx/wS2/Yw1Dw5aj9rD4z2k9z4v8AEyPP4ft78F5bWCbJe9kLcmacMSpPIRickyYWqqqqqr/guXYySab8GdSV12QT63CynqS62JBHt+7P5ivh/fJqngLw3qUaFFu9ItZVRjkqGhU4/Wqqqqqqv21/+Tufi/8A9jhqH/o5qqqqqqqqqqqqv+CHX/I1fFv/ALB+lf8Aoy5qqqq+JPwp+G/xi8Ny+Efif4L0nxLpMuT9nv7cSeW2Mb42+9G/oyEMOxrx/wD8Exfif8EvE0vxW/YL+MOqeG9UjO9vDmpXmEnUHIiWYgpKmekVwrL3L18Ov+CnvjH4WeIbf4X/ALd3we1fwHreNqa/ZWEhtrhQcGVoPmJT1kgaRSTwqivBfjnwZ8R/Dtt4s8A+KdL8Q6NeD9zfaddJcRMe43KThhnlTyDwQKr/AILV/wDJq/hb/soFn/6QX9f8EuP+TEvhj/3Fv/TteVVfFX4neD/gz8PNc+JvjzU1sdE0C0a4uJDjc5HCxoP4pHYqir3ZgO9f8E9PAXir9rP9trxD+1N4t04xaN4f1S41qc7SYvt05cWtqjd/KU789QIUz98Zqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqrm4t7O3lu7u4jhggQvLLIwVUUDJYk8AADJJr9rr9pj4i/8FA/jJYfspfsxpPceDI70Ce8QvHHqrxkb7u4YA7LOI8qCOSA5BYoq/sq/sveAf2Ufhfa+AfB0X2m/uNs+uavKmJdTutuC5/uoOQkY4VfVizNVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVftSftK+Bv2WPhRf/EjxjKJ7k5t9G0tHxLqV4QSkS+ijBZ36KoJ5OAf2Kf2cPHf7dfxu1L9r39pRWv8AwtBqJktbSeIiHVrmM4S3iRsj7HAAFIyQxUId37wgKqqFUAADAAGMVVVVVVV/wXG/5FT4R/8AYQ1X/wBF21fCf/klfg3/ALF+x/8ASdKqqqqqq/bX/wCTufi//wBjhqH/AKOaqqqqqqqqqqqr/gh1/wAjV8W/+wfpX/oy5qqqqr4y/tA/Bz9n7QP+Ei+Lnj7TNAgdSbeCaQvc3RHUQwJmSQ/7qkDuQOa+Lf7Vfxm/4KHwah8E/wBlz9nG1vvCUshjvPFHi2winW1OP9YpkzBayAEkYMkuOUANfsE/seal+x58OdX8O6/4ztfEGs+JL2O9vTZ2zRW9qyR7BHGzHdIOp3sqZz90Y5r/AILV/wDJq/hb/soFn/6QX9f8EuP+TEvhj/3Fv/TteVXirxV4b8EeHdQ8W+MNcstH0bSoDPe315MIooEHVmY8DsPckAcmvjT8Wfi9/wAFUv2gLL4I/BlJ9L+GehXLTpPPEyxpEvyvqN2BzuIJWKI4I3BeGZ2r4B/AvwJ+zl8LtI+Ffw9sBDYacm+4uXUedf3LAebcSnu7kD2UBVGFUAVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX/BQv8AbJ8VfHbxun7Fv7L5m1f+0r3+ztdvLBgW1W53YNlE3AWFCD5smQGwRkIrF/2IP2M/Cf7I3w4WxIt9T8ca5GkniHWVT7zDkW8OeVhjJIHQucscZCrVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVfF34teBfgd8PdX+JvxG1ldO0PRod8r43SSueEiiX+OR2wqr3J5wMkeCPCfxc/wCCtX7TNx458bfbNA+FvheTy3SFiY9Oteq2duxGHupsBpJD90HcRhY4z4U8KeHPA3hnS/BvhHSLfS9F0a1S1sLK3XCQRIMKo79B1PJPJJJzVVVVVVVf8Fv7OCT4bfDC/ZSZodcvYkOeivAhbj6ov5V8BLme9+BXw5vLqQyTT+EtLkkcjG5jaRkn8zVVVVVVX7a//J3Pxf8A+xw1D/0c1VVVVVVVVVVVX/BDr/kavi3/ANg/Sv8A0Zc1VVVfEP8Abw/aq/aN8da58Gf2IPg/e2S6ReyWOo+KtRjjka3ZXKF9z/6PbA7WI3mR2AyoB4r4Kf8ABKfRp/EH/C0P2w/H998VPF90wlmsWu5ms0cdpJnIluAOwPlpjgqwrw/4c8P+EtGtfDnhXQ9P0fSrGPy7WxsLZLeGBR2REAVR7AVVf8Fq/wDk1fwt/wBlAs//AEgv6/4Jcf8AJiXwx/7i3/p2vKvLq3sbSa+upNkNvG0kjYJ2qoyTgc9BX7cf7eGsfta+NofCmnXup+HvhNpN6Ps1nDCJLi8wcG8mjLorybclIi6qoON2SzV+zr/wUh/YP/Zj+G9n8OPhx8MPiqkUeJNQ1CbSdPNzqVzjDTTML3knGAo4UYAAAr/h9X+yv/0IXxV/8FVh/wDJtVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV/wAFMP27L7was/7LvwG1Ca58b62Baa7fWGXl05JgAtpBtGTcyhwCV5jBwPnYFP8AgnV+wjp/7MXhFPiF8QbCG4+J/iC223JJDrots2D9ljIJBckAyOOpwo+VSWqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqvHfjvwh8MvCGqePPHmv2ujaDo1uZ729uWwsaj0A5ZiSFVQCWYgAEkCvGnir4z/wDBWz9o638E+DI7vw/8LPC07SRyywkxabbHIN3cgHEl1KFKxxg4XJUEKJZD8Hfg/wCAvgR8PNJ+GPw30aPTtG0mLCjGZLiU/fmlb+ORzyW+gGAABVVVVVVVf8FubGST4NfDvUlddkHieaJlPUl7ZiCPb92fzFfsz3yap+zf8KNSjjKLd+CNGlVGPKhrKE4/WqqqqqqtV+B3wV13UbnWNa+D/gnUL+8laW5urrw/ayyzOTks7shLMT1JOa/4Z6+AP/RDfh//AOEzaf8Axuv+GevgD/0Q34f/APhM2n/xuv8Ahnr4A/8ARDfh/wD+Ezaf/G6/4Z6+AP8A0Q34f/8AhM2n/wAbr/hnr4A/9EN+H/8A4TNp/wDG6/4Z6+AP/RDfh/8A+Ezaf/G6/wCGevgD/wBEN+H/AP4TNp/8br/hnr4A/wDRDfh//wCEzaf/ABuv+GevgD/0Q34f/wDhM2n/AMbr/hnr4A/9EN+H/wD4TNp/8br/AIZ6+AP/AEQ34f8A/hM2n/xuv+GevgD/ANEN+H//AITNp/8AG6/4Z6+AP/RDfh//AOEzaf8Axuv+GevgD/0Q34f/APhM2n/xuv8Ahnr4A/8ARDfh/wD+Ezaf/G6/4Z6+AP8A0Q34f/8AhM2n/wAbr/hnr4A/9EN+H/8A4TNp/wDG68K/Dj4eeBJLiXwP4D8O+HpLxVW4bStLhtDMFztDmNRuAycZ6ZPrVVVVoPhzw/4W04aR4Z0PT9JslkaQW1lbJBHvY5ZtqADcxJJPUk5NVVV8Q/hd8Ofi3okHhv4neCdH8UaXbXS3cNnqlotxHHOqsgkCsCAwWRxn0Y+teDPBPhD4deGbPwb4D8N6foOh6fv+yadYQCGGHe7SPtReBl3Zj7sT3qeCG6gktriNZIpUKOjDIZSMEEehFf8ADFH7Iv8A0bh8P/8AwRQ//E1/wxR+yL/0bh8P/wDwRQ//ABNf8MUfsi/9G4fD/wD8EUP/AMTVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX/BRT9ujT/2XvBbeBfAd9b3PxN8SWrfYkBWQaNbt8pu5V/v9REpGGYFjlVIb/gmf+wpf+FPJ/al+PlhPdeN9bZrvQrDUQzS2CS5LXk+7k3Mm5iA3KKcn52wlVVVVVVVVVVVVVVVVVVVVVVVVVVVVeLvF/hXwF4cvvF/jXxDp+iaLpkXmXd/fzrDFCvTlmOMkkADqSQBkmvHP/BZD9lXwxqUmmeGNJ8a+LVikK/bbDTo7e3cDPKm4kSQ9usY+tfCb/grN+yr8UvEOneE7k+LfCmpancR21sNY0tXilmchVRXtpJSMsQoLBRk84FU7pGhkkdVVQSzMcAAdTWlftafs1a947g+GOg/Grwvqfie5uvskGn2V4J2kmxkorJlSRgg4PBBB5FVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV4w8YeF/h/wCF9T8aeNNdtNH0PR7dri+vrp9kcMa9ST3J4AAySSAASQK+MXxj+M3/AAVT+O9h8Efg7a3WjfDTSLr7QGuI22RRLlW1G92/xYJEcWeCwUHczNX7O37PPw7/AGZPhnY/DP4c2DJbQt519fTAGfUboqA88pHVjtAAHCqAowBVVVVVVVVX/BbKznk/Z18FX6qDDD41jic56M9lclePojflX7IV5Bf/ALKPwcmtySq+BNGiORj5ksokb9VNVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV+2X+1p4R/ZI+Fc3izU/Jv/Euqh7fw5oxkw15cADLsOohj3KzsPVVHLiv+Cf37I/i/9pz4j3X7Z/7UKz6xZXd+bzR7TUI+NZulbid06C1iKhUjA2sVAwETD1VVVVVVVVVVVVVVVVVVVVVVVVVVVVX/AAVy+LPjD4jftF+Hf2Z9F1WS30PR4bMvaFykVxqV2fllkx94JE8SrnO3dIR94ivgz/wT0/ZU+EXg6x8O3Pwi8M+MNUjgVb/WPEmlxajNdzfxOFmDLECeioAAMZyck3P7CP7KDeO/D/xI0f4N6JoOveGdSi1Gxl0aM2UXnRMHTfBHiJgGCkfJkEDBFV+2F+z7+3F+1F8edU+FXhrxyPDXwPFpaSm6dxBBJvjHnRyJH+9vHEiOQjYjUbMlSdx+Bnwqs/gf/wAFOPCnwl0/WJtVg8L+L0s1vZohE0+ISSxQEhcknjJx6nrVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVXjDxh4X+H/hfU/GnjTXbTR9D0e3a4vr66fZHDGvUk9yeAAMkkgAEkCvjT8afjf/wVN+N9r8EvglYXelfDnSrjzlW4ykSxKdp1C/K55wcRxc4yFGWYk/sz/szfDf8AZY+G9t8P/h/ZeZK+2XVtWnjAudUuQMGWQjoByFQcIOBkkk1VVVVVVVV/wWks57n9lLw9NCoK2vjyzklJOMKbK9TP/fTr+dfsOXkF9+yB8IZrckqvhSziORj5kTY36qaqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqrvxD4fsZ2tL7XdPt548bopbpEZcjIyCcjgg1BPBdQpcW00csUi7kdGDKw9QR1FVVVVUPFHhhpBCviPSzIW2hBeJnPpjPWqqqqqqqqqqqqqqqqqqqqqqqqqqqqvjj8a/An7Pfwz1j4p/EPUTbaXpUXyRR4M15OQfLt4V/ikcjAHAHJJCgkfAf4UfEz/AIKk/tIan8c/jQs1p8ONAuVhlt4mdIvKQ7otLtmGDnawaWQYIDluGkWtK0rTND0y00XRdPtrDT7CBILW1tohFFBEgCqiKuAqgAAAcACqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr/grx+yj43vfFNn+1T8OtNub6ztrCK18SJZhjPYvAT5V4AvPl7CqMw+55ak8Ekfsr/8ABXz4aeJtB03wj+0qJ/C/iS2iSF/EEFu09hqBAA8yRYwXgkbuNrR5ydyAhR4V8X+FPHeh2/ibwT4m0vX9IuxmC/027S5hkx12uhIP58VVf85lf+6gf+0KqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqviT8SfA/wi8F6n8QfiJ4itdF0LSYTJc3Vw2PoiKOXdjwqKCzEgAE18RviX+0H/wAFYvjRD8NvhZpt54d+GOhTJJIlwxEFlGTj7ZelTiSdgGEcIJxghf8AlpIf2c/2bvhj+zB8PbbwB8NtIWP5VfUtTmQG61ScDmWZx16nao+VAcAAVVVVVVVVVVf8FibGW7/Y/wDPiZAtl4qsJZAxIJBSZOPfLj8M1/wT0vo9R/Yt+FE8SMoTRWhIbrmOeRCfoSpNVVVVVVVVVVVVVVVVVVVVVX7Yeq6pof7K/wAVtY0TUrrT7+z8J38ttdWszRSwusLEMjqQVYHoQciv+Ghfj9/0XL4gf+FNef8Axyv2XtR1DWP2Z/hHq2rX9xe3174F0We5ubmUySzyvYws7uzElmYkkk8knJqqqqqqqqqqr40/8FUP2Wvg14j1rwVJdeJfE3iHQL2ewv7LSdKKLBdQuY5I2kuDEp2upBZN44yM1+xz+1Vp/wC1/wDDfWviTpXgy48NWmleIptHhtri+FzJMI7e3m81iqKEJ+0bdg3Y2Z3HOBVVVV8cvixoXwK+EXir4t+IxvsvDOnPc+Tu2m4l4WGEHsZJWRAfVxXwS8GeMv23v2uNL0zxjqMt7feM9afUfEN5kjy7RMyz7efkAiQxxrnC5RR2q5uPCnw58GyXNzLZ6H4b8MaaWdm/dw2NpBHz/uoiJ+AFftDf8FfPjX428Vz+Hf2arSHwpoCzmCyvZtPjvdS1AlsB9kivHEG4xGqMwz94k4Hhn/gpt+3V8F/Fkdj8UNTk1pI8PcaF4p0COykKN3DRxxTISAcEkrnnaea/Zj/aO8EftTfCfT/il4KWS1EkjW2paZPIHl027QAvC5HB4ZWVsDcjqcAkgeItf0XwnoGpeKPEeow2GlaPaS3d9dTHCQQxoXd2x2CqT+Ffs1ftTftXftoftj3un+DfiPqvhj4WWN62p32nQWlufs2lxMFig8xoy3mzkIrENkb5GXhMVVf8FUf2fY/gd+0tL4y8NWrWugfEWN9YtDGNiwXyuBdxqR33skvHTzwB0r/gnn+0nN+0v+zlpOu6/ffaPFnhqT+yPEDO2XmmjUGO4Pc+bEyMT03+YB92qqqqqqqqqqqqqqqqqqqqqqqqqqq8R+IdC8I6BqPinxNqttpmk6TayXV9eXL7I7eGNSzux7AAE14/8V/FL/grN+1Ha+BPApvdG+F3haRnjlmTCWFnuCvezLnDXM2NscfYYXosj18Lfhf4K+DPgDRvhp8PdITTtC0O3ENtCDlmPVpHbqzuxLMx6kk1VVVVVVVVVVVVVVVVVVVVVVVVVVVVVUyqylWAIIwQRXx7/wCCYf7LXxwe71iy8Ly+BvEdyWc6l4cIgjkkPO6S2IMTDPJ2qjNk5bvV2/7R3/BKT9oq20tNdGp6FfhLpoIZGXT/ABHYbirboznyp1wVzy0bYILIw3+DvFWjeO/COh+N/Dlw0+leIdNt9RsJWXaXgnjWSNiOxKsDiq/5zK/91A/9oVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVXx5/aB+F/wCzd4DuPiD8UvECWFkhMdpax4e5v5sZEMEeQXc/gFGSxABIZ/2lv+Cuvxk2KJPCfws8N3JIyDLZ6OpXufl+1Xjr9Nob+BDz8DfgT8Nf2dvAFl8OfhfoMen6dbAPcTNhp76faA0874G+RsDJ4AGAAFAAqqqqqqqqqq/4K2WC3n7FfiK4Mu02WsabMBjO7NwqY9vv5/Cv+CY99JqP7DPwvnlRVZINRhAX0j1K6QH6kKDVVVVVVVVVVVVVVVVVVVVVV+2v/wAmi/F//sT9Q/8ARLVX7J3/ACav8Gv+yf6F/wCkENfFj/gqL+zT8GfiNr3wu8X2HjVtZ8OXZtbw2mlRSRFwAcoxmUkYI5IFeGv25f2dvEHwIb9ou/8AFc/h7wgLyayQ6vAIrma4j6xRwxs7SORyFXJxknABI1r/AILY/AW01lrTQ/hZ441HTUfb9slNtbu4B5ZYvMbjuNzKfUDpX7NX7ZfwK/assbhvhh4huI9YsIhLe6FqkIt722QkDfsBZXTJALRswBIBIJGa8Qf8Fd/2UvDWval4c1LTvHv2vSryW0uNmjwlfMjco2D544yp7V4h/be/Z18IfBXw78dvFnjF9J0LxXafaNHsZ4N2oXnJBVLeMsSQRgnO1cjLDOavf+C2vwLi1Y29h8JPHNxpwfH2mR7WOUjPUReYR0xwXFfs6/tc/Av9qTSpr34U+KzLqFkge+0a/i+zX1oDwC0ZJDL23oXXJxuzxVeLv+Ctn7K/grxXrXg3WLDx01/oOo3FhdGHSImQywyNG+0mcEruU4OBxX7Nn7S/w5/aq8C33xC+GMOsRaXp2rSaXMuqWqwS+ekUUrEKruCu2dOc9c8cV+0X+0V8P/2X/h/H8SfiTDq0uky6jFYKumWyzy+bIrsvys6jbiNsnPpWgf8ABXn9lHxJrum+HdO0/wAeC61S7itYDJo8KqHkcIuT55wMkZr9pb9qP4Z/speE9L8ZfE+DWpbDV9R+wW40u1WdxL5bSfMGdMLtQ8564r46+MtH+I/xu+IXxC8PLcLpfijxVqeqWK3KBJRBcXUkse9QSA21xkAnB7mv+Cc/7ffwL/ZV+CGt/Dz4nWviiXVNQ8VXGqQnS9Pjnj8h7W1iGWaVSG3QPxjpjnnj4DfHHwV+0Z8M9O+K/wAP4tSj0XUpp4oF1GBYZg0UjRvlVZgBuU456V+0T+2l+z3+y+qWfxO8ZE63NF5tvoWmQm6vpU7MUBCxA9mlZA2DgnBqb/gtx8FF1Dyrf4O+Nnsd3Ezz2qy49fLDkZ9t9fsx/t7/AAF/av1q48J/DyTxBp/iKzsmvZtK1fTvLcQIyI8gkiaSLAaRBguD8w461Vf8Fr/ilNonws8D/CGwu9j+KNVm1G+RTyYLRFVFb/ZaScN9YfY1/wAERPhvb3fib4k/Fu7gUy6bZWujWLkZI85mmnx6YEMH/fR/H/grr491PwV+x7e6Xpdy8B8YeILPR53Q4JiKy3LrkdAwtdp9QSOhr/gjT8N9A8XftGa94z1ywt7yTwd4fM2mrMu4wXU0qRiZR0ysYlXPbfkcjI/4LU/D/RNU+A3hP4km0hGtaD4lSxS424ZrW5glZ484yf3kMTAHgfN61/wRD8X6jb/EH4leARM7WF9o1rqflnkJLBMYsj0JW4wfXaM9BX/BYj9qFvDfhPT/ANmLwbqRGqeJVS+8StC3zRWStmG3OOQZZF3EcHbGAcrJX/BOr9l1f2ZvgBYwa9p4h8aeMBHqfiEsuHgYr+5tT/1xRiCP77yEcEVVf8Fi/htD4t/ZXh8dxQA3fgbXra5Mvdbe5P2Z1+hkktz/AMBFf8EZvirceFf2htb+FtxcEWHjnRGeOLPW8s8yxn/vy1z+npVVVVVVVVVVVVVVVVVVVVVVVVVVVft8ftO+NP2v/i5YfsY/s0CbVdHGpLb6ncWkhEesXiNlgzDgWluVLFz8pZC/REY/smfsveCv2UPhPZfD/wAMxx3Wq3G2417VymJNSu9uGb1Ea/dROirzyzMxqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqL4ofDmfxzdfDGPxxov/CW2UEc82iNeIt2I5BlXERO5lI7gHGRnGRVf8Fi/jD4I+KnxW8B/DD4fahb69qng6C9TUptOYTj7TdvAEtQUzukT7Pyo6GXHXIH7O3gbU/hl8A/h38PdcJ/tPw94YsLK9UsG2zxwIsigjsHDAewFV/zmV/7qB/7Qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq1jxV4X8PAnX/Eml6aFAJN5eRw4BOAfmI6nitY/ag/Zq8Pqx1r9oL4cWbKGOyXxRZq529cL5mSR6AVq37f37Geil1vP2hvCshjIB+ySyXXUZ48pWz+HTvWq/8ABUr9hvTCyR/GaW+kSTYyWvh7UWx7hmgCkcdietal/wAFfP2NrFZDa6v4t1ExthRbaEy+YM9R5jJx35wavP8AgtH+yjbTeVB4Q+J92oGfMh0mzC/T57tT+lXH/BbX4ErA7Wnwk8eSTAfIkjWiKT7kSkj8jU//AAW++GKwu1t8C/E8koU7Ek1SBFJ9yFJA98Gv+H4vhT/o3bVf/Ckj/wDjFX//AAXK0uMoNM/ZourgEHeZ/Fyw49MYtHz+lf8AD87/AKtc/wDL3/8AuGo/+C5sbSIsv7L7Km4bmXxruIHcgfYRk+2RX/D8Xwp/0btqv/hSR/8AxirH/guD8P5d/wDafwD8QQYx5fka3DNu9c5jTHb1/CrH/gtx8FJFf+0vg742gIxsEE9rNn1zl1x29a0//gtV+y/cJCmo+Afibayu2JCunWUscYzjOftYYjGCcLn61p3/AAWC/Y7vgftN54zsMMAPtOh7sj1/dyPx+taZ/wAFTP2G9SKxyfGSaxkaQIq3Xh3URnPfcsDKB7kjH0rSP2+f2NtbVWsv2h/CUe5SR9suGtDwcHIlVcfQ9evStG/aI/Z+8SEr4d+Onw91QgkEWXieznwQAT9yQ9iPzqyv7DU7dbvTb63u4GOFlgkDqfoQSKqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr9sT9u74VfslaFJp97cReIfHl3AW03w1bTAOuR8s1ywz5EXpn5n/AIQQGZfgr+y9+0X/AMFK/iIn7QH7Sev3+jeAd+2zZYzCbqANn7Np0LZEcI5BnYHLZP7x95X4ffDzwV8KvB+m+Afh54ds9D0HSYvLtLK1TCoCcliTkszEkszEliSSSTVVVVVVVVVVVf8ABUy0huf2FviPJJFve2fSpIjz8rf2naqT/wB8sw/Gv+CUd5JdfsQeCoHnDi0vtUiReP3YN9M+Pzcnn1qqqqqqqqqqqqqqqqqqqqqq/bX/AOTRfi//ANifqH/olqr9k7/k1f4Nf9k/0L/0ghr/AIKRf8nufFP/ALCFt/6RQV8Cvg38aP2pfE2lfBT4bK18mnC4vVS7nMVlpaOUE1xIwB2hisSkgFmIQAHgV+1H/wAE6Pjr+yt4Ot/iF4ovvD/iHw49wltdXuizSubGV+E81JI0IRj8ocZG4gHaWXPwX+K3ib4H/FLw18VPCV3JDqHh6/juNittFxEDiWFvVJELIw9GNaPqllrukWOt6dIXtdQto7iBiMbkdQyn8iK/aF/5L98TP+xw1b/0rlr9nT9m346ftpeKbfwd4Nu45NP8JafHBPqeqzslno9q0jskY2hjlnaVljRSWYuxwNzD9pn/AIJe/HL9nHwJc/Exdb0Txh4e00A6nJpayR3FihOPNeJx80YJALKxIzkgKCw+E/xS8Y/BX4h6H8TvAOpmy1vQbpZ4GOSkg6PFIARujdSVZe4Y18J/iJo/xd+GXhb4naACtj4n0m3v4oy2Wh8xAzRsR/EjEqfdTX7Qv/JfviZ/2OGrf+lctf8ABFT/AJNX8U/9lAvP/SCwr/gsl/yaLaf9jhY/+ibmvhN/yVTwZ/2MFj/6UJX/AAUD/ZN8Z/tefDTw74J8E+JNF0a60bXf7Qll1UyhHTyJI9q+WrHdlweRjANfEnwRqHwz+Inin4b6td291feFNavNJuZ7fPlyy20zxOybgDtJQkZAOCM1+yl/wTu+J37XHw61D4keCvG/hfR7HTdal0mSDVDOJWljhhlLDy42G0rOo65yDXwA+HOv/sPfsX6lo3jHUdM1y+8A6XrOtTyaeZPJmVPOuQo3KrfdAB469K8c+NvE/wASPF+r+PPGmsT6prmu3b3V9dTNlpJGOTj0UDACjhQABgACvgv/AMEdPC3xM+DXhj4g6r8eruDU/FGkW+oxLp2lR3FrbCaNXCZMgMpUNgkFOQeOK/YG/YC179j3xz488SeJ/FukeJRrVna2WiXlnDJDKsAd3nE0bZCFmWDAV3HynnsKr/gtH4huNR/ad8O+H/Mb7No/g23KoWyBLLc3LOwHbKiMf8Br/gjP4eXSv2UdU1ho8Sa34wvJg5xkokFvEBx2DRv155PbFf8ABT/4Pax8Yv2R/EFt4csJL7VfCd5Br9tbRpueVYA6TbR1yIJpmAHJ2471/wAE0P2mPCH7NPx/m1D4i38ll4W8V6U+l3t4FZ0spfMSSGd1UElQUZCQOBKT0Br/AIK0fthfCP4s+BfDHwX+EXjOx8UeXq41jV7/AE2UyW8IjhkjhhEg+V2YzOxCk7fLXPJr/glhpWm/s9fAv4sfti/E1XsfDwtEstMLgI14kDMZBHn7xkneCFOxdWH0/YW+G3iH9tX9rjxH+1H8Z0im8PeF7/8AtrUWuCRbtd9bS0Usf9TCiBiCSAkKK3D18dP+Cyvwh8A69deGvhB4GvfiC9m7RS6rJfjT7FnGQTC3lyPMoI67UU9VYjBPwh/4LVfDfxNrltonxh+FmoeDrW4cJ/a+n6h/aUMRJ+9LF5cciIB1KeYf9n00TW9H8TaNY+IfD2qWupaZqVulzZ3ltKJIriJ1DI6MOGUgggj1r9uXw+vib9j/AOLumtH5nk+Fby9xwMfZk+0Z59PKz+FfsE+IJ/DP7ZHwl1G2kKPN4jisiQ235blWt2H4rKRjvnFVVVVVVVVVVVVVVVVVVVVVVVVVVf8ABUL9t26+HOlyfszfBvUpZPHPiOFYtburLLS6ZbTAbYI9vIuJlYdPmVGBGC6Ef8E5f2IbP9mHwCPG/jnTopPiX4otlN+zAMdJtjhls0PPzZw0jDqwC8hATVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVfte/8Erz+0N8UdY+NXgr4x3Wi+I9ZeJ7my1Wz8+3DRRJEnlSxlXiUJGnBVz7in/4Jdft738T+HtV/am0eTQJVMckJ8VavKGj9DA0IQj/AGd2K/ZJ/wCCWXwt/Z18SWPxH8b+IH8eeL9OPmWDS2Yt7HT5e0kcJLM8q/wu7YB+YKrAEVX/AA7p8af8Nyf8NY/8LG0T+yf+Ek/tb+yPskvn7PL27N/3c989Kqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq8ReLPCvhCzGoeLPE2k6LanOJ9RvY7ZDjr8zkDivGP8AwUA/Y08DF01r9oPwxctH1XSHk1XnOMD7Kknf/HpXiv8A4LI/sm6EWi0DTfHPiV/4Hs9KjgjPPczyow45+6eo98eJv+C4tgivF4O/Z3nlJHyT6n4kEe3p1jjgbPf+MdqP/BT/APb18fv5Xwt/ZispUc/KbTwvqepy8dgUcL2b+H8sUPit/wAFp/iGmzQ/BWtaDHKNpU+HNO088gD714u5eucgjHPTHA/Zx/4LH+PTv8Q/F7WfDplOWB8bR2mzknpYMwHPHHqB06L/AMEqf22vGG2T4jftO6VJuwSH17U9RZcDjPmRoMgkjgkdSCa0T/ghvcsVk8R/tIxIAfmhsvCpbPHZ3uRjn/ZOQO2eNH/4IjfBeBVHiD4y+Nb0gLuNnbW1rnH3sbkkxnt1x71pP/BGv9kfTihvNU+IOq7CSRd6zAu7I6HyrdOnXjHvmtL/AOCUX7EOn7Ptfwx1PUti4P2rxFfLvPqfKlTn6YFab/wTg/Yk0kxG1+AOkSGEEL9pvry4zxj5vMmbd175qy/Ya/Y/0+EwW/7Ofgd1LbiZ9LSZs4A6vkgcdOn51a/spfsuWTxS2n7N3wuikhIMci+D7EMpHQhvKzn3qH9n/wCA1vKk9v8ABLwDHLEwZHTw1aKysOQQRHwa/wCFTfCv/omnhT/wTW//AMRVj8O/h9pZdtM8C+HrQyABzBpcMe7HTOF5r/hEvCn/AELOk/8AgFH/AIVN4M8H3EL28/hTR5I5FKuj2EZDAjBBGOQR2r/hU3wr/wCiaeFP/BNb/wDxFX/wO+CuqFG1P4P+CbsxghDP4ftZNueuMocdKvv2Yf2adUdJNS/Z4+Gd2yDCNP4SsZCo9sxHFX/7FP7I2pGY3H7OHw+TzwQ3kaFDBj/d8tRt/DFal/wTp/Yo1b/j6/Z/0NPkKf6Nc3Vvwf8ArnKvPPXr71q3/BKP9iDUVIs/hfqOlkptBtPEd82Dn7w82V+e3px0rWf+CNH7JWpsWsNa+Iek5bIW01i3cDjGP3tu5x365961v/giD8L7gY8OfHPxTY8j/j90yC67HP3DF3x/k5F9/wAEUfiX4euDe/D79pTTXuVX5JZ9Gn05uxxuimlIG4D8gab9gz/gqN8OMt4D/aPk1CGPPl2+neOr+IMBnG6OdEjzjtkgFuvU02q/8FsvhZk3NlrevWseflFvpOseYBk5zHumHQ8ZB6DHSm/4KX/8FBPhcpj+Lv7Ndt9ng/1k+o+FdR0yQ4wM+Zv8vHuE/i64xXhL/guF4ZnCRePPgBqdkQPnm0jXUut3TkRyRR478bz09+PBv/BXH9jPxUyR6v4k8S+E2c4A1nQ5Gwe2TamYD65x64HNeB/2rP2afiT5Mfgr46+CdRuJ/wDV2n9sxQ3J/wC2EjLIP++aR0kRZI3VlYZVgcgj1FVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVLJHDG0ssioiKWZmOAoHUk9hX7ZX/BU600G/n+DX7JmzxN4supfscviG3h+0w28rHaI7OMA/aZsnAbBQHGA+eP2Pv+CXura9rafHn9tB7jW9d1Gf7bF4XvpvtDSSNz5uouSfMYnnyM4HAcn5oxb21vZ28VpaQRwwQIEiijQKqKBgKoHAAAAxVVVVVVVVVVVV/wUVspr/8AYp+KsEBXcmkxSnccfLHcwu344U1/wR8vo7v9juC3jRg1l4n1CFye5IifI9sOB+Bqqqqqqqqqqqqqqqqqqqqqq/bX/wCTRfi//wBifqH/AKJaq/ZO/wCTV/g1/wBk/wBC/wDSCGv+CkX/ACe58U/+whbf+kUFf8EM7W2e5+NN60EZuIo9CjjkK/Mqsb8sAewJRSR/sj0r/goPZ2t/+xf8WILuFZUXQ/NVWHR0mjdD9Qyqfwqv2fXeT4CfDWSRyzN4Q0oszHJJ+yRZNftC/wDJfviZ/wBjhq3/AKVy1/wRRsLKP9mXxdqcdsgu7jx3cQyzAfM6R2NkUUn0Bkcj/eNeKvD2l+LfC+seFNbhSXTtZsJ7K8jcZDwyxlHB9irEVX/BLHUL/UP2HvAC3wJFrNqcMDsDloxfzkcnrgkrxxhQO1ftC/8AJfviZ/2OGrf+lctf8EVP+TV/FP8A2UC8/wDSCwr/AILJf8mi2n/Y4WP/AKJua+E3/JVPBn/YwWP/AKUJVftZf8nUfGX/ALKBrv8A6XzV/wAEVP8Ak1fxT/2UC8/9ILCvFXhrSPGfhfWPB+v2/wBo0zXbCewvYv8AnpDNGY5F/FWIr9qz9j/4rfspeNrrRvFui3N34anuHGieI4YSbW/iydgLDiObH3omwQQSNylWPwO/a7/aI/Z1dIPhX8TNSsNLWQu+jXJF1YuSct+4k3IhbPLIFY+tfsH/APBRTQf2sZpvh7410a08NfEKxtjcLb20jG11WFcb3g3EsjrnLRMWO35gzANtqv8AgsBaT237Yt1NKoC3XhrT5IjnOVAkTPtyjV/wSEkjk/Y209I5FZo/EWoq4ByVO5Tg+hwQfxFMqspVgCCMEEV+0N/wSB+C/wAV/EN74w+F3iq6+HGo6hI0txYw2C3unNIeSY4d8bQ7jnIVygzwgAxXwp/4IofD7QNat9W+L3xb1HxXaQSbzpWmab/Zsc2Dwskpkkcqe4UIfRu9f8FR/i/b634q8H/sJ/ATR449J8NyWsd3pOkxbVmv5AFtLJFXg+WjhiOQXlGcNGa/bF8Pn9hf/gnp4U/Z58G36Qa/451AQeJb+2ba14TGZb1lbglSwggHAzFwepz/AMEo/wBlP4fftC/EDxX4x+Kugx634f8ABNtbLBplxuENzeXDOUaQAjeqJC+UPBLrngYP/BWj9lD4a/AnX/BvxE+Evhq38P6V4qF1aajplopW3huYRGySRryE3o7AqMAeUCBljX/BGD4s6v4w+B3in4X6xeyXQ8B6tE+nmR8mG0vFdliA6hRLDOw/66Edq/a3mhg/ZV+MjzSpGp8A62oLtgbmsZgB9SSAB6kV+yFZzX/7VvwcgtwpZfHejSnJx8qXkTt+imqqqqqqqqqqqqqqqqqqqqqqqqqr9uv9sLQf2SfhU+o2clve+OPECvb+G9Mcg/OB81zKvXyosgn+8xVeMll/4Jh/sea54t11v20vj/FPqer6xdPfeGItSy8s8zsS+pyg9yxPlZ95AP8AVtVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVUt5Zw3ENnNdwpPcbvJiaQBpMDJ2jqcDk4qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqp3SNDJI6qqglmY4AA6mviR+2j+yr8JjJD43+OnhaC5hyJLOyujqFwh9GhthI6n6qK+In/AAWl/Z/8P+bbfDnwB4t8XXMZO2W4EWmW0noQ7GST84hU3/BTz9uf44O1p+z3+z3BBbyttW407Q7vWpojnHMvEIAwclo8fSh8Ef8AgsZ8eZFm8X/EPWfB9lc5ObjxHDo8e09mg0/MmO2GTNeHP+CKfjrxDejV/i/+0bbNdOc3C6bpkt/JL7efPJGR9Sh+leDv+CNf7KmgKkvifWPG/iibA3pdalHbQkg/wrBGrjPTlzXhP/gn7+xn4L2HSP2e/C9yUHB1aOTVM855F08gPT+nSvDXw1+HPgtUXwd4A8N6EIxhBpulQ2u3r08tRjqfzqqqqqqqqqqqqqqqqqqqvF/wa+EHxB8w+O/hZ4R8RNKDvbVdEt7pjznrIhOc859ea8Zf8EzP2KfGjSTzfBm30i5kJIm0bULmy2Z9I0k8r/xzj868b/8ABEr4M6oWf4e/F/xd4fdgfl1O1g1NFPbAQQNj2LH61N/wTF/bs+CgNz8Af2hoZreJiVttN1680aWQ/wDXLmIg9w0n51J8e/8AgsD+zwMeO/h9qni7TrTgve+HYtWhVRxl7jTyHwfV5M9Oea8Bf8FtYLe4XS/jN8Bruzlhbbc3OgaiGZWHUC2uFUqR6GWvht/wUz/Y0+JXlW8fxYh8M3smM2viW1fT9mf70zAwf+RK0HxH4e8V6XFrfhbXtO1jTpx+6u7C6S4if/ddCQeo6Gqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr4v8Axp+GPwG8G3Hjv4q+LbPQtKgyqNM26W5kxkRwxjLSucfdUE9zgAkfFr9qT9qL/gpR41uPgb+zf4Wv9A8BucXsfm+UZoM487UblfljiI58hcgnj96wWv2OP+Ce/wAKP2UrGDxHdJF4r+IUsWLjX7qABbTIw0dpGc+UvJBf77AnJCnYKqqqqqqqqqqqq/bhsI9S/ZB+L1vIjuE8J3swC9cxxlwfoCoJ9q/4Is6g11+yx4jspLlHay8dXapGMBo42srJhkDnBYvyff0qqqqqqqqqqqqqqqqqqqqqq/bX/wCTRfi//wBifqH/AKJaq/ZO/wCTV/g1/wBk/wBC/wDSCGv+CkX/ACe58U/+whbf+kUFf8EMf+a2f9wD/wByNft+f8mafFn/ALF+T/0NKr9nr/kgPwz/AOxP0n/0jir9oX/kv3xM/wCxw1b/ANK5a/4JYftu/Db9n6y8QfBr4w6odE0LXtSGqaZrDRNJDb3TRJFLHNsBZVdYodr42gq27AOa/bK/4KV/AnwR8Ite8OfBz4gaf4w8a+ILGWx0/wDseUzQ6f5qFWuZJh8gKBiVQEsW25AG4i3tri8uIrS0gknnmcJFFGhZnYnAVQOSScDFfslfCW8+Bf7Nvw/+FmpxpHqOi6Qh1GNCCEu5mae4UEdQJZZBnvjPev2x/CF34G/ar+K/hy8heIp4sv7mFXHPkXErTwn8YpUOe+a/4Jaftu/DD9nbTvE/wp+Musz6Loet36anpuqC3knhgufLWKVJFjVmXescOGAIG07sDBr/AIKhft4/Cn4++D9G+C3wV1KTXNMtNVTVNV1prWSCJ3jjkSOCFZVV2GZWZm2gfKgUkFq+E3/JVPBn/YwWP/pQlV+2dot3oH7WvxgsL1SJJfGmqXSgrj5J7h5k/wDHZFr/AIJW/tq/BD4CfDzxV8J/jJ4o/wCEa+164dY02/ls5poZzJBFDJETErFGAt0YbgAdx5zxXgr4y/Db4jfDBPjL4K8TJqfg+S3up49TFvLArR20kkczbJVVwFeGQZIAO3IyCCfh3+2x+x5+0H4d+wWvxR8LhdQj8u50HxQ8dlM2cZjaG4wsvb7hdfQnFf8ABUz4P/sceA9L0DxR8Br/AMN6d411PVTHqWg+Hr+OW2NmYnY3DQIzLbkOIlULsVg7fKSpI/4J/Nrq/tmfCY+HFc3f9vKJAoOfs5jcXHTt5Pm59qqv+C2vg2TTvjZ4B8erCUh1zwxJYbgoAeS0uXdjnHLbbuMHPYLX/BFDxZFqPwA8aeDGlDXGieLDdBcjKxXNtEFGOuN9vKcn1PpVV+1j+0Fo37MfwL8Q/FXUjDNfW0X2bRrOVsfbL+QEQx+pGQXbHIRHPav+CTX7Pes/Ff4o+If2wvilFLqH2C/nGkT3aZ+3arNlrm6wRz5ayEA9N8mQQY+P+C2PgrUdX+C3gTx1ZwSy2/h3xBLbXZQZES3UPyu3oN0CrnplwO4r/giL8RPD+m+I/iV8LtRv4INV1uCx1HTInba1ytv5yTquTyVEsTYHONx6A4/4Le+PtDltfhn8MLW9hm1aGa81a8gVsvbxFUihLDsHPnY/65n8f+CI/gLU9I+GHxG+I93bPFaeJdYs7C0ZxjzRZxys7KO67rvbnplWHY4/4KO+LYfB37FnxOvnkIkv9Oi02JVOC7XNxFCQPorsT7A1/wAE0PBkvjX9tP4dQCLdb6PcXGqXDbc+WLe3kdD+MvlLntuFVVVVVVVVVVVVVVVVVVVVVVVVV8a/jF4J+Afwz1v4p+P9Q+zaTosBcohHm3Up4jgiU/ekdsKB05ySACR+zd8JfH3/AAU3/ab1j9oL42QSr8PdAu1SWz3sInVSWg0uA8fIqkNKwwcMScNKDVra2tjaw2VlbxW9vbxrHDDEgRI0UYCqo4AAAAA6VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVXxI8e6P8L/BOqePNftry40/SI1eeKzRXlYM6oNoZlB5YdSOK+Mv7cPxJ8QXPlfDEyeG/Dd9Fus7qeyQX0yjKSbm3yIuHVwCmGwAcjNeCPiz4p8N/FPQ/ifq+tajq19pt6ktxLdXLSyzQk4lj3MSfmRnX2zWn39lqthbapp1wk9reQpNBKnSRGAKsPYgg1VVVVVVVVVVVVVVVVVVVVVVVVVVVfEH4tfC/wCE+m/2t8TPiD4f8MWpUtG+q6hHbmXHUIrEFz7KCa+Kn/BYT9lvwQZ7LwJb+IvH19GSqPYWn2O0LDsZZ9r4z3WNgeoq7/4KVft3/tFXculfszfA0aXbuxRbrTNHl1ma2PTMlxKot1AyOXjAyRmk/YF/4KQftKMt1+0J8ZH0TTblt01hrHiB7zYCc5Sztd1uD3xuTt07fDb/AIIsfAbw95Vz8TPiH4p8X3KY3Q2ix6XbP65VfMk/KUV8Ov2KP2UvhUsLeDfgT4VjuYMGO81Cz/tG4Qjus1yZHU/QioYYbeFLe3iSOOJQqIi7QoAwAAOgx2qqqqqqqqqqqqqqqqqqqqqqqqqq8ffBz4TfFW2Np8Sfhr4Z8Tpt2qdV0uK5dB/ssyllPuCCK+JP/BI39kPxz5tx4a0jxB4Iu3yQ2i6m0kW71MVyJQB/soV9sV4h/wCCSH7S/wAH9Sk8TfszftBRT3KYZVW4uNBvGAyQoeJnRyP9pkHJ6dK/4at/4Kmfsm/ufjV8O7zxToVrw93rWkC7hRB1xf2RA3EAnMrOe+K+E/8AwWh+Bnicw2Pxa8DeIPBN0wAe7tWGqWinoSSgSUeuBG3cZ9fhf8f/AIJ/Gq0S6+FfxR8O+JC0e9razvlNxEvq8BxLH1HDKKqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr9sX/gpN8J/2ZUvfBnhU2/jT4iRhozpVvP8A6Npr9M3cq52kdfJX5zjB2Ahq+EX7IX7UX/BRPxla/HP9qLxXquheC5sNZGSPyZri3PPl6fbMNsEJAH71lw2QwEpJNfCX4OfDT4F+DbXwF8LPCdloOj2oyY4FzJO+OZJZDlpZD3ZiT0HQACqqqqqqqqqqqqq/adsDqn7NfxZ0xZRGbvwNrUIcjO3dYzDOPxr/AIIj3xk+C/xD0zywBb+KIpQ+eu+1QYx7eX+tVVVVVVVVVVVVVVVVVVVVVX7TfgfxF8S/2eviJ8P/AAhaR3Wt+IfDl5ZWEEkqxLJNJGVUFmICgkjknFf8On/23P8Aom+k/wDhRWf/AMcr9n/wnrfgH4DfDbwL4mtkt9Y8OeEdK03UIUkWQR3EFpFHIoZSQwDIwyOD1Fftof8ABOz9qz4y/tPePPiZ4A8D6dfaBrl5DJZXEutWsDOq20UZJR3DL8yMORX/AASz/ZP+N37L3/Czv+FyeG7TSv8AhJf7I/s3yNRhuvM+z/bPNz5TNtx50fXGc8dDX7W3w98U/Fj9m34gfDnwRYx3mu6/pD21jBJMsKySFlOC7kKvAPJNf8On/wBtz/om+k/+FFZ//HK+D/h7VfCPwk8EeE9dgWHUtF8OafZXkSuHCTRW6I4DDIIDKRkcGv2hf+S/fEz/ALHDVv8A0rlr9n79gHx9+0/+zrqPxd+EurWc/iTRfFF1pVxoV9KIFvIEtbWVGhlPyrJuncFXIUjb8y7TutP+CcH7bF5rB0NPgHq8c6nDSzXtpHAOnPnGXyz17Me/oa/Yb/4JY2/wW8Sab8X/AI96hp2teK9MkWfSNEs/31ppso+7NI7AebMpwV2gKjDcC52la/4KH/8ABOy6/advYPix8Jr2wsPH1lai2vbS9cxQaxAmfL+cA7JlBKhmG1htVioUGpf+Ccn7a8Wsf2GfgDrLXG8oJFu7UwZHfzvN8vHHUtivhP8A8EffF+nfCHxn4l+Kc2n3vxBvfD91F4W8OWt2DBZXhT9288/3GkyNoAzGu7cWJwV8Hf8ABLf9trw74u0PxA3w30fGmalb3R3eILNh+7kVuQJckcdBVf8ABRL/AIJt+I/2g/FzfG74I3Onr4snto4Na0e8lEC6n5ShI5opT8qyhFVCr4UqqncpB3N/wTr/AG1RqD6Wf2ftd86PqwuLYxdO0vm7D+Br4I/s3fGPRP8AgnBH+zRqjWHhXx7f6Hq+ny/arhbiG1+2Xty+GkgLg5hmwShbaW74xXjT/gl1+2r4Ou5YYfhRH4gtYs7bzRdWtp0kx/dRnWX80HWvCn/BNH9tbxbeR20XwUu9KhZtr3OrahbWiRD+8Q0m8jn+FWPtwa/YN/4J0aN+yhcTfETx1rlp4k+Id7bG3SW0RhZ6VC2N6QFwGkdsYaVgvy/KFUFi1V/wVx+DFx8Tf2XZPGmkWZm1T4d6iuqHam5zZOPKuVHoAGjlJ9ITX/BG34uQeCf2itX+GWpXqw2fxB0cpboxwJL60LSxDPT/AFLXQHqSB3qq/wCCjHwP/a8/ax+Nfh/wJ4F+GF/B8N/DUiQW+q3F5bxwTXM5UT3joZd5jjXCKNu7CSEZ34r4PfCvwt8Efhh4c+FPgy38rSvDdiltExUBp3HMkz4/jkcu7f7TGvij8M/B3xk+H+ufDLx9pa3+ha/am3uoc4ZeQVdDztdGCurdmUHtXxZ/4JN/tV/Cjxc2s/A6aLxlplrP52m6hpupR6dqFsAfl3pI6YkHrE7Z68dB8LP+CUn7WXxi8Yprvx7vv+ES0+4kEmo6lquqJqeo3KjAwiRyPlyBjMrrgc4OAp+Ffwx8HfBn4e6F8MPAOm/YdC8P2ot7WJjuduSXkduNzu5Z2bAyzE96/wCC2Hxdt9N8BeB/gdYXQ+2a3qD61qEat8yW9urRQhh/deSVyPeA+lf8ETvgvci48c/H/U7TbAYk8P6RIyf6wkrNdMM9httlBHXLjsaqqqqqqqqqqqqqqqqqqqqqqqqvbyz06zn1DULuG1tbWJpZ55pAiRIoyzMx4VQASSeAK+PvxK8e/wDBUb9qjSPgd8H7qe3+G/hu5dobto2EQhUhbjVZk4zkHZEjYOGVflaV6+EXwn8FfA/4c6H8Lvh9pgstF0K2EUIbBkmfq8sjADdI7FmY4GSx4AwBVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVftdRSTfs4+N0iQsws4mIA7CeMk/gATX/Csz4p/Yu0jx7p9uXvvCeuXhm2rktayuiv7na4jb2G8+tV+wt8UV8a/Ck+DNQuS+qeD5BbgMeXtHyYT/AMBw8eOwRfWqqqqqqqqqqqqqqqqqqqqqqqqqr4nfG/4QfBfTxqXxV+JPh/wxE6bok1C9SOWcc/6uLO+ToeFUng18Yf8Ags/8EfC4n0/4N+Cdb8cXigiO9vD/AGXZH0Yb1aZsddpjTPHI7H43/wDBWD9sOQJ8NPC2reCPDd6uY7nSrL+xbbyz0db65PmuQOvkvz2XoB8PP+CMfjDxVqI8TftJ/HiWe8uCGurXQ1e8uJc9d15c4ww/65OMnrxz8Kf+CdH7IHwjNvdaT8ItP1/UoAMX/iRjqcjMOjeXLmFWyM5SNcHkYqzsrPTrSKw0+0htraBAkUMMYRI1HQKo4A9hVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVfFv8AYj/ZZ+NqzzeOfg5oX9oz5LanpkJ0+7Lf3mlgKlz7PuHqK+K3/BFiC1um1/8AZ1+M13pl5A5ktrDxFGTsfqNt3bgMgB4H7pj0545b4h/8FZP2LmKeLdK1fx34WsjzPqEB8QWjIB943MR+0wrgYAkdAOOOlfBn/gtD8H/Eq22mfGzwNq/g29YhZNR04/2lZD1dlAEyD/ZVJD718MvjX8JPjPpn9r/Cv4i6D4nt1XdItheLJJCP+mkX34zyOGUHkVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVfET4k+AvhL4TvPHPxJ8Wad4e0KwXM15fS7Fzg4RR1dzghUUFmPABNfHn/got8ef2sfFrfAb9irwprun2OoloX1K2Tbqd9HnDSBwdtjBzy5YMBgl0yVr9jv/AIJVeBfhK9j8SP2gGtPGvjhWFxFprAyadpkmcgkN/wAfMo67nGwE8KSochVVQqgAAYAAxiqqqqqqqqqqqqqqvidpzax8NfFmkokbte6HewBZB8pLwOuD7c81/wAEN9QSXQvjDpIMm62u9HmIP3cSJdqMc9f3Rzx6fhVVVVVVVVVVVVVVVVVVVVVVVVVVX7Qv/JfviZ/2OGrf+lctf8EVP+TV/FP/AGUC8/8ASCwqqqqqqqqqqqtX0nTde0q80LWbKK8sNRt5La6tpl3JNE6lXRh3BUkEe9an/wAErf2sPhT8cW8afAO80G50zw7riah4Z1C71UQzBEcSRLKhHJX7j9nwezVoVzqt7omn3euaYunalNaxveWazCUW8xUF0DjhgrZG4dcZqqqqqv2tv2Bv21/2ov2gvEHxOu9K8M2ukXM62WiQ3GuKTZadESsQZVU4JBaV1GfnkfGeK+Avwa8M/s/fCLw18I/CeZLLw/ZiN7hkCvdzsS007AdGeRmbHbIHQCqqqqqqqqqqqqqqqqqqqqqqqqv+CpH7XWt+Itai/Ys+BklxqGtazcQ23iaSwO6SaSUjytNjI6liymXHqqE8yLX7C/7IOhfsk/CaPSLhLe78a+IFjufEuooM7pQDst42/wCeUW5gP7zMzcbgBVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV+1L/yb146/wCwU/8A6EtfsKaVZeJP2bNT8PazF9psL7Vb61nhc5DRSRRhl9gQzfnXxP8AAeo/DLx9rfgXVNzS6TdNGkjLt86I/NHIB6MhVvxr9mT4rD4Q/FvSdevbkx6PfH7DqozgCCQgbz/uMEf6KR3pWV1DIwYEZBHeqqqqqqqqqqqqqqqqqqqqqqq8U+LvCngfRZ/EfjTxNpWg6VbD99faneR2sMf+87kKOnrXxt/4K+fs2fDgXGmfDW21T4kavFkKbEGzsQw7NcSruP1jjdT61J+09/wU5/bXkex+B/g++8HeF7ptq3mh25sIgmeC2pXB3FgBz5LIT/d5xXwx/wCCM3iHxLfjxV+058brq8v7pg91ZaEWuJ5Dj+O9uQTu7EeU3s1fB39ir9mH4Ei3uPh/8I9FTU7fBXVtRiN/ehu7LNNuaMn0j2j2FVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVXxn/AGMP2Zvj551z8RfhRpE+qTA51ewQ2N7u7MZoSrPgknD7h7V8T/8AgjZ4u8JaqPGH7LXxtubW+s38y0s9bke0uYCB1jvbYfe6gAxp2y3erT9sT/gpD+xbdRaT+0V4AvfFfh2FhGLrXYPMDDgARanbZVmPH+tMh+bkZ6fA/wD4KxfsufFf7PpnjHU7z4ca1LhWh10BrMtx9y7T5AvP3pRF0PtnStW0rXtNt9Y0PU7TUbC7Tfb3VpMs0Uq/3ldSQw9waqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr9rn/gpV8Gv2aVvPCXhuaHxx4+iDRnSbG5H2fT5Bkf6XMMhCCOYly/YhAQ1fD79mv8AbA/4KX+L7T4tfH/xNe+GfAOd9jLNbmKPyTg7NNsycbSMAzvwcZLSFcV8B/2cfhB+zZ4SXwh8JvCcGmxOFN5fSfvLu/cD788xGXPXA4Vcnaqjiqqqqqqqqqqqqqqqq5t4Ly3ltLmNZIpkKSI3RlIwR+Vf8ESLifSvH3xf8L3TOkxsNOeSNTlN0M1whyfUGXA+pqqqqqqqqqqqqqqqqqqqqqqqqqqqvGX/AAR8/Zn8ceL9c8a6t44+JsV9r+pXGoXMdvqdisaSzSNI4QNZkhQWIAJJxjJPWv2X/wBl7wB+yX4Av/hx8ONX8Qalpuo6xJqssut3EM0yzSQwxFVMUUahNsCEAqTktz0Aqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/4KBfti6f+yh8J3Xw/d283xB8TpJb+HrR8P9nGMSXkinI2R5G0HhnKjBAbH/BK79jm+023/wCGvPjJbT3nijxKJJ/DUd/l5YYZs+ZqEhbkyz7m2k/8s2LZPmDbVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX7UUby/s+eO1jRnI0iQ4UZ4BBJ+gAJr/gnuQfgNcAEHHiC5z7fu4a/4KA/CP8AtLRNO+MGj2ubjS9tlq2xfvQM37qQ/wC67FCep8xey1X7FPxiX4j/AAyTwpq96Zde8JhbaXzGy01sc+TJ74AKH/dBP3hVVVVVVVVVVVVVVVVVVVVVV8dv23P2af2dlns/iD8SbKXWoQw/sLST9tvtw/haNDiInsZSgPrXj3/gq9+0R8ctdf4f/sgfBm7s7m4JWK7eyOr6kUzgSCFAYYAM8lxKo67h1rwr/wAEy/2wv2mNZt/G/wC1r8YbnRo5CGFte3h1a/jQ8lEiVxBbqfQP8uTlOMH4Jf8ABN79k/4ILBe2Pw8i8V61CQ39q+KNt/IGHRkiKiGMg8grGGHHJwKjijhjSGGNURFCoijAUDoAOwqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq7s7TULWWxvrWG5t50KSwzIHSRSMFWU8EEdjXxy/4JefspfGYXOpaX4Sk8Ba7NuYX/hki3iZ+2+1IMJGeTsVGOT83etW/YZ/4KB/sZ6jP4o/Zh+JN34q0WKQyPbaLL5csoB+9Nps5aOUngYQyt16V8If+CxviHwvqY8EftXfCK9sNQs38m71LR4Gt54WHH76xnIIP94q49k7V8Hf2jPgh8f9MOp/CL4kaP4h8tN01rFKYrq3HHMlvIFlQcgZZQD2NVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVfGP44/Cv4BeEpfGvxY8ZWOg6auVhEzFprpwM+XDEuXlfpwoOOpwMmvi7+3r+1D+234sm+Cf7HvgzW9B0S5JSa5tHCajcQk4MlxcghLKE8AhWB7GRg22v2Rv8AglF8NPg+bPxz8dWsfHnjJCssVi0ZbTNOkx2Rublwc/PIAvQhARuoKqqFUAADAAGMVVVVVVVVVVVVVVVVVX/BKg/8I7+238a/BIRofK03Uv3Cnci/Z9UijxnPUebgfU1VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVfGH4s+DPgb8Ntd+KXj7URaaNoNq00uCPMnfpHDGCRukdyqKOOWGSBkj9mb4V+Nv+CmP7U+s/tD/Gm1b/AIQHw7eJ5tiSWhkVDuttKiJHKKpDykYJDEnDSgiGGG3hS3t4kjjiUKiIu0KAMAADoMdqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr9oxWf4DePgikkeH7s4A9Izmv+Cc//ACRHW/8Asarj/wBJbWvEfh/SfFmgah4Z121W50/VLZ7a5ibjcjrg4PY88HqDg18V/hzq/wAKPH2r+BdYBaTT5j5ExXAuIW5jkHsykZ9DkdjXwK+K+ofBn4k6Z4ztC8loreRqVuvWe1cjzFA9RgMv+0q54zWk6rp2u6Xaa1pF5HdWN/Ak9tPGcrJG4BVh7EEVVVVVVVVVVVVVVVVVVVWratpWg6bc6zrmp2mnWFnGZLi7u5lhihQdWd2ICgepOK+Pv/BW79nH4Um40b4bm5+JmuwkqBpknkaejDj5rtgd49DEkin1FH4k/wDBTr/goGxt/Amn3vgrwJesF8/T9+jaeYzwS12xM9yMfeWMuP8AYGa+BH/BGf4UeFjba38fPF97431IfNJpWns9jYBvRnB86X6houvKmvAXw1+H3ws0KPwz8N/Bei+GtLjxi10yyS3Rj/ebaBuY92bJOck1VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV8YP2efgp8fdJ/sf4t/DnR/ESIhSG5nh2XVsD18q4TEsf8AwFhnvXxk/wCCOet+G9V/4Tn9k34r3um6jZyedaaZrFy0E0D/APTC+hAKnBIUMgPTMnU14a/b8/bl/Y31m28D/tX/AA1v/E2lK/lw3Grp9nupFHUwX8YaK5Azk7hIT03r2/Z9/b//AGZ/2jBbaZ4X8bpofiOcAf2Br22zumbH3YiSY5u/EbMeMkCqqqqqqqqqqqqqqqqqqqqqqqqqqqq+vrHTLKfUdSvILS0tY2knnnkEaRIoyzMx4UADJJ4FftUf8FcfA/gme48Bfsz6fB458Su3kf23IrNp1vIeB5Krhrp89Nu2PkEM/Ir4Pf8ABO79pT9r7xZH8av20PGuu6Lp14Q6WV0R/alzHnIjjhI8uxi5OFK5HaMAhq+EnwX+F/wK8JQeCPhT4OsPD+lRYLpboTJcOBjzJpGy8r/7Tknt0qqqqqqqqqqqqqqqqqqr9ipT4W/4KyfGHQ41Mcd9eeJoY0hbCiM36zoG9QFRePUD0zVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVMVVSzMAAOSeMV+198YfGf/BRL9p3Q/wBl34EXpm8F6HqDRi9Uk29zMnFzqMuOsMS7lj67hkqcygD4JfBvwX8AvhhoXwq8BWXkaXoluE811Hm3cx5knlI6yO2WJ6c4GAABVVVVVVVVVVVVVVVVVf8ABV79of47fDn9o7wp4B+EHxP8S+Hre58J21xJZ6NqUkAnuZby6QbkQ8ttjjA74I9q/wCCVf7R/iP49/AXUtG8feJrzXPFvgvVmgvLy9mMs9xa3GZbeR3PJORNGM9oRVftkfGxP2fP2bfG3xKt7pYdVttPa00bJ5N9OfKgIGfm2s4kIH8MbelfsZftT/tQat+1b8LfDXxE+M3jLU9F1/UkWXT9V1aV4bqGaN1QlGbBBOCuepCn0qv2sP8AgoF8E/2UZJfDWum+8ReNjbLPb+HrBCh2sPkeadhsiQ46je3cIa+G3/BU/wDaj+JP7SvgPwPqGleG/Dvh3xV4s0iwm0pNLZpY7O6uIUOJZG3Fmik3B8AHcCAAQKqqqqqqqqqqqqr42wS3XwY8fW0C7pJfC+pIi5xkm1kAr/gnBNE3wy8UW6uDImuhmXHQGCMA/wDjp/Kq/bh+CJ8e+CF+Ivh+yEmueF4mNwqJl7myzlxx1MZJcexfuRVfsG/HVZYW+CXia+G+MPPoEkjcsOWktx9OXX23jsBVVVVVVVVVVVVVVVVVV8QviZ8PfhP4cm8W/ErxlpPhvSIAd11qNysKsf7qAnLseyqCx7A18ef+Cyfhqzu5PCP7MPgK58V6pK/kwazrELw2zSE4Uw2q4mmznjeYjn+EitJ/ZA/4KE/t16lbeJ/2jfG1/wCEfDEjiaG31xTF5SnoYNLi2hWAbGZfLYgcse/wD/4Jm/st/AsW2qTeEj438RQAH+1fEqrcqj8HMVvgQpgjIJVnH980iJGixxqqqowqgYAA9BVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVfE79rT9mr4OXkul/Ef40+GNJ1GA4l08Xf2i6iOcfNBCHkXn1UdD6GtK/wCCmP7Dms3AtbP49WUbkgA3ejaharz/ALUtuq/rXgX4i+APifoaeJfhz400XxNpbnaLvSr6O5jVu6koTtYd1OCO4rxL4X8M+NNEuvDXi/w9puuaTepsubDUbRLmCZfRo3BVh9RX7Qv/AAR2+DXj77Tr3wM1uf4f6w4Zhp0wa702Vyc8KT5kOeeVZlAxhBirf4v/APBSD/gnXcx6T8TNIufGfgG2dYYp9SeTUtPMecAQXq4ltyRwqS4A/wCeZxX7On/BT39mz49va6Dq+sP4A8UzhVGma/MiQTSH+GC6GI35wAH8t2PRTQKsoZWBBHBFVVVVVVVVVVVVVVVVVVVVVVVVV+1H/wAFEPgD+zEt1oN3q3/CXeNIQVXw5o8ys8LjjFzNylvz1B3ScghCOa/4z0/4Kmax38J/C0T/APTS00ePa34vfTLj/aCsP+WYNfssf8E/PgR+y1BbazpWl/8ACUeNETEviXVoFaWNiMN9mj5W3Xr93L4JBdhVVVVVVVVVVVVVVVVVVVV8OCvhf/gtnrWmAKF1TUtS3EHywPN0h7gZH8RJAHuTmqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/4Kr/tjTfC7weP2c/hpqBbxl40tSurzWzbpNO0+T5fLAHIln5UDqE3Hguhr/gm/wDsbw/sw/CkeJfGGmIvxF8YwxzauXGX063+9HZKexXO6THV+MkIpqqqqqqqqqqqqqqqqqqr9s66j8Qf8Fa/hbo8kMd1HYa14UtZYfK3DYbxJWDA8EbZST2wee9fsZ6q/wCyF/wUo8W/ATUp2ttB8SahdaFboxwuGb7RpshBP3mTZGOvNwevWq/4LCfEnVvH/wAQfhn+yZ4Kk+0397dxahd2ytxJeXLm2sozjoQGmJHpKhr4s+E9M+C3/BUD4TeBtFRF03w1qPgzS4JJRtDRJFaQNITnqfmYnPXOe9V4w/ZZ+BPxC+L1h8cfHXgCx17xTpemxWFk9+DLbwpFLJIknkH5GlDSnDsCQAuMYr9pRVX/AILJ6OqgAD4geDAABjH7jTaqqqqqqqqqqqqr4j2q3vw88UWTOVE+i3kZYDkZhYZ/Wv8Agmzcl/C3jey2DEWoWsgbPXdG4x/45+tU6JIhjdQysMFWGQRX7WXwMf4N/EOS50e0K+GPEDPcaYyr8sDZ/eW/tsJBH+yy9SDWkavqWgarZ65ot7JaX9hOk9tPEcNFIhBVh7ggV+z38adL+OHw+tvEULRQ6vZ4t9Xs1OPInA+8B12OPmU/UZypqqqqqqqqqqqqqqqq+L3x4+D/AMBtAbxH8WvH+k+HbXaTFHcy7p7kjtDAuZJTx0RTXxi/4K7fEL4ja4vw4/Y3+FmoTahfsYbbU9RsDe30zetvZRFlBGMgyGTIPKLXw+/4Jh/tQ/tKeIofiT+2R8VdR0lZuTZzXY1DVDHnOxRkwWqHOQBu29DGK+A37Hv7PP7N1rGPhh8PbKDVQm2XW74fa9QlOME+c+SgI6pHsT/Zqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr9uH9uv4r/Gf4tH9kz9ke71BUe+Ol3up6RNsutYuukkMMoI8q3TDB3BG7axJEYy3wP8A+CL3gGz0e31j9obx5q+s67Ogkm0zQpVtrW3Yj5kaZ1aSY5/iHljPY9a1r/gkb+xfqti1pY+FvEmkSt0ubLX5mkX6CbzE/Na/ZQ/ZZ8HfskfDzUPh34O1m+1eDUdZm1KW9v40Wdi6IioxQAEKkajIABJJwM1VXdnaahay2N9aw3NvOhSWGZA6SKRgqynggjsa/aM/4JPfs8/GR7rxB8O43+GviSYM27SoA+nzOe72hICdh+5aMdSQxoX/APwUY/4JoXIjvVl8X/DS2kwN5k1PSBHn+FuJrE9cA+WpY5w9fs3f8FSv2dfjp9k0DxdqH/CufFc2E+w61cL9knc9obvAQ84AEgjYk4APWkdJEWSN1ZWGVYHII9RVVVVVVVVVVVVVVVVVVVVVVX7Q37XnwG/Zh0o3XxQ8ZxR6pJEZLTQrEfaL+7wONsQPyA9nkKJ/tV43/bR/bV/b28R3fww/Za8E6n4W8MOfLupNNl2ziJhgteag21IFIBIRNhPK5k4r9l7/AIJF/C/4cPa+MP2gr2D4geJVIlGlhWGlWz9TuVsNdHPeTCHJyh61Z2Vnp1pDp+nWkNtbW0YjhghjCJGgGAqqOAAOMCqqqqqqqqqqqqqqqqqqqqvi0D4U/wCC1nh3V3YoNY1HSdplXIPm6Ylr8uPUrjnv7VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV+1L+0V4U/Zd+DmsfFLxNtuLiBfs+j6dv2tqF84PlQg9l4LM38KKxwSAD/wTY/Z08V/tJfF/Vv23fj4ZNVii1SSfRhdRfLqOpA4M4U8eTb4CoBwHVQCPJINVVVVVVVVVVVVVVVVVVfFmdvE/wDwWl0uCO6IjsvFWiJHuT7vkWFs7qPq6vz71/wWF+HmrfDT47/D/wDaS8HlrO51aFIpLqJOYdSsHV4ZSf7xjaMD2tzXwq+J+hfFH4R+Gfi5ZXEMGm+INEg1Ny0ny22+MNIjN28ttyn0Kmv2Lra8/bK/4KPeLP2jtXgkl0HwvcTavaiUcIAPs2mwtx95Y1Enbm3J9j/wUckl8Of8FJfBmvSPBtUeHryPexCqI7kj5+mOYz+Heqr9pb/lMro//ZQPBn/ojTaqqqqqqqqqqqqrU7Vr7TbqyRlDXEDxgt0BZSOfbmv+Cad2FufiFYHcTJHp0i+g2m5B/E7h+VVXxj+FehfGTwDqHgrWlVGmXzLK6K5a0uVB2Sj6ZII7qWHevGPhHXfAnifUfCHiaya11LS52hnjPTI6Mp7qwwwPcEHvXwF+M2sfBDx7beJ7HfPp0+INVsgcC4gJ5x/tr95T68dCa8N+ItF8X6BYeJvDt/HeabqUCzW06HhlP8iOhB5BBB6VVVVVVVVVVVVVVfGz9o34K/s76H/bvxc8f6boayIWtbNnMt3d47Q26ZkfngkDaM8kDmvij/wVS+Pnx58Sv8L/ANi/4WajaSXgZIdQlshf6pIuceYsK7obZQDyX8wDruWvg7/wSX+K3xX8Qn4mftn/ABO1H7ZeuJLjS7S/+3ahcf7M925ZIx22x+ZweGQivg3+z58Gv2f9BHh74R+ANL0CFkCz3EUe+5usd5p3zJIf95iB2xVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVftnfFK/8Agv8AstfEf4i6RcPBqOn6M1vYTocNBc3LpbQyD3WSZG/Cv+CJ/wAHtM1C/wDHnx41a1jnvNPeLQ9JkcZMLOvm3TDPRipgUEc4Ljoxqqr9sr/goH8UIvi4v7KX7HGjrq3jprr7FqGrx26XLwXXV7e3R8xhowP3ksmUTDggFCwuP2LP+CqlxY/8Jg/7YE39u7PO/sePxjqKR7jz5e0RiDdzjGNmeM45r9h39vv4q3/xdm/ZP/a5tDb+NY7l7PTdVlgjt5Wuo1JNrciPEbMyjMcq4DnA+YurVU0MNxC9vPEkscilXR1yGBGCCD1BHav2kv8AglT+z18bPtXiDwFbf8K38UzZfz9Jtw1jO/8A01tMhRn1iMfXJ3VB4n/4KIf8EzLpNN1yzl8W/DG0l8uEz79Q0ny84AjlGJbJjzhG2AnJ2PX7Mv8AwUl/Z4/aPNroE2rHwR4vmKoNE1ydEW4kP8NtccJNzwFIRz/c71VVVVVVVVVVVVVVVVVVVV8d/wBqb4Ffs26T/aPxY8fWOm3UkZe10qE+ffXfp5dumXIJ43kBAerCviR/wUo/aj/ar8SS/Cb9jT4barocN4DH9tgjW41Rozx5jy/6myTBwWySpwRKK/Z8/wCCP7X+pj4g/te+NrnxFqt2/nzaFp17I4kkPJN1en95Ix6FY9vI4kYV4K8CeDPhv4ctfCPgDwtpfh/RrJcQWOnWqwRLxy21QMsccscknkkmqqqqqqqqqqqqqqqqqqqqqr9vEjwf/wAFT/gt4kUhFun8NXkpjb5mRdTlhcHPAJWIj0xj3qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqvLu0sLSa+vrqK2traNpJppnCJEijLMzHgAAEkngV8UfE3jL/gqn+2XYfDbwPf3dn8MfCryLBdBDttrFGUXF+yn/AJazsEWNW6AxKQMOa8E+DPDXw68IaP4E8HaVFp2iaFZx2djaxDAjjRQBz1JOMljySSTkkmqqqqqqqqqqqqqqqqqqq8AvJ4r/AOC01yxbLQeMNVH787uLXT5xx+EPy+nHpX/BS74Nt8ZP2RvFtvYWRuNX8JBPEGnKqktutgfOAA5JNu84A7kivgX+1dN4T/4JY/E/wP8A2mya9oWp/wDCPaUFfDJbasWfIPUEKNQII6bB06j/AIJJ/BQfDH9l6DxvqNosesfEa9bU5GK4cWceYrZCe4wJJR/13r/gsvDJof7WHg3xLbW0fzeDbOQEn/WSxX12Tkdfu+WKjkjmjSWGRXR1DIynIYHoQfSq/aW/5TK6P/2UDwZ/6I02q/ab0j9onV4fDkPwCvprfDXK6v5VzbwnB8ryiWlIPGJB8nPPNf8ACAf8FC/+hnvf/Bza/wCNf8IB/wAFC/8AoZ73/wAHNr/jR8K/8FFtOOy11W+ug4yT/aenvtx/10bI/Cj4t/4KJ6Ku+80i/vVA4X+y7CY4Xr/qlyc/me1f8NO/tmeGTnxF8H2njX7z3Xhm7jBA5OHRlXpxnkCrP/gof4q0uYWvi34RWxlA+dYb+S1YdidrxueoPGfxrQ/+Ch/wtvNqa/4Q8Sac5HLQrDcopx3O9Wx2+7Xh/wDbB/Z48RFY4viDDYTNj93qFrNb7c+rsuz/AMerQPGng7xZH5vhbxZo+sJj71hfR3GP++GNVVVVVVfsDgaV8aviH4eUDEdo+Nvyj91dbPu/8D/Cqqv2yP2dh8T/AA0fHvhKxDeKdDgO+KNfm1C2Xkpju68lfXle4wQQdpGCOxr9jz9pL/hV2uL4A8ZX+PCerzZinlbjTbhuN+e0TcBh0Bw3HzZR0kRXjcMrDKkHII9RVVVVVVVVVVVV8Zv2gvg5+z74fPiP4t+PdN0GBkY29vLJvubsjtDAuZJD2+UEDuQOa+K//BU34+/H/wARN8LP2L/hlqtg94SkeoNZC+1WZOhdYl3RWy4PLN5m3g70r4Kf8Ej/AIhfEjXP+Fmftl/ErUZr+/cTXOkWd+by+uG9Lm9csq9MFY9/B4da+FHwU+FPwN8OJ4U+E/gXSvDengDzFtIf3lwwGA0srZeVv9p2J96qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/4Kpabeaj+w949ks1LfY7jTJ5VAJJQX8CnAHpuBOeAAT2r/AIInaxZT/s5eNPD8cgN3ZeNpbqVM9I5rK1VDj3MEn5VVfFbxgfh38LfGPxAUIx8M6BfamA4yD9nt3l5A7fJX7CP7WXw0/Zl+KHiz4t/Frwt4i8U+INasvs9hc2IhkeJpZTJdSOZXX53KxgMMnBcHAPP/AA+0+AP/AESj4gf982n/AMer9rT9qrwV8bP2pPD/AO0T8JvDGs6BPpkOmy3KagIlnmvrSZmWVTEzDHlLbqCTnKemKqqvbKz1K0m0/ULSG6triMxzQTRh0kQjBVlPBBHBBr9p7/gkn8F/i0t34n+C8sPw58TyAt9kgi3aVcvnPzQDmAngZiwo6+WTXhr9pf8Abu/4Jy67a+A/j34YvPGHgcSeTZnUblriKSMH/ly1ABivygYilDbV48tD0/Zt/bT+Af7Umnxr8PPFa23iBYt914c1PFvfwYGWITJEqDu8ZZRkZIPFVVVVVVVVVVVVVVVVVXx9/b0/Zl/Z2jubLxf8QLfVtft1ONA0Ii9vCw/hcKdkJ4/5auleMP8AgoX+2h+2Br0/w8/ZG+Gmo+G9PkbypbrTUFzeqjZAaa8cLDaKQOCNpBziQ18Cf+CPF1rd+PHv7XPxEvtZ1W8cT3Gi6VeNI0jnBP2q9fLyEjhhGBg9JDXw2+FPw3+D3huLwj8L/Bek+G9Jiwfs9hbiPzGAxvkb70j+rOSx7mqqqqqqqqqqqqqqqqqrxR8Zfg/4IZ08Z/FfwdoDRkhxqmu21qVxwc+Y4xzitS/be/ZC0sSG5/aO8Bv5b7G+zaxHcZOcceWWyPcce9ad+3D+yDqqg2v7RvgVMvsH2nVkt+eP+ehXjnr09+DXg74k/Dn4i273fw+8f+G/E8EQHmS6NqsN6qZ6ZMTMBVVVf8FjB/wi/wC0V8HfH7AqsOmffZdy/wCi3olPA5P+t5HuMd6qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv+Csf7WN/o2lW/7I/wquJrrxR4tWMeIhZqXlitZSBDZIF58yckFlHPl4GCJeP2B/2SLD9lD4NQaZqsEMvjfxKI7zxLdLhtku35LVGGcpCGYZHDMzt0YAVVVVVVVVVVVVVVVVVVVX7FpTxR/wAFY/F3iKNYNq+IfFd8ozvwJHuF+Q/9tevpn1q6tba+tZbK8gjmgnjaOWKRdyurDBBHcEHGK8Vfs4+LtK/a41T9j7Rbi6igv/GsNnBHuJBtizG2unB6lLS4aTPYO3XNeG/D2k+EvDmleFNBtVttM0WyhsrKAdIoYkCIo+iqBX/Bb3SxF8VfhrrItyDdeHrq383d97yrgNtxnjHm56fxe3Hwx1Ma38NfCesh4n+36HZT7oTlDvgRsryeOeOar9pb/lMro/8A2UDwZ/6I02qqqqqvdPsNSg+zajY291Eesc8QdfyOa134BfBPxIrDVvhb4ad3B3Sw6ekEhz6vGFb9eK8QfsJ/s/60GOnaXrGhsxyDYakzY+gmEnH+eK8Qf8E6Eib7V4K+KUsUiNmOLULDkeh82Nhg/wDAPyo/Bv8Abp+Fp83wj42utbt4eVjtdZFzGoHpDdgA/QKfxpf2vv2m/hlIsHxV+GMc8CNtaa80yaweQ/7Mq/uzn2Q14S/4KEfCzV2jg8W+G9b0CVsZkQLeQp65Zdr/AJIa8H/G/wCEXj4pH4T+Iei3s8g+S2a4EM5/7ZSbX/Sqqqr9mQjQP23viLo7cC4fWYI/MGWKi8SRTkcZKoDVVVftqfs0t4cvrn4w+BNPP9lXsm7WrOFP+PSZjzOoHSNyfmH8LHPRvlr9jH9qBZlsvg18Q9S/eDEWgahO/wB/oFtXJ79kJ/3P7oNVVVVVVVVVXxE+JfgD4S+Frrxp8SvF2meHdFsx+8u7+cRqWxkIg6u5xwigsx4AJr48/wDBWjxx8RNcPwr/AGK/AWpXuoX7tb2+uXGnNdXlw2PvWlkA23pkNKGODzGuK+DX/BKX4vfGjxAfit+2t8SNXju75lkl0qG+F7qVwOoWa5YukKjoEQPgHAMeK+FHwU+FPwN8OJ4U+E/gXSvDengDzFtIf3lwwGA0srZeVv8Aadifeqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr4wfDfSvjD8LPFfwu1mQxWnijSLiwaYDJgaRCEkA7lG2sB6qK/Yd+OusfsEftP8AiP4WfG23m0rRNVnXSPEYOWWwuInJt7wDHzxAO3I6xzbxnAB0fWdI8RaVa65oGqWmpadfRLLa3dpMssUyEZDI6khgR3FV8VvBx+Inwt8Y/D9WQHxNoF9pgLnAH2i3eLkjt89f8Ek/EHgTwr8dvGvwO+LvhnShq/iW3ji0+LWbGOVor+xeUSWo3g7JGWWQ44yYcddoP/CpvhX/ANE08Kf+Ca3/APiKT4U/C6N1kj+G3hVWU5Vho0AII7j5KqqqvE3hfw1400K78MeL9A07W9Iv4/LurDULVLiCdfRkcFSMgHkda/aT/wCCQ1qmoP8AEj9kHxNP4a1uzk+1Q+Hru9dI1lXkGzu874XyBtWQkbj99AOPgz/wUy+P37NfipPgz+214F1q/SwKxHVJrYQ6rbJnAkbOI7yLA4kBDMMtvk6V8KfjF8Mvjh4Tg8bfCrxnp3iLSZvlaW1k+eB8Z8uWM4eJ8YOxwDgg4wQaqqqqqqqqqqqqqq+Kn7W/7NfwWjmX4jfGbw1p13ADv0+G7F3eD/t3h3y9sZ24r4tf8FqvhxpRm0r4IfC7WfE15u2RX+syixti3ZliTfLIO21vKP8AWQ/8FWf26G8hodU8B+Db7G7Kt4esPLbPPObq5QjsPNXgdOtfAT/gjf8ABjwQbXW/jf4jvfH2qx7XOnW+6x05G64IU+bLggcl0UjOU548J+DfCXgLQrfwv4H8M6XoGkWgxBY6baJbQx+pCIAMnHJ71VVVVVVVVVVVVVVVVe3lnp1pNf6hdw21tbRmSaaaQIkaAZLMx4AA5JNfHP8A4Kr/ALLPwhkuNI8M63dfETW4SV+z+Hdr2qsP712xEZXrzF5v0q6/4KLft/ftN3s2lfsx/Bv+xbEPs+06TpLarLATxiW7uF+zp25Mad+fRf2Av+ClHx/X7X8cvjgdHtJjmTT9Y8TzXezP9y2td8A6dAy9q8K/8EPfCsCo/jf9oDVb5ictHpWhR2u0ccB5JZc9+do6jjjnTf8Agiz+yzaGKS/8a/Ey+dVxIjanZxxucdQFtdw+m7861P8A4It/srXhd7Dxl8S7B/LIRY9Us3QN2YhrUse2RuH4da+J3/BHb4kfDonxx+y98ar261nTQ0trY3pOm3uR0EN5CwXeenzLGM9WGeP2Fv8Agof49ufiHH+yz+1tFNZ+L47s6dpmtX0X2ed7tTtWzu0wB5jH5UlGNx2ggs241Vf8Fx9C83RPhD4mjjUfZbrVrORwoy3mLaugJ68eU+B05PTv8L9bHif4Z+EfEgfd/auhWV3nB58yBHzzz/F3qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr9q/8AaM8Ofst/BXWvijraxXN9Ev2XRdOdiv2++cHyouOQowXc9kRiOcA/8Ev/ANnHxP8AGv4m6v8Atv8AxwM+pStqU8ugtdp/x/agzHzbsA8eXDkpGAMB84x5QFVVVVVVVVVVVVVVVVVVVVab4H8FaNqTazo/hDRLHUH3brq20+KKU7vvZdVB57881R0rS2vhqh021N6BgXBhXzBxj72M9OPpVa74Q8JeKHhk8S+F9I1ZrcEQtfWMc5jBxnbvBxnA6egq3trezt4rS0gjhggQJFFGgVUUDAVQOAAABiquvA/gq+1lfEd54Q0S41ZZEkW/l0+Jpw6Y2MJCu7K7Vwc8YGOlVVVVVVVVSRxyxtFKiujqVZWGQQeoI9K8Xfs4/A/xwj/278NtGEz5zcWUP2OUn1LwlST9c14x/wCCd3gfUPMn8DeN9V0iQglYL+FbyLPoCuxlHuSx+vSm+C37a/wUPm+AfFt1rmnQcrBYah9ojUD/AKdrkAZ7YRSa0f8Abo+LXgO9TRPjJ8LlkmXgsIZNNuMDqxRwyt+AUV4I/bT+A3jMxW914jn8O3cgA8nWIPJUHv8AvVLRge7MK0zVdL1uyj1HRtStb+0lGY57aZZUf6MpINV4VH/CM/8ABRzULAny01C7ut3BTPnWBmHHfLFfqSDVVVXtlZ6lZz6dqFrDdWtzG0U0EyB0kRhhlZTwQQcEGv2qv2bLz4LeID4g8OQSz+DtWmItZDljYynkwOfTglGPUDB5BJR2jZXjcqynKkHBBHcV+yJ+1Qnj62tfhh8Qr4DxLbRbdPv5n/5CSKPusT/y2A/77AJ6g5qqqqqqqqvLu0sLSa+vrqK2traNpJppnCJEijLMzHgAAEkngV+1R/wVx+G3w4e78Ffs8Wdv498TqWhbVnLDS7SToNhXDXRz2QqhyCHPIr4dfsKftfftyeKLb4t/tceONY8NaDL81vBfxgXzQkg7LWy4jtIzzy4U5w2x85PwF/Ze+CP7NWgDRPhN4JtdPmkjC3eqzjz769Ixkyzt8xBIzsXagJO1Vqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/bE/YH+FX7XVlFrGoXEnhjxvYQ+VZeIrO3EheMdIrmIkedGCSR8ysp6MASp0r9i3/AIKh/stTT2PwE8aSarowcuItB8QQi3bPJc2l8UXf2O1WOe5HNLpv/BcDx1H/AGVe6jqemW7HbJcGTQdNKZHXdCFkPT+HOM1+xB8JPj78G/hPqHh39onx5B4r8R6hrc2oxXa6lPfyQxSRxgxSTTKCxDozcZA3kAmv23v+CYX/AAu7xjc/G34C+ILPw144uZFn1GwumaG2v506XEcsYLQTnC5O0q7YYlW3M2kz/wDBbX4eWi+F4dPvdcgtRthurt9H1F2UcAmd2Mj5xn94S3riv2YPhz/wU51T45aB8SP2nfHtnZeEtKW5W+8PNqFsv2lZIHSPbBYKYWZHKMGkbICnqTzVVVVXxo+Afwj/AGhPC58I/FrwVY67ZqGNtLIpS4s3YYLwzLh426dDg4AII4r4s/sBftR/sV+KpvjP+xz4313X9HtQXmtrRVOo28KnOye2A8u9i/3UJ7mMAbq/ZO/4KzfDv4nS2fgL9oW3tPAXi0sIF1QsU0u8kHB3M5zaNntISnH3xkLUM0NxCk9vKkscihkdGyGBGQQR1BFVVVVVVVVVV5e2enWsl7qF3DbW8K7pJppAiIPUk8AV4/8A23v2SvhkZYvFnx88JieEfvLbTrs6lMh9DHaiRgfYgetfEL/gtF+zv4fMlt8PvA/i/wAXXCD5JZY4tOtn9MO7NIPxi/Orr/gp3+3F8eJ20v8AZy+AUNlFISqz6do9zrc8J6ZMpAhUDuWjx71/wyF/wVS/aW/efGX4rXfhrS7rmW01bxF5MTKe4s7ANHn/AGXVPwr4W/8ABFL4R6GYb34u/FDX/FU6gM1npVummW+e6sxMsjr2yrRnvx0r4Sfspfs6fAwQy/C/4ReH9IvYRhNRa3+03vv/AKTMXl59N2Kqqqqqqqqqqqqqqqqqv2uP+ClnwZ/ZokvPB3h5k8cePYQUbSbG4C2+nv2+1TjIUg9YlDPxg7AQ1aX8Kv8AgoF/wUzvYfE/xB1yTwj8N55RJbC7SSy0wIMENa2YO+6I5xLISM5HmjGB8B/+CWP7LfwbS31LxH4ek+ImvxYLXviJQ9urA5/d2g/dAcD/AFnmEf3uSK0/TtP0mxg0zSrG3s7O2QJBb28QjjiUdFVVwAPYVVVVf8FiP2bLHUPBum/tTeEbU2mveG7iCx16aAFDPau4W3nJH8cUpRA3BKygZ+RRX7FXxwl/aH/Zo8F/EjUZxJrMlmbHWjnk3tuTFK57DzNolAHQSAVVf8FqtCN9+zT4U12KMM+meNIEc4Hyxy2lyCcnn7yRjA9c9q/Yt1xvEX7JPwh1F2yyeD9OtmOCCTBCsJJyTz+769+tVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVU00NvC888qRxxKWd3baFAGSST0GK+MvirxP/wAFRv21NL+E/gDUrmH4beFpJYob2MZSGyjYfa9RIPG+UhUjB7GEHGWrwZ4O8N/D7wlpHgbwhpcOnaLoVnHZ2NrEMCKJFCge54ySeSSSeTVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVWsaFoniKybTdf0ex1O0f71veW6zRt9VYEV44/Yk+BHjAST6dot34avHJPm6TcFUJ7ZifcgHsoX61qv7Fnx2+GN2+tfBX4km7YHIiguX0y4cdgRuMbjsdzAH0q2/as/ae+DEsem/GPwC2p26kKs9/aG0kkx2SeIGJ+O+1j79ab43eGvFv7Xvh34xpDJoOn3N9ZC+F3IMW+IlgkYuvVNozkgcdQBWka1o3iGwj1TQNXstSs5PuXFpcLNG30ZSQaqqrxN4Z0LxloF74Y8S6bDf6bqERiuLeVchge49CDggjkEAjBFftGfs9a58CPE4gDy3/hzUWY6ZqJT84pCOBIo/BhyMcgW1zcWdxFd2dxJBPA4eKWNirRsDkMpHIIIByK/ZQ/avtviXbW/w9+IN7HB4sgTbaXTkKuqKB+QmAHI/i6juBVVVVVVX7Uf7dHwK/ZVsJbPxZrf9teLHiLWnhjS5FkunJHytMfu28Z4+Z+SMlVfBFXWu/t2f8FUNdk0/RrY+EPhYlziRVeS20mFVbpJJjffTLgHaAQrYO2IHNfsr/8ABO/4D/swRWmvW+mL4t8bwqC/iPVoAWhfubWHJW3HoRukwSC5HFVVV8Yf+Cm/7L3wT8c6v8OPFF74nvdd0G6+zX8GnaQXWKTGSN8jorAccgnqPfH/AA+p/ZW37P8AhBfipjON39lWGPr/AMfua+H/APwVO/Yz8fXsOmzfEK98L3M7bY18QabJbR5zj5pk3xIO+WcDHU1o+s6P4i0u11vw/q1nqenXsYktryznWaGZD0ZHUlWB9QcVXx7/AGp/gX+zRpcN/wDFzxzbaZc3cZey0yFGuLy7AOMxwpltueN7YQHgsKuP+C1v7NkeomCD4cfEiayVsfaBaWauR6hDc/zYfh0r9m39tL4EftWPfWPwr1rU/wC1tLtxcXul6lp7288ERYKHLDdGw3ED5XJqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq1vXtD8NadJq/iLWbHS7GEfvLm9uFhjT6sxAFeO/27vgX4RaW00S+1DxTeRggLptvthDehlkKgj3QNXwh+JNh8Xvh1o/xC06yNlHqqSFrVpfMMDpI0bKWwM8oew6iqqqqqqqqqqv2tP+Cc3wS/afiu/Etpap4N8eyAsuv6dbjbdvz/x9wAhZs5++CsnT5iBtPhH40/to/wDBLzxVa+APi9oFx4r+Gk0xisopbh5bORAOTp92QTAwAyYHGOpMYLB6/Z5/ai+DX7T/AIWHiX4VeKI7qaBFOoaTdYhvtOY/wzQ5JAzkB1LIxBwxwaqqqqqvEnjDwj4OtBfeLvFWj6HbNws2pX0dsh/4E7AV40/4KE/sZeBA66t8f/Dl9IgOE0UyarvPoGtVkXn1JA9SK8bf8FoP2bNDEsHgvwb428TzoD5cjW0Njbv/AMDeQyD/AL91qf8AwWK/aB+Id7LpHwK/Zt003RACRStd65MueN2y3WHk4OBgjPrjkeIv+Cz/AMemKWGk+IfB2nTLyhsrPw8IQe+6fbc56cAkj061bf8ABJn9rb4u3MOp/tBftIWJZm3E3N/e6/cQ+uRKY0zyeFcjnrXgD/gi7+zp4fMVz498b+MfFs6D54Y5YtOtn+qIrSD8Ja+H37D37JXwwMUnhP4DeFftEI/d3WpWh1KZT/eWS5MjKfcEdT61b21vaW8draQRwwxKFjjjQKqAdAAOAPar6+sdNtJb/UbyC1toF3SzTyCNIx6ljwB9a8b/ALbf7JXw8Mkfif8AaB8HCaEfvILC/GoyofQx23mMDx0xnp614p/4LA/sdeHyy6TfeMfE204B0vQ/L3c4yPtTw/XnHSvEH/BcD4dW24eFvgN4j1HAOz+0NYhs8nA67Elxzkd+x74E/wDwWs+J+vTtB4K/Zs0wuCcRyavPfMAeFz5cMff259qP/BTP/goX4g/5FT9lXTXjf90DH4O1i6YSn+6VmAzgr8pB/HOK/wCGv/8Agrnrnz6f+znquniHhh/wr28h35/67kk9O340Piv/AMFrfEgRdL8H6xpZlPmKT4Z0uDav90/aUOOo4PzcfWl8If8ABbzxSA15q+r6fFOxl8w6todpg+m2Jg6jn7uAP0ofs2/8FjNVJvrz4y6tZyyH5oT41WPbjgfLESgyB2/Hmh+xP/wVmvB9rn/ah1aCSY72iPxK1NdhPJGEG0fRePSn/Zb/AOCwWigCw+O2q6h9jwYseOnk87b0/wBdjdn/AG+vepF/4LbfDQGYtretWseSQp0fV/MGRn5fnlHLegPBxwKtP+Cqf7ZXwXv49P8A2i/2dbVoA4U/a9KvNBuZPXEkm+M8cjEfvkivgJ/wVJ/Ze+Nl1a6Bq+t3fgHxBcsscdl4iCxwTOTjEd0pMfUgDzPLJJ4BpHSRFkjdWVhlWByCPUVVVVVVVVSyRwxtLLIqIilmZjgKB1JPYV+2N/wUT8f/ABg8ZN+zF+xQmoahc6jcNYXfiHSFL3OovjDR2LL/AKuJQG3XHGQpZWVBvb9jb/glb4C+EsNj8RP2gILLxn44YiePTJF87TtKfqBtPFzKD1dxsB+6pKhyFVVCqAABgADGKqqqqr9pDwLb/Ez4AfETwHPAkraz4avoIA44WfyWMT/VZAjD3Ar/AIIheNJr/wCHHxM+HkkpKaLrVnqcSlv+fuF42wPT/Q1/Oqr/AIKx6Gur/sS+LL8jJ0XUtMuxyBjddxwf+1+2P5iv+CXOuLrf7D/w8QtmXTm1G0l4AGUv5yuMH+4ydcc5qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/4Kz/tZy/DX4fxfs6+AtQP/CWeOrc/2s8By9npjEqUAHR5yCg/2BJ03Ka/4JzfslxfswfBKC78S6csfj3xksd9rzOvz2i4Pk2Y9PLViWH/AD0d+SAuKqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqu7S1vraSzvbaK4gmUrJFKgdXB6gg8EV+3D8NvBvg34v8AheTQdDtdD0nWrBPtkdhCsKb1nYSSKowobYyeg4HvWs/sVfG/4ZahJrvwQ+IrXTKcrHFdNpt0w/un5vLcdvmZQfTk1p37Xv7RXwfvY9G+NngB9RiztWW7tTYTyY7pKimKQY9FOfWvh7+2d8DvHnlWl3r8nhq/kwPs+soIU3dOJgTHj/eKn2q1urW+to7yxuYriCZd0csTh1cHuCOCKqvGvgrw18Q/DV74R8W6ZHf6bfJtkjcYKns6nqrA8hhyDX7Qf7PniX4E+Jfs1z5l/wCH752/svUwmBIOvlyY4WQDqOhHI7gW1zcWdxFd2dxJBPA4eKWNirRsDkMpHIIIByK/ZW/a9tPGcVl8OPijqCQeIVURWGqTMFTUMYCpIe03YHo/+8cNVVVVXjbx14N+G3hm88ZePfE2naDomnpuub6/nEUaegyerE8BRkkkAAk1+0L/AMFQPip8cvE5+CP7D3hbWTJqLtAmvRWZfUb0YwxtoSD9mjxk+a/zgYb90Rmv2W/+CSVla38XxP8A2u9XbxR4hupPtTeHY7tpoFlJ3Fry4zuuHz1VTsznLSA1pGj6R4f0u10TQdLs9N06yiEVtZ2kCwxQIOioigBVHoBiqqqr4k+HNB8X/wDBWa58L+KNJtdU0nVPiTaW97ZXUYeK4iZowyOp4KkcEHrUf7JX7KsUaRJ+zR8KyEUKC3g6xY8epMWSfc18b/8Aglj+yr8V9GuF8J+Eh8PdfKH7NqWglliV8cCS1Y+UyZ6hQjH+8K+CHxl+Nf8AwTF/aRuvgz8Vp5rrwTcXkY1exRmlt5baQ4TUrLOMNtySABv2sjgMoKal4j0nTfDN14ua5WfTbWwe+M0J3h4VjL7lx1BUZGOtfslfDmf/AIKMftieIPFvx41e7vdNtrWXW9Sso7hlM0SzRxwWMbDBjhXzFHy4OxCAQzbhbfsafsl2mkR6JD+zb8N2t44vLV5fDdtJPj1MzIZC3+0Wz718E/2Vvgb+ztrfiXXfg/4NXQJfFYtxfwpcSTRqId+0RCQsY1JkYlQdpOOBgVVVVVVVVVVVVVVVVVVVVVVVV3d2ljbtdX11Dbwx43SSuEVcnAyTwOSBWl6tpWu2EWqaJqdpqFlNnyrm1mWWN8EqdrKSDggg+4NV+0r/AMFGfgZ+y147f4bePPD/AI01HXFs4rsJpOnwPEY5Q2075Zo+6kHAOD64ONd/4Lf/AA5t1f8A4Rn4DeI78gHyxfaxDaZ44zsSXHPHfjnnpX7DP7aj/tm6N4v1iX4fR+Em8MXttAlsupm+MiTI5DF/Kj5zGwwF9Kr9vPxf+2Z4Ss/BT/sg6LcalNcy3w1+ODSYbwqqiDyCTKCEGTNwOT/wGll/4LjeJtm0axD5S7hlPD1hkHHX7m48dDkj25r9iXwH/wAFGPDXxem1X9qnxJfXvg6bQ50W2udZs7vZeF4jH8kJJBCiQZHHUZ5qv2vvg/8AED47/AnWfhn8MfFsHhvXtSurR4tRnuprdYkinSRxuhVnyVUjAGCTyRT/APBJf9r/AFFRZax+0/pElnIR5qnVNSm4ByPkZQDyB1Ir4G/8ErfjV8KvjF4M+JOtfH/TdVsPDWtW1/c2KR3Qa4SNwxQbmxkgY54qviP4Uk8efDzxR4HhvVs5PEOi3mnJcNHvEJnheMOVyMgbs4yM461/w458Vf8ARxWk/wDhNyf/AB+v+HHPir/o4rSf/Cbk/wDj9eENCfwx4S0TwzJcrcNpOnW9o0wXaJDFGqFgMnGducVXxf8A+CXX7THi/wCLnjX4h+A/2iNL0qx8WeIb/U47R7y+tmt457h5Y4j5QYNtV8dhxxR/4Jdft66QRN4a/an0lJnG2QnxRq9t8v1SFiee3FfsofDL4j/B74BeF/h38XfFcPiXxdpZuzqWqxX896twZLuaSPEs6rI22J40+ZRjbgZABqv20PCv/BSzUvjSdc/ZT1G9j8GJpdvBHbW2radCGn+cyyNFduOclVz7DHc02uf8Fw/DA8me01e7jtWDugsdAvfMBwcb0VmYc4+U5HTjHH7KX7QX7QXhfwd4u1v9ve9h8MXMF1bjw/BdadDa3F1GUfzvLigGZRuCDgEjnOARXxX/AOCh+v6i02lfB/w+mlQZKrqupqs07DsUh5RD/vF8+grxh488Z/EDUv7W8a+KNR1q6Gdj3lw0gjB7Ip4QeygCq/ZZ/a90X4K+CtR8B+KvDOr6lCLmS90+5s3iKqzhFMDBmBUEqXDAEctnnAPwl+JOl/Fz4f6T490m3NtHqMbeZbNJva3lRiroWwM4IODgZBBwM18Tfih4B+Dfg678f/EvxJBoWgWDxpcXs0buqNI4RBtRWY5ZgOB3rRP24v2QPEGwWH7RvgWLfjH27VkssZGefO2Y/Hvx1rw78YPhJ4wmhtvCXxS8I63LcMFhj07W7e5aQkZAUI5JJHPFVVVVVVVVeM/BPhD4i+Gr7wd468N6fruiajGY7qxv4BLHIOxwehB5DDBBAIIIzX7Rv/BNf4u/s4eKv+F+fsQ+Idbkj0p2uDotrOTqWnKOWEJJP2uEgYMTAuRwRKCcfsSf8FPvDvxu1Cx+Enx0t7Xwr8QGPkWt8P3Nlq0q8bMMf3E5x9w/KzZCkEhKrxX8T/hp4EDnxv8AETwz4eEa7nOq6vBabRjOT5jDAxz9K8Xf8FEv2LvBgkXU/j7oN7IgOE0iKfUt5HYNbo68+pIHvXi//gs3+y5ogeHwv4a8deJJgDseLT4bWE8cZaWUOOcfwHv+Ov8A/BbHxfrd5/ZXwu/Zxt2uZDttzqGsSXkkhx/zxhiQ9ewc/Xnhv2s/+CuPxhCt4A+DN/4aguMCGex8FNBEQe6y6h5iEepzge1L+yj/AMFc/jN8vxA+MOo+GLe4U+ZDf+MzbxEZ6NDp3mKc9gV49q8Of8ETvFWtXo1X4rftGxPcSMDcJp2kSXckvr+/mlUg+5Q/SvBf/BHD9k7w4Y5vE99408WSj/WR3uqLbQsfZbeONwMf7Z+teCv2HP2Rfh8Y28Nfs++DzJCB5cupWX9pSIR3D3RkYH3zmtN0rS9Fso9O0bTbWxtYhiOC2hWJE+iqABVVreuaJ4a0m617xHrFjpWmWMZkur29uFghgQdWd3IVR7kgV8cf+Cu/7Nnwze40j4cw6l8SdYhDANp3+i2CuOzXMgywzj5o45FI70f20f8Agpp+1lI9t+z78NLjw9oly5jju9E0gBMH+F9QvMxK3+0hjI9qs/8AglX+2R8b7uHWf2jv2g7aAs+7bfalda/dQeuEZkiGBwAsuO3ArwV/wRZ/Z10ZUm8bePvG3iWdR8yQywWEDeuUWN3/ACk/OvCn/BOH9irweqGw+BGk30inJk1a6ub/AHHjqs0jL2HAAH5mvDvwB+BPhDyz4U+C3gXR2iwUax8O2sDAgg5yqA5yAc+ozVvbW9pAltaQRwxRjCRxoFVR7AdKqqqqqtS0zTdYsZtL1ewtr6zuUKTW9zEssci+jKwII9jX7SX/AASq/Z4+M1jd6v8ADnSofhv4sYM8VxpMOLCd+yy2gIRVz3i2EZyd2MV+z3+1P8d/+CePxWj/AGbv2r7fULvwJuC2lyWa6/s6Fm2pd2UmMy2hwd0XVecKrqyNpupadrOm2ur6RfW97Y30CT2tzbyCSOaJ1DI6MMhlKkEEcEEVVVVVVUxVVLMwAA5J4xX7cH7afjz9prx//wAMb/sjR3OqWmp3TWGrapp0g3ay4z5kMUgOEtEAYySkgOFPIjBMn7Ev7D3gb9kfwYk0sdprXxA1WEf21r3lZ2Z5NtbZGUhU9TwZCNzYwqpVVVVVVX/BGHGk/Gn4weG7Z9lumnQbYn5b91dSIp9eA5B+oqq/b90Ua9+xp8WbHdjyvD8l197H+odJvf8A559P5V/wRq1v+1f2StQ01m50bxje2wUgAhWgtps8c4zKeT6H0qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqvjh8YfCfwD+FXiH4s+NJ8aboFoZfJVgHupSQsUCf7cjlVHYZyeAa/4J8fB3xb+2P8AtKeIv2zPjbB9s0vRdW+0WUciEw3GpgAwxRg5/dWsflkDsRCOfmqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr/AIKU6SH0/wAB66i8wzX1s59dwhZe/wDsN27/AErwTrA8ReC9A8QCTf8A2npdtdbs5z5kStnv61qWl6ZrNlJpur6fbX1pMuJLe5hWVHHoVYEEfWviH+xD8E/GolutE0658K6g+SJNLf8Aclj6wNlQPZNlXX7NP7VHwHuJNT+D3jOXWLBCXMOnz+UzgckvaSkxseOilzXhP9vLxv4Q1D/hHPjh8OZxcQHbNNawtZ3SdeWgl+Vj24KDivh1+0Z8HPiiIrfwv40tFv5cAaden7NcbvQI+N5/3Cw96rxr4K8NfEPw1e+EfFumR3+m3ybZI3GCp7Op6qwPIYcg1+0T+zd4m+BeuNNGs+peFryTGn6oU5UnJEU2OFkAB54DAZGOVUEggqcEdK/Zg/bPNuLT4e/GXVC0YxFp+vzNkp6Jck9ugEnbjd3YRyRyxrLFIro6gqynIIPQg+lVVV+13/wUJ+Df7KttP4eadfFfjwx5g8O2M4H2ckZVrqXBEK9DtwXOQQuDuHg/4K/tl/8ABUfxbbfEX4t6/N4W+G0Mxaznkt2is4k6FdPtCczP1Bmc44IMhKhK/Z6/Za+C/wCzD4YXw78K/CsVtcSxhb/WLoCW/wBQIxzNNgHGRkIoVAc4UZNVVVVV4m/5TBw/9lQsv/Q4qqv+C3vgHTZfC/w0+KMNvGmoW1/daNPKF+aWKSMTRqT6I0UpH/XRq/YG8QD4m/sQ/DK511ftccugy6TOkmf3kVrNLZ7T35SECtR0r43f8Eqv2rG8RWeiHVNCm8+3sZpyy2viHSZGB8oyqPkmXbGzDBKSIpwyEbvgD/wUf/Ze+PiWmmW3jSPwj4kuAqnRfEbLaO0h42xTE+VKSeAFbeePlGcVVVVVVVVVVVVVVVVVVVVO6RoZJHVVUEszHAAHU146/bX/AGTfhw0kXir4/wDg9J4f9bb2N+NQmjPo0Vt5jg+xGa8bf8Fjf2TfDgkh8L2njPxbKvEb2OlLbQsfdrh43A/4Afp3r4a+OdM+J/w68L/EjRYmisfFOj2up28TsC0aTxLIEYj+JQ2D7g1Xx3/Zt/4Kf/FH4y+NU8E/GLVdA8BXOs3DaGZPFx0+L7G5JRPLtN0mxVOzDrkgAkHk1a/8Ea/j142uEvfi7+0rpL3JOXlihu9YYZxkhp2hJ7+nQevH7K3wAH7MXwW0n4Nx+MZvE0GkXFxLDey2YtTiaVpWURh3wA7seveq8ffsy/s//FXxYnjj4k/CTw54m1yO1S1W71O0FwRChYqm1srgF2PTvWifAL4E+GQB4a+CvgPStvT7D4btIMc5/gjHfn61aWVnYQLa2FpDbQqTtjhjCKM9cAcVVVVVVVVVVVVVrOtaP4d0u41rXtUtdOsLRC89zdSiKONR3ZicCvjl/wAFAVja48OfA+0VyCUfX72HI+sELfhhpB/wDvXiPxL4g8X6vPr/AIo1q81XUbk5lubuYyO3oMnoB2A4A4FRxvM6xRIzu5CqqjJJPQAVf/CDxP4a0iHXPH4j8LwXSbrSzvgRfXY9Y7b74Xj78nloezE8UbWBWOwsy5+XdwcUFVRhQB9BX/BPD4ivFfeIfhXezkxzoNUsFJ4DrtjmA+oMRx/ssa+OXwP8C/tEfD26+GHxGjv5NCvbiGeeOyujbu7RMGQFh23AHHsK1r/gi3+y5fgvpHjP4j6ZJgAKuo2s0fXkkNbbs44+8BwPfPws/wCCQXgn4S/GDwj8VNC+NOtXieFNXg1FdOvNIiY3DRPuC+asi7QSF52HofXivifF4vn+Gni2D4eqW8UyaFeroYEyw5vTA4g+dvlT95s+ZuB1PFC5/wCC3Xw8+R4dZ1a3RsAGPRdU80AkA5XfKBznqD0z0oft5f8ABUf4d4fx1+zfLf2yY8ybUfAmoQqRwTiSB0QHAPqBk5HTGnNevp9q+ppGl4YUNwsf3Vk2jcF5PGc9z9arX/8Agp1+yH4R8d+IPh34x8aaxomqeGdVutLv3n0K5miE9vK8Um1oFkLLuQ4OOQR748A+PvCPxR8HaX4+8BaymraBrMRlsbxInjEyBipIV1VhypHIHSq8TfEDwF4LjMvjHxvoGhIBktqepRWoA45zIw9R+YrxT+3r+xv4OV21b9ojwhceWCT/AGXdNqZ6Z4Fqsmfw+leKf+CwP7HXh8suk33jHxNtOAdL0Py93OMj7U8P15x0rxV/wXB8H24dPA/wB1i/JGEk1XXI7TB9Skccue3G4Z9RX/D0r9uX4nJt+DX7NVjNDMD5clj4d1HWJcYBG1o2CHjPVD1HSvHH7Iv/AAUC/aj+IN18SvFHwCbT9Z1Pb9quGtrLQkYqf9Y6O6Mz88sQzkAZJxVr/wAE5f8Ago58SreK3+J37Ra2dikax/ZdT8Y6hfMigFcLEitHwM/xD73fJrwp/wAEPdPDJcePP2hbm4LcyW+leHxEQe+JZJmz/wB8D9ePCX/BHf8AZD8PeW+unxp4odcF11HWRCjHHOBbRxMBnnG4n3NeEv2D/wBjvwT5Z0T9nnwhM0WNjapaHUyCBgHN0ZCT7+vPXmtA8KeFvCdoLDwr4a0rRrYKFEOn2UdugA6DagAwKqqqqr9sT/gpn8K/2bZL3wN4Ijg8b/EGENHJZQz4s9Mkx/y9SryWBxmFPm4IZo+CfDP7N37eX/BRzU7Xx18dfGN34Q8BTOLixXUIGhgWM9DZacpUt8pGJZSu5cHzHr4D/wDBNj9lf4FxW98vgiPxnr8OGOreJkW8Icd44CPJjweQQm4cfMcZqOKOGNIYY1REUKiKMBQOgA7Cqqqqqqqqqqv24f2TtA/ax+Dd54bFtBD4w0NJbvwvqLAKYrnbzAzdoptqow6AhHwSgr/gkX+0trNzaa1+yN8S5riDXPCRln0CK9DLMkCOVubNg3IaGQ7lU8hWccCMAVVVVVVf8FDv25PEPxH8SyfscfsvNcatqGr3S6ZrmqaXLukvJnO02FsynG3PyyvnB+ZOFDk/sGfsNeGv2TPBK6x4ggs9U+JWtwAaxqiDetohIItLcnpGMDc3BkYZPAULVVVVVVVf8Eiv+Jz+1R8aPE3+p8zT5v3H3sedfh/vcdNmOnOe1VX7RGjnxF+z98TfD6qWOp+DtWtQAwUnzLOVeCenXrX/AARC1cz/AAt+Jegbjiy1+1uQCoAHnW5Xr1/5YdO341VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX/AAUJ+LPi39sj9p7w3+xp8F7gXemaDqptrySNsxT6ngi4mkI/5ZWsQkU9wRN1+Wvgp8IfCXwH+Fvh34UeCrYR6Z4fs1hEpQK9zL1lnkx/HI5Z29244xVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX/AAUL0Y6j8DbPUkTLaXr9vKzY6I0csZ/8edPyr9lvWhr/AOz34EvhJu8rSUtc5z/qGaHHU9PLx/h0qqrxZ4H8H+O9O/srxj4Z03WbXB2x3lusmwnupPKn3BBr4i/8E+vAGt+be/DnxBe+HLk5K2lzm7tvYAkiRfqWf6V9m/bY/ZoUmB7rxL4eteyltUtAg9jiaFB142Cvh1/wUF8D6x5dh8SvDt54euidrXdpm6tvqVGJE+gD/WrPV/hj8afCl1ZWOpaL4q0O9j8u6hjlWZQDyA69UYcEZwwIBGCK/aX/AGT9c+DlzN4p8Krcar4Olf8A1pG6XTySAFmwOVJOFccdjg4zX7Nn7Xuu/Cd7bwf43kuNX8I5CRHO+fTR0zHn70frGenVccg+G/E3h/xjolr4j8L6vbanpt4m+C5t33Kw7+4IPBBwQeCAarxN4m8O+DNAvvFPizXLHR9H0yEzXl9ezrDDBGOrM7EACv2mP+CnPxI+Nnib/hRH7D+hazLLqcjW3/CQW1q51C9PQ/Y4+sEeOTM+HA5xHtyf2RP+CT/h7wlcwfE/9qmS38X+K5pftKaA0v2iytpGO4tcuf8Aj6l3dQf3ec58zIIgggtYI7W1hjihhQJHGihVRQMAADgADjFVVVVVXib/AJTBw/8AZULL/wBDiqq/4LfeM7CDwV8Mvh4lwjXt7ql3qjxDBZI4YliVj3AYzsB67G9K/wCCevhC88EfsX/CrQ76B4pptHbUNjjkLeTy3S/ms4P41458AeCPib4bufCHxB8KaX4i0a7x5tjqNqs8ZI6MAwOGGeGGCDyCK/aW/wCCOPw817SL/wAS/s06rd+HddiRpIvD2pXRuLG6I58uOV8yQsegLs65wDtGWH/BLP8Aa8+JGhfFQfsh/GHUr68tJhc2+hf2izPcaTe2ys0loWbnyikcgCsfkdFVcBiBVVVVVVVVVVVVVVVVr2uaP4Y0PUPEviHUrfT9K0m0lu767uH2R28ESF5JHPZVVSSewFfEb/grT+x54G86DQ/Emu+NLuLKiLQtKcJu/wCutwYkI6ZKlvbPStf/AOC0XjrxTrUeh/Bb9nNLiR5Bs+33st/PMmRnFvbom04z/GwzirK6iv7OC9hV1juIlkQSIVYBhkZB5B56UyqylWAIIwQRXhv/AIIw/s12Gozaj4q8YeNdaDzM8dnDcQ2duiEkhCFjaQ4GBkOK8F/8E9P2MvAixjSvgF4dv5Exl9aEmqFiO5Fy0i/gAB7VpelaXoem22jaJptrp9hZxCK2tbWFYooUUYCoigBVA4AAwKqqqqqqqqqqqqqqqqr45ftG/D/4E6Xv166+363PGWstGtnHnTdgzHkRpn+JuuDgMRivjP8AtA/EX446qbrxVqhh0yGQtZ6RbEpb2/ocfxvj+Nsnk4wOKr4IfsgfE34xrb6zcQf8I34alww1O+iO6dfWCLgyf7xKr/tHpXjzxR8Ef2TEk8IfBrQ7XxB8Qo0KXXiDUQtydNcjnHG0Sf7CAAfx5xtOv+INb8Vavc6/4j1W51LUbxy89zcyF3c/U9uwHQDAHFaB4d13xXq1voHhrSLrU9Rum2w21tEZHc/Qdh1J6Acmvg7/AME/3ljg1v4z6s0ecMNE06UZHtNMOB6FY/wevB3gHwX8PtMGj+CvDOn6Pa4G5baEK0hHd2+8592JNVVVd3dpY27XV9dQ28MeN0krhFXJwMk8DkgV4m/ac/Zx8HAjxP8AHnwBp0g/5YzeIrUSnp0jD7jwwPA7ivE3/BTj9iTwwWil+NUOpTqOItM0m8ud30dYtn5tXif/AILQ/sw6UskXhzwf4/12ZQdjCxt7aFuuPmefeO38HQ/hXiP/AILg3E8ps/A37Oe5nbbDNqPiIszE5AHkxwdenAc+nvXju2+K/wC0jq9/4g8IfsFW9tfaxcyXU+qaDomvSyPLKwd5Gb7SYPmLZyYwBvz3FfDzwB/wWJPgfRfh14JsNe8KeGNBs1tbC3aXS9LMEajgFmKzuSc8ksct2FN/wTn/AOCj3xOOfil+0rHFayH95BqPjHUb5lBznbEqGPGCeNwHP1rwx/wQ9EjC68dftEu7scyQaZ4e5J4yfNkn55z/AAeh9q8K/wDBGb9lfRQkviLxD488RSjG9J9Sht4j16LFCrjOR/GenbmvC/8AwTf/AGKPCQjaw+A+k3sidX1S8ur/AHHjkrNKy/wjgADrxya8K/BD4L+BQn/CE/CPwZoJjxtbTdBtrZgRnnKIDnk8+59aqqqqqqqqr9vz9v7xVqfiqT9k/wDZNmvr/wAV313/AGdrGsaUpedZ2O02VmV58zPDyj7nIUggsv7En/BMHwd8G4LH4n/Hyzs/FfxClxcRafOBPZaLIeeAcrcTjOTI2VVvuDKiRqqqqqqqqqqqqq/4KFeFNb/Y+/bV8Gfta/D60KWHiS8XULmCL92kl5DtS9gYjotxC4JPUmWU9q8JeKdD8c+FdH8aeGb1bzSddsIb6xnXpJDKgdG/FWHFVVVVV/wUT/b91CTUJ/2Uv2Y7261LxZq9yum61q+lMXkhkdgn2C0KctOxIR2X7mSg+ckp/wAE9P2A9N/Ze0BfiH8RILXUPibrNsFlICyR6HCw5t4W5zIQcSSA4P3V+UFnqqqqqqqrq5gsrWW8upAkMEbSSOR91VGSfyFf8ESbafV/H3xh8VXKM0q2OnRyOowgaea4cjHqTEcfQ1Va3pket6Nf6NKyhL+1kt2LJvADqVOV7jnp3r/gh1qRtte+MXh+Xask9rpE+x2w48p7tThfT96M+nHrVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX/BQn9qiD9l74D3t9ol8qeNfFQfTfDkSsA8UhX97dY/uwocg4I3tGDwxI/4JHfsrS+A/AVz+0p46sXbxN46g2aKLkEvbaYW3GXnndcMA+ecxqhB+ds1VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVXN1a2cDXF5cxQRIPmklcKo+pNaz8avhB4f3LrHxP8L20iDmJtVhMn/fAYt29K1X9sP8AZz0nKSfEeG5kHRLWxuJs9OjLHt7+taj+378CbI4tbXxRqHvb6fGv/oyVf8mr/wD4KMeBo1P9l/DrXbhsDAuLmKHnPP3d/b/Per3/AIKQzNldO+EKLhuHn1wtlf8AdEAwenc1/wAN8/FbUgF0P4Q2LtKcw/8AHxPlevRQN3HcY9aP7ZH7Ud+d+kfBWxZFGH/4kV/Ng/VZBj6V/wANQftn6ofM074KMFQYYW3hS/cZ98u3Nf8ADRv7b/8A0RW+/wDCPvv8a+MPxa/at8f/AA51fw14/wDhJfWOhSIk13d/8Ivd2/2dYnWTf5j5VBlOSexNfAr4/wD7R/gv4e22g/DjwANf8O6Rcyp550O6u0jkdvMZGkiYAHMgOOD8w9aP7Yf7VdpifUfgpYrbr94/8I/qEf0+YykDnHaj+3h8ZdPymsfB6xRyMqDFcw8euGzn61af8FIL5CRffCKCUYGPK1wx49esLZqw/wCCjPg2Ugap8NtZtxk58i8jm4xweQnft/8AqrTf+CgHwOvgovNP8U6e2OfPsI2AOO3lyscdulaV+2N+znq+1E+IiWshONl3YXEWP+BGPb+teLdB/ZD+OYabU9c8GXepTAkXllqkVteAnuxVgzd+HBHtXif9hbxr4UvF8V/AT4lSSyxfNbxz3BtblR6JcRfKxyO4QVbftPftHfBiQeGPjv4CbXNNlUwP/adsIXnQjBVZ0BilGM5yHz3NfE/Tvg34sMvjH4PajLoskpL3vhbVF8t4D3a1lyUkTPSPcHHG0EHatfBb4+eO/gfrX27w1efadMuHBvtJuGJguR64/gfHRxz65GQfG/8AwUX/AGePAPwk/wCFl65ql5/aLt5EHhmKPdfTXO3OwfwiPAz5pIXH+1hKjg/bF/4Kz+OvOmP/AAiPwq0q84zvGmaeR2HRr27Cnr0Xd/yyVhX7Mv7Ivwa/ZU8LjRfh1oSy6vcxqNU1+9UPe37Druf+CPI4jTCjrgsSxqqqqqqrxN/ymDh/7KhZf+hxVXx5/aj+CH7N2gT6z8UvHNjZXSQl7XR4ZVlv7044WKAHccnA3HCDIywHNeH9D+KP/BVf9sOTxJqmn3Wm+C7B41vJFz5Wi6PE5K26v0NxLubAHV5HfARTiKHSfDujpBAlvYabplsFRQBHHbwxrgAdlVVH0AFeA/iN4B+KPh+HxT8OfGOkeI9JnA2XWm3aToCR91tpJVvVWwR3Arxb4v8ACvgLw7e+LfGviHT9E0bTYjJdX19OsMUSj1ZjjJ6AdSSAOTX7IkE37R//AAU8ufil4L024g0BPE2q+J5pDHtMFmWlMRkHZneWFSPWQ+lVVVVVVVVVVVVVWt65onhrSrnXfEWsWOlabZJvuby9uFghhXpud3IVRyOSa+KX/BUT9jn4YiW2g+Ik/jG/hzm08MWhvA30nYpAfwkNaf8A8FjvEvxB+J3hzwf8MPgBKdIvdWtotQluZ5dQvWs2mUTNFBbqoRxGWwSzgNgkEcVXiLQdN8U+H9T8MazD51hq9nLZ3Uf9+KVCjj8VYivh1/wTB/Yz+Hawyt8L28UXsRH+leI76S83Y/vQgrCf+/deFfBHgvwJp39j+B/COi+HrHj/AEXStPitIuOnyRqB+leKZJIfDGrywyMkiWE7IynBUiM4IPY1+y/of7Xn7Wvj+/8Ahx8OP2ifEGnalpujyarLLrfi7UYYTDHNDEVUxCVt+6dCAVAwG56A/wDDtn/gpL/0dfpP/hd63/8AI1fsOfsj/tb/AAD+LOreMPj38brHxj4fvPDs1hbWEHiXUdQaO7a4t5Fl8u5hRAAkUq7gdw34xgkj/gsl8UfiX8NvCHw0i+HfxC8R+Fxq19qSX50bVJrI3SpHBtWQxMpZRvb5TxzXwW/4J8/tTfH34W+HfjFH+1bdWUXiqzF3FBd6hfzzIhJUB238t8vbP1r4o+Av+ClP7AVonxFsvjTqfi3wZbTIlxcRalNqlnbgkBVuLS6BMKt93egwMqN4YqK/Yc/a/wBI/bA+Fk3iSXTYNI8V6BOtr4g0uBmaKJ2BMc0RbJ8qQK2ASSrI6knAZqqqqqqqqqqqqqqv2mf209J+H32vwN8LLi31TxKhaK61DiS305hwQO0ko9PuqeuSCta3reseJdWute1/U7jUNRvZDJcXNxIXeRj3JP8AkV4M8D+K/iHr9v4X8GaHc6rqVyfkhhXO0d2ZjwijPLMQB618Af2HfCHw9Ft4m+JYtfEviFcPHalN1lZt7K3+tYf3mGB2XIzX7XH7WH/CFrd/Cv4Z3wGvFfL1PUom/wCQeCOYoiP+WpB5b+DoPm+67tIxd3LMxySxySfWvgv8BvFnxm1J2sZIdJ8P2UirqOuXg2wW2cfKuSN8hyMICOoyVBzXgfxX+xl+zRox0u0+Mfw90y+kRVvLy+8R2hvbw/L94b9235lOxQFGc45JrxJ/wUt/Yk8MBkuvjnY30o+7HpumXl5u6dGjiKd+7CvEn/BZj9lHRw0eiaF4/wBek/ga20uCGM9OplnVh3/hPT8a8Sf8FxNDiVo/CH7PF9ck/cl1LxEsGPcpHA+e4xuHr7UP+Cqv7bPxDKp8I/2Z9MnjnGIzb6DqWrSHO3btMTopPzL/AAnO4etD4p/8Fp/iYAmg+C9b8PRTY/dnw9p+mY6fxXoDr/30O+ehwP2Sv+CuvxKAXxf8ctR8PpLjzIrrx1LBGOn3ksd6nH0PQ/jZ/wDBGP42+MLhLz4q/tJ6T9o/ilhtLrVmGQAcNM8JPQDt0FeGv+CIvwftSn/CY/GnxjqmB839mWdtYZ+nmCfH614a/wCCSH7FuhFf7U8I+IvEW0cjU9fnjz16/ZjF6j8h758M/sNfsg+ESjaR+zt4JlaM5VtR01dQIPPObjee/wCHHoK8O+BvBPg+PyvCPg7Q9ETbt26dp8VsMemEUcVVVVVVVVVVVVVX/BR/9pm8/Zq/Z4vbvwvffZ/F/i+Y6TocisA9qWQma5A/6ZxjAPZ5Iz0r/glF+yDY/Dv4fW/7SXjzT/tHjLxrbmTR/tCktpunPnDjP/LS4GHLcnyygBG5waqqqqqqqqqqqqr9vL9ntv2kv2a/EvgrS7JbjxHpijVvDw25Y3kAYiNfeWMyRemZAe1f8Ee/2k4vFfw51D9mzxVe7Nf8FNJdaNHMdrzae75kjAPJaGZzn0WVAOFNVVVU00NvC888qRxxKWd3baFAGSST0GK/bj/4KOax8Q9Ul/Zl/Y8kvdZv9Zm/s+/8Q6SGklu3Y7WtrDZyQehnHUZ2cfPX7AP/AATt0H9nDTrP4p/FK2t9W+KN5ASqkiWDQUccxxHkNOQSHl9yqfLuZ6qqqqqqqvjjro8L/BT4geJjIU/sjwtqV3uD7NvlWsj5zzj7vXtX/BD3Qjb+Bvir4m8rAv8AVtOsw+08+RFM+M9OPtHTrzz1FVVf8EjidH/ax+M/hmJ1khTTbn52GHPk6gqA9cYIc5/Cqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq+vrLTLG41LUbuK2tLSJpZ55nCJEijLMxPAAAJJPSoE1n/gqd+3k0kq3a/CzwlnjJj8nSIX4HbbNdSH/eUOeoiqzsrTTrODTtPtore2tYlighiQKkaKMKqgcAAAACqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq8S+PvA3gyMyeLfGOjaPhchb6+jhY/RWIJ+grxP+23+z74dLx2viS+1yWMHMemWDtk+geTYh/Bse9eIP8AgozYl/s/g74X3NwznEcuoagIznt+7jVs/wDfYpf2jf20viEufBXw2fT4JF/dz2fh+Rl+vmXBZCf84o/C39vvx982veM7/RY5s7hJrkdopBHOUtM8dsEfhVv/AME/PiN4hlW58c/F6yM2PmeOCe/YEgZwZGjJ6e1aR/wTq+HUAH9vePfEV6R1+yRw22f++lk9v89NK/YY/Z604AXmg6rqmP8An71SVc9f+eRT1H5D3zpv7LX7PmlAC1+FmjybRx9pD3HbH/LRmrT/AISfCrSdp0v4Z+FbQqcgw6NAhzjGchM5x3qx0bR9MCjTNKs7TapCiCBY8DuBgdKqqr4g6D/wlPgLxJ4ZCbzq2kXVoFGeTJEydvrX/BNjXhLpHjjwu74+z3NpeRLzzvWRHPpx5afn+VVeabp2oqEv7C2uQFIAmiD8HqOR0q/+Ffwv1XP9qfDfwteZxn7Ro8EnTp1Q1qX7MP7P+qjF18KtDTIx/o0TW/8A6LK+tat+wz+zzqIIs/D2p6Znp9k1SVsdOnml/T9a1r/gnT8P7gN/wj3xA8QWRP3ftkMN0B0/uiP3/T8bj9gf4seF5Xufh58XbESj7rt9o05m/GIyY6etan4A/b18J2cthJf3XirTXBWaK4u7bVo5R6NHdZdgcf3fyNfETQfGNjI1x4s+EcnhS43fPNFplxZRyHP9xyYwPQRhRVVqel6brVjLpurWUN3azDDxSpuU/hX7Lf7XPw+OgaP8K/FukaT4Nl0+JbXTZbG3S106RR0XauFgY/8AfBOTlcgUrK6hkYMCMgjvVVVVVVVftF/8Eo/ib8Wvjx4q+NXgz41aLo8mv6ib+1gms54prSTAAAkjY9NoO8AHnpxzB/wTA/bgvYmsNa/bIZbIx4Eceu6rODgjAKNtGPx7Dj0+HH/BFT4b6dqS6t8YvjHr/i1vM8x7TTbJdNSQ91kkd5ncHnJUoeeo618NPhX8O/g54TtvA/wv8Iad4d0S0OUtLOPbuYgAu7HLSOQBl3JY4GSa8eeEbL4geCPEHgTUr+9srTxFpdzp1xcWThJ4o542jdo2YMFcKxwSDg4NeIP+CN3xk8Ea5NrH7P37R1palifIbUBc6VcRrnIVprXzN+MDkKvrgVb/APBI39qj4i3trD8dv2nNMu7C2kyhjvr/AFuSMeqLciEKTlh14984r9mH9k74Tfsn+DZfC/w3sJ573UGWTVdZvir3d+6jA3soAVFydsagKuSeSWY1VVVVVVVVVVXxG+Mfwo+EWnnU/if8RfD3hiDyy6DU9QjgeUD/AJ5oTukPHRQTR/4Kv/sv6r8UPDPwy8EyeIPEJ8Q6vb6c2tLY/ZLK1M0gjVyZisrAMwJxGBjJz2qvip8PNG+Lfw18T/DHxA7Jp/ijSrjT55EUFoRKhUSKDxuUkMPdRXwr/wCCTf7Inw6MN34g8O6v461CPB83X74+UG9RBAI0I/2XD14O+H3gL4d6YNF8AeCtC8N2AAH2bSdOitI+PVY1AP1qqqvFv/Iqaz/2D5//AEW1f8EVP+TqPFP/AGT+8/8AS+wqq/4Ljf8AIqfCP/sIar/6Ltq/YD/5M0+Ev/Yvx/8Aob1+0HB4fuPgN8RovFa250c+FNSN954+QRC2kLE/QDOeoxX/AARE/tf/AIWz8SPI3f2X/wAI7bfacZx5/wBo/dZ7Z2+d+vvVVVVVVVVVVVVVav4h8P8Ah+ITa9run6bG33XvLlIQeQOrEdyB9SK/an/bTn1prz4c/BvU2h04botQ12ByHuemUt2HKp1Bfq38OBy1eMviV4T8Am3GvzzzSTOP9FtArTFM8tgkADHTJGT+NeB/+Crngb4N+Hx4X+C/7LoSaYDz9U1bxJ5lxeScgGRY7fJ52kKHAGSABnNeH/8Agpd+1h8S5r21uPAPhXwTo/kMFuRp9016xcEL5Tyy7OAQS3lnkDHXiaWW4leeeVpJZGLO7sSzMepJ7k18Nf2dvjF8ctL1Ob4XWejQtY4jF/rlzJb2iykZC5jjkdyByVVfTJG4Grf/AIIwfGLxbefbfiR+0ZocUruzubPS578ITjOxXeEAYC8DA4A7V4b/AOCInwgtdn/CYfGrxhqePv8A9m2VtY5+nmCbH614a/4JE/sZaFs/tTw94o8R7MZ/tPXpI9/Tr9mEXp2x1Ptiw/Yk/Yb+GlvHd3PwR8B2sMY+WTXh9rU4xnJu3cHtnPr7moPiZ+w38J/3eneOfgj4Vktj8sNlf6baSIVIHypGQ2QQOg4x7VrX/BQ/9izQQ/279oLw/L5e/P2KK4vPu9ceTG+fbH3u2a1f/grD+xHppYWXxH1bVdpGPsnh28XdnrjzY06e/wCtap/wWZ/ZKsCVtNC+I2pYkKg2uj2y5A/iHm3KcH8+elal/wAFtvgbEp/sj4Q+Ork78AXMlrB8nPPyyPz049+vHOo/8FxfC0Ql/sj9nXVbkqf3X2nxLHBu/wB7bbvt/DNX3/BcnWJHQ6Z+zXZwKB8wn8WtMSfYi1XH603/AAWZ+OV9G11pH7N2kG3lBMDm5u5hjtllUBvwx+FN/wAFVP23bofZtO/Zc0k3UvyQg+H9Uky54HyiQFuewIzTf8FIf+ClNwpgh/ZS01ZJBtQp4D1osCeBgG4Iz+Br/huD/grD/wBGr6r/AOGz1X/4qh+1r/wV51fN7Zfs+atZxMcCE/D+4iwR14l+f86/4ai/4LB/9EL1X/whXr/hqL/gsH/0QvVf/CFev+Gov+Cwf/RC9V/8IV6/4ai/4LB/9EL1X/whXo/tgf8ABXLQyDqH7Oerah5w+Qf8K8vJdmOv+oIxnI+9+Hej/wAFC/8Agp54fdoNb/ZYjaR1Dqt/8PtYiZV5GQFmTg4PJz0o/wDBYj9o/wALES/EH9mvRoYcA/cvdP4PHWXeOTjt7V8E/wDgsb8BfiHq0Og/FLwtqnw4uLlgsV7NcjUbEMeAHlREdMnuY9o6llFaTq2la9plrrWhanaajp97Estrd2kyzRTxsMq6OpIZSOQQcGqqqqq/4KSS3f7Qv7fXwv8A2bYbiQ6bp66fYXCITmGS+nElzKPTFsID/wAAz7CwsLLSrC20vTbWK2tLOFYbeCJdqRRqAFVQOgAAAFVVVVVVVVVVVVVVft+/s4/ET9lH452f7bv7OsD22ly6kLzV4reEsml3znEjSIMZtbncwbsGd1JUOgr9kX9tn4U/tZeFLZ9F1O30jxpbQA6t4ZuJQs8LgfNJDn/XQk8hlyVBAYKeKqq+On7SnwW/Zw8P/wDCQfFvxzZaR5qE2lgp828vSM8QwLl2GRgtgKCRuYda+LH7U37VH/BSnxdP8Ev2cvB+o6B4FfAv4Fn8sywk483UbofLHEQCRAuQSCP3rBa/Yw/YG+Gf7JOjprTtF4l+IV7Bs1DxBNDtECsPmgtEOfKj7Fvvv1YgYRaqqqqqqqq/by14+G/2Ofi3qKyMhl8NT2eVJB/0giDHHY+bj0554r/gjL4fOlfspatq8iDfrXjK8mRtuCY0t7aIDryAyOe3U/Wqqv8AgnX/AMSn/gpp8c9L1D91cm38SW4j+9+8XWbYsuRkdEbnpx71VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVf8Fdv2oZfh/8ADiz/AGc/Bl83/CSePofM1bySfMt9LDFdmBzm4kUp/uRyAj5ga/4J6/stRfsv/AOx07W7ER+NPFfl6n4kdkAeGQr+6tcjtChKkZI3tKRw2KqqqqqqqqqqqqqqqqqqqqqqqqqqqqrxB4n8NeFLE6l4o8QabpFoOPPvrpIEz9WIGfavGn7cfwJ8Kl7fStU1DxLcpxs0y1IQH3klKKR7rurU/wBun4v+ObttK+EPwpjEjfKP3M2pzj0IWMKqn2IYV/wrv9vD4xfN4l8SX2gWM/VbnUE06Pb6NDbDeR/vLXhn/gnTBI4u/HvxNnnkc5kh0yzCnPc+bKTn8Urwz+xZ+z34bVWm8JXGtTp0l1O9kk/NEKofxWvDvgLwN4QRU8KeDtF0jauAbGwjgP4lQCfqetVVVVVVVVV+yMT4D/a28d+AJSEimXUbWFMkBmhuVZCPby1c/Q1VVVVVV4i13QPDOhX/AIg8U6rZabpGn27zXt3eyrFDDEo+ZnZuAuPWv2qP2sG/am+KNn8F/wBjD4R2FxPJebR4httGiW/1VkPJTcoENsOpeTkgZOxcg/Dj/gnDrv8AwraxX4ofFFV8cygyXb6bYLJZW4IGIgpKtIw5zJlAc4C8ZPxr/Zw+IfwPvA+u2i6hosrbbfV7NGMLE9FfPMb/AOy3B7E4NV8Bv2u/HXweEGgax5niLwupAFlPKRLaL38hznA/2DlfTbkmvhn8W/APxd0Qa34H12K8VVH2i1c7Li1Y/wAMkfVe4zypwcE1VVVVVVVVVVVVVVVVVVVVWueOvBPhnVdM0HxF4v0XTNT1qZYNNsbu+jhnvZGOAsUbENISQeFB6H0qvj14S8aeOvgz4x8J/DjxPe+HvFWo6RMmjalZ3TW0kF2F3RYlU7owzKFLDkKxIr4c/wDBGLxP4l1Q+J/2lfjpNeXdwwe6ttBD3M8xPXdeXQ4Pb/VN9eOfg/8AsDfsofBJ7W98KfCXTNQ1a0dZI9W1wHUblZF6SIZcrEwwOY1THWqqqqqrxb/yKms/9g+f/wBFtX7G/gL9pH4i/E/U9E/Zc8WXHh/xXBoM1xeXMGrHT2exE8CunmDqDI8B299ue1f8Muf8Fg/+i6ar/wCF09fsHfCj9s74bat4wn/as8fXfiO1v7a0XRUn19tR8mRWl80gH7mQ0fPfHtX/AAXG/wCRU+Ef/YQ1X/0XbV+zf/wVs8E/BH4F+FPhPrHwb1vUb3wtpws1urbVYljucEkNhkymc4x82PevjL+2L+1j/wAFDtPb4NfAf4K6no/ha/mVdSj06R7t7lVIIW6vWSOKGHdtYrhQTgFmHB/YK/Y/t/2Q/hNPomr31rqPjHxLOl34gvLYHylZFIit4yQC0cYZ8MQCWkc4AIAqqqqqqqtT1XS9GtHv9Y1K1sbaMHfNczLEi4GTlmIA4BP4V4p/a6/Zc8GM0XiL9oLwDbzRnD28evQTyqc45jjZmHX09fQ14m/4Kq/sS+HgyWnxPv8AXZU6xaZoN235NLGiH8GNeJ/+C1/7P9gXj8JfDDx3rDqOGu1tbJGPPQiWRsdOqjvxxzrP/BbbxlrFwbD4e/s32aztkRC912W9Z8Z58uKCM+nAY9Dz6N+3p/wVD+JWV8Afs4yafA+RHcad4Gv5go5xulnd48/gASOnWmtP+C23xLyJZ9b0a2fPIk0fSPLBzgfLslPTHcjIzwc1df8ABP8A/wCClPji2m1D4mftPwWFlGjPcLq3jrUZ1iQZJIREaMKNoJ+ZQAc9jjRfgToXw/8AF9/e3ni6Lxpe202y21VYHSJtowXQSEs3PRmwcAYAqv2X/wBkPWfjBPB4y8bRXGl+DYnygwY5tUI/hi44j7NJ+C85K6Z+xv8Asq6VeNqMfwA8EXl2zbjcalpMd/Ju4wd04c5G0Y9PxNeMvEfwX/Z58K6hr1tonhnRZrG1LW+nadbQWs9y3ASNEQA8kqM4wAcngV468a698RfFmpeM/Et15+oanOZJCPuxjoqKOyqoCgegFeI/GvhHwJbW+seNLm9TTmuEjkSwiSW5kBPzCJHZVLBcnlgPerX/AILI/Db4b+GLLwZ8H/2b9SfTNOi8u3bVNfjtm6kl3WOKTczHkncMljzxyf8Agp/+3j8Th5Xwe/Zps3hnH7uSw8Najq0o46hlcJ2J5Q8fQ5/t3/gtf8WW22Wma34dtJDnZ9k0vRhEMj+KbbNxkcZJwOh5pf8Agn7/AMFNviUN/wAQ/wBpE2cDfft9T8cahc7f92OFHj6gZGR681pP/BEXxbqtz9t8cftJWaSsQZTaeH5Lt3AwMb5J0I+UYyQcccGtB/4Ij/Ba32/8JR8ZPGuoYxu+wW9rZ565xvSXHb1xg9c8aL/wR2/Y/wBKVRfHxxrJXGTe62qbsHnPkxR9ehx+GK0n/glz+w3pJSQ/BY3sqEkPd6/qMmcjGCnnhD+K1pf7BH7G+jhBZ/s8eEZNibR9qtWuePfzWbJ9zzWmfsq/sw6MY20v9nX4Z28kabFlTwpZeZj3fy9x/E1p3wi+E+kLCmkfDDwlZC3bdELfRLeLyznOV2oMHPPFWOkaTpe/+y9MtLTzcb/IgWPdjpnAGcZP51VVVVVVVMqspVgCCMEEV+0t+wB+z3+0no1ydQ8K2fhbxSVJtfEmi2aQzrJjjzkXC3CZxlX+bH3WXOa+GPxf/aM/4JWfGBPhH8ZbO78QfDDVJ/MjWAtJbywk/Nd6c74CSKWBkhOMnhsErJXgTx34Q+J3hDS/HngPX7XWdB1m3E9le2zZWRT146qwIKspAKsCCAQRVVVVXg9U8d/8FtdTvr4o6aVql4dpbdk2ujtDHgjHIZEOO23HPWqqqqqqqqqqqqqqq1LTNN1nTrnSNXsLa+sb2Fobm1uYllimjYYZHRgQykEggjBBr9oz/gkQkviJviT+yN4yHhDVopjcpoV5cyxRQyjkNaXSZkhOeiMCATw6AAVZfHv/AIK5/syoNJ8cfDnWfGum2K7RPquhnWogo43NeWLB26ZzJIT6+lL/AMFev2omX+yP+GZdIbWMldotb/72c48nO77vbd7+1XP7RH/BXL9pNP7K+H/wu1HwTp96MfadO8PnSY2TplbvUGYj13RuD6elfB7/AII+eI/FPiD/AIT/APa8+LV3rl/dP511pml3klxPdP8A9PF9N8x4ABCKSe0gxmvhv8Lvh58H/Ctt4J+GPhDTfDmi2pylpYw7AzYALu3LSOQBl3JY45Jqqqqqqqqqq/4K0eIF0b9ijxPppkVTruq6bZAHHzbbpJ8D/wAB88en1r/gmB4ePh79iH4drIm2fURf3svGM+ZfT7D0B/1Yj/ocYqqr9h//AJSw/HD/ALCHiv8A9Oq1VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVXxG8feGfhX4E134jeMb9bTRvDtjJeXkp67EGdqjuzHCqO7EDvX7DXw/wDEv7dP7Y/iX9rH4rWBk0DwxqKX8Ns53RfbBxY2a9NyQRork9zHHuB8w5qqqqqqqqqqqqqqqqqqqqqqqqqqqvE/i/wp4L046r4u8R6bo9oMgTXtysIYjsu4jJ9hzXj79vz4U+HTLaeCtM1LxVdJkLIq/ZLYn/fcF/yTB9af41/tmfHlhH8OPC9zomkzn5J9PtBBGy+93OcZH+wV+leH/wBgn4g+Lr0a38Yvihm4k5dLdpL+4b2aWUgKfoHFeC/2NvgF4MEcreETr11GP9frUxud31jGIv8AxytM0nStEsk07RdMtLC1iHyQWsCxIv0VQAKqqqqqqqqqqqvGZ/4Vv/wUO0rVwfKt9bv7UpkYBF3bi2cn28wuc/4Zqqqqqq+NPxv+Gf7PvgS8+InxT8SQ6VpVqNsan55ryUj5YYIxzJI2Og4AySQoLDxl8R/2pP8AgrD8Tn8AfDnTZvDHwv0m6R5oZZGFpZJn5Z72Rf8AXzkAlIlyAeFHDSH9lz9kT4Sfsn+EBoPgHSxdazeRgav4gu4wbvUGHOCR/q4gfuxL8o6nLEsa1LTNO1mwn0rV7C3vbK6jMc9vcRCSOVT1VlOQR7Gvj5+wjJF9p8VfBLMifNJNoE0nzD1+zueo/wBhjn0Y8LWo6df6TfT6ZqllPZ3lq5jnt54zHJE46qynkH2NeFvFniXwRrUHiLwlrd1pWpWx/d3FtJtbHcEdGU4GVOQe4r4Ift5aHrot/Dvxkhi0i/OEj1i3Q/ZpT0HmoMmInuwynU/IKsr2y1K0hv8ATryC6trhA8M0MgdJFPRlYZBB9RVVVVVVVV8T/jZ8NvhBp/2zxt4jht52XdBYQnzbqf8A3IhzjtuOFHcivjD+3T8QvGpn0f4dxN4U0dwV89WD3sw9TJ0i+icj+8a/Y/8AjYPiz8NotJ1m+aXxJ4ZVLW+MrZe4ixiKck8ksAVYnncpJ+8M1VVVVVVVV8ZvHmqfC34UeK/iRo3haTxJd+GdKn1BdKjuPIa6WJdzqH2tg7Qx+6ScYxzUn7U3/BTf9tWV7L4EeC77wl4ZnYoLzQrb7FEFz/FqVyR8y858lkP+yeK+Av8AwSO+IOm+PdF+L3x3+PM58Q6VqVvqccOhF7q4aeJ1kRnvLgdQyrnEbZ5w3c1VVVVVVVeLf+RU1n/sHz/+i2r/AIIqf8nUeKf+yf3n/pfYVVf8Fxv+RU+Ef/YQ1X/0XbV+wt4D8Da3+yD8JdS1nwZoV/djw/Hi4utOilk/1jn7zKTVvbW9pAltaQRwxRjCRxoFVR7AdKqqqqrx58dPgt8LlYfEb4seEvDkiD/U6lrEMEreyxs25jweACa8df8ABWP9jLwaXi0zxlrfi2eIHdFoWjSnn0D3Hko31DEe9eMv+C3mmNILL4ZfAC9vHkO2OfWdZWI7jwB5MMb7sn/poP14T9uD/gqZ8aFDfCv4Fy6NaTL+5vNM8GzvGB6+feF4ifyHtX/DPf8AwWL+Mpz4z+KWq+FILk/MJ/FcOnRlSOd0Wm7iB/slfwrS/wDgjB8YvF92mo/F39pLSxdlR5slrZXOrP05Aed4TjgDJH4dq8Lf8ET/AIB6eEfxj8UvHOsyL1Wy+zWMbHjqDHK2OvRh1HPHPhn/AIJV/sS+HQjXXwvvdcmQYEup67dt6gkpHIiHr3XsD1rw1+x/+yv4QIfQf2evAEUq/dmm0GC4lXr0eVWYdT39B2FaRoGheH7b7HoGi2Gmwf8APKztkhX8lAHc/nWueI/D3hmzOoeJNe07SrVQcz310kCDGP4nIHcfnXin9tf9krwfJ5Gs/tCeCZJgcGHT9TTUJAc4wUt/MYHPbGa/at/a9l+LO/wL8Nry4tvB4wbm6aN4JdUbqNysAyRDsjAEnlgOAK1PxD8TBq9rafCD4R6z471G2nDTpb6FdahbKVx+7ZYRlzyMgHA6HOSKtfEP/BaP4nQQafoXh7WfCumhFjit49L0zQ0t0GQMGYLKFA42g9AMD1/4d9/8FMfimN/xO/aN+x20g/eW2p+NL+62gjBCxQo0XYAjcB35q3/Z1sv2ePG3iLwzceMrbxXrEEwt7zU7e3aKNXGDJEm5izYfIZjjJXpxk2trc31zDZWcEk9xO4jiijUszuTgKAOSSSBivhp+wd8Crfwhos3xd+G2jeLPFMUJe7m1FWuIoXfkxpGW8vCjC7tuWIz6AeFvg78I/AxRvBPws8IeHzGcodL0O3tNpBzkeWgxzzVVVVVVVVVVVVVVVVX7Qn7P/wAPv2lvhlqXwx+IeniS1ul32d7Gg8/TrkA7LiFj91lzgjoykqchiK+BvxY+LP8AwSz/AGh774H/ABpFzf8Aw1125WWSeKNmiaJsrHqVmCeDwFljGSQpU5ZFNaNrGk+ItIstf0HUbfUNN1K3S5s7u3kDxzxOoZHRhwVKkEEdjVVVV+yMf7a/4K7/ABZvrQFY7HUPEvmiTg/JciE4xn+Ij8Pyqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv8Agthr72n7Pfgrw1G7L/aXjBbh8EjcsNpOMHtjMynB7gHtX7IHh4eF/wBlb4SaKUCSR+DdMllQY+WSW3SRxwSD8ztz361VV+w//wApYfjh/wBhDxX/AOnVaqqqqqqqqqqqqqqp3SNDJI6qqglmY4AA6mtR+JHw60jH9rePvDlllSw+0arDFwOp+ZhxV/8AtN/s2aX5f9p/tC/DS083Pl+f4sso92MZxmUZxkfnV/8Atu/sh6dHLJcftHeAnELbWEGsxTknOPlCElh7jPHPSr//AIKIfsV6cUW4/aC8PuXBx5ENxPjHrsjOPxq8/wCCnf7C9jN9nm+PEDMBnMOganMv/fSWxH61ef8ABVn9h62nMMHxVv7pQBiWHw5fhT9N8Kn9Kv8A/grZ+xXaKht/Gmv3pbORB4fuBt+u8L+ma/4e8/saf9BzxV/4IX/+Kr/h7z+xp/0HPFX/AIIX/wDiq/4e8/saf9BzxV/4IX/+Kr/h7z+xp/0HPFX/AIIX/wDiqs/+CuH7F11N5U/izxHaLtJ8ybQJyufT5Nx/SrL/AIKrfsOXcpin+LN7ZqFyJJ/DeoEE8cfJCxz+GOOtWH/BSz9h7U4xJbfHvT0BfaBcaTfwHP0kgUge/StN/bl/Y/1Z/Ltf2jPA6EMF/wBJ1RLYZPvJtGPU9B3rSP2hvgB4gKjQPjn8PtSLkhRaeJrObOOuNsh6Vp+p6Zq9v9r0nUbW9gzgS28yyLnAPVSR0IP4iqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv8Agrf8fdZ8b+KvDP7F/wALjLqGpX95bXOvW9qSXnuZSBZWRH/AlmYHjLQnjaa/ZX+AOifsz/A7w38J9JaKa6sYPP1a8Rcfbb+QAzy+u3d8qg8hFQdqqqqqqqqqqqqqqqqqqqqqqqqqqiQBkkAAd6+Jn7XPwU+GpmspfEf9vapFkfYdHAuCG9GkyI156jduHoa1P9qz9pH44X8uhfBDwRLpdrnY0tnB9qmTOP8AWXEgEUQPHOFxn73evC/7CvxH8dagPEnxy+I0qXEwBlhina+u2H91pnO1T16bx0r4f/svfBH4bmO40XwVbXt/GBi+1T/S5cj+Ib/lQ+6KtAAAKoAAHAqqqqqqqqqqqqqq/wCChen3Xhv4i+APiRp4K3At3hRwMbXtZllTJ7HMx/I+laXqNtrGmWerWT7re9gSeI+quoYH8iKqqqq/au/a/wDhZ+yV4L/t7xpefb9evo2/sXw9bSAXOoOOM9/LiBI3SMMDoAzYU/C34HftJf8ABU74oD4x/G3Wbzw/8NbKZ47eaCMxwpGG+a106J8gns87BgCvzF2UJXwu+FXw/wDgt4KsPh58MvDNpoehacuIraBSS7HG6SRzlpJGwMuxLHuaqqr4z/s7fDn43WOfEenfZNYij222r2ihJ4/QN2kT/Zbpk4Kk5r40/sy/En4KTyXerWP9qaDvxFrFkhaLnoJF6xN2w3BPQmq+EH7Q/wATPgtdqvhfWTcaSz7p9IvCZLaTnkqucxsf7yEE8ZyOK+DH7XPww+Lv2fSLi7Hh3xFLhRpt/IAszekMvCv7Kdrf7Peqqqqqr4hfFPwB8K9JOr+OfEtppsZUmGFm3TTkdo4xlnP0GB3IFfGD9vfxb4jM2jfCawbw7p5BU6jdKsl5KPVV5SLj/ePcMK1LUtR1m+n1TVr+4vby5cvNcXEpkkkY9SzHJJ+tV8F/ivrPwZ+IGn+NdIBmjizDfWm7AurZiN8ZPY8Ag9mVT7V4S8V6F448N6f4s8M36Xem6nAJbeVfQ9QR2YHII7EEVVVVVVV8WPjf8JfgboDeJfix4+0jw3ZbWMQu5/3txt6iGFcyStyPlRWNePf+CyVlqvj3SPB37PHwb1PxPazanFHcXWoRObm+gDjzUtLSEl97IG2M7cHBaM8iredLq3iuY0kVZUDqskZRgCM4KnkH1B5FSxRzRtDLGro6lWRhkMD1BHpX7Vf7cPwU/ZC0iDTvEUrax4ouIA2neGNLZRMYxwryt92CLjAJBJwdqtg4g/4KB/8ABRn9pC6luP2cfgwumaMrlI7nTNBbUBGeflku7kGAtx2VOnSpfif/AMFsvCEZ1XWfCusaxAnJt18N6TdE46/JaIJDn2/Cv2DP2gfjl+0R8O/EHiD45+ALDwtqehaz/ZUUdvZXFk87pEjyGSCdmZCPMQZBwTu4GMV/wUn/AGsPij+yj4A8Ja58LINDe98Q6pNaXEmqWj3HlokW4GMK6gHJ/iBHHSvA37RP/BYH4v8Ahey+Ifw58M2994d1ZWayuLfSNMjjkCkoxQTNvI3Kwye+cVef8FHP29v2atcs7L9qD4HWd3plw4TzLnTX017gj7whuoS1uzYViQEbrngYr9nP9ov4cftQfDa2+JPw2vZjbNIbe+sLpQlzp9woBaKVQSAcFWBBIKsCDVVVVXi3/kVNZ/7B8/8A6Lav2N/2qtQ/ZA+J+p/EvTfBlv4nl1LQZtINpPfNahBJPBN5m4I+SPs4GMfxZzxX/D8XxV/0btpP/hSSf/GK/ZR/4KqeIP2k/j94W+Ct98GNP0KHxF9s3ahFrb3DQ+RZzXAwhhUHJhC9Rjdntiv+C43/ACKnwj/7CGq/+i7av2A/+TNPhL/2L8f/AKG9VVV8T/2of2ePgyZYviZ8Y/C+iXUBIewe+Wa7GP8Ap2j3Sn/vmvib/wAFnP2dfC/mWvw28JeKPG9ymdkrRrplq/p88u6Uf9+qn/4KWft6fH1msv2dfgLHY20rlEvNL0K41iWE9PmnceQoHq0YGfypv2Sv+CsH7RI8z4s/Fm78M6fcndJa6n4n+zwspHX7JpwdM442sq4yRxk14E/4IieFY2juvir8dtY1F3O6a30LTI7Qg98TTNLu57mMfSvAv/BLn9i3wOY55PhdN4juoxgT67qc9zn6xKyxH8UrwX8JPhV8N4Ug+Hvw18L+GljGF/snSILQ8jBOY1BJOTknrk561Xiz4ifD7wHbm78c+O/D3h2FULmTVtThs1CjqcyMox714x/4KMfsW+CS8epfHnRb+VDgR6PBPqW44zw1vG6fiWA968Y/8Fo/2bNHDw+EPBHjnxFOoyrvbQWUDc9NzSs4/wC/fp+Gtf8ABaf4n+KLw6X8JP2cNPF0QfLS91GfU5GGfveXBHER1HGT160/7TP/AAWB+Lqv/wAIX8JNV8MQzkhHtfBq2aYJ6JJqO/jtkNkc85FP+yZ/wV0+LZZfHfxnvvDkc5JkgvfGzQQ8+sen+YmPbGB2rRf+CKXxA8RXn9p/FL9pGy+0vkytZaTNqDvzn/WzSxkZyTkqee3Oa8Y/s2fAz4NfEA6d8N/Fes+MZ9IDRXGq6h5S27T558iNEz8mCN5dgSSVAABNfse/smDxxLbfFH4m6Ww8PQsr6Vp06YGpMP8AlpID/wAsRgYH8Z/2R8yIkaLHGoVVGFUDAAHpVfH/AOJ8Xwh+FWteMEkjF+sX2fTEfnfdScJx3C8uR6IauLie7nkurqZ5ppnLySO2Wdickk9yTX7B/wAHB4s8aT/FDWrQPpfhltliHHEl8QCCPXy0O7/eZD2qqqqqqqqqqqqqqqqqqr9sL9lHwd+1r8KrnwZrSwWXiDTw9x4d1lkJawusdGxyYnwFdOcjDAblUj/gnp+1N4w/Zl+KN3+xH+0n5mlWcWpPaaLcXkmRpV67ZEBbobacsGRxwGcH7shK1VVX/BNPPiv/AIKJfHDxpcAOWtddnBkOHEk+rwndgcfd3g/UY9qqqqqqqqqqqqqqqqqqqqqqqqqqqqq/4Lk6wNvwe8PRkZzrF1Llen/HmqYP/feR9K8E6IfDXgvQPDhTYdK0u2tCpx8vlxKmPl4/h7cVVV/wTS265/wUW+OHiNUNzC1jr0qXEgyweXWLcq3PIYrv5+uetVVVVVVVVVqmraVolm2oazqdpYWsf3p7qZYkX6sxAHSvFX7ZH7KXgsOviD9oXwGksQ+eC21qG7lXjPMcJdgcEcY5rxR/wVi/Yq8Os8en+PdZ8QvHkFdL0G5AyOMBp1iU/UHHvXiX/gtr8DbPcvhH4R+ONUK/dOoSWtiGP/AJJTj8PwrU/wDgtn481yZrPwD+zZYJPglFutcmvmI5GSkUEZ6leM+ozzkH/goL/wAFNvH52+BP2Zvs8DnEc2n+BdTuduTjLSSSMnUEZwB19K/4Tr/gtn49UJpfh3WdHimxiP8AsfSNOIycj5rkB1xjH3h6HrQ/Z6/4LKeM8LrnxX1jQvNGGJ8Zw22zefm/482ONvt0/hof8E1v+Ci/iwibxn+1LZkOxdlu/Gmr3kgcDAJBi2njvuyAaT/gir8Wdb2yeLP2jdHeVF+QjTbi7wx+8AXkTA6c9/QVY/8ABDXSI1can+0td3BJGwweElhx65zdvn9Ksf8AgiN8F4y/9pfGXxrcAgbBBbW0OPXOUfP6Vp//AARY/ZatjDJfeOPibeOg/eIdTso43OPRbTcB7bvxqx/4I8/se2hc3Efja93AYE+tqNn02Rr+uas/+CS37FNrD5U/gbXbxtxPmTeILkN9PkZRj8Ks/wDglh+wvbQCKb4MT3bAk+ZN4l1MN9PkuFH6VY/8EyP2GdOdnt/gPauXGCJ9c1KcfgHuDj8K/wCHbv7Ef/RAtJ/8GF7/APHq/wCHbv7Ef/RAtJ/8GF7/APHq/wCHbv7Ef/RAtJ/8GF7/APHq/wCHbv7Ef/RAtJ/8GF7/APHqvv8Agmf+w5qEIgn+Atiqq24GDWNQhbOMdUuAcc9OlX3/AASu/YZu41jt/g7c2TK2S8HiTUiT7HfcMMfhWof8Ei/2ML1na28NeJrAMm0Lb69KQhx94eZuOfrke1ap/wAEXf2Vb1S1h4v+JWnuIyEEeqWjpu7MQ9qSe3AI/DrWsf8ABEL4Wzk/2B8cvFNmMjAvNMgucDHP3TH3/L3q/wD+CJXj7RZhd+Cf2lNOkuFGFkuNBmsGHHPzRzyEc4/yMFv2A/8Agp78OMt8P/2knvYFzsg07xzqFuG6/ejmRI8/Mx6nBOc5p/EP/Baz4Rti/wBK1zxJZx8iMWWma0JRnrmDdN+GQcHp0qD/AIK0/tbfCy4jsvjz+zZp8ex9jLPp99oM746gmbzF3fRB9O9fD7/gtL+z54gaO2+IXgLxf4SncjM0CxalbR+pZ1KSflEfwr4X/tgfsx/GaSG0+Hfxr8M6jfXGPK0+e6+xXbn0W3nCSH8Fqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr4+fGTw3+z98IfE3xc8UnfZ+H7MyR24YK11OxCQwr7vIyLntknoDX/BLD4NeJfjp8aPFn7a/xbV7+e21CdNJmmXC3GpzAmeVVP8ABDE4RR0BkGOY+Kqqqqqqqqqqqqqqqqqqqqqqqq1HUtO0ixm1PVtQtrKztkLzXFzKIo4lHUszEAD3NfFX9vP4ceExNpnw8s5PFepKCouMmCzjbp98jdJjr8owezd6TQP2wf2riJtYuJvD3ha5OQs4awsyh5+WIZknGCcFtw7bhXwz/YS+E3g4RX3jGS48X6imCRdDybVT7Qqct/wNmB44FaZpWl6JYxaZoum2thZwLtit7WFYo4x6KqgACqqqqqqqqqqqqqqqr/goT4XOs/BO08Qwx5k0DWIpHbHSKVWiYfi7RflX7LHif/hLv2ffBOptLvkt9NFjJnqDbsYefciMH3znvVVVV+2/+314E/ZM0KTw7oxtPEPxJv4N2n6L5mY7MHGJ7sqQUTByqDDSY42jLj9lD9hj4o/tjeNj+1H+2LqeqT6Hqsi3Fpp10TFc62oyEGFINtaLgbVUAsv3dqkMdH0bSPDuk2mg6Bpdppum6fCsFpZ2kKwxQRqMKiIoAVQAAABgVVVVVcW9vdwSWt1BHNDKpSSORQyup4IIPBBHavjb+wl4U8WGfX/hPPB4c1Vsu2myA/Ypz/s4yYT9AV7bR1rx18O/Gvw01ltA8ceHbvSbwZKCZMpKB/FG4yrr7qSKr4Oftk/FD4XC30fWbj/hKdAiwv2S/lPnQoO0U3JHHAVgyjoAK+En7R3ws+MsKQ+GdcFrq23L6TfkQ3K8c7VyRIPdCcd8VVVVV+3T8GG8b+BoviTodrv1jwpG32pVHzT2JOX+pjOXHopk68VVVX7Iv7ST/CPXv+EN8XXjHwhrEwLSOSf7NnPHmj/YbgOPow6ENFLDcQpPBKkkcihkdGyGBHBBHUEVVVVVWr2M2p6VeabbajcWEt1byRR3dvjzbdmUgSJuBG5ScjIIyBkGvA3/AASN+K/xP8dX3jf9rj433upA3ckYWxu3vb/UIUciNmuJ8rArAbggEhCkD5TkD4K/s0fA79nnSRpfwk+Hel6K7RhLi/EZmvLnH/PS4fMjDOTtztGeAKr9o74z6T+z18EvFvxf1aKO4Hh+wL2ts7FRdXTsI4IiRyA8ropI6Ak9q/YF/Zn1P9uf4y+KP2iP2h7y617QNL1EPexzuQNX1BwGW3JH3YIo9hKLgBTEg+UkVpelaXoem22jaJptrp9hZxCK2tbWFYooUUYCoigBVA4AAwKqv+C3v/JKvhp/2MF1/wCk4r/gm9/yZH8K/wDsH3P/AKWT1+0H8MvCPxg+DHi7wB4106C607UdKn2tJGGNtMsbGOdM/deNwGU+o9M1/wAESfFOsWvxt8feCIXlOlan4VXUJ152Ce2uoo4z9dt1L+Rqqqqqq1XWdH0O3S71vVrPT4JJBEkl1OsKs56KCxAJODgVYazo+qqG0vVrO8BUkGCdZOAcE/KTxniqrU9T0zRbCbVNY1G1sbO2XdNc3MyxRxj1ZmIAH1r4t/8ABTb9j/4Sm4s2+I58X6nbnBsfC0H2/cfafK2/4ebn2rxf/wAFhPjV8SdWbwp+zL+z8GvJgVha8jn1i8kH99Le3CBDyOCZB9c0P2ef+Cs37VgE/wATvHmpeD9EvDlrfVdWXSoSh7GyslLkgcASoCe56mvhh/wRM+GmlGG8+Lvxb13xDKDuaz0W0TToc/3S7mV3HuNh+lfDP9hf9kv4SrFJ4S+B3huW7hIK3urQHU7hW/vLJclyh/3No57VDDDbwpb28SRxxKFREXaFAGAAB0GO1ePf2ifgN8LzJH8QfjF4P0GeI4a2vNYhSfPoIt28/gteOv8Agrf+xt4QWRNE8SeIvGEyHHl6LosiDP8AvXRhUj3BPtmvGP8AwW6e5lOn/Cz9n2WeaRtsE+s6xlmPYeRDGSSfaSm/a5/4K0fGwsPht8Gr3w1azkCK40/wc0MWCOMT6gXQnHJIIxnPGRR/Y7/4KxfGo7/iX8arrw7bzPuktdR8ZSRxDI5Ig08SR5xkAYH4A5rwn/wREuruX7f8TP2hTJNISZoNJ0QuzMep8+aXJ/GOvB3/AAR4/ZD8OeW/iAeMvFbgguupawIEY9wBbJEwH/AifevB37Dn7IfgQRnw/wDs9+DXeIDZJqWnjUpFI6EPcmRs++c1o+gaF4dtBYeHtFsNMtl6QWdskCD/AICoAqq/bZ/agPh+3uvg18PtSI1S4jKa7fQt/wAesbD/AI90I/jYH5j/AAjjqTtr9kD9liT4q6jD8QvHVkyeELCb/R7dxg6pMh5XH/PJSMMf4j8o/iIihht4Ut7eJI441CoiLtCgDAAA6DHaqr/goL8STrPjfSvhlYXQa18P2/2q9RT1uph8oPusW0j/AK6mrW1uL25hsrOF5p53EcUaLlnYnAAHckkCvgl8NbT4TfDHQvBMEaC5tbcSX8i8+bdP80rZ7jcSB/shR2qqqqqqqqqqqqqqqqqqqv8Agpj+xDF+0X4Eb4p/DrSlPxI8J2rFYYk+bWrJcs1sQOsqZZoz3JZP41K/8Ewv22pPjr4P/wCFJfE/VSfiF4StQLW4uX/eazYphd5J5aeLhXzywKvyd5FVVf8ABHlRfftH/GXVrsmW8OnEGVjyd96Wf8yqn8Kqqqqqqqqqqqqqqqqqqqqqqqqqqqqr/gsMB4h/aQ+DXgwAyebpw/dTH90ftF6I+nPXy8Nx0x17VVTzw20ElzcSrHFEhd3Y4CqBkk+2K/4IwRTeJ/jN8YfiBPETI+n26u7Ella6uZJSCRxz5J/LjvVVVVVVr3iHw/4W02TWfE2u6fpFhD/rLu/uUt4k+ruQB07mviH/AMFHf2NPhw0tvqHxq0zWruLIFv4fik1PeR2EkKtF+bivHv8AwW4+GWnmaD4Z/BfxHrbKdsc+s38OnIT/AHtsYnJHscE98do/+CiH/BR343Kq/BT4Ax2dlKcxXuleFbu/wD/enmZoce+1aHwM/wCCyPxs2t4w+JGs+E7W5VgzTeJrfSUKnqGh07LAHptKfhitL/4Iw/GHxjeJqnxh/aT003jYM0lrZXOrOeOQJJ3hPYDJHvjjFeFP+CKX7PemrHJ4w+JXjvXJkA3LavbWMTnHOV8qRwM8gB/xNeF/+CXn7EvhjZIfhC2sXCEYm1TWLyfOCDzGJRGen931HQ4rw1+yl+zJ4PETeHP2f/h9aSwgBLj/AIR22eYYxj96yFzyoPJ6jPWtM0fSNEtRZaLpdnYW6gAQ2sCxIMDA+VQB0AFVVVVVVVVVVVVVVVVXVra3tu9peW8VxBKu14pUDKw9CDwRXxD/AGJP2T/iksreLvgP4Ua4nz5l3p1n/Z1wx/vGW2Mbk/Umvip/wRV+DuvRzXnwi+JXiHwpdkEpa6nEmp2uf7ox5cig9Mlnx1welT/CX/gqZ+wyhvfAXiDUvGvg+wGTb6XM2t2axDnBspl86EY5ZokAGD8+BmvhT/wWs+H0/h6WL46/C7XtL1+1AGfDKR3UF0c4OEnljaEj+6WccH5u1eI/+C33wytQ/wDwiPwK8T6mR9walqsFjn67Fmx29a+LP/BYz9pfxq01n8N9J8O/D+wfIje3g/tG8APYyzjyzx3WFT79Mfsxft0eOfh7+0Xpfxl+PPxL8f8AizS7W2ure7sUm+3NNHLEwCRxzTxxxBZPLf5ePl6V4e/4LIfsw+JvEGmeG7DwN8UEudVvIrSF5tLsQivI4RSxF4SBkjOAfpVVVVVVVVVVVVVVVVVVVVVVVVVVVVX/AAVL+Lfib4+/Hrwd+xZ8KpjeNZX8H9pRRt8k+qXAAiRyM/JBC+5j0BlfP3OPgh8I/DHwI+FHhn4S+EIguneHbFYBIVw1xKctNMw/vSSM7ntluOKqqqqqqqqqqqqqqqqqqqqqqqr+/sNLsptR1O9gtLW3QvNPPII0jUdWZjgAe5r4v/t5eDPDJl0T4U2I8T6oCU+2yho7OJv9no8xz2G1T1DGtK+CX7UH7UV9Fr/xV8QXeg6A7iSJL9DGFXsYLNcc4P3n25HO5q+FH7Knwg+E4gvtP0IaxrUJDDVNUAmkRvWNcbI/YqN3qTVVVVVVVVVVVVVVVVVV+0J4VHjT4I+NPDoi8yWbR5pYE/vTQjzYx/33Gtf8E5/FA1L4Y+IfCUk2+XRdXE6KT9yKeMbR9N8Uh/Gqqq/b+/4KLaH+zlZXXwr+FFxa6z8Tb2ArJKpEsPh8MOHlHIecg5SI9OGfjar/ALD3/BOTX/GevJ+0z+2LBeatrGqXBv7Hw5rAaSWeViGW6v8Acckk8iBh6b/7lBVVQqgAAYAAxiqqqqqqvF/grwn4+0WTw94y0Cz1fT5eTDcx7trYxuU9UbBOGUgj1r4zfsB6pp4n134M6idQtxlm0W+kCzIPSKU4V/o+04H3mNa5oOt+GdUn0TxFpN3puoWzbZba7haKRD7q2DUUssEqTwStHLGwZHRirKw6EEdDXwj/AG3/AIoeAPI0rxg//CX6OmFxeSlbuJf9mfBLfSQMT0yK+FP7Rnwo+MMMcXhfxEkGqMuX0m/xBdKe+FJIkHuhYDviqqqliiniaGaNZEdSrIy5DA9QR6V+1T8Cpfgt8QJP7Kt2/wCEZ1wvcaVJgkQ8/PAT6oSMeqlT1ziqqv2R/wBrEeDTZ/C34l34/sFmEel6nKf+QeSeI5D/AM8iTw38Hf5fuo6SIrxuGVhlSDkEeoqqr4m/Fn4a/BnwxN4y+KPjTS/DekQA5uL2baZGA+5Ggy8r+iIGY9hXxv8A+CsPxN+KniH/AIVX+xP8PdTmvbx2hh1m4003l/cdt1taAMsY775Q/B5RCK/Z3vvizqPwT8H3Hx00RNK8drpqR61brcJMWlTKiVjHlQ8iqsjKOFZyvaqqq/4LSeILvTf2Y/DehWrsiax4yt1uCDw0cdrcvtI/39jf8B96/wCCUXh6w0b9iHwZqNnGqy69f6pe3RC4LSLezW4J9Tst4xn2A7VX/D5L9kX/AKB3xA/8E0P/AMfr9mz9vv4F/tVeOr74efDG18URapp2kyapMdU0+OCPyEliibDLK5Lbp04x0zzxX/Bb3/klXw0/7GC6/wDScV+w1+3b+yf8MP2VPAnw/wDH3xftdG8QeH7OeLULGbTLxmiZrmZ1wyQlXyjKflJ6465Fftmf8FXvh34j+HesfCv9mY6nrWq+JbeSwudfuLF7WG2t5VKP5EcgEryspKgsihNwYZIwP+CTf7JPiz4H+B9b+LfxM0W40nxJ42jhhsdMuo9k1lp6EuDIp5R5XIJRuVWNM4JIFVVVVV/wUKmli+CWlRxvhZvE1ujjHUfZ7k/zUVXh/wD4KK/syfs6/BXwv4X8T+LNS8Q+KrKwP2vRtGtjc3EUjOzESSOViQ5b7pfcB2rxJ/wVP/ay+P2ty+Dv2UPgidOd2wk8Fi+t38Y7O3yiCIdSd6MB/ewCTYf8E3/27/2nL+38QftQ/Gb+xbVn3fZdU1J9VuLcHvFawsLeMYJ4Ei4PbrXwk/4JI/sofDryL7xbpusfEDU4hln1q7MdsH9Vt4dgxj+GQyf4eF/BngD4Y6E2l+DPC2g+FtHt13vBp1lFZQIAPvMECr0HU18Sf27v2R/hT5sPin45+HJ7uHIaz0iZtUmDf3StsH2H/e2++K+I/wDwWy+EukNLbfCz4R+JPEsi5VbjVruLTISezKEEzsPYhCfbrS/t1f8ABTf9oZSnwO+DD6JptyCLe/0jwvJOgU/3ru8LwbvQgL9OM0v7C3/BTv4/OJPjZ8bJdCsJjmay1XxVLOqg5zstbPfBn2yvHfjFeAv+CInw8sgk3xO+N3iDWG6tBoenRaeB7b5TOWHvtX8K8C/8Exf2LfAoilX4SL4gu4sf6TruoT3m/H96IsIT/wB8V4P+GPw2+HluLXwB8PvDfhuIDGzSNKhsx+Uaiqqqqq/a0/aOtvgj4T/sXw/cRSeMNbhYWMZw32OLobhx7HIQHgsO4Uirq6ub65lvby4knuLiRpJZZGLM7scliTySSck1+y1+zjqPx28V/bNVjmt/COjyqdSuQCvntwRbxn+8wxuI+6pz1Kg6VpWm6Fpdpouj2UVpY2MKw21vEu1YkUYVQPQACqrUL+00uwudTv51htrOF5ppG6IiglifYAGvH3i6+8feNdb8Z6iT5+sX0twUJz5as3yoPZVwo9hX7Fnw7Hjv43adqN3b+Zp/heM6lMWXjzVIEI+vmFW+iGqqqqqqqqqqqqqqqqqqqqv+Cj/7Lnir9nn4l2X7bn7OrT6Sqaot1rkdkmP7NvmOPtQUcGGckrIp+Xc5ByJCB+yF+1F4T/aw+ENj4/0QwWetWu218Q6Qr5bT7wL8wAPJif7yN3U4J3KwFV4g1aLQNA1LXZ9vl6bZy3L7jgYjQscnsMCv+CG+kSSap8YNeZCEht9HtkYx/eLtdswDe2xcj/aFVVVVVVVVVVVVVVVVVVVVVVVVVVVVV/wVb/5PS+CX/YPsP/TnJVVX7RHij/hCPgF8SPF6y7JNH8J6ndREEg+YltIUAwQclgAORyeor/giD4Xa0+GPxN8aGIhdV16z08PtPJtYGkIz7faxx7+9VVVe3tnp1pLfahdw21tAheWaaQIkajqWY8Ae5r4of8FCP2QPhP5kGu/GnR9Vvo8j7F4fLapIWHVS0AZEPHR2WviR/wAFs/D6ytpnwV+CWp6pPI+yC78QXi24J6DFvBvZsnGB5imh8Yf+Cvv7UJJ8D+D9X8F6Ldn5WsdKj0ODb/fS5vT5zAeschyc8EjjQv8AgkJ+0X8VNSi8RftH/tFW4un+Z2E1zr13g9VaSdowpPqC4+tfDv8A4I/fsleDxFP4sh8UeN7lSGcapqZtoSR/dS2EbAdOGZvy4rwB+zV+z78LBG/w++DHg/RLiMAC7t9Ii+0HHTMzKZG/FjVVVVVVVVVVVVVVVVVVVVVVVV4//Y9/Zi+KXiuXxx4++CvhvV9dnj2T3kkDRtPz96QIQHftvYFscZxX/DAf7Gn/AEbz4U/78P8A/FV/wwH+xp/0bz4U/wC/D/8AxVWf7Df7IFhD5EH7OfgZl3E5m0pJmz/vPk49qsf2Nf2UdMvbfUtO/Z58B291aSrLBNHokKtG6kFWU44IIBzVVVVVVVVVVVVVVVVVVVVVVVVVVVV+018cdG/Zy+B/in4tauYpJNIsyunWsjY+13snyQReuDIV3Y6KGPav+CRHwM1fxx4u8X/tj/EoSX+o3l5cWejXN0uWnu5jvvbse/ziIMODvmH8NVVVVVVVVVVVVVVVVV+0R8cvDX7N3wg1z4xeLNOvdQ07Q2t1ezsSgnnaa4jhUIHZVJBk3EZ+6relfsr/APBRn4Q/tYfEK7+GnhPwt4k0HVbfTJNQiOrCAJcJG6K6IY5GO8CQNgjoGPaqr9oD/gqd8Ef2ffixrfwi1jwd4r17UtAMSXl1pi2/kLK8ayGMGSRSSocA8YByOxr4Y+PdJ+Kvw48L/EvQoZodP8VaPa6nbQzlTJCk8SyBH2kjeu7acEjIPNfEL4nfDv4TeHpPFXxL8a6P4a0qMlRdaldrArtjOxATl3IBwqgsewrUv+Csn7FFhqf9nwePtZvog203dt4fuvKHOM/OiuR9FNfBr9or4JftB6ZNqvwe+Iul+I0tQDcwQlori2B6GSCQLKgJBALKAcHBOKr43ftkfDr4UyXXh/QWHifxLAWje1tpNsFrIDgiaXkZBzlFycgg7etWHgX9pz9sW9g1vxfqLaF4SZ98DTRNDaovrBb53TH0djjqN/avhB+y78Kfg6Ib/SdI/tXXEUZ1bUQJJVbuY1+7F/wEbscFjVVVVVVVVVVVVVVVVVVVOiSIY5FDKwIKsMgiv2OXf4YftQ+NPhPcOUhnS7tIFJyXe2m3Rtn0MXmH8RVVX/BQb/go1H8H5rz4D/AK5TVPiJdqbbUNTgHnLobPgCONQD5l0c/d6RkjILfKP2AP+Cb0vgi8t/2gP2mtPGq+OLyVb3StGvmM50yRiH+03JbO+7LHO05EZyTlz8lVVVVVVVVfEX4S/Dz4r6Z/ZnjrwzaaiFUrDcFdk9vnvHKuGXnnGcHuDXxc/YG8ZeG/O1f4V6l/wkmnrlv7PuCsV5GPQHhJePTaewU1quk6poeoTaTrWm3Vhe2zbZra6haKSNvRlYAg/Wkd4nWWJ2R0OVZTggjoQa+FX7avxc+HYh07XbxfFukRAKLfUnInjUf3Jxls9vn3gdgK+Fv7W3wb+KIhsodeGg6vJhf7P1crAzN6JJnY+T0AO4/3RVVXxg+Ffh/4yeBL7wTr6hPOHmWd0Fy1pcKDslX6ZII4ypYcZrx34H8RfDjxXqHg3xVZG21HTpSjjqsi9VdD3Vhgg+hqqqv2Wf2v7jwILP4dfE67kuPDgxFYakwLyad0AR+7Qjt3TtlcBbO8s9Rs4L/T7uG6tbmNZIZ4ZA6SIwyGVhwQRggjiqr9on/gmN4q/aT/AGoL74j+MPjTfwfDu5giki09p5Lq+s5Nu2W2tlkBighJUOG+bBcjYcZPwO/Zt+C37OWgf8I/8I/Atjo/mIFur4r5t5eEd5p2y7jPIXO1c/KAKqqqv+Cufw6vvHX7IN7rWmwGWbwVrtprEiquWMOJLaTHfAFyHPsme1f8Ebvi3pniz9nPUPhRLeAav4F1eVlt2cZNndMZkdR1x5puAfTA5+aq/wCGKP2Rf+jcPh//AOCKH/4mv+CKn/J1Hin/ALJ/ef8ApfYV/wAFvf8AklXw0/7GC6/9JxX7Gf8AwTy/ZP8AjR+y98PfiV48+Hl3deIdYsp5NQuodbvIRcMt1Kikosu1cKij5Qo4z15r4U/sb/swfBG/i1f4afBjQNM1KBt0GoTpJfXUJ9Y57hpJE/4Cwqqqqqq+NH7UPwD/AGfbNp/ix8TdH0a6Ee+PTRKZ72YY42W8e6Qg8fNt28jJFfHL/go/4I/amvYPhD8P/h9qdno9tcnUF1rVpkSaV4lZAqW6bgilZGO4vnHG0c1X7Lv/AAT5/Zh+IHhXTvjp4/8AC194q1vX5JZZLLUrwrZWzxSvD8kMW3eD5eSJS4JbOB21vxX8DP2dvCkMOt634M+Hfh+HP2a3eS30yAnHSKMbQx4HCgmviz/wWG/Zh8Dmax+H1n4g+IF/GSqvZ2xsbTI4IM04D/QrEwPXNXP/AAUR/wCCgv7S076d+zT8F/7FsJGKJeaToz6nJEc4xJd3A+zrjgZMaYOeemLf/gnD+37+0ZLFqP7SXxwXSrORgXstW1qXVZYO/wAltCfs649BIvP518NP+CMv7N3hZobv4i+KPFPji5T/AFkDTLptpJ/wCHMo/wC/1fDn9lb9nH4SNFN8PPgt4T0m6hx5d6NOSa6XH/TeTdJ/49VVVVVVVV8YPir4c+DPgS/8b+I5A6248u0tVfa95cMDsiX3OCScHChj2rx9468R/EvxbqPjTxXem51HUpS7kZCxr0WNB2RRhQPQfjXwY+EXiT41+OrPwZ4ejMaP+8vrwpuSztwRukb8wAO7EDvmvAHgPw18M/COneCvCdiLbTtOi2IDy8rfxSOe7sckn1PYYFVVftj+M28G/AHxB5E3l3OubNLhOcZ84/vB+MSy1X/BPzwQuifC3U/Gs8WLjxLqBWJiOsFvlFx/20ab8hVVVVVVVVVVVVVVVVVVVVXiHw/onizQdQ8MeJNLt9S0rVbaS1vbO4TfHPE6lXRh3BBIrxboPxI/4JM/tZW/izwxDf6x8K/FTssUbtkX9huBktXPAF1BuBRzjPytwHdR8PPiD4Q+K3gnR/iH4D1qHVdC121W4srqI/eU9Qw6q6kFWU4KsCDgg1X7YfjGPwF+yx8VfE5mMUkHhS+gt3BxtnniMMR/7+SJX/BFPwc+lfs/+MfGk8JR9f8AFJt4iQP3kNtbx4Yd8b5pR9VP41VVVVVVVVVVVVVVVVVVVVVVVVVVVV/wWRD+Fvj98HPiCEcCHTnw2AwJtbtZSAvr++H1yKjkjmjSWGRXR1DIynIYHoQfSqr/AIKn+Ok8E/sXeMLVJVS68T3NnpFsScZMk6ySDHfMMMo/XtX/AASk8EP4O/Yv8L308HlT+KNQvtWkU9SGmMMbH6xwRkexHvVfFv48/Bz4EaOuufFz4i6P4at5ATDHdzFp7jHXyoEDSS477FOK+Jn/AAWn+Bvh4y2nwu+HfibxhOgIW4vJE0u2Y9iCRJIR9Y1o/t0f8FLv2miYP2fvhFJoOlXBIhv9H8PtOoU8Ye9vN0APuAh4yMYzVl/wTM/bo/aDuIdU/aW+Oy6bAzbjbanrE2t3Fuf9iBGECj2WUCvhf/wRy/Zg8HCG6+IGp+JfHl4mPMjurv7BaMR6R2+JBz2Mrdvx+G/wD+CfwfhSP4YfCvwx4bkRSv2ix02NLhwRg75seY/HHzMeKqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr/gqD8TfE37R37SHgz9jD4Xubr+y7+Fb1FJ8uXVLlRtL4z8kED5Lfw+ZLn7tfCD4X+G/gr8MPDPwq8IxbdL8M6fHaQsV2tMw5kmYD+ORy7t/tOaqqqqqqqqqqqqqqqqr/grtqq6d+xlq1mZEU6pr2nWwDKSWxIZcD0P7rPPYGvhXdah+yL8TP2ef2hUmmbS/EdgdSv5EB+eH7ddWl3CB3Itgh9Mup61BPBdQR3NrNHNDKgeORGDK6kZBBHBBHeviZ4+0T4V/DvxJ8SfEcm3TfDOl3GoXIDbS6xIW2L/ALTEBQO5IFeKPCeu/ED4JeNv2rfGBkm1PXviZa6ZHcjIWWa4tb+8veOnDC0x6AkV+wVqY1f9jj4SXQkjfy/DkNvlOg8otFj6jZg+4NftS/8ABOzRP2rvj1oXxL8ZfEfWdP8AC+m6MtpeaNayF5ZZlkYgwNJuS3RlYb9qnJQHALFqX/gl7+xAugnQv+FM5Jj2/bTrd99p3Y+/v87Gc84xt/2ccV8f/hz4x/4JlftdaPrnwq8S3t1pbQx6po73Em1ruyaQpNY3W0AOMxspIABBRwFb7vhzXbDxT4e0vxNpUhey1eyhu7Zj3jlQOp/JhS/sy/B6T4k6j8Ur/wALx3+r6jKs5husPbQzAfNKsWMF2PzEtu+bkYPNAAAKoAAHAqqqqqqqqqqqqqqqqqqqqr49sPg9+3B4c+ICDyLPV5bK8nccARvm1uPx2KxI/wBr3qq/4KG/8FDJfhjLcfs8/s83ral8RtRYWmo6jYqZm0dpPlEEIXO+7bIGBny8j+PAX/gn3/wTltfg+bH47fHq1GqfEi6BubHTbgiaPQ2fnzHJz5l3ySWyQhJxlhvqqqqqqqqqqqviL8H/AIb/ABXsRZeOvCtpqLIpWG5wY54f9yVcMBnnGcHuDXxY/wCCf/inRTPq3wn1ldds1BZdNvmWG6Ueivwkn47D7GvEPhrxB4T1WbQvE+i3ul6hbn95bXcDROvocHqD2PQ9qr4YftPfGL4UCK00LxO9/pcZH/Er1TNzAAOy5O6MeyMo9a+GX7e3wz8VGHTvH1hceFL9sKZ2JuLRj/vqNyZP95cDu1aLruieJNNi1jw9q9nqdjOMxXNpOs0b/RlJFV+1F+znY/HHwwNQ0ZIbfxdpEZ/s+4Y7RcJ1NvIfQnJUn7rexatS03UNH1C40nVrKa0vbOVori3nQo8TqcMrKeQQR0qqqvgR+1d46+B1rPoy2i+I9BaN2h0u5uTD5EuCQY5drGNS2Nw2sOSQM8n9gv8Aaa/aS/aE8VfEIfGX4ax6V4dsbhG0TUrKER2ts4wGsg7MXuGKMknmDIB3htu6NRVVVVVeJfDeh+MfDup+E/Eumw6hpOs2ctnfWky5SeGRSjow9CpIr4q/BX9o7/gmB8eE+K/wze7vvB5naPT9aMJltby0kYE2F+q42scKOdu4qrxkMPk+En/BYz9mrxfpdunxTsNd8BauFAuQ1m+o2m7uY5IFMhHfDRDHvWv/APBVz9iDRtOkvdO+J+pa7MikrZ2Hhy+SVyOwM8USZPuwr/gip/ydR4p/7J/ef+l9hX/Bb3/klXw0/wCxguv/AEnFf8E3v+TI/hX/ANg+5/8ASyeqqqqppobeF555UjjiUs7u20KAMkknoMV8f/8Agp9+y/8AA43Oj6X4jfx94igJT+zvDjrNFG4yMS3RPlKMjBCl3B6rV9+1l/wUa/bovZ9B/Z68GX/g7wvK7RvdaHm3WNehE2qS7cOOeIjGxx904r4Lf8EZNNlul8U/tOfFG817Up38240rQZGWN3PJ827lHmSZ5yFRDno5r49/s/fAv4J/sw+JND+GHgPw54TwbUwyQxqtxdstxGSrzuTLM2wNjczHj0FVpfi//gof41sLf4K/stQapa+E7WMvPqunWsNoIpJXZnR7+bGxl+8FjZXww4ORXgH/AII3/EPx1qf/AAl37T/x6nuNRusNdQaU0moXUn+/eXPRgOP9W4yepxz8Jf8Agnb+yJ8HjBdaL8JNP13U4QMaj4kJ1OUsOjBJcxI3fKIvNWtra2VtFZ2VvFBBCgSKKJAioo4AAHAGO1VVVVVVVVVd3drYWk19fXMVvb20bSTSysFWNFGWZieAAASSa/am+Pl18c/HzzadPKnhjRi8GkQMCu8Z+edh/efA4PRQo65zomi6r4k1ez0DQrCW91HUJ1gtbeJctLIxwAPxr9nH4E6R8CPAUWiIIbjXdQCz6zfIv+tlxwinr5aAkKPdmwCxqqqv+CjXils+DfBEMvy4uNQuE9T8scR/9HfnVfB/wkPAnwt8K+ETGEl03SoEuBjH74qGlP4uWP41VVVVVVVVVVVVVVVVVVVVX7Q/wC8C/tK/CzVfhX49tj9lvV8yzvY0Bm066UHy7iInoy5II6MpZTwxr9mX46/Ej/gmn8ftU/Zu/aE+0nwBqV2H+0oGkitN5xFqVsACWhkAAkQcjB43xlTpWq6ZrumWmtaLqNtf6ffwJPa3VtKJIp4nAZXRlJDKQQQRwQa/4LN/G+x8MfBrQ/gVp16p1fxpfpfX8Ktkpp9s24bh233Ai2nv5L+lfsO/CO6+CX7K3w98B6naG21VdMF/qcToVeO5umaeRHB/iTzBGf8Acqqqqr4i/ETwZ8JvBOrfET4ga7b6RoGh25mvLuYnCjICqoHLOzFVVQCWZgACSK8Vf8FXf2j/AIy+LLvwr+yF8ApLy1iyEmudMn1a/ZDkLK0UDCOAezeYBj71ah/wUE/4KWfB2Ia98XvgBGdET5p7jVfCF5ZxKvTAnjZUQ8fxZ69OlfB/xtq/xK+FfhL4ha74ZPh6+8SaPb6jLpZuftBtPOjDqhfauTtYZyoIJwRxXizxZ4Z8CeG9Q8X+MtdstG0XSYDPe315KIooEHdmPvgAdSSAMk18ZP8AgsvBJrzeEf2YPhRJ4ouHcxW2qa2koW5ft5VlDiV1PUFnRvVRUX7YH/BXHUVGv2f7PGpLZEeYtqvw/ugrKB0CsfNIOM8HJzx2r9h/9uX9o347/GO9+Cfxu+Cth4evtN0WXUrm9itbrTJbdI3jjHmW9wXLF3kUcMmOSAQMVVVVVVVVVVVVVVVVVVVVf8Frfh1ca98FPBPxLtIfMPhLXpbS4wOY4b2MZc+3mW0K/VxX7HXxQtvjH+zD8OPHcVyJrm40KC1vzuyRd248ifPpmSJyB6EVVf8ABa74gXGpXHwu+A+il7i8vJ5taubRDlmZj9mtMD1JN0P8mvCcXgb9mX4BeGtH8aeJtL0HQvBHh+0sbrUL64WGLdDCqE7j1Z2BwByxbABJxX7Qf/BV3x18SvEH/Cov2JPB+pXl9qEpt4dfl05p7y6J4zZ2hB2Dod8oJwTlEIzXwW/4JLfEn4r6ufil+2Z8SdWTUtTkE9xpNreC71CY8cXF25dE4GNiB+Dwy4xXww/Yv/Zb+DywSeBvgn4ahvIFAS/vrX7fdAj+ITTl3Unr8pAoKqqFUAADAAGMVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV+0V8ZtG/Z8+Cviz4uayscq6BYNJa2zvt+1XTYSCHPX55WRTjkAk9q/wCCRXwb1z4hePfG37ZHxKWS/wBSury4tNJvLgfNNezkvfXIHqFcRgjj97Kvbiqqqqqqqqqqqqqqqqq/4LU6sbT9mLwvpMcjq9/43tiwCjDRx2d2SCe3zFD+FftHfBNPEP8AwSr+B3xN0q0aS78BRiW5dhytnfzMsp+nn/ZsegJr/gmz8Zx8af2SfCN1eXjT6x4TQ+H9TLtubfbBRExJ5Ja3aBiT/EW69a/4LLfGs+DfgXonwb0q82ah4+1Hzb1FPP2C0KuwOOm6doMZ6hHHPOP2svgdH8Fv+CUvwt8I6jaPa6xF4nsdY1NCNrfbLu2u2dJAR1jWYRcY/wBUOvf/AIJg6n/av7DXw0dpI2ktk1G3dU/g2ajchQffYFP41Vf8FY/FP/C7P2u/CvwX+HsK6tq+hWFvo5jgO4vqV3NuEAxxwrwA+jMwOCprwR4Zt/BXgrw/4NtJN8Og6XbWETc/MsMSxg889FHWqqqqqqqqqqqqqqqqqqqqqr/go94N+2+D/C3jyCPL6XfSWM5A5KTJvUn2DQkfV6+CvjAfED4S+E/F7TCWbUNKhNy2c/v1XZL+UiuK/wCCjH/BQub4RNdfs/fAa9a6+Il+gh1PU7YeYdFWQDbFEBnddMGGMf6sEH7xG3/gnX/wT4g+DlnafHn446eb/wCJeqobiysbweZ/YSyclmznN0wPzMeUyVHO4mqqqqqqqqqqqqvGnw+8E/EXSzo3jfwzYaxa4OxbmLLREjBKOMMh91INfFH/AIJ62c5l1P4ReJ/szYLDS9XJZPokyjI9AGU+7CvH3wo+InwwvfsPjrwlf6UWbbHNJHvhlP8AsSrlG+gJqvB3xA8bfD3UP7U8E+KdR0e4JG82s5VZMdA6fdcezAivhv8A8FCvE+mLFYfFHwvBrMS4DahppFvPjuWjPyOfoYxXw5/aK+DvxTEUHhbxnaC/kwBp16fs1zu9Aj43/VCw96/at/ZYtvi3Zy+OPBMEcHjCzhw8RIVNTjUcIx6LIBwrdD91uMFb2yvNNvJ9O1C1ltrq1kaKaCZCjxupwysp5BBGCDVVVfDD4s+OfhB4gXxD4I1hraRsLc20g3wXSD+GROhHXB4IzkEGvgR+1b4B+NEMGj3EyaF4p24fS7mT5Z2A5MDn7477eGHPBA3Gqqqqqv8AT7DVbKfTdTsre8tLmMxzW9xEJI5UIwVZTkEEdjXjX/gnF+xZ48u5tR1T4F6TYXUxyX0a6uNMVT3Iit5EjH/fFaF/wS7/AGINCuFux8GzqEqNlPt+uX0yj22ecEYf7wNeC/hd8IPgxpMsfgLwL4W8HWKJieSwsIbMMOOZHUAtyByxPav2mPH37EnxI0yy0H4zLY+Pho0z3FlY6bJNN5cjLtYiSF0jzjjDPXwO/ay+HOieIvB3wN+GfwvXwt4GFybGzNxfNLJCZWcoApztzM4zl24Y1VVWt65onhrSbrXvEesWOlaZYxmS6vb24WCGBB1Z3chVHuSBX7Rf/BXz4IfDQ3fh34MafL8RteiBQXiObfS4X6Z80jfNjriNdrdpB1qHwB/wUo/4KLyrfeMtSufBfw8vWEiRXYfStMMRPHl2y5mu+ASrSb1/21zX7P8A/wAEnf2bfhCtrrHj60m+JXiGEBml1iIR2MbjqUs1JVh7StJ+FfGn9rv9mX9lzTBo/jfxzpVheWMIS28N6PGJ7xVAG1Ft4v8AVKRjBk2J718QP+Crf7RHx016T4e/sefB29sri4LCG8ey/tbUinQSCFQYYAM87/NUf3hXhH/gnF+2T8WdYi+Mv7TXxbktNS0pGvbXTb+9fV7x2QFxBhX8m3RiAvyM20H7nGKr/gnH4iDWvjXwlLJ9x7W9gTPXIdJDj/gMX51VVVVVVVVVVX7e/wAfxp9n/wAKP8KX2Lm7RZdfljPMcRwyW+fVuGb/AGdo6MRVfsJ/s7r4d0mP40eL9Pxqupw40KGVOba2YYM+OzSA4Hon+/xVVVftyeITrn7QWqWIk3podha2SHOQP3fnED/gUzD65r4QeGx4w+KnhLwy6ho9Q1i2jmBGf3XmAyHH+4Gqqqqqqqqqqqqqqqqqqqqqqr9rz9kbwD+118Ov+ET8Suul67prNNoevRW6yTWEpHzKQcF4nwA6ZGcAggqCPC3x0/bd/wCCYNxd/C/x94NTXvBjyumjnUvNm07zDlt9ldJjarAFmgOCMklEYsa/ZQ/Zf+N/7bfxys/2vP2p7Z18JpNHe6faXUJiXVRGc28FvAc7LJD8xZuJBwN5d3FVVVVf8FtvEWvWPwe+HvhiyeVNJ1XxDPPfFWwGkggxCjeoPmyNjplAewx/wTC8K+AfD/7G/gjVPBNtZ/addjnutbu4lHm3F6JpEcSHrlAojAPRVGOuS6JIhjkVWVgQysMgjvxUcUcMaQwxqiIoVEUYCgdAB2Ff8FKfjl47/aY/aW0r9j74VXTzaTo2qw6a9tHIVj1DWJCA7ykf8s4N2zkfKVlY54x+yr+x/wDCf9lLwVa6L4Q0a2vPEs9uo1nxJPCDdX8uBuwxyY4c/diU4AAJ3MWY0UQuJCqllBAYjkA4z/IflVVVVVVVVVVVVVVVVVVVV8dPhJoHx4+EXin4SeJTssvEmnvbiYLuNtMMNDMB3Mcio4Hcriv2GP2ltZ/YP+Lvif8AZV/aUifRvD9zqpb7dKGaPS73aFE4wPmtp0EZ3gYGEfgFiNM1PTNb0621fRtRtb+wvIlltrq2mWWKaNhlXR1JDKRggg4NeIfEnhzwlpM2veKtf03RtMthma91C6S2hiHqzuQo/E1+2L+03ofij9vO4+Nvg9tN8YaD4MvdPXQUmkZrS7+xxo/JXlojc+Y2BjcD15zXg79lr9tr/gox4ns/iT+0X4ov/CXgkN5tkL62MIWI/wANjYZG0FQB50mNww26TpX7PH7KXwS/Zf8AD39i/CvwnHBeTxql/rV5ia/vyO8suBxnnYgVAeijJqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr/grT8WNe+Lfxd8CfsafDdmvLxb23udSgjfIl1C6xHaQsByPLjcyHIxidT/AA18DfhLoPwJ+EXhX4SeGyHs/DWnJbGbZtNzN96aYjs0kjO5Hq1VVVVVVVVVVVVVVVVVX/BcDU3i+HHwv0UNJtutbvbggNhcxQIoyO5/enB7c+tfBn4V6X8Tf+Cfngj4UazKj2vir4X2Ns0rR/6lrixRkcDnmNnVgfVAeK/4I6/EHV/h38c/iD+zl4qV7SbVLZ50tZWwYdQsJDHNGF/vNG7k/wDXAV8WLQ/tr/8ABVyy+H5X7Z4U8C3sen3ildyC000tLeK49JLkyw5/20r/AIK46e97+xfrlyvl4sNb06Zt/XBm8v5ffMg9OM1/wSL1Mah+xjo9oJI2/s3XdRt8L1XMokw3v+9z9CKr9vb9t/w5+yZ4EbSNDuLe/wDiT4gtW/sPTmXetqhJU3k47RqQdqnl2GBwHK/8Evv2NvE9xrz/ALZHx4hurnXtZeW68N2+pBmnkefJl1KbdzukDN5eeoZn7oaqqqqqqqqqqqqqqqqqqqqqq/aa8Ff8J98CvF+gxQ+bcx6e15aqB8xlgIlUL7ts2/8AAq8Qft9eJP2ef2Y2+Engazul8bavqFzHpOrbQYdLs3CNJIoOd83mO4VcYG7cT8oVv+Ccn/BPy8+HskP7R37RGlve+P8AU2+16PpuokyyaTvyxubjdkm7cnODkx5yfnJ2VVVVVVVVVVVVVVVqOm6dq9lLpurWFte2k67Zbe4iEiSD0ZWyCPrXxM/YV+EfjTzb7wibjwhqLgkfZB5tqzephY/L24RlHtXxJ/Y7+Nnw7aa6h8PjxHpkeSLzR8zEL1y0WBIvHX5So9alikhkaGaNkkjYqyMMFSOoI7Gq+Hv7Ufxt+Gwit9H8ZT6hYRcCw1XN3Dj0G470HsjLXxc+Lnw6+OdqfEOseD38KeOY1Akv7F/Os9SVRwsycPGwAwrgOegbjBSqqqjkkhkSaGRkkjYMjKcFSOhB7GvgZ+3R4m8JC38N/FlLjxBpK4SPU0IN5bL0+fP+uA98P1OW4FeDPHPhH4h6HD4j8F6/aatYSjiWB+UP911PzI3+ywBHpVVVVVf8FFby7svhf4aks7uaBm1/BaNyhI8iX0r+3dc/6DN9/wCBD/40NT1K8Ihu9RuZ067JJWYZ9cE1Vrc3Flcw3lnO8M8DiSKRDhkZTkEHsQQK+DPxEtfit8MtB8cW7IJb+1Au41/5ZXCfLKv0Dg49iD3qtb1zRPDWk3WveI9YsdK0yxjMl1e3twsEMCDqzu5CqPckCv2j/wDgsB8J/AL3Phf4AaMfiDr6kxrqU2+DS4X6fKeJLjB4wgRTkESGtM/Zd/4KD/8ABQTVrPxb8fvFF54O8GySedbQ6vEbaKFOxttMQqS20jEkuwsOfMav2df+Cd37NP7OZs9Z0jwmPE/iq2AYa/r4FzNHIOd0MWPKgIOcMi7wDgua/aL/AOCjn7NP7PCXek3HilfF/iq3LINC8Putw8cg7TTZ8uHBwCCS47IcVqn7Un/BRD9vrUbjw3+z94Qv/BvhCRzHNPo0pto416EXGqSbCWAJykWwsP4Gr4Cf8EaPAeiSw+JP2kfGtz4w1Nn8ybR9JkktbLcTyJJziebPXK+Ucnv1rwB8M/h78KtAj8L/AA28F6P4a0qLpa6bZpArH+820ZdvVmyT3NV8TvDB8F/EXxN4T8vauk6tc28QxjMayEIR7Fdp/Gv2F/FI8PfHuz0ySXbF4g0+4sSD03ACZfxzDgf73vVVVVVVVVVVXx6+L+l/BL4b6h4yvPKlvcfZ9LtXOPtN0wOxfXaMFm/2VPfFa3rWqeI9Yvdf1u+kvNQ1Cd57meU5aSRjlmP4mv2SvgI/xs+ISTa1bOfC2gFLjVG6Cds/u7cH/bIO7HRVbkEiooooYkhhjVI41CoirgKB0AHYVVVVfHTWj4i+M3jbVg+9Jddu1iPrGkrIn/jqrX7EWiDWP2htEuWXcuk2l1dsP+2LRg/99SA/XFVVVVVVVVVVVVVVVVVVVVVVV/wVB+AnxZ/aK+AOgeCfg34T/wCEg1qx8YW2oT2v263tNlulndxs++eRFOHljGAc/NnGASP2XvBfiX4dfs6fDfwH4y03+z9d0Dw3Z2eoWnnJL5M0cQV13xsyNgjGVJHvVVVVVftXfs1eFf2rPg7qPwt8SXbafcmVbzSNTSISNp96gYJLtONykMyMuRlXYAg4I8O+JP25v+CXPiq80u+8PyzeD7u63zQ3ML3mh6k3C+bDMmPJlZQB1R/ub0IAFfs0/wDBV34D/G+/sfCPj61m+HPii8ZY4k1C5WbT7mQnAWO6wuxj6SqgyQAzHrX/AATAWL4qft+XPxB1wma8jtNY15TKuSZ528sk9ecXTnPrVVVVVVVVVVVVVVVVVVVVVVX7V37FPwd/a40WCLxtaT6V4k06Ix6Z4i04KtzbrnPluCNssW452N0ydrIWJMf/AATA/bv+EdxNpXwI/aXt7bQ5WJU6f4k1HQ3Y9y8EQZBnjo7dK03/AIJHftP/ABU1WHU/2jP2k7OdFfcXF7e69dAH7wzceUqscYyGYd+elfs6/wDBOn9mj9nOS11vSvCzeKvFFswdNd8QhbmWFx0MEYURw4OcMq7x3c1VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVXxS+Ivh74R/DnxH8TfFUxTSvDOmzXtwAQGkCKSI1z/E7YVR3ZgK/4JafDrX/2hf2j/HX7ZXxKi+0y6ZezmyfaRG2qXYJk2Dn5Ybd9oX+HzoyPu1VVVVVVVVVVVVVVVVVV/wAFydSWbXPg9o4MebW01ic4b5v3jWa8jsP3XH4+lfA7T/7J+Cfw+0rzFf7F4W02HcF2g7LWNcgdunSv20Yrz9i3/go7o37QHh7S5H0zXJY/EIgiO0SmVXt9QiDf3n/euR0/fjtX/BGn4U39zovxA/aY8VK0+q+LNQbTLG5lX53jRhNdSA9xJM0YP+1Aa/4KgaYdU/Ya+JaJFG8tsmnToX/g2ajbFiPfZuH4+9f8EW9VF7+yz4i015IzJp/ji7VUAwQj2dmwJ+rFx+Fft0ft0eEP2SPCB0rSmtdZ+I2s27HR9HZ8rbKcj7VcgHKxAg4XgyMCBgBmX9i/9kLUv2h/Gjftb/tn+K49Sl1K6W80rRdSuo45dSZSNk1xHwI7ZQAI4AFDADgRgBx4r8JqoVfEukgAYAF5GMfrVtdWt7Al1Z3MU8Lj5JInDK30I4NVVVVVVVVVVVVVVVVVVVVVX/BQL9uDQf2UvAjeHPDctpqPxI8R2zjSbCQLIthEcqbydD/ACCEUj52BH3VbH7E3hnwf4r+OPgC7+L3g0S6kth9u0m31KAxi2vnt1mimMbYBygLJkYy0bDkKaqqqqqqqqqqqqqqqqqviH8DPhR8U43PjTwXYXd0wwL2NDBcr6fvUwxA9CSPaviL/AME77iMSXvwr8ZrKOSun60u1voJoxg98AoO2T3rx98Fvil8MJGHjbwVqOnwK20Xfl+bbsewEyZTPtnPtVVVVVV4H+IXjT4ba0niDwP4iu9JvVwGaFvllUHO2RDlXX/ZYEV8Gv29vC/iBYND+Llkmg6gQFGqWys9pKfV15aIn/gS9SSo4rTtS07WLGHU9I1C2vbO5TfDcW8okjkX1VlJBHuKqqqv+CjsMrfCbw3cKmY08RKrN6E28xA/8dP5VVtjzQPaqr9hH41Wng/xHf/DLxNqUVtpOtg3VlNPKEjt7pE+cEngB41HJ7xqO9ftPf8FZvgp8IDd+F/g+kPxI8UxFozNbTlNLtX9WuAD5+Dg7YsqeR5imtC/Z+/b7/wCCkOqWfi741+JLrwh4BZhcWR1G2a2tVQ9Gs9PUhpTtPEshAZf+Wh6V+zV+wB+zv+zItvq/hvw0fEHiuJRu8Ra2qz3KN3MC42W468oA2Dgs1ftMf8FD/wBnb9mk3ehapr7eKfF8CkL4e0RllkjfsJ5f9XAM4yGJcA5CNWo/GL/god/wUjvp9A+F+i3HhD4ezSNDO1hK9hp6pnBW5vWHmXJHRo48jv5Qr9nH/gkT8Dvheltr/wAZro/EjxCgDG1mjMGl27eiwg7pscjMp2ng+WprStI0rQdNttG0LTLTTrCzjEVtaWkCwxQoOioigBQPQDFVVft1+ET4c+O91q8Ue2DxHp8F6pA43qPJcfXMQY/73vXw98Uy+B/Hfh/xhCWzo2pQXTKvVlRwWX8VBH41BPDdQR3NvKskUqB43U5DKRkEe2Kqqqqqqqqqv2wvja3xg+J89lpF6JfDfhpns9N2NlJ3z+9nHY72AAP91FPc1oGg6t4o1uw8OaDZvd6jqdwlvawJ1eRyAo9uT1PA618EPhNpHwW+HOmeCNMMcs8S+dqN0q4+1XTAeZJ644CqD0VVHaqqqqaWKCF55nCpGpZmPYAcmtTvpdU1G61Kf/WXc7yvk55ZiT/Ov+Cdel+d8S/E2s7c/ZNDEGcdDJPG3/tKqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq+sbHU7ObTtSs4Lu1uEKTQTxiRJFPVWU5BB9DX/AAVJ/YN+Fngj4b3v7SPwe0C28MTaXewJ4h0iyTy7OaKeRYknijHELiV41KoAjBicAg7v+CUvxq8RfGL9li3sfFt/Le6p4H1aXQ1uZmLST2yRRSwMzHqVWXyvXEQJ65P7Gmqwfstf8FIT4F8VH7BZnW9S8LTSSfIFErMtq3XG15Ut+TxtfPYVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVf8ABZf45XVp4Z8J/syeFZnm1LxVcJqmr28HzO1vHJstYdoyT5k4Zsdc26+tfsifAq1/Zy/Z68IfC5YYxqVnZi51mROfNv5vnnOf4grHYp/uotVVVVVVVVVVVVVVVVVVftu/8E+Zv2y/GHhzxQ3xhXwhF4e0x7NLX/hHft5mZ5C5ff8AaYtoxtG3aehOecDR9PXSdIsdKWQSCyto4Q4Xbu2KFzjnHTpX7aX7E3hf9svRPDNjq/i648M6j4Xup5La/gsVumeKZVEkRUunBaOJs5429Oa+BHwf8P8AwC+EPhf4QeGJmuLHw3YiD7S8Qja5lZi80zKOFLyO7kZON3U9a/aH+D8Hx++C3iv4PXGtjR18TWQtxfm1+0/ZmWRXV/L3pvwyDjcPrX7En7IE37G/gjxB4Jb4kjxhFrmqrqCTf2N/Z5gbyljZcedLuyEU5yMelftC/wDBI+4+Pvxn8V/F+5/aMk0lvE179oWwk8KG7NqoRUWMSm8TcAFAHyrgcYr/AIcYf9XR/wDlkf8A3dX/AA4w/wCro/8AyyP/ALur9m74M/8ADPfwR8K/Bv8A4ST+3/8AhGbeWH+0vsf2T7Rvmklz5W99uPMxjceme+Kqqqqqqqqqqqqqqqqqqqqr9sH9q7wZ+yV8K7jxnrjQ32v34e38O6KZCH1C5A6nHKxJkM79hgD5mUH9hL9k/wAZ/tf/ABQvP2yP2omm1fRZtRNzptneL8mtXSNgEoeBZwlQgjxtYqE+4jBv22dMu/hX8fvBfxp0SHH2pYZmUcB7izddwOOgaJolx3AP4aXqVlrWmWesabcLPaX0CT28q9HjdQysPYgg1VVVVVVVVVVVVVVVVVVLFFPE0M0ayI6lWRlyGB6gj0r4hfsifAz4hmS5n8KLod/ISTeaKRatk9SUAMbEnuVz7818QP8Agnt470gyXfw68TWHiC3AJW1vB9kuP90HJjb6lk+leNPhj8Qfh3cfZfG3g7VNIO4qslxbkRSH/YkGUf8A4CTVVVVVfDD42fEn4P3/ANs8EeI5reB2zPYTfvbaf/ejPGeMbhhh2Ir4T/t5/D7xWsGl/Ei0bwtqbYU3K7prORvXcPmjyezAgd2rTtS07WLGHU9I1C2vbO5TfDcW8okjkX1VlJBHuKqq/wCCjAJ+CGi4GceKrfPt/ot1VWME1xMUgjLsqM5VRk7VGScewBJ9ACe1VVxbwXdvJa3MSyxTIUkRxkMpGCCPQiv+Cen7If7K2m+F4/iXYeHf+El8cWV2yXUmulbgaS+coLeHARQV2lZWDPncAwwQP2i/2v8A4E/sv6Qbv4meLo/7VljL2eg2GJ7+69NsQI2Kf78hVffPFeL/ANrv9uH/AIKC+ILv4b/s1+DtR8J+EWcxXb6ZMY2WJh/y+6idqxgrk+VHs3AlcSV+zP8A8EhvhJ8OVtvFHx8v4/iF4iBEn9mqrRaXbP3BXIe5Oe8m1D0MfetN0zTdH0+30nR7C2sbKziWK3traJYo4UUYVURQAqgDAAGBVVVV/wAFEPBZ1HwP4c8dW8IaTRr97SdgORFOuQT7B4gPq/1qv2V/Gw8efAjwtqUs2+70+1/s66yckPb/ALsEn1ZAjf8AAqqqqqqqqqr9sj4v/wDCqvhDd2mmXfla74n36fp+1sNGhX99KP8AdQ4BHRnQ1X/BPr4JpKbr43eILPOwvZ6Crjv92acfrGD/ANdPY1VVVV49vf7N8C+ItRDsv2XSbqXKjkbYmOR78VX/AATesQf+E+1Jk/6B8MbZ/wCvgsMf98VVVVVVVVVVVVVVVVVVVVVVVXxe+Nnws+Avhq28YfFzxfb+HNHvL5bCC6nhllD3DI8ipiNWOSkUhyRj5evSvCHi3w54+8LaV418IapHqWi63aJd2F2iMqzwuMq4DAMAQR1ANVVVVV+1h+2z8Nv2QdT8F2Xj/RtV1OLxdNchjphR5bKGER5mMbFdylpVGNwPDEbtuK8O/wDBRv8AYs8S6ZFqVt8d9Hst6gtb6jb3FpLGe4KvGOR6jI9CRX/BR3/god8N/jl8PH/Z4+AMt94httav7dtX1f7JJDFMsUiyR29vG4EjsZkjJYqB8gC7t2R/wTd/Z3179nL9mfTtC8ZWDWXifxNfy61qtpIo32jSoiRQt3DLFFGWU/ddnFf8FTv2FfE/jrVT+038FNEmv9agt0TxRpVkha4uFiULFeQovLuqKEdRztRGA4Y1+yX/AMFfvDln4c07wB+1Na6jb6hp0SW8fi2zha6W6VRjddwrmQSYAy8YfeTkqvUw/wDBQ39iy408anH+0F4eEJQsFeO4SXA/6ZGMPnjpjNfBj9un9nT9oT4oXPwo+Evie/1rUrXSpdRa8fTpLW2dI5I0ZEMwR2fMoOAmNqsc8c1VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVqF/ZaVYXOp6ldxW1pZwvNcTyttSKNQSzMT0AAJJr9lPT7v9uz/go14i/aC1y0eXwn4OvP7Wto5YztRIj5OlwkH7r/Isx7EwyetVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVXxj+Lvgf4E/DfWvij8Q9U+x6NokBkcKAZbiQ8RwxKSN0jsQqjIGTkkAEj4L/AA1+J3/BVP8AaZ1H4x/FpLvTvhn4dnWOS3jlbyooVO6LTLZsDLsDulkABAYsdpeMHRtG0nw7pFloGg6db2Gm6bbpbWdpbxiOOCJFCoiKOAoUAADoBX7cngJvGnwH1DU7WDzLzwvcx6lHhcsYxlJR7AI5c/7gr9iXx8fHHwE0izuZw974ZlfSpsnnZGA0PHp5Tov1U1VVVVVVVVVVVVVVVVX7b3/BTjwZ8AjffDP4NvY+LPiIuYbiUN5tlor9D5hU/vZgePKU4U/fII2H/gmx8Jf2wNP8R+KPj9+0P4z1GGx8f2yO2gawpe8uplI8q7ZOBaKibkWMDJVsFECIaqqu7S1vraSzvbaK4gmUrJFKgdXB6gg8EV49/Y/+BHj0yTv4SGhXr/8AL1oji1I/7Z4MR+pTPvXj3/gnj4u05Jrz4deMbLWY1BZLLUIjazEf3VcFkY+52CvE/hXxH4M1q48O+K9FutL1K1bEttcxlGHoR6qezDII5BNVVVVfDv4w/En4U3v2vwL4svNPRmzLakiS3m9d0TZUnHGcZHYivhb/AMFBvD2pmHTPiz4ebSJzhTqWmq01uT6tEcug/wB0v9BXhTxp4S8daWus+DvEen6xZsBmW0nEmwkdGA5Vv9lgCPSq/wCCgkMsvwDR403CHXbVnP8AdGyUZ/MgfjVfBbx38Ovhn8RbHxp8WrjyvCNjZ38eq/6O0++OazmhCBEBLFmkVf8AgXYc1ovi/wAJeOILjXfA816+jtdyx2q38ax3KIrHaJVRmVX27SQCRz1qq0T4ifFD4f6Zrn/CqPGlx4a1bWNPa1+1xRrJt5BBw4YA5GA4G5QxIINfsqf8Er4fiZFY/HP9qnx9N4pl19Vvl0ex1B5muS3U3l5nczcYKRkEEY38EV4P8F+Efh94ds/CXgbw3puhaNYJstrGwtlgijHsqgcnuTyTySaqqqqr45+Bj8SPhH4p8HRReZc3unu1ooHJuI8SRD8XRR+NEEHaRgjsa/4J2ePBFeeJ/hldTgCdU1WyQtj5lxFNj1JBhP0U/hVVVVVVVVX7afxS/wCFk/Gm/wBPsLrzdJ8KqdNtAp+VpFP79x2yZMrkdVjWvBHhHVfH3i/R/BeiIGvdYu47aIkcJuPLH/ZUZY+wNeDvCmj+BfCuleDtAg8rT9HtEtoFPUhRjc3qxOST3JJqqqqr40TyWvwd8d3UJHmQ+GdRdMjPItpCP5VX/BOS2jXwn4zuwzb5NRtoyD0wsbEY/wC+j+lVVVVVVVVVVVVVVVVVVVVVVV/wWr/5NX8Lf9lAs/8A0gv6/Yo/5NF+EH/Yn6f/AOiVqqqqqvi58BPg38edIj0T4u/DvR/E1vAGFu93EVntt33vKmQrJFnAzsYZwPStW/4I+/sdajem6s7PxlpcZPFtaa5ujH4yo7e33q+Cf7Cf7LnwA1SHxF8P/hhaNrsBzFq+qzSX9zCw6NGZSVibtmNVPqaqvjJ+wz+yz8d76fWvH3wl006zcHdJqumPJp9zI/8Afd4WUStjvIG/QVF/wR2/Y/jvTdP/AMJxLHuYi2fW1EYBzgZEQbA7fNngZJ5r4IfsZfs2fs7al/b3wq+GVnp2tGFoTqtzcTXl0Eb7wV5nbywQACE25HXqaqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr/gqx8dm+EH7LuoeFNJvvJ134jTHRrcK+HW0K7rxwO6mLER9PPFf8EtPgQvwa/ZZ0fX9Ss/K174hONcvSw+Zbd1xaR/TyQsmOzTNVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVahf2OlWFzqmp3kNrZ2cLzXFxM4RIY1BLOzHgKACSTwAK+OfxI+I3/BUv9qPS/gl8I7i4tPhr4buHeK5ZG8pIUO2fVJ14yxDbIkbGAyqNpkc18H/hH4H+Bfw50X4X/DzSlsdG0WARxg4Mk7nl5pWAG6R2yzH1PAAAArWNJsNe0i90PVIBNZ6jbSW1xGf443Uqw/EEiv2KtYvfhR8evGHwL12fBvXlhhzwHubRnIKj0eIyN7hV/Cqqqqqqqqqqqqqqqr9pD4fePPin8E/FXgX4Y+Pbzwf4m1KyZdP1O2fYQ4IPlM4BaNJAChdMMobIzjB/Yh/4Jk+C/gEth8S/jFFY+KviKAs0ELKJbLRX6jygw/ezA/8ALUjCn7gGN5qqqqr4mfCL4f8Axd0f+x/HGgQ3mxSLe6UbLi2J7xyDlexxypwMg18bP2KPiD8ODca54JE3ivQEJb9xF/plsv8AtxD74H95PqVUUysrFGBDKcEEYINVVVVeHPFPiXwfqces+FNfv9Jvo/uz2dw0L49CVIyPUHg96+Gn/BQDx9oPlWHxJ0W28S2ijBu7fFrdL7kAeW/Hbap96/ab/aJ+EXxo/Z61Oy8K+I3h1eK9tZhpV9CYbg4lAOByrYDEnYzYxzVfDv4Y+GfjP460H4W+Mrq7t9G8RX0VtdyWkqxS7d27CMwYAkqB0PWvjH+xV8MrL4Iw+F/gj4G07w/f+FQ91YQ2kWJL/KjzUmkOXlkdVXDuSdyqMgE0QQcEcjsaqv2SP2l5PhHrS+C/GF47+ENUm/1jEn+zJj/y1Uf88z/Go/3hyCGtrm2vbaK7s7iOeCdA8UsbBldSMhlI4IIwciqqqqqr9prwAfhv8a/Eugwx7bO5uTfWPGB5M/zhR7KSyf8AAK+Anj//AIVj8XPDXjCWby7S2vBFenOB9nlBjlJ+isW+qilZXUMjBgRkEd6qqqqqqqvjr8RI/hV8JvEfjYSKt1Z2bJYq38VzJ8kXHcb2BI9AallknkaaaRpJHYszucliepJ7mv8AgnZ8MhqnifW/itqFtug0WL7BpzMOPtEq5lYe6xkL9Jqqqqqr44/8kV+IH/Yral/6SyVX/BOb/kSfF/8A2FIf/RVVVVVVVVVVVVVVVVVVVVVVVV/wV2+Hnj/4l/s2eG9B+HHgbxB4q1KDxxa3MtlommTX00cK2N6pkZIlZggZ0UsRgFlHcV+yHoeteGv2XfhZ4e8R6Pe6VqmneFbGC8sb63aCe3lWJQySRuAyMDwQQCKqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr9q28n/bh/wCCknhr9n7SLmSfwt4OuxpN20THaiQ5n1OUf3W+RoAehaJPWrW1trK1hsrOCOGC3jWOKJF2qiqMAADoAABiqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/4KiftZeIPGXiO1/Yo+A8tzqGr6vdxWviVtPOZLmaRlEWmxkHuSpl/4ChOPMFfsQ/sj+Hv2SvhHb+HtlveeMdbWO58TapGM+dcAHbDGTz5MQYqvTJLPgFyBVV+2Ppl98HP2jPCfxy0KBtmotFcyqOBJcWxVJUPoHhMY9Tlq0nVLDXNKsta0u4Wezv7dLi3lXo8bqGVh7EEGqqqqqqqqqqqqqqvit8VfAfwT8B6p8SfiTr8Ok6FpEe6edwWZ2JwscaDl3Y4AUckn618K/iRoPxf+Hmg/EvwxZ6pa6Z4gtFubaDU7NrW4jU8YeM9DkHkEqRgqSCCaqqqqqqr4yfspfCz4w+bqVzYHQ9fcEjVNPQK0jf8ATWP7sn1OG/2hXxg/ZV+K3wg83UL7ShrOhoSRqumq0iIvrKmN0XGMkjbngMaqqqqq4GYWGe1V8LPFdr4F+JXhbxjfQyzWujavbXVxHCoZ2iSRS4QEgFtoOMkDOMmvjl8eP20/25/iH/wov4b6FH4E8IakzoLRdVSFLu3zgve3YOXUg5MEY5zjZIRmviB+wz4u/Zy+D+gaoPF8vjH+zovK1ydbbyls2LfuzGMljCMhMtzkA8BsLVV+yb+1lL4Bltfht8SdQaTw1Iwj0/UJWJbTGPRGPeE/+Of7vSKWG4hSeCVJI5FDI6NkMCOCCOoIqqqqqv8Agof8PPtug+HvifZQZl02U6dfMFyfKky8RJ7BXDj6yiq/ZP8AiN/wsr4I6FfXNx5uo6On9mXxLZJkhACsT1JaMxsT6saqqqqqqqv+CjvxCZIfDHwtsrnHmFtVv0BwcDMcAPtnzjj2U1X7Nvw5/wCFXfBfw14XuIPLv2tRd6gCPmFxN87qfdchPogqqqqqvinZf2l8MfF+nCMSfatBvothON26BxjP41X/AATgvfM0jx3pu9j5FzYy7SOBvWYZHv8Au/0FVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX7SvxwHwF+H0fim0sLfUNSvL+K0s7SdyqvnLSE7eQBGjc9mK5z0Pw7/AG/vhD4qENn4ztb/AMJXr4DNMpubbd7SRjcBnuyAD1rQ/FfhfxPpsWseHPEemapYzMFjubS6SWNmPAUMpIznjHWqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/aJ+LunfAb4IeMvi1qJjP/CO6XJNaxSHAmumwlvGf9+Z41/4FX/BGb4Q3+qSePv2n/Fge61DV7ptH066nGXkYss95Lk9dztAu4d1kGeoqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv8AgoD+15Y/spfB6WbQ7mGTx54oSS08OWzEN5LAASXbqf4IgwIBGGcop4LEf8EqP2P77RrFv2tvi/ZzXXinxQskvhtb/LzQ282TJfOW582fc21jz5ZLZPm8VVV+2f8ADgfEL4FavPaweZqPhojVbXA5KxAiYfTymc47lVr9hL4jjxr8FYfDd3cB9Q8IXBsnBPzG3bLwsfbBZB/1yqqqqqqqqqqqqqq/4KK/tBfEX9mn9n+3+IvwvurCDWZPENrYs17ai4TypI5mYbSeuY15r9kPxvr/AO29+yA+o/HSLSdSn1jV5ra5jjsFWB0tp45I8xZwcMgPNf8ABSj9un43/ss/Ebwz4C+FEfhuGz1bQPt89xfae1xMsn2iSPC/OEC7YxxtJ+Y89MaX8Wv+C0viDTLXxFpHgoSafqEK3NsRpOkoGicblwrNvAIIxnmvD3/BVP8Aad+BnjWDwZ+178DFjhdw0rwafLpd8sRODLGrsYbhRg4C7Aefn9Phn8SPBvxf8B6L8Sfh/rMep6Dr1sJ7O4QYOMkMjL1V1YMrKeVZSDyK+L/xF0v4Q/CzxX8T9ZKG18MaRcX7Rs23zmjjJSMH1dgqD3YV8I/+Ctf7S83xM8Hp8VL7w+3gy/1iC31aSLRlgYWzSKszpJnqitu/AA9aqv2jv2nvhL+y14KHjL4o628RuS0em6XaKJbzUpVAJSGMkDABG52KouRlgSAdR/4Kb/trftDeILnRf2VfgcLSxhkwJLXSZdZuoh0DTTECCIHg8oACcbj1J+I//Bbbw/GdZ1Lw3q+pWyjJtf8AhHtHmPr9y3QS9Bj8fXFfsDfHj9oX49eBvE2pftCeAbbwzqfh3Vk023C6ZcafNcOIhJL5sExO0qJIuVwDuPAxzRAYbSAQR0NfFz9jb4TfE7ztS0uy/wCEW1yTJ+2abEBFI3rJBwrepK7WJ6k18WP2TPi/8KjNfT6Kde0aMFv7S0pWlVFHeSPG+PA6kjb/ALRqqqqmGYmAPaqU4YEHoarwb+0X8X/BNjJo1j4um1LR542im0vV1F7bSRsMMm2TJVSOoUr1Pqavp4bu9nureyitI5pGdLeIsUhBOQiliW2joNxJx1JPNVVfswftcaj8K3t/A/j6afUPCTvtgnGZJtLJ7qOrReqdR1Xup0fWNK8Q6Xa63oeo29/YXsQlt7mCQOkqHoQR1qqqqr4n+BrP4lfD7XvAt6yKmr2TwxyOMiKX70b4/wBl1RvwrVNNvtG1K70fU7dre8sZ3guIXGDHIjFWU+4IIr9gv4nDwp8TbnwFqFwEsPFkAWHccBbuIFk+m5DIvudlVVVVVVVX7T/jo/EP45+K9din8y0t702NmQfl8mAeWCvsxUv/AMDNfs3+Al+JPxs8KeGJ4PNs/tour1SuVMEIMjqfZgmz/gQqqqqqqvbSG/s57GfPl3ETRvjrhhg/oavbSawvJ7G5XbNbSNHIuOjKcH9RX/BOnVRD8QPFehbhm70ZLgD/AK5TKv8A7WqqqqqqqqqqqqqqqqqqqqqqqvFPjbwX4Gs4dQ8beL9F8P2s8vlQz6rfxWiSPgnarSMAWwCcDnANadqOn6xp9rq2k39ve2N7Ck9tc28okjnjcBldGUkMrKQQRwQQaqqqqqv+ChXxBPiD4paf4CtLnda+FrENOgPAupwHbPriMQ/TLVVnHex2jzI8iWsz+WwDYWRlwcY/ixkH2yPUV4V+Nnxc8E+Wnhj4i67ZxRY2W/2xpIRj/pk+U/Sv2Uf2n/jL8VPiPaeBPFcmk6nYCznuby9+xeTcIqLhSDGVTl2QH5O/aqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv+Cl/7XnxX/ZP8NeCLn4VQ6CbnxRdX0V1Nqdm9w0QhSIp5YDqoOZSTuDdB758G/H/AP4LEfFTwzYeP/APhaG70DWIvOsJ4NI0yOOVOm5RM2/BIPJqX/gpR+3N+zd4is9K/an+Blpc6fcsAHn059MmnC43GC5jLQOQDkgI3OOVr4AfH74dftK/DWx+J/w01GWbTrmRoLi2uFCXFjcJjfBMgJCuAVPBIKsrAkMDX/BSn9uv44fstfEbw14D+FUfhuK11jQRqE91f6e9xOsn2iSPap8wIF2xjgqTyeemNP8Ai1/wWk17TbbxJpPglX0/UIVurYJpOlANE43LhWbfjaRgH5vxrwl/wVY/aL+C3ji38D/ti/A82kDsPOntdOl0y/iiJwZkilYxXC8HG3YD/erwV408L/EbwlpPjrwVrMGq6Hrdql1YXkOdssbDg4OCD2KkAggggEEVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV/wAFpPjBcw+HPAv7Onh6aSW/8Q3Z1jUreHO94YyYbWPA+8JJWlOP70C1+zB8Hbf4BfAHwT8J4441udD0tBqDJjEl5JmS5YEZyDK745PGKqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq8f+O/C/ww8E618QvGuqJp+h+H7KS7vrh+dsaDOFHVmJwqqOWYgDk18C/BHi/wD4Kifthat8YvihY3EXw38Lyo0tizHy47VWJtdNRh1Z/meVlxkGQ/KXSoIILWCO1tYY4oYUCRxooVUUDAAA4AA4xVVVTwQ3ML21xEskUqFHRxkMpGCCPQivgZPN+zb+2Bq/ws1GVotG1+drC3Ln5WWQiWyk925EfoDI3pVVVVVVVVVVVVVV/wAFkv8Ak0W0/wCxwsf/AETc1/wSG/5M00z/ALGDUf8A0Na/4Laf8l+8C/8AYnj/ANK56+E//JK/Bv8A2L9j/wCk6V/wU4+GXhH4g/sg+NNY8QadA+p+EbddU0e9MYMtrMsiBwrdQsiEow6HIOMqMf8ABErxTrGo/B/4g+ELt5X0/RfEEFzZlskK1xCRIq+37hWx6vnvX/BZb4yp4O+Ami/CDTrvbqHj/VBJdRqf+XGzKyPnuN07W+OxCv6Gv2p/2Urj4c/8E9/gP8QY9NK6rpss02vkLhwmrAXERkH/AEy8uKL2L/l+wP8AGE/G79lHwF4ru7sz6rp9gNI1Uscv9ptP3RZvd0WOX/toOnStZ1fTfD+j32vaxdJbWGm20lzdTP0iijUs7H2CgmvAOieLv+Cpf7bl7qvi++vbPwdZb7u4hRsHTdGhkCw2sXZZZC6gsP4nkkwcYrwL4B8F/DHwrYeCPh/4asNB0PTI/LtbGyiEaIO5PdmJ5LNlmJJJJOaqqqq+Kf7Kvwd+K3nXuo+HxpOsSgn+09LAgkZvV1xsk7ZLLn3FftDfsp+IfgD4fu/HWo+LtDvPClrIqPfXN0llLGWOFVo5GwzE4ACMxPPAxVtc217bpdWdxFPDIMpJG4ZWHqCODVU4BQj2qqBBAI9Kqqqq+An7SnjT4GamsFq7ar4buHzd6RNIQuT1eJufLf8ADDdweCPhf8XPAnxh8Prr/gnWUuVUAXNpJ8lxaOR92ROo74IypwcE1VVVV+3b8Lf+EN+KUfjbTbUppvi6IzOVHypdpgSj23Ao/PUs3pWkatqGg6tZa5pNy1ve6fcJcW0q9Y5EYMrD6ECvhV8QNP8Ail8PdD8dacY1GqWqvNEhz5E4+WWP1+Vww9wAe9VVVVVVfGHxovw7+FvijxoJAsulaZNJbZ7zldsQ/GRkH40zM7FmYsxOSSeSa/4Jv+B/P1bxX8RrmD5bWCPTLRyMgs58ybHoQEh/B6qqqqqq+Omh/wDCOfGXxto6oESHXbpolAHEbyF06f7LLX7EOuDR/wBobRLVn2pq1pdWjEj/AKZNIPzaJR+NVVVVVVVVVVVVVVVVVVVVVVV/wW0/5ID4F/7HAf8ApHPX7J3/ACav8Gv+yf6F/wCkENVVVVVqupWWjaZd6xqVwsNpYQPPcStwEjRSzMfYAE14+8W3vj7xtrnjTUCwn1q/muijHPlh2JVB7KuFHsBVeM9EHhfQfCOiOu25utJGrXakYKyXTkxj6G3S2b6sfrVf8E5fDCy6r4y8ZSx/Nb29vYQPjr5jNJIP/IcX51VVVVVVVVVVVVVVX/BSH4rfHE/ttal8KfB3xj8X6HpFyNItrPT7PXbm2tIJJ7eIF/KjcLktIxJxk5NJ/wAEpP2t7VFurH9r0JexDdHi+1FArjphwxI574/CtR/aD/4KC/8ABOr4gaX4c+NuuXPjjwxeZa3TVb99RtNSiUgP9mvXHnROmfuNjblS0ZUrn4SfFDwp8a/hr4e+Kfgi6efRvEdktzbeYu14zkq8bjs6OrIwBI3KcEjmv+Cknxtu/gd+yf4n1TQ9Vl0/X/Erx6HpE8EpjkjknyZXRlIKssCTsGHIYLX7I/7Rfxv+DP7Tfwt1T4vfEbxbeeGPF6RCW21fWri4gksr0vBHcFZGIwkgEmcdIz71Vft0/wDBSzw9+zZqE/ws+Fen2Pij4iBP9LM7FrPRtwyolCEGSYggiIEYBBYjhT4e/Z6/4Km/th2kXjHx98UtU8G6HfqJba31jVpdJjkTOVKWFomV7YMqISMHJyCX/wCCRH7VGgr/AGh4N/ab0hNSXhC13f2XHX/WRh2HIX+H37V+wF8Lv2ufhfpHjLSf2qPHWo6+Eu7aHw6t3q66kPKRHMsyzHMu1i8ahZCCPLPyjOaqqqqqqqqqqqv+C43/ACKnwj/7CGq/+i7av2A/+TNPhL/2L8f/AKG9ftV/DLwj8XP2e/Hfg/xlp0FzanRLq5tpZIwzWdzFC7xTxk/ddGAOe4yDwSD/AMEQ/FOsQ/EP4leCFeVtKu9FtdQdTkpHPFN5akehZZm+uwenH/BbT/kv3gX/ALE8f+lc9fCf/klfg3/sX7H/ANJ0r/gsxo/gu6/Zi0nWtbitV8QWPia3j0SVgBMfMjk8+NT12FE3MOmUQ9QK/wCCSMHiSH9i/Qv7dVxaPrWoto4cEf6KZucZ7ef5544/Wqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqvh+h/bZ/wCCr2oeK5lW+8J+Ar+S7iyA0Zs9MIitiAcgrLdGOQg9RI30qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv8Agp3+0B4n+Pnxd0H9h/4IO+osmqQxa2tvJhb3UmI8u2ZhwI4Ad8hPAfrjyc1+zD+z74X/AGZPg1oXwp8NLFNLZxedqt+se1tQvnA86du+CQAoOdqKi54qqqqr/goZ8O7qCPw18adAWSG702UWF7PDkMg3GS3kyOm194z6sg7V8F/iLafFf4YeH/HVu6ebqFov2uNT/qrlPlmXHYB1bHsQe9VVVVVVVVVVVX7cP/BRfwZ+yr5XgjwdZWPiz4hT7ZJdOeci30yHIy1yyHcHZc7Yxg8hmwNof4ZeN1+Jfw98P+Pl8OavoH9vWEd3/ZmrW5gurXeM7JEPQj9Rg96/4LJf8mi2n/Y4WP8A6Jua/wCCQ3/Jmmmf9jBqP/oa1/wW0/5L94F/7E8f+lc9fDv/AIKQ/sVaL8L/AA1ban8b7WC503RrW3ubY6NfmVJY4kR12CAk4YEZGQeoOOa/bt/4KJW37UegRfs3fs0+G9d1HTNfvIk1G8exb7VqzJIHit7aBcuELqjEkB2KhdoGd3/BO79mLVf2Xv2fLbw/4uhSPxd4kvG1XW4lYP8AZZGVUjtwy8HZGi5xkb2fBIwa/aokl/bN/wCCnui/Beyke60Dw9qNvoEyocbbe13XGpN6BgftCZ7+WvsB+0r8H7X44/s/+NvhElvD5ut6NJFpysoCRXcYEls3oAs0cZ7cDtX/AARY+LV3ofjDx9+zzrsskH22Ia1p9tKNnlXMJWC6XB53sjQHHYQGv2/9fu/Df7GfxZ1Gyd1km0B7MlWwdlzIkDj6FZWBHcV/wQ98PWEXhD4qeKhGrXtzqWn2Rcrykcccr4B7AmXkd9o9Biqqqqv2sf20PhJ+yT4ZF54vvhqnie+hL6T4as5R9puuoEj9fJhyCDIw5wQoYjFeE/hl+1R/wVQ8fr8S/iv4gPhH4WaZcFbedkMdlapnDQ2ELH9/McYeZjgH7zcLHXg39lH9k/4ffDK3+GGk+DvD50i0DSS3l1cKbyaUqA88lyCH3nAzghQAAAFAA/ao8afs+/C74jL4R+DOo+KfFsULumqS29otxa2b9AkNwCvnkHOcAgcfOxyBc6Rqlnpun6pfWTW8WpwLNAsjrvUEA4dASY255RwGHpRAIxRGDjPSqjIMakHtVVVVVeDvG3iv4f67B4l8G67daVqNv92aBsbh3VlOQ6nAyrAg9xXwJ/bh8JeN0tvDnxRNt4c11iES9ztsro9iSf8AUt7Mdv8AtDOBHJHLGssUiujqCrKcgg9CD6VVVX7SvwpX4wfCXVfDltCH1WzH27Sjjn7TGDhf+BqWT/gee1MrKxRgQynBBGCDX7AXxdXRfEV98I9ZvAtprRN1pe9sBLpV+eMf76AH6x+rVVVVVVV/wUL8Yf2J8HLDwpDNibxJqqLImfvQQDzG/KTyfzqv2L/Bo8Hfs9+HTJFsudcMmqT8Yz5pxGf+/SRVVVVVVV+3L4eGiftBalerHtTW7C1vVGMA/J5JI+rQn8c18IfEa+Efin4S8Su+2LT9ZtpJjnH7vzFDjPupYVVVVVVVVVVVVVVVVVVVVVVVX/BbT/kgPgX/ALHAf+kc9fsnf8mr/Br/ALJ/oX/pBDVVVVVfttePP+EJ+Aer2dvMFvPEsqaXCM87ZMtKcenlI6+xYVXw18GXXxE8f+H/AARZlg+s6hFbs6jmNCw3v9FQM34V+0hq1pq/xv8AFzaeqJaWF9/Z1tGgwqR2qLbqF9gIh/PvVfsHeGxovwFg1ZkG/X9UubsHHO1CIAPpmFj+NVVVVVVVVVVVVVVV/wAFB/8AlJgn/YQ8O/8Aoq3qv+Cx8Hh9/wBkm3n1Zbf7fH4rshpbOPn80xzbwn1iEhI6YHsK/wCCQf8Aa/8AwxtYf2lu+z/8JFqP9n7s48jcmce3m+b075r/AIK5+PNY+L37RXw8/Zc8GObm40wQiS3Qn95qeouiRIwHdYhEQfSdq/4K6/s46d8O/h38G/G/hK2A0/wnpkfg24lWPa3lRR+ZaE46fcuicnqw7mv2Q/jAvx4/Zt8BfEya6E+oahpKQ6oxbJ+2wEw3BPcbpI2YA9mB5zmv2qPjMv7Pv7Pnjb4txpHJeaHppGnxyDKveSusNuGHdfNkQkegNf8ABJr9nO2+N/xK8T/tP/FiN9fHhzUgunG/Xzhd6xL++luZC33niVkYZz88ytwUGaqqqqqqqqqqqqr/AILjf8ip8I/+whqv/ou2r9jf9vj9kf4dfsveAPA/jb4xWuk674d0dbbUbKbS71mikVmJAKwkPwRyhYc468V+2z/wVR8C+O/h3q3wY/Znh1TV7vxVA+nX+v3Fk9uiW0o2SR20TgStJIrFNzKu0ElcsQV/4JT/ALJnin9n/wCGetfEP4laRPpXivx20Hl6ZcptmsLGIMY1kXqkkjSMzIeVCxg4bcB/wW0/5L94F/7E8f8ApXPXh39u3/gpvovgzS9O0f8AZAEmlWGlwxWt+fh7rj7oEiASXeJ9h+UBt2Np64xV58Yj+0r+0Dps3/BQP4k+LdB0TS28sWOn6II0sgWBaHygQ1srYG6URyyHAB6Bl+Gtt4As/h/4es/hW2lt4Qt9Ohj0U6ZKJLf7MqgJ5bAncMDrkknOTnNVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX7aXxhf4E/sw+PviHZXbQapBpbWelOhwy3lyRBC6+6NIH+iGv+CLnwePhr4N+KPjNqNrtu/GeqCysHYdbO0yCynsGnklUj/pitVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVftxftO2H7K/wH1bxpbzQv4o1QHT/AA1ayc+ZeOpxKy90iXMjdjtC5BcV/wAEj/2X9Qt7HUf2vPiZBPc6/wCKGni8PPeZaXyHY/aL0lud8z7kVjzsDnkSA1VVVVXxO8C6d8Tfh/rvgPUyqw6xZvCshGfKk+9HJj1V1Vv+A1+wP461Hwj4w8U/AHxWxtrpLiW4tIJDzHdQny7mIe5VVbH/AEzY96qqqqqqqqqqr/go9+3nq/7NNlafCb4WadM/xB8SWXnx6jPas0GmWzsyCSMEYmnLKwVeVUjLZ4U/sEf8E37vS9Rtv2jv2qLOTV/F2oyDUNM0HU8zPZzO2/7Ve78l7gnDBGzsJJbL8JX/AAWS/wCTRbT/ALHCx/8ARNzX/BIb/kzTTP8AsYNR/wDQ1r/gtp/yX7wL/wBieP8A0rnrwB/wS/8A2Ktf8G+GPEup/Cm7kuL/AEe0uLmJfEOoLHJI8KM5wJsjLEnAIAzxgcV8Jv2avgL8ClJ+E3wr0Dw7cOmx7yC28y6df7rXEhaVh7FsV8cfibp/wY+D3jH4qalsaPwxo9xeRxucCaVUPlR/V5CiD3YV+x/8Fv2wfjL4p8S/GD9nLXE0/XdLnMWo63cXyWskkt1ueQIzKQWO3LYxgOOzV/woL/gsr/0Wsf8AhRw//G60cfF79hf9tzwx4s+NSxR69b6pHqetTW8olS8sr0ul1IrKArEo8/QYDr04r9q7wBcfF39mX4j+BtGi+2XuseG7o6dHGofz7hE82BV7fNIiDPbORX/BFD4s6Xonj3x38GdUu1iuPE9nb6npSu2BJLa+YsyL6sY5VfH92Fj2qqr/AIKBeIfEGj/Evw7BpGu6hYxvoQZktrp4gx8+QZIUjnAFf8Jx41/6HHW//BhL/wDFVN4y8X3MTQXHivWJY2HzI99IwP1Bav7c1r/oMXv/AIEN/jU0stzM1xcStLK+NzuSzNgYGSeelVVVUoxIwHrVQHMK89Kqqqqqq+Hn7RXxj+F0ENh4T8a3S6dCfl0+7VbmADuqo4OwH/Y214C/4KJxsI7T4m+BSpxh73RZc/8AkGU8fXzPwrwL+0V8F/iN5UPhrx9ppu5ThbK8f7LOW9Akm0sf93I96qq/bX+D7fDr4nv4p0mzEeh+LS9zFsXCw3Qx58ftkkOP98gfdNaPq+paBq1lrujXj2t/p9wlxbTocNHIjBlYfQgV8FPilpfxj+HOleN9PCRzTp5V/bK2fs1ymBIn0zhhnkqynvVVVVVV/wAFFvFR1P4paD4Tik3RaHpHmsP7ss7ksP8AviOI/jWj6Xd65q1jotgm+5v7mO3hXHV3YKo/MitA0a08O6Dpvh6wGLbTLSK1hGMYSNAq8fQCqqqqqqv+CjfhlhP4M8Ywx5DJc2E746EFZIx+sv5VXws8Tjxr8NfC/isy75NU0m3nmJOSJTGN4J9Q24fhVVVVVVVVVVVVVVVVVVVVVVVVVVVVX/BRjxydT8d+Hvh/bTZh0Swa7uADx507YAI9Qkakf9dKr9gHwhbXvxH1v4j6pEDY+DtJd1kI+5PMCoI/7ZLP+YrWNSuNa1a91i7OZ765knkJOfmdix/U1XwI8PDwt8GPBWiFAkkOiWzyrtxiSRA7/wDjzNVVVVVVVVVVVVVVVf8ABTLXV8L/APBQfVvEzWxuRpB0S7MIfZ5nlW0D7c4OM7cZwcZq9/4LheBo9NEunfs/67Pf7f8AUT69FFFnHP7wRMcZ77K8SaN+2p/wVS+IWjX994Nl8J/D/TXxZ3E1vLBpenxtjzJld8Ndzlcfcz/CMRqSa+F3w88E/s6/BvRfh/oc62nhzwXpRV7q4wpKoDJPcSY4DOxkkbHGWNfsNSwftQ/8FEPE37RHjC5gg0zRri816Fb2YIFdz5FjBknkxxsGH/Xv+B/bN8DeFPj3+zN47+HFtr+ky6ncaa13pIF3GW+225E0Kjnje0YjJ9HNf8ETPjD5+m+PPgLqN3l7WSPxBpUTPk7G2wXQAPQBhbHA4zIx47/8Fhrm+g/Y9kis1YxXHiiwjutqkgRgSsM+g3qnXvj1r/gj3b6fD+x5DJZCMTT+J9Qe72tkmQCJRuHY7Fj49MHvVeLf+Cpv7TugeKtZ0Gw/ZP8AtdtpuoT20Nx9nvv3yRyMqvwmOQAeOOa/Yx/4KWeP/wBp747wfB/xN8LdE8Pwvp91cyXFtdTPLG8IB2FXGBycHPSqr4x/tgfs2/s/64nhn4u/FKy0DVpLVbpbI2dzcymJiVV9sEbnBIIH0PoauP8Agq3+xBDM8UXxR1G4VTgSR+HL4K3uN0QP5gV4M/4KKfsW+O7mGx0j496JZ3EuBs1iC40tVYjODJcxxp7Z3Yz0Jqwv7DVbKDUtLvre8tLmMSQXEEgkjlQjIZWUkEEdxVfEz9vj9kj4QeJ9Q8FeP/jDa2Ou6VKYbywh0u9u3hkAztYwwuoOCOp71bf8FWv2H7idIZfilqNurHmWXw5flV+u2In8ga+Ev7RfwN+O8Ekvwk+KGheJJIY/Mltba42XMKcDc8DhZUXJAyygZ4qqqqqv+C43/IqfCP8A7CGq/wDou2r9k3/gnR+yP8Wv2bfhx8RvGvw4u7nXda0ZLjUZ4tdvYluZC7ZYosu1cgAYQKPbvXwn/ZC/Zo+B17Hqnwv+DmgaRqMWfK1CSN7y7jz12T3DPIvTswqv+C2n/JfvAv8A2J4/9K56+E//ACSvwb/2L9j/AOk6V/wUX/Z5+H/xm/Zq8beKNb0WzTxL4I0K61nSNYWILcQ/ZY2meHeOWjkRHQocrlg2AVBH/BEz4p+Idb8F/ED4Q6reS3Gl+Gbm01HSFc7hbi684TxjJ+VS8SOFHG55D1PNVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV/wWp+KF7cWnw5/Z60B3mutWun1q9tYid8mCbe0XA+8Gdrjj1RfTj4B/C6y+CnwW8F/CqxVMeGtGgtZ3QYEtwFzPJx/flLufdqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqndI0MkjqqqCWZjgADqa+JOq6x/wAFQv29LHwF4avp/wDhWfhFpIY7mEkpHpkLr9qux2D3MmxEbHRoAR8pNaHomkeGtFsPDugafBYaZpdtHa2drAm1IIY1CoijsAoAA9qqqqqqv2xfDGq/BX47eG/2gfCEGyPUbiOacA7U+2QgB0bHRZYsAjv+8PevCfibSfGnhjS/FmhXAmsNWtI7m3fvtdQcH0I6EdiCKqqqqqqqqqq8SfDvwJ4x1rQfEfirwjpOrap4XuXudGu7u1WWSxlZdrPGxGVJGPxVT1UEVX/BZL/k0W0/7HCx/wDRNzX/AASG/wCTNNM/7GDUf/Q1r/gtp/yX7wL/ANieP/SuevhP/wAkr8G/9i/Y/wDpOlV/wWg+Mp8L/Bnw18FtNuQLrxvqRu79Qf8AlzsyrBSO26d4iP8Ari1f8Ez/AIPD4P8A7IfhCO7tRDqvi9W8Q3+Rgk3IUwZ7jFssAI7HNV/wWv8Ag0+p+E/BHx40uz3S6LcSaHq0ijJ8ibMlux9FWRZl+s4r/gnX8Zf+F2/sleCtZu7jzdW8OwHQNUJbJ860CojE92eAwSHPdz9a/bw/Ys+KX7N3xek/aq/Zstb9fDw1D+1pRpURabw5d53SMUGc2zMWOcFVDMjgLgt8BP8Ags18MNb0S20v9oXwxqPhvXYUCzano9sbuxuSMZfywfNiJ5OwCQf7XatT/wCCqv7DdhZG6tPi5ealKOltbeG9RWQ8eskCL2x97v8AjX7LH7XPw3/a50TxJr/w507V7G38N6otlJFqsccc0qtErpNtR3Cqx8xQCc/Jk4zgftWfsveP/jr4y0jxD4R1fw/aW2n6Z9llTUbiaNy/mu2QEicYww6kc9qvP+CfnxfsLSa+vvGPgO2traNpJpptQuUSJFGWZmNvgAAEkngVcfEjQ9Q+Jp+E/wAOLW/+IWsy3gtLGbw1bma3v5McmFpfLcoOcuUC4Utkr81aR/wT2+Lt/pdpean4j8Nabdzwq81nJNLK1uxHKFkQqSOhKkjPQkc1+1FaeD/2WLxPDHiL4iaN4l8Wuyl9C0MPJJaRkZ33EjhViJGMJyxyDgDmv2W/2cPir+0roI8bXfhyfwN4Wmj3WGoauhaXUM9GhgGCY8f8tGKqeNu7nCf8E4NYLqJPixZBc8kaOxIHsPNr/h23/wBVn/8ALc/+6a/4dwaJ/wBFXvf/AAUL/wDHaX/gnN4L2gN8SNaLY5Is4hz+dftE/Cm1+DHxS1DwLYajPf2kFvBNBcToFdw8alshePvbh9APrVfsl/syfDj47+B9Y1nxRrfiGz1PTNUNuqafPCkflGJGRiHicltxkzyBgD3NePv+CenjjShJd/DvxXYa7CMlbS9T7JP7KGyyMfclBXjX4YfEL4dXJtPG3g/VNIO4qslxbnypCP7kgyj/APASaqqqqqqrwF+0N8ZPhqI4fC3jzUEs4hhbG6f7Tbgegjk3Bf8AgOD714D/AOCil2jR2vxM8CRyrkBrzRZShA/64ykgn/toPp6eBf2mfgj8QxFFofj2wt7yXgWWot9km3f3QJMBz/uFq+O3wp0/4z/DTUvB05jS8ZftGmXDdIbpAfLbP905Kn/ZY1qulahoep3ei6vaSWt9YzPBcQSDDRSIcMpHqCCK/ZF+O5+D3j0aTrt2V8L+InSG+3H5bWUcRzj0Azhv9k552ilZXUMjBgRkEd6qqqqr9qTxL/wlf7QPjfU1m8xIdTayQjpi3UQce37v8evev2S/DJ8VftDeC7Ixho7K9N/ISOF+zo0qn/vtFH1Iqqqqqqqv24fCh8S/AHU76KLzJvD95b6hGAOcBvKc/QJKxP0qv2EPFg8QfAqHRJJt03hzUZ7Tax5EbkTKfpmVgP8Ad9qqqqqqqqqqqqqqqqqqqqqqqqqqqqq+PHjY/EX4w+LfFyzeZDealIlq3rBH+7i/8holV8DNDHw0/Yd8Y+M5lEV74msr6dHbAKqym2hH0yC49d/4VWgaTLr2u6boVucS6jdxWycfxO4UfzqCCG2gjtrdAkcSBEUfwgDAH5VVVVVVVVVVVVVVVX/BQpEk/wCClojkVWVr/wAPBlYZBHlW+eKt/h74BtL86paeB/D8N6WLG5j0yFZMnqdwXOar/gqJ8YR8Jf2RfE1lZXfk6r43kTw/ZhXw2ycFrk464+zpMuegLr64P7Hn/BLVP2mvglY/GHxN8UL3wqurX1zHp1pDpC3Qmt4m8vzSxlQgmVZVxjogOea/4cc+FP8Ao4rVf/Cbj/8Aj9fDzTdT/wCCe/8AwUT0nwrrGtvc6RpOrw6feahJGIUu9Lvo1HnMuSAEWVZCATh4sZ4r9vX4Q33xu/ZQ8feC9FtGudYgsV1LTIkTc8k9q6zCNB/ekVHjH/XSv+CL37QmiaPceKP2b/EepRWtxrF2Na8PCVgouJhGsd1CCf49kULqo6hZD25qv+Cb/wDyke8Qf9x//wBGmqr/AILJf8nc2n/Yn2P/AKOuas/+Caf7D9lbJaxfAXT3WMHDTarfyuec8s05Jr4rf8EiP2UPHOmXC+A9O1j4f6s6kwXWn30t5Ar+rwXDvuX/AGUeP6ivDHj39pb/AIJS/HlPAvi24n1zwTfP9ol02OdzYaxaM203Nru4huBjngEMAr7lIJ8E+MvDnxF8H6L478I6il9ouv2MV7YXCjHmRSKGUkdVODgqeQcg8ivi54L8M/Eb/gqrqXgPxlpv9oaHr3xEtrPULTznh86GQxq6742V1yDjKkEetX3/AATS/Ygv7SS0k+A9hGsgxvg1W+idfcMs4I/zniv25v2R9V/YI+IfhL4v/AbxnrVpompXj/2bcyTj7XpF7GN3lGQACSN0J25ByFdXyOW/Zi+MH/C/vgD4I+LskEMNz4h0xXvYoTlEuo2aK4C/7IljkAB5AwDVVVVX/Bcb/kVPhH/2ENV/9F21fsD67okX7Jvwg8Oy6vZpqs/hhZ4rJp1E0kQkcF1TO4qDwSBiqr/gtp/yX7wL/wBieP8A0rnr4f8A/BTb9iPQPh34b0jU/jQ8d/p2i2sFxbDw5qbMsscKqyBhbbSdwIyDj3xzX7cP/BT/AMOfGzwDf/AX9nHQtbuovFAFpqWs3VsYpJ4iwzb20AJdvMxtZnCnBKhTncP+CXX7KXif9m34Oap4g+IVk9h4t8f3EF1d6dIpV7C1hVxbwyDtLmWV2H8PmBTgqaqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqtIUftd/8Fe574N9q8O/D/VWlBHzKkGkAIhHZkkvQp9CJT9Kqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/wCCrf7UI+CvwPb4WeF9RWPxb8R4pbTMb/PZ6aMC4l9QZAfJUnrukIOUr/gmJ+y4v7PfwDt/FHiTTfJ8afEFItR1MSL89ra4Jtbb2wjGRhwQ8rKfuDFVVVVVX7QXwstvjF8KNb8GmJGvzEbnS3bjy7uMEx89g3KE/wB12r/gn58VZptL1b4IeIXaG+0SR7vTY5RtYRF8TxYPQpId2Ov7xuy1VVVVVVVVX7cH/BTGT4O+NbH4Mfs7WNj4q8a2+pQprUzwtcwQMHH+gRohzJO5+R9p/d52j94f3fgbW9c8TeDNE8QeJvC1x4a1bUbCGe90e4mSaSwmZAXhZ04YqSRkY6dB0FVXxY+Dfwz+OfhZPBXxX8KW/iHRY7tLtbSeWSNRMgYK+Y2VsgOw645r4W/CT4dfBPwlH4F+FvhiDQdCinknSyhlkkUSOcu2ZGZuSB3r4wfsp/s+fH3W7LxJ8XvhrZeJNS060+yWtxPczxGOHcz7AI5FGNzMeRnmtM0yx0bTLTR9LtxBZ2MCQW8QJIjjRQqqCcngADmq+L37Jv7PPx78Q2viv4u/DW08SarZWa2dvcXF5cx+XCrs4QLHIq/edjnGeevAqysrTTrODT7C3jgtrWJYoYkXCxoowqgdgAAKr4j/AA28DfF3wbf/AA++I/h231zw/qnl/a7GdmVZDHIsiHKEMCHRTkEdK+EHwL+DX7P1heeGvhF4TtPDVtrE32q4s4ruWXzpEUKZAsrsRgbQSMds9qr4k/sIfsjfFm9m1Txj8DdA+33BLS3emebpksjnq7m1ePe2ectnPfNaT/wSx/Ye0q4+0v8ACGe/YEFFu/EF+yqR/srMAfo2a+HPwk+GHwg0d9A+F3gHQvC9jKwaaLS7FIDOwGA0jKN0jAHG5iT714y8ZeFPh74X1Hxp431+z0XQ9Igae9vruTZHCg7k9yeAFGSSQACSBX7QH7VPx3/4KOfEr/hnD9mDRtQsPAry/wCkuztCb+FWwbu/kUfubUZUiLnJ25DOURf2Rv2KvhN+xr4Nm1RZrPU/Fs1mW17xXeosW2NRudItx/cW64yRnLbQzk4AX9rr/gqB4g8W6+3wE/YrtrzWdY1Kb7E/iawt2nlnkPBj0+MAlj1HnEepQdJK/Y0/4JYWHhm+t/jF+1f5Xijxdcy/a4fDtxJ9qtrWVjuMl25JF1Nk8rzGDnJkyCqIkaLHGqqqjCqBgAD0FVVV/wAFG9CNp8UvDniBFwmo6F5B68vDM5J/75lQfhVf8E2NdEer+OPDDvn7RbWl3Gp7eW0iOfx8xPyqru0tb62ks722iuIJlKyRSoHVweoIPBFePv2OfgR478y4TwufD97Jz9p0V/s3P/XLBi/8cB968d/8E9PHmlCS6+H/AIr07XolBK214hs5z6KDlkY+5KCvG3wr+I3w5nMHjfwZqmkgNtWaeAmFz6LKuUb8CaqqqqqqrwJ8dvi58NQkXg7x5qdnbRjC2ckgntwPaKQMg+oANfE74j6j8VfEp8Ya5pGm2erXEKpeyWEbRpdOowJWQk4cqACRgHAOAc5r9iX9oxfE+lQ/B/xpqIOsabFjRrmVubu3Uf6kk9XjA49U/wB0k1VVVajfW+mafc6ldMRDaQvLIQOQqgk/oK1fU7nWtWvdYvX3XF/cSTyse7uxZj+ZNf8ABObw4t/8UPEPiaSMMmk6L5KE/wAMk0q4I/4DE4/Gqqqqqqq8a+G7bxl4P1vwldFRFrOnT2bFh93zIyufwzn8KvLS5sLuawvIWiuLaRo5Y2GCjqcEH3BGK/4J3+Ml07x14j8DTy4j1nT0u4Ax/wCWsDYIHuUlY/RPpVVVVVVVVVVVVVVVVVVVVVVX/BabUtR0z4C+B5dN1C5tJG8XhWaCVoyR9kn4JB6V+ynNLcfsufB2eeV5ZZPAOhs7uxLMxsISSSepNVVVVfHjxn/wr74OeLvFqTGKey0uVbZwcbZ5P3UR/wC/jpVafYXWq39tpdjEZbm7mSGFB1Z2ICj8SRX7VtjafDf9klfA2nMqQxR6dpUZXjcImRifqRCSfXJz1qv2bdFGv/HnwNp5UME1iG5II6iE+af/AEXVVVVVVVVVVVVVVVV/wUH/AOUmCf8AYQ8O/wDoq3qq/wCCvnxI1b4qftI+EP2ePCTPev4btoYzaRNkyapqDIVQgdT5X2bGf+ejevPwh+HOl/CH4WeE/hfo2w2vhjSLewWRV2+c0cYV5CPV2DOfdjVf8FsvgyYrzwL8f9NthtnR/D2quqY+Zd89qxx1JBuQSeyqM9MfsR/GVvjx+y/4D8fXt4LjVv7OFhq7Fssby2Pkyu3oX2CTHpIK/be/4JjePLDx1efH/wDZLikeeW8/tK78O2U/2e7srsPva4sWyAV3fP5QIZW+5uBCr4A/4K3ftJfBXy/Av7R3weHiLULBRG016smhakQDjMqmJkcjGOI0zjkk5Nav/wAFxbBrJk8O/s43L3znbGbzxMBGvHUhLclucfLkfUV/wS5+G3xg1P8Aa5n+Nmv/AAw8Q6V4c1Kx1OaTVLjTZYbQS3B3BI5HAD8nAwT71Vf8Fkv+TubT/sT7H/0dc1Vf8Fg/hnpfi/8AZRfx9JZodS8B6za3MFzj50guZFtpYwf7rNJAxHrGvpX/AAR38dXviv8AZKk8OX9y0jeEPE15p9srEkrbukVyv4b55QB2x9K8Tf8AKYOH/sqFl/6HFU00NvC888qRxxKWd3baFAGSST0GK/4KyftJ+CvjnrPgz9nT4M38Xi/UNM1drm+uNKb7RG166+RDawsmRK/zvu2kgEqvJ3Bf2Rvg/f8AwE/Zt8A/CfVin9paJpm7UFR96pdTyPPOqt3CyyuoPcAGqqqqv2hvgx8APit4Yi1n9oLwlY6zpHhKOe6hku7iaIWwZV8wjynUsWCKADnJwBya8T+JrS98QRXXg6wk8PaNo6rbeHtPguHP9mWkZJijRyxbIyWJz95mPev2b/2wviS2rw+E/iA8OvaHaWstxd6vcv5dxYW8SFmkd+koGAMMN7MwAYkgH4N/tIfDD432/leFtWa11aNS0ukXwEVygHVlAJDr7qTjvivjR+yd+z1+0PqdprPxj+G1r4iv7C0NpbXL3lzbSRQ7i2wNDIhxuYn15NQ/8EvP2FIJUmj+BKFo2DKH8R6q4yPVTdEEexGK+GP7Mv7PvwZuEvvhh8IPC+gX0abFv7ewVroLjBHnvukwR1G7nvVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV8e/iTD8Hvgp43+J8siK/hrQrq8tw4yHnWM+SmP9qQov/Aq/wCCKPw1mm0n4k/HbWEknvNUvYtFtLmXJY7B9oujk/e3NJb5Pqh96qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqtZ1jSvDuj33iDXNQgsdN0y2kury6nfZHBDGpZ3YnoqqCSewFfA/RdU/4KUft+6r8WfF1nNN8PfB8yXa208eY1sYJCLGyYdMyvukkXoR5/qKqqqqqqqv2ofDurfs6ftF6H8dfB9sVsNZujdyRp8qm4Hy3UJI6CVGLZ7l3x92vDXiLSfF/h3TfFGg3QuNP1W1jubaT1R1BGR2POCOxyKqqqqqqqv8Agpl+2t4y+AWl6f8ABf4TaLqkXjLxpaEx659kfy7SBm8spanGJbknj5c+WGU/eZcf8E7v+Cd1r8DrW0+Nvxt0+K++JN9H5tjYzYlTQEccknkNdMCdz/wZKqfvMaqqqqqqr4v/ABN0/wCD/gDUfH+qaPf6nbacUV4LMLuy7hFJLEBV3MoJ5xnoa+JH7fXxe8XebY+DILLwjYPkBrcfaLog9jK4wOMcqikHvXwb+M+v+D/jdoHxJ8T+IL/UCLoQ6pc3lw8zvbS5STJYknaGLgeqio5I5o1likV0dQVZTkEHoQfSqr4u/F74e/AvwHqPxI+JviGHSNE01QGkcFnmkP3IokHMkjEcKPc8AEjxR4y/aQ/4K2fGUeDPBltP4X+Fvh65WWSORibbToiSBc3RBAnu2XcEjBwOQuFEkh8E+Bv2av8Agnb8CZ55tQtNB0azVX1XWb077zWbvbgcD5pJGwdsSDCjOABuNfFT9oT9p3/gqB8Q5fgr8AtBvfD/AMOYJVa8ikl8qNoQ3y3GpTrkY4ytuu4ZHAkZQw/ZD/YZ+E37JOhJPosC6941u4BHqfiW7hCyvnG6OBMnyIsj7oJJ43M2Biqqqr/gpH4fNz4J8H+KVjB/s/VJ7Mtnp58Qfp/27/5zVfsFeIV0T9oOy05pNo13S7qywR1IUTj6f6j/ADmqqquLe3u4HtbqCOaGRSrxyIGVh3BB4Irx9+x/8CPHoknbwmNBvZB/x9aK32Ug+vl4MR+uzPvXj7/gnp440oSXfw78V2GuwjJW0vU+yT+yhssjH3JQV44+EnxL+G8pj8b+CtU0pAdonlg3QMemFlXKN+DGqqqqqqq07Ub/AEjULbVdKvZrS9s5Vlt54XKPE6nKspHIIIBzXgP/AIKG+L9NENl8Q/B9jrEKgK13p8htpv8AeKNuRj7DYK8C/tffAbx35UEXjFNEvZB/x660n2UqfTzCTET7B81b3FvdwJc2k8c0Mi7kkjYMrD1BHBFVVftDa2fD3wM8daosmx10K6ijbONryoY1P5uKr/gm7oS2/gTxf4lC83+rQ2mfXyIt/wBf+Xj9aqqqqqqqv2svBh8E/HrxRZxxbLbU7j+0rc4wCs43vj2EhkX8K+AnjYfDz4w+FPFcs3l21tqCRXbZxiCXMcp/BHY/UCqqqqqqqqqqqqqqqqqqqqqqv+C3F95fwW+HumeVnz/FEs2/d93ZauMY9/M/T3r9mKwOl/s1/CbTGlEhtPA2jQlwMbttjCM47dKqqqq/4KJ+LzpHwq0XwhBLtl8QarvkGfvQ267mH/fx4T+FV+yF4NXxr+0F4UtZofMttLnbUp+M7RApdCfbzBGPxr/gonqhh+GPhvRg2Dd675xHqI4JB6/9NR2qv2GdKGo/tC6Vd7c/2ZYXdx06ZiMXr/019/61VVVVVVVVVVVVVVVVVVVVVaro2j67amx1vSrPULc5zDdQLKh4wflYEdK0jwN4J8PTm60DwfoemzEjMlnp8ULcZxyqg9z+Zqqr/gsl/wAnc2n/AGJ9j/6Ouaqv+Cs/iey0D9ifxRpN1Kiy+JdU0zT7YN1aRbpLkge+y2c/QGv+CL/hq70n9l7XtfukZV13xhcyW2ejRRW9vHuH/bRZR/wEV+0j8P734r/8FJ/FXw003XP7GuvE3jiKwh1Dyy/2VpPLUSbQQTjOcAj614r/AOCMfxpk0G6TRf2kNM1u725jsNRsrm2hmI5AZxJLjkDHyHn0r/gnz8QvCP7KH7T1x8EP2gPg9o+l+L7vU/7LtvFNwrSXmk3UgCxx5ZmjW3lDACWIKcSgszIflqqqqr9tL9o1fH+sP8LvBl/v8O6RPm/uIm+W/uVPQEdY4z07M3PIVTVeKLpvA/hp/h/asU1fUik3iJgCGiC4aGyP+4cSSD/nptUjMVWN/faXeQajpl7PaXds4khngkMckTjoysOQR6iv2bZfifd/CDRNQ+LepC81u9Qzxl4RHLHbMB5Sy4+9Jt5JwD8wByQSaqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/4LIfE3/hEf2YbD4fWlyFu/HevQwSxE4LWlsPPkYfSVbYf8Cr/gn78MF+E37IXw48PzWphvtS0saxfbl2uZbxjOA4/vKkkceOwQA1VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVf8ABYL9pA/D/wCEun/AHwzflde+IB83UxG3zwaXE/IOOR50qhB2Kxyg9a/4J8/s3r+zX+zjomg6vYiDxV4jA1fxCWXDx3EqjZAe48qMIhHTeHI+9VVVVVVVV+0B8J7P4zfC3V/Bkkcf28x/aNKlfjybuMExnPYNkoT/AHXav2Bvixd2y6t8AvFsksGoaTLLcaXFOcMgDYuLfB6FXy4X/akP8NVVVVVVVWu+D/Cfii90jUfEfhrS9Uu9AvPtmlTXlokz2NwFKiWIsCUfDEbhg1VVVVVVVV4z8K6b448Jax4O1dc2es2UtrMQOVDqRuHuMgj3ArxV4b1Twd4l1TwnrUPlX+kXktrcLjjejFSR6g4yD3BBqv2Mvij/AMLK+Cmm2t9c+Zq3hgjTLzc2WZUUeS578x7Rk9WRqr4+/tAfDT9m34eXvxF+Jmtpa2tupW0s42BudRnx8sMCEje549lGWYhQSPDvhn9oz/grh8a/+Ep8UTSeF/hd4bufJYwktbaXEcEwW4bie8dcF5CMDKk7V2Ifih8ZP2Z/+CavwS07wfo+nRQSxwOdD8M2cga91ac8NPM55Clh8878DGFDEKleAPg/+0//AMFV/iZ/wtL4r6zceGvhnp1w0dvLGhW3t4wRut9Pib/WSEDDzNkA/eJIVK+DnwV+GvwD8D2fw9+Fvhm30fSbQZfaN0t1JjDSzSH5pJDjlj7AYAAFVVVVftt+HP8AhIP2dPEMsabptImtr6MYz92VVc+2Edz+FV8CfEn/AAiHxl8F+IWmMcVtrVsJ2HaJ3CSf+OM1VVVVVPBDcwvb3ESSxSKVdHXcGB6gg9RXj79kX4E+P/MnuPCCaLfSZ/0vRm+yMCep2AGMnvkoTXj3/gnh4q09ZLv4c+M7LVowMiz1KM20v0V13Ix+uwV45+DnxQ+G0jJ418Eappsakj7S0PmQMR6TJlD+BqqqqqqqvBvxQ+Inw8nWbwV4z1XSAG3GK3uWETn/AGojlG/EGvAv/BQj4h6R5Nr498M6Z4hgXh7i2Js7g+5wGjP0Cr9RXgX9tD4D+NRFb3XiOXw5eyDmDWYvJUHv+9BaMD6sPpVhqFhqtpHf6XfW95bTLmOaCUSI49QwJBFftxaq2m/s3+IoI2IbULi0twR2H2hHP5hCPxqv2ENIGnfs66VeBcf2pqF5c/XEpi9f+mXt/U1VVVVVVX/BRbwOZLTwr8R7aHPkvJpd2+OzZlh/AETf99Cq/Z58d/8ACyPg14W8USzeZdvZLb3hPUzwny5CfTcU3fRhVVVVVVVVVVVVVVVVVVVVVV44+GPw1+J1pbWHxJ+HvhnxZbWUhktodc0mC/SByMFkWVWCkjjIrTtO0/SNPtdJ0mxt7KxsoUgtrW3iEUcEaAKqIi4CqFAAAGAABVVVVX/BQ/xUdX+L+l+F4pt0OgaOm9M/dmmYu35oIqr/AIJueEvO1vxh46lj/wCPW1g06BiOpkYySYPt5UX/AH1X/BSDUCZfAWkoxwq38zjA5z5Cr79m/Oq/4J26aJvin4i1Urn7LoBhByODJPEenXOIz+vrVVVVVf8ABYvxt4z8DfA3wVqHgnxfrXh+6n8WCKWfS7+W0eRPsk52s0bAlcgHB4yBX/BNfxDr/iv9ij4ca/4o13UNY1S6/tTz73ULl7iaXbql2q7nclmwqqoyeAAOgqq+Et5+1h+1/wDHu++EGk/tK+LrK4mN9MJNV8S3xt4oIidyLHGxxkYG0AD1of8ABKX9ray/0rS/2wCt3EMwkXuoxYbt8wclfqAa8az/APBVT9hSJfFeu+PNQ8ceD7Mj7ReTXj+ILBEB4WXzwLi3ToNw8scgBu1fsQft7+Cf2vNHn0K/sYPDfxA0mHzb7RRMXjuYhgG4tmPLJuIDIcshIySCGPxz/YH/AG9/H/xi8ZeNvAP7Smm6P4c1vWrm70vT5PGOr27Wtu7kpGY4rdkQgEDapIHY1+0v+zX+3Z+yr8OY/id8Qv2mrjUdLl1KLT1h0bxnq0s/myK7Kdssca7cRtk7s8jj0/Zy/Ze/bx/af+GcHxV8AftOS2Gkz3k1osOseNNWinDxEBiViikXBzx834Cv2ef2EP27vht8bPB/jv4k/tHabrvhjRdRWfU9Nj8YatdNcxBSCoilgVH5I4YgVVVVV468c+GPht4Xu/GPjC/ez0qx2CaZIHmKl2CKNqAnlmUZxjnkirr9uj9nqAgQ67qtzkdYtLkGPb5sVq37Xvwl0f4eaN8S7qPXG0nXLya0tkjtEaZZIs7t678KOMjnuKs/29f2ebooJtW1q03Lk+dpbnZ7HYW/TNfDf9pH4OfFvWf+Eb8C+K3vtVFu9w1rJp9xCVjUgFtzoE/iXgNnn61VVVVX/BZL/k7m0/7E+x/9HXNV4o8VeGPBOg3fijxj4h07RNIsIzJdX2oXK28MK+rOxAFftv8A7SGv/wDBQD48eF/gb8ANNudU8OaZeNb6MDG0bapeOMS3jgjMcKIpClhlUDu2N21f2ffg9pHwB+C/hL4QaLMs8PhvTlhmuFTYLm4Yl55tvbfK8j45xuxk14m/5TBw/wDZULL/ANDiqv8AgtP8Om8M/GXwH8XdJhe2bxJpElnPcQkqftVlIpVyRyreXPGAfSPjoa+BHxBT4sfBTwL8ShKjyeJPD9ne3G3ok7wqZU+qyb1PuKqqqv2x/wBqaLRLa8+Efw31QNqcytDreowP/wAei94I2H/LQjIc/wAI4+8TtrRI4fBmjxeO9Sjje/nLL4ftJF3bpFOGu3U9Y4yCFB4eQD7yo6meaa5me4uJnlllYtJI7FmZjySSeSSe9fsXfs9N8VPGK+OPE9lu8LeHJlbZImVv7oYKxc8FF4Z/+Arj5iRVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVf8FPbu5+PX7cHws/Zv0ud5IbKOysp1U8wXGoXCmZvYC3W3Yn0HtVpaW1haQWFlAkNvbRrHFEgwEVRgAD0AAFVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVrGraZoGkXuva1exWen6bbSXN3cSnCQxRqWd2PYBQSfpX7M+iat/wUO/4KA638ePF1hK/grwhdJqSWs65SOGJimm2ZHIyxTzXHRvLl/vVVVVVVVVVX7Y3gLWvgz8WtE/aM+H8X2dLy8R7wonyRXyjncB/DNGDkdyHyfmr4b+PdE+KHgfR/HXh+QG01W2EnllstDIOHjb/aRgyn3HpVVVVVV+3v+3poX7JPh238OeE4dO1z4k6zGJLHTbos0Njb5INzcKhDbSQVRAyljk5wpz8A/iRr3xf+DnhP4leJvBN74S1LxBpyXM+k3bBmhJ6MpHOxwA67gG2suQDkVVVVVVVVVVf8FCfhZ/YHjnTfijplrtsvEkX2e+ZV4W7iXAJ9N8QXA9Y2Peq/Yp+LC/DT4x2mk6ldCLRvFirp92WOFSYn9xIfo52ZPAEjHtX7Sf7S/wAMv2Wvh5L4++I+osTIxh0zS7YqbrU58f6uJSRwBgs5wqjqckA/DL4SfH3/AIKtfGyT4v8AxYurnQPhhpFybdWgYiK2hBDfYbFW+/Kw2+ZMRgE7mydkdftT/to/BX9gnwDZ/A74G+HtHuvF9jaCKw0G1B+y6OhHE12ynLSNy2zPmOTucgMGb9lD9gL4m/tWeLx+05+2hqer3GmavILq10m8dobvWQPumQDBtrXHCxqFLLjbsXaW0fRtI8O6TaaDoGl2mm6bp8KwWlnaQrDFBGowqIigBVAAAAGBVVVVVV8R/Df/AAmPw98S+E1UFtY0i6tE4zhpImVSPcEg0ysjFGUhlOCCMEGkd43WSNyrKcqwOCCO4r4f+JE8ZeBPDvixHDf2xpdvdkgYwZI1YjHbBJGKqqqqqqdEkQxuoZWGCrDIIrx9+yn8DPiGJJtR8FW+mXsg/wCPzSP9DkB7kqvyMfdlNeO/+CdmvWokuvhx45tb9QSUs9WhMEgHoJU3Kx+qoK8d/A/4s/DVnbxl4F1Oyt4+t2kXnW//AH9j3J+BOaqqqqqqrwn488a+BLwX3g3xVqmjTbssbO6aMPj+8oOGHsQRXxM/ac+KPxT+H8XgHxxdadqEEV9HdLei0ENwWRXUA7CEx8/93PHWq/ZLjsoP2d/BVrZ3MM3l2BaURyK2x3kdyp29D8/Q8+tVVVVVVVX7RPgJ/iV8GfE/ha1gEt61mbixUDkzwkSIo9CxXZ/wI1X/AATt+IAKeJfhheXHKldUsEJ7cRzgf+QTj3Y1VVVVVVVVVVVVVVVVVVVVVVVVVVX7RfigeMvjn4215ZfMjfV5YIXzndHB+5Qj2KRrVfsIeFV8O/s+6fqTR7ZvEOoXN8+euAwhX8NsII/3vev+Cit4JPiR4X04MmYNDMuAfm+edxyPT93x+NV/wTfslkv/AB9qJI3Qw6fEuV5+czk8/wDABx7j0qqqqqv+C2n/ACQHwL/2OA/9I56/4Jcf8mJfDH/uLf8Ap2vKqv8Agln/AMn6T/8AYP1j+dVdWtrfWstle28Vxb3EbRywyoHSRGGGVlPBBBIINftO+Dbj/gnx+3hpviz4YxSWuiRT2/iLRrJHIH2KZ3juLPJz8hKXEQzzsK555rTtQstW0+11XTblLi0vIUmt5U6SRsAysPYgg1/wWS/5NFtP+xwsf/RNzX/BIb/kzTTP+xg1H/0Nar9qb/grH8OPgv4qvPht8JPCx+IXiWwm+z3t0t55FhazA4MSuqs08gIwQoVQTjeSCo0P/grV8fvBlza6t8fv2Ub3TfDFzIqtf2Npeae6A4+ZPtIZJTjou5M5HzCviL/wVP0bS/jF8Pfh/wDCbwXpfjDw98QbbTZbTW5dWe2aBrq6e3aN4hExVo2QhlJyCCMcVVV+2nPFD+zZ4sjdsNM9kiDHU/bIW/kpqviBcS237M3ww09wwW91rWrhAMYYRmBMnuDliPz9qr/gnVYC4+NGs3zoGW08NzBTnBV2uIAOO/yhqqqqqr/goV/wTy+O37T/AMcrX4n/AA11TwimlpoNvp7walqE0E6yRSSsW2rCy7SJFwQ2eDkDjNl+zh/wWLQLpg+PXlwDcv2ifxQJMDnkt5bSfTuOOlQf8Epf2qvjJqdtf/tOftSJe28beYI0vbzXJY/VUFx5SRnBIyu4D0PSv2Zv2M/gd+ynpUkPw40GS41u8iEd9r+psJr25XgldwAEceQDsQKDgE5IzVaz/wAE9/jzf/t/R/tMwXfhX/hEE8a2+smNtQkF19njZCw8vysbvlPG7HvVf8FC/wBlTxB+1j8FLLwn4In0y38UaHrMV/p0uoTNFEyFWjmjZ1ViAVcN06xrX7B/we+LfwC/Z70/4RfF+TR5r7QNQuhpsumXjXKNZyv5wDFkQhhLJMMYI27eewqqr9q39rm18EwXnw3+GGpJN4kbdDqGoxHK6b6oh6GbtkcJ/vcLLJJPI000jSSOxZmY5LE9ST614S8Maa2mz+OPGJki8NadL5XlxyBJtTuduVtoc/VS74xGhzyxRW8T+JNR8WazNrOpeUjyBUighTZFbRKNscUa/wAKIoCgeg7nJr4NfCXxF8afHdj4K8PJ5YlPmXt2yEpZ26kb5G/PAHGWIHGc14E8D+Hfhx4S03wV4VsxbabpcIjiU8s56s7HuzMSxPck1VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVfsrb/2jf8AgrN44+Ksx+1af4VutVv7eRhlXhgA06057HY8TgdtntVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV/wV6/aH/4Vl8Cbb4OaDfGPXviRI8VwY3w0OmQlTOTjkeYxjiweGUy+lf8ABOb9nYfs8fszaFYatZmLxN4tC65rm9NrxSTIvlQHPI8qIIpXoHMhH3qqqqqqqqqr4leAND+KPgfVvAviCMG01W3MYkC7mgkHKSr/ALSsAw+nvX7IXxB1r4H/ABb1n9nP4iSG2hvr5o7Qu2EhvQBtK5/gnTbg9z5fHzE1VVVVX7a37XPhn9kT4Vt4mu4Y9R8Va35tt4Z0pydtzcKoLSyYIIhi3IXwQTuVQQWBH7AP7Hvi39pT4gT/ALZ37Uv2nWLS+vze6LY6gvOsXKsNtzIh4FrHt2xxYCttAwI0Aeqqqqqqqqqr4+fC63+MXwq1vwSUj+2zQ+dpsj8eVdR/NGc9gT8hP912q6trizuZbO7haKeB2jkjcYZGU4II7EEYpWZGDIxDKcgg4INeAf2YvjZ/wUG/aIuPGvxv8Z3k3w/8MRW8Mt6rqjvEFBFlbRjhHYhmkkxxu3HLMor9sT9vrwj+z1oUH7Kf7G+mWL+IrCIaUbrS4fOt9EJyn2e3Vc+debiMk7grE7tzlgv7DH/BNCXSdRt/2gf2sLV9c8YX8xv7Hw/qLG4FrK53/aL0sT5twSSfLbIQ8tufhKqqqqqqr47+Ff8AhCvjL4y8NJHsitdYnaBcYxDI3mR/+OOtV+w34rHib9nrR7N5d8+gXdxp0pJ54fzEH4RyoPwqqqqqqqqogMNpAII6GvH37MXwR+I3mT634Gs7W9k5N7po+yTbv7xKYDn/AHw1ePf+Cdd5EJLr4ZeO0nABKWWtRbG47edEMEn3jUe/p47/AGe/jJ8N1km8U+AtSjtIs7r22QXMAHqZIywUf72DVVVVVVd/cUA96rwz4q8T+EblNS8KeItS0e72gGaxungYj0JUjI9jXgT9vD4zeF1jtfEw03xVaoQCbyLyJ9o7CSPA/FlY14F/b0+DfifybXxTDqXha7fhjcxfaLcH0EkeWx7siivDXjDwn4zsRqXhLxLpms22BmWxukmCn0O0nB9jzVVVVVVX7S3w7Pwx+M/iLw9Bb+VYXFwb3TwBhfIm+dVX2Ulk+qGvgX8RH+FnxW8O+MzIy2trdCO+A/itpBsl474ViwHqoqOSKeJJoZFkR1DIynIYHoQfSqqqqqqqqqqqqqqqqqqqqq/4K7/F74o/CH4ceANS+F3j/XPC11qGt3EN1NpV49u0yLACFYqRkA84r4H6pqOu/BX4f61rF9NeX+oeFtNuLq5mcs80r2sbO7E8lixJJ9TVVVV4x1+Lwp4Q1zxTMAU0bTbi8YHuIo2c/wDoNTzS3Mz3E8heWVizs3VieSar4V+F18FfDTwt4T8ny30rSLa3lUjB8xY13k+5bcT7mv8AgoNc/aPjfp8Ozb9m8N28ec53ZnnbPt97H4VX/BOO2ZfDHjW83giW/tYwuOQVjc5/8fH5VVVVVX/BbT/kgPgX/scB/wCkc9f8EuP+TEvhj/3Fv/TteVVf8Es/+T9J/wDsH6x/Oqr/AILdS2TfGP4dQRhPtaeGZmlITDeWblgmT3GRJx259a+BcV7B8Efh7BqTObuPwrpizl33t5gtYw2Tk5Oc81/wWS/5NFtP+xwsf/RNzX/BIb/kzTTP+xg1H/0Na/4KKfGrV/gX+yh4t8S+Gr2W013WDFo2mXMTbWgkuTteRT1DLCsrKRyGCnjrX/BHD9mbwnN4R1P9pfxVo0F/rcmpSab4ca4j3ixiiVfOnjB4Eju5Td1URsARvate0HRPFOi3vhzxJpNpqel6lA9veWV3CJYp4mGGR1OQQR2NfEX4O6L8A/8AgozY/Cjw27HSNH+IGjS6ejMWMNvcS21zFEWOSxRJlTJ5O3J61VV+3dfiz/Z/u7bd/wAf2qWkOMZzhi/4f6uq+Krm2+Efwi0psh/7N1O8ZT282/kUEdsFYVOOvX2xX/BNjTjL4j8c6sFyLaxs4M4HHmPI31/5ZVVVVVX7SHxXPwd+E+q+KLK4jTV5wLPSQ6hs3MmcNtPB2KGfBGDswetfDL/goZIoi074teFA4xg6no4wfq8Dn8SVYey14q/bc+BvhifQjHqWoatY63byTfa7C2z9lCvtHmxuVcZIfoCfl6HNeAvjZ8KPicif8IR470vUZ3XItPN8q4A94Xw4HvtxVVVVVEgDJIAA71+1L+2THpwvPhz8HtUD3gJi1HXYGyIeoaKA927GQcD+Hn5ld2kYu7lmY5JY5JPrXgTwDa6zp95428Y3k+leDdGkVL29jTMt1MRlbW2U8PM49flRfmbgAHx744ufG2pQmCxj0vRdNjMGkaTA5MVjBnO0E8s7H5nkPzOxJPYDwt4W17xr4hsPCvhjTZb/AFPUphFbwRrksx7n0UDJJPAAJOAK/Z3+A+h/AfwPHolt5N1rl8Fl1jUFTBnlxwik8+WmSFH1bALGqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr44ePE+Fvwa8cfEZpQj+G/D97fw5/iljhZo1HuXCge5r/giJ4DaPwz8T/irdxtI+oajaaPbzMDwYY2mmGe5P2iAn/dHrVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVMVVSzMAAOSeMV4Yjb/got/wUvn8QzA33w88FziaPIJifS7GTEKZ9Lm4beQTnbM+Pu1VVVVVVVVVV+3h8D59e0KD41eErd01jw4gXU/IyHktVOVmGOd0R5J67SSThBX7LHxxg+N/w1t7y/uYz4k0UJa6xEMAs+PknA7LIAT6Bg4HSqqqqvir8H/hp8b/Ckngj4qeELHxFozzJMLe6UgxyIcq6OpDI3UZUjIJByCQbGxsdMsbfTNMs4LS0tIlht7eCMRxwxqAFRVHCqAAABwAKqqqqqqqqqqr9u74Rx+AviinjbSYFTSvGQe4ZVGBHeJjzhj/a3LJnuXb0qpPiL8a/CXgPxR4T+Cfia70nUvFkENrMLRxHNIA/SKTI8qQqzqHBBAYgEEgj9gj/AIJx+Hf2c7Oz+KPxZtbTW/ifcR740bbNb6BuzlYTyHnwcNN25VOMs9VVVVVVVX/BQjwl/YnxqtPEsMREXiLSIpHcjrNCTEw/BFh/Oq/4JueLwl94x8AzSgmWGDU7ZM9NhMUp/wDH4fy96qqqqqqqqqqr4reAf2UPEHnyfEc+C9OvWyz3Q1CKwuS2M5LIys59m3Z9DXxV+FP7N+itPN8O/wBoOKeVc+Xp97p090GP/XzBHtA5HVD9TTABioYNg8Ed6qqqrv7q/WqT7i/SqrSNa1nw/fR6poGr3mm3sX+ruLSdoZF+jKQRXgX9tz45+DljtdT1e08TWceB5erQ7pAO+JUKuT7sWrwL/wAFA/hhrrR2vjjQtT8MztgNOn+m26+pJQCQf98H6+vg74j+AviDa/a/BXi/S9YQLl1tblWkj/30+8nbhgKqqqq/4KD/AA0Or+EdI+KOnW2640KX7Hfso5NtKfkY+yy8D/rsar9jb4nr8Rvgxp9je3Hmat4XI027DNlmRB+5f15jwuT1ZGqqqqqqqqqqqqqqqqqqqqqv+C3v/JKvhp/2MF1/6Tiv2ev+SA/DP/sT9J/9I4qqqqv2xfEP/COfs6eLpkcCa+hisowWxu86VEcf98Fzj2qvhL4bHjD4o+EvC7x+ZHqWs2sMy/8ATMyrvP4LuNV+3rcNP8e3iKgfZ9GtUB9fvtn/AMeqv+CdNsF+Hnim7D5MmtJHtx02wqc/+PfpVVVVVf8ABbT/AJID4F/7HAf+kc9f8EuP+TEvhj/3Fv8A07XlVX/BLP8A5P0n/wCwfrH86q8u7SwtJr6+uora2to2kmmmcIkSKMszMeAAASSeBXxt1+T/AIKJ/wDBQiw8PeBzcXfhiW8t9FsbpFIKaTasz3N1/sqxa5lXIHDoDgmoIIbaCO2to1jiiQJGijAVQMAAemK/4LJf8mi2n/Y4WP8A6Jua/wCCQ3/Jmmmf9jBqP/oa1/wWR8Oahrn7IlrqdkHMPh/xfY313tUkCNobi3GfQb7iPn1wO9f8Eg/Fuk+IP2P7Lw9ZTKbzwxr19aXke75gZH+0I2OwKzAD3U/hX7WGqafrP/BWKW7026SeKPxx4at2ZTwJIYbGKRfqroyn3FVVf8FFtXWH4eeFdALDdea09yBj/njCyn/0cKr4+KLS58B6IUCPpngbTVdMYKtMJLkgjAwf9I78/wA6r/gm5pXk+CPGOt7f+PvVYLfP/XKIt/7W9fy71VVVV+3L8XF8d/EtfBOkXQk0jwiGgYo2VlvGx5ze+3Cx+xV/WtL0y+1rU7TR9Lt2uLy+nSC3iXq8jkKoH1JArxpdWE2vS2WjzrNp2mILO0lXpKsfDSjuBI++THbfjtSsyMGRiGU5BBwQa8A/tU/HX4dLHbaP48u76yjAAstV/wBMjAHQLvyyD2Vlr9ln44+KPjz4Kv8AxN4l8L2WlNYXv2RJrSZzHcuEDOQjZKBdy/xNnJ6Y5qqrVtW0rQdMuNZ1vUbawsbOMyXFzcyCOOJR1LMeAK/aY/bJ1H4gLd+Bfhfcz6f4bbdFd6hzHPqK9CFHBjiI7feYHnAytV8P/hbZX+gzfEz4kX0+ieBtPkKGdFH2nVZxyLW0Q/ec93PyoASehFfE34n3/wARb20trfTbfRPDujxmHRtDtCfJso+5z1klYjLyN8zH2wBpel6jreo2uj6RZTXl9eyrDb28KF3ldjhVUDkkk1+yv+zFp3wO0L+3fEMdveeMtTiAup1G5bKM4PkRn6j5mH3jx0AzVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV/wVc8cf8Ib+xf4osIrrybjxTqFjpMLDq26YTSKPrFbyA+xNf8ABLLwMvgr9i3wdcyIFufEtzeavcAD/npO0cZ98wxRH8cdqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr/gpZ8e/+FE/steIP7K1D7P4i8aZ0HSdjYdPOU/aJRjkbIBJhh912j9RX/BI/4Aj4W/s6f8LN1mw8rXviVOL0F0w8enR7ltV+j5km64Kyp6VVVVVVVVVVVc29veW8lpdQpNDMhSSN13K6kYII7gjjFeJtO1z9h79oq38Q6LDcT+CNfLFYQ2RLaMw8yA5/5aQkgqT1G3J+ZhWia1pXiPR7LX9DvorzT9QgSe2njOVkjYAqw/A1VV4z8Y+GPh74U1Xxv4z1m30rQ9EtXur68uGwsMajJPqSegUZJJAAJIFfsm/tmfDD9r7RtcvvAdjq2m3/AIcuhHf6fqMGGSKQv5EokXKESKjfLncpVgRjDNVVVVVVVVVVVfG742/Dz9nv4dal8TviZrK2Gk6eNkaKA015OQdkEKZG+R8HA4AALEhVYj4Z2/7S/wDwUt+P2rfGTULmPw54J8K2dzb2xmBaxsUKbo7GMkr5krsI3ll/hADEACOMq0cih4po5UYZV43Dqw7FWBIIPYjg1X7HXxtHxe+F8On6xeCXxJ4ZCWmob2y88eP3M5/3lUqT/eRjxkVVVVVVVVV/wUV8H/2t8MNC8ZQxbptA1QwyNj7sNwmGP/fccQ/Gq/ZA8ZjwV+0D4WuJZgltq0zaZcZOMiddqf8AkXyz+FVVVVVVVrvivwt4Xi8/xL4l0rSY9u4PfXkduMeuXI44rxJ+17+z14aJim+IVvqEwziPTbeW6zj0dFKf+PV4i/4KKfD6zDJ4W8Ca7qjKODeTRWak+xUyHH4fhT/tufH7xu7QfDT4R2zqwwPIsbnUpF57FNq+3KmjN/wUN+IJISPUtGtZCeNlppvlAj1OJT+pH51/wxl+0l43/efEP4u25SXG9LnVLq/ccc5UgLx0wGxWhf8ABOTw1CEbxN8TdSuzgbksdPS2we4BdpPzx+FaL+wl+z9pQUX2ma1rO3r9t1Nkz9fJEf8Ak1o37N3wH0AKNP8AhT4dcr0N3Zi7I/GXca8W/s+fBbxtaLaa98N9Eby02Ry2tsLSVB2AeLa2B6Zx+deO/wDgnZoN2ZLr4ceObqwfBKWerQidCfQSptZR9Vc147/ZQ+OvgAyzX3gi41SziBP2zSD9rQgdTtX94o92UVLFJDI0M0bJJGxVkYYKkdQR2NVdjKKR61UeDEmP7oqqqqtbq6sbiO8sbmW3uIW3RyxOUZCO4I5BrwL+2F8ePAvlwL4u/t60jGPs2tx/as/9tMiX/wAfxXgX/gof4R1AQ2nxD8G32kynh7vTpBcw5/vFG2uo9hvP9PA/xq+FHxIWNfBnjzStQnk+7ambybj/AL8ybX/8dqqrxn4U0rx14T1bwdrUe+y1izktpcAEoGXAZc/xKcMD2IBrxf4W1bwT4o1TwjrsIjv9IupLadR0LKcbl9VIwQe4INfscfFf/hWXxes7DUbzytF8ThbC9DHCpIT+4kPphztyeAsjGqqqqqqqqqqqqqqqqqqqqq+OH7OHwX/aQ0nTND+NHgz/AISKy0e4e4sov7RurTypGXazZt5IycjjBJFeHtA0jwp4f0zwtoFp9l0zR7OKzsoPMZ/KhiQIi7mJZsKoGSSTjkmqqqr/AIKM6+bL4V+HvDscm19U1vzWHHzJDE+R/wB9SIfwqv2HvD4139ovQbh0Dx6RbXV66n2hZFP4PIh/Cq/bv/5OAu/+wVaf+gmq/wCCdn/JMPEn/YeP/oiKqqqqq/4Laf8AJAfAv/Y4D/0jnr9mb/gqp4g/Zt+B/hr4K2PwY0/XofDv2rZqEutvbtN591LcHKCFguDMV6nO3PfFf8PxfFX/AEbtpP8A4Ukn/wAYr9ir9py+/a1+Dk3xS1Hwhb+G5YtauNOFlDeG5UiJIm37yinJ8zGMdutfs5/tDyfsk/tM6h8U9Q8DzeIGtWv7OTTWvfsLgTNjfvMb8gDO3bznqOtX3/BcCxuIRbeG/wBmq9ub+VtkST+JgBkjjhLYljnHy8Z9a8ceNv8Agpb/AMFB4f8AhCtG+Gl74K8BX7bbiJLSXSrGdAetxdXH7y4A7pHlTgHyyQK/Yg/YU8G/sf8Ah251CbUk8ReO9cgWPVdZ8nZHDGDu+z2yn5liyAWJOZCoYhQFVa/4LJf8mi2n/Y4WP/om5r/gkN/yZppn/Ywaj/6GtfFD4b+Fvi/8PPEHwx8aWZudF8R2MlpdopAdQw4dCc4dGCupxwyg9q07wh+2X/wSy+LGr674Y8LS+J/BmpEQ3F6llJPpmr26sTE0vlndbTrubG4gglgN6E7rD/gqF+1J+0ZYL4H/AGZv2WZLPxJqH7j+2mv31SCzJGDJzBDFDjkhpXZAcZB6HUvg94r+DX7e/hL4V+K9fPiLxPbeMfDk2p3yszfab28+yXMoDNy+JLhl3nBbG4gZwKqv+Ci2vi78eeFPC6uCNN0mW6IB6GeXb/KAfp7VaWs99dQ2VrGZJp5FjjQdWZjgD8zX7T08Mnxy8T2Ns4MGkvBpsQAwAttbxwcDtzGfzqv2BdF/sv8AZ8tr7bj+19Xu7oHGM7SsOf8AyDiqqqqv2l/jHB8F/hje61a3CDXNRBtNHjOCfPYcyYP8MYyx4xnaD96ppZbiV555WklkYs7uxLMx6knuTXhkf8In4R1Tx/OoF1dCTSdDDDrM6f6TMP8ArlC+3PUPPGw+6aqooZbiVLe3iaSWRgqIikszHgAAdSfSvgN8OI/hR8JfDngkxqLu0tBJfED71zId8vPcBmKg+iiqqvin8XvAnwd8Ptr/AI11dYAwItbOLD3F2w/hiTIz1GScKMjJFfHj9pPxx8c9RMF9IdL8OwSbrTR4JCUHo8rceY/uQAOwGTmvCfwr8M+BPDlt8Uvj0ZrbTZx5mieGI28u911uoLd4bfpmQjJHTkru+J3xR8R/FTXI9U1oW9pZ2MX2fS9Ks08u1063H3YokHAGAMnqcc9BjRtG1XxFqtpoWhafPfahfSrDbW0CF3ldjgAAV+yn+ynp3wa06Lxh4wggvPGt5Fz0dNLRhzHGehcjhnH+6vGS1VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVf8FwPF/2XwJ8LvAMdwD/aer32pyRBun2aKOJGI/7enAPsfevgL4P/AOFf/A/4feBmhMUmg+GNPspUPXzI7dFcn3LAk+5qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv27tZ1P9sn9vzwb+y34VvJm0bwxdJpdxJCNwimkxPqNwB/0yhQKQR1gbnBzWi6LpXhzRrDw9oVjFZ6bpdrHa2dtEMJBDGoREUegUAD6VVVVVVVVVVVXx2+D2jfG/4eX3gzUWjgu/8AXaZeMmTa3Kg7W9dpyVYd1Y98Gv2NfjHrHw18XXn7NnxR8yykS8ePSjcN/wAet1n57fPTZJ95CONxOM+YMVTFVUszAADknjFftp/tD+M/27vj1o37H/7PE8l34RttUWK9vrbLR6lcIf3t1IRj/RbdQxXsxUt82Y8aV47/AGNv+CZfgTRfgrrvjGfSLy9gOouz6XcXd3q7s3lvcytBEUBym0BiuFQAZxzL/wAFdf2MUmaFPEfieRVYqJF0GUKwz94ZIOO/IB9q+Dv7a37L/wAetSi0L4a/FzSrzWJjiLS71JLC6lYdRHHOqGU45+Td39DVfHv9tH9nz9mfxHp/hP4veK73S9S1Sx+22sUGlz3QaHeybi0asAdyMMHmvh38QvB/xY8D6N8RfAOsx6roGvWwuLG7RWXepJBBVgCrKwKspAKsCCARVftBftYfBH9l7+wP+FyeJLvSv+El+0/2b5GnTXXmfZ/K83PlK23HnR9cZzx0NeGvEOleL/DeleK9CnabTdasYb2zkZCheGVA6MVPIJVgcHkV8Tvi58M/gv4Zk8X/ABT8b6V4a0lG2rPfT7TK+M7I0GXlfAJ2IC2AeK1T/gsB+xzp+pGwtNQ8Y6nCGwL210MrF1xnEro+O/3O/rxXwI/bU/Zt/aR1E6B8LfiJFda6kJmbR761ls7rYv3iqyKBJgcnyy2B1xXxS/4KJ/sp/Brx9q/wz8feOdRsvEGhSrFe28WiXUyxs0auMOiFT8rqeD3r/h7B+xH/ANFI1X/wnb3/AON1/wAPYP2I/wDopGq/+E7e/wDxuvBv/BTT9j7x94v0LwL4Y8f6ncax4j1K303T4X0G7jElxPIscalmjAUFmUZPA6mq+IvxE8GfCbwTq3xE+IGu2+kaBoduZry7mJwoyAqqByzsxVVUAlmYAAkitQ1L4xf8FaP2ipN1xc+FPhL4PPnTNNIBBotjn5pGP3HvJgpPOQAOuyMmvj9+0Vd/FB9D/YD/AOCf2hXcPge2ZtOludMcrL4gfJMxMuQRa5LvJKxHmfMzEJ974n/sW+Iv2W/hT4Mnu/Er+Iy0JttauUjxFaXjEuqRcZ8naSiluSYyfl3BVr9n/wCMGofBL4l6d4vgaWTTnP2fVbVD/r7ViNwA7spAdf8AaUDoTWlapp2uaXaazpF5HdWN/Ak9tPGcrLG6hlYexBBqqqqqqqq+PHgofEP4O+LfCKQ+bPeaZK1qmOtxGPMi/wDIiJVWN7daZfW+pWMzRXNpKssMi8FHUgqR9CBXgbxTZ+OfBmh+MbAAQa1p8N2ig/c3oGK/UEkH3FVVVNLDBE008qRxxqSzu20KB1JPYV4s/aS+BfgsMmt/EzRmlThoLKU3sgPoVhDlT9cV4q/4KG/DPTd8XhPwjrmtyKPle4ZLOJvo3zt+aU37Yf7THxILQfC34XRRwsSFmstLnv3j7cyH92PxWj8Lf29Piiv/ABUvi690O2n6rc6ulkjA9mitMnHsy1oX/BOy/vJ/tvjr4qeZK5zLHY2JkZj6+bIw/VK8OfsJfALQ/LbUtP1jXnTkm/1BkDH6QiPj2/PNeHfgj8H/AAmEPh/4a+HbWSMfLOdPjklH/bRwX/WkRI0WONQqqMKoGAAPSqqqqqrxt8Ifhj8R4mTxr4I0nVJGGPtEkAScD2lXDr+DCvHf/BPLwTqYkuvh74v1DRZiCVtb9Bdw57AMNrqPclzXxl/Zd+K/wb0ptd8S6fY3ejRzLGdRsLoSRhmOFBVtrjPqVx71UBBhX6VVVVVVKzKwZGIZTkEcEGvA37Tfxv8Ah6IodE8e31zZxHiz1E/a4sf3QJMlB/ula8D/APBRVwIrX4keAA3QSXmjT4+pEMp9P+mleB/2pfgV4+8uHSvHtlY3cmB9k1TNnIGP8IMmFY/7rGo5Ip4kmhkWRHUMjKchgehB9K/4KBfCI2mpad8Y9GtP3V6FsdY2L92VR+5lP+8oKE9BsQd6r9kv4zJ8XvhdbJqd2JPEXh4JZ6mGPzSYH7qc/wC+q8n+8r9sVVVVVVVVVVVVVVVVVVVVVVVVVX/BSXWxceLfBfhsSA/YdNubsr6edIqZ/wDIBqv+Cbug/afHfi/xNsB/s/SYrTODwZ5d/wD7b1X7fcUafHeNkQAyaFbMxA6nfKMn8AB+FV/wTnkZvA/i6EyEquqxMEJ4BMXJx74H5VVVVVVVVXdnaX9u1pfWsNxC+N0UqB1bByMg8dQDVjpWl6Wrrpmm2toJCN4ghWPdjpnA5qq1fV9K0DTLnWtc1K2sLCzjMlxc3MojjiUdSzHgCrS7tb+0hvrG5iuLe4jWSGWJgyyIwyrKRwQQQQaqvhz8fPgr8XhIvw1+KHh3X5oJnhltbW+X7RG6EghoSRIBwcErgjkZHNfFP4zfC34J+G7nxX8UfHGk+H7C2iaQC7uFWWfA+7FFnfK5xgKgJPpX7LNt4h/bd/4KTXXx5OjTwaBousjxDdvIpb7HBbKI9OhZunmkxQDGeiSMBhcVVV+2F4lHif8AaE8UPHKHg0x4rCIA52+VGocf9/PMr9nbw1/wlvxx8E6IYzIh1eK4lQD70cH75x9NsZzXj7XF8UeOvEfiVX3DVtWursH18yVnzx/vVX7L+hjw9+z74E08RhDJpEd0VxjmcmbP4+ZVVVVe3lpp1nPqF/cx29taxNLNNK21Y0UZZmJ6AAEk1+0v8a7j43fEe41i1kkXQtMDWujwPxiIHmQjs0hG49wNo/hrwv4b1Pxh4hsPDOjojXeoTCNDI2xIx1Z3Y8KiqCzMeAASelfE/wASaXrOtQaH4YmZ/DfhyD7BpJZdhnRSTJcMvZppC0hHYMq9FFVX7E3wmPxI+MNrrmoWpk0bwiFv7klflecH/R4z9XBfHcRkd6qq/aC/bL8JfDBbnwx4Fa28Q+KVyjlX3Wti3fzGU/O4P/LNT65K4wfGvjjxX8RNfn8T+MtbuNT1G44MsrcIo6KijhFHZQABWj6Nq3iHVLXQ9C064v8AULyQR29tbxl3kY9gBya/sT4f/syW6aj42i07xf8AE8KHtPD6uJbHRGIyr3TDiSUdRGOAe/Rq8aeNvFPxD8RXXivxjrM+p6ndn95NKfugdFVRwqjsoAArwl4Q8S+PPEFn4V8I6PPqeqXz7IbeEcn1JJ4VQOSxIAHJIr9mj9lfw78DNNTXNX+z6r4yuosXF8FJS0VhzFAD0HYvgFvYcVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVf8ABVRT8SP22fgz8HVzNDPZadbtGOf3l9qLxMuMHkpHH2PUelVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV8c/inpPwQ+D/i74sayYzb+GdKmu0ikbaJ5gMQw59ZJSiD3cV/wR1+FeqeNvGvxE/av8bbrzULm6l02xupkyZbudhcXsw9Gw0K5HaWQcd6qqqq1n4r/Czw5rn/CL+IfiX4V0zWcoo0691m3guMuBsHlM4b5gRjjnIxVVVVVVVftt/s8TeMtI/wCFweBrRl8SaDEDqEVuCr3lsnIkXHJlixkdyvGflUV+yP8AtEQ/GvwaNF8QXaDxfoMSrfqTg3kXRblR78BwOjc8BlFV4y8K6V468Jax4L1x7tdO1yxmsrs2ly9vL5UqFG2SIQynBPINfse/sLfDX9j5PEV74d1O58Q65r90yjV7+3RJ7exBzFart44+87ALvbB2gKoH/BbT/kv3gX/sTx/6Vz18Nv2DP2PtV+Gvha+1H4B+Gri5vNDtJZ5pEkLyO8KlmLb85JJOa/4KU/sDfDb9njwdpnx6+Aq3vh+0g1aK11HSTeyTLbvJuaK4t5HYyIQ6bSpY/eUrt2nP/BOv46eJP2gf2WfDvizxpfvf+ItJuZ9I1O9kHzXUkBGyVvVzE8W4923Hvgf8FmtPj1b9pf4baVLcNAl54YihaVY95jDX0ylguRkjOcZGfUV/wTQ+P3iP9m744eIP2KfjPcGztb3WZrbSzMxKWWrIdhjVjj91cKoKnoX2ED96xqv+C5//ADRL/uYP/cdX7PX/ACQH4Z/9ifpP/pHFWqnxV/wUt/b7l8F+IvFl3Y+EbS/vYrBImB/s/SLUtnyUPyiabYm5iD88mTlUC1on/BOX9i3Q9Fi0SL4EaPeJGhVri+nnnnkJHLGRpN2fpgDsBXwP/YZ/Z8/Zz+J2qfFP4S6BqOk32qaW2nNZS373VtbxtIkjNF5u6RWYxqDlyMDAA5r4k/8ABP8A/ZF+L3jfVfiN8Q/hJ/a3iLWpFkvrz+3tRg81lRUU7IrhUX5UUfKo6V/wUI/YU/ZU+Bv7LXiT4jfC34V/2L4isLyxjt7z+29QudiyXMaONk07ocqxHKnGeMV/wS//AGMv2av2ivgDr3jb4x/Db/hINasfGFzp8Fz/AGxfWmy3Szs5FTZbzIpw8shyRn5sZwBjwn/wTh/Yv8DeKtG8beFfgz9i1rw/qEGoabc/8JDqcnkXMEiyRPse5Kth1U4YEHGCCK1nWNJ8O6Re6/ruo29hpum28lzd3dxII44IkUs7ux4ChQSSegFfHf4vfFT/AIKj/tEWPwL+CrT2Pw20G5aZJ5kZIhChCS6pdjI4w2IozggOF4Z2r4zfGW0u9L0n/gnX+wPpl3eaFdXRsNd1u2P+keKb1vln/efw2/ykySHCsi4G2FPn/Yj/AGIvBX7Ingre/wBm1nx9rMCjXNcEfAHB+zW+eUgUgehkIDNjCqnj7wRofxH8G6t4I8RwebYatbNDJgfNGeqyL/tKwVh7gV8RvAWufDHxtq3gXxFFtvdJuDGXCkLMnVJFz/C6lWHsar9gX49CaFvgd4ovlDxB5/D8sjcsvLS2/wBR8zr7bx2Aqqqqqqqqr9oLwP8A8K6+M/i3wnHD5dvb6i8tovYQS4liA+iOo+oNV+wF47Hif4LSeFbiffdeFL94FU9Rby5ljJP+8ZVHsgokAZJAAHevGX7Q/wAFPAQkj8R/EbSEuI8hrW1l+1TAjsY4tzKfqBXjD/goj4HsDJB4I8EarrDrws19MtnGT6gDexHsQp+lL8ef21PjCu34e+C5tJsZ+I57DStiMP8Ar4uSVz7qV/Cof2Of2kfiY63fxY+KEdvG5BMN3qE2oSR/SMERgD0D14U/4J8/CbSQkvinxBruvzKPmRXW0hb/AICoLj/vuvCv7P8A8FfBflt4d+GmhQyxfcnnthczLj0kl3P+tIiRoscahVUYVQMAAelVVVVVVVVVVftdaEfEP7OnjW0RNz21ml2p7r5EqSsR/wABRh9Car9mz9l/w18f/hxqurL4sv8ARdf0rVGt/wDULPbvEYkZCUyrBtxkBO7GAOOteO/2I/jn4OElzpmj2niazjBPmaTNukx7xPtcn2UNWraNq+g3r6brmlXmnXcf37e7gaGRfqrAEdKqqqqqqrwX8WviX8O5EfwX441bS0Q58iK4LQk+8TZRvxU0f22PFXivwhqPgL4t+EtL8S6Vqts0E89sTZ3Ck8rIMBoyyMFYYVeVFMF3EISRngkYNfAD4w6h8E/iNY+KoTJNpsv+j6raof8AX27EbsD+8pAdfdcZwTWi61pXiPR7LXtDvoryw1CBZ7aeI5WRGGQR+FVVVVVVVVVVVVVVVVVVVX/BUb9qX4z/ALMOgfD2/wDg74itdKm8QXl/FfmfT4brzFiSEpgSK23BdunXNfD/AFa+1/wF4b13U5RJeajpFrc3DhQoaR4VZiAOBkk8Cqqqr9vfWRqn7Ql5YiTd/ZGlWlrjOduVM2Opx/rs9uv4mv8Agm9owg+H/i3xCF5vdYjtc4/54whv/a/61X/BRC1ZPjBoN5ldsvhuJAB1ytxOSf8Ax4flVf8ABN+7RrLx9Y4AaOXT5ASeoYXA6e239aqqqqqqqqqq/aa+Dz/Gr4U6h4ZsJ5I9Ws2+26WBKVSSdAcI4zghlLLk/dLBu3P7BXxmu9Y0W9+CXiueRdW8Nq0mmCfIdrYNh4TnnMTkYH91sYwlVeWsN9aTWVwHMVxG0bhHZG2sMHDKQVOD1BBFfEf/AIIo/BjxBfz33w0+KfiTwjHM25LO9tI9Vhg/2Uy0Um3p952PXk548If8EPvAljqMc3jv4+65rVkrZe30rQotNdhxx5kks4Hf+H8q+CvwI+FX7PXg2LwL8JfCVrommhvMnZSZJ7yXGDLNK2Wkc+pOAMAAAACq1PULTSNNutVv5RHbWUDzTOf4URSzH8ga8Sa3deJ/EWqeJL9s3Oq3s13MSc/PI5dv1Y1+yrb/ANk6h48+JcgAXwd4PvriByOlzIhSIfiPMFVbW095cxWdrGZJp3EcaDqzE4A/OvD2kQ+H9A0zQLY5i02zitoz/sxoFH6Cqqqr9ub9oVY45Pgl4OvwXYBvEFzE33R1W2BHc8M//AV7sBV0B8KfhyCzbPFvjyywq/xado7Hkn+7JckYHpCD2lqqVWdgiKWYnAAHJNfsr/B3/hTXwl0/R9QthHruqn7dqxI+ZZXA2xf8AQKuOm7ce9V45+IHg74baFJ4j8ba/a6XYx5CtK3zStjO2NBlnbj7qgmvj5+2r4u+IwufDHw8+0eHPDj7kkmD7by9Q8HewP7tSP4FOT3Yg4qvhn8IPFnxRuLibS1t9P0TThu1PXNQk8mzsEAyS8h4zjooyT7DJGt/GPwZ8IdLufBf7OyzSajOhi1PxxdxbLu5/vJaL1gi/wBr7x47gMZ5prmZ7i4meWWVi0kjsWZmPJJJ5JJ718IPgt46+NniRPD/AIO07dHGQby/mBW3s0P8Tt6+ijLHsOCR8DP2ffA/wI0D+z/D1uLzVrlANQ1eeMCa5PXaOuyMEDCDjjJycmqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqvi2g+In/Babw1oK7Z4/D+qaUUxhgPsunret0B5Vt2c9CDyMZFVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVf8ABaP40PpPw/8AB/7P+iXLtfeKr06rqcMRO42tudsMbKOoknYsPe2/P9kf4LR/s/fs6eCPhc8Cx6hp2mrNquOd19MTLcc5OQJHZQf7qr0AxX7Ynxkb4B/s1+O/iXaXa2+qWOmNb6Qx5IvpyIYCB32ySK5Hoh7c1+zt+3r+1D4S+OHwz1j40fFfxDqXgfxBfJ9ph1ORBb3FjLNLaSXGdo+WKVJGzkfNCaqqqv8AgoP/AMpME/7CHh3/ANFW9VVVVVVVX7R3wp8S/syfE+w+Pvwii+z6JcXm6e3jQ+VZTP8AfhdR/wAsJQTjsCSvHyZ+FHxy8AfFvw1ous6Lrllbahq8cgGkz3SC5SWEKZ0CEhnCb0JYDG11PG4VVf8ABbT/AJL94F/7E8f+lc9fD7/gon+xfoPw58NaXqvx102G707RLSG5hGnXrskkcKhlwsJyQQRxnPav2zP2sde/b/vdJ/Zk/ZL8Ca74g0kajHe6jqcloYTdOgZY+G4gtl3Fmkl2ksF4UL8/7I37P1v+zH8A/DXwlF9FfajZRvc6tdxAhJ7yZt8pTODsUkIpIBKopIBJr/gtHey6Z+0b8PNSt1RpbTwrHKiuCQWW9nIzjtxX/BYH9l+9il0z9rT4fWksN1YmGy8UNaAq8ZUgWt7leQVO2Jmzx+5x0Jr/AIJ8/tYW37U/wQtbrXL2I+OfCqx2HiOEHDTOFxFdgf3ZlUk44DrIBwBn/guf/wA0S/7mD/3HV+z1/wAkB+Gf/Yn6T/6RxV8YPC/xV/4Js/tqr8UdD0Frrw/c6pdXuhSyArbapp85YS2jOAQsiLJsIxlSEkC4K5/Z8/4KF/sy/tDW9lYaR43g8NeJrkBW8P6+62s/mY5WJyfLn5zjYxYgZKr0qq/4Kwf8mR+MP+whpn/pZFX/AARU/wCTV/FP/ZQLz/0gsKr9vX9qrxn+1t8VLL9jH9mEy6tpE2oLa6pdWbfLrN2jZKhx0tINpZnOFYqzH5UVm+KHjOw+CvhaH9gX9j6aXxD4t8R3kdp4+8WaWR5uv37jYdPtmBJS3QsUY7gOHB6ylv2DP2GvDX7JngldY8QQWeqfErW4ANY1RBvW0QkEWluT0jGBubgyMMngKFqv26PgKfH3g4fE7w1ZB9e8MQH7WiL811ZDLN9WjJLj/ZL9TgVWj6vqfh/VrPXdEvpbPUNPnSe1uIjhopEIKsPcECv2efjTpXxy+HVn4ngaKHVrYC31ezU/6i4A5IH9xx8y+xxnINVVVVVVVX/BRzwEbTxH4Z+JNrF+71G2fTbshcASREvGT7sruPpHVfAb4rfFT4b69e6T8J4hcap4niS1FuLT7S7OpLIyJ03DLDkEYY5HcL+zx+2J8aSJfid41l0qwm5e31HUflIP922t8oDz0bb/AErwb/wT1+Gmk7J/GnijWPEEy9YoFWyhP1A3P+TivB/wQ+EfgEI3hP4e6NZTR423BtxNOMf9NZNz/rVVVVVVVVVVVVVV4w0GLxV4S1vwvMV8vV9OuLNtwyMSxshz7fNU8EtvM9vPGUliYo6MMFSOCK/4Ju+JhbeLvGHg55P+Qhp0F9Gp9YJCjY9yJ1/L2qvE3g7wn40sf7N8W+G9M1m27RX1qkwU+o3A4PuOa8d/sFfBzxOZLrwrcal4VunyVW2k+0W4PqY5CW/BXUV47/YP+M/hcSXXhk6b4qtEJ2izl8ifaO5jkwM+ysxrxJ4T8UeD786X4r8PajpF2M4hvbV4WOO4DAZHuOKqqqqqqqvgb+1t4++CelL4Xg0+z13QBM0qWd27pJAW+8IpASEBPJBVhkk8ZNeBf28/g34nEVt4pi1LwrdvwxuYjcQA+0kYLY92RRXhvxf4U8ZWQ1Hwn4k0zWLbAzLZXSTBc9jtJwfY81VVVVVVVVVVVVVVVUviPw8+r/8ACPpr2nNqmwv9hF0hn2jqfLzuwPXFV+1J+1t8NP2RvDmj+J/iXpPiPULbXLt7W1j0W1imfzFUMQ3myxgDaSevY+2f+ChX7dXgn9sXT/BuleC/BOt6HF4WubuaSbVJYi0/nJCAAsZbGDG3fuPw+E//ACSvwb/2L9j/AOk6VVVVftV6o2sftD+ObtmLGPUzbjOf+WMaxDr/ALlV+wbpP9nfs76bebMf2pqV5c5x97EnlZ/8hY/Cq/bp+Enj/wCIXiTwjqHgbwlqOteVZXMNw1rFuWHDoy7j/Dnc2M9ccdDQ/ZJ/aKYAj4YX3Tvc24/9qV+xF8K/if8AC3xd4ntvHfg2+0m31LToWhnlCPGzxyH5dyMRnEhOPY+lVVVVVVVVVVV+1L4e1L9nb9onQPjt4MtDHZaxcG6niT5Ua5X5bmIkdBLG273LvjpWga5pnifQtP8AEei3InsNUtY7m1lAxvjkUMp9uCKqqqq/a88Zf8IZ8AfEs0Mu251iJdMtxnG4znbIP+/Xmn8Krw1af8IP+xL4z8Syfubvx5r1tpts4HMkMDBj+e25XFV+zz4cPiz44+B9E2K6PrUE0qsOGjibzXH4qjCqqqr9qn9omy+CnhU6TodzHL4v1iIiwh4b7LGcg3Dj0BBCg/eb1CtV5d3WoXc1/fXMlxc3MjSTTSuWeR2OWZieSSSSSa+F/hHRms9Q+J/jy3ZvCXhllDQH5Tq16RmGyQ/7RG6QjO2ME8ZBrxb4p1nxt4k1DxX4guBNf6lMZZiq7VXsFVf4UVQFVegAA7VVfsN/A9viJ8QR4/12z36B4SlWRA4+W5vesSe4TiQ+4QHhqd0jQu7hVUZJY4AA718cf23/AAT4CFx4f+HAg8Ua8mUa4VybK1b3cf60+yHHqwIxXj74jeNfifrj+IvG/iC51O7bIjEjYjgU/wAMaD5UX2AHqcnmoYJrmaO2tonlllYJHGilmZjwAAOSSe1ab8KvCHw00+LxP+0DfXFrcSRiWw8G2LgajegjKm4P/LrEe+75yM4AIGfiZ8bPFPxHtrfw/HBa+H/CenH/AIl3h3S18q1twOjMOssncu+TkkjGSKr9nb9jfxf8XmtfFHi0XHh/wkxDrMyYuL9f+mKnop/56MMc8Bu3gnwL4T+HPh228K+C9Et9M021X5Iol5du7ux5djjlmJJqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqvgOF8a/8Fm/GeuufMOg6lre07i23ybVrLqOOjYwenTrVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVaEF/bf/wCCsM+q83/hHwFfmaMNl4/sWlsFjIH/ADzmuyrYPGJzkdqr/gtX8X5pYfAP7O+hySS3F5K2valBESWbG6C0TaOW3Mbg7fVUOOlf8FIP2Uf+FRfspfAbWdOs0Fz4FsR4f194xnfNcp9oMnHRBcrdfjOoya/Yf+MP/C8/2W/APjm5u/P1RNNXTtWZn3P9rtT5MjN6F9gkx6SCq/aQ/bK+Av7LNgjfE3xSz6zcxebZ6BpkYuL+5XkBhHkLGpwQHkZFJBAJPFan/wAFxfC0WqeTo/7Ouq3Wm5P+kXPiWOCbHY+Utu6/hv8Azr9jz9tTwX+2RpPiC98KeC/EPh+48MPbpfrqIieFmnEhQRSo2XIETbgVXGV/vV/wUH/5SYJ/2EPDv/oq3r9pH/gqt8BfgL4qvPAPh7S9S8feIdLmaHUU0yZILSzlU4eJrhs7pFPBCKwBBBYMCB8Lv+C1fwg8Ta1b6X8Uvhbrngq2uHCHUbS/XVoYMn70oEcUgXHUojn/AGT1rQ9c0XxPotj4i8Oara6npepW6XFneWsokinicAq6MOCpBBBFVVVVVXxPuPAlp8O/Ed58T5LOPwnb6bNLrD3ZxGluqkuxI5BAGQR82cY5xXhLX/hf8T/GOm+L9LbxNYaR4X8TRzwzWl39l1S2tVm3LtmjwEkkhTqpGGBwQVzXhbxRoPjXw9YeKvDGpRX+malCJbeeM5DA9iOoYHIKnkEEHBFV/wAFtP8Akv3gX/sTx/6Vz14F/wCCWP7GfiP4e+HtZ1DwDq4vtU0a2nnmj1+7UiWSFWZlXeVHJJxjHtX7WX/BObX/ANk3w7d/tHfso/E3xTZQ+GsTalYyXZS8tICwDTQzwhPMjUkbo3X7uSWYAiv+Ccv7XmqftX/CG9bxqbf/AITfwdcx2msvAgjS7jkVmt7kIOELhJFZRxuiYjAYKP8Agtp/yX7wL/2J4/8ASuevFPhfQPG3hrVPB/inS4NS0fWbSS0vrSdcpNDIpV1I9wT9KsJfHP8AwS0/bf8As1zJe3vhCaTbISP+QxoM78P0AM0RXPGB5sJGdrHP/BbDU9O8VeG/gP4v8O3sWoaNqEOszWt7C2UmjmTT3iZfUMoJFfs9f8kB+Gf/AGJ+k/8ApHFXj/4deBfip4YuvBfxF8Kab4h0S8H72yv4BKmQCAy55Vhk4ZcMOxFftEf8EZfAusWF94g/Zt8U3eg6oimSHw/rMxubOYj/AJZxzn97ET2MhlyeCVByP+Cdn7ZPxc+E/wAbbT9kT4/6jqFxpl1qDaLYJqrmS50LUlJSO3EhJLQu4EQTLBWZCpVd2a/4Kwf8mR+MP+whpn/pZFX/AARU/wCTV/FP/ZQLz/0gsK/4Kf8A7bt/4St5P2VfghfS3PjPxCqW+v3lgS81hDNgLZxBeftEwZQccqjYA3OCmtaif2I/AcnwV+HTG+/aH8fW0dv4s1PT/wB9L4XtZtpj0i0Kg5upAymV4zlSQqkkKy/8E6v2CbD9mbwxH8SfiTYW918T9ctsSIwWRdCgb/l3iYZzKwI8xwcfwL8oLPVUyq6lGUMCMEEda/bB+ATfBvx8dY0CzK+FfEbvNYbR8tpL1ktz6AZyv+ycclTVfs6fG/VPgV8QrfxDG0s+i3u231izQ586DP3lHTehO5encZAY1o2saX4i0mz13Q7+K90+/gWe2uImyssbDKsPYg1VVVVWveI/D3hfT31XxLrthpVnH964vblYEH/AmIFePv28Pg34V8208LJqHiu8TIX7JH5FvuHYyyAHHuqMKk/aU/az+OM72fwj8GPpdi5KCbT7PzNozjD3U/7tT7jYeteMP2N/jrrHg/XfiF8SfHEV9qul2Et3FYy3Ut/cTbBuZC7EKh2g4ClsnA4qvDPiDUfCfiLS/FGjy+XfaReRXVu3o8bBlz7ZA4rwp4k03xj4Y0nxZo8m+y1iyiu4CTyEkUMAfcZwR2Iqqqqqqqqqqqqqqqq/aQ8Kf8IX8dfGugpEIol1aW4gQDhYp8TIB7BZFH4V+x74tHhD9obwpNLNst9VmfTZhnG/z0KRj/v6Yj+FVVVrGh6L4hsX0zXtIstStJB89veW6zRt9VYEGvHX7EHwL8Xh59J0q88MXbZPmaXPiMn3ik3KB7Ltrx1/wT7+KGh+Zc+B9d0vxNAudkMh+xXB9gHJj/EuPpXjD4bePvh/cG18aeD9V0ds4V7q2ZY3/wB1/ut07E1VVVVVVWka1rPh++j1TQNXvNNvYv8AV3FpO0Mi/RlIIrwH+278cfB3l22q6raeKLNMDy9VhzIB3xKm1ifd91eA/wDgoD8LdfEdt440bUvC9yfvTKpvbcf8CQCQfTYfrXhH4g+BvH9n9t8F+LNL1mMLlhaXKu8f++mdyH2YCqqqqqqqqqqvjn+1t+z7+zdd2em/GPx+uh32o2zXFlarp11dSXCAlcr5MbgfMCPmIxxnGa8Vf8Flv2UtE3xeH9E8d+IpRwj22lw28R57tNMrAY/2D+Ffsa/t/eHP2xvF3inwvovw8vPDI8OWMN3FJd6ktw92ruyN8ixgJtITnc2d/bHNft2eCf2p/G/w88PWH7J/ie90jxAmtbdUNtqcVjmyaGTLmR8H5ZBGMId3z5wcZB/4Jkft9/E9fN+Lv7S1o0Mw+eHUPFGo6pIuQMgoU2Y4A4fHHoBX7Hn/AAS/uP2Wvi1p3xgv/jYdfvbO0uLZ9Mt9B+yxOssbJzKZ3JwSrfdHK496r4lfB/4YfGTTbHR/il4H0nxRY6bdfarW21KDzY45tpXftPBO1mHOetfHH/gnB+y/8cdK0PRm8KHwFBoUs0kf/CE2VjpjXJkCAiYm3feBsBHTBLdc14e0S08NeH9M8OWEksltpVnFaQvMQXZI0CKWIABOFGcADPYVVVVfFfUP7X+KXjHVSebzX76bkY+/O7dO3Wq/ZS0waT+zt4GtAgXfppnxgD/WyPJnj135/Gq/4KT/ALWHxR/ZR8AeEtc+FkGhve+IdUmtLiTVLR7jy0SLcDGFdQDk/wAQI46V4G/aJ/4LA/F/wvZfEP4c+Gbe+8O6srNZXFvpGmRxyBSUYoJm3kblYZPfOKvP+Cjn7e37NWuWdl+1B8DrO70y4cJ5lzpr6a9wR94Q3UJa3ZsKxICN1zwMV+zn+0X8OP2oPhtbfEn4bXsxtmkNvfWF0oS50+4UAtFKoJAOCrAgkFWBBr/gpd+2/wDGr9lbxZ4Q8K/CmPw6kOv6XPd3NxqFg1xKjpKFAT5woGPVT161ofxh/wCCz3jHRbHxX4c8HrLpWsW6XdjIuk6UivDIAyMokbeFKkEbucYrSf8AgqX+1h8BPGNt4U/a4+BUYtZm3M0eny6VemPPzSQlmaGcLkYACgkYLjOR8Vvj74q8c/sgaj8c/wBj6J/FWv30FnJoVtFp5upWZryGO5jeAc+ZHGZgy/wlCeQM1/w1F/wWD/6IXqv/AIQr1r//AAUv/wCChXhbxz/wrLxHpOlab4sFxBb/ANjXPhgR3XmzKjRJ5ZOdziRCB33D1r/hqL/gsH/0QvVf/CFev2GPiF+078R/h5r+qftTeELjw94gtdaMOnQT6OdOMlr5MZ3BD94by43e2O1VVVVV+1L8L1+K/wAF9c0O1thLqmnx/wBoaZhct58IJ2r7uhdP+B1/wT6+JreJ/hpffDvULjfeeE7gG23Ny1rMWZR6nbIJB7BkHpVVVVX/AAUV8cCbUfC3w4tpwRbxSandoD/ExMcP4gLN/wB9ClVnYIgLMTgADkmv2wLMfDn4NfCP4MIoSaztJL7UIxxicKoJ/GSW4qv+Cfvhj+2vjq+uPEDH4f0e4uFcj7skm2EAe5WST8Aaqqr49fHPw18C/B763qbR3erXQKaXpgk2vdSDqT3WNcgs30HUgV408ZeIviD4mv8Axf4r1J73U9Rk3zSsMAYGAqjoqgAAAcAAV8L/AIb6v8UfFEegadPFZ2kETXOp6jPxDp9onMk8h9AOgyMkgcZzXxj+IWk+KLyw8H+B4HtPBXhZGt9HgcbXuWJ/e3cvrLKRk/3RtHGDVV4L8Ha98QPFWmeDfDNmbnUtVuFhgTsM9WY9lUAsT2AJq4+JvwQ/ZA+Hmn/DyPWF1LU9Nh+fT7EK9zc3DDLyy84i3Nz8xyBgAHAFfGv9qz4l/GVptLku/wCwvDrkhdKsZCBKv/TaTgy/The+0Hmq8G/DTxB4ytptZElrpHh+yYLe65qcnk2luf7u7rJJjpGgZz2Hem+KnhH4WRPp/wAErF7rWipSbxjqtuv2gHGD9igOVtlPPztukIP8PQX9/f6rezalql9PeXdw5eae4kMkkjHqzMSSSfU14S8HeKPHmu2/hnwdod1q2p3J/d29um447sT0VR3ZiAO5Ffs/fsL+G/BBtvFXxYNr4g1xcPDpoXfZ2jf7QP8ArnHuAg9G4agAAFUAADgVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVf8ABOU/8JV/wUp+N3ixneRPJ8Q3ayxrtRnm1eADII4BVnIHHT2NVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVftefGBfgR+zb49+JkN0INQ0/SXh0tt2D9tnIhtyO5xJIrEDspPGMj/AIIt/BoaD8LfFnxy1S2b7b4t1EabpzyLz9kteZHU+jzOyn3txVeA2X9tz/gq3ceJgy3/AIV8M6w9/GQ2+P8As/S9sduwHTZLOsLEdP37dc4P7VnweT49/s7+OvhWkSPe6xpTtpu/gLewkS2xJ7DzY0BPoTX/AARR+L0tnqvj79n/AFi5ZPNRNe0uCViuyRCsF0oB/iINscdfkY/T43fE/Tfgt8IvF3xW1SHz4PDGkz3qwZx58ir+7jz23uUXPbdX7Bf7PV1+3r8ffGPxt/aFvrjXtG0aeO61WF5Gj/tO9nLeTBlSCkEaRsSqkYCxoPlJx4a8DeCvBmhr4Z8IeEdG0XSUiEQsdPsY7eHYBjbsQAYxx0rw34G8FeDJdRm8H+EdG0N9XnFxfnTrGO2+1TBQvmSbAN7bQBuOTgCv+Co0GrXP7eviG20Dz/7Tlg0dLL7OxWTzjawhNhGCG3YwfXFfso/sgfC/9l3wBpekaL4e0688XNbI2t+I5bcPc3lyRmTbIRuSIEkKgwAME5YsT/wUS/Yx+Hnxt+C/ib4geHfCdhp/xD8LWE2p2epWVqsc2oJCpkltptoHm70VghblX2kHBYN/wRW+N2sa14e8ZfALW717i28PiPWNEV2LGCKVylzGM9EEnlOAP4pZD3qrn9vj4YaP491vwb4j0HVbWz0q/ls4dVtWW5jm8ttrOyfKyqSDjbvyMHvx4G+L3wx+JcKyeBvHGlas5XJt4p9s6jHVoWxIo+qiqqqr/gq7oX7T3xG8M+D/AIRfBzwDqOq+D/EWoxLrl9p7+Y8l2ZQtvbzoOYrdWxIZGyhbZkps+b4Q/sNfDH4cfsy237PuowRXtzcD7bqutRRBZZdUZcG5jJGQE4RAf+WY2nOWz8KviN43/Yu+KV18Kfiiss/hK+mD+dGGdI1bIW8tx3U9HQc/Kf4lwdP1Cx1awttU0u8iurO7hWa3nicMkqMAVZSOoIIOa/4Laf8AJfvAv/Ynj/0rnr4T/wDJK/Bv/Yv2P/pOlf8ABRD43eCPhB+y3460vxHqtqNY8aaFeaJo2mGQeddyXMRhd0TrtiWQuzdBgDqwB/4IheAtbsfD3xP+JV5bSxaXq1zYaZYOwws0kAmkmI9dvnQjI7lh2r/gtp/yX7wL/wBieP8A0rnqv+Ci37Jsf7UHwRnl8N6esnjvwcsl94fZV+e5GB51pn0lVRt/6aInIGa8cfHPU/HP7N3gv4J+LJLltT+GuvXn9lNKuCbC6RS0LZ53RTREDOPllVQPkr9nr/kgPwz/AOxP0n/0jirw/wCJ/Dfi2wbVPCviDTdYs0meBriwuknRZUO10LISAykYKnkHg4qeeC1gkubmaOKGFC8kkjBVRQMkkngADvXirUdM/aS/4Kv2esfCCVbrTbvx1pc0N5acq8WnRwfarpCvVSLWaUN0IIOec1X/AAVg/wCTI/GH/YQ0z/0sir/gml4s1rwD/wAE4Pjh468Nzxw6v4c1PxBqVhJIgdUng0W0kjJU8EBkU4PWvCWqTfsyeEF/aR8fzHVvjd8RIZb3wRa6iPPfSbect5mvXKvnM0hLfZ1fIJzKQw21/wAEy/2F77wyIP2qvjxZT3fjLXN13oFjqAZ5bFJclr2ffybiTcSoP3Fbcfnb5Kqqr4sfDPw/8XvAep+BfEUYEN7HmCcLl7WdeY5U91PbuCQeCa+IHgTxD8NPF+p+CfFFp5GoaXMY3wDtkXqsiE9UZSGB9D2qv2Gf2jh4X1OH4MeM78LpGpTf8SS4lbAtblzzCSeiSMePRz/t5FVVXFxb2kD3N1PHDDEpZ5JGCqoHUkngCviL+2Z8D/APmWlpr7eJtQjyPs2jATID7zEiPH0ZiMdKv/2sf2k/jXeyaJ8FPAj6ZATsaWytzeTRZ/56TuBFGOnO1cZ61oP7D/xd+I2oJ4j+N/xHaCaQ5eIztqN1jPKF2OxPbaXA9K8Bfsi/AnwB5U9v4QTWr6PGLvWm+1sSOhCECMHvkIDUMENvClvbxJFFGoVERdoUDoAB0FOiSIY5FDKwIKsMgivjr8PJPhZ8WfEngkRMltZXrPYk5+a2k+eI57/Iyg+4PpVf8E+/iSvib4W3nw/vbgNe+E7o+QrHk2s5Z169cSCUew2D0qqqqqqqqqqqqqqqq/4KK+D/AOyvifoXjKGLbFr+lGGRsfemt2wx/wC+JIh+FaTqd5omq2Ws6dL5d1YXCXED/wB10YMp/MCvDGv2Xivw1pPijTT/AKLq9jDdwZOfkkQOv6MKqqqqurW1vbd7S8t4p4ZV2vFKgZWHoQeCK8c/shfAXx0Xnm8Gpot2/wDy86K/2Qj1/dgGMn3KGvG//BOzxDah7n4d+O7PUF5K2uqwm3cD0Eibgx+qqK8cfs+fGX4diSXxT8P9UitYvvXlvGLmADsTJEWVc/7RBqqqqqqqrDUL/S7uO/0u+ntLqE5jmgkMbofUMCCK8BftmfHXwN5VtceJE8R2UeB9n1qMztjviUESZx6sR7V4E/4KE/D7WGjtPH/hjUfD0x4NzbN9tgHqSAFkX6BW+teDPid8PfiLbC58E+MdL1gBdzR29wDLGP8AbjOHT/gQFVVV4j8beC/B0Xn+LvF+i6JHt3B9Sv4rYY55y7Djg/lXiz9vX9jjwWJDrP7Q/hGcxfeGl3LamfwFqshP4V8J/it4F+N/gDS/id8NdYbVPDusGYWd01vJAZPKleF8pIFZcPGw5A6Z71X7ZH7Vv7cvw7+OV58J/wBnj4LJr2ipY2txbara+F73U52MqfOGdG8pQHDDleAMk9aMf/Ba74vYVm13w7ZSjrnS9E8kFcdtk5/Ugnt2/YT+Fv7TPwn+HfiDR/2nfHLeKNa1LWzfWNxNrc2qTQRNBGjRtJKvyqGjyEVioyxAGea/aA/Y3+BP7T2v6B4h+MGg3+pzeHYJYLWK3v5LRHSQqzCQxkO2CoI+YYyfU14W/YF/Y18HBBpH7PHhO42dP7Vt31P06/amkz0HX39TXhjwH4G8EQG28F+DNC0CJl2mPS9OitFIznGI1HGSTVVVVVVVVU80VvC9xM4WOJCzsR0AGSau7l726mvJQoeeRnYKMAEnJx7VXwZ08aV8IPA+mBQDbeHLCNvk2ZYW6ZJHYk5J9zVf8Fvf+SVfDT/sYLr/ANJxX7PnxA8TfDD/AIJg+APGPhC7it9TtLfZFJLCsq4fUpVYFWyOQSK8HeIoP20P2YfH/hP4x+HtOlWGCe2+1QwYUSCDzIrhFYnZNG+GyDjIHABIr/giT4p1i1+Nvj7wRC8p0rU/Cq6hOvOwT211FHGfrtupfyNf8Fvf+Sp/DT/sX7r/ANKBX7PX/JAfhn/2J+k/+kcVf8FGPhl4R+JH7Ivj+fxLp0Et34X0uXWNJu2jBktLmAb8ox6b1DRt6q59q/4IfeKdYvfAvxU8F3DynTNI1XTr61DZ2iW5imSXHvi1iz9RVfthf8pam/7HDwp/6S6dVajqOn6RYXGq6tf29lZWcTS3FzcSiOOGNRlndmICqACSTwBXxD/4K6fsg+BNal0TTL/xX4yMDlJLrw9pcbwBh12vcSw7xnjcuQeoJHNfBD9vL9m39oHTNWuvAfi+4i1LQ7CW/vdE1K1NterbxKWeREyVlAA58tmxxnGRn9nD9sH4M/tVz6/B8I7vWbk+GUt21Br7TmtlTzzIIwCSck+S/HtVVVVoCj9mj9uKTRx/o3h/xNcmGNQNqC2vSGiA9FjnCrn0jPPeqqqqv2hPH6/Ez4xeJvFdvcCWykvDb2LA8G3iAjjYem4Lu+rGv2ZfAg+Ifxv8L6HPb+bZ290L29BHHlQDzCG9mKqn/A6/bt8WDxN+0DqGnRSh4fDthb6ehB43bTM/4hpip/3faq/4Ju+FDbeFPF/jWWMZ1C/hsIWK8gQoXfHsTMv/AHz7VVV8avjd4O+CHhd9c8R3Imvp1YadpkbgS3kgxwP7qDI3OeAPUkA/Ev4keKPiv4vvfGXiy8867umxHEpPl20Y+7FGCTtQZ6dySTkkmvBPgnxJ8RPE9j4Q8Jaa97qV/JtjRRwg/idz/Cijkk9BXxp8U+HfhT4Yl/Z1+GGoJdMsobxlrsXDandp/wAuycnEMZ4I7sMdmLVVeFfFHiTwfJc3nhnWLjS7q8gMElxat5cwjJBZVkHzIDgZ2kZHB4JFSO8rtLK7O7klmY5JJ6kmtE0DWfEt+ul6Dps99dMpfy4UztVRlnY9FUDkscADkkUifD7wGPN1l7fxlriH5dPtZ2XTbZh/z2nQhrg/7MJVOh81hkV4v8eeKfHM8EniDUt9vZqUsrGCNYLWzT+5DCgCRjp0HPU5PNV8C/2K/iH8U2ttd8WRzeFfDT4cTXEWLq6Xg/uojyAR0d8DkEBulfDD4QfD/wCD+hroXgXQYrNWUfaLpxvuLph/FLJ1bvxwoycAVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX/BIJW1b9p34zeI7l8XL6dIGVBhcy329jjr1QY+tVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVf8ABar4p3B0P4efADQ53lvNavX1m+tYhl3VMwWq4HJDu8/HrEPQV+zj8KYPgd8CfA/wpiVRL4d0aGG7KkEPdMN9w4x2aZ5G/HvX7cHxh/4UZ+y34+8c2t35GqPpradpLK+1xd3R8mNk9Sm8yY9IzX7Fn7BHxn/aQ8C6p8U/AHxUi8CWcGovpaSMZ0kvCiI7kGIg7AXUc8FgfQ1/w6b/AGqP+jvf/Ji//wDi68O6N4x/4J3ft6eHLTxtr0V8mjahbNqWqRBlivtNvYwtxKA/J2rLJ97+OLOeAa/4KU6bqOs/sPfFG20d285LKzuGMZPMUV9byy9O3lo+e2M54r/giJr2kT/Cj4j+Fopk/tSy8Q295PHjDeTNbhIz7jdBL9Pxqq/4KHTwWv8AwUpNzczRxQw33h55JJGCqiiK3JJJ4AA71XxS13S/C3wy8W+JdamWLT9J0K9urp26LFHA7OfyBr/gibod9cftCeN/Ekat9jsPBj2kpxwJJry2dO/pbyfrXxt8ep8MfhR4n8beaqT6dYP9k3dDcP8AJCPp5jJ+FO7yOZJHLMxJLMckn1qKWWCVJoJWjkjOUdGKlSO4I6V4D/a2+PXw+8mCw8c3Gq2UPAs9YX7YhHpub94o9lcV4E/4KPaROIrX4leALi1fGHvNGmEqk+vkyEFR/wADY+1eBv2kPgj8RRHH4a+Imlm6k4FneSG0nz3ASXaW/wCA5HvVVVfHn4GeF/jv4Nfw9rQFtqNqGk0rUVXL2kxH/jyNgBl7jkYIBHwC+Nni/wDZf8cz/Az43x3EGheeFtp5G3ppxYnE0bfxW79Tj7pycA7gf+Ci/wCwR8Yv2tviH4b8ffC7xB4Qt7TRtA+wTW+rXs8Msr+fJIGj8uGRSpWQcll6Vp37F/8AwVitLG10Oz/aejsbGGJLaJU8bXyLBGAFGNsJYBVA6cjHFeC/+COvjrxn4pi8W/tS/tDXPiGY7ftcWlyT3lzcgc7ftl3hlGSf+WRJz/Ca+H/w/wDB3ws8GaT8PvAGhW+j6BoluILKyhyRGo5JLMSzMSSSzEliSSSSTX/BRH9gH4yftcfE7w341+G/iXwZptjo2g/2fPHrd5cwytL58kmVEVvKCu1wMkg5zx3qq/bE/wCCTPjX4vfGzVPid8B/EHg3Q9M8Rr9q1TTtZuLi38u/YnzXhEMEgKScOckEOz9iMfDfwhqnhD4S+FvAWoahGmo6J4ds9MnurM7lWaK3SJnjLqMjcpI3L6ZHatb/AOCT/wC1b8I/El34j/Zp/aJhAlZm87+07vQ79wcnaxh3pJ1IJLqDnoM4rU/2Bf8Agpt8YbT/AIRb4s/H1BoDt/pFvqvi+6uoZB6mGJGWQjHAfGM8EZNfsZ/sDfDT9kGzutZtNRk8UeNtTh8m8165thD5UWQTDbxAt5SEgFiWZmIGTgBRHLFMu6GRXAYqSpzgg4I+oIIPuK/bc+A3i/8AaV/Z1174ReBdS0ex1jVLqzlhn1aaSK3UQ3CSNuaOORgSqkDCnnHTrXwL+COs/sE/sJ/F/RvjtN4P8SI76rqpsrS8mey1CKbT4LeOzkeSKNg00kRjICniRcHJIH7AP7K3i79rH4pXn7ZX7SqSapow1Bp9Js7qPbHq13GcKwToLS32hFjHylkCfdjZTVVVVX7Y37Oi/GDwkPFvheyVvFvh+FjCiL81/bjloD6sOWT3JH8WQ6PG5jkUqykgqwwQRSsyMGRiGU5BBwQa/Y2/aSX4seG18CeL77Pi7Q4BiWRudSt1wBL7yLwH9eG7nFePvi18N/hhaC68deL9P0ssuY4HffPIPVIly7D3AxXjz/goBe6nd/2B8FvAs93dTtshu9SQu7n/AKZ28RJJ7glvqtW/wC/az/aInj1H4seKLjQ9JkYMINTcpsHrHZRYAYf7ew8da+HP7EHwX8EiG716xn8WajGATJqZxAG77YF+XHs++tP03TtIsotN0mwtrK0gXbFb28QjSMeiquAB9Kqqr/goz8NQ8Xh34sWEAyhOlaiyjsd0kDHH/bVST6oKr9kf4nD4X/G3Rb28uRFpetH+zNQLHChJSNjn0CyCNifQN61VVVVVVVVVVVVVVVft++CT4l+CA8SW0O+48L6jFcswXLCGT904HtueNj7JVfsMeNx4u+Amn6VNNvu/DF1Lp0u48lM+ZEfoEkCj/cNVVVVVVzc21nA91d3EcMMS5eSRwqqPUk8AV43/AGt/gL4GEkVz44g1e7jHFro6/bGY+m9f3YPsXFfG79pn4WfErz49E+AGim4kDAatqTmK5OehItijZHXmRhz065OCcgAe3pWi+HPEHiW5Nl4c0LUNVuFXc0VlavO4HrhATirm2uLOd7W7t5IZojteORCrKfQg8iqqqqqqre4ntJ0ubSeSGaJtySRsVZSO4I5BrwF+2B8dvAXlwJ4tOvWUf/LrrSG6yP8ArpkSj6b8e1fEb/gor+0u9ux+F3gT4eQ/uwGGqi6mlDbcEptkRM55G7gdDnv8AvjVofxd+HPhjVrrXdHHiy60e2l1zSbe5QyWd4YlM6CPJbasm8A8ggdTVfFL/gj7Y/F342eMviprPxwl0nT/ABTrdxqS6ZZ6EJpY/OkLuDM0yjOSf4D17458Kf8ABGb9ljRFjk8R+IvHfiOYAeYk+ow20JIz0WKFXAOe7noPevhB8Hfh78B/Adl8NfhfokmlaBp7ySQ20l5NckPIxaRt8rM3zMScZwCeAKqqqqqqqqqv+Ch37Znjv9jzRvBOpeB/C2g61J4nuryG4XVRKREIViKlPLdeT5hznPQV4K1u48TeDdB8SXcUcU+q6ZbXcscedqNJErkLnJwC3eqqq8eXo03wN4i1HzCn2TSbqXeBkrtiY5x+FVXh2y/s3w9penbGX7LZQxbX+8NqAYPvxVf8Fvf+SVfDT/sYLr/0nFfso/tZ/sSQfsY+DPgl8cviVa293pNtIuq6VNZ6hGyOLuWWPEkEfz8FG+Rj1weciv2nv+ClnwS0X4P6l8B/2NtDuJH16CWxn1j+z5LSC2hmGyRoUkxNLO6kqGdV25ByxAA/4JN/sk+LPgf4H1v4t/EzRbjSfEnjaOGGx0y6j2TWWnoS4MinlHlcglG5VY0zgkgf8Fvf+Sp/DT/sX7r/ANKBXwW/4KLfsY+Gvgl4H0bXfjba2d/ovhrT7O+tX0e/aSKeK2jSRAqwHdhlIyuQcZBI5r9uv/gpJpX7RPhJv2dv2aNB1vUrPxLNHBqepS2LLPfgOrJbWkAzIQ7Ku5mAYgFQuGJr/gm1+y3rf7MfwGNv43tRbeMPF94NS1e2yGNkoQLDbEjgsq5ZvRpGGSADVfthf8pam/7HDwp/6S6dVf8ABYP9o7xVqfjHRf2T/Ad7crayW8F7r8FoxD31xM3+jWrAHJVQFk2YwzSRnqor9l//AIJsfs+/Bn4f6db/ABF+Hfh7x34zu7ZH1i/1ywjv4Y5iMtFbxShkSNCSobbubGSeQo/4KD/sMfD/AMGfDjUP2lP2cNCg8B+LPBKG7vrXQ4/s1te2Z+SYrCuEiZEZmOwBWTerBsrj/ghj/wA1s/7gH/uRqqqq/wCCi/gQyaH4X+KenKyXWmXR065ljyG8twZImyOgV0cZ9ZBXwa8dxfE34WeGfHCSK0up6ejXIXotwvyTKPpIrj8Kqqr9qD4if8Kz+CniHWrefy9Qvof7P08g4PnTAruHuqb3/wCAVX/BPfwLb6Zonij4s6siRLKRp9pNJxsijAknbP8AdJ8oZ/6ZmvH/AInl8beOfEHi+Utu1nU7i7Ct/CJJCwX6AED8Kr9kPwj/AMIf+z34StZIdlxqVs2ozEjBYzsXQ/8Afsxj8Kqvj5+0L4Q+BWgGfUJEv9fu4z/Z2kxuA8h6B5P7kQPVu/IAJzj4i/EXxZ8VPFV14w8Y6ibq+uThVUFY7eMfdjjXJ2oM8D3JJJJJ8J+EvEPjrxDZeFfCuly6hqeoSeXBBGOvqSeiqBkljwACTXj/AFjw/wDsf+B5/hh4F1OO/wDif4jtl/4SDWoSf+JZAwyIoT1UkH5f4sfOcZQAksck5J9aqreHedzDgdK8O+FvEfi/Uk0bwtod7qt7IMiC1haRgO5OOgHcnAHerrw34M8FSFfG+vrq2pRcNoug3CyeWw6rNd4aNMekQlPUEqea13x3q+r2TaLYwWujaMSCdN05DHE5HQysSXmYdjIzEZOMDiq+EnwA+J3xpvhB4N0B/sCPtn1S6BitYfXL4+Yj+6oLe1fA/wDYz+Gnwl+z63rUK+KPEsYDC8vIh5Ns/wD0xh5AI/vNubIyCvSqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv+CH6LceNfi3fTjzLn+ztNHmvy3zS3BbnryVBP0FVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVSqP2wP+CvHlHbfeG/AOq4IxuRLfSRz0+8kl6PoRN3FV/wAFsfjJJcah4D/Z/wBKu8pCj+INViQ5y7boLUHHQgC5OO+9T6Z/ZD+D6/Aj9m3wF8M5rUQahp+kpNqilMH7bOTNcA9ziSRlBPZR0xgV/wAFsvgy11pPgX4+6ZagtZSP4f1Z1XLbH3TWpPoqsLkEnvIo4zz+xV8QNG/am/Yr8O2/jFI9TNxo03hjxJbu2TM0SG3ff/tSwmOQ/wDXWta8IftIf8Epf2hJfGeg6XLrHhC9d7S31CaJmsNcsnbcsEzKP3Nwu0HHDKyEruQnd4f/AOC037NN7oi3niTwL8QNM1NUzLZW1pbXSFvSOUzpuHuyp9K/Y7/bZ8KftkSeNJPCvg/UfD9v4TuLVIxqFwjzXUc6yESFUG1CDEw2hn7EkZxX/BQP4d33xh/4KVXnwt0bUbe0v/EraPp8E8ylkhlks4QpcDkLyM9wDnBr4Qf8FSNW+BFinwP/AG0vhl4t0/xj4UjFnJqtlAkr3iIMJJNHI6ZLKB++jZ1kyGAAOa/a+/4KS6/+1loA/Z4/Zp+HviaCy8TzJBfSzQh9R1NQ2RbRQQM+yNiAWO4lh8pCqWDf8E7v2Rbz9k/4Nz2vi0wP438Xzx3uuiFw62oRSILUMOH8sM5LDjfK4BKhSf8Ago38QPsXhvw38M7O4xLqVw2o3qq2CIohsjBHozs5+sVVVVXgf46fF74cGNPBvxB1exgjGFtWm8+3H/bGTcn/AI7Xgf8A4KMeOtN8u28f+DNL1uIYDXFjI1nNjuSDvRj7AKK8E/txfAHxgI4b/Xrzw3duMeTq9qUXPf8Aexl0A92K/TtWieIdA8TWK6n4b1zT9Vs3+7cWVyk8Z+jISKr9oX9n/wANfHrwi2l3qxWeu2Ss2k6psy0D90bHLRtjBHbqORXwM+PXjb9mjxpJ8DPjylymiQyhLW7lYyf2eDwskbfx2ze33eSAMMtWV7ZanZQajp13DdWt1EssE8Lh0kRhlWVhwQQQQRVVVVVVX7U37akWiG8+HHwb1FZdRG6HUNdibK2x6NHbn+J+oMnRf4cnlf2Vv2o9S+DfiI6D4uvLi88IaxcF7zcWkkspm63CDqQTjevUjkcjB+PP7Zvxl/aQ/aa1Dw/8GPix4h+Gnwv8CW0/9p6zaTzWgSyhZftOoXCKQzs7BI4Im+Ylo0AV5XB+FOk/GP8A4KffHZdC1/xX4stPgn4LNubu2vNTkl228QKwrISSsuoXADl5iDt3SEfKqofD3h/RPCeg6f4Y8NaXb6dpWlWyWtlZ26bI4IkUKqKOwAAFVVVVVX7cn7NreH9QuPjR4J0//iV30udctYk4tp2OPtAA6I5wG9HIP8XFeFfFGveCvEWn+K/DGoyWOqaXOJraeM8qw7EdCpGQVPBBIOQTR/aZ/ah/aLuG8MfCXw//AGSscSi8fSFw6Fhjc9zKcQglWK4KngjLYrwD+wDdalef8JD8bPHE99dzv5k9np0jO0h/6aXEgyT6gL9GrwF8KPh18MbP7H4F8I6fpWV2vNHHumlH+3K2Xbp3Jqqqqqvi/wDD60+Kfw08QeA7oIG1SzZbaRxxFOvzRP8AQSKpPtmtQsLzS7+50vUbd7e7s5nhnhcYaORSQykeoIIoEggqcEdK/Zh+KY+Lvwc0TxDdXSzatZJ9g1Xn5vtEQALH3dSj/wDA6qqqqqqqqqqqqqqrx34Us/HXgrXPBt8VEOtafNaMzLnYXQqG+oJBHuK1PTrvSNRutJ1CIxXVlO8E8Z6o6MVYfgQa/wCCd/j1dE+JWseAbqciHxLYebbqT1uLfLYA94mlJ/3B+FVVVqWp6Zo1lJqOr6jbWNpCuZJ7mZYkQepZiAB9a8fftr/AzwX5ltput3Hie9TjydIi3xg+8zFUI91LfSvHf/BQP4oa75tr4H0PS/DNuwISZx9suV9wzgR/h5Z+teMfiV4/+INx9o8aeMdV1g7srHdXLNGh/wBmPO1fwArwP8Bvi/8AEZY5fCPgDVbq2kxtu5Yvs9uR6iWTah+gJNeCf+Cd3i6/8q5+IHjbTtKjIBa106JrqXH90u21VPuN4rwX+xZ8BPB/lzXPhufxDdRj/XaxcGUZ7/ul2xkfVTWk6Jo2gWSaboOk2WnWkf3Le0t1hRfoqgAV4x+Gfw++IUAg8a+DtJ1jau1JLq2VpIx/syfeX8CK8df8E+fhprQkuPAviHVPDdwclYJv9Ntx6ABiJB9S5+lfGz9lz4gfAnQLvxl4s1Tw6/hqzIEuqDU47dELHCqVmKMXJ4CruJJwM1Z31jqVut3p15BdQP8AdlhkDqfoRxVVVVVVUM01vKk9vM8UsbBkdGKspHQgjoa8B/tcfHbwCsdta+MX1mzjAAtdaT7WuB0AckSAdsBwK8Bf8FDvCuoeXZ/EfwbeaTKeGvNNkFzDn1MbbXUewLmvA3xh+F/xKiVvBPjfS9TlZc/Zlm8udR6mF8OPxWqqqqqqqqqqv+C43/IqfCP/ALCGq/8Aou2r4T/8kr8G/wDYv2P/AKTpVVVfGy4ktPgz49uoSPMh8Mak6ZGeRbSEfyqtOtkvdQtbORiqzzJGSOoBIHFVX/Bb3/klXw0/7GC6/wDScV+zH+xL+xR4o/Y/8CfHH42eEp7e91C0kOqakms30azSfa5YkJjjkIXhUX5FA7nua8LeIv8AgnR+zhcDWPgz8K7LUNbtmJtruDT57m4hf1W5vmLoOOqE/Q1+zr8bo/jx4Ju/FL6XDpl1Z6lLay2aTGTy1AVoyWIGcq45wASD06V/wW9/5Kn8NP8AsX7r/wBKBXwg/wCCZP7GPi/4WeBPGWufC27lv9W8N6deXyp4gv0SeaS2jeRiBNldzMThSAM8Yr4Sfswfs/fAlhP8J/hPoGgXewob6OAzXZU9VNxKWlIPoWxVV+2F/wApam/7HDwp/wCkunVX7UUv2D/grja3njAoLKPx54XlmaZRs+yBLHBPYr5Ywfoc85qv22nRP2RPi+zuqj/hENQAJOOTCwH64Ff8EMf+a2f9wD/3I1VV45/ah+LXhb43eLtY8H+NbsaautTxw6dcP9otHiicxriJshdyoMlNp561+zv+1H4V+OVoNHu449G8WW8e6fTWkytwoHMkDH7y9yp+ZfcDcfj34FHxJ+D3irwfHEJLm7093tF25JuIsSRY+roo/Gv+Cc/jn+0/AviL4f3MwMuh3y3dsGPPkzqQQB6B4yT/ANdBVVVf8FAPiaNf8d6b8NNNuQ1p4bh8+9Cng3coBAP+7Ht/GRh2q0tLnULuCwsoHnuLiRY4YkGWd2OAAPUkgV8TrS1/Z2/Y4vvDFpKi3kOjDTQyHHm3V0dszqfrJK49lqvC3h+88WeJtI8LacCbrV76G0hAGfmkcIOPqa0zTrTR9NtNI0+IR21lAkECf3URQqj8gKr9pL9rXw78IILjwp4Te31fxgyEGINuh04no0xHV+4jHPQnAIz4o8U+IfGuu3fibxTq9xqWp3zl57idssx7AdgoGAFGAAAAABXgXwH4n+JHiO28LeEtOa6vJ+WY/LHbxj70sj9ERR1Y/qSAZPHvwc/ZC+Ht1o3w41zS/F3xR1OHybrUrbE0Vqx6nfyojQgYjB3MwBYAdNV1TUdc1K61jV76a8vr2Zpri4mYs8sjHLMT3JJqq+GPwg8e/F/VZtK8D6L9r+xoJLy5llWGG1Q/xSOxAHQ8ck4OAcGr7wp+z98JSYfGXiyf4ja/ADnSPDkv2fT43H8Mt4Rucdf9UuQeuK8Y/Gzxd4o02TwzpMNh4W8Muf8AkCaFD9mgkHYzMCXnPvIzc8gCq+Hvwr+IHxV1X+x/AXhi81WZSBLJGu2KAHvJI2FQfUjPbNfBj9gDwl4aMGufFvUE8R6iu110y33JZxMOznhpucddq9QVYVp+nafpNjBpmlWNvZ2drGI4Le3iEccSjoqquAAPQVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX/AAQ6/wCRq+Lf/YP0r/0Zc1VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVfHT4k2vwd+DXjT4oXTJ/xTOiXV7CjnAkmSM+VHz3aTYo92Ff8EU/hvd3kXxM+Pmt757rUbqLRbS5kyWdh/pF2ST13M9sfqpqtd+E/ws8U6yPEXib4aeFdX1YBQL+/0a3uJxs+7+8dC3HbniqrxF4Y8NeMNKfQ/Fnh7TNb06VlZ7PUbRLmFmU5UlHBUkHkccV4X8F+DvA9hJpfgrwno3h+zmmM0ltpdhHaRvIQFLlY1ALYVRnrgD0rUNO0/V7GbTNVsbe9s7lCk1vcRCSORT1VlbII9jUv7IX7Kk+onVZv2b/ho9w2d27wtaFGJ6kp5e0nnqRmtA8NeHPCmmpo3hbQNN0ewiJKWmn2iW8SnuQiAAdB2q4+Gnw4uvFI8c3Pw/8ADc3iRXVxrEmkwteBlUIrecV35CgKDngADpXjv4T/AAu+KVsln8Sfhz4Z8UxRKViXWNKhvDED12GRSUPuMGvAPwZ+EXwqWRfhn8MPCvhZpl2zSaRpENo8o/22jUM3QdSelV+1b8Q/+Fk/HPxJqsE4ksdNm/s2xIbI8qDKllPoz+Y4/wB+qqqqqrRdf13w3erqXh3W7/S7tPu3FlcvBIv0ZCDXgv8Abe/aA8IeXDd+JLXxHaxgAQ6xaiQ4/wCuibJCfcsa8F/8FHfCd4sdv8QPAWo6bJjDXOmTrdRk+ux9jKPYFjXxH1b9l39rjwzHo8HxH0ix1+2BOl3lz/olzbueqbJthlQ8ZUZHcEEZr4SfGv4i/si+MP8AhUnxjsrm58Ks5a3kj/e/Z0ZuLi2b/lpCxBJTqDkgBtytoeu6N4n0e01/w9qdvqGnX0Qktrm3kDpIp7gj8sdjwaqqqqqv2zf2T5J5L34w/DDSi7tum17TIF5Pc3MSAfUuo/3gPvVXj7wnq/jLwleeDPD+uWmhxa1qNrc6m7w7EvTCGWL7Q8amRkjErsF+YZ52kgEfsr/BL4bfAL4JeH/AfwvvLbU9NMIurnWISpOr3MijzLpipIO7AAGTtVVXOFqqqqqqmlhgheeeVI441LO7ttCgDkk9hivjT+2b8GvDFhqHhbSbePx1d3EL29xa25BsmVgVZJJiCHUg8hA4PQkV4h8KeILGxj8XSeEdQ0vQNWuZBp00sMnkN/EI45H+/heM5OcH3qvg58XPE3wV8cWfjPw3LvEf7u9s3crHeQEjdG2PoCDztYA84xXw3+Inhj4q+DrDxr4SvhcWN8nKnh4JB9+Jx/CyngjvwRkEE1VVVVVft4fCo+B/iz/wmenWwTS/GKNc/KuAl2mBOP8AgWUkz3Lt6VX7BnxaXwR8UJPAmq3ezSvGCLDFuPyx3qZ8k+28Fo8DqzJ6VVVVVVVVVVVVVVVVfttfD/8A4Qf486te20Gyy8TxLqkBA43vlZufXzVdvo4+tfDXxldfDzx/4f8AG9nuL6NqEVwyL1kQMN6fRk3L+Naff2erafbapp86zWt5Ck0Ei9HRgCrD2IINVXxH/aq+CfwzEttqfi2PVdRi4On6QBdSg+jEEIh9mYGviJ/wUF8fa35tl8OfD9l4dtiCFu7rF3c+zAECNfoVf614u8feN/iBfDUPGfinU9auAf3f2u4aQR57Iv3UHsoArwF+zF8bviMI59D8DXlrZSDIvdSH2SHH94F8M4/3A1eBf+CdenQiK6+JPjyW4cEF7PRofLTjt50gJIP+4p9/TS/hn+y/8BokvLqz8IaBNEA4v9dvo/OB/vCS4YleR/DgV4A+OPwc+LGp6jo3ww+J3hvxXd6RGkl6ujajHeLArlgpLxkryVboe1V+3T+1jrv7IHw10Xx5ovw+tvFJ1nWRpZW41BrZbaRoZJUbCxtvyInGMr079m/4KLf8FHfiqGT4T/s3pb2spIiudN8H6hfugyPvSuxi4GBkoBz7iv2O9T/4Kd+IvjppniD9pjTtSg+Hj2N1HdQ3MWmWSwyGMtCfIh2zMd4C5KnAbk4qv2hPgZ4V/aR+EutfB7xnfX9lpetPbtJdaeUW4haGeOZTGXVlBJj2nKnhjXw9/wCCVP7Jfw+uY7yLT/GGtTxgAvfeI54DJgY+YWnkg+uOntjivHf7BHwe8Sq8/hG61LwpdEfKIZDdQZ9THId35OBXj39hL40eFTJc+Gk0/wAV2a5KmylEM4UdzFIRz7KzV4g8MeJPCd+dK8UaBqGk3ijmC+tngfHqAwBx71VVVVVVHJJDIk0MjJJGwZGU4KkdCD2NeBf2r/jr4AMUNh43uNUs4hgWerj7WhA6Dc37xR7KwrwF/wAFEPDt6IrP4k+C7rTZTgNeaVJ58RPcmN8Mo+jOa8C/Gj4V/EtEHgrxzpeozuMi083yrgfWF8OB74xVVVVVVVVX7c37D037aUHgu2X4rN4Ni8IvfOyf2J/aAvGuBAAcefFsKCBv72fMPTHOgaRBoGhaboNsQYtNtIraMhdo2xoFGBk44HSqqq/aNJHwG8fFT/zL93/6LNV4Ijjm8aaBFKiuj6pbKysMggyrkEelVX/Bb3/klXw0/wCxguv/AEnFfDz/AJRM+Cf+uKf+nSSq/wCCfPj3+xviLq3gC6nxB4jsvOt1J/5eLfLYA7ZjaQn/AHB+H/Bb3/kqfw0/7F+6/wDSgV+z1/yQH4Z/9ifpP/pHFVVX7YX/AClqb/scPCn/AKS6dVf8FeP2RvE3ii5sf2n/AIbaRcX82l2S2fii1tELSxwxkmG9VRywUMUfH3VWNsYDkfs0f8FgfhNfeAtN0D9o0axovirS7ZIbnV7axN3baoVGBNti+eORsZZdm3PIbB2r+2Z+3Xf/ALXHw78UfBz9lzwrrlx4W0vTpNW8Z+Jb23+zoLG2Uy+Uq8lFd0Ubn2s7AIFwWNf8EMf+a2f9wD/3I1VXn2r7JN9gERufLbyRKSE34+XcQCQM4zwa+Kvws8e/CjxNJonj7TGgurjdLDco3mQ3a55eN/4uTyOCM8gVoGv6z4W1qz8Q+HtRmsdR0+US21xC21o2Hp7diOhBIPBr9nf416d8cfh9b+IFEUGsWRFvq9oh4imA+8o67HHzL6cjJKmvg+v/AApD9ufW/Aozb6Zrtxc2cKYwqxTgXNsAPYiNAfc+pqqr4geM9L+HfgnWfG2ssPsuj2bzshbaZWAwkYPq7FVHuwrxN4h1Txd4i1LxRrVwZr/VbqS5uHPd3Yk49AM4A7DAr9iL4Xnx58X4fEl9bCTS/CCC9lLDhrg5EC/UMDJ/2y96/wCCkHjjytP8J/De2m5nlk1S7TOMBQYoeO4Jab/vkfhX7DPgk+Lfj7pupTQh7Xw1azajLuHG4Dy4/wAQ8isP901e3tlptnNqGo3kFra20ZkmnnkEaRoBkszHgADqTX7Rf7cM18LrwV8Fbp4YCGiuvEGCrydiLcHlR/00PP8AdAwGM0stxK888rSSyMWd3YlmY9ST3Jql1zU7HTp9KtdRuIrW7IM9vHIVWYj7u8D7wHbOcZOMZNMxc7mOSaqqXxr4oi8MjwbaazPa6K0hlmsrdvKS4kP8cu3BlIHA35wOBiq8BfDLx78UNWGi+A/C97q9yCPMMKYjhB6GSQ4RB7sRXwg/4J6aHpvkaz8ZNa/tW4GG/sjTnaOBfaSXh3+i7MHuwrw74Z8O+EdJh0HwtollpWn24/d21pAsSL6nC9Se5PJ71VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV/wQ6/5Gr4t/8AYP0r/wBGXNVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX/BY34oDwd+zBZ/D+0utl5481yG3kjBwWtLb9/IfXiVbYf8C/P/gnv8Ll+E37IPw60Oa3Ed9q2mjWr4lcM0t4TOoYeqxvGn/AKqqqqqqqq1K2uLzTrq0s71rSeaF0iuFUMYWKkBwDwSDg49q+IH/BPn4t6C8t34K1fTPFlvklUL/Y7k9+UkJT/wAiZ9q8XeAfG3gK9/s7xp4U1TRZySEF7atEJMdSjEYYe6kiqqqqqqqqbUtRubOHTrjULmW1tyTDA8rMkZPUqpOBnA6V8BP2m/HfwFvJLfTAur6BdHdcaPcylU3/APPSNsExv2JAII6g4BHg3/goX8H9c2QeLtH1rw1McbpDELyBf+BR/Of+/deDvjF8K/iCI18G+P8ARNUmkGVtortVn/GJsOPxWqqqqv2xf2m/D/7KnwV1T4hXzW9zrlwDZ+HdNkbm8vWU7cgc+WgBdzx8q4yCy14R8b6yPCmg+I/i1rdrb6v44vbmbRo2jEct1bK5VrmRVAWONpvMijOPnMUnA2AtX7Nf7U/iX4GajHomqGbVvB1zLm4sN2XtST80sBPQ9yhIVvYndXg3xp4X+IXhy08V+D9Yg1LTLxcxTRHoe6sOqsOhU4IPWqqqqeaG2he4uJkiiiUs7u21UUDJJJ6ADvXxc/bl+GfgXztJ8Dr/AMJfq6ZXfbybLOJvebnf64QEH+8Kt/Cf7Vv7Xs8d/wCI7yTQvCUrB4xOrWllt6gxwj55z1wzZGeN4r4Ufsa/CH4aGLUNS08+KtYQA/a9UiVokbuY4OVX1Bbcw7GviH8O/C/xQ8HX3gfxVYLPp97HtBUAPA4+5JGf4WU8g/gcgkV8ZvhD4m+Cnji78G+I4zIqEyWN6qFY72An5ZF9OmCuTtYEZPU1+zX+0LrXwG8XrcOZrzwzqbqmraepz8vQTRg8CRRn0DD5Tjgr4c8RaJ4u0Gx8TeG9Riv9M1KBZrW4iPDoR+YPYg4IIIOCDVVVVVV+1P8ACYfF/wCDuraJZ2xl1jTB/aGlBRlmnjU/ux670Lpj1YHtRBB2kYI7GrS7ubC6hvrK4eC4tpFkhljba0bqchgR0IIBr4A/FW1+Mvwr0bxojx/bnj+z6nEnHlXcYAkGOwbhwP7rrVVVVVVVVVVVVVVVf8FCPh0PEfwtsPH1nbhrvwreYnYDn7LOVRvriQQ/QFveq/Y1+L+j61+z7bjxTrtpYt4Lc6fd3N5cLEiQD5oGLMQFXYQg5/5Zmvil+3x8PPDBm0z4c6bN4pv0yv2pybe0Q9Mgkb5PoAAezV8Tf2lfjD8V/NtvEfiuW202Xg6Zp2ba3x6MoO5x/vs1eJPGvhTwjD5viLXLWzJGViZt0jD/AGUGWP4CrT9qb4a6ffPJqHg3xDrdtGQEgt7+LTjN6nzWjm2D28s59q/4Jp+Pvht8dfA/iHxfpvwR0XwpqXh/VltYJjM+oTSRNEGEnnzLuB3iQEIFX5RwKr9vb9k79oT9pjXfB7fBn4tReE9J06zu4datrvWbu1imZnjMTLFAjCRuHBLYwAvNeG/+CIV1dzfbviD+0YZJZGJli03w+XZjkc+dLP1xnqnU9a/ZN/YI+Fn7IOs6r4l8D+LPFes6prlgtnenVbiAwbFkDgpHHEpVsgDLM3U9M1UkMMrRvJEjtE26MsuSjYIyPQ4JH0J9aqqqqrXvDnh7xTp76V4l0Ow1azf71ve2yzIf+AsCK8e/sL/BPxaJLnw/bX3ha9bkNYSmSEn3ikyMeyFK8d/sD/F/w2JbnwleaX4qtUBKrDJ9luCB1zHIdvTsHJrxD4c1/wAJ6tPoPibR7vTNRtjiW2uoTG6+hwexHIPQ1VVVVVUrMrBkYhlOQRwQa8BftSfHD4d+VBpHji6v7KLAFlqv+lxbR/CN/wA6D2RlrwH/AMFEtEuVjtPiV4HuLKTGGvNIkE0ZPqYpCGUfR2NeA/jv8I/iYI4/B/jrTbq6kAxZSyeRcZPYRSbWPpkAj3qqqpRIYmEJXzNp2bhxntn2r4LftB+Evi/FPo4I0jxXphaPUtFnky8bodrtGePMTI6gZHcDjNVVVVVX7UjMv7PfjsqxB/slxxx/Euar4WRRzfE7whDKgdH16yVlYZBBnTIqq+MXwA+D37QGmafo/wAYPBFr4ks9JnaezinnmiEMjLtZgY3UnI45zX7Qnwe0Lwj+yZqvw4+FehQ6VonhqFLm206NnkEcEc/nShWYs3d35J6EfSvAfi7UPAPjPRfGelnNzo17HcKmceYFPzIfZlyp9jXxD/Z3/Zy/altfD3jr4keALLxWq6craVcz3M8RS3lAkAAikUc5B5yRXhrS9D0Tw5pWi+GIoY9HsLGG306OGQui26IFjCsSdy7AuDk5HeqqvFf7IP7N3jj4n/8AC5/FXwtsdQ8afa7a7/tZ7q4V/OtljWF9qyBMqsUY+7g7ec81TOilVZ1BY4UE4ycZ4/AGvGf7D/7JPj/XH8R+KPgJ4Vn1GWTzJZra2az8585LSLAyBySeSwOe+a/bw1T4L/sr/sW+MfA/gvwv4f8ADDeM7Z9H0nSNMtI7Y3c0wCzSlUALbItzM5zyFBOWFf8ABFj4dan4e+BPi74i6hbNDF4v15YbHepHnQWkZUyKe6+bLMn1jaqqvjF8JfDXxm8E3nhDxDCiyMpexvAmZLKcD5ZF9uxHG4EjvmvGXhHXPAfinU/B3iS0NvqWlXDQzp2JHIZT3VlIYHuCD3r9l34xSfBv4pWOpXt0U0HViLLV0J+VYmPyy/WNsNnrt3Afer9u/T7vwF8avAvxe0dNsksMbKynG6ezmDgk+6yRj6LWm6ha6vptrqtjJ5lteQJNC2PvI6hlP5EVVf8ABQL4vpc3Wn/BrRLzK2pW+1nYf4yP3MR+ikuR0+aM9RVfso/CVvhL8ItOsdRtvL1rWj/aGphlw0buo2RHuNiBQR03bj3r9rbx4PiB8evE1/b3AlstLmGmWhHQJB8jYPcGTzGB/wBqq/ZC8bfD74B/CvxD8UfHepBL/wARXgttL06EB7m7htwctGueFMkjKWbCgx8noK+O37Tnj344Xj2V1MdI8Nxvm30e2kO04PDTNx5rdOoCjsByTVSyrGuScnsKdi7bmOTVVVfDr4P/ABI+LGoDT/AfhS91IBtstyE2W8HrvlbCLxzjOT2Br4Sf8E9PDWkrDqvxf1s61dDDf2XpztDbL7PJxI/4bPxrw74Z8O+EdJh0HwtollpWn24/d21pAsSL6nC9Se5PJ71VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVf8ERCLHx38XNIu8x3i6dp2YWHI2Szq/wCRZR+NVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX/BTm7uv2gP24/hf+zZpEryRadHZ2NwqnmC41CdXmb2At1t2J9j6VaWltYWkFhZQJDb20axxRIMBFUYAA9AABVVVVVVVVVVqGnafq1nJp+qWNveWsy7ZILiISI49CpyCPrXjv8AYp+APjffPb+F5PDl22f3+iS/Zx/36IaMD6KPrXjr/gnP4500Pc/D7xnpmtxDkW1/GbOb6AjejH3JSvHHwR+LXw4MjeM/AGr6fBFnddeR5tvx/wBNo9yf+PVVVVVVVVAkEFTgjpXg79oX41+AjGvhn4la1DDF9y2uLj7VCPpFLuUfgK8H/wDBRf4k6X5cHjPwfouuxLwZLZ3spm9yfnT8lFeDv2/vgd4hWOHxGNY8M3B4Y3dqZ4gfZ4dzY9yq14S+KHw48eoreDfHOiau7DPlWt6jyr/vR53L+IFVe3lpp1nPqF/dRW9taxNLNNKwVI0UZZmJ4AABJJr9pn43wftu/tGaz4v1vVL2x+DHwytZHR4sqxsEkClow3Aur2by44wR8u+Pd8sTGv8Agnr8ANT/AGvfj/f/ALSnxO0SCHwT4JuYI9J0pIybRriFFFpYxK2cwWsSxEg5JIjDbt7mv2qv2LU1k3nxI+DemJHfYMuo6DBHtW4wOZLdR0f1jHDdR83DTQy28r288TRyxsVdHUqysOCCOx9q+Cvx38d/AzxCNX8KXvnWNww/tDSp2JgvFHqP4XA6OOR7glT8FPj54C+Omhf2j4XvfI1G3QG+0m4YCe1Prj+JM9HHB74OQKqvjL+2V8L/AIXifSdCuF8Va/HlfstjMPIhYcfvZhkDHPyqGORggdaj0r9qn9sadbu/nOheDpHBQPvtbHaOconL3B9GO4A8blr4RfsffCX4WiDUrvTh4m12IA/b9TjDJG/rFDyqc8gncw/vVVV8fPgb4c+O/gmXw3qoS21K13S6VqITL2k2O/co3AZe/B6qCPG3grxH8PPFGoeDvFmnNZappsuyaJuQeMqyn+JWBBB7giq/ZQ/afvPgprY8MeKJ5bjwXqc26dFUu9hKcDz0HUrwN6jqORyMGyvbLU7KDUdOu4bq1uollgnhcOkiMMqysOCCCCCKqqqqqv20fg//AMKu+Llzqml2vl6H4r339ntXCxyk/v4h9HIYAcBZFHaq/YV+Mw8AfEg+AtavBHoni9khj3n5Yb0cRMPTfkxn1JTPC1VVVVVVVVVVVVXir9rX9l/wUzxeJP2gPANrNGSHt116CaZSDjmONmccgjp2NeKv+CrH7E/htXWy+JOo+IJowcxaVoV0cnGcBpUjQ9hw2OfrjwF4z0T4keBvD3xC8NSSvpPibS7bUrEzJsfyZ4lkTcuTtbawyM8HIr4kWPhbVfAWv6T401K0sNEv9Plt7y6upViSFHUruLMQAQSCDkc4q/tUsr64skuYrhYJWjWaI5SQA43L7HGR9atbiVFa2ErCKRgzJuOCwBwSPUZI/E+tfCz9kf4x/FEw3sehf2BpEgz/AGhqymEMPVI8b3yOhA2n+8K+GX7D3wf8DCG+8TW8vi/U0wS+oqFtlb/ZgBwR7OXr4gfsX/svfFTxjF498f8Awf0nWNagsY7GOSWadIlgjLFF8hJBFxuPOzPQZ4FeFP2dfgD4FKv4N+CfgXRpV6TWfh+2ikPOeXCbjyB1PYUiJGixxqqqowqgYAA9BVVVVVVVVVTSwwQvPPKkccalnd22hQBySewxXxT/AG2fhF8PzLp3h26bxfqseQItOkAt0b/anOVx/uB/wr4mfti/Gr4jGWzt9e/4RrTHJAs9HLQsV9Gmz5jcdQCFPpUskk8jTTSNJI7FmZjksT1JPrVeDPgZ8XfiFELjwj8PtXvrcrlblofIhbjPEkhVCcdgfT1rxX4E8Z+BrsWPjHwrqmjTMSEW9tXiD47qSMMPcZFVVVVVVAkEFTgjpXgX9pn43fDwxRaH49v7iziGBZai32uHb/dAkyUH+4VrwL/wUUtnWO1+JXgN424D3miy7gfU+TKcj/v4a8CftEfBj4jrHH4Z8faabuQgCyvH+yz7vQRybS3/AAHI96qv2mtJ1b4W/tI+IdQ8PXtxptxLerqthdW7mN0MyiRmUjpiQyL+Ffs2/tn6T46Fr4J+Kl1baZ4hO2O11I4jt9QboA3aOU+n3WPTBIU1X7bP7cek/sYf8IZ/afw6u/FX/CYfb/L8jUltPs32X7PnOY33bvtA9Mbe+eNKvl1TS7PU1jMYu7dJghOdu5QcZ/Gqqq/al/5N68df9gp//QlqvhR/yVLwb/2H7H/0elVVXlpa6haTWF7Ak9vcRtHLE4yrowwwI9CCRXxh+Ht18LPiVr3ga5VymnXTfZXfrLbt80T59SjLn3yO1V8Iv2oz4O/Zm8U+CbrUPL8Q6On2fw+d2HeO5YglfeEl3+hQdq/Y18ZDxj8AdASSYSXOhNJpc/P3fKOYx/36eKqqqr9uX9mb4iftQfDjRfDPw2+J0Xg/VPD+sDVYGljlVLqZY3jjBmiO+HaJHIYK/J6cZqP4K/8ABZ/wXEuhaF8WTrlrFyt22v2d2W+sl4glP414S/4JX/tG/G7xvbeOP2zfjhJe28LAPaWuoyajeyRBsmJJJFEVshJJGwOBk/KDXgvwZ4X+HfhLSfA3gvRrfStD0O0S1sLOAELFGgwBk5JJ6liSWJJJJJNVVV/wUE+EiXOm6b8YtItgJrMrYavtX70bH9zKfoxKE9TvQdqr4h+IT8Zv2JNI8Q3Unna38PNWhtL6QklzER5SMc84ZZYCT3ZD71+yX4r/AOEw/Z78HX0k2+exszp8wJyVNu5iUH6oqH8RVfFn4kaP8JvAGreOdZYMlhDi3gzg3E7cRxj6tjPoMnoDXibxFq/i/wAQ6j4o167a51HVLl7i4kPd2OTgdgOgHYADtX7H3wgb4p/Fe0vtRtPN0HwwUvr8sMrI4P7mI+u51yR0Kow718ZfHcXwy+Fnibxw0irLpenu1sG6NcN8kK/jIyD8allknkaaaRpJHYszucliepJ7msZ4FICI0QsTtXAB7f5NV4R8BeNPH9//AGZ4L8L6lrNwCN62luziPPd26IOOrECvHn7P9z8G/D0Or/F3xJZ6fq1/GTp3h3TnW6u5iMgtK4Plwxg4+cGTPQAnOHZnbcxyaqq+Fv7N/wAXvjA8U3hPwtLFprnB1W/zb2qj1DkZf3CBiPSvhR+wF8N/CXkap8Rr6TxbqSYb7NtMFnG3psB3SYP94gHutaZpWl6JYQ6Voum2thZW67Yba1hWKOMeiqoAA+gqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv+CQpfSP2ovjN4amVXlTTpd0qnjMN+EOB778/hVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV+xwq/tHf8FT/AB78ZJW+16b4bn1XUrKZhlWjUiwsx6Z8mRWH/XPI6VVVVap4s8K6Jn+2vE2lWG3OftV5HFjHX7xHqPzrU/2gvgZpO4Xvxd8J7kyGSLVopmUjggqjEg+1aj+2P+zZpuVm+J1tKwyAtvYXU2SPQpGR+OcVf/t7fs9WYP2bVNbvsAYEGlsufp5hWr7/AIKL/BmHKWPhXxhcsGxlra3jUj1B84n04IFXn/BSXwgn/IP+GOsTfMQPOv44uPXhW59v1q8/4KWOVdLD4NqDn9282v54z3UW/p70P+Ci3jrUGLaP8IrB0QYcfbJZiD9VQYoft1fH66Hn2HwXsWgf/Vn7DeScf7wYA/lQ/bF/a6mAltvgdYNE43Rn/hG9RbKnpyJueO9f8NR/ts6n/pGnfBCQRr8pFt4Rv3XI9y7HPI71/wANJ/tz/wDREb7/AMI2+/xr/hpP9uf/AKIjff8AhG33+NfEv9tT9qDwHoRX4j6BYeB4NXSS1tL/AFPRZtPYyFDkwtOwUuoO4DBxgEgivhR8KPHmheD5dR0T4V+IvG/h7W0jubLUtQ068u40Xn95b3EGwlWyM/MynAwBzmX4VfFCSRnT4YeKIwzEhF0a5wvsMqTj6k1J4I8aRRtLL4Q1pERSWZtPlAUDqSdvSri1ubSTybq3khkAzskQqfyNVVVVVVVSO8brJG5VlOVYHBBHcV4P/aQ+OXgXy08PfEzWRDHwtveTfbIlHoEmDBR9AK+N/wC1r8avjb8GNa+D9/qOkaOuvIsF7qtjZus8ttnMkJXzNoEmArFQPlLLj5jWsfCfXJPBHhb4P+Eb6xsdJu9TW/8AFOr3TFTc3hZoopHRQW+z20DNsQbmLTXDY+dVX9mZfgD4E+Fvhz4T/BfxromoWGh2ixJHHdoLq4lPzSTyx5DB5HLOflAy2BgAAV+0v+x/4e+MMVx4u8HC30fxiqlncrtg1LA6Sgfdf0kH0YEYK+LfCHiXwLr114Y8XaNc6Zqdm22W3nTBHowPRlPZhkHsa8N+Jdf8H61a+I/C+r3OmanZPvgubd9rIf6g9CDwRweK/Z7/AG6PDvjJbXwn8XpbbQ9cO2OHVR8lpeHp8/aFz7/IeeV4Wvi7+0X8MPg1ZZ8R60t3qkke630qxYS3EmRlSRnEanj5mIB7Z6VqvxI/aW/bA1Gfw94C0uTQvCpYx3CwStDbqp7XFxjMhx/Ao5/uHrXwb/Yl+Gvw7WHV/GKJ4u1xcMGuocWkDf7EJzuP+0+exAWlVVUIgACjAAGMVVVVftQ/s3aV8ePC4utNWCz8XaTE39mXjfKsy5yYJSOqHnB/gY5HBYHWtF1Xw7q13oOu6fNY6hYTNDc206bXidTggiq/ZC/arl+F97b/AA48e3jS+E72fFrdyOSdKkY8nn/liScsP4SSw7gxSw3EKT28qSRyKGR0bIYEcEEdQRVVVVVftPfB2L40fCjUdAtIVOt6eDe6Q5Xnz0B/d59JFLJ6AlT/AA1NDLbyvbzxNHLGxV0dSrKw4II7H2qOR4nWWKRkdCCrKcFSOhBr9lj41RfGv4W2epX9yr+IdHC2esJ0LSqPlmx6SKN3pu3gfdqqqqqqqWSOGNpZZFREUszMcBQOpJ7CvE37QnwE8FiQeLfjZ4E0d4s7o77xFawvkZyArOCT8rDAGcg14f8A23v2UvFvjfSvhx4V+NOjaz4h1u5FrYWmnxT3AmlO7AEiRmMD5TyWA6c8jNfFvxvdfDL4WeL/AIkWWh/21N4V0O81VdO+0GA3f2eFpTGHCPtLBCAdrckcVJ/wV3/aq8efu/hF+zZo0xckLiyv9XYfTyTEM8Ht+HFWnx2/4LLfEa8hl0L4X6zoEBlDPbt4UtdNDpk5TdfqWUEcZBBGAc+oJZQSCDjoe1VpX/BFf9n8apc6j4m+JPja9imneWO008WtlHGpbITmKQ7QOOCO3SvCv/BLD9ibwxskuPhbda7OmNsuq63dyevVEkSM5z3XsK+I37Vfwf8A2fvDtr8M/hJpdnq9xolqtnZWNjJix05EG1UaQZ3bQB8iZPGCVNfE74zfEX4v6n/aPjfxFNdRoxMFlF+7trf/AHIxwDjjccse5NeI/h5418N6Dpvi3XvDF/p+lazI8djc3MJjE7IFJKg84wwIOMNzgnBxp9/eaVf22p6dcNBdWcyzQSr1jdSCrD3BANfCX4gWPxT+HGgePLDao1azV5o1PEU6/LKn/AZFYfhVVVVVVVVVVVfGf9sv4ZfC5ptG0KVfFWvx5VrWymAggYcYlm5AII5VQxGOdtfFf9ov4q/GKV4fFPiFodLLZTSbHMFqvplQSZCPVyxHbFV4O8A+NPiFqQ0fwV4Zv9YuuNy2sJZYwe7t91Bx1YgV8N/+CevibU1iv/il4og0eEjLafpoFxPj0aQ/u0P0Egr4f/sz/BT4bCOXQPBFnc3qci+1Jftc2fVS+Qh4/gC/rValpemazZSabq+n219aTLiS3uYVlRx6FWBBH1rx9+xP8DPGvmXOm6LceGL1+fN0iXy48+8LBkA9lC/WvHf/AAT8+KGhGS58D65pfia3UEpC5+xXB9grkp+O8fSvF/w98ceAbv7D408J6po8pbahu7ZkWQ/7D/dce6kiqqqqqqq8CftCfGT4brFB4V8e6lHaRcLZXLi5gA9BHJuCj/dwfevAv/BRTUIRFa/EnwHFcKBh7zRpvLb6+TISCf8Agaj29P2wvib8MvjFrHhnxv4A1aSe7WylstRtbi2eGWAI++InI2tnzJBlScYqv2bv20NW8DC08E/FW6uNU8PKFitdSIMlxp69AG7yxD05ZR0yAFGkaxpPiDS7bWtC1K2v7C8jElvc28gkjlU91YcEV/wXP/5ol/3MH/uOrwl/yKmi/wDYPg/9FrVVVftS/wDJvXjr/sFP/wChLVfCj/kqXg3/ALD9j/6PSqqq/wCCgfwmbVNC034u6Rbbp9IxZartXkwO37qQ+yuxU/8AXRfSqr/gnT4y8nWfFfw/nl4uraLUrZScYMbeXLj3Iki/75qqqqqqqqqvHvg/TviB4L1rwXqoH2bWLKS3ZiM+WzD5XHurYYe4FavpV9oeq3uianCYbzT7iS3uIz1SRGKsPwIIr9m3VBq1h8QvhFcYePxn4XuTZRHnN9axtNDgfQSH8BX/AATg8Ti98BeKvCDyln0rVI7tFJ6JPHtwPbMBP1PvVftl/HeP4q+N18KeHL0S+GfDMjxxOjZS8uekkwI6qPuKeeNxH3qs7O71G8g0+wtpLi5uZFihhjUs0jscKqgckkkDFfs4/B62+C3wxsPDcscbaxd/6Xq8y877hgMqD3VAAg7HBOAWNf8ABQb4x6LeaLpnwl8Na5bXlyb1rnWkt5Q/keWMRxORwGLMWK9RsXPWqhTfIB6cmvgz+y78TvjSq6lpNlHpOhZIOq6grJG+OoiUDdIevT5cggsDXw5/YS+EPhARXni17vxdfpgn7WTBbBh3EKHJHs7MPavjx8ffAH7M/hFNB8O6bpv9vzQn+ytCtIhFHEDkedKqYCRgg8cFyMDuy+NfGvif4ieJbzxb4w1aXUdUvn3SzScAAdFVRwqgcBRgAVVWVjeaneQadp1pNdXd1IsUEEKF3kdjhVVRySSQABX7NX7GnhLwJoNr4m+KOg2eteK7oLN9mukE0GmDqECcq8g4LOc4IwvQsyIkaLHGoVVGFUDAAHpVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX/BOcf8Iv8A8FLPjf4WdXjXyfEVosMTZjV4tXgIzz0Co4B56+5qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqrUNT03Sbc3eq6jbWcC9ZbiVY1H4sQK+O37S/w18H/AAt8Y3HhT4iaBqXimHQrw6PYWOoRzyS3vkuIE+QnbmTbknGBzX/BM/xz4J/ZQ0Hx3rnxV03VP+Eg8SXFpDaW2nwx3DLbQLIxO/zAo3PLgjP8ANa3/wAFJfCkBI8OfC/Vb0djfahHa/oiye9N+3n8c/FIZfAPwf0+XdkKVtbq/I6/88ygzwe3aj8Tf+Chni4E6R4O1HR1bPy/2BDbYHPH+lAkfz6etf8ACp/+ChHisj+2PHWoaPuOT/xUUdvj/wABSfQdPX61/wAMK/tA+I12+M/jJp8iuMOrX93enGMch1UHjI//AF1pf/BNW2Xa+tfF2R+fmjtdFCY57M0x7f7PB9a03/gnL8I4Nrar4x8WXbLgkRSwQq2OuQYmOD7H8a0/9gv9nizUC50bWb8gYzcapIueev7vb9KsP2Pf2bdOINt8LrNyGyPtF7cz84x/HI3Ht0qx/Z1+A+nBRb/CHwm+1SB5+lRT/wDoYP51Z/C74ZadtOn/AA68MW2xNi+TpECYX0GF6cDirTQtEsPLNho9jbGIfu/Jt1TZ9MDiqqqqv2kpfFH/AAUB/wCCiWnfAHTzqFp4N8D3UlhNujaMwW0JDahd4YfK0jKI426MBAf4q0PRNJ8NaJp/hzQbCKx0zSrWO0srWFcJBDGoREUdgqqAPpVV4m8F+D/GlmbDxd4X0rWoNuBHf2aThfpuBwfccivGv7BfwI8UCSfQrPVPDFy2SDp90ZIt3vHLv49lK140/wCCdPxI0kPP4H8XaP4giXpDco1lMfYAl0P1LCvGnwJ+MPw93t4u+HetWUMX37lbfz4B/wBto9yf+PVVVVVVVVUCQQVOCOleEPj58Z/AgSPwt8StctYYx8ltLcm4hX6RS7kH5V4S/wCCh/xc0cRweK9A0HxDCv35BG1nO/8AwJCUH/fuvG/7Tf7M37Q+hJovxb8F674d1KFSLPVrSNLp7In+7IuHZfVDGVPXggEeN/DGk+GdVMHh/wAYaZ4l0uYFrS/sw8ZdfSSGQCSJ+mQwx6E9arwF4h8O+HvFen6t4y8Mf8JJpFtIDcac100HmqBgfOvPHBx0OMHg18EPi/8ACH4m+G4LX4XXlnaJYwgPoghW2msl9DEONuf4lyue+aqqqqqv2tv2W7X4x6TJ4z8HWscPjTTocKu4IupRL/yycngSAfcY4/unjBW+sbzTLyfTtRtJrW7tZGingmQo8TqcMrKeQQQQQar9j39rJ/A9za/C74masT4blIj0vUJ2z/Zrk8Ru3aE9ifuH/ZztR0kRZI3DKwyrA5BHqKqqqqr9uz4IHwJ48X4k6DZ7ND8Vys1yEXC299jLjjoJADIPVvM9BVfs0/Gq6+B/xMs9fmkkfRNQAtNZgXJ3QMR+8A7uh+Ye24cbjVleWmpWcGoafcxXFtdRLLBNEwZZEYAqykdQQQQaq6urWyga6vLmKCGMfPJK4RV+pPArw5488DeMbq+sfCPjPQtcuNMEf26HTtRiuXtfM3eX5ixsSm7Y+N2M7Tjoar9sz9sbRf2OfC/h/wASaz4D1LxMviG8mtYUtLuO3WF0QN87MCfmBOMKfunOOM6l/wAFn/jJ4uuZLL4T/s26WLjHyR3N9c6s446lYI4T2J+n0zX7Fn7TH7dHxn+NIsvjz8Grvwx4En0m5aO5XwpdabAl0rK0eJbgszDG9MbiOncZqv2vf2dh+1N8DdV+EMXiOPQbm9u7W5t9RktTcLA0Myscxhl3ZTevUYLA9q8N/wDBED4a2oQeMPjv4l1PH3v7N0mCxz9PMabH618If+CW37L3wZ8ZaF4/0E+L9U1zw7ex3ljcanq6kLMhyrMkMcasM84IwfpVSxRzRtDLGro6lWRhkMD1BHpSIkaLHGqqqjCqBgAD0FVVfEP4leCvhZ4fk8S+N9ch0+0XKxKfmknfH3I0HLN7Dp1OBk18ef2xPHPxYe48P+F5J/Dfhd9yG3ikxcXi9P3zr0BH/LNfl5IJbrXwp+CfxD+M2rf2b4K0RpYInC3WoT5jtrXP9+TB5xztGWPYGvgp+xv8NvhYtvrOvwx+KfESAN9qvIR5Fu3/AEyhORkdmbLZ5G3pX7UfwtHxa+DOt6DaW4l1Wwj/ALQ0sbcsZ4gTsX3dC8f/AAOq/wCCdnxUEN1rXwf1S6G24B1LSgx/jACzoPqoRgP9lz3qqqqqq/au8LfGHxr8AfFPhn4Ba7Lo3jy8+x/2TexXxsmi2XkLzYlH3cwrKvvnHev+GXP+Cwf/AEXTVf8Awunr/hlz/gsH/wBF01X/AMLp6tvjV+3nqnxnX4CeG/2iPGeveK31c6VGmmeJZZ4HuFOH2yg4KJhiz/dARjnAzX7SGqeL/h9+x/471Sx8TXcPibQPBNwy6vbTMkouorfmZH+8GLAtnrzX/BHn41/F34x/8Lb/AOFqfEjxB4r/ALI/sT7B/a189x9m837d5mzcTt3eWmcddo9KvrlrOynu0tZrloImcQQKDJIQM7VBIBY4wMkDJ618dv2wPiN8UZrrw7ojT+FvDu5o3soHK3FwBwRPIMH1yi4XnB3YzVeAPhd4++KWqf2R4F8M3eqTKQJZI12xQA95JGwqD6kZ7Zr4Uf8ABP8A8K6KLfVfixrLa7eLhjptizQ2qn0Z+JJPw2D2NeH/AA14e8J6XFovhjRLHSrCAYjtrOBYkHvhQOT3PU1VVVVf6fYaraSafqdjb3ltMuJILiISI49CpyCK8ffsWfArxuJLiy0CXw1euOJtHk8pM9v3LBowPZVU+9fHD9mPTfhELiey+MXhXVfJznTp5vs9/wDQQqXz0xklee3pVVVVVVVVXwG/aS8bfAzUxDYyHU/DtxJuvNInkIQ56vEefLfHfGD3B4x+2n+z/r//AAUi8N/DfxL+z9408JWsPhH+011W38QXU9tNBJdC0KJshhlwR9nfJOAcgqWBJGh2MumaJp+mzsjS2lrHE5QkgsqgHGe3FVVV+0hHHL8BPHyyIrgaDdEBhnkISD+BANV4Cd4/HXhySNyrLq1qVYHBBEq8iqqvER8RLo1y3hNNOk1VVzbJqDOsLnPIZkBZRjPIB57V8dv2l/2ktCXUfh98RPh/4d0W01O3kgZfsMkyXcTDaxSVpWVxz1XBGR0NVX7L/jL/AIQf47eEtVkm2W91eiwuc8DZcAxZPsGZW/4DVVVVVVVVVV+2t4NTwj8fNXubeER23iC3i1KIAYG5wUkP1Mkbn/gVfCjxUfBHxL8MeLDN5cemapBLOScZh3gSA+xQsPxr9jEH4f8A7Tvj34ZyExw+Ve20SEY3PbXICH6eWZDn6V+2n+0bH4H0Sb4VeDb8f8JDq8GNRuIn+bT7Zh90EdJJBwO6qSeNymq/Zruvhh8Mbk/Gn4p6kk0lgWTw9olsomubmccNcbMgKicqrOQC2SDlBXxl/bP+JvxNE+j+HZj4V0CTcv2ezlP2idf+mk3B6fwptHODuqaQyyM5bOT1NV+xJ+zL4N8Z+Gm+K/xC0ltSU3zw6TYT/wDHu6x4DSuv/LT59yhT8vyHIORiGCG2hjtraJIoolCRxou1UUDAAA6ADtX7Tv7V+g/BOyl8MeG2t9U8aXEXyWxOY9PVh8ss2Op6ER9TwTgEZ8R+JNe8X63d+I/E2rXGpanfSb7i5uH3O56fgAAAAOAAAMAVVaZpmo61qNtpGkWU15e3kqxW9vBGXeV2OAqqOSSeMV+yn+yVY/CC2i8b+Ooba98ZXEf7pFO+PSkYcqh6NKQcM44HKqcZZqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr9n0J4Q/4LLeONFISM61qeu7Vx5W4ywNd8L/ABE7c+/LVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVXij4r/DHwWZI/FfxA8P6XLFndBc6hGkvHYR53E+wFeJf26v2ePD4ZLLxDqWvSKOU03Tn6+gaby1P4HFeIv+CkmnhjB4N+FtzcMxxHLqOoCM57fu41bP/fdN+0x+2p8RAR4G+GUmnQOv7uey8PSyAe5knLJn8B06dab4Tf8ABQL4iEHxH41vtEilOSJddjtEIPqlnnj2K/hWnf8ABOzxtrk63nj74v2vn4G829pLesfYPK8Z/HH4V+318DfBv7I3wQ0vxf4b8U6rqvirWtdi062S7WJYFi8qWSWTywu7gRov3+C44Pb9jD9mb4aeIP2b/h949+J3gex1nxV4j0ldTu7q5MhV453aWDEW7ywBC8Q4UZxk5JNaL8LPhl4bVV8P/Dzw1pxQDDW2lQxtx3JC5J9zzQAACqAABwKqqqqqqqqqqpfDXhxfEDeLU0DTRrrWn2NtTFoguTb7t/kmXG/y9wDbM4zzjNVVVVV40+Anwb+ISufFnw60W7mkzuuY7f7POf8AtrFtf9a8Z/8ABOn4a6r5k/gnxdrOgzMCVhuVW9hU+gB2OB9WNeM/2Bfjn4c8ybw9HpHie3UEqLK6EMuB6pNtGfZWavFfw68e+BZfJ8ZeDdZ0Y7tqte2TxI5/2WIw34E1VVVVVVVVVVo+s6v4e1K31nQdUutOv7R98F1azNFJE3qrKQRXwV/4KCappwt9A+NWntqMAwq63YxhZkHrLCMK/wDvJtOB91jXg7xz4P8AiFosXiDwV4istYsJAMS20m4oSPuuv3kb1VgCO4qqqqqv2vP2Trb4lWV18Sfh7YrH4ttYt13ZxLgaqijsP+ewA4P8QG084Imhlt5Xt54mjljYq6OpVlYcEEdj7VX7If7Xj+D3s/hb8UtTLaCxEWlarO2Tp56CKUn/AJY9gx+50Py/cR0kRZI3DKwyrA5BHqKqqqq/a6/ar/Y98O6X4l+BPxm+JP2PXhbp5lha6RdXc9lM0aSwSgxxlFYB43wXGVJHQkVpWrabrunQavo92tzZ3KboZQCNw6dDyDxgg9DVfsF/tAJqenr8EPFd9/plkjSaBNI3MsIyz2+fVOWX/ZyOAgB/bv8Agb+2x8XvH/h0/szfE/UfC/hpdGaHVkh8SSaVH9pEzMGPknzHLIyjhcALjPNW/wDwR0/aI8eXUd38ZP2ltIlmz800YvdadeeSDOYSepPUcmv2K/2EtF/YzuPEt5pXxL1PxRN4pgtortJ7BLSJDA0hVlQM5z+9Ycsep9arxL4L8HeM4rWDxh4T0bXY7GcT2qalYR3QglAwHQSKdrYJGRg81p+mabpNolhpNhbWdtGMJDbxLGi/RVAAqqqqqqqv2hP2o/B/wQs30m2MWs+K5o8waZHJ8sAI4edh9xe4X7zdsD5h4/8AiL42+LHiV/EXjHV59Sv5jshjAwkKk8RxIOFX2HJPJySTX7P/AOw1qviEWvi34yRz6ZprYeHQ1JS5uB1HnHrEp/uj5+udnfQPDuheFdIt9A8NaRaabp1ou2G2tohGiD6DuepPUnk1VftefCn/AIVX8Z9Ugsbby9H17OpadhTtVZCfMjH+7IGAHZSvrXw98a6p8OfG+ieONFY/a9GvEnVN20SqDh4yf7roWU+zGvC/iPSvGHhvS/FWhziaw1a0jubd++x1DAHHQjOCOxBFVVVVVVf8FN/2xx+zp8Lf+FeeBdYWL4h+NYHjtWif95pdicrLdccq7cxxnj5tzA/uyK/4JO/sdf8ACtPBSftH/ELTHXxZ4vtcaHb3CfNp+mvgiXB6ST8NnqI9vTewr9tf/k0X4v8A/Yn6h/6Jav8Aghj/AM1s/wC4B/7kar9tb9nK+0/xPD8TvAOiz3Vv4huhDqVlaQGRo71z8siqoJxKeD/t/wC+K+B37BVzfJbeJfjVcPawsA8eg2smJGHpPKPu/wC4nPP3lIIrw54Y8PeENIg0Dwto1npen2wxHbWsQjQepwOpPcnk96qqqqr4g/G74VfC6Nh418a6fY3KrkWSOZrluOMRJlwD6kAe9fED/golChks/hf4HMmAQl9rUm0Z9RBGckehLj3FeO/2kfjX8RWkj8QePtRjtJMg2Vg/2SDaf4SseN4/39x96qtG8Pa/4jufsXh7Q7/VLgD/AFNnbPO//fKAmtA/ZN/aF8RxCaz+Gd/bRlSc6hLFZkY/2ZWVsnp0rxp8LfiL8O5TF418GarpA3bVmuLY+U5/2ZBlG/Amqqqqqqq+HvxH8YfCzxLB4q8FaxJY3sPyuv3o7iPPMcidHQ46HocEYIBH7Pv7UPg7442KaXMY9H8VQR5uNLkk4mwOZIGP317lfvL3yMMaqq+PMEdz8D/iDFIm8DwvqLAf7S2zkfqBVaDcfZNc067E4i8i7ifzC23Zhwc57YxVVVeKvCHhjxxos/h3xdodnq2nXAw9vcx7hnswPVWHZhgjqCK+Nn7A+p6abjxB8GLx9QtgS7aJeSATRjriGU8SD0VsNjHzMa1TStT0PUJ9I1rTrmwvrV9k9tcwtFJE3oysAQfrUUskEqTwSMkkbBkZTgqR0Ir4a+LofH3w+8O+MoWU/wBr6bDcSKvRJGQb1/Btw/Cqqqqqqqqq/wCCjvh9MeCfFMSDf/pdlMxPUfu3jH/oz86rxX8QNd+FPx9sfiv4dit5bzVNBtNRgWcEoz3mmIsjMoPO2V3bGcZUVrOs6r4i1W71zXNQmvtQvpWluLiZtzyOxySTVVO+yM4PJ4FVbwm5uIrcSRxmRwoeQ4VcnGSewr4Q+MPgtoXgjQPAvg34leGb6PSLGK2RI9SiEsrAfM5jLbgWbcx46k1+1H+2bp3gJLrwF8Kb63v/ABKQY7vUkIkh008ZC9VklxkY6IeuTlRf317ql7PqOpXk13d3MjSTTzOXeRyclmY8kk9zVVo2jar4i1W00LQtPnvtQvpVhtraBC7yuxwAAK/ZZ/ZQ0v4LWaeLfFq2+oeM7qLG9Ruj01GHMcR7ueQ0n4DjJaqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqrx7/wAUB/wWy0fVUUwxa1qdj5WSVz9q0lbZ8E5zl2fpxzjjtVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVapq2laJZvqGs6naWFrH9+e6mWJF+rMQBXi39r79nnwhviuPiJZ6nOo+WHSY3vd/wBHjBj/ADYV4s/4KReGLbfD4H+HGpX55Czandpage+xBISPbctH9on9tr4qjb8P/h9LpNrLxHcWOhttx7zXO5M+4x+FH9mL9s34nkP8RfiS2nW8n+str7XZJQvrtht90WfxH1rwz/wTb8LW4V/GPxK1S+P8Uem2SWuPbc5kz9cCvDf7Ff7OvhwI7eB21adB/rdSvZZs/VAwT/x2vD3gDwL4RRU8K+DND0fYMA2OnxQH81UGqqv+Czuvaj4x+Kvwf+BuiMHvJLaa8WAH/WS3twlvBn6G3kA/3jXhfw9YeEvDOkeFNKUiy0WwhsrYEYxHFGEX9FFVVVVVVVVVVVXgX4z/AAn+J2pavo3w/wDiHoWu6joF5LaalZWd4rz2ssTlH3R53BdwID42tjgmvGfj3wL8OdHXxB8QvGug+F9LaZYFvtZ1GKygMrAlUEkrKu4hWIGcnB9K/wCGsf2V/wDo5j4U/wDhZ2H/AMdr/hrH9lf/AKOY+FP/AIWdh/8AHa8IeN/BXxC0VfEfgDxhoniXSXkaJb/R9QivIC68MokiZlyO4zxVeOPif8NPhja2t78SviJ4Z8J299I0drLrmrwWCTuoyyoZmUMQCCQOleCviD4B+JWkya98OfHHh/xVpkNw1vJe6LqcN9CkyqrNGXiZlDhXQlc5AYHuKqqq4t4LqF7a5hjmikXa8cihlYehB6ivGX7LXwE8db5NX+G+mW1w2T9o01TYvn+8fJKhj/vA18Tf2HPg3oglm0b452nhiRckW/iG5t3QH037oyo4PJDHivH/AMOIvAs5jtfH/hDxNEGwJdD1Lz8DsSGVT+WfrjmqqqqqqqqqrStZ1jQrpb7Q9WvNPuV+7NaztC4+jKQa8K/tf/tDeEhHFbfEW71KBBgxarEl5u+ruDJ+TCvC3/BSHxda7IvGnw60nUR0aXTbqS0b67X8wE/iPwrwr+398CNdVI9dbXPDsvRjeWJmjB9mhLkj6qK8LfGn4SeNgi+FviP4fv5ZPu263yJN/wB+mIcfiKqqr9rn9kaH4gQ3XxM+GenpH4njUyahp0S7RqYA5dB2mx/33/vdZoZbeV7eeJo5Y2KujqVZWHBBHY+1V+yr+2XF8OrBfAPxc1C5l8N2sLHT9TWGS4lsAoz5LIgZ5IyBhQoLKcAAqRt0bV9N8Q6PY69o10Lmw1K2jubWYKVEkUihkYA4IypBwQDzVfGX9uP/AIKHn4ueM/hh8Hf2forqw8O67eafaalp3hK+1CSaGKdkjmaTe0QDKEP3QAW/IaP/AMFrfi6QbvUdb8OWb8BzcaXovlDOOkW2Y9z0Jx+Ffsp+Efi94D+AXhXwh8d9ebW/G+mx3CalqD6lJftcBriR4i00nzOwiaNTnPK8HGKr4s/sAfsx/HP4n3vxZ+Kfg/Udb1nUIIYZ4v7XuLaArFGI0O2BkbO1R/FX7Tf7E3w88CfDGPxJ8BPBUOhp4dMkuo6bavJILi3bBaUb2ZiyYyeeVLegqtG1jVPD2rWeu6Hfy2eoafOs9tcRNhopFOVYfQiv2c/jhpXx1+H1vr8bQw63YhYNZskOPInx95R12OAWX8RklTVVVVVVVVVV+0v+2jZeFDdeBPhFeQ3utKGivNYUB4bJuhWLqJJBzk/dXj7xyF0Dw94z+KHiuPSNDsr7Xte1WYsRuMkkjE5Z3djwO5diAOpIr9nT9kbwv8IIrfxP4qW21zxftDLOV3QaecciAHq3rIRn0C85qqr9tv4Sf8LJ+EFxr2m2+/WPCBe/t8LlpIMf6RH/AN8AP7mMDvVf8E9fi/8A2noeo/BvWbvNxpW6+0je33oGb97GP912DgdT5jdlqqqqqq+N3xi8G/AL4Xa98VvHV55Wl6HbGQRKwEl1MeI4IweryOVUdhnJwASP2SvhD42/4KM/tV63+0F8abYz+DtFv0utThbPkTMvNrpkOesaqql++wckNKCURI0WONVVVGFUDAAHoK/bX/5NF+L/AP2J+of+iWr/AIIY/wDNbP8AuAf+5Gqqqqqq4uLe0gkurqeOGGJS0kkjBVRR1JJ4AxXxQ/bc+D/gLztP8OXUni/VY8qItOcC3Vv9q4OVI90D18TP2yPjT8RRNZWutr4Z0uQkC00gmJ2X0ebPmHjg4KqfSppZbiV555WklkYs7uxLMx6knuTVeAv2cfjP8SRFN4a8CX62cmCL69X7LBtP8QeTG8f7m414H/4J03Uix3PxH8fxxdN9no0G8/8Af6UAA/8AbM/WvB37IvwB8GrG8HgW31e5Trcaw5uy2PVG/d/kgrT9M03SLRLDSdPtrK2jHyQ28Kxov0VQAK8T+L/CngvTjqvi7xHpuj2gyBNe3KwhiOy7iMn2HNfET9vH4N6LFPpnhrSr/wAYyMCjBYvs1q46EF5RuI+kZB9a8TWvi39oTV1v/hv+zzZaTGZD++0GwnVGz2kfIgH12KfetO/YS/aAvtNe+uNM0WwmRcrZ3OpKZX9gYwyA/VhXjn4O/E/4ayMnjbwRqmmRq20XLw+ZAx9pkyh/Bu9VVVVVVjf32l3sGo6ZeTWt3bOJIZ4ZCjxuOQysOQR6iv2bv22LHxCLPwP8YryGz1Q4itdbbEcNyegE/aN/9vhT32nqCGAKkEEcVVfEy2e9+G/iuzjZQ0+iXkalugJgcVVWF19usbe92bPPhWTbnO3cAcZ/Gqqqr4r/AAK+Gvxm0/7L4z0FHvI022+pW2Irq39NsmORyflYMvOcZr41/sb/ABI+FYuNa0KJ/FHh2MFjdWkJ8+3X/prCMkADqy5XHJ29K/YF8ZDX/g3P4Wmm3XHhnUpIkQnlYJv3qH/vsyj8Kqqqqqqqq1/XtG8L6NeeIfEOpQWGnWERlubmdtqxqO5/kAOSSAOTX7WHxNsPjF+z7o3jjTrKSzsW8ZyW2mRy53z28dvKplYdFJcH5RnAIGSQaq/ubi5uI5rm4kmdI0RWkYsQqAKqjPYKAAOwAFVVXL732g8L/OqtkDPvPRRU1xkbIz9TVVVtbyXlzFaxNGHlcIpkcIoJOMliQFHqTgCv2R/gR8Nfhv4eXxNo/iTR/Ffie9gUXmp2NylxHahhzDCVJKr2LHDMRzgYUVVVVVVfFb4n+EPgv8Pda+J/jy8nttB0CFZbyWCBpnUM6ou1F5JLOo/H0rVf+CzX7JenSeXZ6B8R9UG4jfa6PbKMDv8AvblDg/TPrivDH/BYv9kHX7tLbVY/HXhuNn2m41TRUkRBx8xFtNM2Oeyk8Hjpn4efEvwB8WvDFv4z+Gvi7TPEei3JKx3lhOJFDDqrDqjDurAEdxVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX/AAUdI+HP/BRz4M/EtSsdu0ehXs0h+UFrbU5BICcjI8tYwckcHGQKqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq8vLPT7WS9v7uG2t4V3SSzOERB6kngCvGn7XP7P/gjfDefEC01S5Tj7PpCm9JPpvTKA/VhXiz/AIKO2skxsfhv8M7q7kc7YZ9VuQhJ7DyYgxOf+ugoeMv2/vjOB/Yejaj4c0+4GQYLNNKjCn+JZZz5pH+6xrTf2Bfiz40vI9U+LfxchMxGWKvNqc/upeUoAfcFh9a8JfsB/Ajw+Ek12LWvEkw5YXt6YY8+ywhDj2LGvCvwm+GPgfY3hHwDoOlyx/dmt7CNZePWTG49O5qqqqqq+I4Px1/4LNaD4fSNrix8I6tYqmeQi6dafbJN3UAeesgx3yB1OKqqqqqqqqqqqv23Pj4n7OH7Nviz4hWl2sOuTQf2boAzyb+cFY2HY+WA8xB6iIjvX/BGz4Av4Z+G+vftFeIrUnVvG0zWGkyyqS62EMn71wTz+9uFIOf+fdTnmv8Agsl/yaLaf9jhY/8Aom5r9h//AIJp/Cj9qP4DWnxX8XePPFmk6jPqd1aNbaabcRBYmABG+Nmyc881/wAOS/gD/wBFX+IH/fVp/wDGa/Zm/Z28L/su/Cy3+FHhDXNU1bT7e9nu1udS8vzS0pBYHy1UYGOOKr/gpfrGqftL/tvfD39ljwxflYdGa106Zwu9ba7v3SS4mIHVY7cQMR1Gx6/4I5fEu/8AAHxg+IP7NXipjBPqSNd2sLniK+snMVxGP9po2yeOlv271cXFvaQPc3U8cMMSlnkkYKqgdSSeAK8d/ti/AHwG8ltN4zXW7yMc22iR/ayfbzARED7Fwa8X/wDBSO6ZpIPAPw1iQDPl3Or3hcn0zDEBj/v4a8TftvftEeIyyW/i210WFhzFplhGn5O4dx+DVr/xT+JniosfEnxC8RamGGCl1qcsigegUtgD2AxVVVVX2S6+zfbPs0vkZ2+bsO3PpnpVVVVVVVVVV4W+LfxQ8EhE8J/EHX9Mij6QQX8gi49Y87T+Irwv+3r8f9AEceq6ho3iGNMAjUNPCMR/vQGPn3OffNWn/BSqaPSpTf8AwhSfUkX90sOtmKGU/wC0TCzIOvQNXiH/AIKiftGf2/Yx6J+z/wCFLHRnvYlvJZNWl1OWOAyDzCoX7OS2zODtwDgkdq8OeOfBXjGITeEvF2jaymMk2F9HPj67CcfjX7XP7I0PxAhuviZ8M9PSPxPGpk1DTol2jUwBy6DtNj/vv/e6zQy28r288TRyxsVdHUqysOCCOx9qr9k39r6X4c/Zvhx8TLya48MMwSwv2y76WSfut3MP5lOwI4Ftc217bRXlncRTwTxiSKWJwyyKRkMpHBBBBBFVVVTokiGORQysCCrDIIr9r/8AZ6k+DPjU694dsmHhHxBKz2RVfls5jkvbH0A5KZ6rxyVJqvgb8Y9f+CHj208XaOzzWjYh1Kx34W7tyfmX2YdVPYgdsg+DPGPh74geF9O8YeFdQS80zU4RLBKvBHYqw/hZSCpU8ggiqqqqqqqq/v7HS7KbUdSvILW1tozJNPPIESNQMlmY8AAdzX7Tf7ZV942+1+AvhTeTWXh5sx3mqKDHNqA7qmeY4j+DMOuASD8Hvgp42+NniMaF4TsdtvAVN9qMykQWaE9Wbux5wg5OD2BI+C/wK8EfA/w//ZPhi18+/uFH2/VJ0Hn3bD1/uoOyDge5JJqqqpI45o2ilRXR1KsrDIIPUEelftNfCN/g18WtV8OWsDJpF4ftukMRx9nkJwgP+wwZP+Ag96+GXj3Vvhf490Tx5orE3OkXSyGPdtE0Z+WSMn0dCyn614a8RaT4v8O6b4o0G6Fxp+q2sdzbSeqOoIyOx5wR2ORVVVVVftvfHHxj+3v+0voX7MvwJd9Q8M6NqbWlk8bkw394ARcX8hH/ACwiQOFbn5Fdx/rMV+z58DPBv7OXwm0P4T+CLcC00qHNzdMmJL+6bBluJPVnbt0VQqjhQBX7a/8AyaL8X/8AsT9Q/wDRLV/wQx/5rZ/3AP8A3I1VVVVqOpadpFjNqerahbWVnbIXmuLmURRxKOpZmIAHua+LH7evgHwq02k/DXT28U6ggK/bHYw2cbexxvlwf7oUHs1fE348fFL4uzsfGfimeaz37o9Ot/3NrH6YjXhiOzNub3qvCPgTxl4/1IaR4L8M6hrN1xuS0gLiMHu7dEHuxAr4bf8ABPbxTqnlX3xR8TQaLAeWsNOxcXB9mkP7tD7jeK+Hn7NfwZ+GSxTeHfBdpPfxYI1DUF+1XG4fxBnyEP8AuBaqvil8evhd8HrZm8ZeJYkvtm6LTLX99dS+mIwflB7M5VfevEv7Ynxt+MGqS+FfgF4IurBGBBnit/tl3tPAZiR5cIIzyQcdmrwv+w18TviBqK+J/jt8QpobiYAywRzm/uyP7rSsSiY5+7vA/l4C/ZV+Bnw9SKTTfBFtqd7F/wAvurj7ZIT2YBhsU+6qtRxxxRrFEioiKFVVGAoHQAelU6JIhjdQysMFWGQRXjv9lX4F/EEyT6l4HttOvZAf9L0k/Y3BP8RVPkY+7Ka+N37F/h34Z6fNrum/GTRrO1wWgsvEB8iaQDqEaPcZW9hGKdVViquHAOAwBwfcZ5qqqqqv2b/2xdd+GDWvg74gyXOs+FBiOGbO+500dtmfvxD+4eQPunjafDviPQfF2i2viLwzq1tqWm3qb4Lm3fcrj+hB4IPIPBxVarZDUtLvNOIQi6t3iw4yvzKRyPTmqr4bXg1L4c+FdQUAC50SzlA3bsboUPXv161VVVVWhfDvwb4X8R6p4p8N6Fb6Zf62iLqJtV8tLkoSVdkHy7wWf5gATuOc1VVVVVVVa/r2jeF9GvPEPiHUoLDTrCIy3NzO21Y1Hc/yAHJJAHJr9pr9pnWfjhrJ0bRnnsPB9hKTaWhO1rphwJpgO/8AdXooPqSa1X4i+K9Z8DaJ8OLu+jGgaBLLNaWscKqTLI7s0jsBuZvnKjJwBgAVV2MhSPevBn7Cvxm8YaDp/iF77w3pNvqESzRRXl67y+UwDK+IkdeQc43A+uKtv+CdHxAdT9s+IHh6JgflEcM0mfzC4r40fsg+Ifgr4KufG+uePNCurSGSOGKBI5UmuJnOAiAgg/KCxyRwrenJ55Jqi52CNeB396qqRGkO1Rk1FEsa4HJ7mtK1jVtCvY9S0PVbvT7yL/V3FrO0MifRlIIr4e/tvfGvwW8NtrmpW/ivT48BodSTE2P9mdcNn3ffXwR+MmkfHHwd/wAJdo+ialpixTm3mhvEGPMUAt5bjiReQM8HPUCqqodS025upLC31C2luYB+9hSVWdPqoOR261UupabDeR6dNqFsl3Ku6OBpVEjjnkLnJHB/I1X/AAUi/wCTI/in/wBg+2/9LIK/4I9fBX4O/E/4d/EDVviT8K/CXiy8sdat4LWXXNGgvzBGYdxVPNVtoJGeK+Kn/BPj9kf4q+HbnQ7j4L+HPDNzJGRb6l4ZsI9LntnwcOPJVUfGfuyKynuOlfsO+KPHH7IP7fVx+z7qWtNdaRrGuTeGtXhjJEN04LC0ulQ/dbf5ZBPISV171X/BJL4s/FD4q/tbeKNS+JXxD8ReKLhfAN6yNq2pS3IjJv7D7iuxCDthQAKqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr/gt/4cntrz4RePLLfG6DU7GWZMqUZTbyRYYdDzKfXjjvjwT4ji8Y+C9A8XQbfL1vS7a+QKMDE0SuMcnjDetVVVVVVVVVVVVVVVVVVVVVVVVVVVVeMfiZ8Pfh9b/aPG3jPSNGBXKR3d0qSP/upnc34A145/4KFfCTQfNtvBejav4ouEB2S7PsVux/3pAZP/ACHTftNftgfGxmtvhL4EbS7KQ7VudP04yhQezXM+YgfcBT3qy/Yr/aJ+KdwmpfGb4nraKTuEd1eSanNEfZAwjUY/uvXgz9gL4H+HPLn8RnV/E9wvLLd3Jghz7JDtbHsWNeE/hx4A8CQiDwb4M0bRgFwXs7NI3b/ecDc31JNVVVVVVVVf8Ex0Pxh/by+LvxyuN01ukGqX0EgbISW+vl8sZ5+XyRMAPYdhVVVVVVVVVVVV/wAFUfhn+0N8e/iz8LvhX4M+H+tyeBftEcba5Bbefapf3UoiaScxkmJIYwuGk2ffkxkYNfD/AMEaB8NPA2gfD3wtbCDSfDmmwWFnHjny4kCAt6scZJ6kkk9a/wCCyX/Jotp/2OFj/wCibmv+CT3j3wL4e/ZB03Tdf8aaDpt2uvagxt7zUYoZAC64O1mBwa/4Wx8K/wDopnhT/wAHNv8A/F1pup6ZrVjFqej6ja31nOCYri2mWWOQA4JVlJB5BHFeKvEmkeDPC+seMNfufI0zQrCe+vZcf6uGGMySN+CqTX/BL3w9rf7RP7Z/jr9p7xdC0jaIt1qO48rHf6g0kcUY/wBlYPtIAHTatftbW1z+xr/wUx034y6bC8Oj6vqtt4m/dof3kFyWi1FAR1Zm+1cdQJVyDnn9p39obWvgt4G0nxP4O8MQ62mvMY7fU5ZM2lsSgeMsFIZy67ioBUYU89j8RfjR8T/ivdG48deML7UIt26OzD+XbRf7sK4QHoM4zwMk1VeF/gb8YfGgjfw18NfEN5DJ9y4+wvHCf+2jgJ+teHv2BPj/AK0EbU7XQdCB+8L7UQ7AfSASDPtmtD/4Jr37hJPEvxXt4iAN0VjpRkz64d5Fx2/hrR/+CdXwes9r6x4o8Vai46qs8MCH8BGW/wDHq0r9iH9m7TQDN4Hnv3Xo13qlyfX+FXVT17jsK/4KwRfC34B/C/wl4E+FnhTTdA8TeK9SeeS+tAwuYrG1TDgSEll3ySxc5+YI455x/wAEwP2eI/C/7Otn8SPiZpw1nxJ8QJRqMbasn2mS1sANtsi+ZnaHXdLkdRKmfujEenaFo0JuobGwsYraIlpFiSJYkA554woA+lfH39sO98T3snwp+AVnJqt1fsbWXVIYDM07Hgpax4O708wj3UdGr4K/sEaENKfXPjksl7qV7GdmlWt00a2hP8UkqEF5PYHaOfvZ48Xf8E4/AOoK8vgnx1rOjynJEV/Cl7GD6Db5bAe5LH69K8Xf8E//AI5aD5kvh59E8SQryi2t35EpHuswVQfYMa8W/CD4peBQ7+Lvh/rumQxj5riaxfyfwkAKH86qqqqqqqqqo5ZIZFlhkZHQ5VlOCD6g14X/AGhfjf4OZDoHxR8QRpGPkhuLs3US/SOXcv6V488da18R9fl8U+JIbD+1blR9ruLW1W3+0uP+WjomF3nuQBnqcnmqr9lX9rnUfhTc23gX4gXk154MkbbBMVMkmksT95QMs0RJ5TkjqvdW0zU9O1rTrbVtIvoLyyvIllt7iBw6SowyGUjggjvVVVV8Sfh54b+KngvUvA/im282y1CIqHUfPbyDlJUPZlOCO3Y5BIPxY+F3iT4P+N7/AMEeJ4f39q263uFXCXcJJ2Sp7MB07EEHkGq/ZN/aUu/gn4mHh/xHcSzeDdYlH2uMfMbGU8C4QenQOB1XnkqAbK9stTsoNR067hurW6iWWCeFw6SIwyrKw4IIIIIqqqqqqrxX4r8O+B/D954p8V6tBp2l2Ee+e4lJwo6AADJZicAKASSQACTX7SH7U3iT42X0mg6K0+k+D4JP3NkHxJeEHiSfHXkAhPurx1IzX7O/7MPir45aimp3Qm0nwnbSYutSZMNMQfmjgB4Z+oLcqvfJwp8D+BPCvw48N2vhTwdpEOn6faLhUQZaRu7u3VmPcnmqqqqq/ba+Df8Aws34VS+JNJtt+u+EFkvIAq5aa3wPPj/75UOOvKYH3qr/AIJ7/GZLiyvfgnrl0PNtt97ohdvvITmaEfQnzAO+6Q9qqqqq/wCCsP7TniD4E/BbTfh94Me4tNd+Jpu7QalE202dlAIvtOw9RI4njQEdFZyCCFr9hn9sP4Z/seza/wCJtW+Dl74t8W60BbQ6mmqpbLZWYwxhjUxOcu4DO2eQiAAYJP8Aw/F8Kf8ARu2q/wDhSR//ABiv+H4vhT/o3bVf/Ckj/wDjFftM+Ko/Hn7BfjzxxDZNZp4h+HE2opbM+8wiez8wIWwMkbsZwM4r/ghj/wA1s/7gH/uRqqqppYYIXnnlSOONSzu7bQoA5JPYYr4yftx/DzwIJ9F8AJH4t1pNyGWKTbZQMOMtIP8AW+uI+D/eFfFD43/Ev4wXn2jxt4lmuLZH3wafD+6toD22xjjODjc2W96r4a/BD4nfFy5EPgjwtc3Vsr7Zb6UeTbReu6VsDI/ujLe1fCz9gDwVoIh1P4paxJ4jvV5NhaM1vaKfQsMSSc98oPVTWgeG/D3hTTItG8MaJY6VYwj5Lazt1hQep2qAMnue9VXjPxx4S+HuhTeJPGevWulafDwZZ2wXb+6ijLOxxwqgk+lfEH9sD4o/F/XG+H37OnhzULVLnKC8SEPezL0LDqtumD948jg7l6V8LP2DVurkeKvjx4hn1XULh/Nl0y1uWYMx6+fcH5nPqExyPvmvDPhLwx4M0qPRPCWgWGk2Mf3YLSBYlJxjJx1PHJOSe5qqqviN8VfAPwo0c6z458R22nxsD5EBO+a4I/hjjHzMenOMDPJA5r4uft8eMvEnnaR8K9O/4RzT2yv2+4Cy3kg9QOUi49Nx7hhWta5rPiPUZdX8QavealfTnMtzdztLI592Ykmqqq0vwj4r1zaNF8MatqG4ZX7LZSS54zxtB7A1pn7O3x11fAs/hN4nT0Nzpz247f8APQL6/wCcVpf7FX7RmphWl8EQWKN0a61O3X/x1XZh07jvWmf8E9/jNeKr6jr/AIUsFPVWuppHHA7LFj2+92rTP+CcGoyFW1r4sW0OPvJa6O0meezNKuOO+K+Bf7NC/Au8ln0f4ma9qFncr/pOmSxRpayvjG/ZhircDlSDgAEkVVeMbBtL8Xa3pbghrPUbiEhl2nKyMOnbp0qv2dL8an8BvAFwGB2eH7SHgY/1cYj/APZKqqqqqqqqqqqq1/XtG8L6NeeIfEOpQWGnWERlubmdtqxqO5/kAOSSAOTX7TX7TOs/HDWTo2jPPYeD7CUm0tCdrXTDgTTAd/7q9FB9STVW8D3U8VtE0YeVwimSRY1BJxyzEBR7kgDvXwm/4J+i/gtde+Kfi2JreZFlj0/RZA4kU4I3XBGMEdkBznhq/bk+EvgL4b/BzwxH4F8J2WkwQ68IZJIUzJJvgkI8yRsvIf3fVicYr4HXn9o/BbwFeGUSPL4a08uwGPn+zoG/UGq/br+M4+IHxHXwFot2JNF8Hu8LlDxNeniVj6hMCMehD44aqqqr4HfADxx8d/EH9meHLf7Lpds4GoatPGTDaDrjtvcjog5PcgZI8U/8E9vitoiPL4T17Q9fiXO2Nna0nf8A4C4KD/vuvGfwV+LHw+Ej+MPAGsafBFnfcm3MsA/7apuT/wAeqvgd8Htb+Nvj2z8I6WWgtF/falehci0twRub3Y5CqO5I7ZI8KeFtC8E+HNP8J+GrFLPTNLgENvCvYDqSe7Ekkk8kkk9ar/gq1LNbfsW+Kb60uJoLi21PTWhlhkaNkJuo1OCCP4WYfjX7MfiX9qz9rz4Q+GP2SPgx4luPBvhXwZb3U/jLxa88qyXJuby4migMiHewKPtESld+19x2rX7R/wDwS++MP7LPgqb45/Db4vyeKV8N/wClanNY2Umk3+npn5riIrNIXVeCxDKwBJwQCR8Nf28/2rv2nPhh4Q/ZZ+EUtxF8WNSuJ7fWvGbuIvK0qNVKXHmKCYpSGZZJQNw8tSmZJRt+KX/BGv4w6Z4S1Dx94c+OcHjbxpBEbufTbjS5beS7kUbmWK6aaRnlOMLvRNxxkr1H/BJP9sPx18QdV1j9nT4s+I73W73TrA3/AIdv9QlMlwsUTKk1q7t8z7Q6Om7JULIM4Cgf8FIv+TI/in/2D7b/ANLIK/4IveMvB/hj4YfEa38SeK9H0mWbXrZokvr6OBnAgIJAdhkV8X/2yP2bfgl4bufEHjD4s+H55oYi8GlaZfxXl9dtjhY4Y2LckY3NhATywHNfsI/Dbxz+11+23q/7W/iHw4+m+E9H1241mSR0Jie7YMLW0icjEjRZjdmA4EYztLrn9uP9iXxb+1xqvhHUfDPxe/4QtfDVvdRTR/YpJ/tJmaMg/JKmNvlnrn71fsVfsy+IP2rfipqnw78N/EL/AIQ6507w/Nqj332Z5vMRLi3iMW1HQjJnVs5x8nTnj9hz9iXxb+yRqvi7UfEvxe/4TRfEtvaxQx/YpIPsphaQk/PK+d3mDpjpVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX/AAWZ8I/25+yzpPiWGLMvhvxZbSu4XO2GWGaFhnsC7xfkPqP2CvFv/Cb/ALHHwl1oybzB4di04njrZs1qen/XD6+vOaqqqqqqqqqqqqqqqqqqqqqqqqqq8efGX4WfDKJm8ceOdK0yVV3C2ebzLhh6rCmZD+C968ff8FF/Ctkz2Pwz8FXusT52pd6k/wBmiz6rGu53HsShoa7+3l+0CQdMtdQ8L6PcjIeCP+yIAp7iRz57r06Fv514N/4J0G6nGp/Ff4kz3c8hzNb6RGcs3r9omBJ/79g+9eBv2X/gV8PfKm0P4e6dcXcXIvNSU3ku7+8DJkIf90LSIkaLHGoVVGFUDAAHpVVVVVVVVVV+0Z4wX4ffAH4j+NRKqSaN4W1G5gJB5lW3fy149X2j8a/4IheDUtPhp8S/iCYV3atrtrpayHOcWsBlIHoP9MXp14z0FVVVVVVVVVVVVV/wWS/5NFtP+xwsf/RNzX7G/wDwTH0L9qn4J2vxa1D4v3/h2a41G5tDYw6MlyqiJgN28yqec9McV/w458Kf9HFar/4Tcf8A8fr9nf4N2v7PvwY8MfB2z16XWofDVvJCl/LbiBp98zyZKBmC48zHU9K/4K0fGVfhl+yre+D7C5Kat8Q76PSYgpwy2y/vbl/90oixHr/rh9R/wTc/au/Y4/Zl/Z9OifEL4tR6Z4y8RatPqGr2w0DUZ2t1GIoIvMit2RgI4xJwxwZmHXNf8FRv2jP2TP2mvAnhLW/hD8T49a8Z+F9Skh+ynRL+0MthOmZPnmgRCUkihIBOcM+O+f2FPEmgftX/ALB+i+CfF8gubjRLZ/DWosOXgktQv2aVc/xrA1s+T1YHrzWk/sdfHLXPHWreCbHw15Uej3Rgm1e8Jgs2XqsiORlwylWwgZgGGQK+Hv8AwTt8AaQsN58RvE+oeILkcvaWY+x2/upIzIw9wyfSvBnwZ+FPw9WP/hDfAGi6bNF925S1V5+PWZsufxaqqqr9pqG8/bh/4KaW3wg0u6ll0DRdQi8PvLCT/o9pZ7pNQkHYMJPtIB7kIM9K8Q+IvBXwp8HHVNbvLPQ9A0a3SKNQuxI0VQqRRoOpwAFVRnsBXxy/ai8RfH3Wx4L0XVoPCngySXBF5OY/PVefNuWTJIGMiJA3OMB2xXwo+M37I37OWl+Z4eu9a8Y+JJott3q9tpJjZyeqRCcx+XHnsMk45J4rX/8AgpPpyB4/C3wruJuuyXUNTEePQlERs/TcK1v/AIKH/GjUCyaPoXhfS48/KVtZZpB9S0m08f7Nax+2L+0frWVm+JdzbIeiWdnb2+PoyRhu3c1q3xj+LevEjWPif4qvFP8ABLrE5UdOi7sDoO1EljknJPrVVVVVVVVVVVVVeDPjh8Xfh7aRaf4N+IetadZQEmKzW4MkCEnJxE+UGSSTxyTXhv8A4KAfHjRtqaydA19AMMbyw8lz7gwsgB/4CR7V4b/4KS6RKI4vGHwvu7fA+ebTdQWbJ9RHIqY+m814b/bp/Z31/Yl54j1HQ5HXITUtOkGD6FovMUfice9eHfjB8KfFwUeG/iP4c1CRhxFDqURkH1TduH4iqr9pf9n3SPjz4La0hWG28S6WrSaRfOMYY9YnI58t8DP904I6EHXdC1fwzrN54e1/T5rHUdPmaG5tplw0bqcEH/Hoeoqv2Pf2rW+Hd1b/AAx+ImoO/hi7lC6feyvn+y5GP3WJ/wCWLE8/3Dz0JwjpIiyRuGVhlWByCPUVVVVVXxH+JPhH4U+Frnxb4y1RLW0gBEUYIMtzJjiOJc/M5x07DJJABI+PP7QPi747eIPtequ1jolnIx03SY3ykAIxuY4G+Qjqx6ZIGBX7MP7IepfFB7bxx8QYJ9P8JqQ9vb8xzap/u90i9X6t0X+8NK0nS9C0220bRdPt7Gxs4xHb21vGI44kHQKo4AqqqqqqIDDaQCCOhr9rH4MN8G/ite2mnWpj8P64WvdJIGFRGPzwj/rmxwB/dKHvXg3xbrXgPxVpXjHw5c+RqOkXKzwMehK9VYd1YZUjuCR3r4Z/EDRPil4F0fx34fk/0XVbcOYictBIOJI2/wBpWDKfpnoaqqqtd8JeFfFPkf8ACTeGdK1f7Lu8j7fZR3Hlbsbtu8HGdq5x1wPSv+FTfCv/AKJp4U/8E1v/APEV/wAKm+Ff/RNPCn/gmt//AIiv+FTfCv8A6Jp4U/8ABNb/APxFTaRpVxpZ0SfTLSTTjD5Bs3gUxGPGNmzG3bjjGMYrQfDHhDwxLc2/hfw9o+kyzKjXKWFpHAzgbghcIBkffxn/AGsd6qq+NH7UPwy+C8UlhqF//a+vhfk0excNIp/6at92IdOvzYOQpr4x/tNfFD4zzSWmsar/AGZoZY+Xo9gxjhx28w/elPT73GeQFqvhj8FfiR8X9QFl4H8OT3UKPsnv5R5dtbn/AG5TxnHO0ZY9ga+Ef7B/w/8ACHkat8R7r/hK9UTDfZdpjsom/wBz70v1bCnutWNhY6ZZw6fptnBaWtugSGCCMRpGo6BVHAHsKqq/aB/ao8F/BK3k0a0Met+KnTMWmRSfLb5HDTsPuDuF+8eOgO6vBPwP+NX7W2vRfEb4u69daV4ck5tmaPYzxH+C0hPyohH/AC0brwfnOa+HXwu8C/CnQ10DwNoEGnwEAzSAbprhh/FJIfmY9evAzgADiqqqr4w638Y7DR/7P+DXgq21TVLlCDqF9ewxQ2nuI2YNI/sQFHBO7kV8b/DHxh8O+Mnm+NUWoNrV+nmpc3M6zJMmf+WboSm0E/cXG3IGBVvbz3c8draQSTTSsEjjjQszseAABySfSvhV+w38VPHSwan4vMfhDSpAG/0yPfduPaAEbf8AgZUj0NaX/wAE9Pg7aqG1PxJ4qvpAOQLiGJO3YRE+v8Xf8a0v9if9nPTgpm8F3F+6nIa61S4Pf0R1B9OR2rTP2cfgPpCqtp8JvDUm0YH2qwW57Y583dn/ACa0zwb4Q0Uq2jeFdHsChyptrGOLBznjaB35qqqqqq+PmmHSPjd48sAmxV8RXrxqQRhHmZl6/wCywqv2MtTGqfs2eD3L5e2S6t3BzxsuZQOv+zt/OqqviV+0/wDs8fB8zQ/Ej4y+FdFurckSWMuopJdjHX/R4y0p/Ba8df8ABYz9k7wy81v4UtfGPjCVMiGWx0sWsDn3a5eORR/2zJ9q8U/8FxL53aLwT+z1BGoJ2z6r4hLluR1jjhXHGf4zyR6c6z/wWj/ajvn26R4N+HGmxggjGnXcznjkEtc4xnnhR9av/wDgrb+2leKi2/jHw9ZFScmDw/Ad313hv0xX/D2D9tz/AKKRpP8A4Ttn/wDG6t/+Csv7bEMySyfEDRbhVOTFJ4etArex2oD+RFaR/wAFj/2u9NZTeWXgHVdrEkXmiyruyMYPlTpwOv8API4rw5/wW++J1q6Hxd8DPC+pqPvjTdUnsSenQuJsd/XqPTnwX/wWw+BmqtFD47+FfjLw+7jDSWMkGoxRn3JaJsY7hCfbvXw7/wCChv7HHxMMUGjfHHRdMu5Tj7Nrwk0plb+7uuFRGPT7rH061p+o6fq1lDqWlX9ve2lwu6Ge3lEkcg9VZSQR9Kqq+NnwOtfjjp1poet+NNb0rSbZvMex0/y1S4kHRpCyktjsOgPPXGP+Hdfww/6HfxR+cH/xurf/AIJ3/B5Y9t34t8YyPnrHc2yDH0MB/nX/AA7x+Cv/AEM/jb/wNtf/AJHr4Q/BrS/gxpMugeH/ABh4m1TS3OYbPV7mGaO1OckxbIkKg55XO3POMkmv+CjH/JEdE/7Gq3/9Jbqv2cv+SC+Af+xftP8A0WK/ap+NsXwT+GFzfafcqviPWg1poycErIR882PSNTnuNxQHg1JI8ztLLIzu5JZmOSSepJqqqv2bf2TfEvxquIvE3iIzaL4Mgf8AeXrLtlvdv3kgB7djIflHONxBAtPH/wCzv8EvD0HhOy8aeFNBsdNTalhDfRvKD3ZkUtIzHqWIJJ5JrxB+3h+zxopZbHW9X1tkzxp+mOuSPQzeWPb0/DmtY/4KSeDYCw8P/DPWb0DOz7ZfRWufTO0SY/WviV+0t8OPiUsr337N3h6zupGJF7bak8M4z3LxRpvP++CPav2Yf2lpPhtf6d8OfC/w0sLqXxLrEUE19NfMLiVpZRHECQmCEDAAAAE5PG41Vf8ABWD/AJMj8Yf9hDTP/SyKv+CMul6fafsp6tqVtaRpdX3jG7+0SgfNJst7ZVBPoBnA6ck9zXxls7XUfg/450+9hWa3ufDWoxTRsOHRraQMD7EEiv8AgiLY2knxh+IuovAjXUHhqCKOUj5lR7lS6j2JjQn/AHRVfsn2sPh7/gr7qWg6Ogt7GHxr4vtEhVQAIY4dQ2qB2A2L09K/4KRf8mR/FP8A7B9t/wClkFfsC/sCeD/2xPB/irxL4l8f6x4el8PalFaRRWNrHKJQ8e8sS/Q9q/ar/wCCT3ij4B+BL74vfCjx0fGWl+HIxd6ppmoaYkdzBAnLzLgtHMigbmRlXCgn5sEV/wAEzv2sdH/aK+Ek/g658NaN4c8T+A1ht7yx0i0S0tLi2kDeVcQwoAseSjq6KMBgCMBwor/gip/ydR4p/wCyf3n/AKX2FVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV/wAFB/Bo8dfsZfFXSFiDvZ6IdUUkcr9ikS6JHp8sLfgT61/wRv8AGg8Rfso3fheWdTN4U8UXdskXdYZkinVvoXlmH/ATVVVVVVVVVVVVVVVVVVVVVVVVXjT4keAvh1Zfb/HHi7TNGiK5QXVwFeT/AHE+854PCgmviJ/wUR8DaSZLH4aeF73xDcYIS8vSbS3z2IXBkcexCfWjqn7dP7SChtPgvfDGhXPRoc6Rb7Seodj58q9uC49q8Bf8E6dBhdNS+Knjm81W5dt8tppa+TGW7hpXy7g+oVDXgT4IfCX4aJH/AMIV4D0mwniHy3Zh864/7/Puf9aqqqqqqqqqqqq/4Kl+LT4U/Yn8cQwzGO412ax02EhsZ33UbSD8Yo5Bj3r/AIJR+Ej4Y/Yq8J3zoFl8RahqGpOvfm5eFSfqkCH8RVVVVVVVVVVVfG39qj4Gfs6a14a0P4w+NF0CXxWZ/sErWssyKIdgYymNWMakyKAzDbnPIAJFrc297aw3lpMssE8ayRSKchlIyCPYgiq/4LJf8mi2n/Y4WP8A6Jua/wCCQ3/Jmmmf9jBqP/oa1Vf8FR/F2r/tCftoeFf2efB86zt4eFpolsnLKNSv3jeVsjsEa2VvQxtk8cWX/BLL9hi3s4Le5+Cz3c0USpJPJ4k1RWlYDBchbkKCTzgADngCvH3/AASq/Y81PwN4g07wJ8JRo/iS50u4j0fUD4h1KUWt2Y2EMhSS4ZGCvtJDKQRX/BGn4q3Xgn43+MPgRr7vbDxVYG4treYEMl/ZFt8YHYmF5i3/AFxFVVVWueIvD/hmybUvEmu6fpVooO6e+ukgjGOuWcgV4w/bY/Z68Jb4ofFs+vXMecwaPaNNn6SNtjP4PXjL/gpDNLFNa+APhx5TMpEV5qt5uKnsTDGMcdfvmvhOuo/BXxXqfjzwDr+o2viXWIJYL3VppFluJElkWSTBK4Us6KSygHtnBIrxN448Z+NJvtHi7xbrGtSBtym/vZJ9p9gxOOOOKqq0Twd4t8TEL4b8LavqpJwBZWMk+T/wAGtF/ZY/aF18KbH4Ua3Fuxj7ai2fXHXzmTHXv/Q1pH7A37QephftthoOk7sZF5qYbb06+SJP09K0v/gm78QJgv8AbfxE8PWpx8wtYJrjHHbcI88/StN/4Jq6TGVOr/Fy7n9VttGWHt0BaVu/fFWH/BOT4QxKP7S8Y+LrlgB/qZreEE9+DExwfrx6mrT9gL9n63GJrfxDdfNnM2pY/D5FXitb/Zu/Yi8DmQeL/wCx7J1O3ytQ8UzROpHUBRMrE+2DXj2y/wCCc0FmbLTLrVo7mIMN+gyXkrn/AIFcbom56Hn34xXjyD4VR3e74Zal4qmt9+PL1yzgjbb6h4pDn6bR68dKqqqqqqqqqqqq8PfEb4g+Edo8K+Ode0hVIwllqMsK8dMqrAEex4rw9+2n+0Z4fCRnx2upwof9VqNjDNn6uFD/APj1eHv+CkHj612r4p+Hmg6kFGCbG4ls2Pv83mj9K/aN+Nvwk+PdvB4nsfB+seHPGdqojeUGOe3vYR0WVgysrLztYIf7p4wVqv2R/wBr8+Cxa/DH4q6mzaAAI9L1WYljp/pFIeph7A/wdPu/cilhuIUnt5UkjkUMjo2QwI4II6giqqqr4tfF7wb8GfC0nibxdfbd25LOzj5mvJQM7EX8sseFzyelfGP4z+MvjZ4ofxD4qu9sEJZdP0+I/ubKIn7qjuTgbnPLEDsAB+yv+x0+uiy+JXxb05k03CzaZosq4N13WSdT0j6EJ1bqfl4aOKKGJIYY1RI1CoirgKB0AHYVVVVVVVX7UvwWi+NfwsvNJsbdG1/Sd15o8hGCZVHzRZ9JFyvpu2k/dqWKSGRoZo2SSNirI64KkdQR2NfsG/HFfBnjCT4VeIb3Zo/iaUNp7SNhbe+xgKPQSgBf95U9TVVVVVVX7TH7Vnhz4HWUnh7RBDq3jK4izDZlsx2QIGJJ8HIyDkIOW9gc18O/2lPiT4H+K0nxUvtaudZutSkC6zb3EmFvof7mBwhUfcIGEwMDGQfAXjvwz8S/Cen+M/CWoLd6dqEe5G6NGw4ZHH8LKcgj19ua8a+OvCXw70GbxL4z1220vT4eDJM3Lt2VFGWdjj7qgmvjl+3P4s8Ym48O/Cpbjw5o5yj6gxxe3A/2SDiEH/ZJbp8w5FTSy3ErzzytJLIxZ3diWZj1JPcmvB3gfxb8Qdai8O+DNAu9Wv5ekVvHnYOm526IvqzEAetfBn9gbQdHEGu/GK+XV7wAMuj2jslvGf8AppIMNIfYbV/3hWl6TpWh6fBpOiaba2FlbLshtraFYo4x6KqgACqqq/aZ/a7k8OXs/wALfg3Ib/xNJJ9mu9Qgj84WkhIHlQqM+ZNng8EKeOWzt/Z4/Y0WzuI/iP8AHSI6rrtzIbiLSbp/PWNyc+Zck582QnnaSVHfceB+0n+1p8Fv2UfDdvq/xO1uUXl8rf2XounxCa8vtmM+WhICqMgF3KqMgZzgVr3/AAXGlGpOnhf9nRHsEYhJL/xKVlkHqVS3IQ+25vrX7JH/AAU+8F/tS/EWy+FEvwo17wx4i1CCaa3aK9j1C0CxIXfzJdsTJ8q8fuyCSBnJGf2mv+Cqnh/9m343+JPgrffBjUNen8O/Zd2oRa2lus3n2sNwMIYWK4EwXqc7c98V/wAPxfCn/Ru2q/8AhSR//GK/4fi+FP8Ao3bVf/Ckj/8AjFeAf+Czfhjx5478N+B4fgHqdnJ4h1e105LhvEKOITPKsYcr5AyBuzjIzjrX/BUH9sz9pT9nX4/aB4J+DfxJ/wCEf0W98H22oT2v9j2N3vuXu7uNn3zwuwykMYwCB8ucZJJ/aL+CHjH9ofSfhxa6Td2ltHbxSz6pqVwBiASRwnKxrgszFWwowOOSo5r4O/s2/DH4L20c2g6St/rWzEusXqh52PfZ2iX2XHGMk9aqqqq+NX7U/wABP2dtR0XSvjL8QI/Ddz4hSV9OWTT7q4WZYiocloYnVMF1++RnPFVVVVftkaUNJ/aR8ZQquEuJre4Q4xnzLaJm7/3iw/Cq/wCCfOqf2h8BZrMn/kG69cwAexSKT1/6aH0rxL4m8N+DdDu/E3i7X9P0XSNPjMl1f6hcrbwwKP4ndyAo+pr9oP8A4LL/AA28JSXGgfs9+E5fGeoRsyf2zqoe009SOjRx8TTDPBB8odwTXxj/AG7f2qfji09t4w+Lmq2mlzk50nRX/s61Cn+Blhw0q/8AXVnPvwK+FfwZ+Kfxw8RDwn8J/Auq+JtTChpI7OHKQKeA0sjEJEueNzsoz3rw9/wRt/a51uyS61LUfh/oErKCbbUdZmeRT6E28Eq5+jY96+If/BKb9snwFYPqVp4K0rxbBECZP+Ee1NZ5FA7iKURyP9EVj7VZWNxf30GmwKPPuJliRWOPmY4GfTk1/wAOhv2yv+gJ4U/8Hqf/ABNftAfs5/FP9mXxnB4F+K+jwWeoXdkl7bSW04nhniZmXKOOCQysCOoI9xVWP/BJH9sTUbG31C20XwqYrmJZYy2uIDtYAjjHoa1zR73w9rWoeH9SVBd6ZdS2s4RtwEkbFWwe4yDzVV8NvjZ8Xfg7fDUfhd8SfEPhmXfvdNOv5IopT/00iB2SD2ZSK+BP/BZ74n+HJoNI/aA8HWXi/TuFfVtHjSxv0Hdmi4gl9lAi+tfAb9p/4IftK6I2sfCTxxa6pLBGHvNNlBgvbLOB+9gbDAZON4yhPRjVVVVX/BRhlHwS0NNw3HxTAQM88Wt1n+Yr4G6xpfhz9m/wZruu38Vjp9h4Ztp7m4mbasUaxAlj7Yr9oj4z6h8cviRe+KXaaLSbcfZtItJDjyLdTwSOgdzlm9zjOFFVVUDggjHHqK17x9438UW8dn4i8XavqNtCoWG2uLx3iiUcBUjJ2ooHQAACqqq/ZH0EeIv2ivBVmyZS2vXvGJzhfIieUHj/AGkUfUiqr/grB/yZH4w/7CGmf+lkVf8ABG3/AJNFu/8AscL7/wBE21fFn/klfjL/ALF++/8ASd6/4Ih/8lT+Jf8A2L9r/wClBqv2av8AlMrq/wD2UDxn/wCiNSr/AIKRf8mR/FP/ALB9t/6WQV/wRD/5JV8Sv+xgtf8A0nNarpen65pd5ourWqXVjf2729zBIPlljdSrqfYqSPxr/gnHqN/+z5/wUJ1H4PatcvEl/Jq/ha6804DSQO0kTY6Es9qqqf8AppxwTVf8EdtSs/Cf7YOteHvEEy2eoal4Q1DTbaCRgC9yl1aytGOeSEt5Tx/dNVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV4r8PWXi7wtrHhPUcfZda0+eynyu75JY2RuO/DHiv+CLXiW+8MfEX4ufBXWZAl2ttb34tif8AVyWkz285H1M8IP8AujpVVVVVVVVVVVVVVVVVVVVVVV5eWen2st7f3cNtbwIWlmmcIiKOpLHgD3NfEz9uL4KeAjLY6JqM3i3Uo8jytKwYFb/anb5ce6b/AKVcftD/ALXP7RM8mm/CHwpPoelSMUM+mxY2DkYe8lwqt7psPHA614L/AOCfGv8AiC8HiD42/Eeee6nO6e2052nmcn+/czA8+uEb2Pevh3+z38HfhZ5c3g7wNYQ3sfIv7lTc3IPciWTLL9FwPaqqqqqqqqqqqqqqv+C2nilrH4GeBPBscpU6x4pa8YAkbltraRSD6jNypwe4B7V+yd4U/wCEI/Zi+Ffhh4yk1n4R043CnPEz26PL1wcb2aqqqqqqqqqqq+OGP2z/APgqxo3wyjIvfC/gzUIdMuU3ZjNtp+64vg2OAWm86HPf5PYCq/4KdfBv4l/HP9m628FfCjwpP4h1uPxNaXbWkMscbCFIpwz5kZRgF1HXPNfCH4ff8FfvgL4Mi+H3wn8JaroegQ3Elwln9k0S6xJIcu2+cO/JA4zgV/wkv/Bcj/n21X/wUeHf/jdfsSa1+2ZJ4C8Y6h+2Dp9/c67aXavods1rp8Ek0AhJZEFoFQkuABv5yeuK/Yj/AGN/2i5f2zb/APaL/aR+HNzoMUMmo61FJcXUEyz6lcsVVFWORiAqzSuCcAGNfaqr4z/sb/tK/C/9vh/2gPgB8LL7X/DQ8R2/iFHtLy3iyZSGvrcrJIhAdjOMAY2yAZoEMoYZ5HGRiq8d/tA/Br4bGSHxd8QdKtrqMHdZwyG5uB7GKIMwz7gCvHP/AAUc8LWe+2+HfgS/1OQZC3WqTLbRg+ojTezD2JQ141/bW/aB8Zh4YfFcXh+2cEGDRbcQEfSRi0o/BxWsa3rXiG9fUte1i91K7k+/cXlw00jfVmJJqq8I/BT4tePIorjwl8O9d1G2mA8u6SzZIGz6SthP1rwx/wAE/fjrreyTXZNA8Pxn763V750i/QQh1J/4EPrXhr/gm14dhCP4w+Juo3Zzl4tNsUt8DuA8hkz9do+leHP2Hv2ddA2PceE7vWZUPEmpahK35rGUQ/itaB8HfhP4WCHw98NvDVi8eNssWlxCTI6Evt3E/U0qqihEUAKMAAYxVV40/aK+CPgDfH4m+JOjx3EfDWttKbuZT6GOEMy/iBXjH/go14A04yQeB/BGsa1IuQs17KllET/eGPMYj2Kqfp1rxb+398dNf3RaB/YnhyI5Cm0s/Olx7tMXGfcKK8U/Gj4teNd6+KPiP4gv4pPvW737rD+ESkIPyqqqqqqqqqqqqqqqqqqqvh/+1Z8c/hpo1r4c8OeM/M0myUrb2V7aRXCxj+6rMu8KOyhsDsK0P/go18U7TYniDwZ4Z1JF6mATWzt9TvcfktaJ/wAFJfCs4UeJPhfqtn/eNjqEdz9SA6x+3Ga0T9vb9nnVgv2/VNb0bPUX2lu+36+QZP8AIrRv2nP2fteCmx+LXh2PcOPtt19j/PzguOlfEr9oj4W/DXwjL4quvFGn6szgpY2enXaTyXcuOFXaSFHq54H1IB+K/wAVvF3xl8XT+K/Fd3vkfKWlpGT5VpFniONew9T1J5Nfspfsemwaz+Jnxc0pGuCqy6VolwmfKzyJZ1P8XQrGenVueBVVVVVVVVX7d/wNHgfxonxQ8PWezRPFExF4qL8ttfYJb6CUAuP9oP0GBUUstvKk9vK0csbBkdGKsrDoQR0Nfsq/HKD43fDWC61C4Q+JNECWusR9C7Y+ScD0kAJ/3g46Cqqqqqr9rX9mW1+NHh4+KPCtrFF4z0iE+QQAv9oxDnyHP97+4x4BJB4ORdWtzZXMtneW8kFxA7RyxSoUeN1OCrA8ggjBB6V8BP2kvGvwCudRTQ4YNT0zU4mMunXcjCJZ9uEmXachhgBgPvKMZBClfiH8TfG3xW11vEfjfXptRuTkQox2xW6H+CJBwi9OnXqSTzSRvK6xRIzu5wqqMkk9ABXwN/Yb8X+OFtvEXxOkn8NaI4DpZBQL25X/AHTxCD6sC3+zzmvAfw58FfDLQ08PeB/D9rpdooG/yly8zD+KRzlnb3Yn8qqqqv2q/wBqDVLnU5vgf8GZZ7nVrqX7JqV/ZZeTzG4Ntb7eS5JwzDp90c5I/Ze/ZU0v4SWMHjLxpbQX3jS4TcCTvTTFYcpGehkIOGf8F4yWq9062/bN/wCCpV34N+Kd9ctoNz4s1DTRbNMYythpsc5itlwf3fmC2w20g7pXIOTmvCvgzwh4F0KDwx4K8MaVoWk2ybIbHTrNLeFBjHCIAK0r4O/CnQfHMnxL0L4d+H9N8VT2clnNq1np8cE80LsjsrsgG/LRoctkjGAQCc6r8PPAGu38uqa34G8P6hezY825utMhlkfACjLMpJwABz2Ar/gpn8PPAGh/sQfEnVNE8DeH9PvYP7L8q5tdMhhkTOqWinayqCMgkH2Jr/gjN4K8G+JvgT42u/EnhLRtWni8WlI5b6wjnZF+yQHaC6kgZJOPerf4XfDOzuIru0+HfhiGeBw8UsekQKyMDkMpC5BBAOa/4LV/8nUeFv8Asn9n/wCl9/T+OvBHw9+H2j6/498Y6J4b0xbG3U3mr6hFZw7vLHG+RgM+2a+H3x0+C/xZmltPhl8V/Cfii5t0LzW2lavDczRrnG5o1Ysq57kYqqqqr/guN/yNXwj/AOwfqv8A6Mtqqqqrn4j+BrLxb/wgeoeJrKy14wpNFY3b+Q86PkK0RfAl5BHyE4IIOMV/wUG0j+z/AI8Q3ypgapoNtOWx1KvLGe3XEY9e1V+yJ+0R4I+CHwx8ajxXdPLdi/gm03TIOZr2R42UheMKo8tdzk4AI4yVDft8/tM/Eb48+ObDTPE2tSR6Rp0JuLbRraQra2zuSFOz+OQKP9Y2W+cgYHAqwsLvVb620zT4GnuruZYYIl6u7EBVHuSQK/Zg/Zz8F/sw/CPR/ht4Tsbb7XFAkmtakkeJNTvSo82Z26kbshFP3UCr2r4v/wDBZP4PfD/xtqng7wX8Ntc8YxaRdPazamL6OygmkQlWMOVdnTIIDELnGQMYJ8Xf8Fhfgj43+DHjnS9G0LxV4X8a3Gg3cGiR3luk8Ml1JGUiKyxMSCrNvO9VGF65NeEv+Rq0X/sIQf8AoxaYqqlmYAAck8Yr/gpN+yyP2k/gJdX/AIb07z/G3gcS6loflx5kuk2j7RaDufMRQVA6yRxjoTVeEv8AkVNF/wCwfB/6LWvi1/yVTxn/ANjBf/8ApQ9VY6XqWqO0emadc3bIMssELSFR6kAHFVXhDxl4r+H/AIksfF/gjxFf6HremSiS0vrGdoZYm9mHYjgg8EEg5Br/AIJ8/wDBR2x/aK+z/CL4wzWem/EeGI/YbuNRFBryKuWKr0juAASyDhgCy4wVWqq1+Oel63+0B/wpPw8Yrn+zNJnu9Xugc+XOrRhIFx3AclvQlR1DCq/4KSa1BF4T8G+HRMhmudRnujHkbgscYTOOuMykfhXxB+O3j34h+GNA8DX+om08N+HNOtrK0023YqknkRLGJZT/AMtHO3PPC54Ayc1VVXhT4W/Ejx0yjwf4F1zV0bpLa2Mjxj6vjaB9TXhb9gj4++IFjl1ez0Xw7G2CRqF+HcD2WESc47Ej3xXhr/gm1o0YSTxj8T7y4JHzw6ZYLDtPtJIz5/74FeHv2Fv2dtDVTeeHNR1uRRw+o6lJ19SsRjU/litF+AnwT8Pqo0n4U+Fo3T7sj6XFLIP+BuC361YaLo+lKF0rSbOzAUqBb26x4BOSPlA4zzTRRO6SNGrPHnYxXlc8HHpVV+2N8AdY/ab+Aet/B/QvEFnot5qlzaSpeXcTSRoIZ0kIKrzyEx+NfsO/sza7+yd8Fpvhd4h8T2GvXUut3Goi6soXiQLIkShcNzkeWfzrxlocvifwfrnhq3nSGXVtNuLRJXBIRpI2QMQOw3Zr9gX9gXxf+x54v8VeJPEnxA0fxDH4h02G0iisbWSJomSTeWJfqCOKr4V/8E6fGnw9/bjvf2sbz4j6Jd6TdeJNd1ZdIitJVnVNQjulRN5+XKfaVJPQ7Tiv2o/g7qX7QHwD8X/B/R9ZtdKvPEltFDFeXMbPHCUnjkJZV5ORGRx61+wL+x54l/Y88H+KvDXiTxjpniGXxDqUV3FLY27xCIJHsKkP1J61Xxr/AOCa/jfxf+1uP2ofhV8SdC8PyLq9hrC6de2kzH7Zb+XvO5D9yRotxHq7DpVftO/8EkvEfjT4ran8Xv2cPiRpXhq91jUW1GbS9Uae1SyunbfJLb3ECuyguSwTYNpOA2MBf2Ef2Z/j7+ztZeMZPj18YY/HN74iNiLFI9Su75bEQCfzMSXKq3zmZOAoHyZ78VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV8PXH7PP/AAWT1zw/Li103xnq95ESOFddTtxdxKvt9paNcDgEEdqqqqqqqqqqqqqqqqqqqqqrWNVsdC0i91zU5Wjs9OtpLi4dI2kZY0UsxCqCzHAPABJ7CviF/wAFB7jULw+Hfgh4Gn1C7mby4L3Uo2ZpG/6Z20Z3N6gsw91qz/Z2/ay/aNuItW+MHiu50LSnfesOptgoPWOyiwqnt8/ln618NP2Jfgh8PxFealo8nivU0wftGsYkiU/7MAwmOP4gx96trW2s7eO0s7eKCGFQscUaBVQDoABwB7VVVVVVVVVVVVVVVVf8FoL248W/GX4O/C2ykH2gafcSxKSOGvbmOFTzjvbdzjjtVhY2umWFtpljEI7e0hWGFB/CigAD8gKqqqqqqqqqq8T3Wt2PhrVr3wzpi6lrFvYzSafZNIsYubhUJjj3MQq7mCrkkAZySK/4Jafsp/Fv4W/Eb4mfFv4++ENR0bxRdBNNszfBGNz50hnu5ldCUcFkgAdCRneM1VVVVVV8Q/j78IPhaskfjLx1p1tdx8Gxhc3FznsDFHuZc+rAD3r4hf8ABR2JDLZfC7wJ5mMhL/WpcD6iCM5x9ZB7iviB+0n8a/iZ5sHiXx7qC2UuQbCxb7JBtP8ACUjxvH+/uPvVV4K+DvxS+IzIPBXgPWNUic4FxHbFYAfeVsIPxb1rwZ/wTv8AivrYjn8ZeItF8NwsPmjQm9nT6qm1Pykr4wfBL9nP9nHRBZ6xf6n438bXcWbXTJ7r7PbxA9JpUhw6pxwpkyx4GBlh8E7u3+Lf7UHgr4LeGre3u1udTF34guAgaK3s7cGaeIdtzohj9FLgdelvb29pbxWlpBHDDCgSKKNQqooGAABwAAAMVVVJJHFG0ssioiKSzMcBQOpJ9K+If7XfwI+HQkt7vxjFrV9HkfYtFAu3yOoLgiNT2wzg149/4KM+LtREtn8OfBVhpEZ4S81KU3U2P7wRdqKfY7xXjn45/F34kmVPGPxA1e+t5s77RZ/Jtz/2xj2p+lVVVHFJPIsMMbSO5wqquST7CvD3wC+Nfiva2h/C3xJNGxAWWTT3gjOfR5Aq/rxWgfsGftC60qtf6Ro2iBh/zENTRsc9xAJD71on/BNnxROEPiP4o6VZ8fMtlp0lz68Au0ftzjv7c6R/wTg+GkAH9u+PvEt6QOfsiQWwP/fSSVpn7Bv7O1ht+16Jq+pbTz9q1SRd3OefK2fTj/69ad+yV+znpYUWvwr0t9uMfaZZrjp/10ds1Y/Ar4KaYVax+Eng+N0JKudEgZxnr8xUn9asvAvgnTdn9neDtDtfKB2eTp0Sbc9cYXjqfzq40DQruxfTLrRbCa0kbc9vJbI0bHGMlSME4rxR+yt+z74uZ5NT+F+kW8rj/WacrWJB9cQlQT9QfeviN+wh8B9EsH1YfE+/8HwnOJdVuoJLdcdhv8tj/wB9mviV4I8D+Dr42ng74saZ4xVWw7W2nz2xX8XBRv8AgLkVofhvxF4nu/7P8NaDqOrXWP8AUWNq87/98oCa1TRtX0O6NjrWlXmn3K9YbqBonH/AWANVVVVVVVVVVVVVZX99pt3Bf6dez2t1bSLJDPDIY3idTkMrDkEEAgjpWlftW/tEaOVNn8VtXkKnI+1iO6/9Gq1aZ+3l+0RYBRd65o+pbRg/atLjXdxjnytn147+3Fab/wAFHPitCAureCvCl1jqYEngJ495H5z/AJFab/wUqmBVNX+ECN6yW2tlccdlaE559603/go/8NJio1jwF4mtePm+zNBPjjtudO/+e1aZ+31+z7fhTdXmv6dkci60wtjj/pkz/StN/a9/Zv1bAtfilYxn0ubW4t8f9/I19a0v40/B7Wyq6T8VPCd07dI01mDf1x93dkflVlqOnanCLjTb+3u4scPBKJBz7gmqqqr4jeAdA+KHgrVfA3iSHfZapAYy6j5oXHKSJ/tKwDD3HPGa+IngLX/hl4z1TwP4mt/Lv9LnMbMAdsqdUkTPVWUhh7GvgH8Y9W+CHxFsPF9l5k1g/wDo+q2an/j5tmI3Af7QwGU/3lGeCa0DXtH8U6JY+IvD9/Fe6dqMCz2txGcrIjDIPt9DyDwaqqqqrxL4m8P+D9EuvEfijV7bTdNsk3z3Nw+1VHb3JJ4AHJJAAJNftQfELwH8Vvibd+LfAHh2TTrd4xHc3Eh2nUZFP+vMYHyEjA6kkAE4JIqvDllHq+tWOiz6pZ6dHe3CQm7vGKw2+4gb3KgkKM5JxxXwJ/ZR+HXwdgt9adI/EXiTaG/tW5iG2En/AJ4R5IQf7WSx55AOBVVVV+2B+0tceDoH+Efw4upJPFOpKI765tsl7GNxgRx45858jGOVByOWBH7J37MFr8KNKi8ceNLOKfxlqEW5Ecbv7LjYcxr/ANNSD87Dp90cZLVX/BQv9j/4vfBP45Xf7X/wEtb+40q61Qa5eS6ZAZJ9B1AHzJZnQZ3QO4aQuRtBdlcAYLfs6/8ABZfwDrtnY+Hf2kPDNz4b1VQscuv6RC1zYynvJJAMyw/RBKM8/KOB4G8feCfib4atPGPw98VaZ4h0W9B8i+0+4WaNiOqkqeGHQqcEHggGq/4Kj/8AJiXxN/7hP/p2s6/4Il/8kB8df9jgf/SOCq/4LV/8nUeFv+yf2f8A6X39fC79gzx/+3BpGn/tAftffE3xHpdprdoknhfwvobxxnTdOKgwENMkiRhl2tsEZZhhnfcxA/bb/YV8RfsMX/hv4zfBv4g67daBJqQt7e/dxBqGj3oVnjzLDtDK6o+HVUwVKkcjP7Efx/u/2lv2cPC/xK1oRLr22Sw1pYl2qbyBtrOAOAJF2SYHA8zHaqqqr/gr/wDAz4xfGHxJ8Mrn4WfDPxF4qi0ux1JLx9KsHuBbs7wFA+0HBIVsfQ1VVVftm/A8/Fn4Zya5odmZfEnhZXubNY0y9zDgedD6klRuUddygD7xrWPEfiDxClnHruuX2ojT4PItRd3DS+RHknYm4nauSTgcc1VfE3Wf7f8AHmtair7k+1NFGfVI/kUj6hQfxqvD+s3XhzXtN8Q2IU3Gl3kV1CGHBeNwwz7ZAr4TfFHwh8avhzoPxO8DajHeaPr9mlxCQwLRMR88TgfdkRgUZezKa/aj/wCCOknibxDq3jz9nHxhZWMmpXMl1L4Z1oFIY3dizC3uEBKrk/LG68f38dPjH8AvjD8ANfXw38XvAOp+HbuXd9nknQPBdAYyYZkLRygZGdrHGRnFeEv+Rq0X/sIQf+jFrxcqt4T1pWAIOnXAII/6ZtX/AAT6/aGX9o39mjw74h1PUFuPE/h5f7H8QBmy5uIVASZu582IxyE9NzOB901/wVJ/ZcX4CfHeTxx4X00w+DviI8t/aBF+S0vgQbq3/wBkbmEqjgbZCo+4a8Gyw3Hg/Qp7eVJI5NNt2R0bIYGNcEEdQRXxUhluPiz4vt7eJ5ZZPEV8qIikszG4cAADqT6V+xP/AMEt/hr8N/Cul+P/ANoXwzZ+K/HF/DHcf2PqEYlsdGyMiIxH5ZphnDs4ZAeFHG9vHHx9/Zt/Z/Nv4W8a/Erwb4OlCAw6S1zHDIiEcMLeP5lU44O0D3r/AIK8/HL4LfFDwb8N7P4UeJ/Cnii4vtQvLy/1PSZoZ5oFgjjjjildcuu7znOxsf6sHHQ1VaFrmseGNb0/xJ4e1KfT9U0q6jurK7t3KSW80bBkdSOjBgCD6iv2Z/i2Pjv8A/A3xZdES58Q6RHLepGuFS6QmO4VR/dEySAe2Kr9qz9oC1+B3gRo9JnjfxVraPDpUJwTBxhrhgf4UyMA9WIHTdj9hG+vb39o1Ly7vJpri80y9e4lkcs0zNhiWJ5JLcnPfmv2hf24PC3w/W68KfC97XxD4jXMct5u32di3fkf61x/dU7Qep4KnxZ4v8TeO9duPEvi/W7nVdTujmW4uHyfYAdFUdlAAHYCqqvg38AfiL8ctUey8HaakdlbHF1qd4Wjtrc/3SwBLMf7qgnucDmvBH/BOPwdYiK5+IPjrUtWlABa202FbSIHupdt7MOvI2H6V4N/Zw+B3gLy5PDnw20dbiIfJc3cP2uYH1Dylip+mKVVRQiKAFGAAMYqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv+Cuej6h8Jf2mvhF+0doERS4a3jKlTjN3pl0sysT2JSeNfcR+xrRtW0/xBo9jr2k3AnstSto7m2lHR45FDKw+oINVVVVVVVVVVVVVX7ZP/BRvRP2QPifpnw01L4U3vieXUtBi1dbuDV1tQgknnh8vaYnyR9nJzn+LpxWlXy6ppdnqaxmMXdukwQnO3coOM/jVV+01/wUl/Z4/Zr1W58H3V7e+MPF9qSs+j6HscWb/wB24nYhIm4IKDe44yoBzVx/wXI1U3++1/ZstBZBh+7k8VsZCO/zC1wCf90496/Zy/4Kofs7/HjWbTwf4gW9+H3iS9dY7W31mVHtLqRjgRx3S4G4ngCRY8kgDJOKqqZVdSjKGBGCCOteBPhH8NPhjE8fgPwXpmkPLnzJoot0zgnODKxLkexOBVVVVVVVVVVVVVVVVVV+1s6/Ej/grr8M/CLyCWPw/e+HreVC3RI5ftzr14ysp9PvdPWqqqqqqqqqqqqqqqrxJ4p8NeD9Lk1rxVr1hpNjEDuuLy4WJfoCx5PsOTXxL/4KEfDbw752n/DnRrzxTeJkLdSg2loD6gsPMfHptUHs1fEj9rj45fEvzba98Wvo2nSZBsNGBtUx6FwTIw9mYj2pmZ2LMxZickk8k1Xw/wD2bPjX8TPKm8M+AtQWylwRfXy/ZINp/iDyY3j/AHNxrwF/wTgGIrr4m+P+esllokP/ALXlH/tP8a8C/svfAr4eiKXQ/h9p9zeRci81JTeS7v7wMmQh/wB0L/OlVUUIigBRgADGK/aa/aZ0b4IaMdG0Z4L/AMYX8RNpaE7ltVPAmmA7f3V6sfQAmvil4J1zwh+zj47/AGqPjPeXE2q6tam28NWd2T517e3ZEUd047JGrNKicBhFnGwBW/4IlfCKSbUPH/x31C3OyGKPw9pkjJkM7FZ7rB7EBbUcf3z+NV8QPi58NfhbZ/bPHnjDT9KyuY4HffPJ/uRJl2HuBiviX/wUYhQy6f8ACbwaZDyq6lrRwPqsCHPuCzj3WviF8cvix8U5HHjbxxqN9bM2RZI/k2y+mIUwmR0yQT71VVpWj6tr16mm6Hpd3qN3J9y3tYGlkb6KoJNeDP2L/wBoPxmI5j4OGhWzgYm1qcW2M+sfMo/74rwh/wAE24QEm8e/Ep2OPnttIs9uPpLKT/6Lrwp+xT+zx4W2Sv4Nl1q4QcTatdyTZ+salYz/AN814e8EeDPCMQg8KeEtG0ZMY22FjHb5/wC+AKqqqqrxB4m8OeE9NfV/FGvafpNlH964vblYEHtuYgZ9q+In7f8A8IfC/m2fguz1DxbeICFeFTa2270Mkg3HnuqEe9D46/to/H9jD8MfC8+i6VOSFuNOsxDHt97uc43D/YKn2rw/+wD8RPGF8Nd+M/xUzcyD94sDyahcOPRppSAp+gcV4K/Yu/Z98FrHI/hBtfuo8f6RrUxud2PWIbYj/wB8VpWj6RoVkmm6Hpdnp9pF9y3tYFhjX6KoAFalpWl6zamy1fTbW+t26w3MKyof+AsCK8Tfss/s++LC76n8LdGhkfq+no1ic+v7goM/Xr3rxL/wTt+EOpl5fDfiTxHosjA7UaWO5iU+u1lD/wDj9eJP+Cb/AI8tAzeEviHoeqKq5C31tLZsfYbfNH5kfhXiT9jT9orw3udvAD6lCi583TbuK4z9EDb/APx2vEPgbxr4Rcx+KvCGtaOwOMX9hJb8/wDAwKqqqqqqqqqqqqqqqoLi4tZRNazyQyKOHjYqR+IrSvjB8WNDKnR/id4qswp+7DrE6qe/KhsEexrSf2u/2jdFVUtPilfzKoxi7t4LnPTqZEY9v5+prSf+CgPx904AXn/CNapgY/0vTWXP/fp0rSf+Ck3iuHb/AG78LtJvMdfsmoyW2evTcsnt/k8aR/wUh+H04X+3vh34hsz3+yTw3OPpuMftWj/t5/s76nt+261rGk56/bNLkbb9fJ8yv2vdX+Afxx8LReOvAXxH0F/FegxEGCWQ20moWucmICUKS6EllHU5ZcEkYr9hj9oxfC2qR/Brxlf7dJ1Sf/iSXErcWty55hJPRJCePRz/ALZIqqqq+NXx68CfA7RPt/iW8+06ncITY6TA48+5PTOP4Ez1c8cHGTxXxm+O/jz4361/aHim+ENhbuTY6VbkiC1HTIH8T46ueT2wMAAFiFUZJ6CvEf7NHxd8NfDZPitq/hh7XR2dfMgckXMEbY2zPFjKxkkDnkZBIAOar9jT9rFNFFl8IPidqqpp/EWh6pO2Ps5J4t5WP8HZGP3fun5cbaqqr9pr49WXwN8Dm5snim8S6sGh0i2cZCsB80zj+4mRx3YqOhJH7G37P95f3X/DQHxNSa81TUZGuNHiu8sxLHLXj55LMSdmegJbnKkVVV+0D/wTr/Zi/aAtLy7vPA1r4U8TTqTHr3h+IWkok/vSxLiKbJxkuu4jgMvWvgZ4w+Kn/BOP9tU/CPxJrrXXh641e20/X7eFiLXULK42+TeojHCyIsiyDuCHjLYLVX/BUf8A5MS+Jv8A3Cf/AE7Wdf8ABEv/AJID46/7HA/+kcFV/wAFq/8Ak6jwt/2T+z/9L7+vB0UcPhDQ4YY1RE023VEUYCgRrgAdhX/BWSCGb9ibxXJJGrNDqemPGSOUb7XGuR+DEfjX/BFeaWX9lXxLHJK7rF4+vFjVmJCD7DYHA9Bkk49SfWqqqqqqqqv2z/gePhR8Sn1/Q7Py/DnilnubUIuEtp85mh9hkhlHA2tgfdNV4r1hfD/hrVNaLAGztJJEz3YKdo/E4FMS5LMxJJ5Jqq/Zm/bH+N37KWsSXXw21+OfRryUSah4f1JWmsbpsAF9gIMcmAB5iFWIABJAxX7O3/BXX4F/Fi/sPCvxS0m6+HOvXjJDHcXM4utNllJAA88BWhyef3iBFHV+9fFj4SfD345eBNR+HnxI8PWusaLqcRUrIgLwOQQs0L4JjkXOVccg/jXxE+F9/wDBT9onVfhRqVz9pm8L+JhZLcbdvnxrMPLkx23oVbHbdVf8Ey/2n0/Z3/aAtdF8SaobfwZ498rTNWMj7Y7afd/ot02cABHYozE4CSuT0Fftzfs9w/tKfs4eJvAtpYrP4gsIv7U8Otj5lvoFJRFPbzELxH2lz2Br4J3yap8GfAOpRxlFu/DGmyqjHlQ1tGcfrX7J/gvTfH//AAUi8PeHtVgjmto/G2pagY5Pus1mLi6UEYOfmgXjoehr4yeNbn4a/CDxz8RrK3S4uPCvhvUNVihccSPb20koU+xKAV4n8T+IfGviHUfFvizWbrVdZ1a5e5vb26kLyTyscszE+/5VVVX7Bfw71b4WfsgfDDwbrsDwX6aQ19PDICHha8nkutjA8hl88KR2IIr4geOvDvw08H6n428UXYg0/S4DI+MbpG6LGgPV2YhQPUjp1r4t/FDxD8YvHeo+OfEUmJbptltbhsraQKTsiX2APXuST1JrTdY1bRnnl0jU7mykuYGt5nt5TGzxN95CRztIGCO44PFVVV+zp+xD4g8f/ZfF/wAVI7rQ/DrbZILDaUur9eoJB5ijPqfmI6AAhq8N+GfD/g/RLXw54X0i20zTbJNkFtbx7VQf1J6knknkmqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/wCCvnwyPjj9k2XxdaWokvPAmt22ol1XLi3lJtpVH+zmaJz/ANcwegNf8E2fianxQ/Y48A3Utz5t94btn0G8UnJja0bZED9YPIb/AIFVVVVVVVVVVVVVV/wWr/5Oo8Lf9k/s/wD0vv68Jf8AIqaL/wBg+D/0WtV/wU8/ax1T9m74MW/hrwLqRtPG3j15bSwuY2w9haIo+0XKns43oiHjBkLA5TFf8E1P+CengnWPA+nftGfH7w1F4hv/ABAPtPh7RNTjEttFbE/LdTRtkSvJ95A+VCENgswKpoeiR6UNBj0exXTQmwWQt1EIX+7sxtx7Yr/gpP8A8E7PA0/gHVvj/wDAbwra6DrHh6F7vX9E06ERW19aLzLcRxD5Y5Y1y7BQFZAxxuA3f8EnP2r9a+N3wy1L4R/EHWJtR8VeA0jNre3Ll5b7TX+VC7HlniYbCx5KtFnJ3E1VVVVVVVVVVVVVVVVVVVfDsN8Rv+C1uuay4MkWg6nqHmgkjH2TS2tE9Dw4T16Y5HNVVVVVVVVVVVP+1H8BrX41S/s73nxCtbf4gxNCo0aW3mVnaWFZkVZNnlljGytt3ZwelVVVXxN+O/wq+EUDHxv4utLa8Cbo9OhPnXUmRxiJcsAezNhfevin/wAFDvFusGbTPhP4fi0K2OVXUdQVbi5I/vLHzGh6cHzK8WeNPFvjvVG1rxj4k1DWb1s4lvJ2kKg9lBOFX2GAKrwD8IviV8UboWvgTwbqOqgPse4ji2QRH0eZsIv0JFfDn/gnLqt0Ib74q+NYrJDgtp+jJ5kmPQzONqntgI496+Hn7N/wX+F/lT+FvA1ib+IDGoXqm6uMj+IPJnYT/sBR7VVV+0p+0XovwK8NCK0MF/4q1JCNO09m4jHI8+UDkRgjAHBY8DgMV/Zm/Zx1f4p62fj38cDLqC6hP9qsbG7Tm+ftPKuMeUONiAYYAfwABv8Agtp8X/N1HwD8BdNuf3dtFJ4g1OJW4LNugtQQOhAW5OD2dTx3/YC+ES/Bf9kzwB4ZuLRoNT1PTxrGph12v9ou/wB8VYdiiNHH/wBsxXxY/al+DvwhEtnrfiNdS1iPI/srSytxOrDs+CFi7cOQeeAa+Kv7eXxZ8b+dpvgpYfB2luCoNq/m3bj3mIG31+RVI/vGtQ1C/wBWvJdR1S+uLy7nbdLPcSmSSRvVmJJJ+tVXh3wr4l8X6gukeFfD+oavet0gsrZ5nx6kKDge54rwB+wD8Y/FIiuvF1zpvhK0cZK3D/arnHqIozt6dmdT7engT9gr4H+FRHceIodS8VXajJN9OYoQ3qsUW3j2Zmrw54S8K+D7Iab4T8N6Zo1qP+WNhaJApPqQgGT7mqqqqqqvih8evhV8H4GPjbxXbwXuzdHpsH766kHbEa8gHszbV968Wftw/Fv4n6o3hP8AZ/8AAVzayS5CTm2+3XpX++EAMcQ9chwP7wrw9+xP8bfixqUfij49fEGWyZ+TBJcG/uwvdRz5cQ9MFsf3a+HP7JnwM+GohuNO8HQ6tqEOCL/WMXcu4dGCkeWhyM5VQaVVRQiKAFGAAMYqqqqqqdEkQxyKGVgQVYZBFeJPgh8HvF29vEXwz8N3ksn3pzp0aSn/ALaKA3618Xf2dP2JvB0cknirXm8Hz4LfZ7HWHlnPfKwOJWI+i46V8RrH4TWepi3+FWueJdTtlOHk1ayih3+6FGyR/vKprwV+zT8c/iAiTeHfhxqv2Zxlbm9QWcTD1VpioYf7ua8QfsvftA+GQx1P4Ua7IEGSbGEXoH4wF61XQtb0Kf7Lrej3unzAkeXd27wtkdeGANVVVVVVVVVVVVVVVVVSsyMGRiGU5BBwQa0T9rH9onQESOw+KmqSqgwBexxXf5mZGPetG/4KB/HrTSov08NasB977Vp7IT+MToAfwrRv+Ck+uRBF8RfCqwuuzPZao8GOnIVo3z34z6c1o3/BRr4VXQVNd8GeJ7Bz1MCw3Cr9SXQ4/CtG/be/Zw1cKs3jW502Rhwl7plwv5siMo6etfHH9uHwH4R0NbD4U6pZ+JdevYz5c65NtZLyN7k43vkcIPqxAwG8S+J9f8Za3deJPE+r3Gp6nevvnuZ33Mx9PQADgKMADAAArwh4N8T+PtftvC/g/RbjVNTujiOCFeg7sxPCqO7MQB3Nfs8/seeFfhQLXxV4y+z694sQB0YrutrBv+mSkfM4/wCejDI7BepurW2vbaWzvLeKeCeNo5YpUDLIpGCrA8EEHBBr9rT9le8+EGqy+NPBVlNP4KvZOVBLtpcrH/Vuevlk8I5z/dJzgtX7If7YMFvBp3wl+K2oFVQiDSNbnkyAP4YJ2PQD7qv0AwDjGaqq8V+J9F8F+G9R8V+IbxbbTtKtmnuJD1CqOgHdicADuSB3r4ZeF9e/bM+O+ofEPxtBJH4U0Z0MluSdgiUkw2aH35ZyPVjwXFRQwwQpBBEkccahURF2hQBgAAdBjtVVXgr9tH9mHx54n1nwVpHxg0G013QtSudPubDVLkWLySwSFGMJl2rMhKkgxlsjnjnHi/4xfCfwBok/iPxp8SfDejabbqWe4u9TijHHZRuyzeigEk8AV4+1of8ABQj/AIKLWE3wz065l8OXOpWUEdy8TIy6XZ7fPu3BAKBgJGVWwfnjQ4Y1X/BUf/kxL4m/9wn/ANO1nX/BEv8A5ID46/7HA/8ApHBVf8Fq/wDk6jwt/wBk/s//AEvv68Jf8ipov/YPg/8ARa1/wVg/5Mj8Yf8AYQ0z/wBLIq/4Iqf8mr+Kf+ygXn/pBYVVVVeMvi98JfhzqFlpPxC+KPhHwxfaiu6ytdZ1u3spbkZ25jSV1LjOBwDzxVVVVXx2+EumfGn4a6p4JvfLjunXz9NuXH/HtdID5b/Q5Kt/ss1azpGpeH9WvdC1mzktb/Trh7e5gkGGikRirKfcEEVo/wCzb4//AGm/Cvi/wj8Orq1i1TS9KF/FFcnYl66TJttg+cRs43FWPy5TBwCWHijwt4j8E+IdQ8JeLtEvNI1nSp2gvbG8hMUsEi9VZTyP6gg9DVRxSTSJDDGzyOwVEUZLE9AB618Q/wDgjN+0Hollaal8OPFXhvxQstrG9xYXUp0+6gmKAvGN26J1VsqG8xSeDtHOPBn/AASW/bO8SeIIdK8R+CdI8Kae0gE2p6hrlpcRomeWWO2kkkY46AqM8cjkjwh4etfA3grRPCiahJcW/h/S7eyF1cnDSJBEqb3PqQuTX7SnxM0f4xftjeLfiL4dmSbStU8VothMh4nt4WSGOUezpEr/APAqqv8Agmb+1Gf2ivgHbaF4l1MXHjXwEsWm6sXbMl1Bgi2uT3JdEKMTyXjc8bhVnZWmnWcGnafbRW9taxLFBDEgVI0UYVVA4AAAAFfCb4rWPwP/AG5LH4oarK8em6L45u/7QkQEslpLNLDOwA64ikc474x3rU9P0Hxr4ZutKvRb6nouvWDwyhJN0dzbzRlWwynlWRjyD0NfHH/gk5+094C8ZX1t8LfCyeO/C0ty/wDZt9aX9vDcJEeUWeKV0IcDglNykjORkCvjh+yp8dP2cNJ8O6t8ZfBf/CPp4oe5XT4jew3LnyBEX3+S7hMiZcBiCcNxxVV/wTg/4J4a18Wtf0r46/GfQ5bHwHpk6XOk6ZdwlX1+VTlGKn/l1BAJJGJMbRlSxDMqKWZgAoySTjFftkftEN8X/F//AAiXhi+3eEvD0zCBkb5b65GVaf3UAlU9iT/FgVVV4N8E+K/iFr9v4X8GaHc6rqdyfkggXoO7MxwEUd2YgDua/Z3/AGK/CvwuNr4s8fm28Q+KUAeKMputLBu2xT/rHH99gMHoARk1VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVfFz4f2HxY+Fvi34Z6k6pB4o0W605pGGfKMsTIrj3ViGHuBX/BF34g33h7xF8UP2dfEW+3vbWVNYtrRzgxTRN9lvARnrn7KOP7pzVVVVVVVVVVVVVV/wAFq/8Ak6jwt/2T+z/9L7+vCX/IqaL/ANg+D/0WtV/wVi1a/wDiL+23pPw6juSsel6RpmlQKeiSXMjSl8HufPQH2UelaFommeGtD07w5otsttp+lWkVpaQr0iijQIij2CqBVX1jZ6nY3Gm6hbR3FrdRNFPDIu5ZEYYZSO4IJBFf8E3Tc/Cr/gowfhtZ3DtFPJr2gTMvIkjt45ZhnkcbrND35xx3FVVVVVVVVVVVVVVVVVVVX/BNkJ46/wCCivxp+ITFWjMOu3sbLj/WXOqxY6ZGNjSdD6dRmqqqqqqqqqqq/YXH/DRn/BTf4hfHKf8A0nTtCfVtTsJj8yhZJBZ2iZ7n7PIxB/6Z544qqr4u/tRfCL4OCWy13XxqOtRggaRpuJp1Po/IWLt98g45ANfFn9uj4tePzNpnhGVfB2kOSFWxkLXbr/tT8Ff+2YT0yaubi4vJ5Lq7nknnlYtJJIxZnY9SSeST61FDLcSpb28TSSyMFREUlmY8AADqT6V8L/2JvjV8RDDe6rpSeE9KfBNzqylJWH+xAPnz0+9sB9a+GP7DnwW8AiG916wl8X6pGATNqijyA3fbbj5cez7/AK1aWdnp9rFY2FrDbW8CBIoYYwiIo6BVHAHtVVVfHv45eHfgX4NbXdSCXeq3m6PStOD4a5lAGSe4jXILN2yB1Ir9nT4G+Jf2g/GU3x7+NLyXmly3Pm20E6ALqciHaBt6C3TaF24w2NvQNSIkaLHGoVVGFUDAAHpX7Qdxqv7R37aHin4l61pk8/gi28SJaK7OIxc6bZusO2DOcmVImYHBUNISa+LH7ZXxf+Jgl0vTNRHhTQnUoun6U5R2T0km4duOCF2KR/DRJY5JyT61VfDL9kv42/FERXeneFm0fTJDxqGsZtYyPVVIMjj3VSPevhp/wT7+F/hnyb/4gapeeLL1CGMAzaWoP+4p3tg+r4Pda8OeFfDPg/TU0fwp4f0/SLJAMQWVssK5AxkhQMn3PNVVVVVVV8UvjX8N/g5pf9o+OfEUVtK6k29jF+8ubj/cjHOP9o4UdyK8U/tW/tAftA6xN4M+AHhS+0myc7WmtAHu9h4DSznCW4PsQQeN5r4W/wDBPy2lnHiT45+JptVvp282XTbGdgrMeT51wfncnvt28/xGvCXgjwh4C0pdE8GeG9P0eyXGYrSAR7yP4mI5ZvdiT71VVVVVVU0sNvC888qRxxqWd3baFAHJJ7DFfFj9uH4QfDszab4buW8Y6ugIEWmygWyN6Pccr/3wH98UfiT+2d+0+5i8C6bdeHfDsxK+fYA2Nvt6HN053yH1WM9/u14B/wCCdeliVNV+LXjq51O5dt81npYMaM3U7p5AXcHvhUPvXgT4G/CT4ZiN/BXgLSrC4jGFuzD51x/3+k3P+GcVVXVpaX1u1re20VxC4w0cqB1b6g8Vr/7P3wQ8ThzrHwq8MySSZ3yxackEjZ9XjCt+vFa/+wf+zvrW82Oh6torODzYanIdp9QJvMA+mMVr3/BNrwrPvPhj4n6rZ9di39hHc/QEo0f54/CvFn/BPT4keH7G61Ww8eeFbmys42lmkvJJbQrGvJY/IygADJyw71PaSRSvErpKEYgOh4bHcZwcH3FWvw6+IN9o8PiCx8C+IbjS7jd5N9FpkzwSbTtbbIF2nDAg4PB4qe3ntpTBcwvFIv3kkUqRx6H2qqqqqqqqqqqqqqqqqqzjoa8H/ETx38PrqW98EeLdU0WW4ULMbO5aMShTkBlHDAH1Hc+prRv21P2j9GCRnx+t/EgGI73TreTPTq2wOen96tG/4KKfGGyCx6z4Z8Lakg6ssE0Eh/ESFf8Ax2v+HiOh+ItKudA8d/BZLuwvoWhuo4dVEiSowwymN4hxj/arxtJ4LuPEl3c/D+DVLfRJm329tqQUy2+ese5WIdR2Y4JHUZGTX7Lv7ad34OWz+HvxdvprzQ1xFY6w5MktgOyS9S8Q4AP3kHHK4C6Xq2la5ZRajoup2t/azIGjntpllRwRkEMpIIIIP41X7aPxM1n4i+ONJ/Z0+Hxa7k+1Rf2jHEf9fduR5cRPTbGDuYngE842V8Hfhfo3wf8Ah9pngfRwrm1TfeXAXBurhseZIfqeAD0UKO1VVV8W/wDgjd8AfHes3mv+AvGvifwXc3srSyWxK6laqzNklVlIl5yesp7VoP8AwQ48N298knif9o7UtQswRvhsPDCWchHfDvcygdv4TX7Of7J/wT/ZZ0G40b4T+GngutQC/wBoatfS/aL292/dEkmAAo7IgVASTtySar9o/wCB+k/tIfBfxF8F9b1y70ey8Q/ZvNvbWJZJIvIuorgbVbg5MIU57E18IvgN4V/4J5fs9ePG0HxjqevRzTyalFPe2ShkumiSCFNsfG0useScAbjkgV8Kv+ChvhXWTBpfxZ0B9CumIU6lp6tPak+rR8yRj6eZ+FftM/sI/Cb9uvxtpPxih+Md5axWWhQ6TEujRwXUTok00wYsTkNm4IK9to960yxTS9MtNMjkLraQJCrMMFgqgZ/Sv2mvgDo/7Tfwf1X4P674gvNFs9Unt5XvLSJZJEMMqyABW45KAfjX7In7LHh/9kP4a6l8NfDfirUPEFtqWuS6s9zfQJE6PJBBCUATjAFupz6saqqqv+CyH/Jfvg3/ANg9/wD0rWqqqqq/4KA/A77Df2/xt8OWWLe8KWuuqg4SXhYpyPRgAjHpkJ3Ymv8AgnN4P/s34d+IvGs0W2XW9TW2iJHWK3TqPYvK4/4DX7Tn7F3wN/as0kR/EDQGs9ft4tlj4i00LDe24GcKzEESx5P3HBAycbSc1+0P/wAErP2k/gvLc6v4K0ofEnw3GWKXWhwMb2NAePNs+X3Ec/ujKB3Irw3q2ofD3xxpHiC60OKe98OapBdtpupROqSPDIr+VMgKttJXawBBwTyOteAP+C3PwzvLOKP4pfBnxLpV2BiSTQbuC/jY/wB4LM0JUe2Wx6mtb/4LQ/stWFqX0fwj8RdUuCp2RjTbWFARjG5muMgH1Ct0r9q//gqj8Wv2g/D+ofDvwLocXgLwhqUbQX0cN0bi+1CFhho5JtqhI2HVEUEglSzKSDot9Hpms2GpSozpaXUcrKvUhWBIHvxX/D7T4A/9Eo+IH/fNp/8AHqr9jr9pjV/2UvjdpXxLtYLm+0aSNrLXtNgcBryykxuC5IG9GCSLkgbkAJAJr/h9p8Af+iUfED/vm0/+PV411u38T+Mte8SWkUkUGq6nc3cUcmNyLJKzgHGRkBhmv2Qf+CoHxP8A2bNBtPhz4y0NPHPgmyGyxt5Ln7Pe6an9yGXDBoxyRG68cBWUcVpv/BZz9lO7slnv/DXxFsbjHzQNpVtJg47MtwQR9cfQV/wUN/b4+GX7X3hbw14T8BfD/wAQ6WfD2qPeLqesyQxyMjxFGiEMTSAZOxt2/jZjHOR8DP2PP2if2i54X+GXw21G40uRsNrV6n2SwTHX9/JhXIxyqbm6cV+yv/wST+FHwjuLLxl8b722+IXii3YSRWBhK6TZuMf8s2+a5IIPMoCHP+ryAajijhjSGGNURFCoijAUDoAOwr9uX9ov/hEdGk+D3g3UNutatB/xN7iJ+bS1Yf6oY6PIOvon+8CKqq/Z1/Zt1/4/6zMttrFlpeiaa6jULppVecA8hY4QdxJ/vNhBzySNp+Ffwe8A/Bvw+vh/wNoqWwcD7VdyfPcXbD+KWTqe+AMKMnAFVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVXxRRf2N/wDgrPpXjjJtPDHjvUY7+dskIbfU90F0z+yXXmy47bFqqqqqqqqqqqqqq/4LV/8AJ1Hhb/sn9n/6X39eEv8AkVNF/wCwfB/6LWq/4Kv6bf8Aw8/bg03x+lszLqOkaXq1uSPldrd2iK56ZBtxkejA968P67pfijQNN8TaJdLc6dq1nFd2kynIkilQOjD2KsD+NVquqafoml3mtatdpbWOn273FzPJwsUaKWdj7BQTX/BMuzuvi1/wUGvfihHbOI7GPW9flLKf3f2rfAASMDObz0xweO4qqqqqqqqqqqqqqqqqqqtb1SDQ9Gv9auSvk6fayXEm5wg2opY5J4AwOvav+CIemT6n4r+MXjC7cvNHa6XA0hyN7TyXUjHA46wjPpketVVVVVVVVVVfGu+8U6b8HfHF94F0m71PxJD4evjo9narmWe78hxCqjI5L7enPpX/AAR8+Anjf4TeCfiP4l+I/gjWfDOt6zrNtp8dprFhJaT+TbQmTeqSANsZrojcOG2dTiq+L/7UPwj+DSzWOu66NR1uMcaRpuJpwe3mc7Yu33yDg5ANfGH9tD4t/FEz6XpN/wD8IroUmVFnpshWaRfSWfhm44IXap7g0zM7FmYsxOSSeSa0TQdb8TalDo3h3R73VL+c4itbOBppHPsqgk18KP8Agnz448RiDVPinrEfhmxbDGwtts9449CcmOLjv85HQqK+GP7P3wl+EMKHwZ4Sto74Lh9Suh590/HP7xuVB7qm1faqqqqviZ8RvDfwp8GX/jXxRc7LSyXEcSn57iU/ciQd2Y/kMk4AJr4XeAfGX7ZnxWvPib8SDND4T0+YI0SMVQqpyllCeCBg5dxzyTwzg18T/jn8I/gFosFj4h1WC3mgtwtjoenoHuGRRhQsYwETAwGYqvGM18Z/22fil8TjPo/hm4bwloD7l8ixlP2mdDx+8m4PT+FNo5IO6sADA7VVpaXV/cxWVjbS3NxO4SKGJC7ux4AVRySfSvhP+wZ8UvHCwap45mj8HaXIA2y5j828ce0II2enzkEf3TXws/Za+DPwkEV3oXhhNR1aMDGqari5nBHdMgLGfdFU+pNVVVVVVVV/f2Ol2U2o6newWlpbRmSaeeQRpEoHLMx4AA7mvjb+3Pc3moHwD+zzYSapqVxJ5C6x9mMu5zxttocZc543MMeinIavhT+w74s8damPH/7RviC/ae7cSvpguTLdT8cefNk7B22Lk44ypGK8J+DfCvgTRIfDvg7QLLSNOgHyW9rEEBOACzHqzHAyzZJ7k1VVVVVVVfHP9qD4b/A23ay1S5Or+IXTdBo1nIPMGRw0rciJTxycsc5Ctg0kn7U37at6zQyHQPBLSbWwz29goB5H9+5cfiAf7ma+Ev7FHwe+GqwahrWn/wDCW61Fgm61OMGFG/6Z2+SgHQjfvIPQio444o1iiRURFCqqjAUDoAPSqqqqqr9tf9pH/hJb+4+D3gfUM6TYy7dau4n4u5lP+pUjqiEfN6sPRcn9mH9nq/8Ajl4uEuopJb+FdIdW1S5XKmU9RBGf7zdz/CuT1Kg6XpenaJptro+kWUNpZWUKw29vEm1IkUYVQB0AArUtH0jWYRb6vpdnfRjolzAsoH4MD6CtY/Z8+Buu5Oo/CbwszN954dMjgc/Vowp7etav+xH+zdquWi8DTWEhHLWmp3C+n8LOyjp2Hc1q3/BOv4M3gL6V4l8WWDnopuYZUHXsYg3p/F2/GtU/4Jq6dIGbRfi5cQ4B2pdaMJM8cAssq4+uK1P/AIJvfEWIH+xviF4cusZ2/aYpoM+n3VfGe/p718QP2PPiR8M7FtQ8VeK/A1tAFYxmTWxCZcDoiyohYnpgDNTxLDM8KypKEYgOmdrY7jPaqqqqqqqqqqqqqqqqqqra6ubOUT2dzLBKBw8blSPxFaV8Zfi7oeBo/wAUfFdoo/gi1icKevVd2D1PUd68JfFPx34H8Ynx/wCH9dK6+zSM97dW8d07NJ/rGPmq3zNkgt15PPJrS/8AgoB8ftPCi7bw3qeOv2rTSufr5TpWl/8ABSPxxEANa+G+hXZ/6dbuW39P72/3/Mfjpn/BSjw7MQNZ+FOo2vr9m1VJ8c/7Uadv8960z/goh8ELwql/oni2wY43NJZQyIOfVJSTxz0rTf24f2bdQCifxvc2LN0W50m565xyURgPzxWn/tQ/s+aoAbb4s6CmQSPtExt+hx/y0C4/ya0/4s/CvViF0r4meFLwkkAW+s28nI6/dc1Zanpupp5mnajbXS7Q26GVXGD0PBPBqq8ReH9J8V6DqHhrXbRLrT9Ttnt7mFxwyOMH6Hng9jg18Xfhrq3wk+IWseA9X3O2nzn7NOV2i5gbmOUD/aUjI7HI7V4T8a+LfAeqprfgzxJqGj3yYHnWc7RlgDnawHDL/ssCD6V8LP8Agoj4h00Q6Z8XfDkerwDCtqelqsNwPUvEcRuf90xj2NfDj40/DD4tWn2jwJ4ustQlVN0lmW8q5iH+1E2HA98Y9Caqqqq/4LCaPq2pfHr4PS6dpd3dJFp7h2hgZwv+lr1IHFVVVVV4r8L6L418Nal4S8RWgudN1W2e3uIz1KsMZB7MOCD2IB7V8Gvhvb/CP4aaL8Pre7W6/sqOQSXCpt853kZ2bHbJY8dqqvix+zf8CPjnbtB8V/hX4f8AEUrLtF5PaiO7QYxhLiPbKgx2Vx0HpXxM/wCCLXwD8SyTXfwy8f8AijwXNISVt7hU1W1i9lRzHL/31Ka8df8ABF79pHQTJP4I8Z+C/FNupwkbXE1hcP77HQxj/v5Xin/gnR+2n4Rd01D4Ca3eKhOH0ue31AOM4yBBI559CAfYc1rX7PXx98NsU8Q/A/x/phBA/wBM8NXcPJ6fejHXBrUdA13SE8zVdFv7JQ+zdcWzxjdz8vzAc8Hj2qrSyvNQnW1sLSa5mYHbHDGXY464A5rSPhJ8VfEAU6D8MvFepbwdv2TRbibOOuNqHpXhz9h/9r3xU6ppf7OnjmLf906hpL2A/O42CvB3/BIb9sfxOYzrmi+FvCaMfmOra2kpUfS1E3Pt/Kvhz/wRC0eJorn4ufHK8uQceZY+HNMWAj1xcTl8/jCP1wPhH/wTy/ZH+Dbw3ug/CXT9a1SEDGo+IidTl3DoyrLmJG90RTUcUcMaQwxqiIoVEUYCgdAB2FV+0N8bdI+Bfw+ufEtyYrjVrrMGkWTNzPOR1I67E+8x9MDOWFa/r2r+KdavfEWv38t7qOoztPc3Epy0jsck+30HAHAqqqvCmpeK9H1y21HwVeanbavA2YJdOZ1mU+2znHbHQ1+z18fP2iNa+y6H8Uvgr4k1C3YhV1+HTvsbAf3pI5AiOPVkK9Puk1VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVf8Fofg03ij4NeGfjRptruuvBGpGz1B1GP9DvCqhmPfbOkKgf9Nmr9ir40r8fv2ZfA3xBub37Tqx09bHWWYjf9ut/3UzMB0LlRKB/dkX1qqqqqqqqqqqqqv+C1f/J1Hhb/ALJ/Z/8Apff14X/aB+AsPhjSIZfjd4BSRLCBWRvEtoCpEa5BHmcGv+GhfgD/ANFy+H//AIU1p/8AHK/4KZ/suXH7UfwQ0n4jfCqGHW/FHhCNr7TFsnEv9r6dMitLHCVyJGIWOWPBO7awXJcV/wAE2v8Agor4Q8G+ErL9nX9obX00KPRMweHdfviVgWHdxaXLY/dbCTskbC7flJXau5fiR8OpNKGux+PvDjaaU3i8GqwmHb03b923Hvmv+Cj3/BRrwbrfgzUv2eP2d/EMev3fiBDaa/r+nvvt4rdjh7W3cDEzSjKs65UISAWZjs/4Jdfsj6x+zp8J73xx8QNMksfG3jzypbiymXEmm2Ue4wwuP4ZGLtI46jKKQChqqqqqqqqqqqqqqqqqqqq/aS1pvDn7O/xR8QI5V9N8GavcIQQDuSzlYYzxnIGPev8AgiHoywfCf4k+IBt3XviK2tT8xz+5t93ToB+//n6Cqqqqqqqqqqq+LHx5+GPwX083PjXxDGl46brfTLbEt3cem2MHgHGNzFVz3r4z/txfE34jmfRvBrt4Q0JyVAtJibyZenzzDG0H+6gXHQlqkkeZ2llkZ3ckszHJJPUk14d8M+IfF+rQ6D4W0S91bUbg/u7a0gaVz6nC5wB3J4HevhB/wT01zU/I1n4ya1/ZVucN/ZGnOslw3tJLyifRd+QeqmvAHws+Hvwt03+yvAXhWx0mJgBK8SFpZsd3kbLv/wACJqqqqqq8WeHPDnizw7faB4s0iHU9KuoWFxbSRGTeAM/KF+bcOoK/MDgjnFfEf9sw+HNAi+Fn7Omgnwt4f0yM2yajcRf6W+D8xRDkRljkl23OSc/Ka1DUL/Vr2fU9Vvri9vLlzJPcXEpkklc9WZmJJJ9TVWlpdX9zFZWNtLc3E7hIoYkLu7HgBVHJJ9K+Df7BXxC8bLBrPxIuG8I6UxBFq0Ye+mX/AHDxF6fP8w/uV8LvgT8L/g7ZiDwP4Ygguim2bUZx511N67pDyAf7q4X2qqqqqqqqr4k/E3wZ8JvDE/ivxtq6WdnFlYox80tzJjIjiTqzHHToOpIAJGueL/jt+3L4sk8MeErN9F8F2UwMqFyttbrn5XuZB/rZSORGOnYDBavgf+zX8OvgZp6PolkNR16SPbc61dxjznyORGOREn+yvXjJbGaqqqqqqqq8daT4j13wdq+j+EPEJ0PWrq0dLHUBGH+zy44JBB47E4yAcjkCvgV+w0sN8fHX7QNx/bWsTTGYaQ1wZ49+cl7mXJ85iedoO31LZwLa1trO2is7O3igghQJFFGgRUUDAAA4AA7VVVVVVV+2R+0Qfhd4a/4QTwlf7PFOuwHdLG+G0+2PBk45Dtyq+nLdhn4VfDHxH8YPG9h4K8OJ++um3XFy6lktYRjfK/sAenckDqa+HXw+8N/C/wAH6f4K8K2ghsrCPBZh888h+/K57sx5P4AYAAFVVahqGn6TZS6jql9b2dpbrulnuJRGkY9WYkAD618Tf27fg14HaWw8MS3HjHUY8gLp7CO2DD1nYYI90VxXxF/bj+OPjnzbTRtVt/Clg/Ai0lSsxHvO2XB902fSv+Km8Z61/wAxPXdXvX/6aXU8zfqzGvAn7EPx78aiK5vdAtvDVnIM+drM3lOB/wBcVDSA+zKv1rwT/wAE5vAmnBLjx7411TWpQMm3sYls4s+hJ3uw9wVrwf8As6/BDwJ5b+G/hposc8f3Li5g+1TKfUSSlmH4EVqngDwHralNZ8E6DfqRyLrTYpc8Y/iU9iR+Nat+zH+z/rOftfwl8Ox7uv2S1+y+nTytuOn+c14+/ZD/AGS/DOi3HiDxHZyeFtPhX57ka1MoBwcBRKz5Y9lAJJ6DtXxC0b4T6t4mh8O/s/6Z411lppSiPqDRytcHnCwwRxB8dMFjn1FeAf2Afi/4s0r+1fEt9pnhXzFBgtbzdNcHpy6Jwgx2Lbs8FRWr/wDBPL432GW03V/Cupr/AArFeyxv26h4gB1Pc9PwrWP2NP2kNHy0nw4lu4x0ezvrebP/AAFZN36VrHwX+L+gZOsfC/xVaov/AC0k0ifZ+DbcHqO9XNrc2kpt7u3khlXqkiFWH4GqqqqqqqqqqqqqqqqqpWZGDIxDKcgg4INWPjfxppZU6Z4v1q0KklfI1CWPB7kYarH46/GrTAq2Pxc8YRogIWM63cMgz1+UuR+lWX7VH7Q1gFEHxY1p9i4Hnsk3571Ofqa+IXxS8d/FW/tNU8fa6NWvLG3+zwXDWkMTiPcW2s0aKWAJJG7OMnHWqqyvr3TbuK/028mtbqBg0U0EhjeNh0KsOQfcV8Kv27vi14GMOneMWi8Y6WmFP2x/Lu0HtOAd3f74Yn1FfCv9rP4L/FcQWOn+JF0bWJQB/ZmrYt5C3oj52Sc9ArbvYVVVVVVVVVXijxl4P8D6a2s+NfFej6BYLndd6pfR2kQx1y8jAfrXj7/gpv8AsX+ATLA/xcj8QXcWcW2gWM17v/3ZQoh/8iV4x/4Lc/CLT3kTwF8F/Fmt7CQjapfQaaH46/J55Az7dOeOleJP+C3Xxhumf/hD/gv4N0wHOwald3N8V9M+W0Oe/pWs/wDBYL9sTVC5sbvwZpG4MFFnoe7ZnoR50knTtnPvmrz/AIKn/tz3U3mwfGeC0XGPKh8NaYVHv89ux/Wn/wCCkn7bsjl2+PuqZYkkDTrID8AIeKj/AOCkv7bsUiSp8fdTLIwIDabZMMj1BhwR7GrL/gqf+3PazebP8Zre8XaR5c/hrTAufX5LdTn8a0b/AILC/thaWYzfT+CtY2Bci90Mrvx1z5Mkf3u+Me2K8N/8Fu/i5atH/wAJh8FPCGpgY3jTb64sC3rjzDNj9fxrwd/wW3+DuotFH48+Dni7Qy5Ad9Mu7fUlQkcn5zASAfQZx27V4A/4KYfsX/EEx28HxhttBu3PNv4gs5rDZ9ZXXyf/AB/ivDfizwr4z01NZ8H+JtJ12wf7l3pt7HdRN9HjJH61VV4i8QaN4U0K+8SeIL+Kz07TYGnuZ5DgIijJPuewA5JwB1r47fF3xJ+0H8SJ9ejtLprOMm30bTUUu0EAPHyrnLsfmYjucdAK8P8A7PHxy8UBW0f4VeJHjcZSWexa2Rvo8u1T+daB+wL+0DrBX+07LQtDBPP27Ug5H/fgSV4f/wCCbF8+yTxV8VIIsY3w6fphkz64kd1x/wB8GvD/APwT6+BOlbX1a48R624+8tzfLEh+giRSB/wI14f/AGYv2f8AwzsOl/CjQXZPutewG9II75nLnNaZo2j6Jbiz0XSrOwgA4itYFiUY9lAFVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVfGH4a6P8Y/hZ4q+Fuu4Fn4n0qexaQrnyWdCElA9Ufa491Ff8EfviVrPw6+J3xH/ZM8bk2l7HcS39payMQIr61YQXkQHdmRY26DiBjVVVVVVVVVVVVVftd/8E6fBX7XnxJ034k+I/iNregXOm6HFpKW1laRSo6RzzyhyX5yTORjp8or/hyF8Kv+i4eK/wDwXW/+Nf8ADkL4Vf8ARcPFf/gut/8AGvhj4HtPhj8NfCfw1sL6W9tvCeh2Wkw3MyhXnS2gSFXYDgFggJA4ya/aR/4Jrfs4ftHavdeL7vTb7wj4rvGL3GraCyxi7f8AvTwMDG7dcsArknljUf8AwQ10sX4kl/aXumstx/cr4QVZMY4/efayM577f8a/Zv8A+Cb/AOzd+zhqVt4q03RrvxZ4rtTug1nX2WZrVv70EKqscZ9Gwzjs3Jqqqqqqqqqqqqqqqqqqqqr9vrVl0X9jX4tXhlCeZ4ektslS3+udYsYHrvxntnNf8EadIfTf2SdRvGD41XxnfXK7iMEC3tYuMdsxHr3zVVVVVVVVVV4w8beEvAGiS+IvGfiCz0jT4eDNcybdx/uqOrsccKoJPYV8bv2/9c1r7R4e+DFm+kWRBRtZu0BuZB0Jij5WIejHLcg4Q1qmqalreoT6trOo3N9fXTl57m5laSSVj1ZmYkk+5rS9K1PXNQt9I0XTrm/vrp9kFtbQtLJKx7Kq5JPsK+DH/BP7xP4hEGufF/UG0CwYbl0q0ZXvJB/ttykQI7fM3YhTXw++F3gL4V6OND8B+GrTSrcgea8a7pZyP4pJGyzn6k47YFVX7Wnx+1b9mL4Laj8Y9M+H3/CYw6Rd28d9Y/2p9gMUMziPzg/lS5xI0Y27Rw5ORjB/Yr/a/wBG/bH+HereNLPwn/wi+o6JqpsbzSW1H7aUUxq8cwk8uP5X3MMbeDG1V448XaN8P/Bmu+O/EM/laX4d0241C8fusUMbSPj3wpr9ij/goRrX7ZXjzWvClp8Cf+EW0zQNL+2XmrHxIb4LI0ipFCI/ssfzP+8YEsOIm61+07/wVksP2evjB4l+Dmn/AANn8Q3nhyZIZb+bxCLON2eFJAQggkJA8wDBIzg9K/4fSfEKw/0vWP2U9tnFzKf7emhwOg+drUgc46iv2c/+Cq37PXx112y8F+I7bUPh94jv5BFaRavKklncyMcLGlyuAHJ4AkRASQASTiq/br+Bg8BeOF+JXh+z2aF4qmY3KouFtr7GXHsJAC4/2g/QAVX/AAkcFx8TPD/wf0y0uLrxT4h1W006G08soLdrh0VHkJHC7XDcZOOeBzXwj/Z0+FfwXtU/4RPQEm1TbiXVr0Ca6k4wcPjCA/3UCj1B61VVVVVVVVXxk+MnhD4JeEJvFXiq53SNlLCwjYCa9mxwiDsOhZjwo/AHwL8O/ir+294+b4gfEa+uNM8H2MhSJ44ysax7uba0B4J4w0hzjAzk4WvB/g3wz4A8O2fhTwho9vpumWKbYoIVxk92Y9WY9SxySeSaqqqq/wCCYv7X37SHxz/aRufBXxX+Kd94h0WPwzd3a2k9pbxqJklgCvmONWyA7Drjmqr/AIKuftU/Hn9nbVfh9o3wZ8eN4cg8Q2d/JqJj0+2uHlMTwhMNNG5TAdvu4zmvBPw6/wCCwHxO8HaH8R9C+NEi6Z4o06DU7HzddtoWaCdBJGxjWPCZRlO3tnBAORWu/ti/8FLP2L/EOnx/tJ+HYfE3h+8m8uI6la2xhuccssN9ZgBZMZOJN5A52YFfs5ftB+BP2nPhXpvxT8AzSJbXLNBe2M5HnafdIB5kEgHGRlWB/iVlbvXxo+M/w++APw81L4nfEzWf7P0XTQFOxN8txK3CQxJ1eRjwBwOpJCgkal+3Z+3p+2b4w1Dwn+yN4Hn8N6FbNteWyghkmijJO1rm+uB5ULMASFTY3BAL4JOo/BD/AILLeDLaTxZY/EzXdZntwZWsbfxTBdtjqcQTHy3xj7qgk9ADnFfsxf8ABV/xro3jiL4P/tm6Amk3a3Is5fERsTp81hP0xfW2AqqSRl0VAnBKEZYQTwXUEdzazRzQyoHjkRgyupGQQRwQR3qqqqr4xfFTQvg54Cv/ABrrjLIYF8uytd21ru4YHZEv1wST2UMe1eJPEPin4n+M7rXtWkm1PXdeux8kaFmkkchUjRRngfKqqOgAFfsxfAay+CHgVIb6GKTxNq6rNq1wvOw4+WBT/cTJ+rFj0wBVV4i8S+HfCOkza74o1uy0rT7cfvLm8nWJF9BknknsOp7V8XP+ChmgaW0+j/B7Qv7YnUFRq2oq0VuD6pFw7j3Yp9CK+Inxi+JfxXvDd+O/F99qSh90dqX8u3hP+xEuEXjjOMnuTXwx/Zc+NHxYEV1oHhOWx0yXBGp6pm1gKn+JcjdIPdFavhv/AME8fh9oYhvviV4gvfEl0AC1na5tLUHuCQfMf6hk+leEfAHgjwDYjTfBXhTS9Gg2hWFnbLG0mO7sBuc+7Emqqq/aG/a+8F/BdZ/DmhCHxD4tCkfYo5P3NkexuHHQ9/LHzHvtBBrwf8B/j5+1trEHxA+L/iK70fw853Wpni2s0Z522tvwEQjH7xsZ4Pz818Lfgj8NPg5pwsfA3huC2ndNk9/MBJdXA6/PKecZ52jCjsBVVVajpOlavEINV0y0vYx0S4hWQDp2YH0H5CtY/Z++B+vA/wBpfCbwqzMPmki0uKFz06sgDdh3rWf2H/2cNWBNv4MudNkPV7PU7gf+Ou7KPwFa1/wTl+FVyGfQ/GvibTyRwLgw3Cr+ARDjp1NfE7wf4V8H+M7/AMN+DfGZ8T2Fi5jbUTZi3R5ATuCYkfeo4G/gMc4GMEm1kHQqfxrU9C1XRoLKfU7T7ONQg8+3V3Xe0ZOA5TO5VOMgkDcORkc1VVVVVVVVVVVVVVVVVVVV8K/2rvjP8JxDY6V4kOraRCABpmrZuIlUdkOQ8Y9lYDPUGvhV+3j8JvHHk6b40WXwdqkhC5u3820c+04A2ep8xVA/vGrC/sNVsodR0u+t7y1uE3QzwSCRJF9VYEgj3FVVVVVNNDbwvPPKkccSlnd22hQBkkk9Biv2hP8Agqn+zV8FJbrQvCd/N8R/EduSptNCmUWkbjPEl4cpjjB8sSkHqK+Mv/BWD9q74oPNZeFdd0/4e6S4Krb6DBm4Zf8AauZdzhvePy/pXwb+CHx7/bY8a6vp/hrxIPEniTTLL7bdy+ItbYzSQlwhcPKWZsMyg88bh618Vv8Agmj+1N8G/h5rfxO8YaDoLaL4fgE96bLVkmlWMuqlggAyBuyfQAmq+HHw+8T/ABX8d6F8N/Bdkt3rfiK+js7KJ32Lvc4yzfwqBkk9gCal/wCCRX7Y0MbSy6P4SREUszNr6AKB1JOOBXwA/YO+P37TPhHUfHHwps9BvNJ0zV5dKmlutTW3JnjjikbapGSu2aMg+59K/wCHQ37ZX/QE8Kf+D1P/AImv+HQ37ZX/AEBPCn/g9T/4mv8Ah0N+2V/0BPCn/g9T/wCJrxx4P1n4e+NNf8AeI0hTVvDWqXOmX6wyb0We3laKQK38Q3IcHvXwL/4J7/tJftFfD63+Jvw50PR20K6uZbe3lv8AUltmlaJtrsqkHKhsrn1UjtXj3/glx+1p8OfBOu+Ptf8AD3h+TTPDunzX96LTV0llEESF5GVMfMQqk49qrwp418Y+BNUTXPA/izWPD+oxEFLvS76S1lUjkYeMg18Gf+CtH7VHwyeCw8ZanpvxE0iPCmHW4RHdKvH3LmIKxbj70ok6/TH7PX/BUj9mj45S2mg6/q8nw98S3AVRYa/Kq20sh/hiuxiNuSABJ5bMeimkdJEWSN1ZWGVYHII9RWu+HtB8T6edJ8SaLY6rYs6u1re26zxMynKkowIODgjI4IB7VpWh6LoVv9k0PSLLT4cAeVa26wrx04UAVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX/BQnw5rP7If7cngn9rHwTYuNO8RXMeo3EcQ2LJdwBYb2DI6Ca3dSSeWM0noa8MeI9F8ZeG9K8XeHL5LzStasob2xuEGBLDKgdHH1Vgaqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr/gqZqjaZ+w38Qo42kWS+k0y3VkOMZ1G3Zs+xVWH41/wSi0ldO/Yh8F3gRFOqX2qXJKkkti+miyfQ/usfQCqqqqqqqqmZUUszABRkknGK+O37c/gn4f8A2jw78NhbeKdfQFGuVkzZWre7r/rj/socerAjFfEL4m+OviprjeIPHfiO51S65ESyNtjgX+7HGMKi+wAz1OTzQBYhVGSegr4JfsSfEn4nC31vxYj+EvD0gDrLdQk3VwvUeXCcEA/3nwMEEBq+FPwK+GfwZ08Wngjw9HFdOmyfUbjEt1cD/akxwP8AZUKvtVVVfF/4c6X8XvhZ4r+F+s7Ba+J9IuLBpGXd5LSRlUkA9Uba491Ff8EiviBqnwm/am8UfAjxQGspPFVlPZy2snBTU9PZ3Cn3Ef2sY9cVX/BXn4zH4c/swjwBp135eq/EbUUsAFbaws4cTXDD2JEMZHpMa/4JGfBb/hWv7MCePNRs/K1f4jX7aizMMMLKLMVsp9jiWUe01fHD/lMHpv8A2VDwz/6HY06JIhjkVWVgQysMgjvxX/BYH9lj4bfDzRvDf7QXw70Gz8PXer6z/ZGtWVhCIYLmV4ZJorhY1wEcCCUMQPmypOCCW/YA+KeufGP9kb4e+MvE97JeaylnLp97cSks8z2s8kCyMx+8zJGjMe7Ma+J/w80L4q+BdW8CeII/9G1OAqkoUFreUcpKv+0rAH36Hgmvg1+x58K/gvaL4p8UtD4j12yQzyahfxBbez2jJaKIkgYxne2WGMjb0r/gnfpV1+0n/wAFC/Evx11W3eSz0mfVPEh8wZVJbiRoraL2Kicso7eT7VVVVVVVVVXxE+IHhr4X+D9R8beK7zyLDTo8kLy8znhY0HdmOAB+JwASPAXgzx1+3H8Wrrx/46kmsPBukyiIxxk7Y4gdy2cJ4y5By8mOM54yq1o2jaT4e0m00LQtPgsdPsYVhtraBAiRIowAAO1VVVVV/wAEbf8Ak7m7/wCxPvv/AEdbVVf8Fxv+Rq+Ef/YP1X/0ZbV+yd/yav8ABr/sn+hf+kENftx+E/DXjD9kb4sWHii1gmgsfCt9qNqZcDy7u2haaBlJ6N5saDjrkjvX/BDjWdYkT4v+HmeSTSoDpF0ilvlhnf7UrED1dY1z/wBchX/BTn4meK/2jv2vdA/Ze8F3Rex8PX1ppFpBvIjn1W8MfmSvjsgkji5Hy7JD0Y18Cfgn4I/Z6+F2h/CvwFYRw2GkW4Wa4Me2W+uCMy3Ep5y7tknnAGFGFUAV/wAFdv2WtC8cfCaT9ozw1pccPirwZ5aatLCmG1DTncJ84HVoWZWDdkMgOQFx/wAEjvjzqPxY/Zzn+H/iK+a61f4a3iadHI7lnbT5VLWu4/7OyaIDssKe9VVVUkkcUbSyyKiIpLMxwFA6kn0r9q747SfGn4gPDo90x8MaCz2+loOBOc/vLgj1cgY9FC8Ak1+wp8ABKV+NviyyBVS0egQSpnJHyvc49uUX33Hspqqurq1sbaW8vbmK3t4ELyyyuEWNQMlmJ4AA7mvjb+314T8LG48P/CO0i8R6mmUbU5srZQn1QDDTEe21ehDN0r4gfE/x78VdZOt+OvEt5q1zk+UkjbYoAf4Y41wqD2UDPU5PNfCH9in4t/E+KDV9Xtk8KaJKAy3WpRnzpVPeOAYY8cguUB7E18Kf2Rfgx8KlhvLfw+uvaxGATqOrqs7K3rHHjZHz0IG7/aNVVVVftKftfatfavJ8HvgA819q1xKbW61ayQyuXPBitNuct1Bk7c7efmH7OP7FWleD2t/Hnxhhi1rxO7efDp8jCWCxc87nPImlz3OVB6ZIDVVVVVVV+2p+0w2jw3Xwa8A6ji9nTZr17C/MCMObZCP4mH3z2B29S2Ktb+302VbwxRXE8LgxQyx748jnLg8MP9noe/HBv7+91W8m1DUbuW5up2LSyysWZz6kmqqtB8N+IPFWoppHhjQ7/Vr6QfJbWVs88h99qgnFeEf2E/j/AOKI459Q0jTPDsMgBDaregPj/ciDsD7MAfpV7/wTg+JiW6tp3j3wxPPj5knWeJQfZgjE9+1ax+wd+0TpoP2LQtI1bHT7Hqka5+nnbP8AIrWP2Y/2gNDB+2/CXxDLt6/Y7X7X+XklvWtX8GeL/D4J17wprGmhRybywkhx/wB9AVVVVVVVVVVVVVVVVfBjx98cPCuux2Pwa1DXZrqV9z6ZZQNdRTdMl4MMvbG7AIHcV8F/EPxj8ReHBc/GLwNpnh3UFVfLFpe+Y0+epaIbhF248xjz0GK/4KyqL39sP4KaXdEy2r6dZgwsfl+fUnDce4UA/QVVVXxo+NHw8+AHw81H4m/E3Wxp2jacAvyLvluZW+5DCnV5GIOB04JJCgkftdf8FDfjL+1JfXegW19P4S8A7isHh6xnINymeGu5RgzMePk4jGBhc5Y1X/BPb4uP8Gv2tvAWvT3fk6brV7/Ymp7m2qYLvEQLH+6kpik/7Z18QPBul/EXwJ4i+H+tjOn+JNKudNuvlziOeJo2IHqAxNeKvDeqeDvE+seENbh8rUdDv57G8j/uTQyGNx+DKRX/AARv+FA8Z/tKaj8Sby332fw/0WSWJ9uQt5d5gjB7f6r7Ufqo/D/goh8YR8F/2SfHGt2l2IdV161/sLS+cMZrvMblT2ZIfOkHvHX/AAT0/wCChfwW/ZL+C2s/Dj4jeGfGupalqPiifVYpdEsrWaFYZLW1iCsZbiNg+6ByQFIwV56gfs5f8FJfgX+1B8TIPhV4A8K+O7DVp7Oa7WbWLG0igCRAFgWiuZGyc8fLj3FfG34u+GvgL8LNf+LnjCy1O80fw5Ckt1BpsSSXDh5UjGxZHRSdzg8sOM/Sv+H1f7K//QhfFX/wVWH/AMm18avGGn/FP43ePPH/AIas71LHxd4q1HU9Pt7mMC4WO5upJY0dULDftdQQpYZ6E9a+Afw/0z9nb9nDwj4G1BktovB/h1G1STIKicIZbqTPoZWlb8a8D+KPDnxo+FWheMbazWbRPGugwXn2WU7swXMIYxPjuFcqw9c9K+Nnw2vvg78XfGHwu1FZPN8Mazc2SPIMGWJHIik+jx7HHswqqr9kD/go18YP2Y72y8M69eXPjH4ehwk2iXk+6axToWs5WyYyBz5R/dnkYUnePhJ8W/AXxy8AaX8S/hrrseqaHq0ZMUgUo8TqcPFIh5SRSCCp+oyCCaqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr/goT+z6/wC0T+zH4j8O6TYG68SeHh/bWgqiZd7mBW3RKO5liaWMDpuZT2Ff8EfP2gl+IfwQvvgrruoiTW/h5P8A6Esj/PLps7Fo8dyI5PMQ9dqtEOMiqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/4LB6iLL9jue1wx+3+JtPg+U4Ax5knPqP3f54r/AIJz6Z/ZP7E/wqtNkK79LmnxEMD97dTSZ6Dk78n3z161VVVVVVVfFT4peEvg74Qn8aeMp7hLKKQRRx28JlkmlYErGoHAJ2nliF9SK+PH7XnxE+Mz3GiWEz+HfCzkqNNtZTvuF9Z5Bgv/ALgwvTgkZqvhB8AviV8bdS+yeDdFIsYn23OqXWY7W36ZBfB3NyPkUFuc4xzX7N+i69B/wU0sfgzc6gLvQvAV5qDXGLXyvtjwWcirIwYk7ROyFQOMAHn71VVVVV+3bo+o/sn/APBRKx+M2g2kiWOp6jY+LbVIuPOPmBbyInjmSSKbcOPlmGeua0bV9N8Q6PY69o90lzYalbR3NrMnSWKRQyMPYqQfxr/go34k1j9qT9u/w9+z14QujJBoM9p4dtmUF0S7uXV7qYjsEDoj+gtyfevCfhfRvBPhXR/Bnhy1W20rQbCCwsYAAPLhhQIi8eiqBXx9vLPTv+Cu9pqGoXcNra2vxM8NyzzzSBEiRWsizMx4VQASSeAK1b4n/DTQdN/tjXPiJ4Z06w27vtV3q8EMWCM53swGMe9f8FP/ANsLwn+01q/hf9n/AOBEk/inTdK1YXU99ZQvIupaiyNDDDaqBukCiWQbgMO0g25Ay37HvwZvv2f/ANmvwJ8KdXEf9q6Tp5l1MIQwW7uJHnmQMPvBXlZA3cKO1V+3h8RP+FX/ALIfxQ8Txz+XczaHJplqQ2GEt4y2qsvuvnbvbaT2r/gix8MB4e+CHi34p3VuFufF+uLaW7kcta2aEAj0BlmmBx/cHtiqqqqqqqokAZJAAHevjF4s8R/tjfHew+EngC8YeE9EnbNyh3REIcT3rY4IwdkYPXIwR5hrwL4I8O/DnwnpvgvwrZC103TIfLiTqzHqzse7MxLE9yTVVVVVV/wRt/5O5u/+xPvv/R1tVV/wXG/5Gr4R/wDYP1X/ANGW1fAP/gqp+yp8PvgJ4A8DeJJ/F0es+FPCum6XeQRaOJBJPbWscTmNhJtKlkOCSpx1Ar9rP/goh40/bM01P2c/2Zvhr4ji0zxDMiX3mwrLqOqKrAiERQl1hi3AF23tkAZKruDf8E8f2T7/APZS+CJ0rxb5DeM/FN0NQ10QuHW1IXbDbBxw3lrkkjjfI+CVAJ/Zp2+M/wDgrrJd6yC7TePfE13gHO14or6SMDOeFMaAew/Gqr9pzTbLWP2bvippmooGt5/Berq+eMf6HLyPQggEHsRX/BD+8u0+IPxS0+Mn7LNo1hLIMcb0mkCfo71VVVftz/HH/hDPCK/C3w7e7dZ8SQk3zxthrayzgj2MpBX/AHQ/qDX7Pfwbv/jb8RrPwwgki0q2H2nVrpBjybdSMgHs7nCr7nPQGtN02w0bTrXSNKtIrWysoUht4Il2rFGgAVVHYAACq+N37Rvw6+Bem7vEV8b3WZoy1po9owM8vozdo0z/ABN6HAYjFfG39pn4lfHC7eDW9ROnaCr7rfRbNysK4PBkPWVunLcA9Aua+C37L/xQ+Nssd5oumjTNB3Yk1i/UpCcHB8sdZTwRheAeCRXwZ/ZO+FHwbSDUbTTBrniCMAnVtRjDujesScrF9R83qxqqqqqv2sP2mtc1/XJPgB8E3mu7+7m+xapeWWWlllJ2m0hI6ejsPdeMNn9mL9lrQPgfpMWva5Fb6j40u4sXN4BuSzVhzDDnoOzP1bnoOKqqqqqq/aq+P1v8E/BX2TR5438Va2jx6bEcH7OvR7hh6LnCg9Wx1AbF1c3N9cy3l5cST3E7tJLLIxZncnJYk8kknOanlEa7VPzEce1HnknJqq03TNS1q/t9J0ewuL69upBHBbW8Rkklc9FVVyST6Cvgh/wT8ub6G38RfG2+ktEYB00KxlHmY9J5RkL/ALqZPP3gcivB/gXwd8P9JTQ/BXhuw0eyQDMVrCELkd3bq7e7En3qqqq1jwH4G8Qhhr/gzQtT353C806KbdnOc7lPqfzNeAPh34G+PH7W2s2On+FtOt/BOnT3E0ljYRfZoHt4QIYyoj27fMkKOcY4Y1q37CP7Oeogiz8OappmRx9k1WZscDp5pf0/WtW/4Jx/Cy4JOieN/FFmT0FwYLgA89MRocdO/brzX7QP7P3w6+CDtpsHxoXW9eIBXRYdHAkiHHMsgmKxg9cEbsEYBHNVXhP4e+OvHsk0Xgrwfq+ttbjMxsbN5hF6bioIXPvWofCX4qaUCdU+GfiuzwAT9o0W4jxnp1QVd2F9YOIr6zntnOcLNGUPHXg1VVVVVVSI8jrHGpZmOFUDJJ9q+HH7H3x0+JAiuYPCraDp8nP23WybVceojwZGBxwQmOnPNfDf/gnx8MfDgivfiDrF74qvF5NumbO1B9Nqku2PUuAf7teGfCHhXwXpq6P4R8O6do9mv/LCytlhUn1O0cn3PNV/wVb/AOT0vgl/2D7D/wBOclVVV/wVi/aC1n4qftI3/wAMLS/ceF/hvixtrZGISW9ZFa5mYd2DERD0EWRjc2aqo5JIZElhkZJEYMjKcFSOhB7Gv2YfixD8cv2ffAfxSW4SW51zRoWvyp4W8jHlXKj6TRyD8K/4KnfCsfDP9sLxNfWlsYtP8aW0GvWvHBaUFJ+fUzxTN7bhX/BH/wCFH/CC/sst46vLYJf+P9YmvVcrhvssB+zwqfbfHO49pa/4LXfGRdW8b+CvgVpd5uh0C0fWdVjRsj7RP+7gVh2ZIkkb6Tiq/wCCQ3/J5Wm/9i/qP/oC1/wUi/5Mj+Kf/YPtv/SyCq/4J9fCb/hcf7XHw+8PXFv5unaTqH9tajlNyiGzHnBWH915FijP/XSv+Ch/xPX4U/sffEXWIbjy77WdOOiWQBwzSXhEDlT2KxPK+f8AY45r/gkn8U1+IP7JGneF7q6EmoeA9UudJkDH5/JZvPhYj+7tmMY/65Gv+Cy/wj/4RD9obRvipY2hSy8faOouJMcNe2e2J/p+4Nr+tVVV/wAEiv2gdb+Hf7QkXwavdRY+GPiHHJGbaQ5SDUIYmkhlX+6XVGiOPvbkz9wYqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq+KlrL/wTh/4KN2PxE0izlt/h74vka7eGJDs/s67fbeQKB1MEw8xE9Ehzwas7y01Kzg1DT7qK5trmJZYJomDJIjDKspHBBBBB96qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/4LWaisH7MnhPTFkkWS68c274XhWRLK8yD+Loce3tX7FumnSv2SPg/amBIi/g3TZ9q4wfNgWTdx3O/J9zVVXxH0/xJq3w88UaX4Mu2tfEF5ot5DpU6y+UYrpoXWFg/8JDlTu7da/4Zc/4LB/8ARdNV/wDC6ev2ifH3/BQv9l3xPpvhD4r/ALQni+DUdWsPt1stj4oluFMXmNHkkEYO5G4rQ/2dP+Cu/iHRNP1/TfjtrDWmpWsdzbl/HDqxjkUMuR2OCOK/YT+Ef7bXw48WeJ739qn4g3niLSrzToo9Kin8QtqIinEmXYKfu5XjNVVeOPBmgfEPwlqngvxNaC403VrcwzL/ABL3V1PZlYBgexANfED4OeMvAfxQvPhS+l3OpatHcCOxW1gZ2vo25jkjUZJ3LzjnByDyDXwG/YDVfs3if44y5PDx+HrWXgennyqfr8iH0+bqK0nR9J0DTLbRdC0210+ws4xHb2ttCIo4lHZVXAAp4IJJY53hjaWIERuVBZM9cHtnAqqqqqv+CzfwdHi34EaB8XtOtN974D1Xybt1TkWV5tjYkj0nS3AB4HmN0zz+wL+0hpV3+wBbeP8AxVemZvhPpt9YarmTDGKxjMsKL1OTbNAo9SOK/wCCTPgDWPjV+1F41/aY8Zo11LoK3F19oYHD6rqLybmBPXERuMjsXQ59a/a+8D2nxO/4Kda18Nb++lsrbxZ400bSZrmFQzwJcx2kLOoPBIDkgHjIqw/4If8Aw8jvvM1P49+IrizzxDBo0MMmMj+Mu46ZH3evPtX7Ov7Bf7N/7Mt3HrvgTwlNqXiRIyg1/XJhd3aA9fL+VY4SRkExopIOCSKqv+C1vj/+xPgX4L+HME+yfxR4ie7kXPMkFnCdwx6eZcQH8BX7Gnw4X4UfssfDLwQYPJuLbw/Bc3ke3BW5uQbicH6STOPwqqqqqqqqr9tz4xv8M/hW/hvRb3ytd8W77SEocPDbADz5B6HawQHrmTI+7X7FnwTj+Fvwxh8SataBfEPiyOO6uS6/Nb2+Mww+o4O5h/ebB+6Kqqqqqq/4I2/8nc3f/Yn33/o62qq/4Ljf8jV8I/8AsH6r/wCjLav2cP2L/wBlPxd+z58JvGHiT4D+Er/WNR8E6Ld3l1LZc3M72cTvJIAcOzMxLEg5zzmvAvwu+GnwvsG0v4bfD/w74XtZAPMi0fTIrQSEdC/lqNx9zk1XxvuZ/wBkb/gqPdfEHWbaSLSf+ExTxC8u0t51hfkvcug6kjzrhMf3kIHSrG+stTsbfUtNu4rq0u4llgnhcOkqMAVZSOCCCCCPWq/4KVfGDTfhJ+yJ41ilu411Txlat4e023ZsGY3QKT4HXC2/nNnpkAd6/wCCJvwxvdF+GPj34s39uyR+J9Vt9OsC6kborNHZ3X1UvcbfrEfSqqq8Z+LdG8CeFdU8YeILjydP0m2aeZh1IHRVHdmOFA7kiviH44174peOdU8Z62zSX2r3JZIVJYRJ0jiT2VQqj1x6mv2WPgrF8GfhnbW2o2wXxFrYW71ZyvzRsR8kP0jU4/3i570zKilmYAKMkk4xX7SH7c1h4da68FfBe5t9Q1NQY7rXCBJBbHoRAOkjj++fkHbdnizsfGnxO8WC2s4dS8ReIdYnLHlp57hz1Zicn6k8ADnAFfAL9gzQ/Di2/ij4zi31nUwA8Wixnda25/6an/lsw/u/c6/fBzVvbW9pbx2tpBHDDCgSOONAqoo4AAHAGO1VVVVV+2f+0e3wt8Oj4feDb7b4s12A+ZNE3zadbNkFwR0kfBC9wMtxhc/scfszR/C3Qo/iF40sP+Kw1eHMUMq86ZA38GO0jDBcnkfd4+bNVVVVVXi/xXongbwxqfi7xFdi307Srdp53PXA6ADuxOAB3JA718WviZrnxd8d6l4411isl2+22twxK2sC8RxL7AdTxkknvUjrGm4n6CmYudxOSaqvAHw/8VfE7xVZeDfBumNe6jeNwOiRIPvSSN/Cijqfw5JAP7Pn7MXgr4EaUl3FHHqvim4ixeavLHyuRzHAD/q4/wDx5urHoFqqqqr48+N2+HXwf8VeLoJfLubTT3jtGBwVuJcRRH8HdT+Ff8E8vBA03wL4g8e3EQE2tXy2luSOfJgXJIPoXkYH/rmK1DULDSbGfU9UvYLSztY2knuJ5BGkSAZLMx4AA7mv2i/27L/VnufBvwQupbKyGY7jxAVKTTeot1PMa/7Z+Y54C4ybiee7nkurqd5ppnLySSMWZ2JySSeSSe9eBfh540+JmuR+HPA3h661a+fBZIVwsS9NzucKi/7TECvg3/wT78LaGlvrXxg1H+3b8Yb+yrN2jtIj6O/Dyn6bV6ghhWjaJo3hzTINF8P6VZ6bYWy7YbW0gWKOMeiqoAFVNBDcxNBcRJLGw+ZHXcD9Qa1H4d/D/VyTqvgbw9eliSftGmQyZJ6/eU1f/s6/AfUQRcfCHwmuVwfI0qKDp6bAMfWr79jz9m3USTcfC6zXLbv3F7cwc/8AAJRx7dKvP2E/2c7o5g8NanafMTiHVZj+Hzs3FXf/AATx+BVzjydW8X2uCT+6v4T+HzwtxXxd0PwH4Z+IOreHfhvq1/qmiabMYI729kR2nkXh2UoqgpuyAccgZ74FV8Pvgt8UvinKqeBfBWo6lCW2m78vyrdD3zM5CA8HjOeK+HH/AATluJBFffFbxqsQ6tp+irub6GeQYHbICHvg96+HfwG+EnwqRG8FeCdPtLtBj7dKhnuT6/vXywznoCB7VVVXiH4cfDzxdqlrrfivwH4d1rUbFQtreajpcNzNAA24BHdSygMSRgjBOaqqr/gpD8Nda+G37YnxAj1SCQW3iW9/tzTp2HE8FyNxK+yyiWP6xnr1qqqv+CKfxdXWvhp40+CeoXe668Nakmq6cjnk21yuyRVH91JYtx95/wAv+C0vwguvE3hD4a/EzQtOe41Ky1qTQJUiGWmF2u+AH2EkDqOnM3PUV8K/A+l/CD4TeFvh7bzQR2XhLQ7axeYttQ+TEqvIScdSrMSfUk1+038Wp/jr8ffHPxVkld4Ne1iV7EOSSlmmI7ZTn0hSMfh2qv8AgkN/yeVpv/Yv6j/6Atf8FIv+TI/in/2D7b/0sgqv+CI/wl8uz+IXxzvrYZmeHw7psuOQF23F0PoSbT8VNfttfskap+2L4G0HwDb/ABU/4Q3T9I1Q6jcgaKdQ+1yCJo4xjz4tgUSSf3s7h0xz+w7+wnqn7F+peK5o/jN/wl+neKoLZZbI+HvsHkzQM+yUP9olz8srqRgZyDniv+Ctnwi/4WP+yhe+LLCy87VPh/qMOqxso+f7Mx8m4Uf7IWRZD/1xH0NVVf8ABL/4a618Qv2xvBt9p0EhsPCPnazqc6jiGOONkjz/AL00kS49GJ7VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV/wAFIf2Zx+0f+ztqTaDpZuvGPgsPqug+XHulm2qPtFsuOT5sa8KOsiR+lf8ABJD9p0fFf4NS/BLxRqRl8T/DqNUszK+XutJY4hI7nyW/cnjAXyeSWNVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVf8Fu7yVPhB8ONOWQeVN4knlZCOSyWxAPrwHb86/ZrsodN/Zy+FWnWxYxWvgnR4o9xydq2UQGffAqqqq/4Laf8AJfvAv/Ynj/0rnr4T/wDJK/Bv/Yv2P/pOlftS/tAWf7MPwY1b4xX/AIYm8QQ6VcW0JsIboW7SedMsQIcqwGN2enOK/Y3/AGqtP/bA+GGp/EvTfBlx4Zj0zXptINpPfC6Z2jggm8zcETAIuAMY/hznmq+An/BSnQ/jt+0cf2d7T4S32j3K3GoQ/wBqSawsyf6IsjE+WIlPzeXj73Ge9U2iaM+rr4gbSbNtUS3+zremBTMsWd3lh8bguTnbnGaqqqqqqr44/DLT/jP8HvGPwr1IosfibR7izjkccQysh8qX6pIEce6ivBPx08Q/CP8AZ++M37OOoRXVte+M9S02JoSSDZtazSG7BxwC4SKNh3A9M1/wTJ+Cy/Bv9krwvJe2ixax41z4h1AkfNi4Vfs688jFusOV7Mzepqvjh/ymD03/ALKh4Z/9Dsaqqr/gqnfv8Xf22fhj8C7KYyxW9rp2nvEM5S61C8wwGOeY/s9RxRwxpDDGqIihURRgKB0AHYVVVVVVVVVqUR/aq/bUOmTD7V4X8LzGN0xuQ2do3z5x1WWc4z/dkHpQAACqAABwKqqqqqq/YR/aW8J/spfG6b4n+MtB1bV7CXQrnTxb6YIzKHkkiYN+8ZRtAjOec8iv+H2nwB/6JR8QP++bT/49X7PX/BUf4QftG/GDw/8ABnwt8PvGGmap4h+0+RdagLYQx+RbS3DbtkrNysLAYHUjtX/Bcb/kavhH/wBg/Vf/AEZbV+yd/wAmr/Br/sn+hf8ApBDVV/wUU/Yg/wCGsPA9n4k8Dm3t/iL4UicaYZmEcep25+ZrR3OAh3fNG7fKrFgcByy/suf8FFfin+xrGf2ff2kvh3r+p6NoEnk2sUg8jVNHjH/LJUlws8PHyAsu0H5XKhVHiL/gtF+zJYaNLdeG/Bnj/VtT2fuLOayt7VC+OkkvnNsHbKq59jUfhv8Aao/4Ky/Gez8T6tpcvhv4faVI0MN4YnOm6LbFh5iwlsfarptq7sckhd3loFC/DD4beE/g/wDD7Qfhl4G05bLQ/DtmtraRdWIHLOx7u7FnZu7MT3qqqv8AgoB8YftN5Y/BjRLo7LUpfayUbq5GYYT9FPmEHj5oz2r9iD4NL8QfiIfG+t2Ql0TwmyyqHXKzXh5iXHcLgufQhM/erWNY0rw/pV1reuajb2NhYxNLcXNw4RIkA5Yk9BX7T/7Y+sfFCS78DfDm5uNM8JAtFcXAzHPqgzznukR7JwWB+brtHwS+APjv4667/Z3hi0Fvpts4F/q1wh8i1HXH+25HRBz0zgZI+DHwE+H/AMDdE/s7wlp3mahPGBfarcANcXRHPJ/hTPRFwB15OSaqqqqr4h+Lk8A+B9a8ZPpl1qP9kWbzraWsTPJMwHyqAoJAzjLYwoyTwDX7JXwv1v47/E/VP2ifigv221tL8yWiSqdtzejBXaP+eUK7cDpnYOdrCqqqqqqq/b++MTX2rWfwa0S7/wBH08JeawUb70zDMUR9lUhyOmXTutHGOtTyeY+AeB0qq8KeFdf8ceIrDwn4X06S/wBU1KYRW0EY5Zj1JPQKBkkngAEngV+z38APDXwG8IpplkkV5r16itq2p7PmncfwJnlY16Ad+p5Jqqqqqq/4KIeLzp3gLw54Jgm2ya1qL3Uyg8mK3TGD7F5UP1T2NfDWLw18Bv2ffD//AAl2pQaTY6LpMc2oTTZASaX95IABksxkkYBQCScADJxX7Sv7U/ib45anJoulPNpXg61lP2WwDbXuiDxLPg8nuE+6vufmNfs6/sZ+LPi4tt4r8ZNceH/Cb4eNymLq/X/pirDCof8AnowwcjaG5I8B/DrwX8MdAi8NeB9AttLsowCyxLl5mAxvkc/M7e7Emqqqqqr9tP47f8Kp+Hx8K+H74R+JvFEbwwFGw9pbdJZuOQTnYp45JIPyGtA8N+IfFepR6P4Y0O/1a+l+5bWVu00h99qgnHvXw4/4J/8Axa8VeVe+N72x8I2TYJjlIuroj2jQ7Rx/ecEelfDj9i34F/D3yru58PN4m1GPB+1a0wnUHvthAEYGemVJHrUFvBawR21rDHFFEoWOONQqoB0AA6D2qqqqqqqq/bf/AGNPDH7X/wAOo9MN1DpPjPQBJL4e1d0yqOwG6CbAJMMm1c45UgMM4Kt8WPhB8R/gd4zvPAHxR8K3mhazZnmKdcpMmcCSKQZWSM4OHUkde4Iqqr/gmh8Xl+EH7Xvg+a8uvJ0zxaX8PX/OARdFRDntgXCW5JPYGviZ8NPD3xU0XTdD8RJui0rXtN1m3OwNiayu47hAQezeWUP+y5r/AIKN/GE/Bn9kbxrqlndmDVfEkA0DTCDgmW7BSQqezLAJ3BHOUHTqKr/gkN/yeVpv/Yv6j/6Atf8ABSL/AJMj+Kf/AGD7b/0sgqv2EfhKfgv+yf8ADzwhc2oh1K50tdU1NSuHFzdkzsr+rIJFi+kY61+3B/wUz+Ovwg/aR8TfC34O6loMWheGUt7WV7vTVuHlujEskx3EggKz+Xj1jPrXwA/4KxftGeIvjZ4I8OfFLVfDsnhTWNbt7HVDb6SsDxxTOI/MDgkgIXDn2UjvXjjwho3xA8Ga74E8Qw+dpfiLTbjT7xMcmKaNo3x74Y1428Jav4B8Za94F1+IR6p4d1O4068QdFmgkaNwPbcpqq+DvwT+Jvx98aWvgH4V+FbrWtVuCDJ5a7YrWPODLNIfljjH95j1wBkkA/sXfseeD/2QvhufD+nzx6r4r1ry5vEWteXt+0yqDtiiB5WGPcwUHklmY4LYFVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVftVeDvEv/BO39tvQ/2jfhvpjf8ACFeLLyW8+xQDZEd5H2+wOPlUEN5kYxhdy4B8s48D+NfDXxI8HaL498HanHqGia/ZR3ljcp/HHIoIyOqsM4KnlSCDgg1VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVf8FyLq3Tw/8H7FpMTy3mryImDyqJaBjnpwXX8/rXwwtp7P4aeErO5jKTQaFZRyIf4WWBAR+Yqqqq/4Laf8l+8C/wDYnj/0rnr4T/8AJK/Bv/Yv2P8A6TpX/BWD/kyPxh/2ENM/9LIq/wCCKn/Jq/in/soF5/6QWFV/wT4/5SYP/wBhDxF/6KuKqq/ab1t/Df7N/wAVNfin8qXT/BerzQt5vlnzFs5SgDdQS2AMc5Ir/gi9f+I9c+DHjvW/EGv6jqf/ABUkVnb/AGy4km8pYrZGKqWJwP3o4GMfjVVVVVV8Uv8Agkn8A/it8Vtf+KmqeMfFunTeJNUfUbzTLF7dLfzZG3ShSYy4DtuY85yxxVra21law2VnBHDBbxrHFEi7VRVGAAB0AAAxVeLv+CcPws8Y/tOwftTX3jnxVD4gt/ENhra6dEbf7KZrMwlE5jL7G8hc/Nnk4IqqqvCErfHr/gspdak7rcWmi+LrpvMQ5VU0m2aOJh6gyWsYyOpbPTmqqqqqqqqrqBrm1ltkuJYGljZBLEQHjJGNy5BGR1GQRXwC/Zi8N/AHVvEOq6R4gvNYk1tYo4nvIVWS2jQsWXcpw+5mBJ2r90VVVVVVVX/DJv7K/wD0bP8ACn/wjLD/AONV/wAMm/sr/wDRs/wp/wDCMsP/AI1Xhb9nn4A+BtetfFPgn4G/D/w/rVjv+y6jpfhm0tLmDehR9kscYZdyMynBGQxB4Jrxz8IPhL8UJbOf4l/C7wj4tl09XW0fXdEt79rcNgsIzMjbAdq5AxnA9K0nSNK0DSrLQdB0y007TdNt47azsrSBYYbaGNQqRxooCoiqAoUAAAACqqvH3wm+F3xVs47D4l/Dvw34pggB8ldX0yK68nPXYXUlDx1GK0T9h/8AZE8PXqajpv7O/ggzxkFDdaWt0qkdCFl3DPvirKxsdMs4dO02zgtbW3QJDBBGI0jUdFVRwAPQVVVXjzxlpPw98G6x411t8Wej2jzuoODIQPlRf9pmKqPdhXiHW9e+IPi+916/Ml5q+vXzSMqAsXlkbhFHpkhQPTAr4SeDPDn7OvwUs9O8QahaWEemWxvdcvpGwhuHAMjE98HbGuOSFUYya/ae/ai1z45ay+iaHLcad4NspP8ARbMtta8YHiaYDqeMqnRfrk1+zN+yn4g+ON+niDXTPpXg21l2z3YXbLeEdY4Mgg88M54X3IIrwp4S8N+BtAtPC/hLSLfTNMsU2w28C4A9ST1LE8ljkknJNVVVVVVVZWFjptv9l06zgtYd7P5cMYRdzMWY4HGSxJJ7kk1VVVVVV478Xab4B8G6z4z1Y5tdGspLh0BwZCo+VB7s2FHuRXiXxDqni3xDqXifW7kz3+q3UlzcSHu7sSceg5wB2GBVw+yPaDy3Aqqr9if9niP4b+E4/iP4psQPE/iG3BgjkX5rC0bBVcdnfhm7gbV4w2aqqqqqv2p7208e/ta+HPBWp3UUGk6HFZw37zsFjihJNzcSkngAQsM9vkr9qT9pHVPjp4obTtKmltvB+kzN/ZtqQVNwwyDcSDruYZ2g/dU46liUR5HWONSzMcKoGST7V+y3+xRa6fFZ/ET40aWk944WXT9AnXKQDqHuVP3n9IjwP4sn5VAAAVQAAOBVVVVVVWt/sieDfiB8QtQ+I/xb13UPE13dS4tdOjY2lpaW6/6uEBSXbaOpDKGYsSvzV4X8G+E/BGmro/g/w3pujWYxmGytlhDEd22j5j7nJqqqqqqr/gph8Xviz4C/a4+EHhrwL8UfFvh3SNTsbJr3T9J1u4s4Lpm1GRGMkcbqrkqApyDkDHSqqqr40fAT4S/tB+FW8IfFrwXY67ZLuNtJIpSezcjG+GVcPG30OD0II4r9of8A4Iz/ABD8NTXWvfs5eKYfFmmAM66JrEqWuoR+ipNxDN9W8rsMHrXxE+FXxK+EmtN4d+JvgTW/DGognbDqdk8HmgHBaMsMSL/tKSD2NVp2oXuk6ha6rpl1JbXllMk1vNGdrRSIQysp7EEA1o3/AAWz+C40exXX/hV43Opi2jF6bUWpi87aPM2ZlB27s4yAcY4r/goV+3jon7YMfhDQfA3h3WtD0Dw61xc3MGqGMPc3Um1VbEbMNqIrAc5zI3tVV+w/+0R4X/Zd+PFp8V/F+iapq2n2+mXVo1tpoj80tKoAI3sowMc81+1Z/wAFVPg58ff2fPGPwh8N/Dvxlp+peI7WKG3ub4W3kxlJ45CW2Ss2MRkcA8kV4Lu/DNh4x0K+8Z2Vze+H7bUreTVba1AMs9qsimVE3EDcyBgMkDJ60n/BbH9n6NFjj+E3j9VUYVQlmAAPQedXxF8aal8SPH/iT4g6wT9u8S6tdajcAtu2vNK0hGfQbsfhSO8brJG5VlOVYHBBHcV4O/4LVfCSy8I6HZeMPhp42u9ft9Nt49UntRa+TLdLGomePdMG2FwxGQDgjNftd/Fn4f8Ax2+PviX4t/Dfw/qmiaZ4kMNxNZaisYkS5ESpKw8tmGHZN/Xq7cV8N/hB8UfjDrA0D4XeANc8T3oIDpp1k8qw56GRwNsa/wC05A96/Z3/AOCMnjbXZbXxB+0l4sh8Oafw7aBokq3F6/qsk+DFF/wDzcg9VNfCD4IfCv4CeFI/Bnwm8GWGgaauDL5CFpbpwMeZNK2Xlf8A2mJ9BgcVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVftW/s7+Hv2oPglrvwr1ryoLydPtOi37puNhfxg+VKO+OSjAdUdx3zX/AAS5/aQ8Q/Bz4iav+xD8bxLpVzHqU6aAt6+DZX6k+dZZPGyUgyR4wC+7G4yrVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV/wAFy76OTUfgzpqo3mQQa3KzHoQ7WIAHv+7P5itCsptN0PTtOuCpltbSKJ9pyNyoAce2RVVVV/wW0/5L94F/7E8f+lc9fCf/AJJX4N/7F+x/9J0r/gq5BNN+xB40kijZ1hvtLeQgfcX7dCuT7ZYD8RX/AARSurZ/2YfFtikymeHx5cyPH3VWsbIKfxKN+VV/wTzmhuP+ClQnt5UljkvvELI6NkMDFcEEEdQRVVX7fGqDSP2Nvi1d+cke/wAOy2+XGR+9ZYsfU78D3Ir/AII0aUdP/ZJ1G72Sj+0/Gl9cAv0OLe1iyvt+6x9c1VVVVVVVVVV4l12x8LeHNU8Tam+2z0iymu7g5xiOJC7H8lNf8EcdDvPHH7Tnj74raykc89hoEzSPtxtur26Rtw9PlinH0aqqqqqqqqqqqqqqqqqqqqqqqqqqv+ChXxP+zadovwk0y6w92RqOqKp/5ZqSsKH2LB2IP9xD3r9ij4e6f4g+It18R/EskNv4f8B2xvp7i4YLGs+D5ZYngBAskmexjX1r9qr9pzUvjfr7aB4dnuLTwZpkp+y25O030gJH2iQfT7qn7o54JOP2U/2U9R+MuoxeMPGEE1n4Ls5eeqPqbqeY4z1CAjDOP91ecldM0zTtG0620jSLKCzsrOJYre3gjCJEijAVVHAAAxiqqqr9u7xr4r+HX7JXxF8Z+B9eu9F1zTLGBrO/tX2SwM11CjFT2JVmH41+zH8Nf27f23dI13xD4c/ax8Qafp2gXsdvcprHi/U1JmaMspjiiDJjbkE5U89Dk1P/AME9f+Ck/hWBtU8H/tgSXV5HybZPGerQGUDoBuQo3PZ8D3qD9uT/AIKA/sWeMbTwn+1F4Vk8U6NK5Ef9rQxo9zED8zWuoW42SNyCfM80jIBC8Y+Afx7+HX7SPw30/wCJnw01b7VYXX7u5tpcLPp9wAC8EyAna65HsQQwJVgT/wAFUPid8QvhN+zDB4m+GnjDU/DWrTeJ7O1e906cwy+U0U7MgYcgEovT0r9nH9mn9uj9rb4aQfF3RP2utas7GW7msootY8XarJcDyjhs7dwC5Y4AJ/CviV8OP+Cov7EeiSfEy3+OOo+MPCulFWvJIdZm1iC1jLAZmtb1MohJwWjUhc53L1H7Bf7aOnfthfD2/udU0qHR/GnhZ4YddsYCTBIJA3lXEOSSEfy3BQklGUgkgqxqqqqr4ifD7w38UfCV54K8WRXEmm3xQyrBO0TZRgykEdcMoODkcdK8d/8ABOqZVkuvhp49WTk7LLWodpx/12iHXt/qx9a8e/sefHzwTpyazdeEP7WtdhaX+yJRdPb47PGvzdBnKhlA6kVJG8TtFKjI6EqysMFSOoIqv2NPgkvxb+J8Wra1Z+b4c8LlLu9Dr8s82f3MPuCwLEdCqEH7wqqqqqqqtX8O+H/EERg13Q9P1FGjeMrd2ySgoylWX5geCCQR3BNftCfsFyw/afFvwNjMqcyT+HpZPmUdzbux5/65sc+hPC1+xn+yiugQW3xa+J+iOmsFt+jaXdxFTZAHieRGGfMPVQfuj5vvEbaqqqqqqqqqqqqqq/4Kt/8AJ6XwS/7B9h/6c5KqqqqrxF4Y8NeMNKl0LxZ4e0zW9Nm/1tnqNolzC/GOUcFTwT1Hevid/wAEsP2OPiS811Z+A73wbfTnLXPhq/a2AP8AswSCSBR7LGK8f/8ABEDUEd7j4WfHe3lU52WfiDSTGV9Mzws2f+/Q6e/HjH/gkj+2d4XMh0nwr4d8VJGCd+j67Eu4DuBc+Sx+mM8fSvEv7E/7W/hMuNX/AGdfHbiPO97HRpb5FxnJLQBxj5TznHT1GdX+F3xM8PuY9e+HXifTXDbSt5pE8JDYzjDKOcc4qqoKWIVQSScAAVpHwr+J/iFgmgfDjxRqbFioFno88xLAZI+VDyBzj0rw3+xF+114rKDSf2dfHUYc/K1/pElgp6c5uAgxyOeleD/+CRv7ZnicxnV/DfhnwqrjO7WNdjfb9RaiY+/Tv9a8A/8ABECYvFcfFL48IqDHmWegaQST64nmcY/GI18Mf+CXH7G/w0aG6m+Hlz4wvofu3Xie9a8B7/NAgSBu3WP+ZrQfDvh/wtpcOh+F9D0/SNOtxiGzsLVLeGMf7KIAo/AVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVf8FXP2RNS1uxh/a2+EdtNbeJ/CqRv4jWxyk0tvDgxXyFefNgwAxHPlgNkCLn9gH9sHTf2r/hJE2t3kEfj/wAMRx23iK0UBDOcYS7RRxsl2kkAAK4YYA25qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr/AILbE6n8Q/hDoNmC142nX+1TwD5s0Cpz9UNVVVVf8FtP+S/eBf8AsTx/6Vz18J/+SV+Df+xfsf8A0nSv2ivhHafHn4H+M/hFdXCW58SaXJBbTuMrDcKQ8EjAckLKkbEegr9lz9pn4p/8E3vi14p+H/xM+Hl/c6dfukWt6HNL9nmiliLCK6t3IKsCrNyPlkUqQwwDXjb/AIKa/Ez9qSL/AIUZ+xn8H/ENp4o8Sxm3m1vUZY9+mQN8skirHuSIAH/Xu4Cdl3bSP+Cf3w8vPg//AMFLbf4WarqltfXnhibW9NkuYVKJcPFazKWUNyAQpOKqq8b+B/CXxI8K3/gjxzoVrrOhaoipeWFypMcyqwYBgCDwyqfwr4cfDD4f/CHwzH4N+GnhWx8PaJFM8yWVmpWMSOcs2CTycCqqqqqqqqqqv27vF58Dfse/FjXUm8p5fDk+no+4KVa7K2owfXM4x3zjHNf8ER/Bh0/4Q/ETx+0bK2ueIrfTlJGNy2lv5gI9s3jD04PvVVVVVV8bf2p/2n1+O3xLu/Cfx++ItrpWl+KtSmgtbHxVeC0s7YXzJGqRiXYIgXjRVxtwQMYr4FfFLTfjb8HPB/xW0ooIvEukw3UkanIhmK4mi78pIrofdar/AIKy/tE/EHwPa/D34F/BfxZrei+MfFuofbZptD1CSzu/IDeRBCJI2Vgs0zt3wTBzxX/BHH42fFT4ma/8UtG+JnxP8S+Lfs1npk9iuva1cX722HuVkMQmdtgbcm7A5KpnoK/bx8ZeKvh9+yP8R/GHgjX7zRdb06whNpf2chjlgLXUKMUYcg7WYZHPNfsvfCn9t/8Aba0jXvFfh39rDxDYW/h69W2lXWfFmplzJIu/92ke5VTA9R6YxXjD9lP/AIKm/s7aPL468B/tC6x40t9MRrifTtM8SXd7IoUZZvsd4nlzcD7qh2OMbemf+Cd//BRbUv2k9Wk+DnxitLGz8d21o0+n6hap5MesJHzKpi6RzKvzEL8rKGIC7cGv21P2p/2hfHv7T/xOg+CPxY8c6N4W8Axm3e08P+Ibq0gSG0eOC4uSkLqrZuJGO/H3SvJAzX/BNn476h8ef2WNA1LxLr1xq3ifwzPLo+s3V3OZZ5pIyGilkZiWZmgeLLtyzBzknNVVVVVVVVVVVVV+01+yL8XvG3jvWviT4Y1Sw8RJqDhksC32ae3jVQqRoHOxgqqOdyknnbk18Wbjx38IfhlonwNuNA1DQ4dXQavrtzNFs/tKeQKUhVhnMcKCNWXP+sDZAwM/stfs36j8dfFX23Vo5rbwjo8qnUbkAqbhuot4zx8zD7xH3FIPUrnR9H0rw/pVpoeh2EFlYWMKw21tCm1IkUYCgDoMVVVVV/wUi/5Mj+Kf/YPtv/SyCv8AgiH/AMkq+JX/AGMFr/6Tmq+O3wS8DftDfDDWvhZ4/wBOjuLDVYCIbgIDLYXAH7u4hP8ADIjYI7EZU5ViD/wTF8feKv2ef20dY/Z08TXmyz8Q3N9oWpW4c+VHqVl5rRSr7kxSxD184egx/wAFkv8Ak0W0/wCxwsf/AETc1/wSG/5M00z/ALGDUf8A0Na/au+IHgb4a/s7ePvEHxB1G1t9Ln0G8skhmYA3s00DpHbop+87k4A9Mk4AJH/BEXwD4mPiX4jfFCS3ni8Piwg0eKRhhLm6MgmYL6mNFXPp5y+tVVVVVVV8Tf2fPhH8XI5H8Y+ELWS/cYGpWo+z3SnGAfMXBbHo+5favij/AME8PGGj+dqPwp8RQ6/bKCV0/UCttdD/AGVf/Vufc+X9K/Zf+Eh+Dnwh0nw9f2yxa1fD7dq3AyLiQD5CR/cUInBxlSR1qqqqqqqqqqqqqqqqqqqqqqqq+Jn7MvwK+Mni7R/HfxL+HtnrmvaBGkem3s1xPG1uqSGRQBG6qcOS3IPWqqqqqqqqqvtL0zVESPUtOtbtUOUWeFZAp9QCDirzwJ4H1GD7Nf8Ag3Q7mInPlzadE65HfBWrTwN4JsIFtbDwdodvCpO2OLTokUZ64AXFWOmabpkbQ6bYW1ojNuZIIljBPqQB1qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqngguoJLW5hjmhlQpJHIoZXUjBBB4II4xX7Tnwe+IX/BNL9pTS/2j/gPbOfAGuXjhLM58i33ndPpc+M/umUbomPICjGWi3H4C/HPwF+0Z8MdJ+KPw81FZ7DUE23Fs7DzrC4UDzLeZR911J+hBVhlWBNVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX/BZD/kv3wb/AOwe/wD6VrVVVVWueCvBvia4ju/EnhLRdWniTZHLfWEU7Iuc7QXUkDJJx71DDDbwpb28SRxxKFREXaFAGAAB0GO1V40+GPw1+JNvFafET4e+GfFMMH+qi1rSYL5U5zwJVYDkA14S8CeCPAOnf2R4E8G6H4csTjNrpOnRWcXHA+SNVHA9qg8FeDbXWj4ktfCWixauXZzfx2ES3BZgQzeYF3ZIJBOecn1qqqqqqqqqqqqqq/4LD+LP+Ee/ZBbQ0lAbxP4nsbEpnllQSXJOPQG3X8SPWv8Aglt4OHhH9inwPLJEI7nX5b3U5xtxnzLmRYz75iji/wA81VVVVVfsqfDhv2iP+Gm9OtrT7Zd6h4Ju9T0+VIiubqLUIbuFVOMqZGh2444J96/4ItfGceIfhX4r+Bup3jNd+EdQGpaYjtn/AEO6z5iIOwSdGY+9wKr4S3x/bX/4KuX3xEVze+Evh3cPeWDD5oxbaeRFaMhyQN90yXAHfc5AHb/gibfyWnx3+IGgSiOOSfwoJmjfiQGK7iU4HoPNweOCV6d/+CkX/JkfxT/7B9t/6WQV/wAEQ/8AklXxK/7GC1/9JzVeHLW30X/gr6LP4eRRRwRfEy4Ro7cbVjVi4vFAGMAAzgjoMEdK/aW+Llv8B/gL43+LErx+d4f0mWSyWQgLJdviO2Q57NM8Y79ehr/glH+zlb/Fv4afG/xx4yQyReNtNm8IW95KC0n76PzbuTJ5J3PaMD/eQ9xX/BIT4gat8Kv2j/HH7OPi4taT69bzILVzny9T053DoB2JiNwSR18pfTiqqqqqqqqqqv8AgqX+1J8Z/wBmXwv4Cn+DXiS10a68R3d/Fe3Eunw3bbYo4im0TKyjBkJ6HOB2yD8Pb/8A4LKfGbwbpfxO8H+PGm0XxBALixmafRbXzY+gYRBBsBweCAfatf8A20f+Cmf7IGr2T/tLeC7XxBodzMIUl1HT7ZYZsDOyK9sMRrKVBOJN7cEleDX7Mn7SvgD9qn4YW3xK8BtNbgTNbalplyymfTrlQC0b7eCCCrKw4ZWB4OQPiF8OfBvxS8Nz+FPG2iw6hYzDK7hiSB8YEkbjlHGeo+hyCRXgHwH4Z+GfhLT/AAX4RsRbadp0W1AeXkb+KRzxudjkk+p7DAqqqqqv+CkX/JkfxT/7B9t/6WQV/wAEQ/8AklXxK/7GC1/9JzVVdfvv+Cwa/wBg8Y+KCb/K+Xo48/8AlJn159a/4LJf8mi2n/Y4WP8A6Jua/ZC+FP8AwUW8X/Bm21n9mn4lf2L4KbULhIbT+2I7bE6sPNbYyE8nHOa/aV+A37cvhXU7P4iftaeHPF3j3wvpFxunuE8QfbLWOLPzKGiLmzRuBuMaDn1r9gD9pH9nz41fCmDwd8FPCkPgafwjAi3vhEyeY1mrscSpKQDcIzA5lPz7j84BYE1VVXjX/gpTofg39rW3/ZSl+Et9dXdx4m03QxrS6wqIGvDCBL5XlE4Xzx8u7nb1Gaqv2vf2nLD9kv4URfFLUfCE/iSKXV4NOFlDeC1YGVJG37yjcDy8Yx361+yd+0dY/tU/CC2+LWn+FJ/DsVxf3FoLGa7FyymIgFt4VeuemOKr9rH9o6x/ZW+EFz8WtQ8KT+Iore/t7Q2MN2LZiZSRu3lW6Y6Yr9kP9pyw/a1+FEvxS07whP4bii1efTvsU94LliYkjYvvCLwfMxjHbrVVVV+2h+2/4X/YytfCj694F1TxLc+Lvtv2SOzuo7dIvsog3b2YEjJuExhT0Ptm4/4LmW6zOtp+zDJJED8jSeNAjEe6iyOPzNfCz/gs78BPF2q2+k/EnwP4i8DC4YKL8SLqVrDnqZGQLKB7rG3vivD3iLQPF2hWPibwtrVlq2k6nAs9nfWU6zQ3EbDIZHUkMD6ivjX8c/hh+z14Hn+IPxW8TQ6RpUTiKIbTJNdTEErFDGvzO5AJwOgBJIAJHjT/AILg6NBqUlv8PPgDd31gj/Jd6zrq20si+8MUUgU9P+Whr4Bf8FhfCHxa8feH/hv4s+CWt6FqXiPUINOsZ9K1NNSja4lZUUurJCyIWbnG/A555r9s3/gobo37HnjvRPA+pfC698USa1pH9orcQaqtqIh5rx7CpjfJ/d5zkda/4fi+FP8Ao3bVf/Ckj/8AjFf8PxfCn/Ru2q/+FJH/APGK/wCH4vhT/o3bVf8AwpI//jFf8FJP2q/jL8F/gx8L/iB8E/FZ8LXfi65L3Yawtb0mJrZZVT9/E4GC3VQCa/Ym+I/jT4vfsteAPiN8Q9a/tbxFrVnPJfXn2aKDzWW5lRTsiVUX5UUfKo6VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVfEz4beDPi/wCBNZ+G/wAQNFi1TQddtmgu7eQc4PKuh6o6sAysOVZQRyKtL740/wDBIv8AaRksLtLzxP8AC7xS2QASkWq2gbh0/gjvYQQCOhDY+64YfDj4j+Cvi34K0r4hfDzxBbazoOswCW0uoDwR0KsDyjqQVZGAKkEEAiqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr/grB+/8A2yvgnaT/ALyA6fY5iflfm1Jw3HTkAA+uBVVVVVVVVVVX/BST45/EX9nv9m1/HXwr8RvoniGfXrOxgvFsoLoKjiRnBSdHTBWMjO0nOOnWv2M/Hfjr4n/swfD74gfErWl1bxJrumtc316tvFB5pM0mw7IlRFIjCDCqOn41VVVVVVVVVV/wXE8Ttb+FfhR4LjlUi/1DUtQlQNypgjgjQkZ7/aHwcdjz1r9mvwmvgX9nn4Z+EPJ8qTSfCemwTrgjMoto/MJB6EvuOPeqqqqq8aaodD8Ha7rYkdDp+mXFwGQZZdkTNke/Ff8ABD6w8zx/8U9T87H2fR7CHy9vXfNKc57Y8vp7+1fCx3/Yh/4KnXXgpnNh4X1vW5NJCk7UOm6ltktMkcbY5Httx6AxN0xx+3d8aX+A37LPjjxrYXRh1i6sv7K0dlbDrd3X7pXX3jVnl/7Z1/wRn+Cp8H/A3XfjLqdpsv8Ax9qXk2TMP+XC0LRgj03TtcAjuI0PpX/BKaWTw7+3h420KWa3Mk+g6xZMScbyl9budnTn91n6Z+o/4KRf8mR/FP8A7B9t/wClkFf8E+P29/AX7H/hbxX4Y8a+CNf1oeINQiu4Z9LkizFsj2FWWRl69cg/hXxU/wCCxfjH4habP4F/Zl+DGq2viHVlaC01G9cXt1CW4DQ2kKsGk54JZgDjKsOK/wCCaP7BPxB+HPjKX9pj9oKwlsvE88Mw0PSLxvMuoHnBE15cHJ2SsjOgQncBI5YA4Ff8FrPjGmkfD/wZ8C9MvQLrxDfPrGpxo3zC1twUhVh/deV2Ye9v+f7FfwePwK/Zh8A/D65tfI1OLS1vdVVhhheXJM0yt6lGk8seyCv2+dC1L9kv/goNofx68NWzLYa3eWnieGOJQoklRxHfQZ7mQozN04uetaNq+m+IdHsde0e6S5sNSto7m1mTpLFIoZGHsVIP41VVVVVVVVVV/wAFxv8AkVPhH/2ENV/9F21fsB/8mafCX/sX4/8A0N6/aZ8E+F/iH+z98QfCvjCyt7nTbjw7eyMZlH7iSOFnjmUnoyOquD2Kiv8AgiHrusQ/E34leGImlOlXeg2t5MMnYs8U+yPj1KzSfl+X7RP/AAUY/Zq/Zt8Q3ngnxRrGr674psAv2rR9DsvOktyyhlEkkjJEpIION5YAg45FaN/wWx+A13qwtdc+FfjnT9PZ9q3cJtrh1GeGaPzFwOhIDMRzjPf4RfGT4a/HfwVa/EH4V+KrXXdFumMfmxBkeGVcbopY2AaNxkZVgDgg9CCaqqqv+CkX/JkfxT/7B9t/6WQV/wAEQ/8AklXxK/7GC1/9JzVfGT4t+DfgX8NNd+KXjvUUtdJ0K1aVlLAPcSdI4Yweskj7UUep5wMmv+CXfgTxN8f/ANs3Xv2i/E9qz2/h6W+1q+uAh8p9SvjIqRAnviWeQc8eUPWv+CyX/Jotp/2OFj/6Jua/4JDf8maaZ/2MGo/+hrV9Y2OqWNxpmpWcF3aXcTQ3FvPGJI5Y2GGRlOQykEgg8EGv2J9vw0/4KjSeB/hvcOfD/wDwkviLREijkJWfTYlumjBOfmVfIik57xg1VVVfHD/lMHpv/ZUPDP8A6HY1Vf8ABZL/AJNFtP8AscLH/wBE3Nf8Ehv+TNNM/wCxg1H/ANDWq/4K8/8AJmmpf9jBp3/obV/wRt/5NFu/+xwvv/RNtVVVV/wXP/5ol/3MH/uOr9iSGGH9kT4QrDEiBvCNgxCrjLGFST9Sa/4KB/sc/Cf4y/BDxd46tPCemaT458LaTc6rYazZWyQTXP2eNpGt52UDzUdVZRu+4xBBA3A/8ETvi5r2r+HPHvwW1fUJbjTtAe21TRonJb7Ms7SLcIueFQusThRj5nc/xGv+Cm/ibWfjH+3npXwQ1bWJrHQ9Bk0nRrXcf3cDXywzTXAUnG4i4RSeMiFR2r4S/Bf4YfA3wlZ+C/hf4P07Q9PtIVjZoIFE1yQAPMmkxulkOMlmJJrVvg58KNc8YaX8QdV+HXh648T6JcNcWGsNp8YuoJGUqSJQNxyGPBJGcHqAa1zwV4N8TXEd34k8JaLq08SbI5b6winZFznaC6kgZJOPevj78LvhnZ/An4j3dp8O/DEM8HhLVHilj0iBWRhaSEMpC5BBAOa/4IveGPDXif4nfEa38SeHtM1aODQbZokvrRJwjGcgkBwcHFf8Km+Ff/RNPCn/AIJrf/4iv+C28MNv8Jfhjb28SRxxa9cqiIu0KBbgAADoMdq/4J6axpGgfsI/DPWdd1Wz06wtNMuXnurudYYol+2zjLOxAUdOSa8J/tLfs7+PNfXwr4L+OfgTW9Yd9kVjY+ILaaWdueI1VyZOn8Oe3rVVVVVVVVVVVftu/tXfH74QftufCf4QfDrx9/ZPhLxLZ6NJqmnf2VZz/aGuNUuIJT5ksTSLuijRflYYxkYJJqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr43fBH4d/tC/DvUfhl8TNFW/wBKv13RyLhZrOYA7J4Hwdki5OD0IJBBViDofib9oX/gkd8cZPCniCGfxT8KfEl15qhVKwanCMAz25JxBeIuA8ZOGwAcqY5B8IPjH8Ovjx4EsPiN8MPElvrGjX643IdsltIAN0MydY5FyMqfUEZBBNVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVf8FW/+T0vgl/2D7D/05yVVVVVVVVVVV/wWUdF/ZHslZ1BbxjYhQTjJ8i5PH4A1+wIpX9jX4SgjB/4R6Pgj/baqqqqqqqqqqv8Agr/5/jj9qv4S/CmF3b7RotuERc8Pe38kJx15PkL2z068Yhhht4Ut7eJI44lCoiLtCgDAAA6DHaqqqqq/aX1Q6J+zj8VdZQtusfBOsTrtfYcpZSkYPY5HWv8Aghrp5TTfjJqhdSJp9FhVccrsW9JP0PmD8q/4LWfCK4sNX8AftC6HC8TMr6FqVxCChjlQtPaNkc7iDcjPUbFGemP+CgPx81j9qbwX+zR8KvBrrc6v440y11q+tUYlW1K5YWUScdNky3q9OjA8d/hX8PdG+E3w18MfDLw8P+Jf4Y0q30+FyoUyeUgUyNj+JiCxPcsTX7EufC3/AAVb8T+HCttCX1/xVYeXnAHltcPtj56jyfyB+o/4KRf8mR/FP/sH23/pZBX/AARr+HvgHxz8KviKPGvgfw/4g8jxBaiIappkN35f7g/d8xTivDngjwX4OiMHhDwjouhxsu0ppunxWwI9MRqOKr4/adrH7fP/AAUg1P4b+G9cNnp39oPoNlqH2f7SljZWEbmeYIGXepkSeRRuXJkUZGa/4cYf9XR/+WR/93V+11/wS01j9l34M3fxh0v4vHxlBpl/bwahZjw4bD7PBMxjE3mfaZc4lMSbdo/1mc8YP/BKz4xt8V/2StD0XULzztW8A3L6DcbjhvJjAe2IH90QSJGD3MRqqqqqqqqqqr/guN/yKnwj/wCwhqv/AKLtq/ZH/wCCk/7KXwq/Zr8DfDrxx4s1mx17w3pItbu2TRZ5g0isx+R0BUg5HUj8K/bB/wCCo6fHfwfefAX9mDwd4jdvFqnT7/U7q1/0q6gkG17a1t4i7EyAlCzYO0sAnzbh/wAEw/2QfEP7M3wu1XxR8RrFbTxr45khlurEsGbTrSIN5ML4yBITJI7gHjKKeUNal+zP8A9b+Jl/8Ytf+FPh7WPGGoxQxzalqVoLtlEKBIyiSbkjYKAu5QGIABPFfHv4e/sk+PPCGofD74xv8PdNingMMUt5c2lldWDlSEkhdirRuvJGODggggkH/gjn491nwZ+0f4x+C0+rLPpGuaRPL5UT+ZE17ZzKElQjIwYnnBIOG+Tk4FVVVVf8FIv+TI/in/2D7b/0sgr9gb9vzwx+x34Y8UeF/Efw71PxEniHUIrxLiyvkhMOyPZtKOvOeucj6V4i/wCC29jeRf2f8N/2dtRvtSnXbAdR1oAK/P8AyyiiZpBgDgMvU8jHOqfB/wD4KHf8FIvFen6h8VtKuPA/ge1m8y3TUrJ9OsLJT1eC0c+dcybc4dt3p5ig1+zl+zv8P/2YvhhYfDD4fW0jQQMZr6/uAPP1C5YDfNKRxk4AAHCqqgdK/wCCyX/Jotp/2OFj/wCibmv2Kv8Agpl4G/Za+Btr8JvEHwx13XLu31O5u/tdneRRoyysCBtYZBGK+J//AAVu+M3xo025+Hf7MHwT1PSdY1NDCupRyPqt/GrcZggiiCxydAHJfGTgAgMP+CZH7AvjH4Iarc/Hr436eLDxbe2j2ujaM0okk0+KTHmTzlSQJnA2hATtVm3fM2Fqqqvjh/ymD03/ALKh4Z/9Dsaqv+CyX/Jotp/2OFj/AOibmv2K/wDgpl4F/Za+Btp8Jtf+GWu65d2+pXV2buzvIo4yJWBAw3ORiv8Ah958Kv8Aoh/iv/wY2/8AhX7an/BTLwL+1N8Dbr4TeH/hlruh3dxqVtdi7vLyKSMCJiSMLzk5r/gjb/yaLd/9jhff+ibaqqqr/guf/wA0S/7mD/3HV+xNqOnyfsgfCedL+3aO08I2KzuJQREywKWDHPykDqD0r/goX+3L8I/hj8GPFPww8HeMNL8R+OvFumT6VFZaZdpcDTYp0Mcs87oSIysbMVQ/MzFeNuWH/BGD4G694Q+Hfi742eI9Oe1TxpNb2eiLNHteS1tzIZJlz/A8kgUevkk9CDX/AAVR/Ya8cfE3W4f2j/gzo9xrGr2ljHbeINGsoi91OkWfKuoFHzSOqkIyKN21EIBwa/Zl/wCCxN54b0yy8CftReFtR1GWwUW//CTaWgNy23jN1bMV3OMfM6EE4+4Tkn4R/HD4S/Hnw2PFnwj8d6Z4k04ELKbZystsxGQs0LgSRNjna6qcc9Kr9oX/AJID8TP+xP1b/wBI5a/4Ih/8lT+Jf/Yv2v8A6UGq/wCC3v8AySr4af8AYwXX/pOK/ZE/Zc+L37c3wj8M6X8TfiDqfhL4J+A430/RtM0sDzdYuhK8k02HymVaUr5rqwG3Yq5DtX7Z/wDwSl0T4GfCvUvjD8DvGviHV7Xw0guNW0vWjFJOLfIDzxSwpGP3eQzKU+6GIbK4P/BJ/wDaf8UfHn4P6z4E+IOsT6r4k+H1xBEuoXUhea7sZ1fyDIzZLujRSoW6lfLzySTVVVVVVVVVV/wUm/5SS/Ar/sH+Hv8A093dVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV8WvhF8Pfjl4F1D4cfE3w7b6zoeor88UmVeGQfdlicfNHIueGXBGSOhIPxA+D37UX/BKr4lSfFP4Ratc+JvhlqE4juJZojJbzRFvlt9QiXHlyDICzrtBJ+UqWaOv2Tv23/g5+1poSDwvqC6L4utoQ+peGL+YC5hx954TwJ4s/wAajIyN6oSBVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX/BUYLq/7eXwP8NwEpcPp+k/O4+Qebq0yL054KHPHpVVVVVVVVVVX7UP7L3gD9rXwBYfDj4jav4g07TdN1iPVYpdEuIYZjNHDNEFYyxSrs2zuSAoOQvPUH4XfDzRPhJ8OfDfwx8N3V7c6X4X02HT7Oa+dXneOJQqmRkVVLYHJCgewqqqqqqqqqqr9qSIeOP+CwPw68PTZkj0bUvDqlGCgFYiLwjocg7jkHk5I9Kqqqqqv28dTTSP2Ovi3du0YD+Gri3HmNtGZcRD8cuMDucCv+CI2neV8G/iJq25P9J8TQwYC/N+7tlbk+n73+frX7cHwd/4Xn+y34+8C2tp5+qJprajpKqm5/tdqfOjVPQvsMX0kNf8EjfhFe/FL9pKDx9rYmutE+FmlPPbCUlo4rqdpFgjAPQZkuZhjGHTPc1XwvB8I/8ABZi8BjitvP8AHut8O2Qftdtdc9eredkDsWHHav8AgpF/yZH8U/8AsH23/pZBX/BEP/klXxK/7GC1/wDSc1X7V/xfX4Dfs6+PPilFcLFfaRpMi6YSAf8ATZiIbbg9R5siEj0Br/gij8In1LxF4++PurQGX7FEmhabNJlt00pWe6bJ/iCrbjPXErdM818Z/hrpnxk+E/i34WauUW38T6RcWIkZc+RI6ERygeqPtce6iv8Agkb8SNW+Ef7UPib4BeKw1mfFltNZyWsnBi1TTy7BTnp+7+1KfU7aqqqqqqqqqq/4Ljf8ip8I/wDsIar/AOi7av2Pf2Hv2UfiZ+zD8M/HXjj4L6Pqmu6loiTXl489xG08m9vmcJIqseB1FfDb4B/BP4Pbn+F/wr8MeGpnUq9zp+mxxzyKeoaXG9hwOCSKlkjhjaWWRURFLMzHAUDqSewrxZ+0R+0v/wAFJv2h7v4GfAvxpd+DPh3CZZS1rI9v/wAS+Jgr3l4yESSlyyBYchAXRSAd0leEv+CLH7Omm6eg8a/ELx3r+pFcSzW09vYwMeOVi8qR1PXrI3Wv+CYGi2fhz/goDf8Ah3TmkNrpdprVrAZGy2yNti5PGThRmqqqqmVWUqwBBGCCKv8A4eeANU8v+0/A3h+78rPl+fpkMm3OM4ypxnA/KtI8NeHPD6CPQdA03TVC7QtpaJCAM5xhQOM84qqvLKz1CA2t/aQ3MLEZjmjDqcdODxT/AAp+F0jmST4beFWZiSWbRoCSfX7laZpGlaLaLYaNplpYWyfdhtoFiQYAHCqAOgA/AVVVVV8cP+Uwem/9lQ8M/wDodjVV/wAFkv8Ak0W0/wCxwsf/AETc1/wS1+AnwL+IX7Jun+I/H3wX8CeJdWfXL+Jr/WPDlpeTlFdQqmSWNmwOwzgV/wAMm/sr/wDRs/wp/wDCMsP/AI1X/BUr4CfAv4e/sm6h4j8AfBfwJ4a1ZNcsIlv9H8OWllOqM5DKJIo1YA9xnBr/AII2/wDJot3/ANjhff8Aom2qqqq/4Ln/APNEv+5g/wDcdXwW/wCCPdj8Y/hL4J+Ksf7QU+jp4p0W11GbTz4WFyYWlUMyLL9qTIAIAJT39q+DX/BID9mr4capba9471DW/iHe2rh0ttTKW1gWHQtBH8z89VeRlI4KkZzZ2Vnp1nBp2nWkNra2sSxQQQxhEiRRhVVRwqgAAAcAVXx0/Yx/Zv8A2ioZ5viP8NdPfWJlIXXNOX7HfoccMZo8GTGThZQ6+1fFz4f/ABZ/4JcftRaPq/gTxdNqOnTxLfabcsDEmrWPmbZrO7jGVJBUq2M/eSRdrYC+CvFWmePPBug+ONFLf2f4h0y21G0343eVPEsiZxxnawr9oX/kgPxM/wCxP1b/ANI5a/4Ih/8AJU/iX/2L9r/6UGq/4Le/8kq+Gn/YwXX/AKTiv+CbUUcX7EPwsSKNUBsLpiFGOTe3BJ+pJJ/Gv2jIIbr9nr4n21xGskUvgzV0dGGQymzlBBHoRX/BEGWVfid8TbdZWEb6DaMyBjgsJ2AJHqAx/M+tVVVVVVVVVVfGX9in4OfHX4y+F/jp42ufESeJPCMNpFp6WN6kVuVtrmS5j3oY2LHzJWzhhkYHHWqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqrUtM03WdOudI1ewtr6xvYWhubW5iWWKaNhhkdGBDKQSCCMEGv2pf+CUV/pmtn4w/sYazP4e16xm+1r4bF81t5coOd9hc5BhYHkRudvXa6gBK+AP8AwVb+IPwp1xfhB+2x4I1eG+0x1t5tdSwNvf23TBu7QhfMGDnzI8MVAIRycn4bfFX4b/GLw3F4u+F/jXSfEuky4H2iwuBJ5bYzskX70b+qOAw7iqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/wCCk3/KSX4Ff9g/w9/6e7uqqqqqqqqqqqqqqqqqqqqqrTCvjD/guDNOSZI7PU5d7xIcJ9l0EoM56fPEqk+p46iqqqqqq6tbW9ga1vLeKeFxh45UDK31B4NWOmabpkbQ6bYW1ojNuZIIljBPqQB1qqr+w9E+2/2l/Y9j9r3bvP8As6+Zu9d2M5q6tbW9ga1vLeKeFxh45UDK31B4NWOl6ZpaNHpmnWtornLLBCsYY+pAFVeWNjqMH2bULOC6iJyY5ow65HfBqy0+w02H7Np1lb2sW4t5cMQRc9zgd+Kql0DQkvP7Qj0WwW63l/PFsgfcerbsZz71VVVVVVVVVV/wXG/5FT4R/wDYQ1X/ANF21fsB/wDJmnwl/wCxfj/9Deq8R6NH4i8Pap4fkuHgTU7Ka1aVPvRiRCpYe4zmv2P/AIt3P/BPH9q/xDoPx38NajaWsllNomsGCAvJbAyxyRXUanBliPlqcj7ySblyQAfjD/wVc+Cp8PN4Z/ZkGsfEf4ha6BaaFaW2i3MEMV1J8qNIJ0R5NpIIjRW3EAZUEsP+CZGj+IfBv/BQa68JeOyYfEmn2+t2GpRySiRjeRZEw3gkMdyOcg4OMg1VVVVVVVVVVVVVftA6jp+kf8Fc7fVtWv7eysbL4leHJ7m5uZRFHBGhsmd3ZiAqhQSSeABk1/w0L8Af+i5fD/8A8Ka0/wDjlR/tA/AWaRIYfjd4BeR2Coi+JbQliegA8zrX/BZL/k0W0/7HCx/9E3Nf8Ehv+TNNM/7GDUf/AENar/grz/yZpqX/AGMGnf8AobV/wRt/5NFu/wDscL7/ANE21VVVX/Bc/wD5ol/3MH/uOr9ij/k0X4Qf9ifp/wD6JWqrU/2w/wBm7QPi1q/wQ8UfFTR9B8WaK0Cz2+qy/ZIXaaJZEVJ3xEzbXUFdwbJxir74lfDnTNKfXdS8f+G7TTY4/Me8n1WGOFU/vFy20D3zX/BR7466F+2V+0R4M+GvwCR/FEGiRtpVhd2yMF1K+uZV3iLcBmJdkY3n5Th2Hy4Y/CzwUnw2+GHg/wCHUVws6+FdBsdKWUZw4t4Ei3DPPOzPNftC/wDJAfiZ/wBifq3/AKRy1/wRD/5Kn8S/+xftf/Sg1X/Bb3/klXw0/wCxguv/AEnFf8E3v+TI/hX/ANg+5/8ASyev2hf+SA/Ez/sT9W/9I5a/4Ih/8lT+Jf8A2L9r/wClBqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq+O37MnwS/aR0I6J8WfA9lqkscRjtNSjXyb2yzz+5nX51GTnacoT1U18Qf+CXf7TP7OviJ/iN+xn8WdR1MwEstkLwabqQQHIjLZEF0uOobYG6bDnFeCP8Agq/+0Z8Dtaj8A/tc/BO4vLm2GyS7W0bR9RIHWQxOvkzA8YKCJSOcmvhX/wAFLf2O/issUEHxTh8K38gGbLxTEdNKZ9ZmJgPpgSGtF13Q/EunRav4d1mx1WxmH7q6srhZ4n+joSD+dVVVVVVVVVVVVVVVVVVVVVVVVVVVVV/wUm/5SS/Ar/sH+Hv/AE93dVVVVVVVVVVftx/tVah+yB8JdJ+Jem+DLfxPLqXiKHSDaT3zWoQSW9xN5m4I+SPs4GMfxZzxXwN+I8/xh+Dngz4pXOlR6ZL4q0W21F7JJjKtuZUDFA5ALAZxnAqqqqqqqqqqr9ngPrn/AAWS8e38zLG2n6jru1VGQ4SIwD6cHP1qqqqqqqqqqqqqqqqqqqqqqqqqviP8GvhR8YYLG2+KXw80HxVFpbu9kmq2KXAt2cAOUDA4JCrn6CvDHhbw54K8P2PhTwjolnpGj6ZEIrOxs4RFFAg/hRRwByeKqviv+zz8EPjnFBH8WvhhoPiaS1Qpb3N5agXEKnqqTLiRVPXAYDOD2r4Vfsyfs/8AwQuWv/hV8JfDvh++ZChvoLTfdbSMFfPk3SbT3Xdg1pfwD+CuifECX4r6P8LvDdn4ynmmml1yHT0S7eSVSsrGQDcSwZgfXNVVVVVVVVVVVVVXx/8A+CUfw9/aA+MHiT4wax8WPEOlXniSeOWWztrGF44SkKRgKWOTkRg8+tf8OQvhV/0XDxX/AOC63/xrTP8Agih8LdM1K01KP42+KXe0nSVUbToAGKsDjr7V+1h+zNoX7WPwui+F3iHxPf6DaxarDqIurKFJHLRJIoXDcYPmH8q/Za/Z00T9ln4TW/wm8P8AiO91y0t76e7F3eQrHIWlIJGF4wMVX7Uv7OmiftTfCa4+E2v+I73Q7S4voLs3dnCskgaIkgYbjBzX7J/7M2hfsnfC6X4XeHfE9/r1rLqs2om6vYUicNIkalcJxgeWPzqqqq/bL/Yg8Kftmf8ACH/8JN441Xw7/wAIf9u8j7DbRy+f9q8jdu39Nv2dcY/vGvhD8ObH4Q/C7wr8LtN1KfULXwtpUGnQ3U6BHmWJAoZgOATjoKqv2kf+CWvwG/aF8Yar8SItf8SeFPFWsy+dfXNnOLq3uJMAF2hlBIOABhHReOlWv/BDXSE1DzL79pa7mst5/cxeEljl254HmG7YZx32fh2r9mL9gv4A/sq3La94J0m91jxTLCYn8QazKs1yiMPmWJVVUhU8g7VDEHBYiq8d+Fbfx34I8Q+CLu7ktYPEOlXWnSzxqGaJZ4mjLKDwSA2cGv2Pf2BfB/7HniXxB4k8NeP9Y8Qy+IbGO0livrWOIRBH3hlKdSenNV+2H+x54a/bE8NeHvDXiXxjqfh6Lw9fSXcUtjbpK0rOmwgh+gxzXwA+DumfAD4PeGvg/o+s3OrWfhuCSGK8uY1SSUPK8hLKvA5kI49K8d+Fbfx34I8Q+CLu7ktYPEOlXWnSzxqGaJZ4mjLKDwSA2cGv2Pf2BfB/7HniXxB4k8NeP9Y8Qy+IbGO0livrWOIRBH3hlKdSenNVVVVVVVVVVftW/tx/Fn4I/tg/Df8AZ38A+HvCOp6N4wi0n7fLf2lxLeRS3eoS27rG0c6IP3aRlQyNy2TkEAVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVeM/APgb4jaO3h7x/4O0XxJpj5JtNVsI7qPOMZCyAgH3HIr4o/8Eiv2S/HplvPCdhr/AIDvnJYHRr8y25b/AGobgSYX/ZRkHA6d9Y/4I5fHfwBey6r8Bv2krBLg42tcrdaLLgdjJbtNkjJxwB9M0v7KX/BX6xX7Hb/HvVp44SVSQ+PZn3gHrl/mOf8Aa5r/AIZc/wCCwf8A0XTVf/C6ev8Ahlz/AILB/wDRdNV/8Lp6/wCGXP8AgsH/ANF01X/wunr/AIZc/wCCwf8A0XTVf/C6ev8Ahlz/AILB/wDRdNV/8Lp6/wCGXP8AgsH/ANF01X/wunr/AIZc/wCCwf8A0XTVf/C6ev8Ahlz/AILB/wDRdNV/8Lp6/wCGXP8AgsH/ANF01X/wunr/AIZc/wCCwf8A0XTVf/C6ev8Ahlz/AILB/wDRdNV/8Lp6/wCGXP8AgsH/ANF01X/wunr/AIZc/wCCwf8A0XTVf/C6ev8Ahlz/AILB/wDRdNV/8Lp6/wCGXP8AgsH/ANF01X/wunr/AIZc/wCCwf8A0XTVf/C6ev8Ahlz/AILB/wDRdNV/8Lp6/wCGXP8AgsH/ANF01X/wunr/AIZc/wCCwf8A0XTVf/C6ev8Ahlz/AILB/wDRdNV/8Lp6/wCGXP8AgsH/ANF01X/wunr/AIZc/wCCwf8A0XTVf/C6ev8Ahlz/AILB/wDRdNV/8Lp6/wCGXP8AgsH/ANF01X/wunr/AIZc/wCCwf8A0XTVf/C6ev8Ahlz/AILB/wDRdNV/8Lp6/wCGXP8AgsH/ANF01X/wunr/AIZc/wCCwf8A0XTVf/C6ev8Ahlz/AILB/wDRdNV/8Lp6/wCGXP8AgsH/ANF01X/wunr/AIZc/wCCwf8A0XTVf/C6ev8Ahlz/AILB/wDRdNV/8Lp6/wCGXP8AgsH/ANF01X/wunr/AIZc/wCCwf8A0XTVf/C6ev8Ahlz/AILB/wDRdNV/8Lp6/wCGXP8AgsH/ANF01X/wunr/AIZc/wCCwf8A0XTVf/C6ev8Ahlz/AILB/wDRdNV/8Lp6/wCGXP8AgsH/ANF01X/wunr/AIZc/wCCwf8A0XTVf/C6ev8Ahlz/AILB/wDRdNV/8Lp6/wCGXP8AgsH/ANF01X/wunrxd/wTq/4KQ+P/ABdpvj7xt4ostc8SaMkSafql94r824tVikaWMI5GVCyMzDHQsTX/AAy5/wAFg/8Aoumq/wDhdPX/AAy5/wAFg/8Aoumq/wDhdPX/AAy5/wAFg/8Aoumq/wDhdPQ+B3/BZ/Th9htfiTq1zFEcLKfFlm+/33SNvP41/wAKb/4LTf8AQ/6r/wCFRp//AMVX/Cm/+C03/Q/6r/4VGn//ABVf8Kb/AOC03/Q/6r/4VGn/APxVf8Kb/wCC03/Q/wCq/wDhUaf/APFV/wAKb/4LTf8AQ/6r/wCFRp//AMVX/Cm/+C03/Q/6r/4VGn//ABVf8Kb/AOC03/Q/6r/4VGn/APxVf8Kb/wCC03/Q/wCq/wDhUaf/APFV/wAKb/4LTf8AQ/6r/wCFRp//AMVX/Cm/+C03/Q/6r/4VGn//ABVf8Kb/AOC03/Q/6r/4VGn/APxVfFH9jz/gq18a/D9v4V+Kt5L4l0m1vFvYbS+8TWDIk6o6LIMOOQsjj/gRrwt8H/8Ags/8M/DmneBPA8l/DoWjWyW9hBDrehyJDEoAVFM0m/AAxg1/wjP/AAXI/wCfnVf/AAbeHf8A45X/AAjP/Bcj/n51X/wbeHf/AI5X/CM/8FyP+fnVf/Bt4d/+OV/wjP8AwXI/5+dV/wDBt4d/+OV/wjP/AAXI/wCfnVf/AAbeHf8A45X/AAjP/Bcj/n51X/wbeHf/AI5X/CM/8FyP+fnVf/Bt4d/+OV/wjP8AwXI/5+dV/wDBt4d/+OV/wjP/AAXI/wCfnVf/AAbeHf8A45X/AAjP/Bcj/n51X/wbeHf/AI5X/CM/8FyP+fnVf/Bt4d/+OV/wjP8AwXI/5+dV/wDBt4d/+OULv/guRoBNiF1aXcd+4weHbzrx9/D46dM/hzX/AAkv/Bcj/n21X/wUeHf/AI3X/CS/8FyP+fbVf/BR4d/+N14W/Z2/4Kl+CvjDqfx88M/DjVbPx5rMlxJe6tt0iTzWnOZT5LkxLu9kGO2K/wCEl/4Lkf8APtqv/go8O/8Axuv+El/4Lkf8+2q/+Cjw7/8AG6/4SX/guR/z7ar/AOCjw7/8br/hJf8AguR/z7ar/wCCjw7/APG6/wCEl/4Lkf8APtqv/go8O/8Axuv+El/4Lkf8+2q/+Cjw7/8AG6/4SX/guR/z7ar/AOCjw7/8br/hJf8AguR/z7ar/wCCjw7/APG6/wCEl/4Lkf8APtqv/go8O/8Axuv+El/4Lkf8+2q/+Cjw7/8AG6/4SX/guR/z7ar/AOCjw7/8br/hJf8AguR/z7ar/wCCjw7/APG6/wCEl/4Lkf8APtqv/go8O/8Axuv+El/4Lkf8+2q/+Cjw7/8AG6/4SX/guR/z7ar/AOCjw7/8br/hJf8AguR/z7ar/wCCjw7/APG6/wCEl/4Lkf8APtqv/go8O/8Axuv+El/4Lkf8+2q/+Cjw7/8AG6/4SX/guR/z7ar/AOCjw7/8br/hJf8AguR/z7ar/wCCjw7/APG6/wCEl/4Lkf8APtqv/go8O/8Axuv+El/4Lkf8+2q/+Cjw7/8AG6/4SX/guR/z7ar/AOCjw7/8br/hJf8AguR/z7ar/wCCjw7/APG6/wCEl/4Lkf8APtqv/go8O/8Axuv+El/4Lkf8+2q/+Cjw7/8AG6/4SX/guR/z7ar/AOCjw7/8br/hJf8AguR/z7ar/wCCjw7/APG6/wCEl/4Lkf8APtqv/go8O/8Axuv+El/4Lkf8+2q/+Cjw7/8AG6/4SX/guR/z7ar/AOCjw7/8br/hJf8AguR/z7ar/wCCjw7/APG6/wCEl/4Lkf8APtqv/go8O/8Axuv+El/4Lkf8+2q/+Cjw7/8AG6/4SX/guR/z7ar/AOCjw7/8br/hJf8AguR/z7ar/wCCjw7/APG6/wCEl/4Lkf8APtqv/go8O/8Axuv+El/4Lkf8+2q/+Cjw7/8AG6/4SX/guR/z7ar/AOCjw7/8br/hJf8AguR/z7ar/wCCjw7/APG6/wCEl/4Lkf8APtqv/go8O/8Axuv+El/4Lkf8+2q/+Cjw7/8AG6/4SX/guR/z7ar/AOCjw7/8br/hJf8AguR/z7ar/wCCjw7/APG6/wCEl/4Lkf8APtqv/go8O/8Axuv+El/4Lkf8+2q/+Cjw7/8AG6/4SX/guR/z7ar/AOCjw7/8br/hJf8AguR/z7ar/wCCjw7/APG6/wCEl/4Lkf8APtqv/go8O/8Axuv+El/4Lkf8+2q/+Cjw7/8AG6/4SX/guR/z7ar/AOCjw7/8br/hJf8AguR/z7ar/wCCjw7/APG6/wCEl/4Lkf8APtqv/go8O/8Axuv+El/4Lkf8+2q/+Cjw7/8AG6/4SX/guR/z7ar/AOCjw7/8br/hJf8AguR/z7ar/wCCjw7/APG6/wCEl/4Lkf8APtqv/go8O/8Axuv+El/4Lkf8+2q/+Cjw7/8AG6/4SX/guR/z7ar/AOCjw7/8br/hJf8AguR/z7ar/wCCjw7/APG6/wCEl/4Lkf8APtqv/go8O/8Axuv+El/4Lkf8+2q/+Cjw7/8AG6/4SX/guR/z7ar/AOCjw7/8br/hJf8AguR/z7ar/wCCjw7/APG6/wCEl/4Lkf8APtqv/go8O/8Axuv+El/4Lkf8+2q/+Cjw7/8AG6/4SX/guR/z7ar/AOCjw7/8br/hJf8AguR/z7ar/wCCjw7/APG6/wCEl/4Lkf8APtqv/go8O/8Axuv+El/4Lkf8+2q/+Cjw7/8AG6/4SX/guR/z7ar/AOCjw7/8br/hJf8AguR/z7ar/wCCjw7/APG6/wCEl/4Lkf8APtqv/go8O/8Axuv+El/4Lkf8+2q/+Cjw7/8AG6/4SX/guR/z7ar/AOCjw7/8br/hJf8AguR/z7ar/wCCjw7/APG6/wCEl/4Lkf8APtqv/go8O/8Axuv+El/4Lkf8+2q/+Cjw7/8AG6/4SX/guR/z7ar/AOCjw7/8br/hJf8AguR/z7ar/wCCjw7/APG6/wCEl/4Lkf8APtqv/go8O/8Axuv+El/4Lkf8+2q/+Cjw7/8AG6/4SX/guR/z7ar/AOCjw7/8br/hJf8AguR/z7ar/wCCjw7/APG6/wCEl/4Lkf8APtqv/go8O/8Axuv+El/4Lkf8+2q/+Cjw7/8AG6/4SX/guR/z7ar/AOCjw7/8br/hJf8AguR/z7ar/wCCjw7/APG6/wCEl/4Lkf8APtqv/go8O/8Axuv+El/4Lkf8+2q/+Cjw7/8AG6/4SX/guR/z7ar/AOCjw7/8br/hJf8AguR/z7ar/wCCjw7/APG6/wCEl/4Lkf8APtqv/go8O/8Axuv+El/4Lkf8+2q/+Cjw7/8AG6/4SX/guR/z7ar/AOCjw7/8br/hJf8AguR/z7ar/wCCjw7/APG6/wCEl/4Lkf8APtqv/go8O/8Axuv+El/4Lkf8+2q/+Cjw7/8AG6/4SX/guR/z7ar/AOCjw7/8br/hJf8AguR/z7ar/wCCjw7/APG6/wCEl/4Lkf8APtqv/go8O/8Axuv+El/4Lkf8+2q/+Cjw7/8AG6/4SX/guR/z7ar/AOCjw7/8br/hJf8AguR/z7ar/wCCjw7/APG6/wCEl/4Lkf8APtqv/go8O/8Axuv+El/4Lkf8+2q/+Cjw7/8AG6/4SX/guR/z7ar/AOCjw7/8brxz+zt/wVL+JPxY0P44+NPhxquo+N/DclpJpeqbdIh+ztay+dAfKjKxNtkJb5kOehyOK/4SX/guR/z7ar/4KPDv/wAbr/hJf+C5H/Ptqv8A4KPDv/xuv+El/wCC5H/Ptqv/AIKPDv8A8br/AISX/guR/wA+2q/+Cjw7/wDG6/4SX/guR/z7ar/4KPDv/wAbr/hJf+C5H/Ptqv8A4KPDv/xuv+El/wCC5H/Ptqv/AIKPDv8A8br/AISX/guR/wA+2q/+Cjw7/wDG6/4SX/guR/z7ar/4KPDv/wAbr/hJf+C5H/Ptqv8A4KPDv/xuv+El/wCC5H/Ptqv/AIKPDv8A8br/AISX/guR/wA+2q/+Cjw7/wDG6/4SX/guR/z7ar/4KPDv/wAbr/hJf+C5H/Ptqv8A4KPDv/xuv+El/wCC5H/Ptqv/AIKPDv8A8br/AISX/guR/wA+2q/+Cjw7/wDG6/4SX/guR/z7ar/4KPDv/wAbr/hJf+C5H/Ptqv8A4KPDv/xuv+El/wCC5H/Ptqv/AIKPDv8A8br/AISX/guR/wA+2q/+Cjw7/wDG6/4SX/guR/z7ar/4KPDv/wAbr/hJf+C5H/Ptqv8A4KPDv/xuv+El/wCC5H/Ptqv/AIKPDv8A8br/AISX/guR/wA+2q/+Cjw7/wDG6/4SX/guR/z7ar/4KPDv/wAbr/hJf+C5H/Ptqv8A4KPDv/xuv+El/wCC5H/Ptqv/AIKPDv8A8br/AISX/guR/wA+2q/+Cjw7/wDG6/4SX/guR/z7ar/4KPDv/wAbr/hJf+C5H/Ptqv8A4KPDv/xuv+El/wCC5H/Ptqv/AIKPDv8A8br/AISX/guR/wA+2q/+Cjw7/wDG6/4SX/guR/z7ar/4KPDv/wAbr/hJf+C5H/Ptqv8A4KPDv/xuv+El/wCC5H/Ptqv/AIKPDv8A8br/AISX/guR/wA+2q/+Cjw7/wDG6/4SX/guR/z7ar/4KPDv/wAbr/hJf+C5H/Ptqv8A4KPDv/xuv+El/wCC5H/Ptqv/AIKPDv8A8br/AISX/guR/wA+2q/+Cjw7/wDG6/4SX/guR/z7ar/4KPDv/wAbr/hJf+C5H/Ptqv8A4KPDv/xuv+El/wCC5H/Ptqv/AIKPDv8A8br/AISX/guR/wA+2q/+Cjw7/wDG6/4SX/guR/z7ar/4KPDv/wAbr/hJf+C5H/Ptqv8A4KPDv/xuv+El/wCC5H/Ptqv/AIKPDv8A8br/AISX/guR/wA+2q/+Cjw7/wDG6/4SX/guR/z7ar/4KPDv/wAbr/hJf+C5H/Ptqv8A4KPDv/xuv+El/wCC5H/Ptqv/AIKPDv8A8br/AISX/guR/wA+2q/+Cjw7/wDG6/4SX/guR/z7ar/4KPDv/wAbr/hJf+C5H/Ptqv8A4KPDv/xuv+El/wCC5H/Ptqv/AIKPDv8A8br/AISX/guR/wA+2q/+Cjw7/wDG6/4SX/guR/z7ar/4KPDv/wAbr/hJf+C5H/Ptqv8A4KPDv/xuv+El/wCC5H/Ptqv/AIKPDv8A8br/AISX/guR/wA+2q/+Cjw7/wDG6/4SX/guR/z7ar/4KPDv/wAbr/hJf+C5H/Ptqv8A4KPDv/xuv+El/wCC5H/Ptqv/AIKPDv8A8br/AISX/guR/wA+2q/+Cjw7/wDG6/4SX/guR/z7ar/4KPDv/wAbr/hJf+C5H/Ptqv8A4KPDv/xuv+El/wCC5H/Ptqv/AIKPDv8A8br/AISX/guR/wA+2q/+Cjw7/wDG6/4SX/guR/z7ar/4KPDv/wAbr/hJf+C5H/Ptqv8A4KPDv/xuv+El/wCC5H/Ptqv/AIKPDv8A8br/AISX/guR/wA+2q/+Cjw7/wDG6/4SX/guR/z7ar/4KPDv/wAbr/hJf+C5H/Ptqv8A4KPDv/xuv+El/wCC5H/Ptqv/AIKPDv8A8br/AISX/guR/wA+2q/+Cjw7/wDG6/4SX/guR/z7ar/4KPDv/wAbr/hJf+C5H/Ptqv8A4KPDv/xuv+El/wCC5H/Ptqv/AIKPDv8A8br/AISX/guR/wA+2q/+Cjw7/wDG6/4SX/guR/z7ar/4KPDv/wAbr/hJf+C5H/Ptqv8A4KPDv/xuv+El/wCC5H/Ptqv/AIKPDv8A8br/AISX/guR/wA+2q/+Cjw7/wDG6/4SX/guR/z7ar/4KPDv/wAbr/hJf+C5H/Ptqv8A4KPDv/xuv+El/wCC5H/Ptqv/AIKPDv8A8br/AISX/guR/wA+2q/+Cjw7/wDG6/4SX/guR/z7ar/4KPDv/wAbr/hJf+C5H/Ptqv8A4KPDv/xuv+El/wCC5H/Ptqv/AIKPDv8A8br/AISX/guR/wA+2q/+Cjw7/wDG6/4SX/guR/z7ar/4KPDv/wAbr/hJf+C5H/Ptqv8A4KPDv/xuv+El/wCC5H/Ptqv/AIKPDv8A8br/AISX/guR/wA+2q/+Cjw7/wDG6/4SX/guR/z7ar/4KPDv/wAbr/hJf+C5H/Ptqv8A4KPDv/xuv+El/wCC5H/Ptqv/AIKPDv8A8br/AISX/guR/wA+2q/+Cjw7/wDG6/ZE1v8A4KkX3x10W2/aehv1+HzW11/aBm0/Rol8wQP5PzWqCUfvNnQ49eM1VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV+0N/wVO/Zs+B19deGvD97dfELxHasyS2mhSJ9lgkH8Ml03yZyMHyxIQeCARXi/8A4LY/HnUZpF8EfCzwPols3CC/NzqEqf8AA1khUn6p+FaV/wAFnv2q7K4EmoeF/hzqMJI3RyaXdRkDvtZLkYP1BHtX7CX/AAUP1j9sDxjq3gHWvhLF4fvNF0htRm1Oz1Qz27gSxxiPy2QMjEyEj5mGEPSqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv8AgqV+3rrd94g1X9mD4O629npenH7P4s1a1k2yXc/8dlG4PyxpwJCOWbKHCqwevAnwz+InxR1U6F8N/AuveKNQRQz2+kadLdvGp/iYRg7V4PJwK1D9hH9sPTLR7y5/Z18ZvHGOVt7Azv8AgkZZj+Ar/gjF8Hdf8E6B8UvGfjDw1qOj6rcaraaKttqNq9tNB9niM0qmNwGXP2mEnI/hHvVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV+0h8UT8FPgN47+KcJT7V4d0S4nsg4BVrorsgU5zwZWjB+tX17eane3GpahcyXF1dStLPNI25pHY5Zie5JJJNfs8fBjWf2hPjT4T+D+hT/Z5vEV8I57nbn7LbIpknmx3KRI7BeMkAd6+DnwW+G/wD8CWHw7+F/hu30nSrJBvZUBmu5MYaaeTGZJGxyx9gMAACqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv+Cq91d2/7D3jqO1YhLi60uOfC5+T7fA34fMq/y71X/BHr+zv+Gwo/tuzzv+EX1D7JuznzcxZx7+X5n4Zqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/bX+Gt58Xf2U/iZ4E0yFp7660N7qzhVctNPautzEgHqzwqo9zVfs3fGnU/wBnn43+Efi/pcD3P/CP3wa6tVbBubWRTHPEOwLRO4BPAbB7V8Kfiz8P/jb4H074h/DTxJbazoupRho5Ym+eJsfNFKnWORejI2CDVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV/wAFIv2KtY/Zy+Jl78QvBmivJ8NfFd289lJbxEppFw5y9pJj7i7iTGTgFTtGShqvhv8AGH4qfB7U31j4W/EPX/C91Ljzm0y+eBZwM4EiA7ZAMnhgRV5/wUX/AG1762e0m/aA1tUkGCYbW1hcfRkiDDp2Nf8ABHD42+OPiXe/FnRPiP421rxJqyvpuoW91q1/JdylCJ45AGkJIUFYsAcDd271VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVeJ/C/hzxr4fvvCni7Q7HWdH1OEw3lhfQLNDOh/hZGBBHAPsQDXx4/4IueB/Ed7c67+z/wDECTwrJKSw0PWo2u7RSe0c6nzY1HowlPuK1/8A4JDftl6Pctb6doXhXXEVyonsNdREYf3gJxG2PqAfatE/4JDftl6tIsd/oXhXRgXKl77XUcKMfePkCQ47dM8dK/4J/wD/AAT18a/sh+MNY8f+MfifpWsXetaMdNl0fS7KQwITLHIJfPkKsxXy2UL5ajDk56VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV8UvjN8LPgno9pr/AMV/G+m+GdPvrn7LbXF85VZZdpbYMA87VY/hXgLx/wCDPil4SsPHfw+8RWmu6BqfmfY7+1YmOby5GjfaSAeHR1PuDVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVf8FifjF8W/AXxi8G+GvAvxQ8V+HNKu/C32i5stI1mezinl+1Sje6ROodsIoBOcY4rwv/wS+/au8WeGtK8Wn9rZoZNYsor0Rvfag7J5qBwC+8ZPzDJx1rx74l/4KRf8E5tc0zWvF/xHu/Gng28uvLinvb+bWNMuW6mBzPia1cqCQF8vOG2s2Gr9lv8AaJ8L/tSfBrSPiv4Zt2s3uC1rqmnO+9tPvYwPNhLYG4fMrK2BlHUkAkgVVVVVVVVVVVVVVVVVVVVV/wAFtP8AkgPgX/scB/6Rz1/wS4/5MS+GP/cW/wDTteVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX/BbT/kv3gX/sTx/6Vz18J/8Aklfg3/sX7H/0nSv+Cj8Hh+b9in4oHxGtuYI9Oha3Mw+7c/aYhBt9G8woB9fQmv8AgiF/a/8AwrD4m+du/sv+3rT7NnOPP8hvNx2zt8jP4e1fGj/gpTrnwo/bBP7Ltt8JbHUrUa1pGnf2w+sPE+L6G2kL+UIiMp9oIxu529Rniv26P2uNR/Y8+HmgeONN8D23iiTWta/s5rae/a1EQ8mSTeGCPk/u8YwOvWv2Ufjjd/tJ/AHwt8ar7w7DoU/iL7Zu0+K5NwsPkXk1uMOVUnIhDdBjdjtmvjP/AMFANe+E37Y/hf8AZZj+F+n6jZ+JNU0azXWjqzxyRJfSxxsxh8oglCzEDdyAORnj9pb/AILF6H8P/Gt/4H+A3gjT/FqaVK0Fzr+pXTpaTSrwywRx4aRAePMLqGIO0FcMfD3i61vPh3pnj3xHdWemW82ixalfzSSeXBaoYRLIxZj8qKMkkngDk18Sv+Ct9zrfjS4+H/7JPwN1f4l3kBcJqLwXDLcbeN0NpChmePJHzMYz228g1on/AAVu+Lfw38T2mh/tWfsuar4UtL0grNa2tzp9zFH3dba8H74dOjp+PSvAvjnwl8TPCGlePfAmvWus6DrVuJ7G9t2ykqHg9eVYEFWUgFWBBAIIqvjL45vPhf8ACPxr8S9P0uHUp/CXh+91ZbOaYwrcfZoHlKFwrFchCM4PJr4Tf8FV/BniT4EeMPjb8VvBsfhkeHtVh0vS9I0+/N5Pq9zJCZFRNyIFwFJLHhVBJOcA/sOf8FFvGX7X/wAdNd+Heo/DnRfDWh6d4auNWtmgu5bm5Mkd1bRKrudqFds5JwgOVHOOK/ab/as+Ev7KPg2PxV8S9Ume6vy8ek6NZKJLvUZFA3CNSQFRcgs7EKuQMlmVSv8AwVG/a/8AHds/iz4Q/sW6hqHhNSxW8Gm6lqilF+8TcQRpGMYbPBA/Dn9kX/gpv8P/ANozxZH8KvHnhaXwB46lLR2trcXXm2t/KvDRRuyq0c3BxE684wGY8V+3T+1j4r/Y/wDA3h3x/onw4s/Fem6rqjabemfUmtDaymIyQkbY33BhHNnOMFV65r9mL442P7SPwM8LfGOy02PTW16CX7TYpMZfss8UrxSR7iASA0ZIJAyCD3r9r39o+x/ZV+B+p/FmfR4tYvILu3s9O02S4MAu55ZACu8Biu2MSv0P3Md6/YX/AGyrb9snwN4h8QXXhW38Naz4b1RbW502G9N0PIkiDQzbiikbmEy4x/yyznnj4geMtL+HXgTxF4/1tv8AiX+GtKudSuhu25jgiaRgDzyQpFfsI/t0eL/2zNc8V2998J7LwxpHha0t2kvYdVe6MtxMzBIwDGgA2xyEnJxgetftbf8ABRix/Zw+I6/Bfwl8GfEPjbxpLaxXEcSyfZ7ZklUlDGUWSSYjDAqEUZUjd1xrX/BUr9sbwDEnij4mfsaT6P4YkwUnvdM1LTgwJGMXMyGPkH+53H4/sk/ti/DL9r7whd654LiudK1rR2RNY0K9ZWmsy4Ox1ZeJIm2sFcAHKkEKeK/aM+K198Dfgh4u+LemaHb6zceF7D7WthPcmBJwHUMC4Vip2kkcHkAcZyPBH/BVfwRdfsyap8f/AIleDF0fVE8TXOg6J4a0/UDcTarLFbW0xcSMi+Wi/aRvYghQBjczKp/YK/b58Z/ti+PvGPh7XvAWi+HNN0DTYru0W0uJZ5mLylNru2FIAx0Refyr9rj9uH4Sfsh6TbQ+Khc654q1OEy6b4dsHCzSx5I82VzxDFuUruIJJB2q21saV/wVo/ap8TlvFHhX9kyHUvCkbNvltbXULnaq9QbpF8sEcZOz8K/b0/bb+Gn7YP7MfhRvDlhdeH/FOh+Lo21fw/eyiR4Fa0uAssUgAE0WRtLbVKnAZV3KW/4Jcf8AJiXwx/7i3/p2vKqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr/AILaf8l+8C/9ieP/AErnrwd/wWl8C+GPAWi+HpvgTrs+o6RpsFphdaiEUjRRqm7f5e4A7c/dOOnvXxS+LX7Zn/BUbVNN8EfD34U3OgeAbe6E3lRNJ9gSTJAmvL51VZCg3YRFHQkIzV+yf+zloH7LHwU0f4UaLeLf3ULvd6vqIj8v7deyY8yTb2UBURQeQka5JOTX7YX/AClqb/scPCn/AKS6dVf8FtP+SA+Bf+xwH/pHPX/BLj/kxL4Y/wDcW/8ATteV/wAFY7vU7D9t+W+0Se4h1G20fS5LOS3JEqTKCUZMc7gwBGOc4r9kn9lH4efsv/C/RfD2i+G9PPiqSzjfX9bMKvc3l0VzIPNxkRKxZUQHAUdyWJ/4LLfGrVPAfwZ8L/Bzw5dvaP49vJn1EwttJsbQRkwnHRXkli+oiYdCa/YR1/8AY3/Zv/Z+8N6Snx5+E1p4t16wg1DxPczeLLBLl7qRA5gcmXcFhDeWE4AKscAsxP7XvxF/Y6+P37PfjLwJqnx/+E99qH9lT3WiOviywkmt7+KMvA0eJS2S6qpC8srMv8Vf8ESfihqmpeGPiJ8H9RvGls9FubXVtLjZs+X54kjuAMnIXdFCQBxl2PBPNftZf8mr/GX/ALJ/rv8A6QTV/wAExP2dNF/aR+NUtl8Q7Z9T8D+BLVtXudKlYmC6vJSkUUci9CrBCzD+IQhTkEitK0TRtBsbfS9C0iy06ztI/Lt7e0t1hjhTj5VVQAo4HA9BXijxl4A/a4/4KVX9z8efHOi6L8NPC2p3NpENb1WOytXsbAsscCvI6qPtEw3sAQSJXx0FWH7UX7JGlWNvpml/tF/CKzs7SJYre3t/F2nRxwoowqqqygKoAAAHAr/gq1qXwQuvip4E+On7PnxO8H6r4kvhMutSeGNZtrqSG5tXie1un8h2KyHe672xnyU64OP2i9Cb9rX/AIJ13evR2aS6trfgqy8T2ccS8peRQx3TRoOcMdskWP8AbIz3r/gif8TP7Z+Enjn4TXU+6fwzrUWpWoZuRBdx7Sqj0WS2dj7y+4r/AIKv+IdQ+L/xz+C/7Inhy8YyapqEN5qCRnO2W7mFtAzdgY4xcNz0WTJ4xX7E9xF+yz/wUr+JX7OD7rLQfFMt3aaZasSAuz/TbAknr/orSIM9TKPUV/wVo+Kf/CvP2RdU8OWd0I9Q8d6nb6PEFPziEEzzsB/dKQ+WT/01HqCP+CUnwj/4Vj+yPouu3tmYdU8eXk2t3G9fm8lsRW4/3TDEkg/66n1rVYfBvh65u/H2txaLplxDZrb3WtXaxQulsjMyxvO2CIw8jkKTgF2PUmvHn7ff7DGm2l/4b8X/ABy8L6tZXULW95bWlpPq8FxG42sh+zxSI6kEgjkYPNf8E1vFejeEf+Ch0nhf4b6tJd+EPE765pthKqyItzYRRzXNtIVkCuMrbRkblDDOCBk1+35/yZp8Wf8AsX5P/Q0r/glJ+zV4c/aC+IuseIvibpy634O+HKpc2+jXY32txqN38qs8Z+V1Edrll6ErEGyvBjttI0LTgLe1tbGysIW2rFEI0hjAyQABgDjOB6V+zl4Xk/4KG/t96l4q+JQlvNAee51/UrKVjj+z4HSO2s+DwmXt4zg527ucnNafp2n6RYW2laTY29lZWcSw29tbxCOOGNRhURVwFUAAAAYAr/gs3+z54J8Kz+E/j34T0i10vUfEN/LpmvJbxiNb2XyzJDOVHHmYSVWbqw2Z6c/8EuP+TEvhj/3Fv/TteVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVftNftjeJfgP8AEePwRpHg3TNUgfTIrvz7m4dGDOzqVwvGPkH51/w8u8c/9Ez0L/wMlr/h5d45/wCiZ6F/4GS1/wAPLvHP/RM9C/8AAyWv+Hl3jn/omehf+Bktf8PLvHP/AETPQv8AwMlrw7/wUb8a634g0vRpfhxokaX95Fbs63cpKh3Ckj86qqr/AILaf8l+8C/9ieP/AErnr4WfD3wDc/Dvwbq1x4H8Py339gWDfaX0yFpc/Z053Fc5pESNFjjVVVRhVAwAB6Cq/bYmj0P/AIKry6zqjCCzj8T+F7ppWPAiS1sAzfQFG/Kq/wCC213bJ8C/ANi06C4l8WNJHGT8zKlpKGYD0BdAf94V/wAEuP8AkxL4Y/8AcW/9O15X/BU3/k/SD/sH6P8Azqv+C4eg6hH4q+FPiciRrG40/UbMEL8scsckLnJ7FlkHHGdh64OPg9/wSp/Y8+KXwo8H/Eaz1nx1MniTQ7S/LQ6xCFDyxKzrjyDghiykHkEEHpX/AA5t/ZF/6CPxA/8ABzD/APGK/Zl/Yf8Agr+ydrOt698LpvEU15r9rHbXTarfLOFjRiwCBUTBJPJOeg6c5r9rL/k1f4y/9k/13/0gmr/gh1/yNXxb/wCwfpX/AKMuar9mj9nr4dfFr9tnxB8AfjjfaxaRtfaxawmyuFtp5L+2lZtpLq4GUjmOO5A59f8Ahzb+yL/0EfiB/wCDmH/4xX/Dm39kX/oI/ED/AMHMP/xivh38P/Dvww+H3h/4Z+GY520bw3pkOnWgupPNkaKJAgLtgBmIGTwBknAA4r9ia1l/ZZ/4Kc+MPgJKWh0rW31HSLRJMgeTgXtk5/2jDGqj/rqfXNfsk2jftVf8FN/iX+0PO/2vw94Cln/suQ5ZGO02NlgHhd0Mc03sy5681/wVY8P6t8D/ANp/4TftY+FbYhmeBbkxjAe70+ZZFDn/AKawyCPHdYW96/4KieKrj9pH9o74KfAD4f34u7bUdPtrm2kj+752qyoqO49FgiikyeiyNyMmvDfh7SfCXhzSvCmg2q22maLZQ2VlAOkUMSBEUfRVArXvFPxK/wCCpH7ZT/C5vG11o3w5026uprK0gbdFZ6bbHabgRZw9xNlPmbO0ygD5FxXgn/gmR+xb4K06KzPwhi1+4VNsl7rd/PdSyn1K7xGp/wBxFr9kbQtD8L/8Fc38M+GLGCy0fSPGfi2z061g/wBXBbxWuopGi/7IRVA9hX7fn/JmnxZ/7F+T/wBDSv8Agh1/yKnxb/7CGlf+i7mvGuj3HiLwbr3h+zk2T6nplzaxPj7rSRMoPUdyO4r/AII0eI7Dw3+1J4h8KayTa32ueFbm3tI5RtZp4Z4ZHi2nnd5aSt7eWar/AILcajYxfBL4faRJdRreXPiqSeGEn5njjtZFdgPQGWMH/eFf8EuP+TEvhj/3Fv8A07XlVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX/BRH/kvtv/ANi9a/8AoyaqqqrwF/yPPhz/ALC1r/6NWqqq/wCC2n/JfvAv/Ynj/wBK56+E/wDySvwb/wBi/Y/+k6VVf8Fdv2Q/G/jPV9M/aT+GOgXes/YNOFh4msrKNpJ4Y4mZortUXJZQrMkhH3QsbYxvZfhF/wAFovFvhTwFp/hT4lfB0+LfEWmWqW0erWutfZGv2RdqtNGYXxIcDcynDEkhV6V+17oX7Uf7Q/wqvv2xP2gNOPg7w9pt5b6b4O8JvA8bLBO/7yYI/wAyhtiEyON0pAICxqgr/glrNDL+wp8NY4pUdom1ZZFVslG/tW7OD6HBBx6Eetf8FTf+T9IP+wfo/wDOq/bQ/Ze0n9rL4J3/AMPJLm3sNes5hf8Ah/UZlJW2vEBAD4yfLdWZGxnAYNglQK+Cv7Wv7Uf/AATcll+CPxs+Ed7qvhWC6kewsr6ZrYwFmzI1ldqrxyxMSzFAGG4kgoS2dS/4LfeAxaMNA+APiG8vmOIobnWYoEY9sssbnrjotfsjfGzxr+0H8FrD4k+P/h9ceDdWu765i/s6W3miVoVk/dSR+aAzqYyoL9CytjA4FftZf8mr/GX/ALJ/rv8A6QTV/wAEOv8Akavi3/2D9K/9GXNV/wAFEv2HvivYfFmP9rr9l/T7251dJ4r7WNO0lM3lrew423tvGOZQwVd6KC24FsMHbb8PP+C1WqaFpC6B8b/gfcz+ILBRFcXmk3otvOkXgl7aVMxNwScORk8BRXg7/grrrnxd+JvhjwD8Kf2bdYurbUtYtINTu3u3vZ7e0eVVnkW3gi4KoXIZn2jbkjGar/grh4Z1/wCDX7SHw6/aX8DsLO91Wy8pp1UkfbbJhgvjH34Zo06gkRNjGM1/wSR+Dx+G/wCyra+Mb+1MeqfELUZdUcuMMtqn7m3X/dKo8o/67fgP+CnnwgX4tfsheK5rW283VPBZTxDZYXkC3DC498fZnnOPUD0r/glL4T8Q/HL9qo/F3xhJ9ttvhh4WtbW3kdcrHItqthZR4OQMW8UzZ/vR56nIrVNA+Lv/AAS1/a8fx6PBU2r+EXuLqHTpyGjtdY0ucnEInCsIp0AQlSCVeMHDIQW1f/gq341+ONofhv8Asjfs++J9Q8d6vF5UN3fmOWPTC3Bm2R7lYL2aVo0U8tkAqf2PfAfiL4Gf8FQfCngD4h6ol9rulajfwahdROXWa5uNJuDuDNgsC8w+Y4J645xX7fn/ACZp8Wf+xfk/9DSv+CHX/IqfFv8A7CGlf+i7mq/bp/Y++M37OPx7l/az/Zv03UZ9Dm1Q61JJpUBml0G+JLzeZEoObVyXbOCgV2jcAbd/hX/gt/qMXhu3tfFv7Pkeo+I0TY82m68ba2uHxwwjeGR48n+Hc/15wP2tvDH7TH7QXwp1P9s39obTW8IaZY3NnpXg7wv5DRYtpnYyTbHO9QcKd7jdKTkbURAf+CWs0Mv7Cnw1jilR2ibVlkVWyUb+1bs4PocEHHoR61VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVf8FEf+S+2/wD2L1r/AOjJqqqqvAX/ACPPhz/sLWv/AKNWqqqqqqo/B/hKLWG8RQ+F9ITVXJLXy2MYnJPUmTG79aqqqtQ07T9WtJNP1Sxt7y2lGJILiISI491OQa0jwH4G8P3X23QPBmhabcAYE1pp0UL/AJqoNVVVVa34K8G+JZkufEXhLRdVmjBCSXthFOyjjoXUkdBWl6PpGh2i6foml2en2yn5YLWBYkHAHCqAOgA/Cqqqqr/T7DVbOXT9Tsre8tZl2ywTxCRHHoVOQR9a0fQNC8O2v2Hw/othpltnPk2dskCZ9dqgDuaqqqo/BXg2HVz4gh8JaKmqltxvlsIhOW9fM27s8DvVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVf8FEf+S+2//YvWv/oyaqqqrwF/yPPhz/sLWv8A6NWqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/wCCiP8AyX23/wCxetf/AEZNVVVV4C/5Hnw5/wBha1/9GrVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVf8FEf+S+2/wD2L1r/AOjJqqqqvAX/ACPPhz/sLWv/AKNWqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/4KI/8AJfbf/sXrX/0ZNVVVV4C/5Hnw5/2FrX/0atVVV/wVM/aW+NvwL/aA8D6Z8Ofihrvh7Q73w7Dc6hYWMiiOVlvJgz4IzuKAKeeijpXib45/t4ft4+INaH7H6v4C+FOk3b2lvr9xcLYzalInVjPhpQT12QABAwDsTivGPxa/4KSf8E//ABvpVz8VfHms+I9J1OQmAa3qz65p2qKhG+NZZGMsLYI4BjfBzgiv2e/jd4Y/aL+D/hz4veE0eC01y3JmtJHDPZ3CMUmhYjurqwBwNy7WxhhX7bP7Z/hH9j/wDBqM9nHrXjDXd8eg6KZNgkK43zzEcrCmRnHLMQox8zL4D0H/AIKs/tu6OPibpPxel8AeGL9i+luurTaBbzoCf9QlmjzugIwHlyG7M2Djxp8T/wDgpf8A8E9da0nV/it40fx94OvLkQ+ff3z6vZXb4yYvtMqrcwSFQxXdtzgnDhSK/Zs/aH8DftQfCnTvin4EeWKGd2t7+wnIMunXaAGSByOCQGVgw4ZWU4GcCq/as/4KffEHU/iTP+z9+xhoS6zrYu2sG8QRWn2+W5uRkOljBgoyqQcyuHU4YhdoDnTP2Sv+CunjSBfF2t/tLT+HL9k8waXdeNruBgSM7TFZxPbg9sZwD+dfsL+Ff2zfCWm+MdK/a78WHWfs11bReG2aa2uC8YVzPN50QEjKxaJQsuGBRuMMK+Lf7U37Quh/8FOrH4P6R8WNctfBknxB0DT30ZJFEBtp2tBLFjGdrCR88/xGq/4K0/G74sfBD4deAtV+E3jvU/DF3qWtXEN3LYuFaaNYQwU5B4B5r9hTxv4s+JH7Jnw78beOdeudZ13VLKd7y+uSDJMy3UyAsRjoqqPwr9v39v8Au/2b72w+D3we0i31/wCKGuxIyo8TTx6XHIdsRMS8yzyH7kXbhmBBVX079kn/AIKu/FHSYvHXiz9qy68JatexmdNFPie8smiJGQssVjH5EZOB8q7sZ5AORXwR/bj/AGkv2W/jxbfs4/tyXDajpl3JFDBrtwY2ms1lbbDdCdABcWrEEMz/ADryc5Qxmq/4K3/tA/Gz4G+KfhvF8JfiZrXhmHWdPvzew2UiqkzRSQ7GIIPOJGH5V8Vv2pv2zv2wfHt38Lv2JbbWLfwr4WWO31HxFplzFanULhQBJM17IVWKMtu8uONtzqN/zZCr+zVZfEH4W/syeHIf2jNfuR4m8O6bdT+I9S1bVftzoiTSyGWW5LvvUQ7TkscAY4xgeIf2w/2xf27Pitqfwv8A2LFbwd4M0ssLjXpMQSmEkqs9zcsrNbh8ZSKFfM+9kvg7fHH7OH/BVT9nvQrv4q+G/wBpzU/HX9jxtdXumReILvUXEafMzLa3qGKVQNxKj5iBwpr/AIJ+ftv2n7Xfgm+0vxRZW2meP/CyR/2tbW/yw3sLkhLqFSSQCQVdMna23nDqB/wUg/bn1j9lHw7o/g34bW1nP478VQyTQ3N3H5sWl2qHaZjGeHkZsqgPy/IxYHAVvD37K/8AwVq+JuhWfxNv/wBpa+8O3eowLdRaLeeMr6xmVWG5Q1vaxG3jJBHyEgjowXGB+xDpf7WWkfDjWbD9rrWRfeIbXWWt9K3fZnc2SRJiQy2/Egd2fBf5xt564H/BVH9pb45/Aj4/eC9P+GfxO1zQtGvPDUN1eaZZyqkc0i3c4ZjlSdzIFUn0UV4p/aK/ba/bn8Ya34f/AGK4R4L+GOi3T2beLrthaNqDrjLeeys6Z4ZY4F3qrKXI3AD4qaT/AMFUf2J0X4m+IvjFrPi7w7BODeX0etS67YQliAFmhu1DxIxIUMEUAnAZSRn9hn9szQf2wfhzcalPY2+j+M/DzpDr+lROSg3g+XcQ558qTa3ByVZWUkjDN+1J+0x4E/ZU+Fd38SPGha7nZ/s2kaVC4WbUrsqSsak/dUAFmcghVBOCcKfBXjj/AIKef8FBJ9Q8U/DbxuPh54KiuHhgubO+fRrONx1jSaFXup2AxuYblB4+UkCviNb/APBU79hKwh+I/iP4uXHjrwrBMgv7iXVZtfs49zAKk63aLPErEhd6bRkgBwSM/sY/tf8AhD9r/wCG0niXS7MaT4l0Ro7fxDoxk3/ZZWBKSRt1aGTaxUnkFWU/dyf+Ctfxq+LPwS8A+ANb+E3xA1bwxd6hrF1b3bWEir56CFWAbIPQjj6mtH/a9/bK/a08NeFPg3+yDaXMN3oPhXTV8c+ONQEUTyai1sgnxLICsQMokGUUyuQzKFVST8W7f/gqR+xW9t8UPGfxh8Sazoj3SpLenxBJrenpIxOI5YLjPlhskA7FGcAMDtr9iD9rHTv2uvg6njOawt9M8TaPc/YfEGnQMfLinChlliyS3lSKcjPIIdcnbuP/AAUO/bRuv2Rfh3pVv4PsLS+8ceL5JotJW7UvDZRRBfOuXUEFiDJGqIcAsxJyEKnwV+zv/wAFYPj94X074wTftKXvhdNcgW+stNu/Fl7pjyQuN8bfZrKIworKQQrbSAQCBzX7C2gftk+G9A8W6P8AteeIf7Umsb+C38OyPJbzPJCIy0svnRDdIjF4wvmfOCj57Aftl/tb+Ev2QvhePF+rWi6t4g1eRrbw/o3m+WbyZQC7ueSsUYKl2A6sq8Fwa8CeBf8Agqb+2poafFif44v8MPDeqEy6Pa2+oXGjCaEnKtDFaKZDF/ded9zDBBZSGOufG/8A4KA/8E5vHWhQ/H3xS/xM8AavNsE9xfNfpdKv31iupVE8M6qdwWT5G9GwSPh14/8AC/xV8CaD8R/Bd/8AbNE8RWMd5ZSkbW2OM7WX+F1OVZeqsCD0qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr/gtp/yX7wL/ANieP/Suev2dfCGheA/gL8PvCXhqyS10/T/DlkI0UY3M0Ku7n1Z3ZmY9yxPev+Cr/hrSdd/Yj8Y6rqFtHLceHr/TL2xdlyYpWvYbcsD2JjuJB9DX/BFTV728/Zt8W6RPIHg0/wAZzNb5zlRJaWxZfplc/VjX7Zrz/tI/8FObX4VavfSnRo9f0jwxEocgwWx8o3O3PQ+ZLcMMdyOvWtN03T9H0210fSbOG0srGBILa3hQIkMaKFVFUcBQAAB2Ar9pj4X6L8ZfgH46+HOuWkU8eq6JcfZjIP8AUXKIXglHoUlRG/D0r/giP481G1+KHxD+F5unaw1PQItYWEklUltp0hLKOxK3QB6Z2r1wMV+3J8SNT+Ev7JfxM8c6LeNa6hbaP9ktLhAd0Mt1KlqjrjoytOCD2IBr/giZ8I9Burfx78b9RsobjVbS5i0PS5XGWtUKebcFfQuGgXPXCsOjGqr44f8AKYPTf+yoeGf/AEOxqv8Agt7/AMkq+Gn/AGMF1/6Tiv8Agm9/yZH8K/8AsH3P/pZPXhP9p7wR4c/b78RftNfF7Q9V8TWNpr2o3OnWlisbusg3Q2ZxKyjbFEF285BjQ9q/4fafAH/olHxA/wC+bT/49X/BQ39sn4Ufth3HgnVvAngjxBoeq+GkvILy41RYB9pglMTRoDG7E7GSQjOAPMbrnj9lrxLf+Mv2avhZ4n1V3e+1Hwfpkt07nJkl+zRh3/4EwJ/Gq/4Ljf8AI1fCP/sH6r/6Mtq/YC+G2hfDH9kL4ZadoltCj67oVtrt9KiYae4vY1nZnPchXSME/wAMajoBX/BWn4hX/gT9jrVtO026a3l8YazZ6K7qSG8tt88igjsyWzKfVWI71+wx/wAFDfgL+yV8Ex4A1j4beLdR8Rahqc9/q+oWCWxjmZiEiVS8qttWJEGCAAxcjqSf+H2nwB/6JR8QP++bT/49X/BODx7pUn/BRsXfgHTrnS/DfjWbXorSxlCq9vYtHNdwxSBDtyv2eEEDjIGO1f8ABY79mTx/40vfD37RPgnR7nWdP0DRjpWv29qhklsoEmkmjudgyTHmaUOw+5hSflyV/Zr/AOCwfwk8WaPpfhr9oW0u/CHiOOJIbjWYLdrnTbpwAPMITMkBY8lSrIvJ3gdNC17QvFOjWfiHwzrNjq2l6hEJbS9srhZ4Z0PRkdSVYH1Br/gtp/yX7wL/ANieP/Suev2YPh/4e+F/7PXw+8F+GbZIrSz8P2juwQKZ5pIlkllYD+J5GZj7mvE/hvRPGXhvVPCXiXT4b/SdZs5bO+tZl3LNDIhR0I9CpIr/AIJmXuofB/8A4KC3fwmh1B5oL9da8P3JHCzfZBJMrkdM5tOD23EdzX/BZTxrq/i79pXwn8LLS6ZrLw/oELQ27MQou7yZi7enMcduM8/d69h8LPh14f8AhJ8N/Dfwy8LWyQ6Z4a02GygCrtL7FAaRvVnbc7HuzEnrXivwvoPjfwxqvg7xRp0V/pGt2UtlfWsoys0MiFHU/VSa/wCCYuq6r8Hv2+7/AOECag8ltqiaxoN2uflmazEkyuQONwNq2DxgOwzyQf8Agt7/AMkq+Gn/AGMF1/6Tiv8Agl34R0Lwt+xV4Eu9HskiuPEDXmo6jKB81xcNcyRhm+kcUSD2QV+1b4a0nxf+zL8VNA1m2Se2n8Iak4DrnZLHbvJFIB/eSREce6iv+CH2rXsXjv4qaCkg+yXWkafcyIc/6yKWVVI/CZ/0r/gsB+zJ4/8Ait4W8L/GL4d6Pc60/gqC6t9a0+1QyTi1kKOs8cY5YRsr7wMkK4b7qMR+yV/wV0+GEPgzQPhx+0Npl94d1LRrKGxTxBY27XVncxxKEV5Y0zLE5UDO1XUkE/ICFHhPxd4W8eeHbHxb4K8Q6frmi6lH5lpf2Fws8My5wSrKSOCCCOoIIOCK/wCCqfxJ03xF+3DYeFvGYvLvwr4DtNMtb2xtGxI0UwS6ufLyQPMeOZVzkD5EyRirX/gtV+zxY2sNlZfCDx3b29vGscMMUVmiRoowFVRNgAAAADpX7a3/AAUo+Bn7U37P+r/CrRvhr4usdbkvbS80q91FLbybaaKZd7HZIzDMLTIMDq/pmv8Agjn4lv8AXv2QpdKvHcxeHPFt/YWoY8CNo4Lk49t9y/4k1VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV/wW0/5L94F/7E8f+lc9fCf/AJJX4N/7F+x/9J0r/gqP/wAmJfE3/uE/+nazr/giX/yQHx1/2OB/9I4K/brstX/Zo/4KQ2vxjfTJX0y51jSvFNiMcXUcZiFzGD0yZYZlx1AZTxkGvCnirw7458M6X4x8Javb6po2s2qXVjeW77kmicZVgfoenUHg1+2B8bNA+AP7O/jLx7rF9FFd/wBmzWWjwM3zXV/MjJBGo6n5jubHREY9BX/BEf4XanJ4l+IfxoubRo9Pt7CLQLKdl4mlkkWedVP+wIrcn/rov4V+298NNT+L37J/xL8B6JatdaldaMbqzt1GWnmtZEuUjUf3maAKPciv+CLPxz8O6Fq3jH4Ba/qMFnfa/NFq2hLKwX7VKkZS4iBPV9ixOqjkhJD2qq/bLvU+EH/BUs+PfEIZdM07xX4e1/zWjID28aWkjkAdQDFIvHUoelWt1a31rDe2VzFcW9xGskMsTh0kRhkMpHBBBBBFf8FuPiH4XubH4c/Cyz1KG41+0urrVLy2jkBa1gZFjiLjqC537faMnuK/4Jvf8mR/Cv8A7B9z/wClk9fABPCX7MX/AAUp1zwH8Y9G05dButZ1DRt+p2yywwx3L+ZZXBEgICv+4+Y/dWQknANf8Km+Ff8A0TTwp/4Jrf8A+Ir/AIVN8K/+iaeFP/BNb/8AxFWlnaWFpDYWFtFbW1tGsUMMKBEiRRhVVRwAAAABwKr/AILjf8jV8I/+wfqv/oy2r9k7/k1f4Nf9k/0L/wBIIa/4Ky/DnU/iB+x5q9/pNm11N4O1e01x40BLCJBJDK4A7LHcO59FVj2r/gkpr/wd+J/7OA+Hmq+E/DV74r8C6hcR3iXmmwy3E1rcTPPDPuZSWXdJJH148oA4BXP/AAqb4V/9E08Kf+Ca3/8AiK0n4f8AgLQL5NT0LwToGnXkYIS4tNNihkUEYOGVQRkcVX7W3/BPP9nj41eCvEXiXSfB+m+D/GtvZTXVrrekwi1WSZFL/wCkxLiOVWIwzFd+OQwr/giX8T/Fc+t+Pvg7d3c9z4dgsItZs4nJKWdx5gikCenmKyEj1iyOpz/wW0/5L94F/wCxPH/pXPX7LfxG8M/Ff9nrwB408KXsc9pc6Dawyorhmt7iKNY5oXx0ZJEZT9M9CK+IPjrwz8MPBGt/EHxjqUdjovh+yku7ydzjaiDOB6sxwqr1LEAcmv8AglZ4a1z4xftt6/8AG69sdsGhW2paveTqnyR3d+zxJEOOGZZrhh04ib6H/gs38PNa8LftA+Evi9YW8iaf4h0OO2W6AyFvrSViwPYfupICAeuG9DXwH+MHhr49/CTwz8VfC15by22u2Ecs8UT7vslztAmgbuGjk3KQfTPQg18UPiP4U+EPw+174l+NtRSz0bw9ZSXVzIxwWCj5UQd3dsKq9SzAd6/4JUeE9f8AjF+2jr/x1v7HZbaBb6hqt5Oq5RLy/LxJEDjqyS3DDpxGfof+C3v/ACSr4af9jBdf+k4r/gm9/wAmR/Cv/sH3P/pZPX7Qv/JAfiZ/2J+rf+kctf8ABEP/AJKn8S/+xftf/Sg1X7SX/BP39nX9o3SdTur7wZY+G/F91G7W3iTSIBbzrOeQ86JhLgE43bwWIzhlPI/4Ix/EXxfonxu8ZfBJ76S68O3uizam9ujl4oLy3nhi85PQOkhUnjdiPPQV/wAFM9DuPgx+354b+N2qaCL/AEbV30fXFjeMPHeNYtFFNAQ3yk7LePKnjEq5614R8JfAnx54X0rxp4R8E+DdT0XW7SO7sLuHR4Cs0TqGVh8mRweQeQcg4Ir/AIVN8K/+iaeFP/BNb/8AxFaNoGheHLM6f4e0Ww0u1LlzBZWyQJuOMttQAZOBz7VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV/wU6/Yu/aC/aY+LXhXxX8IfCllqmm6X4cFlcyT6pBalJvtEr4CyOCRtdTkcc18P8ASb7QPAXhvQtTiEd5p2kWttcIGDBZI4VVgCODyDyK/br+E/jf45fsqeOPhb8ONNhv/EWtf2f9it5bhIFfytQtpny7kKMRxOeTzjHev+CY37OHxZ/Zn+Eninwn8XtCttL1LVPEZvbWKC+iug0P2eJNxaNmAO5GGDzX7Xf7Inw+/a8+HyeFfFMr6Xrels8uha7BEHlsJWADAqSPMifCh0JGcAgggEeEP2eP+Cr/AOx+l34N+B1/b+IvCbXDPCljd2N3a7m53pBe7ZYWPVgihS2cluCT+wd+31+2L4x0zxF+1/4+j8OaJYkmO3luLeeWCNiCy21paHyI3YAAu7K3C7g+3FfCH4ReA/gX8PdJ+GPw30ZdO0TSItsaE7pJnPLyyv1eR2yWb1PGAABVftdf8EndR8W+N7v4wfsteIrHw/q93dG9utAu5ntY0uclmms50BMTF8Hy2AUEkh1GFGiH/guB4YtR4Ps7O7v/ACkKwXt9NoN24AHX7RK53nv+8JPr6V+w58Pv2wvBlh4x1b9rjxrb67f6/c2sul241L7TJYbFkWVdsaiGNG3RkLEcZViRzmv+Ch37Ar/tZafpvjr4falY6Z8QfD9sbWMXmUg1S13FhDI4BMbozOyNgj52Vhghl+Gfwu/4LM/DrR4fhJ4MuNV0rQbOPyIJL7UtHu4rWHAAEU8zSSIgBACRnKjgKMcfFL/glR8Vb34GXuv3GtL8Svjz4k8Q215qupXOqeVDa2qxyiSOOWdlMxLNGWZwM7UCqoQlv2Kvhp4z+D37L/gT4afELSo9O8Q6Fa3EN7bJcJOqFrqZ1IeMlSCjKeDxnB5Br9ur/gnt4X/a4ht/GXhrWLfwz8QtMtvs8N/NEWttRhGSkNyF+YbSTtlUEqCQVYbQvhbwR/wWY/Z/06LwN4QNz4h0KzTybFnvdM1WKNEwFEbXLedGgGAqNtAHQccfDrwH/wAFfviV8QvC2u/FTxrZ+EvDekazZ313ZTXtlapdQRSI7xNHp6vJIHClSkpAOSDgVVf8FRv2RPjr+09r3w9v/g74Zs9Vh8P2d/FfmfUoLXy2leEpgSMu7IRunTFfs/8AhPW/APwG+G3gXxNbJb6x4c8I6VpuoQpIsgjuILSKORQykhgGRhkcHqK1HTrDV9PudJ1WygvLK9heC5tp4w8c0bgqyMp4ZSCQQeCDXxe/4JbfHf4N/Eh/ix+w947lt1V3ktdKbU/sN/YhjzBHM58ueHHGJWUkYDB+WNp4m/4Lgyouhx+HiJwpjF3NaaApBA+8XJ8snjrjB981+w98OP2xfBUXjDXf2tviBbeILvxE9o+l2S6h9pk00xiUSjaiLDGr74/liJGUJOO/7Y3wr/4KG3nxrtvi3+yn47tU0a10eGwj8PxanHEXZGd3kmt7sfZpCzSEBy24AAcYyfGXh7/gsn8ffD958LvFWg2PhrQ9RiNrqtzHcabZLcRNwyvJFI8pRgCGEIwQSCCrYP7CP7FGkfsd+BNQt9Q1i31zxp4leKTW9RgjKwosYby7eANhvLUu5LEAuWyQAFVf+C2n/JfvAv8A2J4/9K560/8AY7/bV/ZttbL4i/sK/EA33hzxbp1rf3vha7ubffbzSQqzjy7v9zKozhZAyyhcKd2Nx8W/s1/8FU/2x7mz8MfHnUYvD3ha2uEd11C7s7W1Rl6v9mstzzOAxKlxjJIDLzj9lP8AZb8Cfsm/DGL4feDpZL+8uZftOsavcRhJtRuCMbiB91FHyomTtHcksx/aH/Z9+Hv7TPwyv/hh8RbJ3s7hhNZ3kGFn0+5UEJPExzhhkgg8MpZSCCa0T9jf/gpT+xj4g1Qfsw+KLfxN4d1CQPKLG4tRHOBwGlsr04WXAALRFzjHz4yB4g/ZN/4Kd/tm32naX+0j4qtfCvha0nEohvbi1WKMj5TItnZE+bKBu2mYr944ZQ3P7NX7Nfw4/Za+G1t8Ovh5aO+5vP1PU7gA3GpXJADSyEcAYACoOFAx1yT/AMFQP2ZfjB+034C8E6D8H9AtdVvNF1ee5vEnv4rUJG0IUEGRlB57Cv2L/hl4w+DX7MPgP4Z+PtPisvEGhWc8V7bxTpMsbNcyuMOhKn5XU8HvXxg8Par4v+EnjbwnoUCzalrXhzULKziZwgeaW3dEBY8AFmAyeBX/AATA/Y5+Pf7Mnjzxtrvxg8L2el2etaRBbWbwanBdF5FlLMCI2Yjjua/bd+FX7eHiP4keHfiX+yX4+ttPsdB0k2z6LHqa28tzcPKXeSSK4X7NMhURACQ8FTx81eJIf+C0nxY0a5+GuseHbPw7Y38TWuo6lbz6VZs8TDa376OVnAPcwANz6V+wD+wfafsg6DqniDxVrNnrfj3xHCkN9c2at9nsbdTu+zwlgGcFwGZyF3FUAUbcn9q39lb4f/ta/DY+A/Gkkun3tlMbnRtZt4w82m3GMFgCRvjYcPGSAwAOQyqy+Gf2W/8Agqh+x/c3Oh/ALxCniHww07SLDp2oWk9qxY/e+yX2DG7cbjGvbliBmhpP/BbH4rQ/2DqOpx+DdPnJjmvTNpWnsuQOd9uHuFwD1jH6jj4XaN4t8O/DTwp4e8favb6r4l0vRbS11e/gd3S7uo4lWWVWcBiGYFssATnkCqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/4Laf8l+8C/wDYnj/0rnr4T/8AJK/Bv/Yv2P8A6TpVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVXxF/Z/wDgj8XdUtta+J/wq8M+Kb+zt/s9vc6rp0dw8UW4tsUsCQu5icepNWNjZ6ZY2+m6dbR29raRLFBDGu1Y0UAKqjsAABiqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqp54LaF7i5mjiijUs7yMFVQOpJPQVffEX4e6Y6x6l488O2jOMos+qQxlh6gFqv8A9pD9nbS42l1P4+fDm0RG2s1x4ps4wrehJkGD7Vfftk/smafKIJ/2k/hu7FdwMHiW2mGOnVHIB46dfzq6/b7/AGNbQyrL+0P4TYwbg3lTvJnHXbtU7unGM57Vd/8ABSv9h+zt2uZvj5p7ImMiLSb+VuTjhUgJP4Cr3/gqf+wxawiWD4zT3jbseXD4a1MN9fnt1GPxq7/4Kz/sUW0Blh8ea3dOCMRReHroMfpvRR+tN/wV5/Y1CkjW/FZIHAGgvz/49X/D5L9kX/oHfED/AME0P/x+rz/gs1+yXbQebBoHxHu2Bx5UOj2wb6/PcqP1r/h9X+yv/wBCF8Vf/BVYf/Jtf8Pq/wBlf/oQvir/AOCqw/8Ak2v+H1f7K/8A0IXxV/8ABVYf/Jtf8Pq/2V/+hC+Kv/gqsP8A5Nr/AIfV/sr/APQhfFX/AMFVh/8AJtf8Pq/2V/8AoQvir/4KrD/5Nq3/AOCy37JM8CSyaN8RIGYZMUmj25ZfY7bgj8iatv8AgsZ+yFcTpDLB47t1Y8yy6LGVX67ZifyBpP8Agrv+xm7qp1/xSoJwWbQZMD34Oaj/AOCr37EMkiRt8TdTjDMAXbw7e4X3OIicfQVa/wDBT39ha9nS2h+O8Ku+cGXw9qkSjAzyzWwA6dzVj/wUP/Ys1Lf9n/aC8Pp5eM+fFcQdfTfGM9O3T8asP22P2RdSEJt/2j/h+nnkBPtGuQwY/wB7eRt/4FirD9pf9nDVlLaV+0D8NrwK21jb+K7KTBPb5ZTzVj8QvAGqb/7M8c+H7vysb/I1OGTbnpnDHGcH8qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqm8T+GreV7efxFpkcsTFXR7tFZWHBBBPB9qqqqqqqqqqqqqqqqqqqqqqq+PP7Qnws/Zq8H2fjr4ua3caXo99qSabBNBZS3TNcPHJIq7Y1YgbIZDkjHHuK+Dnxj8A/HvwDY/Ez4Z6rNqGgahLLFb3E1rJbszROY3+RwGGGUjkVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVavr+haBALnXdbsNOiIJEl3cpCuBjJyxHTI/MVrn7TX7OHhncviD4/fDuwdN2Y5/E9ojkr94BPM3EjI4AzzWt/8ABQr9i7QCy337QfhyUpuz9iWe86DJx5Mb59sde1ax/wAFY/2JdMYrZfEPWNWAYANZ+HrtQQRkn98icDp659ua1j/gtL+y7ZKU0nwV8SNSkwCpGnWkMfXkEtc7s4/2ce9az/wXA+H0BYeH/gH4hvQN+37ZrUNrnH3c7Y5MZ7+nvV7/AMFuvG2q3AtPCf7NunRzSH91FN4glvGOBluEt4yehPt70f8AgqT+3d4kj3+Cv2XtJkjZSwdfDOq3pCH7pBSVR+OMH0pv2sv+CvfigmLSPgDrGkM2IgR8P7iDaxP3s3WR3wSflGO3NHxL/wAFvvFxMdppusWEU7YWP+ztCsdhUf35VV1Bx3bBzj2ofBT/AILR+KQp1f4g6xpBmYu4Piuxt/LIyAP9FY4B9F45Gcdh+wn/AMFVPFW1df8A2nbm1SZjI0d78RNTZI2GcfJEjqPbaMDPah/wSf8A2zPEAC+Mv2oNIlE5LXP/ABOdTvcsOh/eIm7kDk4x+FQ/8ER/H2qzLdeJf2kNL893AlkTQprpgg4yC86EnHY4+tWP/BDXSI1can+0td3BJGwweElhx65zdvn9Ksf+CInwkjeI6n8bPF1wgX96sFhbwljjqpO/aM84IPHHvVj/AMEUP2aY0Yal8SfiZcNn5DBeWMIA9wbV8/pVr/wRq/ZHt44kl1X4hXJjA3NLrMAMmPXbbgc+2Ktv+CRf7GEE6Sy+GfE1wqnmKXXpQrfXbg/kRVp/wSm/Yetp1lm+Fd/dKAcxTeI78Kf++JlP61a/8Ewf2FrOdLmH4EQs6ZwJfEOqSryMcq1yQfxFf8O3f2I/+iBaT/4ML3/49Q/YB/Y0VQo/Z58K8DAzDIf/AGarT9g39jmynW4h/Z28HM6ggCayMq8/7Lkg/lX/AAxR+yL/ANG4fD//AMEUP/xNf8MUfsi/9G4fD/8A8EUP/wATX/DFH7Iv/RuHw/8A/BFD/wDE1/wxR+yL/wBG4fD/AP8ABFD/APE1/wAMUfsi/wDRuHw//wDBFD/8TX/DFH7Iv/RuHw//APBFD/8AE1P+wR+xvcTPPJ+zx4RDOxJCWrIoPsqsAB7AVd/8E+f2ML23a2m/Z78Mqj4yYhNEwwc8Mjgjp2NS/wDBNr9iGWNom+AelgOpUldSvVPPoRPkH3FTf8Etf2FJYXij+CDxMykLInibVSyHHUZuSMj3BFXf/BJj9ii5gMMPgTW7VyRiWLxDdFh9N7sP0q//AOCPP7Ht4UNvH42stoORBrand9d8bfpitQ/4Irfsu3Cyvp/jz4m2kjnManUbKSOPnpg2gYjHq341qH/BEX4OyvnSfjR4xt12YAuLO2mO7nnKhOOnH688ah/wQ10yTadK/aWuYMA7hceEVl3Htgrdrj9aP/BFj4u6EWk8IftH6PHIy7mJ065s8uuduTHI/HPXtnpR/wCCaX/BRLwsTP4N/amsV2YdRbeMtXs5C/QkAQ7emOSwo/s0/wDBYrwkNmjfGfV9aCAxqf8AhNxcbh13f6WQeoxk8/hS6z/wW+8EnN1p+sajEvzNEbXQtR3ZG0fNFufjrgEep60P20P+CsvgdQ3iX9na+1WOEL5kt38P7xozg7fme1ZFG447jtjANR/8FjP2iPBkkcfxT/Zr0eJuAyr9t0os3OcecJcZKtgc9D1wa8J/8Fu/hNemMeOfgn4s0fP3zpV/b6jt4HTzPIzzn/JxXgn/AIKk/sWeNPLhm+KFx4dupOlvrek3EGPrIivEPxevBHxY+F3xNtlu/h18RvDPiaIruzpOqw3ZUe4jYlT6g4I71VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVf8FuNT1KH4jfDnTINRuUs5tBumkt1mYRu3njkqDgngda+Gf/BGPw946+HPhbxveftAajbTeIdGtdRe3i8OIVh8+JZNgJuMnbuxnjOM4GcD4yfsz/tgf8E2IrX4p/Cf4z3+reC4btI7m704SQQwuzAIL2wdpIjG7NtDZcbiASrMuf2If2p7L9rX4J23ju4s7ew8R6XcnTvEFjBkRx3SqreZGCSfKkVlZck4O5ckqTVVVVVVVVVVVVVVVVVVVVVf8Fq/+TV/C3/ZQLP/ANIL+v8Agk//AMmR+D/+whqf/pZLVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV4h+Nfwa8JBv+Eq+LngvRtg+b+0NetrbbwDzvcY4IP4ivEf7fn7GnhYN/af7Q/hSfZnP9nTPqHTPT7Osmfunp7eoz4h/wCCt/7Fujbhpvi7xFr208f2foE6buR0+0CL1zzjofbPiH/gtn8BbQMvhb4UePNTZfum++y2asee6yynGcdu544wda/4Lf65fTCz8F/s3W6yu22Jr3xG87SE4x+7jt1IPUY3HOR06UP+CmP/AAUH8dFV+HH7L1pJFLzG1n4R1XUJCOTwyybSNv8As9ieKX4xf8FpfHabdC8CazoQbqD4XsLI4xzzeqSM7h6HI4xg0/wL/wCCzvjkMNf+JWsaB5gIbHi60tNv8PH2Fjjjnj69aH/BMX9v/wAbESfEX9qGxeOUkSLdeK9V1CUAnDZDRhTkAHG7ngHHbSf+CH/iK/nF34u/aStI5HbdMLTw29yznPPzvcJyR/EQeexrRP8AgiL8HrcIPEnxo8Y32Mb/ALFZ21pnnnG8S444788+1aJ/wRy/ZC0oKL+fx3rO0cm+1qNN3OefJhj7ccdvfmtH/wCCXn7DmkYb/hS322UZ/eXmvahJwf8AZ88L+ma0f9iD9kLQ2VrH9nLwHKVOR9s0eO75xjpMGyPb8a0j4E/A/wAPBRoHwa8DabsUKos/DtrDtA6AbYxgCrLT7DTYfs2nWVvaxbi3lwxBFz3OB34qqqqqqqqqqqqqqqqqqqqqqqqqpoYbiF7eeJJY5FKujrkMCMEEHqCO1eL/ANmb9nbx+sg8Y/A3wLqssgINxPoNv5wz12yhA6k4HIIrx1/wSf8A2MPGYkfTvBGseFLiUczaFrMy4PqEnMsa/QLj2rxx/wAESvskx1P4O/H65tbiE7re31zTPmDdj9pgYFfqIqb4Sf8ABYH9mNd/g7xhrPjXR7MYC2Oqx69DtH8KW14PPAI7RxjHsTXhX/gsH8cPhrqkfhb9pX9ntBeQ4WdraOfRrxAOrNb3CuGb2BjH0r4Tf8FQf2PvisYbOf4gyeC9RlwBaeKrf7Eo9SbgF4APrID7da0jWdH8Q6bBrGgatZ6lYXK7oLqznWaKUequpII+hqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv+C3v/JU/hp/2L91/wClAr9nr/kgPwz/AOxP0n/0jir9v3VdA0j9jf4rz+I5oUt5/D8tvAJSPmuZGVIAM/xeayY/Ov8Agh1p2rxeFvi5q0xb+y7jUNKgtgRx58cdy0uP+Ayw/pX7Uv8AwUY+N3wQ/a+X9n/wn4X8D3fh43WlRfadRsbqS7xdJE0nzJcomQZDt+TjjOar/goX+1B4/wD2S/gtovxG+HGkeH9S1LUfFEGlSxa3bzTQrDJa3UpZRFLGwfdAgBLEYLcdCP2I/jx4v/aV/Z10H4ueOtO0ex1jVLq8img0mGSK3UQ3DxrtWSSRgSqgnLHn06V+1N+3j8ZPgX+2H4O/Z/8ADOgeC7vw14kbSDPdahZXLXkS3VyYZdrpcKmQFJUlDjPOa+P3/BVf4r+IPiNqXw5/Y2+HkXiOy0h3il1ptLn1Ka9ZeGkghiICRAg4Zw28YOFHB8HeOjb/AAU0D4k/Eu/t9IceGLXVNduLlPs6Wrm2WSdmU/cCktx2xivFf/BTr9oD45+O73wB+wz8Dm1+2sh8+r6nZSTysmSBMYw6R2yE42mZjnuATgap/wAFAf8AgoH+zPqVjf8A7U3wAsLrw1fzCPz0szZtux9yO6heSEPgMdjqWOD0A4+Cnxn8A/tAfDfSfij8N9VN5o+qoflkXZLbSrxJDKnO2RDwRyDwQSpBNeP9a1Hw34E8R+ItHFs1/pWkXV1ai5RniMscTMm9VKkruUZAIJGeR1r9n/8A4K163rPwq+J/xM/aB8O+GYH8JS6Va+HdL8NwT20uq3d2t4TEzTzTAKBahi4ACKHOHJVa/Yq/bc/aq/aW+PlnYeO/h3a6D8ONQ067mtp7LQrhITIiAxL9rlLB2xk8YB5wAOB+13+2V8NP2QfB9tq/iuObV/EOrhxougWkgSW7K43O7nIiiUlQXIJycKrHIGmftcf8FWfjLpcfj74T/s+aXp3he5Uy2ONKH+kRHlWVrqYNMMDho1AOeB0r9l//AIKga74l+KEPwE/aw+H0PgLxdPcrZW1/HDNaxG6YgJBcW8xZ4WckYk3FSWHCg5r9vf4+/F39mb4IJ8WvhNovhrVXsNXgt9Xh1y0nmSO2mDIsieTNGQwm8peSQRJ6jn9hr9pPVP2qv2ftO+J3iSx0uy8QRajdafq9tpkbpbxTRPuTYru7AGGSFiCx5Y9sV+2D8e5v2aP2e/FPxb063sLrV9NSKDSbS+DGK4uppVjRWVWVmUBmdgGB2o3Ir/gnP+3R4w/a+j8Z6N8StI8N6V4g8NtbT2kWi288Mc9rLvVmIlllJZHQAnIGJF46mvE/iLSvCHhrVvFmuXHk6boljNe3kv8AchiQu7c+iqTX/BPf9ub4/wD7YXxL8U6X4y8I+CdJ8I+GtLFxJPpVldLci5llC28ReS4dMFEnYnYCTGMAZOP2v/20/jl8HfipY/A34C/s7XvjfxPqGkx6nFfSwzXUHlO7pxBb4chWjcMzSJjHTBzXif8Aar/4K4fDfS5viB4y+Amlf2BaRGa5h/sHz44IlwWaRYLgzIoGSSxAAyTgDj9hn9vLwt+2JpGo6PeaEvhvxxoECTahpa3Hmw3EBO37RbsQG2BiAysMoXQbm3A1+0b8QvEPwl+BHjr4m+FINOn1fwvodxqFpFqMTyQSPEu7a6o6MQQCOGFfDn/grbqVv+zXr3xb+MPhjw7P4vPiKTSPDOg6Ck1pHe7LeGV5ZWlkmZUQyjcw/vIoGTkfsBftZ/tSftI/E7X4vjR4Gg0DwougG90d7XQLizgll+0Rr8k0zN5nyO3AY9M1+2t+378P/wBkOzt/D0Om/wDCUePNTg8600WOfyktoiSBPcyYJRSQQqgFmwfuj5h4Y/bS/wCCq/xJsYviF4F+ANndeHJ1822jg8Kz/Z7mMAnMbSTebKp/vIxBIwPSv21P251/ad/Zosfhj8QPAl34J+J/hTxxaXGqaRJFIkU0K2d7G8iLJ+8iZHdFaKTkb1IZsnb/AMEn/wDkyPwf/wBhDU//AEslqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqmmht4XnnlSOOJSzu7bQoAySSegxXif8AaI+APgsyJ4t+N3gPR5Ishor3xFaxSZGcgIX3E8HgDPBrxP8A8FJv2JvChaO8+Oum3sq52x6ZYXd9uI9GiiZO3dgK8S/8Flf2TtGDR6Lo/j3X3H3GtdKhhjJ9zNMjAf8AAT9K8S/8FxPD8IaPwf8As86hd54SXUvESW+33KRwSZ+m4fWh/wAFWP21viAMfCb9mbS7iOYfu2g0LUtVfBxgqYnRSfmX+Ejkcc0fjN/wWg+JKFfD3w+1rw5HOPuf8ItZacVBHQG+Xcv3hzkEY6jBo/s0/wDBYr4inHij4wav4bEpJdX8bLaKM5yCNPLDHOMDjkdhx/w6E/al8cEH4r/tLaLc7iS5+13+rc56/vxFnop//Vz4d/4IeeDrfyz4s/aD1m/6b107QI7TuMgF5pe2RnHocdq8O/8ABGr9k3SEQ6vrHj7XZAPn+1atDEhOOwhhQgexJ69a8P8A/BMr9iHw7seD4IW19Mp5k1DVr253cnGUeYp3xwo6DOa0D9kr9l3wuFOh/s8/DuCRBhZn8OW0so4A++6Fu3r71o3hnw34ciMPh7w/pumIRgpZ2iQA856IB3qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq8WeCfBnj7SX0Lxz4S0bxDpsgIaz1Wwju4jnr8kgI/Svi3/AMEmf2S/iQs954Y0PVfAOpyKSs2hXZMG/sWt5t6Benyx+X06jk1rP/BNf9uH9mbUZ/E37LXxkfW7dGLm30vUH0i6nwP+WltK5glGAODI2cD5eleE/wDgqp+1J8Btbh8F/tbfBOa/lU/NcS2L6JqDLwDIFK+TMvcbFQHP3sYr4H/8FDP2VPjyLex0D4jQeH9bnwF0bxIF0+4LcYVWZjFIx/uxyMeOlVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX/AAW9/wCSp/DT/sX7r/0oFfDz/gq98bvBHw+8MfD62/ZdF/c6Ro9pY2V19ouk+0xxwokcnlCIk7goPDYOeK8Z+Gf+CjH/AAUm1nSdL8WeA5fBPgW0n86JLyyl0rTITyPtBExM11IBlRtDhcnAQMxP7Mv7PPhD9l/4QaT8J/CEz3a2jNcahqEsYSTULyTHmzsoztztVVXJ2oiDJxk/8FB/+UmCf9hDw7/6Kt6r/gtX/wAmr+Fv+ygWf/pBf1/wSf8A+TI/B/8A2ENT/wDSyWv+Cxs0tv8AtfWM9vK8csXhGwZHRirKwmuCCCOhr9lT9nvwn+zX8FvDvw98P6PbW+pJYwy67erGolv75lzNJI45YB2ZUBJ2oFUcCv8AgtD8XdU8IfBTwn8J9IvJLf8A4TzVJpdQKMR5tpZCNjEfYyzQN/2z9M1+xD+09+wp+zf+zr4V8FzfGHRLDxLe2cd/4mf+zrsyyX8qhpEd1iIbysiEYOMRgjqSfjr+2p+wV8Zvg74v+GOu/G7Rby38QaTPbxo+nXh2T7cwyLmHhklCOp7FQa/4Ih/EHUk134lfCme7Z7CW0tdatYGJxFKjmGZl7AurwA9z5a+hqviz/wAkr8Zf9i/ff+k71/wTA+A/h749ftLW+n+NbFNR8NeE9OfXb3TpwGgvZonSKCORDw6h5txBGCEZTwxBhhht4Ut7eJI44lCoiLtCgDAAA6DHaviH8U/hf8bP+CmOpeJ/2i/E1vZfDbwnr1xYql7FJNA1rp29IYfLRSxSadN7KR0lfNR/8FHf2H4Y0hh+PWjoiKFRF068AUDoAPJ4Ff8ABVf4w/szfHXUPAvxC+CHj3T9c8UWK3Fhq7WlrPC7Ww2vbsxkRQdjmUDBz+89AMaRplz+15/wT+sNL1uUXer+O/h9Esk0mcHU1gG2U554uow/4dT1r/giX8RZ7LVviZ8EdTLxTeXBrlrbyAq0bRt9nucqeQcvaj2xzX/BYHxhqvj7xl8IP2UfCNyZNS8Q6mmoXNsozmWaT7JZZx7tdZH0NfDvT7D9iv8A4KwjwFpqfYfCPjdY9NtlJwHhv4o2hAHTi9jVP+Amv+CpXxTPwx/Y78U2lrc+VqHjOeHQLU7sErMS84wOoNvFOvp8w+h/4JF/CI/Dz9lWDxnf2hi1L4g6pNqRLrhxax/uIFP+yfLkkHtNXjjx74E+GWgz+LviD4s0fw5pUACy32pXSW6Z52qGYjLHnCjJJPArxZ/wVm/Yt8OzzWdj4x13xGYjtZtL0KbYxzghWnEYI689CBwTxn9hnxx4ci/4KX6drPwvguLbwl4o1zWorCzliFu0dhPDcSQxMilgPLIi4BI/djp2/bX/AOTRfi//ANifqH/olq/4JDfAHwz8YPjJr3jrxxpkOr6T8ObOGez0+6QSQ/2hdsyxStG2Q21LeUgY+8I2z8gp3SNDJI6qqglmY4AA6mv2YPDkP7eP/BQvUPGPxEgOp6JJe3fiO/spkLo1nbsqWtswP/LIF7WMg9UBU9c1HFHDGkMMaoiKFRFGAoHQAdhX/Ba/4ReFB4D8F/HS00+C38Qx62NAvLiNNr3kEtvNNH5h/i8s27hT2EpHIxj/AIJP/wDJkfg//sIan/6WS1VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVWu+JfDfhi1F74l8QabpNvg4mvrtLdOMZ5cgdx+YrxV+2n+yX4MDjXf2h/Au+P78Vlq8d9Ip54KQF2B46Y9PUV4o/4K0/sWeHw40zxpr3iNk426XoFwu48dDcCIdz3xwfbPij/gt38JLRHPgr4J+LtVYA7Bql9b6eDxxny/Pxzj1q4/4LGftDeOLh7X4Q/s1aRJMz7YopDeawwOeARAISTyo7frX/AA0h/wAFi/icufCfwm1fwyko+Ro/BcdmuDg5DagGGMd84+Y+2G/Z0/4LHfE1WHiv4q6x4bSUYdJPGcVkpGDwV09mGDgDGP4hnviP/gj1+0n46lWb4vftLaPO+7czrJfawwbnkGfyueT+Zrwz/wAEQfhjaBB4y+OnifVCD8/9maVBYZ6dPMafHf17V4Y/4JCfsa6AUOq6N4s8S7MZGqa68e7GOv2ZYeuD0x1PTjHhn9gL9jTwkIxpX7PPhSfysbf7TgfUs4x1+0tJn7o69efU58M/Cb4V+ChGPBvwz8KaD5WPL/szRbe12YxjHloMY2r/AN8j0qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq8YeCPBnxC0SXw1478KaR4h0qf8A1llqllHcxMfXY4Iz79RXx1/4I7/AH4gC51f4QaxqPw61eTLLbJm/05m9PKkYSR5PdZNq54Q8CpdC/wCCnn/BPVjPpN3d+N/AFgclYd+t6YsQH8UZxcWajjJXy1z/ABN3/Z9/4LB/A/4im00H4z6Tc/DvWpSEN6WN3psj9M+Yo3wgnH31Krnl8DNeH/EXh/xbo1r4i8K67p+saVfR+Za31hcpcQzL/eR0JVh9DVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV8ZvjL4v8A+L00LQo9Pa3azjmJuIGdtzMwPIYcfKK/4ab+I//PDRv/AVv/i6/wCGm/iP/wA8NG/8BW/+Lr/hpv4j/wDPDRv/AAFb/wCLr/hpv4j/APPDRv8AwFb/AOLr/hpv4j/88NG/8BW/+Lrw7+0Z8QNV8QaZpdzDpHk3d5FDJttmB2s4Bx8/XBqqqq/4Le/8lT+Gn/Yv3X/pQK/Z6/5ID8M/+xP0n/0jiqq/4KVyr4V/4KIjxLrCtHYqdCvw4HJhjSJWYZ94nH4UjpIiyRurKwyrA5BHqK/4LYavpsP7N3gzQJbyNdQvPG8NzBbk/M8UNldrI4HorTRA/wC+PWv+CT//ACZH4P8A+whqf/pZLX/BZL/k7m0/7E+x/wDR1zVf8FxPCd/LYfCbxzBE7WVvNqenXL4+VJJBbyRDp1YRzdT/AAcd6/Zj/YK/YO+Nf7PvgD4mn4PJf3mtaFbPqUy+JdUX/TkQJdKQtyqgidZBgADjgAV/w64/YS/6IX/5c2rf/JVfBH9kD9nP9nHW9Q8SfBn4cr4e1LVLQWl3cHVby7MkIYPsAuJpAo3Kp+UAnAqviz/ySvxl/wBi/ff+k71/wRL/AOS/eOv+xPP/AKVwVXwh+EXwpv8A/go/r3wQ/aG8O/2loOo+KdY06KGa9ntA9wzytaPvhZHIkIQKMgEyrkV/w64/YS/6IX/5c2rf/JVf8OuP2Ev+iF/+XNq3/wAlV4D8C+FPhl4O0j4f+BtIXTNA0K1W10+zWV5fJiXoN8jM7fViSe5rw9p8n7Kf/BYL+x4F+y6D441l44VA2JLBq0e6NF9FS8dUA/6ZYr4D2Z/av/4KueOPi3NIbrw18MJJvsLdYma3X7FahevDP5tyOnKE+1f8FoPhvqOlf8Kz/aQ8Ns8F/od6dHubmNfmjbJubRgRyArpcdeMuvTPP/BST4u3v7Umsfs4fDDwJ5b3XjPRLTWhaxkkLdam6QQxtycGMxSgjqA5J7V4F8H6R8PvBOgeA9Aj2ab4c0y3060XGMRQxrGuffCirmfxJ/wUs/b8fwT4w8WXtl4KsL++WytoJeLPS7UniFTlRNNsTc+Dy+eQirXgP9iv9lH4b6bFpnhn4BeDH8pdoudS0uPUblvXM1wHk59N2K+DFnoWn/8ABX5tO8NWlja6ZZ+PdZgtbeyiWOGFUhuV2Iq4VQMEYHAxX7a//Jovxf8A+xP1D/0S1f8ABDH/AJrZ/wBwD/3I1qdjHqmm3WmyuypdwPCzL1AZSCR781/wSj12H4Sftu33gPxgFstS1jRtS8OBJTsEd5FNFMUOcYJNm6AHkkgdTVf8FsvEOj2/7PPgnwnNexrqmoeM4723tiw3PBBZ3KSuB1wrXEIJ/wBsV/wSf/5Mj8H/APYQ1P8A9LJaqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqrxt8U/hl8Nbb7Z8Q/iH4b8Mw7dwbV9UhtNw9vMYZ6dB1rxz/wVF/Yt8ENJbx/FKXxFdRgkwaHpc9zn6SlViOcf3/rgV4v/AOC3nwqsSy+Avgh4q1nB+U6tqMGm59f9WLjH+elN/wAFbv2tPiMWg+DP7NekzmT5VMWn3+supII48kxjOQcZBHHQ0fir/wAFo/iuNnh7wPrfhmCf/ll/wj1jpOOh+/fASL2/iHcHvX/DE/8AwVY+KoC/ED4/XGjW8p/e22oeOLnyx1B/dWavGTgn88ZHONE/4Il+MNauP7R+JH7SFqLqQAzCx0SW9Zzj/nrLNGevcqenQZ48K/8ABFf9m/S9kvivx/4912VDykVxbWcLc91ELP0wOHHf8PC//BMH9iXwsEkX4OLq1wgA87VNXvLjdwOsZlEfbP3e57cV4Y/Zl/Z08FgN4U+BPgHTJB/y2g8O2wlPXrJs3Hqep71b21vaQJbWkEcMUYwkcaBVUewHSqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv2hv+Cev7M/7RYutV13wYnhzxNcbmOv8Ah9VtLh5D/FMgBjnJOMl1LY4DCvEn7Hf7e/7BuqXHjX9mzx3qXi7wvHIZrmDRY2cyKOpudLcush2jG6PzCoycp1r9m7/gsL4B8XXNt4O/aS8Of8IPre7yW1qyR5dOeQcHzYzmW2OeP+WijksyivD/AIi8P+LdGtfEXhXXdP1jSr6PzLW+sLlLiGZf7yOhKsPoaqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/ab/5KPF/2DIv/Qnqqqq8F/8AI46F/wBhO3/9GLVVVV/wWT+GnxH8d/Ez4d3fgj4f+JPEMFroVyk8ulaTPdrExnBCsY1YKSOcGvgPZXmnfA34dadqFpNa3Vr4T0uKeCaMo8TraRhlZTyrAggg8g1Vf8FUP2JfF/x+07R/jN8I9IbVfFnhmyex1LSYgPO1CxDGRDCMjdJGzSfJ1cSYHKhW+Cn/AAUY/bR+EPhfTvgprHwJufF2p6HbrZacNR0e+i1FI0G1I5UQZl2gKo+VWwOSTzX7Q/7N/wC2P+0B8JPEX7VX7Rmn6vH4ktTaWvhLwHpWnSNJawS3CLM5tl3vCoQk7WzKx+ZyAgDf8EvfD3iDwr+x14X8P+KfD+p6LqdpqOo+dZ6lZSWsybrqRlJSQA4KsCDjHNf8FbfhP8UvGv7VFrrHg34aeKtesF8J2URutM0a4uog4luCV3xoRuAIyM55FV+0p8APCf7TXwf1r4S+LXNvHqCiWxvkj3vp92nMU6jIztJIK5G5WZcjdmvBNx/wUF/4Jm63qfhqz+Hk3i3wHc3RmfyrKfUdKnY4HnxSw4e2kZVAKvtzgEo2Aag/4K8/tBeJVOl+DP2RHutVdf3SpPeXnPTPlRwqxGccbhX7HXxB+P3xN+D58UftIfD9vB/iuTVrhIrA2TWe602o0TiF3aSMZZ0xId2YyTwQTXxRtri8+Gfi20tIJJ55tCvUiijQszsYHAVQOSScDFf8EdPhh8S/A3xy8aah42+Hnibw/az+EzFDPqukT2iSP9qgO1WkUAtgE4HOAar/AIKN/wDBPjxZ8avEcP7QXwA8pPHVnDGuqaWs/wBmk1LyQBDcQSkgLcIqquCVDKi4IZcP4S/4Kbfto/BHT4vBHx4/Z5u9f1HT4/JjvdRsLrSbybbxulPltHKcfxIi54JJzk+D/wDgop+258a/F3h/TPhf+yXNa6LLqlsdTuvsF1dZtRIplUXMgigi3JuG5xxnjB5qv+CufwS+IGpeJvhb8f8A4ReFtX1XxDodw2n3L6Tpz3k8JicXNnIURWO1XFxyRjLKO9f8EmPgRrvwl/Z+1Pxj410K/wBL8TePNZe4uINQt2guI7W33RQrIjgMCX+0ON3USg98n9tX4Pn46fsv+P8A4f2tm1zqcmlve6THGm52vLYiaFVA5y7RiPjs5Hev+CZ/7PPxb8T/ALUXh3xn8W/BHiXS9G+Gmhyy6e+t6VNao8mWjt4UMijcUa4klHp5Y9hVfG/9nn9o79g39qub9ob4K+DLzxF4TfU7m/065tLKS7ghguC3m2N4kfzRgK5UOcAjYytuBC2//BQX9tL9pnSh4B/Z0/ZWu/DWsalF9nn8TXNzLdW9hu+VpleSCKKEjkje0mMcBiK+AX7MHxa/Z1/4KM+DtG13w34k8RabouqxC88Tw6TcGynkutP3PIJypUqss5UszZ+U5wcgfth6Vqmu/sr/ABW0bRNNutQv7zwnfxW1rawtLLM5hYBURQSzE8AAZr/gi58PPH/gL/hcX/Cc+BvEHh37d/YX2X+1tMms/P2fb9+zzFXdt3LnHTcM9RVf8FDP+CfXxPm+KM37UX7MGn3d5qNzcpqGr6RpjbL61v4yG+22gGDJuKqzIuZBJlgGDEJ4M/4Ko/tn6JYW/gPxP+z5B4m8WQDyBPJo95a3U8mMAy20QwXz1CBAewHWv2gP2av2v/jz8IfFH7VX7SGlau3i2IWVn4R8E6Xp7tLZ20lyvms1qm54lCM2FYmQ/MZMbVz/AMEvfD3iDwr+x14X8P8Ainw/qei6naajqPnWepWUlrMm66kZSUkAOCrAg4xzVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVXxI/aN+AvwhWRfiV8XvCugTxA7rO61OP7ScddsCkyN+CmviT/wAFjf2W/CXm23gXTvFPji6XPlyWlj9htmI9XuCsg/CI1qH/AAVb/a8+NN1Lo/7OP7PFvCXfYGtdPutfuYeOPmRUjBxyd0ZGKPwN/wCCwP7RwL+N/iBqvg7TbokGO916PRoSp7Pb2AMhX2eP35614M/4IlXF/cDVfi/+0FPc3MzbrmDRtLLM7HqftM7kk59Yq8C/8Em/2MvBpjl1Pwbrfi2eIfLJrusynn1KW/ko30Kke1eDv2av2evh8kQ8F/BHwPpMkQG2e30K3Exx0JlKl2IPckmkRI0WONVVVGFUDAAHoKqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/aS/YS/Z5/adtp73xh4UXSPE8i/u/EmjKtveBh0804Kzr0GJAxA+6V614k/Z6/by/4Juaxd+N/gn4puvF3gFJWlu/sEL3Fs0Yx817p5JMbY6yxlto/5aLnFfsr/wDBVP4LfHJ7Pwl8TjB8PPGM22NFvLjOnXsh4xFcNjy2J6JLjqAGc0CrKGVgQRwRVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVftN/wDJR4v+wZF/6E9VVVXgv/kcdC/7Cdv/AOjFqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq8bfEn4d/DXTv7W+Ifjvw/4Zs8HbPq+pRWitjspkYZPsOa+J/8AwVr/AGRPABltfDuu6345vozt8vQtOZYg3vNcGNSvum/8a8Qf8Fcf2lfi7qMvhr9mj9nqGO4YBVItrnX71QTwypCsaITjoyuOvXrX/DPP/BW39qE+b8S/iDqXg/SLofNDqetJpUDJ6GzsAWOB2kQH36mvhv8A8ESvBFqY774xfGjW9ZnY75bTQLKOyQN6GabzWcHudiHnt1r4bf8ABPX9jz4XeVNonwR0TVLyLBN3r4fVXZh0bbcF0U9PuKuDz15rT9O0/SbKHTdKsbeztLddsNvbxCOOMeiquAB7Cqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/ar/4JgfA/9oOO88UeCbeH4f8AjeUNJ9u062H2O9k6/wCkW4wMk9ZI9rZOTvxivCXx/wD21/8AgmX4msvhx8a/D1z4s+HfmeTYRXFw0ts8Q76fe4JjIHPkOMAfwJndX7Ov7VXwX/ai8MjX/hb4oSa8gjDX+i3eIr7TzwP3sWT8uSAHUsh6Bs5Aqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/ab/AOSjxf8AYMi/9CeqqqrwX/yOOhf9hO3/APRi1VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVTukaGSR1VVBLMxwAB1NfFb9uj9lD4NGa28Y/GjQpdQhBDadpMh1K4DDPyMluH8tuP4yo6ZIzXxO/4LY+F4p30r4IfBjVdZnd/LgvfEF0tqpbOAVt4fMZwT0HmIenGeAfH/wDwV+/av48M6FrfgPQrvkSWNovhyFFOfmSedvtTDt8jt6+9eCv+CMvxA8YaiPEv7RX7QQmvpyGuYtJil1CeXJ5zd3JXB/7ZuMn8/hh/wTE/Y3+GRhuP+FZHxZfQni78UXTX27/eg+WA/wDfqtB8OeHvCmlxaJ4W0HTtH06AYis7C1S3ij7fKiAAdB0FVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVeM/BPhD4i+Gr7wd468N6fruiajGY7qxv4BLHIOxwehB5DDBBAIIIzX7SP/BMH4mfBHxIfjn+xD4i1lZNLka5XQLe6ZdRscDJFpLnNwmMjyX+cj5f3u4iv2Qf+Crug+MbyD4W/tTxW/hHxZDJ9lj19ovs9ldSqdpS5Q/8esuRyf8AVk5z5eADHJHNGksMiujqGRlOQwPQg+lVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV+03/yUeL/sGRf+hPVVVV4L/wCRx0L/ALCdv/6MWqqqqqqq8aeM/Cvw78Kap448ba5baRoWi2zXF9e3BISGNep4ySTwAoBJJAAJIFfAv9pP4LftKaNqGufBrxrFr0GkTrBfxm2ltprZmBKF45VVgrBWKtjB2sM5UgVXjLXJfDHg/XPElvAk8uk6bcXaROSA7Rxs4BI7Hbiv2Cv+ChHjL9sD4j+IPBHiT4eaL4fg0XRDqMc9jdSytI3nRx7SH4AxIT+FVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVeJfFfhbwZpMuu+MPEulaFpsA/e3mp3kdrCn1eQhR+Jr4s/8FWP2Q/hn59lovizUPHWpwkr9m8OWZki3dv9IlKRFf8AaRn+hrxF/wAFYP2pfjZqs3hf9lz4BrbSOdiyR2U+u3yZ6PhFWKPoSQ6OB68ZpP2Kf+CmX7VrrdftA/FK48O6NcHe9lrWsZTHqlhZ5iVsE8OIzx9K+FX/AARi/Z98LLBd/FLxf4k8c3iY8y3iYaXZv6jZGWm9siYfT0+GP7O/wM+DEEcPwu+FPhrw7JGmz7Va2CfaXGMfPO2ZX/4Exqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr9sX/gnr8Jf2q7G48R2kUXhT4hRxH7Nr9rANt2QMKl3GMeauAAHGHXjBIGw/CL9qL9pz/gmx49tvgV+0r4e1DXfACkrZIsnnGCAHHn6bcNgSRcjMDkAcDETFs/C34sfDz41+C7Hx/8ADDxTZ69ol+uY57dvmjbAzHIhw0cgyMowDDuKqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/ab/5KPF/2DIv/AEJ6qqqvBf8AyOOhf9hO3/8ARi1VVVVVVXx++DOgftC/B7xL8HPE1/dWWn+JLZI2urXHmQSRypLE4B4O2SNCVPBAI71+xF+wv4e/Yx03xKbXx5d+LdZ8Uvbi6vZLAWMUcUHmeWiQiSQg5lYsxc54wBg5/wCClvxW+IXwb/ZY1Txh8MfFNz4f1o6tZ2ovbZUMixSOQ4UsDtJwORgjsa/4JbeNfGPxC/ZRs/FHjvxVq3iLWLnxBqAlvtUvJLmZwHXALuScDsOg7V8Wf+SV+Mv+xfvv/Sd6/wCCJf8AyX7x1/2J5/8ASuCqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqviJ8Xfhb8I9MGr/E74heH/DFqykxtql/HA0uOojVjukPsoJr4uf8ABZD9nDwX9osPhjofiD4g38YPlzRRf2bZM3oZZh5vXuISPepv23P+ClX7V8r2X7PHwtn8O6PMdqXei6TvGCcYkv7zMSn3XyzwfQ14Z/4JMftNfGfVovFX7Unx8EEzEkxtdza7fKCclN8jLHHn/ZZwOOO1fCX/AIJYfsg/C8wXmpeC7vxxqcIB+0+Jrr7RHu7/AOjoEhKk9nRiPXrnw94Z8N+EdJh0Hwn4f03RdNtxiGy060S2hj/3UQBR0HQVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVXxo+B/wx/aB8EXXw/+Knhe21jS7gFomZds1nLjAmgkHzRyDPUdRkEFSQfHvwn/AGof+CUnxNPxM+FusT+Jvhlqc6xzzSxs1rcpnC2+oRLxFMM4SZcAk/KRl46/ZV/a6+Ff7WngoeIvA979h1qxRRrPh66lButOkPfjHmRE52ygAN3CsCoqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv2jfDviDVfiBHc6XoWoXkI06JfMgtXkXIZ8jIBGelf8ACF+Mf+hT1n/wAl/+Jr/hC/GP/Qp6z/4AS/8AxNf8IX4x/wChT1n/AMAJf/ia/wCEL8Y/9CnrP/gBL/8AE1/whfjH/oU9Z/8AACX/AOJrwh4Q8WQeLNEmm8L6tHHHqNuzu9jIAoEi5JOOBiqqqrQ/iX8U/wBm3/gqZH8OvFPxI8VX3hG98Wmyt9P1LWri4tfsWpp/ovyu5XERuIvm7GI56Gq/au/a9+Muo/tbeMPiZ8OfHniWDwj4N8U2thY2tnqk8enu1rlUDojBGWdrWeTBHzKW7CvBvirSPHXhDQ/G2gXCz6Zr+nQX9nIDnfDNGsiH/vlhX/BVP42fFHWv2mPAf7Pvwc8b69pF9b2UEUsOkapNZm41DUJlWOOTymBbCJCRkHHmtjqc/tYWusfC79hbxppmgeJ9VXUvDPg+K2g1dbuRLt3iEaecZQd/mNgktnJJJJ5r/gjJ4+8d+PPDHxTl8ceNte8RSWd/pi2zarqUt2YQ0dxuCGRm2g4GcdcCv+ClH7L37Qvg628c/HPxR8a/7W8Aav4o8yx8M/2reSfZlnlYwjyXHkrsHoeO1fsh/sXftW/HH4M23jz4RftFf8Id4el1C4t00v8AtvULXbLGwDvsgBT5iRz19a8D/Dzxv8KP2Pbj4e/EfxZ/wk3iTRvC+px3+rfaZZ/tTMJ3U75cO2FZV+b09K/Yb+N3xO+DnjvxBpnwQ8Ht4i+IfjvSV0Pw9AYxIlvK06SvO6kgEJHEx+YhR95jtVgfHn/BM/8Abx+IFlJ8QfGH7SdhrvjIg3K6dNrN4qxP97y4ZdojiIPAVVWMHoQOa/4J5/tw/GXwZ8b7b9lT9o3WNU1O3vr6TSLKbWpDJe6NqSEqtu8rEs8bspiCsWKuU2kLkVVftmftb+E/2Q/hd/wl+qWiar4i1eR7Xw9oxl2G8nUAs7kcrFGCpdh/eVRguDXw/wDAv/BRj/govHL8Q9W+K114N8CXEzrat9tm07T5djYK21rB804Qgr5kmeQw8wkEUP8Aglv+2H8NZbXVPg7+11KLmCVS6Je32lsFJ+faEeRX4/hYqG5BPr/wUc1rxF4J/Yc8f6r4a8S6rp+rafFpKQ6na3TW9yCdTtEZhJGQQWUsDjGQxHQ1/wAEdPG3jPx18DfGmoeNvF+teILq38WGKGfVb+W7eNPskB2q0jEhcknA4yTVfsn/ABX+KWtf8FSV8H6x8S/FV/oP/CWeJ4v7LudZuJbXZHbX5jXymcptUopAxgFRjoK+JnxF8KfCPwBrvxL8b6gbPQ/Dtk93eSqu5tq9FRf4nZiqqvdmA714W8S/tt/8FRvGus3vhTx5dfC34U6RcmEiyuJYoIz1WFjEVe9uNpVmDMsa5BATcoPxK/4J0ftW/su+GL34u/AH9pzW/EV7oED3l/ZWsU+l3Dwxjc+yPz5kuAFBJjfG4AgBjwf+CdX7aVx+1n8Or7S/GsdvB4+8IeVHqxgURx6jC4Iju0QYCElGV0UbVYAjAcKtTwR3MEltLu2SoUbaxU4IwcEYIPuOa/ZZ/bl+MPwi0vx48fivxL418a+JbOz0nwjp+qXtxqSxXcsx3TLE7NuZVACqOWdkBBGa/ZB/Y6/be0v9orwZ+0n+0V49hu7S0a+l1DSdS8QT3V/H9osZ4UAiVGgUB5I8oJBtC8D5QK/4KKft26t+zRZ6Z8K/hFbwX/xN8TwiWF3g+0DSrdmKJKIuRJM7hhGhBHyksCNqt4a/4JjftX/HbQrfx5+0P+1XrWh+IdRjEy6VPFPq0loGGQkh+0xJEwz9yMMq5xntUnxT/a5/4JdfGPQvCnxa8aX3xD+GOuAvDHLdvcR3EClRK1q02Xt54tykxZ2NuGc7g4uviJ4StvhvL8WY9UW48MRaG2trewjcJLIQef5ijjIMfzCvCXiL9rz/AIKpfFfxLYaH8TZvA3gPQVWSeyguZY7SwgmZlgiaOIg3U7iOQlpCB8j4KDC18Ov+Cbn7XvwN+JHhTXvh7+1PPdeHV1i1bxDBb3dzZSNaCVTNi3cyQz/IG+VyOccGvi0ZR8KvGZt7qe2lHh6/2TW8hjkib7O+GRhyrDqCOQcGv2R/25Pj78PdG8a+A/Cl34j+IXxH+INzpen+Ek1a+l1BNPkU3ImlVJWILHzYvlOFJAZztQg6r/wTM/ba+Lulnxl8XP2rE/4Sy4VZ49Lnvbu5ht5OSIzIhVItuSMRRsoJO3I5Pw3/AGrf2s/+Ce/xtg+EH7SOo6v4j8Jq8f2uxv7tr4iyc7VvNOuH+YqNrYjyEJVkZUfJTQ9a0nxLomn+I9Bv4r7TNVtY7qyuoTlJ4ZFDo6nuGUgj61+15/wUA+NHxX+M/wDwyv8AsXy3UVwdQbS7nWtPCi71G6UkSJbytxBBGQ26YYJ2FgyoMtF/wST/AGoPEFivijxd+1ag8WyJ5jI8t7dhZOoBumkD9ScsEOO2a+F/7Y/7Un7BnxuX4IftaanqfirwsGjE8t3dPfT29u/CXllcv88sQA/1Tf3WUBHBrTtQsNX0+11XS7yK7s72FJraeFwySxuAyupHBBBBB968E/tm/FD9n79qv4qX7av4r8bMt5rOj+GvDt7q9zdWwvHvwluTEzsSqKrYVRubhQRuyNZ/4J+f8FA/2j9KHxB+NH7Qtppetagn2i28PXmoXPl2O4ZEZjgXybcjgFY1bHck5r4EftYftJ/sJ/tBR/BH9onXNV1PwpBfR22safqV414LOGUDZeWczkkIAwk2g7WXcCFblfOh8nz/ADU8vbu37uMYznPpitZ+NX7UX/BTP9ofVfhV8GfiBc+CvAGlrNcxRw3UlpGlhG6xi4uTF888shdCIidoLYGArPVt/wAEu/2wvhNf2fiD4K/tYO14lxH9qSO5u9NfyywDEKHkSbA5KPtBAxz0pPLsLFTdXhZLaL95cTsqkhRy7EAAdMngD6V8Sf2qf2nv29/jne/AX9j3X7nwp4G0t3+1a/bTvaSXECnY13PcIPMjiYnEcMeGcEFgTwmuf8Ek/wBobwtpkvjD4b/teajf+MreMyxQvBdaYZJAM7Vu1uZGDEgAEoBnGSK/4Jyftx/Efx3451L9lf8AaRlmk8d6MblNN1K7VY7i4e2JE9ncKoAaaMK7B+rKj7skbmqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr4q/tC/BD4IWhuviv8UfD/AIcYJvS1ursG5lX1jt0zLJ/wFTXxh/4LR/Bvw0Z9O+DHgLWvGd2mVS/1Bv7Msz6MoIaZx/sskf1pvjb/AMFYP2xHUfDXwvq3gjw1eAmK40qz/sW3MZ6OL25bzXwD/wAsn5PIXoB8PP8AgjD4s8T6i3iX9pL47S3F9csHubfQVe7nlJ6lry6A+bHH+qbnvxz8Jf8Agn3+yR8G/KufDnwg0vVdSiA/4mPiAHU5iw/iUTZjjb3jRKjijhjSGGNURFCoijAUDoAOwqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq13QtF8T6Le+HPEWl2up6XqVu9veWd1EJIp4nGGR1PBUgkEGv2rf2IPip+xb43H7Un7Hur6rF4f0uV7m7soP31xoSHlwykEXFkRwQ4Yqv39wBev2Iv28vAf7W/hxdG1AWvh/4i6bBu1LQjLlblVwDc2pPLxEnlOWjJwcja7VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV/wWk+G154a+Ifw5+PugGS3nvbZ9LubiIbTDcWr+dbvn+8VlkA9oR6V8TP2lbDw7+xPfftNWM8ccl74Mh1HTgDwt7dxIsCHuMTzIpHUYPpXwN/ZbuvGH/BLj4s/EV7BpNc1rVxr2mSFdxktdILK7DuTh9SX6kepFf8ABJv4tH4lfslaX4bvrsS6l4C1CbRpQx+cwcTW7H/ZEcvlj/rkfTNfsh2L/tbf8FO/FfxynzdaD4av7zWoJSMq0ceLXTkz/eC+VIPXyTX7fn/JmnxZ/wCxfk/9DSv+CHX/ACKnxb/7CGlf+i7mv+CvP/Jmmpf9jBp3/obV/wAEhv8AkzTTP+xg1H/0Na+LP/JK/GX/AGL99/6TvX/BFbRtMvv2j/FmrXdokt1pvg+U2kjDJhL3UCsy+hK5XPoSO5qv247eLwl/wVCGraCv2a4PiDw7qAK8BZ/LtSWGPVlDH3JNVX/BV7XdU+Jf7bGi/Cxr5obPR9O03TLYMMpFLdv5rygepEsYP/XMeleEvCug+BvC2k+DPC2nRWGkaHZRWdjbRrhYoo1Cqo/ADnvVf8FR/wDkxL4m/wDcJ/8ATtZ1/wAES/8AkgPjr/scD/6RwVX7Hv8AylqX/scPFn/pLqNf8FovG1/oX7N3hzwbYXDxJ4n8URi8CniWC3hkk2H280wt9UFfsrf8FLLz9m74G+HvhN4f/ZTv9dj03zpZ9Xi8Qvb/ANoTSys7SlBZPjhlQfM3CKM8U3/BaDxSylW/Y11UgjBB8VSf/K+v+CS83iHT/wBsvV7my8J6ho+j694e1JJLZ4X8u1i86GWNNxUA7SiKDhevbOKqv+CS3grRfGH7Y+j3uswLN/wjOjXuq2kbruUzqEhRiP8AZ88sD2ZVPvVfG/47Pon/AAUt8UfGDXPBc3jZfBXjOeC10MXRtzKdOBt4NriOTYEeFJeEOSp9S1f8PofFX/Rm2q/+FVJ/8r6/ba/bi139sj4e6H4Kk/Zr1PwneaFrI1CLUzqsl+xj8mSN4Qn2WLaGLxsW3H/VgY5yP2IfDlz8Uf8Agnf4R8B+Ore7tU13w1qmhXcc0ZSRLUz3NqpAIH/LEKR7Y69a8D+J/wBpf/gk38adYsvEfgePWPDHiEpBcs6vHY67DEXMM1tcgHy5lDv8rAlRIwZOQR+zF/wUU/Z8/advoPC2kajd+F/F8yjZoWt7Y2uWx8wt5VJSbGPu/K5HOzGa+LP/ACSvxl/2L99/6TvX/BGDwpout/tN694h1K0We78O+E55tOZh/qJZZ4YmkH+15byJ9JGqv+C2/gTR7v4P+APiYYI11XSvEraQsgXDPBc20sxBPcK1opAPTe2Opr9lv4u65pP/AASjf4gi6mOr+EfB+uQWc2TlXtXuUtcHOQFVYV9tpx2r/giP4C0jWviT8SviVfQxzah4a0qysbNpBkx/bZJmkdfQ4tAueuHI6E1X/BbfwFpF38JPAHxQ+zxLqmmeI20cShcO8NzbyzbSe4VrXIB6bzjqa/4Jn+L77xn+xP8ADe71OaSW50y3utMLv/zztrqWKED2EKxr+Ffsr6HpXiL/AIKvw6frFnHdQReO/Ed2iSLkCa3jvZ4X+qyRIw9Coqv+C2+kWFv8avh9rcMKrd3vhiSCZwANyxXLlM/TzX/OvgnPJ4h+BPgG51hvtMmpeEtNe6L/APLUyWkZfP1yfzrVvCf7Rv8AwSk/aIvPHfh/w4Nb8G34ksrfUJ4Xex1bT5JFdYZZF5guVKJ1wQykgOhIb9mn/gp/+z3+0Nqtl4O1N7vwH4tviI7fTtZkRre6lOMRwXK4VmJOArrGzHgA1/wUB8bX/wAPv2Nfin4j0y4eG5fRhp6SRnDJ9snjtSQexxOee3Xiv2E/23ZP2RPCXibTtH/Z5u/G9/4k1GKW51aDWWs9kUUeI4CotZc7WeV87h/renGT/wAPofFX/Rm2q/8AhVSf/K+vAnxT8UfFz/gpD4M+O+h/CzU/CSeJvG+k+fpis90LZZPJtrlzN5Sbg482RjsXG8jPG6qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq+LH7R/wI+Btu03xX+Kvh/w9Kq7hZz3Qku3GM5S2j3SuMf3UPUetfGD/AILUfDPRTNpfwP8Ahpq/im7+7HqOsyCwtg3ZliXfLIOnDeUfy5j8bf8ABWv9sxAvhrTtW8B+F7zLCezh/wCEdtQh4ys8h+1SpzjCM49utfC//gitaXF6uvftCfGq91e6mcSXNj4ehK+Y/U7ru4DM4Pf90p68jPHwi/Yx/Zi+By283w++D2gwajbYMeq30H269Dd2Wefe6E+iFR7CqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqlijmjaGWNXR1KsjDIYHqCPSv25P+Cfvif4P+JZP2sv2Pmu9In0W4/tHUdB0qPZJpjrkvdWYXrFjO+DGFBbAKEon7Av7f/h39qnQU8EeN5bPR/ifpcG65tEPlxavEo5uLcE8MBy8XVfvDK521VVVVVVVVVVVVVVVVVVTukaGSR1VVBLMxwAB1NeHvG3gvxfNf23hPxfouty6VKIr6PTr+K5a0cjIWQIxKMQCQGwaqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv+CmPwnPxZ/Y98Zw2lqJ9R8JrH4hsRtyVNrkzEe/2Z7gfjXiX4/av46/4J+fCf9mbQZXu9cuPHN7YSW6H95LDAY5beMjP3Wk1GMLx1t8fwmvhP8JPD3wx+Cvhr4LxWlvdaZouhRaXcoUyl2fL2zuwPXzGLsfUua+CfxF1D9iPxt+1b8E7/UJLcR+G9Qt9GllODJeQzeTYygf7cV4ZPfA/D/gjR8Hx4P8A2fta+LN9bbb7x/qxW2cjrZWZaJMdxmdrnPrha/b8/wCTNPiz/wBi/J/6Glf8EOv+RU+Lf/YQ0r/0Xc1/wV0t5p/2MdYkijLLBrunPIR/Cvm7cn8WUfjX/BHvVdPv/wBj6KxtLpJJ9N8TX8N1GDzE7COQA/VHU/jXxZ/5JX4y/wCxfvv/AEnev+CJf/JfvHX/AGJ5/wDSuCq/4KD/APKTBP8AsIeHf/RVvVV/wWK+GviHwL+0T4X+OejxyRWXiLTYI0vI0x5OoWbdCR38owFc8na3Za/Zr/aC8FftL/CbRfiV4P1K1klubdF1awjkzJpt4FHmwSL1XDZ2k/eXawyDmq/4KK+FL/xp+xV8VNG02J5JoNKi1AqgydlpdQ3Uh6HgJAxPt6da/wCCJHxM0AaD8Qvg9dX0MOsfbodbs7d2w9xCYxDMVHcIyQ5/66ivHvjvwj8MfB+q+PfHeu2uj6Fotu095eXDbVjUdAB1ZiSFVRksxAAJIFfsAeKYfHX/AAUx8NeNre2e3j8Qa14i1BIX+9GJ9PvpApx3AbFf8FfPhTq3xC/ZYTxVoNk1zc+A9bi1S6VI9ziyaN4ZiO+FMkTtjgLGxPAyP+CUH7QPhb4l/s26T8KX1S3i8W/D8SWl1YO4WSazaVnguEXqUCuI2I6MnONy5qqr/gjb/wAnc3f/AGJ99/6Otqr43Xlz+xb/AMFTh8WfEWnSr4e1DX211Z44c+dYajG8d5JGo+8yNLcjHUtH/tZrQNf0PxXoll4k8M6vZ6ppWpQLPZ3tpMJYp42GVZGUkEEdxVMVVSzMAAOSeMVrugaF4o0m50HxLothq+mXiFLmyvrZJ4Zl7q6OCrD2Ir/gqH8Bfhx+zF8cvCPiT4Gk+GJtetG1RtMsZyn9l3UEy7J4ADuiV25VRwrRttwOBfa1q/iX9mS48R+ILfyNV1XwI11fRbdvlzyWO+RcdsMSMV/wSX+MvhL4TftOyaX4z1CDT7PxrosmkWt5O4SOG782KWIOx4AfymQf7TJ71X/BaT48eHtbl8Ifs7+HNVivr/SL1tZ16KGQMLSUxGK2ibGf3hSWZipwQrIcfOK+An7N99on7AGmfs5+IbdbPVNc8GXttfpOmTa3WoLNK4br80b3GPYpx0Ff8EsfjHZ/s4ftL+KPg18VZx4eXxYo0if7Y4jW21a0mcRRSknC53zxg/32QfxE1X/BZn9oLQPFWq+Ff2cPCWopqN3oN62q68tu+8QXTRmK3gIH/LQJJKzDqBInqQP2LvhJqXwO/Zd+Hvw11y2Nvq2n6WbjUYW+9Dc3Mj3Esbe6PMU/4DxxX7Hv/KWpf+xw8Wf+kuo1X/Bb3/kqfw0/7F+6/wDSgV+z1/yQH4Z/9ifpP/pHFV9Y6fq9hNp2pWdve2d1GUmgnjEkcqEcqynIYEdjxX/BXH9mT4NfBLVPBnxH+FOjWXha+8VXN1DqGjWGIrdzCsbLcQwjiLG/awQBOUIAO4tfeGfHP7UX/BMS20TWIptQ8XeJvh/bXMQmG+S/u4FSaBjn+OZoY2z6yZr/AIIxfH3wz4fj8W/s3+KNRh03VdT1MazoaXDCP7XKYkhuYBux+8CwwsE6keYf4TVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVXxT+PHwZ+CWnjUfiv8S9A8Mo6F4ob68VZ5wOvlQjMkn0RTXxm/4LR/CPw2ZtM+CPgLVvGV2oKrqOpsdNswezKhDTSD1Vli+tR+P/APgrJ+2oWHhLTdV8B+FbrkTWMJ8PWgQ8bluJT9pmXsRGzjrx1r4U/wDBFfS5Lpde/aG+Ml9rN5NIZLmw8PRmNXc8ktdzhncE9f3SH3yePg/+yH+zb8CFgm+Gfwj0LT9QgA26pcQm8vcjuLiYtIuTzhSB7DAxVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV/wAFA/2B9b+GuuT/ALW/7Ki3Gi3WjTnU9b0nSz5T2EiHc19aBeijkyRD7vLAbdwX9gj9urw9+1p4QPh7xGbbSviToFsp1XT1IVL+MfL9rtx/cJxvTrGzAdGUmqqqqqqqqqqqqqqqqq+O3/BQH9lv9n5rnTfFPxEg1rXrbcG0Lw8Bf3SuOqPtIjhbpxK6HmvFn/BWL9pT4261L4L/AGTvgS9vPISsc5s5NavwpOFk8uNRFD77xIo9e9RfsK/8FJf2pXW//aG+LMvh/S7ghnsda1kzYUnOUsbTMCnn7rGM8YOMCv2Jf2HdL/YysfEkdj8S9S8VXXilbX7aJbFLS3jaDzNrRxhnYE+awJLnIA9KqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqtU0yw1rTLvRtUtkubO+geC4hfpJG6lWU+xBI/Gv2Zv+CTHxK+EH7Svhz4oeO/Fvg3VPB/hbVJb+ztrK6uXvZXjDG0Zke3VFKyeU7Yc4KEDPWq/b1/4JnfEz9pf45D4ufCXxH4N0qPUdIt4NXh1y6uYZJbuHcgkXyYJQVMIhXkggxnsePgx8NdN+Dnwl8I/C3SdjQeGNHt7EyIMCaREAkl+rvuc+7Gv2nvhfr/AMa/gB44+FXhW80+11bxLpbWlpNqEjxwI5ZTl2RXYDjsp+lf8E5P2Nvif+yBonjnTfiXrvhfU5fE11ZS2h0O6nmCCFZQ2/zoYsE+YuMZ79K+Nvwj8M/Hj4UeJfhF4vMi6X4lsjbySxAF4HDB4pUB43JIiOM8ZUVof/BNT/goF8CvEmoWfwG+MWn2el6kyrLf6V4hn0wXCKflaeDbwygk4G/GSAxya+H3w++Jlj+zRY/DH4keKLfXPHB8MS6dqOqyXUk6XFy8ToHaV1DuPmXLFcnBODX/AATv/YB+Mn7I/wATvEnjX4keJfBmpWOsaCdPgj0S8uZpVl8+KTLCW3iAXbGRkEnJHHeq/al/4Jz/ABu+OH7Xy/H/AMJ+KfA9p4eW60qX7LqN7dR3eLVIlk+RLd0yTGdvz85GcVVfGv4J/Dr9oP4eah8Mvidoi6jo9/h0ZTsmtJlB2Twvg7JFycHpgkEFWYHV/wDglL+1Z8EfFdz4n/ZV+PdukMhZUY6jcaNfGPOVjk8oNFKOxJZQTg7Rnj/hh3/gp18WjBp3xc/aii0rRSV+0W//AAkNzOSAxxm3gjWKU98O47c+jael7pB0rXFt9QWe28m8DQYjuAV2vlCThW5+Uk8HGTXxl/4JFfF/wZ8RZvH37I/xCtbOxFy0+n2dzqc2n6hpJYn93DcICJEUHAcsj4IB3EFj4H/4JqftQ/GTXdJvv22P2htR1vwvpkyzjw9ba5c38s7DPylnCxwZyQXTexXIBXhh8MP+CdXxE+Fn7eNt+0f4bufAdj8OrC/vHstEtLu5ju7e2msJbZFWP7P5YKtIGI8zBAPOa1HTrDV9PudJ1WygvLK9heC5tp4w8c0bgqyMp4ZSCQQeCDXxo/4JDeLNC8byfEX9j74rDwpOJmmt9Lv724tJNPY5yLa9gDPt5wFdQQOrtVr+zT/wWH1JF0a7/aCWxtmURm6l8VEFQMcl4omlz7jk1+xH+zh8W/2c/B/iTT/jD8Xj471nxNqiajJKXnn+zyCIRufPnbfKWCR8lVxsHXPFV+wJ/wAE5vjd+yr8dJ/id8QvFPgfUdLl0G509YdGvrqWfzZJImU7ZbeNduI2yd2eRx6V+1h+yF8MP2uvBcHhzxws+n6vpRkfRdds0BuLB3ADDB4kibau6M4ztBBUgMNH/wCCe/8AwUa/Z/nn0j4AfHyzk0GSRjHDZa9PZIc9Xe1lQxI59VLHnr1q0/YY/wCCl/xZntrb4y/tVjRtHSRWlgj125u2yrZVvs0KxwyEYB+Zx298fGr4fa58Vfg34s+GOj+LF0DUfE2jTab/AGqLMzCASpskYRh1PzKWX7wI3Z5xWk/shf8ABVz4N2Q8M/C79ovTdU0OAeXaRHXHlEMfYLHeQkRdPuoSBng9a+EP/BKz4leL/irb/GL9tP4s2/jO6gmjmfSrW5mvGvjHjy457iZU2xDABiRSCOAyjr8WFVfhV4xVQAB4evgABjH+jvX/AAT1/Zh8EftZeK/H/wANfGN5c6dLB4XF5pWqWy7pLC6W6hUSbCQHUq7KyEjIbgqQGF9+x1/wVa8FWg8C+Bv2lzqnhuIGG0uo/FE8EkMPRQTKnmxYXGFjZguMKelfsmf8EpbT4a+ObT4xftGeMLXxr4qs7oXtpp1qZJbSO6zuE88soD3DhvmAKqAwyd/av21P+CaPgP8Aai1ab4j+Ddbi8HePpIgt1ctbeZaartGF89FIZZAAF81cnAAKtgYs/wBkD/grXoenL4A0n9odF0RE+zx3cXjGfbFH0GyRovtCqABgKAQOgFfsi/8ABK/wt8FfFlt8Wfjd4ng8e+NLWc3NnbpExsbK4Jz55Mnz3Eob5ldwoUnO0sAwr4Df8E5vjd8Lf24x+0x4g8U+B7jwwNe1vUPslnfXT3vl3sF1HENjW6x7gbhNw8zAAbBPGa/4KLfsH/F79rzxp4Q8R/DXxH4P0228P6XNaXKa5eXMLu7yhwUEMEoIx6kc9q8MeAvEeifAbSfhdD4hg0zX7DwjDo6araxtPHa3SWghE6KShdVcbgDsJAH3c8aJ+xD/AMFOfgHbnw/8Df2jtNv/AA7AdtnaHVpEWJPa2uonii+iMfrXhP8A4JbftBfGv4iWPxD/AG2PjdHrdvbbA+nWF7Ld3M0Kkt9nErKkdtGSST5Qbq2NpO4aVpWm6HpdnomjWUNnYafbpb2ttCgRIYkUKiKB0UKAAOwFftc/8EpfD3xi8Y3nxZ+BXiy28EeLb+5N3fWV1G4sbq5J3GdHiBe3kLfMxVXBY5AU5J039mH/AILA6BFF4e0z4+mWygIjjuZfFjTgL0zvljMuB9MjHAr9i79kr9p/4O/EvVfiz+0T+0D/AMJpe6ror6b/AGWL671ER7po5FkE9xs2FfLZdioR85545qqqqqqqqqqqqqqqqqqqqqqqqqqrx78Tvhz8LdIOvfEjx1oXhnTwDtn1W/jtlcjsu8gs3T5RknI4r4zf8Fkf2ffBXm6f8JfD2s/EK/XIW4wdMsgen+slUytz6RYP96v+F+/8FTv21la1+E/hfUfBnha8HF3o1r/ZFvs9RqFw3mOQDkiFwTnO3oK+F/8AwRh1HXL7/hKP2l/jZd6hqF03mXVl4f3SySNxy97cgljjgjyvo1fBz9jX9mj4DJbzfDn4S6Lb6lbj5dWvoftt9nqSJ5tzpk9kKjpxwKqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqZVZSrAEEYIIr9vX9jrxb+y78Qbf9sn9lYT6Pp2n3y3mqWGnxALoc5IBmjTo1rKWIePBVNxGPLban7Fn7XvhP9rr4Xx+IrNYdO8WaMscHiPR1b/j2nI4ljzyYZMEqeSMFSSVJNVVVVVVVVVVVVVVV+07/AMFLP2e/2dDeeHdO1QeOvGVtlDoujTgx27+lxc4KR+hVd7g9VHWm8d/8FIv+CkEstn4Mtp/B3w6uJGR5LR30rTNnQrJcnM15jjcibwDzsWvgN/wR0+BngNLfV/jRrd98QtXQBms03WGnRt1xsRvNkxwMs4U905xXg7wL4K+HmiReG/AXhLR/DulQ/wCrstLsY7WIH12oAMnuepqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqviz/ySvxl/wBi/ff+k71/wRL/AOS/eOv+xPP/AKVwVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVreu6H4Z0q41zxJrNjpWm2ib7i8vrhYIYV9WdyFUe5NfGr/grB+yr8K/tGm+E9ZvviJrEJKiHQI/9FDg/wAV1JhCv+1EJO1ah+3f/wAFDP2vL2fQ/wBmP4W3HhvSJGMZuNGsxcSRg8FZdRuQsMZ54KiJh614B/4JB/Gb4o6wvjT9q746z/a7hg09vZXMmrX8q5+ZXupzsjb6CUc18Gf2Av2UvgaILrwt8KtP1XVoMEavr6/2lc7h/GpkBSJveNEqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqtQ0+x1WwudL1Ozhu7O8heG4t5ow6SxsCrIyngqQSCDwQa/aO+DPxH/4Jj/tG6V+0N8Coppvh7rN28cVrJIzRRK53TaXcnqY2A3ROcn5Acl4yx+BPxt8EftD/AAv0b4qeAL0y6dqsX7yCQjzbKdeJIJQOjoeD2IwRkEE1VVVVVVVVVVVVXx5/aM+EX7NnhE+MPiz4rh02GQMLKyjHmXeoSKMlIIQcueRk8Ku4FmUHNePP2tP2y/8Agol4ruvhT+zT4U1Hwr4MzsvPsdx5T+SeN9/fcCNSMnyY8bgSuJSAa/Ze/wCCTPwY+EK2fij4ym3+I3iuIiQQXEJGl2jjssDf6/HrLlTwdimre2t7O3itLSCOGCBAkUUaBVRQMBVA4AAAGKqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqmhhuIXt54kljkUq6OuQwIwQQeoI7Vongrwb4ZuHu/DfhLRdKnlTZJLY2EUDMuQdpKKCRkA49qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr41/t/fsq/AgT2fij4oWWr6zBlTo3h8jUboMOqNsPlxN7SuleNv8AgrN+0P8AGrW38C/sk/BGa1u5/lhuZLR9Y1DacDzBCi+VDgnneJVHUkdtE/4Jy/tx/tU6tB4r/as+L0+g2jN5iW+qXh1O6hBycQ2kTCCAcngOm3d93qK+Cv8AwS7/AGTPg+LfUNQ8HS+O9Zhwftvihxcxhh122yhYcZ6b0cjA+bqTp+nafpNjBpmlWNvZ2dsgSC3t4hHHEo6KqrgAewqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqvil8MfBfxl8A6z8NPiDpEepaFrlsYLmFuGXusiN/DIrAMrDkMAa+Dvjnx//AMErf2sdS+EnxNubm++GfieZDLdLGfLmtGYrBqcSjOJI+UlRckhXX5isZqwv7HVbC21TTLyG7s7yFZre4hcOksbAFXVhwVIIII4INVVVVVVVVVVVVftwf8FF/Af7Ldpc+BvBi2fij4lyxfLp3mZttK3DKyXbKc7sEEQghmGCSgIJ/Z9/YL+On7avi5P2i/2x/FOtWeiamyzW1lMfKvtShByqRx422drzwAoJBJVVDB6+H3w48CfCjwrZ+CPhx4V07w9odiuIbKyhCLnABZj1dzgZdiWY8kmqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr4q/HT4PfA/SRrPxY+I2h+GYGQtEl7cgTTgZz5UK5klPB4RSa+NH/BaHwZp80mgfs7/DW/8AFGoSP5UGp62GtbZnJwClumZpQeBgmI5ND4Yf8FTf26wk3jrWNQ8EeDL3kQak7aHZGM44FpEDPOCOVMqsDn74BzXwS/4I4/AHwN9n1T4va/qvxD1OPDNa5OnWAbr/AKuNjK+D/ek2nuvOK8EfDzwF8M9ETw38O/Bmi+GtMTBFppVjHaxlgMbiEA3N6sck9zVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV+3T+yZo/7WfwaufD1rDbw+M9BEl54Xv5MLsuMDdA7dRFMFCt2DBH52YP/BKf9rDWLO8uv2NPjLNPZa94eeaPwyb8lZQIifP05w3IeLazID/CHXgIoNVVVVVVVVVVV+39/wAFHB8Nbm4+AX7N18NX+I13N9i1HUrSL7QNIkYhfIhXBEt2SQuACIzwcvwv7Cn/AATOj8HXdp8d/wBqK1/tvxzczfbbHQr5xcpp0pJbz7piWE9wSQwBysZ5JZ/uVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVf39hpVlPqWp31vaWltGZJri4kEccSAZLMxIAAHc18c/+CqH7K/wfFxpnh3xFN8Q9ciBC2nhwrJbBv8AbuyfK28HmMyEelaj+2v/AMFEf2z7240H9mr4dXnhbQJXMLXOhw5aIHqsupz7Y42HrH5TV8LP+COHivxdqx8a/tWfGi7vtQvHEt3ZaNO91czsf+et7cAnd0BAjbvh+hr4L/smfs8fs+wxn4WfC7R9Nv0Ta2qzRm6vn9c3EpaQA9SqkL7Cqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv+Cq37NGt/D7xXpP7bXwYSbTdR06/tm8RyWS7Ta3SOottQH1YJG/Yt5ZIO5jX7IH7SWh/tT/AAP0X4lWBt4NYVfsev6fE2fsV9GB5igZJCOCsiZ/gdc8g1VVVVVVVVVV/wAFHf2/7r4No/7P/wACr/7V8SNWQRale2oMkmiJKBsSMAHN1IGG0dUBDYyy4/4J3f8ABO61+B1rafG3426fFffEm+j82xsZsSpoCOOSTyGumBO5/wCDJVT95jVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVftW/tFfFb4WfE+Lwx4L122tNPbSobgxyWUUp8xnkBOWUnoo4r/htL9oP/AKGuy/8ABXB/8TX/AA2l+0H/ANDXZf8Agrg/+Jr/AIbS/aD/AOhrsv8AwVwf/E1/w2l+0H/0Ndl/4K4P/ia/4bS/aD/6Guy/8FcH/wATXgb9r74765428PaLqPiezktL/Vba3nQabApZHlVWAIXI4J5qqqqrxP8Atp/sqeC/Emo+D/FXxy8NaZrOkXT2t9Z3EzK9vMhwyN8uAQRikdJEWSN1ZWGVYHII9RVfEb4nfD/4ReGJPGfxL8Waf4d0WKZIWvb2TYgkc4VRjJJPoPQ+lfCr9ob4J/HG41G0+EnxI0jxRNpCRvepYuzGBZCwQtkDqVb8q1rWtG8N6Rea/wCIdWs9M0zToGnu7y8mWGG3iUZZ3diAqgDJJOBXw2+Lnww+Mejz6/8ACzx5oviiwtZzBcTabdrMIZAM7XAOVOMEAgZBBHBzVVVVVVXirXE8L+F9X8SyWxuF0mwnuzCG2mQRRl9oPOM7cZr9jP8A4KG6N+2J471vwPpvwuvfDEmi6R/aLXE+qrdCUeakewKI0wf3mc57VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV8Yv2kPgZ8AtP+3/Fv4maL4fZk3xWcs3m3c6+sdvGGlccYyFIHcivjB/wWVv8AW9QPg79ln4R3mq6jduYbXUtdiaSSVsY/c2UBLMe6lpOwynUVY/sb/wDBRb9tq9h179o/x9e+EvD0jiSO012Ty/KHHMOlwbVRhx/rfKY45Jr4G/8ABKr9lf4QG31TxJoNz8RNciIY3PiPa9srDrstFAjKnjiXzD71p+nafpNjBpmlWNvZ2dsgSC3t4hHHEo6KqrgAewqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq8V+FvD/jfwzqng7xVpcOpaPrVnLZ31pMMrNDIpV1P1BPI5HWvgL4m8Qf8E0P259W+C/jvU5f+FeeLZo7d7y4O2N7SVm+w6gegDRsWjkPRczDnaKqqqqqqqqqv+ChH7aunfsofDcaR4Xube5+I/iiF00O1YCQWMX3XvZV6bVOQin779iqPj/gmb+w/qFrPF+1x+0BZ3F/4t1yRr/w5Z6iN8luJcsdQm3ZLTSbtyZ+4Dv5Zl2VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVft4f8lug/7AVv8A+jJaqqqvhh/yUvwl/wBh2y/9HpVVVVXx18La38S/2gf2hvFWkQyyR+Gtd1jW7uNjudbY6xHbevRTdRk46BSegJH/AATz+NH/AAvH9k7wVr95eC41jQbc6Fq5JJYXFoAisxPVnhMMhPrJVf8ABWzxxqvxX+Lvwn/Y88HXga61HUIL2/RfnAurqT7NaBgOhRDM5B/hmU8V/wAEX3TR/j38UvC0bySKuhKVcnAIhuwmSPU+Z/Ov2mPgv/w0P8CvF3wbXxDJocniS0SOK/jjL+TJHMkyblBBZC0YVhkZVmFf8E9/2HfEX7HOmeL7rxd46std1bxbJaq1vpsci2ttHb+btbL4LuxmbOVG0DAzk1VVVVVV8Wf+SV+Mv+xfvv8A0nev+CJf/JfvHX/Ynn/0rgqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq5uLezt5bu7uI4YIELyyyMFVFAyWJPAAAySa+PP/BUH9lr4KfaNK0nxO/j/X4cqLDw2yzwo3/TS6J8pR2IUuw7rWoftl/8FEf229Rm8Pfs2eA73wj4ekcxvc6Iu0xDuJtUn2qjDOf3XlMccA9/g3/wRrutX1D/AITH9qf4r3er6jdv51zpmhzM7SueT517MCzk9GCoOhw5618IP2dPgh8BNNGmfCT4a6L4fymyW6hg8y6nH/TS4fdK/TozEVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV/wVa/ZhHxs+BbfE3wzYCTxZ8N45b1Qi/PdaaRm5iwOpUKsq56bHA5ev+CYH7TR/aA/Z8tvDPiG/wDO8XfDwRaXqJkk3SXNttP2W5Pf5kRoyTkl4WY/eFVVVVVVVVfHz43eDf2dvhTrvxY8cXBFjo8H7m2RsSXtw3EVvH/tu2BnoBljgKSP2Mfgb42/b+/aI1r9rT9oOD7X4T0rUg0NlIp8i9uY8GGyiU/8u0ClCwP3jtB3F3NBVVQqgAAYAAxiqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv28P8Akt0H/YCt/wD0ZLVVVV8MP+Sl+Ev+w7Zf+j0qqqqr/gm/4G0346ftA/HzQdTlg/s/xf4D1qwkKHeF+2X1vtkQ8/dK7gfXBr/gjt8S9T+Hfxn8f/sz+L2Nncakklzb2sxwYtRsnMdxEB/eaMsT7W/ar69s9MsbjUtQuY7e1tImlnmkbasaKMsxPYAAkmv2E1vf2vP+CiXjf9prWreWTSPDRuNRsVmGfJMoNrp8LA55W3Dt/vQ5r/gk68mgft0fEDQZ2MjSeHdWtWKHC749QtW3Y9MIwH+9X7Zvi/xN4A/Za+JHjLwbrVxpOt6TorzWV7bkCSCQOo3LnPOCa/4JH/Hb4v8Axx8OfEq7+LXj/VPFE2kXunJYvfOGMCyJOXC4A6lVz9BX/BQL4kftgfshfG/R/il4N+K+uan8MvEOorcWmlXjiW2t7hCHm0+XjPluoZk5B2lgOYya+Bnxl8IftAfCvw/8WPBFzv07XLYO0DNmS0mX5ZYJP9tHDKexwCOCDXx6/a9/aE/aa/bB039nz9jn4gX+h6Hps7afLqenlTHdOrZu76R8HMEQXamD820lSTKBWlRWXw2+H8C+KPF1ze23hvS9+pa7q84Mkqwx7prmZugyFZzjCjoAAAK8Q/th/ti/t2fFbU/hf+xYreDvBmllhca9JiCUwklVnubllZrcPjKRQr5n3sl8Hb44/Zw/4Kqfs96Fd/FXw3+05qfjr+x42ur3TIvEF3qLiNPmZltb1DFKoG4lR8xA4U1/wT8/bftP2u/BN9pfiiyttM8f+Fkj/ta2t/lhvYXJCXUKkkgEgq6ZO1tvOHUD9sf9rLwn+yJ8LD411izXVdd1OY2ugaN5vlm9nAyzMwBKxRqdzNjuq9XFfDhv+CpH7ei3PxE8K/Fm98D+FmuHSzuoNWn0CwJUnMcC2qtPMqn5d7B+eC5IOPFnj7/goZ+w9od5p/7Ruor8UPhj4gs5dMudYhvW1BrCSZGQOLiREnVssMLONj/dUhjkf8ES/wDkv3jr/sTz/wClcFVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV4/+JXw++FXh+XxV8SPGej+G9JiODdaldrArN/dXccsx7KuSewr48f8ABZjwHosk3hz9nHwPd+L9TZvKh1fV43tbPeehjgGJpueMN5Rz694PgB/wUu/b3mj1P4weIr7wZ4LumEiW2sFtNtQmePL06IeZIQOVaZRkH/WcmvgR/wAEnf2Y/hIbbVfG1hc/ErXYQC02uRhbJX7lLNSUK/7MrS9a07TNN0ewg0rSLC2sbK1jEcFtbRLFHEg6KqqAFA9AKqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqpoYbiF7eeJJY5FKujrkMCMEEHqCO1ad9q/4Jo/8FGTYO0lt8NfGEgQFiRGNIvJPlbJ72s64J+8Vhb/AJ6UCrKGVgQRwRVVVVVVVV+1V8RvF/8AwUe/a60T9mr4P6gT4G8L3skZv48vAxjO271J8cMiLmOLnDZGCPOxXwu+GfhD4OfD7Qvhl4D0xLDQ/D9ottaxAfM2OWkc/wATuxZ2buzE96qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr9vD/kt0H/YCt/8A0ZLVVVV8MP8AkpfhL/sO2X/o9KqqqvE+rLoPhrVtcMiINOsZrks6khfLQtkgckcV/wAEQtM874ofEzWRCp+y6BaW/mE8r5k5bAHofKz/AMBFfto2V1+x7/wUl0X446NbvDpOtX9p4lKwr/rEkYw6jED/AHpNs5PoJxX/AAU0+OFt8M/2O9em0HVIzffEARaJpksUmd8Nyped1xyVNskq57F19QD/AMEmvgofhb+yxZeL9SszFrHxFvG1eUuuHW0H7u1X/dKK0o6/68/QfsHhfDP/AAVJ8V+Hlt5YgdU8T2IjJIMflySths8nHlY+uK/b8/5M0+LP/Yvyf+hpX/BDr/kVPi3/ANhDSv8A0Xc18dPgz4P/AGgfhXr3wn8b2wfTtbtiiTqgMlnOOYp4/R0cBh2OCDwSDbfGn9oD9gtPjP8AslyXCpJrQ+yfaEmYCxkcJ/p1tjp59mwHZhmJsgx7T/wTP/Y9tP2cfhLF478V2UD+P/HNrHcXsgAc6fZth4rRW9fuvJjgvgciNSf+CtPxCv8AwJ+x1q2nabdNby+MNZs9Fd1JDeW2+eRQR2ZLZlPqrEd6/YY/4KG/AX9kr4JjwBrHw28W6j4i1DU57/V9QsEtjHMzEJEql5VbasSIMEABi5HUk/8AD7T4A/8ARKPiB/3zaf8Ax6v+CcHj3SpP+CjYu/AOnXOl+G/Gs2vRWljKFV7exaOa7hikCHblfs8IIHGQMdq/4K2eJNX+JX7Y+gfCi2vQtvoelWOn2sbZ2x3F5J5jyHPcrJCDjjCDvmvAfgnw78N/BWh+APCVilno/h6whsbKFRjbHGoUE+rHGSTySSTya/aH8KaJ44+A/wAQfCniGzS5sNQ8NXySI6g7SIGZHXPRlZVZT2ZQe1f8ES/+S/eOv+xPP/pXBVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV8Zv2iPgt+z7ow1r4ufELStAWSMvb2ssvmXV0B18m3TMknPGVUgdyK+LX/AAVu+K3xU17/AIVx+xr8KdR+3XpaO21K9sP7Q1GU/wB6Gzj3xpjrmQyDB5VcV8Pv+CX37Tn7R/iCL4k/tj/FjUdK+0EO1lJdjUdTMZ52DkwWq9MKN+3oUXFfAb9jP9nT9nCCGX4a/DuyXWI1w2u6iPteoOSMEiZxmMEcFYwin0qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr/AIK2/s+L8V/2d/8AhZui2Xm+IPhnK99lFy0mnSbVul+iBY5snoIn/vV/wTQ/aAb48/svaHFq995/iPwQRoWql2+eQQoPs8x5yd8BjBY/edJKqqqqqqr/AIKl/tVt8Bfgofh54S1QweNfiFHJa27xPh7GwGBcT8cqzBhEh45dmBzHX/BLr9k6P4BfBeL4i+K9NEfjj4g28V1ciVPnsLA/Nb24yMqzAiSQcfMVUj92DVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV+3h/wAlug/7AVv/AOjJaqqqvhh/yUvwl/2HbL/0elVVVX7RerHQf2fPifrqyOh07wbq9yGRQWXy7OVsgHqeK/4Ia6V/yWTW5IP+gLbwy7v+v1pFxn/rmckfTvX/AAWS+DP/AAm37Puk/FrT7fdf/D3Ux9oIXn7FdlIpOnJxMtsfQDca8Y/E/Xf21bH9lf8AZo0u/llvtJ04aZq0gOTFMbk2/mOTncUsrSOYk5/1rdTmvD+g6T4W0DTfDGg2aWmm6RZxWdlboMLDDEgREA9AqgfhXwTC+F/+CzGpRhJ9tx478SblY4J+0214Qe3y5kBHsB161+35/wAmafFn/sX5P/Q0r/gh1/yKnxb/AOwhpX/ou5r9r/8Aab8NfsqfBjVPiJqrwXOtTg2nh7THPN9fMp2AgHPloAXc8YVSAdzKD8Kf2GPi9+1z8CfiT+1V4i1TUbzxnq15JeeGbaQDdrskbsbskEcBjmKILgB4yMBQK/4JL/tgzeN/DX/DL/xH1Jv+Ej8LWxPhua5bD3dhH962OeTJAPujr5fGAIiT/wAFZfhzqfxA/Y81e/0mza6m8Havaa48aAlhEgkhlcAdljuHc+iqx7V/wSU1/wCDvxP/AGcB8PNV8J+Gr3xX4F1C4jvEvNNhluJrW4meeGfcyksu6SSPrx5QBwCuf+FTfCv/AKJp4U/8E1v/APEVpPw/8BaBfJqeheCdA068jBCXFppsUMigjBwyqCMjiv8Agrr4U1v4cftc+Hvi3YW4WDX9JtLyznZcq13ZPsdCD1KqtuT2w4r4UfEzwv8AGT4ceHvid4Nvo7rSfEVjHdQMj7jGSMPG3o6OGRgQCGUiv2rPiZ4Z+Ef7O/j7xp4pvY4LeHQ7q3to2bDXNzNG0cMKerO7KPYZJ4BNf8ES/wDkv3jr/sTz/wClcFVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVfHH9pn4H/s5aN/a/xa8fafpEkkZe105W869u8f88rdMuwzxuwFBPJFfE7/AIKgftIftHeJm+F/7GHwy1TS/tJKperZrf6pLHnBkK4aG1TB+Zjv29fMWvg3/wAEivH3xC10fEr9sj4o393qN+4mu9Jsb43l7O3924vX3AdMERh+Dw4r4R/An4QfAfQB4a+EfgDSfDdmQBK1tEWnuCOhmmcmSU+7sTVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVqul6drml3miavZxXdjqFu9vdW8q5SWJ1KujDuCpII96/Yj1PUP2L/wDgoV4u/Zk8R3rpoXim6fSbaSdtokcZn0yc/wC1JE/l4H8Vx7VVVVVVVrmtaT4a0TUPEevX8VjpmlWst1e3UxwkEMal3dj2CqpJ+lfAnQNV/wCCln7emr/GHxpp80nw88ITR3TWVyAyJZxMwsLFhyP3rq0ki9CPP6bhVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX7eH/JboP+wFb/8AoyWqqqr4Yf8AJS/CX/Ydsv8A0elVVVX7TnhDxZ8Qf2efiJ4D8C2f2vXvEXh2706xgNwsAkeaMx4LsQoGGOcnBGR3r/gl7+y18Wf2YvBvjrT/AIveHLXStT1/U7WW2EGoRXQkhiiYc+WzBcM7dcE5r4rfD7Sfix8M/FPwz1vAsvE+kXOnyvtyYvNjKhx/tKSGHoQK/wCCdX/BOv4q/s9fGq/+LHxqtNFRtM0qS20KOxvhcn7RMdskpwBt2xB056+afSq1H9iT9oe2/wCCkqftG6X4StLnwIfFkWotfHVrdJBA0KrK3lbt/wArF/lxkhfev2tvh74p+LH7NvxA+HPgixjvNd1/SHtrGCSZYVkkLKcF3IVeAeSa/wCCXP7LXxn/AGYdA+IVh8YvDtrpU3iC8sJLBYNQhuvMWJJg5JiZtuC69eua/ac/Y8/a9/bM/ah0zU/iFoMHhf4TaVeCzsCutW00tnp4OZpxEjsTcT7fQ7cxqSVjzXhbwvoHgrw1pfg/wrpcGnaPotpHaWNpAu1IIY1CqoHsAK/a0/4J5/HPTv2m7D9oj9jiws4bm8uhqt1B9vgtP7O1NHBd1WUqrxzZ3FeRkyAjawFeEH8QeMfhzpy/FTwZaaVq+qaaIte0Npo7yBJGXbNEGUsskTfNjPVWGQDkV8Xv+CW3x3+DfxIf4sfsPeO5bdVd5LXSm1P7Df2IY8wRzOfLnhxxiVlJGAwfljaeJv8AguDKi6HH4eInCmMXc1poCkED7xcnyyeOuMH3zX7D3w4/bF8FReMNd/a2+IFt4gu/ET2j6XZLqH2mTTTGJRKNqIsMavvj+WIkZQk47/tXfsueBv2svhdN8PPF08mn3lvL9p0fV4Ig8unXIBAcKSN6MDtdMjcO4IVh4Y/ZS/4Knfsh6vqGj/ALU59R0S6lJEuj6jZXVlOSP9Z9kvv9XJgAFvLzxjcRg18Lf2Hv2t/2i/Hmh/EH/goF8Q5bvQPDlytza+D1uYJBdyrziSO0xbQoeAzJud1BT5Rhh/wTc/Yx+O/7NHxz8Z+KfiT4OtNL8PatoUtnp08OqwXR3/a4XRCqOzjMasckfw8nJFVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV8Zf2gPg7+z94f8A+Ek+Lnj3TfD9u6sbeCZy9xdkdRDAmZJD0ztUgZ5wOa+K/wDwVE+P/wC0X4ib4SfsWfDTVtOkvyUj1H7MLvVZUHV1Rd0VqmPvOxfaPm3pXwO/4JDeL/G+s/8ACyP2yPiPf3uo3zie50awvzdXc7el1fOW54wVj3ZHSQV8L/g/8MPgr4bj8I/CrwPpPhrS0wWhsYNrTMBgPLIcvK+ONzsze9VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX/BY/4Y6p4M8afDf9qjwYZLS/trhNMvLuIEGG6t2NxZS5H8XE4zx/qkGT2+DPxK0z4yfCfwj8UtHCLb+J9It77ylOfIkdAZIj7o+5D7qaqqqqqv+CwX7SB+H/wl0/4A+Gb8rr3xAPm6mI2+eDS4n5BxyPOlUIOxWOUHrX/BPn9m9f2a/wBnHRNB1exEHirxGBq/iEsuHjuJVGyA9x5UYRCOm8OR96qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv2y/hX8SPGfxdh1fwn4H1nVrJdHgiNxaWjSIHDyErkDqARx71/wz98bv8AolXib/wXSf4V/wAM/fG7/olXib/wXSf4V/wz98bv+iVeJv8AwXSf4V/wz98bv+iVeJv/AAXSf4V/wz98bv8AolXib/wXSf4V8PPgV8ZNO8f+GdQvvhl4jgtrXWLSWaWTT3VY0WZSzE44AAJqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq+InxM+H/wAJPC9z40+JfjDS/Dmi2g+e7v5xGrNgkIg6u5wcIoLN0ANfHj/grX418f6+PhZ+xX4C1C/1G/kMFtrdzpzXV3ctj/l0sgDjpndKG4zmNcZr4Mf8EqfjD8bfEC/Ff9tX4j6vHcX4WV9Kjv8A7Zqc46hJrh9yQKOnlpvIBx+7Ir4T/BP4U/Azw2nhP4T+BtL8OaeAvmi1h/e3DAYDzSnLyt/tOSaqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr9uX4Qx/G79lf4geC4bYS6jDpb6lpeE3N9qtf38ar6F9hj+khr/gjJ8Xn8XfAXxB8JL+5L3fgLVvNtFZvu2V5ukVQPadLknH99fxqqqqq8u7WwtJr6+uY4Le2jaSaWRtqxooyzEnoAASTXwEsbz/goT/wUc1b4wa3ayXHgbwbdDUYYZ48otpbNs0+3I6BpHAlZTkHE1VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVWp6npmi6dc6vrOo2thYWUTS3N1dTLFFDGoyzu7EBVAGSScCv2oP8Agrz4B8Ez3Pgj9mvSovHXiLeYBrU6uNNgkzj90ow902ePl2IcghnHFfDv9gv9r39trxRb/Ff9r3xxrPhrRZsvDb6ggF+Yyc7LezwI7NDj+NVPQ7GzmvgL+y98Ef2atAGifCbwTa6fNJGFu9VnHn316RjJlnb5iCRnYu1ASdqrVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVfsjR/8Mp/8FSfGPwPl/0TRfE09/plnGxwqQyqL6wP+95apGPeQ1VVVVX/AAVU+PLfBv8AZf1DwvpF75Ov/EWVtFtdr4dLUruu5AO48r90fQzqa/4JWfAQfBv9mDTvFOq2Pk+IPiNIus3ZdMOtqRts489SvlHzQD0M7VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX7VX/BRr4EfsyC68NxX48Z+OIgVGgaVOpFq/b7VPysP+5hpOQdmDmrPwz+3r/wVK1eLUvEV6fCPwuW4DRNJHJaaVGoPWGHPmX0owcOxZQ2RvjBxX7L/wCwB8Af2XYbbV9C0T/hJPGMafvfEusRLJOjEfN9nT7luvJHyfPg4Z2qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr/gq7pV38FP2uPhP+0joVuUkuYre4bZx513plyjnP1ilgTHTC/WtN1Gy1jTbXVtOnWe1vYEnglU8OjqGVh7EEGqqqqv2wry6/bW/4KQeGP2dtHuZJfDvhS6TRrloXOEEebjU5hzw6qrRdsmBR162NjZ6ZY2+m6dbR29raRLFBDGu1Y0UAKqjsAABiqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr45ftEfB/9nLwq3iz4teMrTSIHDC0tAfMur5xj5IIV+eQ8jJA2rkFio5r4l/txftcft2+K7n4P/skeC9Z8N+HJhsuZrOQJevCeC93eZEdpGf7qMCTld75Ar9lX/gkp8LvhW9n4z+PVza/EDxUmJE00xn+yrN/9xvmuWHrIAnP+ryA1W9tb2dvFaWkEcMECBIoo0CqigYCqBwAAAMVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX/BZfwAniX9mHSvG8EWbnwf4kgkd9udtvcI8Lj2zIbc/h78fsCePT8R/2O/hbr8twJp7XQ00uclstvsna1+bvuIhDc9d2e9VVVXxu+J2m/Bf4Q+L/irqipJD4Y0ie9SFzgTyqh8qLPq8hRB7sK/4I1/DDVPFniv4kftReMDJeX11O2k2d5N8zTXMzC5vZD/tc243f9NHGeuaqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqrWdUvLO8ENvIoXYDgqDzX9v6n/z2X/vgV/b+p/8APZf++BX9v6n/AM9l/wC+BX9v6n/z2X/vgV/b+p/89l/74FW+uajJcRRtMpDOARsHrVVVV8T/AI+/BT4LNaxfFb4oeHfDE17G0lrb6herHLOikAskf3mAJAyBipv+Cjv7EsErwP8AH/Ry0bFSUsrtxkccEQkEe44r4a/tE/An4xzGz+GHxb8L+I7xAS1nZalG1woGcsYSRIBweduOKqqqqqqqqqqqqqqqqqqqqqqqqqq+IHx1+DPwo1fStA+JXxQ8NeGdR1psWFrqeox27zDON2GPCZBG84XPGc0CrKGVgQRwRVVVVVVVVVVVVWp6npmi6dc6vrOo2thYWUTS3N1dTLFFDGoyzu7EBVAGSScCv2p/+Ct2haNeTfDX9k7Sx4t8Rzym2HiGW3aS0jlJ2hbWHG65fPRiBHnBAkBr4G/8Ezfjj+0l4qX42/tueM9bsl1ArMdJln3apdpkkJITlLOLniNRuAJXbEQDXwy+FPw6+DXhO18D/DDwhp3h7RrUfLbWcW3e2MF5GOWkc4GXclj3NVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVftueCE+If7JHxW8MmFZX/AOEaub2CMjO6W0AuYwPffCuPfFf8EW/Gn9tfs5eJ/BU9wrz+GvFMjxpnmO3uIImXj3kSc5/wqqqq/wCC0HxfPhb4IeG/g7p91tu/HGqm5vUBzmzs9rlSO2Z3gIz/AM82/D9h/wCDv/CjP2W/APgW5tPI1R9NXUdWVk2uLu6PnSK3qU3iL6RiqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqrxH/wAhAf8AXMf1qqqqtP8Aj7g/66L/ADqqqq/4Le/8lT+Gn/Yv3X/pQK+Df/BLz9jrxX8IfBHinX/AerXep614dsL68mOvXUe+aa3SRyFRwoG5jgAcCv24P+CZ1j+zn4Nl/aC/Zy8V69HZeGZ47jUtOurrNzYLvVVubWeMK2EcqWByyglg2FIr/gmZ+1J4i/aY+BU8Xj28+1+LvBV4unaheH719CyBoLh/9tgHRvUxlv4sCqqqqqqqqqqqqqqqqqqqrV/2zP2hdW/4KXaf+z4vjZLHwHYeMFsDpdpYwobmIQ5xLKVMjZJyQGA9qqqqv2c/2zP2hfi7/wAFE7n4Q+LfGyDwXpWp69aQaLZ2MMETLbJOsRdwvmOw8tT8zkZGQBmq/bk/4JreKf2rfjNpfxS8LfFDT9EhbTYdP1K11K2kmMCROxElvsOGyJDmNig3DO75jjwn4eg8JeFdG8KW15cXcWi6fBZRz3LbpZVijVA7nuxC5J9TVVVVVVVVVVVV+1L+3H8Dv2UtNktvF+s/2x4ski32fhjTJFe7fIyrS84gjPHzvyRkqr4IqS6/bj/4KseI/ItlHhL4V21ycjMkGk220/xN9++uAOMchWPSIMTX7LH7B3wM/ZUsodQ8N6T/AG/4wMe258T6pErXOSCGEC8rboQSMJ8xHDM2M1VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVavpdnrelXmi6hH5lrf28lvOn95HUqw/Imv+CLuq3XhT4vfGD4T38q/aGsLe4kjz/HZXMkDkD63Iz+HtVVVV+0sD+1z/AMFVfDXwfVzd+H/Cl7aaXcRhhtMFqrXl/wCwYnzosnuqj0FVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVeI/+QgP+uY/rVVVVaf8fcH/AF0X+dVVVX/Bb3/kqfw0/wCxfuv/AEoFfs9f8kB+Gf8A2J+k/wDpHFX/AAUa+J/hT4a/si+PoPEOpQRXvinTJNG0m0Z8SXdxONhCDvsQtI3YBD6iv+CIfgnVtM+HnxN8f3UEsdhr+q2FhaM4wsjWkczSMvqP9LUZ6ZUjsa/bW/al/aF+Hv7e6/DPwV8WNc0jwub3REOl28iiLbNHAZRggn5izZ+tV/wVb+L/AMTPgr+zt4e8VfCrxnqHhrVrrxpbWU13ZMFd4Gsrx2jOQeC0aH6qK/4Jw/Efxz8WP2TPDHjb4jeJrzX9dvL3UEnvrtgZJFS6kRAcAdFAH4V+3Z+078Yfg5+3X4J8H6D8XtV8PeB7qLRbrV7Lz0S1WJ7t0uGbI4Uxp83Pqa8Y/HT/AIKE/t4+LdY1L9lPTvEvhX4aaXcyW+n3FhqEekGcDjzJbt3RpJWwCY4mIj3KCOrtd/Eay/Z8/Zr07x/8ctVuIJvCnhiz/t2WacXM894sMaPGH3ESyyTHYDuIZmBzg5rw18Yf+CiX/BRXxFqt98D/ABDH8Kfh1p0/krdw3b2ccbddhu40a4nn2kEiMLGvy5CbgT8TPCn/AAVB/Ya0f/hblz8d7j4i+FrKRDqgudVuNZit1Zgo8+G8USJGW2jfE2Rnkrmv2Q/2ofC/7WnwgtPiNodp/Zup20xstc0ppN5sbtVUsFPVo2VlZGxyDg4ZWAq5uLezt5bu7uI4YIELyyyMFVFAyWJPAAAySa+JH7cH7UX7YXxkvfgX+wrAdH8P2Dutx4l2KktxCrbWupJpFItYCcbAg81uOct5Y8Ufsn/8FWfhPpU/xF8LftT6j4z1GwVp5tHtvE17eM+BlvKt7yPyJcc/IQCeAFOcV/wTu/bym/aq0nUfh/8AEe1tdP8AiP4btxcTm3TyotVtdwQzon/LN0ZkWROmXVl4JVP+Clfi39rv9mfx7pHxr+FXxe1//hXms3cSTaU7LLb6bfIN3kupGTBMqFgM9RIvyjZn9mT9oDwr+038G9D+K/hZkiN6nkanYh9zafeoB50Dd+CQVJA3IyN/FX7Yv7ZXx0+LH7Uuifsv/sceL76wn027bTr69011H26/PM25yGxBbIjbmGORKTkKpq58P+Jvh3+ztqeh3nj7V9b8RaP4WuzN4junH2me7EDsbgDkLh+VXkKAo5xk/sv/APBTj4zeC/Cfj2z8c6vrnxQ8d67Npdl4F0m7BlVLhjciZisYDMuWg+RTudtijAJZdV/Z4/4K/wDxe0s+P9b+Oa+E7uZVuIfDtr4ml0mZRztjEdnH5IODyJJM9NxyOPgd/wAFD/2l/wBmH4vp8E/20IdR1LSYblLa/uNThU6hpKtgLcJNGMXUODuJJcsvKMcbWs7y01Kzg1DT7qK5trmJZYJomDJIjDKspHBBBBB96/a3/wCCgPxe134zN+yj+xTpK6l4tS7NjqGuRQR3DrdL/rYbdZMxIsWCJJpAVUh8bQm83v7Fn/BVS30//hMbf9r6e711I/O/saLxlqKozY/1YVoxblscYICZ/iwM1+wd+378SPGHxPu/2Wf2qbQWnj61mmttO1OS3S2eeeAEy2twiYQS4RijqAr4xjJUt488c+Ffhl4N1jx/431iHS9C0K0e6vruU8Roo7AcsxOFVRyzEAAkgV4j/by/bW/bQ+Id18O/2N/C114Z0O3b57i3iia6WAkgTXd3LmO2BxkLHtbIKhpDS/se/wDBW/QrVvE2n/tSHUL98OdLPji+mcEA8BJ4fs49MBscj8PGPgn9vPxN+xh4O0LRfG/2H45i/jn169+021ntt90/7ktEpiJVGt1JXO4qWzzX/CIftMf8Nl/8Ib/wlP8Axev/AISDyf7V+2r/AMf2zO/zsbfu8ZxX7J/wl/4KR+FPjdo+t/tGfE3+2fAsNvdLfWf9tRXG+RoWER2KgJxIVPXiv21tQ/bilvvCPhH9j+y0uO31xLka3rE0MPm6YYzGFJkuCY1R1kbG1GkzGduO+rfsU/8ABVQWb+KIv2xGu9ZjVpBpdt4y1KKNzydigxLDuOcYYBf9rABr9g39vj4zXvxrl/ZS/auJk8Qmaay07U7uBLe7hvock2lxswkgZVbZIBuLAAl/MBX9t7wR+1j438K+GrT9k7xf/YGrW2oSvq0n9oJa+bAY8IMspz83OK+CPhD9pjXP2rb7wp8J/FP2D4tx6hqqXGpfbVi3TxiX7W3mkEfNiTnHOfev2Kvhj+374K+Kmqar+1V8Q/7e8Jy+H5obO2/taO62X5uLco+1VBGIlnGf9r3r/gq3+098f/gr+0R4e8K/Cr4qa14a0m68F217NaWUiqjzteXiNIcg8lY0H/ARXhu4mvPDmlXVzKZJprKF5HbqzFASfzr9sH/gpT8Rp/im37N37G2npqXiMXv9nXOuxWi3kst5na0FnEwKEIQQ0rhhkNgALvI/Y7/4K3avZDxneftQT2WqNH5x0Q+PL6OQPjPl+XFGbXPbAfZ74r9nH/gpD8b/AIL/ABef9n39uS2lxDdLZSa1dW0cN3pMrY2POYgEnt2BU+aBu2tv3OOKBVlDKwII4Ir4P/8ABSH4sfB/4y/E6X4r+NvEnjvTdP0/UbDwvoFxIpSXUhfwrb7mVQVVYlmywycZABJFeIPhD/wWE+Pelf8ACxr/AOId34KF2n2iz8N6d4jbQpY4yMqgigxtOD0nk3jjcQRx+yV/wUM+P3wj+OFv+zx+1zfXuo2U+prpVxd6wii/0S6dgqM8ox50JZl3FyxCsHVto2tVVVVVeK/FvhfwL4evfFnjPxBp+iaNpsRku7+/uFhhhXplmYgDJIAHckAcmv2j/wDgqV8QPi34iPwS/Yh8M6tc3epym2j8Qx2TSX1513fY7cgmJcDPmyDcBk7YyN1fstf8EmPO1OP4rftjavL4k168k+1P4bW9adfOY7i17chi0756ojbSeruCRWkaPpHh/S7XRNB0uz03TrKIRW1naQLDFAg6KiKAFUegGKqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr9kYH4d/8ABXP4n+E4yYo9cvPEUEcYP/LOSYXqLwTwFiXr6c4PSqqvGvirS/Ang3XvHGtybNO8O6Zc6jeNnG2GCJpHP/fKmv8Agj54U1T4l/HP4r/tKeKI/OvAjQLMwyHvNQnaed191WLB9pqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq8R/8hAf9cx/Wqqqq0/4+4P+ui/zqqqq/wCC3v8AyVP4af8AYv3X/pQK+Hmv/wDBZu0+H3hjRfAuiGXQZNHtBod39m0FytkYU+z/ADucf6vZ/rBu/vc5rQv+CaH7YP7S/jKx8Z/tmfF57G0txta2F+uo30cectFAkf8Ao1uGx95SwBOdjV8MPhl4L+DngLR/hr8PdGTTNB0O38m0t1JY9SWdmPLOzFmZjySxNf8ABQf/AJSYJ/2EPDv/AKKt6r/gtX/yav4W/wCygWf/AKQX9f8ABJ//AJMj8H/9hDU//SyWv+CyX/J3Np/2J9j/AOjrmvhD8NtC+D3wv8L/AAv8N20MNh4a0yGzjEabRI6r+8kPqzuWdj1LMSetf8Fu/iFf6b4G+Gvwusrpkttc1K91S+jUkbvsqRxxBvVSbmQ49UB7Cv2cf+CqX7OXwB+B3g34R2Pwo8bvJ4e0uOK+nt0tAlzeMN9zMuZQcPM0jDPOCB2rx7/wWH/Zx8feBvEPgbVvhF48ms/EGl3OnzxyJZlSk0TIc/vj2b0r/giF4lv7f4l/EvwcrubK/wBCtb91z8qyQTmNT7Ei4b8vaq/4KW/ELUPh1+xl4/vdIumgvtahg0eJ1JB2XMyRzDj1gMw/Gv2A/wBvL4G/sf8Aw01vw94m+HfinV/E3iDVjdXmoaclvsNukapDCC8ithT5rdOspr/h9p8Af+iUfED/AL5tP/j1fsofFjQvEH/BTzR/iP8ADnSL3SdB8Z+JtQaPTbgIrxxXkE29GWMlQFkfcACQNq+lfFD4a+EfjD8Ptd+GfjrTVvtE8QWb211EeGXPKuh/hdGCurdmUHtUfxJ+Pf8AwTL+JHxY+BGl363A1/TjBY3hcoiF+bXVIFBO2UQs4K54fhifLFf8ErP2QLf4QfDWH49eNrWGfxp4+sknsC2HOnaZIA8ag8/vJhtkY5+7sXghs/Fn/klfjL/sX77/ANJ3r/gjj8P/AA94v/ai1HxLrlslzN4Q8Nz32mxugYJcvLFCJOe6xySY92ByMc1/wW3+GWjXHw98AfGWK3ij1aw1l9AmlVAHngnhlnjVj1Ija3kIHYzN61+yn8b9W0z/AIJf2fxWuLoyar4J8HatFA0o3ZfTzcR2q+42RQL+nav2Af2t/hT+yl4x8YfEH4m+EPEfiXxBrtpDaaddaf5Ltbxl2kuS7SupLSMIOR/cbPWv+H2nwB/6JR8QP++bT/49Xx5/ac8HfGH9tTwz+0V8KfDer6CY9R0ee4gvhGs0t3bSou/90zAho0iXk5ODniv+C1/xI1Pw/wDCDwJ8MbC8aGDxfrFxd3qKD++iskjwhPTb5lzG2PVFPY1/wTN+Eeg/C39kXwZf6fZQrqvjO2/tvVbpRl7h5iTCCeuEh8tQvQEMerGqr/nMr/3UD/2hVfHP9on4Pfs4eF18V/F3xlbaNbzsyWdsFMtzeuoyVhhTLORkZONq5G4gHNal/wAFjYPEuq3Gl/Az9lvxj4zEeQkk155MgJHysYbeG4OM9t4z6ivEXxa8X/En/goZ4R+LfiD4cXnw98Qal4x8Py3Gi3IkV4Wje2iDESJG2JFjDEFRnceuar9iT/lKzrn/AGMHin+V1Vf8Fq/+TqPC3/ZP7P8A9L7+vjp4+vvhb+yV4u8f6VcNBqOi+CpZbGVSQY7k222FhjniRkP9R1r/AIInfDHSNd8e/EP4v6tAlxqHhyztdO015BuMb3ZlaeQZ6NtgVd3XEjjuar/gtx8L9FXS/h38ZbO0ii1VrqfQ76ZRhriIoZoAf9wrPj/rp7Cv2CPHmo/Ev9j34XeKtXunubw6MbGaaQktIbOaS13MTyWPkZJPU85Oc1+xB4R0Lxl/wUosrPxDZJd2+m+Ita1GKFxlftFutxJCx/3JFRx7oKr/AILReG9L0f8Aaa8N+INNt0gutb8IwSXrIuDLLFcTxrIT3by1jT6RrXw/1W817wF4b1zUHDXWoaRa3E5AwC7wqzEfiTVVVVX7Wn7fvwW/ZTsp9G1C9XxP44aPNt4a0+ceZGSMq1zJyLdMYPILkEEKRkjw38MP20P+CqXi638afETWX8K/DK1uSbaZ4HisLZc4K2VsSDdS9QZWOBggyDAWv2b/ANkn4Kfss+Hf7H+GPhpRqNzGq6hrl7ia+viP78mBtXIz5aBUB5xnJqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqrxGp8Ef8ABbuxvIQwj1LU7cpgBj/pWiCJ8jjA3u/uBzyaqqr/AIKo/En/AIV5+xt4osre48q98YXdrodsc4JEj+ZMMd8wQzD8a/4JSfDM/Dz9jvQNUuYBHe+NNQutbmBHzbHYQw8+higjYf7/ALmqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqvEf/IQH/XMf1qqqqtP+PuD/AK6L/Oqqqr/gt7/yVP4af9i/df8ApQK/Z6/5ID8M/wDsT9J/9I4qqv8AgqRFd+AP294PG99bSSW9xZaPq9umzHmRw4iIU55+a2cZ4547ZrQtc0jxPomn+JPD+ow3+mapax3VldQtuSeGRQyOp9CpB/Gv+C2XxD8LwfCHwR8JxqUL+Ir3xKutC0SQF4rSC2uIS7qOVDPcAKTjdsfGdpx/wSf/AOTI/B//AGENT/8ASyWv+CyX/J3Np/2J9j/6Ouar/gtx8OdT1XwJ8OPilYWbyWvh/ULzTdQkQE+WLpYnhZvRd1u659XA7iv2FdR+CHxz/Zg8DeItP8D+E7rVNI0qDSNcjfSYGlivraNY5C+Vz84VZR6rIPev+FTfCv8A6Jp4U/8ABNb/APxFaH4N8H+GJZJ/DXhTR9JlmULK9jYxwM4HIBKAZHsar/gpT8OdT+Jf7Gvj3TdEs3utQ0aKDV4IkBJK20ySTEAdSIBMceoFf8EefEvwj8efCTxB8JfEvhjw7feLfDeqyX8a32nwyy3FhOqAOrOpLBJVdW/uh4/7wr/hU3wr/wCiaeFP/BNb/wDxFab8Ofh7o19Fqej+BPDtjeQEmK4ttLhikjJGCVZVBHBI49a/aW/aB8H/ALMvwg1r4q+LpVk+xR+TptgH2vqN64PkwJ/vEZY4O1FZui18FP2Pvi9/wUA8MfFv9pvxvrdwviDUDJ/wjBbEcWp6kjK7xYb7tukSi3TBAVnHOImU/wDBJr9rXUbS5n/Y9+LFxPaanpTzf8Is19lJUMZY3GnuG5DIQzop5AEi8bVFfFn/AJJX4y/7F++/9J3r/gjt8RvDPgr9qG+8N+Ir1LSXxl4dm07TJJHCq92s0UyxHPALpHJjnlgqgEsKr/gtl8YNEfw94F+A2n6hDPqf9ot4g1KBCC1qiRPBb7vQv505A9Eyeoz+z5+z3rNr/wAE1bP4GXtiYNc8T+B9SLRTDY0VzqCzTRB+mGQzxqQemzB6V/wSA8TfDvTvih44+CHxR8NaRJrOvxQTaPHq9hHI63FoZRcW48xSQ5WQNt4/1Ld6/wCFTfCv/omnhT/wTW//AMRUXws+GFvKk9v8OPC8csbBkdNHgVlYdCCF4Nf8FrfhpqfiL4N+CPifp9q08Pg3WZ7W+Kj/AFEN6kYEh/2fNt4k+si+9f8ABLn45+Hfiz+yz4c8IwajB/wkfw/h/sjVLHcA6RIzfZpQvUo0W0bum9HHaqq9nh07/gsksl/ILdX+IcaKZPly0kSrGP8AgRZQPXIqv2iZm/ag/wCCokfwx+JOp3cPhqDxZbeHIbV5TH5VnCQHjiOflMz+YQw5zMCOwrwh4L8I/D/w9aeFPA/hvTdC0exQJb2VhbLBEgAA+6oHOAMk8nua/a68UaF4s/4Kp2Fz4f1CO9g0/wAX+HbCaWM5UXEBtkmQH1RwyH/aUiq/YvnhsP8Agq/rFtfSCCWTxN4phRJPlJk23Z24PfCnj2qv+C1f/J1Hhb/sn9n/AOl9/Xxn+Hd38Wv2VfFPw30yISahr/gyW2sELbQbk22YQT2HmBM1/wAEZ/jDpPgL4s+NPgn4ruU0668ZW8EumrdHymN7ZmUPbgHne0czHB/54kdTg1/wWq+Nehaxqngr4A6HeR3WoaNLJrGtJGd32Z5EEdtGcdHKNK5XqA0Z/iFfsY/DDUvg3+y18Nvh5rVq1tqen6Ks9/buu1oLi4driWNh/eV5mU+4PXrX/BPj/lJg/wD2EPEX/oq4qv8Agtp/yX7wL/2J4/8ASuevhP8A8kr8G/8AYv2P/pOlVVV4r8W+F/Avh698WeM/EGn6Jo2mxGS7v7+4WGGFemWZiAMkgAdyQBya/aU/4KefEn41eJh8DP2HdB1maXU5Gthr8Fmxv73qD9kiIzbx45MzgOBziLbk/sl/8EmdI0K8g+KP7Wd1H4s8SzyG5Hhszm4tIZGO4tdy5zdSZ6qD5ec5MoPFpZ2lhaQ2FhbRW1tbRrFDDCgRIkUYVVUcAAAAAcCqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/a9T/hHv8Agrr8JdSSNHOp6l4ZJCfIfnuvs+Se5AXPuABVVVf8FqPGV/4g8X/Cj4FaETNdzJPqklop+aWSeRba14+sdwB/ve1fDnwbY/Dr4e+Gfh/pgUWnhrR7XTYdowCsESxg/iFzVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVXiP/kID/rmP61VVVWn/H3B/wBdF/nVVVV/wVA/Y5+Pf7TfjzwTrvwf8L2eq2ei6RPbXjz6nBalJGlDAASMCeO4r4P+HtV8I/CTwR4T12BYdS0Xw5p9leRK4cJNFbojgMMggMpGRwaqv+CgP7Dlr+194P03U/DGqWmkePfC6yjS7m6UiC9hfBe2mZQWUblDI+G2ncMYckfC74Pf8FjPgtp6fCr4bjU9P0GAlLcT6lpF7aW8ZJyYXuGdol5J2ptOTnbmvGv/AAS2+MPiH4JeLvHPj3xN/wALJ+P/AInnszbvPqhW302FZ0MqrNMVEkhiUqSQFUDYgx8zf8E//g98QfgP+zHoHwx+J2jRaZr+mX1880MV3HcKUluHkRg8bEdHHGc5Ff8ABSH9hj9pD9oz9oW3+IHwo8IWOp6LH4ctbJpp9Wt7ZhNHJMzDZI4bGHXnGOar4l/Dfwd8XvAetfDbx9pCaloOvWrW95bscHB5DKw5V1YKysOVZQR0rWP+Cdn7cH7Kfji+8W/sb/EWTWtNuSAiQX8FldvGDkJc29yRbT7exyc8kIucVBrv/Bb7xOp0iHR49J8xcSXkltoUGxehO5ieef4RuHUYr9jrwD+0B8Ofg+dC/aU8b2/inxfPq1xdi8jvpLto7eRUKxPI6rllYScKNqqVAOBVSxRzRtDLGro6lWRhkMD1BHpXx/8A+CVvxQ8EfEl/jL+xL4xXRrhblrmDQzftYXGnSNnctpcfdaI5I8uQrhflLODgWPiP/guBZwx+H18OvPNEphW6nttBZiRwGMu7YxH945zjJzkk/sR/Df8Abi8MeJ/FXjP9rn4hW2rwa7ZQRWOkjUFneznjdiXWOFBbxKVdgfLJLELnhRX7bP7K37ZH7Y3x90jTJPD1n4e+Eug3iWlhPJrFvIyRMwFxqDwLIWaRlB2pjIVUHykuT8Ofh94V+FHgTQ/hx4I05bHQ/D1klpZwgDO1RyzEAbnY5Zm/iZiT1r9uv/gnl8WPFvx20n9oX9k+xtoNdvbhbzWIVvorJrXUYWVo7yIyEKTJjLjrvTcc7zi31T4laz+zDqd78YfDFvoHjP8A4RS9j1mytrqO4h89YJFaSNo2YbHxvC5yu7ac4yf2Fv2WrL9rDxP418FxeJZ/D2v6L4fXVNB1RNxWC7S5iUCRVIJRldhkYZTtYZwVN9Y/8Fs/AVoPAem3i+JLWEG3tdbgk0i7eRegbzrjbL77plDc5PTj9mb/AIJa/EfXvifH8df20PEsOsal9sW+bQTefb5b64GCpvZ+UKKQP3SFgwABYKCpr9tP/gl5efFbx1c/HP8AZx8SWnhnxnd3Au9Q0y4ke2gurkHJuYJowWgmJAJGNrN825DktpN9/wAFuPA1qnhmPSrvWY7U7Yrq7OjXzsvQEzliz565cluecdK/ZR+HH/BSu6+N2mfEr9qbx/aQ+FbS0uYLvw82oW+ZS8RETJBYr9n3LIEO923Bd2M5NeNvBfhf4jeEtW8C+NdHg1XQ9btXtb6zmB2yxsMEZGCD3DAgggEEEA149/4Jc/tSfs//ABCf4i/safENtRhjZvsUQ1JNN1OBGJzDIZCsE6YwCWZQ3dBjNW8n/BcDxZajw9Kn9kR58ufUZBoNs+MHneuW7feiXPT1r9mfwn8V/AvwN8K+E/jf4jg17xrp1vIuqalFeS3f2gtM7IWllUM7iNkViRgspwSMV/wUJ/4J4fF/4lfGEftH/s3vDda3dpbyalpa3y2N1FeW6qkV1bSuVTOyOPILKwZNwLbzt/Ypb/gptc/FW3f9qdrlfh/a6XPCVvU0lJWuQE8o/wCigTOeDlnypyckmv2+v+Cb/wAQfih8Tj+0V+zZqMEXiy4eGfU9JkvBZSPdQhRFdWsxIVJMImQzJ8y7g2SRWg6V/wAFoPibpMfw917UNN8FWDoba88SXb6dDcFOhO62Mku7B4eJFOR94HJrxp/wSv8AjH8M/jf8N/EfwS09fF+j6Cum6jr+sajqUNtJc6lFePJcMIZJAwTYsW0A9CAWZtzGv2yf+Cdv7Q+n/tCX/wC0p+yhIL241TUv7Xe0tNQjs7/TL8ndNJGZWVJI3fL4DbsuylSOT+wvJ/wUOvPGGu3v7YrOvhv+yQmkJPHpcUn2sSrk7LIBsbA+TIPTHU1/wUz/AGI/2if2lfjxoPjr4ReErHVNHsPCNvps80+q29qy3CXd3Iy7ZHBI2TRnOMc+xrw/Zz2GgabYXShZraziikUHOGVADz9RX7af/BLm6+KHja6+OP7Nuu2fhvxhc3H22/0qeVraG6uwd32m3mQEwTFhkgjazfNuQ5Jgb/gt7a2C+AktblwEECapJJobyBAPvG5LEk7R95svz13V+yP/AMEtNW8LeP4vjn+1d4mtvFvilLv7fb6Ok73kf2stu+0Xc8nM8gb5tgBXcASzjiq/ZC/YY/aQ+En7bTfGnxz4QsbPwobzWJBdR6tbzPtuEmER8tHLcl17cZ5qv+CnX7F37QX7THxa8K+K/hD4UstU03S/DgsrmSfVILUpN9olfAWRwSNrqcjjmvh/pN9oHgLw3oWpxCO807SLW2uEDBgskcKqwBHB5B5FVVftXftxfBj9k3RzH4p1D+2/FtxFvsPDGnzL9plyPleUnIgi/wBthk87VfBFaP4N/bN/4KueMovEXizUG8KfCywvGNvI0TpptnjIK20OQ15cAZBkJwCSC0YIWv2bP2S/gx+yv4aOifDLw/8A8TG7jVdS1y9xLfagR/fkwNqZ5EaBUB5xnJNVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX/AAUeRbT/AIKWfA25tB5Uz2fh2RpE+Vi41q5AbI7gKoz7Cqqq8crH+0V/wWT0nQWAudL8HaxbRgNyqLpdt9pkVvY3KSLjod2D1qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqtesru4vg8Fu7r5YGVXPrX9mah/z5y/8AfBr+zNQ/585f++DX9mah/wA+cv8A3wa/szUP+fOX/vg1/Zmof8+cv/fBq202/W5hZrSUAOCSV6c1VVVVVVVVVVVVVVVVVV8Wf+SV+Mv+xfvv/Sd6/wCCJf8AyX7x1/2J5/8ASuCqqqqqqqqqqqqqqqqqqqrUdR0/SLC41XVr+3srKziaW4ubiURxwxqMs7sxAVQASSeAK/ay/wCCquoarrD/AAW/YxsLjXtcv5vsR8TQ2jXBklb5RHp8GCZXJIAlYYyDtVgQ9fso/wDBKnUtc1dfjP8Ato6jd65rd/MLweGpr1p3klJzv1CfJMrE8mJWx/eZslK0zS9M0XTrbR9G061sLCyiWG2tbaFYooY1GFREUAKoAAAAwKqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr/AIKkY0r9vH4I+ILV83cenaThH5UeVq07KcdeS7Z57Cqqr28tdOs59QvZ1ht7WJpZpG6IijLE+wANf8ElrKf4sftZ/F74/ajbO7C0uJi0gyYrnU70ygkj+LZDMPxNVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV9Y2ep2NxpuoW0dxa3UTRTwyLuWRGGGUjuCCQRXw7/Z/wDgj8I9Uuda+F/wq8M+F7+8t/s9xc6Xp0du8sW4NsYqASu5VOPUCqqqqqqqqqqqqqqqqqqq/aD/AGmPhB+zH4Pfxf8AFTxNHaeYrCw0y3xLe6jIB9yCLILdgWOEXI3Mua8VfEz9sH/gqx42n8C/DrSJPC/wwsLkfaYfNaOwtVBBV72cDNzNjayxKCARlUGGev2Tv2Gfg1+ydo8c/hywGueMZ4dl/wCJ7+IfaJMj5khXJEEX+yvJGNzNgEVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVf8FKFVv+CknwLVgCDp3h4EEf9Ru7qqr9tTxufh3+yf8AFTxTFceRPH4ZurW3lA5Sa5X7PEw9w8y496/4Ir+CF0b9nvxb46miKT+JPFDQISv34LWCMKc/9dJpxj296qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr9tL/gp54A+AJvvh38IhZeMviGm6GYhy9ho8nT98y/62UHjyUIwc7mUjaf2ev8Agn38cf2wPGA/aF/bR8Sa5Z6VqTLPBp1y3l6hqcXVVCcCytueFChiM7VQFXrwL4B8F/DHwrYeCPh/4asNB0PTI/LtbGyiEaIO5PdmJ5LNlmJJJJOaqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr9u0jxD/wVU+Ceizv9piguPC9s8Uf3o4zqskjKdvI4kZsnkA56Yqqr/gsP4x/4Rz9kM+H45iH8VeJrGxZAcFkjElySR6BrdPxIr/gnR4L/AOEF/Yu+F+mSQlJtQ0t9VkJHL/a5pLhCf+2cqAewFVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVXjLxp4S+Hnhm/wDGXjnxHp+haJpkRku76+nEUUS9BknqScAKOSSAASQK/aM/4KC/Gv8Aa68Yv+zv+xV4e1yDStSLQTajaxmLUNUjyA77iQLO255ZirbSC7ICyV+xb/wTD8AfAD7F8Q/i4bHxl8Ql2ywho99jo79f3Kt/rZQf+WzgY42quCzVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVfHFv+Eq/4LP8Ag/R4yWOjajooIiU7h5Vmt182fZskj+H3BNVVf8FwPFEsq/CT4f2bM7yyalqE8KjJZv8AR4ocAc55mHT096+HHhSPwJ8PPC/geHbs8PaLaacm0YGIIUjGB6fLVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVa5q19ZXggtpVVfLBwVB55r/hIdV/57r/3wK/4SHVf+e6/98Cv+Eh1X/nuv/fAr/hIdV/57r/3wK/4SHVf+e6/98Codf1N5o0adSCwBGwVVVVVVVVVrmt6P4a0TUPEfiDUrfT9L0q1lu768uZAkdvBGpeSR2PCqqqSSegFfDj45fBz4wz31t8LPiZ4d8VS6WiPeJpV+lwbdXJCFwpOASrY+hqqqqqqqqqqqqqqqqqqqqqqqqqqr9qH9r34P/sn+Ff7a+IOrm51m8iZtJ8PWTK15fsOMhSf3cQI+aVsKMEDc2FOmeHv2u/+Cs/xBTW9fuz4T+FmkXZETBHGn6eBwVhTg3l1tJy56Z5MakLX7Ov7MHwh/Ze8Hjwn8LvDywSzqp1LVbnEl5qUij70smOnJwi4RcnAGTmqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqvh7j4gf8ABbDWtXIaaLRNT1DzMktj7LpTWq88YAcJ7cAc9aqq/wCCgqn4pf8ABTH4QfDJQJLa0Gg2NxGQCF86/eWZjn/pk68f7PHWqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqrxN/yER/1yH8zVVVVb/8AHxF/vj+dVVVXxn+MXgT4CfDfWPij8RdU+x6Po8W5lQBpbmU8JDEpI3SO2ABkDuSACReftTft9/8ABQPxvqPhb9mq3v8AwR4Qs32yvpl19jFrGT8jXWocP5hAPyQkZGcI2Cauv+CYn7f2nxHxBpn7UGn3GuRDzAIvFeqxyM+OQkxiBz2BO33Ir4bft8/te/sa/EuD4Uftf6Rq/iHRkdRcLqeyXUIIWOPtFtdqSt0o54dnDEbQ6EGvBXjTwv8AEbwlpPjrwVrMGq6Hrdql1YXkOdssbDg4OCD2KkAggggEEV+1Z+1D4M/ZI+Glt8SvG2h6xq9vfarHpdpa6YsZdriSKWVdxkZQqbYHyRk5xwc8fsx/HKD9pT4H+HPjVa+Gn0CHxE94I9OkuxctCsF3NbgmQKoJYQ78Y+XdjJxk/tZf8mr/ABl/7J/rv/pBNX/BDr/kavi3/wBg/Sv/AEZc1VVVVVVVVVVVVVVVVVVVVVVVVVfttf8ABT/wl8E5Lz4XfAiSx8XfEHc1vc3inzrLRpOhU7f9fODx5anarffOQUP7L3/BNz4l/tBeKv8Ahob9tzVtZlGrSrdJoV5My32pf3TckYNvDjAEK7WxgfuwAD4f8PaD4T0Sy8NeGNHs9K0rTYVhs7KzhWGKCNeioi4AH0qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/wCCcJX4i/8ABRr4zfEpz58CRa7ewS5BAe41OMIByePLaQDB6ADOKqq0wD4o/wDBbO5uGYyWmg6nJuAORH9h0jyxjk/8t0Gfdj9KqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqvE3/IRH/XIfzNVVVVv/x8Rf74/nVVVV/wV++LXiH4k/H3wv8As0eGJJJrXw7HbyS2cbf8fGqXoHlhlHUrC8QXuPOf1r9nL4F+Fv2cfg74e+FHhW2hC6ZbK2oXSJta/vGA8+4c9SWYcZ6KFXooAr/gpB+zdonx+/Zw8QanDpkTeLfA9nNq+h3YT97iJd89vnqVljVht6bxGf4a/wCCKfxs1HVvDvjT4A6xdtLFoTJrWiqxyY4pW2XKD0USeU4H96Vz3r/gox+3t/w0PpOo/s8/8Kp/sD/hCvHEs39r/wBu/avtf2Rbq1x5P2dNm/zd/wB9sbcc5yP2XP8Agq3/AMM1fAjwx8E/+FC/8JH/AMI59r/4mf8AwlP2Pz/Pu5rj/VfZJNu3ztv3jnbnjOB8R/iR/wALj/4J9eNfip/Y39kf8JX8JtV1H7B9p+0fZvN02VtnmbV34zjO1c+gr/glV8dPhp+zrpPxn+JPxT14adpNtY6TFCiJ5k95Oz3RSCGMcvI204HAABZiqqWHif8A4LZeJotUXUPCn7OkQ8NNMyQT6prEizXKg9QyRbI24OVHmY9Tiv2Qv24fhV+2Bo95H4Xt7rQfFWjwrLqnh++kV5IoycebDIuBNFuIUthSpI3Ku5d1ftWf8FM/hB+zh4im+HXhzR7rx/44t28u50zT7kQwWUh6RzT7X/eZI/dorEdG2mp/+CuXx08G3EGtfFL9j3UdJ8NXMm1J5JbuybB5XbLNBsc45xhd3qM1+zn+0r8LP2o/ASePfhhqskkcT+TqGm3arHd6dNjISaNSwGQMqwJVhnBOCB+25+19N+xv4G8P+N1+HC+MItb1Y6c0B1n+zzA3lNIGB8mXcCEYYwMcda+Mf/BU34bfCb4ReBvFx8HSat438daBb6xD4Vt9TGzTIZlBU3Fz5fA67QI9zbScKCDX7EP7TWs/tZfBq4+KWueFrLw/PFrtzp6WdrcNMuyNImDFmAJY+YQeAOKqqqqqqqqqqqqqqqqqq8UeKPDfgnw9f+LPF+u2Oj6NpcJmvL69mEMUCDqzM3A7D3JA71+0t/wUE+MP7WvjD/hm/wDYq0LWV03VWe3n1O1QxX2rR9HKsSPsltjlnYqxX7xQblP7Ev8AwTO8Afs6RWHxC+KCWXi34kKBJFIU32WjN6W6sBvkH/PZhkY+QJyWqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq+IHiWPwX4C8SeMZWUJoOkXV+xYZAEMLSH1/u1/wQ/8MvMfi548u90kkj6ZYQysckn/AEiSbJPOTmE9fXPaqqv+CeG74hf8FKvjN8QZmDxwJr95Cx52tNqUaIoycgCN3HfAAHeqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqrxN/yER/1yH8zVVVVb/wDHxF/vj+dVVVXxII1n/gsFbLr4BA+J2kIqy/KCI2txCOeoISPA78etVWqW+n3emXdrqojNlNA6XIkbavllSGyewxnJr/gjI16v7WerLah/LbwTei52rkeX9ptCM+g3hPxxX/Bav/k1fwt/2UCz/wDSC/r/AIJcf8mJfDH/ALi3/p2vK/ay/wCTV/jL/wBk/wBd/wDSCav+CU37Nfgr4/fGjWNd+I1hFqugeBLOC9GkzjdDeXkjssHmr0eNAkrFDwx2g5UsDr3g/wAKeKPDNx4L8R+G9N1LQbu2+zT6Zc2qSW7xYxsMZG3aBjAxxgYr4T6PJ+yZ/wAFSNL+H3hi7mj0y28broUCPJvL2GoERxI553EJcRHJ/iUHgjj9sL4y3PwA/Zr8d/FLTJUXVNM07ydLLANi8ndYIG2n7wV5Vcj0U1/wSf8ACvwJ0y28RftHfGr4g+EV8b3OrSWujr4g1qCO4tVCq892FlcN5kryFfMPICNgje2fEXxj/Zj8XaFfeGPFHxa+Geq6TqUDQXlld+ILKWKeNhgqymTBFfsOeI9P/Z6/4KR6v8IfAvieDU/A/iXUdR0WKeC8WeG4t1WSeykDqSruGSNNwOf3j46kH/gtp/yQHwL/ANjgP/SOev8AglP+yf4L8Z/D3/hpj4v6DbeKtZ1C4/s7w1HqsIuIbGyskFuJFR8qX3RtGCQdiwLtILNVjpmm6XE0GmWFtaRu+9kgiWMFsAZIA64AGfYVVVVVVVVVVVVVVVVVVftEftK/Cf8AZh8DyeNfihr62wkDJp2mwYe71KUDPlwx5yeoyxwq5G4jIq91X9rX/grX8Sm0zSkfwl8K9FuxvUlzp2mjs8h4N5d7Twoxt3ceUrMx/Zo/ZR+EX7Kvg8eGfhto26+ukX+1NbuwHvNRcd3cD5UBztjXCrnpkkmqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv8AgoT4yXwN+xl8VNV88Rve6KdLQFsFzeSJbED1+WZjj0B7Zr/gjl4NPh39kufxHLCRJ4q8UXl3G5/iiiSK3AHsHhk/EmqrWdTt9E0e+1q7YCDT7aS4lLMFAVFLHk8DgdTX/BEXTLnVfFfxi8Z3hLzJbaZA0hXbveaS5kfAHy9YlJ9Mj1qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq8Tf8hEf9ch/M1VVVW//HxF/vj+dVVVX/BUPwp4k+BX7cGk/HXRLUxx69/Z2vaXcBSqfbbHyo5Ez/eBhhkOP+eynvXwv+I/hb4v/Dzw/wDEzwXfLd6N4jsY7u1fI3KGHzIwHR0YMjL2ZSO1V+3L8cNL+Af7MnjTxbPqEdvq2o6fLpWhRk/PLfXCMkZUd9gLSn/ZjNf8ESfhLexyfEH4439o0dvJFDoGmSshHmncJ7nB9BttvxJ6Yr/gtX/yav4W/wCygWf/AKQX9f8ABLj/AJMS+GP/AHFv/TteV+1l/wAmr/GX/sn+u/8ApBNX/BDr/kavi3/2D9K/9GXNV8cP+Uwem/8AZUPDP/odjX/BWHRtR1X9ijxXc2BcppepabdXKp/FF9qSPkZ6BpFPfGM9sj9gP/gnx+zv+1p8Ernx94y8aeObLxFputz6ffWekX9pDFGqpG8bBJLeR/mWTqWwSGAHBr/hyp+yv/0P3xV/8Gth/wDIVfB3/glN+zr8Evid4e+K3hfxd8Q77VvDV39qtLfUtRs3t3k2lRvWO1RiBuzgMOQO3Ff8FtP+SA+Bf+xwH/pHPX/BLj/kxL4Y/wDcW/8ATteVVVVVVVVVVVVVVVVVVftq/wDBQz4b/sp6fP4V0IWvir4kTxf6PoqTfutP3DKy3jLyq4IIiGHcY+4DvH7PX7FPx3/by8cR/tHftc+JNXs/Cl6VktLeT9zdapD1SK1jxttbTkncAC2TtBLmQeCfA/hD4b+FtP8ABPgPw7Y6HoelReVaWNnFsjiXqeOpJOSWOSSSSSSTVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV/wAFqPH6aF+z/wCE/h7BNtufFXiT7Q67vv29pCxcY/66TW5z7e9fsV+Av+FZ/sofCzwg8HkzReHLe7uYiu0pPdA3MoI9RJM4PvVV+0nrf/CNfs7fFHxCsmxtN8G6tOhyAdyWcpUDPGSQAPciv+CIuiC3+D3xF8SCPH27xLDab8HnybZXxnpx9o/XntVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV4m/wCQiP8ArkP5mqqqq3/4+Iv98fzqqqq/a/8A2VvCP7W3wnn8Ba7cLp2sWMhu9A1hY9zWF0FxyOrROPldO4wRhlUj4bfGP9sP/glt4ovfAfj74fS6r4Lvrwv9ivHkOn3T95rC8UFY3dQNwKk4xvjDAYv/APguD4HTSmk0v4A65LqWz5YbjXIo4Q2O8gjZsZ/2Pyq2+HX7Zf8AwVR+Jml+MPG+nSeFvh9Znba30ltJBpun2zEF/skbnddTuMZcEgkKGZFCgfCH4UeDPgf8N9C+FngDT/smi6BaiGEPgyTNnMk0hAAaR3LOxwAWY4AHA/4KdfBHxN8cv2VNU0vwXpU+qa34Y1O31y0sLdC0tyIlkjlVFHLMIp5GCjk7cAEkV+xd/wAFPLf9l34Rp8FfH/w01PXbbRb6eTS7iyvVhkgSWQySQyRyLxiVpGBB/jIIGMn4yeN7X4nfsHeOfiRY2Ulnb+K/hFqGrQ20j7mhS50mSVUJGMkBwCfav+CHX/I1fFv/ALB+lf8Aoy5qvjh/ymD03/sqHhn/ANDsa+IHgbw58TvA+u/DzxdZ/atG8RafNY3sQOGMcilSVPO1hnIPYgHtWlx/tT/8El/jFq1+vhh/Evw91qVYpbhlcafrMCsfJbzVB+y3ShmGGBILN8roQTZf8FuPgtJpscuo/BzxrBqBjzJBDcWssSv6CQupI99g+lfsWft23n7Yfi7xdptv8JL/AML6FoVnBNp+oS3D3S3Ll2WVJJBGsaPgxlUBJwHOTjj/AILaf8kB8C/9jgP/AEjnr/glx/yYl8Mf+4t/6dryqqqqqqqqqqqqqqqqmKqpZmAAHJPGK/bU/wCCn81pqs3wJ/ZDlOv+KL2b7Fc+JLGL7UkUrHaIbBVB8+Uk480AqONu8ncv7FH/AAS9XR7+3+OP7WkX/CQeLLyb7bbeHL2X7VHbysdxmvmYnz5iTnYSUB5YuThQqqoVQAAMAAYxVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX/BSyeb9oT9vL4Z/s36U7yQ6alhp92qnmGa+mWWd/YLbeQx78GoYYbeFLe3iSOOJQqIi7QoAwAAOgx2qq/b+1f8AsT9jT4s3gkK+Z4fe2yF3f650ixj38zGe3Wv+CN2kDTf2Rbq8CAf2r4wvrnO7O7ENvFnHb/U4x7Z71VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVeILG7uL8SQW0ki+WBlVzzzX9laj/AM+U3/fBr+ytR/58pv8Avg1/ZWo/8+U3/fBr+ytR/wCfKb/vg1/ZWo/8+U3/AHwag0zUFnjZrKYAOMkofWqqqqr6xsdTtJdP1Gzgu7addssE8YkRx6FTkEexqx+CfwZ0y/Gq6Z8I/BdpehtwuYNAto5N2c53BM5yAfrVVWr/AAl+FXiDWT4i134ZeE9S1ZiM393otvNOcdP3jIW/WmtbVrU2TW8RtzH5ZhKDZsxjbt6YxxirHR9I0su2maXZ2hkADmCBY92OmcDmqk0HQ5bwajLo1i92HDidrZTJuHQ7sZyMDmquba3vLeS1u4I54ZVKyRyIGVweoIPBHtQ+BPwPW8/tFfg34GF3v3+ePDtr5m713eXnPvVta2tlbx2llbxQQRLtjiiQIqD0AHAFX2mabqkaw6lYW12iNuVJ4lkAPqAQeatbO0sbdLSxtYbeGPOyKJAirk5OAOBySaqqqqqqqqqqqqqqqvF3i/wv4C8Nah4w8aa9ZaNoulQNNeX15KI44UHUkn8gBySQBkmv2jv21Pjd+3j45b9mj9kbQdVt/Ct+Xiu7iLMFzq0I4eW4kOBbWmCAUJBYEBydwjH7Ff8AwT6+G/7KOl2/ibVha+J/iRcwbbvW5Isx2W4fPFZqwyi4JUyEB3Gc7Qdgqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq6uraytZb28njhgt42klkkbaqKoyWJ7AAZr/AIJ+QXH7Tn/BRH4gftJ3kby6ZobX+pWbyA/u2uWa2s4iDzkWxkx6GIVVV/wVO1R9N/Yb+IMUTSK99LpluGRtu0HUbdmz7FUKke9f8EqNMWx/Yd8C3S+XnUbvVJztXB41CePn1P7v8selVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVXx3+P8A8L/2cPAdz8Qfil4ij0+yiylrbJh7nUJscQwR5Bdz+SjLMVUEjVNd/an/AOCt3xVOh6FDL4U+FOg3m50ZmNjpi9pJmGPtd4VOFQfd3HAjUu5/Zt/Zf+FP7LXgaPwZ8NdHAnmVW1TV7hQ13qcoH35X9Bk7UGFXJwMkk1VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX/AAUf+Mx+Cv7JPjHUbK6EWr+KIh4f0z59reZdBllZT1DLbrO4x3UdOo/4JBfB4fD79mF/iBf2oj1P4iapJehiMN9jgJhgUj03LPIPUSiqqv8AgsDqCWX7HVzbMZM3/iXT4F29MjzJPm56YjP44+tf8E7NPfTf2KfhTbOyEvpMkw2jjElzLIPxwwz71VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX7Xn7afwu/ZG8KfavEVzHrHi3UIS2j+GracLPc8482Q8+TCCDl2HJBChiCB8F/2b/2hv+CnHxK/4X1+0Trt/ovw+jlZLV4YzCJog3NrpsT5CRA5DTtu+YHJkcNj4d/DjwP8JvB+neAfhz4as9C0HS49ltZ2qYA9WYnLO7HlnYlmPJJNVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVf8FS/GGt/tC/tUfDz9kXwHOZZdLmgiuVHKrqN+U5cA/dit/LfPGBJJ0xmvBPhHRfh/4N0LwJ4cgMOleHdNt9Psoz1WGGNY0z6naoyfWqqv+C0t5NbfsqeHbeGUKLrx5ZxyrgHcosr1vw+ZVPH9a/Yls0sf2RPhBAluYQ3hCwl2lSMl4g5b8S2fxqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr9uz/gop4O/Zh0+68AeAZLPxF8TriLC2hbfb6KGXKy3WDy+CCsIwSMFiqkbv2Qf+CfHj39o3xV/w09+2ld6rfQaxML2z0S/cpc6uThkluAMeTbYOFhUKSAOFQAPYafYaVY2+maXZW9nZ2kSxW9vbxCOOFFGFVVXAVQAAAOBVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVXxQ+Ifh74S/DrxH8S/FVwItK8NabNfXPzAFxGpIRc9Wc4VR3ZgO9f8Eqvh94g+PX7Rvj/9sT4hw/aJbC6uBZyMh2HU7zLS+WTnAht2KbewnT0qqqv+C2skY+AvgOEyL5jeL9yoTyQLSbJA9BkfmK/ZWt5rT9l/4P2lzGY5ofAWhpIjdVYWEII/MVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVft6f8FKV8A3t18A/wBma6Gs+PbmX7FqGs2kf2hdLmZtn2e3UAia6J+XgFUJx8z5CfsIf8E0j4WvrT4//tSWra141u5RfafoN+/2hbCVju+0XhbPm3WfmCkkRnk7nxsqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv+Cxvx8vF0fwv+yt4JuJbjVvE88Wo63bW3zO8Ik22lvgdTJMC+3r+6jPRq/ZD+Atr+zZ+z74U+FgSI6paW32nWpo+RNfzfPOcj7wVj5an+5GtVVV/wW9/5JV8NP+xguv8A0nFfs9f8kB+Gf/Yn6T/6RxVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVUxVVLMwAA5J4xX7dX/BRDxJ4z8SP+y5+x9dXOq6rqs407UfEOjsZJriZjtNrYMh/4C0w9whGN9fsE/8ABOfw7+zdZW3xN+KcFnrnxPuYyUIImttDVv4ICR80xHDTdslUwNzPVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVfETx54b+F3gXXviJ4vvltNG8O2Et7eSnrsjUnao7sxAVV6liAOtfsEeBPEn7a37ZXir9rz4mWW7SPDOpC+t7eQl4xesCtlbITjcttEitnqDHFn79VVVX/AAXCvjH4B+FemCMET6xqExfPTZDEMY9/M/SvgzZwad8HvAun2wIitfDWnRRhjk7VtowP0FVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVTFVUszAADknjFft5ft4+Kfjd4pP7I/wCyO93q41e5Onatq2lHdJq8h4a0tWHSAc+ZLkBwCARGGZ/2Dv2CfCv7KPhiPxR4ohs9a+JuqwAahqQTemmow5trUnkL2eTgufRQAKqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv8AgrX+0HrPjbxP4e/Yt+FZl1DUtSvLWfX4LU7muLiRl+x2WPXJWVh6mI9jX7KH7P2jfsyfAvw58KdNEEt9aRG51m8iTb9tv5cGaT1IBARc8hI0Haqqqr/guN/yKnwj/wCwhqv/AKLtq+E//JK/Bv8A2L9j/wCk6VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVft/8A7cvi34v+Lz+x/wDsnTXGsPq0/wDZ2tanpDF5dTmbIeyt2HAhAB82UHDDIyEDF/2DP2DPC/7J/hdfE3iZLTWPiZrFuF1HUlXfHp8bYJtbYnovTe/BcjsoAFVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX7Vf7RHhz9l/wCCmufFTXPKuLu3T7No2nyNt+338gPkw8c7cgu5HIRHIzjFf8Eqv2fPEfxV+IOv/tu/GMS6leXV/cjQJrpB/pd9IzC6vAMY2plokxwGMmMGMVVVVV/wXG/5FT4R/wDYQ1X/ANF21fCf/klfg3/sX7H/ANJ0qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv8AgoX+3fr/AIz19v2Rv2VLy71bWdVuTp2vanpAMkk8jHabC1ZeSSciV16fdB+/X7AP7B+g/sqeEU8V+MbWz1L4n63BjUL1cSJpcTYP2SBvwG9x99uMlVWqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqp54LWCS5uZo4oYULySSMFVFAySSeAAO9fHDxf4p/4Ki/tl6R8IPhrfTxfDfwpJIkd8g+SO0RlF3qJB/ikIVIgeo8oHbuavA/grw18OPB2i+AvB2mR6fomgWUdnY2yD7kcahRk9SxxkseSSSckmqqqqv8AguBZQyfDn4X6ixbzYNbvokGeNrwxk8euY1/WvgReTaj8Dvh3qFyVM1z4U0yWQgYG5rWMnA+pqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr/go3+35qeiX037Ln7Nl/JqPjHVXNhrup6YDNLZPIQn2G12ZLXLk7WZcmPO0fvCTH/wTv/YD0v8AZl8PQfE34i2sV78T9bswsinDx6FA4Ba3iIyDKeBJIPdFO3cz1VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV/wVZ/a/u9A0tf2TPhJezXXizxSiReI2scvLb2s2BHZKFyfNuAw3KORGQMHzRj/gn1+yFa/spfB2OLX7aGTx54qWO78RTqAxtyB+7s1YdViDHJBIZ2cg424qqqqr/gt7/wAkq+Gn/YwXX/pOK/Z6/wCSA/DP/sT9J/8ASOKqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr/AIKQft+t8E7Gb4D/AAU1Jrj4laxCqXl9aESHQopOAFxn/SnGNq4yisH4JTP/AATh/wCCf5+Ctlb/AB5+NenvcfErVomksrG7+c6HFIOS2c5unUncTygYrwS+aqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/bf/a38PfslfCO48QiS3vPGOtrJa+GNLkOfOuABumkA58mIMGbpklUyC4I/4Jf/ALJWv+OvEs/7bHx7W41TVNUu5brw1HqK7pLm4dj5mpSA/wC1uEXb7zgDEbVVVVVVf8Fvf+SVfDT/ALGC6/8AScV+z1/yQH4Z/wDYn6T/AOkcVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV/wAFDP279L/Za8It4I8CXltefE/X7YmxiIWVNHgJwbqZT1Y8iNCMMw3HKqQ3/BN39g3U7S/h/av/AGjrGe/8WavK1/oOl6om+S2eRt/9oXAbk3DElkVvuA7z85XZVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVXxp+MngT4B/DfWPij8RNVWz0jSIs7FIMt1KeI4IlJG6RzwB9SSACR8B/hh8Sf+Co/wC05qXxy+MME1p8N/D1yscttHIREsSHdBpdu3BJIIaVwAcMx+VpEFadp1hpGn2uk6VZQWdlZQpBbW0EYjjhjQBVRVHCqAAABwAKqqqqqv8AgtrDC3wG8BzmJDKni4qrlfmUG0myAfQ7VyPYelfspSSTfst/ByWWRnkfwBobMzHJYmwhySfWqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv23f2yPCv7Inw3OpEW+p+NdcR4/DujOxAkcYDTy45EMe4E9C5woIyWX/gn3+xp4q/aC8bSftn/ALU/n61Fqd4b7RLHUkDHWJ+QLuZCMC3TaoijwFbapwI1UPVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVXirxT4d8EeG9S8YeLdYttK0bR7V7q+vbltqQRIMsxP0HQck8Dk18RPGXxa/wCCsv7TNp8PPAH2zRfhf4XmaSKWaPMdha7tr31wowGuJQMRxZ+XO0EDzHPwm+FPgf4I/D7R/hl8O9HTTtD0SARQx8F5W6vLIwA3yO2WZu5J6dKqqqqqq/4LZWs7/s8eCbxYyYYvGiRu/ozWdyVH4hG/Kv2RLq3vf2U/g3NbSb0XwHosZOCMMllErDn0ZSKqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv2n/ANpTwH+yz8LL74keNpxPNzBpGlJJtl1O7KkpChwdo4JZyMKoJ5OAf2Sv2cviJ/wUP+NmpftUftKvJceCre92x2pBji1N4uEsoFBzHaxDG9h945XJdpGW3trezt4rS0gjhggQJFFGgVUUDAVQOAAABiqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr6+sdMsbjUtSvILS0tImluLieQRxwxqMs7MeFUAEkngAV+1Z+0h8Sv+Civxo0/9lz9mmC4k8EW93ulnY+WmptE3z31w3VLWPgop5JwxBdkRf2Xv2ZfAH7K3wvs/h34JgW4umxNrGryRBJ9UusfNI+M7VH3UTJCrxkksxqqqqqqr/gtX/yav4W/7KBZ/wDpBf1+xR/yaL8IP+xP0/8A9ErVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVfFj4q+B/gn8PtY+JnxD1iPTtE0S3MszkjfK3RIo1JG+R2wqr3JH1rwL4T+Ln/BWj9pq48c+N/tmh/C3wvL5bpExMen2udy2UDYw9zLgGST+EHcRhY4z4T8J+G/AnhnTPBvg/RrbStF0a1S1sbK3XakESDCqO59ycknJJJJNVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVXNxb2dvLd3dxHDBAheWWRgqooGSxJ4AAGSTX7aH7Xfjz9tX4i2v7Iv7JcF5qfh+6uzDqF/aOUGuOh+Ylv4LKPBYs3DkBj8oXd+xl+x34K/ZF+HCaFpwttU8XaqiyeIdeEW1rqQciKPPKwJnCrxk5YgFiBVVVVVVVf8FlER/2R7JmVSU8Y2JUkdD5FyMj8CR+NfsCu8n7GvwlLuWI8PRgEnPAdgP0FVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVa5rej+GtGvvEPiHVLXTtM0y3e5vLy6lEcVvEilnd2PCqACSTXxh+JHxU/wCCrH7Sen/CL4TG70v4ZeHJ2ljmmjIiggDbJNSuU+XdIwO2KInIDbRgtI1fBf4NeAvgF8OdJ+F/w40lbLSNKjxuYAy3Up+/PMwA3yORknHoAAAAKqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqrm4t7O3lu7u4jhggQvLLIwVUUDJYk8AADJJr9tT9tTx7+1749i/ZJ/ZJgvdS0PUro2l9f2JKvr7j76q3AjskAJZyQHALEhB837Ef7EXgr9kTwVvf7NrPj7WYFGua4I+AOD9mt88pApA9DIQGbGFVKqqqqqqqv+CvsEM37G99JLGrtD4j094yRyjbnXI98MR+Jr/gnFNLP+xL8K3mleRhptwoLsSQFu5wB9AAAB2AFVVVVVVVVVVVVVoX7Vf7Ovib4hD4T6H8XNBuvF/2yay/scSss4uId3mxYYD5l8twR6jFVXxS/aV+A3wS1a00H4rfFLQ/DOo39t9ptrW+mKvJFuK7wAD8u5WGT3B9K/4b8/Y0/wCjh/Cn/f5//ia/4b8/Y0/6OH8Kf9/n/wDia/4b8/Y0/wCjh/Cn/f5//ia+Hv7Wv7NvxY8U23gj4c/GHQNf128SR4LC0lYySKilnIBUdFBP4VXhz9tL9lTxf4m0/wAG+Gvjp4Y1DWtVu0s7KzhuGLzzu21UXjBJYgDnkkVVVVVVVVVVVVVWqanpmiabdaxrOo2thYWMLz3V1dTLFFBGgyzu7EBVABJJIAAr4afGP4VfGXTbrVvhV8QdD8U2thN5N0+mXizeQ/OA4ByucEgkYYDIyKqqqqqqr4gfFT4Z/CnSxrXxL8faB4Ys2zsl1XUI7bzCOyByC59lBNah/wAFNf2GtNuzY3Hx5tXkU4LW+h6lOnXH347dl7etfDb9rf8AZn+L19DpPw8+NvhXVdSuH2Qae16La6mbOMJBNskf8FPb1qq+IH7XP7NPwr8V3ngb4h/GTw9oWv6esZurC6mYSQiRFdNwAOMoysPYitF1nSfEmjWHiHQdQhvtN1S1jurK6hbck8MihkdT3VlIIPoar4n/ALQnwP8AgtPb2fxV+Knhvwzd3cPnW9pf3ypPLHu271i5cruBG4DHB9DiX/go3+xNFO1u3x/0YujlSVsrtlyDjhhFgj3BxXw1+PPwW+Mav/wq34peGfE0sK7pbfT9RjlmiX1aLO9RyOSAKqviT8U/h38HvDX/AAmHxO8Xaf4c0YXCW/2y9cqhlfO1BgE5O08exr4U/tAfBf45HU0+EnxG0fxQ2jCI362MpY24k3eWWBAOG8t8f7pqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr9vX9qjxr+1z8V7H9jH9mJpdV0iTUBbanc2cgCa1docsN46WkG0szk7WKlj8qKx/ZG/ZW8FfsmfCu28DeHRFfa1ehJ/EGtGILJqNyB19REmWWND90EnlmYmqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr9on9vn9nv9mHxbH4E+JWoa6+uy2KXq2mm6Y0/wC5dmVTvYqmSUbjd/DzjIzcf8Fpv2VIZnii8FfFG4VTgSR6TYhW9xuvAfzArwZ/wVv/AGNfFlzDaan4j8SeFmmwA2taK+xSR0ZrdpgOeMnj3xzXgvx14K+I+gQeKfAHizSfEWkXHEd7pl4lzEWwCV3ISAwBGVPIzyBVfGT47/CT9n/wwPF3xd8b2Hh7T5GZLcTFnmunAyUhiQF5GxjIVTjOTgc1rP8AwWo/ZosdRNppPgL4h6lao2DdCztYQ3PVFa43EY/vbT7V+zl/wUH/AGc/2nvEkfgnwHqWu6d4lmhkmh0nWdMMMsiRgl2DxNJFwBnG/Ppmqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq1bVtK0HS7vW9b1K10/T7CFprq6upRFFBGoyzuzEBVABJJ4FftYftjfFP9ur4gQ/st/skaZqs3he6maK9uYA0MmtqrbWlmbjybJQQcPjdkFx91B+xj+xR8Pv2RvBUcFnFbax441OADXPELRYeQnBMEGeUgUgYHViNzc4C1VVVVVVVV/wAFY4YZf2JfFjyRI7RalpjRllyUb7XGMj0OCR9CfWv+CXczz/sKfDJpJTIyjVVyzZIA1W7AH4DAx6Yqqqqqqqqqqqqqv+CgvhrVP2Wv2/rP4x+Frcwwave2Xi2xEfAadZALqMnoS8sUjMPSYZ4NeG/EGk+LvDmleKtBuludM1myhvbKYDiWGVA6MPqrA/jVfGOR/wBun/gpvH4LsLg3fh1Ndj0WNlOVXS9PDNduhHZ/LunU9zIvrWsf8Ef/ANkvWtWvdYnuvHMMt9cyTvFb6rCkaM7FiEXyPlUZwB2GK/4KSfsOfBb9k7wP4O8Q/C668SS3WvarPa3Q1W+SdQiRBhtCxpg5+tfsg/8ABMX9m745/s3eCPiv41vvGSa34htJpbtbHVIooQyXEsY2qYWIG1F7nnNfAb/gm9+z1+zn8S9O+K/w/vPF0mt6ZDPFAuo6lHNDtljaN8qsSknaxxz1r9u34wj4H/sq+PfGNrdiHVLrTm0vSiDhvtV1+5Vl90V2l/7Zn6VdfCnxl8Mfg78NP2ndNuri3Ov+JL+PTpVTAtnsGgMEgb+80ouf+/H1r4NfErSvjJ8KPCXxS0YKlt4n0i3vhEG3eQ7oDJET3KPuQ+6mqqqqqqq/4KPfH34k/s4/s6r49+FWpWlhrlzr1tp4ubi0S58qOSOVmKo4K7sxrywI68V/wTr+J/j34x/sq+HPiB8S/Ek+u+INR1DURc3s6IjOEu5FUBUCqoCgAAAAAVVVX/BUX9pr4vfs0/DTwlqXwe1210fUPEOrTWtzeS2MV08caQ7hsWUMgOT1Kn8K/Y18Y+J/iF+y78OPGvjPWZ9W1zWNGWe+vZ8b55C7ZY4AHYdBX7QfwlT47/BXxf8ACJtem0Y+J9Ne1S/iQuYHyGVioI3JuUBlyNykjIzmv+CfP7B/iX9jubxdrnjDx9Y67qfieO3t0tdMjkS2hihZ2DsXwXkJfA+UBQDydxxVVVVVV/wUS/bvi/ZT8N2ngjwB9lvPiR4jtmmtBMgkj0m1yUF1Ih+8xYMI0IKkoxbhdrfs8/8ABNf4x/tbGL49ftWfEnX9Ls/ECLPbRSnz9Wv4DyrlpcpbREHKAq3HRFUgnTf+CSP7FljZLaXXg/xDqMqjm5udfuFkPHcRlF/8drwL/wAEpPgB8MfjT4Q+MHgbxD4mhXwtqH246NqEsd3DPKiN5RV9qvGUkKPkl87McZyK1XVNP0TS7zWtWu0trHT7d7i5nk4WKNFLOx9goJrxD4a8Xftbaz+0H+0wguBF4aC628ATdhLi+SKKEnnCx2ombrnEH1r/AIJP/GYfFL9lLTfC1/dGTV/h5ePo04dss1t/rLZ8dlEb+UP+uBqv+C2n/JfvAv8A2J4/9K56+H3/AASx/Yz174e+G9Y1TwHq8t9qOjWtxcTjX7pS0kkKszBQ+0cknGMe1ft2/sGJ+xfa6F+0D+z5438RW2mW+rJBKs11i80i5YM0UsM8YQmMlSnI3KSvLbjt/YR/aB1j9pX9mvw58RPFDRN4igkm03WXiQIstzA2PMCjAUvGY5CoAALkAYAqv+CzPxJ1Dxb43+Gn7M/hdjc3bP8A2rdWqEnzLm4c21mnHRgBPx1xKp4zz+xPeXv7Gn/BRnWfgFr+oySaXrd1P4aaeUbPOLlZtPnK/wB5yIlHp9oaqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr/gp9+29e+EbeT9lb4IXst1408Qqttr93YkvLYQzABLSLbz9omDDOOVRgAC0gKf8E7P2HtP/Zc8BL4y8b2EE3xN8TWynUpDh/7JgOGWzjYcZGAZGHDOAASqKTVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX/AAWS/wCTubT/ALE+x/8AR1zVn+yF+ylY2yWkH7NXwvZIwQpm8JWUr9e7PGWP4mvit/wTc/Y++Kul3Fo/wl0/wpfyqRDqPhdRpslux/iWJB5LfR42H0rxVoP7Qf8AwSb/AGibK+0HXW1nwtrQMluxDR2fiGyRgJIZo8kR3Ee4cgkoWVgSrkH4XfEfwz8X/h14d+J3g+5abR/Emnx3lqXADoGHKOBnDo2UYdmUitfi1j/gob/wUan8B+MfE17beF49ZvrKzSJ8Gz0qxEjbIVYYSSURZJIOHlJOQMV4d/Yo/ZH8L6JD4f039nH4f3FtCpVZNR0GC/uD7tPOrysfctXwx/Y6/Z2+DHxMuviz8Lfh3a+HdcvdNk0+VbSZ/s4jeRHZkhYlY2JjUZTaMZGOTVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVXj/4geC/hZ4Q1Lx58QfElloeg6TCZbu9u5NqoOygdWdjhVRQWZiAASQK+N/7RXx6/4KdfFKP4Afs+aJfaV8O4bhZLgT5jEsaP/wAfuoSLkJEOCsIJ+YDG99uP2Sf2QPhn+yR4FXQPCdsuoeItQiQ694gmjxPqEo5wBk+XCpJ2xjgdSWYljVVVVVVVVVf8FQLea6/YV+J8VvGXZU0xyB2VdUtGY/gAT+Ff8Embq3uP2J/C0MMm57XVNTjlGCNrG6dse/ysp49aqqqqqqqqqqqqr/gsz8Il8Xfs/wChfFixtGkvvAesCO5kVfu2V5tjcn6TJbAem4+tf8ErPi6fil+yLoGkX16J9U8CXUug3AJ+YRR4e24/uiCSNAen7s+hr9rP4xQ/AX9nTx18TxdLDfaZpUkWlk9TfTYitsDviV0J/wBkE9q/4IpfBmS81jxz+0Jq9uzrbRroGlSyLndK+2a6YE/xBRbjPpIwyOc1/wAFvf8AklXw0/7GC6/9JxX/AATe/wCTI/hX/wBg+5/9LJ6r/gtD8VL/AMQ+LPh3+zf4aeS6mUHWb6zhyxmuZmNvZpgfxAC4464lXpnn9rf9k6ytf+CbkXwm0e1juNU+FWi2urW0iL/rLi1QteyADnMkcl02P7zCv+CMvxlTxb8DNe+Deo3m+/8AAmqGezjZjxY3ZZwFB/uzrcE46eYvTPNV+2l+1n8XvgZ+39oHh+3+LOt6Z8Ore60K91fSY3HkfZS6falwFLbWRHJAyfmOKm+LP/BRD9vjU77XP2apF+FPwqhuWh0zU725+wyXwRiu9p0SSZ2yDuWAeWuNpLMMn4ieJv8Agp9+wLqll4s8efErVPF3hia5EJvb7VJNe0y5fk+VI1xieAsOh/dE4+UnBA/ZE/ak8JftafCS2+IWgWw03VbWX7JrmkNJvawugASA2BujZSGRscg4OGVgP+CtX7R3xy+CHxE8BaV8JviZrHhi01PRbia7hsXVRNIswUMcg8gcV8E9Y1PxD8GPAOv61eyXmo6l4Y025u7iQ5aaWS2jZ3PuWJJ+tft2eN/Fnw3/AGTPiJ428C69c6NrulWUD2d9bECSBmuoUJUnPVWYfjX/AASW+N3xY+N/w68ear8WPHep+J7vTNat4bSW+cM0MbQliowBwTzX/BSL4eftr6HbeLfFfxZ8c/2h8Ir7xxOfD2mf2mkvkRySztZjygoK7Ycjk8dK/ZP+Ev8AwUj8WfBHR9b/AGc/ib/Y/gWa4ulsbP8AtmK32SLMwlOxkJGZAx685r4Q6T8aPDv7PeiaR491Kz1T4nWehut1c3k3nwz34DbDIybSybtmduDjOCKg+DX/AAV4/aKi/wCEj8X/ABltPhTYXWXg0uDUn0yWFCMqFSxR5B24mk3jvyMV8TvFX/BSz/gnnqmj+K/HfxYk8feEb28+zrcX+ozaxZXD8v8AZ5muFWeB2VWIKlc7SFYhSK/Z9+Nfhv8AaI+D3hr4veFozBa69a7prR3DvZ3CMUmhY8ZKSKwzgbhhsAEV+3r8PP21/B2k6Nf/ALT/AI5/tzw9eaxcroMH9ppc+TJtyTtVQV/d4HNfAH4M/wDBUnxB8GfCWs/Bv4r/ANneCrvT1fRbT+3YofJg3HA2FCV5zxmvjbrnxG+HP7FPifXrrX57bx14e8ANLcanBIGdNQitB5kqtjBPmBjnGDX/AASI+P3xl+On/C2P+FufEPVfFH9if2L/AGf9vdW+zeb9u8zbgD73lR5/3RX7T/7Tf7Sf7J/7d8Wk+IPit4hvvhrcavaazDpU7hopdJuH/fwqNucRsJ415OPLU8mre4t7y3iu7S4jngmQPFLGwZXUjIZSOCCMHNf8FTP2mPE/7PXwN0rTfh34juNF8X+MdWWCyvLYgS29tb4kuJEJzzkwxnjpKa/4J4S/GDW/2Z9D8ffG7xvq/iPXvGM0mp2zaiwLWtk2Et0UAAYdU87PORMOeK/ai/b++NfxA+Ncn7Kn7DumLe67DdPZX/iGKGOd3nTiZIPNzFFDFhg87g8glSoUO+p/sYf8FVdI09vGmnftd3Oqa5Agn/sWHxjqGyRgP9WiSxi3LcDhgqE557n9gH9v3x18TvHt7+zL+0vaLZ/EbTmnisdQa3W2a9ktwfPtp4lwq3ChHYFQFZVYYBUF6r4YadF+2/8A8FOrrU/GKrf+H38Q3d/LbyjKtpunqRbQFcYKsIoI2HGQ7HqaCqqhVAAAwABjFVVf8FS/jG3wl/ZI8Qabp975Gr+Op00C02sQ3lSgtcnA52m3jlQnoDIvrg/8Ewf2cdIi/Yn1qPxjp5I+MhvDeoyYf+zzG1rEv0IEsin0mBr/AIJbeMtV/Z8/bI8Xfs5+MrjyG8Q/atHmjyVQ6np8kjRt6YKLcqPUyLg84Nf8FtP+S/eBf+xPH/pXPXw2/aq/Zg0v4Y+FbXUf2jvhjb3FnoNmk8Eni2xEkbrAgZSnm7twII24zniv+Ci/7ZPhX9qLTtF/ZY/Zks73xzd6jq8VxfX+n2jss8kYYR29upXdJ8zb2kACgIMFgWK/sQ/s93v7Mv7OXhv4aa5JDJr7GTUNbMJDIt3OdzIpH3hGoSPdzu2ZHBADukaGSR1VVBLMxwAB1Nfs2Gb9tL/gqLqnxdui954d8M6lPrtszKSq2tkVg04DPCtv+zSY7lXPqa/4LC/DrU/hn+0B4F/aL8JFrOfXbZFe6iHMepae6GOUnsTE0IH/AFxNfB34k6R8Y/hV4U+KOhlRaeJtJgvVjBz5LugLxn3R9yH3U1VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV/wUC/bR0v9k/4anT/Dlxb3XxF8TQvHoVm2H+yJ91r2Vf7iHIUHh3GOQHx/wTD/AGLNVa7X9sD492s9/wCJtdke98N22pZeVPNJL6jNu5Msm4mPPQHf1ZStVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV/wWS/5O5tP+xPsf/R1zVV/wV28A6b4r/Y81TxXcW8ZvPBesWN/bTFfnUTTLauoPoRcKSOhKKewI/wCCNXi+78Qfsp6j4cvJmYeGPFl3bWqnOEglihnAH/bSWY4Hr71+1x8I/i/+wX+14v7R/wAOtPLeG9S16bVtD1DymktkefcbjT7jGCgIklQDILRMCrbg239nn/gqh+zV8Z7Wy0rxpra/DnxRKoWWz1yULZvJgZ8q8wI9uenm+Wx9D1q3uLe8t4ru0uI54JkDxSxsGV1IyGUjggjBzVVVVVVVVVVVVVVVVVVVVVVVVV9qOn6Xb/atSv7e0hBx5k8ojXP1Jx2rWf2hvgB4dOPEHxz+H2mHIGLzxNZwden3pB1xWsft7/sbaGhe9/aI8IygKGxZ3LXZwTjpCrc+3UDnpzWrf8FT/wBhzTHMUPxguNQdWZWFp4e1AgEf7TQKCD2IJFan/wAFif2P7Fc2o8c6idhOLbREXkdv3kqcn8vcVff8FsP2b43QaZ8M/iTcLj5jPa2UJB9gLl8/pV9/wXA+HUcIOm/AbxHPLuwVn1iGIAY65CNz04x+NXv/AAXLsI5gunfsyzzxbfvT+MRE2e/As24981df8Fx/ELzs1n+zhp0UJPyJL4oeRh9WFsoP5Ch/wWn+LWsHzfDX7N+kyQxjbJ/xMri5w31SNQOO2K/4fIfH7/o2nSv+/t5/8TX/AA+Q+P3/AEbTpX/f28/+Jr/h8h8fv+jadK/7+3n/AMTR/wCC1HxZ0dw/iT9nHSI45AREDqVxbEsMZ5eNs8dsd6tP+C5GvJOrX/7N+nzQgHKQ+KniY+mGNswH5VY/8FytLkLjU/2aLqAADYYPFyzZ9c5tFx+tab/wW8+Fcwi/tj4H+K7bOfN+zajbz7PTbuCbu3XFaX/wWo/Zfu/Lj1PwJ8S7GRiQ7DT7OWNPQ5F0GP8A3z/jWk/8Fdf2MdSVTeeI/E+lEoSRd6DKxBzjH7ovz39PftWj/wDBSL9iLXTix+PulR8kf6Zp17Z9P+u0K/8A160L9q39mHxOVj0P9oX4dXUrH5YR4ltUlPT+BnDY5HOK0jxBoHiGA3Oga5p+pwgDMlncpMoznHKkjsfyNVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV+0v+1V8I/wBlbwYfFPxJ1rN7cqw0rRLUh7zUpB1EaZ4UZG6RsKuRzkqDYaJ+1Z/wVv8AieNX1edvCXwq0G8KxvsZrDTB3SJeDeXhQ/MxwF3cmNWVT8BP2e/hb+zX4Dt/h/8ACzQFsrRSHvLuXD3WoTYwZp5MDex+gVRwoUcVVVVVVVVVVV/wURsX1H9iv4rW8cioU0hJckdo7iJyPxC4/Gv+CPV99s/Y9jt/K2fYfFGoQ53Z35ET59vv4x7e9VVVVVVVVVVVVV8aPhrpvxj+Evi74W6rsEHifR7ixEjjIhkdCI5fqj7XHuor/gjv8Q9R+HH7RHjT4B+JhJaS+JLF8Wsjcx6jp7vuTGeD5T3GSP8AnkPTj/gtn8YTZ6B4D+A+nXZEmoTya/qkYOD5Ue6G2B7lWdrg46ZiU89v2LfguvwD/Zm8DfDy5sxb6rHpy3usKR83264/ezKx7lC/lg+kYqv+C3v/ACSr4af9jBdf+k4r/gm9/wAmR/Cv/sH3P/pZPTukaGSR1VVBLMxwAB1ND9o/4YePf+Cik37R/wAXtQvP+EJ0zxE17YrbW7XLvDZps08KgxjLRQSMOn3upPN7/wAFbf2KtRs59PvtV8Tz21zE0U0UmgOVkRhhlIzyCCRX/BPT4u+FPg5+3Xb6d4V1iZ/A3jS+u/D1pNdAxM9vNLusWdG6OZI7dTnkb25Peq/4Kx2H9q/tvy6X53lfa9H0uHft3bNwIzjjOM9K8E+DfD3w88H6L4E8KWKWej6BYxWVlAoA2RRqFXOAMkgZJ7kk96+NPwy0b4zfCbxZ8LNet4pbTxLpU9pmVAwilZcxSgH+KOQI6nsyA9q/4IpeNr/S/j74y8Ai4caf4g8LG8eIHhri1uIxGSPZLifn396/4Le/8lT+Gn/Yv3X/AKUCv2ev+SA/DP8A7E/Sf/SOKv8AgpF/yZH8U/8AsH23/pZBX/BEP/klXxK/7GC1/wDSc1/wWS/5NFtP+xwsf/RNzX/BJ/8A5Mj8H/8AYQ1P/wBLJalkjhjaWWRURFLMzHAUDqSewr43/wDBWr9mz4WaxceF/BMOrfEfWLaQxu2i7I7FZBkFRcufnOccxJIpz97jFftift4/FT9oz9n/AFzwXrX7IWv+EfDd5NaTr4iu7i4uI7OSO5Qq29rWNPn/ANV94cydT0P/AARf1G8vv2T9ctbmYvHp/jm9gt1J+4htLOQgf8Dkc/jX/Bb3/klXw0/7GC6/9JxX7Af/ACZp8Jf+xfj/APQ3r9tf/k0X4v8A/Yn6h/6Jav8Aghj/AM1s/wC4B/7ka/4LN/BA+Lfg54e+N2kWRe/8D3/2TUnRefsF0VUMx9EnWIAdvOY1/wAE1/jMPjP+yR4RuLy6M2reEUPh7Udz7m3WqqImJ6ktbtAxJ7k8mv25rrU/2xP+Cinhv9nXw7cyNpfh+e30J5YBnys/6RqFwAf4o0LKRx/x7j6n9oLxPB8C/wBmDxx4i8IRJpv/AAiPhG5TRY4gQttJHAY7YDvhW8sfQV/wT5/a/wDhB+yLqnjDxV8QPBHiPxB4g8QRW9rZXWmrA32W3Us8ykyOpzI5iJ6/6pa/4fafAH/olHxA/wC+bT/49Xj79o/wl8Vv2/8Awp+0N8K/Duq6FBeeJNEmns70RrLJNE8MUvETFcSIgB5ydzE9aqv+CTMyeEP23pvDGtSbb+40PVNNRTkbp4mSRxzz92CQ/hVfttfBD9tz4o+P9C1f9mD4v/8ACJaDaaP5GoWv/CQT2Hm3XnSNv2Rowb5CgyTnjHav2lvGn/BQT9lLxXpfg34n/tLeJpb/AFfTvt9udL8T3M6CLzGj+YsEIbch4x0xXhmea68N6Tc3ErSSy2MLu7HJZigJJ9ya/wCCxvxig8b/ALQXh74NW2peVpngOwVr9xlgl5ebJHJUfe2QLAR3yzDjnPgj/gqV+w58P/Bmg+BPDureKotL8O6bb6fZp/YL5WKGNY0zz1wor9q/9oX4Ya5+2Np37TP7OV9fOqz6fq1zHeWrWjDUbZgHG05yjpFESe5d8j18HeK9G8eeEdE8beHLpbjS9f0+C/spQQd8M0YdDx/ssK/4Laf8l+8C/wDYnj/0rnrwb/wRt/Zn8T+BdD8RT+PfibDfatpNvdOE1GxMaSyRKxwv2TO0Fum7OO/evjt+xL8f/wDgnrDP+0P+zV8YNTv9D08pHqxWBYbq1hZwF8+LLRXUG8qDlRtLA7MAsP2EP2ubX9rv4Qv4l1OytdN8XeH7kWXiCxtyfLDld0U8QJJEcig4BOQyOvIAY/8ABQn4zD4IfsneNvENpdeTq2t2v9h6SVba32i7BjLKezJF50o946/4Jn/tW/s0fsp+FfGOpfFLU9Yj8V+J7+GJFstLe5WKygQlPnBGC0ksuQOyISfT9vj9ub9kf9qP9njUfAXhfVtfbxTp9/b6loTXWjPFGJ0bZIrPngNDJMvpuKk9K/4Iy/GNfFvwJ174Pajeh77wJqpms42fkWN3ucBQeyzrcEkcDzF6Z5qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv2kf2g/A/7Mvwo1X4peOLkNHaL5WnWCuFl1K7YHy7eP3YgknB2qGY8Ka/Y4/Z98df8ABQH49at+1b+0bHJeeD7HUA0dpKpEOpTx/wCqsolPS1hG3f8A3vunJZyI4o4Y0hhjVERQqIowFA6ADsKqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv+CyX/J3Np/2J9j/AOjrmqr/AIK4+M7Dw1+xnrfh+4uEW48W6xp+n28ZxucxzrdMQOuAtscn3HrX/BGnwhd6D+ypqXiK8hdB4m8WXdzasRgPBFDBACP+2kUwz7e1a/4f0HxXo134c8T6LY6vpV/EYruxvrdZ4Z0PVXRgVYexFftCf8EgPgR8Q7K91n4KXVz8PvETKzw2oke50yaTrh423SRAnjMbbVBPyHgV+wv+0f8AGP8AZF/aVg/ZQ+Ml3dHw3f60ui3WmXUxmXSL2VgsE9s3O2J3aPcAdhSTzMZGTVVVVVVVVVVVVVVVVVVVa/4l8N+FbBtU8UeINN0eyQHdc392lvGuOuWcgCvG3/BQX9jTwEJF1j4/eG72SNSRHozSaqWI7A2qyLnPqQPXA5rxj/wWd/Zh0QvB4U8K+OfEkyn5ZEsobSBhj+9JLvHb/ln6/jrv/BbPxfrVydN+Gn7OFt9pfIgN/rcl47nHXyoYUPXsGPTqM8f8Nyf8FUPiZx4D/Z3m0uCXiK40/wAC3rIAemZblnjzhgew4zjGaOh/8FtfiUvm3Wo63o1tL0b7Xo+k+WMHA2xFZejdcZ4GTkV/w77/AOCm/wAQTu8eftMfZoXPzw6j451K42jP8McUbJ1VTjIHTvVl/wAETfiJr1wLzx1+0ppqXDDDywaJNft0z96SeMn5if5+1aL/AMEQfhhBj/hIvjp4ovuTn7FpcFrxjj7zS98n6ce9aL/wRo/ZL0zY2oa38Q9XIOWF1q8CKeMYxFboQM89c+9aV/wSk/Yf05At58LdR1QhNu678R36kn+9+6mTn9PatO/4J4/sWaWpW1/Z98PPnGftMlxcdPTzJGx/WrH9jv8AZP05XW3/AGbPhm+8jPn+F7SfGPTfGcfhVj8B/gdpcgm0z4M+BbSQLtDweHbWM49MiPpwOKsvAXgXTYTb6b4L0G1jZtxSDTYowWwBnAXrgD8qtbO0sbdLSxtYbeGPOyKJAirk5OAOBySaqqqr3T7DUoRbajZW91EG3COeIOuR3wc81ffDv4faoUbUvAvh67MYIQz6XDJtz1xlTjpWpfs6fs+aysi6v8Cfh5fCZt0gufC9nLvbOcndGcnPrWrfsT/si60ZDd/s4fD+PzCN32TQ4bXGMdPKC7enbGe/WtY/4Jo/sP62Wa6+A9jCzMWBs9Wv7XaT6COdRj2xj2rWv+CQn7Guqgix0bxZo+cYNlrrtjH/AF2WTrWv/wDBEn4F3KOPC/xd8d6exHyG/S0vADjuEjizzz244961n/giL4r0uf7d4D/aRs3nQt5QvfD8loyZyMeZFO56HBIUdelN+wL/AMFPvhrmT4e/tIPfwx5Mdtp3jjUIA/Xhop0SPOWJ5JGTnNHxN/wWt+ExK6ho2t+I7SEnEf2DTNZ8wA9d0AabkDpkHB6A9F/4KsftofC4pbfG39mywiWEfO15ouoaLMwz1YyFlB6jhAK8Gf8ABbr4UX5jT4g/BTxVopJwzaRfwakB7/vPs5x0/wDr14H/AOCn37Fnjgw2/wDwtn+wLuUf8e+uabcWmz/el2GIf9914M+Kfwx+I9uLn4e/EXwz4mixndpGrQ3ePr5bHH41VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV+2x/wU28Afs8JffDz4VPY+L/iKoaOULJvsdGfpm4ZT+8lB/wCWKnIwd7LgBv2bf+Cffxm/a08ZL+0b+2trusppmpss8Ol3bmK/1WPqilRgWdtz8qKFYjO1UBVz4V8J+GfA3hzT/CPg3QrHRtF0qEQ2VhZQiKKBB2VRwOSST1JJJ5NVVVVVVVVVVV+27YR6j+yH8XreSBpgnhG+mCrnIMcRcNx2BUH0454r/gizftc/sseJLKS5V2s/Hd2qR5GY0aysmHHXBYvyff0qqqqqqqqqqqqqq/a20+f9kb/gpzo/xds0+y6LrWsWXiXeFwGguGMWoLnpuZhck+gkXI551HS/+G0/+Cs11YsgvvC/hLWgtyMF4hYaSArg/wDTOa5Tbnp/pHvVV/wW9/5JV8NP+xguv/ScV/wTe/5Mj+Ff/YPuf/Syev8AgoL8Y1+CX7JvjnxFa3og1bWLP+xNJIfa5uLvMZZD/eSIyyj/AK51/wAEpf2NPhX8VvhT4o+K3xp+H2n+JYNS1ZbDQ4tQQlYY7dMzSoAR995Qhz3hPTv/AMMB/saf9G8+FP8Avw//AMVX/BVD9mvwx+zZ8UvA3xA+DPhyHwvoOt2TLFDYAqtrqVpIGMinJKlkkhI942I74+AnxRsfjZ8F/BnxVsGjA8S6PBdTRxnIhuCuJ4/+ASq6fVar/grHff2X+2/Lqfleb9j0fS5vL3bd20E4zzjOK8B+OPDPxM8FaJ8QPB2pJf6L4gsYryynUj5o3UEAgZ2sPusvVWBB5Br9ov4waJ8Bfgp4u+KetahDbf2LpkrWKyEZuLxlK28Kg9WeUouPck8Amv8AgiZ8NdR1D4p+O/i5NaP/AGbo2hro8Mzr8rXFzMkrbT3ZUt+cdBKM/eFf8FwvD17F4s+FXirY7Wd1p2o2W4JwkkckL4J9SJeAcfdPXnH7HnjfRfiH+y38LfEmh3cU8X/CLWNpcCNsiK5t4Vhnj+qyxuvPpX/BU/4h+F/Bf7HPi7w/rOpQx6p4ua20/SLMyASXMi3MUshVeu1I0ZicYB2gkFhX/BEP/klXxK/7GC1/9JzX/BZCKST9kS3aONmEfi+xZyBnaPKuBk+gyQPxFf8ABJm7trn9ifwtDBOkj2uqanHMqnmNzdO2D6Ha6n6EV/wVm+JXiT4dfsh39r4Zurm1l8X61baHd3FuSrJbSRzSyqSOiusHlH1EhHev+CQHwI+GGnfAaL44NolhqXjTXNUuoG1C4iWWXToYX8tYIic+Vux5jEYZt65JCrj/AIKv+KNC0H9iXxjouq6hHBe+JL3TbLTIWPzXEyX0Nwyge0VvK3/Afev+CKn/ACav4p/7KBef+kFhX/BbyKRvhN8NpVRii+IrlWbHAJt+Bn3wfyNf8E+ru2vf2MPhPNaTpKi6H5ZZTkB0lkRx9QykH3Fftr/8mi/F/wD7E/UP/RLV/wAEMf8Amtn/AHAP/cjXxV+HeifFv4aeJ/hj4iQHT/E+lT2EzbQxi8xCokUH+JGIYHsVBr/gnF8c7r9kv4hfG/4V/E1zbw6Lol/qMtm8mM6lpRcPFGD/ABSoXGe/lJX/AAR7+GGrfED4hfEf9rTxypvNQuLuWwsrqRf9be3LfaL2UDswVolBHaZxX7TXw+vfir+zz8Rfh5pUHnajrnhu9gsI8433PlMYR+MgQV/wRz8VfDKfxV44+CPxE8O6Lca5qoh1DRV1SwjkeRoA6XMKmRSQwUxuEHJCyHHymv8AhU3wr/6Jp4U/8E1v/wDEVb/C74Z2dxFd2nw78MQzwOHilj0iBWRgchlIXIIIBzVV+2p4K8a/sSft2W3x18H6ft0rW9ZPibQ3GUindmBvrNiB8vzvIpUdI5k9cD4A/tBfDT9pP4eWPxE+Gmtx3VvOii9sXcC502fHzQzoDlGBBwejD5lJBBqv+C2n/JfvAv8A2J4/9K56std0nwt8NoPE2u3iWmm6Roa3l5cSHCwwxQB3c+wVSfwr9iP4cWP7cf7bni34lfFTw+mreHEN/r2q2F0S0TtO5jtbZiOcJ5gZRxxb49j/AMMB/saf9G8+FP8Avw//AMVX/BS/9hz4MeC/2a7n4l/BT4Y6X4b1Twnqdvcai2nxsDcWMhMLggk52ySRPnsFbtyP+CRPxh/4WL+y3H4Fv7vzdU+HepSacVZ8ubSUma3Y+g+aWID0hr/gtp/yX7wL/wBieP8A0rnr4T/8kr8G/wDYv2P/AKTpX7fXxG8IfDj9kj4lz+LLu3Vte8PXejabbOw33F3dRNDEEU8sVZxIcdFQnjGa/wCCHfhjWobD4teMpFmj0i7m0uwhODsnuIhcSSc5xuRZo+3Sbt3/AOC0vxZn8R+P/Af7P2gTPcnSbdtWv7eE7y93cnyreMqP41jRyB6XA9a+FX/BOf8AZd8NfDPwtoHjj4LeHda8R2OkW0er6hcRs73N2Ix5z5DYwX3Y9sVL+wB+xpLG0R/Z78LLuUjKxSAj6Hdwa/Yj1jUv2Qv+Cidx8IfEV28djqGqXfhG7klbaJhJIDZy46ZeVLcg8HbKfXFVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV4o8TeH/BfhzUvFvivV7bS9H0e1kur69uX2xwRIMszH0AFa5qPxK/4K1/taRaNov2/RvhX4SJ2yMnGm6eW+aZxypu7krhV5wABysTtXgPwL4V+GXg3R/AHgjR4dL0LQrRLWxtIhxGijuerMTksx5ZiSSSSaqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/4LJf8nc2n/Yn2P/o65qvij8ZPhZ8FPDsvin4qeO9I8N6fGpKte3AWSYj+GKMZeVv9lFJ9q/aQ+MvxI/4KeftJeH/hh8GtDvY/C2lyPDo0FyhQRRsV+0ajebchBhRgc7VVVALuQ3wd+F3h74KfC3wx8KfCwLab4Z06OzilZArTsozJKwHAaRyztjuxrwX8UPhz8R21GLwH440XXpdIu5bTUIrG8SWS0mjco6SIDuQhlPUDPBGQQa1LUtN0fT7jVdX1C2sbKziaW4ubmVYo4UUZZmZiAqgDJJ4FfFHV9N/a/wD+CoFpc/CPN/pV/wCJ9Mgjv7dciW2sI4VubsEdUCQSurd0VPXFVVVVVVVVVVVVVVVVV8QvjX8H/hNbNc/Ev4neGfDIVNwj1PU4oJXH+xGzb3PsoJr4k/8ABX79krwX5tt4RufEvjm6TKodK0w20G4f3pLkxnHuqP7ZHNax/wAFfP2jvilqUnh79nb9nKzN252pGYrrXrrB4DCOBYgGPoVYduetDwj/AMFlv2h8Nq2u+IPBel3HAZ7628OiIHHBS323JH1U9x7VoH/BGP4reMr9db+OX7R9o964BnNjaXGqyyHuvn3DxEfUqfpXgr/gjd+yl4dEcvinUvGniuYD94l3qaWsJPstvGjj8XNeD/2Ff2P/AAMsY0H9nnwdK0YGx9Usf7TdSOhDXRkOffOa0Hwx4a8LWgsPDHh7TNItgABBYWiW6YHT5UAHGT+dVVVVVVVVVVVVVVVVVVMqspVgCCMEEV40/Zk/Z2+Im9vGnwP8EatM5JNxPocAnyepEoUOM98H0rxx/wAElf2M/F4Z9I8J6/4SmYHMmi61KefXbc+co+gAFeM/+CJM1lM2p/CT9oKa3uISWtrfWdJKsp7E3MEgI+oir/hQP/BXz9nJN/gD4iar4u02zGEisdfj1aFUHZLbUAGwfRI881Y/8FVf2w/gjfRaN+0l+z1bz7X27rzTbrQLqb1w7q8TDAyCseD15FfDP/gsV+y14x8q18dWfifwLdnAeS9svttsCf7slvuc/UxrXw4+OPwb+L9sLn4YfFDwz4m+Xc0OnalHLNEP9uIHeh9mUGqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqvG3jjwf8N/DF94z8eeJLDQtE0yPzLq+vZhHHGO3J6kngKMkkgAEmv2iP+Ch/xq/ax8X/APDPn7E/h3Xbaw1PfBLqdvGY9S1JOjOrZxZ2+DkuxDYwWaPla/Ys/wCCX/gX4DPYfEf4xix8YfEGMia3iKmSw0iTqDErAebKDz5rDAP3VBG41VVVVVVVVVVVftPWL6p+zT8WtMjkVGu/A2tQqzDhS1jMM/rX/BEe+8z4LfELTPKx9n8URzb93XfaoMY7Y8v9faqqqqqqqqqqqqqr/gp7+x546/ai8H+DtX+EujWmoeLfDGoTQvDPdx2oksZ0y53uQpKyRRYUno7kd8/8Ew/2LPiH+zOnjbxr8ZtGtbHxVrzw2NjHFexXRjs0/eSNvjLAeZIyggnP7kHHPNV/wVA/Zl+MH7TfgLwToPwf0C11W80XV57m8Se/itQkbQhQQZGUHnsK/Yv+GXjD4Nfsw+A/hn4+0+Ky8QaFZzxXtvFOkyxs1zK4w6EqfldTwe9f8FQP2dP2l/2nB4I8HfBvwna6h4b0P7Rf6jLPq1vaiW8fEca7JHBPlxiQ7sY/fEdjX7NHwig+A/wE8D/CeNIhceH9IijvmixtkvHzJcuMdmmeQ/Q1X/BQb9mzV/2n/wBna+8G+ELC3uvFuk6hb6loKzTLCGmVtkqGRiAoaGSXqcbgmelf8E1/hB8e/gH8F9U+FPxx8OQacNO1h7rQni1KG7DQTqDJHiJm27ZVZ+cZMxx0NV/wVYtLe/8A26hY3UfmQ3Gl6THImSNytkEZHI4Nar+yX/wUG/Y81nVbT9i7x8fFHw+1C4kuINEu5rRprLcejQ3mIy+No8yFgz7fmUcCta/Y6/4KVftn+ItMX9pvxJD4a8O6fNvj/tC5tfLtx0Z4bKyOHlxkBpNhIP38Gv2fvgN4E/Zt+F2l/Cr4e20gsLDMlxdT4M19cvjzJ5SAAXbAHTAVVUcKK/bA/Zc8Nfta/B+5+HGsX40vU7WcXuiaqIvMNldKpUFl4LRsrMrLkcEEcqK+Hn7M3/BWL9lfUb3wl8EIrsaVe3BYNp+qabeafO/AMqxXpxExAXLNGjEAZ6cad/wTl/aE+L/h3xr8V/2vfHR8afEWXwtqNr4P8OrqAMFnfyWzi3eSRdkMe2UqwjjHlhsMzHBWv+CX/wCzP8Y/2ZfA/jfw98YfDdvpV1rGq291ZeRqEN0JUWIq3MTNtIIHXGc8d6/aj+AulftL/A7xL8H9Svxp8urQpJYXxj3/AGS6icSRSFe67lCsByVZgMZzXw0/Zw/4Ky/sq3mp+CPgfYzxaVqN15ryafe6Xe2FxJgL5yLe/wCqJCqCzJGxCrngCvGH7PmuftJ/sh6V8G/2hb023jG90OybU9St1jke01aJFJnAT5G/eBgwXCsrOFKggj4Xfs7f8FUf2NNY1Lwj8ENH0/xB4d1W68wiC9sriwnmwFEwjuXjkgfaFBbCAgDJbaMeK/2A/wBrz9pLwj4k+JX7V3xBh1jxvZ6JPF4L8I2N1FFbWl02NplaIJAnTGELbjsLyELtP/BMn9n74wfs2fBfxJ8P/jB4dtdKvbzxRJqdmsF7FdeZHJa28RJaN2AwYOhwee+eP23f2Xov2tPgZd/Dmy1S30zX7C8j1PQry5UmKO7jV02S7QWEbpJIhIB2llbDbdp+FnwT/wCCv/7PWny/Cr4R2F9YaCbl5IlW90e8tFLZLPE90WMKtyxUbCScldxr47eBvHnxJ/ZW8X/D20tILrxj4g8HzWPk+ckaSXslvtYbyQoBcnknFf8ABLP9k/43fsvf8LO/4XJ4btNK/wCEl/sj+zfI1GG68z7P9s83PlM23HnR9cZzx0NV+3//AME4/jd8V/2idQ+KvwD8M2N9pvibT4pdVV9ThszDfKpikAWRxlXjSNiR1Znz6n9kz4Iw/s7/ALPXgz4UtFCuo6ZYCXVnjORJfTEyXB3c7gJHZQf7qr0AAFftjf8ABLfWPHfxBuvjt+y/4ntvDPiu6u/t97pM072iPeZ3G5tbiPmGVm+YqQFLEsGTpWlap/wW68IW8fhldGudWW2bZFdXa6Ldsynpmfdlh3y5LDPJ4wP2Q/hx/wAFHV+MsXxL/as+IFqfDi6VcWkvh9tQgYmSTaY3SCyT7OGVkGXLbgCQM7jVV8c/gP8ADL9ozwDdfDn4p6Cuo6ZOwkgljby7iymAIWaCTBKOMkZ5BBKsCpIPir/gld+1t8BvF0vjH9lL4sLqSoSLd7bVG0XUghP+rkyRDIuODmQBucoM4q1vP+C5FgY9DhTVX8s+WryweHZuvczsDnr94tx61qf/AATb/b+/ab8UW3i39o7x/o2n3UNuIBPrGqLez28WS3lxQ2itEAGZiQHUZJOTX7Zfw++MPjb9lbWvhJ8EtJi1XxHrttbaVM73sdosdplftD7pGAIZEMe3OcS+xr/gmb+yV4w/Za+FviT/AIWdpVrZeMfFOrrJcxQXEdwI7OBNsCF0JBO5524P8YHUGq+IXgjRPiX4D8Q/DzxJD5ul+JNMuNPu1AyRHNGUJX0Ybsg9iAa/4Jw/sh/tW/sq/HHWrrx/4TsY/BXiLSpLO9ubfWbeUCaJw9vN5SuXPSRAMZHnEnGDX/BUL9jH9oX9pH4o+F/GXwc8H2uuWGk+HPsVyr6rbWkgmFxK+AJnQEbXXnOOtaNoX/BcfStKsfDOnj7Hp1nbx2kAaXwufJiVQi/Ny5woHPLcZ5NW3/BMr9s39o7xXZeJf2tvjjbwWduMeW1++qXcCnBaOCFQtvCDjBKtgHna3f4N/B3wH8Bvh1pPwv8AhvpP2HRdIjITeQ0txIeXmlcAb5HPJOB6AAAAaJ+wx+0h8Tf+Cgf/AA0R8avCFjY+C18UvqyM2rW90/2e1X/iXwmNHJP+qtlbjGA30qq/4KG/sB/HD4uftC6d8bf2d9BtLi4vdPt21WRtThs5Ib+2bbHMvmMucxLCBjODFz1FeD7rxFf+EdEvfGGlx6br0+nQPqllHKsq210Y1MsaupIYK+4Ag4IGaqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/4KD/ALTHi/8Aav8Ai/pn7FX7OLyappy6qtrqs1rJiPVr9GyULDj7Nb7WZmPylkZukasf2UP2ZfB/7Knwi0/4c+GljutRkxc65quza+o3hUB39QgxtRf4VA7liaqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/bk/4Js+L/2sfjDbfFPw38T9H0KOLRoNOeyvrCSQgxPI28Ojc58zGMDGOpzVl/wTB/bkg26f/wANi/Z9PG5dsOv6qcKc9I8Ac9xnua8H/wDBFay1HV11z43ftDazr7yEG4g0rT/Klc/9fM7ykjr/AMsx9a+Bv7OPwa/Zw8Nt4Z+EXgq00eKfaby7OZbq9YDhppmy79yFztXJ2gZp1LoUVypIIDADI9xmviF/wRn8Y6Z4nuPFXwF/aDe0mkkeSFNdilguYS3XN3bZL5yefLX8ak/4JQ/toePQmj/F39qDR7zRg6sUbW9T1fb6sIZ44lyMDHzD6jFfsjfsK/CL9kTT7m88Mvc+IPFmpQiK/wDEOoRKspjByYoEXIhiJAJUFixA3M21QKqvE/j/AMC+CovN8YeMtF0VduQL+/jgJ+gYgn8K8Q/tyfs3aCZIofGtxq00ecx6fp0z5I9HZVQ59mxWp/8ABSX4Rwbl0jwV4suyvAM8dvACc+0rHGOen4Cvhx+3v4e+JXxA0jwJpnw51G0/tV2QXU+oIfLKozn5ApyPlx170viq3IG60kH0YGk8Tac+AyTJ9VB/kah1jTJzhLxAfR8r/OlZHUMjhh2IOaqqqqqqqWSOGNpZZFREUszMcBQOpJ7Cvi7+3/8Asl/Bcz2fiX4vaXqeqQEqdM0EnUp946o3k5SNvaRkr4i/8FpdY1u/Ph39nb4Ez3t5cOUsrrxBM00kp7Ys7U5JwCcCY0fDv/BYD9rNf+JpqGueAvD91xtnmTw1CisPutHGBdyKR/eVxXw7/wCCJ8V5crrHx1+Ot5f3MzB7m18PWmGZv4j9quNxbPqYRXw2/wCCbn7G3wz8qey+Dth4gvYsZuvEcr6kXI7mKUmEfhGK0XQdD8N6dFo/h3RrHS7GEYitbK2WCJPoiAAflVVVVVVVVVVVVVVVVVVVVVVVVVahp2n6tZS6bqtjb3tpcLtlt7iISRyD0ZWyCPY18Tv+Cd37HvxV86fV/gxpOjX03IvfDxbS3Vu7bISsTE9y6NknPXmviR/wRSis7v8Atv4B/HK9066gffbWniG2yyMDwRd221lx7RE+9NJ/wV4/ZAUvI2t+PvDVgOSxHiW3dB+d3Eij/rmAPYV8LP8AgtVoguo9C+P/AMGtQ0W7hby7m/8AD83nKjjg7rWfa8YB6jzHPXjtXwi/bF/Zm+OjQWvw3+MGg3uozgbNLupTZXhJ6gQThHfBOCVBHvyKqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv2s/28fgx+yhpkthrd8viLxpLFusvDGnzr5xyPle4fkW8Z45YFiPuq2Djwz8Mf2xf+Cq/jODxx8RNXfwr8MbG5b7JMYGSxtRnDLZW5YG5lxkGVjgYILjASv2ev2Yvg9+zD4RHhP4VeGltWnVf7Q1S5IlvdRdejTS4GepwihUXJ2qMmqqqqqqqqqqqqq+KunNrHwv8YaQlutwb3QL6AQtjEheB12nPGDnHNf8ABDjUVl8OfF7SBNITbXukT+WSdq+Yl0uR7nysH6Cqqqqqqqqqqqqqqqqqqqqqr/gqb/yfpB/2D9H/AJ1VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX/BT/8Abak+Bng//hSPwv1Yr8QvFlsRdXNs/wC80axf5S4I5WeXlUxyq7n+U7Cf+CZn7Eo/Z18Bf8LR+IukqnxH8W2y7oZU+fRrFsMtuPSV8K8noQqYGwlqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq1vXtD8NaZNrPiLWLLTLC3XMtzeTrDGg92YgCvip/wUQ+HXhozaZ8M9GufFV8hKi8mJtbNT6gkeZJg9tqg9mr4hfti/H74hvJHP41l0KyfIFnoYNmoB6jeCZGGOMM5FXVzc3s73V5cyTzynLySuWZj6knk1Vfsrf8nAeDv+vmX/0RJVVUNxPbtugmeM/7LEVa+Jb2HC3CrOo7n5W/MVZ61YXuEWXy5D/BJwfwqqqqr40ft8fsq/AoXFn4s+Ken6lq9uSraPoJ/tG6D4ztYRkpEcf89GTqPWviD/wWE+LPxG1lvBv7K3wLma7uAVtp9Rt5NUv5Rx8yWlv8qMCR1aUdOKj/AGPf+CmX7YUqX3x++IV34Y0C5YO1nruoeTHt9U020GxWHTEixnjr6/CP/gjf+zj4LNvf/E7Xtf8AiDfRqPMglk/s2yZh3EUJ838DMR6ivhz8G/hP8IdP/sz4X/Dnw74XhZAkh0zT44HmAx/rHUbpDwOWJPHWqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqvip+zp8CvjdbvD8VPhV4c8RSumwXl1ZKt0i+iXCYlToPusK+MH/BFr4Ua+ZtT+CXxF1nwhdkl49P1RP7RtM9lV8rLGP9pmlPtT+Ev+Csf7E779A1LVvH3hLTxnZayHxDZGJeSohkH2qBAOSUWMDn5upr4O/wDBaTwneTpoP7Qvwuv/AA5fRuY59S0Im4gRx1320hEsQBGMBpD7V8Jf2hvgj8d9PGofCb4maH4jwm+S1t7nbdQj1kt3xLH/AMCUVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV4o8U+GvBWgXvinxfr1ho2j6bEZbu+vrhYYYEHUs7EAV+0v/AMFS/HPxT8Q/8KR/Yi0DVbu81OU2yeIYrFpL28PORZW5BMa4GfNcbgMkKmA1fsn/APBJ2K31OH4tftgagfEviK5m+1/8I010bmISk7i99Pkm5fcclFOzI+ZpAxAsbGx0yyg03TbOC0tLWNYoLeCMRpEijAVVHCgAAADgVVVVVVVVVVVVVVXtnBqFnPYXSlobmJopFBxlWGDz24Nf8EQL2aw8X/GDw5cSGKWWy0uWS3ZMNuhkuUJPHG3zcY/2vbiqqqqqqqqqqqqqqqqqqqqq8afs5fAX4j+Jh4z8efCHwrr+vKkaDUdQ0yOacLH9wb2GcL29Kqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/a2/aa8K/sp/B7UviNrphu9VkBttB0pn2tqF6wOxOORGuNzt2UHHJUH/gm/8AsyeKf2j/AIpaj+29+0MZdWiOqvc6Gl5HxqWoK2Dc7TwILcqEjUfLvQAYEWDVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVUSAMkjgd6+Ov7Z/hX4fR3mgfDmC28T+IoUYGTzD9jtnHUMy8ysP7qEDPG4HiviX8XPiH8XdZOtePfE1zqUgYmC3J2QWwP8McQ+VRjjOMnuSeaqqqv2Vv8Ak4Dwd/18y/8AoiSqqqqrDXbyyIR286IfwseR9DVjqNrqEe63k+YD5kPBWq+K/wAefg38DdLGrfFr4kaH4ahdN0UV5cjz5xnH7qBcyS9D9xT0r4zf8FovC1pct4e/Zz+GF74lv5H8qDU9eDW8DyEgL5dtETLKDnGGaJs9jQ+EX/BU39uUiX4ia3qXgjwdfD5rbVJTotmYjjK/Yoh583HKmVCD/fGa+C//AARs+AXgtbfUvi94j1n4g6igBe1Vjpthn02RMZWwe5lAPdecV4B+GHw5+FejL4f+G3gbQ/DOnqADBpdjHbByO77QC7erNkknJNVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVfGT9lj9n34/W8ifFX4WaJrN46bF1IQ/Z72MDoFuYisoA/u7tvqDXxc/4IyT6Vff8JX+zB8X7zStQtX8210/X3ZGjcf8APK8t1DIeoAMZPIy/Um2/ar/4KUfsP3MOlfH7wZfeL/CsDCJbrXI/tUbLk48vU4CfnbjAmMhA/hHb4E/8FYf2Yvi21vpPjTULn4ba5LhTFrrhrJm/2bxfkA95RF0rTNT0zWtPg1XRtRtb+yukDwXNtMsscqnoyspII9xVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV+1h/wAFBfgh+yza3Oh3eop4q8chP3PhrTZwXhYjg3UoytuvQ4OXIIIQjkaJ8P8A9tr/AIKpeKYfE/jjVn8I/C+3uC9rK9u8Wm24HH+i2+4NeTdQZGYgfMC68LX7NH7IPwV/ZV8Pf2X8N/Dwl1e5iCaj4gvwJb6+6EhnwNkeQMRoFXgEgnLGqqqqqqqqqqqqqqq/4JNkeH/2xPjV4LDGPytOvf3Mo/ef6PqSR8/TzMH3Iqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq8U+KPD3gnw3qfi/xZq9vpejaNayXV9eXDbUgiRdzMfoB0HJ6CreHx//AMFZ/wBsI3Ew1DS/hX4RI3DO3+ztM3/dB5UXd0y+5GP4lhxXhnwzoHgzw7pvhLwrpNvpmj6Pax2tjZ26bY4IY1Cqij0AAqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq8Q+IdD8KaLd+IfEeqQafp1jHvnuJ22qg6fiScAAckkAcmv2g/2uvEXxOa58K+CHuNF8LHKSMG2XOoL38wj7iH+4DyPvE5wASpBU4I6H0rxBp4trgXUKAQzknAHCN3H07j/61VVVX7K3/JwHg7/r5l/9ESVVVVVUUskLrLDIVdehBxiv23b39v7W20nR/wBlzVdOj0LVWW0vE0uOO21WGVs5d7id9iQ8D54vLZScHIOa+FH/AARy8V+MNUPjb9q74yXt5qN64lurHR7hrq5mY/8APa9uAct0BARvZ+9fBf8AZU/Z9/Z9tYovhV8L9H0m8RNj6pJF9ovpcjndcybpMHGdoYKOwFVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV1a2t9bSWd7bxXEEyFJIpUDq6nggg8EEdjXx3/wCCXn7LHxqNzquleGJPAOvz5YX/AIbCwQs+OC9qQYiM8nYqMf71an+xj/wUN/Yj1GfxJ+zV48vvFvh2OVpZLXQ2LtKMYzPpc25ZHIGP3XmsOxHb4Kf8FlIrPUB4N/am+GF1oOo2z+TcatokLgROOD59lKfMjIxklGY5PCCvhZ8avhP8btCHiP4T+P8AR/E1iAPMNlcAyQE9BLEcSRN/suqn2qqqqqqqqqqqqqqqqqqqqqqqqqqqvin8Xvhn8E/Ck/jX4qeM9N8O6RBkCa7kw0zf3Ioxl5X/ANhAW9q+Nn/BSL9oL9qnxS/wS/Yo8Fa7pdtfho21G3jB1a7j6NIGB2WMXPL7tw4PmJkrX7KX/BJTwn4PuoPiL+1FfW/jbxPLJ9oGghzLp9vITuLXDt812+cEg4jzuBEgINWlnaWFpDYWFtFbW1tGsUMMKBEiRRhVVRwAAAABwKqqqqqqqqqqqqqqqq/YVB8Kf8FVPjR4fDtCl1P4mtY0kGWkQanHKnPusYbPHT3qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/4KT/ALS3iz9of4q6Z+w/+z80mqRnVI7bXDaScalqIYFbYsOBDbkF5CTtDqSceTk/sl/sz+Ff2Vfg5pfw20EQ3WpsBc67qiphtQvWUb39Qi4CIp6Kozkliaqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqrxb4s0DwP4dvvFPibUY7PTtPiMk0rn8lUd2JwABySQK+P/7QniT4464okV9O8O2Lk6fpgfOD082UjhpCPwUHA6ktVXNtHe2z2kpAWQcMR91h0P8Antmp4ZLeV4JV2vGxVh7iqqq/ZW/5OA8Hf9fMv/oiSqqqqqr6GtE1wShbK9f5+kbn+L2PvVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVXxs/Zd+A37RGnmz+LHw40vWLkJsh1JYzBewDtsuI8SAcD5clTjkGvir/wAElPi/8Itc/wCFjfsb/FzUZb2zJeDTry9/s7UYgMHbFdx7Y5ckD5XEQx1LV8PP+Cof7TH7OXiGL4aftkfCfUdV+ykRvetajTtTVAcGQAgQXS4xgrs3dTIc18B/2tfgD+0lYLP8K/iDY3uoBczaPdH7NfwYGTugfDEDH3k3L/tVVVVVVVVVVVVVVVVVVVVVVVVVe3lnp1pNf6hdw21tbRmSaaaQIkaAZLMx4AA5JNftT/8ABXH4cfD5rrwR+znZ2/j3xOcwjWGJOl2sh4Gwrhrps9k2ocgh25Wvhd+wR+1b+214sg+Mf7YHjbWvD2iTgPBb3ygahLCxz5dta4EdlEeeWUEHBEbZJr4K/AH4R/s8+FV8IfCTwZZaJZsFNzMil7i9cDG+eZsvI3XqcDOAAOKqqqqqqqqqqqqqqqqqvg7nwp/wWn8T6OMw/wBsajq+UGH3+bp73XJ7Z27vbp7VVVVVVVVVVVVVVVVVVVVVVVVVJJHFG0ssioiDLMxwAPUmvE3x1+G/hoNEda/tO4X/AJY6evnf+P5Cf+PZrXf2qdVlJj8NeGLW3HQSXsrSk++1duPzNap8ePilqjEHxM1qh6R2sCR4+hxu/Wrzxx401DIvfF+szg9Vkv5CPXpuxWhapqlx4q0FptRupWbWbEMWmYkj7RGOefShNMv3ZnH0Yik1G/j+5ezDHYuT+lReIdUiwGmWQDs6D+lW/ipThbq1I94zn9DVrqun3gAhuV3f3G+U/lVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV/wUj/bHj/Zi+FJ8L+DdTVPiL4zgkh0nYcvptv92W9I/hIztjz1fnkIwr/glZ+xtL8LfB4/aL+Jmnu3jXxlbbtJgulJk03T5Pm3tnkTT8Me4TaOCziqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/vrLS7G41LUbuK2tLSJpZ5pW2rGijLMxPQAAnNftNftBX3xp8TnT9Jmlh8KaTKw0+3I2m4ccG4cdckfdB+6vuWzVVXiGwEsQv4h88YCyj1Xsfw6fl6VVVX7K3/JwHg7/r5l/9ESVVVVVVV9DWg6ubtBZ3D5mQfKx/iH+NVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV8RPhd8Ofi34ek8K/EzwTo/ibSpMkW2pWizCNiMb0JGY3x0ZSGHYivjz/wAEcbKC+PjT9lH4gXXh7VLaXz4NF1m6cxxODkfZ7xAZYyMDAkDknq4rwj+33+2v+xj4ktvh1+1v4A1LxPpSHy4Z9UAivGjXgvb3yBo7sDOSX3knA3rX7PX7Z37Pf7TdnGnw28bwprZj3TeH9Txa6hDgZP7okiQDu0RdR3NVVVVVVVVVVVVVVVVVVVVVVftQ/wDBSL9n79m1bzw9b6oPG3jS3LJ/YOjzqVt5B2ubjlIeeCoDyD+5jmvs37fX/BUjVRJMT4R+FjTkpuElnpCKrcYHMl9KCvX5lVh/yyBr9lv/AIJ4/AP9mBLbXbLS/wDhLfGkQBbxHrECs8LjvbQ8rbjryMvg4LkVVVVVVVVVVVVVVVVVVVXjfHhT/gtzpl/CRGmpalY7VhbYR5+ipA2fqxYkdw3vVVVVVVVVVVVVVVVVVVVVVVVXd3aWFrLe311Fb28CF5ZZnCJGoHJJPAAHc18W/wBuTwZ4Ukl0b4a2SeJtRTKteSMUs4zjsR80vP8Ad2j0Y1pnx7+IXxO165svHHiSW5juR5ltaxgRQRFeqrGvHTucn5evNVVV4U/5G3QP+wtaf+j0qqqqs9av7PCrN5iD+B+fy7irDXbK+IjZvKlP8Lnr9DVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV8Zvi54O+BPwy174qeO73yNI0G1MrqhHmXEh4jhjBIBkdyqKOmWGcDJr9kT4ReMv+Cin7UWuftP8AxzsDN4K0LUFdrJwTbzzIM22mxZ4aKJdjSf3gRu5mJoKqqFUAADAAGMVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVftvfHh3lf4L+FbzEaBZNdnjb7x4ZLcEenDN/wEdmFVVVRCsCrKGUjBUjgjuK1OxNhdNCCSh+aNj3X/ADxVVX7K3/JwHg7/AK+Zf/RElVVVVVVVHI8UiyxOVdDlSOxrTL9NRtVmXAccOvoaqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqvGngbwZ8R/Dtz4S8e+FtL8Q6NeDE1jqVqlxEx7HawOGGeGHIPIIr9oX/gjpoF3dyeNf2VvGU3hXVoJPPh0LVLiR7dXByPs90MywkYGA/mcn7yCvBH7en7aH7FfiS0+Gv7XHgLVvEuiodkVxqmFvTGvBe2vlzHdqOp3lyTgb1r9nz9rv4CftO6b9p+FnjWKbUoow91od+v2a/tfXdCT8wHd4y6f7VVVVVVVVVVVVVVVVVVVVX7R/7fH7OX7M6XOl+KvFy634ohBC+HNDK3N2r+kxyEg7Z8xlbByFavEH7SX7fH/BRbWLvwV8CfCt54Q8CTSmC6bTpWggijPUXmosAWO0kmKILuHHltX7MP8AwSS+DXwmNr4o+NNxD8RvE0ZDizmg2aVat6CFuZyDn5pflIx+7BGat7a3s7eK0tII4YIECRRRoFVFAwFUDgAAAYqqqqqqqqqqqqqqqqqqqq/bVP8AwiP/AAVj+EGvy4SLULvwzO7yodojN81u7DHJwI259R3xiqqqqqqqqqqqqqqqqqqqqqqvi98b/AnwX0b7f4o1DzL6dCbLTICGnuT04H8K56u2APc4B+Mn7RfxD+M928Gr3x07Q1fMGj2jkRLg8GQ9ZWHHLcA9AuarT72bTb6C/tmxLbyB1+oP8q0+9g1Oxt7+2bMdxGHX2yKqq8Kf8jboH/YWtP8A0elVVVVVpuvXFniK4Jmh9CfmX6f4Vb3MF3CJreQOp7jtVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX7aPxg8X/wDBQD9qHQP2U/gXfG58JaHqLQteREtb3N0oIub+QjgwwJvVD3+cqT5qivgp8HvBvwE+F+gfCjwJaGLStCthGskgHmXMp+aSeQjq7uWY9snAAAAFVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV8dPijafCD4ban4ukaNr4L9n0yF+RLdODsBHcDBYj+6prUtRvtY1C51XU7uS6vLyZ5rieVtzSSMcsxPckkmqqqqtWsPt9oURcyx/NH7nuPx/niiO1VX7K3/JwHg7/r5l/9ESVVVVVVVVaLqBsLsbm/dSfK49PQ/hXBHBqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqrx18P/BHxO8NXPg74heFNL8RaLeD99Y6jbLNGSOjAN91hnhhgg8giv2h/wDgkDLpuot8RP2P/Gd1oOq2TfaLfw/f3roY5Acj7Jeg7426ALJnnkyKK+E//BTP9oz9mXxQnwj/AG1Ph7rOrJZsEbUJLdbfVYI+gcdIryPjh8qWyT5j9K+Dfx6+EP7QHhpfFXwj8c6d4gs1AE8cLlJ7Rj/DNC2JIm9mUZ6jI5qqqqqqqqqqqqqqqqpiqqWZgAByTxiv2g/+Ckf7MHwBW50qXxevjLxLASn9i+HGW6aNwcESz58qLBGGUsXH9w1rH7Uv/BQb/goHqF34R/Z+8I3fg/weXaK5m0eVrZEQ/wANzqcm3LbScxxbCw/gav2cf+CPHwy8FPbeKP2h9ePjvWwRI2kWjPBpsT9TvbiW455yfLU8goa8P+HPD/hLRrXw54V0PT9H0qxj8u1sbC2S3hgUdkRAFUewFVVVVVVVVVVVVVVVVVVVVV/wVrYeF/2vvgx4+VlRrfTLQ71fDj7NqLyjrwAPN4P1z2qqqqqqqqqqqqqqqqqqqqqq/aO/aa0P4MWDaDooh1LxddQ7oLUnMdoD0kmwc9OQg5PsDmvFPirxD421y68SeKdWn1HUrxt0s8zZJ9AAOFUDgKAABwBVVXwo1sXFjPoM0mXtj5kIP9wnkD6Nz/wKqqvCn/I26B/2FrT/ANHpVVVVVVY39xp8olgfj+JD0arC/g1GASwnBHDIeqmqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/4Knftgj4F/DH/AIU94F1Xy/HPjq1dJZYmw+maacpJNn+F5MNGh6gCRgQVXP8AwTE/Y9H7O3woHxD8baX5XxA8c28ct0kq/PplicNDa4PKueJJBx821SP3YNVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVftvfFFvGHxJTwPp1zu0zwopicKeJLtwDKffaNqexD+tVVVVV9DWv2AgnF3EP3cx+YY+63f8+v51Vfsrf8AJwHg7/r5l/8ARElVVVVVVVVaDe/bLBVdsyQ/I39D+VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVXxb+CXwq+O/hh/B/wAWfBGm+I9NIbyhdR4ltmIALwyrh4X4HzIwPbpXxm/4Jh/Hf9nTxKfi/wDsUePtav8A7DukXTFuRBqluuclEYYju48DlGCk8Da9fs2f8FdBbamnw1/bC8M3HhzWrST7PL4htbB4lSQHBF5aY3wt6tGCM/wKMmvDniXw74x0Oz8S+E9e0/WdJ1CIS2l9YXCzwzoejI6Ehh9DVVVVVVVVVVVVVV8VP2mfgB8Eo5D8Uvi34b0G4jUk2Ut4JbsgdSttHulbqOiHqPWvjD/wWm+G+kedpPwK+GureKb0nZFqOsv9htd2eGWJd0sg7YPlHJ9uW8G/8FRf2/lH/CUXN74J8C3wBMV4H0PTmiPIAgUG4ulI5BcSL0+YcGv2f/8AgkV+z58Lvs2t/FS4uPiVrsQDGO9j+zadE3+zbKxMncfvXZTwdorStI0rQdNttG0LTLTTrCzjEVtaWkCwxQoOioigBQPQDFVVVVVVVVVVVVVVVc3FvZ28t3d3EcMECF5ZZGCqigZLEngAAZJNfGr/AIKc/sl/BqSfTIvG8njXWYNymw8LxreKrDoGuCywjng4dmGD8vavEH/BXT9o/wCLWqyeGf2Zv2eYRcMMKGtrnXb0A9HWOARqhz/eVx/Ok8J/8FoPjoi3Opa/r3hCwuFIVm1Cy0AxAjoUt9twD7spI9aX/glx+3Z40Bn+I37UGnubg/vVn8SapqMmCcndvjUE7ucbiOc5r/hzf8fv+jl9K/79Xn/xVXn/AATv/wCCjvwgiGtfCT9oH+1ZLVRsstK8WXlnLIF+6vlzhYWXAA2s+O2Mc1+zZ/wU5+Knw5+Iy/An9ufRZdMvEuBbt4hurEWVxYO2Nn2uJFCNCcjEyKMAhjuBLCCeC6gjubWaOaGVA8ciMGV1IyCCOCCO9VX/AAXI0X/kj3iGNv8AoMWsoLf9ejJgf995/CvBmtjxN4O0LxIr7xqumW92GIAz5kavnA4/i7cVVVVVVVVVVVVVVVVVVVVV+01+0RYfBbw8NK0WWG58W6pETYwMA62qZwZ5B6dQoP3mHoGrV9W1PX9Tuta1q/mvb69laW4uJnLPI56kmqqq8Naw+g63a6krHZG+JQO6Hhv0/XFI6SIrxsGVhlSO4qq8Kf8AI26B/wBha0/9HpVVVVVVVneT2M6zwPgjqD0I9DVjfQ6hbieE+zKeqmqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr43/GHwh8A/hb4g+K/je68vTNBtTJ5StiS6mPyxQR/wC3I5VR2GcnABI/YT+DPjD9uP8AaY179r/46Wxu/D2iaqJ7e3kQtBeX6gGC1QNnMFtH5bEHriJTu3PVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVXxD8XWvgHwNrnjK7CsmkWMs6o3AkcL8if8Cbav41qeo3usajdatqVw093ezvPcSueZJHYszH3JJNVVVVVVzbx3du9vKPlcYz6Hsangkt5nglGHQ4NV+yt/wAnAeDv+vmX/wBESVVVVVVVVV4duvs+oCJm+WYbfx7f4fjVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVftK/sW/Af9qbS5F+IPhhbXxAkWy08SaYFgv7fH3QXwRKg5+SQMoycbTyPEHwd/bi/wCCX/iK78b/AAu1uTxb8Nml829kt4HnsJUAxm+tM7rdwAP3yHAGB5nJWv2TP+CkfwT/AGmVs/C2r3Efgnx5KAg0TULgGK9f/p1nIAkzj/VsFfngMAWqqqqqqqqqqrx9+1L+zj8L45D47+N3g7S5ogd1q2qxy3PHXEEZaQ/gtfEL/gsR+yh4UEsHg+LxX41uFOI3sNN+yQMfd7lo3A+iGta/4K5ftNfFvUZPD37N37Otslw52qBb3Wv3ag9GVYVjVSfRlcDpz1o/A7/grv8AtQkD4g+N9V8HaNdn5k1DV49Gg2/3WtbEeYQB2kj5PU5ya+Fv/BE/wLYPFqXxq+L2r69OWDy2OhWq2UW7urTS+Y7gnuFjP0618If2Q/2bfgSIZ/hl8ItB06/hxs1OeE3l6CO4uJi8i5POFYD24GKqqqqqqqqqqqqqqqv2tP8Agp38Gf2dJr3wZ4N8vx546t8pJY2VwBZ6fJ0xcXAyNwPWJMsCCGKdasvhp/wUU/4KUSw69441lvB/w5uZA9ul2JNO00x9d0FouZbogZ2ySbhnjzB2+CX/AAST/Zf+GEVvf+O7S/8AiRrUYBabV2MFmH45S1ibGOD8srS9fpjw34V8L+DNIh8P+D/Del6Fplv/AKqy02zjtYY/92OMBR0HQVVVX/BR/wDZM0b9o/4I6l4k0bS4h498FWct9o11GgEl1EgLy2bH+JXUMUB+7JtPAZs/8EiP2hr34rfAe9+FfibVGutc+G08VtbNK2XfTJVJtxk8ny2SWL0VFiHeqr/gtlobXX7PvgjxGoz/AGf4wW2OCcgTWk7ZxjGMwgZPqPWv2RtcHiT9ln4R6uZfMkl8F6Ukr8fNIlrGjn5eB8ytx29qqqqqqqqqqqqqqqqqqqqvjF8VND+DvgW98YawVllQeVYWm7a13cEHZGPQcEseygnnpXjDxdr/AI88SX3izxPftealqEpkmkbgDsFUfwqAAAOwAqqqqr4a62NV8PrZyvmfTyIiD1KfwH8uPwqq8KDPi3QAP+gtaf8Ao9KqqqqqqrTNQk064EqnKNw6+o/xqKSOaNZYnDKwypFVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV+3H8WfFv7d37U/h79kf4K3ZuPDfh/VWt5bqJt8E96oIur18HDQ28YkVT3/eFSfMUV8GPhH4P+BPwx0D4VeBbPydJ0C1EKOygPcSHmSaQjq7uWZj6txgYFVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVft8eMn0b4Z6V4Ot5dsviLUN0wz96C3AZhj/AK6NCfwqqqqqqqvEFh5sQvYl+eMYcAdR6/h/npVfsrf8nAeDv+vmX/0RJVVVVVVVVUbtFIsiH5kII+oqGRZ4UmQ8OoYfiKqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqWKOaNoZY1dHUqyMMhgeoI9K/a3/4JO/DT4tm98cfAV7PwJ4wctLJp4Urpd+/JPyKCbZycfNGCnHKZJavgz+35+0p+xd4yj+Bf7ZPhPXNa0ay2xw3d1h9RtIc4EsM5Oy9h6gZYnjAcbdlfCz4t/Df42eELXx18LfF1h4g0a64E9s/zRPjmORDho3GRlHAIyOOaqqqqrUNR0/SrSS/1S/t7O2hGZJriURog9SxIArxr+2d+yj8PjJH4n/aB8FRzQnbLb2eqJfTRn0aK33uD9RXjT/gsL+yJ4aLx+HJPGHi1wSEbTdH8iNvcm6eJgP8AgJPtXij/AILY+JdZu00j4Sfs7RvdysVhfVNWe6eU9gIII1OcDs5/Tlv2if8AgsF8eAIvAnww1XwnaXZ+R7LwvHpkTqf7txqJYgf7SuOnXrQ/4J0f8FE/jkxl+Onx8Sxsp+JrPVvE91qLKD1CW8IMAHqA6jJ9ya8A/wDBEn4TaYIpviX8Y/E/iCRSC0WkWUOmRk/3Tv8APYjtkFSfbt8Pf+Cdf7G3w28mXSvgho+q3cWCbnXnk1RnYfxFJ2aMH/dQD2rRdB0Pw3p0Wj+HdGsdLsYRiK1srZYIk+iIAB+VeKvHXgjwLZHUvG/jLQ/D1oBkz6rqMVpGB67pGArxj/wUa/Ys8EyPBqXx40e/mTomj29xqQY+z28bp+bAV4j/AOCzf7KuklotE8N/EHXJMDa8GmW8MR6ZyZZ1YcE/wnkenNaz/wAFxfC0Bx4e/Z11W95H/H54ljtfr923k9v/AK1L/wAFofjF4hIi8G/s2aVNIT5YAv7m8/eH7oxHGvt8vU+tH/gp5/wUC1dRN4a/ZU0p4U+WQ/8ACJaxc/N9UnUDjtQ/bO/4K1a+Fg0z9mvVLEn94JV+HV/GCvoGmJXHIPrx9aHxZ/4LWeJwv9leDdY0rzWMi58MaZb7V5+T/SUOBz0PzcDnrk6P/wAFxtZja987WIxcZyv27w9akdjhNylOnoPUV/wpv/gtN/0P+q/+FRp//wAVTeBf+C3egKYbHXNW1CPHmGQa3ok+D/d/fPu6AcAY59San/aB/wCCynw1VrrxX8NNY8RRQ5Lg+Dra8XHByTp6qcAHrnjnPQ48I/8ABaDx/wCF9UGg/Hz9nmKC6jIFw2kXE1hNCO5+zXIcn6GRfrX7Pn7b/wCzj+0uYtP+HXjlINfdCzeH9Xj+x364GTtQkrLgckxM4Hc1VVVVVVVX19Y6ZY3GpaleQWlpaRNLcXE8gjjhjUZZ2Y8KoAJJPAAr9qr9vj4tftTePT+zF+xTZarLp2oytaXWsaepju9YA4fy34+zWgHLSEqWXliiblb9j3/gln8LvgdbWHjb4yW9j458eIFlWKVPM03S39IYmA85x/z0kHUAqqEZIVVUKoAAGAAMYqqqqqv2EoF+Bf8AwU8+JPwZsSsOlam2tadaW4OB5cUwurcgD+IQxEY7Bmqq/wCCueh/2v8AsYa1fiLf/Yuuadd55+TdN5GeP+u+OeOfXFf8E0NeHiH9iH4Y3LShpLS1vLN13AlPJvZ41Bx0+RVOPQiqqqqqqqqqqqqqqqqqqqkeOKNpJHVUQEszHAAHUmv2ofjTL8YviFK2m3LHw7oZe20tM8Sc/PP9XIGP9lV75qqqqqvh/rf9jeIoRI+ILv8Acy56DP3T+Bx+BNVXgv8A5HHQv+wnb/8AoxaqqqqqqqvDeo7H/s+VvlbmPPY9xVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX/BTP8Aa4/4Zw+DT+EfB2reR4+8cxyWumtE+JNPtek937EA+Wh4O99wz5ZFf8EqP2Rv+FJ/Cz/hc3jXTDF418e2qNbxyoVfTtLJDRxEHo8pCyv7eUMAq2aqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/b08RtqnxcsPD8cmYtE0iNWTPSWVmdj+KeV+VVVVVVVUyqylWAIIwQR1rU7I2N00Q+4eUJ9K/ZW/5OA8Hf9fMv/oiSqqqqqqqqq0CbztLiBPMeVP4H/DFVVVVVVVVVVVVVVVVV4p+Kvwv8Dh/+E1+JPhbw/5YO/8AtTWILTbgEnPmOMcAn8DXw7+M3wn+Lzamvwt+Ieg+K10Z40vpNJvVuUgZ92wF0JXnY2MHtVfto/ttaB+xnZeFbnW/h9qvidvFZvFt/sl0lvHA1uIciR2VsbhNxgH7jVq//BcHXr2b7J4T/ZttY5HcpCbvxM9w0hJwvyJbJyf7oJ5PX1/YX/bd+Nv7UfxK1zwz8RfhHZeFNDstDe/sbu3sbtfOkWeKPYZpT5Z+WRjgAE4yOFNV+3FY/tVah8JNLg/ZAmnTxmPEMJvfJns4WOn/AGe48wbrsiP/AFvkdDu444zS/BX/AILR+JQF1T4g6vpX2li0hPiuxg8ojn/l2Y4BwOF459M1+yH+zl/wUJ8E/HbR/Hn7RHxm1HWfCNpb3UV5pE3jG5vVmZoGWJ/s/wDqmw7KcnkEZ9Kr9rD4O+Jfj/8AADxT8I/CHiWDQNW182Yg1GYuFgEN5BM+fL+blImUAdSwzgZr/hzf8fv+jl9K/wC/V5/8VXwM/wCCRnjf4XfGLwZ8XvEXx20zVbjwzrVvqNxZx6RKWuBC4bYJXlyCQAMlePfFV8X/AANL8UPhL42+GlvqSafL4t8O3+kJdvEZBbtc27wiQqCNwXfnGRnHUV/w458Vf9HFaT/4Tcn/AMfrTP8Agin4+8P6nZ6rpP7QujyyQXCNJG2iTQB4wQWXcsrdQMYIwc1VXH/BNP8A4KKw3Mw0r9qrRo7eSVpFVPGOsW4yxy37tLcqvJPAND9hz/gq5pMezTv2odRlS0H7mKH4kaoFfb0Cq6gYOOjYHPOK+FmkeJ9A+GHg/QfG+py6l4j03QbG21e8mm817m7jgRZpGf8AiLSBiW7k5qvjV8UP+Ct/hz4veNYPhv4M1S98Fw67eHw+E0HTrsHT1lZYdrIu8hkUNh8yfMM8037Zf/BWvw4WbVv2cNTv12+YWk+Hl9IiKM5y0BAA9cnP0r9lv4g/Ev4rfAXwp8QPjB4XtfDvi7WI7h9Q0y2sp7RLfbcypEPKnZpEJiSNiGY8kkcECq/aO/4Kl+A/2bvjfr/wW1/4Xa3rM2gJaGW/sb+NQxnt459vluoxhZV5DEH2rR/+C037L97iPVvBHxI05+ct/Z9pNGMdOVud2T/u/jX7Pf7RXw3/AGnfA9z8QfhbNqcukWmpSadI1/Zm2czJHHI2FJOVxKgyOM5Haq1/4ifD7wpqdvonijx14e0fUbuIy29nqGpw280qDOWVHYMy/K3IGOD6Vp+p6Zq9v9r0nUbW9gzgS28yyLnAPVSR0IP4iqqqqqqq+NfwF+FH7Q3g+TwT8WfCNprVh8zW0rjZcWUhGPNglHzRv05BwejAjIPxU/ZZ/ar/AOCbXjK4+NP7N3izUvEPgNWDXzrD5pjhXJ8rUrVflkjAJHnoABkn90xFfsXft9fDf9rfSf7Dlji8NfECxh332gTT7hcKB809q5x5kfqv3075GGbX/FHhnwraC+8UeI9L0e2IOJr+7S3Tjr8zkDjivF37cn7IXggyJrv7Q3gx3iJDx6dqA1F1I4IK23mHPtjNeLv+CwP7Hnh0yLol74w8UlSQh0vRDEr+h/0p4SB+GfbtXi7/AILg6QjPbfD/APZ9vbotkRXGr68sJHpmGKJ859BIMepo/wDBQr/gpP8AF0mD4S/s7LZW0uRFdaZ4PvrwpkAgtNMzQ8DBBKqOe+RQ+Gf/AAWg+MxEuv8AjTXPCdrOVKynXLLRdgIx9yxIlXA65XP1Oa0//gjp+0J8QLuPUfjd+0npZuCCWliF3rcq+26doe3Xnj3rwZ/wRW/Z50jZN42+InjfxFMhBMdvJBYQP6gqI3fn2cY/WvBX/BOf9i/wKI3034E6LqUyHJl1uWbU9590uHdO3QKB7cmvC/gbwT4Gs/7P8E+DtD8P2uMeRpenxWif98xqBVV8UvjN8KvgnoB8TfFfx7o/hnT8N5b31wFecqMlYoxl5Wx/CisfavjN/wAFovCNjcyeH/2d/hjfeJr1n8qDVNdLW1u7k4Ux20ZMsoPGAzRN7VHf/wDBX/8Aa0UTWa694F8PXgJUoqeGoI0buCcXciEe78ex58J/8EWfHvia6/tz44/tDwHUJcGePSrGXUJJP+3m4eM/nGa8G/8ABHv9kHw2I28QweL/ABY6kF11PWfIRiOoAtUiIB/3ifevDH7Bv7HPhFETSf2d/B04QYH9qWR1I9Mcm5MmfxrRPhF8J/DKLH4b+GHhLSlQEKtjolvABkYONiDsBQVVUKoAAGAAMYqqqqqvHHw3+H3xN0htB+IngnQ/EunsCPs2q2Ed0i57qHB2n3GCOoNftTf8EifDt1bT/EP9kq+ufD3iKxb7TH4bnvWNvO6/MPss7nfby7hwHYpnGDGBmv8Agnz/AMFAvFHiLxNH+y3+1FNd2fjuxnax0nVtTTyZryWP5TZ3YbBFyCCFc8yHhv3mDJVVVVVVc3FvZ28t3d3EcMECF5ZZGCqigZLEngAAZJNftc/tY/E79ur4ox/skfsnW9zc+FZroxahfwuY11ny2G+aWQf6uxjODz9/gkElEr9j79jj4d/sjeA00bQoYdU8V6jEp13xDJCFmu34Jjj7xwKR8qZ7bmyxJqqqqqqrUgND/wCC46CwkMfmalDuLYOfP8Or5g59fMYe2eKqv+CiuhHxF+xT8VdPWLzDFpMV3tCk4+z3MM+ePTys+gxzxX/BHrXBq/7HseniTd/YvijULTHmbtu4RT4x/D/r849896qqqqqqqqqqqqqqqqqqr9tH4sv4B+Gv/CKaReeXrHisvbgqfmitQB5z+24FUH++xHIqqqqqqBxyDyOleDtbGv8Ah+2vHfdOg8ufPXevU/iMH8arwX/yOOhf9hO3/wDRi1VVVVVVVIzIyujYZTkEdjWn3a31nHcLjJGGHoR1qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqvHPjXw18OPB2s+PfGOpx6fomgWUt5fXMn8EcaknA6ljjAUckkAZJFfs5eB/E3/BS79tDWvjv8TtMl/wCFf+F7mOeSxn+eIQRsfsWmA42sDgvLgAMPMJwZBQVVUKoAAGAAMYqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv2oNWOtfH3xpeMeY78W3Ix/qY0i/9p1VVVVVVVatYi+tSqj95Hyn19Pxr9lYEftA+DgRyLqbg/wDXCSqqqqqqqqq8LPm1miz92TP5j/61VVVVVVVVVVVVVXx4/bo/Zp/Zw8QP4P8Aid44ubfxElulwNKs9KubiUxuDtbcqeWM47uK8af8Ft/g9pxdPh98G/FuvMhIVtVvINMR8dwU884PuAfYV+wp+1xqv7YHw/8AEHjLW/BMHhm40fWfskNtBO86SwGJGV/MZV3Nv81TgYG0VX7d/wCxF8Z/2q/H3h7Wfh/8WbTwx4fsdGNlqVjd312FllEzusi28SmNjtcgszA/Ko6AV4X/AOCHeiRKkvjX9oW9uSfvQaX4eWDbz2kkmfPGP4Bj3r9k39ij4Y/sdw+Il+HXiPxPqsvipbX+0W1meCRc2/m+WYxFEhUfvpMgls5HpVeJvAngjxq9nJ4x8G6Hrzac5ezbU9OiujbMcZaPzFOwnauSMdB6VpPh/QNAi8jQdE0/To9oXZaWyQjA6DCgccmqqqqqqqqqqqqqqqrxT8H/AISeObuW/wDGvwt8I+ILqdNks2qaHb3buu0LhmkQkjaAMHsAK1j9iP8AZD1xi97+zj4BjJYNiz0WK0GQMdIQvGO3QnnrzXwn+Dfwz+BnhZvBXwo8KW/h7RXu3u2tIZZJFMzhQz5kZmyQijrjiq/bG/4JzeGv2v8A4h6f8RtW+KWq+G73TdFi0qK3g06O5iMcc00u8gup3EzkdcfKK1D/AIIleP8ARJftfgf9pPTZblBhHuNCmsD0OfmjnlI5wP17YP7Bv7MXxt/Zo0fxjpvxm+KFv4xfWrq1bSjBqV3draxRLJv4uFXyyzSchM52gk9hX7e37RH7UnwGu/BUv7OPwlm8Y2t9Heya/K/hu81OG32tAtupa2ZTEWLS9SN3GOhr9hX9tr46/tL/ABI1zwD8WfhBpvhSPQ9Da+e8tra6tmefzoUSPy5y2AVeVvvZ+Ue5r4yftKfB74AXWlW/xX8R3WjprCSvbzppdzdRgRlQ28wo+3llABHOfrXwu+Mvwx+Nej3OvfC3xfa+ILCzn8ieaCORPKkxnaQ6qQcYPToQe9a54n8NeGLb7X4l8RaZpMGCfNvrtLdcDqcuQPSvFP7an7JXg5X/ALb/AGiPApePIeKy1iO+kUjOQUgLsDweMenqK8U/8FaP2LPDysNM8ba74kZP4dL0C4XJzjg3AiH644+leKv+C3nwosw//CEfBHxZqxH3BquoW+nZ69fL8/Hb16n05m/4K/ftO/ECSS3+DX7NmjzPuKhfs99rLocA/wDLDyuQMnp3HHrJ8ZP+CzfxZBt/D3w51nwzDc/KYT4XtNMGCSMFtQG5RyOdw4HXrnwh/wAEnf23Nc1s+J7658J+CtSlnadrifXPLkid8ligsUkC9SMLgc4HHTQP+CJni/Wrkal8Tf2jrb7S/My2GiyXjucf89ppkPXuUPTtnjwn/wAEXv2ZNHCy+KPGHjzxDMM7kN7BaQn/AICkO8Hr/H+FeE/+CcP7FXg7a+n/AAI0m+lA+Z9WurnUNx9Ss8jKPwUV4T+FPwt8BBF8C/Dbwr4dCZ2jSdGgs9ueDjy0XHQVVVVVc3FvZ28t3d3EcMECF5ZZGCqigZLEngAAZJNftW/8FYFtNZk+Ef7HuljxX4kuJvsg8Rram7h80/KEsYACbl9x4dgUyOFkDZHwj/4JcfHz9ojX4/iz+2d8S9X01tQIkk043Iu9WlTOQjO26K0XB4QByo+Uonb4K/so/s+fs92yJ8KvhjpOl3oTa+qSxm5vpPXNxIWkAP8AdBC+gFVVVVVVVVVVf8Fa/wBlFbzRIP2t/hlZtY+JPDUkK+JGs/3bz26sBDeDbz5sL7FLddhUkgRV+wl+0rF+1F+z3ovjPUriM+KNJP8AZfiSJcA/bIlH74KMYWVCknAwCzKM7TVVVVVMVVSzMAAOSeMV+3f+2Z4w/ad8exfsa/sprPq1jqV8LHVNS0+b/kNzD70Ebg7RaJgmSQna+0nIjUl/2J/2MPBn7Ivw9SzjjtdU8c6xCp8Q66qE+Y3UW8JPKwIcYHBcjcwHCrVVVVVVXjv/AInP/Bb3TbG7OI7HUNP8rZwfk0OOYZ/4Gfyqq/an0ZfEH7MvxZ0YqrNdeCdXWPdnAk+xylDxzwwB/Cv+CJGtG4+CHxA8OeZkWPipLoJvHHnWsaZx1GfI698e1VVVVVVVVVVVVVVVVVVV+038SD8TvjDrWq21x5um6a/9n6dg5HkxEgsPZ3Lv9GHpVVVVVVXws1v7Fq76RNJiK+X5AegkXp+YyPyqvBf/ACOOhf8AYTt//Ri1VVVVVVVV4Yu9k72TNxINyj3HX9P5VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX/BVj9obXvij480H9iP4PGXUb261C2OvRWrZ+1XsjL9lssg4whZZXzwGMfIKGv2Uv2d/D37L/AMEtC+Fei+VPeQJ9p1m/RNpv7+QDzpT3xwqKD0REHbNVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVXxcuWvfit4zu3ADTeIb9yM9M3DnFVVVVVVVVfs6WAg/aH8HXUa/JLdTZA7N5En86qqqqqqqqrwofmuVHov8AWqqqqqqqqqqq/an/AOCmXwn/AGXfHd/8LdU8CeK9f8T2FvDOyQJDb2bLLGrpiZ3LHhsEiMgEEdqvP+Cq37YvxqupdI/Zz/Z2tot5IzZ6bd6/cw+h3oEjGB1LRkd+K/Yd1z9qDX/hDe3v7WWi3dh4vOuTmza5htoGlsTHEY/3VvgR7X81cMoOADznNV8ZP2I/2cP2gPiNafE/4t+CJ9d1ez06KwjjOpT20DRxySOpZIXQs2ZWHJIIABGBXgn9ln9m74crEfBfwM8E6bPEfkul0WGS4H/bZ1aQ9O7UiJGixxqqqowqgYAA9BVVVVVVVVVVVVVVVVVVVVVVVTzwW0L3FzNHFFGpZ3kYKqgdSSegrxN+0T+z/wCDQ6+LPjh4D0h485jvPEVrE+RngIX3E8HgDPFeJf8AgpR+xL4XDJd/HXT72Vekem6dd3u4+zRRMv5kCvE3/BZf9lHRi8Wh6H4+19xwj22lQwxnp1M06MB1/hPSvE3/AAXF0qMPF4N/Z3u7jP3J9T8RrDt6dY44Hz/EPvjseegP/BTv9vn4jAx/Cj9mW0khcfI9j4W1PVJRwejK4X+Fj9zt7Gn8Wf8ABa34qBoNO0DWtBtZQcxf2bpekbQcg/PcbZehPG7PAPXBrTv+Cf3/AAU08dmVfF3x0h8P2124lube58Y3exm458m0Roy2MDtwoGcAVon/AARH8V6vcf2h8Q/2kLVbiQAzCx0KS7ZzjH+tlnQ+nJU9Ogrwv/wRY/Zu0xkm8U+PvHuuSKeY47m2tIm57qIWfpgcOO/rx4V/4Jg/sS+Fgkg+Dq6vcJ/y21XV7u4z06x+aI+2fudz2rwr+zZ+zz4H2N4R+BvgPSpY+VmtvD1skuc5BMmzcevc8VDDDbwpb28SRxxKFREXaFAGAAB0GO1VVVVVVVTukaGSR1VVBLMxwAB1NftdftVfFn9ub4u/8Mk/sqC4uPCD3Bt766tz5a6wY2HmXE0oz5dlGQCOz8MQxZFH7Hf7CHwq/ZL0GK+tbeDxF49uotuo+Jbm3Adcj5orZTnyYuxwdz9WJG1VqqqqqqqqqqqvEfh7RvFvh/U/CviKwivtK1izls761lGVmhlQo6H2Kkj8a/YY13V/2Kv29fFv7LPjLUJBonim7/sq3mmIUSzrmXTbggcAyxSbNo/inUfw1VVVUxVVLMwAA5J4xX7eX7ePin43eKT+yP8Asjvd6uNXuTp2ratpR3SavIeGtLVh0gHPmS5AcAgERhmf9gn9hfw9+yX4OOt+IhZ6t8SNdgUarqSIGSxjOD9kt2IzsBALNwZGAJ4VQKqqqqqqrwoP+Et/4Le397IDNHp2o3e/zTtI+z6G0K4x1w6pj2FVXjXSTr3g3XtDVZGOoaZc2wWPAY74mX5c9+a/4IbaqDH8Y9DklXKtotxEm3k5+2q5z6cR/nVVVVVVVVVVVVVVVVVVXx+8cn4dfCDxN4ngmMd3HZmCzYdRPKRGjD/dLBvopqqqqqqqra4ms7mK7t32ywuHRh2IORWjanDrOlW2pwEbZ4wxH909x+ByK8F/8jjoX/YTt/8A0YtVVVVVVVVW07W1xHOh5jYGlZXQOhyCMg1VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVfteftG6J+y58Ddc+Jt+YJtUC/Y9BsZT/AMfmoSKfKQjIJVcNI+OdiNjnFf8ABJf9m/WfFniDW/20fiwst/qmqXdzH4dmvBukuJ5GYXl+fcszxKfebj7pqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr4of8AJS/Fv/Ydvf8A0e9VQBJAAyewq08LeJ9QwLDw5qlyS20CKzkfn04HWrT4RfFS/Aa1+HHiV1K5DHS5lU9uCVxVp+zn8bL3Ah+HuoLkgDznji6/77D/AOtVn+yZ8cLrAm8M2lpnP+u1GA4/74ZqtP2Lvi3cgGfUPDdrxkiW8kJ+nyRtzVp+w74wkIF/430aEZ/5YwSScfjtqz/YWXAa/wDiYScHKQ6RjB7fMZfT2q0/Ye8Eof8ATvGetzdOIo4o/r1Df/W96s/2MPhDbEGa78RXeCeJr2MZ4/2I1rwn+zn8JvBGt2fiLQtBuBqViSYJ5r2V9jFSpbbu2k4JHIxzVVVVVVVVVeFP9Zcf7q/1qqqqqqqqr9pj/go18AP2YfFF38P/ABRD4k1vxXZwxyy6ZpmnFVjEiB4y00xSMgqwOUL45BGQQPEH/BWn9p74x6lL4a/Zk/Z9jhmbhWS0uNevVHZgsSoif8CRwPXvX7D2s/tPa38G7i6/ax0W6sfGH9tTm1e5jtonnsWSNo2aK34jKu0qbWVThVPfNV4p/Z7+BvjrxqnxF8bfCfwv4h8RxWsdrHf6rpsd2yRRszIFWQMoILt8wGeevAqysbHTbSLT9Ns4LW2gXbFBBGI0jHoqjgD2FVVVVVVVVVVVVVVVVVVVVVd3dpY27XV9dQ28MeN0krhFXJwMk8DkgV4j/aV/Z18Iqf8AhJvjv8P9NcDiO48R2qSHp0TzNx6joK8R/wDBSv8AYk8NB1uvjpYXki9I9O028vNx9mjiK/mQK8Sf8Flf2TdH3x6NpHj7XnAOxrTSYYkJ7ZM06MB/wE/SvE3/AAXE8NQB4/B37Pep3uchJdT8Qpa464JSOGTPbjcOp59T/wAFXf2zviENnwi/Zn0udJ+Img0XUdWfB6bTE6KTyv8ACR7c03xo/wCCz/xJAXw98Ptb8NxzgjYPC1np+Ac/xXylk4Yc5BGBznNH9l//AILBfE0lPF/xj1Tw4lw372OfxuLWMZzkFNPLjbyeACOnHAqD/gjj+0J41mS4+LP7Smjyyltzui3mrsp7kGcxEnlvT9Tjw1/wRB+GFqEHjH46eKNUxjf/AGbpcFhnpnHmNPjv69R6c+GP+CQv7GugCMarovivxJsIz/amuvHv6dfsyw+h6Y+8fbHhn9gb9jbwksa6V+zx4Sn8oAL/AGnbNqWcY6/aWkz0HXPf1NeGfhh8NPBSxr4N+HnhnQREMRjTNIgtduPTy1GKqqqqqqqqqqqqr/gq/wDtE3XwX/Z2/wCEG8OX7W3iL4lTSabFJG+14bFFBu3X3KvHD9JycggV/wAExv2V9L+AfwH03x1rWmR/8Jv8QbSLUNQnkj/eWlm4329qpPKgIVdxwS7EHOxcVVVVVVVVVVVVV/wWG/Z71WSx8N/tX+A4riHVPC7xWGuTWuVeGESbrS6BH3THKxQt1/eR9lr9jH9pfRf2p/gZo3j2C4gTxDZotj4kskIBtr5FG9gvZJBiRO2G25ypqqqte17Q/C+jXniLxLrFnpel6dC013e3k6ww28ajLO7sQFAHc1+1x+3v8SP2tPFY/Zh/Y40zV7nSNYdrW71GzRorvW1wRIiZx5Fpt5Zm2llB3bU3Bv2FP2CPCX7Jfh3/AISPX5LTXviTq0ATUNVWPMdjGcE21rnkJn7z8NIQMgABRVVVVVVVV+xOp8c/8FX/AIu+J1jVk0u58S3UZ2gFYxfJbIcE5B2yqDj1PQZqqr/gjGyaN8bfjB4WhkEcaabCVgkPzkQ3ToDzzwJMH3YZ7VVVVVVVVVVVVVVVVVVV/wAFBfFbWnhXwx4KglIOo3st7OoOPlhUKoPsTKT9U9qqqqqqqqvhNreVudAmfp++gB/Jh/I4+teC/wDkcdC/7Cdv/wCjFqqqqqqqqqtDnE+lwknJQbD+H/1sVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVftNeLvEX/BSD9t/RPgD8N9UlPgTwncyWi30A3RJHGQb/UT2P3RFHk7W2x4wZDnwb4Q8O/D/wAJ6P4H8I6ZFp+jaFZRWdjaxjAiijUKo9zgck8k5J5NVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVaz+zp8EtV8Q6pr1z4OgvLrUL2eaeU3s5VpHkLPhRJtX5iRgAY6VafAr4O2RDQ/DjQmIII861EvT/fzVp8Ovh9p+37B4F8PW23O3ydLhTGeuMLVrp9hYqFsbK3twFCgRRBOB0HHaqqqqqpmAHB57Uee9VVVVVVVVV4UT5Ll/UqBx9aqqqqqviV8Z/hL8HNN/tX4pfEbw/4YgKboxqN8kUkw/wCmcZO+Q8HhQTwa+L3/AAWX/Z88HmWx+E/hfXviBeJwlwynSrJvcPKpmP08kD3r9hP9qDxp+1Z8MNX8b+O/Aa+F7yz1l4bKO3tp0t7myaNGikSSXPmtuMisV4+UHAziq+IX7JP7OnxY+IsPxV+JXwr0nxJ4jt7GOxjm1AySw+TGzsoa33eU5zIRudScADOBXh/w14c8J6XFofhXQNN0bToP9VZ6faJbQp/uogCj8BVVVVVVVVVVVVVVVVVVWs+IfD/h23F14g13T9MhPSS9uUgU8gdWIHUgfiPWvEX7YH7KvhUMNa/aJ+Hscifehh8QW9xKvTGY4mZhwQeleI/+Cpn7EXh8tHD8W59XmQcx6doV7J6dHaJUP4N9cV4k/wCC0/7NOnF4vDfgP4g6zIoO13tLW1ib0wTOW/NK8Q/8Fw5ZWa18Gfs5ZdjiKbUfEpYk8gZijt/XaeH9R703/BS3/goV8Qsx/DH9mS28ibPlSWHhHU9RlHX+MSbDxj+DtnocU3jj/gth8SsxaT4b1vQ4ZcnyhpOlaVjr/HchXHQ/xenqMt+xv/wVp+I3yeL/AI/XmjRSk74bzx5cxxde8dmrqegI4OMj3xa/8EYfjR4tuUuvid+0ppRmB5lisrrVHXgDgzSRHoAO3QV4b/4Ih/CS1Kf8Jf8AGzxdqYB+YabYW9jnpnG/zsd/XqPTnw3/AMEh/wBjPQzGdT0HxT4h2Y3DUtekTfjHX7MIuuD0x94+2Jv2dv8AgmP8G12+IPDPwg0k245TxHrMVy/AU4xdzOWOAvHJOf8AaOW/bF/4JnfCAmPw34n+H2nS25xGnhnws0nI6BWtbcqPujByBwOela//AMFjP2QtGdk06Dx3roU4DWGixoDzjI8+aI44zz6+vFaz/wAFvfhZBx4f+B3iq95H/H5qVva/X7ok9v8A61ah/wAFvvFupSLa+Ff2a7CG4kAWNLjxJJdktnnCpbRk8cY7HnJ6Uf8Agqn+3B4lIbwN+y7pEyOS6D/hHtVvsoODzHKmee/T2pP2zf8AgrT4mVX0f9nDUrJCvmrJH8PL6NHQ9MNOSCORjBzS/FT/AILX+I0RNM8I6vppcearHw1pUBC4+6ftCEDqOD83H1o6J/wXE8QqbiW61e3S7yrH7d4ftCg+6TsVlZPXgA9xR+AP/BZjXHFvf/FPVrBIwWWT/hMLeIE8DH7glj+IxxX/AAxt/wAFbtbP2m//AGlNWsHQbBH/AMLFvotw65xCCvfqef0of8E7f+Cm2sk3+q/tWWy3DHDC8+IGsySYHAyVgYYx71/w64/by1c/aPEf7U+kvcKNiN/wk+rXPydfvPCpHJPGP503/BJ79sm5U217+1DpLW8o2SqdY1J8oeG+UoAeM8HrX/Dm/wCP3/Ry+lf9+rz/AOKr/hzf8fv+jl9K/wC/V5/8VX/Dm/4/f9HL6V/36vP/AIqv+HN/x+/6OX0r/v1ef/FU3/BKT9tDJC/tSaURngnWtTH/ALJX/Dur/gpl4fEVxoP7U1sZIj5ca2Pj7WIXjXGON0KgDAxgHv6VL8Gv+CzfwrjOoaB8Rdc8TJaDOF8S2uqlgueRHeks/TptJORwecfs/f8ABWXxV4Y8W/8ACpv21vBE3h3U7aYW82uwadJaS2j8f8ftmRkDnO+IDAx+7Iy1aRq+k+INLtNb0HU7XUdOvoVmtbu1mWWKeNhlXR1JDKRyCOKqqqq/4KQI/wAcP+ChXwp+A8jFtMgTStOuY9wOw3l2XuJPb9wYuOvye4qOKOGNIYY1REUKiKMBQOgA7Cqqqqqqqqqqqqq8S+G9B8Y+HdS8J+KNLt9S0jWLSS0vrO4Xck8MilXRh6FSRXj/AOHP7Qf/AASh+Oc/xQ+FsF34g+FGuTiFmnJa3uoCcraXpUfup0LN5c2Bu5K5zJGP2ff+CiX7Mf7QFhaQWfji18J+JJlxLoPiGZbSYOOoilY+XMDyRsbdjkqvSo5I5o0lhkV0dQyMpyGB6EH0q+vrHTbSW/1G8gtbaBd0s08gjSMepY8AfWvj5/wU+/Zb+ClrcWWi+LE+IPiFFIj03w3Ks8QboPMuuYkGeDtLsP7tSL+3B/wVY8TxB4D4R+Fdtc/e2yQ6TbbTyefnvrgDtyFJH+qDE1+yx+x58Iv2TPCp0jwJpxvtevYgur+IryMG7viMErkf6qIEAiJeBgE7myxqqqqqqqqnnhtoJLm4lWOKJC7uxwFUDJJ9sV/wRot5fFvx1+MPxMmiO9tNjRyRjaby7eYjgY/5d/UdOh7VVf8ABK0i0/bg+Nul3WYrsadqeYXGGGzVYg3HsWUH61VVVVVVVVVVVVVVVVVV+3jrzan8ZbbR1kJj0fR4YymeBI7PIx/FWQfgKqqqqqqqtE1SXRdWtdThyTBICVH8S9GH4jIrwJPFdeKvDtzA4eOXULV0YdwZFIP5VVVVVVVVVXhWXdbTw5+44b8x/wDWqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv8Agp3+1If2efgLN4Y8Mal5HjT4gLLp2mGN8SWlrtAurkehVWEanIIeVWH3DX/BKP8AZaX4LfBMfFjxTpoi8XfEaGO5USJ89npg5t4vYyZ85sYyGjBGUqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq1HUdP0iwuNV1a/t7Kys4mluLm4lEccMajLO7MQFUAEkngCv2uP+Cknjf4y+JT+zh+xNa6nf3GrTmym8Q6dExur9jw0dkOsUeM7pzg4BIKKN5/ZX+Fvj74L/s++DPhr8Trq1uPEmj2kgu3trgzqvmTySIm8gbiiOqEjIypwSMVVVVVVVRIAyTRJY5Jqqqqqqqqqq8MxbNPaQjl5CR9AAP8aqqqviJ8Vvhn8JNGPiD4m+PND8MWGG2S6nepB5pAyVjVjukb/ZUEn0r4D/tO/Bn9pe28QXvwd8UPrNv4avUtbyR7SS2yXTcjqsgVijYcAkDJRuMDJr9qn/gnD8Ov2r/jDo/xT8VeN9X0KOz0pNP1Gx0u1jEt8I5GaNxM+4RsFdlOY2yAvTHPwi/YO/ZR+CiwT+Evg/o97qcGCNV1uP8AtK63j+NWm3CJu37tU/U0FVVCqAABgADGKqqqqqqqqqqqqqqqvEvjPwd4MtPt3jDxZo2hW2P9dqV/Hap/31IwHY14w/4KD/sY+CC66v8AtBeG7tkz8ujmXVMnpwbVJB+uPwrxb/wWV/ZT0LzYfDmi+OvEkq5Eb22mRW8LYPUtNKrgEf7BPqBXiX/guBLNL9j8Cfs6l2c7YptS8QkszHIA8mOD/d6P6j3of8FFv+CkHxMwvww/Zqjit5MbJ9O8G6jekA4ILSM5jAwRztA+b3FJdf8ABbT4nqpii1vRLVwCA0Wj6P5Y467tkp5A45IyRwCRQ/YD/wCCnvxIw3xA/aSeygbG+DUvHOoThen3Y4UePPyqeoyRnOa0X/giN4q1W4+3ePf2kLRJ2x5ostAku2fAxjzJZ0I4A5Knp0rw5/wRP/Z9sij+Kfif491Vk5K2j2tmjn3BikbHXowPvXhv/glH+xNoAja9+G+p67JGQQ+pa/d8kY5KwyRqenQjHJ4rw3+xP+yP4TVBo/7OvgN2jA2PfaNFfOpGMENOHOflHOc9fU5uvG/7MfwYjMN34t+GPgcQgjy2vrHTSuAAQFyp6FRj3FeK/wDgpL+xR4RZ4b3466bfzKcCPSrG6v8AcfZ4YmT82FeKv+Cz37L+kK8Xhrwn491+YA7GWwgtoScd2km3jnA+4e/4+Iv+C3erX0/2D4e/s5oZXbEMmo6+0zPz/wA8YoAc4z0c9fbn/hvH/gqH8VAE+G/7O76bbycR3Wm+Cb2ZV6j5prhnizn2A+X60fA//Bar4sr5mr+Jdc8O20w4k/tbTdH2ggHGy1KyjoOq56980f8AglN+2j8S28z4zftMadcRzH94LrW9R1iUDPOVlVFJ5Y43Hr1GTXhX/gh94GtgjeN/j5rupcfOmlaLFZY46BpJJu+Ocdugzx4Z/wCCQX7G+ghBquleLvEm08/2nrrR7unX7MsPp2x1rw//AME8/wBi3wyFGm/s++HZtgwP7Qee/wCxHP2iR89e/t6CtE/Zz/Z88NBR4d+BXw+0wqSQbTwzaRHJGCcrGCSQACe9adpGk6RCbfSNMtLKM4ylvAsQ4GBwoHaqqqqqqqqqqq/a0/Y1+FX7Wvg99L8VWMemeKLKFl0XxJbQg3Nk3UK/I82Ek8xMcckqVbDD9nX9oz4vf8E1fjDcfs0ftKWt1c/D+4uPMtriLdMtikjHbfWZxmS2cgmSIDIYMQBIHR9I1fS/EGk2Wu6HqFvf6dqNulxaXVu4eOeJ1DI6sOCpBBB96qqqteU+MP8Agt/a2qh5Y7HU7fy0dtuz7NoSyNj2Dxs2O/44qqqqqqqqqqqqqqrWdG0fxFpV1oXiDSrPU9NvomhurO8gWaGeNhhkdGBVlI6gjFfGn/gjt+zx8QrubV/hhr2r/Di+nYs1vbx/2hYgk5JEEjK6/RZQo7KKj/4JL/th+BS1t8KP2mtFtLUOQgXVtS0nKHqSsCSAHheMn68cw/8ABID9pPx7cRN8Zv2ntNuE3BndJL7WWU9CV+0GHJwBzkV8EP8Agk/+y38Jp7fWPFOmX/xE1mEhhJr7L9kRx3W1QBCOPuymTrVnZWenWkOn6daQ21tbRiOGCGMIkaAYCqo4AA4wKqqqqqqqqr4++ID4S+BXxG8VJKUbR/CeqXisCQQYrWRxjGDnK9ua/wCCH/h4W3w8+KPioIP+JjrVjZbuOfs8Mj465/5efTv35xVV/wAE2f8AlJL8dP8AsH+Iv/T3aVVVVVVVVVVVVVVVVVVV+1jqB1P9oPxhMZN4iuIYF4Ix5dvGmMH3U/jz3qqqqqqqqr9nrWxf6vomlTPmaw1S2C5PWNpQR+RyPyqqqqqqqqqvCr4uJ4weqA/kf/r1VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVqmp6doumXes6vfQWdhYQPPdXM7hI4YkUs7sx4ChQSSegFeAtN1L/AIKfft+3vjXW7aeX4Y+EXSVoJ0wqaVA5+zWzL2a5k3M65yA8uD8gpESNFjjVVVRhVAwAB6Cqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq+JfxM8C/B/wXqXxB+I/iSz0TQtKiL3F1cvjJ/hRF6vIx4VFBZiQACa+L37Qn7SP/AAU/+Jj/AAO/Z/0O90T4dW0wa5SRzEjwg/LdanMuQF+UskC55GAJHAI/ZF/Yo+Ff7JHhYW/h23XWfF19CF1fxJdQgT3HQmOIc+TCCBhAcnALFiAR4nhw8FwB1Uqfw5H8zVVVVVVRxjrTNuPH4VVVVVVVVVVVpkH2fT4ISMEICR7nk/zqqqv+CgviL9uPTZfCHhj9kfS7i4svEiXMGq3emackl3ZzLtK77iUmK3idGIV8KQyN84yor4ef8Egvi98T9Y/4Tj9rH433IvrvDXFtY3L6pqEg/uyXc+URhz90SjnrX7O/7J/wR/Zb0u/0/wCEPhq4sp9XEQ1O+u72S5nvTHu2Fyx2jG9sBFUfMeOTVVVVVVVVVVVVVVVajqWm6RZSajq2oW1lawLuknuZViRB6lmIAH1r4jf8FB/2O/hiZYNc+OWhajdxAj7LoRfVXLDqu63DorezMuO+K+IX/BbT4RaUJYPhj8IfE/iKVTtWbVruHTIif7w2eexHQ4KqT7daH/BRX/gop8dXWH4E/ASOxsp+YbzS/DNzqDKD0LXExMGOnJRRzTfs5/8ABYL46Ev47+KOp+FbW7OXS78Ux6bCwP8Aeg03dgf7JT8K8Nf8ETvFWt3n9q/Fj9ouJ7mRgZ003SZLt5fX9/PKpBx3KGvB/wDwRv8A2T9AjRvEmo+NfFE4/wBYLvVEtoif9lYI0YD6se/Pp4U/YC/Y08Gbf7H/AGefClxt6f2tA+qfn9qaTPTvXhzwH4G8HxiLwj4M0LREAIC6dp0VsB68IorxB4q8L+E7P+0PFXiTS9GtQCfP1C8jtkwOp3OQOO9eL/27P2PvA5lXXf2hvB8rQg700y8OpsCOo22okOfbrnivF3/BYb9kLw8JE0J/Gfih1B8s6dowhRj2ybmSIge+0n2NeLf+C4elRiSHwH+z5d3GciO41fX1h29MExRwvnvxvHbk9m/4KM/8FG/iqzQfCf8AZyjt7eXJin03whf37oMjlpXcxccDJQDn3FDRP+C1nxdIN5qeu+HbOQjDm60zRPKG4/wxbZuOexOAOvFL/wAEtv25vigm/wCMv7SdlJDMB5kV/wCItR1iXGOdyuoQ9h989/x8Kf8ABD3wfbqr+Ofj/rGoMeWj0rQ47Pbx0DSSy5577R9BXhb/AIJA/sb+Hyh1bTPF/ibYckarrrR7ueh+ypD9OMf1rwn+wl+x74KKHRP2ePB0zRgbW1Oy/tIjHfN0ZDn3614d8G+D/CFv9l8JeFNH0WHGPL06xjtlxx2QAdh+Qqqqqqqqqqqqqqqqqqv2wf2SfA37W/wyn8J67HBYeI9PVpfD2u+Vuk0+c4ypxy0L4CuncYYYZVI/YL/an8b/ALJPxVu/2L/2nVm0rSY9QNtpN1dtldHu3bKrv6G0n3BlcZVWYN912K1VVXwuI8Rf8Fsdd1JnBOmajqmPJ5X93pL2/wA3Xsef9r06VVVVVVVVVVVVVVVVVVVVVVVVVVVX/BQbXf8AhHf2MPixqAcL5uh/Y8kgf8fEscHf/rr9fTmv+CN+hrpX7ItzqAC51nxdfXWQST8sVvD+H+p6c/qaqq/4Jbj+1v28Pjf4iuWIun07VsqnCfvdWgZuOvVBjnuaqqqqqqqqqqqqqqqqqq/aFlkm+OHjh5HLMNbuFBPoHIH6ACqqqqqqqqvg3r0fh34neG7+5nWK1bUreO5d22qsZlXLE9gMA/hW1vSirY6UVPp+lYPpRFVVVVVXhcn7fIoP/LE/zFVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVf8Fe/2mpPh78MLL9nvwjfsviLx+hk1XyjloNKVipT1BnkGwY6pHKD1Ff8ABPf9mKP9mP8AZ80zSNZ08Q+MPE+zVPEbOgEkUzqPLtieuIUwuM43mQj71VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV+0R+0h8Lf2YfAM/j74naz5MZ3Jp+m25VrvU5gB+6gjJG48jLEhVByxArSNA/af/wCCt3xdGua/PJ4V+Fnh+6KqyAtZ6WhxmOBTj7Vdso+ZzwuedilEPwN+A3wx/Z08BWnw6+Fnh9NO063w88zkPcX02AGnnkwC8jY64AAwFCqABWt232nTpAoy0fzj8Ov6Zqqqqqqp2z8oNVVVVVVVVVVafbG8vYbcDIZvm+g6/pWBjAq9vLPTrSa/1C7htra2jMk000gRI0AyWZjwABySa/aL/wCCsv7PXwgE+h/DR2+JviOMlSumXAi0+Funz3ZDB/UCJXB6Flr9gr9pH9tf9oj4r6n4y+Kvw/az+FGp6Y62E0OnpY21lcKwaN4GlPnXSuu9GYFwDtOVwQaqqqqqqqqqqqqqqr4rfthfsy/BQSw/ET4zeHLG+hyH062uftt4pHY28AeRckYyygZzzwa+KX/Ba/4WaN59l8IfhXr3iWdcql5rFwmm25PZ1RfNkdenDCMn2pv2ov8Agq/+1Apj+EXw1vvCmj3Y2JcaToS2ULqfS9vyQDjBLRuv4ZxWmf8ABKH9rb4030WuftJftDW8Ls24rc311r93F6jDska+g2yECvh1/wAEcP2WvCojn8cal4r8bXAI8yO6v/sNu2PRLcLIM/8AXU/hXgH9lf8AZv8AhcsbeBPgj4O0ueIALdjSo5bnj/pvIGkPTu1X9/YaXZy6hqd9b2lrAu6WeeQRog9SxIAH1rxx+2p+yd8OvOTxV8f/AAbHNb/622sdQGoTofQxW3mOD7YzXjX/AILFfsk+G/Mi8NReMvFkitiN9P0kW8Te5Ny8bAf8AJ9u9eJv+C2niPV7r+yvhX+zpE1zIzLbvqesvdPJ6fuIYlOcAkgSH9OR+1v/AMFbvjJg/D34N3/hyC42+VcWHgpoYSD0Kzah5iH3OeOvFH9lL/grr8ZV2+Pfi/qPhqC4U+ZBf+NDbQkE9Gi07zF98FePbpWg/wDBE/xtr11/afxR/aQtftTnMosNHlvnk/7bTTRkck9VP614R/4Iw/sw6Lsm8UeK/HfiOYfeje+gtYTx/dji3jnJ+/6e+fCP/BOb9izwZsfTfgNot9IvV9XuLjUdxxgkrcSOv4AAZ5xXhP4U/C3wEEXwL8NvCvh0JnaNJ0aCz254OPLRcdBVVVVVVVVVVVVVVVVVVVVVVX/BRv8AYhs/2n/AJ8b+BtOij+Jfha2Y2DKAp1a2GWazc/3s5aNj0YleA5I/4Jd/ts3fxJ0Zf2afjDqUsfjrwxC0ejXN8xWbU7WIYaBw3JuIQpBz8zIMkZR2NVVfsXg6r/wVq+M13fMZJbPUfFPkt93bt1ARLwOuEOP161VVVVVVVVVVVVVVVVVVVVVVVVVVX/BWHVzpv7Efi+yEhH9q6jpltjbndi8ilx7f6rOfbHev+CXekHSf2HPhzvQrLenUblxuzndqFxtI9PkCcVVMVVSzMAAOSeMV/wAEYc+IfjV8YPGTKXMumwZkmOZf9Iunk5POc+Xzz1AqqqqqqqWSOGNpZZFREUszMcBQOpJ7CvHX7W/7MXw1LxeM/jv4LsbiIHfaR6tHc3C/WGItJ/47XjL/AIK//seeGg66BqHi3xayj5DpWiNCrH63TQkD8PwNeLf+C4FozG1+Hv7PdxO7nEc+ra8EOe37mKJs8/8ATQfrx/w8H/4KWfFcbfhX+zn9jtZB+7udN8G313tBGQWmlZounIJUA570dC/4LY/FRt17qet6BaycBvtek6P5YJPG2HbLwCecE8DnIFH/AIJtf8FE/iMPO+Jv7TVssM5/ew3/AIv1O/kXIGRsEZjwAWGA+OMdDmv2T/gv4h/Z6+Avhv4Q+KPFcHiO/wBCa6L6jFC8Yl8+5lnxh2LHaZSuT1AHAqqqqqqqv2lbMWXx48awCMpu1R5ME/3wHz+O7P41VVVVVVVVX7MHxTHxF8AR6Zqdzv1vw+Ftrrcfmlix+6l98gFSf7yk9xVVVEA1gegoqp7D8qKqewravoKKKe1bFxiti14YjAvJXBPEWP1H+FVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVXjDxZ4f8B+FNY8beKtRjsdH0Gxlvb65fpFDEhZ2x3OAeByTxX7HvhLXv2/P25fEX7THxE06aTwl4Svk1CG1uDujR1O3TrIdiEVBK4HDGM7h+95qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr9rj9sH4a/sjeBhr3iqX+0vEWpJIuhaBBIBNfSKPvMf8AlnCpxukIOM4AYkCvgR+zd8ef+CmXxQk/aA/aN1u+0z4fwzlIRCGhFyi/8umnRtkRwjo8xzk55d9xXwV4I8I/Djwrp3gjwJ4estE0LSYfKs7Czi2RxLnJwO5JJJY5JJJJJJNVTKGXaRkEYINX9qbO7ktz0U/L9O1VVVVU7ADaDzVVVVVVVVVVVeF7PPmXrj/YTP6/0qv2xP2cF/ap+B2o/CiHxG+h30l5b3ljfHe0ccsT8iWNSPMQxtINp4DFW6qK/Z0/4Jj/ALNXwEjtdY1bQh4/8VQFXOra/ArxRSDvBa8xxjIBBbzHB6PQVVUKoAAGAAMYqqqqqqqqqqqqqmKqpZmAAHJPGK+MP7fH7KPwRM9p4r+Lel6hqsBKnStCP9pXIcfwsIcrEf8AroydvUV8Q/8Agsj498bamfCX7L/wJnnvrnK2tzrCPfXUmf7lnbHAYD/po4yeRxz/AMM+f8FWv2wP3/xU8Z6j4M8OX/L2mr6gNJg2D+E2FqPMJHQeagJPVupr4Vf8EVfg9oCw3vxe+JOv+LbpcM1ppkS6Za9sqxzJKw6jIZCfQV8L/wBlL9nD4MCF/hr8GvDGkXVvjyr42Yubxcf9PM2+X/x+viJ+0z+z38JvNj+Inxm8I6Lcwkh7OfVI2usjriBCZDjvheMj1r4gf8FiP2TvCnmweEIvFnjSdciN7DTPssDH3e5aNwPcIevSta/4LFfHn4h30uifAL9nGxN2wwi3ButbnAPG4R26xYPpncM+tLff8Fn/AI+qjQW2v+DdMn7GGz8OmAHud+26/Dkj0q0/4JF/tSfFK5TU/j3+0pp7Tu+9nkub3Xp0J6kmcxDd9GPXrXgr/giv+zxo22bxv8QfGviSZcfu4JILCBvXKhHf8nGPfrXgv/gnb+xh4FMb6X8BdB1CVCCX1p5tU3Eeq3Lun4BQPavDPgjwX4KtTY+DfCOi6DbkYMOmafFap/3zGoFVVVVVVVVVVVVVVVVVVVVVVVVVVV/wU9/ZL1/4d+Krf9tr4ALPpWpadfRXfiWPTl2vaXKsPL1JFHGC2BLx1Icg7pGH7Ev7Wvhz9rf4R2/iaJ7ez8W6KI7XxNpUZx9nuCDtlQHnyZQrMh5wQy5JQmqqv+CaGNe/4KKfG/xKsZnhax16RZpcF1eXWLcqeedxUPk/UHrzVVVVVVVVVVVVVVVVVVVVVVVVVVX/AAWMup7f9kGOGGTalz4ssI5RgHcojnbHt8yKePSv+CfNnBZfsX/CeC2UqraH5hBOfmeaR2P/AH0xqq+NHiFPCPwc8d+K3cIui+GdRvSxHQRW0j55/wB2v+CHfh57fwf8WPFbIdmoalplkjEHGYI53IHGP+Xhc8+nA4zVVVX19Y6baS3+o3kFrbQLulmnkEaRj1LHgD618Tv+ChX7H/wp86DW/jVo+rX0RK/YvD5bVJCw6qWgDIhGMEOy4PHWviN/wW28H27vY/B74J6xq0jHbFd6/fJZgHsfJhEpcH03qf5U37TP/BW79o9VT4X/AAy1Dwrpt3ny5tM8Opp8Mi9sXeolh6fMjr/So/8AgmL+3T8cmS9/aD/aCt7eCVgz22pa5d6zND9IhiEYwMBZMfSvA3/BE/4HaQIpfiD8VPGHiSZB8yWEMGmQue+VImfHsHB968F/8E3f2LfA6xtZfA/TNVuEGGn1q6n1AyH1KTO0Y/BQK8J/DH4a+AYxF4E+Hvhnw4ijAXSdJgswBjHSNV7cVVVVVVVVVVftn6T/AGZ+0Drs6x7V1G3tblRjAP7hEJH/AAKM/jmqqqqqqqqq+C/xJufhZ4+sPEiu7WLn7PqMS8+ZbsRu47lSAw91HrVpdW1/aQ31lOk1vcRrJFIhyrqwyGB7ggg1VVVVVVVeGI8m4k9AoH6//Wqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/4LCftF3Gm+GND/ZW8ETyz654ukivdbhtgWcWgkxb2+ByWlmUNt64iHGHFfsU/s52v7MH7Pvh/4dTRQnXrgHUfEM8Yz5t/MBvGR1EahIge4jB7mqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv22v23fBH7IfgxVC22t+PdYhY6JoJkIGOn2m42nKQKQR2ZyCq4wzJ+yV+xZ8S/22vHz/tXftcX17c+GNWnNxa2EzPDNrYXiNUVceRZKOF2kFguFwDvrStK0zQ9MtNF0XT7aw0+wgSC1tbaIRRQRIAqoirgKoAAAHAAqqqvEdl5sK3ka5aPhvp/n+dVVVVMwAwDz2o896qqqqqqqqqq8beMfD3w98Iax458V36WekaFZS3l5Mx+7HGpJwO7HGAvUkgDk14G+K3/BRj9uv4j6N4l+E4fwT4D8OatFeWjkyWekIYpNwFxIP3l+xKgPGu5cn7kYORD5vkp5+zzdo37M43Y5xntVVVVVVVVVVVVVV8Yf2lPgT8A7I3Xxa+J2i6BKY98dlJMZbuZfVLeMNK49wuPevi7/AMFnYb29Phf9mb4Q3mtahcP5VrqPiBW/ePxgR2cBLyZ5xmRT0yvOAvwF/wCCpP7bbLdfFbxTqPgvwpeYJtNYuDpFt5Z5x/Z9uvmSEA4BmTJzgv1NfBr/AII2/s++CvK1D4teItZ+IV+uC1vk6ZZA/wDXOJzK3PrLg/3aDfsy/so+GBCH8A/DHSHXIUm3077SR37NM/8A30xr4qf8Ff8A9lbwMZrPwQfEPj6+QEIdNsjaWu70aW42Nj3RHHpmtR/4KiftpfHu9k0T9mf4BxWMbvtE1lpk+u3UBx/FKVWBR3O6L05Hf/hkL/gql+0t+8+MvxWu/DWl3XMtpq3iLyYmU9xZ2AaPP+y6p+FfDr/giT8M9NWG4+Kvxj8Q67KCGe20Syi06PP90vJ5zMPcBD9K+H3/AATv/Y2+G4ik0n4HaLql1Gcm414yaozt67LhnQdB91QP1rRNA0Lw1p0ekeHNFsNKsYv9Xa2VskESduEQADgDtVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVWo6dYavp9zpOq2UF5ZXsLwXNtPGHjmjcFWRlPDKQSCDwQa+MvgL4if8Erv2rNP+L3wttri++GfiaZ1gtXkPlzWzENPpkzckPHgPE5ySFRvmKyCvhX8UPBfxo+H+i/Ez4fasuoaHrtsJraXGHQ9Gjdf4ZEYFWXsQRVaxqlpoekXutX7lbbT7aS4mYY4RFLMecDoD1r/gilpV54g+Inxg+JN/FmX7JZW7ylThpLmaaVwCck8wKSMnque1VVVVVVVVVVVVVVVVVVVVVVVVVVV/wWIsZLv9j4zxuoFl4qsJXB6kFZkwPfLg/ga/4J5X8ep/sWfCi4iUqE0ZoSCc8xzyxk/mhqq/4KPeMk8E/sW/E2+E22bUtPi0uFQcFzdTxwsB/2zdyfYGv+CPvg5vDX7H0GvPFhvFniS/1BHK4LJHstRz3Aa1b8Saqvid+13+zL8HRNF8QvjZ4X0+7gBMlhDeC8u1x628G+UdMfd5r4nf8ABab4G+HlltPhZ8PPE3i+6QEJcXrppdqx7EE+ZKR6gxr/AIN+3B/wUz/aeIt/gF8JpvD+lXGFivdG0EyJtPTfe3m6FSf7y7OmeOtWP/BMP9t34/3cOr/tLfHqPT4Xfe0Go6rPrlzbH/ZhVlgUc8BJQPpXwy/4I5/sueDzBd+PNQ8T+OrtAPMiu7z7DasR3EdvtkA9jK1fDn9nv4GfCJIx8NPhL4V8PTRjAurPTI1uG/3piDI34saqqqqqqqqqqqqr/goR4ba38W+FfFqRfJfadLZO4/vQybxn8Jj9ce1VVVVVVVVVX7HfxUGveHZvhtrF1uv9FTzbAueZLUnlR7oxx/usoHC1VVVVVVVeHYdlgZCOZHJH0HH9DVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVeN/GOgfDzwbrnjzxTefZdH8PafPf302MlIYkLvgdzhTgdzgd6/YK8F+If21f21PFX7WfxJsTJpHhi/GowQSfPGt63y2FspPVbeJA+eoMUWfv1VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV+3R+3R4Q/ZI8IHStKa11n4jazbsdH0dnytspyPtVyAcrECDheDIwIGAGZf2Hv2GPGH7TXjB/wBrX9rtrzV9O1i4+3abpepj95rrn7s8y8bbRQAEiAAcAcCIAPb21vZ28VpaQRwwQIEiijQKqKBgKoHAAAAxVVVU6JIhjcBlYYIPetSsWsLloScr1QnuKqqpiAM5oncck1VVVVVVVVVVXiX4OeC/i5oMOifEfSTq2hLdx3EmlSyFbe9aM5QToMeZGH2tsPylkUkEDFafp2n6RYW+l6TY29lZ2kYit7a3iEccSKMBVVcBQBwAOKqqqqqqqqqqqqubi3s7eW7u7iOGCBC8ssjBVRQMliTwAAMkmvj7/wAFSP2X/gobnR9A1+T4heIoMqLDw86yW6P6SXZ/dAdQfL8xgRyoq8/az/4KPftyXk2ifs9eCr7wh4WmlaNrrQ1+zpGuekuqTbQHUdRCY2OD8p6V8HP+CMyX98PFv7UHxYvNa1G5k8650zQpGPmOTk+deTAvJnoQqKeuHPWtI8Hfsc/sR+GRf21l4H+HFs0RX7fezIL28CgAqJZC085/2QW6nA5r4y/8FnPgj4Te40z4N+DNZ8dXkeVjv7onTLEn+8u9WmbHXBjTPHPcH42/8FYf2wyP+FaeF9V8EeGr0fu7jSrMaLb+Wejre3Lec+B1ML8novQDwB/wRi8XeKdQPiT9o/49S3N7cENcQaIj3k8pPXdd3WMH/tk31r4Wf8E4f2PPhQsE+nfCKx8RahCBm+8SudTdyOjeVJ+5U+6xr+grT9O0/SbKHTdKsbeztLddsNvbxCOOMeiquAB7Cqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr45/BTwN+0L8MNZ+FXxBsPP0zVovkmQDzrKdeY7iFjnbIjcg9CMqcqxB/Z2+L/AMSP+CYX7R2q/s+/HN7if4d63dLI91FEzRKr/LDqlsOpQqoWVBk/KRy8QB0zU9N1vTLTWdG1C3vrC/gS4tLq3kEkc8TqGR0YZDKykEEcEEGv+ChPxWs/hF+yJ8QtYluQl7rmmvoWnJu2tJPeAwnb7pG0sn0jNf8ABHH4bTeEf2Xb3xxe25Sfxv4gnubdiMbrW3VbdP8AyKlwc+hH1NVVVVVVVVVVVVVVVVVVVVVVVVVVf8FIvBlx45/Yr+Jmn2UTPcabYQ6om3+FbS4inlOPTyo5P59q/wCCQPju38Vfsg2nhZZlM/gzXr6waPoQkri6VvoTcOM+qn0qq/4LZfEeLSvhJ4D+Fdvc4ufEOuS6nOinnyLSIoA3oC9ypHqY/Y1+yr8PX+E/7Nnw38BXcBt7rSfDdp9ujYY2XLxiScf9/Xev2qf+CqHwT+BMl54R+G5i+InjKAtHJFZXIXT7GQcHzrgZDsD1jiDcghmQitM+Hv8AwUs/4KIomueLfEdz4P8Ah/qBDwpdu+k6a8LDI8m1jBlulwTteQODnHmY6fDP/gij8HNDEV18Vvij4k8VTrgm20y3j0u3J7q2TLIw7ZDIe/HSvhl+xh+y18H/ACZfAvwS8M295AAEvr21+33SkdxNcF3Un2IoKqqFUAADAAGMVVVVVVVVVVVVVVVX7bng5/E/wTn1a1h33Hhy+ivflGW8o5jkH0xIGP8AuVVVVVVVVVVeBvGGqeAvFmmeLdHkIudOnD7M4EqdHQ+zKSp+teFvEmleMfDun+J9En82y1KBZoieoz1U+hByCOxBqqqqqqrGeBVlB9mtIYO6IAfr3qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv+Cxn7Q13ofg7Qf2Y/CE8r6v4yZL/WY4AWf7Ekm2CEAckyzoTgc4gx0av2K/2fbX9mn9nbwv8OpbWOPXJYBqHiF1wTJqEyqZQSPvCMBYge6xKaqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv26/wBuHwt+yL4LSx01LfWPiFr0DnRNJdspAv3TdXGDkRKeAvBkYFRgBmX9hv8AYb8ZftLeMj+1z+1y95q1hq1yL/TNM1Nf3muycbJ5k4C2igAJEAA4A4EQAeOKOGNIYY1REUKiKMBQOgA7Cqqqqq1TT01C2KDAkTmNj2Pp9DUkbxOY5FKspwQexqqOMdaY7j/Kqqqqqqqqqqq0nTG1G4CnIiTmRv6D60qIiCNFAVRgADpVVVVVVVVVVVV4n8U+GfBWh3Xibxh4h03RNIsU33N/qN0lvDCvqzuQB+Jr9oX/AILF/CXwT9p8PfALw/N491lfkTVLtXtNNjb1AIE0+DxgBFOchzVt8E/+Ck3/AAUIuI9V+KuuXfgvwLcsskcGqK+mWWzqDDYJ+8nODlXlGDn/AFlfAf8A4JVfswfBeGHWvG2nv8R9dt1DyXniCNVsoyOSUswTHt4ziUy455r41/8ABSj9kr9n20bw3oniOLxdqtgnlQ6N4SSOaGDbwEacEQRgEYKqzMuD8vatU/bt/wCCgX7YuoT+G/2XvhhdeGNHeTy3udHg8+WIE8LNqNwFhiOMcqIj15xXw0/4I7/EXx/q48a/tXfG67mvrtg91aaXcPqF9Nn/AJ6XtxkKw9kkHP3q+DP7EH7L/wAB/s134D+E+kvq1tgrrGqp9vvA4/jWSXd5R/65hB7VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX7YX7Ivgb9rr4bN4V15o9M8RaYXm8P66kIaSxmIwVbu8L4AdM84VhhlUj9nz9sD45/8ABO3xrN+zf+034T1bVPCFlIfsUcRDz2EZY4nsXbC3Fs5BPlkgA5wVYMp+P/xm+Jn/AAVP+Ovhn4MfBfwtqmkeC9EmMudQQZgDELLqN4EJWMIh2pGGY8kAlpMV8NvAHh34VfD/AMPfDbwnbmHSPDWnQ2NorcsUjULuY92Ygsx7kk96qqqqqqqqqqqqqqqqqqqqqqqqqqrX9D0vxPoWo+GtbtEutO1a0ltLyBxxLDIhR1PsVYj8a/ZJ+It7/wAE6f2yfF/wB+MF9JZeDvEVwlm+pXC7YlwxawvyegjdJGRyOF8wliPKIFvcW95bxXdpcRzwTIHiljYMrqRkMpHBBGDmq/4KU/Ffwxr37f8Apmn+OTd33g/4crpdrqFpZKsjzxZW7uUjVmVfMcTeVkkAFRk8V4v/AGjf23P+ClfiG7+HHwK8LXXhPwCX8m/S0uWigWM9ft9/gb8qSfJjA3L/AMs3Iyf2Uf8Agl38FPgALLxZ4/it/iD43hIkW7vrYfYbFxyPs9u2QWU4xJJlsgFQnSqqqqqqqqqqqqqqqqqrxLoFh4q8O6n4Z1RC1nqtnLazgddkilTj3wfzrxV4c1Hwh4l1TwrqyBbzSbuS2mAHBZGKkj2OMj2Iqqqqqqqqqq/Y1+Kgsr+4+FmsXX7m8LXGklzwsoGZIh/vAbgOmVbu1VVVVVVaPa/ar+NSMqh3t+H/ANfFVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV4g13SPC2gal4n1+9js9M0izlvL24k+7DDEheRz7BVJ/Cv2M9A1n9ur9vnxN+0z4ztGl8NeDr1dTgtrhcrE43R6XbYzgGNY/NJHBaAk/fqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/bV/bJ8GfsifDs6ndiLU/GWtxSx+HNFyf30igAzTY5WBCV3dCxwq9SV/Ye/Y18dftYfEKb9r39rcXWsaNqFx9r0uw1EYOuSgjZI8eMLZIBhIwAr4UAeWMPHFHDGkMMaoiKFRFGAoHQAdhVVVVVVWtaSLxftNuB5yjkf3x/jRBBwRyOxqnYHgGqqqqqqqqqqqsbCfUJxBAP95iOFFWlpDZW628C4C9SepPqaqqqqqqqqqqrxb4x8JeAdAuvFPjfxLpmg6PZLuuL7UbpLeGMe7uQMnsOp7V+0L/AMFhvCelXcngr9lzwjN4z1uaTyIdZ1C3kjtPMJwPItxiack8DPljOMbhXhP9h/8Abl/bi1+28f8A7VHjrVPCegM/mQW+sRkXKIe1tpqlUt+MgmTy26Ha9fBL9ij9lT9kfRh4vsdC006lpEXm3Xi/xRPG81vjGZBI+2K2HbKBODgk5NftD/8ABYP4KfDs3WgfBLSZviHrcTNH9uZmtNMiYdxIR5k2D2RQrdn6VD8N/wDgpd/wUOlW78d6nd+C/AF4wYRXyvpGmmMnI8u0XM11xkq0gYdvMGa+An/BJX9mz4ULbav8Q4Ln4l69EFYvqyeTYI46lLRCQw68TNIPYVpOj6RoGmW2i6Dpdnpun2cYjtrS0gWGKFB0VEUAKPYCqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq+PHwm8C/Fv4dazo/jHwDpHiee30+5fTI72wS4lguPLOxoWILI+4KMqQT0r9i34ZW/wv8A2c/B+j3XgeDw1r9xYLLrkJsFtria53Nlp8AMz4wMtkgYHSqqqqqqqqqqqqq1DUtO0m1a91S/t7O3T70txKI1H4k4rxF+0X8OdEZoLG5utXmXjFpFhAf99yAR7jNfGr9u74geEL+2sfBvgvRLeC7gLLPqDyXLBgxBwEMYHG08561rv7bH7Smulk/4WGbCI/8ALKx0+3hx9G2F/wDx6r/9on486lJ5tx8Y/GCtknEGszQDn2RgO34V+w3478b+LfC3ie88VeM9c1m4h1CJI5dQ1GW4dF8vOAXYkDPal1TUU+7fTH6uTUev6pGMG4DgdmQVD4qmGBcWqN7oSv6c1b+INNuMK0pib0kGP16UrJIoZHDAjgg5qqqqqqqqr9s79iP4fftg+FbePUroaD4x0aNl0fxBFB5jRoTkwTJkeZCSScZBViSpGWDaF8M/+Ct/7IkLeB/hu2o+JfCtllbJdPNtrNpsHP7mOcefCvH3AqDJ6HNaj4f/AOCzP7QaHQNautd8KaVN8k0guLLQEVSBnc0G24dSDggbh1GOtfs//wDBG3wXoF7D4q/aU8ayeM9UEvnSaNpMkkFi79W82dgJpwSc8CL3yCRXhTwh4V8CeH7Pwp4K8O6boWjaemy1sNPtlt4YR1O1FAAycknuSSetVVVVVVVVVVVVVVVVVVV+3l8Lzovi2w+KGm25+ya8gtr8heEuo1whP+/GOnrGT3qqqqqqqqqq0vU77RdTtNY0u5a3vLGZJreVeqOhBUj6ECvhX8QLD4neBtN8W2WxJJ49l3Cpz5E68SJ9M8j1BB71VVVVVeHrPyLU3Lj55umeyjpVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVf8Fgv2gz8OvgfY/BfQb/y9a+IkxF4I3w8WmwMrSZx08yTy4/8AaUSjnmv+Cev7PX/DOv7M3h3QdV0823ibxGv9s68JE2yR3EyrshbuPKiEaEdNwc/xGqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/aq/ae8B/spfC27+IHjCZbm/m3QaHo8cgWbU7rblUX+6g4LvjCr6sVVv2Sf2Z/iP/wAFBvjLf/tV/tOPPP4LjvP3Nm6vFHqzRkhLS2GcpaRHhiD8xyoJYyOtjY2OmWNvpmmWcFpaWkSw29vBGI44Y1ACoqjhVAAAA4AFVVVVVVVWr6Mt4DcWwCzAcjoH/wDr1MrxsYnUqy8EEYIqqqqqqqqqqqtP0241GURwrhR99yOF/wA+lWVjb2EAggX/AHmPVj61VVVVVVVVVVq+saR4f0u51vXtVs9N06yiMtzd3k6wxQIOrO7EBVHck4r9pf8A4K+/DvwXNc+Df2btGXx34h3eSmr3KOmmRueP3agiW5OePl2KeCHYV4Q/Yo/bd/bw1+2+JX7UPjnUfCfh2R/MtYNWhInSM9rTTl2rApHG6TYTw2JOp+Bf7Iv7NP7Ivh59Z8LeH9PtL2xty1/4s16ZHu9gHzs07gLAhHVYwieo71+0n/wV/wDhV4DF14W/Z80k+P8AxCCYk1SdXh0uB+mV6SXJB4wmxTkESGvDv7KX7fv7f+rW/jH9ofxlqPg/wdLIJreLWImhVE7G10xCuDtPEkuzcDne9fs7f8E9f2av2cVt9U0PwiviXxNCQw1/xAqXVxG4OQYU2iODB6FFDerGqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqrxD4m0Dwpp7an4h1WCyt16NI3Ln0VRyx9gCa8bftP31x5lj4F00WqZIF9dqGc+6x8gf8Cz9BWueI9e8TXX23X9Yur+bna08pbb7KOij2GBVfH3Sxc+GrHVUXL2V1tJx0Vxz+qrVV/wT7/5E3xX/ANhOH/0VVVVW15c2jb7adkPoDwfwqx8TqcR38eD/AM9EH8xUM0NxGJYJVdT0INVVVVVVVVVVVVVVVVVVVVVVVVVVVVV8Vfh5pfxT8Bat4J1QIov4D9nmZc/Z515jkH0YDI7jI714i8P6t4U12/8ADWu2jW2oaZcPBcRN/C6nBx6juD0III61VVVVVVVVVfsnfFMeCfG48J6rc7dI8SOsQLHiG6HEbewbOw/VSeFqqqqqtOsmvrpIAPlHLn0FKqooRAAFGAPSqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqndI0MkjqqqCWZjgADqa8IRt/wUT/4KZXHiW4/0/wCHvgmf7RHld0T6XYSYgTByCLm4YOy9ds0hH3aqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr4yfF/wN8B/hxrPxR+Imqiy0bRoC7BcGW4kPCQxKSN0jthVGQMnJIAJHwu+H/xX/wCCr/7St98UviY13pHwv8NziGSKBmEdrbj5k0+2YjDTuCrSydgxbjMaHw34a0Dwd4f07wp4V0i10vSNJtktrKytYxHHbxIMKqqOgAFVVVVVVVVVqWkW2oplgElA+WQDn8fUVfaddae+ydOD91x0NVVVVVVVVVjPArTfD891ia6zFF2GPmb/AAqCCG2iWGCMIijgCqqqqqqqqqqv2qf+CnfwL/Z6N54W8JXMfj/xrBujOnaZcj7LZSY/5eLkZUEHgxpucEYOzrWkfCv9vX/gp3q1v4n+IetyeEfhsZhJbNcwva6dGnZrW0B33TgZxI5x1HmDpX7NH7Av7PP7MENvq3h3w6uv+K4VBk8Sa2iTXKNjkwDGy3HX7gDYOGZq/ak/4Km/Av4EG98LfD6aL4ieMoC0bW2n3AFjZyDI/fXIBDFT1jj3HIIJQ81pHwh/b6/4Ka6raeKvibrcvhD4cSS+baPdW72unxoRkNaWYIe6OOkrkg8jzeMV+zV/wT7/AGdv2Z0s9Y0Lw0PEfi63UFvEetIs06SY5aBPuW4znBQb8HBdutVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVfG34u+GvgL8LNf8Ai54wstTvNH8OQpLdQabEklw4eVIxsWR0Unc4PLDjP0r9nL9oXwV+0/8ADOD4q+ANM1uw0me8mtFh1iCKKcPEQGJWKSRcHPHzfgKqqqqqqqqqqr4ofH/R/CTS6L4W8nVNWXKvJnMNsfcj7zf7I6dzxivEPiXXfFeovqviDU5r25fgNIeFHoqjhR7AAVVV8QtL/tnwVq9iq7n+zNIgA5LJ84A+pUCqr/gn3/yJviv/ALCcP/oqqqqqrW9ubKTzLaUqe47H6itM1u3vwIpMRT/3SeG+n+FVVVVVVVVVVVVVVVVVVVVVVVVVVVVV+258CJNcsD8YvC1pvvdPhEetQRrzNAvCzjHJKDCt/sYPAQ5qqqqqqqqqgSpDKxBB4I4xX7O/xRX4o/D22ub6ffrOlbbXUgx5dwPll/4Goz/vBh2qqqqCliFUZJ4AArSNPFhbAOB5snLn09qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr/gpZ8e/wDhRP7LXiD+ytQ+z+IvGmdB0nY2HTzlP2iUY5GyASYYfddo/UV/wSP+AY+Fn7OP/CytZsPJ174lXAvgXXDpp8W5LVfo2ZZgR1WZfSqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqtb1rSPDejX3iHX9TttP0zTLaS5vLu5kEcVvDGpZ5HY8KqqCST0Ar4rePfid/wVc/agsPhV8M5LnSvhj4ameSKeZGEdtaqdsuozrxmaQHbFGcY3KvGZHr4QfCTwN8DPh3o3ww+HekrYaLosHlxqeZJnPLzSN/FI7ZZj6njAwBVVVVVVVVVVJFHMhjlRXUjlWGavvDEbkyWEgQ/wDPN+n4HtVzZXVm2y5gZD2JHB+hqqqqqqqtDtB4itItR0m6t7iylz5dzHKHR8Eg7Suc4II+oqw0OysSJAvmyj+Nx0+g7VVVVVVVVVVV+0L+1P8ABT9mHw6Nc+Kvi2K1uZ4y1ho9qBNfX5Gf9VCDnbkEb22oDwWFeOP2pv20v+Cjfii9+F/7OnhbUPCvgYnyb5bOcxDyWON1/f4G0Muf3MeNw3Ltlxmv2Vf+CVHwY+CK2Xiz4qi3+InjKILIBdQf8S2xkBz+5gb/AFpB43y5zgEIhr9pH9sf4Efsq6MH+IXiVJdaeHdY+HNM2zX1wOiny8gRIcffkKrwQCSMV4k+OP7c/wDwU11678DfCHw9ceEvh0ZTDex2tw8FlHGf4b69wDO20j9ygwRz5Zxmv2Wv+CW3wL+Ay2Xijx7BF8Q/GcIVxdajbgWNnJ1/cWxJBIPR5NxyAQE6UFVVCqAABgADGKqqqqqqqqqqqqqqqqqqqqqqqqqqpiqqWZgAByTxirf4g+Abu/OlWnjjw/Nehipto9ThaQEdRtDZz+FVVVVp+3P+zfffG0/s823jC+bxwuryaSbE6TcBPtSEhl83Zsx8p+bOKqqqqqqqqqqr/gpF/wAmR/FP/sH23/pZBX/BIb/kzTTP+xg1H/0NaqqqqqqqqqppYYIXnnlSOONSzu7bQoA5JPYYr4w/Hu51eS48L+B7pobBSUuL+NsPceqxn+FPfq3sOtVVU6K6lGAIIwQR1rXtNbR9bv8ASWBzaXMkQz3CsQD+WKr/AIJ9/wDIm+K/+wnD/wCiqqqqqoEggg8jpWj68XK2l+/PRJD/ACP+NVVVVVVVVVVVVVVVVVVVVVVVVVVVVSxRTxPBPGkkcilXR1yGB4II7jFftV/s6z/CXxC3inwvaO/hHVpT5QUE/wBnzHkwsf7p5KE9vlPIyaqqqqqqqqr4AfFB/hb8QbTUruZhpGoYtdSQHgRseJMeqNhvXG4d6R0kRZI3VlYZVgcgj1FVVVoWlFdt9cLzj92pHT3qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/bp1jVP2zv2/fCH7LfhS9kbRfC1yul3EsJLCKVwJtSuAPWOKMJgj70B5wa0bR9M8PaPY+H9Eso7TT9Mto7W0t4xhYYo1Coi+wUAfhVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX/AAUd/al8WftC/Eq0/Yh/ZzE+rLLqaWevPZNn+075Wz9lDDgQwMpaRjgbkJOFjJP7Hn7KnhL9kv4S2ngfRxb3viC/CXPiPWFjw1/dY6AkAiGPJWNTjAySNzsTVVVVVVVVVVVU6JIhSRVZSOQwyKuvD2nXHMaGFvVDx+VT+FrpObe4jkHowKn+tS6LqcXDWbn/AHfm/lT2tzHw9vIuP7yEUyleGBB9CKEbsBtRj6YFeKf2hfgN4JnuLPxZ8ZvBWl3drI0Utpc67brOrqQGTyt+/cCQCuMjvXxZ/wCCh37Mnwe1S68OeIPEOu3uvWQU3Gk2ehXCTx70V0yZ1iQblZSPm6EHoa+Jf/BY+8lhks/g/wDCGOCQg7L/AMR3nmY/7d4Mf+jfwr4wftjftG/HJZrPxz8TdR/sqcENpOnMLK0K4+60cWPMH/XQsfev+CMfxw/4Sv4QeIfgZq17uvvBF79s0xGPJsbpmZlUd9k4kJ9PPWqqqqqqqqqrVtW0rQdMuta1zU7TTtPsYmlubu7mWGKCNRlnd2ICqBySSAK/ao/4Kyh9Tl+E37HekS+JfEF5J9kTxL9ja4TzW422NttJnfJGHcbcjhHBBr9nn/gln8SPjH4i/wCF0/tu+LNWkuNTkFxJoBvWk1C7z0F1Pk+QuMfuo8sAcZjIxSR/Br9mz4abUHh3wD4J8Pw8klLS3hH1/ikY/V3Y9ya/aH/4KofEP4t+I/8AhTH7EPhTVZ7rU5Gtotf+wNLf3mQQTaWxB8lcZPmyDcBztjK5r9mn/gkpqOt6svxW/bN8R3et6zfTfapfDkN+0zSyHkm+uwS0jE9UibHA/eEErXhnwv4b8F6DZeF/CGg6fo2kadEIrSwsLdYIYEHZUUAAVVVVVVVVVVVVVVVVVVVVVVVVVVVVeM/Fuh+AfCGt+OfEt0bfSfD2nT6hfSgZKQwxl3IHc7VOB3NQ+Kv2rv8Agq/8Y9Z8LeH/ABPJ4V+H2lqJp9P+0Oun6XbEkQ+cseDdXDlWxu6kPjYi8X//AAQ308aYV0z9pC4Ooqgw0/hYCJ2A54FzuUE+7Y96/Ye/ZZ+Kv7Ovwl8X/D34qfEU+IbrVdRkj0kQX89zaWdkIAqGOOUKYmZ3kLoBj5V5OM1/w458Vf8ARxWk/wDhNyf/AB+v21f2KtU/Y01Twppmp/EG18UnxTb3UyPBpzWnkeS0YIIMj7s+Z7YxXws/4I5eJfih8MPB/wAS7f486Zp8fi3QbHV0tH8PvI1uLmBJhGWE43FQ+M4GcZwK/Yc/4Jx61+yB8WdW+JepfFax8Tx6l4dm0hbSDSGtWRpLi3m8zcZXyALcjGP4s54rw1/ymDm/7Khe/wDoctVVVVVVVVVVV+2P8LPFnxs/Zo8c/C3wLDay67r9pDFZpczCGMstzFIdznp8qNX/AAT8+BPj/wDZz/Zzsvhp8S7axh1uDVry5dLO5E8flyMCvzDvgdKqqqqqqqqpiAMkgADkntXxy+MzeKJ5fCPha6ZdIgcrdXCN/wAfjA9AR/yzBH/AuvQCqqqqq+Nel/2d46nuFUBL+COdcDvjYf1Qn8ar/gn3/wAib4r/AOwnD/6KqqqqqqrQtZxtsLt+Okbk9PY1VVVVVVVVVVVVVVVVVVVVVVVVVVVVr+gaP4p0W88O+INPivtO1CFobi3lXKup/ke4I5BAIwRX7RP7Nuv/AAU1VtTsBLqPhO8mK2d8RloCeRFNjo3o3RsZ4OQKqqqqqqqqr9kj4p/8Jh4MPgrVrrfq3hxFSIsfmmtOiH/gB+Q+g2etVVaPopmK3V2mEHKIR973PtWBjAqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqvjv8VdK+B/wc8X/ABZ1gxtD4Z0qW6jic4E8+NsMXUcvK0adRy1f8Ec/hRqnjDxX8Qv2rvGzSXuo3l1JpdjdzjLTXMzC4vZ/975oV3D+/IPWqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv+Cmf7aZ/Zx+HY+Gvw/wBUCfEXxjauIJon+fR7Ikq916iRsMkXoQz5+QA/8Evv2J3+CHgxfjj8TdKI+IHi61BtLa5T95o1g+GCEHlZ5eGfPKrtT5T5gNVVVVVVVVVVVVVVVVfELwpN478C6/4LtvEep6BLrenT2ceqaZJ5dzZNIhUSxN2Zc5H9K/Yy/wCCYPgz9n/WD8SPi9f2Hjjx1DdM+muI2ay04BvkmRZBl5zw29x8hICjI3n/AILTfA86F8QfCnx90mz22nie1Okas6JwLy3BaF2P954SVHtbVVfsGfHVf2e/2n/CHjO/vjb6FqM/9ka6xOFFnckKzt/sxyCKY/8AXL8Kqqqqqqqqr9p39sP4L/speHjqHxB10XOu3UDSaZ4dsWD3t71AO3pHHkEGR8LwQNxG2rvVv21/+Cr3i1tP023/AOET+FlldDfGHePSrMqcgyPgNe3IGOAMKTkLErE1+yx+w38Ef2UdLin8J6QNZ8WyRbLzxPqUKtdSZGGWEci3jPPyJyRjcz4Br9rr/gpD8Gv2Y0uvCmizReNfHyKyjRbG4HlWD44N3MMiP/rmuZDxkKCGrwb8AP20f+CnHii0+JXxq8S3Phb4eLIXsJri2aK2jjJOV0+yyPMOPlM7nkDl3K7a/Z3/AGVPgp+y/wCHP7C+FfhSO3u54wl/rN3ia/v8f89ZsA7cjIRQqA8hRk1VVVVVVVVVVVVVVVVVVVVVVVVVVVVX7V3gTX/id+zX8SfAfhWOSXWNX8OXcVjDGcNPMELLEP8AfKhP+BV/wSP/AGmfhz8EPG/jD4X/ABS1a28PR+NGtG07U70+VDHdW5lUwTOeI94myrNtUFCCcsoqGaG4hSe3lSWORQyOjZDAjIII6giqr/guN/yNXwj/AOwfqv8A6Mtq/ZO/5NX+DX/ZP9C/9IIaq68W+HPAX/BVrW/Gvi/VodM0XQ/iLqN3f3cxO2GKMyszYHJOBwACScAAk144/wCCjP7ZfxhmvdX/AGQf2aNbl8EWkjJBrdz4cudTuLvYeSPLPkoSBgxL5rDI+bJxX7LP/BW7xFrfxBs/hP8AtTeE9N0Oe8vPsMevWUL2YsrjdtCXkEjHYN3ymRSoQ43Jjcwqqqqqqqqqqqqqqqqqqqv2h/is2nRP4A8PXW25mT/iZTIeY0I4iB9WHJ9iB3OKqqqqr9oPSw9lpOtIv+qle3c467huX/0Fvzqv+Cff/Im+K/8AsJw/+iqqqqqqq+hrQdVN5F9lnbM0Y4J/iH+NVVVVVVVVVVVVVVVVVVVVVVVVVVVVaxo2leIdLudE1vT4L6wvIzHcW86B0kU9QQa/aH/Y+1zwC914v+G9vPq3hsFpJrNcvcaevU8dZIx/e5YD73ALVVVVVVVVVXwy8eah8NfGum+LtPLP9kkxcQg48+FuHT8R09CAe1aNq+neINIs9c0i5W4sr+BJoJV/iRhkH24PTtSqzsERSzHgADmtL0ERlbi+UFuqxnt9f8KwMYFVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX/AAWi+M9xYeDfBn7O/h+eR77xPd/2tqkEIJdreFvLt4yoHzCSYuwA53W49a/ZS+C9v+z9+z34J+FawRpe6VpqPqjIAfMvpcy3DZH3h5rsAf7oUdAKqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqrUtV0rRrV77WNTtLG2jUs81zMsSKB1JZiABXxO/a9+CHw98K6vrWn+NtJ8TarYWkklnpOkXi3El7MF+SJXTKrubALMcLyT0r9ny30b4mftU6n+0l+21q4Vba6XULPSlt2vI7y6BxBFti3hbeBUXCMfm2xg7hvrwF8XfA/wASvDUHi3wpfzz6bcSSRxSS2zRlijFWO08gZB60uvaS4wLsA+hQj+lR39jNxFeQsfTeM/lXBHBqqqqqqqqqqqqqq8ZeMvCnw98L6j408b6/Z6LoekQNPe313JsjhQdye5PACjJJIABJAr9mL9sX4PftZWniCX4aXt5Dd+HL1obix1CNYp5ICSIrpEBJ8qQDvgqQQwHGar9tr4Fr+0P+zT4y+HtpaefrSWh1DQwFywvrf95Eq+8mGiJ9JTTo8bmORSrKSGVhggjrxVV/wTr+Py/tA/sveGtU1K/W48R+F0Gh62GbLtNAqiOVsnJMkJict0LFwOhqqqqqqqpZI4Y2llkVERSzMxwFA6knsK/bC/4KsaV4avLj4S/soJF4p8Vzym1k8RRw/abW1lJ2hLSMA/apc8BuYwcY8zJA/Zh/4Jd+Nvir4gHx1/be1nVby91WUXZ8OXF4zXl4T0N9MDujXAH7lCGAwCUwUrxT4u+EH7OPw2Gq+I9T0PwT4O0CARQIEW3hiUD5YoYkGWY84RAWY9ATXx1/4KGftA/te+L3+A/7FvhPXdN06/LRSX9quzU76LhWkaQHbYwcjLbg2MbnXJWv2Rf+CUHw9+FRsvHv7QL2fjnxirCaPTCpfTNPfqMqwzcuD1ZwEyeEOAxREjRY41VVUYVQMAAegqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr9r3/AIJafDH9ofW7/wCI3w81hfAvjW/dpb1ktvNsNSlPV5YlIMcjHlpEPJJLIzEmtRt/+CiX/BNG6t7uXVby58Cx3CohWc6roU5PRCrYa1LdOkLtjgnFfsV/tm+D/wBsPwHc6tYaf/YnivQDHFr2itL5ghZwdk0LcFon2tjIypUqc8M1f8Fxv+Rq+Ef/AGD9V/8ARltX7J3/ACav8Gv+yf6F/wCkENV42+Guj/GH/gqBr3w08RO40rXvidcQX4jJVntxcM0iKR90sisoPYnPatD0LRfDGi2Phzw5pdrpul6Zbpb2dnaxCKK3iQAKiKOFUAAACv8Agtf8MfD/AIb+KXgH4oaTZW9tf+M9NvLXUzEu03ElkYAkr+reXcImeu2NR2Ffsw+LNQ8efs4/DDxhq88k2oat4S02e8lkOWknNsnmOSSTy4Y8nPPNVVVVVVVVV8Uvib4P+DXgHV/iZ4+1CWy0DQolkvbiK3eZo1Z1jBCICx+Z1HA718Ffjd8Ov2hPA0XxG+Furz6loU1zLbJPNaSWzGSMgONkgDcEjnFVVVVVVVVfGH4o6L8J/CD65qd5DFdXUn2bT45TxJOwJGf9lQCxPA4xnkVdXs+p3Muo3Vybia6cyySscmRmOS2fcnNVVVVVXxZ0oar4D1JFXMlqouE46bDlv/Hd1V/wT7/5E3xX/wBhOH/0VVVVVVVVQTSW0yTwth0OQasruO+tUuYj94cj0PcVVVVVVVVVVVVVVVVVVVVVVVVVVVVVXxx/Y78GfEx5/EPg54PDXiKQl5GSL/Rbtj3kRfuMT/GvqSVYnNfEf4S+PvhRqf8AZnjXw/NZh2IgulG+C4x3SQcHjnHBHcCqqqqqqqqv2JpPHeq+DbvQtX0O9i0G0k83StSnTbGwYnzIkzywDfMCoIyzAkcCrDSrWwXMY3yY5dhz+HpVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVfD4j9uL/AIKsXni53+3+D/Ad895bnrGbLTWEdsVzkFZboxyFT1WV+KqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqvFnjXwj4E0w6v4w8R2GkWnIWS6mCbyByEXq7eygmr39tTwXq2o3Wj/D/SLrU5LePct3eA28T84JVPvkD5eu0814g+OnxM8Q7kbxC2nwt0isF8jH0YfP/wCPVrn2nxBa3cGpXcty93A8TyTSF2IYEck896ljeKRoZFw6EqwPYjrVfsZf8m/aH/183f8A6PeqqG7urcjyLiRMdlYgflVt4lvosCdUmHuNp/T/AAq017T7rCNJ5L+knH69KBBGQc+lVVVVVVVVVVV8Xvi74B+Bfw/1T4l/ErXI9M0TSo8yORukmkPCRRIOXkY8BR9TgAkeJvF/7Sf/AAVs+MQ8IeD7Wfwv8K/D90JHSQsbXT4txAuLkg4nvGQnbEDgchdq75D+zj+zP8Lf2XfAMHgX4a6OsbSBX1PVJwGutTnC4Msr/nhBhVycAZOaqv8AgpT8Aj8B/wBqPxAul2PkeHfGedd0javyJ5zHz4h2GycSYXsjR9ARVV/wSX/aFX4R/tDr8Ntdv1h8PfEqNLA+Y5CxagmTav6Zcs8OO5lXniqqqqqqviz8Xvhz8DvBN78Qfih4otNE0WxGDLM2XnkwSsUSD5pJGwcIoJOCegJHxb/ai/af/wCClnjmb4F/s4+Gb/QPAZb/AE2IzeX50BOBPqVwuRHFgEiBcgnj96wXH7Hn/BPb4SfsqWFv4huIofFfxBkj/wBI8QXluALUkcpaRnPkrjgvzI2TkhTsH7YX/BSD4RfsxQ3nhLw9Nb+M/iHGCg0W0n/c6e/reSjOzHXylzIeMhAQ1fDX9l/9rj/gpP4ws/jD+0V4pv8Aw34CLb7KSWExB4GOTHptoeFQjaPPfg8HMpUivgh+z/8ACb9nXwhH4L+E3hK10iz4a5uMb7m9kA/1k8p+aRvrwOigDiqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqvGnhjwx408I6x4T8aafa32hatZS22o29yB5bwspD7ienHOe2M8Yr/gj9canY/tianpvh64kudKl8Magl4/IDW6TQmOQjI58zyh043n1qv+C43/ACNXwj/7B+q/+jLav2Tv+TV/g1/2T/Qv/SCGq8Nf8pg5v+yoXv8A6HLVf8Fz/wDmiX/cwf8AuOr9ij/k0X4Qf9ifp/8A6JWqqqqqqqqq/wCCkX/JkfxT/wCwfbf+lkFf8Ehv+TNNM/7GDUf/AENaqqqqqqqq/a3+Lb/E/wCKN1p+nXfmaF4aZ7KxCn5ZHB/fSj13MMA91RT3rw5411nw4yxQTefa55t5Tlf+Anqv4ce1eHfGWi+JECWs3lXWPmt5ThvfH94fT9Kqqqqq9tIr+znsZxmO4iaNx7MMH9DV3ay2V1NZTjEkEjRuPQqcH+Vf8E+/+RN8V/8AYTh/9FVVVVVVVVeHL7yLn7I7fJN0z2b/AOv0/KqqqqqqqqqqqqqqqqqqqqqqqqqqqqqrVtH0nXtPl0rW9MtNQspxiW2uoVljce6sCDXxL/YT+H/iVptR+H+pz+GL1yWFs6m4tGPoFJ3pk+jEDsteOv2UPjf4E8yefwk+s2cef9K0dvtSkDqdgAkAxzkoBVxbXFpM9tdQSQzRnDxyIVZT6EHkVVVVVpOiaz4gvE03QdIvdSu3+5b2lu00jfRVBNeAP2K/jJ4xaO41yytvC1i2CZNRbMxH+zCmWz7Psr4Y/scfCT4f+VfavZN4q1WPB+0akgMKsO6QDKjoPvbz70kccUaxRIqogwqqMAAdgKqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr9tv4zN8Bv2YPHfj+zuxDqw082GkHdhvtlyRDGy+pTeZMekZr/AIIwfBxfC/wT8R/GXULRVvfG+qfZbGRlyfsVnuTKntunecEDr5S56cVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVazrWj+HdMuNa17VLXT7C1TdNc3MojjjHqWPA7D618ZP27dpuNA+DViDjKNrd9F+sMJ/DDSe/yd68T+LfE3jXVZNc8Wa9e6tfS8Ga6lLkD0XPCr6KMAeleE9X/sTxBZ37PiIPsl/3G4P5dfwrgjg1Xj3T/wCzPGes2YUALdu6D0VjuH6MKr9jL/k37Q/+vm7/APR71VVVWWrXthhYZdyD/lm/I/8ArVp+t2l/iMnypv7jHr9D3qqqqqqqqqqr4jfDnwX8WvBOrfDv4g6Db6xoGtQGG7tJxwwzkMpHKurAMrDBVgCCCK+Evwi+HvwO8C6f8OPhj4dt9H0PTlOyKPLPLIcbpZXPzSSNgZZuTwOgAFVVf8FYf2fR8YP2bZ/HujWCzeIfhpI+pxMq/O9gwAvIwfQKqTH/AK4Y71VadqF9pGoWuq6XeS2t7ZTJNbTwuUeKRCGV1I5BBAIPqK/ZG+PVh+0n+z/4U+KMM8Tanc2ottbhjAXyNQiAWddo+6C3zqP7kietVVVVV+15+2/8J/2SPD23Xrldb8ZX0BfSvDNpMBNKOglmbnyIcgjcwy2CFDYbHww+AH7UH/BUb4hR/GT46eIbzw78ObaVktJo4jHEIgw3W+mQMSOwDTtkZHJkZdteD/BPwG/ZA+EkljokOjeCfB2hxedfX13ME818AGWeVvmllY4Azkk7VUdFr9of/gpB8Zf2m/FZ+AX7EPhvW4YNSZoW1m2iKalfqDhmi5As4O5kYh8YJMXIP7Hn/BKjwV8LZbP4k/tEvZ+NfGhPnx6S487TtOkPOXDD/SpR/eYbAc4ViA9BVVQqgAAYAAxiqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr9qa2+Md9+z/4zsPgBbyS+PbqxEOkeVeJayRlpEEjxyOQqyLEZCpJHzBeRWj/APBRf/goP8DLceHPjd8CZdbktvkF7rPh260+4kxwD5sIWGQdPmCc+pzmviD+2N/wUE/bK0Gf4W/Cr4I6hoOja3E1tqEuiaXcbriJwUeOW9nIjhiblSRs7gsQSK/4J4fsOt+yV4P1HxB41u7W++IHiqONdRNsd8OnW6ElbaN/4ySdztwCQoGQgZq/4LN/Dj4h+PPE3wsl8D+A/EXiJLOw1NbhtK0ua7EJZ7faHMattJwcZ64NfsvadqGkfsz/AAj0nVrG4sr6y8C6LBc2tzEYpYJUsYVdHRsFWBBBBGQRg1Xh34T/ABSi/wCCr8vjKX4a+Kk8Pn4k3lwNVbRrgWvklpMSeds2bDkfNnHNV/wWj+Hnj/x7/wAKd/4QbwN4g8RfYf7d+1f2Tpk135G/7Bs3+Wrbd2xsZxnafQ1+x5pWqaH+yv8ACnRta0260+/s/CdhFc2t1C0UsLrCoKujAFWB4IIyKqqqqqqqqr/goJ4G8cfEr9kH4geB/hvoV7rPiLVE09LSxswDLMq6hbPKBkjjylcn2Br/AIJu/DDxn8If2T/Dvgz4g+Fbvw9r8WoX813ZXShZBvuHKMQCeqBPwxVVVVVVVV+0n8SW+F3wh1rXrO58nU7tPsOmkHDCeUEBh7ood/8AgFVSO8brJE5V1OVZTgg+orw18ULyy2Wmvq11COBOv+sX6/3v5/WtO1PT9XtVvNNu454m7qeh9COoPsaqqqq+K+ljSvHmpxouEuXFwnvvALH/AL63V/wT7/5E3xX/ANhOH/0VVVVVVVVUrMjB0bDKcgjsasblb2ziuRjLryPQ9/1qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqvEvgTwT4yiEXizwlpGsADCm9skmK/7pYEj8K1/9jL4A65I80Hhe60qRzlmsL+RBn2Vyyj6AAVqP/BPr4dTOTpPjjxFbL2W4WGbHHqFTv8A571P/wAE79NaIrbfFe5STs0mjK4H4CYfzq3/AOCd+lKiC6+Kt1I4PzGPR1QH6AynHH1rTP8Agn/8K7fa2q+K/E94w/hikhhU/UeWx9O9eH/2SfgD4dZZYvAcN/Mp/wBZqFzLcZ+qM2z/AMdrRvD+g+HbX7D4e0Sw0y34/c2dskCcdPlUAVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX/BaH4lalrmrfDP9m7wyXuby/nbWbqzjOWlldjbWaj3JNzwfVa+Cvw20/wCD3wj8H/C7TFTyfDOjW9kzp0llRAJJPq77nPuxqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq+Mvxx8FfBPQxqPiO4NxqFyp+w6ZAw865I4zz91AerngdsnAPxf+Ofjv40aub3xNqBh0+FybPSrditvbjscfxPg8u3PPGBgCqrwNq/9s+GrWZ33TQDyZcnncvc/UYP41Xxz08Wnjc3SqMXtpHITjuMp/JBVfsZf8m/aH/183f8A6PeqqqqunIPIrS/ELwkQX7F4+gk6lfr6ikdJUDxuGVhkEHOaqqqqqqqv2oP2pvhl+yj8Pn8bfEC8e4vLotFo+i2zr9q1OcD7qA/dRcgvIeFBHViqt+wR+1T46/ay+F2qeNfHPw4PhubT9Ue2tb22DCy1KMliPI3kvuiGI3PKk4IOSyJVVVVe2VpqVnPp1/bRXNtcxNFNDKgZJEYYZWB4IIJBB9a/bA+Al3+zZ+0H4r+FxSRtLt7n7Xok8gP76wm+eE5/iKgmNj03xtVV/wAEhP2kf+Fa/GW6+B3iPUPL0D4ibfsHmNhYNUjU+Xj085MxnuWWIVVVVV+3V/wU00D4IvffCP4DzWniL4inNvdX6qJ7TQ5CcFccia4HaPlUbG/JBjP7IH/BNHxb8T/EQ/aJ/bZl1PUL7VJlvoPDWpTM1zeueVk1An5lXAXFvwcAB9oBjP7T37Y3wQ/Y58IwWmvSwXWuC0C6J4S0orHNKijamQBtt4BgDewxgEKrEba0HwN+2H/wVe8cx+KvGepP4S+Fmn3R+zy+U66fZgEgrawkg3dxjIaRjgcgsg2pX7Ov7Lvwd/Zd8Jf8It8LfDiwTXCr/aOr3WJb7UWHQzS4GQMnCKAi5OFGSTVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV+3745Op+NNE8AWs2YdEtDdXKg8efN90EeqxopH/XQ1VVWl6vqWi3Iu9Mu3gkHXaeGHoR0I+teGvibp2pbLTWgllcngSZ/dOfr/D+PHvQKuoZWBBHBFVVV+0Hpey/0nWkX/XQvA5x02ncv/oTflX/AAT7/wCRN8V/9hOH/wBFVVVVVVVVV4WucpNZs3Kneo/Q/wBKqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr4XD/hr/8A4K26t40f/TfDngTUZr2Df8yC30wLb2rJ2Aa58qUf77HrVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV+0N+0VoHwP0dbWKOPUfE+oRFrDTy2Ai8jzpcchAQQB1YggYwSPFvi3xH461+68T+KtWn1HUrx90s0rZx6Ko6KoHAUYAHAqqqvhRq/2bVZ9Hkf5LxN0YP8AfX/Fc/kKr9oXT90Ojaoo+60sLnHqFK/yaq/Yy/5N+0P/AK+bv/0e9VVVVVWlaxLp7iNyXgJ5X+77iopYriJZoXDowyCKqqqqqq/aS+O2ifs2/B3Xvi7ruh6lrEOkRqsVlYwszTTOwWMO4BEMe4jdI3Cjpliqn4A/s4fGn/gpl8Wp/wBo/wDaK1S6sPh/DdeVDFEGiF5FGxxY2Ck/u4EOQ8vUsWwWkLsvhnwz4e8GeHtP8KeFNHtNK0fSbdLeysrWIRxQRKMKqqOgxVVVVVf8Fgv2dE+IfwYsvjl4f04ya98Pn2X5iTLTaXKwD57nypCrj+6ryn3qq0nVdS0HVbLXNGvpbPUNOuI7i0uYW2vDKjBkdT2IYAg+or9kf9oLSf2m/gN4b+KVlJEuozQ/ZNctU4+y6hEAJkx2UkiRf9iRPWqqrm4t7O3lu7u4jhggQvLLIwVUUDJYk8AADJJr9sf/AIKO+Mvi74n/AOGav2KRqOoXOqz/AGG68R6Wp+0ag54aKxI5SPH3rjgnBKlVG9v2FP8Agmt4W/Z4isfip8YY7HxD8SSomgjOJbTQSef3RPEk47zdFPCdN7ftn/8ABU7RvA13c/CL9lx7fxT4zmk+yT69FH9ptLCVjtCWyjIup8nAIzGDj/WHKj9k/wD4Jf8Air4ja/8A8L7/AG2r7U9Q1LVJhep4avLlnubtzgiTUJM7lGAP3CkEDAYrgx1pOkaVoOmWui6Fplpp2n2MSw2tpaQLDFBGowqIigBVA4AAAFVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVePvH/gz4XeEr7x38QfEVpoegaZ5f2u/u2Ijh8yRY0yQCeXdF+rCvh78R/A3xY8K23jb4c+JrPX9Cu3kSC+tGJjkZGKOASAeGBH4VVVVVVVVfG/xW3jj4ueLPEpl8yO51OVIGznMMZ8uL/wAhotVVVVeG/G+s+HGWGKX7RaA828p4H+6f4f5e1eHvF+jeJIwtnP5dwB81vJw49ceo9x+lVVfGzS/7Q8CzXKrl9PnjmGBzjOw/hh8/hX/BPv8A5E3xX/2E4f8A0VVVVVVVVVWhziDU4STw52H8en64qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr9qL4p/wDCk/2efH/xQinWK60PRJmsWY4H2uQeVbD8ZpIx+Nf8EUPhWdL+HHjv4zahb/6R4i1SLSbF3X5hBbJ5krKf7ryThT7w+1VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV8efjVonwS8GPrd2EudWvN0WlWJbBnlxyx7hFyCx9wOpFeKvFWv+N9fvPFHijU5b/Ur+QvNNIevoAOiqBgBRwAABVVVVpt9LpmoW+oW5/eW8gcD1wen49KtbmG8tYru3fdHNGHQ+oIyK+Nun/bfAc84XJsriOYYHvsP6PVfsZf8m/aH/183f8A6PeqqqqqqtG1ZrCXypWzA55H90+ooFXUMrAgjgiqqqqqrVdK0zXdMu9F1rT7a/0+/geC6tbmISRTxOCrI6tkMpBIIPBBrRND0bw1o9l4e8O6VaabpmmwJb2dnaQrFFbxIMKiIuAqgAAAVVVVVVa5omk+JdE1Dw5r1hFfaZqtrLaXtrMuUnhkUo6MO4ZSQfrX7WHwB1T9mf47eJfhRfNNNZWU/wBo0e7lXBu7CX5oZM9CwGUbHG9HHaqr/gk9+06Pg38b/wDhU/ijUjF4V+I7x2sfmPhLXVBxbyc9BJkwnHUvGTwtVXibxN4e8GeHtQ8V+K9ZtNK0fSbd7i9vbuURxQRKMszMegxX7SP7XHxq/wCChPxGH7Mv7KmkahD4LuJCLqXmB9ViVhuubtyM29opwRGeWyNwLssa/s3/ALLHwE/4J8fC2/8AH/jTxDpn9ux2e7xF4w1FRGEU4Jt7ZTlkjLAAIuXlbbnJ2qvx+/bP+P8A+3z45l/Z0/ZL8PatYeEr1mjupoz5FzqMAOHlupc4trX/AGMgsDhixYRj9jD/AIJ1fDD9lm0tfFviAWvi74jvH+91maD9zpxI+ZLNG+5xlTKfnYZ+4rFKqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq1HU9N0i1a91XULezt0HzSzyiNR+JNeIf2ivhxopaGxvLnV5l422cPyg/774BHuM1rH7VesSMV0DwpZwKOjXkzTE/gu3H5mr/APaI+Kd4xMGs21mD/DBZxn/0MMal+MvxQmfe3jO+BPZdqj8gMV8IvE3iTU/h/p1/q2tXV3dSy3O+WaQuxAuJAuSfRQB+FJrmqoMLeE/7yg/0qPxNqCYDrFIPdcH9Kg8UwNgXFq6e6Hd/hVtqlhd4EFyhb+63yn8jVVVVVVVVVf8ABUf/AJMS+Jv/AHCf/TtZ1/wSf/5Mj8H/APYQ1P8A9LJaqqqqqqr4ha63hfwF4j8SRyBH0rSbq5Q5x80cTMPxyBRJY5JyfeqqqqqjkkidZYZGR0OVZTggjuDXhr4o3dpss/ECm5iHAuFH7xfqP4v5/WrDUbHVbZbvTrqOeFujIf0PofY1XiDTF1nQtQ0kgE3dtJEueMEqQD+eK/4J+qy+D/FiMCGGpxAgjBB8uqqqqqqqqonMUiSqeUYEfhSkMoZTwRxVVpX/AAUb/wCCgnxs+JV78NvgxpXhubVxNcfZ7HS9FjLCKJzubdcyOAAuAST+vNX/AMb/APgs34Fs5fEPiH4WvqtlbLukgTw/ZXZwOSfLs280jHXHav2Pv+Cq2gfG/wAZWHwj+NXhO28G+LtTnFtp17aSt9hvLg8CBkkJe3kZhtUFnDMduQSAfip4o1DwP8MPF/jXSYbea+8P6DfahbR3Klo3lhgeRQ4UglSVAIBBx3HWvhx+33/wUr/aQ1m+0X4J6Tod/eabCtxdwaXotsogjPyglrp2GC3vnPtxWt/tL/8ABX34O6dJ4u+IXwnj1XR7T95cmbw7BcxxRqMsZDYuHRMdWJAHqK/Yn/4KTeB/2qtWT4ceKdAXwj4+8h5be0W48211NEGXNu5wwkChmMTAnapIZgG2/tNfH/8A4KX+Efjr4s8N/BH4S6hq3giyuYl0e8j8JNdrMhgjZyJR9/8AeFxn2x2rxx+2/wD8FT/hp4cn8XfED4aP4e0W1dEmv9R8GmCGNnYKoLNwCWIA9zXw8/bz/wCCn3xb0SbxH8MfAKeKNLtrprSa80vwgbiOOdVV2jLKSAwWRDj0YetfCH9o7/gqnrvxZ8E6J8Q/g1qVl4V1DxFYW+uXL+DHgEFi9wi3DmT+ACMud3bGaqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr/gtN8T28OfAvwp8LbO42XHjLXGubhQfv2tmgZlI95prc5/2Py/Yv8AhgPg9+yz8NvAkluIbu30OK7vkxyt1c5uJge5IklZfoBVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVXijxLo3g7w7qHinxBeLbadpkDTXEh7KOwHck4AHckDvXxl+K2t/GPxzeeLtXZooSfKsLMtlbS3BO1B78kse7En0Aqqqqr4Xav9v0A6fI+ZbB9oB67G5X/ANmH4V4y0/8AtXwnq+nqu5pbOTYMfxBSV/UCq/Yy/wCTftD/AOvm7/8AR71VVVVVVXhzU+mnTv8A9cif5VVVVV+0p+0d8Pv2Xvhhf/Enx7dh/LBi0zTIpAtxqd0R8sMQP5s2CFUFj0wf+CdH7V/xy/aq0fxn4g+KHgOzsNCsdSzous2aGGF95JayVWyZTEAuZQf4gG55NVVVVVVV/wAFcP2Y/wDha/wYi+NPhfTTN4n+HSPJdiJcvc6UxzMD6+S2JR6L53UmqqGaW3lSe3leOWJgyOjFWVhyCCOhr9gL9p6H9qH9n/S9f1e/jl8YeHdumeJYhwzXCL8lxj0mTD5A27/MUfdr4j/EfwV8JPBWq/EL4heILbRtB0aAy3V1OeAOgVQOXdiQqoASxIABJr4ofFr9oP8A4KtfGNPhJ8ItOudB+GWj3Pn7boFIbeMcC8v3TIaU8+XCpOM4XJDyVbp+yx/wSu+BQSe48zVNQUFyqq+q+JrxVxwufkjBzgZEcYbqWb5tN0T9q/8A4K2fFBtY1e6fwn8K9EvCiMFY6fpi/wByJMj7ZeFDlmOMbhkxqyLX7P8A+zj8J/2ZvBEPgb4V+Hls4SFa+v5yJLvUZQMGWeXALN1woAVc4VVHFVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVXifxh4a8G2P2/wAR6vBZxnOxWOXkI7KoyW/AV41/ad1a8aSy8D6eljDjC3l0oeU+6pyq/ju/CtZ1/W/EV0b3XdWur+c9HnlL4+meg9hVVVfBf/km2lf9dLn/ANKJKqqq01i/s8LHOXQfwPyP/rVY+IrO6IjuB5En+0flP49vxoEEZBz6VVVVVVVX/BSnw7r/AIr/AGKPiNoHhbQ9Q1jVLr+y/IsrC1e4ml26paM22NAWbCqzHA4AJ6Cv+CYHhnxH4R/Y48J6F4r8P6loupQ3+pNJZ6jaPbTIGu5CpKOAwBBBGRyDVVVVVVVftTai2lfs/wDjO5SQIXskgyc9JZo4yOPUPj8aqqqqqqrSda1PRLkXWmXbwv8AxAchh6EdCK8NfEzTdU2WmsBLK5PAcn925+v8P48e9AhgCpBBHBr9jrShozfEOwVdqrrkbovorxb1/RhVVVVVVVVVWD+ZY275yTEufyqq/wCCWf8AyfpP/wBg/WP51X/BYn4d+Gfh7+0b4b8eeDbVNK1LxXo/23URaqI995DMVFzx0kZdmT3KbupJrxr4gv8Axb+xrr3irVYzHfaz8Mrm9uUZcFZJdMZ2GO3LGv8AgiH/AMlT+Jf/AGL9r/6UGq/bK8N6T+zp/wAFHbPV/hVYrpqprGka5b2FnGESG4kZGljRV6K7qx24x+8IAxgVX/BWD/kyPxh/2ENM/wDSyKv+CKn/ACav4p/7KBef+kFhVa9r2h+F9GvPEXiXWLPS9L06Fpru9vJ1hht41GWd3YgKAO5rxt/wWB/ZC8Ja1Jo+lv4z8WRxOUa+0TSI/IyDg4NzNCzD3CkHqCa+F/7d/wCzX8YPAPijx74L8ZzSJ4L0a41fW9JubVodQtbWCMySOIT/AKwADG6Msu4gZBNfs5/tVfCX9qfS9a1j4TXOrT2ugXEVvdyX9g1t+8kUsAuSd3C8+mR61VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV+3b/xkj/wUv8Ah98BIv8ASdM0N9L0u/hHzKFlf7XePjuRbyKCP+meDjmgqqoVQAAMAAYxVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX7dPxlk1TWYfg9oN3/oemlbjWGQ/62cjMcR9QikMR/eYd1qqqqqq+HOr/wBl+JYYZHxFejyWz6n7p/MAfjTKrKVYAgjBBFa1YHS9YvtMIwbS5kix/usR/Sv2Mv8Ak37Q/wDr5u//AEe9VVVVVVUrMjK6NhlOQR2NaZfLqFmk4I3j5XHoRVVVfELxrpvw38C694+1ew1K+svD+nzXs9tptq1zcSpGpYrHGvJY474A6kgAkfDvwD8aP+Cs/wC0Pc/Ef4jy3fh74ZeHH8lvsxJisYMhlsbUsMPcyAhpJSOB8xAHlRnwf4P8L/D/AML6Z4K8FaHaaPoej2629jY2seyOGNegA7k8kk5JJJJJJNVVVVVVVV7ZWmpWc+nX9tFc21zE0U0MqBkkRhhlYHgggkEH1r9uT9mq7/Ze/aA1vwRa28v/AAjWpMdS8NzsCQ9lKxxHnu0TBoj3OwNgBhVV+wd+1XcfsofG+18UarLdS+D9ci+weJbSBd7NATmOdV7vE+GHcqXUffrW9c/aH/4K3fHZdA0BL3wv8KPD9zv2yBmtdLhHHnTYIWe9dSQqA8biAVQO5+MH7QP7Of8AwS9+EVv8F/g9o9prHjaSIywaS8weV5nUf6dqUqYIz8pCDazABUCINy/s1/sU/Gj9u3xyP2mP2u9b1eHwtqDLNaWspNvcavCOUjt0GPs1njoygFgcp97zB4V8J+GfA3hzT/CPg3QrHRtF0qEQ2VhZQiKKBB2VRwOSST1JJJ5NVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVUzKqlmYAAZJPGK+Jv7Rdho7TaJ4F8q+vVJSS/YboYj/sD+M+/3f96tZ1vVvEN/Jqmt6jPe3cv3pZm3H6D0HsOBVVVVXwX/AOSbaV/10uf/AEokqqqqrT9Yu9PIRW8yLvGx/l6VY6ja6hHvgk+YD5kPVaqqqqqqqqqqqqqq/bPlkj/Z68QIjkCS4s1ceo+0Rn+YFVVVVVVVVeG/HWs+HSsCyfarQdYJW+7/ALp6r/L2r9lPWNG1+w8SanpU2Jpp7f7TbuMPGwRgCR6EDAP+yaKnHT9KwTwBWD6Vg+lVVVVVVaQS2mWxJ/5ZgVVfsG/GP4b/AAK/bGuPHnxV8SLoegrDqdq141rNcBZZGwgKxI7AEjrjA7kCvGf/AAVP/Yq8JaZLe2PxOuvEt2ibo7DR9Gunll9g8qRxA/7zil0/4v8A/BV39rOHxJ/wjN1o/grTWhs7q4jBaHQ9JjdnKGUgK9zJvkIHVmfoEQ7fjxY2mm/s6fETTdPgSC2tPBWpxQRIMCNFspAqj2AAFf8ABKn9oj4N/s9fEfxrqXxi8aR+HLTXNHgt7GeWznnWSVJSzKTCj7Pl5y2B2zmviZ/wVj/Y/wDA+i3Nz4V8Xaj441ZIz9n07StMnhDvj5Q006IirnAJUsR/dJGK/ZS+FXxb/b8/bCP7TvxI0KS18I6brUOp6jeCEpau1sF+y6fblv8AWY8uJXxkhAxYhnXdX/BWD/kyPxh/2ENM/wDSyKv+CKn/ACav4p/7KBef+kFhVf8ABVn47eN/i38etF/Y8+HV5M1hY3NlBe2UEhX+09XuyhhjfBwyRrJDtXs7uTkquP2fv+Ccv7M3wX8DWOieIvhp4b8d+ImgB1TWfEOlx3/nzHBbyopgyQxgjChQDj7xYkk/8FJP2I/BPwn+HeoftJ/s56WPBd3pcTaf4m0rSsxWt1p96ptZHjiHEZxMEdEARkdmIBUlv+CIf/JKviV/2MFr/wCk5qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqYqqlmYAAck8Yr/gnZGf2g/wDgoh8S/wBoO6zPZ6UNT1KwlK58t7ub7PboPQC1aYevy/Wqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqviv4/sPhf8Pda8bX7ITp9sTbxN/y2nb5Yk9eXKg+gye1arqd/rep3es6pdPc3t9O89xM5+aSR2LMx9ySTVVVVVUjvE6yRuVdDlSOoI71oOqJrWjWmpoRmeIFgOzDhh+YNfFvTxp/j7VFUALOyTLx13ICf/Hs1+xl/yb9of/Xzd/8Ao96qqqqqqqvD179mvRAzfJP8vPZu3+H41VVVeHfC/hrwhpx0fwpoGnaPYtcS3BtrC1SCMyyuXkfagA3M7MxPckmqqqqqqqqq/wCCkP7LB/aW+At1c+G9N+0eN/BYk1HQhGmZLpdo8+0Hr5iKCo/vonYmmUqSrAgg4IIxiqr4F/8ABSjxN8BP2UF+BPw68D2cXjhNTuBYa6tvH5UdvMd3nPEBme6DEou4bdqpu3bdrfsSf8E1dZ1/W4v2i/2xYbrV9d1Gf7fZeG9VdpZHlJJ+0ahuyXYnDCE8DjfnlAiJGixxqqqowqgYAA9BVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX9/ZaXZTajqN1Fb21uheWWRtqoo6kmvi38ctR8aSS6F4bllstDBKsfuyXfu3onovfv6Cqqqqq+C//ACTbSv8Arpc/+lElVVVVVDPLbSLNBIUdehFaTrMWoIIpSEnA5Xs3uP8ACqqqqqrVtZ0fQLF9T13VrPTrOMgPcXc6wxqScDLMQBk4FaXq2la7YRapomp2moWU2fKubWZZY3wSp2spIOCCD7g1VVVVVVX7ZFslx+zr4okYtut3s3UD1+1xLz7YY1VVVVVVVVXwn+Jer/Crxja+JtNLSwf6q+tQ2Bcwn7yn36EHsQO2RXh3xDpHizQrLxHoN4l1YX8IlglXuD1BHYgggjqCCO1VVUQD2ogHtWB6CiqnsPyoqp7Ctq4wAK2r6CiikYxWlqF063UD/lmOtVX7CvwW+G3x+/a+vvh38VfD7azoM9rqs7Wy3k1sRLGco4eF1bIJzjOD3Brw5/wTB/Yi8N3iX8fwZXUZo2DJ/aWsXtygx2MbS7GHsymvC3hHwr4G0O28M+C/Del6FpNmu23sdNtEtoYh/sogAH5V+0L/AMkB+Jn/AGJ+rf8ApHLX/BLv9mr4N/tL+OPHfh74x+FpdZtdJ0a3uLHytQuLRoJGm2swMLruyoxhsj2zzXhD/gmp+xR4MvI9RsvghY6lcxNkNrF/dX6H2MM0jREfVK0vStL0PTrfR9E0210+xtIxHb2trCsUUKDoqooAUewFV/wVg/5Mj8Yf9hDTP/SyKv8Agip/yav4p/7KBef+kFhVXUv2H/gsIr+Lii7vieiw+cox87AWmPf5ocd8471X/BSR0T9iL4ps7qo+wWoBJxyb23A/XAr/AIIh/wDJKviV/wBjBa/+k5qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/a5+IDfC79mP4m+OIZvKubDw3dpZv/duZkMMB/7+yJxX/BE/4f8A9j/BTxx8SJ4dk3ibxDHYxEjl4bOEEMPbzLmUfVT7VVVVVVVVVVVVVVVVVVVVVVVVVVVVftY/tLeHv2UPhBcfFbX9Fl1phf29hZabFci3a7mlJJUOQ23bGkr9DnZjjOR+xz/wUf8ACH7XXxD1P4bW/wAPbvwnqVlpTahbG51NboXapIiSIuI0ww8xWxzkbj2NVVVWh/trftN3v/BSOT4BXPxN3+A18eXWljSf7GsRi1RnCx+cIfN4Cj5t+73qqqq/b/8AiI0t3oPwtsZ/khU6nfgHqxykKn6DzCR/tKe1VVVVVVV8JNX8yC70OV+Yz50QPoeG/Xb+dftA6f5WvaZqYGBc2jRdOpRs/wAnFfsZf8m/aH/183f/AKPeqqqqqqqoEqQVOCOh9KsLkXlnFcA8svP1HB/Wqvr6x0yxuNS1K8gtLS0iaW4uJ5BHHDGoyzsx4VQASSeABXwZ/wCClE/x2/a/uvgn8N/htPr3w/e3aO0161ylxE8WTLfShyFW1bKoqnD/AHTyziMVVVVVVVVVVV/wVQ/ZSHwK+NDfE3whpYh8F/EKaS5jSJMR2Oo/euIMDhVfPmoOB8zqABHVV8J/iPrXwf8AiV4a+J/hyC2n1Hw1qMV7BDcxh45Sh5RgezDIyORnIIIBr4K/F3wj8ePhd4e+K3gi683S9ftFmWNmBe2lHEkEmOA8bhkYdMrxwQaqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq8u7WwtZb29uI4LeBC8ssjbVRQOST2FfGL4uXnxC1I6dpkskOg2r/uIiNpnYf8ALRx/Idh7k1VVVVVV8F/+SbaV/wBdLn/0okqqqqqqR2jYPGxVlOQRwQa0bV11CPyZiBOg5HTcPUVVVVVX/BWD/kyPxh/2ENM/9LIq/wCCXH/JiXwx/wC4t/6dryqqqqqqq/aO0v8Atf4FeN7QLu8vSJbjH/XLEuf/AByqqqqqqqqqr9lX41nwTro8CeI7zGhavKPs0kjcWdweAfZH4B7A4PHzZqqqqqqqqto/LtoYv7kYH5Cqr/gln/yfpP8A9g/WP51VftC/8kB+Jn/Yn6t/6Ry1/wAEQ/8AkqfxL/7F+1/9KDVVX/BWD/kyPxh/2ENM/wDSyKv+CKn/ACav4p/7KBef+kFhVf8ABWb9lXxnoXxAtf2vvhbYXUtuUt/+EkezQtLpt1bhVgvcDkRlEjUtjCNGCT8/HwG/4LEfAzX/AAXY2vx3i1fwv4rs7dEvp7bT2u7O+kAAMkXlAvGWOWKMoC9AzV+3J+2rrv7Yvw28Q+CP2fPCGtxfDTwgkWqeL/EOoQiEXQWVFghCjOxfNdGCk73Kg7VWNif+CIf/ACSr4lf9jBa/+k5qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/4LFeOT4Z/ZLj8KwzkS+MPElnZvGDjdDCHuWJ9g8EX4kV/wTv8AAyeAf2Mvhfpnk7JtS0k6tKxXBkN5I9wpPr8kqAH0Aqqqqqqqqqqqqqqqqqqqqqqqqqqqq/4LC/EDU/iT8Z/hr+zD4PlW5u7bZcz26E/Pf30iw20bYz8you4YHS479viZoVv/AME9v+Ci3hTVtD3WfhS2XTLkNtIEumzwC1vWx0PzLdMBzhgp6ikdJEWSN1ZWGVYHII9RVX9/YaXZTajqd9b2lrboXmnuJBGkajqWYkAD3NS/tb/srwamdHm/aP8Ahol0uQyt4pswqkHBUt5m0NkY2k59q0PxBoHijTIta8M65p+rafOP3V3Y3KTxSf7roSD+Brw1/wApg5v+yoXv/octePPi78KfhbFHL8SviX4X8LCZC8I1jV4bNpQM52LIwL9DwoPNeAvi78KfipBJcfDP4leGPFSwKGmGj6rDdtCD03rGxKfRgKqq+OvjNviB8XPFHihZA8NxfvFakHjyIsRxn8URT9Saqqqqqqrwpq/9ia/Z37PiNX2S/wC43B/IHP4V8ftPFx4ZsdRQZa1u9pOOiupyfzVfzr9jL/k37Q/+vm7/APR71VVVVVVVV4WuN0M1qTyh3KPr1/l+tV/wUl/a68VfFvxun7En7OIu9Tu72+Ww8RzaeSXvrrdj+z0YdI0IzM2QMqVJCo+79iP9j/wx+yN8Lo9EjFvqHjLW1SfxJq6p/rZQPlgjPUQx5IUH7xLMcbsCqqqqqqqqqrx7478I/DHwfqvj3x3rtro+haLbtPeXlw21Y1HQAdWYkhVUZLMQACSBXjXxP8cv+Ct/x/8A+EU8DxyeGfhx4VVpYTeFmt9MgJKi6uQp2yXUvIWNTwMgHaskh1/Rrnw7ruo+Hry4tJ7jS7uW1lltLhZ4XeNypaORCVdCRkMpIIwRwaqv+CU/7Xg+C3xNPwT8cassXgrx1dKLSWd9sem6oQEjkz2SYBYmzwCI2yArZqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr9oL4rHXr2TwPoF0f7Ns5MXsqHi4lU/d91Uj8WGewNVVVVVVV8GkKfDfSVYjO64P53EhqqqqqqOMda+N37VFp4Ink8OfDq4hvdcibE17gPDakdh2dvboK+A/xl0f42eB4PENn5cGqWuIdVsgebebHUDrsbqp+o6qaqqqq/wCCsH/JkfjD/sIaZ/6WRV/wS4/5MS+GP/cW/wDTteVVVVVVVWuaXBrmi3+iXJHlahay28mRn5XUqf0NX9jc6ZfXGm3sZjuLSVopUPVXU4YfmDVVVVVVVVVV+yx8bB460FfA/iS8Da/o8I8iSRvmvLccBs93XgHuRg8/NiqqqqqqrWLz7mKEc73A/Wqqvht+yD+zd8IPGp+Ivw3+FtjoniNo5YzfxXVxIxWX/WDa8jLz9Kqte0PSvE2h6h4b12zW703VbSW0vLdiVEsMiFHQkEEAqxHBB5r4PfsufAP4AalqGsfB/wCHNn4bvNVgWC8lguZ5TLGrbgpEjsBg88Yqqr4m/C3wD8ZfB914A+JnhyHXdAvZI5LixmkkjV2jcOhzGytwyg9e1fCL4JfCz4C+Grnwf8IvCFv4c0e8vnvp7WCaWUPcMiRtJmRmOSkUYxnHy/Wql8ll8mbYRLlQrfxccjHfgHivEv7CX7H/AIu1s+Ida/Z+8KG+eQyO1rbtaJI5xlmjhZEYkjJypySfU1/wUy8SfCT9nr9jLU/gx4K8P6D4duPG9zBaaToul2sdsoWOeKa4uDEgGQEiCF+u+RMk1/wR6+HWp+Df2UZvE2rWzxP408RXOo2gdSpNrGkduhwfV4ZWB7hgfeqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr/gtt4kutU1/4RfDDTCXnZL++eAPjzHleCGHjoOUmGT/ePTBrwb4btvB3g/QvCFk2bfQ9Nt7GI+qwxqg/RRVVVVVVVVVVVVVVVVVVVVVVVVVVVXV1bWVrLe3k8cMFvG0kskjbVRVGSxPYADNfsYWtx+2F/wAFKPEXx61WBp9F0G8uvEEXmxnCohFvp0RJ6OimJx/1wPSv+C1PwcOu/DHwf8btMtN1z4W1B9L1J0Xn7LcgGNm/2UmjCj3uO/b/AIJ7fGY/HD9k7wR4hvLvz9X0S2/sPVSW3N9otAI1Zj3Z4fJkPvJXxI8f+GvhV4C1/wCI/jC8Nto3hzT5b28cDLbI1ztUd2Y4VV7sQO9aPL+05/wVq+NOqaXP4sk8M/D3Q3E8lt8zWGjQMxEK+UpX7TdOAx3OQTh+UUBRp3/BFv8AZfg0UWOqeNfiLd6gy/PfR39rDhuPuR/Z2AXjo248nnpj9kL9k3wv+yD4B1jwJ4a8R3evLrGtSajJe3lusUoUxxxpEQpIIURk5GMljwK+Nfj3xV8MP+Ch/jrx14F02PUPEelePdQbSbZ4jIJLp5HjiGwcv87qdv8AFjHGa+H3/BJDS/iDZv8AET9rj4seL/EfxA19RcajHp19Gkdq7Afu2lkjkaVk+7ldiDGFUgA1+2B+w34+/YJ1bQ/j18CviHrd14ft9QSKLUSBFf6NcHOwTNGAkkUgBXdtVSTsZfmG79h79phP2qvgFpXxC1CK3g8R2E76Z4ht4FKol5EFJdAeiSRvHIB/CXK5O3NV8XvEreD/AIW+K/EsUvlzWOkXD27ekxQiP/x8rVVVVVVVVWtT/wDCVfBy8LHfcWUI3nuDEwbP4oM/jX7GX/Jv2h/9fN3/AOj3qqqqqqqqrw9N5WpohPEilT/P+lVZ/AP4Paf8Xbj48WPgHS4PHd1Ymym1iOMrI8ZxlioO3zCAFMmN5X5S23iqqqqqqqqqqtd1zRfDGi3viLxFqtrpul6ZbvcXl5dSiKK3iQZZ3Y8KoAJJNfHX4w/Fj/gqR+0BYfAz4LRXGn/DXQ7rz1mni2okSHZLqd3z0AYiOLIOGA++7V8V/jbDYaRp3/BPv/gn7p95qGl3kzWOv+IrJF+2+K7xuJtsoxtgwpDy5ClFwCsK5f8Aag/4Jb618A/2Y9J+Kui65N4g8UaEzTeNreBC0EcEm0K9sMbtkDDDs3LB2kOxV2iqBKkMrEEHgjjFf8Ey/wBsAftH/CYeBfGmqeb8QPA8EcF88z5k1Kz+7Fd+rMOEkPPzBWJ/eAVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVfHf4jN4I8Mf2bpdxt1fVg0cBU4MMf8cnsecD3Oe1E55Jyfeqqqqqqq+EH/JOtI/7bf8Ao56qqqqq+vrLTLObUNQuora2t0LyyysFVFHUknoK+O/7Ut54jNz4R+HFzJa6XzHcaiuVkufUJ3VPfqfYUSWOSck9Sa+AXxg1D4L/ABAtPEUbSS6Vc4t9VtV582AnlgP76n5l+hHQmrC+stVsLbU9Ouo7m0u4VmgmjbcsiMAVZT3BBB/Gqqqqqqqqqqqqv2qfB7+Dfjp4mthHtt9UuP7StztwGWcb2x7CTzF/4DVVVVVVVVVV4N8byaJ4gi8QeENbgOo6JdAM0EofypAASjgHuDgqexIr4U/EnR/ir4OtPE+lsscxHl3truybacAbkPt0IPcEe4qqqqqqrw/B52oCQjiJSfx6f1qqqqqqqqqq/bu/ZX+LX7TOieEZ/hD8Wo/Bms+DLy4vbaOQzQLczyIqpJ9phJeFowrgFUbPmt0wKX4Of8Fo/DiLoGmfFQ6naRjat4ddsZyQeCfMuEEx9cnmvhz/AMEofjL8VfHcXxG/bU+McmtHcpm0+z1Ga+u7lFPEL3MgCwR9RtjDYHCleo0HQdG8L6Hp/hrw5plvp2l6VbR2tlZ28YSO3hjUKiKo6AKAAKqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv24Ub4o/8FUvhR4AJMsGly+H7K4iGTtjN211KcAHny5TzjsM8Diqqqqqqqqqqqqqqqqqqqqqqqqqqq/4KWfGV/g1+yP4tuNPvfs+r+LQvh7TiDht1yGExB6gi3WcgjoQtfsU/H/8AaT/Zn8L65qnwc/ZsufGFp4xnieXWZtC1C5WRLfzEWON4CEKq7S56ncSD0wPjP+2x+2t8c/hZ4k+Evi/9jiddK8S2TW00sHhjVhJC2QySpuJG5HVHGQRlRmv+CK/xhbQvHnjb4BazO8Sa7arq+mRSEqEurf8Ad3CAf3njdGPtbmv+CvWu6jo/7G2o2FgziLWvEOn2d3tJH7oM0wz7b4Y/xxX/AARy8P6Tpf7JU+sWUa/bNZ8U3st7JwW3RpFGi+oAVAQPVie9VWi2NpqH/BX6S3vYEmjT4q3Mqq4yA8czujfUMqke4FV+3romk6/+xz8WrLWYUkgh8OTXUYbHE0BWaIjPcSRoa/4Ib6hqUuhfGHSpQ32C2u9HmgJBx5siXayYPTO2KLOOeme1V+2vrB0v4A6raLIVOqXtrajHfEgkI/KI1VVVVVVVV8OrtLmTUfC9w48rVbV0UH+9tIP5qT+Qr9jWN4vgFosUi7XS7vAwPYi4fNVVVVVVVVapr9r4U0u88UXsU0ttpFtJdzRwgF3SNS7BQSBkhSBkgZ7ivhv8RvBvxc8DaP8AEf4fa0mq+H9dt/PsbtEZN6hirAqwDKysrKVIBBUg9KqqqqqqqqqqlkjhjaWWRURFLMzHAUDqSewr9tf9qXx5+298WrL9kD9l3ztR8MnUBBfXls5Eeszxt80ruuQLGHG/djDFd/ICV8QviDafDHw7F+wV+xTFceIdc1+5Fn438XaagN14mvsFXtLZ15Wzj+dS2cFd3O0yPJ+wv+wv4Q/ZI8IDVNUFrrPxG1m3A1jWFTK2ynB+y22RlYgQMtwZGAJwAqrqOnWGr6fc6TqtlBeWV7C8FzbTxh45o3BVkZTwykEgg8EGv28P2UNR/ZR+NV3oFhBPL4N18ve+GbyQFswZ+e3Zu8kJIU9ypRuN+KqvgN8avF/7PPxW0D4s+CZ/9P0W4DS2zuVjvYG4lt5MfwOhI9jhhyoI+C3xe8G/Hn4Y6D8VfAl75+la7bCRUYjzLaUcSQyAdHRwykdMjIyCDVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV3dW1jaTXt5MsUFvG0ksjHARVGST7ACviL4xuPHfi6+8Qyl1hkfZaxt1jhXhB7HHJ9yaqqqqqqqvhB/yTrSP+23/o56qqqq8ZeNPDfgLQ5vEPijUo7S0hHGeXkbsqL1Zj6V8avj/wCJPixdtp9uZNN8PRPmGxV+ZcdHlI+8fboPfrVVX7CnxafxD4XvPhZrN5vvtAXz9NLnl7RjhkHr5bkfhIoHC1VVVVVVVVVVVV/wUA+H73mjaB8S7KIs2nyHTr4gdI3JeJvYBw4+sgqqqqqqqqqrq2hvLWWzuFLRTxtG4BIJBGDyORxXwfmuvhh8adU+Hd7Ofst+zQIXONzKC8D/AFKEj6vXwJ+Lt78JPGMd/I8kmi35WHU7decpniRR/eTJI9QSO9WF/ZarY2+p6bdR3NrdRLLBNG2VkRhkMD6EEVVVVVVeHbXyrMzsPmmbj6D/ACaqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqtCCfEX/gtveXcy+Zb6Pqc+4HHyGz0YxIQDkf62ND2PJPB6VVVVVVVVVVVVVVVVVVVVVVVVVVVX/BY/4pX3xA+N3gr9njwt5l3L4dtlnuLWI8y6hfFBFGR3ZYljIP/Twa+B3wy0/4MfB7wd8K9NKNH4Y0e3s3kUYE0qoPNl+ryF3Pu1V8d4Z/2IP+CmQ8eWUMltoY8QxeIIhGCPM02+yLuNAOw33UQGD9wcGv20PgvL+0t+y94t8A+GXiudUvbOPUdCdXG2W5gZZolVumJApjz0xJn3r/AIJc/tl+Fv2ddV8QfAD44X0nh3RdX1M3Njf3yNGml6gFEU0NyCMxK4jjG4gBGQ7sBiy2firwvqOiDxJp/iTS7nSCu4X8N5G8GDjB8wHbjkd+4rwl458F+P7G61PwN4r0nxBZ2V29nPc6bdpcxRzoFLxl0JG4BlyM8Zr4v+O9f+Ff/BRbxv8AFLw5o82qXHg74g3mqz2sSFvMt4bgtMCQDtUx7gW/hBz2r4NfH74SfH3whbeM/hf400/VbSaAS3FsJlW5sTj5kuIc7omU8HPHcEggn/gqz+2Z8PdP+Emo/s6fDzxTYa54m8TyxR602n3KyppdpG6yMjuuR5sjIqeXnIQuTjK7v+CUfwC1n4Mfs1jxH4r02Sy1z4gX39qtbyqVkhswgS2Vx2JXfL64mAOCCKr/AIKB3ip8LfD+n7kzNr6ygE8/JbzDj2+f+VVVVVVVVVabfS6ZqFvqFuf3lvIHA9cHp+PSv2Y7eG3+E9stsR5MmpXssOBj5HuHdf8Ax1hVVVVVVVVV7Z2+oWc9hdJvhuYmikXOMqwwR+Rrw14b0Hwf4d0zwn4W0u303R9HtI7Sws4F2pBDGoVEUegUAVVVVVVVVVVV/wAFL/21tc1nWX/Y3/Z2uLjUdf1eddP8S3emEvK0jkKNNhK872JAlI6f6vu4HiPU1/Zj8Ky/srfAj/ioPjP44Edh488RaSfOls3dsf2DYOmSSGws8iH5myvbEf8AwT//AGD9E/ZU8JDxZ4ztrHUvidrcP+nXiDzF0uFgD9kgbp1GXkGN545VQTVfti/syeH/ANqz4K6p8PL4W9trlsDeeHdRkXmzvUU7ckc+W4JRxz8rZxlVx4s8K+IPA3ibVPBvizSp9N1nRbuS0vrSdcPDNGxVlP4jqOD1HFVX/BM79s5v2b/iV/wrzx3q5j+HXjO5Rbp5nOzSb04WO7HZUIASX/ZCMT+7wQVZQysCCOCKqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv2kvGB0LwbH4dtJcXWuOUfB5WFMF/zJVfcE1VVVVVVVV8IP8AknWkf9tv/Rz1VVVfFr4z+E/hJpX2jVphdanOpNppsTjzJT/eP9xM9WP4ZNfEb4m+K/ijrbaz4mvi4UkW1rGSIrZf7qL/ADJ5NVVV8IvH918L/iPoXjW3ZzHYXQ+1RoeZbdvllX6lC2M9Dg9qs7u11C0gv7KdJre5jWSGRTkOjDII9iCK13xD4f8AC+ntq3iXXdP0mxjOGub65S3jU47u5AHAP5V4o/bc/ZG8Hs8etftEeB2kiyHjsdVS+ZSOCCtvvIOR0618M/8AgoP+yz8ZPilo/wAIfhn45vtb13XDOLRl0a5t4CYYGmfLzIhHyRvjjkrjuM+Jdbi8M+HNV8ST2F9fR6TZTXb2thAZricRoXKRRj78jBcKvckDvX/BQL9pzxN8R/2WvEmlaP8AAL4seDtNlurGQ+IPEemDTI4ttzGygAOzHeRtGPUGv+CYV3d3/wCwz8NLq+upbid/7V3SSuXZsardgZJ56ACqqqqqqqr4j+CdO+I/gbWvBOqACHVbVolcj/VSDmNx7q4VvwrW9H1Dw9rF9oOrW5gvdOuJLe4iYcpIjFWH5g1VVVVVVVVX7RsMnhX4m+G/G9ohDMiSHA+9JbyAn/x1kFRyRzRJLE4ZHUMpHcHoa/ZH+NZ0+6i+FPia8P2a5cnRppG/1UhOTB9GOSv+1kfxDFVVVVW1u91PHbx9XbH09TUcaRRLFGMKigAfSqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr9hfd4y/4Kq/GXxIS7LZTeJbuPfhGWM6lHAgIHBIWVR+uTVVVVVVVVVVVVVVVVVVVVVVVVVVVUf+CbfgfUP2qP+GqPFPxK1zW9VXxD/bSaTPZxLAkif8e8YbltsO2Lb3/diqr9sX9grwD+2LqvhrXvEXi3VPDmo+HLee2E9hbxym6hkZWVXD9NjK5GP+ejZ7V8KvA8vwy+Gvhn4dS+IbrXf+Ea0uDTo9QuowktxHCgRGcDjdtVQT3Iz3r9pr/gnJ+zx+03q83jDWLC/wDC3i2df3+s6G6RtdsAApuInVkkIA+8ArkYBbAAFv8A8EN9CXUDJd/tJX8tjxtgj8KpHKOmf3humHTP8HcenP7Kv7LHgj9kj4f3fw+8C6/rurWuo6g2oXc2qzRsTcNGkbMixooQFY0GOT8oyTXg7/gnj8PvDH7TXiT9pXVfGWo6/deJp9UkutBvbCE2Zjv1kSSJupdBHIyYYfMOtfFr/gi58G/GGu3etfC34k6z4Ehu5C/9mzWC6pbW5J5WINJHIFx0DO2PXHFfs+/8EkvgF8G/EVl4y8bazqPxE1jTpVltItRtktrCKRTlXNspYyEHs7sv+zVV/wAFCQT4M8JkDpqc3/oqqqqqqqqqvAHwz8Y/E7VhpHhLSXuCuPPuH+SG3B7u/QfTknHANfCjwDL8M/Amm+D59UGoTWgZpJxHsBZmLEAZJwM4BPYDp0qqqqqqqqqo/wDVp/uiqqqqqqqqqr/gpF+3FB+zP4H/AOFefD3U4m+Jfii2b7MykMdGtWypu2H984ZYweNwLHITDaZf3X7JnhY6nGZLz9oHx/Z5ticy3Hg3T7pT+8zncup3KuCvG6KJ85DSYH/BNz9ghPgNosPxs+LunCf4l65blrW1uAWOg28g5Qg/8vLg/vG6qDsH8Zaqqv8Agrd+xo3izRZP2pPhxpO/WNGgWPxbawJ811aIAqXYA5LxABX/AOme08CM5qq/4JR/tq/8LI8Lwfs2fEvVw3ijw3a48N3c7/NqVgg/1BJ6ywqAB3aMDjMbE1VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVXx98St4i+I9/DHLut9JAs4gDwCn3/wAd5YfgKqqqqqqqq+EH/JOtI/7bf+jnqqqvjp+0ro3w5Sbw34VaDU/EmCrjO6GxPq+PvP8A7A/Ejode17WfE+q3Gua/qU19f3TbpZ5myzH+gHQAcAdKqqqq/Y48dnxt8EtNs7mYve+G5W0ybceSiANEQPTy2Rfqhr9rT9mrSP2r/hI/wp1fxNPoEf8AaVvfR30NoLho3i3DAQso5V2Gc8Zrwv8A8EWv2aNLMc3ibxx4/wBclQ/NGt3bWsL9OqrCXHQjh+/418If+Cfn7KPwP8U6b468BfDaWLxJpLs9nql3q93cSRMUKEhGk8vJVmH3P4vpiv8AgrNcwQfsT+KYpZAr3GqaZHECPvMLqNsf98qx/Cv+CX9vNbfsK/DCKeMoxTU3AP8AdbVLtlP4gg/jVVVVVVVVX7dvwgl03W7b4vaLaD7HqQS11YIP9XcKMRyEejKApPqg7tVVVVVVVVVftZaesvhTRNU2jdb6g0IPp5kZJ/8ARYr4e3ral4D8O3znLzaXbl+P4vLXP65qKWWCVJoJWjkjYMjqcFSOhBHQ1+zj8Z4vip4UGn6vcIPEmjoqXqE4NwnRZwPfo2OjegYVVVVV4dsSiNfSLyw2xgjt3NVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVXinxZ4V8DaDdeKfG3ibSfD+i2Oz7TqOq3sdpbwb3VE3yyEKu52VRkjJYDqa8E/Ej4dfE3T59W+G/j7w54rsbWbyJ7nRNVhvo4pcBtjNEzBW2kHB5wQe9VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX/AASMQa5+1f8AGbxa0aRs+m3A2N8zL5+oK5Ab0/d8+vFVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVft/2TzfCXRL1CSLfxBGrKF6BoJuSe3KgfjVVVVVVVIjyOscalmY4VQMkn2FfBz9kbW/EvkeIPiSJ9I0w4aPT1+W5nH+1/zyU+/wA3svBrw94b0HwnpMGheG9Kt9PsbcYjggTaPcnuSe5OSe5qQAgEVVVVVVVVUi72VAeScDiqqqqqqqqqr9rP9pzwf+yn8ItQ+IviJorvU5AbbQdIMm19RvCPlT1Ea/edv4VB6kqDa+MtVjv9R/bL+O7x+I/GHia/ll8GaTfRBor+8jba1/NEePsNoQEjj+7JLGsf3IpQP+CaH7FGs6/q8f7ZH7RFvcanrusTtqHhqz1QF5HeQknU5w3VmJJiBHAxIOsZFVVVdWtrfWstle28Vxb3EbRywyoHSRGGGVlPBBBIINf8FE/2N7n9lj4rNq3hWzkb4eeLpZJ9EkALCwkzmSyZjzlM5QnlkI5LK1VXhPxX4i8C+J9L8ZeEdWn0zWtFu47qxvIG2vDKhBVh+I6Hgjg8Gv2Kv2r/AA7+1p8ILXxZA0Fp4p0kJa+JNLRv+Pa5xxIg6+VIAWQ9vmXJKmqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqtc1OHRNFv9YnI8uxtpJ2z6IpP9Kurme9uZby5kMk07s8jHqzE5J/OqqqqqqqqvhB/wAk60j/ALbf+jnqqmlhgheeeVI441LO7ttCgDkk9hivjv8AtVtMLjwh8LL0qnMd1rKHBb1WD0Hbf/3z2apHeV2kkcu7ElmY5JPcmqqqqq/4J++LXsvGviLwVLLiLVdPS7iVj/y1gfaQB6lZST/ue1VVV4w8D+CviForeG/H3hDRPEukvIsjWGsafFeQF1+6xjlVlyOxxxXhvwx4a8GaHaeGPB/h7TND0ewUra6fptolrbwAsWISOMBVBYk4AHJJ71VVVVVVVV4v8KaL458Mal4R8Q2wn0/VLdoZ06EA9GU9mBAYHsQDXxX+GeufCXxvqHgvXV3tbNvtrgLhbqBvuSL9RwR2YEdqqqqqqqqq/aegWb4YGQnmDUIXHHfDL/7NXwRmM/wp8OOQARalcD/Zdh/Sq8A+ONa+HXiqx8WaDLi4s3+eJiQs8Z+9G2P4WHHtwRyBXgbxponxC8LWHivQJw9tex5KE/NC4+9Gw7Mp4P5jgg1VVWm2D6hciIcIvLt6CkRI0WNAAqjAA7VVVVVVVVVVVVVVVVVVVXxV/aS/aC/Zd/4KIyeGfGfxf8V6h8P4vFUN1/Zt9qcj2n9kXpBwEYkEQpMyjPRoOoxmqr9vD9p/49+Lf21rD9n39nv4o+IfDsVm9hoPlaRfvDHc6jcOGkkfYednnJGc/d8pvetY/ZZ/4K4Ratew6J8fNbn05LmRbSWfxu6vJEGOxmA6MVwSPWv2i3/4KV/ss6JpGv8AxY/aA8UQWmuXT2tobHxbJcsZEXccgEYGO9fCX4X/APBVf42fDrRfij4F+PWvS6Fr8Ty2b3PjOSGQqkjRnch5HzI1fsifAr/got4G+Oui+I/2hvinf634It7a6W9spvFLXqvI0DrEfKPXEhU57da+OPxN0/4MfB7xj8VNS2NH4Y0e4vI43OBNKqHyo/q8hRB7sK8Dfts/te+DPEfg/wCKviz4v+ONX8Lt4g/eWt3qcj22pratBJdW+wnBHlzxgjgDzBjGONJ1XTde0qy1zR7yO7sNQt47i1uIzlZYnUMjr7FSD+NVVVVftc/tp/DT9jrS/D91480LX9YvfFJuhpdrpUMZDG38rzDI8jqEX9/HjAYnJ44r4WeOYfih8MPB/wAS7fTn0+PxboNjq6Wjy+Y1uLmBJhGWAG4qHxnAzjOBX/BUf/kxL4m/9wn/ANO1nX/BEv8A5ID46/7HA/8ApHBVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVTzw20ElzcSrHFEhd3Y4CqBkk+2K/4IfQvc+L/i7qU0RllWw0tWuGXcwLyXJILf7WzOO+32qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr9szRH1j4Aa5NEm99MntrpR7CVUY/grsfw/Cqqqqqqvh38LPGfxR1X+zPCeltKkZAuLuXKQWwPd3/9lGWPYGvhD+zl4L+FkcWpzRrrPiADLahcRjEJ9IU5Cf73LHnkA4qqYZXA9Kqqqqqqqq1TxF4c8JWL+I/Fuu2GjaPYFXvL+/uFggt03AbndiFUZI5JxzWh6/oXifS4Nb8Na3Yatp10u6C8sblJ4ZR6q6EqR9DVVVVVVVVXjLxh4Z+H3hTVfG/jLWLfS9E0S0e6vryc4WGJBknjknsAMkkgAEkCvjv8c7v9tD4zaz8avibLf6T8JfAwEFlp0cgWXyWLGCyh6r9su2jLO3OxEd/mSACv2Ev2WNc/bP8AixJ+0D8Y9Fjt/hv4Zmjt9M0iOIpa3ZgAEFhAh6WsChQ/Xd90lmaRhHFHDGkMMaoiKFRFGAoHQAdhVVVVXx/+Bvgv9o34U638KPHNvmy1WLNvdIgaWwuV5iuIs9HRvwILKeGIr43/AAZ8bfs//E7W/hV4/sfI1TRpyqyoD5V3CeY54ifvRuuGHcZIIBBAqv2WP2k/GX7LHxd0z4l+FWa5tARb61pTSFY9SsmI8yI+jDAZG/hdVOCMqfhd8TvBfxl8A6N8Svh9rEepaFrlsJraYcMvZo3X+GRWBVlPIYEVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVfHnVDpfwt1go+JLoR26e+913f8Ajoaqqqqqqqqq+EH/ACTrSP8Att/6Oeq8ReI9D8JaNca/4j1OCwsLRN0s0rYA9AB1JPQAck8Cvjl+0hrnxNlm8P8Ah1ptM8MqcGLO2W8x3lI6L6IOPXPGKqqqqqv2WNebw98fPCFyJdqXV21m4J4YTRtGAf8AgTKfqBVVVVVVVVVVVVVftIfAfT/jb4PMVmsUHiXS1aTSrphgMe8Ln+42Ov8ACcH1B1fSNT0HU7rRdasJrK+spWiuLeZCrxuOCCKqqqqqqqv2kERvhPqRZQSs9uVyOh81Rx+Br9n12k+EHh8uxYgXAGfQXEgH6Cqr9mz40P8AC7xSNJ1q6b/hG9YkVboMeLWXgLOB7DhvVeedoqOSOWNZYpFdHUFWU5BB6EH0qqggkuZVghXc7HAFafYR2FuIUwWPLtjqaqqqqqqqqqqqqqqqqqqqv+C2vweDQ+Afj1p1qMoZPD2qyAc4O6e1/AYugSfVRX7DXxbX42fsq/D3xpPcmbUYtKTTdTLNlzdWn7iRm9C/liT6SCviN440f4ZeAPEfxE8QOF07wzpVxqNyN20skMbOVB/vHbgDuSBX/BKrwDrPx6/a78T/ALQ/jRWvD4YW51W4uGBZX1S/aRUzn0Rrlx1wUX61X/Bb3/klXw0/7GC6/wDScV/wTe/5Mj+Ff/YPuf8A0snqv+C0HxlHhf4MeG/grptxi88b6kbu/UN0srMqwVh/tTvCQf8Api34ftLfskT+D/8Aglx8MdYj0sp4h8Ezx67rA2EyLHqhHnow9UaS0UkcAQH61/wSy+MI+K37I3h7Sr27Euq+BJ5NAugTz5UQDWxx/dEEkSZ6Ext6YFV8avjl8Mf2fPBFx8QPir4mh0jSon8qIEF5rqYglYoYx80jkAnA6AEkgAkeJ/8AgtbFe6tLZ/Cn9nHVNYsYTkXOpav5MzL2JhhikCf9/DX7P3/BXv4JfFbxFZ+Dvid4Xvfhvqd9IIYLy5vVvNPMhOAsk4VGiye7JsH8TDrWqagmm6Td6rs81bW3ebaGxvCqTgH3xX7e37cek/tof8IL/Zfw6u/Cv/CHf2jv8/Ulu/tP2r7NjGI027fs59c7u2OfhD/wWN8NfDD4S+CfhpP8B9T1CXwl4dsNIe7TxAkYuGtrdITIFMJ2htmcZOM9TX/BTK+XVP8Agn98QtTWMxi7t9GmCE527tUsjjP41/wTY/ac+Ev7Lf7Jvjjxn8UtbeH7V4zePTdMtUEt3qMq2duWSGPIGACCzMQq5GSCwBk/4LjwDX9sX7OTnRA+3e3icC5K5+/t+z7QcfwZ/wCBV+zh+018Kv2pvAv/AAnPwu1Sd0tpRBqOm3sQiu9OmIyElQEjkchlLKecHIYCv2g/+Csfwa+E/iuf4f8Awv8ACuofFHxDaTGCc6ddi3sllBwY0nCSNKwOc7EK543Zzjw9/wAFmJtE8QW2lfHD9mnXfC1jdYYXVpftJMiZwWEE8MW8ZzyHHToa+HHxH8FfFvwVpXxC+HniC21nQdZgEtpdQHgjoVYHlHUgqyMAVIIIBFftn/8ABQHTP2OPF3h7wtqnwtu/E6+INNe8iuYNWW18spIUZCpjfP8ACc57+1ftdf8ABT/4W/syeKpfhv4c8MT+PfF1mB/aNrb362lrpzEZEck2yQmXBBKKhx0LKeK/Z5+Ktx8cfgn4P+LV1osekTeJ9OW7exjnMywEsRtDkAt064FV+03/AMFAdM/Zp+PPhn4G6l8LbvXJfE9lZXUGpQ6ssCxLcXMtvgxmNidrQsevII6V+1F/wVZ+DnwA8U33w98GeHbr4heJtLcxagLW9W1sbOUcNE1xtcvIv8SohAOVLBgQPg3/AMFp/h34p8R22g/GT4X3fgyyupAi6zYaidRhgJPWaLykdUHdk8w/7PerC/sdVsLbVNMvIbuzvIVmt7iFw6SxsAVdWHBUgggjgg18Vviz8Pfgj4Iv/iJ8TvE1romh6eAJJ5iS0jn7scaDLSSNg4RQScHsCRr3/BZK913Vrq0+CP7MHiHxRp1q+De3d80chXnG6CCGUJng8yH6V+zF/wAFVvhB8efFtl8N/HHhi9+HfinUZvIskvLxbmyuZugiE+2MpIx4CugBJChixANfsx/8FKdD/aW+OT/BKx+Et9oM8dvdzf2hLrC3C/6P1HliJT8314qv2yf+CjeifsgfE/TPhpqXwpvfE8upaDFq63cGrrahBJPPD5e0xPkj7OTnP8XTitKvl1TS7PU1jMYu7dJghOdu5QcZ/Gvg7/wUk8HfE342eNfhD4k8Bt4OtfAlnqd1qevXutJLAkdjMsUjFfLXaDktnPGO+a8K/wDBXHwn8TP2hPC3wa+GnwpvLrRPEWvW+mL4h1LUvs8hWRwhlS1EbEDnI3SAkdQp4Hjfxx4R+G3hTUvHHjvxDZ6JoWkQma8vruTZHEvQe5JJACgEsSAASQK+If8AwWo8G2+ty6J8FvglrXi2NHKx32o332HzQOrJAkcrlSAcbihxyQOlfCn/AILQfDjXNfh8O/Gv4U6v4G8yURPqVpef2jDAT/FLGY45UUd9okPfHpoeuaL4n0Wx8ReHNVtdT0vUrdLizvLWUSRTxOAVdGHBUgggiv20/wBvPTf2Nda8L6Tqnwxu/FCeKLW4nimg1VbXyTCyKylTG+f9YpzxX7T3/BT34Mfs53UPhWx0a88aeM3tY57nSLC6SGHT96BljuLkhtjkEfKqOQOWC5Gfhr/wW38J6t4hg034rfBa88P6TPIFbU9J1b7e1uDxl4GijLKOpKsTjopPB8KeKvDnjrwzpfjHwjrFvqui6zapdWF5btlJ4nGVYdxwehwQeCARX7Q/7SHws/Zg8Bt4++KWsSwW8kvkWNjaxiW61CbGfLhjJAJwMlmKqo6kZGb/AP4LZazcXk1/4a/ZfnutBtnIee48RMsjAcksUtmSM7SOMtjrk5r9kH9rTw1+2B4A1Dxz4b8G614eGkah/Z93DftHJG0/lrIRFIh+cBXTOVQjcOOaqqqqqqqqqqqqqqqqqvFv/Iqaz/2D5/8A0W1f8EMf+a2f9wD/ANyNVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVfEfwwPGngDxF4TCgvq2l3FvFn+GRoyEb8G2n8KdGRjG6lWU4KkYINVVVVRRSzypBBG0kkjBURVyWJ4AAHU18HP2Q9V10QeIfieJtM084ePS0O24mH/TQ/wDLJT6fe/3a0Lw/onhjS4dE8O6XbafY24xHBbxhFHqeOpPcnknrVVVMMHAqqqqqqqq/4Kr+Ok8KfsqXfhuO42XHi/WbTTwgPzGONjcufp+4UH/eA718PPi58UfhLqJ1b4Y/ELxB4XunIMjaXqEluJcdnVSFcezAivhN/wAFj/2kvBSw2PxK0Tw98QbKMANLNF/Zt42PSWEeX+JhJ96+E/8AwV6/ZW8ftDY+NZNd+H9/JhSdVtDc2pY9lmg3ED/adEFeBvib8Ofifpa618OPHmgeJ7EgEz6TqMV0qezbGO09iDgg8Gqqqqqqv+CmH7UusftA/E2H9kb4M6rHJ4f0W7Y+I74TiO3u7uEF5PNk6LbWio7yMflDI7HiJWr4E/ArVf2wfixofwE+GFxd2Xwz8HM1xqesmDYzqxUXGoSIeBcXBRUijOdiJGhLCN3PgLwJ4T+GPgzR/h/4G0aDStC0K1W2sbSEYEaL3J6sxJLMx5ZiSSSSaqqqqqv+Civ7GFp+1P8ADD/hIPCNjEnxH8IwSS6NKAEOow/eeydv9rloyeFfuqu5q8s7uwu5rC/tZba5tpGimhmQo8TqcMrKeQQQQQehqq/4JwftvXP7Mnj5fAXjzUXb4aeKrpRelyW/se6OFW8Qf3MALKo6qAwyUAa3uLe8t4ru0uI54JkDxSxsGV1IyGUjggjBzVVVVVVVVVVVVVVVVVVVVVVVVVVVVVftSXnleB9OslYBrjU1Yj1Cxvn9StVVVVVVVVV8IP8AknWkf9tv/Rz18TPin4T+FWhnV/El5maQEWllEQZrlx2Udh0yx4H1IB+K3xf8V/FvWPt2uXHkWMDH7Hp0THyrcev+05HVjyfYYAqqqqqqrwDqJ0fx34c1YSbDY6tazhskbdkqtnjntVVVf8FSvG/jT4e/sm6h4j8AeMNb8NasmuWES3+j6hLZTqjOQyiSJlYA9xnBr9gLxP4l8Z/sffDbxN4w8RanrusX1jcNdahqV291cTsLuZQXkkJZiFUDk9AB2qqqqqqqqqq/ad/ZjsPi7YSeLfCcMNr4wtIsAnCJqSKOI5D0DgDCufZTxgrqOnX+kX9xpWqWc1peWkrRTwTIUeJ1OCrA8ggjpVVVVVVVftJyiP4U3yEZMtzAo9vnB/pXwChMHwi8PRlgSUnb/vqeQ/1qqr9kj41/2xYx/CzxNeZvrKM/2RNI3M0KjJhP+0gGV/2eP4eaihknkWGFCzscACtL0uPT4tzENMw+ZvT2HtVVVVVVVVVVVVVVVVVVVVX7cHwd/wCF5/st+PvAtraefqiaa2o6Sqpuf7Xanzo1T0L7DF9JDX/BEn4uGS0+IHwJv7xf3LReINLhLfNhtsF1gegItendj61/wWJ+MZ8Cfs32Hwy069MWpfEPVVhlRX2sbG12zTEY5wZDbIR0IdgfQ/8ABK74Lr8Jv2TdE1y+tFj1fx/O2u3LFfm8iQBbVc9SvkokgHQGVvWq/wCC3v8AySr4af8AYwXX/pOK/wCCb3/Jkfwr/wCwfc/+lk9V8fGH7a//AAVL0v4Y2zC+8NeHdUh0WUKNyfYrDdPf57DdILlA3Q5TrwK+JvgHRvij8OPEvw11xF/s7xLpNxp0+Fz5ayxlNy+hXIYehAr/AIJNeOdY+Cn7VnjT9m/xjIbWXX4riye2Yn5dU053OAD0zELoHudq1VftO694n/bu/wCCiFp8DLHWJ4/Dmka/J4d09IW3La29sSdQukB+VnPkzuD3VIxyADXwt+E/w9+C/g2w8BfDTwvY6Ho9hGqrFbxBWmYAAySt1kkbGWdsknqa/wCCs/7Jfg3xb8INQ/aK8JaBa6f4v8JPHJq81rCIzqtlJIsbmUAfPJGXVw552Bwc/Lt/4JKfHXWPi9+zdN4K8VXj3mqfDu+GmRzyvveWxdN9tuJ7riSIf7MSdTmv+C4GkaTpf/Clv7L0y0tPN/t/f5ECx7sf2fjOAM4yfzr9lPwx4auP2XPg7PP4e0yWWTwDobO72iFmY2EJJJI5Jr/gqMqr+wh8TVUAADSQABjH/E2s6/4JE/sreA/izd+IfjT8TNJttesvCeoJZaHpN4gltlvGjDy3EkTZVmVDCEyCMknGVUj4wfA/4Z/HPwBf/Dj4g+FrG/0y7tWhgY26+bYNtwstu2MxOnBBHpg5GRX/AAS01/X/AIQftz3/AMG7rUCYtag1TRNQhXOyWeyEkyyAdAV+zyAE9A7DvX/BUf49av8AA/8AZfvrTwrqMllr/ji9XRLW4hYrJbwujvcSKR0PloYwRyDKCORkf8Et/hR8CPhB8EtK+LXiXxJ4TX4g+MUkuJbm91KAT6dZlmWKCMMwMe5AJHwAWL4JIVcfHvSP2df2hvhdrfwx8c+N/Bd1b6lbSCzuZNUtnk0+52kR3MRLZV0Y5yOoypyGIP8AwRX+KWt6Z4+8f/ALUb3zNOm0/wDtu0hMgZYbmCWOCbZjg71liJx2hB9a/wCC3v8AyVP4af8AYv3X/pQK/wCCf/7G/wAPNM+FGk/Hr4u+FdN8YfEj4jx/29ealrdot0bVLo+bGI0kBVHZWDvIFDFpGXO0AVpul6Zo1jFpej6da2NnACIre2hWKOME5O1VAA5JPHrVf8Fpppbf9q3wnPbyvHLF4BsmR0YqysNQvyCCOhr9ij9k/wCH37N3wk0B7PQbG58bavYRXev67LEslzNcSoGeJJT8ywoSVVRgHG4jczE/8FhP2b/Ad58ILf8AaD8P+H7LTPE+hapBbapdWsCxHULWclB523G90kMe1zk4ZhyMY/4JKfETVPH37Hum6dq129zL4N1u70SKSQkt5KLHPGuT1Cpcqg9FUDtX/BQnxxfftOftz+Fv2XW8UrpPg/w1f2thdTvOIoop51WW8umLfKXjiby13DAMbY4c5+Gt5+zj8IPBmnfD/wCHHijwTomh6XEEgtrfVrcZIABd235eRsZZ2JZjySa/4LCeAvg3rnhXw38ePh/rvhqTxdDq66Zq50q9haa+t3ido5pQjZZomhCBuuJQCcKuP2Ivivqnxt/ZV+HfxD167+06td6Y1rqEzNlpbi1le3eRv9pzDvP+/wClf8Es/wDk/Sf/ALB+sfzqv+C1f/J1Hhb/ALJ/Z/8Apff14S/5FTRf+wfB/wCi1qy+HviX4x/tveLPgt4c1qfTE8eeOdS0zU5ol3EWS6g1xMSv8WwW4k28AmMAkDNfCz9mX4C/BjQdP0D4efCzw7p66cUeO8ewjmu5JUORLJcODI8gPIYnjgDAAA/4K5/GPxZ8TPj94d/ZY8I3EkllogtGnsI5MfbNVvADEHHQ7YpIguehmc9xX7MX7Mfw4/Zd+G2m+CfBej2h1IW6f2zrRgUXOp3GMu7vjds3Z2J0VcAdyf2/f2S/Bv7SHwV1/U4NAtU8e+HNPlvtB1SKELPI8SFzau4GXjkAK7TwrFWHTn/giv8AHfWtUtvF/wCztrd69zaaTbDXdDV3ybaMyLFdRjPRC8kLgDgM8h/ir/guN/yNXwj/AOwfqv8A6Mtq/wCCbn7KngT4W/A3wx8XtY0a01f4gePtPh1u71u8jE80ENyvmwxQu2TGPLdGcg5dySxIChf+Cqv7N/gP4gfs5+IPi9beH7K18Z+CVivIdSggVJrq2MqpNBK45dAjtIM5IZBjAZs/8EWviJqniP4C+K/h7qV288XhDX1exDknyYLqPf5Y7bfNjmfHrIfav+C3UPiwfGH4dz3ayf8ACMnw1Munkk7ftouW+1Y7Z8s2me/TPav2ZfiP8BviL8I9BP7P+paM3huwsIoU0qzKJLpuFAMM8I+aOQHOdw+Y/MCQcnw/4V8MeE7e4s/Cvh3TNHgvLl7q4isLRLdZZ2ADSMqAAuQq5Y8nAzVVVVVVVVVVVVVVVVVV4t/5FTWf+wfP/wCi2r/ghj/zWz/uAf8AuRqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv2kPBJ8A/GnxPokcWy1nuze2gA48qf94APZSxT/gNVVVXw2+EvjX4q6n9g8LaYWgjYC5vpspBb/7zeuP4Rkn0r4R/s8eCPhVDFfpAura8F+fUrmMZjOOREnIjHvyx7nHAqqqqkHII9Kqqqqqqqv8Agr78UovEXxd8MfCjT7sSQ+ENLa6vUXjZdXZU7W9SIY4WHoJT6mqqtB8Q6/4V1SHW/C+u6hpGo25zDeWF09vNGfVXQhh+Br4U/wDBUP8AbC+F3lWtx8QovGenxY/0TxRbC8Y465nUpOSR6yH1xXwr/wCC2fgPUhDZfGb4RavokvCvfaBdJfRE92MMvlug9g0hr4W/txfso/GNYYvBnxs8PLfTYVdP1WY6bclz/CsdwELn/c3D3NI6SIskbqysMqwOQR6iqqq/4KWftip+zV8KD4M8F6oI/iH41geHTTG/z6Za/dlvCOobqkfT5yW58thVj4W8R6fDp/wZ8HaXc6n8R/iDNbxapDEMy20UrK8Gngn7sjnZNOSflxFGShSZT+xt+yx4a/ZN+Dtj4F0/7PeeIb7bdeI9VjTBvbsjkKTz5UYOxBxwCxAZ2zVVVVVVX/BWL9h85vf2q/hVo56bvGmnW6dOgGoIo/AS49pD/wAtGqqr/glT+3Z5Dad+yx8XdadkYiHwZqlzJnZ6ae7Ht/zyz0/1fTYKqqqqqqqqqqqqqqqqqqqqqqqqqtX1XT9C0q81vVrkW9lp9vJcXMzKSI40Us7YGScAE8V4r/4Ku/sU+GtyWHxC1XxFKnWPStCuevs0yxqfwJHNeLP+C3XwhsvMHgb4LeL9YKj5Dqt7b6cGOO/lmfAzx345x2rTdQtNX0211WwlElteQJNC4/iR1DKfyIqvC3wl/bC8AeKNb139pn4kanr+kalI8Hh+zutda9EYSRiZfKBMcRKMnTkg4OMYqqqqqqqqpmVVLMwAA5JOMVdftGeGPhj8LdL03R5rfWPEUqT+Vaxybo7fMz4eYg8DuFHJ9gc14r8W+IfG+tz+IfE+qS319cHl3PCDsqr0VR2A4FVVVVVVVUE8ltPHcwsBJE4dCRnBByKqqr/grz/yZpqX/Ywad/6G1f8ABN7/AJMj+Ff/AGD7n/0snqqqqqqqqqqr9oX9mPw38abJtZ0swaT4sgTEN9sIS5AHEcwHUdAHwWXjqOK8b+BPFfw61+fwz4x0afTr+DkLIMrIvZ0YcOpx1HH4g1VVVVVV+1BP5PwyEYK/v9ShTnr0duP++a+CkAt/hV4cjBPNpv5H952b+tVVabqN/o+oW2q6XdyW15ZyrLBNGcNG6nIYH1BFfAT4nQfGnwjDqECxx6xZbYdUt1OBFJjhx/sOBkenI5KmtO0uDT48L88pHzOR/L0FVVVVVVVVVVVVVVVVVVVVVXgq2H7F/wDwVYXw5Ggs/D194maxiQjEY07VVBgGf7sRniyfWE56Gv2859T/AGtf+Civhv8AZ60C7kfT9Fms9A3wruEJf/SL64A9Y0dg3b/R/wAa0rS9P0TS7PRdJtUtrHT7dLe2gQYWKNFCqo9goAqv+C3v/JKvhp/2MF1/6Tiv+Cb3/Jkfwr/7B9z/AOlk9ftC/FW0+CHwQ8a/Fe6eMN4b0ee4tUkHyy3JGy3jP+/M0a/8Cr9i39iD4hftkr4u8bad8TP+ESj0S7iik1GazkuXvriYO8gBWRCCoCFiTz5q+9f8OZvip/0dh/5R7j/5Jr45fBj4hf8ABO39pvwVrM3ixfEt5ZNa+ILHVIrdrdbsLOyywOGZjk+WytycrIPWvDfiDSfF3hzSvFWg3S3OmazZQ3tlMBxLDKgdGH1Vgfxqv2PrpPh1/wAFUZdH8XzeVcnxX4h0t5ZW/wCXl1ukTk9d77VB7lx61X7fniTSfC/7HHxWvdYnjjju/D8tjCGYAvNcEQxgep3ODj2Nf8EOtA1G28J/FvxRLv8AsOoajpdnBkfL5sEdy8mDnrtuYs8en4f8Fz/+aJf9zB/7jq/ZO/5NX+DX/ZP9C/8ASCGv+Co//JiXxN/7hP8A6drOv+CJf/JAfHX/AGOB/wDSOCq/Y9/5S1L/ANjh4s/9JdRr/gt9pWozfDH4Za1E7/YLTXry3nUE7TLLArRk++2GXH1Nfsz/APBKn4B/Hv4DeC/i5P8AFLxpDdeI9NWa8gs2tfKhuFYpNGuYicLIjDkk8c1/w5L+AP8A0Vf4gf8AfVp/8Zr9ln/gnF8Kv2UPiVP8TvB/jbxTrOoT6VNpwg1NoPKRJHjcuPLjU7v3QHJx8x9q/wCC3v8AyVP4af8AYv3X/pQK/Z6/5ID8M/8AsT9J/wDSOKqr/gtX/wAnUeFv+yf2f/pff14S/wCRU0X/ALB8H/ota/4Kwf8AJkfjD/sIaZ/6WRV/wRU/5NX8U/8AZQLz/wBILCvjJ8JdD+I3/BT7xN8Jfibr1/pWm+K/GzwSXtsyiaNbmMPahDICvJkhUZB4PHav+HJfwB/6Kv8AED/vq0/+M1/w5L+AP/RV/iB/31af/Ga/Zy+A3hn9mn4SaT8H/COq6jqenaTLcSpdagUM0jTTPK27YqrwXwMAcAd6/wCCYEiab+3/ACWF+TBcSWutQJE4IYyKGYrj1ARj+FV/wWomhl/aq8NRxSo7ReAbNZFVslG+3XxwfQ4IOPQj1rwl/wAipov/AGD4P/Ra1+x7/wApal/7HDxZ/wCkuo1X7XN0nw4/4Kuw+MPFs2zTLPxl4a1d55WwptI0s2Y5PQKI3Xn+4e1V4u8SaT4N8Kaz4u16eOHTdE0+e9vJJGCqkUUZdySe21TX/BFvQNRv/wBpzxJ4gh3rZaV4NuEncDhnlubYIh54yEdu/wDq/wAa/wCC43/I1fCP/sH6r/6Mtq/ZO/5NX+DX/ZP9C/8ASCGv2/P+TNPiz/2L8n/oaV/wQ6/5FT4t/wDYQ0r/ANF3NfG74FfDH9ojwJc/Dz4qeHI9V0uZvMhcN5c9nMAQs0Mg5Rxk89CCQQVJB+Pf/BLX4x/s222ofGj9mL4rarqcHh6F7t7eORtP1e0gT5nMcsJCz7VXcQBGSBgKx4P/AATE/bY8W/tOeGtd+H/xVnhuvGXhGKKdNTjiWI6nZuSm90XAEqOAGYABhInGQxNVVVVVVVVVVVVVVVVV4t/5FTWf+wfP/wCi2r/ghj/zWz/uAf8AuRqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv2/vh015pGh/FCwti0mnsdO1BlGSInJaJj6APvX6yLVVUFvPdTR21rC800rBY40UszMeAAByT7V8HP2Qb/VfI8Q/FQSWNpw8ekRttmlH/AE1Yf6sf7I+b1KmtF0PR/DmmQaNoOm29hY2y7YoIIwiqPoO57nqTyaqqqqpxleO1VVVVVVV4i1/SPCnh/UvFGv3qWmmaRZy3d5cP0ihjQu7H2CqTXxo+JepfGT4r+Kvifqqsk/iPU5bpImOTBETiKLPokYRB7LVVVVVfDL9pT4+/Bp4/+FY/F7xPoMEXSyg1B3tTjput3LRN+Knr718MP+Czf7RPhbybX4l+EvDHjm1THmTKh0u7kx1+eINEM+0P+FfDL/gsX+y54x8m08eWPifwJdsB5kl5ZfbrVT6LJb7pD9TEor4cfHv4KfF+KN/hj8VfC/iSR03fZrDU45J0GM/PDnzEOOcMoNfFb4neEPgz8O9e+J/jrURZ6J4es2ublxjc+OEjQEjdI7lUVc8swHeviH8YPEfxo+IviX9q34pQxTvLfm38OaXKN8DXSKDBbqrcNBaxtHJICCHZolYfvyw/4JP/ALJV5pOlzftcfFe1mu/FHisSP4bN6S8sVtKT5t6xbnzZySFY8+Xk5Il4qqqqqqqryztNQtJrC+torm2uY2imhmQOkiMMMrKeCCCQQeDmv+Cjf7El5+zB8QT4z8EabI/w08VXLHTnUFhpVyQWazc9hgFoyfvKCOSjGqqGaW3lSe3leOWJgyOjFWVhyCCOhr/gmv8At1wftGeEY/hR8TNVjX4leHLXieVwDrtqnAnX1mUYEi9/9YOCwSqqqqqqqqqqqqqqqqqqqmmht4XnnlSOOJSzu7bQoAySSegxXir9pL9nnwPvTxd8cvAelSx8GC58Q2yS5zggR79x69hxWgft+/sjeLfHmi/DTwn8YbPWfEOv3qWdjb2mn3TRvK+doMxiEQyRj72cketV8bvFnizwF8HvGfjjwLpFvquv+HtDutQsLCeJ5UuZIY2k8sojKzFgpACnJJGM9KH7V3/BXj4ssG8DfBjUPDcc2BHNaeCGt4epGVk1HzFPPB5IGOcc14N+CH/BYPxn4v0PxL46+JOp6HZ2Oo211PaXXiyG0iuI0cMyPDYFlKsq4KMuDu5HWq1TTbPWdMu9I1CES2t7A8E8bDIZHUqw/EEivCn/AAS1/Ym8LNHNL8KJtbuIiCsurazdzZ5zzGsixnsOV7fXPhP9mP8AZz8ChD4R+BfgTTJUxieHQLfzjjpmQoXPU9T3PrSIkaLHGqqqjCqBgAD0FV+1ZCraDoNxtJKXcqg+mUB/9l/Sqqqqqq4uba1j825uI4kHVpGCgfiav/H/AITsMq2qrO46LApkz+I4/Wr/AOL1soK6Zo8j+jTyBcfgM/zq/wDiZ4qvQVhuYbRT2hiGfzbJq91TUtRbdf6hcXBz/wAtZC2Ppmqqqqqqqqqs7Y3l5BZq4UzyKgYjOMnGaqqr4p/CT4dfGzwlJ4F+KPhiDX9ClnjneymlkjUyIco2Y2VuCfWvAHw/8H/CzwfpvgDwBokWkaBo8bR2VjE7usKs7OQC5LHLMx5J61VVVVVVVVVVXxG+F3gj4raG2g+NdEivYgD5E4Gya2Y/xRuOVPA46HGCCOK+M/7HHj/4cm41rwkknijQE3MXt4v9KtkHP7yIfeAH8SZHBJCiiCDtI5HY1VVVVX7WFyE8FaRZ7hmXVBJjHPyxOP8A2evhtamz+Hnhm3IIZdJtiwbsTEpP6mqqC3nu547W1heaaVgsccalmZjwAAOSfavgv+xL4w8Ym3134ltP4b0ckMLLbi9uF/3TxCD6sC3+zzmvBfgbwp8PNBg8NeDtFt9NsIBxHGMs7d2dj8zse7MSaqqqqqqqqqqqqqqqqqqqqqq/4LU/Ci50nXPh5+0RoMbQSndoeoXMS7THNGWuLRsj+Ig3IyecRqPp/wAEkvBGs/GL4/8AxL/ay8cxLcXkEs0cExXCnUb92lndPTZECmM8LOB9Kr/gt7/ySr4af9jBdf8ApOK/4Jvf8mR/Cv8A7B9z/wClk9f8FqPjK+gfDDwj8D9Lvdtx4rv31PU40PP2S1wI0b/ZeZww97ft3/4Jy/B4/Bn9kbwTpV3amDVfEkB1/UgRgmW7AaMEdmWAQIQecoenQV/wWS+DR8bfs+6R8WdOtw9/8PdUH2lgOfsN2Uik6cnEy2x9AC5r/gk78ZW+KX7KeneFtRuxLq3w9vX0aUMTvNrgSWzH/ZCOYh/1xP41/wAFMv2I/iRH8R1/a2/Z70y+vL5ZILrXbHSkY3lndwbfLv4EXlhhEL7fmVl34ILFfg7/AMFqPBFr4Os9M+Pvw38UHxRZRiG6vfDkNvNDesvBlMU0sRhY90G4ZyRgHaP2gP2jfjz/AMFP/GukfBn4FfDfUbDwfp10tw9tJJuBlIKi6v5lGyKNFLbUBPJOC7FQP2VP2d9A/Zd+Ceh/CbRblL25td1zq2oCPyzf3smDLLjsOFRQeQkaAkkE1/wWZ+DHifx78HfCPxO8M6TPqK+AdQuhqUdujO8FpdpHvnKjqivbxBj1UPnoGI/Zi/4K7eB/hX8EPCvwu+JXw48RX+q+FNPj0y3vtMmhaG4toVCQlg7KyFYwqEANnbkddo/4Kj/8mJfE3/uE/wDp2s6/4Il/8kB8df8AY4H/ANI4Kr9j3/lLUv8A2OHiz/0l1Gv2r/2edF/ag+B2v/CbU7iO0vLpVudIv3XcLK+iyYpCBk7Tko2OdjuByQa+AH7VP7QH/BNDxHqPwN+OHwvvtQ8Lz3b3ENjLMYXhckB7ixnw0c0LgAlOhbBDId+65/4LXfs0LYGSz+G/xNlvAg2wy2VjHGW7jeLpiB152/gK/Yx/ayi/a+8B6947h8B3fhWHS9bewtbee4Nx9ogEUbLL5mxFLFmkUqoO3aOTnNf8Fvf+Sp/DT/sX7r/0oFfs9f8AJAfhn/2J+k/+kcVVX/Bav/k6jwt/2T+z/wDS+/rwl/yKmi/9g+D/ANFrX/BWD/kyPxh/2ENM/wDSyKv+CKn/ACav4p/7KBef+kFhX/BUX9iTx18QtdsP2mvgRpV3feJ9Kgji13TtPyLudYTmC8gC/M8qABGVfmKrGVHytn4Jf8Fn7DRPD1v4Z/aS+GWuz69pafZ7jVfDyxFrt04LS2szxCKTj5trkFskKo+UeLv+C2HwkW3W2+GPwX8a63qc+Ehh1eW3sFLscADyXnZu3AAJPHHWtNv4NV0211O2SVYruBJo1ljKOFZQQGU8g4PIPSv2v/hf8Vv2Fv2y/wDhoLwJpcq6BqWvya3oGomJmti8+5rmwlI+79+aPbkFomBB64f/AILa+CbvwzHHo3wG8R3Hi+4QRw2B1GI2hmbgASqpkYZxgCIE9OOtftp+Bv2g4dd8PfHT9pN0s/FXxUS5u4tG8oxPpdrb+UkMTIf9V8rjEZyyhQXJdmA8IOknhLRJI3VlbTrcqwOQR5a8iv2Pf+UtS/8AY4eLP/SXUar/AIKh/sP+If2h9E074v8Awm01b3xt4XtGtbzTEwsmq2IYuqx9jLGzOQpxuV2AJIVT+zR/wVquvg94Ot/hJ+1H4A8V3+q+FkFlDqenwxi+aOMBUiure4eL94gAUybssMbl3As37U3/AAUL8f8A7atgn7OX7L3wx8Swaf4hcLqHmRrJqOpRAg+V5cJZIIc4LsXbIABKruDf8E+f2PD+yT8Jri18Sy2t1448Vyx3evTQYZLYIpEVojj7yx7nJboXkfGVC1/wXG/5Gr4R/wDYP1X/ANGW1fsnf8mr/Br/ALJ/oX/pBDX7fn/JmnxZ/wCxfk/9DSv+CHX/ACKnxb/7CGlf+i7mv2pv+Ckmj/sqfHu0+F3ij4W6trPh2XRobq41Wzl8qdLh2fKwpIBHOgQRZO9cMxGeMV8Zv+Cy3wXuvhzq2mfCPwV4tvfE2p2MkFo2sWkFtbWbuu3fIUldnK5zsUYYjG4A5r/gj3+zB46+G+l+Jfjr8QdDu9FbxTZRafoVldxtFNJah/MkuGjPKo7LEEyASFZvuspNVVVVVVVVVVVVVVVVVPBDdQSW1xGskUqFHRhkMpGCCPQiv+CHk8kHiv4u6c8zRl7DS3aBmxkpJcjJX1XeRntu96qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqrx14Q0vx/4O1fwZrCg2urWjwM2MmNiPlce6sFYe4FeK/DGreC/EupeE9dt/J1DSrl7edB03KcZB7qRgg9wQar4ZfB7xt8V9R+y+GtO22kTYudQuAUgg9i38TdPlXJ57DmvhN+z94H+FEMd5aQf2nrmzEmp3KDcCRyIl5EY+mTzyTVVVVVVRAIxRHaqqqqqqv8AgpbcfGe6/Z4l8I/CHwD4g16LxDd+Vr93pFm9y1nYx4dlZYwXAkbYCQMbFkB4NXVrc2VxJZ3tvLBcQuUkilQoyMOCCDyD7Gqqqqqqqhmlt5Unt5XjliYMjoxVlYcggjoa8QfHb40+LfBKfDjxV8VvFWteGYrlLiPS9R1Wa5hSRAQpVXY4A3HCj5cnOM814b8c6BN4t8GzfEjw7Jq3hDww0Mc2h6dP9lNzAr+ZKvmMGIeVyzOx5+bauxVQL8Kf+CsP7HHiyys9J1TVdX+H8kUawxWmraS32eMKMBEktvMRUAAALbAAO1eBPi18Lfilafbvhv8AEbw34nhC7mOk6pFdFB/tBGJU+xAIqqqqqqqq+KXwx8F/GXwDrPw0+IOkR6loWuWxguYW4Ze6yI38MisAysOQwBr9rH9l/wAa/so/Fe8+H3idJLvTJ91xoWsCLbHqVoTww7CReFdOqt6qysarwV408U/DnxbpPjrwTrVxpOu6HdJdWF5AcNFIp4ODwQeQVIIYEgggkV+xH+2J4U/a5+GMerI9vp/jTRESHxHoyvzFJjAuIh1MMmCQf4TuUk7cmqqqqqqqqqqqqqrVdZ0fQ7U3ut6tZ6fbrnM11OsKDAyfmYgdBXhv4u/Cjxn4gm8JeDvid4T13W7W2a5n03TdZt7q4ihV1RpGijcsqB3VSxGMsB3qv2tP2l9L/ZP+FK/FPWPCGoeI7dtThsPstnOsJRpEkZXZmBAXMYXoTlhxWr/8FrfiD4iu/wCzPhl+zbZC6c4hS71ibUHfnAPlwwxkZyvAJ57nNfsn/tc/t6/Gj48+GtO+KvwKuPD/AMONQFyuoXtp4RvbOCEi2kaFvPuGfgypGDhv4/oKr9qf4Fp+0p8B/E/wY/tuLR5NfW2MOoSWxnFs8NzFMG2BlJz5W3qPvGvCn/BEL4U2aoPHHxw8V6uQfmOladb6dnp08z7Rjv69q+Ff/BL/APZL+EvifRfGmj+G/EGqa54evob7Tr3UtcmzDcQyCSN9kJjRsMF+VlKnGCCCc1VVVVVV+0/Ym5+H9rdoObTU42b/AHWR1/mVqqqr3VNN05d1/qFvbjH/AC1kC/zq/wDiX4Vs8rDcy3bDtDEcZ+rYFX/xeuGBXTNHjT0aeQt+gx/Or/4geK7/ACraq0C9lgUR4/Ec/rVxc3F3J5t1cSTP/ekcsfzNVVVVVVVVVVVXwt0t9b+JnhPR0QsbzW7OIgehmUE/lmqqqqqqqqqqqqqqqqvil+zJ8Jviu0t9q2h/2bq8gJOp6biGVm9XGCsnblgT6EV8Q/2GPij4YeW78F3Vp4qsV5VIyLa5A943O0/8BYk+g6Vr/hnxF4VvTpvibQdQ0q7Gf3N7bPC/1AYDI96qqqv2uLzEfhjT1bqbmVhz28sD+bVo9n/Z2kWOnlQDa20cWAMY2qB2+leGfBfi7xpeDT/CXhrUtXnzylnbNLs92IGFHucCvh3+wb8QNdeK8+IWr2nhyzJBa2gIurojuPlPlrkd9zY/u18MP2fvhb8I0Sfwr4eR9SC7W1O9PnXLcYOGIwgI6hAoPpVVVVVVVVVVVVVVVVVVVVVVVVrWgaF4ksxp/iHRbDVLUOHEF7bJOgYdG2sCM8nn3rRPDvh/w1aNYeHND0/SrZ5DI8NlapAjOQAWKoACcKBn2HpVVrnhjw14nhit/Enh7TNWjgYtEl9aJOEYjBIDg4OK03S9M0axi0vR9OtbGzgBEVvbQrFHGCcnaqgAcknj1rW/BPgzxNcpe+I/COi6rcRx+Wkt9p8U7qgJO0F1JAySce5qOKOGNIYY1REUKiKMBQOgA7Cq1DTtP1ayl03VbG3vbS4XbLb3EQkjkHoytkEexrQvCnhbwusy+GfDWlaSLnb5wsbKO38zbnG7YBnGTjPTJ9aqvFXwD+BXjrUW1fxt8FvAfiG/dizXWq+HLS7lYnqS8kZOfxrQPDXhzwppqaN4W0DTdHsIiSlpp9olvEp7kIgAHQdqplVlKsAQRggil/Z0/Z8XW28Sp8Cfh4NYaTzG1AeF7MXBf+8ZPL3Z4HOa1XSNK12wl0rW9MtNQspsebbXUCyxvggjKsCDggHkdQDWieG/Dvhm3e08N6BpulQSvvkisbRIFZsY3EIACcADPtVWfgTwRp2r/wBv6f4N0O21Te7/AG2HTokn3OCHbzAu7LBiCc85OetV4i8LeGfF+nHSPFnh3S9asWbcbXUbNLmIkd9jgjP4Vpv7MX7NejXaX+j/ALPPwzsbqM/JPbeErGJ1wQeGWIEcgH8Kt7a3tLeO1tII4YYlCxxxoFVAOgAHAHtWueDfB/ieWKfxJ4U0fVpYFKxPfWMc7IDyQC4OB7Vb21vZ28VpaQRwwQIEiijQKqKBgKoHAAAAxVVrfgfwV4mukvvEfhDRNVuY4xEk19p8U7qgJIUM6kgZJOOmSfWkRI0WONVVVGFUDAAHoK1bRtH1+xfTNd0qz1GzkIL293As0bEHIJVgQcEA1onh3w/4atGsPDmh6fpVs8hkeGytUgRnIALFUABOFAz7D0qvGPwU+DXxEuxf+P8A4SeC/E1yCMTaxoFtevwMD5pUY9OK8IfBz4RfD2YXHgH4V+D/AA1KBgPo+h29k3fvEinufzNVrGjaP4h02bR9e0qz1KwuV2zWt5As0Uo9GRgQR7EV4T+CHwW8BaidY8C/CDwT4dv2JJutJ8P2tnKSep3xop/Wtd8IeEvFDwyeJfC+kas1uCIWvrGOcxg4zt3g4zgdPQVDDDbwpb28SRxxKFREXaFAGAAB0GO1WfgTwRp2r/2/p/g3Q7bVN7v9th06JJ9zgh28wLuywYgnPOTnrVV4z+D/AMJPiPMlx8Q/hb4R8USxrtR9a0O3vWUDsDKjECvCHw/8BfD6xOmeAfBOgeGrNsbrfSNNis4zjp8sSqKrXfCHhLxQ8MniXwvpGrNbgiFr6xjnMYOM7d4OM4HT0FWlnaWFpDYWFtFbW1tGsUMMKBEiRRhVVRwAAAABwK1DTtP1ayl03VbG3vbS4XbLb3EQkjkHoytkEexrQ/Cvhfwuk0fhnw5pekrcEGZbGzjtxIRnBbYBnGT19a8Q+GPDXi3T20jxV4e0zWrFjlrXULRLiMnGMlHBHQntXh34BfAnwfqn9ueEvgp4D0TUt4f7Zp3hu0tptwyQ29Iw2ck8571VVVVVVVVVVVVVVVVVVX/BIjOj/tTfGfwywEpj06bMwO3/AFN+E+77789eMd81VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVftw/A261lbT4s+E9NkuLxPLs9Xt7eLc8qkhYZQo5ZgSIz1OCnZTXwe/Y+ub4QeIPisXtoDho9HhfEjj/ps4+4P9lefUr0rSdI0rQdOg0jRNPt7Gytk2w28EYREHsBVVVVVVVU4AbAqqqqqqhhkuJUhiXc7nCirG0jsbWO2jwdo5OOp7mvip+zn8CvjfA0XxV+FXh3xFKybBd3NmFukXGMJcJtlQY/usOgr4t/8ABFr4L+JBPf8Awf8AiBr3g67YEx2WoINTtAeyjJSVR2yXfHXB6H4sf8Epf2vPhoZ7rRPCmneOtNhBYXPh28Eku3tm3lCSlvZFf6nrXibwl4q8F6pJoXjHwzquhalD/rLPU7KS1mTtykgDDkHtVVVVVVVVZX17pt3Ff6beT2t1A26KaCQxvG3qrDkH6V8Nv+CgH7X/AMLPKh8P/HDXdQtIsAWmuMuqxlR/CDcB2Qf7jLivht/wW2+IuneVa/Fn4OaDriDCvd6HeyafIB/eMcnnKx9gUH0r4cf8Fcf2QvHBitvEWs+IfBF0527db0tni3e0lsZQB7tt98V4C+M3wi+KkKzfDb4neF/E2U3FNL1WG4kQf7SIxZT7EAiqqqqqr9rL9l7wT+1f8KLz4f8AidI7TVLctcaDrAj3SabdhcBh3aNvuunRl54ZVZfir8LfGvwW+IGs/DL4haQ+na7oVwYbiI8q4xlJI2/ijdSrK3dWBqq+A/xz8ffs6fEzSvij8OtSNvqOnvtnt5GPk31u2PMt5lBG6NgB7ghWGGUEfs3/ALRHgH9p74W6d8TvAN3iOb9zqOnyODNpl2qgyW8nTkbgQ2AGUqw4NVVVVVVVVVp+saRqxnXStVs702r+XOLedZPKfAO1tpO04IOD2Iqv2uP28fhv+x9qeiaH408G+KdavvENpLc2TaZFAINsbbWV5JJFYNkrwEYYbOe1eI/+C395cz/YvAf7OYd5H2wy6j4hLs5Jwo8mKDqeOA55OPc/sF/td/Hb9p3WvF9v8XvhNbeE9N0y0tZ9FubXS7y3S4LM4mVpZ3ZZCB5RAQLgE5zkYr9vD9mz41/tJeFvCui/BX4lweD7rStQmfU5LjVbuzS5t3RRjFureYysikBwAMnBHfSv+CJnjzXLhb/4hftJWCXL4MzWuiTX7N0yA8s0Z6lhkj0OOcD9kj/gm74H/ZL+IT/EzRfib4h1/VptLl06WGe2ht7d45GjZjsXc2Q0SkfP+dV4k8KeFvGWnDR/F/hrStcsRKsotdSso7mIOvRtjgjcMnBxkZrSdD0TQbYWWhaPY6dbjpFaW6wr+SgCqqqqqqqqqqvjVpJ1j4X6/bqCWgthcKR28pg5/RTV/wCI9B0zIvtXtYmHVDIC3/fI5q/+Kvhy2ytlFc3jDoVTYp/Fuf0q/wDi3q8wK6dp9tbA9C5MjD+Q/Sr/AMY+J9SyLnWrgKeqRt5Y/JcU7NIxd2LMepJyTVVVVVVVVVVVVVVX7IXh1/EXx98OHy98OmedezHH3RHG20/9/GQfjVVVf8FLvjh8Uf2ff2dLfx38IfFP9g67J4ktLNrr7FBdZheKZmXZOjpyUXnGeOvJr9kLx74s+KX7M/w8+IPjvVv7T1/XNHS4v7vyI4fOkLMC2yNVReAOFUCqqqqqqqqqqqqqrVdG0fXrNtO1zSrPUbV/vQXcCzI31VgRXib9kX4B+Jg7nwSulzuOJtMuJLfb9EBMf/jtaz/wT48B3BY+H/Hmu2Ofui7giucf98iOr/8A4J463GcaX8UbG4GR/r9KeHjHPSR+9f8ADvbxl/0UPRf/AAElqx/4J4anIVOp/FS1hGPmEGkNJz6AmVe3f9K/bb+AmleEf2v/AIW/AnTdduda/t2HTEuJXgEJWS81CSHy1Ubv4EQ555b8K8L/ALKfwG8K7JLbwDa6hOhz5upu93uPujkp+SirHT7DS7WOx0yyt7S2iGI4YIhGiD2UcCqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr9gj/ikv+CpXxp8NyYBuW8TWUfmoULquqRSqVHusQPpg/Sqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqp0SRDG6hlYYIPetU09rC4KAExtzGT/KqqqqqqqqpBkAiqqqqqrw9pfkRi+nX944+RSPuj1/Gqqq8Y+AvA3xD0o6H498HaJ4k085/0XVrCO7jGRgkLICAfevir/wSY/ZG+IrT3vhzQdY8CahLlvM0G+JhL+pgnEiBenyx7Onbk18U/wDgit8aPD5mvPhL8R/Dvi+1Qkpa6jE+l3RHZV5kiYjpkugOM4GcD4pfsnftIfBdpm+JHwa8S6Xa2+fMv0szdWgx1P2iHfF0/wBqqqqqqqqqre4uLSeO6tLiSGaJg0ckbFWRh0II5Br4c/t0ftb/AAraJPCnx38TyW0WAtpq1wNUgVR/Csd0JAgx/dxjqMHmvh1/wWs+NmhrDbfEz4Y+FvFMKEBp9Pml0u4cdyT+9jJ+iKP518O/+Cx/7LXioRW/jjTPFngq4Y4klurAX1snuHty0h/79D8a+Hn7UX7OnxXMUXw++NXhHV7mYgJZpqkcVyc9P3EhWQfitVVVX/BQX9iLSf2r/AH9veFra2tPiV4bt2OjXbkRi/iBLNZTN/dYklGP3HPUKz51vRdX8N6xfeHtf0250/U9MuZLa8s7mMxy280bFXR1PKsrAgg9CKqv2Q/2rvG37JXxRg8aeHjJf6Hf7IPEGiGTamoWwPbOQsqZJR+xJByrMD8Lfih4J+M/gHR/iX8O9aj1TQdbg822nUYZSDh43XqrowKsp5BBFVVVVfFv9t/9ln4G+IL3wj8S/i3Y6Zr2nbRc6ZFZXN3PEWjWRAywxvt3I6EZwMMDmvFn/BZX9lLQzJD4e0bx34kkUkRyWulxW8TYPUmaZHAI5HyH3xX7Jf7Tvh79rX4XT/E/w34fudEgttXn0yWyubhZpEeNI3yxUADKyocds1X7b/wv/wCChfj34wWdv+zB431TSfAk+hQm5kt9ct9NW3vhLKJEyGE7ZRYW3AbfmIB60f8AglN+2j8S28z4zftMadcRzH94LrW9R1iUDPOVlVFJ5Y43Hr1GTX7D37G0/wCxt4S8ReGZfiW3i7/hIr2G8YjSfsKWsiIUYKPNkLbht5JH3RxzVeO/gz8I/ile6dqPxL+GXhfxXcaQsi2L61pMN79nEm3eFEqsBnap+oBrw54I8F+DovI8I+EdF0SPbt2adp8VsMccYRRxwPyFVVVVVVVVVVVVVVVWqabaaxpl3pF9Hvtr2B4Jk/vI6lWH5E14m0K78L+I9U8NX4IudKvZrWUEY+aNyp/lVVVVVVVVVVVVVVVVVf8ABPjwe0l/4q8fTR/LDDHpts2OpY+ZKPwCw/8AfVVVV/wWS/5NFtP+xwsf/RNzX7Af/Jmnwl/7F+P/ANDeqqqqqqqqqqqqqqqqqr4gY+K3/BanQ9Fh2zQeHdVsfLIxx9h00Xb56dJUcc5PH0FVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV8N2PgL/gtdrujyKYI9b1PUQ+SU/4+dLa6XgjJ3Ps9B8wI4qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr/gpt+1T48/Z++KXwVsPhtdvPeia9v8AVdIMhEepW7mKGOFwOob9/g4JVgrDkVNa3VqVivIhHLtBZVbcM98HjI96qqqqqqqnAK4qqqqqtC0Y3LLeXKfulPyKR98/4VgYwKqqqqqvih+x1+zB8ZTNN8QPgp4ZvbyfPmX9ta/Ybtj6me3KSHn1Y/rXxR/4Ip/B7XRNefCX4neIvCty2WW11SFNTtgeyrjypFHbLM5789K+J/8AwSP/AGuPARlufDGj6F46sozkPouorHMF9TDceWSf9lC/4148+FXxN+F19/ZnxI+H3iHwxc7tqx6tpstrvP8Asl1AYY5yMgiqqqqqqqqq+Hf7TP7Qnwm8qP4dfGbxbotvCMJZw6pI1tx0zA5MZx7rxn3r4ff8Fif2rvCgjt/GEHhPxrAMCSS/037JcED+69s0aA/WM18Pv+C2vwn1RYoPid8H/E3h+UgB5tIvIdSiB/vEP5LAewDEZ79a+Hn/AAUO/Y3+JSxx6R8ctD0u5c4Ntr+/SmRvTdcKiN/wFiO2c1o+uaJ4j0+PVfD2sWOp2Uv+rubO4WaN/oykg1X/AAU5/YGX4waPeftAfB7Rd3jnSbcvremW0ZLa5bRqPnRR1uI1XgAZkX5eWVQWUqSrAgg4IIxiqr9gr9t3xB+yT48On63Jdal8OfEE6jXNMT53tnxtW7twTgSKMBl6SKMHlUZfBPjTwv8AEfwho/jzwVq8eqaFr1nHeafeRoyCaF1BVtrgMpweVYBlOQQCCKr9oL9vv9t7w78aPGXwh+DP7P8AZ6tb+HtWls7a/tfDeoapPPEMNG4EThAxRkJG0jmjr/8AwWs+LrGOx0rXPDlnJ1T7HpmiiIZwSGm2zenAJOB061+yf4d+NXhP4CeGfDn7QuqnU/Hdj9pXUbx7/wC2PMrXEjQl5f4mETIvf7vWq+L3/BOz9mn46/FjUvjF8S9I13UtY1aKCO5totUa2tm8mFYkOIgr52IuTv7enFeFP2Av2NPBm3+x/wBnnwpcben9rQPqn5/amkz0714Z8I+E/BWljRPBnhjSdB04OZBZ6ZZR2sQc4y2yMAZOBzjsKqqqqqqqqqqqqqqqqqqqqr9uDwC3hX4vHxNbW+yx8VWq3KsFwvnxgJKo9+I3PvJVVVVVVVVVVVVVVVVVfsxeAf8AhXfwW8P6VPCY77UIf7QvQy7WEswDAEdiqbEP+7VVVfH79n34c/tLeBY/h18ULW/n0eK/jvlSyujbv5sauqncM8Ykbivhh8OPDPwh+H+h/DTwbFcR6J4etRa2SXEplkEYJPzMep5PNVVVVVVVVVVVVVVVVVX7A5Pxc/4KdfFb4osPMttOOu39rJv3BRLeLbwrnv8AuZG9Pu/hVVVVVVVUzKil3YAKMkk4xXiv9oj4G+CRIviL4peH4ZYiRJbwXYuZlI9Yotzj8q1z/goL+zzpJK6fc+ItawcA2OmbM+/79o6u/wDgpX8OEk22Hw78STR4+9NNDEfyDN/OvgT+0rpvxy8P6j4gsvClzpMWn3xtRHLdLKz/ACK27hRjhsY5+tL4pszw9tMPpg1H4i0uTAaV4/8AeQ/0zUN9ZXOBBdROewDc/lVVVVVVVVVVVVVVVVVVVVVVVVVVTukaF3cKqjJLHAFXnjbwZYErfeLdGtyO0t/Gp/ItS/FD4dSSxQReNNJkknlSKJEuVZnd2CqABzkkgfjQ1TTScC+g/FwKS6tZcCK5ic9trg1VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX7S7D4cf8FifAHipm8lfEWo6A7ODt+WZBYMSc9MIQc9h0I61VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVftQBfjp/wV28C/DsqLq08L3mjWc0f3leGBTqM6kf7skin6H0q/sINQh8qUYYfcYDlTV5ZT2Mvkzrz/AAkdD9KqqqqqqogEYojBxmqqqrRtDa6K3V2pEI5VTwX/APrUFVVCqAABgADGKqqqqqqqq/4K7fGCP4d/stv4GspoxqnxD1KPT0U43JaxETXEig/7sUZ9pq/4Jp/sDeAv2lPB/i34k/GrS9Tl0BZV0zQFtbp7VmuFw89wGX72wGNBkMhLyZGVGPiX/wAERPD9wZrv4P8AxtvrLkmKw8RactwD6A3EJQgf9sjXxM/4JZ/tkfDlpprT4fWnjCxhzm78N36XO76QyeXOfwjP8q8WeB/GngLUjo3jnwhrfh3UBnNpq2ny2cox1+SRVP6VVVVVVVVVXhbxr4y8Dah/a3gnxdrPh++4/wBJ0u/ltJeOnzxsDxXw/wD+Cl/7Z3w9McUHxgufEFpGADbeILSK/wB+PWV1878pBnP0q3/4LZ/HuPRfs1z8KfAU2qhcC7H2tISfUw+aT+Tiv2ufiH+zx8Z/E9v8XPg1oms+FfEGund4r8PXlkkdr9sIy93aSRyONrtnejBTuIYA7mxVV/wTj/b5vP2cPEMHwo+J+oz3Pwz1q6Ajldtx8P3EjczqOpgYnMiDpy6jO4PY31jqljb6lpl5Bd2l3EstvcQSCSOWNgCrqwyGUgggjgg1VVVVVVVVVVVVVVVVVVVVVVVVVftd/DF/iP8ACG9n06283VvDjHULQKPmdVBEyD1ymSB3ZFqqqqqqqqqqqqqqqqr9nD4Zt8VPi1o2gXFsZdMtH+26nxlRbxEEqfZ22J/wOgAAFUDgcCqqqqqqqqqqqqqqqqqqqqq+K3jCP4efC7xh4+kk2L4a0G91IttBx5EDydD1+70r/giB4PkNh8VviNdIzm4uNP0y3lYd0E0swz3J8yA/hz1FVVVVVVfFL48fCv4OWvm+O/Flta3TJuh0+H99dS+m2JckA/3mwvvXxN/4KP8AibUHlsPhP4SttKt8kLqGrfv52HqsSnYh+pkFeOfjP8VviVI7eN/H2sarE5ybaS4KW4PtCmI1/BR2qqr9gH/kmviH/sOH/wBER1VVb6rqFrgQ3T4H8LHcPyNWnihWIS9gx/tx/wCFW91bXcfmW0yyD2PT6jtVVVVVVVVVVVVVVVVVVVVVVXjf4l+AfhxZC+8beKrDSkZS0cc0mZZQP7ka5d/+Ag1q37Wvh+9s0ufA2hzX8cykx3N63kp6ZCDLH6Eqa1349/E7XCyrrw0+I/8ALKxiEWPo3L/+PVqOs6xrEnm6tq15evnO65naQ59csTVeHv8AkaPD/wD2GrD/ANKY6qoru6g/1FzInsrEVb+I9RhwJGSYejrg/mKtfE1lNhbhGhb1+8KiminQSQyq6noVOaqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/4LDQzeBf2kfg38YIY33Q6aqxsgOS1he+fgHI5H2kenXrUE8N1BHc20qyRSoHjdTkMpGQR7EVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVfsJ4+Nf/AAVB+J3xadlmtdGfXNRspc7gFkuBaQKD6+RK3PTCmqurSC8iMM6Bh29QfUVqOjXFiTIgMkPZgOR9aqqqqqqqQc8elVVJG8riOJCzE4CgZJrS/DqxFbi/AZx92LqB9fWsDGBVVVVVVVVVf8FSfHGt/H79s3Svgl4KVtQfwzHa6Bp9tG+Vl1K6ZXlI9DmSGJvQw+1fAv4S6F8CvhD4U+EvhwK1p4b06O2aYLtNxN96aYjsZJGdz7tVVr3h3w/4p02TRvE2h6fq9hN/rLS/tUuIn+qOCD+Ir4k/8E1P2NPiX5s9x8IbTw5eyZ23Xhud9N8vP92GM+R+cZxXxJ/4IhaTL5t18H/jhdwYz5Vj4l01Zt3puuICmPwhP4V8Rv8Aglj+2Z8PjLLa/Dyy8XWcIObrw5qUdxn/AHYZPLnbPtHXi/wB46+H2o/2R498F674cvgSPs2radLZycdflkUGqqqqqqqqqqqr/gm5/wAFFYvhB9n+Bfx516U+CX+XQtanJf8AsRuSYZOrG3Y/dPPlnjGxvk8DfG/4NfE5V/4V38VvCXiR2H+q0zWILiRfYojFlPsQDVVVVVVVVVVVVVVVVVVVVVVVVVEBhtIB46Gv2oPhC/wk+J15a2Fvs0PWS15pRC4VEY/PEP8AcY4x/dKnvVVVVVVVVVVVVVVVV+xz8Hn+G/w5HiPWbXy9c8UhLiVXXDQW4B8mP2JDFz7sAfu1VVVf8FCP2iviB+y/8BYPiT8NYdJl1eXxBbWDLqds08XlSRzM3yq6ndmNec+tfstfEvxH8ZP2evAnxQ8XJZprPiPSlurxbOIxxByzA7FJYgYA4JNVVVVVVVVVVVVVVVVV/wAFN/HC+B/2LPH7xTtHda7HbaTbBTjf59wglH08kTfXp71/wSQ8Df8ACIfsa6NrLxlJfF+tX+quGHOBILVfwK2qkfXPeqqqqq8beOfCXw68PXHirxrrttpWmWoG+eYnknoqqMszHsqgk18c/wDgoB4v8WNc+HvhBBL4b0kkodTlwb2ceq4ysIPtluh3L0q+vr3U7uXUNSvZ7u6nYvLPPIZHkY9SzHJJ9zVVVV+wD/yTXxD/ANhw/wDoiOqqqqKaWBxJBKyOOhU4rT/EoOIdQXnoJFH8x/hSOkqB43DKRwQcg1VVVVVVVVVVVVVVVVVVVeO/iH4M+GmiNr/jXXrfTbQEiPzCWeZv7saDLOfYA46nA5r4tft0+LvEMk2k/CyyPh/TiCv2+5VZLyUeoHKRd+m49CGFapqmp63fzaprOo3N/e3Dbpri5laWSRvVmYkk/WvhLq/n2Fzosj5a2bzYgT/C3UD6Ef8Aj1VVeFQD4s0AEA/8Tez6/wDXdKqqqoLm4tXElvMyN7GrDxMrkRX6BT2kUcfiKjkjmQSROrqw4IOc1VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX/Bbnwk198IPh145WAN/Y3iOfTzJgZQXVuX+uCbQe2QM9q/Zh8Wjx5+zj8MfFxmWWXU/CenSXDA5xMLdBKM+zhh+FVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV8VPFq+APhh4v8eM+0eG9BvtSLYBx5EDyZweP4O9f8EQPCD/ANmfFf4iXUZdrq50/TLeVl5BRZpZhu758yEkew9aqiARg8+xq/8AD9vc5ktSIZPTHyn8O1Xdhd2bbZ4SB2Ycg/jVVVVVVIMgEdqoAsQqjJ7CrHw9eXRDzjyI/Vh8x+g/xqy020sE228fzEcu3JNVVVVVVVVVXxG8caP8MvAHiP4ieIHC6d4Z0q41G5G7aWSGNnKg/wB47cAdyQK/4JU/DTWfj3+1D4x/aj8eQi6/4R+ee9ErLhZdXv2c5UHqEjM7Y/hLRH0qqvL2z0+2e8v7uG2giHzyzSBFXnHJPA5xWj/GP4R+IvE0fgrw98UfCeq+IJI3kXSrHWYLi62IMs3lI5YKO5IxVftT/Ho/sy/BLW/jM3hCXxLFoctskthHefZSwmmSFWMmx9oDyJ/D3q5/4LC/tI+Pp5LX4Mfs1aTJJuIWN0vdadORgEQCHJwcdOpB9q+BvxZ/4KW/Gj4weErT41fBq6tPhhdagE8Q6df+Dre0ga2YMpZheKZsKWVvlOSF6HJB+Iv/AATo/Y3+JfnTan8FNJ0e8lyRdeH3k0tkPqI4WWIn/eQiviL/AMERfAd8JLj4T/GvW9IcElLXXtPjv1b/AGfNiMJQe+1j/OviJ/wSN/a/8FebP4d0Xw741tkJIbRdVWOTb6mO5ERz7Lu/GviB8EPjF8KZGj+JXwu8U+GlV9gm1PSpoInP+zIy7GHupIqqqqqqqqqqBKkMrEEHgjjFeBf2qP2kfhmYk8D/ABy8Z6ZBAAI7QavLLbjHT9xIWjP4rXgb/grz+2H4UMUfiDV/C/jGJDhhq+irC5X0DWph5x0JB9814J/4Lg6fII7f4kfAK4hx/rLvRNbEmfXEMsa4/wC/hrwT/wAFaf2MvFxVNV8W6/4TlYDamt6JKefQtbeco+pIHvXgr9pX9nr4jhE8D/G3wTrEzAEW1vrcBnGemYiwdfxUVVVVVVVVVVVVVVVVVVVVVVV+0H8HrL40fDy68PhY49Xs83Ok3DceXOB90n+44+U+mQcEqK1LTb/R9QudJ1S0ltbyzlaG4glXa8TqcMpHYgjFVVVVVVVVVVVVVVfsl/A1vix43XXddtC3hnw7Ist1vX5bqbqkHuONzf7IwcbwaAAAVQOBwKqqqv8Agsl/yaLaf9jhY/8Aom5r9gP/AJM0+Ev/AGL8f/ob1VVVVVVVVVVVVVVVVX/Bbrx8bH4dfDj4YQXHzaxrFzqs8annbaxCJM+xN03HqvtX7OngJ/hf8A/h58PZ4RFc6D4asba7UDH+kLCvnH8ZC5/Gqqqqvib8V9B+HFjsnZbvVpkzbWKNgn/ac/wr79T274+IfiTVvilJcnxjc/a4biNoxbj5Y4UPZF/h7c9cgHNeMvC134Q16fRrrLIp3QSkY82M9G/ofcGqqqqv2Af+Sa+If+w4f/REdVVVVVp2q3OnOBG2+Mn5oyePw9DVlfW9/CJrd8/3lPVT71VVVVVVVVVVVVVVVVVV+0F+1l4Z+EwuPC/hdYda8Vqu1ot2YLEnvKw6t/0zHPqV4z418d+LfiJrkviLxlrtzqd9KThpW+WMf3UQfKi/7KgCqrwXq/8AYviO0umfbE7eVLnptbjn6HB/CqqvCn/I26B/2FrT/wBHpVVVVVWOpXWnvugkypPzI3Q1p+p22pR5ibbIo+aM9R/9aqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv+Cpvgw+MP2KvGk8MIlufD09lqkK7MkeXcokhB7YillOfbHev+CUnjQ+L/wBi3wrYSTiWfwxqF/pUp7jbO0yA/SOeMfQD61VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX/AAUi8WHwf+xT8Tr6OTEt/YQ6bGuM7vtNzFC4/wC+Hc/h36V/wR/8Ijw7+x3a62YdreKfEeoagGKkFwhS16nqM2xHHHXvmqqqZVddrAEEcgirrQbC4JaNTC3+x0/Krjw3eR8wSJKOw+6am0+9gGZbWQAd9uR+dEdqqqSN5DtjRmPoBmotF1G4GFtmUer/AC1beFc4N3df8BjH9T/hX/BYH4s6tp+ofDr9nT4b3d3FrOp3A1i9SxmZJ5GLGCzjDKc/M5nO3PVUPpXwk8Jax4C+F3hTwX4i8RX2vato2j21rf6nfXLzzXlwkYEsrO5LHc+4jJOAQO1VVeOPjt8HfhwJE8ZfEXRbCeL79qJ/OuB/2xj3Sf8AjteJ/wDgor8E9Icw+HtI8R684PEkdslvER65kYP/AOOVq/8AwUz1SRyug/CO0hUdGvNXaUn8FiXH5mrv/gpJ8ZHcGx8G+DIlxyJbe5kP5ida+GHxP1/xr8O/Dni3VbPT4rzV9NhuZ47eN1jV3UEhQzEgfUmk8VXA+/aRt9GI/wAai8U2zYE1rIv+6Q3+FQa1plwQqXSqfRxt/nSsrqGRgQehFVVf8FhvjaPAv7P+nfCHSb5I9V+I2oCO4UNhl0+2ZZJT6rulMC+6lx6iv+CdvibxL8G/g14O+Fej/sxfEue51/VBe+IPE11pi2Nhm5lRRcxu53SxR23k4IALCMkdar9uL4SfHn4y/CPT/C37Ovj+68JeJotdinubyHWp9MWWy8mZZInkgyxUu8R24Odv1q1/4I3/AB/8dXaXvxj/AGltJe4yS00Ud5rLgk5ODO0JOefT9a/ZZ/4Jd+BP2Y/ijo/xdtfiv4i1/W9HiuI44GsobW1lE0LxNvT53wFfIw4+ZRnIyKrVNK0vW7GTTNZ021v7OXHmW91Cssb7SGGVYEHBAI9CAat7a3tIEtrSCOGKMYSONAqqPYDpVV4z+Jvw3+HNt9s+IHxA8OeGodpYPq+qQ2gIHp5jDNeOP+CnX7FnggPEfi4uvXUfS30PTri73fSQIIv/AB+vGv8AwW0+DdrHNb+B/gt4s19SGXGrXdvpySDGP4PPOD7jp27V8b/24vAnxiFxs/Yu+D2mS3KkNePb3P2sEjlvPtJLZi2eQWz9DV1LHcXMs8NrFbJJIzLBEWKRAnhVLFmwOgySfUnrVVVVVVVVVVVVXgv43/Gb4bhF+H/xZ8X+HUQACLTNbuLZCB0BRHCkexGK8F/8FSv21PBxSKb4o2/iG2jIIg1rSLefPrmRESU56ff+mK8F/wDBbz4mWRiT4h/BDwzrADYkfRtRn0449Qson579efbrXgv/AILR/s360I4PGXgjxv4bnYfNIlvBfQJ/wJJFkP8A377V4O/4KJ/sXeN/LTS/j3oVjK5AKazHNpmw+ha4RF/EEj3rwr4+8CeO7YXngjxtoPiGApvEulalFdqVzjOY2YYz3qqqqqqqqqqqqqqqqqqqv2yf2dH8T2k/xb8E2IbVbKHOsWka83UKD/XKO7oowR3UDuuDVVVVVVVVVVVVVfC34Y+Jfi54wtPCHhiDMsx33Fy6ny7SEfekc+g7DuSAOTXw98BeH/hn4Q07wZ4ZtvLs7CPBdh888h+/I57sxyT27DAAAqqqq/aw/Zm0L9rH4XRfC7xD4nv9BtYtVh1EXVlCkjlokkULhuMHzD+VfBH4V6f8EPhN4Y+E2larcanaeGLFbSG7uECSTAEncwXgHntVVVVVVVVVVVVVVVVVftxEftD/APBT34efBKE/adP0GTSdMvoVXcArub26b3PkSAHt+7x61VVVV8X/AIu2Hw70/wCwae0VzrtymYICciBT/wAtHHp6Dv8ATNapqeoa1fz6pqt5JdXdy5eWWRssx/z26Cq+JvgiPxnoLJbIv9pWYMlox43H+JCfRsD8QKlikhkaGVCjoSrKwwVI6giqqqr9gH/kmviH/sOH/wBER1VVVVVWd5PYzrPbvgjqD0I9DVhfwajAJYThhwyHqpqqqqqqqqqqqqqqqqqv2ov2uF0Y3fw5+E+qA6ipaHU9YhIItuzRQnu/UFx93oPm5WWWWeV555WkkkYs7ucliepJ7mqqq8Hav/bfhyzvHfdKqeXLnrvXgn8eD+NVXhT/AJG3QP8AsLWn/o9KqqqqqoZpbeRZoJCrqeCK0jWItRQRSELcKOV/ve4qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq+N3gcfEz4NeOfh4q7n8SeHb7T4uOVklgdEI9wxB/Cv+CIXjkTeF/if8MLicq9hqFnq9vCxxnzo3hlYDrx5EIP8AvCqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/4LQeKv7I/Zh0DwzDIBLr/i63V1OOYYbeeRsc5+/5PbHrjIz+wt4V/wCEN/Y++EujGPY0vhm2vmXGCGugbk5GTzmY/wD1ulVVVVVUkMMoxJEj/wC8uabTrB+tnDn/AK5gV/Zmnf8APnD/AN8UtlZIcpaQg+0YFBVUbVAHsBiqr4H2z/tnf8FVte+J1wPtXhjwNqEupW5ZfkMFgVtrHbn7paYRTY68P7kVrWuaL4b0yfWvEOr2em2Fqu6a6u51hjjHqWYgCvix/wAFEfAfht59J+FuizeJ71MqL+5zb2an1Uf6yTv2QdCGNfEj9qb45fFFpYNf8c3dpYSk/wDEu0s/Y4Ap/hIT5nHP8bN2qqqvgF/yRLwP/wBgO2/9Fiqqre7ubVt1tO6H0B4P4VZ+J5Vwl7EHH99OD+XT+VWt9aXq7radXwOV6EfhVftOalcftuf8FL9L+EmmTyXPh7R9Xh8OIIiSFtbRml1GUAHGdwuiGGMqiZPFQQQWsEdrawxxQwoEjjRQqooGAABwABxiqqqrxp8QvAXw30r+3PiD410Pw1p/OLnVtQjtI2IHIDSMAT7Dmvij/wAFb/2R/ABls/DGr6747voyVCaJp5SEN7zXBjBX/aQOOR15x8S/+C1/xe1kzWvwp+FXhrwzAxKpcatcS6ncAdmUL5SKx9CrgdOetfET9uv9rn4ovKvij48eJ4reXIa10m4Glwlf7pS1EYYf72T6knmry9vNRupL3ULua6uJm3STTSF3c+pJ5JqqqqqrLT77Upvs2nWU91KFLeXDEXbA6nAycVp3wn+KesCE6R8NPFV755xF9n0a4k8z/d2oc9O1WP7M/wC0fqkjQ6b+z98SbuRF3MkHhS9kIHqQIjxXjL4QfFn4cxrL8Qvhd4t8MI2NrazolxZA56YMqL6iqqqqqqqqqqqt7i4tJkubS4khmjOUkjYqyn1BHIrwj+1h+014EZP+EV+PnjuzjT7tu2uzzQ/9+pGZP0rwh/wVj/bS8LtGupeOdF8TxRniLWNCtxkYxgtbiJz65LZ968Jf8FvviXZiMeO/gX4Z1bA+c6Tqk+nZ46gSLPjnt/8Arrwp/wAFr/gFqISPxj8L/HOiyMOWsxbX0anA6kyRNjqOFPb148Lf8FRv2JfFDJC/xck0a4c4EWq6Ndw45xzII2jHbq39a8MftR/s1+NAi+F/j54Av5ZBlYI/ENss34xlw4/EVa3dpf26XVjdQ3EL52SROHVsHBwRx1BFVVVVVVVVVVVVVVV+1f8AsqNozXvxQ+GWnFtPYtNq2lQrk2x6tNEB/wAs+7KPu9R8uQtVVVVVVVVVVVXgHwB4o+Jnia08J+EdNe7vblhuODsgTIDSSN/CgyMn8BkkCvgn8FPC/wAEvCq6Joqi51C5CvqWoumJLqQD/wAdQZIVe2T1JJNVVVVVVVVVVVVVVVVVVVVVUskcMbSyyKiIpZmY4CgdST2Ff8E9BJ+0R/wUW+JH7Qs4MtjpI1PUbKVhnyzdS/ZraP2/0ZpRn/Y96qqqvin8SLD4ceHjfOEn1G6yljbMfvuByzd9q8Z+oHetV1TUNc1G41bVbp7m7upC8sr9WY/yHt0FVVfG7wILSc+MdLh/czsFvUUfcc8CT6HoffHrVVVV+wD/AMk18Q/9hw/+iI6qqqqqqsr2awnWeBuRwynow9KtLqG9t0uIGyrDoeoPoaqqqqqqqqqqqqqqqv2vf2mJfCUc/wALPh9qWzWZo9urX0R5skYf6pD2kYHk/wAIIx8xytVVVXwl1cRXl1okj/LOvmxA/wB5eG/MY/75qq8Kf8jboH/YWtP/AEelVVVVVVRyPFIssTlXQ5Ujsa0jVE1KDDYEyD519fce1VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX7HO39nz/gqp8QPhDIFtdP8R3GsafZwk4VYmYX9r+PkxBR7vjvVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVf8ABb7xBPeaj8IPAViXkkK6neyQqCd7O1tFDgevyyj8a8GeHYfCHg7QvCduF8rRdMt7KML02xRqgxwOML6D6CqqqqqqqqqqrLwl4V03Xb3xTp3hnSbXWtSjWK91GGyjjuLlFOVWSUDc4BJIBJAzX7Qf7ZngD4M/aPDmgmLxL4sQFTZQS4gs2/6byDoR/wA81y3HO3INfFX43/Er4z6p/aXjvxHLdRRsTb2EX7q2tv8AciHAOONxyx7k1VVVV8Av+SJeB/8AsB23/osVVVVRySROJIpCrL0IOCK8d+NPGOk/DrxNfeDdFGr+KLXR7qTRLUOiC4vBExhRi5CgF9oJJxX/AAR9+BPiX/hd/wAQvix8QtFv7PUvB9v/AGSsWowtHMuoXTF5mYN8wdYo8HPa4754qq8aePfA/wAONEk8R/EDxho3hzS4uGvNVvY7WPOM4DOQC2BwBya+NH/BYj9nTwELjTfhdpOs/ETVIiVSWBTp9hkdczSqZD/wGJgcdehr4wf8FX/2svieZrHw34h07wBpUhIEHh+2xOV7brmXfIGH96Py/pXiXxV4n8Z6tLr/AIw8SapruqT/AOtvdSvJLqaT/ekkJY9T1Peqqq8C/sz/ALQvxN2P4D+CvjPWYHxi6g0aYW4z0zMyiMZ92rwZ/wAEkv2zvFYRtW8KeHfCkbgFX1rXIm4Pqtr5zD3BAPtmvCf/AAQ98YXOx/HXx+0bT8DLxaTocl5nnoHkkixx32n6V4Y/4Iqfs56diTxV8RPHutSL/BBPbWcR65yvku3p0YdO+ePDv/BK/wDYi0Aq8/woutYlUfLJqOu3r+ufkSVUPB7r2BGDzWifsSfsieH1VLD9nHwDKFBA+3aJFenpjkzB89K0n4P/AAk0Bg2g/C3wjpxVy4NpodvCQxGCflQc44zVvbW9pAltaQRwxRjCRxoFVR7AdKqpYo5o2hljV0dSrIwyGB6gj0rx9+x3+y18Tlc+MvgP4Oup5M77q201bK4bPrNBskP4txXjz/gjd+yr4meS48Ian4x8HynPlw2mord26/VbhHkP/fwV47/4Ii/E6wEsvw1+NfhrWwCTHDrOnzaa2P7u6MzgnHGcAE+nbx5/wTR/bQ8BGSSf4OXWu2qZ23Gg3kN/vx6RI/m/mg/nXi34f+PPAN39g8deCdf8OXQYr5GrabNZvuHUbZFU59qqqqqqqqqqqqq0TxL4j8NT/afDniHUtKmPWSyu3gb80IPc14c/bI/au8JqkeiftEePljj+5Fc65NdIv0SVmUDnpjrzXh3/AIKo/tuaCVS5+KtnrMSjCx6joNk3r/FHEjnqDyx6DtkHQf8AgtL+05p5Eeu+B/h3q0Y/iFjdW8meO63BXGAf4ep68YrQ/wDguNr0IRPEv7Oen3ZzhnsfEz2+OeoV7eTPHbI57jtov/Bbr4Q3AX/hI/gr4vsTj5hZXtvdY57bzFnjntzx71ov/BY/9kPVQPt1p490fIJxe6LE2MHGP3M8nXr/APX4rRP+Cnv7D2uFY0+NkdlMQT5d9ol/BjnHLmDZ3HG7p9DWg/tlfsneJSi6V+0Z8Pd8hwkdzr8Fq7EnAAWVlJJPbGenrWgePfAvizYPC3jXQdZLjKf2fqMVxuGCcjYx7An8KqqqqqqqqogMNpAPHQ1+0j+xumrvd+O/hBYxx3jEy3uhoAizHqz2/ZW7mPgH+HBwpurW5sbmWyvbeW3uIHKSwyoUeNgcFWU8gg8YNVVVVVVVVVVfCD4JeN/jTrg0vwvZeXZQMPt2pzqRBar7n+JyOiDk+wBI+D/wY8HfBfw2uh+GbYS3UwDX2ozIPPu3HdiOijJ2oOB7kkmqqqqq/bw/an1/9kP4QaR8SfDnhXT/ABBc6l4kg0l7a+neJESS2uZS4Kc5BtwMdPmNfAb4jXvxf+C3gn4o6lpsGn3XinRLbUZrWByyQtKgYqpPJAz3qqqqqqqqqqqqqqqqr9uL4nr8If2UPiT4xjuPKvG0WTT7FgcMLm7It42X3VpQ/wBFPav+CMHwwHhj9n3xF8Trq22XXjbXTHA5TG+0s1MaEHv++kuR+FVVVrOr6foGlXWtarcLDaWcRklc9gPT1J6AdyQK+IHjbUvH/iW516/YqjHZawE5EEQPyqPfnJPckmqqqvbO11G0msL2FZoJ0KSRsOGUjBFeOvCF14M1+XTJQz27/Paykf6yM9PxHQ//AFxVVVfsA/8AJNfEP/YcP/oiOqqqqqqq0TUjYXOyRv3Mhw3+yfWuCODVVVVVVVVVVVVVVX7TXxzt/gv4HZtMmjfxLrAaHSoWAby8ffnYf3UBGAerFR0zi8u7rULua/vrmW4ubmRpJppXLPI7HLMxPJJJJJNVVVVaJqUmj6taanGTm3lDEDuvRh+IyKjkjmiSaJwyOoZSO4PQ1XhT/kbdA/7C1p/6PSqqqqqqqtbqWznS4hbDKenqO4q0uor23S4hPDDp6HuKqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr/go7FL+z9/wUF+GP7RFtC0dhqK6bqN3KqkedJZTiG5j46/6MIAfZ8fWGaG4hSe3lSWORQyOjZDAjIII6giqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv+Ciq/wDCxv8AgpJ8Gvhuq+bBGmg2M0YXdtM+pSNIxGDgeW6E9QAuaqqqqqqqqqquta0axmltr3V7K3mgtjcyxyzqjRwjOZCCchBg/MeODzX7UX7cl7rE138P/glqb2unpuivtfiO2S5PQrbnqif9NOrZ+XAGWd3kcySOWdiSzMcknuaqqqqr4Bf8kS8D/wDYDtv/AEWKqqqqrSbwaRcSz20EY+0sGnAUAyEAAEnqSAAM+gFWl5b30Imt5Nw7g9VPuK+Pf7cv7NP7OQuLHx58Qre7123yDoGjAXt9uH8LIpCwnj/lqyD3r46/8FlvjJ4xNzo/wO8Laf4E05sqmo3gXUNQYf3gGHkxZH8OyQjs1eO/iP4/+KGtv4k+IvjTWfEupvkfatUvXuHUH+FSxO1f9kYA9Kqq+FX7KX7RvxtMMnwy+D3iPV7Scjy9Qa1+zWZz/wBPMxSLpz96vhj/AMEVfjX4gWK7+KnxJ8NeEYHALW1hE+qXK+qsMxxg9sh2FfDn/gj7+yb4OWC48Xp4n8cXaAGQalqRtbdm/wBmO2EbBfZnb3JHFfD/APZ1+A3wqEL/AA6+D/hHQZ4DlLq00mJbjPqZipkJ9y1VVVVVVVVVVX+n2Gq2cun6nZW95azLtlgniEiOPQqcgj6146/Yf/ZI+JBeTxT8APCJmlB8yfTrL+zZXPqZLUxsT7k5rxz/AMEZv2YfEIefwb4k8aeFJyfkjjvY7y3Uf7kqGQ/9/Pzrx1/wRE+JtiZJPht8bPDOtLnMcWtafNpzY9N0RnBOO+ACfTt45/4JhftpeBzLJ/wqX+37WIf8fOh6lb3e/wD3Yt4mP/fFeM/hL8VPhzI0XxB+Gvinw0yEZGr6PPZ9enMijg5GD3yKqqqqqqqqqqqqqqqvD3xc+K/hEofCfxP8WaKU+7/Z+tXFtt6dNjjH3V/IeleH/wBu39sPwyFGm/tFeM5tgwP7Qvzf9iOftAfPXv7egrw7/wAFZv21tEKjUfHmia+FPTUfD9qmR6HyFiPt614c/wCC2Xx7swqeK/hV4D1RVwCbL7VZMw46lpZRnhugA5HHHPhz/guJ4cmCJ4u/Z51G0I4eTTvESXGfcI8EeO/G4/WvDf8AwWV/ZO1kJHrWj+PdAf8Aja70qGaMH2MMzsR/wEfSvDf/AAUo/Yl8UhEs/jrp9nK3WPUtOu7LafdpYlX8iRXhv9o/9nvxkEXwr8c/AOqu/SK18R2skg9igfcD7EVb3FveQJc2lxHNFIMpJGwZWHqCOtVVVVfG39mbwD8aIn1K6iOkeIlj2xaraoNz4GAJk4EijgdmGAAwHFfFf4CfEf4PXjJ4o0Yy6cX2waraAyW0vp82Mof9lwD6ZHNVVVVVVVVWVjealdxWGnWk11czsEighjLvIx6BVGST7Cvgp+w9ruvm38Q/FuWXR9POHTSIj/pUw9JG6RA8cctyQdhrw54Z8P8AhDRrbw94Y0i203TrRdsNvbx7VX1PqSTySeSeSSaqqqqqq/4LV/8AJq/hb/soFn/6QX9fsUf8mi/CD/sT9P8A/RK1VVVVVVVVVVVVVVVV/wAFr/inJaeDvh/8D9LlL3GuahLrF9FHy3lwL5UCkDkh3mlOPWIV+zX8LY/gp8A/AXwtWNUn8P6Hbw3u0YDXbLvuGA9GmeQ/j1PWqqq/aY+IBuryL4f6ZOfKtis2oFT96QjKRn2AO4+5XuKqqqq+IngyDxroD2ahVvrfMlnIez45U+zdD+B7VcW81rPJa3MTRzQuUdGGCrDggiqqv2Af+Sa+If8AsOH/ANER1VVVVVVVeHdQN1bfZZGzJAMAnuvb8un5VVVVVVVVVVVVVVavq2naDpV5rer3aW1jp8Dz3MznAjjRSzMfoAa+NPxS1T4w/EDUPGGob47d28nT7Vmz9mtlJ2J9eSzerMaqqqqqvhtq/wDafhqK3kfMti3ktnrtHKn8uPwqvBihvGGgggH/AImdt1H/AE1Wqqqqqqqrw/qH2S5+zSN+6mOOezdj/Sqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/4LN/DBvFX7OmhfEq0g33PgbXVE77f9XaXgETnPvMtqK/YQ+KA+L37Jfw38VzXXn31tpCaZfljl/tFoTbszehbyg/0cHvVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVXiYp8R/+C2un2ChZbfRtUt/LDAHabLRxM3qOJY3x0PToaqqqqqqqqqppobeF555UjiiUs7u20KAMkknoMV+134u8O/Gb9pbUvibpN+2o6fpGnw6PoUrIUVIYwzSyKD/fllmwTzsK9MkVVVVVVV8Av+SJeB/+wHbf+ixVVVVVV8Wvh+/xU+HGu/D5PFuu+Gm1m1MK6no149tPA3UHKEb1zwyE4ZSRxnI+PvwI8e/s6/Ea++HfxAtFF1EPOtL2LJh1C3YkJPGT1BwQQeVIIPIqq+DX7Kn7Qfx/mjHwq+Fus6vZO+xtTeL7NZRnODm4lKx5HdQxbjoa+Df/AARN1O5W31P49/FmOzU4MmkeGIPMkx3BuphtU9sCJh78c/CD9hH9lT4IrDP4P+EOkXepwj/kK60n9pXW7+8rTbhGe37tUH5mgqqoVQAAMAAYxVVVVVVVVVVVVVVVVLFHNG0MsaujqVZGGQwPUEeleN/2UP2Z/iP5j+MvgV4J1CeYYe6GjxQ3B/7bRhZP/Hq8bf8ABIr9jfxWZX0TQfE3hGSQkg6PrTuqn2W6EwxnsMe2K8af8EPbBzJP8O/j/PENx8u11rQhJx2zNFKvP/bPn2rxp/wR4/a68NrJJ4dXwd4tRVJjTTdY8iRvbF0kSg/8Cx714z/Yl/a18AiR/En7PvjMRxHDzWGnNqEa+5e28xQPfOM8Vqmkarol42nazpl3YXUf34LqBonX6qwBHSqqqqqqqqqqqqqqqqq0HxX4p8K3Au/C/iXVdInB3CSwvJLdsjvlCDXhn9tL9rTwgY/7E/aJ8dlYseXFe6zLexrjGAEnLrjgcYx19TXhj/gq/wDtr+HvLW/+Iek+II4wAE1TQLUZAxwWgSNj06k5O45PTHhn/gtn8dbLYvi/4TeBtWVRgmwe6sWbpyS0koz16DHPQV4Z/wCC4Pge5Cr4y+AOuacRwzaZrkV7n3Akjh9uM/jXhr/gsN+x/roH9qyeNfDp7/2joiyY6/8APvJL6D8x748Nf8FC/wBi3xYFOlftB+HYNxwP7TSfTvXr9pjjx0/l6ivDfxp+DfjIL/wiHxa8Ga5v+7/ZuvW1znr08tz6H8qvLO01C0lsb+1hubadCksM0YdJFIwVZTwQR2NfFH9h74ceL/M1HwLcP4T1FiWMUaGa0kP/AFzJBT0+QhR/dNfEH9lj40/Dx3lu/CkusWKZxe6QDdJgdSVA8xB7soHvToyMY3UqynBUjBBqqqqqvDPg3xZ40vRp3hLw3qOr3GeUs7ZpdvuxAwo9zgV8OP2DPHOtvFe/EfWbbw9ZnBa0tiLm6b1BIPlpx3y+PSvhl8Dfhn8JLYJ4O8OxR3hTbLqNz++upR3zIfug91UKvtX7U/xb8ffAf4T3nxc8D+CrPxbbeGplude0mSZ4Jn0/BEssMqhgjRkq7bkYbFfpjNfs/f8ABRX9mD9oQ22laT40Xwv4knwo0PxGVs5nc8bYpNximJPRUcuRyVFVVVVVVftx/sq6h+2B8JdJ+Gmm+NLfwzLpniKHVzdz2LXQcR29xD5e0OmCftAOc/w9Oa+Bvw4n+D/wc8GfCy51aPVJfCui22nPeJCYluDEgUuEJJUHHTJ+tVVVVVVVVVVVVVVVVau6/tn/APBW23s7Ytd+F/AWpJGzH50W10nLSfWOW83KD0xOPrVVVeLvEVp4S8Naj4jvCCljAzhScb36Kv4sQPxrU9RvNY1G51XUJjLc3crSyuf4mY5J/Wqqqqq+N3gQY/4TLSoPRb9FH4CTH6H8D6mqqv2Af+Sa+If+w4f/AERHVVVVVVVVp12bG8juAflBw49QetAqyhlIII4NVVVVVVVVVVVVV+3j8VX0Xw3p/wALNIu9l1rY+1alsbBW2RsIh9ncE/SMjvVVVVVVXwv1cWHiD7BI+Ir9NmD/AHxyv9R+NV4L/wCRx0L/ALCdv/6MWqqqqqqqr6GtHvft1ikjNmRPlf6jv+NVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVftCfDGD40fA/xx8LZURpPEeiXNral8YS52EwPzx8sqxt+Ff8ABFT4oXCaR8RfgDrckkV3pF5HrVlbSkh0D4gulweVCvHb8esh/Gqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr9kJj4//wCCuPxT8TqpmTRbzxHLFIpLARx3As0bIAABWRevHIHJwaqqqqqqqqqv26v2nm1G4u/gf4B1Mi1t3MfiK8hbHmyDraqR/Cp+/wCp+XoGBqqqqqqr4Bf8kS8D/wDYDtv/AEWKqqqqqq/ay/Zh8KftSfDC58IaqsNnr9iHn8P6uy/NZXOOjEcmJ8BXXnjBA3KpHwW/4JGftLfEfU3f4gtpfw+0OC4eKS7vJFvLicI20vBBE2GUkHBd4wRyMgjPwL/4Jh/sq/BWO21C/wDCB8d69CAW1HxMFuUVx1MdtgQqM8jcrMMD5u9W9tb2dvFaWkEcMECBIoo0CqigYCqBwAAAMVVVVVVVVVVVVVVVVVVVVVVa94Z8N+KbI6d4n8P6bq9qQQYL+0S4Q56/K4I7V4x/YQ/Y98dmQ69+z14RiaUku+l2h0tiT1O61MZz7+vNeMf+COn7I/iMvJ4el8aeFnJyiafq6zxrx0IuY5WI7/eB968Xf8EO4SGm8B/tBuDn5LbV/D+ePeWKYf8Aovv+fiz/AII2ftYaEJJfD+q+BfEsY/1aWmqSwSt9VniRQf8AgZ+teKv+CeH7aPg7edU+AHiC6VM86U8GpZHHIFtJIe44xn8jXir4UfFHwKXHjf4beKfDxj++NV0a4tNvUc+Ygx0P5GqqtI+H/jzxCiyaB4J17UlfG1rPTZpgc9MbVPoas/2dPjrfAeR8KfEi5TcPOsWh4/4Hjnnp1rT/ANjT9qTV9LGs6X8EPEt3ZkPtkht1fdsYqwABySCpGAM5Fav+y5+0noa79U+AXxBhjCgmQeHLp0GTgZZUIBz2znp61qui6xoV0bHW9JvNPuQMmG7gaFwPXawBqqqqqqqqqqqqqvDXxM+JHgsIPBvxC8S6F5Z+QaZq09rt+nlsMV4b/bu/bE8KhRpf7RXjOfZ0/tK//tH16/aBJnr39vSvDv8AwVv/AG0dEULqfi7w74gIHJ1HQIEz1/59xF6/oPerz/grF4h8Zqf+FtfszfDHxLK33p7NLixmxz0kZ5XXjb37H1GH/bW/Zk8QOX1H4CeN/CTN0GjeKotSjT/gNxAjEYzxvH171a/tD/sr6qqm38d+OdHcgbk1bwnEyqcDOHt7uQsOvOwHjpzWleM/2dvEDJHpX7Tvgu3kZiDHq2narY7RkgEs1oU5Azw3GRmtB+G3w/8AEwX+xv2rfgRMxGRE/jDypMYBzseIN0PpxXhf9h/UvFDM1h8YvB95Co5fS5GvCMdcgFQOcDr3/PQ/+CevhOBgfEvxG1a+HcWNlHaH83Mv8q0n9jj4A6Xplzp0nhKa/e6geF7q7vpWlUMpBKFSFRhnIZVBBwQeKl1L4zf8EoP2qpptUn1XxT8MvF8nzzTMzf2rZBuCGPyi9t93I4znssoI+HfxC8HfFfwTpHxD8Aa5Bq+ga5bieyu4sgMucEEHlWVgVZSAVYEEAg1Wo6dYavp9zpOqWkV3Z3sLwXFvMgZJY3BVlYHgggkEe9ftqfs5337MH7QHiD4dLC50O4f+0fDs7ZPm6fMzeWMnq0ZVomPdoyehFfs/f8FC/wBpz9ngWul6D42bxH4btsKNB8Qlry3RB/DE5IkhHJwEcLnkqelfs+/8FbP2d/iuttovxNM3w08QSYU/2lL52nSN/s3SgeWOpPmqgHHzGtM1PTNa0+DVdG1G1v7K6QPBc20yyxyqejKykgj3FVVVVVVVVVVVVVVVVVVVVVcf8FHvDsPwy/aKfULqztPGXwl1nVLDRYc4/tKFrt7bT51UgZ2yGNZFGcBQx+9x/wAEXPg3PZeEPGv7QuvQvJfeJbwaRpk8xJdreEiS5kBP3hJMyKSed1uaqqr9qXxSYLHS/B1tLhrljd3IBx8i/KgPsTuP1UVVVVVVXFvBd28lrcxLLFKhR0YZDKRggj0xXxE8GT+C9fezUM1lcZks5D3Tup916H8D3qq/YB/5Jr4h/wCw4f8A0RHVVVVVVVVXh+6+06ciMctCdh+nb9Kqqqqqqqqqqqqmmit4XnnlWOOJSzuxwFAGSSfTFfGfx/L8T/idr/jMuxt726K2at/BboAkQx2OxVJ9yT3qqqqqqq2uJbO5iu7d9ssLh0PoQcj+VaZfw6pp1tqMB+S4iDgemRyPw6V4L/5HHQv+wnb/APoxaqqqqqqqq8N3fkXv2dm+WcY5/vDp/UVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVWt7v2LP8AgrRFqsh+w+FPHup+czMxjjaz1UlZGJPAjivNzegEA6dBVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV/wAEiwPF/wC1b8ZviUAZPP024/e5Mg/0vUFm+/0OfIzyMnGexqqqqqqqqqv2wvj9/wAKS+HX2PQboL4q8Rh7fTMHm2QAeZcEf7IIC/7TA8gGpZZJ5HmmlaSSRizu5yWJ6knuaqqqqqqr4Bf8kS8D/wDYDtv/AEWKqqqqqqrw7qXkTfYZn/dyH5Cf4W/+vVVVVVVVVVVVVVVVVVVVVVVVVVVVXt7ZadbPeaheQW1vEMvLNIEVR6kngV8UPEf7LutmS31n4P8AhPxzcgkE3mgW0seeP+WksZPYcqD0HPFePPgH+zb43ufPg/Z98JaEqsGUaUJ7Y8diI5FUj/gIrw38Lvhv4OVB4W8C6Fpjp0kt7CNZD7l8bifcmqr4Hf8AJMdL/wCvi9/9K5qrVtF0fX7J9N13SrPUbST79vdwLNG3GOVYEHgkfjXxA/YN/ZO+I8U39q/BvRtLupQcXWhq2mujf3gICqMf95WHtXxV/wCCOdhIkt98E/itLDIATHp3iWAOpPYfaYFBUduYm+tfGf8AZO+P3wDeSb4j/DvULbTUbC6vaAXVk2Tgfvo8qhPZX2t7VVVVVVVVVVVVVVVVVHJJDIksMjJIjBkZTgqR0IPY1ofxo+MXhcxnwz8WfGWkmIjyzY69c2+zByMbHGOefrWiftx/tfeHwgsP2jfHMuzGPt2qve9Dnnzt+fx+nSvih+2t+0j8bPBD/Dv4t+PLXxVohlWaGO+0OxE1tMpyJYp44VlR8ErkNyrFTwcV+zB+218cv2S5tRh+G+oabqOkaqubjRNciluLMSjpMiRyRskmOCVYBhjcGwuNJ/4Lb/G2FVGvfB7wPeEKu42k11bZP8RG55MA9h2960r/AILk3ClY9c/ZrjcF+ZbTxaV2rjsjWpyc5/iFfty/t1/Cn9snwFoVlH8JNZ8NeL/Dd68ljqD38VzEbeQATQsQqsQ22NwccMmOjE1VfBD9qT48fs66it78J/iLqWlWxkDz6XI/n2Nxzzvt3yhJ5G4AMMnBFfs+/wDBZ3wL4gFroP7Rng6XwxekBX13REe6smbuzwHM0Q/3TL+ArwB8Svh98VvD8Xir4beM9H8S6TKcC6027WdVburbTlGHdWwR3FVVV+19+1l4Y/ZA8A6R478S+F9R19NY1hdNitLGVYnUmGSQyFn4wPLAx1+Yehr4VeO4fin8MvCnxLttKn02DxXo1rqsFpPIrvDHcRLKgYrwTtcVVVVVXjv41/B74XozfEb4p+FPDTKMiPVNXhtpG/3UdgzH2AJrxj/wVM/Yp8Is8EHxRufEFxHndDo+j3M3T0kdEjOfZz74rxN/wWy+A1k0kfhL4U+OdWKZCtem1slcjOCNskpAOByRnB6cYrXP+C4+rSF08Nfs5WcAAYJJfeJ2mz6EoluuPcbj9av/APgtt8c5ZGOl/CLwJbptwq3D3cxDepIkTI9sD61/w+r/AGqP+hC+FP8A4Kr/AP8Ak2v+H1f7VH/QhfCn/wAFV/8A/JtWP/BbD9pCNGGpfDP4a3DZ+QwWt7CAPcG5fP6VpX/BcHx/C4OufALw/dqNuRa63Nbn/a5aOTr29PevD/8AwXB8A3JUeKvgFr+nDHzf2frcN7jr03xw57enU+nPhT/gsB+x14iMa6zf+MPDG8gMdU0MyBOcZP2V5jjvwM4PrxXgf9s79lL4jGKLwn8fvBs082BFbXmpLYzuT2EVxscn2xUE8F1ClxbTRyxSLuR0YMrD1BHUVVV4215vFHjPxB4l80t/bGp3N2WOfm8yVn5zz371/wAE5/E37dfxa8K+HfBfgTxBpPgL4O+EFNtJrf8Awj0Ekt385aSK381W82ZnZy0mNiFiWycI0aMkaRtK0jKoBdgAWPqcADP0Aqqr4z+IP+Ej+JOtXaPuhtpvssI7BYhsOPYsGP41VVVVVV478IWvjPQJdMlCrcp89rKR/q5AOPwPQ/X2FXtnc6fdzWF7C0U9u5SSNhgqw4Iqv2Af+Sa+If8AsOH/ANER1VVVVVVVV4YuPLvHtyeJU4+o/wDrZqqqqqqqqqqqqr9qrxqfA/wN8RXcExS71SIabbFTg7pvlfB7ER+Yfw/Gqqqqqqqr4Uav9q0mfR5Hy9m+6ME/wN6fRs/mK8F/8jjoX/YTt/8A0YtVVVVVVVVRSNFIkqH5kYEfUVFIk8KTIfldQR+NVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVf8Fofgq3iP4U+F/jppFoTeeDb82GpyIDn7HdECN2Pok6oo95zX7GXxtT9oP9mvwT8R7i8WfVpbBbLWjuywv7f91MWHbey+YB/dkWqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq8c6u3h/wT4g16NmVtN0q5uVKnBBjiZuD26V/wQ30dBD8YtfkVCxbRrWIhjuUAXjPkdMHMfPXg9O9VVVVVVVV3dW1jazXt3MsUFvG0ksjnARVGST7ACv2itBj+OfjG88YJqE1pdIn2exSQl41gQnYpX+EkksSO7Hg14l8I6/4TuvsutWDRBiRHKvzRyf7rdPw6juBVVVVVVVfAL/kiXgf/ALAdt/6LFVVVVVVUDjkHkdK0e+F/ZK7NmRPlf6+v41VVVVVVVVVVVVVVVVVVVVVVVVU80NtC9xcTJFFEpZ3dtqqB1JJ6CviD+0npGkNJpngeCPVLpSVa8lyIEP8AsgYL/Xge5rxP4z8T+Mrv7Z4j1m4vGBykbNiOP/dQfKv4Cqqqr4Hf8kx0v/r4vf8A0rmqqqp4IbmF7a4iSWKVCjxuu5WUjBBB6gjtXjD/AIJ9fseeOtWudb1v4M2lpe3bbpZdI1C509QeclYYpBCCSck7OTWqf8Edf2SfEqNL4e8T/EHRZQOIodTt5VXp1WWBmI4PRhyfoK8Rf8EOvD0yM/hL9ofUbVgPkj1Hw4lwG46FknjxzjnafpXiX/gib8fLJXfwn8VPAerBASFvftVk7YzwAsUoycL1IHJ5GMnxN/wSd/bW0AO2n+ANH8QJHnLaZr9qMgZ5AneNj06YzyOK8TfsSftc+Eg51f8AZ28cused72OkSXyqBnJ3QBxjg89PzrxD4I8Z+EZDD4r8I61osgbaU1HT5bYg88YdRzwfyqqqqqqqqqqqqqqqqqqqq+HnxR+Ivwk8QR+Kvhl421jw1qseB9p027aEyKDnY4BxImeqMCp7iv2fv+C0HjHRBbaF+0f4Jj8R2q4Vte0BEtrwDu0lsSIpST/cMIA7Gvgp+018Cv2h9NF/8JPiNpetTLHvn0/eYb22HGfMt3xIoBON2NpPQmqr/gtp/wAkB8C/9jgP/SOev2Tv+TV/g1/2T/Qv/SCGqqq+On7W/wCz3+zjAV+KvxH0+w1IxeZFo9tm6vpRj5cQR5ZQ3Zn2r/tcGvjJ/wAFsdcu2uNM+AnwotrGLlYtW8TzGaUj1FrCwVGxzzK4yeRxz8T/ANuD9q74vPKvjH43+I1tJgQ1hpdx/ZtsV/umK2CK4/39x9TUskk0jSyyM8jsWZmOSxPUk+teH/DniHxbq9v4e8KaDqOs6rdlhb2On2r3E821SzbI0BZsKrE4HABPatL/AGKv2uNY8v7J+zh8QY/NJC/atBntcY9fNVdvTvitR/YU/bC0yD7Rc/s6+NXXOMW+nG4bpn7sZY9vSvF/w/8AHnw+vV0zx74J1/w1eNnbb6xps1nIcdcLKqmqq1tbi+uYbKzgea4nkWOKJF3M7McAADqSSBiv+GKP2uf+jcPiB/4Ipv8A4mviP8HPir8ILmysvil8Pde8Kz6lG8lnHqti9sZ1QgMU3AZAJGcdMj1qq+H3xq+L/wAKJ1uPhp8TvE/hkhtxj0zVJbeNz/txq21x7MCK+Ev/AAWF/ae8DG3sfiFaeH/iFp8eBI17bCxvCo6BZoAEz7vE5Pc5ya+BX/BVX9lz4xS2+j+JNZufh5rkxCi38QlVtXY8fJdqTGB05k8v6V8S/EtrpXwk8V+L9PvIp7ey8O3t7DPCwkR1S3dwylT8wIGeDzX7B3/BMLXfjN/Z3xa+P1he6L4EcJPp2jtugu9cXOQzdGht2H8Qw7g5XaCHrw/4e0Hwnoll4a8MaPZ6VpWmwrDZ2VnCsMUEa9FRFwAPpVVWt6kmjaLf6vIAVsbWSdgeOEUsf5VNLJcTPPM5eSRizsepJ6mqqqqqqq+N3gT7XAfGOlw5mgULfIo5dBwJPqOh9selV+wD/wAk18Q/9hw/+iI6qqqqqqqq0ybyNQt5AekgB+h4P6Gqqqqqqqqqqqqv+ChXidxF4Q8FwyfKzT386Z7jEcZ/WWqqqqqqqrwJq/8AY/iW1kd8RXB8mXPo3T8jg14L/wCRx0L/ALCdv/6MWqqqqqqqqq8Pzedpcak5MZKH/P0Iqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr4wfDXRvjH8LfFPwt18AWXibSp7F5MZMLOpCSgf3kfa491Ff8EiPibrPwt+LXxD/AGQ/HzNZ3zXc13ZW0j4EWoWh8m8iUHqzxojdOluT3qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv2qNVXQ/2Y/i1q2+MNbeCNYaPzAdpf7HKEBxzyxA/Gv+CJGlGH4H+P8AW9r4u/FaW4YphT5VrE3B7n99yO2R61VVVVVVVV+0n4xbQvCEXhuzl23WtuVkx1WBMF/pklR7jdVX1hZanavZahaxXMEgw8cqBlP4GvGXwKVg9/4NmwcEmxmbr7I5/k351qGnX+lXb2GpWcttcRHDRyoVI/8Are9VVVVVXwC/5Il4H/7Adt/6LFVVVVVVVXh+8NrfLEzfJP8AKR79j/n1qqqqqqqqqqqqqqqqqqqqqqqqrxr468O+AtKOqa/eBN2RBbpzLOw7Iv5ZPQZ5NfEb4v8Aif4hzvbzzGy0lWzFYQt8px0Ln+M9OvA7AVVVVVXwO/5Jjpf/AF8Xv/pXNVVVVUcjxOJInKsp4IOCK0rxCk223v2Cv0WToD9fSuCODVVLFHNG0MsaujqVZGGQwPUEelft7/tV/sg/BoX/AIEsfgn8O/iV8SCGSW0vNDtp7bSXOfmupdhbzAST5SEP13NHkE+INYl8Q63fa5NY6fZPfXDzG20+1S2t4dxzsjiQBUQdAB2qqqqqqqqqqqqqqqqqqqq0nWNW0DUrbWtB1W703ULNxJbXdpO0MsLjoyOpDKfcGv2fP+CuX7QPwtNrofxVig+JWgRbUMl7J9n1KJPVblQRKec/vVZjgDetfs6/t7/s3/tLy2ui+DvGB0nxRcLx4d1tBa3bMFyRFyUmwAxxGzHAJIAr/goj+yj8RP2uPhh4b8FfDfWfDmm32ja9/aE8mt3E0MbR+RJHhTFFKS2XHBAGM818EPBOq/DT4LeAPhxrtxaT6l4V8L6bpV5LaOzQyTW1rHE7RllVihZCQSqnGMgdKqvGfjTwn8O/C+o+NfHHiCy0XQ9IgM15fXkojjiQe/ck4AUZLEgAEkCv2uP+Ct3xC+Id1feCf2b5Lrwd4YDNE2vMNmqXwH8UZ/5dUPbb+8xg7kyVGo6jqGr30+qarf3F7e3UjST3FxKZJJXJyWZmJLEnqTVVX/BLj/k+34Y/9xb/ANNN5XxE8d+H/hf4F174ieK5Z49H8OWEt9fNBEZHWKNSzFVHU4HStH/4K/fscanqK2N7qfi/SYS2DeXmhFohz1xC8j47/drQPEnwU/aW+HZv9EvfDPxA8H6oDHKjxx3kDNjlJYnB2OARlHAYZ5Ar/go5/wAE4NG+CmiXPx4+BFrOnhCOdRrmgu7SnSd7BVnhZiWaAsVVlYlkLAglCdlf8E4/hKfi/wDte+BdMubUTab4cuTr2oBk3Ksdph49w6FWn8hCDxh/wrWfFeg6Bq+gaFql8sV94mvZLPTIT1mljt5bhwPYRQSHPsPWv+CxvwmHjX9mmw+JFnbh774fazHNI+zJFndbYJQPT979mYn0Q1VVVfs6/tm/G39myZtM8K66ms+EbvKaj4U1oG6067ibIkURt/qiwJBZMZ43BhxX7JH7afwm/a38MmfwnP8A2N4o06ENqvhq8lU3FqBgGSMjAlhyQA4AxkBgpIFVVV8ab9tO+FviG4RsF7YQ9O0jrGf0aqqqqqqqqWKOaNoZUV0dSrKwyCD1BFfE3wQ/gzXmS2Rjp15mS0Y87fVCfVcj8CK/YB/5Jr4h/wCw4f8A0RHVVVVVVVVQJBBU8jpUbCSNXH8Sg1VVVVVVVVVVVftx60+qfHa408yhl0fS7a2VR/DuUzHv1/e+3aqqqqqqqoHHIPI6V8ItWGt6l4W1Avuka/tll/31lUN+oz+NVVVVVVVVV4VkzFcQ5+6wYD6//qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv+Ck/g3xB+y1+2B4G/bA8AWbLba5dxXN0qDahv7UKk0TEDCrPbbQe7Hzj614F8aeH/iP4L0Px94Uvlu9H8Q6fDfWUw/iilQMuR2YA4I6ggg8iqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv2+9VGjfsa/Fm7Mwj8zw9Jbbiu7PnOkWMe+/Ge2c8V/wAEatL/ALP/AGSL+78p1/tLxlfXGWPD4gtosj2/dY+oNVVVVVVVVfHLxOfFHxG1J45d9tpzfY7cA5AEfDEfVy5+mKqq8ReFdB8VWn2PW7BJwAfLk+68Z9VYcj6dPWvGXwW1zQvMvdAL6pZLk7FX99GPdR976r+QplZWKMCGBwQRgg1VVVV8Av8AkiXgf/sB23/osVVVVVVVVAkEFTgjoasbkXlnDcAjLrz9e/61VVVVVVVVVVVVVVVVVVVVVVXxM+J+ifDfSxNdkXOpXCn7JZK2Gc/3m/uoD3/LNeKPFWueMtXl1rX75ri4k4A6LGvZUXso9PxOSSaqqqqqvgsqr8NdKVQB+9ujj3NzLmqqqqqrR9da2K2t45aLornqn/1qBVlDKwII4IqaaG3heeeVI44lLO7ttCgDJJJ6DFft7f8ABU64vn1L4Nfsu64YbUb7fV/GVs5DynOGjsWH3V4IM45Of3eAA7SySTSNLLIzyOxZmY5LE9ST61VV8HP2efjT+0BrH9i/CL4d6t4heOQJPcwxbLW1JGR51w+I4+OcMwJ7Zr4J/wDBFB5YYNV/aD+KTRMQC+jeF4wSvs11MpGexCxfRu9fDb9gX9kL4WJE/h34HeH767iA/wBM1uJtUlLf3gbguEP+4FHoBUXhbwxb6X/Ydv4d0uPTiebNLNFh6Y+4Bt6ADpXib9lb9mfxjvbxJ8Afh/eyyZ3Tt4dtkm5zn94qBx1J69ea8Tf8Ewv2I/E2+RvgymmTvnEumaxe2+3OeiCUx9/7vYduK8Sf8EX/ANlzVQ8mgeLfiDokpJ2ImoW9xEv/AAF4Cx/77rxH/wAEObBw0nhH9oqeIg/LDqXhoSZ6dZEuFx3/AIDn2rxF/wAEU/2jLEl/DPxH+H2rRqPu3E93aSN06DyHXrnqw4A9cDxD/wAEpf22tDV3s/hnputIgJLafr9nnAGThZZEY9OgBJzwK1/9h/8Aa98NO0eo/s5eO5ipwfsGjyXwznHBgDgjjr+PQ1r/AMMviR4VVj4o+H3iXRwilmN/pM9vtAGSTvUYGOfpVVVVVVVVVVXwS/Yh/ac/aA+z3fgD4Xaimjz4I1rVV+wWW0/xrJLjzR7RBz7V8E/+CKnhDSvI1X4//Eu51y4XBfSPDqG2tge4a4kHmSKePupER618Jf2ePgj8CdPGn/Cb4Z6H4dymyS5t7bddTD0kuH3Syf8AAmNVVV/wVO/a11r4z/GPUfgv4c1Mx+BvAF89sYomwL/UowUnmfHURtviQcgBWb+PAqqqv+CXH/J9vwx/7i3/AKabyv21/wDk0X4v/wDYn6h/6Jaq/wCCa37QXiH4JftOeFtCj1WZfDHjzUIdF1ixaQ+VI87eXbzYzgPHM6HeRwhkHAYmvGPhXRfHfhLWvBPiO0W60rXtPnsL2FhkPDKhRx/3yxrxNoV14X8R6r4ZvmBudIvprOYgY+eJyjcfVTX/AARK+ExtPDfxA+N9/a/PqN1DoOmyMMEJEomuceqs0luM+sRHrj/gpB+0xF8KP2t/2e7C3vttv4JvDrutJG/W3upVt2U9g3kRXOB6Sgngiviz4A0v4u/CvxV8NdRljFp4q0W508zbdwj86Iqsg91JVh7gVrej6j4e1m/8P6vbNb3+mXUlrdQsMGOWNirqfowIqqqq+GXxL8Z/B7x3o3xI+H2tS6Zruh3Kz2s6E4bH3kdf443XKsh4ZSQeDX7OXxp0j9of4J+FPjBo1uLZPEFlvuLXdu+zXMbGOeLPcLKjgE9Rg96qq/bI1y/0H4GajdabL5c8t9axh8A7R5gJ4IIP3cfjUXxL8XRjD30UvH8cCj+QFRfFjxInD29hJ/vRMD+jVF8XtSGPO0i2f/ddl/xqP4woQBNoBHHVbnP6baj+LujNgTaZerz/AA7W/qKi+KnheQfOt7Hx/HCP6E1F8R/B8vB1RkPYPA/9BUXjbwnNgJrtsP8AfJX+dR+IvD8wHla7p7HHQXKZ/LNR3dpPgQ3UMnOBtcHmqrxl4Ws/GGgT6PdYV2G6CUj/AFUg6N9Ox9ia/YT0280fwL4p0vUITFc23iBkkQ9iII+nt3z3qqrGelbSeAP0raxHArYx6CtjEV5ZPcV5eeN1eWMYzXlr615a4xVid1jbn/pkv8hVVVVVVVVVVVV+1Hevf/H7xnPISSl8sQy2eI4kQfoo47VVVVVVVVV+zhq4XxrpuhyPw+pW08QPqJFDfpt/I1VVVVVVVVXhZiLqZAeDHn8j/wDXqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv22f2e4v2mP2dPE/wAOrS3ifXoYhqPh+RwB5d/AC0agn7okUvCT2WUntX/BHL9oK717wT4g/Zo8XXTx6v4Lka+0aG4ysv2KSQieLaeR5M7AkHn/AEgDotVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVf8FQdSOm/sM/Epo50jluF02BA2Pn36lbBgB67N35Zr/gkzpy2X7E/ha5VJAb/VNTnJccMRdPHlfb92B9QaqqqqqqqvFutp4a8MaprzkZsLSSVAe7BTtH4nA/GpJHmkaWVy7uSWZjkknqaqqqvGHwx8NeL1eeaD7HfkcXcC4Yn/aHRvx59xXi/wCG/iTwczS3lt9oss4W7gBKe24dVPTrx6E1VVVfAL/kiXgf/sB23/osVVVVVVVVV4Xn32stsTzG2R9D/wDqqqqqqqqqqqqqqqqqqqqqqqr4i+PtK+Hfh6TWdQIlnc7LS1DYaeT09lHUnsPcgHxL4j1fxbrNxr2uXRnu7lsscYCgdFUdlA4Aqqqqqqr4L/8AJNtK/wCulz/6USVVVVVVVomtraD7LeyqsAB2yMcBPqfT+Vf8FG/+CjVx8YrjUPgX8C9Zkh8CQO0Os6zA5VteYHmOMjkWoI/7a/7n3qqvBvgrxb8RPEth4N8C+HL/AF3XNTl8u0sLGAyyyt3wB0AAJJPAAJJABNfsp/8ABHbR9MjsvGn7VF+NRvcCRPCOmXBEER7C5uEOZD6pEQoI++4JFeFvCfhjwPoFn4V8G+H9P0TR9PjEdrY2FssEMKjsqKAB/WqqqqqqqrX/AIbfDrxYGHinwD4c1neSW/tDSobjccgknepzyAfqBWufsW/sk+ImL6j+zl8P1Y9WtNChtCTknJMKrk88nvWtf8EvP2HtZw3/AApf7DIMDfZa7fxcDtt88r364z71rP8AwRu/ZF1QYsdQ8f6RwP8Ajz1mFun/AF2gk6/5xWtf8ERPhBcBh4d+NXjCyyPk+22VtdYOe+wR5447c8+1ax/wQ31BGZ/D/wC0lbzAltsd54VaMqOwLLdNn3O0fStW/wCCJXx4hVv7C+LXgG8IUbRdi7tstnkHbFJgY78/StW/4I4ftd6cWFneeAdU2sADaa1Ku4Y6jzYE47c81+0J+yJ8av2XodLm+L+l6Rpw1uaWPT0ttXguZLgRgb5BGjFwg3L8zAcsB1NVVfDD4I/F341ap/Y/wp+HWu+J7hWCytYWbPFBnHMsvCRDkcuwHIr4J/8ABFv4m+IVg1X47eP9O8J2rAM2laQov7z3V5MiGM+6mUV8E/2Av2VvgN5F54V+GNnq2swYI1nxBjUbrcP4l3jy4mwcZiRKqq/aW/aj+Gf7KXhPS/GXxPg1qWw1fUfsFuNLtVncS+W0nzBnTC7UPOeuK8BeMtH+I/gXw58QvDy3C6X4o0m11SxW5QJKILiJZY96gkBtrjIBOD3NVXxc0rWtD+K3jPRfEisNWsPEF9BfBxg+ek7rJn/gQNVVVX/BLj/k+34Y/wDcW/8ATTeV+2v/AMmi/F//ALE/UP8A0S1V+yr4ev8AxV+0z8KtC02NmmuPGOmElV3bES5jeR8eiorMfYVXxd1218U/Fjxr4msWVrfV/EV/eQlTkFJbh3GD6YYV+xR8JT8E/wBlr4eeA7i1aDUY9IS+1NHXDrd3RM8yt6lGkMf0Qelfti/8E4f2r/2j/wBorxd8WNJv/BMekajNFDpUN1q8qSRWsMSRx7lEJCk7S5GTguea+BWheOvC3wa8F+FviZLZS+KNF0S2sdTls5jLFNLDGI/MDEKTvChjwOWPFf8ABUT4S/8ACq/2v/FN1aW5j07xrHH4gtPl4LT5W457k3EczfRh9TVVVV/wSC0zVrD9jeyutSVxb6l4j1CfTyxyDCCkRI9B5sUo+uaqq/bh/wCSDXf/AGE7X/0I1VVVVVVUV3dQDEF1LHgY+VyP5VF4h1+H/U65fp7C5cfpmovGviuHGzXbk4/vsG/nmvh7+0b8QPh0tzFp8el38N5KJJ0u7XBZgoUHdGVOcADJz0FeHf24NCmCReLPA15akDDS6fcrOD77HCYHtuNeHP2lvgr4k2RxeMYNPmYcxalE1tt+rsNn5NWm6rpOs2wvNH1O0voD0ltpllU/ipIqqqqqqqtPA+wWwH/PFf5Cqqqqqqqqqqqr9oH/AJLd45/7Dt1/6MNVVVVVVVVfs5/Bv4h694w0Lx1aacbDRdNvYrh7y7BQXCKwLLEvV8jI3fdB78Yoo3bFFTjpRHaqqqqqqq8L/wDIQkAP/LE/zFVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX7bvhTxH+wx+3B4b/AGqPhzYyJ4e8W3rajcW8Q2xyXHC6jaN2AmR/NBP8UrED93x4J8ZeHPiL4P0Xx34R1FL7RdfsYr2wuFGPMikUMpI6qcHBU8g5B5FVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX/BW6+Np+xZ4gt1jDfbdZ02EknG3FwHz7/cx+Nf8Ey7D+zv2Gvhdb+d5m+3v5s7duPM1G5fH4bse+O1VVVVVVVX7U/jHTfB/wpkOp3TQR6tfwWasqFjnmU9MnGIj/k1aeLPDN9gW+uWhJ6K8gQn8GwaSSOVA8Uiup6FTkVVVVOiSIY5FVlYEMrDII78V4y+CWjazvvvDbppl2eTCR+5c/Qcp+HHtWv8AhrW/DN39i1rT5Ld+djEZVx6qw4Iqqr4Bf8kS8D/9gO2/9FiqqqqqqqrBPAFeGpGj1AxnpJGRj3HP+NVVVVVVVVVVVVVVVVVVVVVWq6nYaLptzq2p3KwWtpE0k0jdFUDn/wDV3r4k+PtR+IfiWbWLpmjtY8x2VsTxDGOn/Aj1J9fYDFVVVVVVfBf/AJJtpX/XS5/9KJKqqqqqqr/gpp+yBD8JvFh+OPw80pYfCHie6K6naQJhNLv25JUD7sUvLAdFcMOAUFVX7K/7IXxW/a08Y/2B4FsRZaLYyL/bHiG7jb7Lp6Htx/rJSM7YlOT1JVcsP2Y/2Qvg5+yl4XGjfDzRBPrF1Eq6p4gvVD3t+w65b/lnHnpGmFHBOWyxqqqqqqqqqqqqqvE/iTQvBvhzU/FvifU4NO0jRrSW7vrudtqQQxqWdyfQAE1+1/8AtJa7+1P8b9a+JWoNPBpCt9j8P6fI3/HlYRk+WpGSA7ZMj443u2OAMaF4f17xTqtvoPhjRL/V9Tu22W9lYWz3E0rHsqICzH2Ar4J/8Elv2o/ikLbU/Gdhp/w50ebDGTW38y8KHHK2seWDc/dlaI8H2z8E/wDgkv8Ast/C0W+peNNPv/iPrMRDGXW38u0Vx3S1jIUqf7splHP5aF4f0DwtpNvoPhjRNP0jTbRNlvZWFslvDCo7KiAKo9gKqqqv+C2n/JAfAv8A2OA/9I56/ZO/5NX+DX/ZP9C/9IIaqv8Agq9+w7rsuvX37U/wp0eW/tLuIN4w063jzJbOihReoo5aMqAJccqV3nIZylVVV/wS4/5Pt+GP/cW/9NN5X7ZVje6n+yj8WdO02znu7q48JX6QwQRmR5GMLYVVGSSfQVoHwD+OXirUotH8OfBzxrqV7KcLDb6DcufqcJwB3JwB1Jr/AIJtf8E6fFXwM8TL8d/jnaWtt4pjtXh0LRI5VmbTPNUrJPM6Er5pjLIFUkKrvk7iAv8AwUF/aX0r9m39nrXLy21KOPxd4ptpdL8OWyviTzpF2yXAxyFhRi+em7Yp+9X7H/wlX44/tMfD34bXFsJ7DUNZjn1KMjhrO3BnuFPpuiidQT3YdelfEjxtpvw1+Hvib4iawN1l4Y0i61K4XdgskETSFR7nbge5Ff8AD8XxV/0btpP/AIUkn/xiv2EP234/2zNH8Wy33g238Mat4VubZXs4b43QlgnV9kgYohzuikBGDjC881/wWq+Eo174TeDfjJYW5a58Kaq+m3xVf+XW7UFXY+iywoo95jVVVV+yp+yx8Qv2r/iVb+CPBtq9tpdqyS67rUkZMGmWxPLH+9IwBCR9WI7KGZfh18P/AAv8KvAmg/DjwXYfY9E8O2MdnZRE5bYgxuY/xOxyzN1LEk9aqq/bJ0/7f+zz4kkVQz2ctpMo2ZPFzGpI9PlYnPpn1qqqqqqqqqqqrDUtR0q4W70rULmznX7stvK0bD6FSDXh79pT40eHAscHja5vol6x6ii3W76s4L/k1eHf24dfgCxeLPBNheDoZbC4a3I99r7wT7ZFeHf2wfg/rOyPU7nU9Ekbg/bLQugPs0Rfj3IFeH/iD4F8WBR4b8X6RqLt0jgu0Zx9UzuH4iqqqqrVNlrCmPuxqMfhVVVVVVVVVVVV+03Zix+PfjSAKi7tRMuF/wBtFfP1+bn3qqqqqqqvAvw38Z/EnUxpfhDRJrxlI82cjZDAPV3PC/TqewNfCj9kvwd4LEGr+MzD4i1hMMI3T/RYG/2UP+sI9X46fKKVERAiKFVRgADAAqqIBoqp7CjGpGAaMfHDUUbHHNFSO1EdqqqrwsgN3K/pFj8yP8Kqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv2v/ANnXSf2ofgRr/wAMLvyotUKfbdCu5B/x66hED5TZ7K2Wjb/Zkbviv+CSP7R2q+HNW1v9jL4pNNYavo91czeHoLs4eKWNmN5Y47MrB5VH/Xb0FVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX/AAWQunt/2RLeFJygufF9jGy7seYBFcNj35UHHtX/AAT9tYbT9jL4TQwRCNW0FZCoGPmeV2Y/iST+NVVVVVVVX/BQ3VHh8P8AgvRATtu7y7uCMd4kjUf+jjVQXV1atvtbmWFvWNyp/SrTxx4rs8CLW53A7TYk/wDQgatPix4ghwt1a2dwB1O0ox/EHH6VafF6wcAX2jzx8cmKQP8AocVafEjwldYVtQeBj0E0TD9RkfrVprmi34AstWtJieipMpP5ZzVVqelabrVm9hqtlFdQOOY5FyPqPQ+4rxl8CriDffeDpjMnU2UzYcf7jHg/Q4Puau7O6sLh7O9tpYJ4jh4pUKsp9weRVfAL/kiXgf8A7Adt/wCixVVVVjPAoKxHAryye4oRjua2L9aCqOgFVpD7NSt2B/ix+YxVVVVVVVVVVVVVVVVVVVVVV+0t8QzdXSfD7S5v3VuVl1FlP3n6pH9AMMfcj0qqqqqqqq+C/wDyTbSv+ulz/wClElVVVVVVV8Rvh/4Y+KvgXW/h14xsRd6Pr9m9rdRnqA3R1PZ1YKyt2ZQe1fHP4Q+IfgR8VfEPwt8SqzXOiXTJDceWUW7gb5oplHo6FTjsSR1Br9iP9iLxr+13413v9p0bwDo06jXNcEfJPB+zW+eHnYEeojBDNnKq/wAMfhf4D+DfgnTvh58NvDlromg6WhW3tYATkk5Z3Y5Z3Y8lmJJPU1VVVVVVVVVVVVVV/wAFB/BH7TP7Rdhp37OHwG8KPa+Hr1kvPFniXUrkWVmVBzDaKxy8q7h5knlo+CIhn74r4Kf8EWvhh4eNvqvx18f6j4tu1wz6VpCnT7MHurS5M0gx3UxGvhj8EvhH8F9KGjfCr4d6F4Ztym2Q2FmqSzAf89JeXkPA5dieKqqqqqv+Co/7PXxg/aN+EHhPwt8GfCH/AAkOqaZ4kF5dQf2hbWnlw/ZpU3briSNT8zqMAk89MV+z14W17wN8Afhn4J8U2H2LWvD/AIP0nT9RtfNSTyLmCzijlTehKth1YZUkHGQSOaqmVWUqwBBGCCK/a4/4JJeAvincX3jz9nu7sfBPiadjLPossZXS71z1KBATasc/who+ANi5LV8ZP2e/jN+z/rh0D4t/D/VNAlLlYLmWLfbXXXmGdMxycDPysSO4FVVfsE/FHwJ8F/2s/AnxK+JevDRvDejnUBe3ptpbjyvN0+5hj+SJXc5kkReFOM5OACRZf8FCv2LtQhM1v+0H4cVVbaROs8LZwP4XjBxz1xitb/4KO/sT6BA0158fNHn2puCWVnd3bHnGAIom5z2/E8c18Zv+C0nwn0GyuNP+B3gLWPFWqY2xX2sAWFkhIOG2AtNJg4ypEef7wr43fHT4m/tEeOrn4h/FTxHJquqTL5cKBfLhs4QSVhhjHCIMnjqSSSSSSf8Agl78YP2ePgH8UfFHxO+Ovj1PD9zFo6afocTaXeXhmaaTdPIPs8UmwosKL82MiU4zg4/b+/4KIfs6fE79mPxD8Nfgf8S313xB4muba0nhTR760MNoJBLM++4hRcERiMgEn950wCRX/BNf9pXwl+zP+0FJrnxH16TSvB/iDRriw1S6FtLcLA64lgkMcKu7HfH5Y2qceaScDJH7Tf7d37B/x0/Z/wDHfwoHxvSS51/RpY7ASeGtVCi8jxLbMSbXAAnjiPbp1FVVaTo+reINSttF0HS7vUtQvHEdtaWkDTSzOeioigsx9gK/Zh/4JD/Fr4kT2Xij4+XMngLw0xDtpihX1W6X+7s5S3B9Xy4/5585r4S/B34bfAzwXaeAPhZ4Us9C0a058qBSXnkwAZZZDlpZDgZZiTgAdAAKqq+OWht4j+DvjPSI4/Mll0W5aJAM7pEjLoPxZRVVVVVVVVVVVVVUCQQQeR0rw78X/ih4U2LoPjvWII0+7C9yZYh/2zfcv6V4d/bP+J+mFI9e07R9aiH3maE28p+jIdo/74rw7+214EvtkfiXwxq2lSNwXgZLqNfqfkbH0U14d+PXwf8AFIRdL8faWkjjiK8kNo+fQCULk/TNQzQXMSz20ySxuMq6MGB+hFQxmaVIl6uwUfjWBjAqqqqqqqqqqqq/bc0dtM+Pmo3rIQNW0+1uVJHUCPyc/wDkHHbpVVVVVVpelanrl/DpWjafcX15cNtht7eIyO59Ao5NfCj9jWe5EGtfFa6MMZAZdItJPnPtLIPu/wC6nPP3hjFaF4f0PwxpkWjeHdKtdOsoB+7gt4gij1OB1J7k8nvVVVVVVVEA0UUjGKManoaMfofzoo3Qc14ViIS4lI6lVH65/pVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX/BU/wDZ28SfCX4iaH+278FhNpt3BqFudfktEx9jvoyot73A42ybRHJn5SwTOTK1fsoftH+Gf2pvgvo/xP0Iw29+6/Zdc01H3HTr5APNi552nIdCeqOpODkCqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv+C1s8K/su+E7ZpVEr+PrV0QnkqthfAkD0BZfzFfsWwywfskfB+OaJ42Pg3TWCupBw0CkH6EEEexFVVVVVVVX/BQ26D+IvBlkN2YrK6k56fM8Y49/k/lVVVVVWmsatYACx1S6gA6COZlH5A1afEXxba4B1ITqP4Zolb9cZ/WrT4vakgAvtJtpv8Ark7R/wA91WnxZ0KbC3dldwHHUAOB+oP6Vrlx8MfHduINVu4PNC4jnYGGWL6MR09jkV4p+FeqaOj3+gXUWt6eBnzLYhpEH+0oJ49xkeuK+AX/ACRLwP8A9gO2/wDRYraT0H6VsYivLyOWry1xjNbVxgAVgYwKqqqqsSFvbds9JV/mKqqqqqqqqqqqqqqqqqqqqrxt4otfBnhbUfEl0Aws4SY4z/y0kPCL+LED2HNahf3eq31xqV/OZrm6laWaRurOxyT+Zqqqqqqqq+C//JNtK/66XP8A6USVVVVVVVVftnfsKw/taeL/AABrWh6naaJqFhe/Yde1B1+dtKIaQlBj55EcEIpwMzsScCvhf8MfBPwb8B6P8Nvh5osel6DokHk2tupLE85Z3Y8s7MSzMeSSTVVVVVVVVVVVVVVVVVVVVVVVVVVWv+HfD/ivSbjQfFGh6frGmXS7Z7K/tUuIZR6Mjgqw9iK+Mf8AwSS/ZW+JbT6j4OsNV+HuqyksH0WfzLUt/tW025Qv+zGYx+ufif8A8EZv2jvCpluvht4o8L+OrVCfLh806Zdv/wBs5iYh/wB/q+IP7JH7TXwtMreOfgb4vsIICRJdx6Y91bLj/pvDvj/8eqWKSGRoZY2SRGKsjDBUjqCOxqqqqqqrwZ8LPib8R5xa/D34d+JfE0ucbdI0ma7I+vlqcfjXw2/4JWftkfEJ4pb/AMCWHg+ykI/0rxFqKQkDvmGLzJh+KCvhN/wRP+H2ktBqHxp+K2q+IJVIZ9O0K2Wxgzx8rSvvd168qIz9K+EH7OPwM+Alj9i+Efwy0Tw8zJslu4YPMupl44kuJC0rjjozEZ+tVVVVSxxzRtDKgdHUqysMgg9RXjnw1P4N8Z654TuQfM0jUJ7XJ/iCOVDD2IAP41VVVVVVVVVVVVVVVVofirxP4Ym8/wAN+ItS0uTPLWd08JP12kZr9k/4wfF7x58VNN8J6z4i/tPSYbea5vDcWyGRI0TCkOAG/wBY0Y5J61VVVVVVVVVVVV/wUK8NOmo+EPGEUeVlgnsJnx0KsJIx+O+T8qqqqqr4U/ss+OfiB5Oq69G/h7RHwRLcxnz5l/6ZxHBwf7zYHORur4efCfwN8L9P+xeFNHjimdcT3so33E/+8/XH+yMKOwqqqqqqqqqqqq0CHy9NViOZGLf0/pVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVXjLwf4b+IPhPVvBHi/SodT0XXLOS0vrSYZWWJ1ww9QecgjkHBBBFeBPEHjn/AIJRftiXngnxZcXmofC/xWyebcBCVvNPLkQ3iAf8t7clldB1+cDh0atI1fS/EGk2Wu6HqFvf6dqNulxaXVu4eOeJ1DI6sOCpBBB96qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/4Laug+AngSMuu4+L8hSeSBaT5OPxH51+yd/wAmr/Br/sn+hf8ApBDVVVVVVVV/wUMtmXxP4Ou94IksLlAuOQVkQ5/8eFVVVVVVVVKzIwZHKsOhBwRXwSJb4ReD2Y5Y6Pbkk9SdgqqqqqqqqqyXdeW6g9ZVH6iqqqqqqqqqqqqqqqqqqqqq/al8VmS40zwXay/LEpu7oA9WOVjB+g3nH+0Kqqqqqqqq+D6qvw50kKAOZzj6zyZqqqqqqqqikeGVJozhkYFfqKt5kuYI54zw6gj2qqqqqqqqqqqqqqqqqqqqqqr44ftH/Bf9m/SdM1v40eM/+EestYuHt7KX+zrq782RV3MuLeOQrxzkgCvD2v6R4r8P6Z4o0C7+1aZrFnFeWU/lsnmwyoHRtrAMuVYHBAIzyBVVVVVV8StK/Z11gPF8YdO+HF9x86+JYbKToMcicHoBivGfwK/4JV63Iya34c+G8bMxB/sbU5bUZGOhs5FAHA6cHn1OdV/Yd/4Jd61ZjUNM8YX+lW8sjRRyWXiKZhvTazAecsnOHTOexFP/AMEyv+CfeuH/AIkH7SviO0llcMkdx4i00qATwiq9urZ6Dkk+uag/4Iy/s7eIrUTeEfj34pugD80sbWV2mP8AgCrjt3q2/wCCInwhWdGvPjX4vlhB+ZI7K2jY/RiGA/I1af8ABE39nVJ1a/8Aih8RpoQDlIZ7KJj6YY27AflWk/8ABG79kTTlQXl94/1UoCCbvWYl3ZPU+VAnTpxj3zXh7/gmD+xD4ddZ0+C6ajOhyH1HWb64B5BwUM2w8j+73I6HFeEv2Xf2bvAjRS+EfgR4D02eEYS5j0C3M4wMf61kLnj1Pc+pqGGG3hS3t4kjjiUKiIu0KAMAADoMdqqqqqqqr9ufwK3hv4uR+K4IgLTxTZrLkDA8+ELHIP8AvkRN9XNVVVVVVVVVVVVVVVVV/wAE/PAxg0zxJ8RrqIhrqRdNs2Ix8qYklPuCTEPqhqqqqqqqqqqqqr9sDwW/jP4F609vHvudBZNUhGO0WRKfbETSH8Kqqqvhv8HPHnxTvBD4Y0k/ZEbE2oXOY7eH1y2PmP8AsqCfavhT+zJ4D+G/k6pfxDXtcjwwvLqMeXC3/TKPkL/vHLehHSqqqqqqqqqqqqkRpHWNBlmOAPeoIVggjgU8RqFH4VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX7Z37KPhn9rb4RXPgy+aCx8SaYXuvDerSJk2d1jlGI58mQAK4Gf4WwSi1/wTp/a58T/s/wDj65/Yr/aUefSY7TUWstCuL98HSrwtzaO3QwSsQ0bg4DN1KuCtVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX/Bb3/klXw0/7GC6/9JxX7PX/ACQH4Z/9ifpP/pHFVVVVVVVV/wAFD9M32PgfWUUfupb23c467hCy/wDoDfnVVVVVVVVVfBH/AJJB4O/7A1v/AOgCqqqqqqqqtLQSajbr6OD+XP8ASqqqqqqqqqqqqqqqqqqqqq+K3xQ1fxb8V/EvjPStWnjhvb9xa7X+UwJ+7iyvQ/Iq1pPxZ1GALFrFjHcqOskR8tvy6H9K0rx34Y1fakWorBKf+WdwPLP59D+BoEMAVIII4NVVVVVVfCD/AJJ1pH/bb/0c9VVVVVVVVeGbjzbJrcnJhbj6H/6+aqqqqqqqqqqqqqqqqqqqqqq/4Le/8kq+Gn/YwXX/AKTiv2ev+SA/DP8A7E/Sf/SOKqqqubm2tIGubu4jhijGWkkYKq/UnpXxL/bu+B3gRpLLQdQuPF+oJx5elAeQp/2p2wpHum+vHP8AwUQ+M3iF5IfB2naN4VtjnY0cP2y4H1eUbD+EYrxX8Zfix45DJ4s+I3iHUomJJt5tQk8kZ64jBCD8BVVo3/JP9O/7DF9/6JtKqGea2lWe2meKRDlXRipU+xFeFv2hvjX4N8tND+I+seVHwsF3N9rjA9AkoYAfTFeCf+CgPimyMVr8QPBtjqcQwr3WnSG2lA7sUbcrH2BQV8Ov2lvg78Tnis9D8Ux2epS4C6dqa/ZpiT0Vcna59kZqqqqqqqqqr9sj4bnx58HbzVLKDfqXhdzqEGBy0SjE6/TZ8/1jFVVVVVVVVVVVVVVVVWtrcX1zDZWcDzXE8ixxRIuWdmOAAO5JIGK+EXgK2+GXw30DwTAE8zTrRRcuvSS4b5pW+hdmx7YHaqqqqqqqqqqqqr+xtNTsbjTb+BZra6iaKaJujowwwPsQSK+Jvgm7+HPj7XfBN4GLaVePHE7DmSI/NG//AAJCrfjVV4c8MeIPF+qxaJ4Z0i51K+m+7DAm4gdyT0VR3JwB3NfCj9jjTNNMGt/FG5TULlSGXSrdiIUP/TRxy5/2RheOrA1YafYaVZQ6bpdlBaWtugSGCCMRpGo7Ko4AqqqqqqqqqqqqqtBtTcXyyEZWEbj9e3+faqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv8Agov+wdZftO+FT8Rfh1ZQ2/xO8P2pWBRiNdbt15+zSMSAJByY3PGSVb5SGT/gnL+33f63dQfsu/tJajPp3jTSZf7P0LUdTQwyXhj+T7FclsFblSNqlgDJjaf3gG+qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/4Lg3yx/D74WaZ5ZJn1m/mD56bIYhjHv5n6V8DrF9L+Cfw+0ySRXa08LabCzAYDFbWMEj8qqqqqqqqv27dBOq/BSPVY48to2sQTsw7I6vEfwLSJ+Qqqqqqqqqqvgj/AMkg8Hf9ga3/APQBVVVVVVVVXh+LzNRVsf6tCf6f1qqqqqqqqqqqqqqqqqqqqr44eKH8GfCLxb4jhcrPa6VMtuw/hlkHlxn8HdTVVWleJNc0Ugabqc0Sj/lmTuT/AL5ORWlfFyZdsWtaasg7y25wf++Tx+orSvGPhzWdqWepxiU/8spf3bZ9gev4ZqqqqqvhB/yTrSP+23/o56qqqqqqqq8MzeXfmEniWMgD3HP8s1VVVVVVVVVVVVVVVVVVVVVV/wAFd/hD8Ufi/wDDjwBpvwu8A654qutP1u4luodKsnuGhQwABmCg4BPFfA/StR0P4K/D/RNYsprO/wBP8Labb3VtMhR4ZUtY1dGB5DBgQR2IqqZlVSzMAAMknjFfGT9tXwN4Dkn0LwJFH4p1qMlWlSTFnA3vIOZCOOE4/wBoEYr4g/G34nfE++W88W+LLuaOKXzILSBvJggYdCka4GR2Y5b3rxfYxW2rm8toljt9RT7TEigAIWJDqAOihw4A/uhfWqqq0ghfAWmxE/MdVvXA/wBkxWwB/NWH4VVVVfCn9q74rfC4w6edU/4SDRY8D+z9TcvsX0jl+8nsOVH92vg/+0j8N/jHDHaaRqP9m65szJpF6wWXIHPlnpKvB5XkDkgVVVVVVVVSxRTxPBPGkkcilXR1yGB4II7jFftCfCub4RfE/VPDUUDrpc7fatKkYHDWzklVz3KHKH3XPeqqqqqqqqqqqqqqqr9iT4Ty+MviL/wnupWobSPChEkZdeJbxgfLA/3BmTjoQnrVVVVVVVVVVVVVV+3x8LDLDpfxb0m1JMQFhqxVf4ScwyH8SyEn1QVHFJNIkMMbSSOwVEUZLE9AB618Kf2RPFfivydY8fSS+H9LbDC12j7XMP8AdPEQ/wB7Lf7POa8F+AfCPw90oaP4Q0S3sIODIyDMkzD+J3OWY/U8dsVVVVVVVVVVVVVVVolmLWyVmH7yX5mz29B+X86qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr/goT/wAE9LD9oaxn+LvwftLfTfijp0QaWFGEMevog+VHbgJcKAAkpwDgIxxtZP2G/wDgpPqOi6nH+zj+2BeXWka/pMxsLLxLq5aOTzEO37NqBflJARtEzdf48HLsjpIiyRurKwyrA5BHqKqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/wCC43/IqfCP/sIar/6Ltq+E/wDySvwb/wBi/Y/+k6VVVVVVVVfGnwm3jj4T+KvC8MXmT3mmS/Z0xnMyDfGP++1Wqqqqqqqqq+CP/JIPB3/YGt//AEAVVVVVVVVV4ZgxHNcEdSFX8Ov9Kqqqqqqqqqqqqqqqqqqqq/bl1p9L+BM1gj4Gsata2rDnkLum/nCKqqqq0nxf4i0UKljqcnlL0ik+dPoAen4YrSfi5E22PW9NKHHMtucj/vk9PzNaV4m0HWwBp2pwyOf+WTHa/wD3yeaqqr4Qf8k60j/tt/6OeqqqqqqqqtJk8rUrZgf+WgH58f1qqqqqqqqqqqqqqqqqqqqqqqqrx1498KfDbw7P4o8YavFY2MAwCxy8z44SNerMccAfXgAmvjx+1b4y+Lss+h6I8+g+FslRZRyYluh6zsOoP9wfKO+4jNVWr2p1HQpoVUtNYubiIA/wkASgDvwEbPYRn3qqqtM/5E3S/wDr6uf/AGnVVVVVvcXFpPHdWk8kM8LB45I2KsjDoQRyCPWvgB+2vd2klr4Q+Mtybi2wI7fXguZI+wFwB94f9NByOrbslhaXlpqFpDf6fdQ3NtcRiSGaFw6SIRkMrDggjBBFVVVVVVV+1d8Fj8XPh493o1qJPEfh8PcaeFX5p0x+8g/4EACP9pVHc0ysrFGBDA4IIwQaqqqqqqqqqqqqqq8O+HtW8Wa7YeGtBtGutQ1KdYLaJeNzscDJ7DuSeAMmvhD8M9K+EngDTPBWlssj2yb7y4Awbi4bmST6E8AdlCjtVVVVVVVVVVVVVV4r8M6R4z8N6l4U163E9hqls9vOnQ7WGMg9mBwQexANfDj9nfwJ8H5A9jYnUNZRcPql4oaQ/wDXMdIx9OccEmqqqqqqqqqqqqqqq0awN7dBnXMUXLZHX0FVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVftq/wDBP/4c/taaY/iOwkg8M/EWzg2WeuJDlLsKPlhu1Xl0wMBx86dtwGw/BX9sL9pP/gnf4xi+AP7UnhLVda8G2vyWO6TzJ7SAHAlsLgnbcW+OPKYjbwAYyGU/CP4z/DH47+D7fxz8KvF9jr+kzYV3gbElu+MmOaM4eJwCMqwB79CDVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV/wXG/5FT4R/wDYQ1X/ANF21fCf/klfg3/sX7H/ANJ0qqqqqqqqr9oPwI/w6+MHiXw4kIjtDdtdWQVcL5E37xAP90Ns+qmqqqqqqqqvgj/ySDwd/wBga3/9AFVVVVVVVVaXbG1sIoSPm25b6mqqqqqqqqqqqqqqqqqqqqv+ChN6I/BnhPTvMH77U5pdm3rsixnPt5n61VVVVVA45B5HStK8deJtI2pDqLTxL0iuP3g/M8j8CK0r4tafMBHrFhJbt/z0hO9fqR1H61puu6PrKBtM1KC4OM7VbDD6qeR+IqvhB/yTrSP+23/o56qqqqqqqqt22XETg/dcH9aqqqqqqqqqqqqqqqqqqqqqq8cfE74a/DG0tr/4k/EPwz4Ttr2QxW02uatBYJO4GSqNKyhiBzgVp2o6frGn2uraTf297Y3sKT21zbyiSOeNwGV0ZSQyspBBHBBBr4t/Fvwp8HPCkvibxNcbnbKWVlGwEt5LjhEHYdMt0UfgD8Wfi94w+MniV/EPiq9/dx5WysYmIgs0P8KKe5wMseSevQAVVQTeRMkoRXCn5kYZDjoVI7gjII7g1q9gum6lPZI5dEbMTHqyEZUn6qQcVVVpn/Im6X/19XP/ALTqqqqqq/Zy/ag134O3kXhzxC0+qeEZ5Pnt92ZLEk8yQ56jqTH0PUYJOdD1zR/E2j2mv+H9Rgv9PvohLb3ELZWRT3H8iDyDkHmqqqqqqq/bP/Z9bwtqs3xa8IWJ/sfU5v8AibQRrxZ3DH/WgDokhPPYOf8AaAFVVVVVVVVVVVVVX7Gf7PreDdJT4peL9P2a5qkONNt5kw1lbsOXIPR5AfqF4/iYVVVVVVVVVVVVVVVWt6b9tg86FczRDgf3h6UR2qqqqqqqqqqqqqqhhkuJVhhXc7nAFWFlHY2ywIQT1ZsdTVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVfGH4JfC74+eD5/A3xX8IWWvaXLlohMu2W1kxgSQyrh4nHTcpHGQcgkH4pf8ABPT9qf8AY88V3Pxg/Yx8c61rumQAs9lZlRqcMQOfLlt8eXfRjjhVJJ/5Z8bq/Z7/AOCxegXV1H4L/ap8Gz+FtWt38ibXdKtpJLcOvB+0WhzLCRjnZ5mSfuqK8AfEv4e/FbQI/FHw28aaP4l0qTgXWm3aTqp/uttOUbjlWwR3FVVVVVVVVVVVVVVVVVVVVVVVVVVVVf8ABcb/AJFT4R/9hDVf/RdtXwn/AOSV+Df+xfsf/SdKqqqqqqqq/b6+Gh1DQtI+Kmnw7pdKYWGokD/lg7ExMfZZCy/WUelVVVVVVVVfBH/kkHg7/sDW/wD6AKqqqqqqqtJtPtl7HGRlE+Z/oKqqqqqqqqqqqqqqqqqqqqr/AIKH/wDIG8Ef9fV5/wCgxVVVVVVVVI7xuskTlXU5VlOCD7VpPxD8T6WAjXovIh/Bcjef++uv618Av2h/AGo+HtP8G67fromrQGQL9rYLBMWkZhsk6A4YDDY56ZoDcoZSCCOCOaI7VVVVVVVUf+sXHqKqqqqqqqqqqqqqqqqqqqqqq+KnwQ+E3xv0+x0r4seBNM8T2mmTNNaQ3yFhDIw2lhgjkjivF3ivwR8DfhyNSvUj0/RNBtI7WwsoeCwRAsUEQJ5OFAA7AEngE18WPip4n+MHi+58WeJZ8bvktLNHJjs4R0jQH8ye5JPeqqqrxNbme0gu0UlrbMbn/YJyvHbDFuf9oVVVpjKfB+loGG4XNySM8j/V/wCBqqqqqqr9lv8AaNuvhBrq+GvEtxJN4R1SYecOWNhKcDz0H93++o6gZHIwba5tr22ivLOeOeCeMPFLG25XUjIYEdQRg5qqqqqqrU9M0/WtOudI1Wziu7O8iaKeCVdyyIwwykdwRX7S37OmpfBbXv7V0hJLrwnqczfYbjBY2rHnyJD6gfdY/eA9QQKqqqqqqqqqqqq/ZG/ZjfxRc2fxU+IFgV0a2kEmlafMmPtzjpM4P/LIHkD+Mj+6Pmqqqqqqqqqqqqqqqq13SSpN9bLkHmRQOnvVVVVVVVVVVVVVBSxCqMk8AAVo2lCyj8+ZR57jp/cHpVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVfHz9jn9nj9pO2kb4m/D6zl1dkCx67YD7JqEWOB++QZcAdFkDqPSvHv/BJT49fB7XJfHH7InxwupLiAEwWtxevpGoqv/PNbiIiKXPGd3lKeePWP9t7/AIKWfsrbbD9oL4TzeItJtjse+1rRigwOyX9mRCx/2m8w/wA68Cf8FtPg1qcSR/Ef4R+LdAnIwW0q4g1OIH1JcwMB16KT0q3/AOCsv7E08CSyfEDWYGYZMUnh67LL7HahH5E1/wAPYP2I/wDopGq/+E7e/wDxuv8Ah7B+xH/0UjVf/Cdvf/jdf8PYP2I/+ikar/4Tt7/8br/h7B+xH/0UjVf/AAnb3/43X/D2D9iP/opGq/8AhO3v/wAbr/h7B+xH/wBFI1X/AMJ29/8Ajdf8PYP2I/8AopGq/wDhO3v/AMbr/h7B+xH/ANFI1X/wnb3/AON1/wAPYP2I/wDopGq/+E7e/wDxuv8Ah7B+xH/0UjVf/Cdvf/jdf8PYP2I/+ikar/4Tt7/8br/h7B+xH/0UjVf/AAnb3/43X/D2D9iP/opGq/8AhO3v/wAbr/h7B+xH/wBFI1X/AMJ29/8Ajdf8PYP2I/8AopGq/wDhO3v/AMbr/h7B+xH/ANFI1X/wnb3/AON1/wAPYP2I/wDopGq/+E7e/wDxuv8Ah7B+xH/0UjVf/Cdvf/jdf8PYP2I/+ikar/4Tt7/8br/h7B+xH/0UjVf/AAnb3/43X/D2D9iP/opGq/8AhO3v/wAbr/h7B+xH/wBFI1X/AMJ29/8Ajdf8PYP2I/8AopGq/wDhO3v/AMbr/h7B+xH/ANFI1X/wnb3/AON1/wAPYP2I/wDopGq/+E7e/wDxuv8Ah7B+xH/0UjVf/Cdvf/jdf8PYP2I/+ikar/4Tt7/8br/h7B+xH/0UjVf/AAnb3/43X/D2D9iP/opGq/8AhO3v/wAbr/h7B+xH/wBFI1X/AMJ29/8Ajdf8PYP2I/8AopGq/wDhO3v/AMbr/h7B+xH/ANFI1X/wnb3/AON1/wAPYP2I/wDopGq/+E7e/wDxuv8Ah7B+xH/0UjVf/Cdvf/jdf8PYP2I/+ikar/4Tt7/8br/h7B+xH/0UjVf/AAnb3/43X/D2D9iP/opGq/8AhO3v/wAbr/h7B+xH/wBFI1X/AMJ29/8Ajdf8PYP2I/8AopGq/wDhO3v/AMbr/gqN+138Cv2ntA+Hth8HfE93qs3h+8v5L9Z9NntfLWVIQhBkVd2SjdOmK+H/APwVK/Yx0DwH4b0LU/iJqkd5p2kWttcIPD94wWSOFVYAiPB5B5Ff8PYP2I/+ikar/wCE7e//ABuv+HsH7Ef/AEUjVf8Awnb3/wCN1/w9g/Yj/wCikar/AOE7e/8Axuv+HsH7Ef8A0UjVf/Cdvf8A43X/AA9g/Yj/AOikar/4Tt7/APG6/wCHsH7Ef/RSNV/8J29/+N1/w9g/Yj/6KRqv/hO3v/xuv+HsH7Ef/RSNV/8ACdvf/jdf8PYP2I/+ikar/wCE7e//ABurH/gqn+w1eK5uPjBdWRUjAn8N6kd302QN+uK/4ej/ALCX/Rdf/LZ1b/5Fr/h6P+wl/wBF1/8ALZ1b/wCRa8W/8FIP+CfnjXwxqfhPXPjf5tjqtq9vMo8MatkBhjcP9F4YHBB7EA1a6r4c1uN9R8Ja/DrekPLItnqEULxLcorFQ4SQB0zjO1gCOhFVVVVVVVXwR/5JB4O/7A1v/wCgCqqqqqqqqX9vX9jnwVquo+HfEPxz0e21fTrqS1vYBaXUvkyRsVZNyRFThgeQSK/4eQ/sR/8ARftJ/wDBfe//ABmv+HkP7Ef/AEX7Sf8AwX3v/wAZr/h5D+xH/wBF+0n/AMF97/8AGa/4eQ/sR/8ARftJ/wDBfe//ABmv+HkP7Ef/AEX7Sf8AwX3v/wAZr/h5D+xH/wBF+0n/AMF97/8AGa/4eQ/sR/8ARftJ/wDBfe//ABmv+HkP7Ef/AEX7Sf8AwX3v/wAZr/h5D+xH/wBF+0n/AMF97/8AGa/4eQ/sR/8ARftJ/wDBfe//ABmv+HkP7Ef/AEX7Sf8AwX3v/wAZr/h5D+xH/wBF+0n/AMF97/8AGa/4eQ/sR/8ARftJ/wDBfe//ABmv+HkP7Ef/AEX7Sf8AwX3v/wAZr/h5D+xH/wBF+0n/AMF97/8AGa/4eQ/sR/8ARftJ/wDBfe//ABmv+HkP7Ef/AEX7Sf8AwX3v/wAZr/h5D+xH/wBF+0n/AMF97/8AGa/4eQ/sR/8ARftJ/wDBfe//ABmv+HkP7Ef/AEX7Sf8AwX3v/wAZr/h5D+xH/wBF+0n/AMF97/8AGa/4eQ/sR/8ARftJ/wDBfe//ABmv+HkP7Ef/AEX7Sf8AwX3v/wAZr/h5D+xH/wBF+0n/AMF97/8AGa/4eQ/sR/8ARftJ/wDBfe//ABmv+HkP7Ef/AEX7Sf8AwX3v/wAZr/h5D+xH/wBF+0n/AMF97/8AGa/4eQ/sR/8ARftJ/wDBfe//ABmv+HkP7Ef/AEX7Sf8AwX3v/wAZr/h5D+xH/wBF+0n/AMF97/8AGa/4eQ/sR/8ARftJ/wDBfe//ABmv2q/2nv2ff2gdM8O6Z8IPiVaeJNR0eeeW6ggtp4vKhdUG4+ZGoPzBRwe9VVVVVVVVVXw2+P3xI+GRitdJ1c32locHTb7MsQHonO6P/gJAz1Br4Z/tRfDn4gLDYandDw9rDgA2t9IBG7f9M5eFP0baT2BrajAEYPpijGD0NGM44INFSOMUR2qqqq1TV9L8P6Zd69rd6lpp2mwPc3dw4O2GKNSzucc4Cgn8K/4eQ/sR/wDRftJ/8F97/wDGa/4eQ/sR/wDRftJ/8F97/wDGa/4eQ/sR/wDRftJ/8F97/wDGa/4eQ/sR/wDRftJ/8F97/wDGa/4eQ/sR/wDRftJ/8F97/wDGa/4eQ/sR/wDRftJ/8F97/wDGa/4eQ/sR/wDRftJ/8F97/wDGa/4eQ/sR/wDRftJ/8F97/wDGa/4eQ/sR/wDRftJ/8F97/wDGa/4eQ/sR/wDRftJ/8F97/wDGa/4eQ/sR/wDRftJ/8F97/wDGa/4eQ/sR/wDRftJ/8F97/wDGa/4eQ/sR/wDRftJ/8F97/wDGa/4eQ/sR/wDRftJ/8F97/wDGa/4eQ/sR/wDRftJ/8F97/wDGa/4eQ/sR/wDRftJ/8F97/wDGa/4eQ/sR/wDRftJ/8F97/wDGa/4eQ/sR/wDRftJ/8F97/wDGa/4eQ/sR/wDRftJ/8F97/wDGa/4eQ/sR/wDRftJ/8F97/wDGa/4eQ/sR/wDRftJ/8F97/wDGa/4eQ/sR/wDRftJ/8F97/wDGa/4eQ/sR/wDRftJ/8F97/wDGa/4eQ/sR/wDRftJ/8F97/wDGa/4eQ/sR/wDRftJ/8F97/wDGa/4eQ/sR/wDRftJ/8F97/wDGa/4eQ/sR/wDRftJ/8F97/wDGa/4eQ/sR/wDRftJ/8F97/wDGa/4eQ/sR/wDRftJ/8F97/wDGa/4eQ/sR/wDRftJ/8F97/wDGa/4eQ/sR/wDRftJ/8F97/wDGa/4eQ/sR/wDRftJ/8F97/wDGa/4eQ/sR/wDRftJ/8F97/wDGa/aw/be/Zo+J/wAOdI8N/C/4x2uq6xH4q0u6e3tLW7jf7PFOGlbLRrkBe2cn0NftM/Ha6+NPjRhpk8ieGNIZotLgYFfN7NOwP8T44B+6uB13ZqqqqmhjuYXtpCNsqlSTnj0PHocH8KZSrFSRkHHBzVVpP/IuWP8A10m/mKqqqqqqr9mX9r3wr8L/AAneeFfjT4l/s7QNIi83TdUlhln+zqWAaBhGrNt5ypxhRuBONoH/AA8h/Yj/AOi/aT/4L73/AOM1/wAPIf2I/wDov2k/+C+9/wDjNf8ADyH9iP8A6L9pP/gvvf8A4zX/AA8h/Yj/AOi/aT/4L73/AOM1/wAPIf2I/wDov2k/+C+9/wDjNf8ADyH9iP8A6L9pP/gvvf8A4zX/AA8h/Yj/AOi/aT/4L73/AOM1/wAPIf2I/wDov2k/+C+9/wDjNf8ADyH9iP8A6L9pP/gvvf8A4zX/AA8h/Yj/AOi/aT/4L73/AOM14j/b/wD2B/F2iXfhvxJ8a9D1DTb+Mx3FvNp16VcfhDkEHBBGCCAQQRXj3xZ+z/N43bSfgt8XtN8V6beh5bSERTQ3FuBjMbiVEDkA8Fc5AJIGKqqqqqqqqqqqvgT8RP2QLPxFLrPx1+M2i2sOmTFItDEFzMbiVf4pWijZfLB/hDEsQc4Aw0X/AAUc/YegiSCD49aPHHGoVEXTbwBQOAABDwMV/wAPIf2I/wDov2k/+C+9/wDjNf8ADyH9iP8A6L9pP/gvvf8A4zX/AA8h/Yj/AOi/aT/4L73/AOM1/wAPIf2I/wDov2k/+C+9/wDjNf8ADyH9iP8A6L9pP/gvvf8A4zX/AA8h/Yj/AOi/aT/4L73/AOM1/wAPIf2I/wDov2k/+C+9/wDjNf8ADyH9iP8A6L9pP/gvvf8A4zX/AA8h/Yj/AOi/aT/4L73/AOM1/wAPIf2I/wDov2k/+C+9/wDjNf8ADyH9iP8A6L9pP/gvvf8A4zX/AA8h/Yj/AOi/aT/4L73/AOM1pmpWOtaZaaxpdwJ7O+gSe3lAIEkbqGVgDzyCDzVVVVVVVUQCMVrGimHN1ZqTH1dB/D7j2qqqqqqqqqqqqVWdgiAlicAAda0fRhagXNyAZsfKv9z/AOvVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVMqspVgCCMEEV47/ZR/Zq+Jjyz+N/gZ4L1K6nz5l5/ZEUNy31mjCyf+PVN/wS8/YUnleZ/gSgaRixCeI9VQAn0UXQAHsBgV/wAOuP2Ev+iF/wDlzat/8lV/w64/YS/6IX/5c2rf/JVf8OuP2Ev+iF/+XNq3/wAlV/w64/YS/wCiF/8Alzat/wDJVf8ADrj9hL/ohf8A5c2rf/JVf8OuP2Ev+iF/+XNq3/yVX/Drj9hL/ohf/lzat/8AJVf8OuP2Ev8Aohf/AJc2rf8AyVX/AA64/YS/6IX/AOXNq3/yVX/Drj9hL/ohf/lzat/8lV/w64/YS/6IX/5c2rf/ACVX/Drj9hL/AKIX/wCXNq3/AMlV/wAOuP2Ev+iF/wDlzat/8lV/w64/YS/6IX/5c2rf/JVf8OuP2Ev+iF/+XNq3/wAlV/w64/YS/wCiF/8Alzat/wDJVf8ADrj9hL/ohf8A5c2rf/JVf8OuP2Ev+iF/+XNq3/yVX/Drj9hL/ohf/lzat/8AJVf8OuP2Ev8Aohf/AJc2rf8AyVX/AA64/YS/6IX/AOXNq3/yVX/Drj9hL/ohf/lzat/8lV/w64/YS/6IX/5c2rf/ACVX/Drj9hL/AKIX/wCXNq3/AMlV/wAOuP2Ev+iF/wDlzat/8lV/w64/YS/6IX/5c2rf/JVf8OuP2Ev+iF/+XNq3/wAlV/w64/YS/wCiF/8Alzat/wDJVf8ADrj9hL/ohf8A5c2rf/JVf8OuP2Ev+iF/+XNq3/yVX/Drj9hL/ohf/lzat/8AJVf8OuP2Ev8Aohf/AJc2rf8AyVX/AA64/YS/6IX/AOXNq3/yVX/Drj9hL/ohf/lzat/8lV/w64/YS/6IX/5c2rf/ACVX/Drj9hL/AKIX/wCXNq3/AMlV/wAOuP2Ev+iF/wDlzat/8lV/w64/YS/6IX/5c2rf/JVf8OuP2Ev+iF/+XNq3/wAlV/w64/YS/wCiF/8Alzat/wDJVf8ADrj9hL/ohf8A5c2rf/JVf8OuP2Ev+iF/+XNq3/yVX/Drj9hL/ohf/lzat/8AJVf8OuP2Ev8Aohf/AJc2rf8AyVX/AA64/YS/6IX/AOXNq3/yVX/Drj9hL/ohf/lzat/8lV/w64/YS/6IX/5c2rf/ACVX/Drj9hL/AKIX/wCXNq3/AMlV/wAOuP2Ev+iF/wDlzat/8lV/w64/YS/6IX/5c2rf/JVX3/BKr9hu8dGt/hJeWQUYKweJNRIb3O+dj+WK/wCHT/7Ef/RN9V/8KK9/+OV/w6f/AGI/+ib6r/4UV7/8cr/h0/8AsR/9E31X/wAKK9/+OV8Yv2C/ht8IPhlPrXwD0PULNNLle61Kwn1Ca782EqA8ieYzFSgUMQCAV3HqBVVVVVVVV8Ef+SQeDv8AsDW//oAqqqqqqqh4eHiaxvdKlu7q0hubeSF7i1k8uWLepXdG3O1xnIOOCBT/APBIj9jaRi76L4sZmOSW16Qkn16V/wAOhv2NP+gH4q/8Hr/4V/w6G/Y0/wCgH4q/8Hr/AOFf8Ohv2NP+gH4q/wDB6/8AhX/Dob9jT/oB+Kv/AAev/hX/AA6G/Y0/6Afir/wev/hX/Dob9jT/AKAfir/wev8A4V/w6G/Y0/6Afir/AMHr/wCFf8Ohv2NP+gH4q/8AB6/+Ff8ADob9jT/oB+Kv/B6/+Ff8Ohv2NP8AoB+Kv/B6/wDhX/Dob9jT/oB+Kv8Awev/AIV/w6G/Y0/6Afir/wAHr/4V/wAOhv2NP+gH4q/8Hr/4V/w6G/Y0/wCgH4q/8Hr/AOFf8Ohv2NP+gH4q/wDB6/8AhX/Dob9jT/oB+Kv/AAev/hX/AA6G/Y0/6Afir/wev/hX/Dob9jT/AKAfir/wev8A4V/w6G/Y0/6Afir/AMHr/wCFf8Ohv2NP+gH4q/8AB6/+Ff8ADob9jT/oB+Kv/B6/+Ff8Ohv2NP8AoB+Kv/B6/wDhX/Dob9jT/oB+Kv8Awev/AIV/w6G/Y0/6Afir/wAHr/4V/wAOhv2NP+gH4q/8Hr/4V/w6G/Y0/wCgH4q/8Hr/AOFf8Ohv2NP+gH4q/wDB6/8AhX/Dob9jT/oB+Kv/AAev/hX/AA6G/Y0/6Afir/wev/hX/Dob9jT/AKAfir/wev8A4V/w6G/Y0/6Afir/AMHr/wCFfGb9gD4Cfs4eALz4j/CfTtdg1SOaG1uTeam1xGYJHAPykcHeI+fqO9VVVVVVVVVVV8M/2hviN8M3htLPUzqmkR8HTb5i6KvpG33o/wADjPUGvhr+058NviH5VhcX39gau+B9j1Bwqu3pHLwregB2sfSqogEYNFFPajGD0NGM9iKKt6UR2rXvCOl+P9B1TwLrhuBp3iCxn0+8NvKYpBDNGySbHHKttY4I6HBr/h0N+xp/0A/FX/g9f/Cv+HQ37Gn/AEA/FX/g9f8Awr/h0N+xp/0A/FX/AIPX/wAK/wCHQ37Gn/QD8Vf+D1/8K/4dDfsaf9APxV/4PX/wr/h0N+xp/wBAPxV/4PX/AMK/4dDfsaf9APxV/wCD1/8ACv8Ah0N+xp/0A/FX/g9f/Cv+HQ37Gn/QD8Vf+D1/8K/4dDfsaf8AQD8Vf+D1/wDCv+HQ37Gn/QD8Vf8Ag9f/AAr/AIdDfsaf9APxV/4PX/wr/h0N+xp/0A/FX/g9f/Cv+HQ37Gn/AEA/FX/g9f8Awr/h0N+xp/0A/FX/AIPX/wAK/wCHQ37Gn/QD8Vf+D1/8K/4dDfsaf9APxV/4PX/wr/h0N+xp/wBAPxV/4PX/AMK/4dDfsaf9APxV/wCD1/8ACv8Ah0N+xp/0A/FX/g9f/Cv+HQ37Gn/QD8Vf+D1/8K/4dDfsaf8AQD8Vf+D1/wDCv+HQ37Gn/QD8Vf8Ag9f/AAr/AIdDfsaf9APxV/4PX/wr/h0N+xp/0A/FX/g9f/Cv+HQ37Gn/AEA/FX/g9f8Awr/h0N+xp/0A/FX/AIPX/wAK/wCHQ37Gn/QD8Vf+D1/8K/4dDfsaf9APxV/4PX/wr/h0N+xp/wBAPxV/4PX/AMK/4dDfsaf9APxV/wCD1/8ACv8Ah0N+xp/0A/FX/g9f/Cv+HQ37Gn/QD8Vf+D1/8K/aN/Y5/ZY/Z48RaBZfDPRNXl8UKTeTPfaq9xHaxYKp8mAN7NkjP3Qmccg1VVVVVa/Zrb3KzRgBZsk85+bv/MVVaT/yLlj/ANdJv5iqqqqqqquba3vbaWzuoUmhnRo5I3XIdSMEEehHFfAT/gnF+xR8Zvh3ZeKF0PxLFqcP+j6pbR69IBDcKBkgYJCsCGXrwcZyDX/Dob9jT/oB+Kv/AAev/hX/AA6G/Y0/6Afir/wev/hX/Dob9jT/AKAfir/wev8A4V/w6G/Y0/6Afir/AMHr/wCFf8Ohv2NP+gH4q/8AB6/+Ff8ADob9jT/oB+Kv/B6/+Ff8Ohv2NP8AoB+Kv/B6/wDhX/Dob9jT/oB+Kv8Awev/AIV/w6G/Y0/6Afir/wAHr/4V/wAOhv2NP+gH4q/8Hr/4VD/wSM/Y6t5UuLfSPFsUsbBkdNfkVlI6EEDg18Xv2D20bR49T+DV/e6i1pEFm0vUp1eaYAfejlwAzdMq3Xkg54rUtM1HR76fS9WsLiyvLVyk1vcRGOSJh1DKcEH2NVVVVVVVVV4S8G+KPHmtw+HfCGiXOqahNysMCZ2juzHoqjIyzEAeteE/+CeHgvWfBz6f8X9d1a41C/AM0Gi3ptUt1xzH5gG5ye5G0dgCMk/8Ohv2NP8AoB+Kv/B6/wDhX/Dob9jT/oB+Kv8Awev/AIV/w6G/Y0/6Afir/wAHr/4V/wAOhv2NP+gH4q/8Hr/4V/w6G/Y0/wCgH4q/8Hr/AOFf8Ohv2NP+gH4q/wDB6/8AhX/Dob9jT/oB+Kv/AAev/hX/AA6G/Y0/6Afir/wev/hX/Dob9jT/AKAfir/wev8A4V/w6G/Y0/6Afir/AMHr/wCFf8Ohv2NP+gH4q/8AB6/+Ff8ADob9jT/oB+Kv/B6/+Ff8Ohv2NP8AoB+Kv/B6/wDhX/Dob9jT/oB+Kv8Awev/AIVoej2Xh7RNP0DTVcWmmWsdtbh23MI41Crk9zgDmqqqqqqqqq1TQVmJuLIBX6mPoD9PSpI3jYxyKVZeCCMEVVVVVVVVVVW1rPeSCG3jLN39vc1pmjw2CiRyJJsctj7v0qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqdEkQxuoZWGCrDIIr9rH9nt/hP4kPirwzZn/hE9amPkqi8WE5yTCfRTglPYEfw5NVVVVVVfBH/AJJB4O/7A1v/AOgCqqqqqqra2lu5lt4VyzH8verO0isrdbeLt1J7nuaqqqqqqqqqqqqqqqqqqqqqvj94Zbxf8F/GGgxxCWWTS5JoUxndJDiVAPfdGv41VVVVVVVVVVVV8Mv2kfiN8NVh0+O+Gs6PHgCwvmLBF9I3+8n05X/Zr4Z/tJ/Df4kGHT1vzo2sSkKLC/YLvY9o5Puvz0HDH+7VVVUQCMV4dgEmoeaB/qkJ/E8f1qqqqqqqqqqqqqqqqqqqqqq8Sa/pnhTw/qPiXWZxDY6XayXFw57IiknHqTjAHckV8QPGmq/ETxnq/jTWXzdarctKUzkRL0SMeyqFUewFVVVVVWsWn2uxcKMvH8y/h1/SqrSf+Rcsf+uk38xVVVVVVVV+xx8VW+HnxUg0HULgro/ispZTgniOfP7iT/volPQCQntVVVVVVVVXxU+BPw2+MNrs8XaGv2+OMpBqVqfKuYR2+cfeA/uuGAyeK+Kf7FHxO8ENNqPg8L4s0lAWH2VNl1GPRocnd/wAsT1wKu7S6sbmSyvraW3uIWKyRSoUdGHUEHkH2NVVVVVVaH4e13xRqMWj+HNGvdTvpThLe0gaVz74UE496+FP7CHinWmh1X4q6mNCs8gnTrR1lupB6M4ykf4bz6gV4E+HHgn4Z6Ouh+CPD1rplvwZGjXMkzD+KRzlnPuScdBgVVVVVVVVVVVVVVVVVVVVVe6da36bZk+YD5XHBFX2h3doS8a+dGP4lHI+oojtVVVVVVVKjSMERSzHoAMmrHw7NNiS8byl/uD7x/wq3toLWMRW8YRR2Heqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq8T+GdD8ZaBfeGPEmnxXum6jCYriCQdQe4PUMDggjkEAjBFftAfATX/AII+JmgkSW78PX0jHS9RIzuHXy5CBgSAdR0I5HcCqqqqqr4I/wDJIPB3/YGt/wD0AVVVVVVRQyTyLDChZ2OABWl6bHp8ODhpWHzt/Qe1VVVVVVVVVVVVVVVVVVVVVVOiSIY3UMrDBVhkEV8WPBkvw++JPiPwbIm1dMv5EgyMZhY7om/GNkP41VVVVVVVVVVVVXw1/ac+JPw88qwuL7+39JjwPseoOWZF9I5eWX0AO5R6V8Mv2h/hz8Tlis7PUhpervwdNv2COzf9M2ztkHsPm9QKqqrw3b7LV7gjmRsD6D/6+aqqqqqqqqqqqqqqqqqqqqqv29PiL/YngXTfh1Yz4ufEU/n3YB6W0JBAP+9Jsx/1zaqqqqqqq1W0+yXrxKPlb5k47H/OKrSf+Rcsf+uk38xVVVVVVVVHJJDIssMjJIhDIynBUjoQe1fA74gD4n/Cvw/4vklV7y4tRFfADGLiM7JOO2WUsB6MKqqqqqqqqrxz8KPhz8Srf7P428IafqjAYWd49kyD0WVMOo9ga8af8E/vB+oM9z4E8ZahpDkki2vohdRfRWBVlHudxrxN+xD8c9CdzpWn6Vr8SgkNY3yo2P8Adm2HPsM/jWr/AAQ+MOhMy6n8MfE0aoPmkTTJZIx/wNAV/WrvRNZsA/27SL238v7/AJ1uybfrkcVVl4Y8Samyppvh/UrtmAKiC0eQnPTGAetaL+z38bvEDomnfC7xCBIPle6smtVI453S7Rjkc5rwv+wh8YtYeN/EN5ougQn76y3JuJV+ixAqf++xXgj9g34YaCyXPjLWNS8TTqeYv+PO3P1VCX/8f/CvC3gvwl4I04aV4Q8OafpFqOsdpbrHvI7sRyx9zk1VVVVVVVVVVVVVVVVVVVVVVV3pVje5aWEBz/GvBq58NTL81rOsg/usNpqfT7234ltZBjuBkfmKqqqgCxCqMnsMVBpOoXGNlswHq42/zq28Mjhru4z/ALMY/qatrK1tF228Kr6nGSfxqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqvF3hDw3478P3XhfxXpUOoabeLiSGUdD2ZSOVYHkMMEGv2g/2YvE/wAGb6XWNKS41fwlK/7m/CbntcnhJwBhT0AfAVuOhO0VVVVVXwR/5JB4O/7A1v8A+gCqqqqqtrWe8lENuhZj19B7mtN0uDT4+MPKR8zkfoPaqqqqqqqqqqqqqqqqqqqqqqqr9v34eNYeItE+JllCfI1SH+z71gOBPGC0ZPuybh9IqqqqqqqqqqqqqqgSCCDyOlfDT9qL4j+ADDp+o3n/AAkOkx4H2W+cmSNf+mcvLD6NuA7AV8Nvj98N/icsVrpOrix1Vxg6bfYimJ9E52yf8BJOOoFUiNI6xoMsxwB71a2621tHbp/AuPqe5qqqqqqqqqqqqqqqqqqqqqq/a48bt43+OOuiOUtaaERpdsCfu+TnzP8AyKZPwxVVVVVVVeILTzrUXCDLQnn6Hr/Sq0kqfDtioYZEkuR+Iqqqqqqqqr/gnz40Z4fFHw8uJ8iMx6naIT0BxHN+GfJ/M1VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVUtrbT/AOut43P+0gNPo2mSdbRfwJH8jR0DTD0hb6bzQ0HSweYCfYuf8aTSNNj+7aIf97J/nUcMMQxFEicdFXFVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV1a2t9bS2V7bxXFvOhSWGVA6upGCrKeCCOMGvjr+w/HObnxR8GNqOSZJtBmfCn1+zuen+4xx6MOFrWNF1bw9qdxouu6bc6ff2j7Jra5iMckbehU8iqqqqvgj/ySDwd/wBga3/9AFVVVVafolze4kkBii/vEcn6CrWzt7OIQ26ADuT1J96qqqqqqqqqqqqqqqqqqqqqqqq+NPw4tviv8Nda8FyiMXNzB5ljK4/1VynzRtnsNw2n/ZY+tX1ld6beT6df27wXNrK0U0Ugw0bqcMpHYggiqqqqqqqqqqqqqqgSpDKxBB4I4xXwx/aq+IXgQRabrkp8S6SmAIryU+fEP9iXk49mDD0xXwP+JHhj4waZL4k8OLeJHYuIriG6gKNFKRnbu5VuOflJxkZxmqqqqqqqqqqqqqqqqqqqqqrxFrEHh3w/qfiC5x5OmWU11Jn+7GhY/oK1K/utW1C61S+l8y5vJnmmc/xOxJY/mTVVVVVVVSIkiGNwCrDBBq7t2tbiS3bqhwM9x2P5Vojo+lwKjAlNwYeh3E/yIqqqqqqqqr9kfxOfDHx78Ns8u2DVGk0+YZxu81CEH/fwR/lVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV8SPg98O/izYCy8beHYLySNSsF4n7u4gz/ckXkDPO05U9wa+KH7CPjbw+82pfDTU4vEdgoLLZ3DLBdoPQdEk4HUFSey1r3hzX/C+oPpPiTRb3S72I4aC7t2icfgwHHvVVVfBH/kkHg7/ALA1v/6AKqqxngVaaLfXeGEflp/efj9OtWWh2dph2XzpB/E44H0FVVVVVVVVVVVVVVVVVVVVVVVVVVftx/Bp/D3iaP4r6FZ403XWEeprGvEN2Bw5A6CRR/30rE8sKqqqqqqqqqqqqqqrQdC1XxPrVj4e0Kye71DUZ1gtoEHLuxwB7D1J4A5NfB/4a6b8Jfh7pXgnTzHJJax77y4VcfaLhuZH9cZ4GeihR2qqqqqqqqqqqqqqqqqqqqqq/ak1p9B+APjK8il2NNYrajHUieRIiPyc1VVVVVVVV4jtMiO8Qf7D/wBDXhz/AI8X/wCup/kKqqqqqqqq8Ia03hvxZoniJG2tpWo290DzwY5Fftz/AA1VVVVVVVVUskcMbSyyKiIpZmY4CgdST2Far8Zvg9oUnka38WPBunybiuy7122hO4dRhnHI9K8OfFD4Z+MZxaeEfiL4Y1yckgRadq8FyxIAJ4jYngEH8aq5uLezt5bu7uI4YIELyyyMFVFAyWJPAAAySa0r4g+Adevo9L0Pxx4f1G9lB8u2tNThmkfAycKrEnABP0FVWu+L/CXhh4Y/EvinSNJa4BMK317HbmQDGSocjOMjp6itJ1nR9fsU1PQtWs9Rs5CQlxaTrNGxBwcMpIOCMV/wtj4V/wDRTPCn/g5t/wD4uv8AhbHwr/6KZ4U/8HNv/wDF1/wtj4V/9FM8Kf8Ag5t//i60bx14I8R3h07w94y0PVLoIXMFlqMU7hRjLbUYnHI596qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqrxF4V8M+LrA6Z4p8P6dq1qQcQ3tssygnuAwOD7jmvGP7DvwW8Rs8+hR6p4buGyQLK582LPqUl3HHsrKK8Sf8E+PGVs7t4S8e6PqCDJVb+3ktG+nyeYCffj8O2r/sYftBaY7LbeE7TUkQHMlpqcGCB6CRlY59MZqb9mP49wyGJ/hhq5ZepQI4/AhiDXwj8L+INH+GHhfSdU0i4tby00uCKeCVNrRuqgEEetJompv0tSP95gP61H4bvX/wBZJEg+pJqHwzbLgz3DyH0UbR/Wrews7UD7PbopH8WMn86qqqqqqqqqqqqqqqqqqqqqqqqqqqvGXhLRPHnhfUvCHiK2E+n6pbtDMuBlc9GUnOGUgMD2IBr4tfDDXvhD43vvBmurvMJ8y0uQuFurdidkq/XBBHZgR2qqqqqqqqqqqqqqr9if4CNodhH8YvFdntv7+ErokEi8wwMMNOR2ZxkL/sEnnfxVVVVVVVVVVVVVVVVVVVVVV+3JefZvgRPAGcfa9VtYsL0OCz8+3yfniqqqqqqqqrq3S6t3t36OuAfQ9jWgRNFZyRuMMJmBB7EAD+lVVVVVVVVV4Pvf7T8I6JqJdXN1ptvLuXod0anI/Oqqqqqqqqv+CkX/ACZH8U/+wfbf+lkFf8E+P2CPh9+2B4T8WeJvG3jXxDop8P6jFZwQ6UsI8zfHvLMZFb6YAr4of8ETtLtNAn1L4I/GLU2120TzLay8QW8fl3LjkKJ4QpiJOMMUYZ646j/gmt+2d8XtD+M5/ZJ/aA1jUtSE81xY6XNq8hlvNK1C33l7WSRss8bBHUBidrKgUhTgftaaqdE/Zb+L2pJK0ckXgfVxE6rkrI1nKqHH+8RXwG8YeJfgJ8Qfh5+0Xa2sx0zTfE8kAZAR9oW3SA3kAPTLW94q+3mVpuo2Gs6ba6vpV3FdWV9Ak9tPE25JY3UMrKe4IIIPvVf8FGvHes/tH/tVeP4/DlwJvD/wh0drIsWO1Et5kjuXwONxvLjy891VOeAK/wCCQeqDUP2NrC0Ezv8A2b4i1G32sMBMskuB7fvc/Umv+HHPhT/o4rVf/Cbj/wDj9ft7fsOaT+xh/wAIL/ZfxFu/FX/CYf2jv8/TVtPs32X7NjGJH3bvtB9Mbe+ePgb/AMEgPDfxh+Dngz4pXPx01LTJfFWi22ovZJoCSrbmVAxQOZgWAzjOBX7If/BNXQ/2S/ivL8UtO+Ld94kll0ifTvsU+jraqBK8bF94lY5Hl4xjv1qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/aC+Buj/G/wc2myeVba7p4aTSb5l/1bkcxuevlvgA+hAPbB8R+HNc8I65eeG/EmmzWGpWEpjuLeZcMjD9CCMEEcEEEEg1VVVVVVVVVVVVX7KP7Ns/xO1iHxx4x0908J6dLmKORMDU5VP3BnrGCPmPQ/dH8RVI44o1iiRVRBhVUYAA7AVVVVVVVVVVVVVVVVVVVVVVV+3iyr8EbZSeW163A4/6Zy/4VVVVVVVVVSRJGWKD77bj9cf8A1qqqqqqqqqr4Yf8AJNPCX/YCsv8A0QlVVVVVVVVf8FIv+TI/in/2D7b/ANLIK/4Ih/8AJKviV/2MFr/6Tmr6+sdMsbjUtSvILS0tImluLieQRxwxqMs7MeFUAEkngAV8EIov2lP+Cq8njv4aQzS+Hrfxhc6+97GhVRZWxOJ24yqzOI1GRnM6g8k1/wAFCNTfSP2L/ivdxtIC+ifZ/kbacSzRxH8MPyO4yK8KfBy48a/8EpvEnjm2tg974L+KU2rRFBuY2klnZWs647DcySH2i9M1/wAEuPjMfi7+yV4e06+u/N1bwJK3h+7DN83lwqrWxweceQ8SZ6Zjb0wP2mvjBafAP4C+NvizcSIJtB0qRrBH6S3kmI7ZD14aZ4weDgEntX7Onwe1G7/4J/ftJ/H3WInuNT8SRwadZXE/DtBb3UNxdSBj1DyMgP8AtQGv+CK2qfav2YfFGlvM7vY+OLkqpHCRvZ2ZAH/Alc/jVf8ABc//AJol/wBzB/7jq/Yo/wCTRfhB/wBifp//AKJWq/bs/wCCjXhv9lS5Hw68D6TaeJviLPAJpba5dha6TG4BR7jaQzuwIKxKQdvzFlBXd4b+KH/Bavx3Yw/EXw54Vki0O4Xz4dOuNJ0ezDxkZG2G523RUggjnJ4wTXjz/gq/8YNH+DWqeHNa8JWfgP45+GddtrbUNPvdNc293ZsknmOkMp3RSKwiyjE8MrKSGIX9jL4oeMfjT+zL4G+KHj+8tbnX9ftria7ktbYQRnbdTIgVASBhEUe5Ge9VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX7Rn7OWifG7RhfWTxaf4p0+JhY3hUBZh18mbAyUznB6qSTyCQfFHhbxB4L1278M+KNLm0/U7F9k1vMMFT1BBHBBGCCMgggg1VVVVVVVVVVVfs1fsuap8XLuLxT4tiudP8IQNkOBsk1FgeUjPZMghn/Ac5K6XpenaJplro2kWcVpZWUKw29vEu1YkUYVQOwAAqqqqqqqqqqqqqqqqqqqqqqqv25rNrr4EyzLuxaatbSthc8HcnPoMuP09aqqqqqqqqqqqqqqqqqq8E2H9l+DNA0v/AJ89LtoeTn7sSjr+FVVVVVVVVf8ABSL/AJMj+Kf/AGD7b/0sgr9iHw3+3vrnhTxLL+x/r/2DR4tQiXWF+1WMW648v5Di5BJ+T+7xX7Xdn/wUa8M6HbW37V+s+MZ/Bl1PHHK9lfwPp8xzkJJ9k/diThtomGcjIBAzX/BNPw3+yzZ/A0eJ/wBm21vXuNQlWDxLc6yyPqi3ca58mcqAoRQ+5BGAhDZ+8Wr/AIKr6klh+w547tGCE6jdaXAu5sEEahbycDucRnj0ye1f8EwPh1o3jj/gn9qfgnxBbSf2b41v9ZtbvIzujlUW7MmfQJx6Fa/4JQ+M9Y+B37Wfjf8AZr8YyC2m12O5sHhJOP7T015CAPYxfaue+Fr/AILS/Fu9OkeAP2dfDzyzXmu3TazqFtCCXkRCYLRAByweRpzj+9Ev4fE34I2XwW/4JjeJ/g9EqrN4e8ATvfPCBiW9CG4uHHXhpi578Hr3r/giFqgm+GHxN0UTOfsmvWlx5ZHC+bAy5HufJ5+gqv8Aguf/AM0S/wC5g/8AcdX7FH/Jovwg/wCxP0//ANErWq6laaNpd5rF/KI7axt3nmckDaiKWY88dAetf8E9fCo/a1/bs1b4q/E60i1JdOF54ru7a4Akie6aZEgjwf4Y3mVlGMAQqMY4qv8Agtx8N/DFtb/Dr4sWdjHBr15Pc6TezIgBuoVRZIt57lD5gHtIR2Ff8E3v+TI/hX/2D7n/ANLJ6qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr9tfxB8XfBv7NHjPxz8DvEsmi+KvDFqupxzpYwXe+2hcNcqUnR1x5PmPnbnKDkAmv8Agln+1/8AEf8AaX8M+NvDnxk8Tprnirw3ewXVvefYoLQyWU6FQmyBEQ7JImJbGf3yjPAqvi/8RdL+EPws8V/E/WShtfDGkXF+0bNt85o4yUjB9XYKg92Ff8Ewf2lP2tf2nvib4p1P4rfFE6t4L8K6Ynm2SaJYWolvbhyIF3xQK+1UjnYgN1CZyDX7bv7b/wC1/wCCP2wfEX7PvwS8dW+l2Ud3pdlpFpHpNjJLLNd2Vs+1pbmNsbpZzySoHHIAJr/hDv8Agt9p6fb7fxUbiWEblg+3aHIWPptcbT9DxXw8/wCCpn7RnwL+IafDL9tT4as6RMou7uLTfsGpWyMcCcRriG4jwDjYEzyQx6VoGv6L4r0LT/E3hzU7fUdK1W1jurK7gfdHPDIoZHU9wVIP41VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVfGj4E+Cvjbon2HX4Psup26EWOqwRgzWx9D/AH0yeUPHOQQeR8Xfgf48+DGr/YPFOn+ZYzMRZ6nbgtb3I9j/AAt6o2CPcYJqqqqqqqqqre2uLyeO0tIJJ55mCRxRoWZ2PAAA5JPpX7P37FNxcvaeMfjJbGGFSJLbQD96TuDcEfdHT92OezEcqbe2t7O3itLSCOGCBAkUUaBVRQMBVA4AAAGKqqqqqqqqqqqqqqqqqqqqqqqv2q9GbXf2f/GFrGgZ4LWO6UkcqIZkkYj0+VGH0Jqqqqqqqqqqqqqqqqqq8LaM/iPxPo/h6MEvqd/BaqFGTmSRV4/OgAAFUDgcCqqqqqqqqv8AgpF/yZH8U/8AsH23/pZBX/BEP/klXxK/7GC1/wDSc18Vvhz4f+L3w18S/DHxRbRzaZ4k02aymDpu8suuFkUdmRtrqeoZQRgiv+CQPjrV/ht+1J4o+CeuStAniXTLi3ltSTxqFg5cce0X2odM8iv+Cx+rLp37IdvZmRFOqeLrC2AZSS2IriXA9D+6zz2Br/glnpn9nfsNfDyRklSS9k1Od1kGMZ1G5VSBjoVVT75z0Nf8FC9Avv2Wv2/fB/7R2g25j03xBdWeuFYlwHntnSK9h/4HGEZj3Nwa8DWVt+2f/wAFZNX8YxyLqPg/4bXQuYpEJeJo9O2xW+1uhV7wiUDoyluo5r9pyw/tX9mz4saX53lfa/A+sw79u7ZusZhnHGcZ6V/wQ11TEvxk0WSZzuXRbiKPHAx9tVz9TmP8qr/guf8A80S/7mD/ANx1fsUf8mi/CD/sT9P/APRK14n0SLxN4a1bw3NKY49VsZrR3A5USIUJH51/wSi8YQfBn9snVfht48VdMv8AxBpl54e2XDBDDqMM8cgiYnuTBLGF6l2UDk4Nf8FvvGegHQvhl8PIr+KTWheXmpy2yuC0NvsWJHYdQHYuFPfy39K/4Jvf8mR/Cv8A7B9z/wClk9VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVqul6frml3mi6tapdWN/bvb3MEg+WWN1Kup9ipI/Gv2L7++/ZB/wCCj0/wk168aKwvdVvPCVxLIceekrg2UmOP9ZKlqR04kP0qv+CzPxkbwf8AAjQfhBpl5svvHuqebeIrcmxs9sjAjqN07W+D32MOecf8EuvgufhD+yV4ev7+0MOseOpX1+9DphgkwVbZeeceQkT44wZG9cn9sL/lLU3/AGOHhT/0l06q/wCC03gnwvqH7Pfhjx/dWVuviDR/FEVjZ3e0CRreeCdpYc9SpaJHx2KH1Nf8Eptd1jXP2KfCEWrNK40y+1CztJJCSXgW5dlwfRS7IPQKB2xVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVWtaHo3iTS59F1/S7XUbC6TbNbXMQkRx7g/wCRXxh/YQJa4134OagoGSx0S+kPHtDMf0WT/vvtXiXwp4l8G6m+i+K9BvtJvoxkwXcDRMR/eGeqnsRwfWqqqqqqqr4UfsvfFT4rywXdro76LosgDHVNSjaNGX1iT70uR0IG3PVhXwb/AGa/h18GoUvNMs/7V10riTV71AZRxyIl6RLyenJB5JqqqqqqqqqqqqqqqqqqqqqqqqqvFOhw+J/DGr+GrhgItWsJ7RywyAskZQ/o1XtncafeT2F3EY57aRopUbqrKcEfmKqqqqqqqqqqqqqqqqr9lDwwPFPx78LQSRF4dOne/lOPueShdD/38EY/Gqqqqqqqqq/b18IeKPH37IvxG8IeCtAvtb1vUbG3S0sLGEyzTsLqFiFQckhVJ+gNf8EiPhD8UfhD8OPH+m/FHwDrnha61DW7eW1h1Wye3aZFgILKGAyAeKr4nfs5/H34I/8ABSI/G74ZfCHxX4h8JN4rh1x73R9LknRre7wb+IFQQG/e3K49Cp74r/grf8Nvil8Wfgj4Q8IfC3wLrvim5j8VLfXdtpVg1y0SR2s8YdtoJUZmwOxz7V+w54J134dfsm/Dbwb4n0a/0nV9P0pvttjfRGOa3leaSRlZSARgv0PbFf8ABUb9m3xN+0L8AbKb4e+HZtZ8X+EdYju7C0t1BmuLeX91cRJkj1jkPtDX/BKn9lvxb+z78KvEfiX4m+F5tE8X+LtTCtZ3Sr5tvZW4KxA4J2lpHmYjuNhr4gaTPr/gLxJoVqu6bUdIuraNdu7LPCyjjvyelf8ABIj4JfG74O/Ejx8fif8AC7xV4WsNW0O3+zzapp0lvDNLFP8AdBYDL7ZCQPTdVf8ABYb4J/F34x/8Kj/4VX8OPEHiv+yP7b+3/wBk2L3H2bzfsPl79oO3d5b4z12n0r9krw7rvhL9mL4X+GPE+k3Wl6vpfhaxt72yuojHLbyrEoZHU8hgRgg1X7e//BNXxT8VfHcn7QP7Nl5bWPjGd0uNU0d7oWf2u5jxsurWbhYpztXcGKqSN+4Nnd4O+NX/AAWU0qzj+Hg+EFzq9/CnlR6vrOhwhwBgAm58xLdyOPmfcTySTya/bp/Zi+Knwn+Hvhz4zftF/EOTxX8U/HutyR6hsm8yHT7aODKwKQApILDIRRGmAqDHzN/wTt0+4039ir4U210oDvpEkwA/uyXMsi/+OsKqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv+Cwfw21L4Z/tH+Evj94YL2j+KbKJzdRrzHqWnlFD57HyTbYB/55t1xx8G/iNp/xf+E/hD4oaZ5aw+KNGtr8xo2RC8kYLx9+Ufch91NfteXl9+2f/wAFKNP+C+jXTzaPpOqW/hmMxP8A6qG3Jl1GUAfxI32n6iJfw07T7HSdPtdK0y1jtrOyhSG3gjXasUaAKqqOwAAFft2+JNM8G/8ABULWvF+tNINP0LxD4bvrsxJucRQ2NhI+0dztU4Fal/wVy/YvstMF9a+KfEuoT7QfsVtoEyygkZxmTYmR0+919ua/aI+Pvxr/AOConxR0L4VfBP4d6ja+FdFuDNa2crbvLd/kN9fyrlIVVcqoydu5gC7OBX7PXwb0f9n74LeEvg9olx9ph8OWAimudu37TcOxknlx2DyvIwHOAwGeKqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqvFngrwl470w6P4w8O2Gr2hztjuoQ+wnuh6ofdSDXxB/YE8Has8l78OfE11oUpyRZXqm6g+ivkSIPcl68Y/si/Hbwg8jDwgdbto84uNIlFzv+kfEv8A45WqaPq+iXTWOtaXd2Fyn3obqBonH1VgDVVVV4U+D/xR8cFT4V8B6zfxscCdbVkhz7yNhB+JrwL+wP8AEHWfLufHfiHTvD0B5a3gH2y4HsdpEY+oZvpXw5/ZV+DXw4aG8tPDg1nU4SCL/ViLhww7qmBGpB6ELketVVVVVVVVVVVVVVVVVVVVVVVVVVV+1X4LbwT8cvEdtHFtttWm/tO2OMZWf5nwPQSeYv8AwGqqqqqqqqqqqqqqqqq/4J8+Cma48UfEO4gO1Ej0y0kK8EnEkwB9QBD/AN9VVVVVVVVVVVVVVVVVVV8Vf28/2cPgj8Yn+C3xR8VXeh6pHpsF61+1i89ohlL7YXaLc6PtVX5QLiRfm6gaj+3b+x3pmlvq9z+0V4LeCOPeUt7/AO0TEe0UYaQn2C59q/a8+OXiL/gpL+0X4R+EvwC0O/uvD+jtJbaS9xCYzO8rJ9pv5l6xQqsaY3cqqknDOUHw28DaT8Mfh54Z+HGhHdp/hfSLbTbZiu0ukESxhiOfmO3J9yeTVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV/wVQ+ECfFP9kXxDq1pbCTVPAlxHr1oQpz5ceUuRkfw+RJI5HTMa56ZH/BNv9p+y8HfsG/EW5126jluPgyby5toHb/WQXEbT20fXrJc+fGOg6c9cf8ABGn4V3/jT4n+Pf2kfFQa8m02M6dZ3Uy5Mt9dt5tzKD/fWMKD7XJqv229H0zxD/wVT1Dw/rVlHeafqXifwxa3dvIMrNFJZaerofYqSD9atP8Agnl+xbZX51KD9n3w60xYttmeeWPJOf8AVvIUA9sYxx0rwh4G8FfD7R4/D3gPwjo3h3TI+Us9KsY7SIHGM7IwBn3qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqrUNM03VrY2eqafbXkDdYriJZFP4EEVq/7PnwQ1sN9u+FvhxS33mtrFbYn3zFtOeetXn7Hv7O14HI+H/kO/wDFDqd0u36Dzdo/Kov2MP2eo5Fd/B91KAeUfVbkA/k4Naf+yr+z7pjI1t8NLByh4FxcT3A/ESO2fxrQvht8O/C7rJ4b8C+H9MkTpJaabFE+R33KuSfeqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/b6+Hg1Pwpo/wASrGDNxos32O9ZR1t5T8jH2WTge8tVVVVVVVVVVVVVVVVAFiFUZPYV+z78PG+GHwj0DwtdQhL8Qfab8EYP2iU73U+65CfRBVVVVVVVVVVVVVVVVVVXxw/Yw/Zq/aJvn1r4ofDGxvdbZFU6xZyyWV4wUALvlhZTIAoCgSbgBgDGBVv/AMEef2PYb/7ZJH42uItxP2WTW1EeD2ysQfA/3s+9fBr9nb4Kfs+6RJo3wf8Ah5pfh2O4UC5niVpbm5x082eQtJIAc4DMQM8YqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqrX9D0vxPoWo+GtbtEutO1a0ltLyBxxLDIhR1PsVYj8a8ZXvjn9nDV/jH+zY8zi11a+j0nVSwKGdbG8E0EoHo4UEeqyfgf2B/guPgV+yr4I8J3dmbfV9SsxrGsKwAcXV0BIVbHdEMcX/bMcmq/bC/5S1N/2OHhT/wBJdOqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqrxh4X0vxt4W1XwlrMYez1a0kt5eMlQwwGH+0Dgg9iAa8Z+FNW8DeKtU8Ia5F5d9pNy8EoxgNg8MP9lhhh7EVVVVVVVVVVVVVVVVfskfCpviZ8V7O8v7cvo3horf3pI+V3U/uYj/vOMkd1RqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqvF37Lv7Ofj/wAT3fjTxt8EvBuua7fsjXWoX2kRTTTFEVFLMRk4VFHPYCqrX/2cfgL4q8c/8LM8R/CHwrqXiz7RBcf2zc6ZHJdebAqLE/mEZ3II0APbaPSqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq+1PTNLRJNS1G1tFc4Rp5ljDH2yatrq1vYEurO5inhcfJJE4ZW+hHBqqstd0PUpvs2nazY3UoUt5cNwrtgdTgEnFVVrruh39x9ksdZsbifB/dRXCu3HXgHNVVVVVVVVVVVVVVVVVVVVVVVVVVVVft0/Bh9SsIPjF4ftN09ii22spGnLxZxHMcddpOwn0K9lqqqqqqqqqqqqqqqoIJrqaO2toXlmlYJHGilmdicAADkknjFfs3fCCH4O/DWz0i6hUa3qQF3q0nU+cw4jB9EXC+mdx/iqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/4Le/8kq+Gn/YwXX/pOK/4Jvf8mR/Cv/sH3P8A6WT1Vf8ABG3/AJO5u/8AsT77/wBHW1VX/BLP/k/Sf/sH6x/OqqqqqqqqqqqqqqqqqqqqqqqqqqqqtS06w1jT7nSdTtYrqzvIXhuIJV3LJGwwykdwQSK/aJ+CeofBXx1NpkUUsmg6iWm0i6YZ3R942P8AfQkA+o2t/FVVVVVVVVVVVVVVX7E/wDk1fUofjH4rsSLCxcjQ4ZF/184ODPg9VQ5C/wC3z/BzVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV/wAFvf8AklXw0/7GC6/9JxX/AATe/wCTI/hX/wBg+5/9LJ6qv+CNv/J3N3/2J99/6Otqqv8Agln/AMn6T/8AYP1j+dVVVVVVVVVVVVVVVVVVVVVVVVVVVVV8Wvhb4d+L/gu78H+IE2eYPMtLpVy9pOB8si/TOCO4JHfNfEf4d+Jfhb4svPB/im08q7tWzHIoPl3EZ+7LGSBlTj8DkHBBFVVVVVVVVVVVVV+zV+z5qHxs8Tfa9TSW28K6VIp1C5AKmduogjP94jqR90HPUqDpum2Gj6dbaTpVpFa2dnCsNvBEu1YkUYVQOwAAFVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVf8ABb3/AJJV8NP+xguv/ScV/wAE3v8AkyP4V/8AYPuf/Syeqr9k/wDZm139rH4oy/C7w94nsNBuotKm1E3V7C8iFYnjUrhOcnzB+Vf8OQvin/0XHwp/4Lbj/Gv+HIXxT/6Lj4U/8Ftx/jX/AASZsX0v9t2DTJJFdrTRtUhZgMBiqgEj8qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqvjZ8E/C3xt8LNoutILbUbYM2m6kiZktZCP/HkOBuXv14IBHxF+G3i74WeJJ/C/jDTGtbmMkxSDmK5jzgSRt/Ep/MdCAQRVVVVVVVVVVVV+z7+zh4l+NurLeziXTfC9pIBeaiyYMpB5ihzwz46novU84U+FfCnh/wT4fsvC/hfTIrDTbCPZDDGOnqSepYnJJPJJJNVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV/wAFvf8AklXw0/7GC6/9JxX/AATe/wCTI/hX/wBg+5/9LJ6qv+CNv/J3N3/2J99/6Otqqv8Agln/AMn6T/8AYP1j+dVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVfE34V+DPi54cfw34y0wTxjJt7mPCz2jn+OJ8HaeBkcg4wQRXxw/Zr8c/Be8kvJ4H1bw474t9Xt4jtXJwFmXnym6dflOeCeQKqqqqqqqqoAsQqjJ7CvgB+xjrfiyW18V/Fa2n0nRMB4dLOY7q79N46xIf++j6Lwa0fRtJ8P6XbaLoenW9jYWcYjt7a3jCJGo7ADpVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV/wWF+GPxJ+J3w1+H1h8Nfh74l8WXNlrlzJcw6HpM9+8CGAAM6wqxUE8ZPGa+Dn7RH/BTj4GfDPQvhR4K/Za8RyaL4eheK0a++HOqSzFXkaQ7mG0E7nPYcYr/huD/grD/wBGr6r/AOGz1X/4qv2N/iN8bvip8E7Xxb+0F4Kn8LeL5NRuYZdOm0efTGWFGAjbyZiXGRnnoe1f8EoPgV8bvhz+1FdeIPiF8HPHHhfS28K3sC32s+HbqygMrS25VBJLGq7iFYgZycH0qq/4JufAr43eBf21pvFXjf4OeOPD2iGx1VRqWq+Hbq0tizn5B5skYXLdhnntVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVXVra31tLZXtvFcW86FJYZUDq6kYKsp4II4wa+Mn7DPh3xAZ9d+E93Fol+2XbS7gk2sp6ny25aI+3K9AAorxx8OfG/w21Q6P428N3mlXBJ8sypmOYDqY5BlXHupNVVVVVVVXwr/Z4+J/xdnSTw7obWulk/Pqt+rQ24Hfa2CZD7ID74618Fv2UPh58JfI1i9iXxD4jjww1C7iAS3b/pjHyE/wB45b0Iziqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqtb0HQ/EunyaT4h0ey1Oyl+/bXkCzRt9VYEV8Qf2E/hl4ld73wVqV74WumyfKUG6tif9xyGX8HwPSvGf7Fvxv8ACzyS6XpFp4jtE5Eum3A34/65vtfPsoate8KeKPC832bxL4c1PSpQxXZe2jwHP/AgKqqqvC3wt+I/jYI3hPwPrWpxOcCeCycxD6yY2j8TXgj9g74pa8YrjxjqumeGrdsb4i32u4X/AICh2f8Aj9fDr9kD4M+AHhvrjRn8R6lFyLnVyJUVvVYQAg9shiPWkjjijWKJFVEGFVRgADsBVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVXFtb3kD2t1BHNDIMPHIgZWHoQeDWsfA/4O6+S2qfDLw1LIx+aRNNjjc/VkAb9f51efsk/s73zB5vhtbqQScQ391EOfZJR/wDWr/hjz9nH/onX/lXvf/j1WH7Kv7PumlTbfDSwfYOPtE88/fPPmO2fxrQ/hh8N/DLB/D3gHw9p0g/5aW2mxRufqwXJ/Oqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr/hqf4Y/wDPDWv/AAEX/wCLrwB8SNA+JVndX/h63vkgs5RE73MIQMxGcLgnOBjP1FeI9ctPDOh3viC+hnlt7CEyzLAgZ9g6kAkdBz9BX/DU/wAMf+eGtf8AgIv/AMXX/DU/wx/54a1/4CL/APF1/wANT/DH/nhrX/gIv/xdf8NT/DH/AJ4a1/4CL/8AF1/w1P8ADH/nhrX/AICL/wDF1/w1P8Mf+eGtf+Ai/wDxdf8ADU/wx/54a1/4CL/8XX/DU/wx/wCeGtf+Ai//ABdf8NT/AAx/54a1/wCAi/8AxdV4p8aeF/BVtBd+KNWSwhuHKRO8bsGYDOPlB7Vpuo2Osafbarptys9rdxLLBKoIDowyDzz0PetX1bTdC0y41jV7tLaztELzSuDhF9eOa8L+NfC3jW3nufC2sRX8ds4SUorLsJGRwwB6VWj/ABC8GeINcuPDWja7Fd6lab/OgSN8psba3JGOCQOveqqqqqrWfjRpejfEq2+Gsui3UlzczQxC5WRQgMgBBx14zVVVVVXt3bafZz395KI4LaJpJXIJ2qoyTx7CvC3jjwp42juJfC2sx6gtoVWYojLsLZx94Drg1Xir4geDvBMlvF4p1yKwe7VjCHjdt4XGfug+o/Oo5EmjWWNtyuoKn1Bqq+IHjS1+H/he58UXllLdxWzopiiYKx3MF6n618P/ABpa/EHwvbeKLKyltIrl5FEUrBmGxip5H0qqi8c+EJ/Ej+D4fENm2sxg7rQP82QMkehIHJGciqr4geNLX4f+F7nxReWUt3FbOimKJgrHcwXqfrXw/wDGlr8QfC9t4osrKW0iuXkURSsGYbGKnkfSqqq8ReJdC8J6Y2seItRSys0cIZXViAx6DCgmvC/jXwt41t57rwvrMV/HbOElKKy7GIyMhgD0qvEHiLRPCulya14g1COzsoSqvK4JALHAGACTya8LeOPCnjaK4m8LazFqCWjKsxRGXYWzj7wHXBqqqqqqqqr4i+OrT4deGn8S3thLeRpMkXlRMFbLHGcmvAni628eeE7HxXZ2clrFfeZthkYMy7JGQ5I9SufxqqrVfFfhbQpBDrniXStPkPRLu8jhJ/BiK03VtJ1m3+1aPqlpfQ5x5ltMsq5+qkiqu7u0sbd7q9uoreGMZeSVwir9SeBWl+KPDOuyNDoniLTNQkTO5LW7SUjHXIUmqoappjXn9nLqNqbvp5HnLv6Z+7nPTmqutZ0exl+z3urWdvKBnZLOqNj1wTX/AAkvhz/oYNN/8C0/xqLxBoE8iQQa5p8kjsFRFuUJYnoAAeTVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVfAnwwPC3wx0i3kTbcX8f22cEYO6XBXjsQmwfhV7Z22o2c+n3cYkguYmilQ9GVhgj8jXijQrjwx4i1Lw9dZMmnXUkJYjG4KcBvoRg/jVVVVVVX7R+gf238LL+eNN0ulTR3aDHOAdrf+OOx/Cv2cNc/tr4V6fA7hpdLmltH/AAO5f/HXUfhX7UfiD+yvh0mjxSYl1i8SMgHH7tPnY/mqD8a/ZxF14O+JPiX4f6hJh2g3AHjc8TcYHqUkJ/CvFWtReG/DWqa/KVxp9nLMAe5VSQPxIA/Gv2TdBkkg1/xndhnluZVtIpG6nHzyH3yWj/Krq7tLGE3F7dQ28Q4LyuEUfia0/XtC1djHpOt2F6yjlbe5SQj/AL5Jp2RELuwVVGSScACvih4+8R2nxu8LeG9M8SzQ6NLc2DTwQShEfdcYcMy8kEAAgnGKivLO4by4LuGRsZ2o4J/Sri5trSEz3VxHDGvV5GCgfiasPEPh/VpTb6Xrun3kqjlLe6SRh+Ck1Xjn/k6jS/8Ar9sP/QErUNX0jSUEmq6rZ2anobidYwf++iKt7m2vIFuLO4inicfLJG4ZT9CKq/1LTdLh+0anqFtaRdN88qxr+ZNafqulavEZtK1O1vY1OC9vMsgB+qk+lVXjf/kS9f8A+wXc/wDopq/ZD/5Bvib/AK72/wD6DJVfte/8hHwz/wBcLn/0KOtN/wCQba/9cE/9BFSyxQRtNPKkaIMsznAH1NWviPw7fzfZrHXtOuJg23y4rpHbPpgHOa/aT/5JHqn/AF3t/wD0atfs4yRw/CDTZZZFREmuSzMcADzW5JrT9d0PVpHi0rWrG8eP7629wshX6hScVVj8E/Cdn8UJPiBFrk73TXL3K2BZcLO2Sxz1IySdvb1xxVahrWjaSUXVdXsrMyfcFxOse76biM1+0ZNDc/B7Up7eZJY3mtyjo24MPNXkEV+zb/ySPS/+u9x/6Nary9srCE3F/eQW0Q6vNIEUfia07XdD1gsuk6zY3pT7wt7hZMfXaTVV+1Rc+R8Moog+PtGqQx4x1wkjfh92vgJeXHgH4mr4Rv5CkHiLSreVA3QytCsyflukT6mq/aJv5vFPijwp8J9OmO6+uknuwv8ACGOxCfovmMR9DX7I0jwv4s06VlDxPakL3z+9DfyFVVOyIhd2CqoySTgAVa+JfDd9c/YbHxBptxcZwIYrtHfP+6DmiQBknp61+0T4+8R6L4g0DSfDfiWa0s7qMtcLaShSx3gcsvzAY7ZxUd9ZTOIobyB3PRVkBJ/Cqq88S+G7Cf7Lf+INNtps48ua7RGz9Cc1HLFPGssMiyIwyrKcgj1Br9p3/klU/wD1+wfzNfs7/wDJHPD/AP28/wDpTLVV430vXNb8KalpfhrVm03U54Cttcq20o2QcZwSAQCMjkZyOa0j9lTwmLbzfFHiDVtS1CT5ppYpFiTd3wCGY/Unn0FeOvCWufs6eLNM8ReD9buJtPvSwVZjgtsxuilC4DAggg4HfABGa0/V7LUNEttfWQR2tzarchnONqMgbJP0NaTb6x+0v8QL6bVNRurTwpo5zHbxtjCkkIAOm98EliDgDHpXxS/Z/wBI8LeHpfGXw6ub+wv9FX7Q8YuGYsi/edG+8rKPm64wD0r4I/ECb4h+B4b/AFF1bU7GQ216QMb2ABD4/wBpSCe2d2K+LGnfFvXb7T9B+HuoQ6dp11C5vr0tseIgjjdywyDxsGcg84r4c+Gn8H/tKWXhyXUXv5LOScPcuu0ys1m7MxGT3Y9zVfED4BeGviJ4ifxJqes6lbTvCkRjgKbcKMA8qTX/AAyT4K/6GTW/zi/+Ir4beANM1b43rYeFru5uNG8N3i3L3c2Cz+Swx0AGGkAA/wBnJ7VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV4G8OSeLvGGkeG41LC+u0STHBEYOXP4IGP4Uxt7O2ySkUEEf0CKo/QACvAPjC08eeFbPxNZp5a3JkUxnqhVypH6A/jX7VPhgaX43tPEcEeItatf3hx1miwrf+OGP9aqqqqqq13SoNd0S/wBEuceVf2slu+R0DqVP86/ZS1KfT9T8T+C707ZoWSdYz1VkYxyfqYxXxdz44+O3hHwLGfNt9O2TXKY6bj5kg/79Rp+deOyfBH7TeieIQfLg1jyPMb+HDqbd8/QAMfrmv2nfEA0j4ZvpkcmJdYu47cAddinzGP0+RR/wKvgr4cHhj4ZaFYPHtnnt/tU2Rg75TvwfcAqv4V4s+Ami+OfF914l8TeI9WntpAgt9Pjl2pBhQGALZwCQTgAcnrXxZ+A+keBPDj+OfAOpajZ3WkujyI1xk7SwXcjABlYEg9emfx+HGuD4tfCVU1uZxNf2s+n6hJFhW3YKM68EAlSG6YBPTFeO/hX4e8L/ABc0LwDp95qMmn6pJaLNJNIhlUSzFG2kKAOBxkHn1rwB8CvCPw411vEGh6jq89y1u0BS7mjZNrEEnCxqc/KO9eOPgdpXxC8VHXvEPiPVTYrAiR6fDJhUcZBYFsgAjHAA5yc818T/ANnjw94Z8L3fi3wPe6hZ32jx/aCjz7gyLyxU4DKwHIIPb8a+A/ji98d/D+C91WYzahp8zWlzK3WUqFKufcqy5Pcg18Z73VdN/aB+26FCs2pRSWZtI2XIaXy0CDHf5iOKsP2atD1W3bUviB4g1bWNcu13XFwtxsVGPZAQSQOmTx7DpWt2XiX9mfx3Z3Gj6nc33h7UTvMMhwLhAQHRh08xQQQ4A6jsSK8dePtP8H+A7nxrEUuE8hGs0JwJnkx5Y+nIJ9ga+H3wiu/i9af8LF+KWtX9ydQZjZ2sUmwLGDjPfauQcKoHAByc18TfhbcfBZLb4i/DTWb62S2mWO6glk37Qx4PQbkJwCrZ5I/DwV4mt/GXhPS/E1sgRb+3DtGDnY/R1z7MCPwqvG//ACJev/8AYLuf/RTV+yH/AMg3xN/13t//AEGSq/a9/wCQj4Z/64XP/oUdar4hsPCng2XxHqb4ttPshK4HViFGFHuTgD3Irwp4c8YftJatd+IvF2u3Fj4fs5vLitrc/KGxnZGDwCARl2BPI69r39lf4b3FsIrK71i0mUfLMtwr5PqQVx+WK/aMhit/g7qFvAu2OKS2VF9AJVAr4R/DvxB8VPBlrp2ua9dab4T0uaRIba0+V7yUsXdyTkYXcACQRkHAHJr4r/BWT4SWdr448D6/qJjtbhUkMrATW7H7rh0CgqTwRjuOuePhb4ul8deA9J8SXKqtzcRFLgKMDzUYoxA7Alc47ZrQP+Tt5f8Ar9u//SWSvjt8SZ/h14SV9KdRq2qOYbNmGfKAGXkweuAQB7sO2a+HP7PVv4v0mLxn8S9V1G6vdWUTxwLPhgjDKtI5BJYjBwMYB59vjL8LdR+F3hqZvCuu3tx4Z1SZI7yxuWDmGUEMjjAAwSuM4BHAJOeP2bf+SR6X/wBd7j/0a1eJf2f9C8Z+Lr3xL4o8RatdQTMDbWKS7VgG1Qy7juO0kE4XbjIr4ufA/TfhzoP/AAn3gDVNRsp9LljMyNPkqGYKHRgAwIZhkZIwe2OfhR4vm8deAdK8Q3ZX7XJGYrrAxmVGKscDpnG7HbdVftbXBXwfotmC37zUi+O3yxsP/Zq+N+j3nhe1+H3xD0sgT2NnbWzyBcDzIlWSIn1z8/4LWj6zY63odnr9pKBa3tslwjMcbVZQ3PpgHmvg6H+JXxp8RfEudWa007KWW4H5d4McePpErZHq1fswObXx14v0wyKTsyRjrslYZH/fX61VVqV9r37RHxEvvCdhrE2n+ENFY+c0B/14DbQx7MzkHbnhVGcE5zr/AOyz4Ll0iRfDF9qNlqkSbreaafejOBxvGOAT3XGOvPSv2evHur+LdH1TwT4ruJZdS0YbBLI2ZHiOVIY92QjGT13DrzXxo+Ffh74c+IdF0jQ7zUZ4dRjLStdyIzKd4X5dqqBx6g14M/Z58FeBvEtn4p0jVNbmu7Hf5aXM0TId6MhyFjB6Me/XFV448Mz+MPDF54etdaudKkugoF1bkhkAYEjAIyCAQRnvVv8AssfDaOz+z3NzrE85Xm4NyqsD6gBcfmD+NfDe81r4QfGaT4XXWqy3ejX8myFXPygum+KRV/hYnCNjrk9cCv2nf+SVT/8AX7B/M1+zv/yRzw//ANvP/pTLVVVftRawPEOveHvh5oqfatRWQySRpyQ8u1Y09iRk/Qqe9fEKI+DvghqWm28v/Hhoi2SuB22LFkfga/ZQsIoPh9f3wA8261RwTjnascYA/Mt+dahZxajYXOnzgGO5heJwRnhgQf0Nfsh3rrqHiTTSxKyQ28oGOAVLg/8AoQ/KqsP+Tvm/67y/+kLVVfHPx5/wgfgO6ntJ9mp6lm1ssHDKzD5nH+6uTn12+tfs5+BV8JeA4tVu4duo69tuZdwwVix+6X/vklvq59Kqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/ZQ8Mfb/FOpeKZkBj0q2EMJI/5ay9x9EVh/wIV8dvEn/CMfC/WbiOXbcXsYs4MHBLS/K2D2ITefwr9knxL5thrXhCaXmCRby3U9cMNkn0AKp/30a/aS8Lf8JF8Nbm/gi3XOiSrdpgc7PuyD6bW3H/cFVVVVVVVYxjwN+1TLAB5drrruQem7z49//o4Y/CvgzH/wmfxo8Z/EBx5kFo7QWrn/AGm2oR9I4sf8Cr9rLRpG0XQPFVtlZNPu3gZ16jeoZT+BjP8A31Xxc1UfFLxp8O/C9s26DULOC6mCH7guCpfPuqR59s0iJGixxqqqowqgYAA9BV98UviL8WPGN14P+E9zBpenWYbztRkALOgbaZMkHaCcbVUbj1J64+IPwd8YaT4H1bX9f+Lutas9pbmR7RzJ5TjI+XmQ8fh+Ffssf8kxk/7Ck3/oEdfF7/k5Lwh/1303/wBKTVeJviv498e+Orj4c/CVoLNbNnW51GUAkhCA7gkEKgY7RgEnIIxnFeLfgx44tvCGs654i+MetX8lnp1xO9oPM8l9sZJTmTG1sYPyj6V+yV/yJWsf9hQ/+ikrx2FP7U+lhgD/AKdYdfXalV+1w9sPCuhRsV886gxQHrtEZ3fqV/SvjIl/D+zz4FhmyF/0MSDoR/or7AR9P1FeDfh98cdQ8JaPfaB8UrW106exie2tw7jyUKjCHCHkdD7ita+DHxz8RaZNo2t/E6yvLK42+bBI0hVtrBhn5OxAP4Vovwn16x+EUPw5/wCEtOnX8czONQsldtgMxkwo3IeQcHkdT1r/AIZ28a/9F01v/vxL/wDJFWPhLUtC+GF14Ru9cn1q8Wwuo/tsyEPMZN5GQWY8BgvU9O1fshXcBh8TWBfE262kCnuuJASPocfmKr9rSVbzxH4a0i2Be6FtK3ljv5jqq/mUav2jYri2+Dt3bQMWSOW2SZsY+UOvOP8AeC1+zebQ/CLSRbbfMEtx5+Ou/wA58Z99uz9Kr9pP/kkeqf8AXe3/APRq1+zb/wAkj0v/AK73H/o1q/aI/wCSOeIP+3b/ANKYq/Zi/wCSVW//AF+z/wAxWgf8nby/9ft3/wCkslftefaDq/hsMD5H2afZ/vbk3fptqy+y/Y4PsW37P5S+Tt6bMcY9sYr9pMqPhJqYLDJntwAf+uq1+zeyr8IdMZmACzXBJPGP3rVJ8TfiV8YvFV34b+Fl1Do2k2QJlv5AN7JnAckgld3ZVGfU9cfEv4P+LND8Car4i174ta1rLWsas9pL5nlPl1G35pDwCc9O3QV+zF/ySq3/AOv2f+Yqv2vrjbbeFrMc73unPPTaIh0/4Efyr4teD/8AhI/hTqOhxRK1zZWizWyoP+WkIzhfqAV/4FXhX4lvYfs2axYm5xf2MzaZb8/MUn5Bz2wrS4/3BX7P3hL/AIRX4aaeZ4tl3qxN7PkcjeBsH/fATjsSa+A/+gfHfxjphCoAl6gT/aS6QYH4Zqq1lrhdIvmtP9ettIYscfNtOP1xXwO8MfELxHHrH/CA+MoNEaAw/a0csDLnfsPCngYb86/4Vl+0R/0Vy2/77f8A+Ir4QfBnxT4A8Yaj4p8Q6/Y351C0kicQF9zSPIjlzkAfwn86/am/5HXwr/1wP/o0VVfGP4pQfC7w/Fdw2qXWp37tHZQOSFyoG52xztXK8DkkgcdRofgb44fESwt9f8T/ABOuvD9veIssNpYKUcRsARkRsgXIPcsfXmrnwrd+Df2gvDmiXviG71qVby0f7VdZ3kM3TljwPrX7Tv8AySqf/r9g/ma/Z3/5I54f/wC3n/0plqqr4wfF7TPhppPkWrRXeu3S4tLMtnYDn944HIUY6dWPA7kfA34U6pa3snxO8fCSXXtQLSW0U4+eAOOZGHZ2BIC/wj3OB8YNOfVfhh4ltIk3sNPklC9z5fz8f981+yhfxXHgDUNPDDzbTVHJUf3XjTB/MN+VatfxaVpV5qkzAR2dvJM5PQBFJP8AKv2Q9Ocz+JdWaP5FS3gRj3JLsw/RfzFVYf8AJ3zf9d5f/SFqqvEzP8cfjxb+Hbd/O0Hw+xWYj7pjjYGZv+BviMEdtppURECIoVVGAoGABVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX7O3hgeHPhhp88sW241dmvZcjnDcR/hsVT+Jr9rfxJ5l3ofhCGTiKN72dfdiUj/ICT8xXwI8S/wDCMfE/R55JQlvfObKfJwCsnC59g+w/hWoWNrqdhcabexiS3uoXilQ/xIwww/I14j0S48N6/qPh+7OZdOupIGOPvbWIyPY4z+NVVVVVVftTWF3ouu+FvH2lnZcWzGHzcfceNhJF+pf8q/Zm8PDRvhlBqEiYm1i5kuTkc7Qdij6YTP8AwKvjdoJ8RfC3X7NE3S29t9qjwOQYiHOPqqkfjX7Mum3Xibx/J4i1A+Ynh/TFhhJH3GZfLQf98CSpY1miaJicOpU4ODzXgLxDffs7eOtV0jxlo11JZ3qCMTQIMuEbKSx5IDKQTkZBGfUEV8QvirqfxZ8J6roXw88O366Xb2r3Oq6leII1WKIeYY1wT8zbQMZyemMZI/ZVljk+GlxGj5aLVZlYY6HZGf5EV+0jpOt+H/Hvh/4mafYtPa2SwBn2kqk0MpdQ3oGBGPoa+G/xw8NfE3V5NC0jS9StbqCza5lNyqbAAyKQpViTy46gVo2p6j+z18WNTn8SaPdXGl6gJY0miXJliZw6yITgMwwAVyMZPtXi/wCL998VdA1Dwl8MPDuoutxbONQ1G7QRx28IXLqMFuWUFecdTgHOR+yTLG3g/WoFfLpqQYrjoDGoB/Q18VdD1jxN+0TLpPhu6jh1QRRTWrscbZYrbzVGexOwAZ9RVj+05pekWx03x/4V1jTtctV23EMMClXYcEqHZSoPXByPc1La+Lf2l/G9nqEulzab4V047BI3RI8gvhv4pHwBxwuBnpk/ELwFY+OfBF14RUpbfu1Nk+PlgkT7hwO3G0+xNeCPit4l+CEP/CCfEjwxfvZWzt9knhALKpJJCFiFkTJJBDDGSPYXn7VHgllEGh6Brmo3bjEUPkpGGPpncT+SmrC6+3WFte+TJD9ohWTy5FKsm4A4IPIIzgiqrxJ4b8YfAL4hz+OPC2lvf+Hrtn3ogJRY3OWhkwMptIBVunA68ipf2tfB/wBhD2/hjWXvdvELGNY93pvDE4z3218O/BXjL4p/EVPir48097LT7WVZbSCRCnmFP9UqKedinDFj94+uTjxZ4bsfF/hvUPDWokiC/gMZYDJQ9VYD1DAH8K8KeJfHP7OGq3egeKfD817oV3NvWSI4QtjHmRP0JKgZQ4PA6Y5u/wBqvwS0KJovh/XL68k4S3aJI+fQkMx/IGv2jZVn+D2ozIGCySWzAMuCMyr1Hav2bf8Akkel/wDXe4/9GtX7RH/JHPEH/bt/6UxV+zF/ySq3/wCv2f8AmK0D/k7eX/r9u/8A0lkr47fDW4+I3hJU0pVOraXIZrRWOPNBGHjz23AAj3UdASa8A/tET+BNKi8GfEbw7qn2jSkEMUkUYEoRRhVeNyvIGBuzyMcdz8TPFvi34y+FL3VtK0G40nwloS/aWlufv3sudqgY4woYnAyBySc4Ffs9W0d/8F7OxaQqJmuo2Knld0jD+Rr4b+LLz9nvxdrOgeN9Eu/s98EVpIEBJMZbZIm4gMhDt3z07giviV8UNV+LXhDVNK8BeHr6LRLOE3OqaleKI1KRfP5a4JGSQp65OOgGTX7L8scnwsjRHyYr+dWGOh+U/wAiKr9qMi+8aeEdJAjJKNw3+3Kq8+3y/wA6IBGCM+xq/wDhlqi/GCf4Z20M6aNeask5VVOzyArOGz6rE7r9c1HHHDGsMSKqIoVVAwAB0Ar4c40/9p/xFbAbPtM19w3U7m8zj8s1+0P8S/FPgc6HpXg2/Fvfai0rSEQJK21SgUAOCOST27VoNvqVrodhb6zdm61CO2QXUxAHmS7RvOBgAbs8Cq1nQPGH7Pnj278YeGdIfUvDF+W82JASscbHPluQCUKn7rkEYx6kVD+1d8OntRLPpeuxzAfNCsEbc+x8wAj64r4bfEOL4kaVdatBoV7psMFz5UIuR/rk2ghwcY6kjAJxgc81+1Z4Y1e5i0TxhpltJNDpokiunRd3k5KsjEf3chgT0HHrXw5/aB8PfEHV7Hw1Fo+oWuqXMLNIWVDCrIu5trbtxHBxlRVftS+CNa8Q6LpXiLRrSW7GjmVbqGJSzKj7cOFHUApzj1B6AkeHv2pfC39gWdrqGhavJrMcKRNBbRIyTSAAZU7sgE9sZGe/e8uPFVx8efC/iLxtYDTbnWry0uILQkk28JlMcaNxw3yc59cnGSB+07/ySqf/AK/YP5mv2d/+SOeH/wDt5/8ASmWvAHxo+IPiT4vyeEdV06JdPaa4je1W32vZrGGwxbr1Cg54JPGMgVXxg+Ib/DTwc2u29mLm7uJxa2qt9xZGVmDP32gITgdTgcZyPBvxE0fS/Fk/jfxxot14l1ZpfMgaS5VEjf8AvlSpyRxtHAXHA6Y/4a803/oRrn/wPX/4ivBfiW38e+D7HxGdP+zw6nE+baRhJgBmUgnAznHp3q2XxB+zP49vbiXS7q/8I6qdokiGcqCSnzHgSJkjBxuBPrkfEH44zfE/SW8BfDDw/qs9zqoEdzJJEqsI8jKqFJwD0ZmIAGfXI+EXw/T4ceC7bQ5WR76ZjcXzqcgysACB7KAq++M96qw/5O+b/rvL/wCkLVXxt8eDwF4Eu7y1n2alfZtbEA8q7Dlx/urk/XHrX7M/gb/hHPBZ8S3seL7xARKu4crAufLH/AslvcFfSq/aE+Jfi74d2WkDwvFFEL+SQTXckPmBCu3CAHgEgseR24718P8AXdS8T+C9H1/WLMWt7e2qyTRhSoz/AHgDyAwAYD0IqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqrW1t7K1hsrWMRw28axxoOiqowB+QrxD8Kvh/wCLNTfWfEPhuG9vZFVWleWQEhRgDAYDpUXwL+FEEqTw+DrdJI2DIwnlBUjoR89V4i+C3wz8Vatc67rnhkT393gyzLdzR7iFCg7VcLnAHbnvXib9lTwXqFu7+GNSvdKugp8tZX8+In0IPzD67uPQ14w8H674G1ybw/4gtRFcxDcrKcpKh6Op7g4/ocEEVVVVVVVVVd2VnfxeRe2kNxHnOyWMOM/Q1DBDbxLBbxJHGgwqIu0D6AVTokiFHUMrDBUjIIq1sLGxUrZWcEAY5YRRhMn8KubS1vYTb3ltFPGeqSoGB/A1b21taQrBaQRwxr91I0CgfQCqq70+wvwgvrK3uNhygliD7T6jPSlVVUIgACjAAGMVU8EFzEYLiFJY2GGR1DA/gatNO0+wXbY2Nvbg9RFEE/lVVVR6RpMNwbuHTLRJz1lWBQ354z2FVUkccqGORFdSMFWGQfwq10zTbEk2On21uTnPlRKmfyHsKqqqqutN06+ZHvbC2uGj+4ZYg5X6Z6UFVVCqAABgADpVXdjY38YhvrOC5QHIWaMOAfXBqKGKCNYYI0jRBhUVcAfQCqqqqq1n4eeI/GPx1s/FOtaZ5fhzQYEFpK8qfv5E+cYQEsP3jk5IAIT3FVVDR9IFwbtdLsxOesvkLuP44z2qiARgjPsat9O0+zdpLOxt4Xf77RxBS31x16VVR6ZpsNy17Dp9slw2d0qwqHPrk4zVVSWlrHO91FbRLNKAHkVAGYDpk9TVVVU6JIhjkVWVhgqwyDVrY2NhH5VjZwW6HqsUYQfkKqqqqq5tbW8i8i7t4p48g7JEDDI6HBoAAAAdOgqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqvHXiq38E+EdT8UXEQlFhBuSItt8xyQqLntliB+Nf8Neaj/wBCNbf+B7f/ABFfBv4yXPxVudUt7jQYtO/s6OJgUuDJv3lh3UYxt/Wvjd4m1zwh8Or/AF3w7ffZL6CWFY5fKWTAaRQeHBHQntVj+0t8WbR0a41ezvQvVZ7GMBvrsCn24xXwd+LVp8UtIneW0Wz1TTyourdWypDZ2umecHBGD0I+hP7T/hK21rwD/wAJIkY+2aFKrBwOWikYI6/TJVv+An1qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv2svEwtPDuk+FIJB5moXDXEwB5EcYwAR6Fmz/wAAqv2Q/wDkI+Jv+uFt/wChSV+0n/ySPVP+u9v/AOjVqv2R7e6bxNrt0it9njsERz23s4K/orV8criG1+E/iOSdgFa1CD/eZ1UfqRVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVftDeJf+Ek+KGpJFIGg0lVsYsdimd//kRnH4Cq/ZD/AOQj4m/64W3/AKFJXxA8F2vxB8L3Phe8vpbSK5dGMsShmGxg3Q/Srb9kjwkkoa78U6tLGOqxpHGfzIP8q8G+B/DfgLSRo3hmwFvCW3yOzFnlb+8zHkn9B2xX7TvxP07UYY/h7oN2k/kzCXUpY2yqsv3Ys9zk5PoQB6gVVVVVXxe+O/jDS/Ft/o/gK5ijsNEQR3s7QJKGmLYPLA4AJC49Qx6dPBmsS+IfCGia5O4aa/0+CaUgY+dkBbj6k1VVVVVVVVXxO+J3xFj+KUHw3+Hl1ZxTPDGD5sCsfNZS5yWyANm09K1OX9qrwraPrFzqOma5BbjfLBBbxMwUdTtVEY/RSTXwb+L1r8UtNnjubRLLV7AL9pgRso6no6Z5xkEEHOOOTmqqqqqr4y+ML/wN8PtQ17SJljv1eKK2ZkDAMzgHg8H5d1fs+/FzXPH0uqaH4suopdRtlS4tmWJYy8JwGGFAHBKnP+3VL8SfFup/tBHwBpmoRLoVoP8ASEECljtg3t82Mj5yFqqqqqqqqqqqrqSWK1llt4fOlSNikecb2A4Ge2elfA/x78SvGeq61F400n7PaWwBhc2hg8qTcQYhn73GTzkjAyeaqq8S+ItK8J6FeeIdZn8q0soy7kDJY9AoHck4AHqa+FXx/wBW+IvjJPDV14dtLKB4JJVkSZmYbRkDnimKquSQAByT2rXP2gdY1jxBL4X+EfhI6/cQZD3UoYxHHBYKpHyZwNzMoP5GtG/aF17Q/EMPhv4ueERock+Nt1CGVEycBirE5TOcsrHGOh5pWSRA6MGVhkEHIIqvib8YdT8Ga9b+EPDngu91vV7q3E8YUHZsJIyoQMzYKkEfLj1rUvjl8a/C6Lqniv4YW8GmZG5hBNFtz0Bk3MFJ91r4d/EPQ/iVoI1vRhJE0b+Xc20uN8D4zg46gjkHv7HIHxX+JPjHwNqFhaeGPBUuuRXMLPLIkUr+WwOAPkB7etf8NA/Ff/okNz/4DXH/AMTV5+0948066isNQ+HUVrcz48qGfzY3fJwMAjJyeK8X+JLbwf4av/Et3Z3F1Dp8XmPFAAXYZA4yQMDOSewB69KHxq+NniCD+0vCPwmC2JG6N7mGabeuOqkGPd26A18PP2iV8QeIY/B/jbw+2iarNL5UTDcEMh6I6N8yE8Adck9qqqr4h/Hmz8Na5/whvhDRJfEOvlvLaGInZE/935QS7Duoxjucgir343/F7wcYtR8f/DGGHS5XAMttvQoD0Bbc6hvY7c+1eE/Fei+NdCtvEOgXJmtbgcBhtaNh1Rh2YH/62QQar4x/GG0+F9naW9paRX+rXp3R2zuVCRD7zsRzjPAHfn0NfCHx/efErwm3iK+0+GzkW7eARxMWGFCnOT/vVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVd3U99dTXt1KZJp5Gkkc9WZjkn8zVfsh/wDIR8Tf9cLb/wBCkr4u+Pr34b+GbbxFZWUF2DqEUM8UuRmJgxbaQRhsLwTkD0NeH9e0vxRotnr+i3Ins72IPE3Q+4I7EHII7EGv2h/EXxZ8Na8+mTeJrlPD+oBmsmtUEGV/ijdlAYkZAIJwQQcckVVVVVVfFrx1F8PfBF7riMpvXHkWKN/FM2cHHooBY+y1b/Du40v9nDXfEWoxs+q628F+zOMsIVlXbk9eVZ3z/tjPSv2fdQGo/CTQmLAvbrLAw9NsrAf+O7a+Knj4/Dbwk/iRLBb2QTxwxwNJ5YYtnPOD0AJ6Vf8Ax9Labpdn4b8KXOueJdRsY7uTT7MtIlorqGG9gpOcEHGO/JGRXh79o3XrbxZb+F/iP4OGi/apFRZAHiaHccKzK/VSf4gR681Xhz9pTR9Q0zXNY17SxZW+mzRw2UUE3my3zvvwFUgYwEBJzgbvz139ob4l6GY9W1D4US2GjSOAjXiyozA9B5m0KpPuv514K8XaZ478M2XibSdyw3SndG/3onBwyn3BH4jB714r+MVzoPxR0r4b6foUV4b8wCa4acqYTIxz8oU5wgDdR1qv+FxXM/xj/wCFV2OhRSxI2JL4zkFcQea3y7ex+Xr1qqr4ZkeLv2lfEXiHIePTvtTRPjOQpECY+qE/hmtX1XTtC0y51jVruO2tLSMyTSucBVH9ewHc8V+y3pV3f+LvEfjKG2aDT2ha3jXou+SRZNo7HaqD6bh618R/ip4Z+Glik2ryvcXtwCbaxh5klxxk/wB1c9z+GelXvxz+L1lZHxHcfCJ4NFUb2aUS71T+8WwMDH8RTFaN8VrHxR8NdQ8f+HtPaSXTbeZ5rGZ9rJJGm5k3AHOVwQQOcjgHIHwh+JZ+KPh251mXTEsJrW8a3aFZTJwEVg2cDruIx7V408VWHgnwvqHibUBujsYSyx7tplc8IgPuxA9s5r4SeP8AWviToM3iLUfD0Wl2vneVa7ZzIZ9v325UYAPHfkN6VVfta6o0fhvQfD8RYvfXzzbVH3hGm3H5yjivGNrL8FPjL4e8RRDbp9zaQLPtBwUWMQzD3OAH+rCkdJEWSN1ZWGVYHII9RX7PhPij4r+NPGxYOn7xUI5AE0xZcf8AAYsfSviD8SPDPw30oajr1yzSzZFtaRDMs7DrgdgOMseB9SAbj45fF17JvEdl8IpI9EVPM3yiVn8sDO7cAPlxzu24xXwx+JWk/E/QG1fT7d7W4t5PKurWRtxifGRg91I6HA6Hjiv+FxXM/wAY/wDhVdjoUUsSNiS+M5BXEHmt8u3sfl69arwr8YrnxV8UdV+H1roUS22lGcPeickkRMEJ27cDLEDrXxh+K83wvt9JNro0epT6pLIiRPP5eAgXJGAc8so/GvF/7QV7B4hl8KfDzwu2vXdo/l3Nzh2iR84IAQZKg5BYkDjuOa8X+MtA8C6JJrviS+EMCfKqquXmfsqL3J/TqSACai+O3xO8TRvqXgX4Tz3OloTtnnDyGQDrgrtGfYbsV8L/AI6ad491N/C+s6VJouvRBsW0jErKV+8FyAQwwSVI6A8nBx4q8UaN4N0G68Ra7cmK0tFy20ZZyTgKo7knAx/Ic1B8dfin4mjbVfBPwokudKRiFllEkjSY67Su0E8HgbsV4E8Qat4p8L2eua3oL6Nd3G8PZyMSybWK5IKgjOM4I4BFfEv43eFfhw/9myq+p6uy5Wxt2AKZ6eY3OzPYYJ9sc03xw+M5T+00+DlwNOxuwba4L7fXdjp77cV4l8az+Gfh1N441HShFcwWUc72LS/ckfaBGWx2ZgM4r4V+Obv4i+Eo/E93pSaf5s8kccSSmQFVwN2SB3yPwqqrxpqd78eviTb/AA+0Cd/+EY0OXzNRuYz8srKcMwPQ90T3LNyOnwztoLP9pfWbG1jWOC2N3DDGvREQBVUewAA/Cv2g/Es/hn4X6i9pL5dxqTLZRsDggPnfj/gCuK/Zt8LWug/Da11UQqLzWna4mfHJUMVjXPoFGcerGv2mfDVrrXw1n1gxKbvRZkmifADbWYI659CGBx6qK+BXixdW+ElhqOq3QB0iOS3uJnP3Ui6E/SPb+VfD341eEPiTq15ouiRXsFzaxmVFuo1UTRggFlwx6Ejg4PI98eVH5nmhF3ldpbHOB2z6V4+1PQNJ8HatdeJpoV09rSSOVZf+Wm5SAgHcnoBX7I2k6lBp/iHWpVZLG7kghhyOJHQOWI+gdR+PtVTzw20ElzcSrHFEhd3c4CqBkkn0xXw2gl+Mnxr1L4h30TPpGiODZrIMAEZEC49QA0h9G+tSxRzRtDKiujqVZWGQwPUEelV8Z0svF/x00XSvBzpPqSCCG7lgGQkyyMSWI6lEwSewGO3FVXivVZNC8L6xrcQy+n2E9woIzykZYfyr9k/SLe+m8Q+Lr4faNRMqQpNJ8zLuy0hz6sduT7e9a3o9h4g0i80TU4Fmtb2FopUYZ4Ixke46g9jg1+yrqd3p3iHxJ4Jnm3JGn2hUzwrxv5bkD33Jn/dFeL/FWleCvDt54k1mXbb2iZCA/NIx4VF9ycD9elWnh/W/GPhHxn8bfF6kT3dlJFpURBARD8jMo7KFyg9csT61+yx/yTGT/sKTf+gR1VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVfsh/8AIR8Tf9cLb/0KSv2p/wDkmMX/AGFIf/QHr9n34tHwTrI8M67dY0PU5Bh3PFpMeA/sp4DenB7HPjvwVpHxB8M3PhzVkG2UboJguWglA+WRfp+oJHevFPhnVvB+vXfhzWoPLurOQq2PuuP4XU91IwR9aqqqqr4teN/Dfjb4u2Ph/wARawLXwr4fmMdy6o7iWQcy4CAtyVEYPbBPevHPxq+D+v8AgbWvDth4pQy3WnSw20Q0+4Ub9h2DmPAG7b9Pav2UNR+0+AL/AE9ny1nqj7R6K8aEfqGr9rnVSmieHtAQ5N1dy3BUdf3ahR/6NNfCX4e2Pw88I2unpAn9pXMay6hNt+Z5SPu5/urnaB+Pc1+1Lbw6p428KaPaAC+nhKFgOcPKFT9Q/wCdePtW/sLwPr2rqwD2unTvHk4+fYdo/FsCv2Wvh5Zag1z4/wBWt1mNpN5GnI65CSAAvJj1GVAPY57gV8eJbSH4S+ImvFVlMCKgYZ+cyIFI/wCBEGv2Y7KW1+FVtPIxK3l7PLGD2AYJ/NDXhMjxf+1RquqBjJHpL3BHHGIoxbj8NxB+tV8B8eK/jX4s8YlvMhjFw8TenmzYT/xxWFVWualHouiahrEpXZYWss7buBhFLHP5V8F/Dvxf1ODVPEnw41mwsfNmEF3LdKjNIwG/A3Rvj7wJxjqK8Vz+NB4207w18fdb1RtKmdWDWUkaQkE4DgKoUgHg8bgM/jpWj+H/AAXoI0/SLKKw02wiZ9iA4UAZZiepPck8mvAXjTwb4n+J+qfEb4na1Dbi3YHS7W4jZwpydmAoIxGoH/Amz1zXj39pDwJLoN7o/hQ3WtX1/bvbwhbZ441LqVy28Bj16Ac9OOtfAH4dan4d+Hl/aeJrZ7ebX3ZntZFw0URTYAw7MeTjtkZ5zX7J1zLZ3Xivw3ckCSCSFwo7MpdH/ktfHLVtQ+JXxA0j4PeGrjMcEwe+deVWQjJLeojjyfqxHUCtD0XT/DujWehaVCIrWxhWKJcdgOp9SepPckmqr4xk+Kfj74P8KITJDaeQ0yDtulLyf+Q0Wv2kvBw8TfDyXVbaHdeaC/2pCBz5XSUfTbhj/uCvht8RW1D4B6jqdxPm/wDDlhPbOWPLFI8wn8VKDPqDX7KOkCz8C6jrEi4a/wBQKqT3SNFAP/fTPWj+NPBvjj4xX3jP4i6zDDo+nZGl208bOrhWxGCoB4xlzn+IjqK8V/tNfDzT9Omj8OTXGtXzoVhjS2eOPcRxuLgHH0BzX7MngTW/Cfh3UtY121ltJdakiMVtKpV1jjDYYg8jcXPB7AetfAcjxX8bPFfjEsZIkFw8LY6ebMAn/jisKvryDT7G4v7ltsVtE0sh9FUZP6Cv2U7ObU9Z8VeLrvmWUxxBj3aRmd/1VPzr9p+e81z4g+GfCellmu1twYQCRiWaXavTof3a/pXgbwVo/gLw7baBpECjy1Bnm24a4lx8zsfUn8hgdBXiwS/Gb4/xeDriZ20PQZHjljUkDbGAZunQs4Eeewx6VbW1vZ28VpaQRwwwoEjjjUKqKBgAAdAB2r43W8fh348+GNb0dNl5d/ZZpkTgyOJimTj+8qhffFftNeHdY1/4dLJpELzjTL1Lq4hRckxhHUsAOTt3An2ye1fAj4q+E/EPhjSPByzpY6zp1olv9lk+X7RsXG+M9GJAyR1BzxgZrx/4pj8FeDdV8Tuqs1jbkxI3RpCQqA+xYqK/Zx8Cx64Lv4r+Kl+36je3TizecbtpB+eXn+ItlR6bT68V+1Pq5sPhvFpqON+p6hHGy55KIGcn/vpU/OvhDpA0P4Y+G9PK7W+wJMy46NL+8P6uaqv2gPiXP4f06LwJ4bmzruugRllfabeFjtznszcqPQZPHFfCjw74P+GvhWLSY9f0qTULjEt/cC6T95JjoDn7q9APqepNeANR06H9pfXr6W/tktnmvdszSgIcnjDZxX7VdxHe/DzSLqyuY57VtXX543DKW8qUDBHXowr4SFD8MfDHllSP7Mh6eu0Z/WvjWyr8KvEpZgP9CI5453Cv2W7Rbv4W6jaX0Ylt7nVZ08tzkMhhiDDHoTu4rwH8GfBfw61W71nw/HePc3SGMNczB/JQkEqmAOMgcnJ461440LVvE3he90TQ9fn0W+ufL8q+hLB4dsiscFSDyFK8Eda8cfAb4mWUa+IbrxAPGyWA3vaXk05kdR1AXfkjA6K4Jxxmvgj8TdA8eaAdM07SYNHu9JRVksIcCNUPR4/9knOR1B65yCa/aa8dnw94RTwnp82L/X8o4U8pbj7/AP30cL7gt6V8H/Aw8AeBLHRpowL6cfaL44/5bOBkf8BAVf8AgNV47+DPxF8T6tqeoaX8Vb22s7yQtDpryzLDGMfd+VyAPovevh5rn/ChvF6+EvHnhGwt3vSFGtwFmcoxwG3MSDHnAIUKRjJBIrxX4v0HwVox1/xBdtDZK6oZEjaQ5bpwuTX/AA0l8I/+g/c/+AMv/wATX/DSXwj/AOg/c/8AgDL/APE14a8Z+D/ilo+oxaDdyXdmFNtch4WiPzqcj5gOxrw5qfib9mjxlf6fr+kXF9oOosFE8QwJVUnZIhPy7gGIKEjr14Bq7/aLg8TJ/Yvwv8K6rqmt3C7YjcQqkUGf43IY8Drzgepr9lazuJ/HXiHWricTeRYGKWYHKs0kqtuzjv5ZNeKNbt/jx8S4vDSavDaeDtAffPM84jFywOGZST1blV9Fy3fFfFfU/DMPwl13SdJ1PTQkWneVb28E6fKq4AVVB6ADpX7L+q6Xa/DprO51K1infVJdsUkyqxyqAYBOeaqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv2Q/+Qj4m/wCuFt/6FJX7U/8AyTGL/sKQ/wDoD1X7OHxaHiDTk8B6/dZ1Kwi/0GVzzcQqPu+7KPzX6Gvjv8Jo/iHoX9qaTAo17TIybcgYNxH1MR/mvoc+pqSN4naKVGR0JDKwwQR1BFVVVXxn8eD4f+BbzUraYLqN3/o1gO4lYH5/+AqC3pkAd6+Bnwb8PN4Ji17xnoFpqN/rLfaEF5CJDFCfuYz3YZYnvuHpUvwj+GUkbRjwNoy7lI3LZoCPccV+yZcyWWpeKvD1wf3iCFwvTBRpFftnuv5V8YwPFf7QPhfwuqh4rX7MkyE+shkf/wAcx+VV4kP/AAl37VemacGLxaO8IB6hfJjM5/8AHyR9a/aX1b+zPhTeWwbDaldQ2ynOP4vMP6RkV8CNI/sb4UaBAyASXEDXLn+95jl1P/fJUfhX7Vmrmy+H9npUcmH1HUUDLnqiKzH/AMe2V8LtI/sP4deHNNK4ePTonkUjo7rvYf8AfTGv2WLiK+8Z+KL++fGozwBwjnDENKTJx14bZn618Y/iFY+AfCF3Ktwp1W9heLT4AfnLkYMmOu1B8xPsB3r9knSBB4X1vXCmGvL5IASOoiTPHtmU/lSePfBsviU+D4/Edm2sjINoH+bIGSuem7A+7nPtVfHzVxo/wn12RWxJdRJbIOmfMcK3/jpav2bdI/sv4UafOy7X1Gea5YY/2tgP4rGpr9rmxjk8O6BqJRd8N7JEGPUB0yR9PkH6VL9r8Q/B1vsTl7rUvDJ8kjOS8ltwR36kV+zR4X8AeLtM1ay8R6JY6hqlpcLLGLhcsISoHAzyAwOfqPWtJ8HeD/Dh8/RPDWlae6g5lgtEjbHfLAZ/WqsPE1v8JfjP4/vblRtNpcvbxNwHlkeOWJfXB3Y+hz2r9m7wTd2+nXnxK8Qq0mq+IXZoXkHzCEtln+rtz9Ap714m8X+GfB1nHfeJtZt9PhlfZGZScu3oFGSffjirG+s9TsoNR066juLa5jEkMsbbldSMgg+mKrwWB4s/aj17WSSyaMs4U9gY1W2A/Un8zVzbwXdvJaXMayRTIUkRhwykYIPtitUmu/hfN8QfhtPI/l30cSWxP8e2dGRj9YXfPvivhRoE+nfBfSNJgIS4u9MeVW6ENPucE/TePyr9mzwx4I8TXmu6J4u0W1vNRg8qS3iuVO4ICyyYGexKZ+orSfBHgzw/KLnRfCuk2My8iWGzRHH/AAIDNeJbuaz8NarfWZLTQWM0kW3klghIx+IFfshrZDTvEjJIv2tprcOmeQgV9px6ZLV+0L8QrHwr4Mu/DtrcK+r61A0KQqctHC3EkjDsNuVHufY1+y7pA0/4Yi/KfNqd/NMGI6quIx+GUb8zVuB4v/avkkI8y30iRsj+55MO3/0bj86r4W3MfhX9pDxHpmtP5U2oyXkEDSHbuZ5llQ8/3lXj13D1q8vLTT7SW+vrmOC3t0LyyyNtVFAySSegxXh0y/G74+nxTbQu2haA6PHIykDZET5Q+ryZfae270rIPAPSv2mfCfh3wjqmi+KfDCJpmp3k0jSR2x2Asm0iVVH3SCeSOpIPXJPx1TV9U+BTXk8ZFyI7Se9QLggkrv49mIPtiv2dNW07UfhTpVpZSxmbT2lhuYlPMbmRmGR7hg341X7Uk0uueLfCXgu1f97KCwX1aaRY1P5ofzrUdS0XwtozX2p3kFhp1jGFaSVtqoowAP5ADvXh/wASaF4r01dX8O6pDfWjMVEsRPDDqCDyD04PqKrxn8B/A/jvX5/Emty6mLu4RFcQTqi4VQowCp7Cv+GWPhj/AM99a/8AApf/AIivCfww8Na38atV+H95JeDTLKS5WIpKBJiM/LlsY/Svif8AC6zsvgfP4R8NJcTJoZN5arK292w7PJyAMna8mMD0Ffs1eLbPXvh1b6H56m+0N2hmjJ+by2YtG2PTBK/VTX7T3iy00f4ft4bWdftutzIixg/MIkYOzfTKqv8AwL618D/DM3hL4ZaRY30Riup0a6uFcYKmQ7gCOxCbQR6g14d8c+EPFtxdWnhvxBaahNZHEyQvkr2yPUe4yPeoru0nmmtoLqKSa2IWaNXBaIkAgMByMgg89jWpalYaPYT6pql3FbWlrGXmmkbCoo6k1+zXHNq/xU8R+ItNgeHTTbzErjhfNmVo0PvhWP8AwGppYoInnmkVI41LOzHAUAckn0xXgxJPjh8c7rxdeRtJomgkPbo442oSIFI7bmzIR7MKq5u7SzVGu7qKESOI0MjhdzHooz1J9Kr9rDWdM1TV/D/hrTmW51OzErTJH8zR+YUCJx/EdpOOo49a03Q4J/DGm6N4hs7e+aC0hSZLiJZFMioATg5Gcg1/whHgv/oUNE/8F8X/AMTX/CEeC/8AoUNE/wDBfF/8TWnaNpGjxvFpGlWdkkhy620CxBj6kKBmpoIbmJoLiJJY3GGR13A/UGvjV8V/DvgrQLvwd4Qe2bW79GhaOzUAWitwzNt4D4yAvUE5PbPwP+GMng/4fzWWuQtFqGvAyXkeMNChXasefUKSfYsR2r/hlj4Y/wDPfWv/AAKX/wCIr4i/s7+APCvgfWPEOlzaqbqwtjJEJbhWXOR1AUcV+z98IfCfi3w9b+M9UkvxqFjqh8oRTBU/d7GXI2k9TzzVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX/DK/xN/5+NE/8Cn/APiK/wCGV/ib/wA/Gif+BT//ABFfAP4TeKfhpd6zP4iksXW/ihWL7NKXIKls5yox94V8bvAutfEPwYmgaA9styt7HMTcSFF2qrA8gHn5hX/DK/xN/wCfjRP/AAKf/wCIrTP2afi3o+oW2q6ZqOj293aSCSGVLtwUYHIP3K0V9Xl0m0fX4LeHUfKAuUt3LxhxwSpIBweuD0zivjH+ztfeLfEP/CS+CZLG3lvATfQXDmNTJ/z0XAPJ7j157mv+GV/ib/z8aJ/4FP8A/EV/wyv8Tf8An40T/wACn/8AiK/4ZX+Jv/Pxon/gU/8A8RX/AAyv8Tf+fjRP/Ap//iKr4kfBmT4l+JtM1bVvFLRaVpoUDTEs87xuzJ+83jBYADO3gAUkccUaxRIqIi4VVGAAOgA9KrwF8Gh4E8dax4xtvEn2mDVlmUWJsvL8oPKHHzhznbjH3RnParT4NCL4ty/FW88R/aXZmaOyNnt2Zi8pfn3nOF/2fyqvCvwcPh34l6r8R7vxJ9vl1EzmO1+x+X5HmOCPn3ndtUFegznNfF34WTfFTTtP01fEf9lxWUzSsPsnn+YxXA/jXGBu9etaPpsOjaRY6PbnMVjbRwR8Y+VFCjj6Cvi58H3+K0+kmTxMdNg0wSfuhZ+b5hcrk53rjhAOh60kaRxrFGoCoMKB2ArxV+zXcz+KZvFHgPxfJoctxK0piCMphZuWMbowIBJ+7jj1xxVh+z5ZRaTqj6x4qvNX8R6pZvbf2tfIZvs6sMNsRmzkqSMls4PGMkH4Z+A4vhx4Th8MR6iL5o5ZJHuPI8reWOfu7mxgYHXtWmfs8x2HxTfx/J4jMtoL576K18kiTzGYttL5+6Cc5xkgYPrVfFj4dTfE/wAOweHo9e/stIrtbh5Psvnb9qsAuN64+9n8BXhXQIfC3hrS/DkE3nLptpHB5mzb5hVQC2MnGTk4z3r4sfDeP4o+G4PD7av/AGa1veLcrN9n87lUddu3cvUP69q8I6FL4Y8MaX4cmv8A7a2m2qW4n8vy94QYB25OOAB1NeLv2cYLzxC/irwD4on8N30jmRo41barnqUZWDID3HI54wOKP7P3jLxAFg8dfF7Vr+zH37WHeQ34uxA7dVNRp5cax72faoG5jknHc18QP2ebDx/43/4S658RtawyiIXNolpuMoQAHEm8bcqAPunHvVtbW9nbRWlrCkUMCBI40GAigYAA7AAV8Z/g9J8VYtLktdbXT7jTGkAEkRkR1fbngEYI2D614N8M2/g3wvpvhi1uXuE0+AR+a4wXOclsZOMknjt0qvhf8If+FdaxreuXXiL+1rrWSpLG08jy/mZm/jbOSw9Mbffiviz8BbP4n63a69D4g/sm5ht/ImxZ+f5wBJU/fXBGSO+Rj0qwsoNNsLbTrYYitYVijGMYVQAP0FePP2d7HxB4hfxd4P8AEM/h3VZJDLIYkJRpD1dSpDIxzyRkH05NH4B+PdcjW18ZfGPVLuz6PbReYwcfVnx+amrDToLDS7bSVZ5oba3WAGbDM6qoX5uMEkDnir/9mPVNM8QS6t4A8e3GiwTMdqKHWWBSeVV0YFh6Zx2yT1qT9nTSm8LappTeJbq51zWPLF1rV7D58m1XVyqKWBUEqM/MT6kjivA3hWLwT4S03wrFdi5GnxFDMI/L8wlixbbk4ySeMmvAHwbHgrxtrPje68Sf2ndauJQY/sfk+UZJRIxzvbPIA6Cq+KfwQ0L4kzR6vFfSaVrMKhVu403iQDoHXIzjsQQR78Cv+GevHGuJHp3jP4uaje6XER/oyGSTfjp998A+5DYrwl4O8PeB9Gj0Pw3Yrb26HcxJy8r92dupJx/QYFeNvg14r1rxXeeMPCXxL1DR7m9VFkgAZUwqhQAyMOMDoVPPOa8Nfs8xrr8Pij4i+LLrxTewYMcU6nywR03FmYuoPRflHqCOKv7Gz1Oyn03ULdJ7a6iaKaJxkOjDBB9iDU/7Net6Bqsuo/Df4i3mjxzHmF96so54Low3jngFfxNeA/hH4k8OeJYvFfir4jahr91DC8aQShiihhg8szH8gK8QfBseIfilp/xIuvEeI9OaEx2H2PP+r5H7zf8A3ju+7/jXxT8Aj4keEZvDa6ibKUypNDKU3KGXswyMggke3B7Yr4SfDZPhh4YfQ21P7dcXNy1xPKEKLuKquFXJ4AUc9/0FVXh34M/2B8UdQ+JP/CSef9uedvsX2PZs8z/b3nOP90Z9qIBGCM+xrxH+zmE16TxN8N/F1z4ZupSS0EQYICeoRlYFVP8Ad5H0HFeF/wBna2g16PxR8QfFF14o1CJlaNJw2wEdNxZmLgdhwPY1qdhDqum3Wl3DusV3A8LlDhgGUgkH1wa+EXwHb4ZeIr3xBc+IRftLbtb28ccJjARmViz8nJ+UDHT+njP9nh9d8S3ni/w1461HSNRvZTJLlS4BPZWRlZRwOOab9m3xh4gkjTxv8V7/AFC2jbPlEyTk/QyPhTyecH9a8GeCPDvgLRk0Pw3ZeTCDukdzukmfuzt3PH0HYAV478N3/i/wrfeGtP1v+ynv08uS5Fv5xEZPzKF3L94cZz0Jr4WfDax+GHhxtDtr37dPNO01xdGHyjITwo25bACgDGTzk96r4l/DrTfiboCaFqWo3dmsM4nieAj74UqNykcjDHjj60v7PXxIsUGnaT8ZdQj0/GPLDzxAD02LIR+tfDv9nrwx4J1KPxBql/NrurxNvimnj2RxP/fVMklvdieeRg81VVXjn4L+OvGfie/1CP4n3mn6RdMpisVaWRYgFUEBN6ryQTxXw++AXgnwFdR6qFl1bU4+Y7m7AxEfVEHAPuckdiKqvGvhr/hMPCmpeGPtv2T+0YDF5/leZ5fI525GenqK+Fnw9/4Vn4Ybw5/bH9pbrp5/O+z+TjcFGNu5v7vXPeqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq//aAAgBAhEBPwCwHUOh0DcsWBgAADRYsAWY9hgdA6G6DcBgwA3RuAB0A6FtALAdQA3AYIPIC10MAGblzoboLoubs2BgABYBhoWuFjqADAAAHsMGgGjoAwsWLAWAA6gDAAuHQGDAAYWLaFgeoAAAAgOh0GgAANwAewi1xBuWsHkAAABoDAACxugGdAB7D1ALGhbQ6BYdjoAaM0NBBYtdFgGAw2Nw6FjdG4bgAA9gsbg9Q6BoaG4dDQ2AAGgsW0LGo9iwywAAdAuXGHoMFqdQAAABgbgbhY0YWNCyLAAD2ANWMDqG5ZMYHQudAN0DAAA0DQOhfQ0saHQ3DoPYA0BFzoFzQuDaAAOgw0udAuIvoAAAAD2A0Bm4BoxnQAAAZqHQ6HQCwAaBuBYHYNDQYWRoFjcN0AADLGgBoD2OgAAaMAewxliw1c6A0WNw0NGBoAAwAC1wB2NCyNB2sNIaBg9jSxoaADGaGhuMCwbltCw0DDQsjQEi2g1oWBoewAAD2OgaGhoX0OgjQ0LIsaBobgbgAFgA0DQNAAHYsbgaDDcAAAAZudDcBgAGlgAA0CxYABDOhuaB0AAGAABbQOhbQexY0AsM3OgABuHQAsWCwXNB2NLDdzQA3LBYNA3AAAYwsDsW0LFhoGi1yxYAAGFzcvoXuXNAYMAsBuGgrGhox2LaAWLBoAAAzcdmdDoAG4ADQAD2LFjoFkFyxoaGw7M6Fi2h5AFg0A0DQ0NAZoGliysdDRjDQB2aADcYMCwWLDJ1FbQu2NjbOZ9C8m9zc5mhVPIVSLAGAAAAAaDNDqaFtA0AaA6AADADoWOhoFhgAABZGha4W0OgADAAGAasNwALDQAAPYDUAA3GLYZuBoADQBYLAdTodCxuMWwDAewAAGgAE3aItQ3LXGiw0NG4QnbQ0Z0LDAAAAHsdTSxYDRlg3B7m6GIQHUAd7HU1A6jAtobgWLXANA6AbhYbNzcAAABh0AOgx7G4bosdAAAex0GgDc3GixYOpuBYsbgDAHqB1A3NAZ0Nxgag9gGdAbQ5pI9o2y9xNpbkZXQDsVWpaCtsi/QQAFiwQlZidzctcAAAALDSNAtodDc3AsWAADcBgbltANywPUsAAPU6lixbQBmp0DoFgYADAtc3QWCxYsaBoAkAD2OhqM1A6DEWDQLIY0AAwY0DNwsasA3DodDU1AewMCc7aI3DoG6I7C2Ct8NiysaGgg3A6AMIysbnQOh0DoAB1B7FtDc3LaAdA3ANQAAGAw8g6FzoFw3LGyAAA6AHQAAABgFy9y4PYtdHQAAAHsAdQA3C4G4HQOgDAuMNLB0AGg3Og9hgXHKyHK6HJimxzkwAuCYpHNoSaaNGNKxZ3FuAOzOgXNGMIS0sboAAAOoPYA6AAGhcAAAZudQsDRuNAwCxYCxYsWZZhY6FgsNaFgGboBbANBuFkaFixoMsW0LaGjLaAbgD2CwWDQ0sOzBbFg6Ay2gwYDLFixYLFjUsbh0GFhjdkOQpXRcepuMC4bj1DcNwZublg0AdgBnQE7MU7OxudALAAAFgsNM1CxYsWABAAPcsFhlixYtoLY3C1zcAAYkADDoBoAMAGaBobDsbljdDLAFgNwB7Aw0A0A3DodA6BbQDoDAALgdA6ANpIchyFqaGlh7Aw0BFixY0A6GpZDsgsI0Dce4dB2DQYU3dWAAAAexcZqXN0BpYAADcAGAbgAAAAAthgy4AFkzcAADQ0LFhq4bnQsBuABuA9gsGhoaMsO1h6jGPY0A6AANFhlgZYsFg3LHQlLsN3HsWuWAYNDtcLAbgFxsNywzc3A6jZuAAQlZl00FkAAA9gBlgB7AAaGhuADsWLAWCxbQ6GgMugAC2gWBjA3A0LAG4A0WLaBYHuWNLABfQAAHsB0LXLXRYsWC10XAOg9i4AMDSwAwOgaE5JIbCxYAbui9wAAAAAsHQFuAADCNrHQAsAA9gANwuXGy50ADRgBa5YsmGgBuAy4wANBisGgzoBsAdDQLAA7BYsW0LAdAOhcNwAB7FjQ0AQFgtoOzR0DQLAAwAADoOS6FwAG0y6YWAALnULBBtrUAqUpQtfqAwsGgAAEGgAAAYOxuMANLGhZWGWuDLFkADLCNzQsM0AA0AANi6DSx0ANANCwAAAFtDoDAAAAB7Frg0NFtALaGpqMAAAGOxdF0aDkkc6ZdMCb0sAA2AD1N0aGhoAtSpTcJOL6AyUlFXNzTYBu6QAzRnQBlgLAyOjI6o6AAAPYLgahuHQ3Q7Ay2huAAOxbSxYaNTUsA9gAOgFhjOhYOh0NB6mgbm6AAAADYAaAAAAep0DoaGjNzoXNwOgMAAGSvcbGy7sFhOwppnMiTTYFx7AAwADc0ItJpmJxEq9WVSW7dwJRUlZgwBnQ0sWLDNwAA0NCDOgAAD2BgGhoADA6G4ADAANzodDdFg1CwG4dBj2LdhjRYsbjCzA1AAYw3B3NQZ0C2gdADQAewdDobhYDcAeoaBYYWQxjYwDQY9hMACwzcLGlhmhpYGFgNzcYAAHQsMANwAIs3QWYAAaG5oaGgF1YckhzQpJlzQAABhc3OgdAOpubgDNCwMCx0N0WADcAAAerDcLHQGdAepbQsFgdmWAGrgW0Dc6hublgsdAY0WB7EpJHMrEmPUYB1ANAA6AdADSwdBhYHsAMALFixYsM3BnQ6AFwIsAAB7GlgtcOg2hzRzXN0FjmaOcUk0AAMuBe4AGgBYDoA0Fi2gWLag9ywB0AABgBuXOhuBsFjTYtoG4A9gDYuBYNzdFroLHQNCxJpIerEtBjYAxBoMAABB0ANDcJwaS8zQ0AGFgLMsMCwbgABYEXubgAAbjYMeoeQ9C4xjIOzLnQAYMOh0DoAdAeodAGMAAADqHQC9wAADcGbgDQAAA9gexoGhcAAGMk7Dm7ildFTV2LIGgAtdADADQCwaFkAASnKSV3sacrAaHuMLB0EaAMa1HqAAWuC1BC0NwAB7AMexJjAbbDqDVzbYjJsWobmljcDcADcGFixYLAOxugOg9jQZoWNw3BgAAaM0HqaAwAAAAHsAAAGhugJSshyuboV7ktWdBajAHsMNxHUANA6mghbDGM0NCKg6bvvpYewzoAFgLFgCwM6AFiwGgjciNXIoAOgPYBkmNjGNgFg0YRlc0sXEA7AaFtCxZAzdHQ3AALILGgaWOgAAHQAAZc0A3LhY0GtC49gAAHsMGwAAOg7IlURKTkCQhq40dANweoxhZIAGhDOobEZxk2l0LDCwFkWAsBYsDRYEG50DQAA3NwIgdAAGASNwLagAzcE7EWdABmljQep0A6G4BYAAYNBuMQXOgy4zQ3AAGCLgANBYAAA3BpWLXGgAcke0Q6p7Rscm0AloIGNpgMBh0A6m5uElogluN2kicklcxFJQk4opUuRt99Qle6sSmkkIDoMA3OggGaGgAwdgAEWLAIAA3BsB7EtWBuhhvoABa4mRegAzcYwLDBltCwWLAAW0BosmAABug0YADRY0ZoFg0OhYsAAAB1HsEi6CU0hzuDRYsBYLlzcA3DQDoB2OoWHsh2JatMm1dDabsUqsalNSTKtF0+W/VXNDoVKcZ2v0dxIsAGhuWAsBYtctqWGdBjABbi1RoArCQjQsaDZKaQ6yue1HUbOdkJXJMYAw3GA1c0EMixO4DNA3CysHQ6lhgAG40AM6D1QNAFgCw9QAZYsWLFixbQsAAAAwsPYvqVHZl2JkmAAblgNwB9i2gw3GPYNBq49Gbg/hGVIWjB91+rX6FTYjGxQoRpQ5U9CNZVG/LQsFgAC1wsG4zc0ZYBG4HUYAdDoG4kaAtR6IciUiTuC1G9BMTsN3HqMYA9gAFZgtCL0Nwe4AzQOhubm6GjoboBgMYOxbQNLAGjCxbQZYLADRYtqNFixYAAAGANpInLli2SlenchrNMrfEgGAFxsumXED2CxowLm5Y6D2DfQWrGIZe6sEtrmJi50nFdSLuxJWIRsrAABuAB1GjQ3LCR5AbgMAQblgSEF7EpaEndk2PULXOgi9xbgx7BYGADDctcTsRZYYbm5qbj1LM6AdANwA3NzcB6lrltCwWLaAblgAGWCxYDoFgAAB7E60YqxXu6aJv90hW9iQXvIrfEADeoAzdgXRobgM6GgDH8KFqPRi0Q2jcIq6ZpcsnDUaTZFJIsB0ANLFjQauGhoKxoOxYtoDDc0BltB6oA6m4WuJB0FqPVE3ZWHsPVAK1hao6kdRDY2ADsxKwBoaCFsEWhbDRZAbjOgB0AauAAwsbjOha4dAsbhYauAAWAEDALG4WOhow6EppLUnNzYopDk2rXLtqxzO1hXT0G29wBnUsHQeqBj1QhBe4xjLlgls0WtTibpDV9CnKU5TXZj+B3KbUo6ElZKxlkqV6kajteL772028ySeyMNXjDCVKfWVhojseQG6AAABosAdDS4AMhBSZKEbEY8zsj2cbbE0lJ2AWogew9hbEnZE3dj1N2blhBcRpYZYLAFhbAAxARY9QLnQAuaA9Dcep0Gg2QFzcLI0AZoGhox6osAAbhY6lgYG6A3ACrWUVZbnNKe4loBdXLly5c1YMYxo6AtzRj2E1expY0ED6EmaMZuh4J/UXiOzt/X/AAxSUpYSnKW5HVE3y2HFXdiUedcvco0VRgoRNb2E3G5WnTcoyh2X39QsWEHQtoBYWw9gt0BI0A3LaWLaB1BlhjKexd9ykrIlJt7m4wQXuXFqhMqv3Rq7GrnU06Ghpc6CFa4DACxYe5YADQIuxuDNw3LANCAGBubjRoB0GFtBm4HQ6AAXQMRuaGhoXQ2jQZoXNC6BtJFWq27ISFYsMbG9C10jc6HcQPYZK2h2NDqaC3HsRtzWHYdti+orNktyfwkIJy5uuwx6MqVHzcqKdCCy3ne99PvGtFYqp3SJq8nYa00OxugLXQ9gLXNAsJXEaGhuXVhWZYDdAAaBoD1KbSdicE1cj8AXOgbl7ILlyLKkk0NjepdF1YumaHQW4gG9BsuAND1FsAG4MViOoBa50NCxZFixYLJlgsAG5qG4AAXC6AAAANDQ0NACc1FEqrloKKLDLjdzRAJ6jY1cg7oHqhorSagmldjZbUfxHRD3HqrCfvIZJ6i0Y9JXG9UTqQVBw5db7/pYpaOxqyEqamvaPTqV/dxVWdN80X8PS/8AXmU6sqmHUnpa2gqqei3Kym5UZ9G2vmkyaSlZGyE9xeRbUauWAsBuAFixYLly9wAexoAAU22inZxsS0dh7FzcRcuXL3OhPcLXA1Oo9wxFeNGHNLZFhsLiZuADDQAZcWwhGgBoAAABuAi4dAuD2DyNw0ZoBuAMANC1yxYAnUSRJuTLdBl9BvURbQZuHUa0PspkCxKVlcfwijeBJXRbRD3RZ8qHeyZucyjYk7ovYcryRK1jFVnTgpLuV6yVJyKM07PuTTiml1Mfy4qMFtaKT+SsSw6UoNfZVjL+bE4eq4/Yu36XsYag6cRTbgodncq6VGSld8payLaCScHfcsAFhrQ6AWuWGrGg97Fy9x6m4dANxq4wAhFNajtGOhTaTsOEXuTik2iwWOgblwk/eG7G5Ya0EILmJoxrQ5HsXu7DdjdHQ2NwAYCAGhGwmrCaYwsWLBYBq4AAGjQOwbgxgdDSxZAHQ0ANDQ0NA0GM0CdRJaDd0W1uIbGywkWbQ1ce1x7D1Vhaj6oT1ugeug9Sn1Q9xtNWQ7rUlWUqUaaW1/xOjFK5UjOCnJvTc5043RzaFeuoKMu7sRmpqyK6nLl5ejJ0ueFnsUormSMbh3Srzg+n+Q0My+aw0KkIaKaaf5/mNKS0KlJ0ZuEt0VYXSk+qOVc11uJAy2gFiwHQLaCRYbRcAC5oXE9C4AaAm1sNthzSta4xhcGCG1sS3JRstRbWQaFrhuFrlyT0EtBoYtUAD1RY3FYA6AzmvYUhNMsWR0CwAAAAdDQdrDC2owsblixsi10AAAWDcAnO2iJaosNWG7F2zSw9UdERla9hbl0oWL6CeolZE00xaJCeiHuhtXsQ0kSSTZTpuEbPz/MqVYSwsaf2k3+NieIjTrRi+p1NLIzWm6UZwfb81cwSvQi/IrYqEGk3qU5RxFGPk7nLT9nFwettSPYmlZmFaumuy/X/ACHH2lOVVjaZJPkTHZ6CWhK8nqTwkpYGNbs2vyGveEABYGMOgths3AAAC4AMsAzcHsW0GgdjoCLakt0V4qFRxR0HqhlwC92glqjQGhaFtQAYAaAbltDcTsJ3QFg3BhYYAB6Fh7D2LCRYsWLFgLAABoGgM6Fhk5WQ3djGSskN6egtjS51sdLEXdEtyTd9Tq2J2YndE3ezKsuxRnzRTLqyJ7oT1JxuYrBOlRhN9Vcb9246VOpJOSu1sVq0acop9SFeDuuqMTN1pNS6q36GBVKjTcJK+jS9baFfD06s9d0YagqUVFGHqxlHQg/esT2MHGqsTKT21S22vcU5ctkOCsct9RLUtZFh42r9X+r39zt56fPoNanQDoW0NywMHoWGwADcAeoAbgwBjHG0bj1B2HsSaLovoIvoSdzm0N0DEdDcaOgxjPI6D0FsAD2Og1poAGgbhchIRYsAAAG4LcHsdB2LHQAC4ADAA3AB7E3dgPRG5JaidnYauO9rola6YnrYnrEnqrkpe7dEqiirsUtCavErTjGWphmnT0KfM1K/c0simpOr5GaYanh5QUXe8U/m/wDhiVarXhyfyrT+vmP3Yso1ItuxVoqpKL7EMM41ZS6MqqTqxa2sym1OMmu7PZS9s5dLItfYpUlCNkRtzFSKV0RppIj0La2AtcW4WCwHQBjLDSY9UMAALBpYABFjQB2HqWN6foD3Ogx6nQtoal3YnJRi2U5RqQUlsI6BbsaHQFa5JaDRY6FrjTsIBgaWsAPYAEMWxB3RqPUNQdw1NQDQ6FrnQ3C1yysFga0LAzoaAAMAACWwDsBJNrQl3ItNGqJK6aFqky90Jw9jJPdPQTbi12NW0umpGTcPkUKqq0790YmF0l0MLJJ8qKUHOElEq87py5fiWxQm3BOe5gMQ8Rg50Yq+t/uTR9YdCSUXq9CXtI4l/wAthUlGo5LrYtdGzHHqSjcjBJFtTR2I0nJNrZFenQVClKHxWfNvvd2/C2wo6MjZosuhYstjU0LB0A6AwsWuySGPUAC1wANADoLYGaAzcexT1TQXGPUnNQTbHFqMW1ui6bZ0sU6inG6Lq9hPQVg3FuWHuboSHsNbGljRliyLHQLgW1AAHsG49QTEw6AAAADszQFsHQauWAAA6AWCxYsBYJ6RGD1egSdmMcdbEN7Ejc6tFN3RVklVa7ohXSqyh5EJWTMljUxMqdJbt2/Eo07SjGK6mY03RXLLdGHqWqxb6mXYqFD2nN9qLX3ntmsVKn5X/EclymR4p4GNXm+1dL5r5lZJrmfQlbQmtrBJ2sbosaJDtcpyk8ROHlcUmtF1KivBfP8AQW5FaMsOxYWqEAFhnQYkDQ2PUAsWQaGg0WAYWAtc6B1Bg9UU3aSJKzaGN3JMlS9pHUrTukuxGDU5S6EncwtRTi7dLllzXHHUtohpICw9i2gPVD2N0NAC2ALAhgdQH2LaA3cRCV9ANRhuBugDcZuDOha4AjodQLAAAABOVhyuEtjoDavynSw72Kl2rl7xuc/QqK0kRbTsY1qM4S6spTtXcn3scr9qmU8U6c047ooY2Ua049tUYzHzxsJVp72b/rRFeX7xSRdVsLpo2YeLVvRDXNYxk2qba3KcPbYdKfVCglFLsV5clKUuxQn7SmpGOnKNBuO5Sk504t7sauhxRa6EjG87o+5vdGDj7SE0+1/u/wCHGhIex1LHQsAdAZo0WQrF1sbIeoBZDSsHQ0OgNFgY9wAGbjAewnZplZe8xoqz5Yto1lEp1aioSqdVf8CFCnUoRxMvit/X9WPbPn20tcT54aEasZNpdCOx1EO1jRltDyLDB7DWh0LaAdAAAex0B7gPUQ9WDsyOjFqboAAAA6FtCxbQLAHQ3BgABYA0BtJD1AeoSdkNJtMcdFJDsTSRCS1iVanJJPz/AEKrurk6z9ql3ROPtIpdh4KUqEpfy/qVKkvZpxKbUqrMJNVItrfY5YqHItiGHi20yFFKnZFJaNE/dVxwVRoslZIaJRUlZkFZtDhpqUZKd/IYy2iJaWMNU9ommtmUa3snp1TX3j2LjCxYLG4FhrQhGySB7EY9R6h0Cwbo6Fiw0G40CWgAAWHsA7WGyceaCmN2FFttMjG8ClPmUk1azaJtOHKtjD0ld3d+hP8AdU6aW0tjG4Knh6nLB3TSf3oSsjcpQUnY3LWLaBoFh7WGtCOyABi1QBYBbA0J3Q9h6AhNMCEuhoXRoAAAaAMHqdAQD2OgAAAkAFR3dgGboaNxaJidlYjBOEib01FJRaZVXM1JHtU1yGN92Kl1Rha9GrGnRtq077lCo8G6lF/bTX4/MxNo0m0YapGcr9SnTUYvlErpMpx1uR2FZSsSSaFobjZO1xXdd22sWbFZIaRuW1MTzqUOXvqKKOhZNFiyZ0LCGDOozcB3bGro2QwNg6C2AYWRYexYoUJVVK3RXLACYADGVJqJJ/uV5CqKcuVbjpVKWKqU57rQdflpRg+jb+9JfoYmVOlTlprfT0KF5ya8ylGOHr+zn9oxNGMXFN7MSnOaXfRHK05XIa3aFYsbgADV0R1QG4MADdAEtgbu0jc3ABaEZJoAZ0AAsBqB1HsAB0DQAAACTSRe4PQYSTa0N1oM8uo9rlenOUm1tYrwqKlbqUqjqUk12FC8oVH1Wn9fIrKMnyvqUafI1boY2h78anloVJRlh2yhOm6y5R1FFqPfQpuSk6clsJWXoRld6FT3ZJ/Ir140qbv1aL3RGzGmkVWuYnCo5wcPn6FjnSqcpfQS1LalncSLFhrQ6DAYHUsFg5V1FYkAkG4zoDLDQMGYTEujSqW+0rfl/kQk5LUNA0vYGAxkmTXM0j4qUomGoSjLme+pPmlV11uV5XkkieHp1pyjF3WhT0reyW9nb5IxLoyxCqzlaMU+/b7zGKclGou6MNOamktbdSezKMHGNvM+Ger3NQtoAbm49hKysdCwD1AAAtcewK25sjQ0ACnKzFYNAAA3Z6AWOgMuAA9QDoAABUd3YDcsPYT1JJJjWjJzcV5oi+aNzntKxiW6dKTe6ZhqiurGNqQpSp0ae0VdfPUxEZVJRnFl7WZicRCeChTtrBv8Xcp0J3dJ7f5kcDGEl5f13IRvYULyQlqK3ToYqbjTdtynH6zRTmRVlYilcepi6koVoJbMg7oSTROjzVFLsW0BoF2Oohi1GBbQBbliw7WLq49R2QWLXANzQYzoAG5CrTjgp02tW1+AAdQAdgk0kU5x1b66GEjN1JX22KM063L2Mb7uMfs9ispSqabodN04yqLZ/qOLjVUo72/NFTDJzlzbMzDE05VW4KyvYhUlTm/S4pqcU+4kr3HFOx0ADoGg1dEdjdBoxDR0FsAADVw0uN6WFuPYANiEroYAMCwWGJFgAsAbgABoaAD2G9QbJRaQyUX0FcUozhYvf3epicPKdaFRdCNeEJKDer2JPQxscPVUeR3VlffciqFKrN1XZNWW/wAXTbyuu2tzEUlyOa3sZfSk6b7M9rGVJ23WnzRGcK1JTXUUHPD3X2Xb7zlla9ilTcafM1uKE/aN9DVIhRq/WZT6MqSUY80hWaujlLdCWpOmnEwUaqUlNbEdiwlcSSQjdDWo0PYkDAaugLXCTSEubY5DoPUDoGhoFx6gG4A1akAFgYNDJPQcozjdCqR5rLcWNrU6vsFHS6b1X9bEIRp4mrfv/wAH9RUmoe0ktG2l+H+ZPCuE5wm7SVtO/wDloyjVgo06M5e4vLp+Zi8FKVb2dB8zT220XW7MZi44qEKiVrRS+5bjjTrwjJarcrQjbX0KStBdLD0ZYTsmg6BYLFroewjQYA1qIBgAx2LDsKxa4PQ1AjKzFqgsWGAG5YsAgLaHQsWNA0sdAABgFT4S1wsN30BlS/Qo14ybgt0SaaU0TMfG1anIclye0KLTMTQUndkFGpSaRCmo2j5FPDuEpP8AmMBRtS9n2MFD2c60n1j+N0NpxdipXcoxh0QruNhLQUd2SpqS5WRjbQtcLLYtoTfKuYVug9hBuD2BAjoB0CxYscnNNHUY2AFg3AAAENrkQwAA6DY7WIJcjRhOWWKknslYxSUrTXQxFNyqRrdLK5WqwlUTWzFldWDlUf8AS77kowjUXL1V/wASnWUsX9btpf8Ar+rDgp1ZpbSenzJYN0qPsYvYxFFySIrSw1cWoAARHsINwOg0AAAMY7CQAJ3Apu6sGo9UAFwuM0LgG50DQ6B0AGBoDAqPQsPYvc3HuLcqK6IYSMazqLdmHpKCcejv+I/dTj2KlKFSzn0I04xjy9DEy9jOLXV2NJRRSoRpxaiY+NSLhKBUjyxujCNurFLqY+KwlaVNvZtf1uUKc54txjtpb1K0HBtPdHNe1hSioXZC10VYpT90aBajSCvTdSjKC6qxQg4Uoxe6SX4G6AB7HQBABoJIS0OhGyTfUsMepuCsAMAAFsBZcp5AFlc0AJyjHVsnaw3bRblSEXGc4721/r7zDUFUw6g+pioSpUYqS1jdEp+0iqj3ZXzNUqcJUle8FH0avfdFGm4xc+7KUK7o2i/ce603Wt+5ialVVacaa0vqPC1fb38noYicpxir/CaDaSI3sADAQFzRg9QNC2oAdAaGhrUtqDDQ3Y1qLYIuzFqgAA0DQDoGgG4WDcEg3AYwNyoA1cuboHrsSiLcx0ZU4Kv23MNKLld7NfoMqwU2rmFUmnfoxqydjDSlUptS3TaJQ5oW7GGrR53Fbor01VSuUG6clLsYlupU5u5hakaicV00JU5Va1Sn0f8AmU4ckEmN6ktWrEQZa51LNsAHsHQBm4PohgkX1Iu47Ax7AxbG4xgwuABcWrRJ6j1AAGPYxMOaFipWSlGBQlFVG32aMVhYYek6UHdTs362/wCCYZeyjHyJy5qcr+TMRKNSXu9l+Biqqm4KOyil803/AMAcaH1CEH8S9e7I1VDA1IrcqUkkmuxhsyqUacox6/12Kl2rkrONyjVVWCkgDoACHsCGJ3DQDoaWAAGDQMJXeiOhbcu2dApu8QBgGgbmhoMB6mgBfQAOgAAVFoaEYOTsiV0aNGzL9OpTm4SUluiNWVWtUbVk27LsrjRUjTq8sZO8ZJJ9Nev9IxWClhZeye36C1umWTVhKzLX0FFK5BK9jEwVGvLkWrTfzRhJyqUYuW7Jx92y3Groo0YRXNFa9Rx1GndDVxLSwFrlgPIex0FsBsPQGJAkrklZCWgtIssixpYsSWgwkm9C2gA9i5a5ZFkzltsaXsO6L7GjQeYM0NAGMlcsmkOkoyuKFtSlifbehUipYLmvrF2+++v4GHs5ThbdKw7Kmmt22vuS/wAyVBxp+89dGvnuSqUvb16dvhvbfdMjOXJZjqU4c0bX5tvIWiJWehWmqcUylFRhZHQAsgA3QDsIAZuWFewB0AsD3NwvcuRNxaOwQdmWGeQMHY0NGaFkM6AwbSOZHNqXDcAewBYtcsE1eJYp1XCV0N3HFPYb6MmmncUrlouHMt0VItLQhK1jG4ueKtOb1SsS93U0LA46hOlGaU+pCKjZItcsmYSnGdeMJbMnFXYtgQAACQBa49h7m5JqxYpavUilKWo99DcCwkTGFtRgbjEgADdFndB6CNwOoNjfQb0J6QcuxLETlXim+44RjTjKXVSXzVv8yiuWpG3mU4c0Fd6NmIofVcZyXMPQpqlU51vsYrEVJU4pP4dCM3LEOr/Ne4p80iSvbyJ11CDlLa5VmoxuTpqcbMS0QwBjQMTGMAWobjEMA6AxoBlkyzsC1YLYaOh5id0AABc3Bu45IcxybDmHIuxSZGaZe4AAAPYYGzJRhKkm9yrFxi2tiliHWpc0XZ9ChOUIxb3tqVYqFn0ZOm43ZTetjMMNDD1eWO1jDqopSjJ6X0OVHLYabWha+hhKPtaip99PmYijKjNwluiOotGTjoU5qpG6Iqyt2FYejBLQAHY6HkAD1kyxuOPUcVdHaxYsrBa5y3EtAqbg9iKHqBctcAAANCyC5KSSuJXaYPVEtSVynhY18ZBS2ul95PCyp4ha/DcxMaapUWldtO/rfT8Dlam0YPE3oRw8+qdvnsVYO6dZ6pafL0KuNdWn7NPRfqODhJsp0aVGMIx67/16FOPJKVnpd/noRV4sxFCNaEoPyMXT9pBU3s/0Kd+VJ9Bl+gADC2huhjACw1oFiwBuWGA7D2NQVgegK3QKT0BgGgGlgbHIlqBK7QAN2RCetjmTOa5cACwPYkdC42KV1Y50tGVKcI2jHqYqnUwtGM6ytrb5/K5gqKxdRUG9XsVK1L6o6TXvp/15EpWehVxSqza6oikxqwxDVmQbhNSXQx2IeIrSqvdkXYasPXQpRUVyokrNMSJK9mLVHWwwOgdBoC4yEbXBK5Poh6ljcSbOQURItc3Km41oNCVkNAWAAAAB9xslHnViO1gdiWqKk9beYsQ6ai4L3rp/cPERnXcpbsdTmcY/IxLVKpOUnqjCYl1sRFpbWMxjaq1zXtcwdNThNt7DV7Jk5WlFFJQjUXM9CEtUSlpYhRdVaCjZliKlzO4NADL6m6OgyxYBbBYtqBYYWBg9Bj2GXudbD12HrYRYKdky4WAAY2SlYvoMOgBuAJnMc+gpCdwAJbD1BpDimi8eblTHdFPl9pCp1g7r5GfY/wDtTDSpctlo977fJGHqST16GJrvEP2kt+vm+5OmmYLLlXx172bVv61I0fZNx8xo0tYsWuhJyjctZWLWdhLmRazJLlaY1eInpcVmiJYko8itubm5uAboetjdgkWuxIkrsErnKiyANy5VB2FsMAAsABYsBUdkjkva5GKitDqPcexXUqVOFZd/yKtZzrS7u7K/u0lLyue0naKXUw1H2tRxT1Sb+4mo1qsk/wCt/wDIy6caMldXWo9XKC9TA1JRSbMdShhlHl1XKn94qsJbEpxlBRtqhys0O17GAuqjs7WT/Jl7tgAAPctoADQaFgAGCGA2mdBm7Dct0GWNwi9Tc3ALFh2GNm7A0ALl7l7huAXsKVhSTACSuhg2htj5VPmI1E7E482kNxVmqihbcjHVp7ojFp2JR7CbjK6N3qNWGrsTuhaFKuqcJx/mRLVaEtdURaGh+9GxC+w7J2ErHLqCqxc3BPVGgJXGhi1RZtD1IxYkxCWodSzZGNgsdDoaMsFQYxbDAASGhpWLFgsWQ1oMKkb2FEYncZuirdyV3ohYSVS0IbvQcKkWqaV7aPyJ0EpruiFd09fVfeUbwv63/r7yPuqRiFVcueC0aMFRq05pvawn7Sk1P0FhOWrKqnoy14ltDaw5spRagkAANDLe8boACwNMGFgAYFrIsPVDDS6Q0IdkIInQ3OgA9hjaHqdAYwvoACWhoMGgIvUWqACSsw5Tk0HTTRLDv7I3OlH3jL8bLEU5JvZkZKXN3MHjpUqVSl0kn9/cUkqbT6lrlug10LFrMfYSuiOj5S1haOxa6Fox3jO/clqjR2aHqhaipJVHPqx6j1CW4xIaOpHoPY2EtBijcSA3AtoWCoMEMAEriCyaLFi2gD2ALFglKyuJMr1FHUx9dRwq5d2Qm3FSfxblTmqVVV8rFSlTdCMvtt/hb7tyjQjOUVPe+vojE8rxL5Hov0KteNWo1DZlOHLFX3JySikU3B02upUcFSio7lKonPXoYuT9ouTv+Ba9hbAAG4W1LbBuGg9iyuDBnQABgD1Y1oWL7F0G4tRAtyNmgAGSY9QuMv0GzcDSxYsmaGgwBhB3QAVNJGg/Id2OlJRUraEY3dyymnGWwsrrYWsnS1i3rt+rK8lOStvbUruUYPl3E20joWugaueRuFSLvdCd0SStoRZJXJax0IO6EugtjrYexuBLct7wWGhJJICzHsK4BYLXDoaE7WGMQwASQThKDtLcvoXLmjOoNgAwlZvlHeL1MSo1V7PqzF0XTxcaXRIxOGjTkkt0tfUp0Z1ko01dswylLENPaKb+49tGtiI1JaXsVqk3NqL7inUpqFtyFeUqlpdyVVOty+QnroLW7Iu7S7EakaklyspzirpiVkbgAaGhvY0B3DoCWoMGaACA3Y9h7CWg9UNHQ6gWEEUR2GgAk7olsIGDsCRZA9x2EPsFlYVrD1QG5T1QBuVVsMJT0sNylFRFTdzHU60GvZ9H5GSypVMVGFbbX8n2Myy+i6KlSfvK669PVl1JHkeQmNA1oG6NLC92Vi2thM0sbSsPSRLozqO1iT0FsdBKyDS4dhrU8joRJaIeotjoA3c6Ddw6BLYGDA3EO1gnJyd2AJXLDNwjd3uDMJTjUrRhLZsrLlqOK2KlJ+2U12Jy0KVSNKrGb6NfmYqSniJ1FuytV0v5FDFtUuVdHcxNKyUkQu7In7lVJ9Tks0iqkkmiUHKon5MovmgJ2TRKrGOphcMqcL9biVmhnQNALA90RjZWLDGAxbBbQHsACHqPUSGb2AaHG6NCyEhbCBENgAZLY3BgW7GmwMEy1xhuWCxa40EHZmhoaHQmroVmNK25ZCSuXSk2jLKtKNaPtvhur77fLUx9CmsTU9ntzO33jVdTioTtr2TIS9pBS7i7Cs3YsXHqhamhsE4qUS6kidmroiTRP346bkJXVmaMVmhRvEjqhj+ELPmQxq41oW0EtRWQ3qWLFxNMAuhF0PYnblBgwNy+gACiDNyw0+hYBkpOTuxyfO4lWcJ8sOqJct7NmX0FjK3ItE1L8E35ditSipW7GHXRlWspUvZR2Tvf5Moy1aIqMpKUio1HEtX0HBzjJEZtxiYWFSOJlG+mpUpNvfZlfCy9jTlT2T19BYRrD+2W17G47DYABKxe7uBYGbgWGAB0B2sbjFuA9joWTAtdFtCwQZ0AGSDoPUQ9ToBa5Y3NyxYsFkNaFh6o3CDuB0CWkh6q49iKHG7FNukodmYbD0XGHtH8V+/byMVyyjTa3cl+ZKHK2kOKZRrynXlSf2ToNC2CyGr6ANcrGlexEWqHZOxPSSaFJNl9BJRViOqGNXVi3vEraI6Ceg1oLY3EkiR0Eh6iQjQsJKxuW0LE1oFgAAASBsAAGbk2OCdRSK9STqtrpcw03CvDz0+/QzjDxo1XGLuidOMW2hWUWkUU2pEZKyiYqlKNeMu6/Ui7FOnJ112KcEqtz2CppO/xXf4jbUbDrXl7Neo9Ni10UqfJJmwAHUtogA3AAALXGbggsANBYa0EARZHUAYwLAdDc6Ba4kAWCxugdjqCdmbm5oFRJoVkh6F3cTdhzdiVeUopPoUaypVlUSu0cynJsasxU4qfOlqx2A3BrQ6C1Gk0U48/u31RKzVxSsiSukyesdBxvJJHRrsNXRFaD1WhYaN0htIjHQbdyGHg8P7Vy1va1une/wChK1xOw3zaD0FtYsWEbBuFwnqgLAABuWBu5uB1LBuDYT7HtX7VRt3MVhnVk+V2aJwlGMZsws1jZqlJ2um7+ib8uxhqUasJNu1rlanTVCEo77MikkkjlcpK25ODnFrsVlyQvbYpU4sWjTfoYity1Yw6svrocsVK63FsX0EtAAGSaSudR7Axo6AABYsdBliyDcCwPY0LD2IoAIM6ASYy2hsbm5uWVg6FtADQAaGrg0NBYg7oAauh2sMoxvNXM0q0niZOl8PTfsu46kvZ3W5dGkk4mGpexi4+ZZOLLKwM6FjctpcZ1GSXVDtuSXLIvoLW6Ev3pZ3YtYkHpYSsbo3QkJX0NW9CNNXKlGVOEXL7Wxjans6Tl1RCcatNSjszDQnGNpCDoB0DoAS2GAG4Ba4kdDRjAk0lcWxuaGEoxq1owb3JK0mi6GTTSuYd+0tVXVCtJaEYqdV0pbJMpUlGTS6EKM6EpwZiFFtcmxyShJMnBRs+q/yJzs79xyjJ3KFJ1KsY92TimkuxiUlXpy6Wt87CauR1R0JbC2ALHUYmpyaXQ6j2FsWAADoACA6DtYANxsvoMR1AIsi00BJjAVjcsWLFtCxuFhosWDoGgxpG5Yho0BoE10BNrVEakFZSjffqWtsS7DpulKdWKu2tiDnKMXJWfVdhLUYNaHQuM5vdsuppsNEXZjaTt0JRSRJ3u+xTu43YlZi1dxrQi7osr2J6CskaXsKLbSR7JpEoRpNRk9ZbCSSJUauJlSjN6Lb+v8zOMvjha0qTd7L0/UjSUIqMdEugkFzoIC2hYLBLYe4DQaBZcoLYfkX0NwBpNlgZe2qHqx7D2KtVeytbY+r+zS5Xa3QpU/ZQ9mmTpKcn72vK38+xyuMObruVsQ61T1X5EIvlb7CbnC0uh7Zzpa/1Yxaas1siKSg79DCYupGr7T+VpolOk4ttau76mOhTquKpaKyffW2u/n8iMFCCj2DrYYloAJN6IVNuVjFwqRV4uxliqODlPdu4rEmLUAOgMAaVgZYLIsrFrosNDQW1GkOzQwNw3NCD0AYM3OgIsAHQCxY0NCxYY0OwkdBO6AKuxuaWGkyVkkWOb3UjCYGvjalqcbv5fq0WtoWAa0LiOtho3Q3fY+KI/ejYqQtFi91ITukJWQtdCPYe5KyZF30LxVzCw5qqUuhUqQdJRj0ZjKkZShUnvHRfcKSlsfWZRpwXYx1VVKradzcLi1FsdLm4LsWGrmlh2JbD3AaACyGABuWAYDSG17OxWsotlGUX70tzWVW6JRlGsrehWknUUfL8ijRftXNmJq05zahsVKqpRu9tii7QbKtpUmmazp2Ki5Fy9SrQp6crvp+I4tWfcqNL5GCq+1oRn3HQfsvadL2HudBRbTNLBGVpJkan7xyZmNWnWrNw2ZCmorlQrNkrIWwGjASGFg3CwWEgBgDepcvcBbWECIPodA3A6DFYQAAAhCNLAx7AgTugYVFdDtoNaG2hKLc0+w3ZamDxVKnP3o30fXrYp5xiMPUlDdz06f5EXOrh/ayjbW29xgaMauFhtMQ0lsLQV1IsregneRukzoJWYrDRLTV9BR1ROCm7llFowtCVfm5eiv8AJGZZZBqPMrrRr177/mKHI2upOTZLYOhXw06UYSl9pXRFiGJ3LAHQ3JmhZBuD2BD2HqBYZK7TsDDcJXYr3HqyRJalWo1TuldlKjKg6jk7t6oxafNGVJ7r8fmUU5z5pdhcsMPbq3+RVouVbmv0KSpuhrvf9CtRU6fIRUXQvfVf5FOk1S57aX/E+rKM+ZrVlKoo35tbovLS7K1NVYcpF6CqO3K9jRyJJJDs4iimizsSkoq4tYkoe8mJ2TI7EthANBoBoNl0XQ7WCwdDU6BdMAsrjVwbFuWYrJAhMvoGhuBudBaBoAaAzQ0LFjRnQaHubiYiL1DcJbHUe4KyepWpSjNxfQhTvI+qqU4z6orUJxwym9m3942mWAWpJaAwaurC3sP4lYmve9SG40dB7Fky5Gk60lCO7MHhp4mo6cehCheooedjF04wqSindRb/AAMPWcE7ehWzF1PZxltBWX9WJu7uOSasNXJRSsMqVW1eXREJKSUl1EwSsw3LJosW0NCo+gthgWDQ0NwAldqyAY2A3dkqijKK7ha6JNXsSnaDZjqsZ1XKC0svyJ4epRi4dUKUeWFui1MTV5VFHK20UpqLaY2ndIhScaav9r/hivH2eDVPomr+uv6P0JawTFew3pYvbUUbI6CfvaEuxoiOiKFL2l49bFSjzsbUWo9yNROKfmSYh7C2AYAMAAFsWAtoMB7g2N6B1A0uEdUAbjADQAAuHQ6APUYCOgtUHQepJWZsi2grt3ZOV22xvoYGjCrVSk+j/JkpO1jFYmdOUFFbvUE7G6BPoSiLsIJqzKllZkldJltWy97FrIWwjluzAYepGupLTl1MVCjgI8kdZTbfX18yvKpiFzP+rIw1CnKcPau0Xv5L5ajjGMG763Iz5pNdhtq6FaxGLsKN2T0dmOzQoqMUl0ExG5a4dACWrLDRbQsWGtDQYALVh0HqdAHuLVnQdrCs9SqtBNN26lWai25GJw2HeGhBy1S7MhRgqsop6K5VtKTZTmuVpFCnSlOTn/K7b79DDpuPM9ytiJOnTpt7Xt82aNWEny3K9KMMLTknrK/+Qppy5RWszYeyF8Q7sdRc/L5EbNC0kivU9n83YrJTcW94jelz4iO7ALDVw0BgBoAkFrhY0sWBhKxqzewACI6oBgCAAAtcSC1zqWBjtcBCIlgsVIklocoSSasWISaehicNUoyiprdXMTRlPlt0aOgDHqJ2Vhl9LD0Q76E1eJHWNhpXE+qJ2aVhtJk1szC4iFaajD7L1KznVpTqQ26kKNWtiXXa92Kt83+JUlNScLntGotdyb0KdG8HNdP1G7MSdzL/AGEaVT2u7Wm+/wAv1KklF26mc42Sartb2X4JGrEESwbgx7E3ZHQGBa6JOyFd2ZsFwGGhcYwr35Hbch8KFaxa6sbLQkuhRi1UbZiIKai29rf5FWU5xcY7ohVlCtKDXclLQp30SJKKehGovZS/rqVYc0o1HsrlJ8+qI12sM6Vt3f7ialJpt6Iw1BqpKq+un3EXdkkloTWpCKbGlY5Umr7k42EuaSRWjFzFUjKo4p7Ekmii+SPohJaljqGhY6BoNHQ0AsJBpYtoFjc3NGOyGMktDdAgSTQBC1gGIDcAAAtcYwB7D3NGWEC0N0A1dD7DNkB8OpVrzq25nsMnUjGSi+oaND2CyZ5DRG6uhq8CGqEkrosRVm0W00Lt6dyMG9FuZbg+SPm9SlSbwVVPoQrujNS72fzMbJzqOfV/qTinG63PYynG6Ww706Vns9SUm3YoyiruXYq1H0KFSVS7lumVEpRTLDaQyJuAdRvQlqy2gzoA3ZDQAD2LFwBq6Gbo0MdOcaS5N7lNpQRBqUborJJlebjTco7jipU5ryOWcFJrqcvtMQo9UrlSd5JEIcqTIxV9RSSVu7KrjC0n3SHJRSQ5XdiNOKot9RXUSL3PaLTuSsimlKSR9rQtdpleSlO6He5e8jlSbaHqhO0SyTYO1xgwGSehuAb6Ahgw3BljoNDfQLC3Oh0NwtdENgGJi2Op0ADcA6AboAexLcsCEWTRB3R0AnFXGPUsJDd9DUa8yrh4zlF31TuaWOh0LXF5D1d0NpjbdmhaaEU02PctdWIyd7MquS+EwFHmak1exWopqVWCskZSqCwzq1Xqtlr+g8ynTcmlvfT+kVJcyuYzGU50IwXxr8vyKUOZNvoUsfKEZwS3Vv60K2JqVeSm/hin+aHJIU3cx2XyjTpTUtXe6t0aTTvfrr+pVcKUbt7EXB21HrsSinYrOSg3HcixWOhodDoTfQtoWRY0NGPYbuOxugZbQtceobhrck3pYuVqnJTcuxCXNFMLX3KPJKqoNkIRovljqkVKqrJztbclVXKl1a0KTbqOHcxFaFSEVHfr9xOdOCi4/Fe3y/IjT99s5LppFSC+q80Vdp/gezVRcy6FWlz07dncpLmvHsJxm7xL+6OSsVpJRTj1KdCXt3NvRDdyScXYi4pXZSdLnXO/d6mMlRlVl7F3j03/AF1FZ9SyS0LaWHFIklypGlwdrmjLXAHY0Eg6AAbmgFgHohljRmha5oAIha4EthFtA3FsbltAALBYewdAJLULAthGzE7oCSTQxlrjCxFJK7K0YTq+0W9rFgZY3NEyslF3RFJ3QrDWp9lA1dFGleaMBGnyRUlot/vf6GKp8keWOxSqODsTwc6sY1v60ILljr8KZHDqtXfq/wAScnH3Uc1palSNmSim7FRuEG0YW1VwjLrYxlFOMqfS5FajVhotcWgFgk9BDWh0Bj0G22dAaAB7FrlgNCWxTVN0J8y97S336iBrQjFRba3ZzFSSUlFFKlTlNOXS5hJuFaFZborUrXUfUrVLu66NCk1UfoYNSmoqPW7+5NsnXkk1H7SsUqkaMZJinCU2+jRCMYq3cVNRjoUqXtZcsSrGzcShFKivLT7iMrQv3KV5GIqQnTgluifvPTsLSNhWcLF/duWurE6DjThJ9blk0N/gTtfQGJ9AAlsFwuaB0A0ALG4AWLGhYLAkR3IgwQC1AAYBYNw3Bq6LDBK4jdEdgCaLXBlkxIbuOVotm6TLBa412NGJKULELp27HLqSRuhJMUZN6EY06dKm4u8ne/ltYpY10aE4W+K3yMZUdLErD1F7zV/lbyHRk9tzD1a+KjHDUn7z22/X/MblUwsmntujLcf9TnN8t200tbWffZ3JtvceTvEU41E9rP8AFLv5kKkaqvF3RzbEneWgm7aGKxirQhHls0rXvv8A5GjFGk8O3f3r7eQ0WEbi2NDoTd9Cy6DDobom2wvcZ0DQC/Q3LaHQ6j0JXvY2G1cclFXOZpoxHuNW6mIq+znBdypRiqinfcrwlLlcXs/vJVFGUYlerGbuivXjTk0+pQn7SXs0zlVKpGDlZxdnptcxTk+WrGNo7b31WpScKspyqaJL738id40edrZEKsZxTW7OfR3MJivYS5rXdn/kVJp3fcow5IWM3k8NTopPWVn8rkZJRsupTvd3GrCVzdaDjdC8ipJza1K+GhTklCXNom9LWb3Xy7nJ7tyUbIWo0NXNUR1AluXNLlgS0LHQAAADyAOhoFjoROgx7HUQB0HsAG4FtBAA9RgLYBaG4E43QWHqWGtAaug3GDYJ6ktGO1rkiK0EUIK4o3XoYOEOV1n9kxUvbV3Ve5SrNXt6FalPBYh23i7f1uQr8ra6Mc0lpuiEo1aCqxI1qntYJbalNWk4jjaZiarpxc10MLGMqDnIryk5x5duouwnoPca0QhbCBuyEdAOhJ8qHqMOgwADoDNxxuyMW9hrUqt80UthqTasN8ySFTjOouboV4R52lsVIN07IqUpyvrrYjOXtFDyMRFPFcjIpU6kZonzOpOq95O5GvOphlTeyd/nsSpKEXB/11JyU8PKPkZZFwqNS6FRa6HtG5uPYh70UR1VzMKUKtTkXSKRC8Kqj2RTldJknoiOskctkJXZfVIjaLRGtzzcESWlh7WHsDsC2Amx7BcSHqX1QdCx0NwZYsD1CwDWhYYFuhEtdDRuNWAewG6AANwYDBokgYtC4Ca2AJKxcAGRTk7Lc2Y7MAsug1dEUrDjoQaehTwtStdwWi3KMFRVnK7t2sYqpCnSU4+87bbfiRqOrNRprV20+RzqnXjGS0vZiqU44rmjtcxVOjiaHPze+lro9X+SKkehKfu37FGrH2S5dtyM7TUkVpOb5u5RglVjWkrxjo1te6779CdSDm4rcVRqFivUVOUP7xGzG7MehzNpLsEdhBUeliw0JgSaYDOgwsABa5q2C2MLXVCqp2va+m26t+pJq7aJalFxjJ3V9BK4pVnirW91Ly3/ADIUvaNGIoRpOUVK9m1tbbr/AMAnTcpolRUmruxjY0VhZVFG81117/cU7NJSK80pcopWp6Ilz1MO5bNik4U1pe+hG1JpnK9X0OePtHbcU+WSiiK1TK1Re0TuQXMuZoTNGizbuPYTVyri1HFRo21ZgcDPFT5Yv+vvHgpU6sqa3Td/kO3snJm5y3QMtqdDoEhaoLK9w0C5voV8HOlSjUe0joINDQAGixa6LAFhK4kdCWwxtMT1EWANLAAwOpYAbGrjCwCCL0AqLUBgFKp7OakuhKnU+u+1ezX4gMBqyF2HqhK1QwFadB3j1sVYLmaRsjDV54epGpHeLK16k3Lq7ii1a5Cuk0jERcpOXckuhQjaVh22JU7rQSKyTXmJaamN5nT9zcgrxGNLkcuwrMIiWox6sfkXAk9AewdAAQAA9Q6DWhKajUUX1Lpsg7q5b3myjyexlrqVG2VKEuZtdEYiLtLl6XLuVNwZVU5SjOP2U/yJRc6sGvP8ildtpkIQlh+ZPW9ivBSlTium40lJoqYqCwbprfm/Qw8L4iUn1t+ByJTuc1ivCcqsGtkUKiqXXYSs0hXukbMctDBpNuUtkTw69r7R7mEqqlhJ97r7rEKvPUcmVK9OWGhT63ZCleLJwadhoSYgGzcWglqBYYJamKx062Hp0W9I3t8zdh0GAA9gQAbkUIJ7A9jcAAuaAGgPY3FsA7BJAwEX0GJ3QVFewgALJsC1wuLQ0G9UyO+g4pySZCo4STfQo03VjJrcxUoUHTu/iT/CxK9tChOnGHM90yvUVSUpLqLXUqTh9XVviu/uKkdLoo3bOdyuuzMD7KVWKqrR3XXto9Ozsyj7KniF7TWKZWSu2tipJyUb9FZfn+pJW2MLh6c6FSUpWcdtNxmJpupSlBPcjaEEuxL3oisIk9BD1AbQ9ywx7Ggw6DNDoLVD2FsBY0ehFN6FrErSNIq5B7Ek29BxUbNdUSlZkLyq8jegpKztqi75nfuzB1OZvmWhiKnsat6f9aHLyLXccvZq/cdWMYO3qUKjlTTK1WElePcw0JTrW2dkVqbU7PoRpym0kV5KnBLu7FOEU3YstF2I0XKV49CbvIrc/PG23Uw8pyej919O9tv1+8m+Z3fkvuJ4icYKlHZtXHOFOLMPRU6ySXRt+T1IZheE6Pf9PkOTTtclu0dABgMWwFlc6CN4ltRbBYsbjAAASERQIlqxj1A6HQADcANy4AMGixY6CAg7oJq8SIHQHsW0GJFtA3AktyHwpkYqTTKVGEsDVrS3i1b53MFUqLmv1Y+V1Iynryilzy0JPmTSForEdXYjSvTc+xyu1imrTsYT2SlP2nVO2+/TYw2HqVasaVNXk3oVJpzt1JWcGVYuKV+qv/X3FrqxzSULIUp1qfu73a+52MRTdOo4vdE4XVhERE+wNB0JOwNjLXBlgFqaWA6BoFkmUZcruVaiUku4rczJw5lYhGzSFZ7EW3ErTSaT66FNXi2KD0iicErrvcwtSPJVj10PZqc0+xj4RdRJdkY+lafKuy/Ixl4tJEJqFHQaVmjJIxlXhVlsY6cZ4mco7XKU+TXqYiq5VlSXRX+Y5qFr9dBdyOKlRi4x+1p8hx5noVYuSaRh6bjBeSIrUkryuUKEJRvLYynFRoV/f2ej9C8KuKny9GUq1OpUlBboqxW6GjcsM2GNWQ1dFroZohK7LGhpZiRYLDB7FtCwAFiwkJCGSRLVDSC4xbHQ0DodDodTQANwGA1cNAWpsMvZhuWDQY9jdgxm9iw0gS0GQaTIpJ3MRh3RhCLe6uKhKdFzW0d/mSu1YxOHhVSjJXRZW0He2goK6UhwqVo2vtr9xlc6CxEViI80dbrVdPLXcUW1ccnGpoU68oVXVp6NXt+RVutepq6eu5Um20mX0aJJX02ErT0K9Tli5SMI4wxPtKivFK1tvxPeS0I7CJasR0AnK7sFg6F9x7CB7AHQBgmmihB1JqJKNm0cqUm1uyFKMsNOp1TS+9P/ACI4i+I9l2Gn9kvZDfNp0IKyRzOEk09SvHlS8yhQhUi3D4r/AIWI1HGSa76mNotSipPZ3ROreq12Papuy6DqJvVmtnLsUG1TKcXOy6k4uM+RkIJSuTcUr9ii6Dwk3L4tOXfzv5ff8iSvJWGnbQdFxoqp3dvu/wCHOX3j2UINtbvV/l+hCKbVzGxhVXPBcqvtuZPRw9XC1KslrfR6mPwFLD0qLkrrr/VyeXRoYeNe29/63KtnR5etxvl2LASsaWuPUNLDsG5a6Oh0EjUNwAYAAkRQXNxjWhsAtToaDQAblgsHQ6C1QPYCxJD1BNC1Og9haoJKzC4yWxbQ6HQdj+zauKpTlCduTXa99dt9C10dS1xOxpYhqyL1R7aM6Hs5LXoynXlCnKC6/oVNrnKmOk1Dm6EdGiSOe0NDL4QnUcW+jG7TcR8q1IYpRbS2IuMolaoloh+9qhroZXRpuVqj01HNuu4rYklLRjS6C0REN2CdyxJ6DLaBYsWHsG49i6sAANCRGbUtCWpIdROlyoUI8/MtyKu2N6MWjMFhliKc43s4pv7k2SWl0Yin+6hK+6MtaoTqP+6/x0IcsKig+piqrqM+ruMVJ9SK5Zy8ynBubJTkm0UfehG5gatOniouWqRiKqr1XUXVspOydyMFJNGGrJ81N/ZIS5veQ6qd/IWJU8OoN7N/kj2ym1Bdb/oRpwpYKFNu87t38vyKzinFJbIquNfDQUY2te5CUKNBxjstTFOpjcLTt1X6fInRq0qEYVlsvvXyMbh4NKa2ev3lWCV0mPQlUSkkx6lkO1hbBuiw9ToFhAbnQAGdAOhucl0KJYsWLD1Y9ySDoIOgACNLAIBB1CxY3GtCxYE0x7WIbD1JK6GDaDobjEroc+VryMdio4lU2lblil626k9FchLmimNWYI57Qd+hgqkZYac0r6fm0Ymo5xguysSbS0Kjg1Fx7Ff2SwkYx3bffyKeGck0ilFSmovYrKEa/K/hvb5X+8q+zhUl7J3jd29PmTqVfrDh9lL8fzKinJxsybs7DSa9Sokox8ild38hq7KicYpEOjL3Ny7RElsA9TcnqzcLAMaFqzqPYsAADQ1qdCWrLX0Jtx1Qti3Ua1RTm1qiat8xxUkOipOxOK515E6vJfyI1HNxvsV2lJog+WTIvVpk4OMaa8rnKr8xSqyvLsig26aHDk07kcPH2vtHu1Yo6trujDU4WqSTvbf5mIhKVFNdEyUuSnFy7FWioUKVXpNfqZlFJ2i7paL0/ruYWfssJ7Kb0b19OphKrnVqRg/dhJ2/r/MeZOVN0HHbz/4BjJrE0/deibf5/mZe6fvUKnwyt8vu1MzwMsLJRk73/rux7k4ptMdki9y1zoboQDZbQSuINDQYAFtASbFAUEi2gXN0HQdrktxq6Blzc3QAHQ0Nw0sdAALdA0sDWg0AQfQ3CUboNxGgJXGPUwlFVa0KcnZNpX7GMoxo150ou6i2r7Xs7XEktDNKVGniHGlLmjZWdmr6Lo9Rdi2pVpWgvNGFnGhShFbbP8DEtTm5RWg9U0crUdGYOlCvVhCUrK+9rlSq6DTXVP7tUVLrVGJqKVKXM7Iw1P2NNQvexjK0aeGlNatEK/PThOPUxNOpUl7vQjG0Vcr11FK5lThXcpSdrJr5os1uTfMkQtawoT9vJv4baEXaFis5OnJR+LoRsT2AJWSHqMBoexa50EbMQbnQ6AAPYsmkNXGE3yqx5EKFSSlUS91FOvHncL6oxMOSdlqilSUMKq7/AJrW7mYyiqsJR6r7ibb1IUZy5m9iMlblKmyZGCc7FaKhUkjNZQoxp0U7vlT+Wv8AkO3La+tkcnKmjDxShuSneauJpuwqqU1FPUweHkoVakdY3V/0MRB0qFG+0rszDL6ftFCcrKUeZfPpuUptUPZy+ytPwKDVSrThJ6GKrVqznGkrtt9lp8zD8lKlGUVre7Mv/e13UlH3Xdb/ANMoZnCtilRpPdtL1b03Xcr0quHnKFTSXy/QxVadSEU+l/xJJct+o9VocylJpbo6D1E9ELUBalrs5XYtcL3BgFrgoNnImKCRQp05KXM7WQ5nMczOdiaaA6DJLUHqBoINALCCwdAAZuhjHsAMAhLobhKF9jYujc3Og0BbQG7osWJpyw8X8iKXNG5XpwhUahsSb5rkXzCaSMVjFUqQg+isvkUKMKkdd7ozXLYVaU4dF/XcnduxPms7EYOEW+pR+JFVLmaMRRn7WMWtHqQ/dV+aX2tPu/4cbuPue2vTUOxUkpNWJKzsLmc79BOyJbCBIm09BpJFlYsbgHQtcsJWQrWGaMA6G4y90XTsSaS1G7RuhaolvYp8rlZ7Cm1HlWzFQiqrn1Y0pbjrv2Hsul7lWzRhpR9rZ9CFV0pziutypPlSSKlRysXd/kcnNHm6lTG1sVaVR3a06bGBoyqQlJfZVyU2pJPqRlZX7HxbijZXRKUYx5jJZTrfuo7Sf+X+Ri66VeNOW70XyK8qk6ic+it9xKMajlN7u1irhpUm/Iw1WVB80Xq/yZWqwlQjybowWMlCEot2snb1K2GWHxsqa3UlL7rMqYx16vtJPVfoSqbkndEZ810jlUUS1ZJu2hFNRV9xLQGEF1GzqdDqAAmNnNcOYGBugTsXTLEo3Wg0MsAFg0A3QjqdA3AR0OgAAACkKSaFJMcExwsCN0PYOp0HqgeqQrPQVeTy2NHpz3+dmidOyjJdSuoxow11d/kSWpSouV7dCMIOMuZ2f5ns4uV3uRm4vQqYiUk0+pXqNTaS1sfFoKKsYeg6j06Eo80tCpOc5U6S8ytQUqkeZaxGclSU42enUhTcG03c0bHsJJIQ9UdQvZGjZLY8i2ghodrDVwWxbQaErIabLBuAPUs0R+IzDBOhyQk73Sf3pM0SsQqxnJxT1RJDUrqxUUo0+ZdBfETlH2jimSadhp3RGKjVbfVDfM5Mqrr2JTTkmOcZwVilR9py8vdr7iNNFKpyXUepUjF9Dlai1Yi7q6KrlGnz9CVCpVjyvZmCn7GnFQ3iVIqpUjUlurlbF1KtClSts3f+v62KOJjKl7uuuny0Ep1cLWm93b7jldrrsQrU40mm/euZpKpRqOUY9b2uYeUsRV9rJayPZOnzK241c5bllEnNRV2zEVlCClfcgtLgN62N0JalrIs2FjQGA7IbGLRC2DQLl0NgJ2LpoHoMGg6AbgMDoAAx2Gy+gF9RsudDQsHQsbF2RndBOCtcRcewI3Qw3QtxS/dpFKjOpHTZGIi9F2HJsVSUX6kpPncewrtonh+WkmzFUZQSXlcpvmir7nI4xckexlyxkupSSw7lL+YrqCmuTYlTSq81PoionOTaErtXKVNOLl2KtRRldvQaZJWdmWVhbF7mgSsHQLFmtxoZYewkWNwaN0AxNMHsdSLs0VZXm2JXFFJtk6Eo0lN7Pb8iEf3N/kTmvY8nmJImtXYejsSkkVvcjd9dirNKN0Q1TuOHvWMHZVFcklSfLB7EbuV0UoKpVV+rKivVa6IUZJKT2J2SfKe056ah2KMW4psjaMbHNdJIpVIqMlLs0YWn7OmoohimqVSKfRCm4wce5Kh7WpDXYxU41nzvciorYnHmXmU4qT1HCLIU073JUKclqjGYWi6cLbq/fysLZIdkhu7sO3QIIeqGhasaGXGCsSd2OxctcS0EjoNj1AuczLidgktCSESNwWpuAAgNwZ1B7h0NCxoaBoMep0Aewxbis0NaDRuhbDQO1jRoYy1xU24LzMFjo4ajKjJbu/3J/wCZWm2rntY1aScGN323RGMGnJmKdKg4SpvmVk3016ozDFQq+9F7t6drsnWdRLmd7aEiGEp/2dCcNZJ6+lv62MRm1HCUrNXbvbfr8nsSrrF0V7N6FGEXTs3sYesoyce+hVwtKnhlKMryvqrPTt946FScrQWopa6GNwXtoKN7dSW2m5UlzMkKwi5uh7AeR0JtMeo9SUoumopaltDobg9jcAFFIOg1dD3RVvytkL8qb3HFpIdWU6ai3otvzFW/d8vQTU1ddRNJpXJRvByXQm0mr9SpGTTZi6qr0YJL4EVrSSsRdo26knpoUtHcdpQ5jDOPKQjezT2Gk37piKsfqbpr4rnsnFLm9TCU3JSkupCvHSNx3lYnBxSKerRCEqs+SBUwkadaopPdL8kY+Hs1C2qaTIyTmpRehT9nKM1N6209SFNwVikraGHc3Wqcy00sQb5yq5c9kyo2oJkrznoPDNW1KtNp2LWFqIgtBo3QLWOpKwmM6DZ1JGghAPUABM0YNXJK6C2pIANwsCBiHqeQHUEA1cbuFxlwAauWIO6GSWpa6GtCyErjtcNzcgk2iUJLD030X6k3zNX6FaK5UzE0J1VGMOjuKGt0YjmVByjuYmh7KpKO9rfkiUuZWFFxVylV55SXYoc0qMrdv1Rh6VKSm59nbfcdGNOXLEvYkm6qa2KWOnCModHb8CVSVmo7mGqQmrr+rDqXl6GPm403JFKanC5KNhieqQ0dBktjoLWwG4PUEblhoWwNINwBkloPoiTSY3oi6tY9moq3cirWSGloE2rEpX0MRWcaE4LqS0ih2vYtqRVxzvSUF0f+X+RSScUuxhoKc4wb0Y42ghO7t3MZWapSj1sYCfscLzW+yzB0WoOT3kYelzJ+RShCc3GTtZN/Mo037KU+2hhqzpT51uUGpt159Gl+BjpNucTLcM3QjzE0loV8Sq7UrbKxaKbZhpubk/MjTSdzDTo4bDVK0ldy91bq2j1/4BVd6aZQV5DqRclFfP1/4Yru0i+gITL3Rcuhy0HqLUtoM6WJW2JFjoFx6gPYuARYDWhJajRKwD0QmhhuAAAFzpoBuMAOgA0ENzoT3LEkkNBuWGdCDSZCs54eNBO12iuvZyafQb5qehQl7J3W5SrRpt3jdvzMPye3i27K5jMVSpSrwpu8XdJ7f8EWMgqkV0fX/gErq6FKKdilRnOMWnomvzMfVw9KjGEH779dl+Bip0IUINPXW+/y/pCs9DkUFZbDXYjTacpX0ZTio00o9GQasWTTuUOWLslYxOIlXm6k3qzS4viQ7co7jWhuGlzqA9ToPY6FgEgGWES2sDO41oTrqE7SFJTipR2KU3K7ts7DVzzItPQqLSxOHusjFOnYlytvUqNe6iorJeY0kxKxGF4uxS0e/QjaOu5e8ddyCW5LCSq11JvRCUHQcX1sQkpVmuiItxXqUoKVS72Mxq0pTfstn6/qVIWjHQjWbjyIu/aSX9bEW76l1yyuYa7lPtfT7iFWc5uMlon95Gzm2iVSTdiTdyp/BXyMLVbrShbZIin7Ur25hlStCLs2J3RqX0AAHqOzGrs3ZYGXAvcNAAFqElclYsNHQ0LAaWANBhodAe40biEWGG4aWCx0AsCehPY0HqrGglcSGNhHYkmoQMVT9rRklvYy+rLkcZbrQkrou704r+ZfdYxlOEcRJQd0ezUG0urb+8q4anyR8hb2IYCNerRcHeTTVv6ZSqunB0u6KyUpkoqUHFmFo+0nGmupiIuCd+hTu7SJSai4mHaglGXmSjaTSJ1FB6lpVXCUejHpZF9bENWOD5UxKKiirBON0UYpw2FG07FWPvpIUIxVhRjztHLHsV0lax0HsWOoFjQAGrg9jQloivT5lZdSnBQjyroPRENrFrISsiXcrK7THpTKmjb8ys1zDldkr3OVOK8ildqyKdJpscdkhxXJ5lNXmku5CC5WylXlV5odmj2TpOXe5iK6nFLolYoUH9WlPzS/Mw13F37j5JQUHvp+ROi6fvdHt8hx55q3UraCi56Iw0HCkovdCdig7tm0mMqfwkUWozuWinfqYj4hsr4aFVpvoQjbQNBB0GDWgzoNXQkboAGOwHQBaiQ9AerJRTYDSLBuWCwAFgGdDRINRjSGG4DADQg7olqjoNj2COuhJakaylVdNdBass1YVCM8C6j3VjDU3N2RmODjTwtGVrP3v01+f6CTlojAxjTrNVFcxcHCo7LR7ErRSMybq4iNSjG0Oqvfp56lKmqkkm7XKcalPEwrRl8PQxjg69ScPhbdkONxVLyceqOblimtx1OZaknHljFfMsosxuGvVpzfm/0/QpuE6UZxerv8rFbC0q9OTkvftZPXT5bCwX1elTje/uoktVYt71+onoX90qfw0bUtSnZQJx/eJk/4qK6d0XF/CL6B0FqFwPIDdD2AJbEispe64sckVV7WGjFshp2TJyTiklqSWhUV+uhFO1mzFct2kV0tGiK5kVYwbSiYanRdGpzq7SVt+/8AW5Q00RCDVNNvRk6SUHNPYjTlUg+VamBcaFX2jV7J/lYx1OjSceTayvvv1IW53Yi00TindPZmXyXw230RKMKc0t0jFYmDqq3bRdkV8RP3IN3SvbyIVGppxM4waoVY3d7xT+8hVdKL5dCGJVST5ClKq5y5tugrahZDbloQjdqJTptSuyu05FkSE1YaTHqdAC10HUtcsbgCC4B5AA1cBpDQM6CQPY6DsCAA6FiwxnkNAaAdAALXI6MeqNBrQsgwEIzxMIvZtGdRpQxko09lp+Gv4kacVNyW7HTcJWe4nfQoTjLBuBHERcnyP4dH6lfFTr0oxl0Ifu9uh9bjLE8r+Jq5Vl7ajKUn8O3zG1KQ4/utCGr0GrS8io9C6ZVopS5kJ3sitT5YK3Uqy5KMpdirVo1qNOdPsr779TiCtVpxpeS/Df8AUwd4umltKN/mVa0lKUY9B1nKMfSxi41aUXLpFN/cmLVJkbNj2E0oq5VV43TI/wAJkveUWVnaaE4ziVI8srC/hFtDcED2NDoLVgBsMY9VYktCo7RKUGmyKtobIk9FYtfUlZrQd27E1aFysoyROmmkOCS0FZyRSbkjB4LnimVMHU5XJbX0LSeGbtoYWbprTqQouOD9pLeTsVJuVPlbIQSVxR0F700ijzSkqdPcw+GpwxEoVNUo3+ZjFQnUjKn28yVGE4RturijCKffoRdWfM6nfT0+QoXbbIQjShZdC+hFPmd9h2HoYmEITSi+i+8hLldx11bQVmaWLISTQFiwCRLUtcOgFixoBuFhWOgA9RgNDTAsMsAG5bQtcLDGW6j2GPYAA6BJpvQAQdDqPYjByTa6DRCSpq63HqRepi8RLEVfaS3ZCUVzcyvpp5My2MqsnTXVP8Ff9Chl0pQqOGlk5fdZdyM5OduhUSuThGDU+uxQrQlQlTau3t5Cw6pznJddfwsKo+WxSUalRRS3MZepiY0Kekl8ydKpD93Ud5Lf+kO6uUHGdlIqxtLl7ChicRhnOctY6LREoXi0yEXh4tN+72/rUzC83eWtjAtQgo9yNPqiVJe0jNfZMTKVaL5t2LVEUMn/AA0Q1gR/hMpO8bFbWZTbUlYr7oj/AAgHsIC6si4g0sK1jQsaDV0NaEtiVraijazRTfM9S+ljQw9OM07vZHNGM9VoNJq427PUrXlEjTbpuQ6b27lKDc35FF2dkYTEOMeRLYxGIU8PZPUqTUcLCPcxdb2sk4qy2sYmlyUIdraEIrlu1oiVB/V41b7uw2+W4ktDDYqOHxcLbv8Ar9SGHlUqydV2itf6tqToqVScYbJaFCTjF27GMxSw6TtoU3WqTbXw28v+HMfRnh68aT1uk/vVzEUZVXHXRO5Rp+zgoiQi4JAlYG9ReQAWui1yFk9TGww65XS7K61367jdyx0A0ZuPYsAFtQAHuDsNDV0INwDobi1EbgG5bULFhgbgbgBfQ3Ajqga0KNbkjKNt0RS6knzO4JDd0NpyMPOphq8ZJaO/5GFjLFTVOPXT7ytk7oVFB7/15mKoypzcXuZniY0lFrbRfMU1Tg5diNaNaCqR2ZNqLKU25xkvssxUn9aVeStKSuvS9v0MViHWrOq+pjIVaVtNXb7mUoXWhWd2Sq054eEE/ei3807fdaw5Su7lSCq07MdBypumu5Uw84pcu+hT93DyduliTSlYlZxFdXQgSTiipJRjZEf4TKMvesTt7RHLFalSXNLQj/CDQlsWGA9TcLXZYaBjQya0JXkiPMiMklcWquLUhLRpFa9nbcp4hONuxskThF02inUh9WcOopxXs79DmSqyt1KELuyMKlCLlLYxF5L3RNuybKUFJoxeJVanFLoidS1NxIN2s9irU56cILoxU5Suo7pN/cYWrKpapJbOxHFfXZKne1tP62K84QqPlehFtNoq0oy0kLGxjSWHite5Ouqk2+q0LltC4jQ0Ogbj1Gri1RYbtYsDubnUbuy1i1xhfUDcA3AABm4HLa4xuzSNGdALDsIT6AFkx7DHsWuML3AGHQA3BBBktGPYG9Cm04nQFqKN2htIweIdGpGS6MzesqmAjXJV5VakpS6GOo+2pyhfcr0lUoSp9zBYdKnCkumhm2FnCpKh11RSpVIR5ZbolO9k+hGmpNu+hLEOVdxa2RRnZNdzGTShz9iM1o+5U96LZTqU3h1paT/Iqy5WmupFcy1JVZQhyo+sWqKk1q+pWioUoyXfYZ0DpYkAXNzoG49joWGAA0bgOwo2bfcmrkmoqxBJ9Spyt+7sQVlYjK6LJJFW7Y4WbKU+RO3VE17vKS91NDqReCUet/0Ie82U90xKSw1yjBOkpvqirTSSfkYWg409xu6sRSk3c5dj2bbtfYw2Gcqzadm1YhG0+V9DFTp1cXONPuYijFyS7FCybl2K7jzJLdJX/Ml7zuzEYiNNq5CopO3Y6G4kC2OgtQktDoYfm97m7sEWSEkwsFgew0DRYsBuWEOwAyyLIaTLF77AS1kFrFiwALYDcsdBhY3AGAbgAAtyVmyWwNJmCm+apB9Gw3KNOVSShFXb2FTcZNS3RWwVSdKE0tLr79SpRlSq8s90Y/GxWW8iet1+TFTvRcuzS++/+Q4Sk3YaTRCMqbV99zG4n61XdWe7MQpKo7jR7VKHL5lDCOtzzj9lXJScJLsy/tcPKNtLr9RRTmooq05QVnuSwk+WNa2mq+63+ZGtClX5qvw7fP5EpyjayJ76EcO6tSPKru6sYqcY2v1dhaoAZZG6LG4x7Fg3Q1oLYsAAdB7EJRkrpjs1YktD1K6utCfYpJpWISs7ClzMltYrT54xiuhKzQuVJIrSamVZqTuRd4MowZQpylKPcjByio26DpLljFdDE03GoqfUnTnSqypyVmtGOXuuxCCdJu+pGTtyoTfMVab5rrsYdycm2YfBOliXUm7KdvwuRhCs5UqTvK/p+ZCEqMHCW6b/ADFTcXKTe+pQqxqc1ns7GZYWjOhBwleW7Vno10MJLmjzNWbDZC1NxJWEkW0KcOZmI5XK0RrQihi2C2gFgAsbgWAACw0AMSsyXQGtDdDLXLAGgBYBq7BoaDoWTADoABuhBui2hubiSTubi1MJUdKrGcd1qcznNy6ixclhvY9Fr8yjKE4zVTfS39ehicI4Q5o6plVtNJFFNU249Ec8nXilsKm6yk76xRUXUjPnqM2Q3fQpycYns7xUkV3Z8oqiVislioKk3voKi6FoXvbT7irTtK43dElUhW02bItRjc3OhYGhpXOR8vMNXLDDc6WDodQB6hYLBLbQhBQjZFrktiWw1dmI5nD3dxtoqVUrpFJ6p9LEmmxmIqOnF+SKFV1FdGIgYqz93sQ1pxMLZ1En3IxUJza2VjFOUbRiYtOc0yhFVMZHne9l8v8AhjF6V5Rjtcdk+Ubu7F9U10PerVdd2UMXHDUquFtdyaV/R9v+CZxKlTnTlTfTXfyHUqVaMalR/DovRu4sUlQjCCs027972JO6KkFUhykUkrDs5Glwegle5y2Z0OzLXRCTjoiSuWTLIGWuBoWuDBjQPU6AB0ABlgsSRYBrQGDsAAtUdDc0GMCxY6APUYdAGMtoAK1h7BoC2EZLgo4vFKlJ2Xf5HDUFiajVbT4vw22K8oU4Pmdun3kruSKOOnSpcu/kSg3IajRoxUXdyTv5a2/QxeWQpUY1oO/fyf3638iNSUG5Jk5OUdDD4KUnKUVotzER5W0hRvUvfZCamnBblNqELdCtSc4XiYVzl/E3Tf8AWhh5zcpX6PT7kU6cqqfcxDc6Me8fyOZJXI2kCRu0CsOwk5NIqtX5V0N2G4Gh0LaHQ6HQAALD10RayHsS10JX6GyFTbhzJaFHDyqSl2IcqtFFGNWNeXN8L2HezfYqRc00+pRgoJWMW7NEo80rdyskq3LHZFCK5lIoJOgnvfUlzW8zGJ8j5TSNOE7+9Yk7id2c1vmUVyxae5Qbcrp7FKlTnCVVv3r2/C9ytBS0ZPDSnhmr2S1+/wD4ZlOcJSdJbxKrUXqaIdKUYpvqSko6tiIq4N2GaNGhhYKpVjBvRmY4RYWs6V72OodTQ3B7Fhm4AdCwdBlrosAAbgdAsWCa93Qs7CBoAsHQ0NAOhoWVjoaWBgHQA0NLAAN3GtBAbmHbvaO7MJQlGnOa+z+rsV8HP6oqzWjdvmRpP2kY9zHUFRqJd0TozhTUpFDEuVeVOX2TF4iCpRq9db/e/wBDFVadXDUqlPqrsbUY3MLJKVn1MRT560lHp+hmtOcWqkPslCaTR7W1Fw80/wA/8z2j5bHLBtrtqJ+9GxRqunsY6SnUfLs/8i0mkktFoQSSRNRTsiUbIWqIq7sSWp0F7sb9SQthoYD1QIsAMADoDGTeuhJpF24sTklylaTpysupUqtO66FOd0icmk/Qukil7y1MZJt2RSfLdmFh7WdigrysUqvMlFdBXe5ioqpJKO9yrNNOxGxKPQdNTqRk+hXc78sN7ojVjBqHVlKDaaWxmNOlTnem7qy779SrWnUa5n5fcTkk9ChReIqKEdzERVJuL3Wg22kTpKpNN9AWiGJqT0JJisjuJ2ZOTa1LoDdhoDHsB0OgD2LDAABnQGBYNGD2GPQ8hoaLDOhuBuBuBY0Y0dBj2AAAZoW0AtcSLFtAyqXJioTSu4tO3pqUKk8T7WS0u7tfNFbAtU4YdSu5a2tbVL1MRl3LKKfuu6Xf9TOMxlKfOlfW3/B2Mwxka+Dpx6x/yIucMVKUvhdkhpSo6+hO0IqK2RQrRxFHm6EaiU0UatSnF3erv+JiOVJXV72X3tIlR5LIpcsa/LU2a0+4moqEnfYWNhOc4weqX/DmBxMayuhWViq0yFSMU0inCpWiorRt/qVcPOjNwqK0k7Nf8MNaXE7IgrajIx5mVJXenQYPVCHqDCwbnQ6A9gA3FfW4yaTQ1qSkr6nOtkcvUxSulLsTVR2cdupGo3ZJHtVOVkOzZF6mKleVyo7pJGFTh7xRqcsuYwLVS89rv9EVE+ZJMkkp1G3bTQV2WVyTvLRkqc24yi9E/wBCVSnCrGH2pfoSwbnCNWCuorX5lT2krRjK3fTf/IlQdbDc1NfD8T9W7f0jFJOnCMHqnf8A4BNalatOilKC966sT5pS5pPViV0KxYSHuZbRqRp8s92zNcMsPWjTT+yn95a5P4LowNSpUlPn6PQ3KdSUsTOm9kkWNAdjcOgMaGAMNwAANzY3AewMuS1QthrQsSGraAACAGMGAWBoAADoAAWRg6sqVVTjuv8AhjDqNOgoLq238yhOdShBR+KPNb5GYY2WJinLdMnVUqrpvo1+JPSNnuYbCU5T999LmaUVh5Qguqv95iFzvlKcFFKKHDllzEqCrw5WVqfLFJkh6tMpqM6kKcn8TsSwkKNWbizAxd1LvcjJPQbuiEJe1a6WIRTaRiFLnbZKn+7RGLlKxTvZrsJ6Hwx8wZ1OgNAwYAAwANBjHDUtoS0Whyc0Xctol5EqahEoJRg7EJXkXsrldpxsimlJpHLalYhFXMLHlhG3Ym911sPmki1pCaaQ43tYyvBrE1Wm9EU7RxTkulypio08KpyWsrfgyCpyrJX0ZOooOdNbP9DGKVOkprqnYhJyhFvew1zSsNbEEuWVzQLaCVyK5ZRZiJurJze7HJ3HqhQSYKKvcDc6B1Gbgw6AGgAAAFi2gMsrGgMa1CyGiRuNAB1C5e4dABm5c6DAANAABBuYPCOtQqVE/ht+LSKaUpxhcwjoRUXfWKl33a/yMfJ0XVqp3Vtv+CZvTccLSqT+3yNeVl/wSMI1ZyqSlZrW1t/8irNuWo8VCrBS3srdrEYwlCTlKzS0KNalKKs9TEU4RgordHPSw3LVrfC3r6bdNTGzjKfIndRuk/IluQipNIvGCkpK/by/pHLSnT5oEKnMpWW2hCo/Zc1texCUZtXK0PZqMk90n96L2lcwcViq0YT8kY6EcNTmkrrZGW1qFOq51ldJPvvbTbu9PxIO6uQjdk3zMHqhgSpWoqd+tgYbgDLXN0IABEtiT1I0L0fbPvYGyvN20KVNyi3fUxEk7IclGOhQneTbHqmVOXZGHppv0JU6coU4Ld3v+n6mIw0KLir7lCScE0Si5J8u5im42p9iMVYp0ZKDb1RhqlKkpOcebTTVrXv8uxVrwTUaUeXvre/37E5uLWm7JzlKOu3QVGpWlGEHbX7yc7V+X7v1KtWU4Qg9op/iS1kLRjdzobB0KTSlcr8ru4knZFroYtx+RfWxY0LFrlrBYsMNwsFWhKFOM3swYbgABZAdAYDLANDWglYbAB7ACAAA3LAMewWAADcMFjJUouivt/pr+hLmjK5gJp1LSMxwVJ4Syd3ceDeIw7h1Vis/YqT7DlHEU6c09GilShSw/s492/vFCV2pfL7v8xZfOhFy7t/1uYOVOUEpvsY1QqWpp6LYlyyw06ifvJpJfPX8CdeMZqDerI14qaj1MdBVKaUd7EYcmHSXQWKaSkvhkrr1KUVUajHsQptYjn6Wt+ZUqudvIjSc4NroYByUJVI9GjiHEJUPd3f+RiYydCUYbsw0ZRpxjLclaMeXqboewASqe4ogw3ANx7AzcGArMlsON2OXu8o9h6vQqJKNzVxv5nM5yKtTlgyjNapEK3NKyFF3TMLTu0ihTgsTZdLfkTSnUd+n6lL3IqJ9ZUW0jEzVS0iN2tD2zVJ0+5GSTRTjzS06k4+9bsTalZIwlaNCLkviWxKgp1I1OquQoyqO0ehJpOw1cPQQS0shEWtWx6myGr6CsmNAFhItctoAG4WCtiJVKEKb2iWBAAFtQ3QBdBoaGg9hg1ckhpPUsMAeoAFzcLaHQaDodBgWCxoAACV2ZZg6GIxMPayslfWzetvIr0qaqNOVkrvbd9vmXjJv2Ttdf1uYdylScZSvYqReArRqQldNbWt/mVqVSWIqOb3tp95UwKeUylB6xf8AXUwcJ1MRKKlpbsVLqdmYmCq4blb11/IniZUmoxVyhKrKq4z00TWxiIOkpeV2UlSxVH2slZrYpUpRUru76EcReMYy3sRc5OouX3Vbr3HQk6sIR0hFaet/vIuMKcm+hQdoJ+Q9EQqOMLLqUXUco0ou12Z9Sd467NfgRSaKcU3cfvO5LUaNjmAlrYuPcRcexdG4AMbHqASa2JNFtB2Q4JrQcHazJ8rXLbSxGcYO6MQ10WpRjo7kEoplL3p27GHpKclBGHwijUUr6Iqq0lb+rf8ADlk5alZOFSyJy5Y2RTtFXfYldjd7EJcqTRJSbckRtoiUZe05UtxzcW0tyriYxa9kuXSz637vUkr6l9BoB7G5a+wpxULMY9TQsmJ2EPWwiwbjAYdC4EtkgsABY3AexuWAeqLaHQaLDLBND2LXJK6sAywAAbjLD1LAAB0A6AG6OhFmBbUlJdNSnQliXFSeq1fyX+ZhKNSm2pb6shgFOnGXf/geZiMthKlGbfw/13KspYms6sdtU/wMPKpKlUox2kvyKVD2MpNboqTUptmKVacYxp738v1M6wUsLWpt7Kz+9FWDco1eyt+hg5wrUpSlreL/ACOWFKCj0uYpqhRc+xKn7SSmuxhsSuSVNvXS/wCJhnJ1Z32urfcTgnGxpGPKhJyiriuYCzxEGzNo875uiuU1pYqNxtFHSxGak2uxLZA1odTQBgItqNdgGgLagKzB6skro5bRKiTsiKSRPYqtFZqMku5Vle5f3bGyuygkrMy6nKdVW6EaTUrrYqRk3GxysrU05Rl/WhCXNF3G/dQ1bQUVYdraCq/u+VMatJMc3KSl1IxTbk1qcsX0KiS2LG4dB7Ckm7CY4qeg1qSQ0hbiF2MVKUaTcNxLQ6AzoG4HQYD1QMAAAAC4NgWNR7DVySY4kth6o6BoywwABDGAMAAsAaHQIq7Mpw0ZKrJ/Zi38zB4WNFR5d5Ihl9WpJabf13I5fCnKEGtGnf8Aq5iIKjalS210/wCHMllSVV+2X9fIrU60J1pQ2bfbZ2MdFxatvoJKTfMyrGdKKjB7u/3meQpY9pNdF36fcVU3BoqxqeykoPWzsSoRrU48yvb8zE4114wh1grfiV5z95xepGtWqRTT95en/DFFNRvLcg7xSZOmrmFwvsnN3+J3Hd6FSq6c4zp9CeNlyJLZFJKEeZiu1qdBRSehLYTuiWuxozc6D2GIvcV7BuPUWwWvoNWACWiNx7ia5bE1duwopbE17piKijTu+hUd7MqNWsN30HGTikylSU4RVtUZfh/YxbfUbsl2JcsU5dbEJOcFLujGJ8sUkU1d2N2T3YvhIu5T6lZXSE1ew9UrFS9tBJzaQ6Oq1JKzsdDQsjD0uRy7tsaIKwwluLmdRroW0KUeeSj3MTQ9hVlSvezsaFroaN9joWLFrha6DcLHQ3NkWAAAQWDqW0OhYtcsGj0BoJJMtfQs7sUk20AWAauPYAugAGDAYB0Ap25kYGpGHxOy6lOq3ivd2RhEuW73v+jJyWHpTUndvRfMUlCLjP4kx4ZVKijB7mLUaGX3e/8Aw5mmJTw0JpdX+hyqzknuaxvcw9WNVSfb/hjExhTpKHUoRjUrKktyvh1RlydzC4Snz1ZSla6/Ep1W61VLoUk/r7iirGU4rk7ojrsTg9V2Erotd3MXKUOVJbsrys+Upz5oRTXQVoozLDLD1VTTvon94lZFroemw1oLY6AWsIYtjodCFKKWo4R9okezh2Jwimrdyvug3AkWHuXsPUKj9xmIgpJLuSrfu15F24tlJJtFODm2+iMqpxqNxfQjT5I2Y1zRsVWpJplOCjBRRjpuKj5lONkkK7kSQknEei0KezOXmsJe82O5UV1YpRtchNybRU+Jlrj3ELdDVyceXQ0sdQtqMV0Xb1YFgsAdDoMsD2BoCwAAWuFjoW0AsD2AsDRJHUkrkoEldEXJzafQLDQAAABuPUB6gAg6ENWYROcVGO5gYKnGNOW7jzP1KL9lhorrr+NxYONOlVcHrOz+4yzD1PYSdR63/IwUlhsfyN+5y3+d38ziC2Jwq9k/iVv6uVITpU1B7Iowck3F+6k/vJVYpq+5hYJScY9ShyKoo1NrlBQoTWIXxLT5Mw2Olimp1d7v+tDHYWKcqd7plajSpYOLi9VdfKyt+pl+GftnUXUvHnst+v4mGnGVOpL+V2/AouUqak+xyNFCKdVIxOHc6k2tipTbqKT6FGPKtDDJc15FWTlK7HblQ1qNaj1VhdjYYMvoPUjsbkFeSK99C7ve5TbcHqRbcldlSnzyRODi7BuPaw9WNaG5zK9jVs0bJzbg7j/hlZK4l7ti1k2YKVsHXV9+W336mSJqo5fL8htWaOZJWZWfPXjJdCNWN9OqJ4VYiUYr1JP3mdOYl72ov4ZF80Smrpo5l0Eupj6dCnJKi7q3nv8AMhT52kYvDKhJQTvomUV7xP4mACH7zGNXRFNMYJa3FYWozcW7GaAWA3GNDAA6AA9QLFixoPVFiwxhuA0mOKsNElcabbJR5lYYFgAAsAD2DQ6Fh7AB0FozKsTLD1lURCUJ041F2MG7ws+rb+SMqrKbqKT15mkU6qrU3JPVXRjMQlXVOT3uQxCjDlT0SMyqxrUIpqz0/IymcKHtsM9XLb5u5j8NCnOUXun+Rl+YueJlyLSFn/WhmFCNSlzRfxF24+RWoTWKpuL92zv80TUpSUm9v8jFUFKgrdTCYKvHFwhTV4pO+3r6jgoSvbfX9CpFSpzgtLlBOnS5ShONWPI+lxydOsn5kqijl82t1/mV5yhTjHz/ADMLJSgmijJQnd9CeqG0i+poxJWHbcVmi1waGC1QbCanEqQ5XYpa02Q+JFSpytIrrRG5oNpsWo9iV1JMVncjoNNlS7jZ7ntVyuBKMroopNNPsVIe7ZGHjeKiZfh3Tp67mIhJxTj3KzvSbZKbU4pGCbdKVV7EsQoRtHexFtu437o/hH8BR2sR+FlJrlZRlzIcryJT5NRVOaWrEop6FTWTLHU0C1wkiNlFoS94sDFsLSwGX03Xxdan0UL/ADTX6XOgAWuAx7AAMAA3AtcsANHQGaAA0h6krN2GtQd9LAdAABDQMGAAAWEYWSTMvqyrKMeiHgKapXi/fSb/AK1sZVeliVbdtv5lFyw1BuO93+gqlSm5y6t/gZbXp0acro4gcamKU4PRpffr+ljK6EZYhN9F/wAExU/b1HIbhRaj0ZPE/u1TIuq8Zy/Zt/XmYij7KUPNGPhChRTtra7MlSxa12sVKNOcU6fRNf1clhvZ1XLul+FyMbLkfcxOKWHScldFKraWm6JQWKq0l1lZfMxeHdKkqTZVrwjR5bXcotelykuSKijGSqRpxcO6+65jKPspW+ZN+5ch70bk9CL90qtNq3YsA0WDdnQWpCTUlYxC2KXwMj8SMRuiv8KLDGkVG4xujdWOViTjHUUndk7uI5e6YhO10SfNIw0IpXJr3kYBKTS8zDyi6tSMepXqTppQvvuY2upU4wj13+RKi6Kae9vz/wCGMLVthH6/iQl70m+xGCUbon8J9lHS5h6Eqzaj0Tf3K49gV7C0G7kYc2iKcGtWSd27D2GANanQikkNErlgtohotoCdthWcWBudQ3QA9QNwAAaVgCxuWOgBugsjQNCyGkNXHBNpjjdj20HeysSV1ZDc+VaCabt1ACyLDQNAW0DcYDAAouxkFRqpp2J1/Y49wk7LW7+RlsqLlNyXvN6bmMxUVC3XsZjhqsqEZpfFcy6inVhSnL8PxMdKnTo1JXvy7feUsVTo1FzztzXS0vrsinRh7aMJuyb9bGNw6qpK+2pipzVSLRia8KWB9rH4kU8fUxVeEZbKOn9WJVY1Kc61daNcq9fl5J+RgHKMvZUna/6a9SNZ0sLO2jbsUuapGMZP3l/wRwV2fVYV5LnenYo0eZSnFa6X9EYXFQm4Tg7paoqTlVpcz6jw8Ve+6IJNEcO50ZSX2Vf8Uv1MVz16brPpZGLbjR93cw0r00nuSim9RK2hc3QtRrQsXL9C2oiK95HJG97FaScrFL+GyHxIxCV0V/hQx6Ia0I07xRZcy7FdxU3y7FriWrkVPeRibxdolSUuVK5GN3YoUlL3UONzDUY06PN1KdapSV+p7Wdabk+g3KUlbfoYluLcJ/FZFSMacPZwd7pP0fYw0udzT9BWUbXJPUdrFlaxTrSpfA91Ydmh7m6Ha40rIjPlZKrdWQNaFixa4thprQjBz0iPQkW0Eh7EbNpGZww8KqVB3Vl33sr7+ZpYewkNpC1AC1g6DADcACwBuB0A6DEM0ANRoGiyGkNaDRGNnK44tNtdSVragbuxY5bjQ46DQ1oWBjAN0dAAi7My/MJ0ZKUXr/XkV69CvgXVk/f26/8ADHOlCHLv1KanX92T6fkYuFWnh4c/w9CoqMsCqq+KN+/VletH3oy6lfBynUhN7atff/wCpW59yVirQjJWIUZKbvvoiglCtOKVtrfcY51qmEhQT0U0+nZlKo3Z9UYqNSvNQT8zDUpUYOb7k6MnD2nS5SUI1FzLQw1qTcXs01+GhhJ8qil0bX3MwzqVKSttqLL4Roc8uif3/eQTbMtpVMRVlh4bzVvu979CUn9XcPMxdJ1IKK6GGdSFZJ9SWxuxIvZCNxs0DRo0QtRyla1y2hdrYG7jk2boeoqbbSFK7sibUVdl1JXI2ZJ8rsTklYnFcrb3JN8xSp3kUo6OxyK6SMIlObh2VydNucddytGVFuJSr8lTmfQxcpV6jmyc3FOb6O34GSuNSnUlLuy6shu90fV2qcZdzMsLHCz9n9pb+o9EVqinNSHudC3Q3OgkMeoXBD11Kc+Ruw9WHQHsU3zNMnJtsepCHPJRGnF2KkFK1xDCwbmh0HqAHQYDANA6m4M6FgdjcDc0ALD1LDjoculiUdT2au2WaZUhzRaF2LXQo6FtCwNXHHQ6D2DQsbnQCxYBDdkQnYpV7xUbiq2kkUsSpRXSz/MzXF0ZYWNJO7079mVswovCexUbStrq3106dhUXUqr0JY1Yflg1dtNL53/zKftZqdW3W1u1t9epiMVCSpxpw1ej1/H/AIBKSjKJVw/tZOdPd6+hTcpJSktVozEV4KPNN6XRRftcTCK+F3/Qy2tVw+MdWcbpPvYdRShOL2kyr7KGAjFSvzO60a20HQklz20KkuaCIYWpPEQUVo3+IsK0uVbNL8DHunHDunbp+hhIzjF82phKsqNdTi9SpdaEld2Pq6UuYkNMe4tUhLoMaLXLIQxbD0N0APUbsezmkpNaMlWcqjn5WsQThv1KjjOEn2RhJupRujVJsqOcmpJbEoynKLWxjeRQVnqQ3uULMowbjyE5qM3HsYasqFT2j7GDrUlUc57JO3rpYrV1Vq3UdXfT1JxlSk4SWqLq0W9jMaVOcUqPw/13Moh7OjKL7jSWgmt0RrSUUn0K1aVWblJ3bJI+zc5lcdKSgpvZm7LG4OzA6CVzCYGtied01flTb22W/US1sNaiabaRYDdER6hFtSTQ02PUpyjNXQAwtc6AAwOhoAzQ3DcYAjQ3EO1gLGg9SxYsBZDico43OW6OVHIJAA1ckrMYDWgAG4FyWwtiLsyM7u5QqWg/UxmJU6icehXqKSfdmHqKMbPoYqpHEVeZL4UvwRjalqM4x3epl2GXJCUtm7FanTlPTZMwUGoSn01RPC1Y1XN7Mr0nWw0ovqZemuVdtCth3Tk4sxMFg4QhJ3dr/i/U+tSqUKcVsrv77EqilDlRJcq1MtqQUlzGJre0bhEzOsvYKa6jipUXNeSJVY+0UUVNUipuNXiSRayJEVZWAlqLYdrhoE7taHRADRLexKu5qMOwvdVxzvqKd58goKmnYS0sQ5WTsmYp3k7EVfQwVPllZ9DD29pcrxXtZMpNSnFPqVq0fYSiu9zBThSqKpJXKlZ1Kjb6k5+415kW+W3kYVKMUOV3YQNXY9Uxr3bDV2Oo3BR7Glx2L3B7HQAwOMnhnLl+0mvv0FuPViVmMEruw9Q6B0GNpopQUE0jS4hj0EgNwACwAFkbgaGh0DdBZHkWuWBoA6AWAtcLAaBoaGhJaDBFkWRuBudACWwJopRbg5LoRmuWyJOxJ3dxbHM7io8tadRPfoZFWXtXh6nwQf5+mpUhH27cX7t/1KeKbXsrehja3t8JTSWq/wAiEKlHD3eq2+8wMnHli+5mFRvDzqRet19xiZU5uEp79CEU43RgcMqvu3szNKSp1HFO9iHvQ5EYPEunRk2ryey/rQx9NU4qEZX+VhzfslHsPCKWJVZPZWHfl5jE1OWnKS6IwtT2tFTta5LUa0HZsQmErWNLGghjR0HsWSp3AZd7EqS9opX1SIO6uyLUloW3Kmitci9ComldFSerHWjTg4takIOMFcwy5pJsp0FCqovXoVIpVpR8/wAi6m0l0MVVjKnBJbaClyRu9z2TjSVdPraxW96ELdWVakVHk5ba7/Iw91BRNG9DYjqyck2kgeqHqNaghnUOg7G4N2NxWY9UdDQZpYGjSxoLUqycIXSuLoy1ixuPY6G47WEG47MLXNzoaAFgAOh0LHQ0Oha50LGgbnQAAADcAGSV0MaC2g1cYDDqBLVCtYRRq8tOUe4pWJyTDQN0UKcadV367mZUX7ZVI/DYhKVWNOUd4JX/AK+Z7CU6fN02fzJxTy9UlupXfpaxXfsFdmXc2IoVoS6Wt+JjcvlPDwq9Iv8AT1IKyjbsYWvCjCUo7tK3qV6kqk5Se7uZdh6sK0nPZpf8EjNwqtvZt2KUnKryR3at95By9kr7kNzVaCjdns3FcpKLWg1Ya1GdBDWo1Ysw3DdDKmkYxL2GcpCPNoYP2cZVHPdxaX3p/oVG5zUYdLkFaNhtRfqTp8zTPrEZzcV0NHB3KkbSKyvMhedNeRhI2lcxGJdoz6oqVYclRveW33mEw0qsHJbIr2u4R2TucvMhxcqaiU5KUo9kYyso1U33KV3BPqQ1ZIi0j7Q7XFsPVCvYsDOgzca0OoPVD0QhOw9TqOxY6ljoLU7DcUpen43X6FSSirgDH0NwOgG4AdA6AbBpY0DodC1xggAALFgAAsbgBLYGKxa4MBjLFhAnoWEHQaQJXRoLdFWDlFGCqPCYtV4b6Jruk07dbapE8xeKV7WS/H8DG4uNFuT6tGZ0VisPCzsmn+hlFZ0lKnfdW+49rbLZwk95fkv+CWtCxRrSTRWbVSUvmYiEZUlWirKS/HqVo2jFsm/Z2qediblytopRhKjUcnqlp96v+F9x25blJ2mmzNJUJ1VKh2V99/mSaaJbDSuNiCW5ozyJbCehqEVd2K+s9CxZkYEYt6Ix0ozgo0o6pf8AD7kadSlGKT9TmTVkV7upG3RjkmKlFTcvO5Ke/mVqrtqJwVKd99LGEi21HuYatGN42vfQcIzkk9jFzX1jl6K6Rg0lg3Ubsk9v1v8AMrUVCTV9GVKijyKK2KNdxnfok195G0ZXvuNPEJ2drMpq6V3qULc6ctitJSnJrYWwty1xPQZfQWpewwfkX0ACT0EPYAvdWAsIY37qJRUlZhbU0NC2gncA3GLYA0A0LINyxYGjoMaGtCx1LXEWBgPYALABuFgsDWhJEtg3CQB0ACnDmkSikhxLI3LDRFpI3DmdidRUoTqT6XKcm8DGrD7X/DmawlVpKK7ojWTw0FHz/Qo4OawNCv8Azcyfylp+BVbdorYltcSd9CpeUI3Fiqk6aodFqYr4uTsSd4KLHGPs33KdROvCl/Nf8FcaTINXaL+62VIoltYktCSR1NCW5uhrQ3Q9joKzKSvNE9WyzvYkrLTcy+nB4uHtNrpf1YcfZVrdjMYRp1oSp9Vf8Cc4xwtWT3SVv/Akis4xw8ZLdsp2tzMnNKOhzOU5eRK7VzEWSIe8ihLlvIpQXsFNbsx1owjbsRh7Xmk/s6/ic7lBpPRIxH8CHdjVmipTjSwtOXWV/wA7HsE5JFCEZKSXQpv3rMW6Q0rC1RfU6lgsM3Q0PVDLaljdAM3QLQbi0PYQLYirIsM6WDodC2pZMJOzsHUex0NA6gFgtcADQADoWGg6AAAAAM6B0Dobk9hlkJjGAAG5Tbi9B6olHTQ5eaxQw7quVuibGhm+g4Ow0gqRUoNMUWoKC2I4R1Jqe6j+tzNsUsNB0qH2o2+5rTXzH7eFC6+K3luYL2rwkJVN3uVpxho+rsQbUtBSfK4P1MHRdbEKC3eiKkHGbjLdafcZbSnipyjL7N/uSuSTUUyhUlRn7SDs9V96s/wLqxZJ3JW5S2g0iyuyotTluxbD3NyyLXQk0aDKWzYlpdmrXuiSQqTcHLojF1VUcbdkjF1XU5W+isZhFzpWj1aFGMYScmKU9Wn5oxUJc0Kl+/6EUmrrcnNKCRiZNooSSpcpSTk0r6lKXs05dRV2lcd3MnNJNI51ysilGi29yk+b3pPSxGS9rcjNJXRTSlqhL3kbkdh7i1GXRbUaVjdDRoD2GR1aRWpuErFky2hYqPljcpyUoKSFqJLl1EjoWItJM30NzyFsWuVKbgl5nQbG9NDUYaMFqBbQAsFhgbh0LHQDoalzcN0AAdAAsBYLMNToSWh0GjqPUYaAHQ3RoKlFQi09yFB8vN0I0FOagnuRprkduhRbTdidHTQcSxzNqw1pcLXiYOjLEVFTjvZ/gr/oUpuCcV1ORVKq5u5XjFTUV0HyvBRSWtyvTi5amDwqqK7ZmFOlTowkt3f8CFez03KsG5JmGqNVqkV2/Qn70Euxypuw7LfoilUjVgpLqWTReysSWlxpWJK5YN5HQZYaB7lODdGUl0LucEyjBxpJPco4KeJrRUXtuKo1Hl6Mqyelism46Gk24kqd6bjcjFRio9ifvpIppSkkYqynJJ6In7zsUVdpFKi7Ka6FRc60J8sX8ySUqjaKybk2Q1gjEU0nGn1tcpRbhfsWtPl6/wDAX+YqTk0QgopEtrj3LD1Qti6N2XHroaJErNDsMlsJELKRXbrU1U7aG6GWuVFzxcSlFRppLoIV2AbAdbgtybckl2AtoAWA0ZYLFgBXDQ3BgGgbHQ3QtS4wYABZWBlrmjDoG5uW0LDSsSWo9hCGAFtQNzB04SjK/Rfqh4uo6MaF/cve39alXE/XK01CNlBaa9PwMRClGqlBbpX3/r7hOjByurprTcoYb2tB1F/WxLCRjTlfdFSKT0Ntis6ji+V2ZF3QrWMHiJ0KnPB2dmvvVv1I7ktGRxE54qNO2lrsoYqUKDpPa4kpw5UZdhoLCSnPe7t8ipzKUuZ31f5l3cjWTw7g1dtrXt308zWMiGrZiZaxXVIrQ54y16P8TBUnSw8IJ7IwFejTnL2kbpprdqztoyrWjBpy66CV4jV0SRONySHuJFk0D1FF3KWDc5qHVlSkoN0n0/QnCk0pte8vyMNzXXtHdX9NL7fcYmrBzn7NWi3ojCUuWDS31ZG9F06nZ3+5mKqKpOUl1dypK0lbqV694pRR9XXLBrd/8Aabn6FSooMnK8RL3iELWZg581l0K8+Wcl2K3s/q14rV6kK0YwlC12+vYrtwdnuhVbQcUtX+RQpq9+5h6SjDyYoRbvbUhBIGro3Q1cTELY6mpF21IxumwQNXGIi7RJaII2bJWewlY3RE6HQBljcFuPQNwOgwBWAN0PUtoWCxYAsWC2gweoG50GtAACxYZubjQWOgWLDQ7Mkh7G4PUB7AXubsRhotuxCrTVCWu6sjKszVKu7LdW+63kYurzVG0PHYaOHkqkrS0S0et/T9TAOSg1HqtDC4SOYYf2zdm9O+z+RmGElSxEqa1sVVO/uoUXJD00I2sRaSIpktylZTb6kn7qKVCvUqqEI3vG+62WhXxChhqdNdE/xdytK8h2RQmk7Mg4c1pFOLdRLuVbykU6bS16mEnCm5c60aa+ZiJShBuCu+xi8C6/LGWlrP+tSEbaChYnG7Fqxoa7DQn0LamiRhKE69eNOCu2YjG3xKqdVZfcUabq1HfqVrXfke0pqjy9bjbTsU58kZJdSpVcoxXYle6JfHoKKaHPla8itPnlzmIi3JFWd7WKNNzkrH1WdGTjNf1sL93h4rrqYirSk0k/XfclOHsYLrqKm0lJGYVo1KzcXcpyu0ilFJ6bkI3iiyb0FqOwtRWsPUbL6CaE9R6tBfQ6huIe5a6NtDcaLaWBsQhK4MYbBpYWgdCwWGG4eQxIC1ywWuJAAFjoG4amp0NQ1AAZYsdANxha5YtcRY6D1RJWGgALAixuYe10Yel7VWvoiEYzrRp9DErlk0KHt5e8tn+Gn/AATJ61KlVak7NfqYJxVOUk9m/wATHVJTxck+r/4BWjGFd04u5TvCd2Yqm6eKlRa2PZpxaKeFqLDz5H8OvyucsnKLuSjZMjHZkPeRlNWrOc4xfdehjKictOmhI5LxuUGvaxvtcjHmjcwlf2GIp1Wr8rTt6FWlKWJnVvpIwGFVKDjHa9ypFKJKS2JSdSzfoJXkRi72HGxJK49hotoLU5ew9UkYatKhNVIPVDnzy1FVaG7smr7GJrxox5pGGpyjFyb1eolK7bZKaSuTlCUrpE5xhTJ1uaxVa5XJMdkiUXHlfcovlqIq1JVVC/T/ADMXH2VJW6oocr5m97DfuvyIzsvW35lZLnbIU3OSS3MJCUKkpdxaLUgtBG4kIGtS49EIdjpYaNx6C2EC2JFxD2GNXEjQL3QOwW6HQsFxnQZYYWGWGLYYAAWLBa4dDcYWOoFugWLFhoLDQ0WZYsNFixYGrhoMLE43HsNAFgsdDSx0sU37pliamrq61+5J3/C5gqkHiotrS5mUfZ4qUH0Kcm6ifQxmEhFxrU3rJJv5XJY6ccDz095P8kiq6sMRL2i1Vvyv0K8qcsbF03sig/bS5SnSliqc6r1lFaj5nF8ivK2i8xyqwVtrooRTdpdTE+zjVfK7ocHdeZh0pT5UUqccupylDXmb/wCD3MwpuhWdKW6t+KT/AFHa43fQjbmRh5RVOSfVGKws6UYSe0ldHsZuo6aXvdjDqVONpdCVGpLD+1S0vy/Npv8AQ5GpNPdEU3HQox1uNJbblaFlZk4jjoNLoSjoKi3Z9hRaZJJIV+hBvUnZRu9iULWJaIr5fOrQ9rOPuvRP+nctZJE2kitCTh5mGhy0m5v3uhV5ZQsTstR3lKxUS2XQi51GovpsUacak4KL16kKTlT5+2hjcS6kFfdaFOlKmk31RTgnCfp+I5RWjK0IVaspU9tzC1HTndb6/kYOL69R2aSFtoLUsIY7c1xiVtzcS1LXQxg7NCWhbQsRTbRVacrLoWEtB3bSQwGAPUNLGgWuNMex0C+gdBhc3QG4bgDWgajsdDcBgzqFjoAAAdDQHqag9QewaAaDsSiSGXAACEmoyXf/ADMHiakLqL0s196a/Uoz5ZKxWjCtiOaez/yI0tbRMQpwwiqy+Fafr6lVJ4KMYvZt/fYdZynGPkYzCTg410tEU26Sj5ooPmwkm3ZybRhMFKLlOfRO36GIptUk5fIpUJutzN6JfiQwrlXdSWzViK6mCxFOjSqRcbylazvtZ6+pjcdHEyjFbJfj1Ma+erKQ9RanJUlWhGO2tyNOVn2HOU4wg3toiVZ+19otz926S73ZJuyitt/mVpR5VO+t3f8AC36ifu6GEjz+6KmtLmIq03QVO3vK5VpSg1fYe9xuKdiEUpRTKsn0MLVnKF57jnGauiOqUkRs0r7NkJ4WnOpTSvHpuv8AglHCyrU5Tj0HFJEswvg1huzv/Wn6kmrD953LylNp7E4pxJJc2g6baaIJxcmydzCVIQqKUijGGk13Kk1CKgnu7mMtJt+bHU2i+mgptRaJLldyFRp6GHVKdGcpfFpbfvqYePvMdrWOggtcaLDSY0PRC1Y9haoIzTm12JOyITUo6bDWpToucHJdCatBFinJSTt0Bg0HQBh0OgAzRh0GgsC2LaiQbhYBK6LdixYLAbDLAwAAAsWGjqMDSxYsPYsNDQywWHG6JxGABFEtxlCKv72xR5owkrbr9TB0XVrRgkSw8lBvorkZcsrRRXarUVSl8JSdJzlTT02W5C+8VdJ2b7Mw1Pnp1Ke6aaXTb+upiqLglLuhexSh7Z2Ta79/IdSlJQcO2vqY2nCeHg09eUp4dyoNt6piV24roVKDp0oSfVfcSilJns7LQqU+ZXRKm+hGk9hwjClFLSWtyVWXsuXo3f7iDhNKUXsOy66mHwVSpRqVo7Rt+OhUpTho9yVPmjZkHBUHF/FdfqUZOKTJO+iK6aloZvjKVenS5FsrPcs7MkouS7r/ACLy51d7E8O401KW7JRdn5qxRo8sbLZElh1g7395PbX/AIYqNKyKUk0yli5UJuMepzNq5GlJVXNvRji3oNpLlQ1qu5VlJS02G29UKdnYmoyt5E2u+gk7NlK6jZHOpKyK1p0o27u423NibkrIqpvUirvQwWHqSpOdvh3+f/BKEbvQauwpK8WyQloND2JND2HdqyEGg2KKT03MwnJUJKO/QwNF0sPGEt7BRrclGVPv+hJuQtjDU5U4NN7u4PY3Re4ywPULaDRuWdtDcdmFgHqM6HkILXHqILAdDoAAdAC4zyGAB0GDuHQubhuG5c3AAHsNEo2YAEXZlSPLJm7Oe8UjDuMsP8zJqsKc5c+2v5H1xzw86fS9xStex7ByptGFwU6GIlCXdswOKoV/aR5fibe76afoYWPPUlBbNMzWEVQjDqrL8EU6Tbin0bJ1YtQUfsoyqpH2kebZp/kUZQxNRxtpYjQvWrdkv1SJX+qzv0sSXPFNEo66Ek9LGIfs7NddDktOxUk5SuWXLYyjLmv3MfN/h6k46LuQqunp8/uMdiVXbml0X4E4PRdynQ5oOXZkU5Ky6Eou69DEO02u6KqXMvITSfKRheVylL9/b5lO9m2RTdSMI7scoLCVaMlaS1X+RmVBxhT5XulcqLllZmL5ni4xj1ViUlFpCqwWG9m1rcxEXKLSOZx9SLcoolNKxiFqSRrcVpTS7lemoT5SlFSbiyhJOjPvoUZNTj6k3y1bdEPWRpGZNXkylC5Rje6Rho2RBJysypTjDYpRTieyiz2URK7sVKaUboY4R9nfqyNCNtSdNRmkipSUVdEKalFtj1IzUqjj2JRTkW0OoDFsEtgOgEoOMVcaBiOjLXFqWOhY6gw3YtUCGIdhAG4aGhugNDQ2DQ0DcAALnQHqPU3AALB5ADVx7E43Q0MLoexclUfKkQvzNltCnUklZDqudBQ2tf8AEwaVOk6S+y/zMPSUmiFKnOlQg3qk/uOIvY0abe8/n2VvIyWo8PgYU56tt3+//gmIpxoRduuyMbVp0sPBVFeTb/rQnJwpx5nrojAJSqe89Fzf+kv9TL8HTr4eLfX/ACJz+pUnUitbpW8vxMFWnWjKUlo2vusZo5RrtdCg4puLIxjJtEsD7GnCTeslf8Wu/wDVz2UnO3Qq4aEcPzx1d2QlGajpZsUbq/YpVZwkpQ0aKi5rWJJuURxcY+pKq4yiluio2oNoyqqq2HVS1mVaSjThNvWV9PS3+ZWtOmlbVEk3KzJYaLle2ph8O5xklq0m/uKWDrzxcGlpe3Q+o81SpFS+BPpvb56GYxlFQnF7MklVoRU1729/+AU5xVVOWtrjruc5uRSkqnvkaUJUp3+Lp5f15lWq8PSjzK7VvImnJdiMIcxO/NYm7NX6Fabq1pu1kNJR9m1qtSUm58yJq09OpXbqb7k46XRhIqUIQ6tmKlGlWShurp/e/wBCSlKLXW5PC8tKNS+/6CgpNTZKK9nzW6lKmopNdSgrpFLcVkysrxTKekBSbdipJp6FNXkS1ixk0koRK0pKyRzOU02TXNFopfAyMbvQlR5feK7jKWitt+RfQHsjBYRYiE9dYxcvu1Gm9EJNKwfV5ypSmtkhP3UdBgirUc9zoW0Gix0R1AALXN0Ba6AASAABBodA0AAOgHQA6gAaC2L6HUEAAMqR6lgLGgXSaKkUopoiyfvQsYem5SVjLVGnBOXn+hTqSeK93oZhTp4tyd+35Iw+Ift5JdGjGYn97zPsZy3VwXOnqnp8zG0KlGMYvpYyWftYKK3kn99r/oZbhnToNv4rDwkp0IQqb9f6Rg6ThScX0MclOyXmcjT9DCwu2iC9u+eOz2/I9hGEHJ9DEzh7GyK8FKUX20JRTh7pKi502uxQgvYJFKnT+rTT3urE4NWuYzDqnUaXSxBSaaRB0qeCUY/FfzMXXlToOUd0UJqpQjLqOldu5UVmkUYTjUlLyFOdPFxttuQlLmlPqZhhKVbCqpF9bdexUqyVPX7K/IqRUpc8equRpydSz2sVKnspRhEjJRV7lSPPJoa5tENXloVIpy1MRTak0NuS5UYeDqLXfX8idPldmU6SlLlK2GjCDXZpfeiUbTaXQUuWOm57W8tBwlQlFy+0r/JmDotzUJbMo+zcJKXZ29RuCwFVdbq336iinFWKUEtinY3Y9aYtIIitSqrspJXuQ1uNN6FbSat0GozicrjNIv79iKtzIoQvtuY6nOlN0ZLVbjLCsMw1eUJu3aw9bsbuCqyjTlBbMWwdDQG9QeowaHqgHsdA3C2thbFrgOwbha50DUuBuFhhuMLIDc0AHsdAAAsD2LFhq40WRoxosWLDGronBINxtt6isdCcXOLinuYbDylRmr/BFP11S+W4yHw6GVVJQ96qubX00+RhK050eVLRfqOLpTVWK1bt+Gpi4UVFVIKzV9Ndu57dYeTUVpPf5FvbxcW9ehSwsK2HUnq1pb5mNrxTdO92nbsZLOpSUJQ3T0KVdR95Q38+5Ktz1VBaL8+33GNxslT9mn1sR9lpJa2MRQhUm5RVrmDouCk7GBVOdHlhoo3X6v8AMlh6VSmk1dGLowVNwa0bJQjGTi1oRvGVkYlwhBcqs+vqQfI4w6EI1KCUHrfUq1vaxhGp0vb8O3oTw0pSbT3MLajU5ktRU5c1x01K1yNH2NOLS/pFepzyc7FRKcYonNRpezjs7XJUotXFdUpOPwy0t2t1uRwcnhZWei1t5/0yUHLTuU1BSUUrPZE8O9ZWMZhYwUHJXuVIqVFwS1KUUorn1f5EYXUmnsiTtLQko+0inqV222kKk4vme5Kly0lK2rKNN13OTduVXKV3OWupVbUWr6MhBuTV9CvZKyKVoVEypXdSLU5baL0EpxglLqvwZhU3PyKzU3OLVrdNyhQUUktinHQitLFtSnrTsS2SOqKu5TWgklsct6liq7zZTqSjKxVXvRKjtNMa3MPNwSmt1r9xiqs69Rzm9WQulZu4xrobFRUI4NWXv389vyKkKKwcWvjv57fkaPQY9gtfQlDlSLDLXJRcXZl7huMsMAOpoPUBpHQYWNwA0NAtoDNzdA0WujoAFgCw0DAuFzqbgdAAC6GWJRTJRsywtxWGk2U8Ovq8q3ZpfeYvFewp3vvp+JFKVkU4u1kYOhKSUEtWZVOpRnOhUVpb/LpsPGr6x7GXQqYenUsivgXOq4Ja6/h8yphmqalDe1zB4ZRpPGN6R/r+tDD4Cli8XVxC2cW+vSPqZXQeGpQT3ujN5uFKMV1d/wAilTlPFKa2ZUqPFYmajsrmFu3Yjh4p37lOhFJpFHLlh21F6N3/AK1JRjTp+hXo89mVqTlJtFaNldGLrxhTcp9BpOopPoQxDVJzn2K1VSjFvdGHxMoxuxYuDxCj3RTw9KrBPyMTg1TimVKidosxFOMMNHu9SjFVOVLcdJu0WRjH2Mn1KF5p22KdR06M6b2ZhaMPaa+ZUwtNU3U6xf5/8MTh+7aXX/gle04KL6Hs4Ri5R3IUFy67sWGjTpuF9zEQhTlZPVCnyuXNvsUcLJxSa3MPhnKcpVFov66E6MpUpN9L/gRk4ttdVYpvlrq5jKdNOVujIRtsVqSUfMqxcamphKSqySkVr1afK90kvusRpuErRJRlVqTqT3ZCOtiKRGOhZFHZoauy95Iqq7ErUynpIt+8ElKR7GN7lRpySK61QneBRd1YqwSjcsrDWhHceqLc1kVH0LWZYY0U0nNJmYqnHENU3orfkbjSLD97USLFix1sFgLXLdQsA1YHqg3Gi10NFkwB2N0dCwBuWLAWCww1AOpoaWA0NA0NzoDDoPYLXNw6E43QAJcysV6EKllNXsJKyIS5Wr9Xb+vuMJUlSacN0TjGNRT8jEtqr7QljXUwySeqK0akFGtfW2vzWpTjUhHmvoUq/Ph6tKSumrfiU1VVFU6Ls1f7vmQ9l9Xpyj8SfmQca1J1Eru1vwJVI0KftJ6vZfc7lGPNXfL1MJh/ZYiKJ4aM1JLqUaLg3d6WSFaUNzEKU42RThywXNsjFRjOLaK1GUlyLbqOnRjGUWvzMRS9koqXUiqdKLlUlaPXT/L5FemuVSTumR0jvuU6CqTSjuUeen+7b2IYlwduhL2c/esZnUprC05t+6tPwVijGE3yp6q5LEU4V5Nbf5kaEKk+anpYxGFr0aqcdvkUXFUPaS3vb8DFYf3FOHUll0a8lUit9/VW8zH0atelzX0J05SVl0Kabje+g24xnK+xXqVJwvJ6GLikvMjGTfJ1KNfla5tinKnOktfMxcU6bQsMpSaS2K1O8m4LQrVHUu29TD0lGfMOnqYmg3NFDDydRLoPDWVmVcPKnh41GtHp8xRvF2ORLVkI2QkdCndIkxaO43ccnaxdp6CqPqHtJ23IyTm12G3JXFOSVkxNpqxKbkgSumbHQp86quTenQvcbuPUdrEb3Ymk7jldljc3NzQAtqWuABoaMNAYFi2h0Ny3Y6Fi2hbQ3LFixYsWuho8ixY3LaAbgDRa40W0LFiwFjoMNwAAexuSVmDd9BGW5XVxkZey1atp6+rRWjex1MNFScb9HcwWFVOi6tr32K+Nc2oLdDqRtcpzi3yFKpGtS5JMp4Z0cE4SdynXp4aM5z2ehQzSNCsqclq01/Whh8Suf2ae2ph8VKOHqRS6X/C5iMXKrGMe2phfdqKRQk6ldyFGTg+XV9BKXKubR9SjWi7xXR/on+o3zak1dW6GLXJTR9YUo8r0K9JWszF0ZSqqo3paxOjeLXcqUpzhCK2ufV3JNx2RTko+8ilUc15krNq5WpuEIO26K3LisL9XfV/oe7QxCvsnqSoLEqpUjokn+Cv+hQryp3S6mGqQlT5anUr4RU1yX0ZLmo4eTf2df0MPzJJxWiv/AJlWo6lKPLs0Qy5yvr5/1qfUHyeyWj/rzMThHZ07/wBIxGAn7BWe39dzExlUd+phGnU87HsvaVpRvtcp4Vxil2RUpczirn1blu09XoVk6TcXsyolzMpNXscrepWpX1ISjThKT7M9nKVNT6MxHMsPChPaLYqcNddiUr2FYSEriVkN3FqdAaBq6NkKyfmU5JU5R7lugPCuOFWIezbX3Jf5jSuJ2Q7CdxQvTb7FhbEYNvQqRaaTGB0CwWALAAG4WHsboGWOgdRAg3AAZuGoAwexug3CxY6gAMNS4NgHQ3QDHsFWN0IOhCTjsXU4JdiUbaGCl76iurKso0sPTiuzJSjKTk11MU3Zi99XKUW4819Ue1lUwztqzNcu9rhZJaN2/rcWCnCrz3/qxGEaSeKk9la39f5EakqkbNlPCrljNvQr0LJKJh4WaZKUeQo1vaU1JrcrRlNx5XsTd1YjdxtcxVlNsrckneBSs6c23sOEXqitT99uJUpOMY363f3CmuX3SGDcoqyMPlzUV/X6ksra2/r8TB4CrUjHnezf3XfmLLOS6v8A195UwbhPyKtNVIvl6/5WPZqCkmihLktFGGxMa8eSotv66GKw7aSNItLsTrS9ryvZlaPvaMh7WnieZu6MzpylUbj1Kk529l1FGDbhLuRpKFSMn5/lYdGN27adS+iVyMIxkXuzGwUoNW6FRpteRSjztlOCSRUjdNEaamuWS0KeLVXDezmtlp8lZfoSrfWZyjN6O7+e5ye6aNiIq4o2JbAdBj1A6GmwwSuSfMrHQGuwzEYpVMPTpJW5V97JwcYK/U6EXYnLmaOlgNywx7GgA9RA9gAsDGOx0NywDegtrADRZAGg9SwHQB7B0AGMDcOgWAGHQ1LB0AHsW0GroaswS0Nik0jEUeZ8yMKpQrQkt00ypW9pKTgrpW/J3JShKq7MxU1J6bFKSincw7jKjJPdGAqqi79Cpj54uMqcVpH+uxTasoyMZS5qbT+FL+vMoTckmuxRkqlGEI76kJttRa2ViFO8ddilFQauyK0Y5Rgrt6DgtyonzXRidZpIlCUJNJaainyRk32MGniKU0t7afLX8iNRxlzMx9ejUivZ7Ri+/Vf5lJ+5Foji9kjC1lKMdeh7WNON7kqi1lHYdZOHM9iSp1noyhTUKKimV3Gc3F76lWjUp201Rhqjle/QjOEoPW6KqXLoKSnH3ehNpMqNRnqYi0mTjapzPYxtKSqRmloNTdVXRW5nflOROxZta7jkrFZJ07lam079CjHWyEvdGn0IpWKk3Ck2tSnh6kpxSWrKy5Xy9SwloQWgyWwAxg1dA0tyjJyi211Buy0OgGhUi4OzOSXs1LoVJucYp9NCSfMiKTi2xgwSOgwHqiw7MHYNGgSAuvZ26jWgPVDLXLaA2dAsy2gDA6HQYBcudAN0MuA9jcYBcGbhdlzcsAAD2BlWNnc3LOxZWILVFCrReVzp399td9vyErTRl+LcVyPYzOKdTmXUtoK3MmU4qUb2MJGN4tdEQwtWhXrPo2/zJKM4pChKmropyU3sZZShTqc6fR/iexSdiMnCXl/wCK9pJyPaaWJxhNWexzJsqfEY6lKtKKTMRh3yJMdOXK2yXNyKK6EqE50+dGCpyeErS62Ri5TrYi/LayS37JalGDpu6MFWnO0X1K+JcU0kUcZLkauUsZGKlF7NEqyklFE8XyYZQ6o9uqrXnoYtQnNSXS34FLDQpzk4bf8AAHBRjzLcVO/LUT2K01FJj5U3bUrrmrxT2MYueCsYtt0FEU1GnZk4RjUWpPW9ivFyjZlGLhBR7EtWU3fQqfxZFKNoitYlaxHW5gq3sa0Z72FUcI3RK8pXFsQiBPYLliw0DVxaoLD2HsRaldIsrFhGLlNRjy91+ZVxMp0o0ui2HpoNG6sWOh0AARY3OgWR0AAuXuFho2HqhLQEiyACxoDQ7BuDLBe4MNAeoBY3C2gbgAAwYGjB7ATV42EXSp26i1LpPQpVHzEWucwVK8k+hmmEpwp3sSik9Cmk5JFGUXFlGPO7LexioS+qxUlrpcs5VJJLYy+g6s1B9SlhKNmrd09ynRjSw8JJ6tijemmtxynKagiSdGDkuhh260YyJJydhw5YpdiEarxD5vh0t/W5UhdSa3JR5qScnqVYVpSVlpfyJYaTV0jBOmsLytakEqNdU+kithp2Tt3KWDppXaIYeCWiMTh4sw+DhroTy9af1+p9XdOS8iphnOE5vqVqE6MIzhquvkYmpVpNX6lLFSUVHuSvUiknqXcVGBWpc632X9fkQjOD5b3Kii0m+hWlGyszEzk9EShzPlJ0uaopM1ckcsU7X3LNMaIxvFjot1WaLTsIkldFknoJ9Bo5bsUbiSQE1oB6FxjsHQ6HQauOb9ryeVyMVHbqdbBpYaTYiFpSJqzsdQAugsaAD2KNWNSF0FKnKpNQjuyUXGTi90HQEgsWGdB2GMtqBpYAuD1GOxYYDsaGhZBoAAwAAALXLI2LIABgFRWkSa9krCV0ENBw91SMtqSUo8pjXCpQlKo9EYuioTcEQ1ehQT1MHTlWqe0h0KEJVIJMxGXwnO6X9feSk6UnFb9ChOSqRXW4sGo15yl5EpQcbLZFGEeZsupJopxVkcqRJsqQd010FOOvkKMqlKVWn9qz/wAzEO1PzMDTVSrFPbX8mRrVoP2d9L+RLEVJJWJ4uspK7/Io4t1a3JJ7HO4UeYeMTl7xQxkWnYljeYo14xqXew0lH2aK82oWexXlKaS7Eor3dOhCjJP3epTTU3zdNiMWlbuY2LjB23HXqJpPyPaKSlzLZ2MSlKouU9o5xfcpuptIjewlpqPUceZjheLRZOTsOKHvYSuaWElc3IxuKKQBLYAAACyudBjLanUaumYPLpSw8qsVpFf5Lv5kpJpBa5bQA0sDDqBCCgrIKVSVOanHdE25Sbe5Y0sJALYGh7FrlrD3ALBYBh0C4DNw0OgPUAB7gBfQvcsFxmgaHUYI6APYsFitG6uJ3VihCMk7saJWVijR9vl8pxWsHr6Nf5owdKf1Z14fZaFOeZ1o0Y6U46vz/rp95mGEg37u5UgqWi3KSnKUVb1MLVWFi4wV3Zvt/mUMam1HZu5Og4025ff/AMAxEKc67T0kiVJxqpkKqndyXQundEGk+VIjNJ2Q+ZpOPc0a0MzxVSmqMabs5TS+XUlOSna5XhL2UrbshaFFKK0sYuL9klbqUZTi5K9nZpHs5QjG76FB8rcWipg06Try+HoYpwhyVY76L8yo1LDxMRC12kQwypUXU5t0undX/Ui3HqU6sFfXVbEatS6mVqqnBu5Si21ZmMpOMoq2hDnT5r6DaUXfcoufI77k7XtInCLSZNNVZvutPuKkm2raSJczm12VyLnLobbDcb8oooSsVnyxYlZvTUcWQhzOxOhKm1dbnIrns2yNO4kkgAktBoYLYGBY0BrQ6G+hNOTVjcw+EqYhy5F8KbfotWaGHr1JU3Sjs1f7tf0Ju70FqU9mrdDcGJADB7BYFuMSLaBYsiwFtC1kPUtctcSuWRuWB6gAy5c6AB0A0CwDACwGgPYNzc6AWQGg0bgSV1YTKdTlTIxciV72MBjJUadSktpKxTxdWGHlho7Sav8AIyqgsPyLuY7DN1XNbWMdheaSZSozhVXYpUFDEQl0aMPScZNdrsxWNjQpqVtzG4tzxHtF6mErKtUSZUjGK5SEG5PsU4Weg42q69SPuRs+hT9yKRjbTx+Gh25n9y0JpNomvaKUehGEYUkjF1I06TkTqJ02ys3TtfoUa96kZmIxL+pqPe6/FMqYOMqVNSJNOjy9jMKbhBPvoPEv2EovZL8iolB69CFaKp2tqSmnDmRJXRGTU0kUn9ZqNy6IqSVBu5KLVKMx1KkG+wm6zaRK12n9kdOzcuj1+8nhueqmThZ6ntG5NIlNwrW7nw1LinfYSTRVs4tM9koslTvocll6E6rnGKfRWFE5SwADRJD2GhAwATsypPmlcsFi5GbjF26m5CTiz3OR9zS5sgAsdQOgzcANx7WAOgdAASuMLDAADoA7AbhbQY9UG4AAAaGgaGhYsaAAMNwKkbSI6mWYZVptX2TK6tNop3IyVzD46aUYvoZfUjiaTjLcxWTKVb2sZaW2t/wStg224LfT8h4Nzw6pzeumvZ97GGwslRtzavr6md1JxrU4bxSMXVjVrtpWXYow5XozCYj2kOWW5QrKSafQU+eSS3Ks7Jytclimrabnt0kmUZe1zVN9Ifi3/kJtJIcWouzITThtqYhRnRcZIjCNOm7v4rL01RjOWvONNdNPU+rKmuVH1SCwSnfVFCtz0oq+u/yKlS0EjHT9pT16skqdnDurFS8iK5luVKEIYVVb69ikpVpqEVuyMbOzI1VTpprd3I4mVSTur2/UgqdSldPUVKE4MoYSFJyV+pUhzXUtii4yprtsKkuW6K2Fk3e+xVw8Yx5uo1fVlSCcRQ9nG6ZTm4u5Um3Jjg2h67j2sbi0AAdgJoGhbj2HYAvc8jRgMIR5tCVloC1Q2uaxd7HkNo3E7hbQHqM0AAG7ltAAAFEGWuDVwA3DoaBYNGWNiyLBYLDGtBoACwHQLBY6FrljYsW0B7ABW6FPRXMLiHSndE5e0loSfKrIpNN6kfiMBj3Rdl1/rsYXHxrNJMdNKdluVZxpR5pGMziMWkn/AF9xisd7WbfcgnJ6lOFtCKVOSUhV3yWMJVcJX6orYn2svdKkEox/rsKqnBmWVE8XVk/JfgU5xlYnPlqJPZlSUZIrSTdn1ZiUlFxZhYxlNR6kKMKmI33ZiaUYUORMwWFcm77ojBKEnfVuxiv3jXZMdk2j2kKbdypLl0RVxDdD2ctihTpvDyqJ6q34koJQS6onVi4qPYpzgoTj3S/BlGMqc0+jJOy5WfFqv60Q6kKk5wv8LsKEYtw6InBqcWtjFRckrFdRcT2WhZNjirNiV42RUTjK5GvGzTK0GpWBLqJAAATV0WGrhuNIAsAAtBsbTH2Iys7l02X0Ro2PcIrQAGCAsC3OggALXLFkAFrlgNzSwB0GgsFg3LBqADA0B2NDQNDcGehoAAAG5VjeI6nucpFXdkaQXmPYTsQlcwcpRnJyd1fT0MsxsI1GpLXoPOIyqQit1v8A1YzLH0qtF8r1T8yrUVXV9DrYo3dWzVkuoopJWHKU5OU3qK9kyNbnbb0MPX9nexzRkm5OxKTa5UjLZW5pLdyZRnzMpy9ouVP0Ky9mvdJtSUUu+hiqLak73sYatGlO7WtrfkYmtJYmGm1hYyNSLqqOsXbcopqLlAqycJxaelrmNcI1JRT/AK/4YbjFlRUakZKX6iqWVxypurHm1jdX9Oq7koxUpeyVo3dl+W+pUcUotb21I8zV9mQcYxWl3r+g61KHKoLbX70v8jD8lWfMlZrb9TT2ri+w6Kq1pJUrd3e97eRTw0rJ9El89NRUnJWb0KtByndPRmIjdciepNVOS3XqyV1G6JV1DWxUzFxkuSOvr/wCpXq1Lqw6Moy5WS2ELsCsMNAaDQauNag0geoAAMHqbjSC1xpCI2UlcxXs/bS9nt0BbG5pY0DoaWCxSpqc1FuxVhyScQSDQASuciYkDsWAsAbgAPYDcZ0FuAW0ALDAAAAHsB0NwDoMAaurDVmUY6XKifMGlyL0KUlYhdNNbmGm1iIyfczJ0/eUOrv95Com2VI9SEruxFpIjfmuXbTS2ZCaVOUOun4FOTjzLuht2SKt4K5l0uSlF/Mo14+zb6lFzjUVupOtGSXNuYflnUT7Fbn5pNbEYylNp7kJpYhOp5f1oS9yc0tijVlGnP0EpVU1/KjFRcop9Wv+B+hOOiXVD2Q07EkmlYTlaxVVOy5GS1VyFRRd3sVnGo4qHcyyMYyeupVg3Wbj2/4cxMKiqOUehDExV49EKSc2l6lWUVBvt+hVo+0qRlEnGSkrkKL57GIo04OVytFX5o9BVXfmfUrVXVkmSR0EhgAAElqFg6DHcAAAHYuPUEOzWgLUCwWAE9UTk5MsW0AYlc5QC2gWLAAAAAAFy55AG4GgAAAdTcsBcYD2BgDppyue097yJNORTop4eVQ0YrEXbYpXcU0jDcjxEObq1cxzj7eTWy0QqEadWTS3dyqoyhZblOPvEnykdKTb30FF0/dl1FShTipJ6scV7BSe93+gqfNHToZhWjKk2l0sYWKfLC3RDi4y5ezJ1G0kjESU5c19yk3yuzKmI9k/aJXW34WIY22KVR/1oYipGo5Se/QpVI1Ywpz0UU/mTx2GnRcab961nuYfFclTmWpmFe6jF7r+vzu/nboUJ0FTqOo/e6b/ANfeSbaOZKNiNWyFUViMm9LkU7ak4xaLOOqZlsJ3c7XJ4iaqOnF7L7u5WxMnLktvb8h0VUjKrHS5hpwV5TjqvMqKU4OMd5X+VylTklrt0JUbSk5bdCvzKMdd3YzDD021fdf8MVI+5yodJJJLclBxeo0mJXOgzQDQOgdDoTQAncYdALGlgAGWDcBhYaLaFhACRa5ZCC1xRQAIuA2AAAAAWYWEdLAg6HQGAAWNC2gHQsB0LA9gewywW0LE9Y2Kjj7NJBGbdLlLHQjJWMNUSkr7FNttSXcqVVJWJw5kyqrLQotc6MVVg7tFapGrShy77EptpX6EZNxKLU6Dv30LuEVJehjNaT83YpxtUt12HFqN33KtdN6FKfNZHPa6K1eMqXL3FeNS7KXLKpF93YrYRyryUNlb8R4ajgHNOXM5K60atq159hVE3p1Ksm43ZOauc6kyrBRpwn3E02y1kNNNEajjqXjWjaJGXP7nUwtWdHCyv1aRRx0VWbktypiKLlG+/wAyceSMKcUVVGnFQ3uOvHDVJSbur/qYrH06FKy1f9eRLEylG76nPGdKC6rUxqVWXL1s39yuTV0iTsOo5OxylgLDQAAPUYwGbjAGADGMDoG4AdAASBha4lYAsiwWRYsWDqWADoAAAAAFwAABgWA6GgAaAwOgE1Z2A6BHUqWi427EKjasmSV07DnJaofvIjorlJKdovYg0tOxHlnPyFDZIhZLlOaEIuLV77eRiornpx8/yLtaoUajp819ESunYik6atuVJ2asU6HtlFdyvUhKDUXoOMVEwuOlh6UuV/0kxVKkowrVNeb+uhgZqnXcr2tqYvEVMRP2k3d7E2rCmraEmnYRzpounsc13qRZGqpRs+hUlKdCDvormHxFSkrwe+hi8a6U4xkr3MTOVCjGPUnJ1UpSfkzFQlRlvdFajGviFbRNmP5cPKE4R93Z7lOk4QUk9zEV7xSaMTXdV3XRWHoiwDN0dBhuAASVhgDAAAOgwJNJXN7G49wL3YACVy2g0WuWANwDoAPYC5c6BYsHQAsWCwA2MA3GABYAADQ6Gh0OgaD2LDSsDSCrHS4UYqU0mzG04U6zjB3S67AnYnNyehRnaVmTl7qsXaZBXOe75SN0xLRn1CFLC05c13K729PMw9NSdmQVPllfcmloOLq4qMY9E2VuW6Uey+/qKo6cVF+pdVJIlypvlehOnCrTck/gjf7ilKay5St7z29PyKbprDwUNTFQjLDxqw1S0fk9f8uhgsW60ZtrZ2K8aypUlONlZ21Xr+qK9RyjYlcbbRbR26Deghbjt0G4rRiu9iL90o1OVqBCHLNxkZfgaOI+LW3r/mY6NS6Uo+7te/6blWMUpRT2bMJOhyP2z9N/0MPOhVgnJ8su2rIYSjWjKE3o1qKrOinh5PbZ+RVqudJRa2Jy5WN3ALHQsdQAAJJWB2YDVw0ACwA9we1iUU1YFYAsAFhLQSuWLFgDqWLBuBqPUAOgAG4BuBcEblw6AwAAAYAwubmgg6FtBA9hag1dWHGzE9DoGhe5FajbsmJ8ysVsQp0oQtshqzMPhpVYTmvsq/6EFdpFGDb5SS5qTlfUV5JO5KSctSlVTxs3Ha1vv1MXH2dRx7FSSat5GhQoc7STJp8yXYxOLqQjKD62/Ao62jHbX8Fcwcvav2Deknf5q/8AmfV6cakoT6X/ACMTjKtaMVJ6Lby0S/RbmKoypQp1G9JXt8nYci7kSHZnMmznsOVxu7G7Cl7pQadmU2mmpK6MLhYShVg9k7L8/wBSnVeIrKgtkjEYdU6snHs/8jA89R1FbSBOc4pJ7WKElVfspbMzVU02lvF6MdX2kUT1YaFiw1oW0N0MAAAaLIsW0LA2A0AAwALDi1ow3ALFrEVdFiwwANwOgwZY6jNywAAAAGgAGhuiwFgDoAaGg7BoaGhZBZBuIsW0OgFWPUR1C5FNsUPcu9x1G4qPYUpLYVVshLmRRrTp3jF6PRlNKT0MPJ32uYjDxpRjaWrRGpJSZCaS94wkHKpUqW0vYxMqk5u61Gm4slomYevKEfMm2kYqs6nvMpySj7u5GThJSRRTr06tST1SXz1sVrqMUVKrdNKT22IUZ1ISmlpHclJJaDkNsuTbUdCnUUoLue0HLQTTRQavuYetFTSRiMY1Vc1pFW/yv38iMIRm6lJe80v61K9ChUkpQV2/X1MX7PCVJcq1l/XmYuopR5Y6uO5h8XGDk5au2nrsYmlHEJ87sN8r0HqADFsIsB0GBY0NBpMaLA9jcsbg0AD2GAMCpPnd2iwFhIsiMblhIGAAAAHQ0AAAAAADc0YWRYDQANAALg2aMdhhuPYQO1i6GHQnHmjYas7AxJyaSKdJxXMSqpwcSwmaEXZnNexTm4u5gHGSb6qzRWrObd9y6UirOMpuaKf7tOD6u/3jrTTbvuUqjhPmJa3sOWisYqEI0YuO7K8nFW7inGLt5CdkKu1omYurGUYNb2G+YjU5aLs9yUrgAo2loOzegn0Ein8RSnaVijOShOk/tNfgl+phMReqovZJE5NVXFLS1zF8s5RjPzMJTdPCzjU3d/8AgbGIklLToSqN6jd2NADLaHQQwANg6ADihoGhoaDcYBa4wNyxYsWLMa0LIsKNxRRYsWsDAsAFzoBc6AAaAWA6FgCxoaBYDcQMAA0OgGljQCw7FrhYQwAAsVY2dwjFydkUvdl6HPenYrUnC1+pVw0IYWNS+r6GgnYV3G4pK1iMtEYas4NuJOtFy3KtaEpXirEpxurmJxCqzUoq2hKvBwStszEayc4qyb0EmkOLTSFK0Y8z2ZiZKTbRJtrQctCMOWT13Ziargr2LpDYAF9BOwncjK6sc/Krop1NUx1G2mjDxdalJx3VrmCxNRVHGo9P67GIzDnUuXz/AK2JY7lg09WVJqY2hhYDc6AAAaGgaADjcasMsWLC2LFhoBltDcAVhRuKGhyqw4XFFILWDQBgWANA6AOxYDQADdAAAWALFi1y2hYA3AAACyC10WLFiwwAAJxurDi0yi0rjeop3SRWnzpeQ6jcVELFOStYvZiehCdhSg4NdSnNXV+5iHFT93YVW23QlUTs0SrudBQ87nM0rDqMctERUXJ83b9BRTpt9iS5lp0LqxGKr1Yqe2xWUVNpbILAPUHsGwthajlqRqJso4hxTt1MRmSnThGK2ST/AKsVMXfRFSo5HNoHQYG49QYAAbBbQNwBq44jQNFhrQGBYtoWAQkRWgBuC2ABgMA0DoWVjQN0NFiwAHQAAB6gG47G5siwAAAAABsjcsMAANwqRuhgdAgky6FY3FsJlxTuj2jaRzJx3L3RGqowcS90zmZzNpamIcZW5exdJEJ8kW+4mN3Y0JAA1odCwkJ3IbDQtERlZaEpXALADLm5oOzNCwBoG4INgNDcsNKw4g9S1yxZWAABIigARpYABgA0B0NLBbQshHkMHqAAAADAsAdDWwwDoAABYLFhghoewAAaAypEEdCwdABEdYiu2dQvZCehfQ1Y3Zkpati1HK4W0GgANywhq5a2gh6sAAAC4BoAAMNC1wtcYFhosNFg6DHqWuWCwhCVy2gNFgAAGAMeoABozc6AdAAAAA6ABubAAAB0AGjQDdBoB0DcAALjSaJQtsNAAAU3dEN2J3kX94k7DkXT0Odjkw6GliGo0WucqOVFi1y2hYaLXLFkIAANwQxamgwsAGgABoAWABpDihoaHsDLaCSLFr6CVgZcuG4bgGgwGMDcYAM6G4AAAAAADGFxgAABYAOgFzc6AAAANE4AwLG4blN62IqzZB3kWW5N3egJl9BK5bQsjlQAFkwGtBnKWCxoBuANAhiLajVywbgAAdACwWudAOhuWuNFrBa5bQsgjG4kkG4wABgDANCxbQNCxZDA6AAAB0OgBuFhgG4AAAAAWLFtC2gxDGDdwC4APUHZjgiUQ3ACn8SJaJlP4ip8QK5qxRuWAAAALaAFy6sbhuAAC1AGAAAGgHQ0AOgA0mWQ4nIcpyCikG5bQuXDc0AABgAdA6FiyLItcYAAAdAsHQDcOgdAAaLABY3CwG4HQRug0DoAAFiwNFixYcRwuOLTNwBaMnOLjZMg0pE7SYooViyEB0GbjQWGhgD2LgDVywAAIdgGdDoWQFjcACxYANwaAtoW0AAAAseRY6lhAAwsblhltAsbD1LAAAAHQuXGdBly50LlxgAw0A6AaDDcAEbgD2AADQLBuBcGkyVNdDlYoXRyIlEsXuXEbDQgAABhoAaAAwABAWQ7FixZDA0AOhuhhcC50AAYAAFrgM0YGhoAaDEDCxobhcA3AA0LHQDodALBYCwAbgABYsAzc0C+gBobg9gsAWLBYsFrjQWNwtcsiyHsS1C1xIsNXOgFgA3HqdAAAA0sAFgsAAA9S1wA3HZnQA0NABoAewAAB0Bo0LBYOoFh2LFg6A9TcLBYHqWBgAdANxjADoGhoBYGAADB6hcAC4xbD2AAADcA3AtqPYaGkOJyCiWQHUADcDoM0G0BoAwAAZuHQLaFgAYFtDUuBcNg3GbgBYsh7AABoBuWLXCwWABgBbQsWCxqHQ3AGrmhoBoPYOgbg7FkItoWRoaAaMAAAAAsjQHYsWB7ABYAALFgLFixoWQwAAALIsBYsWANCwAAaFgsaNFgLG6AYDOgbgas1OgbgAA9gALBoFix6AdAABhYsmGhYDctoAGgMAtoBYAADQ0NLHQRsDAAAB6gI3RbQSLAPYAAAA6B0AsWublrjAAACwAAwAAAAGNHQ6FwDoD1LFrmo0G5YBnQOhYAAaDQGaGhbQ0NCx0LaFgsHQsAPcDQGG5Y6Azc3GG4ywFjQsmGgFgsaHQ3AOgAbhYsWLFixYGixYtoWHY0saB0LAWAsBuFiwywWNDQ0sXHqAAGjNA6GhZAMYyxYsWudCxYsWLFix0LaFgOhYGXRoy6A3ALFhmgMsWAC5dGg7WB6m4AMLD2Og1dAaAwLjADQ3AAAAC2gFh2OhudAsi4FzQeoAzSxoWAALAAA9joGxYaHqFiwjQtoHQsmAkWAYAAAABoBYaADoABcuAX0A3AewwLAB0AudAuXOhuAPYANAABgHQ3HqHQ3QwAAAC4AAFkWLFixYsWLFrlixYsFhoLFjoWCxYsWuWRYAAACxYsWCxYAAAACxYsWLFixYsWLFixYsWRYsWLaFiyLFixYtoWLFixYtoWOhYsWAsWRYsWLFiyLFixYsWLaFixYsWLFgGixYsWLFixYsWLFixYsWLFixYLFixYsWLFixYsWLFixYsW0LFiwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAF0wAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAM1oYmjlMsdSaspKHnqr3XQ4erOcakZO7vf7/APhgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAMfTVXhKvH/p4n90b/AKGQ1eTF8v8AMmv1/QAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAACnFS4eqp/8APxf+klFvDYtX+y/yeoAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAFH/AJJ+r/18X5Gd0fZ4yT76/wBfMy+t7bCwn1t+WgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAYf/AJEFb/HH8jiOjeEKq6aHDtfmpTpPo7/eAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAGE1yHEf4omZ0PbYScVva6+Wpk1f2WLj2lp9//AAQAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAwOuR4vycPzDGUXh8TKC6PT80Yasq9GNRdUAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAZbrk2OX/Xv/0oOIsPaUKy66P9Dh7Ec1OVF9NV6P8Ar8QAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADHUszlXk6L93S2q7f5nsM5Wqf4o+uZtR+OF/l/kUuIo3tVhb0/yMPj8NiNKc1fts/uADKNcsx0fKP4NhmOG+sYaUFvuvVGXYn6viYze2z9GAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAVsNRrK1SKZiOH6M9aUuV/ev8z22ZYD41zR+/8AHdfMweb4fEWjfll2f6MDI9cHjY/3PyYGa4b2GKkls9V8zKMV7fCxu9Y6P+vQAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADGZLh6+sfdl5bfcLEY7LWo1VzQ/rZ9PRmEx1DFRvTevbqcPe9Txcf8Ap1L8LAZ9hfa0FUW8fyMkxXscSoPaWnz6f5fMAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAlFSVmroxeSWl7TCvlkun+Xb8vQ4HzGtUxdfDYiNpeynrt23/qwmpJNO6CcYzi4vZmLw8sNXlT7f0jLsWsTh4z67P1AAAAAAAAAAAAAADKskxeZStSVoreT2X+b8kYvDvD150W7uLa+52AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADhelCpmSi1q4yX3occZlUrr3qf9fc/wMJjaOKhzU3r26oOIMHz01XjutH6f8D9TJsZ7Cvyyfuy0+fRgAAAAAAAAAAAAAHDvDU8e1WraUl98vTy7v5Lyo0adGmqdNWitkjiOvCOf4jDJar3vvSf6gAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABwrLlzej8/wD0llemlOUGurRjMonTn7bCOzXT/L/IwGbxqv2Vb3Z/n/k/IqQjUg4y2ZjMNLD1pU3029DKcb9ZoJSfvR0f+fz/ADAAAAAAAAAAAAADh3IpZlX5p6U47+fkv17L5EKcKcFCCslsuxxr4ivDTeDyyS5vtT3t5R8/Pbt3WXYyrUzGNWrJylK923du67gAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABw/LlzSg/7y/HQzKHJja0e0pfmwx+V0sWr7S7/5lDH18DNUMUrro/63X4oznCwxOHVelq1+K/4H+Zl+MeFrqfTr6EZRnFSi7pgAAAAAAAAAAAAZfgKuOxMaFLd/gur+RgcFSweHjQpLRfj3fzOO+L/YUZ4bDS8m+77LyXXvt6+/Vn3b/MyzK4YWKlLWb/DyQAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADskVszwlL4qi+Wv5FTiHDr4It/gPiGpJ2hT/G5/a2Yz+Gl/wCSv/MyXM8zWZ4dyp6c8b+69uZXOJ8fj8PnOJhGneKm7aPZ69xcQ1Yu06f5r/Mp8Q4eXxxa/ElisBjYezlJP10fyuTpYnK5c0Pepv8ArXt67GJVL2jlSfuv8PL5GQ4//mHm/T/IAAAAAAAAAAAADhTJ/qeF9tUX7yf4Lov1f/AOK88+p0fq9F/vJL7l39X0+8zrF+3xDinpHT59TIsAkvrE1r0/zAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADFZjhsPpOWvZav+vUnnWJry5cNT/V/5IWVY/Fa152X3/gtClw/hY/G3L8Py/zKeX4Wn8NNfdf8xRSVkgoz5KsZdmjiqHLm9Zej/wDJUNJqzKmAwtT4qa+7/IrcP4afwNxf3/n/AJksszDDJ+xnzLt/wHoV4yjNqUeV9tiMnGSlF6oy3HRxdFS+0t/68wAAAAAAAAAAADhfKfr2MUpr3IavzfRfP8kZjjqWBw0q9TZfi+i+ZneZ1Zxq4qo7yf5vb5L8jB4Z4mvGmuu/p1IQjCKjFaIAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADGZhQwq9969luPFZhmD5aK5Y/11/yMLkFCnrVfM/uRCnCEeWCsgADiv3sdGf80Iv8P+AAFbD0q8eWpFNGZ5MqEHVpO6W67GCxc8LWU4/Nd0UasK1NTg7pgAAAAAAAAAAAk27IyHLFl+BjTa956y9X/lscYZt9axX1em/ch+Muv3bfecRV7uFFer/JfqcPYa0JVn10X6/15AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABOcYRcpOyRic2rYifscGr+f9berMHkcIvnxD5pfh/wRRSVkAABxF79PCVO9KK+7/hwAGk1Zma5e8LVvH4Xt/kZTmX1afJN+4/wff/MTTV0ABm+aqinRpP3ur7f8H8jAV/b4aE+tvxWjAAAAAAAAA4RyxYvG+1kvdp6/Pp/n8jiDM1l+BlNP33pH1fX5bjbbuzMqzrYubXey+WhhKCoUIU+y/HqAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAYvF0sNT56j+XVkaeKzWfNP3aa/r5vz2MNhaWHhyU1YAAAM29/KcFU8pr7mAAYnDU8RSdOez/q5isLPDVXTn0/HzMmzTltQqvTo/0/yADNs0WHj7Om/ff4f8Ebbd3ucO4i8JUX01X6/wBeYAAAAAAAAHDmW/UcvhBr3pav1fT5LQ4tzL63jnTi/dp6L16v79PkYmr7KjKfZMyyj7bFwT73+7UAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAzDMKeEhd6yey/roYTLquLn9YxfyX9bLyFFJJIAAACv+84epS/lqNfergABmOAhi6VvtLZ/wBdCpTlTm4TVmjJ8150qFV69H38vUAz2h7PFuS2kr/oGX4n6viYz6bP0NwAAAAk1FXbE1JXTAAAOG8v+u5hCLXux95/L/N2Rn2YrAYCdVP3tl6v/Lf5D1M8qcmDa7tL9f0OHaV6s6nZW+//AIYAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADMcwhhKd95PZf10Mvy2dSf1nFayeyfT+ui6AAAABhf3mQYiP8ALOL++yAAAzXLFioc8PjX4+X+RKMoys9GjKc29rajWfvdH3/4P5gcQYfnw6qLeL/B/wBIDJsX7fDKLfvR0/yAAAADNP8Ac6nocO/x5+n6oAAA4Ly/2GCdeS1m/wAFovxv+BxrmHtsXHDRekN/V/5K34hxHU92nD1Zw/T5cK5d3/wAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAx2MhhaTnLfou7MuwM61T63idW9l28/wDL7wAAAAMk9/AY2l/dT/8AAW2AAAZtlSxC9pT+Nfj/AMEacXZ7oyrOFK1Gu9ej/wA/8wr0o1aUqb2asVKcqc3CW60DLMZ9WxCk9noxNNaAAAAGZK+DqejOHX/tMl/d/VAAAYTDTxNeFGG8mkVZ0cvwbl9mnH8lovmV6069WVSbu5Nt/MOIZ3xMY9l/mZXT5MHTXlf79QAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAxGFpV+XnV7O4AAAABwt7+Jq0f56cl+QAAAZrlMcQvaU9J/n/wAElFxbjJWaMszl0rUqzvHo+3/AIyUkmndGf4X2ddVVtL80BkeO9rT9hJ6x29P+AAAAAY6PNhai/uv8jIHbF27p/oAABwTgfa4yWIa0gtPV6flc44x3s8NDDRes3d+i/wA3+QGcy5sbPyt+SKMOSlGPZJAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAcMVfZ5tRfdtfemjF0vZYipT/lbX3MAAAMyyqGKjzR0n37+pVozpTcJqzRl+aVMK+V6x7f5GJjSzHCP2bv28mv6sNNOzCjVnRqKcHqjB4uGKoqpH5+TAAxWMo4aN6kvl1Mvx6xkJSStZ2ACtHmpyj3TMllbGw+f5MAADhTBfVssg2tZ+8/nt+FjiXHfW8yqST0j7q9F/m7sDFfvcfJd5W/GwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABl1X2WMpVO0k/xRxHR9lmlePnf79f1AAAAx2X0sXC0tGtmYvBVcLPlqL0fRmGxVXDz56bs/wAzGVoVqntIqze68/L1Ay7HywlW/wBl7ohONSClF3TAOEKcJZVBtJ6v8zxUjGOepJW9yP5yOHatqk6fdX+7/hwAMJ+6x0F2lb8bAABgcK8ViadBfaaX+ZmmKjgMvqVI6cqsvXZfjYeoGH/eZhH/ABfqAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHFa58ZCsvtwjL8LfoAAAAV6FOvBwqK6Mxyephrzh70PxXr/AJgBlOZvDS9nUfuP8P8AgdxNNXQHB1nlUfV/meKytnsf+vcfzkZVW9li4Po9Pv0AAxv7rHTfaV/xuboAA4JwntcfKs9oL8Xp+VzjrGctGnhk/id36LRfi/wAZla5sbT9f0AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADOf3uW4Ksv5XH/wABdl+oAAAAGYZHCpedDR9uj/y/Iq0p0pOE1ZoAynNvYWpVX7vR9v8AgCaaug4N/wCRVH1Z4sJ/25T/AOvcf/SpCbTujC1lXoRqLqv+HADPIcuMk+6T/C36GEn7TDwl3S/IAA4Jwvs8BKq95v8ABafnc4qxf1jNKlnpH3V8t/xuBUdoNmTK+Oh8/wAmAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAD/fcOrvTqfg1/mwAAAADFYKjiYctRfPqjMcsqYRp3vF9f8AMAyvNpYdqnU1h+X/AAPIhOM4qUXdM4Ls8s/7ef6Hi3h74yjVXSKX4yDh7Ec1OVF7rVej/wCD+YAcR07VYT7q33f8OZNU58FDyuvxAAMspxwWV01L7Mbv7rsq1JVakpy3bb+8CvpSl6MyRXxsfn+QAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAZN++y3G0P7qkv8At13f6AAAAABWowrU3CaumY/AzwlXke3R9wDL8yq4SVlrHt/kcAYqnicpc6b05n+SPEjDe3qxh1cNPVNtBl+J+r4iM+mz9DRrQA4ip3oRn2f5r/gHDtS9GcOzv96/4AAGCofWMTTpfzNL72cTV1Qyqq11VvvdvyAK/wDCl6P8jI/99j6P8gAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA4Uknj3Re1SMo/hf8AQlFxk4vdAAAAABi8JTxNJ05/8MYvCVMNUdOf/DgHhJK+TVV/08f/AKTA4/xFKnjaEJOzcXb7zNsN7DFSS2eq+f8AwQyXF+3w6g370dPl0AM3p+0wU121+44eqcuIlDuvyAA4Voe1zWl2V39yf62OOq/LhKVL+aV/uX/BQBVV6cl5MyV2xsPn+TAAAAAAADBcNV8bl/1mi7yu/d7pdn3vcqU5U5uE1ZrdPdAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABk9f2GYUanaS+69n+BntD2GZVof3m/v1/UAAAAADG4KniqfJPfo+xi8JUw1Tkmv8AggeEUr5biI9p/wDtq/yPGCH77CS8p/g4/wCZVxNSrCMZu/Lt39Ay7FvC11Pps/QTUkmgKtNVKcoPqmjLajpYyDfe336AAcC0ebGVKnaNvva/yOOq3Ni6VLtG/wB7/wCAANXVjKvdxtP1/QAAAAAAAOHaHscroR7q/wB+v6niRj5/6xzVKVnCMY6enN89zKcVVxOH56m97AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAACdtjipKpiaWJW1SEX8/wCrAAAAAAGKwlLE0+Sov+AY7L6uEnaWq6MPCCd8Nio9nF/en/keMMLwwcuzn/7Z/kAZFj1KP1eb1W3p2+QBmNN0cZNLvf79SlUVSnGa6q4AcCUbUK1Tu0vuX/BOLavtM2qLtZfgv1ADD+5mEV2lb8QAAAAAACMXJqK3ZRpKlSjTWySX3HFuJ+sZ5iql/ttfJaL8EZLDlwUPO/5gAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABmH7/I8NW6wcoP56r8EAAAAAAFeFKpBwqWafczHLo4d81OScfXVHg9P3sZD/AAP/ANLPF6F8DhpdpNfev+AAQnKElKLs0Zdjo4ukpfaW6/roBxFR5a8Z91+X/DmSVvaYOK6xuv6+QAcG0uTK0/5pN/p+hnFX2uYVp95P8wAxX7vHyfaV/wAbgAAAAAAGTUfbZhRh/eX4O7MRVVKjKo/spv7kYybqYipN9W3+Jl0eXCU15IAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAMs/f5Ni6HWNpr9fwQAAAAAZ3JxwbcXZ3Q61Vr4n945Sas2B4Q1LY7Ew7xT+5/8E8WqfNktKXaov8A0mQAYfEVaE+em7MwGc0sRaFT3Zfg/T/IM/o8+F5l9l/8A4drWqTpPqr/AHABw9FUcoo3/lv992Tk5ycnuwAzmPLjZ/L8kUpc9OMu6AAAAAAA4Qo+0zWD/lTf4W/U4jrexyqvLyt9+n6knzNsoR5aMI9kvyAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADhSpH6+6MtqkZRf3X/Qq05U5uEt07fcAAAAAZlhZ4nDunBq+m5/q7iv5o/j/kLh2v1mvxFw5PrUX3C4bXWr+H/BPDjLIYPNJyjJu8Gvxizj7A08Zk0oTeilF6fd59zH5TWwvvLWPf/MADAZzVw9oVPej+K9P8j2lHG4eSpyumrehgqzw+KjJ9Hr+TACX+z5L/AIaf5RAA4ghbFJ90v1Mulz4Sm/JfhoAAAAAABwJRviatTtFL73/wDj2v7LJ5Lu/yTf6EY8zSAAAAAAAAAlJLdgAAAAAAAAAAAAAAAAAAAAAAAAYHEPDYqnWX2Wn9zOJsMqGaVbbS95fPX87gAAAA00AAcHT5c1iu6a/C/wChxTDnyisvR/dJDSasY/IoVLzoaPt0+Xb8vQq0p0pOE1ZoAKNepRmp05WZWqOrUc2tWZViPb4WLb1Wj+QEYuTSRxE1Syitbtb72kABxJD3qcvVfkZHPmwUV2uvxuAAAAAABwJSthqtTvJL7l/wTxMr8uChTXVSf4JfqYaPNXgvNfmAAAAAAAABxFiOWEKSer1+7+vwMhw+Mx2WTxbjeNN2b6vTf5dfW/cAAAAAAAAAAAAAAAAAAAAAAAADPf8AaMBhMWt+XlfrH/PUAAAA4W4eo46LxFd3inbl7vR6+WuxxvhI0q9GcFZONtPJ/wDBAAOGJ8mbUX5tfemjPafPlldf3W/uVwMVgqOJhy1F8+qMflNbDXkvej3/AMwAMuzKeDk7K8Xuv8jC4yjiYc1N38uqDAw58VSj3kl+KOL58uUzXdpfin+gAHEUL4eMuz/Q4dnejOHZ/mv+AAAAAAABwdS5Mqi/5m3+Nv0PE6teTh2h+bf/AADL1fF015oAAAAAAAADM8T9YxUpLZaL5HBeUrL8io0Zr3pLml6y11+VkZiqKxdRUFaF3b0uAAAAAAAAAAAAAAAAAAAAAAAAGB/2rIsRQ602pr02f4X+8AAAA4DxGlai/Jr8n+hxvhvaZfGot4y/B6fnYAAyafJmNCX96P5oxlP2mGqQ7xa/AAauY/I6dW86Gku3R/5fkVqNSjNwmrMAKdWdKSlB2aMBnsJ2hX0ffp8+35ehkqU8xw9v5o/mjjaVstS7yX5MADOafPgp+Vn+Jw7UtWnDuvy/4cAAAAAADh6n7PK6EfK/36/qeI9XnxFdduVfkZUr4yn6gAAAFScacHOWy1Mpr080xkcJh37zvvotP67GM4XzLC0pVZxTjHVtNf8ADgAABmuK+r4aTT1eiOE8oea5vRw7Xu3vL/CtX9+3zM8xqwWX1Ki0drL1ei+7f5AAAAAAAAAAAAAAAAAAAAAAAAAcK1oxx/sZ/DUi4v5r/gW+ZiKMqFaVKW8W192gAAAHB+I9lmkY30kmv1/QzrDfWcvrUuri7eq1X4oAAoT9nVjPs0/uNGivT9nVlDs2vuADE4SjiYctRX/Nehj8nrYe8o+9H8V6gAcC4utHO8NRTvFy2+V9DjqSWCpR7y/JMADE0/aUZw7pr8DJqnJjYed1+AAAAAAAGAp+zwlKHaKX3JHHFX2lXEv+/b7nb9DJlfHU/n+TAAAAMf8A7rU/wv8AI8No83EVL0l+TOInbK6/oAAAGdYxV8Ryxfux0+fU8KMk9jhamYVFrP3Y/wCFbv5v8jjnH806eEi9vefrsvwv94AAAAAAAAAAAAAAAAAAAAAAAAGHrSoVoVY7xaa+RxVQjHHe3h8NSKkvmv6fzAAAAwOIeHxNOsvstP7mJprQzPC/VsZVo/yt29On4AAGDqe1w1OfeKf3ozql7PMa8f7z/F3AAMfklKvedL3Zfg/8ivh6tCfJUVmAcBpPiPC+r/8ASWeLdSUMFhnF2fO/yMDn20MR9/8AmiE4zipRd0AE/wDZsY/7svyYAAAAAAUoe0qRh3diySsjiep7SjOfeV/xZkivjYfP8gAAADMXbCVPRnhfDm4gi+0ZflY4oly5RW9F/wClIAxVX2VCc+yZw3UnnNdYWFlUs35Oy/pGKwlfC1XSrRcZLv8A1sAZrjPq2HbT956L/P5GV5dVzHG08LS+KbS/4JhMNQy7BRow0hTj+CWr/UzHGSxmKqV5faf4dF8kAAAAAAAAAAAAAAAAAAAAAAAAAYn/AGzIKVX7VGTi/R7fogAAADIcV9Zy2jUvraz9Vp+hxthPZY9VVtNfitPysAAcP1PaZXQf91L7tP0OLKXJm1Xzs/wQAAYunh507V7W89DGUaFOpajPmX5f5geH3/JSYX1l/wCkSPF//dcL/il+SDCY+vhZXg9O3QwOaUMUrJ2l2f6dwDPKXJjJPuk/0/QwNX2uGpy8kAAAAAAZTT9pj6Ee8o/mjEz9nQnPsm/uRn/+6fNGRq+Nj6P8gAAADNnbBVPT9Twop82czl2g/wA0cXytlNRd3H80AZ9W5MJy/wAzt+p4R4Lnx2IxLXwxUfnJ3/8AbTH5dhsdS9nXjdfivR9DNcHDCYypQhK6i7Xf9fIJNJXbMyxjxVdyWy0Xp/wTws4e5YSzOqtXpH9X+hxnmXsMIsPF+9P8lv8Aft94AAAAAAAAAAAAAAAAAAAAAAAAAcLyjWlWwM3pVi7eq2/V/InCUJOMt0AAABwLiubD1KDfwu69H/w34nG2E9rgI1lvB/g9H+NgADg6rz5VGP8AK2vxv+pxzS5cdCfeP5N/8AAJzjCN5OyMTnuGpaQ95+W33/8ADn17MsZ/BjZf11f6FPIatR82Iqa/e/vZRyfB0vs3fnr/AMD8DHZNQxCvD3Zfh80YrCVsNLlqK35P0OA2lxHhfV/+ks8Xo3wGGf8Aef5AbbGBz2dO0K/vLv1/4P5lGtTrQU6bug4jpfw6i81/l+pw/V58M4fyv8/6YAAAAABw5DnzWgvO/wBybM6nyZdXf92X5HEH+6L1X6mR/wC+R9H+QAAABnkrYKS72/M8IqV8fiJ9oJfe/wDgHGsuXLEu8l+Tf6AHENbmrxpr7K/F/wDAseF2A+r5J7ZrWpJv5LRfkx2SMbX+sYmpV/mbf3sM+x/LH6vB6vf07fP+tzh/JqubY+nhqa3evkuphcNRwOFjShpCC/Bdf1ZneYvMMbOt02Xotvv39WAAAAAAAAAAAAAAAAAAAAAAAAAGBxUsLiadaO8Wn/XqcUYWNLHupD4aiUl89/x1+YAAAHCOL9hmcYt6TTj+q/FW+Zj8LHFYWpQf2k1/kSi4ycXugAOBKt8NVp9pJ/erfoceUbwoVOza++z/AEAr4mjQjzVJWK+e1KkuTCwu/wCuhHKcZimpYmdvLd/dsjD5ThKGqjd93r/wAAq0adaDhNXRw7lDw2fYWrSd48606q+nzPFunfJ6M+1RfjGX+QAYfFVcPPmpyszA53RrWjV92X4f8D5/eZ1R9rg5W6a/18jh+tyYh0/5l+K/4FwAAAAADhGHNm1N9lL8mv1OJp8mU1n5Jfe0jP8A/c/mjI/99j6P8gAAjCU5KMVdsaadnuAcQythoru/0Z4QUvdxVT/CvzOOp2wdKPeX5J/5gNpIrzlisU3HVyen5IyjAxwOAo4aP2IpfO2v4md4n6vl1ap1UXb1ei/Fhj8ZHC0XN79F5kpTq1LvWT/M8PeGVlmB+sVV+8qfgv8Ag/1uzjLNvYYdYWm/env5R/4O3pcAAAAAAAAAAAAAAAAAAAAAAAAAArf7fkEJ7zoOz/wv+l9zAAAAoVZUasakd4tNfIoVo16UasdpJNfM4pwX1bM6llpL3l89/wAbgAcCVbYqrT7xv9z/AOCcaUfaZZzfyyT/AE/Ur4ilQhzVJWRWziviJezwkPn1/wAl8yhkU6kufFTu+3+b/r1KOHpUY8tOKSAADK58mOoy7Sj+aPEHARxmSyg3a0ov9P1MZgK2Flaa079AAMLmdajB05e9B6Wf6f1Yw1Z0a0ai6MTUkmgAAAAAOCoc2ZN9ov8ANI4wly5TNd2vzT/Qz1Xwb9UZK/8AbofP8mAAGW4r6ri6Vb+Vp/Lr+BxJhVh8zqKO0veXz1/O4BxJLSnH1/Q8JqPLllWf80vyOPZ+7Qh/if5AZxiPY4SVt3ovn/wDgHK/r+e0U1eMPefy2/GwccYr2eChRT1m/wAF/wAFoqVIU4OcnZIzDGyxdZyey2R4ecKvMcT9brx/dw/F/wBf1sYvFUsJh5VqjtGK/pfPYzDG1MbiZ16m8vwXRfIAAAAAAAAAAAAAAAAAAAAAAAAAA4WxMI4uWGqfBVTi/Xp+q+Zi8NPDV50Z7xbQAAAHB+M9vlqpt6wdvluv8vkcc4Lnw1PExWsXZ+j/AOD+YAHCVb2ea01/Mmvwv+aOKKVSrk2JVNXkotr1Wv6FDJq1eXtcXLXt1/4HoijQpUY8tONkAAAUp8k4y7O5xPBVMorJdk/uaZOEZxcZK6Zj8h3nh/u/y/4JOEoScZKzQAGW51GnCNKtstE/8yE4zipRd0AAAAAHAsb42o/7v6r/ACONpWyxLvJfkzNqM6uElGCu9PzRluXYqjioTnCyX+QAAGef7RgMHiuri4v1j/nqAcQzviYx7L9WeGlD2WQxfeTf4I47nfFUodot/e/+ABn+J9pXVJPSP5s8J8p9lg6uNktZuy9Fv97/ACDjPF+2zH2aekEl83q/zS+RnOZe3l7Km/dX4v8AyOHchr5xjo4emtOr7Iy3LqGX4WGGoq0Yr7/M4uzr61W+q0n7kN/OX+S/O/kAAAAAAAAAAAAAAAAAAAAAAAAAAFOcqc1OLs1qvVHE1OOJp0cyprSorPykv6t8gAAAOC8b7HHOi3pNfitV+FzMcJHF4SpQf2k18+j+8nCUJOMlqgDG5jRwsfed5duv/AMhr46vmmHxUny04zi/lfX106lSEakHB7NWKtN05uD3Tt9wAAAFZfWclf8Aep/nEDGZfQxUbTWvfqY7Kq+F13j3X69gAMHj62FleD07dDA5pQxSsnaXZ/p3AAAAAOA43rVn5L82cdS/2GnH+9+jAAACh/tHD9WHWlNS+T0/zAM4nz42flZfgcG4f2OSUI+T/Nr9DjSpz5nbtFL83+oYmvGhRlUl0KVOpi8QoLWUn+LMmy6GX5fSwsfspL59fxK1WFKnKpJ6RTb+RxHncq9apGD1k25P16L+vIwWDrYyvChRjeUnZI4T4Zo5JglTWtR/E/Pt6I4qzxYKh7Ck/wB5Jfcu/q+n3gAAAAAAAAAAAAAAAAAAAAAAAAAZlw5jMFSjWtzQaTbXT1X67egBkDWPwVbLZPVrmh6r+l8rkouLae6AAADC4iWHrwrQ3i0/uKFaFalGpB6SSa+ZxbgPq2Yykl7s/e+fX8dfmDtYxubylL2OEXNLv/l/nsYLJVF+1xL5pdv8+/5G2xg66r4anV/mSf3o4iw/sM0rR7u/36/qAAABw9NV8oo37W+66/QqQcJuD3WgDSasY7IqdW8qPuvt0/4BXw9WhPkqKzABNp3RgM9lC0K+q79fn3/P1KdWFWClB3TAAAADgKP+8P8Aw/8Atxx4/wDZ6K83+QSxdSOYKh9lxv8APUAADhv97HFYb+em7eq2/MAX+0Y1f3pfmzJKXs8toR/ur8Vc4oqe0zas+zS+5JBxBi02qEXtq/0PDPJfrmaLETXu09fn0/r1DxKziWAyZ0qcrTqvlXe28n92nzIQlUkoxV29kcCcGxyigsTiVevJf+ArsvPv9xnGa0suwzqz1fRd3/l3MViauKrSrVXeTAAAAAAAAAAAAAAAAAAAAAAAAADCUfb4inS/maX3uxZWsZxwfh8Tephfcn2+y/8AL5aeRjcBiMHU9nXg4v8AP0fUMBjJ4PEwrw3i/wDh181ocUYSnDExxVH4Kq5l69f8/mABbQADgzHe3wHsW9abt8nqv1XyOMsB9YwHtYr3qbv8no/0fyKtWnSg5zdkipiMTmk3To+7T6v/AD/yMHgKOFhaC16vqBwrX9tlVPvG6+56fhY44w/JjoVV9qP4p/5WAAAA4Lq8+Wcv8smvyf6md0fZZjXh/ef4u4A7JGaZrhZQdKMVN/gvn/l95GMpNKKuxq2gAYXGVsNLmpv5dGYHNqOKtF6S7f5AAAAHAa/dV35x/U49fuUF/i/9tD2FP2vtbe9a1/IAADhit7LNaXZ3X3pr8zF0fY4idL+Vtfc7Bi6ns6E59kzJKXtMbDyu/wACjTVOlGC6JL7jNantcdWn3lL8zFYiOHoyqS6EnUxFW+8pP8WcC5NHLsphde9PV+nT/P5h4l5pLMM7+rU9VSXKv8T1l+i+RwHwLHL4xx2Njeq/hX8vn/i/L1MbjaODoSrVnZL+rLzZm+a1sxxDq1Nui7L+twAAAAAAAAAAAAAAAAAAAAAAAAAOGKHtc1ors2/uTf5gYvB0MXTdOtFSXn+nYzfgurSvUwb5l/K916d/z9ScJQk4yVmuj3Mo5czyypl8vjh70P1X9d/IlFp2e4AYDhlPJp0pr97Nc3o18K/z9WSi4txe6ADhPH/VcxjFv3Z+6/Xp+OnzKtONSDhJXTVn8zNMmxkszq0MS7QpyaXmujXqrO/mUqUKUFCCskAcCYi9GtR7NP71b9DjnDc+Dp1VvGVvk1/mkAAABwHWvCtS7NP77r9EcY0fZ5rKX8yT/C36AYzHUcLG83r26mOzSvitL2j2X69zBZNXxFpS92Pnv8kYTAUMNG1Na9+pjsqoYpX2l3/z7mMwFbCytNad+gAGAzypStCt7y79V/mUq1OtBTpu6AAADgRL6vWfmvyOPXrh1/i/9tDGZlh8LpN69jC1/b0Y1O6AAM7xVShCm6bs7/oZLmEZzo4mPRpvyaaucTUfZZrWXd3+9J/qGeVeTBtd2l+v6HBOF9tmMfWK+96/kVJqEHJ9FclJybb3ZnmO9tV9jF+7H8/+AcFZMswzGnGW1/y1b+S09WiMYxioxWiMzxiweDqV7XcU2l3fRfN2RwfwUsNUeY4/3q0nfXo3q2/P+nrosTiKWHpSq1ZWit2Z9ndXM699oLZfq/N/hsAAAAAAAAAAAAAAAAAAAAAAAAAAcDUObG1Kn8sfxbX+TADM8lweYRtWjr0a0a+f6O6KuQZhk+JjisP+8jF9N7dU16dVfvocUYGnGrHG0P4dXX0fX79/W4AcMZksbl8eZ+9D3X8tn81+NzjHK/q2L+sQXu1Pwl1+/f7wAxmYQwcFNv3ui63MgzNZnllHF2s5LVdns/x28jjnL7Shi4rf3X+j/NfJABwZifZZl7P+dNfNa/ozP8N9YyytBb2uvVa/oAAABwTW5MwlD+aL+9NP8rnHlC1SjV7pr7rP9WGY53GlenQ1l36L/P8AIpUcRjKvu+9J7v8AzMDktGhaU/el+C9ACcIzi4yV0zH5DvPD/d/l/wAEnCUW4yVmgAw2LrYaXNTdvyZgM3o4m0Ze7Lt39P8AIAAA4E/3Sr/i/Q49/iUPSX6BxD/vS/wr82ZHU58Gl2bX6/qAAcRUpypwmtle/wA7HD2I5asqL2eq9V/wPyOKPfr0a389OL+YcR1dadNeb/T/ADPDfCc2LpSfdv7lp+JndX2WXV5f3X+KsZtj1hqNov3nt5eY9TwryN0cHPMaq96p7sfKKev3y/K/UKlGFS3Or2d/n/X46mIxFLD0pVasrRW7M/z+rmdXljpTWy/V+f5AAAAAAAAAAAAAAAAAAAAAAAAAABwtneHy6c4VlpO2va1+nbUo1qdamqlOScXs0ABmOW0cbhZYeasn+D7/ANbmacNY7AXk1zQ7r9Vuvy8wDhfNFgcclJ+5PR/o/l+TZm+XQzDCToS3ez7NbP8Az8itSnSqSpzVpJ2a80Bj8dTwlPmlq3su5gcBUxNT6zitb7L+unZfecD5haU8JJ7+8vyf6P5MzPAxxuEnQl1Wnk+j+8qU5U5uElZrR+qAMsxP1bGUqv8AK193X8BpNWZjsM8NiqlH+VtfcwAAA4dr+xzShLu7ffp+pxvRUstVR/Ykn8ndf5GaZy6t6VF2j1ff/gGXZRUxVpy0h+L9P8yhh6VCChTVkAAY3LqGKj76179THZbWwj97WPR/1sABgM7qUbQq+9H8V/mUa9KvBTpu6Aw2IjiKSnH/AIZ9QA4F/wByqf4v0Rx5/Go+j/NBxDFrExfdfqzhypeNSn6P+vwNGgAK9GNalKnLZow83hsVFveL1/JmcfvMrwVXylH7mkgzit7XGTtstPu/4J4dYPkra/Zhb5tr/gnG+MhhcmqSm7JtL8b/AKGLxU8TVdSX/DIyXKquaY+lg6W83a/ZdX8ldmFwtLC0IUKStGCSS8krBisTRw1J1asrRRn2f1czqcq0prZfq/P8vxAAAAAAAAAAAAAAAAAAAAAAAAAAAMtzfF5fPmoS06p7P5fruZRxPhMfaEnyVOz6+j/TcAAzThXA428orkn3W3zW35PzMz4dx2AvKceaH8y1Xz6r5/eBwvm317BKM378NH59n8/zRxnk1/8AbqS8pfo/0fyDG4ynhaXPLfou5gcFUxVT61idey/rp2+8MBi54TEwrw3i7/5r5rQo1YVqcakHdNXXozjPLfq+MVeK92p+a3+/R/eAGTYn6zl9Gr1aV/VaP8UcZ4T2OZe0W00n81o/yX3gAABCr7GSqX+HX7jimhHMOHsQoaqUHJedlzL8kZXk3NarXWnRf5/5CSSsgAAxuOpYWnzT36LuYzGVcVPmm/RdEUaFStLkpxuypTlTk4yVmgAw+Jq4efNTlZmAzmliLQqe7L8H6f5BgsX9Vx06Un7rb+TvowA4F/3Kp/i/RHHn8aj6P80HEdK9OE+za+//AIYy7FfV6kpX3i187XX4owM+fDU5eS/IAAzqj7PGSt11/r5mHq/WeFaFTrGdvvjd/iVqipU5TfRXMHTeIxcE9bvX82cCULYetV7tL7lf9TxexXLg8Nh7/FJy/wDAVb/24p05VJKMFds8NOFngaMsfX+OatHyXX72vw8wx2OoYKi61aVkvvfkvMzrPK+Z1by0gto9vN92AAAAAAAAAAAAAAAAAAAAAAAAABQy3GYj+FSlL0Tt95R4SzWpvTUfVr9LspcC4t/xKsV6Xf8AkR4EpRV6lf8A8lt+rP8AVPJ4fxMQ/wDwKK/Rn+r/AA7HR4lX/wAcT+weHP8AoJX/AIHEwOIwGHp8kMUppd5xdvnv95CtSqfBJP0YAG5mfCmBxl5QXJPutvmtvusZnw3j8DeUo80F1Wq+fVfl5mS5nLLsXGsttmu6/rVeYnSxNHS0oSXyaZxPl6yWrJz/AIb1i+/l6r/gmDw08fW+s4he70X9dPzYBwXmPtsK8NJ+9Db0f+T/ADRn2WrH4GdJfEtV6r/Pb5jVtADgfFe0wc6DesXf5P8A4KZxxhPaYKFdbwf4P/g2AAADOcz9q3RpP3Vv5/8AAOD67xPD+GdRa8vK7/3fd/FIxVB0K86T3i2vudgAAxuMp4Wlzy36LuYnE1MRUc5u7Mvyqpi3zPSPf/Iw+FpYeHJTVkYzAUcVG01r36ox2V1sK7vWPf8Az7AAYHOauHtCfvR/Fen+RmFWnVxMp03o/wDIybHfWKPJJ+9H8V3/AMwDgX/cqn+L9EcefxqPo/zQZvR9rg5rqtfu/wCAGSz5sFDyuvxAAOI6fvU5+qOFqntuFcRD+Saf3sz6v7PC8i3k/wAFv+hw9Q5q0qj6K3zZwhSUMppyX2m3+NvyR4n+0xudUcJT+zC78rt3/BI4e4fhUxMMPSWr3fl1f/A9CjRhRpxpwVklZfIk0ldszzNamYYuU2/cWkV2X/B3YAAAAAAAAAAAAAAAAAAAAAAAAGFy7F4t2oU3L0Wn37GE4IxtSzrSUF97/DT8T/V7IcD/AL1Wu+zdvwWp/rDkOC0w1G77qKX4vUxHHeIl/BpJerb/ACsVuLM1q7VOX0S/4LKubY+r8daT+b/InOU3eTuwApValKanTk011WjMq41qQtTxq5l/Mt/muvyt8zC4zD4qmqlCakvL+tPmAAZnwvgMbeSjyT7r9Vs/wfmZFg8bl18LW96nvGS6d01uu66eeqM+yLC5zg3hsQvNPqmuv6PujF4GrgqzoVI2cf608gDJMxeAx0K19Nn6Pf8Az+QmpJNM4tyz6pjnUivdnqvXqv1+YBwdi/Y5kqbek01891+VvmZlhFi8JUofzJr59PxGmnZ7gAAZ3mPsoexpv3nv5L/gmTZYqrVeovdWy7+focDYjnwdSl/LL8Gv80zivDexzWpbaVn961/FMAAeiMdjJ4qq5y+S7GVZU8S/aVPgX4/8AjGMYqMVZIBpNWZmuUUIQdWnJR8nt8v8vyAAMJiZ4etGpHoUasK1NTg9GBwL/ulX/F+iOPf4lD0l+gZxicTQp3pxTi9H5Bw7O+HlHs/zQABxBDmwqfZ/5nAU+fK8xo/3Yy+65nmJ9rinFbR0+fX/AC+RHG/VsGqNJ+9LVvt5etvuMmwf1PLqGH6whFfNJX/E4jcKmcYiot20v/AUl+n3nB2U/V8N9Zmvent5R/4O/wBwcYZosLg/YQfv1NPRdfv2+8AAAAAAAAAAAAAAAAAAAAAAAAoYatiJ8lGLk+yVzA8E4uraWIkoLtu/8l9/yPYcN5V8bVSa/wC3n9y91fMxfHE7cmEpKK7v/JaL72YvPMxxX8Sq7dlovuVgAAADCYzEYSoqlCbi/L9e/wAzK+Nac7QxsbP+ZbfNbr5X9EUa9KvBTpSUk+q1AADiDIqeZ0Lx0qR2f6Py/J/MrUalGo6dRWktGgDhHMvreBVKT96np8un+XyOIss+v4GUIr346x9V0+a0+4Aw1eWHrQqx3i0/uKVWNWnGpF6NJr5nE+C+q5lUSWkveXz3/G4AAY3FRw1F1JfLzZgcLPH4lym9N2/0IxjGKjFaI4IxPJjp0ntKP4r/AIFzjzDe9Rrrzi/zX6gAA8ndXHyT0hv9/REYxjFRirJAGYZxSw14Q96f4L1/yMRiauInzVJXZGLk0oq7KuX4mlT9pODS/rft8wAMix3JP2E3o9vX/g/n6gcCP/Zqq/vL8jj1e9h3/i/9tCcIzi4yV0zM8ulhKmmsXt/kcOTtOpDyT+7/AIcAAzmPNgp/L80eH1Rr6/BbujL70Sbk23ucO4L67m2Gw9rqU439L3f4XKlSNODnJ2SV38jIsLHO8wbTvG7lJrtfb57fj0FFJJLYnKMIuUnZIzrMnmGMnW6bLyS2/wA/VgAAAAAAAAAAAAAAAAAAAAAABgMpxmOlahBtd9kvnt+ph+E8Dgqaq5jVXpey/wA38rGI4twWDh7LLqKt3asvu3fzsY/PMfjbqrUduy0X3L9QAAAAAMDmOKwU+ehNr8n6rZmRZ9iMclGtQaf8yT5fx2/H5AABj8pweOjavBN99mvnv+hmPA9WF5YSfMuz0f37P8DE4Svhp8laDi/NBw/mf1DHRqN+69Jej6/Lc0a0OLMr+p45zgvcnqvXqv1+YBwhjfrGWxg3rB2+W6/DT5HHGB9phoYmK1i7P0f/AAfzAADNsXLF4hUqeqWi82YHBxwtBU1v19QyTE/Vsxo1OzSfo9H+DOLcL7fK5tbxtL7tH+DYAAAO1jM86bvSw79X/l/mPUwOS1a6Uqnux/FmGwVDDRtTjbz6mOzKhhY2lrLt/WxHAYjGydSnTUU/kvl/VitRqUZuE1ZoATad0ZZjViqCb+JaP/P5hwG/3NZea/JnHq0w7/xf+2gYihTr03TmrpmXYaeDzD2ctmnZ9+v6AAGb1oqhKl1ab+SaPDt82cul/PCa/wDJf+AHhbgfb557ZrSnFv5v3V+DZxnjHhchxM1u48q9Ze7+pwPw4smyyMZr97P3peXaP/bq/G4cZ51ToUlgYSXPNXa68v8AwWreiYAAAAAAAAAAAAAAAAAAAAAABgMrxePny0IX8+i9WUeHcsyumq2Y1FJ9unyW8vy7ox/GclH2WAgoRXVpX+S2X4mIxNbET56s3J+YAAAAACTbsjKuDcTiUqmJfJHt9r/gfPXyMDkGX4NL2dNN93q/x2+VgBNPYAAxGGo4iHJVipLs1czHgjD1LywsuR9nqv8ANfiZhkeOwN3Wp6d1qvv6fOxwlmf1vAqnJ+/T0+XR/p8jP8rWYYGVNfEtY+q/z2GmnZgcF472OOdFvSa/Far8LmOwsMVhp0JbSTX/AAfkVqU6VSVOas02n6oADOcb9XockX70vy6syHA3/wBokvJfq/0AMNOOYZdFy2nHX5qzKlOVObhLdafcAAA2krszXN3XvSpP3e/f/gEKcqklGCu2ZdksKKU62su3Rf5sMxzRwl7DD6zf4f8AB/IwOTRg/a4j3pv7v+CwxOEo4mHLUV/zXoZjgo4WryKd/wA16gGX4x4Wup9Nn6EZRnFSi7pnATvHEL/D/wC3HHivRoPzf5IAnThJxbWq2AAlJRi23ojF46datOa2lp8v6R4dz5eJcN58y/8AJJD4cxHM05JfeeGORywOFr15u7m0l6RX+b/Ax2Ap4x0lU1jCSlbu1e33N39Ugx2No4LDTxFZ2jBNv5f1p5mdZvXzTMKmNqO0pPTyS2S9F/mZPmlSvL2NTV9H/mAAAAAAAAAAAAAAAAAAAAAAUMPVxFRU6UXKT6IwXCmHwtP6xmc0kvs30+b6+i+8zDi+NOHsMugoxXW35Lp8/uK9erXm51ZOUn1YAAAAAAcHZRhvq6xkvem27f3bafe+/b5hj+Jcuwd4ynzS7R1f+S+bMXx1Wlph6SXm9fwVv1MRxNmtbes16WX5albF4irrUqN+rbMqzjH/AF91cJVcKce20vVbP7tF6mW8b052hjI2fdbfNbr5X9DD4mjiaanRmpLyAAGk0UsrwlGu69KHLJqzton6rb57hxflX1XF+3gvcqa+kuv37/eBh686FaNWG8WmvkYXEQxNCFaG0kn95xnl3sMYq8V7s/zW/wB6s/vACcowi5SeiH7TM8bps/wS/r7ynTjTgoRWiAOC8V7XLnSe8G18nr+dzinCfV80qW2l7y+e/wCNwAAM3zX2zdGk/d6vv/wPzKFCpXmoU1dsy/LaeEh3k93/AJeQZpj5UUqNHWpL8P66GW5bHDR5paze7/yAzbNFh17Km/ff4f8ABJScndu7CMJTdoq7KWU4yptBr10/MpcO1n8c0vTX/IweG+rUVT5r27nAb9+uvKP6nHcb4Sk/736AAAGfYv2dFUYvWX5f8Etc4HyyGFzShUlrNv7rp7f5mMVsRUXm/wAzIcJ9Wy2jT62u/V6/qB4lYyti6Sy/DSVlrPz7L5bvzt2K1CpRly1FZnD2GtTlWfXRfr/XkAAAAAAAAAAAAAAAAAAAAABk3DGJzC1SfuU+73fov129TEZzlmS03QwEVOfV/wCb6+i09DHZhicbU9pXnd/gvRdAAAAAAADhPOqGCoVoV5WS95efRpeexnHE2Lx7cIvkp9l19X19NgDF0MbjGsJgqbnOW9k9F5vpfzMo8P8AE06MYVpqCXzfn5fizCcH5ZQ1lFzfm/0VvxuUMLQw8eWjBRXkkvyAAAMVhKGKp+zrQUo9n/Whj+B8PUvLCz5X2eq+/dfiY7h3McHdzp3Xdar/ADXzSDgnMPa4aWFk9Yar0f8Ak/zRxBlv1/ATppe8tV6r/NafMAM/xnLBYeL1er9DJMF7Gh7SS96X5dAA4IxXs8bOi3pNfiv+Bc47wl40sQul4v8ANfqAA1YM6zPfD0n6v9P8yjQnWqKnBXbMvy+nhKdlrJ7v+ugY3FQw1F1JfLzZlWDld4qtrOX4L+vuQGIlVjSfsleXQxEKsaso1PivqYLIFOClXbTfRfqUspwdPaF/XX8yMIwVoqwBwJL/AGmrH+6vzOOY3y+D7TX5SAAAMXk9PE1XUnN3+R/q7hbfFL8P8jhaiqOYYemm2k+u5Rwn1vN/Yd5u/pdt/gWDOMyhl+ElWe+yXd9P+D5GHyDNcxm6rg1zaty0369/uQ/DqniKfLiqqfotvRt/oZ1w7RyRUqFKbkmnuv8ALuAAAAAAAAAAAAAAAAAAAAAUMPVr1FTpR5pPZIweQ4HKaSxWZSTl0jur+n2n+C/EznifE469On7lPst36v8ATb1AAAAAAAAAy7JcbmErUIad3ol8/wBFdmXcF4SjaWJfPLtsv83/AFoUqNKjBQpxUUuiVgxebYHC6VqqT7X1+5a/gYnjfAU9KUZTf3L8dfwK/HeJl/CpRXq2/wArEeKM7xU+Shv2jG/53Mklnj/31R5fPSX/AJLp+TAAAMdkWX4y7q01futH963+ZguFpZfjY4jC1bx2alvZ76r79lqtw4syv6njXUgvcnqvXqv1+YFetCjTdSWyMFRlmGMc57bv9F+noABleK+q4ylW/lav6dfwOIcIsXllWC1aV16rX8dvmAGUZbPMMXChHbdvslv/AJLzscaZVCh7LEUo2jblfy2/DT5IzjMvq8PZ0377/BEISqSUYq7ZluXQwlPXWT3f6IBr+0Mbb/l3T/F/1+HqAEcujLGyxE9drfctQADgaVsfOP8Acf5o40jfK79pL9QAAAOHf+RpQ9ThbBc+Z4jEtaRbS9W/8vzAnRpzlGUoptbeXoBx1O+Opx7R/Nv/ACAAAAAAAAAAAAAAAAAAAAAyrJ8TmVXkpLRbt7L/AIPkVsbl3D1J0cMues93/n2/wr59zGY2vjKrq15c0v627IAAAAAAAAMJgsRjKip0IuT/AK37fMyngyhRtUxb55dvsr/P8F5MjCMIqMVZLoYrF4fDQ5601FeZj+OKELxwsOZ93ovu3f4GN4hzHF3U6rS7LRfhv87hg8DiMZU9nQg5P8vV7L5mWcEU4pTxkuZ/yrb5vd/K3qYbCUMNDkowUV5KwAAAAZhluGx9L2VeN1uuln8jGcCU3rhqtvKSv+K/yMXwtmmH/wCXfMu8dfw3/A4krzpyWFaaa1aenoZVg/q2HSa956v/AC+QABw/i1i8spTerSs/Vafjv8zNcG8JjalD+Vu3puvwsAZLj3gcdTrdNn6Pf/P1M4wSx2AqUY2ba09d18r/AIGJdWVaXtfivr6mT5YqEPa1F77/AAX+fcDNcU6GHfL8UtEZfhFhsPGHXr6gAAAcIUqlHM486tzQbXmtHocQ4GpjcBKjT3bj+au/ktTN6FKhjalGntHT7kk383qAAAYPByxCqPpCLk/lt+NjhiDlm1Feb/BMweDpYWl7Omu7fm3uwMy4hwGBvGc7y7LV/wCS+ZjuNcZVusPFQX3v8dPw+ZicfisS71qjl6t/kbgAAAAAAAAAAAAAAAAAAABkGRf2lOU5y5acd+/y7erM24kpYel9TyxcsVo5L9P/AJLd9O4227sAAAAAAAAMk4VxGOtVre5T/F+i/V/K5gsBh8FSVKhGy/F+r6mJxVHDU3UrSUYrqziHj/6vQm8HHbaT6vpZf5/cYSeKrx+sYuo51J6tt3aXReS8ugWuZLwfVxCVXF+7Ht9p/wCS/H03MLg6GFpKnQiopdv61+YO1jG8S5ZhbqVTmfaOv/A/ExXHc3ph6Xzk/wBF/mYjivNa3/LzlXkkv+D+JVzHGVv4lWT9W/8AMweaYzCS5qFRr8vmnocPZ9jcw0q0dP5lovue/wAn8j21J1PZ8y5u19fuAAAMTg8PiY8temprs0n+ZiOFcqrf8u+V+Ta/Db8CvwJQf8Gs16pP8rGI4IzCH8OUZfNp/irfiYjIcyofHRfyV/yuSi4uzVmBwLjNKuFb/vL8n+hxzguSvTxMVpJWfqtvvX5AE5RjFyb0RwLn0M2yz+9TfK/T7L+a09Uzi7hinhc7ljEvcqe8l/e+1+Ovq/IAivreYt/Zp/n/AF+QAAABwZiIV8D7OSvKm3byT1/zQZhV9ri6tTvJv72AAAZPgFSyDE15LWadvRf8G/4HCmR/UqHt6q/eT/BdvV9fkgx2NpYPDyr1Xov6svUzXivGYy8Kb5Idlu/V/orfMAAAAAAAAAAAAAAAAAAAAACnia1OlKlCVoytdd7bfmAAAAAAAAFOnOpNQgrt7IyLhGnQSrYxc0+kei9e7/D1DOuLMPg70qHv1PwXq+vovm0Y7MMTjqntK87v8F6LoYn/AG3HRoL4Iav1/rT7ww2FrYmqqVGPNJ9DIuF6GBSq1rSqfgvTz8/usEpRjFyk7JGacZ4WheGGXPLv9n/N/LTzMwzvHY5v21TTstF93+dwCnTnUkoQV29ktzL+GcNg6X1rNJJJfZ/ztu/JfiZpxfVqR9jgV7OC0v1+XRfn5o9pNz5+Z83frf1MBxbmOFtGUuePaW/37/fcwPGWAxFo1b0356r71+qRSrU60FOnJSXdO4AAAFfCYfEK1WCl6pMxPCWVVtoOL8n+juvwMVwJLfD1vlJfqv8AIyvJM0yvMKdV07xvZtO+j0em+m+3Q4kwP1zLakEvej7y9V/mrr5gGfY//mHg/X/I8MsyqYTN/Z/8u6i5ZeT+y/W+no2cS5b9ewEoxV5R1Xy3XzX42AxldUKEqnZfj0Mpw7o4WN95av5/8AAAAA4LxfsswdJvSat81qvwuYmp7KjOfZN/cjcAAChRnWqxpQ3k0l8zD4SnRw8aCV4xSX3AcV519dxHsKT/AHcPxfV+i2X39QAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAweDrYusqNGN5P+rvsjI+HqGWw5n71R7v9F2X4v8AAxOJo4ak6tWVorqzPOK62MvRw/u0/wAX/kvL7wxmIWHoSqPp+fQynDOlQ55fFPVmX5dXx9dUaKu/wS7syfJcPltLlp6ye8ur/wAl5Bm+fYTLY++7z6RW/wA+y/pXM1z3GZjL95K0ekVt/wAH5/gAGDwVbGVlRoxvJ/1d+Qll/DdG79/ENf16L8X+WY5licfV9pXld9F0Xov68wAw2LxGGlzUZuL8nY4bx+b4uKliIr2f8z0b9EtH9y9SWf5ZGs6LrLmX3fft+JCcZxUou6AAAADF8J5XiNVDlf8Addvw1X4GK4EqLXD1U/KSt+Kv+Rm2RZrgaMp+wcmtuVc35XdvVIlTqzrcsk+dvZ73MvwkcJRUI77t+ZkmYrH4GFa/vbP1W/37/M4nyt4HHS5V7k9V+q+T/CwZp++q0sMuru/Rf0wAAAAwWJeGxNOsvstP7jPK8Y5VWnF6OL/HT9QAADgzA+3x7rNaU1f5vRfq/kBxbnf1Sh9WpP8AeTWvkv8AN7L7+wAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAZfl1fH11Roq7/BLuzKMmw+W0eSnrJ7vq/wDgdkZrm2Gy6j7Sq9ei6v8ArqzNs4xOZVeeq9Fsui/4PmBjv9rxkMMvhjrL+v63Mty2vj66o0Vr1fRLuzK8qw+XUFSpLXq+rf8AWy6Bn/FsKF6GDd59ZdF6d3+C8ypUnVm5zd2+rADCYSri60aNFXk/6+4xGJw3DmG+r4e0q8lq+39dF8351qtStN1KjvJ7tgBa+iMryDDYGgsbmbsukX+q6vy+/wAs64nxGOvSpe5T7dX6/wCW3qGFx2Jwsr0Kjj6P9Opg+NsbSsq0VNfc/wANPwIcf5NFxjiZOm33Ta+9X/FIwuOw2Lgp4eopp9ncAAAAMTl+ExVnXpRk13SbXo+hiuDcsrawTg/J/o7/AKGR5HWyupOKqc1OXS1mn+PTfbocQ5SsxwbhFe/HWPr2+e33MlCUW4yVmjB/v8dVrdI+6v1/rzADC0XXrwpL7TS+92M94frZbUuvepvZ/o/P8wAAMTmHtuFopvW6g/k7r8EgAADhHA/VsujNr3p6/Lp+GvzDM8fDA4SdeSvbp59DF4qriq0q1V3lIAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAy/Lq+PrqjRV3+CXdmVZTQy6gqVJa9X1b/AK2XQzzPaGWUu9R7L9X2X5/ljcbXxlZ1q0ryf4eS7ICrUjTpuctkrnDWW4jHVOaCvOo7+i/y/wCAZRlNDLcOqVPV9X1b/wAuyG0ldnEfFLr3w2EdobOXfyXl+fpuACi5NJaspqnw5gOeSTxFRbdl/kuvd+SKtWdWbnN3k92ABkWXUMDhv7Uxq0XwLu+jt3fT7/MzbNsRmVb2lV6LZdEv63YAYlvH5jyLa9vkt/1IN02uR2ttYw3EmaYeyjWbXnr+d2YfjrEx0rUlL0bX+ZguMcJiZxp+zkpPolf8nf8AA3QAAAAYjCYfEK1WCl6pMnwXkzTVOnyX191vf0d1+BiOA4vWjW+TX6r/ACK/BmZ0/gSl6P8AzsV8lzCj8dGX3Nr70Si4uzVmQnKE1KL1WpSlSxuFjKSTjNJ2fmjiHhieBbrUNaX4x9fLz+/uwABYuawjw3RyUvmk1+N19wAAGX4SWLxVOhH7Tt8ur+SIQjCKjFWS0QVY05Qanble99rGe4LCYXFOOFqKUX2d7eV/68wAAAAAAAAAAAAAAAAAAAAAAAAAAAAAMFgq2Mrxo0VeT/q78jJ8oo5bh1ThrJ7vu/8ALsjP8/pZbS5Y61Hsu3m/L8/vaxOIq4mq6tWV5PdgEoRnFxezOHcho5VhVFK82ld/ovJfjuNpK7OJuJXi28Nhn+7W7/m/4H5+gABw1gqVGnPM8T8FP4fOX9aLz9DMcfVx2IlXqvV/guiQAFGUI1IymrpNXXddUcQZw8xxHuaU46RX6/P8EAGY4j2GGlNb7L1Zw9htJVn6L9f0AyjJcTmVXlpK0VvJ7L/N+RlmUYPLKVqa16ye7+fReWxm/GdKleng1zS/me3y7/l6lfNMbXq+1nVfN62t6W2+RguI83jONOnUc29Emr3/AF/ExmeLLcLB4yzqv7Mf+C3ou/V7GG4wyutZSk4PzX6q5h8Xh8Qr0ZqXo0wAAAAqUadVWnFP1VyrkWWVfioR+St+VjD4elh6UaVJWitl/wAOSipKz2OIuFHSvicGrx6x7ea8vLp6bAAAAAcEYJSrVMVLaKsvV7/cvzMbxPlmFunU5n2jr+O34mN44xE/dw0FFd3q/u2X4mMzLF4x3r1HLy6fdt+AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAFKjOtUVOmryeiRkGR08toa61Jbv9F5L8d/TPs8pZZR71Hsv1fl+f5YnEVcRVlVqyvJ7sADh/GfWstpVG9UrP1Wn47nFXEftW8Hhpe6viffyXl37+m4AGDwtTFYiFCnvJ2/4PyOJ8ZTg4Zdh3+7pb+cv+B+bYAAABxDXcpwoR9f0X9eZg8OqFCFPsvx6hkOQVczqXelNbv8ARef5FatgcowivaMFsurfl3b/AOCzOuIsTmUnG/LT6R/z7/kgMpoUcmwP9o4hXqS+Bev+fV9F62MZi62LrSrVXeT/AK08gjKUWnF2ZheJMzw9lGq2uz1/PUq+JTwcIvFUea/8rs/Wzv8AmjIONMrzup7HDOSna9nF7equvxAAAAAOIuFVWvicGrS6x7+a8/Lr67yi4tprVAAAAB7ap7P2fM+Xe19L97AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAcLZB9TprE11+8lsv5V/m+vbbuZxm1HLcO6s9W9l3f8Al3ZjMZWxlaVas7yf9WXkAAYTPMRhcBUwlLTmd79UrWdvXT8QAA4dUcHha+ZTWsVyx/xP+l8rk5SnJyk7tgAAAGFX13M3U6J3+S2/QMiySpmdfl2gvif6Lzf4bmOx2DybBrSyWkYrd/11f6mZZniMwrOrWfouiXZAcO5ZHHYu9T+HDWX+Xz/K5n2bSzHFua+COkV5d/n/AMDoAGc4n2+KaW0dP8zw0yBYDK/rVRfvK2vpH7K+e/zXYKk4U4uU3ZLqzE8a5fSq8kE5Luv0va/4GF4oyvEWSq8r/vafjt+JTq06seaEk15agAAAcaYXL42qKVqz6Lqu77evXbXoAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABwhkccRP65V1jF6Lu11fp+ZjsZRwdCVaq7Jf1ZebM1zOtmOJdap8l2Xb/MAAAAA4gf1TB4bL1vFc0v8T/y1/AAAAAzSv7HCTl1tZfPQ4eoctGVV9Xb7jL8BVx2IjQpLV/gurfof7HkWX/3Y/fJ/wCb/BeSMzzKvmGIdaq/RdEuyAMY/wCysmhho6VK2svKPb9PvADHYhUMPKp1S09ehwvk7zfNqOFfwt3l/hWr+/ZebRGMYxUYqyRKUYxbbskcR8QTzCq6VJ2pLbz83+i/UClWqUpc1OTT8nb8jI+Ls4VOU5VnJczspWenrv8AiZLxVi8diI0HRTb3abVl31v+YTzDCQqOlKrFSXRtJ/cJqSumABnvF0KN6ODd5dZdF6d3+HqVas6s3Obu3u2AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAcEY+MPa4ebsrcy+Wj/AE+44jzyWZV+WD/dx28/N/p2XzAAAAAMjwv1rMaNLpe79Fq/wRneL+tZhVq30vZei0X4IAAAA4iraQpL1/RfqYOh7HDwp9l+PU4bymGWYN162k5K7v0Xb9X5+hxBnM8yxN18EdIr9fV/8AAyHAfXcwp0mvdvd+i1/Hb5mfY/69j6lVP3dl6L/Pf5gBxFXtGFFddf8jwkylQoVswktZPkj6LV/e7fcHGmbOjSWDpvWWr9O3z/AC9QDET9nRlPsm/wMopNYSnFLV/qzh3Jo5dhUpL95LWX+Xy/M4l4hWAh7Gi71X/5Ku/r2XzfnOcpycpO7ZSr1aTvTm16Nr8ilxFmlL4a7+ev53KXGeaQ3cZeq/yaMLOrOhCVVWk0rpdHYxWLoYWk6taXLFf1/SM94prY69Kh7tP8X6+Xl99wAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAIzlG/K7X0AAAAADhn908Riv8An3B29XsAAAAFb/as2Ueidvu3/E4Ryj63ivb1F7kPxfRfLd/LucZ5z/zA0n5y/Rfq/l5gBk044TLMVi0/edoLur7/AOfyAAzet7XGTt00+7/gnDWWLLcpw+FtZxir/wCJ6y/Fsq1IU4OcnZJXfyMxxs8bip15faf3LovkgDNp8mCqPyt97scC5N7SUcRNe7TSS/xW/T87GdZpTy7COq9ZbJd3/l3K9epXqyq1HeT1YBkmD+t5hSpPa936LV/lYzjPcLlsPfd5vaK3+fZf0jM82xOY1eetLToui9P89wAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAMNmE6GErYeK0qW16qzuAAAAVJqEHJ7JXOHaE6+JlNK7ei9WxulkGT/wB5L75P+vuRVqTqzc5u7er9QAu7WvoAFSahByfQ4WwLzHPMPRlqnK79F7z/AAQcY476vl/s4vWbt8t3+i+YAZvFzowprVykkZTgI4HBU6C3S19ev4nEmbPMMY3F+5HSP6v5/lYAMtzOpgJTqUl77Vk307u3cq1Z1Zuc3eT3b3AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADOKvs8HPz0+//gHhdlHtW8VNaRd167L7rN/ccY5n9Zxn1eD92n/6V1+7b7wAAAq041IOEtmrHA2Cw2WZ17atP3XFqLfSTa3+V1f77G5xri/a49UU9IL8Xq/wsACbTTW6K/FSrZLKN7Vn7r+e8vmvuYAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAGb4OpiaCjTeqd7dzIvZZLwvTqRacuW/rJ9Pk9H6MnKU5OUndsAAAA4f4png7UMS+an0fWP+a8unTsZnifrOMq1ukm2vTp+AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA8bXeFWF5vcve3nawAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAZLl0MfivYylZWb+4/1Lw/8Az9f3I/1Lw/8Az9f3InwZh4xb9q9PJAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHCP/ACMf+3X+gFX+HL0YAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABwj/yMf+3X+gFX+HL0YAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABwj/yMf8At1/oBV/hy9GAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAcI/wDIx/7df6AVf4cvRgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHCP/ACMf+3X+gFX+HL0YAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABwj/yMf+3X+gFX+HL0YAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAGT4Kli8T7OptZvQ/1ZwXeX3/8A/1ZwXeX3/8AAKnDeCjBtN6Lv/wAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADhr/fvk/0ArfwpejAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAOGv9++T/QCt/Cl6MAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA4a/375P9AK38KXowAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADhr/fvk/0ArfwpejAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAOGv9++T/QCt/Cl6MAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA4eqQp4y82krPfQ+uYb/n5H70fXMN/wA/I/eitjMM6ckqi2fVAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABnWLq4XC+0pOzuj/WHMP519yP8AWHMP519yKWf4+U4pzW/ZAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHEv8AuPzQFH+LH1QAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABxL/uPzQFH+LH1QAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABxL/uPzQFH+LH1QAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABxL/uPzQFH+LH1QAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABxL/ALj80BR/ix9UAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAZ/QqVsHy043d1sf2Xjf8An1L7j+y8b/z6l9xSyzGKpFuk910AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAOKsxxGAy/22HdpXS2v37n+uuc/wDPxf8AgK/yP9dc5/5+L/wFf5FDjPOJ1YxdRWbX2V/kAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAcd/8AIp/7eX6gYX+PD1X5gAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHHf8AyKf+3l+oGF/jw9V+YAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABx3/yKf8At5fqBhf48PVfmAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAcd/8in/t5fqBhf48PVfmAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAcd/8AIp/7eX6gYX+PD1X5gAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHGWEr4rLPZ0IOUuZaJXfU/wBXs1/6B5fcz/V7Nf8AoHl9zMPkGaRrQbw8rJro+4AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABiJyhC8T61V7n1qr3Fiare4AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABi/wCGAt0AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAYv+GAt0AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAYv8AhgLdAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAGL/hgLdAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAGL/AIYC3QAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABiouVOyR7Gp/Kz2NT+Vio1Lr3QAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAClRqVZqFOLbfRGG4Sx9VJztD1ev4X/Mz3BPKq8aMpczavpp1a/QhiacutjcAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAMnyWtmNTTSC3f6Lu/yMBluGwVPkoxt3fV+r/pBx/wD8jCn/AIF+cgpV5U3psU6kakboAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABV8NTr0qdeVlOSj97tf5GHw9LD0o0qatFbAcf8A/Iwp/wCBfnICnUdOV0QkpxTQAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABOUYRcpOyRmWYTxeIdTZLbyRw9mSzLKqGKvrKKv6rR/imBx//wAjCn/gX5yAMLV5Zcr2YAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABxLjvZ0lh4vWW/p/wAH9A8Jcz9rgK2Ck9abuvSX/BT+8Dj/AP5GFP8AwL85ABQqc8F3AAAAAAAAAAAAAAAAAAAAAAAAAAAAwWAr4ufLSXq+iMHw5haSTq++/uX3f5lXDUKdCahBLR7JdhTktmU8XJaS1ITjNXiwAAAAAAAAAAAAAAAAAAAAAAG0ldmYYp4rEzq9Ht6dA8Ocz+pZ9Ti37tROD+eq/FJfMDj/AP5GFP8AwL85ABhZ8s7dwAAAAAAAAAAAAAAAAAAAAAAAAAAAzTM6eCpXesnsv66HAePeNyaMpfFGUk/W9/yaQYr+BP0f5AU6koO6KdSNSN0AAAAAAAAAAAAAAAAAAAAAAGe4n2GClbeWn37/AIAUK06FWNWDtKLTXqtUZdjIY3B0sTDacU/vVw4//wCRhT/wL85AAnZ6EJKUUwAAAAAAAAAAAAAAAAAAAAAAAAAAMViaeHoyqzeiMZi6mKrOrN6v8F2PCvGf7xhW+0l+T/QMV/An6P8AIApVXTldCakroAAAAAAAAAAAAAAAAAAAAAA4pxHNWhRXRX+//hvxAPC7M/rWSvDt+9Sk18nqvxuvkHH/APyMKf8AgX5yAAwcrwt2AAAAAAAAAAAAAAAAAAAAAAAAAAA4jx/ta3sIP3Y7+v8AwPzuHh9jPq+e04t6TTj+F1+KQYr+BP0f5ABhKv2GAAAAAAAAAAAAAAAAAAAAAAZvW9tjakvO33afoAeFmZ/Vs4eGk9KsWvnHVfhdfMOP/wDkYU/8C/OQAGDladu4AAAAAAAAAAAAAAAAAAAAAAAAAAZjilhcNOr1W3r0JScm23qwy3FvCYyliF9iSf3O4mmroxX8Cfo/yACMnFpojJSimgAAAAAAAAAAAAAAAAAAAAAqzVOnKb6Jv7iTcndgGWY6eBxlLFQ3hJP7nt89ijVhWpxqQd1JJr0Zx/8A8jCn/gX5yAAou1SLAAAAAAAAAAAAAAAAAAAAAAAAAADijFXnCgntq/0/rzAOGMZ9byfDVb68qT9Y+6/xRiv4E/R/kABg53i49gAAAAAAAAAAAAAAAAAAAAAzmpyYCq/K336fqAB4d5n9dyCkm7yp3g/lt/5K0cf/APIwp/4F+cgAFoLVAAAAAAAAAAAAAAAAAAAAAAAAAAGYYj6xiZ1O709Nl+AB4ZYz2uVToN6wk/uav+dzF/7vP0f5AAYeXLUQAAAAAAAAAAAAAAAAAAAAAcST5cC13aX6/oAB4S5n7PG1sFJ6TXMvWO/3p/gcf/8AIwp/4F+cgACk704+gAAAAAAAAAAAAAAAAAAAAAAAAAGaV/Y4OpPra3zegAeF2M9nmFbDt6Tjf5xf+TZi/wDd5+j/ACAAIy5opgAAAAAAAAAAAAAAAAAAAAHFL/2WC/vfowAOG8y/s3NsPir2UZK/+F6S/Bs4/s8wp/4F+cgADD60kAAAAAAAAAAAAAAAAAAAAAAAAABxRV5cNGmur/L+kAHCGM+q53hp30cuV/8Ab3u/qYv/AHefo/yAAMLK9JeQAAAAAAAAAAAAAAAAAAAABxU/3NNeb/IAAxuZ/wBo4PB1m7yVNRfrGUl+Nr/MAAw38JAAAAAAAAAAAAAAAAAAAAAAAAAAcU1L16cOyv8Ae/8AgABTqSpzU4vVO6+QsRHE5f7aO0oXXzVwADBPRoAAAAAAAAAAAAAAAAAAAAA4q/hU/VhKUYrVk8bh47zRPNsMtm38v8yedR+zD8SecVn8KSOHYVll8JVXrLX0T2/DX5gAGG/hIAAAAAAAAAAAAAAAAAAAAAAAAADiGfNj5Lskvwv+oAHB+M+s8NQu9YqUX8r2/CwABg377XkAAAAAAAAAAAAAAAAAAAAAce1qtLCUpU3b3mvwJ4qvPeb+8bb3AMswbxmLp0F1evp1/AjFRiorZAAGGX7pAAAAAAAAAAAAAAAAAAAAAAAAAAZvLmx1V+f5aAAeG2M5sBi8M3t7y+aaf5IAAwr/AHqAAAAAAAAAAAAAAAAAAAAAOP6fNlkJdpr8mABwXgdJ4qS/ur83+n4gABQVqcQAAAAAAAAAAAAAAAAAAAAAAAAAMe+bF1X/AHn+YAHAeM9hmcqbelSEo/NLm/T8QADD/wAVAAAAAAAAAAAAAAAAAAAAAHGdL2mTVX2s/wAV+gARg5yUYq7Zl2DjhMJToL7K/Hr+IAAQVopAAAAAAAAAAAAAAAAAAAAAAAAAAYvXEVPV/mABl+LeExVOuvstfd1/AhnuWz2rR+bt+ZDH4Sp8FWL9Gv8AMTUldO4BQ/iRAAAAAAAAAAAAAAAAAAAAAM6oe3y6vTW7i7ettPxADhTA/WMeqjXuw1+fT/P5AABCPNJIAAAAAAAAAAAAAAAAAAAAAAAAAAxqtiai/vP8wADF0vZ15R8wUnF6MhmGLh8NWS9JP/Mhn2ZQ2rP56/mQ4qzSO9S/ql+iMg4nx2KzClQqKLUn2d9n5/oAAAAAAAAAAAAAAAAAAAAANJqzMdh3h8VUo/ytr7nYA4WwP1bARk1709fl0/DX5gABho81RAAAAAAAAAAAAAAAAAAAAAAAAAAGZR5cZVX95/mAAZzStUjNdf0AA4PhzZ1R8ub/ANJYAAAAAAAAAAAAAAAAAAAAAcbYT2GbTklpNKX6P8UBleCeMxdOgtm9fRb/AICiopJbIAAMFHVyAAAAAAAAAAAAAAAAAAAAAAAAAAM8hy4+ov61SAAM0pc+Hb6rUADgSlz5tf8Ali3+S/UAAAAAAAAAAAAAAAAAAAAAPEHA+0wtPExWsXZ+j/4K/EDgvAWjPFSX91fm/wBPxAAAw8OWmgAAAAAAAAAAAAAAAAAAAAAAAAADiany41S7pfqgACcFODi+pOLjJxe6ADw7o3xFer2SX3u/6AAAAAAAAAAAAAAAAAAAAABmuBjjsFUw7+0tPXo/k7E4ShJxkrNaEISnJRirt6GX4OOEwtOgvsr8ev4gABThzzUSwAAAAAAAAAAAAAAAAAAAAAAAAABxVS0pVF5r+vxAADM6XJiH56gB4f4fky+dV/al+CS/W4AAAAAAAAAAAAAAAAAAAAAHG+V/Vcw9tFe7U1+fX/P5nCeA+sY5VGvdhr8+n+fyAAAwdPRyAAAAAAAAAAAAAAAAAIxlJqMVdsyzhaVaSeJlyrst/wDgfiUMgy2irRpJ+uv53OJf3Oa1oU9EmrJaJaLoQxc1vqU6sKi91gAAAAAABxDR9pgZPrFp/p+TAADOaV4RmumgAcN4X6tlNCn1tf8A8C1/UAAAAAAAAAAAAAAAAAAAAADiHKVmeAnRXxLWPqv89vmcM5dLB4Fc6tOWr7+S+78wAAhBzkoojFRikgAAAAAAAAAAAAAAAAKVKdWahBXbMqyeng4KUtZvr28l/WpRdqiDiv8A5HFf1X5IIycXdFCuqit1AAAAAAAK9JVqUqb2aaJwcJOL3QABjaXtMPKPl+QBl2EeLxdOgvtNL/P8BRUUktkAAAAAAAAAAAAAAAAAAAAAAYujb30AAGGo8keZ7sAAAAAAAAAAAAAAAAAyHK1h6ftqi9+X4L/NhB2kmHFf/I4r+q/JARk4tNFKoqkboAAAAAAA4gwvscbJraWv+f4gABiaXsq0odmBwFgPbY6WIa0prT1en5XAAAAAAAAAAAAAAAAAAAAAABpNWMRh3B3WwAGHw/2pAAAAAAAAAAAAAAAAAGRZesViOaS92Or9eiAE7pHFf/I4r+q/JAGHq8k/JgAAAAAAHEmE9thfaLeH5Pf9GAAGcUuWqp91+QHCeW/UcsgpL3p+8/nt9yt8wAAAAAAAAAAAAAAAAAAAAAAGk0VcInrAlRnHdFiFCpPZFLCxhq9WAAAAAAAAAAAAAAAAAGU4L6rhIwfxPV+r/wAtgCk7wRxX/wAjiv6r8kAGGqc9Nd0AAAAAABOEZxcXqmZhg5YXESpPpt6dAADNqXPh79g4Yyl5jmEYSXuR1l6Lp83p6XAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAMlwixGMinstX8v8Ag2ADDu9NHFf/ACOK/qvyQAYWfLO3cAAAAAAAz/LvrND2kF70fxXVAAFWCqQcH1RyScuRLXY4YyVZZglGS/eS1l+i+X53AAAAAAAAAAAAAAAAAAADC5disV/BpuX5ffsZhlGLwFFVsRGybtunr8vQjXpy2YAAAAAAAAAAAAAAAAAAAAAHCkqElVSknNWuuqXT7/0ADCu8Gjiv/kcV/VfkgATs7oi1KKaAAAAAAAM+yh0ZOvSXuvddn/kwADhvhlRxTx9decV/7c/0+/sAAAAAAAAAAAAAAAAAAAVKkKcHObskcLYPBY/CrGfFdtWeys7bdb7699jBpKLiuhx7/wAiuP8AjX5SCnXnDzRTqRqK6AAAAAAAAAAAAAAAAAAAAAKGf4nA5tLG0Hre1ujjtZ+qXyeplGbYbNMJHE0Ho911T6p+a/4OwBhHujiv/kcV/VfkgAMLLmp27AAAAAAAEoqSaaujNeHp026mGV49uq9O6/EatoBGMpNRirsynh6zVXEr0j/n/l94AAAAAAAAAAAAAAAAAABKUYxbbskZxm0sZPkg7QW3n5v9DwtxXPgK9C/wyT/8CX/2rML8TRx7/wAiuP8AjX5SAhOUJXRTqKpG6AAAAAAAAAAAAAAAAAAAADG1vY4adTsmHCvEtXJcXzb0paSX6rzX47eaw2IpYmjGtRlzRkrp+QGFfvtHFf8AyOK/qvyQAGClq0AAAAAAABi8qwmK1qQ17rR/16lThWk37lRr1V/8inwrRT9+o36K3+Zhcuw2FX7qFn36/eAAAAAAAAAAAAAAAAAAAHEmZf8AMLTf+L/L9X8g8L8V7PM6tBvScL/OLX6NmG+M49/5Fcf8a/KQBQq+zl5GjQAAAAAAAAAAAAAAAAAAABxDU5MBJd2l+N/0A4G4s/s2ssHiZfuZPR/yt9fR9e2/c0a0DDv94jiv/kcV/VfkgAMM7VUAAAAAAAAAAAAAAAAAAAAAAAAAAAAY7FxwuHlVfTb16FScqk3OTu2HBuK+rZ7hpX3fL/4EnH82UP4iOPf+RXH/ABr8pABhKnNHlfQAAAAAAAAAAAAAAAAAAAA4qnahTh3d/uX/AAQDw/4t51HLMXLVaQb/APSX/wC2/d2Ck7TRxX/yOK/qvyQAFJ2mmAAAAAAAAAAAAAAAAAAAAAAAAAAAAcT4zmqRw8XotX6vb8PzAw9aVCtCrHeLTXydzDVI1OScXo7NfM49/wCRXH/GvykAFKfJNMAAAAAAAAAAAAAAjCU2oxV2zBcM1KiUsRLlXZb/AOS/EpZFgKatyX9bmZ1JU8dWhB2SlJJeV2Qxj2kiM4zV4sAAAA4rledKPZP9P8gCMnFpxdmjgnitZrQ+r4h/voL/AMCXf17/AH9dE7NHFX/I4r+q/JAALRgAAAAAAAAAAAAAAAAAAAAAAAAAAATnGEXJ7IxNeVetKo927gHCOL+s5Nhal/spf+A+7+hx7/yK4/41+UgAMNPmpryAAAAAAAAAAAAACjRnWqKnTV2zK8opYOCb1m93+i/rUDN/+RhX/wAcvzYQqSg7xKVWNSN0AAABxS74qC/u/qwAweLrYSvCvQlyzi7p/wBficM8RUM6waqx0nHSUez7ryfT7t0cUO+bVvl/6SgACOsUAAAAAAAAAAAAAAAAAAAAAAAAAAAHEGI9lgZJby0/z/AAPDPFe1yiVJvWE39zSf53OPNcrh/jX5SAAwcrScQAAAAAAAAAAAAAyPKlhaXtJr35fgu3+YBm/wDyMK/+OX5sCnUcJXRCSnFNAAABxP8A75H/AAr82ABkmdYjKcZHE0Hto10a6p/1o9TNuIcuxmNnWhViua2jaTWi0auRxuFltUi/miM4y+F3ACHwIAAAAAAAAAAAAAAAAAAAAAAAAAAAOKq16lOkuib+/T9AA8K8Vy4nEYe+8VL7nZ/+lI451ymH+JflIACjLlqJgAAAAAAAAAAAAHD2XrEV/azXux/F9Pu3+4AM3/5GFf8Axy/NgGFq8suV9QAAA4oX+2R/wr82AAZzRtKNRddAI16sfhk182RzHGR+GtJf9vP/ADI51mMdq8vvbI8RZpHas/wf6GClKeFpyk9XFX+4AAAAAAAAAAAAAAAAAAAAAAAAAAAM9q+0x8/LT7l/mAHAGK9hn1JX0mnH71dfikcba5RD/GvykAARfNFMAAAAAAAAAAAAFFyaS3MuwiwuGjSW639eoAZv/wAjCv8A45fmwApT54JgAAHFMf8AaIPy/UAAx1H21CUevT5AAGHhyUYR7JL8AAAAAAAAAAAAAAAAAAAAAAAAAAAAxVT2lec+7b/EAMqxX1XHUa9/glF/c0caO+UQ/wAa/KQABh3ekgAAAAAAAAAAAA4fwnt8YpPaOvz6f5/IADN9cwr/AOOX5sAMnzb2+MnSv7rXu/Ld/P8AJLzAAAOK4a0pev6AABjqPsq8o9OgAYan7WtCHdpfewAAAAAAAAAAAAAAAAAAAAAAAAAAAxE+SjKXZP8AIAAzbF/W+FsNWvrJwv68rv8AiAAYN3pgAAAAAAAAAAABw1hvZ4R1HvJ/gtP8wAM2/wB/r/45fmwDOsXyU/YRest/Jf8AB2+/sYOu8PXhVXR/8P8AgJpq6AAA4op82FjLs/zTAADO8NJKM2rNaP57f15gBw9R9tmuHj/eT+7X9AAAAAAAAAAAAAAAAAAAAAAAAAAAAzOXLgqr/uv8gADLcX7fg6nC+sKnL+En+TAAMF8LQAAAAAAAAAAAAk5NJbmGoqjRhTXRJAAZt/v9f/HL82BXrQo03UnsitVnWqOpPd/1b5Bk1f22Bpvqlb7tAAAM8pe0wFRdtfuf+QAAnZ3RxNRjXqcy/wCXkU/n/SQ007MA4Gw/tc2U/wCSLf6fqAAAAAAAAAAAAAAAAAAAAAAAAAAAGcu2Aq+gABw7i75ViMM39uEl90k/yQABgvtAAAAAAAAAAAAGU0fa42nHzv8Adr+gABm3+/1/8cvzYGc4v2lT2MXpHf1/4H5+gHC1a9KpS7O/3/8ADAAAVqaq05QfVNfeSi4ycXugADMvfwWFn5NfczM6Ps8Q2tnqAeHmFtCviH1aivlq/wA0AAAAAAAAAAAAAAAAAAAAAAAAAAAGd/8AIvqen6oAAyXGxjjvq6erTv8AKzAAMF9oAAAAAAAAAAAA4YpKWLcuy/yAAM2/3+v/AI5fmwzDFrDUW18T0Xr/AMAeoHDNXlxbh3T/AAsAAAZ3h/Y46a6PX7/+DcAAxHvZVRfaUl+pm9Hnoqa3iAcJ4P6rlFJNay95/PVfhYAAAAAAAAAAAAAAAAAAAAAAAAAAAM4V8BV9AAMbio4enfq9jh6u45rSm3q2196aAAMF9oAAAAAAAAAAAA4Vp+7Vn6L8wADNv9/r/wCOX5sbSV2Zhiniazktlov68wDJJ8mPpvzt96sAAAcU4a8IV100f6fr94ABU1yiHlN/kVIKcHF9SpBwk4vdBluCljcZTw8ftNL5dX8kRjGMVGOyAAAAAAAAAAAAAAAAAAAAAAAAAAADMo82Dqr+6/yACrUjTg5yeiMTiJV6jnL/AIYwVX2WJp1O0k/uYABgl7rAAAAAAAAAAAAOF4Wwkn3k/wAkAAZt/v8AX/xy/Nmc4vkh7GL1e/p/wQAwMuXFUpdpL8wAADHYVYnDTpPqvx6fiSi4txe6AAl/yJ1/18/9tDN6PJWU11/NB4f5bz1p4yS0j7q9Xv8ActPmAAAAAAAAAAAAAAAAAAAAAAAAAAABWhz05R7poAMxxnt58kX7q/HzAwlX2uHp1O6T+9AAYRWpgAAAAAAAAAAAHDkbYBebYABntaNHGYictlKX5srVZVqjnLdgBSdqkX5gAABxHgfY4j2sVpP8+v37/eABL/kTr/r5/wC2hmdH2mHbW61/zKVKdWcacFdt2S82ZPl0MvwVPDx3S182939/4AAAAAAAAAAAAAAAAAAAAAAAAAAAAGNp+zxNSHZv8wM0xnKvYwevX/IA4eq+1yyi/K33O36AAYdWpRAAAAAAAAAAAAOH1/wnw+f5sABtJXZxXjo18xrQpu8VKTutnqwAFoAAAGYYOOLw8qT+XkyrSnSm4TVmgAqaZRD/ABv8gaTVmcG8PtYmWMqrSLaj5vv8lp6+gAAAAAAAAAAAAAAAAAAAAAAAAAAAAcQUfZ4+XnZ/h/mgx2LWHp6fE9iTcndvUA4Pq8+Xcv8ALJr8n+oAWvoRVkkAAAAAAAAAAAAcPO+Xw+f5gFSpCnBym0kur0SM68Rstwd6eF/ez8tIr59fkreZnPFWaZq2q9S0P5Y6R+7r82wABK7sAAABn+UuvH29Je8t13X+aADEaZVRXeUgyvLZ42tyrSK3f9dWU6UKcFCCslsAAAAAAAAAAAAAAAAAAAAAAAAAAAAHFVDWnVXmv1X6larClBzk9EYivKvUc5ABwRVvCtT7NP77r9AAoR5qiQAAAAAAAAAAAAcNyvgUuzYYjE0cPTdStNRiurdkZ14l4ShengI+0l3d1H/N/h6mbcQZjmkr4qq2ui2ivRLT57+YAAFCPNVivNAAAAGcZCqzdbDr3uq7+nmThKEnGSs0BjNMswy7835mW5PWxsk9od/8u5hcLSw1JU6Ssl/VwAAAAAAAAAAAAAAAAAAAAAAAAAAAAM9w/tsDPvHX7t/wuZhjPbz5Y/Cv6uABwXV5cbOHeP5NABg4auQAAAAAAAAAAAAZJmWFweAnPE1FCKlu3botu/ojOvE6nG9PLoXf80tF8o7/AH29DMs3x2Y1PaYqq5vz2Xotl8kAAAGXw58XSj/eX5gAAABi8uw2LVqsbvv1+8rcKxb/AHVT71+q/wAiPCtRv3qq+7/glPKMMqVOFRc3Je1/PyEklZAAAAAAAAAAAAAAAAAAAAAAAAAAAAAEoxlFxeqZm2BlgcbUw7+y3b06P5qwAHDFX2eaUvO6/B/qAFCHLTSAAAAAAAAAAAADiun7tKfqvyAAAAMip8+Pp+V3+AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAeIGV608bBf3Zfo/zX3AAZZV9ljaM+0l+YBQp880gAAAAAAAAAAAAOJaXPgub+Vp/p+oAAABwvS5sTOfZfm/+AAAAAAAAGGweIxU+SjByfl+vb5lDg7GzV6kox/F/hp+JmSWCxc8PLXldrkK9OeiYAAAAAAWuRy/FSjzKm7ejFUhLZgAAAAAAAAAAAAAAAAAABj8FTxuFnh6m0lb07P5PUxmEq4TEToVVaUXZ/12fQAE2ndFGoqlOM11Sf3gYWlyQu92AAAAAAAAAAAABmNH22EqQ6tP7+gAAABwvR5cNOp3f5f8OwAAAAAADI+G54xKtW0p9O7/AMl5/d3MPhqOHpqnSiopdEHFH/I3r+v6IKOJlHSWqE1JXQAAAAAZTh6WPxjwymk4q7XW10v1MJluFwq/dx17vf7zE/wZ+j/IKdecNmUq0ai03AAAAAAAAAAAAAAAAAAAON8heJpfXaK9+K95d49/Vfl6AAZHV9rltGX91L7tP0DD0eeV3sgAAAAAAAAAAAAAx+H9hiZ0+z/Dp+AAAAGWYf2GDp0+tvxerAAAAAAA4cyP67U9tVX7uP4vt6d/uEkkkgOKP+RvX9f0QFCs6bs9jRoAAAADO83WFj7Kk/ff4L/Pt954f4x0s/pqT+NST+6/5pBiv4E/R/kAm07ooVlUj5gAAAAAAAAAAAAAAAAAANJo4s4WlhJvF4WN6b3S+y//AJH8vQAOEqvPlkY/ytr9f1KVKVSVkQgoRSQAAAAAAAAAAAAAcUYblrRrL7St81/wPyAAADLcN9ZxcKfS+votWAAAAAABgsJPFV40o9fwXVmXU4UKaowVktgDij/kb1/X9EAYSrdcjAAAADMsdDB4d1Hvsl3ZVqzqzc5u7Zk2K+q5jQr3+GUX8rq/4Biv4E/R/kAQm4STRCanFNAAAAAAAAAAAAAAAAAAASipKzM94GjVk62Asn/K9vk+no9PNGLwOJwk+SvBxfmvy7hCnKpJRgrt9EcHZRjMPhZLER5U3dJ77W26bdSMIxVkGGwlbE1FToxcn/X3GE4MqySdeoo+S1/H/hzifJ6OVYenUoybcnZ3t28kiON/mRCpCa91gAAAAAAAAAAAZzhPrODlFbrVeq/4GgAAAcL4SyniGvJfr+gAAAAAAHDeB9lQdeS1lt6f8H/Ipy5ZJgHFH/I3r+v6IAjJxaaISU4poAAAAz3H/WsS4xfux0X6sDKMV9ay+hXv8UYv5tK/4mK/gT9H+QAYSpyy5X1AAAAAAAAAAAAAAAAAAACpSp1Y8s0muz1JZBlcnd4eH/gKMPgsNh1ajTjH0SX5AZNktXMammkFu/0Xn+RgsDh8HSVOjGy/F+oeIP8AudH/ABfoCk4u6ZQxCno9wAAAAAAAAAAAM6wP1XFNL4Xqv8vkAABRpSq1I04K7ehhcPDD0Y0o7JAAAAAABg8M8TXhSXV/8P8AgQhGEVGOyCjLmggOKP8Akb1/X9EAGDno4AAAAZxjPquElJP3novV/wDAAPD/ABXt8hpK+sHKP43X4NGK/gT9H+QALTYpzU4JgAAAAAAAAAAAAAAAAAAABleW1MfiFSht1fZf1sYXC0sNRjSpK0UB4g/7nR/xfoBtsYet7SOu6AAAAAAAAAAADOcvWMwzUfijqv8AL5jTTswAA4ay63+1TXlH9X+n3gAAAAAAcL4XmnOu+mi/X+vMDCys3EDij/kb1/X9EAFKfJNMAAADibFe0xEaKekV+L/4FgDwrxXNh8Rh77NS+9Wf/pKMV/An6P8AIADBz3iAAAAAAAAAAAAAAAAAAAAJNuyMhytYHCpNe/LV/wCXy/O4B4g/7nR/xfoAU5uEk0RkpJNAAAAAAAAAAABxDlLTeJpLT7S/X/P7wADKcslja1n8C3f6erIQjCKjFWSAAAAAAAyTD+xwMF1ev3/8CwEJcskzcOKP+RvX9f0QAFGXNTTAAAJSUYtvZGKrOvWlUfVtgHhnivZZxKk3pODXzVn+SZi/93n6P8gAKMuWomAAAAAAAAAAAAAAAAAAAAcLZd9ZxntZL3Ya/Pp/n8gA8Qf9zo/4v0ADB1PsMAAAAAAAAAAAGk1Zmb5BKm3VwyvHquq9PL8gAy3KK2NlfaHf/Iw2GpYekqdNWSAAAAAAAo03VqRgt20vvIxUYqK2QBQlzU0HFH/I3r+v6IADByvFoAAAzqv7LA1Gt2rffp+QAcKYr6tnWGqX+0l/4F7r/Mxf+7z9H+QABTlzQTAAAAAAAAAAAAAAAAAAAA4dwP1XAQTXvS1fz2/CwAeIP+50f8X6ABCXLJNCaaugAAAAAACnSnUfLCLb8lcxOEr4amqlaDim7a9/6RGcZbO4AAAGNybCYt80laXdf1qVeFayf7uon63X5XIcLYlv3pxX3v8ARGE4bwtFp1Hzvz0X3f5sUUkktgAAAAAADI6XtMfTXbX7l/mAGFlaTiHFH/I3r+v6IADBu07AAAHFVW1GnT7tv7l/wQAp1JU5qcd07r5Eq0a+BdWO0o3+9XAAMI707dgAAAAAAAAAAAAAAAAAAAyrCfW8bTo9G9fRav8AAsrAB4g/7nR/xfoABhZ81O3YAAAAAAFFyaSV2Zbw2mlPFf8AgP8An/kilRp0o8tOKS8jjT/kXQ/xr/0mQUsVKOktURnGaumAAAAAAAAAAABwvTviZy7L82v8gAhLlkmaNHFH/I3r+v6IACg7VIgAAHFFTmxUYdl+bAA4XxX1nhqlK+qg4/8AgN4/kgADBPdAAAAAAAAAAAAAAAAAAABwlTtXnWtsrff/AMN+JGSkroAPEH/c6P8Ai/QADBytNruAAAAAAJNuyMlyeOGgqtVe+/w/4PcDjT/kXQ/xr/0mQFKrKnK6ISU4poAAAAAAAAAAAOFIe7Vl6L8wAMPLmpryOKP+RvX9f0QAEHaSYAABn8+bMJ+Vl+CAA8O8V7TJMRRb1g39zj/mmAAYN/vLeQAAAAAAAAAAAAAAAAAAAcOUeTAqX8zb/T9CjV5HboaNaAHiD/udH/F+gAFGXLUTAAAAAAOHMsU5fWai0W3r3+X5+gBxp/yLof41/wCkyAMNW5JWezAAAAAAAAAAADhaP+yzf979EABhpWlbucUf8jev6/ogADdAAAZrLmxtV+b/AA0AA8N8Vy1sVh7/ABQv/wCA6f8AtwABhf4qAAAAAAAAAAAAAAAAAAADAUvZYWnDsl+QUK3L7r2APEH/AHOj/i/QAAi+aKYAAAAAYPCyxVeNKPX+mylShSpxpwVktADjT/kXw/xr/wBJkAGGqc8LPdAAAAAAAAAAAGP4qzTKsR7HC1EoWvZpPXvtfp3MP4o5lDSrShL0un+b/Iw/iphZfxsPJejT/NRMP4i5FVtzTlD1i/8A23mMPxPk2I/h4qHzaT+52ZTq06kbwkmvLUi+VpnE9nm9f1/RBVxVCl/Emo+rSK3EOV0t6yfpd/lcrcZZfD4FKXyt+b/QrcbvanR+9/ol+pV4xzGfwqMfRf5tmFm50ISe7S/IAADHO+KqP+8/zAA4QxX1fNqeuklKP3xdvxsAAYf+KgAAAAAAAAAAAAAAAAAACjT9pUjDu0iysBh61/dYHiD/ALnR/wAX6AAGHd6SAAAAADhjB2hLES3ei9Ov9eQAcaf8i+H+Nf8ApMgAoT5JpgAAAAAAAAAAHE3++r/Cv1ACnVnTlzQk0/LQw/E2c4f+HiZ/OTa+53MzzbNsbN1XiHzPfRK/3IxlbME7V6kmvVtAAYH/AHWl/hX5IAADF/7xP1f5gALHRwU4129YtNeqMHxJl2KslU5X2lp+O34iaaugAofxIgAAAAAAAAAAAAAAAAAAFDE0cLVhXrO0ItNvfRPsjAZrgcfDmwtVTXk9V6rdfNAFCtzrle4eIP8AudH/ABfoABVq06Ueackl3ehlWNoYug5UZcyTa+ej/UAAAAAjBzkox3ZhcPHD0IUo9FYAONP+RfD/ABr/ANJkABQnz00wAAAAAAAAAAOKFbGR/wAK/NgAA0mrMxGVUamsPdf4fcYjAV6O6uu6AMPHlowj2S/IAADGq2JqL+8/zACpUjTg5SeiMZipYid3t0QYTM8XhH+5qNeXT7tjB8aVo2WIpqXmtH9235GD4hy7FWUaln2lo/8AL7maNBh/4sQAAAAAAAAAAAAAAAAAADPXbL6ny/NFOpOnJThJprqtGZZ4gZzg7RnNVY9pb/8AgS1++5lniVleJtHEp0peesfvWv3pephcZhsXT9ph6inHumn+Qm07oo1VNeZ4g/7nR/xfoBKSirt6GN4ny7DXSnzvtHX8dvxMbxli6t1QioL73/l+BiMVXxEuatNyfmzw9lfAVY9p/ov8gAAAADIMP7bHRb2jr92342AA40/5F8P8a/8ASZAAYOW8QAAAAAAAAAAOKo/vqcvL9f8AggAABiMuoVtbWfkYjLK1LWPvLy/yKVNzqRh3dgAADNI8uNqr+8/zAG0ldmYY115csfhX4+YAGEzXG4T+DUaXbdfc9DB8a1FZYmnfzjo/uf8AmjJs6wOOrKNGfva6PR7f1sAAAAAAAAAAAAAAAAAAAcQP/hPn8vzQBhsXXw01UoTcZd02n+BlniRmuGtHEJVY+ekvvWn3pmVeImT4ppTk6Uv7233rT77HG+Ko4nL6NSlJSTlundbd0Y3iDL8LdTqXfZav/JfNoxvGlaWmGgo+b1f3bL8TF5jisW71qjl+X3bAHh4/9mrr+8vyAAAAAOFqFqdSr3dvu1f5oADjT/kXw/xr/wBJkABh5ctRAAAAAAAAAAAcVwvClLs2vvt/kAAAAYPA0cRjKXNHXmX4MAAAz6HLmFTzt+SAMzx3O/ZU3p18/wDgAAAcBUufNHL+WL/NL9QAAAAAAAAAAAAAAAAAADiFXy+fy/NAAGb1qkaChGTSb1V9Hp2AAPDtf7NXfmvyAAAAAMipezwFPu7v73/kABxr/wAi+H+NflIABOzE00mAAAAAAAUMsxldXp0pNd7O33mIxVDD15YetNRnHRpu1iMoyV4u6AADiSlz4Hm/laf6fqAAAAZBS58fDyu/wAAAOJ6fLi4y7r8rgZnjuROlTevXyAAAPDuheder2UV+bf5IAAAAAAAAAAAAAAAAAAAz6N8vqfL80AAZz/Cj6gAHh7D/AGGrL+9+SX+YAAAABkfHGNyyvJSftKLbfK+l39l9PTb03MozvBZrQ9rhZ37rqvVf0n0YAca/8i+H+NflIAAoO9KIAAAAAAZZlVfMKvJS2W76L+uiMu4fwWCSajzS7v8ARdPz8w45/wCShxX+L9EYfF18PLmpSaf9dDLOIKeIap1vdl36P/IAAzKj7bB1IeT/AA1AAAAOFaN6tSp2SX3/APDAAAHFVK9OnU7Nr7/+GDMMaqEeWPxP8PMbbd2AAAcB4b2eVuo/tyb+SsvzTAAAAAAAAAAAAAAAAAAAM2jzYGqvJ/hqAAZz/Cj6gAHAVPlypvvJv8Ev0AAAAAxM+ShOXZN/gGAzDE4Cuq+Gm4yXVfr3Xk9Dhnj7C5hy0MXanV7/AGZenZ+T+T6AHGv/ACL4f41+UgADCO9IAAAAAAzPMaeCo871b2Xf/gdzwszWpUzXEUqsrupG/wA4vZfKTA45s+IcV/i/RAZFnLk1h6716P8AR/oAA1cxlD2GInT7NgAAAcOUPZ4JSe8m3+n6AAAGe0Pa4Cdt1r92/wCFzGYuOHp369EVJyqScpO7YAAAZHhPquW0aPVRV/V6v8WAAAAAAAAAAAAAAAAAAAYmn7SjOHdNfgAAZz/Cj6gAHCNH2eTUV1d397b/ACAAAAAzWXLgqr/uv/IA4Z49xWXctDF3qUv/ACaPp3Xk/k1sYDMcLj6Cr4aalF9vya3T8mHGv/Ivh/jX5SAAMH8DQAAAAAFWrClTc5uyRmOOnjK7qS26LsjgjG/VM/ws76OXL/4EuX82Bxv/AMlDiv8AF+iATtsZJmX1yhaT9+O/n5/11AA4mw3JilUW0l+K0/KwAAAQg5yUY7sw9GNGjGmuiSAAAKkIzg4vZ6GZKrHFVIVfii2vudgAAAyTBfXcwo0LXTav6LV/ggAAAAAAAAAAAAAAAAAAAMZS9liJw7Nr8QAM5/hR9QADLcP9XwVGj1jFL7kgAAAAM7dsvqen6oAMpzrG5VXVbCzs+q6Pya6/mujRw3xpgs3SpS9yt/K9n/hfX039dzjX/kXw/wAa/KQABgtmAAAAABxNj9sNB+b/AEX6/cFGrKlUjUg9YtNeqMJiIYnD060NpJNejVw43/5KHFf4v0QBl2NlhMRGotuvoRlGcVKLumAHEGF9vg3Jbx1/z/D8gAAA4ewvtsYpPaGvz6f5/IAAADjrL/q+YqtFaVFf5rR/o/mAAAHh9l/PWqYuS0iuVer1f3K33gAAAAAAAAAAAAAAAAAAAcQUfZ46T/ms/wBPzQAGc/wo+oAGU4X61jqNH+aSXyvr+AAAAABnv/IvqfL80AAm4tNPVD41xVXAxwuM99Raal9rRNWfffff1MJmGFxcb0JqX5/dugAMFtIAAAAAK1WNKnKpLZK5iK8q9WVSW7dwOAMb9a4ew7b1inF/9uuy/Cwcb68Q4v8AxfogA4cxntsN7JvWH5dP8gAlFSVmZjhHhcTKl0W3p0AAAMgwf1fCKT+KWv8Al/XmAAABxfln13LJOKvKHvL5br7vxSAAAEm3ZbmQZasvy+nQfxWu/V7/AHbfIAAAAAAAAAAAAAAAAAAADiqhdU6y81+q/UADOf4UfUADgTB+2zP2r2gm/m9F+DYAAAABnivl9T0X5oAAMdgpJ+2oaSXb80YHizHYe0aj5157/f8A53MDxRl+KspS5Jdpf57ffYTUldMDBLSQAAAAAcTYr2eGVJbyf4L/AINgDwixvNhMThW/hkpL/t5Wf/pK+8ONf+Sgxf8Aif5IAMixXsMbG70lo/nt+NgAOI8v9tRVaC96O/p/wN/vAAAyfAfW8Sk17q1f+XzLAAAAbnE2Uf2bj5QivclrH07fJ6fcAABwXlDxmOVea9ynr6y6L9fl5gAAAAAAAAAAAAAAAAAAAGcYb6xgpxW6V18tf+AABnP8KPqABwHgfY5fKu1rUf4LRfjcAAAAAzaPNgaq8mAABmOXXvVpLXqv1QYLNcZg3+5qNLtuvu2MDxpB2jioW81/l/wWYTH4XFx5qE1L8/u3Rg17jAAAAADiHEe1xrito6f5/iAeFuN9hnvsW9KkWvmve/JMONf+Sgxf+J/kgATad0YSuq+HhUXVAA0mrGdZY8HWvFe5Lby8gACnTlUmoQV2zK8vjg8Oofae78/+AAAAAHEuSrNME4R+OOsfXt6P87PoThKEnGSs1pYAAw2Gq4mtGjSV5SdkjJsrp5bg4YeGrWrfd9X/AJeVgAAAAAAAAAAAAAAAAAAAAzTCfVsVOmtuno/6sAGc/wAKPqAGGw88RWhRgryk0l8zB4WGFw8KENopL7gAAAADFQ56E4901+AAABmOXXvVpL1X6gQnOnJSg7NdUcHV69fK41K0uZtvV9loAAAAATkoxcnsitUdWpKb3bb+8A4dxv1LNsNiL6RnG/pez/C4ca/8lBi/8T/JAAcNV+fB8n8rf46/5gAYrDUsTSdKorpmY5bVwVTllqns+/8AwfIAIwlOSjFXbMlyZYSPtams3+H/AAe4AAAABxhww8QnjcLH318SXXzXn3XX13ACMZSkoxV2zhPhpZfT+sV1+9kv/AV29X1+7vcAAAAAAAAAAAAAAAAAAAAOJsF7Siq8VrHR+n/Af5gBnP8ACj6gBwFlftsTLGTXuw0X+J/5L80AAAAABiKfs6sodm19wAABmOXWvVpL1X6gcL0PY5PQj3V/vbf6gAAAAGc1fZYGo+6t9+gAGRY369lmHxN7uUIt+ttfxuca/wDJQYv/ABP8kABwtVtWqU+6v93/AA4ABWoU69NwqK6ZjuGakW5YZ3XZ7/f/AMMVsJXou1SDXqiMJTdoq7MLkWMrtXjyru9Pw3MuyjD4NXjrLu/07AAAAAAcRcG0sa3Xwlo1Oq6S/wAn+D69zGYHE4Op7OvBxl5/p3+QGX5Xi8fU5MPByf4L1eyOHuE6GW2rVXz1e/Ren+f5AAAAAAAAAAAAAAAAAAAAAFSnGpBwkrp6GYYKWExEqT26eaAM5/hR9QDDYapia0aNJXlJ2RlWXU8vwcMPDotX3fVgAAAAAZ5S9nj6i76/egAAAzHL7Xq0lp1X6kIOclGO7MNRVGjCktopL7lYAAAAA4nqcuEjDu/yX/DAAeGGN+sZBGk3rTlKP3+8v/SvwONf+Sgxf+J/kgAMhqcmPh53X4AAAAAAAAAAFfDUcRDkqwUl2auvxKvB+TVHf2Nn5OS/C9ihwjk1F3VFN+bb/Bu34FOlTpQUKcUkuiVkAAAAAAAAAAAAAAAAAAAAABnOWLG0Pd+OO3+XzJRcW4tWaAzn+FH1AOC+Hnhaf12uvfkvdXZd/V/gvVgAAAAAHFNC1WnVXVW+7/hwAAAMsyZVs1oyh8N7temv3O1gAAAAA4rnrSh6v8gAPCHG2r4nCN7pSXydn+aONF/xkGL/AMbAAy+fJi6Uv7y/MAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAM6yRYle1o/H+f/AAScJQk4yVmgzn+FH1C1zhbhCTccXjY6bxi/zf6L7wAAAAAA4hw/tcC2t46/o/wAAAA4Ww96k6z6Ky+e/wDXmAAAAAHFMr4qEf7v6sADw9xv1XiGhd6TvF/NafjY42/5KDF/4v0QAFOXLNS7AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAY/KsNjF76tLut/+CYnhrF03+6akvuf4/5ma5DmVSEYwotu5g+CM1rte0ioLzf6K/42Mm4RwOXNVH79RdX09F0/F+YAAAAAAFSnGpBwls1YxFGVGrKnLdOwAAAZLhfq+ChF7vV/P/gWAAAAADiZ3xq/wr9QAMFiZYXE068d4SUl8nc4vqxrZ5iZxejlp+AABB3gmAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAcT4PkqxrxWktH6r/NfkAAAZVg/rWKjT6bv0X+ewAAAAAHEv8Av3yQABVqyqy5pO70/BWAAKP8KPogAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADH4SOKw8qT67evQqQlCThJWa0AAA4dwHsMP7WS96f5dPv3+4AAAAADiZWxq9F+oAAAAU1aCXkAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHEmW/wDMVTX+L/P9GAAGTZa8ZXXMvcjv/l8yySsgAAAAAOKY2xUH/d/VgAAAEI80lHuAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABKMZRcWrpmcZTLBz5oawe3l5P8AQADBYKri6qp016vt5mDwdPC0VSp9PxfcAAAAAA4rhrSl6r8gAAADAQ58VSj/AHl+YAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABUpwqQcJq6ZmXDlSm3PDe9Ht1X+f5kouLs1ZgYDI8Timm1yx7v9F/SMHgqOEp8lJf5v1AAAAAADiinzYWMuz/ADTAAAAMhp8+Pp+V3+AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAFfB4euv3sE/VD4fy9u/J+L/wAyhlmDoO9Omk/vf43AAAAAAAM7pe1wFRdlf7tQAAADhalzV51Oyt97/wCAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABUgpwcHs1YqU3Tm4PdO33AAAAcM0OTCOf8z/BafncAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAz/D+xx0mtpa/5/iAAAFr6IwWHVDDQpdl+PX8QAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADifC89CNZbxevo/+Db7wAAAyXC/WMbBdFq/l/wAGwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAYijGvSlTls1Yr0ZUakqct07AAAHDWD9nh3We8vyX9fkAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABxNgbSWJit9H+j/T7gAAMBhJYvERpR67+S6kIRhBQitFoAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABXoQr0pU5q6ZjsHPCV3Sn02813AADIst+q0eea9+X4Lt/mAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAGa5ZDG0rbSWz/T0ZWozozdOas0ABkOTOTWIrLTov19OwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABmWVUcbD3tJLZ/1ujG5XicI/fjp3W3/AAPmBRoVK0uSnFt+RlnDsabVTE6vt0+ff029QAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAGk1YrZRgarvKmvlp+ViORZfF3VP8AF/5lKjTpR5acUl5KwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADi7CVzlZys5WcrNwGgBK4AFgtoFjcYbhbQAAtoG4WAsAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABLYiAC3Le8S1ZLcGtEEQAluPZB9kG7i3HuC3Jbn2QBrRAfZBOyJaovZ2NEyWpzMkAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAEhbgAdTsX1HqhvQUhLUTvoLRD1QS3HqkN20Qth6oau0Xu7F7MeqFuS2HsCN0N6Be59kDoh7ktx9BK7HqwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAerH8QAD2FshJDs0PZGiOrFuaWG1YbTJbnYe59k+ydULctqPVEdxao3uCtYurD2QldmnQ+yCVxu7HuS3H0EAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAtZEtwAH8Ithaqw1oNXQ1d3OpawtVYasDVxvUaufZPsj3RbU3uhqyI7kRbMaVhW2HGyLXQtxKx9kkkhLQcbFr6j1Y+h9kSuxgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA9TmYAAAAAAAAAwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABRui2tiUVYjsAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAR2H8Q9iOwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAth7jd0JaACSYwACyGrACVxqzGgAAGlcEroSuNWErjStoWTW40OKA5UNFtAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAauxJXABbFrs5RK40rDWg1YJbjV2OKGtbDQtyWqQ1d2GlYasW1sNDiJXRa7GkNB0sfZN4jVj7ISI7gS3JK7HsAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAD+IlowAs9D7QtWLY+ySJbho2ItZHUEbo6stZEtz7Qu5Hc3iLYW49xK7N7j2Psjd2PVCV2PUVmrFmyzQ0nqdT7IAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAP4iSuiOqASuxvU6iPsj+EauyWrFufaOjNx7FtBbETozcavI6sWwiO49iO5YTsJtm6PslhbMWwR3ASuxvUvoAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAcyLq5zITszmRzITsFy5fSw9Uc2hcRfUTG7jd0N3Vi+gnYTsN3OYuJ2G7l1YTsXL6WBMTsN3ObUbui+ljYctNAL6WC+lgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAP/aAAgBAxEBPwDobh0NLGgaAXNGHQAADoXNDQ0NGaGjNDQLHQYBoCA6G5oGhpYsGjNA0uGgAAW0A3DoADGBuG49QNLG4WQdRq5YsW0LXLG5YB6gBbQEMYDOhYYCSsAAAWBnQ0AY9QsGgMB6gaAaWNGaGhoxoALBY3AtcNAC1wAAAAAHsNaGgbsBh0C2gWOgFgEPU6D2DoboGAFjcsNDLHQsWDcaNGDQdAaGCN2MDRgdAAtcAZYLaHQZZjALgW0LDVywPUex0AAAAAsPYsWNkWCw9QLHU6G50AARpYuDtY0BltB6gG4NAzQsWANDQZpYAOgAAA9joPUCwbAFxg0ACNANADoaAWLAW0ANgQG6OgdACxYsWLGpYtcALAag0WLFrljVjQ9SxZBuix0AtoWLaBYaGixuWGWHYdgAAAsCNEMGFtC1xo0sABoMRZB1AaOgNaBYZ5FgDoADAOgXCxoAAC2PQ6Gg7WHsNFixoWudCxYtoHQAAC6FawBpYCwAWADoG4AHQDUDU3NzoPVAIGBYANbAAaAdAFsBqam4zqFwDQ6AAAGoWLDGamoG4asOgwDodAAC2huDABgBY0YFtA6AAAAC2OhYLFhD2LFi3QAOgMAN0HQLaDAaLFgaNABjA6ABYA0ALAABZMCwW0A3ANA3LXDlbHBjTLMs9z2crDTAC4xnQCwdAAAC+huINAC4dDoDAAS0CxYsWLAABY6AMNRnQNWAHQLXAAAtcsrA0WTGFg0GAAAAbm4BYGWubgAG4PUNbFhnQA1CwMDcAsNBYAGHQAHsAlfRGFwc2+Z9DlitjlTFCNjkjsP2fLaxpclThMng09mTwtSK01BgbgAAAAA0bo3B6gBoPUGAWBBZAWPI6gbgAA9QNxgAMAAABWOhoGgaWAANANwANA0GgdrGjNGaM0Bo0AsAAgAYdBnQAZoMGALcDoAFrig2hxadgeqAwlLnmipanCyGguXLiEHSxicOp6xJQcXZhdGljQZ0AAADoAbo6BYGdAsMdgOgGhodB2ZoGlgZudRh0LgMHsdDqdBgFkWADcAALFlYsBuAMaB2sDA0AEaHQdrGhZFiyNAYboNA6AGgFgA6FtAexYZZWNGWCyNA0GAARpylsQwjb1FhoRQkoqyHShJ6orUVGVkNdDoJN6IwcXSin1KrnJKT6ko2sSN0ABcvoVqUakbMqUnB2Y7AAAAAdDcHqXNwDcAAAA8jQ3NA6DGDA0ABiHsdBzQmmFkaGhZGgwurGha4AFkNIBgdDcL6D1AYPUAEOw0iyNCx0CyHsD1LHQ6CDoAC2GdABmhoaDsWRZGhoyyGaGli2gaXLXOVjVi1zD4e+rHFLYYxj0K0btEo6komXw/eqfYk25NkZOUWuxZ2TZNasGg3BghNFWkqkWSVnZlkaWLDBgAdAALmhoaDSNB2HY0NA0AAEgDcsHUtctqMsWC5uWQAT2LEdyxZBYsW0LaFgBoAEHQZbQsWDQDQOg7GgAC1AN0boaOhZMsi2hY0HY6BoaG5oAtgsWAAsbh0LAAJXFSViNDmasQoRiKEb7FTDU5PRFLDUYq7WvzLq1kOw0WuWQSSHTTPYpuxThyuy2BTepLllTS6r8iasW0LAgaLXLAmYqjzO6JRcXZj2GAAA7WADcAGDVyxYLAwAEHQsDB2A3DoaABoXQOzG0O1y6ZoG4AAG4wAYHQuAMGFw0GAIOgAAM3GdTyNwABbGgaGgaGljSwDNGzQAjBzdkLDRtYeHSehyWYtDcsFy+g9RK40kNDVjoNFhajWg0kki66D1VwLaC1OgWuASjzIqYbnptrdD0GBfQDoAA9jQdjQ0NDQ0NC4aWDoWGAAtgYdAbRcGGhsdBgAEnZG4dAHsWTZY0NQ0DQAC2g0NG4aAGjA0AABABYtoFiyLAdQ3BgAtixa4dALFiwBYsRpyk9CFBJIjBWsgYJABrYa0FsPXYlboMNy1yyFe2hCDm9yzaSb02HazQ0WAWwFiwF2tjG00pc0QsWADcC1wsWGkWGiwWAsWAaOhYLAItcDQBjNC2gaBYsWBha5PcitCW4kNDR0AYNG4dAAsAwLGww0CyC2gFgDqAahsXNQ3EdSwaG4WAEXQBuaAXQDEruxTwytqKKWwkbDAW4LU6CYuw2hGyBlrlhI5dDlew9HdF9iTT1GB0LFgtctoDFqVqXPFolHlk0AAAAaGlg0YaBYaLFixYsWGgA6AAGgG4dCwFi1wGyerFJJWNxbAPYAaDcsDAEPY3DyGdAGW0NAsWAAEAB0AYW0LAPULAWBbAAAM6lrlmzDUne7EgBpBYSOhYNi6bHqJJijccVYaBqwkWLFh6bD1YwsPYLFi2h0Nxg3oYmL5rhuAABYaAsrFiw0WLFiwWuWN0FgALjZuAMGh7FrlgAN0BJXRytjixAOxuDPIZ0AAOgM0ZoMZYNwsNFhosWABAa2DdDVx6oGwDdhYAAWwAABZtlKg0tTkiaWsAFiwbnQLIsi2g7PYSvoOOgtg5bsxFOEZ2i7osJDasRkpXS6DceVaiauWA3shotcsmhos7Fhoa0NzEQbiMLDAAA1AtoFgBgAbgbgi4wBAywAAFkWN0WGFrjR0DoAPUAADodDoBoaAAAAAaHQudC4aM0HYdjQ0DQYAAtiNOUloKhJodGR7OXYjhpSQ8JJInSlEDC0/tABa5YaCxYSRZMshDXYauNJlkU6kKkFOL0YoplijRlVmoR6kuxJ3Fe1jlG7MlrsNj1A6gdC3Y1QtC2pLcJK6K8eWR1A6AAFiyLIsmWQAdQ6GgHQBgBc3NB2NBgbjAAN0GlgAYdDUewAAaAW6Fg3QbmhYYBoAAAdQC2gWLaFi2hoWAAGBThe1ynFKNkKKQkKCLJlrqxUipIq4WV/dR9VmU4csUgY1csJDRJFixY6gki10O2wNEkmrGEw1PDYeFCC0ikl8lY1Rq2ijUlTkpReqG01ruNaHQexJiaTNGPUCNgsJXNR3YkIYdDEwurgAAADHax0BoBgGgwABbB6FgeqNAZuGhoOx0ADQEdQQHQ2LXRpY0A0AAS0Nyx0NzQ0OhZAAB0ABBYDQNB6mgG5sABuC10KcbpEUkhINy12OLsSRThzSSJx1sdA3LaFkw0aRLUR5DSFuMsG5bQe6NWh3Yri0sJj10NbA3cbHqD1OpcWwjc0Oh0GWuaIYTimrE48rsGgaWAALaGpZmpZscGkKLbsRwze59VSJ4Z9CUHHcsDQAAkmjRlkGh0LI3RodAGdAQHSx0DQHqDGABYAAFsdA3CyNwGdA0Z0AAAOhoaFjSwWC4DAA6lPDzn0HhmmrEYpJCEJXZytiR0JbaC0JasLFk0WuxKzEi2g0rGlhjEtUWux2aLK4lZmo9rWGkxbFx7XLqw2hu6C5pYC4PUWwrG+w7GgrPRjRzXVgauWGYmnfVBoDAAALXIU2hUGxUkrF0GhKCkrEqCkrFWjKm7Ma0AALaAWAsAwNzoaGgAjodBs3DoaHQDSwAbgAtToPUsDLAWuPY3Gi4M3AAB7FrlgYA9xgMtodAoU+eSOSKpk2mxaliKuQQxbDHZocdSUdC2g1odBaj0RoWHYsmhxRy3QlqiS0LXQloNbmuhdSbXYt0Og2mN3G9C6C90XLjszoAJgMY9h6oJWAKiTuicbOw0dAAALBRhfUgrA9QAjqYiClElGzsNDWgALY0LXDc6lgsdSwPU0LXGlY0saD2GNILFiwWGtAtoFgGgDcLXBlgCwzoCGAAAAFrlgAaDRlgp0nN6CwsLFWk4MwsUqdxts3NLFi1kRk7GhfUfYSLXErjtsNFtBFmxWbJLQFa1ixYS1Ry6aji9LDXQSWh9k5VFtotc66l7HQY9dDQeodRh0DQWwaF0MsPYdhgwqWsTgpolFp2AAEG5YoxUpakIosFrnIor3h8r6HJdaDTBq5WoRaJxcWWGADOgWCw9AsAHQGGrDoNI3CzAeoAFgAC1xosFjdHQCwW0A6BYAEAAAG4BRo87I01FGxUSlFoppKNjcaEiw43WhZIhdsiupa5ZWJJIurFkNFgWiGtbIabihqxYmtWWuJXaIpMd7jvsPm50ltZiV2eRLdg7F7ouPYAADoIBaiuPQkAxlirsRlZlZps6AdBgAUIbCQthK5GnFK7JWkzl6HLYWq1JRsNFrlaldE4tOxYZYENXCw9y+h0Nwe5uaD2DodDQALAMYAWAA6AHQcUopljoPYZYALAAA0i2giwWRYsixYaLaFixa5GEpOyKeDutSFFU1oSdxk43E7ITTNBLQitSTsWexC6FsQXQtoSk5I3Q1c5U0OKSNth7EbIesUxq6Iw5mkVsPOkoymviV0MVkxob01GzSxzWRzN9Ru7JPohu71DcuaDsGgBYAFuPUauNIYMHsVdiQbgDACKu7FKNogQ02HJSjylrCsS11LWYve0GrA0mVqSaGrOwAthoBltAsMYNAFgBBudAsNBYtqWLFtBoLAALY6DLGg0dDcsbnQAAAG2mPYjJtkmkiL5kAKN9iOGkyOCnIjlra1X9fefUIRS/r9RYdRlYdlohtAwUb6CjYIrREGhtS0FGyEeZfUb0NRq6HHYaZJbMS1LDirIUL02+xToyqTUVu3Yw9S9pGZ4p4hUoPaCshxuUvZunJPfSxSoSqN8vRX+4kro0HZodk0XT0N0MAYCAALFixYUb7DTY9xgD1Kgy2gBYBItccWUo3ZBWVhK40iKsRSTuS3ErK47SSQ0JaEoXAlFNFenZ3AEHQ0HqaBoADLlwHqAGgbnQsHQ0GGwAAIAHqdBjsAaGg7BoAD1Y/gI7k4PcpQcloOnIjSlIp4Vt6kKCiQSQpqw6mpKRN6WHsNaDGrliwrNFhJ2VhaLU1voJMdmkDudDlRYteKT6DRJ6DjcjHUVuWxh43hP0FFwgmjMcvq4XFTo1I2a/J6rr2IV1WjJLo2vudizuK3PoYPG1MMqig/ii4v0dr/kTlaKQ5JsclbQbuhgD2AAA3RqAtxFhbDGMB6ktipIY0GtiNKUiGDnIhlj5btf195LArt/X3n1RJ7EsNFIrUkloUoq4iKRK2wloReo48y0NGrELcthx1JLQhJKLTJRshhVgmrFSHK7AICwBodA0NLBoGhoBuD2AsWAYPUNwAAN0XHqXC5cuAAABodWPWKQviORum2YGN4P1HTSRKMU9CGjG7sQmmhDuSu0PYsct2ezdjTQepbQ6EErIauxIextYFZ7nWwtxSUU2LZkg0ewo3ZB8zkuzt+Cf6mGi20u+hVlfRbIzTM6mPxLrzVpNL8FbsjF4OdDkurKaUl8xvVHNeWg5XRLVDatoXL6AMGwAAewNmgWuhbG6Bu6GAwqMqO5a5LZIpx5nYp0Y2KVDmMNQpwXvdiUYyjZdCaFHmkThrYqQuRhZgjRoYU3oNWRdmjRZuxe2hdNWJJoGrlencYLYOgdCwW1ABnUewWLD2DodBAwNDQGBoAAB0NA0DQNA0DQAHsBTpuc2kYek6tWMF1ZSouOMVP+tjExUMJOC7MynSg/X9ET1YxCsJCjc5DkSPZpodPYcUPbQabVhxucqZa5oR+FA0mPobiSdhxVzQRNJJIiiSuhLUUdCzuRSasZLONHGRqyV1Eqw5IqP9aE3qrGIrObchu6HuOSY2OzHqzoMGAAAXuMuFtALBLUYMJOyJSuVNxK0UOmpLQo0VHRmHppR0KMUtSVPkitdyE0ouJNpNllzIqRTROOhJ6WLiI7j1QJ2ZG0kWa0G1ohq5JtjWo3zIYTjdFam0wWwM0NGOxoaBZWLIdg0LGlgCwzoDEM3BgdB6hcABFywHQYxmgAABFNuyMNl1So7taf15mVKMMZO/S/5mFglj5W6N/wCRJv8AtK/9bGNd6EvRmVv/AGd+v6IlqwILUUVY5UkX0Ga3NlY6DsSjdDi9iKGOyIWcR2GiVmkRTb0Iq7S6k1d6Ekk7EYtrTcqLmkQg2WdmNGisNNq5CVnoKTUWupVm5NIc1q3vdjkaNO5uxjNLA2MuPUAYD3DoDAWrR1LHSw3fYHsDJMm9B6uxNXkSRF8rRh4OpJMprVItqVGpJW6FlqiV27EklLQ5U0iqraMnFWOW5ykdBotqWISsydpLQ6mrGtLj2NbklcYVIXROLTEG5Y2NAsNG4MDcAtoHQ6AD0Og9QuPYHYAdgC6AHsCVwDoAzcBlHDTqySRhcLDDw13K1ZzZTpU4S5oqzZyRUrpHsabn7Tl1KkYzjyshShBWgrIkrnIKmRikyK0N0JC1RazJLSwkJW1JbklY6EtGNkEnESvYa2JakIXaKaV4tlbWtKK6Mas2imvfT8n+RjcNSoUcPOL1km2U4uVeCXdGNozpVnGas7jimziF4ulSozwyu1ON9tm7N69l8zeRjcFUrZjhq8doc1/mrF73Kyi5KxUtq+49jQTs0PTYZoWNy2hYAB67GgXOh0NzqBiq7pQ03ZSxFVzSve5VqqnByZLFVZO9yhJypRctwaCaJJsjF3IxuyRSjzSsYenywQtBbXLJIbJq5ZXQ0rWRVSaZNaCV0WG7FupayLBCSuTWiYnYlaQ7JtD2EyQNXK8LosDSLXDQ3QWNxrQtcADcZ0LAdAAepuiwG4AAaB0GABuWAOha7MBlkqklKa0/rzFRpUF7q1+ZOWo5cxC9yw4nI2hU3YcFaw0jdFtRaIki10hRu7EkiSuhppCV7Jk4twTY020W0ZNXsyUXYpxbgRV7IcHuJXZSilOKKePpSzGGCe7XN8r27fqZvTo0c1r06LvBS0eq0+epXXvv1MJDnbXk/wAmOq5JJ9ChN0pqot4tP7jNczqZliJYirpJpfh6JDs4XtsaVFEpqpHnjN9Xb0voTa5XYlJq3clJNkm2BcvoMGG4AMuErXC1x7G4ywdDHa1Ip7HsqbtZbGPk3KMUUsPCEErCSikl0EtDluSjoSVzkRyIcEmThZGDpXqEZJRt1IO24muWyL2Wo7vQexazRLWzKkfdKidiMdkctxU7os72GhotqIi+aNn0JJm40bjQ0mtAKkbxKkLAdAsFtAuMNB6m50OgAzQNzyNy5Y0NCysAwAsWN0MYwsWLFjdFixbQsWIQcpJGBwEIw5pL+vvKk3eyJNtXHJsUXIhTsyFNMtGN7FrFhq9kSFFNCiJPlOVuJb3SKZUTshxdrDTcSKbaJxfs/mJPQgm02ODcSUWoXKa9xlBJyKlaXI6X2b3+Y1exQjzTS8zD4VThKb72RisfUqcQzw0PhjFN+rV15/cxxTk7mESSnL+6/wAdCySTIzXNeRo011Jtxdhy0HqybUZWErktR7D2HcB7D2LXNTZFgYwGFixa6LFiyGGNpOUVJdDC4hxkoPYxGuJS9AUb6CVtBRd7FSNolky3QcRwJRRhKL5rii7Cg+U9m7DjqSTQ7mt0TtYm09GSV0dBU0yyHFbj1Yxl7oplRasW44uxJK6sND0GrltCxXUeUZ0NzoM1C7AADcN0BoWRoWLICxbQexYsWLAAbo6BbQCw7gah0DDYV1GilgIU7Sb2/ruTqt7DYtdBU76kYq5yuSGtNCy2OVWIx6CunsYmmoTa7CXusXQUtFEy+jTq1XCpPlVnra+yutF32KcU00hNNIeseUjFXabLJwId30KkEqUlfZiSS+8pxuiycLdiylS5LbakabdOSSMNh5SrqXNpZq1uve5iVz8skraJDpqKu+uxh8Piq01DCQ56j2V0rvtd6L5mDdOWX4bC4mn7KpTl+815uXWz20dnf4W7maYWlh8xqqlrdvXXVdHZ7aFTBSheUtFexgsNhnlmMje9SCg0tdnOKb7dfUabjfocqkyfupNE5d+pJqSFKKVycuZ3LkrMugB2LXSLaGlrCV1qOyG7m5YAsaMYAAGNpxhNOPUxl41VIi1JJopxsxQLKxKKaFSiezTORNE6aaOVc2pQilT0ErnRCb3B2GtDdWHaysZNlFbNcSsNRV5yTtt0TfVpdO5dNiimWtoOOpOJJWHsNWBOzI2khx1sW5hxuKzJIUXIskrEtycVJFSNmaltAOgABYALBZDWhYsNXLDQDRbQsAAAHQAYbm4G5hcE5u7KNONOCZKdw5bspwTjcl0XYbtYiunYcbMtr5i+Gxf3iaUargYjUjqrGHpOrNRXn+CuK1xu0rIw0lGoJq7RLsSiuYjFXaFpoewnV50uqv8AdqUqd5W7EKLm3H+tCNG9GT7W/Mgnzcvc4eytY3EzpSdlyv8AD5oy3K5zx9PDreTt/WpjsK6cuX+W6+5sopVJQnLaOn5nD+NqZXXq1Yu8ueUl5Xk2u+ytuPOXWoYiNRa1Hd+r1ey/yOKqUctxuGpVN6qVvVxv0v8AoZ5mcMfiGkrXf5fJEZunUnJ9VYj70ERptU+foSmpWJzu0NtT02/4YvdskDOgBa5axq0J2JScgtpcsWFohlgsWubgAGMqzhUXK+hHnxFRc2pjKTqQut0QxNWnHlizCzc6MW9y7aGWuOI1qSVhq5KF2UV+7SI03KyRKLXyLiV9h7EuxZNHKtjI80q5ZiliKPxK6+9W6p9zkSjchG7RJJPQWjJK5PYY9QKbb0JK6Iu8SYnsSSbJaIvfQa0Gk0VaTnLQnBx0AB6nQ6AdAABlgCwWuWA8gNwDcewWuFh3ZqABYSuYXBub1IQjT2OZcvKSRCHMKkhP3bI3dzS4nyu5yLm5RJqVxWjK/QqRtqjdqROLUbPpoQ1diN4Xaeq/U1i0S3TRRilJFak6dVohSjNWW5LD8lT2nSX6EoJVF5nskpcpgp4fEyw1CnG0uVxbu9b36dNxYScavK9NT6uozjbXo/u3MpyepiFXp20UHL/wFrzMZhpYecXLqrmUuhRjVVTXmjbqrO6fT0MNjZYfEKdN+8ndfcY2X7t1F1bb9epgcwWIwcJxVlLX+tBT1duoqiTtujPF/bNShUxTvKg4uPS1lbpa9lprcblSnZ9B1liFzx6kZpXiug6j9lyN6X/EqNPYeugnZ6s5nqTk29WXG9Bu4CV2DRJ20DlucoouSSHEsNFi2hY5bjRYA3AnCM1Zq5CnGKtFWLKwqFKTu4q5FJWsLVCV9hR00LNuwlpYtdEkKD1a6FC7jYhzQlzRepUlzVHKSOR2sLmS0G7MtcaVhpNF1FXRb3WUIXmiT94jJMvdW7FSKRYkk0W2JJJkHZ3LpqxsiSurD0I2JK6Gkx6ojBslBRjYqUlJWKtJwGagNgAAAzoFtQEg6HQZc2Bm4AAAMANzDYVuzZTtTHK7Iu9rEY8yZyqOhZ3I3Ukzl1aFFP5DSkrIacoqS6aE4e966/eVE1CL/rQlFv3SlyuKKtNTlK3VX/EcbN+RJ3uRi3DmHZq453il2MxrQryjKP8ALFfdFIwtOdPGyqP4bL703f8ANFPJa+My2viKSuqT122t5tfqSUpU1LtoSTU3c4PtUzHDVo7c6X3SscSJUs0rRt9p/mZZw/icZRlUpRukr7r9WjHqvleJlRW9rPbZ/eYyVac5KrsnZbbfL9TEu1mv60KLcaql6GcRnyyTWnNJfhF/qSqKnVp4ePW/4WPZySv3KbUZLuhS5tSUryuxSVlboU82i85ngX/KpL8fL9Rv93Yk1y2RJpIk7jY2DBFro0NIWbJO7ErnLZFtDodC+lh2NNiyuNaFgsWViyaHYB6gCWhFEY3dzqhItoRV2zl1sNaOxOI37phtXYp+/TUnuNuTIycJX7C2GtR9hq9iUbIcbpiV0ygrST9SWqLW9SL1Jq6uOXutDGxhTd0SWiCe4tyauiw0Qmou5OXNK40TipKxVp8jNw3DYAAALBoAMuABoaAABqbmoG4bmHoczTIJRjoXb0FEhFt6EEnbzJ6uw03Gw9Y3ErNTWz0JwcZ2Kcfddt0UqScXD+ZX+4VFzoxS3VyrT5qDXVajh7vMmU46O3QwVO6XMtNV8jMsP7KvKPcdNN38ikrwaOXSzFoYPM6eMrVKUd6b5X9yfZdxU1Gqovsf2jjcHSqUsPPljU0londW807fKxlWWVcZha0oPSCv/WqMTlOIjOE2rRmrp6du1zLIfU4Qq0nrGV/ne/W/Uz+tiMxxSrxlytyTez0urrputL9NzK87x2AwyVN2g35apbrZv5meZtLMK8q810/LbojOMDWpTSl1Sa22a9SpC9OMu+n3GGp++ovc4kr5bUyOjh6bvO8ZS+L4lFRflrZbaaElT5rr5H1mpJq78hz0aHL3V+I2pS3E09Lksqwbx7x3L+9ta93trpa9uvYvoOWl7lSzZe42r6DabNDobiRuinFNXKk1LYUblklYuNl2XLhe4O7VjqXHcep0sPQLAwSb0RGKSbZTnGVR01uiKaRYWjFFtkINIcWcjuVIto5NUYfR3sSoc0ml2uXafN2G27sSSZO0ncsaF7jZC9mkR2+TFqtC7eoKz1KkbO47DGFLyHGzsRknuVEhJXE7qwthQ5kSetjZjdzlsV6SnEkrOwzcAADoAPYewMAACwAAAAAQjzSsUKajAu9BXuiCuxLks0QklG3YqRTSaItKxGyvFlFNwnD5/d/w5UV4qf8AWhQsqmuzMM0kk94uxh4ONWdPt+pSwsqz9mlrqiNN25OupS9yq0+tzLMFVrUo8i1VvzS/UzqhKOMalv8A5GNp01SoShvbX1IU3zSiv66lZ0Fh4r7Xz2Mgx1XH4Wc5qzjOUfknv0FQw2Dcai3qTs992vn0XkiUFWqwkvQzHB1IQimtbL8NH+Rl2YzwdKrSi/iVvl9zMXnlPEZfh6PWmrf5dP8AMwVWj/Z1aD+Jyi1vtfX+mY2nLD1aSezjF/erlTH0nlcMP9pSb67NehKShDlZjsfVrzu3skvkiWmFXqyhUbkpdTE4xura+lrE7XG7RTN16jY2ktB6LQuki9x2WxKXMdBsaTeg9DoC2EjnaVkW1ErDasMsboAFsN3LA0W0I6EtwNAIuzE+aNi3LjFbqvyF8LQl7okRsxIUHdFm5NnJfclBc1zCUXVqxpx3bS+b0X4mPw9bB4mdKeklo9t/xROyfKPyG7l0O9xvUvqVL2KbXMxapobstC93ZD7IjJJ2JpOIx6gimrMldtMvdlRaXGUdhpXuXSgRjdjeliS1KceZMaMTSt7xoaWHaxoGhoaGgADAYdToDNDoG5YOgbgFkwC1yyLIMOk5oWqOhCJolYa6lJwUk33KMYPmh9xVpODsOKauUXCEoz6rccEpTp9Hqjl5X6CozeKhNbSjr6pFaFOE0+sl+NiNOCpzm3794tfdr5feKhFV2rbSZmGWzwmLdO9+V/11ZklZ05Oa+K34meYarVj7eXVlacabi5adPvMHTwzxtCdb+G2lLf57a/cZnRpLF8tF3im0vT56me5TTwGZ08TV0bjyrr8Vn0b7dilln1qjNyV1C0vu07+ZD6lPJaf/AD9jJ3320t5FbGyq4OFGT+Hm/F3HNUndPUa56crvRMjiFyuHRbGHxKgkpPUr4uUpvleg5twbOVpNPex7eMKPJN6tq33O5hK2KniMRCovdUly7bcsb+fxX31+VipJtplWLjK3dXHfS5N2XmOUtWupJRkO6epKQ7NgM0GgsEEmxyWti1y1kNstcsDdy7RcCwizaGmW0DQaQLaxpYYldWI3S0IJIxaUZ059nb7wimJaEIOTsjC4KVSUYxHFTrzhF3SbR7FwtpofV25pruZjgpYdqMt9yEJezU+mxUp89rdionyoHZWJWaL9BWsiVloSs9CPxCejQ7t6iJMU7CldFSNnYkEVdpHKuVy7F7wVi1pFouLJWsR2JfAhEF79hxu7EoNFOajqSbZKN1YqQ5ZNFtC2oAwAOgPUsG6GMA6BpYZYAADQ0LodjQDCxvMQlcjG0TZlKHNFoiim7xut0ZhBOEZrp+pBJwa8yyicnNGFT5f195i6fLPlS31MLTnVwUKiXwtr8jGZROeAoYrpdr7nbv5djE0dYtbXa+5nHscLltavio6QUFLr0ivV/gYuum5Smzh2SxdRShs7P8UZzl/Nl1WnBfB+SaXcz3KZ41UoU5WcZRlt0i7tbonlalk9PGJ7TcLf9u3vv5dvmQpN1DjnAUc/qYT2UrckYt6N6xb7tFCbjN0o9VYUpSulsUG+WSkS0iijHmhKPlf9SceWSXXccrp2Hdv1KbfLYx2HprI8Pire85Sg/kk7/j2+ZyKVNt9NV+X6lOT1JX5bdipNNp9jnujdajZOyQ5e7clZpA7XLpj1QCJtO1g6CVyKsh7CRYbNSwzoXNLgQs1Ycd2OxuaWHa47FgaI7iRFNGPhzYeVumpS/eRjJdUQp3jcdPl0MDh24ohWdCpzR6IwlBRnKS66letCVCnTS1jdP5spw5bme4KWHlBSd20n945tUOXzHP8AdxXky93LyI6oY9VoS1Vupu7DeqJ7Ii0pkdx3ItE3dId9COquVlfUdhkdGmOadNRE1eyKytYjqrE1qQSsRSlTHYi7SuQWjYp2k7k4pFrIZiqSa5ixYsi10BYsWAsWRY6AbjN0dCw0M6AAAAWCwGGo82rFBRVkRWhSjfUtdvyErCpyac+xyL3Z9yGk04mE0Si3uKmoYhwezJ4Nb/P7jB2lQdxxhOKktzIqNTEYPEUlK0Y2drfIzHBqeTUqEOkeZ+triqU3llSm1s01+RVyt1qLlJe7t9/zuY3I6c8rpYh/bvF/1cyrhqGQYyjllJ6KUYr0bS6t/mZVRTwTw899U/yJU/7Lzdc/vckrdv8AMzPE058ygrPmk/vtYjVVO91fQyCEK2JhCp8N/wCttTGYpZbm054Z/DJ2/p3K9eVWvOUt5GVUXiMbRpfztR+/QzrCRw2LqUk72bX3Oxwng6GIzaFOu/cfNffs7ba7mY4anQxdSlTd1GTS/K4nZ6CnoKXLIqT5uVvocKQw39oNYj4XGS69vLUzOrTw8oyT0cuX7/8Ahje5KWiOtjRpF7pEndnMrXJO6GPY3GJXJJp2Bnsrwch6sUdC4MuC1OoNDE7CegtCAkmiUdRrQTRKzY0JIYtRCimiKJ0/aQlHumZU+bDRfVafcQhaNjCYONWpGMupShGE7GJweFqZjRw6+BqN992k359Stja+FxNTLabvTu+3R2Xd9O5XyxU8LCopXcnbbt8yVL2NX3lqlsYrAVKdODl1/ruYqSVSyNHDzHHqJvmGrG7NdxyvqIbui15EXqS+IvZ3L8w2lsR+IcNCStoMI21F8SL3TIq1kVUkyPYhHmpeg0RV9CnC1kyqrkVdcoE7ShylSPLKwBbQAAYGjNA0DQ0sdAGhmgAMAAClT55EYOMdh7kFchG0RLRlOPM7EZyVN07blKSV4S67epG6fL1MNOc3ZboxdB80Ki3MJgo4uhNc1nytrTqmlb7mYCk43jL0MPldOWWSqJ6wl+DXr5GHxH1Zvk+1+hS4howzCNLpNWW/2Uk+hgcBh/rtWjVlZLmTVn+jMfhqtLK6U1tJu/yOIsLPBVqdGT9211/WrJYipPELES1knf7v67GMzqq6cZxlq9dv+AYjHyniLz/r8DH1LVItddTD3qtRsPE/VKbXe6/rcnOTfNe7E22pIp4idGXPFaoxNT2lONS97rUeJcZrk0aMyw8sO6bvdyim/JtCfYjraw5XbsUY+0jNeV/6+8z/AACwkqMov4oKX9aszHBQxlOEJPRSUvu+4le40mtTdHMkKVopkpaadBfA2PYHZosaEWk0yvP2lSUu40RjzSsVaia5ULcZa4x6nQDoAGg7bFJpsur2Q1ccU+pyolBoa0GtDcSIq7IrQS1IQ6mUwjGVWnfVS09GU8OpNMxtOjQUfZSvp2e/zIzSq3ZjsHGEqbpyu3FPa1nbYhQ95SfxMzLFScIQUbWu/vZChVxGJxPOvfpK8l2t57fdco5xWxtByq6OLklt0fkkVpc01bsJXKkmkrDL3SQ5e8Nu1jVo8hvRia57lO3MrlZrndhvSw4koq4rC1RVhaQ0WQaXRy32HdNFVbEdGUKi5bFem1KyKesiNprckna/QqJWTRVhZJ9y1xwaVzFUrrmQ0WLDLAAW0AtoG4Fiwy2gW1CxYAAGboC1zCUVGN2SepZMhHS5G19BOyLWL3aJLmdxza5JdjD01zxdNbkqM61NwS1RgY8lKVOW48DOH75bRt+Pz8jhGaqV50X8Mlf5r/hzM8FjMI6+L3hFqKWm/Xu+vYx2GjnMaOPitaLS++MV5duzMjvWzKMan2nr/SM/wFbC0OT7Kb7dde7ZjsbOvOCqu/LZL+kVU4ylDqYmslTt2MRO8799CtHmoxn02/r7ijUkptS7FaftE77j0jbuU2lFMw6lOCaHTpQymDl8fO/usvkNxi7oq3nNvuRk2jSKuKeljJXhfZYj27s+R8u/xXXby76FWtOVnJ7aEr3JSdxu6t1OZoSvLQ57uxPRXFsy10bjXuDGK47WI8vK779BScXoWuJDLjsy1wYzc6AXEFTEU6HI5v4nyr1af+RK490JXVxpXJwSjoSWg1oNaC00EtiKIRu2ZfhXWbXRamEpKGczh/PFP5p2K1CWGoOrJaK7+5Cr4fFZRha9L7cW3vvfz8ijl3tMTOtHdxS+5yff+8zLcNisZjIQT0trtuvu/wAjHxVKEH1t+bMVTnjcEsVR+wrP77dbfkYfHzxCqVrazjr+BjPZQotrdXbJVozVOMd0Yyb92D3RUVhyurD0HvoN6Gysh6FlYktbEPiRXXLNiSi7MaJR6FkhaonG+o0NKwmrWBq1miLsyaXLZBQV0ieskQpqMXLqSjyRSKkWopInG1kS96MSKdxwu0ipFNNIrUnB2AsAAbjA0ZoaM6FrljoA9R6lhgAA1cAw9H2kiUVBJG7IQ5kJWikJX1IRUrMklezIJtWW4o3jzrbqU7N8r2f9IyjFYaGEjCp8cZrvtrfbTe3mZXiMHPMlJ/C3br109dzGYOODzKdN7J/g/vMVCpRoVsOlomr/AHXX4NGWzr0qca9HTl0e3r1MwzKWIjJ9Ju7Xn9xlGaShTnh47OSv8vl1RhqVWhnipx3Uvzehm+Bxiymaq6tavbZPTZ9ilg5V6dWovsLmMwpYedGniqMruS1Vno1vq9/krE5wm1G+90YnDOFNS6p7GFg61GdPpa/zX/AuZNl1bMMRyw3UW+my+aOS1RpvYrXSijmUtEYCMnQsulzC4vBQwuKpYl3k0+Tf4r+X66Emnt1Hh5SwftUtIuz+Y4q7XYm04WHdLQcly6lWWrSHL3UybWhBq6J2vYTtJG97EneJFqzOiPIt7jG9Bq5fSyGLa49SKuA2WAYHQGAldGMy2GMqUHN/w583zSa7ruYqlGnPlg7qy12EtBXaTLsld079CfYvoLUSRHYiUafNLQwVH2cG+uxi6aoZlg670Tbi/wDt5afqZnjaNWDowV42Xfs7+ZRhSjh3yuyWy8vUy/DxjByl03MPj8ThFGq9JO/Z/wCZjEpYVYmfw3V/m7ev4GWwxTwUsHQjedRx6paJ676beaMm+rwdXDT6xl33SuZ3RoShJ09pX0129SjrUSMzxMK9VTj/ACr7y3tsLaMdY6t+WxpshkmubQNhl07ErEPiRValK4rNkndstcktSG9iS0JasjG7SJKzsJNq4pPlJq0jpcKOtkhq8V3KkWvdZrKZNSXTQlqyyUFqXs7ohZweupyNK3UxmHvBPqMAOgAW0BlgsMGi1wAAADcOgWuYOnyxRPVkI3YkoxsjRpIS1sKytYlG8dCMndW3uYXCxrTTa9yV180m/UqUXTq8nZlKjFwv1ZlVGjisfRjTVk0r77r+vQzvATjSeI/vf118jJ6E8fl88wlvWlZ/9u+6u3bokZNVpYSlWwtVav16adL/AJih7Rzj2MBgcTTzeda/uTSdtN1p3v8AoZhmGGUaWYU/4nXfp8rfgYniutiaFSEno1bp1+RVqOKk1syeJ5YSRiqrcU49Co5ySfRq5kdGFfFxU37rdn6NWfn9xja6yPHzhhHorpejXncrz55KS6lSTaQrJnDmAoYjLMRUk/ejtvv95j4ctT+uhNvmKGPdPA1MP0lZ/d8hy1PJid1YtfQmrJMt7pLVEdiejLaElaRKNrobTloPaw7D/hoGyzbFT9xyLi1Hogb0Gbm5ojoLYBgHQVCrLH06il7qTTXm7ddyo7uw+xuhvWxJvk8hu4xIir2ZFEI3MBhudjw8moxj01ZxfTpwyqlVh8UJKX3O36mIpylhI1Y7NaGSUFUymPt/jT1/padDL40oxs17srpfcVpKpiI4d/Et/kKqqmGlRlsrv7mYXOHh6VP2T96Df4+qMqwVeOGtN3dnLpsYqlDEUElvzW+8xFCVCvKD3WhzNwsQqSgpKPXRkloWGveEh6M5dbE1YZF2kislGWhblehrY6Ek3qhNIZKOpLcL6WE9CUWKzY9yDsizHFzTfYUby5ibtGxFLmsNJpIlFJss7XOqJtN6mKo8juuoWuW0Gg3ABmgzQsHQADYAAC2hYYUouU0inF8jsalODUdRJMVtCE42XNuiUUvmeznhqnvonS9nNVYr3W7/APA7mQ5zRwuW4nBTV3USa30t8nf70VMnxOLovF0o3jD4ndaX06u7+SKEVz3vdMyvE4zDScp+7JSdtndd/wDhxVMwzDLKdDCw55wnzvVL3LNNa2W7T016WsZdmdR4qlhqj0v+voca46g8dFQfvRVnv3b/AFFl9bD5hTU1ZTtJbaxl13/4JjsNVwOMnQk78jav31+ZVmqddOX21dfLT9CU6blZMxU+afJfYnjMLLB06dvfTd99vyJOFVuN7Ir5lgHkNHDL+JGW+uzvfpYwdKpXqyw9JXbTf3J9yrzQ9ye/+RzboUk9SFl6mGxk6dSNnpfU4tq5bNUJYR3binL4tH13/TQq3UmyU1bl6k2loOTk7jaeiG+Via5WJxsRtexTV9OxPVA2pai1uRlaSsNjuN2WhuYfDzry5YE0oVFBdCpVTvGOqCKQrNnU6huBa4kaEkNK9hrQYFLWasSjqStax0shq7G/csOwrNHKQVkRV2ihTcpqJhsLPDvknufU5ql7R7bGacP4XFZLKvKp70o1ElZ7paLful0MvrVMZkeBaenJ+K9xv5uJeE6ipUpXcUnLT1TWvp07lOqp0qNek70nfXaz2ej1eqt+JmmXVlWrYylC9Wa2utX2u3ZfcZHmWHjg1icx/dRlFK+srTl0tFa2vva3oYfKqmX46vh3K/PNtaWtrtu/xJTxOXV6lKppLVNaej2uUMTUU3ydGn9z3Mzk5Yibve+vYjeUdBsd7jV7s66m6SGtHoaJE5XSHJRauRVpFZapjixRd+400xu47XE043Kl2tCWw007BHR2uNNrcT0JEZWsQ96KuRk02ugk0rMq31Ze1iWltSMfaLc5Y8trk97roOzbK8FUhYmnGVmMGB0DQsipyuMWh6mhoNFkaHUe5uAbsNwAADCr3jmSjykU27G6sLRXLJ6ItqYOUfhlsY/J8RRhHE1Phnqtv0d/vRHDO0qD+Rg4pvle6OEJ3yrGUXu4pffdfqU6D+svC9V/w/8AWpjacopMyrM54ZNQ6q33/IxPtMLjE5brX+tyviJYhyqPdszHOaWJw+HjFWlSVn56Ly/zOIsesVjp4lbSt+SXby7GeYipXpYCNNfBOTfo1/W2pCm1VV+1xUI80qnV2/ArOEarfSxUrOU2irW+GHYpY2VCoqsN0mvvTTMRU9o+YUgcnuNrmRho+3k6S3e3qTempu7klew3oOySaJ6xTIaux1XkPSRqm0S2aLXQJ2Oo2eQ17qDDTVLDTkt9vvJNvXqLUSuW2LDehoXNywlcSvocuhyjVxxuWAKWkhtNj1QS1F8Fhi2IqzQld2KULmFjaqj4qsWZtF0sow/Lu5N/gjK4+0qewl9p6fc2cIVFQwOJy/7VOc4r0/4e5hMLKnhpc29ihxvhcfSjh4K0YzcOvxXu/sruvLzMw9tWotT3ilF7dt/x/wCCTwUsLlEMsvdrb+v+CVKsqeHoVJ/HGKb9Vr6dCtnkMXjXi6i0d7/NeS/QwmMjRrTn3TX3orv3nLuLTQatYtcY9yxvoStJIlZorJWQlqipqjsX7FtBq6JXIPmJK8SUb7ErvU2I7kl7txOzGrpsWxSd4LyKSvFlr3fcqJPQnFSWg1qRT9noON2ipCyRKIYulZ8xZGgDsBbQGixa5YLDRYaBoaGgAsbgWLaFjQwcE2T7FFa3GlYaViLSj5jd0rFGVtCrxHVxGWwwFRXjHZ/pt+pmGMli3TrL4oxin8klf8Bw5qqqrZ7/AD3MDjcThFNYaVrqz22+dzF4ipVqe3b9/f8AroZNhlmWHqRcrOEXP1s0n273/QU/ZzavsZhmVTFzjOq7yStfbb0XmcK1sFXpYiliXZNK271Xp+ph5qpU5WZhZUZOXRP8DJb5nhVVoq6cVL5NJ9bGKr4ank1KpNfvLyTevwp3Wm33a9zC1VVhFrVMdFu93qtSdCf1hRirt9PL1Kt5NtClenqKSat0G1Yk9LF7qwnd3MpxkMJmVHFSV1TkpW72d/O33GcYqGLzDEV4aKU5yXzk2vwY2rkneImrWFLT0HqiLV0Sad0TtZDZK3LdD1NlY3Nle+pZ2uLUbUpabIcHa/cr1Eqapx2QEdB7EVdk4WAEriihRvoTjcSTRZInFWGhjSLFNtDdxMvrqSaexzNwaLltEcq3ILXQwWFnWfLDcw1FupzPoUaV9W9DC+2hLDUqusHK8dvJPbXtv/mY7MHhMxlWhpyPR9vwZkEqGJ4hxVODvTrKM1vr9mXZ7tkqCoV/q9PSMXyr0svn95w9wXCrSxNDGys44idRK26fLbWMtNv80ZpU9pyRTslG33KxmWKy+EqdOov36fuP3tnurbd9WZdDBTyqvLEa1FFqG/Z9tPvKmY4aeW8ltbpqWuz0ta3dmFp4eNOrzq8pK6330/QqQcqafYUHOaS9SpytvUew7NJHUTFZDtZFlsTWwtWNpqyKkFoSi073NbaEdHqNtLQV4sabE0oNE7uyRK8nqR1dh6WR7qlYmrRaL6FKdkylVjyWOZKnZE7bvqQjoSjrdEW/hOVKApKUGrFVe/7o1Z2K8OeDRJWdmaAaAAAAdAYdANxmljyAsGh0FozCK6HbYgm9iEu5JapolFaM03KcUxWuUmnRfc4ehDGzlgPtSvy+qV/JdOrHJ4Wsl1i9f1HUU20ZfXlQpz5Xq9PyOIJ0Iypum9HFX33KFRzmk+5nuGpYXFUVQd4zgpPfRu/e/YhiFTqKS6mdZVVhhadeS9yorr7l53/IyzHPAwl7PqnH7/vMa1VvT6NGBnGlRcY9Fp8jPsHWws6daS0muZbf8Ep4ujgMswONXxQck99nFrzXXsY2vGviJTjs3cspRKdlFpk7bGjEkXsmN3joXSSRJJai20I2uWvFoibIno7E02kK71I22Q3bQ3RCNk5MRN6olshPlhYxCUbJdEWugQtSWpTjdMr7pFjQWpCNyMbIsiSTsSiuhKN0OK1HEkjlGhRdkWLjaZfQtoCScSK00KavJGRVPY1Obsvz0MNgZOlOt0X6/MrwnKhyx7r8DKc1xWZVqeIraSoOUI7bXi+iX8vW/qY5qvWqN9f+ASprD59g5R0U4zp/cuZfizBU3QopSet3+LuYDAyw2FqXfvOpKXyaj590/MrYnFw4hnUj8E7rp0ivmY/L/b53hcQ/s3/J+fcjiJNcvqjMshpVsTGrL7O34Pv5Flz8qMMv3yi/QzPA1MFiZUZ7r+u7HZRGtDQ0crCRo2Wsi12yotC/vG/yJJSiiBOI1oSu1caskbpErti1a8iWjBNtaE1Yk04pFtCCTZF6HPY0cSNkuUlo0UlG/MxauxGpFKMX1uKMY3iSilKzHZJIxsLVLh0DQsAwsWDoWALFuwWHY6HQCwbhubhgnpYkmtCE7JtsioT8h80WK0lpucq3S0HBTi4/Ix2Ew9DCYWFN3nyRUnqrtJJu2yv2WhSlKOi3MnxGKwdWdZe7OE5OL0enTTVbd9e5TzXD5hKWKpO+rvvvfXdL8rFf3FGpB6MU5Rd+5UqylBeRGo4qMlun/kYitJ8srlWS5It7/wDBMmrvMMqoQxcrwpzjFq32X5qz6ebOJMNQwWZ16VFWipOy10121uYWv+9UparqRko1b9E9DNsyxVZqhWlzRirR0Ssvkr/ee2cqKT6Da5X3OfTQm/euhtNDsX10ZKd9i/vWTHvuSXuXuQeqQ9GSl0L7eQ9RR502xJNW8hvREW7kl1ErQS7k2uSy/plOKcjFU1FpF3zJ9iU3J6jdwbsiEroibop8qV3qOXM2wtoQjdbkUkJXJSUSVW5Gqluc6kveHGSjzRexCcZWT0J0YtOxKmk2OnoJJRsycbDSAS0uOK0IJLUirrQoQi5q5hY6u2yI1FOlFX11uhSUqigttF82V8nhhKeju9X+XmykpwzNQ5bqabvfa1la3W99+lji+EcPg8HjoO0qVVOXmm1r5drdepK8qzpt6JJ+t2/0S+8xWKw9VSlhVorxa10te2r3+RPBYmOAweKk/jtd6drPT/gGNUbucd9f+CexrVqdOsp2VO91Ze9pZenyMY3KtGK2JXhO66My7CzxuIkk9bN/cr+XRGOrTrYmUpO41qNjtbQ0bG7q3Y3LW0JW0sStZIkkpFrrcimRi7FSNkSimlYSWyCDXUa1Oe0UkV+V1HbYuQlZMk3JjLkNy9nsboVuWxbS5y39RRVrjit0jERbs+w007sq+9FStqJ2dzFQVSm32NDoWDc0NSwdAAOUVJtn1Y9jFRHSViUGjQ0HYAA0NAwsrVEhu6HTUo2ZGWiIzbRGKaujDqDi4y7H1dJ6bPYm5pxT7aGFnFzfN0RFqc3Dvcy/KaeWRq0qfWTl829erIU41LUn129TlunF7rYVSzfkO7uihJVIOD6LQtdJEMwq4enKlF6X/roYivKqpSm7sU7JWKqcJWMXWk8LKcdZRRGWiRK17DsmipZPToOzRoOzaNLWNjW5dNWFuTabTN43JRW41dXFJpKxTg+ewoe62cvQiuadjmX3FeK5+VDm4RTRXqOdr7huMk7Iq1eiKFnBGhGxU0hoJBGKtckkkQitEVJ8uiNWDLu2hGbSsNpoknF6EakJQae/Qcroi09ycVLYnCzLJbkVd2LRVNDldkIJpIpU0yEEpKSMFBOUY97FDLqUcBWko66d+6v1MNSUqtXl3hKFvR81/wD0krx5qM3LdtP5bP8ANFaooTm6avKMXb7rles874TxFVbqN36xd+y/l6Ir43EYmnl9ei+kXLbqtd/PsZdldChXq1W9Kru99LJLv/kY1cmAjhHtBq35f1dmLp8lL0/zMPUiqU4dyOVyxOIjSoq8tH+vVkcPKrW5Fv8A5GGxdTDVPaQeuq+9WKsk22i4xbFtTctsPRCjoVIvSxqxHRWFpdjd1qTVmJ2dhop6lTuON43JqLjdBC1iTRK2thbEWXuRaeglpYu0mjmi2iUd0iUUoE9Uok0nKwp20KvxMsmrFWHLNxLB5BY6FhlixyXFTI0G9SOGXUVKCWxZdBwFAcUTpKSKuG7EqbidAAACk2poV2iKuzdkbOJTnJSt0MCqWIlGL0loupjslp5XjVRrLnho2trr1TbXyMZh6VarU9krQTfLvor6b67d/mUafNdPdFGXNOKMbL20faR3juYHE1MVRU7Wd/yM6lgqtGhVoK0+W0t9++un3aHMN3Kc1CRL3UpR6mOxip0ZVZdFr6IweKp4qjGvB3THHkuTtKNjD1lGT81YxuGnhKzhPf8Ar1K1RzkpvqSvfUj70Ny9+pJ6ssuU3Hoa7EZWY1zNtFrwv2I66GjdmSSucrk/QSUacb7vX5E6jsX0Y5JLljuyT9n7q6EZ/FfsPUd2aEmkSqRRUrN6IMNrA3KaTepiGnaKYhK7SEklYcdCdRJWW49dywWHcLOwm0WuJtHP7tiMyyk0xwuUaLnLlW7KlWKhKnbW5FXZBWehRpX0MNh4qytdlPMamDyarTpu0mm79rK+1n2KONhVwM4yV3KzT29dDKKmKqVsW6krKE4qOi+Fp329Ouo6iqUVNbfqZ9ldaNd4nCSs1KLbsnpbVWb9NlfsQjTjRlSoRvGompPtdLo97pmSZBVy+hRliYWqWa36KTa622ZPFwr4eNJLVN/pZfIx+OrY+VWpN7Wtt2+XUzeq61KDSs1GF/XlV383d26EHeon0ZlePq4DEwrQ3Sl+Ka8zI8VPCYh4yk7Tje3XfTrdbPsZhGCqycFZN7Cuhq+pJmrGDW1hqx0JPWw3aViGqI6fM02HZuxJaXHqx2kQbTsxx5loTklHlQ3qdCGrsiSWw7G4gTbaIxcmKN5WZaLViWqsyV2kNpxQouVyVkVYvmuzlRjKdp8yNwsAxlhJkYpIVO5GldEUkrCV9C10UeVN8xybXGkMepCHNJGIwycbo+rSuPDtHsxpoAApfGiGpFakbNkY32HFxMPGTipx6MyiFXN4Sm371Nbd1+FjAYWnmuKrUMA+Zpc3b3ej963TpuZliJYKEq8VdR39Px2RTpVljVWT9zl8t/zMOm5Svs07kMqq4ejCTXuyvb+rmJcoPlfQjNPQvqTsQlzxUSpGNWEqT9DJsuhl2DhhIu6irfL73+ZU39BSu/QfuWaMwqzxElUlu0U3GVJx6rUqO6Kck7x7kvdbGrpNFugnqNXdjZ3PInK2xCSty9y1mySukyNNJ3ObSxiq3Ny27Frob3IJKPP2G7tsuNkpJK5LEpbEsQmTm5BpYwutM3ZSdkVXeQtWQ1YtSbsh2ZubF0w3C+gzdF0ITuym02oigrEK3sZKS1ZVinPnEr2ILVGXUrsw2FUVd9EU8vhWpTVWVouLjtfV+j7FLJ6kcNThHaKfzsvUnhGqc1H1fyuZNReLwUaEFu27/db79TPsEsFllXW8rN/l5s4exVargISqR5XOzaunbp08kZzWs6Sp67/gKvaLkujIU+ejUaZj5VatBqmryt+RKny0/Qek7/1qTrvDSfMYipze8OS6FedOVKCjvrf8C90XdjdiLM3LXih2Uia95kJHNZaHPohtbj2J7l09CL945rRdholFKwykm5aEknqTVgjZhSjzNFOKs/I3ZypFSHKJXjoU4XbTHB01IqLS5L3gxceanfsWGtADoCVyMCNK5CCZyigrXLaFi1kbFiUNBxsJEndDihxJUU2OimrFWhyq6AYdDDpOohaFyML9SGlrMlhp+w9t0bsUIK6a6GGrVaNCrRpu3tYtP0f9eRwHldThbF0q6nzSjFxva26fS8l1MwwlK14dbXMNg5YVSp3vFP3fJdu7+ZTxEoN266GO4inQyBYJxvGM+ffya7eff5GPxn1uUankhO3yLt+8OSkkmOTUrlRxUr9WSd3zdhtSipEpco0pQSRRftIOn2FJQqJE6aUrdCSdOXoVovRnM2uUjzKTbJaMUmmmhrWxcbukascWp2IJJ/I0jTfdkrJkpNoeiuPVCl7ljclKyJ17bE6jkrGgzdDRYwfwWLFIluLUhuLRFSV9CxfQb0G7ly5udBsbObQQnqYWnzXHiJQUuXtYr151ZR5uish6RKcbRKSMojCvUnh5L7O/m1YwOChTwtNX2tFFDDtYqNHpf8e5SoYdxn2gtN92tTMcasLRjVa0clH73bzMDUqYLBqonpfRfdfucQKpi4qdKVndX0vp8xJNRqS6NGb0qMZNQ0tZ9TIqtXNKs1W92UpzXf4W0trLVFTC1aTtLtuQw1SNWUm9G9F2FQUk/MryaizNmpU7NXu4rt1RVjaKQh6JD0egaXN2O5u7Csok1dktXYWiL6F7oucqepPR2Je8iFm15HSyJEujJPmdyG5LVonJsEixRhyxTHFxk0Qs7sbvFeQ0pEtDnafN1JS5r3JWTZL3b+Y/ebK0VKDQG4HQIU2tSMdURjcirI6EbtEY3Q43Ra41dnJZIcdLEoXHEaGkxxGhwZKnzIrUXF3QAUXaoi10JKxGm07kKcWtHqU1Xlh/YJaJ36DpcrcZ6P7zCyUIqdT4Vpf/AICKeQ/W8M61OV3FpNW7/MxtL2bUU79vMqyTi0S2uSlzxUGVGnFW6EZKS03ITVrdyas7ErS23RUwyq1Kc7/Bf8f+GNIuzKNk+RlWLUdSErKxBulUUzFRT95CbnBSXQraq4qvNFLsK17FbL68MJTxTj7k20npq42vpe/3ofvRv2G1YnsmJpoja0UVLKWjFNXb8ilZRuVpJrQnZpWG7ob0SN0dCUkitWTdkBuDDoYS3JYKb0JKzOghNJMeoEmdBFy6sPYGwi9LCTZg6ipqRVrJpqPUjFyJQaRBXKcLuyMBGMaLil7z/Qw2NoUKzq137kFfrul5akquHm5YmUuVS1jo3fr8vmjCZhKNBq10/wCuxWy+ljKUotaJp/c790YxRrwgtklb5/0itGNWcEnqtGZXUwlHDOlXfvc2i1/TTuZ3jcHiaM4U3719N9V+Ri4qFeE6Lvpf57Pf0JZuq+Ap4WS96N2v6t+o6nJVcWTuppJ7E7T3P3XM5J7f5mPqwqYiUo7FjdDTvodBaMepe6sIvaJLSfoOz1NOUbsRaFJDa0JLUhJWsbsivdJKzJO6sEF1JqzGJXEtCKux1YyUY9hyu3Yg2mmL4bdxQ5nZbk4SUJSe2xHS1yabZGDqFRWVi6sTJL3rAB0Ka1IxuRiR0L3ERWhy3NCyuNW0HY0HaxLYldjC+o7MauyrTvG5UjysAW5TalBM66CmmKo20QqyjK6MLmSStU1/ryRhuSvJOi9f67nEGS0MuUIxVnKK7/5vqZvTrKdOrUd3NXvpsZ/k0sbicNWg7OnNN+nVbr9SUfeXkSfcbT1FJbl01p1JPmj6EbXuX5ZFZcyU0N3ikuhUSqQv1G2mvIfvU0kQlGphuTrG/wBxSfLJxfUUWk4spNKTROOiK2NqVcJDDt+7Btr57kHY5Xaw78thfDYjFWT7Ik7u4pq7fZf8Ag1y8v8AXQqvlk0aD1GOxKdkVJNoaNAsDAwm1joQ0J7iI7Enohq4SYwC5cZuxCsJpDmKzFskhxbRRpc8lFdTDu0m0ZZhpVnyLd6GQZXUr5paW0d/16mIoqVaUF8N2v66lGVOhhpYVLeV/wCv+HMDia39oVaMdKUYX6b3Xz2v5GMzCVClVnTd4qDt01fy8uplMalXLIVK38SV7/e1006Ip4aVCl+9+JN/d8jGV1J+7slf72yhB1JufRGKVZTjb4fluKnUliHKWz2+4jQpqm/aO90zLKMPYTdbdJtevyG0nJepU1kxpMOpbRCWoWdh6pMlL3WS3kW1Y9UWV7nLdEFZkn+7t5k1dkHYdiMU4tFSNnoMIK7JaDEhK0CCTZo3dEbKLvuU5XkSkpRuuguo430LJRQ2lC3claCVib96446ktisrTYAJXIRuRiiMRwdrkVqRirHKrFh7WNiTuPyGx3Hqh3FFsemghxswlqjEw0ADB2lR9BqxBu4tSEot8vUXw2MFUlStODs0S4nwGc5fOOKdqkY6b7/JJffoc01QSm7qN1HpZf13MvVOpioxq7P/AC8iq4qTsSab1L20IqL9BNJ2JWWqHpsX5tt0YepDl5WSioSsym7Oz6lSNmmQlysilCrHsYykoS93YlNS5ZFSKvoW5oJ9iGjSY1ZtD2UkOC1Fdovahp/Wp0QnuQ7roVG5tyA5kXvoVLJDtYnEOg7M6BuGFvzhHclqIRJ6I3Q2PUUXLYUVJXRy6lizLDsbsUUW1HG6uNLoKKRTV2SWpQjOMVUXRmGgpIyWVTDVFiVtB3f6f1YyTHRrZfPEWtNydvnden4GXY2piI1OZ6OTa9OhPERw/NOu7RXX122HUpf2euXeckn87GIwc8HhZ0aWtrvt+dyFOgqFOrLdWT37NlaGDr1q3O/ds7b9v63Mbh6P1aKo9I676O77+VjD4SSy9zWzbXzVv0aJwV5RfYq2iorvqYim4UpTXV6GMw9TD0Jc6t/wSspS1RJpsaSHYOhoK1x6FnsypSlTpq631Q72TLXQuXlY9UhXUrG7sTdoWHpohdhptFN3WrJaoYRutSbuWTRFaFRWgmUrXIJuROd3YhJp3ISS0ZHcaVmy0WlYei1L6WZO1ibJ7GJWqYAQVyEbMgtbC7CuRh3NhA2O9geiEyWiG1y3JTTY5O49Sw3dC1OW5irRJ2ZZD1OhgHo0WuiCs9CFNPUThB36ixLil2scM4jA1ozhXWvK7b7/ACOMFi8Jldavg/iSTW3l3ujIc7xcswq0MT8D5Wtutuyv1MVB0qjiiabH0ZKKZHTUVmJ3Y12HdfIldSuiqnOmpLoJXimTVnYaeqJR5qV+xGLnQfkyhqpQY0+RrsQvdohFuSuTVpJDfNNR7DnzNIFdUWv63BR5k0iDsnYteLLE9EU1zMSsipfmJbjjdDRYZYZbUZhv4g1oJEldCQg2GJXIrsK1rIdhK9yyukN20NxLQIxuYjliopdhu7RBJp37GJryoYeVRdEUZKrRjPul+KMDjoLLJ4RrWUk/uVv61MFBzlY9hPE0KlGL+JNfgYOE8PgaWG/ls/mZThfaV1GWzdjMMth7fmturf1qZbioRST3en5GIajBsi3XwVaUd4pfn/wCvVdSPN5GEm5Xg+qa+4o1VChKPmjMocmI9UvyRVXNyW6FTCVJx5H5/kZ9mzxdSz2cUvmkvIlrCUuw7JjV2SSSNxkUrXHYshK8WzG1XJRT6IeisIUE2OK1SGkRjdXKju7EonUimyL5WJXQ43NyXw2HuJaFJLQnK6sQ2OVRi2jRsvoRlcp1GmajfYm7qxUVncqbkou5JGK1kAEYq5FWIxOXUhHuM2Ohq2NoUk0W01JXTv0JT10Lt6Du0aWGxlwiyC6mJpc8WSg0GoYWpy1ETjd6Ck1sRbYlaKOeUoKEnojO6GJq4OosK7VOV8u29tN9PvOGcZi1llD6zrNU4822/KubbTe+2nYyz+zcUp+3o80rNpc0ldrXdGbUpYXGVKE9eVtfd95Xi0udLRk01G6G9LihcStJ9iaSOll8xvm9ULX0ZRq+zqWe3UlCVKaT2ZRTT5ZblZaeaKTu/Io/uqqU9nuYug6c+eO3+exK6ZNPn0JS5Zq/YxGmqKdua5T/AIpFXiScFh5R66fmJJwYp66C3aRfUbXKVE3qynFKKsNob1JwTWhKLSJ7gWGWDDJ+0QxaFk47h0sW0JCV2WS0RpaxJK9kJdDTlsSktjcSNDRkJRUddtfvHJyepoyKViF7lkk7FOlH6vGqnre1jLaNanCVVLR6dOqMDzxTnFbGcVqmW4X21T3pc0PL4mlfrtfYwGMaUZrd2b/MzKcKnvx6iwLhinWmrScbW7JtPe/kY1OOq6orzqU+enTerR7tbLlO1pK1/wASnXhSqU52vbRkYRp1ZoznEYaeX0Z8tppK++q/LYWIjBWtfmW/YwWY0faYmGKfvODcd9ZfJW276GKzKE8Z9VfxcvN8tPK34k9mS1SS3FC63JLVIeqvcS1E76C1LFPzKzuNaaiVkNo0sJX3JaRdhRumSVgVmhK6sWb1LXQrKPmSbsbsUPcuUYNFVJELWsVNKI9RJtXINaIjuOejQtY26jfMiTaaTJtOTsPe5JFeHMh7mgFNEEh6PQteRaw2i92XdhuyJSSQpNDnfY1sPQuXL3GwLhTasSjdWMTTvEATs7mGkqlNEopSRHV2I6o2QqajVlPukvuMwx9eLksOvglFPbZtLqZO1SoYty+GFOX5GKqe1mpvqKpy37GZZNTw+UUMdB3VRtelr+fl2Ra8iLtclqxO6sxvqJvdE0t0K7XmJ+2p26oheaT6oqa/mS0dkJOVNPsULVKE4vdW/EnRaXK+g4JtR7IqVHUlf5FdW0I7ojLkqcw7SoRSKd5wlLtb/IW9iSskiNrklroE05aEFdWLdSb1QlZXJSTRUXUGMZoGF+MLmyGhklZ2HsCEbhOXQtcSAhG7HZLQEEVew1oUNYKJha0vqjh0vf8AQyqhTjh1Ce0rP8SWGhicFXi90m18ldHCOMnjKS9otYya+5/Ir15+zjTa1uTm51oORiprSK8/0MVFt+0XoYDFU6uVTpdU/wBENKSaKteisrl/O359LfLqYiu6uES7OxXx6xMnFK3slGP3RX9dT2sas4vuVsAoYT6093p/Wv6Gkrol7skY7FqvSppLWK1G+ZCV9DdCWx0NGiKvHQrv336i10GeRLY0RUYk2iTuCFexbS4ldaFt/Im7kVd2PZ6oUbIr2crIbSsVJ+64jIuxdRaaIy0JClZEJe8YhWZoyQ0rXJx0K8OVnQCCIQbIK41doZI1FdjlZD1C42bjeg9jQALMhoy10Vad7lSPLKxYDLajcnEnG8iMb6kbcl+pN66EYtsoYZNvl6mPwlTE5VVwrlZVItPT8f6aMTS5bLyN9yeLrTw6w7l7i1S039dxvTQhqaO5OL0IyvoR+LlLLZkouOpGTptNdSr7j51s9yF1LlKlLmtYh7rlAo29onLsKtD2Dc1qtvxJRd41P5kKTTsV5XlYi+Work2nBdyFR2t2G7SaWzKcOe+pO0pJkKKUX57Eq0oVnQ5dGr81+q6W877+RyO5KF0KPLqaNE97nPocy3KruzcsBZBhtJm5oc1yKurI5bnUtfU9BJDaG0iTuWuWARGmuRyuSd2WuJWIwQo2sLVWMEk1djwNKGDlV59Vy6WfW1+vT8TJM0hhUo1I88W9Ve3lulcy+vHETqUFo0Zhg/7JTxNP3vehG22s5xje+u172t0tpuZnWdHFQS62/QwVWrWxlanONoxs0/PW+n+Zi3JTcu343Ks0qTu9FqYatFax6mFhGvWhTbsmV8RNRcUx2lCUU+lzB4GVbA1a19Fb7/vKdNwXK1oVsdOrRjTey1J/HoNa2Jyd2kW1Hvoa21NWNJJFuhRpc1SEejZVty/eJXY1c3uJpob10JK+pblRI3Ey4nscyJT0Hq7FOny2ZdbnN7hN3ZNOyJu6GN3QtUU9iWo3chbmMTJS2EtNSSTNCW9jEU1KI1qBGNyKsrCVmJNvUd7aEr3LjbAY9QBjWh0A6HQjqxjepi4W1AKE+SaY05Ig9LFRxdOTW5keDxEsCva/Eld7d32I4WksYqcnaLS79iDat6EK0qcISto9jOsdHHzhVW6ik/l9w4p0nLqiTaZv6kVuSXYTUlYat8iOqFroP3oELXsyk9HCRG/wv5FGKqU230FT181cnaEYvqOS+pNd3/me0TpxX8pV92pcxULy5l2Jtzce5KPLUSZH93NNlSPK7Gq1KMVy2fUlN8qRCtGvOaj9jRmTYKWMxkKEev8AXkYnB18DjJYeurSg7NafpdGdVcNVxHNQejv37vv5WJbEldlkFSwaDsHQKFudGgzQWwtGVIHKk0WV9BjaQ9S10JCQFCk5zUUV2tkFrmKqOjQlUXQguaCY4NkDAKDlyvraxmlOWFlLBveLf4/f+ZSTpVUn0KCVLBLFQ3k0vv8A67FfGPEOMp90/u1X5GJx9LMPZ14O+/4fJFOc6cvf3SK1WFSm13KlV2lHo/8AMwlPVxJRlHTszE11QoTm+iRh6rTcls1+BgE55diIfaTTt5XX6GIptxt1MXBRqtInFKdhIqR95DsTVkiS0FtYt7qZTpuSujFU5YenTfeP+ZUbcWiBLcvoJq1iV90RSehN6WHqCOoJjd0U1eSJ2vZFtBxukiV2xJtktRmlyMdbCj7tyEeZ2OT3rECok0mWXImPVElZj3JxuirG0mgKULl7OyElYZJtbEm0hu43cudRg9joXNkAHQQmyMrxKl0zERU4MLF2GBrOtS16Fr6GiQp1o05ezna9k9FsXcvi2KUmkmYKvHM6OGwFWfLGMtHa/wAT7K3fuZjh4YbFVqFOXNGLaTta6T3t0KvuxsWukhxTIyfMSi3LQilLQUrpXOVKV+qObqtzn0uiolZOPQf7yLmt0U6rqta6rYpUI8sKfWV393T5mM5YVXGG2xKpz00uxW+Hlv5kZPmt30K0bSaJVJ8q13RQ9/lbdrE25S16E03FSfU5Yypcz3ReLdmUqMq1OU4K8Ybvtdpeo1ezIV8NgqeIdJWc9Zb9Ld79uhw9nTx+EhiYR5bu61vs9Hsu3YxWYTxVWrWrrmqSd+bb10StqS1EtCUESUbk4JK9ypsAM6GhR+NHQCDVx26DndWHK4+g3clqIsgBIU/Zx06jdwjDVEIq9jlsmRSbIx1aMBRcsTFX6o+ue3cnVV5S15tvwWhWrvFV1iHu/wDhjA4hxw/Ly2i5KO/yv3+Q1GGJ9nF3Wpgsso4DD8tKNopvq3vvu2ZjWUasYp3UlbsYi1Oa5OuhiMIlVutrP9DKYwn7kn7ze/yKvNOquTrYzbKMLXwXsJ6qopJrVde9/wArGGw9eEoU4P3IpRS02SS9djKsVWwqqOt7zbkui927S2v0+ZUqyr1Z1LWu3p21/wAivFwqcvYfw3LXJu7H0HLmsn0Hdosi/LEyGeGlV9nVhzX21as++m/oeIMsDDF06GF+GnHl673d9/XzJXcfQpQT3JxS0Le6LuWdkkcttie4HQeo9QuUopal7q5uyT901crknZ6D1GIUm1ZEpvlUSLe6ZFdRLc5dLCjeLXYSUWTV0ySGtDFQ6gUVYaViK0OhKXQk7j1L6Fy5uMNwAAWuwmQbtYqO47FaDjJoAyiT5nEl7pJu5CbUR2kjmbRTcozunsZtmmFy+Lq1pWjfez/RMr2nqia10HqKSaszm5ZJPqThpdF09EPWPMJajaixQUbKWzGnRmkU17Kpf5mGrOVaLfZ/kSftJN+Y4crkipLmcScbamIu2peRf3UijCU4tLpqV4Waa6pFKhOpQTtojFwhTpXh10KWGmsRKUuqX3mVU68qdfCYfVTs3tsmu/nbZlalKlLle60PqkJVKrf2l/mZZTdHDQjJWauvxJaMZYkSimzRvlZUjoDHqBqUF76EtAEX0Q0mi1h6jZuKwOx0HsNWQ3ctchC5GFyKu7DWtiKdj7bsZapTrRit7lXn5lCG2tylBRwzXVtfgYTE054CcJ9NV6oyiPtaU6j6PT5mZ4mlLDQpx36lDCVoUITqbp26foUMO8ZV5I7q7/rYxzVWUYparQw8nRxKkujOeNHEKfS5hv3nNN6qzRg8fXXNGorWk7bbdPwKsozcl2Rhbyait20vvOJ8CsFmNWh/K/66s9vH23sutrkY3Q1dkkk0ONmPVDhzRKkFyKC3sZRh62GoJy+JenqY2vLEVOZ7srXppRfXUpa3RUfveZf3Ru7shXsN2RJ3ZuGgWuctkKN2RXQsdCpNKyLq7Za9kJJpjV3oNaEXYumRg+W5ytoUUtiTbEnYqK0roqau41oSRiI3iMSuR0RZCVkPexLyGMZ0HqaDAsrGgw3ARCN2bRsiSGYiDUrgGUTtNxJps0ctC6TuylWiqE4Nau34EcO1OKWt1cxWW1a1NKnUsrrpfS+vXsYjgrLM1wtOrf8A3e02/e0aW9uZX321XkZjGhhce8FCrzvlUr2a0e3f8yUL6jaHsPVEJWeo4+9oLZpns3bck0mU5uorPck1Kye6KkVKm090iDlOTbfxK3ztdfkShy00+tyUeWbj31HFuWhWanTVuhyuULvoKVrMoR54qMN3dGjjz21RQnOHU/iU2l0MxxdPB8qqP4mor1e21zh7iWvQqVvq8+WVnCeifuu11qutlqtV3Ktf2tm+1iMUlZC6E3qww+LpV51IwesHyv1sn+pON0TiiNrlaLSGWBhbQwkbji7IepLYHqIGWuJBZGgk27FGEFOPPtcnqxK5FJpHLZ2ItKLIK6ZJK2hTTasimrSuUZNJNdbGX4enUxCjJ2RPEUccsLSirKNlLd6vr0+5GXRoKnVoV1qpW6/Dez2/4cr1Z4amqNHSLd/8t9Ss6tbGaO0Yrb1ae+/QjjlSwSpJfau/usTrV4YySpv3ZR8u6/NfcYPGTw9Z1I9Grfc1+pPmWLs1pLr53MVWgqyoqXvWvt0Q8dOdD2UXom/0MXScuXkdrMn7OV1FamBxEsHUVVbr+vMxFJttv1JU4pX6k5XoaCXM0hpplRWa0GlexSpSqXilqfDOz6FPF8tKcFomvxFUjPboYnlbVuxSsm0VEnLVFkk0KKauhFR6WGrjVxLQjFsjCwqbQ4sjdO5HyG7D0Ju8iKi02cqUtTlSi0up7J8pUSTshKxF3Oa8EhCjfQjC5JqMVYk1d26lS8nckhomrxsShZjJJ2IJtoeiJoegxlzSxYsbljyH2OhYsAzcpqyuNE0SK0eamwDATUayKgla5JqTRTi5OyIYmHsotPQlXbj5GHzOph6dSmnpNWf9WI4qhVx84L44xV99nt5dCS5USjGw1oK1iKs7MpySeo0kR190ktXcpNqaRVj9pdCEYzpSb3t/kUZt0LdYtP8ANfqYj4Wl/WgnzxT6oXxW9BJOTSIScbo9m5NxMPL6qnWlsl/XczWpSyzBwxFR6Tsl87JbX7lXEKNJ1F2uZXXqVsPTnOPK5JXW9m1tfqZhhI1prmW2ph8lhQVadN+9Uab+S9f8jbQUdjoWcrjjZlLDJz5YLWT/ABMRRlRqSpT3i2n6rQlEehVtOAwZoMZgYOyZLUktCSTRJIBD1EtALCi3oiiowfNLoRbGiNk0QWhdXZboJWVjD4R1aNSS+yrlLlQmlO5RoycOZGHoc1WEE9zIcNOjQjTqPW/6kcdg8ZXjVhs7336v5GMpTU6n+LT0ZlWGVWFaT3SRiZws7en9fcOi5RjKPT/InBxafcnVjUr2j9m1yDdXM3NvWzsvLT9V6iko1HHuVkr8vUdH3k0TjKSaZKtzSdyoo89vIlaOHVyKV00SWqj1RVnzyuYiuqSU3tcw2NjQTaWrIUZ1YTqLpa/zMVg6lOpKn1Sv96v+ph1dMrJ3VynJcyKjT1QpaCfNohK0WTabuGhGOxGIo6HLoNXQ1ZClZDdxtWJ7in0Gk4svzMpz97UlG2gRRH4bCV9SFO7VhR1aJ6xsuhZKCsSjZ3JxWjQ1oFde+xoILQeiJa6j1GhjRYsrG6A6APU6gCKWyJapkkNBXSU3YCL5WmU2p04snBN6Cp8q82ScaUVYpxcrRjua2MZWnQotxV9V+aT/AAIwTS1MmybDYyjiXVnZwg5R0e6a7NdL76EtNEWuSik7JikxNuWhd2sct3fqVNSVnK/chLmjqYL95F0/JlOVpyj5Em3CMe6/zKUeVOLH8ZKK9o2mVYqzsUuZ01/WhmWLoPBui3fnVluYLFY/P6ntqulKiuRLR/D7vSz6efqYWlQwM3Tg9W2+v2nfz7mZYqvQw9aWHV6kU+VaK7S0Wui176EZSqSgmul/noYnCzpU4VOkv0OVSSuJNKxUtoJ21I2qU1KLuRUotW6FaUqk5Tlu9fvGtCqkJ2TJbh1Airuxh42idB6oaRLVDVnYtqcrexySsWdhFmKDZZQiuXdjgmiUNULYsnK4lpYSViOqsWaRHdIoRk5WQk1LkZg23HzscrSUuljB4bnUYQMmx2ZrMa1aWsJ/Cvd0+e+3cxeNq1sNCc1rKy+9pfqUJSVOMH5mNpuTUnvqYmvXp0qcaf8ANG+2z3M0a9u6a23MLl8I1q1aC1lq/VJLuOTTbe9idam68ad/eavYw+JqV8yrU3HSFknfe6T+W5Uoyp0fa20ZUTdr9GVIpu1+py3lJmIgnRv5oVNKTj5EsFJ4dVb6J2+8ktbLocqcU+5gcE8TKWtlFN/dYwmJqUKVWnF6VLJ/L+vISfO4x9CUOR2MZTUaFN97l9UyprFlrC7C0SJPVjI6lNXYo9hbFtx7IfvaEtBPUnIbQnqRk27GnLZF0pMlaTRJEIiirFPZCVkXcW2cqaJRTSsSWnKcrkrsnqrdhoxKtINCnsdCV0SVx7APUZY3R0DoOzBgCIXasiaT2HYY9jERUo3NAMsrc0eRmid0e0skTld3KVZ05KS3RGe43zuzMHjKGIhJwlflbWz3WhkmPwlChiIVd5waW++nZFRLm0G7qxbQik2QbTsyLUhrSz3RTpqUrjgpTaFKMldGFnGNWzdipFe0UvUUrwt20JxvLlbKkeWRCDq03HqjDwvGUWYzJ6+Cw0a1dWUldbO6tps3bTuYX6vRxNGjU36LX+vvK+PwGDyVZfTl++nLmtr8Ke97Nb+dynTpyiqltWeyjJp21TIWbdjEYh88aTfe3yJRvE5lKKXUzuni6tWh9X2jJOW21vP9NSNFzicD8Lwrx/s1VLcqnK9vNyta/wAtxpRTXUrJcqRJaFVJobTGtSwwMLT5pXIaIew2NpqxISTIUeaxJU4RlFEveWg1dWFTsiyVizeglojSxyuyEtCK0IiSsZaoPEQU9v8AgGIVqskUnrYjLkq39RWnUeurZSlFK6ZWkpYZRTMlrOlOrG26l+CbMrVGhVjKromnbfqnbYxWGo1sFTrQlduytZ7o9nouV7MxHLK8rjU1C7W1j2EpYqLa0d0//AXr+RSrqlTq0FvJq33GJiqatIq5fCWYRxaesY2Xzb8/0KM6cabgl7zdzM8wU8JSwi+xr838v1MVD3SKcmmUneLsTlJ07diV4S1JTqSpyS2KclZopuzl6EKlSMGl6GJwE6GEp1Zr49V6J2fXuupSbhNSRi5e1nfu7mYP9xSR1JWcLm6DVoe50Cloi13YsaMe1x2ZK1rE0S0ZcTFJJWIxSiyS1YnYjBy2IoTKejEr2HroVHHlSQ0TnroSbdkWvoWMUhIaViOiHYltYYxpBY0sAGjDSxoA7BSehJWVh6kkEo8yaJR5XYDCz5KqZUTauh6RL3ExshFSaIUKdG6gtG22R8uhTwNStRqVIL4Vd+l0u/mbjFuRak0mczvykWnGwpNaorKN+ZF+Ssuz0MQuWV0SblKMu6LtrlROScFLqtBNysnsyNBRSkuhUcVOFSWzOIs6eIagn7sVZeSXy/MxdaCzjC1E+j/EqUFiqfL2TS9DLqSp0o0+kV+RCTTaYsRTpzs3uLlq17xfvRVvvt/kTirKxVpylaMX1KFGO3czTCUsK4QpO6lFN/O/e4pON11Jy0uezc4totfRFRDJLUGi2hCHNJIowUYofYshpMdktBpMpwu0N3aGlscqOVWLdBoR0LItpYsJKxC19CKukQXvFX3uWRwzhcNXxj+sP3Yxb67rbYxlPmryUe5UoypV+WfT/Iwd5aLqZbh4VaqpT2MBUlTrUpLuV3RqU6b3cbdyjzUsplUT0ckn6XX62KVFUoSuupVqc0pLyK1STirblaM1JLql/kUFUxClTi9k5fdY9m6t31Q4uNPm7lavN4mMVtrf9CUlKqYmF1GxDCu+q01HNyVib5YMqpummyE2k49bP8ijBxp6lJqNOT6lSPJFInWnOMKfa/4sUbTs+iOVyaSMbUc7eSLC1pWIxXUdiCvcYRV2UYaHQOxy30LaXJbE1ZEhq7CLE7ocdBraxJ66EN7itZWIQ91Mlq0J6snZw8xrWw1zbEkuWxux7mJWxax0EStYl2J6ItcauaG5YNDcaNDQAGblPUbTGlbQkOxfVGKp2d0AnZmCqe1oW6oqr3tDodA5mQaUbshUTeiMLm1bDUalOK0qR5Xttp5enYejsPctc2QpJrUpyVmnuRjv6Dpxs4t62uiSuuYrcsoxsN3ivIUlpJdCtCDp80NV/VzK6NKrFuo7JfP8jNq0acZU6Ts3p/wdTB15whDD1Jcztva22+xxHHGTxsKGHXuuzk9NFfXR6/cSyCjilT5pWcVdOz0+V/zKMFCcV2MqySvDFTr816Um+iVn993oYmootQXVlfKY1J0qkpXcW3t3+ZDBYfD06mIi/wB5OUdNdknd9tNPvOVzi/Ip04uzb3MizzB1K2Jpzp80LRUJ3a95SmprltfS0d9HfTZlGjUxdZUqKu27JbfmVYyT8yXYpTcFKK66GX06M8TFVXaHV6v8tSa0GiasxgYSgt2JJMBtM3IQ5hNRI3bVhx1Q1dHkM2QJX0HGysy1l5nupb6lCnSk58zs7aepGKS03MswixWKhQX2nYrUPY1nTvqnb7iaSlysjNQtyPcxUasMO6/Lu7LXtYxLljKkqknrL8NPkUMJLCzjRvdpLX5LpqYbCylOUlsnZlSEKWH9r0jfXzZgsHUpxm6mzs196/rUoLEypVYSfuKN0tPiTT9enoV8U61OKatZMlUcJxlIw1OdbMXFv3eV29br9CtXlTnyy6rX7yliZUK/Mna6aK0uW1RacyK0J0qahLcUW5WsQot1NDCYdylKNXRpv8PQxWOi8thRivek7t+nkKEX8ipTSsn1KsJSlyIxlLEewqexV52dtt7ab6feZdDELC0liVadtdt/loVk6fuIqylKaXYUmp8y6HM5J9xc0Z3XQrJyp3S2EmimrxaKkWi6sxJoZpYoQ5pISstBu5a499Bstc5bk4IlCNmhxsNEVfQVnuOVxJNlrO5GKcmkRirabmr0HFoV9LEp2diWpe2qJq7GlHUlqYj4R7ESGrGtBlRdgY0WLaG6B2GM3DQCwUy2hKyJaoZJDipRsypDklYDA1nTqJdCt8WgHQik2Sk2UknIeKqrCxwzfuxba+f4/iSaGhF7o2KbbdjCtztF7lRtRTW60JJos/ZvyFZzkhRfK7FOTUkiVVul5ozh4mFatKnL3m046LT3Y3/G+/5FDFvEVfaS3f8AkkV6Uai5jD5pRwqqYbq1br1+RO06qt8bj/XkVca8Pg4O9rxSfyt+qKcFJuUtyOHdSk1Htf7iE3JO3Q9o4u6MHThVxMIT2Zj2sLGpKG0btGV5hKKp4mGkrL8V6foVZJm9kJaEm0iWqJWsVTQKEFKdhJR0NxWueRo9CKu7E1CMFFb9SyuR+JWE7q4Kw9UWVhLUi7M3L6BF3ZTbbRaccVTlB6a3+52/Ed27k029Sn7yRVqSnTjTeyf5kYWVzD0pSpub2IYqdNcsOrV/vM0w8cThZ4Se14v5p/11Msx7nOPttbLl/DyRQw65OR7VIt/cr/1sKnH2EZPdSSM4pQo87qbxt/5M0v1I4OE5U+beDujFYWeKnHlfRlSnONJJPWLS/MrzlKS8ipiJ1KtrmJxUMJDnqd/z+8w9RS5Z+dzHyqSxF/5tfvK1G9RQX2TExhCWhh6VX285Sej2HanFSe9yvUb1WzIL3+foSadSxTkozUmOrGVWcF9n9SHNz6dCcXzMtdWJRRDQqQUlcsr6lkkNXLamEindmlhIVrWGtjrYuOyY3ZDSaZOOnoNMWgCYtbWJRSWgkrX6jTSHIcrNMbROSsN3sOVy2lx6lZXiVrXshNWZHVIauiSViSRJDWg0dAC47G6LAA7BBiaa1FZjSGOxsYmOtwE7O5QqKrBEtx6m43ZWQ7MuU4OrJQW7Jq0nF9CVhHQVmRTuUqnLK5iqcbc8dpr7iTUoK/oUZKWhJNSZGo07fIVODSu7Nfj/AJDnVlXqxkrRVrPvvfz+8xOUfWsbSq89lBvS29166GBy5YjKpZhQd6cHyt+d30bv+Fj6xCPu9O/qSp4DL6ssbjf4cfi3/wDbbv7kSUaOOUbX5r2f4/1czvLf7SoU48/LyyTel7rqt1b13OWK2MDxY8trVaPLdzjKNu9032dvh/4JjKFXC1uSorSVrlWG7KdpQTKkFexgssqYadeo6nMpO6VrcuiVr311Td33sSh5n754hJL3bb6b+m4rlXbcdrE1Yq9AMLRtG7LIsOKuNFihGMU5PUepy2Ixaehy30Ra2pJWQ1d6G6OT3bktNDm1uSjZ2LJwTFZpWKcVKHN1RGTsmiMbp/Io0eefKyhFTjLTYwEIVoS5nblTfr5GVZe8VSryv8CTX9XKOLqyoPDNaRv+JllanSVSNSF3JWWtuV9/P0MLR9rTnUbs1f5vt5GBoVKdoswGEr4yEJU/sPy/4BjMDLDUXiJqyu7r+n+hVxU61CdZQ5vaK8Xe12n/AFvYw9GnCM6LnzTspNWtZS6dna2/UrRqYeNL2GspNprayS89zCRdfGrDxes2V8JOjOcZfZuh4ZuouXqZzk0Mzpexm9E1+GvdFHDXSS6IzLFfW68Zp2skjg1Us1xeMitqXMn6pfL9UVKF6rnLaJi+TkTi9ytKLjysrPlhbuyr7jivK/3kWkiPRMhBcraRhMXUrQk6lPl95pa3ulonptfs9itrLl6l7OxVjGzsxMjJIcItXK6UY+p1HqzCK0DlR0NCT0JNN3Q3ujlu0iZKLY43RNNCjcsWOhTdmJuSE7ik7XJNDYSabSGSVldjldWGVX7rJasjsU3oPWKJLQkk0SV0NDNwB7AA3dAAiD0HawxjGtColJWJx5XYDC1eV2JO4JWVy4EJuN7F7u5IRvoaOJFalOMVFX3ZH3qfL2JLlbiU/dmYhpTaQo8xWm3FMlNbvqZvWrRccNT3nez0026GAqSw2BeEh8MtX6/15mJwesbvbUoYqlmmCj/LJX+T+4xGG50pLeBGm5Np7MxmGnhMVLC1N0r/ANWv+ZQoYb6lXlNe/pbfpv5bGMtNqae6RJ80EZRhY4nERot2UtDMlOjivYw1sYKhQnhqrm/eSTW/9feTelzltqcrKlrD3Ku5VegUKfPNIUOWNgY0WKVJzkkWsrERK7Oothu5LZIa6CiSJ6pDN2J7oS5WkRqWjYbSIyvBeRgKdN06lST1SVvmUlCNOd9+hC0L2KVepSw8vZ7yVjB1pqlGU/ie/qYeuo1bsy/EUqdope65X6n1SlKhOunu9PuMFKcMr9tT7NP8iXPicLUot6LX9CFWLw9HDpaQTX3mJwcMPmU60Xq4qPy3J1nUmpx/roYPmw2YUq3RNM4nqU61JVKW822/v8zC2sr7s+rRjTVTu2YhOE2l2JJqSRkHPgqU6q+3Ucn979TEqNfBSqdXL8LGJjyuUCrBtt9ivZUbeZWqe0ncWxGPUjRc4tmMy54XCUqr+0r/AItd/IhNcyZWunci76FgRiZXsCVzCaxFa9i2g1oTloRlocqUWSeiRLctoPXQrJ6AAmRk4u4p7H2TpY3OfVl00R5XF33Oa1hhW2ZLSQmKOiIu6sNJolG4k2NXLDVmMOh0C6AOgwOhF2AlFjRbQlYrUnJXQEXZoTTimWQ2hjAuo6jjzJtFmHQWhzXUfIhNcyv1Krk5K61Q9HqVoXcX3I4ujhZxjVlq9lZ6231WxmFWGLknThypN2V79F/XzMowEsXivYVpezg2vetzW7vlTu7djEYeGFU3VleMW1e26TdnZd1bToUMOsRhZzhKzUbrTfsvL5k6dSrgnTm/et+NjLJ4nA4p0lG9O9o6rRJL1b+epSq6pvXoUqN6vsk/isvR3MwoVPrcqdXWauv6toSilRlTktXoUVCk0rXSW2xi5qphamEg+WpOzjLeyi9dNnfmS1enQhg6tKhCq/hel9OhKnGUmzLsFLFUqzi9YJP9CtHlSKVNTi7u1lcilJXKsFdsa1KxVWhuYGm78w3c6HQvdCVyKcVZCWhYSsJXZFJoavohNNDWqG9EaaG6sWTVhuyse6o83U+JJj3MZhXiaHs1Llu1ra+zT/Qo007xuYeLhFpldSq048jtqc0USp4BZImpXrOXZ6R29HtfvqVsR7GDdr2uZdjJ4iMJyhyuUU2r3s30v1t3KOMjTw7fUw2MdPfVGS1cVWzGGGc7U5dLJ6tfeVeZOpyPyf3mFoSlByauluV6PPiE27J/gOnQpYtK/NGPqr/5HsY16spJ2Ude/oiUXioyV7NXf5FJKUIxvqQw9T6tHm2e3yKtJOMpyWtibiqbjfVmDofuHCxiY8sfZ05X67WKsU1sVoOEmu6J1IqMotbiWlhU9NzL8gdbJamYc+kHa1t9ut/PsZznVHK6HtZq6V9Nf8mV84hjMPSqt6NRt5Jr0XXuKnKVZQju7leLsimkmSp2jbsOOhzJRuTabNGxK7MNflNEx22HLSxN30QtEc1xrYaTdzD5jh8RXqUKb96G+/X5WNLlZ+6NgHQsKT0RCdy6JOzuXTCUklYdi42kipK6JpNkSOsSKJLQZJ2ViepJahYAsMYzpYLAlZHQRGSGr2sS1Q0EldBWovmuh6hhpJ00BoaDsHslUjyvqPHYCfDjwcF+9jPmvr8PLb0387jSJLoI3I3IS5rE/e97uSV4ok28Ko9UzNMDSx1Jwn2a69vVdjCylGlG+6t+Q5qX3mZUKWNp1aUvhkkuvaz7GE5aMYx6Ky+4m4ycuXZk8FKUZSWyKDSpxj1Qm5rnW5xBWlPC+0TtK6FTrqPtUnbvr+ZlePdVclR6r8TNK8qlb2Udl+ZkrrQqyg20rbeenQk29EcPyoU8YlXdotPvv02KrfOWuKDlJQW7Kmt0xq+hV6FV3iRXNJIoQ5aZ0Dc6aEFrcs2K1yI/iQrXNDQRLVDabGiw9h6jV9SKaRurG1l1I05RkmYfDSr4WVaP2Wr/AD/4YVPlivMrw5ZpIWtNRMV7WWLpJL3bO/4W8/uMKoxcShioOEYPq2vuMHOk6kFPq13MPaji4VYbLX5mFlSpqth6j1qOP4O/Ty9CnNRw1am3vy2/8CT/AEMdUi0nHdaGIqzhjFTe3Lf7v+HMJW9nh8SnvPl5fRJ3/PqRvKCa3KGU1P7RhWl8Kg/vbfmY/EL+zadNbxv+JUk5UlHrZfgSpuS0MDUowwVdSfvtWW//AA33mNpyoKEu6uObmmzE8qi5dkSs2KN2Yy8acYR3lr8luU8zcMveGi9P1+4zLBzxebYeX2Yxlf1dree3yK2EdOnCC7KxRyrFQzKpiY/CoR7er6369ivUSkkRUWkyErqSMROK91FWSSsh7EUrlOF0RXLHQ0cUOWliaSVh20sfFdEULVXJbGDyylh8TWxEV71S19+m3X9EW93zK0tECRyOx7N2HBnIzlaIaMk7ak22JpGjVxu4wqNcpNe6SSdiIpNKxEewxxu9yUWSSJILDWgdADc0Gi2gLYZB2YtyzZJajHsPQtcqQ5ZWDBv3WhgBpYvaNu4nYTs9Se5uW1Fq7kNOpFNXVxptMgpKLceg6SqU5crte/4rUr1lRnBfZZgcDUx9Kq6K1g4r5ST11a2t87ijyTcZbrQxdOrOooJ6NamGoulTjC97EkrehTpz+tPm+Gy+/r5lFx50nsZ/GCwbXVNfcYClClhqXVTjr97Rm1PE4SvOODdpKStt8N02tdNY3V+m61K06kqknF8sns9HZv10KPwpPexCCg3bq7v1sl+gpupyvqtzF4qtDFUacIXjO93dK1l263ErNGS4uODx9LESV1F3ttt9/wCRiU8RXnNK123+ooqnUUr/AA6/cVtiolymFhzVENWVkW6D2LXQo3LaJXL6CTSI/FYloiCvr1GnYlFXRHViTdtRx1sTtcjFcxUWrS2HqWTNZJK4k46iSi012HKUle/Yw14Ri2jWpJQ7FaGrSKUoqHvbnvTTT3TKdJySt0GoQwyqJXl2Hh2pRU1ZpEVDlhyvovv6szWivc5JayW5luGliKTVTTa/3rsVaidW8XdNtfcSgq1+6I4OdatFfIxmBjRxHslut/y7kMLVjJQqdIq3pdvoZjUhQwfM9U7+XkYarCVK6W9/xKteNCLk15mAoPE1pyv8Kcn8jF1JSpq/Q9pLlcu5i8bShCMan2nZb779PQsrGX06LoVXNXnpy7/Py+8zLC4WlSipQvVhy2ld6Xu5K2zvaGr2tpuyFNxVltq/vMLgqTc8TPWUIu3r/XdFDDVsRWUb6/Ir4iEcscpStLnUUrbqyvr0t5mK4fSq0cZLXlvb579fzRVoxUuWIoP2bkloVYoaLXEikrLUjZqyClSU5pFdJTdjeNkXlE1kriWtrklqXSYnaD1KzvI2CNR2aIVHoKztY5bxshq44q5JKxzdjUYx7DKr6F4vcnZ7EFdIgrkFYemhKxZtjTvbsPckru5JIasPYBmgFgaBbARehd2uS11GhkjoYuGt0GEladiYAJXdhlro7FRWSJb2C4loU5Npd0Vfi5Uc9uZInXrLN8PhKa92cZN7aWtb8+jM/wANh4unGnvGOu+5Tq1IYadKk7c+j67evqVIqjT957FNqM1IaT1RL4blSpHmUOopJszj3sHfzRJ4ieAoKjumr7bX13/4czfGUcNip4iq7QjZt67JK+2pjcDNwVeCun/VzKsVP2ipN6dCnJTbS6O34f8ABKcuWZGnCnU97pt8zE4TD4DGxjV2dOL67ygn0v3MPUjVhGUXoylWUG5GIuvmVtiolymCirJjFYYlsRjc3Ek0x25UK249UQSaHrax1sa3JLlSLtsldiuNN2Ook2xvl0QnzRSJJz5V5GHw0p0qk1tC346EOeVGn21/D/hyFT2c0ypLmcpeo73SY1aVjA4eVVTlH7KbJ1FGSXS5UxXtIub3v+CKM5TcZLpYzHCTSw8+lpFLFSp4WVP+a353/QwFWcaXvd2ZJipVaLlPe7/NnD9KE4VHLdbetiVKdTGe9vqRhJTjKXaxxlGUcDVw9P4rWXqZbRqUsBRozfvJa/1sYuj7ay6MwGWxhl8sZJ2u+X5ff+gsO6znGHRX+SK0lrFGPyqnjHCc/wDl3JSXqvmu/oPszLbU68ZS2Rm+KhWrzttKV/wJJyhdeaFKSpuMSviqtOp+73M+yupjcEvZuzjqvX70V4VMJlNCNT/l5+mnS/6GY5ViMLhaeImrKe23+f5ou1RcESjoOJoQV2RXNHToQuoid5WIScZJoqytJkE7ehKXNoWaRr0Gm0W0K0rRRLVjVxiEilfmSFLXQWyiSViotRovdWNGhjsSaRVloyRoQZRauLQa0GtDoTV2OCsNW0LLYa1sWGdAYAGh0Ai7EdTexOKQ9B2CceaLQQbi00PUANEkBHV2QvglcexfQ3RCWthytKyIvmWvQ502k+jRQzGnjq9apTVuWTj91u6Q8VCniY0pP3pLTztuQ5Y2aMuzWvgXKpRladnZ2Tt96aHXnKb52QSe4qnLeUemxQxeGwd4wj/Ei4vV9fv/AEM/o4t4Wawc+SpdWdk+qvo9NVp5GbNfV7ehl/s5YCKntZ/qUcFSzGv7Gsrxlvur29GmUY0/aci1UdP6+RTjCOatU9rv8j2aim4iWqt2P4lGLe6vf06FSrzwSe5gsK69dU4GYyhLK/q9B8tSUr82+naz0JKjKonNaJPvvbT8Ssrk1eJhYWp3LaFhxRFalOmlTcmWsJdyKQ1qcmi8yyTJK47ISvIS6DVlcS11Hq2xaFkb7iW1jlcWyc1CHM/61FP3YkcQ5Uo02/djdr52/wAkSxEo46jRS0lGTf8A2642/wDSmTyrly6OMv8AE2vut5/oRmm3zdiPv3RH3Ib6tEpRlJrsRSqQcX1ME/at6baGKxVailGs/dSt0307amFw8MRelJWbTt6rX+rmU4+FWnOdNaSVn8n6eQsJTWChNLVt/cj6o40VJ9dvkKhN2aiOjGcoRj1M1S9s79dStOKnOXRXRGSlS9rHpsYmtKpRVO+i1MPGfNZbtW+RjoY1Z3QhSdqVp8+2rsuXfXe+3zOVqm0+4oxdr7lKvGWJdBPZX++9vyY5r2TTWpVx1WvThTm9IKy9G2/zb3KtTflMoxLw1dxqrmbil21aWun5HGWbZjhc+y/Lou1NxXOvd6rva+mmzOFs5nmmLx84Ts/iird212X4i4gqY7MZ5en8KT9NUuy/MpqpTqv+WS/rzKkUysrNIcdLkYtpJFKD5SN02kJJIk9bIb5mNWViSTsNe6Ik3e6JSs7lapd6DacUNIaaQ0uTzLCsiTVtCLTRd6oqDSGtA0sNlaSsS1Y4Kw9BLQpStI5W1clayY1ckbkkLUGh2sA0W0ACWjNBbFjQIMTsiaTiSiNaWLG7KsbTaClJSgmAJXDcIRbehT4ho5fWpQqUedVHyv3mraaPRO/poS5btdhx0E7EUmiSSaKjS27CUZaLclhHRxnt4P3dmvNve+/TYr4WnVqwrPeN7fMpWvynNdFTlqT5eoov2fmiLvEs/a6mauapc8V1RmS58JdLW6ISr1IqlFtrtr+Rl2Syp07z0nv6IzCGJo4qcmmrvTzRkmVVb+3qqyeiv+ZODg+ViasZ7i68qblh4+8+VWuvJPcqYOnDAKtf3r2t6WI1JQfNF6jd4X7FdKWnYqq6OS9kU1amrBIvchG7IK6UehVilN2DpYTHJid2i1ycXZkIdRp3GhlxWuPQIu6HI5Lw1EtiDSu2U4Wqqb+Xz/4YWInKio30KjSjHzIJaMs5K76GaYx4OvRmldTlGL6Wu0m+phpJy5X1djAV3HF1afSDX4md/wC3YOlG9mppv5NPy7E4VJ4b61DaLS/r/hjLsDSw9JRi+9/v9Stio1F7OG0f1Jr2lCmltHQquP1aDtbT/gfoUMMoxTfb8zO4KMm46tK/4Ga5ZXxGT1IRdnLro932uYXC/U6EMPJ3aSX3L5mNptuKguiMTUcFB26GaYNxjTrR3qfo1/XQxOH9mo038RDL5uMW1v8A13JZLOji51IK94x/N+fqVMsdGjKvLVRcbr1u+/8AdK9Wpjc5q14rlp8sUlvqt3fR7W8ihSbjJ33kZdhKmDzKtOdTmcnorWt5X1v8yrhq2aY2nN/xJaLb/gL7zKMpnkOf432r0Uml/wCBO2zf9bn17CYjHTxOD+3Jxe+lk+++q7GSYqouWlV1dN8vql1029CUlOV7WuVqepTwUqtJyj9nchAXw2RFX2Jb2HdyFBKRzJnUk1dI2aJOxWlobgDWiFqzRF9SHwjtzWHaV2NKJGPNoOFr3JVEiddE58wmOXQeorWRApyumWQ0S7FhxuxxLe7oNXJIaGM3AsD1LC2NwLEJu6Lpsew4XVxxEupiYe9cKNVxdhaoB7AbjfREYpxfmZdl31NVlzXU5yktNuZ3tu7+pQjz1IxvuYqi6VadN9G19zI6IdmkGGpudaKW8tPv0M6wlWGYUaLlbknd6XvZSVvLfcwdFU51H/M7/hYpxTlbuU41IympdzD/AFh4+U5P3VFW23u7+fYr42nTs5PdpfeVajpw547/AKmGhUrUOZfHa/zt92/yMLKrUo01XjadldXvZ9tNCOBwX9lQrJ/vnNprXa2nW25hpUKdOrGpG91purO61/rQwkPaRu9kUZNVbdVqvlsUqjqOblu/zMdBKEEuqOhSjGpL0Kl5RcfkTo8rsyTS0JxU73KsSkkyKSghkHZu4rFBJRQkrlRJpm4jrYSTTIu7SJWUbE07EN9STvYatKx5Cjdmlh6oSuyxFlNpxZBXgmRXLBohNtPyMPFVrQbG+aTj2FK65SnZRld9DEU4uKi+xhaikl3Woqri0UsTKnHnXRkJy9i0no7MwOF9tJP1/Alh401KSeqMJBTpJoxFNww8Gu6/IjSThdFWp7atU9WipX/dOnHe5mGDpuFOS3e/3mYUlGu/JEpOqot7Iq5nWeCWFi/djK627en6kmqbUmr7/imv1K9WtTw8OZdNNtldv8+pgq9JY5Rm92m/QpQlicTOjR2lL/Pv6lOq5YvE4b7VN2/D+u5w7OpKMXUVpauXrd9vLsY/L44zOIY2jG84Rbi79b6KzdvvRm+U08PRwnO7VK1OPM+3yvbTysYThif1/wDtP2ms1HS2zsr9f0OGcv8A7AapyjedSCTd9tul2tF6XOJsJiJxoY/Cv95TbdtNVbXfRfczJM5p5nh/awVrW/rZFen7qaKNVwjKKejQk7Ch0HJKKSOW7uWSZNJ6okrFOKtcjC60G1dorNKJJgh6uwkN3EMaI7obXMSfuslJJEsRylTFykTm5A0WHsMh8JAg0tRNM0Y7NDsDS2EtB7DSJJDsWLFtCwAywW1AFKxGorDaaJWZZWMRG60JQ0ChW1UWaND2AG0kKxF6mOryo4epUhG7im0trtLa/S5gMROvhaVaceWUoptXvZtJtX622uOTk+Z9TJsRia+EU8TDkneStdO1m0tVpsaMjaxl+K5anMt4P8UZnSqZjXr1ZO0pap9nZ37dzBQdOjGnN3aW/cg7STZKp7So79TMsTVwmGqVKUOaVmkrpfi7kMLHFw5JfZab8nZMoWm1HuZZQnTxtN0ldp3ttt6mc4x43G1MTazk7/1t+RkeBePzGlhZPlU2lfe2va6v96MflCoYqvQm7OF/wfqZLjMHhaf7x3bumte290VqsZ1JOHR6ehleAliE3Ba2b+S1fXyOLMHPBwp0qfvc3LK+2jV332JJPZlOKSfmKo5VLGIr4ZZfSjHWrGTutdunlr5Fa0qjaMvpYdY6lLEO1JNcz1dl10Wu3bUmmtGUIXlYmrJI5nciU43diK5Y2LG47IWhFpsbskK0pNokrnLzRaRJKxZjS+Y1Z6CY02izSOlxDSRS30KL9zlJtRaRSk7bb/oUXzS03MNSVaTt0Tf3Fklcr4mjSVOi378r2Wv/AAxiMDU+rwrJe7K6Xqt+pltVVKLb0ehXxM6uZSwS2cG79ntt/wAE4fjWnhq0aq0jLR6arvpt8ylGCiorex9aoRpwhB+8lruVKKlPmvoygkmopaWKlXmpxbfcjJSoRlfo/wAjhPNqua1sbXcbKFWUN73t12VvT8T2clK7WibKlT2kYya/pGYSl7daEsO1h2orVanspRT06XKWXzlT9rJe7t/WpxNnFJVsLhajtJxait7231S007sw+IhiMZiVBXcOVP7mzhriKSh9Zw65nCbi+lrbvVee1vmZhSc8d7ekr+0kr+lnrr59rFanKnha04K7Rw/gcHhcRSxWKq+zjSje3K5a/J3/AAZnEq2Lx1TnekYuK27t/kziqtHLcqVONT95FRdrb6pb6oxnC+Kw+T1cfiI6JKV7rSMVeTsnrZJ6Wv2MNjcLj6MK+Gd6bWj1WluzszBYKhh6s5U/tW79LlScnp0IpX1KeHlTpqpNWi9hySTSLLQqw5W4diceVpDikmShsSi4rQlU5IXHiVKRUlcCxCFxxSiu47XIxTKk7OxdWJ10lZbk6810HiZNWsc1RtabioJrU9jAdKI6EWirh3FXRYGLsQnbYhsLRIGx6jLaBew7DWjGhlh6oABbB0AASbEkNKw4plan2JUluSVmUcQrWZuWLBYB6gtiyb0I6aDV0YWrBYqcE9bJtfgXajPl82ihWqVaSdT4iyUUTjypeZJcz0MFlMqeHqYhbc2vq9uvZdCviKlOVo/C0/vOF+IK2Bx9LERfvK9ttmmn0fRiioq8djCVIRaUtiviI4mrGnF6bFdNXRSk0kzKq+HjgpVFL346Ws9vy6mJtisrdKkv4bcm/KVl1/wlSHLLQpxi049yrRdOt7Tq9CpzRg09hvmVyq6Sw3Lb3r+e1vu3KyvJkd2TfMjqK5Qg1qL3nYqcyYrKNiWm5bQjvc3QviReyN0Sd9CSdyKWpNajsrnMPY02H2I7EIe+vMoKVmydOUmlHdkIqVTlewptTaWyKErJ26lXmUeaG6Jwi5873jsVcfVlgoYe/uwba+e/Qofu46bshh08V9ZW6Vv6/wCGMMpOVkYylU+q88H7zTt6ow+F+s06cp7pL7ylh1OdntYpYZRi2jDe9GT6t2JJXUOlj+xcLllJ06S0k3Lru3fq2ZviIUqsKcvttr7iFGM6UnH7KKmHcpOK6lSg4pW7FCj9Yq+xfVfkU6FStVVCPyOP8vw9BwxNZXnQWm+7uns7bPszKcqniMBVxUF7sEm/n87/AIFKFKFNwp97/eUqlSlGFOL0V2/vMHjKdWlFp77Gc4ChmdL2VZXjdPrvHVbNf5GEo4qWYVfau8Jbbad9tfvM0yvnxdKrFXcnFP8Awpp9/wDglLNnieG6OIltOEqf3prt2/4cp5VHLsJHDU1ZWuvn82O8mvI9i0k+5PCqklKXUlWlUlzMhBNNdSlSTmubZGJlGVRuGzJttm7IK6sSd9TFVW2oCiOPuJhugUrI31LEdCpTu1YjQckSw1rFRX3FTVywwAqUlNFSm4sITSldiaKc1awpJpF9BrQaGhvQexvEcbIa0sMYwAHsGgABG1htWsNjY7MnTUlYq0H0ROhKKuiGJcdGU60Z7MtdaD1OpuKzNLDsLcT1EtWSTsU8spwzyeOv70qfJby5k77+Xb5lOom5076q34/8MYV1J16t42UbJa76b+RG0olSrGLim99CpOopQUFdN6+S7jrVGnTg/dtdkoRkrMjQjG3L0MuwUa9BSlK0U7Py0J0pQbfYc+XZ6MxlRUopz+1t6kJWWpg6VL2NbFN2tb/L9OxhsznTwtSEH7tRWfy26f5E43aRGphaVGrpeb+HfRdfL7zGYiGJjFqNrJLe+qVr/Mm26Pyt9w0+pWblYrRtISFsj0IpkItRsyDsybcm5HM27dirZkXcim3oKSimiLu0SV2OSLpsqOLat2ItbDaaJNPQ2RYlHRD1ZHRXYpJpd0ihL93aRw9m88bTq1lG3LJxWv8AK7dkRV5NsxOAnhqdOpNWjUV16Xa7+XkUtZIjKCU7vXoYZwqV/Zye9/8AgElaGu5Spy+rxnJWuU6fLGyKDUr23MPJ1KFuz/Mwz9nNRWxhIQnePVlOg+VpFLDSp1HpoYvGKgp36JP73b9DHVW1zt7bGJw3tYqrPda+hQ543aloe0jOSkmYpOknCW9jJZQljFTWsmh4jCYHEOq37yWm/XR9+hxFN47FzdXaf9dLEMZLDYSpgqT0ny3+V+/+ZgMlw9PG4nFyq2clFRjyvV63V76W8118jF5VVo1uSorO2q9UmtU+zMS8LhsxwNCnK1ozVrN3ae9+lk0uw3Ccmr9SOGrzxd4r3LavTftbc4UwOExkPY1attEk+V6vbZNW+ZxFCGCwkcDTd4Urq/ovm/xZisXDEQhyu/KrEYpppLUq1dOVbIcpT96Rh8NUqzSpxuZbls8TiJ01HWKbeu1t+piXBRlG+o9TR7CotR5iL5UrlaVojfPK7NNi+gAlcsX0IQdRaCprchBWJpNpFaCGrAWucuhylixUpqasTg4OwJ3SsQV2WSRF3RdM0uNFjS1htWsh6j7jGgauB0ASALXLaFiwagFy1zlTVithuqFJxZh8Q20iWpa4i6HsCEKKtc0cbCi3Ir4ilSqWb1f6GCqpU5r+a34EKPRLc9hGaa/l/Q+rcuHhVezNI8z7lPE89ZxizKq9KpVu3pdr7tDHRhSrtU3oQqqpJUqm+x7qlKm+mxjP9vpUqUV70E/66GGVSUHzrVNlHFWwvs6u8nb+rFOXslFdCp8DaMVV5ZxilqYfByrUeWKu1+g5RqKUV1F7yZLVoxCcndDRGWhHVlKN9xO6Qx3aaJJWKltkRaI2aL6JMWk7IlLTQbTQ9LMbuyOzZJWZYfYim1YqU2mkWI7kHdI5lOKSKFFRoKMTERtsSrSnBJ7R2KNenUnKnDeO/q0n+TRWbeJUVvuU8PbGe2X8tv1KjurFF+5GL6EUpRujD0nJWW5h17ZcsHqtzBU+ednuv+ASlCnaxF3p3Ri01RbW9hy+s0lOot/66GJhH2Ti+rMwqOhQaWvKiElHAxl1asc1KTdOO6T/ACKrlWqx59yjhFh63tnvsZ1UiqdSpF9kjESdSopLoiFC0pTl1HhpTqwdPpJMzTEzxdedSfxWX4JJfgSyyVXE0pSV3d2/XqVKUZ1YtdLmCx1PCYaupR96S0/rUwdaeAm6cXov68zPMyxXOoqdk7voYXM61Ka5pXiZxi6uHpQVGdr3+exRzPE05p8112ZnOPqKNKVGVlJX/Ip51mFN+5VaOFs8xnLWc5Xbsru2t736ehWnecmuok5OyI0mo8zIJvcqSul5GMnpYSsLUl8JGSaLaCi2RpysPQtKTsihS5Iii3ojlJuyK87vQkFrli2g0ctxoaKtNSRoym1ZIotXG1YptWNhq5LY6A0tB21JLQ0sMGdBgMVrAG41cCwWOoArMkrorRtNopStNC1Rpcmo302DoWOoQbcWizTFVSm/IzHI62PxMcVSfwJq2nWUXe7a2SeltTB0486h8iOEqYHHcuIVnFar5aaq/Qn8Tk9m2z285wjRSMtpYjHzqUq8fZ+9JLVSulonpbfez1RleX1aK5ZK9klfvZLW19BYeNJe6rX1Iu/QqZrianEFalWjanNLk1T97W60V9tddDJuEMXnFa0XaMbNvR2ttpzJu7svK9x4Crk2LaxEfeS017rybK9TmqynFb3MVhHVpRqL7Lv89ShjsRWxcqU4csElZ3Tu9bq26tpvvcp4ujSpXquy2vr+hWpvl5XuZHnsMurVKrhz3i42vbVqz1szDQtU97ZHsnBtEVHVFSLtoVYcu44uxDRpMjF7Mih9BRfK5LoW1JIskiKe5Zublcb99D1dug9HoNuWhBO49iWuhtoOxHcnUlNps0JWUiEmmYeTTdjBRjKcIydk9zHUoczUHdJsp21XYo0IUpuaXvT1fnZW/JCwj9tFta6IcXTk4vdaDpzlGUraIpNQqRhL7V/6+4owlKMnHZGDlBOKvqZZhp4LETcndzd106a9zDxcZN9NCUoy1a0sUUuTQqJNWK1/aqG5mdOp7Sy2X+RWk7tNbs5WqXvmAw045ssTUf7u1vndfP8AAeMhOs+RXey6GeVqcKkaTVne35GYZbXdHnlGyuuq/wA+xPkpppa2uUasaqmrdiVR0p3XkOrBN4mu7L/geX+RLO5zweG9hC7hKWt+7d9Gum3mZNOWJrVYzjZwlyrrfRa+XoYxOnRlTnC00+/9IxVStzU/ZRvrrqtFbfUzmXNXp/11OIaN6Mar3vb8H/kcQUsNHJ8vlTlebi+ZWem1t9Hp2M1pQll8ZW2St+CZldKhUwFN1IpvXdX0uzA0adTMakHFNe9pbTcao4ShJ2SS7EM9pz5k4W001Mtx8akFV5bbolJzil0OXlTTdmicdNDEzfOris7Itq0O5UclV90ppy0PZ2egloidO7dmUqaUlYWyRTg01qNuy1K0mk0S7DCK1GrMsNGiZoyUbjTQJ2IPUgyK0ZDVFtbDSY1oWLaklZD2sPYYNXAA6Aw3EBbQsNGjLAD1MXHW5sUneCDqCV3YsWuLYgrNktNSlVhLE1Yrdv8AJFCfsac0vtIwzfO49jK8zoYJVquIV3KDit1q1ZPRMnWTgosyZU/7RhTqbJ/p5GFxSxEI1Fo23/6U7fhY5HFletDEqMVumzG4OOHhTaeslcrVYUsVSjJayk7f+At/oYrFYmDpqi/tJvbb5mNxn1iinLeyROOuhhnThg/ZyWr1Mfw7SxVWjiL+9Tbtv1tfr5dmYKnGlWhUqK6TM4wdWhU5Z7rX79SdBKHr+pw1hoV8ZGlN6S0MzoSpV0mXTd47ElqkVqfuNkZbeRFc1RC1ZFWIR5pkqahCpD0HuSs0x7CWhFLlNLjWiQ9C6TISJ7sewth6o5UmPUtcuLVWMLJe0SMJO8W0TpymiNN8/KKLcvRn1l1Zqp2SX3KxVk5Nt9SDsnHuRSkkupRi7qxh4Ne8ZJhoVMTSqSeyf5FCXNUl5MppqCFHQlsVMPfFe1vukvubf6mNbU5Po2Yqo4UnNLVEJqpNx6LQxVJQba6GTYGNTFxn0Tv9yv3M6hHG524X0U0/u18jibM41akaVP4Ya/OzXbzMZVjTsu+n3mZV6+Gw0alFXbnFdNm9dzNIqOLp0/5lf7rf5mLoLEYd03sjNajhUWAp7zi36Wf9dUZRRgqtKfmr/ecY5jShmNRUtdX/AFqjDy9pdmb4N4OrFd7v8fmY7Mq2OUKSja3TuzibCxw+Hw0E72X5JGLzSeIoQoKNkvxMVhMXiMRg8JB8sYXnLZ3206Nb7rtsZdpmlT/t78zP6zjhVT7v8v6RUy3kwXtm9dH8nYyaN8MvVluhJ3Gk4sxFFXLFhq6I09SlSUY3JpKxKproUoiSbuiC3ZTV38iT2XYqybY02NaCjcsWFFNHIlYcdSSEycUmWGtCO5F3RSs42Ke1hnkNXJI1tuS0SB7jALAC2LFgAAGgAK8VKA9zDSvTsN6aEdUG5ZWBFO/Miom0kSoQpYmeJevKm/w/roYP/aKalHqrkY8lZX6GMh9ZioT1RisNKqlyzsldbb7f5GKVR4ScYq7tp0voZPluIxNHB1sRHkqJRk43T3S0unYqcP1qmFnNL95B2cdNn1ve2n4kIRbjd7ipVZrmS0QsTQoTnGrG7cZJbrXldnp2dtDJ6WNr4yc6kP3UVo7rdtJaLX9DB0cXWxdeG6duVaaWTb1/z+QoyScuxCtLEJOT12KFZw9yfyKuJjUp04Jaq7/IxdSdeu51Xq0vy8irSabJSlBRt0MbCVWCnJ3dl5dDC4KnhKMaNNWil/Xca5ooxCj7F23FfmMMo3ItNokkloKVhtqLGlyseisSXQjHQirSLJzSHfQbT0LJ9TZ2JvSxq2WsLYdrEijBN8zexa5Cye5T3RhZe8rbK5Sy+piqSlS3vb5282V6UqNVwqKzT1/pGLwkaap8kr3V3p1bf+RD3I2fUuloypHl1sYaaaV+hRq3aXmczjV5vUwalaOnUw1K/O7GHSnqJtonqrDqpSSZmFK0LR3b/UjTnVqeyenfqSp8k3GGzuY6T+HsRzenhcBKjGPvS0vfp32/A9pUhioyTu1f53Myw/sstg5K06jv8l+G9uzJxhWk0pfD5eQ5Sp4Nxi9bO3zMmhi6WHhTxU+aUb62S69kUK805w7mKwsYVFVkrsxjpxwdCUN3dtejaX5DUZrzI0ZKcGtupxkoOeFj15Xf/wACdvwHkWBwNKFWh702nv0/T8DiK/sqSb11/QwmVYWjGFTlu2k1d7fIlZx0MB/yM6nrL8zijBQhleHxClq5STVuyVnfz1+4xkk8svfov0MkdsKvVlzB5XVr0+aMb2/L7ytT5JuJOKaZ7GNycItJIlTi1oQppD1QtxQTiRVhXTFKyZJpR8yc3YactSEdNScOxypFhU1Zs5TdDGtCe4rG41ZGxB9CldpnMiMk02XQ0DVx7j3GNIANAFsDQx7G4tgAGMAdmitG0jCvVo0sR0dzcY9UbliNrX7EtJFPllKa7aGU11hcZSnLZNHE2AhTrRq0n7slf+rsTs7GDjFqvKXSm7et1YwdWU8PF1FaRDFvExi57wjGK9Iqy/BGX5pXVSq/5vTr8jEwjbmRPMKmFweJpyX7uPLJPzsr6Wb6sxVBVqirLdP8DDVJU6TttdfgYetOGIjUhujHYlUoTqy6asy61ecHExcIxhKit09DCe9NNvVf8Ax1N1k5Q2svyFL3FJlPBuuk4rZkHTwUa9Opu4tL1+Vycfcv8j2TUHJbXMXZQ06nPHnceqJyrTqyjFt6va5lmMqKsqVR3T79GZxXqRxVoyaVl1KtbnwTqRerjf8AAyus/qkp1Hezf5IrYmtXm5tv/Iq1arwMJOTvf/M9vV/mf3syec5qopO+36l7kdWiT0sNLlNxtNkndjvZErpaDLDsyLdtC2hDcjsjCQ5qiZluJ9i1NvbX5oxsnXqSqPdv9SkuaVl0MSvf06EdZXOa71KSXKoow1kmutiOta3kZfG/J6XMFGXLK/YwyvAjaxKVrlWMYzcn2MXiVKmkiDcpOfdlWUvrOnw2b+ZmVRU6Mpze6ZiKrVVLo2YnKqOE9jVvrNNvfT8WZjjo46rTaeiVvu+4yjLpYfGaa88+b8vMzHMYVMzjR7Jv8UuxntClTqxVJ3XKm/6YlU5vbw6J3+VrfhbY9tHE3i3rFa+tkStCPvdL/mzB0uaXkOpGi02caVoVsyjUhs9vvKjvB36HE1nTpW8/0HNSpQ8or8iSSikjL1fNKi85fmZzSlXwvLHXl1X6/gKpXqUXTv7sdTIKbnhtPMlBq1zK84rYSnKnDaSt0/yZjK6qtS6jd5I1Q1oWOpHsPaxGyaZa9hO7sPlTJzvZFtTRjsi61JJNosmxRuaIVhpXJRuyokMlJkfejcYtrlOrOCai99CLuJ30E7uw7Mtcb1sOz0JJXHG6uABbQ6Ai1xLmdg0NxCsWLDWo7AxgaGKgtzD6TRZWEgsPQTK+WzpYGnim9JtpfL5/oJ6EpJoWJlTzJUo7STv62MXNU4qRlGd1cTmeLwzd1BQae26ldbdLJ3vrfyG1DVmZ15VsPF0na39dTL6inSjzO7W5Ccpt22OH6tDDZbVp4qfNWT912a0v2Wmm2piKzhTk4q7X4lDGYeeU18FUj700rO7+63n6lGcnhKFKovfikm+9utlovkQqOOhPDOnSjVTvF7mk5uL2KUHTkkijFy5py1elvLe/9MlD2jTitepw5mjpYXEUEr8zUb7W0T7a6NGY0alGpKnOOitZ97oyfOMTl9anCDvTUuZx0V2ttbNr5GNzqOaYvEVHG3vvTe13e17K9tr9SpJSpSW+w58tNx6fqVIc1Sz2JQXtDL1/tsvmTtUzJcvdfgjM4OeLsu36GDqqeW1YPeN/x/pmG/5FtS3f/IyedP2Mo9Tkila2hVS/tCK9Dls9ELUStJJk1qrMtc5W3oNaaCTsdbmlri2HZIBdiL1IPoYVXcbbmAlRiqkKqvdabqz+X6joXhexg4xwmIvJX3Xb/Mrt87TFOCvFdijCV3JvTojD6vRGGdk290Tspc0TJOZpc+//AAUYK9WM4y30MPBwppXuUnKz5jN8Vi6OIw6ofDKTUttrb66/JGIk52b3RjKsYylHqtV6GHx1L6xDDT+OV2t9l+HTuZjmeEwDh9YnyqTstG9XstEziWFXNcMqKny+9HW19FJNq2m9reW5k9fFYrmjUWvM1FabbL+mYqVXkjzy122JwknZdTC13SmqkX78dv62M0pS5XV5tUrvz6/IhOpVpOTdm1+ZQp4uOBklK8b2b01f/DdjBYGjOhXmlaeje+vTvZWsfVeeLUn0OGs0eJws+WNnGco79vkVsOq9Rc+tvlqzG5OqNODxEU3e2qTtZ231Mzp4NYej7H47Pm3/AF0+4mlNRuk/VJ/mbobclYp0YQftElf0MZXVGm6j6GYZhQqUOSm7t76WMhpOGFi3po/xKtaV2ig72uV4tNpEXyovexKKv6DWhLexfVDlysveKsRilBNDqJOxKbaLRS03GlZiSstBpEhroWVxaaHkPUe40nsSSGjmbKctLD1OliMtLEXoU3e7IvRF1sNrYlZoslYe43oNIaDQ0Y9gdjoCNw3Bo3GzzGrjDoAytFSgyl8aFsIDMJyp4Wc47pM4beJnltKWJd5tXe3fTbTYqYupPDwot+7G9iMlKN11HFFehKGNjPuYjL6tOlTdZaTXMvTVdH5dbGCwFLDYqpVhvK19+l/MxL9u3dbqxHJ6kMs+sRXuRko/O1+9/wCtzD4dUMTHk/5eb/L+vI9m6cPmKadXXcklYpxisOn9q5KN5DjoYbFv2fsnsytRSu4mCn7WTT+yjLaKxGLp0P5mYbA4vBY7F0sR0qScdvhsktm+qe+p4c5dhsXUxSl1kn13SUX17RRn1JThib706nKvS7X6eZg8JTapTk9JOxicsjRrzVtXq/6uZf8A2fio0sNf36s4RW+zkk/LbzRWTjJwe6dvuMT7sOY3ZOnOdaSgru7MrnGliOSS1enp/wAOYjTM4fIo/uZ1qL6p/h/wDKoKeFlF7Nv8itSq4Wra9mtmYSv7ekpfeVP+RjH5fkKTvcslC4m3JEotjelyL94nfZDv0Kl+U5dDQYrM0eoiFiCTsYN2nYw8XKZiavPTirbK35mItLXqN+0qMpJSbb6EXZKPYowlFq5TSSKPvVDKedVGYepNc1uhCWiuPSNjEJJ69EZ/n1PAK83v/XZmGzajialGnfVxu9+sVJdPMVLDLMIN/HbTfpv5dzijLqeOnS5/+Xb5l6r5rt5mLx8K2eywlP4KcLv10+f42MPCNCsqsV8LuYmu5ydj2r5htRi2iUUqDr1/g0X59tTNsViMZlkKlB2k5pLZ6Xt1sUquLwdKphavR36dvK/5mHxNWFWcW9HYUptpPbW5i4YOlGksL1iubf4uu/6aFaqlBQS1WpWxNbGYhuTvd36bvXyJpc7T7mJlT9hTUd9b/hYgtEKN2/Iw1SpOm3UWt391zGYf6xRdO9rlLI3z+/LTyJzcKfJHYbbkRdkiq3Gw1dFktDRuw3qS1VjlsiUii1y6k20tDUaVzSw2rF9DmuWuaaltLj3Lm5JO2glsPW6GlcexTYiTEiMnYouOqYczZzIe1iV1sMkhjBlrjvYBWsdBbiEAIt2DRjVy1iwwsNXQlaqIW5KSTS7l9CcOda7A1oYHDU8LSVKGyK0KsnTcJWSeul7qz08tbO67W6mbYinh6NOrU2i0r/4mo9PUxmdL2+HVd8yuoLpZO7Wy1/rUrYehHDqcV71332KTk4lGtWq0fYX9297ab7X77GLw1eljaVWE+WKvdWTuumvS3kYvMp4yjQpy+wuX5Xb7LuTw1pXaHTqKm5b8u7MrpQw2SyzGr+8pVNF9nz6a9LbIqV8NXnKphoctNvRXbsvV6v5kEqjUTEwdOUkt0YSuoq7WjK7ynBZjTjhaXuTi21eXxJK+ru936dinVdKqqkXs7oq4iGZ1IzjG1W7bd907dNEuv3nCdWGFxCglbmXrr+JxNCWIxMq0OjaZWqqacb7bDzSc8PUo1NZTad/T0Rg/Z4apB09OVpr1TTX5FRtz5r3MXPmhyoSVzDWeMl8/zMXaOMjy76GJf/ClB+hmcXTrqa+0v+A/wMpaWEbb6v8AJGPpRlh5c3RXMov7KXqVl/wopen5GrL30IWckTWug0riV2NXbsKJONhpbD3JXbC7SLtIhPUg72MCn7RNFJt6JnOpJxkYhKCXYlGzdinFN+hi686dktW2U/a1aS112FJqajcShJJJamWR5Kuoq0YTUVuRmuReRVqRSil1MVTTTZnXD+Fx9T2uIjzO6turW9GjLcnxWHzyVdyvTaello+mt77WMNga1TiLEYqfwxiox27a9et+qMNQeFjOEpXk3KTdu7b2MFjKU8diIw+JNc2+9lb+kVKtSdS19WUsZbGzwzWiSd/XyLRdVwT8xStJxHlM8yymvCcrQhra3WWndPojGZnQw+EpUcIuaom4vdWsk+qs9yeIcKVKdX4pNJ+vyK9GLnFve5kuSzzSq6cZWl6f8FGLwuDwuHjd3nzNden3oy7E0MZgauJjpyycVv0bT7duxl+Z0sJSqvlvKS5U77fKzuZrjFi8TKraze5eNmdCrqxrS5Pc0KtRzio9hRIxUoknzvXctfTsPUbujlui+pKVh2cWZc8T7OUa+rTdnptfTbyGtzmWw7bGxyou9hpJkWtRdUdA3He2hzaD6sViUEEWJrlQLUT0sJpDs43IvobMbukx6otoPXQa0GMe4zoWDoaFx2aEG4PU2E9BaltBq+hYsNAV42qJi2CrS55Rd9h36D0ikF9Bauwv4aMThsLj8vmm9YJaa6+8uuljH4ilgKUq9RXUVf7vS5g+KKeYYT29P4X+m/RGCrQq01JM4YyieMlOCeurXoreaHhJYutGhbV6GJwNTBV54ap8UHZ/L0bKUPbU49yU4YajOk1fnSX6mX17ZTLLqcr06UlF+tvv2836mEw8aNGNGP2UZRTw2Lc3KpyqHN0b1jfT8N9irO7SZSjaNuxTpVaeJlWavCSVttGr383e69LeZSpwnGCT1sYapPCYi8Ve2n9bjzSNKtHEPT3Ul8vkYPMcPW5lUdk1Lvv8kV17XGQV7a3/AOAQp87chxdOV5dCryeyhy72dyvO7LWJVJU60pRet2Zdh6mIxCqS2Wt/MxStmUF6Gb0lKgpLo/z/AKRhuZ5fNRV3d/oOpVmlTu35GBouhRUXvuV/+Rivl+RFXegym0pId2xLoNXdiSsJ2H5j20OZcom9jcumhaqxGSKdnGLMvkk2UUou5NRZiKbm7FRRT5XuOLirIrUdVJmBivd5nZXMVgOSomnvqv6uYehKtJqPRX+4weImq8XbexUp1J4uEumxKjKSq8r3sKlelTTeqMRKybfY4rzOpTxWGwFJe9Vf3W9V+qMPT5WuZ66fgYrljCU4rqZrjJQnNGEwKw2KqzTvzP8ArqUKS9sp9k0YhQ5rpamDoqnVq1JO91Zet0OdKCjOo7JtR+926GbYGGDqfVqVS6nFPb/NszDIJZCqtSD55SfMltuku77GEo1MTSipr3rXt2KkabipGDxdSk+aDs2V8pquf12pPRvRW+e6f6EsI6FKMYqylr9+pJ2i49iUryuRV0iTLajeg3qLclGzLEHyq5Ncsr/MvdEKTldeTf3FzRIVtSXVCs1ZCu0NqWhzIiJakY6F1a5Jcy0LGiZYsPQbZYeqEnYSAhqhqzFozYlVU1FdlYWxTpKcJNu1i7WhF6WHogew0hoaGFhgthHQNywC3NyysMvoOziDVmNDQYqN43IO8Ubm4ouTSMZHlqKNtkvyGCbTOa8LEISmtOmr/L9TMsFHE0p031TX3nB9B4TiHEZalpv91vXv3HhaVGhShDdpt773t+RkOYvAYqnWSva/9bMy7HvD4ynXW6d/60M+x0q+MrYuW8m5f1/wxwfjsNOnTxr1UWnbXX59PuM2q4etW5qLvGST69depTpNxk+mrJT5Ukt2xZZH+zoVFLWbfTtr3/yMTh3pfp/kYHDyq11QT1bsj6vL2Th1jf8AMwqUasUnu0jE4ev9dlad4Qutkrv80UKPt6TUuhL9zUfK9yjhqVSs6j03JZbKph546EvdjurdO97/AKGGft69SEtFZu/5KxJ2ikyerGlYUYt6ow6siW5pdMtocqvdISWiGlJpIjJIdrEUuZWJK7uTsyPce4m7jd1ce1kNA7WLaMaskL3vdQ6ynTjC2sTB1Fy8pTvNplRtpLoRcoxXNq0YqCm+YqwUZaO45NtowsUoeYqt+VdinV9nOV+qaMCnKpzIwTU7Mp0sTHO5z5v3bilbTf13K6sopeZUajCxjadCpxJRct0rLffV/r1Mxz32ecRwCXd3838v1MViXLmT6P8AQz3MVicTzRWjsvu07CjyyU+pN8qg13JSXL6MnWpwg3FaSS/P/gGYZzCjlrpShzRTXW2raSexiqknR5o6XT/IwVLF08qoVMVq2nbbo/Iy/E1KUZpfajb77P8AQzCEo8tNaPT7jDQk4c0tm3b5O36FWvyxUDLcvnjacnDda/d80YzCToxjJ7MlYjshpXsWuSQ0yolFL0Fdsik3Zkkm2kZ17C9GNHpCN997K+/mRabSKktiWrSJN2HqrjlcbfU9reKLpkU00Jpote5FqxdWsOV0haolaw3YctLD20NbEkcqsmh6kboCnITuSjZXYxEbKnr1Bvmdxbl7oNGiw7NjNyx0HqHQsMGFrnQLJm4y6LXQNXGk+o0E1zRsUo2jbsLcsUm00zizDU1TwmKp7VKcb/4lo1+HZIsFSpCnBzm7JK7IVIzSlF6GW5vSoVq9NvVwaa10V077eWxh8VSxOH9rSd4vbz+84b4bVXiV4qWi5Gn96ff9DFVKaxUYRerTf3W/zFKMbKT1KUmpKxiJxqxaT01RlWFWBwqoUtl/XW5hpxdKLjsQqWSjcjQ5p819l+JXx31ZUqcnu7L1sUcMsXRnZ6pX+7/gGKw7wmNpz5rS5XZW8469du3n5EalSOE9p1en9fcYWcas046rr5MWa02p4NS1upP53S6eT6lPC1MVl3scPrNNt+mnfQp4enVUnKVnG+limr77NWYsfChh5KUrR5Wnp/wLmBw8sRTnGGvLFt/IxVldJkvhGkRtzIoq6JPQ3kTlayE2k0U0nqL4rjbN0LR6F/eJavQvZWQbMjqbktUNaliK1uYmhOjPkmrNEVJO66FFpyKKu9DAS0uynZ6lWSm7orUedFSjyR5ShBOVzC01T5p3ve5BWY+aVzL5R9nZbmU+5Hl62J6TiVpxa3MdiadOnOU3oiONjicyliKTuoJyj0vdtddtuqM3o4qrjqeYwjdxXM1pv2/TYWYxr5ZHEydm1r5StqttbO6vaz3WhSq0sRhKeIpu6eq9P68j2SlOOuxWrP6woLYlSi5czZJxcFHyMLVjGDjNdfy2M1p06UYxg77P0b6GZZ0sflFPDUVeWHclbb4rPql+pOTy+nTxuN9yny3b362+zd9H0M3xdDHYt16D9ySjbf8AlXez+8r1lV9jTivhSXzvd/izO8BPC+z51ZyipL0e3U4azbEYbFVIzjaLvFO6d01a9rXW+xxJT9lV9kpXjHRaWJIppMmrOxFXaRUTUnEb0Ju8tCUko3KMny+95/mcyTKtkiLsip8RLRE2rl3c0Kk00kjocyuX5mxO6HK5qkeha1jSyRLsSasNaHlck+iLrTU22LqwmhliLsxO6RJ3iinbUWxF3sdQ20E7ssDGNFh6nQBbAbh5CbWwnoPc8hq5axINGX0HuPcZufaFqw2ROo5RUX0DoV6Ma9OVOWzVmcqhTjBbJWI4GDxft/tPT5feV4VYVKfsX7t2mvKztvrvbYec0KsaNCS5ZQTXV31v2sjBRpzpST3WpiVGVeMZ7NksJRjl9WTfvK1t/wCvvJV1QlBNaSb/ADIPoV6XssDC+7N2Qi0rlWSqVU0vIo4j2VV05ehFOpJOK/pH1Zyg5dDLqrwFb6ytXF3t6ff+RisYsZWqYhKym27dr6+RhsSnTai9bWZThzycXsRnha+CaatKC89Xf8CLbqJdxycYqJJN3RJtqxNWdiOskRfuWJYqPtVR62uU5WiyUuZi1Qkh6Ftbj0RGzkjTlGgI7WFohybSXY6D2NyFm9ehisTLET55bidpFBe8jDxi5p9Sk+WFjL/ZKsnWV1/XYhGLfkQot2fYxcXyuPW7FDkVikrxSZgcN9YrRpr7TsY7CKhUcJO7Rl1VRcm+z/I4dfLS5l1JO8mZjJ08PKS6JmZwePy2FF6Smn8r/cZRgKNDLoub1hzJb+nfyOHqEoZfJyd+aUpL5tmfyeFwM1SV2uaXzbb6+ZgsRTlk0Yz+NRS/G78iUuaPN1IQtHne44WTv1HOGGoK20UZhkssdisHmd7RpKUkvOStvdfkzhChVxVLFRrrVNW9Lu+3y3MZRoYPF1MHhvt2m99Wlbrfo+9vIxeCqvESlVldOMUla1rXb1W+rIxWph8S8NiFUW6MVWdSXMyLUaW47uJJCkvZco9Eh1pOCjfb9RpykkTbV4jRJc2opaitdEm5O5FMcryRKpqSOW+pJ3Y2K9hLsavUhZo00ElYjdO5uWvYsNbG5JrnGrmmxze8XumJXAKLVhvoXsrF1YjPUvcskgiXutAZJWQxkkiwHQANLgdToItcaubAmNXHsPYsNaoS1GjoMZ0OJc2lleXyxMY8zTirXtvJLez79jj9vL4r6iubSm+3xcvN8V9rv16GX4XEYudqMb8sXJ6pWUVdvX8tym4Om+muhjsoo4utGpL4lez1026X12KM0qas+liDnXxE5VFZQat56J3/ABMDn+JxOPqYOrGyVnB3XvLrolpZ93qV8PTqxjBx21+e5hFSjWXtdv67GZZjDlpQlLV3srdl6du5hrNRlLZlNXwMko313vt8iFGpSnGpJab/AHGJbr1nUe7dzC4j2dVRkZxhqEKaWF1puMW91r1Wuv6Gc4WjTjQdF35o3e+/NJdfJLbQnWhSlFdDLfZUq9RWvzrz0e35JeRDDuUnFbp2/wAzF0pU22th1NFJPUrtSkOK9nJk42ZPUgtUVqsKNFyl0Rl9GXK60/il+X9foPSJrcVkxK5ZcpZvQ05tRWckNWZo2kP4iTV7IS1OXTzLFvdQ0rI2RJ6WQtLSexfmldlK10YdJJSW5RtypyLKUtWSqqMlC+r2MTiYUqVKy1tZ+t3+litzTTkY+eFqYCl7P+JFa7/8N9xRhFyjHuUqqpSUqe62MVVlWblN+8ZZH2ikvRf5mV80IaLRIwvM6ClPdmMd4uPkY3GuOZrDJWaT+UdNfvurXvoVKdGlSbqS9yKXR9PTUyenFOMm/dlql2T19TNFKpjKtO3up2T/AK1JJ3stkezkovQVLn91fZWpj5xqNOCsjEqPJaS3KmMxlKvHBcv7rkUk7rXW1rb/ADuYXFTwyvDqihmMIY6NRxvJpre22/TzRjsNWp0KWLqL3Kl7PTpo/P70iinO8YrpcrVPaS06DqxlJxT2KdCdVqMFdk3okVdCNrISujoSurDTuYqo6NCU4q9lcynMFjsJDELRS/rsiclypIUtC65WS2VjRPUYhuydjclY0sIiuZiVkNNtWLo0sOzYtR2ucytYdt0PVmqGrq44q+gotCbS8zQ0sEGSkSkmiMrootKaucy5nYHsJ6WEHQeqJLQB7DOgAB5HoAthIavsOJylgCxZMsD2BjWhWjBxvPZamYYunUxFKnf41p8lf9DLs2pf2pLBxfvKN36P5frcq1IulKaexl2IeJhfs0Qr4arVnGm7uNu/nf8AIzjLKNPJ6GJg9Z6Pf/Py7GW5bVqYmWGirapL7k+/fuYLCVMHjK+Hr7wsvw12uRpynPliYynKUPQp1FSw0Ofdv8zgmvhpxnhKy0qRcVvvby/zRj8NL332f6lTDfvlUXZr8V/kKhH4vIp16kIU2vtPl+Wr/QrYZTw1WUnrG1vxK1JVFZ9DKpTpQjd6/wDBZF0leTer1K8Z1KsorZkVK2u6IxcnYlTSVu5VjaFya0KUVzoxP+1YlUF8MdWRiml5FRJuxoRSFaw+5F6mjdySvqic9bEtUjdWLNsjG7sO1zcsNXJI30LXZFXRBXaSMFSjyXZSgpJjpwjNMbpuSlbYoWxULv7K/IwuGjUag+pXw/LKS+RCEXy+pZznZGLl7PVGQ0oyfM0YemrWXWxj6ioUbr0MTNKhzNmOy+nCNXGLeS/J+v6FDGVM4Uql7wjO33Rj6Pcw+IWCwkZVekUl/ia8r/5DjJz5n11ZiW+R2MPJNO5HEuhhalNfatf5GU4ehK1Wv8PLJ9d0tNvMlgKlbDzxKfuwtf5vTr+hXnFyi+trL5GRVsXXo2xPxKUkttr6beRTwqp0m4rZ/n/wwo83xbGLxMMJhnWl8KTv/WphP9pUZx+Fq6/rc0TZhswnhcPUhT3n+XXoyN7om22Uo3su5UouEEpLUTQ7ydySTcfQ9nzRdiEFey2HFuJynKnsNJQNZMabRLRaDdxG6NxDersJuw272RHcchO+hK7WhZ2EtDVnIOKG1sRtcSuxp2Ebm5exzXC5SV7EdEXGLYWpfUTLMlF2GhoAALBuAK1iKOotSVuhYtdgCHuOyQ3cewGbUVWwNWlKVlKLje17XVtvmV8HRwc8NTlryqyeq+y9ba/iYDP8LUxOKzBw5Y0kk5Xbum/5baW9GYLOo1Kcpx9+HK2ul+na63Rwhw1Tr3pynb3HLa+q1tv1vv07GTZFPA5xia/PeNWzStta+l7u/wCBONDFZRSoQf7ynzOW+2lvLq9icpUsWpQ3smvuIuriKzqvVtq/rZIzLKa2W432Kdpr00ur92irhpLDylYzKjQxmIvSh7qs0rvRpLW7t1Mro1as3Gk7OKk7/wCGLk9+6VjE43nk+V6Mq05Ty+NSk7uLtLyTem/z2MJGLqqDWjX3EeHsRgsJQrV4/u5O17rd3tom3/WpxJldfL5+zntby/zZKLc3ylODkly7oVCrKUHPRPVMxP1XA1+de/FRTe61cU33ej+8p4qjjaca+HfNCaTT1V09dnZq/mU1Gcly7oxEVfyMTCUZKJJXZVrqhBy69DAYd0qN5fFLVkXoXurnK00jZGiQ9VZCOW0RtWJKL16jVtxJdyS94Td73Ek3oaWVjRs/UaGrElBqNu2otNihZOxhqjlTaXQoQl7PR6jotq9z2lnyMyvSo6KdlIwjwdONSFXe1o77/L9dB0Ek23vqSw0qdNSlpfbzIp7FWjGULMyLDJKMGtWrlFWbZjUqi5DF4T2uHlRT3TX3nEOJjl1GhgWuZtct9tr621/MyPL62V1aVK/NCSk2tFq9ureyW2hxBSqYzDYOUdFGom1/hdv06FVxUnFGIl7tupSpuEE2tzDYjDfV8RGsvecVy7780e3lfco4bEVcDVrXtCnbtu2l6/mf2l7FVMLUlbneitvbXe2n3mAxGGoU5+2o87asnzNcvnZb389j67SwePjTrPWpfkXflSvqr9+tvIyvFcs63tldSjZLaz76fkVZJxtEyvLqOYc9PEu1JRk29enkmnqyVeMIKFJWSVu/5jSvuSdxWWtyole6ZDSK7nGGNwlbGc+G+CMEuu6Wr11Mhx8sfhHWat70kvRPfZbl7KxTgnVSexxVgsFg6WG+qPWUE5b6u7739NO1+o04xv3MZgKNPJcPi4u8pSkmtdLbdf0JyFdRNHGxeyHKybQ5aolboW0uLYWr0ExJcrQtUzdju1oP4SKstR2VrE35jWj1E7Cd+o029BpJWIJMasiymjoFjqBYoSSaJrW6IVLslMjsJ8zRfUi7kXceqJRuOJJFjQAEMANw6mx0Gi2o46FhbnU6AAYvDwr4d0pbO34NMx1eeKxrrPpGMV/26rGKy2jRxGKU1+7qKnffed13v0W1vkZFk9LLmqcPha/y82YbBw/syOKpP3mpaeS5V19ew6bdRyWxisyqwpXpq7vb8PQyKv8AXaVSt1hJx+aS9O5ldRUqLqMxWKnUquo+p7ZToKmUcbLAVVVp9mvvVvMwmM5azqx3RVn7WTk+o21H2a9X8ilTq/Vq1enr7OPN06tJfj6n9q18bg8PTq7RXl+iRxJOmlKindKzXzWpXpqO3QVJRlG3UxVam8JGX2k7fLQjiZRTbMJ7KpTUYdNCiuSo2ys0qd+xmklKrzdypFXZTisTil/LH8yeha7sTXuktWcuhui1yKQ0R1Ha2hYR0sJaGwoXOvoPQdmi1yxbQprVGEjFqxGT5bDla5Skpy1IVlGceUlK2vmQqe2kzG1Z4irBvokirR5aVyFNOSRlGHksQpPohrljcbUq9ypO8XY4pxFRZym/hir/AJr9DJ66qqjX6Tcv/JdP62M7qQglFdCprST6lWO9+hSqWWvYz7NFgMErK8m7L1e3RmJcngfZ37X/AAMNl9avjZ0KD/hRl23av1fl5mJrV6OBnO15xV0tN0vuKNN4mnRxE1ql/wClWv8AkZV7KvivZN6JpP5mOpwWIqcnwptIdV8li+jFJxWg1oyS90erQrKxUtVU+bt/wDDYaNCnGlDZIjRvBX6idmTqykkm9iZVqy5OXoi2hZPQaSFqTaNOQltYa0RBJiEjdivqa3sSWmgth6nVFlZjV2OKEtENu9kWckRVnYWqADcBiZCbcLCumczehB9BEotWuJidi+h2GlZjVy1yxYtoBbQaNyyGgQhu+grN2Gkroeg2mkNNli2o0dAdhjVwxmZU8PiqOGa1qXt8k328vIk3CnOdr6GbV8x5KkYK8Jzp3+H4VJWffR9FZ6nDtKjmMsLguWzc0+bV2VtradUtbnhvjKGKxeMo0Icio+2i9W+bmtrr8NrbK97+RWxVTD0/YRhdSdm77ab26mHw37nmi9mPKsRh5+zfuOTUujureulxupGcIRjdN2euxjMBiKc3eOitbVdvUp1ajxM5NWV3ZdrOxSweJzOFXC4R/vFGTjtuotrey1emrsYDDSpxc2rSlZtedv62LRlCyeo5tXkQqTXI6LspWUl37avbWz0J1sTTrKlU6WXTS6b6GLwfLODlLdX/ABa/QrYSH1h0lLTvbyXQdGcYvlfkiaXtalJ68knG/ezavbpfew4rlSK05YLDzqQ6Xdvx8zK8RPNaVCtJcjerW+3S+hxDgsXXw8aeFlablDonpzLm302v59tTMKXK+XqjMK3sqVo/E9EYGgqFLl69fUk22jZkdUS3SNoka8XXdF7pX/Fr9BK+5GKQ0XSH2NOUaTvYSvYWqaZONmkhuysWuJFlew0kdLDvcpaysYJc1NsqY2KxCwy3av8ALY9pGcmuwrN6GApR15mVKqUlG2hhI8qbRGnKdVa7maYeNKnTgvUpRb5TJ4z5XKRiaijFeZlmYYh43MJ1FaFNxUdtdG5bK/bf5GXZlLHUXUSsl/wfJGe0q9LF4ipUV4uy6bfLUwNClSkpy0jBtJeTSu77733/ACMXVpYhqvT1UkYmrLmUV0MXioVJwjFWdl53tu/K5jaGIrygqdTktJN6J3Seq8rrS+6KWGqPmnXnzXldK1uVdtN7d2YbC0alKTc/hje1n6JfiU4wjUtHd7v5Cx+Dy2nWr4iHN7uju1yt9dE7+nmU6NOpl0asHdu7fltb1vr6W1KdKlRnUnFWc2r/ACVirK9H0ZLYSHoWvoOOlrlveSHHmjYoc0eWMyEfaVHYnJxlp0FtYcWo6m6J00oc9+pKXu2Fd6DZOXKiLU2kTSTsh66mg0krIik2LQ925d9DRtalPE06lWdKLu47/Mjqjl5vkNI5Uk7ErFk9RxVi9tiLa0N2XaLu+hYLBYsWCMrKwmK1yEhMnLmS8ixHU0FZotdWOW44JjhdFtBosWDSw1oWEroaBF7mjNBalro3Ru7HLdjRZDtYGrhoytkbxklirXdFN+l9O679mU+WdPkMdGSp3gtdjIc1xdDMp+0Voq6W3l2X5lLOKeXY2nK9vaX/AA36Pv5GHisdUpw/mdvx+QsJPLquIw8l70Xov6uZpjquLxir1OkYxXpFeQq1KdOnKD1SfN63f6W2Kmf0MwcKK+KMV36adkZ3TrSxU6kFu5Pp1dzJ69bBy9utJNa7dreZCnVhmVGhNfupRk5Ps+VuOm+srLTbrpchlFapSlWgvdjvt/mVsqqfV5Vor3U0n6u7XXyZlFVYaqpT2v8AlqYjEutmcpS2na33JGJyWHvRqP8Ae05KLX92173Ttu9tzEt4R80v5ml/WpXxtJ5VGj9rnb+TSX6HJCEHbqKtCFRKWz0KGBhiMXRoS2ab/D1OAqVXGYicKqtGnv8Afrtb9TLa1KnmFOrW2i038nfoZ7OFXE1K0Pgd7f1uUb4rEOs/hjov6/roJC3GnqdBxuzluRopzclvaw9iCGr6DVlYtdWGlYtrdG1rCeo9i10NWRbQik9jrc0ZJW0KCvIo11Ck49SnBuSn1sU4JK/cUUoa7lCTlKxS5VJR8iNFQgreRgMH7SqkZpg2+ScuqSJ5Z7KMW9tPzXmYWMVRaRja6UJPsjOI1q+T1I0l700+3dLr5IyuhLB5dSorezv8n8zE1Vj8VXoPZJP72/TsUclq+0qSn8MkrfdbvceGWEjHCr7Ct933/mV7Kep9Xg8RGt1SsSpucZJblaXLTu9kiNTlpKXcoXhGUn3M2w7x0lCXwvf+tH9xRzOVHCTwkfhk4/gnbp59yvXhTV5PQn7+orKNh9ipsh2sJ3aRTi5NyEyetkiD5LSRK85OxBtak4ylTv0vf9P0G9LFTVWHDUehdcpVmpbEZ2G7ltCNidmROlhK7FJXJLVGHwNOjialaO87X+St3ObRiRUVrCvZnLpcsrIktCxyOwnokIl5DAsFtDU1BNnMQlYg7ifQbIpydkNq4mmJpj2HFWJK2w4qw0WLDAKb6DirKxKIIWwKRzaWOboboSuiUbEo22GiysFrj0RjuI8flOV4j6nT55TSTV0rq6b1adtjD160qKlGF5SaW60XV+dt7GGUY1YfW1dXflutNuxm0KVTGRlhqfKnpa7eqW+vfsYGpPOsJVoVaXI4Sa3vonvpbft0KGMwtLLsNRox1puTvd635ej7W+ZheIW+K6Tq/b0+70X+Rn31fD5VTk6dpqUne72b0020KcqboRUFbq/UyrELCZt9ZUb6JNX3V299d79inlUMSnUlLl1V9L2T676mPwuDjg4VKMuZ3alo1bfv+hgV9cqQu/itFf1oYxYnKMXLBRfNF2utFfT52t5MxeKjUjCnBcsW1zLe7SaXn1exicsblOdLVX/rdlfBYeOEw1b2v7x3urPTlfe9mVc1pxwVWvV97EValpPb3FFW0S5d77WfyPZyxeJhCL3aM0g3VnS6pv8ABlNe1bitGVKEXV11szhyrQw0K2Lqq6jHz0t6f5HCWJo1suq1aUbKpGTvdvR7aP8A4BjFJKy66GeYybhHDQfvS0+X9fqU6EaNONJdCEbEdUcq0PZ6E0WKWjfoONokLWHaw4pai1dxwdrsa0sJLQdmh62NbWNxRumLTYsraCSb1G27IwsG3cp03K9iNopRS1sU4tx3IYhqfK1exGrFyvHQoOcPeTvdkoznJXMnpqpK8n7qMfJSnHldxQ+sRh2X6FowpuyMfiVh6E60topv7jGZi6uFSSs57dbX17GT16tejXhWlflk4rTo4rt56/MwWa1cFnmIp35lK3lor+TIY2nicDTqrRSWi7GITqV3KW5i/fbSetyFk7EFyxfdlSmnzIjOm0qb3Krte2yL03QU27kqPPCM3sUsLJwl7aXNrdaWt2Wm46nQ5U1YnZPQeug0Lcj8NhWvqQpuq5Weyuc2hTlKLui3UUpR0JJaak1qWtF3epVWxUla6GtGW0HsO60Ero0aFZHNc03ErDlazLpISu7jn2Je8tS1np0JbWQpLlsK71Y5JtaCbuXsJOR1sMA0ADoCZRlpoLWQUp8klJG7FsRZdWGk0NJqw0r2LXJKysWGiwXLCknFEloSVgTN0dBEjVIi72HZ6Mki2hJJFhjjfUxsVKDj30K2Pw+CliJwjdTVob9Wl5/Zvv8AmZziMPiJRlR+GyXXdLXco56qVWsnq6a/NNdjKOKHLGVcPT3aV/nddj6tSy7CfVanxtRa32s77XXbdmMpUY4qhipbwd1v1Vv60MZmf1+nGMttvu+SIUuSMUvP7jh2lgpYmcsU7JRdt916HB+cQzDCYmmtW04r1i7dl2MPXUYVMKvtTb+93/XuZxQq4LFQpw05ZR/Bp+ZJ4jF4jmjrJK726IyyjUzDGUsPHeTf4Jvy7FOs8PQnRktbr8L3/QzTL1yUq0V7mtvwv1vv3M1jR+p0FFe8k1LfdydvwtsYKq4VOZPVD/fT9q93+pWl7DET5fP7hLmV+piMPUnk+IoQ3ascG2w2T08HL4+VL5XXy/EzCtGC9pJ6K7MBS+tSqY2pu3aK7L+tPvJ6yTMThZUoQm9pbEEtTSxGVxpONxNDGnykdi1x2aJNLYcrpIvpqLRnQT0LKw7dBv3Uh7lySaSOhho2ikUZe/ZDqNzKEmk2NNSbIfGkjCJta9DB0nUpSn/Kv+AZbh405Qh3KlFe2iUoKMVGPQxCcVymcVKcMPKNTZ6feYvERrUvYQ+JNf19xPF4fB4arVl3897L1MFlVWtW+tz3nJpel35+vQrVY0qTw1P7Fvx1MwjKOI90ml7Rsc09Y9DNcbilipQhNpaWS9PIwGa4inXUK0m4t2d9159zPFVoypzi7Xv+hlOJlXwjU3eV2jEYqrGvKlGb5b7dDOcdiKVSNGnUaile19L9/wAEf2hirW9o/vMBKVTDwcnd2HLew9x6j0YkmydGUaSn02LJowtV0JN90196L6EZWfoJ3TQ1ohpNElfUyPD0MRjqdPEO0Xe+/by1Kr6jd5aktlYstjTmJWsbIsD0LrRD1Grqw9WaJWNFEltYTui+ly1yVkXvc2SuJN7CloNu9zQ3GjcsFhoCyFYpysiE0o2FNMhZspr30miSV7I0VxOxo0i5LU5WajTsbjQyxEgyV1sO9g0uPsdDQ0LpsTVhO6JXaJDAg7Mz3MJ4eWHpwXx1Ixfknu9v8jMsX/aDmq0dKclZ37PfS3bzKmcYahDkb1ltv0V+3kVc5r1sHiJ0Zcs4Ncrsnre/VW+8wOLjjKH13EK03bme93teysldJKyVjif61LDL6o/e+X6mIxGBxOHwMJ/FTpxTWvxJyu/ufoZTKFVO/wAOprBxUFvb7jL8Th8VX9tWW0Ulv0SXQ4Zx2JyBupRlZ3bWi0cr33v3MLKKqpsweJwzxVGeIV48yb3Wl/LX7jDZlVwuKnOhLlcrra+j0a1XbS+5gsko4N1KyjaNaXMt+is+r/Qy+lRvSjUXut679121KuBwGFxMoyj+4mknq9t+7luvIxzjOtyUtIvRfe+5ioRpzly7fqYbEyppq+hn2cLMFh42t7KChfuk3boraNLrtdspNRaaOHaVKrTrQxOqmmlv+hhMmSpVL7y1X9XOJas8Zi45fS3erfaK/q/3dytSp0qcYU9lp9w1crVnVhFPoKKbKitsUVHldyej0GrOw+4ktiFnY3JK2xNLUbfcejE7Mbu7lmtxtxV7kakZx913J62E2kbj1ZBKUkyF0ULcvmKLcm+xTklTRzudrlFWqL1MowTxGIUF1MLScL03vdfgYSlJTc+4qcpSjJ9CLSTaK9VwnJt6M434iWDoQnHdP8PuZRoVcxyydWLtObTXor+iMoqR9jSy/Faybvbzbavdenczfky/MKNCnpFczt59O5lFOLw3tpv3mtTMZcrcnuyduVkYe6tCvG2cxj6fkZ/TUalNrdp/hscRtSdFev6HD01T9rGXT/g/5FSEnBVn1b/Cz/Uq0Kd3zJP1SMlpwnKfMk9t/mYivTwtNya06IhnScJXjr67lCs6tKM2rXRLR6Gr0E3ZWM0xzxXsbbQhGPzW5FsqyTskXtoLRkJWTRUVL6nCS+K7T3HuipP2dNy7GExaxWFhiErcyT+9FaWlh6G8bG25UUbabmhuboZy6k7aJFk2czaG07IlvoW8xJt6sbbSVycdB3N7IaTtqa2Lrl0Grsd0hnQGdC6Bs8gLCfQjIS0Is5nFqRUnGUI8u+typTkoRn0d/wALCZewndm6HZocS+lhbk49UWRyssQeo0noNdBoBWbOpuXIaaFrq6Gm7Dty2GPcbGV8urZhJ08PG9SzcVdLVJvq0tl1JYZfUlGatKS19TM8Q44+pS/lipelmk/W9zB055piafsFaEdX58tr72fXoVKDxFWnWofwnGV/W8bb69GYjG1MPh51HG7Sdlff/IyqrVzDN4wSsmn9+mnT/I4cyiSzGrQnLaK0t2u316mLqNqEFG3LG34so1FKVNQeiWpm+GqYepGMutmvuuYONWri/bX0Stbzv3MTCpRwf1lr3dvn+f4GEryrR9qtkYnH4rFQwuGoxvyy01Wz33XSxmWW+wweGu7N3uu23mY3l/1ahKe9/wBWUKkaFWTqbcsrer22Hd6S7EJJRXmX1KduWxwzhIYihWnKVuSN9jAQUsPz32TRWyz6rja+IlLmc5O3lHt+nyRiZczVu5k2YPHYedRq3vNfd8kWurHNZitJFO3NYrr3mNJMbsiOqFZvQdtxK6J2b1HFXsSWpiswq1aj5JWXQhiqzwMpuWqdr/cPH4l/bZhcTWqwqqcr2izJf4UvUkiS6GxbUpQIWS1KUbRRGPOrrqSi+UjGyRho804syupKhLnj0IZaniWk93f8fUVKMUoLoSukkirUjBKPVnHP1mWV1Fhvjdrbd03vpsjNac5YPDyxD+w0/W976focN5m6MJxe3Lp+XYlh61OpSxvLd6X1Wijzffe/YyTEPH+3xlZ3/eTt6K3a3fqjLHCtgny7x3MzlzVZdidlBkZapozKtOlmDqR3VvyRhXPH4yMqsttfu6L+u5nyvWopf1qiNVYeviV5P87L8zEK2BoLzk/yX6E5xkr9zIvim/QzublOEEY/AwoUoyj6MwOuFgvIbLuw9BStFkZEXfUmrO5JrlRLVJl700iOq1GlKJVcV7kdiok9SxbdDWiJWaLaDQldGxNbFrmzLaXIq8mN3dx6rQi9CKs3cnqiw9Ilh2dkNO9hWRoxljcA0C2gDLouRnoU3dNm61IVHDVFHEO8U+iZSklJ9mYinSWHpzg/ed7799PwFqRlZClcZJDWgnoTbk7nLoOKRotBNWJxViUbDQOxdM0RoQV2O2xZMlGxJDWo9Ror1adCXtKvw3X5+WpnGMnPGYrF03+7pS5EvJv7+nVP1Mvw8sRXqVJO0Xa3XVNfP7x599Zx+Cw9daUYyj8pNJ7LsvNnGWY5ZRxkMFg3aKg19reXqr/iZth3iOGIzofx3Plf+FRj393e/meFFB5VmUqmJjb2c3J67vTte34mKxtPMcfUxMd5P8lby/Ixfso4Ne1Vqrkrb7WXbTUoYGrJScFddf6uZzVnUpwnVfwpL7lboYlVZ0nKlrJrT+mVq1bE4d4CT9x+90+Jfj+NjiLKaGBqOlg9Y8sX130vu2zKMxqQcK8NHH+uxgcZiMXnNV1dYy5JLb4nzX2+Xl26nEmZqWBhhp/ZS/H5ehUhUlh483w62/C/6bmZ4CpCth4J35483a2rXfyMwoQhiJ0lpZtL7zmjJuxVuqWm9jJ8eqFGjSk9W7/d8ihXprBuKerX/AM5lGtJc3QzWXuKMNyjTUIqK6G82PVCiuVPuR9ySkivr7w9S1xqysQSY0RSsVuXm0GrIxLaozS7P8jJo0nGd17w6NP2fLyq29jMKVOOKhGMUk7dPNmIpU6dCpyRS0ey8jBY6OGoS0u2zCYqGJp8yVvI6MklaxFNO5TilFMo23YoWasfV7RWpJKMEkQjpfuYelGMlYgrzijK3Jx87our3JO8kjNaNSrnuCkvhgqjfq4tLz6nEkVOnBJ6p3/Br9Su1Wo4yLWkUr+mpg8RQeZQoUY68iad389GUY2wkovdr9DD4R4LF1IVH7itJ+cpSn2u1ol5fO5QqRyrCV5PXmqP72k/Mr00k7jacuToxtRio/IxMIzzhRkrrT8jGUo4PM4OCsnZ/ozOpWxtF+n5mbU3SxdRP7Wv36mZ0/Z4ahHy/RGR1cVicM6mJVmrro9E3bbysZTUUFN+hVxjxNSNVq1m19zM5knQivP9GYL/AHaHoMlqyWgti9i6UVYSuhdjFShOgmlqtCKTiJXJtKNiV27EoIcfdLJLQnG8IjWo9VdigrCTPI1uN3LXE9LCdrF7uxLViskrEryE7G4/IvpYWj0FbqSVzZWGADLGgG6OhYQtCE3F3I1bkZX0EyDhFJ3voUp8klK17C2uKxzWQnckmK3Ua6D3sXshWaLXYiMlZEoko6ANKwkaEHqhbis7omrRJajHuR1ZnGX08dhHQns/0d11XY+puliq+Ek9JtNL56a/8E41vCrRjh9oqPN5PrvvprocV+wdLA4qgvddK7evddHr32Rj+H6bzGNZu0Xyy+7531MqyurVwlXFU43VPlb16O9uvl0TMWnXquq42cne1/kZFQnhsS5Rd0r36W1ZxfhcRmFPB5ktIwsns7qKS8mtF2MhzCrONNx+GSV15P5HEPDP1fLqU60tarmmrbW809fwMpxUsLi1GS0gckU/MweNoSyXEU6q/eOUeXfZPXbTbuYOpShh6lO13JWXlqn+SMrxHscW3f4dfuM1zbAV8mr1sRO1Wbjyqzd7Wi9lb7yvU9rhoxjK/Irbdd3+hl2InRxNHENXcdLbf19xmzVbEuqjE03Sn7SPW1zA0IVqbv8Ayv77GEwbr8TYCEno23b0iUcPTrOc49Y6fI4mjKnjJU+xiourFQW7ZGCUnbazHrYqR0t6FeFnZE73sVHzbEl0NUPch5FNXLMluiS1JJSun1KtOrha1k7NbMwWJWIop9dmZl/vkPRfmzFa4eb7p/kYLArEQk27W29TJpO84+hKLSSOW5CLsQitzDxT1uUYwlTa6lmkkYiCa0KTi0kjDwippp6GBwTdWlP5lGnGN7d/1KujTRGV3cxMkm31sZ9j41qc69GV4yUUum0rPf1MPisBR5o15WVRKPV3e3TbfyMlyHEYbiSeJt+6tyxemuna9180OmlSlzHEeFo4jFUcEvibUnvstL9t/O5g8rrvBy+srerN9Nk7LZ+Rj7bHKnVsRUVVd9lcqP8A4V0/NfkcSRaqU6ne/wCH/DmayVTG0LdVH8WZ9QlUxNLlWslb8f8AgnEUeWNONrWuvyKcJLDxfkvxRksOdzXp+pm2EvSUoL4ScqtSkpSd0tF/XyMC7YeHoS1ZvoRs9CTXKrE3y2Glyoi9STvO5VaVJEHGzJOzHO7sOK5xpNkopaFlczmFLDcP4PELWU6zi/Jcrt+NunzsT30Ja6C2RK6Grq5K5uhK5Fq2pLV2EtUWLK2hLsaNCskbIVuo2rD1Q9ES1LBoG5YALXEWEPYZFkHcgyjzKLqR6HM3FruWVimo637fiSg9hLQXkWvoJaFkSaRfQuOwmkRkmi1yasG6AiRa3L9iWqGugwZVhzQOIKtPLMJWxK+KXKl5e8r91qr9DDcY4upT5ai/cVJQSWmyvbVR5t/Q4qrf2jlypVdYwikl5J36Wf3mOmswzilhr+5aLflZP0fYxlChXhSpQ+GOttfiVrO+/fTYzvB1sVWglKySd9EcAQngsm+q1I2lGpPW+6bXa/W/W5xFmdSnls1Dq9Py7GAi8JRjyvVf15kfa4ynKpvJK/8AWxSy7nqSrvqtf6uYlYOORX/5e8z77fkYKu6+HqJbRdvzMijVx2NVO+jaS/LyOKYSynEypKXvXaXy+8o1cRRlKOId22mtl08v1KuaKvgKVHrByf3qPl5GJq87VaG6SX3XMoymWaxcacrTs3e19t+qRi8E4rXZ3t8jCV3l1KvU3UU38vxMozhKpLGUlstPvt1XbyKuHxFWvKpzWjTkpLb7K+8x9R4ipKpPe5kVLDVcRUhXeihK2+/K7bf8MZbiViKTt5ooUlLExg9tjGU4QruC6GGp812Yqny1WxqSi79/8hts3Za+hHRWEMtoSSuSSMVRhVouMl0+4yRtKfy/UzNf7ZD0X5sxX+71F5P8jJEnSl6/oZLb20v66l2/UsnZEFcwGHjOaUttfyIRUWhTirWKjjKfu7FSEVylKKi9DD005IymStZ9CilyIxU23ykXeBmjcaU2uz/IxWGVLDUG/sP/ADMNl+Extb67FXaslv0v5rv2MFgFLFvET3p3S/7etfr6bpkMfDHuE6fw3a+61+3cx+Bg80hW+1GNl6fecRSao0oRW7f6GNqc82nuW5Z3Qt36EYwfRXMdiKWHinVV05JfOTS8+rGo6aLTyE+Wk2VFGbvJXL86t0t+QoRi7RVjFYuGHtKSunoZljaddKFPZFCHJSjHskRV5FkyKTFZuxKN9WNLlG1clLmdxMjZ3RKyRZMWsrHKmyWsmS7CfMrE4pJIfkLWwk7E1yxVh66D901sbLQsr3HtYeothtWLXZJXNFAsnEasXtoKyQpO4wZuAPU3LB0GD1RrYuRm0iNTQhWkotJ7nPZWIpOSTZTUJSfM9EUppSi5EKNCVaUVLS6to+6/QqU3GKfRvQS5tEXQ2lsSdzcLo6CkRnqSV0dbAwiLYjFMtoT1Y7DSLXLqSOP1fLZOWqUl+f8AnZEcEsXkUZUYc0ly8sb21TXV/qZnQxM/ZqMrRUXdWW9r/hboZFkOJhm3toaxkrN6Kzve1r32+RleZ4OOOnQ5rypWbWq3vbW3Wz2ucQVZww1XFUadmtUr+Sdrv87GR4atmGNpUYrl57t7O1l8rjyjFY6g3Qpc3s7TkuZK0d3u1+F35E61SWFlUpR5mlotrv1exlGYrCXcd2uV/PTszKqNF4WpTlu159zLMsnjM7jhZ/BJ2v8A00/xMdkWEy7Lqkqe/tGpb7J2T3f3IwOX1IYilh8HKzi+Z6bRvvq9dWtN/kZ9GNaX1nELmlFvXbfRvT8rFLCRxWPpN6xik+2ia+Zn1KjRxdSeHVqb237Lvr948S1TjFLYWbVcBRaoKzfXsnvo073/AAMRmKcKVGrKybajp9qTvbRdW+uhWymtRlVw2KjZ2akrp/in+TMqo4LC4p4NLZJ212/Hr5mcZxUpwVOC0lp/WhjOZSsVMbGniIQvrJtL5Rbf4JmB9nhKqorreX5epkdKFXHfvNlr/VjOsK44iVSC91lOs1G0ehXkqtNSb2KkVOn/AF2GlsTtfQjqxW6Ci2NX1Y2uVNbm5inajNrezPrlf2fs+bT+vmZVh5UqLlJav8jMrPGU/l+bMX/Aqej/ACMjb9nJeZk1vazE02R97UwtFSYpKDsvIt+7v1RhYSlTg5rXUlJpaFROyXcpRasZRRjN3n0MBRpyqOfLZevUbSRiKltWKVrGJqurX5On/AK2FpV049Nuphcuo5fhY0KS323317tklCMGmtOv9fIy2hRp0ovDK0OZ993vvr0IVKlWoq1SNrScd+nfT/hziyUqU8PGHZv8TEpym5W1JwtHmI2baKXuyRiMHDEpxqK9mmvVO6enmSi1PyKfIqbU3vb8yooqTsxP3WSerMbhFiIqLdmjD5TGnNSnK9uhoc1ncTTuNpbEWr3HayLpbFRxik2xasgknqOau7FSVxLXUs76GSSxksO3ilaXNLttzPl202t599TVy0EtSSaEnONl0KmmiLJpD0ijd2HqStoi9xJNiaaL7Mk0noO2iG03Y1T0Ha2pK3QiruxIbux6nIx6g9jcADoPY3Qw6FjQTE0RlpYuXaQpXYpX0KtSE1S5FZW/Ip14SUKdTVR2+/XbyI3vaO7uNaD1TGrRuSkkOoKpqRmpAmRlfQkg3CJBaD00Y9yUUxo3BaWMzymljKUqdRXi/wCujRhqOLwubww9Nfukr9OlvmRp1JVqvP8ADpb8b+Y8THL6katNapr8VbzMJPC4jH1vZfxGlffu++nXoU6uNjncsLN3pzUWtt0reu3mYHB1octag7ODd9tvmZTndDD4TE0I/wAVqMZPXa17bW2e6IYZ0JO2zZGPNFu+xhMxnTej06/1YnjITowUfhfM/uVzNfaVMFRnUleTcm/S6tscNVcHhsyr42UbuVGUFq+sof5dvmY7BwUnbVNXMmxeHyqjLETWlmuvW68/yM5xscyxEYL+RW+X3HtYe09l1tf9DFutUwklRdpaW26epmMamIhCdP4oSjJf9uvXyKuFWKlUqz1lOEZr/t6Kf9fkZriMDlMnWrO7sl1+e1zMeJPr1VKi/ikmvTrul/mYuybbOIcTh8DhY42rtSfN16+50v0l2f6lGjGdZVfL87GSYynh6kpPqrfiZxDCYnAydLTlS77vfcgtdCMV7FW3uSkoslFORURJW2Ixsi7Olhm40r2PqtHm5nBX9BO+hKEJNNpNocVJWZCnCMbRViFGEVeKSElexRpOcrEOWELp+ROnZJsoUnOTSJR5LJ7k1sUoKqvQw9KUnbojA01v/VzCU+WlH7yrVsiq9Vc5rpnEGZLAQpTa+Oaj9/yZHEQhSnKWihqYHGUsbRjXjtbTf/gGdZc8Vl86Md5Wv6J37mUYWGAwUaMNrP8ArqU2m6dG2sk393/DniK508ZhoU+kV92piYNzaHB2FiV7adNdEcN5lLMqXt38Lenp9yf3kHzTcSjQ9jTlBd2/vG7xsSVlcckyW9h/EXuxu6LF9LGrQrOxbQa0JwUoq49ESTOXUbukR0kVIcjcewn7tiFkr9iTUYuRpIw1d0VLl3at+RUV9haQHqhyukkNWQxQurnLZijqbRQ3d2LNjfvF3ctzElysWtzlGldDaFJ3BmlgtdBYexYGWAYdAuQdkc6aIzTdkOpqRqJoni3KEY9iVWm6fKt7GCxfsa0Z2vb/ACsT0lzdDnipXsTqJMnUTZKpc9oU6iQqkWwi7Mlqg1eiGiO4pW2Gmxtl9xrS40W0KcOeSQ0tSpQSk5I9nem2uxVwaU7p3ur/AHGRYDExzWVeUbQalrdbtxsrXvsm7/Jkckx8cxeJVTmp8ytolpyq63v8XVq/bQljFhsI49W/zsv0MFw7WzOcqtGfKueLbsnflST0bVtFbQzjB4bDVaWCveSjpLX3r6rTpo11MvyWqvrNXEVeVQs0uW/lbR9X1KVDmpVV1Sv8tjLsxWGjTp4j4Y6LzurPZXRxBRpU3GFF+7NXj/T1+8yjAVlW9nThd8rS1S108yphoUMpxFao/fTil6+9ddul7+VupxlhsBmuRxweGnyTkopuzeq1vrZfK5CH1evh6m8oR1W1+n6FPFPEZ3XtGzp2Ute/vL8O1x4iLfs09ShHlqPUyXHYeGVYic5Xklo9dkrf11OK8U8Vh3ed3CUpJ23u3ZW6W2ucIYWvGFHFTqXV00rJWvpa/wDwDPqmHqyh7FW0s93r8zMsPRxeFlRmrq1nvqUpJxsuiKUnG0uz/wAytmcp0HSSt3/q3kcvKvUozpqNmOjeMXYqK0nF9CXcv0KcrovZWNx2shDHHmWnQk0noadTlubCVo2KVGU37pRhSjemn7y33IYWMKUaT2ve/fW+xX5ar912sYWnKFaEOsn93T5mdYVUMZ7O+yX+f6kYp2Rg40YU5RnKza0dno9P+CYStRp0qkZat2s9fmZXTnKUV0uS0jyroVtivNRfMQhzQUr7mZZZDMKKpS3Tun2afqjifLcVicveFwrtKo1FvTSNnd2b12ta6fYwOWU8vwMMMpe7BLW3Rb6XZ9Yp4ujGVGV4tNX+fnr0Z7OUqMqcdJJWT87aOxkmIxGCoqGM96qrpPRXV+yujjbEfWsdSqR25fybMRNzakvQknzctyth4c111VjBYWFKChDSKRGdp6dSrRXtFFMnSbV1tsOtGT9knql/X5FtESd2P4jRjdnYulEklyoT10EkY/MsNg1T9vK3NJRWjd29EtE/x0JO+hurE6UowjJ7PYky1xpJWZZ9GTbb1EtNzmsrEneLixSVmkQRiqE6E1Ga3V/kzkvEatdGiGrq44WROVlY6+g03JIbTVhWHaxFJvUe5F2ZN3ZZpj0RuOLauINASuSSSLAxmwzcOhc3C4J6EXcvY57kKisKqmrCqJOyHUR7RtWQ6qJ1LjdwE7EZ2KU+ZCHqhlzpYQmrl0Ia3Gr6IktEJFH4wveNjlXI4mJh769DLsM6dNxl1MunGk1zbK5mlSNaV49UvvMpnLA0vZt355P/AMmfzMniq+Oo1qnwx0+7TocS46ftq9Givfir28+m+hhamJpwcWtZRs9vJszmLdSjC+rs/u/4BUzHC4zLIYan8dO19+tvKxgcR9SzSnOP2LN+j0ffucUzTc3HaT5vz/zMLjVXVKotv8m1+hQrPPsXiKlFWUZKP/kqfW3VslltChisTKXxNJdd035kaHLU53uQfPJWMVhsXPDzdDs+36me5dUp08PiHs039z9TKsuWCxEsH/Ir/r3f5lXESlio0n0Tf42/UhhKjw0qj2uUopcyKEF7N33NU2K7XoQipSsuxSvJcpirSm5IsNKxT2Hq0RV1YVmkNp7CinFlGzlZ7Ma1ZoLawlcSvojA0U6an2IYP2Up1f5hp1eWPYp0lFpMlR5aCr362+4qVJVpq+5K1m0PmSXyMPFvRmS024RTJ7GOnzQ066GK/h8phpfuYIxEnCjOS6K5Sw1T63Gq3py2+ZxDg62Y4Gphaejfpt+H5mFy6OEwVOknqlb7l6spQUakfQzKmnVT7szh89V36XRiKdoKxXdqlyT5oIoSXKyckppkpOUnI9o1BxHTSm59zWw22rDVhalrsaG7pES2rMfl1LGQgp/ZkpL1TuuqJrUtoVKjlGMew02DVyN02X0FudRiEmmzH4mWIlGT6JL7iTajclrFFinHmSRPVaCj7uo9TSyY9LkVZDSYrXOhoh7kloh7j2AATsPUOoB0BgGlg0Ny45aG5e6ObQjPuOb2PaO1htm5ZgBh5bIOgwEKxe4XuONtTldiy6FFWmh2TN0SqRU1F7slC8rsjZJCqLk5SUru6Iuws3WIwVDCtWdNvXe93fa2n3nHuHpyoUs2w6vWrQV/klbfTy2RgsRJ4RRnpNKT+bi1bt13MVw41Rji3U9dNvx1+4yCmsvzXERgvdm079/e2trYxeJwuYZmnFcraTtq/ht6HEMKdec6tPTlSVvN3/yOEcHSrZpSw1WN4tS0v13W2pSw1fCVa9ChrFr3ttNXbe9/kYiTVSz3Rm+ZLCUpVZK8Unf+rM4eryxeDp17WUkn3tfoZfi5UpR032OJMkp47NaMJVOWhGMW3a+ujel1IoZjPHVp161PkvJq176LRPRLdfcOEXXc+5Tzx08pngXDdp3v8trfqaXUHvuZXg/b4ulRbspO1/W3QzjBrA42ph+bm5Xa+34alNWXKUtWmuhN8kPIqU17Pl6okrWGm0U7jXvaBbUsn6l727DXUUexJylj+VPSMfzZyuxy7FKjKVihSgo3vuUa0vqs4KPutrW/X0K+HcG4xZUg4OxF/u1FlJXfN2JQ1sYdKT5ZGFpc6Vu5lWHlUkqkZaLyKlRSk0uhi3ywaRUzalWwEsXD4UnL7t1t+hg6yr4SnVj1V/wFFU47mDwsqVSrKc73d7WtZdvMklVdo7JCxaq4hYbl6N3vt8v+CYaUY4mqqmlkvwPY1K3M4z5rLtbq3+tvlczWMZYhzvoY73YR16EpOUpItaKiQpuNNvcsnMm1FWLbov7thNNEttDmurEdSSsJFjldhK7RGN0NO49rCTU07lTWTsNNI1bHds5dHqcy0GncTuS03MJShiK3JOXKn1tfpfbz2KtlddiUuZJDTsJpIirCjd37Ek0rFrNxRtFHLcs0iN0mh3G7Rt1L6WJJ3uNa7j0Y1qNjdyw9hpWLFgYM6AD1GAAdADQAAAofEI3GAkjRhGzSLJiXumqQ42KLtUVxq0mdCpT5qkZdjoW0sFy60FNJkswqVsJGnfSK0+evY4LzOmsunRxH8WUkvv8ARWMVWw+EwuKjUelZyUd+ib6X6LrYw2JpLGJRd7NP7mR5FxDVxk3pKm0v8V0/yT3VjLqEsc+SGr/r0OIVSyzE4OtRl7z51Lyatbe/nsZXxCqeNrYe+tWNvubfbz8jEvmq1E97szbByxdSlF/Cm+b5/j9xgaVOlRhSjskvwM8zTBV8JQp0PijKTe+zem6/rqZq6eLw0YUd4QV/kknuYxxjS9vP4YtN/L/hjGU6X1yXI/du7FazQ1GcF3R7Vxp6dCtVjVmp33ISTuUWrNF3Kml0JxTk302KkUpElpZFNrksKSkkhtWRFX1RZX0GvduR0dyxgUqlavV87fcNXRGlJrUp20itkS5Y2ZmEa9SGFjR+GM+aW38rj1169DDUfZUnOpu7f1oYmLlU87FOnKqku2pCainEq5ZOlQp1JL4tvl8/MndVVYwNnB9zKbLDaE0ozfmY+rFJQb1exgsrpwpTw6+Fr8/mUMPJVKMafww0f3Neu5jsdGjNU+rTKF3CM57tWJSdOPqOvSo1XWk9Fp95jr0sPUfWS/AyKlKpQqJbtL83cxbgq04dL/qzNmlCNhT1Kk5OxSlaHL5Ch7jlHoVLSkxOzJ2baWw3qTsK2x1HG4rEZK9htOKEm7WE3axBJtJk0ktBkkrkUWuX1Q7crOg9XYlFIla1jlk+W3f8LP8AUw+HdeTit7Ma90VzlVtBWaZBXiydkrGmjHHQS1uLaxfSx5FtbllcclZF1ZDeo7ltB26G6NwGbmjLAWGWBosbgD2FYHsAAAD1KHxC2EMBGltDRog7i3Oaysc3MiaWgVY637nQ5dTRD1ZYa0G09i5GSWxgMTChN2V2ZpKWcZNLLartpJxlvyynFwbsrX91tWbtrfR6mXZBHKpyhz8znK+1reW7uZRkNfMuWEdGot9Oi9VvscGNZdiqzlDnkpRsr2tpNPut7HF+WqtUjWcbcrbt6/PoPCw/tmnWjvCP5vVf+Sok+au2VcPGSb+Rg6UJU4Q+X6GErOniJYeTvKD/AAvp+HzKFZyc1F9DBQeKcsJf7Lf3K/8AWph6VJ14Rlt1K0qlPF0oxXutvm9LO3nvbb8iFOUanK+v6mKUVTahvqZDSx9PDS+ua3nLlemqVui7X66kI62KTV7IlJqny9SUIuK12RV5XKyJKyRS+ESsMpJe0jFdf1KkHGTi3sTtyqzJqKsVWoRlLolcyel/sib3d2U43tfYpwctUJcsdEOcU1d6GQc6rpYifuuS6bK67bmMlh8VWqtrbbful+XcdGSkqklrYwDgqdTmWrWnrp/wSFCyv9xiMdWnRjTvolZbdWQpxlZPoYHBqUrL+rGGpVUqXK7RV7ru+noYqaUXLsZlln1jEUcTz2dO7tbe6t30+5lbEyw9KUoq8rNpd7K9r9NjLKb+qc6VpSak+urs2Y7GVJZzHCKlfmXx82ytdrl+W9yjX9pTTtqTp81OftXft/SMwwlKvh3QmtG03v8AZd0Y286DaV+VL8SpX/s2ahNc3Mtelr7972sV5qLairozn+A1DdNlJOMYpu7KuskvL9BpWT8jnskkSaTaa1Gr6MUeiJpJIcboSu0WstCOtrlSLTepuajv3GlYpq7ZNMerJK+wk0i2hZ73Em0xtmiSfUetzdCVtiNWVOTlF6kkpKxpy26kruKEnoN390nDRWGupa3UtfYfuq1yCv1JfHuO7juSutEzoKzGPUZuHQAuAbhbQHsB0AAAAAOgwpuzKck0I0GgjYSuiztYvZWOo1dklqT0SHrsKd4a9DSS0IRuTg0PQT0G0yyZJ22FZop2vcwuGqYzEUcLQ1c7L5v1t+ZD2NLP6uFr/wDLm6e+9munmu7OEK1PD4iVRy3jK3rp/Wpl8nhM3rSq7XX6vpfyMdnznxDj8NUXu0uRw81KDvsr723bIct5Te7F8ZGmptR7lP8Ad1pqPQWW0VN4r7clZ79Pnb8DBSvCVTuUG41pVIvuKpL2sde5LAzll9bFdKfL/wCTSSXXz7Mc3Fq71RiFenGXdkpNyjB7K9vna/5EW2UY8z0LWqib5XFDhaLY1fQpL3Vc2E7ilZolrK5NNSsNpysZtL2WCn5q336GEpezpQS6Jfhoe66d+5h1Z+SM8xFZZTVWHXv2k1tulpvoQqLEYe6e6uZDVqVcLV9tvGTS+9djD4OpiMww8I7Nu/yhJ/oYelOrXnCS+FfiTp3ShHe6ORc/oVaSWGprq2/u0/4JKMVLlRk656yLKMUkV48yUSvOTxLg9kZdGVWrU9otE9PR6dBzVJRivtafgQoQVRTa96+hQdT65XVvdSVvXW/mVJXi092PFSr46rD7MGl+F2YzHS+qNp76Ga1KtKNKU3fnRXTdNNehi3z0fO4tZ8pUny1H9xThd28iSurIVrMnq1YvoRV3d9CWgtWdTofFDXcTaujW4lcdrKxZk7pkkuW47WdiOi1LpaErGnKS5XJFVxlLQdrEbOKEjS47aWGlck00huyFZpy6lKHNFt9C10kW9xCVmWuzltsSukSleNh3JL3Lj0G9C9xtFk0gsPYDRliyLIC4bnUCwAFix0BjPIGrjQBhn0EIkl0BK4hpWRbUd2tB6Cd1YauiUbIpRTkkxU+R2Kd27dyX7uEr9V+pi68aSjfq7FuhezN0aPqJ6l+hgasqeIpzi9U0/uZTq05VJVp6ybv8xZ1DDYSdFO0p2t8pJ9jgnASzSpHF43aE9fmp6+610i1a3XyKkctxWaODd4X1+La339jiSGDhm9elhvgjbl32+ev3mCwlTFXlDaKu/QmnZMnBOcai3tYzPGQwOBeIlqo6tbXtrvqU8RCrShWp/DOKkvSSul30ucXVsLlFGhVo6+05F13m1Hrfq/JehF05tq+iMVCOIw0qE9Yuza84yTXno0v10JRcanyZKTkuQjfn0JvWxT0SfQqy7GEqRcW2VpxjB23ZJJMpKPLqTStdEW+o3ZkuWVktiF+W3QVJpXaM3XPUo0V1lf5LchG7siMYxV3q0ObnFJMlXiqqp31dzLcO6KmpO95N/ezKoKjGcOkm2vV9Th/FwoS5qmqSf3v0JOVetFQWlrfPX/gFPC0Zz5JR2Vnq/P8AXsZXOlOhWoyWrtbySbMVWSfI9uhSpynUdjh/DR9pe2xXh+85vIqyUIt9CvD2rUOm46Cb07EWlBFKm3Z72K8XyaFa9XExh9m+pmiVBunD4rmIlKWF5GypBzspO7X/AAxmleVGLjfUpVnOE0/UvYm1KoRi52HFw2FJWSfQnF8zNOWw27jSSE3ctsbi0aHFXLJsp1IzV10IuyJSVxSuyhQdapGC3bMXQlRrypS3i7DsdhtbjtYvorjVj3baFkkiStFSXUk77CSSIyjJtLoNpMWsrEI6+8aWYk1sxJWHFjROyWg77mq1JayRK6GrjsNqw7dBnQLFgA6AaFkOx0GMLAAAbgDNDQdrBRlaSED2BCVyPYV7C8xa2Q172ha1kSdylOUqkotaIqYlOXLfU+sSp03O2yuYrmh7NS+2rmJpxcY36MU07pko2N1YskxRUkJXKXuzT7GZYuGEoOrLZNfi7efcrtVeWT6JlLGVMPh5Rhppf10f+Zgq1WVGU9mypzLMZuT6W/P/ADMFXqUqcox+0vwMZiHGooRRgamI+u16FRaQatt1ipdPUnFVYWlqjC1Yxoulbbb5dDNcJRllWEqy11a6q1m7ELU5t9yc2oX7lOnKrD3VdtpL5tGYYWeExEqc1qtP63FKzZyXd/K5Td7R8hp3uUpWl5Fe6diUGQsqDfmWUtRxsx7EJO4paFC04NGI5Z51Sot7R/F/8A9hGlUcXumY6tCeJm4bXMRm1HBYebkr323/AMmSoKU1PqkYWnFRm5P0MvlHm16EOZQjLo3Yw1ZRxUalvl6a/oYzFuVabit9TD/uItvexXTcreVzCQvGK6vUyqnyNFd6NmIrq/s+5TtF2ZTvJfIp3hTUWUbciRir2aQqloyn0TMzxPNWtfVklzYfnvpb8W2v0K2LjSg2Y/Gyr1JSMLNRq8vc3RGV7EJKEiclJSXccW9ESXu3HT0EhxvoWszW4IlK5HVFKCpVPZd9R6Fk0K1zDVnRrRmt0ZjWlVxUqkt5ajSucqLJqw0aaIk00WVtBK6N0kNaMkrJC0bsStZGjY3aTN0WF0HrqNaDXQmrRSG+ayG7slYkzRnKmJJjsdB7BYHqBuG6AZbQQWAAsgBnQYaB0NwTsynLmimIGLUWugvMfZDI/CrC1Q+xOKTMTUnFwUer/Rn9m0XXni0v33K4p66X8r23t0MuymrlOEpSxFXnnWm23a2tlfRNrr5Ili8VVwrVSV+Tm5dErf8AD+ZCGIr0qbUrSur6J6W1X/DGYZgsLmkMHNfEr3+/pZ/mSrzk6bWzT/AVNNDhcwEsPGqvbR5l2u1+RUjFS0ILQzXBUcZR9jVV43i+u8WmtvNIkrQSKVpWTKmBw9HJZ4hP3ue0d+yb6+fVGIyyliMYsTb34xaT9WvO34E+akud69DE4xvM6dKD0SV163/rQznHUsTUhKlDlSjFWu38MUuve1xTZh8PiKmOhWjUtTjF80bLVvZ33Vuy36kVGVIxEFHltsYKPNCV9pO/oYCs8PKgkvtp/c0cR4pYzMq+Itbmlf8ArYzjDYqtSg8PU5XGSb0Tur6rXa667mEy6piabVLeKu/6bE7T1F7sV/XUg1zIvd6kZNRfmU7OHK+he6JRRKKsiybsJ+6f2lTo0nUTuv8AL5GU4l169XGQ3dkvu/4boUcRXUZUVL3HZvRb/mZjClLm+rx5W1Zat62s3r3ettuhl2Cqww9FYmXNNRSbta76uy0VzNcS69aLntaMUvRJL8iq3i41sOnvFx9Lqxl2HlRpU6b1cUlcwUOWlJy6ber0MtwMacalao7y1S+7yYsTzV5QS0X/AAf8jE8tKjG2vNqYLCSr02+xTpRVaK30MGnyXKsr3RjafK5Nb2MPC8IvujCVak8dOnL4YaLzuirh6s69KalaMb3Vt9GlrutbbGHlCceaOzMW1G077X+/uZvmNWUfZ30T/roZji3OukviRXxMoU2r6GYZjOtLfT+vIbutCMrSiybtNpEdNRvmjuTb6Mny8+xUjek5X2G00kStqio9EWuNWC9lYWpdoteXoLVkk7EXoczvcnLmZtYfYZY3Vy12JWRoiW7GtBpOKHYa3RZq7QWXNoW1QotN2Eruw9S1x3e/QatqWTjcYwbLjsAABcDoMGdQewB0A0EkGhcdg6hoXEIw81ohWYNaC0NLlriV2jd3FZRGrFnZLqSTZVsoq5j8BiZ4ujGCtyzTlttZ+fe22pxLwrJ5bCpOWujSt/Mm77/3TBUFGjFSZSybNa+PprCUueD5nJ80Vy2Sto3d3u9uxmlGlXrQlU3i7X+S7HFmbT4ZzBYBx5lH3r3tZSStpaX5mWV6GJy+niHKznZpWfX+uxQwtGafPO2mmj37FbDypTtI5HYpwb2KlOTlbqVFeOnQpxbirGKcnRhTT0/r/Ip3VVrq9CvSw0MqeJqVLONRQtZ7yV915W6ddyjl8aOYYitPd2j/AOAq3cqbHKyhq7JlSM3Bcu9zETTpN9jDRUIWZj8RGNdKKty6f8EzT2mJppU3qpJ/K+pltCjWxEI158sOrs3bzstXqZLn39mSqyprmU04rppqr7MqT505LqOTlZibTdyXuoWrsRs7tidiavYbshXbsY7F0cJhZ1qsrRXXX8ldmBymNHLJUW7p8zv669zLpLD5fBpdX/X3GCb5F5ksPXljI1PspPt/w5GMZRTuV6SquE5dEYfDRjVnP+Z/oQUeWST8ylBywvL5oxU500qb3Sv9+hSprk5r6spxXJGktdNOhldSFOjOHWy/W5gMNFuUnui8aNDXoLF068VKm7/1/kNuripvorfkYajWim5LTS2239dyFOarzl00FUTbgZXRqwwcVVVpdt/xRmMEqV+lmY+cnyp7XMXiIQxdkZhiOSLj3KvcjeWxJWSJvZi0iRFBtpIkpX1J25EiKtFj3HF31LrlsfEkPexJq1xfCITu7oT0uKaUbMjJSm7FnZ2Erq5KOg1dIdhtcpFXQok2kx6xLEtx+9Yaux6akpK1iO1xLVEbNI0ZokmXWthq8OZEXdE2rF042Q1oSHsMvoPUAsWVgNA0LKxoO1jQ0NDQNADoAG4AM6D2OgQdpEJcyEx7CiFhLQ59EboaCOhVTktDFYhUrO2r/wCAYvEOOF9pb31/wPkYVKcEzB42OWYdyoSu5Q7bNtp73vpby/E43yTF4rKIzpq6m79Ft8zPq8q+Jhg2tXGF33srbfK25gVGOX07L4V+rf6lD2lXCRrNWVlf1sVJRrU+S+pl0KeNyanmVOWkna1v107djLsUsJi6da13Fp/d95isbhsTmVKVbSM1JdXqotrbzXkhTpRoVYW3tb71+hRXtJxgnbWxik1KUbknaSZxRhMDhMtw9atDWVpWu9Xqlte2i9Dlbd31u38ySux6spqXLLl3sVfdqOKMzwccXgq2Gi7c8Wr9rqxQxtCOU4fCqPvQe93r8v8AgnE+byx2KVeoveso/d6JFObnNkY21fQjFUU1Hq7/AHkGlBE2pU+bsTkqiVtxJ2sJX0IKLhyshC0kyrB8jikNc25BWaMVh4YmDhUV0zM/3WDdu1vv0KOFjGhCmt0iC9yK7FJ733Mpy2pjqyoUnZv+urRmlSk6iow2hePXW2l/Ik4WppLYpUm5RiurKWGq0o+87pFOlUr4rlvun+RTwqjOXWysjL8O5T1KdNtowUYuMnuyu+am0UMNTw0qs19rV/JJfkjJMxhmNaq4rSErLz/BGI5lyRWxy+8vMcb/ACGnKlYrVk4TT7NfeZ7iYVaNKj/ImVXGpi210MxmpVWuxJJoSsVHeyHZxRf3bkXeBZaEU6lTVk9iKbTQnqOd0a3Lu5L4ire3KR2sJm6YleJiJJqxR3ZurF0lYk3ZC2BQSsIT0Y739BtsWjuh3GvdSOVNWFr7pNO5y6D10JN8uhFNWLti7jdtBL3rIl1TFJKQ5bj1QPYABmmxuAaHQ6ADAuAzQ6GjC5pY0C4A1obgYeothEXdWIxujlsyTuxSHG6I3EctxXve2xBrnSM/m44aTp/Fol6tpL8bb6d9DMoTjl8ruzUbt+a3MjdLG5PSxNGV1JaPVX6dbfkezVPDSpte9d/kjh3iXFOhUweLp2VNyjHVap8uui8no2zC8OYfF8Rc1fSMYK2+us30fnb5XK1PDTw1ONCV9HffdNrr5JeRgsJiVw7U543Tmuq0VjG4d4VKduxOdLLKlDAUlaE3ot9d+t397KNGj7WDxE+SndKUrN2XV2Wrt2RVo4avTU078srLdatO33pMxEJtNUldpNv0XqZbTq1KXvq0vkYhrlbvqjEXhCMmcR4+PEjwtCcuX2EU317pfy9F5+Zl+Np4/CrEUneLbS/7dbT3t1XYk9RaGEi73MRJRqxkv60MDjaWInUpwesJWa7M+sU1RhUb9297/wDA3M2qU61bmg7pkMVQjjfqrl7/AC89rP4U0m77btaXv5E5xlFO+jJzXOkSndKKZRUnFroynFyu0RSltuaRlzLoU4OUG3uU18I8VBRlDdtfkV4pptEYXHBR3M0gp+wp/wA0vwW/5mFoucrR1drlKd0yj70rGX51SwWLWHoVP3qXM1Z7erTQnzOUrbkU5ySRgqkFXXVIzXExr1lSoK0F/XXUoxqqs6lulvv0/UweEqSajHV9SWEVKC8ynL+bqUacKUHJddzEVnTpTk9la34FfEU1VVCT1mm/wMoyqhgnKFBWi9ev6tk6sKjcYvVFWbU4JdX/AMOcjb0Kaq08NGNV6/12MSoOEm3dWv8AdYzytGVRuOjRGso15S6lb353ZO2wnckr7Cb5LEG3Boh8JJp2syWi1J2a0F7raE0aNaDWg0+a5LVpsfxCav6GhGL5bsu1FWK7TZh4xcJyb1VrFrqxZ7ErSsWbQ0rqxLUafQXupXHc6lnaw7pHN7orbiXvXQ2nJF7uw4p7DaaVhwbegmrFklZscU0P3Wn1JLqyVm3YlsW00CwAzSwaDSsMLAdAAAOgAanQLIaLG6Cxa4W0CDaZTqKSKbRTV4jikONhXb0OiRpYiuomrEdWrnInKL7f8MZll+FxFDmqL31KLW/SUZd7fZRVpe0i0+phas8NlypUN4vT7/MqYlyblN6mVRjisz+pw+Nrm/T0/Ey2uqONquoveaUV8m77evUlgoKhUq31UrJeTSf5mQZzQnRq5ZKVpVE0tHuvlb72itCli51Lr4fxsVYqjnVNU1zKmlLt0Tf9amfZ1h6lKGHw8d2r7/PdGDr3m6cX1/HUqZnh6WXezgv3rk03r8LTXa36mIzOEcrhhaa9+MpO/k1FLpbo+pXlzK19XuZtl2KxeMwlSnO0abk5Ky1uklre6tZ7XMuySeXc8qnxybv6N3Wza2f+Zg6Sp0o0+g48srHK27GWQw1LBVqtT4rJR33fpp95PEU3OEXuSp04Va1eC1k7v1MNhY+xjRfwrQrqr9Yl/LZL8SMVfn62t8jDQre3lT+wlG22/vX8+2/y6jglJX3MVP2UlLzsKtdWRg8HiKePniOb3JJaWW638zD1KVWD5dx027RZQw8/ZXtorXK0pSjJx6GGoJtXM1wOHp1XDDarS2+/zZUwlSjJKS1K1FxnZmKUqmOtD/l3Bter0/yK+DzLGQwOKUuWcfiVov5dF9yMXjIYerCm/tIhU2XYp5B/wpyzJbuPL93z/QhTk3oRh7NcnW46EKdGEqfxdSniKiqpWMO3Cjapu/66FKpCEU11MT79KCj0v+hBQlJW6GJoznhnCO5XlL4H2KWFjiMT9aa96Kcfxf8AWxg04pLrZHskryXUVOLkmfFFpGIUvZ2ZnFbFQxtOlB/uuSXNtvdW8++2hmuJhKHmiDu5S7kkYmSTSKbTdhytdEHfQg0k11INpEbWQo87t3JaISW5UVpWNEyeGcaEKj+1f8CNJzlyrcxGHnRlyTVmi2tytVhCpCD3lf8ABC1djqjFYedCEebqrlSXM9Cla9iSObQl8KQ7WGSV2bEtSw1cvqavQe2gbId7jE7ojeyOdO0ew37tiWruKdouJJ3aJyVmhuzG9LBcG9QNAHY3NzoMDoaHQ3AA3QB0FsDBgAAFKpyspVFJFF3VhonHTQSsWT1Ykmi6LW0ItX8iq3y+7uYi05Q12f6MxVVUKEqknsQx1N1kovVpE6calK8nZ3/pmXSngca8RS1qK6T8vR3X4GPo4nlpYhqzbTa0fr9/oYiny1o0qsuWpKPPFWveL1TutF6PXyMfSUcRQxMdJxacutm91237IweIg5Om49TK6mJhia08LDnlCE76pWXK7vXsunUpUKydRVd7u223Tb9TJKtaGPxKnt7Ty0WtvUxWK/2yEYq6ad35q1vv19CeHqUqUK81pK9vUoYhV6tSC3i/v0FVaUWlfVfgYrGfWK0qkvtEajjZNFSHM7op0nuylCftpyesdLeWmvnv3I4Sm8Uqn2krL52MVh8ThpSo1oWcl3X6FJTS5mtOn6ma5xhqGNoYWbtOrey1+yrvpbbu0QrUqkU47fmynXdOacdyUJyxKmvhSf6W/UqwU3KNvMoSas2UHGULN23OGcnxeErYh1Z3UpNpWS0+Tf4kkpNeRhHUhQknrGS/Jp+vQnRpypXhHRvXXuQxtKpXnSp/Ztf57FLlTVl8LT+4zjFqtXlPq3f8Cm8RUxqjy+609brt2+RlFN1cXia3S6jf0MXQnTUYvrqYjK4YqlCU+n+fqfV/Z1OR9l+KuPEUnhI0YL3k9d9SnUhCPN17FCg6k1OT13MNg3KLnLa5h8JRqUbLSXfUWFp3Sk7srUYu1n/wxGTalDvsYajOHN7t3obxRVs5anI09zB0q1PG15TejtyrTZJX/G+4orlQoqKuQ0Vr6mIlGMLzOIcbRdVwjLSUdNHq1v6aGazUaMpMpv8Ad3Jq6MwnarGK7FJ2ZOSuU5LbqU2m7shtYW6IuMWm/Maumi6vYkuaPoKPMSqSnGK6LY4Tw1GrmtF1naEXd77L01+44szGnj85r4il8HM0vRaX1Setr26XsNpsxOFhUr0619Yp/irCtuaNmf42ni5xlT0UYpfchlGN5oa95ocdbXLDtYklZMs1ZkrX0GR1R0saMaI3SsPQ0buySbsyTSIK3qLyEkncsnqTV4uxfUdTsjYlqWugCxa4aFjQCwWC1wOgwA6nQNGaBY0LDRa40WGhgAFKpZmGmpITurEoko+7ZDVzodRSckhxaSFDW5jIzhilbt/kcRUKuIoxVF62X5ruSylUcfGu/icEn8vn5k6d1Eo42MK6kntf+vwM7zijmmX0qlFWtBR67pvXVIznh/GZZSwftal5wS6L7T5uja2Z9ZpUE6u84OHfp+BkeIqYqo8Y9p3n8pSbX4NdPVGMx9On7R018cEvmkr9PIw1GcJVpSf8Rq3lfT+tjjLD1KeXVnR+NOP/AKUl10MXTnl+Fp1G9ev9aksZOlLAYa97/wCUmT5fr0XHre5GXI3FkNFqQacbMyzB/WXON/hTf3EGvZpopQSp6EpP2imcXcSvEz+uVd1yr8Uui8+wsRzwjT6JMxGHhWfO91dfJ7mU5b9RiqLd9W/v+bIVI3k30Y8YqcuRbzT/AAsXUEm+pSqRSb82UcN/ssK1+rX3f8OYVtQae5Uws3SjWXXQU+Wm49mY7AqOVxqrrZP+rmPhBVIxj/ViFOMsJiMRLalFyfyt/XUweGqVc7wmPpv3JLlkvJ2d+/TZK5wniFPFYn2y+BzS/Tb1Muwrw9JqLvdtmSuiskrVau8JXW/Zaaepg8PKvTqT6JalXAzlmKxCenLZr5vz/QyflhXg57Jn1SNW1tm7lDAxpSt/X5ksOnG3YwSTjoU5WfKuhOmnE9ly0r9iinVgmitLkjdFeNq0PK5i6SqUpx7ogualfqyC01Oa6SFK0bma4n2b12aZmVZ+zTe6M+qfu9O5iJThh3KO6TZluOqYpS57XRmVeUa3uksbVhJxVj+0K3kSqOEHPyMFj6lSqoStYhumRxNV42VK/uxX+X+Zis4q+0apbL8TCY6pVwsqklqr/grmAzGpXqOE7bXRjcbUpVYQjazI2VythHSwdOt/O3+GhTr8mFaW5dXFuWTZKJGOxidI2LXZh4OU9CWzLXY1sJaD1Vi6voJ7FilpoxpMvdWNEh2sOSeiLK9h2egmnEtqJIbT0LJRTKjSZJJtIas7FiSFdjsAPRAFiw1oBuWuhoOgNHQACwFgNwYWLBYGAGFq2dilUuhWY0WbHHWwkRir37lacHQpxW6vc0JQjNq/QwVFUscqy1as7f4dTiSSxmIp47Z1Vt25XbfS999kYuryxaW6K1Spl9bH4pK8W07ba21119dup4UUcZjY0KblaDT0sukpNvo9rHiTg4ZxnVevhlaMKcXFese7ts113OEqWJqY6dWptHeWmtttFtoYaNPF5tiKeBlanTp001bdafza6fM+rwr42rGivd1a+WvUzlKnhk6cLN8ltf70U3913YzjNa2Cx0ktUuum915Mz6Es2rwwtSdlKMpXt1VrbW/MjTjRxChB/BTVv8Tdvx+5GQUqawMI7y1187t7fMxia5ZIqSlyxMPmixdWtTUbezlbfe8Yyvtpvb5X6lOrFU1LrsYXETq4z2M9FZa769dDG4WVCdSMZc8YW1tb8NypNc3L3MRhKdelOnUXMmvT8ik404u5BqFOTJtTmmuiIYeFSnKTejMLGMq0IN9UvvOKsvll2Z1MJzcyi9Ha1/ld/mYHMPbYuvhlG3s+V378y7W8vMwc5Qm1ze6+liKgo+6yOc1o4VUVLRa7L/IxueUKEqE6isqlSMXv9p76Lp209TOM9ynDZHWpyneShfaS2s+3ZXKvEOH+r4erPaq0o76X26eXWxwl7LEfWcPOF+aLW/TS5TqTwuIqqhL3NY2t593r0OSpKlKMNOa3y1KuWwpU6Eab1a19b+pjsO8LbDdtX6tf11I42WHxNPlf7vTmXe34r5GByyGaYurGnPli+ZrS9km7dU3oZfhYxm4vVK5rCC8ynOMkpW1KUYzhaC1aMNg5YKlRvPm76W327mHXtJuvGV1JWtbbzIQgqfK9bEfeh6FKCpxsi99DGtw56m6S2MCp4jDc9VaSUWl2vFPp5keVNPyMNj/a4uthnC3s7a33ur7dLFWq0pU46sqVJuTpX1sZzUbhGMnqZjJxk03sZvJ8sfmP34W6GUTdOrOD7fkYx82JsVsPTVNytqYXD06kLyRjHy0WigvZ1qb7/roU7PQw0nKWJqrpe3yuZRhaNZSdRXJUKdHCzhDazf4GFm6VWFTpf/h/zM1VsTT/AK6lapGlByk9DD5pTxEo0U3ptp/X4lNtaN33/MlGzJR0F8JmGYwwU6PMtJzjD5ydl0fX/hzRzujFvnldCjfQwdeFPEU4t6yZVh+9kvMlo2eQ9hWaIU4w2Wm/3knzO4rtidzRokt30HqtBaxQlqy+iLWiWvHQ0S9C9k9RxSGtLF7rUluN3Q9joct1c5GSBjYDBoYPyLGp0GB0DQAHsHQN0DFbqWGWOgM6HQA6ETC1bqwnctoW0JXEhO6Qk5aLc11XYinYy+o6VbmXZr79P1MdUUYtX22M8pSxVR04vW8X91zE4an9RtU+1p8/kZZi8Rkn1eCXuRjJPbfmk/N7NGYYN0sLSqP7UXb5Nr9DKINYN4aK0bv9xwHyYXP4U2vdqJ3+SuvPftYyythcXi61aD0m5Lrs9DjylHCYt8z92nOK+Tkoeffz/UzrMoVcdRpL4G739GulrnFmPis4eIwusLe76aX3V/vRgcapVmr6ya/My9Sgm30HKMoWZXn7sbddCvhll6dGrpUW/wA0mtrrZoeIrVq0KdLW712/UwtCp9dcuqTXzMPXnTpVIX+J3Ym1VtIw2Njhq8ZPqmvvVn+ZmU5fXZya/rQxlWqsZT5fhs7/AKf0ilVTdk9Uv6/IwGJ9vQUu9/wZNQTTb2KkcTXzGU5fAku2/XzOHspp43MYUZ6KTs36fNGZYaWEzCrR6Rk19za8x12mrdUUZ21MRicNVwdLD/aTb6+diWGw2LyGq6j99aW12enp1MRh6PLSoLa2m5wxmtfJ8w9hH7MG1ttdLszB4aNTEWj9uX5scfq69jU0tJr5qz/JkqsY4GLj8Sf4GEwcsww1avU+JJW/rQlgp1IOMVqZdehTj3KMIU4ojdUrFKC5PQwbjJRl1sOEXZsrTjRXLHZWKdTnjdbGOxscNh3We2n5pfqQzJzr06f86k//AAFpdvMnJ8q7spv2l10HRUYWKWIp4t1KcN6bs/XczfHU8JRliFuml8728zH1K8JwdPe6vtt1JTqzzrDfyOEr+tn8/uMzryVeUWYyo2rmZ3fKJ+4iNqWYST6t/jqP38RJlZ/umvIwP8L5mYS9xIxa5HTfZfkRko+8ZXBSw8k/tN/kQqVsHWdt1+J9ZjXwkpx7P77Hs+bB83aX5pGJqe19hLy/Jmb4i7jC+i1MgoUGo4pTvGUXbR9fx/Aik5WJO7sO8rMjqzEYSFWMebvdfIfwlVJxErRKVKEpRn1RWbc2xosrIkhWRKzQtGhJFtCVuUk/dsRstCVrI0aOo9WWWxJWRtC4/hUiUkmibV9Bx0LaCLXdhx5dCXYejZo2NDsWOhuAG4WLADGAA7BoaWHawFwGMC5qB0BakZOLujDV+ZaiZK6Vycr6ITY4IwlaNGpCrJX5WnbvYzTHUpYuEuW3tZNJXbt7spfPRW6EUUIJuzZxdhMLiLUsBP2b5Vd2cve66S+7tqY7K6FDGrEzl70ko7Po797dex7OOY0Z4OvKzjFzWnVStHa3TXfrqjKJY+U5UsRPmi1F3slZ21Wnn8uw8HPMsLHnnrQT5VbdNttdOt97mH9nha0JSjeCd5K9rrt3XyKWOp0M5eFvany8ze+ltrb/AImTZfXrxjWtyRautndr53Rx3hsNiq9elVV4yTTWuv3foYrFyjl1P2crNS5bW6R2++xic2nWw7qOPvJaa7aq/TW6TWu179DIshpU2sTa8pRTe/8An+hNVnFwvvfsYadSnTjGbv8AgYhxlKEb21ucR4+pRxjq4lXnNKV9rpe4tFotIpfnrqYLPsWsROrTqcrT0Vk/0/My3FVp4iOIUveSs9Frff0u/IdadVKpzWfXQq04yhd9DAfWKuKmqkrx0cdFovlr95iubFUKle+ul380ZhUw2LbrU48qj7ttXtu7vvfYoYSOHrVa9FazUU/lzW39XsUsXCEUrbGOpvEUXFvTcVSLhyrZFLGVaE/3b2ZisbLGYipKbu27t+vloUKTpxVLmu0UpSpym++hh8K6uKWLk/eiml6N3/TsRxE4zSvoyoqdfE0py+Ok20+6as1bZd76ksxoxxkIzj70tE7vazdrWt0MNUjRaknazVvUxEKlbBSlUfMotSfTV6Lb9OxCMfZJQlddrfqZBmFZOqqVWyha6svzaMsc44v2s5e67rb+n1MRh6bqNQ01+/X8DEVsNhalKlNWdRtJa9Pv/QoRcoXvp/kQj+6m+q2MLCMYpy3JVoTXLHVGFxlWrj61NrlhG1ut2769103MfjaeAp0k17sny+l7+pW5Y042WhRs5JtarYx9XkUbLUy7Fy5Z6akZOtSZCgqGtOOr1evUWZKpUkqcfeTfXqvkZxWjGhGX2nYdWWHpUKkJczavzWt+BmuZTxM5TnrJ/wBdjEVbRdzFS5m5EZ+6jHrkxql3t/kUPecpE1elIwP8L5mPd5RRUnUklzO463+xc3l/wDAw5cLH0uY/B0q9Hm+0kzLZfuK0fL9GYGn7TB1If1sQldwj2f8AkZjRhVxXsp/C7J+j0ZgcNSwlKFGkrRirJa7fO7MXOnUquVKPKnsr3t83uRdyDvYkr6FBYuWYNuX7rl2st77332+Qp4h46cX8CSttv+ZKPujjoU1aKRKTY7sunsN6iE7sTT1Q7rUvZh0G7Ii7rUstbDtbUjqK7dvmPSDZd2SI6RsTsSWhqWOVNGo5NktCS0LNltBgWGblkM6jFqM8g6Ax6gHQuboNLFixoWTLaDNwC2ha5uFtCwi5Rq8hSrcyRKTa0G9RO6IyaWpPHP6/DDfzRb+6xkuTzzTEqnTV3FOXbaL80JtplKSSRjasY81WT0WrOO5YfHYDC5jl7vSUuRvVe8rN6Ss9mulvMqU6NTBxxNB++klLf9dPuRhOKatG/tdr2/JdEUc1jhsE68no7X+fyMurxrZgqNT4ZSUfm/TU4gxcKWZU8mjH97VtFa9+nb8UYnPcZk2WYbLaz1hUint1mr7J9PM4nxdPNsyqzjpCV/u19GZHUoTzJwjqoKSS1WtmVa9BYN0Ptv176eRhcvllOWUY1370kvufo2YlPcniXJWT2MbmbpuPO9v835Ga8UTzSrKnNawXKvz7Lv1MrpV6lTlitb+RgcUoJwe5TqJQWpheWbUZvuzJssrY7Fww9BXlLbbor9Wux7WdPCSoR+20n8mYjAqWI9hT/m/TXqUsPyTlFbP9DFYKMqiih4BrLJ1b+9GSXyafn5EsXWoN33bfb5GFzB1ZtN/19xRpyjFyXUwuI9rj5qL0i7P13/UxkvY83ZlHEcibROtP6zFdNPzJVFFpvcq0YYjE0q63i/0t+pmWLqywqUHqpL7uv5FLNMfCvSoNfuasW76bwsu1/teXzMBilOvOMX8Lt96Rl1GdOrLk3lv8ilVqzcYSdknf8v8AI9qnW5lsmn9xi6UcdiqWIe9N3XzXy/UwLqVaaf2WVKPM4tPS9zG57h6Eq13/AA0r79dO3+ZnuYJ4VUKD95x5vkmm912MHmdKrSpVYbSt36/LyMVhoYikoT6NP7tTMaTxOAqQg9WtP6+RlUsR7Gmp7212M8rOnTTZgcd77UnoZXjIVMJzRexi8xVKCfVvT+rFassDXjXjtJyf3t+vczPM1XpR/r9CtmShh6OHpvSKS/z6FaatcxtdqLsV6qbSPa2RmjcpRkUtKZOHLQkYH+F8yp7+LS7WMcr0fQdW+BUfO36kpOlhml0j+g8zruk6enr1MDTccJUl3T/BGTv91JeZWp+zxTj5mb02pxqL0Mvx9SvWjTkjmdyEtbDqWaSHPXQhVsmyNS6uSm2i90rDk0iMklcqSag2tzLp1pYWLqr3nft3fbyJzsczZccrMk2xzvI57oc+he8bl+iFMbbdkRk20hSe5OblGyG7xRKWhe6L6CZe4tGNp3RLRWEy7E2DVyyYFkWQ7GgAdDoaGgOxoaGhoaFtC2hYsAABuG49w0sBoaNmGrWdhO8SV7kHoJ9CD5dTB5lisHJzw0+VyTi9E9GrNap7r5kXd3Zg8LKtTqOP2IuT9E0v1RVo4evTn7eXLB+Tf5a6lXM3h8vrYOS0lUuvkrX2811MjrqMJReid/zI5LQhXhKesZSffyfe/UoYjCYlfUIrS10tdlt+Fut+5icTQ9pCHXpvuY3DRWZYLHJ/vKcue/nbtt+D9CWNwVfNZ4zGw5oyd7Xa97prHz8rE54mWY1qU4+5y6O63v23M7wM8HjXTvrN3S+fqzJ8lr4jMIwqK0YrmezvZq2zvvr8rMxmJdWjFz2jp/X3GMxUcVls6tPpt8jA5+6UKVaXxRv/AFsY/FvF4qpNrWV38+5zwVb2lGOrS6+Vupllanh5KU1eNrryf/AK2M+sTf1d6yWnzfmZM8RQp06dZ3aWu36FOuoy9rLd6IpVsTzxlB6pPt1VjDV/rHO49NP8zEVMTOXLh4c09LK6Xrq9NrlCcud02rSjuvzKjUpJW2KuJdGhKUtiq4V4qra17P8AAnglUTaepCNSEeU4WyrEVs/xNKjG85R5ktF1d9W7asxTqwip7p2uYPC4qrltKNbWfXbp6aDxEqdHkq63v+XkYLGYetR5JfEvUxCcsX7GG1r/AIowOLUpunN6x/L+kf6wyw+Dnl9SWsNdtlJPy1+84WlQy6P1Vu8lu9dW7u/Xo+5gqrjKM5O+uhKTjWcF8TKPPOrTp2u3fyMFTowrcsI6rffcy/EqvJuEvdTfTs9TmioqpfRfoZnk1LEU5KC1lKLe+qjJPvpouhXniqePlDlvDlstVvb7zIcO6OZTa2cUvRpv/NjxLhFXe+nzKFVKCjUd2/66FCkqSiktDirGqpR9kugsT7quZJm1Knh5Qa31/rQxWa0JYd2fvL1/yMVm7rQt939WMDi4ZhmDwsZe+tdun4L8Ss4wmk3fUqVnyNJ6Ixtdze5Uk9y92YmMJe6yjSTWmxOMZRsynTjBWiKjTU+e2o4xnHlexUwkJJRhor3Er6Pqf2fhua/L+I8MlQTto9Cnh6VKLUFYnhKNSXPJa/MlThODUldMpYOjSneK1Gm1cktFYXvLzRdXKlTDvBwpwXv3u3r91tvuGtLITdhWRe9yp7NwjZepYcNLok0rXL2Qr2sdB3vqcqegvIex73La5ey8z1Gm3oSslZEk09C+4r7HNrceuw1oKzLbCVyz3RzJ7jabYG49DoAzoGjL3ADoDAOgAMuF0aF0MB7i2LFixbQLFtAaFuRdncw8+eGhON2Q2G9TPuJsNk9WjHEaRnfXXT5JNlKzV0JKw606dOXK/iTXydjE4yOIxSwbekUr/N6fl0Zm2X0MThJYql8Kk116Jd3+h7OpTSjHb+mUpyVF1XqovYnXp4aSxkFeSTVvJO2/y7Gb1Xmud4fGUI8sFZtXvZpWers/wKuW4vNa9HD4RXmtbabJa7tLZGL4VqYvAzxFN+7Tab+WvV/oY/LprCLF8vu35fmvnf8AAzjBQnmeDrN3bk42+WpSwUKcp1Fu7IxS5qSgY3BRw+URoRem9/V9jE1cHhZxpV5WjHd2ezv0XoVsNCVZ1aUrwu+XTddHrqvnqYrLJYSjSqX+JX/8ma7vsUKyhGNF6v8AzSMNXp05xjH4loZbV9tVlfp+g8HKMuZapGFrtO6epg8XShlywyj7/M3e72flt+JTxSjWUk7NGHr0qFWrOWraa67seJhBxUnq9irTcrQK+HVPXpoQ5lF2MFiqdWpVpxldwlZ+Tsv8zIqlTJ8ylm1ON+SPK+m7+f5GJjUxmWThSdpSjp6203t+JUzOeAhhqMldylCL6W5mot7Pa+xicLCrCMr63f5GPpVFWjOi9rX/AKZgsxVZqqlqrr+tDDqlPFJt25lbr5v9Co6Ua2IxFSWsuVbPaOi273MvhSVWdR76fkYriyngqUHNac3L+vZmXZo6fNi6u3M2vR2tsv0MtzlQxNOva97pfP5GE4sw0s7lgNpSTfX/ACt+JlFGGChKDd1KUmvm2/PuZ9jqeAwUq1R2imr/AD082RxKjhYVFqnYebqs5Spq+v8Aw/QyTFfV5VoPVv3l6XS/UeY1Kjprl63euy+7vYoyVeMZx3RGfLRUn0M8m6rlJbXFJRjYwmMUHyrqiFOpiMRClT3k0iS5asqfWLt+NjBqFHHVMZTfvSSXy/ryKmLvyWW7KtRcjSKz3Ks/dSJOyJPnZCKirDVxIQuxHUhNKSH7z0Lya30RVi5ThJPYbbVzdEMXGWMlhl8Sipfe2l+TItuIpEVZk43tYjJKSv1NVI02GkkrkEnF2LXQmXtdE4tyQ9Uhuxe5uxrcclyoSTiN/gPVNmliMlyoY90mNXdhzsvMloJrlZe40iNmhK6fkNtuw462LDTsG49QYPYtoAdDoaMLIZYtcGDA6HQHsFkxhZGhZGhZFkaFjQGB1DA1bSsTV0SjYbujR6MjHkk33N2ToqWEqVm7Kmrv00X6mSRdfMcTiGrO8fO6Sdn5fcZtN88I03aLUm/wMt5ZTsxThh24tXuzCY107pK6qWS8raf1sZbh/q2Z06uI+Dl1+99tThvP4YHOqdRq6jzabbprezMXxJQqYCVDk3vfV9X6H12eOw0Mnpx95zcr37q22n5lSjGlUbXTQrYxuUqS3szKsQqnM27tafMzqvH6vGD1s7mY0HjayV7a2/rYxmXTo4udGErqNktPL1MLGnRpOjVV+ZJdtvQy6dCjJyUdYWtqzFumqkKijdycXv8AMyxuVBQtqv8AM9lUoS9nV/r7jENxrU1Fbv8ARkak7cr1ISTpWkUcTSqTmovWNk/mTotTtMnjkpPXYxebKb1e39dinncG1zb3/rocRZnhMLWmsMtHCL678ib3Xe/l8iOde2jBpbpP+tCjjoygu5hYU6FbmkveV/xlzHtXU5JRexiYc7cmY7BTwk1XoPdar5+dzKMw9nUb7rT+rHI6tKX9691+f3/gYHL6tOjiKsZW5bRirdLN737vqYDExll9OljIczcu9tV10Ks8PiMreHXX1MjlClh6feFl9w8NhKNVZty3nGLW7Wn5fgYvFYyGJoSoO0WveWn66/cY3DUswy+phpq8Xvv/AMD8zKlLCYSlhG7uKsn5L7/xZHCVnn7rRVoJO+2uj+aM1x6+qVG9o2/NWX3or46dbEUJLRxd/k1YyTFuvGNSLtZ2ZVtGg/Mzb3JJt3MTVbm0u7KE7OLvrcji50JKrTlaUWmvVfgVqMaGZrFU95X5vNuXM35Xvsl6EaawOHp+xjtyx3+zout9vv8AMlKEJ89tSvNaFZ6FSaSKtXm0Kau0gLahYViySaLJbDi3G4rKNup1Ho7sUnF6CVpXGlcUrog00UMI6WJq1pO7m18klaxGopSsuhsKXMhaJknd3OuhK7HoMSuJXY00O97IabQldm5J8zG7oUU3sWSJNojeLJWuSd9B7Al1I63HrqhpLUuDbtYSux2Y7IN0G4AaAMNDQNOgdADoAAA0AzQ0GGhpY0BBTlyyTKclKCaJXaEWTZKN1oUKqcVHqYahTxWAxuGqSsp0mv8AyaL/AEMMqeU4CgsbLlnVUuVWvdRkkvhulp3sZpWdVxqU17qVv66mW4eUYvm0aMTTc5JIVarQr0uVX1M9wazbAU6dKXvOKW3+djHcLxyapRxM5XlNWtb/AIL/ACMTOXtJSj3MmqVKeIhKgr1JSSivN6ddPvMfhfY1ZU6mkk3defyKsHTxM6k9FoYPBxpRnUg/id/vsZ/jIqmoR3v+RKnLllWkrasUpKcJJXvqY2qvaK/xR3XrqUqc3B1V9rUwjcsI1Ja2VvuMs0jK/S34mYVI1/flvZfkiVH2soqO6f5GZ1HgsTh5y+BTtLyvFpd38TS0+elyvTjUp+zju9jJcnr05OFbSVSqn06STWz7ehmdBUcVOne6VvyHgdW+/wDXczDDShKVl1JYSeIkopWepRwjajRm7SQ8NyVVSi/e6fItUow1jqYzGzqZvyNaW/HRmFvCjGSemhSxFGpzPm0l5MxMEkl1J0a3tIO1pJ7abGBqNVbvYcXCfvO10ZlKrXq0qVJX5dX8/Uw2GxMMXGpb91y6vTSWult9reRw3UrYJVPrisptuO2q+V7fMxWJisFBrum/KO/9dTh/MsLisNJQne+2j/yKfslhmovyKDoqpFSfvW039CjXqzpyVtLJX9dziahOpgfY0dYu2vmpJ7PyuV8zjTcpydmrJfL5HBWNdehyra6ZWqxVK3ZGa105Nt9SpWtiXroRq03q3qYiq2049jL4wr42EZOy/wCB/mSxVONOTb92O/ke1jOk3ElLQrSRiZ30IrUpfEAFhFJ8r1JyFJ8vKZlQp0asVTlfRX0tr1Q9UUaXtZKDY7czRZtWOtiKbKXvq8Ryg5ct9SEYwba66lJ0/Yzu9bK336/gOztYatoNXRFdQqPSyNhbG2w73IX3HYj7o/IUfeLNNDbRZt2HeSTKo9iN+cHZ7FrIptK5KceRRQ3dli2lzdjHsNoDQ3QzQCyOgWLBYsMsWNxh0LG6LBZMsWViyLIsiyLI6DDQAsLcDLqqceUlohpbEdHobIxNHFR4gp1V/CUXfbfRLz/QpSunfqZpktLFRTktU9P6uYXDSwlSdF9P68zn5ahU1g0VKzjUUUx5/Uo4eVKavzNq/wDSM9zrC5jl+BhH4owV9+y8kOUuZlSUZTTW6K05QVmzOpyrYf2LWnMn9xSx0o0rx6GPk6k/ZS1cnp6L/hzFNUf9nfa/3v8A4BRkquJ9k9mjG2jdN7FC/sVCWz1+8wEeahF9kjJMZRwlOopLVvTf/glDMYVpzlBWSdv60KVWHuwiQpQ55Slrc9tQw+JdOburefXQzyUJZng6a2UpO/ko2t+JlOAp4DA8rnzOcpS2ta723f36FWSq2T9DNMLTpKU47rUw2BUmpN/195ictpyqxnbVP+upjMvliJ06vWL0/q4sPKF5tlLIoVM3eNT1/Tl9f0PY+wjb+VXMhxk62Fcu7aMHm+KxNeVDErWNrbbXt0XmU6kqk1F7EcQlQqYKS1nr939dzLFUxDqUofYRSTlaVRWaSXfv29SdKLynERfTcwWDp0HhsLUXMtbPbTfb59zCyq4yGKoSWkNF8tPLt1OE8lqYep7jsml97WvXuYHG4irhZxlq1fXTp5EcdCDotq/M7emhRrUqCc5aGd5vUxOMdSGnK01+XYzGTnKTlu3/AJnB2YVaWIjTjs9H/VjMZL6hTXdGbVW5ySK1+fmXQjBy2K81CEbdUYqh7ak4LqVFGUVEUrQ5VsVpct0Ymd0S95sSsUFuFhm4mbiq2VvIlBKSHs0SldIpSdxbsq0ZU1GT6q/9fcczUroctLDte5w/QwtWvUeIdkoya33SbW3nby7lLDQhVdVfE1ZmkncSsjqrF9bj1HoTd3YaujZIkeQ9Vcb2JOw1d2HbRDk0S1+QtdCWrRNWjYbJPRISuPsgFI3I+60ypPmk2gkN6DZYANwA6AFhoN0WGBY3RoBoDsMGWLFixYtoFixYEG5hanJURNKxKPvX6CaSJSvYqQUoalnyJLcxWN5YuKfvLX5oljK2InGUnq277a7fcKTe+9yu2oXRiaVRTi0+/wCZUbUFCSum3YypQqYh1Kc7rVJWMThY08HSqqV5Svpbs2ZrinhqbqR3RisXXqSUltZNbfP8B4urWzKpRlH3YxVter/EnJRqShFatbHNCVOVWXxR0JKGOrQje0npf+rFfBww2JcL3a0+4WHhiZSi3qmyHtJ4aMVvF3+5/wCRHE4Z5VFU/wCIr33+XkYOfLXg6msVZv7vIhT9li6ip/DJ3X9bmXSwkI1PaS97ldtHvYo42nHnjOW+i063RnWHrVMyjJS93roujTXn0MRJYrBOtH4oXXzOHcwdWlJVpXfTT/JGJx81aMJbXMTmlRYiFOTupJ/p5GXY1yWi07mbZjOMIOD15lf79ehDMYyenr/Wh7eNaEkno/wI4hUqlOlHotTA5jRxdWpQqLV8yW/T0XUy3DYOopRo6KLa67/MeXU3WlUtqkvz9TDwVOcnJadCFGM5zrbtael0ZBm9XJ8XVhiFZylC3mnfsnYzCOEzGlz8vLyu27eq+7uKFaVR4TpVTvt0/rujAZPjKVelDl5owg0tUvJdb7JGTZZQUOdxtffV7r5/kZNmWHnh5ulLaUls+ja6owdWGEwzpp35m/x+8y7GYavWtDWEYt9VZxaXr+hmMpVcJJ20Vvxat9xjcPPCY33tUqf43+ZWl+8UZr3rnBNGEswm1toZ5jowwkLef6GK5qjbW7Ky5naOxGpJQdmXlKKT6CilBslUXLuSq+6ytUuVKjkxNIumjDSVwLaFhIVyzTFe+pH3rktJWRdLRCwkP7P+tX+3y2+V/wCtCvXqVIpSd+VWXoaONxxk7E1rYUmopDTa0JxlCNmtSMuaIkmroatsbNDWpYlBu1iSTsXWw02aMbRe5GS5kZhgamCxCp1FZtJ/J7bNkmtLlWpGEHOT0RG3IprVMSKj96xJ3dyQ9C+gwDoW0Ogx32N2SlYeoWQG4XNwAYWB7AAtgAHqAHQ6DNy50GD2OhoAnbVGDqOdHUlFqRsRVzqQm+ZxM0hTlTmp9FclBUfZwpbtt/kYbEOouZ+ZW0V31MTKN0kY90aVCOHrLV2tvt8itW+pYxTT933v+B3ZhM2qSw8U5X0dv6sOj7WClLbqYmKVGT6JP8CWbU8XltCnQ3XN/WqMDXnLEVnLeLX43/yMwj7SkoooNxjK+8XoW9qva+Rh6badt3dnIoQi12Zg8XCMnGW0rMpwdXESpxe+hiq8MNiMPhauqpycX83pt/mzBx/e6bGf4qeFwVSpDdWt/wCBJeZmuHwtaUcRGNpOKW73/ryI4KjGLv2v1MNgMNGlKy39e/qVsr9jg1iILRtr7kr9fNFT2dfMPq/925/Z7jC1PS239XMXlcnZS7/11I5cqa/r/MzHLniaCit1/XcheUvayMJhKcZuUVq2395gsNClOTWnM7v1Katzq+7sOrFK0ujK/PVglB2V7v5J/wCfQxSp4hwk9l+jJweYYmjTg/dctfuXozC5bh/Y2T1imuvTTuVcV7DMsJhqcvdnTvt1W/n+SMmThhnz+f8AwDBZFhMBXdSmtJtt79X6sx7w0IScN7XW/QqYulVwVV0VaV5Jb/qYXGVqVGlKu9OV9t0m+noYmUa9FVKeraX3Xv8AoYij9ZqXitUzhSCw1WcktVa/zvYziLVOMZdVcxs7N2FaK5luVZWY7pk3LlQ9CtW1divWb0RuBh9wAI6j1iRd4vyEny8w1bUtqjmly2IvmI/yi1Re6uTpWpKfchLllFozLOYRx9KlUfv1W0vVJyeytsn2EmbD2NxvoD6F7Ow7MVmaMaVrjSaQ9EYmvOvJTk7tJL5IckyvRp1qTpy2a1KcVGCS2SJS0Km47uxGzepuh7gDehuL4Rg2iUuxe4bltCwDLILXLaHQGAAB0LA9SxYshhYCw0dCxuMGCNAMuqpPkJRuVpuNrbkHqXSk0yvifq+bwpSelSLt6xf6p9bLTzMfiKLxqwdXVzTtv0tfb1XUr04YeLqJ3aVk7bf1Yo1Z0qnNa8Xt/W5Sl9Y32RX9lCE23tqjLcNTzmsnVqcvK4paXvd+VrWOIOGva0azpO6gvS6enV6GUY+s8Z9Xi7xirdN2vQwleaw0esWt9rfIupUHF7Wsf2e8Lhowoy2vbTv6swUKbi3b3nu/Nf1+JWpS51Jz79CplyjScm7/ANepThTpSnTnty6eTKLnCqope6uvb5bmGwVP97KqvdUW/n3J0EsNGa2Vl95lmIhDM4Nax5kv666kqXtMRepLW93oZfGX1udTnvGySjbZrfUq0qVSVOUo80FJOSu1dJ3tff7jG4iOJr1pxVk5Npb2Vlp5+piFzpNO5Szil9bjl8JfvNW1Z7aeVuq6mQYepjqdXAyfuLmnf5RVuj6b3+RhU6OdVYzXvOLsv7t9H/w+ph5KVtbks4WLx0cJCGqlJN3/AJHbt1t308ytFSbVitSquUElo3qSpwUHD7ihGVOrGLiVandHD2ZzxcK7mrOMrd9/l5fiYmVCVKMFG0le+r1va3psyMZznC3wu9/63MTTw/toKCtHtr89SOGaftsOuu3b5sw+Kk5zhH+ur/Mr0aNelgZc3LKm3d6vRu609O1zCUYQU0/eh326dtz61h6EKftJazailZ7/AC/UzODp42jWpVOZOai1a1lfXV7mKwmJo1nTw8LqN5PVK90++xi8NCjQ9riNIctl1s3o1prsVMjjQpXoS6aL8erMLlCo4aUpr35L7vxsZbg4UoxSVnbX5X/zM3fPBe9dJdjGTg00tSpVUY76kqkakW76oliIpWe5PFwtZalbFtaE6rkDQFB2kLVCRa49wh3Qr8rEnexraxd8qHa43Z2ZpGWmxLTUo+zjGTlrdaepZRiu7MVj6WHVNVJW5morfd6Lb9Rq7K+GoqtHETV3F2W+7939RR01JxSROLaWo1poOzFbQm7XfYaurjs+omLUVj3boavoOSSsWTQ9EQsvQlLUlu77ktCTVhgwNwJE520HJsaa3LtjRYGWGixYsWGrli1y2gywywDBbAAMsAMsWBgdB7AgLGHnyVExq6RUpKVhbEY31M1y2GJq0az3pu6/Xr/mZnhqVfG06sfjimvS9r/kZm1Ww1SlHdfqVMf9WwNPCy3T/r+rmV45qn95jMXRrYZtb28+iOH5Oph6yW8GpfJHE+OVPL5V4/E1FP70ZDktXGVHO9rpS/LzRl+AjSwvsb7af1qYum6dCVjDV5zjeW5UxEaHvLz/AEK2I9tCLltYVRPDq32TGKpiKjlS66f1cdH2r53vHR/InzfUa766RXzepKtOMp0n0S/AwtZ4PFU629mn/W5TzGriMbO/dv736GU4Spi6/s4vo39yIUZKuoJ6a/gm/wBClFYqTS6uxjsB7OlKl8v61MFkVKOeSxX2rL8rd/0MFntajicRTw26Svts733XkU6E1mLrye609DK8TTrVnTW61+9v/IwuVxq4+ly/FJtL5v1Ic1VadSWGnKtGTfupii1U5GKfLIcVKm2zEQeBoQjS3clf9d7lG+JprlMuzClias6dtm193yKODo4hxs9V6lWH1Cmm+36v1KMJ0X9ZW1SP/Af9WRXx1PE4WnGK1ikn6owObPCZXrqkn+HyZhsD9fdLEwenNGa/B91+XyI4OnRoqU/70vnqyuoV26Efi9m3/wCTJHs1muWyoy0cXb7vuHRUIQUvT7kTlHljbv8A5FWpySUkZpj/AG0bLp/XYrVL6i96Vu5VSoym1u9fwt+hWrNu46u5KXMeYAQdpIg7oRF6EkFLRmwtrmlhP3UfEhR93lFLZHkxzco2Em16DhGUlfoNDs1YXNzK2xJ3Viw1ZXLLQVrE11E/dLJx8xvcWxa7SOV3sT0dhq7sLsWWpP4S5N3YwGAxFxsqVFFpCd2aDldEbMAAAAAEB0GiwAAABudAOgbosCCxYMJX56S7komZ4pYekn3aX3uxQb5bmOxSo0/7z2MJg3Thzy+J7mJy+mm5x6/13OMcL7HkcN5P/Mw+e1cNh3hZ09b3vf8AC1v1KWaKE4zktF0/pGXcTyyzM/rtGHuXd433V9uZptetrnE+ef2nVpezjZNfDvsrb2RwLgqMcnrynK1VyaWj2Tt6feZZhamFwUKdSfPJLV2tf5IqVFJrmXUxWGXtVKL0b18jMsBThJKLvd/p6lSMaVNzk9Fp95RSbjBOztexHBRkrJ2aPqHxxWzd39yXfyM0otYHkg7Wf5FSlHELm2ur/PsQhNuEais7/gLK40K0Ffmt12/C5l7qUMXGrSlun0/zMVOrXqRklrDmdu75Xp5GUOphaTxEtG/et28r9duxicfLEzdV7sWaVlnX1Vx0ns7rSy10t19Svlqo46dZLRx5X6733MuwlSWJrVpuyk7KPblSV7+fZ7WMqwcaOJc4LVRt+Lf6mGnVjUjWjo4u62KMoRUk1fTTyJSUbKxQzL2+ZzwcY7K979/K36mInDD0nUm9lc9rzpNaC5MRiKlJP4Gl9+v6DoxoqMYSs2u3b/hyUcXRzNKP8JxatppK9793ftsrFbG4rDOEYu8r2b0V9exmOfV8fSo1PZ2SiotXXRvXZb+RkdWtWw9WnWWrm0tvh0tt/wAOVnOjKrQjGzgk733ur/IpZtRxUYxW04q+/VK/QyzOKVLkoUI3SaXyWnVdPvMx4kryowgtOb0fy26r7jAYhw/2io9Gl+DXYp5rRp4mMU9Hq9/l0PbxxUnG2hjMJF0VTp6NO/8AmYyi1SWu9zF4izSKmqkkV6vKVJtu5N3ZN3dgsAF9Si7pAtWSVwV2PW1iNmhwXKSVrD1V0OLRoSSmRtuXuh6ai3TOXmdh6N2OW9Pm+QnYdnFC1RomiceUurDaehJ2iKyQrDvcl7xbUk7MVmyUubQ0sbnUB2AGSaSKlZJWQnzS1E1YUlc0ehFqwdBgB0NwA0YBc6HQRubgAG4BoF0aGgbj1B6gAZW1domkrIxOGjXhyvv+pKUaFPmlsjB0pV6v1ir8l/X9dSovdfKNXiZtlEMWouS+F3X9XMyyZUoSbWv9eZiMLCtQlXi+r/DQwOWV8wlGjFbPy2+9GXcJRdR1ZLWOi/rmMFlyoU4xXT+u5XcIpcvYqzUlddRT9pB8mpOlebbMxw0MVQcHs/0ZhcHKhTSluUqjblb+tyVP30jGUXOFkV8POm5O2gsOq9GcY7p/oYShOjP8DCQlGnzreMb/AHGGcm1NGauo8NJ9JJr70VMTXwmU6K8ox8ulvXuZTialXEKvWVml+fp6GfZpSoKlKnqpWv66+XoVcRKWNhSS0Ueb5tmX11Qo1YLeaS+7+vIhNuK9D2NStZQIK7uUcHCOO+sQ+K1v6/4YxWJrxzGnhnH3ZJt6rp+JSqN1W+j0XqYTAKlVq1o7z1fy+ZXws6mIoVesHL8VYq1IVacknqirmWJoV5ua3V1t3aXT1Mjp1KWGbrPWXvL+kYdSw+CoYqPw1It/NaP+rI+svEYaFZfFNRT+7T8DKKcaGDqYftt6rcwWP+qKNtpJr+tGYirVlypLVSi/kv8AgEM4Twyg9k/09DE4GniKCqN6tL8Gn3KNOdGk1R15pJ/lffyMbXrxrwVPZvXYrwjWoOC3T/AxmVz0klt/XcnUj7Ftef52/Qxc1sVJdCrK2iA3AAw0ug9RCu0MTErkkkk0RepzMh70Wi7tYlC70FZIdkX0IS1Fe1iMk9TkS0HFqArO6Olh6WHbcqSvoNKyRZPUaLI0EVLpD1Q03E0a0EubQltZDHsNA9xjWhKaROtqSm5DE7DZFnMkznAAABmmxdD2LHQsjQdjQGW0GiwAAB0DcHqAG5uWLGyMBU5Kq8xxV7jslzMd8bWt/wAu4/iKy0Qm7k42VkZssPOhSjRXLLl956u7u9ddtLbaGc4CpOknF6Lf0+/8iHCyo0azT92Wq/q5w/k+IwuKTl8Liu2jXzZCn7KyWt/zN1dbmJp044NTjO827cttl3vt8hzcm/Lb9SKhShGMFoOSu4joqEElr5GLw3tGrjjKMkoxu/XYpU0pczZUi5JmKg0mlv3MUnh26rje6uzDt1knPqQ5qbnN6WTv8tTB4qn7sHpczLDPF0FT5tLp7dtbGDw0auCrRjL4uazts7H9iVaTeBq1eZVFzX5bWs1pZP06oxeIoRlDD1XrZW39Ft+pSarUp+77ydrehk1WriMPCpOHK9rXvtpe/mRi5RuYSvXw1WE6b1XoezvKyWxVhXWGq+xdqlpcr0dnZ2euj1s7PTuYSrWnh6TxTvUsuZ6K7trotNfIw7lKpNSVkpaea/QnOEbrdFTmlNtOy0t+NzLMrrR9rOctZ7aLRJvz1vcz3LZTp03Kesb9N79N9Nt/MxGKjh8ujXatdpW+Xf0MTi/7HyuhClX9tCUm1Hl5OTmt1d3LVPf8DNc2x9Or9XoS1Uo62XT1RLOKGErcnLabk3u3e79GkVaVPGOnVqS5HRstr3112ta3zKtCo66d7qdr9OW2nz+RDL8E41cSp+cdH7vRrfW6vujArD15QhN7LzKWDjNKEZWXoU+H6EJ1sRVn9ldHpZt30f4WIUMLHlkpXe6dmZlmUJxc4dbr7vkYvFWvFlR3dyWiuNuTbGa2B6lzcsFJ2dyL5lcIyaGEXdWLISvZdiW4rLU3vY10GrsSItJWItojJuwmKSSR7WTRJpO58UWr6lH2ipRjUfvLT+rHkSSvoNpIY72PQ3ZZ3tcqbD7FSco05Na26EJXgpLqJ9RgwZuVKiihYnTYnNyALIGzQBh0A3AAHsMexugtcY7WLi2AAAA3QG5a4WRoWRoAFOVpJkZqUVYx1aVWf1al8/6/MpUoUqahHYTTBNsrxuypyyTizGUozwcoR3cbL7jJoV1GPtOkbPbdadCtRcIx81cpSurFenZE4tvQkk428hqPut7oqYeSrQm9kn+Nv8irFTUfJkIKzZGScR0+anbuYihN1FFbGLVOdJqXQoYdxb5drOxmc5ww0ox3lp96KPsVSp83xJWW5WlBUlJPQxNOX1KSw+9m1628yhatRpSl8S0f9bGNw1Gdak5b38+hiKlPD8sl9qWv3f8AARgpQhdLZO36/qQnr5MjJXZGaUvUg2n7xKNPlUuphnWU5Kotr222I3ehKlKUbR3MsnHDuc6y+y7evyOJ6zqUYyW0dX8jC4zC5hk/JVlqpu+/ZW2t5/qcNVKXsqWFxGqlFv7m7bf5+pPLKk4xqy+J/wBdzOMuUqMMTL4o6fr3/QoYedSDw1PWUot/OS/4PoYLH1MDgZwxL95NN+nXa/QwWc5di8C1TfxLT4t1vuvzMPVre2VaHwpW6GUZ1VxkI8m0bp+qt5LuLGe1pyot/ErMxVJ06SjB/CrFaTpRcbmLnzSJFWXQirs3AAC1olJpxQD1NyOjuRcHv2NlcvqWi4lrbCWhFEt2RepZjdkLsNC0RuWukS1kEtCSvGwzdDTE+473uSaY7Nj0TRpawNjYdCUkkSxCSsiUnJgAXNDcYABoAWA3GWLGljSxYsmAdAOgDBgdCwzQexYsAgAI4x06XLH4uhgsIqNG8vie5JbFSqlXjTXUW4ndElcr0lZu+hmUq0MBV9kveUXbbe3n59zIvavA0lX+OUU5erWu2m/YxWPq4uhRU5X5IqK9Fol/X3mHcoVG/IrzvC/cglNFTWslHs/v6Daq2cXs9Svja2KqJT2iklttqRadecV0Sf33/wAh1eWVpbFKlKnCd31/MqU3TwkayfdW9LCadOM0/iVyNCKvJ62MLDkioJWsVoK8U1/w5TyqOIwssLOWru/TW66/qVcrX1B4eO2n33XmYanKjy01tbUrYWWFnWxFLV1JR02tZJPff8CWR4+GMjUxcLQUuaDutU1rondWstzM8qVeg6E9E7f59zIMNV/ezm9JSf8AwPwsvlfqYxYp16EaKvFv3ttl66/cQUYu67Eo80+YlRTHS18iUUpcxOUX8JSqSjLcU4VFZnGGYYWjhpUJStJrRWf528jh7K8H/ZVOvyWcnrq9el99PRGX5Fh8NBVqb1hFrr1lJ9+5l2bzo/V8HifevBNvbWyu9F+FzifDrFYd0ada0G19nbt5sy6eFoYlVa6uo8qvrra3bYzTB0avM4L30mm9dU122XyMkpexwFLDUNKkZNt+Xo9DJa+HVZprZXe+r/4fscOY2WGo1IUpXjOTf32766WMPiU67lKWnTQxWPbvzPQx2MjOHukpNu5UqqKa6l7yI/EMAAirsqPWxhpJqwBOKTaTErkGN+8kx2ZezIytaxd3Ro4l2mRSGrot7xDUbTSQ1zKw1ZWLtDtYWiGJ3QmmrC1G9iTTRzNj7G5N6WLjHqhq42kipXs7EpOQFh6jQFh2NAAAABmg9i6GGg7Ghoyxa50AABmgdDS5oMsrDsaBoGgmmBzXdgwqSrJsjdtklclBc/MXI6hiaUpQaXcrxsnB9inRcX+BCqotIovmdjFR/dNGX4efNGD3bt95h8LUpYmcJ7Pr21SJUVSnKKJRUXoVVKGIVuqLKpJxfTUq29mu/wDX+RUnKVG3TcjUi7Rj0RSoPlafUrxUdUcqlZswmElTmrvYrunUoQUN7GMUoYeoo/Eotr7mLPI4TLadbFaSaem+3omPiPGcUYTB1IU+SNKTi9U7q0X2jbfpcq05cqcn5fcUYJStEp02kkOnyxKcnKrOn/K7ElZIbu0QSfyHRU1yos6EnzDp+zXtOhnOS0MwzKhUk9YJu2uzsu67FXKZfVI06Tsl/XVmDwGKo0akE7p37Ld37mBxLr4vF1amnJov6+7cw8nmuFqVaTtzLRea0626/Ip5FiM4y2jho6VIxj2d5WS7pLX5GCyHE4nFrmdkrLp/mUlRotQpq8oqz39ChgqlLMJ1b+7NNW89P8jA1o4LDObfuxcV/wCBNRXd7sp1bXkjE1Z8lmVpNlSsSlzO5H4kPSQzcACCtqPVlF2diLug6G4bK5FpxLpoUUxaMvdG43csok7WFqxaMWqL2SY2mh6tob90dy/MhaRuLVaByoe6JXTsWB6jYMlNIqVLvQbuABuAw6AAAAAABuNXLXLFjoMAAAN0DGMsWBk9iHYBfEC7mFnz0kzdDWlhbHQUuVlKLnGTe9/8itSXNd9SOjTkezg0kyKcJaMmruyZVnKk+eO5VvJ3vqyrzwjvroOakrlSDupdzlqTmpxlZLdW3JPmaOWOkSUqKr8ltbeZCzjcm2ptPW5Spqadyvivq3PK17K/4ow1KtQqxnUVpLXoVK0qtW5j+H8NmuJowrxuk/NbtdmuxXp0I1quFoaKC8+77+nczHDvFYVU0uZt2tt8zK8uw2Ao/V6EbRu3a7er1e7b1ZFNy0JU3fcpxeo1dsVN2TQk4uzHBKFoko3Y6DjLTZlOMaeIqRtq/wDJGMwNHFNKqr2aa33XozJeHY5hha1anLldNXel/wBV+plsKWOxlSpF6aJrX18u5Qw9PCOcaUdFdperu979TLazq04zj7rV331RgMTPDYWXO7tRb7a2FjsTHeV/uKmLw9SK5Y/iyhh+WpKpF72X3X/zMJQhhabivtNv+tzGYjVpFeo2zcI7ofxjWotjRo0ARZKDAoTvGwALUdrGmiJpN6DIvUZq2iyViyasNJOwtDQ1ehSpSqS5VuS0TTPIjG8WWsW0FFKKLDtY0sbIm10JMbFJMlOyJ1X0HJsA3A0CyYWQyxYtcOgAABodDRlrjLBYHqCAABgdAsGpY1HqWCWrSF8RJ2RFtsvZkZO4ZdV97kLXMQ3Gm5JapGDnUq0YyqRs30vc8iSRh0ktWV6alTdinaUrS7HLGSsiraK8z2fLHmuTScSTV0PNKmJzCrDksoWW++j8vIrzcY3jqVJVZOCWyv8Ajb/Ipa3tuScack5Moxlyty7u3pfQdGNWpzrpoSi6UGyHPOKclqYaVSOLpUbX9rUjD0Ter+S1tpfuYrLlW4nUKcr0oXTdt3b1urbmb08R9fqrEK0lZW0e3mtDA1JQxs8PV0ck5R80rJvTbV9dTiLJqeW18PCnPmvGMnpa1/VswmJwVXG4qFGpzVE1zaNWve2+j2e3z6GW2o1ebyIpWS6iSU7DfvK/UtZ6DV9EiWsSN3uhKTV4k0kP4loYiCnDnS1RTr0Z0JSb1tpvqzPM9xOWUZVIPlT0ez0+5/gZJUwilKVGp+8d5Ws9V01ei0s/mYetNuMn1S+VzNcJjamJpzw32d1pqvV7fIzPC1MJiZRw79pTdve+HV7qzu/mZvja1OknRjeV0kr92kZe/wB4oy3FTjRrTlGV72e1tSdXmp3K9W7Y3c0HZoW6JW50N6nQAIq5U0sgKErMWqAEX0H0GiWjOoRexo/kIVmtCKSWokLe5TqOm+aO5LUmtR6bFtB7HMuVDkc5KSsOVxsbJzS0JTSQ5toAN0aDHY2NDQdjQtcABgAbgdAshq4yxYLXLXNAAAA6BYAsgauMaHbmLpyKnQ2EryJqzQUp8k0yjU54p9Sa1uxLU6DTaI3sKo2rCjG9iyjK5hMJVp4mtVc78z2tsc11oYvF06FSnTlvN2Xra/6FSVovyKzXLddSMfZ1VC2lhpK6QqbUbolQcopSesTAy9rRU+5SppO67krvV7lfEezi2UqkY027e9t8noYDCU51YV49L/joZlNzqOrUd5O34u3QzW2GisVFXlFW+Umr/kuhUxNedGMqDs3bt313MtybCYKpVq0o+9P4nrrq2t20t3sZbjqeJrV6MFrSaT+aT/J+Zyq1y0Y2NEhz0IWVrHs7+6Rgm3EinGKRFKTd+hyJy0MTGyaRi1UcV7KXLLva/wCZneM5sXg4VHzS5eZvbduPT/CPJ6GQZfDM7Xk5SSeq637vo0tuhlWcPNaPM1tUX6Psu5xBDD4bDYWbneVVdnp/XyOaE5OSeqZjnKjH6xDVxvYwOHxOJpQrQXS7Wm+/9aEKqlS51ue3ajyk5XYMbFuTfvD3OgBa5FWJu7AT1Kc9EJ3QAtVYUbxb7G49hSdy92X1uiysJ3RYiixZMdrFlyjta5fm1HJX0HK6IvQ2HN2GxvQbuVaiiObkSlc6ABuMYzqIeojcexa4AbgDCyLFlYewaD1DcsDDY6AdAAAdzU3OgampcC3NIlGxPoXTE1zErOWg7BltVSaj1Kjux7FtC2g1q+xKVpJIpxjzuXcnKEtBUoxXqVIKOqMThaddKclrHVepUulqYiEdLOxhMbUxNSfNHROyd9/8ipCk6cOXfrv/AFsVIOT9x2OeKXK3q/0MLGlTpqz0+ZdJifM0YnC05zWuiEoya10MDho0Y8kfkYiLlK8/QnTVSPJNGNqLCVsPRhH45Nb7aN+fYoKLlK/oUcPCnWlKmrc2/wAlb+rFXFUaNWFKTs53S31srv8ApjVtydR3OZtEZu5huSdSPM7JsxFB0a0o9j2d9DlW1izUroxKlZIxGGlLmbdnbQwXDkMRTpxfvVm5NLbRa23tu2/nYzzEYx5bTwdZ80IylfZWbem2r0t19TAY7Ncsm8PbR6te7o1pvr2sZXOtn2EpOpU0p7aLRp36W7eZQws6KjVqaKpdrrs7P8fQxmEqV1TVN2Sd2u6/4e2xlWazyyopUld2a7bq3VMxC9lVbXwyGzc0Haw0LdFRe+S3OgBBXLj1AITsRqdBNNaAHQtZXGxMRojpdlkrWI6sVk9B6MsWuJ3iTWjRzOKsRslYbVhSHK5zaDY3cq1lHREqjluNgtgCxdWDoaFhrQ0sPUsiwAAAAAADAYbgAFgLAWQWLBa5YaIppk1dKw43jYUWmODZGLubhhavsqqkRaqLmQ0jSxKyTJTTbj1I0vfUvIvy6ibbuSm2X5lYcFqirBTVjM4zXLH7Lun9xhKCpwSjsi3NAoU5QpqmyeEpwSlSd9Pu1Z9WpyUVbYrUVVp8pFWHBa36GAr4ieYVaNRe7Hbb9NTL6cKk030T/JnspTj8/wAiom5FTC3UZSXmjB0pRlOL2voaKxVpqVeLktErr1KktbIAhNpCr81Ple4pu12XTVyTTKrXIytFtpsxXs+fD4mlpKnGSfq5SfXya6GeZZCpgm3q5Xa9U7Lr5GAksTTpVKvxRaX9WMlw8sFRaw2iVtPnd736HFOPWMz+jPCK1KKS+dlfez3MMm4+pK0FYrzU42IyewAblRXkiW50ASuJKxLSJuACm0U6xCaaAvoJ2GjoX0EXvY8iL0I6ofQViWwpK1iUla5KTbuXG9C5cb0J14x0KmIk1ZDdy5ugWwABYsFi1yxYYAGhYDQsBuGgDDc3BljodAAAAA6AA9RAABuZbiIyh7N7ji0iVkrsmuaJKly1NNSlUjJO3Q+tzniZ0eXRdb/oK9kOQpWG7u99SVmjE0FNJSZh6TUdjD0KkYWk7vU9lLl93cwmDdKk1KV9ewsLUVSUk9GjCLkiqcpXlFWbta/n8+xJ8z0Qp88W7DTlKXItWtzBxdNJNmEnThUantaX38rt+I48zXkYrFqtClHltyxtvvq3f8TA4ZYiThzWsm9uw5JKzKtbm0HdgFynK8khyU9EOdnYdm73KdOFSahJ2T69jFUuWVou6WhFJJxe1jHV4YfE0qc9YyUmvl/w5nWWwqUaVTCQvNNaX3Xq3Ylkn9nYmnTqSu0o308k+jZLDxqTVtFe4puGlypUuN3JLqhNNABJXUWNanQtcsJI0KlrAABSrJaEJpoTTA0YxvQuRaE1dsjrqLYWjsXWw3dehzKxOadxk5qKJ4lt6IdZ3PrLS2J15SHqPUZYBbAB0AAsDYywBYCwG4AAMDdDZcZ0GOwAB0AAGBcEAABh6vs5plPExqR0Jasel7DvYilG9upUSUtOp0sS2Grj20OboVIpojGcZxa2syspKMuXexg1OdCKn8VtR0lZxfUjRabTFQUcS5vtY5YtMUEzlvdFRyVOPJvdfmh1HGtGPe5zKFr9SpJvQqTlhsNOpDVq7KdeVWnGb3aX5G4ACeuhGQncbOZJE+hWfKrIxVOE1BvdGXZVPD1q1Scr883JabJ9Nx3lLmk9SLSZKSY3cCySA0HYVnD0C1yxoNpIuN3HqaGgAFOpYjVQpJgXNB6kX0DZEZXVhzs0OZUldaHNoPUk7Ir1LuwtWOwxs3GAC2AOgAywanQAAtcCwADGrhYACxa4dBg0AAAFgsG7LDEFgCwwMFUSfKyElKOhJEnoX0G1KQ2WUi7Yy9hy0siFSHwcyuNRSu3ZEfYtOUZL1uLEUXUSUl95y8tRroTpOdVSTHG0lY5EtCUIJuyMM5Qi+bvoOV5R1MQlNqz2dxXehOVo8qLjYAI3LnNsdCbXMVKvMl5FSd5Ik73L6DAABoaLEHyji+hokXViw9QGDDcOgF9CFRpkKqaLm5c0Op6CegRldnMrk5XLjK80kPUjuaEthosMAFsBugNwBrQYMFsWFYYFjcAAAAA0NBLQtdAAMAABmlhjsHUALAAnZ3MFiE1YeqsTV0TVmRdrjFoXEElcrLkxUX6GMly4eRCKWXy83+qFhKro+1S0MsqurRtJ6rQtZ6DhdjtypjkraEneKiXHboPQ5m2J3QAzcB6kWuo6iuipPUbTjYmxu6HqMGBpYBosW0LBugvoAbg9QGaWGPYuaEZWZComKSexuXBGw5e5Y2Q5aDegTkkipLmkF0XHaxoAALYALFrgWuP4TobBYaDRgHQ0sADOhbQLAAbnQOh0AGBYAHqDLFtB3DcAAewARk4u6MPinKKuQlzImlykVdDL9B6mnQY7WMxjZwkv6sZjO9CPn/kVYcuBS9PzMLJLA6+Zk9+WfyNLXObmKtmiotkAkYmfLZITHU5Ue3kxVpPcU7xFU7kqmugppiaZccj2jbJu+pKXQlqMAAm7IjqibaCDbLFwADcAAsPYDQE7FOo7CqdBO6DS50FYurjCcklcrVHJgW0LAWALAtgFsABbQdrBa5uADWgWAAAAA0CxYaR0AAAADQGHQsHUAAACM3HYw+IuSknApq8WSjqW1LiLaDtYzCN6MX2ZiZOdOlHy/wCAY6yw7Xp+gp1JRVNPTsYOi6NNRe7Lu+g7om72KkkiVTU9ux4iTViUnJ6ik0N3NwUnYcmKRGVxMdXsSqts5xu6L3NxgBuE9iOxUFsQOgdAAAA0A3NBhoCdiM7kZimmXQAVKiS0KlRtD1LAMAABbAA9hm5ui4AaGlgAAAAOg9S6DQGdBmlgHsADN0BYZ0DRgW0HYAsWLFixYNwsEZOLuiniOZWZSrWVhxU1oSp2HEOgzFx5qDRRfPUhHt/ncx2tBv0/MwNvZRLrqOa6EqisTqJFSo29xtsACLSHJdCDJhdo9o7mrFAcUz2aa3PYtMcEhxGWCxYJrQSuia0LsiCADVgAAA9gsDsBGTQqoqx7YdZEsQktB15PYk+o3cGrliw0WLDAGC2AAaAuAbgbgAAPU0DQ0saWHYsg30LBZWLaB0NxgAaF7gGjLXLXGG6Cx1AADQ0A0LovYhWaKeJaVynWjNK5OnYcRxGiceam49zCYWpCpzSVrGLi50XGO5hoyhTSe5KbJSk3oXk0TdkMANwudCOxLUCFO7LJDZdIchVHclNDlcYAE9iOxPYsrEFc0AAYAGgG4bgXLAdQuczN2F9Bu6NwAsXOgxgAtgC50LgbgxgAMBm5YsWLHQsWLDVwaLFiwWuWOgAAdAAGbjLFgFuBugHqBYACyYm1sQrNFLFyatcdXXQqV3e57a5GbaLtjQ0VXobiloVUuW6ACxYLFiNrEnc1ErHO7jk2N3ADoDSACexzMlqkdCAABYAAZYGiw0WAAAAAAAGACHsdDSww3OgWLANAAdAYGjAAugBnQsDQAAAABYsW0LXBgAAM3AeoADFNrYVaRKs2QuymvdFaxJoqSSJS1IysySQno0ABuALYYg6jsAAdDc3LoB2HFDSHYsuhoaGh0AAAaLHQtcDcABAAAADADoALY0ZdBodDoWRoG5oaAwAGBuaMsjc0aCwDWpYLHQGHQALDAsgsNFixYsrFkWNBiABgDDQNA6HQN0IITaI4hNH1hEq6JVbscrsWxvEfceoBYLBY1LFtAGtAADoAAAx6oGHQLDsaFkaGliwbg0WLItcNBgdQACwAAA3oAC2ANA0Nx2NDQdjQdgDYAAAAYx7hqPY6HQDoAAAAAMBh1AABgABoXLlwudDQuCOg2SaaAOgAGgNnQW4wAAAewAHQAWw9gsWGaG4aGhobj2NwAAAANQAGAPUtcBAwAsAbg9Q6HQ0ALhYA0DodALFjc3GbnQ6AGgBZAFgaRYGDDqBuAAAWDcLDA6HQNDQSuxkwGGgG4D1Qi+oD1Cw0HQB6gdAOpZM0sFix0EF0OwNGhc6HQ6G4MYAADDodANbBcYBcAA6G4dAtc0HawWLaA0CLD0LXBhcDcAGABYCxYNB2DQLobLoLo0L6F1YvcuXQNAAAGg9QtYC4AFmWLHQAg4q9xyjYlZ7AD1OpqMtoFgNBtF0MNC6LoLoGaFwLAFixZlhoLFg3OgwAYAFixYsFiwNDAsAB0N0FrlixYWwABuB0DyLFgLGgwALFiw9QsNDWhYOgPUA3HqdAaBgNAACA3A6BoDAA3BgHQGAAAAAdDQ3NA0YwsFkWGWLFiyCx0NLDSCxYWwAFkBYsWLXCxbQtcAEADAAYtgAsgOoFgAAsNDBoHY0YaGljQOc5znOc5znOcc7nOc5znOc+mxznMc5znMc5znOc5zHOc5znOc5znMc5zHMcxzHMcxzHMcxzjnc59DnOc5zn0OY5jmOc5jmOY5jm0OY5jmOY5jnOc9oe0Oc5znOc5znOc5znOc5znOc5znOc5znOc5znOc5znOfyOddjnOc5znOc5znOc5zmOc5znOfQ5znOc5znOccznOc5znOc5xzuc5zjnc5zmOY5z2hznOe0Oc5znOc5znOc5znOc5/I5znHO5znOc5znOc5z+RznOc5z+RznOc/kc5znOc5znOc5znOc5zAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABKEopNrfb8v0AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA8NMBlWccUwyzHxb/AHbqJLRNxaVn1ta7srbbnjflNLDVsFXowUY8soWSskotNKy/xOwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHAWP8AqPiLgKrejjy/+Btx/U8Zsu+s8NOslrSnGXyfuv8AGS+4AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAD2s6XENGpB2ag2vVSuZnRhn/DdSNNX9vSvH1lG8fudhoAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAK3/I9p/wDXt/meE2Z/XuF6MW7ypOUH8ndf+StI44yr+zOIcXhkrJSbX+GXvL7k7AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABiP+R5R/wS/M8C815cRisvk/iSmvl7svvvH7jxyyj2ePw2YRWk4uL9Yu6+9P8AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAxX/ACO8P/hkcAZv/ZfEeFrt2i5csvSXuu/kr3+R4p5N/aXDNblV5UrVF/278X/krkAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAGN0znC+k/yNtjhbM6eeZBh8TU972kLS82vdl+KZxDlM8pzWvgZ/8u5NLzXR/NWfzAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAMx0zfBP/AB/+kh4G53z4fEZZN6xfPH0ekvudv/AjxwyL2ONoZnBaVFyy/wAUdn846f8AboAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABwjmXAFDJqMM1pqWI97mvGb+07apW+G2353P7W8KavuypRS78lRfkrn+qvhpmvu4TEqm3tao0/uqX/IzPwNrOHtMtxamuimrf8Ak0bp/cjPODM9ya7xmHkor7S96P8A4ErpfOzADNtMxwT85fkg4Ezz+xs/w+Kk7Qvyy/wy0f3b/I47yH+28gr4WKvNLmh/ijqreusfmNW0AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAyzPczyyfNgsRKn6NpP1Wz+aMk8a80w9qeZUo1o9Wvdl+Huv0svU/szgLjTXCS9hiH0VoSv/AIPhl58uvmcUeGGeZInVUfbUl9qCen+KO69dUu4GdaYrBv8Av2+9AeHGf/2zw9RqSd6kPcl6x2fzjZ+rPE7h3+x+IKqgrU6vvx+fxL5Svp2sAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAACbT0OF/FfOso5aWIl7ekukn7y9Jav5O67WKuScH8eU3Wy+fsMVu1ZJ3/vQ2ku8ou/d9DiXg7NuH63JjKfuvaa1i/R9H5Oz8jP8ASeEl/wBPY/jcDwa4h+o5xLAVJe5XVl5TjqvvV15ux4tcN/2rkUsRTV6lC8l5x+2vu97/ALdAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAKVapRqKpSk4yWqadmn3TWxw14tc9H+z+IaarUpac1k3b+9HaS8173X3meLvAmWYbLMNnOSVuejOtBKK1s3e3K999HF6p9eirUalGo6dSLjJaNNWafZroGHxFXD1oVqUrSi00+zTumcN51Rz3KKONilacdV2e0l99/kcecNPIM7q4aK/dv3of4Xsvk7x+V+oAAAAAAAAAAAAAAZnnOFy+F6rvLolu/8AJebMNXVehCqvtJP71cAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAM+zHE4XLWqc3yKcJON9G4yVm13XffzI1eGPEagoz/cY5Lyu/Tb2kfLSS8lq+J+Esz4exPscZDR/DJaxl6Pv3Ts12DwV4m+r4yeUVpe7U96HlNLVf9vRX3x8zxW4V/tnJnXoxvWoXku7j9qP3arzVuoAAAAAAAAAAAAABxBxDDAx9jR1qP8A8l8359l8351qtStNzqSvJ9WZJw1i48F4TPJyXs5ycEuvuuSu+2sWkvn2uAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAcTx5sqq/L/0pGGrTioVYStLRprRp90zhnxOwuPw39lcTQVSnLRTav6c6WvpNard9ZHGnhfXy6m8wymXtsM1fTWUV30+KP95bLdWVzB4uthMRTxFGVpwaafZp3Rwxn1HPMqo46l9partJaSXye3lZ9TxM4T/sHOJSpRtQq3lDsv5o/J7f3WvMAAAAAAAAAAAAAOIM6jl9HlhrUlsu3m/07v5k5zqTcpO7Z4MeBNLOKMc5z+D9j9inque32ptaqPaKs292lo/EXI8LHg2vhsLSjCFJKUYxSSSjJN2S0WlwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAz6PNltZf3X+Bl8ufB0pd4x/JBwX4hZjw5UVNP2lB7wb284v7L/B9VfVZ1wVk3F+ElmvDk1Gr9qnsm+zX2Jefwy37yPCviDEZFnM8mx6cI1Xa0tOWp0/8AAlp5vl6HGvC9LiHKKmEek1rB9pLb5PZ+TvuYjD1cPWlRqx5ZxbTT3TWjQAAAAAAAAAAAAGPxtLB4eVao9F+L6L5mNxlXGV5Vqj1f4eSPB/w2ln+bUniY+4veflFdfWWij2vfpp/s2BwvSFOnH0UYxX4JJHiF4hYjiCu8Nhm44WL0Wzm19qX6Lpu9dgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAjFyajFXbMt4A4lzBJ0MHKz6yXIvX3mr/IwXgfndWzxFanBeV5P7rJfiU/A7AUI82Lx7t5RUfxcpH/ABDPgXD/AMfMXdb3q0kv/Sb/AInE/A3AMMjxksNj06ipVHFe2pu8lF2VkrvW2hwHwJwdmvDOBxFXHOFaVOPNFVKekrWa5XFtel/8yfghluIjfB49284xl+TiY7wOzmkr4avTqet4v8mvxKPDnGfCuKWLoUJxcesffi12ly3VvX8GYfMMg8QsOqGLX1fHwWjW91/L/Mr6uD95a2f2jIJ5h9TjSzBfvoe62tp22mv8S1a0ad1bY8ZODP8Amd4WPZVEvuU/0l8n3YAAAAAAAAAAAAHFGbfXMT7Gm/ch+L6v9F/wThjJFjKvt6q9yP4v/Jdfu7nhTwysoyKNeov3te035R+wvud/JyaPGTjNzqf2JhZaKzqNdXuofLd+dl0YAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAcP8CZ7nlpYWg1B/bl7sfk3v/wBupmE8J8gyeksRn+MT8rqEfS796Xy5X5EvEfgzII+zybCc8l1UeVP1nJOb+aZmfjVn+JusLGFJdLLml98rr/yUx3GvEWOv7fGzafRScV90bL8CrVqVZc05Nvu9WFaHPTlHumjhmfPlVL5r7myE505KUHZrsYHjPiDBW9hjKiS6OTa+53X4GV+NWf4ZpYqMKy81yy++On/kpS8QeC+IJRWa4f2VTpJrZ9LVIWmvWyRlNbD1cJGWHr+2h0ldSv6taO3ffvd6mIw9LEUZUasVKMk009mno0cecIVeHM0dFa0Z3cH3XZ+cdn8n1AAAAAAAAAAAAOJc0+pYRxg/fnovLu/66sy/A1MbiY0Ke7/BdWcAcMU8xzbC5bCP7tO8v8MdZfftfuzinPKWRZPWxsl8C91d5PSK9L2v2V2YnE1cTWnXrS5pybbb3berYAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHDHA+ccQz/2Snan1nLSK+fV+STfexT4c4K4KgquaVFXxC1UWk3f+7T2Xk5v0aOIfGbNsZellsVQh30lP73ovkrruYvG4nGVXWxFRzm93Jtv72AAcMaYKUP5ZSX4gBlWd5jlVZVsDWlTl5PR+q2a8mmjgDxVlnGJjl2ZRUasvhktFJ9mujfSzs3oktL8W8MYbiLLJ4OtpLeMusZLZ+nRrqn8zNMsxWWYypg8VHlqQdmv1XdNap9VqAAAAAAAAAAADaSuzPMxePxkqifurSPov89zhTKvq2G+sTXvz/BdPv3+48DMltTxOZzW9qcflaUv/bfuZ44Z86mJoZVTekFzy9XpFfJXf/byAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADDYWviq0aFCDlOTskldt+SRkXhplmS4ZZnxRVSS2p30v2dtZP+7HTza0OKPF3E14fU8kh7CitFKyUmvJLSC9NfNbFWpOrNznJuT1berb7tgAAZB7tTFU+1ST+/8A4YACnUnTmpwdpLVNaNNdUeHXGsOIsu5artiKdlNd+015Pr2fk0eJnAcc/wAJ9awsf9pprT+/H+V+f8r76bO6nCUJOMlZrRp9AAPDPw3eazjmWYx/cL4Yv7bXV/3F/wCTPTa5xllH9lZ9isIlaMZNx/wy96P4NAAAAAAAAAcV5j9Vwfsov3p6fLr/AJfMyHLXj8ZGDXurV+i6fPYSSVkcB5XHKuGsLRlo+Xml6y9539L2+RxNm8s3zfEY1vScm1/h2ivlFJAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHDfDGY8QYtYbBQv3k/hiu7f5Ld9EVsbw74c4Z0MMlXx0lq3uvX+SPaK956XfVZ9xFmOeYp4nHVOaXRbKK7RWyX4vdtsAAAMr9zNMZT84v70AAZBnuLyTMKeNwrtKPTo11T8n/wVqkcO8QYPPcvhjcK9JbrrGXWL81+Ks1ozxW8PPrCnnGXQ99a1Irqv50u6+0uu+97gHht4dTzuqsdjo2w0XotnUa6L+6ur67Lq1TpwpwUIKySsktkl0R45ZI4YnD5nBaSXJL1WsfvTa/7dAAAAAAAAA4gzD67jpST92Oi9F1+b1OFsu+q4FTkvenq/Tovu1+ZkWXPMc0w+DX/AC8nGPybSb+SPELM1lnDOKqRdm48kfWXu6eibfyAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAOC+CMdxLiuSn7tKPxz6LyXeT7dN3ZHEvHGXcNYR5Jw0kpLSdTez62f2p95bR2X92rUnVm5zk3J6tvVt92wAAAKPuZ9Uj/ADQT+52AADgXjPEcNZgqmsqM7Kce67r+8unfbrdYHHYbHYaGJw01KE1dNdV/W63T0Z4o+G/1VzzbLIfu3rOC+z3lFfy919nfbYDwezVYzhuNBv3qMnH5P3l+dvkHG+QLO8ir4SKvO14/4o6r7/h9GNNOz3AAAAClRqVpqnTi5SeySu36JFWlUpScKkWmujVmAAAcRY/6ngJyT96Wi9X/AJK5kmX/AF7GwpP4Vq/Rf57fMsrHhDgFiuKKU2rqnGUvw5V+MkeOmY+zy/C4JP45OT9Iq35y/AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADgfgnFcS4zlXu0YfHPt5LvJ/hu+z4146wmCwv8AYHD1oUY6SnH7XdRfVP7Ut5dNNWAAAAYr3M8oS/mhJfdqAAB4ccf1OH8SsLim3hpvX+4/5l5fzL5rVWdKrSxFKNSnJSjJXTWqafVd00eJnhm8C55plcP3W84L7H96K/l7r7P+HYPBTOfquc1MDJ+7Wjp/ihdr/wAl5vwA8V+GXlGeSr01+6r3kvKX2197v6SSAAAADw+inxRgk/51+p452/sXDf8AXz/22QAABxhjvbYxUIvSC/F7/hb8Tg/AeywrxElrPb0X+b/QPAjBKVfG4prZRivm23/6Sjxrx3t+IYUE9KdNL5ttv8GgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA4R4VxfEeYxwlDSK1lLpGPfzb2S6vsrtcc8XYTKsF/q3kL5acNKk1u31jfq39t9fhVkmAAAABnPuY7B1O0mv8AwJJAAAHht4jSyWosBj5N4aT0e7pt9V/dfVdN11Tp1KdampwalFq6a1TT6rumeI/hY6XPmWTwvHedNdO8oLt3j06aaIynMauW4+jjKXxU5KS+T29HszBYujjMNTxNF3hNKSfk1dBx/wALx4gyWpQgv3sPeh/iXT0ktPWz6E4ShJxkrNdAAAADgKXLxNgX/wBPI/joeOUL5BQl2qr/ANIn/kAABisRDD0J1ZbRTf3FKFXH4tR+1N/m9WUaMKNKNOG0VZfIPBHCKnkFWs951H9yjFfnc8RMX9a4oxs77T5f/AEo/oAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAGTcQY/KPbfU58rqxcG+qTad12elk+ibtZ6oAAAADib3cNSq/wAk4sAAAPDrxJq5JOOBxzcsM9nu6bfVd49103XVOhXpYilGrSkpRkrprVNPqjxC8K6eYc+YZTFRrbyhsp+a6KXfo/J3brUalGpKlUi4yi7NNWaa3TXQ8GOIVjconl1R+/Qen+CWq+53XkmgPGHg94HG/wBrYaP7uq/et9mff0lv/iv3QAAABwlU9nxBgZ9qtP8A9KR40Uufhnm/lqRf4SX6gAAcZYz2eEjQT1m/wX/BscGYH2mIniGtIqy9X/wPzA8K8MqPCeF7y5n985W/CxmuJ+tY+vX/AJ5yl97bAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA4kpe0yusuyT+5pmFqe1oQqd0n96AAADgLxFxXDtRYeveeGb1j1j5x/WOz8nqZdmWEzHCwxWEqKdOWzX9aNdU9U9zjjw6wHEdN1oWp4hLSdtH5TXVee681oZFWzHgfiam8dBwXwy7Sg93F9UtJK3VWdtUQnGcVKLunsGaZZhszwdTB4mPNCas1+q7NPVPo9TirhvFcP5nPBV9UtYvpKL2a/Jro7oADh7hXNc+r+ywNJyS3k9Ix9X+mrfRM444MnwxiKFCVTnc4czdrLmu00vJK2r37IAMsrewxtGr/LKL+5pnivQ9rwlirbx5H9043/C4AAHFGM+sZjNJ6Q91fLf8bnD2C+q5dTi1q/efq/+BZAcPf7Dwbh5/wAtBS/8k5gAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADH0va4SrT7xa/AyCr7XLKL8rfdp+gAAAHCPGmY8N4nnw75qb+KD2fn5S7NfO60OGuKst4hwixGDnqvii/ii+zX5NaPozP+HcuzzCPDY6nzLo9nF94vo/wfVNHDOWYvK8BHA16ntFT0hLq4dFJd47aaNJPe6Qcd8G0OJcudLRVoXcJdn1T/uy2fbR9DGYOvg8RPD4iLjODaae6aAOLKk45nJJ9F+R9GWTlwVK7v++n/wCkwPHXL+fAYTGJfBJxf/byuv8A0n8QAOJ1/aHB+InvzUXL7o836AABjcTHDYadZ/ZTZluGljsdCnLXmd36bv8AAsrAZ3/snBdaP8uHcf8AynYAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAOGPdwk6P8k5R/X9QAAADKc4x2U4qOKwVRwmuq6+TWzT6p6HBHidgc+UcNirUsT2+zL/C31/uvXs3rYA8S/D6Oe0HjcHG2JgvTnS6P+8vsv5PSzVSnOnNwmrNaNPRprdNAcWr/hTl6L8j6MUlLgqaXSvP/wBJgeI+Vf2jwziqcVeUY869Ye87eqTXzAA4VazLhHDQf2qKh90eV/kSi4uz3QABxlivZ4GNJPWb/Ba/nY4KwnNVqYh9FZer3/L8QEm3ZHiK1Q4SxiXSCX3tIAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAMo/d5jjKPmpf8AgSu/0AAAABNp3RwV4vYjAqOEzdupS2U95x9f5l/5MvPYwGYYXMMPHEYWopwls07r/h+63XUA8SvDWObxlmOXRtiF8UdlNL/2789mVKc6c3Cas1o09Gmt00HF/wDyM36I+i9KL4NrJPVV5/8ApFMnCM4uMldM4iymWU5tiMDL/l3Jpea+y/mrMAPCLFe34VowvrCU4/8Akzl+UjifCfVM6xeH6RqTS9OZ2/AAA4yxPtMdGknpBfi9fyscMYX2GW0+8ve+/b8LAYKHPiace8kvxPFafLwji/PkX/lSAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAL91n77Tp/in/kgAAAADh3irNMgxHtsDUsnvF6xl6r9VZro0cD+IOC4mpuny+zrxV3G97rvF9V36q/XcA8QvDShnkZY3ApQxK36Kfk+0u0vk9LNYvCV8JXlQrwcZxdmmrNM4wX/AApf9ur9T6LGbqnHGZfJ/G+desVFO3qn+AeN+Rexx1HM4LSouWX+KOz+cdP+3QA8CsZz5bi8Nf4ZqX/gUbf+2Hivgvq3FWIa2nyyXzik/wAUwADMpvGZnU5ftSsvvsilTjTpxhHZK33AZTFSx9BP+eP5o8XJNcJ4hd3D/wBLiAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAGb/uswwdbzcf8AwJWQAAAAAZXmeKyzGU8XhZ8tSDun+j7prRrqjg7i3CcSZcsRS0qLScesX/k+j6+qYBxrwDl/EtHml7leK92aX4SXVfiunVPxQ4fx2R599TxsbTUU+6avJJp9U7fo7M8Gs6/sjMKWLbtGNT3v8Mkoy/Bv5iaa0ON+H1nmR18IleduaH+KOq+/4fRslFxbi1ZoA8Dcb7POMRhn9unf5xkv0kzx0wPJmeFxSXxwcf8AwF3/APbgAMZWVDD1Kv8AKm/uRw5Q9vmdJPo7/cr/AJ2AMo/5GOH/AMcf/SkeLv8AySdf1h/6WgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA4oi1gVVW8JRl+Nv1IyUoprZgAAAABw1xJjcgx8cZhXqtGntJdU/0fR6nDfEeBz/ARxmElo9GnvF9U/wCrNaoA+lNC3FuFl3w8V91Sr/meHuQZjjcoxWNw1Pnp05JStq43V7tb2036dbHhtnqzfh2hOTvOn7kvWOz+cbP1YeLPDH9k528RSjalXvJeUvtr73zf9vW6AHhjjvqnFWEk3pJuL/7ei0vxseN+A9tklHEpa052+Uk0/wAVEADiet7LK6lt3Zfe1+lzgqhzYqpU/ljb73/wADLpcmMoy7Sj+aPFmHNwjin2cP8A05FfqAAAAAAAGM4ioYPH/VqqtGy17N912tYhKM4KcXdPZ9H6AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABm1D2+Bq0+8X96V0ZLX9tl1Gf8AdS+7T9AAAAAAOFeKsdw7jlicM7p6Si9pLs/Ps+j8rp8N8S4DP8DHFYOV+8XvF9mvyez3QH0qqVs/wM+9Jr7pv/M+ihUvg80p9pUn96n/AJGX5BgcvxVbEYSHJ7W3NFfC2vtW6PXW1k+qvqHHPDEOIcmqYVfxF70H2ktvk9n5O/Qq0p0qkqdRWlFtNPdNaNAZdjJYPGUcTHenKMl/260/0OPcFHM+FsXCGvuc6/7dtNW9bAAca1eXCU6feV/uX/BOCqNsLUqd5W+5f8EAjJxkpLdHiRBV+EsZb+VP7pRf6AAAAAAABxBW9tmdaXZ2+7T9DwD4ZwOK8PKUcbSVSNadSdpK9teTTt8HSx4lcPZfkWd/VcDdQcVKzd7Nt6J72slvd+YAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA0mrHDPuYeph3vTnJfL+rgAAAAAHDvEePyHGxxWDnZ9U9pLs12/Fbqxwhxpl/EmF9pQfLUj8UHvHz849n99noH0r6NsfllTvCovucf8z6J1W1bNqfdUX9zq/5gHjHwa6Ff+2sLH3J6VEuktlL0ls/71usgDgbGQzThfCTnreHI/8At33Hf1sZngp4HG1sLPenKUf/AAFtfoAHG9W9ajT7Jv73b9Dhal7PK6fd3f4v9AAzv/bOC60v5sO5f+U7gAAAAAAEpKMW30K1R1akpvdtv7zwmwP1LgjLKNrfuov5y95/meLGI9txZiV0ioL/AMki3+LYAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAYH9xnWJpdJpSXy0f4sAAAAAAMpxWY4LEwxWBcozi9HFP7vNPqno+pwNxvWzukqGNw8qVdLV8slCXmm1o/7rfo30+ljRvTymr2dZff7J/oz6KlZLOswpd6cX90rfqAYzCUMZh54evFShNNNPqmcc8H1+G8ydF3dKV3CXddn/AHls/k9mB4H5l7bKK+Db1pzuvSa/zi/vPFvK/qXE9WaVo1VGa+as/wDyaLfzADi+pz5k4/ypL9f1MqpeywFGHaK/IAOHV9e4Nw8N+ago/wDknKAAAAAAAZvW9lgK0/7r/FWKFJ1asaa6tL7zhvDLDZLhKCVuWnBfdFI46r+34lx0u1SS/wDAXy/oAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAGZfuM2wtfpK8H+n4sAAAAAPCWhSr8T0oVYqS5Z6NXXw9mLKcvTTVCF/8K/yIYejB3jBJ+iA+lZQ5sly+r2qSX3xv/7afRbr8nF2Ip/zUJfeqlP9LgBneRZfnOEeFx1NTg9fNPumtU/+G2OM/CjMcn5sRgb1qHkvfiv7yW6/vL5pB4MZp9V4heGk9K0Gv+3o+8vwUl8zxzyn2mCw2YRWsJOD9JK6+ScX94AZ9J1s1q23vb7kkQioxUV0ADwrxHtuE8J3jzL7pyt+FjMsP9Wxtah/JKS+5tAAAAAAAcWVeTK5r+Zpfjf9Dh3DvEZrQprfm/LX9ClTVOnGC2St9xnVf2+ZYir/ADTm/vk2AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHE9N/UVWjvTkpL5O36lOcakFOOzVwAAAADgPiHC5DnUMdiYylBKStFJvVW6tL8R+OPD1tKNb/wGH/ywn455Qo+7hqjfnyr9WVPHfCJ+5gpP1ml+jKnjzUduTAJetS//ALYjxm8RcTxPkdLDVaEYKFRSum2/hmra+v4HhVxRi+HOIVjMLGMpOEo2ldqzs+jT6dzg3xIyziGKoyfssR/I3v8A4H19N12tqABxl4U5bnXNicHajXfZe5J/3ktm/wCZerTPqOacJZ3QqYyk4SpyUl2kk9eV7NNafPWzOLcqhnnD+Iw1P3ueF4+bXvR+9pfJjQBH9/nC/vVPzkAB4KYn2vDk6d9YVJL5NRf6s47w31fiXHQ/6eSf/gT5v1AAAAAAA43q2w1Kn3bf3L/gnhlhPrPEuGh/ej+Mor9StUVKlKb6Jv7iTcnd7gAAAAAAABTo1Krapxbsm3ZXslu/RdQAAAAAAAAAAAAAAAAAAAAAAAAMbh1iMNUpfzJr8DhzEe2y2nfePuv5aflYAAAALpgAHFsObK5Ps1+dv1OGZ8ua0vmvwZGUoyUouzRwb4w4nBqOFzi9Snspr41/i/mXn8X+Iy/McJmOHjicJUU4S2a/rR909V1ADNsmwGbYZ4bG0lOD6Pp5p7p+aaZlmAhgMHSwkJOUaaUU3vZaK/orI8R8h/sfiKvSirQm+ePpLWy9HdfICTSVzIIurmtK/e/3JsADwIxV6ONw99nCX3qSf5I8XsJ7HiqtP+eMJf8Akqj/AO2gAAAAAAcb1b4ilT7Jv73/AMA8E8L7XiXDy7VKf4Su/wAjiCr7HKMVU/lpzf3RYAAAAAAAAHgfkqrYnE5hUjeMY8ivs3LWX3JJekjxWy3KeH+JYYHCyt7aPOo9IttrlT7O10um3YAAAAAAAAAAAAAAAAAAAAAAAADJf3GOxWF6KXMvSX9IAAAA4mz+tgpLD0VaTV79ltp56HBmKlVoVYSd2pX181/wAADiSHPldZeS/BpmSz5MxoP+8l97sBw5xVmmQYj22BqWT3i9Yy9V+qs10aOD/ErKuIFGjN+yxH8jejf919fTR+TWoAHHXAeF4noQbnyVoX5ZWuteklu1p0d1v3T4i4VzTIMR7HHU7X2ktYy9H+js11SDGy5MNUl2i/yOFIc2aQfZP8rfqAB4H4v2ed16Dfx02/nGUf0bPHXCcmZ4XE/zQcf/AAGV/wD24AAAAAADi6pz5nKP8qS/C/6ngHhubPcK+9R/+SwucbVPZ8N45/8ATqa++LX6gAAAAAAAAeH+Q/2Nw9h8PJWnJc8v8UtbP0Vo/I8WeJv7f4xxuNpyvBS5IP8Auw91Netub5mA9s8LT9s7zsr+tgAAAAAAAAAAAAAAAAAAAAAAAAxv+zZ1QrdKicH67r9AAAADjehrRrLzX6r9Tg2vyY6VP+aL+9a/lcAAzeHPl9aP92X5GEn7PEU59mn9zAE2mmnqcG+L+MwHLhc2vVpbKf24+v8AMvX3vN7GWZpgszw0cTg6inB7Nfk+qfdOzXUAMwy7CZhh5YfFU1OEt01df8B9nuuhxj4OYjC82Jya9SG7pv4l/hf2l5fF/iZnalSwOIjJWajJWe6dmjg2N8xb7Rf5oADwtxf1bivC3ekuaP3xdvxseOmC58qw2J/km4/+BRb/APbUAAAAAABn9T2mZ1n52+7T9D6P+HtnWAf/AF8f/kk1/keI8+ThXGv+7b72kAAAAYLB1sbiaeGoK85tRirpXbdkrvTfucV8E5vwxlTzPMYpU01Gyak7u/RaW079jCcSZdiakaUJPmeiTTAAADw34ced5/ShNXp0/fn6R2X/AG87K3a76HizxUuGuEsXjYytUceSH+Oeia9FeXyMlwbxmPp03qr3fotX9+3zAAAAAAAAAAAAAAAAAAAAAAAAA4noylgfaw+Km1JfL+r/ACKFaNalGpHaST+8AAADizD+1yyUlvFp/p+pk+I+r4+lU7Nfc9H+DAAK8PaUpQ7poKM/aUoz7pMAMg4kzLIsSsRganK+q3jJdpLr+a6NHBvihlme8uHxFqOI/lb92T/uv/2169r7gAeOPC2VYnhXHZlOny16cLqS0b1StLuteuq6NI4Jpyli6s0tFH7rtf5AAZBjVgs1w2Jb0hOEvkpJv8DxVwX1rhXE2WsOWS+Ulf8A8luAAAAAABjqntMVUn3k3+J4DUOXOsGu1OT++P8AwTxUny8I4x/4F99SCAAAAOD4uXEWAS/5+0//AEtH0kZqPAs/OpBfmZBG+Z0V5gAAB4UcLvJ8lVetG1WvaT7qP2V9zu/N2ex9JzjFY7OKOR0JXhh1zT/xyW3/AG7G3zkzgvA2hPFSW/ur83+n3AAAAAAAAAAAAAAAAAAAAAAAAAYijGtSlSltJNfecM1pSwXsZ/FTbi/l/VvkAAABjcOsRhqlL+ZNfehpp2MuxP1nCU6vVpff1/EAAxlP2eIqQ7Nr7mZPU9pl9GX91fgrAAHBvi1j8q5cNmN61FaX+3FeTfxLyevZpaGT53l+cYZYnA1VOD7bp9mt0/JoA8bpOPAGZNfyx/8AS4n0XMJQxOa5lTrwUoulFNNXTTls09HscY+DSfNiskfm6bf/AKRJ/lL7+hisLXwtaVGvBxnHRpppp+aYAYe2e8LRT1dejb5yhZ/cxpp2YAAAAABVmoQcuyHqeCVHk4ipx/lpy/JI8W5W4SxC7uH/AKXEAAAA4Ghz8S4Ff9PIv7nc+k3U5eCYx71oflNnDUebNaK9f/SWAcO5asyzbDYNq6nOKfo3r+FzxR4JlwfTePg3PCykkn9qLeya0v1s121Sdr4bFUcTTVSjJSiwDw44Vef5zCNSN6NO0p9mukf+3n+Fzi/iTDcOZJiMzr7U4tpd5bRXzdvlqZlj8Vm+Y1MVWfNVqybfm5MwGEjhMLChH7K/Hq/mwAAAAAAAAAAAAAAAAAAAAAAAADD/AOyZ5UpfZqx5l6rf9WAAAAZ5hvq+Y1YdL3Xo9f1ODsT7XAOk3rB/g9fzuAAZ9T9nmVZebf36/qcL1OfKqfldfi/0AADhnG53hMaqmT87qdopyuuzik016o4WzLN8dglPNML7CovNNS80rtx9JfewPHf/AJN7mXpD/wBOwPoo/wDIyzL/AAQ/9KkHE/BmU8Q0eXF07TW01pJfPqvJ3Xz1OL/DrNuHpOpJe0odJxW3+Jbx/FdncA8I8w+t8LUYN3dOUoP7+ZfhJHGOXf2fn+Mw1rJTlb0buvwaAAAAAAM0nyYGtLtGX5Mw0PaVoQ7tL8TwWinxK32py/OJ4vytwpWXeUP/AEpAAAAHhrS9rxXgo/3m/ui3+h9KSvy8MYWn/NV/KL/zOFI3zSD7J/kwDwayz61xGq7WlGMpfN+6vwbfyPpVZyqeVYDLIvWc5TfpCPKvvc39xgcwxGCqe0oSs/wfqjLMVPF4OnWmrOS2ClSnVnGnTV5N2SW7b2S9TgPhWHD2Tww8l+9l703/AHn09IrRfN9T6SvHv13HQ4fws/cpazt1n0X/AG7+dzhDLvb4p4iS92G3q/8ALf7gAAAAAAAAAAAAAAAAAAAAAAAAA4li6UaONhvSkr+j0f6L5kZRnFSi7pgAAAca4Xlr066+0rfNf8P+BwdivZ450m9Jr8Vr+VwADi2lyZnJ/wAyT/C36HBdXmwM4dpfmkAGHw1bE1FSoQc5PZJNt+iWpkXg9n+YWnibUIP+bWX/AICvyk4n+p/APDCvmmIVWquknd3/AOvcNf8AwK68zG+M2XYGl9XyXBJRWzdoR/8AAY7/AHozbxQ4nzG6eI9nHtTXL+Pxf+THCPipmuTSVLFt16PZv3l5xk/yena25kHEuWZ7hvb4GqpLqtpR8pLdfk+jZ43wc+AMyS/lj+E4s+ipJLOswj/07j+Ev+CBKMZRcZK6Zxj4PYPHc2Jyi1Kpu4fYfp/K/wDyXyW5mmUY7K8S8NjKThNdH+aezXmroPAnMdMZgW/5Zr8Yy/8AbTxry36vn8MSlpVgv/Ao+6/w5QAAAAADiGfJllZ+VvvaRlEOfMKC/vL8zwV/5KWX/XuX5xPGD/klav8Aih/6UAAAAeEND2nFdGX8sZv/AMla/U+ldiLZfltBdZzf3KK/U4OjzZi32i/0QB4IZT7DKK2OktasrL/DDT/0py+4+kfnizHjWeGi7xw8Iw+b9+X/AKVb5Fr6IwdD2GGp0v5Ul9yDwc4M9vW/trFR92GlNPrLZy9I7L+9frE8SuNaHCeQVcbJ/vGmoLvJ9fRb+tl1MZisRmGLnXqvmnN3fdtmT5esDg4Ueu79Xv8A5fIAAAAAAAAAAAAAAAAAAAAAAAAAMbho4nDzoy+0mjhrEyq4FU5/FTbi/lt+GnyAAAA4rwvt8tlJbwaf6P8ABmBxLw2Jp1l9lp/5/gRkpRTT0YAHG9K2IpVO6a+5/wDBOCKtp1qfkn911+qAyfh/M84rexwFF1H1stF6t6L5tGVeD2DwNH63xBilGK3imor0c3+SS8mYnxK4W4dpPD5BhVN7cyXLF+snecvmvRme+JHEWb3jUruEH9mHur5te815NtD1AMtzPGZbiI4nB1HCa2a/J913T0fU4j8U6Wd8F5hl2ZQ5a0qb5ZJe7JrXVbxenmvTY+izX5eK8VS/moSf3VKf+YAZ5w9lud4Z4fHUlOPTun3i90/6d0cXeEWZZXzYjLr1qK6W9+K80vi9Y6/3UjwpzJ4DiijGTsqicH81df8AkyR42ZT9ZyOnjIrWjPX/AAz0f/k3KAAAAAAcVy5cqqLu1+af6HDkOfNKK83+CbPBhtcTf9w5foeLqT4Tr+sP/SkAAEpRjFyk7JCaaugDwPw/Pn1ar/LSf3uUf0ufSsxV8xy/D32hKX3yt+hwVC+LqS7R/Nr/ACAjCU5KMVdsyXBUeH8gpUajtGjTvJ+aXNN/fdnEWb1M3zfE5hUfvVZyl97b/AyfD/WMfRp+a+5av8EHBfClfiPNIYWGlNazl2j/AJvZeeuyZQoYXLsHGnTShSpx9Eopfot38zxq8Q58VZ5KFGX+z0rxivzfq+v3dEcI5X7eu8VNe7Dbzl/wPzsAAAAAAAAAAAAAAAAAAAAAAAAAAUv9hzyUPs1ldf4lv+v3oAAACtSjWpSpy2aa+8r0ZUasqct4tr7jhrF/Wcthd6x91/Lb8LAAcbUr4alU7St96/4BwfV5My5f5otfr+hk+RZjnGIVDA0nOXlsvNvZLzbRlnhZk2SYdY7iXEqy+ym1G/a/xTflFL5oznxgw2Do/U+HsMoQWik1ZesYL85fOJm2d5jm1b22OrSqS83ovRbJeSSQAAZnDnwVaPeMvyZ4Q8V1uGuJoYynBTvCUWnpdOzdn0ei1s/Q4Y4zyniGjzYSp763g9JL5dV5q6+YAHEXh/lma4iGOor2WJg1JTit2ndcy2lr10l520M+yuGa5XXwU/8Al5Fr0dtH8nZlajOjUlTmrSi2muzWjAAAAAA4xlbLku8l+Tf6HCcObNIPsn+VjwdklxTTXeE/yPFiPNwjivLk/wDTkQAAzHDfWcJUoreSa+fQ4exLxGXU294+6/lp+VgDwHw15Y6u+nJFf+TN/kj6TmM9txdTpX/h00vv979TgeF5V5f4V+YHhfkf9q8SUeZXhS/eS/7d2++Vvlc8fOJFk3BWIhCVqle1OPf3vif/AICmvmHBmG58XOs9or8X/wABMwOBxGOxMMNh4805uyS6t/18tzgrhKhw5lkcNHWpLWcu8v8AJbL792z6QviastwjyHAT/ezXvtdF0j8938ltzIwuGq4uvGjT1lJ/0zA4Ong8PGhT2X4vq/mAAAAAAAAAAAAAAAAAAAAAAAAAAcTYecsLHE0/jpNSXp1/z+RhcRDE0IVobSVwAAAOLcH7HMHNbTV/ns/8/mcF4zkxE8O3pJXXqv8AgfkABxVR9plc3/LZ/jb8mcCPBf6y4GOOdqMqkYyd7WjJ8rbfRK+r7GceKeWZPh/7P4ZoRUV9tq0b90t5P+9L7mjNM3x2aV3iMbVdSb6t7eSWyXkkkAAAVYKcHHurHDk/Z5rS9WvvTRhcVXwtaNahNxnHVNOzT8mjg7xlT5cLna8lUS/9Livzivl1MNiaOJoxrUJqUJK6ad015NAAcf8AhNWxuJq5llUrzm3KVN6XfVxe13vZ9bu/QxWFr4WtKjXg4zjo01Zr1TAAAAADjaX+x04/3v0ZwbG+Yt9ov80eGmZ4XLuJKGIxVRQppTu3svcdvvehx3xvw7mnDuKwmFxUZVJJWVpK9pRel0l0AAAyVewxuLwvRSUl/wBvf0gDwQwns8irVnvOo/uUY/q2ePGN+t8c4uV/htH/AMB939DgmFsLVn3lb7l/wQPBjIPqWTSx9Re/Xen+CN0vvd35qx9J3ihY7PqOU0pXjh43f+Oev4RsvJ3DhDCeyy9TtrNt/LZfl+J4WcBf2Ph1mONj/tE1on9iL6eUn17LTueJvH+F4PyWeKm060rqnHu+9uy/Oy81m+a4rNMbUxmKk5Tm2235nCuT/VqP1mqvfktPJf8AB/y8wAAAAAAAAAAAAAAAAAAAAAAAAAAqQjUg4SV09Dh2csPOtl9R603df4X/AFf5gAAAcY4P22BVZbwf4PR/jYwGKeFxVOsvsv8ADr96IyjOKlF3TAOE+Bc14jq/7PHlpLeb+FeS/mfkvm1ucZYbhHhfh3G5RQXtsXVpyg5aOUW07NvaCT15Vq7K/chNwmpLdalOcakFOOzVwAAAKP8As+cJfy1LfdIDhfjTNuHqvNhKl4PeD1i/l0fmrPvdaHCPiLlHEMVSjL2dfrCT3/wvaS+591YADijgvKeIqPLi4WmlpNaSXz6ryd16PU4v8O824dk6k17Sh0nFaf8Aby3i/XTs2AAAAAcbv9zRXm/yRwVG+MqP+7+qAAACv+4z2lPpUg4/Na/5AHhdhPq3CmFT3kpSfzk2vwscf4769xJi8Re/NJv79f1OD6fLlt+8m/yX6BkGT1s4zOjgaW9RpX7Lq/krv5GNxODyHJ51WuWlQhovKK0X4JepxFnNbOc2xGYVneVWTl97KVKVWpGnHduy+Z4Q+Gip06OaY+HuxS9nFre202u3WPd+9ta+f57gcjy6rmGNny06au/Psl5vZHiNx5jeMM5nja7tBaQj0jFbfPv537s4ZyX65W9tVX7uP4vt6d/uAAAAAAAAAAAAAAAAAAAAAAAAAAy/iDCYyq6SfLNNqz6+j/TcAz1PBYyjmMdl7svR/wBP52E1JJp6AAABiaEcRRnSltJNfeVqMqNSVOW8W0/kcLY36zl8Yt+9D3fl0/DT5BGLk0krs4V8L6NHDf2rxJL2VGOvI3Zv/G91fpFe83po9Hxb4q1K1L+z8hj7GhFW5krSa/upfAv/ACb01RJuTu9TGUfYYipS/lbX3MyGv7bLaMuyt92n6AAAAZ/B0c1q23vf70mQmpwUl1AjJxacXZo4Q8YcdgOXD5qnWp7c3216/wAy9bPzexlGd5fm+GWIwNVTh5bp9mt0/JpABOEKkXCSuno0+pxl4O4bF82Kya1Oe7pv4X/hf2X5fD/hRmGXYrAYiWHxVNwnHdNWf/Ddns+gAAAAccvSgv8AF/7acERvXqvyX5hQ4awVbgqrm+qrQq8u+ji1HS3dN3uAABxD+7lhsR/JNfc9/wAgCqv7F4Ta2dGhb5xh+rM6qe0zCtL+8/wdjhunyZXRXk397bDwT4ZdOlUzitHWXuQ9E/el82uVej7n0leMFgMlhk9GXv1tZf4Vsvm0/uXcPo+8HUeIOK1WxVNTo4aLm09nJ6QT+d5W2fLrpoYnEUMLQlWrSUYQTbb0SS3b7JI8Z/FerxfmH1XByawdJ+6tud/zv/21dF5tmU5XVzDEKlDRLd9l/WxhsNSw1GNKkrRWwAAAAAAAAAAAAAAAAAAAAAAAAAYut7HDzqfypv7kXd7mU8WV8Paniffj3+0v8/nr5mExuHxdP2lCSkvy9V0DHYSGLw06E9pK3+T+TOGsVOeGlhqvx0nyv06f5fIADQADi/BexxqqpaTV/mtH+j+ZwljvYY72Tek1b5rb9V8zLcsxeZYqGFwlNzqS2S/rRLq3oupgMjyHw+wkcdmklVxjXuxXR/3U9vOb+S6PivjLM+I8R7TFStBfDBfDH/N929fRaAcUUPZZnUts7P71r+Nzgyvz4KdPrGX4Nf53AAAA4wpcmY8380U/zX6GTVfa5fRl/dX4KwBGLk1GKu2eHfhzxBSxMMwq1pYWPZfHJdnF3SX+JN/3epXr0qFN1KslGK3bdkvVvREZRlFOLumAHEfCuV5/hvY42ndraS0lH0f6O6fVM4x8NM0yDmrwXtaH8yWsf8S6euq81sAAAAccP95RXlL9DgdXlXf+H9Q/tfG/2esu9o/Y83Py6W5rJXfV6LZuy6AAAcSUfa5ZVturP7mn+RhaqrUIVP5kn96DhrA/X85wuGtpOcU/S6v+FzxYxywvCmIV7ObjFfOSb/8AJUytU9pUlPu2zLKfs8FRh2ivyRw7kdfO8zo4GjvN6vslq38l/l1KNLB5PlqhH3KNGP3Rit38ld92eKXFdXiTiSvipfCnZLslpb5JJPzV+ofR64eo8P8ABbzTFNQliG6jb0tTjpC/la8l5SPGrxpqcSVJZRlMnHBxfvPZ1Gvygui67volg8HVxdaNGkrt/wBXfkZXllHL8OqVPV9X3f8AWwAAAAAAAAAAAAAAAAAAAAAAAAAHElb2WV1X3VvvaX5AYXF18LUVSjJxf9ff8zKuL6VW1PFrlfdbfPt+XoQnGcVKLumZsnl2Y08fH4Je7P8Az+78vMTUkmgAx3Eb/teFSD/dwdvVP4n/AJeiIyUkmndMAOKcD9Zy+Ukveh7y/X8PyKdSVOanF2ad16oyTjjh3hzhjD1smiqmLxEFKTevK3upP+7K6UFva7et3mWZYvMsTLFYuo51Jbt/1ol0S0XQA43w9qtKsuqa+7X9TgvEcmLnS/mV/mn/AJNgAAAcb0bTo1PJr7rf5nCVb2mWRj/K2v1/UDhjg7NeIq/Jg4e4t5vSMfn1fkrv5anCPhzlHD0VUjH2lf8AnktV/hW0V977uxxd4pZTknNQoP21dacsXpF/3pforvo7HEvGOb8QVebGVfdW0FpFei6vzd35nCPiNm3DzjST9pQ/kk9v8L+z+K8upwxxllPENHmwdT31vB6SXy6rzV18wAaUlZnGXhBg8w5sTlNqVXdx+xL0/lfp7vktzM8qxmWYmWGxlNwmuj/NdGuzWjAAADjd/v6K8n+ZwOtK7/w/+3BwxwHnPEKVTCQSpXs5tpJNWb0+J6NbI4jyj+yM1r4Hm5vZyavtddHbzQAB4ScO4HOcRjKWNpqcORLzTct0+j00a/K5xvwvXybG4jLauqs+V/zRadn+j7NNHDtX2uV0X2VvubX6B4QZf9a4op1GtKUZS/DlX4yR9IfNFQyajhk9+ef/AIDGy+/mZTg5zUVu9CMVFJLoeEPCH9m5d/aWIj+9rLTvGG6/8C3fly+Z488W1cuyOpg8Nu0nN9Em7Qi/OUtbb8sW7WuSk5Ntu7Zk2Bp47MKOGqS5YyklJ72jf3nbrZXdjxQ8W6+e045Pla9lgqSUYxXVRVle29kvRdL6t4fD1cRVjSpK8nsjJMlp5dRtvN7v9F5fnuAAAAAAAAAAAAAAAAAAAAAAAAAAcaVuXBwpr7UvyX/BQAZdnGLwEr0Zadns/l/lYpZ7gM3w0sLX9yUlbXa/Rp+vR2+Zw3jZypSwdb+JS09V0+7b0t3ADiXLvqeOlyr3Z6r9V8n+FjhLMvrGE9hJ+9D8un3bfcAHCXB2N4mxn1egrU18cmtIr9W+i6+Su1xrw9Hh/iDF5ZGanGlNqMt7x3V7aXtZSXSV10OC8feM8JJ7ar9f0+9gBxhh/aZdzr7LT+/T9UZFiPq+Y0Z30vb79P1AAAA4yo8+AjNfZkvxTX+RwRWvTrUuzT+/T9EHAvhLXzBRxmbJwpbqG0pev8sX/wCBPy0ZmGaZLwvl0XVcaVKOkYpavyjFat9/vb6nGHitmec82Hwd6NB6WT9+S/vNbL+6tOjbAMLiq+FrRrUJuM46pp2afk0cHeMqly4XO9HsqiX/AKXFfnH5pasw+Io4ilGrRmpRkrpp3TXdNABn/DWWZ7hvYY6kpLo9pR84vdfk+qZxn4YZlkPNiKH73Dr7SXvRX95f+3LTvbYAAA42/wB5pf4f1OB1+7resf1DwS/5Jyp/19l/6TA8YcF9X4pqT/5+RjL8OX/20AA8DMxwlHF4rCTlarUUXHzUOa6Xnre3ZN9DxuyJV8uo5nBe9SfLL/DLa/pLRf4mcNe5Qq0f5JyQeBWW2pYvHNbuMF8vel+cT6R2be0x1ShF/wAOEY/OT5n/AOStfcZNS9rmFGP95fg7nhpwa8/zNVK8f9npWcv7z6R+fXy7XQkkrI+kzxhTxebU8hwrXJR9+pbrVkrK/dwhZeXM100KdWdO/I7XVvkUKFSvUjSpK8nsjI8ipZdSu9aj3f6Ly/MAAAAAAAAAAAAAAAAAAAAAAAAAAOJsmr5hThKi9YX0fW9uvy/4JWo1KM3CpG0l0YAGAzCrg8TGvB3a/Fdv62Mt4iwWOtFPln2f6Pr+fkAcS5Z9dwTcV78NV+q+a/GxlWYTwOKjWjst13XX+u5Sqwq04zg7pq6A4O4OxvEuN9jR92nH459Ir9W+i+b0uzjDjPBZDg/9X+HfdUdJ1Fvfqk+sn9qXTaNraca4DmjDFx3Wj/R/n96MtxksHioV10evp1/AhONSClF3T1QBmWH+sYOpS7p/f0/ETad0YLELEYanVX2kn+AAAAZ/Q9tllaPZX+7X9Dgh1J5mqFNOUpqyS1bd1ZJdWeHvhZSy5Qx+axUq+8YbqHm+8vwj0u9Vxz4l4Hh9Sw2HtVxP8vSPnJr/ANJWr6tJpmcZ3j84xTxONquc332S7JbJeSAAOFeOM24dqr6tPmpveEtYv07PzXzutDhHjvK+JKdsO+WqleUHuvNPaSv1XldK4ANXOM/CPA5pzYnLLUa27X2JfJfC/NK3da3M3ybHZTiXhsbScJro+vmns15rQFa+pxDkdfJswqYOtrbVP+aL1jJeq+53XQAONv8Ae6f+H9WcEJexreq/Jh4IV4TyCtSW8ajv84xt+TPHbAWq4PGJbqUH8rNfnIlFxbTWqAAyjM62WY+jjKL96nJNedt16NaPyZnuEpZ9w9WpUtVWp3j6tc0H99mZV+7zLGUvOL+9ah4X5X9Q4XwyatKpeb/7ed1/5Lyni9nCzHHV66d1Uqtr/CrpfhY8OsixWdcQ0MHhY3m7+i0s2+yV7s4a4ewuQ5bTwOH2ju+spPeT9fwVl0OMOJcLw3keJzXEfDSi2l/NLaMV/ik0vnczTMcRmWNrY3Ey5qlWUpSfdyd3+LDDYariaqpUo3kzJMipZdTu9aj3f6Ly/MAAAAAAAAAAAAAAAAAAAAAAAAAAAzHKcLj4ctaOvR9V6P8ATYzbhrFYG84+/Dut16r+kAAZbxPjcHaMnzx7Pf5Pf80Zbn+CxyUYStLs9H8u/wAgOJcr+pYtygvcnqvLuv66M4Qzf/mCqPzj+q/VfMOE+FMZxHmEcLh1aK1lLpFd/V9F1fZXa4y4swXD+B/1c4fduXSpNb36pNbyf2n0+FWtoY7CQxeGnQltJW/yfyZWpSpVJU5qzTafqjhHMPb4N0JP3ofk9vu2+4AM4w31fH1aXRN29HqvwZwjifa5cqbesG18nqvzAAABYaWK/cQi5OeiS1bb0sl3Z4aY2eR8d5fOsuVwrKEr9OZunK/pzM8RPFVYdzy7Jp3ntKoundQ8+8tl011U5SnJyk7tgABwnwhj+JMZ7DDK0V8U3tFfq30XXyV2uGeFcu4ewiw+Dhq/ik/ik+7f5LZdOpmucYDKsO8RjaqpwXV/klu35JNmExmHxlCNfDzU4S2ad0/mgAzzh/Lc6wzw+OpKcendPvF7p/09DjLwozHJubEYK9aguy9+K/vJbr+8vVpBxbwx/rHwhg8xw8b4inSi/OUeVc0fNp3a87pbgBxt/vlP/D+rOCP4Nb1X5MPArMOTG4vBt/FGMl/267P/ANKX3HHXDv8AbmAo0Ur8tWnL5c3LL7oyb+RxfhPquf42j0VSdvRybX4NAAHhTmf17hegm7yptwfyd1/5K0cU5ess48x+FSsndr05vd/8lMrwFTMMdRwlP4qklFfN2OKMbTyXhvEVaeip0+WPk7csfxaON6169Kl2Tf3u36H0U8tVTNswx1vgpxh/4HJv/wBxmMxuHwdCWIxE1CEVdtuyR49+K0OJa0MowCaw9J8zb0c5WstOiSbsnq73dmrIwWBr4ysqVFXb/DzfkZPktDLqVo6ze7/y7IAAAAAAAAAAAAAAAAAAAAAAAAACvmGEofxasV6tX+4rcVZXT2qc3on/AMBFXjXCL+HTk/Wy/VkuNqknanQ/G/5JH+tGbT/h4df+Ayf6n9u8QS1WH/8AJJH9t8Qf9A7/APAJGOoY3EVOeWGcW+0ZK/y2J0akF70WvVAAGXcUY3CWjN88ez3+T3++5l3EGBx1oxlyy7PR/LozOMthmGElRe+6fZ/1oxqrh61vhlF/NNHAtDE8V1qeFwkb1X8XaK6yfaP66K7OKM+wfBuWf6v5LL9+1+8qdU3v/wBvPp/JG1tXdPUDjHL/AGWJjiIrSe/qv81+pkeYfUcbCo37r0fo/wDLc0aAONMLyYqFZbSVvmv+A0cGYr2eLnRb0mvxX/AuAAAHhZ4erLqUc1x8P30l7kX9hPq/7zX3LTdu3irh6GF42zJYWacXVlJNbXl7zt6SbXyMNWVejCqvtJP70AAHCXCmM4jzBYXD6RWspdIrv6vZLq/K7WRZFgclwMMHg4WhH72+rb6t/wDAVkkjjjxEwPDlN0YWqYhrSHRecn0Xlu/JamecQZjneKeJx1Rzl07JdorZL/h3dnDHGOa8PV+fBz9x7wesZfLo/NWfy0OEfEPKeIoqnB+zr9YSev8A26/tL7n3SAA4x8KstzrmxGEtRrvW6Xuyf95LZv8AmWvVpnBOX4zLshw+DxkbVKacXrdaSdmmujjZ/nY8VeDv7FzP63h42oVm2u0ZbuPknvHyulsAcbf75T/w/qzgj+DW9V+TDwzzJYDijCyb0m3B/wDbysv/ACaweLOE9hxXiGtpqMv/ACVJ/imAAeBOOvRxmEb2cZL5pp/lE8bsEsL4h0qtv41K/wBya/8AbTwayb67n7xUl7tCLf8A29L3Y/hzP5Hjhm6o5XQwEXrVlzP/AAwX6ya+44w9pHNp06iacVFWfmk/1Po/47B8NcEYzOcY/wCJV5UlvLlirJed3L0SbPEDxFzDN4TxGKly04/BTWyeyv3fdvzsktCtVnVqSqTd23d+ok5NJbmS5ZTwGFjBL3mvefd/8DZAAAAAAAAAAAAAAAAAAAAAAAAAYrH4XCq9aoo+r1+7cxXGWCp6UYub+5fjr+B/b2d43/daVl3Sv+L0P7BzzGa4mtZdnJv8FoUOCKC/i1W/RJfnco8LZXT/AOXfM/Nv/hvwKWVYGl8FGK+SIwjFWirABVpU6sHCaTT6MzTg6nO88G7P+V7fJ9Pn+BisJXw1T2daLi/P+tQADLuJcdg7RcuePZ/o91+XkZ3i8FmFsTR92ptKL6+afXs+vlucHcZZrwrmP17LqnLJpxkuji+jXk7Ndmk/Io5hHMIfWYz5ufVt7363877gGc5esdgp0eu69Vt/l8ySadnucLZj9awKhJ+9DR+nR/dp8gDi7C+2y5zW8Gn8tn+d/kZfinhcVTrL7LX3dfwE01dAAAeEvAyzLELNcZG9Gm/dT+1Jdf8ADH8Xp0aPFbxAeApyyjAT/eyXvyX2U/sr+81v2Xm9ONcPyYuFVfaVvmn/AJNHC+I9tllPvG6+56fhYAAhFSko3tfucI8L4Th7LYYWhrJ6yl1lLv6dEui87t+I/iNDIqbwWCaliZLXqqafV/3n0Xzelk8RXq4irKrVk5Sk7tvVtvdtgQnKElKDs1s1umeHXiZmuLxEMtxtKVe+04q8orvPo13lo115mAAcSZBhs8yyrga+0lo+sZLaS9H96uupmmW4jLcZUweJjacG01+q8mtU+qdwONl/tVL/AA/qcEfw63qv1Dwv4fyLOMa4YyrONeDUoRTSUktd7N3W7Stpqr62PHLCcmc4euvtU7fOMn+kkAAeCeK9lxDUpX0nTkvmnF/lc+kdhlTzvJ8X354fl/8AJHhHkLy3h6NeatOu+d/4dor7ve/7eK/CX9vcTSzPMI/uKFoU4P7TjrKTX8qk2l/NZX93fi/Nf7W4gxuPTuqtWcl6OTa+5WRkuMxMsiweDm/cpqTS85ycm35u6V+yS6HF2afWMQsNB+7Dfzl/wNvvDhPLPrOL9vJe7D8+n3b/AHAAAAAAAAAAAAAAAAAAAAAAAAFfEUaEOerJRXm7GN4xwlK8cPFzf3L/AD/D5ntuIc0+BckH/wBur7/ifyMLwXBvmxVVyfZf5u7/AARhclwGGt7Okr93q/vdwAAADFYOhiqfs60VJef6djM+DZxvPByuv5Xv8n/nb1K1CpRm4VItNdHoAAGQ53PLq1pa03uv1Xn+f3FKrTrU1Om7p7MA4ry5YbG+0ivdnr8+v+fzMgzH6jjYyb916P0fX5bgGJoRr0ZUpbSTX3lWlKlUlTlum0/kcOYz61l1Nt6x91/L/gWAADhXh2vn+aU8DR0T1k/5Yrd/ou7aXU4w4hwfBuQwoYRJTtyUo+i1k+6W77tq+9yvXq4irKrVk5Sk223u292/U4zw/Pgo1FvF/g/+DY4IxF41aD8mvyf6AABT8UKeB4Mw9SElLFNOml2cNOeXys/Nvte2IxFXEVZVq0nKcm22923u2AcEeF2Oz3lxWKvSw/f7Uv8ACnsv7z07J62yXIcuybDLD4GkoR692+7e7fr8tCtXpUKbqVZKMVu27Jerexl/G3D+YY14LC4qMqi6a2f+FvSX/brYAHjHwd9cwqzjDR/eU1aaXWHf1j1/u/4QON0vrFL0f5nA792uv8P6hhcVWwtaFehJxnFpprdNdTw/44o8SYG1RqOIgvfj3/vLyfX+V6dm/HfCc2EwWI/llKP/AIEk/wD20AA8Kq3suLcL2fOvvhL9bH0gsBSxM8jdT4frMYu29pct7fcUqUKVONOmrRikkuyWiRx9nH9kcL5hjk7OFKbX+Jxaj/5M0UKFSvVjSpq8pNJLu3okcTUMZw1g1TxNKVOduWKkmr6Wur7pb9vvJNyd29SMXOSjFXbMny+OBwcKPXd+r3/yAAAAAAAAAAAAAAAAAAAAAAADHZphMFG9eaT7dX8t/wBCvxRjcZN0svpP1td/dsvncocK4vFz9rj6uvbd/fsvldGCyXA4O3sqav3er+9/oAAAAABjcvw2MhyV4J/mvR7ozvI8Pgm5Uayf91tc34b/AIAABgc0xeCd6E2l26fcZfxnSnaOKjyvutV926/Ew+KoYmHPRmpLyDPstWOwUqaXvLVeq/z2GraHDGZLF4JQk/fho/To/wBPkAcV4P2GYOa2mr/PZ/5/M4MxvJiJ4dvSSuvVf5r8gAA8N+GqPDWRyx+N92pUXPNv7MErpfdq/N26I4y4nrcQ5tUxk9IbQX8sVt83u/N9gznDfWMBVp9bO3qtV+KOFsT7HMoLpJNfqvxSAAACMXJpJXZ4feE8aahmGcwvLeNN9Ozn5/3en2uySSVkcY+K+W5M5YbBWrV1po/ci/Nrd+S9G0ziDivNs9q8+OrOS6R2ivSK0+e/dnCHAebcQ1FOguSknrUeyt/L1k/JaLq0YnjPJOFcNTwGNxkq9WOj2lP1layVuzfNbuZZmmCzPDRxODqKcHs1+T6p907NdQCcIzi4yV09GmeIXCMuHc2lTgv3NS8oPy6x9Y7eln1DjhfvaPo/0OB3rXX+H/24DJs4xmUY2njMJLlnF/JrqmuqezRxzn+E4p4HWOoaSpzg5R6xlrG3o+bR9V53SAA8M8orTzXD5j9iFWMF5ylGTf3RV36o8fIqHCSxNtaVajL/AMnS/UPpLZz9S4L+qxfvYipGPyjebf3xivmeEWUrM+Nsuoy+GM1N32tTTqO/l7tjxn8QHxfxHOdCV8NRvCl2av70/wDt9q/+FRT2Dg/hvF4im81lSk6EHyqVny89k7X2uk0/VoAAAAAAAAAAAAAAAAAAAAAAAx2ZYXBQ5q87eXV+iK2f5jmU3Ry+m0u/X5vaP5+ZgeEIuXtcbPnk+iv+L3f4FDD0aEFClFRXZKwAAAAADaSM04uw+Hbp4dc8u/2V/n8tPMxue4/F39pUaXZaL8N/ncAaaAAKGIrUJ89KTi/LQwHGVenaOJjzLutH/k/wMDnOCxqSpTV+z0f3f5HFOW/VcY6kV7s9fn1X6/MyLMvqGMjUb916P0f+W4mmroDjDBe2wSrJawf4PR/oYPEyw2IhWjvF3/r1KVSFWnGcHdNXXzAA8KuEf7azZYmvG9GhZvs5fZj+r8lZ7njPxdZLJMNLe0qn5xj+Un/275gNXMTCWAzCSjvCWnyd0QnGcFKOz1AAAhTnUmoQV29Elu32SPDjwzp5VGGYZlFPEPWMelP/ADl/6T011MZjMPg6EsRiJqEIq7b0SOO/FbE5q5YPLG6dDZy2lP8A+Ri+27672DgXw8pYuh/bGdS9nhI6pN250urfSP4y2Xc4w8U62Jh/Z+SL2OHirXS5ZNLtb4I+S17taobbZkHEmZ5Hifb4Gq4vqt4y8mtn+a6NHA3FdXiPLvrNTDum1pf7Mn1cHvbvpptd6gHG/C1LiLKJ4V2VRe9B9pL9Hs/W+6RiMPVw9WVGrHllFtNPdNaNHHK1oP8Axf8AtpwQ/wB7WXkv1AMPjsRQpVaVOVo1FaS6NJpq/o0mn0+bAAo0alerGlTV5SaSXVt6JfM4c4QwmV5VhMJNXlRfPfvUcWm/Ne80r9Eux4+Ufa+HmYLqvZv7qsP0K3jhksIpUqNSb72ik/Na3t6peh49eIlHirFYOhh6coQoxk2nbWU2l07KKt6sybPK+VRxDw+k6tN079Yxk1zW83FOHpJ9bBk2UYvOMwo5fg481WrJRivN9+yW7fRanCXCWB4eyChk1GKlCEbSuvjb1lJr+823botNkeKXh3gspof2pgGoQbSlDzfWH6x6brTRAAAAAAAAAAAAAAAAAAAAAAV69KhTc6kkkurMbxPXxVT6vlsG2+ttfkunq/uMDwnKpP22YTcpPpf83/l95RoUqMFCnFJLogAAAAAA4tzXEe3eDj7sElf+9f8ATy7hgeHcwxaTjDlj3lp/wfwMNwTRSTr1W32Wn53/AEKHDmWUVpSTfnd/noUcHRg1GlTSfkl+hxRwvkmRcJQwWY4eNXG4j3tV71JeTWqtto7OTe6jYzHgypG8sJK67Pf5Pb77epiMNWw8+SrFxfZgABVzLFVqCoVZ80Vqr6tej3+WwcKZn9awnsZv3oafLp9233dwMRRhXpSpT2kmvvMTh54etKlPeLa+44Qx/t8G6En70Pye36r7gAw2Gq4qvChRjzTk0ku7bskU44PgLhS7s5QWv9+pL8bX+6K8jG4yvjcTPE15c05ttvu3uAcY4X2ePVRbTSfzWn5WOGsV9Yyyn3j7v3bfhYAAEm3Zbnhl4cRyyEczzGH796xi/sLu/wC8/wDyXbe9s3zfB5Tg54vGTUYR6/ol1b6I4449xvEuI5dYUIv3Yf8At0u8vwWy6tnh5wXRzOU80zP3cHR1beik1rb/AAr7X3Ld24948rcQV/q+H9zCw0jHa9vtSX5LaK87sDw18OZZ3NY/HK2Gi9Fs5tdP8K6vrsurVGjTo0406cVGMVZJKySXRLsGIxNDDw5601GK6tpL72Zj4k8L4JNTxcZPtC8/ximvvZmPjnllO6weGnN/3moL8OZ/gjinP/7dzKeOdGNNySuo31a6u/W1k7W273OOI3p0H5y/Q4Jf+01V/d/UAAA8GuGPr2ZyzOtH93Q+Hzm9v/AVr5NxZJpJs8Y/ETFcQYDEYPD3hho9NnJprWXkukem7u7Wwrvh4PyX5GeYn6zmFWael7L0Wn6AeA2OynIMylm2Z05NtctOS15L6Slbd3XuprVLm0dzLM2wOaYdYjBVVUg+qf4Nbp+Tszxwzz2uLoZXB6QXPL/E9I/crv8A7eAAAAAAAAAAAAAAAAAAAAAAzfiPDYG9OPv1Oy6er/Tf0KOT5jnE1Wx0nGHRdfkunq9fUwWAw2Dp+zoRsvxfqwAAAAAADinJ62MrUZ0I3b91+XVN+W5lXDmEwKUmuefd9PRdPzAOAaGT5fOWfZ3WjToUL8ik0nOole0U9ZOOjsvtOPmcYeKMc5zOrjeVylJ6dFGK2S66LyV3r1MVxZmNbSElBeS/V3/Qr4mtXlzVZuT822AAAYbFV8NU9pRk4vyMFxnXhaOJgpLutH92z/AwWf5fi7KFSz7PR/8AB+Vw4ywHs8RHExWk9H6r/NfkZFmH1LHQqN+69H6P/LcAPBbhX2+IlnNePuwvGF+sre9L5LRebfVHi3xX/a2a/UaEr0aF15Of2n8vhXza0YAcZ4X2mCjVW8H+D0/OxwTiveq4dvtJfk/0AA0YeFHh6koZzmENd6cX/wCltf8ApP8A4F2M3zbB5Vg54zFz5YQWv6Jd29kjjXjXGcS4znn7tKPwQ7eb7yfV9NloHCvDmI4gzSngqOiesn/LFbv9F3bSPEjijDqEOHsq93DUNJW+1Jfmk939qV29kwMko5fVx9OOYVOSjf3mk27Lokk9Xtfpv5GS4rLq2W0auAa9hyrltoklp12tazvs9zizxnlhsVLD5PCM4x0c5Xab/upNaLu9+itq8w8S+KcbdSxbiu0LQt84pP72YnFV8TPnrTc5d22397AON43w1J+b/I4LlbHzX91/mgAADhrxRx2QZdDA4XD03FNtt8123u3ZryXokS8ceIG/do0rf4Z//JnHOazzWnisbUpxhKau1FNRvpd2bb13eu7KuKWFyr23VQVvWyS/Gw9QynLp4/FRorbdvsuv+XqYjPMswEFSU0+VWSjrt+C+bMr8VcfkuKWIyxOMvN6NdpR1TXr8tSHGGL4pxuIzDFQUZyl0vbbRK/RJJIAAAAAAAAAAAAAAAAAAAAAr1qdGm6lSSUV1Zi87xmaVXhsui1HrLZ/f0X4/kZRw3hsFapU9+p3ey9F+u/oAAAAAAAABmGb4PAx/fT17LV/d/nZGYcYYqteOHXIu+7/yX9alatUrS5qknJ927hhcrxuKt7Gk2u9tPvehh+DMdU1qSjH8X+Gn4lHgjDr+LVb9El/mS4aybDQ5623eUmvysZzHJV/ublzf+S/jr+aAAAMFnWOwllSqO3Z6r7nt8rGM4ljj8HKhiadpbpruvJ/du9w4XzJYvBKEn78NH6dH+nyAynK8RmmOpYLDq86jSX6t+SWr8kcW5phuC+Fo4XBu07clPvd/FP1Wsu3M13G23dgBmWFWKwdSj3Tt69PxMhxX1bMaUns3Z/PT/ggBm2YQwOElWe60S7vp/wAHyOEMznXVShVleV+ZfPf8dfmeF/Af9t4n69jI/wCz03s/tyXT/Cvtd9u9sXisPgsNKvXko04K7b2SRx9xxiOJcbaN44eD9yP/ALdLzf8A5KtF1bCnNcEcJKa0xuMWneELfg0n680uvKPUAr8cVqXCmHyPCNr4/aPyc5NQXk07y73S7oAA41jfAwfaS/JnB8rZlbvF/oAAABn/APyLK3ocT4xQy7D4dPWSTfol/n+QEK1SEXGMmk9/P1A4KhbBTl3l+SQAAAAAAAAAAAAAAAAAAAAGaZthsvpc1V6vZdX/AMDzKWDx+f1FWxD5KK2X+X/yT+XYwmDoYSkqVGNkv617gAAAAAAABi8ZQwtN1K0lFf1t3M14vrVr08IuWPfq/wDL8/NE5SnJyk7tmGwdfEy5aMHJ+S/qxgeC607SxM+Vdlq/v2X4mDyHL8KlyU033er/AB2+VgxeNw+Ep89aaiv62W7Mx4yqSvDCRsu73+S2XzuYjFVsRPnqzcn5gAAABgcwxGCqe0oSs9jC8bTVliKV/OOn4P8AzMNxLlley9pyvtLT8dvxPBDh2E4VM7nZp3hDr/il/wC2r/t7ueJPFH9vZ3OVKV6NL3YdnbeX/bz/AASAAM+wv1XMakFte6+ev/AMsxSxeDp1urWvrs/xAM4wCxuCnR62uvVbf5GR4ung8yo1azagpJStvy3tKy72vbzMpw2CwuX0aWCSVFRXLba1rp3633b63ueKPHzznEPL8FL/AGeD1a+3Jdf8K6d373aweHHDsM4zqLr/AMGiued9rR2T9Xv5XONeJJ5/nNXF39z4YLtBbffu/NsAAADiupTrZbLkd3GST8nqjIcbTweNVapslL8np83oZVWqV8HTq1N5a/e7r7loAAAYvFxoezXWUlFfr+BxJJRyqt6L80YzF1MVV9pUeuiXklsgMvyDHY2zhG0e70Xy6v5IwXB2DpJOvJzf3L8NfxKGBwuHVqVNR9EiyAAAAAAAAAAAAAAAAAAAADPc7WXwjCEeapLZdP68jLOHquIqfW8yfNJ7Rf6//I7Lr2EklZAAAAAAAABnPE1DBXp0vfqfgvX/AC++xjMdiMZU9pWld/l6LoYfDVsTUVOjFyk+iOAfC2pnmbUMJiHpJ3kl9mK+Jt+mit1sr6nHuMy1Y/8AszKaUaWFw94xUEknL7Unbdtq13dtK99QZnHFlLD3pYW0pd+i/wA/yMVi6+KqOpWk5Pz/AE7Ba5g+HMxxVnGnyrvLT/g/gYbgiFk69X5RX6v/ACKHC+V0v+XfM/Nt/ht+BSy7B0vgpRXokYvLMJi48tamn+a9GtTP8jwWBV6dbX+V6v71t818x0ans/acr5e9tPvAAAMDmeOwE3PCVpU5PrGTi/vTRQ4nzOjp7TmXmk/x3/Eo8bVl/FpJ+ja/O5Q4ywE9KkZRfpdfg7/gUM7y6v8ABWXz0/OwmpK6dwONsH/CxKX91/mv1OC8Zz0J4dvWLuvR7/j+YBh8PVxFaFGjHmnJpJLdt6JI8WeAMTwjnFOnVd41oKaa25tpxX+GX4NHDPizi8TwNSyGDarU7wlL/p19hL5e4+0Y9ebQCtL/AFa4GhSWmIx7u+6pr9Gmv/A32AAAAOL6EqON54v3aiV/NrT/ACDA0vZYWlT7RS+5AAAGb432ueYehF6Qa+9/8CxxRnP1yt7Ck/cj+L7+i2X3hgsHVxleNGktX+Hn6GV8L4PCJSqLnn3ey9F/n+AAAAAAAAAAAAAAAAAAAAAAFTDUalSNSUU5RvZ9r7/kAAAAAAAAFSpCnBzm7JdWZ5xVOvejhHaPWXV+nZfj6BlHC1fF2qV/ch+L9F09X9xgsBhsHT9nQjZfi/VmQ/8AGKcH1s2lpicV7lPuo/zL8ZdtIdwxGIpYek6lWVoozviWtjW6VH3af4v18vL7wjGUmoxV2zLeEMTXSniHyR7fa/4Hz+4wOTYLBJeygr93q/v/AMrAFScKcXKTsl1Zj+I8RjKv1bLItv8Amt+XZeb/AAMt4TpQftcY+eb6dPn1f5Hs4cnJZW2t0+4x3C2X4m7jHkl5bfdt91jHcI46hd0rTXlo/uf6NlWjUpS5KkWmujVmAAABRxVeg70puL8m1+Rh+Kszo6OakvNfqrP8TDcbRelel80/0f8AmZlnOW5lgKlKNS0rXSatqtd9tdt+pw9jfqmYU5N6P3X6P/g2YB4N8GK39t4qPdU0/uc//bV832Z9JbLMBi+FqdSo7YinPmprq1a1ReS5bSb7xiupw7mH1LHRcn7stH8+vyf4XA4XyWWc5xh8CtpyV/KK1k//AAFM8Ss6jmef1Y0v4VH93BLa0dHb1le3lYAAAA4wwntcAqq3g/wej/Qw9P2taEO7S+9gAAFarCjSlUk9Ipt/Ir4qpWxEq7fvNt/eBwxk/wBTw/tqi/eT/Bdvnu/+AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAGLxdHC0XVrStFf1oZ1n1bMZ8q92mtl+r8/wAF+Jh8NVxNRUqUbyfQyXhejhLVa/vVPwXp3fn9wcLZFUzzN6GBhtJ6vtFayf3bedkeJmfU8wzj6rhtKGGXs4JbaaSa+asvKKMfj6GCourWdl+LfZGbZxXzGrzTdorZdF/m/MMqyPFZjL3FaPWT2+Xd/wBOxlmSYTL4/u1eX8z3/wCB8gAxeLo4Si6taVkv608xvHcRVbL3KCf9er/BfngMuw+Bpezoxt3fV+rADEYShiYctaCkvNHEOAynCtxw8n7T+Vapet9V979BZFmMqKqqi7fj92/4E4Sg+WSswAAAAw3FOZULJz5l/eV/x0f4mG42pvSvSa807/g7fmcHYvJc7zWjha+KjSg37zm1DRbpOVlzPZWvqyNfAYHL1VhKMaFOOjXwqMV0t0SRxpxRW4jzSeKnpBe7CPaPT5vd+b7JGdZe8DjJ0um69Ht/l8jhvMljcFFN+9DR/o/mvxuHh5bKsszHP5b0oezh/jnb8vd+TY227vcAAAAxmHjicPOi/tJoyWg5ZnRg1qpL8Nf0AAAOL8b7HAqknrN2+S1f6L5gcLZN9arfWKq9yO3m/wDJdf8AhwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAx+PoYKg6tV6L72+yM2zavmNbnm7RWy6L/g92ZZlWIzCryUlot30X9djK8pw+X0uSkter6v8A4HkBwjbhzhbF5/LSrW/dUu/m1803/wBueZmOY0cFQdaq/RdW+yMzzOvmFb2lV+i6Jf1uwyPhaVdKvi1aPSPV+vZfj6FOnCnBQgrJbIAMViqWFoyq1XaKKGHxHEGI9vX92hHZd/66v5LypUqdKChTVktkADtYzPPMRja31LLdX1kv0fRef3eeUcNYfBWqVPfqd+i9P83r6BicFhsTHlrQUvVf1YxfBuCq3dGTg/vX46/iZb4R8UZtSq1csoqtGna9pKL1vspNXemybe3czDLMZl9eVDF0nCcW01JNNNaNfIAAAAMJm+YYOnKlh68oRlulJpP1Sdn80Ybi7MaVlNqa81/lYzrOqOZ04N0+WcfO6a/B+nzMhzR4DFqbfuPSXp3+X+ZGUZRTi7o4o/4SuD8tytaTrXrT+fw3+Tt6xADFVVRoTqfypv7kZJntHMadnpUW6/VeX5fiAABh8B7LiV2WlnJfNWf4tgAAHFWN+sZhKCekNPn1/HT5BluBljcVChF2v+XUwuGpYajGjSVoxAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAMfj6GCoOrVei+9vsjNM0r5hXdSo9Oi6Jf1uzJcjrZjU00gt3+i8/y6mEwdDCUVSoxsl/V35gZdgK2PxlLCUVedSSivVu33dzxezzB4CpRymlK1DBwUfWbSv6u1l683mZrmlbMK7qT2Wy7L+t2JNuyOH+GY0UsRileXSPbzfn+XrsAEmkrtlT2nEGN5Iu2Hpv73/m+nZebKVKFKChBWS0SAAzvH18biP7Nwb1+0+3denf7jK8qoZfRVOkter6t/wBbIAMjhDg3gV4morVXHna7znZRT9PdT9GytJ15SlU95y1d+re9zE8PZbiPipJPy0/IxHBOGl/BqteqT/yMZwjisPB1PaRcV3dvz0/EaAAAAChiq9B3pTcfRtGL8QOIMbUhUxlf2rjFRXMlpFbLSz6lDjeW1aj9z/R/5lDi7LanxNx9V/lco5vgK3wVo/fZ/c9RSjJXTuThGcHGWz0KsauDxMoxdpQbV/QyHiOGNSo1nap+EvTz8vu8gAB4WDxSxHVRcfvaf6fiAABj8VHC4adZ/ZV/n0+9k5ynJyk7thTlOM04P3ltbe5kuLxWJwyeJpuMl3Vr+dvz/AAAAAAAAAAAAAAAAAAAAAAAAAAAAAADGYujhKMq1V2S/qy8zN81rZjX9pPSK2XZf592ZHkVTMal3pTW7/Ref5FDD0sPSVKkrRWyAMPiauGrQrUZcs4tNNbprVNHFHEWJzvHTr1ZNptvXq27tvzf4bCTbsjhzh1YWKxGIX7zov5f+D+QABxFjKtacMtwz9+e/kv638vUwGBpYLDxo09l+L6sACtGUqcowdm07Ps+5kWUrAYf39aktZP9Pl+YAcC5F/bOf4fCyV4X5pf4Y6v7/h+Z4457eWHymm9vfl+Kiv8A0p280wM1zfD5dT5qjvJ7Lq/8l5mY5ri8yqrnenSK2Xy6syrhCpVSqYt8q/lW/wA+35+hRyzB0aXsoU1y+l/vvv8AMxnD+Uyg51Kail1Wlv0/AwmSvMcTNYO6pL7Uv+AuvbotzE8J5lS+GKmvJ/o7FfCV6DtVg4+qaAAAACnVqU3eEmn5OxSzvMafw1pfN3/O5iMRUxFR1ajvJ7/0hNppp6mQcTqrbD4t+90l38n5+fX13AAAAA4zxjjRhhYbyd36Lb73+Rg+G8xxNmocq7y0/Df8DB8F4eFniJuT7LRf5/kYXLsJhVajTUfz+/cAAAAAAAAAAAAAAAAAAAAAAAAAAAAACrVhSpuc3ZLdme51PMa2mlOOy/V+b/AyPJamY1u0Fu/0Xn+RQw9KhSjSpK0VsgAM+wn1bMKsFte69Hr/AMA4Z4fVJLF4he99ldvN+fbt67AAYvE08NQnWntFX/r12OG8JOanmFf46m3lH/g/kkAAAAeCGTRo4XE5tV0v7kW+iWsn6N2/8BZxTnMs5zjEY5vScnbyitIr/wABSDPM8pZdTstaj2X6vy/P8qVHG5tinb3pvdvZf5L+kZPkGGy+Kl8VTu/07fmBmtarm+N/s/Du0I/G/T/Lt1fpcwuFpYWjGjSVooJRjJWaujE8PZbiPipJPy0/LT8DhbwVr8UYirTwOI9moRu3NXV27JXVrX16PRM478IuI+DaCxOYKEqTlyqUJp3dm0uV8s9k/s2XVgAAAAGQcTulbD4t3j0l28n5efT02TUkmndAAAAB7Gnz+05VzbXtr94AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHE+e/W6n1ei/wB3Hd93/kun39jKcqq5jXVOGiW77L/PsjCYSjhaMaNJWiv6v6gAGLyWhicbTxVTXlVrd3fS/pr+AAAZ+5YzE0cug/ifNL0X9P52IxjGKjFWSAAAASbdkcRyXCnAEcHHSpKKp/8Ab07uf4c1vkGd5xTy6hfeb2X6vyRg8Fi84xb1u3rKT6f10X6GX5bQwNFUqK9X1b7sDiDMpYPC2p/xJ6R9e/y/OxkmVxwGFUH8b1k/P/gf8HqAHhXw+sq4epzmrVK3vv0fwr/wGzt0bZ9Injb+3OJnl1CV6OEvHydR/G/lpD1i+4QpyqSUYK7fQw/B2Oq0+abUX2f62vb8TE8M5nQ/5d8y8tfw3/AqUp05cs4tPzVgAAAOEMTj5XptXpLq+j7Lv6dN9OoAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHFmcyoQ+qUnaUlq+y7L1/L1MHg6uLrxo0ldv+r+hlmW0cBh1Sp/N933AAAAAMiX1rF4jHvZvlj6L/PQAAAA8Pcn/tTiTC0Wrxi+eXpD3tfVpL5njhnHtsxoZfB6U48z/wAUtvuik/8At4x+NpYPDyrVHovxfZepbF53j/7z+6K/yX4vzZl2XUcDQVGkvV9W+7AMIv7TzeeIetOjpH17/r9wAcJZI85zrD4K2kpe9/hWsvwT+djxI4qp8KcLYrMY2U4x5aa/vy92OnZPVrsmVKk6k3ObvJ6tvdt7sjFyajFXbOH8hhgKSqVFeq9/LyX6/wCQFWjTqx5ZxTXmrniXwNkGCxuGwmHw6pyjRhzuN1ebTu2trtW6Gc8MYTBYeVZVmktk0nd9tLfkEcBip01UjTk497Ow4tOz3ADcyThSda1bFq0eker9ey/H0KdOFOChBWS2SAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAOM8C5qlXgru/L9+q/X7zh/JY5fQvP+JLfy8l+vd/IAAAAAznE/VsBVqJ6pWXq9F+Zk+FWGwFKl1S19XqwAAADwLypc2KzGS2Sgn/AOTS/KJxVmzzbOsTjb3U5O3+FaR/8lSOIc0nmOLVGjrCLsvN7X/Rf8EyLJ4Zdh7PWb3f6ei/4IBneN+p4GpUT961l6vT8NzJMD9TwMKTXvbv1e/3bfIAPAzJlOticykvhShH1esvuSj959Kbil1cZhMipS92C9rNf3pXjBeqipP0kg4QypVarxdRaR0Xr3+X5+gBkuDWMzPDYb+ecY/fJI8S8asRxTjajekZcvpyxUX+Rn+byzDEtxfuR0j/AJ/P8rHDuQPHT9tWX7tfi+3p3+70jCMIqMVZIq0KVVWqRT9Vcq5BllX4qK+Wn5WKvB+Wz+FSj6P/ADuYqFOFacaTvFNpX7GGwtbE1VSoxvJmS8M0cFarW96p+C9PPz+6wAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAEoRla6vYAAAAAOI17WNDDfzzV/RbgAAABlb/ANXfDWVfadSDl86rtF/KLj9xxVmv1XC+xg/fn+C6v57fecIZQrfXaq8o/q/0XzADOISxWY4bC291Xm+2m3+XzAA8MsrWX8L4ZNWlNc7/AO3tV/5LY8Q+InxDxTjsyTvGc3y/4I+7D/yVIp05VJqEFdvRepgMHDCYaFCP2V+PV/NgHhthfrPFWCh2k5f+Axcv0PEnPvaYvEKD96tOcn/hcm/x/K5k+WTzDFKktFu32X9aIo0KdGnGnTVorRIAznF/VcBVqp62svV6IynI8TmM/cVoLeT2+Xd/0zLsrw2ApclFa9X1fr/lsAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABiMBCviqNdvWney6O6sAAAAYLCTxeJp4en8U5KK9W7I8bcdSy/JcJl0HaN7+kacbK/lqvuLVM8zb+6390V/X3sp04U4KEFZLRABypu/UAMFhZ4rE08PD4pyUV6t2PEjNocO8E47EUny8lJwh5OVqcfubX3BwlglXx/tGtIK/z2X+fyADwulGjm1fFt29jRqz9LJL9TNcbLG4ydbu9PRbfgcP5WsBhEpL35av8ARfL87gBmOW08dGFOq/cTu137a9inShSgoQVktkgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA8L8u+u8U4VNXUG5v/t1Nr/yax9JXiHnzn6jB6xhGL/9Lf38yXyOEst+r4T28l70/wAun37/AHAAABgMbVwOLpYqjbnpyUldX1TutDxX8Q63FXB8cDSpctdVIymlqpQipfD1+LlfK+2jfRo4OwvssC6r3m/wWn53ABSaTs9018no16NaMo8MeyzeLtekveXy+z9/3oAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADwy4nwGQZw6+NT5Zx5OZa8t2ndrdrTW2q7M45xtTiXjjF1PsyqSS/wJ6P5x1XqiMYxioxWiAAAAM+4Zhi71sPpU6rpL/J+f39zLsN9XwlOl1ikvn1/EAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA+p0VifrCXv2tfyAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADF13Rp8yVz+1p/wAqP7Wn/KhZrNtLlQAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABmn8D5gR+JAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAGafwPmBH4kAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAZp/A+YEfiQAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABmn8D5gR+JAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAGafwPmBH4kAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAZp/A+YEfiQAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAGcYyphMN7Sna90tT/WbG9o/d/wAE/wBZsb2j93/BKfEmMlNRaWr7f8EAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA4l/wBx+aAo/wAWPqgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADiX/cfmgKP8WPqgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADiX/cfmgKP8WPqgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADiX/cfmgKP8WPqgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADiX/AHH5oCj/ABY+qAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAOIac6mC5YK7utj6lif8An3L7mfUsT/z7l9zKODxCqR/dvddGAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHh5keCznOfquMjzQ5ZO12tVbqj/AIhVwv8A8+X/AOBy/wAz/iFXC/8Az5f/AIHL/MxfhdwzTw9ScaLuk2vfl0XqAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAeEP8AyUa/wS/QDMP90q/4ZfkwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADwh/5KNf4JfoBmH+6Vf8MvyYAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB4Q/8lGv8Ev0AzD/AHSr/hl+TAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPCH/AJKNf4JfoBmH+6Vf8MvyYAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB4Q/8lGv8Ev0AzD/dKv8Ahl+TAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPDDMcJgM99tiqihDkkrydlfQ/1z4d/wCg2n/4Ej/XPh3/AKDaf/gSMbxjw/PC1IxxlNtxf2l2AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAMNTjOdmfVKXY+qUuxLCUrbAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAGD/AInyAl8LAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAMH/E+QEvhYAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABg/4nyAl8LAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAMH/E+QEvhYAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABg/4nyAl8LAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAMLKMal2z29L+ZHt6X8yJVqbT95AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAEUmzkickTkQAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABDcB7AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAENwHsAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAQ3AewAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABDcB7AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAENwHsAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAARsmcy7nMu45LuAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAASlGKu3ZFTM6EdFqYSssRByStZ2HBgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAYrFwoR7vsVq9StK8mGTfwZev6IHFMasAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABgcmx2YUMRVwsOZUYSnL0im7ebdtF1Kk5VJOUndgZN/Bl6/ogGk0NW0AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAKFCpXqxpUleUmkl1beiXzOEuF6GR5THB2UpS1m/wCaTWvyWy8td2ziLJ5ZPm2Jy+X/AC6m4rzjf3X842fzAyb+DL1/RAE43QAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB4QcNrFYyeaVV7tLSPnJrV/wDbq/Fp9A8eMl+rZ3Rx8F7teFn/AIoWT/8AJXFL0YGTfwZev6IAJKzAAAAAAAAAAAAAAAAAAAAAAAAAAAAxuPoYSHNVfour9DGcR4mq7UvdX4/f/kQxNepWjzzb1W7Y0mOCew1YAAAAAAAAAAAAAAAAAAAAAACEJTkoxV29EjhbJY5PlFDBreK97zk9ZP79vKweNGSf2hwvOtFXnQkpr0+GXys+Z/4UBk38GXr+iACaugAAAAAAAAAAAAAAAAAAAAAAAAAAA4N4PxPEWM5I+7Sj8cu3ku8n07bvz8ZeHaOR8UzoUFalKEJRV27K3K9X3lFv1YYf+ND1X5gNJoasAAAAAAAAAAAAAAAAAAAAAAHhpk6zLiGjzK8aXvv/ALd2/wDJmgMdg6WNwtTDVleE4uLXlJWf4MzHA1cBjKuEq/HTlKL9Ytp/kGTfwZev6IABqzAAAAAAAAAAAAAAAAAAAAAAAAAAAybKMTm2Op4PDK8pu3kl1b8ktWZDkeFyXAQweGWkd31k+rfm/wAFZbI+ktlX+4ZjFfz03+Eo/wDtwYf+ND1X5gEldDAAAAAAAAAAAAAAAAAAAAAAPBjK1TwGIx0lrOSivSKu/vb/AAAPGzJfqHEzxEV7teKl/wBvL3ZL8E3/AIgyb+DL1/RAATWoAAAAAAAAAAAAAAAAAAAAAAAAAAHhLwusFgHmVaP7yt8PlD/7Z6+nL5h46ZT9f4MrzSvKjKM18nyv/wAlk2GH/jQ9V+YATXUAAAAAAAAAAAAAAAAAAAAAA4Hy5YDh7CUbauKk/WXvP87AHjrkv1vIKeOiveoS1/wztF/+TcgZN/Bl6/ogAJrQAAAAAAAAAAAAAAAAAAAAAAAAAAOFcjlnWb0cEvhk7y8orWX4aLzaKdKFKnGnBWilZLskHEGWRzTKcTgX/wAvYSj/AOBRaRKLjJxas0Yf+ND1X5gA1dDVgAAAAAAAAAAAAAAAAAAAAAwOFeKxVLDx3nJR+9pEKcacFCKslovkAZ9lVPNcrxGBntUjKPo2tH8nZ/IrUZ0akqdRWlFtNdmtGZN/Bl6/ogAJK6AAAAAAAAAAAAAAAAAAAAAAAAAADwYyVQw9fMprWT5I+i1l97sv+3QDxGyn+y+K8fhUrJVJNek/fj+EkYf+ND1X5gATQAAAAAAAAAAAAAAAAAAAAAcA4VYniXBw7S5v/AU5foAB4tZL/ZnFWI5VaNW1Rf8Ab3xf+TqRk38GXr+iAAAAAAAAAAAAAAAAAAAAAAAAAAAAEr6I4YypZXk+HwdtYxV/8T1l+LYB9IrKvq3EtLGRWlamr/4oNp/+S8phv48PVfmABJXQAAAAAAAAAAAAAAAAAAAAAeElD2nEkZfyQk/yj+oAHj7kvtcvw2ZQWtOThL0lqm/JNW9ZGTfwZev6IAAluAAAAAAAAAAAAAAAAAAAAAAAAABwXlqzHP8AC4dq65k36R95/elYAPpIZT7fI8LjorWlUcX5Ka1/GMV8zDfx4eq/MAAa1AAAAAAAAAAAAAAAAAAAAAPBelfOK8+1O33yj/kABxnkqznIMVgUrylF8v8AiXvR/wDJkjJ1ajJef6IAAluAAAAAAAAAAFtAAAAAAAAAAAAAAADwZwHtc3rYlrSnC3zk1+iYAeKmU/2nwdj6KV3GDmvWm1PT15bfMw38eHqvzAAJrUAAAAAAAAAAAAAAAAAAAAA8FIp43Fv+7H83/kAAcWZL/ZGf4zDxVoublH/DNKSt5K9vkAAT3AAAAAAAAAAAAAAAAAAAAAAAAAA8FsHyZZicR/NNR/8AAY3/APbmAGIoU69KVKorxkmmvJqzMXgJ5fms8HU+KnUcX6xlZ/kAAVFsAAAAAAAAAAAAAAAAAAAAAeCf+94v/DH82FbEUaEearNRXm0vzMTxfkOH+PFw+T5v/SbmJ8T+H6XwSlP0j/8AJOJifF6gv4GFb/xSS/JP8zE+LGb1NKNKEfk2/wA0vwOIc9xmc4z6xipJyS5dElorvp6vcAAnuAAAAAAAAAAAAAAAAAAAAAAAAAB4WYb2PDFCXWbk/wDyZr8kAB4v5V/Z/HWISVo1HGov+3kub/yZSAAJ7AAAAAAAAAAAAAAAAAAAAAHDme5hlVSo8FU5HJJPRPReqZieJs5xOlXFTa7czS+5OxOcpy5pO78wCpJRi2PUAAnuAAAAAAAAAAAAAAAAAAAAAAAAABwNRVLhvBx/uJ/fr+oAH0jMq5cyy7MIr4k4P/t2Skvv5pfcAAT+EAAAAAAAAAAAAAAAAAAAAAwT/eP0AAxU9ogABLcAAAAAAAAAAAAAAAAAAAAAAAAADhmHJkmDj2pU/wD0lAAeOuVfXOFVXS1oVIT+TfI//Sk36AAEvhAAAAAAAAAAAAAAAAAAAAAMK7VUADtYqS5pNgAA9WAAAAAAAAAAAAAAAAAAAAAAAAABkiSyvDJfyQ/9JQAHFGWRzPJcVg2vjhJL1to/k7MdGa6DhJdAAlsAAAAAAAAAAAAAAAAAAAAAUnapFgBiJ8sLdwAAewAAAAAAAAAAAAAAAAAAAAAAAAABkEufKMLLvTh/6SgADijLv7PzjEYdKyUm16PVfg0FrjhF9B0ab6Dw9N9Cvh4RptoAAAAAAAAAAAAAAAAAAAAAIvmimAYifNO3YAAJ7AAAAAAAAAAAAAAAAAAAAAAAAAAHCNT2mQYKX/TuC+6KQAB4tZd7PHUcXFaTi4v1j/mmvuAAxX8FgAAAAAAAAAAAAAAAAAAAABhJc1JLsBUlyxbHqAAFTawAAAAAAAAAAAAAAAAAAAAAAAAAB4c11W4Xwkuya+6TX6AAHiVl31vIZzS96k1Jemz/AAd/kABjHakAAAAAAAAAAAAAAAAAAAAAGCnaTiBip7RAAAnqwAAAAAAAAAAAAAAAAAAAAAAAAADwfxSq8POnfWE5L70pfqwADG4WGKw1TDz+GacX6NWMTh54etOjP4otp+qdmAGOfupAAAAAAAAAAAAAAAAAAAAABTnyTUjRodrE5c0mwAAbsgAAAAAAAAAAAAAAAAAAAAAAAAAA8E8ar4vCt/yyX4p/+2gAB4i5d9Tz+q0tKiU189H/AOTJgBjXeaXYAAAAAAAAAAAAAAAAAAAAADCVOanbqjEz5YW7gAATfQAAAAAAAAAAAAADoAAASlGMW5OyRU4gpSq+yoK/n0+Xf8CeNrz+192hgbzw0XLVjgnsOLQAAAAAAAeFWYfVeJKcG9KkZQ/DmX4xS+YAAeLeXc+FoYyK1i3F+jV19zT+8AMRLmqNgAAAAAAAAAAAAAAAAAAAAAUKns5pmImpz06AAA3ZD1AAAAAAAAAAAAAAAAAq1IUoOc3ZIzbOKmMlyx0gunfzf+XQwbtXiGX/AO6w/rqDSaJRsAAAAAAAZbjZ4HG0cVDenJS+53KFanXpRq03eMkmn5NXQABxfl39oZJiKCV3ytr1j7y++1vmATkoxb7D1AAAAAAAAAAAAAAAAAAAAAAIPoAAE3fQAAAAAAAAAAAAAAAAAz/NHiKnsab9yP4v/Jf8EKT5akX5hl/+6w/rqA0miSswAAAAAADwvzhZhw9Sg371H3H6L4f/ACVpfJgAG5xDl39n5rXwttIydvR6r8GgMbO0FHuAAAAAAAAAAAAAAAAAAAAAABGVwAJS6IAAAAAAAAAAAAAAAAAz7MPquG5Yv3paLyXVgEXzRTMv/wB1h/XUAlG6AAAAAAAPCTPVgc5eDm/crq3/AG8tY/fqvVoAAPFjLvY5nSxUVpUjZ+sdPya+4DE1Oeo/IAAAAAAAAAAAAAAAAAAAAAAAU+5dMHJIlJsAAAAAAAAAAAAAAAAAM3xn1rFSmnotF6L/AD3AMM+ajF+Rl/8AusP66gBNWYAAAAAAFGtUo1Y1abtKLTT7NapnDGe0s7yqljYbtWku0luvv28mmAAeJ2XfWsjdZLWlJS+T91/mn8gxFT2dNvqAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAZ3ivq+Dk09Xovn/AMAAMA74eJl/+6w/rqAE1dAAAAAAAHhfxWspzD6piJWo1ml5Rlsn5J7P5N6IAAzHBwxuDq4ae04uP3qxWpSpTlTmrOLafqtzEVfaT02QAAAAAAAAAAAAAAAAAAAVK9Kn8UkijiqVaXLB3Y4tAAAAAAAAAAAAAAAAAAAAAAcXYXGU6WHqzptUp83LLo2nZpea/UAMsd6TXmZf/usP66gA9RoAAAAAAA8MuOY5hQjlmMl++gvdb+3FdP8AEl9613uAAeIUsPTz/FU8NJOLld26Nq8l8pXv93cAAAAAAAAAAAAAAAAAAAwuEr4uvGhQg5Tk7JLVtnHdHM8gzKWWVbRkoxba1+JJ6Py2duq0djLJuXPd3ehk/wDvD9P8gcUxxaAAAAAAAAAAAAAAAAAAAAAHwHl+acH0clxsdORO/WE3q5LzUm/VXT0bOKuF8fw3mlTLsbG0o7PpKPSUfJ/g7p6pgGVv4l6GX/7rD+uoAE1qAAAAAAAU6s6U1OnJqS1TWjTXVHBXilQxUI4PNpKFTZT2jL/F/K/P4X5bCakk07oCrVp0qbqVJKMVq23ZL1Zxx4pxnGWCyeXlKp+ah/8AJfd0Y3cAAAAAAAAAAAAAAAAAAClSqVqkadNXk3ZJbtvZL1OAuBqOQ4ZVq6UsTNav+Vfyr9X1flY+kllnss6weNS0qU3H5wlf8poyx+/JeRk/+8P0/wAgGk0NWYAAAAAAAAAAAAAAAAAAAAcPYBY/NsNhWrqc4p+l1f8AC4eJnh7huL8rdNWjiKd3Tl59Yv8Auy69nZrazzDAYnAYqphcVBwqQbUk901/Xo90Blj/AHrXkZf/ALrD+uoAFRaAAAAAAAAZHxrneTJQwtZ8n8svej8k9vlYw3jXjYxSr4SMn5ScfzUjF+NWYTjbDYWEX/ebl+XKZ3xXnGcv/bK7lH+VaRX/AG6rL5u78wAAAAAAAAAAAAAAAAAAA8JOEU1/bOJj3VNP7nL9F832YfSOyz2/D2HxaWtKpb0U4u/4qJlztW+Rk/8AvD9P8gCUboAAAAAAAAAAAAAAAAAAAAPCrB/WOJqUv+fcZS/Dl/OSA8ZvDBcQYV5pl8P9rprVL/l5FdP8Ufs917v8tmmnZ7hgHaujL/8AdYf11AAmrxAAAAAAAAAAAAAAAAAAAAAAAAAAAAOGsjq51mlLBQ05nq+0Vq3923nZGGw1LDUIUKMbQikkuyWiDxay3+0ODMfTSu4w51/241N/hFmBdq8f66GT/wC8P0/yACa1uAAAAAAAAAAAAAAAAAAAAeCuGUswxVf+WCj/AOBSv/7aAeOPhf7Jz4hyyHuvWtFdH/z8S7P7fZ+9tzNGGdq0fUy//dYf11AAeqAAAAAAAAAAAAAAAAAAAAAAAAAAAAPBzIVRwdXM6i96o+WP+Fbv5y0/7dAx+DhjMJVw0/hnFxfpJWf5kaM8PjPZVFaUZWfqnZmT/wC8P0/yACSurAAAAAAAAAAAAAAEpRjFuTskY3ialTbjh1zPu9v83+BVz3H1H/Et6JIy+88HSlJ3bjG/rZDgug1YAAAA8E6KWFxlXvKC+5N/qAThCpBwkrp6NPZo8YfDKXDON+vYGN8JVen/AE7k/svye8X20equ4u0kzL/91gAAAAAAAAAAAAAAAAAAAAAAAAAAAAABh8PUxFaFGmryk0kvNuyMoy6nl2Ao4OntTil62Wr+b1APErLv7N4yx9G1l7RyXpO01+EjJ/8AeH6f5AATVmAAAAAAAAAAAAAFatTo03ObskZrm9TGy5VpBbL9X/WgGWf7jR/wx/JA0miUbAAAAeC9NLJa8+9Rr7ox/wAwAzXK8JmmCqYLGQU6dRWafb9Gt0907NaniLwFi+Ec1eGneVGd3Tn/ADR7P+9HaS9Hs0ZU+bBU35AAD3AAAAAAAAAAAAAAAAAAAAAAAAAAADwuytY7iOlKSvGknN/LRf8AkzT+QAfSIy36vxTTxKWlWnF/OLcX+CiZM71r+X+QAE1oAAAAAAAAAAAAAGeZo8VV9nB+5H8X3/yAMs/3Gj/hj+SAaTQ1YAAAPBtJcP1P+vsv/SYAAcY8I4DijKp5fi1vrGXWEltJfququupVyDG5JN5dio+/T0dtU+qa8mrNddddTkl2LAA9wAAAAAAAAAAAAAAAAAAAAAAAAAAA8Fcu5cLisa18UlFf9uq7/wDSl9wAfSVy3ny/AY5L4Jyg/wDt9KS/9IZkDvOL/u/5AASV0AAAAAAAAAAAAAcQ5h9Xoeyg/el+C6/ft94AZZ/uNH/DH8kATV0AAAHg1K+QVV2qy/8ASYAAHi1lPJiKOPgtJLll6rVferr5ANJjhB9B0oPoOhTfQmkpOwAAAAAAAAAAAAAAAAAAAAAAAAAAAeGuBWF4Zwy6zvJ/9vN2/CwAeN+W/XeCsU0ruk4zXykk/wDyVs4bd2vR/mAANAAAAAAAAAAAABJpK7MxxbxWJlV6dPToAGWf7jR/wx/JABJWYAAB4LVL5ViIdql/viv8gADjHKf7TyWvQSvJLmj6x1/HVfMAAlq2AAAAAAAAAAAAAAAAAAAAAAAAAAAGT4RYTLcPh/5IRj90UgA4ly3+08lxeCtd1Kc4r1cWl+Jwu7u3ZP8ANAAE9wAAAAAAAAAAAA4gxXsMG4p6y0+XX8NPmABln+5Uf8MfyQAZ3wJWy/hmjmc0/aOXvLtGXw/NPfzlboAAAeCVdOONpf4H/wClJ/oAAHGOU/2ZnNeglaLfNH0lqvu2+QASdk2AAAAAAAAAAAAAAAAAAAAAAAAAAAGWYdYjHUaP80or72kAAYjLv7N4lzDB2soVKiX+Hm938LAAE9wAAAAAAAAAAAA4lxPtMWqaekV+L1/yAAy3/cqP+GP5IA8NeFf7XzL6xWjejSs35y6R/V+WnUz3K4ZnllfBS+3FpeT6P5OzJwlCTjJWa0AAA8GcSoZzWov7VN/epL9GwAA8VMvp4nC0MzoNSUW4NrXTW2vk016sAK7tSl6AAAAAAAAAAAAAAAAAAAAAAAAAAABwdS9rxBgo/wDTyD+6Sf6AAHibl31TjrEzSsqtOE/w5H+MGAAVAAAAAAAAAAAABtJXZiazrVp1H1bYAGW/7lR/wx/JAYLBVsbiYYahG85tJLzf9anDeRUMly2ng6Wtt33k93/l2VkHiDlqwHEeJpxVoyfMv+3ld/i2vkAAB4b41YXibCtvSTcf/Ak0vxsAATgpxcXszhHLvr3DGLyao9aU6lNX6NPmT/8AAn+BOEoScJKzWjAMY7UrdwAAAAAAAAAAAAAAAAAAAAAAAAAAA8P4c3E2DX978k2AAeNeXWzDBY2K3hOD+TjKP5yAAKnQAAAAAAAAAAAAzat7LBVJeVvv0/UAAy3/AHKj/hj+SA8JeFfZ03m+IjrK6p+S2cvnsvK/cDxqwHJjMLi0vii4v/t13X/pX4ANAAYDFywmLpYiG8JKS9U0/wBClVhVpxqQd00mvRgAHCf7niDOMN2nCf8A4HFt/oeImU/UM8qSivdq++vV/F/5Mm/mgDHS1jEAAAAAAAAAAAAAAAAAAAAAAAAAAAPDr/kqMJ6v/wBJkAAeMOcYWpRpZfDWpGXM3/Lo0l6u9/JJdwACp0AAAAAAAAAAAAOJ6nLg1FdX+X9IAAy3/cqP+GP5IOEOHKme5pDCrSC1m+0Vv83svN9rlChToUo0qStGKSSXRLRIDxiwftshhWW9Oa+5pr87AAAHh1max/DmGle8oLkf/bui/wDJbP5gAGVL2fGuYR/mp039yseKeU/Wsqji4r3qT1/wy0f42f3gGJlzVWAAAAAAAAAAAAAAAAAAAAAAAAAAAHAMlHiXBv8AvfmmgAOMOJqWR4B1FrVlpBefd+S6/JdTHVquJc6tWXNKTu292+rAAKnQAAAAAAAAAAAA4rnrSh6v8gADLf8AcqP+GP5IUXJpLc4A4XWR5WlUX76paU/LtH/t1fi2AeImGWI4Yxce0U//AAGSf6AAAHgvnCjWxGWyfxJTj6rSX3rl+5gAGGXLx3XXfDxf/k6RjsHTxmFqYep8M00/mrGNwlTCYmph6i96DafqnYKklCDl2HqAAAAAAAAAAAAAAAAAAAAAAAAAAAHCNVUs/wAFL/p5D8ZJABmWY4fL8JPFYiVoRV3/AJLzey8ziHPcRnOOniq3XSK/lj0S/Xu7smrxaAAKgAAAAAAAAAAABxRK+LjHtH9WAAZb/uVH/DH8keFnCv8AaGO/tGvH91SennPp/wCA7+tvMAOJKKrZNi6fenNf+SsAAA4aziWUZrQxsdoPXzi9JL7m/mU6kKtOM4O8Wrp90+oAENOPpJf9Av8A7lQeKWU/Vc2jior3aq/8mjo/ws/vDG1LRUEAAAAAAAAAAAAAAAAAAAAAAAAAAABga/1fFUq38sk/uaZo0A2krs4+4teb4r6vh5fuKb0/vP8Am9OkfLXqBJWk0ABPcAAAAAAAAAAAA4jd8c12SAAOF8rxGZ/VcHh1ec1FLy0V2/JLV+Rk2U4fKsBSwdBe7BW9X1b829WAGNp+0w1SHeLX4AAAB4T8R/2hlX1Gq/3lDRecH8P3fD5K3cACl73Hs32wyX/lRMPEXKfr+RznFe9S99ei+L8Lv5IbSVyrP2k3IAAAAAAAAAAAAAAAAAAAAAAAAAAAAyDF/W8pw2I/mhF/NxV/xA8SuLvYQeV4WXvSXvtdE/s+r6+WnXQCurVGABLVgAAAAAAAAAAAHEOuYT+X5IABRcmktWeDnCFTLcqp4/Fwca04pJNWcYpLdPVOT/BLuwAJJSVmAAAHC+f1cjzSnjKeqWkl3i91+q80jB4yhjMPDEUJc0JpNPyf9fIAML73HVd9sPFf+TphUpxqQcJK6as0cUYKWWZhWwL+zJr5bp/NWYAAAAAAAAAAAAAAAAAAAAAAAAAAAAeF+O+tcM0Fe7puUX8ndfg0HGfFFPI8C3B3rTuoL85PyX4uy7latUrVJVKkryk7tvdt7sAxKtMAB6gAAAAAAAAAAAHEK/4UJ+i/JAFGhVr1I0qUXKT0SSu2/JLVnCXgFn+a8tbMn9WpPo1eo/8At37P/bzTX8rOE/DPhvhpKWDoKVVf8vJ+9P5PaP8A26l5gAE3yxb7AAAAeGPHEctqrLMbL9zN+639iT6PtGX3J67NsAMs97jXHP8AlpU19+ocZ8W4bh7AOrKzqy0hHu+7/urr927MZi62MxE8RXlzTm2231bAAAAAAAAAAAAAAAAAAAAAAAAAAAADwUzFOGKwLfaa/wDSZf8AtpmuaYbLMHPFYh2jFfNvol5vZGe51ic4x08VXer2XSK6Jf1q9eoAYtbMAJbAAAAAAAAAAAABxJG2OfmkGBy/F4+vHD4SlKpUltGKbb+S1OEfo85pjeWtnVT2EP5I2lUfq9Yx/wDJn3SOGOBcg4bpqOXYdRlazk9Zv1k9fkrLskAABmNRU8HVn2jJ/cmAAAAcB+JrwUYYDNJN01pGe7j5S6uPZ7rbVbUK9LEUo1aMlKL1TTun6NAZB7/F+by/lVFffC/6HF3HeXcP0nBvnrvaCf4yf2V+L6Lqs6zrGZxjJYvFz5pv7kuiS6Jf8F6tsAAAAAAAAAAAAAAAAAAAAAAAAAAAADw5zeOW8RUJzlaE7wf/AG9t/wCTKJx3xZLOsX7Ki/3FN+7/AHn1k/07L1YAGKV4IAKm1gAAAAAAAAAAAAzLIcyzfMqdDL6Eqs3HaKvbV6vol5uyOEfo6V6nLXz6tyr/AJ9w1fpKey81FPykjIOFsmyDD+wy3DxprrZe8/8AFJ3lL5tgAABxVXVDIsZU7U5/fytL8QAAAAyLirNsklfBVmo9YvWL+T/NWfmYDxrqqKWMwib7xlb/AMlaf/pRW8bMIo/u8JJvzkl+Sf5GK47zWWLxeIwsvZfWGnK265VZJS3W+6syc5VJOUndvdvcAAAAAAAAAAAAAAAAAAAAAAAAAAAAAKc1OCkABiFemwAk7sAAAAAAAAAAAAPBLE2njKD7QkvlzJ/mgAAADxLxKocMYl9ZcsV85K/4XAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADBVN4AAVFeDQBJ2QAAAAAAAAAAAAHhDjFQ4h9k3/ABISXzVpflFgAAAHjPjPZ5PRw6es53+UU/1aAAAAAAACpVhTV5OxPNKK+FNlCXtaamuo4tAAAAAAASzDCRfK6iv6oswAAAAAAAAAAAAAAAAAAAhNwkpIjJSimgAGrOwE3dgAAAAAAAAAAAAcJ49YDPMLiG7KM1f0bs/wbAAAAPGfH+1zWhhU9KcL/OT/AMooAAAAAAAxmPjS9yGsvyKlSVSXNJ3YZf8A7tD0CUE9hoAAAAAM5oY3Lcphmc6L9lOXJFvROVm/W2j12vpfcxmZYnFP95LTtsvu/wAyh/Fj6oHFMlFoAAAAAAAAAAAAAAAAAAAwlblfIwAKytNhN2QAAAAAAAAAAAAAcM5mszyfDYu93KKv/iWkvxTAAAA4yzNZln2KxKd05NL0j7q+9K4AAAAAAGPxipR5Iv3n+A9QMv8A92h6ASjdAAAAAHh1wE86q/XcYrYeL2/na6f4V1fyXVrxxymGI4Irezjb2LhJJLZJqLt2tGT+QYf+ND1X5gbko2AAAAAAAAAAAAAAAAAAAMNiFNcstwAxKtUG0kN3AAAAAAAAAAAAADwazdVsurYCT1py5l/hl/k0/vAAADi/N1lWSYjFJ2kotR/xPRfi7gAAAAAAY3FQwtCVWXT8X2MHi51q03Ud3LUAy/8A3aHoATj1AAAADhLhutn+ZwwkNI7yfaK3+b2Xm+1zB4Ohg8PDD0I8sIKyS6JHFuW/2lkONwaV3UpzS9XF2/GwYf8AjQ9V+YA0mhqzsAAAAAAAAAAAAAAAAAAAUcZZWmRnGSvF3B2SMVVg5Lldxu4VKkKcbydkVc1gtIRuYDFzxE3GSWiHT7DTQDQAAAAAAAAAAHh/niyjPqNWTtCfuS9Jdfk7P5AAAB4z54pSoZXTe3vy/KK+67t5pgAAAAAAcSY72tdUIvSO/r/wP8yhU9nUjLsbgZf/ALtD0AGroatoAAAAeG3DKyfJ41KsbVqtpS7pfZj8lq/NsDirLf7NzzGYNKyp1JxXopNL8LGH/jQ9V+YATV0AAAAAAAAAAAAAAAAAAACbWx7ap/MyU5S3dwMVi4UI932K1adWXNJ3DJv4svQGrko22AAAAAAAAAAAA8PuIlnWS05zd6tP3Z+q2f8A28rP1v2AAAx2Mo4LDVMTWdoQTbfkjO81rZrmFXG1vim7+i2S+SsvkAAAAAAGMxMcNh51X0X49CcnOTlJ6sMHU9pRXdaAZf8A7tD0ACa6gAAAcAZAs5zylRmr04e/L0j0+bsvRsA8c8t+p8a4mSVlVUJr5xUX/wCTRZh/40PVfmAA1Z2AAAAAAAAAAAAAAAAAAAADE140abkypUlUk5SeoGTfxZegBKNgAAAAAAAAAAA4A4o/sLNYyqP9zU92fkukv+3X+DZGUZRTi7pgAB4vcVJpZNh5dnU/OMf/AG5/9u+YAAAAAAHFOJtThQT31fy2/ryAyypaTh3Ay/8A3aHoAEldWAAAA8HslWGymeOmverOy/wx0/GV/uQB9JbLeXG4DHJfFGcH/wButSX/AKWzD/xoeq/MACouoAAAAAAAAAAAAAAAAAAAAzG4j21TTZbAGTfxZegA0mrDQAAAAAAAAAAAeFnG8alOOT42XvLSm31X8j819nutOiuABxxxhQ4ewLlFp156Qj/7c/Jfi9O7WIr1cRVlWqy5pSbbb3be7AAAAAAAzzEe2x0+y0+7/g3Ao1PZ1FLsaNaBl/8Au0PQACSswAAClSnVnGnBXbdkvNmUZdDLsvo4SG1OKj62Wr+b1APpD5b9Z4UhiEtaVSL+Uk4v8XEw38aHqvzAAkroAAAAAAAAAAAAAAAAAAAAzGv7Olyrd/0wAyb+LL0ACa6gAAAAAAAAAABGUoyUouzRwL4n0sVCOBzafLUWim9FLyl2fns/J76NAHGHHeA4fpOF+eu1pBPbzl2X4vp3Wb5vi82xc8Xi5805fcl0SXRLov1AAAAAAArVFTpym+ib+4nJzk5PdgGCqe0orutAy/8A3aHoABUQAAB4eZcsdxJhYNXUXzv/ALdV1+KSADxMy3+0eEMwoWu/ZykvWHvr8YmG/jw9V+YAA1ZgAAAAAAAAAAAAAAAAAAAY+t7Ws7bLQAMm/iy9AAaurDAAAAAAAKlWnTjeckl56FHG4avP2dKak7X0GmgAAAOHvEHO8liqdOpz019meqXo916J28jBeNWAlFfWsLKL/utS/PlK/jRk8Y/ucPUk/PlX4qUvyM88W85x8XTwiVCL7ay/8CdrfJJ+ZUqTqTc5u7e7erbAAAAAAAM8q+zwFTz0+9/5ABltS03DuGX/AO7Q9AAJrQAAA8FcDz4/E4p/Yio/+BO//toAYihCvRlSmrxkmn6NWZWwc8HmUsNP4oTcX6xlZ/kAATWoAAAAAAAAAAAAAAAAAAAYmp7OlKXYADJv4svQACa1AAAAAACTUVdvRGZcSNN08L/4F/kv8yrWqVpc1STb8zhT/fpf4H+cQcE9hqwAAAAAAAAAAAHFFS2FhDu/yQAUpuE1JdBNNXRl/wDu0PQACWqAAAPBnCezyWtWe86jXyjFfq2AB4o5b9Q46xlNLSVRTX/b6U3+MmAAVOgAAAAAAAAAAAAAAAAAAAcQ410I04Lq7v0X/D/gQnGpFSi9AAyb+LL0AAmtAAAAAABtJGd5w8TJ0qT9xfj/AMDt94HCn+/S/wAD/OIDSaGrAAAAAAAAAAABxXO8qUfX9AAMDU56KXVaGX/7tD0AAewAAB4Y4dUeF8N3lzP75P8ASwAH0hMt9hxThMWlpVgl84Saf4OIABPYAAAAAAAAAAAAAAAAAAAOJK3tMa4/ypL9f1MJinRlZ7MTTV0AZN/Fl6AASV0AAAAAAcR5m4R+rU3q9/Tt8+vl6gHCn+/S/wAD/OIBON0AAAAAAAAAAAHFMr4qEf7v6sADLanLUcO5l/8Au0PQAAAAA4LpKlw7go/9O4v71f8AUAD6RWW+1y3AY1LWnV5flNX/ADggACfwgAAAAAAAAAAAAAAAAAABmFX2uKqT7thgcX7N8knp+QBk38WXoAANAAAAABjMVDDUJVZdPzK1Wdao6k3dsA4U/wB+l/hf5xACaswAAAAAAAAAADgfwz4b4lyZ4rMaLlVU3FSUpRaSSsrJ8u7b1VzH/Rv4eq3eFxVWD8+WS+7li/xMf9GnMIf7nj4T/wAcJQ/JzMf4CcaYa/s6UKtv5Zpf+l8hj/DnizBX9tl1XTqoOS++N0V8NWw8+StBxfZpp/iU5uE1JdDLmpYWDXYFFvZCoVH0FhZsWE7sWGgiSSk0AABw5BQybCRXSnD/ANJQAHi9lv17hDEpL3oOE1/27NN/+S3AAJfCAAAAAAAAAAAAAAAAAAAVpqFOU+ybHqBgcXe1Ob9P8gMm/iy9AACW4AAAAAcUYzmnHDxe2r9en4fmAHCn++y/wv8AOIASV0AAAAAAAAAAAeD/APyTsv8Ar5L8ogBXw1HEQ5KsFJdmk19zMf4dcJ46/tsupXfVQUX98bP8TNfB3LPZ/wDCU/ZW2jJuUfk23JfPm+Rm/DGY5RO2KouK6SWsX6NafJ6+QAE/iYAAGS/8izDf4If+koADjLHYLC5JiFi37s4ygl1bkmrL+tNydCcegAEtgAAAAAAAAAAAAAAAAAACGWYvM28Fg481Womoq6V213dkvmZ3wznGSVPZ5jhp0n05k0n6S2fybAMFi/aLkluvxDJv4svQABJvRFWDjKzAAAAAJyjGLk3ojF15YivKq+rADhT/AH2X+F/nEACSswAAAAAAAAAAPBqfNkFSParL/wBJgAAFSnCpBwmk0+j2M88Msqx154X9zPy1j/4D0+TS8jPOCs4ym8qtPmgvtR1Xz6r5pIAlq2AABw9LnyfCS704f+koAMfj8PgcNPE4iXLCKu3/AF1eyXVnFPEuIz3GutPSC0jHsv8AN9X8tkglTjLdEsKn8LJUJx6AS2AAAAAAAAAAAAAAAAAAAPDamqnFOEXnJ/dCT/QxGHo4ik6VaClF7ppNP1T0OIvA3hLNk50aTw9R9absv/AHeNvKKj6nEf0fOI8BeeXzjiYLovcn/wCAydvuk35GZZTj8srOhjaMqU10lFxf47rz2Itxaa3MJiVWjrujJv4svQC1yOHnLyI4WK31FGMVZIxy99egAAAABn+I9jgZJby0+/f8AAOFP99l/hf5xAAqIAAAAAAAAAADwVqp5diqfaaf3xt+gAAAZ5wDk2aXnyezqP7UdPvWz89E/Mzzw5zjLrzpR9tBdY7/ADjv9115kk43T3QAABwbVVXh7BP/AKdwX3JL9ACpUhTg5zdkldt7JLdnHHGE86xPsaLtQg9F/M/5n+i6LzbAAlThLdE8Kn8LK1KcI6gAAAAAAAAAAAAAAAAAAHhZG/FOHfZT/wDSJAGYZZgsxouhjKMakH0lFSX3O5xF9H7hnMbzwMpYab7e9D/wGTv90kvIz7wN4tyeTq4WCxEF1g/et5wdn8o8xlFOtTrzp1oOE4qzjJNNPzT1I0Zy2RHCr7TIwjHZAGO+KIAAAABxVXvOnSXRN/fp+jAA4U/32X+F/nEACSugAAAAAAAAAAPBPEJYjGUO6g/ucl/7cgAAADjvIsqr5TicXXpL2kISaktHdLS7W6vbR3AAAPDWuq3C+FfVKS+6Ul+VgDxD40+uTeXYKX7tfE19prov7q/F+S1AADGu1IAAAAAAAAAAAAAAAAAAA8LHbiigu6n/AOkMAA8XaVN4HDz5VzczV7a2s9L9gADHfFEAAAAAz2r7THz8tPuX+YAHCn++y/wv84gADAAAAAAAJ4ilDSUkYbhjOcVhI4vD4ac6ctnGLd+nS7/ArUalGbp1IuMlumrNfJgAB4RYtUOI1T/5+QlH7rS/9tAAAAPFDF/V+GK6T1m4xXzkm/wTAAAPBzFKrkVSj1hUf3NJ/ncDxE40+rQllmCl77+OS+yv5V5vr2Xm9AAAMc9IoAAAAAAAAAAAAAAAAAAA8M6nJxThPPmX3wkAAeLn/Iuw/wDjf/pLAAMd8aXkAAAAAcV+DGUZ/ltJQSo4qEIxU0tJcsUrTXXb4viXdpWOKeD834Zxn1XMaXK/syWsZLvF9fTddUgA4U/32X+F/mgACS1AAAAAADEV4UIc0uu3mV8dVraXsuweFv8AySGB/wAL/wDSpGbZHl2bUvZY2iprzWq9GtV8mjjLwtxOWQli8ubqUlq4/aivl8S9NV1TV2AAcIY76ln2Er30U0n6P3X+DYAAAB4147lwWFwifxScv/AVZf8ApQAAB4KY3lxOLwjfxRjJf9utp/8ApSDjrjCOTYf6vh3evNaf3V/M/Psu+uy1nOVSTlJ3b1be7YAABjZXqW7AAAAAAAAAAAAAAAAAAAHA9b2XEeCl/fS+/T9QADxc/wCRdh/8b/8ASWAAY13qfIAAAADKMOsTmOHofzTivvkkGdZFl2dYOWDzCkqlOXR/mnumujTTR4ieCGY5Fz43Kr18MtWvtwXml8SX8yV11SSuAcKf77L/AAv80AAT3AAAAAAOEOFMTxFj1Qp+7TjrOXZf5vovnsmeNPC+EwHD+Dng6ajGjNx+U1dtvq7xV2+rA8Lf+SQwP+F/+lSA8S/D6NKM81y6FlvUiuneSXb+ZdN9rgAJtO6MhzFZjleHxafxxi3621Xyd0AAAB4t5isVxC6MXpSio/N+8/zSfoAAAeGuYfU+JcPd2U7wf/by0/8AJrHFnE9DIsE6ktaktIR7vu/Jdfu6mNxtfG4ieIry5pyd2/6/BdAAAArS5qjYAAAAAAAAAAAAAAAAAABlOJWGzChX/knF/c0wADxc/wCRdh/8b/8ASWAAYp3qsAAAADgqkqvEWCj/ANPIv7nf9ADxE8Estz7nxuWWoYl6tbQm/wC8l8Lf8yWvVNu5nmQ5jkmMlg8wpOnUj0fVd09mn0aug4U/32X+F/mgACpuAAAAABgsFXxuJhhqEeac2kl5s4W4cw+Q5bDCUtZbyf8ANLq/TouyseJ2Wf2hwnjaaWsY86/7cak/wTXzA8Lv+SQwP+F/+lSAlFSTTV0eIvCP9hZjz0F+4q3cf7r6x+XTy72YAHg/mqxOSzwkn71GX/kstV+PMAAAFetToUpVajtGKbfoldma4+eYY6ti571JOX3u9vlsAAAYbETw9aFam7Si016p3RnWd4nOcXLF4h6vZdEuiXkvxer1AAACtPkptgAAAAAAAAAAAAAAAAAAAZHjFjMrw+Iv8cIv70rgAeLn/Iuw/wDjf/pLAAKj5ptgAAAAeHUOfijCLzf4RkwA4o4RyjiXBvC5jSUl0e0ovvF7p/g+qaPEHwhzfhaUsRSTrYX+dLWP+NdP8S91903Y4U/32X+F/mgACp0AAAAAA8HuGk1POKy7xh/7dL/21f8AbwYnD08RRnRqK8ZJp+jVmY7CTwmKqYap8UJOL9U7P8g8Lv8AkkMD/hf/AKVIA4s4fpZ7lVTBy+LeL7SWz/R+TZWo1KNSVKorSi2mn0a0aADwtzr+z8/hSm/crLkfrvH8dPmAAAHipnSy/IJ0Yu06z5F6by+VtH/iAAAAwc+anbsAAAGOn7qiAAAAAAAAAAAAAAAAAAAB4XY/63w1RjfWm5Qfyd1+DQAHi5/yLsP/AI3/AOksACrLlg2AAAAAeGv/ACVOE9Zf+kSAAnCM4uMldPocY+DmCo1qma5JDklZ81JfC9m3BdHp8K0fS1rOdOUdGgAKnQAAAAAMvwNXHYunhaKvKbUV6t2+7uZXl1HLsFSwlH4YJJfLr6vd+bA8Vst+o8W4uKWk2pr/ALeSb/8AJrh4Xq3CGB/wv/0qQAeLeQ/Uc3WNpr3K6v8A9vKyl96s/VsAKdSVOanB2a1T7NHC2eQzrKKOMW8laS7SWkvx1Xk0AAAeJ+frNM8lSpu9Oj7i839p/fp6JAAAAYWpyVF5gAABXqe0m2AAAAAAAAAAAAAAAAAAAB4K5laeKwDe9pr5e7L84gAeLn/Iuw/+N/8ApLAAxkrU7dwAAAADw5ny8UYR+bX3xkgAA4+4DVdSzDL4e9vOC695Jd+66777zw0JbaE8POO2oBU6AAAAAB4PZMsVms8dNe7RWn+KWi+6PN96APpAZbyY/B45L44yg/8At13X/pT+4PDL/kkcD/g/VgB4lZMsy4ercqvOl76/7d+L/wAlv87AAeE3E6wGYPLq8rU623lPp/4EtPXlAAA4+4mjkWUTqQlarP3YevWX/bq19bLqNtu7AAAAMPU9pBPqAABi6vJCy3YAAAAAAAAAAAAAAAAAAABwFmyyziDD1pO0ZPll6S0/B2fyAA8XP+Rdh/8AG/8A0lgAY2d5qPYAAAAA4Iq+z4jwT/vxX36fqAABx9wI6nNmOXx13nBde8orv3XXda3uTpxnuieF/lZKEo7oqbgAAAAB4WZUsFw7TqNe9Vbm/TZfgk/mAeOOW/WuGPbpa0Zxl8neD/GS+4PDL/kkcD/g/VgBOEZxcZK6ZnuWyy3M6+Df2JNL0vo/mrMAIycWnF2aPD7jCGfZeoVpf7RTVpLuukl69ez7JoAAxWKo4WhOvWkowim230SOM+KKvEGZyxD0px0gu0e/q9392yQAAAAUKvs5+Ro1oAASkoq7K1R1J3AAAAAAAAAAAAAAAAAAAANtjg7OlnGS0MU3eTVpf4o6P79/RoAPFz/kXYf/ABv/ANJYASaim2Tk5SbYAAAABktdYfM8NWv8M4P7pJgAAHH3Ad+fMcvj5zgvxlH9V80A0mrGLUVUsgAAAAChQnXqxpQV5SaS9XoYLCQwmFp4en8MIqK9ErfoAcX5b/aWQYzCJXcqcrf4krx/FIPDL/kkcD/g/VgAeMGXfV8+jiEtKsE/nH3X+CiABk+cYvKcZDF4WVpx+5rqmuqfX/M4U4twPEOEVWi7VF8UHvF/qn0f32d0AFetSoU5VKslGMVdt6JLu2eIXH0s7qPB4NtYeL32c2ur/urovm9bJAAAAAYXEJe5IAB2sYmv7R8sdgAAAAAAAAAAAAAAAAAAAAPB7iBYfG1Msqy92rrH/ElqvnH/ANJQAeLn/Iuw/wDjf/pLADG1LR5F1AAAAADbYy3FLF4KjiF9uMZfekwAADj7gNQUsxy+Om84Lp3lFdu66brS9gxLvVYAAAAB4f4H65xLhIdIy5v/AABOX5pAAcUZb/ZudYvBpWUJyS9Lu34WPDL/AJJHA/4P1YAHjVgVPLsNiktYTcflJX/9tAAMvzHFZfiI4nC1HCcdmv61XdPR9ThvxgwlaMaWbQ5JfzxTcX6rdfK69DA53lmPipYXEQn6STf3XuvmVq9GjHmqzUV3bt+ZnfiVw/lsWo1faz/lhr98vhX338jivjvM+IJclR8lFbQW3rJ/af4LokAAAAABQxTiuWWqIzjNXi7gTqQgryZXxLqaLRAAAAAAAAAAAAAAAAAAAAAGFxNXC14V6MrTg00+zWqOF8/o55ldPGU93pJfyyW6/VeTTAPFz/kXYf8Axv8A9JYBKSirsqzdSTkwAAAAAPDnGrF8M4WV9Ypxf/braX4JAAABx9wJ7HmzHL4e7vOK6f3ort3XTdaXs7WJPmbYAAAAB4N4RVc7q1n9im/vbS/K4AHjblv1XiqVZLStCMvmvcf/AKTf5nhmv+MSwP8Ag/VgAeJ2EWI4YxHePLJfKSv+DYAAAAAAAAAApOLumLFVUtyWJqtbjbb1AAAAAAAAAAAAAAAAAAAAAA4A4vlkGYWqu9CpZTXbtJenXuvOxSq061ONSnJOLV01s09mgPFz/kXYf/G//SWAYuvzPkjsAAAAAAeC2YqeBxODb1hJSXpJWf4x/EAAANzxL4Np5fSnmWDsqbdpR2s3pePk306dNNgAAAADwSw65MZW/wAC/wDSm/0AA+kDlnNhcHjkvhlKD/7eSkv/AEl/eeG//JJ4D/AvzYAHFOH+sZHjKXV0529eV2/EAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPD7xDllDWBx7bw72e7g/wBY9103XVFCvSr0o1aUlKLV01qmu6YeLn/Iuw/+N/8ApLAxOKVuWDAAAAAADwrzRYLiKnTk7Rqpw+e6/FJfMAAADxpzRQwuHwEXrJub9Iqy+9t/cAAAAAHgvSSyavU71GvujH/MADxbyz69wlibK8qdpr/t1q//AJK2eG3/ACSeA/wL82ABiqSrUJ0/5k196sAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABwzxtmuQSth581PrCWsfl1T8187mU+L2R4qKWLUqMut05R+Tjr98UeJXE+T5ngKEcJiIzaldpXulZk8XSWzuVcTOpotEAAAAAABhcTUwteFek7Sg016p3RleYUswwNLF0vhnFSXldbfLZgAAB4h5ys04grzi7wh7kfSO/3yu/mAAAAAHg/C3DrfepL8or9AAMywUMdgq2FntUjKL9JJr9Tw/w86HDGCpTVpRgk/W7uAAYqHJXnHs3+YAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB4O58q+BqZZUl71J80f8ADJ6/dL/0pAAAHGmerJslrYlO02uWH+J6L7tZeiG7gAAAAB4RL/jHF/jl+gABQoU6MOSCstX97v8AqAAZh/vlX/FL82AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAcMZ5UyXNaONhqovVd4vRr7tvOzMPiKWJowrUpXjJJp901dMAADxY4lWY5msDRd6dC6fnN/F/4D8PrfuAAAAAB4QSUuHWu1SX5RYAAAAYufPiKku7f5gAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB4ScXJx/sbEy11dNv73H9V812AADxA4shkOWv2b/AH9S6gu3eXpHp3dvMlJybbd2AAAAAB4MVVLJa9PtUb++Mf8AIAAAAxFVUqM6n8qb+5AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAUatSjUjUpytKLumt01s0cBccUc/wyo1mo4iC1X8y/mj+q6PyaAA4h4gwWR4KWKxUtFsusn2X9aLV6HEGfYrO8fPGYl6vZdIrol5L8Xd7sAAAAAA8EsQnTxlHs4P7+ZP8kAAAAcT4j6vkmLq31VOf38rt+IAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABhcVXwlaNehNxnF3TWjTOEvFjCYuEcPmzVOptz/AGZev8r/APJfNbFOrTqwU6ck4vZrVP0YHFHiLlOSxlThJVay+zF7P+89UvTV+RxBxFj88xTxGMnd9Etorsl/TfVsAAAAAAPBnFKnnNag/t0398ZL9GwAAADxMxSw/DGJ11lyxXzkr/hcAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADLc+zPLH/ALHiJQ8k3Z+q2fzRDxR4pjDl+sp+bhC//pP5mZcY59mMXDE4qTi90nyp+qjZP7gAAAAAADw7xv1TiXCyvpJuP/gScV+LQAAAB4041U8rw+FT1nNv5RVvzkgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAMLiJ4evCtD4otNeqd0YXEU8Th4V6b92aTXo1dAAAAeMWY/WM7hhk9KUF98tX+HKAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAeGGarH8OUYt3lSvB/LVf+StIAAAJNJXbOIszeZ5tiMXfScm16bR/BIAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA8G859hmVXATelVXX+KP8AnG/3IAAAPETOVlnD9eadpzXJH1lo/ujd/IAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAMrzCrl2NpYul8VOSa+XT0ezMvxtHHYSniqLvGaTXo1/V/MAAA8X8++t5pDL6b92itf8UrN/crLybYAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHg9xIqlGeUVpaxvKHo/iXyevzfYAADiXPaOS5XVxtT7K0XeT2X379ld9DE4iria061WV5SbbfdvVsAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADLcxxGXYyni8PK04O6/yfk1o11TOHc+w2d5dTxlB77rrGS3T9PxVn1AADxM4uWc49YbDyvQpXS7Sls5enSPld9QAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA4I4wrcO43mfvUZ2U4/8Aty81+K081gcdhsdhoYnDTUoSV01/X3rdPRgAeJnH0aMJ5Tl87zek5LousV5v7XZab3sAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABwlxrmHD1b90+ek/ig3o/Nfyvz69Uzh3jPKM9pr6tVtPrCWkl8uvqroDH5hhMBRdbFVFCC6t2/p+W5xl4rzxMZYTKLxi9HUejf8AhW8V5vXskN3AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAACMnFpp6owXHfEeDio0sXKy/mtL/ANKTKviTxTVhyyxb+UYJ/eopmNzDF46p7TFVZTl3k23+IAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAF7jLoui6LoAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAEMAHsdDodABgAtgOoD2OgPYR1AAOoNXC1zdCLIQAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAIewAHQLaBYaHsWHqAtgtce4FtC10D2AGFgtY6gHQWwM6AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB0OgAAxnUNzoPYZYsLYOh1Op0HsdDqPYYDLAHUC2h0FsDAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAewtgAOo9xm4eR0Nxm4bHQudTqF9AHsMYMvc6j2NzqDL3PIDqPYAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAANyxZAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA2dBMYAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAzoIYAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAzoWGAPYAANTcAZ0AAAAYzcYMLgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHQewAM6FxgAdA6A9g6AdAuM6AB1OpuAMBbC2AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA6C1QAaHQewdQ6B0GbnQGdTobnQ6DHsA9gexoHUtoD2A0NDyOh1AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADoIYBY6DOodDoM6DAGMsHQ6DGPY6j2Bq40HUBg9gGW0LagAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAFi2hYauixYeoWALFgGrlixYGrjVyxYGrliw1csANXLFtCwFgLAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB/9k=)
![[[@Watabe-Uchida2017-gi]](https://doi.org/10.1146/annurev-neuro-072116-031109)](data:image/jpeg;base64,/9j/4AAQSkZJRgABAQAASABIAAD/4QB0RXhpZgAATU0AKgAAAAgABAEaAAUAAAABAAAAPgEbAAUAAAABAAAARgEoAAMAAAABAAIAAIdpAAQAAAABAAAATgAAAAAAAABIAAAAAQAAAEgAAAABAAKgAgAEAAAAAQAABK+gAwAEAAAAAQAAAaAAAAAA/+0AOFBob3Rvc2hvcCAzLjAAOEJJTQQEAAAAAAAAOEJJTQQlAAAAAAAQ1B2M2Y8AsgTpgAmY7PhCfv/iG5BJQ0NfUFJPRklMRQABAQAAG4BhcHBsAhAAAG1udHJSR0IgWFlaIAfjAAYABwANADUAEGFjc3BBUFBMAAAAAEFQUEwAAAAAAAAAAAAAAAAAAAAAAAD21gABAAAAANMtYXBwbAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAEWRlc2MAAAFQAAAAYmRzY20AAAG0AAAEhGNwcnQAAAY4AAAAI3d0cHQAAAZcAAAAFHJYWVoAAAZwAAAAFGdYWVoAAAaEAAAAFGJYWVoAAAaYAAAAFHJUUkMAAAasAAAIDGFhcmcAAA64AAAAIHZjZ3QAAA7YAAAGEm5kaW4AABTsAAAGPmNoYWQAABssAAAALG1tb2QAABtYAAAAKGJUUkMAAAasAAAIDGdUUkMAAAasAAAIDGFhYmcAAA64AAAAIGFhZ2cAAA64AAAAIGRlc2MAAAAAAAAACERpc3BsYXkAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABtbHVjAAAAAAAAACYAAAAMaHJIUgAAABQAAAHYa29LUgAAAAwAAAHsbmJOTwAAABIAAAH4aWQAAAAAABIAAAIKaHVIVQAAABQAAAIcY3NDWgAAABYAAAIwZGFESwAAABwAAAJGbmxOTAAAABYAAAJiZmlGSQAAABAAAAJ4aXRJVAAAABQAAAKIZXNFUwAAABIAAAKccm9STwAAABIAAAKcZnJDQQAAABYAAAKuYXIAAAAAABQAAALEdWtVQQAAABwAAALYaGVJTAAAABYAAAL0emhUVwAAAAwAAAMKdmlWTgAAAA4AAAMWc2tTSwAAABYAAAMkemhDTgAAAAwAAAMKcnVSVQAAACQAAAM6ZW5HQgAAABQAAANeZnJGUgAAABYAAANybXMAAAAAABIAAAOIaGlJTgAAABIAAAOadGhUSAAAAAwAAAOsY2FFUwAAABgAAAO4ZW5BVQAAABQAAANeZXNYTAAAABIAAAKcZGVERQAAABAAAAPQZW5VUwAAABIAAAPgcHRCUgAAABgAAAPycGxQTAAAABIAAAQKZWxHUgAAACIAAAQcc3ZTRQAAABAAAAQ+dHJUUgAAABQAAAROcHRQVAAAABYAAARiamFKUAAAAAwAAAR4AEwAQwBEACAAdQAgAGIAbwBqAGnO7LfsACAATABDAEQARgBhAHIAZwBlAC0ATABDAEQATABDAEQAIABXAGEAcgBuAGEAUwB6AO0AbgBlAHMAIABMAEMARABCAGEAcgBlAHYAbgD9ACAATABDAEQATABDAEQALQBmAGEAcgB2AGUAcwBrAOYAcgBtAEsAbABlAHUAcgBlAG4ALQBMAEMARABWAOQAcgBpAC0ATABDAEQATABDAEQAIABjAG8AbABvAHIAaQBMAEMARAAgAGMAbwBsAG8AcgBBAEMATAAgAGMAbwB1AGwAZQB1AHIgDwBMAEMARAAgBkUGRAZIBkYGKQQaBD4EOwRMBD4EQAQ+BDIEOAQ5ACAATABDAEQgDwBMAEMARAAgBeYF0QXiBdUF4AXZX2mCcgAgAEwAQwBEAEwAQwBEACAATQDgAHUARgBhAHIAZQBiAG4A/QAgAEwAQwBEBCYEMgQ1BEIEPQQ+BDkAIAQWBBoALQQ0BDgEQQQ/BDsENQQ5AEMAbwBsAG8AdQByACAATABDAEQATABDAEQAIABjAG8AdQBsAGUAdQByAFcAYQByAG4AYQAgAEwAQwBECTAJAgkXCUAJKAAgAEwAQwBEAEwAQwBEACAOKg41AEwAQwBEACAAZQBuACAAYwBvAGwAbwByAEYAYQByAGIALQBMAEMARABDAG8AbABvAHIAIABMAEMARABMAEMARAAgAEMAbwBsAG8AcgBpAGQAbwBLAG8AbABvAHIAIABMAEMARAOIA7MDxwPBA8kDvAO3ACADvwO4A8wDvQO3ACAATABDAEQARgDkAHIAZwAtAEwAQwBEAFIAZQBuAGsAbABpACAATABDAEQATABDAEQAIABhACAAQwBvAHIAZQBzMKsw6TD8AEwAQwBEdGV4dAAAAABDb3B5cmlnaHQgQXBwbGUgSW5jLiwgMjAxOQAAWFlaIAAAAAAAAPNSAAEAAAABFs9YWVogAAAAAAAAYWwAADgyAAAKQlhZWiAAAAAAAABvHAAArpUAABaZWFlaIAAAAAAAACZOAAAZOQAAslJjdXJ2AAAAAAAABAAAAAAFAAoADwAUABkAHgAjACgALQAyADYAOwBAAEUASgBPAFQAWQBeAGMAaABtAHIAdwB8AIEAhgCLAJAAlQCaAJ8AowCoAK0AsgC3ALwAwQDGAMsA0ADVANsA4ADlAOsA8AD2APsBAQEHAQ0BEwEZAR8BJQErATIBOAE+AUUBTAFSAVkBYAFnAW4BdQF8AYMBiwGSAZoBoQGpAbEBuQHBAckB0QHZAeEB6QHyAfoCAwIMAhQCHQImAi8COAJBAksCVAJdAmcCcQJ6AoQCjgKYAqICrAK2AsECywLVAuAC6wL1AwADCwMWAyEDLQM4A0MDTwNaA2YDcgN+A4oDlgOiA64DugPHA9MD4APsA/kEBgQTBCAELQQ7BEgEVQRjBHEEfgSMBJoEqAS2BMQE0wThBPAE/gUNBRwFKwU6BUkFWAVnBXcFhgWWBaYFtQXFBdUF5QX2BgYGFgYnBjcGSAZZBmoGewaMBp0GrwbABtEG4wb1BwcHGQcrBz0HTwdhB3QHhgeZB6wHvwfSB+UH+AgLCB8IMghGCFoIbgiCCJYIqgi+CNII5wj7CRAJJQk6CU8JZAl5CY8JpAm6Cc8J5Qn7ChEKJwo9ClQKagqBCpgKrgrFCtwK8wsLCyILOQtRC2kLgAuYC7ALyAvhC/kMEgwqDEMMXAx1DI4MpwzADNkM8w0NDSYNQA1aDXQNjg2pDcMN3g34DhMOLg5JDmQOfw6bDrYO0g7uDwkPJQ9BD14Peg+WD7MPzw/sEAkQJhBDEGEQfhCbELkQ1xD1ERMRMRFPEW0RjBGqEckR6BIHEiYSRRJkEoQSoxLDEuMTAxMjE0MTYxODE6QTxRPlFAYUJxRJFGoUixStFM4U8BUSFTQVVhV4FZsVvRXgFgMWJhZJFmwWjxayFtYW+hcdF0EXZReJF64X0hf3GBsYQBhlGIoYrxjVGPoZIBlFGWsZkRm3Gd0aBBoqGlEadxqeGsUa7BsUGzsbYxuKG7Ib2hwCHCocUhx7HKMczBz1HR4dRx1wHZkdwx3sHhYeQB5qHpQevh7pHxMfPh9pH5Qfvx/qIBUgQSBsIJggxCDwIRwhSCF1IaEhziH7IiciVSKCIq8i3SMKIzgjZiOUI8Ij8CQfJE0kfCSrJNolCSU4JWgllyXHJfcmJyZXJocmtyboJxgnSSd6J6sn3CgNKD8ocSiiKNQpBik4KWspnSnQKgIqNSpoKpsqzysCKzYraSudK9EsBSw5LG4soizXLQwtQS12Last4S4WLkwugi63Lu4vJC9aL5Evxy/+MDUwbDCkMNsxEjFKMYIxujHyMioyYzKbMtQzDTNGM38zuDPxNCs0ZTSeNNg1EzVNNYc1wjX9Njc2cjauNuk3JDdgN5w31zgUOFA4jDjIOQU5Qjl/Obw5+To2OnQ6sjrvOy07azuqO+g8JzxlPKQ84z0iPWE9oT3gPiA+YD6gPuA/IT9hP6I/4kAjQGRApkDnQSlBakGsQe5CMEJyQrVC90M6Q31DwEQDREdEikTORRJFVUWaRd5GIkZnRqtG8Ec1R3tHwEgFSEtIkUjXSR1JY0mpSfBKN0p9SsRLDEtTS5pL4kwqTHJMuk0CTUpNk03cTiVObk63TwBPSU+TT91QJ1BxULtRBlFQUZtR5lIxUnxSx1MTU19TqlP2VEJUj1TbVShVdVXCVg9WXFapVvdXRFeSV+BYL1h9WMtZGllpWbhaB1pWWqZa9VtFW5Vb5Vw1XIZc1l0nXXhdyV4aXmxevV8PX2Ffs2AFYFdgqmD8YU9homH1YklinGLwY0Njl2PrZEBklGTpZT1lkmXnZj1mkmboZz1nk2fpaD9olmjsaUNpmmnxakhqn2r3a09rp2v/bFdsr20IbWBtuW4SbmtuxG8eb3hv0XArcIZw4HE6cZVx8HJLcqZzAXNdc7h0FHRwdMx1KHWFdeF2Pnabdvh3VnezeBF4bnjMeSp5iXnnekZ6pXsEe2N7wnwhfIF84X1BfaF+AX5ifsJ/I3+Ef+WAR4CogQqBa4HNgjCCkoL0g1eDuoQdhICE44VHhauGDoZyhteHO4efiASIaYjOiTOJmYn+imSKyoswi5aL/IxjjMqNMY2Yjf+OZo7OjzaPnpAGkG6Q1pE/kaiSEZJ6kuOTTZO2lCCUipT0lV+VyZY0lp+XCpd1l+CYTJi4mSSZkJn8mmia1ZtCm6+cHJyJnPedZJ3SnkCerp8dn4uf+qBpoNihR6G2oiailqMGo3aj5qRWpMelOKWpphqmi6b9p26n4KhSqMSpN6mpqhyqj6sCq3Wr6axcrNCtRK24ri2uoa8Wr4uwALB1sOqxYLHWskuywrM4s660JbSctRO1irYBtnm28Ldot+C4WbjRuUq5wro7urW7LrunvCG8m70VvY++Cr6Evv+/er/1wHDA7MFnwePCX8Lbw1jD1MRRxM7FS8XIxkbGw8dBx7/IPci8yTrJuco4yrfLNsu2zDXMtc01zbXONs62zzfPuNA50LrRPNG+0j/SwdNE08bUSdTL1U7V0dZV1tjXXNfg2GTY6Nls2fHadtr724DcBdyK3RDdlt4c3qLfKd+v4DbgveFE4cziU+Lb42Pj6+Rz5PzlhOYN5pbnH+ep6DLovOlG6dDqW+rl63Dr++yG7RHtnO4o7rTvQO/M8Fjw5fFy8f/yjPMZ86f0NPTC9VD13vZt9vv3ivgZ+Kj5OPnH+lf65/t3/Af8mP0p/br+S/7c/23//3BhcmEAAAAAAAMAAAACZmYAAPKnAAANWQAAE9AAAApbdmNndAAAAAAAAAAAAAMBAAACAAAAVgEuAesCxAODBFMFQAYnBxoIGAkfCj0LTgxrDZAOuQ/sES0SZROjFOMWKhd5GL8aDxtXHKQd8x88IIQhzSMRJFIljSbLKAApMypmK5csyi33LygwUTGAMqsz1TUCNjE3XTiKObY64TwNPTg+YD+JQLFB3EMIRDZFY0aQR7hI3koBSyRMR01qTo9PtFDYUfhTFlQzVU9WbVeNWK5Zy1rhW/Bc9137XwBgCmEZYi1jQWRSZV9mZWdhaFNpPGogawFr4Gy/baBug29ocFBxOHIgcwhz73TWdb52qHeTeH55bHpZe0l8PX03fjZ/OYBBgUuCV4NlhHSFhYaUh6SIs4nAisyL1ozfjeeO7Y/ykPWR9pL1k/WU+pYElxWYLJlHmmObf5yYna6ev5/LoNSh2qLgo+mk9KYDpxWoKqlCqlyrd6yUrbKu0a/xsRGyM7NTtHO1kLapt7q4wbnDusC7u7y1va++qb+jwJ3BlcKKw3zEasVTxjbHGMf5yODJz8rGy8PMwc28zq/PmNBz0UPSDdLT05vUZtU11gXW19eo2HbZQNoG2sfbgNw13Ofdkd473uPfjeA74Ovhn+JS4wPjr+RV5PnllOYs5r/nUufk6IXpL+nk6qjrbuwz7PXtr+5j7xHvtfBQ8ObxfPIX8sbzj/Ru9WH2ZveK+OT6xf0u//8AAABWASMBsAJqAzID6ASxBYAGWAc9CC0JJwonCyIMMA06DlEPYxB+EZ8SvhPpFRAWOBdhGIgZtBreHAsdMx5bH4AgoSG/Itsj9SULJh8nNShJKWAqeSuTLLEtzC7oMAQxITI8M1o0dDWLNqQ3uTjNOeI69DwGPRs+MT9NQGxBj0KwQ9BE7UYIRx1IMklFSlVLY0xuTXhOfk+DUIVRhlKIU4pUjFWKVoRXeVhoWVRaRFs/XEZdWl53X5VgsGHCYspjxWS1ZZxme2dXaDBpDGnsatBrumypbZtuj2+CcHJxXHJAcx9z+3TVda12hndieEV5MHokex18GX0VfhB/Bn/2gOKByoKxg5WEd4VdhkqHRIhMiWGKgYuljMaN3o7uj/WQ95H1kvOT8JTtleyW7pf4mRCaOZtunKadzp7jn+mg5aHfotmj0aTIpb6mvKfKqO2qIatdrJyt2a8UsE6xiLLCs/m1LrZkt5u40roFuzS8X72Fvqa/vMC+warCkMN9xHfFgMaPx53Io8meyo7LdcxTzS3OBM7az7LQi9Fq0lHTRdRF1VHWYtd12ITZh9p922bcQ90a3ezeud+H4FjhLeII4ubjw+Sd5XHmQOcM59Too+l/6mnrZOxo7W3ucO9w8HDxcvJ283n0d/Vp9kf3D/ez+EX4y/la+e360PwM/dr//wAAACsAxAFLAdgCZQMGA48EKwTPBXoGLwbpB6UIaAkvCfkKygufDHcNUg4yDw4P9hDZEcASqROTFH0VZxZSFz0YKRkVGfwa6BvSHL4drh6jH5sgliGUIo8jhiR2JWEmRScgJ/gozCmfKnMrSCwhLQIt6S7XL8gwujGqMpgzgDRgNTk2BjbPN5U4WDkZOdk6nTtkPDI9Bj3ePrY/j0BoQURCJkMQRABE80XnRtZHu0iWSWVKJUrgS5hMT00HTcNOg09JUBhQ6lG+UpNTaFQ7VQ1V3latV3xYSlkVWd9aqFtvXDVc+V29XoBfQ2AIYNFhnGJrYzxkDGTaZadmcWc4Z/xov2mAakBrAGvAbIFtQ24EbsVvhXBBcPhxq3JZcwFzqXRPdPR1mHY8dt53gHgdeL15YHoWetp7q3yDfVh+KH72f8OAk4FngkSDKoQYhQuGAIb0h+OIzYmvioyLYYw0jQeN2o6wj4qQa5FHkhKSxpNbk+yUepVBlkKXiJkImoqb5J0ZnjKfOaA5oTWiL6MopCGlGaYRpwqoCakPqiGrPaxgrYeura/TsPyyKLNatJK10bcUuFm5nLrXvAW9Kb5Hv2LAf8Gewr7D38T/xiHHQ8hnyYzKscvWzP3OKc9b0JfR3tMy1I3V7tdT2LnaIduV3S/fDuF35Ivoee1O8vb5Uf//AABuZGluAAAAAAAABjYAAJOVAABWjQAAVukAAJG3AAAmuwAAFwoAAFANAABUOQACh64AAkeuAAFrhQADAQAAAgAAAAEABAAIAA8AFgAfACkANABAAEwAWgBpAHgAiQCaAKwAvgDSAOYA/AESASkBQAFZAXMBjQGpAcUB4wIBAiECQgJkAocCrALSAvoDIwNNA3oDqAPXBAgEOgRtBKME2QUSBUsFhgXDBgEGQAaABsIHBQdKB5AH2AghCGwIuQkHCVcJqAn6Ck0KoQr3C04LqAwEDGIMww0lDYkN7g5UDrwPJQ+REAAQcRDkEVkRzxJGEr0TOBO2FDoUwxVSFeMWdRcGF5YYJhi4GU4Z6BqIGzEb4xyfHWIeKx74H8YglSFkIjQjBiPbJLMljyZsJ0soKykNKfAq1Su7LJ4tfi5cLzkwFDDwMc0yqjOJNGg1SjYuNxU3/zjtOd460jvKPMU9xD7HP85A2kHoQvZEAEUGRghHCEgISQlKDEsUTCFNNE5OT25QlFG8UuVUDVU0VllXfVigWcRa51wLXTBeVl98YKRhzmL4ZCRlU2aFZ7xo+mpDa5ls+W5jb9JxRnK8dDR1r3cteLF6PHvQfW9/HIDWgpSETIX2h5SJLIrFjGWOFY/fkc2T3ZX/mCCaOZxMnl+geaKgpNqnLamgrDCu4LGhtF63C7muvF2/K8IfxUbIoswMzyjR9tSR1yjZ0dye35fi0uZg6fntEu+s8gD0MPY59//5ivq4+9T8u/2j/nD/N///AAAAAQAFAAoAEgAbACYAMgA/AE0AXABtAH4AkQCkALkAzwDlAP0BFgEvAUoBZgGDAaIBwQHiAgQCJwJMAnICmgLDAu8DHANLA3wDrwPkBBoEUgSLBMUFAQU9BXsFuwX8Bj8GgwbJBxAHWQelB/IIQQiTCOYJOwmTCewKRgqhCvwLWAu0DBIMcgzUDTkNoA4LDngO5w9ZD84QRhDAET4RvxJDEsoTUxPdFGoU+hWPFioWyxdxGBYYthlRGeYaeRsNG6QcQBzkHZEeSh8OH90gsyGOImkjQiQYJOslvSaQJ2YoQiklKhIrCCwFLQguDi8SMBExCDH5Mugz1zTKNcE2wTfKONs58jsOPDA9UT5pP3RAckFoQlhDR0Q5RTRGOEdHSF9JfkqhS8dM8E4bT0VQa1GGUpRTl1SVVZRWoFe/WPFaNVuAXM1eHl90YM9iI2NmZJVltWbOZ+Ro+2oWazNsVG12bppvwHDqchdzRnR2dad22ngQeU16j3vYfSZ+fX/jgWeDEITKhnSIBomHiwSMiI4bj8ORgpNYlUOXO5k7mzudOZ8poQSixqR2phunvaliqxWs3a7FsM+y9bUxt3y5y7wPvkTAcsKmxOnHQsmyzDHOnNDd0vTU8dbn2OPa6Nz03wHhCuMU5STnSOmW7DfvivOm92z5zvuE/MP99P75//8AAAACAAgAEQAeAC0APgBRAGYAfQCWALAAzADqAQkBKwFNAXIBmAHAAeoCFgJEAnQCpgLaAxEDSQOEA8AD/QQ7BHoEugT8BUEFigXXBikGgQbdBz4HoggICG8I1Ak5CZ0KAgpqCtULRAu7DDkMwQ1QDeYOgg8hD8AQXBD2EZASLRLNE24UDRSoFUIV3BZ5Fx4XzhiOGWIaQhsqHBQc+x3dHrgfkiBtIUwiMCMYJAYk+SXxJu8n8yj9Kg4rJCxALWAugC+dMLgx0zLzNBo1STaCN8M5DDpcO7A9Bz5gP75BIkKTRBZFrkdaSRFK0UyaTm9QVVIyU9pVWlbMWEhZ01tmXPheg2ABYXBi1WQ3ZZxnCWiBagpro21NbwBwt3JvdB51w3d1eWR77n6kgHmB1oL8hAKFCIYQhzOIc4nUi1WM6Y6IkC6R2ZOIlTuW8Zipml6cCp2nnzegvaI/o8ClRKbKqFCp1KtUrNCuRq+5sSiyl7QItX22/LiJuia70L2CvzbA6cKcxE/GBMe7yXPLK8zkzp7QWNIU09LVj9dK2P/arNxP3ejfdeD94oLkA+WC5wLog+oE63/s7u5G73nwnfGN8nXzRPP39Kr1TvXb9mj29vd29+n4XPjP+UP5q/oN+nD60vs1+5j78fxJ/KL8+/1U/a3+A/5Y/qz/Af9V/6r//wAAc2YzMgAAAAAAAQxCAAAF3v//8yYAAAeSAAD9kf//+6L///2jAAAD3AAAwGxtbW9kAAAAAAAABhAAAJzfAAAAAMoq1YAAAAAAAAAAAAAAAAAAAAAA/8AAEQgBoASvAwEiAAIRAQMRAf/EAB8AAAEFAQEBAQEBAAAAAAAAAAABAgMEBQYHCAkKC//EALUQAAIBAwMCBAMFBQQEAAABfQECAwAEEQUSITFBBhNRYQcicRQygZGhCCNCscEVUtHwJDNicoIJChYXGBkaJSYnKCkqNDU2Nzg5OkNERUZHSElKU1RVVldYWVpjZGVmZ2hpanN0dXZ3eHl6g4SFhoeIiYqSk5SVlpeYmZqio6Slpqeoqaqys7S1tre4ubrCw8TFxsfIycrS09TV1tfY2drh4uPk5ebn6Onq8fLz9PX29/j5+v/EAB8BAAMBAQEBAQEBAQEAAAAAAAABAgMEBQYHCAkKC//EALURAAIBAgQEAwQHBQQEAAECdwABAgMRBAUhMQYSQVEHYXETIjKBCBRCkaGxwQkjM1LwFWJy0QoWJDThJfEXGBkaJicoKSo1Njc4OTpDREVGR0hJSlNUVVZXWFlaY2RlZmdoaWpzdHV2d3h5eoKDhIWGh4iJipKTlJWWl5iZmqKjpKWmp6ipqrKztLW2t7i5usLDxMXGx8jJytLT1NXW19jZ2uLj5OXm5+jp6vLz9PX29/j5+v/bAEMAAgICAgICAwICAwUDAwMFBgUFBQUGCAYGBgYGCAoICAgICAgKCgoKCgoKCgwMDAwMDA4ODg4ODw8PDw8PDw8PD//bAEMBAgICBAQEBwQEBxALCQsQEBAQEBAQEBAQEBAQEBAQEBAQEBAQEBAQEBAQEBAQEBAQEBAQEBAQEBAQEBAQEBAQEP/dAAQAS//aAAwDAQACEQMRAD8A/fyiiigAooooAKKKKACiiigAooooAKKKKACiiigAooooAKKKKACiiigAooooAKKKKACiiigAooooAKKKKACiiigAooooAKKKKACiiigAooooAKKKKACiiigAooooAKKKKACiiigAooooAKKKKACiiigAooooAKKKKACiiigAooooAKKKKACiiigAooooAKKKKACiiigAooooAKKKKACiiigAooooAKKKKACiiigAooooAKKKKACiiigAooooAKKKKACiiigAooooAKKKKACiiigAooooAKKKKACiiigAooooAKKKKACiiigAooooAKKKKACiiigAooooAKKKKACiiigAooooAKKKKACiiigAooooAKKKKACiiigAooooAKKKKACiiigAooooAKKKKAP/0P38ooooAKK+J/29P2nvFX7JvwZ0/wCJfhDSbLWb281q20xob7zBEI5oLiUuPKZG3AwgDnGCa5n4H/FT9vXxZ4zsbf41fCnQPB/gy6tbiaXVbTUYrqWFxAz258lL2RmVn2hht6E8jrQB9/UV8RfsmfFrWZ/gTr/xA+OPxh8NePYdI1W7E3iDS/LtNOs7VY4XSCZzFbJ5i79x+QYEirucjJ7nwx+2z+yj4z1vTvDfhn4m6Rfapq11FZWlssrLLPczOI440V1Uku7AL2OeKAPqSivI9F+PXwb8RWfjC/0PxdYXtv8AD8SnX3jlyumiASNJ5/Hy7RDIT/umqP8Aw0Z8DR4N0H4hnxrpo8NeKL3+zdMv/O/cXd5vePyY2xy+6Nxj1U0Ae1UV4ppn7R3wI1jT/Fmsab460qbTfA0iw61d/aFW1spH3BVedsRkkqQArHnjrgVf+FXx6+Dfxwt725+E3i/T/E405gtytpLulh3Z2l42AdVbB2sVwcHBOKAPXKK8m+Ofxj8KfAL4VeIvix4ykxp+g2xlEQYLJczt8sNvHn+OWQqg7DOTwCa+Xf2Gf2xvFH7WaePIfFng+Lwfe+Cbq0tXgjuHndpLjz/MWQOiFGjaHGPUnPSgD77oorwr9pj4o678FvgV4u+KHhm0tr7VNAtkmt4bzf8AZ3d5kjxJsZWxh88EUAe60V8s2H7SlnJ+yg/7R9/YCK8tdGluLjTRnK6zblraTT15Lbvtym3XuTir/wCzn8c734k/s46L8bPit/Zvhi4nTUG1MpL5NhaixvZ7Vm8yZ22oBECWZ8Zz2oA+l6K8v+G3xs+EXxijvJfhb4w0vxSNOYLcrp91HO8O4kKXRSWUNg7WIw2OCa5zxB+05+zv4Veyj8SfEjQNNfUJ5raBZ9RgRnltpmt5hgtkCOVGjdjgKwKk5FAHudFcf41+IPgb4ceHJfF/j7X7Hw9okJUNeX1wkEG5/uqHcgFm/hAyT2FVvAfxN+HnxQ8Pf8JZ8OvEdh4j0fcyNdWNwk8SOgyyOVJ2MAQSrYIHUUAdzRXhvhr9pr9nrxlfarpnhT4i6Fq11okE11dx21/DI0dvbAtNNgN80cYBLOuVHc16De/EPwJp3ggfEu/8QWMHhNrWO+GqvcItkbWYKY5hMTs2OGXa2cHIx1oA7GivI/GPx8+Cfw9TUX8ceONH0I6S9vFdJeXkUMkUl1GZoEKMwbfJGC6qASVBIGATXSH4mfDkeB4/iY/ijTE8Iywi5XV3u4VsDC3Af7QzCPBPGd3XjrQB3FFeYfDX41/CT4xw30/ws8X6Z4pXTHVLr+z7lJ2hLZ271U5UNg7SRhsHBODXKan+1L+zfo11p1lqvxN8PWs+rPJHapJqVupkaKV4H6v8oEsbxktgb1K5yCKAPeqK+T/i7+2D8Jfgz8XvBvwr8Xa5pVkfEcV9PqF3dajFb/2TFbwLLbNNGwORdMSke5k6Ejd0r0nw74s1XWfjDqul23i7RNR8Of2HY31ppFspOqwtcSPi8lk8wq1tMoxHhBkjg8EsAez0V40f2iPgQPiAPhUfH+if8Je0v2caX9vh+1eeekOzd/rT/wA8/v8AtU/xt1/xB4b8FR6l4a8UaN4QvG1HT4Te67GZLQxzXCRtAArxnzZgdkfPLEDgnIAPXqK828Z/GP4T/DqS9h8e+L9K8PSadaw3twl9dxW7R21xI8UMhDsDtkkjdEx95lIGSKNO+MXwq1b4fP8AFjTfFumXPg2JGkfV0uozZIqNsYvLnau1vlYEgg8HmgD0mivIdC/aA+B/ijxzP8M/DnjzRdT8VW28Sabb30MtyGiz5ihFYlmTB3qMlcHcBXe6T4t8Ma9q+saBouq219qXh+WOHUbeGVXltJZoxLGkyg5RnQhlBxkHNAHQ0V8o/HH9r74T/B/4SWXxXsdb0vX7bWL2Cz0yJNRjhS+/06OzvXhlAk3CzVnkl2qceWVYrnI968LfEbwD438J/wDCeeD/ABDY6x4c2zN/aNrcJLa7bckSnzVJXCFTu54wc0AdpRXyf8Af2wPhN8fde8SeFvD+t6WmsaPrOo2FjaQahHdT6jYWOzbqMSAKfKl3EjG5QBnea9L0X9oj4EeI/HMvwz0Hx/omoeKoXeNtNgv4XuTJHnfGqBvmdMHci5ZcHIGKAPZaKK/P/wDaR/bD8DaX4SbS/gd8SdGu/Gdl4h0Oyura0uLS9nW2udRhtrpRE/mA/I5RmUEoT1U4oA/QCivMfiT8afhL8Hbe0uvin4v0zwsmoMUtv7Quo4GmK43eWrEMwXI3EDC5GcVtT/EfwBbeDE+I03iPTx4VljilTVRcxmyaOZ1jjYThthVnYKCDgkgUAdpRXkPhf4//AAP8beMrr4eeEPHmi6z4lst/m6faX0M1wPK/1gCIxLFP4wuSv8WKo+LP2kvgB4F8VReB/GPxE0LR9fmdY/sNzqEEc6O/3RIhbMe7PG/bntQB7ZRXHeKviH4D8DPbR+NPENhoRvIrmeH7dcx24eKzQSXDgyMo2xIQznooOTXnC/tRfs5v4Ik+JC/EnQD4ZiuTZNfjUIfJF0F3+RndnzdvzBMbivzAbeaAPeKK8I+InxBa88C+GPF3wz8c+H9Ksdc1bS44dS1Ai6sr+1uZQpt7Zo5EBnnztiIY88YBOR1HjP41/CH4dHUU8eeMtJ0B9Jit5rpL28igeKO7MggJV2B/emKQIAMsVbAODQB6fRXI+GfH3gjxn4Sh8e+FNestV8OXETzJqNvOklqY4siRjKDtAQqQ2SNpBBwQa5X4d/Hf4MfFy+v9M+GPjbSPFF3pg3XMOn3kVxJGhO0OVRiShPAcZUnjNAHrFFcx4k8beD/Bpsh4t1qz0b+0nkjtjeTpAJXhieeQKXIB2RRvI3oqkngGuY+HHxp+EnxfXUG+Fvi/S/FQ0l1ju/7Ouo7jyWfO0sEJwrbTtb7rYOCcGgD06ivH3/aB+B0fxBHwofx5oq+MGkEI0o30P2rziMiLy92fNI5Ef3/au/tfFvhi+8S33gyz1W2m13TIIbm6sUlU3EEFwWEUkkYO5Vcq20kYODigDoaK4Jvin8N00LV/FD+JtPXSPD99Jpuo3ZuY/ItL2GVYJLeZ84SRZWVCpwQxA61l2fxu+D2oW/ii9svGmkS2vgmTytbuBeReRp0nPyXEu7ZGwKkEFsggg4IxQB6jRXmfw2+M3wn+MNpdX3wt8W6Z4phsXCXB0+5jnaFmzt8xVO5d2DtJABxxmuW1b9p79nXQZdKh1n4leH7OTW5JI7NZdRgUzNFM9u5GW4VZo3jLHCh1K5yCKAPdaK81+JPxk+FXwesrPUfil4r07wvBqDtHbG/uEhM7KAWEasdzbQQWIBC5GcZrr/DfiXw74x0Oz8T+E9TttZ0jUU822vLOZJ7eZM43JIhKsMgjg9RigDboriPiX470n4XfDvxN8SNdVn0/wxpt1qU6J990tYmlKL/tNt2j3IrA+EUvjPTfhHoGp/F7U45vEcll9u1adhHDDby3GbiSEFQqiK2D+UrHkogZiWJNAHq1FfMOrfHb4d/GHRPFXw6/Z6+KOiXXxEj025l0/wCw3dteNHcRr+7cod6PGJNqyFc7VJ5U4NepfBn4kWnxf+FXhb4mWdubNfENhDdSW7fegnIxNCT3McgZM98ZoA9Mor4x8AftPa54h/a7+IP7OPiTS7Wy0vQ4Yn0K+i3iW7mhtLO5vYZtzMpdFvY2Taq/IrZzjNJa/tPa5qv7aDfs36RpdrJ4ZstIuZbrUjvNx/a1rHb3MlvGQ3l7EgvLctlS25iMjGKAPs+ivHPGf7Q3wK+HXii18E+O/H2iaBr14EMdle30MM2JPuFlZgUDfwlsA9s123iPx94I8HvYp4s1+x0Y6mtw1sby4jgEy2sLXE7IXIBEUKtI5zhUBY8DNAHW0V5r8N/jH8KvjBa3178LfFmm+KodMlEN02nXMdx5LtnaHCEkBsHaTw2DgnBrET9oj4ESfEA/CpPH+iN4vEnkf2WL+H7V5/8Azx2bv9b/ANM/v+1AHstFeV/En44/B/4PNYp8UvGOl+F31MsLZL+6jheYLjcyox3FVyNzY2r3IrodG+I3gHxHq9voHh/xFp+p6jd6dHq8EFtcxzPLp0r+Wl3GEY7oGcbRIuVJ4zmgDs6K4PUvil8ONGuNatNW8TafaTeHJLGHUkluY0NnJqbKlmk+T8jXDMoiDYLZGOorC8J/Hj4LeO/F9/4B8F+ONH1zxFpiu1zYWd7FPPGI22uSqMc7GOHxnaeGwaAPWaK8T8WftJfADwL4qi8D+MfiJoWj6/M6x/YbnUII50d/uiRC2Y92eN+3Pauy8Y/E/wCHHw8aBfHvifTfDpuoLi6iOoXUVqHgtNnnyKZGUFY/NTce25fUUAd1RXhcf7SvwMv/AIaa58W9B8caPqfhjw+CLu9ivovIhmYhY4ZXz+7eR2RUDAFiy4ByK8d8LftWeG/jd+zXqXxL+G3jPw54T8T2ekWd7qA1C6W/tPD893yEv9hiYD5JFBZV+YZ2nBUgH2tRXmvjD4q+APhT4Ts/E3xX8V6XoFpMsafa7qZLWGecpkiBZGLNu5YICzY9cZrovBvjbwf8Q/D1t4s8Ca1Z+INGvM+TeWM6XEDlThgHQkZUjBHUHg80AdRRXgvhf4jLo/8AwsnX/iN468P3WgeGtXeFJLX/AEf+xrZYoz9m1GR5GXzwXDE/LwwPQgL0Oj/Hj4LeIPH9x8K9C8caPqHi+08zzdLgvYpLpWhyZE8tWJ3xgEug+ZQCWAAoA9Zorxvxr+0R8CfhxPfWvjzx9oug3OmzxWtzDd30UU0U80SzpG0ZbfuMTrJjH3CG6c11k/xM+Hdt4Y0zxrN4l05fD+syW8VlqH2mM2tzJdsEgWKUMUcysQqAE5PAoA7iiua1bxl4T0LW9M8Na1q9rY6rrUd1NZWs0qpNcx2Sq9y8aE5ZYlZWcj7oIJr5v+Bf7Y/wg+Ofi3xR4K0LXtJTVdH1q707TLeHUYrmbVrK1toZzfwIAp8pi8i/LvH7onceQAD61orxrwb+0R8CPiH4quPA/gbx9ouu6/beZusrO+hmnIi/1hRVY+YEx8xTIHeuN/aO/ae8B/s3weEj4rvLFbnxTrdhpohur1LR4bK4nSK7v8MrForRH3ycAdAWXOaAPpeivJNV+PfwU0PwDZ/FPV/HGj2nhHUSVtdUkvIha3DBmUrDJuxI2VYbUyeDxwa7DwV478F/Ejw/B4s8Aa5ZeItGuSwju7CdLiFmU4Zd6EgMp4KnkHqKAOror518K/GHxBe/tK+OPgP4osra1g0zR9M1/QJ4Q4lvLC4Z7e8M25iu6G6UKCgA2sMjPXgZfi58d/iJ8S/iX4I+CFp4bisPh7eaVp327XFvHjuru5tXub6NTauMNb77dQNvVnyelAH2RRX59fAz4vfthfFvxD4pt7u28CWeleBPFtx4Z1XYmqfaJvsKwSXEtrmRk+aObEfmY+YfMMV1fxL/AGqdc8EftJeH/hZp+k2t34MSfSNO8R6m5cXFjqPiMXa6XFHhhHtL26eaWBwJkxigD7aoorxOw/aS/Z/1XxzH8M9L+Img3nimWRoU06HUIJJ2mXrEFVjmQYOU+9x0oA9sorzDxn8a/hD8OjqKePPGWk6A+kxW810l7eRQPFHdmQQEq7A/vTFIEAGWKtgHBroPDXj/AMD+MfCMPj7wrr1jqvhueJ5k1G3uEktTHFkSMZQdoCFSHyRtIIOMGgDr6K+B/Hn7XHhDxN8SfhB4a+A3xD0nXbbW/Fn9l69Bp81teO1mbK4lVWyHdFaSMESR4zjAbGRX0j+0P8YrP4AfBfxV8X77T31VPDlssqWiOIzNNNIkEKFyDsQySLvfB2rlsHGKAPZ6K+W/htq37YP/AAkOjv8AFbSPBd74c1VHNzJoNzfRXWmHyzJGT9qEkd2pYCMmMxnLBgNoJPF/DH9q3UPE/wC0948/Z/8AGOlW+l2ul3ktp4bv4i4/tCWxtre6vreXcxHnRx3McqbQA0YfjKHIB9sUV4B8KPi3rXj34mfFzwTqlpb21n8PdXstPtJYt4kmjubCG7ZptzEbg0hA2hRgDIzzW34T/aH+BPjvxhP8P/Bnj/RNb8R23mb7C0voZpz5WfM2KrHfswd+3O3HOKAPZKK5rxd4y8JeAPD914r8caxaaDo1iAZ7y+mS3gj3HA3O5AySQAOpJwOa+SPAX7SVl8Vf2t7fwZ8NfGNj4l+HkvgGfVylg1vOg1aLVo7Ys0yqZkdYXwYmYDBDFc4NAH25RXjWqftEfAjQ/Hkfww1jx/oln4sldYhpst/ClyJXxsjKFvlkbI2ocM2RgGum8ZfFX4Z/DuQRePfFWmeHXNrLegahdxW2baB0jklHmMuVV5EUn1ZR1IoA7+ivPfAnxZ+GPxP8NzeL/h74p07xBotszrNd2dzHLFCyLuZZWB/dlVIYhsEDnpXN+Cv2ivgP8SPFE/grwB8QNE8Q67bo0jWdjfQ3EpRPvsgRjvC/xFc474oA9morxvxf+0P8CfABuU8bePtE0N7O8/s+ZLu+hieO78tJTCys24MI5Udhj5VYMcA5qa+/aA+B2m+LNH8CX3j3RIvEHiBIZNPsTfwGe5S5AaBo1DciYEGL/npn5M0AevUV5DJr/iD/AIXxB4WTxRoy6KfDj3raAYz/AG01wLsRfblffj7IFIiI2f6w9eRjIvP2n/2dNPvNF0+9+JXh+G58REf2fG2owbrgGRogVG77pkVkDHALAqDkYoA91orhviB8Tfh58KdD/wCEl+JXiOx8NaWXESz386QI8jAkIm4gu5AJCrk4B4rT8H+NPCPxB8PWvizwLrNpr+jXoJhvLGZLiCTacMA6EjKkEEdQeDzQB01FeVeM/jl8G/h3/ag8d+NdI0FtF+zfbEvLyKF4TeK7W4ZGYNumWKQxqASwRiAcHHU6J488E+JPCEPxA0HXrG/8MzwNdJqcNxG1mYI875DMDsCptIYk/KQQcEGgDrKK8r+HHxx+Dvxflvrf4XeM9K8UTaZj7THp93HO8QY4VmVSTsYj5WxtPYmtWL4q/DWbwC3xUh8T6c/g5I3mbVxcxmxEcbmJ3M2dmFcFSc8EYoA7+iuB8U/FT4aeCFDeMvFGm6IpspdSBvbqKAGzhaNJJwXYfu1eWNS3Tc6jqRXK+Jf2jPgR4N8L6H4z8WePNH0jRfE0ST6Zc3V3HEt5E4DCSEMQzrhgSwGACM4oA9oorjJviN4Bt/BkfxFl8RaePC0scUqaoLmM2TRzOscbLOCUId2Cgg8kgVzPh348fBbxd44vPhr4W8caPq3inTxIZ9Ntb2Ka5TyjiQFFYktGeHUZK/xAUAes0V41r37RPwI8LeOI/hp4k8f6JpnimVo0GnXF9DHcB5sGNGVmG13BBRWwzAjAORXpfiTX9N8J+HdU8U6y5i0/RrWe9uXAyVht4zJIQO+FU0AbVFeAfBLxf4pl+BFh8Uvi3e7bvVrW58RTx7UVNOsLsveQWgKKu77LbMkbMxLMykk9MeI/spftW+E/Gvw88F6b8W/iLosvxN8ZLLfppJubSG5igvZ3lsbUQxBPnFq0QUMPMf73OaAPu2ivlzxl8R/GU37WXw8+DXhO/FrpEWg6x4j8SQ+VE5ntQ0dlp6B3Vnj/ANJd3OwqW2YJK5FezwfFH4cXXgWb4n2/iXT5PCNuk0smrLcxmyVLd2ilYzZ2YR0ZWOeCCOtAHeUVwfjP4o/Dn4deGY/GXjzxLp+gaJNs8u7vbhIIpDINyLGXI3sw5CrknsK0fBfjrwZ8RvDlt4w8Ba3Z+INEvN3k3llMk8DlCVYB0JGVIIYHkHgjNAHV0V4pov7SPwA8SeNx8N/D3xE0LU/E7MyLp9tqEEs7SJkvGoRiGdQDuQEsMHI4rc8Z/Gv4Q/Do6injzxlpOgPpMVvNdJe3kUDxR3ZkEBKuwP70xSBABlirYBwaAPT6K5Dw14/8D+MfCMPj7wrr1jqvhueJ5k1G3uEktTHFkSMZQdoCFSHyRtIIOMGvjbx5+1x4Q8TfEn4QeGvgN8Q9J1221vxZ/ZevQafNbXjtZmyuJVVsh3RWkjBEkeM4wGxkUAffFFFeVaV8cvg5rviDR/Cmh+NNJ1HWfECXEthZ213HNPcR2pkWaRI0JbYjRSKWI25RhnIIoA9VoryDw5+0D8DvF/jaf4b+FvHmi6t4ntvM36da30MtxmLPmKFVjuaPB3qMlcHcBUnj749/BT4V6vY6B8SPHOj+GtS1LBt7a/vYoJXVjgPsdgVQnje2FzxmgD1uivln9lP4k+Mfi94f8b/EHxDqAvdCvPFusWnhpUiijSPRdOlFpEQ0ahpDJNFK5Z2Y8gA4FfU1ABRRRQAUUUUAFFFFABRRRQB//9H9/KKKKAPyP/4LP/8AJqOh/wDY22H/AKRX1fQH7Of7MfxH+DU194x8XfG7xL8R7DUdBktYtK1iWV7W2klMUqzxh7iUb0WMxj5R8rnnsfsLxp8P/AfxI0lNB+InhvTfFOmRTLcJa6rZw30CzIrKsgjnR1DhWYBsZAJGeTXUrBAsAtVjUQhdgQAbQuMbcdMY4xQB/LV8ONO0y/8A+CQ3xJl1LUZtP+x+P0mhWKMyC5m+z6ciQyYI2odxbcTgFQcE4B+1P2Tvhp4c+J37VOh/GnxbcfDTSB4a8P22m6d4d8M6tYatPd3tqMpqBhhJETxjLK3+sTZGP4S1fbP7Vf7JNx8Qf2cr/wCDP7N1rofw/F3qltqd1p8FounabqaQj95BcCyjBXzCsTFwpJ8pFJC8j5K8Bf8ABPn4j+Ivjj8Pvin4z8J+CvhJpXgC+j1J7Xwg93LdapcQSpLGsplxFHEGjAyDuCsy85BUA+S/hv8AGH4efCG1/bv+HPxE1UaP4i8XTazY6RZyRyNLe3Mx1S2WOIIrZO+eLJOBtbdnaCRwXxZz/wAOkPgNtYqf+Ez1DBBwR+/1fkGv6RL34JfBvUvGE/xC1HwNod14nuYzFLqcunW0l5IjJ5RDTMhc5j+Q5PK/L93imXfwN+Cl/wCEbDwBffD7w9c+F9Kma5s9Jk0m0ewtp3LlpYbYxGKNyZHJZVBO5ueTkA/Hj/gpz8D/AAx8Dv2fPh5ofwf8NWmg+B7TxDbnXvIt2KXEkUJjtJtQMWJJxjzAzOSxZgAQxWum/Ym8BWeoftYv8VPDfxJ+Hepsvhl7a/0PwLbTWMclozoIppLfy1jDJJ5YfJDDCZUE5P7Q6xo2keIdLutD1+xg1PTr2MxXFtdRLNBNG3BSSNwVZT3BBFcN4B+DHwi+Fcl1N8NPBWjeFZb3id9MsILR5RnIV2iRSyg9ATgdhQB+fP7f3we/ae+MnxI+HFv8NfBll41+HfhKRNYvtMu9Rt7KG+1VJHVI7lZZUZ4o4guAowRJIpPPHgv/AAS9174wXX7Rv7QsOv8Ahi0srHUteuLnxFLHdK7adq4urspaQqHPmxlmmBkAYDYvPzc/uXXI+G/h/wCA/Bl/q+q+D/Dem6Fe+IJzdalPYWcNrLfXBZmMty8SK00m52O5yTlic8mgDrq+Nv8AgoMSP2NPikRwRpsf/pTFX2TWN4h8OeHvF2jXXhzxXpdrrWk3y7LizvYEubeZQQ22SKQMjDIBwQeRQB+cd54A8VD9pyf9nlNMnPw81fxFB8UpLrYTbCOFT5+ns33dz61HDdbO6OxwRXzvpJsx+w5+ztc+MMH4fw/EQP4p83m1Gn/2xqIia8z8n2UXZg83f8udueM1+3tctY+B/BWmeFm8D6b4f0+08OOksbaZFaQx2JS4ZnlU26qI9sjOzONuGLEnJJoA+JPHl74T1P8Abl+CEnwsntLnW4NI8QnxK+msj48PtaoLIXhi48s3vl/Zw3G7JHFeI/CnwF4Kv/2L/wBpHVbzQrKe+1LU/iC1xcPAjTSNayXJgLSEFv3RUNHz8rfMuDzX6Y+AfhL8LvhXBd23wz8I6T4Uiv2D3C6XYwWYmYZ2mTyVXdtydueg4GBWxZ+B/BWnaHqHhjT/AA/p9to+rtcve2UVpCltdNeEm5aaJVCSGYsfNLA78ndnJoA/MXTL/Sz42/Y01r4sTRHwnL4JY2U18QLMeJpNOs2haZpP3fnND5v2cvzv3bPmrtLeP4da/wDtX/HWLRtUttO+HN58Pra08a6haTpBaQ6zLNcqJXnH7pbiPT2Yu55UY3c5r791n4c/D/xF4Qj+H2veGdM1HwvDFFBHpVxZwy2KRQALEiW7KY1WMABAFAXAxjFHhT4d+AfAnhs+DvBXhvTtB0Jt+6wsbSK3tm8wYctFGqqxcfeJBLd80AfB3wf8R658G/iD8Lv2avFNz4Z+I3hbxDpN/Y+GNd0mFINRtbLTrUSul/bK0sLwzRIE8+F1V3A3pk5rxrRdOvJtR8P/APBO++LS2/hrxu1/MJCXE3gXTgut2IkPcNPJBZE9MxkfT9LvCvwT+EnwwutS8RfC3wBoXh/WryJw0un2FvYvOfvCN5YYwVRmAyMEZ5wTXlXwY+FHxDf4s+KP2iPjZbaXYeLdc0610LTdN0qZ7uHTNHtZHuGje6kihM008773IjCqFQL6AA8x+EnhLwvrn7c37SGta1pNrf31hYeD7e3muIUleKG70+fz0QsDtWXy03gfe2jPSvAPhX8SNH+EX7Iw0tvD2ma3FqHxO1Pw1o9pq+1NIsZrjW52tp7pij+XBbMpkyq5DBQpUkEfrDY+GfDema1qfiXTdJtLTV9bEAv7yGCOO5uxaqUgE8qgPKIlYrHvJ2gkLgGsGf4YfDW58MX3gm48J6TJ4e1OWWe7042MBs7iad/Mlklg2eW7u/zsxUkt8xOeaAPz2+Fd3rkX/BRCS18QeLtB8TazcfDi4XUk8PWX2G2tnh1S2MEMyG6u3eRFclXkcNsYfKqkV5r8CNZ/Z60b9hb4qSeOJtCSSa98XLrUUjW/2uadry6Fokq/6wzFPKFuCN33Nnav1N8J/Cj4XeA2s38EeENI0B9Pgltbd7CwgtnhgndJJY0aNFKpI8aM6g4ZlUtkgGvmT9nT9kfwZ4J8EaYPi54M8Pa14z0jVtXvbbUXs4LyeGK81Ge7t9lxLEHDIsgIH8DZ2+tAHzP4fsLvw544/YX/AOFqpHa6iPC/iCyvXvwFb7S2kWXkQztJ/wAtcKVwxyXBA54r2O7OuSftgfGc+B2B1hvhfpf9mGMjH2kzX32cqRx9/bjFfbHjz4a/Dz4paQmgfEnw1p3ijTYpRMlvqVrFdRpKoIDqsqsFbBIyMHBI6E1oaT4J8G6DqP8AbGh6DYadf/Y4NO+0W9rFFN9its+RbeYihvJiydkedq5OAKAPin9inXfgFY/stfDS3jvtGtb8LZx38d7JAl5/wlDuEuBKsxEn2xrssE3fvDldvG2ui/4KAf8AJBLP/sa/C3/p2tq+iR8DvguPHH/CzB4D0IeLt/m/2v8A2bbfbvN/56efs8zfjjfu3Y4ziu38QeGfDfiywGleKtKtNZsllinEF7BHcRCaBxJFIEkDLvjcBkbGVYAggigD4juPDPh3Xv8AgpPPfa3ptvqFxpPwvsZrN7iJZTbyvrN3GZI9wO19jFdw5wSM4Jz8neLrGz0b9lL9tvQdKgS006y8Z6sYLeJQkUXnW+nu4RRwoLEnA4yTX7Gr4Z8Nr4jfxiuk2g1+S0WwbURBH9sazSQyrbmfb5hiEjFwm7aGJOMkmvn39pL4G/8ACwv2f/iZ8Ofhjpem6Xr/AI6gd5JBGlpHdXzmMNPcyRoS8hSMKZGDMQAM4AoA+WP2hbr4Sah4G/Z58NfBuTR5vEr+L/Dc3huPRWgZ4bWBvMvpoxb8i1W3D+eeE6bucV3Hwf8AH3gj4eftNftSJ4716y0AxahoGpn7bcRwZszo8QMy7yCyBhtJXOGwDyQK+tPA/wAEvhT4E1VvFvhvwXouj+JryBY7zUbKwt4LqYkDzN0yIrsHYZbJ+Y8nJrS8QfB/4T+LPFen+O/FPg3R9X8R6UFFpqV3YQT3cARtyeXM6F12MSVwflJJGCTQB+Ptt9gb/glH4P1u8jX7LYeJobmWWVMeTB/wmEm923DKjB+bOPev200qTRLvTkbQ2t5rCQHabco0LBuTjZ8pzmsYeAfAo8KXHgRfDmnL4au1nWbSxaQixkFy7SzB7cL5beZI7O+V+ZmLHJJNL4J8BeCfhtoEXhX4faDZeHNHgZ5EtNPt0toA8hy7bIwBuY8k9TQB+Z3wn+2p8EP2udC+Hpii8fW/jH4gDTre32rfo80Ze18hVxIC45hI4LdOhxyPxW1H4J3f/BOzwHpnwgm05/E7R+HF8IQ2DR/2kniUXFvu8lU/ei6D+Z9oP3vv7+tfqTF8Jfhdb+PX+KkHhHSo/GUkZibWFsoRfsjL5ZBuAvmHKfLnOdvy9OKo6V8Efg3oXjOf4jaL4F0Ow8VXLO0mqwadbR3rNJnexnVA+58nc2ct3JoA4T9rR/F8f7LvxQfwYZP7dXw5qBhNvu80HyG8wxY+bfs3bMc5xjmvgn496v8As9r+xt8HNJ+Hd1ohjfWPCB0WGBoGugwu7c3LIo/eCXbv+0Nw24sJOSQf2Brx7Tv2efgJpF3qN9pfw38OWtxq8kct5JHpNorTyRSrPG0hEfzFZkWVc9JFDj5hmgD5e8H3vhbTP28fjBL8U5rW21afQPD3/CKPqLImdFWGYaitoZeMfbMmYL7E8Zr408bf2bJ+w3+1bc+Ddn/CuZ/GU7eGzF/x6G2+22Au2s8fJ9lN0JfL8v5M79vFfsd4++FHww+Klra2PxM8JaV4qgsnMlumqWUN4IXOMmPzlbaTgA4xkcHivHP2pfgnqfxP/Zh8UfBP4W2ljpl1qFtZ22n25AtLOFLe6hl2ARoQihIyFCrjoKAPnj9o+5+E+p61+zZ4e+CsmkT+IIvG+j3OjR6IYGMOg26SPqTx/Zuln9mGJAMI3y+nHlfx0+Jn/C0vhL+0ZJpd34N8AeEdAuNd0G+hvrJbrXdc1XT4DEHY+fbR27zShVtmaO4mOFYbSAK/TTwr8G/hJ4F8Qah4t8FeC9G0HW9V3fa72w0+3trmcOQzB5Y0ViGYbmBOCeTzzUN/8Evg1qniq48dan4E0K88RXcTwzajNpttJdyRyKUdXmaMuwZCVbJ5U7TxxQB+fGo6Xpnjvxn+whD4ztItaivNF1K8mju0EySXEXh2C4jkZXBBZZlWQE9GAPUV6V8Kfh74Fl/bw+Pd9L4fsZJotF8MPGzW8bbGv4blbplBGAZxDGJCOX2jOa+408DeCop9AuY/D+npN4UjeHR3FpCG02KSIQOlmduYFaICMiPaCg2njir1p4Z8N2Gu6h4osNKtLbWdWjhivL2OCNLq5jtgwhSaZQHkWMM2wMSFycYyaAPxf0WKOz/Y68A6RaqIrPS/jXb2ttEowkUEfiabbGo7KMnAr7C0Twv4c1z/AIKJ+N9U1rTLa/u9L8CaIbSWeJZWtzNeXSyNGWB2syjBI5xkdCa+wl+Gfw3TSYdATwppK6Xb3w1SK0FjAIE1ASmcXSx7Non80mTzQN+87s55rch8M+G7fxBc+LbfSrSLXLyCO1nv1gjW7lt4mLRxPMB5jRozMVUkgEkgcmgD8WfFFlqml/sm/tF6N4WijtdA0f4x6hHfQCGR7aDQkvrGW7QwW7I5tlVmMscbLmLeMgcj7D8JeAvG/i/40/Cr4m634/8AAdxbeFbTUlsbTwtZT2s2paZfWnkmJS9/cK9vE/lSqFTapUYxmvt7SvCfhbQotTg0PRrLTo9auZr2/W3t44Vu7q4AE084RQJJZAAHd8s2BknFcX4L+BnwW+HGsXPiH4feA9C8NapdgrLdadpttazsrdV8yJFbaTyVBxnnGaAPmH9s3QNE8T/ED9nPQvEdjDqenXXjtPOtrhBLDIE0+6cB0bKsNwBwRg96zdR0qTRf2+9dh8D2sFhqGp/CKSYLEixpNewav5VtJIFADMoIQE87cDoBX3Tq3hnw3r93pt/rulWmo3OjT/arGW5gjmktLjaU82BnBMcm1mXcuDgkZwTR/wAIz4b/AOEkHjL+ybT+3xafYP7R8iP7Z9jMnm/Z/Px5nleZ8/l7tu75sZ5oA/Fv4P8AgLx78Xf2NtN+HEfj7wJ4YspJoxqL3enXcfiDTvEMF4JJJLqeTUVA1D7Umd7RAuCNq7WAr7B0DxT4c8Fft7/E1fGer2mjHVfBPh+5tnvJktknitZ7tZ3QyMAQh+9gnA69K+ptT+A3wR1rxkvxE1jwBoF94oR1kGpz6ZbSXnmJja/nNGXLrgbWJyOxFafjX4Q/Cn4kX+nar8QvB2j+JrzSCTZzalYQXclvuOT5bSoxUEgHA4yAeoFAH48+IdZ03xH/AME+v2pPEehXIudO1X4ia9eWlxHlRJBcazaSRSLnBwyMGH1r62/ay8IeBvh9pXwG8P3Wm2uk/CTRvGWnQ61DsCWMccdvMmnm9z8ptxdFPMaXKlipc5Oa+4bv4b/Du/0PVfC994W0q40bXbh7vUbKSxge2vLmRldpriEoUlkZkVi7gsSoJOQK6LWdF0fxFpV1oXiCxg1PTb6Nori1uokmgmjbhkkjcFWU9wQQaAPhDxJeeGNT/b5+F83wrmtbjUoPDOvDxfJp7Ky/2Uwt/wCy1umj+XP2rcYg3OMkcYr5u+HfgLwVcf8ABNX4zazPoVlJf38fjq8nuGgQzS3FldXv2aRnI3FovLTYc/LjjFfqr4C+Ffwz+Fdlcad8M/CmleFba8cSTx6XZw2ayuOjSCJV3EA4BOcDgcVoW3gHwLZeGLvwRZ+HNNg8O6gtwtzpkdnCtlOt4Wa4EluEEbiYuxkDKd5Yls5NAHwjqHhK916X4Q/FX4eeP9B0b4r+HvA9sv8AZXiMLcWuoaXfRRPJI8aSJc2582Ij7VEG6FGVhkV7h8Af2jPAnjX4OeD/ABn4hj03wJL4k1O50K0sRcxC1udUguZoDHYSYRZ1neJni2rls45PJ9b8YfBH4N/ELTtO0jx14G0TX7LR0WKxhvtOt7hLWNQAEgDofLQBQNq4GBjGK3Lj4b/D27sNB0q68MaZLZeFriG70iBrOExafcW4IhltU27YXjBIRkAK9sUAeDftxwTz/smfE0wqXSDSjcToOr2tvKktyn/AoVcfjWx+0l8YtI+HXhDQdFGlWPiK/wDiVqUPhzTrPU5Vh0uVr6Ny73sjJIPsyxKxdQjNJkIB82R794g0HSPFWg6l4Y8QWy3ul6vbTWd1A/KSwXCGORG9mViDXj3hP4IaNP8ABfw/8IPjTaad8QrfQIUtg+o2kdzHPHaFo7SV45gw88QbBI46vuIODQB8VeBI9c8eft2eENN1PxP4d8RL8KPCuq3048Mad9hs9PuNXkjso9PdzdXTyOI1aUBim0c7FLYr6b/YgDH9mvw7crxbXl/r11aL/ds7nWLyW2XPcCJlx7V6V4p+EOlWvgHxHoHwb0/SfA3iDWNJOk2uo2lhFCbWICQQ4ECoSsBlkeJM7VdiccnPoHgjwdoPw98HaJ4E8LW4tdI8P2cFjaR91ht0EaZPdsDJPUnJPNAH5ZeMbDU9H+LPx5+Nnh2zmvtZ+E3jjw3rwgtl33FxpjaDa2urW8Y7l7KWVgO7xp6Vu/BvwjrPhf8AaL+CGqeLITB4o8Z+GvGfibWlb7yX+s3VjcvC3/XvG0duB2WICv0/07w54e0jUdT1jSdLtbK/1qSOa/uIIEjmu5IoxEjzuoDSMkaqilySFAUcACkuPDfh2712z8U3Wl2s2tafDLb2188EbXUEM5UyxxzEb0SQopdVIDbRkHAoA/Nn9nbUvgxpvw1+PFn8fp9Mh18+KfER8ZpqjRi4ktGkY2hKynzGtjaGMW23K9fL+bNeN+CPDOpeI/AP7CmgfF3T/wC0pZL3U3e21GPzd1tDp1xNp/mJIDkpCsBAI6gV+qviv4JfBvx34itPF/jbwNoev65YBRBfX+nW9zcxhDlAssiM2FPKjPynkYNdnqXhjw3rOo6XrGr6TaX1/ocjzafcTwRyzWckiGN3gdlLRMyEqxQglSQeKAPgPV9K1HRP20virZfDa2jsNY1b4TWt3CsCLGJtUS+vIbaVguMuMKoY84GK+ebnUfgQf+CWlvpNrJZf8JD/AGNDFHbIV/tb/hNdyg7UH+kfbPt+SeN+zn7lfsGnhnw3H4jk8YJpNouvTWq2L6gIIxeNaI5kWBp9vmGJXYsELbQxJAyTXFL8Evg2njk/E5PAuhr4uL+YdXGnW/2/zCMeZ9o2eZvxxu3bscZxQB8ieOfB/im0+IekfGHwV468M2nxb0PwXZ6X4h0DxIySWlxYl3uTIZIpFuLItMZf3oV43AG5cLk8ZqfxI0jWPA/wB/bq0LRR4S0nS2Gi6/ZAqIbXw9rMgsZCXRVDW9neRwTxEKo8vL4AOK+8fHvwV+D3xTubW9+JXgjRfFNxYjbBLqen293JEuc7VeVGIUnkrnB7ivG/2jfhn8Uvin4Vi+A3w/sNI0PwD4gtYrLWtVkmZbqzsFkAltbCwSExl3hXYjtIqx5OFyAaAPhL4oaG+vf8E+/iR8YPENuft/xm8UaT4jkSUHeumXOu2EGmQEnnathHCQOxc19R/Gvwp4X8IftPfsst4U0i00f7PqHiHToxaQJAq2Z0aYiABAB5YIBC9AeQM19n3ngTwVqXhO38B6toVlqPh22jtoY9Pu7eO4tQlmUa3HlShlPlNGjJkfKygjkA1o6l4Z8N6zqel63q+k2l9qOhySS6fczwRyzWckyGOR4JGUtEzoSjFCCVJB44oA/J346fEz/haXwl/aMk0u78G+APCOgXGu6DfQ31kt1ruuarp8BiDsfPto7d5pQq2zNHcTHCsNpAFaWqX/g/UfiT+wxqnxQmsJbe48JavOJdVMXlNfvpWltC2Z/l80ynKfxb8bfmxX6OX/wS+DWqeKrjx1qfgTQrzxFdxPDNqM2m20l3JHIpR1eZoy7BkJVsnlTtPHFeK/FT9nO08ffGX4PawdC0i7+H3gbTPEWm6hpVzDG1v5Oo21rDaRRWhjaJo0MBBXgIAuB6AHz7LD4f8WftYfHa6+GMdvqGhr8Nrey8QNZBZLefxGZrl7VXKZR7hLTg9WXgHBBrznUNQ8Nah/wSDuG0Ge1nni8D2UV39nKF1liKIyy7eQyurD5u4Nfqn4J8AeBvhtoa+Gvh74fsPDWko5kFrp1tHaw+Y2NzlIlUFjgZYjJ7muT0v4B/A7RbbxDZaT8PtAtLbxZgaxDHplssWoBWLAXKeXtlAZi2GBGST1JNAHx9e3Xhqx/bu8F3HxTlt4bFvh2i+EXvyq2o1X7Z/wATEW5k+QXhg8nO35vKwB3r7x8Ial4I1PTJ38A3Wn3Wnw3VxHKdNeJ4Vu95a4VvJJUS72JkB+bcSW5NU/Gvw1+HfxI0NPDPxB8M6b4k0mJg8drqNpFdQxuo2qyJKrBWAOAVwQOla/hjwp4X8E6JbeGvBuj2eg6RZgiGzsII7a3iBOTsiiVVXJ5OByaAPyA+I3/Juv7c3/Y2Xn/pJp9fQnx+8GeEfBV1+ytN4R0az0Z9P8daVp8DWkCQmO1utOu/OiUqAdsm0Fh3Iyea+5rn4b/Du807XdHu/C2lT2HiiY3Gr272MDQ6jMwVTJdoU2zuQigtIGJCgZ4Fa2q+FfC+unSzrej2eoHRLmO8sPtFvHL9juolZI5oN6nypEVmVXTDAEgHBNAHwB8JL74V6Z+2t+1BfeMbnR7TWo28NeXJfvbx3C2H9iQ/aNjSkMId23zMfLnbu7V8nlrPTv2LdV8W6Yv2P4dwfGVNW0F8FLa38Nf2/EIpIwcBIPM8wr0GD+J+/tG/ZV8NeIfjf8Y/HXxj8KaF4r0Hxlf6Fd6LHqFtFfSQnT9MjtJyyTRkRFpEwNpO5evpX1ff+EvCuqeGpPBepaNZ3fh+W3Fo+nS28b2bW4G0QmBlMZjAAAXbjHGKAPhf4k+OvBXi/wDbp/Z70nwvrVnrNxpmleMJrtbSZJ1hjvbO18guyFlHmCJyoznAzjBBOP8Asyf8K9tpP2oPDfi69s9H2eP/ABHLdF5Y7aa20u5061zOHODHH5aSsH6AKx7Gvtnwp8HfhL4Fj02LwX4L0bQxo7zyWRstPt4GtnukWOd4mRAUaVFVZGBBcABiQBS3Hwg+FN346HxPufB2kS+LxE0P9rtYwG+8t4zCymcp5hBiJj5P3CV+6SKAPhf4O+I9Y/Z98Z/CP4BXupeG/iT4J8QRXdh4T1vSkjg1mwhtbVp911DG0sM8LxJsa6hZCzfNIuTz6B+3oNMtPDPwi13WFijsdL+J3hKe7uJgvlwWouz5jSM3Cx9N2ePWvpvwb8Dfgv8ADvXLrxP4B8B6F4c1e9UpNd6dpttazurcspeJFbaxGSAcE8nmu08UeFfDHjbQrvwv4y0m01zR79QlxZ30CXFvKoIYB45AytggEZHBAI5FAHwv40u/CFp+3l8PL/xxNZx+Fm8FajH4UlnZBZL4hGoJ9tEDH92Lk2mwDHO3cBzitD9nCXRLv9q/9oK++GbQyeCH/sFbp7Mg2D+Ilgl+2mAp+6MoiMX2krzv27/mzX1NcfBD4N3fgK1+Ft34H0WfwfZEtb6RJp8D2MLlmcvHAUKK5Z2bcAG3MTnJJrr/AAr4Q8KeBdDt/DPgnRrPQNHtM+TZ2FvHa28e45O2OJVUZPJwOTyaAPjb9rDVtN+DPxP+Ev7UGoyfZtI0G8vPDXiCVR/zC9bhJhkkP9yC9hiPPdz9R3X7FXhjU9H+AmmeLvEkXleIviPd3njDVAev2jXJTcopHYx25ijx22Y46V9I+KfCfhXxxoVz4X8a6NZeINGvdnn2OoW8d3ay+W4kTfDMrI211DDIOCARyBW1b28FpBHa2sawwwqEREAVUVRgKoHAAHAA6UAfDn7Gt3a6fP8AtH399KsFvbfFbxFLLI5wqIlpYszMT0AAyTXxzpXw2/ab/aD+APxO8f8AhWx8NLpnxn1S48T2El9PfJrsEOnyRro6RCOJoQ0cdnC0OWwd/wAxGSB+w9r4H8FWVjremWXh/T7ez8SzT3Oqwx2kSRahPdII55bpAoEzyooWRpAxZQAxIGK2NJ0nStA0uz0PQrKHTtN06GO3trW2jWGCCGJQkcccaAKiIoAVVAAAAAxQB5J8EPi/p/xa+A3hX4yxKWTWdIjvrmOFdxjuI0IuolUdTHMjpjuRX5MfGj4m3/xF/Zj8H/FOx1Hwb4J0HUPEui6h4b8L6VaLJq6TJqkY8xrwTxpHcRx73mSG0+UblZzkmv208N+FvDHg3Sk0LwhpFnoemo8kq2tjbx20AkmcySMI4lVQzuxZjjJYknk1wFr+z/8AAmyu9Uv7T4deHYbnW2R76RdJtA100cizIZT5fz7ZUWQZ/jUP94ZoA+Y9E8L+HNc/4KJ+N9U1rTLa/u9L8CaIbSWeJZWtzNeXSyNGWB2syjBI5xkdCa+NfFNsNI/ZR/aG0Szie18Iab8Z7231WC0VkEHh86hYPeRxrFgrEFdtyqMBC3GM1+1EPhnw3b+ILnxbb6VaRa5eQR2s9+sEa3ctvExaOJ5gPMaNGZiqkkAkkDk1WsPBvhDS7TVrDTNDsbO21+4nu9RihtYo4724ugFnmuFVQJZJQAHd8lgAGJoA/Pj9oHV/g9dfGv8AZa03wHdaJNew+JS1nHprW7PHpX9nzgeX5OStuW2YAwhIGM44+2/jJ4g+FPh34datP8bLiyt/Bl8sdhfnUF3WrJeuIFSXggKzOAWOAo+YkAEiv4X+AvwP8EmI+D/h94f0VoLtdQjNnpdrAY7xEeNJ0KRgrIiSOisOVV2UEAkV6bqGn2GrWNxpmqW0V5Z3cbRTQTIskUsbjDI6MCGUjggjBFAH5Y2/9lfsy/GD4T+Ff2cfiXP4r8IfEDWk0u48EXGoJrUVjpzwySPqGnTlnntoLXYCyF2RsnnjibTfhNqnxY1n9pKLwjcrp3jfwt8QLXXPDN83At9Ws9Ks2iVz/wA8p13QTA5Bjdsg4FfoB4I+BvwX+Gmq3Ou/DvwJoXhnUrwFZrnTdNtrSZ0PJUvEittJGducZ5xmu50rw14c0K81LUNE0q00+61mcXN9NbwRxSXc4RYxLOyAGR9iqu5iTtAGcAUAfi7o/j/xX8V/gL+2V48+H9ldab4g1Y2DT2Sh1urSeLRrWLVLXAw3mQFLiLjkleK/TP4Oa/8As5nwR8PLL4Z32grYTWSx+HYYZLYXBX7PulSBM+Z5ojVjOAN/Db+c17PovhDwn4avNU1Hw7otlpV3rlwbrUJrS2igkvLgjBmuGjVTLJjjc5Jx3rj/AAz8Dvgx4L8T3PjXwf4E0LRPEF5v83ULLTba3un8z7+ZY0V/n6tz8x5OTQB8qftdS6FH8eP2bP8AhZLQr8PP7e1c3pvSBYf2yNPP9j+fv+TPm+b5e7+L2zWPo2qeDdV/4KSau/gC50251CH4WTR372ZjfF6NYgKLctFyZBGY+GO4IV7Yr7z8V+EPCnjvQrnwv420az1/R7zAms7+3jubeTady7o5VZTggEZHB5HNYXhH4UfC/wAANbSeBvCGkeHpLK2ks4XsLCC2eO2lkWWSJWiRWCPIqu65wzAMckZoA/Kr4c6j8D7b/gmr4y0v4my2C+JBZ6+niaC+aP8AtVvFZmuMecrfvTeGfy/JJ+bGzbwBXZlbG3+Pn7JI+P7Wa69D4Bv/ALQdZMYYa2Lex3cz/wDLzv34x827OK/RHUPgj8G9X8aw/EnVfAuh3niyBlePVptOtnvlePGxxOyGTcgACtnKjgEV4/8AFX9n8/FH9oXwN448S6Tpmu+CtF0HW9M1Ky1FEuBLLfvbPBi3kRkdR5RJJPBxgegB8xr4/wDA/g34/wD7Tnxb8KadD4h8C+HfBmnHxFb2QSS01HXLdbmR4SwDRPILUiOc4bZu+fuK898ZeIvEeo/En9ljxNrfiPwdb3eoeJbVtM8P+F7TbLYaZfWMwmzeNcM00JBSNwltDGWI44Ffq94W+HvgPwR4Z/4Qvwb4d07Q9Aw4On2VrFb2pEoxJuijUId/8RI+bvmuZ8O/Aj4I+ECp8K/D/wAP6QUvU1FTaaXawlb2IOI7hSkYxKgkcI4+ZQzBSATQB8h/AvwJ4L8SfHj9qq+8QaHZ6jc3Ot2WnyS3ECSubR9JgZocsCQjE5KjgnGc4FfIcPhLwvpn/BILQ/E+n6Ta2+sR/wBj6kt6kKC5F6uuwRLP5uN+8Rfuw2chPkHy8V+2Om+F/DOjX2qanpGkWljea5Ks+oTQW8cUt5KiCNZLh1UGVggChnJIUAdOKxj8NfhyfBifDg+FdKPhKNUVdH+wwf2cqxyCZALXZ5ICyAOBt4YBhzzQB8szf8pH7T/sk9x/6foa+KfBngPwU/8AwSQ8Yay2h2R1C/sfEd/cXPkJ50t1aapdpBM0mNxeJI0VDnKqoA4r9lz4Z8NnxIPGR0m0OvraGwGo+RH9sFmZBKbcT48zyjIA5j3bdw3YzzWTB8O/h/beD5fh5beGdMi8KzJLHJpCWUC6e8dw7SSq1sE8orI7MzgrhmJJySaAPzZ+ImpfEBf2q/g7qkWs+H9JS8+HzjQrvxXaXF5ZNrTzxtfrbeTc2oS9e28khixJjDqBzX1R+zH8ONd8D6n8S9d1nxToWvv4u11NQntfDsMlvY6ffC1iiuV8uS4uWWSYLHLIC+ctnGGGPfPFfw5+H/jvw4vg/wAbeGtN17Qk2bLG+tIri2TyxtQrFIrKpQcKQAV7Yqx4N8CeCfh1oqeG/AGgWHhvSo2Li0021itIN7Y3N5cSqu445OMnvQB8YeAfCvhnW/2/fjPrGs6Va397pvhrwvHazTwpK8CXK3QmEZYHbvCKGx1Ax0r4t1621PSv2QPiFo+grb2nhTQfjXqFvqcM8Ek1hbeH4NbVpUnt4GjdrRHKGaNGXMe4ZHUftja+GfDdjrt/4pstJtLfWtUjhhu76OCNLq5it93kpLMFDuse5tgYkLk4xk1BpnhDwlotlqOmaNoljYWerz3F1ew29tFFHdXF2c3EsyIoEkkxOZGYEv8AxE0AfGHgvwD478TfHv4f/FTxJ498EXZ8PaRqdtBZ+F7Oe1n1LS72OJQGMl/ch7eCZYZEKrtUng5YV8P2XxC8DaN/wSH1LwZqeu2dv4gistW0ptOaZPtgvl1Wd2gMAPmBgn7wjbxH85wvNfsB4H+CHwZ+GWo3WsfDnwLofhi/vQVmuNM063tJXUnO0vEittyM7c4zziqx+AfwNOs634hf4e+H31PxJHLFqdy2l2rS3sc/+tS4Yx5kWT/loGyH/izQB8hfEbwl4c8X/tmfs9Wviawi1K3sPCmvXsUMw3xefD9j8t2Q/K2wncoYEBgrD5lBGl4MufCej/t3fF7/AIWdJaWmpyeH/Dw8JNflIx/YiQzDUVszJhQovM+cF9sjGa+5pPCvhebWrDxJNo9m+r6VBJbWd41vGbm2gm2+ZFDKV3xo+1dyqQDgZHArn/H3wo+GHxUtbWx+JnhLSvFUFk5kt01SyhvBC5xkx+crbScAHGMjg8UAfjn4z/s+X9h39qy48Fbf+Fd3HjOdvDRh/wCPRrf7bYC6ayx8n2U3Yl8vy/kzv219lftBeDfCXgj4xfsunwdo1nov9neKLnTbf7HAkHlWUul3IeBdgGIzgZXpX2re+AvA2p+Ev+EA1Hw7pt14X8qOD+ypbOF7DyoiGjj+zMpi2oVBVduAQCOlaOq+GfDevXmmajrmlWmo3eizm5sJrmCOaS0nKlDLAzgmN9jFdyEHBIzgmgD8yPH2vQ/swH4i/Gj4fa94Y+JHw61vxP8A2l4k8PXTRHWbXVpJ4redLC7jZ0mljlQMlrPEGQrtRwev2p+1TZ6lqn7MPxZstHYpeXHhPWlj4+Yk2Uvyj0LD5R6E10958CPglqHjWP4j3/gHQbjxVFIJl1WTTbZr0SrjbJ55j3l1xw5O4djXqF1a219azWV7Es9vcI0ckbgMjo4wysDwQQcEGgD4D/a4+IOneHf2D7ibQ7yHS08a6RpOg2Es0gjijj1sRW7s0jEBVjtnkcsSAApPFcT8dPCfwm1iy+DH7KXwPtdMn1O38Q6RrRXS1hZtJ0bR3E11qUrw8RvNhYkdiDM8hwSc19gfDD4IaT4S+GOjfC7xxDY+LtO8KXdx/YpvbZbg21ikkqWCHzgw863tJBbmRcZUHsxz3PgP4T/C/wCFkN3b/DTwjpPhWO/cPcLpdjBZiZhnBk8lF3YycZ6DgYFAH566j4/n0vXf2xP2lrNjJN4MsI/CWjH+OGXRrBp5x6hWvrsEn0X1BrjfG0/w/j/ZP+Bf7HPgzXrLVtT8fz+F9Lv4bGdJ3XT96X+p3TeWSER/KkxuI37mAzhiP1Ftvhp8O7MeI1tfDGmxL4wdn1pVtIgupO6GNjdrtxMWUkNvByCc5yayPCnwU+DvgWOwi8GeBtD0NdLne6tDZ6dbQNBcSxtC80bIgKyNEzRs4O4oSpOCRQB8CX+pfEPxj+2t8SrrSPEPhPSLv4ZWGk2GiWniqzubo29nf2q3d1fWSQ3lqqNLK3lSygMwSNE3KMhvqH4EeC7n4K/DTxv418Raxp/iU+KNU1HxlNLoFu0Vi0d1bRNss42mnLK6w71PmHezk5Oc16z44+CXwc+Jup2es/EbwNonii/09dlvcanp1veSxoCTsV5kY7cknb0zzjNejWlnaafaQ2FhAltbW6LHFFGoSNEUYVVUYAUDgADAFAH4h/FTx/qfjb4M/Bj4jx6n4L8I6RqXi/w1qPhvwpodoG1K1V71Mlrzz0USRxsROkNmiqcozHv9maJ4X8Oa5/wUT8b6prWmW1/d6X4E0Q2ks8Sytbma8ulkaMsDtZlGCRzjI6E19O6Z8APgTo1xf3Wk/Drw7ZzapLFPdPFpNojTywSLNE7kR/M0cqLIpP3XAcYbmvQYfDPhu38QXPi230q0i1y8gjtZ79YI1u5beJi0cTzAeY0aMzFVJIBJIHJoA/FfxTbDSP2Uf2htEs4ntfCGm/Ge9t9VgtFZBB4fOoWD3kcaxYKxBXbcqjAQtxjNfTH7QOr/AAeuvjX+y1pvgO60Sa9h8SlrOPTWt2ePSv7PnA8vyclbctswBhCQMZxx+g9h4N8IaXaatYaZodjZ22v3E93qMUNrFHHe3F0As81wqqBLJKAA7vksAAxNcX4X+AvwP8EmI+D/AIfeH9FaC7XUIzZ6XawGO8RHjSdCkYKyIkjorDlVdlBAJFAFL9oj4jL8IvgV48+JQkEc3h/Rry5tye90Iituv/Apii/jX55/Ej4aR/sxfsDeGPBPhRYPCus+KpvD2keJ9flDh7X+15Yo9RurmdWWURLuaH767I22oVIUj9VvEfhnw34x0a48O+LtKtNb0m7KGazvoI7m3kMbrIm+KUMjbXVWXI4YAjkCp9a0TRfEmk3egeIrC31XTL+Nori1uokngmjbhkkjcFWU9wQQaAPgzRfhD431zxR8NbbVfE/gQaL8Kbo65ZaV4S0u4tb4wCzntEijMl/cBbeQzAthMOVXJNfPXwp+Lnwm8K/sO+Nv2lfHWp6PqfxE+I1trd5qf2ySGa5utSZpobLSxFLufyYEWKNYMbVQF9oUk1+pHgH4RfCr4VQ3EHwz8H6R4VS8IM/9mWMFoZcdPMMSKWx23E47VhR/s8/AOPUtX1hPhv4c+3a+SdQmOk2hku9ziRvOYx5fc4DtuzlhuPPNAHn/AOzJp/g34N/Cv4bfs63WrWdr4x07wza3s2ktMi3j7ubu4WEneY/tLSDdjGc+hr6hrxa9+EFvrHx40z416xLavJ4d0i40vS4orRUuVa9dXuJLi6LFpFUIFhiUKqb5WO5nG32mgAooooAKKKKACiiigAooooA//9L9/KKKKACivj39sz4g/E/4feFPh6fhHqUGma94l8b6JoQe6jEts8eo+dHsmXBby9+xm2FXwuAwzXiPxi+IXjP4NeJPAvwB1n4p+IHk8VJqWua34pi0dNR1eO0tvJiSz0+1sLGaGBZZnJ8xrdxCgIyWZaAP0wor8wfDHxi/aC1T4afHLwz8I7vV/HOq+ELfTrnwfreuaNJp+oXkV+jfaoXguLWyiuLiyMUhjPlASlo1YHPPd/Br4j6dfeBfiHqPhP4x6/4y1jQ9DnnuNG8UWNrY6xot/HDI5lkgFpazKjMAAjrJECp2uwNAH6B0V+SFx4n/AGmdA/ZA8P8A7Ys/xcu9U17TtD03W7nQZNPsINFvLJliM9vKqwm48+SMljOsw/eZ2RxqQq+kfGf9omTXfjdD8Jj4z174eeFtL8P2OtXt34d0afVNWvbrU2fyLcOljfpbQxRx73YxhpGbYCArUAfpPVe7u7WwtJr++mS3trZGkllkYIiIg3MzMeAABkk8AV+Wtj+0z8bLr4Qab4Q0m+l/4THxJ8Q/+EF0XxPq+kSWLy6XJF9rTWJLCeG3Vpkt90fl7FRplzt25B9z+Jvwg+JfhH4B/FOK6+M3iPXUl0C6uYZL220s3NvLbQSy3CpJHZoPJuVCxlChaNclHDEFQD6R8Z/F7wR4I+HKfFa7uJtW8NzJZywXGkQSakbiK+dEglhW2EhkjfzFYMuRtOQelenV+S9lqHxO+BX/AATh8PfEbw38QNUv9TvtF8CSaeLuGxMek293PYQS21sI7dC0ZhmMeZjI4AB3bsk/Wlt8S/Evgb9qnxV4A8faxv8ACHiLwzD4j0BpljjisW0pvs+rW6uqhnyHhuTuLFQzY44AB9aVjeI9ctPDHh/U/EmoK72uk2s93KsYBcxwIZGCgkAsQpxkgZ7ivyzvv2mfirZfB74bap4h8RXfh/Ufjx4h1XULXUIdKbU7rQfDEayXNvDaWdtbytNM1ssOx5Ipdpld3yFyOu8C/FPxJqdx8WPhjbeI/EPj3wSngq61bT9b8R6NPpd7a3gWWC5sJJJLGxScFWjmjYR7lBdSWA4APv8A+Gfj/Rvir8PfDnxK8OxTwaX4nsLfUbWO6VUnSG5QSIJFRnUMAeQGYZ6E1U+KXxI0b4S+Dp/HXiOCeXSbK4s4ruSAK32WC6uI7drmUMy/uYPM8yUrkhFYgHFfAfwe8deM/hf8Kf2SPEq6kw8AeJNG07wxrdmUjKRXuo2aNpV2H271/wBIjMD/ADbcSrxnGfbfDuq+Ifjxrf7QunaxfiX4eWYk8GaZZyJGIWubazYavcO23c2Z5xCMsVHlNxnOAD7SBBGRXI+PPHfhP4ZeEdT8eeOtQXStB0aMTXd06u6xRlguSsasx5YDgGvLv2UvEd74u/Zm+FfiPUi7Xl74Z0lp2kzueVbWNXck8newLA9wa8x/4KGf8mXfFb/sFp/6URUAfZMUsc8STRHckgDKfUEZBqSvhH4ueJ/i9e/tBfCr4OfDzxifCGk+KvDWsXWozpZwXk6mya0KSW4nVkWcBmRWcPGquxMbMFxx/g34h/FvwRp37UPww8QeMrrxXffCbS4tS0PW72C3S+VNS0ia8jSYQxpFI0EsXDFPmzyMYUAH6PUV+TfiXxX+0d8O/wBlfwv+2XffFi813UrXStD1rUvDslhY2+i3en34t/OtlCRG4S4CTbvP887nB2oisFX610L4h+L739tDxJ8M5dSd/C1n4H0zVoLIxoFS8uL+4ikl3bRJlo0VcFsDHTOaAPp/V9X0rQNJvde128h07TdNgkubq5uJFihgghUvJLJI5CoiKCzMSAACTxU9neWmoWkF/YTJc21yiyxSxsHSSNxuVlYZBUg5BHBFflJ8S/FvxG+J/wCyp+1mNY8W3lp/whvijxVptp9nitRv0izsYSunPvhb9y/mtucYl54kFdh4osvjr8LP2b/h9L4X8aeJ9f0nVLrSpfEOp2On2N7rWjaE9j866fbQ2oV0WUIGcxTSohZgGxwAfppRXz7+zX4k8P8Air4fSap4X+J1x8VdMN7KkOoXsdvFe2qqqZs7pbeC2PmxnLHzYkkwwDDABPzF4o+OfxQ8Fj9ov4c3Ostc+K9K1XSE8GzyRRB4ofGKxWmnRxIEAlFpeGTlwxO35yRQB+j9Ffm94R+OfxQ8W6P8Afh0utND421LxNquneLLhYot81t4NFxFqSyJsKx/a5lt8lApAlAXbkY8Y1P9pjxX8RoPH3j3SviX4l8J6voWo6rZeFfD2j+F7rUNJmGlyPDEdTuBplz57XksZ3rHcRiFWAGGBNAH7EUV+b/iz4p/Hn4j/Er4FeDvBmuP8O4/ib4NvdX1yN7SOa60+WNbKd/IjuY2xcxl3gXzQUUOzsjMqiuV8a/GTUI/jJrnwF1/4peK/CuifDHS9Igm1HRdEk1bWNb1S+t/tJnu7iDTLyCCKOIx/ulijMrs5ztXaAD9Sq8u+HXxb8NfE3WPGuiaBb3UE/gTWZNDvjcoiJJcxwxTloSjuWj2yqAWCnOflxyfOv2TviN42+Jnwhi1b4hRzvrWmajqGmNez2MumtqUFnOyW98LWaOJ4vtEOxyuxQGLYC/dHhPwGv20uf8Aau1FPENv4Sa28b6jINXu0SW3sCumWhFxIkrIjJH94qzAEDBIoA/QyvLvhj8W/DXxYk8YReHLe6gPgnxBe+G737UiJvvLFInkeHY77oiJV2s21jzlRxn4C+G/xs8S6J+0f8MfCugfEbxT8SfB/wASk1e2vLnxFoa6dZC6sbNryC40m4Wxslkjby2R1j8yPawYMTiq3wF+F3xR+IWtftBzeD/irqXgC0tfiZ4iFpBpdlZSl78RWpM93JdxTNLDtKKII/K6MWdtwCgH6r1jeI9ctPDHh/U/EmoK72uk2s93KsYBcxwIZGCgkAsQpxkgZ7ivy5uP2yfHXiD9nv4KXV/qZ8K+J/idqOp6Xq+s6dpsuqSWcWgNcR31xZWUMM5ea4aBRH+6dI/MZmXCEjqPAvxT8SancfFj4Y23iPxD498Ep4KutW0/W/EejT6Xe2t4FlgubCSSSxsUnBVo5o2Ee5QXUlgOAD7/APhn4/0b4q/D3w58SvDsU8Gl+J7C31G1julVJ0huUEiCRUZ1DAHkBmGehNdxX5G6f8cdW+HH7Mv7LPwz0bXLrwm3xB0GA3ut2Omy6ve2Wn6Vp8Ms62lpFBcZnmeWONZGidY1LOVOAR1/hf8Aae8TfDPRvjHd/wBq678RvCXgnw1H4g0PVvEWkXGlXZu/3sU2m3Ej2ViswEgikWRYgwR2BLEfKAfePxf+Lnhr4K+E4fGPiu3u7myn1Gw0xUs0SSXztRuEtoiRI8Y2B3BY7sgZwCeK9Ku7u1sLSa/vpkt7a2RpJZZGCIiINzMzHgAAZJPAFfj/APtD/D3412H7PnhH4hfEr4rX/iW/1zxB4UudV0d7Kxj0mKS51G3kSOx8qFLiHyGIXe0snmqCWUFgV/Q79qHTNW1T9nr4hRaLrd14fuoNEv7gXNosLylYIHkaIieORdkoUo5ChgpJVlbBABsfE745eBfhb8JX+NWoSya34Y/4lzQzaUYrk3EWq3ENtbSwsZEjeNmnR9wfBTJXPAPsVfi/458IeJ/DX/BLDTNRvPF9/wCJP7a0b4fXNja6lHbC20xTe6ay28H2aGGRohuVf3ru+FHzZyT9PXVz8bvgv+0l8JNG8T/E288caL8UX1jT9UsLuxs7W2tLqysXvIJrBbeNZIkDJsKSSSsVOWd25AB+gVFfnr8KdZ+NfxV+MHxWl1/4hXOjeCPhh4zkhtbHT7a2+0X0UVvbXDWl1LJC7C0Rc4EW2VzLJukASMV574b8Q/tKfFH9nfUv2xNG+JtxoGqS2moa7o/hVLOyl0GPTLFpGjsrzfCbmaWeKL551nQoz/KoC0AfqZRXwrpvx68U+N/jP+zlLod0+m+GPid4R1fXr7TSqOGlFrZz24aRk35hMzD5SobuDxWno3xY+Ja/Gr9pvQbJn16D4f6N4fvPD2k+WuPtd1pt3cSRKUUSN9omjQEFjjouKAPtevLvCHxb8NeNfiH49+GelW91Hqnw7m06DUJJkRYJG1O0W8hMDK7MwEbAPuVMNwMjmvzA0Hxl8UvF9v8AAXVvDn7Q+r634g+LF3GPEOj6Wujk6fC1lLc3ElrA1lI1tHYTIsMwmDFs4JV69P8ADHw/+I3xD/a0/ab0zwh8Q7z4f6fFeeGDNLpVpbTX1xcNocPlZmu0lRIY8EsiRh3Jx5igEMAfqDWNYeI/D+q6pqmh6ZqVtd6jojxR39tFMjzWjzxiaJZ0Uloy8bK6hgCVIYcEGvnb9jT4leMPiv8As7+G/FnxBukv/Ecc2padfXUcaxLcy6ZfT2Ym2JhVMixKzAADcTgAYr5p/Zd+HvjSP9qP48arffErW76LQvEOmpd20kOnrFqofSYvLN1stVZfJUqieS0Ywi7snJIB+nFFfnb+zRqvxx+LcXiT4lePPiZNpmheEvEfiXTLDT7e2tEguIbS5uI0n1OV4g7LAGVY443jXbCrOzMzY8it/jt4m8H/ABM+Fms+Efi54k+Kuj+MvE9p4b1qS+0COz8NTDUEkWOXTLyKygjEkUqhgsVxPvTO5ztOQD9b6K/PO2f46fGn9oX46fDPTPile+CPC3gi60NbBdLsbOS+EmoaRDOU8+5jkAgEu6RlCeY7NgSog2ngLH40/tFeNP2L/A3jnS7u/n13/hIZdJ8XatoGnw3Wqx6Pp17d2dze2NmyOjTP5MRfbG5QM7JGcDAB+pdFfnZdeP8Axze/st+KvEn7M3xK1P4uarBqEEUV3LZWcuu6XZ+ZCt7CLRbe2E17DEZZI47iFXO5RtbC7jwp4h8ReOfgV8S/+GefjdrXjPxdplrHNYW+uWVnFrGk6jbBpmtbmA2dtIFvCnlATQ/KNxjY9QAfonRXw0fj/rXxi1v9nrTfhTqDaZD49hk8V66YlSV4dG023UTWcm9W2+bfTxW7MMMCjgEc14fpWpftJ/FjwF8cPHFn8X73wuvw58U+LrLQ7XT9OsWEiaPPJJAl9LPFI0sSptiVI/Lwo3SNIx4AP1Vor4V1z4vS+Lf2b/hR8RvFHxNn+F1x4007Tru7/sjT4r/U9RuLuzWV7TToZIbpw3mEt+7t5X2jGB1rxfwH8ffifb/Cb9qTw9L4g1nUtQ+EmkTah4f1jX9NXTdaWK90ia8txd2rwQgvBLESrPCpdSCy4IAAP1SrGh8R+H7nXrrwtb6lbS6zYwRXVxZLMhuYYJ2dYpXiB3qjtG4ViMMVYA8GvzL8a+Jf2hvhH8B/Cf7W2qfFW88RyougXmt+HZdPsoNIuNO1aa3gkgtUji+0xzx/aAyzNO5YqflAIStXwP8AD3xpqH/BQH4p3S/ErW7aDS9O8Oag9vHDp/l3NnNcXcyabLutSwtohujVkKzFWJaVmwwAPvvwb8RvD3jrVPE2j6JHeJP4T1BtMvDdWk1sjTqiyEwPKqiVMMMOmQfoQT3tfmbrX7T3xK8CfD39pDxaZ01zVfCPjaPw74at7pFS3tzfJYwW6yGNVLRxS3LStuO5gCu4ZGPq34VfCT4p+B9Zh1zxt8XtY8cfabRkv7G9s9PhsmvGKFZrQW9vHLbxoA4ERkcNuBJyvIB9C0V8V/tU+Nvi9ofxF+C3gT4SeIIvD0/jrV9T069mnt47lFgTT5JPOCOp3SQYMsS5CtIqrJlCwrxf4ofFHXfA3xS0L9mbXPil4o0vS9H0Btd1fxJYaOdV8QanPfXk0dtaKbPTriC1iiVGZpBbgsBHGpB3kgH6R+IPEfh/wnpFx4g8U6lbaPpdpt866vJkggj3sEXfJIQq7mYKMnkkDqa2a/En47+NPiX8Tv2K/iYmu+KdaNv4I8UadYWepXGlLpcviLSbq/0/7NLdw3VnC6yQNMwZoUiDSRAsCrFT9YfHa/8Ajr8Irf4O/D/wH8RrrVtb8ZeLJdKutW1u1s5X+y3FjcN80VtBDG32cqJYlCrvdFV2KlqAP0Gor8+9cPxcuvjD4Y/ZH0D4navarbeHbvxb4h8USQ2L61dwS3xtLaztf9G+y24EhYs4gYhEVRg5LP8AC/xa+Mvw9uvjv8IdYvW+IviT4ZaDH4g8NX01vHHealBeWs8kNpeRWqxxtNFPB5e6NVMqsDtDUAfoDRX5BeGfFHxK8S+JPgJ/wi/7ROseKb34nrJdeI7DTho7i0jjsWunmgiFi5tbeGcC3kSXLEsFDCTNe9/D/UPjh8Xv2iPix4cuviDc+H/BHw58SWAtbTT7e2+13glsra4azmnlhfZaAhidn71zKwLqqKCAfoFRX5A/HH49eJ/hjBqnxS8B/GfxD8QNQ8La3bR6hptn4fiPhFLKW+jt5rGW9hszHHOiPtEn215fMXG1Swx9PfEDxD8Tfi7+0vdfADwT4xu/AHhrwfoNrrWs3+lxW76pe3WoTSR21rE93FNHDCiRGR3CMzEhOnNAH0d4D+Lnhr4h+LPHPg7RLe7hvfh/qMOmX73CIsUk01ulypgKuxZNkgBLKhznjHNepV+P/hL4leKv2aPCv7Xnj/UtR/4S/wAReGvEOmQw3l1brB9puruys7W1kuIbUBflaaMyiJV37WKhNwA7vwD8Y/GGg/FP4d2Gg/ETxZ8UYPFd82neJrTWPC11pdnZ+dbu8V/YyHTLQW8UU6KjRPNLlHz1BcAH6Wa/4j8P+FNLk1zxRqVtpGnRPFG9zdzJBCrzyLDEpdyFBkkdUUZyzMFHJArZr8+v+Ckuia3qvwCtLjS/El7olvDr+hRT29qlu0d15+qWqxNIZopHBt3Akj2MoLD5wy8Unxrv/jv8M9f+B3wn8B/Ea5v9U8aazq9hf6xrNpZzTPb/AGGWYSGKCCKIyWoy8ChVV3RFl3IWyAfoNRXwT8V1uPAEfhj4d+If2gPFdtqD2dxIllouk2up+J9XlM7uLlxbWFy6QRBhEBHbRphfmkJzXkvhj9q74ut+yVbasJkvfiVeeNT8PLHUdRs/sqm8l1D7LDe3dooURyJCd0kWAPNXBGCRQB+nutata6Do9/rl8JGttOgluZRDG00hSFC7BI0BZ2wOFUEk8AZrI8D+MdI+IPg/R/G+gpcR6drdtHdQLdQPbTiOQZHmRSAMh9iPpxzXgFn8Pvih8I/B/jDxPqvxb1vxtJHoF7KsWq2unIkGoxRNItzbm2t4mjQbSBCxdec5OK+a9Q+Mfxz1X4K/sp33hnxYmm+JPifcWFpq+oXFnFcrKLrSZppJWgARS6uBIgUqu9VDZTcpAP09or4X+FGr/FT4d/tWa78BfF3jq/8AiFompeD4vFVlcapBaQ3dpcpfmymgVrSKFGikyrqCoC9BjBLfHSfFX4y+M/hl4I+JcXxv1PTPiN4y8XQaJqPgzTE0tfsEMuoNbT2dvaT2k1xFcWcC+a805cEKzMNpU0AftbRXz7+0V4g8T/DX9mLx/wCJfDerzDX/AA14Zvri11GRYmnN1a2rFJ2XYIi5ZdxAQLnooHFePfF34q+P/Dnws/Z71/RdXe11Dxj4u8Gadq0ojiY3Vpqa5u42DIQol7lApH8JFAH3HXBP8RvD0fxNi+EzR3n9uTaU2sq4tJjZ/ZVn+zkG62+UJd5/1e7djnGCM/A3xN+IvjjRf2gPE+k/F74o698HPDyz2MXgu7tdOs5PDd7HJChmN9e3NrOhnM+9WimlgCqAUJyDX0SfiD4vH7blr8LU1Nm8KyfDqbWjZ7I9ragurxWyz7wu/PlMVwG285xnmgD6tor8atM8b/tS+Iv2Lbj9rGT4v3VprHhmPUr610qDTbEWN3Bpd/PBImoFomlmklWIgGN4kRdoCF9zt9O/Gb4l+K/EXxU+F3w6k8bT/Cnwb4x0G91m41i1a1iury+g8gxabBc3kU0UTLHK0zYTe6rgEYNAH3xRXxF+xHq3xB8U+GfGfifxr4+1Hx7Yw+I9S0jR725jsks7rT9NnaOK9tTa20JbztxSRi8iFoiY8A1zWq6h8cPil+1f8R/g1oXxBufB3grw7pHh/UWk0+3tn1JZrpbgGG3kuIZUSOYrvmdlZx5aLHtDuaAP0Cry7xN8W/DXhT4o+CfhJqNvdSax49h1WewliRDbRrpEcUs4nZnV1LCZdm1GyQc7eM/nv+058WfE/gxfiF4r+Hvxs8Q6t4p8Eedew6Dovh+K98P6bHbKJWs9XuIrOYBigId5byN1zuCLjFdN8cZfFXxR/aF/ZW1DwbrreDb/AMR6F4ruzfRW8V5PbQ3Fjp8siwJODF5pUlFeRHVSdxRsYoA/SDWtWtdB0e/1y+EjW2nQS3MohjaaQpChdgkaAs7YHCqCSeAM1keB/GOkfEHwfo/jfQUuI9O1u2juoFuoHtpxHIMjzIpAGQ+xH045r4c0Txx8V/hN8Ufih8BfEnjO78a29j4Ffxn4f1bUIbZdRswsk1pLbXDW8UUU2Jo1kjYxqQMqdw6eea9+0743tPgb+zdo+p+KLvRNc+Lmlre6z4js9KfVL+C2sbGK4uja2Vvbzr9ouJJo0VzCyRKWfZwCAD9UKK/ND4e/F74zXMPxc8C/CrVNa+I50jw2mreEdX8S6PLpd1/abiWKTT52ms7CO4KuscsTeWuQ5VmIBK97+yR45tvE+sXGn6h8XvEXiTxTb6aj634T8VafaabqFhesyb7iKBbS2mSFTuj2o0sXzL8+QMgH3lXBfEv4jeHvhR4PuvG/imO8l06zkgidbG0mvZ91xKkKbYYFdyNzjJA4FeKePvHfjL4f/tRfDnTr/UnfwD8RNO1DRRasiCK11+zH222m83bvzc26zRBN20lAQAevy38Rvjz8X3+A/wAbfjx4W8RyWGmQ+MNL0PwmEhgeOLTrDVbPS724jLxneLyZrkEvu2qBs20AfqZRXwP8SdZ+Nnjj9sGf4DeC/HU3gzwk3gey12+ntLe3mv45TqV3bMLN7iKRI3mAjDu6uFSP5FDtuHoPwi8ceNNR/ag+Mvwy1vWZtS0LwfpfhJtOimSINHLe21y11IXRFZmmaNWbJIBHygDigD63or8t4fi7+0Z4l/Zp8Q+KPCt/qOo6rZfETVNK1G80uxgu9WsfDNrqUkMz6falNk08USqqAo7bSzYLAGvq39l7xT4a8V+FNUvfCfxWv/ilp8N2I1fVobeDU9MZUG+1ukitrSQPuyw86FXAOOQMkA+m6+UdC/bi/ZR8RX9rp2n/ABFsYpL6XyLd7yO4sYJZd2zYk91FFEx3DGA3Xivq6vz2/YZ8G+FfiD+wL4T8F+N9Ph1TQ9Xg1u3u7e4UNG0b6reZPPQr1VhypAIIIBoA/QgEMAynIPIIpkssUETzzuI441LMzHCqoGSST0AFfl38B/iHqX/DAPwv17xZ8VJvh6fNl00aulpDf6hfWdpd3VpaW1nDNHNm4lhiiKFYZpNqnCEncuZ4C+IvxG8YW3x8+CL+OfFEll4f0Gx1rQtc1zSYtN1+KC4SY3EEsNzZwrJHI0G1JHtlfY77cMFZQD9RtC13RPE+j2fiLw3f2+q6VqMSz213ayrNBPE4yrxyISrKw5BBINatfmd+zPpWt/D39hnQPGOtfGW78N2OpeHNGnt7/VLXT5rXw/EoG+O0j8mLzDKriNPtDTEMEIDHKvN8A/jD4qtf2m7b4Up4x8UeOPBvinw1data3XizRjpV1Ff2VxEjtaMbOxMtrLHL08ohWC7WwTkA+z/jT8ZPD3wQ8I2/inXrG+1eXUdQtNJ0/TtNjSW9vtQvn8uC3gSR40LMck7nACgn2rufCOu3vibw3p+vajot54dub2MPJp9/5X2q2bJBSXyJJY88Z+V2GCPoPgz9vvwnr3iDV/gHJpXi3UPD6T/EXSLFEs47VhFcTwXbJeqZ4ZD50IQqikmIh23Ix2kSapJ8efGP7Tmsfs9aJ8Tb3QfC+ieD9D1G81KG1s31ea6Mtzbu0LPAYI2uioed/LIXywsSIHYgA/RKivz/ANCn+MH7SXxF+Jlv4d+Jep/Dvwv8NNXPhnTYtJtrKW5vNTtbeKa5u797yCbzIt8yqkMYjVgCS2ea8t1f9pX44+IfgL8O77SNWs9D+IMnxPtfAOs3iWonsLiWK5uLOWbyGKkxSbY5WRHQ5yqsooA/VKivzUNl+0foP7S0f7OkXxnv9Q0PxZ4Zk8Ry6reaZpzatYPZ3S208NgY4EtkWcyoQZYJRGqkIu4769o/ZG8Y/EHUh8Tvhr8R/EEnizUPhv4om0i21aeGKG4urF7aC5g89YQqGVPNZWYAZAHfJIB03xJ+JOtf21caHodw1nbWbGN3jO2R5F+983UAHjA9Mn28ivviBrul25u9T8SXNpACAZJrx40BPQbmYDmrni//AJGzW/8Ar+uf/RrV4N8cNI/tn4VeIrYLlobf7SPUfZ2Ep/RTW+FpKdSMJOybSMMVVcKUpxV2k2e2N8QtbS6hsH8S3C3NypeKI3jh5FAySq7skAdxS2vxA1y+knisvElzcPauY5VjvHcxuOquAx2n2PNfngniW81Pxr4V+ICSf6J4dTQtOnPYfb4JGnbP+zuKn6iup8Ha/eaJ8OL/AMWWetw6HdeKNfuZVmktmu5ZUb5QkMK/efeD1BHJyRxXt1cgcYp82unTq76aXe2p4NHiFSk1y6a9eitrrZb6H3e/jPxTEhkk129RFGSTdSAAe53UqeMvFUih01y9ZWAIIuZCCD3HzV+fmoePPHPi34VeONL1e923Xh6eFJpZLQQS3FrOXTy3i48p9wBJAyB8vvXe2+s+K/CPwo8M/a/F1vZ3GpLbC0YaebicWzW6eXBDCmfMdWxlmHOeoOM41MjnFWclzXtbXsnfbs+xvTz6EndQfLa99O7Vt+67n1lrnxN1bw1pk2ta74ku7Oyg275XuZtq72CrnBJ5JArQt/G/ia7t4rq2129eKZVdGFzLgqwyD97uK/P/AFrx54j8Y/BDxvYeKW82/wBEvba3MrQ/Z5HU3KY8yLA2OCpyMDHQjIJrufDHjLx/4R8YeFvDPim/t9U0vX9P82KOCARtaiOIsoBHzPwoBLE5yemKqeRSVNu65k3p3SSemnZ9bE08/hKolyvlaWvZtta6910ufZv/AAl/iz/oN33/AIEy/wDxVIfGHitQWbW70Ackm5l/+Kr4H0P43fEPWL+08RWxe4sbm+EL6VFpztGlqW2lxdgcyDuM4zzjHy19Mp480fU9du/CFtb3gvoxMhZ7aRYCY1OcSkbTntzzXLisorUXaSv6dLb3/qx14TOaNZXi7dr9b7W/q/kemxfErU57B9Vg8VTSWUed063zGJdvXLh9ox35q7b+N/Et1BHc2uv3c0MqhkdLqRlZTyCCGwQexFfB/giWMfssa/lgNou1PsxZcD9RXSReMfF9rpXw2+Gvg6eHTb/WtJgnkvJ4xL5caQ7gERvlJIRuoPYcda6amSvmlGMtpNa9kr3OWlnicYynHeKem927JH1rqvxVv9DvdP07VvE13bXOqyeTao1zNmWQEDaME8/MOvrVrWfiRrPh7S7nWtZ8RXltZWi75ZDcTEKvTopJPJ6AZr44+Ldl4ttdV+GFlealBfa+NQmVbtofLhMpeLYzRoei8ZA64pV+IPjm18O/ErQvEl5BqWo+GBEYbj7NGI3SYkFWi27CMAcEHqck0QybmhCcZXvv/wCBct1p+f3BPOuWpOE42tt/4DzWeu++33n2bp3xA8QatYW2qad4gvJ7W7jWWKRbmXDI43KRls8g96ztF+Kt/wCImvE0XxNd3Z0+ZrecJczfu5V6qckcivlO48feOtYu/AHgbwvewaTfa5pEGoXV40CSAAxM21IiNgH7tuAO4xtANcx8Ntav/DXg/wCIWpXGsWulX0Osur3c0LSQiQsA5SJSWJPOxeecZGM0f2K+SUm9dLLfd210/L7hf24ueMUtNbvbaN7LX89PM+8P+Ev8Wf8AQbvv/AmX/wCKo/4S/wAWf9Bu+/8AAmX/AOKr5J+D3xF8Wa5401jwb4lu31KO3tVvLa5ms/sEpQlAQYcD5TvBUnkgZzzxQ+JvxR8VWnj2bwV4bvG0mPT7RbiSaOxa/lmlcAqmwA7Y8MAWxwc9eBWCyar7Z0bq9r31tb7r/gdDzuiqKr2dm7W0vf77fifYMnjXxPEVEuvXiFztXddSDJPYZbk1J/wl/iz/AKDd9/4Ey/8AxVfn5431rx14wT4X6tdzjR7vUL5YxA9tjyb6GfyxcFX5KsCpCHjrzzX1hrV7qPh7wPcXmoarbw6hZWY8y+njIg85VAMjRocgM3RVz1wAehnE5Y6SheSbk2rejt8y8LmqqudotKKTvp1V/kepHxh4sAJOt32B/wBPMv8A8VWJ4f8AinqHirThq3h7xNd3toWZBIlzNjcvUckHivkz4cfE/wAXX/xATwprepNrWm6pYvdW1xJYfYGyoJDRqQC8ZCsAxzk9MYIrpP2Xv+SWR/8AX5cf+y1picpdGnKU3quW1uzv3SfQywucRrVYRprR81773Vuza6n1S/jLxTGjSSa5eqiAkk3MgAA6kndWFpnxYvdY0ODxLZeKLr+zLkExzyXMsSkAlT/rCpHIPWvKvjZqNxpfwr8R3dsWWQ23lZXqBM6xN+jGsXxJ4Q+G9v4E0a+8cxB9L8N2aCNfNdYyWjVfuKw3uxHy9yT71z0MLCUIylfV20V9l2+aOnEYucakoQS0jfV23ffXaz6dj6JTxt4mlhFxHr148TDcHF1IVK9cg7sYqK08e+IL+Iz2PiK6uYwSu6O7d1yOoyGPNfn1Lb+IfDv7OrQTmWxs9f1ZfIjdjvg06cghST0DFCfcN7mvUvCug6Z4O+OGpeDPDUbWuj6joazzQI7EJKriMOCSSDt7+rZrtq5PGMZtTva9tOkbX676/gcNLOpSlBOFk+W+uzle3TbTy3PrZPHPiSSZ7aPxBdtNFgugu5Cy56ZG7IzUQ+IGum8/s8eJLk3QGfK+2P5mPXbuzj8K+Q/g5oVh4Z+L/jzQ9M3/AGa1jtQnmOZH+Ybjlm5JyT1rC8e+CfCdp478O+Hvh9BIPF0upDUbu5E0kjQW2S7mZmY4ySCo64H+0N0rK6ftnS53sne3lfXXRfeU81qexVXkXxNNX7O2mmrfyPsy0+J+q32s3/h+18R3r6hpixNcRefMCgmBZDkkBgQP4ScdDzXRWnjrxhZTrcRaxdOynOJJWlU/VXJB/KvmTWcab8e/DtxbDB1nSru3nC9xbnzUZh9eAfwr3CvOxNBQ5HHZpP8AR/ij08LXc3NS3i2v1X4M+4fA3if/AIS3w9DqjqEnVjFMo6CRcZx7EEH8cV19eM/A3/kU7v8A6/pP/RUdezVynUFFFFAH/9P9/KKKKAPEfjd8G/8AhckPgaL+1/7I/wCEL8WaR4oz9n+0faf7Kdn+zf6yPy/M3Y8z5tv9xqyvjT8DtQ+I2veFviH4G8TP4M8eeC2uRp2pfZVvreS2vVVbm0u7VnjE0MoRSMSIyOodGBHP0HRQB4DY+A/j1c+BdZ0nxD8UbePxVfzQzWOp6XoUMFtp6xFWMItLma5M6SbSJC8oYhjsKHBHimo/AbxxpF38Qfjv8W/GFp4n8UN4I1Hw7aR6XpX9k2cNgQ107SI9xdSSyvIinJkCoMhV5zX3TTJI0lRopVDo4IZSMgg9QR6UAflP8Cf2cPib8Z/2WPhV4P8AHHxOE3wq1HRdGvLzQ4NIig1G5gjjjnXT5NRSbH2UOqglbcSsg2M5JLn6/wDiV8BvFGp/Emz+NHwa8XR+CvGEWmjR71buwGpaZqenxyGaGK4thLbuskMjExzRyqwBZSGU4H0tb29vaQR21rGsMMShURFCqqjoABwAPSpqAPl3x1+z34m+K/wqsPCPxH8cvP4z0TVodd0nxHpmnx2J07UrRy1s8No0kytHGrNGySSMZEZgWBIYbPhb4XfFzUtG8S6F8cviDbeLdO8QabJpa2mmaNHpEMMcyvHLOWae6leaRGxjesa9kJ5r6JooA+C4f2RviTqv7Nd5+zN41+JVpqWj2S6FbaHfQ6IYbm1s9EvIrlY7lftjLOzpBHEGXy9gG4hycV6d+1L+zR/w0foWiWOneKJfBuraNLexjUYLYXUj6dqtnLZahZ7DJEFFxFIPnydjIrBSRX1NRQB8+fFv9n/S/iF4Z8Kaf4S1aXwTr3w+uYbzw3qdpEk/2CSGE2/lvBIQs1vJCxjkiLLuXHzDFXvCfgD4t3eieJNG+NHjex8TQ69ZtYxQ6Vo/9lRWqOkkckgL3N1JJJIHGcuFXaNqjkn3WigD80/jNo3g/wCA37EkX7OfxN8WQa54nGkNpnhWKwt/s2palfWEiHSTa2fmzO00MwtvMZWKhvmO0HFeojwv4r+An7F48HWFo+s+PdR0/wCxuIgXNx4l8RzbZpnZAcRLeXTSO/RYlJyAK+0JdPsJ7uG/ntopLm2DCKVkUyRh/vbWIyucc461boA+fofhD4z8PeBPhX4C+HXjVvDNj4Ak0mHUdtjFc/2vpmm24gksz5jDyPPwCZFyy9gTW1+0N8Iv+F9fBfxX8IP7W/sP/hJ7UW323yPtPkYkSTd5PmRb/u4xvXr1r2eigDwnV/gp/avxv8DfGT+2fK/4QzRtS0n7D9n3faf7RMB83zvMHl+X5P3djbt33hjnyT4o/Bv/AIQvRP2nfi7/AGv9s/4WP4WA+xfZ/L+x/wBjaRc23+t8xvN83fu+4m3GPm619o1HLFFPE8E6CSOQFWVgCrKRggg8EEUAfmL8IP2bviX8YP2evhD4b+JHxNXUvhY2ieHNTl0GHR4re9u44LaC5gsLm/WYq9rG6p923SR1UBn3Zc/RvxP/AGffH+ufGux+Onwh8fReCtcfRf8AhHtUiu9KXVbe7sUna5hdEM8BjmikdsNlgQQCMAhvqyGGG2hjt7eNYoolCIiAKqqowAAOAAOgqWgD5B8D/skab4Y+EPxX+EPiDxXe+JLb4rajq2oXeoXESRXkT6tZw20pJUmOSRXiMoYJGmWCiMKvLvDPwd/ac8NfD7RPC9t8Y9O/tTwyY4LW4/4RpWtbyxhgEKRX8LXhlaTKhhJBNDg5yr19eUUAfPXwF+CGpfCafxl4m8VeIE8S+LPH+prqmq3VvZjTrMSRQpBFHb2okl2KqJ8zNIzuxLMc8VzPxG/ZesfiB+0R4K+PDa+1hD4Ziijv9JW1Ei6o1k802nu0/mL5Rtp52k/1b7sAZXGa+q6KAPlDwD+y3Z+Bf2j/ABf8fE8Qte2niGO4NjorWoRNNudQFmdQnS481jIbl7NGx5abcsMnJJyLf9nX4s+APEPiST4DfEu28LeGPFuoT6tc6VqeiDVvsF9eHddTafKLq22LM/z+TKsiK5ZlGCVr7GooA8I1b4LXGsfG3wL8Z7vXt0/g3RtS0qS1NqM3j6iYC0/mLIBFtMOdgjYHd1GOeS8c/AXxuvxTvvjP8D/GsPg3xBr1nbWOuWuoab/aumamlluFrM8Sz20sdxCrsgdJcMmFK9TX1JRQByngjSvFWieGLLTfG2vJ4m1uLebnUI7RLBJmd2YBbdHcRqikIo3scKCzMxJPyj4j/Y8HijRfjn4Lv/FbQ+G/jLe2+qokNpi70zUI0hWVjK0pSeF2t4mEZRMDepJBzX2zRQB8a2P7NvxU134o/Dn4rfFj4oQ+IL34c3F9JbWFhocenWU0d7ZS2bs3+kTSicl1cvvMYClFiXcXrBt/2XfjZ4QvvH7/AAn+Mkfh2x+I+v6jrl7FPoEV3JYyahtUtYzfaY2WZURVLS70JAZY0IIb7oooA+RtV/ZD8KWvwk8A/Dj4b63d+EtW+FsyXnh3W1RLq4gvNrrcSXMT7EnS682Q3EfyB9xwVwMeieE/AHxbu9E8SaN8aPG9j4mh16zaxih0rR/7KitUdJI5JAXubqSSSQOM5cKu0bVHJPutFAHxJp/7I+u2nwe+GXgyLxz/AGf46+EDMPD3iaz09VVYBG1t5NzYyzOssc1qVjnUSruZQysvSvVbD4S+P/GPgXxf4F/aE8W2XjDT/F1k2ntb6VpX9kQ2sEkbxymMvcXUjyPvDbmfClV2qOSfoaigD89tV/Y8+M/i3wHofwu8c/Gv+0vDPhS80u501YtAiiup00q4ilhTUZzcsZ8RxmMGIQkvtlcvgxt91+KvDmn+MPC+seEtW3fYdbs7ixuNhw3lXMbRPtJzg7WOK3qKAPguH9kb4k6r+zXefszeNfiVaalo9kuhW2h30OiGG5tbPRLyK5WO5X7Yyzs6QRxBl8vYBuIcnFfQ3xH+Dn/CwPid8L/iP/a/2D/hW99qF79l+z+b9t+32T2ezzPMTytm/fna+cYwOte3UUAeI/CP4Nj4V+IPiRrh1f8AtT/hYPiKXX/L+z+T9k823hg8jd5j+bjyd2/CdcbeMn5xb9jf4g6X4P1b4HeCfivJoXwd1mW6DaQNJjm1aysL6RpLnT7PUmnCpBJ5kiq0lvJJGjbQzda+/KKAPkz4rfs0anr2ofDHxP8ABbxPF4A174Uw3FhpbTWA1Oyk0y7git5bSWBpYWICQpsYSZXB7ncOJX4N6x+z94d+PPxv8TeLNU8f6x4w8PwXF6ltbR2N4JtGs7tSLMxF1jDrMqwoIyYQgLNKxLV900UAfiV4K8d+Nv2evgz4R8Z/Crx58M/iFql3aaPp0GhaZoq22u6yZfJiNol1aXhlkuI1OS0lsMFC0iLyB+o/w++Dn/CC/Fv4pfFP+1/tv/Cy7jSbj7H9n8v7F/Zditlt83zG83zNu/OxNudvzda9B0nwD4F0HVZtd0Lw5punalc7jLdW1nDDPJu+9ukRQxz3yea62gDxH9nr4Of8KG+GNt8OP7X/ALc+z32p3v2r7P8AZs/2jezXmzy/Mlx5fm7M7vmxnAzgcRofwH8c+C/2gfFHxZ8EeMra18M+PZrK613Q7vTDcStc2NsLZXtLtbiPyfMRVLh4pMEHHUBfqWigD5e8Gfsy6f4b+Cfj74Katr0uoWXj678RTz3cEH2WW3i8QvKzogMkoLQiUgMThiMlQOK8rt/2Rvilq9j8MtF8ffFmLU9I+E+s6RqelWdloEVklymjjZGLxjcyu0zRfIrxGONMsxikO3b960UAeI/D/wCDn/CC/Fv4pfFP+1/t3/Cy7jSbj7H9n8r7F/Zditlt83zG83zNu/OxNudvzda8y8E/s6/Ej4VfB/SPhz8MPiUmlarpGranqbXtxo0V1aXseo3Vxcm2uLVpxIFQzAB4rhGJUMeDsr67ooA+OPCX7NvxN8LW/jnxdZ/E5LP4leP9SsNQv9WtNFgGmKmmw+RDaLp00sjGJo8iR/tCzOcHeuMG/wCCPhlcfB7xd4y/aS+P/j7Tb3WNT02z065vIbNdD0mw0+zkdkDLLcTs7tJLzLLLnG1FAA5+uKrXllZ6hbSWWoQR3NvKMPHKodGHXBVsg0Afnf8AsNfD/R5/FvxP+OXh155fB2u6pc6b4ME6GNI9EF3Nf3UlqpAItrnULmZoiRkpGnQYr6G8Bfs9f8IR4C+KHgj+3/tv/Cydc8R6z9o+y+X9i/t8sfK2ea3m+Rn725N/91a+kURIkWONQiIAFAGAAOgAp1AHxJd/sl+JtJ8O/BNvh/47i0rxZ8FNKl0i0vrzSvtljfwXVrDa3BmsxcxvGzLCGQpPlclSWBrgfiN8Adc+FXwV/am+JHifxpL4x1n4jeDrqS7eSxishDLpmkXcGIlhdl8oq6hEI3IqAM8rEvX6M1FPBDdQyW1zGssMqlHRwGVlYYIIPBBHUGgD85vhz+zd8S/ip8L/AIV6Z8U/iaut/DbTLLQ9XTQ4NHitLu8e1hintba+vUmZZYIXCkhLeNpNo3ncNx958QfAfxzb/tDH47/DTxlbaEmt2FhpfiDTL7TDfx3ltYTPJG9vKtxA0ExR2jyQ6gHdtJGD9RRRRwxpDCgjjjAVVUYCgcAADoBT6APkWb9kXwxrnhP4zeCfGOsS6jpfxf1x9bf7PF9ln02TyrdYfKkLyB5IZbZJUcqo3YBQgHPYfC34eftB+GdatJfid8V7fxfo2m2z28dtb6DFp0945ACT3twbicl0APEKxKxOWz92voqigDxH4lfBz/hYfxI+GHxB/tf+z/8AhXGo3t/9m+z+b9t+2Wclps8zzE8rZv352vnGMDOa5n4rfArxJ4k+Iuk/Gn4T+LV8GeOdM0+TR55bmyGpafqWmPL56211bebA+Y5svFJHKrLuYHcDgfSlFAHy/wCP/gF4u+MX7Pvib4M/Fnxuupat4jKyLq1hpq2MdnJBNFc2witfOlLJFLCrHfMWcZG5eMUr34DfEjxs/wAMtZ+Knjix1TXfh34ibWzPp+jtZw3sQtZLZYDG13J5b5kMjSgkE/KIwOa+raKAPm74vfAnXfGHjvQPjD8L/FQ8F+PvD1pcaYLqazGo2N/ply6yvZ3lt5kLMqyqJI3SVGRsn5uAPH/E/wALrz4D/BP45/FzxpreoePPHfi7RrmXVtQ0+FdOlENpaSQWtvp8KGYW0VsjswYmR9xaRixwK+8aCM8GgD8Z/Cfjnxt+z54F8G6z8LfHfwy+JN3q76NpSaLoWirZa1rEcrxQOsU9peOzSQRkuzy26qoQs6qflr9KPhn8GR8O/HfxO8anWDqA+I+q2+pfZxAYDZ+RaR2vl+YJHMm7y927amM4xxmvRNF8BeBfDeoT6v4d8OabpV9c5824tbOGCWTdyd7xqGbPfJrrKAPzZuf2EviHd/A2b9mhvjB5Hw8tHDadDHoMP2/al2LyOO+uTc4nRJBn91HA7EKWcjKN7/8AE39n/wAZav8AFu0+Ofwa8bR+CvFbaYNE1NLzTRqun6jYJKZovMgE9uyTQuzFJFkyR8hG3NfU9FAHxf4W/Y50yLwp8XfCXxQ8WXPjaH4w3EV3qNw1rFYz286W0cJaExFoxsljEsHyDywERvMKl29A+Gvw7/aJ8L6xpkPjz4r2firw5pKPH5K6AtpqN+DGUjN5d/apU3ISGJihjLsMnAJU/SFFAHif7Q/wZt/j78JtX+GcmrSaDcXslrc2moRRiZrW7sbiO5gl8tiodRJGAy7hlSQCDgjhZvgT8QPFviD4UeNPid40stS174a6nqGoStp+ktZwX6XtpJaJEEe6lMJQPvL5cMRgIvWvqaigD5T8dfAH4gXnxruPjd8KPHsHhTU9X0a30PU4L/SV1aJ7a1mkmiltibi3MMqtIQQd8Z6lCRg8jpH7Fehw/A3xR8F/EXi6/wBWl13xJd+KLXXFiittRstRmuVu4JhszG0sUq5ZlSNXBYBEBwPtqigD5k8N/Cr49Xema9o3xa+Kdp4lstT0e60qCGw0CLTVEtyuz7bcH7RO8kqjIEcbRR8tkE7SuBYfsr/YvCHwG8Kf8JPv/wCFI3Npced9ix/af2Wxkstu3zz9n3eZvzmXGNuDnI+uqKAPAtW+FX9mfHy6/aV/tCS5+xeDZvDv9jw2u+WTbe/b/OSbzOWO3yxF5fJOd/avym+FnjjxB8KPgBB8bvh98Svhzfaslpc3kOjappCz+JpZ7qWScaRcX9tdw3M96zv5TH7MC0pJI2/Mf3WrkoPAPgW219/Fdt4c02LW5CWa/SzhW6YnqTMF3knPrQBzWseGx8Zvgtd+E/HdhLo3/CbaC9pqVorZms21G12TxhiPvxFyAcdRnFfJ9h+x58WdW0r4caH8S/jGde0/4Wa1ouqaVb2uiR2UdwmjSqyC9IuHeWZol8pHVlSPLO0crEEfoFRQB8efGT4AfG34sWXirwI/xWtbfwB4y3xXVjc+H4bjUbO0lAElvZ3iTwx4Izsea3kkjJzuYgV6Do3wDs9A+OulfGPTtXYWmj+Cx4Oh014S7GNLyO6W5a5MmSQIwhTy+c7t/avoKigD480T9k/+x/2P9S/ZR/4Snzv7QstWs/7Y+w7dn9qXc91v+y+ec+X523HnDdtzlc4Hzv8AtE6Tay/tBfDP4ZTeLdB8LzeH/Bd20dz4w0+HUNE1Xzrq2tzBb2k8sKC7j+zeY7rPvWNlUKys5H6l1geIfCnhbxdZrp/izR7PWrVG3rDe28dzGG6bgsisAfegD5L/AGVfiz4/8aeKvHnw78Q3fh7xP4f8Df2dBp3iLwtbSWmm3Es8btNZeW01xF5tqFTf5MhVA4VgG4r2Xwt8HP8AhGvjr46+Nf8Aa/2n/hNdO0iw+wfZ9n2b+yhMN/neYfM8zzunlrtx1bPHsGm6ZpujWMOmaRaRWNnbjbHDBGsUSDrhUUAAfQVeoA/P3Uf2MviFJ4P+Ifwk0D4sf2T8PvH11rF9JbDRIp9Uhm1ku80LXrz7ZLfzGyw8lZmTKCVDhx6D44/Zh8Ua5q3wj8T+CvH58Ma78I9Iv9NtJ20uO9hvXvoLS3Z54pJl2xbLZt0atvO8FZUKZb7BooA+SvA/7NOu2Nx8QPGnxO8ZDxX4++IOl/2LLqUVgLKz07TkjkWK2s7QSyMEV5GkctMWkbBO05JoXP7JrxfCX4VeDvDvjCbRvGvwet7ePRPEkNmjjelt9kuFnspJGV7e5i4ki80HpiTjn7EooA8C0PwL8e5fCviPTvGnxPtn13VYok0290fQobOLS3j3EyLDdTXn2gyEjeJGAwMIEJLHi/BXwA+IjfGTSvjV8ZvHFl4p1bwzpl3pWkw6Vo/9kQpHfMhnmuS1zdPM7CMBUDLGhJIGTX1jRQB4f+0D8HJvjd4Ci8NaXrreFte0rUrDWNH1hLcXT6ff6fMsscohLxiTcm+NlLgFXP0rzrxV+ylpOt/sqab+y5omtnSbTTYNHiGpNa+e0kml3lveyyvD5sfz3MkLFv3nys5b5sYP1rRQB4jB8HPJ/aOvP2gf7Xz9r8K2/hn+zfs/3fIvpb37R9o8znPm7PL8vjG7fzgeZeNf2dviPL8btU+Mvwe+I6eCZfFem2em69az6RHqgnFgZPs9xbtJNGIpkSQoNyuncqfun67ooA+P/An7NPj34SfC6bwH8LvidNp+qf8ACR6jrq6jf6ZDqAuIb+WSQ2l7E8iNLjeC00UsDs6hgFBKVF4V/Zw+JOgWfxV8XS/EaKH4ofFGCzifWrDSUt7HTf7NheG18ixkmmMhUSPveSYs2RjbtFfYtFAGToFlqem6Fp2n61qB1bULW2hiubwxrCbmZECyTGNPlTzGBbaOFzgcV8BeBv2LPjB4U+Fdr8B7n45zR/D+IXMM1tpGgQ6fqU1teXElxPCb+W6umQOZWXckasFOM1+idFAHyl8R/wBmG31bQvhlafB/WYvA2ofCK483QfNs/wC0bHyzataPDcW5liZ90bf6wSBw2WyWOaj+Hv7NOuaB8RfG/wAS/iJ44fxjqHj7Q7TRr+JdPj0+GFbZp8/ZhHI5SIxyhQj+Y4YMzSvuwv1jRQB+fWmfsafEUfBC0+BWv/FCG70zwjeaVe+E72LRRHc2L6RcGeAXytdPFeJjahQLFwu7OTgen+Ef2ePiFF8bdA+PPxO+IkfiXWtG0i/0j7DaaQmn2CxXrwyK0A8+aVWVo28wyPL5m5dvlBcN9bUUAeI/Gn4Of8Lfufh5cf2v/ZP/AAgfizT/ABRj7P5/2v7DFPF9m/1kfl7/AD8+Z8+NuNhzkGj/AAc/sr9oLxH8dv7X83/hINB0/RP7P+z7fJ+wzzTed5/mHdv83GzyxjGdxzge3UUAfIPiL9nb4laJ8QfFXjz4A/EaHwMvjx4p9bsb/R01e2+3RRCEX1mDcW5hnaNVEit5kchAZhxioG/Y98PWHw0+Gvw28Oa/cW0fgDxdp3i+5vbuEXdzq15aXElzcecweII9zJKxMg3BBgBGAFfYtFAHiN98HPtv7ROk/Hz+19n9l+Grvw9/Z32fO/7VdxXXn+f5gxt8rbs8s5zncMYJ8Kfg5/wrHxf8SvFX9r/2l/wsPXF1nyfs/k/Y9trFbeTu8x/N/wBVu3YTrjbxk+3UUAfF/wAS9AvdF8V3006HyL+V54pMfKwkO4jPqpJBFeZ6jY2+qafc6bdAmC7ieGQDrskUqf0NfondWdpfQm3voEuIj1SRQ6n8DkVjf8Ih4T/6Alj/AOA0X/xNNNp3Qmk1Zn5Taf8AAfwZpvgrU/AsNxePZ6pPHPJM8kZuFaIoVCMIwoA2d1PU/hZvPgn4WufCuieFYLu9tF8PStPZ3UMqpco7OXY7gm3JY5yFBBAxX6of8Ih4T/6Alj/4DRf/ABNH/CIeE/8AoCWP/gNF/wDE12vM8Q3fne9/na35aHCsqw9rci2t8r3/AD1Py20j4MeEtI0/xFpvnXl7D4oWNbw3MwkctGG+dX2htxZixLE89MDisY/AbQX0bTdIk13V2fRrgz2Nz9oXz7ZWVFMUbbMKmEUgAcHkYGRX6xf8Ih4T/wCgJY/+A0X/AMTR/wAIh4T/AOgJY/8AgNF/8TTWZ4hO/O/6Vvy0E8qw7SXIrf8ABv8AnqflRH8CvCMfh7XvDYvNQa38RTQz3MjzI8wkhcOCrtGfvNyxbcTntWj4X+Dnhjwv4jHiiO6vtRu4IjBai9n89bWIjGyLIyAASoyTgHHqT+o3/CIeE/8AoCWP/gNF/wDE0f8ACIeE/wDoCWP/AIDRf/E0pZlXacXN2e/5fkhxyvDpxkoK62/F/mz8sNL+C+g6FrY1TQdW1XTbP7SLo6dBdbLIyg55j25KnHK59ugAr2Gvu7/hEPCf/QEsf/AaL/4mj/hEPCf/AEBLH/wGi/8Aiawr4mpVadR3OihhadJNU1Y/Je//AGdvA99qNzcrd6hbWF5N9on06G42Wckmc8ptyB9G4/hIFdj4z+FnhrxpFpvntPpd1o/Fnc2LiGaFcAbVOCNvAxxkdiMmv03/AOEQ8J/9ASx/8Bov/iaP+EQ8J/8AQEsf/AaL/wCJrd5liG4yc3dbHOsrw6UoqCs9z8vn+E2iTjw017qWo3k3he4e6t5Z5lkklkkdXPnMyEsMrxjbgd6bd/CDw1ezeKppbm7DeL1jW72vHhBEcjysxnHvu3V+of8AwiHhP/oCWP8A4DRf/E0f8Ih4T/6Alj/4DRf/ABNSswrfzf1e/wCepTy6h1j/AFa35aH5Z638F/DOs2WgQJe32n3fhuBLW1vbSZYrnykUKAzhcdOcgDBJxwSKor8BfBaeGNU8Kie9a11S6W8aR5VaWKZBgFGKc9879xOTzX6tf8Ih4T/6Alj/AOA0X/xNH/CIeE/+gJY/+A0X/wATVLM8Qkkpv+nf8yZZXh223Ba6fhb8j8wfCXwp0Twh4jn8V2+o6hqOpXVr9lmlvZxOZF3K28kqG3fIAMHaAOneneL/AIV6J4s1mDxJHfX2iavDH5JutOn8iR4s/cc4bI/I+uQAK/Tz/hEPCf8A0BLH/wABov8A4mj/AIRDwn/0BLH/AMBov/iaj6/W5/ac2u3y7F/2fR5PZ8ul7/PufmD4k+FGh+J/DulaBf318smiuslrerNm7V1/iMjKcnpzjPAxjFdDq3gvTde8GP4J1u4uL21lgSB55HBuHMeCsjPjBfcobJHJ6g1+jv8AwiHhP/oCWP8A4DRf/E0f8Ih4T/6Alj/4DRf/ABNT9cq6e9s7ryZX1Kldvl3Vn5o/LLwx8GPD/hjXrDxMmp6lqN/p8DWyNeTrKpiZSoXGwYVATtCkAd81nQ/AXwtb+H5fC8Wo6imnPfpqEcazqPJkQEbEIT7pz35yAc5Ga/V7/hEPCf8A0BLH/wABov8A4mj/AIRDwn/0BLH/AMBov/ia2eZ4i9+d9Pw2/MxWVYe3LyK2v47/AJH52eMfDsfi3wtqnhuV/LGoW7xB/wC65Hyt+DYNeU3XwmPxG8G+GrT4hT32mX2k24jkt7SeLyzIoCeY3ySKWKrkEHjcRX60/wDCIeE/+gJY/wDgNF/8TR/wiHhP/oCWP/gNF/8AE1nQx1SkrQdrO9/lb8TTEYClVbdRXurNdHrf8D8urP4QaDF4c1Xwvq2palrdlq2wub+5E0kRj5UxMFXaQcHvkgZ44qfwx8KdF8Lrqk0Oo6hfajq0H2aS/up/MukiC7VWN9oC7eCDg8gdgAP0+/4RDwn/ANASx/8AAaL/AOJo/wCEQ8J/9ASx/wDAaL/4mnLH1mmnLR7/ANfJCjl1BNNR1W39fN/eflL4c+BmieGfEa+KLPxBrc95vV5fPu0ZbjaMBZsRhnA9CayU/Z18Ow6hd6rbeJdft7u+YvPLFeRo8hJz8zCHJ59a/XD/AIRDwn/0BLH/AMBov/iaP+EQ8J/9ASx/8Bov/ia1/tbEXb5/Iy/sjDWS5Ntfmz8vbXwtq918VpfFOpR+XpukadHZWDF1ZpnmO+aQgHIK428gZ6ivVQCxCqMk9BX3b/wiHhP/AKAlj/4DRf8AxNWLXw34esZluLLS7W3lXo8cCIw+hAzXJWrudr9Fb+vzOyjQjTvbq7/1+R4Lp3w3+JNx4d8Inwt4ufwcbLXodW1aD7FHdHUtORdslgxkIMPmgKTIuWX0zX0pRRWJsFFFFAH/1P38ooooA+ff2n/G+v8Agj4O6k3g6UW/ibxHdWHh3SJT/wAsb/XLqKwhn46+QZvNx32Yrjv2pfGkv7PX7IHjDWdG1O5W/wBB0FNL06+uLhmu/tlwqWFtO87nc0okkWRnJySCxPU0v7X4Fl4Q8BeKbg7bHw3498KXt4W4Rbd9RjtS7+0bTq+exUGvJf26fHPhPS/E3wR8C+N7waf4cvPFUfiHWrl4pJYIdO8PqJB9qEatsgkvJ7ZGdwEBI3EDJoA1tA+Mvw6+I/giw/Za0vxlr+n+ONb8NPa6Rr19Z6nYyapc2NqBLfWd9dRxm7ZHXznZWYSLuOWUmvpT9n34kXfxd+C/hH4ianAtrqOr2KG9hX7sd7CTDdKv+yJkcD2xmvl608SaJ+1H+054C8ffDuX+0fh18GrfVrufX1RlsdQ1jVIPskdraSuAJltot8s0kZKBiik54r1P9iGJj+zR4X1PkRa1c6zq0C/wrb6nqt3eQBf9ny5Vx7UAeQzeP/jd8Z/2jfiP8J/BnxQs/hTD8O2sYbPTBpNpqWo6ut1bLcPeyi8YEWylgiiAAkcs6nG76k1X4lW3wT+E9l4s/aI8Q6fBfWqxW95d2FvMkN3eSMVjjtLXM07yScYjTcxOSBgcfIH7Rvj79hHx74h1fwz+0Y7+DfGHhGR4rPUru2utK1cRocx3Ol3kCb54SxJjCM4JBJj9fEbLWvihY/CP9mb48/H231TVPD3gXxHqlxrE9/bPJqEOmXMVxbaPq2oQoGYNApjkkfBYBg5yxJoA+lf2i/2irLxf+yT8XvEvwxuPEHhXxB4Y0xGJvdPv9C1G1ed1MUkRuI4Xw6hiGjJ464r1jwF+138E/Fnizw78NbDWLyTVtegYabeT6feQ6dqk1vHumWzv5YlguCApO6N2Vv4WYkZ8N/az+Ofwm+M37HXxsj+FniS38TRaNo8f2q4sg8lqjXEgKItxtETvhSWRGLICNwG5c9J+0/DDbTfsvG3RYvK+IWiRJtG3bG2m3gKjHQEADFAHunxG/ae+FPwy8S3PhHWZdT1TVtNt0vNSh0bSb3VjplpICVnvjZwyi3RgCw3kMV+YDbzWh4x/aV+CfgLRvCfiPxN4nhg0nxwkkmjXcSSTw3ipbm6AjMSsS0keBEmN0jlY0DOwU/ntDq03we+PHxn0X4n/ABru/g43inX217TJbjTtKksNY02a1gjjMN5qFpMXktvLMLwCQbcAqmWYnqz4D8IfD7Wf2MPBvgvVp/EXh2z1jWp7C9vIhFLNDcaZdXUTGIJGE2bwFXYpUAcAigD6XtP24/2fL2x1NoNQ1Q61pM/2abQTouoDXd5Qyhl00wfaTGYxuMuzy1HDsp4r0rTv2j/g9qvwft/jvYa40/g65IRLiO1uZJ/Oab7N5H2VI2uPO87935YjLFuADkV4d4Htrdf+CgnxRuFjUSnwT4ey+Bu5uroHnrztXP0HpXzH4L+Lfin4Lfs5eONd8JNFpyXfxg13SLrVri2e7ttB0671R1n1GWBCu9IF6AkLuZS3GQQD728E/tQfCzx4fE1ppo1iw1bwlYnU77StR0a/stU+w4YrPBZTQrPOj7CEESMxbC43MoPgnwU/a90/48fCXx3eazNqfhLUdJtvEtx/atrol9FbWWm2E8sMFxFNdRyQS3kURSR4NxYurAxgKyjyb4Ga7ofiL9vKx1nwz8Q9T+J9pP8ADrUYJdcvIbeO0lmi1azd4bJ7O1tbeSOPeCfL8zYzbS5bIGV8D/H3g7T/ANlP43/ArUNVgtfH/h2P4gTXuiyt5d7HA8tzIswibDNEyzRkOODnigD7it/jh8N/hd8CfA/jzxx4wuNZ0/W9P0yPT9QmtJG1LW57y3V4THZW8ZleedTvMaR5XnOACa9C+F3xa8LfFzSr3U/DVvqdi+mXH2W7tdW0270u7t59iyBHhu4425R1YFcjBHNfm42o2nw60H9i346eOIZD8P8Awr4RFhqV8Immh0m71XRrOO0vJwgYrExR4jKRtjLDJG6v0i+Gfxl+GXxktdS1D4X69D4jsdKnW2nu7VZGtTKyh9sU5URy4BG7y2YKeGweKAPPfGH7Vnwi8F+NNS8C3z6vqV/oIhOry6Vo2oanaaV9pUPEL2e0gkSEshDYJyFOSAK9Stfib4OvfiVe/CS3u3PifT9Lg1ma2MMgVbK4leCN/MK7CS6MNudwxkjFfm18ffiX8P8A4L+OfiF8RfgZ8U/+Ea+JhnhfVvA2p2b3Fr4kv4IkigW2tJUjuhLcIEjW5s5DG2ATn5jXZ678XPCfwc/bWj8ffGy5XwVp3jf4b6dBaS3gcwf2jaX80tzY+aqkGaNZVO3jdkY5IBAPr67/AGjPhLY+DPHfxAutWkTQ/hrqV1pOuTfZpiba8sxEZkVAm6QL5yfMgYHPB4Nefr+2/wDs1r4gfQL3xRJYK9vPc2d9dWF5Bp+px23+t/s67eERXhB4UQM/mEjyt9fn5f6smv8A7En7ZGvxWlzZQ6p431+7iivIHtrhYriHTZIzJDIA8bFGB2sAwzggHIr63/aIsrP/AIT/APZSTyE2weK1WMbRhAuk3BAUdsFVIx0IHpQB9C/C/wDaS+FfxcbxNbeFbq9t7/wfsbVLHU9PutNvbeKVWeKU291HHIUkVGKkL2wQDgV5rYft5fsxajJoMtv4omXSvETQRQavJp95HpMVzcKHjtri/eIW8M+D80buDGciTaeK4K/Aj/bg+J6x/KJfhXYO+P4mW+vVBPqQOPpXzHr1naP/AMEjfAkTQIU8vws20qMbn1y13HHq2459cn1oA/RL4d/tUfBn4o+O3+HHhXUbway9o2oWa3unXdjFqNmjBXuLKW5ijW4jBI+ZCQR8y5XLVjeNP2xPgh4C1y/0jxDdaqtro12LHU9Vh0XUZdI065LBNl1qCQG3TDMATvIUn5iK4T43qqfto/sxsg2kweN0JHGV/s63OPpkA4r4o+Pfx2uvip8LfjroPxA+Il74Z13ST4g0vTfh9pNhD9pmtrJHWC5v2ktri7mhmRfOklieGBI8ktgZoA/Ub4m/tFfCP4Qarp+iePNZazv9Xs5r6xghtp7qW7jglhhKQJAjtLKzzxhI0BdgSQNqsRqeAfjZ4G+I2v3vhPRHvLPXdN07TtVuLDUbKexuY7TVEZrdyk6KSfkZZAMmNxtcA8V8V6VaWup/tMfss3WowpczW3w81a4ieRQzJMbaxQyKT0ba7DI5wT612v7XviA/s9eM/Cf7X1navc2eiWV94a8QxRKWaewv0M+nscdotRjjQd/35oA+wPBHxI8I/ESXxFF4Su2vB4W1W40S+cxPGiX9oEM8SM4Ak8suFZkyu7K5yCB5R8Rv2r/g38MfFF94O1y61HUdU0aCO61aPR9KvdVXSbaVd6TX72cMq26svzAOQ235sbSDWl+y78NtQ+FPwK8K+FtfJfxDNA2pazIw+eTVtSdru9Zj3InlZQT/AAgDtivkr4VfGP4afsv+PPjp4W/aA1aPwrrWueMNR8TafcXkb41fR7+KH7ILN1U/aHgCGEwx7mVhjbkmgDof22v2hNJ034R/D638C+INVj034pa/o1r/AGz4bhu5pxo01xG101pc2iPsuJIyEijBEsmWEasVYD618G6l4O+HvwVtfEkF5qx8MaPpUuqGfWmvLnU0s1RrpzcfbN10XRCRskG9QAmBgCvzB0/wj4k8Gfssfs46f4m0ybQpL74x6FqVrplwnly2FjqGs3Nza27p/AyxSKSh5UttIBGK/Tn9or/k334nf9ivrX/pFLQB5ZpP7b/7O2tajoVtYazfHTPEk8NpY6zJpV9Fost5OB5dqNReAW3nEnaV34VsqxDAiu7+J/7SHw3+FGtnwzrsWsaprCWq301po2jX+qyW9mzMouJ2tIZEijyj8uwJ2tgHBr8z/GHxi+Gnxc/YK8C/swfDjUF134n+JdC8LaNa6JDBKLy0urYWkkt1OjIrQwQLG0v2hsRsoDKxUhq+kvix8fLu0+P/AIk+Evj74lXHwi8M+H9O0y401rGygl1XxPNeo7T/AGSa5t7pSsDhYvJtoDM7k4YDigD6j1r9pf4I+H/hv4a+L2q+KbeDwb4tuLe20/Uyr/Z3kuVd08w7cxACNw5kC7CCrYPFU/hf+078Jfi54y1H4f8AhW61C28QadaLqH2TVNMvNLluLFmCC6gS8iiaSLcyjcB3HGCDX5d+ALbH7Gv7MunXcUuyH4z2UJiu02yqq+INRGyWNgNrDoykDByMCvurx4An7ffwokT5Wl8G+JUYjqyrcWZAPqAeaAPQNf8A2vfgd4b8S6h4e1HUr57fRbwadqer2+l3s+iadesyp9nu9Sjha1hkDMFfdJhCcOVPFc3L4o8Sf8N82vgxdWuv+Efb4ZT6gdP89/sZvBrUUQuPJzs80RkoHxu2nGcV+dPw2tfD2gfBPXP2avj/APHvUPh7qlvNq2l6z4YuNK0rzLxL66mb7RYyS2Ml3dpdrIJUlid33NhSNq19rWNpYeBv2+/BOg398THd/CefR9OnuWAe/ubHU4ZpEUjAeUQIZXVf4QWxgUAfdeva5pPhjRNQ8Sa/dJZaZpVvLdXU8mdkUEKl5HbGThVBJrwXwR+1X8I/H3j+y+GulPq+n6zq9tPd6YNW0XUNLi1KC2wZntHvIIhKEBBOO3IyM1g/tbfHbUvgZ4R8M3Gl3Npoz+K9dtdFl1zUomm07RYJ1d5Ly4RWQMVVNsas6KXYFm2gg/HfgzxXoGv/ALWH/CztX+IetePfB/wa8C6nr1zrN/Zww2qXGqP5TnT4bOzthLB9ltpSHQS72XarsVagD9Z72K5nsriCzn+y3EkbrHNtD+W5BCvtPDbTzg8GvzV8VJ+1p4d/aD8CfBOP47me38Y6Xq+ovenwxpivA2meTtRY+QwfzeSWGMd6/QvwR4y8P/ETwdonjzwpO11o3iGzgv7OVo3iZ4LhBJGxRwrKSpHBAIr5I+Jv/J+fwR/7FnxX/wC2lAHF/GH4m698CPjl+z1YfE/x/d3umT6d4vGsTQwPbpq9zHFZizB02y3iaZXl2wxojtuJKjJNfVXwi/aB+Gnxsttek8GXdzDdeF7gW2q2Wp2c+m3tjI6l08+3uUjdFdAWViMHB7ggeFfGuKKT9tz9moyIHKaf44ZcjOG+x2QyPQ15D4o8KeI/Gvxt/bG8F+Cz5eueIvAmgWloAwTzLqaw1GKIFjgKWJCbv4Qc9qAPqTwr+118EfGXiXTPDui6hfiDXrl7LSdWuNKvbfRtUuk3ZhstRlhW2nc7G27Xw+PkLVU/bF/aAuv2bPgTr3xI0i0N5rUCCLTkks7m7tPtLEbTdG32+VFjPzvJGpbChtxAPwn8Mrj4bfFfwn8MPhn4v/aB1O31fRb7RGi8E3GkaVZalp+q6O8bx2rxRWCXcSwyR7DIWClOS5U5r7H/AOCg2iar4h/Y0+KemaLayXl1/ZiTCKJS7mO3uIppSAOTtjRmPsKAPOP2kv2idPm+Hfwt+JPhfWNX8H6GvxL8PWWrT6hb3uhO2nqzyXaTx3SQSNatH9/cpjYAjnBr6O+GH7Tvwl+LnjLUfh/4Vur+28QadaLqH2TVNMvNLluLFmCC6gS8iiaSLcyjcB3HGCDXy7+0j8VPhr8bPBHwN8VfDbXLPxLor/FrwhBJLbOsqJLvaRoZR/DIqSKWRhkZ5FekePAE/b7+FEiDa0vg3xKjkdWVbizIB9QDzQB2/wAKfiz4C8M/AfUfij4q+JVx4k8M6XfaobnXtYt1s3j8m9kgNv5UcaEiKQeTGFQtIcBAcqKwfEf7SPhX4j/B34oP4CGvaHrWieFtT1KB9R0nUNGm8s2s3kXVtJcxRbhvXKMjbgRnAr4N0XSNUuP2JvAvi6DTJ9a0PwN8V7nxDr1lbRG4lm0ex129+0EQKCZREXSVkAPyoSeATX3L49/aN+Cfxm+CfxV0T4V+K7bxRc2ng3WLyc2KySRQRPaSKoll2eWkhJ4iZhJgE7cAkAHUfB/4z+H9G+DvwH0vxxq9xqHi34heH9HFrGVmvb29uP7OimubmUqHYImS808hCqSCzZYZk8Z/tj/AnwRrPiHwzqGpX+o654VnMGpadpml3uoXVuFhjnaeSO3ifbbrHKp85iIydyhiykD4T/Zns/GX7P118GvjV8YdTt/EHh74q+FdE8Lx3wh2L4WZYEk0e2jc5K215GQly2ADdhXY7SK+sf2YYov+F/8A7Tkuxd7+KdMVmwMlV0m3IBPoMnA9zQB77e/H74RWHwmsvjfceIof+EN1OGCa0vESR2uftJCwxRQKpmeZ2OwQqhk3ZXbkEVQ8A/tEfDL4grr0dpcXug33hi3W81Ox16wudHvLWzcMy3Lw3scTeQwRsSDK8EEgjFfk14X0jxJF+xF+zd8QNO1G+0nw/wCBPFepX2r3unWcWoT6ZaNealbR6gLWaKdJEtGcF/3TlEYuoyuR9XfD21+B/jjxj4x+K2rfGm5+NJ0/wXeaJq8FrYWjxJot5KLiSMjR7SNpJcxuFjBaUB2wozQB9SfC/wDac+GPxe1i30jwhFrSrqNvJeadeX2h6jY2Oo2se3dNa3VxAkMifMpHzAkEEAg1S8C/tW/CL4keJrTw54PfV76DU557Ww1YaLqC6NfT2wczJb6gYPs77BG/zbwp2naTXyB8CPi7p3g/4seCvgx8FvicPjJ8LtQ0zUGuLSW3F3qfhS0062323mXdsiNJFI4FukFzH5wO0KWxiue+FfxS8J/C34g/Dr4cfssfEn/hPvBXifWGsbrwNe25l1Hw3YyrLNPdxT7I7q1htHADQXitw21SG5oA/Tz4h/ETwX8KfCGoePPiDqsWjaHpiq09xLuOC7BEREQM7u7EKiIpZmIABJrzT4dftM/Cz4mavqPhrR5NT0zxBpdmdRk0rV9KvdM1CSxztFzDb3USPNETgbow2CQCASAfJP237C/t/D3wy+IcthPqnhv4feONJ17XoLeFriSPToEmia78pAzyLaySpKyqpIUF8fLXA3fxE8E/tGftd/BvX/gXqUXifTfh1aa/eeIdYsQXs7e31OzW2tbJrjAR5pZsSeSCWQIWIGDgAxf2OfiPYfHD40fFDx7rGseLptY07xBrFnpdjdLqlnoNrolm0NtBEbaVY7QXhJMkkUg89GJJUbSa+z/ih8d/h/8ACW+0zRPET32o67rSySWWk6Pp9zqupTxQ482Vba0jkkESZG6RgFB4znivCP2Iv+Qd8bv+yseMf/Spa8R+OUup/DX9sW88e+L/AIk3nwq8MeLvC1hpum+IEsbC6sRc2NxNJPp1xPf21wlsziQTpzGsnzAlmUAAH2m37SfwdHwztfi1FrE1x4fvbo2EQgsbue9a+DNG1obGOJrlbhXRlaIxBlIOQBzV74TfHz4efGW41zTPCsl9Zax4aeFNS0zVrC50vULQXCl4XktrpI3CSKpKMAQcHnIxXwTP48074C/CO38UfCT4lnXdE+K/j+WbXfHV/p8ElppBurcJdXMcFvFBbYkktlVJWUwB5Gdt3APHeB7WX4j/AB7+NukfDDx1qnju58S/Cg2ljr+pJDAl1fyXF1BG9rJb21pbvbxyMFWSJCgbeu8lTgA+9vC37XHwW8beLrXwp4TutU1WK+u30+21e30e/k0S4vI92+CLUlgNszDa3zB9nBwxr5l+Hv7WfhX4Y+OPjzp3xT13WdZk0fxndfZLK0tb7WJNO0mOztSZDHAkotbRZC5BbYhbftyQ2O1/ZS/aX+BOnfCj4Z/BWXVF0Hx1ptjYeH7rwxLbTJqdtqdrEsNwJbZYy6p5itI07AJtO9mHNaX7K1tbyeNP2mzJErGbx9eRuSBlkGm2mFPqBuPHufWgD7L8JeK/DvjrwxpXjPwlfR6nout20V5Z3MWdksEyh0YAgEZB5BAIPBAIIr8+fAvxF1Y/s+/tD+JPHPxC1bw3b+H/AB94stIddjDajdaVY2t8iQx20Ugl/dxqdixquFUnbtOCPWP+CeDs/wCxb8KmcliNMkHPoLmYAfgK+JvEd7aR/sa/tleG5JlXVNP8b+Kri4ticSxQ3t9HLbSMvULKgLI3RgDjoaAP0m8ZftHfCT4T6jo/g3xxr87a7qWlDULOFLKe5u9RiSSOA+RDaxMZZ3dw3kxpu27n2hEYja+Fv7QHwu+L/hHV/GnhTVHgsPDs89tqyalBJp9xp01sgklW6iuVRotqEMSRtx34OPnCWKKT9u74ZvIgZo/hnqjISMlW+3Wi5HocEj6E14H4+8E+LviDL+3X4F+H8Ty61qR0D7Nbw/fnf+xoJZYUHd50Vkx3LYoA+4/An7V3wc+InifTPC2g3WpW8viASto11qGk32n2OsLApkkOn3VzDHFcbUBfCNkqCygqM1r/ALRuv2Phv4dw6jqPjW+8Awtq2lQDUtPtlu5mee7jjS3MbxyDZOzCNmx8oOT8uQfh74fan8LPjh4j+F2m6h+0PqHiHVvD+r2msWHhSbStJsb6zvtMifdb3UVpYwz26xxmSF9xVDnaCSVr3/8A4KAf8kEs/wDsa/C3/p2tqAPV/iB+1F8HPhp4tvPAHiPUrqbxTaWtteLpNhp93f31zDdGURm3gtopHlx5L7yoxGMFyoZc7fhH9oX4QeNfhPc/G3SPEMUPg+wW4a8u7tHtTaNaMVnjuIpVWSORGGChXJyNucjPhPhGKM/8FCPiJMUBkTwFoKhscgNfXRIB9DgZ+gr4x8R+Hta1z9nD9o5dH0ybW7PRfjZf6pqWm20fmyXemWV7YXN5GsY+98gaRl7hTQB754//AGrdA+JHxW+Ami/DLU/EGixax4q3XMN5p+o6NFq2ltY3BWRPtEcSXVv5mw4BYA7SQMrX1j4A8UaRL8Wfi7p8njq71lvD82lG80q8t0t7PQFlsROBBPsTzVnjIlkYuwQjHB3Z+Ofi5+0H8IPjb8YP2cI/hL4gj8Tw2njFbi5ns4pWt7US6bdCKKeUoEincBisLESYRiVAU1m+J/CHirx94n/bu8GeCA0mu6xZaDb2kaHDTSP4dT9ypPGZRmMf71AH2L4A/aw+D3xO8T2XhrwZLq17Hqzzxafqh0XUI9Iv5LZXeVba/eBbeTYqMch9pA+Ums2b9sf4G2/ie18OXF5qkNteaiNIj1iXRdQi0U6iZDEtt/aMkC25cyAoCH25GN1cz+z9+1P+z94n8L+Bfhr4U1iO08Tmwh08eGhbzJqGnTWFtia3ubfZm3WHyym+TbGSAAx3Ln8zvjt8eL343/sueItc8a/Ei+h8cz3ttI/w90uwiih0VbPVov8AkJZtpL0GKNA/nyzwxNJtCLkgEA/RrXf2yNL8PftV3HwJvdL1KbR7XQo7l5bbQtTuLn+05b3yFw0UbIbLyus4Qx78jzcgrXqXjz9q74OfDvxLqfhbWrrUr248PpHJrU+maTfalaaPHKu9G1C4tYZI7fKfPh23BfmIC814D498feDvhJ/wUFsfFPxL1WDw3oniX4bR6Vp99et5NrPqFtrMk8lssrfL5vlyqwUnkH1IB+bfD17a/DLxP8Zfhp8ZfjlefCWfWfFOuaqthdaZpL2utaXq774bu1uL2zme4LxEQvEkjNGybNowBQB+lXxD/aN+GHw4k8P2V9cXuval4qge70yw0GwudYvLq0jCs9ykNnHI3kqHU+YcKc/Lk5r0rwn420Dxt4Rs/G/h9p5NLvommjM1rPbT7UJVg1vMiTKwKkbWQHPQdK/Orxx4b/Z++FGi/Cuwg+M198L/ABj4S8KpaeHvE17Aq2+oaMwT/RryO7hW0uBmNHNtujmU7WG3Ir65/ZV+JvjX4v8AwN0Dx74/so7TV75rqMyQQyW8F7Bb3EkUF7FDNmSNLmNVlVW7NkfKRQB5P+zj+2Fp3xx8Y+N/B11peo2s2l+Ir/T9IlGh6lawf2faWkEo+3XE8flQXRkaXMUhifGweWCQW8O/ZN/be+GWg/s3/DaH4u+K9V1XXLmMWuqa1cWt/qFpbXdzcyCCO/1Py3hjkZWQbXl3KpBYKuDXoH7Kvj7wd4Q+MHx3+C/ijVYNL8ba38Q9X1rT9LuG8u5vNPu7C2ljngRsGRNkEjErnAGT2r5o062t/wDhyxIgiUKfDs0hGBguNUZt31yM59aAP1K+KXxx8A/CKXStO8Ty3l7rOvGUadpWlWNxqepXYgAaVorW1SSQpGCC7kBFyAWyQD0Hw2+Jvgz4teGV8W+Br5ryx86W1lWWGW2uLa5t22zW9xBMqSwyxtwyOoI4PQgn87vjpLrHgD9qzQPih4q+Id78MPCniPwTb6La+I47Gxu7SLULa8kuZbK6lvre4jtvPSRJUkOzzChXcdgFfT37JvhvwLYaF4x8Z+A/iDP8SYPGWuy6he6o8FvbwyXyQQwSmFbWGCFlKxpudFKs2eSc0Adr8SP2lPhh8MPEh8H6w2p6vrkNsL66s9E0q91eays2JAuLtbKKXyIzg4L4LAEqCAa3LP49fCrUdW8EaTp+uJc/8LHtZrvw/cRxyNa36W8YlkRJ9vlrKIzuEblXIBwMqQPzu1LVLr4QftM/GKP4j/GW8+DcPjK/stY0i9l0/S5LDV7KOyitzGl5qFpPia0ZDG0HmKcMGVDuYmfxd4D8K3nwL+En7KHwG8RTa94k1/UZPE+heI54vJn0Kxtr6S/n1jy4Vg8sK8xtbdNqLJ5m0fKCKAPeP2gP23/CPw38MRal4AS41y8j8WW/hq4lGk393ZK8F1bx6kiTQIqNNHHMViAc+ZMCiCQo6j3LWP2mfhboPgrQPG2qNqsI8VTTW2k6UdIvv7bvZrdmWRItM8n7WduwsSYwoQqxO1lJ/OTxV4h0ez/Yg8M+HzozaDqfwQ8Y+Hrfxjp6bp5LKbTdRjkvL5jy8kV1v+1rNyHWQkkkMa9C+PHjjRdV+O3w0/aM8NfE5/DvwwvtA1XQI/Fuk21lqljYak15E5S5e7guYoEuRGYvN2qQ8YUsqlsgH25a/tNfB+5+FviH4wPqlxa6B4SeSHWUubK5t77Tp4tu+G5spI1uI5F3qdpTlSGGQQaxPDf7XfwL8WfETTPhrour3T3+vecNKvJdPu4dM1N7dS0q2V9JEtvcbVUkNG5Rh9xmyM/CHxU0HwDN+yV+018UPBfxPm+Kl14ttLOLU9SMFpBbi502OOJPJ+xQQQSZhdFd0DAlACdwavo/9p+GG2m/ZeNuixeV8QtEiTaNu2NtNvAVGOgIAGKANbwR+2RpfiX9pbx38ErzS9SGn6HJo1npk0Ohan5n2y7+0C7N7KYzHDBuSPyJXWNHXe6u68r9T/Ej4l+B/hH4Su/HPxD1WPSNHsyiNK6vI7ySNtjiiijDSSyOxwqIrMx6Cvi3wf4+8HfDD9vL4x6F8QdVg0C8+I9h4M/4R5btvKGpvbRXNpJHbs3DyCaRU2A5JPTg11n7aUFxo1z8IPitqdnNqHhD4e+L7fVNeSCFrg2tq1tNBHqDRIGdks5ZFdiqkqCWxwaAPaPhv+0j8MPihqGraDocmo6dr+iWovrrSNW0y80zUxZscLcJa3MSSSxMeA0YYZIU4JAPyL+xz8R7D44fGj4oePdY1jxdNrGneINYs9LsbpdUs9BtdEs2htoIjbSrHaC8JJkkikHnoxJKjaTXQWvjzwf+0N+2V8M/GPwOv4/EWh/DzRNfHiLWrIF7FhqscEdlYfaANkkokRpjGCSgGTgk13H7EX/IO+N3/ZWPGP8A6VLQB7Fba/Yn9pq98K/8JrfSXyeE4L4+FzbKLGOF754l1EXPl7jMzI0Xl7+F+Yj7uPLJ/wBvv9mOHTbPXI/EN5c6PcTLBc6jBpV9LY6dI8hiVL+5WExWzFh9yRg4UhyoRgxoWP8Aykc1j/slWn/+ny6r5O8FW1uf+CP/AIn/AHS/PpHiqRuBy66ve4Y+pGBg+woA/Rv4n/tGfCT4Pa7pPhnx3q0trq2v2091ptrb2lzez3ot3jRo7eO2jkaSUmVdsagsRuYDarEZ3hD9p74Q+NfB3jHxppuoXVnD8PoZZ/EFlf2FzZalpqQwNcEz2c8azDdEjMmFIfBCkkED4++InxG8G/DP9pX9nXxj8Q9SXStJj8D6zHLezI7wwPKliFeZ1VhEhPBkfCgkAsMisViP2h/Gf7TfxR+D8MmqeE9Y+HreEtPvoYmWDW9Yjt7xna2JAMwg81IN44JOFJHNAH6Caz8bvhzoHgfwv8RdU1F4tC8ZT6XbaZMIJWaaXWSoswUCl0371yWAC/xYr52139sjS/D37Vdx8Cb3S9Sm0e10KO5eW20LU7i5/tOW98hcNFGyGy8rrOEMe/I83IK18aeJv2gPhf8AEf8AZl/Z98AeCNSk1jXtC13wBHq9tBbyn+yXsri3tpVvXZQkLef+7RWO6Q8oCoLD6f8AHvj7wd8JP+Cgtj4p+JeqweG9E8S/DaPStPvr1vJtZ9QttZknktllb5fN8uVWCk8g+pAIB77qf7Vvwi03xzfeAo31fUrnSL2LTNRvdO0XUL7TLG/m2hLa5vbeB4Y5cuoILYUkBypyK3v2lvihJ8G/gR44+Ilqk732kaTeyWXkW0t0ReCBzA0iwo5SISBTJIwCIuWchQTX59fFT4r+DPgb4v8AGPi/9mb4kMfGeo+Ig+r/AAx1G0eb+2tVnmS3uHsoJI4722lmUeZ58ReB8BsbcGvvn9qwk/stfGEkYJ8GeIePT/iXT0AfMX7Nvxr+Dnwm+A3hTxp8QPGfiZrrxiYYr2+8WNqkpm1aHTft101st4p2QOofyjCPLlIVELvgV9Fat+1b8JtD8IaB4t1X+2LeTxXPcW+j6S2jX39t6g9qxEhg00Qm6KADdvMYUKVJI3DPyZ48sLHU/CP7EdrqNvHcw/2zoUmyRQ674tDlkRsHIyrqGB7EA16L8X/EeifBn9tDwr8avipL/ZvgPVPBl14btdXmQtZabrH9oLdEXEoBW3+0wYRJHwGKFc9aAPpXwh+0J8KvHHgDxB8SNA1WRtK8JrdHWIp7We2vtPeyjMs8VzZzIk8cioM7SmWH3cgivL7X9ur9mS51HSbZ/FT2una2hNprFzY3dvossqRGZ4BqUsS2vnIgO9PM+VgYziQba+Y7HUrL4lX37W/x18Bq7+ANf8Hx6XZaiI2jg1m+0vTLtbm6gLhTJHCJFgEoG1yCFJC5qD4i2VnJ+xT+ylBJBG0f9tfDI7SoK5dYA3HT5gxz65PrQB9vfC79pv4S/F/xfqfgPwjeX0OvaZapqBtNS0280yWexkYIt3Al3FE0kJYgbgO44wQT3vxM+Kngf4Q+HU8T+PL82VrPcRWdtHFDLc3V3dz58q3treBXlmmfB2oik4BPQEj5m8ZAJ/wUD+G0ifK0vgXxAjkdWVby0IB9QDzXGftwWGv6N48+CvxaGv33hTwn4Q1DV4dY1mwsoNQfSm1OzWG1vJYLiGeMQqyvFJKYz5YlzkHFAH1D4K/aG+GHjjQfEuu2l3d6SfBkfm65Z6vY3Om3+mx+UZg9xa3MaSqjRqzI4UqwB2kkEDnvAf7Vfwi+IXjax+H+lSatpur6xBNc6UNY0a/0qPVIbdQ0z2T3kEQm2KdzAc7cnGATXyZpHirQfCmkfGD9p34R/Eyb49eNtE8MW9lJFHa2aWe2CSW4ttx0y3gSZo90jSBWMgj+UlcpXkb+PNG8ffHf9nDxDpvxevfirqn/AAkE/wBue2sre10XTXutLuD5EQtrZDFM4Vh5M9xLNsViw4zQB+hXjz9rL4NfD3xbqHgnU7rUtV1XQ445tXXRtJvtVj0mGVd6PfSWcMqwAr82GO7b823BBrwnxv8AtB6b8M/21bnT/Feu6m3hy++HOn3On6JaRXl7JdalNqt0C9vpturyPOYY8O4jyqL85CiuP+Bnxp+F/wCzN4i+MfgD4+axH4Q8UXvjXWfEMEt9HIp1rTdTdXs7izZVb7QQg8oxRbmRl2bc8V6holxZ6p/wURvdWt0LJP8ACXTpYmkjaOQJLrdy3KuA6EjGVIBHQjNAH018IfjH4A+OXhAeNvh1fve6elzNZzrNDJbXFtd25xLBPDKqvHIhIyrDoQRkEGuU+J/7SHw3+FGtnwzrsWsaprCWq301po2jX+qyW9mzMouJ2tIZEijyj8uwJ2tgHBrx79jwCPxh+0ZDGNqD4l6k+0dNz2NkWOPUnrXCfFv4/XNv+0F4m+E/jz4kT/CDwxoGnabc6YbGzhl1XxNNeq7Ti0mure6BW3cLF5NvAZ3cnDDpQB9Sat+0n8FNE+HXhn4t6j4ngi8H+Lp7e20/Uyr+Q73KO6eYduYgBG+8yBdhBDYPFZEn7Uvwktvh/B8SdQl1Sw0vUNQ/szToLnSL6HUdTumAaNbKwaEXU/mqdyFYuVBJwASPy78FaZbXv7Ff7NWg6xavPaN8ZbO1ltr6LDmP+39RVo5onAwcZDoVHOQR2r61/bUtNd8O/Fz4MfF248TX3g3wh4cOuWGo69ZWNvfjR7jVIYVtrieK5guESGXyngeYx4i3D5lDnIB9SeGv2kPhP4l8H+JPGi6hdaVa+DjjW7XU7C6stQ01igkUXFlNEs43oQyEIQ4+6TzVH4cftOfC34neMX8AaMdV0nXzaNqFtaa3pF9pEl7ZIwRri1F7DF5qKzAHbyM5xjJr4ql8b6T8NPA3xl/ae+DPxMk+Nni8WOi6be3BsrVrG2SG4aOO4ZNKggSf7LDcSyyBCzbECsVGCOZ8O+K9B8Yftlfs/wCr+HfivqHxcdIPFUd7qP2a2g0i2eXTFkS3tGs7aGIMdhaSJpZpUCoXIyCwB9+ftP8AjfX/AAR8HdSbwdKLfxN4jurDw7pEp/5Y3+uXUVhDPx18gzebjvsxUHi344fCH9ndPC3w08b69enU7nTCumRvBd6lf6ith5UDBfJjllnuXLqSoBd/mfGFYjlv2vwLLwh4C8U3B22Phvx74Uvbwtwi276jHal39o2nV89ioNePftDfEPwV8L/21Pg34t+IGoppGjQ+GvE0Ul5KjtDA0sloqtK6KwiTPymR8ICQCRkUAfVfwt/aA+GPxf0/Xbzwtfz2lx4WkEWsWWq2s2mX2msyGRftVvdJG8asgLK5G0gHB+U45LwZ+1p8F/HnibS/DWh3uoxL4gkki0fULzSb6z0vV5IlZnSwvp4UgnbarMoR/nAJTcK+G/Emmar+0/4h/aY+IXwE33vh/W/Att4V07UIkMdvruq2/wBomnFs7hfNVEcWxk+7lsAkDNSfDe++GHxkT4VeE9f/AGg9SvdX8PatpGo2ng2fSdKstQsdT0bDi1uIbewjuYFh2vC7EqhXI3EEZAPuzUf2p/g7Y+O5/hhb317qfiu01SPSp9OsNOurueGWRIJPOlEUbLHbKtxGWnciMEkbiVYDP1b9rn4LaV40vfA8N1qerXOk3iadqN5pej3+oadp97IwRbe6vLaCSGOTcwVlLfITh9pzXmP7MFhYp+0V+09qi28YvJfE2lwvMFHmNFHpUDIhbqVUuxA6Ak+teQ/si/Hb4T/s9/CSL4E/GfWF8MfEnw1q+pW+padcwTNfanc3l/NLDd2kcaNJdpcpIhR4g3p90A0AfYPxG/ae+FPwy8S3PhHWZdT1TVtNt0vNSh0bSb3VjplpICVnvjZwyi3RgCw3kMV+YDbzVvxl+038D/AWneE9Z8S+J4otP8c2811otxDHLcx30cMSS4h8lHLu6yIIowC8jMFRWbivz7h1ab4PfHj4z6L8T/jXd/BxvFOvtr2mS3GnaVJYaxps1rBHGYbzULSYvJbeWYXgEg24BVMsxPVaN4C8H/D74mfsVeDvBWrT+IvDtmvjKewvbyIRSzQ3GlPcxsYwkYTb5gCrsUqABgEUAfY3gn9qj4SeP9E8XaxoEmq/aPAxjGr6ZPo99Fq1uJgTCfsDQ/aXEoBKbEJODwCDXlH7M37Z3hz4ufBH/hZfj+K40O60qzuL/VbhdI1C30xYEuZIozbTypItw2xV3LFJI27IwOgseBgI/wBv34piMbRL4L8OO+P4mFzeKCfUgcfSvk39n/8AaMi+D/8AwT6k0DwRJb3/AMU/hnbXNvqehTJJLdaYW1lreW6ubSPEzRQJN5x2/eC4yPmIAP0L+HH7Tnwt+J3jF/AGjHVdJ182jahbWmt6RfaRJe2SMEa4tRewxeaiswB28jOcYya4zxv+218B/Aep6zZarc6vfWnh29GmalqWnaLf3+m2moMwQWcl3BC8Xn7mClFYkMQpw3FfG/h3xXoPjD9sr9n/AFfw78V9Q+LjpB4qjvdR+zW0GkWzy6YsiW9o1nbQxBjsLSRNLNKgVC5GQW8wTxN4K1HRPi1+y94o+JvhL4e+CtX8bayL2DXpHh8SW9q9+J5liWUx2zrcOpa3uGZjHE6kh2AwAfqj4q/aT+Gng7RPDeqaqurSX/i6N5NK0a30i+n1u5WIBpT/AGakJuUEYILmRFC5AJBIB774bfE3wZ8WvDK+LfA1815Y+dLayrLDLbXFtc27bZre4gmVJYZY24ZHUEcHoQT+eHx31GTwj+1foXxev/iXcfDvwH4r8FQaTpnim0tbDUNOF1DeyXTWk1xeQXMUC3EUiSxS5QSlCu47QK+nP2TfDfgWw0Lxj4z8B/EGf4kweMtdl1C91R4Le3hkvkghglMK2sMELKVjTc6KVZs8k5oA6f4k/tTfCL4W+KJ/Bet3GpaprVhbJe39roulXurPp1o/Kz3v2OKUQIQMjeQxHIBHNeDftpftHaVon7LkHi74W6/elfH0+n2mm61ocNxOYbW5u4UuZY5oEbyZfJZ0i3FHMh2pmQYrl/h58W/hz+zF8Y/jtpP7QOrx+FdQ8VeIj4j0nUb5HWLVdIks4IoYbWQKfNe0eN4zCuXBb5VOa8G1fwzr3h39gP7VrOlz6Da+IviJZa1pemXMfkzWOmaj4lhmtYXiP+rJRt+3tvwcHigD9Kfhvqvgn4e/A+28S6PL4m1Dw7aQy3YOsRapqOuMrSNuVoLlHvmYN92PZkLjC4ryD9lr9sLQPjV8Gl+IXjpZPD93plldajq13Lp15YaPFbQ3EqK8N3dAwy7YkUvslcht2cdB9tV+G/ga9tvH3/BNG4/Z98JyDVviN4FXzte8KR/8hT7Ppevi4u4Gtz858yJfkAB8wkIMkkUAfpn8PP2tPgx8TPFGneENDvNR0/UddhkuNH/tbSb7S4tXgiXe8lhLdwxJcBV+YhDu2/NjaCa8y+DX7ZGl/E/45ePfhZLpepRWek6laWWizDQtThOw2CT3J1CaWPy7dvO3iLzBFuTaQGyGPk/xb+M3ww/af8TfA/wb8ANUTxNr2l+MtI8SXbWkTg6JpOmCRrxr0lR9mZ1byBDJtZ2bbg4rofhJ8R/Cnw2/ax/aD8C+LL6PT/E3irVNI1LQtPnYQy6vD/ZMaAWZfAlO+NkIXOGBB6GgD3S0/bH+Bt54p0vw0t5qkMGt366VY6tcaLqFvo1zqDsUS2i1CaBIGkdgVXDlWPAY1i+FvFHiW5/bh8e+ELjVbqXQrLwXol3BYNO5tYria8ulklSEnYsjqqhmAyQACeK/Mv4ufHG4+Nn7Pth4n8WfE28vfHM+uaLc3HgfSrCKCy0Dy9XgUrfj7M94jRDCrNcXKI8pUIvzAH9DvD97aaV/wUP8YafqUy21xrvgHR5LBJDta5W1vroTGLP3vLyNwHIBz0oA+5aKKKACiiigAooooAKKKKACiiigD//V/fyiiigDjviF4G0H4m+Bte+HviiLztK8Q2U9jcAcMEnQruQ9nQncjdQwBHIrW8O6Vd6R4c0zRNUv5NYurK0ht57yZQsl1JHGEeZ1XgNIQWYDjJrnfHvjDWPB0Ohy6P4X1DxS2raraadMmn+VmyguSQ97N5roPJhwN+3LcjjGSO9oA4X4jeCR4++HmvfD631GbQotdsZtPa6swomghnQxuYsjCtsJAP8ADnI5ArptE0XSvDei2Hh3Q7ZLLTdLt4rW1gjGEiggQJGijsFUAD2FalFAEMtvBOUM8ayGJt6bgDtYdxnofepqK43xp8QPCXw9ttJu/F999hi1vVLHRrM+XJL5t/qMohtosRqxG9yBubCr1Ygc0AdbDBBbRiG3jWKNeioAoGeeAKloryXx38UD4C8d+BPDeq6eP7F8bXV1pg1LzdottTWHz7OBo9uCtykc6htwxIqLg7+AD1Wa3guAonjWQIwZdwBww6EZ6EdjUtFeT/Gv4nr8I/h/d+K7ex/tfVpp7bT9J04SeU1/qd/MtvaW4fDFQ8rjewVtqBmwQpoA9YpGUMCrDIPBB70iFyimQBXwMgHIB74OBn8hXk/jD4nnwX8TPBHgrVtPC6T43+3WkGpebgRarbRrcQWjoVx/pECXDI2/O6LZt+YGgD1hESJFjjUIiAAADAAHQAVGLa3WdrpYlEzgKzhRuKjoCeuBXP8AjXxZpfgPwbr3jnW1kbTvDthdajciFQ0pgtImmkCKSoLbVOASMnuK+bfA/wC1zp/j+TQJdD+FXj5NM8SG1NtqU+ixpYiC82+XcPKLhsQhXDs4BwvODQB9bkBgVYZB4INMhhit4lggRY40GFVQAoA7ADpXDeB/GGseLZvEcWreF9Q8NLoeq3GnW73/AJWNRghClb238t3Pky7js3YbjkA5APiP4w1jwP4dTW9D8L6h4vuWu7W3NlpvleeEnlWN5j5rouyIHc3PQdhkgA7d7e3kljnkiVpIs7GKgsueuD1Ge+KWSGGYoZY1cxtuXcAdrDuM9D71LRQAUUUUAFFFFABTBFGJDMEHmEBS2OSBkgZ64GTXIS/EDwlD8Qbf4WSX2PE93pc2sx2nlyHNhbzx28kvmbfLGJZUXaW3HOQCASDQPiB4S8UeJ/E/g7Q777Tq/g6e2t9Vh8uRfs8t5bpdQrvZQj7oZFbKFgM4OGBFAHZV80fFv4KeNPjN4z0Ww8T+JrW3+F2kXdlqdxodvZN9s1O8sZPOijurt5in2VZljkMaw5Yrhmxgj6XooAKikggmZGmjVzE25CwBKt0yM9Dz1qWigAriviV4Tl8e/DnxV4FguRZyeItKvtNWdlLrE15A8IcqCCQpbOMjOOtdrRQB518JfAn/AArT4YeDfANxPHfXXhfRNN0iS7SPy/P+wW6QlwCSQrFSwUk4zXoTRRO6SOgZo8lSRkqSMHB7cV5f8bviro/wP+E3in4s69A13Z+GLGS7MCMEaeQYWKJWIIUySMqAkHGc4NXPhZrvxK8R+FE1X4q+F7TwjrUshIsLPUTqarAVVkLzGC3xICWVkCkArkMQcAA9HooooAie3gklSd41aWLOxiAWXd1weoz3pXhikdJJEVniJKEgEqSMEg9uCR9KkooAZLFFNG0UyCRG4KsMg/UGnEAggjINLRQA1VVFCIAqqMADgACnUUUAFFFFAEIt4BO1yI1EzKFL4G4qOQCeuPapqKKAIYLa3tY/KtYlhTJO1FCjJOScD1NTUV5VY/FzQPFPws1H4q/DS2ufGNlax6j9mtbOMxXN7cabNLbywRLcCP5zPC8ak4DYyuQQSAeq1FDBBbhlgjWMMxYhQBlm5JOO57msvw5ql1rnh7S9avtOn0e51C1guJbK52+favKgdoZdhZd8ZO1trEZBwSK2aACiuC8N+MNY13xh4r8M33hfUNHsvDslolrqd15X2XVRcxeY7Wux2fELfI+9RyRjuB3tABUMNvBbJ5dvGsSElsIAoyTknA7k9amooAhjt7eF5JIYlRpjucqoBY9MkjqcetC29ukz3CRKssgAZwoDMF6AnqcdqmrjdE+IHhLxF4u8SeBNHvvtGt+Efsf9qW/lyL9n+3xGa3+dlCPvjBb5GbHRsGgDsqihght08u3jWJSS2FAAyxyTgdyeTUtcF8QvGGseCtLsNQ0XwvqHiyW81C0spLfTvK8yCK4fa91J5roPKhHL4OfwyQAd7UU0MNzE0FxGssbjDKwDKR6EHg1LRQBG0UTxGB0DRsNpUjKlTxgjpjFPAAAAGAKWigCIQQCY3IjUTMoUvgbio5Az1x7VLRXiekfGNdV/aA8R/Ar+yDEfD+hWGt/2h5+4S/bp5ofJ8nyxt2eVndvOc42jGSAe2VGsMSSPMiKryY3MAAWx0ye+O1SUUAFFFcF4j8YaxofjLwp4XsfC+oavZeInvFutUtvK+yaULaISI11vdXxO3yJsVuQc9gQDuFt4EledI1WWTAdwAGbb0yepx2qWiigAorxP46/GNfgp4f8AD2utpB1j+3vEWkaB5Yn+z+V/atwIPP3eXJu8vO7Zgbum4da9soAjihhgUrCixhiWIUAAsxyTx3J61JRRQBEsECSvcJGqyyABnAAZgvQE9TjtTliiR3kRAryY3EDBbHAye+KfRQBDNbW9xs+0RLL5bB13KG2sOhGehHrRLbwTNG80ayNE25CwBKt0yM9D714v8dvjIvwU0Hw5rbaQdY/4SDxHpHh/yxP9n8r+1bgQefu8uTd5ed2zA3dNw612Nl4w1i6+I+peB5fC+oW2mWNhBeR64/lfYLmWV2VraPDmTzEA3NlQMeny7gDt57e3uo/KuYllTIO11DDIOQcH0NTV5Tp/xc0DxX8Jrj4u/DK2ufGOnG0u7mxtrOMw3N89o0kZhiS4EZDtJGyLuAz1GQRnvfDmqXWueHtL1q+06fR7nULWC4lsrnb59q8qB2hl2Fl3xk7W2sRkHBIoA0zbW5nF0YlMyqVD7RuCnkgHrj2qaiigCOWGKeJoZ0WSNwQysAVIPUEHrTlVUUIgCqowAOAAK4SLxhrEnxMn8BN4X1BNLh0pNRXXj5X9nvO0xiNmuH8zzgv7z7m3b36Z72gCGa3guVCXEayqpDAMAwDLyDz3HapqKKAEZVZSrDIPBB6EVDHa20VuLSKFEgC7RGFATb6bemPap6KAGRRRwxrFCgjRAAqqMAAdAAKfRRQBDJbW80kcssSu8JJRmUEqSMEqT049KmoooAihggtoxFbxrEgJO1QFGScngepqWvlT4YftK6t8T/gt4G+L+ieANQvD4x1h9LmsLC4iuW02CO9uLN72eWUW4aFPI3vtXcN4GCAWr6roAKKKKAPCPE/wcufEP7QPgr40jUo4rXwppGr6XJZNEWac6m0BDh92FCeUcgqc56ivdI444Y1ihUIiABVUYAA6AAdBT6KAIo4YYd/kxqnmMXbaANzHqTjqfekmtre42faIll8tg67lDbWHQjPQj1rxT9n/APaE+Hv7Sngm48efDlroWNpfTafPFeRLDcRXEAVyGRXcYZHR1O7lWB9qT4RftCfD343eIPHPh3wK11LN8P8AU20nUJpolSCS4VnVjbuHbzEDRsNxC5xxkc0Ae2Nb27TrctEpmQFVcqNwB6gHqAamoooAKjliinjaGZBJG4IZWGQQeoIPUVJRQAiqqKEQBVUYAHAAFLRRQAUdeDRRQBFBBBbRLBbRrFGvRUAVR9AOKdHFHCuyJAikk4UYGWOSeO5JyafRQBFJBBK8cksau0R3IWAJU4xkHscVLXG+PviB4S+GHhibxj43vv7O0i3ntLd5vLkmxLe3EdrAuyJXc75pUXIGBnJwoJFTxV4w1jw94k8LaFp3hfUNctfENzNBdX1p5X2fS0iiMiy3O91bY5G1doPPvtBAO9pjRRO6SOgZo8lSRkqSMHB7cU+szWr+40rR7/U7Wyl1KezglmjtYNvnXDxoWEUe8qu9yNq7iBk8kDmgDTpCAQQRkGuW8D+IdR8W+D9H8Tatol14bvNTto55dNvtn2m0dxkxS7Cy7l74P1wcgdVQBFDBDbRLBbxrFGgwqoAqgewHAp0UUUKCKFBGi9FUYA+gFPooA474heBtB+JvgbXvh74oi87SvENlPY3AHDBJ0K7kPZ0J3I3UMARyK8fk+BOrT/F/4bfFHUfEX9pN4F8O6hol0J4f3+oy3y24NyzKwRCTCWZdpBLcYxX0ReXdvp9pPf3b+XBbI0sjHnaiDcx454Arn/BHjTw38RvCGj+PPB139v0PX7WK8srgxvF5sEy7kbZKquuQejKCO4oA6aOOOGNYoVCIgAVVGAAOgAHQUxbeBZnuVjUSuArOANxA6AnqQO1TVxsvxA8JQ/EG3+Fkl9jxPd6XNrMdp5chzYW88dvJL5m3yxiWVF2ltxzkAgEgA7KomggeZLh41aWMEK5ALKG6gHqM96looAimt4LgKJ41kCMGXcAcMOhGehHY1LUcr+VE8mM7ATj6V4z+zr8YV+P3wW8LfGBdJ/sIeJrd5/sRn+0+TsmeLb5uyLdnZnOwdce9AHtVQpbW8c0lxHEqyy43uFAZtvTJ6nHbNTUUAMiiihQRQoI0XoqjAH0Ar4J0D9nT4/eDPCOs/CXQNf8ACGq+FtTudSlh1XWdJuLnWIo9VuJLib7RCswt7uZDKwWRmQNgF1PIr75ooA80+E3wu0D4RfCzw38JtFeS90rw5YRWCPdYeSZY1wzyDG3LnJIAwM4AxXpKqqKEQBVUYAHAAFOqpf3MtnY3N5DbvdyQRvIsMW3zJWVSQibiq7m6DJAz1IoAmlghn2edGsnlsHXcAdrDoRnofepa434feKNU8aeC9J8U614fvPCt7qUIll0vUNn2q1bJGyXy2Zc8Z4OcEZAOQOyoAKhS2t45pLiOJVllxvcKAzbemT1OO2amooAijghiaR4o1RpTucqACzYxk46nApGtrd5kuXiVpYgQjlQWUN1weoz3qaigBixRozuiBWkILEDBYgYBPrwMUjQwvIkzIpkjBCsQMqG64PbOBmpKKACiiigAooooAKKKKACiiigAooooA//W/fyiiigD45/bG17XdBtfgw2hajc6cdQ+J/hWzuTbTPD59rNNIJYZNhG+NwMMjZVu4ry268EXvxn/AGwvi98PvFfi3xBaeD9J0Lw1cDSdM1SfT4ZZ7lLtd7SW7JMqgAlkjdUkYqZQ+xMfY3xS+Efhv4uR+FIvElzd2w8H+IdO8S2n2R4033mmOzwpL5kcmYmLHeF2sezLRoPwj8N+Hfix4s+MVlc3b614xstOsbuGR4zaxxaYJRCYlEYcM3mtvLOwOBgLzkA/LzWPHHxF0b9gP4nabaeLtVOr+APHz+GdO1p7ln1IWdl4ltIIjJOfmdhE5jO7IZPlIK8V7n8TfAc37NHxO+DHjXwB4t8R6ld+M/F1p4X1+01fWLrU4dVttSt52a6khndo4pbZoVdDAkSKCV27flK/tXfAGx8F/shfE7wh8P4tU1ufxj4rs/ENxCP39yLjUtesri6WAW0aOsUYDFeCyKCS5xmvfvCf7LGgaJ8Q9I+JPi/xn4l8f6h4YWZNCh1+8hnt9LNwnlySxLDBCZZymU86cyPt77huoA87/Zz8SeIdVtv2lf7V1S6vG0jx7r1tZedO8htbeOwtGjih3E+XGrMSqrgAkkDmvizxZ4c/4Wd+yD+yN4k8ZazrF5qWpeLfCmnXM66reRPNHf3bCWRzHKN1wNg8uc/vYjkoy5NfoHrX7HvgzVviD4o8Z2vizxLo+k+OZEn1/wAPaffpb6VqU6RrCzygRG4Tzo1CzCKZPMHBOMg6mpfslfDvUP2fvDv7PC6nq1tpPhFrKbSdUiniXVbS60+Xzbe5WURCLzFbP/LLaQTwDyAD59+PXwj8TeEvE3g2STSPGHj34L+HdHure60rQNevhrVvqslz5qahORdQ3eoIsRMaIJ3eM5IQjAO38YPEngrxB+xl4W8dfC/XLvxNpuma34Tu9Gv7+aW4vri5tfEFnEEnknAlM2/fE+8Bg2Qehr2rU/2cvE1/babNbfGjxvp+s2lobK6voLmwIvozNJKGltJLJ7WOVBIUSWCKN9gUOXKg1x2t/ACDw/pHwe/Z8+HmnXKeBPD+sjxDq9/cSec7f2PML+GKV2wXnvdRljlcgY2JLgAYFAH2jXyb+0Mxvfi7+zzoN2NunT+L7y7kc9PtFjoWoyWyH/eckj3X1xXs8Pw0hh+Ltx8Xf+Eh1d5bjRk0b+x2us6OqpP5/wBqW228XJ+55m77mRjnNef/ALTPg7xL4g8FaP4u8EWb6l4m+HetWXiXT7JMbr02geK6tVzj557OaeOPkfvCtAHnnxt8SeLPiJ8efCv7LXhHXbvwtYXmi3XinxLqOnv5GoNpcNwlnb2dpOMmFp7hj5si4kVE+RgSTXn/AMcPgx4V+CXw80bU/A99rN1KfHvgu8tbPVNWvNVRbkatBbulub2WV4zPHNIZMN830AA+gPit+zhoXxR8b6F8VdJ8Ta34E8aaHZyafFqmiTQRzTWE7iR7W4iuobiKSMP86gplX+bJwK5LxV8IvEN5r/wk+GNrdarrfhjw1qs3irWNc1a5+13U9zpreZY2zytgbpbu4EwVEVEityiBRtFAHpv7TP8Aybf8Vv8AsU9d/wDSCavAf2R5P2oD8MPhQviS38HL4D/4RvSsPaTaidX+y/2en2YlZIxB5udnmDdtHzbSeK+w/HPhHTfiB4J8QeA9Zklh0/xJp93ptw9uVWZIbyFoZGjLq6hwrkqSrAHGQRxSeBPCGm/D3wR4e8A6LLNNp/hrTrTTLaS4ZWmeGzhWGNpGRUUuVQFiFUE5wAOKAPy08T+O/G9v+yp+2HrkHiDUI9R0Dx14ht9OuVu5RNZQRLYeXFbybt0SJubaqEAZOBya9A+PPgrWPg7+zzH4ysPF+ual4x8V+IfCD6xqs+o3IMzvqUQkS2gEnlWluwmdBBCqr5e1G3bQa+mdR/ZR+Hep/Df4o/C+fUdVXSvi3rF9reqyrNALiC51AQiVbVjAUSMeQu0SJIwycseMeg/FP4M+F/i74Fs/h94kury206yvNNvkktHjSYyaXPHcQgtJHIu1mjAcBckZwVPIAPlDx/4Z1X4pftyX3wv1jxZruleDk+HVhqdzpmk6jNp6XdwNWvIRvlgZZYwVPz+S8bSBVV2KDafHdG8UfFjwN+yn+034W8DeJb+91L4W+JNW07RNR1C6+0X1ppEcNpcyKLic5eSCGWcwszbs7QOgr9GovhH4bh+NVz8d1ubs6/deH4fDbQF4/sYs4LuS8Vwnl+Z5pkkIJ8zbtwNoOSflb9pD4K2XgT9nf41r4H03VfE1z8TNWTVtUgjH2m4tftclrb3VxYw2yxSP9lgi8+OIsxZ0wzFSVoA8Y+FXgb4c+K/2kvAWofAnXfF+u+DtJ0i61DxJJfaxry2ceqRSW7ac8xuZkWS7lYSie1YFNgJeIV9EftxX3iaDwz8K9J8L+IdQ8MT+IPiL4c0me90ybyblba+aaKUKSGQ8HIDqy5AJU4rwfwx408ezfED4fWP7Pnxj8WfFO2u9ZtY/EFjr2iwixtdBG77XNPe/2dZvb3CKB5SmQySOQNhGa9z/AG6PBmp+PPDHwo8P6dBqEkcnxI8NNdTaYZUubS18yVZblZofnh8oNuEuRsODkUAcbN4Sk/Zz/au+EHhvwB4i1y90L4pw6/Z63pur6td6ujy6XZC8hv4jeyytFLvGyUqwVlYAKO/y38cfGFhpHgXU/wBpH4Ct8Qdcn8O63a3CeM9R1x00K5hbU47aezh0+a6QXNm29oVMNiE4B8xgGJ/SH4efszaB4M8fyfFLxT4q174heKY7J9NsrzxDPBKNPspWDSR20NtBbxK0uFEshQu4GNwBIPkdx/wT/wDhzeeAJ/hFd+OPF7eAUfzNP0JdRgS009vPFwPLYW3mzKkmSiXLzIuchdwVgAeaeNvgr4T8Zf8ABRmC21fUNcijvfh3cau4tNb1C0K3EOr2tuqRmCdPLgKctAuImf52UtzWP+z38KvCvhX9oL9ovxvBda9fXfgTWbWazhk1vUJVuN2jpKy3MbzlLo5YqnnB9g2hcBQB9lfFb9nXS/iX480D4o6R4r1rwR4t0C0n05NQ0WS2D3FhcuskltNHdwXEbLvUMp2gq3PPGEt/2dNK0n436r8bvC/ivWtDuPEptX1zSLaS2bS9UksofIhkmjmgklRhHgMYpE3YHfOQD8xvCus/Gfx/8GNJ+Ofgrwr8SNZ+M2sfZ9ZtNZXUYE8NSB51kawTT/7VEIsDDuhAa183Pzt89fU/ifwfrXxk/bU8X/DbxD4s1vTPA9n4P0TUbnSNN1Cex+03TXN3Gm6a3kWSNACxlWIr5rCPcxVAD7T4f/ZL0vwVqJtvh58RPFnhTwib1r8eGtPvLUabFK8vnPHA0ttJdQW8jlmeGKdU+YgbQcV7Dpvwj8N6X8Yda+Ntvc3ba5ruk2ejTws8ZtFt7KWSaNkQRiQSFpWDEyEEYwo5JAPiXw/8Mpfjh+0t+0L4J8aeMfEsHhPwxdeHY7DSdN1e50+CKW70SBnlLwMsrbWXcsZfyi5Z3R2II4Twgnx8+L/7DngQeH9R1TxRqOh+JLq28QRW2pnTdY13QdJ1C8tHtor8shWZ0SEuTIjSBGG8Fuf0V8H/AAj8N+CfiJ4++JmlXN3LqnxFm06fUI5nja3ibTLRbOIW6rGrKGjUF97vluQQOK8y0r9lrw/4Y+GWlfDTwR408T+GU0TU77VbXUbC+hju/P1Cea4lSZDbm1nh3TsFjlgbAAOdw3UAUf2UfEPwt1Lw74i0D4cP4k0640TUFTU9B8V3F5PqWizSwoyW+L2WZ1hkUebGUlkjYsxViOB5HF4Yk/aZ/aV+LnhX4h67rNn4W+F39j6bpWj6Vqd3pKtcahZi9m1C5ezliklk3MEg3NsVVJ2kkmvp34PfA3Q/hDP4j1pNZ1LxT4m8YXMN1rGs6vJE93dvbRCGBNsEUMMcUKfLGiRgAE5zXMfET9mjRfGXjub4n+FPFuv/AA88U6haR6fqV74fngj/ALRtYSfKW5huoLiJpIgzCKUIJEBxuKgAAH5s/Gy98QeOf2B/jv4U8ea7qOv3Xwb8XXmhafqj3UsU+oW1lcWwgN+YmRbl0jujHIJFKs8ayEGQbq/YnwH4I0f4deF7XwjoM99c2VmZCkmo3txqNyTK5kbfcXTySvgscbmOBgDAAFeJ6j+yX8Kr79nnVf2arZtQsfDWthmvbyK4WTVLm4kuFuprqW5nSUSTyyrl2ZCMHChQFA+nKAPwrs9E8aSf8E/X/atl+I/iqT4g+GHvb3Spjq9wLS2jsNZltvs7WisIbiOVEPmtOsjtnZu8tVQfVfiDwZq3wR/aA+BHinRfGniLWbz4hahfaT4jTVNTmubXUVfT5bqOUWjH7PbmKVAY1t440VflCgZz9Fw/snfDqH9mi7/ZXTUdVPhO9iu4nujNB/aIW8vHvnxJ5HlZEkhC5hOFwDk816L4z+Dnhjxz4j8A+J9WuryG6+HV++oaesDxrHLLJbPakXAaNiy7HJAQod2DnHBAPjjwD4Ck/av8afGHxX8SPFPiHT4/CHizUfCmgWOjavd6TFpUWlRxD7aqWsiCW5neQybpxIu3aAu3geM3XxS+LHxK/Z5/Z9F/4xv9K8Q6j8UB4U1HXNLZbea/tbUapZtOAQ0ZMqRK53KyiUbwuVGPtzxp+yjoPiHxjr3jXwb438TfDu68XrGNeh8O3cEEGovGnliZknt5/JuNgCmaAxuQMnLfNXS3X7MfwtPhP4aeBtFgudD0T4U6zZ63o9vZSKA1zZRzRqtw0qSNIj+e7yEFZHc7i+c5APnXwJ4dm+BP7Zsnw68La7rureF/FHgS71y50/VdUutWYanYX8MInhku5JJFeWORlcBsMcHgAAeMeC/BniH4s/slal+2FrPxB8Qaf8Tr/TdV8SWN7Z6tcw6ZpQsmmkg0+PTg/wBka1VYVjmWWJ2f5yWya/SS7+Efhu8+M2nfHKW5uxr2maJcaBHAHj+yG1ubiO5d2TyzIZQ8YAIkC7c5UnkfP2p/sPeAL5NV8NWHi7xPpHw9168lvdQ8H2V9FFo00k8nmzxJ+4N1DbzOWaSCGdEJY4CjIoA8V+JcvxP+L/hj4L/GvxD4b8QeKfhxqXhaLUNe8OeEtSm03UYdV1CGCeK7EUNxbTXkMSl0WFJt6k7wrcg7N1450Hw3p3wI/aC+FPizWNc+HEWoS+FNdGp3l1NKbXWJjbQT3wuTvE1hqCxxM8g3hXK7iuDX1X4++Bs3iu/07UvB3j3xH8O5dNsl05YdBuLcWL2qElFayu7e4t1dASEljRHAwNxVQB84/HHwf4G+DP7M15+zB4F8M654y1fxtY6la6XBFb3F5NcaldyeY9/f36xiC3KXMyztJIyAbTsXC4AB5D8YvGXjfX/g3+1d8dNI1/ULCwspB4b8NC3upYkt4vD7LDeXdvsYKrT3rzKZF+YrEBnHXsPHvgfVvgl8SPgL8TtE8beJNX1zxr4otdA8Qf2lqk1xZ6jb6nZXMrE2RItoPKkjBiSCONE7DIBr6W039mLwzdfsrWv7MHia+uv7PuNISw1K9snVLma5kYTXdwjzJIoea4LyEsjfePFeiePvgz4X+IreBG1u6vID8PdatNdsPs7xr5t1ZwywRpPvjfdGVlYsE2NkDDAZBAPh/wDaKg8JfEPxV8Q7XwVH8QfGPivw1a7ZJ9D12TSdD8MXsdoJE2F7uzgknA2zSIEupPmwwUbVHK6h408f/GLwd+xw2oeLdW0C7+IC3K63daPdGznuwujSPLllBAMjITuC7oy26Mo4Vl+tNY/ZF8J6j4t8V69pnjDxNoGj+O7g3evaFpl9Hb6ff3LRLC8hfyTdQmVFAlEM8e/jOAMV4D8Y/wBnaHRNW/Zb+Dngy71+HQPCWoajaf2vZS7NQsoodKlEE8lzDEsSMZAo+aMRvnYyMrFSAamieBo/hp+1RN+zz4e8Q6/d+A/iH4KvtTurC51m+urnTb20u44PtVpeyzNdQCdJSrbZM+YAwIwAPBP2cPAth4K/4Jx+PPi14d1XWbfxHc+GPHSCQ6vevBA1reagIpbe3aYxQTAxKxljVXLbmLZZif0U+E37O/h/4X+Kta+ImpeIdY8b+NNegis7jWtdmiluY7GFi8dpbpbxQQwwByXKpGNzncxPGOH8OfsfeFfC/hnx78O9M8Y+IR4F8eWWr2baA01o1ppn9tljdS2LtbGZXBd/LEjyIu5iVYnIAPB9X1jxX8Utb/Zz+AN94l1PRPD/AIo8HnxFr1zYXctrqGqmxtbVY7MXsbCZFeSUyXBRg7qMbhkmvur4Y/C7QPhNolz4c8M32qXenTXLXEUeq6jc6m1qGRV8mGW7eSVYgV3BS7YZmOcEAeceN/2YfA3jTwp4H0FNU1bQtX+G8UUWga/plwkGrWYjgW3f94Y2idZ40CzRvEUcfwjjHofwv+HN18ONJvLLUfF2t+M77Ubj7TPfa5cRzShtipshjgjhghiAUYSONRkljliTQB8H+L/GHi6C9/buWDW76MeG9Cs5NKC3Mo+wSN4XMxa1w37ljL85Me07/m681xvibwhr3w18C/s+/tCWPjvxNqnjHX/EHhGz1eS91WeWyvrHXNkdzamwBFrHGof93sjDAjcxd/mr7r1T9mzwNq0vxjmuL7UVb43WcVjrWyWEC3ji046Yps8wnY3kncTJ5o3842/LWh4p/Z+8G+LvAHgj4c6leX8em+Ar7RdQsZIZIlnkl0EqbYTs0TKyvsHmBVQnnaVoA+Wp/Asvxn/bH+LfgXxj4p8QQ+EtF0Pw1cJo+m6tdadayz3KXQLu1q8cwxtzsSRUckGRXKpt+fPGvjv4heFP2RP2i/BVt4t1W/b4XeNLPRtG1qW7kOorp8l/psot5bsMJJHhWZ4mcnJQhTxxXvtx8F/FHj39sz4ua3Y+JvE3gJF0HwzFaapozpDFdZW6E8TLdQT20xTCkHZviJ+Vl3MG+ibP9lD4SWvwH1z9nqSG8u/D/iX7RLql3cXJl1O9vbpxLLfT3LL89yZVV95XaCqgLtAWgDjv2hte13Sv2gP2cdL0zUbmzs9W1/V4ryCGZ44rmOPSZ3RJkUhZFVwGAYEAjI5rwj4A/BXwqn7aP7QepLqOuNceFdR8Mz2fma3qDxu97pInkF1GZ9tyiux8tJtyxr8iBVAFfQXhn9j3wxpPi7wZ8QPFfjnxV418SeBLiSbTbzWb+GbZFLbSWrQGKOBI9jLJvdwBM7oheQqu2ur1D9m3SH+NOpfGvw34t13wzfeI4rOHXdP0+eBbLVl09DHbmYSQSTRusbbN8MkbbehU5JAPjr4C2Z+FPxI8M6F+0yPGOkfFXVdRu4IvEUus3l54W8TzzeaYbeNY5mtYf3fzQ2stvCylAFLHKn6P/bV17XfD3gXwHc6BqNzps1z498K20r20zwtJBNfoskTFCCyOvDKeGHBBrT0j9kvSofE3hnWfF/xC8WeNdL8F3qalo2k61eW09tbXsSMkM8ksdtHc3Dwhj5Rnmcjqdx5r2H4s/CPw38Y9H0bRPE9zd2sGh61p2uwtZvGjtc6ZMJ4UcyRyAxsww4ADEdGU80AfIHiTwjqPxd/bT8bfDbxF4s16w8HWfg7RL6XStL1KfT4ri4e6u4wzy27LMi7c71idPMIXzNwQCu9/Yf1LXx4L+IfgXWtZvdet/h94717w5pt1qM7XN4dPs2ikgSaZ/mkZBKV3HsABgAAfQWm/CPw3pfxh1r4229zdtrmu6TZ6NPCzxm0W3spZJo2RBGJBIWlYMTIQRjCjkk+F/wAI/Dfwlbxe3hy5u7n/AITTxDfeJbz7W8b+XeagIxKkPlxx7Yh5Y2htzDnLHsAfJPx6sPA3xL+Kmt+B9LtfH3jrxVpFjarcad4a1t9G0rQXuEaS3uJpWurK3FzKuJArtcOVCkRhTz63+w94/wDFXxP/AGU/h5408bXj6hrl3ZTQXVzId0k72VzLaiSRv4ndYgWbuxJ71d8Sfss+H9Z+JHiH4kaF4y8S+EpvGa2q6/Y6NexW9tqf2KIQQszNC88DiIbGe2licj+INzXp/wAF/hH4X+BPw00f4U+C5bqbRdD+0C2a8kWWfbc3ElwwZ0RAQrSkL8udoGSTkkA+Kfgn8PW/arh8Z/GL4oeK/EdvqCeJtW0rRLLSNZvNJg0G00q4a2iCQWsiRvcvtLyyTLJu3AbQBz6N4RaK2/b9+I7yPsji8BaAWZ26Kt7d5JY+3Umus1n9kbQJPFniHxL4D8eeKvh/a+Mblr3W9M0C+ggs7y7cAS3KCa3lltp5gAJZLeSNmwDw3zV6PJ8CfDLfGCT4xxahfJe32gL4c1CxLxyWd/aRSPJE829DL5qGRxuWQbgcMD3APy7+J/iy00Hw7pH7RHwKHxC1WPTvEmmb/GOta440jVbO61FLWa1TTZ7pTLayCQrG0VhGgwrK+Bmve/2jfD+uaD8ZvEfxG+Nlh4w1n4UnTLIaNqPhDVbu2/4ReWBH+3XF3Y2M8M0hdsS/aAk6oihWTA49OH7BHgCbwVbfDLUfHfjC88G6TPb3GkaQ+owLbac1rcJcRqm22DzqrKUVblplRG+UK4Vx6d8S/wBmuT4jarrlzH8TfF/hzSfFEQg1XSdPvrdrGeLyhC6wi6tp5LXzEAD/AGeSMHk4DEtQB83/AB90fW/EPjPwh8Ubuy8U/Eb4Ft4Xh8iHwhq9zb31vqEknnJqs0FtcW1xfJJbNHsKO7IQzeWS2TtXXj7S9Y+Ln7Ib/DDxbqOueDfEMHicfaZ7ueR9TitdGzE975m1pZUkUlvNXcsgPAYV7tr37M1vJDpdl8OviH4q+HWn6XpVroqWOi3sD2f2GzQxwhYb63ulilVDt86LY54LFiARc8O/stfDDwldfCefw2b6xh+DkepR6PAsyPHMdWtzb3L3ZeNnkdtzSAoyfvCScj5aAPmr4U+AW/au134n/EX4n+KvENq+heLdW8N6Dp+j6xeaTDotvo7iBJljtJED3crZkkeYSDlVC7Riuk+PFh4A8dfE6b4eWtv498feJtH0q08/RvDety6Vp2lCYyNb3l5cm6soRczAEqJJZnKoGWLBYt6j4j/ZM0K+8aeIPGfgTx14o+Hb+MJVn1yz8P3kENrf3AXY1zsnt52t7h1Ch5YGjZsZPzfNVnXf2VPD+oePdQ+IPhrxt4o8I33iCzsbHW49Kv40XVYtOjMVu08s8M06TLGdhmhkjkIz8wY7qAPg7/hOfF3xD/YZ+BfiDx3ezajrsPxG0LT7i5uHEk839neIJbSNpXGd8nlxLvfJ3Nls81+iP7WPxP8AEHwZ/Zx8f/EzwogfWdD0ySSzLIHWOeRliSUqeGEZfeQeCF54rk2/Y6+G1t8BbD9n7RNU1fTdI0bVTrWmX6TxS6hZXy376jE6SSwsjiOVyoEkbEpwzFsvX0j4j8LaH4x8L6j4N8WWqatpOr2sllewTD5biGZCkittxjcCemMdsUAfKPhf4HaP8DvB83xnh8T+JvFvi3RdEvL+/nvNcvrm11mcWrSHzLJ5Wtgm/mFYkTZxgnFfKc/grxPZ/sbx/tmR/EXX2+LH9gx+MGv21W5OmMXjF2dLOl+Z9i+y7D5AQRBg3zbs8V9t/DP9mKH4Z6npZg+JPi/XfDugRyQ6boOp38MunwQyRtCI5fLt457mOJG2xJPLIqYBwWUEcOv7C/w6W1Twc3ivxM/w0jvBer4Ja+iOhhhL54gP7j7UbUS/OLc3Hl7u2PloA838ZyeIfjJ+1H8NPCt14k1zwv4c8TfDq71fUtO0q+lsWnb7ZaMImkjIkiILgNJEyS7QU3hHcHzGH4leM/2aPhv+1n4b8J6xf61bfCm4sJfDj6tcSajPZf21p8EuzzZyzvFbyyb1VyRgHOcnP6NXfwh8M3fxi0n42vPdJrejaLc6DDbo8Ysza3U8c7syeXv8wNEoUiQKBkFScEc8n7O3w6k1b4p6lq8dxq8HxgW2TXLK6dDbeXa2QsVSARokiBohliXZg/zKV4AAPz9+P/7O+kfC/wAAfCXxVH4u8R+IfEF3468IDVrvUNZu7221Saa9jZpntZ5HgjKsMxGFEKr8uSM5+kfJvvG/7ZHxO+Gmtavqcfh648C6I4t7S/ubTyZZby6DywNBIjQysFALxlWIGM4q1b/sN+E5rLw5o/ij4jeM/Emk+DNQ0/UdDsr/AFG3MFi+mTJLApEdqhnACeVmcyMsZIQo3z19Fab8I/Del/GHWvjbb3N22ua7pNno08LPGbRbeylkmjZEEYkEhaVgxMhBGMKOSQD8wP2aPBln4J/4JjeJfiv4c1XV7bxFqfgXxLKZDqt48NvLbi9MclrA0pitpAVDb4VRs85zzXrhv/Efxl8a/Av4Aa94h1TS/C998PI/FmtPY3s1nea1PEtrbR2sl5C6zhA0rTShHVn43Gve/B/7H3hXwX4B8afCLTPGPiGTwD4v0/VNNi0SWa0a30mLVzIbg2MhtfNDAyN5YleRVycqxOa6nxl+zF4L8V+H/A+n2Gsav4b134cWyWmha/pk8UWp28KwpBJG5eJ4JY5kRfNjeIo2OAtAHzh4av8AxH8JviF8cf2ftK8QanrHhfSPBkfibRJL+8mvLzSJLpLmCW0F5MzTsm6JZYQ7FkGQD3r541Pwz4n8Jfsb/C39rJfiB4p1H4g2EfhK6M1zq9w1nLa311aWslnJZBhbtEYpcOzIZZWBeV3ZiT+kPgH9mzwb4G0TxlaS6rqviLXfiBG0Wua9qk8c2p3a+S0EahkjSGOOCNiIY0iCL6HnMWs/syeAtc/Z80f9my7v9TTwzokGk28NyksIv2TRp4biAvIYTESzQKJMRDIJ2hTggA4G31/XT+33rHhk6jcnR4/hjZXi2XnP9mW5bWbmMzCLOwSFAFL43bQBnAr88NO8NeL5P+CdGk/tW3fxG8V3XxG8PadHqWnXT6xci1gW0vvJFu1orCGZJI1IleZXlcn5n24UfsInwi8Nx/Gm6+Ooubv+3rvw/D4beDfH9jFnDdSXiuE8vzPNMkhBPmFduBtB5Pm0P7KHw7g/ZhP7J6ajqp8Imxew+1GaD+0vKec3BPmeR5O7ccZ8nGO2eaAPnr9pXxH4evP2gfAvhz45eIdS0L4U6n4Wv76C30y9vbNbnXYrmFW+2Sac6XHlR28qmHcRGZWxncQp9H/YV8E6x4c+GWs+Itcn165bX9Z1BtNm8Q31/cXk2hwXUw0yaS2vZH+zO8D5YIkZddjOCcGvJv2i1tNG/ac0DU/E/inxH8KtF0/wibHT/E+iWpuo9RuZ73dNYXbvbXdvEIUhSWNWiVnMhPmYVVPs/wCyV4g+Leu6l4//AOEt1/VPF3gG1urGPwprWuaZHpWo3imFjf5iSG2MkCS7BDM0Kb/nwWUA0AeaT+BZfjP+2P8AFvwL4x8U+IIfCWi6H4auE0fTdWutOtZZ7lLoF3a1eOYY252JIqOSDIrlU28X4F8eWPhz4UfF34XfFPxx4muNE8E+O/8AhGND1LTLqaTxHex3At57bSoriMNPNcFpGgZlIfyzwybdy9dcfBfxR49/bM+Lmt2PibxN4CRdB8MxWmqaM6QxXWVuhPEy3UE9tMUwpB2b4iflZdzBvbpv2N/hP/wqOx+EthdatYjTdZTxJDrkV5u1o68khkOpyXMiMslwzE7i8ZXB2hQAoAB85/s6ah4j8Dftd3nwwsNE8T+E/BviTwVJrsekeKNYXWLhb601CK3+1xt9svmg82OYiSJpsllBKjAAwf2UdFsrv9mJP2mPjj8SfEPn2tn4ggivm1O4SHRtPhurqzZ4oAXjnufkMizTRyybiiKNqKtfY3gD9mTw94I+KMXxo1HxZ4h8WeMRpFxos15rF3DLHLa3E8NxgQQwRRQ+W8I2CFY1wzl1djuFjQ/2Xfhno37O9x+zHO19qng+6gvoJGupk+2Fb+6lu3bzIY41DJLKTGQnAVc7uSQD4S8Nanqfw9/aD+CuueANF8eaB4a8fX19pWpzeMNae9GuRvYyXEE5sbi9up7eWN4w6s0VudpKlMEivW/hp4Im/al+Inxd8dfEXxT4gsovBfi6/wDCnh/T9H1e70qDS49ISIG8CWsiCW4uHkLkzCRdu0bduFHs+j/sh+H4PFHg3xn4t8feLPGGr+Abw3Wkvqt9btFEhgktzC0EFtFE4ZHBeVl89mRf3oXcjW/EH7JvhvUfiBr/AI98J+NPE/gj/hMWjk1/TtCvoraz1KWJPL85g8EksEzoAsktvJE7AZyHy1AHwH8Jdc1/w7+xT+y2NF1S6snvvipYWV08EzQtc202v6iJYpfLKh45APnQja3cV6r8cdZ8F6n+0j8RPC/7RPivX9H0TTfD+k3Pgy00K81G3Ae4W4F3cBNMYGW++0RbYY5wdyL8isucfVuj/sifDHQvhh8PvhJpt7qseh/DXxHB4n0tjPC073dveT3qRTu0JDweZOykBVcoF+fdlj8m/EO8stH/AGpPiXq/jLx94q+C8l/b6NbaZNo1gbux8QWtrbM7XLST2d7EZ4pZXg8qMRMqqOHZyQAczqHhf4qeBv2bv2ZvDGqeIvEXhLxf418c6ND4guTqd1c6ipv9OvEmQteyT+WSiIxhI8uKYFljDCu2s/gxLpf7XWo/s8af498XR/DvXPBcXim+059evJria/h1FrJlW/kka8hhlWQSTLDKhkdQGYp8lep/BX4e+Pfjp4A0DWf2gdT1a5k8DeO5PEHhS9uLODSNQ1HTbCN4bCTUrPyR5fmedMSoSGRlEbHaSQfqM/CPw2fjYPjx9pu/7fXw8fDXkb4/sf2M3YvN+zy/M83zBjPmbdvG3PNAHz1+xHe65a6B8Ufh1q2s32u2Xw98d6voWl3GpTtdXa6dFFbXEMUk75eQxmZlDMc7cDoAK+16+b5f2ZvC8fg34neD9E8R67op+KesXGuX9/Y3ccF7Z3dykCOLOVIh5ce23UYcOxDMCxBGPoPTLEaZptpponluRaRRxebO/mTSeWoXfI/8TtjLHueaAPwx/Z4+Ieqfs0fB7R/Gekxed/wt3wldxaVD1EvjHR7+WxsYcdM3dvcwr6kWx9K9U+Gz6l+yZ4Y/ayXwli/1T4c6T4a8iV1Mgmvo/DkLyXMin7we4d55AeuWr9D/AAP+zf8AC7wT8PPBXw1axfX9O+H98NS0ebUyktzBerJLKk+6JIk3oZnCkIABjgkZrW0z4G+B7DxP8S/E12s2qf8AC1/si61Z3hjktDHZ2K6eI4kWNWCSQr+8Ds+WJwQOAAfA/wATfAesfs4/Abw9+094U8d+Ita8daZPoV1rMl9q91eWXiKLVLq3t7m2exlka2jQi4LW/kxo0e1QD3r0K68EXvxn/bC+L3w+8V+LfEFp4P0nQvDVwNJ0zVJ9PhlnuUu13tJbskyqACWSN1SRiplD7Ex6f4e/Yr8FaReeHbLWvGXijxP4Q8HXUN5onhnVb6KfS7Ka2ObbO2BLidLbjyEnmkVAACGr3fQfhH4b8O/FjxZ8YrK5u31rxjZadY3cMjxm1ji0wSiExKIw4ZvNbeWdgcDAXnIB+aOkfFX4o+F/2Vr/AMA6T4rv/wC1U+Kk/wANbTxFdSfaL+y0yTVfs63LzSZ3SxwkojtkqSpHQY9s8SeDX/ZY+Mvwdl+G3iLXb7R/iDrMnh3XNJ1fVrvV0ujLaSzx6hH9skkaGaKSLMjR7VZWwVFfQcP7K3wqb4ceNvhZq8d5q+hePNb1DxBei5mUTQ32oXAuS1tJCkRjEMqq0J5ZSBlmql4D/Zd0Twt450v4jeMfGfiP4ja74dgmttFl8RXMEyaZHcLsmeGO2gt1aeVBseaQO7Lxkc5APkfULN/hd8atd8WftS/8JjZx6n4rE/hzxjpWs3n/AAjtrpc86Cx0y8tLWdUs0ORBL59sySFixlGQ1d9q3xf1r9mbxV+0H4f8T31zqlvFpp8eeE1u5XuGdb4fY59PjLk7Ui1JYljiXhVuFAAGK9f8T/siaP4zvJ9P8V/ETxdq/gu71FdTuPDN3fwT6fNMlwLlImme3N6bZZQGEP2naMADCgCvJvjJ4a0r9pn9pb4ceF9H8P6p/Z3wq1O51DxHrNzZ3Fnp7xwNDNb6ZE8yIl7515BBK4TdGqxbskmgDgdb8AfETRfHH7K3wG1/x94hsZdW0DxT/wAJNc2WpSpd6hcxQWN1MrXBLOv79nVJFIkjjJWJ0zmvQPgc1t8HfjF+0N8Kb7xXqi+APCmmaLr1rNqmoTX0ukR31rdNfNHc3Jlk8seSJAGLbSCeSTn6+8T/AAj8N+K/il4I+Lmo3N3HrHgKHVYLGKJ4xbSLrEcUU5nVo2diohXy9jpgk7g3GOa1H9nb4f6z4o+JXijWTd3x+Kuj2uh6xaSSoLb7HaQzQKINiLIjuk77mMjc4K7ccgH5vtrs/gjx78Evih8JrD4g22jeL/FumaFfa74s1qS4tvENhq8UoDf2bcXckyHgTQyfZbdVxwvK4940/wCHx+OH7XHx58H+OfFPiJfCvhm38Kva6Pp2sXem2hmv7CUvIxtXjl+UxAhFkEZZizozBSvqWm/sVeE4m8Fr4l8feLvE0Hw71Kw1LQLe/v7f7PZnTT+5iMUNtEswK4RpJg8wXISRNzZ8n074IeKPHn7Xnx+1208VeKPh9G9t4SjtNQ0WRIIb6M2Ey3CFbqCeCYxFVw6r5kTE4YbmDAHyf8UbLVfGX7LHjXwn408R6xrw+E/xWj8KaXqEmo3CT3enf2lp+wXrROguZoFmKLJICUZQyFW5r7d+I+hv8JPi3+zT8P8AwbrOrro154h1xbpLzVLu9kukOl3EwS4luJZJJlV/mRXZguBtAwK9buf2SPhHP+z9d/s4xJfQeHr5vtE16tyW1STUPtC3Zv3unVt9yZ1EhZlK8bduzCjSsv2dbGa++HmueM/GWu+K9a+G9/e6hY3t89mslxJe272rR3KwWsasiRudmwI27lmbpQB82fDzwM/7V/jz4veLPid4m1+0tPBvi3UPCmg6Xo+r3mkQabHpUcQN7izljMtzO8hk3Tb1C7QBt4H2t8REn0L4Q+J0sbufztO0K9EVw8rNcbobV9shl+8ZMjJbrnnrXj3iv9lPRNW8d658QfAvjjxN8Ob/AMV+X/bkXh66t4rfUZIk8tZ2jube4EVxsAUzQ7GIGTljur3678Iafe+B5vAVxc3UtjPpzaY88kxmu2iaHyS7TS7y8pU5LvuJbk55oA/L+81zx54k+BP7Gun2njPWNFvfGN3ptrqmo2d0wvbiGfRp2m3ySbw7tyQ7hir4kX51BHs3wq8OT/Bj9s7Xvg94Z1/WdR8J6/4Ej8Staaxqdzqhg1OHUzZvLDLdvJIoljbLjdywz0ChferT9mTwFZ+G/hJ4Xiv9TNr8Gp7e40djLD5k7W1q9mguz5IDgxuSfLEfzYIIHFdrL8IvDjfGg/Hj7Td/8JAPDreGhBvj+x/Yzdi837PL8zzvMGM+Zt28bc80AflR8Ofh1rPiL/gnzB+0xqvxK8XD4jaH4c1DW9P1N9du/JtX0ozPFbi0Di3lilEISUzRySSbjljwB9VeHfiL4s8W/tOfAy/1C7uLG28VfDW+1a905JXS1N5K1lJlod20vH5jKpIJAyM14z+yb+x6njr9lX4faN8T/FPjCw0G6gabVfBs10tvp08kV3Iwjljkg+2RwvtDvAkyIxO4AZOfuD4w/s4eHfixrfhLxbY+IdY8DeI/BXnxadqXh+aG3mW1ulRZ7Z0mhmieJgi4Up8pHHGQQD5muYr/AMe+Pf2yfBXiHWNTOkabD4fezit9QubY2m3Qo7plt3hkVolklQGVUIEgLBwQxB8y+FXwP8cD9gXwhefAnUNZl8ReKdN8OXuq2Z166hku9Ptj5l3ZaZNcStFpzzxuyhohGuMK3AGPu34U/s1/D/4R33jTUNFutT1abx/9lbWG1a6+2NcS20LQtKXKq+6YOzSZYrk4jVEAUcB4O/ZBtPA3gVvhx4e+KfjSz0Sxngm0VYb+2il0dYDKRFBIlqDPEwlKtHdCZCqp8uVDUAR/sna78LJD4r8I+BofFPh3WtIktG1Xwz4tvL26u9LMqN5TwfbJrkGG4wzB4ZnjcjPHArwnxt8FfCfjL/gozBbavqGuRR3vw7uNXcWmt6haFbiHV7W3VIzBOnlwFOWgXETP87KW5r7E+EnwF0n4Wa/4h8bX3iLVvGXi3xUttFf6vrMkDTtBZhhBBFHbQwQxRIXY7UjBJJLE8Yzfit+zrpfxL8eaB8UdI8V614I8W6BaT6cmoaLJbB7iwuXWSS2mju4LiNl3qGU7QVbnnjAB84eEfBLftV/FT4xan8SfEmvWGm+A/EL+GNC0vR9Wu9ISxFpawyvqBFpJGZp55JiUaXeqquAuK9n/AGMfH3i/x58HJovHWotrer+FNc1jw6+qOoVtRj0q7eCK6bHBZ4wocj7zAnvV/wAc/staJ4m8b6v8QvBvjXxJ8OdY8Twwwa23h26ghj1NYE8uKSVLm3nCTonyLNFscL1JOCPZ/hn8NvB3wh8D6V8OvAVj/Z+iaNGY4IyzSOS7F5JJHYlnkkdmd2JyzEmgD4S+FHgW4/af174pfFX4h+LPEVldeH/Fmr+HfD9lpGr3WmQaNbaK4hSUQW7rHLczNmSUzrKpBC7doxXhHwj8TeKv+GMv2WfhH4a1q68MQfE7Vn0nUtUsX8m7h0+F7y6mhtphzDNceWI0kHKjdjmvu3V/2RfC1z428R+LPC/jPxP4PsPGs/2vX9G0a+jttP1G5KhJJzuhee3lmUATPbSxM+Achua0ov2R/hXH8B/D/wCz75upf2L4UlS60jUVuVi1bT7yGZ54bq3uYo0CTRNIwVtmCvysGBOQDx/StDn/AGcP2qvh18M/Aut6tf8Agz4o6Xrpu9J1XUbrVVsL3RY4bhL23mvJJZo/OEpjkTfsJwcZxjwD4R/Ce8+K/wCy34n+LXi34i+LYvFOkXXiqXR72PXryCHSTpt9d+SUgikSOVcplzOJGKHy1ZUVFX71+GX7OOieAfG1x8TvEXijW/H/AIxls/7Nh1TXpoJHs7EuHaC1htYbeGIOwDSMELuRy2OK+KP2a/2TLn4hfA250vx14u8ZeG9G1vXNf/tbw1BdLaWV9bnVLjarLNA1zDFcRbRIsEsayqS2MsWIB1Nz8RPDvxr+FvwWX4iJ4w8S+N/FfheLV5fC3hC/k0tb5JUhWW/vpIrizSK3WU4jMt1GmZCArsBt8gsviF8S9O/YV/aS0u/1PWNM1X4b6/q2k6VLeagbnVbGzQWlxDBJfRSOZZIfPZBIJGyoADECv0A8b/sv+FPEnirw/wCN/B3iHWfh3rnhzSB4fguPD0ttEH0cOJFs3iure4i2Iy5jKoGU8g8DGFafsbfC+y+EvxF+DcOqa5Jo/wAT76XUdVuZ71bm/W5nigileK4nickv9nVyZRIdzNzjaqgHy/8AFP4YeJvh/wDEb4KX/hj4neLoNT+KWpy6J4lupNVeYXkE9lJdNJDbyBrazkQxlYWtooxCG+QAjNd38ONKv/g7+0b8Wfgz4d1/WNQ8LT+C7LxNawarqVzqUtlfSzXVtMYJ7l3lAlCK7AufmGa+vvGfwZ8L+OdZ8Aa3q11eQz/DnUP7S09YHjVJZvsz2u24DRsWTZIThCh3YOccGKf4KeFbj4qa58Xnur0az4g8PxeG54Q8f2ZbOGaWdXjXy94lLSsCS5XAHy5ySAfm4vib4ha9+yP+yKLPxnq+kat4v8V6Jp+oarbXJa+lgu7PUFlDyTeYJCcAjzA4DBWwSor3XwJ4dm+BP7Zsnw68La7rureF/FHgS71y50/VdUutWYanYX8MIuIZLuSSRXljkZXUNhjg8AAD3ex/ZW+Hun/D/wCFXw3h1DVG0z4Q6rZaxpMjTQGee4sI5oo1u2EAV4yJ23CNYySBhhyD6Ld/CPw3efGbTvjlLc3Y17TNEuNAjgDx/ZDa3NxHcu7J5ZkMoeMAESBducqTyAD8h/CXiH4wfF34Kx/Hbwt4X+I+p/GHXWk1TSdZs9Rgj8NwFbgmLT4rD+1EiNiI08mQS2rSMdztlq/WT416rq1t+zz481uIy6XqcXhbVLhTE5SW3nWxkcbXQ5DI3RgeCMg15jp/7JWleGdVvB8OviH4s8FeGNQvZNQm8PaVd2q6cs80nmyi2M1rLcWscshZnjhmRcsdoSvo/wAaeFdP8deDtd8E6vJLDY+IbC6064eAqsqxXcTQu0ZZWUMFYlSVIz1B6UAfkzrfhXxF4F/Z6+Bv7UMHj3xPqXjq8uvBkl7Jd6tO9ldWestbRz2TWIYWwhCS4BEYkYje7u5LV3Hj7xTqnxX/AGlPif4O8Y+FvHnirw38PP7K0/S9P8IagmmQQT3lml5Le3jLqVhNLOzSBYMlo1RM43Ma+1PEH7OPgjxJ8GvCfwOvr7UU0Lwd/Yf2SeOWEXcn9gNE1t5rmExneYV83bGucnbs4xn+P/2btI8WePZfin4P8W678PPFl7axWOoXugzW4XUbaAnyVu7e8guYJHiDMIpdgkUHG4qAAAfHbeJfjno3gf8AZm034oXGr6Rr8nxH/su8N5Osd5qGmJFf/Y/t32aV45HkgWIyKWYNIpY5PNfRs2va6P8AgoHaeFxqNz/YzfDC4vDZec/2U3Q1uGMTeTnZ5gQld+N204ziu4+If7NXhz4l/C7Qvht4j8T6+954Zv7fVtN8QreJ/bVvqNqztHdCcxeWXAkdMeUF2HChcAiD4b/sweE/h18Tm+MkniPXvE/jG40ebRLu/wBYu47g3NvLcQ3ALIkMaRmNoVWNIRHGFLEozsXoA+H/AIHP4p0f9mjxl+1l4o8Ta14s8T+BF8aS6BY3N/c/2daRWU98irNbLIEuWLFi0kwZkjEaJtESmpfiB4L1/wCCP7LGlftdaB4+8Q6r8SdMtNH13Urq81a6uNO1pb+WD7RYyaezm1S3ZZ2WBYYkaMhNrZGa/RH4XfBLwZ8KfhxcfCzSTPq2hXdxqU86akYpmlGqzy3FxG+yONGjLSsoUr93AJJyT4do/wCxF4D006JoOpeL/E+u+A/DN3Fe6X4S1C+im0i2kt3326NiBbqeCBgpihmndF2gEMMCgDg/HfhvVfit+3DefDPWPFWvaR4OHw50/VLnS9L1GbT1urj+1ruEeZJAyyxja3z+S6M+1Vdig2nznwDovxu139nr9oD4JfCnxRqN54g8FeM7zRvD15fagw1AaWgsrp7Nb+TcySmKSaKKZvuFl5UKCP0Ai+EfhuH41XPx3W5uzr914fh8NtAXj+xizgu5LxXCeX5nmmSQgnzNu3A2g5J4eL9mvwtZaV4807RPEfiDRbj4ga+fEl1e6ffLa3dpfbIUAtXjiA8nEC5jmWVWywfcCAADyr9lDWPhfZeIvEngXw9YeLvB3i60tLW51Hwv4t1C9vmgi3On22ye5uLqKWKaQlHlglKkquVQnB+36+ffhd+z5pnw88a6p8TNd8Va1468YapYxaWdT1qS23wafDIZltoIrSC3hRDId7nYWZuSe1fQVABRRRQAUUUUAf/X/fyiiigAorwz4q/GG5+F/jz4Y+HbvSluNE+IGrz6JPqBmKNZXjWsk9koj2kSfaHiaM5ZdvBGelcF4F/ak0zxb8cfiX8KNR0saXpfgW3Nzaas025b9LLYmqHy9o8sWk8iRkhm3ZJOOlAH1hRXw9o37V/jLxd4B+Fsvg/wPDcfEH4tWl1qmn6TdX7QWNjpVuwb7de3awO4QxSQkIkLM7ybF4BarVz+1V4m8M/D34x3njvwnbWXjv4MWMd9f6daXzz6dfQXVu1xaT2108EcgjlCOjBodyMjA54NAH2vRXxS37UPxB0j4en4p+Kvh6thpviifS7DwXpCX+7WdXvdVcrAl2jRLDZB12yHMkhjj3lxuXaez8CfGn4l2/xWsfg38cvC2m+H9Z8Q6Zc6to13o2oTahZXMdk8aXVrIZ7a2dLiESxvwrI6kkEEYIB9E3/iTw7per6Z4f1PVLW01TWzMLC0mnjjuLs26eZMIImIaUxp8z7Adq8nArar4j+PX/J4f7Ln/Xx4y/8ATKa6L44fGT9oL4ZQeJvFvh34eaJeeDfCUJupbnUvEH2S+1OGKETSiyt47WVEZfmVRPKpdlOFwVJAPqC/8SeHdL1fTPD+p6pa2mqa2ZhYWk08cdxdm3TzJhBExDSmNPmfYDtXk4FbVfmV8dviTqXif4xfsjfFL4WaC3ia58R2/iPUNMsJbmOyEkeo6JG6GeZw4iSKOTfKQrthSER2wp+gvhx+0L4tb4keJPg78evDlj4R8TaFoo8SwXOm3z6hpl7owlMM06SSQwSxvBLhJEdMnIYcEUAfWdYtl4k8O6lrOpeHNO1S1utW0UQG+s4p43uLQXKl4DPEpLxiVVJTcBuAJGQK+Io/2tfibb+DtJ+PmufD200/4Na1dWUcV22pyNr0On6jOlvbanNYi28gQOZI3Ma3BkWNt3OCKl+F2uaV4Y/a3/as8Sa7cC003SdO8GXd1MwJWKCDTLuSRzjJwqqTxQB95UV8I2f7VXxUs/D/AIY+MnjP4d2mi/Cfxde6fa29z/ajy65ZW2rSrDY317Zi2ECxStJFvjS4aSMOM7iCtdXq37QHxY8WfFDxV4A+APgXT/Etj8P5YbbXNS1fVX0yGW+liExsbER21wXlSNlLvJtRWO09iQD6p8P+JfDnizTRrPhXVbTWdPaSWIXFlPHcQmSFzHIm+Msu5HUqwzlWBBwRRYeJPDuq6tqmg6XqlreanojRJf2sM8ck9m06CWITxqS0RkjIdA4G5eRkc18Zf8E6Jprj9l7Tbi5t2tZZdc8SM8LMrNGzavdEoWUlSVPBIJB7cV5R4R8R/GnS/wBrr9pjTvg34P07xBczXvhea5u9Y1NtOs4BHocOyJRDBcTSSy5OPkWNQMs/IBAP0+or4b8Oftkya1oPw+8S6p4SbRrXXvFl14I8TQzXQebw/rsW+KGIlU2TxSXCCPzMpgSIcbiVr3ex+Llzq/7Qeq/BXSdLWex8O6Bbavqmp+ac291f3Dx2ln5W3BaSKKSYsWGAFABzkAHttYuseJPDvh19Pj8Qapa6Y+rXSWNkLqeOE3N3KGZIIQ5HmSsqsVRcsQpIHBr57+LPx08XaF8TdH+Bfwc8MW/irx1qmmya1cnULxrDTNL0tJfIW4upo4ppGMswMccUUZY4ZiQBz83/ALWHij4mR+EP2f8AxF408Gxw+LrX4paZjRtK1BLuO5kitdRWHybqZLdVWfCtmRV8tW+bkEUAfpdRXyV8L/jx8T9S+ON/8CPjP4LsPC+sS6H/AMJHpdxpeptqVrcWSXC200MjSQW7LNE7rnCbWGSOACeO+KP7S3xs+DAi8efEj4eaPpPw7XVYNOuGXX/P1xLe5uVto74Wq2wt2QF1doknaQKeSMNgA+5qK+R/iJ+0D8R9J+PUv7Pnww8CQeJdafw1a+IY7281IWFjbpNeXFpJ9qYRTSBVMSFBEkjOz4IRVL1xNp+2Hr9t+zf8T/i74n8Gx6f4t+Euo3ukavoqXvmW8l5ZGE7oboR5MUkc6MpMeQcjnGaAPu6iviUftLfFvQPiN4AsPiR8Nrfw74I+Jl9/ZelXi6qt1qlreSQSXFst/apEIUM6RkFYppRGertjB6HXfjr8W/E/jfxb4W/Z/wDBGm+JrL4fzLaavfavqkmmx3OomFbh9PsFitrnfLHHIm+SUpGrMF55NAH0/qXiTw7o+o6Zo+r6pa2N/rUjw2FvPPHFNdyRoZHSBGIaRlQFmCAkKCTxW1X5s+Mfihofxp+IP7H3xS8ORS29h4i1fWbhYJxiaBxpFwksMg6b4pFZGxwSvHFetaN+0Z8UvHXxp8ZfCD4d+Aba4h8Ba1aWerazqGoPb2iWFzBBOGhRIHeW7YPLiEYRQis8g8wCgD6y0TxJ4d8SpeSeHNUtdVTTrqaxujaTxziC7tztmglMZbZLG3Do2GU8ECtqvyH+DfxV+OXw78LfHzW/hf8ADqz8TaR4a+IvjTUtQutR1YWBnSK7LyW9jDHDM0kiRruZ5TEmSFXec4+vvFf7U2fCXwquPhZ4bbxN4t+M9ql5oOmXNyLOGG1Fql3c3N7cBJfLito5FDhUZ2chUBJyAD66rH1/xDoHhTR7rxF4p1O20fSrFPMuLu8mS3t4UzjdJLIVRRkgZJAr5w+G37QXiC48ceJfhJ8c9AtfB/i/wzpKa/5ljePf6XqGjlzFJd20zxQyr5Mq7JY5IwwJUgsDx8QftN/HX42fGf8AY0+IHxB074cWWnfC7xDpr/YLq41Yrrhs/OURX8lkbfyRFLgERi48wKdw3cAgH7BxyRyxrLEwdHAZWU5BB5BBHY0+vkvxv8cfGOi+L/C/wL+Dfhm18T+ONS0Ndbum1K8ew0zTNMRhbpPcyxwzyO00wKRxRxljtZiVUc5elftUaoPhn8YtW8W+Exovj/4KWF1d6vogu/Ptpwlk99ZzW92I1LW93Gh2s0YdCGDJkDIB9kUV8AXn7Wfxi8L+FvCvxo+IPwws9F+FfiObTIp7qLWftWsabDqrJHb3tzarbiDyGklQFI53kUMCRuyg7PXf2hfi1rfxq8afCT4NeDdF1qT4dQ2EupjWdafTbu+bUIBcIthDHa3A2IjBTNMVQvlR03UAfYLX9il6mmvcRrdyI0qwlwJGjUgM4TOSoJAJxgEirdfK2r+IfCo/bC8JeF9Q8IWkvii48G6jfRa8ZSbi1t4ryCN7NF2AMjtIW3kgjBAGGNeAH9tH436v8H9Y+OvhP4RWlx4S8IXGprq73etiG5uINLupoZ5NPiS3feqRRh2aZozv3oiOFDsAfpRRWP4e1yw8TaBpviTSmLWWrW0N3AWGCYp0EiEjnB2sK+Rf2i/gJ8MtT0Dx18YPin4u8Q2/9nWEl3ZXEWs3Vjb6CtpB8rWVvaSQRGRpBvLTCSR2bZu24WgD7Sorwv8AZj1Xx9rn7PXw71n4omRvFV7ollNftKNszyvECGlGBiRlwZBgYYniub+Hf7QN1458HfFrxVJoiWbfDPxD4h0NIhOXF4uhqGWVm2DyzLnlQG2+poA+mKqXl/Y6fGkt/cR2ySOkStK4QNJIdqICxGWYnAHUngV8Q6p+1x4luvh98D9S8I+GdP8A+Er+OFuJrKLVdSaz0nTylot3Ks10sMkkjncEhjSLdI2cYxg3Pjj4y1Cy+FHgy/8Aj98NtJ1DU7vxrommrp0WoveWcE896IYNQhnNvE5Kg+YsbxqcHax5NAH27RXxz4o/aF+LF18avGXwK+E3w/tNe1fwxp+l6gNR1DVPsOnpHqCTFhPthlm8zdGqxJGjBwXLvGFG7E0/9srH7NuofGfWvB80PinSdak8L3Hh2G6V92vJfDT1t0uioXy2kZWMhT5VJ4YgbgD7hor4o8I/HL9ot/2gNM+BXxE8D+GtOe80d9fkv7DWrudTYRzrbSpDHJYJvnilkjDq7RqVbcrHkD0b9pT4+3nwA0jwZqVh4Zn8WXHi/wAR23h6KytZBHcGW7trmaIx7gVLNJAsfzFVAcsWAXkA+kaxfEXiTw74Q0a58R+LNUtdF0myCme8vp47a3iDMEUvLKVRQWIAyRkkDqa+fL/xn+1bD4c0sWnw68N/8JBfG6kvHn8QyppWmQxlfIWSUWTXE8sisd3lwrGuxsuMqD8d/tGfHOX46f8ABPv4zapqmkx6Lr3hTUh4f1eztrlb23W9sdQs2Zre4Cp5sUkciOjFR1wemSAfrDVS3v7G6muLa1uI5pbRgkyI4ZonKhgrgHKkqQQD2INfHF1+0V8Y/AnjbwXbfGn4c2Xhvwh8QdVh0TT7uy1f+0L7T9RuwxtIdRhFvHEPPKld0EsqRtwXbIJp+FPjF8Lfh7rv7SXje+8LweGoPAWpQS63qFoxluNYddOinSRoyFCyfvPKRAxBY5JBY0Afb1FfGmk/tB/GPw94j8Dn42fD6w8LeG/iRfppemy2WqyX1/p1/cQvNa2+pwNawxqZhGyboZHVJMK3B3VF4J/aG+L3xT+IHimy+HPgvRLrwl4M8STeGtRN7rb22tF7WURT3iWi2skSQrkvGkkqvKi5XBIFAH2hRXhn7Q3xgu/gT8P4fiSdJXVdHsdU06DWGMxiaz028uFt5rxAEfeYDIrFDtBXcSwxmn+Kvi9PpHxw8D/BXQ9LTU7jxLY6nq2pXBmKf2bp1gI445igVvMM9xKkSjKgfMcnGCAey2V/Y6nbLe6dcR3Vu5YLJE4dCVJVgGUkHBBB9CMVbr81vAX7S3hL4S/sceA/iz4H+GyaVomteI20geHtKlaVoBc6vc2kslvlAZZXZDIsZCgs2zcAAa9tl+PXxY8Ja78NPDnxT8Fabo+pfEnxBdaZFb2OqPe/YLOKxa7jklkNvGsk+9HR0QbAMMrnOKAPryivnXWPj0ND+POu/CC70ZpbLQvBK+MHvYXLzyf6ZNbNbJb7QCdsO5W35JO3HeuP/Z2+N3xm+Num6B8QtQ8H6BZeAfFVs93a3Nhrr3uo2K7d0Ud5AbSOIyMfldY5SYmyGGQaAPrqiivzoXwPo/7Wv7SnxR0b4tPcap8P/hTJpujaZoC3M0Flc6hdWou7u9vI4nQzOnmLFErEoFBO3JNAH6L0V8A/CfSn/Z5/awuv2ePDF9dTfDvxf4Yk8RaTp93cS3X9kahZXa29zb2rzM8i28ySCXYzEK4O3AJzzmgfth/H/wAX/BeT9ofw18H7CXwVpCXs9/HNr23Urq306eWK6lsIEtWQrEImOJ5I3cqVVMbWYA/SGivzI+JHxu+OXiL9pf4MTfBbTdP1Lwt4n8MatrGlwXmtXNhBqsFzb2Uvm30UVrKsb224eQMS53ucxnIPvnin48/FTWfihrXwg+BXg3TfEOseDLOyufEV9q+pyafp9pPfx+db2MDRW08s08kQ37tioildxycAA+u6K+Jpf2y7E/A2X4k23hK6/wCEwh8RJ4Ml8MS3CLJD4me5W1+xvdBWTytzrIJlU5jIIXcdtReFP2gfjtafHGf4PfFvwh4a0S203QW8TXuqWOs3dwn9mB5IWaCOSwTe8MyqsqyPH8rb13DigD7eor4Ij/a1+Jtv4O0n4+a58PbTT/g1rV1ZRxXbanI2vQ6fqM6W9tqc1iLbyBA5kjcxrcGRY23c4IrsfFH7QvxYuvjV4y+BXwm+H9pr2r+GNP0vUBqOoap9h09I9QSYsJ9sMs3mbo1WJI0YOC5d4wo3AH2NRXxZ4c/bCtrv9n25+LXiTwrPaeKbHW5fCknhy2nSeWbxEl59hjs4JyFUrLKVbeQNqEkhtvOppXxy+Nei+OLP4XfFPwRo2meJvFmkalqPhh9N1ie60+6u9NRXk0+9mks4nt5AJEYypHKhTeVBK7SAfXc00NtC9xcOsUUSlndiFVVUZJJPAAHU023uLe8t4ru0lWeCdVeORGDI6MMqysOCCOQR1r8tv2b/AInfGTxR+zV8TPEvxm8Oad4i8IW0HjSeRrjWbi7uruSC6uN+mPDLbAJaCMSQo4lJCKgEQBwvttt+0FbeCPg78DPD3wm8CxXnij4oaNYt4e8NxXX2aysbWKwiurh57to2K29nG6gsIzJISoVck4APuKivjjx/+0H8Wfgx8Ib3xn8WvBOk23iL+2LDRdOi0/WWk0q5fUXjijuri7ntontLeN3IkMkZI2ejA17j8LNa+L2rWd8nxc8P6Po9zA0RtLjRNTl1G0vIpFLMwE9tbyRFDgYIYNnIOKAPVqK+ZpP2i7XRfiD8X/BnjHTF0u1+GGjWXiGG6E246hplxbTSzS7Co8swSwPEeWB4ORnA8v0P9sTVNY/Z80T4oS+EFs/GmseJ7bwifDsl2cQ6rcagLQo1wI8kLbn7Tny+V46fNQB900V8ja78dfi34n8b+LfC37P/AII03xNZfD+ZbTV77V9Uk02O51EwrcPp9gsVtc75Y45E3ySlI1ZgvPJrkfEX7acY8I/Brxb8PfBl14nPxikurW0sPOFtc217DbO6wyllZAq3CeVPITtiQPKAwXaQD7nor4+1T44fGyHXfD/wd0XwZot98VtQ0yfW9WhbVZ49B0jTRcvb28kl39la4mknZdqRpADuWQkhFBb1D4F/F+8+LGk+IbXxFow8O+KvBurT6JrVgk/2mGO6hSOVZLecpGZIJopUkjYopwSCMigD3GivCvhl8Y5/iD8UPip8O5dKWxT4cahp9ilwJjIbsXtkl2XZCq+XtL7cAtnGcjpXzwP21NTuv2cPAXxlsfCtrHrnxF8RSeGrCyu9R+z6da3IvLu2We7vTESkW20LHEZYswRQfvUAfflFfMmt/Gzx18KfhJrvj/45+FrO01TS7qGzsbPw/fvqKavPeNFDaLbmeC3eNpZ5RFtkX5cFySuDWT4U+N/xY0j4oeFvhl8d/BumeHZ/H0F7Lolzo2qS6nGtxp8QuJ7K8821ttkwh3Orxl432MB0BIB9S6jqOn6Pp91q2rXUVlY2UTz3FxO6xxQxRqWeSR2IVVVQSzEgADJpul6ppmt6Zaa1ot3Df6ffwx3Ftc28iywzwyqHjkjkQlXR1IZWBIIIIOK/Nr4lftDfF74x/BH4reK/hf8AD6xvvhXb6Z4g0uPVLzVza6nqMdtBNbXN9ZWv2d4jBE6vtWWZHkCHG0/LX1Z+zhLqcH7J/wALptFt47vUI/BOiNbQzSGGKSYabCY0eQK5RWbALBGIHO09KAPoeivzV/ZG+NPxkbwd8TPHvxqtbGHwV4f17xbeXmpHV7m+u7I2N08klnDbyWyg2luiusTCRTtVQIlzgdUP2t/i54f8H6J8dfiP8MrbQvhFr8tnm6j1Vp9b0ux1CRY7W/vrL7OsIicyRmSOKd5Ig3IYgigD76ubm3s7eW8vJUgggVnkkkYKiIoyzMx4AA5JPSnQzRXEST27rJFIoZHUhlZSMggjggjoa8h/aG1TRdC+AnxH17xJosXiPS9L8O6rd3OmTuY4r2K3tZJWgdwCVEgXaWAJGcjkV4Zqvx9uvBngn4OfD/4N+CYNS8XfEPR7ebRtFa6Nppul6daWcUk0tzciN2WC2R0jQLGWkOFUZ4oA+u4fEvhy41+58KW+q2kut2cEdzPYLPG11FBKSscrwg71RypCsQASCAeK2q/NH4TeMPFmm/tjfFvxT8atJsvCt5oXgPSJr42N62oWRtba4u5WuYZWihl2FM5R4ldSpGCMMfQLP9qr4qWfh/wx8ZPGfw7tNF+E/i690+1t7n+1Hl1yyttWlWGxvr2zFsIFilaSLfGlw0kYcZ3EFaAPu6ivjLXv2h/jBq3xi+IXwT+D/wAO7PXdU8DRaXcPqOpar9hsBHqVqZ1WQJDLMZmZSkaIpQhWZ5I8AN88fH/9pz4oeOv2Fbv4vfD3R/8AhFtX/tGLStbUarJBd6TdW2rw2Mi2ssER84SSjYTujxE5b5iNhAP1Jub+xs5LeG7uI4JLp/KhV3CmSTBbagJ+ZsAnA5wCa47xB8U/hp4T8T6R4J8UeK9K0nxDr7Kun6ddXsMN3dl22KIYXYO+5htG0HLcDnivl34h+L9Vt9c+CEfx0+HWiz+J9Z8YPZ2BtNRlvItJkW0mljvIZZLaBnkIj2lCigEg7iQK+fPD2u/G9f2yv2iPE9l8P9C8S3vhfSdES2jn1SX7TBbRWl9c6fFZ/wChNmS+lVTOuUELkbWlxmgD9YaK+S9V/alsJ/gl8Nfir4J0pdY1D4pX+i6bpWnPOYwt1qbD7QssqoxAtI0naQhesRHAORU/bw8YfEPwL+yz468R/DZVj1CCxlSe7F7LY3FjbSIytc2rxRuzzo5QIm6PqW8wFQGAPqXWvEnh3w2LFvEWqWuljU7qGwtDdTxwfaLu4OIreLzCN8shBCIuWbsDW1X5ZftIeJ/i03wb+CfiP4keF7UeILP4n+EpbPT9G1E3738SKXi/fXUVoqTzSbkKt8i8MZOTj6T+Hvx7+Kk3xzX4IfGrwRYeFr3WdGm1zRrrTNUbU4ZYrWWOKe2mL29uVmj8xWyq7CM47EgH11RXwTqX7WnxMuPB2v8Ax48GfD6z1T4PeGZrsS3supvDrWoWOnytDd6hY2Ytnh8mMxyMizTo8iJu+TIFdR4k/aR+IWv/ABfu/hJ8CPDWi67c6Roljr1zPrury6X9tt9R3mGPT44rW4eTCpl5WARGYK2DgkA+zqK+Y/iD8bfHemeK/DHwi+HPhO01b4jeINKfWby2v79rbTNHsYmSJ5bm6iglkl3Tv5USxREuQzHYo56X4HfGDU/icninw74v0RPDnjLwLqf9laxYw3Bu7be8MdxBcW07RxM8E8Mium6NGHIK8AkA9sjv7GW8m0+K4je6t1R5IQ4MiLJnYzKDkBsHBPXBx0q3XyP4d8YeErf9pj4x2OneD7W08ReHfD2iXl5rSSn7RqUU0dy0UEi7cIIRDgMCSwIyPlFeFRftu/GD/hTnhn9pS/8AhPa2Hwxvv7P/ALUml1kNqkUd7NHbPdW9qkBRreOZ8LvlWSRcMUjXkAH6W0V8mfEz4+/EfQvjvZ/AD4YeBrbxNrOo+Gz4gW8vdT/s+ztY1vDaObkiGaQop248pHdmYDaq5ccE/wC2D4m8P/DD4yap478GQ6Z8QPgpFFNqekQXxnsbuG7i86zuLa78pW8qdA3DR7kKkMM0AfeFFfO3xP8Ajpc/Dyw+E97Fo6Xx+JfiXStAdWnMf2NdStp7gzKdjeYU8naFO3Oc5GK+bvCfxG/aSv8A9uL4h+D4NG0258M6XY6CklvNrtyIrXTJrq6Iv7e3+yGNryaHJlh+QAoiec4+YAH6NVUvb+x022a81G4jtbdCoaSVwiAuQqgsxA5YgD1JxXx18Hv2hvi98a9buPEPhDwXon/Cv7XXLvRZ2m1t0162WyneCS6nshatEmSm5YGmEm0g5IINdX+2d4g8I+Fv2eNf17xz4StfG+kW95o8cmlXkhihma51O2t0cuqsQYmkEgGPm27TwTQB9S0V8nfEv9oDx/4e+Otp8Afht4Gi8Ua3qXhv+34rq51EWFnbKt4bWQ3TeVK4iX5SDEkjs7KuwLl16P8AZy+NviH4v6d4s0nx34di8LeMPAmsy6Lq1lb3P2y2MixRzxTwTbEJjljkBAZQQQc0AfR1FfH3jj4//E+b446t8Dfg34V0bVtU8NaTaavfvr2ryaWbqO8aQRw2EcVtctIVEf7yVgI0YhWwcE9L8Qfjb470zxX4Y+EXw58J2mrfEbxBpT6zeW1/ftbaZo9jEyRPLc3UUEsku6d/KiWKIlyGY7FHIB9OUV8K337YWueG/Cmoar408Hxabq3gnxfpnhnxjbx3zS2+nWmqeX5Oq20xhQzwMJ4WCukbYZs42jd7d4v+N0Xhr426B8I7exjngm0HU/EmuX8kxjGl6bZMkMEmwI3mGeZmXBZdqozDdjFAHvlFfBEf7WvxNt/B2k/HzXPh7aaf8GtaurKOK7bU5G16HT9RnS3ttTmsRbeQIHMkbmNbgyLG27nBFdj4o/aF+LF18avGXwK+E3w/tNe1fwxp+l6gNR1DVPsOnpHqCTFhPthlm8zdGqxJGjBwXLvGFG4A+xqxbDxJ4d1XVtU0HS9UtbzU9EaJL+1hnjkns2nQSxCeNSWiMkZDoHA3LyMjmvH/ANnH40XPx0+HL+KtX0Q+HNd0vUr/AEXVtO84XK2uoabO0E6JMAu9CQGU4HBxzjJ+O/CPiP406X+11+0xp3wb8H6d4guZr3wvNc3esam2nWcAj0OHZEohguJpJZcnHyLGoGWfkAgH6fUV8I6j+2zb6b+zjJ8dLrwfcxapo3iKHwzrugCYS3NjqC36WN3FE6Jido92+IAL5mVB2knHqMXjr9qafwpFqKfC/RYte1O9ZILObxCUg0+w8kOs1/OtozNN5mUMVtHIBwd+MmgD6Xubm3s7eW8vJUgggVnkkkYKiIoyzMx4AA5JPSnQzRXEST27rJFIoZHUhlZSMggjggjoa+Cf+GjF8d/BX4/6L8WfBunXOu/CjTL5dc0i0vzeaVqds+nPeQmG68pHWO4jVkYNHvjIORu4G3q/7Q+ofDnQ/gL4M+G3w7TV5fido8i6ZpdvepaRaeLHToLmKHzZEKi3jjYh3xuVEJWORiEIB9v0V8t/A/45ePPGfxF8bfB34ueFbPwv4t8Hw2N8p02+a/sb3T9R8wRSxSSRQyKyNEVcMg5wQByB6T8U/EXxg0ZNOs/hF4S07xBcXvnG5vNX1Q6bY2CxhShkEcFxPKZMnAjjwNpLMvyggHrdFfB9r+2XqMX7PPxS+K2s+F7aLxT8JL2503VNKttQ+0Wc88HlMslveCIEwyxyqykxhgQVI4yej1H9pX4keHPBY8ceKfh4lofGGo6dpfgfREv86rqdzqPmsg1HdCsNjiJBM4DS+UgkDEuoDAH2bRXy98P/AI0/Ec/FCX4L/Gfwvp2i+JrvR5dd0qfRtQlvtPvbaCZYJ4S9xb2zxzwvJHuBUqytuBGCK8+1X9pT40fDzxl4Hg+NHw/0fw54b8faza6DbLZa9/aGrWF5fbltmuoPs0ULRu4COYZXEZOdzDGQD7ior4wu/wBob4u+KfjB44+HHwd8GaLrFv8ADiWzg1NdX1t9O1G8ku4EuM2VulrOqxKrgCWZlV2yF4Ga9x+PPxG1v4RfBrxZ8UdC0VfEF54WsH1FrFpjAssNvh7j94EcjZEHcfKclccZzQB69WL4i8SeHfCGjXPiPxZqlrouk2QUz3l9PHbW8QZgil5ZSqKCxAGSMkgdTXy/46/av0fwr8U/hN4F0jSv7W0f4kQx3NzqYm8sadBflIdLfy9reZ9ruG8sfMu3BI3dK+cf21fi5P8AEP8AZz/aZ8JW2liLRvh9PoelR6gkpc3l7LLZ3V3H5e0BPs/mxp95txJ6YGQD9SKxb7xJ4d0yx1TU9S1S1tLPRI3lv5pp444rOOOPzXed2IESrH85LkALyeOa+SbT9ov4waD8XfA/g74sfDe18L+GviTLc2mk3UOrC9v7S8gge5SHUIFhWJHljQjEMsqo3G9uTXk/j/xB4R1f4Z/tq6b4e8JWvh/UdG0zU7fUr+CQvJq0z6A0yzyqVAQoJNgUEg8t1JoA/R2wv7HVLG31PTLiO7s7uNJoZoXEkUsUgDI6OpIZWBBBBwRyKt18SaR8ZNe8FfDD4I/DH4caBF4n8d+MvDdlLZ211dGysbWysbK3NzeXlwsczpEhkRVVI2eR2CqByR6z8FfjD4h8ea/4y+HHxC0KDw9438BTWS6jBY3L3thcW2oxGa0urWeSKBykgSRSrxhkZGBzwaAPoKiiigAooooAKKKKAP/Q/fyiiigD5s/aw+HHif4lfB25tfANoL3xh4d1HTNf0OIyRwltQ0m7juUQSSsqIZEV49zMAN+SQK+KfiX+zJ+0A3wJ8AS+AdIjm+JOpW/iTT/FcX2u3iMEPjlJbnUpWneRUmNnd+UVEbMWC/u8jmv1pooA/PT9o39luHU9f+F/jjwt4CsfiTo/w902fQLvwveSwwSXGmSpEIJrOS4KwC4tmiB2yMokRmUMpxmjqHwOGt/s0fGTw38LvgZF8Ltf8Y6RNp9nYG505bvUyIX8kztbTPBDteR1UPNxkk7c4r9GaKAPib9or9nnxB8X/wBnjwX4Ss9NsdR8Q+BrvRdaj0nU3AsdQm02IxT2M8i7wqzRSSoHGQG2nIHzCD9nn4Y+EtK8cjxFZfs2xfCW7sLOVRqs1xpksxnkKo9vAtjNO7RshYmVtgOAApydv3BRQB8mfHTwJ431T49/AT4n+GNGk1rS/Bmp6xBqywSwxy21vrVj9jS62zOm+OFzukVNz7eVVjxXy144+B3xZ8Q+MPi7B4y+ENt8TNd8VXV+PCnibVNSsm0vRtIuLZYra3S3uWee1mt23MTb2xMznLSAfOP1YooA/Mo/B/48+DtG/ZL8QeHvBS6/qfwf0G8tte006laWrxzXGjQ2Bhimd2jkk379m0mMlcNIisHr0Lwn8KfiP8bfi/4y+Mnxg8MyeAdJ1HwfN4I0jSJbq3u9Qa0vpjcXl7ctavJDEzNsSJFkY4BLY43fedFAH5JfCb9mGx8FaX4f+F3xF/Zd0vxRqujPb2M3i21udMXTb2zhZU/tCRJ5heLP5Y3PD5DFpAcMFOR9Er8EvFes/Hf9ojTfEWlzQ+DfjP4Y0qyttZhkhaOGS0sp9Pubd4vMEwlIufMjOzYVVgWBwD9yUUAflF8Hf2cLHwpB4V+H3xC/Za0vUtc0SS0tbvxZa3emHS7iG2ZUOpKskovPOYL5hgNvuL8ZVTuHsXh3Q/jx+z18V/ifD4M+HL/EPwr8SdaPiTT7y01OzsWsdQubeKG6t75bt0cRboQ6SQrL8vG0sdo++6KAPlP9i/4bePfhP8B7LwZ8TLCLTfEEWq63czQwTpcQ7LzUri4ieORCco6OrLuwwBwyq2VHk9rY/H74OftCfG74jaB8Krjxz4e8f3ehvpr2Orafa3O/TtKhtmaSG6ljCwmXchctvUoW8pkIav0EooA/Prw9+zFrjfsl/EbwZ8VL200vxf8AEK/1nxfqE8Ev+iaTrF3MLy2ZJjjK2bwwln6ZRsHbg11f7CkXiTxb8LdR/aE8eWq23ij4wX39tToP+WVjBClnp8SnA+TyIRKv/XU19CfFr4M+Cvjbott4a8f/AG+fR4ZTJNZ2moXNjDeKRtMV0LaSMzQnvG5Kk9q9K0/T7HSbC20rS7eO0srOJIYIYlCRxRRqFREVcBVVQAAOAKAPjf4o+Dfit8Pf2j7P9o/4ZeFm8eaZq3h1fDeu6PbXVvaahCLa5a6tr21a7eOGX77RyRGRDjay7jnFT4qeGfjP8adN+C3iG88Cnw5feGfiLp+t6jp0mpWlzLaaRa295F9olkRliaQmVMwwtKw3cFsHH2/RQB8s6x8OPGt1+2V4f+K9rp4Phex8D6jo8t6ZYsJfz6hbzxxeTvEpzGjNuC7eMFgSBX5zeKP2Yvjv4u+A2v8AgDXPgrb638Xry5SbUvHWpatp1xJfiLUUugdNllke5jDxKIxAy20UabhkthW/b+igD5atfhz4zj/bV1L4tvp+PCdx8P7PREvfNi51CLVZ7l4fK3+aMROrbymw5wGzkV82+MP2evjBqnwO/ar8H2Ggebq/xJ8S3uoeH4PtVqv222mt7JEfe0oSLLRONsrIwxyMEZ/TeigD5I+Pnwx8ceNZPgS3hnTftg8G+NNJ1bVv30Mf2aytrK6ill/eOvmbXkQbY9zHOQpAJHzT4o/Zjt/Bfxl+IHifXfgPa/Gvw/491I61Y3tvNp8epabdzxIlxZ3CahNbq1u0ieZHJG7bNzBlyRX6m0UAfBnjL4P+JdLvP2cPEXw7+Hsej6Z4B1qebVfD+nXFrjS4dXs5oJnjdmijmSCeYvJ5fzMCWVTzXp3wJ+HPjPwb8Xfjn4o8Saf9j0zxj4gsr7SpvNik+028OnQQO+2N2ZMSIy4kCk4yBjBr6looA+I/hJ8IfiJ4Y+Efx48Ma5pP2bU/GfinxpqWkxefA/2m01ZmNnJuSRlj83I+WQqy/wAYWvNbD4HfGfwB4G/Zt+I/hjw7HrHjL4Q+H/7F1vw215bwS3Vrf2UEFyttdM5tvPtpYVdAXEcg3DePlz+k1FAHwd4b+E/xG+Nvxa8YfGP4seHX+H+maj4On8E6RpM1zBeagba/lNxd3t0bV3hjYtsSKJZHOAS204z8/eJ/A37XOrfsdaj+yCnwnS51jSdFTRIdfGs2MelX1nYbRbyW6NJ9oE80cSr5c0cSKx3PIowK/XKigD4e8b+Bvit8M/jro/7Qnw88Kt45s9Q8LQ+GNe0a2u7e11CE2k5uba8tWunjglwXeKSMyIcbWXdyK8n8VfDr4iW3wX/ax+PXxU0qPw1rfxI8KXsNtoiXEd4+n6bo2kXMNuJ54SYmnmaR5JBGWRRtAY8gfpzXM+NPCOh+P/B+u+BPE0TT6P4jsbnTr2NHaNntruJoZVDrhlJRiARyOooA/M9tE+Pf7Rv7Pnw6/Z7u/hw3hfwtf2fhuTWvEs+qWdzZyaTp/wBnulFlDE32pp7jyUG2WFBHkhiQQ1d1+1Z4B8bfE++1vTNP+BM2qeK7WIw+EfG+ka3ZWM9jK0eY57idpba9gEEzMzQqk0bgYBJY4/QPw5oGmeFPD2l+F9FjMWn6PawWdsjMXKw26COMFjySFUcnk1s0AfG2kfCv4p2/7TXw6+IniRF1Wy0P4fXWiatqySRIkusS3NrI22EsJcS+XI4Ij2joSDgV554H+BvxS0f9hL4ifBrUdE8nxjrtr4zjsrH7TbN5r6tcXj2Y85ZTCvmrKh+Zxtz8+0g4/Qyqt7fWenWz3l/MlvBGMs7kKo/E0AfM2n/svfCTxx4J8Df8Ll8D6frXiPw94f03S3e7VZng+zQjdCHRipCyF+VJBJJBIr55/aF039pD4k/Giz0fVPg7f+K/g54TlhvLexs9Z0a2XxBqUe145r9bq7RxaW7cx2xT944DyEgKg+52+KvgFWKnVRkekMxH5hKb/wALX8Af9BT/AMgTf/G6AL/w98Q+KvFHha21nxn4Vn8F6rK0gk0y5ura8liVHKoxmtHkhbeoDAKxIBwcHIr4F0XwV+0l8Obr44fCXwv8Oo9c0z4leIda13SvEkuqWtvp9vFrsKCSO5ty32oyQODtVIyJCcbkUb6+6/8Aha/gD/oKf+QJv/jdH/C1/AH/AEFP/IE3/wAboA+PrP4f+MfDX7Lfwq+EXxC+CUXxR0/TtBtLPX9MW9083en3lrBGiNBHdOlvMQfMBkiuVdCAU3AkjzHTv2e/j3cfATQ/CN5o9yhsvifpXiDStEvdVhvrrQ/C9reQyLbS3ksxWVoFSRwiSSMFYIpYjA/Q/wD4Wv4A/wCgp/5Am/8AjdH/AAtfwB/0FP8AyBN/8boA8e8BfDnxnov7WHxX+Jep6f5PhvxLo3hy00+782JvPmsBdfaF8tXMibPMXl1UHPyk4OPj34h/Djxp4C/ZS+LvhDxXosY1jx98TLy40SwM8TXGowatrcEtqLOSEyiG7kiVmgaTBhcB5AgUkfpH/wALX8Af9BT/AMgTf/G684+KZ+Bnxk8KHwf45u5Z7NLiC8t5bf7Xa3VreWr74Lm3nhVZIpYmGVZT6g5BIIB8y/CLxhoXwx+MGiD4qfD/AMd6N4v+ICjw9p3iHxVf2Osx7YVe6XTklsbmQQK7Iz/6oGRhukY4yPoD9pr4c+M/iFqfwZuPCGn/AG+Pwp8QNK1vUz5sUX2fT7a2u45Zv3rpv2tKg2Juc54UgHHPeEfA3wL8NeKdM8b614r13xprmhrImmXPiG8vNQ+wecpSR7eIosKSuh2tLsMhXjfgnP0F/wALX8Af9BT/AMgTf/G6APlL9pP4dfEDxP8AGnwr4k1L4fy/Fz4Y2Wj3Fu/hyK+tLeOHXHuFeO+ura+mgt7qLyR5ahy/lks4TPX5/k/Zd+Ndz+yn+0N8M7XwVp/h/XviD4sGq6JpFhe2xsUspP7NYKko8lEWIQyKQyRklDtQgpu/S3/ha/gD/oKf+QJv/jdH/C1/AH/QU/8AIE3/AMboA+R/Guh/Hr9o3xx8OvC/jD4cN8P/AAt4G8RWXibV9RudUs79b650rc9raaelszSPFJKwZ5Z0hIRcbQ3ynM1b9l7xl8RNL/am8GeIol0Sy+K2oWs2h3ryxypJ9n0+3RJXSJ2dEW5iwyuqsVBIBBBr7L/4Wv4A/wCgp/5Am/8AjdH/AAtfwB/0FP8AyBN/8boA+H/gr8GNFsfFnhePxN+ypp3hPXdFmSa88RW93pcmnwXNspaO6sRHM105eVVKI0MbRhvmbK4bD+Nvwz+K3xK8bRar4L+C0ngf4n2ur232Tx/p2s2SWQ02C6QvLd+VJFd3QktlMbWs1q3LYBKjJ+/P+Fr+AP8AoKf+QJv/AI3R/wALX8Af9BT/AMgTf/G6ANP4heCNF+JfgPxD8PfESb9M8SWFzp9wMZIjuY2jZl/2l3ZU9iAa+Q/2O/hR8ZfD+pa78RP2htPSy8WR6Xo/hPTgLmG7L6VokGXuw8LuF+3XUskrI2GG1dyjv9U/8LX8Af8AQU/8gTf/ABuj/ha/gD/oKf8AkCb/AON0Afnz4f8A2cvjLY/sefCP4WXXh7Z4o8MeObLWdRs/tdofIsIvEFxfPL5omMT4t3V9qOznO0LuyK+iP2svA/xJ1a8+F/xQ+Fvh/wD4S3Vfht4jGpz6OtzFaS3dlcWs1tP5Ms5WPzE8wMFYjIzg5wD9UaN4i0TxDC0+jXiXSp94LkMv1U4YfiK2qAPgvwT4Y/aD1b9pnxH+0B4l8CW3h+0u/AS6LpOnz6pBLJ9str+S5igvpbfzRG8pdmLwpLGkZUbmcMtcN4D+FnxAuf2jPCfxF8E/CC5+CNtbteyeM5F1Sxk03Wllt2WGCKzsJ5EnkE7LKLl4YWXB3ZbC1+ltFAHmnwy8SfEXxLa6/L8RvCS+EZrDWbyz06Nb2O9+3aZCV+z3xMYHlGbLfum+ZdvPUV8z+JfB3xj+B/x58W/GX4TeE/8AhYfhf4kwWLa5o1ve29lqNlqWmxG3ju7U3TRwyxSwBVkj3h94DDIyK+46KAPjX4N/Dr4q+K/jlrf7TPxq0aLwlfNoyeG/D/h+O7jvpbHT/P8AtNxPdzw5ha4nlVNojZgiDaWJNfFP7OWvftD+If2IrP4O/DX4Zfb18Ux+IdNsfEsuq2kWm2sF9qd5FcXF1C7LdpJbl32xxxSiTapDcsi/tBXAfC/4ZeEfg74H0/4d+BLZ7TRNLa5eCKSV5mU3U8lzLl3JY5klYjJ4BwOBQB8Y+M/g78RPg14w/Z/8X/C/wvcfELSPhb4evvC2oWdpcWtpfNDPa20MF1ELuWKJvntz5i+YCNwIyMkbjaH8afgh8bfHnxO8FfDy48feG/itFpd9cWNlf2NpqOkavY2otHjlF3NHDLBMoRjJHKxRg3ylcE/eNFAH5J/Ej4UeN/Av7LniK38c2NtdeP8A4xfEWy1yXRdPucTx3V9fW7rp+mXpUKl3DaWuVuWCpGyyPu2qGPY+BL3wnpnjif4cfFnwJ460Dxb8ZdNu/D1t4h8UXljq3m21vbSymxinsbmRLdVVpJFURKJHyzsSAR93/Ff4R+C/jP4ag8MeNoJ2isryDUbK5tLiW0vLK+tSTBdW1xCyvHLHk7WB6EgggkVx/gv9nTwX4T8WWPjzVdU1vxj4j0mGWDT77xBqUt+9jHOu2b7NGdsMTyrhXkEfmMvyliCQQD4O+E37MNj4K0vw/wDC74i/su6X4o1XRnt7Gbxba3OmLpt7Zwsqf2hIk8wvFn8sbnh8hi0gOGCnI+yPAXw58Z6L+1h8V/iXqen+T4b8S6N4ctNPu/Nibz5rAXX2hfLVzImzzF5dVBz8pODj6looA/LjUv2SPiJ4y+AvxE8GavpNnDrz/FDVfGui2WpSxzWOpWwv/tEEVyYGk2R3cBdCrYZdw3qOa9b/AGf/AIXeE9N8fWniGD9mWH4U3+l2sx/tiW60uZ47mRRE0NqLKaaR0eN3BlYRjGBtO47fuyigD82PhZ8PPjZ4Q+Dnxc/Zr1fwDcst5F4vn0TX4r2xNhqI1h5pLSHY06zxTOZyG3xhF2/MwBBN+8+Cvxm8FeFP2bfid4N8PRa54v8AhD4ci0TW/Dj3kFvLd217p1tbXS290zG38+2mgDJlxHINw3/dz+jFFAHyprHjT4v+N/hxq0+vfAUX9pc3cVs/hvVdX017u+0x0JmmaP8Ae2QdX2hYJLgb1yS6EBTwn7I3wx8b+AvFfj7U08K3vw3+G+r/AGD+wfCt/qEV/JaXMSyfbLmNIJriK0imLIBCkpGVJwo2ivuaigD89v2t/wBn/wCJXxI+KPhTXvhvpq3ml+KdNbwh4xla4hh+z6DJqdnfNKEldGlIjS6i2oGb970xnMM/7PvxLk/bIg1xdLT/AIVEmqjxwZxcQhR4iXSTpAg+z7/Ozwtzv2bM8bs8V+h9FAH5ZeKP2Y7fwX8ZfiB4n134D2vxr8P+PdSOtWN7bzafHqWm3c8SJcWdwmoTW6tbtInmRyRu2zcwZckV7PrPwT8Qf8JR+zXfeB/A1v4V0HwJqOp3ur6ZaXNsYNIW+06dCiEMnnZuZSCYlbJJYgDJr7mooA/PD9pH9nSTWvjzpnx4f4Y2fxg0S40JdC1TQ5JbaG+tXt7h7i3v7M3jxwSZEjxSxGRDjay7jkV9Ifs6+CdB8HeD71tF+FsHwlOpXjyyaXHLazSyqqKkc87WbSRB2AxsDvtAGW5wPf6KAPgG20T9oD4P/tG/FjXPA/w5HjTQfilJpN/p+pDVLWyt9Ou7OySzmjv0lPn+XlBIGhjkbb8qqzZ25Pwo+GPxO+Ff7HnhP4TeOvhPafEm4ivdWj17Qze2IP2S41K8uoZ7dblmtZyQ8TiN542UHr5i7R+ilFAH5MWX7JPxN8efA74neBU0Q+CtAv8AWtK1jwR4T1+/GqR6edM8uSeC5eGW4WK2vpFYCBJXEIYnHY+3fAX4V+FbDx/peuH9l63+F2p6RDM7a093pc4gunjMZjs/sc0ssqyI7qZGSIBeCMkgffNFAH5YeFfAH7TXwt/Z98V/sgaF8M18RWottd03Q/E41ezttOl03VGnkje6id/tSXEXnlPLWFlZgP3gTMlfe3wD8M634K+BXw58G+Jrb7HrGg+G9IsL2DeknlXNrZxRSpvjZkba6kZVipxkEjmvWaKAPzg+G/wc+JieE/jT+y7468I3Vl4c8daj4pvNP8WW9zZzWX2XXSzw77fzluVnjaQ5TytuV5bbgnnvFHgz9qH4wfAnSv2SfFvw6Tw6kkWm6Tr3i0apZz6W2m6fJEZLiwgRzdvNcpDhYpIYxGX+ZgADX6gUUAeNftCeEtc8a/s+/EjwL4StPtmsa74Z1fTrC33pH5tzc2csMMe+VlRdzsBudgo6kgc18r+IvhV8Y/h/qXwJ+M3gjwt/wlWr+AfDB8NeIfD8d5b2929tc21vuktJpnFu0lvPCSymTEinCnvX6G0UAfnh4d+EHxa+LHxg+Lniv4q+Ef8AhB/DXxJ8C2/hq2UX9tf3UOXuopVuFgcqswSXzMIXi2soErOHC+Y/B39nCx8KQeFfh98Qv2WtL1LXNEktLW78WWt3ph0u4htmVDqSrJKLzzmC+YYDb7i/GVU7h+rtFAHy18K/hz4z8N/tPfHT4ia1p/2fw94yh8KppVz5sT/aW02zuIroeWjmRPLd1H7xV3ZyuRk18x6h+zT8YdV/Yk+Jvwfj0mK18Yaz4m1PWtNtJrmApcRLrianbKZY5GjQzxxAAOy7Sw37ecfqDRQB8N+OdG+LXxt1j4G+Npfh9feFH8JeMDf6tZaheWEk1tZrYTw/aA0Fw6yIZZAqqpMhHzFAK7/4XfDnxn4c/ah+OXxF1nT/ALP4e8Y2vhSPSrrzYn+0vplrdR3Q8tHMieW0ij94q7s5XIBI+pay9c0mDX9E1DQrmae2h1G3ltnltZnt7iNZkKFopoyHjkAOVdSGU4IIIoA/M74IfDXUof2wvE3w+Rop/APwWvNS1/R1jbIt9Q8axRSraMn3V+yL9uZADkLcITjIz9mftQ/DbXPjB+z14/8Ahp4YMY1fX9JuLezErbEa4xujRmPChmULuPAzk11fwq+D/gL4L+H5/DngGxktob25kvbye4uJbu7vLuUAPPc3M7PLLIwAGWY4AAGAAK9OoA/Pfxl4f+Nnxr+HXweS/wDhve+FNY8E+P8AwvqOp2d5fadJjT9L5u7yF4bl1eJWYhU/1rYyqEEV7H4v+HPjPVP2uvh38T7HT/N8M6F4b12wvbvzYl8q5vJbZoE8ouJW3iNuVQqMckZGfqWigD8fvA37J1l8ItOuvhf4l/Zn0/4qGzvLr+yvE0F3psMV5YTzNLCNSF5LHPDNAjiN9kUoYKNm45z73+0x8PdR8Xxy+DYf2fH8WxaTYRweE/EOj6tYWF3ps/k4VWkmltbmzWCQZBhaaN1AyuTtr9BqKAPyz8ffsuePpNX+F3xZ+JHgux+Our6L4Pg8L+KdLmmgjunnhcXKajYyXjRQzSLI0qSCR4y6tuGWOB9h/s4+BvD3g/w1ql1onwmh+ETand7m09ZbSWe4iijURTzmyeWJHyzqEEjlQAScnaPomigD5A0f4W+O7X9o342ePLjTNuheLvDeh2Gl3HnQn7Rc2cV2syeWH8xNhlT5nVVOeCcHHh+vfs/fF29/4Jo2H7P1toO/x9Do+l2r6b9qtRia3v4Z5V+0GX7P8saM2fMwcYBJwK/S+igD84Pih4o8a+EP+CgWm6z4I8HzeN51+F8yXNha3lvZ3S2x1tCZYDdMkMjK4QGNpI8qSQxKhWqv+zn8XPix4A/aM8XeOtMt/CnjD412Fvp+l6K11HdDT7TSLV4bFbq4h3R+bPI7PL5ZdUBGCSCB91N8MvCLfFNPjKbZ/wDhKI9GbQBP5r+X9ge5W6KeVnZu81Qd2M44ziu/oA/MvV9C/ag+MsXwMtfEPwqHg2y+HHivRdU1hrrV7G5uJvsUE1vJLapBIym3QOzNvYSsWQRxsoZq9XvfDnxW+HP7YWufEvQ/BNz4u8JfEPRtG0ye8sbuzhk0q506eZXe4iupomeHypfM3R7242hc4B+3aKAPy/8AFnwz+Kvjj4t+FvGPhP4LSfC3x/Za9aXGteL7HWbI6ZeaRDLm6jlS2kS4vzcxDasdxaK6kjLKoJP03+2f8OfGfxY/Z28QeBPh/p/9q65fXuiSw2/mxQbktNVtLmY753jQbYonbBYE4wMkgH6looA+WpPhz4zb9te3+Lg0/wD4pOP4ezaG175sXGovq0d0IfK3+bzEpbfs2dt27ij4C/Dnxn4L+Knxy8SeJtP+x6d4x8SW1/pUvmxSfabaPT4IGfbG7MmJEZcSBW4zjGDX1LRQB8DftVeDdf8AiLe6hoV78A5fHE9rbAeGfE+laxY2N9Y3ckecySzy2t1ZiKb5sxNMjgAsuSVryzx9+y54+k1f4XfFn4keC7H466vovg+Dwv4p0uaaCO6eeFxcpqNjJeNFDNIsjSpIJHjLq24ZY4H6mUUAfGfgr4A+B/FnwM8e/DxvhNB8IbTx1DcWUtislpPcSRtBsgu5vsTyxJLG7EogkcrtBJBO0cd+zd8Evivrnhn4g+JP2mdPGk+MPG+lWHhZ447mG8MelabpwtmmSWF3VTdXU1zcFM5G5Q3I4+/aKAPyS+E37MNj4K0vw/8AC74i/su6X4o1XRnt7Gbxba3OmLpt7Zwsqf2hIk8wvFn8sbnh8hi0gOGCnI+yPAXw58Z6L+1h8V/iXqen+T4b8S6N4ctNPu/Nibz5rAXX2hfLVzImzzF5dVBz8pODj6looA+Wv2VPhz4z+G2h/EWz8a6f/Z02u+PPEutWS+bFN5thqF15lvNmJ3C7052Nh16MoNeQWtj8fvg5+0J8bviNoHwquPHPh7x/d6G+mvY6tp9rc79O0qG2ZpIbqWMLCZdyFy29ShbymQhq/QSigD8wdT/Zh+MCfsvXnhy4sLfUfiF4z8fWvjbWrO0uIlt7WS51aG7uIopp2jVxb28YBwcsykJu+XPtH7WngH4h+M/Enw5vtM8KT/ET4faRPqLeJfDFrfQ2Ul+80KJYSutzLBDcxQP5heCSQKdwJDYwPtaigD8s/Cv7O3xUtvC/7Vsdp8ONP8ERfFPw3a2fhnQ9MurIwCaLSLuz8gmHyoYZTI6GQkLEHc7ZJFBc+y/8Kc+I/wDwnn7KGs/2R/ofw00vVLbxBJ9og/0KW50NLOJdvmbpd04KZhDgfeJC819z0UAfLXhb4c+M9O/a/wDHnxSvdP8AL8Maz4W0XTrS782I+ZdWlxcvNH5QcyrtWRTuZApzwSQceeftP/Dz4geK/in4J12fwLN8Vvhpp1jexX/hmC+trVTqsskRtry5gvZoLe7iRAyBJGYIWL7a+6KKAPyVX9mb4zN+z5+014MsvANh4c1P4j38d14e0XTLyz+yCBrW1URJIDBFGY2Rkfesa71YpuQqzfT37V3wB1n4zfCrwfbaPpFjr+teBdW0/Wk0bU3CWeqpbRPBc2EkuGWMyxStskIKiRVzhSSPsyigD4C+FvwotrZPFGqfDn9n+D4J+Kf7CvbTTNbu59NllGoTptRESwmuW8lXCu0jFdwUAIc8fMlh+zd8VNX8O/CzT4PgXDoni/wp4p8Nar4q8V6nq2nXmqaqdPuY2vZ7e5Es1xNG/wA07LNJFtA8uKN2wB+zFFAH5q/tTfDv4hfFHUtZs9B+Bk3/AAndqfJ8KePdK1uysmtBgGK4up/Mt76MQsSXtvKmRhkISTkfoCnh6TVvBI8KeNJE1V73TxZai4XYlyZIfLnIXsJMscdga6migD8j/h5+yr8ef+GfviNovjnTkh8d6RYaLong0m6t5TNbeDHN7pMyyJIUgFzdOQVkZGUD5woruPFX7PPxf1/9g7x/4Ebw+D8UviNqd5r9/pgu7Yhb2/1dLryftJlFufJtURM+ZtOzC9hX6cUUAfLX7QHw58Z+N/iT8D9f8Maf9tsPB/imTUdVk82KP7PatYXEIk2yOrP+8dRtjDNznGATXk8nwI+JWo6V+1vpEmmran4rLcxeH5HnhZLrztDWyVmCOxiAnyp8wKeM4xzX37RQB+Wvjj9nHxZ4v8LfAz4ma38LrfxhqPgPQDoGv+DdUuLRbiWBoooxPaTtI1p50EsPmKGkAkicruRuK+q/2a/AXhjwpaa9q+gfBiL4OvqckEZhMtlJd30MCsUedbGSaOPY0jhVMjHkk4zivp+igAooooAKKKKACiiigD//0f38ooooA57xF4t8L+EYrKfxTqtrpMWo3UdlbPdSrCs1zKGKRIXIBdgrEDqcGvL9K/aa/Z31zRNd8SaR8SvD13pXhkoNTuo9St2htPMYrGZH34AkYFYz0cjCknivAv29dG0rxF4c+Dfh/XbSK/03U/ij4Vtrq2mUPFPBNLKkkcinhldSVYHgg4NYPiH4e+BZv+Ci/g6eTQLFmt/h5f3UY+zx7VuLXUYIYJQuMb445XRGxlVYgYFAH0z4h+K/h7xd8D9e+JXwg8c6FFaR2VxJa69cyLc6Vaywj5nuSjrhYyPnBIK9weh6LXPil4I+GvgDTPGPxY8XaTpNnLBbiTUZZltbS5nkjDE24kckiQ5ZEBZtvrgmvzH1uxs9F+Bf7dWgaRAlnptrq+qSxW8ShIo3uNItnlKquANzcnHevT1ufDll+1t8Ebn4qyQReH/+FalfC735VbRfEfnw/a/KMnyC5Nns299uQOdtAH3dp/xn+Eeq+AJfirp3jLSbjwdArNLq63sP2KPYQrB5t2xWBIBUkEEgYzVv4dfFj4Z/F3SZtd+GHifT/E9hbyeTNLYXCTiKTGdkgU5RscgMASOelfA/7YOs6Pca38GtQ8B6t4Zh8H2Pja7TWrm9iF7odtrj2TtYtqUdpND8wmcnLyLtlaN3yOD7Z8HPh343g/aE8SfFrxj408LarqGp+HbPTL3TPDVrNa7/ACrl5bO9ukmvLpiwQzRI5xleAflNAH0T4z+Lnwt+HU0lt4+8W6X4dlis2v2S/u4rZhaJIsJmxIy5TzHVMj+Jgo5IFXfAHxK+H/xV8Pr4q+G3iGx8S6QztF9psJ0njWRQCyMUJ2uAQSrYIyOOa+Kvim/gKL/goX8MpfHracip4K1c6c2omIAX/wBug2GAy8ed5fmbdvzbd2OM14b4jbVNc8Sfto6z+zuRNYTeGtOtDLpfMM/iCKyuDe/ZWi+U3KQMqyGP5vNK5+agD9HvC37QPwO8b+Mbr4feD/Hmi6z4ks/M8ywtL6GW4Bi/1gVFY7in8YXO3+LFWte+Ovwa8L6iuj+IfGuk2GoPqUejrbS3caztqMixutsI87zIUmjYqBkB1JwCK/P3wB4G8SfFLwP8C7vRviB8PNP8KeFtV0bVtETRdOurXUGFov76xR5dRkAkmhMkVxGYyxJbcMg16v8As0+DvDF3+1F+0r4yvNOhuNZt/EOm2kNzKu94YP7Mt5GWLdnZubBcrgttXcTtXABmfs5ftBr8f/j58QtSsfi7avoHhbVLuw0nwnZDT5Ir3SrS2gV9Vkn8trp43uJso6Sqg27TkEivrLwR+0L8C/iV4kuvB/w/8e6L4h1uyDNJZ2N9DPNtT77KqMS6r/Ey5A7mvzA17S9bH7K/7aDeCLV11FPiHrkchtExONMVdLa/RCvO37J52R0xniva/wBonUvg7qfhn9nu0+As+lzeIB4u0BvCSaQ0Zlj0xSTfECL51sxab/tAOE6B/mxQB9w+Lfjl8HPAc95aeMvGmk6NcafcWtpPDc3cccqXF6nmW8RjJ3b5UBdVAJKgt0BNZniv9oz4CeBjKvjH4g6HozQX8mmSLdX8MTJewrG8sLKzAho1ljZ8j5A6lsAivmb4ReD/AAzrP7en7RHirVtOhvNU0Ww8IQWU0q7zbpeWEpn8sHIUyeRGCwGcLjOCQeP+C/gTwZr+rftdalreh2d/d3vijU7CaaeBJJHtU0y3kWEswJ2B3ZsdMnPXFAH2vdfHr4J2XjjTvhrdeOtFj8U6ssbWmmm+h+0zCZQ8W1A2cyKQYx1cHK5rR+JHxi+FfwfsbXUvij4r03wvb3zmO3a/uUgMzLywjVjufaCC2AcDk4r8mrzwl4X0b/gmD8K/E2laTa2urW174S1OO7jhRZxey6tbI8/mY3eYyMVLZzt+XoAK9w8U3XxJs/29fFyaZrfhfQ9Su/C2kL4bfxTY3N2JbBXn/tCPT2hu7VUcXBBnX5mZdhwFU5AP0Nk+IngGHwV/wsmXxHpyeE/s4u/7WN1ELD7Oekv2jd5ew9jux2r5K0v9qDRviR+1B8PPB3wg8bad4j8FaxoWvXGpRWD29yPtti9t5G+QAyxMqyMQu5QwOSDwa+UPG/w7tfhf4M+FOk/EjxLo3iP4bav8XLnVNXbS42h0Gy+2LNJb2TpJNOqWkWohy6SOUViFfBFfQev6t8PtT/4KI/DOPwjdabdapb+DteXUDYtE8qoXt/syztHz0DmNWPAJIGDyAfdni/xr4P8Ah/oU/ifx1rdl4e0i2wJLvULiO2gUt0BklZVyewzk9q5/4b/F74X/ABh0u41r4XeKdP8AFFlZy+TPJYXCTiGTGQsgU5QkcjcBkcjivHP2qPidbeANM8DaJFoekaxrPjHxHbaXpc2v4Gl6fdmKWUXczbS25ERljVCruzYDqMmvnP8AZ5vNTj/bv+JtjqXizRvFV/deDtIm1WbQbMWFol7BeTRpFJF9pu2MsUTAbnlL7WAIGKAPov40ftcfC34HfFPwL8M/GWsaZYv4qe8a/uLvUI7Y6TawWss0FxNGwOUuJoxAm5kG45BJG0+geM/2j/gH8PLTS77xx8QdE0WLW4Irqx+030KNc28wBSaNN25omBBDgbcd6+dP2mf7C079qv8AZf1rxALeDTzqfie1mnuAoi8y40aRLeN2bjLyHCA9W6c1D40+Kf8Awkvxx8b/AA98FXHg3wRD4M0TTH1vxL4itFvbi6stQjlnhhtIRPaI1rChO+SWdo1aQqIzkkgH134k+K/wy8IeCoviR4n8VabpvhWdIpItTmuoltJVnGYjHLu2v5g5QKSW7Zr5o/aN/aPsov2W9d+MX7PXjCw1Kaz1HRrSLULFre+jjNzqtnbXETo4kQOYZmUq67l3Bhg4NfIf7PXgnQ/iP+xv8ELZPiBp3hXxb4X8Sa1qHhSfUUhntL2ex1C/iEJspZEM0Ygk4Ebh4htZDxgy/H/4x+IPiB+x98b9D17w/pMPi7wL4o8O2WozeH5t+k6vdvqemTJNBLIodZCpWOVJC7RkAMx6AA/Wq68Y+FLHxRYeCLvVrWLxBqcEt1bae0q/apbeAgSSrFncUUkAtjGTjOa6Svzw/Znl8V+Dv2g/HPhz9oq3tD8V/G9nba3pup2zM9rPokaKkmkWRk+4NMuC29Vw0wkWdgTkj7n8J+N/B/jywudU8Fa1aa5Z2d1NZTS2cyTpHc27bZYmZCQHQ9QeeQehFAHyTrnxg/aQ8SftC+PPhB8HrDwn9g8C2GjXkj68b+Oa4OqxyvtWS1MiqFMR5MR4I4PNem/A79oA/Em38X6B8QNGXwR41+HNxHbeItOluUmt4Emi8+C8hucIHtbiMM6OwUjawYcZPiHgXxf4T8IftvftAXni3WrLRLd9E8HESX1zFbIQkN5uIaVlHGRn0yK8m0X4q+Gl1v8Aal/bAg0Y+KPhoNK0vRrBNgNt4gOk280d4wDqyvbeZOsIl2sjJvOCFIoA+9vAn7Q3wK+KHiG68J/Dnx7oviTWLNDJJa2F9DcSeWpAZ1CMd6qSAWXIGRk15T8Gv2zfg38ZviL4o+HGheIdI/tDStTax0hYNSiuJdat47NLuW6t41A+RCZUO0uMRs27qB8feJNY8TS/tFfsv6t4j8U+EJdTutSvRaaL4VtChsNNu9Mm3hrt7mR54W+VQRBBGzDKrxXs37OOueEfBXxN/aek1KOBNR0bxZNqK2aKn2s2n9kwSqYY+GIdUfbt4PNAH1Tov7SPwA8SeNx8N/D3xE0LU/E7MyLp9tqEEs7SJkvGoRiGdQDuQEsMHI4q58e/ivpHwR+D3i34navd21qdD026ntVupFjS4vEiZre3UsRueaQKiqDlicDmvyZ+Knj/AFPxt8Gfgx8R49T8F+EdI1Lxf4a1Hw34U0O0Dalaq96mS1556KJI42InSGzRVOUZj3/UP9rOGGf9lr4wrMiyBfB+vsAwBwy6fOQRnuDyDQB5T+yt8W9Lv/g3ofj74jfGmz8b3viuS2hklmOnWltYau9n9ruNNg+yxxfNEm5ikpZwqFuBmvc2/aI+BSeAn+KUnj3Rk8IJcPaDVWvYhZvcRsVaOOUnbI2QcBCSe2a+CPjR4T8OeJP2f/2PvC2s2EVxpOr+LPBsd5bY2x3EcmjXQkSQLjcrj5XB+8pIOQa9a+KsPhbQf22/g+fiDHa2Pg6Hw3rMXh77QqRWMXiTz4MgbsRLMbMMIc4OchOaAPsLw18V/hn4y8Fz/EXwn4o07V/DFrHNLNqVrcxy20SW675TJIpITy1GXDYKjriud079oX4Fav4vtvAGl+P9DuvEd7AtzDp8WoQNcSRPF5ysqBskmL94B12fPjbzXwLrM2h3vxS/a4vvhg8MvhE+AFTWJLMg2b+JRaXhfYU/dmYWpQT7ed2N/wA1cd8Q/hl8PW/Yl/ZctP8AhHLBYr3W/ADXAW3RfOOqrEL4yEDLfaRI/nZ+/n5s0Afp54A+N3wf+K2oappXw08Z6T4ou9GIF5Fp15FctCGOAxEbH5SQQGGVzxnNeb/HTU7ptWsNG3EW6QeftHQu7MuT9AvH1NeU61omjeG/+CgXw/bw9YQab/aPgHWre4FtEsQlhtby0MKMEABEf8PoOBxXovxy/wCRstD/ANOUf/o2SgDxSZ2ihkkRDIyKSFHViB0/Gvm6T4+eJItcj8NSeAL5dUmi89LYzr5jRDI3AeX0+U/lX0tXzjq3/Jz+jf8AYCf/ANDmr1crhSlzqpC9k3u+nozyc1nVioOnPlvJLZPf1R6VoviXxFqPi3+zL23srTT206K68nzw2oRTvtLJJFnhFyRnaBnHPOB1Vl4l8Oaneyabpuq2l1dxAl4Yp45JFA65VSSPxFfO1xdNZftBeKrxbU3xg8MvILcDJmK+UfLHB+9jHTvXhPg/UrK98e+AtZ0eO0sZXvHgntrCzlgW3DEKqSTuT5zuhYnngdznj0Y5NGpFzWnup6LrZvX7t/8AhjzJ53KlJQevvNavpdLT79v+HPvqfxZ4WtAWutZsoQsrQEvcRriZcbozlvvjIyvUelb4IIyOlfn5eeG9D1Lw78X9cvrNJr+w1aUW8zZLRZuTnb6Z7+o4r6U1VPEGsfs/RLopeXVLrQ7Vhs/1j7oUMgXHJZl3AAckniuXFZTCHKoz3aTv0uk7/idWEzidTmcobRbVutm1b8D1S18S+HL77V9i1W0uPsQLXHlzxv5IXqZME7QO+cVzHgT4leHfiAl7/ZMgjmsriWHyXkQyOkW398qqSfLJYAH1r5V+Dej+Htc8T6GsOpWQntbC4gvtOi06aF5onjMckd1IxKSNuOcnrj6Adt+zavhjSb/XvDd3bJbeLLa7usq0RWUWQ8obQ+MbQ4ztznvjHNdGKyilShUs23Gz2t1a+7TfzOfCZzVqzpXSSk2t73sk/v128j6d1TxH4e0R44ta1S1sHl+4LidIi30DkZ/CuF8ZeP7zw14x8H+HLO3iuLfxJLNHJIxO5BHswUxwc7+9fO/x01TTb7xrrPh6806zsZ49MBgu5bKS7u72TbvSOArhYgCxUyEZG05JAAFJNQNppvwQ17VXYWls9xDJMQWVPnjSNSRnsvHsD6VeFyWPJCpLXmT0/wC3W1b7jPFZ5JznSjpyta/9vJO/yf8AW590V51deMb5viVaeCdMhSS3gsZb7UJGDFkViEhWPBxuLZJBByvTFafhnxxo/ivVNc0jTY50m8P3H2a4M0exWf5hmM5OR8p64PQ4wQT57oMqWPx08ZG+YAzaZYzxE9oYhskx6jf1rycPhrOoqi1Ub29bfo7ns4jFXVN03o5Wv6X/AFVh9j8cLGbxFp+jap4f1LSbTWJjbWV5dReWksu7aAVPK5OAOp5GQBk1L4s+N2keGNcvtIh0m91SHRhE2pXNsoMVoJvu7snk+o4785Bx4be/F/wb4++Jmm32v3rWPh7w5MJLCHyZHlu7okbZX2qQiKQCATnpxy2JfEmqweC9R+KfhjXIpft/isq+lqsTv9qE4dQEKgj5C469wQMkYr3VlFNTip02nba7/mS38k7vovvPn5Z1UcJOFVNXetl/K3t5tWXV/ce/+PfjDoHgTSbHV5bS61KDUY1mhe2TMXltjazyNgLkEYHU+lb/AIt+IOjeDvDMHiTUo5ZBeGJLe3iG6aaWUZWNR646/T6A/OHxe1fTvC/wS0j4b6xOY9fexsGEGxiMRMu8bwNo2lSOT2q38RtVsvF/gzwT4z8P3lx/Y+h30aXtzbxN5tsFVVaZVdc/uyDhsEZI61z0spptU207OUlfult9+x01s4qRdWKkrqMXbs3v92564/xF8T6j4K8Qa5p3hq50nVdFj8xYNSjIjlCje2wqVLEID6c469K9L8P61aeI9DsNfsM/Z9QgjnQHqA6g4PuOh96+dPhn4gvbuw8eudVvNd8I2MRNle6gzNI+InM6h3AJUY7gY4OBk16d8EILi3+FHhuO6OXNsXGf7juzJ/46RXHmGEjTjKys0136xvbXtb5XO3LsZKpON3dOL7dJWvpprf52Poj4fandaX4w0t7ZiouJ44HHZklYKQfzz9RX27Xwj4P58W6Jj/n9tv8A0atfY3hbxv4P8cRajP4O1q01qPSL2fTbxrOZJhb3ttgTQSFCdsiZG5TyMj1rxT3D5o/a6/aI8Y/AjQtCi+Guh2viTxNqx1K+azuzIqDSNDspL3UJlMZDeYAsccY6F5FBB6Un7Tnxq1rwt+z9ofxW+FWrraLrmqeGvIuzFFMr6fqt7bo/yzK6ASQSEbsZXOQQea8M1A/Gj4vftZ/EDxr8IrPw3qWh/DvTI/A3/FRS3awPdXgjv9TMC2sb5cboYJd2OFAGcmvl7xJqc2j/ALCWt/Ab4rvaf2j8HfHei+HdUEjl7Z9L/tS3ubSUmYKTbtaShFLgZSM7gOaAP158D/Hv4J/EzxDf+E/h7450bxHrGmBmuLSwvYbiVFUhWYKjEsqkgFlyATgnNfM/w8/az8KeG/Fvxd034+fEDStDttC8aXGj6FHqEttZutlFZ2suxAAjyhXlYtI+4jdhmxiub+N118PdX/aH/Zp0n4RS6bP4m0/V7u6zpJib7P4ZWwlW6DmDhLeQtGsYbCs3CZOayP2dNT+C+j/FT9qO/wDHlzoVpqK+Lrn7c+otbLN/ZAsrfAl835jbb/N4PyFt3fNAH39qPxC8CaRpmka1qniGwtdO1+RItOuZLmNYLt5IXuEEMhba+6KN5BtJyqlugrj1/aB+CD+AJfioPHWjjwfDO9qdWN5ELIzxtsMaTE7XbcMAKTu7Zr8kNF8JWfiX9lH9l/w14nsHm8La58XENhZXQcF9CuZNWe1icMQxikgIwp4MTAYwcV9S/tTR+IdA/aa+BX9j3fh/w9oUVprUOlTeILOWfR4dcKw+UpjgntVS5a3Drasz9d6oCxGAD7X0n44fB3XfClj460bxnpN54e1K8i063v4ryJrd72ZwkdvvDYErMQoQ4bJAxzXZ6z4s8M+Hr/SdK13VLawvdeuDa6fDNIqSXc6o0hjhUnLsEVmIGcAE9K/Paf8AZj8UfEPwx+0HZeIfF/h7WNc+I1tpksVr4cgltLfTdd0mF3tbp0lurlklldbdnO5SwjB75o/Z6+IT/tY/G3wn8VruPFl8LPB8CXERG3yfFniEbb+PHrbW9uYyDyPO6DPIB9X/AA1+I0dl8K7nxz8VPH3hzVbSzvb9ZtbsHWy0uKGG5eNIneaVlWSLHlyZb74I5IydTwp+0T8CPHPh/W/FfhHx9ouq6P4aQyandQXsTRWUYBPmTtu/doQpIdsKQDgnBr87vhP8TtN+GP7FWhTX2g6b4hufEvxB1HRLGLWSq6Xb3t5r90Ybq8cpJsit2j8zcF3blUKQSCOx+GE2rt/wUMn0/wAT+K/D/irVbv4b3KapFoFj9itbdodUtjDBNG11dvJIiOSrSOG2MPlCkUAe2/DH9sDwT8ffgr4k8T+BfFWg+F/F+m6frV09veXiXselQWFxNbwX96q+U4t3RI52JAAVwMsOT9CN8S/C3gX4WaP48+Kvi7SLOzaxs3utYMq2mnXM80SkyW/mOfkmbLRJuZtpABPWvza+Ctx4fX/glz8RdJt3threm+HviFb3sKlBcwst1qThJVHzjCOhwexX2rqpptEs/ib+yFffE54Y/Bq+EJ0057wgWKeJGsLQ2/ml/wB2JjAJBb7+d+7Z81AH6H+Dviv8M/iD4Tm8deB/FGna54fthIZr60uY5oIfKXdIJGUnYUXlg2CByRXKwftI/AC68UaN4KtviHoU2u+IYoJtPsk1CBprmO6RZLcooY8zIytEOrggqDkV8W3k+hX37Tf7QV78L5IJfDX/AAraNPEkliQbV/Eoe6MG8p8huBZcP3AwG5zXkfirwZ4R0D/glv8ADfX9E0azsdStE8F6rFcwwIky31xqNkJbjeBu8xxIwZs5IOOlAH6e/ET4+fBT4R6lY6P8TvG+keGL7Ul8y3gv7yOCR487fM2sQRHuBG84XIIzkV3S+L/CbXmkacutWRuvEETz6bCLmLzL2GNBI8lsu7MqKjBmZAQFIJOCK+F/jTo114I+K3jz47/CHxj4WvtftdDtbTxZ4W8RuhSSzso3ng8u4jfzrF5IpCAskbwykqzDjNezeBfFnwI+KX/CmfidrOmadovjbXNElvvCtjeNGmo21vc2iNeRWifKXVISAxVcbOQACaAPdvH3jPR/hz4G8QeP/ED7NM8OWFzqNyc4PlWsTSsB7kLgDueK8B/ZO+OvjP40+Fdbt/ilo1r4c8b+Gbu3h1DT7QyeXHBf2cF/ZyASszDfFNtOT99GxxivP/27LrxF4u8L+C/2c/BC2s3iD4ra3HA0N67x2x0rSF/tG/MzRKziMiKOJ9oJKyEDriuA8K3fxl+FX7aWia18ZYNAs7L406JLog/4R+W6e3Oq+Hw11ayXBu40IkktpZokC5DbQOCPmAPsbxJ+0R8CPB/jSD4deKfH+iaV4muWRE0+5v4Y7gPLjy1ZGb5GfI2hsFsjGc18/wBl+1Bovgb9ob42+GPjT4307w94Q8Kp4WXRI9Qe3tdk2pWU810sb4WWZnZFYgl9gHG0Zrzj9h7XPhJo/wCz3qY+Kd9o9l42g8Qas/jc6vJBHcjWl1GZle6+0EHIXyjCTxgDb8wNanw2vPhlp/7ef7RN54xn0m21qPTPCf2GS/eBLgWn2CX7Z5JlIby8+V523j7m7tQB9++G/Evh3xjoVl4n8J6nbazpGoxiW2vLOVJ4Joz/ABJIhKsM8cHrxVzVdU03Q9LvNa1i5jsrDT4ZLi4nlYJHFDEpd3djwFVQSSegFfnr+xr42+H/AIB+F3xO8c6jrFj4c+GWp/EbXG8M3VzMltp50+4lhhj+ys5VFgkuhN5YXAHPbp9bftFf8m+/E7/sV9a/9IpaAGWv7RXwGv8Axjp/w9sfiBodx4l1aOOW10+O/he4mWZBLHtQMTudCHRfvMpBAIr0W18W+GL7xLfeDLPVbabXdMghubqxSVTcQQXBYRSSRg7lVyrbSRg4OK/Kbx34M8I+G/2BfgBrWgaNZ6ff2d98PtTiuIIEjlW9vJrRrifeBu8yVnYu2csTzXv2h+KfDPgr9vr4njxfq1pog1bwVoFzaNezpbrNDaz3azOjSFQRGfvY6dTQB9o6X478F61p+r6vpWuWV1Y6Bc3VlqM6ToYrO5sji5infO2N4T/rAxG3vXmvh79ob4O/E3TPEcXwk8daR4l1TRLOeeSPT7uG5kiCKdshRScpuwN2CpPGa/KG91nTvEX7KfxK8SwTPe/D7U/jpeXviCWAPtm8Lz6vDLO5CgN5To0bNx9wnPGa/WmfWfhJf6Rdab4IvtEuNUl8P3JsYdOktnnbTFUDMKwncbcOyD5RsyV74oA4L9nH4yy6z+yf4I+NHxo8RWtrNe6PFe6pql4YLK3DsxBd9ojiTJwAAAM8AZr1nwZ8afhJ8RfDGoeNPAvi/TNc0PSQ5vbu1uo5IrURJ5j+cQf3eEG47gOOelfkHbXmtRfsu/saX8F/pFl4Xtr8LqFzrkEl3o0Go/ZJ101r6OGaA7Vn3BS0iqkuxm4GD9YXHwv+IF14g+MHxK8SeL/DGt6xrnw+uNHudJ8MWk9s0sirPJY3dwk15dMz7WlhRsDcvA+6aAPuW7+JHgCx8Jaf48vPENjB4c1YWhs9RedBa3Aviq2xjlJ2t5xdQmD82RjrXK/ED9oH4IfCnWrLw58SfHWj+GtU1BBJDbX95FBKYySokKswKoWBAdsKSCAcg1+YHjn4nfD3Uf8Agnl8C/DOn+IrG61e5bwJapZwzpJcedYXNmtyrRoSy+UUKuWAAbCk5ZQfoH402E3wy+InxU+P/wAJvFfhXXLo6VaR+NfCfiF4yXg0y2LRLBdRuZLSSW3cARTRPFIzBjjOQAforDd2txapfQTJJbSIJElVgyMhGQwYcFSOQemK8d8N/tIfALxl4zPw78J/EPQtY8SDeBY2l/DNMxjBLqgRiHZACWVSSoBJAwat6b8VNFvPgJbfGx9JuLfSZvDSeIDpxQG4S3Nn9rNvs6FwnyY6E+1flT8RPGmseKvCn7NXjqfV/Bnhuw1bxx4UvtA8KeHbQG9s7S6nVZA95567vLjk8q4SK0jjDEIScDIB+snxJ+Nvwh+DyWb/ABS8Y6X4W/tAkWy6hdRwPNtIDFEY7mVcjcwGBnkijxR8bfg/4K8HWXxB8V+NNI0zw1qYVrPUJr2EW10HXcvkPuxLleQEzkc9K+Rfhrf+D9O/bg+OrfFKe0tvE0ljoJ8OvqLImfDosz54sTL/AMs/tfmfaAnG/wC9WM2sfC9v26vB2p61eaU3hF/h+0fgSdXhOki/TUGW/WxYHyRciARD92c+UMDvQB9t6V8Y/hRruj6B4g0XxfpV9pvim6FjpVzDdxSRXt3hm8iFlYh5QEbKD5htORwa6g+LfDA8Vr4FOq2w8RNZHUhp/mr9qNksghNx5Wd3liQhN2MbjjrXwX+1P4j+HNld/ATxV4c1LS4/DOkfFK1jv7uzlg+x21zNbXolM8kZ8tGMrfvCxBDNluTWponjHwr4t/4KRyp4X1W21b+yfhfcWt21rIJkhuP7atpfKZ1yu8I6MVzkBhkDIoA+k7v9pb9nuwv9H0u9+I+gQXevySRWET6jAr3DxzPbMEBftPG8WTgGRSg+YYrb0n44/BzXfiBdfCrRfGukX3jCyDmbSobyKS7Qxf6xTGrE70xl1+8o5YAV+UXh/wAC+DZf+CYvxe1uTRLNtRvLnxZeTXJgQzSXFnqlwtvIZCN26JUUIc/KBx3r6B+K/hHwt4Qb9kS+8LaTa6VcW3ivTbOOS2hSJxBeaVctcJuUAkSsAz5PzHk5PNAHouj/ALU3h/wT8d/jl4Z+N/jrTfD/AIZ8Lah4etdBi1CS3tCn23SY7q5SNsLLMTK287i+wHjavFfX6+O/BL+Dx8Qk1+wPhc232z+1ftMX2H7NjPnfaN3l7Mc7t2K+DfhJffCvTP21v2oL7xjc6PaayjeGvLkv3t47hbD+xIftGxpSGEO7b5mPlzt3dq+NJ11OD9mLRdZ8NvY6f8KLr433t7YvqlvLNosPhmS7nGnvdwRyQ504X+1ipdVyUJ4zkA/Zz4cfGb4T/GC0vL34XeLdN8UQ6e4S5NhcpOYWbO3zFU7lDYJUkYYA4Jrln/ae/Z1TVNC0VviT4f8At/iZYn02EajAXuknO2JkAbpIeEJxuPC5NeKfDr4fePL79oi2+L3jDxx4Q1G+i8MXGlS6f4atJ7WW7s5biKaCeYTXt1uSF1IjYKAPMIzzXwno/gbwbbf8Ea7jV7bRbOLUJ9IfUpLlIEE7XsOpERzmTG4yKqhVbOQoCjjigD9ePiT8cfg/8HmsU+KXjHS/C76mWFsl/dRwvMFxuZUY7iq5G5sbV7kV1Vt468E3v9h/Y/EGnz/8JOjyaTsu4m/tBI0812tcN++Cp85Me7C8nivjT4seFtQX4wQ/Gr4T+O/DNp8RdJ8KQ6bq2geJWR7W50l5pLqOQvHItxZMzmT96FeNwBuTC89Z8KPiF8A/jP4S+CHxW8U6JpXhjxRqdteHwlp15JEtzbSqnkXkWnj5PMj2IMFEAKbG2qSBQB3n7V3xf8T/AAL+COr/ABH8HWdnfavaXel2sEV+JDbE6hfwWhLiJkf5VlLDDdQOvSvN7j4zftA/Cv4ieA/C3xx0jwzqei/ELVDotpeeG5ryO6tL0wyTxtNa3YfzIWEZDOkg2dWHQHL/AOCj8Uc/7JfiOCaVoI5NU8Oq0iOY2QHWbMFlccqR1B7da8B+KugfDX4MfEz4a+Pfhj8SdR8e/EV9esNItdA1rWx4lln07U5ljv3to7kzT2TpCC/2qJkVQCr7lcigD6P0j9q3UIv2wvE37OHizSrey0CFbK30TVoy4aXU5rCO/ezudzFQ0sTSNAQF3eUy/MxGPTPCfxq1PV/jV8Y/h3rFnDFo/wANLLQbu3ngV2uZhqlrcXE4kBYqdhhAQKo6nOTivk+9+EcXxr+OX7Vfg2G8Olazbz+CNR0TUl/1mnaxZaW0tndIRkjZIoDY5ZCy/wAVVv2OPi2fGHxi/aG+JHj22/4R/VNL0vwnF4jtXVsWN/pNtqNvfqAMkxh4WeMjO6MqRkmgD0f4SfGj9r34yeFNE+NnhPwz4MbwL4jlE1po0t7epra6eZjEZJL1Ve0EwUFzEIsADaW38VL+0f8AF79rL4LPB4i0S28EX3hvWfEGn6JpyXCamb5Bqc4ghe4KSLF8pOX2fgK+b/i6fg/8Cvh3fftM/sZ/FSHQJrmeG7j8KWd8l/oOvz3MyK9smlMzSQXEm/H7gI0fPyJgsv05+3LcXF38Ivh7dXUBtZ5vHXg95IWOTG7X8ZKEjqVPFAH078Lh8aRpt7/wupvD7ah5o+y/8I+t2sPk7efN+1ktv3dNvGPevIv2pvir4l+FU3whudC1eLRrDxF4703SdYlmSExvpc1reTTozzKREuYVYyKVZQvDAE5+g7rxz4NsfF9l8P7zW7ODxLqVtJeWumvOi3c1vCdryxxE7mVT1IHY+hx8U/t/DwibL4EDx79i/wCEe/4Whon27+0fL+x+T9jv93n+b+72eu7j14oA+rfhz8cPg98Xpb+D4XeM9J8Uy6WQLpNOu4rhogxIVmVGJ2sQdrfdOOCapX37QPwO0zx/H8K9Q8eaLb+LpXWJdLe+hW6818FIihbIkbI2ocM2eAa+UfGV14b1z9vD4V/8Khlsp9Z0jwt4jPiWWxKMiaZMluulxXTRcbftQLRoTkDkDBBr5H+C/gf4hfFb9j7UPhnqvjnwJ4ajvLq9i8RnU9Pu08Q2Gvreu8s95cNqMai9WdQ6SNEMrs2jbgUAfr58SPjJ8Kfg9Z2l/wDFLxbpvhaC/cx251C5jgMzDG4RqxBbbkbiAQoOTivE/wBlr4wa58XtY+L9ze67Br+iaB4vm0/Q57YQGEaZ9itZ4gksCgSqWlZg7FmIP3sAV5D4XuPD2lft6+NB8Yr20/tq18F6Evhae/Kxo1mGnGrPa+adodrgKX2ndtyD8ua0v2Erzwnf6z+0Fe+BZLOXQpviJfNavYbPsrobK0y0Rj+QqWycrweo60AfWfjP41/CH4dHUU8eeMtJ0B9Jit5rpL28igeKO7MggJV2B/emKQIAMsVbAODU8Pxh+Fdx8Pk+K8HizTH8GyBCNXF1GbL95KIFBmztBMrCPBOQ52nnivk7RPC/hzXP+CifjfVNa0y2v7vS/AmiG0lniWVrczXl0sjRlgdrMowSOcZHQmvjjxpp8WnfsSftY6NpFsIdO0T4paiYLeBMR29rBqumXEixoowqIN7YAwBntQB+z+reLfDGg6ro+ha1qltY6j4glkg063mlVJbuWKMyukKk5dlRSxA6AZrgviF8f/gj8JtUs9E+JnjrRvDOoagA0Fvf3sUErITgOUZgVTII3sAue9fJ3xz+I3gLxd+09+zFpfhTxDY61dLrGrXrJZTpcbbabSLgRSMYywVZCDsyRvwxXIU4p/s26j8OtP8Aij+0lF8XbjTbbxtJ4vvTef2u8SSv4XNvD/ZWDOQDaCDdx90HOeNtAHuEPxR8U3n7Zlj8LrHVUn8F3fw7k8Qpbxxwukl9/asVulws4UyFTCxAUPsOd2M4Nel337QPwO0zx/H8K9Q8eaLb+LpXWJdLe+hW6818FIihbIkbI2ocM2eAa+aY76wu/wBv/SL7wzcW0tnf/B+dtNliZWtplOswvEYih2smwhhsz8vI4r4v+C/gf4hfFb9j7UPhnqvjnwJ4ajvLq9i8RnU9Pu08Q2Gvreu8s95cNqMai9WdQ6SNEMrs2jbgUAftLF4t8MT+KZ/A8Oq2z+ILW0jvpbASqblLWV2jSZo87hGzqyhsYJBFZUfxK+Hsmk+INeXxJp40zwpcz2er3TXMaw2FxaqrTRXMhIWJ41dSwYggEZ618YWGvaT4H/b+1aLxzrNrYS6j8MdL8q4upUto7p7TUrn7QUMjAEr98qCSF5PHNeH/AAv0b4XfHP4C/tIaJ4i8ZW3h7wx4v+K2pPYa08kK25mJ024sXUXBWOZJZUjxGTiVDgHkNQB+kvw2+Nnwj+MUN3N8LfF+meKBp5UXK2Fykzw787TIincobB2kjDYOCa0fiJ8Vfht8I9FTxD8TvE2n+GNOlfyo5r+4SBZJMZ2RhiC7YGdqgnHOK+bPgP8AFbxpN8afEfwL+J1r4f1bxNpOhWWrR+I/DStHBeWBne3igvIHLtbXCMS6xiV0KuzJtHXlvFNzoGn/APBQvQrv4nSQQWL+BHTwlJfFVthqY1BjqQtzJ8guzB5OdvzeTx0zQB7v8TP2ivh14S/Z+8R/Hnw74k0vVdH0/TLu40+6ju4pLW8vIonMFskittaSSZRGEB3bjtxmuO/Yz8V6v4/+D1j461v4pD4n6hrEdtLfPFFYxW+lX728ctxYxiyijI8syKCspZxweMkV8eXC6BqGg/t33nw+WCf4evocptmtQrae2vx6Dc/2s9vt/d+YG8jziv8Ay05zmv0T/Z1tba1+Afw4W2iSESeHNHdgihdzGyiyxx1J9aACf9on4EW3xA/4VVceP9Ei8Xeatv8A2Y19CLnz2+7DsLf609o/vn0ryv4YftmfBv4n/F7xd8I9K8Q6QLzRL62stIeLUoppNa8yzW5uHt4gBxbyF4mCs/KEkjoPmVtfh/ZfZ/FXgfXvDPxS+E3jDxqtxJYbom1+w1XV78I7WVxE0kd8be4ctskRZkVcCT5d1esfs3N4a079qT9pTRLr7JbaquvaLcW8D7Em8mbSYdrxqcHaxB5Xv1oA+ol+OXwhvNb0zwjpnjbSJdf16S8t9Os1uo5Z55rFpI7gLCjbyIXikV+gBRlyCDXKfD/4t6LoXwM0P4jfGL4ieGr6GdH8/wAQ2UyWWjXLGaQR/Z2mkIztAQgMSzqxAHQfP/7Ang/wzYeCPiH4uttOhGtap458ULcXhXdMyQX8qRxhzkqijJCLhcszYyzE/PnwG+Jsfgv9jT9mbwdYaJoupeIfGl7c2ulXniIZ0vS7i3e7la6kIG8y7N0cKI0bOzlRIozkA/Uf4c/Ff4a/F7RZPEPww8TWHifToZTDJNYTpOscoAOxwpyjYIOGAOCD0rxX40ftcfC34HfFPwL8M/GWsaZYv4qe8a/uLvUI7Y6TawWss0FxNGwOUuJoxAm5kG45BJG0/PX7Ld5fD9s7442d34q0nxXdXGieHZtRudDtBY2X22NrmMoYRcXR82NCFZmlZj0bBGK7r9pn+wtO/ar/AGX9a8QC3g086n4ntZp7gKIvMuNGkS3jdm4y8hwgPVunNAH0z40+PnwU+HWi6R4i8deN9I0PTfEEay6dNdXkUa3cTqHEkOWy6bWViygqAQSQDXT3XxJ+Htjo+jeIbvxLp0WleInSLTbtrqIW947xPOohl3bHzFG7jaT8qk9Aa/Oie7+IkH7cvxMj8P694S0W9vdA8P8A/CON4nsbm8a40QQSfaV0x4Ly1VYxeeYZ1UNuJQ9BXn/xC+Cun+HPhJ8CPhp4n1nS/Geg618Z459mko8elpa3y6lJNYQoZpj5McjSRlDIeMo2TnIB+l2mfH/4Iazotl4j03x1o0+k6lJexWt4L2IW876dG0135cpYIwhjRndgSAoLZxzWNB+1D+znc+DLz4h2/wASdAk8N2FwLSe+GoQGGO5ILLCTuz5jKCypjLLyoI5r56/al8JeF9X+On7MXhnU9ItbrSP+Ej1VvsckKNbkwaVLJFmPG3COisBjGQKoeCfh/wCB2/4KE/E6+fQbF508H6HcKzW8Z2zXE1zDLIARgO8caozdSowTgmgD6P8AiL8SI9S+Fem+PPhV488PaVZape6eLbWdRIu9NngnuVieKNo5EBllyY0+b7/y8Hkdn4z+Mfwn+HUl7D498X6V4ek061hvbhL67it2jtriR4oZCHYHbJJG6Jj7zKQMkV+Rdzbwad+xt4t0KwjWDT9I+NT2lpAgCxwW8fiaErGijhVGTgDivsa48M+Hde/4KTz32t6bb6hcaT8L7Gaze4iWU28r6zdxmSPcDtfYxXcOcEjOCcgH134e+KPw38WeCh8SPDnifTtQ8K7HkOqRXUZs0SIkSF5t2xNhBDbiNp64rn/h18ffgp8XNSv9G+GHjjSPFF9pi77iDT7yK4kSPIXzNqEkpkgbxlckDOTX56eE/H/hb4EfCT9q7xBfeGrTXNA0b4jXkUWjTLGlg76lHp0CrKGR447cTTB5DsIVQxAzwbbX3ieH9ub4Ar4o8XeF9X1u70rxPDcaf4XszapY2BsVmt45pXup5Z43kQtCzpEuUcogycAHtPgH4yfErxJ8Gfjf4m1PxlpWhap4R8beJdH0vVtat4007T7DTrxYrdbhYfK3KiErvYliSCxbGD9JeNvjn8JvhFp+jH4ueN9H8P3WqxKYWubhLYXLKo8ySKN2LeWCck5IUEZavzA8QOn/AAxB+2DBuHmR+PfGW5c/Mu7UIiuR1GRyPUV9Q/EnwjeW3xSsfjH8IvHPhmz+IWi+ELbS9U8P+JWR7W40hpHuY3Mkci3FkS5kHmhXjcAbkwvIB9q23jrwTe/2H9j8QafP/wAJOjyaTsu4m/tBI0812tcN++Cp85Me7C8niuqr42+FHxC+Afxn8JfBD4reKdE0rwx4o1O2vD4S068kiW5tpVTyLyLTx8nmR7EGCiAFNjbVJAr7JoAKKKKACiiigD//0v38ooooAxNb8M+G/EwsV8SaTaasNLu4b+0F3BHP9nvLckw3EXmBtksZJKSLhl7EUSeGfDcviKLxfLpVo+vW9s9lHqBgjN2lrI4keBZ8eYImdQxQNtLAEjIFbTMqqWY4A5JPQCuC8MfFb4XeNtVudC8G+MdG17UrIEz2thqFvdTwgYyZI4nZlxkdQKAL03w98A3Fnr2nz+GtMltfFTM+sRNZwtHqTsgjZrtSmJyUUKTIGyoA6DFQ+LPhr8PPHnhuPwd428M6Zr2hQ7PLsb60huLaPyxtQpFIrKpQcKVAKjpiu2ooA4Ww+F/w10rwZJ8OdM8J6Ta+FJUaN9IjsYFsHRzlla2CeUwY8nK8nk1X8A/CX4W/Cq3ubX4Z+EdJ8KxXjBpxpdlDaecRnBkMSqXxnjdnHQV6FRQB8lfET9nmL4l/tKaH8QfGOjaX4g8DWvhLUNEu7LUY0ufMu7i9guIj9nkRkZQsRO7OVbGB3H0j4S8G+EfAOhW/hfwNotl4f0e0z5Vnp9vHa26FjliscSqoLHknGSeTzXSUUAeT6V8BvgjoXjCT4haL4A0Cw8Tyu0janBpltHeGR87385Yw4dsncwOW7k13ul+GfDeiX+p6roulWlhe63Ks9/PbwRxS3cyII1kndAGldUAUM5JCgDOBW0WUEAkAt0HrS0AYejeGPDXhw6i3h7SbTSzrF3Jf3xtYI4PtV5MFWS4m2KPMlcIoZ2yxCgE8CuM8I/BP4OeANevPFPgXwNofh7WdQDC4vNP063tbiQOQWDSRIrYYjLDOCeTzzXp9cT4k+Ivg3wj4j8M+EvEWoi01bxjcT2mlQeVK/wBpmt4WuJV3IjIm2NS2XKg4wCTxQBt2Phnw3pmtan4l03SbS01fWxAL+8hgjjubsWqlIBPKoDyiJWKx7ydoJC4BqHT/AAl4U0k6s2laLZWZ1+Z7nUTDbRx/bZ5EEby3G1R5rsihSz5JUAE4FdDRQBxsnw6+H0vhS18By+GNLfwzZeT5GltZQGxh+zuJIfLtynlL5bqGTCjawBGCAarePfhf8Nvinp0OkfEvwtpnimyt3MkUWp2cN2kTkYLRiVW2MRwSuDjiu7ooA4mP4afDmHwY3w4h8LaUnhN4zC2kLYwDTzGTuKG2CeUVJ5xtxnms3wj8HPhJ4B+wHwP4K0XQG0sXAtXsdPt7d4BdhBceW0aBl87y08zB+fYu7O0Y9IooA5jxf4J8HfEHQ5vDPjvQ7LxDpFwQZLTULeO6gYjoxjlVlyOxxkdqoeGPhr8OvBMkM3g3wtpWgyW1oLCJ7CygtmS0WRphbqYkUiISMz7B8u5i2Mkmu2rifFfxG8GeCNW8N6H4o1EWV94vvv7N0uPypZPtF2I3m8vdGjKnyIx3OVXjGc4FAEnjn4d+Avidon/CNfEbw7p/ibSvMWYWuo20d1EJUBCuqyqwDAEgMOQCeea5zU/gV8E9ZutDvdX8A6DfXHhmGK20uSfTLaRrGCAYiity0Z8tIwBsVcBcDAGK9VrifEnxF8G+EfEfhnwl4i1EWmreMbie00qDypX+0zW8LXEq7kRkTbGpbLlQcYBJ4oAztZ+D3wl8Q+Fv+EH13wXot/4d8+W5GnT6fbvaC4ndpZJhCU2CR5HZ2cDcWYsTkk1JYfCL4U6Z4MHw50/wbo1v4U8xJf7JTT7cWBkjkWZHNvs8susiK4YrkOobOQDXodFAGDqfhbwxrWraVr+saRZ32qaE8smn3c9vHLPZvOnlytbyMpaIunysUI3LweKi8M+D/CfgqyuNO8H6NZ6Ja3dzNeTRWUEduklzcNulmdYwAXc8sx5NWfEPiTw74S0mbX/FeqWui6ZbFRLdXs8dtBGXYIu6SQqo3MQoyeSQByazvC/jvwP43hkuPBfiHTtfiixvfT7uG6Vc+piZgKAOW8TfAv4JeNfEJ8W+Mvh94e17XCEU39/pNpdXeIxhB50sTSYUcL83A6V6D/Yujf2OfD32C3/sowG2Np5SfZ/IK7DF5WNmzb8u3GMcYxVbX/E3hvwpZx6h4o1a00e1mlSBJbyeO3jaaThI1aRlBdv4VBye1blAHlnhn4G/BXwX5H/CIeAdB0U2139viNnplrAY7wI8YnQpGCsoSR0DjkKzKDtJFaN58JPhbqHjy1+KV94R0qfxjZJ5cOsPZQtfxrsMeFuCvmDCEqPm4UkDgmvQq4n4h/EbwZ8KPCdz448f6iNK0SzltoZbgxSzbZLudLaEbIUdzulkRchcDOTgAmgDmdM+AHwJ0a4v7rSfh14ds5tUlinuni0m0Rp5YJFmidyI/maOVFkUn7rgOMNzXpmraTpevaXeaHrlnDqOm6jDJbXVrcxrNBPBMpSSKWNwVdHUlWVgQQSCMVoUUAcxc+CfBl7Z6Lp15oGnz2nhuaC40uGS1iaOwmtkMcMlqpUiF4kYqjRhSqkgEA03xj4H8GfETQ5fDHj7QbHxHpE7Kz2mo20d1AzL91vLlVl3L2OMjsa6migDjNB+HPw+8LeEpPAPhrwzpml+GZY5YpNMtrOGGyeOcFZVeBFEbCQEhwVO7JzmrNx4E8EXei6T4bu/D2nTaRoL2smnWb2kLW1k9jj7K1vEV2RNBgeUUAKYG3GK6qigDEn8M+G7nxBa+LLnSbSXXLKCS1gv3gja7ht5iGkijmK71RyqllDAEgEjgVxnxI8At4xtobmwkWLULUELvztkQ87SR0weQfr9R0PjTx54M+HOjL4i8d6za6Fpjzw2ouLuQRRme4cRxJuPdmOB+fQE11tAHxq3wn8fKxA0wMB3E8OD+bim/wDCqPH/AP0C/wDyPD/8cr7MooA+M/8AhVHj/wD6Bf8A5Hh/+OUi/Cbx6owukgZ54mg6n/gdfZtef658WPhZ4Z1v/hGvEnjLRtJ1fCn7Fd6jbQXOHGVPlSOr/MORxz2oA+c/+FUeP/8AoF/+R4f/AI5R/wAKo8f/APQL/wDI8P8A8cr6/fUtOj05tXkuolsEiM5uC6iEQhdxkL527NvO7OMc1wvh/wCMXwi8W6imj+FfHGh6zfy/ct7LU7W4mbHPCRyMx49qAPngfCbx6CWGkgFup86Dn/x+gfCXx4HLjSRuIwT50GcfXfX2dRQB8ZH4TePSwY6SMjofOgzz/wADpG+Evjxxh9JDDrzNAeR/wOvpi4+I3gy0+IVn8KrjUQninUNOm1aCz8qUl7K3lWGSXzQnlDbI6jaXDHOQCMmrPg3x54M+IenXWreB9ZtdcsrK7nsJprSQSpHdWzbZYmI6Mh6j3BHBBoA+Xx8J/HwyRpXXr+/g/wDjlczefs9+LLzxXp3jFtNki1DToJrcbLi3CyxS4O2QFiSFPzLgjBOea+86KuFSUdYsidOMlaS8z4z/AOFUeP8A/oF/+R4f/jlIfhN49JBOlAkdP30HH/kSvs2ioLPjP/hVHj//AKBf/keH/wCOUf8ACqPH/wD0C/8AyPD/APHK+zKKAPhLxR8A/GXivw7eeGr3T5be1vUEbtBcW6OFyCQCWIwcYPHQkVtWvwe8b2VtDZ2mjiKCBFjjRZoAFRRhQP3nQAV9O698RfBvhnxh4X8Ba5qItte8ZteJpNt5Ur/aW0+H7RcjeiFE2RfN+8Zc9FyeKPH/AMRfBvwv0FPE/jrURpemSXVrZLMYpZs3F7KsECbYUdvnkdVzjAzkkDJq3Uly8t9CFTjzc1tdjzX4efCrUdK1SLXfEexHtjuigVg539mYjjjqACef19d8N+D/AAn4Oiv4PCWjWeix6peTahdrZwR24uLy4IMtxKIwN8smBudsscDJ4ro6Kgsw9C8MeGvC6XsfhnSbTSE1K7mv7oWkEduLi8uDma4lEarvlkIy7tlmPUmsS9+Gfw41K/1LVdS8KaTd3usTWVxfTy2MDy3U+m4NnJO7IWke2wPJZiTH/ARXb0UAec+Cvg/8J/htf6hqvw88G6P4ZvdWObubTbCC0kn5ziRokUsM84PGcnqTWV4k+AXwL8ZXBu/Fnw88PaxO129+0l3pVrM73cqosk7F4yWkdY0DMclgqg52jHS+OviN4M+Glhp2p+NtRGm22rajaaTauYpZfMvb+TyreLESORvc43MAo6sQK7agDA1Twn4W1xdLTWtGstQXQ7mK808XFvHKLO6hVkjnt96nypEVmVXTDKCQDgmo/Ffg/wAJeO9Dn8M+N9FsvEGkXWPNs9Qt47q3facqWjlVlJB5BxweRXR1434w+Pnww8DfETQ/hRruoXD+KvEKRS21jZ2F3eskE032eOe4a2hkS3haX5PMlZFyCScAkAHX+Bfhv8PfhhpLaD8OPDWm+F9OdvMe30y0itI3fGN7rEqhmx/Ecn3q/wCGvBXg7wWl/H4O0Gw0JdVupL68FhaxWoububHmXEwiVfMlfA3O2WOBkmjxV418G+BbGLVPG+vWHh6ynkEMc+o3UVpE8pUsEV5mVSxVScA5wCe1N8L+N/Bfji2kvfBev6fr9vCQryafdRXSIT0DNEzAE+9AGdL8MPhrN4SuPAMvhPSW8M3jySTaWbGD7DI80hlkZ7fZ5ZZ5CXYlcliWPPNQ+E/hR8LvAbWb+CPCGkaA+nwS2tu9hYQWzwwTukksaNGilUkeNGdQcMyqWyQDXf0UAeWwfA74MWut+IPElv4E0OPVPFltNZ6xcjTrfzdRtrkgzxXTbP3ySkAyK+Q+Buziuj134feA/FHhQeBPEnhzTtU8NrHHCumXVpDNZLHCAI1WB1MYCADYAvy4GMYrr6KAOL8L/Df4e+CfDD+CvB3hnTdE8PyiRX0+ztIYLVxKNsm+JFCNvHDEg7u+amuPh/4Cu/Cdv4Cu/DemTeGbRYEh0p7OFrCJLVleBUtinlKImRWjAX5SoK4IFddRQB5Z41+BvwX+JOr22v8AxB8CaF4l1OzCiK61HTba6nRV5CiSVGbaCchc4zziusuvBXg6+13R/FF7odjPrHh6OaLTbx7eNriyjuEEcy28hXdEJEAVghGRweKwbf4v/CW71yXwxa+NtEm1mCVoJLJNStmuUmRtjRtCJN4dWG0qRkHjGa6vxD4k8O+EtJm1/wAV6pa6LplsVEt1ezx20EZdgi7pJCqjcxCjJ5JAHJoAbdeGPDV7r9j4rvNJtJ9b0yKWC0vpII3ureKfHmxxTFd6LJtG9VIDYGc4FN1rwt4Y8STabceItIs9Vl0a6S+sXureOdrS7jBVJ4DIpMcqhiA64YAkA8msfwt8S/hx45mlt/BPivSfEEsC7pE06+gu2Rc4ywhdiBnua7agDy3XPgd8F/E3i+D4geI/Aehap4mtijR6ndabbTXitHjY3nPGXymBsOcr2xS+L/gh8GfiBLdT+OfAmheIJr6WGa4kvtNtriSaW3Ty4nd5I2ZmRPkUknC/KPl4r1GuJ8I/EbwZ48v/ABHpnhTURf3PhLUX0nVE8qWP7PexxpK0WZEUPhJFO5Cy84zkGgDz/wCM/wAD9G+LPwti+DUEWn6T4XuZbaG8g+wpKE063+Yw2UeVjt5SVRY5dreUu4ou8KR7Rf6Zpuq6bc6NqlpFeafeRPbz280ayQywyKUeN42BVkZSQVIwQcEYq9RQByt34E8EX/h2w8IX3h7TrjQtL+zfZNPktIXtLb7EVNt5UBUxp5JVfK2qNmBtxgVieNfhD8KfiRf6dqvxC8HaP4mvNIJNnNqVhBdyW+45PltKjFQSAcDjIB6gV6LRQBz9j4S8K6XZ6lp2m6NZWlprE01xfQw28ccd1PcjE0k6qoEjyDh2YEt3JrlPAnwZ+EXwuuby9+GvgrRfCtxqPFxJpen29m8oByFdoUUlQeQvQdhWr4r+Ivg3wRqvhvRfFGoiyvfF1/8A2ZpcflSyfaLvy3m8vMaME+SNjucqvGM5wK7agDgtO+Ffwy0jwleeAdM8J6VbeGNQeZ7jSksoRYzNO2+Xfb7PKbe3LArgmovAPwl+Fvwqt7m1+GfhHSfCsV4wacaXZQ2nnEZwZDEql8Z43Zx0FehUUAeRaV+z/wDAnQ31OTR/h14ds21qWOa+MWlWifaZIZVuI2lxH8xSZFlXP3ZFDjDDNWPGHwK+CnxB1638U+O/AWheIdZtdvl3l/pttc3ChPujzJEZiF7AnAPI5r1WuJPxF8Gr8Rk+Ep1Ef8JY+lNri2XlS5OnJOLYzebs8riVgu3fv77cc0Adl5UXleTsHl427cfLt6Yx6YryjRfgF8C/Dk9xc6B8O/DunTXVzDeyvb6VaRs9zbSCWGYlYx88UgDo3VX+ZcHmvW6KAPO/H3wi+FXxVS0j+Jng/SPFa2DFrf8AtSxgvPJLYLeWZkbbuwNwHB75p3iX4S/C3xn4WtfA/i3whpGseHrEILbT7qxgmtYPLXanlRMhSMqvClQMDgV6FXA+IPit8LvCesJ4e8VeMNG0bVXVXW0vdQt7e4KvnawikdXIbBwcc0ARTfCL4VXHgY/DCbwdo7eD8Bf7H+wQf2eAG3jFts8sEN8wIXIbnrzVrwv8MPht4Iazk8GeFNJ0J9OtZLG2axsYLZobWWQTSQRtGilY3kVXZBwzgMQSM12VrdW17bRXllMlxBOoeOSNg6OrDIZWHBBHQiuFtPi18Kr/AMWS+ArHxnotz4mhYpJpceo2z36MuchrZZDKCMHIK9qANCL4efD+Hwrd+BYfDOmR+G7/AM/7Rpa2UAsZvtLmSbzLcJ5Tea7Mz5U7mJJySav3/hDwnqo0gapotleDQJkudNE1tFJ9iniQxpLb7lPlOqMVVkwQpIBwTXRUUAeVeLfgX8FPH09xdeN/AOg6/PeTx3U8t9pltcSSzxRrCkkjyRlmdYkWMEknYoT7oArvZPDvh+bQT4Wm0y1fRWt/spsWhQ2pt9uzyfJI2eXt+XbjGOMYrBs/iN4M1Dx/qXwus9REnifSLG31K6s/KlBjtLp3jik8woIjuZGG1XLDHIAxXbUAeaeAvgz8IvhZJdzfDTwVo3hWW/4uH0ywgs3lXOQrtEillB6KTgdhWuvw3+HaeCv+FbJ4W0pfCPlGH+xhYwDTfKLbyn2XZ5O0t823ZjPPWu0ooA8v8c/BL4N/E6azufiN4G0TxPNp6hLeTUtOt7t4UBzsRpUYqmeqg4PcVznjX4J6V408cfDnXLlLG10P4b3Et/ZWcVkn2j7X5Bt4EjnziC2jVt7RRoC7pFlgqFW9zrifH/xF8G/C/QU8T+OtRGl6ZJdWtksxilmzcXsqwQJthR2+eR1XOMDOSQMmgDU8VeEPCfjrQ7jwx430Wy8Q6NdlDNZajbRXdtKY2DoXhmVkba6hlyOCARyK5XwZ8Fvg58Ob19S+HvgTQfC93IpRptL0u1spGU9QXgjQkHuM16ZRQBi2Phrw5pesan4h0zSrS01XW/JN/dwwRx3F39mTy4fPlUB5fLT5U3k7V4GBUGl+EfCeia3q/iXRdFsrDV/EBhbUr23toorm9NspSE3MqKHlMakqm8naCQMCugVlYblII9RXJ+OPHngz4a+HJ/F3j/WbXQNFtXijlu7yQRQo0zrHGCx4+Z2AH1oA46x/Z9+BGl+L18f6b8O/D1p4lVzKupRaVapdrKSSZBKsYYSHJy4O4+teha/4Y8NeK7WCy8UaTaaxb2txDdwxXkEdwkdzbtvimRZFYLJGw3I4+ZTyCDW5SMyqNzEAepoA8X1v4QW/iP45eHPjDq8tqy+D9NurXTIo7RRdi5v8pcSTXZYs0Sw4WKJVUBnkZix2BfSPEXhHwp4vgtrbxbotlrcNlMbiBL62juVimMbxGRBKrBX8uR03DB2sy5wxB6GigDgfAfwq+GPwstruz+GnhPSvCsF/IJbhNLsobNZnGcGQQqu7GSFznA4GBWLrvwG+CPifxbH498SeANA1XxJEVZdSutMtprvcmNjGZ4y5ZMDaScr2xXrFFAHn/jz4T/C/4pxWcHxL8I6T4qj05zJbLqljBeCFjjcY/ORtu7A3Y6gYORW/4f8ACPhTwmt0nhXRbLRlvpFluBZW0duJpEjSFXk8tV3MscaICckKqqOABXQ1wfir4p/DHwLfRaZ438X6P4evJ4xNHBqOoW9pK8RJUOqTOpK5UjIGMgjtQB0MPhnw3b+ILnxbb6VaRa5eQR2s9+sEa3ctvExaOJ5gPMaNGZiqkkAkkDk1RtvA/gqz0/WdJtPD+nwWPiOee51OCO0iWK/nulCzy3KBdszyqAJGcEuAAxNP8K+NfBvjqwk1XwTr1h4hsoZTDJPp11FdxJKFDFGeFmUMFZTgnOCD3FWtE8TeG/EwvW8N6taaqNNuZLO6NpPHP5FzFjzIZfLY7JEyNyNhhkZFAHF+Ffgj8GvA0NnB4N8C6HoiafdNe232PTbaAw3TxtE08ZRAVlMbtHvHzbGK52nFO8b/AAU+DvxM1Kz1n4i+B9E8T3+ngLb3Gp6db3csSgk7VeVGIXJztzjPOM16dRQByd34D8E33iDRfFd5oFjNrXhtJYtMvWtozcWUc8ZikSCXbujV0JVlUgEdRXJ678Bvgj4n8Wx+PfEngDQNV8SRFWXUrrTLaa73JjYxmeMuWTA2knK9sV6xSMyoCzEADqTQB5745+Efwr+J02n3HxH8H6R4ol0li9o2p2MF4YC2N3lmZG2g4GQODgZ6CtGX4c/D2fR9X8PT+F9Lk0rX5muNStGsoDb3szKiNJcxFNkrlY0Us4JIVRnAGOyooA4DwB8Kfhj8KrO50/4ZeE9L8K2164knTTLOG0EzjOGk8pV3EAkDOcDgcVc8c/DrwB8TdGHh74jeG9O8T6YHEi22pWsV3EsgBAdVlVgrAE4YYIz1rs6KAOSsPAPgXSvCD/D7TPDunWnhaS3ltG0mK0hSwa3nBWWI2yqIjHIGYOpXDAnIOTXQ6fp9hpFhbaVpVtFZWVlEkEEECLHFFFGoVERFAVVVQAqgAADArlfBfxG8GfENtfTwdqI1A+F9WudD1LEUsXkajZhGmh/eom/YJF+ZNyHPDHBrtqAPKLf4D/BK08a/8LJtfAOgw+K/MMv9qpplst75rZzJ54Tf5hzy+dx9a1dT+Evwu1rxvYfEvV/COlXvi3S1C2urTWUL30KqCFCTlTINoY7eeMnGMmup8R+INH8JeHtU8VeIbj7JpWjWs97dzbWfyre3QySvtQMzbUUnCgk9gTUfhfxLonjTwzpHjHwzc/bdH12zt7+yn2PH5ttdRrLE+yQK67kYHDKCM4IB4oAXQfDHhvwtaTWHhjSbTSLW5nmupYrOCO3jkuLhi80rLGqgySMSzsRlickk1ymofB74S6t4Jg+GupeC9FuPCVpzBpD6fbmwhOWIMdts8tCCzEFVBBJPc16PRQBx3hn4d/D/AMFyLN4O8M6ZoUiWsdirWFlBasLSF3kjgBiRT5SPI7Kn3QzMQMk5Txz8O/AXxO0T/hGviN4d0/xNpXmLMLXUbaO6iEqAhXVZVYBgCQGHIBPPNdlRQB5h4o+Cfwd8beHtK8JeMPA+i61ouhRpFp9neafbzwWccahES3jdCsShFCgIANoA6cVtwfDf4d22laJoVv4W0qLTfDVwl5pVqtjAsGn3MQYJNaxhNsMih2CvGFYBmweTXaUUAYmpeGfDetalpes6xpNpfahokjzWFxPBHLNZyyoY3eCRlLRMyEqxQglSQeKIPDPhu28QXXiy20m0i1y+gjtri/SCNbuaCElo4pJgu9kQsxVSxAJJA5NO8R+INH8JeHtU8VeIbj7JpWjWs97dzbWfyre3QySvtQMzbUUnCgk9gTUfhfxLonjTwzpHjHwzc/bdH12zt7+yn2PH5ttdRrLE+yQK67kYHDKCM4IB4oAxX+Gfw3k0m40CTwppLaXd3x1Sa0NjAYJdQMonN08eza05lAk80jfvAbOea3F8M+G18Rv4xXSbQa/JaLYNqIgj+2NZpIZVtzPt8wxCRi4TdtDEnGSTUXizxToPgbwtrHjXxTdfYdF0CzuNQvrjY8nk21rG0ssmyNWdtqKThVLHGACeKtaDrml+JtD07xJoc/2nTdWtobu2l2snmQToJI32uFYblYHDAEdwDQBkr4C8DJZ63pyeHNNFr4lme41WEWkPl6hNKqo8l0uzEzsqqrNIGJAAJwBXP+FPgt8HvAn2E+CvA+h6C2lyzTWjWOnW1u1vLcJ5U0kbRoCjyR/I7AgsvykkcV6ZRQB5/J8KPhfOniSKfwjpM0XjFkfW43sYHj1N0GFa7RkKzEergms/xx8Efg18TZbOf4i+BtE8TS6coS2fUdOt7p4UByERpUYqn+yPlPcVRn/aD+AltNJbXPxK8MxSxMUdH1myVlZTgggy5BB4INdD4l+Knww8GS2sHjDxho+hSX0Ymt1v9Qt7VpoicB4xK6llz3GRQBxPjX4J6V408cfDnXLlLG10P4b3Et/ZWcVkn2j7X5Bt4EjnziC2jVt7RRoC7pFlgqFW9zrK0XXdE8SadFrHh7ULfVLCfPl3FrKk8L4ODtdCVOD6GtWgAooooAKKKKAP/9P9/KKKKAPiD9tu6vtd0z4X/Bhbuaw0f4oeMLHRtakgdonl0qOKa6uLVZVIZDceUsZIOShZehNe/wCjfs+fA3w5qega14d8BaLpWoeFyx024s7GG3mtt8TQsFeNVYhkdgQxIJ+b7wBrH/aI+Ca/HLwLa6HYau/h3xHoGpWmuaDqyR+abDVbBi0MpjJAkQqzxuhPKOe+K47wTF+2dqXibRoviVJ4K0PQNNk36jLozX19daoAhAjijuY4EtELMGZi8rgrgZUnIB8seJf20PFGu/8ACd+NfBPxJ8A+FNM8E3mo2em+Gdcmjk1fxAdKLJM8khvrdrNbmRGW1CwSEja7ZBxX6NfDXxvp/wATPh34X+I2lRNBZ+KdLstUhjcgvHHewpMqMRxuUNg+4r430b4C/G34O614n0T4UaJ4M8XeEfEWq3mr2DeIpLi0vtHm1CQzXEDeRa3C3VsspZohuikG4qSQA1ew+I/H3xF8PfHv4WfC3w9/Ylxoer6bqE3iGyhjm+3WUdtbk293DtOyK1a4UQKJBli2FyVYoAS/tF/GLxf8PZvBXw/+F9hZ3/jz4k6m+maUdRL/AGG0jt4Wubu8uFjKySJBEufLQgsSOR34X4g/FX4w/sy/B3xN45+Met6B4yvEuLCz0KS1tJNCje7v5RAFvjNc3EaQxu6uZFcYiVyRnFd7+0V8GfFnxIfwZ46+GWqWmkePfhzqbanpD6ijvYXKzwtb3Vnc+VmRIp4mwXQFlIBANcP41+Efxw/aG+FniTwZ8Z28PeEL2SbT7zw++hyXOqNZahp032hbi6kuo7dJUaREXyliGE35clhtAPP/AIY/tOeIYfjZ4M+Fnif4l+Cfilb/ABAh1AJL4U2Qy6Pf2Fv9r8qVFvrszW00SyLHIwjfeoBHzYriYfj1+1740+EPxD+MXg+48J6Pp3w31HxHGLW6srq6uNYg0K4mJUss8aWv7hAgx5jPIC2Y1IA+pvhRoXx4t/EdrL8TfB/gXRLGygdZLzQJ7qe7ubkgBXhjmtIBbxEFtytJK3OAccnkfAf7O/jXwv8As6fFL4Rahe6fJrHje78Wz2UsUkxto11552thMzRK6lBIPN2o2OdpbjIB86fGbxN8Xvib8bv2TvGHw81vSdAt/GNnqmq6XFfadNemzuZtBknna4KXUPnI0E3lxqojKPlyzg7R7d4/+K37Rcf7QOhfs4/Dx9C+3al4MTXL/W721maGxnhvDbXFwlqs6tKrnakUHmDazhnkZUIan4p/Z3+L2leE/wBnjXPh5No1/wCNfgdZx2k9nf3E8Gn38c+lDTbwRXCQPIh43RM0PI5YAjB9X0f4T+Pbj9prSfj74ibTba2XwB/wjl7Z2s800iapJqEV7IYS8MYe2UKyrIxRycZjGTgA8e/aE+K3xr+EttdXlx8V/h/4Xk07To59O07VLOaTUNeuobcNOGQ3sRt45JgyxiCOYquCz5JA5LxR8SIvjB4y/Yw+KUVr9h/4Sm91DUGt87hDJPoUzPGG7hGJUHuBmutn+AHx98P/ABK+K+q+Cn8K3th8Vbgy/wBv6sbptY0q2a1S2NmttFFsuIYtpaBftMSgklgc4Mejfsw/E/wt4G/ZostPudJvtc+C10i6pE1xPHbXNrcWcllcvay+QXaSMPvjV40D4IJXigD7U8eXPiuz8H6rdeB5dNg1uKEtbS6w0i6fGQQWe4MOH2Km5sAjJABZRkj4d+DX7Tfii/8A2i7H4I+I/iB4U+KFh4i0q9vrfUfDFubU6dfaeyGW0mAu7uOSNo3JRt4cMpDCvor9qX4ReI/jl8F9X+HXhXULaw1C8nsbhVvg7WN0tndR3DWt0IwX8icIUfaCcHoRkHyLw38Fvjtrfx3+G3xg8fQ+FfDujeB9M1jTU0PQ5LqYwrqMUKo6TyQQpJlo8GMRRCJVG1pSx2AHisv7Rf7W+vfB34g/HTw0fClho3wy1TxBFJY3Npdz3GsWmiXMwkzIs6LaFYECrgSmSRSxKKQteg3Px2/aL8M6/wDCH4ieMV8PDwB8W9YsNFXRLOG4bUdLOr28k9jK1+8gS4cbAJwII0GdqA/6yut8O/s0+OtI/Ze+LfwTub7TW1zx7ceLpbGZJZjaRrr7zNbee5hDqUEg83bG+Odu/jPR+P8A4A+MfFXgH4EeFdPvLCO7+F/iPw1q+ptLJKI5rfRraSGdbYrExZ2ZwYw4QEZ3Mp4oAl8MftCanoGsfHLQvi+La2m+FEjaxbyWyNCtz4burVrq0lw7tulQxTQysuFLoMKM4ryu2+Nv7RviO1+CHw0jk0bwt45+KGi3/iDVdTubGW5ttPt7VYpltLWzNxEZbkLcRrJumwoVn2nI28t+2L4Bs/iL+0B8LfA/hXUol1bx7BPofiqyj+eSXwnY3UGqTSSYP7oLJC1ujOPm+0uq5Oa2f2tvD+r/ABN+Pnww+GnhDS9L8S6joek6zr1xo+sXtzpNt5DvbWcN1De2Ya4W4jYyIiJGy7GdnKkJkA5/Qv2pPjn4J+GfxY8afEmfRvFur+C/FB8FaNp2l6fJp6X2rvJbxwSvcPdzgxv9pQyRhFaIo67mODWX8VtJ/aI0r4w/s4f8Lq8Q+HvEEF14x81Ro+nXGnvZ3f8AZ11uhUy3E4uINpO2RhE4IGVO75ZLXwrqnxh8J+N/2Ln8G6V8JvE3gWHSPEmk3uhXv9raXHdvdNc20kxaC3lWZ5YSZBIrSOjNJknGfVdW+E37VfxW8e/Cnxh8U28H6HZfDrXRqVxZ6RcX11Jeg2s8EkySzwRiPDOvlwFTlWZmmBQKwBZ/ab+LnxY+F17q+qab8T/AXguys7U3Oi6PrFvNdaprMkUW54pD9rtzErSBkQwQykDDFgSVXgfFHxIi+MHjL9jD4pRWv2H/AISm91DUGt87hDJPoUzPGG7hGJUHuBmutn+AHx98P/Er4r6r4Kfwre2HxVuDL/b+rG6bWNKtmtUtjZrbRRbLiGLaWgX7TEoJJYHODHo37MPxP8LeBv2aLLT7nSb7XPgtdIuqRNcTx21za3FnJZXL2svkF2kjD741eNA+CCV4oAd+0/8AGr4ufCG41/XLH4m+AfC9vpds95ovh3U7ea51bWlhh3vHK/2uFoi7hkTyYJAPlLNyQvu/h/4kfE/4i+HPg/8AEH4b6Jpr+GPGlpb6l4g+3Tut1YWd5ZrPCLUJ8skgkba24EcDgAll+drn9nP9oLRvEHxk07whL4TutP8Ai5e3103iTUzdtrNha3tstuLD7NHFtmigVcW5+0oqAlijH5T1tpd/Ff8AZ/8ACf7NfwR8P3Oh6lqWNP0LxBZn7RLcTWVhZIl3eWBAjKxW5Uu8kyKPmjU4ZwrAD/8AgotFFP8Asp69BOiyRyav4cVlYAqynWrMEEHggjqK4D9sf4Y+Avgh4N0r9pb4SaDZeEfGfgnWtHJm0i3jsjqdjf30Nnc2F1HAEWeORJd2HBZSo2kc5+jv2tvhH4v+OPwM1f4deA57G21y7vNKurd9SklhtP8AQL+C7ZZHhimcbliIGEPJGcDkeaat8E/j/wDHPxD4bX9ovU/DmleC/DOowawdB8NG8ujqd7aNvthe3d4kH7iJ8OYkh+cgbjwCADyf9siDwV8cPjBo/wCzn4u8QWGh6TofhfWfEVzJf3UVvF/at/C+l6PgyMv7yAyXFwByRtU+mfrP9lT4rv8AGz9nzwT8RLuQSane2Cwalgg41GzJtrvpngzRuR/skHkHNcN4G/Ze0S9+IHxN+Jnx10HQPF2s+MtbEmm+dapqCWWiWVvHb2MH+lQL5cpCu8wjBUs3DNjNeOeJPCvxQ/Y++G/xr1r4UXPhy203WtdttZ8F6VcpcP8A6XfLEt9pa2kCRgPPLGRZRwuwLPlhGASAD7s+I3jjSPhl8P8AxL8RteDNp3hjTbvU7hY8b2itImlZUzwWYLhR3JFfk/8AtM6t+1J40/ZCk+J3xJ1Dw3aeHPFVz4bvX8PWljcre6ZbXGrWctpt1Bp2W4mGY1nVoEXlijfKA36qePPBFr8U/hhr/wAO/FQNrB4q0m50288k7jF9sgaKQxsQMlCxKkjqAa+BPF/7PX7Y/wAQfgLZ/s6+JNT8GRaXoR0mJNZjlv2utVt9IuoJYBJB5Gy0kKQq0rq04ZxtVVVt6gHuvjf4nfGLx58c9X+A3wLvtK8ODwfpdnqWv65qtlJqRjm1IyfY7O2tI57cFnSJpJJHkwFwAuevmWtftXfE7w9+zx8atc17TtMtvih8Fp2srxYVlk0u8Z1imtbuKJnEqxTwy58tn3KwI3EV6n45+E3xh8I/G7U/jv8AANtF1K48V6ZaaZr+i67PPZwztp7P9jvLe6t4Z2WSNZGjeN49rJyCGr5t+PHwb8S/DL9i39oXxb8RdTtdW8c/ENZNX1iWwR0sYGXyYLe0tfMxI0NvEgVXcB2JJIHSgD1TxX8W/wBpn4PXvgb4jfE+68Nap4K8Yazp2jajpOl2lzFc6N/azeXbTx3005F2I5GUTFoItwOURc8M/af+NXxc+ENxr+uWPxN8A+F7fS7Z7zRfDup281zq2tLDDveOV/tcLRF3DInkwSAfKWbkhZrn4Q/tGfGu9+H2ifGKfwzpngPwdqOn69M+iyXkt9rdzpwD2cbwXEapaQ78PMolmYlQqtjLVj3P7Of7QWjeIPjJp3hCXwndaf8AFy9vrpvEmpm7bWbC1vbZbcWH2aOLbNFAq4tz9pRUBLFGPykA6Pxz+0v8Sr/Sv2etR+EOmaX9o+NkbSNDq5lMVos2lG+jYvCQx8hjudQMyhSgaMtvXmvD3xI/bE1f4r+Mv2b5NU8JDXdAsrPWYvFP9n3Qh+wX3mJHB/Zf2k5n81CN5udioMlZGIrtNE/Zp8dabpv7L9nPf6az/BS3EWslZZiLhv7HOn/6HmEbx5p3fvPK+Tnr8ter+HfhH4k0j9p/xl8arm5tG0PxD4d0nSLeFHkN2s9jNPJI0iGMIEIlXaRISTnKjjIB8sX/AO074+1T9lRPHninRtFufGGh+NrXwpqsL2zz6bJcW2tR2MlxBFI+5SUIkjJY7H5HAAr134hfFP486j+07cfs9fCd9C0yzPg+08Qyarq1vPdPZvJf3NpJst4ZYhcFgkYRWkjVDuZmfhDwF9+yP8SLn4H+J/hpHqWkjVNa+I8njCGQzT+QunvrEeoCN28jcJ/KQrtCFN+Bvx81fRNv8I/EkP7WN/8AHdrm0OgXXgq18NrAHk+2C8g1Ka8Zynl+X5RjkAB8zduyNoGCQD530L9r3xl4J/Z8+Kvjb4wWenah4v8AhR4juPC7/YWNlY6ndM1sllN++ZjBHIbpPNJY7FV2xjioPh9+1D4msPi94E8B+K/ib4F+Jtr8Q3urV08JlI59Ev4LZrmNTi+umuLWURvGJHSNg+0ng7a6m6/ZC1bxT4D+O/gTxXq9tZL8T/Fb+I9HvLPdPJYNElm9o88cqRqXS4tQXjVmVkO3f8xx6T8KvD3x/tfEenr8TPCXgPTdPsI3FxqOhy3Ut5dzbMI8EEtpCtspblw00hAyq5+9QB638LL/AOK+oaJqMvxg0vTNK1VNTvI7SPS5pJ4X01XxaySNJyJWTlwOOhwpJVfjzwl8PvAPxA/be+Ptl498NaZ4kt4NE8HiOPUrOG8RPMhu9+1ZkcDdtGcdcDPSvf8A9mn4jfED4meHvFmseOn0q9trDxLqNjouoaMsq2eoaVB5flToZSd5WRpIWdDsYxkqSPmbynWvhR+0z4U/aL+IHxd+EKeELzSvHVhotmV1281CK4tm0qOVS3k2tq6OGMpwPOXgDkUAeQ+CtNi+FeuftUfs8eGJJB4H0Dw9Brmi2TO0qaWdX0+6+12sTOSUhMkQkiiGAgLY6mvkbV5/C/iH9iP4f+HLT4Kal4X1P+yNAZ/iJc6PbLZaQIBBJLq63enyT3+AqsyuYkzuzKVXdX6feA/2bfEXhrwL8UrvxX4gh8SfEz4tW8/9rap5TWtkji0e1srWCMeY6WtqrkITuc5ZjyQo8a0L4J/tpj4CaT+zZd3ngPw/4fg0CPw3eaxbT6nqmoNZC2FrJJBbS21nCsrJnG+RlBPTigD9F9Ou7W/0+1vrG4W7t7iJJIplIZZUdQVcFeCGByCOPSvnD9o/4weMvh9J4J+H3wssrO88d/EnVW0zS21HebKzit4Wuby8nSMq8iwRJkRqwLEjnjBxNO1nxd8NPjR8Lf2ePAs2nal4H0/wtLHqcMvmyazZRadEsFldSSIREsUzKsShwC7eYVzsO3a/aP8Ag340+Ilx4G+IHwq1Cx0/x38NtVfUtMGqCT7Bdw3MLW13aXDQhpI1mibiRFYqR05yAD5w8Jab8XNI/b88L6f8XNb0nxHeR/D3Vza3+l2MummSI6jablntpJrhVdWzhkkwykZVSOfT/g98UPiL43+BnjPxR4XTwx4b1zS/FWu2Ec97BLbaTFbWWoNE9zcpDIGeXygzsd6CST7zICSLPgf4SfH7V/2k9I/aC+LUvhzT7Wy8M3+grpOjzXVxJbtcXVvcRubieGITltjh8JEEwgUSZZh5jqX7IHxOm+A118O7fUdFudWi+IVz4zWyumnk0jVLJtQe8j06/Pkhwjhl8wCORQ6gfMOaAOi/Z3/aW8ReLvj5rHwL1/xx4a+JtqNAOv2Wu+Grc2sMbw3SWtzZTILm6jYqZUdHST7ud3PTx3Wf2k/2pNZ+C0H7TfhfW/DeieGtV8RR6Nb+HrjSJr2+trSXVv7KWV5xewma8D/vGt1RF25GQQa+ivAvwi+NMf7RNj+0H8Un8OaTpeneELzw8mj6I91MbMPeW91GfOlhiE4KxybiI4tnyKqPy9fD3wp0jxj4Y+GVx+1hB8MPCPj3SIbvWfF9vrN/r9zp+p3ETXVxOl3JYNb3On298sXyqVk3JhUyrKQAD7av/iP+0B8QPizrHwL+Fes6Hol18OdK0qbxR4hv9Mmu1udU1OEzRW1lYLdReVE0aGR2ed2QOqDJG5vP/En7X3xM8Pfs1fETxpfaJpsfxM+FniWx8N6raIZDpt3LNqFnD9otyzeYkVzbXW6PeS0b/eDBeeq0vwr8YpvGA/ao/Z4tNJmg+MPh/RLnW/D3iee4sZIbi3tv9DuIrm2huMPHDL5U0TR4O3IfOAMLX/2P/iDrX7N/j7wLd67pl98SPiZ4ksvE2r3rCa30yOeDULO4a2t8JLMIYbe1EURZSzt8zbQx2gHVwfE79pD4d/H/AOHHgn4v3nhzVfDvxT/tS2ig0e0ubebR73T7VrxE8+aZ/tUbopQuY4iW+YIg4PKftQ/Hj4vfBZ/EniWD4leANETRIJL7SfCl7bzT6trNtBH5jpJL9rieKR8MqeVbugO3c+CcfRHxZ+EfiTx38Yfg38QdIubSHTvh5qWqXmoRzvIs0sd9p8lpGIFWNlZg7gsHZAFyQSeK+Yr39mX9ofT7X41+CvC03hGbTvi7e6zdt4l1BrttZhtdWiMS2ElvHDtdYF/dwyfaAka/MIWI2EA3vHviy18e/tFfsc+OrGJoLbxFbeJ9SijY5ZEvPD6zKpPcgNg10P8AwURe/i/Z0STS4o571PE3hkwRyuY43lGq2+xXcKxVS2ASFJA5welXZ/2e/iJpt3+zJrmjzaZd3vwYt/7N1iGWeWKKe2vNLTTbqa0cQsXeLaZI0kWMSDglCePSf2qvg/4t+OHwoXwN4J1Cy0vVV1jR9QS41ASPbxpp97FcuSkQ3O21DtTKBjgF0B3AA8tuPib+0T8Hvi18PtD+NOo+HvEfhb4m38ujo+j2FxYTaPqnkPPbxq01xP8AaoJRGyF2Ebg/NgD5T5n+0v8AtKfFv4BTa94svfiJ4BzoU/2i18ECGZ9Y1DSxKAWe5N2JIrgwneNtqYg3BZgMn1q3+Enx9+Kfxd8FeNPjy3h3SfDvw1uJ9Q0/T9AnuruTUtUlha3jubh7mGHyIoUdmjiXe28ncxABPg2o/sl/tJJ8Ivid8CNBufBwsfHVzq95J4pumvG1fUP7QmeeKG8gSDasi5ERn86UJHjZC5GKAPeviN8Z/jJefHvwl8HPg6miw2ni3wnda6b/AFiKeX7EYbqBBMIYZEM/7uTaId8fzsHMm1Cjcf4X/an8b/DvwD8dZfj3HYazr/wNnjElzpET2cGqQX1pHdWI8qV5PKkdpBG+GKjIwDgk+wWHwR8V2v7Qngj4sSXdkdI8NeCrnw3cxCST7S95NcW8qvGvl7DEFhYEl1bJHy9SOD1/9k3UfHN7+0jpvi3UraDRPjWNLGnyWxeW5s2sNOjtvMnjdETK3EYdVR2DKOWUnAAPm79oaH9qOfwP8KfEnxn1bw3NYaz488Izz6Rpthc21xpMz30bRxJdvcSrchQSsu6KP5uVYgYP6O/HD4raT8D/AISeKvizrcDXdp4YsZLswIwVp5BhYogx4XzJGVc4OM5welfHXiz4L/tjfFrw54G8IfEe98G2Nr4I8QaFq9xeWU9/NcawNJuY3dtr26Jalowzbf3u+XC7o0+avsr40/CzRfjb8KfFHwn8QTPbWPiexltGmjAZ4XbmOVVPBMbhXAPBxigD5K1n4s/tSfBPS/CfxV+N8/hzVfCXiHUdPsNa0rS7G4tLrw//AGrIsMEsd3LcyrdrDK6pOGijJzlOK5r4c6X8X5v+ChfxWun8RaQdMtNI0E3MJ0yYzPpcr3jWttDKbrEU0ZyZZSrLIeRGldXqfwV/ag+L+n+E/hn8db/wxB4M8NahYahqd/pEt3NqHiA6VIsttG9vPDHFaJLIivcYlkO4YjwpzXuvhD4R+JNA/aV+IfxkvLm0fRfFuj6Jp9pDG8hukl003BlMqmMIFbzV2FXYnByBxkA+f/8AgoBcWdonwJub/QJvFNtH8RtOaTS7eGK4mvFFje5iSKdkict2DsAe5rzv4c3XhjUP22vDGt6T8O7r4Gxjw/qdklpqOnRadceLLiQpKY0FiZrORbCOIyndN53zAhCgJX6k/al+E3xM+J0Pw31b4VNpB1jwH4rtPEJh1q4uLW2njtra4h8sSW1vcOGLTL/BjAPOcA8tZfCD9oL4m/FXwX49+PmoeGtJ0X4e3c+paZpPho3l1Jc309u9sr3d5eJBhIkkYqkcI3E/MSAKAOS+Fnxy+PHxE1r4g+LdbuNE8N/Dn4VeKPEemX0gtJbnUNVtdHmlwsX79UthFAELyEOZJN21UUc8i3x6/att/glH+1/cQeHm8EPaprbeD1tJ/wC0l8PvhxONUNx5Zu/s5ExQ24jx8uc19HfBL4Ean4H8JfFHwj48ltL+z+IHizxLrKpaSSMBp2uSkpFKZI4ysoQkOF3KD0ZutfPDfs5ftSz/AAaT9kq413w3/wAK5S3XRz4mV7r+3W0BCFFt9g8n7MLkwgQGXzymz5tu+gD1nx18bPiP46+K3h/4Ifs9XGm6bc6h4dj8Vap4g1a2kvYbLTbqXybSO3tI5YfNuJnDMd8iqiLn5iRjs/FmpfHvwj8KLS617xn4K0TXbW6kXVfEGpWtzb6VDp4L+XNHbPdJ/pDAR7ke5WMEsQzBQG5Dx98DPiR4T+K3h/42/s7PpL6hp3h9PC2paHrcs1taXul28vn2rw3UEczw3EDllBaN1dGwduOeb+KPwb/aK+JSfDPx7r0PhDVfEvgPV9Q1Cfw3LJeLoV3FdwGC2JuZIZZGurQEvHK1sF3sSEXAyAR/s1ftG+N/itdfFn4fR634f8e6/wDD+Ozm0jXdKR7PS9XTU7aWS3EqCW48sxzQtHK0bsu0grnGT9e/Dm68eX3gXQ7v4oWVnpviyW1jbU7bT5GltIrkj51iZyWKj3Jx03N1Pyv8Pvhp8Xfhp46+M/x++KOveGLK68YaLpRtzbtcpp2lyaNDdqftTzhGe3QSRs0wZC/zny4gFB9y/Z08X+PvH3wS8JeMvifZ29j4l1a0866jtI5YYGBkYRSpHN+8QSxBJNrAEbsYHSgD40/Zb+Bfwb+Lfgr4w23xJ8F6T4ha7+JHjCB57u0ie5VPtpA8u42+dGVydrI6sp5Ug189+OvEus3X/BO34heEvF99ceIY/ht4+j8LRXlwDcXF7YaT4itEhZzgmVvKYR8DLbQME9fqjwF8G/2yvhhH488LeAr/AMD6fpPi3xTrmvW2rXUuo3t/aR6tcGRP9CFtDA0sa4ODMybuDuXr03jT9j64h/ZLH7Onw41SK41RtS0/U7rVNXd0N9dxapDqN7cTtEkreZLsfYNpA+VSwA3UAfPOpah8Kfjd8ffh3pP7MPw3uPDHi/4f+IdM1rxHrcmiDw41poDJMs1pMJkguZvtinYkflMpGWyFBNfrjXzJ8Tfgx4qv/jb4E+PPwsurKx13R9+j+IILx5IodU8O3J3vFuijlJntpgJbcEBSxYMwFXPhH8RviB4x+L/xa8M62+lX/hHwreafb6Ne6asu8TSRSG9srp3Jjee2ZYy/l8IZNrfMCqAHC+Ofih8ZvHXx51L4C/Am80nw7H4Q0uz1TxBrmrWcmpFJdRZ/sdlbWkc9uCzpGzySPJwvCjI5+avgv8XdY+Anw3/ao+LHxGtbbVNa8O+OLlp7fTPMitru+ltLC3gSLztzxpNNIgwxcoGPL4yfpXxt8JPjZ4Q+Peq/HT4CNoWpr4y0uz0zX9I164ubJDNpzOLS8t7i2huDlUlZHjZMFRkEkjbwvhv9j/xj4g+E3xw+HXxl17T5b34u65JrUd7o0cqx2czQWphIhnw3+j3VuCq+Y29FUllZiFALmq/FD9p34G3vgjxb8dbzw5r3hbxlq9jouqWmj2M9ncaBdamfLtpI7iW5mW7t0lwkxZI35DICMitCf4q/tE+Pv2kfiN8DvhvJomhaH4GXQ7ubWb+1lvJki1G1842qW6TxB5JnVyJSwWJEIKOzKRVv/g/+038ZdQ8E+G/j9deGLHwp4M1Wy1q9m0Oa7nvNfvNMO+1VobiCKO0gMmJJlDysSoRSFO6vaPhz8I/EnhD9oT4w/FjUrm0l0j4gxeHI7CKF5DcxHR7WaCfz1aNUUM0gMex3yM7tp4IB4B8Kfjr8Yfip8X/EWgN4y8LeF7jw54hudOl8D6hp0/8AbT6Ray7ftq3Ju42eSaL97G0du8GCoLdTXXftKfE74qfDrUb270z4oeA/AGmx2om0mx1y3lutS1a5RCXicfa7cRxlxtUwxzPjk88Dkvit8Cv2ifjdrPh/RvHGneCrK38Pa7aapbeL9OkvF1u3tLK6E6xWtpJAwhmkVRHIftjx4JbBOANu4+B3x68MfHD4kePPAZ8LajZ/Ek2gTWNZN0dU0SG3tUtmt4reKIrcw5BkjjFxANxO85OSAeW+Lfir/wALw0j9jr4staCwl8TeKobqa3UlkinOmXizIhPJUSKwUnkjGea+tP2svi14g+BX7Pni74seFoILnU/D8dpJFFcKXicS3cMLgqrKclHbHIwcH2r510P9lP4peEvgt+z14S0270jUPEnwb1yHUL1HuJ4rO7tmS5hmEEvkF/MVZ1ZQ8agkEEjgn6L/AGsvhH4k+O/7PfjD4T+Ebm0s9X8QRW0cEt88kdsphuoZ28xoo5XAKxkDCHnHbkAHI3upftQaN8PtQ8W+LvFPgrwrfX1xbzsNRtrj+z/D2nlH86OSf7VGL24VjGu9mt4id7AY2qfO/wBnP9prxH498Q/FL4e6z4n0Hx7P4GsbTU9N8RaDA1vZ6hbXcUpKSQ+fOqyQyxbSY5WVlYYwQc+n/tP/AAa8c/FKb4f+IvA66Tqtx4E1ptWk0PXmkTTNTBt5IUEjxRzFJYGfzIWMTKH5I4ri/Bvwd+MGg/Ez4n/G34p3mhMfFnha006Kx0Y3JWyfT/tJMe+4RfOTZIpE2IyzFh5SKoLAHiWl/tAfth3f7MWl/tdTy+E5NGstKj1a98NQ2V3595YRY+1TLftcAQTFFeSOIQOijCs8hGTseMNX+IXi39ubw3qnwSn0yzutf+E/ni/1iKWeG0sZ9Yjl85bWB4zPKTsVYzNGo3FyxC7G8q+A/wANv2mvjP8AsVeAvg3b3fhnR/h54n0a3gvNajkuzrUelO+ZraOyaNrczuuY/OM+zYS3lq2BX3jpnwP1bRv2n9M+L2ly2kPhTS/AR8JRWu+T7Ys66hFdRsF8vy/JEUe3d5m7dgbcc0AeGeHf2s/Gnw/+DPxo8R/HG2sNV8T/AAX1VtKkk0sNZ2mqG5jt2099szP5Ble4RJOWCD5gD0rH8FftU+KtI+Jfw/8ADvi/4m+AviNafES9bTLiw8KOgutCvnt3mgKsL65a6tWaMxPI8cRDMrDrsPf6l+yPf+M9I/aF8K+M9Tt7bTPjBqtvqGmz2ZeWeyNra26wyTRyJGu+O5t1k2K7BlGNwJ47T4V+Gf2hrDxBpNr8SvCvgKDTtNVhd6vpElyb69dYyI5ILV7SJLZi+1pMzSADKoOQwAPZvh1f/Fe9ufFS/FDTNM02C31m5i0I6dNJK1xo6hfs81yH+7Ox3bwuAOgHGW+QbHwH4H8e/t6fFDT/ABz4d07xFaxeC/DxSLUrSG8jUtcXYbCzKwGQOcda96+A3xF+IHj/AMSfFC38UvpV/wCH/DviOXTtB1DSllEdxaJEjvFI0hKyTWzt5Uzx/J5yyIOUIHmHi34UftI6F+0n4m+NPweTwnead4m0LTNIePXry/hmgewkmkMgitbWRXU+bwDKp47UAcP8HfCsvw4+Pnxt/Zl+HN9LovhG90Cw1/RYI2Z4tBvdTE9tcLbAn93G0iLOkQIVTnaACa8S+D8PwU+B/h7wb8CP2tPg3a+Dte0q7torLxbJYxXujavqcUoeG7GrxL5kNxO6h2S42YB2sQvyj7W+FHwB8b+C7b4geOvE/i+G/wDiz8R0T7VrUFl/oOmi2haKxt7Ozlc7oLXcWxI+ZWJL9a8X+Jvwb/bF+P3gV/gX8WrvwVYeEdUktV1jXNKa+k1G6tbaeOfFtZTwiGCeQxAMxmdU3ZQZAFAH6K18pfHn4tfEfS/iH4H+A/wXTT4PF/jaO+vp9T1SN7i00rS9OCedObeN42mlkeRY4U3qu7JY4rd0n4h+P7v9qXV/hdbPpd/4G0zwzDfTNbiZtR07VXuEWKC7kJMWLmBnkjT7+2PccAgvg/Hj4QfErWfiR4H+O3wUutMHi7wZFfWE+n6y8sNjqemaiEMsTTwJJJDLG8avEwRl3fe4oA+b/h54g8ZfDT9rL4z+Kvjfqen6tN4W+HumX9xfaRayWcc9jaTXkxka2mll8qXarBlErIcAgjJVdO9+PH7VfhX4MWH7XHimLw9P4IuYLTVrzwjbWk41K00K8ZCk0eptcbJbuOGRZJI2t0jOGUEHFej+Dv2d/il4j+KnxH+IHx4udDk034keErfw3Np2iPcsbSNXuFli824iXzgYpQ3nYQl2ZfKVUDNwF3+zp+1J4l+EFh+yl4s1vw2Ph1axWul3XiS1e7/ty80OzZfLtlsWi8iG5kijWKSY3DqFJYKW4oA9E+JXxm+N2pftGaR8BfgoNGitNf8ACA8RHWNThluF09FvTA84hjliNxuUxpHFuTDvvZiqlTxOvfHD43z/AB71n4IweOfCngTUfDtjpTaXHr+lzufF9zdW6vczWz/bIFihSbMQjgM0qsCWzjFfQkHwZ1y0/aqsvjZZzWcXhmz8Cv4WS1Dyfa1uTqUd2jBPL8vyREm3Pmbt2BtxzXlv7Sfwo/aB+M2jeKfhivh/wLr/AIT8QRtDpmo6rJeQ6hovmxBGn+ziC4SaeJ8vDJHND820FQASQDvf2gvGXxE8GR6fd2PxK8GfDHQHtyJ9S8Q273NxPeg8RW0D3drEEIxkl5HycBe5+LviR8cdV+P/AOxFpvjLxClmutaf470bSL6TTt/2KefT9dgi8+3EhLiOZNsgDEkbiM8V9Ban+z78bfB/xi8P/FHwDP4f8cSaf4SsPC/meKZrm3ubCayZmkv7Z4ILncbnIMyfI7MAPM29OOl/ZC+Lcf7NPjD4ZT6zo2peMtU8cSeMbS4Xz7XT7g/2nDfiKUFJZIDII2XavmhCQN7jLUAfcHxf8XeIPAPws8W+N/CmjP4i1nQtLu7yz06MMWup4ImdIwEyxyRyFBYjhRnFfMfwC8f/ABp+L3gu78TaD8WPBHi3+0dNWSM2Gj3CPo2qyFWEFzANQZ5IlTeCsvkzbgD0yK+svEy+PLnwVd/8Ic+naf4se2U2329ZbmwjucAlZPKaKRo85XcuCOG2nG0/J3w6+B/xYvf2jrb4/wDxE0fwx4Lm0/R7zS54PDNzcXc2uS3ckbCa/lmtrUeXD5eYlKu+5uWwAKAPmL9l3x18bfhd/wAE7r74ytrOj6lYaT4c1XUtEsDp0yTQXUV3cOxu5/tRWdGbJCpHEQCBk4yfY/if8Vfj/wCF/wBmTX/jr8QLbw3Mmqt4bl0nw41g91Fp8N/qdtE/22d59tzP5UysNiRpHKuV3AAmn4M/Zm+P+g/sreNf2RtSbw5Low0jVdN8O60l7dLPcfbp5JYBfW32UiERrIQ7RvIcgAK3LV738e/gZ4t+KX7L4+Cnh+7sbfXAmgL511JKlpnSr20uZvnSJ3+ZIGCfu+SRnaMkAFv9ojxn4/8ABrWV3pXxI8HfC/w5JCwl1HxJA91dTXgb5YbeFrq0h2bcZO93JOAnc/Afxp+OHxP+P/7D9l4ws9Q0bTru08XWOh675FpPcW17c2Ot2qWtxZM06NFCzrHM6OHLKSgK/er7M+Ivwc+MsP7SMfx4+G1t4a8RpPoEOiR2/iOa5t5NHkinkma5s3gt7jcsocCVP3bMVA3henmDfsefFOf9mHx58JL/AMRaRN4z17xhc+LNPv44549Pac6jFqESzxFWkiDmMq6qZfL3DDSY5APXfFnxL+Mnwt8TfBPwN4t1LSNe1Hx34gvdN1a7tLCWziNtFaT3MX2eJ7mYxuCihmZ3B5wBnjsoviz4nf8Aa21D4HmO2/4R618DW3iNH8tvtP22bUp7RgX3bfL8uNcLtzuyd2OK8y+LXwv/AGifij4U+Hfj23tPDeifFD4d6/8A2vDp7Xt1daPdW7QyW0sL3It45keSOTcCImCkYyc7hr/DP4N/GyH9ozVP2gPizqOhs2reE4vD6adpBuGWxaG+a5RFknjQzoVZmaZvLbe2wRBFDEA+U7L9q/8Aa1T9lrw/+2NqcHhVvDNuLRtQ8Pw2919tv7aW9WxmnjvDNstpN7Zji8qRVX5ndz8g+kLL4o/tG+Av2g/h14D+L8/h3UNA+KserRW9vpFtcRTaNe6Zam8ERuJpW+1xvGrIZDFES/zBEX5TyR/ZJ+I5/wCCfFt+yf8A2lpP/CXQ2kFubrzp/wCzd8WprekiTyPOx5a4H7n73GMc19D/ABT+EfiTxx8bfgz8SdJubSLTPh3e61c6hHO8izypqOnPZxC3VY2ViJGBfeyALyCTxQB33gK/+K95rXi+L4j6Zpmn6Xbam0fh+SwmklludN2KVkulfhJd+QQuB2xgBm/P/wCO2qeH9J/b7sLjxH8MtQ+Kdu/wyCrp2nafZ6lLA/8AbTkXDRXs0MaqACm5WLZcDGCSPrz4KfEX4geOPiF8V9H8RPpV/wCGPDGsxWOh3+lrKA48rddW07yEo89q+1JTH8okLIcFSq+c/FH4U/tCwftNWvx9+C0fha+t/wDhEB4ZntfEF7fWjhzqDXplT7JaXAIwEUZYHO7jgEgHSax8UfB/wa/Zf8V/GnRvAcnw5g0yyvb4aLe2Nrp9wb5M29uJobR5It1xIsSqQ5JVlzjoPlP9j4eGfgd8bNM+Eem+I7HXB8S/Btlq97Ja3cV1nxXpBK6pnymbDXEU6yZPXyj6c/QHjv4SftC/Hnw/4T8I/Ge28Jafolj4pstV1u10m8vryK/0rTl8+K0KXVnEHMt2E8xW2qI1yCSdta/xV/ZV0CfV/AHjj4CeHvDvg7xX4J8R2mpvNFaJp63emlJIL+zeS0gZz50ErbcqRuAyQCTQB9jV8BfFr9pzUR8bte+C3hf4jeD/AIWQ+DbOxuNS1TxS0c815eaghnhtbO0e8sx5ccO155i7EGRVVR94+3fBT4i/EDxx8Qvivo/iJ9Kv/DHhjWYrHQ7/AEtZQHHlbrq2neQlHntX2pKY/lEhZDgqVXzrx58DfiV4d+NmvfGv4Pad4c8Tx+NrOxt9c0XxI8tsBc6ajQ293Z3cVvdFCYWEcsTRYYKGDZwAAee6T+11481L4I+Hf2gI7TS7/QPC+vXujePY9N33UYtLac2rarpc6SNugibZcshEm6B2w3yb28c/aq+Jfxg+Ln7EXxa+Kmmz6do3gTU7hY9Ft5rKV76+8PJNHbfajL56CN7yY+dFmIhbcKNpL7l+u/GXwY+KHxV8M+BvAPjmPw7o/hb+0H1HxlY6M1x5d/HayebZafbrJCoe3kcRm7eQxs4TaqbXIXzzVf2SfHV9+zz8Vv2XrfXLJfBurlm8FzySTPc6dBK63X2C7Qx4+z29wuyF0kkfyWwVBRQQC/8AFr45/FD4E6N4K8A+OfGHhNfHHxA1O8it9ev7d9I0LTNNsYEluJ54Zrx2lmUsqRIJ0EjyKCVCndz3gn9qzxHJafGDwdP4t8NfETWfAHhWTxNpOv8Ah8ILG8i8mcGG7t4rq5EU0E8I3Ks3zxupwnfqvGvwa+PnxW0XwP8AEXxPa+F9G+Kvw41G6ksbbz7jUdC1KxvbZLe7hume3ilgM5G9CkcvlFEPzEnb3tr4C+L/AIx+GHxE8L+NfDnhLwlqPibRbrTNMj0W4uLlVkubeaJnvLh7W3Oze6FVjiYqAxJYkAAHzrL+0F+1P4f+Hvwq/aH8Wv4YHgvxndeH7fUNCtLW6a9htNeMUaXa3zzBTMGlDmEQhEB2b5CN59n8V/E/42/Ef48+JPgl8Cr7R/DNl8PrHT7rX9Z1exl1N3vNURprWztrWO4tgF8lN8srSZG4BVHBaLxr+zj438Sfsr/Db4HWN9pya74O/wCER+1zySzC0k/sB7ZrnynEJkO8Qt5W6Nc5G7ZziXxP8JPjp4E+Pfif40fAVtA1a0+Idjp1trul6/cXNn5V5pSNBa3ltPbQXG5fJbZJEyAnaCGOQFAPl34NfGzWP2f/AIGftCfETxhb6dd+J4/ivq9iIY52tdMl1W9Fhbqxmn+aK1WRjI7OcrErckjNeyfDH9pzxDD8bPBnws8T/EvwT8Urf4gQ6gEl8KbIZdHv7C3+1+VKi312ZraaJZFjkYRvvUAj5sVHoX7G3jnVfgn8UPh58R/EunDxP418aT+M9O1XS7eRre0vT9kmgZra452rPbsrR73zEfv7ice6fCjQvjxb+I7WX4m+D/AuiWNlA6yXmgT3U93c3JACvDHNaQC3iILblaSVucA45IB8m+Jvif8AtHftBfs7/E340+D77w7ofwzudL8RW2l6ReWNxcahqWlWkc9tNdy3sdwi280hjkaGNYJEA2hiQdx+2P2Wv+TZPhF/2J+gf+m+CvlDSP2cv2n/AAH8KPFf7MXgDUfCsvw71RNXt9I1jUHvP7UsNP1ZpZHtJLOKLypZEaZ1Sbz1AXDGM4EdfbXwa8Gap8PPg34G+HmtTRSal4Z0HTNKuZbZmaFp7K1jgkaIuqsULISpZASMZUdKAPg745/tSfFT4B6w3iPxT8RfAWprp+qW0N34I0+CZtX/ALKubpIPPF092ZBcRxuJCHtUi4YAtgZ9v+JvxU+Oc/7TFn+z38JW0Owg1Dwg3iCXU9WgnuWsmj1D7I7pBDJF9oJDIqxl4wCS5chfLb5ln/Y6/aYk/Zu1j9mG0ufBdrp9zdC6n8RF75tS1x0v1vke8jEGIJiVAkmMlyTt2qgB3r9vyfCPxI37WkHx4Fzaf2BF4Il8NGDfJ9s+2Pqcd4HCeX5fleWpBPmbt3G3HNAHyj4Z+M37Z3j7wb8RrfRp/CGj678H9S1HTNQvZLS7uYtdubFFuEWG386P7HG1u6b3Z5mZ2IVYwMn1fwf+034r17W/gL4m1WxtbPwT8adCdGwj+bYeIxbreww+eW2tDPEs8calNxdAd3OD23w0+Bvizwba/HGDVLuxlb4meIdS1bTfJklYRQXljBbRrcbol2uHjJYJvAXGGJyB4D8XPACfBH/gn9onhXxrrFla+L/hhp+lXukXNszypJ4g0iRJbOO1DrHJIbiVfIxsDFZG4HNAHY+NP2q/Gnhbwr+0J8UNN0m11Xwx8Lby20TRIsPHJdarEkSahJcy7seRBcXCIdoXCxSZbP3adl4y/au8L/tCfDv4U+MvG3hjWNL8Z2F7q9w9joE0csS6W9ubm1Q/2gdkciTYhuXVjvGDGawfiL4Rv/gr/wAE7tX8Ia7fCx8U69poh1C4KKyza94muh9pSV2ZVWOS5umieUsPLi+ccqBXP+DJfE/7KXjXwavin4MeF9Jh+IGq2HhJdU8PeILvUb63NyzGJUg1C1ST7GjLveKKULGo3bTtoAo+Jvif+0d+0F+zv8TfjT4PvvDuh/DO50vxFbaXpF5Y3FxqGpaVaRz2013Lex3CLbzSGORoY1gkQDaGJB3H2Hwp4j8c+F/2Q/gtqnhbxj4X8A6YvhPQxqes+J0eaO3j/s2DyxbxCe1iaRjnPmTKABwGPTh9I/Zy/af8B/CjxX+zF4A1HwrL8O9UTV7fSNY1B7z+1LDT9WaWR7SSzii8qWRGmdUm89QFwxjOBHW/rX7NPxe0my/Z/wBa8MJ4d8T6p8IPDy6NeaRrU9xFpst21pbWxvrSdLaZlliaFtjPBkxscbGJFAHntv8AtA618a/2S/2nfD3ibVNK8Ral4E0XXbBda0SN4bDVLO40iSe3uUid5TG5BZXUOy5XKkg171/wkfjnwv8As1/C7VPC3jHwv4B0xdA0sanrPidHmjt4/sMXli3iE9rE0jHOfMmUADgMenNeHP2ZPiteaD+0bbfETWtGk1X44WQgt5NMW4SCzd9JbTysiTKW2xMQA4ZjIq7ysZby1x9X/Z8+Oslv8CvGFpY+GNZ1/wCFekT6VeaFql3c/wBlyTSRQ28d/aXS2kjrOiQ5G+3yFdlUg8sAaPwI/aL+K/xd+HPxV0nwpP4e8aePvh9emy0vUbXzbXRdZS4gSe0nZfMdoyQWWRVkK7l+VwDkfcugSa3NoWmzeJYYbfV3toWvIrZi8EdyUBlWJmALIHyFJAJGCa+EtM8FfGT4A+H/ANoP43+MPEnhe38Q+MhZapp8+y6XTbW4tLFbVYJ43HmsPMCxRFGZ5jhtiM/lD7L+GOseKvEPw38Ka/45sE0rxHqelWNzqdpGGVLe9mgR54lVyWASQsoBJIxgk9aAPyb/AGN9Y+H7fC/TtB1/9nnWPGd1ca/rUUniKLQtJvLFxLq1wA7XFxcpOUgUhXzHldpCggDPrf7QXiT4PeCf24vBl78YrGG58Oj4e6hBDDJpU2rItwdTgMe22t4J2XCK4DbAB0yMgHq/gr8L/wBtr4D+BF+G/hnT/h5q2mWt/qd3BcXmravFOy6heTXeHSPTioK+bt4J6da9+Hwj8a3v7Tvhv47anPp8en6f4LutAvLaGWZpv7QubyC5LQhogrQARsNzOr5x8nUgA8V/ZM8P2Mnxq+K3xE+F/hW78F/CjxBBpMWn2txp8mkQ3+qWyyi7v7WwlSJ4Y2Ro0ZzEnmsM4JU1+gVfOH7RPxG+IHgBfAFv8NH0q41XxD4o03TrjTtQWV7i802eTZeGzEJyslvG3nySOCkcSOzZOFb6PoAKKKKACiiigD//1P38ooooAKK4f4i/EjwX8KPCtz408fakul6VbNHGXKPLJJLKwSOGGKJWkllkYhUjRWZj0FcT8PP2gvh18SW1u00ptR0rVfDkCXV/pms6bd6VqMNtIGMc/wBmu445GifY210DLkEZzxQB7dUYhhWZrhUUSuqqz4G4qpJUE9SAWOB2yfWvj+z/AG8f2adQbw/PYeILu40vxFLa28Oqppd8dLgubwKYbe5vfJ8iGYlgGjZ90Z4kCYNYPxR/bI0v4Z/tLaT8Gb7S9SutJ/sa8vdRks9C1O+uvtQa2a1FqbaNllh8uV/OdEdUfajOjAqQD7ior51+JP7U3wi+F3iifwZrdxqWqa1YWyXt/a6LpV7qz6daOMrPe/Y4pRAhAyN5DEcgEc1seKf2kvgn4O+Hnh/4s674pto/B/ie4t7Ww1SMPLbySXKu8eWRSUGI2BLAbSNrYPFAHuVFfPvw2/ac+E/xR8Yz/D/QZ9R03xDHa/b4bLWdKvdImu7Ldt+02yXsMRliyeSvI6kAc1keLP2uvgj4N8San4d1bUL+aLQJ1tdY1O00u9u9I0m4fAEV9qEML28DDcN4d/kz8+2gD6Zorxb4rfH/AOGnwcbRbPxXd3N3qviRnXS9M0qzuNT1G+8pQ8jQWtqkkjIikFnwFGRzkgV8qfA79o7RNV+If7TnxM1vxFqNx4A8ISaJdwR3cd2p023h0gPfRx2MyiWBxMj+bGI1YyA5BNAH6KUV5d4h+MngDwtoXhDxJrV+8On+OtQ03S9JcQSuZ7vVhm0RlVSUD92cAL/ERXC+PP2qvgz8O/Fer+Atc1G8uvFOjQ2txJpOnadd6hfSxXaSOjww20UjSIqxkyOPkiyokZS65APouivGPDv7Qfwf8T/CBvjvp/iSCHwRFDLNPf3Ie3Fv5DmKVJY5FWRJFkBTYV3FsBQcjPNeC/2pfhX45tNavtPj1vTYNE0yXWpH1TQ9R05Z9NhGWubZrmBBMnoFJY/3aAPoyivHdT+PPwy0f4W6D8Zb/UpI/CniVdMaxuRbzM8g1ho0tMxBfMXeZUzuUbc/NjBryKz8UeJW/b31TwY2q3R8Px/DWxv108zv9kW8fWLmJrgQ52CUxqEL43FQBnAoA+gPDfwq+G3hDxRrfjfwx4Z0/TPEPiRzJqWoQW6Jd3bE7j5swG9hnnGcZ5xmsz4lfBP4U/F/+z3+JHhq11ufSWZ7O4kDR3NsXxu8m4iKSxhsDcFYA4Gegryy0/bH+Bt54p0vw0t5qkMGt366VY6tcaLqFvo1zqDsUS2i1CaBIGkdgVXDlWPAY103xK/ab+EHwo8Uv4F8W6lcnxKbG3v7fTLKxub68u4rqWaGIW0NvG7SuWt5NyqCUVdz7V5oA9D8AfC/4e/CvS5tH+Hnh+00G1upTPOLaMK9xMesk0hy8r443OzHHGcV3lef+N/ib4R+HPw6v/ip4znl03w/pdot7dSPBI0sUT7fvQqpk3DcAVxkGvJ/Df7XfwL8WfETTPhrour3T3+vecNKvJdPu4dM1N7dS0q2V9JEtvcbVUkNG5Rh9xmyMgH0xRXBeF/iX4P8ZeKvFngvw/dtcat4JuLe11WIxOghmuoFuIgrsoV8xsDlCQOh5rhYv2lvgu/wbT4+XHiFbTwTK80cd5PDNG8ksFy9o0ccBTznkaaNkRFQs/8ACCDmgD3eozDC0y3DIplRWVXwNwViCwB6gEqMjvgelfGXxJ/aF0Tx7+zf8adW+Hx17w5r/hbwnq14P7Q0y/0S9tmexuHtbmFrmKJuWiLRvGSQVzwcV22i/G/wn8NfgN8NvE/xJ1S6utS8Q6RpUVtDBBcalqmp301mkjLBbwLJPPK3LOQpxyzEDmgD6bor5/sf2nPhDe/DbxP8U5NRurHSfBRddct7ywurXUdNkjVXMdxZSxrcIxVlZRs+YHKkiu48U/FnwP4Mt/CN14hvXt4/HOp2mj6SVhkfzr29ikmhRgqnYGSNiWfCjGCckUAekVHJDDKUaVFcxNuQsAdrYIyPQ4JGfevnLxH+1b8IvDfjfUfAcz6vqV5ok0Ntql1pmi6hqNhps9wFaOK7urWCSKJyGUkEnbn5sV7x4j1yy8MeHtU8S6kSLTSbWe7mI6+XAhkfGfZTQBs0V8B6Br+s6d+zf8P/AB38RPiHqnhHWfiH4h0vWZbi0iN9iTxBcie20ZElSQRWwjkjtyVAClC5PzNu9x8eftVfBn4d+K9X8Ba5qN5deKdGhtbiTSdO0671C+liu0kdHhhtopGkRVjJkcfJFlRIyl1yAfRdc/4q8K+G/HHh6+8J+L9Ng1fRtTj8q6tLlBJDNHkHa6ngjIFfHPxy/bT8I+E/2Ur79ov4STf2+LtHh0oy6feSwLepIY3jvkjCPbKjKysZWjG7ADEsufaj+0r8KrX4by/FPW7jUdE0VLxdPjj1LSr6yvri8fb5cNvYzQrczvIWwgjjbcQcfdbAB7xDFHbxJBCoSONQqqOAABgAfSpK8F8J/tJfC7xdoPivW4JtQ0mXwPate61p+q6bd6dqVlbCJ5llezuY0lKOkbFGRWDbSAcgiuWtf2x/gRf+HvEHjLT9Uvbrw14Y05NSvtXj0y8bT1jcoDFHceVslnQyL5kUZZkOQwDKwAB9R0V8z+G/2u/gX4s+ImmfDXRdXunv9e84aVeS6fdw6Zqb26lpVsr6SJbe42qpIaNyjD7jNkZm+JX7V3wi+Fet6noHiNtYup9ARJdWl03RNR1C10yGSMSrJd3NvA8MS+WwcgvuCkEjBFAH0lRXxd+0B8QNQfxl+zjqPgXxBOmieLvFkQlexuHSDUbCbTbmeNZNhAlibCuA2VJAOOlfTPxH+I/gz4S+DtQ8e+P9RGl6LpoTzZijysXlcRxxxxxqzySSOyqiIpZiQAKAO4orxr4f/HbwN8Q9O1rUbWHVPD48ORJcahH4g0u80aS3t5Fd0mb7bFEpjKxudykgBTuxXKeBP2rvg58RPE+meFtButSt5fEAlbRrrUNJvtPsdYWBTJIdPurmGOK42oC+EbJUFlBUZoA+jIYYbaFLe3RYoolCoigKqqBgAAcAAdBUlfJ3j/8AbT+BXw41XXdO1q61a9t/C1wtnrGoabo19f6dp12+NttcXdvC8SzfMoKBiVJCthuK6LxB+1f8F/CUPh5vFN/f6Rd+KrK4vtMsbnSr6PULpLaaOB4kszD9oM5eRdsPl+Yy5cLtBIAPo+ivjsft3fs5y+G5vEFnqupXlxZyzw3mlW+j38ur2JtApna7skhM1vHGHBaWVVjPRWLcV7LqPx6+Eul/Cmy+Nl34ih/4Q7UoYJ7S8RJJGuftRAhjhgRTM80jEKsSoZN3y7cggAHrYhhWZrhUUSuqqzgDcVUkqCepALHA7ZPrUleI/Dz9oL4dfEltbtNKbUdK1Xw5Al1f6ZrOm3elajDbSBjHP9mu445GifY210DLkEZzxXlNn+3j+zTqDeH57DxBd3Gl+IpbW3h1VNLvjpcFzeBTDb3N75PkQzEsA0bPujPEgTBoA+xKK+Hfij+2Rpfwz/aW0n4M32l6ldaT/Y15e6jJZ6Fqd9dfag1s1qLU20bLLD5cr+c6I6o+1GdGBU/ZXiDxBofhTQr/AMT+Jb6HTNJ0qCS5urq4cRxQwxKWd3Y8BVAJJoA2K+cV/ZE/ZoXWZtcHw70rzri4+1yQeUTZNcBt3mmz3fZt+7nPl5zz1qD4eftafBj4meKNO8IaHeajp+o67DJcaP8A2tpN9pcWrwRLveSwlu4YkuAq/MQh3bfmxtBNfLPw6+KGmfGT9ufx5o2s6v4wt7bwTJpdn4d0+3j1bT9IV0s5bm/l1GNUjgJlk+WH7ThZkVTHvDKaAP06AAAAGAKWvjP9pD9q62+BHxM+GngT+zry9TxTeyHU2g0i/v2XT1trkr9ka1Qq9z58Sboh5jrFl2QKVavN/Fv7R2k+DP2vvD9/4h1vV7Xwhr/wxF/Y6IttfST3WpXOqxmLZpMaNMbwQBww8nzI0Dh9qq2AD9FKK8X+G37QXwp+KvgDVPiX4Y1jyND0GW6g1R9QiksJdOmslD3CXcdwqNEY0IZtwxtOc4rnfh7+1N8IviX4m0/wpoM+p2d5rkMtxpEmqaTfabb6vBAu+STT5ruGNLgKh3kId2z5gNuTQB9FUV578Tvin4H+D3hdvF/j7UDY2BnhtIVjikuLi5urhtsNvbwQq8s00jcKiKSeT0BI8R8P/tX/AA4+Jl/4j8AeFbXxFp/ifSLGeW7gv9C1HTzYMYHli+0TTQrHC0iqWi3OC+PlzQB9YUV8Sfs6fG3RvB/7E3w8+Lnxr8TSlJNHtnu9QvpJbu6ubiZyqKPvzTzSNgKqhnY9BXtPwt/aI+Gnxc1rUPCvh2W/03xFpcCXVxpGs6ddaTqC2sjbUuFt7uON3hZuN6AgEgHBIFAHuVFflp+zT+2j4C8I/B1x8XfEGuazqOm65rseqal9g1HVrfS4P7UuFthfXscUscCiLZsR3yse07QhBr9A9R+LfgHTfFngvwXNqYl1P4gw3txoghRpYbqKwhS4mcTIDGqiKRWUsw3Z+XNAHpFFebR/FzwA/jDxV4Gk1QQar4KsLbU9WEyPHDbWl2sjxytMwEZG2Jy2GO0DnFcJ8L/2m/hj8XtYt9H8Hxa2q6jbyXmnXd9oeo2NjqNrHt3TWt1cQJFInzKR8wJBBAINAH0JRXkvxY+OXww+B9ro998T9ZGi22vXTWVpI0UsokuFhefy8RI53MsZCDGXbCKCzAHB+F37SHwv+LniLVPB3hybUNP8RaRbpeT6XrGm3ek3xs5G2JcxwXkUTvCzYG9QcEgNgkZAPeKK+W9F/bG+BuveKtG8MWl5qkEfiS7FhpWqXejahaaRf3rZ229vfTwJBI74OzDYfopORnB+HnijxLfftkfGfwte6tdz6LpOgeF5rOykndra2luFu/OeKInYjSbV3lQC2BnoKAPsKivzK+AvxG1XVP2PP2e/E3j34laxomseIvEGnWb34Q6ldaxcT3tzHHp91JOsrLFcBQrSkjaFHzY4P1x8Tf2lfhZ8KvFNn4F16bUdU8SXdsb7+zNF0y81e8hsg2w3U8VlFK0UIbI3NjODtBwaAPfaK/O39mf9o3QLH4QfEn4t/EXxRean4bX4ka3YadeT/aLspZXd/FBp8MSENIkI81AibQqA9FANfaviL4l+D/Cvjbwl8Pdbu2h1zxw98mlQiJ3WZtOg+03G51BVNsXI3kZ6DJoA7maGG4he3uEWWKVSrowDKysMEEHggjqKkr5uf9q34Rf8J3d+ArJ9X1GfTtSXRbzULLRdQu9KtdUd1jFnNfQwPAkwdlVgWwrHDEHiqngTXNb8MftJ+P8A4V6rfTahpuvafZeL9IM8rStaCRjp99aKXJIiWWGKaNR8q+cwGBgUAfTlFcF8PPif4C+K+k3uu/DzWItasNOvrnTLiWJXUR3lo22aIh1UkqSOQCpBBUkEGvmPxb8QfjP8YPjj4m+CPwP1+28C6N8PbeyfxD4hlsY9SvJL/UYjPBZWVvORAAsO2SWVw+GIUKMHcAfbNRxQwwKUgRY1LMxCgAFnJZjx3JJJPcnNfK2pH48fCH4E/FHXfHPjiz8ZaroGjanqOh6omlx2FyptrGSVftcMbNbOyzKNuxFUqPmBzgfMmr/Ef9qX4Vfsz6H+1hq3xKsfGWnro+ka3qfh6/0O2sllt9RWAzQ217aOjpKnnYiLxuGYAFecUAfqRRUcMomhSZQVEihgGGGGRnBB6H2r5q+Jn7W3wb+FWv6v4b1+fVNQvPDdtHea0dJ0m91OHSLeVd8cl9LbROkAdfmAZt235iAvNAH0zRXwL8Ufja/jXXP2dPE/w31HWNK0LxT40NrcRzwXmlNfWg0+5kCy286xPJAzBXTcpRsBhng17h8Sf2pvhF8LfFE/gvW7jUtU1qwtkvb+10XSr3Vn060flZ737HFKIEIGRvIYjkAjmgD6Kor4I/av+OWjab8M/g38WfBHjI2XhLVvH3hp7vU7G6eK3uNHkeVrlJmjILQsiHzY2GDjay5GK92+G/7Tnwo+KPjCfwBoU+o6b4hjtft8NlrOlXukT3dlu2/abZL2GIyxZIyV5HUgCgD6BoryfQ/jb8OfEXgvxT8QNJ1B5dE8GXOqWmqTGCVWhm0bcLxQjKGfZtOCoIb+HNct4o/ab+E/hXwp4L8Vz3N9qa/ES3ju/D9hpmnXV/qWowvAlyXis4I2mwkTq0hZQEyAxBIBAPoGobi3hu7eS1uUEkMysjowyGVhggj0Irk/APjzw/8AEnwxbeLvDP2oWN00sYW9s7ixuEkgdopUkguY45UZHVlIKjkcZrwfxL+2b8CvDWs+IfDbXuqavq/hO7ktNWs9K0e/1Gax8lEd55xbwOI4AHGJWIViHCFijAAH0Z4X8L+HfBXh+w8KeEtOh0nR9MiENraW6COGGNeioo4AHpW9Xh+s/tGfCPRvh7oHxQTV5dW0HxUUTSH0uyutRnv5JEaRY4be1ikmL7UcspQFNrbtuDjzzXf2gfCPxR+AXxZ174a6hf6brfhLRdWju7a7trnS9V0u9SxkmiMkE6xzRPjDxuBg4ypyDgA+s6K/Ni48e+OV8AfsW3q+IdQFx4rutDXWJPtUu/UVm0GSaQXbbszh5QHbzN2W+Y8819TeAvFOjv8AFj4vWM3ju71g+HptKN5pV5AlvZ6AstiJwIJ9ieas8ZEsjMzBCMcHdkA99hhht4xFAixoM4VQABk5PA96kr8tf2k/2zfA/i/4baZb/B3Xtd0+6v8AxPoVvZavFp+padY6nCNShS7is9QaKOGdTHuDqr/Om4gMuTX6G23xL8H3fxLvvhFBds3ijTtLh1ia38pwq2VxK8Eb+bt2El0YbQ24YyRigDvaK8Kvf2kvg7pvhHxp451HXDbaN4A1ebQdWle3m3R6lAYlNvFGELzMzTRqnlK29mwuea3Phf8AGXwr8WRqMWg2OsaVeaT5JubTWtIvdJuES4DGJwl3FHvVwjYKFunOKAPVUhhiaR40VGmbc5AALMAFy3qcADJ7ACpK+ZPitrmt6/8AHP4XfB7Q76bT7V/t3izWJLeVopJbLRWhht7UlCCY57y7iaQHhkiZDkMRWj8Qv2pvhF8NfEt/4T1q41LUdQ0SGO51f+yNJvtUi0i3lXekt/JaQypbqyDeA5DbPmxt5oA+iqK8ttfjR8OL3xn4e8B2urLLqfi3SX1rR3VHNrqFlHtLtb3IHlSOqurmMNv8sh8bea+VPjJ+3Z4M8HWngfUfh/Fd67Y+IfGCaDdXSaNqN1byWdvJLFfNYywoEmmEkZWEIZPM2uURwpIAPv2ivn/xL+0x8MPCug+G9Y1EatNe+L0kk0rRrbSL6fW7pIRmVhpqQm5RYxgu0iKq5GSCRmyv7SvwcPwrn+M764YfC1neRaddzS208U9neS3Udl5FzbSIs0MiTyorrIg2A7j8vNAHu9FfMbftf/Am38Mf8JzqesXOl+F5NVg0e31a+sLq0sbqe4jeRJLeaaNBJbkRt/pC/uuOGIIJ2vhf+078Jfi54y1H4f8AhW61C28QadaLqH2TVNMvNLluLFmCC6gS8iiaSLcyjcB3HGCDQB9BUV8tav8Atj/A3Q/Eh0DUrzVIbWPUf7Im1htF1BdFg1DzfIFvJqLQC2Deb8hO8qGyCwwaxdf8UeJYf26fCHg+HVbpNBuvAmrXkunrO4tJLmLULZEmaEHYZFVmVXIyASAcGgD6/oryv4r/ABm8CfBnS9O1HxnPctPrN0LHTrGwtJ7+/vroqX8q3trZHkchVLEgYUDkjIzlWf7QPwvm+Gep/FzVb+58P+HdEd4b99YsLrTbi2mjKqYntrmJJjIWdVRVRi7MFTcTigD2mivDvhn+0L8Ofinrl54W0X+09J12ytVv207W9LvNIu5LF32LdRRXkURkhLYUumQpIDYJAPk19+3h+z/Z3Fr5Emv6jp2p3r6dp2pWPh7U7vT9SvIyweCyuIbdluHBVv8AV7g2CVJANAH2VRXzT4y/a2+CngbXtR8I6xf38/iTTbWzvZNIstLvbzUHt72N5UkS3hidyqJGTM2AsRKiQqWUHkL/APb1/ZgstD03xRB4nn1HRb9YZJr6y029ubXTUncxp/aMscJWzYsCDHMVkHXZt5oA+xaK8o+Jvxq+H/wm07S73xTdzz3GvSmDS7DTbWbUb/UZQu8ra2tqkksu1fmZgu1RgsRkVysH7T/wbl+F+t/F6fVprLQPDNytlq63dlc217p10ZI4vJurOWNZ4nDSoSGT7rBuV5oA99ihhgUpAixqWZiFAA3OSzHjuSSSe5Oakr5q8H/tb/A/xz8RNP8AhloGq3Z1TWop5tKnuNOvLWw1VLZS0xsLuaJIbkKoLbo2ZWUbkLDmvMvhl+2Rpfjn9ojx98HLnS9SSy0S70uy0mZNC1NG824tmlujfyvGY4E8wDyGkWJXT5lLqQ1AH3FRXCfEj4l+B/hH4Su/HPxD1WPSNHsyiNK6vI7ySNtjiiijDSSyOxwqIrMx6CvO/h/+0v8AC34j3OtaTo8mp6drnh+zOo3ej6ppV7p+qCy523EVncRJLNGxGFMSt8xC/eIBAPe4oYYFKQIsalmYhQANzksx47kkknuTmpK/NL9iP4gWfxq+IPxE+Jms6x4sute/tvWbe1sr7+07bQbPR4rlYbSCK3mVLP7SioDIuPOVmfcqnNel67+2Rpfh79qu4+BN7pepTaPa6FHcvLbaFqdxc/2nLe+QuGijZDZeV1nCGPfkebkFaAPuKivze0n9p3Qvhf8AtL/tC+H/AIia1rGq2+n3fh59I0axtr3V5ba2/saGa7lhtLZJfIg3uHlkIRC55JY19bN+0T8HF+EFj8d/+EjibwVqccb2t4scrPO8r+UkMVuqGZ52kzGIVQybwV25BoA9rorxv4X/AB28BfFq/wBT0Tw//aGma5oyRS3elazp11pOoRQT58qb7Pdxxu0Um0hXUFcggkHip/ir8cPAPweOj2niuW8utW8RSyQ6Zpel2VxqWo3zwqHl8m2tkeRljUgu5AVQRkjIyAeu0V+b/wAef2r/AAx8S/2U/jD4h+Cmo6/pGreFdOkVr99O1DR5LW7jmSOSKO4mjixPEciREben8QGa+s/HPxw8A/B7wr4d1Dx7fztfa8sVvp9jZ20+oalqFz5Ydkt7W3SSaVgOWIXC5BYjIyAe20V8qeIvj14O+J37PnxT8T/C/V7q11Xwzomrx3MMsNxpuqaXfRWUsiCaCdY54JAQGRiBnGVJxmvKvgD+2d8JJfhd8J9C8Ya9qc+q6xo+i6dPrt5p9++mXGtvaRLLbyatJF9ne5abcHJlPz5DNuyKAP0Aorgrb4l+D7v4l33wigu2bxRp2lw6xNb+U4VbK4leCN/N27CS6MNobcMZIxXD3v7SXwd03wj408c6jrhttG8AavNoOrSvbzbo9SgMSm3ijCF5mZpo1TylbezYXPNAHutFeTfC/wCMvhX4sjUYtBsdY0q80nyTc2mtaRe6TcIlwGMThLuKPerhGwULdOcVzvxK/aX+Efwn8XW/gDxfqN1/wk19YjULPTLGwur+8vIWlaEC3htYpGlfcjZVRlVBZsKCaAPe64DxH8K/ht4v8VaL438VeGdP1jXvDm46be3duk01mWYMWhZwdjbgCGHIIyDXGeCf2jvhF4+8Ba98R9F1h4NJ8KNPHrKXtrPaXmmSWq75UurWVFmjZV5xs5H3c1kfD79qT4UfEjxpF8P9JbVtK1u9tpL2wh1rR7/SP7RtYceZLZm9hi85U3AsB8wHOMAmgD2vxP4X8N+NdAvvCvi/TLbWdG1KMxXNndxLNBMhOdro4IPIBHHBAI5FeWeAv2a/gX8MtZg8ReCvB1lYarZxvFbXT+ZczW0cgw6W7ztI0KsOCIyoI4PFfEc3xF+IA/Zo/bC18eJdSGp+F/Fni220m6+1zefp8FtaWrQxWsm7dCkTMxRUICkkgDNfUdn4piHjj4F6bqvjvUdP1LX9D1CdNEW3E9trjQWVu80t1csjNG1sZBIvzqXZu/zAgH1VRXyb8Uf2tvg/4R1HxR4DGpajdavoVpINSutL0y+vbPR5ZYiYvtt5awvFbN0J3MNnV9o5rlf2WvjZptn+zr8B4viZrt3f+J/iJZJa2lxdeddz3l4kUk7+bMd5B8tCd0jDOMZzQB9uUVwB+J/gtfiXcfCN74p4mtNGTX5YGjcRrp0k72yymYjy/wDWxsCu7cAMkY5rzD4d/tU/Cb4p+IrPQPBv9sXMOqtOmnanLouoQ6TqBtlZpfst/JAtvIFVGOd4DYO0mgD6KlhhnUJOiyKGVgGAI3IQynnuCAQexGakr5Q+BPjafwlpfxQ8A/EHV5LlPhHrF0raleSvNK+iXdsmrWkk8jZd3gt5zC7HJbyc8kmvoXwR428LfEjwlpXjvwTqCapoWtwLc2d0isiyxP0ba4V19CGUEHggGgDqqK/MrWv2k/i5Z/sk/H34rW+qxL4k8CeKvEOl6TP9lhKw2thqEcECNGU2OVjYgs4JPUkmuu8d+Kv2j/2dbTwh8QfFHxEs/iN4b1rW9J0fUdLutEg068SPVplgWazns3AaWJmDeW8bB13cggGgD9A2hhaVJ2RTJGCFYgblDYyAeozgZ+lSV+ef7Sf7SfxD+HXxksbbwZcwx+Bvh3baTqPjvdAkzta6/qKWMCK7DdE1vCk10dpGVxu+Xp7X+0f8XvGvgu88EfCz4SQWk3j/AOJ19cWWmT36s9nYWtlCbi+v5UQgyeRHt2R5G9mHOAQQD6hor53+GngP9ofwh4qWbx98UrXx94cuLaXzoZ9Dg027gugV8o20lo4QxEb9yyq7DC4Y5JH0RQAUUUUAf//V/fyiiigD4G/by0jxImmfC74h6fqN9pWg+B/FUWoazfadZxahPpttJbTW6aj9mminSRLV5AX/AHTlFYuBlcjnfhVafDbxx8QfFPxY8P8Axyl+LviLTPB93o0ojttOitoLG6lW5QPJp9tChkWSI7UZi6hm4ANfo3UMNvBbJ5dvGsSElsIAoyTknA7k9aAPxt8SQxRf8EbNK8pFTboeiSjaAMP/AGtbvu477uc+vNfR3x18c+E/hB+2r8MviN8S9Ti8O+F77wlrukpqd2fLtFvftFtcLC8p+VGaNSV3EZ6Cv0KqGe2t7pBHcxLMgIba6hhkcg4PcHpQB+a/w4+L/wANf2aPi/8AHPT/AI96zD4YvPF3iH/hJdH1O9VhDq2jS2cEcMVpKqsJntWR4zEuXBb5VOc184ajZ3nw6/Zf+EPivxnpl1oGjX3xtsfElppkttI1zYaNd6jdXUERtY1aTd5WZBEqlvmwBniv24lghn2edGsnlsHXcAdrDoRnofevDvjt8Hrv4ww+AorTU00z/hDfF+jeJ3LxGTz49KkZ2gXDLtaTdgMcgehoA+WL74j+CP2kP2wPg1qvwQ1KPxPpvwztfEGoa/q9krPZ20WqWQtLWzM+NjTSy/OYgdyqm4jg4+V/An9ieDvh745/Z++PXx4v/hrqDalr9vqmgXOlaU39qWmrXU7i7spbixlubtbuKXgxyPIrZVQoCCv3BihhgDLDGsYZixCgDLNyScdz3NI9vBLJHNJGryRZKMQCVzwcHtkelAH5j6hqfgv9mH9q/wAH+JfipqzWfgm5+Gll4V0XxHqK7bdL7TroyzQTzKuyGW4h2S7m2h9pUdAK87sdQi+NFh+3TqXw7huNSh8TaVp40sG3lhkvdvh5okaGORVdlmZD5LAYkUqykqwJ/X2aGG4jMU6LIhxlWAI4ORwfepKAPxv8Y/tCfC34t/DX9mHwt8OtTfXdR0nxt4Dk1RbeCUppjQOtu0V5IyhIpWlbakZO58MygqC1fWPwviiP7d/xymKAyJ4c8JKGx8wVheEgH0OBn6CvtqKGGAMIY1jDsWO0AZY9Scdz3NS0Afijc+Hda1b9mv4i3+laVPruleEPjpqetatpVrEZpLvR9O1lZbqFIR98Kv7wqOymvvi++P3wc/aJ8A+NPAHwV8UW/i3W9Q8MalKkNiksgjE8DQxrM+wJDIzyALFIyyH5iFwrY+tqiiggg3+RGsfmMXbaANzHqTjqT60Afh14g+Pvwz8ZfsM/Cn4VeF9Ql1PxV4cm8E2Ws2MNtMX0mTTb6zt5jfFkCwKZlEabjl3ZQoIyR9s6lq0Ph/8A4KI3k10uZ9W+FcUenQFgjX1xZ6tczywQk8NIqEMR1CnJ45r7sjhhiLtFGqGRtzlQBubGMnHU4HWlaGJ5EldFZ487WIBK54OD2zQB+CHxc+ONx8bP2fbDxP4s+Jt5e+OZ9c0W5uPA+lWEUFloHl6vApW/H2Z7xGiGFWa4uUR5SoRfmAP6GW9hY3H/AAUm1C+nt45Lm0+FVl5MjKC8fma3dB9hPK7hwcdRxX3IsUaM7ogVpCCxAwWIGAT68DFPoA+PP+CgH/JmnxX/AOwO3/o2OuK/afhhtpv2XjbosXlfELRIk2jbtjbTbwFRjoCABivviigD8zvB/wAZ/h58Bf2rv2g9C+KOovpGo+K7zQdV0O2+zzTTavB/Zkduy2McSM08omQx+WmWLcAHBI+bvAEF9oP7If7NPxS1/S7q68KfDvxzq2qeIrZIGnlsoP7S1SFL6WGMMxWymcO+0MV5YDjNft60MLypO8atJHkKxALKG64PUZ71LQB8UfFr43fCz48/s3/G7Qvg34hh8X3Vl4N1fzTpyyTQh7uwuBFEJwvlNKSpzGrF143KMjPyP4v1OS0t/wBl/wCOdr44ufCngGy8HzaLP4l06ztdRg0m/ube1VWuRdQXEcMc5he3eXYPLdQrMoY5/YqGCC2j8q3jWJASdqgKMk5JwPU0iW8EUP2eONUi5GwABeevHTmgD8nPE/gPwv44/Z+/aY8b/C74mXPxl8QeL9CgtL+4itrRIml0e3laFIPsFvDFNIYpNp2bidqqeRiq/wAQv2hvhX8bLX9mHSvhfqj+IJNN8d+GrrUJLeCXyNPK2dzAILqVlCxzu7nZETuIR2xtAJ/W2GGK3iWCBFjjQYVVACgDsAOlJFDDApSCNY1JLEKABljknjuTyaAPyV+NfxS8DfA/xl498d/s7/E46b8QLjVkfV/h1qVm9xD4g1QeXbk2lrJHHeRS3CKuLm2Z4ZMKSMc1+iHxp0jVPGPwD8d6DaxPaajrnhnVLZI1O6SKe5spECgjqys2MjvXrLW9u8yXDxK0sYIVyoLKG6gHqM96moA/Mb4265Y+Jv2SP2d/EOm7Bbaj4i+Hc6BDlVDzQHaPTb0I7EYr1r4XxRH9u/45TFAZE8OeElDY+YKwvCQD6HAz9BX0p8Kvhnpvwl8N3XhHQ7uWfSTqN/fWUEgULZQ307XJtYsdYonkby88qpC9AK9LoA/FrX9D1TXf2Cf2qtF0K1ku5ovHfi144IlLEQ22rRTSbVHZURmOOwNeuftM/Ebw38VtO+Dv7QPwp8byN4C8FeIboa5rWiW0GqSaO97pxjhup7aeKeMC2MqibfEzRLLuADAEfqTUMFtb2sfk20Swx5J2ooUZJyTgepoA/LWPT/hz4w0H42fFvw38a5fi7r9t8OtS0S6eO30+K1gtJIp7qAGTT7aCN5VcScFmZQ53AArVn4mWFjpv/BJCG10+3jtoT8OtGlKRqEUyTW9tJI2B/E7szMepYknk1+oUEEFtGIbaNYoxkhUAUZJyeB6mpaAPgf8Aafhhtpv2XjbosXlfELRIk2jbtjbTbwFRjoCABivIfiv8drrxd43+Lnwz+IHxEvvh5H4eaTTND8K6RYQy6v4iims1dbkPNbXU08VxI7IsdokZRVy785r9VqYYozIJigMigqGxyAcZAPocD8qAPx10PUbfSP2fP2G/E+t3C2ul2Gt6TFPdTNtihM+mXUUIdjwoZsKueM4FfoJ+1LZ/CHUfhBfaf8b9Qu9F8Mz3Vn/xM7ITCbTruOZZbW7WaGOTyPKlRW82QeWvR+Dg/QssMM6GKdFkQkHawBGQcjg+hGakoA/HnWPF/wAWvjz8IPj78BfAXjKL4yaVpvhy2l0fxTY20dvNcXMzs0+jzSW+La5uDBEMPCq58zDjc3HWfD7U/hZ8cPEfwu03UP2h9Q8Q6t4f1e01iw8KTaVpNjfWd9pkT7re6itLGGe3WOMyQvuKoc7QSStfqpBbwWsQgto1hjXoqAKoz6AcUq28CSvOkarLJgO4ADNt6ZPU47UAfiLqXivwqtz8e/2bda+JXhP4deFfFfjPV2v4/EbSR+IIItQMUl3LbI5jtpY7lt7WsrsTHGVZt5wF+xPEUnh6/wD20/gJd+HLqHU9KHg3xI1jdQyrcRyQ/wChqkkcqlg4ZDwwJyD15rQ0H4A/H7wHY+I/A3hHXPCGseGtd1PU9Riv9e0q4udVtzqs73EyzxpKsN6Yy5CM7R5UKrDaAB9LfBD4TaJ8C/hP4Z+Evh65mvbHw3ai3WefAkmdmaSSQheF3SMxCjhQQBwKAPnH9na1tk/ar/alZIkBfVvDIYhRkg6LExB9eST9Sa+DfC2k+JI/2G/2ZPiDp+o32laB4E8R3d9q97p1nFqE+m2rXGoW0eoC2minSRLRnBf925RGLqMrkfurRQB+cnwqtPht44+IPin4seH/AI5S/F3xFpng+70aUR22nRW0FjdSrcoHk0+2hQyLJEdqMxdQzcAGvAvEkMUX/BGzSvKRU26Hoko2gDD/ANrW77uO+7nPrzX7JQ28Fsnl28axISWwgCjJOScDuT1qagD89fjr458J/CD9tX4ZfEb4l6nF4d8L33hLXdJTU7s+XaLe/aLa4WF5T8qM0akruIz0Femftz+EvEfxB/ZZ8W6T4MsJNduAdN1B7C2G+S/s7G+gu7iCMDl2khjbaoyXOFAO7FfXE9tb3SCO5iWZAQ211DDI5Bwe4PSpqAPzF+Lfxm+GH7T/AIm+B/g34AaonibXtL8ZaR4ku2tInB0TSdMEjXjXpKj7Mzq3kCGTazs23BxXr/wC/wCTv/2pf+v3wf8A+mNK+1o4IYmkeKNUaU7nKgAs2MZOOpwKloA+Cf2y9e0rwH8R/wBnz4n+LJxp3hbw74ruU1LUJAfIsxfadPBC8zDhEMhwWOAO9Jo+t+H/ABf/AMFBdC8U+HbuDVdL1D4Qy3VldwMskUsU2twlJI3GQQytwR2NfeU0MNzE0FxGssTjDKwDKR6EHg09VVFCIAqqMADgACgD8efHPgjxf4/8Afts+Dfh/bS3Wr3PibT7iG1t1DSXCwWlhc3MUaEEO80UboFwd5IGDnFeh/DvVPhX8cviD8NJZP2i77xprXh3UP7Y03w/JpmlWV5b3FvbSpLFeR2djDPbqsTujpIyqWwBltlfqLUSW8Eckk0carJLjewABbHAyepwKAPhn9vvULPw78Pvh/44mu4dLu/C/jnQ7+01C+R5NLsZR50bTaisYLi2Mbum5MMJXjIZeTXO/AnXPBnxJ8a/E34jRfEnwr4x8eeK9FttPGj+F7sSxWdhpyTeWcSP587yS3B3ysiqo2oAO/0N8f8A4R+L/iX/AMIb4g8Ba1Z6Xr/gXV/7XtLfVbZ7zS7x2t5bZo7mKN0cFVlLRyKS0bDIGeRxvhH4CeNdW+M2gfHL4u3WhQar4Ssb2y0uw8OWcsEWdRVEnmu7qdjLPhE2xRhEVMlvmJoA+D/B1/axfsjfspfFJ0fWPCXws1y2uPE8VtE1y+n+TFcWxupoUDP/AKDM6vIApZAd2MDNfS1r488H/tDftlfDPxj8Dr+PxFofw80TXx4i1qyBexYarHBHZWH2gDZJKJEaYxgkoBk4JNfofHDDCGEKKgdix2gDLNyScdyeppIYILaMRW8axICTtUBRknJ4HqaAPxp+Cv7Rnwa+H37H/i74c+KtQEnivUrzxdb2egfZpmvdXk1HUbyK3jtItn+krKzCIvHuRWDK5BVgOq1i3k/Zsl/Ys8S/Ge5Oj6R4I0nVtE1vUJgzwafe6hpEEVvFM6Bgq+ZE0QY/L8uScZNfra0MLyJM6K0kedrEAlc8HB7Z70k0MNxG0NxGssbdVYBgfqDQB+Y/w28dfDn4kftEftH+MLu3u9T8B6j4N0BZZPsVyf7R05Y9QiuJLaFE8+aJwJERo0zJgmPIKsan7PnxXtPC/wAY/AnwQ+B3xQX4xfDTVdPvhLazxJNqPhO2sLfdaebewLHuhkcC3WK6QSr8uGOMV+pFQx29vC8kkMSo0x3OVUAsemSR1OPWgD4H/be8UaD4K8Y/s9+K/E8rW+laX44E9zKsUk/lRrp11mQpErvtT7zEKdoBY4AJrkv+FlaB8bf2tdI+KfwI/wCKw0T4X+Dtbj1PUtPBa1vb3UDE9npcVxjbLKPLMuFyF3cndkV9a/F/4PXfxO8XfDDxNbamlgnw+8Qf21JG8RkN0v2Wa38pSGGw5l3biD0xivcooYbdPKgjWNMk4UADJOScD1PNAH4PfET40N8Z/hb8PPFviH4p3ev+NZ/Ffhi+vfCWlWENvpnh1jqUSNHdhbd7qN4XYRLJc3Q8xyNiYOK+/fAF5a6b+3x8YtHv5lt73XPCnhm7sYXO17mC0a7inkjB+8sbuqsR0JGa+5UijjZ2jQKZDuYgYLHAGT6nAApDDE0qzsimRAVViBuAbGQD1AOBn6CgD8RPh5fWc/7Fn7J2lwzK95onxU8O6ffQg/vLa7h1K6MkMq9UdQykqecEHoRX0xofxR+H/wCzx+178cLn456nF4X/AOE9TQdQ8P6nfKyW9/Y2NiLaa2glAK+ZBPnMIO9t4YKc1+kccMUO7ykVN7Fm2gDLHqTjqfeiSGGYoZUV/LYMu4A7WHQjPQ+9AH4f3ctz4j/Ye+OvizSdPvJLeD4sX+vvbvbSxXa2MOuWd5Kz27qsiNHBl3VlDKFOQCK+jfEvxy+Gnxv/AGvP2atQ+FOqHxHpWmT+KvP1G3hlFistzokjJbid1VWnCoWeNctGMbwpYA/pvUcMMNvGsMCLHGnAVQFA+gFAH5FeMvix4N+CfjTWtd/Ze+I7ah4g13xWP7W+F2o2jSy6hqN9drDfS2MUkcV9ZOwLz78vbtjdt2kCvseYDU/25bR7ZuPD/wAO7hbnb/e1XVoTAr+mBZSlR15Jr6sNvbmcXRiUzKu0PtG4KecZ649q868FfDPTvB3ivxr41+1y6hq/ja+guriaYKDDb2lulvbWke3jyoQrsO5eR2PJoAzvg98Tfhb8T9K167+FUsb2uhaze6XqKJavZmPU4GDXAeN0Qlm3q+/BDbs5znHxxc+MPDn7Lv7RvxZPxlmn0DwD8Yjp+p6V4iCzC0gvobP7HeWVxcxKfs8x2CWFmKrtPDbhivt74afDjTvhro9/ZW99c6vqOtX02qanqF4U+0Xl9OFV5WWJUjQBI0jREUKiIqgcZPoUsUc0bQzIJI3BDKwyCD1BB6igD8gPBGoaJqfg39rif4aeKdd8Y/DSHwZFDpGoarqGoanaterpupNqCWlzfMwfaWi8zyyRymT0r3P9nT9kv4Z+JPgx8J/FfjrU/EXjCNNC0LU4NK1nXL270e3ufskUqGPTzILfbEx/do6MqgAAcV+hqIkaLHGoVVAAAGAAOgAoYFlKglSRjI6j354oA8z+FHxh+H/xs8P3vif4c6i2o2Gnahc6XcF4Jbd4ru0YCWNo5lRgRlSOOQR9B+Ynj/4haf4A+Pn7Qfw1tvHfhT4f2Xjo6d9sfxf5y3O+70qOGa904LsiuINhCLFLIMTI+SE+U/qN8Mvhzpvwy8OSaJZ3txqt5fXdxqGoaheFDcX19dtvmnkEapGu44CoiqiIFVQABXzjH8Evjp4I8b+O9W+HWreFtb0Tx5qr6zIviWxupL2xuJYIoDEslvIFuLdFiXy0byyinaGwM0AeM/FcfD3wb4Z/ZFm8H+IbbWfAnhbxTpulQa0lzHPbSRRaXc2cDvPGTHl3TaSDgP8ALx0q58PPi38Of2YvjH8dtJ/aB1ePwrqHirxEfEek6jfI6xarpElnBFDDayBT5r2jxvGYVy4LfKpzX1x8AfgdpPwL+GsfgCK7XWJZr271O9uDbpbQy3t9O1xKYbZCyQRKxxHGpOxQOSck+3SwQz7POjWTy2DruAO1h0Iz0PvQB+Jd5pmpeAf2YvhF4n8WaReaJpF98bLPxNaaW9pI91YaJcahdXcUZtYleTIhBl8tVLANjGeK+nL74j+CP2kP2wPg1qvwQ1KPxPpvwztfEGoa/q9krPZ20WqWQtLWzNxjY00svzmIHcqpuI4OP0dqOKGGAMIUWMMxYhQBlm5JOO57mgD8cPCvxl8B/Dj4P/tJ/AbxPdTR/EO68Q+NzZ6FFbTS315BqwkktbiGNVO6B433tLkKiAs5VcE+hXD/AAXt/gH+zTrPjr4jXPwl8aaP4N06Xw74jRQltGJtNtEurad7mJ7KVJQEJglYOwXKEYJP6meTCJjcCNfNK7S+Bu2g5xnrjPaieCC5iaC5jWWN+GVwGU/UHigD5j/ZF+Kvjb4wfCaXxP44MF9NaatqGnWWr2ls9na65YWkuyDU4YJCxjS4GeASuVJX5SAPlP4N/tHfBP4H/Ev9o+2+KniWDw7cy+OLu8hW4jlBuo0sbRSluwQiaUEcwoWkG5SVwwJ/UwAAAAYArwb4P/BiX4Z+I/iVrl/qEWpr488TS+IYUEJQ2ge2ggEZLFtzAxE7hjrjFAH52+A/HfxF/Z7/AGefg94W1m8Hwm0b4ja74gvNQ1vULNJh4Z0+5mnv7G1MUw8iGe5EiqpnVkiO7chIwG/C6S18TeKP2xH8IeIdX8dWuteC9INpqmpwpHPqbnTtThEkIit7WOSIspjieOIIwX5SwwT+xcsUU0bRTIJEbgqwyCPcGn0AfkdP4n8PP8EP2G/GiajAdC03VdBsrq9DjyILlNFns2ikforLcI0TA4w4KnmtTxb4W8VeN/EX7ePhLwTG8+uarY6DBawxf6yd28OpmFPVpVygHctX6q/Y7Tyfs3kJ5JO7ZtG3du3Zx0zu5+vNWKAPxq+Nf7RnwR+In7LXw+8AfDfURNrFjqvg6O60aC3l8/Rlsr+1ikjvkCYtRHIBCpkKh3KqhbIr3rx38TvA3wJ/byvfFnxZ1VPDeheLPh/Z2enX9yri2nvLHUp5JrZXVSPNCSKwXqQQBkkA/oukMMbvJGiq0py5AALEDGSe/AxSSQwzFDLGrmNty7gDtYdxnofegD8ifhndfB7xj8EPj/f/ABqm1Twx4T1v4s6hdxXv2e5tb/TZpItPubG7fZG72jRsEffMoRMgSgZK19F/sofGTxZ40+IXjT4cDxvbfF3wb4as7G4sPGFtax27G4uWkWTTrmS3xbXE0SIsnmQqvDfONxGPvCoYLeC1iEFtGsMa9FQBVGfQDigD5T1kDSP23/C19eHEPiLwFq9lalun2jT9SsriRE93jn3EekY9K+JNJ1MfCT4p/Grwd8Wfjdd/CCTxD4p1HXbOC603SZLHWNK1FI/Ilt7q/s5mmeNB9nkhWQ7CgAUEmv1C+IHwz03x7qfhHXnu5dN1XwXq8Wq2V1AFL48t4Lm3bP8AyyuYJXjce4YcqK9Glt4JyhnjWQxsHXcAdrDoRnofegD8kfin8MbPxT8M/g9+yN+zrrs114s0TSz4k07xTdB4LnRdIjgkjgll8sROjX7yraLEV+5vZkzGKoeO/iH4Jn/Zs/Z78VafpH/CI6N8JPiP4fs/FOnbS3/COyaXFc2d2tyRltkcsqN5jcusiuclq/YKmSRxzRtFKodHBDKwyCD1BBoA/KH4v+L9Nt/2oNA+PFn8VpPBHw48ZeDU0bS/F2n2thqGmfbbTUJpprOa4vLe5igEwZXSQ7BI0ZTcdgFcd8afCfw+j/Yk+MHijwb49l+J1l448V6Fe6jqMtvbQ2814NU0u3m8pLWGCBlZETcyKVZs8k5r9iza2xtvsZhQ2+3Z5e0bNvTG3pjHapVVUUIgCqowAOAAKAPhz9tmztI9J+BdgkCLap8VfCEYiCgRhBNIAoXptA4xjGKs+PAE/b7+FEiDa0vg3xKjkdWVbizIB9QDzX25RQB+Bf7RPx3vPjX+y/8AEi58e/EW+03xl5l5Evw40uwhT+zIbC/wramWtprxgsMaztO00MAYgKM4B/QXxTqFtpP7fPwuvNYnW2TXPAetWFm8p2i5vEu7W4eGMn70giVn29doJr7sWKJZGlVAHfAZgOSB0ye+M8UjwxSsjyIrtEdyEgEq2CMj0OCR9DQB8hftaR/BfyfBl78UPG938M9b02/nuvDniW2zEljeiHy5UlnliktFSaKQq0VxtEqhgvIOPibxx46+MHxx/Zzi8V3WqyeIdM+FvxL066PinQdLXdrOgacMyanb6dMs0Uz200wd1RHhYwkqMKTX7LSxRTxNDOiyRuCGVgCpB6gg9aciJGgjjUKqjAAGAAOwFAH5y/CSz+FPxh+MGm+NLL4+XHxd1vw/oWpwJZ29rp1usen6n5Uc6zyafawFXLKhSOVw4IJVcB6+T/gF4/8AAPjP4b/CH4WfEL4s+DPC/hr4f61Zatb6bczG18TGbSbl5LKwvVuDFBBKjlRcSRgmQqVULuLH9q/EPh2DXPDeseHIJpNMGsW1xbtcWuI5omuIyhlRh0kXO4N6gV8HP+yv8cNe+DNv+zZ4t1zwePBcFhBo76vZ6PP/AGxLp8CiMFIJZTbQ3TovMwZwrEuqbgKAOy+FscT/ALd3xyn2qzr4c8JBXwMhWF4SAfQ4GfXAr5n8GWVpH/wTo/aEijhRU+2fEFsBQBlLm5Cn6gKuPTA9K/WbRtKs9B0ix0PTwVtdOgitoQzFmEcKhFBJ5JwBz3rSoA/I34njXPBvj/4DfGDXvGt98P8AwdJ4DGhv4itrG0v4dN1K4+yzqLv7bBcRwR3aLsE20HcgUuFLZ5D446B4Bn/ZB/aP+J/gz4nzfFS58XSaHFqepGC0htvtGmz20SeT9igggkzC6K7oGBKAE7gwr9n3RJEaORQysCCCMgg9QRSRRRwxrFCgjRAAqqMAAdAAKAPiP9qWKK3+MX7M7QIIzF4zmjTaMbUbS7oFRjoCABiuQ+Gnj7wd8O/25vjz4T8darBoWq+PpPCNx4fhu28o6okWmfZJBalsCRkmUoVHOfocfodUL21vLLHPJEryw52MVBZd3B2nqM98UAfEn7aUFxo1z8IPitqdnNqHhD4e+L7fVNeSCFrg2tq1tNBHqDRIGdks5ZFdiqkqCWxwa4218eeD/wBob9sr4Z+Mfgdfx+ItD+Hmia+PEWtWQL2LDVY4I7Kw+0AbJJRIjTGMElAMnBJr9E6ihggtoxFbxrEgJO1QFGScngepoA+Kf2EP+Sa+Ov8AsoHi3/04yVxnj3x94O+En/BQWx8U/EvVYPDeieJfhtHpWn3163k2s+oW2syTyWyyt8vm+XKrBSeQfUgH9Dqhmtre42faIll8tg67lDbWHQjPQj1oA+Jv2foov+Gu/wBqWbYPMOoeElLY+baNDiOM+lfBGiaP4jg/ZL+DHjzTNQvtI8O+AfiJ4gvtWvNOs4tQm0u0fUdSgTUBazRTo8dq8m5/3TlFYuBlcj91qKAPgj9nm0+G3jj433vxY8P/AByl+LviLTPDp0aUR22nRW0FjdXUdygeTT7aFDIskR2ozF1DNwAa5r9sLxna/Cr4/wDwY+JEGr6V4d1BbPxFYG/8RtLHojW8iWrG2eaFWeG5d8NC/ClEkDbuBX6LQ28Fsnl28axISWwgCjJOScDuT1r5i+Lfwf8AiXrHxY8P/Gf4XavpDajpOk3WiT6T4htpZ7CW3u547hpoJIGEkFxuiVWba4dMAgbRkA+KvGVn4P1X9hf9om/8FeOtF+IviLxVLf8AiTXW8PSo9paXNyIR5MUKySSJHHBbDDSnfIVZz1wPUfHHjfwn4H/aC+Df7SPi6+jl+F2o+C7vRrXXFBl0/TdSvZLe4huJpVBESXUCtCshwueCQK+ivg38CNZ8H/EjxX8avHt9ptx4s8W2lpp722iWbWWnWtnZM7IoEjvLPM7OTJNIQcBVVFUc/Sy21stuLRYkEAXZ5YUbNuMbdvTGO1AH5VXWs6Z8VvFf7T/xz+G5Nx4Auvh4dCTVEjZLXWNUsra8klmt3YDzlto3WAyD5SeFJAzXmvjb4ufDr4p/sH+CP2WfAF7/AGx8U/EWh+F9Et9CS3mS9sry1Fo81zcxuitBDAsTS+ewCEAMrFSGr9pERI0WONQqKAAAMAAdABTRDCJjcBF80qFL4G4qDkDPXGT0oA/Obxx8TfA/wH/bvu/FXxb1dPD2h+K/h9Z2WnahdI4t7i9sNRnea3DqpHnBJFcL3DADkgHyH4Z3Xwe8Y/BD4/3/AMaptU8MeE9b+LOoXcV79nubW/02aSLT7mxu32Ru9o0bBH3zKETIEoGStfrtJDDMUMsauY23LuAO1h3Geh96loA+D/2UPjJ4s8afELxp8OB43tvi74N8NWdjcWHjC2tY7djcXLSLJp1zJb4triaJEWTzIVXhvnG4jHI/E34o/D/4Sf8ABQfSvEvxJ1eLQtJuPhhNaLe3Cv8AZ4ppNbRl82VVKwqwQgPIVUthc7mUH9GILeC1iEFtGsMa9FQBVGfQDivDJ/g1PcftLR/HqXUYnsl8Hy+F205oiWZ5NQS984yE7du1dmzb3zntQB8JaP8AFfxFo6ftV/tifBnRJNd0HVI9BtvDzvbyiHVJ9Gt2trzUEiAWSW3iMw+dMCRYWAbjjmdO8ZaD42/av/Zx1vQvi5ffFy6jvdfW9vIrS2ttGsmuNFnZYLf7LbRrHI4QloZZ5plVQXxwT+xaqqKEQAKowAOAAKbHFFCnlwoEUZOFGBknJ4HqaAPyEx9q/Z2/bm8GW/7zXbbxP4q1CSxXm4W0vbCB7aYx9dkywyFD/FsbHQ16sNb0jxH8ff2ONb0G8iv7C78LeKminhYPG4Gm6eDgj0IIPoeK/SRYYlledUUSSABmAG5gucAnqcZOPrRDDDbxrDAixxoMKqgAAewFAH5SfBL4x/Dr4H+Dvi58FvinetpHxKl8T+JLv+zmtppL/XRq0ryWV1ZRRoz3XnxlEXYCRtwwUYNeZaHrum+AP2af2Lfi14pka08K+E9Sh/tW92M8dnHe2NzbxSy7AxWPzCFLYwCQO4r9qDDC0qzsimRAQrEDcA3UA9RnAzRLFFPG0M6LJG4IZWAIIPYg9aAPzY8EfE3wL8UP24PFfjfw202r+Eo/hZFai8S1maDUEg1aeSdrMBd1zGBJ5e6NSGdWVdxHPC/Bj4p+HPh18TPhz8J/2Z/iZ/ws3wB4iup7Wfwrdw+dqXhexSGSb7QLsJHPDBBIqxGC9UuAwVTu5H6yABQABgDtUSW9vHLJPHEqyS43sFAZscDJ6nFAHx58MtR0u3+N/wC0t461Ft2g6dcaJYXLIhmRm0zRkubo7EDF2VblUZQCTsC4zX0H8JPHfgD4mfDfQPHHwtljm8K6pb7rAxQG2RYo2MRQQsqlNjIUK7RgjjisTwN8I4/h98ONS8EeH9cuotV1ebUr651vy4WvH1LU5ZJpbwpIjxF1d/kVlZQqquNoxXZeAfA+g/DbwdpXgbwwkiabo8IhiMzmSVzks8kjnlpJHJd27sSaAPyC8Sf8mB/tW/8AY9eLf/TtFX3z4O/ZR+Fuh6joPj3xZq/iHxlfeHQl9p7eJtdvNTtdPnVP+PiG3nk8hJEGdrlCV6gggEfWdFAH5GfDD4J/Fj9qH4f/ABN+LFj8Q4fC/hz49Xuo50640GK/mfRrffpun5uJJ43Q/Zog6bVG0tvByeMfQPHfiu48B/s7/tP69pt1q1x8ErjXfCPjq2tI3uby0V4Rp1xfNEgZmEL20c8oQFtkmcYBx+xdFAHzl4A/aw+BfxY8YWXgn4WeID4uvrqCW5ll022mmtbKKNcg3c+wRwFydqK5DFuMV3/wo+MPw/8AjZ4fvfE/w51FtRsNO1C50u4LwS27xXdowEsbRzKjAjKkccgj6D0hLeKJHS3UQ+YSxKAD5m6t0wT9a4L4ZfDnTfhl4ck0SzvbjVby+u7jUNQ1C8KG4vr67bfNPII1SNdxwFRFVEQKqgACgD0OiiigD//W/fyiiigDgvHvjDWPB0Ohy6P4X1DxS2raraadMmn+VmyguSQ97N5roPJhwN+3LcjjGSO9r45/bG17XdBtfgw2hajc6cdQ+J/hWzuTbTPD59rNNIJYZNhG+NwMMjZVu4ry268EXvxn/bC+L3w+8V+LfEFp4P0nQvDVwNJ0zVJ9PhlnuUu13tJbskyqACWSN1SRiplD7EwAfc3xB8f+E/hZ4L1f4heOr7+zdA0KE3F5ceXJN5cYIGfLiV3bkgYVSfauyr8N/inbar4n/wCCcXxg0bxZr+raxP8ACvxXrWi6ddz38/n3FrYaoltCt6yOv2oLFKVCyhlyFOMqpH0p+0LbXPwe1D4Tfs9eA/8AhMdT8PeP9T1i+1ddL1aa71+6g0u0jl+xW99qF5HJDFKzK8uydGEcbhCCzZAP00or8oGk+KngT4a/tEwaBoHjDwb8PIvAV/qOhf8ACS363N9puswWl0twllcrfXs6xMoilXdL8kgbYF79knwP+LWqfspr4w+H3i7W9f8Ain430fRLvULm41ia0M9oWhubix07c5t7BmhLxRyogbJ3SPli4AP0U8R+INI8J+HtU8VeIJ/sul6Naz3t3NtZ/Lt7dDJI+1AWO1VJwASewzXBeJPig1n8PNJ+IvgTw7qHji11s6fJa2+mqiXD2moFCtwVuGi2okbh2BwwHUDkj859et/g/wCIP2aPj74G8PR+N/BviPw74XvNVv8Awz4i1jUzc2cltZXTwzwSSXU6zWty6kS+XM8Mu0B0GcG14q8Lr8Hf2GfAeteAta1qzvfEF34DnuZZdXvbhl+0XFmssUBmmbyYXVipij2oVONuOKAP0yl+IHhKH4g2/wALJL7Hie70ubWY7Ty5Dmwt547eSXzNvljEsqLtLbjnIBAJFTw34w1jXfGHivwzfeF9Q0ey8OyWiWup3XlfZdVFzF5jta7HZ8Qt8j71HJGO4HwD42+CvhPxl/wUZgttX1DXIo734d3GruLTW9QtCtxDq9rbqkZgnTy4CnLQLiJn+dlLc16R4J8b/YPjZ+1q/jfxRd6V4Z8Nf2BItwbhiulWz6CktxNbKwdYmBzJ8qHc/JVjQB9514n8MfjGvxH8e/EzwONIOnn4darbaYbjz/N+2faLSO68zZ5aeVt8zbt3PnGcjOK/OPTtYvfAXxf+CPjr4Z6V4+0rw/4315NE1DUfF+tSXcXiC2v7SaSOQ6fcXk88LoYxLG5gt8DI24IFfVv7MX/Je/2mv+xr07/0021AH23RX59/E3QNY+JX7cNh8LL7xZrmjeEpPh1Jql7YaTqEtgLyWHVxCqvLCVljBEnzNC0cjBQhfYWU/P8Ae634v+Hn7NH7W3w60jxPq9zB8NdQuIdBvrq+mm1GztbmytrlYVui3mkRO7bCW3AHGaAP2Eor4U/aG8S+IdM0T9miXStVurRtY8feG7a9aGd4zdW82n3jyRTFSPMjdlBZWyCQCRxXNaJ4Uk/ai+PfxetPiT4g1q18N/DbULPQ9G0TStUu9IjSR7SO6m1C4aylikmld5AIS7bEVSNpJzQB9SeI/jGugfHjwb8Ef7IM58W6Vqmp/b/P2/Z/7MaFfL8nyzv8zzvvb1246HPHtlfBXiPRU8OftqfAnw/HeXWoLpvgzxJbi5vZjcXU/lGyXzJpW5eRsZZj1JJr5RtPEHjr47ab8RPiDceGviVqfjGPWNb0/wAMX3h7UYbLR9C/s2aS2s40tRqlssrh4w90bi3cyElQNuMgH7R0V+fnh3xf8Tb39p74Haf49ludJ1PVvhvqF5relLKVthqyvZeduiRjEzRuzqp5wCQDisR3+Mfjb4h/ti+B/hl4knsPEVunh238PNPcP5NjNcaHG7iDO4QGRiTvQDDkOeRmgD9HqK+Af2Ub/wCHfh/xve/D+TTPGfgf4hjSVnvtA8Vateapb3kccqrLqFlPLcXFtP8AvCEaWBkODgxrzjY+OD6x8VP2nfBP7OF9rWoaD4Nk8Oaj4n1VNMu5bC41aS3uYbSGyN1AyTJEhkMsixspcYBOAKAPuKaaK3hkuJ3EcUSlmZjgKqjJJPoBXzd+zZ8bPG/x98NP8RtR8HQeGfBurAy6BcHUjdXt7biWSPzLi2FvGtvuCB1Akfhsdsn528LaHeeH/id8Y/2ULnxBrOseBk8L2PiDTHn1O6fUdK+1PNDNZLqPmfajGzQrJGGkLKhZckE57D/gnF4H0fwv+yh4F13TZ76W48R6bBc3K3V7cXUKOhdQLeKZ2S3TB5SIKpPJGaAPrzRPiB4S8ReLvEngTR777RrfhH7H/alv5ci/Z/t8Rmt/nZQj74wW+Rmx0bBrhfgP8Y1+N3hXWPEy6QdG/snXdW0TyjP9o8z+y7lrfzt3lx48zbu24O3ONx618Yfs/fBXwmn7af7QWpnUNcebwrqXhie0D63qDRu17pInkFyhnK3KB2IjSYOsa4VAqgCvDPAvwZTxR+zH8bPidL4x8SaZrHhjXfHN/okemarPp9pp9zp9zcTpIILYos7ySL87T7zsO1NgGaAP2uor8pviN8YPG3xBT9nnwHqtj4l1fSvHHhAeKNftfCc0VlqWpzJbWu23Fw1zZtFbiSdpJxFKrsNij5d1fQn7Jtr8SNF8R/EPw7rHh7xLoHw9il0258LweKrmO81CAzRSLf2wnF3eSPAkkaPF5kpKiQr0FAH2rRX56eP/AApefFb9ua9+Gev+KNe0/wAHw/Dqw1O40rStUuNOhu7n+1ruAGV7ZklUbD83lOhfaocsq7a8H174r/Er4BfD/wDaV+GPg7xLe6lD8P8AWfDdh4c1fVpmv7jS4fFa2wljeeYs0i2JmYxGQsQSoYkcAA/YOivza+Ivw+m/ZP8AEfwj8bfDnxZ4i1KXxL4u0nwt4gtNZ1e71WLWYdY3xNdPFcyOsV1C6iVGgEa4DKV28VyPxR064+Hnxm8YfEH9ptfGQ8JXepWs3hzxb4c1i9i0rQNPCRILa9sbKdGg/fBvNnkgmjl3jJU5yAfpDY/EDwlqPjvVfhrZ33meI9EsrXULu18uQeVbXrypA/mFfLO5oXG1WLDGSACM9lX5g+APg14P1n9v74p6xJqetyHS9O8N67bGLXNQWJ7i5nup2R1WcJLahlGy3cGFFJVUCkivCdH8Q/Ej46+AvFnxbtfDPxKv/iNf32sr4Z1LRdRgtdD0ZrC6mt7K2js/7UgSWNTEou/tFq7SEv8AeG0kA/bSuCsvGGsXXxH1LwPL4X1C20yxsILyPXH8r7BcyyuytbR4cyeYgG5sqBj0+Xd8F+MrL4j/ABX/AGlvhZ8PfF3iHVvB9lr3w5uNR8TaVpd5JaNLcR3VqZbdZYZMwsJmCtLE28xB4g4WQmupsfEuqfDD9pP4n6JpFzfX+heBfhfpN3YaZPdz3KGSze7AP712LSyLEqvISXfqzE0AfoRRX5CT+CvE9n+xvH+2ZH8Rdfb4sf2DH4wa/bVbk6YxeMXZ0s6X5n2L7LsPkBBEGDfNuzxXqXj3/hKfjV+1B8MvB8nibWPCvhjxT8PLrV9Y07TbuW0kuF+12riESxsrwvvdVeWPEhiDxBgshNAH2z8bPiUPg58I/F3xUbTv7WHhXTbjUfsgl8jz/s6F/L83ZJs3Yxu2tj0NVo/iVrE978P4bDwjqF/Y+NreS4ur+2aJ7XRwtstwgumdkciVm8tCiHJByBwD49+13odh4Z/Yo+JnhvS/M+xaV4Ru7SDzpXnl8qC28tN8spZ5GwBud2LMeSSTmvOvEGva7p3xm/ZA0TT9RubbTtXsNeF7bRTOkN0INAWSITRqQsmx/mXcDtPIwaAPv+ivyn0H4b6p8bdU/aTuPFnjzxTa2/hLxVqUOh22naxc2MOnzR6fbTrOvkMrSFW27I5C0SYYogZ3Y+ZX0Pja2/ZP+F37Y9/4+8R3/wARftfhqed31KWPTprW+voLOazbT4itr5TxPl28vzHfLO5zgAH6c6R8Y11X9oDxH8Cv7IMR8P6FYa3/AGh5+4S/bp5ofJ8nyxt2eVndvOc42jGT7ZX5q6/8KtI+LP7evjrRPEer6vY6Rb+B9DkuLTStQn0z7YTeXaqs89q0dwUTkhEkVWJG4NgCvUf2QtV8RaV4j+MXwY1LW73xLo/w48SR2mj32oztdXa2d7aRXX2SS4cl5fszuUDOS2CATgAAA+2a8s+N3xV0f4H/AAm8U/FnXoGu7PwxYyXZgRgjTyDCxRKxBCmSRlQEg4znBr5Ji8MSftM/tK/Fzwr8Q9d1mz8LfC7+x9N0rR9K1O70lWuNQsxezahcvZyxSSybmCQbm2Kqk7SSTXyf8bL3xB45/YH+O/hTx5ruo6/dfBvxdeaFp+qPdSxT6hbWVxbCA35iZFuXSO6McgkUqzxrIQZBuoA/XL4Wa78SvEfhRNV+Kvhe08I61LISLCz1E6mqwFVZC8xgt8SAllZApAK5DEHAp2nxe8Ja/wDDvxD8R/Bk/wDbVh4e/tWOQbZLffc6Q0sdxCDKgIxLEybwpU/eXcMZvaB8MPDvhz4eTfDLT7vVH0meC6tzPcapeXGohLvfvK38srXKuu8+W4kDR8bCMDH5t/se/C3w7oP7LXxC8f2d5qs2p3KeNdOeO51W8ubPyYr24AcWksrQCY+Uu6YJ5jEsSx3tkA/R74M/EUfF34TeEPiiNP8A7KHivS7TU/snm+f5H2qJZPL8zam/bnG7aufQV6XX4iQ/D/Ufhh+xF8JP2lvD/jXxIfGGhweEZYVbVJ1037De3Vpavp/9noVtfIEMu0ny/McjdI7Emvc/if4s1T4n/tQ/EH4d+LPDHjjxX4U+HVlo0Nlpvg++TTYfteqW7Xcl7eyLqNhPI4+WKBQzRr5bkruOSAfqRRXzT+ydJ8Vl+EMWmfF+01K21bS9R1C1s5NZaJ9SuNKjnY2Et20MkqGfyGVJCHYllJJYksfib9pe48P+KbT4t+L/AIWH4h+JvFHgaPUJW16w16TTtA8N6hplv5slvBFJd20U/klf38UVvcPyylw2MAH630hIUFmOAOpr8xvGmu+NPjB8Qf2T9Ln8Wat4Zs/iJ4U1zUNcXRbprJ7o/wBn6bd7A6cx/OzASJiRFZhG6Fs1c+GngoaL8Rv2jv2b/wDhIdeu/Bem6ZoOraak+sXr31hJqEFw9zHBfGX7SsbyQI2PM6bgSdzZAPv34fePvCnxS8GaT8QfA17/AGjoOuQi4s7gxSQ+bESQG8uVUdeQeGUGuxr8T/hrLf8AwE/4JzeFviT8Nb3Wo/E/juHRdJkmF9NerZC+v/s7y2NrdzfZbeUJIwj2+WnmFCxwK+hfhJofj3wl8dfBDfDfwT8QtA8H6jDqNt4ubxhqcepW87C2Mtlex79TvXjuftCbHMSIrLJ04GAD9J5naKF5UjaVkUkIuNzEDoMkDJ7ZIHvXF/Dbxbq3jrwTpfivXfDV94QvtQV2k0rUvL+12xSRkAk8pnX5gocYOdrDIByB8LfsceCdW8f/APCQfFv4g+Ltb1278N+MPE9joli+o3MdhZW8V7cRnzIEkCXMh8xgrTBhGgjRFXZk+K/D34h/EbxN+yN+zF4AHivU9OvPizrD6XrOvJcudSFjA13cSxQ3blnjnnESxJJksozjnFAH7F0V+f8ApWhz/s4ftVfDr4Z+Bdb1a/8ABnxR0vXTd6Tquo3WqrYXuixw3CXtvNeSSzR+cJTHIm/YTg4zjGD+wx4I1bxv4TtPjX8QvFut+ItX0vW/EdnpFtPqNytjZ2iX93blXt1k8u5kYs5EkwYogiRAoiU0Afo9RXwv8cpdc+Kf7Tngj9m2513UPD3g6Xw9qHifVhpd1LYXWrNb3MVpDZfaoGSZIlMhlkWNlLjgnpjnPjh4b1D4VaN8Mf2evhx4p17RtH+J3i77Be6nPqdxeahY6cttJczWdle3LyTQmbyQkTbmZNzkHJ4AP0Lor8/9K0Of9nD9qr4dfDPwLrerX/gz4o6Xrpu9J1XUbrVVsL3RY4bhL23mvJJZo/OEpjkTfsJwcZxj5CtPDPi7XP2J/GP7Rl78RvFf/CZ+DL3xFd6HJHrFxFbWSabqtyFhNujCO4WQAhzcCRthEalY1VQAft7RX5geI/DGufCb4h/s9/FjTfGniHVtc+Ieu2+j+Ikv9Smmsb+HUtPnuDixyLaAQyIDCkMaKgxwTydXSfhtdfHr9pr9onwh418aeJrTw34bvPDo07TdK1a406GCe80WB3nDQMrkqy70jLeTvLO0bMQQAfpVXBXfjDWLb4kaf4Gi8L6hcaZe6fNeya4nlfYLeWKQItrJlxJ5rg7hhSMevzbfxte3+IGpf8E/LL9rvWviP4mvfiV4UiE+n3H9qTQ2UcWl6obDyZLOIrDcCeKMmeSdZJZHY7nK4Wv0E13xDrx/bq8G+HrfUbiLSbvwBq929mJX+zNcLqFqqSvCGCM6qxAYjIBIBGaAPsuivyq+AtmfhT8SPDOhftMjxjpHxV1XUbuCLxFLrN5eeFvE883mmG3jWOZrWH9380NrLbwspQBSxypTS7N/hZ8Z7vWf2pP+ExsNW1bxYz6D4xsNZvH8My2Vzcf8S/TLi1t5xFZoylYXjuLba5y3nYIagD9VqK+Nvglr+u6j+1r+0noeoalc3OnaTJ4Q+xWsszvDaifSS8vkxsSsfmONzbQNzcnJr4sPjf4n3f7F/g3VNE8Yahp3iW/+Ko06PVHmkuJY438RXECJIHb97Ci7QYXOxlGwjbxQB+zlFfmF8fNNu/gvqPwq/Z/8FTeM9d0j4j6rrGpa/Np2qvdeI9RGl2UTtbRXt7dQmBJ2KvMIZYiI45BGF3Nnqfgz4O+KN9rXxM+G+iWHjX4cfDjX9Cg/sW61+9jvNT0fV3MsFz9gnN7fS+WUMUyh5flcNs2ZyQD9FK8y+FHxY8L/ABk8N3finwklzHZ2WpX+lOLqNY3+0adO1vMQFZxsLqdpzkjqB0rrfCei3fhvwto3h2/1S41u50uzt7WW/uyDc3bwRqjTzFQAZJCNzEDqTXyD+wH/AMkY8Q/9jp4q/wDTnNQB6H8R/wBqTwv8OviafhKvhHxR4q1+LSodamXQNNF+sNlPNJAruBKsmd8RBCoeoxknFbHh79pb4feNfh1rXxC8AWmq+J38PTraX+iWVjIuu210ZFjaCWwuPKkSRN25g2BsVmUkCvJNH/5SMeJf+yY6Z/6eLqqHgkWyf8FEfiWvhjb9ifwLoba95XT+1/tc4tPNxx5n2LG3d82zGOKALum/tz+G9Y8Saz4P0v4UfEO61vw6tq+o2cehxtNaLeqz25lX7VlRKqMUz1ANfbkUnmxJLtZN6htrDDDIzgj1HeviP4I/8nuftN/9eXgX/wBIbyvgTxGZfiL4O8VW154n8U6h+0m/jO406DTtN1LVobOO1g1YRfZoUs5UtI9PWxUtLOCrRtuLOHXBAP3brxP4r/GNfhf4q+G/hk6QdS/4WFrv9iCXz/J+yf6NNcedt8t/M/1W3blOud3GD6p4e0Kx8MaFp/h3TGne00yCO3ia5nlupykShVMk87PLI2By7szE8kk18eftYf8AJVf2bP8Asex/6bbugD7brjfFvxA8JeBbrw9Z+KL77HN4r1OPR9NXy5JPPvpopZki/dq2zMcLnc+FGME5IB+Ev2gLHwb8S/iH4v8ADHhqD4geNfFmhWcKTReH9cfR9G8N3EkHmQyGRruyt2uWUiVkIuX5GUAIFfNWv2kv7QXwt/Y1+IHxE1XVZtZ8S6zZ6ZqEtrqd3ZiURWt7uuAtvLGqXLsmTOgEmCVDBTigD9tKK/LD4oahc3/7QS/s93mheOfFfgHwB4X066j0zw3qjQ3d7d3ssqC51G+m1Czu544khCxqJmzIWaTJ255Px1rv7QXhP9kbXLbU7nX/AAfqem/EHSrPwvea1Mk+rDQ7jUbNrUXrw3EwuPLMkkLh5W81EAfOTQB+vdFfm58TvAV34H8bfCH9mfwz4v8AEVrpvxY1XWNR8S63Nq1zNq96ui6ekxtYrp3LWy3Tbd62/lgIrbNoJB1vD899+zZ+0rd/Cfwrqmqa54G13wVf+JYtK1K+uNTm0y+0qdI2+z3F08k6wXKSYMbOwEoypGSKAPrv43fEsfBr4ReLviq2nf2uPCunT6h9jEvkef5C7tnm7JNmfXa2PQ12/hnWf+Ei8N6V4gEP2f8AtO0guvL3b9nnRh9u7AzjOM4GfSvxr8S/DCf4kfsA+K/2nPHfjLxBqfjzxX4bvtZnkg1e6h02KCYOV06PThIbT7MkeEZWiZ92WDq2CPo3Uo9T+MPxo+Gv7Oes61qOjeCNP+HUHiu/t9LvJtOm1e4eeOwitpbm2eOYQQjMjJG67mZd2RjAB+kVFfmRp+t+K/hlJ+078B7DxHqer6D4M8JLrugXd7dy3WoaX/aNhdl7Q3sjGZ1jkhEluXYuikjccA1D4U+C3xc8R/sc2/xB8B+Nda1j4v8Aj3wpo032271ae3RbWb7NdSWdkhZre0kNuGhS5CeYZGMrvuZmAB+ntFfnn8DvGHwT8JaT8RNBluPGPwu1fRdFF/rmkeKdSvLybTbRFlV9U0+W6mvIpFZsgzQO6lkTMak4bwPTtYvfAXxf+CPjr4Z6V4+0rw/4315NE1DUfF+tSXcXiC2v7SaSOQ6fcXk88LoYxLG5gt8DI24IFAH6OfDH4xr8R/HvxM8DjSDp5+HWq22mG48/zftn2i0juvM2eWnlbfM27dz5xnIziu6vfiB4S07x3pfw0vL7Z4j1qyutRtLXy5D5lrZPFHO/mBTGu1pkGGYE5+UHBx8rfsxf8l7/AGmv+xr07/0021eP/F/4K+E/Hf8AwUB8K2utahrkMeteDNUvZhY63qFl5ctrcWluggNvOnkRsgzJHHtSR/ncFuaAP07or8oPFPibVPjD8e/iro/jTwt8QfE+ieBL620bRrPwfqKaXbWL/ZI7iW8nK6lYyzXUjy5jLq8aRhQASWqj4u8T/Fe0+FX7POjftP6prHhjQNS1bVtO8ZSQXD2up3YtIrk6P9pk06RplinjhEt20LkA8uQvzAA/W2ivyE8I+GJ/DPwa/ap+KHgfV/E7eF7bRdZt/BmoanqurM4tI9IWWd7WG8mOI4bxGFtdBBIVDASFar+NvAvjHwX4H/Z6+LPhz4n+LIfGXxG1zw5oWtX02pvcQTWXiC0cTCOwlDWUbQHBtysPysN7b3+agD9g6K/OPwrp11+z1+1vrngXwxreu654Y1j4d3fiebT9W1S51V/7V06/jg82GS7eSRWmjkIcBsFsHGAAPljwrrPxn8f/AAY0n45+CvCvxI1n4zax9n1m01ldRgTw1IHnWRrBNP8A7VEIsDDuhAa183Pzt89AH7h1yvjjxr4a+HHg/WPHnjK8+waHoNrLeXtxseXyoIV3O2yNWdsAdFUk9hXwt4h8Jal8X/20PGvw58R+K9f03wfaeDdEvpdI0vUp9PjnuJLm7jy8tuyyooUnesToJCF8zcEAr5i8a6dqOv8A7BP7Q/gzxN4g1jV4fhN4t8Q6XpFzPqNx9qks7Axi3hvJUdTdRqlyylJdyHCnb8i4AP2os7u3v7SC+tW3wXCLJG2CMo4yDg8jIPesbxdrw8K+FNa8UGD7V/Y9lc3nlbtnmfZ4mk2bsNt3bcZwcehr4p8YeFPhR8JPgn4W8EahrvjZx4r1K2ltNO0XWdQutb1e8e0G+yinkmM8VrsTzHVZoI025Z1DMG8W+D+q+KfDXiT9or4PXmm67oHhmy8J22taXo/iHVV1i7043dtdxTqlytzd4jlMav5ZnfYwPTJoA/RX4M/EUfF34TeEPiiNP/soeK9LtNT+yeb5/kfaolk8vzNqb9ucbtq59BXpdfM/7GP/ACaX8IP+xW0n/wBJkr4d8N/DXVvjF8Mv2hPGni7x/wCK1vvCHjDxvB4fistZubK30w6dPJNA6JAy+cQxCgTF1SMBI1TkkA/XqivzM0f4m+NvFWtfsQ6/qesXKzeM9O1C51lY5Wihvpz4d87dPGhCP++zIoYEK3I5r0LU/iD4j8K/tY/GC5W5ub/TvDXw40/VrfTDM5tzcxTXrsUiztDyBFUsACRgZ4FAH3jXifwj+Ma/FTxB8R9CGkHTP+FfeIpdAMnn+d9r8u3hn8/b5aeXnztuzLdM7ucD5t/Zd+DEXj74e+AP2jfHnjTxHrPjzxJb2niC5uYdavILBRdqJxp66fHILQWsasI2j8rJKk7ua8btvid4g+Dfwx/bO+JfhNA2s6H4xvpLNmUOsc8tjYxJKVPDCIvvIPBC88UAfrFRX5UeCNB+JXhjx18Ndd+Gfg/4lxahPqtvB4v1HxVqcF5p2q6VdRslzcyQDVbqOKaGQpNCbeFAqhk5U4PqfwL8UeJdS+Ff7TV5qOr3d1c6P448cW1lJLcSO9rBbxKYYoWZiY0jzlFXAXsBQB+glFflRLonxf8AiH+yv+zl4otItd8beGbTSbW88Y6PpGrSafrWsRTWCiCRbrzoZJvJmJkkh89GlJXkkcfQfwy0bwh8Zvgvp2k/BXx/4t8M2nh3xGHvPt1zcya1aT2MokuNGvTfl5ljAdUKMzfLt5dSQwB9XeNfFml+A/BuveOdbWRtO8O2F1qNyIVDSmC0iaaQIpKgttU4BIye4pPBPi3S/H3gzQPHehrIum+I9PtdSthMoSUQXkSzRh1BYBtrDIBOD3NecftM/wDJt/xW/wCxT13/ANIJqZ+zF/ybZ8Jv+xS0H/0ghoA6T4d/Fjwv8TtR8Y6Z4cS5SbwPrU2g3/2iNUVruCKKVjEVZt0e2VcMcHOePVPAPxZ8L/EfXvGnhzw+lyl14E1X+yNQM8aojXPkxz5iIZiybZByQpznivz5+Blr+0zP8TP2hG+CuqeELHRx8Q9QE6eIbDULq5Nz9ktMmN7S7gQR7NmAVLbtxzggD079hpPGEfjH9oRPH81hceIR43/02TTIpYbJpf7PtuYY53kkVcY4Z2Oe9AH6DV4l+zr8Y1+P3wf0P4sLpB0IazJfx/YzP9p8v7Fez2efN8uLdv8AJ3/cGM45xk+21+VX7PHiHwZ4f/4J1+EP+E413WtBs9S1TUbGBvDsksesXd1P4gvBFZWZgVpTJckeWRHtbaWIdMbwAfqrXG+AfiB4S+J/heDxn4Hvv7R0e5mu7eOfy5It0tjcSWk42yqrfLNE65xg4ypKkE/m18GLDV9P/ac1v4GtpPjDwZ4D8b+BLjVf7L13xA97qEd5BfxWxvLa5gvruazaWOZg6i4D71DYXAA8d+Ckk37Pv/BOTXP2h/AV5qjeL5o9X0+M3GoXN5Z25n8Ry2KXMVjcTG1SSIESlwg3MGLk7nyAftzRX5deAfD/AMRfCPxT+HepfDLwZ8SrCKe+a18Y3Xi/VIb6y1Gwnt3Bu5Izql2I7mKcRunkQxjaWX7uFO7+zv4Su/iJ8RPjB8RviX4z1m60n4e/EXW7fR9N/tK4ttPshaCG4eaZY5FE6hZAiRy5iiVW2rl2NAH6UV4n4X+Ma+Jfjp45+Co0g258F6dpF+b7z9/2n+1RMdnk+WNnl+T13tuz0GOfy/8AF/iaLwyngH46/BOP4h31nN4r0a1uPFviLW3Ona7p2o3Yt5YV0y4ut7QSq+YnSxgReGQ4xX2j8Lv+T6/jp/2LvhD+V7QB7f4z+Ma+EPjZ8N/g4dIN2fiFb65cC+8/Z9k/sWGCXb5Xlt5nm+djO9Nu3PzZwPbK/PL9qTwjJ46/a2/Z08LjWtQ0GC8sPGonuNKn+y3jQrbWLPFHcAF4fMxtZ4ysgUnYykhhxngz4a+IX8ZftMfsreEfF+uW+kRaToeo+Hbi61S6uLvSL/U7e4ZvKu5Ha48rz4I3KlzxuGTuJIB+oFFfmxoXxk8T/tED9nPwhp95c6Rq99Lc+IvGUdrK0EsA8K/6Jc2cwjIPlXGquiFDwyKQQQcHH+D3weX9oXV/jndfETxt4s8vQ/iFr2k6Pb2Gu3lhBpiQpbypNClu6b5FaXCiYyRoFARFy24A/T+ivyE8IfGH4p/GT4Ifs1/DvW/FF7plz8UdY1nS9d8QWEhtry6sPDn2vCxXC/NFJfrboGkXD/e2nmvZrfwWvwl/aS0j9nzw7r+u3fw++KfhjV559OudYvrm60q801oR9ps76WZru3WdJihCy8SAMpBoA+otb+PfhFPgJ4q+P/gn/ipNG8N6brV9HH+8tPtMmh+ek8IaWPcmZrd49/lkfxAMuM+j+AvFA8b+BvDvjQW32Ia/p1pqHkb/ADPK+1QrLs34Xdt3YztGcZwOlfkR8Avhn4f0P/gml8RviHaXeqTapq/hPx7azR3Op3dxZhIbrUUVorSWVoI5CIlLOiBmJYsSWYnqNU+H+o/B74afs5/G3w9408SXXiXUtb8HaZqIutUnewutM1hY4ZrMaeCLWKJEYCLy4lII3MXf5qAP17rxP4y/GNfhFe/DyzbSDqv/AAnviqx8MhvP8j7Kb2Geb7Rjy38zZ5GPL+XO7O4Y5+Y/j1YeBviX8VNb8D6Xa+PvHXirSLG1W407w1rb6NpWgvcI0lvcTStdWVuLmVcSBXa4cqFIjCnn5+0vx/4q+J/7Nv7GnjTxtePqGuXfxI0mC6uZDukneyTVbUSSN/E7rECzd2JPegD9iaK/Pf49WHgb4l/FTW/A+l2vj7x14q0ixtVuNO8Na2+jaVoL3CNJb3E0rXVlbi5lXEgV2uHKhSIwp5+fLf4mfFP4m/smfspaveeMNT0bxB4t8cWOianqtjMEvJ7dI9Us5C7MGR3dYVcl1YeYA+CwBoA/Yuivy3s/gxLpf7XWo/s8af498XR/DvXPBcXim+059evJria/h1FrJlW/kka8hhlWQSTLDKhkdQGYp8leYa94q8c+D/2If2nfC9h4p1a4m+GXizUdF0TUpr2V9Sg0+OSwmijN3uErGPz3UMWztwOgxQB+y9FfGP7SniDXdK+IH7OttpGpXNnBq3jNILxIJnjS5hOm3T+XMFIEibgG2tkZAOMivnr4o6dcfDz4zeMPiD+02vjIeErvUrWbw54t8OaxexaVoGnhIkFte2NlOjQfvg3mzyQTRy7xkqc5AP1TorzDS/A2qw/FPU/icPGGo3ukarpdtZQ6Ezq2mW8kTmQ3cIAz5kgOCc8jOSRtC+n0AFFFFAH/1/38ooooA8t+KXwj8N/FyPwpF4kubu2Hg/xDp3iW0+yPGm+80x2eFJfMjkzExY7wu1j2ZaNB+Efhvw78WPFnxisrm7fWvGNlp1jdwyPGbWOLTBKITEojDhm81t5Z2BwMBec+pUUAfNyfss/DNvhd8QfhDfyX9/oPxJ1TVdX1Lzpo/OjudXl8+T7O6RoEWKQBogyuRgbi9ZF3+yro2vfDzTPBPjbxx4k8Q6p4e1BdS0bxHLcW1vrelTRxCFBbT21vGhUICHE0cnmF237vlC/VFFAHzvafs+vdfD7xt8PvHfxB8S+MoPHemTaVd3OozWayWtvPDLA32OO3tYoInKyklmicswUtuCgV1Ws/B3TdT+F+jfC7TPEWt+HoNAgsYbLUtKvBa6in2BFjiZnCGKTcq/vEkiaJ+6dMeieIvEnh3who1z4j8Wapa6LpNkFM95fTx21vEGYIpeWUqigsQBkjJIHU1tUAfM3hD9l3wno0vjLVPG+v6v8AEDW/Hel/2Jqeoa1LAJTpQWRfscEdpDbxQxHzXY7U3FjknpXM6Z+x74fh+E8HwX8Q+O/E3iHw3p15pNzpi3k1l59hFosyTW1tFJHaLujJRVfzAzFQArJjNfX9FAHzt8Vv2ddL+JfjzQPijpHivWvBHi3QLSfTk1DRZLYPcWFy6ySW00d3BcRsu9QynaCrc88YsX/7NXw51e++LV1rLXl/D8Z4bSDW7eSVFiRLOyFgn2Uxxq8ZMYDEs7nfyuBxX0DRQB8e6X+xt4Zi1PwXq/inx94t8V3Hw91G3v8ARF1K+tzDbC2jeJIWhgtoklDIwDyyBp/lAWVVLq3sXg34NaJ4F+J/jj4naHqd6H8ftaT6hp0hiazS7s4VtxcQ4jEqs8aKHBkZSRkAdvYKxdA8SeHfFenDWPC2qWusWBklhFxZzx3EPmQuY5U3xll3I6lWGcqwIOCKAOHb4R+G2+Nkfx4Nzd/2/F4efw0IN8f2P7G92t4XKeX5nm+YoAPmbdvG3PNcpH+zf8O2tvirYaibvUbL4wyvLrUE8qbF32i2ZW2McaNGPLQEFmdg/IYcAe+0UAfFvh/9iPwdp0/gu88UeO/F3i+5+Hmo2l/oTatqEMiWi2e4JAI47eNHR1KiR3UzEIoWRFyp7bx3+y/onif4hX3xT8GeMfEPw68S63bw2mr3Hh+4t0j1OG3G2E3MN3b3EZmiQlY5lVXVTjJGK+gtV8SeHdCutOsdb1S10+51if7LYxXE8cL3VxtL+VArkGSTarNtXJwCcYFbVAHgNz+z14fl8XfDbxxBr+srrPw0tbqwguJrlLqXUrS8jjSaO/kuI3eUsYlferIwbPODgclq37Kelr4r1/xP8O/H3ij4dw+Lblr3WdP0K5tVsru7kAEt0iXVtcNbzygDzJIGQtgE/N81fUFlf2Op2y3unXEd1buWCyROHQlSVYBlJBwQQfQjFW6APnH4tfs26F8VNV8HeKYfFWveEvFHghZobHWdHuYkvHt7pUS4hnNxDNHKsgjVjuThhnuQcjwr+yV4C8Gaf48tNA8Q+JLe8+IjWE2pakuquuord2C4W6hulUSLLI3zyBi0ZPyBFi/d19S1wfh34p/DTxf4k1bwd4U8V6VrOu6CcahY2d7DcXNoQ2wiaKNiyEN8p3AYPB5oA8u+HX7OWn+C/iC3xT8UeMte8feKItPfSrO71yS1xZWMkiySRwxWdvbR75GRfMkZWdto5AyDu/GD4D+Gvi9d6D4gl1XUvC3irwrJNJpOuaNMkF/ai4ULPF+9jlikhmAAkjkjZTjjB5r3CigDwf4Xfs+eFPhfbeJrpNU1PxJ4j8aFW1nXdXnSfUbzy4zFChaOOOKOKFGKxRxxqig9Ccmux+EPww0D4L/DTw98K/C1xc3Wk+GrVbS2lvHR7h41JIMjRpGhbn+FFHtXo9FAHztP+zrpcHxxvfjr4X8V634c1HXUsU1zTrKS2bT9XGmqUtjcJPBLIpVDsLRSISnAIJJN/wAN/s7+CvC/wr8Y/CLT73UJNH8bza3PeyyyQm5jbXmka5ELLEqKEMh8rcjY43Fuc+9Vi6x4k8O+HX0+PxBqlrpj6tdJY2Qup44Tc3coZkghDkeZKyqxVFyxCkgcGgDwbxL+y54E8QeB/APhK11TV9E1L4Y2tva+H9f0+5jh1ezSC3S1b955TQyLPEgWeN4TG/8AcGBj0f4Z/D3U/h/p97BrPjPW/G17fyrNJda1Jbs0ZVQuyCK1gt4Yk4ztVMk8kk16ZRQB+dvxB+EXiT4hft2Xuq6br3iHwXBa/Dexjt9a0VliQ3I1e6L20huIZrab5GDmJ0LD5WXacGvoTwt+yx8JvDvws8TfCXVLe68Uad43luLnxBeavObjUNVurkAPPPOoTEihV8sxhBGVBQKRmvo6igD5Q8Hfsl+HtA8WeHfFnjDxv4n+ILeDCzaBaeILyCe206Rk8sThYLeAz3CoSqS3DSMoORhvmqPx/wDsmad8RrnWtO174keMP+EP8S3LXOp+GxfwSafPvcO8CSy273kFu5zuhiuFTBwoUcV9Z0UAfOHiD9m3RNQ+L+nfGjwp4o1nwbrFvZ2mm39vpT232PU7CylMsMFzFcQTYCklN0ZRthKgjgjnr39k7SrTxHrusfDv4g+Kvh/pvim7kv8AVdJ0O7tUsp7ubmaeEXNrPLayTHmRreRMnkbTzX1hRQB5TcfB/wAN3Pxf0j41zXd62u6LolzoMUXmR/ZntrqaOd3kXy/MMu6IYYSBcE5UnBC2Pwf8L2Pxd134zrNdS6z4h0i00S5t5GjazFtZyySoVTy9+9jKwYs5UjGFHJPqtFAHxOv7C/w6W1Twc3ivxM/w0jvBer4Ja+iOhhhL54gP7j7UbUS/OLc3Hl7u2Plr6Eu/hD4Zu/jFpPxtee6TW9G0W50GG3R4xZm1up453Zk8vf5gaJQpEgUDIKk4I9UooA4z4i+BNB+KHgLxD8OPFAkOk+JrC40668lgkqxXMZjZo2IYB1BypIIBAJB6V5lYfs8+HodV+E/iDV9d1TVdV+EFvfW+mzzPbr9rF/Z/YpGvFSFQxWIDZ5fl/Ny27pX0BRQB474P+CPhTwU/xCk0q7vZT8StTuNV1HzpI28qe5t47Z1t9sa7UCRggPvO7OSRwOMu/wBln4fXv7P2hfs3S6hqg8M+H/7N8i4WWD7c39l3Md1D5khgMR3PGA+IhlcgbTgj3u48SeHbPXbPwvd6pawazqMUs9tYvPGt1PFBgSyRwk73RNy7mUELkZxkVtUAfMXjz9l/RvGHxL1H4w6F428S+DfFuo6baaUbnR7uBIVtbR5XCG3nt5opN7S5YyBiu1TGYzuLeifBr4MeEPgd4Um8LeE3u7x7+8n1HUdQ1Gf7Tf6jf3JBmurqbC75XwASFVQAAAAK9ZooA+bPiJ+zRovjLx3N8T/Cni3X/h54p1C0j0/Ur3w/PBH/AGjawk+UtzDdQXETSRBmEUoQSIDjcVAAraj+yX8Kr79nnVf2arZtQsfDWthmvbyK4WTVLm4kuFuprqW5nSUSTyyrl2ZCMHChQFA+nKKACvl3wV+yzoHgG+8aW3h7xdr0fhTxr/ack3h55bV9PtLnV23XM9qWtjOjZyUUysilmypyMfRHiHxJ4d8JaTLr3ivVLXRdMgaNJLq9njtoEaZ1ijDSSFVBeRlRQTyxAHJAraoA+dNZ/Zk8Ba5+z5o/7Nl3f6mnhnRINJt4blJYRfsmjTw3EBeQwmIlmgUSYiGQTtCnBDviP+zno3jbxzF8UfC/inW/h/4xFoun3Oo6FLbr9us0YvHFd293BcQS+WWYxuYw65xu24FfRNFAHKeCPC0ngvwxZeG5da1HxFJab91/q0yz3s7SO0haV0SNeC2FCoqqoCqAABXzDq/7FXgjVJfGmmW/jPxTpXhPx9d3uoat4esb+KDT5b3UQftMquIDcoshO5oRP5LHhkZCUP2TRQB4Jp37O3grTNd+FHiGC91Brn4PaTd6NpCtJCUnt7y1gtHa7AiBeQJboVMZjAYsSpBAHTaF8H/DGgfFLxl8W7ae6m1bxxZ6bY30EzxtapFpayrF5SCMOCwmbfudgeMBec+q0UAfIfgv9jX4f+FPhz4g+C2p69rXif4c6zE0FtoGqTQPbaZG0xuALSaKGK5DJIQY2eVim0EfMNx774afA7Wvh9rUGqan8UvFvjG1soHtrWx1m6tJLdEcAbpTb2sEtxKuPlkmdyPrkn36qkd/Yy3k2nxXEb3VuqPJCHBkRZM7GZQcgNg4J64OOlAHm3wm+EXhv4N+HdT8NeGLm7urXVdW1HWJWvHjeRbjU52nmVTHHGBGrMQgIJA6sx5ry+1/ZE+Flt8A/Dv7PLXGqSaN4SkjudI1P7SkWr2N7DM88N3Bcwxxqk8byNtYR7dp2srAkH6kooA+dfhl+zjongHxtcfE7xF4o1vx/wCMZbP+zYdU16aCR7OxLh2gtYbWG3hiDsA0jBC7kctjiu4+Dnwj8N/BHwSngPwpc3d3p6Xt/fB7145JvN1C5kupRmOONdoeQhRtyFwCSeT6lRQB4X8Y/gF4X+MV5oHiGfVNT8LeKvCkksmk67os6W9/ai4ULNFmRJYpIZQAJI5I2UgcY61zWpfsu+F/E/w3l+H3j3xT4i8U3banHrVvrt7fqurWGpQBRBcWEkEccVqYdvyJHEE5bcrb2z9M0UAfOvwy/Zx0TwD42uPid4i8Ua34/wDGMtn/AGbDqmvTQSPZ2JcO0FrDaw28MQdgGkYIXcjlscVXsf2XPh/YfALxB+znDqGpnw34k/tT7RcNLB9uT+1riS5m8txAIhteVgmYmwuM7jkn6SooA8a8XfA7wn4zT4dx6pd3sQ+Gmp2urab5MkS+bPaW8lsi3G6JtyFJCWCbDuxhgMg6fg/4R+G/BPxE8ffEzSrm7l1T4izadPqEczxtbxNplotnELdVjVlDRqC+93y3IIHFepUUAfLq/slfDlf2ZLj9lIajq3/CJXMU8LXXnQf2iFuLxr5iJPI8nIkYgfufu8HJ5r0bU/gx4X1X4sad8Yri7vk1rTNCu/D0UUUyx25tLyaOZ3O1BKJg0Y2usigDPy5wR65RQB8maR+yXpUPibwzrPi/4heLPGul+C71NS0bSdavLae2tr2JGSGeSWO2jubh4Qx8ozzOR1O481Dqv7Imj+ItRtrbxX8RPF3iDwha6lFqy+G9Qv4Liwa5gnFxCkk7W/22SCOQKyxPcsAVHJAxX1zRQB8teNv2VPDviz4p6j8V9G8ZeJfB2oeIbO2sdcttCvo7W31WGzDLAZy0LyJIiMUEkLo4TIVlJLGLTv2PvhZpHwr0D4O6XdapbeH/AA14kj8T2YWeEzLdRX76gkDO0JBtxI+zG3zNgAMm7LH6fa/sUvU017iNbuRGlWEuBI0akBnCZyVBIBOMAkVboA8i+MPwV8JfGnR9OsPEM97pWo6FeJqGlatpc/2XUdOvEUr5tvLtZeVYq6Oro6nDKcDHnupfCXxN4T+FfjrTbrxl4y+I2r+I9Pazjze2NnfQsyPEn9nvDDaW9q+ZdzSsCflDMWCha+n6KAPNPgz4Y8VeCvhJ4N8I+OdVfXPEWj6TZWuo3skjTNcXcUKrM5kfDPlwcO3zMOW5Jr5x0T9iiz8Kf2lB4J+MnxA8N6fqeoXmpPZWF/piW0dxfTNPN5avprsAXY4yxPuetfbNFAHyp45/ZR0nxp8SIvitZfEPxd4V18aJbaBcS6NeWcAurO1mknUymWzlfzGeQlmRk7YA7+n/AAg+CPgL4JaRf6b4MguJbrWblr3U9Sv7iS81HUbpuDNdXMpLyNjgDhVH3QMmvXKKAPK/C3wh8NeEfil46+Lmm3N3LrHxBi0mK/imeM20S6PFLDB5CrGrqWWVjJvd8kDbt5B/JjwNr2reF/h74gj034neO/h18SpL/Wr5fAVloQ1KC11K8vZ54ra0iudPuHnglZ1bzFufLbeXLRgkL+3dFAHxZ4M8I/HXxL8UvhH4/wDHkuo6NeaD4RceK4Yb2NdFvdSvowoto7ONm3XEMwaWWXCooWNUL5+X3P4ufBrRPi6fCd3qGp3ujal4L1q31zTrywMQkW4hR4mjcTRyo0UkcjK67QehBGK9gooA+WNc/ZQ8M6n4+8S+N9H8YeJfDVt43eKXX9J0q+jtrLUpYYlgDs3ktcwM8a7ZDbzxMw7g9aOofsdfDyf4J+CvgppWt61pMHw6vINR0HWLeeEapaXlu0hSTeYfJfiV1ZWi2lT0yAa+taKAPmLxV+zFp3ia78OeLIPHHiHRvH/hywbTR4psZbSLUL60dvMaC+hNs1nPEZPnCG3GxuVIOc6viD9nLQ/GPwtt/hb418V+IPEEMOqWmrvqV5c276hLcWV0l3GpYW4hWLegXYkSgJkLtPNfRFFAHkPxi+CnhL416Npun+Ibi90rUdCvE1HSdW0uf7NqOnXkYKiW3lKuvKsVdHR0cHDKcDHO/Cr9nnw98NfE2seP9V17VvHHjPXbaKxuda12WKW4WxhYulpBHBFDBBBvJcokY3OdzliBj6AooA+Frz9gf4eXPhXXPhlbeOPF2n/DnWRdFPC9rqMMWm2T3ZZ2+zn7O04jSRvNSB5Xh3/fR1JWvX/iB+zX4U8cR+EtQ07W9X8J+J/A9sbPSte0ieKK/jtnjWOSCYSxSQTxSbFLRyREbhldpJz9FUUAfOXhH9mPwL4T8EeOfCb6lqutaj8SYriLxBr2oXCT6teme3a1UmXy1iQQxNtgjSIRxjohyc9RL8FdGT4OaH8F9I1/W9EsPDljp1hZalp179l1ONdMSNIHMsaCN2IjHmK0RifJBjxxXstFAHydpv7Ifgq5XxjdfEvxHrfxD1bxtop8O3l/rE1uk0Oklmf7NbJZQW0UQ8xvMLBCxcBs9qztL/Y28Mxan4L1fxT4+8W+K7j4e6jb3+iLqV9bmG2FtG8SQtDBbRJKGRgHlkDT/KAsqqXVvsKigDx/wb8GtE8C/E/xx8TtD1O9D+P2tJ9Q06QxNZpd2cK24uIcRiVWeNFDgyMpIyAO3M/GD9njSfiv4s8NfEHT/FOs+CfFfhWK6trXU9Ekt1le0vdvnwSpdQTxOhKArlMq3IPavoaigD5o8Z/szaXr/ju6+Jvgvxp4g+H3ifVraC01a60Oa1CarHbDbC11b3dtcQNNEhKxyqiuoOMlQAPnH9qXQD4S8QfBGw1zxJ4p0Pw74ZbVpJ/Hdkj6lqlpdtarBBBdssE0Sx3glfzJZLdlzGqLsDMw/SWigD81/hbp3j74yeJPiH8Kb7x54h8f/BHxF4SuNOm1zVdMt9MvItWv2a3ki064FnbrcoLVnZnMEkaSbBk5K19aeI/2fvBvifwl8OPBt/eX8dl8MNU0bVtNeKSISzT6HGY7dbktEysjA/vAioSfula90ooA8suPhF4aufjRafHSS4u/7es9An8OrBvj+xm0uLqO7Z2Ty/MMu+MAHzAu3IKk8jx/w/8Asl6X4K1E23w8+Inizwp4RN61+PDWn3lqNNileXznjgaW2kuoLeRyzPDFOqfMQNoOK+s6KAPLdN+EfhvS/jDrXxtt7m7bXNd0mz0aeFnjNotvZSyTRsiCMSCQtKwYmQgjGFHJPGWf7M3w4t/A3xN+Hd497qGjfFfVdV1fV0nlTfHPq6Iky2zRxpsRPLUx7t7KeSzV9C0UAfHkn7H2n3nhHw1oOrfEnxXfa14JvzfeH9feexXU9NBtha/Z0K2ghlhaIYkE0bs5J+YA4rrvAn7LngjwR4n8V+Mp9b1zxNrHjfSodI1ifWbxLk3MUBlxIAkUYiYrKU2RbIgqrsjU7i3vei+JfDniQ348ParaaodKu5LC8+yzxz/ZryEAyW82wny5UDLuRsMMjI5FbVAHmHwZ+F1j8Fvhpofwu0nVr3WtO8OxG2s59QMTXK2qsfJiZoY4kYRIRGp2AkKM5OTWB4R+Ang/wZ4R8eeDNLvL+Wy+Ieq61q+oPNJE0sVxrpJuFgKxKqomf3YdXI/iZq9uooA+VfFH7JHgfxB8Lvh58NdN1/W9AuPhYLX/AIR/XLC4hj1W2a1t/s2XcwmFxLFxKvlBW7AdK3vhl+zL4K+GPjvWviVbaxrPiHxF4k0uDS9UudZukuzeLbyySrNIBEgWQiQRbU2wiNEVY1IJb6MooA+SfBf7Iuh/DrVbKHwR8QfF2j+DdOvhqFt4Vg1CJdKhkEvneSj+R9rFqXyTb/aNjZIOVJU91o37OvgbStZ+J1zcy3OqaJ8WXWbWNFuzE9iJ2t/s1xJDtjWVftESoJA0jAFcrtr3yigD5n+G/wCzfc/DTVdJbT/ih4w1Tw7oAK6fod/eWstlFGUaNYpJFtUup44lOIklnYLhSclQRymr/sYeB9U8W+MNag8W+JtL8PePrl77XPDdjfpBpN7eSxiOaVwIfPXzgo81EmVZMAOCnyV9h0UAfND/ALNNnpvgLwP4G8CePvFHg/8A4QDT102wvNOu7cyXFusaRgXtvNbyWlwf3akMYFZTnYVBIPk/j/8AZk1Pwv8AB5Pht8HdY8SN4m8TeK7bW7/xGupRwXhvWdZLm+1GVRGHgEMQQQQxYZxEgVRlx930UAct458I6b8QPBPiDwHrMksOn+JNPu9NuHtyqzJDeQtDI0ZdXUOFclSVYA4yCOK+b/h9+ybcfDiTw5Bo/wAZvH93pHhj7IlvpV1faa9jJbWe0R20qJpyOYSihGCurbejA819c0UAeWfDX4ReG/hbqXjXVPD9zd3EvjvXJ/EF8Lp43WO6niihZIAkaFYgsSkBi7ZJ+YjAHmuu/BLSPBPg34w3vg4a9qmq/FCa4vrmDT7y3tb1Ly5tks1FhcMsS2+0KrB5WfYctkgba+naKAPNPgz4Y8VeCvhJ4N8I+OdVfXPEWj6TZWuo3skjTNcXcUKrM5kfDPlwcO3zMOW5Jrw2D9jP4ewfCSL4Nx69rcWk6R4hfxLoN1FPBHfaHemZrlBZyrCFKRzSSsomSQ4kYEkBcfXtFAHzX4A/Zk8PeCPijF8aNR8WeIfFnjEaRcaLNeaxdwyxy2txPDcYEEMEUUPlvCNghWNcM5dXY7hjeCf2RfA3gvTvF3gv+3NX1r4eeLk1BJPCeoSW8ml2h1ObzrhrZkgS6T5i3lgzkJuZhl8MPq2igD5w+Gv7PWofDfWNMul+KXi/xBo2iI8djpGqXdpLaJG0ZiCzPHax3FwI1P7vzpm2kAnLAGus8D/A7wX4E07x7pFm91qNl8R9a1HXNUivHRl8/VIo4Z4YvKSMrDsjAUMWcZOXPGPY6KAPh63/AGE/A/8AwjWieCdY8e+MNW8NeFLuwu9D0241C3Ftpx06ZJoUVY7VDOo2GIfaDKUjYiPY4Dj6BsPg1omlfG7Vfjnpmp3tvqWvaTb6TqNgGiNjcrZyM9tOQY/NWaIO6ArIFKtyuRmvYKKAPLfE/wAI/Dfiv4peCPi5qNzdx6x4Ch1WCxiieMW0i6xHFFOZ1aNnYqIV8vY6YJO4NxjnrrwV4G+E/jP4gftF31zqUl14h0/TotThjia8jjg0hZViNtbW0LXDOwmbeAZCcDao5z7nRQB8G/sifDPd8RPiz+0jc+H77wzbfEbU1XRNO1ON4LuDTYRvnuWgfDQHULx5Lho2AbG0nrXlXwc/Zs174gat8eJ9T8aeMfAFprXxF1+Oa00q4S0t9S06SO3KSotzbylC+5k+0W5RnUbSzbV2/qPRQB8+eKf2Y/hT4k+Fnh34SWdpceHtK8GvbTaDdaVObe/0q5swVhuLadg580AtuaQOH3NvDEmk+Fv7O2hfDnxhqHxK1vxHrPjvxpqNomnHWNdmhkmt7BH8wWttFbQwQwxM/wA7hY9ztyzHpX0JRQB8i+HP2PvCvhfwz49+HemeMfEI8C+PLLV7NtAaa0a00z+2yxupbF2tjMrgu/liR5EXcxKsTkej+Kf2fvBvi7wB4I+HOpXl/HpvgK+0XULGSGSJZ5JdBKm2E7NEysr7B5gVUJ52la9Sg8XeFLq91nTbbWrKW78O7P7ThS5jaSx82Pzk+0oGzDuj+dd4GV+Ycc1uwzRXEST27rJFIoZHUhlZSMggjggjoaAPmPxJ+yz4f1n4keIfiRoXjLxL4Sm8Zrarr9jo17Fb22p/YohBCzM0LzwOIhsZ7aWJyP4g3NZyfsffDyw+D/gf4OaBq+r6XY/DnWo9e0S/SWCS9gvI555l3NJA0Tp/pMibTHkoRklhuP09rWt6N4b0m71/xFf2+l6ZYRtNc3V1KkEEESDLPJJIQqKBySxAFXLa5tr22ivLOVJ7edFkjkjYMjowyrKw4IIOQRwRQB8zeJP2WfD+s/EjxD8SNC8ZeJfCU3jNbVdfsdGvYre21P7FEIIWZmheeBxENjPbSxOR/EG5pmifsj/DPw/8Pvhp8NNNvtUTSPhV4gXxHpJaeFppLpHunWK4Yw4eEG7cYUI+Avz5BLfUdFAHlp+Efhs/GwfHj7Td/wBvr4ePhryN8f2P7GbsXm/Z5fmeb5gxnzNu3jbnmuJT9mL4at4P+KPgXUWvdR0j4uanfatq8c8qbo57+GKFxbNHGhRUEKNHu3sG5LEYA+iKKAPjjw7+xh4U03WPBXiTxV478WeMtX+H99HeaPcavfwyiCOOGSH7OY47eONo5FdWkcr5zNGn70KCh1PH/wCyZp3xGuda07XviR4w/wCEP8S3LXOp+GxfwSafPvcO8CSy273kFu5zuhiuFTBwoUcV9Z0UAfM3h/wD4xg/ai1jx7aT6rp3gqy8MwaKLG4vFbTLu+86OeOeys1LGIW8QeKSR9pd3Kou1WZvpmiigAooooA//9D9/KKKKAPif9uS5uLbwf8AC9raVoi/xJ8IoxRiuVa9AKnHUHuK9l/aY+KF/wDBf4A+PPijpKJJqHh7Srie0Eg3J9qI2QFh3USMpI7jjjrXm/7ZvgT4ieOvh/4SPwy8PP4o1bw34w0HXZLCO5trR5bbTbgzShZbuSKIEgADLZyenWmX0vxK/aO8JeLPg38WvhFqfw88P+JdGu7VtUn1fSb8LLKAiKkVlcSybwWMikrs+TBPIBAPMfD37Bvw81r4aWWteJNU1af4u6hYx3k3jQandjVIdWlQS+dCVlEaQxynCQhdmwAEE/NXun7HfxV8Q/Gr9m3wT8Q/F2067eW89rqDoABJd6dcy2U0uAAB5jwlyAMDdgcV4ho/i/8Abf8ADHw6tvg9F8LLbUvGGn2a6VbeMBrNmmhyLGohj1KW3Y/bA4XEjwCAlmB24U4HX6VDqv7GPwl+Cvwj8J6Ivi7T7nWdP8OandtdpazRSapKzTXsULhjNm4keUxg5WMEk4BcAEf/AAUc/wCTLPiZ/wBe9j/6cLavevir4k+M+jPp9j8IvCOl6690kz3V/rOrHTbGxEW3YHWKC4nlaTcSNiBQFO5h8oPCftm/DTxZ8YP2Y/Hnw68C2y3uv6taQ/Y7dpEiE0lvcxT+WHkKopcRkAsQMkZIFeK/HbwV8SvH3xN8C+OfEPwvu/iN8O/+EeZbjwe9/YwtYeILiVJVuL63ubiO0u1jizDjfKsbbnUHIJAPJ/j5+1P8TfG37EfxM8beCtLi8M+KfB+qXXhvxBJZ6xIG06a2lhU3Om3UEIa4EgmiKZMWFdvmJXDfVPiT46fFrwRovhjw14k8EabdfFHxzqdxY6Lo2natJLYNb2sAnnvLq+ltImiigQMZAsDsSUVAxb5flWw/ZT+Luq/ssftG/CyfwtpvhLWfiF4iutX0LTLG6gksRbmCwkhgjkjWNYxvgaH544wGG7aEIY918a/hh8Qv2jtG+HPxc8SfB9ZNc+Heo30d54L169sWXVrDUbaKO5e2uYZpYFkjlRWt/OaPd5ZLbQV3AHsul/tH+M9Ft/id4X+Kfhay0vxx8N/D58SLBp19JdabquntFO0ctvPLBDKmJbd4pFeLKnBUsDxH8Mvjv8efil8PR8V9B+FdtHomraXaXWh2NzrAt9Rv552jDSSh4PJt7TYzyRvukleNQwjDOEHDeE/g7pdz8Lvitb+BPgHH8J9Z8QeHL3SrESXGm/bNSkubaZfKcWk8scMay7ApklG4sSQoXJueN/hV8bW/Yn8AfC/wdaSJ4p0PS/C9rrelQagljcXlpYRQJqWnw30bhInlVGTzFkCkZAbDcgHc/DP4/wDxBvPjfP8AAP4zeG9I0TX7nR31zTrjQ9UfUrWWCGZYZreYTW9tLHPGXVvubXXJBGOed/4J8f8AJt8H/YxeJ/8A083deZfCH4IeKNN/aa8H/FLw/wDBiy+E/gjTfDmrabLDFcaeb97q4ltpI5LyKyZ1JkCsEYSzP8rGUoSoPu37G/gTxv8AC/4X6t8PfHejSaXd6T4j1yS3mMsMsN/aX17JeQ3MJidyqlZgpWQK4ZSCtAGbrvx/+Lnir4h+LfA/7PPgXTvE9v8AD6SO21nUNY1V9Mgm1CSITGwshHbXBeVI2XfJJsjVmAPrWF4i/bW0XTf2f/DXx50bwtfX66r4gs/D9/oxIGo2V3JdtaXcKoARLNDIjBE+USfLyu7jK03RPjj+zj8SfibceA/h1N8S/CvxI1dvElk1hqNlY3FhqlxBFDc294t7JEPJdoVdJYt+wZDKTiuBuv2XfitpP7PXg7wn9mt9Z8Z3XxH07xr4hS0mjitrczap9tvPJa4aPelvGQoAyzlSVU5AoAzv2jfFfxsOo/AHxL408DWUHilfiAzadoWm6qLjzIW0u48pLi8mihjSQOziUorIqruXeTtr6U+Hnx3+I5+Mi/Av45+ELDw1rmraXNrGi3mkajJqNhewW0ix3FuzTW9tIlxDvRjlNrKSQRwDn/tV+Bfij4o8QfCDxX8LvDsfiW48D+J21i9tpb2GwBtksbiPAklz87s6ogCsNzDeVTc64ngjwp8Wvi1+0jpXx2+JXg6T4faD4G0a90rRtNvLy1vNQu7zU3jN1dS/Y5JoYoUjiEaJvLMfn+7gUAfJ/jH4s+J7D9gLwj4w/Z38H2PgHStX8QR20tvZarPavYFPEggVYHjg3SLeSRss+WTZHIwAkHy19neL/j58V/CC+Cvh1L4I03Ufi746kvjbaVa6rI2k2llYYaa9ur97VZFjVGT5FgLO7bEz1r50tv2bfjOn/BPHTPg4miRx/EHQtTbVotLlu7fbObbxDJqSQ/aEkeENLBgqS+AxAYrzj1LxppPxp1/xx8Ov2qPCfw4u4Nd8N22p6LrHhK/v7GLULjTL8xss9rcJNJaiSKWIOEkkQyISMo2MgHovgT48fEnxBrfjP4ReKfB1ho/xY8K6fBqdrYrqUkmj6tZXTNHDc2979mE0cYlRo5A9vuRgMBsnHw5+yx8bfFPwB/YW0L4xeKPBFjf2V3qNpHFc6fdPLquqjVtXlivZ7iP7OuJo2cGKMSSebwNycV9nfB3wP8TPFvx+8SftL/FDw7/whRuNAtfC+iaLLcw3d6tjFcvezXN49s0kCvLM4CRo7FVHzEHr8+6V+zl8Zbb9gH4c/BOfw9t8aaDq2kXV7Yfa7Q+VDa62LyVvOE3kttg+fCyEnoAW4oA+gPEn7RHxZ8B+FdGPjT4e2i+PfHesrpfhbw3Zar57SI0HnPLqF20CxwCBVdpzEsqqAApbOR0fw2+OfjyX4rP8DPjl4WsvDPiu60x9Y0q50m+fUNL1K0hkWO4WOSaC3ljngZ13xunKncDioP2lvhp4+1/W/hv8YfhXYQ634o+F2q3F6mkzzrajUrHULV7S8t4p5P3cc5jYNE0mEyMMRmuU8FeFPiv8W/2i9F+PHxH8HyfD7QvA+jX2maPpt5d213qN3d6o8Zubmb7HJLDFCkcQjRDIXZsscDAoA8ji/bf+NWp/A0ftI6N8IbU+BNIadtWefWwl9LBa3T21xLp8C27K8cWw5aZ42ZgyrHtAdtr4/wDxR+PVn+1B8DtF+FGnWF9oniCx1+8tILnWrmxg1UxWMLyC9jitZVQW4kV4DiXcxPEeMmr4X+AXxa07/gm/rfwEvNC8vx3eaZ4ht4tO+1Wx3S32o3U9uvniUwDfHIjZMgAzhsEEDsfi/wCAvivoPjL9nv4veDfCM3jBvhnaarp+saRaXVrBe+Xq2nwW/mwNcyxQv5MkPzL5nIPHGSAD1r4mePv2ktAMj+A/AGgXdlp9hFdXV9q/iBrKKa4Kky2tqkdrK2VIwJZzEhyOOuPmT4s/FvR/jv8ADz9k/wCLmg20llaeJfiVoVx9nlYM8Eq2uoxzRFgAG8uRGXdgZxnAzirXi74b/FHX/jz4k8a/EH4MJ8U9I1mz0k+FINU1LTzpnhyRLUC9hvLW4kkVJDcsztc20Ny7KMJwAtcj4c/Z4+NPgz9lr9njwrP4YN/4m+FfjiDXdV0u1urUyNYpc6iGaCRpVhchLqOQLvB25BAYEAA/Qn4z/Fjw18Dvhjr/AMU/FqzS6boMKuYbdd89xNK6wwQRLwC80rpGuSBlhkgZNeZ+CPiZ8eoI7vxP8dPAujeDPCMOl3GqS3NprUl/d6ctuokMF5CbSJWfZuYvC7KuwjkkZ1v2p/g/rHx0+BviH4e+GryLT9dmNpe6ZPcAmBb7TrmO7gWXaCRG7xBHIBIViQCRiuIstc+M3x18Ma58Jvih8JbrwDpniHQ9Q0zVtUm1ewu4kmu7cwbbGK1kllnU7y2+TydoGDluAAeaH9r/AOLWl+AdP/aJ8VfDC3034Nak1vMbldUaXxBaaXdyKkGp3FgLfyTCwdHaKO4aREbcc4Neh/Ej9o74g6T8dh+z58KfA8HinXb3w3a+Iba9ur82enwQzXdxbStdyLDKyxp5KbPLV3kaQKAoBcfPmr+Cv2qfG37O1j+xxq/w8i0mT7DZeHdQ8ZDU7SXSP7Is9kbXdvbK/wBse4mgi2iF4UCyNlmCivpbQ/hT4q0b9sS4+I9tppTwZH8OrDw9BeGaIk31tqc85h8rf53ELK28psOcBs5FAHhmlfEj9p+6/bg13whHoeky6TZeFtIaexbXrpbSK0l1C436hDF9jKNeOCyGMqvyIgMxHC+ia3+038V9UtvGvjr4SfD+x8Q/D/4e3N9a313e6pJZ6jqkmlZF/wD2XbJazRssLK6K00qCV1IXA5qTxV4c+KvgH9sFvjB4Z8E3PjPwt4u8MWOgXUlhd2cE+m3dpeyzCWWO7mh3wNHLklCzAqRjoG+bdP8A2WLb4X+IvF3h/wAQ/s52XxistX1i+1TRddtrnTYZRBqErT/Y9RW/mhkRrdmZRNGJVdNuFDDaQD6w1z9rvwn4Q13wxq3jK3TSvht458Mvr+ieJHlISSaC3+2S2VxCV/dStanzYfnPmYZANwwek8P/ABm+K2qeAfh14xufhbdtd+OtUt4byxgvE8zQ9Kuy7R314ZEUsyRBGliUAqzFNxZRu8W+Mf7M2v8A7Ql1o3wb1bSD4E+EvgnQ0n037FPCxm8QPA1vZiKOJ9622koSwEixiaRlADIua7a8+NXx3+Gfwk+HmrfErwHDe+LNR8R6Z4Z12OHUraNGS8uPsi6nZLGZPNEzmOQW5CSIGbeFVGYAHSftwTzW37IvxZnt5Gilj8P3hV0JVgdvUEcivefh6zP4B8NO5JZtMsySeSSYUry39q3wP4o+JX7OHxF8BeCrL+0de13R7m1srbzI4fNmkXCr5kzJGufVmA96434T/En4+l/DPgvxb8CtU8O6dBBBaXWqy67otzDAIYgpkMNvdPMwJXGFUnn60AfnL8J9M/Yvl+F3jzxV8bvFdlofjq38S+J9txBr81nrsQiv5vsptraC5WVnUAGJRGwbA4IzXtreIvjN8TPh5+y9+z18SdW1HQdX+KNlfX3i27hke11ObTdEtVm+yvIu143vBLEJ2GHHzA9WB+mP2SfgRP8ADjwZq7/EPwnY2Xie48T69qENw6Wtxcm1u76Sa2cTxGQjMZBC7gy9CAeK3v2lvhT498Uap4C+MPwgW2uvHfwuv7m7s7C9l8i31Ox1CD7Pf2JlwRFJNGF8uRgVV1G7AO4AHzp8afhF4M/Y3Pg/45/AKGfwta2viDTNL8R6VHd3M1hq2l6pOtrIZYZpJB9pid1kjmHzZ3bt2eP02r8/fFXh/wCPf7U3iPwb4a8e/DyT4X/D/wAMaxaa7q7X+pWd9e6rcaa3mW9lbx2TyqkHnYeSWRlLKo2Acg/Rmk/FjxLeftFa98FL/wANJBpGn6Ba63aaxHeJK0xmmMDwzWwXdCdwbyyx+cRuR7AHjXxG/wCT8/g3/wBip4p/9GWdT/HL4/8Ax6+Dem+IviJP8ONHk8A+FZGad7jxB5esX9nGR5lxaWyWzQKQuWWKWcOwGMKTiuh+JngTxvL+1Z8I/iroejSat4f0vTdd0fU5YZYUawa/Fu8E7pK6M8ZMTK3lhmU4yORXxV48/Z6+NHirw/8AGPw34o+Dlt47+IHiqbXP7G8Z6nqdhJaW+m3QcWMNklw73FnNFHtjSGOGOPfhpJlUl6APYvi/8W/j7cftWfCXTPhJpmnaj4f1jw/rWo2Fvda3c2NvqscsNm7S3sUdrKqPbE/6OMS53scxkkH3DXPjj8XfEnj7xF8PvgV4L0vX5/AyWy69e6xqsun2gv7mFbhdPsjDa3DSyiJlZ5HCRpuUHJJA8p8a/D74w+BvF/7P3xY8LeC5/GJ8CeHbzQdc0myu7SG9ha9tLVFlha5mjglCSwsrgSdCCMjJHn/jD9mwaB8avGnxF8R/Ae3+MGg/ESS11OIRz6cmq6JqAt0huLWZb2eGKWCRlDrJFKdh3gqQQ1AHsXiD9taOD4afCb4i+DvBV5r0vxS1SbRY9KEyw3drqMMVyrQMxUodl3bmGSRiqom6XkLtPpmqeM/2rLfw9pMdj8PfDX9vXUdzNqE9z4hmi0nT1jkIhiDrZPczyPHhmIhSNSCN3TPlviP4Ia/Pb/s7p4A+H9v4P0vwZ4obWNW0e1ubUxaVBPa3QkIZWRZiZphu8kMSWJAIyaq/tD/Dj4geJPj3ofijWvhxJ8YPhnb6CbWDQVvrOG3tNeN00hvLu0v5oYLhGg2Irt5hjwcR5OSAeQftCfHAfHn9grxr4lu9LXRNZ0TxNpWh6tZR3C3cMGoad4hsY5RDcKFEsTAq6OFGQ3fGT+o2tagdJ0e/1VU802cEswTON3loWxnnGcV+VA/Zi+NUf7IPxq+HC+D7LSfEvibx0/iXSdF067t2tGskvtPvVgtpf3SIoS3kjjEixHKjKKCK/TjXbyfUPh7qF/dWkunzXOlyyvbTFDLAzwFjG5jZk3ITtO1iMjgkc0AfBth+2Z8crv4I6V+05P8ACKzg+G/2OG91JBrnmayLViFuLu2tltvJaGH5mVZJkkdBuKx5wPXPiJ+014w074xaN8FPhH4Mi8Yat4m8Mp4i0+8mvWs7GKJrloWkvHEMjRwKgVgyqzvI6RhRu3D4r+EB/aI+K37Cng79nvwf8N/sVl4u0GLTG8WT6paPplvpVwSs87W4Zbw3HlFkEIixuO4SMoNfbugfBzxP4Z/a20Pxvpunk+CNF+GY8Lx3rTQ5+2xanDNHCYt/m5MCFt4TZ23Z4oAl1b44fGTXfHOpfC34Q+DdI1rxB4OsbCfxPd6nqk1jplrfX0InjsLR4rWeaeVo/n3siIiMm47m2jn5f2xk1j4c+AtW8CeEZtU8ffEbUb3RrDw5cXK24tr7SmkXUzd3WxgsFmYmLOsZZwV2p8x2+QfFD9mmLRf2g/GnxT1f4LWvxq8NeP0sLgLFLZRano9/ZwLayJsv5YI5ba4VEfcku5HDAptwa7DxJ8DfFvh7Tfg78XPgt8NrLw7rvwzvdWnn8Ex3ltEs1lrsRgvI4rtCLb7UNscq5YRltyl+hYA9p+Fnxy8a6n8UdQ+BXxp8MWnhjxrb6Yut2Mul3r6hpep6d5oglkgllhgkjkhlIWSKRA2CGBKmvn7QP2w/j/4v+C8n7Q/hr4P2EvgrSEvZ7+ObXtupXVvp08sV1LYQJashWIRMcTyRu5UqqY2s3p/w58IfFP4lftGL+0P8SfCj+AtJ8O+H5tB0TSLq6t7vUbiS9nSe6vLk2jywxIFjWKOISOx+Zm28CvjH9nLXv2h/EP7EVn8Hfhr8Mvt6+KY/EOm2PiWXVbSLTbWC+1O8iuLi6hdlu0kty77Y44pRJtUhuWRQD2n4kfG745eIv2l/gxN8FtN0/UvC3ifwxq2saXBea1c2EGqwXNvZS+bfRRWsqxvbbh5AxLne5zGcg+7eGfHvgPSf2jfjC+r+GLLQdV8K+G9D1HWfEEcrSS3VrJFcSGORdgwlssJwwyXGMgbQK8z8Z/B34ifBrxh+z/4v+F/he4+IWkfC3w9feFtQs7S4tbS+aGe1toYLqIXcsUTfPbnzF8wEbgRkZI61PgZ4u8a/GP48aj4r02TRvC3xS8HaPodvcmaCWQS/ZryG7Ty4pWYND568kBWP3GYZNAHQ/CL42fH/AOLh8PeP7H4Z6dpXw08TsslrNd6yy64mnyqXhv5LNbVoNkq7WWFbguAwOcZr6H+IeteONB8NS33w78Nx+K9caSOOKynvk06EB2w0ss7pJtSMfMwVHcjhVJr5a+AviH9pXwL4b8F/BHxr8JjM3hiG00i68TW+sWQ0iXT7NBCt5FEWN4ZXjQHyDAvz9WRTkdf+1/4M+J3jf4faDp3w5sJtctLTX7C78QaNa3w0251fRIRJ9ps4rlnjUF3MbFDIgdVKlgDggDfg/wDH3xp4j+LXiX4E/FrQNL0bxdoelwa5BJompPqVjdWE8rQNlpYYJYpoZVCujpzuDDAxnzn4MftgeMvib8K/+GhvFPgi38H/AAt07R7vUL6+uL95r+WezUmUWVqsCh4FZGjEkkitI4+VNuGOB8Bvgn4t8OftP3XxUsvhLYfCjwPP4HbRLaxs57Az/bF1FLjddw2JKLNImeUaZdqLul3nYN34Qfs2eLL3/gn5Y/sy/EeH/hHPEN7oF9plwvmRXItLieaZ4XLQO8bhSyMQrnI4yDQB0Gl/tI/F/Rr7wN4h+Lfw8sfDXgv4kalaaTp0ltqsl5qunXWpKTYjUrY2scKidgEbypn8p2CtnrUnxy+P/wAevg3pviL4iT/DjR5PAPhWRmne48QeXrF/ZxkeZcWlsls0CkLllilnDsBjCk4rxv4LfAzStI1vwlpHjL9lXTdH8Q6FNbte+JrW70x9MSe0+ZL+zAlN4zPIgdYmt0ZM/M3GT5f48/Z6+NHirw/8Y/Dfij4OW3jv4geKptc/sbxnqep2Elpb6bdBxYw2SXDvcWc0Ue2NIY4Y49+GkmVSXoA/Su5+JevXHjHwHpXhjwld634W8Y2d1eXOvxzRx2+mJHAs1sJYmG9zcltq4I2nnnnHzF/wUU8O6j4y+DvhHwVpOoS6VeeJPGugaZDdQu0bwy3srwxyBl5+R2DfhXplv8QPG3wlv/gH8Irrwql9beLLNtI1K7W+iWfTbvTtM+0DbbgN58Z8mQPIrbUwOSWQNr/tN/D7xf8AELTfhxb+D7D7fJoPjzw1rV6PNii8rT9PuxLczfvXTdsTnYuXboqk0AfIH7Qnxs1v4vfsa+FtG0ySTTfFHxDtLtdbSE+XLZReGYJbrX1BH3cTWhtPrMvHNe5x+E7rxz+wZ4PhXxLrPhyey8D6bqH2rRr1rK6leDSAwjkmAZzG7HLgEEkDnrnM1f8AZL1fSfEPxz8b6JfLqq+NtC1e28NaP8sf2C91y3Dap+8kKoPtt3DC4OQF+bccHNdVqNl8X/Af7Kfg34ZeHfh1d+LvEUnhK20PULa11HTrX+z7iPTUt2Mkl1cRxyqJMr+6Z+mRwQaAO/8A2QNQv9W/ZZ+E+p6pcy3l5deGdLkmmmdpJZHa3QszuxJZieSScmvo2viD9nbU/jd8NP2Xk8K+L/hjLoPiH4beGbW3so7/AFjTTa6xc2dq4YLPazzrbRhol3PPtADg9FbH0v8AB3x5dfFL4UeEPiTe6W2iT+KNKs9Sayd/MMBuoll2b8LuA3cEqpIwSAeAAfGeh/tbftB+Nfhlq3xi8D/CHT7rw14ZuNVS+S514xXt9HpVxNFM2nQpayKdqRZPntGWfciIwCu3eeOP2vbqzvPg7ZfCjwdL40l+NWk3+o6QjXIsjE1rFaToLlikgjiEdw7TPyU8shVdmAr5K/Z38V/tCTfsv658Nfhd8MDrb+IdV8U2Wna+2rWcGn2pu9Tu45Zr2GVluUNuzMwSKOXzVAwwYlV+nNF/Zy8V+AviV+zDaeHrf+0/Dfwn8O6/pOraj5kUW2a6srOGB/JdxI3nyRSHCKwT+IgYJAOp03xMkv7UXgTw58QvBGlRfEO68E6jfS6zZXMk4s44r2GKSygMkUZkicyb97BWHIC4Y14437bvxgufhJqPx60n4SW8vgfwre31vrUsutLHeSw2N7JbSzafAIGEixoiu5meIl98aqVUSH6G1v4c+M7v9snwt8VrfT9/hbTvBmp6TPeebENl7cX1vNHF5RfzTuRGO4IVGMFgSBXgGifAL4tWf7AvxA+ClxoWzxnrf/CUfY7D7VbHzf7R1C4ntv3wl8lfMjdW+aQbc4bByKAPSrT9p34n6d8R/h/Y+PPh1BoHgT4pXLWGi341RbjU4LloHubYX9osQjj+0RocJHNJ5Z+8xORXXfGv4rftCeAD4g13wZ8PtEv/AAr4Zt/tb3mra+bK41JUi82WOzgitZlR1OUBnkTcwOF24Y4Pxb+FPj7xPD+zwmh6X9pPgXxPpeo6z+/hT7La22nXEEsnzuvmbZHVdse5jnIBAJrwnxv8GPilrnxW+J1z41+Edt8U7nxHMy+Edd1PUbM6RoenPapEsLWtyzzW0kUu52e3tneYt94AZAB7T41/bCOleGvgx4m8AeD7jxWPjQrjT7NZ1t54ZnsTdQJISrIq+ZhJ5CcRIHkw4Xae18PfGv4h6R8V/Bfwj+MXhrTtE1Dxro2pX1rd6ZfS3lsdR02fMliplghZj9idJ95AyRIoXChj8+eA/gF8WtF8OfsiWGp6F5M/wv8AtX/CRr9qtm+w+ZpUtsvKykTZlYL+539cnjmvX/24YRoPwZj+NOnvFDrvwk1Oy8TaeZX8pZzbyCG4sy4GcXlvLJAFHV2TgnFAHrPwt+Llz8TvGvxJ0Wy0tYNB8C6vHodvqAlLNfXsVuk18vl7QEW3kkWIEM25g3TGK8d8QftEfGHVPjD8Qvgr8Hvh1Z6/qfgSLS7iTUNR1b7BYmPUrU3CowWGWXzmZWSNVXYQGZ5EwFb1D9l74d6n8Mvgb4Z0HxGM+JL6KTVtbcj531bVZGvL0sepKzSsgJ/hUCuZ+Ffw58Z+G/2nvjp8RNa0/wCz+HvGUPhVNKufNif7S2m2dxFdDy0cyJ5buo/eKu7OVyMmgD52f9ub4q6l8FI/2jfD/wAIlTwLony+IPtusJHqCvBMbe+/s+GOF0mitZAw8yZ4WkKkLEAN1fR/xT+PfiLSfG/hn4Q/Bjw7b+LfG3ijTptZAvrtrDTdO0mFlj+2XcyRTSESSusccccbMzZyVAGfnPSf2f8A4uW3/BPjxt8Dp9B2+NtXHiMWun/arU+Z9v1O4uLf9+JfIXfE6t80g25w2DkD0Pxn8PPiv8O/i74L/aG+H/hg+NJLbwivhHxBocN1b2t8tuky3kNzZvcOkEjRzb0kRpFypBUsegBy/wAQP2lPijqnwQ+OPh5PC0Hhn4p/DHSLiXVLZdUk+zJYXNjNPDqmm3scAeU7I2aONkjZZE2OyH5qv+Df2ifih8Nf2Z/BHiv4oeFoNY8S+Jo9A0fwvZafqsl3da7fajaqYmu57m3iFs7BWlmYmUKoY7mOAYdK+BfxW+KFl8f/AIh/EDSofB+v/F3w43hjRtHe5ju5NPsYLO4gjkvJ7ctEZZ5pzIyxlxGgC7mOa5Q/D349+PvgL8MbZvh/J4V+IXwJ1DQb+zstTv7N7LXn0u1e0u4ILi1mn8lJomJjklVcOVH3QzAA+gPBXx3+JWmfFnRvgx8ffCOn+GtX8XWd3eaBf6NqMmo6fetYKJLqzczW9vLFcQxMJOVKOoYqwIwcX4TftGfFL4z+PfEOgeFPANtZeGvBPifVvD+s6xfag6CT+zriSFBYQJAxmlZAkku9kSPeFDO2cY+k+G/jB8cv2gPAHxU8feBpvhx4Z+FsGqTWttf3tpeX+papqlv9jbC2Uk0cdvBFvIZpAzsRhNvI7/8AZX+HPjP4b6Z8Tbfxpp/9nSeIfiB4l1uxHmxTedp+oXIktpv3Tvt3rzsbDr/EoNAG98dfjjqPww1Lwj4E8E+HT4t8e+Prm4t9H01rgWduIrKMS3d3dXJSTy4LdGUttRnYsqqpJyOK8V/H/wCKPwl+HY1b4teBbJ/Gera3aeH/AA9pWhao11b61fXyhods9xbwNbIMSeYZUO1Y2bkFQW/tE/Dz4lH4l/DT9oD4TaPF4o1f4ftqdnfaG9zHZy6hpurxRxym3nmIiWeB4ldFkZVcFgWBwDxXxV8PfHT47eEPD/jzTfh//wAId4p+Gniiw8RaHpGr6nayS6xDbRPHc2872hmhtGlSZ1iJkkwyqX2AmgD0HwV8d/iVpnxZ0b4MfH3wjp/hrV/F1nd3mgX+jajJqOn3rWCiS6s3M1vbyxXEMTCTlSjqGKsCMHkPAf7SXxr+L3iLxlpHwx+G9gbDwJr+uaDe6hq2rSWkF1caZLLFbx2oitZnZpNsbzuwCQh8L5rAgM0nw38YPjl+0B4A+Knj7wNN8OPDPwtg1Sa1tr+9tLy/1LVNUt/sbYWykmjjt4It5DNIGdiMJt5Hf/sr/Dnxn8N9M+Jtv400/wDs6TxD8QPEut2I82KbztP1C5EltN+6d9u9edjYdf4lBoA+Z/2U/wBoL4q6H8C/GXxb+PFranwV4fuvE18+prqtxqGpm4t9SkAsUtpoEUQoN8UDCbosY2KGO33LSf2g/jH4e8R+Bz8bPh9YeFvDfxIv00vTZbLVZL6/06/uIXmtbfU4GtYY1MwjZN0MjqkmFbg7q8E8I/AH4q638DPip+x1408KT6Pp2r3Ov3mkeLFurSbTbgXt+b2yDwpN9qR97fvFMOAqHnJGeh+C3wZ0Wx8WeF4/E37KmneFNe0WVJrzxFb3elyafBc2ylo7qwEczXTl5VUojQxmMN8zZXDAHfXn7YWq6fJq/wAN7nwlH/wuGz8SweHrLw4LxvKvYbvM9vqiXHlF1svsSSTyP5RMbRtGwzgn6a03xp4uu/irqvgK88HXVr4fsNMt72DxEZoza3VzK5R7RIR86tGBuLEnvkD5S3wvqnwP+P8AqHjZv2x49IK/FbTtX+zWHhV7y18o+DUZrdtMacSm2W7mV3vfO80ospEecZFfXdl8W/E8v7RcvwVvvC4t9HfwyNft9WF5HJL5qXKW8ltNaqCY+ZMo+8h9j7d2DtAPkD/goZH4xfxf8E7rwDNKniDRr7XtcsoYnZRcz6Hpx1NbZgPvLOLcxEHg7680/wCCifxSHxR+DHhvTPh3qMqaNPo0Pjy/ngcxudPeWCz0yJmB6T3N35m3uLdvQ192fFr4feLvE/x++BfjXRLD7Tovg698QTatP5sSfZ477SZrWA7HcPJvlYLiNWIzlsLzXyb4y/Yn8Y+F/wBnf4z+CfBUy+K9c8X3NnbeHbQPHb/YdAsdRF5aad5lw0cY+zma5OSwDDaByAKAPsT4tfs+eA/ivq8viP4na5rUui2Nj5UemwavcaXptq6FnkvSLN4HebBA3TSOqBcqqkknhf2F9c8WeIv2d9K1LxNqd3rlr/aGqxaLqOoFmvLzRYbyWOwnnZ/mZnhUYY5LJtbJzk+d/tgWH7RvxD8Q6T8LfBvw51DX/hTMi3HiO40zV9M0+81cAkjS0N3dQyQWzYH2hwpaRT5alV3M3e+KPjd8Tfhf+zh4m+JUnwbPhe68DxQi08PXesacsc2nQeUrvDNZPPFH5cZYRw43syBEUllBAPpT4jeNLP4b/D7xN8Q9Rtpr218MaZeanLBbjdNLHZwtMyRg8bmC4GeM9eK8P+BPxa+MnxH0nTvHHjnwv4d0rwZr2l/2raX+k66+oyWysEZbe6R7SFGfYzFpIpGRSjKecZ938Q6j4kTwXe6r4Z0SPU9c+xtLbaZeXAtUmnKZEEs4SUJk/KW2sM+3Nfn18NPgn4rvvjpH4i8K/Cq8+CHgm/0nVbTxdYNqVlJZ61c3kax232Wx0+4ngR4G3ObnZEWUlcHkUAdnN+198RofAiftFSfD+0i+CbXEQF7JqUg19tMluFt11UWAtjCIMsJPKNx5vlfPx92vT/HXx4+Id78WtQ+CPwD8JWPibX/Dtjbahrt/rF/Jp2mact7uNrbloYLiaW4nRGcBUCquCWPIHxr8L/2VrH4e6Jpnwm+IP7MOmePNQ0qUWa+LLa50yOw1CxEmFu7lLiZbqKdYiPMiEMm91JVsHI+kta8LfF/4HftE+Nvi78PPA8vxF8LfE+y0ldQs7C8tLPUdN1HRoWtYmVL2WGKW3mhZd22TcrAnbtGWAOr8W/tAfFHT9a8HfCTwt4Es7v4s+J9Mm1e+0641UrpOi2NvIIXuLm9it3klV5mEcaxQ7mOc7ccp4Y/ab1u48MfFiy8d+FU0H4g/CHT5L/UdKivDc2V3A1pJdWlxaXflIWhuBEyndGHjYFWXI54jxP4e+O2j/Fvwl+1Zo3w+Gp6rdeGrjwz4j8KW+qWrXtra/bTe2k9rdSmG2nlRsiaPeo+bEbPjdWbpHwf+L3jWz+P3xc8a+Hk8P+Jvib4cOgaF4dF3BcT21pZ2dxHD9quY2+z+fczzFiFcpEu0FydxABFF+2D8XbXw/wDDn4teJfhbbaR8MfHl1pFk102sCXVrU6z5a2929qkHlfZjJINo84ylCGZY2Owes+Ovjz8R7z4wX/wQ+AvhCx8Ta14bsbbUdfv9X1CTTtO09b0sbW1DQ29xLJcTojOMIEVcEk8geZfED4JfE7W/2NfhP8KdL0bzvFPhn/hCP7Qs/tFuvkf2PJaNe/vWkET+UIn+47bsfJuyM6mr+HPjN8FP2lPHnxW8BeA5viN4Y+KtlpH2qKxv7OzvdN1PR4WtI9y3ssKPbzRMpLq5ZGU5XGNwBxf7HPiPx5beAf2gPEmn+EPtni0/E7xBJ/YP9oQxqLp4rEPD9tYeXsQknzNvKjIXJC17B4B/aB+I6fHK1+BHxr8MaPoera7pVxq2k3Gh6tJqcLC0dFntblZra2kjmVZA4YKUYZwcivnXTPgl+1DB+zx8a9HttJj0Hxr468f3XiKOwsNWRPtWkXjWL3Ntb6ghQwvLHHNAHcRNwThVZWrV+GnwN8T2/wC0j8M/ib4X+CVn8J/Bvh7TNdtL1Rdac2pTXF7FCIpbxLR5PMDFSsb+dNJneZRHldwB678Nf2j/AIv/ABk8X+KtD8BfD2zg0XwP4j1nQNT1XUtSeGOaTTZ5IYVs4o7d2kdwsckxYqkYfapkbOLK/tcJJ+yn/wANCR+Gz/wkIkGlnw4bjEo183w03+zzJszn7ScbtmdnzbcV1P7K/wAOfGfw30z4m2/jTT/7Ok8Q/EDxLrdiPNim87T9QuRJbTfunfbvXnY2HX+JQa+X3+GupT/t1S/CnT2im8Ax3tv8Wr2BG5ttZ8mXTEgcdvOulS+UE8sjEDjIAPuj40/Eq++D/wAFPFnxUl06PUbzwtpM+otZiUxxyyQR7zGJdhIUkYDbM47V8sa3+138ZfCOpeCdW8U/B4p4X+Jkyaf4fW11iGXVv7QuIHns4r2B40t4VuAvJWeQQrkuSRtr6K/ak8F+JviN+zp8RvAfg2z/ALQ13XtDvbOyt/Mji82eWMqib5WSNcnuzADua8q+LXwn+IHifRf2drTQ9K+0y+BfFeialrK+fAn2W0s9PuIJpMvIok2ySKu2PcxzkAgEgA1vhr+0P43n+KPiv4RfHfwtYeDtZ0HQ4vE9vcadqLalZT6Q8rwSM0jwQOskEiFW+TDckADG7y7/AIbB+LMfw9T9o65+F8EfwXcrP9o/tRj4iXSWk2DVDp/2fyfKKkSeULjzBH83SvQ/FnwV8T+Mf2q9f8W6lYsngjXvhhL4Ulv0mi3C9uNSkleIRb/NyIH3h9mztuzxXz7J4K/asuv2bo/2MJPh9DHMNMTws3jT+0rQ6L/YqKLf7Ytt5n203Bths8gxD9582/ZQB03xP+KX7Q4/bM8I+G/hbpel6toF14S1G/sLa6125s7TUYZJLPfdXMcdpKqTQvlIBtk3Rszb0LFK9f134/8Axc8VfELxb4H/AGefAuneJ7f4fSR22s6hrGqvpkE2oSRCY2FiI7a4LypGy75JNkaswB9a5D4mfDz4kfDD42/C34r/AAw8G3Xj7Q/C/hi+8KXljZ3dpbX0McjW7206/bJYY5FJh2vh8r1xioNI0f45/s6fET4lXngb4bz/ABH8L/ErVj4ls/sWpWNldadqtzbxQ3Vtei8ljXyXaJWSWEybRkFScUAafiP9trS7D4IeCvjN4e8I32qS+J/Etv4YutD3BdRs9Qd54ZrdVwVkmSeHy0BKq+4NuUV9TfDTUviZq3hs3/xW0PT/AA7rMk7lLLTb19QjjtyFMYkneGDdKMkPtTbxlSQa+DE/Zd+K+ifCL4VaJJbW+seK4vitp3jvxP8AZJo47a1Sa9murwwmdozIlurqgC5dyCVU5xX6aUAfFXg79oj41/GC8ufFvwW+Hem6r8N7TU5dNi1LVNZexvtUS2nNvc3djbrazRiCN1fb50itJtOAp4rR134//FzxV8Q/Fvgf9nnwLp3ie3+H0kdtrOoaxqr6ZBNqEkQmNhZCO2uC8qRsu+STZGrMAfWvNPgdpv7RX7MnhGP9nzS/hY/jfRtEv7tdB8QW2r2VnZPpt3dPcL9vSd/tMM0AlKsI4ZQ+Bs3ck7Om6J8cf2cfiT8TbjwH8OpviX4V+JGrt4ksmsNRsrG4sNUuIIobm3vFvZIh5LtCrpLFv2DIZScUAX/Ef7bWl2HwQ8FfGbw94RvtUl8T+JbfwxdaHuC6jZ6g7zwzW6rgrJMk8PloCVV9wbcoryf9oL4o/teaTrPwTtZfC2iaBe654yiX7JZeJLvy7nFpdGOwvZEslDQlTvlYB18xEAjIwwtp+y78V9E+EXwq0SS2t9Y8VxfFbTvHfif7JNHHbWqTXs11eGEztGZEt1dUAXLuQSqnOK93/a38DfEPxDF8MvHnw10E+KdR+HXiy11u40qO4htp7qyEE9vMIJJ2SPzV80MqswBAPOcAgF/xV8bvirbeKPDvwc8E+ENL1P4m6how1zV47nU5YdE0e08zyA73SWrzzmWcNHCiQBmCs7bAOeOuP2tfEmmfDDX/AB9rHgfy7/4b+Iho3jfTre7adtPs0EbzajYuIVN1CkM0c4VkjJjL5IKc+YfGn4F6t44+LHhz9pDW/gzH4/sNU8Opo2s+E7+4sF1bTJYLh7i3u7Z5JzZzNtkeKaMTjjYVLHcB1s/ww8Zj4JXXw6+Cfwkh+EE/xD1UWGsNHdae8mmaNIgS51GRbeVke6eENDDFE0hRmV2YBTQB7J4X/aD1L4g3fxK1T4a+Gx4o8L+Bbcwafe2t2u7XtZjgM81nZ5XyxHHmOEzFyDKxABCE17n4F13WfFHgzQ/EfiHRJvDWqanZQXN1plw6yy2U0qBngd1wGaMkqTgZxyAeB8o/BX4efEj9mXS/H/wq8J+H11r4faLb3OteCZpr6CDZLc+ZNNot08jmZAlxl4rl0dPLkO99y7a+gvgZ8R774vfCDwj8TtR0g6FceJtOhvms/NE4i80ZGyUAB0YYZGwMqQaAPzy/Yr8X6j4K+OPxT8OeI79zoHxA8QeLdT06S4kPlwaj4f1ea3voULHADWc1rIBxxE57Va/Yu1/X/Gn7U/xP+Jutzz+X8QfDuma/ZW0rNtg02a/vbfTQqngb7CC3kbAHzMa6ab9j/wAf+PPg/wCIfCV/e/8ACEeJj8QfE2t6dfq8c7/2Prlxc29wFaFn2/adPuZNqnDK+zcFIyPobwJ8J9f8JftR+KfGdnpC2PgqbwdoOiabKksW3zdOnui8CxBzKojjePBZApzgEkHAB+Vfwhs/2QLT9jGDxt4q8XW+jfGFbTWXt5tN8QTw+IhqSXlytgIrS3uRI0hxEFQxFSpBIwc19Z/F3SfiV+0R4K+C/wCzL4su59L8Y654Un8VeKpImNvPbXNlpotrXzdm3Zv1W5R2UDGYWHQc/Qv7FHwIn+EXwB8I6J8QfCdjpPjjTjqBu5NlrPcr51/cSxZuYDIGzC6EYc4HBwQQOTs/2VtP+MXxx+JvxY/aF8OTPDLcWWjeFYV1OWFk0bT4dz3GdPuVwt1cyyOI5SHXaMoueQD3/wDZl+KE/wAZPgR4N+IGoApqt9YrDqcZGGj1KzY216hHbbcRuAPTFep+Mdebwr4R1zxOkIuW0exubwRFtokNvE0m3dg4ztxnBx6V8BeHdE+In7FHgb4/XPg/whHq3gPQrqLxP4WiutWigjkhuYIzq1s0szyzR/ZmjeVPMU+aWwrFmr7X8Tpq3jf4QatFZ6c9tqmv6FOI7KVlWSOe6tW2wuzFVDKzbSSQM+goA+Iz+278WdO+HfgT466/8IRB8OPGB022ZrfV45taS41JVWKWO0MSQtbvO3lxbp1kZSsjrHnZXtHgf4/fFT/hedh8GfjP4DsvCkninSrrVtCubDVf7SDixaNbi1uswQhZkWUOWTcnYFvvV5L4j+BHxWv/ANiX4TfCK00Pf4t8My+D21Gy+02w8gaVc28l3+9Mohfy1Rj8jtux8u4kV7l8Qfhz4z1z9q74R/ErS9P87w34Y0jxLa6jdebEvkTaglqLdfLZxI+8xtyisFx8xGRkA841j9qj4pX3h/xZ8Xvhv8PrLXPhZ4Jnv4rm8udUe21XVItJdo7+4021W2kiaOFo5AgmmRpShxtyK7Lx1+0l4gvfGPhL4Yfs/eHLbxh4n8V6IviYzaldvp2madokjBIbm4kSGaVmnkOyOKOMtwzHAHPyB4b/AGU7X4Tza54F8Q/s2WHxchk1K+utF8RQXWmwedaXs7zxwamL2aOaKS33mMyRxyqyBdoJGD9EeMfh78Sfg78fNC+Nvwg8B/8ACW+Gbrwlb+EtR0HT7u1s7vTksJzPZz232uSKGSJVcwtGHUgAMMjNAHg3gvxNqMGoftveLPi54MtXudO0/TbnVPD/ANt+1Wlytn4dLNEt0sakxXCIGVjGrqrjcocEV9pWXiv4jWvwe+HU3wJ8AaZcx6to9i6wX2qHTtN0a1+yRPCjMkE88qqD5aLHF0XLFeM/NFh8Hfj14l8Pftbax4p8HxaJrHxg0aOHQdPi1G2uvMkTQ5LBIXmDIiSh9iyF9sYcnY7xjebvxP8AhF8VrjRPgVp+r+Bp/iJ4K8KaELPxJ4TttQtbcyamLW3htppkuJ4rW8igZZFMbSFATvww6AGZ8T/jtqXxj/Y//aM0HxZpFtofi3wFYalpWqW9hefb7KQtaCaG4trjZGWjlR+FZAykEMK+/fhj/wAk18J/9giw/wDSdK/Obw9+zP8AFab4bftS+H08C6b4Hb4n2cA8N6Rp91atZrs0ryBBmERpE/mgLISixiQsUZ48O36H/CX+2R8L/CkXiHSZtC1ODTLSG5sbh4nlt5YolR0ZoXeM8rwVYjFAHoVFFFABRRRQAUUUUAFFFFABRRRQB//R/fyiiigAorm/E3jLwn4LtrS98X6xaaLb31ylnBJeTJAklxIGZIlZyAWYKxA9Aa4z4efHX4M/Fu/1DS/hj420jxReaVzcw6deRXDxqTtDlUYkoTwHGVJ4zQB6vWFf+F/Dmq63pfiXU9Mt7vVdEE62NzLGry2ouQqzeSxBKF1QBiuCRx0Jrh2+OXwcXxBp3hMeM9JbW9WvrrTbSxW7ja5mvLJ2juYViBLboXUrJx8rDBOeKqW/7QPwOu/iC3wptfHmiy+MEcxHS1voTdecoy0WwNkyADJj+8O4oA9goryz4k/G/wCD/wAHhZf8LS8ZaV4WbUSRbJqF1HA820gMURiGKqSNzAYHciuivPiH4D0/w1Y+M7zxFp8Wgam9tHa6gbmP7JO146x24jm3bG812VUwfmJGKAOxorx/wb+0F8DfiJrepeG/Anj3Rdf1TSEeW6t7K+hnkjjjwHkwjHKKSAzLlQeCc15V8C/2xvhD8dPFvijwToOvaUmraPrV3pul28GoxXM2r2VrbQzm/gjUKfKYvIvy7x+6J38kAA+taK8K+FPj9k+C9t4/+J3jnw9rkMH22S71/TGFpo5ihuZIwQ0sjhfKCiOQl/vqR9eo+G3xo+Evxitry7+Fni7TPFMenMEuf7PuY52hLZ2+YqncobB2kjDYOCaAPTaK8o+JHx1+DXwea2j+KPjXSfC814N0EV/eRQzSrnBZImO9lB4LAYHc100nxD8BxeGtN8ZP4hsP7B1h7WOyvxcRm1uXvXWO3WKUHa5ldlVMH5iQBQB2NFeR2nx8+Cd/4Z1zxrZ+OdGl8PeGrk2epait7D9jtrlQpMTz7vL3/OvAYkkgdeK1/BHxd+F3xJ8M3HjLwH4r03XdDsy63F5a3UckMBjXc4mYH92VX5iHwQOelAHotFeOeBP2hvgV8UPEN14T+HPj3RfEmsWaGSS1sL6G4k8tSAzqEY71UkAsuQMjJr0vS/EXh/XLnULPRdTtdQn0mc217HbzJK9tOFDGKZUJMb7WB2tg4IOMGgDYorzTxr8ZvhN8N3vYvH/i/SvD0mn2sN7Ol9dxQOlvcSPFDIVdgSJJI3RMD5mUgZIrl9X/AGnP2dtB0PQ/Eus/Enw/aaV4m3f2ZcyajAIrsI3lu0Tb/mVHG126I3DEHigD3Oivk39ob4xfE/wX40+FXw/+EEGiXOo/Em91G2FzrIuJLWFLGzN4GX7K6MdwUjuOR9aw/C/7QPxL8EfFRvhR+03p/h7R/tmh3/iGx17Rbyf+zjaaWyfbEu471Ee3aJHEpfe0e3POQaAPs6ivlj9m79rT4Y/tIWt/b+HNV06LX7G61NTpUF/Hd3R0+xvDaxXpUKjCOdTHIPlIHmKNzcE/U9ABRXlTfHL4OL4g07wmPGektrerX11ptpYrdxtczXlk7R3MKxAlt0LqVk4+VhgnPFVLf9oH4HXfxBb4U2vjzRZfGCOYjpa30JuvOUZaLYGyZABkx/eHcUAewUV8jab+2Z8G7z9oXXvgDdeIdItLvSksLe3uG1KIveavd3E1tLpscGB+/geNAyhy26QKVUjn1f4WeJdb1ZvHM/iTxZoviG30jxDfWludKQxf2ZbQJGwsr5mkcG6hDZlPyjDA4AOAAexUV434I/aH+BPxJ1u+8N+APH2ieIdU01HlntrK+hnkWOP78gVGO5FyMsuVHc10r/FX4axeAB8VZPE+nL4OaJZhq5uY/sJidxGr+fnZtLkKDnrxQB39FfLmt/EfxlZ/tj+F/hRb34TwtqXgzU9WntPKiJe9t763hjk80r5o2pIw2hgpzkgkA129z+0l+z/aeOovhncfETQk8VTTC2XTv7QgNx9oJwISoY7ZSTgRnDE8AUAe2UV5/wCMPit8M/h89xF458U6boD2lk2pSrfXUduUs0lSAzkOR8nmyJHnu7Ko5IFYPxD+M3gvwL8FtU+Nr6raPoFvpX9p2d1JMsdvdCaIPaKjsQD9oZkWPuxYAcmgD16sLUvC/hzWdW0vXdX0y3vdQ0RpZLCeaNZJLV5l2SPCzA7GZflLLg4JGcE5+L/2TfjPY6l8AR8cPiz8abLxTFf21lc6tJcHTrGw8P3s0KzS2Aa3SIqyedGjJM7PuUYwWIP1X8Ofix8M/i9pEuvfDDxRp/iiwgfypZdPuEnEUmM7JApJRiOQGAOOelAHoVFeQWf7QHwQ1HU7DRdP8daPdX+p2dxqNtBDeRSSSWdpv86farEiNPLfLHA+RuflNQ2H7RPwG1XxTpfgjTfiDoV1r+tW8V1ZWMeoQNPcQ3EYmiaNA+T5kTCRB1ZDuAI5oA9loryzwL8cPg78Ttb1Tw38O/GmkeJNU0Xm8t9PvIriSFc7dxEbHKhvlLDIDcE54rG1X9pL9n/RPG0Xw31b4iaFa+KJphbjTn1CAXAnYgLE6bvkkYkBUbDEkYFAHtlYWmeF/Dmi6rq2uaTplvaajr0sc1/cRRqst1JDGsMbSuBlykahVyeAOO9fMfxE/bM+Dnwv+O2nfBXxh4h0jS/M0y6vtRvrvUooP7OnjNu1tayxMOHuYpjKm51O1chWDZHrfhHX/EGo/Frx9oN94o0bVNK0iPSTaaRZxkanpZuYXaRr997Ai4K74MIvyg+hJAPXqK8a0X9oj4EeI/HMvwz0Hx/omoeKoXeNtNgv4XuTJHnfGqBvmdMHci5ZcHIGK1vGfxr+EPw6Oop488ZaToD6TFbzXSXt5FA8Ud2ZBASrsD+9MUgQAZYq2AcGgD0+iuB034qfDPWPA0fxO07xVpc3hGVN41f7ZELELv8AL5nLBFw/yEEghvlPPFdrdT+TZzXUeG8uNnHocDIoAs0V+ZXgT9pT9r3xD+zxZ/tQz+GvBereGP7MuNYudJtZ9QsdT+x2hk84RSy/aIDKFjZlVgAemc4FfWepftT/ALP/AId8K+GfFvjPxxpXhi38W6faapYQaldxQXUlteRrLG/kli2MMAzAFQe9AH0FVW+s7fUbK40+6BaC6jeKQA4JRwVIyOnBqHSdW0rXtLtNb0O8h1DTr+JJ7e5t5FlhmikG5HjdCVZWByCDgiq/iHxDoXhPQ77xN4nv4dL0nTIXuLq6uXEUMESDLO7tgKoHJJoAwvhz8P8Awx8KvA2i/DrwZA9roegW62tpFJI0rJEmcAu5LMeepNdrXjuj/tC/ArxD45Pw00Lx/omoeKhkf2bBfwyXJZQWZFRWOXUAlkGWAGSAKb8Qf2h/gT8KNVh0L4k+PtF8N6lcBStre30MM+1/uu0ZbcqH+8wC+9AHslFchrPxA8C+HbbTL3X/ABBYadbazvNlNPcxxxXAjge5cxuzBWCwI8pIOAilugzXmmoftSfs5aV4SsfHepfEnQbbQNUeaOzu5L+JY7prdzHKIMtmXY6kEoCBQB71XAfC/wCGXhH4O+B9P+HfgS2e00TS2uXgikleZlN1PJcy5dyWOZJWIyeAcDgU2y+LHwx1LTPDmtad4q0250/xhN9n0e4juo3i1CfazeVbuGIkfCN8qnPykYyDXQ33izwzpviLTPCOoapbW+t61HcS2Vk8ircXEdqFM7xxk7mWMOu4gYGRnrQB0FFeP6Z+0D8Dta8ey/C7SPHmi3ni2F3jbTIr6F7rzI8l4wgYkyIAdyD5lwcgVN40+PXwV+HX9pjx3440fQX0eSCG7S8vIopIZbqMywoUZg2+SMF1UAkqCQMAmgD1qivlv48/tYfDP4LfBNPjRa6rpviCz1L7P/Y8SahHEmqedNHG/wBnmAk3+WjtI21WwqnOOSPTdK+O3wX1zQrDxRpPjjRrrR9UnuLa1vEvoTbzTWkTzzokm7aWiijd3GflVSTwDQB6vRXzpZfFzTPHnjr4e3/wy+IPh298H6/HrQltB+/vdXewVU3adMsgXbayHM52sNpA7gjqPG/7Q3wK+G3iS18H+P8Ax9onh7W7wKY7O9voYJtsn3GdWYFFb+FmwD2NAHsdFc5Z+MPCuo+I73whYata3Gt6bbw3VzZRyq1xDBclhDI6A5CSbW2k8HBxXy7+1l8dLvwh+y98SPiV8F/Elsdd8IzpYG6txBeC0vor6C3uYJEkWSPzEDsrK6kqT0BxQB9UP4X8OS+JI/GMumW765DbGyjvmjU3CWzPvaJJCNyozYLAEAkDOcDG7XmXxI+NHwl+D9rBefFLxfpfhaO7z5A1C7jgebb18pGId8d9oOO9aMXxR+HE/gaL4nQeJ9Ok8JTojpqy3MbWTLJIIlImB2cyEJ1+98vXigDvKK4fwl8TPh9491LXNH8E+IrHXbzw1cC11KOynSc2lw2791KUJCuNrArnIIIOCCK+TfCX7U/h7wl8S/jTpnx28daZoOheGvEdnpuhJqElvaFYpNOguJY4yAskxEkhZi28qD1C4oA+0PEXhvQPF2jXPhzxRp8Gq6VehVntbmNZYZVVgwV0YFWXIGQQQe9a8cccMaxRKERAFVVGAAOAAB0Arj7T4i+AtQ1DQ9KsPENhc3fia0e/0qOO4jdr60iCs81vgnzY1V1JZMgBgehpNY+I3gLw/f6ppeueILGxvND0xtZv4pp0R7XTVLg3cwJykIMbjecD5T6GgCp8M/hl4R+EfhZfBvgi2e00tbq7vAkkrzN519O9zMdzknBkkYgdAOBxXf14xpf7RnwE1vxrD8ONH+IWhXvie4AMWnQ6hBJcOSu8KqKxy+35tg+bHOMVqfEn43/B/wCDwsv+FpeMtK8LNqJItk1C6jgebaQGKIxDFVJG5gMDuRQB6nRXzB+0H+1V8N/gR8LdP+Jkuqabq8euXFjHpUI1CKFNQhubmCKeeCUCQOlvDKZ2KqRtXBKg7hwvxc/aQsYtR+CfiP4V+MtPufBvi7xVNp+q39u9tcWc1jb2V1LMpuHDCMRyQgs6MpG0gnGaAPtmivMfhz8avhJ8Xl1F/hd4w0vxSNJdY7v+zrqO48ktnaX2E4Vtp2t91sHBODWFov7SPwA8SeNx8N/D3xE0LU/E7MyLp9tqEEs7SJkvGoRiGdQDuQEsMHI4oA9rrx3x/wDAn4d/FHxXoPizx3b3eqt4ceOa0sHvbhdLNxC/mRTzWKuIJpY25VpEbHHoK898O/EjxjfftgeNPhZeagH8L6R4T0jVLa08qIeXd3V1cRyyeaFEh3LGo2sxUYyACTXc6L+0j8APEnjcfDfw98RNC1PxOzMi6fbahBLO0iZLxqEYhnUA7kBLDByOKAPa6K8u8a/G34QfDg6gvj3xlpPh99Kjt5rpL27igeJLsyCAlWYN+9MUgQAZbY2M4NYGs/tMfs8+HrHQNU1r4keH7O08ULv0uWTUrcJeR7ihkibfgoHBUv8AdDDaSDxQB7hRXkHinxBr9l8W/Aehaf4p0bTtJ1eDVXutIuoy2qaobeKNo3sXEgAW3Zw03yN8rAdwRH43/aG+BXw28SWvg/x/4+0Tw9rd4FMdne30ME22T7jOrMCit/CzYB7GgD2OisrWdd0bw9od94l12+hsNJ023ku7m6mcJDDbwoZJJXc8KioCxYnAAzXmdj+0J8DNU8c2/wAM9N8e6Jd+KrpQ0WmxX0L3L5TzAoRWJ3FPm2fe284xzQB7DRXPWHi3wxqniLVfCOm6rbXOtaElvJf2UcqtPapdhmgaWMHcglCMUJHIBx0qrpPjvwXr2jX/AIj0bXLK80rS5rmC7u450MFvLZsVuElkztQxFSHyRtwc0AdXRXzl/wANIfCTx/4M8a3Hwa8eaR4i1rw5pN7dlNPuobqSFoYWKSmMFtyB8fNgoTxk5xVf4J/GK2m/Za8B/GX4y+IrSwbUfD+nX+qanevDZW5nuIUZ3bHlxJvduFUAZICjoKAPpWivMfCvxp+EvjjwbqHxC8IeL9M1fw3pCTSXt/b3UbwWq26eZKZ2B/dbE+Zg+CF56Vu33xC8DaZoOk+KdQ16yttH16S0i0+7knRYLqS/x9lWJycOZtw8sD72RigDsaK8g+IH7QPwQ+FOtWXhz4k+OtH8NapqCCSG2v7yKCUxklRIVZgVQsCA7YUkEA5Br0FPFXheXVbPQYtYs31PULY3ttai4jM89qpAM8ce7c8QLAF1BXkc80Ab1YWn+F/Dmla1qviPTdMt7bVdcMLX13HGqz3Jt4xFF5sgG5hGg2qCcDnHU5+fv2pfi949+EXh/wAD/wDCt7TTLrW/Gvi3S/DMZ1ZZmtYRqSzYlYQOj/KyL0J4zwTiuX0T40fG7wN8YPBvwo+PuiaBJb/EIX0Wkav4cuLny47zT4Dcvb3VteLuUSRKxSRJGG4bSvOQAfY1FfEug/tU65q37VV38HZtJtV8Cy3N/oOn6upf7RN4i0qztdQurd8t5YjEM8iLhdxkhcZrsPi/8cfHlh8TdK+AnwJ0Ox13x3f6e2sX1zq00kOlaNpiyeSk1z5IMssk0mUihjwxwWJCjkA+qqwte8L+HPFMVpb+JdMt9UisLmK9gS5jWVI7mAkxSqrAjehOVOMqeRyAa8s+El5+0V9u1bSvjrp/htooEhksNT8Oy3SR3BcuJIpLS73yRtGFU7xMysGAAyGx7hQAUV4b4q/aa/Z58DzNbeL/AIj6DpE6Xk1g8dzqEEciXVuQs0bIW3KYyQHyMLkZIr0vW/G3g7w34Wl8c+INcstO8OwwrcvqNxcRx2ghcApJ5zEJtbI2nODkY6igDp6K8z8EfGb4T/Erw3feL/APi3TNe0bTN/2u6tLlJY7by13t52DmPC/N8wHHPSvPfG3x8+H2p+F/Emk/DX4leG7LxVZ+HzrsNxd3CXVrZWUqRvDfXSRuCIGWWNgxIyrqwBUjIB9HUV494j+NHw2+Ffg7Rdd+L/jjRdGN/bRMLqa4S1hvZfLVne1jkcuyMTuVVLkKRknrXb+F/HPgzxr4Xg8b+EdcstY8P3UbSx39rOktsyISHPmKSo2kENk/KQQcEGgDqqK8K8MftPfs6+NNTGjeE/iToGrX7XcVikFtqEEkklzNny441DZcvtIUrkHBwa9U0nxb4Y17V9Y0DRdVtr7UvD8scOo28MqvLaSzRiWNJlByjOhDKDjIOaAOhorirD4k/D7U/Bb/ABHsPEmnzeFY0mkfVRcx/YlS2dopmactsAjdGViTwQQelZPw3+Mvwo+MNreXvwt8W6b4oi09wlydPuUnMLNnb5iqdyhsHaSMNg4JoA9IkQSxvGSVDgjKnBGe4I6GvI/hN8DPh58F4tWbwZbXUmoa/Kk+pajqN5cajqF68S7Y/Purp5JWEakhF3bVycAEkn5v/ay/a88AeAPhR8RtK+GPxI0W2+KHhyweW2skubW4uoriN13r9nl3o8iLuLRlSyjJKjFfXuleNfDs+qab4Pu9Wtj4mvNMXVBYGRRcvaqyxvcCIc+WJGClgMAkCgDsqK53SPF3hfXtS1jR9F1W2vr7w/MlvqMEMqvJaTOglVJlByjFGDAHHBBrzDRv2mv2ePEWval4X0P4kaBfarpEU891bw6jAzxRWql53OGwViVS0hBOwAlsAUAe40V8l/AT9sb4QfHZvElnpmv6TZ6hoOpavClqmpRTyz6Vps/lJqeMJsgmTbIDyqhh87da9R8CftDfAr4oeIbrwn8OfHui+JNYs0MklrYX0NxJ5akBnUIx3qpIBZcgZGTQB7HRXy5+zp8R/GXjzxh8atK8VX4vbXwj4zn0nS08qKLyLJLO2lWPMaqXw8jHc5ZucZwBXq3xJ+M/wn+DtraXnxS8Wab4Yj1BmW2+3XCQtMUAL+WhO5goILEDC5GSM0Aem0V454b/AGhvgX4w0qfXPCvj3RdW061u7WwluLa+hliju75xHbQsysQJJnIVF6seBVOT9pb9nyHw/qXiyb4jaCmi6Tetp1zetqEAt0vUUO1uJC215ArA7VJPPSgD2+ivJ9K+O3wZ1vwDJ8UtJ8a6TdeEYZVgl1VLuM2kUzusaxySZwjl3RdrYOWXjkZ7jX/Ffhnwp/Z3/CS6pb6Z/bF7Dp1l9okWP7Re3GfKgi3EbpH2nao5OD6UAdBRWPo3iLw/4jS7k8Pana6olhcy2dwbWZJxDdQHEsMhQnZIhOGQ4Ze4FfHetfF79o7xR+0L48+D3whtfCcOn+BrDRrySbXVv/OnbVY5X2q1q+0BTEeSnQjrQB9uUV+fuu/tgeN9D/Z5+L/jrVfDFjpnxD+Dt2dO1HT/ALRJd6ZPP+4eKeCUCGUwTRTBlVgrqQQ3TJ2tZ+Nv7R3wg1fwLe/GfSPCur+GPHGuaf4eE3h+a9gv7O71QlbeTyLsSLPGGH7wI6uB8wBAIoA+0PEHh/Q/Fei3nhzxLYw6npWoxtDc2twgkhmib7yOjcMp7g8GtO3t4LSCO1tY1hhhUIiIAqqqjAVQOAAOABUtcL4y+J/w5+HbQJ498T6b4dNzBcXMX9oXUVqHgtNnnyKZWUFY/NTce25fUUAd1RXnXw++Lvwv+K+h3HiX4beKtN8SaXZuY57ixuY5khdRuKy7T8h284bHHPSuc8I/tG/ATx94vk8A+CfiDoeu+IY1dvsVlfwzzMI8l9gRjvKAEsFJKgEnAoA9ooryrxL8cvg54Nv5tJ8U+M9J0u/t763017ae7jScXt3GksMHlk7zI8ciOFAztYN05rIuf2kv2f7Tx1F8M7j4iaEniqaYWy6d/aEBuPtBOBCVDHbKScCM4YngCgD0/wAQ+F/Dni2yi03xRplvq1pDPDdJDdRrLGJ7dxJFJtYEbkcBlPYgEcit2vjDx1+0Gvw0/a1Hg7x94ssvDvw6h+H8mtyi/a3t4Rqf9rR2qP8AaJAJNxiJRYw+1jyFLc19FaB8Xfhd4q8CXPxP8N+K9N1PwlZRTz3GqQXUclpBHbKXmaWQHCeWoJcNgqOoFAHotFeQj4//AAPN1rdmfHuiLJ4at4bvVC1/AqWME/8AqnuHLBI9+RgMQeRxyM3/AIa/Gv4SfGOG+n+Fni/TPFK6Y6pdf2fcpO0JbO3eqnKhsHaSMNg4JwaAPT6K8avf2iPgRp3j6P4WX/j/AESDxdJKsA0x7+EXPntjbCU3ZErZG2M/Mc8Cvn/Rv2oNE8F/HH46aD8bvG+neH/CvhPUPD1poSag9vabDe6THdXMcbYWWdmkYuQxcqOmF4oA+5aK8L+IPj2W58PeBfEfw58b+H9L07xHrulwreX+Lq31Wyui2bSxeORQbm448lgWHBGOcjU+Iv7QXwO+Ed9b6V8TfHejeGr66AaO2vr2KGdkY4D+UW3hM/xEbfegD2CivlD9on4r+I/CcnwRvPh3rES6d448d6RpN5NCsNzFeaVe2l3MyxuyuAshjRhJGQ2B8rYJz9X0AFFFFABRRRQAUUUUAFFFFABRRRQB/9L9/KKKKAPgL/goQPCx8F/CQeOPsn/COn4m+F/7S+37Psf2PzJvP+0eb8nlbM79/wAu3OeKo+P7rwbrX7cfwFg+F0tlc6xpGl+I5tffTTG3k6DJZrHaJdGHjy2uynkKxwGyVHPPuf7Tfwg8QfGLT/hzp2hx2s0PhrxvoWvajHdthJNOsHc3CKu1g7MrYCHAbkE16t4D+Evwt+Fq3qfDXwhpHhUak4e5/suxgs/OZc7fM8lF3bcnaDwM8YoA+Ev2WfBscXg/9pfxj4N0+L/hOr/x544tbS9Khp/Mt55DaRK55VFmcsFXA3MW6nNeBfCHwH40+Ln7IvgfwRpfj/wD4b0mCbSpi5027h12w8Q2l1HK/nTSaiANQe6QpIxiBkLnaoVwK/ZHQ/DPhvwwl7H4a0m00lNSu5r+6W0gjgFxeXLbpriURqu+WRuXdssx5JNcFc/Ab4I3njUfEi78AaBP4qEgm/tV9MtmvfNHSTzzHv8AMGOHzuHTNAHyh8OL7whpf7avx6b4qT2dt4gnsdBbQn1JkQN4cFmROLMzH/VC7837SE48zlu1fFniMeHX/Yr+JE0AgHwtn+L6P4e8zA0//hHzrFp5pg3fILMz/aCuPlxu7V+y/j74R/Cv4qR2kXxM8H6R4rXT2L241SxgvPJLY3eX5yNt3YG4Dg9DmvIv2m/gtqvxP+Ddh8Nvh7a2Nl9h1jQrqO3fFvax2em30M8kaKilRiKMhFC46DgUAeD/ABvufh3rH7Q37NWkfCOTTbjxLYavd3P/ABKTEwt/DK6fKt2HMHCW8haNYw2FZuFyc1vfsYP4bsfGn7QegyG1t9ah+J2szR252LcLa3FnZmJlX73luEcrjg4PvX1p4J+D3wn+Gt/qGqfDzwZo/hm81U5u5tNsILSSfnOJGiRSwzzg8ZyepNSP8Jfhc/j+P4rP4R0o+M4kMaaz9ih/tAIYzER9o2+Z/qyUzuzsJXocUAfjf4PfSov2Qv2Xp/H+z/hW0fxA1A+IjP8A8eYH9p6n9hN7n5PsoutnmeZ+7zt3V+znh/Wvhzf6/f2nhW+0q41pba1e8Sykge6Fr84tmlEZL+X98RFvl+9t71Pa/D7wFZeEX8AWfhvTIPC8qSxvpUdnClgyTu0kqm2CeURI7Mzjb8zEk5JNZ/gH4UfDD4V2lzY/DPwlpXhWC9cSTppdlDZiZhnBk8pV3EZwM5wOBxQB8rfEX4o3ur/tCeIfhl4Pfwn4Ov8Awv4fs7vWPE3iW1F3cyWV68xS2tIBNab4YsM0sklx5aNJjyySSfgjT7ax17/gmlaeHpbtNR0j/hYcVjDJbZgiezfxMAPJCNmNGDkoFb5QRg8A1+zfi/4Q/Cn4g6tp+veO/Buj+ItS0rH2S51Gwt7qaDBJAjeVGZQCSQAcA89ea0Jfhv8ADufRpPDk3hbSpNJmvTqT2bWMBt2vjN9oNy0RTYZjN+98wjdv+bO7mgD4I/bD0e48G+Pf2dbHwuNA8M+C9O1nUoi2s2Ty6Ba6r9hCaSbiC3mtgGGJ1gYuAshU9qdqser/AAYm+Nv7SfxR1jwx45jl8KWkGseG/DlpJBDdNbtMLaa9jubm8/10cjwszLtMSn5TtYH9D/EXhvw74u0a68OeK9Ltda0m+XZcWd7AlxbzLnO2SKQMjDIzgg1yXg/4O/Cb4e6LfeHPAvgzR/D+l6opW8trGwgt4rpWBUidUQCQFSR8+eDjpQB+ZniTWPE0v7RX7L+reI/FPhCXU7rUr0Wmi+FbQobDTbvTJt4a7e5keeFvlUEQQRswyq8V+j3w1tPgraeI/HP/AAqs6V/bc2qmTxMunyI841NkGTdqpJWQr2IHO7+LdVnwz8Dfgr4L8j/hEPAOg6Kba7+3xGz0y1gMd4EeMToUjBWUJI6BxyFZlB2kisn4X/CC38A+LvH3j67ltZ9b8fakl1cmytFtIUtrRTFaRlQzNJKEJaaV2JeR2wFQKoAPnG48M+Hde/4KTXF7rem2+oT6V8LrKa0e4iWUwSvrN3Gzx7gdrFGK7hztJGcE58O+Dvwz+Hv/AApP9r9f+EcsP+Rs8b2Ck20ZKWltbLNBAhx8kcUkjvGi4CMxZQDzX6mr4Z8Nr4jfxiuk2g1+S0WwbURBH9sazSQyrbmfb5hiEjFwm7aGJOMkmqln4K8G6dZaxpun6DYWtn4hnuLnU4YrWJI76e7ULPLcoqgTPKoAkZwS4GGJoA/JzVNH1vxx4W/YN0qx8R33hvUtQ0piNVsRC95Cy+GldmT7THNES4Uq25G4Y45wR7H+zH4Qj8K/tLfErwf8b9X1Dxh8UdOtvN0LWdXn8xbzwffOpC2lsoSCIxXCmO62RjL4x8pIr75h8A+BbceH1g8OabGPCSGPRgtnCP7MQxeQVs8J/o48r93iPaNny9OKvXfhTwtf+IbDxdfaNZXGu6VHLDZ6hJbxvd20U+BKkMxUyIsmBvCsA3fNAHw1+wLq/hDRPgbr8d7d2Vjf6B4k8VDUg7xxy2kUOpzyN52cFERCrEtwBivubwv4p8N+NfD9j4r8Ianb6zo2pxia1vLSVZoJozxuR1JB5BB9CCDzXn2p/Bb4f2uo+JfG/gvwn4f03x14gsbm1k1afTIpGnadel2Y/LkmiZgplXzAXAwTwMafwc+F+i/Bj4ZaB8MfD7+bZaFAYxJ5aQ+ZJI7SzSCOMBEDyOzBFGFzgcCgD4c/ZZ8GxxeD/wBpfxj4N0+L/hOr/wAeeOLW0vSoafzLeeQ2kSueVRZnLBVwNzFupzXgXwh8B+NPi5+yL4H8EaX4/wDAPhvSYJtKmLnTbuHXbDxDaXUcr+dNJqIA1B7pCkjGIGQudqhXAr9kdD8M+G/DCXsfhrSbTSU1K7mv7pbSCOAXF5ctumuJRGq75ZG5d2yzHkk1wVz8BvgjeeNR8SLvwBoE/ioSCb+1X0y2a980dJPPMe/zBjh87h0zQB8ueB28NaX/AMFAfixZav8AZLW71Lwx4ZubFJtiPMUmukkki3YLMJCoJXndjvXyj4xXW2/ZT/a8/scXBgHxW1Q6mLTd550gT6WdR27OcfZfM8z/AKZ7q/WfxV8Jfhd461/SPFXjTwjpWu6zoDrJp97e2UNxc2rI3mKYZZFLJtcBhg8NhhzzXSaR4V8MeH11JNB0ez01dZupb6+Ftbxwi7u5wFlnn2KPMlkCqHdsswABJwKAPLfD2r/A2/0TRNK+G194fkku9Hni0GGwktvMaw8tSy2qod/kjCFwo2jA3cgV+SutfEnwDF/wR9sPCcmv2a65Np0Gmrp/nKbv7Zbaosk0JgBMgaNEZ2yo2oNxwOa/YrwX8Efg38ONYu/EPw/8DaH4a1O/BWe607Tre0mkViCVLxIrbSRkrnBPOM81Sj/Z/wDgTFqGt6qnw68O/bPEqumqSnSrQvfLIwd1uCY/3gdgGYNkMw3HJ5oA+f8AxQ6Rf8FC/AhlYIJ/h7rcce443uuo2jFV9SF5IHbmvgf9pD4q3HxY/Y28c/EnSb/wf4A8JXupXD6VoUNmJPEN5qFnqeDPNcefClvdu6NcMkdtLIEJLSHJNftdq/gnwfr+vaN4q1vRLO/1nw60r6bezQI9zZtOnly+RKw3x70OGCkbhjOcCuLl+APwLn1fV9fuPh34dl1LxBG8Wo3L6VaNLeRy/wCsWdjGTIH/AIw2d38WaAPlTx14W8PeMv8AgoF8MZfE9hFqQ0zwHquoW6TLujW6W+t0WQoflYqJGK7gdrYYYZVI+ov2hLa2f9n/AOI9u8SNEnhrV9qFQVG2zlK4HTggY9K9Hbwv4ZfxBb+LH0izbXLS2eyhvzbxm7itZGDvAk23esTMqsUDbSQCRkCtHUNPsNWsLnStVtor2yvYnhngmRZIpYpFKujowKsrKSGUggg4NAH5ImHw9p3g39hO98ex28Hw6Gm2r6k1yFWx/tt/D8I0d7kn5N3m+b5bP/HjnrXv2jXXh7Uf+Chc918MJbeeKLwK6eL5LFla3Ny18h0xbkx/J9r2CUru/eeTn+HFfaN74E8Eal4QX4fah4e0658Lpbx2a6VLaQvYLbQgLHCLYqYhGgVQqbdqgDA4FVfAvw3+Hvww0htA+HHhrTfC+nSOZXt9NtIrSN5CAC7LEqhmwANxyfegD88P2MvB/hnR/wBgDVfE2nadDFq2v2fiue+utuZp3hur22j3OcnCQxIiqOABwMkk+N+Lvhx4CH/BN34GlPD9kjXF94LuZHSBFd5tQurdLqRnADFplkYOc5IODxX7CaR4N8IeH/Dn/CHaDodjpugBZk/s62toobPZcMzzL5CKI8SM7M42/MWJOSTVWb4f+Arjw3YeDbjw3pkvh/Sjbmz05rOFrO2NoQ1uYYCnlp5LKDHtUbCAVxgUAfHXjfw9oPhX9un4Hnwzp1tpQvvC/imymFrCkIe2tBZvBEQgA2RsSUXovavjj9ov4nt8W/2VPi54w0688HeAfBrahqdvb6Q1kJvEGp6nY3hXzppfPgS2uppkMqqsE0u0hzJyTX7P3fhnw3f67YeKL7SrS41nSo5orO+kgje6to7jaJkhmKl41k2rvCkBsDOcCuDufgP8ELzXtV8U3nw+8P3Gsa5FLBqF5Jpdq893FOCsyTyNGWkWVWIkDE7xw2RQB8caxeaFD+2h8G9a8YPaxLrfw71RBPd7FW4u1ms5WG5+GcJlvUCud1mPxhN8V/24Yvh75p8Sv4V8NjTfs+fPN0dFvvK8nbz5u7Hl4/ixX3543+EXwr+Jel2GifEPwhpPiSw0tg1pBqFlDcx25AC/ulkVgnygAhcZHB4rpdN8KeF9G1bUdf0fR7Ox1PV1t0vbqC3jinuktFKW6zSKoaQQoxWMMTsBIXANAH5J/FbUfgnd/wDBOzwHpnwgm05/E7R+HF8IQ2DR/wBpJ4lFxb7vJVP3oug/mfaD977+/rX0hoHhfw/rn/BQ/wAa6pr2mWt/faV4D0Q20s0SyGB5ry6WRot4O0sBgkc4yM4Jr6r0r4I/BvQvGc/xG0XwLodh4quWdpNVg062jvWaTO9jOqB9z5O5s5buTXaw+GfDdv4gufFtvpVpFrl5BHaz36wRrdy28TFo4nmA8xo0ZmKqSQCSQOTQB+cfwE0f4MRfCL9onwz8ZYtLtfhtp3xR8QwzwaiyQadbwCazkhTkqsai4ZSgGMMQB2r9H5jZNoDtppja0NqTCYsGMx7Pl2beNuMYxxjpXkfxV+CGheO/hj4r+GXhqz0rw9beOZ3k1iYabDN5zXRX7VdeV8iPesqgxzyh9sio7K+0LXsOlaPp2i6PZ6Bp0IisLCCO1hi6hYYkCKvPUBQBQB+TP7H37O3jb4u/sd+A9H8WfFzW7fwBrmmSx3Hh3TbPT7MPatPKsls1+IGuzFJyHAcFlYrnHT3PxJ46tW+Mfi74OfDxfB3w7sPhx4f0mDVtd8RWYu7iXTrmGR4LW0g8+0U2tvGCHkmnZFZyojOST92eHfDfh3whotp4b8J6Xa6JpFgpS3s7GCO2toUJLFY4ogqKMknAA5Ncp4p+EHwo8ca/p/irxn4M0bXtZ0rH2S9v7C3ubiDady+XLIjMuDyMHg8jnmgD4x/YG+IPg7wX+xj8PLjxn4kstNsrvVtW0fTri7mS2juJX1e8S2gh3tgsyL+7QE4UYHAr3D9tv/k0X4u/9i1qP/ok10XxQ+AHhj4h+GNB+H1lY6XovhWx1yDWr21i06ItK9vcG8ItiNqW8k85PnzBGdkeUDDSbxq/tI+ANe+KvwE8ffDfwuYRq/iTR7uxtTcOY4vOnjKrvYBiFyeTg0AfA/xzuvhJe/s2fA3wx8JJdHl8W3Ou+FH8KxaQ0DSxXUU8L3c0Yt+VRIVl89+FBPznJFej/EH4mDxz4++M/g7Rrvwb8P8ARPB1vb6f4k1fX7Jb3UtVFxZCUAQme0RLdI3EaSTPNuIIVAMV9X/DL4DfC/4ff2d4n03wXomm+MRYQW17qlnYQR3csixKkpNwqLI28j5iTluN2a6nWfhB8KPEfi6z8f8AiDwZo2p+JtP2/Z9TudPt5r2HZjYUndDICuPlIPy9sc0AfkV4Y0bTfiD+zf8AsPaF4ygGq6fdeIoYZoJiWSWG2s70RxOP4o9saqUPysvykFSRX298VPiLPaftBaT8FPh7Z+FvDOu2Phh9cm8ReIbXzxa6bLdG3NtY28cls0jF4zJLm4SNAFJDHgfVNt8OPh5Z2Wh6bZ+F9Lgs/DEpuNJhjsoFj0+YhlMlqgQCB8Ow3RhThjzyao+N/hN8LfiZLYz/ABF8H6R4ok0xt9q2p2EF4YGPXyzMjbc9wODQB+SPwt8Iaj45/Y/+Md74E1K113Wfh18TNa8TeF7ywiWK0ludHlgv4zZwxs6JFcq0qxorFQsuASOa918L+I9X/aO1v40ftJ/CtpJzpPgoeFfBUkRyx1C5sP7Vu5ExyJBcz20GV5zEw7V+j2ieF/DXhmO8h8N6TaaVHqFw93craQRwCe4kVVeaQRgB5GVVBZskgAE8Cq3hLwZ4P8A6NH4c8C6FYeHNJid5Es9NtYrO3V5DudhFCqICx5Jxknk0Afkj8JvAfiv4s/szfCnw7ofj/wCH2g+H9LutBvLFrXTbqDWbTWLKeKRoWkk1Er9vkmDxTExbpGd8L8wr6W+EnhLwvrn7c37SGta1pNrf31hYeD7e3muIUleKG70+fz0QsDtWXy03gfe2jPSvqUfAf4Ir41PxJHgDQB4rMnn/ANq/2ZbfbfOznzfP8vf5n+3nd713tj4Z8N6ZrWp+JdN0m0tNX1sQC/vIYI47m7FqpSATyqA8oiVise8naCQuAaAPxV16ys7T/gmN8TLCK3RbDw941vY7aNUBW1tLfxZC7BAB8qIm4kDtmvrX9rvQvAvj3xT+zlobQ2ereHdQ8dxmSKBke2nQadcyhW8s7XR8DcpyrqcMCpIP3Jb+AfAtp4fv/CVr4c02HQ9Va4e8sEtIVtLlrslrhpoQuxzMWJkLA7ySWzmuf8OfBb4QeENN0zR/C/gnRdLstFvH1Gxht9Pt40tb2RSjXMICfu5ihKmRcNt+XOOKAPmj4vWtrZftp/s02tnCkEEem+OQqRqEVR9ksuABgCvOv2SdX+FWlaB8aE+L93pNp4zXxhrreLf7ZeFJDbeeTZNL9pOTZfZdnkZ/dY3be9fobe+GfDepa3pvibUdJtLrWNGWdLG9lgje5tFugqzrBKyl4xKFUSBCNwADZwK4nxZ8D/gz498Q2vi3xv4E0PX9bsgohvr/AE22ublAnKASyIzYU8qM4B5HNAHyh4d8XeFPCX7enxLm8UazZ6RDrXgnw/d2Ul1cRwR3FvbT3ayyRu7BWVM8kHgc9K+NvEGr6d4l/wCCfX7UviLQpxeabqnxD1+9tpkB2y202sWkscoyAdrRkNkjpzX7B+M/g/8ACj4jX2m6n8QPBuj+JbvRyTZS6lYQXb2+eSI2lRioyAcDjIB6gVsSfD/wJLouseG5PDmmtpPiCSabUrP7JD9nvZbnHnSXEe3bK8mBvZwS2BkmgD5V+LPxUvLn9oXSvhJ4ETwvoniCHww+vz+KPEdv9r8jTZro2/2exhjltnmZmjMkhNwkaqFJDHgfnkbuKf8A4Jx/tMW+navbaxp9n491WOyvNPRYLOWBtSsJVktI43dY4XaRpI1R2UBuCRzX7Ja98Dfgx4q0zQ9G8UeBND1mx8MxRwaXBe6db3MdlFCoSOOBZUYRoqqAFXAwBxwK3J/hl8N7rQdV8K3XhTSZtF12c3OoWL2MDWt5OdhMtxCU2Sv+7T5nBPyrzwKANLwj4P8ADHgPw9Y+FfB+mQaTpWmwpbwQQIEVY4xhR6k+pJJJySSSTXxB+zt4I8Ha7+0Z+09q+t6HZahezeINPsHmubeOZzaPpVuzQZcH92xJLL0bvnAr9BKxNL8M+G9Ev9T1XRdKtLC91uVZ7+e3gjilu5kQRrJO6ANK6oAoZySFAGcCgD8fPhfpd/o37BXwV/aA0WN59X+CeqahqxCZaWTQV1K7stVtgf7v2El/rCteo3Ih8f8A7PP7Wn7SO9bi38e6T4i07RplO5H0Hw9p9xY20kZ7LPOLif0YOp9K+r/jN4C+I8fw2T4O/s3+HvDvh/Sdet76wvLqc/ZLbRre7/1k1rp9vAyXEj+bK+0tGvmYLFgzY0fF/wAD4rP9lDxB+zt8N1jjz4PvfDmmfaG2IZJLF7aJ5nUHG52DSNg8knBoA+CPjBc/CS8/YT+E/hf4XSaPJ40v7jwgPCsOltA1ymuLc2r3MsQg+ZZFQTee/BBJDkMa99+HF94Q0v8AbV+PTfFSeztvEE9joLaE+pMiBvDgsyJxZmY/6oXfm/aQnHmct2r6E+Dv7PPw0+HGk+GtePgvQ7Px3YaRZWV9q9rYwLdyTQ2yQzH7SI1kbeQQWJBYYz6V6B4++Efwr+KkdpF8TPB+keK109i9uNUsYLzyS2N3l+cjbd2BuA4PQ5oA/G/W4tJX9iz4m65oscY+HVn8W0uvDMoXNrFoP9tWQeS1P3VtDcefs2/LjPavs39rfRvBfjDxj+zfo8tpY6xoN547QvAFjmtJQun3UgDKMow3AHByD3r7ku/CHhPUPDD+Cb7RbK48OyW4tG02S3jazNsBtEJgK+X5YHG3bjHauZ8P/Bz4S+FdL0jRPDfgzR9NsPD9099p0MFhAiWd3IGV7iABP3crKzAyLhiCRnBoA+Q9Qm0T4f8A7feu67ZWC29ovwik1G9is4lVrhrHV8IxRcb5BHlFJ5xhc4Ar4++Knj/U/G3wZ+DHxHj1PwX4R0jUvF/hrUfDfhTQ7QNqVqr3qZLXnnookjjYidIbNFU5RmPf9qT4Z8NnxJ/wmJ0m0Ov/AGT7B/aPkR/bPsZk837P5+3zPJ8z5/L3bd3zYzzXn+mfAD4E6NcX91pPw68O2c2qSxT3TxaTaI08sEizRO5EfzNHKiyKT91wHGG5oA+ZbXXbTwt+3N8Vdd1BHeKx+HGkXpiQZllitru9Z/LXqx4xx34r4w+Knj/U/G3wZ+DHxHj1PwX4R0jUvF/hrUfDfhTQ7QNqVqr3qZLXnnookjjYidIbNFU5RmPf9nbnwT4Ou/Ftp49utEs5fEtjbPZQak0CG8jtZG3PCs2N4jLc7c4ySccmuL0z4AfAnRri/utJ+HXh2zm1SWKe6eLSbRGnlgkWaJ3Ij+Zo5UWRSfuuA4w3NAHzFovhfw3rv/BRHxxqWt6XbX91pvgPRfsslxEsrQefeXayGPcDtLKNpI5xkdCa+cvht8Nfh/B+x/8AtYiLw9YqYdb+IdvG32ePdHDpwma0jRsZVYG+aJRgI2WXBJNfrXD4Z8N2/iC58W2+lWkWuXkEdrPfrBGt3LbxMWjieYDzGjRmYqpJAJJA5NULfwL4Is9H1fw9aeHtOg0rX5LqbUrRLSFbe9kvs/anuIguyVp8nzS4JfJ3ZoA/O3Qbq4vvi7+xDe3chlnuPBGvySOxyWd9H0wsSfUk1qfsk6v8KtK0D40J8X7vSbTxmvjDXW8W/wBsvCkhtvPJsml+0nJsvsuzyM/usbtvev0Ai8D+CoLrQ76Dw/p8dz4YgktdJlW0iD6dbyosckVowXMEboiKyxlQVVQRgCuW8WfA/wCDPj3xDa+LfG/gTQ9f1uyCiG+v9Ntrm5QJygEsiM2FPKjOAeRzQBxP7V8sM/7KPxgnt3WSKTwZr7IykFWU6dMQQRwQR0NfFnxQ8F+EfC37MX7MF/4b0az0y5sPFngG4hlt4EjdZbt4vtD7lAJMxYmQk/OTlsmv1J1XR9I13SbzQNcsYNR0vUIJLa5tLiJZoJ4JVKSRSRuCro6kqysCCCQRisq+8FeDdT0rTdC1LQbC703RpbaextZrWKSC0ls8G2eCNlKxtDgeWygFMDbigD4l8C+L/CngX9uH4/jxprNnoQ1TQfCN9bNfXEdssttZw3kc8qGRlBWNmAYjgd6+Mf7TtNZ/Yf1jWy8lx4EvPi7c3PiRolkHmeGpNe33JZQA/llShcEfdzkda/Yjxf8ACD4UfEHWdN8RePPBuj+ItU0fizutQsILqa3Gd2I3lRmUBvmABwG568109j4T8LaXYX+laZo1laWWqzXFxeQQ28ccVzPeMXuJJkVQsjzMxMjMCXJJYkmgDxTxprPwkv8A4f8AiPTPBF9olxqkvhPUTYw6dJbPO2mLDjMKwksbcOyD5RsyV7kV+bF7ea/F8G/2H9Qs9R0aw8NQWgSe78QQS3eiw61/ZSJphuo4Z4PnDC4WFmkASbB7V+sngb4LfB/4ZTX1z8OfBGi+F5tTG26fTNPt7Rplzna5iRSyg9FPA7Cr1n8KPhhp/gqX4bWXhLSYvCU5lMmjiyh/s9vOkMsgNtt8rDSEsRtxnmgD4L8T/C7x4NF/aM+IWv8Ai3w3ruseKPh7dadd6T4YtZrYG4gtLz7JdXEc13dOZZEd4lPG5VAH3TnzT4k/E74e61+yZ+zF4e0fxFY32q3ms/D7y7SCdJZx9jktkuC8aEsgif5HLABXIQ/MQD+n/gP4U/DH4W2Fxpfw18J6V4WtLtg08emWUNoszDIBk8pV3kAkAtnA4rC0P4AfArwz9r/4R34d+HtN+33MN3P9n0q1j824tpBNDK22MZaKVRJGf4HG5cHmgD4++NNhN8MviJ8VPj/8JvFfhXXLo6VaR+NfCfiF4yXg0y2LRLBdRuZLSSW3cARTRPFIzBjjOR9FeCPEfwK+IHiT4d/Emez07S/iN4j8KxXuj2l00aavDpF2iTyRJHkMUjLYYquBh8cbq77xh8Cvgp8Qdet/FPjvwFoXiHWbXb5d5f6bbXNwoT7o8yRGYhewJwDyOaydY+D1p4g+OHhr4vapJaeX4N026tNLhitFW7W4v8x3Dy3RYsYVhAWKFFUBnkZi3yBQD5y/b+sNV1Xw98FtM0LVG0TUbr4o+Gora+SKOd7WZxchJlilBjcocMFcFTjBGK29c+Ftv8Gl1H9qP46fEDVfiRq/w20fU7nTPtsNnp9lYrJARObe1soYlNxOg8ne5ckNtA6V9g634Y8N+JvsH/CSaTaar/ZV3Ff2f2uCOf7NeQZ8q4h8xW8uaPJ2SLhlycEZo8SeGfDfjLRLrw14v0m01zSL4KtxZX0EdzbTBWDqJIpVZGAZQwyDggHqKAPxY1T4bftY+Av2VvDXxL1jT/C87eB9VT4n3UkM18NdmuppZL++SZWi8jcYLiWGVQ2Ni4BJAJ+qdJ+Ing/4f/tm3fxB8V6nDYeEvjd4O0aTw5rN26xWb3OnNI72fnMQqPLDOkyhiA/QHdgH9D76wsNTsLjS9Sto7uyu43hmgmRZIpYpFKujowKsrKSCCMEcGuT1L4Z/DfWfB8Hw81fwppV74VtY44odJmsYJLCKOEYjRLZkMSqg4UBQF7YoA+O/hJ408Y6X+1vrfwsvvizN8RvDMPg5tbkSdNMX+z719RWFUL2MMRAEHIEhJIbcexr7U8HeNfCHxD8O2vi7wJrNpr+iX2/yL2xmS4t5PLcxuFkQlSVdSpHYgg81z3hX4V+A/hj4fv8ARvhF4X0bwobqNiEs7GK3t5JwpEbTpAI2kAJ5y24jIBFZfwN+Eek/BH4c2XgHSpIpjFPeXtzLBbJZwyXV/cSXU5it48pDEHkKxRKSEjCrk4yQD4f/AGZ9W+BOial+1HffEa50G2ul+IHiD+1jqBt/PbRxFBsWdZPna3L+dsBBUtvABbNfMOkQeOtF/Zj/AGR7jWriw0nwxH4i1GZ5fE1vNdaZbJcG8fw419Gk0DGMRunlM0iojGItkDB/QL4Vfsk+D4NV+I2r/Gvwd4f8UXWu+O9X8R6NNd2kN/JBY30duI1LTxExvuiYsgyucHJNfYGteHtA8SaLc+G/EWm22qaTeR+TPZ3UKT28sf8AceJwUZfYgigD4Hi+HfjeDxZ8Wfi14x8aeFtV1DU/AU2mXumeGrWa13+V58tne3STXl0xYIZokc4yvAPymvL9L8H+GfCv/BJS8l0DTobOXU/h297dSov72e4urQSSPI5+ZiWOBk4VQqjCqAP0W8GfBn4RfDrTL/RvAXgrRvD1hqqlLyCw0+3t47pWBBWYRoBIMEjDZGDjpXRt4I8GN4R/4QBtA08+F/sosv7KNrF9g+yBdgg+zbfK8rbxs27ccYxQB8Qa18Tr+Pxr8LPg94NtfDek+KX8Eprh8SeJYDcJY2GYLeSCyhSS3kmmkZQ8ifaIkVEDNu4A+I4L7UH/AGJv2o7fQdai1/SIPiPP9svdFi8i3m0maXTJNTe0iheQJA8TTMyq7LsZzk5zX7O+L/g/8J/iDZ6bp3jvwZo3iK10fH2KLUNPt7qO2CgACFZUYIMKBhcDAA6V0Ok+DfCGg22oWehaHY6db6tIZbyO2tooUuZDEsJeZUUCRvKRI8tk7FVegAoA/Ov9pPV/ghP4m/ZY0r4cXWhSzx+O9IfS4tNa3aRNHFtcBjCIsstvvMOcYUttzyBXWfB/x94I+Hn7TX7UieO9estAMWoaBqZ+23EcGbM6PEDMu8gsgYbSVzhsA8kCu2+Jn7JnhNdW+GN98D/CGgeFE8OeOdN8R6ybK0h09p7Ozt7qNgPIiHmOrTjYhwACcEV9IeIPg/8ACfxZ4r0/x34p8G6Pq/iPSgotNSu7CCe7gCNuTy5nQuuxiSuD8pJIwSaAPxn06fV/+HfPwA1C3nsbXw1H8QJZdbm1e3ln0uK1Os6n9nfUoUkhZrRbswGQM6gNs3cAivtGH4f/ABdvfibrXxfbxx4S1HxXD4G1TS7TTvDFpPbT3YkKTWU8omvbossM6gRsFAHmEZ5FfdFp4L8HWHhyXwdY6FYW+gTidZNOjtYks3F07SThoAojIld2Z8r8zMxOSTXNeAvgz8I/hXLdz/DPwVo3hWW/4uH0uwgs3lGchXaJFLKD0UnA7CgD8h/FOsfAW3/4JJfYdPu9HGp3Wh23nRTNCb1/EhlQ3hZH/em6E3mHJG8Jz90A19canruieEv27/h9qnie/g0mz1b4X31jaTXUqwxzXUepWs7RIzkAuIwWwDnHNfVk37PvwHudc1bxNc/Dnw7Nq2vRyxahdSaVaPNdpOCJVmdoyZBICQ+7O8fezW944+FHwv8AiZZWOm/EXwlpPie00t/MtItSsobtIHwBmNZUYLkAA46jg8UAflLrHiLU/GXwz/bl1/4N3x1Oa7v4fstzYsX82CPS7eO6MDJy2Yll2smQ3BUkEE/od8INb/Zvl8C/D7TvhveaD/Z1zYrF4et45Lbz2T7MWmSFM+YZfKVvPAG/hvM717XonhLwp4amvLjw5otlpUuoeT9pe0to4Gn+zRLBD5hjVS/lxIsabs7UUKMKAK47wz8Dvgx4L8T3PjXwf4E0LRPEF5v83ULLTba3un8z7+ZY0V/n6tz8x5OTQB+evwO+I/gH4R/sz/HzxR4g0i116bwp4s8Z/b9GKxGaeJrxgLeVGVsQyK4DFlZQhJwQCKwfEmseJpf2iv2X9W8R+KfCEup3WpXotNF8K2hQ2Gm3emTbw129zI88LfKoIggjZhlV4r9L/wDhTHwh/wCEu1Hx9/whWjHxJq8Elre6j9gg+1XMEqhJEml2bnV1AVgxO5Rg5HFV/DPwN+CvgvyP+EQ8A6Doptrv7fEbPTLWAx3gR4xOhSMFZQkjoHHIVmUHaSKAPnX9kZ0X4j/tI2zMBMvxCnkKE/MEfT7PaxHXDYOD3xxXCfFvXbzwV+3JpviTw7otr4w1a9+H0lpJpt5qFtpjWUA1NmSe2muz5cv2hi6TRqNypGGJIIRvvHTfBPg/RvEureMtI0Szstd15YE1G+hgSO5vFtgVh8+RQGk8sMQpYnA4HFfF/wAcvgh4s8T/ABwk+IGu/DPQ/jT4RudHtdOstM1W5ht59DuYZpJLiaCO7ieCRbrdH5jhllHlqo+UYYA+ffjP8HLr4dfsFab8P/FUtpNLqvjDS7q5g0yV2s7WPVfECz/Y7WYbHMcCy+WrjacgkY4x9UfHLxfpPw08XfCb4LfDzw14asNa8TyakdHv9btwumaNFpVshlaGKLy3ed0kVI445Ytw3bnAHMH7OH7Llh4CsfHMvjjwvoum6P401ez1Wz8IWn/Ex0nRTYxIsbxmeNENzJKvnSNHEqq4Xy+FBr6k8cfDj4ffE3S00P4jeGtN8UafFIJUt9TtIryJJB0ZVmVgG9xzjigD80v2ffBGkfF7x7+158JfEfiPTfFek+I20SC6vtGtUtbA3l5pssdxNBbrNcKksUsahj5rMZY9zHdnGj8BfFfiH9oL4o/Czwl41BfU/gBpOoTeKUJLqfFCzS6HZmTPG5oYLq7Q+kiMOor9AdQ8KWvw98OavqfwZ8F6OPEMlvbwxWqCLSYbpbX5IIpriGGQqkMbMIxsYKPlUAE157+zl8HvEXw2s/FvjD4iTWNz48+IuryazrL6cG+xwYRYbazgZ1V3jt4UADuoZnZ2PWgDtfhFafBaytvE8PwVOleQ2u3z62ulSJIF1tiv2sXGwnbPwu5TggY4xiviS18E/Efxn+3H8cI/h/8AEW7+H/2bRvCX2prTTrG/a6V4brYP9OilEZjw2Co53c5wK+zvg18ILf4WDxbqtxLa3Gt+ONam1vU5LG0WytRNJHHCkcUIZzhY4lLuzM8krSSscvgem2nhjw3p+u6h4osNJtLbWdWSGK9vooI0urqO2DCFZplUPIsYZggYkLk4xk0AfnZ+1J8FfDnwT/YP+M+l6PfX2t6lr0T6rq2q6nKs17qF/PPArzSsioigKiqiIqqqgADqT5Z8dvDfww+DuleEfit4A+J2p+M/iZoV/po8P+G9a14eJF1Ca8nitporeyummkil8p32zwbHjI+9jIP60eIvDfh3xdot34b8WaXa61pF+uy4s72BLm2mTIO2SKUMjjIBwQeRXC+EPgX8Evh9qQ1rwF8PfD3hrUACoudM0m0s5gGGCPMhiRsEdeaAOqs/HPg3UPFuoeAbHW7O48SaTBFdXempOjXcEE/+rkkiB3KrdiR3HqM/D/7TZ8Cr+2R+zI/xCOnrpyw+MTCdTMQgF2Lex+z7fO+XzN+PL77sbecV9Q6F8ILew+OPiP44anLay6lqemW2i2SW1osEkVhEwmkNzNuZ7iV5uh+VUjRFVc72Pnnxn+As3xY+O/wp8Y6zpmm6z4O8K2Pia11iy1FFnWb+1oLZLdRBIjpIu6Fi27GOCM0AfOUnjfwX4Y/ax+O3xV8G2sWs+EPCnw2gm8Vx6cEkhvNbtJrm5jhcrmN5/sKlGznaDhsc14t488V69r15+yx4v1DXfBukrq3jXw/caN4Z8M2mLqx0y+idZt16bjMkao6xTLHawxb2UckLn9cPB/w68A/D7w8fCXgTw5p3h7RGLs1lYWsVtbs0gw7NHGqqxYDDEgk981zGgfAX4HeFZGm8NfD3w9pUj3kWoFrbSrWJvtluxeGfKRgiSJiTG/VCSVIoA+WPgX4P8M6j+23+0z4v1HTobrV9OufC9ra3Eqh2t4p9FgeURbshDIUTeQAW2qCcAV8VftIfFW4+LH7G3jn4k6Tf+D/AHhK91K4fStChsxJ4hvNQs9TwZ5rjz4Ut7t3RrhkjtpZAhJaQ5Jr9utP8M+G9J1fVfEGlaTaWWqa60L6hdwwRx3F41vGIomuJFUNKY4wEQuTtUYGBxXn8vwB+Bc+r6vr9x8O/DsupeII3i1G5fSrRpbyOX/WLOxjJkD/xhs7v4s0AfIvii58Bj/go34SvPHsmmxzj4ZTPpj6gYVI1A6sObYy9JvJ83Gz5tm/tmvF7yXQ9T0r9vbxB8OzDJ4GuvD5gils8Gxm1u30G6/tRoSnyFwWhExX7z8k55P6eeLvhL8LPiAJv+E68H6P4gM9stm7X9hBcs1tG5lSLdIjHYsh3qucBvmGDzWjp3w98A6P4Of4d6V4b02z8KyQS2r6TFZwpYNbzgiWJrYL5RSQMQ6lcNk5ByaAPhe917wP+z/8AsdfCi20fwfo+pSeLG8M6bbRaqFTTV1XVFjnGoajMySNtjlVpnkwXLhcEEgjzvwdd+M4/279YtD4u0DxL4suPhbfrcLoFl9htbW6h1C3NpDOjXV27um/KvI4bYw+VVIr9M9W8D+C9f8LHwNrmgWGoeHDClv8A2ZcWsUtl5MQAjj8hlMexABtXbgYGMYrJ8J/Cj4XeA2s38EeENI0B9Pgltbd7CwgtnhgndJJY0aNFKpI8aM6g4ZlUtkgGgD4r/Yb134G6d+yz4Nh1i+0i01uO7C66uqSQJfDxQbthJ9qE5En2s3OPKL/PjZs421qfBHwd4T1r9sr9qLW9Z0azv7+K48L2aT3ECSyLbT6HCZYlLg4SQgb1HDYGc4FfVtz8DvgveeN1+Jd34D0KbxajiQas+m2zXwkXAWTzynmbwAAGzuA4BxXb6f4Z8N6Tq+q+INK0m0stU11oX1C7hgjjuLxreMRRNcSKoaUxxgIhcnaowMDigD8YPBSLb/sl/s36dCNtvYfG6ztoEHSOGLxBqARB7KOAOw4r638cfE9vFnxn+KXgDQbrwb4BsfA9hp0PiXX/ABJZLe3t/Bf2puUjjgM9ogtYo32mSeWRdzMFjxyfsyH4Z/De30rT9Ct/CmkxabpF+NUsrVbGAQWuoLI0wu4Ywm2OcSOziVQH3MWzkk1R8Q/CD4UeLfE9j418VeDNG1jxBpe37LqF5p9vPdwbTlfLmkQuu08rg8HkYoA/Iz4a3s0/7IH7Ft3fTmRLb4l6fD5rnAWKGTV4YgSegCqqr+Ar9uK4DUPhR8MNU8KQ+BNQ8JaTP4ctpvtMOmtYwfY4pxIZvNSHZsV/MZn3KAdxJzkmu/oAKKKKACiiigAooooAKKKKACiiigD/0/38ooooAKK+Qv2xPHvxJ8CeHPhwnws1iLRNY8T+OtC0F57i3F1B5GomWNhLESpZA21iFdGO3AYZzXF+DL/4t/Cj9rHRvhN4t+Il/wDEHw7458NahqijU7WzgmstQ02eBWMLWcMKiGRJSPLKnaQOSckgH3jRX53eA/jN8TNW/Yi+LfxT1HXJJvFPh1PHJsL4xRBoDpUl2LPCBBGfKEaY3Kc4+bPNTa38Uviz4+ufgT8EvB3iZvDOrePvDA8R+IfEMdvDNex2lrbW++OzjmRoFmuZ5sF2QiNQSqk4FAH118X/AIt+Gvgr4Rj8Z+K7e6ubGXULDTQlmiSS+dqNwltESJHjGwPICx3ZC5wCeK9Rr83P2svBXifwH+zQmieJvGuo+OSfGfhiS1u9VitY7uG3OqWgEMj2kUKSkMGbzCgYhgD93JX41/HqfVP2htc+DN7451/4eeGvBml6fdXNx4Z0WfVNS1HUNT8ySNGmSw1CO3t4IkUkGNWldiAxVWAAP0ior8sov2ifjLP8F/Aep3epXdvqq/FnSvCM+qXGmNp0uuaHLehUuWtLmCMwm5t3QSbEXDhtu3oPqPxv8QPGGlftefDD4cWGotD4c17w/wCILy9swiFZriza1EDlipcFBI2ArAHPIPFAH1VRX5j/AAo+I3je6+Mp0H47/FPXvAnj6fX7yO18K3enWcHhzVNKSdxaw6ZcyWpNw0kOwmRLrz9+V2HGT92+CtG+KGm+KvGV9448RWusaDqN5DJoFpBaC3l061WILJFNIOZmaT5gTkjnnBCqAcx8T/2kvgt8GtesfC/xH8Rf2VqupWzXlvbraXd0726P5bSf6NDKAobg5xWn4W+Pvwb8b+BtZ+JPhHxZZat4d8OxTy6lc27M7Wa2sZllE8IHmxsqKW2sgYjoDxXx98ZfHtz8Ov29vCOu2nhTWvGDyfDzUoPsWgwQ3F2ofVIG81kmmgURjbgndnLLx1IwdX8HePdW0L9qH9oXxX4Tm8Aad4x8DzabY6RdyW76hcDTdPvN19fLavLFHI3miONPMZgi/NjjIB9PeFf20P2YfGWsaZoOiePbRb3WnSOxW8hubBbqSX/VpDJdxRI7PkBFViWJAGSai8Z/tp/sz/D3xBf+F/GXjH+zNS025NpPG+nag6rODt2CRLdkYk8Daxz2r4lkv/iL+0h+xl4L/Zy8FfCbXrSfV/D3h2zbxHriWVpo9lFbRWzNqEDC5kuJiAm6FUiDHIJxyK+mv28IzF8LPAcTOZCnj7wiCzfebGoJycdzQB9K/C74zfDf4z6be6v8NtWbVrTT5RBO7Wtza7JGXcBtuY4mPBzkAj3r1Cvi/wDah8deLtK+Ivwm+Gdl4uk+HXhPxvc6qmr+IYBbpcJLZWyy2lhBcXccsEEl0xchmQswjKpySDh/sb6j4+8Q678Tr3xH8StU+Ifh7w7r0+h6Ld3C2H2K5t4YoJWmR7W2iaSaGR3gkkEhiYqdqgg4APuyiivyR+HOo/tQfEn9kmf9pVvjRfWPiLR7XW7/AE/TYtN09dMmTRrq6QxagHhaWZpxbld6SRLGpGELKWYA/W6ivy38dftYX/jrWvhX4QbxJq3w30jxf4Hs/G2r3ugaTPq+qP8A2gVjt7C28uzvRboHEryTvCxIVFUqWyakP7U3xV8KfA74i/2dc3fibWtJ8R6Z4e8I6/r2lTaU1/FrrQpbzXdvJb2u6SzeSVZGWFFkMa/L82WAP0B+IPxc8NfDbxD4I8M67b3c11491U6PYNbojRx3AgkuN05d0KptjIyoY5xxjmvUq/LD4rfDD4i/D34w/s4SeMPijqvxAjvPGP7+PVLSxhEd4NOui0tq1pDC0UTDcPJcygfKQwwd31H8DviD4w8VfHb4+eE/EGpNeaT4R1jSLbSoCiKLWG50yKeVVZVDNukYtlySM4GBxQB9WUV+TGl/H348at+yL8DvGuk+KUi8aeN/iDF4futRu7SKaKS2uNW1CzRZYEEQZESOLIQoxCffBJNe7eDL/wCLfwo/ax0b4TeLfiJf/EHw7458NahqijU7WzgmstQ02eBWMLWcMKiGRJSPLKnaQOSckgH3jRX55fAV/jb+094Wtf2i5PinqXgzS9a1K5fQ/D+l2dhJYw6VZ3j24S++028k0886xMWcSIELDaoxtHsPwx+IPjDXf2rvjd8PtW1Jrjw/4WsPCs2m2hRAtvJqFvdPckMqh28xo1J3McY4xzQB638X/i34a+CvhGPxn4rt7q5sZdQsNNCWaJJL52o3CW0RIkeMbA8gLHdkLnAJ4r1Gvx5+InxA8YfEf9kPWtY8a6i2p3lj8WYdOhkZEQpaWXiWKKCPEaqCEQAAkZPck19V+PfEPxJ+Lv7SV/8AADwZ4wvPAHhzwhoNrrOsX+lxW76pe3WozSR21tDJdxTxwwxpEZHdYyzMQnAyaAPtuivyz8YfHv40eAvgh+0p4J1bxL/aHjn4KCybT/EaW0MUt1ZarBHdWbzwBPI+0RoXjkKoFbAO0Hk9h8RtS+PHwEX4e/GXV/ijeeMLTxD4g0jSPEGhXFhZ22mfZtYkEIfTkhi+0Qvbu6lQ88rSAfOx5yAfo5RXwXDf/Fj9o34zfEzw54c+Ieo/Djwl8Mby20a1XRoLN7u/1R7ZLme4upLyCceRF5iokKBRIMszDivINb/aT+OV1+z3Y3Fnq9nY/EXwz8UbXwDqmoLahrG/e31BbZ5mtsgrFcRuhkSN1YHcEZONoB+qdY2k+I/D+vyajDoWpW2oyaPdNY3q28qStbXSIkjQTBCSkgSRGKNhgGU4wRXw7pVz8Y/hH+1L4L+Gnif4l6h460L4n6JrczrqFnZQPp+oaQsEnnWv2WGMLG6ylREwYDqSx5rh/wBhrwB4xtPHXxn1/UviNrWq22jfEXXLC5sJ4rBbfUpksrNReXBitUkWbDLxC8cfyL8n3sgH3v8ADT4jeHviv4Os/HPhaO8i02+edI1vrSayuM28rwvuhnVXUbkOMjkYNd5X5MaX8ffjxq37InwN8baT4pSLxp43+IEPh+61G7tIpopLa41XULNVlgQRhkRI48hCjEJ98Ek17t4Mv/i38KP2sdG+E3i34iX/AMQfDvjnw1qGqKNTtbOCay1DTZ4FYwtZwwqIZElI8sqdpA5JySAfeNFfitF+1X45+IHw61n47+GviJ4lsPFvm3t14d8HWHha7u9Bks7Sd0gsru5GmSm4muo4/nnS7jWN3wuwIa/VXxRJrPjj4O3V9o+pXvg7UdW0pLuO4t0ia8smeNZigS5ikj3AZRt8eRk4AbBAB3fh7xH4f8W6LaeJPCupW2saTqCeZbXdnMk9vMmcbo5IyVYZHUEitmvxq+C3jfxr+zh/wT18L/FPTvFt9rV54ktNK0vR7DULWGew0ae/vDb+dFHZ263UyormQxu0jOUCLy1e1fCT4ueJtN+OvgjwZ4c8d+LPih4d8XQ6jDrkniLw3caWul3ltbG5t7q2m/syxRIZmjeEwsz7SyEHOSQD9K6K/FbVvi18ZPFXwrtvirH8adS0L4k6r4xTQX8CacumKbWJtX+wtp8NtPaTXAu47YfaDNKXGASVK4NfUviif42eI/2pk/Z28PfE6/8AD/hyy+Hum6te38dnZTarcXa6hc2kk0UkkJghluAqGVhCygLiJIy24AH6CUV+S3hnVP2pvH3wf+Leq3nxludK1L4J6v4i0ewubHS7FJdYm0RftEdxqhljkUq8bpEIoFiAwXcyseO0+JH7VPiPUPBP7P2lDXLrwVP8XtDXXta1fSdLm1a9s7aCyt55YLG1jt7rEk81wEErxOsSKzEbitAH6a0V+UQ+P/xM0P4UftB2fhvxPrviHT/BPh2PV/C/ijXNGk07UBLLFMJ7aYXFlaRTvbyRq6OIOVkw2cV13iLXf2gPhfqXwP8Aizr3xPuPEUHxF8R6NoOsaAbC0t9Jig1yF2RrMIhuUa3dR80kztJ1JA+QgH6LQ+I/D9zr114Wt9StpdZsYIrq4slmQ3MME7OsUrxA71R2jcKxGGKsAeDWzX5j+B/h7401D/goD8U7pfiVrdtBpeneHNQe3jh0/wAu5s5ri7mTTZd1qWFtEN0ashWYqxLSs2GHGftL/GjxV8P7bx348+HXxq8QeJfEvgi7kuP7E0rw/Fc+GLGCKUM+n6ndQ2cgWQRZR5JL1JQ3zBF4UAH6NXvxb8NWHxj0v4ITW90de1bRrrXIplRDaC2tJ47d1Zy4cSFpVKgIRjOWB4PqNfCFzqaa1+3n8OtYjQxpffDPVZwp5KiXULNgPwzXZftiePfiT4E8OfDhPhZrEWiax4n8daFoLz3FuLqDyNRMsbCWIlSyBtrEK6MduAwzmgD69or4O8GX/wAW/hR+1jo3wm8W/ES/+IPh3xz4a1DVFGp2tnBNZahps8CsYWs4YVEMiSkeWVO0gck5J4bwl8cPinqP/BOfxb8ab3XpJfGmnWXimW31ExQh0ew1C8htiIwgjPlxxooynOOcnNAH6VV5d8V/i34a+Duj6Nrfie3uriDXNZ07Q4BaIjutzqcwghZxI8YEYY5cgkgdFJ4r5Y8X/EP4qfETx/8AC39n3wR4nk8Iz+IPCh8V+Itdt4IJr/7HEYLdYLNZ0eCOSaeUl3MbbFGVHrzX7UXg/wAS+B/g94A0LxP4wv8AxvKnxM8KPBfanDbRXaW7X8eyGRrWKGOQoQT5mwMc89BQB+jtFfm78a/j1Pqn7Q2ufBm98c6/8PPDXgzS9Purm48M6LPqmpajqGp+ZJGjTJYahHb28ESKSDGrSuxAYqrAcpF+0T8ZZ/gv4D1O71K7t9VX4s6V4Rn1S40xtOl1zQ5b0Kly1pcwRmE3Nu6CTYi4cNt29AAfqbRXyr43+IHjDSv2vPhh8OLDUWh8Oa94f8QXl7ZhEKzXFm1qIHLFS4KCRsBWAOeQeK+cfgP8RvG+pfFK00P48fFPXvB/xNn1S+E3g2/06zttA1CxjklFvFpE72uZ18ry282K7aViGDLzmgD9OK8H+Jn7TPwQ+D3ia28G/ETxJ/ZetXdmNQjtls7u6c2pkaISn7NDKAu9GXkjkfSut8BaN8UNK1rxfc/EHxFa65puoam02hQW9oLZ9P04ooFvM4/1rhsneck9c4IVfiT4nfEK6+G//BQi11i08I654ze4+F32f7JoFvDcXMYOtl/NdZ54FEY27SdxO5l4xkgA+3PAnxj+FnxN8KXXjjwH4psNY0KwMi3V3DOoS1aFd8i3AbBhZF+ZhIFIXBPHNeX+CP2yP2aPiN4otfBvhDx1a3eqahK0NmskFzbQ3kqcFLW4uIo4bhuOBE757ZrwLwr8AfG3xkP7RHifxxos/wAMbH426dY6RY6dJJBNfwpY2k0Bv71LZ3iEkzTD90spOxCrtyDVjwT8ZPH/AMD5/Anwa/ap+H1pYactxY6JoHi/Q2W70OW9VPItFmgdVn0+V1G1SVKbiQu1ASAD1PUf28f2UtJ1D+y9S8cG3uzK8IjbTNS3NJHncq/6L8xGD93PHNdkn7UfwnvYPAl74fuLzWbL4h61JoOnXEFq8KJdxQyTOZ0uvJkWMLEw3KjEnGBjkeU/tO/8nD/sxf8AYz6t/wCmmesL9tyz8Saj4y/Z7sPCOpRaPq9x438u3vZoPtSWzNpt2DL5JZA7IuSisdpbG7K5BAPv2ivhn4Tat8Uvh7+1frnwD8Y+O774gaFqfhCLxVZ3OqQWkV3Z3CX5sp4Q1pFCjRPkOo2DbjAA5Lfc1ABRX5zeBrX47/Hrx38adIHxf1Pwdo3gvxbc6ZpEOlWFg06FbW2mQXE1zDKZLdC/yxKEYkvvlYFQjvCHx38ReO/2UdB8afEj4lp8M9dt9ZutD1bVdPsIbufUp9PuZ7Qxafbyxyjz7oxrIBHBKRh1SPoVAP0Xor8yvgl8bPH66x8c/hvdeI9f1+x8IeH7fXfD+peJ9L/svWYlube4DpPC9tamRFlhV4neBSQSCCMVv/BfU/jBrH7Nmn/tEfF741/8I3c674WtJRI1lY/2PpMbiBhfNHJGjTXkqoS2+TyhJMyJEVVAQD9FqK/Mn4LfGTxRpn7Tfhv4bWfjfxT4+8GeO9E1K7jufFWi/wBlPHf6aYpDNp8v2Kx862kjkKlRGyqdpVzmm/Cj4jeN7r4ynQfjv8U9e8CePp9fvI7Xwrd6dZweHNU0pJ3FrDplzJak3DSQ7CZEuvP35XYcZIB+nFeXfFH4t+GvhJH4Vl8S291cDxf4g07w3afZUR9l5qbskLy73TEQKney7mHZTXkX7PXxB8Y+MviL8ddF8S6k19ZeE/Fa6fpcbIii2tTYW8vlgooLDe7HLEnnrivhLxD42+JHxe/Yw/Ze8Z61ryyeNNf+Ifhw/wBq3FskqpdfbbuKKZ7ePykcR4UlAVDAYyM5oA/ZuuNuPH3hRPDWveKtN1CHVrLw19tS++wyxztFPpwb7TbnDYE0ZUqyMQVbhsV8W3s3xi+DHx/8A/DDVfifqvjLw/8AGGz1yyM2pWunpfaPqmnWZu0u7M21tFGY2UMvkyRuiEKTuBIrxf8AZV8EeK7L4JfGzxVqnj7V9YtF1bx3Zy6Zcx2S2s1ys0oe+dordJhPIQWYLII8scIBgAA/TT4Z+P8ARvir8PfDnxK8OxTwaX4nsLfUbWO6VUnSG5QSIJFRnUMAeQGYZ6E13FfjB4Xuvjt8G/2N/hB+0JpvxNuZLLR7bwtC/hiOwtE0mXSL+a2sjA7MjXLXGyUO0xmxuz5aIMAfZv7SHiSC28Z6d4ab4xeIPCNxdWHm23hzwho8eq6zcyrIw+1ybbS+nW3+6mPLijyrZkOcKAfQPxX+Lfhr4O6Po2t+J7e6uINc1nTtDgFoiO63OpzCCFnEjxgRhjlyCSB0UnivUa/HjXPit4u+MX7IHwm8UePZTc+IbL4oaFpV7cNALV7iTTtba3WZ4AAIpHRFMiAAB92ABxX1R+0Z4kWPx1B4Yi+MviLwvfS6es1p4a8HaJFq2qvKHcfbLnbaXswtz8qhWWGP5WzIc/KAfb9FflQP2lvjZ4m/Ys+FHxS0jVodM8da74u0zw/e3UtmpgnKavLp0pmtvl2rOIg0qRlGGWVGTgjrdTtP2kvCP7R+hfAiD4zXup6R8QdCvdVn1G90vT21DTJtMliWYacIoUgQTiVVUTRzLGMkBn5IB9/+OvGWj/Dzwfq/jjX0uJNO0S2e6nW0ge5nMcYydkUYLMfYD3OBk1taPqlrrmk2WtWQkW31CCK4iEsbRSBJVDrvjcBkbB5VgCDwQDX5kp8W/jN4B+Cv7UfhPVfGNx4g8R/BvzTo2v3FvbpeGC706O9txMiIIXkhZiNxT5u4wAK9Q8YfEP4qfET4gfC79n3wP4nk8Iz6/wCFD4r8Ra7bwQTX/wBjiMFusFms6PBHJNPKS7mNtijKj1APveivPfhp4P8AEvgfw9JoXifxhf8AjeVLiR4L7U4baK7S3YLshka1ihjkKEE+ZsDHPPQV8WaGnx7+Ofxb+OHgmw+K974J8P8AgrW7a10waZYWcl4HuNOt5gkk1xHIDbo5L7FVZHLsDKFCrQB+itFfl1oP7SHxZ8Tfs4fsyfEG41YWuu+NvHuleH9elhhiC3tss19a3KlShVBObZXPlhSD93aOK+m9d+IHjC0/bP8ACnwxt9RZPDGoeC9U1Sey2IVe8t763ijlLld4Ko7DAYDnkZoA+qq8u8LfFvw14u+J3jn4UaZb3Uer/D9NKkv5JURbeQavDJPB5DK7OxVYyH3IuDjG4cj5D+Ar/G39p7wta/tFyfFPUvBml61qVy+h+H9Ls7CSxh0qzvHtwl99pt5Jp551iYs4kQIWG1RjaGeGPGdt8Of2j/2xPiFewNc2/hfQvCmqSRKcNIllpF7OyA4OCwTGcGgD9EKK/HDw9+0h8Rk8O+DPi1YfEHxP4u8Ya3e6VNrHhFPCl5B4fGnahKi3FvYznTFZHtIpd6XLXjiXyyTuDBa+4/gd8QfGHir47fHzwn4g1JrzSfCOsaRbaVAURRaw3OmRTyqrKoZt0jFsuSRnAwOKAPqyivyr0P4v/tFeLv2Ifh/8RtH1PVb7VtU8RXcHijVdHsILzWLbQINTvoJZrK08sxvLGsUKZWJ2VNzhGYEj6A+HuoeJviT8BvEcX7O3xybxZrct4sem63rtjbTXOkmNoTNaXttFb2zGXaJMefCrr5i5UhQSAfaMkiRRtLIdqICxPoBya+SNL/by/ZI1Z7IQfEa0t4tRcR29xeW93ZWsjE4wLi5gjhH1L4r6nvFmTRp1uXEkot2Dsq7QzbOSFycAntk49a/DL4TfGnxrcfsSfDX9m+1+F05PxS0+98LaT4g1q8tINBe5vPtBWQmNp59y/MY43iR5HTEeTigD9mvEvxc+HPg/xj4V8AeJdchsNf8AG7XC6NbOrk3jWqq8oR1UoCAy43Mu4kBcnil+I3xa+HXwktNIvfiLrcWiw6/qMGlWJkWRzPe3AYxxKsascttPJAUdyOK/MT9p34QeJrDxD8Afh74ZuzqXjD4feB9butIu9p3z6r4aGj3EDAckfaHtvLYf3ZCK5b9qLx7pv7Rs/hz4k+HX83wn4I/4QmW2yf8AmMeK9Vsbtwcfx21hFEpBwV+0sMdaAP0Q+If7Yn7OXwp8Raj4V+IPi7+x9S0lo0ukewvpEjaZVdB5sVu8Z3B1xhjycdeK734V/HX4WfGxdTf4Z6y2rjRzCLrdaXVr5ZuN/l/8fMUW7Plt93OMc4yM/Pf/AAUc/wCTO/Gv/X54d/8AT7YVu/tZ+P8Axj4Vufhp4T0PxKfAeheNtfOmaz4mVYS+nwLbSzRQxPcpJDFLdyIIkkkRgp6DJBoA+w6K+Bf2VL3x5q3xk+LGnX/xT1f4ieDfBdzZabpNxcf2fJZzyXVtHcXCSTWtrG0t1Zv+7Zll2YkAZA44++qACivzI+DfxD+M+t+CviB+0N8SvHV3N4Z+Fuq+Lxa6DYQW0S6lbaRJdMPt85haQlF2xxJEUAESu5dmYVj6z4h/aY8Dfs3WP7Zeo/EyfV9XGn2XiPUfCjWlnH4ffSrvy5XsbcrD9qimigk+W4M7lnX5gQaAP1PrGu/Efh/T9a07w3falbW+rauk8llaSTItxcpahDO0MZIZxEJE3lQQu5c4yK/Pf9oT4m+MdL+LGl3fjHxx4j+F/wAH7vQba607XtB0+C5t21iaVt6atcTWt2beNIjGY0ZEjfLFpMggcT8TvBXjDxn+1v8AA0aP8YtWaPV/CetXFrqumQ6WUHlQWSzT2oNrLEUvjiR93mAcCIqvFAH6r0V+e97efHP4mftT/EL4KaN8Rbvwp4N8NaN4ev5J7G2tX1Tz7lLhWigkmhkjRJ2XzJnKMwMaLHtV3qTw3cfGf9pzxj8UNX8OfE3UfhvoHgHxFe+FtFs9JtLKczXulogubzUDeQStPG8smEgQxAIvJJO4gH6CUV+WGp/tK/G7xv8AAr4Ia94b1W08LeNvE3xCXwbrk6Wou7JngXUbaeRIXKkozwJOqh1wwCFtuc9HZ2H7Rmn/ALS2o/syD4z6jeaBq/haLxWdZudN0463ZlL17Ke0s3SBLZFmdkffJbyeWq7I1VjvoA/Syivy40z9pr4o/Cv9mT4yat4r1ZfF3iz4YeMrnwfpmp3VttN2JpbOOzubqG1X52iF5udY1BkEe0ZZsm/8M/i/4q0b4zeAfD3h74geLPijpPi2S6s/Eaa94YudLh0+dbZpre9s5DplmsMRlTymheSQYcEHIzQB+j3iXXrPwt4d1XxPqKO9rpFpPeTLEAZDHbxmRgoJALEKcZIGe4ryXw9+0L4F8SaD8MvEFpbanHB8V4Um0hTZPKYle3+0gXbwebFAdnGWcgnOCQCR8UaZd/Gr9pL4N/ET4/j4mX3hPQpB4httA8P2FnZS2B03TGntGOoefC880twYXLFJYvLBGwjoLXwm+IHjDwp8PP2JfCvh/UWs9K8XaetrqsARGFzDb6E08aEspZdsihsqQeMHjigD9Qa8u8W/Fvw14M+JHgP4Xapb3UmrfER9Sj0+SFEa3jOlWv2ubz2Z1ZQycJtV8twdo5r4h/ac+IOuaLrPjmXwX8bfEaeKfDVk19YeG/DOgQ6lZaa8VsJFXWZks7k7JWBYmeeAKjDap27mkvvHV38T/i/+xF8SNQhW3u/FOl+INTmijzsSW88OJM6rnJ2hmIHtQB+iXiDxH4f8J6RceIPFOpW2j6XabfOuryZIII97BF3ySEKu5mCjJ5JA6mtmvz//AOCmOkaxf/so+INQ0vX7vR4rC80jz7e2SBo71LjVLSAJMZY3cCMv5i+WyEsoDblypzfj38RvGXwFtfhr8GJ/iNrU974/1DU5L3xXLpMeqaraabpsKTSxWtnp9kYzNI8scaSNbuI1LuwbAIAP0Sor8qdP/aK+K/g7wp8a7TwBqGvfEvT/AAn4Yi13w5q/iHRLjT7yO6Jkiu7S4D2Vit15AVLlCsQJQlCzY+Wbwpqfjy7+PHwg8LeD/wBofWfiDpHivTb7WvECWh0iaOEWSQSwynyrImCyupHNv5RKuCRsk3BsAH6oUV+ZFjfftE/F7U/j6NI+LN34QsPhz4jv7XRYrDT7KSUvBZW9wkd1LcRSb7ZCfljRUcl3LyMuxV4O6+Kf7R0P7PXw3/bI1L4jP5mo3eg/b/C9rp9rFo82n6ldw2UqZZXuftDb/MMvnbVYlY0VQKAP1zor80vi78fJ9e/aH8X/AAi1Lx94i+HPhzwBZ6Zufwxoc+q3+pajqUP2rdLcJp9/HBbwRGMCLYrSszEsVXA5nU/2g/j9qH7P3w01PTtUOj+MdU+Jdl4Ol1S/0mS3i1OxmuJ4Ib2SwnSCREni8qV0XyzuDBSg4AB+qdY2jeI/D/iL7d/YGpW2pf2ZdS2N39mmSb7Pdw48yCXYTskTcNyNhhkZFfDvhu4+MPwy/ak074OeJfiZqXjXQvH/AIW1LUYpr+0sYbnTdS0+WFDJbG2gjjETJKSImRgCBnPOfPP2IPBPi/TJfjN4n1H4m6rPa6T498XWE1vew6eLKae3kRP7SuNlvE4l4DuqSRw8cIooA/T6vLvCHxb8NeNfiH49+GelW91Hqnw7m06DUJJkRYJG1O0W8hMDK7MwEbAPuVMNwMjmvzht/jt4m8H/ABM+Fms+Efi54k+Kuj+MvE9p4b1qS+0COz8NTDUEkWOXTLyKygjEkUqhgsVxPvTO5ztOfpT9n7/k7f8Aal/7CPhL/wBMUVAH23RXw78XvEXxg1z9q7wt8FPAnjV/B+g614Q1HU7+WGzt7q5V7a+gjElqbhHSOYq2wNIsiBGc+WX2svkVn8VvjJ4B+E/7VPgnWPGVx4j1/wCDdvJPomv3Nvbpe+Ve6St/biZY4xFI8DkjcU+buMYAAP0/or4c+LvxV8f+HPhZ+z3r+i6u9rqHjHxd4M07VpRHExurTU1zdxsGQhRL3KBSP4SKq3erfFT9oL9oD4ifDnw348v/AIdeDfhYNNs3fRYbRtR1PVNQtheNJJNeQTrHbwIyoI1jzIxYlsYFAH054m+Lfhrwp8UfBPwk1G3upNY8ew6rPYSxIhto10iOKWcTszq6lhMuzajZIOdvGfUa+BPHumazov7W37LOk+ItYfxBqlpo3jWK51CSGO3e6lWysQ8pihAjQuedqjA6V4Jrn7R/iH4ja98Stbtvid4l8DXXhLWNT0bwzo+h+GbnU9Pnk0pjD9o1O4GmXYn+0zq2YUlj8qPH8ZJoA/Xaivgb4f8Ax0+JnjX42/BTTdbWbQbLxl8P77W9X0WSAR+XqkUlqpz5qecnll3VVJHB5BNWY/ib8cNU8eftW+GfAk66xq/geDRh4S0+dIhFDdXeiLclAcLv8yc7sSMRn5cqtAH3hRXwP+yh43XxVqF5pk3xh8Q6/wCL4NKDax4W8Waba6ff6dfuUzcx26WtrMsCMSm1HliO5f3mcZ+s/hNpHxI0H4faRpPxd1+28UeLbdZRfajZ2ws4Jy0rtHthXAXbGVU4AyQTgZxQB5x8SP2tP2ePhL4qk8E+PfGMNhrkEST3FrDb3V49rE4yr3JtYZVt1YEEGUpkEHoQa9Ruvij8NrHwVB8Sb7xTpdt4Uuolni1WW8hjsZInGVZZ2YIQQOMHmvi3U/DPx2/Zf+J/xE+JfgDwbD8VPAfxC1Fda1Kzsplt/EmnTrAsUohSUeXewAJmKFWVwWKgdS30Bo+u/B74/wD7Ott4s8MaTZ6t4N1DTZ57C1u7KLy7d7ZZISn2d1KRyQSK8ZC/dIIUkckA9t8KeK/Dnjjw7p/i7wjqEOraNqsQntbuBt0U0bdGQ9wa6Cvkf9gv/kzr4Tf9gOD+bV9cUAFFFFABRRRQAUUUUAf/1P38ooooA8R+N3wb/wCFyQ+Bov7X/sj/AIQvxZpHijP2f7R9p/sp2f7N/rI/L8zdjzPm2/3Go1/4Of258fPCXxx/tfyP+EW0fU9J/s/7Pu8/+0XhfzfP8wbPL8rG3y23Z6jHPt1FAH58XP7FvxCXQPiJ8LND+LcmkfDHx5c6tef2XDpET39rJq+957cXzTfNamV8snlK7xkxiRMlz6f4x/Zf1DU9E+GGo+BfGL+GPHvwosE0/TdaFkt1b3Vs1vHb3UF3ZNIoeGcRqwUSho2wyvkc/XFFAHyD48/Z5+KPxV+C2s/D/wCI3xHg1HxNe6nYatYajb6MlrY6fLp08FxFAlos7SSxF4TvMlwXO8kEABa1vG3wF8eT/ESH4z/CXxtb+EvGl5plvpWtpdaadR0nVobYs8LSWwuIJYpYmd/LkSYkIdhBHNfVFFAHyz8Tv2fvGPxc+Ctp4B8YePN3jTS9TtdbsPEdtpkcEdtqNjc/aLZlsPNZTHGP3e1pSxXlmLZJxvCP7N/xCj+Mvhb46/FD4knxTr/h7TNS0xrS30tLCw8m+EJQ28azSNGyNG7SO7SGXcoHlrGFP1/RQB8UeI/2aPjB8Q9Q8PaL8VPirb+I/BvhvW7PXYoU0GKz1i4msJfOtopr2KfyVVWADvDaxu65Hy5Jr0T4PXnxQ1H4ufFu78Ravd6n4FS+sbfw/Heaetiba4gjlTUordsLJcW6uIgszDa7h9hIBZvpOigDxC++DX2z9ovSvj9/bGz+zPDN14d/s37Pnf8AabuK68/z/M42+Xt2eWc5zuGMHuPiZ4N/4WL8N/Ffw++2f2f/AMJPpN9pf2ny/N8j7bA8HmeXuTfs37tu5c4xkda7eigDh/hj4M/4Vx8NfCfw8+2f2j/wi+kWGl/afL8nz/sVukHmeXufZv2btu5sZxk9a4f49/Bn/hd/hjQ/Dn9sf2L/AGL4g0nXfN+z/afN/su4WfydvmR7fMxt35O3rtbpXuFFAH5xftmTWmufGv4QeDLrxHpHg9kttd1CO+8U2kV/oF25jgtvsTWszwpJdssheNvOjMcYkADmQLXM+HfjJ8XGsPin8ItJ8Q6LrNh4V8OQS6T4n+H+iySLY6vezNBDpgsBNdQSXB+WQLG4EatmQqOV/SnXvDfh3xVp50nxRpdrrFixDGC8gjuIiw6EpIGXI+lT6Rouj+H7CPStBsINNsoc7ILaJIYkzydqIAo/AUAc18MY/GkXw38KRfEiVJ/FqaVYjWJI1RUfUBAn2kgR/IAZd33cL6ADivyt/ZT+Bnxh+Lv7IGheErT4q/2B8PPFEutwajp0Ojwyamlq2rXcVzbWmoeaqpHcgMWaSCV0LsFYrhV/Yqq9paWlhAtrYwJbwpnakahFG45OAMDkkk+9AHzD47/Ztnl1rwf44+CHiJfh/wCKfBGlHQLSR7Majp91ovyFbC7tmkidkjeNWjdJVZGyfmzxZ8W/ALxN8XPgprvws+NvjIazqer3Ed3b6ppOnrpX9mzW7xy2ptojLOx8maLfuklZn3MpIXAH07RQB8Sf8MzfGTxb4y+HnjT4v/FuLxDP8OdWXUbW1sdBi062uh9nlgdp8XMr/aH3qQ6sI0AdRCS4ddTW/wBmv4k2nxn8YfEv4V/E4+DtK+Isdj/b1gdJivbgT2EAtkuLC5klVYJGhAUl4pVDfOVbgL9j0UAfEehfsawaB8GfhX8Hrbxc8sPww8Y2/itLySy3Perb6jc34tWTz/kYi48szbn5Xfs+baPbtf8Ag5/bnx88JfHH+1/I/wCEW0fU9J/s/wCz7vP/ALReF/N8/wAwbPL8rG3y23Z6jHPt1FAHxR4X/Zl+LfwtuL/wt8F/iunhr4eX+oz6hFpVzokWoXumfa5jPcW9hdyTrGsLuzlRNbymPP8AESTW541/Z2+I8vxu1T4y/B74jp4Jl8V6bZ6br1rPpEeqCcWBk+z3Fu0k0YimRJCg3K6dyp+6fruigD4Uf9is2v7Omu/ArTPGs0t5e+JZPE1hrF7aefLDcDUk1GJLmMSr9oIKbJJA0e8kuFXha9F+IXwD8bat8QdM+NXwt8aQeEPH0WkrouqPPp39oaXqtmr+ciy2pnikjeKYs0UiTblUlG3Ka+pqKAPzE/aN+CI+EP7Ffx61fXNem8W+MfGls2pa7rM0KW32mdTFDDHDboWWGCCJQkUe5iozljmvXdJ/Zu+JfjjV/AupfGf4mr4s8I+Cbm11nTNJt9Hi06a51C2T/RJtRuY5nWb7OWLBIooUd8MyjG2vte6tbW+t3tL2FLiCUYeORQ6MPQqeDUwAUBVGAOABQB8leK/2eviJpvxM8SfFD4CfEGHwReeN47ZddstQ0ldXs57m0j8mG9t08+3aG4WL5WG5o5MKWXIycmb9jnRIfhD4Y+Fuk+JbhLjRvFdj4v1HVryBbm51bUYLz7bcvKqvEqNcP8oYZEa4G1sc/ZlFAHiPjD4Of8JZ8cPh18Zf7X+y/wDCA2mt2v2H7Pv+1/2xHBHu87zF8vyvJzjY+7d1XHPEfDr4D+Ofhb8YvF/i3wt4ytm8B+OdWn1/UdCudML3aalcWywSNb3y3ChEd445GDQMcLsGMlq+paKAPiPQf2OP7E+DHwq+EP8Awl/nf8Ky8YW/iz7b/Z+37b9n1C5v/svlfaD5W77R5fmb3xt3bDnaPbtf+Dn9ufHzwl8cf7X8j/hFtH1PSf7P+z7vP/tF4X83z/MGzy/Kxt8tt2eoxz7dRQB8Y+Hf2cfjB8Ln1Dwv8EfilbeHPAV9e3N9b6XqGhJqd1pJu5WnnhsLn7VCghaR3ZFmhl8vP8dfY1xbRXdrLZ3S745kaNx0yrDBH4irFFAHwh4L/Y21zTfgjqn7N3jzx0Ne8CWyIvh2W10/7BrOlvBc/a7aZrsTyRyyW8gXYRCgO35uDtr2z4d+Dv2g/DWqJe/E/wCJVj4v0jT7aSNbWw8PLp11eSYG2aeX7XOPMGDhIY41JPPYD6DooA/C/wCFnjjWPAXwXvvjz4I+JXw8h1ONtY1GLR/EGkJceJo5rq7uJxpV3fwXdvdS3Ts/ln/RgxZsKGXBP6Z/Db4ca74h+KGjftTeKGbQ9Z8Q+AtL0S98OvAd1jc/aG1CUm4Lgko8xhMZiBG3cW52j3AeAfAq+ID4sXw5po1xjuN+LOH7WW9fO278/jXW0AfM/g79nX/hE/BXxf8AB/8AwkH2r/ha2t69rPn/AGTy/sH9uQJD5WzzW87ydud26PfnG1etcrqH7Kdynw5+EmieEvGUuheOfgzp8Fjo/iFLJZopoxaR2d3Hc2LyYeC6SNS0YlDIyqVkyvP2FRQB8ua/8Ffix8SPgz4++GHxY+IFlqV5410+XT4LnTtE+x22mpLGyMywNdyyTkkhjvmXpgBec7PxF+AX/Cf+GPhb4c/t37B/wrXxDoWveb9l837b/YqMvk7fNTyvN3Z35fZ/davoqigD5a8QfAfxzb/tDH47/DTxlbaEmt2FhpfiDTL7TDfx3ltYTPJG9vKtxA0ExR2jyQ6gHdtJGD4lqX7EnxCuPhj4x+A2k/Fv+zPh14mm1O5ht10OKXVIpNTmkuWinvXuNs0CzPuYLFHK65TzUGDX6JUUAfN118BdRi+Mvw8+Lui+II4H8IaFdeHdQtJrQyC/srjypFaKQSr5EiTQq3KyBlJXjrXR/G74N/8AC5IfA0X9r/2R/wAIX4s0jxRn7P8AaPtP9lOz/Zv9ZH5fmbseZ823+41e3UUAeI6/8HP7c+PnhL44/wBr+R/wi2j6npP9n/Z93n/2i8L+b5/mDZ5flY2+W27PUY5+Vrn9h/4hN8PvG3wM0v4vSaZ8L/E0mqz2Wmw6NEb6zbU3kn8iS9M+ZbVJ5N7IqRvKuYzIikk/ovRQB8pfEH9m3V9bufAPjP4c+MG8JeP/AIe2B0u11NrJb20vrGWONJ7a8s2kjLxu0aum2UNG3zAk1nePf2efih8TvhIPCfjj4jwah4ws9f0/xFp+ppoyW9hZz6bNHNDbLZJP5jw5RtzSXDSEuTnAVB9fUUAfLvj74DeNbn4mD40/BzxnD4O8XX2nQ6XrEV5p39paVqtvbMzwNLbie3kjmhZ22SpLnYdhBHNR/E79n7xj8XPgraeAfGHjzd400vU7XW7DxHbaZHBHbajY3P2i2ZbDzWUxxj93taUsV5Zi2SfqaigD4+8Kfs2/ENfjF4X+OfxO+JbeJvEGgaZqWltbW2lx2FgIr8Q7DbRiaQxujRu0jyGUy7lGI1jCnL1L9mr4v+ONf8GxfF34p2/ijwx4G1y08Q2ccOgxWGq3V5p7FrVbq7juGhCoSC5htojJyDtBr7VooA+bPgfefFHUfH/xYvvFmr3eqeD/AO20h8OLe6eunyQLBGUvYohhZJrdJsRxzOB5hR2XcpDN0n/Cmv8AjJIftCf2x08JHwv/AGZ9n9b/AO3faftHmf8AAPL8v/a39q9vooA474geFLzxv4N1Xwrp+vX/AIXutQi2RanpbrHeWrhgyvEzqy9RhgRypI4zmvk+5/Zh+MvxA1vwzD8fPi7H4w8J+E9TtdYh02w0GHSZdQvbFt9s99Ok8oZI3OWijRFcgE4wK+4aKAPD/id8Gf8AhY/xD+GHj3+2P7O/4Vxqd3qP2b7P532z7VaSWvl+Z5ieVt37t2184xgdad8WPg5/wtDxV8NvE39r/wBmf8K917+2/K+z+d9s/wBGmt/J3eYnlf63dvw/TG3nI9uooA8KuvhHFa/tDv8AtGT6wRHb+EH8NNpq2xY7RfC/NyJlcknA2eUIiT13fw1j/ssSfFW4+E0V58XtWutb1S51HUZLK7v7FNNvZdKNy4snuLRABDI8QD7CA6qyq4DhgPo2igD8r/hB4H+N/in4k/tGSfCX4lW/gmCfx3d2tzFdaLHquxjYWhF1bOZoGSYBtuJPMjIVSEUhi3u+pfsb2uh+Afhb4a+EPimTw7rfwkvbjUNNv9RtRqkV5cXscqXj30HmQF2naZ3Lo6lCTt7Y+0YLO0tWle2gSFrh/MkKKFLuRjc2OpwByeasUAfH3hT9mDxRpvjD4h+PvGnxBbxLrfxG8OwaFct/ZkdnBaNB9oVZLeOOYkRBZlAidmfcrM0zb8LZ1z9lWDXP2V/Dn7Nb+KJbafwxaaPFa6zFaqc3WiyRSwzPaPIysjPEC0TSHg/eyAa+uKKAPkLw/wDs6/Em6+M3g344/FL4kR+ItX8JWWqWKWFlo6afYeXqUcaFogbiaZZAybpGkkkD/KFWLad2L4j/AGaPjB8Q9Q8PaL8VPirb+I/BvhvW7PXYoU0GKz1i4msJfOtopr2KfyVVWADvDaxu65Hy5Jr7XooA+Mr79mX4kab8WfG3jP4afFF/CXhn4kS291remppcVzeR3UMCWzzWF68oFu8saDJeGXY3zAHjbit+xdLB+z38MfgVpHjqbTJ/hlrtprlrrEOnxvLJLZTXE8KiCaWSNSrTLy5kU7OUO7A+56KAPlDwR+z547f4uab8Z/jj47i8baz4Ys7qx0C1sdLGk2Oni9wt1cmMz3LyXM0aqhYuFVcgKcgjI8Dfs0eOfAN58RvDWkeObaX4e+PbnWtRTTJtKLX1he62D5uy8W5VXgR2Z1QwhjnbuHLH7GooA+RfEP7K/wDb37KHh/8AZg/4SfyP7CttCtv7W+xbvN/sS5t7jd9m88bfO8jbjzTs3Zy2MGz4t/Z++If/AAunWvjJ8JviDD4UufFmm2Omavb3ukJqvyaeZDDNZu08IhkAkIKuskZJ3FSRivrGigD4QH7Fl3p/7P0fwa0fxw8uraZ4rPi3TtZvbBZSLtNROoRJc28csayjkpIyNGGJ3qi/crtdb/Z5+Jdp8YfFHxY+F/xHi8LP49tNNtdcguNHTUnDaXE0UM1hJJPGsDbHI2yxzJuO4q3SvrqigD4f0n9jKPR/gN4M+B0XjGS4Twf4sh8TrqM1iGluRDqsupi3eNZlCs3meWZQcZG8RjOwe5a/8HP7c+PnhL44/wBr+R/wi2j6npP9n/Z93n/2i8L+b5/mDZ5flY2+W27PUY59uooA+QPEv7KX/CRad8frD/hKfs//AAvJIE3fYd/9l+Tp6WOceePtGdu/rFjO3nrWl8Qf2bdX1y58BeNPhz4wbwl4/wDh7YHS7XU2slvbS9sZY0Se2vLNpIy8btGrptlDRt8wJNfVtFAHnPwy8PfETw7odxD8T/FsXjDWbq5efz7fT49MtreJlVVt4IFeV9ilS26SV3JY8gYUcv8ADD4Of8K48ffE3xx/a/8AaP8AwsbVbbU/s/2fyfsf2e0jtfL3+Y/m7vL3btqYzjBxmvbqKAPhQfsXXFn+zN4N+Bej+N3s/EHw/wBaXxDo3iBdPUrFqMN9PeRNJZPMwdALho2Uy/N97/ZrtPAn7OXjrS/jbpfx8+JXxFbxZ4gs9DvdEktYdMSwsUiupoJkNtGs0jRhDE28uZWlLg7kVFSvreigD4o8L/sy/Fv4W3F/4W+C/wAV08NfDy/1GfUItKudEi1C90z7XMZ7i3sLuSdY1hd2cqJreUx5/iJJr0jT/gAlp8aviV8RbzU4dQ8NfFTRrDTdX0aa2bzPO0+N7aORLkS48qS3lkSSMx53bWDjBB+j6KAPkP4Z/Ar46/CyPQvBGh/FqC9+Hnh2SFLS0vdCSbWP7Ot2Gywe/FysRQRgRiX7KJAvAwQGqprf7NfxJtPjP4w+Jfwr+Jx8HaV8RY7H+3rA6TFe3AnsIBbJcWFzJKqwSNCApLxSqG+cq3AX7HooA+RPAf7OXxB+EHwM8LfCT4T/ABJ/sq/8K3t/ci+vNJjvbe/hvbq5uRb3Vu8yy4TzwPMiuEYsm48HYPM/FXwe+KPwg+FXxf8AH+i+NLrUfjF8S5rGWK+0LQ1ES3lmiW9la21g7zqiSKDFNcTykKrmV2UJmv0HooAwtKtdYfwzZ2XiGdJdVazjju5Yl2xtcGMCV0XspfJA9K+U9P8A2PdET9kbTP2WNZ197uTRbULY69DbfZ57XUIJ2ubW9hhErlHhlIO0S/MAV3AMa+yaKAPn8/BfXNS+JXwy+KXibxOmoav4C0bU9MuxHYeQmpz6olqslyo89/s4DW5by/3md+Nw288Drf7HHw/i+FF18Jvh1cHwtYX3iq08VzyNF9r3XNvfQ3jRBd8WEKwrDH8x8tAvDYwfr6igDw/9o74M/wDDQPwd1r4Tf2x/YP8AbE2nTfbPs/2ry/sF9Be48rzIt2/yNmd427t3OMH5o/blvLS88T/Bfwhd6/p/hIajreo3i6rr9vHeeH1a006VBa3ttM0cUstx9oxbh5Y9pV2Ulwqn9B6y9Z0PRPEenyaR4h0+31Sxmxvt7qJJ4mxyNyOCpx7igD83vAXxd+I+h+J/F/wStvEnhrXvDej+EbzUk1/wFocu/QNQeTyreB7CKe8gmmk3NNFAhDtsJZdpzX2z8BU+JSfBnwaPjDObjxo2mW7aq7xxxSfaGXJEiRZjEighZNh27wccYr0TQvD2geF7BdK8NaZa6TZISywWkKQRAnqQkYVc8elbFAHz98IPgFpfw0+Hfif4ca3fr4l07xTq2t6jch7c2ymDW53lktiokkJCrIUL7hu64XpXgI/Y0+IF/wCCdP8AgN4n+LEuq/BnTXt0XSDpUcWsXOnWkgkg0241RZ9pt02IrMlukjxjZuA5r7/ooA+cPif8NPjt4i1PUP8AhW3xMsfD+gatZrZy6Xqfh+LVI7UbSjy2siXFq2XU8xziZM9MD5a8ovP2O9W8K6V8H5fgn41Hh7X/AIQafd6VbXOq2H9p2+oWeoRotwtxCk1uVbfGHQo4C8rjGCPuaigDwjwV8GLrwr8avGfxmvtdXULnxlpei6dLaJaeQsL6SswaVXMsm4TGbITaNmMbmzmvMNZ/Zw+J/h3xp4v8S/AD4lx+BtP+IFz9v1ewvNHTVkh1J41ilvrBjcQeVLKqKXWQSoWG7A+7X2NRQB8hH9kPw1pngL4QfDzwlrU2n6f8KfE9p4mM1zF9ruNUnhW688TPvi2SXEt00rSAMFPATBGPTz8G8/tIL+0F/a/3fCZ8L/2b9n9b8Xv2n7R5nts8vy/9rf2r26igD5Jh/ZJ8M6h4I+MPw/8AF+sTanpnxb8R3XiFnt4vsk+myzJbeSIX3yh5LeW1SVJCFBbAKYBz2Hw18B/tD+Hdasj8R/ilYeK9D06J4hBb+H1sLy9JTakl1cfapkDIeSIYowx5OBla+hqKAPhOH9kj4j+HLbxd4A+HHxWbw58MvGN1f3kulHR4rvUNPOplmu4LG+kmCRwyO7MA9u7R5O07jvrs9D/ZX/sbTfgBp/8Awk/nf8KMjaPd9i2/2pu05tPzjzz9n+95n/LX+7/tV9c0UAfDt5+yf8QrW/8AiVong34o/wBheCvinqN9q2qWZ0eK51OG51GFYblLa+km2LE6qAA9u7Io2oyn5h0Nr+y1qOjWf7PjaF4qjXUvgTCtkZp7EtFqllNYLp14vlicGCSSJd0bb5BG3UOK+waKAPGv2g/g5YfH/wCDviT4R6jqMujx69FCI72FBI9tPbTx3MEoQldwSWJCV3LkAjIzkeWeJf2fPiX4/wDC/hbU/GXxBt7b4o+B9Rlv9F8SaTpIt4IVmh+zzQXFjLcSrPFPGWEwEse7I27AvP1xRQB8DftD23xZ8D/so/F3VPjD4xj8UvqWkCyhXQdLbR49OS5b7LJOW868mdB5wluGJIWKNgiDknxbw38QfHXwT1zwHpvw+8afDX4lR+N9Z0/TbrTPC+ipp2qXVpK22a/R7G9njK20e6SR5IhEqg5K5Ar9YHRJUaORQ6OCCCMgg9QRXL+H/AngjwnPPdeFfD2naNNdf62SytIbdpOc/OY1UtyO9AHk3w/+BH/CCyfFiT+3Ptv/AAtDWrrV8fZvL+w/arSK18r/AFredt8rdu+TOcbRjJ88v/2S/t37KXhn9mL/AISrZ/wjn9j/APE2+w587+ybyK7/AOPbz/l83ytn+tO3Ofmxg/Y1FAHy345+AvjdfinffGf4H+NYfBviDXrO2sdctdQ03+1dM1NLLcLWZ4lntpY7iFXZA6S4ZMKV6mrfjf4DeKviT4S+HOkeNvGqX2t+BvFel+KbnUI9MSBL9tNnklFstuk+IFZXEavvkKhQWDsSa+maKAPEdf8Ag5/bnx88JfHH+1/I/wCEW0fU9J/s/wCz7vP/ALReF/N8/wAwbPL8rG3y23Z6jHPiun/soeKdMvfix4TtfHMY+GfxduNbv7/Sjpx/tO0vddtzDctbX4uAgj3nzArwN/d7lj9sUUAfBVv+yN8UtXsfhlovj74sxanpHwn1nSNT0qzstAiskuU0cbIxeMbmV2maL5FeIxxplmMUh27fefBHwZ1HwR8dviP8WrPW47jSviPFpclzpr2xE1ve6XbraJIlwJMNG8K/MhjBDchscH3yigDxHUvg5/aH7Q2ifHn+1/L/ALH8O3ugf2d9nz5n2y5iuPP8/wAwbdvlbdnlnOc7hjB40/syaTf6t8drjXtZe80344wW1tc20cAhksIodMGmvslLuJWYfvFbYm04GGxmvqCigD8/bD9jz4s6tpXw40P4l/GM69p/ws1rRdU0q3tdEjso7hNGlVkF6RcO8szRL5SOrKkeWdo5WII9P8Zfs9ePbX4s618ZfgT48h8F6v4utrW21+z1DSxq2n3zWK+VbXSxi4tpIriKIlMiQo6hQyggk/WdFAHy/rXwE8X6z42+DvxHvfGw1DxH8MRqMN9c3WnRqur2+rxRxXQEUEkSWzjylMRUOF6MG5J5+T9nn4qeCPGHinW/gJ8R7XwponjfUJdW1HStV0UavFbalcgfabqxkW6tWjM5UO8UnmJvJYYztr7AooA+UPiz+z5498ZeLvh/8UfAHj5PDXjrwTaXOnTX11paX9rqdpfLELhZrVZYNjF4hImxwASV9CtT4f8A7M3jDwDc/FPxBa/E++uvFXxOOnTvrD6fbedZXlhb+QXSF98DxNwEhMa+XEAm5mHmV9dUUAfH2gfAzx9YfFC2+PXxu8eWviPUfCGjX+n6Ymj6G2mpFDdbHuLidPtF5LcSlYhsiXCKc7VJau+/Zc/4Ws3wX0eX4zanc6v4ilnv3W6vbNNPvJbE3cv2F7m1j+WGZrby2aP7yZ2Pl1Yn6Bd1jUu5CqoJJJwAB3NCOkqLJGwdHAIYHIIPQg0AfIniX4KftKxa3rcnw1+O0mlaHr1zLci11jQoNXudNM53Otjc+db4jU8RRSo6xrwCcVoaf+zp4n+HfwL8PfBD4E+No/C1vpCTx3V/quljWpr4XZkkuHZftNqEkkmlaQsCQM7QuMY+raKAPhzwB8IPin+zd+zprvgLUvic2t2WiaIbDw/caR4YI1LT5sOkciwLdXZvZC7ptTagyOTgkr9MfBlPiDH8JPBq/FiUT+M/7JsjrDhUTN8YVM+RH8m4PkNswpOSoAwK9LooAKKKKACiiigAooooA//V/fyiiigDxP46fHXwz8AfD2ieJPFGnahqcGu6zaaJBDpkH2q5NzehzFthBDvkptCoCxJAANcX4F/aZj8Q/E6z+Enj7wJrnw61/XLOe/0VdY+xyRanb2uDOsUlncXCLPErB5IWIZVOTXlv7feqTaJ4X+D2s2+n3Gqy2HxP8LXC2dmEa5uDFJM4ihV2RTI+NqBmUEkAkdazotY139pD9qH4X+NPDfhHXvD3hL4TQ61eX2o6/ps+kNdX+q2n2GKytoblUmkMYLyTOF8sYC7skZAPVPAf7Vml/FH4gap4A8A+C9c1U+G9c1DQ9d1Hbbw2GlyWE8kG+SV5g0plMe9I4VdwjBnCZAPl/wANv2j/AIQ/CX9mDwD8QNLs/E994X8T6/caJZpfSPq+sLdT315GTJ+8kklUywOI0jLvhkAUnNd9+x14Y8ReF9J+LcfiTSbvSH1P4l+K7+2F3BJbm4tLm6DQ3EYkVd8UijKSLlWHQkV8f+C/hn8SLX9kr9mrw3deFNWh1bQvipYajqNm9jOtzZ2Sa7qEzXNxEU3xQiN1cyOAoVlbOCCQD6ptv20bceL7z4Z618KvF+l+O3tYr/SdCkt7OW41SzlZk89J4bp7WBYiv783E0YjyFBZztr0z4W/tHaL8T/h34o8bW3hjWtN1TwVd3mn6v4fktlm1WC9so1leCKKF2WZnR1MRRsPkAc5FcXrPhjxHL+3h4Z8YRaTdvoUHw+1Oyk1AQSG0S6k1O2kSBp9vliVkVmCFtxUEgYBrw0aZ8evBHhr9rTxF8K/D97F4t1PxIt34eMloQbuM2NnFLcWazKEnKKJDHjcrSKF5ORQB9G+Df2mptY+Jui/C34gfDvXvh5qXiy3u7nQZNWaxlj1BbFBJcRZsrm48mdIzvMcmDtBOegNPwt+1dpnj74j6x8M/APgrXNbvfC+u3Gi65dotvDY6YsD+WLiWaWZd4lIYpFEry7VLMqjbu+PfDXhRPEX7SfwF8b+BPC3xE1Cy0S91r+3fEXjFNTyslzpE6RoYL5wIAJAVZ4beK33lY0ZmIUfWP7IvhfxH4au/jk/iLSbvSv7X+JuvX9mbqCSD7VZzQ2giuIfMVfMhcqwWRcqcHB4NAHYaL+1B4D1T4A6/wDtB3tpeaVpfhePVBqen3IjF/bXWkyyQz2rorsnnM8eEG/B3LyM16Rr3xR0Lwf8Jbr4w+NLe40XStO0j+1723lQPdW0YhEzwsiEgyr9zarEFuAcc1+dfxW8Aa5D+07J+zhplq0ngb4261pfjjUAuPJt00EF9ZhdfvYvZrfT+cYLSuM1+kXxQl0+H4eeITq/hm58Z6e9nLHc6NZxRXE99BINksKRTPGkhZCfkLAsMgZJAIB538KfjX4l+IUpk8T/AAz13wJp09h/adpf6pLp8lrLbErgSNa3UzQTFWD+VIoO0Mc/KRXjy/ty+FjpafESTwL4jj+E0l0LRfGrRWw07mXyBdG28/7aLMyfKLgwbe+Mc14d8KfDWv6v461b4WfA8eNtJ+D+seFtVstStfF9nfW9tomozIsNlFpMmpIt2SoaTzYld4lUAg5K4wJ/EnxIvP2QI/2Ml+FniJPiZ/YUfg9t2myjQVRIxanVDq2Pspt/KHnAhzIZPk2Z5oA+1/iR+03ovgL4m2nwf0nwprXjDxXqmijW7C10iKGRJ4TcNbkNNNLFFCqldzSSsseCArM7Kh8mt/27tJ1fwlrHiDwr8LPGGtaj4QluovE+mx2tvFLob2bFZkmlmnSKaQqDIkdu0rlMM4jBGdHwt8PPEvhj9tDw3eGxvLrQdH+Eq6I2rNBJ9le8h1aAiJp8eX5zRoZPL3btuWxjmqfwW8H+K9KsP2n49U0S9sz4g8Xa1c6cJraSP7dBNpltGktuGUeajurKrJkFgQDkGgD3zVfjjZv8NfDHxL+H/hbWvHtt4xitptNtdIhi84x3cBnjkuHuZYYbeMKMM8jgBsLyxAPjWp/tY6hqXwr+K2o6X4G1vR/H/wAM7bGpaDObF7q0NzbtPbXiy/aGtprbYGk3K5JEbAIxwG+d7RfjD4E/Zm/Zt8KXmmeK9D8JpYLb+N/+Eds7j/hIbJIrP/RIfKija8hje44neFBKiqAGXNS/Br4V6/qHi39p1PC/gvxH4Z0Px14V0iz0J/EzXT3N9N9j1G3d3nvZZpd3mOpaOWQSxqy70TcBQB7f8JP2sL2x/Ze0H4w/HXw9rGkSpo+hbbqSOzmm8SX+owIFbTbaylY7riUgpG6REBxlVVW2+neAf2lY/EPxB0/4WfEXwPrXw28S69az3ujQawbSWHU4LYBplgns550E8KEPJA5V1U5wRnHxhZQ+NfiL+yf8IdK8MeCdfj8Yfs86j4P1PVdC1LTbjTpdRbQoWtrm3spLlEhuH2K0qbHbkRg/Myg+y3mva3+01+0D8Jde8JeEPEHh7wx8MrjUdY1XVNf0u40dmubmze0gsLaO6VJJnLOzTlV8tVUDcSQKAPW/Cn7UFz488WXNh4J+GviHWvCNnrMugS+JIDYi0F5bzeRM4t5LpbpraKQEPMIiBgkAgZq540/aXOjeMtd8FfDvwDrvxGu/CCRya/Lo5s44dPMsfnLAHvLiD7RcmMh/Ih3MARnBIFfHfiyS70r4mQa5+zL4P8feBfiPq/iW3fWtGuNNuU8K6jbvchL+9vnbzdNUPAGdbi1mErNjguThmo/D2P4TfGr4ot8Tbf4kjRfG2uP4g0XU/BN1rUllN9rgijltLm20hiYrmKSPajyJ+8j2gMAgFAH134m/bA+GGg+Hfhp4r0qz1TxNp3xW84aL/ZVr9onkljtmuFiaHcriR2HlYPCPnzWRFZ1u/DT9p/R/GeveM/CHjbwrq3w78QeB7KPVbyy1gQSF9LmEhW7hktJZ43UeWwcKx2twCxzjwS8+E8HhrxF+yfp/ww8K63ZeFfDeq6xdzxX0M81zpcV/ptzJjUJCZPKYzSlP3j43EKCeK9B1DQfEmn/tqeMPHbeF7/V9B/4Vha2qNFb5gvbuLU7qVrGOabZbtOyEfu2kHDAthTmgDvvg5+0VrPxhn0jUrX4X+ItD8J+JbZrzSddvGsGtri32eZG80MN1JcW/nLgxb4/myORXC61+2podmPEPiXw54A8SeJ/h74Qup7PV/FOnxWrWMD2jbbmSCB51urqC3IbzpYYmVdrFdwGa8J+DkV34e+OXgvR/2adC8c+GPA12183i3w94l069tdC0mEwM8Jsmv1Pk3BuSuI7OV4mUk4CAms34d+JvHX7P/wCzrrP7KOqfDPxN4h8baWutaZo0thpktzpOtRalcTy2122ogfZoI8XC/afPdHjwxK5OKAPr34oftX/D/wCGmo+AdMh0/UfFc/xNsry88ProsSXLXxtVtnSOMM6DMy3KujkiNUV2dkUZrpPCPx2i1v4j6d8JvFfhjUPCXifU/Dw8QxQXj28sZiS4+zz26y28sitPAxRnVcja4YEjOPlrwz8FfGnw5+J/7IXhq5sbnVYPh/4X8Sadq2o28Mktna3L6fYxIrzhdsayOjrDvKlguAOCK9L/AG02m+Hvhzwr+1BpFubnUfg9qYvbmJCFkudF1ICy1K3XdhclHSZcn70QxzigD6C+HvxZ0X4leIvHGhaBZ3KweBdWGi3F7IEFvc3qQRzzpBhix8jzVjcsF+fIGQM15b4y/aeTS/H+sfDT4Z+Atd+Jes+Fo4ZdcOjmzit9N89fMjhea9uIEkuXj+dYY9zY64PFb37Knw+1X4cfAvw3pvicf8VPrCy63rjsMO+ravK17d7u5KSSmME/wqK+c/B/ifXP2Wvi18XdL8b+C/Eeu6H8QPEDeJ9E1fQNJuNZSZru2hhlsJ1tVd4JYnhxEJAFZSSGAFAHuvjX9pmDQfEWjeAPBvgbXPGfjjVdLj1qbQ7MWttPpljIdoe/mup4oIHL5RY/MZmZSAMYJq/8Nc/Dw/Anxb8dP7O1OGDwLJPba3o08KQ6rYXtsyrLayxNJ5YkXerAiQqykEMa8aPiLxF8GP2ifEfx28S+B/Ed74Q+LXh3QgWsNOk1PUNE1HS0lU2V7aWXnSRrIkwPmJvTzQVJ71454x+GnxP8Xfs0ftRfER/CGqadqnxeulu9G8OG2eTVBZ2cNvawPLaRB3S4uBG0jxYLKMbuc4APSf2jf2wviHpP7Pvib4lfCrwJ4g0qwWfS49H8SXEenC3ube8vI4vtaWtxM0ywzKQkRkh3N5yPsVdzL9Ga5+0m3hfwvoF54j8Aa9ZeMfFl9Np+j+FAbKbVbySBPMklDRXLW0cEcY3ySyTKsa/ewSoPGftm+BvF3jf9jfxP4U8GaRPq2uR2uj3UWnwITPN/Zt9a3csUaAZMnlwsFUDJOAAScV4h8fI7n4za78KP2jvDOgeOG8LeFJNY0rWrHS4dS0PxJbQ6nFblbu2tl8i5mjiliCTrFu3qcoHCGgD6Lj/av06Dwb491vW/A+uad4l+GKW8+v8Ahw/ZJ9Qhs7lfMS7geGd7e4h8pXkykmcRuu3cAD6HoHx88F+MPiNpvw88ELN4g+26DF4jn1K02NY2ljdnFl5rlg3m3XzNFGqltisxwMZ+WPB954Q8AeE/iD8Zvg14L+IPi7xbf2ljpNsniiHV5p9VlLSC0RF1LMyWkEkrNPMypGiFiGNM/ZG+DvjD9kzx3qHwc1jTpdf0PxxZQa5D4js7JvItdWtYI4b7TbmSJNkFuCPMsA+xQheIFnFAH2h8I/ijo3xk8CWXxA0DTdS0myvpLiNLfV7RrK8U20zwsXhYnAZkJU55BGcHIH59+KvEkfjj9qzx18NPjn8XfEXwoTTzYL4K07StS/sKz1KzlgDTXQuShS8nM5KeU7EJgqEfnb92/BD4x6d8cPBk/i/T9D1Pw29nqF7plzp+rwrBeQXNjKYpFdFZ16jseOQeQa+cfjd8WPBOtSeIfhd8f/gH4o8S6XBO6aa9loZ8QWepwkYjmtp7XP2W4bn5HMbpgHfnoAa3xy8W/FP4Y+Bfhp8EvBvit9T+IXxD1aLQIvEt9bQGa3to43uLzUDbxqsTzRQJhF2hS5DN3z6H8PP2eNa+HHivT/Etl8WfGXiCBElTUdP17UI9Ttb7ehCsqyRA2zI+HHkFAQNpXBNfGPhv4FfH/wAOfszfBnxiukTaj4/+EWu3Gt2vh65ullvH0G6eeJtK+0uQpuI7KVAm44BQR4OAK+xPAv7TEvxI8WaT4X8N/DDxnYpcFzqd7rekSaPa6WiRlgHe6KieVn2KI4N/DF84XBAPqCvlfxT+05cad8RvEnw78A/DnxB4+m8ELaNr9zpRsY47Jr2EXEMUSXdzA9zMYiHKRKcAgZLcV6P8KPjHp3xWv/G2lW2h6noF74F1yfQ7uLU4VhaZ4445o7iHazBoZopUkjJwSrBsYKk/EH7UA0W38ZeK/E/w68K/EPwl8Z4LZINI1jw3plzPp/iGWGEG0S7aAT2E9ujkRyfbBHJGinooWgD7jg+MWhzfG/8A4UQbC6i1r/hGk8TmdhH9nW2e8Nn5JIct5ocZOF24/izxXm+uftYeCdB8E/Fzx1daTqD2fwd1KTTNSjRYfNuZYooZS1tmQAridQN5U5B46Z8C8Ta38QfhN+0/4K+NvxF8G6zrdr4h+HEPhvUn8M6dNqws9eivlvZI3itw7rFIXZY35BIHbJHgfjrS/G0H7J/7YviPxr4V1Dwi/ifXJ9Ts7XUE2yNaTWlh5bh1zG54Kv5bOqyBk3EqaAPuC0/a706HxV4X03xh8PvEvhLwz44vYtN0PxBqkNtFa3V5cAm3imgSd7q1Nxj9z58SMx4ZV5x0/jf9o59E8c6p8Ovhz4E1r4j614bghudbXSGs4YdNS4XzIYnlvbi3WS5kj+dII9zlcE4yM/MnxI8V+I/2mbb4ZfBvwf4C8T6LLpviDRNa8QahrWly6fZ6ZZaO4uHWO6fMVxPMyqkItnkVgS24KM1yvin4eD4X/tA/E3WviRb/ABF/4Rrx7fW2saPqngi61l7Yy/ZY7eezvbXSGLxzI8Y8mR4yrxkDeNuKAPqfxR+2b8JPDfw++HnxPhi1HV9D+JV01lpn2K28y5+1iCaRbd4CwfzmlhNsEXJ84hThcsMt/iDo/iP40/BWbx34Z8S+D/GGvW3ihtM0yW/h+yRQ2UUQmbUoLWeSGV5I3R4AN+wk7sMBjyPXPhNaaJpv7MGmfCvwpr9t4c0XxnPqtzb6lDcT32nxXlvfTyy37OZGhzPMcmRhhmAznivZfi34Z8Sal+1/+z/4m07Sbu60fRrDxkl9exQSPbWjXVrZrAs8qqUjMpVhGHI3EELnBoAZ4m/a+sNP1rxTB4G+HviPx5oHgK4ktPEOs6QlobWzuYFDXEMKT3EU13LbgjzlgRtnTJPFbvj79rv4WeBdD+HfiaCO+8SaX8UfNXRJNJgFxJcSi2NxBCImZH824bbDGpAxKwEmwBmHzt8L/HHiz9lTRfHXwi8S/DnxP4n1ZvEGs6r4dudH0ufULLW7fVp2uYBJdxKY7aZGcpOLgoEVQ4LKa5/wR8A/iH8LtL/Y58E6rpdxqNz4Kv8AWJ9bmtYnuLbT3vNPupNssyBkREllEKMSFYgBScigD9HPBPijVvEfg608T+KvD9z4Ou5llefTtQlgkntUjkZQZZLeSSH5kUSfK5wGwcEEV8/eAv2qpPihq9heeAvhp4m1fwHqd6bC38VIlnHYysHMZuUt5Llbs2gYHM/k44zivonx34ck8Y+B/EPhGG4NpJrmnXdiswzmI3MLRBxjn5d2a+Kf2ZvjBrvw8+HfgL9nnxx8MvFdj4z8MQWXh+5+y6RLNpLR2oW3Gorqfy2htmRRK7eZvzuUIzYBAO+8TftfWGn614pg8DfD3xH480DwFcSWniHWdIS0NrZ3MChriGFJ7iKa7ltwR5ywI2zpkniuD+MHjLw74/8AjH+yZ428HX6ajomvaxq13aXEeQssM2izspwcEHnlSAVOQQCCK4/4X+OPFn7Kmi+OvhF4l+HPifxPqzeINZ1Xw7c6Ppc+oWWt2+rTtcwCS7iUx20yM5ScXBQIqhwWU1yWgfBXx58D/CH7H2k+IdMu9TbwLrF7FrjadBJfCwl1q0uVTf5CviCKecRNL/q1ABJ24NAHf+G77SdP+L/7bF/4iN5/ZVna6JLdfYJTBeC3Tw0Gl+zyhlMcu0HYwYbWwcjrXsFr+0B8PvhX8DPhNeaLp+ueIrnxxpmmw+GNERlvdd1ASWaTgSvLIEzDCQ1xM8gjTqW5UHxfUvDHiu2+JP7ZOktoGpSP438PWN3oksdlPJBqCwaAbGSKCZUKPOLj5PJB3nOQpFYMvhL4hfDjw5+yf8bv+EQ1XXrX4b+FV0XxFo9navLq9iuqaVaQm4isiBK728sGyeNQZQp4U7WwAfTut/tNzeCvhdqXxK+Jnw717wlJpOqWGlS6fdGymllk1CeGCOa3mt7iSGWFWnXcwcEEMuCRivTvGPxe0LwX8Tvh78LNQs7mfUfiNJqkdlNEE8mA6Vafa5fOLMGG5Bhdqtz1wOa+S/2lvEev/tHfst+Mpvhn4J8R/atD1DR7+0tdT0yXTrnVU0++t7yf7HbT7bhsRowXdGpdxtQNnNZeoeNfE3x0/af/AGffHfhj4eeKNI8JeF5fEn27UtZ0ubTxHNf6PIiI0Mg8xEVlCec4WJ5GVI2c5wAM+E3xP0n4NWP7WXxT8Q21xfad4Y8cXt5NDbbTM8cen2WVjDsq557sBX1348+N/h34f2fw8vNTsbq4T4j65p+g2QhCZguNRglnjkm3OMIqwkNt3HJGAetfCmq/DDx34j8DftmfCmz0K+j1zxTqd1q2j+bbSR22owXmnQCAW1y6iGR2kt3jZVYlGwGxmm+KvHnjL462/wCzvYeGfhf4t0y38HeM9Av9dudV0mexSxa2tri2dAkg3yorOzSTovlRhVDOGcCgD6q1v9p+5j+IPiTwP4B+G3iHx1B4Jmgttdv9LNikVpcXESzCGGK6uYZbmRI3VnWJTtzjk8V6/B8U9GuPi5P8Gxpmppqtvoya2161ow00wPP9nEK3WdpnDclMfdyQTggfn7+0qbKz8ZeLPFPwe8J/ELwf8blMcGnX2h6ZcS6R4klgRRatetH5+mzWwBCu9z5c0aDnG0CvtjSfjLPb/FjQfgb4s8P39r4i1Tw0ddOpRRBtIkltpI4by2im3FvMieVGIK4CumTllyAeGftyeO/FHgrRPAUI8R6l4I8AazrotfFniPR0JvtNsjC5gCyhJDbpNPtR5wuU45+bDdn8M/Ctp4U+H/inxd4A+L2s/EbwtqOlSy6c2o6lDrH2W4hjkLywaiq+c4YbBsd2CFSerYHovxn+KerfDFNKmf4ea1448N6l58OpzaJAl/cWI2jyt9gD508cuXDmMHZgZB3CviD4MeCTqXxo+J3xG+Cvw91f4d/C/WfB81jcaffafLpKax4iaUvHd2emOAyKlvmMyBEDs+ApO40Abv7Dfxy8fa98Kl+G3xf1efVPF/8AwjVt4p0fVbhyZtU0fUYdzMXJJaWxui9vITzt8pu5Nd98Fv2jLLwD+xX8L/in8Vr7UfEet+ILKys7eGFTe6rrGqXbMsNvArEGWaTB+8wAAJJAFea6T8GPHsX7HPwY8beG9EurP4pfCbRba5g02aF4Lu7tngEWp6RLE4Dq11BkKrLkTJGeOa5XRPhp8UfDf7K/7MXjm08IalqesfCG8tdS1fw4IGi1R7OSCe2uDDazBGa6gEgkSI4ZuQOcZAP0K+FfxK8WePX1O18X/DrWvAF3pwhdV1R7KeG5jn37TDNZXE6Fl2fvEOCmVz1FerXl3bafaT395IIre2RpZHPRUQbmJ9gBmvnGw/aX0/WvBvi/xroXgDxhNZ+ErD7Z5d3os2nT6hIA5e2s4bwxSyyR7PnOzZyArMcivW/CnjrTvHfwx0j4l6Bp91c2PiDSINWtbJ0RLqSK6txPHCVdxGJGVguC+3d/FjmgDxj4P/tKal8ZLrSNU0b4ZeItP8F+JVlk0rxFcmw+yzxRKzLLNBHdNdW6Shf3Rki+YlQQMiuj8L/tEeFfFfw08efFCy069i0/4f3uu2N5DII/OmfQC4naEBypV9h8vcyn+8BXxV8MY7nw98afBGn/ALMPhvx14O8PahfXD+L/AAz4g028tfDmm2LwySPJbG8VooLoz7PLSxlaNwTxsHOJoOsePvh58PP2gf2dT8NPFGseKfEmueLbvR57TTZDpV3Ya6JHguP7QP7hdiud8eTIWGxFeQ7QAd9+0F8XdO1TxT+yV8Z9K0bVNQstevLvVLbTbOAXGoyrqOhu8MIjVtnmZlUMS4jTlmcICw+kvAf7UGjeIPFPiLwF8RPC2q/DfxP4c0ptelsdY+zzCfSFYo93bzWU08UixsNsihtysQOecfKWt6H498I+Dv2MfEK+CNe1k+A9PifW7PT7CSW8sVXw6LeQTQtsKOjkjy2IkZgY0VpCqH0Lw7f+IfjN+0lffH+w8Da5aeCvA/g6/wBFtIdVsG02+17UL6dLiWK3tL3ymMKRxCMNNsRpXxnAJAB7B8Mv2lvEHxPsrTxFpfwj8T2nhnW7CbUdG1SV9NMd/DHEZYg0S3hlt2uRgQ+cqqxYZZRzXk37K37TfxP+JGi+PNV8e+BvEEtlout+JHt71ItPkEVvYXLCDSY4LSYzzXUSfuwRGwkZf9axIJ84+Dkd34e+OXgvR/2adC8c+GPA12183i3w94l069tdC0mEwM8Jsmv1Pk3BuSuI7OV4mUk4CAmqXw98SfGL4VfDL47fCLwR4Q1qH4oW+ueKdc0CeTSp5NMvbe9uPPt57e8KG1eQo52Qs+5pF2FScgAH1b4W/agn1D4keHfhx8QPhx4h+H03jRLptButXNjJFevZxedLBILO5na3m8sF1jlwWAPOcA4+v/teQQeLPG3gLwF8NvE/jnxB4Bu1t9Sh0yC3WBI3t4rlZRcTzRoxdZCqQrumZkb92Fwx+PdL8LTeKvjH8BPF3hHwt8R9Wn0TXZX8Ra/4uj1NRC8un3CGP7NduI4VEgIeS2t0t1O1N5ZlU/Yv7OvhnxJonxs/aH1XWtKu7Cy1vxLYT2E9xBJFFdwppdvG0kDuAsqK4KlkJAYEZyKAPKPj5+2BrafAL4a/F/4EaRql/ZePtY0mNbmKOyDW0T30Mc1jPHdS4E9yPNt0KBlVlYmRBtY+q63480HX/iv8BpPHXh3xL4T8WeIJvEn9maY19CkFu1naM0/9qRWlxJDOHhQPb7TJtLc7SWFfHWm/DT4maZ/wTn+HOnnwhq0uv+C/E9trt5ogs5U1RrSy8RT3DiO1dRIzmFhIq7csvKg5FfSvi3UtQ+Lf7QP7MnxR8JeHtbXw9YXHi83s17pV3ZNZCXS2giN0k8aNAs0g2xGQASZG0nNAHofxU/af1b4XTa/qdx8JvFeq+E/CWX1fXYI7KK2igRQ8k9vDPcpc3UUSnLvHFtADcnacfT2j6vp3iDSLHXtHnW6sNSgiubeZfuyQzKHRxnsykEV+RHxah+IPxCsvjh4U+KfhX4g+IvGE02uWXhHStJS/t/DK6S0DR6dd+bavBZXDEfPMLqSSVmzHHCSQh/Sr9n3TdS0b4C/DbSNZtZrHULHw1o8Fzb3CNFNDNFZRLJHIjgMrqwIZSAQQQeaAPlo/8FBfC0/w9h+L2lfDXxbfeBLWVotX1lLa2S307ZO0Dna84luRGVzI8CPGoO0vvDIvtPifxF4Ef9pz4b6XcSavJ4h1PQdZutOa2vGTSGtYzB5xubcOBJIfMUxNsOOSTkLj5E8I/Dvx/bf8Er/EHw8ufDOpxeKptJ8TRx6Q9lOuoPJPql5JCq2xTzS0iMrIAuWUgjIIr2k+EfFh/ac+Auv/ANi339maP4I1m0vrv7NL5FrcypZbIZ5Nu2OR9jbUYhjtOBwaAIfFv7demaDo+r+OdA+GviHxJ4A0bVTokviK2l06C0kvluRaHyo7m6ilNuJyIzcMqx7u+Oa9C8cftX6V4C8U+Gfh3qngjXrrxt4u0RtW0/RLRLWe5eaOVI5LJ5Fn8hZIwzSSSGTyVjjc+YTtVvzi8M+IvBP/AAinirwD8Udc8VaF8KLfxfrV9daLpfhe61LSPstpq8twIotftYrgNZPLGJJwuGDGSMOqivvCFJfHP7ZPw8+LnhK2m1bwXffDvUvI1mCGR7Bnu721mgTz9uxZJI8sqEhiAeODQB23jP8AaL8b+E7Uz2fwV8Wa2dP0631HVWtv7PSGzEsImlgikmuo/tk8PKutsrruGAxNe7fD3x34b+J/gbQviJ4QuDc6L4is4b60kZdrGKZQwDr/AAsucMvYgivz6+JF74x8Q/G34heGfi94e8e63o6C1h8F6V4ZF9a6JfQPbKZpLu9sZLeLzvtDMJBfTrFGuMAjk/Q/7DXhvxH4P/ZM+Gvhnxbpl1o2sadprRXNnewvb3ELieQ7XjkAZTgg4I6YoA5Kb9tJNQ/4S+XwH8KPFvi608Aatqela3c2UNnHDA+lTNFKYDcXMZunYL5ixQBnCkb9rELXrF/+0HZaj4B8J+PvhT4S1r4kW3jWAXGnJpEcEapGUD7rue8mghtsZ2kO+7eCoUkHHw78Av2gZPhTpvxw0W18AeKfFGrT/ErxjLpY0jSZr6zvbh7whYGuYdyW7KwBkM/lgIdy7gDitYeBfi58A/hP8Avg74wTxRH4FstN1FvFk3gqK5udQTVJ5BPa2ckunq93HbB5pEL2xG5lUbguMgH2b4a/an8Ma58KviL8R9V8P6poOofCpNQ/4SDQrtYft9vLp9r9rZEaORoZFliw0UiybGz1GDXJx/to+GW+Huo/F1/BXiCLwWiWSaRfvFbq+u3l/MtvFb2FuZhKQZHULLKI42GWVimGPyr4I+Gviu2+GP7ZNtongPxHoVl4v0mT/hH7HVo7m61G/EmgPEux5JLiWeWSU8p5jyozCNwrgoPrvWtH8O2v7I3hPwp8Ufh9q3jPSH0PRLHVNF0+zae/g2QRBpfsweK4DW8igkQ5mRgCi5XgA9k+F/xE8WeORqNv4x+HuseAb3TvJYJqUtlcw3CThiDBPZXE6MU2/vFO0oSvXNcj8Vf2hdP+H3jPS/hb4V8Man4+8eavaSahHo2k/Z0aCwjfyzdXdxdSwwQQmT5EZnyz8AGvEf2Sbvx0vj3xno+jXHiy/wDg9aWdidFn8a21xBqMWos0gube1e9jjvJrRIxHhpwdrfKjEbiU8c3PiH4E/tY6p8bdW8L6x4l8EeOPDVlpE15oljLqlzpN9ps8sirNa26vMLaeOTPmIrASDDAZBoA9V8TftPWHhXw14Sm1HwP4g/4TbxvcXNppXhERW39ryy2Rb7Q7nz/syW8SjzGnabZ5bKwzuAra+Gf7ROi+P5fFXh/WvD+peD/GPgmFLjVdC1QQm5SCVGeKeCWCSWC4hk2sFkjcgMMMFOM/O/j/AMQ+KJfit8Lf2t9N8A+Jbnwxpljreg6rpb6c39uWNveyRm31FNNUtM0bND+8QATLEwYx8FRd8C2Hir4vfHn4gftFQeGdV8N+Fx4Kj8I6RFq1pJY32qyrcTXs10LOUCaONS6xRGRQXySoxmgCS2/4KDeBpPCPhr4pX3gLxVYfDfxA1rBJ4lns4RZ2dzdEIqPCsxuZI0kPlvNHE0ZcYjMler+B/wBqOz8UfFew+E/ifwJ4h8D3viKxudR0G41qGCOPVILPZ54VIZpXhlRXVzFKquEOXCMQp+RdZ+HHxBl/4JfeC/AEfhfVH8UWkXhkS6ULKc38Rt9ZtZZd9ts81fLjVnfK/KoJOACa+oPjH4Z8Sap+1t+zt4l0zSbu70nRI/GAv7yGCSS2tPtWnwJAJ5VBSLzXUqm8jcQQMkUAYk37aSah/wAJfL4D+FHi3xdaeANW1PStbubKGzjhgfSpmilMBuLmM3TsF8xYoAzhSN+1iFr16++P+nah8OfC3xH+F3hbWfiLaeM40l06HR4oVISSPzN11LdywQ2yrgqxkcEONmC3FfC/wC/aBk+FOm/HDRbXwB4p8UatP8SvGMuljSNJmvrO9uHvCFga5h3JbsrAGQz+WAh3LuAOCDwL8YPgB8Ef2fPhDry+JIvBtrb6kfG9x4MinudUt7qYfabO1D2KvdJbefNJHJJajd8igMqkEgH2p8Pv2nPB3ivwx451zxnp154A1D4ZSMniXTtX8ozaeogFyku+3eWOWKWE7o3Rjv5wOmcn4f8A7SfiX4hNBqdh8HvFtj4d1azuL3SdTuV09FvUhiMsYeD7X51sbgACAzKoYsu4qDkfCehfs9+MPiN4c/a18JeD/Cev+DrDx/p/h0+HB4ke5NxevaWkpbfc3cs8hMsqAOskpkiWRVlVD8g+7fhB+0a/jq60DwTffDbxd4b19oNmqJfaLNa6bpcsERLBr2XZBLG7rshMDSM4KttC7iADyL9lb9pv4n/EjRfHmq+PfA3iCWy0XW/Ej296kWnyCK3sLlhBpMcFpMZ5rqJP3YIjYSMv+tYkE+s+Fv2oJ9Q+JHh34cfED4ceIfh9N40S6bQbrVzYyRXr2cXnSwSCzuZ2t5vLBdY5cFgDznAPyl8PfEnxi+FXwy+O3wi8EeENah+KFvrninXNAnk0qeTTL23vbjz7ee3vChtXkKOdkLPuaRdhUnIHJaX4Wm8VfGP4CeLvCPhb4j6tPomuyv4i1/xdHqaiF5dPuEMf2a7cRwqJAQ8ltbpbqdqbyzKpAP068FfFLRvHXirxl4R03TdTsrnwTeQ2V1Ne2jW9tcvNEJQ9pISRMgBwxwMHBxtKk/En7UHjeWH9pTwr8Oviv8RdZ+FHwq1HQGubLU9Iu20lNR8Q/amR7S61RVJhSK2AdYyyKxbJc8LX2L4H+MeneN/iX49+Fy6Hqek6j4BawMs97CsdtfQaikjQz2jqzb03QyKcgcr9Qvmfxt+L+meF9ZvPAnxQ+EWu+LfA2o2kbpqWnaUPEFjPKSRLb3NnCJJYdvybGeMq5JwRtyQC3ZfAG+1z4f2Xhi9+MXjDU7OC8kvbDVrLU4rW/ezmQCO3mu7eIfao1OWWRxuORkkAV84fsZ/DvxL42sNW+IHiz4peN9VuvDHjHW9MgtLjW5JLGe10u7aGFLiFkPmZUfPyA3tXon7CvgXxF4N0H4gXieHNR8D+Atd8QSXnhTw7q25LzT7ExIJWaB2ZrZJ5gzrASCnPHOT1P7FHhjxL4U+HvjOy8UaTd6PcXXjjxPdwx3kElu8ltcXzvFMiyKpaORTuRx8rDkEigD5W+B/7RV38H9D+Ok9j8PPEnja00L4keMdQ1a70qGBbaxtBeEt891NCZ5VRS5igDlVwXKZGf1L8IeKtF8deE9F8beG5vtOk+ILK21CzlIKl7e6jWWJiDyMowOO1fEHwV8GeL9L+Cf7Ruk6roV9Z3uueMPHtzYQTWssct5b3jP8AZ5YEZQ0sc2f3bICH/hJrofhD8Sp/gV8BP2cfA3jbwrra3viiw0Xw9MyWhA0q+a0jVFv0kKSQ7pPkwVyCGyBjBAPof40fGDw/8EfBY8X69Z3eqSXV7aaZYafYIr3d9qF/KIbe3iDsiBnc9XZQACc9j55qP7RupeEvhT40+KPxO+HGu+D18EWwuriyuZLC5a7iK7s2k1rcywyY6MGZCp4NdD+0jbeCtQ+GNxpHxF8Dar4+8OX9zBFeWejWz3d5bKCXS7SOGRLj9y6qd1vumUkMqkAkfBln4T+MPxN+CX7Qvwy8IHxVrvgG70OOLwc/jK3mt9XmvzFK91awteJFdS22ViWJ7hQQx2hiAxoA/Qbx58b/AA78P7P4eXmp2N1cJ8R9c0/QbIQhMwXGowSzxyTbnGEVYSG27jkjAPWvjHQ/idr/AIB/bQ/aKsvC3gXWvHuq6ja+DpI7XShbxRQx2+nz73nuryaCCPdvARNxkkOdqkKxXA8VePPGXx1t/wBnew8M/C/xbplv4O8Z6Bf67c6rpM9ili1tbXFs6BJBvlRWdmknRfKjCqGcM4Fd7Z+NNX+CH7V3x38beKvAvirUvDXiyDwrHp+paPo1xqUEs+n2EqyRBbcNISWlCh1QxhgVd0baCAUfjv8Ato6tN+zZb/FD4I6HrEGqT6/aaJfiWCzS50O9t9StobuxvoLqQgTTIzQJsWQAuHLIMMKX7Q/xR1KH4h/s0fELxP4L1zQ7qLxFrinQvLhvdUkc6ZPDCiJZzTRMZWK7T5gVQd0jIoYjzXxF8H/i5e/sh/E7xjP4RvovEnjn4gR+PovDSJ5mpQafHqFk627RISDdfZrYytGuWydmN42j6F8e6zffGX4rfs2/EbwZ4c11dF0zxBrT3z32kXllJZRnSpolkuY54laJGkIRHfCu3Ck0Ae0/B79oq3+KHjPxN8NfEHg/V/Ani7wxBb3sum6v9ndp7G7LrDcQy2s00TruQq4DfK3GTzjjpP2tZNJ8ReHbfxv8LvFPhDwv4q1ODRrDXNVjs44vt10xS3W4tI7l7q3SZgFR5Yl5IDBecY2oeHvH1p+2p4x8b+HtDnmt2+GFraWF3NC6WE2qRaldSpbG4IEe/BQsu7cEIYgDmvgvxHoXjn4nfDbwRda74S+JXiX4s2XiPw9feJLjVrfUYNK0tor+FrpbSz3R2LwrnKG1gkZYv3srqAzUAfeXwvdv+G6PjoOSF8O+ESB/wG96VtyftayaT4i8O2/jf4XeKfCHhfxVqcGjWGuarHZxxfbrpilutxaR3L3VukzAKjyxLyQGC845jStP8a+GP2w/jFq2naHdSy+KfB2jT6FdSW8v9m3F1pf2mOS3kugvlJIJJY8ozhih3AYGa+GfEeheOfid8NvBF1rvhL4leJfizZeI/D194kuNWt9Rg0rS2iv4WultLPdHYvCucobWCRli/eyuoDNQB+kPjX9qiy8O/EvxJ8HfCngPxD418XeHLOxv2tdLigEMltepI3mG4nmiijEflhSrsHdnURI+HK+Xzf8ABQHwdcfD4fFbw38O/Fus+E9MyPEN8llBCuiPG5jnhmSadHnlgIzKLcSIikEuDxXefDjwz4ksf2zvjN4pvdJu7fRdU0HwtDaX0kEiWtzLbi785IpioR2j3LvCklcjOMivn3wr8PvHdv8AsHfHPwdceGtSi13Vrnx21lpzWcy3dyLu4uWtjDAU8yQTBgY9qneCCucigD6j8beKPh/N+0L8F7e5m1efWfEFh4huNGksrto9JeCG2t3na9t96iYskqmAlG2tknBCmsaX9q9dY8U61pHwv+G/iT4gaJ4Xv20zVta0pbNbSG8jYLNFbrc3MMt20BOJfJQhexNecSeDfGB+N37J2rf2Hfmy8OeF/ENvqlx9ml8qwmn02wjijun27YXd0ZVVypZlIAJBrlPgF478Qfss+FNU+Afjz4eeLNZ1XSda1WfSL7RNIm1K01uz1K9lu4Zhdx4ghlUS7ZVuJI/LCglsdAD7p+K/xF0r4RfDTxL8T9dtp7vT/C9hPqE8NsFM0kcClmVA7Ku4gcZYD3r55039sfRbvxf4N0rVfAPiXQ/C3xBu00/QvEeoW8ENld3k0bSwRmDzjdRLOqnyWliQv1C7csO3/a90TW/FH7LfxR0Dw5p1zqmq6j4evobaztYmnuJpXiIWOOKMMzuTwAoJJ6V478evB/i3WPh3+zhZaRol9fXOheOfBt3qEcFtLK9nbWsUgnmnVVJijiziR3wq5+YigD6Z8HfF/QvGnxQ+IXwq0+zuYNR+HLaUt7NKE8ic6ta/aovJKsWOxBh9yrz0yOa81sv2t/hk/wADbv4+arDfafolvqF3pUVoYlmv7q9tr17FILaGJm8ySeZP3ag9DltoDEeHQ+KfEXwI/a4+Mes6x4D8S+I9O+J9t4cutDudC0yS+t5Z9Lsns57aeZcRW77sMDMyoEyzMowT4n4c+D/xjT9kDwRqC+Drz/hLfht8SrjxlP4ckHl3V7bWesXjyQwGQIsjNFN5sDfdk2qyZyoIB9YeNPiv4w8ffA34tWHiv4Za/wCAHt/COrXUE2qvZSw3EclnMFVXs7mfZMuMvE4BUY5qH4b/ABl0L4RfspfAtr2wvfEGu+JfDfh7T9H0bTESS+1C7OmRSMsYkeONEjjVnllkdUjUZZskA6+s/Fr/AIaA+F/xF8FeC/BXirTrm68L6nEk2t6NPpMLXlxbvClmn2rZJJNubny0aMAcvkgH5J8Q/DrxX4h+CP7LPxPg0LxY1p8MNJbSPEWk6Kb3SfEluk+nxafPPbxIYblzb3Frho0+aWJyUDK2aAPsa3/aw0Ky8KfEbVPHHhTVfCviP4XaZ/a+r6DdNaTXT2TRSSxTWs1vPJbzxyeU6BhINrqVcLxnuvGHx98M+DPCnw78XX+n3k9t8SNY0XRrJIhH5kE2uDMLzbnACJ/HtLH0Br49h+Engj4m/C3403vwo0vxzdeKvEPg+68P29941bVUa9EsU8kFtbf2uwl2xzH5m2BFMnyk5bHDa7418cfGPwD+zx4Q8O/C3xdYT+CPGHgy68Qz6npE9lFYf2dIsEwUSDfOiks7SxqY4413SOhIUgH2R4p/acuNO+I3iT4d+Afhz4g8fTeCFtG1+50o2Mcdk17CLiGKJLu5ge5mMRDlIlOAQMluK9ZHxV0cfFi1+Dz6VqiatdaIddF2bQnTkhWfyDA9yCVFxu52YI287uQD8HftQDRbfxl4r8T/AA68K/EPwl8Z4LZINI1jw3plzPp/iGWGEG0S7aAT2E9ujkRyfbBHJGinooWvrvQ/jPqmmeP/AAL8FviD4evrfxX4m8N/2tPqNtEraOt9aKi3toswct5kbHdjaV2snzZYUAWPjh8JX+IFg2uw+N/FHhOXR7O4KR6BqjafFM23eGmVUbeQRgHjAyK+PP2Zp7zwT+yzon7YPxH8feNPFt1ZeGrvWdR0y61c3VlMIkkLiO2lCrvCrlN0gAbnNfpJ4nhluPDWrQQI0kslpOqqoJZmMbAAAckk9BXwT8MvhZ451f8A4JmxfCKXR7nTfFt/4I1HTk0+9ia1uFvJoZliikjlCtGWYqPmAxnNAH074w+PvhnwZ4U+Hfi6/wBPvJ7b4kaxoujWSRCPzIJtcGYXm3OAET+PaWPoDXutfkfrvjXxx8Y/AP7PHhDw78LfF1hP4I8YeDLrxDPqekT2UVh/Z0iwTBRIN86KSztLGpjjjXdI6EhT+h+m/GPTr/44ax8C5dD1Oz1HS9Ih1qLUZoVGnXlvJIInWCUMSXjdlVgVHO70yQDK+MXx40f4T6j4f8J2Wh6h4w8ZeLHmXStD0oRfaZ0tlDTzySTyRQwQRAjfJI4GSAATnHj/AIm+O+i/Fr4FfG7w3No2oeEfGHhDw5qkWr6HqqxLd2v2jT5nglV4JJYpoZlBMcsbkHBzg1V+N1l4m+GX7S3gz9pKHw7qXirwrD4d1Dwxq8WkWz397pv2i5iu4L1LSIGWWNmjMcpjUsi4JBFeaWnhTx18a/E3x7+Ndh4X1Pw9pPifwOfCPh6z1W2exv8AU5IorqR7qS0kAliQyzLHD5gDMuWwBxQB4r8bfEmrD9kj9k34dt4R1LxRoHi6XwTBqsFpJbJFqMEWnif+y286eJvMuWjVhkCLajBpFJUH7Rvfi/ffBrwxaeGfh78BPE82ieHNLgvLq306PTrW006GWLz5LaDzbtBdTw5YOlsJF3ggMTXgVz4f8WeJ/wBnb9jq30rw1q/2vwr4q8Irq9tLp1zFc6euk6ddWd3LdQvGHhijmXaZXAQgqwYhlJt/Ei98Y+Ifjb8QvDPxe8PePdb0dBaw+C9K8Mi+tdEvoHtlM0l3e2MlvF532hmEgvp1ijXGARyQD6m8TftS/DHQPhH4V+L9l9t12z8dm0i8P6dp8Hm6lqd1fJvitoYWZQJAAd+9lVNp3MMc+H/H39p74ueEv2bPiD44sPhf4j8FeKNDtxHBLdHSrqK1+0xSNHfiRLmaGWKBkVZUCu6u6Dy2XcR4F4I+G3xU8B/s8fss/E0+DdV1TUfhBcXkmueHEtnXVRZ6lDcWkssNpJteSe33rIkQAZgflr6O+NvivUv2of2Yvi74R+G/g7xHa3T6K8dkdZ0qbSv7RuGDSGC0iutk7svlhSTGqFmUKzHdgA7r/hqJfCvwo0nxr8SfBOu6Lrus31vpGk6EVs7nVNavJ4UkRrVLW4khCPlyTJJHsCMWCjbu6v4XftDWnjrxvffCvxj4U1T4feOLGyXU10rVjbS/arBn8s3NrcWk00MyI5CSAMGRjgjvXyz4w8R+IPiHpXwU/aG8E+CfEl6PhBqsset+H7vSbmy1Rre/0z7JczWltcpGbp7VpAVEWd5DhTkGu88JXWu/H39qrwz8ZNF8Maz4c8FfD7QNTsFvdbsJtKuNUv8AVnhzFDa3KpM0EEcW4yMoUyHaucZoA6bwN+2RYfFCzv8AVfhv8O/Eeu6Zoa6smqXiJaww2t3pfn7bJTJOGnuLjyUKLEGVRPEZHUlgvoOrftN+BLD4D+Hvj7p1td6tpfisaUmmWNsIze3Nzq80cENsqs4TzVkkxIN+F2NycVxf7DHhjxF4Q+AUWj+KtJu9F1D+3vEUxt72CS2m8ufVbmSJykiq2142VlbGGUgjIIr5i+HXgDXLb9qa0/ZkmtWPgL4Va3qXxFsG48gQavGF0uyAGSDbX1zfyKDgbYkIzQB+r1FFFABRRRQB/9b9/KKKKAPJPi38IdG+L0XhCLWb24sh4O8SaZ4mt/s+z97caW7PHFJvB/duW+bGD6EV63Xlvxc+Lvhb4MeHbHxL4sWaS31DU9P0qJLfyzIZtRuY7aNsSPGNiNIGkOchQSATgH0aLUdPnEJguopBcbvK2up8zZ97bg84746UAXKKxNc1ddN0jVLu1eOS60+1ln8tjnBVGZdygg4JHtmvGv2bvjDdfF/4J+AvH/i17Gw8Q+LtMS+ks7ZjGm4khvJjlkeTYMd2bHrQB9AUVTv9Q0/SrV77VLmKztovvyzOsaL9WYgCpPtlp5Edz56eTLt2PuG1t/3cHod2eMdaALFFUodS064NyLe6ilNmxSfa6t5Tgbir4PykAg4OOK8F/aO+Mt58Kv2efG3xi8AtYaxe+G7Fri2EzGe0eVXVSsnkyIxA3cgODnvQBa+GnwEs/A3jzW/ip4l8T6n438YazbLp66hqnkILLTUkMy2dpDbRRRxRFyHc4LOwBY8V77WLb6zaxaDb65q88VnC0EcsskjCOJN6gnLMcAZPc1qwTwXUKXFtIssUgDI6EMrKehBHBBoAloryz43/ABHvfhB8JfFPxRsdEfxG/hexk1CSxjmEDywQYaYiQo4GyIM/3TnbjjORz/xP+PPhf4cfALUv2gkibVtGt9Lg1KzhjYI959sCC0iVsNtMzyxoDhsbuh6UAe50V8aa7+0t8U0+K958IPAvwm/4SXWdI0PTda1HdrtvYrb/ANoF18lfOhxIUeNhuBAPXAr1n4JfHCw+Mdv4h0+50K98J+KPB98NO1vRtQMTz2dw0azRlZYHeKWGWNg0ciNhhzgUAe5UVC9xbxzR28kqrLNnYhYBm28naOpx3xSm4gWZbZpFEzAsEyNxUdSB1wKAJaKz4NW0u6vZ9NtryGa7tcGaFJFaSMHpvQHK57ZFMvNa0fT4Hur+/gtoY5BEzyyoirIcYQliAGORx1oA06K8s+Kvxf8ACvwgsvD174pEzp4l1zTdAthB5ZZbnVJxBHI/mOgESE7pGBJCgkKTxXpEeoWEqxPFcxus4YxlXUhwv3iuDzjvjpQBborGl8R+HodNj1ibVLWOwmOEuGnQQsScYDk7Sc8cHrXh/wC0h8XvEHwf8K+Fde8L29peS694p0LQ5RdK7oLXVLpYJXTy3Q+YqnKEkqD1UjigD6JopksscMbzTOI44wWZmOAAOSST0AqkuraU99/ZaXkLXuwS+QJFMvlno+zO7b74xQBoV4F8V/gJZ/GXxLotz4y8Tam3hDSXtrmbwxB5Eem6hd2kxnhlvH8o3EqK2wmHzBGTGpK9c+2ajrGk6OkcurXsFkkrBEaeRYgzHooLEZJ9K8I+P/xc8RfCnUPhXaeH7W1uU8c+NdO8N3pulkYx2l5b3MrvDsdMShoVClty4Jyp4IAPoiivmvw78YfF2uftBfFH4RJY2T2PgvRdF1DT3zJFNPcaklyXSeUs6CMGFQpWMFQSTu4r1v4ea14s1fwFpOufEWws9E16e382+trK6+12kDgnIS4woZdoBJ6A5GSBkgHc0V81TfG3VJv2nvDnwc0cWF74Z1rwnf6815Hukn+02l5DbKkciSeV5ZWQlhsLZAwwHB+ixfWTWxvVuIzbrkmQONg2nBy3TgjmgC1RVdrq1QorzIpkUuoLAblXBJHqBkZNUTr2hrpg1ttRthpxGftJmTycE4/1mdvXjr1oA1qhuYBc28tszvGJUZC0bFHXcMZVhyCOxHSpFZXUOhDKwyCOQQa8T+Pvxng+Avgqy8f6npL6no39r6bp+pSpMIfsFpqFwtsbxsq29YndMp8uQfvDFAHoXgjwR4a+HXhm08I+ErU2mm2ZkZVaR5pJJZpGlmlllkLSSyyyO0kkjsWd2LMSSTXWV4t8c/jHB8GPDmiaomlPruqeJdd0zw/ptikwgM93qUwQZkKvtWOMPKx2nhD06j56+JP7VHx1+GnijQPDeq/Azzh4u1htG0aZfE9mBdT7JJULKID5QeOMt8546E5oA+7qK+bfEXxy8XfD74Ia18Vvih4FOgazp0qW9toUOpw3z3k9zLHb2caXMSBFaeaVUxtJXqQeld98Evipp/xr+FugfEzTrN9NGsRP59nK4eS0uoJGgubd2AG5oZo3QnAzjOB0oA6LwR4B8MfDzTbrTPDNu8a6hdzX93NNNJcXF1dz48yaeaVmeRyFVQSflVVRQFVQOypGZUUu5CqoySeAAKhF1atbfbFmQ2+3f5gYbNoGd27pjHegCevL/jV8K9J+N3wr8SfCjXbyewsPE1qbWae22+dGpYNlN4Zc/L3Brvr3WdI023iu9RvoLWCdlWOSWVUR2f7oVmIBJ7AdauyTwxFBLIqGVtqbiBubBOBnqcAnFAEVjapYWVvYxkstvGkYJ6kIAAT+VWqz7TVtLv8A7R9hvIbj7I5jm8uRX8px1V8E7SO4NebfC34z+Dfi5Z69e+GXeJfD+s6nosy3BiVnl0qcwSzRhHfMLMMoxwSuCVHSgD1miqMep6bLdJYxXcT3EkXnrEJFLtFkDzAuclMkDd0pljrGk6o88WmXsF29q2yZYZVkMbf3XCk7T7GgDRor53/Z0+LniL4vaf4/u/Edra2r+FPGuv8Ahu1FosiB7TSrgRQvL5jvmVl5crtUnoq9K6+P4zeDZPjHffBEu6a7YaNba1JKxiFsYrq5ltkhDb95mDQliuzG1lIYk4AB6zRWfc6tpdnd2+n3d5DBdXeRDE8irJKR12KTlse1LqWq6Xo1qb7WLyGxtlIBlnkWJAT0G5iBk0AX6KzYdZ0i5iknt76CWOLbvZZVZV3cLkg4Ge3rU0Wo6fcXU9jBcxSXNrt82JXUvHuGV3qDlcjpnrQBcoqlYalp2qwG60u6ivIQzIXhdZF3KcMuVJGQeo7UyLVtLnv5tKgvIZL23UNJAsimVFPQsgO4A9iRQBoUVnXmr6Tp0M1zqF7BbQ2xAleWRUWMtjAYsQFzkYzXnnxf+L/hT4K+BJfiD4qEtxp6XNharHa+W00j6hdRWkZQSOilVaYO53cIGIycAgHqlFZ0msaRDYPqst9AllGCWnaVREoHBy5O0fnXknx++J2ofDD4CeNviz4RW01G88O6PdajZifdLayvDGXTf5TozISOdrg+hoA9srjbTwD4YtPHN98RxbvN4gvrRLA3E00kohtEIfyYEZikKO4DyCNR5jAFydq4v+DtYufEPhHRNfvFRLjU7G2uZFjBCB5oldgoJJwCeMknHetL+2NJ/tL+xvtsH9obd/2fzF87Z/e8vO7HvigDRoqjd6npthHPNfXcNvHbKHlaSRUEaHgMxJG0HHU0y81jSdP+z/b72C2+1sEh82RU81j0VNxG4+woA0aKq3t9ZabayX2o3EdrbxDLyyuERR6lmIA/Gn211bXtvHd2cqTwSgMkkbBkZT0IYZBH0oA57xp4Q0Xx94Yv/B/iITtpeqKsdylvcS2zyxBgzRGSFkcJIBskAYbkLKeCa3rCxstLsbfTNNgS1tLSNIYYYlCRxxxgKiKo4CqAAAOgqaVnSJ2iTzHAJVc43HsM9s18B/FD9rT4/fB/wynivxx8BBb2k93bWFvHD4qs7i5ury8kEUEEEEduzyyOx4VQTgEngEgA/QCivB7f41zD436B8DtU0BrPU9Z8KzeJZpxcrIls0FxFbPa7VT5yGlz5gYA7fu817xQAUVlz65olrM1vdahbwyp95HlRWH1BOakvdX0rTLMajqN7BaWp24mlkVIzu+78zEDntzzQBoUVA9zbRW/2uSVFgADeYWATB6Hd0xUCanpsmoSaTHdwtfRIJHgEimVUPAYpncFPrjFAF6is6+1jSdLeCLU72C0e6bZEs0ixmR/7qBiNx9hWjQAUV5x8O/EPjbVtEvb34k6Zp+hXsepXVtbR2V99siktEl2W8jSFU2yyD70eMg+hO0dzb6ppt3dXFjaXcM1zaECaJJFZ4i3I3qDlc9s0AXqKz7vVtKsLm3s769ht7i7bbDHJIqPKw7IpILH2FLqGq6XpEaTareQ2UcjiNWnkWMM7dFBYgEnsKAL9FRSTwxFBLIqGVtqbiBubBOBnqcAnFeSeNfjj4E8DeIvBvhrUrn7VceN9Um0m1e2eF4oJ7e3luJGuGaRSqAQlDtDEOQCOpAB4ToX7JXi/wn4PuPhJ4T+MGtaX8OJVuIItMWxsJb+1s7p3eW2g1KSJpAn7xlRnjeRFwFcEA19Y+DfCPh/wB4S0bwP4TtRY6NoFpBY2cAJby4LdBHGu5iWYhQMkkknknNaN7rei6baxX2o39va20xURyyyoiOX+6FZiAc9sda0wQQCDkGgBaKKKAPJPhF8IdG+D9p4qtNFvbi+XxZ4j1XxJObjZmK41WXzZIk2Afu0PC5ycdSa9boooAKKKKACiiigAooooAKKKKAPJPhF8IdG+D9p4qtNFvbi+XxZ4j1XxJObjZmK41WXzZIk2Afu0PC5ycdSa9boooAKKKKACiiigDjfC3gHwz4P1HXtZ0a3f+0fE1417qF1PNJPNPJjbGu+RmKxQp8kUS4SNRhVGTnsqKKACiiigArjdd8A+GfE3ibw/4s1y3e7vfDDTy6ejTSfZ4p50EZnMAby2mRNyRyMpZFdwpG412VFABRRRQAUUUUAFFFFABRRRQAUUUUAFFFFABRRRQAUUUUAFFFFABRRRQAUUUUAFcbH4B8Mp48l+JUlu8/iB7H+zY55ZpJFt7QuJJI4ImYxxea6o0pRQZCibydi47KigAooooAK43SPAPhnRfF+uePLS3d9d8QrbxXNzNNJMywWq7YoIQ7MIYVJaTy4wqmR3cgsxNdlRQAUUUUAFFFFABRRRQAUUUUAUNVtbq+0y8srK8fT7i4hkjiuY1R3gd1IWRVcMhZCdwDAgkcgivH/g18DtJ+ETeINZm1vUPFnirxbcRXOsa3qrRG7u2t08qCMLBHFFFDCmRFGiALk9c17fRQAUUUUAFFFFAH//1/38ooooA/PX/gov4G8FeKfhx4D1XxVotnqU1h408P2iTXUKSGK01C/hju4wzD5Y5lRVkHRgAD0qT4ieDfCHw5/at/ZU8JeCdItPD+g2SeOo7azs4lgt4nn0+KZgiIAoLsXc4HJya+zPiV8NPBHxe8Gah8PviJpaavoOqBPPt2d4zuicSRukkTJIjo6hlZGDAjg15X4g/ZR+CvivwBoXw48Tadf6np3hm7a+025uNW1B9Stbl2Zmkj1Az/agTuIx5m3GABhVwAeD+Gbq1uf2v/2lEtpkla38I+G45AjBijiC+ba2OhwQcHnBFfHek/Az4I6P/wAEw7L40y6Pb6f450/w3Fq9j4jDt/asGqQP/oawXbMZY080JEsKMEAOAua/WHwF+zx8G/hhcatd+BPDcelT69ZQ6ffyLPcSPdQQNM6ea0kjF5S08haY/vXLfO7YXHm2jfsO/sweH7zRJ9K8HGK08PPDNZ6c+o38umJcW4AjuHsZLhraScYyZZI2dm+dyz/NQB8ofEqbx/8AFD9pzwZ4J8ZfD/SfiHBpnw9s9eh8Pa9qQ07Tjqt3ctDfXhgazuo7mWAKkSo6fug5cYL1xHxa+FnxN+G37Kvivwt4otIPBmman8RNAvfDlho+om+/sS0u9Qs90NtO0EARYrgSSwqIwE3YAwK/T34qfAz4ZfGZNMk8e6U9xfaG7y6df2l1cafqFk8q7XMF3aSRToHAG5Q+1sDcDgVkR/s3/B5fAC/DK40ee88P/wBowau8d1qN9cTyX9tKk0c8l1LO1w7CSNDhpCpxgjbxQB8f/H74SfDXwNq/wP8A2e9A0mLwz8MviB4puF8Sx2rGD+1ri1sHltLa9nBEkxvJo1WTexaTaASa6P8Aa9+Dfwp+Ev7HPxrPww8K6d4VGsaRG13FptulrFKYHVY2MUeEBAY5IUE9ya+0viT8MPAXxf8AClx4J+JGjQ65o1yySGGUsjJLGcpLFLGVkikU/dkjZWHYivONA/Za+Cfh7wn4p8FxaLcajp/jW2Wz1l9S1G+1C6vLdFdUia5uZ5JlSMO2wI6hSSy4Y5oA+Ofjkvibxz+1D8O/hjd+CdM+InhzT/A765ZaFrepjTtNudTN0tvNO6Na3SXcttAE2ROhCCVpBgivRPhh+zPqt58M/in8KPjPo9t4M+HfizVYNQ0nRdF1l5U0qHbDJcQw3KwWvlQtdQ+asSoEAdlxgkn6S8V/s3/CPxv4P8NeDPFOl3F9b+Do0TSL37ddxapZFIxFvi1CKVLoMyqA58358DfuwK88+KH7MPhvUv2ePFfwI+H2lR3MHjFx9pl1rU7+6KTyNHm/lnlkmuZpbfykkSLzFVzGqFkQkgA+or7RNM1PRLjw5qMP2nTru3e0mikJcSQyIY2ViSSdykgknJr8f/h3LqHjDRvhB+xTrMzXV78PPG+pw67u583RPBbC708yJ/zyuftOnqpPDYOOhA/X/QNHg8PaFpugWsjzQ6ZbQ2qPK26RlhQIC7HqxAyT3NcBo3wR+Fvh74r678cNH0GO28b+JrSKx1HUhLMWnt4RGETymcxLxDHkogZto3E4oA+I9Xg+Nk/7enxFX4LXvh6yuv8AhDvD/wBsbxBbXd1H5f2i62eStpPAdwOc7mwRjGK+qPgT8G7z4UXXinxH428Sp4o8d/EO/XUtWvUt1soXNtAlvBb2tvvkZYbeJQq7nZjkljzgeo2Pw48GaZ8QNV+KVlp3l+J9bsrbTry786U+ba2jO8MflFzEu1pGO5UDHPJOBXlXxH+DE3xC+O/wv+JM1rbW1t8N21C8S/FxML2aS/t3tTZJCoEQgOVmkkdmYlFjVQrSGgDz39tTTrjw74Q8J/tCaREz6l8HNdttbm8sZlk0aY/ZNXhHoGtZWkb/AK5j6H5+8e+KvE2uT/tPftQ+A5TLceBPDsvgzwvdQ/MYzYw/bdVuo+oJW5lUBh1+z4PTA/TrXtC0nxRoeo+GtftlvdM1a2mtLqB87ZYJ0MciNjBwysQcHvXI/Dj4T/D34SeBLX4Z/D/R00zw1Z+fstGkluQftMjSzeZJcPJJIXd2JLs3XHQAUAfmp8OfgJ4/ng+D3jX4U/Cvwz4Om0fUdK1W88WWXiM3upazpFwo/tBbtxpsUl39shkaT95M2JNpXGK6P4R/s+/CP40+O/2lLn4p+H4vEpt/Gt/aWsd6zyw2Yk0+0aSW2iLbIZ3+UNMgEhCIN2FAr678H/slfArwD4ks/EvhLRrzT20yd7qysU1bUW0u0nk3bnt9Oa4NpGTuOAsQVf4QK9Y8KfDbwV4IvfE2oeF9O+xXHjDUH1XVW86WT7ReSRpC0uJHYJlI1G2PavGcZJJAPxtv/Dnhn4m/sW/sh+JfiLpdrr19/wAJh4W0aW5volmc6ZNfPbzW7u+T5MkcUayKThgozX09+0b8G/h/e/HL9mD4L2GmJovgu4n8WxTaZpv+iQSWsenLdSWpWLH7id02zIMB0ZlPDGvri7/Zp+Cl78GrT9n+fw4D4F09UFrY/abnfbtFKZo3iuTL9oV0kO5XEm4dM44rT0b4DfDTRbjwRfpZXd9f/Do6g2iXd9qN7eXMDaojR3bSSzzO05kRio87eEGAgXAwAfJHxC8BfBrxN8V/+FJfDb4HeHPGeteCtDthcnW5VsdA0Ox1CWaSCKGBYLom4lYSOTDbKcbd0wwoHyx4Zv8AVZ/2I/hXo+qTi4/4R74uabpVuUleZEtrPxA8cMaSSfO0ca4SMtzsCjtX6jfEH9mb4MfE/wAXJ468XaJM+t/Zlsp7iz1C9083dohLLb3a2k0K3EQJztlDDt93io7j9l/4Gy/C65+DNr4ZWw8I3F+2qLZ2dzcW5t71rj7UJraWORZICk3zIsTKqD5VUJ8tAEX7Wv8Ayav8Y/8AsTfEH/pvmr88vi38G/hZ8L/2PPh38bPh3okGmfFa1l8I3ulazEzNq2oanfzWqTQy3LFprhJoZJQYnZkCDAXauK/SX9onwvrXiz9nb4l+C/C9q+oatrHhfWLCytww8ye4uLKWKJNzkDc7sBlj1PJrxv4IfsdfBvwbo3gHxdqvhaSPxV4f06xlS2ur+7ubLT9T+zoLia3sZJ3tIZvMyd8UYw3zKQckgHnPxb8N/Dv45fGrxp4L8O/B/QfH/iPwvY2Njr2ueKr1rew05b2BriCCzRYLuUzeU29nhjhAJG6XcAB83+Btf1nxL+yl+xXqevXb314nxI0u1M0jF2aOyGrW0QLHk7Y41Gfav0j8ZfstfA3x942u/iD4n8PPNrGqRQw6iYb68tbfUordQsSX9tBNHBdqijaFnRxt+UgrxVm4/Zm+C03w30H4TQaA1l4Z8Lagmq6VBbXl1DLY3yTSTrNDcJKJlYPLJgb8AMVA24AAPnnw/wD8ne/tIf8AYn+Gv/RN/Xyt4Vgg8Xfs6fsY/BvxXM8XgTxzcmLXI1kMUd79hs5rmzsZmUgtHcTgBkz8+0Cv1auPg94AuPiLe/FVrCRPEeqaV/Yl5NHczpFc2IcuiSwK4iZkJbZIV3qGIDYOK5vU/wBmz4J6z8JNM+BmpeGY5/BeiCEWFm1xceZaNbkmKSG68z7SkqZOJFlD4JGcEigD5O8P/C34ZfCr/goX4c0v4ZeH7HwzbX/w91W4uLPTo1t7fzBqNqgkFvHiNGZVwWVQW285Irxnwde2dn/wSX8epeTpAYrTxlbv5jBds0mrXqpGc/xMWUAdSSAOSK/Qz4efsxfBT4XeKYfHfhDQZI/E0dpPZPqt1fXl9ez29w0TyJPNczSNLzCmzfnywCI9oZgeOvv2Iv2Y9U1fxJrGo+DRcf8ACWPdS39q19e/YWnvkaO4uI7QTiCGd1c4mjRZEPzIytzQB81/FDwP4b+I/wAa/wBknwd4xtmv9Dv/AAz4lN3Z+Y8cN0kVhpsqxThCvmQl1UvG2UfGGBXIOX8IP2ZPgdrn7S37Qfwv1rwrbX3gjw1L4evNK8Pyl20mxuta01mvbiCz3eUk0hiXbIF3RjIjKAkV+iM3wm+H9x4k8H+LptK3at4Ctbqy0Sbz5x9kt72KOGdNgk2Sb0iQZlVyMZUgkk3tC+HPgzw14z8T/ELRNP8As3iDxmLEatc+dK/2kadE0Nt+7dzHH5cbEfu1XdnLZPNAHwl+yb8O/DHxh/Ys+HHhv4i3t5JZ+Gdcu3spI72S2l36LrV1DYxNIrAsqoiRhM8gLjDBSPtT41/DWw+MXwj8X/C7Udoi8TaZdWSuwyIpZYyIpfrHJtce4rwv4tfspeEvEnwW0T4B/D/RLbTfDkWvQ6k0s97dmXTB9qkvri6tiWeSad5HdER5FRTLu+6gQ/YlAH5V/A3x3f8A7TXxG+AsGt7nm+FXhe813xFG/wA2PEZkfQYY5RztkR4L2ZQeQMH0J90/a5/5KZ+zb/2UCH/033dfRfw9+CPwt+FXiDxd4p+H+gx6Rqnju9/tHWpklmkN3dbpH3lZXdYxulc7Ywq5YnGTW74u+G/gvx3qfhrWPFenfbrzwhqA1XS386WL7PeLG8QkxG6h/kdhtkDLznGQDQB8Y/tbeJvF3iH40/CD4Q+APC8njW60a7k8davpkN5BYsbXSSINPLzXJEYQ3swfBySYeB3EP7HvibxT4a+LXxh+DXj/AMNv4Lu7y/XxxpWmTXcF4UsdbZkvPLlt2MZjS9idsDG0y4Nfalj8N/BenfEHVPinaadjxTrNjb6bc3rTSuWs7VneKFI3cxxqHdmPlqu4nLZIFcp8QvhJ4Q8Sanf/ABFXw3DrXjGHQL/Q7YT3txZw3VleDe9lO0RZPKkkA+donaPll56gHrQNpqFpkFLm1uU9njkjcfiGUg/Qivx3ll1XRPg54h/YAsriSLWZ/HMfg+wZWLTJ4R1ctrJuT/eWPTRcQHsCgHtX6Wfs9/Chfgd8GPCvwrFwt02gWpjd4zIYvMlkeZ1i81mk8pGcrHuJbYBnmtC4+CXwuuvi5bfHafQY28dWdh/ZsWpebMCtr8/y+SH8kviR1EhjMgViu7bxQB+c2o+F9Y+I37VHxZ8K6h8JfD3xO03wDa6FpOiaZ4g1ZbO20nS7mwWYyWlo9jdxs1xKXDXAw6+WsYIC1n/ET4VeOvDnwP8A2cPhF8YJnSdPipaWG2y1KaaWPSLiHVPs9r9tRYJSY7VxAWUKdo4xX6G/Ev8AZz+EvxZ1+z8W+LNMuYfENhAbWLVNL1C80m/+ys28wPcWM0MkkWckI5ZQSSoBJNarfAn4VyaB4O8MSaK0mneAdSh1jRY2u7pmtr+AShJ2kMvmTN++kyJmcMWywJAIAPjjw98Ifhl8Lv25E8AfD/w3ZaF4W8a/DW/Or6TaxCOxu5LbUYIUkkgHyF/KleNjjLKxzkkmvPP2UvhN8LvD/wACPjb4w0Lwtpth4hsdY8d6Ql7DbRpcxWMMsgjtRIBuWNVVcJnAAHHSv0vn+HPgy5+Ilp8V59P3eKrHTZtHhvPNlGyxnlSeSLyg/lHdIituKFxjAYAkV5zpn7Mnwc0Xxl4n8caRpV1ZX3jJbr+1oIdSvY7C6kvV2XEzWSzi3E0i/ekWMNnLAhiSQD4i+D3g34A/Bv8AYQ0X40/ETw9Lr954i8I6Xa6tdqzNq+oRagLaC202O5DxukAfyIYo1dEREUt0Zjm+EvBmq/Cf9sz4Jtpnw48OfCW08a6X4msbvS/D16bmW7t7G0iuojqIjtbaDzY5QCrRmU8sDIwAr9F7n4I/Cy++EkXwJv8Aw/Fd+BILGDTU0yeSWVBa223yU8x3MpMZRSrl94YBt24ZrjPBn7KnwL8CeKdG8daHoM83ibQPOFnqt9qV/qF8iXELW7xtPdTyu8QjZgkbEohJZFViTQB5Z+w//wAgX4zf9lV8Zf8ApYK8p0v4J/B3V/8Agop40l1jwbpN1PF4S0jX4TNaRMw1N9SuPMvVyM+cSq7pBzkDJr7s8EfCrwR8OdY8U634OspLCfxlqDarqafaJpIJL6QYknSGR2SJ5OsnlqocgEgkCuY8e/s9fCv4keOdF+JfiTT7qPxPoMQt7e/sNQvNOma2EnnfZ5jaTRedD5nzbJNwyT2YggH5deD/AIeeOP2h9C+Leu6t8JfD/jPxbqniPX9J/wCEh1TxB9m1XQpLGZ4bKCziOnTvZizTy5IxFMpkJ3sfmr1fx/oN7F8avg7p37SHhmX4t/YPh1Ml/olhaDWktNeimtYrrVpbN1VZI7jeYY5NpZWydoGWX7N8Z/sn/A3x34rv/GusaNd2mrawqLqb6XquoaVHqSxjaovIrG4hjuCBxmRSxGFJK8V81/tDfC77d8e9A8ReMfAXiTxD4D0nwyumaReeDLmW31Gxv3ud1wl4bS4t7ySBoUiEWGeNTvym5twAPnnx18ER8Ef+Ccfxguj4Ug8Fal4j1JtRgtY0hXULXTH1uOXTLa8ngz5k1vFIcZd/LDbAeCK9R+NX7M3wa8H/ALQnwC0LwpobaRbeNLvXdJ8RPa3M8U+uWcdg16Y9SnV/NuvMmiVpTKzGQEq5IOK9S+CP7P8Aq3i/w58UvB/xgsPEEnwl8YXGmroXhzxRq895qttDaLvuZXuIriSaCOe4EbRwm4LqEJYLvKn7N8R/DnwZ4t8T+F/GXiHT/teseDJ7i50mfzZY/s0t3C1vM2xHVJN0TFcSKwGcgA80AfmkbFf2dfiR+1bovwG0yPQbXT/Aen+I9P02yQLbW+qrbXymeC3GUUny0ZkVQHKgY6VzHwy+BXxE1zwX8I/HHwr+F3hrw9rFjeaNrsvjWHxIbvVtWtZSj6h9tf8As5Jbj7bC8m+N5yqsRj7or9UrH4beCtO8d6z8TLPTQniTxBZ22n310ZZWE1rZlzDGYmcxLtMjcqgJz8xOBjyPwz+yN8BvBviW18TeF9Eu9NNhd/b7XT4dW1FNJt7ssX82LTRcfY0O45AEW0HkKDQB8xeCfgh8LfjJ+1z+0uvxU0CHxTaaZe+GktbO/LTWcL3OhQCWZLcnyxOVRVE23zFAwjLk5+Vdc8K+GvE3/BNTwdP4t0+DWJfCHi5dKsLm+QTy2tgPFBszGsj5KobdVhbnBQBTxX7S+H/hz4M8LeLvFPjvQdP+y6541ktJdXuPNlf7S9jALa3Ox3ZI9kShcRqoPVsnmuM/4Z2+DR+EuofAt/DaSeB9Te4kuNOknuJA0l1ctdyOJXkMyt57GRSrgo2Nm3AwAfNXxg8FfAzSPFHgP9nrwZ8GNG8Z67Ha6lrGmaLdPHp3h/TbMyxpc3l2nlzRsXlZVQLazSZL42gsT8oeH7XVfDP7JX7ZHw2vLWx0yy8LX+sJaadpk0k+n6eLrT4Z5be0aVI2EKyMxC7EAYthRX6IXn7H3wK1LSNB0vUNO1S4m8NGcWGoHXdVXU4o7ogzQ/b1uhcmFwoHlGQoB91RXXeG/wBm74IeEPCnizwN4a8KW9hoPjlGTWbOOSbyrsPbLaOSDIdjPEoDNGVZmzISZCWIB1/w9vLbT/hV4av72QRW9totnLI56KiWyMxPsAM1+NXxV0/w7cfCDw7+0t8F/hNongPR4PE+jalpPiW5v3bxVqAudTjhMjxpDITDcK7bo571z5ZOYwQBX7W+EPB2h+B/COmeBtDSVtJ0i2S0gS6mkupPIQbVV5Zmd3wOPmY8cV86QfsNfsvw2racfBzTacsyXFtZy6lqElpYSpKs4axga4MdoTIuWMCoSCyHKMVIB4ZrPwj8C/F79vzxnpfxI09de0TTPBehXY0q6/eafPdfa7yOKa4gPyytCrSCMOCql2ON20jyiw8J+IPjT8bPjmvin4P+HvijNoGt/wDCP2aa9rYsn0fSEs4XtVsrRtPuxAJ97zfaI3R3ckDHl5P6j2fw58GWHxB1H4p2mn+X4o1awt9MurzzZT5lpau8kUflFzEu1pGO5UDHOCSAK86+If7Mvwd+J3ikeOPEelXVp4haBbWbUNJ1O+0i5ubZekNxJYTwGZB0Ak3FRwpAoA+Nj4bng8KfBr9nX4z/AA/074xfGDStIv7qO31XUM6NaafazpbC8vp5YZBO+wwRqRaSyFt+NoLM1b9l34a6T4u+H/7RH7O3xEit9G8KWHiye3Njol9MlnpsVzbW11LBZ3DrE6QrLk7SiqCWBTBIr7O8WfsvfBPxpY+GLHV9DmgbwbbGy0m5sNRvrC8trRgqtALq1ninaNlUBld2B69ea4fxx+yr8O7D4KeP/hf8IvB2m2f/AAsERJeW9zdXUNm022K3F2+wu+6BEExWPYZnT5mDu0lAH1VpthbaRptrpdoW+z2UKQxmR2kbZGoVdzsSzHA5JJJ6mvh34bg/tTfHSX46Xv7/AOGnw0uLnTfBsZ5h1PVlzDf6zjo8cRBt7RuRxJIu1q+tNH+H2j6f8MbH4VX7y6hpVto8Wiyu0skc09uluLZiZI2Eiu6DllYMCcg55r5x0/8AYB/ZY0iyh03SvDmq2Vpbrtjhh8T+II40X0VF1AAD2AoAy9b/AOUivhX/ALJpqv8A6drWvtiG4guA5t5FlEbFG2kNtZeCpx0I7ivPdP8AhH8P9L8YaR4+s9MYeINC0Y+H7O7e5uJGj0wukhhZXkZZCXjUmRw0hx9/k586/Z3+DE3wlTx7rF5a22l3fj/xLd+IZdPsria5trM3EcUW0STBS8kpjM0pCqoeQog2IpIB8m/tIfA/4O/tEfHi2+CfhzwVpD65KbXW/HviZLKI31jpiY+zWSXO3ct5f+WFU7t0durOAQRXSQfDj4f/ABY/bK8YfDj4r6JZ6x4b+HHhTQh4T8P30SzactvemZLy8S0cGN3jaOOAOVOxcAYOK9x1j9ir9nXXPFWu+Nb3Q9Ti1nxLdNe6jPa+ItbsxcXDfxNHb30cYwOFVVCqMKoAAFdd44/Zk+DXxCs/D1v4j0i5F14UtVsdN1C01K+s9TgtVUIYTf286XMqMo+YSyPuOWOWO6gD80PGQHhz9lf9sP4V+GZ5J/AvgXVGs9ADSGVLJZobae7sIWYkiO1ndlVMnZuxXsvxU+C/w2+DXjn9m74i/DrR00rxTqvjSy0rUtVDySX2pW2qafdNdfbbh2Mly8jIGLSljnOCBxX29H+zx8GYPhDefAe28Mw2/gbUInhudPilniMwkYO7SXCSCdpHYZaQyF2PVjXV+Kfhj4H8anww3ibTfth8G6jBq2k/vpo/s17bRvFFL+7dfM2pI42yblOclSQCAD87fjh4Z+Hnx18Q/GL/AIQz4QaB4nvvDUEul694s8S3zQfZLy2sVkMOlxpBdTK9vGyMzJ9lTzMncxLMfr/9j7xFrHiz9lr4V+IfEFy97qN54d08zzyMWkldYVTe7HlmbGWJ5JyTUniT9k74B+LvGWpeOtf8Mm41DW2STUoFvbyLT9QkiUIj3lhHMtrcMqjGZYmz1OTg17D4G8EeGPht4P0jwF4Ksv7O0LQrdLSytvMkm8qGMYVfMlZ5GwO7MT6mgD8a9Ytzd/sSeKrVZ5rYzfGOZBLbyNDNHu8UIN8ciEMjr1Vgcg4I5r6a174QfDX4Fftl/s/t8I9At/C//CWWnizT9X+xhk/tCG0sI7qE3JJJmkWYb/Mcl2JyzHAx9bv+zl8GpPBdz8PH8PZ8P3esnxDLbfa7v5tUN0L0z+Z53mD/AEgB9gbZ/Dt28V2+vfDnwZ4m8ZeF/iBren/adf8ABhvTpNz50qfZjqMIguf3aOscm+Mbf3itt6rg80Afn9+zX8G/hN+0RYfEz4k/Hbw9ZeMvGlz4v1vSrttUQTzaPbabcGK0sbQnm1WKEK4aLY7F9xY8Y5PXtE8RfFj9rX4w6Nr/AMKtA+Ktv4Lh0XTtJsPEWrrZwaZp97YrcPNa2j2N3Gz3MxffcDDr5YjBAWvs/wAbfsifAH4g+LNQ8aeIfDsyanrQRdU+w6jfafBqaxjCi+t7SeKG5wOCZUYsPlYleK3/AIifs2fB/wCJ+saf4k8R6TPa63pdt9it9R0m/vNIvVs87vszXFhNBI8Oc4jcsqkkqASTQB+ePxM+E/xA0D4Efs5/B34s3klndn4pWunBtP1CS4nh0e5h1QQWy3uyJyyWjiDeFVgoyMHp237Rv7Nn7PmifGP9m7w5p/w90K00e+13UNNubdbGERT2y6fc3EcEg24dfPYyANnLkt945r7yb4E/CuTQPB3hiTRWk07wDqUOsaLG13dM1tfwCUJO0hl8yZv30mRMzhi2WBIBFr4tfBj4d/G7QbTw78RdOe9g067jv7OWC5ns7q0u4gQk0FxbPHLG4DEZVhkHBzQB8b/EbwV8J/iJ8W9S+EXgH4KeHfG+s+AND0vTr+78RXP2TRtDsbqOR7G1tIFgu380xKWzBBHhQgaYbVA7z/gnpr19e/sgeB38RX4nlsJtU0yOaSUuDFZ6pc2ttGrvywWNEjTPJAHevRtY/ZC+AevanZ6zquh3c17bWFvpk0o1bUkbULO0XbDDqJS5H25VHH+k+YSOGJHFbc37L3wLl+HejfChfC6Q+E/D+rDXLCwiuLmOO3v1uZLvzFZZQ+3zZXPl7vLAO0KFAAAPfaKKKACiiigAooooAKKKKACiiigAooooAKKKKACiiigAooooAKKKKACiiigAooooAKKKKACiiigAooooAKKKKACiiigAooooAKKKKACiiigAooooAKKKKACiiigAooooAKKKKACiiigAooooAKKKKACiiigAooooAKKKKACiiigAooooA//Q/fyiiigAooooAKKKKACiiigAooooAKKKKACiiigAooooAKKKKACiiigAooooAKKKKACiiigAooooAKKKKACiiigAooooAKKKKACiiigAooooAKKKKACiiigAooooAKKKKACiiigAooooAKKKKACiiigAooooAKKKKACiiigAooooAKKKKACiiigAooooAKKKKACiiigAooooAKKKKACiiigAooooAKKKKACiiigAooooAKKKKACiiigAooooAKKKKACiiigAooooAKKKKACiiigAooooAKKKKACiiigAooooAKKKKACiiigAooooAKKKKACiiigAooooAKKKKACiiigAooooAKKKKACiiigAooooAKKKKACiiigAooooAKKKKACiiigAooooAKKKKAP/2Q==)
Reward & Aversion Networks
![[[@Watabe-Uchida2017-gi]](https://doi.org/10.1146/annurev-neuro-072116-031109)](data:image/jpeg; charset=UTF-8;base64,/9j/4AAQSkZJRgABAQECWAJYAAD//gAuSGFuZG1hZGUgU29mdHdhcmUsIEluYy4gSW1hZ2UgQWxjaGVteSB2MS4xNAr/2wCEAAMCAgICAgMCAgIDAwMDBAcFBAQEBAkGBwUHCgkLCwoJCgoLDRAOCwwPDAoKDhMODxEREhISCw4UFhQSFRASEhIBAwMDBAQECAUFCBIMCgwSEhISEhISEhISEhISEhISEhISEhISEhISEhISEhISEhISEhISEhISEhISEhISEhISEv/CABEIB20OGQMBIQACEQEDEQH/xAGiAAABBQEBAQEBAQAAAAAAAAAAAQIDBAUGBwgJCgsBAAMBAQEBAQEBAQEAAAAAAAABAgMEBQYHCAkKCxAAAgEDAwIEAwUFBAQAAAF9AQIDAAQRBRIhMUEGE1FhByJxFDKBkaEII0KxwRVS0fAkM2JyggkKFhcYGRolJicoKSo0NTY3ODk6Q0RFRkdISUpTVFVWV1hZWmNkZWZnaGlqc3R1dnd4eXqDhIWGh4iJipKTlJWWl5iZmqKjpKWmp6ipqrKztLW2t7i5usLDxMXGx8jJytLT1NXW19jZ2uHi4+Tl5ufo6erx8vP09fb3+Pn6EQACAQIEBAMEBwUEBAABAncAAQIDEQQFITEGEkFRB2FxEyIygQgUQpGhscEJIzNS8BVictEKFiQ04SXxFxgZGiYnKCkqNTY3ODk6Q0RFRkdISUpTVFVWV1hZWmNkZWZnaGlqc3R1dnd4eXqCg4SFhoeIiYqSk5SVlpeYmZqio6Slpqeoqaqys7S1tre4ubrCw8TFxsfIycrS09TV1tfY2dri4+Tl5ufo6ery8/T19vf4+fr/2gAMAwEAAhEDEQAAAP1VAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADn+PDlOSDlOXDmecDn8IOXxgyuaCty4HKAZQZQBy4AAAAAAB1AAAAS9KAAADw6gAAOoB4D+oAA6sDT2AsdEFvoA1tUOm1Q2+oDoOgDpt0AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAACl54Hnnn4eecIHn/Ehw/CBxnHhS20dV1CfUdLL6PoJe90MvpNRGz0Eux1CZ0qH6I9cQdEiUGAACrQsQucGAABX58AaACIOcUAQvLADUHPAMZzQ1lZbKeGKliMyM2lzWUzG5lrB56lRlZ1HVh1vXB2HYB2fbB3fah6B3wdxYAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPP/KQ8q8sDy7zQPNPNA4XWR6L28v0HtM677ss32XczXX9an1t+WAAAA4AAAAAAAAAAABWKrAAAAAAAAFAUQogAAAAAAAAAAAAAAAAAAKHNtcvy7XL8ja5Tj6XJcPa4rjaXn3FXMHds9I9ID1b2YPbvcw70AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAACv5UHjfjgeP+Qh5J5IEPpUv0/03OvSvRsn6L6XnXoHSzQ4FJAAKAYAAAAqgAAAAAAAAAACqxRgAAAAAAKIVUAAAAAAAAAAAAAAAAAAAAAAAAAAAAACDERnG8HS8787ufLfGLnxjx/Wev+gGvob6JD6L9YAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAqeah5z5yHKc+GVkho7QbfVh3HoYeobYAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAFQPMvJA8k8kDxbxQPH9JHrfredeueq436v6rlp6BLAAgAGCscSAAMCgAAABVAAAAAAAAAAAFWhQAAAAABQFWQAAAAAAAAAAAAAAAEoAgAMAAAAAAAAAAAAAAERgjAAps8a8HufBPnrWPAeg1j6Y+ng+pvosLYAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAZQfNnzMHzt4OHkOgj0fuM66zoorR1ZM7JDE5e1xHB6Tx/otr2b3gPoH6WD2uUAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAKXkgeM+Ph494+Hj/lwafqcv1T1PKvXvYMa9f77KxUPUoAAkAAAoVyAQDAoAAAAHAAAAAAAAAACgOoAAAAABQUQogAAAAAAAAAUEK0ACtAAAAAAAAIACaCBBgMAAAAAAAAQBGIMAACDw6l84fNGsfMPM7Z/Uf1yz6792AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPH/AI7D40+Vg9D+j8b+gfbufT2TtcLBMAAGkg0EKMTyi58c8T3jwP596M5fqbSfr77GD1MAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAKQeDfOQfN3zMHjW8j2D1rK/WfV8r9Y9WyrtCAAAABXjUKAgAAAYOYpIABQDAAAFBQAAAAAAAABQVaAAAAAAUFWQAAAAAAABQSjSjQAAAAAAAAAAAAAAAAAACAImImAwAAAAAAQYiMAAAAPJWvln5P3j5X6nXP7L+3GfVdgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA4oPzg+BA8++n86+vfrDj29JTG2gAAAAACJIANSgAACIrxj5g3z+VPmHrw9y+5NJ+8fUAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAMzz8OSALodh3YWAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADl/BA+fvAg+e/Cg6/33Kvob6H5tfeOuysVNwwCQEAAAoPUoBAIAAAocsgADAoAAAAcAAAAAAAAAKCrQAAAAAKqFEAAAAAAAqtCuQAAAAAAAAAAAAAAAAAAAAAAAAAAQTREwGAAAAACAiUAAAAAFb5oa+PPjfoyw/t7Wfvz6YAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADKD8xvzID6X/Qjk2+n7nLoCYAjQAAAAABqAQjWAwAEG0GYrPlL4k6sPlD6l6sf0Z+0WWAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPn/wCQQ+Yfn4PMtpHWvllAOShpem+0s97+jg+o/QAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABnkQfP3gQeB+BB5h67L9x91w094915tO7M6BgUK5ANASAAAArxqDAkAAAoVyAQDAoAAAAVQAAAAAAAAFBVoAAAAAHKgEAAAAACqIWpAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAQTREwGAAAAAIjEGAAAAIiPn34e1j4e7Tpy/Rf9A2dGAAAAAAAAAAAAAAAAAAAAAAAAAAAAB5J+EAbv6vcuv0Ic2wQAAANBAAAAAABpIIDSgAAGoUAAHJfD+mX5/J6GH6b/oNS2AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADlA/PT87g4T6wyr6V9449fXuhwsBgBx7XmHjW0+HfPHTn4F7B0Z/X32gH1TYAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA8c+bw+dvnQPnxEe5e6437n7lz6e2bGNgmAAAUADxgEgIAABQepQCAQADBWOJAAKAYAAA4AAAAAAAAABXUAAAACgqyAAAAAAolGlGgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAEARNBUAAAAAIjEGAAAANRFb5Ba/O7wPsx/Rb9N6XagAAAAAAAAAAAAAAAAAAAAAAAAABwQfzv/Zudfqm/z+lBgSAgAAGoAAAAAANQCGpQAAAwYDAAmYvknSfza879Hn/Vn9BrVsAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAKQfmh+XIe+/oJx6/V9zj1AYAAAAAAAGP833Pyb8g92OX9w7x+gf0cAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABUD50+Vw+XvmYMn3+K9/wDoHm0939Y59HEsAeAAAAMCgchRoCQAAAFeNQYEgAAFCuQCAYFAAAADgAAAAAAAAFWhQAAAFBVkAAAAAFVyK0AAAAAAAAAAAAAwABgAAAAAAAAACAQAAAAAAAAAAAAg0SWAwAAAERiDAAAAGog8FF+YXy/2Y/qJ+mlq6AAAAAAAAAAAAAAAAAAAAAAAAAfhgGh+w/FsJjq0QABIAAA0EAAAAAEBCQYwGAAg2gwAEgSPku5/LqX0+f8AYn6xpAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHg/wCIQWv1a49fos49gAAAAAAAAAAAADwD4f6c/iX23uw/VH7BYAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAeZfGYfH3yoGl9N419M/SPLr7TPjoAAAAOBQAAACgAcrAJAQAACg9SgEAgAGDmKSAAUAwAAFUAAAAAAAAFVijAAAFBVkAAAAAFVyo0AAAAAAAAAAMBgMAAAAAAAAAAAAAAAABAACAQAAAAAAAAgCTSAwAAAERiDAAAEBEkPnGl+RsfZj+3/ALnSAAAAAAAAAAAAAAAAAAAAAAAA8J/nqR/RT1XndAKkamDAkBAAIDQAAAAABpIIDSgAAGoUAAIJQBB8Gax+Xv2z6XP+v3VsAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA+HA/Fn9IML/RyfzugAAAAAAAAAAAAAAADH+DNo/Ob1X0ef9dfqBgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAeTh8LfDYeEfTsV9Y/VXDr7IYaOKAAAAABXAAAAAFg5CjQEgAAAK8agwJAAAKHKgEAwKAAAHAAAAAAAAAKxwwAABQVZAAAABRKtIBAAAAAAAAAMCgAAAAAAAAAAAAAAAAAAAAAAAAEAgEAAAAAAAAiJoKgAAAERiDAAAEBpJW/MbSfzp/cvsx+uwAAAAAAAAAAAAAAAAAAAAAAA/KHyPN/sEcG4DEKaIAAkAABGgAAAAADUAhqUAAAMGAwAJkAAOG/JHpy8P/AG978fqAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA/Ob8lEfsx9TeX0hFAAAAAAAAAAAAAAAAAGV+fu0fm19gejz/r76KwAAAAAAAAAAAAAAAAAAAAAAAAAAMr4bD88fm9H1l9ncuv1vu8uryGKClAAAAAoOAAAAGBQrkA5AQAACg9SgEAgAGDmKSAAWAAAAOAAAAAAAAUFWgAAAByyAAAACiVakAAAAAAAABgUAAAAAAACFABCgAAAAAIAAxAAYAAAAAAAIBAIAAAAAAERNBUAAAAiMQYAAA0EJPjP8durL98/pDaAAAAAAAAAAAAAAAAAAAAAAA/BD7Qxv7nOLUAApGpgwIAAAGggAAAAAgISDGAwAEG0GAApBAAAfBn5V+hz/q/+l3RAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB8Mh+L/7ecuv0IcOwAAAAAAAAAAAAAAAAAAAHG/l10Z/HP66+hh+hbwAAAAAAAAAAAAAAAAAAAAAAAA4r80w/Ov0GX98favBv0hlY5CiBzAYAAAAPAAAAAKBQcMAgAAABXjUGBIAABQ5UAgKAYAAA4AAAAAAAAFdQAAAA5ZAAAABRKtSAAAAAAADAYDAAAAASglBAAAAAAAAAAAAAAAACDEGAwAAAAABASAAAAAACImgqAAAARtAAAACNQfnvU/nn/Rv242wAAAAAAAAAAAAAAAAAAAAAD+cr9XcL+oDj1AAGNVIMCQEAAA1AAAAAAGkggNKAAAahQACQJAAAeGUvxY+pPU5v2c6CkAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHzZ+BIftF9Q+V0hFAAAAAAAAAAAAAAAAAAAAAHzH+Qfbj75+1vXl2oAAAAAAAAAAAAAAAAAAAAABxn5fB+cf1VnX6L/RnB0BDAFcgEOKAAAAABygAAAMCgegGgJAAAAe4oAAkABg5ikgMCgAABQUAAAAAAAFWhQAAAVUKIAAAUSrUgAAAAAABQDAAAAFEKIBGYzy7zjWeI5LRYGPaqgDw19hG/wBFL6jqofZd3m94hgAAAAAAACDQKAAAAAAJAQAAAAAgm0VAAAANSgAABARkn89f699+H2SNAAAAAAAAAAAAAAAAAAAAAH84H6989/RhyagAABbUQABIAACNAAAAAAGoBDUoAAAYUAAATIAAAHJfjp05ecft134/RYAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABwgfzq/pLhf30ed0AAAAAAAAAAAAAAAAAAAAAAAYAvyW+a/U5/3K+mNJAAAAAAAAAAAAAAAAAAAAqfnIH5S/TWdfp37N5/U4lAAA5ZBQUoAAAAFBwAAABQAOVgEgIAAB4OCwIAAAKHKgEBQDAAAVQAAAAAAAVjhgAADkKIAAAUSrUgAAAAAABQDAAAUSggEVGeUeJaz4r5J0z2vpEnd9jlXQbEO4IIGZmG1g87RyvJ6Lj+OudP0hP0z1fGvXO6xoJAAAAAAAEGgUAAAABICAAAAAERNBUAAAgNKAAAGh+QvrfXj+phcgAAAAAAAAAAAAAAAAAAAAH89H6U4X9bHHqAAAFI1MGBICAAQGgABmVPH53fhLpybu5w7qyaBgAINoMACZAAAAGh+dv5lejz/pB+mm8d2AAeZeWBU0w9j7UAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABn4NhvfsH524YaAAAAAAAAAAAAAAAAAAAAAAAAHwbrP5V/sN6XN+gwwAAAAAAAAAAAAAAAAAPBA/FXKl/rJ9Ied1j4AEADgUkHMBgAAACuAAAAALByFGgJAAAAc8YDAkABg5ikgMCgAABQUAAAAAAFBVoAAAUHEgAACiVakAAAAAAYFAAAAKpICODo8n8n2ny3yzeeq9rg9p9u576UzYAAAAAAAAARM4rza15d5PtPkO1rPuX0FhfudnCgQAAAAAACIUAwAAAQEgAAAACImgqAAAaxBgAAfjD9MdmH6YlIAAAAAAAAAAAAAAAAAAAAA/ED3bKv0ROHYAAAGIU0QABIAADUAAy/J+7niOvEALsvses83qcZWAADUKAASBIAAAA+Tbn8UbHq83sncAePhw/qUO9jp+YetaT9t/oeHroAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfnt+VUv9+93yekBgAAAAAAAAAAAAAAAAAAAAAAAAHzp+Kvdj+j/6s9WTwAAAAAAAAAAAAAAAAPzb/JkP0d/R7z+qyY25RAIAVyAQ4oAAAAAHKAAAAwKFcgHICAAAcDwoBAIAChyoBAUAwAAHAAAAAAAAq0KAAADlkAAAActSAgAAAAGBQAACiUJqeWUePeQdEeR0NZ9Z9Wzr1b1nCurM2AAAADAAABgAAAAIAIfI6PBfnbpjH+nhfW/UcmgIAAAAAAAQaBQAAAICQAAAABEVIJgAAjWAw5dn85f9BnZz+9jAAAAAAAAAAAAAAAAAAAAAD8wfnKH+yZ5+4AAAAFNRMGBAAAAwADzvkvU5A1gAAC9L9A3vM6wzoARpQAABMgAAAH4u9Gfpf6XI8O5lM7cPoCzjQDzPmy5+JfjD0MPtv9Z9Z9TAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAOPD+bL9kebT6mODcAAAAAAAAAAAAAAAAAAAAAAAAAADylr8RPsP1eb9kX0gAAAAAAAAAAAAAAyvxaD5g/Znl296OPcUHBIAAOWQUFKAAAABQcAAAAUCg4YBAAAACvGoMCQAGDmKSAwKAAAHAAAAAAAArHDAAAVyAQAACq5UaAAAAAYFAAArCeH8t0nybyXefLuq0n172DGvXvR8LlJAAABgMBgAAAMAAAAAAQAAAjyD5P3nwX6yufsy3yaAgAAAAAAABEKAYAAgJAAAAAEbNAMAAGpQfjNrFH90unIAAAAAAAAAAAAAAAAAAAAAAPmz8JUf0aL5vQAAAABQ1NBgSAgAEBoHm3L+pxhrIAAAC9fjfdO83qBMaNAYAEyAAABx/863Zj/RF2/JqCYAAAAAcG1+aHxL6XP8Aqb+les6YAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAH5AB4t+0Xn7hz6AAAAAAAAAAAAAAAAAAAAAAAAAAAAeffiL15fVv7I9uLwAAAAAAAAAAAADlw/B2KH+0HU+b1gmOBQkAFBxIOYDAAAAFcAAAAAWDkKNASAAAoPUoBAIAChXIBAUAwAAHAAAAAAAArqAAAHApIAACq5UaAAAAAYFABhM8X8S3z8t873iX1JHqnq+FevdVjQIAAAGBQAAMBgADAAAAAAAAAAEAgDzv4N2jK/SlHRGFAgAAAAAAABEKAYAAgJAAAAEBEmgGAEYflX8VduP8AQ73FyAFQOK4wOfxwADpQ3utDtpQAAAAAAAAAAAAi/mSD9sforzuhCQAAAGIU0QABIAADUDD8s7+Zp1ZAAAAAG76TwdExy7ADAoABIEgAA+P/AM3O3H94zi2AAAAAAAAPAKX5N8n6fN+v/wBrWgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA88/myD95fZfJ6glgAAAAAAAAAAAAAAAAAAAAAAAAAAAHGtfhZ9ierzfsOUgAAAAAAAAAAAOX/A0O8/Yrz97xjsAK4QCAB6AQq0AAAAADlAAAAYFDlACQEAAA8bgYEgAALQ4kAAsAAAcAAAAAAADmKMAAByyAAAOEpUgAAAAMCjGZ474ztHi3mvRn6h7fm/WfScK7Rc2AAAADAoAYDAYAAACggQACggAAACNgAAAEQvhDybqy/UK3y2CAQCAAAAAAARCgGAAICQAAABBtJoAw/wAftY+df3Z68ua+dg8E8XDyTzwLPWo6HZlgM5kWDytKLu2eneph7B7MHtHuYdWAAAAAAAB+Pwch+vnFvGYsAAAACkamDAgAAAYBzmkedVvU5QaAAAANWK9Os+V1gm1CgAEgSAAD85PHu3H9fDi2AAAAAAAAAG/E+k/ld9K+nzfsp6QwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA/JjwjOv2YPL6QAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA88/CXsx/R39RuzIAAAAAAAAAADMD8C+mzr9lJ/M6QdADwkAAHLIAOKAAAABQcAAAAUCg4YBAAAAD3FAAEgAMVwAgKAYAADgAAAAAAUFWgAAFcgEAAoKtQAAAAAzDb8g8Z2nxjzPoz9G9nh+0e089bBmwAAAAYDAYBWVyVGZG1odCn0k0AJKCBAAAAAAAAAAI2AINoNxfmDrH0N9eywgAQICQAAAAAABpQDAAEBIAAACImnza3+N3kHbj7r4413f0LFe7ez5X6x6XnXWJDQQAAAByHB0vN/K7nyXyPWfFvK9Y9O+gGfRP0WH0j2oAAAAHhgfzy/0Zc2vTM5bAAAAAY1UgwJAQACA0CvxfRlyVf0OcGgAAANP1Ph6Jjk2GFAAAEyAAH5cN7Mf1JOPYAAAAAAAAAAOe/LXoz+MP2F9DD77AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAzw/md/aHm0+izg3AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA8dpfhL+zPq8v3SMAAAAAAAAAD8Sw8d/cLg3vHPqDYoOCQAUHEg5gMAAAAVwAAAABYOQo0BIAADgeFAIBAAUPJAALAAAHAAAAAAACqxRgAKDiQAAFVyo0AAByrPHfHd14/5zvHo/s0P2j2TnrdM2AAAAABQMZg5lrnMDacni9I5IuSEdSiirvJ9R1GVdT0eb1YJrqOphgAAAAAACAAEAgRs8Xo/Oj9cN5umGgCAQCCQEAAAAACIUAwABASAAGR8iWviT4p6I7r6rT+sfqPn09HbnSFAAANSQAAAAAAGgGMHiPgeseAfP20eH+uaz9MfcwfYNgAAA/BX3rK/09Zw7NBAAAAAWwQABIAADUAAi5baOPzPR5QpAAAbMV6fJ5XWIqaDAAmQAA/Kra7Mf07OPYAAAAAAAAAAAD5c/Hzuw+sP2Z6s+jAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAD4xD8hf6AuHYOXUAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAD5c/F3vw/oN9+6cwAAAAAAAAD86/wArpf729l5XSDsAHgBIAOQohVoAAAAAHKAAAAwKHKAEgIAAFeNQYEgAMHMUkBgUAAA4AAAAAAFBxQAADlkAAAcJSpAG+e0eP+Pbz4/ym0eq+wZ37J6/z3srAAgAAADlGcfkbTlZO0YeXrNvZC/q5Xt9ri5iWQsqUrVGhSz6NqpTpZuTS5+IOm6bG+29YwvVJYAAAAAAAAIBH5XfZHTP0QnPog2AASAgkBAAAAAAiFAMAAkBfMnwBtPxfidEfc36S89+6u59QAARtAAADQQkAAAAABoAAAAY3z8L5Y+HOrLB+2dp+zvqsOgA+bPwPR/Q52fndTEUgAAAAUjUwYEAAAI0AAAOc0jisb0uYLkAAOm9G87pDn1ahQACQJAH5mcn24/q+cWwAAAAAAAAAAAByTX4+ebenz/uf7hpIAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfiaHe/pf5vQGNgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAH50ax8H/0Uenz9KAAAAAAAB5CH86n7a82n0ccG4VQA5RAIAVyAQ4oAAAAFBwAAABQKDhgEFcPDm6TwOkz3ns89ASAQAFDlQCAoBgAKCgAAAAACuoAAAchRAAKCrUUvK7PH/IN48iZtPrvsGN+w+q4aXSGqkgIAAOF8m1Xm+XvFHZ2jo+gDpuhx03tvK9yfJqSgYFAAAAAAAFFnNcxrPJ8dvnznB2q29jfpntmN+kmbAAAAAAAA+G595+3TmoQaDaDYCAkEBIAAAAAIhQDGB8S/mb0Y+Em0/Wf6rYX74YUiJoKgAQGlAAANSQAAAAAAGgAAAEgxHzt8b9EfJHjXTn9DfSdL6J/OAPbf194t0jxoBAAAAxqpBgSAgAGggAAAHPaTwGd6fIFIAPQOfTrDzupGlAAJAkAfn9839uP7InFsAAAAAAAAAAAAAN/NXaPzu/br0uf7JAAAAAAAAAAAAAAAAAAAAAAAAAAAAAi/mPD9uvdfJ6glgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADQ/Fjs/U5v28NJAAAAAAA/BAPVv1T83oDLQGwB4SAADlkFBSgAAAAeAAAAAWDkKxrxH5d2z+WvmfpyntaLvskXmH11F/rJ6bwbAgAFocSAwKAAAVQAAAAABQcUAAA5ZAAAwfJNI8f8AHemPLer0n1/1/GvYfQsLkJsAHCAkQPPvE9V4v430Rqer6z6H1wu77Lm16qfHVuc1nbhFvLlz3WAAAAAAAAAAAACBzHnu0edcj058J4lFdZ9XRX0tc57siAAAAAD4I3+iPtg57BASIlNB0AgECQSAAAAAGRxeyw/xvZ83m+f1n+omF/RxhQAANSaAYAjWAwAGghIAAAAAIgAAACAkEY3i6Pm3501jwjwveOE/daK+oW8ezAkAAAAC2CAAJAAAYAAAABH5505cqehzABIj1bS8ntGzSAwAJkA+Pfzt7cf3UOLYAAAAAAAAAAAAAAPkH8e+/D9Z/wBIenMAAAAAAAAAAAAAAAAAAAAAAAAAAADwf+fpH9GrvI6wAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA5L8A+zH9gfu7syAAAAAAD40/FtH9BW/wCR1A6BscChIAKDiQcwGAAAAK4AAABgUc58n3HyP8odeOb9A7R9CfQjXp40Act8Ap/Ln7x8m3sZy6AxXACAoBgAKCgAAAAACuoAAAchRHH+OaT4/wCPdOfA+l6L2H17CvYupwoEAWACiXnXPlflG0+TeTbyz2IO4sNdd6eHpG3ybGaPkOX3xwpNY0vVsL3fK5riLvTl0u1je9vZW8lgAAAAAAAAAA3nKOZ8b6M+h+Tpfk329rHpPU5Vn9vjfQksAwvmnRfEv6jaT6Oc9gAIBAg0SqAAJBIJAAiZyHEdOeLwWk/jxwI/pz9UcNPpgwsCQAAEG0mgAAalAAANSQAAAAAEBAAAACQENSgJCAPxL5Lux/eGfh6GNJAAAAApGpgwIAAARoAAAAAHm3L+pyBrABqeq8PQ9OTdoMACZAPCvxT7cf6ITi2AAAAAAAAAAAAAAA8HpfiV+h/q8v6wvYAAAAAAAAAAAAAAAAAAAAAAAAAAH54/Cmb/AGzPL6gAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPku5/H/APo/9Xm7sAAAAAYH87H3Vhf30ed0BVAA8QCAByFEKtAAAAAoOAAAAq0vkX4X6svkz1nqy+mfpKp9jkaAAAAAPz48Bzv9/JfN6AocsgAFgAADgAAAAABQcUAAocP4zc+OeOdOfOer0vYvYsK9d1MaBAABQDj88peQ+Rb5+Q8VtHo/q8P1j1zC+18RqeI9i3n06Xk3AcPge2PTdWLo9HHSKNk7kMwqXN4+0c90dT61nc21DXasksAAAAAAAAAj+TeiPozqsa53y5k1XbPyv451SdCnJSRz/rY/qr3fM3jn0AAAACQQaJTAYEgkZIcNxfXjq9xFdf8AgbnXGfrVR9gHPqAASAgAARs0AwAaxBgANBCQAAAAARAAAAEBIMYAAjn/AMF+iPpz9bItGZNAQAAAMaqQYEgIABoIAAAAAzzLpy549DmADr8b708zrahQACQJc3/OD2Y/0m6vHqAwAAAAAAAAAAAAAA8u/D/sx+w/2R7MngAAAAAAAAAAAAAAAAAAAAAAAAAH48TY1+nZ5vSAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB+PV/1Ob9vDSQAAAD48D8Xf6EeLa8cmoVQA5RAIAV0gA4oAAAAHAoAAYTX5/fnf3YWvsXoz+sO7cgwAAAAAAIPxWl/q59oef0jpakgMCgAAHAAAAAACuoAON8W1nxbxrozxfYBezewc2nq9vKlHIAAAPI8lt+ReR75+Ty7R657BjXrvq2FaBDOfo+dem7sfdr3Hucizzn13TPai57weV3joOtivNPO+jKTcpadrOtvt8tIctGlzDPMs7qxa4VzRT9vXj3zNRgIUAAADxTSdn1FMSGMDx+fpy7Dn8NG2mdhqQ/Itvoz9COfQAAAAABAiESqAacpWfCYPTl2PdYXtGF+IfFOq/TPSzaIqAYCAkAABAaTYAANSgAAGpIAAAAADQAAAACAQGlASHnTPwc+vu3L9YLHDuwJAAAAELaIAAkABAaAAAAAQi8qyfW5AuRUemb/AJfYjc9AAEgSP51OvL9uPc+bQJYAAAAAAAAAAAAAAHnjX4j/AFb6vN+yL6QAAAAAAAAAAAAAAAAAAAAAAAAB+Cv2Lz39vnn9AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHG/z69mP7qfV3ZkAAAAfhKHun6QeZ0BnoDYA8JAAByyCsUYAAAArgACAPg/8v+7D0X7x6cvot1SAAAAAAAAAHw0n5f8Atjxbq7CwQFAMABwAAAAAAoYniu0+JeK9GfL+vUva/ZuXT1OfGwBy1AAAcR4/qvIfIt5889AteveuYV6932NKSAABw/z/ANmXrfruWpiRXl9Dq5/R+p5tOG4feDrWu03ebUE6GDaoVLm33kVzvj2sU7Guerrxe32mN1qKfT1pfz/a68Om6XK+j0sdAQAAAAIAgY/hvRn9DO59EAAON4Hoz9wOfQAAAAAAAQCKfD7HDP6Me67Xm1sGTBAAAANoUAASAgAARqoEwAaxBgANBCQAAAAAaAAAASAhG0BIAedfih055f3lZ6R1mFdOSwDnIqQFbXq8PqiKCQAAGoAAAAAgUfJezCuduABc9a4t5m8fQgMACZPw76c/ur7ayoM6AAAAAAAAAAAAAAAA438NuvL6t/ZHtxeAAAAAAAAAAAAAAAAAAAAAAAAB/OB+s/Np9JHBuAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfAGsfB/9Fvp89gAAAML+YQP6D/Q/I6gKBscChIAKDiQcwGAAAAOUADgvxY6suF/Rntw+kSpAAAAAAAAAAA8gR+TP9IXJu45dAYFAAAKoAAAAR+VUvDPD+rPzL0/WfbvaufT1y5z2CAAc0sbnznxvVeO+OdEZXrDPX/X8NPWNrGnCkAAAA861XhnvXRHfQ8u/m3FdXP6F6Bz3x3l28dH3010q4WAAAAAAAoNoMyMXSMP2EbPOEb3Qp2eeRzPN758/Z0n2jd5NuZipbl6K5TqmnEsAPmvfP6LsY6CIRAg+fd8/ctbHQEAAAAAABhcJrHHdJtHedLzbqSwJCQEAAANGBQABICABAaTYAANSgAAGpIAAAAADQAAAEBINYgAIKofkr+fnfjo+t0ueEAHQWEAPnPOKXvv6M4afaBx7AgARoAAAAADeauPNT1uQAOvxvvU8zsaDAAmT8qton/U4AxsAAAAAAAAAAAAAAAA4pr8I/tv1eb9fCkAAAAAAAAAAAAAAAAAAAAAAAB/MP8Auvy6etnDuAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAVg/Ab9P/W5f0JKQAAfI/wCN8v8AoCPJ6gqgAeAEgA5CiFWgAAAAUHAHMNfgx7H6nL+llvSFUFUFGKgCIRARAAAAoB+Cf9MHHvtHLoDAoABQUAAMYPDPCOnPw2/0R7h7phftW3hoCHLIAAHAeJ75+K+MdEP9hD2X2LC/TZ8tAQqkgIAAAZw/z/2ZfQ3Zc+x541z/AKNUUOvispoa5LAAAAAABVBygAAABQ851jn8TfPZ6iK6npMNDEDWnl+W8705YvrQfM/2AO4c2gBmeC9Gf0YvPoAACeTaTL6gAZ0AAAAQhyHCdGeV21T3epy7CS0SqAQEhAAAAAiFAMBASAACNTBUADWIMAERCCAAAABAQAAAAgEBpQEh8BaT+UX6d9Mff+9xbMBAAAAznh/Ff5kdePvv7FY30Rz6AA0EAAAAAREeY8963GGkjketaHk9rCbAASPie4+DP3L1kObQAAAAAAAAAAAAAAAAA88/B7sx/ST9RuzIAAAAAAAAAAAAAAAAAAAAAAAD+V3+ifk17U4tgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAD4//Kj0MP6StPozAAD8y/mjKv2FPM6QqgBXCAQArkAhxQAAAAOBQD8bt8+D/Vj0eZ7mnOY5wKqABEBrWNahqAgAB+Bf9HnHv25y6BQDAAcAByfgGs+BeP8AXl6f7xnXu3pvNoomADkKIxfENJ8M8R6s8j2ml7R7ZzX6K7JgUAxRKEgAADDEs+YffOnPuOPw17inkvPOG7Meu9ax0DOgDE22c75RrGfPrOtrQ+j7fG1JYAAAAAAFdnNctrHNe7BL5vL2tdF6zNZL2vkf7G2iU59ADzXE6M/ZTn0AAAxWeJ/QO0PbhaAABhtcTxfTlrd1NdhNz6AASCDaVQEgkEgAACDQKAAJAQAIDSbAABhQACA0kAAAAAEQAAACQEI2gJPkD8a+zL9y/ojl1CajQkAAAACm8yH408d3YfuP0vn7gmAMAAAAAGozvI+7nadmIB03o3ndKN59wAEjysn8Cf6QerKycmwAAAAAAAAAAAAAAAAAeO0vwo/YT1eX77GAAAAAAAAAAAAAAAAAAAAAAB/Kr/R9x69McewAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfg/05foV+jnfiAAH44bWN/peeb0BVADwkAAHLIKClAAAACuAA+dvw47sP226rrwfIx70OUPzZ3ZriP0Wa/OLdT6X9AmvhTzccv3+LoGtAH4Cf0pce/UHNoFAAAOYHzx8rdWXlntOs/QHvPJr0hnSuoAAB3EJeA+BdWfkfp2s+7e4c9esy5UCAGBQA4JAQAABQnzRvPYeqj8c0Ncu58th9D6VNa5jYBD4FtNb6MDkuGFuV2ZWRc6fuGV8H4LrG7vM6DeyrotvKwQAAAAAQeX6zh5e+VIpHoEV5H9YZ2GFnM0vGfcNo3Dn0AAAAgnAARoZ/GaTxef0Zdn2+Gm2ZWAAAAEiI6QbAkJQIAAAaUAwEBIAAI1MFQAjWAwARqAQAAAANAAAAQEg1icqz+eX9furL7GOPUCkjEAgAAApGpwfih0ZwftxJKc+gA1AAAAAEBPOts+VPT5QB/sHHvZZxdIAABM/zpfth24++nFsAAAAAAAAAAAAAAAAAAHzb+Jfdh+8n1R1ZgAAAAAAAAAAAAAAAAAAAAAB/Kr/AEfcevTHHsAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHyXc/k5/Sb6vNoAAH4cfRWF/e55/SDYoOCQAUHEg5gMAAAAcoAH4k+m+py/fkm2fgXmsvu8gMf7MD8r/1SD5n5cPm39UAT5yDyr7aBkTUbGM49H4ef02cW8phoDAArh84/I3XlW+ug+idfk1ABQcUAHC/OmsfPHD9WfuPvuF+8dHz0oIAAAKAY4QEgAADAp+e+L9WX014tm+v2w8s9XueuObUAPPLXh+91ZfRNzj2WmHBXtY0tCK0rcuHAawMDWcHn9YsfTON874azc6lrs9zDQEAAAAJXZN4PpM3uoBlXFeU9OXq/aY6BDAAAAMClamDj+O2jA6m57Lq+fWQhgAAAAAAEiNdAwCQkBAAANCwACQEACA0mwAQGlAANBCQAAAAAaAAAAQCA38rt58a/bYQYWBQwTQQAAAMaqTEa/A/7t68f0eOHcAGAAAAAANzBeRns8QAdzhp2bfN60BgATP47+x9uP6SnFsAAAAAAABDg6xZ2pakUAAAAAAB8SaT+VH9DXqc3rwAAAAAAAAAAAAAAAAAAAAAB/Kv/Rrya9acWwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAND8CP1G9bl++SkAfhR9Vc9/c5wdINjgUJAByFEKtAAAAAoOAPP2v53v3K9fk7KRz+SH6iJ/mZ98h8v/WwfBX0In86/o+z8xPZkeh8G19c+jtRsCOIIvngf51/0K8O4Y2DDHD5g+UezLe+v0fQk3JqAACuoOfZ84/NfTl5971c/Q/u3HrfIpy1AAAAMCgBwQAAAMCgC/mf07pwyLIvW+Qw0vdCASwDzvwnqy9973DRXw1xWeG5/XjnZ1KjNS1diK2+jzrqOrxuwQ8/idJwOR2jkNPWfSPZ+XUM2DAAA838H6c/qXb5tKPlWk877BS6U59AAAAABvjOsc9F0Z9N1+V9faw0EQ1oAAAAAAACBE0SqAQKQkAAAaMCgEBIAAI1UCYAjaAAAakgAAAAAiAAAASAjF/nG6s/3S9+57CWAxBsQkAAAAQtojwX8JOzH9/fWOPUJY0EAAAAAREeR5nr8QWi7L9eXyO1gUABM/EvwB24/uycWwAAAAAAFGl5hT9fjNhP0a35HWCYAAAAAAfl78oelz/0L9BtAAAAAAAAAAAAAAAAAAAAAB/L/wDvTy6eonDuAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHw3+f/AKHP/RUdEAH4UfVXPf3OcPUAK4QCAFcgEOKAAAABwKAfnprHxX+vnp8skrX5Y/qqn+Z/6KD+VPRw+MP0bF3czX5R/qyqT4zFvfUoMjaijCL8+x5v7E8HQGdHItfKHzD25er/AFvlXty82gAAAC+Jar5e8I68fZPpXK/ftPk0ABwlKkAAAKAY4kBAAAFAOjMZ8qfS3Rh5P73LsGGgAAAHyj7l1Z987l0UAAAACPPZkY9zhc5rPK420dj6Ljfo1rCwFFx1rG9PZxvl9z2PSy9NkuLoUecalrseAk817HafT7fPoCYJggATibXm7+nHqPSsNbRlQAACA1oAAAAAAACBqaFUBISgQAADQsAAkBAAgNJsAGsQYADUIIAAAAGgAAAICT4k/Nnry/f45dQQAAU1hIAAAAFNRM/KnaPE/wBwhBz6ADAAAAABqOF4r0+QN4APSui8rsamegACXJfzp9eH9FnW8moJgAAAAB5d1ZY56PMAXPUeHe4cW4AAAAAARfip0Z9V+5PoYPAAAAAAAAAAAAAAAAAAAAA/m1/YPl0+gTh3AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAACj/O315fux9QduIB+FH1Vhf3OcHUAOUQCAHLIKClAAAACuAAPw/wCjP6I+2/Q5pZGvyu/VpV+ZX6MC+U/SA+LP1hAGvye/V9NuW1+V/pM1Y+32t6JqL8U0/wBRftTz+k89R8i/Pndl799c4X6Uc9gAAAZfzLrPyrU68vqD6a5tetdjQIAHLUgIAAYFADggAAAYFMCncG48h9r6M6Hf415p01rpTGwAjDzf021IQwAAAAAAAAAy6XAeb9OXNeq0ev3OWxRgGN5zpPHc7tFFaO2+iuezFh+QJ0Z+sdDz6IJgAACDl2/LW9GXpXY8+qkMAAAAAEBrQAAAAAAAJEa6BgKQkAAAaMCgEBIAAI2aAYA1KAAQGkgAAAAA0AAAAg/Fr3frz/Ss5tAkAAKRgkBAAABSNTOfa/nv/Wzqx+xDi3AGoAAAAAgYnlXdzB15AG76f5/SjebcABI/Cjoy++vt3KgzoAAAAAzvJ+/nDsxADRh+py+P2AMAAAAAOWa/BX9JfV5v00KQAAAAAAAAAAAAAAAAAAAH8+X6E82n18cG4AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAH5e7Ryf7aelzgH4a/R+Gn3qcHSAPCQAAeSDmAwAAABygAVf5tOvH9rvUO3nllZnbQ8/VFWnCprgAZGoDGgziUS9IOKNr5+/JGK/o8z+Dp+IvAe7H6a+tua+wXGwAAAOY+UNp+XPTOrL6w975NpSG5ZAAUTipAAACgGOJAQAABQDochfNKjgfonVBjXkmn0Z+kHPoAK4HAAAAAAAAAAwHIMAOY8W2jlvbLXpr+awGAAAAAcL5Vvn6N6PnTiKAEAQBMlry/nujP0vuufRxnYAAAAAAAA0GoAAAAAAACS2lUBISgQAADQsAAkBAAg2k0ADGAwAaiAQAAAAiAAAASfzbfuf1Z+8JzaBIAAMaNgSAAADGqkGfDP5m9eH9BN/i2BNGgAAAAAR+NdeFc7sABfW+TbQbw9KAwA/Ny8vE/2PtBz6AAAAAGB5r6PMHVkAAHS+ieb0hzagAAAAAB4Z+Ffdj++v0x1ZAAAAAAAAAAAAAAAAAAAAfiN7Rhf6FnndAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHlbX4Df00evy9WB+K3p+N/ooef1Cg4JABQcSDmAwAABQcAAeD/hr24fvZL2YSyjmkFJIDnA5QAEaDWhHGEcQRYSf4off+OnvfxkP3f7XyrpzC3AAABzHyJtPzB7t05/XvqPJqEscCkgArnINAADAoAcEAAADApilKsweb6rzr3zSau3jeD4tvn7to4aDkOUPHMvpz9f2OfQEAAAABSBoAVQUAA4DxXeLP0ejRMbAAAAr+T6TzHslLozGwAAACgLzbh+iO89IyqyRQAE0CAAAAAAAGA0AAAAAABA0pBsCAgAAAaMCgEBIAANSaAYgNKAAaCEgAAAA0AAAPIxfz/f0q7RYZz6AAIAAKjQkAAAAQtoiL8DOjL7w/QjKgysaCAAAAAJ5n0Zc2ejzAB2ON92nmdjQYAeGEfhz/Rp05ynJqAAAAAY/l3ocwdeQAAHf8+nVHmdQAAAAAAHwLrP5xf0YenzdaAAAAAAAAAAAAAAAAAAAH5U+WZV+sZ5nSAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB+GX6Bery/oiWvx+lyv9NjzuocChIAOQohxQAAAAOBQAD4I+KfQ5/18m6cpZxSShI8JHg5QAEaDGBHGEUYUPyBVed/Z8UfdOV+xnLoKCgAEPybpPxt7z15/Y3onHqomAK5AIAHLUAAABQDHEgIAAAsAtXKQEcla+fffOjPz33TOl8qls9ZB650AYviO8cD0HRn1/R4X3W1hpj0aVejc5ONc9h3mdeR+c7Rbtsu+s5vqDKyIXz/g9UfT8nLYDADkWvKNzfP125z6AMAADBa4Hi947L0rOtQzpEBAAAJYJgAAAAAAjAQAAAAAACRqVYCBSEgAANCwACQEAIiaCoARtAAIDSQAAAAAaAAHxXS/PD946kTG2jAkAAKawQCAAACmomfJ9z+PX9D+saxy6iA0AAAAA47WPPz1OQALcv2BfI7WBQANmf51v2q7cfoE4tgAAAACLx3uwaduAAAEiPUdTx+wJoAAAAAAPxk1/U5v3KNJAAAAAAAAAAAAAAAAAAD4W/NWH+5p5XUAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB8G/Gvo8/9ARvH5UebZX+rR53UrhAIAVyAQ4oAAAAVwAAB+Ss/pc36Lzb5zyimkCWQHvB6gAIwGsCOEI8wPyV4ya9z+zs7+qJuLYAHAAHj1L4Xd25fe/qHFqrhgAoOJAAc0o5AAACwBwQAAAMCqFG4mAoIg+R/p3pz8n7UXoebz6+E+7756xz6AABW4y1yfO75+qdthp4JxHVlPda1tiH6B2eF+b/P/RF/1YPVrvNYDAIvlzWPU/RZegRVLlaXAZ2+fqPbYaBLAADK4q54yjtHbeg5XqGdAAAICIAACYSwAAAAAAY0AAAAAAEDU0KoCAgAAAaMCgEBIAIDSbABrEGACNQCAAAAEQAA/MPSeG/XVAY0woCQAAojEgIAAAKRqYM/Bb7X6sP0SOLcAagAAAAGTS8kPX4gAA9O3vK7GpnoAAl+Pnq/Zh+lhxbAAAAAB5p05YJ6XMAAABc9S4d7ZxbgAAAAABy34D9eX6yfoH24gAAAAAAAAAAAAAAAAAB4p/PGj+j93kdYAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHKfzqdmP9K/ovZl+a3yxnX7FHm9blEAgByyCgpQAABAHzv4dvnQ1KXsX0Djf4hfbXo8/1vPrnNMEs4SSBJxXXx5Ho++Fw8302tBjAijDA/KVNPpKL+8eq4dwQAOAOMa+JfFO7L7L+p+TSZc6cUAA5CiABy1ICAAGBQ4QEgAAFgFq6ZBoGAHz/AKvVn6j4nJ6p2uV+b5esevHPoAAAADAqQBXAAYWq1n8LS9NlhgMAADzKl5BtdOb1RD0CO27nDSUlgBWFxHEbRjdda7XqsNHksAAAAAAEQEABMJYAAAAAIDEAAAAAACRqVQACkJAAAaFgAEAAAjZoBgDCgAGghIAAAANAAPxs1j3r9GYYZUjaAkAAGNGwJAAABjVSDPkr8i+rD+h+1x7ghGgAAAANDxeD2OIGgDrPQPO6kbz7AAJfEHwj2YfuacWwAAAAAc/pPmx6/GAAAAakP1CXx+wBgAAAAB8x3P4vf0V+rzesAAAAAAAAAAAAAAAAAAEX8vgfvP635HUCYAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAH4i9Gf3P+jPoYfIP5By/30PL7HhIAAPJBzAYAAHxX+V/bhne39WWtjh5RwScH7ONfRE9TNME8oTOD4p8B/cfwWfc9Lyv0F7v8H/oRGcfXgee+14XqniPi+7+a/Zl/dH1H5+wTQAKC0w+QPkrtx+lvs3C90xscxRgA5ZAAUTipAAACgG4JAQADAqhRuJgKAAA5z5l6c/phca8e7vfP0l3NtJQC+IAAAKkazkPO9o9et5V555tvPKfSYuzOfQAAAAAPG7nlPoyigZu9ImAABnC854foje7qX2E+NAMAAAAAAAAARAQAJYJgAAAACNBoAAAAAIGpoVQEBAAAA0oBgSAgAamgqAEbQACA0kAAAAEBAD8H9o/RT7MzoMmNYgAIAAKjQkAAAAQtohv8/HRl+mn2rjYZ0MAAAAAA8twPV4w0kAu+v8PQM5NwGAeffz2dfP8A0ebHFqAwAAABovH4Pa4waAAAA6X0TzekObUAAAAAAPyo8d9Pm/fx+sgAAAAAAAAAAAAAAAAB+BofY33N5fSGdAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfAesfKv72enz+f8A8zwf0laXldrhSACg4kVWAwAPze1j83/0r9Lm+o3VKgHEo+BflNV+p/2U1NO1NOEnjPq+R8L+3ftP4Dr+RYadn9ffE/pXifi/v/K+afYvXxfNvxp+Jf0B9K/o7hp66cmoMABweY/nz05733cHpBzaACuoABQcSAA5yo0AAMChRKEgAAFgFq6ZBoGAAAeT6T5R9S6TD5XL9av5Xk+E759/6RnUpnQVJVa4fgN4l6kPOuS0XX+j516NcwoBjQy42gChz9LzXj+iPVvUsa2zKwAAEDzvzXfPsPRYraM6AAAAAAAAAAAAAEQEACWCYAAAAAxoAAAAAASNSqAAmQQACDQKAQEgAA1JsAGsQYANRAIAAAAaAfzwbx+y30TlQZMRjRgSAAFNYSAAAABTUTPz50j4y/dOkHPoNBAAAAAOF4j0+UNswA9d5Nr7eHpQGAH8+3Rh+tv05hYRQAAAAB5vz3rcYayAAAAelcuu+ed0gAAAAABmfgX1ZfpT+kndiAAAAAAAAAAAAAAAAAH5Zh4L+wHnbhhoAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB541/Ph/T76/Lt/wA0ofsh9DeX2KSgAchRDigAAPlX8Xu7D9j/AEzqwc9j3AqAz5wH+Wn2zL/UCxU2JQn+YPf+d+Zvev1b8X6Lx7h7uL+8fG+m+KfNvvPzMTu83Ot+Z7Hf+p/NfT9Du+H7/SdN8/8AR9X13zP1eh87cPpfBn2P1ZfWb+LYAFBxQAD0AgActSAgACgGqkgIABgUxSnEwFAAAAA8Pn3I68vo+5y6lIPn+72YZfey/Vzm1BrzPzzozy8fSfqe7ya+b+gUtIigDD5G54/znecCncgGJwqfhPqEP3H2MWr20vstbGwTj8b0nK9fa3TKwAAAAAAAAAAAAAAAEaAAE0CAAAABoMAAAAABA1NCqAgyJKHTACEQoBgSAgBGqgTAGFAANBCQAAAAaFf+Zvoz/ou77DQIAQ1KAkACBvIu3NwzYAAAUjUzmGv5zP6B+nL1M49hAaAAAABzvmHocwdOQAej8+vSp53S0KAD8iNcvRv0xQGNgAAAAHHcP6nKG+YAAABoS/WF8btAAAAAAA+ean8Rf6LvW5vWAAAAAAAAAAAAAAAAAD5kD8Lv6JuLYOTUAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAD8BP179bl+xvw7pe2for53SGVACuQCHFAAZjX8/f2/wCpy/Zc2kTSA9wCA2MPPPyKVe8/r4K3YaseXeh5/wCd3sP7h/PO75Djp7H98fmH7JF83cHb8LfOX2/597j9xfC/oPVYfsfPYOB7ng8xzX0PzXKc/wDR/MdDzXm+t6h9JfDfoHoXo3wH6Hrni+8OYowAcsgAKJxUgAABYA4IAAAKAdq6ZBoGAAAKDnBF887Rh/RtLaOfShyWk856vS53j6l/O0cOus9t3uNd1NjQDMsXmnlfTHHeO2vIfN5dn1JHofSstZYuE8dT+buiT/SX61S5PRtdBczeAlHt9jnsBoAgCAAAAAAAAAAAAAADUAEBNAAAAACMBAAAAAAkbxNV8F/Ie+fgXNXBaZ7B9HZv6U+lsdPYzOwEBIAINpNACNoABAaSAAAADQ8U/CHpz/pJObQEBIjaAkAMTyXtxgm6MvUt/wA3qCGAAMYqBn4ye09WP6ZHFsANQAAAAKXjfdzh14gB22OnbHmdTUKAD4WvP4s/by5Dn0AAAAAOf829HmDqyAAAAAPRek8jrDOwAAAAAA/KXy30+b98jWQAAAAAAAAAAAAAAACuH8w/7mcuntxw7gAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAflRvenz/r/+b2sfGn7U8O4c+gA5ZBQUoAD8yto+Qf2H9LltztSzA9wxAbGKHjkfjn9Xqv1SsuZ5x/KvyJ+yfidD2D1/G/Qve/Cv6Bb5Nht/P1+m3XwS/o15HqBhYUAB5Xrn+af3D9d8hhef/Y/BedcT918B03oPl+r6F6L8L+hei9/8J+h2l8z1XEgAOcqNAADAocICQAGBTFKcTAUAAAAK8FAGh4p5Z2Y+y+o8+lpkVxfm/Rljem0puZl+hdXlSiYBlcBc+dcH0R89+Nh1P0XJ9Gde1DEyCAK/ND8aUPUfmwPzg+v86/ZDGxrlOd1lu3LjpSygymBo2gL6LesjQ10aIMAAAAAAAAAEBEAEwlgAAACAxAAAAAK6PzQ/Mnoj3v6u2j27vqm7yiPJPDR+DeHS/QP0Rx0/QXS59QUAAA1JsAGsQYANSQAAAAA/Pr5C6c/3AObQEBINYgAI5DzLvwDfOX1rl12Dj2AAGIU0D40/Mbqx/oEOLYARoAAAAHi3ZhXO3AAOqyv0I8vrRpQAePufwe/o/wCjJxyagAAAAGV5X6HOHXiAAAAAbfp3ndIcuoAAAAAAYX8/vXl+vX3X24gAAAAAAAAAAAAAAAAH4h+n43+k55vQAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB82/jV6GP9MXinRl/Pn/R1x7SnLqAOWQcwGAHJ/zt9eP7Be4d2FiwKaYb3iEBkY4YBec/irNfrJ9rsnsisMZU37hzcdYvgKj4h9Zzr9HPVuDUBgwAPzg3j1L7PlhlRSuOC8++t+M8888+9/PPOuf+0+F7z0v5v6j0r0388/SvStT4n7xyzYCAAKAaqSAgAKAdK5AEgwAABQeoAAAcfS8V5Tsx7a1Jj4lrrfobDSwZUAGO15153vHFeG6KDqJPpL1gQjGxhFAEFcOD+BiqaBs+4h4t5Un6NTCjaDV1wuX0FtO+Bn1QM5qrjMzKoWt0N/oZOo6WX1nXydd2kvsLEsAAAAaCABLBMAAABGAgAAAYv4faT5d+onTn7LI5eoKAjQZRD5m+BlWf+zHPr9JnPQCAEaqBMQGlAAjUAgAAAD8UfpDpz/Rw5tAAIBGNGBJl+L9uIdORdl+wWvM6gQAAWwRk/wA3fVj/AEE+o8uoSxgAAAAHlXVlhnfzAB0WdelHldiNKACJL+b/APezsw9ZOLYAAAAAo+Sd/OHZiAAAAAL69xbWjh6AAAAAAAPjrSfyl/pI9Tm3QAAAAAAAAAAAAAAAAPhb8tpf72Hk9QAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADf5+OuP15+0+3n/AJjf285tPdji3FBxIOYDAD82fk/0eb9eLXRlZsDsTCkeAgRxhBXHX+Wkflh/Qgq7ey5sTlWZpHNCH8mA8y6ya/W53nbBQDADg/y/6s/1l2eXQAABWOGGBvz+ceb/AG3wXmXmX6H+acX3Xt+B6l6p+e/pXqno/wCc/p055nqAWKJQkAAAtgU5ZkKQAAACvBQAAA5Ty7eOFo9Gfdem5VkdpD6MzoAZ53S8s8/6c/nfjQ9f+tUehStSvQrRshaigCvXD4k8ZK7uNzwXfqvadAX5ffcudfptfzsEAAACAAEABTFiYLMHmKOa5elyvI0cbxtLL69nd99D9B9Ah+jdnA4TRoAARQAAAAgMQAAYH4c8D1Z/rhtaZySA9wKAjQZGEVcPin87Zr9dvvPj2CAAGjQmgBG0AA0EJAAAIA/mx/eXpz9uObQACQEMKAkPEOrKkdmIB1XpvBuGGgABTUTPxM3z+q/0IzYZWNBAAAAPPOjPkT0eUAOizr0o8rsGjQGAfhf9/wDXz/aJxbAAAAAFLyPv5w7MQAAAAA9E5telPN6QAAAAAAAPw3+tfV5f1ZLQAAAAAAAAAAAAAAAHP/zFB+/fqnkdQJgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAANn517T83/vV6PL+Hvs+N/owef0DgUkVaAAA/no/R71eT6kt6RashYnCV4K0I4QgrBWqD/I72aX+sNoU9p1ZnkexkH4uIy/aor9MDz9gsAAD5F846sv0DOXUAABzFGADyQAyPMvc+f8v8r/SPzDyrI+8/PPVPWPjfufXfWvzD9Z1HeF74EgAMCqHIUJBgAACg9QAAA8Y0nyb1Hpz9C6nmuVU1AAGeXXPjvjnTHyC+X92fTYrM7XlHsycvmY+80heXdqHyZ9bhWrNfAvnZXq0NT5R6/NfUTSfFvyHmv6IPW8bBMAAAAAAAQCAAAEBoABg8Q1wXBWcB5zovPnUvSfTofp/qub9KtwwTCWAAAAjAQA/LO18HftL1Y78zUsgPcNQEaEcYooAh8DH+SH7B4afchy2ACA0mwAYwGADUkAAA+aqX4t/0cawhhoAAgJEbQEh5pvHHHfzgB61zabpxbgAFI1M/PfWPln9rmg59BAaAAABx+0efHp8gAdFnXpR5XYINoMA/KDoOvn/T84tgAAAACl5H384dmIAAAAAdZjfenldYAAAAAAAHhP4Z92H9JHoHVmAAAAAAAAAAAAAAAAfh2Hqn6Veb0BjYAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADZyVP8Ang/oy9Ti+U/gmX+355nUPQCHFAAeEUvww/ef1ePSt0W7QrU4TPBWhHAFesFSoPxb8hOf0f6C+xvmntaY2p5JoQg/F4PMPteK++jg2CgAHB+cnvnVn9PnLoAACuoABwKSADlqQEcd18fk/kX6L+aeRcH+lflnpHsfy/1ntHrv5T+waJ4fvBQDtyzIUgAAAFeCgAAByHzj1Z/Ue1hQoOAADk2vn7wrrz+Gpof6W+8ubVplm0H5DfsSqn/MVL7w9NH+X/3Kz8xv12FWrNfNXxyV0DBYH2YP0iMk/NhO7+2mOgSwAAAAAAAAAJAAAEBEAAAABnFNeZeY2eXeWaLlvTWvWfYM3693OGgIAAAQGIHyF+Km8fs56rtlPMyWUcjxKA1oRxDhhFXgD54/JdV+9fv/AA7BIAI2aAYgNKABqIBAAfl3pPnf7ENNMrAAQEgxgAc5U+THp8wAb8V6weZ0gADGKjxO1+Ev9IOucpy6gDUAAADB0nyw9XjADbzr1E8rsEBoUAfBXyb18/7TnFsAAAAAUvI+/nDsxAAAAADazr088jsAAAAAAAAPxr6M/Wf2B9DAAAAAAAAAAAAAAAAA+MQ/Hv8AoM4dg5dQAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAKYD/IbuPS5P0y/m/uf6P9Py+pXSwQ4oAD829Y+cf1g9PluXWW7YrVlFiQbmDhriq1WqnI4eh8l8v+X/ANmzfUno/K/VVj9F/k+zPI9jIfyOR8w/pdFfYpwbBQADg/Kr756s/XDl0AAB5QAD0AgUHFQAAAFmJth5F43+i/mPjHD/AKd+Ueq+2/H/AG3t/pf5V+wqvneq4mAoAAAUHqAAAAHilbpy90MbVwAAB5V899OfwV4AH6afVrVu01btBatp/kF+yCdj8vx/RPoAfnx+kYvyo9vDyj9Hmu8+WWeAyD+gvaxVIBHHI/En+iHPT2AzoAAAAAAAAAAAQCAAQGgAAAAAABzPk9LyLyDVeSa1r2f2rGvafQsbBAAmKH86X6HdeP19YubEw55RSOBQEYEcQQQjr1xVvg4fyn/QZya3TGgAamgqAGpQADUIIAD8Fto/Q77bimkMAAAgGsQAajxCp6vKDQB7HqeX1BLAY1UjWfzcfvF04+znHsANQAAAMy58hPW4wA2869RPK7ABgUAfNH5LdeH9ABxagAAAAFLyPv5w7MQAAAAAuy/XDxu0AAAAAAAA8apfg1/Sx63L3AAAAAAAAAAAAAAAAFIP5n/2V5tPpI4NwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAKoA8U/CXt5/wCmb+dfpy/W36d8zpcuVCgpQAH4tdGft33V6HNdutXbgW7admcHRor1mU6bVP43+d/WOL6z82/ecvlNez9BvUP1v+P55ujxXNCH8yGfJP6gZV9MHFsDABVD8gv1I6s+1OXQAFY4YAOWQAHNKOQABgUOEBJzPTzeK+Kfpv5V4jN+i/mPuPvH5p+q+2aP5r+phGgAADwcAAAAB4Zc6svZ3YWoAAeQXPzV+X3RH0N+jaN+2y3bFbthbtp/kB+yqqz+XgdF2KPi/wDWNn5Sfsg15T8JC/Seo1UqDq1BU4AQD8vvfc7/AFeMrAAAAAAAAAAAABAIABoIAAAAAAAAgV/MGvFfFtTxrW0n3H3TG/c9/GvjjwDafi/9mdsrNhliwKaYJHg4BrAihCGuFesFbPD8Xf0Tx1/RQ5LABAaTYAjWAwQGkgBT/mg6M/6K/QMNBqAAACQRDSgJPN+L9HmDWQDuO/8AP6AxsAELaI/D37c6sfuY49gBGgAABRpeOHr8QAbedeonldgA1CgDz7+ersw/pUdw6gAAAAFLyPv5w7MQAAAAAkT9lPF7QAAAAAAAAD8W/pb1Ob9VDSQAAAAAAAAAAAAAAAPyO8nyr9gzzOkAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAACqAHfhZ0ZfoN8893Pq/p/wCf0OXnscwGAB/P305fpT9Jd/PeutXrgXrqdydU6NlakKlSFR/P35T9v6D034L9G5fzevU6vrt/O3Nr1flN7d9T5jf+UPZ+S+S/1E9D5X28yyAABwH42fr11Z7Jy6AA9gMFBxIAK5yDQABQDVSQEABQzy71PN8N8H/WvxXg/avd+f8AoD3f8a/c9w+a+pAHuAAAAAGeT8D1Y/S7ufQAMJrxHxTqz/OT71T+k7bm3bHbtit2wuW0/wAf/wBmFVz8uUff/pwflb91M/Jz9jReKfJbX6GVGqlQdSoKpWBAPnb875r+jUw0AEAEAAAAAAAAAABAIAEQEAAAAAAABGgCIvJivDvCt48n9WtZHjwvpH62c2rDKm4KxMErwcA2MIoAgrhWrDrVRfMH5tTX9GNvh3BIAak0AxrEGADUkAPCPw16c/6QTm0GgAAASAhqUBJh+P8AbgHTmAavsXFuHNoAAU1E/wAtcnrx/WA4tgAaCAABXa8WPY4gA2Ir1Q8nsAEaUAN/mr68f6Gu847BMAAACl5H384dmIAAAAAAe0nidwAAAAAAAAB8m/ll34f0inTmAAAAAAAAAAAAAAAB5F/OOj+iLrfJ6gTAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAACqAH/AArpn8K/pZ6XN+Z37tcO7jl0HMBgAfzh9WX7M+u9/P8AEfCy8n6aD6z/ABGH9G/VKPrD8mAs9Qz9KaDmj8WfO/rPIdr+b/uuHyO3X9k7f3f88ZeRx+lz+J5X1PPct5H1vPXPN+h6PpvS+Z6bqPZ+J6zrPc+C6677Hxn42fr12c2ycugArqAAcCkgA5agAAALAHBAADApilOJjj/Avsfi/n/yz9i/EPZvor4j773zW/Kf2B5nqAAAAFTU+Yt43vYgTmpfAecbx8j+Lp/pj2Tm5aC3bHbtiuWgu2x/kV0E16Z6ij607IPhH6dZ8T8SLmv0aa1uZ2w8l0vpPmfQKvy31lSsCAUPwemv6U+5x0BAAAACIAAAAAAAAAIBAAjQAAAAAAAQGgBNAjC+ebXwLW6s7f1+10HxyHN9kP7m9SIlkB4DIgigCvWCtVHVqCp/iEn+vf2Dw7hIAI1UCYgNKABqIBHxh+d/Tn+75zaCAgAACAkRtASHiXVlQOzEAX23l1snHsAAUjU/h74X68f3JOLYAGggAAQNeKnscQAasP1c8ntABhQAfz1dWX7KfQPKwmgAAAKXkffzh2YgAAAAAB7SeJ3AAAAAAAAAFP8AnS68v3s+k+3EAAAAAAAAAAAAAAAA/BgPoH9IPM6AysAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAKoUHYTn+dH94PU5PwO/pF5ddo4tlWgAAD+bfqy/bKz3c/xX+nAXvx8D9b/AM2R/rvfT/KD7+R6ZQpZ9Bqjw3P6fx/z35d/ZsXsvo/L/bk36P8AyPI9NUQzz6l+TP6l8Xo7fM/Pfa8vy/i/c8rynh/d890fH7XA/S/0P596B2vv/m3aXPc+DcumYArkAgUTipAAGBQ4QEgAMCqVyAJBhy/TzfP/AM5frn4v5x7z9F8x9K+3/iP75IeN7gADAqXUxeCcp1Zu3Q6v8LYr3P8ASi5svat2wt2guWwt2wuW07tody1LfCFSoynTap1HPlXwj+o/k8lL7f8AP/R/r/8AKv2GvX+M+3QA/F9P9k/qnDQEwAAAAAARAAAAAAAAEAgQEQAAAAAAARoAgJo+UrX56dh05/RuzS+TLIb3Pi1P1DDyD4nl2/ah/aPyg18zX0ffPzE16L68L5a9pHt/nqzK/Zjj3DIABqaCoAalAAMQCPz++WOjP9qDn0AGgAAICQYwAPO9Y4c9DnAD1jn03zh3AAGjVXgX4rdeP9ExxbAAgNAABrXiB7HEAGnD9ZPJ7QAaNAZ+EPTl+lX1pzsIoAAACl5H384dmIAAAAAAe0nidwAAAAAAAAAfmPtHi37z+lzgAAAAAAAAAAAAAAAHyCH4rf0McW1k5NQAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABsGxQcE/kJ6P6XN8efqXlX1ecG6rQAAB/PH1Zfq1yPdzyfYQXvzWD7W/IxP66+w0/zz/ZgKNGln0RUaLWdxXN6/5i/T98v6Fya8kjnKoDcof45frFz6+lnJqAKoUfNeP2vzt+yPE/ROL4j5f9M57reP2u9776b8w77vvqfym+vZ4AAOaUcgADApVJAQAFAO3LMhSAAAUON+b/tfhPmzP/XvxT6V+nPyT9n7g+H+/CkOaUAAAPzg/O3af2Wk1zHhZthbtjt2xW7QW7ady0nYez5Q8M/UvyXq/rfwfoumq/F/b06jX595/wC6/wA9er5nNHhf1V530np0H5J+yIAfkkn+k33Nz6IJgCDEAAAAARAAAAAAAAJAAaCAAAAAAAgNACaIkfza/q52Y/QPD0vgb0APS/M6Xnv6oxfzdML7I/PpGn5eH3F52183eeJ8r+rDX5y/Ygdz4Oz8pv6RuXYMGACA0mwBG0AA0EJPz8+WunP9qDm0AEQAAACAaxADn6XkZ6fKAHo2F9icHQAAMaq5L+cDrx/pfuceoJgDAAAA8OPZ4QAuy/XzyO0AEG0Gfi30Z/WX3lkgysAAADM8p9DnDrxAAAAAByPZzxe4AAAAAAAAAOZF/PJ++Pr8v0sMAAAAAAAAAAAAAAAYH85f6L89/bR5/QAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAANg2OBQn5u/Gbuw+3qtz+rJ5nS4oAAA/D3oz+ovffQ5vk39KAvfkOH6wfm6n+t9gf5CfpoHd0WqVEVKi1Soj/MH06X+mMjT1GqAjA/F/9W8NPWzk1AHAHyzpPzd+m9oMLzOG4PouF4D5n9T8/wCX+Z/Ue19G9X5D0f0r678c6h3s/Eg0AAUA3BAAABbFKcTAUAAAPBwCeP8Aq+R8yfPn7f8AgPrH1P8AJ/Ze9Tflf6+5VQAAAH4c/NG8foP9Z3NxzVi2Fu0O3aFbtBatp2bQT/Ff03y3zJ3X69+H4EMdP6Z2fwL+jKlaL/Nvtv3j+dPXsTxvb8B9o0Pdofxz9wAD8kE/0j+68NATAAQIAjEAAAAQEAAAAAAAkAEaAAAAAAAI0ATPnJP8Lv3g68NChR+b2+PqOVapfrCn8nTSfZkTf43enSeteNUvpn43F7zxI+Q+xRdzktfgX/R1zbehHPYCAGpNAMaxBggNJPjr80+nP98Tm0ABoAAAEgiGlBHJ4XH6vIDDusb708/oAAGIU3+aXpx/oR9J5dQlgDUAAEDw89nhACwn7IeP2gAgNCj8kt8/SP0lzQZWAAABmeU+hzh14gAAAABZT9gPF7QAAAAAAAAAA/Mzwz0uf97DaAAAAAAAAAAAAAAAAPz+/MSX++jvJ6gAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABsGx4ASfz49OX2F8f8AXl/QeeT1OKAAAPze1j5m/W70+b88ZEUvdmvrv80x/rPOjzr8nR+sbwv0Qp1NOkFKiP8ALX2uX+jclS9wKgNYH4sfpthr9IHHsAOA4L8xOrP9DPbOewlq6gAMvPfz3zj5n9P8387+S/Ydj03q8z1L1H7T8P6c9n4gCxRKEgAMCqHoAkGAAA54AAAY2uPzT8ufs34bm/U/RzfUW9+K/vAY7gAAz40a/LfzraftX7IueuttW7Q7VoVq0FmyFmdVP+T36F+b+penfVfB4Pz53cf217d+O/utat5Hs/GPkH7d+C+p4vT4/l1r1ePpvV/k/rfVd38o/Zz8VeXr/Z/6qw0BMAAAAAEQiMAAAQEAAAAABAIEBoAAAAAAIDQD8uR/P36odWNu4z5++QBkw/vP6OS+QJ0/tHnUfk32zPXPlgX6RfnY1+jv5cBufc4dzUa/Ev8AY/n2+oDm0AkARqoExAaUADURwP8AOlvn/SN0vPoADQAAAJAQ1KAk8exPT5QtHaZV6Ged0gAAIW3+efoy/ZT6E57CKAEaAAQNeKnscQAWE/ZU8ftAABgUfmdtHH/rCpDCwAAAKXkffzh2YgAAAABoS/WTxu0AAAAAAAAAAOZ/nk7Mf33+kOzIAAAAAAAAAAAAAAADP/myD9Y/rTyukIoAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAACqAFcIBH5t6x4F8rejz/AL4+u+T1KDAAA80pfz4/u76vJ3EVLU1RWrSLEhTmC5q+DKrVOmFKkP8AJb6yl/eTqlzgAGoH4offGWv26cOwoLw7X5o/UXZl9iHFsAOWgAHoBBEq4LzD5/8ARvLPN/jP27ofWO3w/WPUfuPwm670fmgJAAoB05ZQUgAAFB6gAAAAJc+J/KH6j+R+PfSv1vxn1r234Z/QQcnaAACfJbX5w/KG0/TH2La+nNVzZtDtWRWLAWJ05fyx/QPzr0P0/wCu/P8AD+fO3k+zfePx79zrVvG9zE+PvsvifH9f9Q/KfpD0/wDGv23yvyf7L4Hzjs/tfgO+/LP8o/Z/6R+n+K+7BAAAAAAACIRGAAANAAAAAACQARoAAAAAAA1APwUpfVv2R0Z3LjLeKLl/SE+rvD+Ps5P6h/LxH398eM+5/NWvBfHkfo9y7n8Rv2cVd1Tqfx8/R7DX7fOXUCQAamgqAGFAAjUcn+HG0fX36awwzoQEAAAQEiNoCTzHkfS5g0k7TKvQzzukAAAKb+CW+f6dfWWNhnQAMAAKdLxo9fiACVP2lvjdoMAGoUfAes/Mv7LPMOfQAAACHxzuwDtwAAAAADazr088jsAAAAAAAAAAA/MLaPMP3S9LnAAAAAAAAAAAAAAAAPzeD8+v3g4Ng5tQAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAKoAcogEefNfz1fS/q8v0/8ApL53QGVgAAKH4r9f6fL+ltzaLdoVqcJ5EK0IYWVqoVaY6lMX4sfpnNfVj6lXAqAjQ/Bv7GjT9TDz9xweTfnh1ZfSX2tlYZUKDigAVyAQOaUclVaebeUfMfqnk/IfGftfqHsH0X5v7B0f2n4gGvGFgpbiYCgAAB4OAAAAGC1LgPPvkr7/APN/nL3n9G/MPr3038K/oUPP9IcCgHmLXwX8I6rz36rufrL6Ya3LLVicJ5Qk+XPoPnPkzsf2X8Lw9XDryvo/zO76Uj/FP32tWKrVRVqgRIFHzT3/AJr5Htfrn4n7L9SfM/V/R978k/aAjQAAAAAAAQiMQAARAAAAAAAkEBoAAAAAANCH+Y7Sf2v9D2z8F5R16d9JC0LoXL6fnfxyn1P1uj0L53Z6fM184qL2NzXzZ7CneqNfjb+oGGv2WcuwEAAgNJsAalAAfPrX4rfRnTl8efujD+jjm1AGgAAICRG0Ajz3hfQ5w1g7XKvQjzukAAAKb+Gu0fe/2lnQZUADQQAzKnyE9fjACwn7M3xu0GACNKPjPSfgX9xrzDm0AAAAh8c7sA7cAAAAAA6XK/RDyesAAAAAAAAAADimv56P6MvW5fYBgAAAAAAAAAAAAAAGOH81v6782n1acG4AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAFUAOUQCAPxDx/T5cX98+XUObUAABQ8f/AAK7cf2m9m68LdlqxOErwVAjhCvWCrVHxHK+L+g/kJ+0/p/HerP385ygAI0P54/rqb/TL2Dg38s+Xdp+fvtnWfqU4tQAVjhgA5ZAAe5BoAAo5vLt8i8f+L/bPL+5+f8A0X2b2n7v8B6s+h/PhyahIMAABzwAAAACkOaUAA5j5O+0+E+WvW/1T8f+yvWPwn+hnL5vqgAUAHz78W0virybSfpP6ca+mPZ3NiVj3B4f5B+gfnXVfRPie7yvxZ9N8dofYvN1bdf89/UK1YVWsDQD5YT/ADL/AKAEuf8Alf8AV/xnhvrH1PI+sNr8S/fQy2AAAAAAAENRgAANAAAAACQARoABQDGyGsnKZjZzOG/KTRfo35ZpPn14M/6ID7eupXLw7l1OygVaQU6LVOk5p0wp0wp1A/D/APajn2+kTm1AkBADUmwAYw4Cj82vz/6sf1M/RPl0+Dfyc3j9zvonm0BNEAAAAgGsQDgeB7+cNoO9xvujz+gAAAKT8T9o+w/uuKDGgAQGgGFpPlZ6vGAFhP2dnjdoMAEaUfMNr8p/321yDl0AAAAh8c7sA7cAAAAAA7HC+4PL6wAAAAAAAAAAAPyG770+b9gTWQAAAAAAAAAAAAAAAPzb/PiH+8bvK6gAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAKoAeEgAofGH5fd3P55/QEq9VOLYAABwL+Zu0fDv7U+jy9BZonmFK8BAjiCGsFb558X9D8XsfnH9VUPqr6H809ld+gfy6qgqAND+eT9Q8tfD6mk9X9D5V9adbx6gmADmKMFBxIAOWpAQABYolCc7yPy/rPFvI/gP6H7T2z1Plvbun+6/Aw6OEAABQeoAAAABcuUAAAAMb5Q+u+L+U/U/1j8a+y/Wfwv8AoQPN9UKAAAPKGvkL5LpfL0Nr3v3Vr3D15r1W+JsbIqFR8peJ/uH89/W/p35H+2163k+1WjBPGEfDvzzNfuJ9W5WCfj/xr+m/k/kv1z9N8l9b6f4h/QCJnqAAAAAAAIajAAQEAzOUa5zmaOa58XOc8zn8KjBw2sSqy5rhra6NfRT4Px1r3vyyl23ZueF5JV+qm8TcuKrdpOdEV6zKdNrgflFrn/Yx+3UxU8sX4Df0h823cnPqBICAEbNAMy2fmb+b3Xj9k/ppnfshz2fnzc/l7+8Ws+zHPoNAAAAkEBoHnnDehzhrB6Vhp1ycG4AAAM/GbafpD9BpYYWACA0A5fWPNj1OQALkv2NvjdqFAAMGeGUfix/Q1vkHJoAAABD453YB24AAAAAB2uGnaHl9QAAAAAAAAAAAHgv4e92H9Nur1ZgAAAAAAAAAAAAAAAZ/82Qfqv8AYHl9IZ0AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAFUKDgkAFCh/OZ04979rbz+mZ5/QAAA8CAPxJx/T5f1z1Ns55RyPEIDIghgHy3x58T/RPbbXxH23I5vb7H3bL+v/xSquVAagfzz/0I8u/pWJz6dAIAAAAV1AAOBSQActQAAMChwgJACga+Q+J/Gftnk3onzf6V7j7X+hfzvqnsfGAA8HAAAADQtJwAAAAABnfJ31fxvyP6/wDrf4x9p+l/hX9EBweiAACqAA3xUXzd8+WvCvE6XE9wz0Hrmup3Qk839nwvSPZP1j8Y8y5/8P8A6CwPPlXkUif3f+nudehEsAAPJPij9O/JvOvtL3PnvqCX8W/eERUgAAAADeXa4rjWchyNLkOTo5fkqWZvB0vSo6bope/0km/vS9zbRtbcl0GAAH5rfKe8foj8OaLu+9qfO+Mmv1N6Em5aTtWE5EbjrpVOVa/MTPbnzA+s/fxVPFEvyI/pV5tmmGoCAkAGJgq8f/E7oz3f2AZ7iYaNSQA/NT8+OvL9+e55NRoAAAEgiGlHl/J+hzBpJ67za7acWwAAAM/HXafdf0SlhhYAAMAOK2z4M9PlADRl+vJ43a0oABo08ro/Bf8Ao86MQ49QAAAK3j/dzh24gAAAAB1mN96eV1gAAAAAAAAAAAH4HdWX6nfdPdiAAAAAAAAAAAAAAAAfn2H5d/vzw7WTl1AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAGwbFBwSADkL+Su0fLUffh/Qm7y+oAAFcABhfhh05an6y9/PvytSOGIDIwigFwHzH8F/S3pPY/Kep5959X1/3df/AGP+LHLeIAlcP52f6VeTfpTn0cAAAAA5aAAegECicVIAAwKVSQEABbFKzfFfG+v8N86/Pv6J9d92+n/L/XJvsvxZZHIAAAAUh7QAAAAAKDgMX5G+w+J+T/oL9S/IvtHsPwn+iA5escAAKDhAMAOa8ua824tnHcw02uDj9M/J/nP2zi9DvfR/gP0n3T3aSwJgAAAAHhXw3+tfi9r7yx6PXT8r/YDBa4HhaOE4WlxPE0carXY9ijsOul9f1Uvq+sR0lxMEAAAEg0EAAAAA/JPj94/Q/wCH9J47RHl+3h91WibdlOxOEiJx12q/zEP4p9BpdD5qGl9/oqfDKPHP2/5NUTLQBgQADRp82j/Dn9EOrL9Pp+XYARqAQfjrrPkX7vNWUxtAAAJAQ1KPGMr0uUKR7ny7Ts49QAAAZ+O20+6folLDGwQANQDzzoz5E9HlADXivWE8fsaUAAg2+ZU/wL/pB6MQ49AAAAKXkffzh2YgAAAABdl+ry+N2AMAAAAAAAAAAD869Y+f/wB2vT5wAAAAAAAAAAAAAAACuH86H6A4X94HndAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA2DY4FCQAeg8Ba/DjV9bl/X7608zpCKAFcAAYjX4v8Akfpc36uev65vcwQGxhFCGX8k/IfvOnqfCfquFtejf2E79Z/i9VAATjA/Bb+nHk2F59FAAAAActAArkAgc0o5AAKAbgkBAwKY5NQk5vLt8K8J+A/oPL92w7vf+z/RP5yDo84AAAuXKAAAAAArgAAo5f4v+5/PfnT6l/QPzb7D2/wz+hHGO4ArkAwAAABVBQAAzdM/z+8x/d/52/SD0H8R/fg4u8AAAAAAKvm1z8I+M/pv5R7jg/nn6fwGHlv2fdI7ruZfb9tL7fppHCYA1AABoAAAABIANQAAAAPyL57eP0Hq6R888sV6R9OC1bQrFlE8hTxpkSdf5na+LPRGdF5kGr98IqfkSj7b/RXi2GxYDAkBCB85fhRvP65feGNtRUADEAgzA/Av7F6sv1COXVoAACAkbXs8LT0uQGaPtfF0JHzaAIAAD8hNl6r+kcsMbBAAjQPLenLAPQ5gA3869QPI7GFAAINvmVP8C/6QejEOPQAAADN8o9DnDrxAAAAAADa9F8/otHJsAAAAAAAAAAB5k1/P7/T76/LsAAAAAAAAAAAAAAAAAfGn4to/oP2vI6gGAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA2DY4FCQBXIBH8/va+ty4/7z8moc2oA5QAACD8194/Nb7e7+b7z6K5EBrAjjBPOPM+r+Wek/Of627X6R+v/AAronfVfiaqAAniiPyX/AKMOTcdhoAAAAKDigAcsgAPcg0AAUA3BAADAqlckDQAAea+BfJfrvino3yf659D+y/o/82SnpfMgwWpcAAAAAAOUAAKADz74n/RfzDx77J+q+P8Aqqz+Jfvg6bUAAAABVBQAAAAG/G/1HyPyR+jH3H5/7Sfj/wC3AABB5615l5ta8y84s4Xfa9F9Dl8x4Z+k/lH3F9afE/e2jxvcEBEAAAAARDRgA0AAAAQCBgAAAAfkrw+8fohb0i1k4ejU7rLtv2ezwbE4TSocDZEKHnWfmvmFWcsPrH3cXKfhMn/RX6ZwbDUwKAkBHmbP5+v1S3n78OXVGsBg0EJADxL8C+nL99va+fRomAAgJE5rWfJj0eYA6r1Lg6SPC0BAAAfk7qdN+nIBhYIAEaB47140Du5wA6bK/STyetqUAAAzzKq/Av8ApB6MQ49AAAAMXzD0eYOrIAAAAAACdPsur8zplMNAAAAAAAAAAA/n/wD2I9bl+vikAAAAAAAAAAAAAAAAfgKHv36beb0BjYAAAAAAAAAAAAAAAAAAAAAAAAAAAAADYNjxAIAV0gofnR8t+lz+H/r7L+rzh3AHgAAAFnjP5U9GXzP9e9mH1d7fSsIJAKXz58r+oeI/f3d4em73Pi3OBwCoB8jJ/If7s8e47GwAAABzFGADyQAV1SAgACxRKEgAUA6csoKQAADwzfB/F+2+es/8/wD6H+hvoL73+f8AZPf+BHNKAAAAAOBQAYDAAA8d+HP1X8eyvubo5veF/If2oGAAAOBQAAAAAAA+cPgX9p/B/vL3380/VvL/ACrxPofLvL7Xm+tS9N9Ml+kejw/S9+QEz5r93574V+iP178T+5dH8I/okM9BAaAAAAAjQABoAAAABIIDQAAA/Nv5D2j9S59Z8E8n/M/7BdtcP0H1/wBj+pfxjYm6PLlkBQGsCGEOO+aR890A3+8C+LvAIr94Dh1EGgUAgJr/AIJbT9EfrVnYZ0MYDBAaSAB+VOk+Pftu00ysAAAgPN944g7+cA9C7rzuoZlTQQAAH5O6rpv05TDKwkABgN8S7cGnZgAHX436AeV1tSgAAGeMU/w9/op6Mg49AAAAOY899LmDpyAAAAAAAALCfV9Z5vTaOfQAAAAAAAAA/Jf030+b9YTWQAAAAAAAAAAAAAAADwEP57f6BeTX0U4tgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAKoAVwgEAOWQVnEfzuduP6a/GXRn++LvL6gB4AAABYrkeF/BnRl8ccd14+ufQ9L0LhRfOP25+f/AL59Obf334W9+nM5QcAAH5yp9B+s3D0KudAAAADmKMFBxIAOWoAAGBQ4QEgAUA7eTAUAAAsgACeU8PvfOXln5t/S/u30j9l+Ndi76j8tAAAAAFcAAFAAADgGNfM3xR+0fhHqf3X819T3B+d/p4AKDgAAAAAAAzPKKXkfzH6Hn+ZMo9C9S4u/1L1CH6h1kAJgAAAAHJ/n5+ifmPJfon3ed6Sflv6+ADQQAAABENGADQAAABAIRoAAB8r/AIuaz+2nEYe9839t+Sf11p8VHH2H17+k/g0/GfRflHn/AKAP0JBJGEUQQVwr5w/zqgK8u/QfF/bRyaADQsBAT+bnwd0x/QFf5dQTGsQYAMJAA5MP5yv6A+nP11ObQAAAgPEenLOO3EA9W6bzOobFsCQAAPyd1XTfpymGVhIADQy/Ie/nDqxADuMNO3PL6kawGADfmS3+Tn7+7ZhyWAAABw/HepyBvAAAAAAAAAASI6zs/N6pjn0AAAAAAAA+GfkH0Of96jogAAAAAAAAAAAAAAAAA/Gbz7Ov2UPL6QAAAAAAAAAAAAAAAAAAAAAAAAAAAAAKoAVwgEAOWQcw/E7pz+o/z+7sP0v+4PN6Rc6cAAAwKHKAEHmrXzn8z9EedfIG8foN9BfAf0B7ov6L/PbnAqscAqAfjjL/AEm+3eDocRQAZweR8Bc5PX0em+kyxyakgA8qAABgUqkgIAC2KNwpBgAAPcAAAHHc/o/N/hP5t/THrv0t9J+a+jn2H48AAAA8AACgAAHAADsrTH4x+W/3b+efrr7B/MP1q6fKfZOUAAAABrPO/HqXkHkFrzfuqXrXq8PuPhf9H/K/pH7f8D6Nx8l9mAAAAAAAARtfIPxx+5fz592fTv5X+wB4H0gAjQAAAARoAA0AAAAAkGggABlfzPaT+x2b5v3HlHpv5f8A0h0vmD8nJ+3vv/zP52+SPqfxeaEf1b9TEtaKKIIa44Kwvkbw4rd+wcr+upuSwQiFABxLn+dP90d4+kW82iCoQGlAA1JAAA/Gb2vpy/S5vNqAAASYnjfZgHTmAez6vl9Yk1GEgAAfk7qun/ThMMrCQAGhzfmPocwdOQAehYadaeX1DGAwAT4Z0PjD9s9JDmsEgAA8xxPX4w0kAAAAAAAAAACwn3HU+V1BlYAAAAAAeAfjf3Yf0qnVmAAAAAAAAAAAAAAAAAcJ/N6H7N/TnldIRQAAAAAAAAAAAAAAAAAAAAAAAAAAAAVQA5RAIAeSDmHx5rP5ffqv6XN+S/8AQhx7Su5dVAAAYDHKwCAAAX8It5+kvqb4r9560/RP59FAUFAAOX/DKa/of9S4d3EV4H8l6x8kfO3Tnv8AdUtLmRed9en9ffffPr9GmFiicVIAAwKVSQEDApjk1CQYAChIAAAANGHHT86fPH5t/SvcfTvrfJetr9x+GgA4FABgMABQUAcCgHm/f53wXzv7p/PP3n7b+Jfvynk+yAFVnknjFLxnx7SYvYQ9f9fivXN2AVM47q4/zM9w/bPwT70X8K/oYEwAQEAAAAAPGvU8j87foT9l/C/um1+Ef0QE2IDQAAAEQ0YICAAAAEggNAAD8eHUPpf5j/UeF2HB9pp8ZHH3f0p+lfz7+b+79B+YdVzTnmv0pVbbCY4hwwigoh8DVlcf6W4160ctgA0oBn5OaZ+Z/tewTKmk0AMKABGoBAB+f3yh05/tg3m0AAAJPOeG9DnDWCRHur/K6wKjQkAAD8kdp739LpoM7CAAEDiOD9LlDfMAPSufXpTzOkYwGACflftMX6pjDCgSABovHI/a4gYAAAAAAAAAAAAb2deiT+T1gmAAAAAcR/O524/1QHXkAAAAAAAAAAAAAAAAAH51h+X/AO+3FtsHJqAAAAAAAAAAAAAAAAAAAAAAAAAAAAFUAOUQCFBxIOYU6X86n7n+py/ix+p+Gn2A7h3UAABgMeMAgAAF+V+P3vmr7B837uQ+0/GwAAAAD4e+cJr9v+m4t/hz83dY81+l+jP6M+iGuvGgCv40n8r/AB8n9A/rfy6+2uxtRyAAUA3BAADAqlckDQAADgeAAAAUh7RQjX59+bfzr+jdr6i7PI9mX7z8EVwAAUAAAOAAcqAYAN+XfofmfiP6C/XfxT7nufg/9F+QeJRr4l5FpPT+0D9m9mzfosksVAoAABy/5ofpf5T6h+gfk+1IfFfeAAAACAgAAAct+cn6R+W3P0Xz06E/O/04ARoAAAAI0ABEAAAAQCGoAByXxH8Z+z/Ov0r0YzeNcP6TD1/n/UfT1f8AT/40/PXu9eT1Dzip88/Qaa7ZomRCihDyP8w5cf7DZa8r8v0v1Pm5LBA0YFc3/N5vl++XuuVCSNJsAalAAjUAgA+Lvz46M/3YTn0QAACTxfI9PlCkdBFeuHmdIFsaQAAB+M+0/SP6CxSEWEAAIHm/KepyBrAAepc2u6eb0jWIMAD8GenP9GfsrLQMmCQAZ9LyY9niAAAAAAAAAAAAAADT9P4Oic49gAAAACD+aLsx/qC6jsyAAAAAAAAAAAAAAAAABgfgH67lX6uHmdIAAAAAAAAAAAAAAAAAAAAAAAAAAAAVQA8JABQcSKtAB+a3zJ6HP9qfFtz+4T/M6gAACgUHDQEgAA75G8H9E/O76O9TzPsM+g+DAAAAMwPxO/TWK9j/ACdzvJ+9dY+prdSKAAIAgJkI+EfiGa/VL9G+TYIAAChQUJAAoB05ZQUgAAJAUAAAAqVcwAK6rwL5j/Ov6M2PqTq8z2dfvPwQACgAAHACg4AAABGeCfnT9j8RJm+J9F1vtnn+n7b7Vm+zIYAKCgAAAAHO78/5lel/uH8/foRJ+E/0OCoAAAAQEAACncfAfkf7t/O/6S9/+J/vgcXeIDQAAAGoQYNAAAAAJBgHjvn/AEHxh9B/Af0D8y/A36p/Lv7A9Dbq7S2mk04/gbGK38QVz9DQlYJkYeWfByfyP+juV/qDpc+0X5ZbR9SfV2QGbEGgV+e/w90ZfvSc9ADUmwBG0AA0EJADyvxa5+vUikAACqjwtPT5AYeiY33B5/QBTWEgAAfhVvH6CfZ+eiEUEAAAeTYnrcYXIAes8uuunndI0EKADO/B7pz/AHG6nn2CUCQAc3pHnR6/IAAAAAAAAAAAAAAB0mV+ink9YAAAAAfzN9mP9P8A1fZkAAAAAAAAAAAAAAAAAAHl/wDOYH7L/U3ldIRQAAAAAAAAAAAAAAAAAAAAAAAAAABVADwkAFBxIq0AGF/PP15fsL+K/Xj/AEF+oeX1BLAAoFBw0BIAARfnd8h+09j+Sv2f5J+zfe+j4wMAAA/NpPivrzHT8ufvrWPuW3UvcxyoUBEBrWNYDfFpPyk+2MtP1xfyaAAFiiUJAAoB28mAoAABzwAAAAC5eAAABXVeBfMf51/RnQ/WPf4XqR9x+HAwAHAAK4AAA5Dwil4N4dqrHugel/Jn6H+W+6fdnn+jvnwv6GACgoAAAAAAAc7+ZX6V+V+wfe3je4p8d9wAAAAACAgAA341+p+R+Tf0g+0+C9ePyL9rAEaAAAADUAAaAAAASCB8x/M35n/Tf2d7L93+DRfi16fznzN+qO8etrUySM4f5GVc33A/rHuyE4NHy58nqvFPvGK/TT03m0GKjwu1+f36z3F456AbQr8EfvPoy++znoAak0AxjAYNBCQAAABoAAczU+THo8wAe28ut849gKSMQCAD+efoj9jvonHRpNBICADxSt7HEDQAev8ALtop5/QIhpQADRoDAAmQA856cucPS5gAAAAAAAAAAAAAAFR67b8btBMAAACt/NJ2Y/1C9L2ZAAAAAAAAAAAAAAAAAAAfBIfkN+/PHr2Bx7AAAAAAAAAAAAAAAAAAAAAAAAAAAFUKDgkAHIUQ4oAD88dY+LPVfR5/Wv018/oDGwAKBwKNASAAHlnyz8R+9ff35T/efgnxX+uffz96VIARh8AfIsX93fnnFfrX71vnJIk54K4AAQGtBjAjjZwn4+y/uL9XuPYM2BY4QEgAUCu3EwFAAAPcAAAAwWpcAAAAAVpv53+Yvzj+k/SPrL3PiO3Pr/yAAVQAHKEXjbXgPgeq899oa9694zr0x0s53bn+DPHf3n+dfvX6C/Gf3UXxfdUAAAAAABgAc7vz/l/7v+1/gf3Wfhv9CAgAAAAAEBAAD5i+FP2v8E/Q73r8s/Xw8f3BAaAAAAwABEAAAARD8dfOfovlP3P5P1vbn3H4aMD8yvzB2n6A+rrn2ryKl5N9zj6DFc+PfP8AL+afHpr6n+ycq+3upxsAGy0Kf5c/Ue+X1GctgDeMd/zc/wBJe+XXHPQAjVQJjWIMEBpIAAAA0AAPP9I4A7+cA1PaOLYObUCkjEAgA/ml6c/6DvS+fZomEgIM+l42evxAAAex8m15vB0CIaUAA0aAwAJkAp+S93Ow7cQAAAAAAAAAAAAAAAPS97yOwM6AAAA4P+eftx/qaOvIAAAAAAAAAAAAAAAAAAAD8SPOIr9tneV0gAAAAAAAAAAAAAAAAAAAAAAAAANg2KDgkAHIUQ4oACi1/PP9e+py/LH9CfHsHNqABQOBRoCQAUPk3X+J/evp9v234L+Vf599+P2v9IbZ7FUPG/jBO99sY6fmx+vu2fuMlTJID3A5RqCEG1omRhHGEXnTPxl/Xrn1+4jl0AocSAgALY5NQkGAAoPUAAABodSUAAAABXAZUbfMnzn+Z/07719U/W/kmyfSfm7gDHF4P896LwTU0Xv/AL7m/cdSGCYAAHzr7fgfBvu/7F+Hfd2z+D/0UGWwAAAwGAwAAOS/L79P/I/p77Q+Y+tD5b68EAgAAAAaAAeBez4f58feX6h+R/SR+LfvAAjQAAAQGgDQAAApRp8Q4f5f/UP3Tt/pn8xhrzABwIvhTxfZY5qvPvJpOb5RHrn0xD+tPr3G+lJYAACS2lP5s+ROjL9TjksAT4ft/nd++FyGLABioExrEGCA0kAAAARAADyfbPmDtwAOv9N4OgMLAZGCAgA/mN6c/wCkrqOfZhLBAIOV1jzU9TkAAA9q49p28PQCGFAAINoMACZAPO+a9bkDWAAAAAAAAAAAAAAAAPVtPxuwJoAAAD5t/JDvw/pHOnMAAAAAAAAAAAAAAAAAAADn/wCd0PtL9KvN6AxsAAAAAAAAAAAAAAAAAAAAAAAAGwbHAoSAD0AhxQAAHwt+bnfz8P8AulL91OLcACwegHICABQ/Pf7H+J/evRT7b8FPnel+e3zz2ZZdil6/9o5V7z+EDP1m+jOnKaRqV4SPG5whQRAY0bIwjiFFEHzT+V81/Q96Bw7gqVSQEDApj0wJBgAOB4AAABSVzQAAAAA8AGHK5dvyb5N+Xf1H9VfQ/wCgfz9zfz37Xx/zv4rpPp30Kn9A+n5MVNwAAAAABzvwH9/+b+YfoN7vzvtJ+R/tQADAoAAAAADg+zh/L37V/XfxL6pPxb97AAQEgAAADQADxT85v2H8Q+8PpT87/Tg8L6EQGgAAA1AAGgAGX8L/ACX61d+4qxus+p/KwAAAD56pfnJW6J+5es5tOs9Nh7ggYDQAAACRhV0vyH3y/T30jnoJE/IXQ6b9UxhmgAamgqEBpQAMJAAAAEQAA8R6Ms468QD0PufO6AysBjAaCAD+Y3pz/pK6jn2YSwQCDzvoz489HlAAA9q49528O4IYUAAg2gwAJk5HaOEPV5AAAXpsr3bnDvMQyBlWjrGdl9Oecm2YMAAA14r1I8fsAAAAAPg35d9Hn/do3gAAAAAAAAAAAAAAAAAAAA8MD+ez9lubT6qODcAAAAAAAAAAAAAAAAAAAAAAAAGwbHAoSAD0AhxQAAFb+f3qy8r/AEp2j9OTz+gACxXIByAgA4zn9P4e/Rb5n9UefXfjAALQ4kKtH4DfS/oc/wCg02kSyjllE94PUFUEaDWAyMIohwxCh/LJPrf2V5NwytwSAgYFUrkgaAAAkBQAAACpcrAAAABQcADAYeA8Xt/B9XyvseS9v+o/M/ov6EzXWEsAVwAAAAAAA4D5i+FP2v8ABPpv7f8Ahf0K8fH/AHIUAAAAAAAAeV/mV+t/in6E/QH57+nB43ugAICQAABoAB5N6Xl/mv8Adv6t+N/Sh+L/ALwANQAAARoADQDLjb4P3vzH+nft6x+nfy8I5aAAAHz9+eW8/oz8kyfpQYWCYAIEYAAAAhqOz8+NM+4+z5AyZ+Auy/Sj7QiwiQAaNCaAGFAA1JAAAABoAEfg3Ti06sgD1Hq/N6QzoBjBtCQA+WqX03am2omABIeRdWOQd/OAAB7LybXG8HQCGpQACA0KAAmcjzD0Odh14gAqPRej8nrDOwAAAACHG1jFyOzHNg6cyYOh7rzuiwc2oAAAAB+S/qHp836vGsgAAAAAAAAAAAAAAAAAAAAHw9+LyP3f9d8rqCWAAAAAAAAAAAAAAAAAAAAAAANg2OBQkAVyAQ4oAAA+MNJ/GL6I7+f95zzOoACxXIByAgA+fvK+v86+y+P2w+h/NQAWhxIfnrrP52fth6HPfmZNOKSUJHg9wKANBjAjjCKIIIRw8Il+G/8AQvz7esnPq4IAAYFUrpQUgAAV4KAAAAXLwAAAABwKAFHL/ONr5v8AKNJ9z9s5+3xbpPyn+pPrn0/9K/moO3xgBygAAAAADgADj+rk/PnmP3b+dv0O9P8AxH+gg4PSAABQFAAAA8I9fxPzp/Tn9E/L/Uj8k/awAABASAAAiAAeTfmv+u/in3b9Kfn/AOlB4X0IwAAAGggA0M/4L+S/W977e247B9N+YAhgAAeJNfmz+iWy+gcTCtgTCWAAEaAAAAIRpR87XPyP+oNSGFH8ze8/0E+s46AkAhBtJoAi5rWWdIEiZ0CAAAGgAZXivXiHRkAeq9P5vSGdAMaNgSAAAFNRMACSPxHtwadmAAAHrPLrsJwdKEjUoAABgUAEKjyqj7HGFIAOnyv0I8nrAAAAAAAAACMTZgAYAAAAAAfz/wD7C+ty/YJSAAAAAAAAAAAAAAAAAAAAAD8qA+B/3c4tutOTUAAAAAAAAAAAAAAAAAAAAAAAbBseAEgCukFBSgAABofgV4j6nL/SlreZ0gmFiuQDkBAB8afPfpXqPv8A2+IHqfIAA6hSTL/nU6s/1F+nevGedk8wpZQ+FfF5rQ+/2vyi+gk+J/V5ojajiCGIIIBwQi/LD1vLT9YDj3cEAAUA6csoKQAAOeAAAAwWpcAAAAArgMYXzt8z7rxv2ul9LfQmVbZLg+cvF+0+XPXvgv3v626b9R/lwdtxqAAAAAAOAAABnyJ9P8n8ZfZH6b+T/YD/AMP/AKDFTVQAAAAAAD5T+Mf2j8F/UvrfyD9vQ5usAAAkBAANAAPHPU8n82f0E/U/x/6FPxj92EBoAAAMAKyr4Q0PzH+nPt6x+nfzGBIIagAHlP5j7T9//R+bGy0EBNAAAxoAAACEaUc7S/Ir9hdY0jmrz3+cboj+m63hsCkBCA0mwDybkfQ5i1S9J67h6AzoAABoAHMeT9mAb5gHqvT+b0hnQDEHGEgAABSNTAAkxPJu/mDqyAAA9E59OtTzupCRqUAAAwKADzrfHmz0uUAAD1Dk12Tz+kAAAAAAAAAAAAAAAAAADytr8BP6ffX5dgAAAAAAAAAAAAAAAAAAAAAAGB+I3j0P9wtXyukBgAAAAAAAAAAAAAAAAAAAAANg2PEAgBXSCgpQAAAofKP4gd2H76+9cewJhQ5QAgAAD88Pk/2P7e7b6P8AMQ6fKAB1Cknwh+bndj+2ljoznsCnmCfyJP49/S4Hva/IT9fU/wAvfusPJPqcPkD61CCFqCAK8A/nL8u4f9G68PQoSABQDt5MBQAAD3AAAANDqSgAAAAReFNfMHgHRPqn0zL+jOizYqagK4MT5Q+a/S/CvqT5f9Q+lbP6Z/MwOQAAABwAAADAo8z/ADv/AFH8g6n9DeD0uoX89/TQAAAAAAAD4O+u+J8f/UH1fGvp+efp6AAAAEAAANAAPD/zk/Y/w39IPa/zH9aDyvYagAAA1Ab8R/NfpWB93Y9t0+r/ACgACRGgcK1+Wf21svqwwoQGiAmgAA8F/PrSflfyPaKeiz2T6Hh/Q/1hjp6oS/yc/QTfP185b+WvyM6J/oWOfUCQEAMJspNeDJ6XKAHY516RY4OkAABoAHC+dd3OGsAHpfY+f0hjQAIVGEgAABSNTAAk43z30uUN8wAAOnzr0pPK7EJEbQAADUKAOW8/9HkDoyAAA9l4t5Dh3AAAAAAAAAAAAAAAAAAA/MzaPNv3C9LnAAAAAAAAAAAAAAAAAAAAAAAAz/wTDS/Z3z95zn0AAAAAAAAAAAAAAAAAAAAAAqgBXCAQA5ZBWKMAAAUHfjTvn+mXsmVgmFDlACAAA5b4P+K/df0bf9p+Gg4AB1Ckn4P/AFX6XP8AbtjSbFgU9gJvitP0X6hCV4/w1+rE/MP1CF+ZX6qtfkB+uYQQNV64QVx5X8/8v+iv0zh6AUgAUCu3EwFAAASKAAAA0PpAAAAebfL2i+Z9nWfpv6ayrtyWADgAHKAB5z8cfEft8H2dN+kn3P4aAAAA4AAAAoBhRvP4Q8D/AHX+dv0Q9s/HP3UPK9gAAAAAACJz+aXRftv4D+iB+I/0AiAgAAEgIABoAB86e58/8Dfp599+cekH5L+zo0AAAGB8leP/AJp/S/6Aaf6R/NwXiABIMDB/LXWfo37blhDAGABFAGR+Rui+Efsboy+nvW6WlkC8l8ST8P8AnZP6k/VrDT5a95pfUZy38J/DnRP7mnPqAgJAGpNGNR4iejygABZ7TK+w1OLcExAQDyXmvQ5guQDuPRODoDCwACo0JAAABo1UABJ5jzPq8YaSAABKj2G35Ha0ho1gMAGoUZdry6P1eAGAAAezcW7zh3AAAAAAAAAAAAAAAAAACj/PJ15ft59Z9uIAAAAAAAAAAAAAAAAAAAAAAABy4fgB65lX7Bu8zpAAAAAAAAAAAAAAAAAAAAAAqgBXCAQA5ZBzAYAAAoOAAAYDHKwCAAA8Z8G+K/d/uQ+1/CQEDB4HGh/OF+93p83XWaVicVmcJ/n9P5w/SQc0ovxt/XpPZ5Zr8t/1iH+Ov69igrtV64V64V/xRVfqN9m8WwQAAWxRuFIMABQkAAAAKSuaADJF82fLey87+kaPqL2HBgMGACqAoOAABg/m75U/M/6Y+hvrD6z8nuH0H58ACgoAADAoAFcB80e7878FfZf6n+PfX7vw/wDoIBgAAAAABhflj+l/lH1f9efMfWh839U1AAAAIAAQEAAPkL5O/bfwL9R+n/Gv3Qbz9KAAB8+eT9V8ofoP87+j9On3H4cAAASFUPzI6non9E3c9AAA1ACaKCPwo5Hqy/VnutIleDgEjCPmA+Ffhua+vftSL+3Djv8AOPwXon9lDn1AQEgDUmjMZ4WelygAAAGtL6fpuXbbdhonK3Plad/MDADW9m49w5dAAC40IAAABjFQAJJ4tV9jiBoAAADf9R8/pQ5NUawGADUKreWdmFI7eYAAAA9H5teiPN6QAAAAAAAAAAAAAAAAAA/Pn4k9Hn/onN4AAAAAAAAAAAAAAAAAAAAAAAAAOH/AIPoT9afO6HGFgAAAAAAAAAAAAAAAAAAABVACuEAgByyDmAwAABwKAAAwGPGAQAAHzHX+K/dvqk+1/CgEDB4HxhS/Mj9rvS57NlzYshYsBZsJ/l3kp9N99D/MP9lQbGH4f+6C+Yv3LZWrua9YK9YK35VJ/Qv6Zce4ZgAFscmsKMTD4vexsnzvos3P4/aoUeT1oKmPbWM+kAJnOhm+n83wXzh9x+Mze/8Azv6D9R9Uubb3/S+cunX5gAAOAHAoAAOBeTw7vi7nvy/+oftb0j9M/mcOvyBwAAAFAMFBwAB5r+cP6l+P+r/oD4nv6B8R+ggAAAAAAeZd/nfl9+k/6h+Q+zn5B+3jQQAACQEACIAAfA/2Pw/l36cen5Oin53+nNAAPNvgj8//AKB+8vS/tPxcb2eMAAAIPgSl5V+oDLpmwAARoBNN/F614T+wfTjuztSyDcA2MIoBV/Ew/Pz7DjT9JTh0/MLhOiP12OfYBASAI2aGj+f4vS5AYAAAAASaMuCiwGgAAPX+fTdOHcAApjSQAAAYxUDDIJ8iPW4wAAAAA7PG+8PK6xjAYAJlU/O831eIKkAAAAuy/VZ/H7ATAAAAAAAAAAAAAAAA8++A98/z2/eT0cPqgAAAAAAAAAAAAAAAAAAAAAAAAAAOHD8AvoLKv1mn8zpAAAAAAAAAAAAAAAAAAAACqAFcIBADyQcwGAAAOBQAAGAx40BIAAfGPsHxf7r7mfafhYCBiuA/K/SeA/Tr0MLNlzYshZshasJ2q6a7zcqyRwMp5ArVdqrWarVwrVgq/n2nx/7KcmxBjpyPJeR9jynK+L95zXN+L9pzuJ5X1tvbvn2tX0fndTR7PF1LvX5Ni1txWjXlCpz/AJqb8gtfWfmP0b0/k/SGV8b+wZWRw+7iZXn/AEOh0W/B0vSex8f1HWe38V13Xe78H0T/AE/mgAAHgAN+bvH+w+T/AKb+I/b/AKkk/T/5hcCAAYFAArgAAAwtsPzoxf3T+eP0k6r8N/oQOfqAAAAAAD5i+I/2X8J/VPpvyH9tDDpRoAAICQAGgAFep/Nfe/b/AOf/ANEHfh/9AtRMws+n88fqT88/oX6FT9H/AJxGgAAAI+W/h/af1Y6nBgMAAEaAS/zk/NTeP2g7HXOzO3PIJwDYwhrjrVheCfBar9GPs3i1/NbxnWf2OTn1AAJAEaqBPxnnvQ5QtAAAAAAAAAAAAG3D9gXzOoAApjSQAAAY1Ugw4TTPhj0uUAAAAAAOu73zOqRmGgMMa55Hm/R5kNYAAAAAADX9J4OiycewAAAAAAAAB5h8vbR8y/PvZlR3amyJnENct9jaL9XPeAAAAAAAAAAAAAAAAAAAAAAAAAAAAA4f8Ag9j/X7z97xz6AAAAAAAAAAAAAAAAAAAFUAOUQCAHkg5gMAAAcCgAAMFY4aAkAAPgL69+L/AHb0U+0/CgEDFcB+J20e+/cHdhastWLIWbQWrKduyqs2kTqyKsipXZVqOa9RqtXCrXDmPmbw/tPnT7F+Q/VeK5X5v9I1ew6PO6vq/e+D6bpfY+L6ToPY+O0jr8gAJAUAAAOJ+Tdo+X++2n6v+kcKuksAVwFabwOd4foub5jx/seX5XxPteS5HwfvLPaac/cdz9J+b95330/5h0K+p8uAAHD/ABF8V+2af29tx9MfX/j4AFAMFBwAAAAEPwf9h8P8/fpR9N8l6iv5T+ygAAgCIAAfnz9l8Lyv6W9HI8+F/RRoIAABICBAQAAw/wAvP0r8q+k/sz536dG/L/XJ8G/MfqHQfaevIjfovzoQEAADy9r8uf0wteyJjQAAAAwDjk/5xv1z6svoO1StWRTyg4BsQQVwq1B+D/EyfzZ/RlzafCvyuz9r059AACQAYqBPgPN+7nDWAAAAAAAAAAAAADve/wDO6AysCmsJAAABjVSDDxvoxzzu5wAAAAAALcvpdHj2Zl0YNPqxCkAAAAAAAAWpfcdN5fUplYAAAAAAAAAY4vnb596Y8O8a6M/J8zaPWPe2e1+9h9F+zAAAAAAAAAAAAAAAAAAAAAAAAAAABxQfg1uQ/wBntryukBgAAAAAAAAAAAAAAAAAVQA5RAIAeSDmAwAABwKAABQKDhoCQAiVfnF+h3xv7jsn2f4aAgYrgPwQ2j72+pe7C1Zas2Qs2QtWk7dlVatog0Z1r1qyqVmfOnq3g/ea+L7nw/nvAfKfqXDcb8Z+xQ7Ge/yv+of035f23Z/Qfm2seh88FUrkgaAABXgoAB4p8j7z4X9HaT9b+q4MEwAAVwAAUAA4CMfM8V4/1vEcN85+k8Dw/wAx+m7HofV5no3pP1v5J6Vr/UfmBUrP5F8O/Lv6h+4vVv0X+dw7fDCwAVwAAAAACqHyP8Xftv4B+l/rf5H+1B53qAADEBEAzdM/yz+mv2T8K+xD8X/eQBqAABICABoAAefdvB+Xv6B/rH419AM/Gv3T5n+evzj+j/0N0P0X+dUbWQA0AAy/yn0n6q+ws2jZYAAAA0E/NBnxt+t3Tl84fXYWvjwPJnp/YVhr4UYmz7ba9bps8D8ZH8h/amNezfmZD/fhOfUBgQADEwVZvhfVih05AAAAAAAAAAAAAC+k4adkcG4FJGIBAADGqkGYPlPbzB05AAAAAAAAAAAAAAAAAAAAAAAXJfTdFwdGqcuoAAAAAAAAAAADfPGvKPJt48a8R6I8C3t8/sH7rZ9eSgAAAAAAAAAAAAAAAAAAAAAAAAAYn4YB5p+2PFt3xyagAAAAAAAAAAAAAAAAAVQA5RAIUHEg5gMAAAeAAABQKDhoCQA5nHu+DP0e+X/WQ+u/GQEDHKB/PzvH6RfSHbhZstWLQWbAW7Cduyn5D8Y/E/0fzHpnhfpH3R1X6Z/HWT8ffPfeeJ++/GfsPm/H8Xs9T2/rfK9j1n2n45t4/v8Awn8+H9N3PpsnPQMCqV0oKQAALIBVD5u+Q+iOa+rqX1V0uDcDAAAAcoAwGAA4AB4AAQquG878D7/zrzf5L9a4TsfI+u9R9S+w/H/VfLPb+J+Nfqv4z9n+mT9N/mUKABygAAAAOBQD5e9/5r4U/TP73849bPyP9pABAEKEG0DzT8vP1r8W/Tn1X81/Vg8/0xgAAICQARAAADwT89P2T8M/UOb8f/d/z5+/vzz969CPv/wEaCANAAPz15faf0wdz0jQAAAAGgn859r9E/ozfP8APX9RgufkoH6/0g/Ij9T0++/Hxn641XNOkHzr5OV0f50QfuZ+AON/0wt59QGBAANTQVHju+fNHZgAAAAAAAAAAAAACo7fv+DolMdUjEAgABjVSUqXl+R6XGFoAAAAAAAAAAAAAAAAAAAAAAAAAudDz6dHsef0KRQAAAAAAAAAAAABB4TS+QfiLvw1P1f3j7rAAAAAAAAAAAAAAAAAAAAAAAAAK4fjt8Qo/ar3/wArqCWAAAAAAAAAAAAAAAAFUAPBAIUHEirQAAAAPAAAAoFBw0BIAeacft/K/wB8eH9+H1H5MAgY5QP5+d4/SD6Q7cLNhqzZCxZCzYTs5mfX+bu1+Q/2rtctzcfX/cH33458UcT8B+unqnRwe19x+n/zRZret8tXrBVrj/n1/oq5tPSTnsGBVK6UFIAAFymfK3yR0Rv/AF+j6RtYWOqVAAAAAcoAwGACqAoOAABwKAAZ/mfnfReY+V/Gfs/mvTfP/oG55lePvf2h99+DWj6b81HAoAAAADlAcSAvlr4Y/avwb9H/AG38s/YQ8r2AQCGhSINPkj6P5b5T/VP6P5XQPzn9TEBoAAEAANAAAD40+o+R+bvSPN+79w+ofj/qA9r4oaCAIgHztS/P/wDWS10phQwAAAABA+cPwo1n946u0fnl+paL35HB+xNkPlPkk/tr8cmfrxSJpUmfK3MlfTX4Mo/oE/APnv8Af/2PHQJoCAAamgqOepeMno8oAAAAAAAAAAAAAAF2X3vXcHS+PKgEAAMoN8xxnbz0ToyAAAAAAAAAAAAAAAAAAAAAAAAAAAALKe7tce23e49glgAAAAAAAAAAAEQfF35ad+H3N+wPTmAAAAAAAAAAAAAAAAAAAAAAAAAH5zB+R36pc9/eR5/QAAAAAAAAAAAAAAAAFUAOUkAFBxIq0AAAADwAAAKBzFBASAHinnfT+Mfa/m/Th9H+ZAIKHLIfghvH3Z9Vd2Fiy1YshYsBPZTn8m4fpPif0z8l/q/svPD53zr9APrPC5Hk+j5Dl4fG+u+pV/TP5vgrVnXrBXrD/n6/oo5tPSTnsGBVOWUFIAOYZ8k/K3Tn6v8AYMV7mYUDQ6koAAAADwAYDABVAcCgAA4FAAAAAr+Y8Hv+S+QfDfuXn3deJ9t757R+ifz32rvovzwAAAAcCqJQkKD5u9zwPz9/Qr9I/Kvok/HP3QACQRNAprBfmZ6H+0/g33kfi370ANQAAJAQNAAABrX5xYXzP7P+mkv0v40AhAaAiBzn5M6z+gf0NDDNjQQAAABA/JryneP1HytJ/PD9UEXvyLD9kbg/ibok/rj8aA/X2g5o8818ddkV7z+O6X6M/EHPr9afohlYRQEgIaNCaEDy3i/Q5guQAAAAAAAAAAAAAALSfV9Bx7a1znsROhz+0cpiduKFyAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAW5e1t8m29b4tgTAAAAAAAAAAA88a/Bn9ZvW5f0JGAAAAAAAAAAAAAAAAAAAAAAAAAHzD+HAfS/61+fvunPoAAAAAAAAAAAAAAAVQA8JABQcSKtAAAAA8AAACgcCjQEgB87eT9jy31vh6Qe7+eAAUOWQ/F3aPWvurvws2GWLArE4TzhPyXP6v5+91+Qf2Tvcfj5Nf9G/0P8AI4ovqfxWGu5hgCtAFesOvji/n4/pu5ttk56BgVTllBSOOZ8e/MfVn739j5V6kY0AA0OpKAAAAoOABgMAHADgUAAHgAAAAKCgAGF8nfN/pnzwfmv9Idt7X9J+de5+ifoX8+h1+UAOUFUkKQMAPC/zl/XvxD7a+tPgv0gPnvpwAJERUjR8d+Uv6p+OfpD7V+f/AKWHl+wjQAAkBAgIAAHIYd/52c+/rvqv60+q/GQ+b+pAGADQ/Oh+0/ooY0Eg0EAAAAA/ng/QHoz+q8u1+X/6Jo7D81h/rh5an+aP7XIs/jAz9gM9z8h/J7cHcj+zfzgU/QHY8uvxF+8CYZaASAhBtJoQIvJ+jPljrwAAAAAAAAAAAAAAAAAB6JYBoNAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABqQ+g6Dg6NU5tQAAAAAAAAAA+avxU78P6O/S+nMAAAAAAAAAAAAAAAAAAAAAAAAAOHD8U/DJf7EfRPl9ATQAAAAAAAAAAAAAFUAPCQAUHEirQAAAArgAAAKBwKNASAHzB4v3L/qEoPa+EAAocsh+Xes+U/px6HPYnZYsCmsBNMEsofMXzn+b/ANcTu8n7L7N9Z/T/AOOY4u/5uKEIYAggCvXHX8YS/IP+lDm2DCgoB25Zk4S18ffNfVn9GfY2NehLlQAAA0PpAAAACg4AKAAAcAPAAAFcAAAAA4AAACz51+PfzL+mPZNvXl8V0fD+19y95+8/Cu+d9R+YqolHIUAAAeW9/m/mj9O/sP4b9xL+H/0GAwQJLRCm/LXv/NfG/wCrP1Xx+ofm36qNBAAQEgA0AAPg/wCZ/Tev9u9f578yf0v/AG3+Z/Vz8h/bABgB4HS/PD9Z7XRGVBINBAAAAOYD+Zr97+nPq30vkK0j1K0Pxn1JP7Q2AofLAfQvjVT+cHTj0eVF6X4An9M/fnHr/Nz/AEQaL1E59QJAQg2k0AMbZ5tw3ZzhpAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAF/pubXp7vndAJgAAAAAAAH5Db57f7SejgAAAAAAAAAAAAAAAAAAAAAAAAAADPzkD8kvvfGv022PN6AGAAAAAAAAAAAAFUAPCQAUHEirQAAAArgAAAKBwKNASAHyf4X6B0/0dtyB6/xQAFDyQ+K9F+aX7M+jzT2Gp5wnmCaYJJAk5Hn9Tlu/5Pc22+n8eyMIoghhCGuEEA4PihHz1+53JqGVBQDt/DKPjT5z7M/pn7FxrsjKpFAAAAaH0gAAABwKAFAAAOAVwAAA5QAAAAHAAAwKDzXj9j4W9q/N/wCjPsDzD9D/AJ88H8Q/Of6J6f37v8L37pP0P+ew34wYAAABxP5n/pn5N6r+hHj+5ZPi/vQAJGiprA/NH0T9m/CPuw/Gf3gaCAAgJABoAB498V/n39B/pDqfffgXyh8kftf4F+pex+OfuIZbNQKX5M6T9ofUMsJYIEQ0AAAD5na/Ev8AcvpzvaDV7QC/oJ6F5VcEVKLM75Pa+OPTKXYeQiqfPc3+lP21xafjN1Wy/W05tQJAQg2k0AMKDmbnzCh284NAAAAAAB2NTn05k6MwAAAAAAAAAAAAAAAAAAAAAAAAAAJ09q1z6rXRQzd82FyAAAAAAAAAAAAAAAAALv5V2Gz5nUEUAAAAAAB5k1/P5/S36/L3YAAAAAAAAAAAAAAAAAAAAAAAAAAAeOfjMHkX6v8ALp9aHDuAAAAAAAAAAAAFUAPCQAcCkirQAAAArgAAAKBzFBASAHyL4P6J3f0J0+YHq/HAAUPJDz6j+dT93/S5ulnqZ5xzyillCWQHOBwCNBsYRxBFEEEI4IBQfkQn9t/orxbBDCg4+q+O/mnpx+lvsbKuxMqAJFAAAApK5oAAAAcCgBQAADgFcAAAOUAAAABwAAMCgcHPfCnx37Bb+6bx2a31v5R418//AA/7b5B6381+kfRPtH6b/NMx6nzQAAABgbYfmtqft/8APv6RaX4b/QoRoCBs0jR8T+U/6t+N/pt6x+dfqAnneo0AACAARAIR/nZ9Jfnn9DfQ5+h/zw384vufz7R/QvPUPivvGoHxj4FtP6juyoEAgRDQAAAPz4a+Ff1t6c795l+8K9fTu5WXb49f8D9R9md9F+T0vE6x/PHrW9XjRd1+c81/QJ7Lxa+K/wA/e0f0WegYbApAQg2k0AMKACt55rHEt6+cGAAAAAHs+x5fVy/mPZjWOjMAAAAAAAAAAAAAAAAAAAAAAAAA67uPO63nPqCQxTk4fVnz+X3YBcgAAAAAAAAAAAAAAAGxFdlv+X1BnQAAAAAAfiZ9q+py/pIaIAAAAAAAAAAAAAAAAAAAAAAAAAABgfAP5HB7z+qfDt7AcuoAAAAAAAAAABVADwkAHApIq0AAAAK4AAACgcCjQEgB8Z/PfpXr/uXf4Ael8oABQ8kBn4IfWPo8/wBnz6RNME0o5pBSPB6goA0GsCOMIohwwihgDzD8U5r+j3uOHYEHP2fG/wAvdUfRH2hi+0M6ABQeoAAAFJXNAAAADgUAKAAUFAVwAAA5QAAAAVQAAYFA8CpN/Fnm/wCW/wBRfeXafpn81POjzsLwDwfu/nXM/O/6G+ifov8AQfwLpz6T85AAGAZ/5x/dfnXPfpn0c20fnv6aCBssYUfIP0nyvzV+pvvfOTH57+msAAAgAEQD598v6r5m/RXxfs5j678iOe3w/K37c/X/AMR+lD8Z/dU5MX5J/p9qvVjKgAECIaAAAB+W9Lw39MujO9eZevCu3A5D4V+J/omJnyX7D0v3p9v+C2Kf1n4f8ZfNBS9MP1j8z5X9JbuDY/GTVSfsqMMmAhAaTYAwoAGojD8v6cco6cgAAAC2n7mvldRm2vJcz0eYKQAAAAAAAAAAAAAAAAAAAAAAO9L5td9vn9SAwAJkAKNLnsHuwx4+rEGAAAAAAAAAAAAAAAaUPrOl83qmMbAAAAAPhb4y9Dn/AH8OiAAAAAAAAAAAAAAAAAAAAAAAAAAAADlw/LP84g+1/wBLPP39IOfQAAAAAAAAACqAHhIAOBSRVoAAAAVwAAAFA4FGgJAD4z+e/SvYPce/wA9L5QACh5IDPhf8vu3H9rrXRlNMEsoSyDkeJzgUBEBrAjjHHEKKEIoA/Kb17LT9fTj0MxnyL8h9efu32rGnoJzgUAAoSAAAAFJXNAAAADgUAKAAUFAcoAAA8AAAABVAACgGK4ABPlTw/t/m77s+a/SPVF/Qf5/UqU8p4fc+cfFfzH+mPY/pj6j8x9UX7X8acCEGlWo/PXzT9x/nv9POi/Ev34MegSGjSkhc/lb9Q/sf4Z9bn43+6tQAAIAAaFf83Pjv2P7B9q+g/Pg9H50YHiX51/sX4f8Aqj2H5P8Asqfn5z9Wf+iDEIYACAQwAzI247kvE+25r5S8j7HjvaeL33SZ9Utlxp7Pb43lfL+P+nd7D5fm+dfSf1Hn/QFT9H/lOpx4c90g/wA0uoyv9fzj0PP2/wCeD9seiPqw5rAQAwmwBqUADUQMRwvB9vPXNoAAA7nK/Rjzugayp5D1ZZZ14gAAAAAAAAAAAAAAAAAAAAABp+s8HQN5d0BgATIAAADMjScjM68aNbeKSaQDAAAAAAAAAAAAB6NvoeTbo5+HcEwAA4b+ertx/o89Y68gAAAAAAAAAAAAAAAAAAAAAAAAAAAAA8vD8pvhQPtf9F+Db2A5tQAAAAAAAAqhQcEgA4FJFWgAAABXAAAAUDgUcgIAPiT5v9P979l9T5YPQ+ZAAoeSAyr/AD5dMfYX232YTTClkCV4PeNzgUEiDawGMFHEEUQQ/Ho/z5/oZ5de0hxr5a+MuqPSPt6X6kuOriYCgAFCQAAAApK5oAAAAeAAFAAKCgOUAAAeAAAAA4AACgGK4AAA8B+OPzX+kPtb3T7j8SD1PmQYc7j1/OPzj+Z/0n0/073+F7xL+hfgAOUaOL88/s/gvO/099DzNg/PP08JGoraB47+a367+JfrJ0/5V+yNTn6QAJAQND51+d/z3+hv0Qd+hfzyADQT4V+v+K82/THu87hPyc+G/RP1o7SGgmAAip5v5/v+Z+b/AB/7FwHO/M/p3T9b3eB0/l3tfGfL33D63zlufXksKrrZ/D7Xi8/zH7L6dTx+S8t9o9zp+lKv6b/JlWmOnyCX4Z/vXhr7+c1iD+Dfyv6Y/oO7PnsEgBhNgDUoAGogEFVrieK7MKZtABf9l5drZx6jWJRa8dp+nzA0AAAAAAAAAAAAAAAAAAAAAXvXuHoG8m6AwAJkAAAAAAACLOuc7G6ssKn2YhSAAAAAAAAAAAAf0mN9pe8zqCWAH4+b5+sfrj6OAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHk4fmN8BB9J/d3Hr9ZavHqAwAAAAAKoUHBIAOBSRxQAAAArgAAAKBwKOQEAHxn89+lewe49/gB6XygAFDyQGHzVovwy/YLv5/Y5KmWUJHg94K4AAQGtBjAjjCKNnyX+a0V+5H05xb+CfD3Rna+4Ee5mFiu3EwFAAKEgAAABSVzQAAAA8AAKAAUFAcoAAA8AAAABwAAFAMVwAAAeScHu/DP1p8F+7/AEkfp380AwAK86eBfMX51/Qmf9P59X0Vc/Sf5xG1CROfzm5z9v8A59/Tiz+I/wBAhnTUVtAPz2+y+EX9CARvxv3YAEgIK4/zg+uPz/8AoH20/QP59ABoJn6Z/ll9Y/sf4b47c/Hf3T7/ADKmgHMY9njPjvw37l5f0XzX6X6h6N9X+Teg9v8AVflNw7fFPHxfzxfun0xtXKm3bHasC8H+ePzj+r+qg+a+l5j7J+x+A7ut93/NVaoFT8tU7f7b8moZUINPxi1Xnv7qObZjQAwmwBqUACNQCAAYHN8v0ZZN7WPRdDj3DNjWIBh+RdmLDpyAAAAAAAAAAAAAAAAAAAAAD1vk20W8PSgMACZAAAAAAAAAAAzann+f78MlOnIGAAAAAAAAAABL6Ly67553SAHzfc/jb/TH6vNKAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABxgfnt8GB4v8AUOdfV/1XxberGFgAAABVCg4JABwKSOKAAAAFcAAABQOBRyAgA+M/nv0r2D3Hv8APS+UAAoeSAwKPzw/Lvsx/XX2DbJXskex7kOUFARARoMYyNiOB/PFP5/8A2z59fVvhsfnf2tc/Tb+SwoFduJgKAAUJAAAACkrmgAAAB4AAUAAoKA5QABQcAAAAA4AACgGK4AAAHB538DfAfvf0p9WfS/nAe78QADhqgvFvlb4D955L6b876H6Vv/pv81tLxqfmP97+b9v+hnH3B8Z900VsAOW6OX8nP00/VPx31dv5H+2AASAg8L836X5T/Rvw/uHn135EADQQDyX80v1X8exv1r/L/wBf7IyrH8M8X7Xwfz/85/ov1j2T6P8AOPXun+1/EmG3GAAAB+AH0n0R9rWLi3ZCzYG/5/8AG+/8it/D/wBH+9et/oH8uQwej8pWwUvzc+aJv+gzuuPUJBBpiuvwX936cf2LOWwBhNgDUoAEagEAAANAAAAgGsQA4TSPPj0ecAAAAAAAAAADfzrtk4ejnuU68a5vmAAAAAHf4ada3zetAYAEyAAAAAAAAAAAAAVee3z53F9DnaaSAAAAAAAAAFmX6+vjdoAND+dj91/X5PpgYAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAeVfGYfIPyiHW/S+N/SP09x6+qGNgAVQoOCQAcCkjigAAABXAAAAUDgUcgIAPkDwP0X0X3/AK/JD1PkAAKHkgMCg/OHSfy8/RPtw+y5KlVBXA5WCgiAjQRqPK/iFP5I+58r/TL5lzv5r+t9Z+v9PksEBQK7cTAUAAoSAAAAFJXNAAAADwAAoABQUBygACg4AAAABwAAFAMcoAAAOBeP5+78+vd/zf8ApD6/X9T/AJeABw1BAHk/yf8AC/uHB/UHk/V/Stz9O/mg53o4/wArPpn9n/B/tw/EP6AaK2AAHxz4F+x/hf6gH45+5gMJAQfnp8r+rfQH0T6nyoet8mADQQA/NDu4uR/VnXDzv5x/Pv3DxL0r5D9f959n/Rv5z1D1vkAQwAAAAD5c/C3Wf2R9O1znsssThLINwJsY4swXx7+ek17Z+zOGvo5gwQINArhPwF3z+8/1CgDNsJsAalAAjUAgAABoAAAEA1iACI8gw/T5QtAAAAAAAAAGl69w9Dk5NiBrhuQ9LlabQAAAAbUV6k3ye1AYAEyAAAAAAAAAAAAAABUa5nmPQ5846sgAAAAAAAAD2SXxe0EwD8f/AKA9Pm/UY1kAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAACkHzZ8th8ufLIfX/wCtPn775z6BVCg4JABwKSOKAAAAFcAAABQOBRyAgA+UPD+/6X6P15Q9f4oACh5IDAoOJ0n5a/MHqyh+47n6m13IMAFAEA8uR81/KyrzD7pzr9HvNMb+K/beift7tePVRSABQK7cTAUAAoSAAAAFJXNAAAADwAAoABQUBygACg4AAAABwAAFAMVwAAAOBXE8z+fPxP7X699m+v8AKqfRfno4aggAA8n+Tvhf3Divqnzfo/oqx+lfzdwn5gfqX4/9VfaHyn2LU+X+waAAUPyf/SPyr7k+iPkftg8X3wkBHnfwN8D++fpbe+8/BwrMAGggGN+Pus9N61wffYH0F+cfs30P1n6V/NYb8IIBDAAAAAE/Mml+af6f7x9HPqZ5wleCgZ/zsn8w/JSfpv6d46fapjoEyCBBoFHjjn8D/wBSejP9CjmthNgDUoAEagEAAANAAAAgGsQAEZnjXZjGdWQAAAAAAAAB6L1/l9bUysAMe583zvU5ApAAAAep6/k9yEUABMgAAAAAAAAAAAAAAAAGVzfVlz1Dv5wpAAAAABoS/WTxu0AAPgP5t9Hn/dQ3gAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAOED8U/Nof7jdJ5fUDoUHBIAOBSRxQAAAArgAAAKBwKOQEAHyx4n3mt9LXmHsfDgAUPJAYFB55rCd6Hw9+eOk+Ee3bx7L6Q11FxqjzIee+Xp+LVU/qb7Qwv7N4LOvg6v15/dftXJsLAoSABQK7cTAUAAoSAAAAFJXNAAAADwAAoABQUBygAADwAAAAHAAAUAxXAAAA4FcI5nPq+BfUvzT+kvs1f1L+YRRuBAAANH458j/AfvGb9cY9nufC/qH83fmB9Z/r34f8AYqfif7+0AAPmj3fnfi79Xfqfj7B+c/qQSAj4yufAf0B9fn3/APP4AADQQD5M+Tfif2/ruE9nh/UPrPofxxpnQACAQwAAAABGh8Ifklquo+l9I9j6pp/Lh5j4Wn89+ny/rr7cxv6RbjaFMJkECDQKAnwX8GejP9Vv0KyphloANSgAaiAQAAA0AAACAaxAAQcHrPn56PMAAAAAAAAAexaXkdrUlgABHwHRlyJ6PMAAABqep8HS45dwAJkAAAAAAAAAAAAAAAAAADPpYeN2Y5VHrxQpAABJ6Tya7h5/SAAHzH+Vnfh/RqdOYAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAV/wAQA8p/cLh31Tn2AHhIAOBSRVoAAAAVwAAAFA4FHICAD5i8X7k+oiw9r4MACh5IDAoPL9o2e5lmdnXwb+W/Xlx13WPVNsVwZ45zSf6Ofq9ybWMTN/Enzp3Y/Zf1lzXOZ2KChIAFArtxMBQAChIAAFSdOexOD6LIyuD382px+xnGHfRhx636V4lzXmuaXV5utsd/hbW96PzuwdnjOUAAAeAAAAAqgABQDFcAAACg5RKVOH+ffxn7L6l9m+l84p9F+eq4YCAGoMQXgnyV+cf0T0X2H2+LJ+YX9HfxZ9qfV3xP6A0+e+nABjX5X/Un7B+HfVh+PfuQQFGdPzL/AEH+E/du5PvfwUAABocn8h/J/rXh/wBQeB999Vfnx+//AMW7X3tDQ+O+5AAQCGAAAAANQDLmvkD5T0nxHlbk6sPXvoKa+q/XMqBA2WhTCZBAg0CgJPnOl+Ff6rbx+g5ybADUoAGogEAAANAAAAgGsQAEDR+N5PqcgUgAAAAAAAPbZ/H7WomAAABzvCd3PlnXiAAB1mWnenl9gAEyAAAAAAAAAAAAAAAAAAAAAEIszN6c6VfaC6HS3uHcEwAA8W/Ebuw/poOrMAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADPD8GLWdftBP5vWAPCQAcCkirQAAAArgAAAKBwKNASAHzp5H2XNfWuPoh7v54ABQ8kBgUHkG+fV+Iy/iP4t3jo/qXWPpD25q2NAEPzIn+eP1Lnf3x8RzXsf3TB1hhQFCgoSABQDt5MBQAYPF+b9Nx/I/PfonJct4f3fN5Pk/Xae7v5+zrej89p3OzxtI38+3Z25Y8+NVpZdVLN5/Tycjh9zFxPO+in6bTn6jq/a+K7Dsff+B7Tsvofz2ydXlgAAAKoAAMCgeAAACg5RKOTDjp/Pv1L8y/pX7NX9S/mEUYoCDQJACpN/NHyx+Yf017X7P9r+RfnV98/v38mfRx+NfuwAAeEfn5+vfiP603vyT9qDHQ8G8z6f5p/Q3x/sA+r/KAAAh+ZPF+1+Xeu+G/c/Dv17/U/wCXZ+P6fO/Kr9LP1X8e9VPyH9sAAQCGAAAAANQAmgQAEYIAAAA1NBsJkECDQKAkBfOf4XdEfqB+jWGgRTUoAGogEAAANAAADy9L8gP3F2mcwsEAPF8f7cWnTiAAAAAAAB7Ho+R2tSWAAAANRg8V24YZ24AAB2Xb+b2BhoEyAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB4/8Ahr3Yf04HVmAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABz/APPkH0z+p3n9QY28JABwKSKtAAAACuAAAAoHMUEBIAeHeb9T5J9o+f8ARh9F+ZgAUPJAYFB+a3Rn+b97oz+p/rap9fGgAAADxwfzH5sq7r9IMa94OWwAChQUJAAoB2/Aw6POfOvmv1LgOE+V/Vsvsuf0ew7L6D8963qPd+D6XpfY+MtHX5auaAAAAHAoAUABEPD5nzvouY5Pw/tuQ4/wfvuHzPC+67fvvZ+N9F9F+r/KPQbn0f50DhVAABgUDgUAAAHKDhwFHP8A55/EftXtH2J63ygfQ/AAAAAAAAc/j1fIviH5V/UvoPzb/T38LfpP7d8d9IHleyAM/Mb7z8594+wfI9sPl/qz4R+Z/TfWPpTu8IPY+PADh+f0vimH8v8A6f8As/5E/UP5itfefV5TUl/IHz/+y/hf6du/G/3MEwQIhoAAAADUAIoAACMEAAAAaqQYEyCBBoFASAjw1n4S/eHTH6rP49mFAA1EAgAABoAAAfl3U/A/9HOshzaAgBnne0cOehzAAAAAAAAej9b5fW1MrAAAAGoERi8P3YYZ24AB1GV97J5fYEoAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAD5o/J/vw/o9OnMAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA8pD+dr9d+fT6+OHpeEgAoOJFWgAAABXAAAAUDgUaAkAPLuL3fmP7y8X7sPp/ycACh5IDAo/K/wCBOzH9Pfo/bMVoUDz0PJNUr23ZJ5/4dKz/AEUPz+/bPnv6lOawAChRqEABz2fZ5X5d8l+xeZc98n+t976H7PxPoPffV/k/ZTe38NIrkAAApK5oAAAAcCgBQAADgFcAAYGXVwfA/Ofo3nfnHyf6xzfoXH7Pp/qv2X436jrfV/lAXiDAoHAoAAAK4FUkLRzOPZ+en0B+a/0d9Yn6h/MoAAAAAABInmPF7Xxl5j+e/wBGZX6Yf0T/ABB6UfCfeCA3xf1PH/Of9bfuPz3TPzD9ay/zK+H/AHD9Jei+z/Gg15AD55+Q/wA2/pT6X+ofrfyR348/QfCfpZ6/DaidT8qP0n8s+0PpD5P7IPC+hBAiGgAAAANQAigAARgIAAAAxUDAmQQINAoCQEIPyr8NNp9i/ZuXvGdgDUQCAAARAAADkP54N8+v/ocGHNYIAZAHkGT6nIFIAAAAAANL2Dh6BOTYAAABEQIhpRh8L24ZJ2YAGh3vHvuHBuAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB+engno8/7mG8AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHxN+LqP387ny+14pABQcSKtAAAACuAAAAoHAo0BIAcvh3/CP6N/M/rAfW/jAAFDyQGHwbrP5Ufsf3Yd4tS5zFcHkHyMqvQhq/b4vkXzoq314cZ4bmf0DnFqAAUA3Ri828g+X/AFTyXkvjP270j1L3vgvUfQfsfxW0d/hAABIoAAADQ+kAAAAKDgAoAABwA8AAAVwAGB5j5P1vlvlfxX7Vw3o3lfVevexfc/hnozvp/wAxCwUHAAAAOBXCCpBnHfnl8F+6/U/0v9J+dB7Px4AAAAAEDQuv81+L9l8oL85+sfpF0v3387jVKAfmP7r+wfh32Gfjn7j4T81/Bfvn6EH3v4GAVPjb5v8ASPJft7xvs/T2/e/gnif526z+vjsqagHzx7fgfD/6rfX/ABN8/Nf1UJBAaBy3lLXn/HMr5TNTpg9V9xyu0IAEjAAAAAYqBoIQAINAoCQEINvMzX4yeMdmP7QfQvPqEtqSAAAAIgABQD8RuL7MfPP6PMqDm0BADIwyvH+zBh1ZAAAAAAAeh4adenm9QAAACIgRDSgQOX4b0OaqdOQBtdfxb7zuHYBgAAAAAAAAAAAAAAAAAAAAAAAAAAAAH4X/AKC+ry/oiWgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA/FMOF/aDh6XnPQAKDiRVoAAAAVwAAAFA4FGgJABif5y/oD8Z+6dCfafhQAFDlkBnG/zt9Wf6sfQHVjI+k+RDlD4X50r0LMqfP/oCa73rGugEqH4eTX7g/TfFsEgFR+X8Pv8Ainj3wH9C2vYLx9b9R+7/AAO4el865ZQUgAAe4AAABofSAAAAFBwAMBgA4AeAAAK4AAAAMTyfyfrvH/H/AIH96PZ3Pt3rX6L/AD1Mej885QAAAUHAnBIWHn/J6n58fZ3wH737yfpX84AAAAAgbLGlIBy/xd8j+t8R9peZ9J68z9B/AADxf85/138S/XK9+RftPxL2/wCf/v8A9Qn3v4IYefT8Jz/l/wDUH3Fv/qH8vtTTm+Eaus/fBlTABofmN79+yfhv12fjf7kEnjXxzS+W/nnRcB3FLtukatXQo84Hmucn9k/o3jf0aZUkYAAACBhQNBCABowKAkBCDaTTPzl0n8tf0A6c/wBPOo5dmpAAAAAiAAec/jjvGD+3Mn84X9JdrqDl0BAFIwScDtnwx6HOAAAAAABYT9fteT2BLAABEQIhpQAFZvh+V9PkQ1zALG5jevo8e13Qw0CWAAAAAAAAAAAAAAAAAAAAAAAAAHy9c/jT/S16vN0YAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAcoH83v63c+n14cO4AKDiRVoAAAAHgAAAUDgUaAkAAPhj6P+K/dvXT7X8JAAocsgM/Lb5r78f1hk3yllQ9wPUPgjJK9SxanzP6VmvoOMmKJjfzhl9N+wPHsGb4Xwj5r9S8TufFfuHtftP2v4b3y/S/mANBVOWUFIAAHuAAAAaHUlAAAAUHAAwGADgBwKAADwAAAAFBWh5z4n8v+o+J4Xw37l7Z7r9l+O+tyfZfjwAAACuAcSDQUeU/BH51/Q33n619t+LB3+EAAAQNCkaMAK9T+dnn/AMt+1fRX19Xz+gfS/m5+X/3n5v8ARn1n4f0H5j/ev4//AFF6Efe/gvJ/BPxP7Z6b9m+n81YPo/zhgH5L/dus+9plTQBoeQfnH+v/AIl+sN/8m/Z/hP8AOKNPE/pC5+ifbqXq+k05QRAYwG+Wh8i/Fk19ofr/AM+m+zOgAABCNKBoIQANGBQEgIQbSaEDyP8AJboj5y/Q/SPvv0Pk2BAAAJUD55+JNo+EPuDaP1h2eDb8CP0d7Mvtc49QACkYJGC8owvT5guQAAAAAA6b0vz+kObUAAESQaCFAAMKMW58+o+pxBSAAAuy+k6nz+i+c2oAAAAAAAAAAAAAAAAAAAAAAAHjX4d9uP60/oN15AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfBn5KS/6Bdby+oAFBxIq0AAAADwAAAKBQcNASAAHyrofFfu/wBPn2v4SAgocshEz+cr9PfT5voeepkkCR4OUPnb5YK2K4RffQdDGTHCEXz6P8wP6N+Lq8Q8B/Pv6A5T2zzvpPc/Rv0P+dJDr8MABgVTllBSAABzwAAAGZuXz+jmZ/H7UOdz98Zn0Chaua8ty/0+fqa/d4t06PPVQFBwAAOBQAAAAHAABwfP6fg/hP53/Q8Hvqv6D7T9I/nIOjzwAcCgnBIUHhPmfTfF36G/J/qvdH6B+CAASDUxCmgAABhfl195+e1fQPwH+1ftn1D7X8FPB/ovi/gj9KPp/mvg39NPw7+pn8f9X+UfAPu35x/R/wBbO/Tv5hEBvLs/ID9ltJ0UypoCIB+cH3P57U/P7j9HG+8Pmfrfq3YqZZQe8FUEQGxhHCEXHh+YfO53+8nZ82oIAAQjSgaCEADSgGBAANTQVCA0o+c/zZ2z+Q/f9o+m/ZML6aKX594Ta+ffINo6b7El/of7Tyahm/zo+ROzL9zzj1AAKSMQCpePdeNY68gAAAAAAPT+bXojz+kAAESQaCFAAMKAIuB6cuZPQ5AAAABUb3YcW+0cW4AAAAAAAAAAAAAAAGfktHTIAYBx/wAYbR+Xf6m+jz/p4MAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAZ/P2H0z+lfndAZWKDiQcwGAAAPAAAAoFBw0BIAAeNeFfFfu32+fa/hQCBjlA+eKX4d/un6XNalamkCR4OcNPAQ8Z1B/SHdCjjFFEEHM4d/5t/TH5Z/TnoHv3ofL+02vuPwkHLgkBAwKpXSgpABXV8jyfj/AGXJ8t4X3nNc74/2PPY3l/VV9LPo09Ds8ezodHnSGnOAVauXVRo8/pZVLj9e5ub8W/0vq/K9L1Hs/G9f2Hu/CdhY9n40Vy4AAAABVAABglHlnz98d+weH+jfIfrf0V7h+l/zZYPV+XAFcAolCQZ8u/N35n/S36Kb/wCj/wA5hpyiDQKRoAAAAHm3d5/5efRv3fxfnP0l/Mn9y/Vcn6f/AC/+VPa/pH5/3X3N+Jf0Ty/58+b9L719ae18UHu/CiA35spfI/6l0Dc2gCIB8Lej5vyL0P6N+X/Y/W/nf6jPNy9c0oSPG4ENGyMI4RQ1wqfmYn5r++3PrqmVAAIRpQNBCABpQDAgAGpoKhAaUAHIfKTn5x8n3jnyl3PtkP6F9oxv0sigQEnM/wA6/Vn+1X1Dz2EsCkjEAjltJ8xPS5gAAAAAALsv1+XyewEwBEkGsQYADCgAEuW4L0eVh05AAAAAG33HDvrHFuAAAAAAAB5J4FtHknAbxylW5zvP6XFcnpNQoAOv9IzfXeSJ+e/Rus/rP9XAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHzyH8/X9BnLr3hx7Cg4kHMBgAADwAAAKBQcNASAAHL/AAh8V+7fo077X8LAkGOUD819J+Zv1U9DnmmallB8g3uEADQZGNkQouQ4/X8g4X8q/qLyH6x+s/JPuE+k/KQGBSqSAgYFUrpWR555/wBN5/wfyv6tw/JfN/pWv2PR5/W9T7vwfT9D7PxnQ7nq/KbE3f4LhoAAABygAFZ3jYPB7uBzvlfVcxy3i/aclx3g/eY/W8/o9x330n5v6D6H9V+V9Qev8gAAA4AAGBQAZ3gvi/Z/OeB+cf0X9GfR36B+Bb59D+fCgqgAlUR8WfO/ofA/oNx+vaT678lApEBAAAAAEaHzp8E/sn4V+jvyj/N39p2vuDHTxD4n/RvyP7V+gPjfs/z59i+P/Yfrw/TP5lABoJ8FWr33OAyGANA8U/nz0n7w9e/ZvwrlvtD5f6+xxfwv6HzXqQb/ACot3UCHjAtc4EUI+7jar0A/H36ny0/Wg5tAAENR2CkJQANKAYEAA1NBUIDSgQGkgAAADQAAEBIn586z+dP7/wBztnPoBSRiAQB5lyvpcwaSAAAAAAHa955vUGGgAiSRPYgwARpQACRk1Hm9T1uQKQAAAAqOk7nz+m2cuoAAAAAcyL+cHqPX5fQfRsq9I9KxvuJIrIoNcyUudlZu7slL5htfN/1H05/pn9gsAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAD8TYM6/XY83pAHkg5gMAAAcCgAAUCg4aAkAAHfnX8l+yfcXa/S/l4dHlgxXAfinrHsf3F34TTtSyjleJzwUBGhz+1z90XFZdfiHIflX9Reqeofe/hX50ek+/8Anv6sHJoAwKVSQEDOa8t8f7fzTzX4z9p5Ts/O+k9B7/6j8s7nt/pvzDXk9D51QAAAC5eAAAAArgAAoABwADlDJ4jz/oOF4H5n9M8+4H5X9U1PSunzvS/VPsvxv0Sb6X81HCAAAKAYOBfKuH2/mrx38x/pr3r6c+x/Hu2Pq/ysUFUAArqvgPpPzL+l/uA/Tv5mRAQAAAAAAagHwv8AWfF+P/pr6XlfKXgf82/3b9mfE3t/IfZfz90edvfbvV4bj6T83AEaB+WWi+wfoyRGywBoH4mc99X8h9Deb/sP4bp/ob8H+j2PyP8Azf8AVfsv4zT/AFU/OJr2H78D5b/O1P8ASH55a+WvrFP7enpVqovKfxcVf0U+m8ewICRqVYKAkAEQoBgQADEwVDWIMEBpIAAAA0AABASJHR+GnZdeX7Mu4tQKawQCAIfI+nLKO3EAAAAAAJPYOTa6cPQAN/OjSOH/AFNYhlYAjSgAEgSp+Y9uFA7sAAAAAAlR2fX+X1OMdAAAAACjeEAwAAAAAAAADkvhjaPzt9l9Hn/Zf3BgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB5f/ADiB+8/sPl9QS3kg5gMAAAcCgAAMFY4aAkAAHfGfz36X6h7/ANvhB6nyIMVwH89G8fpf9D9vPPM3NMKSQHuBQEoRt8f/AEp8J+4+DcZ83+iew+tfoH4JPB9F+f8Axb4Hnf7gHFqAwKVSa/l3B9D5H5N8N+8816Vw+/6d6X9j+L+i3vpfzILyAB4OAAAALlygAAAAPAACgAFBQFcAAALGPhPNfnv0Hy/y34z9myfT+f0PXPXvuvwvvD6L87AAYFAoOAORw7vmX5+/L/6b9l+ovp/zT0I+w/IQBVAAxfzr+K/aPpH6b9z4gPZ+OAAAAABoIAV3P5ies/tP4J91eJ/gP9P/ABdkfk/9ZVvTvuPyb9Abn2H4ODgAagN/GbSf1g7uG1ExAQD+Yn9hO3l8V4X+hP5jsfpF8D+k2vyg/NP1j9cPz3D6r+FwofrWH5Z82n+s/K1P5pfqEnxn54B0XUj+xvyjF9HfqZy7BDCRhVgoCQARCgACQAYqBMaxBggNJAAAAGgAAICRqUch+CG+f2h+p2dBlo1hIAAFJnkVL0+UKQAAAAAdjjfoB5nWAN8UF+Dn9E/Xl0hybACNKAASBIqNeXUvY4wpAAAAABoy+83PJ6wigAAAAAAAAAAAAAAAADK/NnaPz0/Z70uf7uAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPyIDw/wDaTh3Dn0eSDmAwAABwKAAAwGPGAQAADvn7yvsPNvs3j9kPofzYGK4Iv5lujP8Adju+3GecJphSShyHO+R933W96XyLflT5z9A8tufmf9Ie1ey/qX8x2a/0X5/DXF86/m3Ff0NnDsAymtvH/Hfjv2fyLovlv1X1v1z7L8W9KtfVflI5yoSDAAcDwAAACpcrAAAABwKAFAAAOAHgAAA8AAA5DyjwvuvIvJvhP3bR9l6OD2j1j9A/n+wej88FAArgADAy6fmf5w/Mf6a9a+sfo/zruz7D8hAAA4Hl9P8APj7r+F/cvWj9E/noAAAAAagAAI4/8rv1P8e/QH6A+D/RPGvg/wDH/wCqz776uD1g+9/AAAAYBxf5J6z+zzsqYANA5Vn8yn9Ae+UvyJ93+c+Y/pf6fk3/AMdfzH9c9k86T/Y/8m2dv9pB8Nckj9buRa/Mz9Rw/Mb7lDX/AC8H+pnyoL4Q/oQ5tQzoSW0qgUBIANCwEBIAjVQJjGAwaCEgAAADQAAEBI1KDyZL8I/0E7M/0xOLdjSQAAAeX5N2Y1TqxAAAAACdP2d/kdgJtJPxR6b0MP2FOHcARpQACQJAGd5b388J2YgAAAAAAdFlXdXfK6wTAAAAAAAAAAAAAAAAAPl78We/D9v/ALO6cwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADjA/my/bjm0+hTi3csg5gMAAAUHAAAMBjxgEAAAvJ8/pfCv6KfM/qsx9d+MAweHA/zn9Wf9CNjrxsTCnlCf5n8X9D8DsfnH9Vs+vfpPy7H+bfG2Lvg/c/THpf7B/JEMHqfNV64/PPw/i/6azh0Z5Xw+/wCF+Pfnn9Idh7V6PzfsnYfc/gob+cFscmoSDAAUJAAAACkrmgAAABQcAFAAAOAHAoAAOBQAAAIx+ZeNfK/qni3P/D/uHtHuv1/5B69N9p+NhUigqgAGJn0fMfzb+X/057b9YfU/l/WH1f5WAAHhPxx+cf0X+ivUfoP4AG3GAACICAAASHz77PhfAP6p/dfnnlXxZ/MH98dhDz+r91d/+k/zMHV5YgNA8O+D9Z/WFMqaANAhZ/Lh+9m2fWSXn8jfYf235/v/AJK/D/of6/8A5Sh+o/5bj/Q78lk/1T+AA/Xjj2vzE/UwX5WfoSF/8tA/U7h2vwl/p259rpjQktpTCZBAA0LAQEgCNmgGMYDBoISAAAANAAAkBDUoCTyb8I+rP7a/VnLRG5AAAAMw6PK4PT5AaAAAAAPU97yuwM6aScZ/Px2Y/sF9a4aBFNQoABIEgA5vSPOj1+QAAAAAAAB/W4adjY8zpBMAAAAAAAAAAAAAAAA+P9J/I3+jv1ObvQAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPy2+M4f7pHmdTlkHMBgAACg4AABgMcrAIAABZB/nz9cfE/vHpx9t+DAwez59/Erqy/d+x1ZWJxTzhyvwn8P/Rnc7nxH2/JZvd7H05r/Zfz5vbPt/G6kfp/NQQNV64+H/Ln5X9P/YnzjLLwaX4T999z9v8AuPwjsHfQ/nAEgAUxXThQDAAUHqAAABSHtAAAACg4AGAwAHAKDgAAAeAAAAAAO4Pm9Pw3wz8+/oLG9y5/R+gPSP0n+cA7PIHAoAAc7l1fK3gP5b/UH0b9T/a/jGqfRfngAfH3gffeY/oPwe3YPrvyYAEBoAACAkAPgvzz9s/A/XM7+TP9DPpT82P2z+We4+2fxb+n/bD9E/nYYAB8qeBaz+lDcqQBEAz2fy9fvrtn0/A+l5fzfm/rf477v83/AJn+p/qB8q/P/R9h8yp/p/8Ajaj9k/zHZ+imI1+fP6Gi8t/PAPRvFU/1Lp1P8/P9H/Pr6AY2NloUwmQQANCwEBIA1JoBo1gMGghIAAAgIAABICGFASHmDPw0+ie3L9fr3DqAAADGKsbyvt54DqxAAAAAO/7LzOsMbaSJ8Maz+af9AW8Xzl1ahQAEyAAAB57zHrcgawAAAAAAABL1mGnVXvN6QlgAAAAAAAAAAAAAAAfjn3fqc37LmkgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAYX82Qfrl9a+Z0uXKhWKMAAAUHAAAMChygBAAAEg/lqD4n94+rT7b8GAB1HyV+WHVj+2NnqzsWBWJg8u+O/gP6a9M7T5X0vO/Oq+w/Saz+yfxTFDWXM6GPoug382vx/J6ny3yH47/XfRey9Xk+/eofov82PO3xBRKEgAUA7eTAUAABIoAAADQ6koAAAADlAGAwAHAAOUAAAVwAAAAACqAB5z4H8r+q+D9B8T+1/Qvvv6P8AzvsHufDqoKAAHF8/ofInmX5Z/UP1l9D/AKJ/Pbz1/kiBV+ffpX5r/SP2Cfpf82CAiAAAASAgAofmD9Z8tW+wfwb+xfpP8s/6O/hj1Xzj8Z/rD6g+nuz41G+z8YAfE/O6z98tyoAaAAfzO/sNvn3Hwr+oflPaaf2H53579V+N9T9YXfxn9wu3VV2dENJlKk1SpC838rDw6wP7Jpuf58/6S+fXuDGxqaDYTIIAbQoCQEANSbAEbQADEAgAABAQAACQEMKAkAOO/EjonT/asXSnPYABSNTMa58uq+nyA0AAAB3fbeZ1Bjo0kSNn4G/fPdh99nFu1CgAJkAAACHyftwqHdgAAAAAAAAAGrFdH0fn9F459AAAAAAAAAAAAAAA8m/Avux/pj7DqyAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPzrD80P324trbuTUUFKAAAFBwAADAocoASAgAkH598Y/E/vP6Gv+1/BgaHUfDfwT2Y/sZZ6MrFkJ7AZv59/Hfvuvu/B/qnK9f6s/ebP1f+MuG+LvkP3nkbvyf7j9Oey/pP8hcr8lfJ/pGWnxH7V9ZfRX6x/IgdvggUKChIAFAO3LMhSAABzwAAAGC1LqYUNkHAACuAACgAFBQFcAAADgUAAAAAUFAAK/i/mfTfO3mX5t/Snuv0n9l+Od8fW/lCqCgAB5Dwe98fXfzL+lfsr0j9Q/mMOzyOX/On4T9y+1Pafqvy9E9H50AAAQEgAAN+C/mP0/yD7X8D9A98+Wv2f+S/mj9A/U8/4O9e/nn+2PsWT9R/l0BfAHX6T9nJnQgIAAfhJ9CbR6v4X+zfhvvG0sPIsL1fM/Sy7+D/ANFXbiq5MiKoynSapUhc98zhv+1BT8xa/EP+nHm2lMaGFA0EoECDQKAkBADCbAGpQAI1AIAAAaAAABIIhpQEgAZBX41+L93P+3/p/FqCYFI1MAzfLuvDOOzAAAAD0vpvL6xMrQkRtHx5+VXbh/QnLxbsCgAJkAAAA53SfOD1+MAAAAAAAAAAADW6Tl16O55/QCYAAAAAAAAAAAB+JXRn9dfqD6GAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABUD+c39AcL++nef0CoUoAAAAeAAAAWK5AOQEADxr+fXyv699aepe7+eh2+ELR+dXzF2Y/rTZ6MrFkLM4WfLeH6P5J5b81/r/0z7O+t/Cuoj+q/EvgDmPyf+0evx/Jw5P7n+6/PeB58+bi+PMO76L/AEC9b4Z56vxwFANwQAAwKpyygpRckzjuRtcjz1rlsClh4lLMy2s2swGAABp20bGujb6BPo+jR1vWy+w7ZHQEscCgAAoOAAAAAAHAAAAcdh3/ADl89/mn9LeifTPtfG+yL93+GKAqgAQfNPifafK3v/wn7p9aaf6n/LqeK8PufFf6L/LfqHSn3X4aAAASAgAGh5r8I/A/vt3L+w+G/Tel6n5p+RH6m/qn4/v/AAf/ADp/ZPQfcHR590+p/Lfzj9q1X1eZMaAAAfnZ8Faz+inzr+yfiHvOpt5nkMvdh+itv8K/oa5ZTnkbZAitTaqUxVKbVSmOp8Vo+Wv3n5tQypGlA0EIAGjAoCQEIDSbAGpQAI1AIAAAaAAABIIDQAQAxB1vyz1z+Ef2z2j6MOTUCkamAFdLzbnPU5A0kALCfsc3kdgiaEjUoi/nx6sv1Z+usNGEWABMgAAAAHm3P+vxhpIAAAAAAAAAAAC7GddJ0fn9Fkw0AAAAAAAAAAA+Tbn8pf6T/V5ngAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfHn4lI/oP6nyeoE3FAAAADlAAAAsVyAcgIAHA/5m8b7nlPsTHtD3/zkWj8u/PO3D9QbO+dqwFm0FidOxMm8TZE1+W/ffjX9uekcqvi/L/q36fi9TrfoP8xVfyQVfX36G8WoQwKAbggABmV5xo/OfO9I4HgtZ4uzS7Pr5fWdTD6Xooe9sy9XWReuNOtBo4wNAyqrWTjMxeeo53maXK8fRx3JUtXvEehehy/S/S4fevTBDlAAAAAAHAAAAAZy1+evmj85/o3T+od/P90l/Qv5+BpQADlfjf5H9b4L7K8v6f2Q/Qv59+N+J/NP6T+/Hfpf82AASAgAAGgfDna/n/8AQXz3yH6B+X/Qv259X+N/n50/2fwv23m/j/7j8OZv5f8A1B927H6f/MP5qfQ/RzfUJlSIAAAch/Njc/st86/o35tV0P0T8s5j7m+U+39tt/kv7DYsDmVCRMr1wqVmqlQKtMdT8YUfoR+hPJsENqOgEEIAGlAMCQEINpNADCgAaiAQAAA0AAACAaxAAQAxo2BPwZ+Tvbl+2f1Fy6BFtRMACQ5LWOCq+nyg0vonPp1h53UICEjUoPzg1j5z/auxhz6gATIAAAABX8x7Mcw7+cAAAAAAAAAAAAAHI19jm11NLl10HYWAwAAAAAAIA/nU/eL1+T6WGAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAH4Oh3n6zed0BhbigAAABygAAAWD0A5AQAOB/N49/wAA/oX8n+w659l+HhR+R+r3Yfoxa2ztWQs207FoLEqqQZHEj858n8l/tHssXzODiPuj7j4bpav2/wDOdb8sk/bv0449gzYDApabnyvyXWPK/LN54jsrXpPomVehd3nXedJDBA8HAAAAFSrmAAAACg4AGAwrM43g2vPvOrXm/mdKv6iHq3rsP1/vYaiYAAAAAqgAAAARN+D/AC/+e/0HX+pZ097l/Rf54BoAAPBvjz83/o32f7A+i/PbP53cnq/QP056/wAiHsfGggAABoHP5dX5s/pL8V+1fnB9udnH+bv6R/0d/FMX56Rf62Sfkf7TX+LPmf0zhfvLh9z83vrv7j8X+hjKmgAAAfkSLwr9XO3m8Z8e/U/yP236X+J+7sz/AD30tiZN7gSMIYBV67KtUKtUfyn+ZkP+jbX5NgkalWCgJAAaUAwIABqaCoQGlAA1JAAAAEQAAACAaxAAQAxg2hJ8YWvx5/ZXpz+wDj3aiYIBABDzGsYp1ZdNvcmwZ0NQCGpQefNfz0/0V9WfUnJuABMgAAAABXa8zyvX5AuQAAAAAAAAAAAAAAAHacVtdHw765y6gAAAAAB+RfrPp836wmsgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHkgfzn/uby6e/HDu4oAAAAcoAAAFA8AcgIAFCQK+KvSPhv336NPufwAKPxg9x78Pu21rnatBatJ2raJ7DqYkir0eX/C3xP9D46/L/ALJ9I/Qv6R/JlWr6/wAJW/NNPo/1g49gzZnN+OeMbLxry3oz631uT1T1TCvT9WLHJqEgwAFCQAAAApD2gAAAABygABQACgoA8OH8speSeSaLyTUpeze2w/cPSM2CYAACgoAAAAA1nhnyv+f/ANBH1a59vX9B/n8AADn8ur4188/LP6j+19X7/wDA/wA+/wBC/mP0vuj738DAAAQEAPmnx/sPKPu3h938rP0+8v6j5t+Pf6h/hP8AVT8qfD+h++PcPi/vBnke18eeSfmH9P8AA/a36b/Nf0MlciAAAAcbK/nN/Tzpj6v8A+l+U437M7vOsT/HfdTSA9QRgRxBBXCvVCtwyPxQ/W/LX7mOexJbSqBQEgAiFAMCAAYmCoQGlAAwkAAAARAAAAIBjAAEAMjBAQB8j/jT2Zfrj9u82zDOgQCABGgAAAANQCGFAB+FH3h24/chxdAAEyAAAAAAEXnfVlz56PMAAAAAAAAAAAAAAAAAAGn23Ht0BwdAAAAAB8a/nx6GH9C50ZgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAH5g/AUv8Ae+55PS4pgAAAK4AAACgcCjQEgABIpXlvyL8P+9/oLP8Ab/ggXP4Tfa/oc/2Ha0i1ZC5ZTuWk7Voc4iGqyrkq/Ot/j+h0a3f8xVqhV/PVPmf2G5NvNvCGeGeQdEd77PL9l9k576czAALYo3CkGAABIoAAADQ6koAAAACuAACgAAHACg4AAi8ua8Y8T0XiOpa959+zfuu1DBMAHAAAAAADQ8B+Vfzv+id7687vG9QPufw8ABB8++T9V8gfRnwf7xZ8W6PO/Q+b9H/nAHIA0AAPz9+W/VPon3n1vkvys/T75j9O0/zc/dP5I6no/ofmOS9Xx6Pqw/Kv2FPk3wfu/lj7U8b7b6Db9v8AhwAAAgEfE/4yaz+0t32PF+WPv3674meb8/8A0yWQHgDAZEEMAV64cx+PCf0Z+yPNsGbEltKYTIIAGhYCAkARqoExrEGCA0kAAAAaAAAgJEbQEgABSMEgIAPl21+LH6mdUffRw7ggEADUAAAAAagRDSgA/Oz5t7Mf2iOPoAAmQAAAAAAAOP4n0+Vh0ZgAAAAAAAAAAAAAAAAAB2PceX1BhoAAABwf88nbj/UVp9eQAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABU/nwD6S/UjzehVzsAAABXAAAAUDgUaAkAAJFKT4J+X/AFf6G9x9b44PT+X/AJ5f1W9Ln+hrVxashctJ3LQ7dxOwqK9MKlWlT8AD5XgZ9De6Cp/CiPE/R5vH9yR7r7fzX1RnoOEBIAFArtxMBQAAD3AAAAMFqXAAAAADgUAKAAAHAAOUAAAFBnkbXg3gGq8y9oa+jfoXOusIYAqgAAAAAFZX83/LH5p/Svr31z9D+e9SfV/lIAAcj8P/ABn7Mcr8r+rfR/1T+hfzyHt/EogAABl/mR8P+5fpV0P2f4v+Vn6d/FftlKv97+F/lT7l+/fzT879j6vj/rKfzT/ViND88sT8y/pz74679G/nIN+AACQQG/lLS+Dv107+P5K/Rf8ARPy6aX81/WZXicoI0GRhDEODzxL8j/Yo0/bW/wAugANTQbCZBAA0LAQEgCNmgGMYDBoISAAAANAAAQEiNoCQAApIxAIAAPnr8ReuP0K/TLGwwsEADUAAAAAagRDSgA8h/BLsx/o8ucnQCAmQAAAAAAADOpcJheryBpIAAAAAAAAAAAAAAAAAes8e2gcHQAAAAfzs9eX7yfRHbiAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHjIfzv/tHzafUi8W4AAAA8AAACgUHDQEgAA9xR4x530nzH9/eN9xK36v8h/mb/eH0+b0e1U8P+Syf7c5ofnv+oSry/wDGsP6AHor1GU6rXiP5wM13ixfsAfy3zydP9Q8K9gn57BAFiiUJAAoB05ZQUgAAWQAAAAAuXKAAAAAoOABgMABQUAcCgAAA5QAOU+e7Xzr4hpPsf0tL+kN/NgmqgAAAAAGNn0fJ/gf5f/UX0/8ATv3H4bZPc+HACuq+Sfmn8r/qmx+iXr/Jd03778BAAAA8b+U/g/3j7o+FOnzfP+w+M/ZvsSh+sfyh8Q+W/v38xG99H8t9l++/zb/VHZCf5z8z433Or9+eB9v1Z9x+HgASDQRgfkZ8K7x9m/oB+q/jurL+Xfr8jgcAINkYo64fHf5uTX3f+tvNrYIYIGFA0EoECDQKAkBADUmgGjWAwaCEgAAADQAAEBI1KAkAAKawkAAAGeON/ht9e9uH6yz+f0AgAagAAAAIiBENKAA/nk6s/wBgfpfDUIYTIAAAAAAAAAYdzx2H6nKGkgAAAAAAAAAAAAAAAHrPHtoHB0AAAAH4i9Gf6Bff3oYAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB8Gfj0j97fTfL6gTAAAcCgAAMFY4aAkAAHPGNH8C/Rfwv8AQHuPmv3n8+/zy/0M+lzXLVT59+WCr6n+6g/M/wDW5P8ANfng+0fpNHkfwwH0P9LtfB/zrR6ZcqfLfT5r6c8aS/Pv9/8Ak1UyoAAoBuCAAGBVK6UFIAACQFAAAAKlXMAAAAAHKAAFAAAOAFBwAAADgUAAMBr50+ZNF5B75S+p/fMqlJaqAAAAAAefcvq/G+P+Yf099l+rfpn80B2+GAHifnfRfFXb/mX9N/oJJ+vfyGAgAA+VMX4T93+xfi72vjPn3nPzj+jPgzL/AG3+LfX/AKL+5/PamL+o/kHz15P/ADt/UHt/3LLs+ws3/E/y7+m/vfpf0b+cg14wQjQAD882vnih+t/jf3b1n5d+vyOBwAgMzQ+Vvg6apfrzjf16Y2AAhGugEEIAGjAoCQEAMJsARtAAMQCAAAEBAAAJAQ1KAkAAKY0kAAAGNVeefiJ1Zelfstm9059ABgAAAACJINBCgAPyK3ntf09TDCgSAAAAAAAAAAAz6XMc/wChz5p05AAAAAAAAAAAAAAB2fbeX1BhoAAAAflHvHon6vejzgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfkMHx3+7fHt0hy6gAAOBQAAGAx4wCAAAWQYB458t/C/0B9//ACX95+Afln+33p8dqy15/wDFCfC/oiP4R/WtH5A/qqP81P1gR+Kn7EsfZF8R/MNHolip839nmvsrwFL8qf6KuTZTJgAFANwQAAwKoegHIAADgeAAAANDqSgAAAADwAAoAAAcAArkAwAAFcAAAAHH/L9r5ZztZ+qvqvKuyIYoKAAAAA3548n635K9u+B/ffrvV/Uf5bEvFAOM+DPiP3Duvtnu8HTPqfy0AAPhr1v4T93vflP9t+Le+XvzL+mPVPav1r+U5cDp5fhb0T92/nX7BxfwD+lPm/44Vct9jj/UP5R+Z/QvKvvnzPptw+x/GgQ1AADM0y/H/mP1P8j+Pfp78x/Xfo71il09EPP/ACBHhvzwn67+h+V/el/CwAACRqVYKAkABpQDAkBCA0mwBqUACNQCAAAGgAAASAhhQEgABcaEAAAAxio59r8ZPO+7D9sfTuDcSW0AAAABpINYgwAT4z0Pzm/ePWA5dASAAAAAAAAAAAAAKmNtGPgduFE6MwAAAAAAAAAA0u349ugODoAAAAAD88NY8j/bH0+cAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAACuH4ceew/272PN6QTABQcAAAwKHKAEAAASDUGnwp8z+p+w/Ifq/NeYfqV9N8LZtNef/FKf3p+bA9r9BUflP9CJ/M37NM/Or9VAr1XPB/mcOqrE+/Q7jyYX4s/0rcmzjFgADApVJAQAFscmoSDAAAkUAAABgtS4AAAAAUHAAwGAAA4AUHAAAAKDgAAAAG+KUvkz581n336+h+0GdAAoAAAAc98bfKfq3nP2h5X1fr5+gfz8AHlfF7fwD3H5R/V33d1n67/Igb8IB+dvgvw37r8DfeP6B+De9+nfE/t1Tq/e+Q649r8+8H+e/wBh/D/TdPy/d902vzn9L8D+Cufqx/3Kyv5v4z8u/p3710P0n+aBLyaAAHm/f535sfrf93+d+T/Dv5v+qfLvjui5HQa9F90l/Rv1flfsbIaAAAAJLaVQKAkAEQoBgQADRoTQAwoAGogEAAANAAAAkEQ0oCQAEKYhIAAANGqgCD8sNI+Ef2Z3z+q04t2gAAAANJBjAYAJyTr+c/8Ao66sOhOPYEgAAAAAAAAAAAAAAAKONvnk5nXlmwdGYNAAAALoQ9vouLfbOTYAAAAAAD4u0n4i/fT1OYAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADP/AAzDhf204tukObUAFBwAADAocoASAgAeDgo885PW+J/kn579V+p/tv8AQfwi1Za8/wDitP8AS/4GDy/sE/tj2gPzw95T+A/ocPXfpqlX4oXznVb9z6omrxY/wM/qA5NnmLAAGBQ4kBAAWxSnEwFAAAOeAAAAAXLlAAAAAAcoAAUAAAqgAK5AMAABVBQAAAADlGvk75N1Xa/YyPo+TOgQAAAAAeDfHv5z/Rvuf119J+b3j6L86APijwb8m/rPa+6vb+J9MPvvwI5dr85PkD4f91/Sj1f7/wDBfEPBvzn+srNbxfu/fPff0n+TvF/j391/l7sKH03xdP7q+B/TuJ+R/wA6/TfPvA4f7u/PHyX6lF9zxc7fqPy0AAD5o9/5z5x/SX3Pnkb8J+iAmEsAAGsAAAAAbLQphMggAaFgICQAYmCoQGlAA1JAAAABoAAAEA1iAAgBiDjCQAAApGpgASfFX5D92H6Efpzz6WTGwAAAGkiNYDABqFfz5/r/ANmH0acWwJAAAAAAAAAAAAAAAAAAAQNU6uswx3I4J7cVelxoBgAAAAAAAHzDc/lp/Rf6vMAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAZ4fiX8+S/2y9X83pCWAPAAAALFcgHICABwPCgJ+Rfhr4X+h/qf6D++/EOts+l85JEjbeyuqroZSYK47/modnMOtA5rVgr1gq5w/wCer+kHg9Db6nn2DbgAALHCAkACgHblmQpAAArwUAAAAqVcwAAAAAVwAAUAAAKCgCg4AAAABygAAAAAFFr5h+ONUz7TD6cnyoEAAAAHN5dfxVzf5d/UX276B+mfzMHV5eB+cHw/7j7T4r4P3n2l7b+i/wA6+Xfk3yez6x7R8Z+y+gdl+j/z18PdL+Wf151XPcvh8V90/dfA/Ek/9U/56et0uDq8F+hsvQ9d/PL8e/b/ANHPzmS+Vv305+j5S7X8y/pf7KX9L/mUAAD8+/tfg/RPsDh72p8x9aE0CAAQGIAAAAgYUDQSgQANCwEBIAjVQJjWIMEBpIAAAAiAAAAQDWIACAGNGwJAAACmomABIeRUvxkud/P+xHsPB0BLABAQkRrAYANQr8NP0E7MPsc4dQAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA8Mpfi9/S163KAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAM/KYPzY/WXn0+4Dh3AHKAAABYPQDkBAAoSBQE5H5rfH/ALX0fuOfsewWf0r+cbFkJ7ATzoleU4aaxOOAmCuzgeO8T9D7XZ9X4ytU14Pxr+z/AMv/AKY/Q7V+4/AIqgVjPpJB6C7ceqbeeAAwKpXSgpAAAOB4AAAA0OpKAefC4Tu2egiYoOABgMAABVAAVyAYAAAoKoAAAAABGz5t+NdJyftNn05PlQIAAAAb8yeN9n8tfWvx37H9Gn6V/NJ81eP9h4F9YfN/pPxF9OcXt/kb9Q+18b0Nv4P90+yKP7Z/G3w/6R+Qf2P6BxU/I+Y/af1/ifGep/WH+c3sGd5fpeBe/G/0P+bH47+4/dPUk/md4xF/vJ8M/Cftfvf1B7Pxge18UAZP5K/qX5D+ofefnP6e04+8IoAABoMAAAAEI10AghAAg0CgJAQA1JoBjGAwaCEgAAADQAAEBIMoBAIAZGCAgAAApqJgASAFL8yNo/PD9IOrL9H9Hg3BMQEJGpQAADAr8WPsDsw+5zh1AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADzJr8EP6gvX5QAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA+SfxYD1/9XOPb2Q5dXKAAABQOBRyAgAAkUoaL5m+WPzn+m9X0/L0fo6z+w/yRYnFYnCaUJpEKDSMUULIfl/xf0PwOz+cf1VB9M/Q/mvW/P3R8RyB8B+7ABaqLRryESqgY9hbrPT1evx9be9D5/c6T1flej6r1/juq6L2viA24wAUHqAAAAAXMX5f0vg/zDWew49mz7sj1D0aX9X+25vvLqApgAAKCgCg4QDAAAFBwAAAAAANZ87/AA7pMX3EH0a7OgQAAAB5t8PfB/vHpv2b63ymhD9N+Z/nR9Mfnn9Dd5+c+3H5t9jd/jeIeq/Lfp/0fL+v/wAk/Euf+Sf2t0uD5+cP3t+gfj3z78kf0d/EXfZ3u/J8v+gHxf6Npfnx+afqW77GP278i0vWv118/wBX4A+wfgf3z3A/RP5zAPkLxL9j/D/0qb+OfuCCAmgAARgIAAAASMKsFASAA0YFASAgBhNgCNoABoISAAAANAAAQEiNoCQAApGCQEAAAFsEAgEACNDw/wDIvry4z9UKn7FdxbjUAhqUAAAwK/EP7w7MPsw4dQAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA4Zr+eT+pr1+UAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADnA/LP84kfpj+i3n9Fh2VgAAFA4FGgJAAB7igFyv4r+V9t9VbX59/Rn1znfs/8AH3B6/ifofqFz3/zGWVzI8FUGsBkIcn8IfE/0T2+/8N9zyeJ2+x9Q8N9n/P8A8peofDftf0t9Ofd/z7Oe58IAAMaVn0uf0M3K5PXx8Hz/AKPC57zPqeZ5fx/tOVm8v6jr+39r4ruu++m/MPQNP6L84c+8AAAAAqfxz+U94/R33rSbsbXnHk6fE+ap+CeZp524Ho3uUv69+55e6SwBVAAVQUAAAAAUFUAAAAAAjZ8x/EWs9V94I9jMqAAAAMeN/iPmvy3+o/unsP07+ZPG/jD4X91g+GPuvw/7k9F/OP6L81+nfS+e76b9F/nvmvlb5P8AcuE7T5b9o+sO8/S/4+Pnv6T4nw3W/Rfzb6S7X8t/WPz38/4vQvZwe2/RCX4l/sPlf0N8PfnP9Dfefof234ic36XgflZ+i/6z+N+vJ+P/ALaEsEwAAawAAAABJbSmCkJAAaUAwIABBtJoAalAAjUAgAABAQAABASNSgJAACkjEAgAAELaAEgIAGoA0Ph/8u+7Dv8A9M5f1kcWwIYUAAg2g/56vtHv5v1EPO3AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADmRfzg/1K+xy6oAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHzyH4naUP8AYb2TzukJYAFAoOGgJAABzxgHL/zYdWX73+HfLfr3rPzHh9msHzP6n3n2V97/ADlbk+l/IZHgoCMBkYebfG/wX9Kemdr8n7HnvnNfX/bcn7J/Ff8APj/SbzaUfRPN9dx2+KAAMChxICAAoB2czzdnFcX4P6HwnBfM/p/Hdb5n03pPqH1n5J6l1H135CG3CAHyDa/Ff9penLrSpAAAAAMbk0cX4+P5S4qa/Yr7LyoExQUAAcqAYAAAAKCqAAAAAAUWvkT4w1Xvn3dL64zYAAAEfyn4X3Xzr9v/ADf6T6x8Eez8Z8EffXpeH6180eH9sz1Tg+r8Sby/e+6fSP3n8xzSex8G9QRgRxBD5oH519uV2nBOeS/RhV8wfmpB/Rn4z4H2XzN+gvg/e7n5zft/8pb/AN94dDT4z7oIoAABAYgAAACBhQMCZBAA0LAQEgA1NBUAMKABqIBAAADQAAAJAQwoCQAApjSQAAAY1UgwJAQANQAAzGvgL86+3G5+gwfd3Q8GzRsAGjTm2/5yNH1OL99vVPJ6gTAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAP5m/1V9jk/VUAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADM/LEPzR/SDG/0j0eDcEwYDHjAIAABXjUHB/Mf1Y/vZ0nZj8o5f5L/ZncUfK8/wA++p/rvD9gk/Qf5ekcCgIgRxBl/CnyH7zpbnwX6zzOv6Z9Z87+s/xb+CH9OvJvdOfRy1AAAAWOEBIAFAOnLKCkAZ/nnn/RebeYfH/sfm/ReB+g+tew/Zfi/qVn6z8kb/PBtl9lfYfRkDQAAAAAAAB8uJ/mV+lEX+pRlQAOAAFBVQDAAAAABVBQAAAADmWvhv5v2n7T+wMqnJYAICIHhvxn+c/0b9afnd9P+aep/WXk/XQ6X135R8rcb+Y/111LuDDlfq76r5T3yX9C/ll6jRgRQkweWs/Oruyu587c8f8Ao4ql/IJL6r/Vfn0+Hcv84/pLZ8I/rX/PD9P9P8T/AHlDLUJoAABgNAAAAEI0oGglAgAaFgICQBGqgTEBpQANSQAAAAaAAABIIhpQEgABUaEgAAAxqpBgQAAAwAAACAPkX88evHwH7q1n729o4dwljRp+f2s/EHufVjifsRy2GGgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABRF/NT+3/scv3WAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAeCfjGHF/rfy6/UJx7AwKHKAEAAASDUGfzS9OP7SelduHyFo/kH9o+m43N8x5d759AveZP0j+VnOBQEQGRhH55x+9808v8Amv8AX3b/AEV9Z+H7Pn31X4n+JX9NnJuHPornINAAFANwQAAwKoegCQYAAEPm3B7/AJD498L+84HrvJ60f5D/AKT/ADl+2ln2flgBVBVBwxUBEBGoEDzT8hJr9O/0lysJYA4AAFBwgGAAAAAAoCgAKAAHkLX5/V9p/Qn13GkE0AADznk9b4V+Wfzz+ifrKSo2/pn678o+YfPflP3D02x5Hn+X+g+p6v2rN+qfyBIokjCGEqtUc/nH5+VoZge3fXgqviQvx6/frp4fBvP+f6/C/RqvP7ROHkCWCYAAJGAAAAASMKsFASAAg0CgJAQA1JoBjWIMEBpIAAAAiAAAAQDWIACAGIOMJAAABjFQMCAAEBoAAAADSTw/8+erP4n9c68frj6E5de/ys9Pmz81erH9pfZ+Pb8Bv0V68P0Zd5/QAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAZ4v5q/2/9jl+6wAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAiD88PyWD6z/Vfh37gw0ChXABICABwPCj+e3oy/R76C7sPBfL/wAy/rrq6XgetzX2V9p+d9jJ9x/OTngoCMBrAiiHFAKGAIPCg/MP+i3j3DDQc0o5AAKAbggAAAtijcKQYAACvBQOM8T+d/R/lab4b959I9F+7/Dujd9d+USPY9yFUABGg1jGMBnkMn48fvznp9CmdAAoKAAKoKIBgAAAAAAAAAAAEXyLS+I/qjVfcWjiwTAAD8P/AFP439p6uj8R+19x9Yfov89fPPingfr/AKFL87v537h9BP19Y/Tf5HkcqbGKvXar1Rc98st8J6OP6DaKpWF+RvqH6N+W7nov2fw+x9m/xz/ov7CfoX88BFAAAARoAAAAAktpTBSEgANKAYEgIAYTYAjaAAaCEgAAADQAAACAYwABADGjYEgAABSNTAAkABoIAAAAA0kGMx/kzWflbwnrx4y5ovav0P5tfpU5NfFvxT68PSv1FR9DHJsAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABAH80X61exyfqAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABwQfkL8ao/UD7/APP6JybVyAcgIAFCQKPxL3z9t+0O/nb4N4P6d53pfE/0P736r+ifylI/0Pl5FEoCIDIxx5wTxChgCD5BH8z/ALscW4Y2OEpUgADApVJAQAFBA78O8crOh6BR712cMEDgeAND+ar9RD0drz/8z/pLc9T9f5T0Wf8AQfwFzqlygAI0GsCOMI4mfA/jed/vkY2AAA4AAAVQUQDAAAAAAAAAAAAOH/PTSeQ/Q5nshlQAeMtfzw/uVWvi/I/jn9fdN9i/b/i1D4s09bFX4v8Adtn75+y/Duvn+u/CJnMbCEFUmrVCrVaq+cdvB815X6l+T+t+w/nX6V8d+b/s/wCCn0HD+k/kz+N/9IP0I6X7j8IDTkAAAaDAAAABAwoGBMggARCgGBAAINpNADUoABiAQAAAA0AABASI2gJAABkYICAAACmomABIADQQAAAABpIjWAwARpQACRiqfzo/O/0cPpP9Lcb+gTk1AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADj2v50P6pfX5QAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA+VA/G2xL/AFs+juDoHxQOQEAAEilH5hax4b+mnoc0k1KeVEsgSSA5RqAiA3mg+NPNStv6nF65A5j/ADbl+n/q1xdCmVADyoAAGBQ4QEgBl0fmt+d/TFL1a40OIZ5b9IzX6I/bPPcpLe4Dxhn8+n7vdmNlFfnvlf5x/Q1T13Lq9Xtfpn83OWoAGoDGCZEEcQYf4Zqv3v8AoXCwTAAAcAAACgqoBgAAAAAAAAAADQ+TPhvafrj7Zh2CGfk5S8K/UvbN/wA6/J/qvL/WPnfSae59t+M+aTeN9/7Bve7+a2rVY2JER12VaoqdZqnVa4n88/0z8t6jR+z/ADngvXeL2pfin573/wBIPof5H7D0H5b/ADf9u8+Zwe55xF9X+VaHooe8+6Z1NGgAAAAJGpVAIIQAANCwEBIANTQVADCgARqAQAAAgIAAAgJGpQEgABSRiAQAAAWwQCAQAI0AAAABAQkRtAAANQoACZADmRfnl+ePo4e3/fWdfY+jxagMAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADx+l+Gv9OHrcoAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAUvzbD8rfrTOv1Q9G4OhQAJAAB7ij5cqfxw/bb0uVZKUswSShI4HOBQEaDfh0fn3RhmY4/u7fJZ+JMv9jfrDi6XGbAHLUgIAAsAcEAHlf4bbzb/Qvoz99sVMjw5/5NT+DO9iv2Y9u5tVehfiql+YX7F9eM0rU0ifDeMfA/u+D7R5/wBB3ng31PxGN6H5n2XujvuP5vbG5jjFFCEX5iJ+vfrNjoEsAAABVAAABQUQoAMAAAAAAAAADz9r85pdl+jPdYNv84tH6N/S2+dqll2fI/O/lH9qp7N3eJ9faH6b/HXz6iPqTgB/nAxHEdgH6G+is/G79emio5+X/nT9j/EPddY5vIsX0eH7N/Jn8K/of7e+uOfT83PiWq+4/O/zj+jbNj9M/m2h5yHjTk/s/wC+cb+lDKgAAEltKoFASAAg0CgJAQAjVQJjWIMAGpIAAAANAAAAkBDCgJAACmsJAAAAELaAEgIAEaAAAAA1AIalAAAMCgAJkAADCF8O/BXfj5l9kXP199R8O+wZ0AAAAAAAAAAAAAAAAAAAAAAAAAAAAAHzLc/lb/Rl6vMAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABwX5Oh8H/oflf6QdRxbgpAABZBlAP5uf1+9Tl9bWpfIyWRD3g5wKAiAz82h9T601xPmw/qr2BHzd+bEP8AoytcXS5YABXOQaAAKAbgk5xr+fP6M7cv0WuXFicJXgIFL4BT+C/1fy0/Q53Pb/zVtfLn6ndWM0zU0o5ZBcH4R8B+8cS34/8AbNHB6J9p+lP0z+YmM9v86iiHFCKH5ST+Jv6B8NATAAAAAFUAAAABQUQoAADAAAAAQIFb4ftfKf6DaLO/D5H7f6+mdr538P8ASfL+2/MP6Mg4e9frb3f9W/lLnvy59D5T9ifzZH9bfR6PxT/ZoKXyGz4V+1hfUVWp+ZPmj9i/EfeNW+Lx+l3c32R8I/hn9C+M+N519D/btz6P8u/nv799Q9T9v+Lsh7PIq+Lh8ifHc19zfr/zahDABqaDYKQkABpQDAkBADUmwAYwGCA0kAAAAGgAAASCA0YEgABcaEAAAAxqpBgSAgAagAAAADUAhhQACDaDAAmQAAAADxP4w6M/j/ybtx+jvprG/pL6H49dMigAAAAAAAAAAAAAAAAAAAAAAAAAAPiLSfjb97PU5gAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADwf8kw+Z/0dxv9Cd7i1AAAkGoP8guf9Dm/SE2zHA97JHocoKAiAnwUPL7NmLx6f2zvJfjR+mWOn6BHLsrkAgcJSpAAGBQ4k/JnSfm39buzG5baszjmeDmCZCPwD8l5f2p+vHNrL+UgcB+kvVjNMyeYUkoSeS+f9T8s93+R/wBW9b5/Xh8x+gP3v5vGz638NiiCGEcHlovxe/psw0DOgoAAAAABQUAAAAAAUAEKAIAMAEAAPBfzr1m1n2v0AtuLXyX81+v1PUfzv9epeWPp9p+sP03+fbX43e1+e/tD+NKf7ZSI/Fn9mAqfkWz9L/y8a/Vuq55f88/0X80taH3v5lx31j8z9p6h86fkf7L+Q/66y/qe1Uz+NeJ9p5N6Z8r+wp6p9X+WX6vtfntPz0Px5/TXHT9Fjn0BCNKBoJQIAGhYABAAINpNACNoABoISAAAANAAAAIBrEABADEHGEgAAAxioGBAAADAAAAAERAiGlAANGgMACZAAAAAAA8+a+Wvmzry+Z+B6s/pP6v57+vPUeLYEwAAAAAAAAAAAAAAAAAAAAAAAD80NowP2U9LnAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAD5Z/KMPBf0Yxr9BOo49gQOB4V4v8AgV14ftl1/ViDFUHOY5QUBEBPMx/IFdVHth9k/lap9L/bXk3cZWPQCBQcVAAAAWKLnf5tt8/2N9h6sZOQT72RlWRLo1ZiXAi86T/GT7Azv9f/AMk8bzv0U6sZrDJphTShJ5ZwfS/IPo/5L/WPQcFPkZH6A/oH5dHF9X+IxQhDAEPKj/AX+pXCwzoGAwAAAAAAFUAAAAAAAAAAAAAAA5X4A0Xz0az9r+mk2vnrw/0byzuPzH+joOFrX639z/V/5QtfBHofLa/BJ/qE9H4wfsozG/CQPqD5RD9v56ipye/P88436N+aetd98H+gVPNfO9P8R/1gmvsu1U2eU5vV+Hrv5N/a5LpzfYuv+r/xbSo7+d4F+TU1/Rp2fJsEDCrAgIAABtCgEBIANTQVADCgARqAQAAAA0AABASDKAQCAGNGwJAAACkamABIACA0AAAABEkGghQADRoDAAmQAAAAAAAAA81a+V/krsy+V/VujP7S+1uHb1Ew0AAAAAAAAAAAAAAAAAAAAAAA/Gvoz+rv0s9DAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAD5U/MAPlz76yr9G/QOLYVEgUfjhtnwP6o9/ODQAKCgKAgCIGXwQ5fjNPzX0zOv2J0ePcAFBxIAOWpAQABQD+Bvzo6sP2R8K3j479ql+lfPDNnVR+iujU/iD95zX1pTqeA/Gia+0qeOlL9F98rE7U84TShKyb+FuW/Kf7UvY/H6H1B9Bfp38nxxet8RDAEMAV+WH/AD/f1KYWpnQADAYAAAAAAAoCgAAAAAAAAAACAct4RS/NPodp7j7kcvfn0fLXD/l/9mr7b0eX9Y3P0v8Aj63yI/wM/eOX64tH40/sgj465sX2f8NM3fqsVWq1Uqjq1RVObD8I/wBXJr7duVNn5T8L9K8j9B/Mv6EXzwPXfov9P/nWhQ9v87ofkYn93/odybDYpBsFISAA0YFASAgBGzQDEBpQANRAIAAAQEAAAQEiNoCQAAojEgIAAAKaiYAEgANBAAAAAGkgxgMABhQACQJAAAAAAAAAAAAZXybpPxh8jd+Hvv3njf2Zp8WoDAAAAAAAAAAAAAAAAAAAAP58erL9rvqbuxAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPB/y4D4q+3c6/R/2Dh1e5Wcg1/P8AfYPpc33mXAAAAAAAHnSPnH5tVfPH01Fe2fX019Uu5NVGADyQAHOVGgACgH+GHvXbz/b/AOV9z+tV9q7+XSf6Z9YDvHQ+RPJk/wBWKVTS8yT/ABQ66K+q/wBJrmew1YnCeUJpkUfA/H/QML1Dz/qPWY/qPxiGJqCEK8AV+UH/AD8/1JYWrs6AAAGAwAAAAAAAAAAFAAAAAQAAAAD+Zb9g98/KvGKK/qgfZ5h36fR1lZs68dmwn4X9BBK5Pxn18XlHZt3ueJ4H0QKtVlWqOrVFVrB+BX6yzX3Bdqbnxx8/+o+b+n/m37ovlwei/Uv6X+D5+f735lnfCA+E/a3j2RsWDQSgQAIhQDAgAAGpNgAxgMEBpIAAAANAAAAkBDUoCQAApIxAIAAALYIBAIABgAAAAADSRGsBgAjSgAEgSAAAAAAAAAAAAADmxfFnwP3Y+b/bei+zvong2lIoAAAAAAAAAAAAAAAAAAPPP57u3H+n/b68gAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA8k/MkPgD6qh/o/wDSnD0BJ4h+F/Vl9KfoR149ONABSDjfOU/IfF0/CufT+oPrTC/r70/n08v/ADR6s/0C9959HCYArkAgUTipAAGBX83f6yd/N7h+XLX6y3mr35aJ7mun+nf5ds+hPglH663qmhQD44/KSa+6v1HasWGrFgJ7CJpSpXkq0GRDirhBA1XrBXqh/On/AE04adSZ05QAAAGAwAAAAAAAAAAAAAAAAAADFZ/MJ+2e2fxxvUX+aF7P9ohZsitWAnsInkKkVpI0RwBXgarVgrVgq1h1aoq/PB+Ef7LzX1xoOb/jnn/VfE/T/lX9cpzLf2R6L+r/AMf52d3fN53zIP4J/oN5NmkaAQEAAANCwEBIAINpNADUoABoISAAAAIgAAAEgiGlASAAFMaSAAAAIW0AJAQAI0AAAABAQkRtAAANQoABIEgAAAAAAAAAAAAAAAPAPh3py+Os7ty+ofqbl0+nPVOTUEwAAAAAAAAAAAAAAAPz8+XPR5/3sN4AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA4f82w/OP1uX+jn2Tw7y+dSfkr8kd+HuvbaTy/jgc7QT3fVZftHv8AlX0n9A4aXyWAHjFL87fvjqz+g15dQAchRAA5agAAAL/mf/aTv5vUfyWa/XPpmr/5YJ/qN2Scn4kj+4fARep/YLWfQayvwnl/Rn63Ms2WrFgLNhOewEr1SoEcBMEDdau5rVwrVg/nN/ppw06kzoUFUAAABgMAAAAAAAAAAAAAAAAADztn83f68bZ/N/pVrrvIwX9FU7NkmxYCeVP578Q+C/p219H7+b7Sz7b+dIoWoYAr1wrVh1qwqtUIvMQ/FH95Jr3LTa0r6fgHzV8R/R59BVl7Bm/b/wA552YGd4cH5W/0dc26GVgpCQAGjAoCQEAI1MFQAwoAEagEAAAANAAAAIBrEABAAIUxCQAAAY1UgwJAQANQAAAABqAQ1KAAAYFAATIAAAAAAAAAAAAAAAAAAN8OpfLPy92Y/N/ZdEfVv19x6/UNzl0AYAAAAAAAAAAAAB+B36fety/fJSAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAMT8/g/NGgj9G/u3h23vNYr5l47eOm94l9v1Gb2xsAFBQAAA8Y/Ojry+6fpjGwihQcSACucg0AAV/N3+uXfz+1eVNfBHZJ/SHkzOP05f2z8+B+hnMUvy2/T0WdntZ/5cJ8T+zYWLTVmyFmynZsJzStqxEUIVqzmvUCvWaq1g/nK/ppw06kzoAFBVAAAAAKAAAAAAAAAAAAAAAADzNn84/7L7Z/InYUa/Etd191J4nhQm++B3nhnl/ZfL/Tfln9XR83V/b/AKJ+s/xwyLs8KGAIK4V6461YVWuDPmhP8rv6KU/Pe9a9AyVXoOejgtmlmZrnMzQzvnMf5wf0P824ZMJQIAEQoBgSAgBqTQDGsQYANSQAAAAQEAAAQEiNoBAIAY0bAkAAAGMVAwIAAAYAAAAANQIhpQACDaDAAmQAAAAAAAAAAAAAAAAAAAACj88XPyb8f9uP0J+t3Pp8rfX2dBnQAAAAAAAAAAHyNc/kt/Sx6vNoAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABF8XB+ZHhKP0C/Qvj29DMqVwAgLAAAcAAAB5JS/OH7H68vrU5NgByyAA5pRyAAP8AnS/Ujuw9+0bnR0A0tJPS01VqkjNy6WdnE5uezO+E0/jz9zwtWGrNtFn8niu1zE/1g/LhH6qxhX8KZ+X37NOatYKtVqtWD+dH+k3DTuzOgAAFBVAAAAAACgAAAAAAAAAAAAAMVH8wn709Gfy55JQ/UH9sbJPwbTG4D9Avn/5z9X5z0v8AOP2it5g9/aPpv9P/AJ5ZH7X57DCENcIKw69cVWIE+AE/H/bU/c+sZ9C/hOn/AEN1w/Kv9WBZmbU5uaGd8Pj8T/cHl3DICQAAGhYCAkAEBpNgCNYDBAaSAAAADQAAAJAQ1KAkAAKIxICAAACkamABIACA0AAAABEkEBpQADRoDAAmQAAAAAAAAAAAAAAAAAAAAAAAMzm2v55P1k78Pux3n9AAAAAAAAAAAc3+BfXl+sv3z24gAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB84h+ZfxaH2b+kHJr7o7G1JAoBgAqgAAAB5z+bvVn9C/cMVKZUrkAgAeVAAAfgp9j9uX15o6RoaIaGinxXi/1vx/W/QPm+m3O8T3s7Oc52eGf8Vp/nV/QUFqy1Ztp2fx9H+zH5OJ/qF+TCP2phCr+WAcb+hbXplZqpWaq1g/nm/ofx09TMqAAAAAFUFAAAAAAAYAAwAAAAEAgAAP5qP1c6c/YeJqcn0cev8AMgeB9+FvzxnsPU/PfqPB+g/mn7zR8+fV9A/Qn6n/ADKxnr/CRQhDAOvAKvWCBoH46p/cvzqn+mvsLXn/AObCfR/p0P8ALb9QGvzXgR93aVLN/IdH2l+j/JuGYCAAaMCgEBIANTQVADUoABoISAAAAIgAAAEgIYUBIAAUkYgEAAAFNRMACQAGggAAAADSQaxBgAMGAwAJkAAAAAAAAAAAAAAAAAAAAAAAAA/Ij4S9Pm+gf02xv6tOLYAAAAAAAAzhfip2fq837nloAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADywPzZ/PAPob9IOfT6vfyaAwKAAcAAAAByTX5y9X25foPscmw5CkgArqkBAH5F4PZn+k+hrnf0A0PGvW8j4P6H9x/n/L9A4vR/RqL8G/obPoIz88M/4jT/ADE/osT2rNTZtp2fxVH7d4jL/c/8Zw/amFlb8bkfoz8WNfpDVaq1mqtYP55v6H8dPUzKgAAAAAAFBVAAAAAAAAAAAAAAAAAA/EHu94+6TSCQJ/lofjHp7NDykPR/rXk9j48d+U/2dDraY/a2r+sfxQxmnLFCEUAQ1wrwBGB4Z+Tk1+9lNr8/erT+6Ph9h9Fh8j+hI+hvaXOfns+ffyDmv6Lu949ggAABEKAYEgIARs0AxAaUACNQCAAAAGgAAAEA1iAAgAAtjSAAAABC2gBICAAYAAAAAA0kRrAYAI0oABIEgAAAAAAAAAAAAAAAAAAAAAAAADyv+f3tx/SD4R68u3/QTl0+zN3h2AYAAAAB5v8AjX15dR+7/bjtgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHLh+d35rB0/6Tc9/bdrl1CgAFBQAAAAM38/t489/RTReiHNo8kABzSjkA+PPyQ6o/cK/vlfvhd/Oj7n4HT9S+5/MszwPpx+3/bfxv8AeaVHyPYo54UPziT+Cv3dT9ns1Nm2iz+PxX7K/Bqfqvwoj9oYw+dvy5Z7v86C/beq1VrNVa4fgR+6uOnvhlQAAAAAAAAAoCgKAAAAAAAAAAAAAiB8V/kLrP7W2dsxQfwDfxZqpyYofZHpSXlHi35Z/WC/UfV5HpyfpX8ytjCOIcMQoIArsBIQ/HP62mv0Bv1N38kk/wBOfiMP0e/JUOi6NP7B9Kaoefh+Lf6e5X+hpzaBIAANoUAgJABAaTYAMYDABqSAAAACAgAACAkGMAAQAxBxhIAAAMaqQYEgIAEaAAAAAgISNSgAAGoUAAkCQAAAAAAAAAAAAAAAAAAAAAAAAAfkbvn0n7A+jh8H/nWHg31VlX1h9E8W3r1nGgGcyL5q+MuvP47/AET7Mf1v0GAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAZ4fCn5gBzP6Q4X98a/NqAADgAAAABGfH/yL24/df0nzaK7OgQKDioAM/wDnE3n9TfpXpyvXmrn5sff/AJ36H619V8JhfP3bx/V30H+S/ttSn4fvUaIUPxyT8D/YVP66s1NmyFn8aJr7f+Jw/Wn8fR/WWgHmH6BB6H+XDn7V9XCtWarVg/A3908dPfDKgAAAAAAAAAAABQAFAAAAAABAAEAAIv5+LXtH6G75g0rw+c/ByvOPKU/NY8tfpv6B/Nv6Nt+JqtT6R/Tv5q+kN9qONqKIIIGMRB8BJ/LX7RBt/BgZnZp/Ynyi195fLIeJfZofmv3Cf1V+TQfVf7Bc2oZMEAA0oBgSAgAamgqAGpQADQQkAAAAGgAAASAhqUBIAAMYNoSAAAFDEwYEAAANQAAAABqAQ1KAAAYFAATIAAAAAAAAAAAAAAAAAAAAAAAAAAHklL8EP6XfW5ezA8h+KQ+SfmwOd9Bl9JHD4zi9J94+zmfoP6wAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAMD40/LkPGv0eyr9Dd7k2AFUAAAAAB3i2i/P32jry+7tji2UkAFdUgI/N61+f/7T9uOrcat/J30/ynhvo/6z+KYnFVf6K7/4L/RVanz9NOkHnf4hKvrH2SH+jFnSLFgLPnKeL6+G5wKdF6en19TXwB1dImtXZWrh+CH7pY6e+mVAAAAAAAAAAAAAAAAAAAAAAAAAAAeSi/n9+ruiP0Iv1JTZ8QfJSuH9Gcn9r+m5Pz74O+C/ffvP52+r/KPnH5L9f5jwT3m19QfVlT28LUTQSAPgv4cmv2P9Sqbd0Ldoc8olQI64VvnZP8n/ALRzv9dp+XQJAAAaFgAEAAAjZoBiA0oAEagEAAAAIgAAAEgIYUBIAAUjBICAAACmomABIACA0AAAABEQIhpQACDaDAAmQAAAAAAAAAAAAAAAAAAAAAAAAAAAPxH+yvV5f00LQAAcF5iHPgdx6wG2AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfKn5Zh87fojlX6L9RybDgAAAAAFdRz3wNvHmf3/c+1HHqADmlHJSZ+AHvnZl+kFvSLStfL/kH7B+MdB9Q+H73MerfN/R3Kvz31FSmL84quV/QHzon+sdjbOxYCecJ5VUj2hBMiVRQCgrtQVgr1w/n2/oRx09bXKlAAAAAAADyr5kpeVchS1esD0v3SX7oSwAAAAAAAAAAAADyr8XLnxj3facuUPd/pOK+47WNAHxV8vfAf0H8W6X6N/PP1F9u517v8hxXxp8mbR9u/UWkd7yIfPfxYnxPpSf6s9/UPtMtWAmlGqCj8ET+C/nCa/V/9BebQM2CAAQaBQCAkAEBpNgA1iDABqSAAAAADQAAACAaxAAQAAU1hIAAAAFsEAASAA0EAAAAAaSDQQoABo0BgATIAAAAAAAAAAAAAAAAAAAAAAAAAAAHx7+ZXoYf0enRmAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHzT+VIfNv6R5V+hu9ybOAAAAAFY4Z8z/DnXl9C/b+FbRjYA5agA8o/APeft/8AQLpysztTPGrQ+R/E/wB5/nD0/wBe+V+x9H8y/M/1r8av335enwT4Oa/Y6xtlPYCaYJZUSOGINsYoYmoYQrwBXrj/AJ4f6SsL7szpVBQAAAAPk/8ALXSfnb3LSfTuoor84jz7xhHpf2DFfR3vkP1CxLAAAAAAAAAAAYHzZ4HS8y4nZfq33WDBM5Dn7/zz+XPkP2X6g+sPv/wj5E+AQ639Ncq+3fn/ACv84fljfPzzuaX0R91Z39g/lFL+Afo3fP3zuam6wMDARk8MP5xy0/0A/STn17MzAkAABEKAYEgIARE0FQA1KAAQGkgAAAAgIAAAgJBjAAEAMQbEJAAABiFNACQEAAwAAAAABpIMYDAAYUAAIJQAAAAAAAAAAAAAAAAAAAAAAAAAAAFH+dHry/fz6G7cQAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPmT8mA8Q/TLK/wBAr/JqAAAAAOWg5Rnwt5F2Y/b/ANJ8ejiKVzkGjw2j8Nfb+zL9LOwqJHsVA5H4T/V/xhu79j8N4h81fgX9Jfpv+nHk+v8AL/406R+7c++M8wTyhJID3BRs3m5kaRxBFAEMAV+eH/Pv/UBz63zNg0qgoABS/Iml8L/oDvH19sVMkzJXokph84fOKfhXkkvO9QT9V9al+0e8y/onfgAYAAAAAAHCtfll98bL6PMKEBPi35z9H1fAtuf53/dz6X89KQfBf5ZhJ+mudfdV/CwBGg0DzD4utfPXn+sxWBdT1ae/1sv6Y+m40uEoJQIABBoFAICQAAak0AxAaUAA1CCAAAABoAAAEgIalASAADGjYEgAAAxqpBgQAAA1AAAAAEBCRGsBgAjSgAEgSAAAAAAAAAAAAAAAAAAAAAAAAAAAAPyG3z9e/WH0cAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAD5E/JMOT/VTC/s93NqAAAArqAA8FtfDHU9WP3T6lxbDpFKkA4z8iNZ+QftPqz+ufYKl7QzPin7T4Py3pP1P8c8x/Wr8l/ZveD4T9C5z+aTaf3163oxnmFLKErwxvib7j4LyTT/AE/8q+sfoj8U/c44vD9+KIIYAg8oH+MX9MPNqGVBSBoUFgD8RvGd4/WHsLh81E8qJZAe8HOGAcb5mjzjzMPKfFU/NfpeX91/deVbhLAAAAAOc/LfWfpD7blokNAPPPg74H9//RbF+3/DP5vP6Q+/g9TAIg+Gfy9Dif0jzr9Bt7C2IAAAAAACEaUDAUhIAAA0oBgSAgAQbSaABjAYANSQAAAAAaAAAAQCA0oCQAApGCQEAAAFI1MACQAAGAAAAADUAhqUAAA1CgAFIIAAAAAAAAAAAAAAAAAAAAAAAAAAAAPkG5/NT+i/1eYAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAGB8Gfk2Hof628+nvhz6AAAKxwwAp/JWk/H/0Z0Z/afS8WzlcgIA+dvzw2n48zOiO4ha8/wDojv8AN+ufq7bi8T8B/ZPw73L6C+O+39q/CH81/WPs77HsmmalkCR4fFvmf7h+BeqYm3keT/fvzf3XeR/lv61FEEMI4PnBL85/6HOfYMqAAaCkfldS+Gv2F3z3pKU84TTBLIOR4PUSoCNCNg4ohcZ8sp/GXnc1+rH6D4U8TAAA5sX5i+udC+/znoAGp/AXs355/RX0yfoP87fjJv8ATn+y5SABgfH/AOYAfP8A+hkP9FvQMNAQAAAAJLaVQEhKBAAA2hQABAAAI1UCYA1KAAQGkgAAAAgIAAAgJBrEABAABTWEgAAAAU1EwAJAAQGgAAAAIiBENKAAQGhQAEyAAAAAAAAAAAAAAAAAAAAAAAAAAAAAGKL+cP8ApX9jl9FAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADHD8y/zAD7f/U/k168ysAAHLQAChynxNrHgP13tH1tq8eijQAAM8udee2LXqvdwgQcp1cvzl4H+w/ifmOX9l8Rc+gvkftt3Y8H6PUs8HpaGrw+h8T+ifq/4563h+J7vgH0Y9PoOP8f/AGyKMcMQofi1Pwb9x+bUM6AAAGfPX4HbZ/sv6ZtnJPR8cfa0ub5ZD6G7Fnyl9EI674+Dq/pgEY0yMIohwwir/Nif5l+8xX7Zb+FAMQOA/NLWfbvveXKQwTPnXx/sPnj9CPI+wsn1v5B5h/O3tH9Bf0HrIAAAHzf+YofG323L/R33vn0BAAEiNdAwFISAAANKAYCAkAEBpNgAgNKABGoBAAAAA0AAABASI2gJAABiDYhIAAAMQpoASAgAGggAAAADSQaCFAANGgMACZAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAD8Mv0d9Xl+9i0AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB5kH5BfJyP1e+7ePZxFACuYDAByyeWfEnTn5p9j6T9TX+WwQAAUAxwgJAADN8v9rxPxl+nvvPgPQsz3vnaUfd5tHq5vDzdV67kYZ+F/ZHl/T+rM/Jv2KOIUcIRfm+n3/6y8uwQwAAAPwH9r6svvl+kWLAflx+qyc/53B9r+ltfnL9tp+lflaGf+sgNa1HGEUIQQjgri5/8pE2/vLhpskNPCmvzu+st19przUCoEcf+fnw37v92ej/AF35AHX45+Ydr40/oa6M7AAAAAHlX5sB+e3vKf6LfZnPdohjU0KoCQlAgAAGhYABAAADU0FQAjWAwAakgAAAAA0AAAAkBDUoCQAAYwGggAAAY1UgwJAQAI0AAAAABpIMYDAAYUAAATIAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfmZy/pc/wC0JtAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB8fB+N3pkv9hvUuLYExQcUAA5CiPGfjLpz85+uNZ+qtrksEAAFgDgkBAAM/MT4q6s/2TsbZyPB7g4f4t/TfyuvW+8/PPXfr38k/Zms+R+xZGEUQQ/kSn+iH3pybBLAAA+XaPw3/b/qx2Zqm1YD8s/1bVT/AJ1i8/6FPwL9g2ZHzSLyz9LA6FjUcQQwhBAOvAKtlB+SPsWd/tHUwv4n+Veif0B+jOewhgBWV/AnrX53/Rv1Ufov83gBSD+fX7s6sv1GGAAAAAHOfn8H5ycsj9Af0E59PQUnQBApCQAABpQDAQEgAA1JoBiA0oABoISAAAACAgAAABAIxowJAACkjEAgAAApGpgwIAAAagAAAACAhIjaAAARpQACQJAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB8vflP34f0fnTmAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAYn5PB+en6j5V+hc/JsAK6gAAHkgB4z8g9Ofj/ANObR9X95yaBIBQDHCAkAAMv+fTafqP7968pHie4FoXn5/te74Hat+f+jawGRBHGDfwol/vx7lybggAAD8RdFv8A6EdOMllqzYD8sf1fTsfnOH276e1+bn3Gn8M4qfIfSofdDKmKIIIQgrhXrDr1hch+Kar9UfLM66H9CIruTGgAAPjn5/8AQ+O+8uf0pT6j8qAADwj8Bto/fX6W1kAAAAAAGfKQfnB8cB9X/f2Wn15axYSAgAAbQoAAkBAA1NBUADWIMAEagEAAAADQAAAEBIjaAQCAAC2NIAAAAApqJgASAAgNAAAAAGoBDCgAAGoUAAkCQAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAYX83/AGY/07db2ZAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB80/i8En6+c+n0Sc+g5ijABXIBAB5x8mdGfzl6/0R9S+/cl2CGBYAqkgIAPFaPwL/SDtx+uH3L3IVQAEaDWMYxCIHmH47TX9MU/Hs5QAADlP5rtp/an0HfGe0yzZD8gf0CVdD8sC+4fUGvzY+50/zG/YxOp+PgfsXG5ihZBAFeuFbAHerC4f4zHx333np9bv5bAAAA+cPm/86/pH9Ad79A/ngNeUARoJ+eVL8zv6JenPuAAAAAAAAPPw+Afz2Dm/2eyr6qOawAAGlAMBASAACNmgGANSgAEBpIAAAAANAAAAJAQ1KAkAAGNGwJAAABiFNACQEAA0EAAAAARJBAaUAAg2gwAJkAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA/ATqy/YD7B7sQAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAKQfmn+WYfaX6n8mvbLlbigAFBxIADhYfzJvHy/zvVH0j9K89eqGGgDFEoSAAHyx+HfTH6KfZm+aq0qgIwRCICAAHwRxmd/tycuooOcAB8b/k/wBEfspJtlashZsj+ZYkdR34em7rXlHoSfm3ug1+fhe+xjg5knUYytXD5nyivS/nQXlXt4/z8/UbG/uc57AAA8O836b5B+8vlv1TvT7v8FAGoDQAD8ZPM+nP999CkAAAAAAAAAfBn42B+v8A9ycmgQxBoFAAEgIAEG0mgAaxBgANBCQAAAAQEAAAAIBGNGBIAAURiQEAAADGqkGBAAACNAAAAAAaSDWIMABo0BgATIAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB+Ru+fs/wCqPo4AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHm4fkL8go/UX9BOPa05UAACuQCABy1J5LS+afnLrz6b6Wl/RHc82oS1UkBAB86UfiF7v2Zfo909QDAAAAAA+ek/yl/d3HT6RMLABQVwfmN82dOf6dTa52rQWbI7VkVqwFmyixMVPIJUTj8na/P/jCuh+3GvXPjcXzdpFWPdxe86BPxL8653++By6gAB458Y/n39Efb/qX2n4mJ2+G1AQAAAM38GtJ9j/bHeHgAAAAAAAAHycH4Ufu/lX0Mc1ohQDAQEgAAjZoBgCNoAABqSAAAAAA0AAABASDGAAIAALawgAAAACkamABIAADUAAAAAQEJEawGACNKAAEEoAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA+DdI+Xf3a9TnAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPOuT1uo3duINeYAPncPBvaQ+mAAAAAAAAAAAA+PvpEOwAAAAAAAAAAAA+Vw/HKKX+t/wBPcezgYAOBSQAHCUqSmzwf5t6J8I9D2n373vm09NkxtwQAAHFfkZtPx79t9Of2p17kGAAAcIj86fn2b/XL7w5tAlgAAAfjpb6svvKxpFu0Fm0O1ZFashatJ2LITyKnIyD8ngxOkqcKqn6p4uMuB+gXRk1aguT/AAymv6Q/QOTYEHhHmfUfIf2/8n+t+pp9/wDz41AAAAAADjGfgR9ndOX6+DAAAAAAAAAD8tvleX+5zePUCgACAAAEG0mgAQGlAANBCQAAAAAaAAAASAhqUBIAAMQbEJAAAAAtggACQAEBoAAAAA1AIalAAANQoABIEgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAD55qfx5/pS9bmAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAK9HLqCdDznj9b4F95/Nf6P+k/Sv1H+Yw7PIPxTD9VPzuD64+sQAAAAAAAAAAD8qP0BD1YAAAAAAAAAAAACp+cwflN9WQ/1h9F5dwQAOWQAAHLUgIAy/DNF4H4V0w32wfs/sXPXoj8mAAHzj+aO8/Inte8e9+wVPddYHI/PYeD+ATX3Z+quGnbGbAAAAAPwv8Aorqy+ubOkW7IW7Q7VoVm0FuynbtJzztvZJxX5IUd96jced+cJ/e/Vo5vmQ1PQwq1RVfyCT/RX755NhJr5l+avzX+mfuD0z77+f0OzxQAAAAAAAPN/wAAdp/SL9RNYAAAAAAAAAPJ/wCcEP6b3cPQAASAgAARs0AwBG0AAA1EAgAAAAaAAAAgJBGNGBIAAUME0EAAADGqkGBICAAaCAAAAAIkggNKAAAYFAATIAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAco1/OX/Ux6/LdAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA+e/iP8w/pkPmv0YAAF+qvd+J+tF/V/5WPxTD9rPzTD6J8kDj+RD6B7UOX86D7D+Lg7v58D136cD8uPskPHKgfK/7XB6sAAAAAAAAAAAAAAcJ+TYfCX6aZ1+j1/m2BA4FJAAcJSpAAAjH5p4nsvGvIujOv6oHqfpmNeleg41ZJOK+RdF82eL7T5ryNz1P0tNfS/2VhXopLBgAAAAB/P8AfcPXl9KFxqWgtWwr/NY+E9OH9X66Vy0naspyMDO/H1k3WXPN88n+n+qTUrBUqjq1RVfgROj+y/Ltn/HHy/6f5/8AcPn/AEXZn2v4eAAAAAAAAAHiyPwE/avry+2RgAAAAAAADP5WA/pc6ni6ASAkAABBtJoAEBpQACA0kAAAAAEQAAABASI2gEAgAAprCQAAAAKRqYMCAAAEaAAAAAA0kGsQYADRoDAAmQAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA/mw/pK9fk9LGAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAeX8Xs/AX0H+af0h9Jdx+n/wA0B0+afimHqHggful/P0H298mB+snw8FjFD1r6cDlPnQPlv9Ug+Mf0bD8s/wBfw/Jj9EQ9WAAAAAAAAAAAAAAAD5x/HMOO/XHG/q859QByyAAArnINAAABQ0fCeVaz5f5ntn5xzus956FD7/u8q7frofX3ZoChROJkGAwAAAD+ez9A+vH6E8Dtec+5D9q2RfBXmw9DND3P9DEXbQ7lqSeEKvzcz89alDvtEX0hVJqVgqVR1aoqnysP5H/aryvc+L+w/NP6V+y9f9O/l0NOcAAAAAAAAJBo/kL8WemP6FfcNIAAAAAAAApfyxB/TNv8XQCkBAAAjZoBgCNYDABGoBAAAAAgIAAAASAhqUBIAAMQbEJAAAAApqJgASAAA1AAAAAEBCRGsBgAMKAAAJkAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAP54v3j9fl+gBoAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADhOX0+p0t+ILxAD8Uw/az84w+hPzeD9tngfj8HoF0PIv2RD8Rv2sD8Rf1HD5T/QcPyF/Z8Pxo/UEPVgAAAAAAAAAAAAAAAIg/Pb8lQ+oP1n59fRjGxwKSAAonFSAAAABYAOEYTnzjz3VcDwuk8VyGqw9xrpd+XvbUvXvy9N8muADDLCGu18Kek6z3mLS82B3veRfPXbh3PCueI/WVV1tpO5akljCrTZzPl7XYbQqtVqpVCrVHUrCqeEzr8gfR/5P/Uv1N9Ffa/hqnr/HgAAAAAAABINGhVH5hXHxR/RF0ZaoAAAAAAAcF/M8H9PJw7gSAAA1NBUACA0oABAaSAAAAAA0AAABASDWIMCQAAoYJoIAAAGIU0AJAQADQQAAAAAagENSgAAGoUAAkCQAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfgT+yPrcv1WUgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAD88w/QzycPHN0PjH0sP0h+HA9z2g+V/v4PzGiDrfsEPFPrsPhHxcN39BA7UAAAAAAAAAAAAAAAAA48PyT+FEfqD+iPNtaM6csgAAKrlRoAAAGBQAqkgIAAM/k6OdwdFhYdrKot50LWYogQaDLUyPkzQ1n6Gzqn5w9EK+nO1J/Oztx95wVTxP6yTXVWZLVgcjEV6rK1NzVqBVrNVKoVao6tNHm3kH5t/RfAfp53/ADfXH1v4+AAAAAAAIBCI2g6ARAT+Dn0505/riUgAAAAAA8P/AJ/0f0lnFqAAAgIk0AwBG0AAAiIQQAAAAANAAAAJAQjaAQCAACmsJAAAAAoYmDAkBAAMAAAAAARJBAaUAAAwKAAmQAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPwL/AGP9Xl+qy0AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB8zh+Mrk/wBhfoDk2HS1JAAActQAAAABQDAHBICAAAACgGwKFBVmQEDPy+8A6s/0z851ib10LVoPhHzEejmh7x+gCLVlKeZ09iIoArVGvO6HmfY9lF6fx1Wq5q0lXmnjn51/QcFhV+cX9Hn338/BlQAAAAAEgAiOkGwEBISeSfz37x/Q37ntIAAAAAAfP/4Ro/o3OLUAAERNBUACA0oABARJAAAAAAaAAAAAQADCgJAABiDYhIAAAAFNRMACQAAGoAAAAAA0kGsQYADRoDAAmQAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPwL/Y/1eX6rLQAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAFT8yQ/LX9Bor9RtTl2V0gIABwlKkAAAACwAUShIAAAAAFANgMBqqSkz8gfkn1R+ztjfK1aCzaHF86hw3p4/p/TSnshPIqcgRwE5Pw383+w+br8L/RvWfY/2v8/3+e+k/HPJvJvzb+jpfVujz+88R/Rv53/NP+irDQMqAAAEAgQaJTAYCAkJAQflDovKf3X6swAAAAAA+f8A8IEf0dHFqACA0mwAARtAAAIiEEAAAAADQAAABASDWIACAAChgmggAAAYhTRAAEgAIDQAAAABAQkRrAYADCgAACZAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA/nu/cf1uX6QKQAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB45+MAeZ/sljp9Irz6OJAAAVzkGgAAAYFAAolCQAAAAAKAYBQDFFzX81Wuf7gdn042rIWrI7VkVmwFmwFiZVNISqDjiD5u+YPzn+rO7m+W9Xhe89Xok47jx7z07638p7Ct91+JQ/NaPiX9/ObUM6BAIBCJVINgAEgIJAQAHGM/nH/ou7cvYAAAAAADzT+bEP6cX8OoAjZoBgA1iDAAQESQAAAAAEBAAAACQENSgJAAAAtrCAAAAAKGpoMCQEAA0EAAAAAGoBDUoAABqFAAJAkAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB/Ob/Qb6/J7WMAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADNp9PLvHN1AAAAfL/ABwfaIAAAAAAAGOHxT93AAAAAAAAAAAAAAAAAAAAAAAAAMD85PyXD9A/1H5dtJ2VAgAFEq1IAAAADAoAFEoSAAAAAAwKAGBR+F2keyfcnThZtNWbI7NkViyE84TzokcU5AZGL5B8n/L/AOufUbvkeb5ZZf1Puvf/AKZ/MU9b2/i6tYI/ixPy39tOXYMmCEG0SqAAEAgSBAIAAAA/GT1/rz/V8tAAAAADP5eA/oH9e4tUSWgqAABqUAAAjUAgAAAAARAAAAEBIIxowJAABiDjCQAAAAKaiYAEgAAI0AAAAABpIIDSgAAGBQAEyAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAUxfzU/09+xy9aAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADfzE+6/O+w+rujl9fPz39OAAD8eOfD9p+HDuKgc7uh5f66GbRDF0g431oPn/wCgw5rhw9dygqeaB7Q8AAAAAAAAAAAAAAAAAAAADxwPxQ5pP9o/aePZywwAAHNKOQAAAAKAYAOEBIAAAAADAYDZ8c0fjv8Atr1c+rapWLIWLI7E4rEwTShI8FQGRh4b8ofnv9S9pN8l9JxPu/03j+7V/wBJ/k+tWFWgBv5rJ+2fqXx7CTYjYAAASAgQEgAAAAAAfGv5hdMf0VGsgAAAAB/PwH6F/a3Hq0zsAAERiDAAQGkgAAAAAICAAAAEgIRtAIBAABSMEgIAAAGIU0QABIAADAAAAABAQkGMBgANGgMACZAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPJKX4N/wBQvrcoAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB4j8kfpX5QH0PzBxfTyeWn1HyPqH1p8D+k/UrvyH9tOXH+W3Rh94fAIfenhIUvl4Prb5uD68/MYP0+/OEPs77xD8WP05D84few4n6hD82/pMM39TgAAAAAAAAAAAAAAAAAAAAAK/5ch+YP6v53+gTuXVSQAAFVyo0AAAABQDABwgJAAAAAAAGDaPwF9v6sv0FsaRZshYnHYsCmmCWUHuAAawIflzw/0fw5n59/VHq/0z95/Ma1vofyyvWCtGB+Maf6xfY3J0hLAAEBIIBAIAAAAAABoVyP8ANb0R/UVt7ZgAAAAB+QYU/wBYOPUM7ABAaUAAA1EAgAAAAARAAAAEBIIDSgJAABgNjSQAAAAKGJgwJAQADQQAAAAAagENSgAAEaUAAATIAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB8daT+e39CnqcwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABwfjf1HyAel5J8/ex4nzr1f6F+Y/U/1T+N/uvYHx/3J8DB8ibwekfeIfBfEh9v/AJ0B9J+fh9g+Qh+hP5HB0H6mB+LHswfcHswfid+gweU/oCH4p/tYAAAAAAAAAAAAAAAAAAAAAAAfMn4eh9G/sPza77sbBAAKCrUAAAAAAFAMAFEoSAAAAAAAM8V/AHeP0/8AqTpzszisWAnnCaUJZAe4FQEYEUYUYM+l8GnPXrir1ghQOM/EWa/pW6Pj6AEAgAJAQAAAAAAADaFAB/Njvn/QH7hvmAAAAAHxYH5Pf0GcuoY2ANSgAABoISAAAAAAgIAAAASAgaxAAQAAUNBgSAAAABTUTAAkAABGgAAAAAIkggNKAAAahQACQJAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAH5cbRJ+xPpc4AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAGb8A/a/A/Q30H4nvzni++AH4oB+0V0Pw2/ckPxg9OD9HPyID9dIg8R8pD758yD82P0cD8s/ukPEfsMPzZ+6Q8p/QEPxT/awAAAAAAAAAAAAAAAAAAAAAAAOPD8S/HU/239l49lWGAAAolWpAAAAAAKAYAKJQkAAAAAAD4+pfin+ufbl7vYqZ5wnmHPKKR4PcAA1oRxhFCEMAQQBXgBgHwRwed/t+cuoIBAIAAAAAAAAEGgUAAB/PXtn+x31X0ZgAAAABhB/MP8A0K4X6wc2o1iDAAAakgAAAAAAIgAAAABAANSgJAAAAprCQAAAAGIU0AEBIAADAAAAAABpINYgwAEG0GABMgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB+C/Tl+qH2934gAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB8sh9TAfMP0iHh3dh6H4YHyd9KB6hz4ekfBIel/YYfL/ANQB8iebh918uHP+wB8v/UAAAAAAAAAAAAAAAAAAAAAAAAAM/LEPzL/ZnK/sF3NqpIAAA4SlSAAAAAAUAwAUSggEAAAAAfAP5N9Wf6u/Qe8WJhTTBNIOV4nOAAa0GRhFEOGEUEAQMBMJH4jfuBhr9RHPYIAAAAAAAAABBoFAAABJ/N3vH73e97wAAAAAAH4e+pZ1+pCcezSgAAERCCAAAAAAQEAAAAEBINYgwJAABiDYhIAAAAFI1MGBICAAaCAAAAADUAhGsBgANGgMACZAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAOB/nt7cf6buo68gAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAD4u/E0P04/SDl2V2VAgAAFVyo0AAAAAAUAwAAVSQEAAAAfCNr8gvsjsy/RDXqZpgklCR43KCgmtGxgRRCihCCFjEQ38tE939vOTUMqAAAAAAAAAEQoBgAABJ5g1+TXyx15/0t9LSAAAAAAD5X/DRH9GerxdAAACA0kAAAAAABEAAAAAIABqUBIAAAFIwSAgAAAALaiAAJAAARoAAAAACIgEMKAAAahQAAglAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAH5w+E+jz/ALtG8AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB4IH4MfasV+rsvJs5ZAAAAFVyo0AAAAAAUAwAAFEoIBAAB5N+NO8+UfcnTn9U+gEyyMe8HKAAjQYwI4wigCJoJgI/M7zSL/ffsOPUEAAAAAAAINAoAAAACSo1+bX5kdMffv6v6T2AAAAAAAAH89AfaP6N8mwRQA1EAgAAAAAEBAAAABASDWIMCQAAYDY0kAAAABjVSDAkBAAAMAAAAAAGkg0EKAAAYFAAJAkAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAVg/nw/ZX1uX7JKQAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABlaASgAAAAAAAAAAAAAAAAAABibYAAAAAAAAAAAAAAB5T+Dwd5+ynPr3i42qyAAAAOHKjQAAAAAAwKAAAAFEoSADQ+Nvz36I+XOp2n1z3hr6J9dc9CrFrBw3NJtYDYmSd+Kx5kj5f8AiJV9UfsbzadaY0AAAAACDEGAwAAAQCAXgv427za/Z/ePogAAAAAAAAA+Vw/Cf+iDDTtTDREQggAAAAAAGgAAAABAANSgEAgAAoYJoIAAAACkamDAgAAAaCAAAAACIQQjWAwAGjQGABMgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB8LaT+eP9HXqczwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAD56DK+kAtAAB+UYfq4B+Uf6uAAAAB+VH6Ah6sAAfPn0GAAAAAAAAH5bfp4F0AAAAAA/FP9rAAAAAAAAAAAAAAAAwg/ID4bR+p337y7WnRSkgAAADlqQEAAAAAAMBgMAAAAUSggFm+G0eG/K+sfNPl206Vphghs9qgEwDL4Vo7APqf9Dsb+ojCgAAABBoDAYAAAACAkApfmRovzd/Ufqz/AE5tgAAAAAAAAAAH4l8tL/bJvHuhIAAAAAAA0AAAAAkBA1iDAkAAGBTWEAAAAAMQpogACQAAEaAAAAAAiIBDUoAABGlAAAEyAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAYH8/HXl+zP2n24gAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABwHizAA5MPhLrGfWv1ojrBB+KYftYB+Kf7WB8yeLh9B/QAeSfIIeGfpsGt8Zh9U8kH5Bfr6HqHxmH1T6GHhOEHS+fhu2w88+6Q+X/qAPmzxkPZvpMPJPkwPsn0UPkzgQ+P/ANsQAAAAAAAAAAAAAAAAPk4Px7wUfpv9y8u11c6UQAAAA4cqNAAAAAAAAwKAAAAAAEoAojNa4iuw6kOoJAAAMtlXeAEACAxBgMAAAAAAQCAQAeC0fjVr9eX7Ze1sAAAAAAAAAAAA5L+dIPrb9VuTXoDKwAAAAAAaAAAAAEAA1KAQCAAChoMCQAAAAKGJgwJAQACA0AAAAAAaSCA0oAABgUAAkCQAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB+SnE+nzfvOayAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHhfx+wGFQPn0Z6D9ho+u3SH4ph+1gH4p/tYH89f7TB+SP7oh+If7CB+Mf7Eh+Xn67B+PP66B+NH7OB+S367B+PP6nB+MH7Sh+SH6jB+Y33CHg/1sH51/tYH8/f7HB+T37WB+UP6zB+QP6DB8jfegfjL+/QAAAAAAAAAAAAAAAAAEXwyH5a8Wj9Avvbm29HXKlEAAAAKrlRoAAAAAAAACgGAAAAAAAAAgBQAAAAEAAYAAAAAAAAABICAAAymfmH+dXVn+ov6d2rYAAAAAAAAAAAAB5OH42/PCP1K+8OPayRQAAACIAAAAASAgaxBgSAAABbWEAAAAAAFtRAAEgAADQQAAAAAagEI1gMABo0BgATIAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfGOk/k9/Rb6nN6QAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHzOHy59es97egAPxoD9f9gPxW/akPxT/awPxT/WEPzm/WQPym+6Q/GD7jA/RkPxm/ZEPwL+4wPsQPhD9Vw/FP9rA/Kj9AQ+fwPin9rA/FP9rA/H77mD8u/rMNOuH0n9HB+KH7XgAAAAAAAAAAAAAAAAAADPk8Pzj+OA+mfuTHT7J6fm0UQAAAAquVGgAAAAAAAAACgGAAAAAAAAAAAAAAAAAAAAAAIBAIAAAAA+UPyG6I9A/aLafawAAAAAAAAAAAAAAAPjYPyRw0fpZ90ce2iRQAICAAAACAkAGpQCAQAAUNBgSAAAAAxqpBgSAgAAGoAAAAAAiIBDUoAABGlAAAEyAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfO34g92H7qfXPVmAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABRC8AAAB8kh8TfQweL/riH4p/tYH4p/tSH4ffbIfnX+1Ifl59QB032mH4MfryH5qfUAdN76H5/fquH4p/tYH5UfoCHz+B8U/tYH4p/tYH4/fqcH47faYdj6aH5ffUwfDf71AAAAAAAAAAAAAAAAAAAAAcUHwt8Fh88fVsv7Z+wOXXqzOgAAABRKtSAAAAAAAAAAADAYAAwAAAAAAAAAAAAEAgABAAAAAAB45+Um0/NX61dEfoW8AAAAAAAAAAAAAAAAACL4bD8w/LkffX3vy6+rmVtAAAAACQEDWIMCQAAALRhKAgAAAAKRqYMCAAABAaAAAAAANJBoIUAAA1CgAEgSAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPnS5/Er9gPU5v0GGAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHBcuHsoHFdqHFdqGZ5OHpGwE3igdx3AcFKHZ+KB3HWhz/VhxXahynQBngYvahxXahx/VhX8UD1DqA4ekGx1oAAAAAAAAAAAAAAAAAAAAAAB5f8Nh8SfOAfRv1njf1T9C8+lglgAAKJRpRoAAAAAAAAAAAAAAYAAAAAAAACAAAAAAAAAAAA8P/Nrafiv9FeiP1M60AAAAAAAAAAAAAAAAAAAAD5fD86/iMPdPtrC/sX03n0BMABASADUoCQAAAKEBiEgAAAAMQpogACQAAEaAAAAAANQCEawGAA0aAwAJkAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA+LPyO9DD9hv0E6MwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADig+PvkQPkvmEfSn0llf0b9B4adcQwAAVRC1IAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAByzPjr4M3j57/Qbaf009QAAAAAAAAAAAAAAAAAAAAAAADmg+K/hkPkn1NP6d+hsL9y9cxvoTJgCNYAAgAAALRhKAgAAAAKRqYMCQEAAAwAAAAAARJABhQAACNKAAAJkAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADF/K7fP44/b/0cPrkAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADxwPlz5kD5r8PD0j3fOvb/Z8q9n9byvpiWAKrkVoAAAAAAAAAAAAAAAAAAAAAAAAAADmWfOfzbrPyx847T9Kfd1L706gAAAAAAAAAAAAAAAAAAAAAAAAAAAx/lkPlz52DwzzIOv8ASIffeh512focvvvSsq9S6/OgQBQgMQkAAAABiFNEAASAAANBAAAAABAQkGsQYAADUKAASBIAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAD440n8rPbvS5/2n9LpAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAYgeD+CB4N4kHjPmIdb6jL9O9Ezr0Hv4fcdnD7DsZfTKIAAAAAAAAAAAAAAAAAAADD84o848uteQ+NaT4b5fpPtf08z6p+uA7sAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADKDynzIPPPPA4HzwPN/Lw806lHt/0Njf1D9G823QsgAQAAAAFI1MGBICAAAagAAAAAA1AIRtAAADRoDAAmQAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAi+SdJ/NriPR5/1s+7rTwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAMwPJ/KQ8s83Dz/AM3DieKDieaA6VHQ7iep0yH9Qg0E9kAAv7KAAAADGCgM5ugznuXa5TjKWT2DPS/VQ9h9rD3P3sOgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAApeMh4D80h8meKB6r9FZV9MfTvNp6yZUAAMQpogACQAABoIAAAAAANJBAaUAAAjSgAACZAAAAAAAAAIPBanw7y3pjlPW2vqH27l1lJYAAAAAAAAAAAAAAAAB5X8U75/Ck3fj+mn6E0tgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAACIOX5YOawgzOUBnLgZgY4ABlZIAAAAG2GgB0egHR9QHXdqHcaYAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHOB89fMYfK/y8HVfU+VfTX0LzaewS5aDU0GBICAAARoAAAAAAgISDGAwAAGBQACQJAAAAAAAZwvl74x68/kHf7Mfo72NnTeDB8o0EfQ30Bzae++3c2npbsrAAAAAAAAAAAA5L520n5l+VOzHyv7O6I/QH7BCUAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAACoHzV8lh8w/OgZ/vEP3r3jn19z9mxu+ZUAAAgNAAAAAAERAIalAAADRoDAAmQAAAAAA8y+Wto+T/lntx9M+49Z+5PewAABnigfMXzmHzz4UDvWYfq3puF9/3OVdd0+dafQywDKx2ctx9zwfmWs+TeL7xwHvGs/VP1gH1zsAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAeSfO4fO3z+HgPEB7B7Xjfs/s+F+u+tY3qtigAAAAAAaSCA0oAABqFAAAEyAABRF4n8+7x87fNXXnxv1BtH2D9lh7MAAAAAAABEHlnjIeSeWB51wIcZy4ZPOgAauwHXdqHofqIewe4B7tsAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABwgeFeEh4d4kHi3CB6F6vnXqPpuN+leiZV6D3uddORQ1AIRrAYACDaDAASi4gnzXyvafKPHN8/GvHd47z6OtfS31CH0hoAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAcuHkPkQeTeVB5d5uHnPChe7GX13Vw+n6KHq6qcvRJgihjMgwGc7zdrj+MqOKtaL031kPZfZg9z95DvQAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAACoHD8UHE8oHL8uGPkhV58AC7uhN0odB1Idh6AHcd2AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfB2eAAAeFethtAAAAAcf8ApiABx/MgdCHa+aBUA7vbAAAAA8/DJA7HoA8uAPSwtgAABlfOweo+mgAVPnQPbuoAAPMg9NAADPD5/wDcA6AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPz+/QEAAA/FP7GD7mAAAAA/P79AQAMT8Zg5n9ug6Dz8P52v1PD9CboAAAAHJB+BX12H6qaAeGfzlh/QZ9MhKAAHLh+HfYB8g/roH6DZgfjB+mQfnt+kwex5Qfnd+aAf0qgAHxN84h+lXegAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB+f36AgAGeH8xv1gH7pAAAAAfn9+gIAB+VAcr+wIAH86X7BB9VgAAAAAH4UfawfoCAfyqh/T/ANWAAAHyYH0xth8qfjQH9IX5zBx/6kh87/laH7tAcf8Azrh/SwAfFIfCP7iAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfn9+gIAB8kh+BtsP6curAAAAD8/v0BAAPyoDlf2BAA/nS/YIPc/zDD7E+kA/MH6QD7OAAPwo+1g/QEA/lVD+n/qwAAAAAPPPxQD98vwgD7++4Qyv5fw/qSeHKfzpB/SwBmfzUB+4XOB9MdmAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAH5/foCAAfBIfnR8xB+1P6AgAAAB+f36AgAH5UB8pfbAAH5jft0H1X+PwfQH6Ah+VHVh+lgAB+FG2H0gAfkCH9P8A1YAAAAAH51+6B9QfzpB+wX1WB/KqH9P/AFYcp/OkH9LAHyZ+JQftR5yH5Tf0SB3YAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfn9+gIAGf8PByn44h9h/vOAAAAH5/foCAAflQHK/sCAB/Ol+wQfVf4/B9AfoCH5UdWH6WAAH4UfawfoCAfyqh/T/1YAAAAHl4fJ36Agfz6h+qP1yGP/LkH9TF0OU/nSD+lgD8/vhwP3kA/n1++Q/REAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPz+/QEAD4++gQP5lwP6aA9AAAAD8/v0BAAPyoDlf2BAA/nS/YIPqv8fg+gP0BD8qOrD9LAAD8KPtYP0BAP5VQ/p/6sAAAAOH+QQ+9gK/5Jh7B+igeL/iQH9D4HKfzpB/SwB8v/kgH9B4H4ofXIfewAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfn9+gIAH5/foCB/P6Hyv+xwfo4AAAH5/foCAAfkwHK/siAB/OB+wgfXf5gh5J+hwfmB3wfrJ1AAH4MfawfoiBmfy0B/Tf3Yflz9YB9LAHCB+M31QB5YH6XvD8hf3HD8wfaw+0wOM/nSD+mABn4Ch+xXrAfhR+6Qef/nGH6+AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAH5/foCBnh8KffYH5xh+OP1GH9A4AAAfn9+gIAHm/wAchX+wQ9V+dw+W/Rg+vnh+ZHuwdbMH1g8DxQPkfqA+w+wD5P8ACw9m+tg+DvpEPcAD5BDxEA6sP0BA8E+ZA9j+swAPkfwsPqz6AA5oPgPQD7M9FDzr4YD9LAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAD8/v0BA+OQ8Z9WA8/D8k4Q/ZL0APobsAAPz+/QEAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAD8/v0BA/OMPvED+YAD+qqIPOfzeD8zv0nD9cZQ/P79AQAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPz+/QEM/8/g/RED+VUD+qoAA/DP4hD9if0hD8/v0BAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA/P79AQ+OfcQ9VA/lVA/qqAAPzu/GYPsP95w/P79AQAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPz+/QEPxz/YwAD+VUD+qoAA/P78Vg+uP3yD8/v0BAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA/On0sPHvVg9W+gA/lVA/qqAAPxe/PYP2v++w/P79AQAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPz+/QEAAA/lVA/qqAz/wA+Q/Nj9Ug+6wPz+/QEAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAD8/v0BAAAP5VQP2pqh8s8yH6L/XYSgfn9+gIAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAH5/foCAAAfyqgf1VAH5xh+OP6PB+xgH5/foCAAAAH5zfoyAAAAAAAAAAAAAAAAB5+Hwd54Hs3Hh9XfSAAAAAAAAAAAAfPgfDP08H2WAfKIfLn38HqYAAAAAAAAAAAAAAAAAAAfDPxsH7XgAAAeUh+WlQP1R9bA8ED5Z5oP0B9FAAAPgb4sD279Qw22B+flIJ/0MA+FA+aQD7OD6zAAABn5sh8s/ZYfoIB51+WAcv8AooH1QAAeH/JIfpMAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAH5/foCAAAfyqgf1VAHNB/L04P6XfSw/P79AQAAA8JD+dP+lUPTQAAAAAAAAAAAAAAAAPzL8HD9rPLw/nS/dIPqsAAAAAAAAAAAPjT8NQ/oK+iwAPyW/WkAAAAAAAAAAAAAAAAAAAAOc4oPWAAAAZ+dwfanwmHxp++4cV+HAfvz50H4Rf0NBbAPlQPAfpYPym9ID9pPzODrv0DD8nvSw/Qv8kQ+npQ+PP0DD6WAAAD5J6sOg/BMP3h92D8kfsgPJ/x1D+ir14DMD8I/Qw/awAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPz+/QEAAA/lVA/qqAAP5d+dD92ftEPz+/QEAAAPyhDw73kP1WAOcDFohy/uYHlQeleFB6H3AeXawegeaBN6KAAAAH5p/P4ftYB/PV9nh+owFT50D0H1UPNKgdN2AeX6od2AAAHz/APlwHzJ/Q+HoYH5U/qsByQeE+/Bq+ch0Pah5Tth3fKBV80D3iwHlXngfRGgB5IHGenB6AB5p5eH0XoBX8vD1gAAAAA8/D8oP2fD8+flkP2sA/n7/AFRD6w8KD3jy8PUAPDPxYD+hr+c0P2y+gA+fPxND+h3QCwB+OH7GA8AAAAA/F4P1k7UPP/SAPw1D9B/sMD80w6P5cD9rAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA/P79AQAAD+VUD+qoAA/nN8HD9ef0xD8/v0BAAAOM/PEPp78Qw/pXsB55+Gwe4ewB+e37VBY/AAP1n5kPz2/pND42/NMP6AvlcPx6/otAAAAD80/n8P2U+FA/Kj+gQPVuaD80PuQPx1/RkO5/AoP6deoD8y/tEPYwAAA+f/KQ+dvkcP6AtUPyp/VYPmHxwPY/xYD9+viQPnn9rA+AfiYP2H/CgPo7wYP10+aQ9O7MPyV/eAPHvm8Pp74AD9j/AM/A6iiH5k/0Dh+Q2IH7WAAAAAcv+UYfpj6mH58/nyH9BgH4MfdIfWP864f0y6AAHyj8vh+p385ofqj9wh4z/PKH9QABzQflh+ugAAAADA+PPDw/TYAAPwoD9sulD5UYHL/FIftYAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAH5/foCAAAfyqgf1VAAH4cfDgfoz+yIfn9+gIAAB+Zf3uHe/zhB+t/2cB+Pwe8foSH5PdwH6U/zTh/RL2YfzkfvIHJfj6H9AXlIfhR/RaAAAAH5p/Bwe4/HIf0j9WB+bweVe6h87sD9jPwYD9DvtkPzm/RkAAAAPn/AMpD7T/DYNj9tA/Kn9Vg/Bj7mA+GQ/QmwHxT+1gfKn5ph+6/4ph9Z/fIcP8Az+h/R0B+QF4Pc/jMP2Pwg9F/BYP38rh8HffIfFPxSH7WAAAAAfKn5vB4d/RwHKfzwB+kPrAfjH+54fUYAAAfkd+rgav5+B+Zf64h8o/Jwf0SgHwoHpH1GAAAAB5oHxD+Uwft/wDYYAHzP5SH3dx4fBf6SB8U/FIftYAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAH5/foCAAAfyqgf1VAAH5s/j2H1n++4fn9+gIABSD8VfosD5yDmf3fA/H4PoD9AQ/Kjqw/Sz+acP6LerD+dL91w5X8fQ/oC8pD8KP6LQAAAA/NP5/D9pPwFD3b9jAPx+D6m+2wAD4+D8wf10CL6QAAAAD5/8pD7WxA/n1+6Qxf1WD+bX+jcNMA+KQ+Kf2sD5U/NMP3X/ABTD7W+1g+WfxoD+j0D89vjwP3E/L4Pze/TQPuP8Iw/ovAKgfGnxSH7WAAAAAEQfzW/0FB6bhB8/Afih/SKFgAAPhQPqD0sADx/nw/Nv3sP0pAPxwD9ipQ/NMAAAP0sAPzODzf8AXkDhA+P/AL7A/LsPgm6HL8uH6Q/q4AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfn9+gIAHw/zwcoB1YH6AgeafzZB2H9NAfn9+gIAB8KB7R9BgceH8zv8AR+Hqf4/B9AfoCH5UdWH6WfzTh/Rb1YfzpfuuHP8A40B/Qb4eH4hf0WhUyg6AAA/NP5/D9rPOg/nU/aIPs786Q+C/3tDa+Sg+uGB/N79iB+wIEXPh0oAB8/8AlIfawHnX88gfc/68h+Cf02H6o8oHjXCB8J/tuHxz+dwfuv8AimH2t9rBw/8ANOH9LHcB+a0QfUf0WHD/AM7of0ZfzTh+7H1QHyJ9HB8rfFIftZUDK6AAAAAPwT/eQLAFf8Kw/Tz6gA5QOrA+MfcQ9d48Iu1A+dA/J793wugY/wCTYfr4AAAAAAH58hsfdwcP8iB95geM+zAAfFPxSH7WAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB+d30iAAAAAAAHwSH5l/s+HM/piAB4z+FwftP9IAeTh/Pr9EB+uX48B7d+qIfjR0AfpZ/PgH7n+0B/PR+wIfa385AfWPvAfln++QfKnymH7pAHn4fkB5CH7geuh8efiAH7CfaIfhl8/h7X+1gexgfnDth+gYHwJ+fwfv4Ac+H51+fh+onYAfOgfEn6zB8/wD4NBp/TIftdxQfzy/aYd38KB+1P5AB9E/rIHQfnmHyD+ggfJ36xB+d3mIfVfx+H7E/Awfjd6wH6LfooH5TfKIfu/8ABIfKf7pAAcJ+IAfaOeH039PBU8MD4k+ww+lgPL/5tw/qI/PEPyg6UDRD+kLmQ+T/AAMP082wAPiHqw+swAAAAPyWxA945IP024cPwF5wDPD9V/0RAA+KfikP2sAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADz/AM/AAAAAAAAAA6D2AAAAAAAAAAAAAAAAMcMfpQpbAVMoOgAAAAAAAA86DtdAADPeF0CLnw6UAAAAAAytUCoHn/pYAHONDQsBdAAADn8IO9AzK4cr6KABzUIdWAAFQMroAADjAyvRQeBlcuHegABygdWAAABwgdHsAAAHKdMEoAAAAFLzYO72wAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA//aAAgBAQABPwCqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqrVfiF4A0O/l0rW/HPh/T72DHm211qcMMiZAIyrMCMgg8joQavP2pv2YbC7msL/APaP+F9tc20jRTQzeMLFHidThlZTLkEEEEHoavP28/2ObK6mspv2ifB7SW8jRuYbwyoSpwSroCrDjhlJB6gkVN/wVC/YUgleB/jshaNipKeG9VcZHHBFsQR7jipv+Cu37GUcrxJ4i8USqjELImgygMB3GSDg+4Bq8/4LS/sp213NbQeDfihdRxSMiTw6TZBJQDgOoe8VgD1G4A88gHitV/4LcfBWG/li0T4OeNbuyGPKmuri1t5G4Gcxq8gXnI+8cgA8ZwNc/wCC43hq3liXw1+znqd/GVPmtfeJ0tGVuwUJby5GO5I+lax/wXJ1qeyMegfs2WNnd7hiW88VtcxgdxsW1jOffdx6Gv8Ah958U/8Aoh/hT/wY3H+FTf8ABaz9qJpXNv8AD/4WJGWOxX02/ZlHYEi8GTjvgfQVrv8AwWS/a51fyP7P074f6J5W7f8AYdGmfzs4xu8+eTGMHG3HU5zxj/h7z+2V/wBBzwp/4IU/+Kq8/wCCpv7dNzdzXEHxqhtY5ZGZLeHw1phSIE5CqXt2bA6DcSeOSTzU3/BUL9uueF4H+OzhZFKkp4b0pGAIxwRbAg+45Ff8N+ftlf8ARw3ir/v8n/xNf8N+ftlf9HDeKv8Av8n/AMTX/DWP7VH/AEcx8Vf/AAs7/wD+O1/w1j+1R/0cx8Vf/Czv/wD47Wu/tC/H7xT5H/CTfHL4gav9l3eR9v8AE15ceVuxu275DjO1c464HpX/AAtn4qf9FN8V/wDg5uP/AIuv+Fs/FT/opviv/wAHNx/8XX/C2fip/wBFN8V/+Dm4/wDi6mmluJXnnleWWRizu7EszHkkk9Saqqqqqqhmlt5Unt5XjliYMjoxVlYcggjoa/4Wz8VP+im+K/8Awc3H/wAXX/C2fip/0U3xX/4Obj/4uv8AhbPxU/6Kb4r/APBzcf8Axdf8LZ+Kn/RTfFf/AIObj/4uv+Fs/FT/AKKb4r/8HNx/8XX/AAtn4qf9FN8V/wDg5uP/AIurT4yfF+wuob6w+K3jC2ubaRZIZodduUeJ1OVZWD5BBAII6Gv+Gsf2qP8Ao5j4q/8AhZ3/AP8AHa/4ax/ao/6OY+Kv/hZ3/wD8dr/hrH9qj/o5j4q/+Fnf/wDx2v8AhrH9qj/o5j4q/wDhZ3//AMdr/hrH9qj/AKOY+Kv/AIWd/wD/AB2v+Gsf2qP+jmPir/4Wd/8A/Hah/a2/aqglSaP9pf4plo2DKH8YXzjI9VMpBHsRiv8Ahvz9sr/o4bxV/wB/k/8Aia/4b8/bK/6OG8Vf9/k/+Jr/AIb8/bK/6OG8Vf8Af5P/AImv+G/P2yv+jhvFX/f5P/ia/wCG/P2yv+jhvFX/AH+T/wCJqH/gqF+3XBCkEfx2crGoVS/hvSnbAGOSbYkn3PNf8PR/27f+i6/+WzpP/wAi1/w9H/bt/wCi6/8Als6T/wDItQ/8FSv2645Ukf43pKqMCY38M6UA4HY4tgcHpwQfev8Ah7z+2V/0HPCn/ghT/wCKr/h7z+2V/wBBzwp/4IU/+Kr/AIe8/tlf9Bzwp/4IU/8Aiq/4fV/tUf8AQhfCn/wVX/8A8m1/w+r/AGqP+hC+FP8A4Kr/AP8Ak2tD/wCC2f7QdvLKfEvwr+Hl/GVAiWxjvLRlbuSXnlyMdsD61/w+8+Kf/RD/AAp/4Mbj/CtP/wCC4HxEjvYX1X4C+HLi0Vv3sVvrM0Mjj0V2Rwp9yp+lf8Pzv+rXP/L3/wDuGtK/4LlaTNfxRa3+zTd2lkc+bNa+LluJF4ONsbWkYbJwDlhgEnnGD/w+8+FX/RD/ABX/AODG3/wrQv8Agtp8Abjz/wDhJvhT8QLDbt8j7Ctnd7+u7dvmi24+XGN2cnpjn/h9X+yv/wBCF8Vf/BVYf/JtWn/BX/8AY4ubSC4m1HxhaySxqz282hEvESMlWKOy5HQ7SRkcEjmtE/4KzfsT6tdNbX3j7WtGjWMsJ77w9dMjEEDaBCkjZOSeQBwec4B/4ej/ALCX/Rdf/LZ1b/5FqH/goD+xlPCk0f7QvhcLIoZQ8kiMAR3UoCD7EZFaR+2T+yZrlkL+y/aT+G8cZYqFvPEtraSZHX93M6tj3xg1of7Rf7PniiWW38NfHf4eatLAoaVLHxRZzsgPAJCSHA+taR498C+Ib0aboHjTQdTuypYW9nqMU0hA6narE4FVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVaxrmieHrI6jr+sWOm2isFM95cLDGGPQbmIGT6V4g/al/Zo8KyXdt4h/aD+HNjc2MZkntZfE9p56jbuGIhJvJKkEKAScjAORWuf8FHf2JfD0MU1/8f8AR5VlYqosbK7vWBA7rBC5Ue5AFa3/AMFdv2MtKa8Ww8ReKNZFsrGI2WgyILogZAj84x4J6DftGepA5rXf+C2nwBt/I/4Rn4U/EC/3bvP+3rZ2ezpt27Jpd2ec524wOueNW/4LlWiT3MWhfs0yzQjItp7vxcI2PHBeNbRgOewc/Wtb/wCC23xzuLRE8N/CLwJY3IkBeS+e7u0KYOQESSIg5287iOCMc5Gt/wDBYz9r3VrpLiwh8CaNGsYQwWWiyMjEEneTNNI2TkDggcDjOSdR/wCCpH7c2oS3LR/GtLSG5ZysFv4c01RCrZ+VGNuXAAOASxbgck81rH7cv7YGu2RsL39ozxxHGWDbrPVHtJMj/ppDtbHtnBrXf2hfj94p8j/hJvjl8QNX+y7vI+3+Jry48rdjdt3yHGdq5x1wPStb8Q+IPE12l/4j13UNVuY4xEk19dPO6oCSFDOSQMknHTJPrVaZ4A8d63YxanovgrXr+zmz5Vxa6bLLG+CQcMqkHBBH1Bqw/Zp/aN1Wzi1DS/2f/iReWs67op4PCl7Ikg9VYREEfSrD9iH9rbUrOK+tv2f/ABekcy7lW4sfIcD/AGkchlPsQDVh/wAE3P21tSs4r63+CE6RzLuVbjXNOgcD/aR7gMp9iAa0z/glZ+2JqFjFd3XhHQdNlfO61utdgaSPBI5MRdOQAeGPBGcHIGjf8Eg/2qNUtWuL3W/h9pEiyFRBe6tcM7DAO4GG3kXHJHJByDxjBOi/8Ea/j1cSyL4i+J3gGxjC/umspLu6LHuCHhiwMd8n6VYf8EXvH8t5FHqfxy8P29qWxLLBpE0zoPVULoGPsWH1of8ABFAn/m5j/wAsv/7tq1/4IvfDpLWFL743+I5rlY1E0kWlQxo74+YqpZioJzgFjj1PWtG/4IzfBOC6Z/EPxX8b3tv5ZCR2aWtq4fIwSzRyAjGRjaOSDnjB/wCHOH7MX/Q9fFD/AMGdj/8AIdWH/BJb9kizs4rW4tfF99JGuGuLjWtryH1YRoq5+igV/wAOn/2Qv+gL4o/8Hb/4V/w7N/Yh/wCiJf8Alyap/wDJNf8ADs39iH/oiX/lyap/8k1F+wf+yBDEkKfAPwyVjUKC8cjHAHclySfc81/wwn+yF/0QLwv/AN+X/wDiq0X9jL9lDQJZJrD9nvwLK0qhWF7o8V4AB6CYOFPuMGv+GWf2Yv8Ao3L4X/8AhH2P/wAar/hln9mL/o3L4X/+EfY//Gq/4ZZ/Zi/6Ny+F/wD4R9j/APGqtfhD8JrG1hsbH4X+Ere3t41jhhi0S3RI0UYVVUJgAAAADgV/wqz4Yf8AROPC/wD4J4P/AImv+FWfDD/onHhf/wAE8H/xNf8ACrPhh/0Tjwv/AOCeD/4mv+FWfDD/AKJx4X/8E8H/AMTX/CrPhh/0Tjwv/wCCeD/4mv8AhVnww/6Jx4X/APBPB/8AE1/wqv4Ynp8OPC//AIJ4P/ia/wCFVfDD/onPhf8A8E8H/wATX/Cq/hhjA+HHhf8A8E8H/wATV38IfhNf2s1jffC/wlcW9xG0c0MuiW7pIjDDKylMEEEgg8Gv+GWP2Yf+jcfhf/4R9j/8ar/hlj9mH/o3H4X/APhH2P8A8ar/AIZY/Zh/6Nx+F/8A4R9j/wDGq/4ZY/Zh/wCjcfhf/wCEfY//ABqv+GWP2Yf+jcfhf/4R9j/8ar/hlj9mH/o3H4X/APhH2P8A8aq//ZL/AGW9Ss5bG4/Z0+GyRzLtZoPC9pA4H+y8cYZT7gg1/wAMJfsg/wDRAfC//fl//iq/4YS/ZB/6ID4X/wC/L/8AxVf8MJfsg/8ARAfC/wD35f8A+Kr/AIYS/ZB/6ID4X/78v/8AFV/wwl+yD/0QHwv/AN+X/wDiq/4YS/ZB/wCiA+F/+/L/APxVXf7BP7H17azWc3wD8OKk8bRsYhLE4BGCVdHDKeeGBBB5BFf8Oy/2ITx/wpL/AMuTVf8A5Jr/AIdlfsQn/miX/lyar/8AJNf8Oyv2IMY/4Ul/5cmq/wDyTX/Dsv8AYgxj/hSX/lyar/8AJNf8Oy/2IP8AoiP/AJcmq/8AyTX/AA6c/ZA/6Avij/weP/hX/Dpz9kD/AKAvij/weP8A4V/w6c/ZA/6Avij/AMHj/wCFX/8AwST/AGRryzltba18YWMki4W4g1rLxn1USIy5+qkV/wAObf2Yf+h6+KH/AIM7H/5Dr/hzb+zD/wBD18UP/BnY/wDyHU3/AARs/ZmaJ1g8e/E5JCpCM+o2LAHHBIFoMj2yPqK/4cufDD/otXij/wAF0H+Nf8OXPhh/0WrxR/4LoP8AGtc/4IqeF7jyP+Eb/aC1Sx27vO+3eHY7vf027dk8W3HOc7s5HTHP/Dkz/q5r/wAsv/7trV/+CKOsw2ZfQf2i7K8u9wAiu/CzWyY7netzIc+238RX/Dlz4n/9Fq8L/wDgun/xrV/+CMPxohsy+g/FvwVeXe4ARXcV1bJjud6pIc+238RX/Dm39p7/AKHr4Xf+DO//APkOtb/4JD/tYaVLFHYXfgXWVdSWey1iVBGfQ+dDGc/QEV/w6c/a+/6Avhf/AMHif4VN/wAEyP23Y5XjT4LpKqsQHTxHpgDAdxm5BwfcA1rf/BOr9tHQIopr74FajKsrFVFlqVleEEDuIZnKj3OBU37C37XkMTzP8APFZWNSxCWyscAdgGJJ9hzX/DK/7T3/AEbh8UP/AAj7/wD+NVqnwW+MeiX0ml6z8JvGVheQ48y3utBuYpEyARlWQEZBB57EGtb8NeI/DUsUHiLQNR0qSZS0SXto8BcDgkBwMiq0/UdQ0m9h1LSr+4s7u3bdDPbytHJG3qrKQQfpVp8ZPi/YXUN9YfFbxhbXNtIskM0Ou3KPE6nKsrB8gggEEdDVp+13+1ZZXcF5B+0t8UGkt5FkRZvFt7KhKnI3I8hVhxyrAg9CCK0//goV+2jpl7Df237QfiN5YG3ItwsE8ZP+1HJGysPYgitK/wCCqf7cmn38V5d/F+01OKPO60u/DenLHJkEDJigR+CQeGHIGcjIOif8FjP2vdKumuL6HwJrMbRlBBfaLIiKcg7wYZo2zwRySOTxnBGhf8FtPj9b+f8A8JN8KPh/f7tvkfYVvLPZ13bt80u7Py4xtxg9c8aR/wAFydagshHr/wCzZY3t3uOZbPxW1tHjsNjWshz77vwFad/wW9+GEsNqdW+Bnii2lZU+0rb6pBOImIG8IWCbwOcEhc8ZC540f/gs3+yZqd6LW98PfEjSYypP2m80e2aMEdsQ3MjZP+7j3FaH/wAFXf2IdWhkkv8A4nanorRsAsd94cvnaQY6jyIpBj6kGtE/b3/Y28QNZpYftEeEYjfKrRfbrlrIKGGR5hmVPKOOofaQeDg1oX7QvwB8U+f/AMIx8cvh/q/2Xb5/2HxNaXHlbs7d2yQ7c7Wxnrg+lafqOn6tZQ6lpV/b3tpcLuhnt5RJHIPVWUkEfSqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqrXf2hfgD4W8j/hJvjl8P9I+07vI+3+JrS383bjdt3yDONy5x0yPWtd/4KQ/sR+HfI/tD4/aTN9o3bPsOn3t9jbjO7yIX2dRjdjPOM4ONd/4LJfsi6R5H9n6d8QNb83dv+w6NCnk4xjd588fXJxtz0OccZ1z/guJ4Pt4Yj4a/Z51i/lLESrfeIo7QKMcFSkEuT7ED614h/4Lb/G65h2+FPg/4H06XzshtQmur0eXg/LhJIfm+782ccH5eePEv/BXb9szXUul0vxF4X8Om4YGNtN0GNzbAEEhPtJlyCAR8+44J5BwR4j/AOChX7aPilp21P8AaD8Rwm5hMLjTlg08BSMZUW8abGwfvLhs85zzWuftAfHnxPLFP4k+N3j3VpYFKxPfeJLudkB5IBeQ4HtU00txK888ryyyMWd3YlmY8kknqTWjeC/GHiO1a98PeE9Z1S3SQxtNZ2Es6K4AJUsqkA4IOPcetaN+yB+1Pr101nY/s8/ECORIy5N74fuLNMAgcPMqqTyPlBz1OMA1on/BOP8AbP1+CO6tfgndW0LybC19q1jasnOCTHJMHx77ee2a0b/gkf8AtYapctb303gjSEWMsJ73WJGRjkDaBDFI2eSeQBweegOk/wDBGf4zT2YfXfi14LtLrcQYrSK5uUA7HeyRnPtt49TWmf8ABFXS4r6KTWf2jLq6sxnzIbXwotvI3Bxh2unA5weVOQCOM5Gif8EavgTB53/CR/FHx5fbtvk/YmtLTZ13bt8Mu7PGMbcYPXPGmf8ABJX9kqwsYrS6i8Y6lLHnddXWshZJMknkRRonAIHCjgDOTk1Yf8E3P2KNOvIr63+CFu8kLblW41zUZ0J/2ke4ZWHsQRWjfsXfsn6FdNd2P7PngiWR4zGVvdJS8TBIOQkwdQeByBnqM4JrRPgH8CvDPnf8I38FfAelfadvnfYvDdpB5m3O3dsjGcZOM9Mn1rRtA0Hw5atY+HtFsNMt3kMjQ2dskCM5ABYqoAzgAZ9hVVVVQGeBQUVVVVVVVVVVVVVgnpW0etYGMCqqqqqqqrGeBWCegrbnjNbRjGa2jsKwMYFVVVVVVVVVVVVVVVVVVVWteH9B8SWqWPiHRLDVLeOQSJDe2yTorgEBgrAgHBIz7n1rVPgt8HNbsZNL1n4TeDL+zmx5lvdaDbSxvggjKshBwQDz3ANap+yN+yzq9jJp11+zp8OEilxua18M2ttIMEHiSJFdeQOhGRkHgkVqn7AH7HGr2MmnXXwF0FIpcbmtZZ7aQYIPEkUiuvIHQjIyDwSK1T/gl/8AsUahYyWlp8JrrTZXxturXxFqDSR4IJwJZ3TkDHKngnGDgjWv+CRX7JmqWqW9jN430iRZAxnstZRnYAEbSJopFxyDwAcgc4yDrf8AwRm+AdxFEvhz4n+P7GQMfMa9ktLoMMcABIIsH3yfpWtf8EUtBnukfw7+0Pf2VuIwGjvfDCXTl8nJDJcxgDGBjaeQTnnA1T/giz48hvpI9F+Oug3VmMeVLdaNLbyNwM5RZHA5yPvHIAPGcDVf+CO37T1lBcXFh4t+HOo+VkxQRandxyzDPGA9sEBx2LY961v/AIJZftmaVFFJYeAdI1lnYho7LX7RDGMdT5zxjH0JNat/wT8/bI0W8NjefAfW5JAobdaT290mD/txSMufbORWr/s5ftC6BZnUNd+A/wARNOtVYKZ7vwveQoCeg3NGBk1rXh/XvDl0lj4h0S/0u5eMSJDe2zwOyEkBgrAHGQRnpkGtP1HUNJvYdS0q/uLO7t23Qz28rRyRt6qykEH6Vof7Rf7QfhiGS38NfHf4h6THMwaVLHxReQB2AwCQkgycetaF/wAFD/20/Ds8Vzp/7QXiCZ4o/LUX8VvfKRjGSs8bhjx94gn3rw3/AMFdv2zNCS0XVPEXhfxEbdiZG1LQY4zcgknD/ZjFgAED5NpwBkk5J8J/8Fu/izZtCfHXwS8I6uqs3nDSb+400uMHaF8z7RtIOCSc5HGBnI8Hf8FtfgnqKRp48+EPjPQ5XVdx0ye31KNGJwcl2gbaBzkLn/Zrwh/wVH/Yn8XSLav8WJdDuXkZEi1jR7qAEKu7eZRG0Sg8gbnBJGMcjPgf4/fAz4mNFD8PvjF4M8QzyrGRbadrlvPOvmAlA0SuXRiAflYA8EY4NVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVWoajp+k2Uuparf29naW67pZ7iURxxj1ZmIAH1rxX+17+y14JZ4fEf7QfgGCeKbyZLaLXYLieN8E4aKJmdRgdSABkDPIrxR/wAFXv2J/DuE0/4iar4hkHmB00vQLobCuMDdOkSndzgqSODkgYJ8V/8ABbn4N2aOfA/wZ8Z6uwhyg1W6ttOBlycqTG0+Fxg7uTkkbeMnxZ/wW7+LN60x8C/BLwjpCll8katf3GolAANwby/s+4k5IIxgEDB6nxN/wVs/bT14SDSvGXh/w5vuDKv9meH7d9inP7ofaRN8gyOTlvlHzdc+Iv24P2vfFAUal+0b47h2yNIP7O1eTT+T1/49ymR6L0HYCtd8W+KvFPkf8JN4n1XV/su7yPt97JceVuxu27ydudq5x1wPStG0DXfEd01j4e0W/wBUuUjMjQ2ds87qgIBYqoJAyQM9MketeHP2S/2nfFnltonwC8dyRS24ninn0Ke2ikjOMMkkqqjZDAgAkkcjgE14c/4Jj/tl+IPLkuPhha6NBLbiaOXUtbs0znGEKRyPIrYOcMoxgg4PFeH/APgjp+0PftBJ4i8feAdKglhDusNzdXU0TkA7CvkKhIPBIcjjjNeHv+CLUQa1n8WftBMy7T9ptdO8N7TnBwEme4PAODkx8jIwOo8P/wDBHT9nixSCTxF4+8farPFMHdYbm1tYZVBB2FfIZwCOCQ4PPGK0X/gl5+xjpcUkd98N9R1hnYFXvfEF6hjGOg8mWMY+oJrRf2M/2UdBlkmsP2fPA0rSqFYXujx3gAHoJg4U+4wa8PfDL4beEGtX8JfD3w1orWKlbU6dpMFsYAQQQmxRtBBI4xwT61WM8CgoHeqqqqqqqqqxngUFGOTWBjAqqqqqqqqqqqsZ4Fbc96wB0qqqqqqqsZ4FbSa2+9bRWBjAqqqqqqqqqqqqqqqqqqqqqqqqqqqqqiARiioNFQehoqe1Edq1z4L/AAc8T28lr4k+E3g3VoZZPMkjvtBtp1d853EOhBOe9a5+xJ+yT4hnkuL/APZ98GxPLH5bCx04WSgYxkLBsCn3AB961r/glx+xfqsUUdh8NtR0do2JZ7LxBesZBjofOlkGPoAa8S/8Ecv2dNS86bwz458eaNNJICkcl1bXUES91CmFXP1Mh/GvFH/BFe/Rnm8FfH63lVpsJbap4fMZjjwckyxzNubOOPLUYJ5GOfFP/BJL9rXw/AZdIi8G+JnEZYRaXrJjYnP3f9KjhGT164968X/sU/tY+B2kXXfgF4vkWJmV5NNsDqSLtGSxe2Mi7cfxZx71q+iazoF4dO13SbzTrtVDGC7t2hcA9DtYA4NeCPjx8bfhq8DfD/4u+MPD6W2wRw6frU8MW1W3BGjVtjJn+BgVPcc14I/4Kq/tp+DngS9+I2n+J7W32BbbXNGt5AwVskNJCscrbhwSXzg8EHmvAf8AwW/8VW/lW/xQ+BOlX+7aJLrQdWks9nXcRDMsu7Py4BkXGDyc8eBP+Cwn7IvioQReKZfFvgyZlHnPqekG5hRtmTta0aV2XcNoJRScglQM48BftY/sz/E9ooPA/wAdPBmo3UzbYrJtWjgunOFPEEpWQj5gM7cZyOoIFVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVWq6tpWh2Euqa3qdpp9lBjzbm6mWGNMkKMsxAGSQOe5FeKf2rv2Y/BX2qPxN+0D8P7O4stnn2n/AAkNtJcpu27f3COZDkMp4Xoc9Oa1v/gpX+w/4fu0sr/4+afLI8YkBstJv7xACSOXhgdQeD8pOehxgitV/wCCwv7Hun2Et5ZzeNdTmjxttLXRAskmSAdplkROAcnLDgHGTgHVf+C1/wCzZDYSyaJ8NfiXd3ox5UN1aWVvG3Izl1upCvGT905IA4zkf8PvPhV/0Q/xX/4Mbf8AwrXf+C4vhS38j/hGP2dtVv8Adu8/7f4kjs9nTbt2QS7s/NnO3GB1zx/w/O/6tc/8vf8A+4a0T/guP4fuLtk8R/s36hY2wjJSSy8UpduXyMAo9tEAMZ53HkAY5yP+H3nwq/6If4r/APBjb/4Von/BbH9na4tGfxH8L/iLY3IkISOygs7pCmBglnuIiDnIxtI4BzzgaJ/wWL/ZB1a0a4v4vHejSLIUFvfaLGzsAAd4MM0i4OSOSDkHjGCdE/4Kj/sOa0lmj/GdtOubxlQW99oGoRmFmOAJJBAY1Hq2/aBySOa0L9tf9kbxH5/9n/tH/D+L7Pt3/b9dhsc7s42+eyb+hztzjjOMjPh7xv4K8X5/4RPxhomtbYxIf7O1CK5wh6N8jHg9j0qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq1XVtK0Owl1TW9TtNPsoMebc3UywxpkhRlmIAySBz3IrxZ+1l+zD4HaaHxR+0B4Bs7m3ZVltBr1vNcIWAK5hjZpACCDnbjBB714r/AOCpf7Evhdngh+K9xrlzFN5bw6Vol3KAME7xI8axsvAHyuTkjjqR4n/4LWfs76dK0HhT4b+PdZMczo0tzFa2cboOA6HznYg9cMqkA84PFeJ/+C4niW4haLwX+zzpljKGcLNqniJ7sMMfKTHHBFtPQkbz6Z714w/4K/8A7Y3iaJo9F1Hwf4TZo1QPo+hCQqQ2S4+1vOMkfKcjGOgB5rxb+2/+1142Eq67+0P41RJpHeSPTtSbTkbfncu228sbOSAmNo7AVrfiHxB4mu0v/Eeu6hqtzHGIkmvrp53VASQoZySBkk46ZJ9a8KfDf4h+PZFh8DeAvEXiKR5GjVNJ0qa7JdV3MoEatyF+YjsOeleGP2Cv2wPF6o2k/AfxBAHhMoGqNDppC5AwRcvHhuR8p+bGTjANeGv+CSH7U+t/NrF94J8Pquwst7qskrkNndtEEUgJXHIJAORgnkjwv/wRev5GSbxp8fIIlWY77bTNAMhkjwORLJMu1s548thgDnnjwt/wR6/Z00pobjxP4z8c67LGzb4hdW9pBICCACqxGQEZzkSDJA4xkHw3/wAE3f2M/DXkyx/B6LUrmKMo0+pareXPmZ6lo2l8vPuEGK8Ofs4fs+eEPLbwx8D/AAHp0sduLfz4PD1sszxjHDSbN75KqSWJJIycnmrW0tbG1hsbG2it7e3jWOGGJAiRoowqqo4AAAAA4FVVVVVQXNYGMCqqqqqqqqqC5HWsADAqqqqqqqqqqqtpI4oKPWsDGBVVVVVVWCRxW3I61tFYGMCqqqqqqqqqqqqqrGRxWD6Vg+lbSRwK2mtpraaKnFYPpWD6UQcdKqqqqqqqqqqqqqiARiioNFfQ0QaqqrV9F0bX7M6druk2WpWpYMYLu3WZCR0O1gRkV4y/Yg/ZK8eGVtf+AnhSN5lAkk0y1OmO2H37t1qYzuJPLZyRwSRxXjL/AII//sz68Zbjwr4h8a+GZyoEUUN9FdW6HfkkpLGZCdp2/wCsAGAeec+Nf+CL3xFs3Zvh18afDmrKcFU1rTptOK5JyC0RnzhdvOBkk8DHPjr/AIJwftieBLeS+n+Ek+t2kS5M2hXkN8xO0sQIUbzicKRwmCSAMkgHxV4F8beBLwad438Ha34eumZlEGq6dLaOSuNw2yKDkblz6ZHrXw/+O/xq+FLwt8Nviv4r8ORwZ2W+n6tNDCQWLENEG2MNxJwVIyc9a+HP/BXH9sHwNDFaa9rfhzxvbxqEUa9pIWVUAQcSWrQszYU/M+8kuxbdxj4f/wDBbzwVdRwwfFT4H63pzrkS3Ph/UYr0N8pwVimEO35sDBkPBJyeh+HP/BRv9jT4l7IdP+Nmk6HdnO+38Ro+leX97GZZwsJyEJ+VzjcoOCwFaXq2la7YRapomp2moWU2fKubWZZY3wSp2spIOCCD7g1VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVeJvFnhXwVpUmu+MvE2k6DpsWfMvNTvY7WFMKWOXkIUYVWPXopPY18Rv+Cm/wCxl8OZZLJ/iuniW9iYg2/hyzkvwQNwyJwBARlMcSZO5T907q8e/wDBb/wrbiSD4X/AnVb/AHbhHda9q0dns6bSYYVl3Z+bI8xcYHJycePP+Cwn7XPiozReFpfCPgyFmPkPpmkC5mRd+Rua6aVGbaNpIRQckhQcY8aftq/tZ+P/ADk8SftCeNTFcecJrex1R9PhkWX76NFbeWjIRwEI2qCQAATWsa5rXiK9Opa/rF7qd2VCme8uGmkIHQbmJOB6V4d8BeOfGPl/8Ij4L13W/NnFun9nabLc7pjjEY2Kcsdy/L1+YetaN+xv+1Vr0Uk1j+z944iWJtrC90eWzJPsJghYe4yK0v8A4Jsftoap9jk/4U79kgu9h82613T4/JVsfM6eeZFwDkrt3DBGM8Va/wDBKL9rS4uobeWw8J2ySSKrTy60CkQJwWYKjNgdTgE+gPSv+HPX7TP/AEPHwx/8Gd9/8h1o3/BHP473Esi+IfiZ4Csowv7trKS7uix7ghoY8DHfJ+lH/gjZ8TOn/C5vDH/gvn/xrWv+COfxtgijPh34peB76QsRIt6Lq1CjsQUikyfbA+tf8OfP2mP+h4+GP/gzvv8A5DrVv+CTf7VunXZtrM+DNUjCgi4tNYZUJPbEsSNkf7uK1b/gmV+2Zp12baz+F9nqkYUEXFp4gsVQk9sSzI2R/u4rXf2JP2s/Dsd1Lf8AwE8Vyizk2SCxtBelju2/IIC5kGe6ZGOc45rXvgr8Y/C0vkeJvhL4z0iQQ+dsv9Bubc+XyN+HQfL8rc9OD6VXhj9pT9ofwXKsvhT46+PdLAmWZorfxFdLHI69C8e/a44xhgQRwRivCf8AwVE/bZ8KJDbt8XU1q1gVgsGraNaTlixJy0ojErEEnGXwBgdAAPCH/Bbr4vWUinx98FfB+sxiRiy6Pe3OmEptwoBlNxghuScEEcYB+avA/wDwWj/Zx1yNYfG/gjxr4YuTkl44Ib+3ACj+NHWTJbcMeXjABJGcD4f/ALbv7JfxQeGDwf8AHzwpJcXGfJttQujps0hDFcLFdCNycg8AZxg9CDVpd2moWkN9YXUVzbXMayQzQuHSRGGVZWHBBBBBHBzVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVaxrmieHrI6jr+sWOm2isFM95cLDGGPQbmIGT6V4u/bQ/ZO8DyvbeIf2hfA6zxNKksFnq8d7LE0fDq6QF2RgeNrAEkEAHBx4r/wCCt/7F3h1HbR/FviPxQUh8xV0rQJ4yzZI8sfahD82ADzhcEc5yB4s/4Le/DCzaYeBfgZ4o1dVZRCdW1SDTS4wNxYRi42kHIAGcjByM4Hif/gtv8bruUnwX8H/A+lx+cxVdUmur9hH/AArmOSD5gMZbGD/dFeI/+Crf7bOvQxwWXxJ0zQ1VXWQ6doFpulDADkzRyFSMHBTaeT14x4r/AGvf2pfGyPD4j/aD8e3EEkPkyW0Ouz20EiAk/NFEyoxyepBPQZwBjWNc1rxFenUtf1i91O7KhTPeXDTSEDoNzEnA9K8MfDzx/wCN2RPBfgbxBr7STGFBpmmTXRaQAEoPLU5bBBx1wRXhf9hv9rjxg0KaT8A/FUBmZlT+1LZdMwVBJ3G6aPaMDgnAJwBkmvC//BKL9rjxCkLatpfhXw0ZFYsuqa0shiIJADfZVm5IGRjIwRnByK8Mf8EZPH12qHxn8cPD+lsYSXGmaRNf4kyPlHmPBlcZO7g5AG3nI8K/8EdPgZp0qTeL/iT4z1oxtGwitPs9jG+376uDHIxVuB8rKQM8kkEeFP8Agm1+xz4UCyj4U/2xcruHnatqlzcZB7GPzBHx2OzPvXhX4A/AzwPEkXg/4O+C9IKrGDJaaFbxyP5f3C7hNzsMn5mJOSTnJNVQXPJoL3NVVVVVVVQX1rAxgVVVVVVVVUBngUFAFVVVVVVVVVVYzwK2kjrW0dqqqqqqqwT0raMdawO1VVVVVVVVVWM8CsE9BW0kVtyOtbRjGa2ito7VgegrAxgVVVVVVVVVRAIxWB6CioNFQRgUVBGAaK8cGipxxRB9KI7VVVVVVVVVVEA0VBHBoqaqqqqv9PsNVs5dO1Oyt7y1nXbLBcRCRJB6MpyCPrXj39hX9kj4jgvr/wAC/DlrN187Rom0l93zfMTatGHOXJO4HJAznaMePv8AgjX8FtZR5fh18TvFXhm4diQmoQw6pBGMrgKgEL4wGHzSE5YHtg+P/wDgkP8AtM+GYmuvBeq+FPGUYX5YLa9NlcE45G24Cx49D5n1Ar4i/s5fHj4TebJ8RfhH4o0S3hzuvJ9OdrbjridAYz+DV4O+Ifj/AOHV+2qfD7xz4g8MXr43XOjanNZSHAZRlomU9HcfRj6mvhp/wVb/AGxfh9LDFq/jPTPGthE3/Hr4g02N2wd24edD5cpJLZBZ2xtUAbQVPwu/4LafDvVHisfjF8Idb0BztQ32g3aahEWLAF2ik8p40AJJ2mRuOAc8fCz9uP8AZS+McdvH4N+Nfh9L6ePcNO1Wc6bcghcsojuAhcqAc7NwwCQSOaqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqviT+1/+zD8I/Nj8ffHHwpYXMGfNsoL0Xt0mPW3t98o/755r4w/8FqPhVoDTab8FPhxrHi25Rtq6jq0o020xjO9EAeVxnA2ssR6nPAz8VP8Agqj+2D8TGlt9M8bWPgnT5GOLXw3ZCBgMAAefIZJgRgnKuuSx4xtC+JvFnirxrqsmu+MvE2q69qUufMvNTvZLqZssWOXkJY5ZmPXqxPc18Pf2UP2kPir5T+Bvgx4nvrebHl3k9kbO2f6Tz7Iz/wB9cV4H/wCCRH7RWvtFN4z8TeEfC1sWxJG1zJe3K8A5CRp5ZHJH+tByp7EE+Dv+COHwi0+ND49+LPizXJl5P9mW0Gmxsd2eVcTtjbgEBgc5ORnA8I/8E4P2PfCKll+FCazcMrKZ9X1G4uTtJBxsLiMEYGGCBsZGeTXhf4IfBjwR5R8G/CXwdojwyJIj2Gh28DiRMbX3KgJYYHzE596qqqqYZHBqqph3FVXiTwR4L8YxGDxd4Q0TXI2haEpqWnxXIMbcMmHU/KR1HQ14o/Yf/ZK8YLKmrfAXwtAJlVW/su3bTCNpBG02rR7TxyRgkcHINeL/APgkx+y74g82Xw9d+MPDEp80wpZams8Ss33Ay3EcjsqEcAOpIJyxOCPG3/BGvWIhNcfDj43WdySp8m01vSWgwQnAaeJ3yC3cRDaD/ERz43/4JjftdeDvMksfBWmeKLeLcWm0TVYn4GMERzGKRs5OAqE/KcgcZ8dfCb4n/DC5Nn8Rfh74i8NyeZ5aHU9Nlt0kOWA2OyhXB2PgqSDtJBIFfD/4xfFj4T3QvPhl8SvEvheTzPMddK1SW2SU5UneiMFcHy0yrAg7QCCBXwz/AOCuX7XfgX7Pa+JtY8P+OrKLajJremLHN5Y2DCy2xiJfCnDyBzl2LbuMfCv/AILUfBXxE0Nj8Wvhx4j8HXMjBTd6fKmq2iDB3O5AjlUZAAVY5D83J4yfhT+1J+zx8cIY2+F3xf8ADmtzyNtWxF19nvM4GM20wSYA56lME5HY1VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVaxrmieHrI6jr+sWOm2isFM95cLDGGPQbmIGT6V44/wCCgn7Gnw/jZtY/aA8M37jAWPQ5H1csSpIANosgH3SMkgAkAkZFfEL/AILVfAjQmktvhz8NfFviuaNTiW8eLS7d2wpAVsyyY5YEmMYK8Bgcjx9/wWk/aI164li8AeBPBnhWxZsxmeKbUbpAGJAMjOkZG0qD+6ByCQRnA8e/tvftb/EqSR/FP7QPjDy5o2ilt9MvjpcEqMoVlaG0EcbAqoGCpHLHqzZ1XVtV12/l1XW9Uu9QvZ8ebc3czSyPgBRlmJJwABz2Arwv8KPil45SF/BXw18U+IFnVmiOl6NcXYkCkhivlocgEEHHQivDf/BPf9sXxUN1h8EdTtUXZufUry2sdofODtmlVjjByFBI4yORnwx/wSH/AGl9YZJPEHiPwPoMAmKSLLfzXEwXAO9VjhKEZOMF1PB9s+Fv+CMEWyGfxr8e2ZirCW10vQMbTk7Ss0kxyMYJBjHJxnjJ8Of8Ehv2YtJ8uXW/EHjvXJfs4SVJ9SghiMnGZEWKFXXkHCl2ADc5ODXhr/gnt+xz4W+aw+CGl3cjbNz6neXN9uKZwds0rKucnIUAHjIwBjw38FPg14NlWfwh8JPBehyLMsyvpugW1qRIv3XBjQfMMDB6iqqqqqoLnr+VBRVVVVVVVYzwKCgd6qqqqqqqqxngUFyOTWBjAqqqqqqqqqqxngVtyOtAADAqqqqqqwT0rbnvWABxVVVVVVVVWM9K2k1tHrW0VgYwBVVVVVVVVVVVVVVVVVVVVVEA9qKgjAoqD0NFTjiiDVVVVVVVVRANFQRwaIIqqqqqq+Iv7Jn7NfxX82Tx18FvC99czZ8y8gshZ3T/AFng2S/+PcV8Sv8Agjp8EfEbtc/DPx94j8HTNn9xdRrqtsnI+6rGOTpuHMp5KnsQ3xO/4JO/tQ+B0nvfCMOg+ObKGPeBpd55FzgLlswz7ATnOFRnLDGBk7R4++FHxN+Fl+NM+JPw/wDEHhm4ZmWJdU06W2E2CQTGzgCRcq2GUkHHBNfCn9qT9of4ISxt8Lvi/wCI9FgiXatiLr7RZ44xm2mDwk8dSmQCR0Jr4T/8FqPi/wCH0tNO+MHw30HxdbxsFmv9NlbTbtlJ5dlw8TsAeFVYwcAZHLV8H/8AgqB+yF8W0gtLrx+3gfVJFy1l4riFkq4OD/pILW+OhGZASDnAwQNP1HT9WsodS0q/t720uF3Qz28okjkHqrKSCPpVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVfFj41fCj4GeG28WfFrx5pPhnTfmET3k37y4ZRkpDEuZJnAOdkas2OcV8ev+C1EIS50T9m/4dszFXjXXfE64CtnG+K1iY5GMlTI45I3R8FT8Yf2tv2jvjy00XxP+Lmu6pYTtltLhmFpYjAwP9GhCxEgZGSpPJySSSfCXgLx18QL1tM8B+C9d8SXifet9I02W8kGQx5WJWPRGP0U+hr4Yf8ABLj9qn4gmO517Q9J8EWBbmXXb0eayhwrbYYBI+7GSBJsBx1AIJ+Fv/BIH4LeG3t774p+Ndd8Z3Me0yWlso0y0c5JZWCFpSMbVysiHhjxkBfhr+zb8Bfg+kf/AArf4UeHdGuItu28SzE118qgDNxJulPAB5bqSepJNMMjg0R2qqqqqqph3FVRAIxRGOBVVVUQCMZqaGKeJ4J40kjkUq6OuQwPBBHcYr4hfsY/su/E/wA2TxT8FvDq3MuS13psB06dm/vM9sULH/ez75HFfEn/AII9/DLV0kuvhT8Tdd8O3HzMLXV7dNRgJ2nCKy+U8YJ25YmQgZ4PGPih/wAExf2pvh4t1e6L4d07xrp1uxKzaDdh5imcAm3kCSFumVQPjnkgZrxP4S8VeCtVfQvGXhnVtB1KNdz2ep2UlrMoyRkpIAwGQR07Gvg//wAFAP2svgo0Fv4a+Lep6tpcTZOleIT/AGnbkYxtUy5kiXgHETpz9Tn4O/8ABbLw3ffZ9M+PPwnu9LmOxJNV8Mz/AGiEseGc20xDxoDzxJI2CeCR83wc/ac+Anx/tEn+E/xQ0TXLkx+ZJpqz+TewrgEl7aTbKoGQN23bnIycGqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqtc8T+GvDEMU/iTxFpmkxzMVie+u0gDsByAXIycdq1H9qv9l/SZbm21L9o34Y289kzpPA/i2yEsbJkMpTzd24EEbcZzxiv+G/P2NP+jh/Cn/f5/wD4mv8Ahvz9jT/o4fwp/wB/n/8Aia/4b8/Y0/6OH8Kf9/n/APia0j9uX9j/AFy9FhZftGeB45SpYNeaolpHgdf3k21c+2cmtD/aL/Z88USy2/hr47/DzVpYFDSpY+KLOdkB4BISQ4H1rT9R0/VrKHUtKv7e9tLhd0M9vKJI5B6qykgj6VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVXjP4p/DD4cor/EL4j+F/C6sqsp1nWILIEMSFI8114JVgPUg+leNf+Cmf7FPgkywT/Ge31m7iUMtvounXN75gL7TtlSPycjBOC4OBkZyufGv/AAW3+D2mmVPh78G/FuvMijy21W8g0xJG34blPPIXZhgSMk8FV6142/4LT/tDa1E9t4I+H3gnw3G/SaeOe/nTDAjazOkf3RtOYznJI2nGPGv/AAUJ/bM8emVdX+P/AIjsUlUIE0Ux6TsUPuAVrVY2BzxuzuI4JI4rxD4n8S+LtQ/tbxV4i1PWr4rt+06hdvcy4yWxuck4yzHHqSe9eCf2efjt8SBDJ4G+D/i7WYJ1DJdW+kTG3wU3qTMVEYBXkZYZyMZyK8Gf8Erf2ufFaLJq2geHfCatnada1lGJG0EHbaiYjOcYIBBByBwT4J/4Iy6NEYbj4j/G+8uQWHnWmiaQsG0B+Qs8rvklMcmIbSf4gOfA3/BND9kLwSiNcfD268S3SRhPtGu6lLOT8uCTGhSLJxnOzgnjA4rwx8HvhJ4JZH8F/C3whoDRzGZDpmh29qVkwAXHloMNgAZ64AqqC5rAxgVVVVVVVQWgoA96qqqqqqoDPAoAAYFVVVVVVVVAZ4FAADiqqqqqqqqqrGeBQXPU1gAYFVVVVVjPAraSOtbRVVVVVVVVgnpW3Peto9awO1VVVVVVVVVVVVVVVVVVVVVVVVVVVRANFQelFfQ0QfSqqqqqqqoqDRX0NEdqqqqqogEYrUNNsNVs5dO1Oyt7y1nXbLBcRCRJB6MpyCPY18Uf+Cb/AOyR8UIpGPw2TwnfO2Re+F5fsBQY5Ahw0GOh5iyMccZB+LX/AARt8b6Y02ofBT4nabrlvuymm69CbOdFxyBNGHSRs46rEMH25+LX7Kv7QvwOaaT4lfCrW9NsYW2tqcUP2qy5GR/pEJaMEjJwWB4OQCCB8L/jt8ZPgpfjUPhT8TPEHhlzIZJIbG9ZYJmIxmWEkxy8AffU9B6DHwY/4LQfGPwy1vpnxt8E6P40sUVVfUNOH9m3xIBBdgoMLknB2qkY688jHwQ/4KLfso/HZksNE+IaeGtYdgq6T4oVdOnbIyNjlmhkPBGEkYjHIGRmqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqrxV4r8MeBvD174s8Z+IdP0TRdNj8y7v8AULhYIYFyACzsQBkkAepIA5NftP8A/BZO1tpbnwn+ytoaXYClH8V63asqbueba1bDEYwQ8wHOQYsAE/ED4i/EL4x+MLjxl8Q/E+peJfEOosqPdXbmSRuyxoo4VRnARQAM8AV8BP8AgmN+0H8Y4INc8V20Xw80Cbay3GswM13KhIy0doCG6c/vDHnjB5zXwe/4Jp/st/Cjy73UvCcvjnVBGoa58TlLqIN1bZbBViwTj76uwAxu5OdH0XR/D2mW+i+H9Js9M0+1XbBaWcCwxRDOcKigBRkk8DvVVVUwBGRVVVVVVRAIxTDHSqogEYojtVVVUwBGRVVXizwR4L8e6Z/YvjnwhoniLT9wb7Jq2nxXcWQQQdkisM5APTsK+K//AASx/Zo8frNeeDbbVfAepOvyNpdwZ7UvnlngmLcY42xvGOAfXPxc/wCCWv7Snw7S41LwfbaZ4+0uHLBtJl8q7CBcktbS4JORjbG0hOR741TR/FfgTX/sGtaXq3h7XNNmDeTdQSWlzbSI3B2sA6MGHXggivgX/wAFQ/2qPgvHDpOp+KIfHuhxYAsfEoaeWNQu3EdyCJRwF4dnUY4Ayc/AX/grJ+zN8W2ttH8d3tx8M9euGVBDrUnmWLORztvFUIig5G6YRDp64rT9R0/VrKHUtKv7e9tLhd0M9vKJI5B6qykgj6VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVfFX9vX9kv4O+db+KfjRol5qEOAdO0Rzqc+47vlYQBhGflOd5XGVzjcufiZ/wW58JWcs1l8IPgrqeqqGxHf8AiDUUswANvPkQiQsD+8AzIpGFJzkqPHH/AAVx/bK8XxtFo3iPwz4PVsBhoeiIxI2kEBrszkZyDkEEFRgjkHxh+1H+0j4+kZ/F3x38d6jG0iyC3fXrhIEdV2hlhVxGp2kjIUZyT1JzNNLcSvPPK8ssjFnd2JZmPJJJ6k1pvgLxzrVjFqejeDNdv7ObPl3Ftp0ssb4JBwyqQcEEfUGov2ZP2kp4kng/Z7+JUkcihkdfCd6QwPQgiLkYr/hl/wDaX/6N3+Jv/hI33/xqv+GX/wBpf/o3f4m/+Ejff/Gq1j9n349eHrVb3X/gh4+0y3eQRpNeeGruFGcgkKC0YGcAnHsa1rw54h8OSxweIdB1HS5JlLRpe2rwFwOCQGAyK0rVtV0K/i1XQ9Uu9PvYM+Vc2szQyR5BU7WUgjIJB9ia8J/tZ/tPeB0hh8L/ALQHj2zt7dWWK0bXria3QMSWxDIzRgkknO3OST3rwZ/wVd/bT8JeTFqHj/SfFEEHkhIda0S3Pyx9VZ4FikbeMBmZixxkMCST4I/4Lf8AjS2eCL4kfAnRNRQ7Fnn0TVpbIp83zOscqzbvl6IXHP8AEM8eBP8AgsJ+yL4qEEXimXxb4MmZR5z6npBuYUbZk7WtGldl3DaCUUnIJUDOPhz+0D8Dfi9sj+GXxb8KeI7h8/6JY6rFJcLjd96Dd5i5EbkZUZCkjjmqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqaaG3heeeVI44lLO7ttCgDJJJ6DFeNv2wP2W/h3I9t4t+Pngm1uYv9Zaw6tHdTx/KGG6KEu65UgjI5yMZzXjb/AILB/sgeF5Hg8O3HjDxgRwkulaL5EZO0EEm7eFgMnaTtJyCcEYJ8a/8ABcLUJBLbfDn9n+3hIYeVea3rpl3DZyGgiiXBDkciU5A6At8vjP8A4K7ftmeKGVtD8ReF/CADKSujaDHKCACCP9MNxwcgnvkDBAyD4w/at/aY8e7k8WfHvx3fQvt3W39uzxQZXofJRlTPvtzVjp99qt5Fp+mWU93dTttiggiMjufRVGST9K8GfscftS+P2iHhv4EeLjHMsbRT6hp50+GRXBKsstz5aMpAzuBwMjJ5GfBn/BJb9qXxIkU3iSfwh4UjLR+bFqGqNcTKpJ3bVtkkRmUDOC6g5GD1I8Gf8EZfDsKxTfEP436jeMWjMlto2kJbbACd6iWV5NxIxhig2nOVavBH/BMH9kLwcI5b7wTqfim5i27Jtc1aV+RnJMcJijbORkMhHyjAHOfBHwH+Cnw18tvAPwn8J6FNHtxcWWkQxzsVztLShd7EbmwSSRk+tVVVVAZ71gYwKqqqqqqoLQUAe9VVVVVVUBnvWBjAqqqqqqqqoLnrWBjAqqqqqqqqqsZ4FBc9TWABgVVVVVWM8Cto7msDGBVVVVVVWM8CtpPeto9awMYFVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVUQCMUVBHBoqexojtVVVVVVRAIxRUHoaIIqqqqqiARiivoaI7V8X/ANg39lv40wXMmv8AwvsNG1WeMqur+HlGnXMbZ/1mIx5cjc4zKj8Y9Bj4v/8ABG/xzpCXOqfBH4kWHiGFMtHpWuQ/YrnaF+6s6bo5HJAxuWJcHk8c/FT4EfGH4Jaj/ZnxV+Hes+HZDIY45rmDdBMwzxHOm6OT7rfdY8DPSvgX+2n+0n+zq8Nt8OPibqA0aLAOhamft1gVDZKrDJkQ5PVoijH+9XwG/wCCz/w28RW9no/7Qngy78J6kcJPrGio95YMcjLmHmeIcn5R5x+Xqc4HgT4jeAfijoEHij4c+MtH8SaVOoKXWmXiXCDPZtpO1hggq2CCCCAQRVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV+0r+078Lf2VvAJ8dfEvUZS1zIYdM0qzCvealMMZWJGIGFBBZ2IVQRk5ZQf2pf2wvi/wDtY+KW1Xx1qzWegWkzNpPhy0lYWdiuW2sV4Es21ypmYbiMgbVwo/Zh/YQ+M/7TckWs6baL4Z8HiZVn8QanC211/i+zRcNcMB6FUzwXU1+zn+w18Bf2bPJ1fwxoUut+KVj2yeINYYTXAJ6+SgAjgGc42KHwcMzVVVVVVUw9KqqqqqqIBGKIxxVUwyOKqqqu2AaYY6VVVVfEX4R/DD4uaUdF+JfgPRPEdrtIQX9msjw5BGY5Mb42wT8yEEZPNfGv/gkN4E137XrHwI8bXPhm8fc8Ojazuu7LdxhEnH76Jep3MJzXxn/ZW+PPwClkk+JPw81Gz01GVV1i2T7TYsW4UefHlFYnorFW9q+CH7VPx8/Z1vUn+E/xI1PSrPzhLNpUjC4sbg9Dvt5MpkgkFgA3cMCAR+z3/wAFlvhh4ths9A/aF8LXHg3WHZY31jSka70yQ45doyTPAM4AUCYdy46V4S8Y+EvH2gWvinwR4l0zXtHvV3W99p10lxDIO+HQkZHcdR3qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr4k/FX4b/B7w3L4u+J/jXSfDekxZH2i/uBH5jAZ2Rr96R/REBY9hXx//wCC0fhLQrq40H9nDwH/AMJNJHlV8QeIPMtbRmBHMdqu2aRCO7tCwP8ACRzXxq/bD/aQ/aD8y1+J3xU1W+0xpGddItCtnZLnoDBCFV9o4DSbmAJ5OST4A+E/xM+K2oHS/ht4C13xJcJ/rBp1i8ywjjl2UbUHI5Ygcj1r4df8EmP2kPFSx3XjrUvDfgmAsRJDc3f266XBxkJb7oiCBn/Wg8j3x4F/4JAfBTRWjn8f/EPxT4mlRRmK0SLTYHOGBLLiR8cqQBIMFeSwOB4T/YJ/ZD8GtHJpnwQ0S8kRlYtq0k2pbmAxkrcO64PdQAue1eHPhz8PfBwhHhDwJ4d0P7NGY4f7N0uG28tD1VfLUYHsKqqqq8RfBv4Q+MA48W/CrwfrfmTm4f8AtHQra53SnOZDvQ5Y7m+br8x9a8R/sD/sf+KpTNqfwM0SFjK0pGnT3Gnjc3UAW8iAL6L90dgK8Xf8Ei/2dNZUSeFvFHjLw9Oqqu1buK7hOCSWKyR79xBA4cAYHHXPjP8A4I3/ABCtHLfD34y+HdVU4KprOnzaeV5ORuiM+cLt5wMkngY58df8E+P2uvASS3F38Ib7WbSL7s+hTxagZPlBO2KJjNxnHKDJBxkc1reg654a1CTSPEWi32lX0P8ArLW9tnglT6o4BHT0r4aftr/tV/CSWE+DPjl4oW2gbKWGpXh1G1A+bIENxvRQS7E7QCSQc5AI+FX/AAWw+JWkeTY/GT4U6J4jhXKvfaJcvp84Hy4Zo38xHPDZA8sHK9Np3fCT/gp/+x78WXSzf4hS+CtQfJFp4ugFgAA2Mm4DPbjPBwZc4PTg40/UdP1ayh1LSr+3vbS4XdDPbyiSOQeqspII+lVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVePP2xf2WPhoJk8YfHvwZbT2ylpbS21NL25QBN/MEG+TJUggbctkYzkV48/4LJ/sseGjNbeDtH8Z+L50YiGW209LO2cB8ZLzusigrlh+6J6A7cnHjj/gt58Sb12T4a/A/wzpCjAV9c1GfUi2GOSVhFvtyu3jJwQeTnjxx/wAFNv20/HMbW0vxhl0O1OP3Gh6fb2RBClSRKqebyGPG/GcEAEDHjH4kfET4iXP234gePvEfia43BvO1jVZr19wUKDmVmOdoA+gArwp8OPiH48kWHwN4D8ReIpHkaNU0rS5rsl1XcygRq3IX5iOw56V4P/4J0ftheMhFND8Ip9ItZGKtPrF/b2flkJuGYnfzcHIGQhGTg4wceD/+CN/xY1ARP47+LnhXRFdjvXTLSfUXRdmRw/kAtu+UjOAOQT0rwd/wR++AmkSJceMvHfjHxDIn/LGF4bGB/lwdyhHfqdww4xgA5Gc+Cf2B/wBkTwGIX0z4JaJqU8SgNLrZk1TzW2bSzJcM6ZOScBQoPIAwMeG/CHhTwdYrpXhDwxpOh2aqqrbabZR20YCjaoCxgDAAAHoKwMYFVVVVVVAZ4FYGMCqqqqqqqC5oKB3qqqqqqqguawMYFVVVVVVVWM8CgoAqqqqqqqqqqCk0AAMCqqqqqwT0raMVgYwKqqqqqsZ4FbcjrW0VgYwKqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqogEYoqCKK+hojtVVVVVVRUHpRUiqqqqqIBGKK+hqtU0rS9c0640fWtNtdQsbuMx3FrdQrLFMh6qyMCGB9CMV8b/+CWf7NfxVF5qvguxuvh1rtxudJtFUPZGQ4wXs2OwIAPuQtCPevjr/AME3f2mPglFdazbeGE8a+HrZdz6n4ezO8a4yTJbcTKAASzBWRRyWFfDb4q/Ej4PeJIvF3wv8a6r4b1aLA+0WFwY/MUc7JF+7Inqjgqe4r9m7/gs3rum/ZPDH7T3hX+2IBhP+En0GFIrhf9qe0+WN+TktEY8AcRsa+FHxq+FHx08Nr4s+EnjzSfE2m/KJXs5v3lszDISaJsSQuQM7JFVsc4qqqqqqqqqqqqqqqqqqqqqqqqqqqq+Onxp8F/s9/C3XPiv48uXXTNFh3LBEy+ddzMcRwRBiAXdiABnjknABI/aO/aF8e/tN/FLVPib47v5S1zI0emad5u+LSrQMTHbRcAYUHlsAuxLHljX7A/8AwTjg8Z2Nh8a/2hdFlXRpvLuNA8NzqUN8oIZbi5Xr5LAfLEf9YDub5MB7W0tbG1hsbG2it7e3jWOGGJAiRoowFVRwAAAAB0ojHQ1VVVVVUQCMURjgGqqqqqqmAI461VUw7iqqqogEYzRHaqqqqrq1tb61lsb23iuLe4jaOWGVA6SIwwysp4IIJBBr49f8EzP2fvjDPPrnhS2l+H2vzbi1xo0CtaSuRw0loSF68/uzHnnJ5zX7Qf7Cf7QH7PH2nVtc8Nf8JB4Xgy3/AAkGiK1xBGg7zpgSQYGMl1CZOA7V8G/2gfjH+z9r48RfCPx9qegTuwNxBFJvtroDtNA+Y5B/vKSOxHWv2ZP+CwXwz+I09l4R/aB0SLwHrs2EGtW0hk0mZ8dX3EyWuTgAMZFHJaRelaVq2la9pttrOhanaajYXkYktru0mWaKZD0ZHUkMD6g4qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq1vXNE8NaPeeIPEesWOlaXp0LT3l9fXCwQW8ajLPJI5CooHJJIAr9qX/gsboOiC88HfsuaPFrd4Y2jbxXqsTpbwOcgm3tmAaUjgh5CqggZSRTXxJ+KvxI+MXiSXxd8T/Guq+JdWlyPtF/cGTy167I1+7GnoiAKOwr4Df8E4v2ivjbBBrmoaNF4G8Oy7WW/wDECPFNOhIyYbYDzG4IYFxGjDo5r4Nf8Ew/2avhgLTUvFWk3Pj/AFq2k8zz9bOLXd2AtEPlsuP4ZfM5JPoBpGjaP4e0yDRdA0qz03T7VdsFpZwLDFEM5wqKAFGSTwO9VVVVVVVVVVeKPBvg/wAb6cdI8Z+FNH1+xOc2uqWMd1Fz1+SQEfpXxH/4Jo/sm/EN2uLXwZfeEbt/vT+HL024PIPEUgkhXgEcIOGPcKR8Tf8Agj38Q9K828+EvxO0fX4RgpZaxbvYTgc5UOnmI54Xk7AcnptG74nfsxftAfBuJ7r4kfCfX9Is4m2vfC3+0Wqn0NxCXizgE/e5wfSvhf8AHb4yfBS/GofCn4meIPDLmQySQ2N6ywTMRjMsJJjl4A++p6D0GPgr/wAFofi94Z8vTPjj4G0nxrZrGqDUdNYaZehh953UBoZMgfdVIsEk5xgV8EP+Cgv7Knx8lg0zwv8AEu30XXJ1Tbo3iNRp1yWbgRoXPlTPnjbFI54z05qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq1XVtK0Owl1TW9TtNPsoMebc3UywxpkhRlmIAySBz3Ir4jf8ABRv9jT4ab4dQ+Nmk65drjZb+HEfVfM+5nEsAaEYDg/M46MBkqRXxC/4Le+D7Znt/hT8DNY1JWUiO78Q6pHZGNsLgmCETbhneMeYvAU55IHxE/wCCtX7YvjhpodB8TaD4KtJVZDDoekRs2wgj/W3PnOrYOdyFSDyMV4/+Nvxj+Kru3xK+KnivxOjSGQQ6pq81xFGS275I2YogyeFUADsBXhP4e+PvH1z9i8C+B9f8R3G4r5Wk6ZNePkKWIxGrHO0E/QE14H/4Jt/tfeN1iuG+GSeH7SVcrca5qENqQcgYMQZpgcHPMeMKe+AfBP8AwRp8aXRhl+I/xp0TTQrAzQaJpst7vAflVklMO0lMfMUOCfukDJ8G/wDBJr9ljw6kD+Iz4s8VTLtMwv8AVPs8TsFwwVbZY2VCecbiRwNxrwT+yX+zP8OxC3hP4H+Ebee3ULFdXOmpeXCYTZxNPvkyVJBO7LZOc5NRQxQRJBBGkccahURFwFA4AAHQYqqrGeBQAAxVVVVVVWM8CgABgVVVVVVVQWgoA96qqqqqqoLnrVVVVVVVVQXNYAGBVVVVVVVVVAZ6UFAqqqqqgpNBQKqqqqqsZ4FbSe9bRWBjAqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqKgiivHBojtVVVVVUQDRU9qqqqqogEUVwKqv2g/wBiT9n79pCKe98ZeEk07xFIpCeINIC214DjguQCsw9pFbA6Eda/aI/4Jf8Ax8+Df2rXfA1t/wALE8Mw5bz9Ktyt9Ag/56WuSx+sRf1O2vAHxF+IXwd8YW/jH4eeJ9S8NeIdNZkS6tXMci9mjdTwynGGRgQe4r9lP/gsRpGri38G/tU6fFpd3wsPizS7djBL0AFzbICYz1JkjypyBsQAk6JrmieJtHs/EHhvWLHVdL1CFZ7O+srhZ4LiNhlXjkQlWUjkEEg1VVVVVVVVVVVVVVVVVVVVVVVVVV/wVR/aql+Ofxqf4YeFdYSfwT8PJpLeA28pMd9qBAFxOcHa4QgxIecASEHEhr/gmT+x3Z/GHxU3xu+JGkJdeD/DFz5enWF1DmLVr4DOWBGHhiyGI6M+1TkB1NUQCMURjiqqqqqqiARiiO1VVVVVVMB1FVRAIxRGOM1VVVMMjIqqqqqqr9o//gm/8DPjjFca34S0+DwD4skYudQ0q1Atrluf9dagqhJJJLpsck5Yt0r9ob9jv44fs1XJn8deG/tmgySFLfxBpm6eyc5AAd8AwscjCyBS3O3dgmv2a/2zfjx+yzqiP8OfFbz6BJMZb3w3qRafT7okYZvLyDE5wv7yIox2qGLKNp/ZR/4KOfAz9pyOw8MXN9/wh3j2fEbaBqco23cm0E/ZJ/uzA8gKdsnB+TGGNVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX7Vf7ZXwg/ZJ8MjUfHGoPqPiK+hL6R4bsXU3d6ckB2zxDDuBBlbjhgodhtP7Uf7Zvxm/au8RPeeN9aaw8OW8ztpnhuykK2lohbKlxx50oGAZXGeuAoO0fsz/sS/Gf9pu5i1Hw/pf8AYfhISFZ/EmpRlbfgkFYE4a4YFWBCfKpGGZcjP7Of7CXwH/Z1gs9U07w/F4l8XW+GbxFq8IkmR85BgjOUt8cgFBvwcF2qqqqqqqqqqqqqqqvi/wDsJ/sx/GeKWXXPhvZ6JqcikDVPDyLp9wDz8xCL5cjc9ZEfoOwxXxl/4JDfETQvterfBDxtYeKLSOPfFpWrYsr1j02JJ/qZD3y5iHOO2T8SvhJ8TPg7rY8OfE7wRqvhy/dS0KX1uVWdRjLRSDKSKCcFkJAPGc1+zx/wUR/aZ/Z3nt7DTPGcvivwzFtVtA8RyveQogAAEEhbzbfC5wqMEycsjdK/Zu/4Kf8A7N/x9+yaDr+rf8K78WT4X+y9euFW3nkPaC8wsb8kKFkETsThUPWqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqvit+01+z/8AA8SR/FT4t+HNAuol3NYTXYlvCNhfIto90xBUcYTklQOWAPxU/wCC1HwV8OtLY/CT4ceI/GNzGxUXeoSppVowwMOhxJKwySCrRxn5evOR8Uf+Ctv7XPxBSey8N6zoPgWxkYhU0LTg0/l5bAaa4MjBsMoLIEzsGAuWB8c/FL4mfE+9GofEf4heI/FFyGLJJq+pzXZQ/wCz5jHaMcYGABxXgD4E/Gf4q7H+HXwt8T+IIHz/AKVZaZK9uuN33pseWvKOBlhkqQOeK8Af8Emv2nPFOyfxjP4Y8GQc+ZHe6h9ruB94DalsHjPKrnMgwHB5IKjwH/wR4+DWkLHL8QviX4p8S3EbA7LCGLTIHGWyGQiZ8YKDiRTkE98DwF+xL+yp8N3t5/DfwS8Oy3Vtgx3OqwtqUquCrBw1yX2sCoIK4284wCRVjp9hpdnFp+mWUFpawLtiggiEaRj0VRwB9KqqqqqgOKqqqqqqxngUAAMZqqqqqqqgtBQB71VVVVVVQXuaqqqqqqqqC561gYwKqqqqqqqqxngUFHc1gYwKqqqqCk0FAqqqqqqwT0raMdawMYFVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVUQCMUVB6GipqqqqqqIBoqe1VVVVUVB6GiMcVVftLfsG/Aj9paK71jV9DTw74xnUbPEulRiOd2AAHnxghLgYCrlxvCgBXUV+05+xP8af2Xb+S68VaWmseFXm2WniTTUZrZ9xOxZQRugkIAyjcZJCs+M1+yx+2h8ZP2TvEiXvgrVP7U8OXMgOp+GtQlZrS6XPzMgBzDLjpInfG4OuVP7LX7YXwg/ax8LLqvgTVks9ftIVbVvDl5KovLFsLuYLwZYdzhRMo2k4B2tlRVVVVVVVVVVVVVVVVVVVVVVVVX7bfx4/4Zz/Zr8XfEOxvPI1x7f+ztBwm4m/uPkjYDp8g3SnPaI9TgH4c+Bda+J3j7w78O/Dke7UvEepQWFuSpZUaVwu9schFBLMeygntXwt+G/hn4QfDvw/8ADPwfZpb6T4esktoAECmQjl5Wx1eRyzse7Ox71VUQCMZojHFVVVVVVMARx1ojtVVVVVUQCMURjiqogEYBojtVVVfSmAHSqqqqqqr+wsNVsLjS9UsoLyzvIXhuLa4iEkc0bDayOpyGUgkEHgg1+03/AMErPAnjsT+K/wBn25tfB2uNlpdGuGY6bcnkkx4DPbsSRwN0eAAFXkn4g/DP4j/BrxVJ4X+IfhbU/DetWUhKx3CbNxU8SRSKSsi5HDoxU9QTX7If/BWT4j/CZrHwN+0AL3xz4PiURQaopU6rpqgAL8zEC6QYORIRJ82RIQoQ/DL4qfDr4z+ELTx38LvF+n+I9DvOI7qzkJ2PtDGORCA8UgDLmNwrLkZAqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr9vj/go94d/ZthvvhV8Lmt9a+JlxakSS5WS28PlwNrzDo8xUlli6D5Wf5SFfxF4h8ffGHx3eeItfvdU8U+LPEl4ZJpdjT3F3M3AVUUZ6AKqKMKAFAAAFfshf8EuLDTFsviH+05YJd3ysstn4SSYNDCQWwbxkJEp+4wiVtvZ9wJQWFhYaVYW2l6XZQWdnZwrDbW0EQjjhjUBVRFXAVQAAAOABVVVVVVVVVVVVVVVVVeLPB3hLx5osvhzxr4a0zXdLn/1lnqNolxETgjO1wQDgkZ6jNfH7/gkl4L8TSza9+z94lTwtfSMWOi6u8k9iSf7koDSxD2IkHYbRXxn/AGf/AIt/s/8AiAeHPin4PudJllybW6BE1tdr/eimQlW4GSuQy5G4KeK/Zo/4KD/tFfszy2Oj6N4nbxN4OtmVX8Na5I08CRDqtvJ/rLY43YCHywx3Mj9K/Zf/AOCh37P37TvkaFp+s/8ACJ+MJNq/8I9rc8cUlxIe1rJkLcdD8q4fHJQCqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq+M37Vf7Pf7P0Lj4r/ABS0fR7xF3DTUdrm9fIGMW0QaTB3LyVAwckgZNfGP/gtl4bsftGmfAb4T3eqTL5iR6r4mn+zwhhwri2hLPIhPPMkTYAyASdvxh/4KAftZfGxprfxJ8W9T0nS5WyNK8Pn+zLdRjG0mLEki9TiV35x6DGgeG/EPizVYNC8K6DqOs6lcsFgstPtXuJpSSAAqICxOSBwO9fDD/gmX+1Z8RttzqXhSw8GWJwRceIrvyXYd8QxCSUEDHDqoORz1x8Nv+CO/wANtKaG8+KvxR1rxFIF3PZ6Tapp0IbC/KXYyu6g7+R5ZOV4XBB+Hf7Hn7MnwsihHhD4MeGluYVwl7qFoL+5B+YEia43upIdgdpAIIHQACsZ4FAADFVVVVVVAZ5qqqqqqqgMDrVVVVVVVBc8/lQUAe9VVVVVVYzwKCjFVVVVVVVWM8CgoHeqqqqqqqqqC5rAAwKqqqqAz0oKBVVVVVYzwKC56msADiqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqIBFFeODRHaqqqqqIBoqRVVVVRAIxRXjg1VappWl65p1xo+taba6hY3cZjuLW6hWWKZD1VkYEMD6EYr9sn/glckMV98Sv2X7JisavNfeDySxwAzO9m5JJP3QLcjPXa3RK8JeMPiJ8GPHUHiTwhrer+E/Ffh+6ZBLCWt7i2lUlZIpFPbgo8bggjKsCCRX7Cv/BQDwZ+1fosHg7xGIdD+J2nWZk1DTQpWDUFTAa4tWJPHILRE7kycblG+qqqqqqqqqqqqqqqqqqqqqqqqv8Agt58TpX1D4bfBizvWWOKG51/ULcOcOzH7PauR6gJdjP+2cY5z/wSD+F0Xin46+IPide2qSweBtHCWzlsGK8vC0aMBnJHkR3a9MfMOnGaqqogEcGiO1VVVVVVMO4qqqqqqogEYzRHaqph3FVVVRAIxRHaqqqqqqq+MHwS+GPx48Kv4P8Aih4VtdYsvma3kcbJ7OQjHmQyD5o29xwcYII4r9q3/gnJ8TvgILvxf4CFz408DxYZrmGHN7YKevnxL95R/wA9UG3uwSvgH+0p8Yf2afFa+LPhP4rlsHk4vNPnzLZ3y/3ZoScN7MMMv8LCv2Pf2+/hL+1lpsWjQSxeF/HsMZa78NXd0GaXG4l7VyF+0IFQswChk/iGMMaqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/4KO/8ABR2H4Uw6l8BfgLraS+NpFaDXddtnyNCBGGhhYdbojgsP9T/10/1fgbwL49+NHj2y8H+D9MvNf8S6/cnapYu8jsSzyyOx4UcszscAZJNfsbfsKeCP2Y9KtvE/iBLPxB8SJ4XW61hAzQ2SuAGhtVYDC4GDKVDtub7qtsFVVVVVVVVVVVVVVVVVVV4p8JeF/HGh3Phnxj4e07W9Ku12z2V/bLPFJ9VYEZ9D1FftIf8ABJnw7rvneJv2btZi0K9+Z5fDurTO9rL3/cT/ADPEeCNr71JYfNGBz8Qfhn8R/g14qk8L/EPwtqfhvWrKQlY7hNm4qeJIpFJWRcjh0YqeoJr9lP8A4KufGH4Ki38J/GBbr4jeEVwqTXNyf7VsRwB5c7kiZAAf3cvOSMSIBg/A/wDaD+EP7RvhMeMPhF4ztNbtI9q3duMxXNi5zhJ4Ww8ZJVsEjawUlSy81VVVVVVVVVVVVVVVVVVVVVVVVVVVeKPFnhXwRos/iPxp4m0nQNJtsedf6pex2kEWem6SQhRn3NfHT/gsN+z38PkuNK+Eelan8SNYibYs0QbT9OUgkNmeRTI5GARsiKMOjjrXxw/4KU/tYfHD7Zp1x49/4RDQbuPym0jwwhskKdw0+WnbcOGBk2HkbQCRWg+HPFfjrXE0bwvoWreIdZvWZktLC1ku7iZurEIgLMe54r4Uf8Er/wBpfx+0N54yt9J8Baa7fM2qXAnuiuOGSCEtznjbI8Z4J9M/Cb/glJ+zn4FFvfePbnWPH+pRbGYXsps7PevOVghIbBPVZJHGAB658H+A/BPw80hNA8B+EdH8PadH0tdMso7aPPqVQAEkkkk8kkk9aAzwDWBjAqqAB5qqqqqqoDPFYGMCqqqqqqgO9VVVVVVUFzyaA71VVVVVVjPAoKAPeqqqqqqqoDPAoAAYFVVVVVVVVYzwKCgCqqqqgM8CgoxVVVVVYzwKC+prAAwKqqqqqqqqqqqqqqqqqqwfStpraSOtbcjrW0etbR61tFbRW0VtFYHoKwPQVgegoqDW0VtFFQelbR60VB6GivHBrafUVtNEHpiiO1VVVVVVVVVVVVVVVVVRAIxRUEcGiMcVVVVVRUHoaIxxVVVVRUGiMcVVftzf8E+/C37RGl3/AMRvhrYWuj/E+CPe0m7y4NcVFwIZhnasuAAk3HQK5K7WTSdW+I3wQ+I0Oq6Vc6r4T8Z+EtQO1tphuLK4jJVlZWH1VkYEMCQQQSK/YZ/a80L9rb4Sw63cy2ln420LbbeJNLiITZLj5biJNzMIJOdpPRldeduTVVVVVVVVVVVVVVVVVVVVVVVf8FWvGUviz9tXxdp/2xLm28M2On6Xask5lVQLZJ5EHJClZp5QVHRg2eSa/wCCQvhOPRv2bNZ8TSJEbnxD4puGV0difIhhhjRWU8AhxMeOoYZPYVVVVMMjiqqqqqqiARimGOlVVVVVVMARkVVEAjFEY4qqqqYAjIqqqqqqqqr9r/8A4Jk+E/iZFd+Pv2frLTvDHisKZLnRAPIsNTI6+WB8ttKRwMARscbghLSVq2k+OvhR40m0rVLbV/C/inw/c4dCXtbqzmXBDKwIZTgghgeQQQSCDX7DH/BVbT/HEth8Jf2ntSsdK1wqsWm+LnYQW2oNwBHdKAFgl7iXIjboQhAL1VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX/BSD9vqP8AZs0IfCr4U6haXPxK1y3YzThlkHh63YDbM6cgzuGzGh4AG9hjYr+APAHjf4weN9P8D+B9Hutb8Qa1OVhhU5LE8vI7HhVAyzOxAABJNfsf/sfeDP2VvBnkwmDVvGmrQr/beueX97ofs8GeUgUgehcgM38KpVVVVVVVVVVVVVVVVVVVVVfFT4O/DP42eGpPCfxO8Hadr1gysITcwjzbVmGC8Mo+eJ8fxIQe3Tiv2ov+CW3j34cC78X/AAFlvvGvh1cu+jtFu1SzHHCKnF0M5PyKrgYG1sFj8PPiX8R/gz4ui8X/AA38War4Y1+z3RG4s5TE+3ILRSKeHQlRlHBUkDI4r9j3/grJ4H+KTWHw/wD2hhY+EPFcrJBba0h2abqDYxmQk/6M5I/iPlknhl4WoZobiFJ7eVJY5FDI6NkMCMggjqCKqqqqqqqqqqqqqqqqqqqqqqqqr4sfGr4UfAzw23iz4tePNJ8M6b8wie8m/eXDKMlIYlzJM4BzsjVmxziv2g/+C0d/dfbvDn7NfgP7HH9yLxL4hw8h6gtFZrlV7FWkdsg/NGDxXxX+N/xc+OWvSeJfiz8QdY8S3ztlBeT/ALmDrxDCuI4V5Pyxqo5PHJr4JfsQ/tHfHuC11Xwf4El0/QrvBi1vWmNlaMhI+dCQXlXnrEj9D3GK+Df/AASN+EPhdU1D4zeJ9R8a3pUZsLNn06zQ55yUbznOAACHQdeDxjwN8Nfh78MdKGh/DvwTonhyy2qHh0yxS38zaMAuVALtyfmbJJJJOSaqqAAFVQGeKwMYFVVVVVWM8CgABgGqqqqqqgM96qqqqqqoLmgoxmqqqqqqsZ4FBQB71VVVVVVVBc1gYwKqqqqqqqqC561gYwKqqqsZ4FBR3NYGMCqqqqsZ4FBR3NYGMCqqqqqqqqqqqqqqqrBPAFbSRW0etbRWBjAFYGMCqqqqqqqqqqqqqqqqiARjFFQRgUVB6GipxwaIPpRHaqqqqqqqqqqqqqqqIBGKKgjg0RjiqqqqiARiioPQ0R2qqqqIBGKK8cGqr/gqJ+xrB8QPDV1+0b8ONHz4q0K3z4jtreIl9UskVVE55/1kCLzwS0fBP7tRX7IH7R+ufsufHPQviTp80smjtILPxBYqTtvLCQgSjaCMunEif7ca5yMg6Tqum69pVlrmjX0V5Yajbx3FpcwtuSaJ1DI6nuCpBB96qqqqqqqqqqqqqqqqqqqqqqr9u/ULLU/2xPi3c2Fyk8aeJriFmXoJIsRyL9VdGU+4r/gmhDFH+xP8O3jjRWlbVGchcFj/AGndDJ9TgAfQCqqqqqYDqDVVVVVVRAIxRGOKqqqqqqYY6VVEAjFEdqqqqmAHANVVVVVVVVftSfsh/DH9qfw2LPxPB/ZfiaxgaPSPEVtFunsssG2um5RNFkHMbHjcxUoWLV8fv2d/id+zd4zbwd8SNG8jz976bqEDb7bUYVbb5kTj8CUbDruXcoyM/sP/APBTzxn8BHsvht8aJtQ8WfD75YbW53h77QxuJ3ISMzxc4MTNlQBsIC+W3gL4geCfil4T0/xz8PPFGn+INB1OPfa31jMJEb1U91dTkMjAMrAhgCCKqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv26P2wdA/ZI+FT6pbSW954319ZIPDOmSLvVpQBuuJlDA+THkZwcszKo6krrGr+O/jF8QZtW1a6v8AxL4u8W6mNzbfMnvruZwFVVUdWZgqqoAHAAAAFfsMfsdaV+zB4GGs+IIUufiF4itk/tq4EgdLNM7ltIscbVONzDO9xnO0KBVVVVVVVVVVVVVVVVVVVVVVVftUfsEfCP8AaWiuPEUEKeFPHTKNmvWNuCLorwFuoQQJhjjflZBtQbiq7D8ef2bvi5+zf4lHhz4n+Gmto52f7BqlsTLZagqnBaGXAycYJRgrqGUsq5Ffscf8FLPit+zXJY+CvG01341+HMWI1024l3XmloF2j7HKx4QYT9y5KYXC+WWZj8IvjN8M/jx4MtfHvwq8WWmu6RdZBeLKSQOCQY5YmAeJwQflYDIwRkEE1VVVVVVVVVVVVVVVVVVVVVV8VvjJ8Lfgd4Wk8afFnxxpnhrSEbYk15Id874J8uGJQZJnwCdkas2ATjANftP/APBZDxB4ghufCX7L+g3Hh+2LFH8UaxFG91KuCD5FsdyRDPIdyzEH7iEV4j8T+NviZ4ok1zxVrur+JvEGqzBWubyeS6uLh2OFUFiWJyQAo9gBXwJ/4JfftAfFKW01b4g2ifDvw/MqyGTU4999Ip7LaAhkbjkTGMjrg18Df2Bf2bvgSLfUdJ8H/wDCSa/BIJF1rxCVu542HQxptEUWMnBVA3TLHANVVVQGeaqsZ4FAADFVVVVVVAcVVVVVVUBngVgYwKqqqqqqAz3oKCKqqqqqqxngUFAHvVVVVVVVUFyOtYGMCqqqqqqqrGeBQUDvVVVVQXNAADAqqqqrGeBQUdzWBjAqqqqqqqqqqqqqqxngVtJrb71tGKwMYFVVa94o8M+FbYXnifxHpekW7dJb+7S3U8gdXIHVlH4j1rxF+13+z/4dYwv47TUZh/yz061luMjJGd4XZ26bs4we4rW/+Cg/w7tt6+HvBHiDUGVG2m6eK1VmH3eVaQ7T64yPQ1q//BQ/xNPu/sD4aaXZ/uiF+2ahJc4k5wx2rHleny8Hg888al+3n8cL7y/str4Y0/Znd9msJG35x18yR+mO2OpznjF/+2f+0TeXHnW/ja3skwAIYNJtSo9/njY/rWo/tU/tBapMs9z8TdQRlTaBbwQ264yT92NFBPPXGenpX/DTHx6/6KjrX/fxf8K/4aY+PX/RUda/7+L/AIV/w0x8ev8AoqOtf9/F/wAKg/ae+PtvNHPH8UNWLRMGUSbHUkHPKspBHsRg1/w2L+0f/wBFH/8AKRZf/GaT9uL49pGqNqmjuVGCzaamT7nBxVl/wUC+LsUsQvvDHhK4iTAkCW1xG7/RvOIB/wCA49q07/gojrkTudW+F1hcoR8gt9UeAg+5aN8/kK0z/goZ4Mm8gaz8O9atd3+v+y3UVxs/3d2zd+O2tL/bv+BmoZ+1p4j0zDBR9q09WyP737qR+B+ftWlftWfs96y7R2fxNsIyhUH7Xbz2o56YMqKD05x074zWi/EDwF4kKr4d8b6BqhaTy1FlqUU+XwDtGxjzgg496qqqqqqqqogEYoqDRUHoaKntRHaqqqqqqqqqqqqqogEYoqD0NEY4qqqqogEYorxwaqqqqKg96IxxU0MVxE8E8aSRyKVdHXIYEYII7jFftj/BiD4CftHeMvh3psHlaRDdi70gAkgWlwoljQEgE7A5jPvGevU/8Eo/jDdfFT9knSdD1fUHudU8AX0ugyGWRTIbZFWS1O0HIRYZVhUkDPkEc4JNVVVVVVVVVVVVVVVVVVVVVVftuaHD4e/a7+L2nwXDzLL4uv7ss4wQ1xKZ2XjsDIQPYCv+CeOqxa1+xn8M7yGxS0WOyurYxq2cmG9niL9By5QuR2LdT1NVVVVEAjFEY6VVVVVVUwyOKqqqqqqIBGM0R2qqYDqKqqqiARjNEdqqqqqqqqq+Kfwo+H/xq8GXfgH4leHINZ0a7ZXMMhZHikX7skbqQ0bjJ+ZSDgkHIJB/a9/4J/8AxC/ZtlufF/hl5/FPw+3ZGppH/pGnA7QFu0AAUFm2iRcqcc7CQtfsz/tcfGb9lPxM+t/DPXUfTbxh/aeg36mWx1AAYBdAQUccYkQq4xjJUsp/ZU/bo+Cf7WGmx2nhjU/7D8Xxxs934X1GVRcoF+88TDAnj77l5A+8q1VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV8b/jN4J/Z/8AhhrnxV8fX/kaXotuXWFCPNvJjxFbxA9ZJGwozgDOSQoJH7Qvx58cftJ/FTV/it49uUN7qDeXa2kXEVhaqT5VvHwPlQHqeWJLHlia/wCCb37En/Cs9ItPj18V9G2+LtVgJ0TS7u3w+jW7ZHmuG+7PIuMDAMaNg/M7KtVVVVVVVVVVVVVVVVVVVVVVVVeO/APgz4neFr3wT4+8O2euaJqCgXFldJuVsEFSCMFWBAIYEEEAgiv2wP8Agmv4y+DjX/xA+DKXninwTGrz3NmV3X2jooLNuA/18IAJ8xQGUcMvyl2+Av7RXxZ/Zr8Zp42+FHieXTrhsLeWUpMlpqCAMAlxDkCQDc2CfmUnKlTzX7HH7dvwt/a60P7DYFPDvjuwhMup+GbiYyMsYbHn28hVRNEcjOAGQnDAAqzVVVVVVVVVVVVVVVVVVVVVq2raVoOm3Os65qdpp1hZxmS4u7uZYYoUHVndiAoHqTiv2r/+Cv8A4P8ABj3/AII/ZlsrTxXrMeYpPE16j/2fbOGw3kR/K1yRg4fKx52sPNXg/ET4ofEz40eKG8T/ABJ8Y6x4p1q5bYkt7M0pXJ4SJB8sa56IgAHYV+zj/wAEy/jT8YpbfXviNDP8PfC7qHEt9bbr66HHyx2xZTGCM/PJtxwQr9K+A/7I3wH/AGcrWN/h34NibWRHsm17UiLnUJcjDfvSAIwRjKRKiHAJXPNKM85o896qqrGeBWBjAqqUZ5qqqqqqoAGqqqqqqsZ4FAADAqqqqqqoDPApVBGKwMYFVVVVVWM8CgoA96qqqqqqqgoxzVVVVVVVVQXNYAGBVVVVjPAoKB3qqqqrGeBQX1NYGMCqqqqqqqqqqqqqwSOK2571tFYGMCqqtU1fSNEtftutapaWFuDjzrqdYkzgnGWIHQE/ga8YftbfAbwfHKr+NotZuYwStvo8ZujJjssg/dfm4rxd/wAFDP8AWQeAvh36+Vdavdf7wGYYv+Ak/vPUejV4y/a4+PHjJnRvGb6LbtnFvo0YtduSTxIMydCB9/sO+Sb291PW797zULy6v765Yb5ZpGlklboMk5JPQV4d+APxp8VKj6L8M9feOQqEluLQ20bbjgEPLtUjjk5wO+K0X9hX47aqqm+g0DRyY9xF7qO7acj5T5Kyc9/TjrWnf8E7dZl8z+1/inZW2MeX9m0lp93XOd0qY7eucnpjnT/+Ce3w5jmLar458R3MWwhUt1hgYNkc5ZH4xnjHcc1p/wCwh8DLKAxXMniO/YsSJLjUFVgMDgeWijHGemeetad+xZ+zzZRulz4Qu9QLHIa51W4Ur7Dy3Ufnmv8Ahjn9m/8A6Jz/AOVe9/8Aj1f8Mc/s3/8AROf/ACr3v/x6v+GOf2b/APonP/lXvf8A49V7+xl+zrdWz28Hgaa0dsYmh1a6Lpg9g8jLz05B6/jX/DDfwE/6B2tf+DJv8Kk/4J+fBwxsIvE/jJXwdpa7tiAfceQM1e/8E7/DMls6ad8TNThuDjZJNp6SqOecqGUnjPcf0rUf+CduuROg0j4pWFyhHzm50p4CD7BZHz+lar+wL8ZbJZJNO1nwtqKLIRGkd3LHIy54Yh4go46jcfxrWP2OP2hdI8x08EJfxRJuMlnqFu+fYIXDk/Ra1f4E/GfQ9x1L4W+J1SOIyvLFpks0aKM5LOgKjGCTk8DmtR0rU9ImW21XTrqymZN6x3ELRsVyRkBgDjIIz7Vofjzxz4XRY/DPjTXdJVUKKtjqMtuApO4gBGHBPOPXmtD/AGvP2gtCdQnj17+IMWaG+s4ZgxIx94pvAHBwGAz9Tnw//wAFCfHVoEXxP4D0PUgpO42U8tmzDbgfe80A55JxgjjA614a/b2+D+rypb6/pev6EzH5ppbdbiFRkDrGxfoSfudAepwD4T+Ofwf8cbE8M/ETRbmaT7lvLcfZ5m+kUm1/0qqqqqqqqogEYoqDRX0NFTVVVVVVVVVVVVVEAjFFQehojtVVVVRUEcURjiqqqogEYorjoa/4LLaBYWnxu8EeJIAwutS8Lm3nGAFIhuZCjcDO798wJJPCqOMc/wDBDXVb+bSfjJoclxusrS40W4hi2AbZJVvVkbOMnIhjGCcDbxjJzVVVVVVVVVVVVVVVVVVVVVVX/BSLQv8AhHf23Pinp/2v7R5uoW13v8vZj7RZQT7cZP3fN2574zgZwP8Agltr/wDbP7HPhrTtsQ/sLUtRs8o2Sd1y8+WHY/v+npg96qqqqqiARiiMcVVVVVVUw7iqqqrWfEOmaHGGvJt0jD5Yk5Y/h2HuavfiNqczEWNpBAnYvl2/oP0o+NvE5ORqYHt5KcfpUPjzxHFjfcxTYOfniA/D5ccVa/EibIF9piNxy0Tlf0OfbvVh4z8P3+F+1m3f+7cDZ+vI/WlZXUMjBgRkEd6IBGKIxxmqqqph3FVVVVVVVVVLFFPE8E8aSRyKVdHXIYHggjuMV+1t/wAEtNK8Ttc+PP2ZoLPSNT2s9z4UlcRWt0QM5tZGOIHJGPLbEZ3ZDRhcNqOneOPhd4vew1K11nwt4n0O4BaNxJZ3dnKACpHRkOCCCMcEEcGv2Tv+CwmqaFDaeB/2prC41azjURweLdOiBuYwAABdW4A80YyTKmH4GUcksPhf8dfg38a7Aah8KfiX4f8AEyCMySQ2N6rTwKDjMsJIki5I++o6j1FVVV41/ad/Zz+HRli8bfHLwPpNzCoZrObXYDc4L7MiBWMjDcCDhTjac9DjVf8Agpp+w1o9/Lpt38ebSSaHG5rXRNRuozkAjbJFbsjcEZwTg5B5BFf8PR/2Ev8Aouv/AJbOrf8AyLWh/wDBSb9iHxFLLBp/x90yJoVDMb7TL6yBHT5TPAgY+wya8E/G74M/EuRIPh58WfB/iWeT7sGla3b3UgO0tgojlgQoJIIyADnpVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVU00NvC888qRxxKWd3baFAGSST0GK/4KUftj/8ADTnxWXwr4I1WWT4c+C5Hh0va+E1O6PE16VHUHGyLOcICw2mV1r/gmr+x7F8YvFY+NHxD0tJvBnhm522FncR5TVb5cEBlZSrwRg5bnl9q8gOKqqqqqqqqqqqqqqqqqqqqqqqqqqr9sj/gmj4a+J63vxG+AVjZ6B4wkYy3mjBhDZaqxPzMmflt5SOcjCMRyFLM5K+O/hN44Usur+FvFnhm9V1yHtbqxuIyCpHRlYHBFfsE/wDBTjRvjWNP+EXx7vrTSPH3llLLW28u3tNcZeisBtWC4I/hACOQdu0lY6qqqqqqqqqqqqqqqqqqqv2qP20Pg3+yd4ba+8aap/aniS5jJ0zw1p8qteXTY4ZwTiGLOMyP2ztDthT+1P8Atz/HH9q7UJbTxbqyaL4Sjm32fhjTHZLVNpOx5iTunlAIy7cZyVVM4r9nD9jf41ftNagkng7Q/wCzfDkcm268R6mjRWceM7ljIBM8gxjZGDgld5QHdX7OH7BnwL/ZyEGtWGk/8JR4rj2t/b2swJJJbuO9tHgrb9TyuXwcFyKrGeBWABgVVVVADFVQGeAawMYFVVVVVQGeBWBjAqqqqqqoAAdaqqqqqqxngUqjoKwMYFVVVVVWM8CgoA96qqqqqqrGeBQUVVVVVVVVYzwKC+tYGMCqqqoLnrWBjAqqqqoLnvWABgVVVVVVVVVVVVVtJ6VtHrWB2qqrxJ8d/g14SEi678S9AiliBLwQ3i3Eq/Lu5jj3PyMYGOcjGc14m/b0+DukM0Ph/T9d15wxAkithbxEA4zmQhuRyBs+uK8Rf8FCvG94pTwr4B0XTM8br65kvCODnG3yhnOCMgjjoc8eJP2tPj94mEkU3j+40+BwQItNgjtdmVwcOi+Z78twTxjitX1vWfEF62pa9q97qV2+d095cNNIckk5ZiT1JP1J9a8L/BH4u+NGi/4Rv4da7dRTf6u4ezaGA8gf62Tancd+mT0BNeE/2Afibq2yXxZ4l0XQYm+9HHuvJl+qrtT8nrwh+wd8HtDKT+J7zWPEsoBDxzXH2WE8dQsWHHr989q8J/DT4e+BF/4o7wXo+kSEYaa1s0SVxknDSY3N1PUnGcdKqqqqqqqqqqqrq0tb63e0vLaK4hlGHilQOrD0IPBrXfgV8GvEqyDV/hj4bkeVtzzRaekErHBGTJGFY8H19D2Fa/8AsM/AbWSDp1lreh4IOLDUS+cA5H78S9c5/AdOa8Tf8E8LlYXm8HfEqKWUD5LfU7AxgnB6yxs2OcD7nAJPOMHxP+xj8ffDjO1v4Ytdbgj6zaZeo+eQOEcpIevZeOpxWveFvE3hW5Fl4n8O6npFw3SG/s5LdzwD91wD0ZT+I9a8J/Fv4neBdieEvHmtadFH923ju2MPHrExKH8RXgv9vr4m6KEg8Z6BpXiWFRzIhNjOx245ZAydRniPuegxjwL+2t8E/GBjtdW1K78M3jnaI9UhxETjORKm5QPd9vI+mdM1TTNb0+DVdF1K1v7K5TfBc2syyxyr6qykgj3FVVVVVVVVEAjFFQehoqccUR2qqqqqqqqqqqqIBGKK8cGiO1VVVRAIxRX0qqqqr/gr146tvEv7Tlh4Qsbtnj8IeG7a3uoWRQIrqd3nYgjkgwyW3XoQcDkk/wDBEDwfJZfDX4n+Pikvl6zrlnpisZFKE2kDykBR8wP+mjJPByuOQ1VVVVVVVVVVVVVVVVVVVVVVV/wWR8Iy6D+1jZ+IVjcweJvCtnciX7OUXzYpJoGTf0dgsUbE8ECRRjGCf+CNHi5L34W/ELwEFTdo2v2+pEhCCRd2/lctnBH+hHjAIyck5GKqqqqqiARgUR2qqqqqqIBGKIxxVVXiTWo9D0x7oFWmb5YUJ6se/wBB1q5uZ7yd7m5laSWQ5Zm6k1VVVadrOp6U++wvHj9U6qfqp4rSPiDbT7YdYh8l+nmxglT9R1H61DPb3cKz20ySow+VkbINVVVRAIxRGOM1VVVVVVVVVf8ABRvwr+z03wH1Tx78ZPC8V3rVjGLLw1eWkn2a+a9cOYoUmAOY/wDWSMjhkAV227gDVhYX2q31tpel2U95e3kqw21tBEZJJpGIVURVyWYkgADkk0ra94V1gMjX+j6rYScEb7ee3cfkyn8q8KftyfteeCoYLfQ/2hvGbx20yyxJqGoHUApUKAuLjzMphR+7PydeOTn/AIej/t2/9F1/8tnSf/kWv+Ho/wC3b/0XX/y2dJ/+Ra+JP7S37QHxg82P4k/GLxXrttNnfZT6lItrz1xboREM+yitB8L+JvFVybHwv4d1PWLhcZhsLOS4cZOB8qAnk8Vo/wCxr+1Vrtq13Y/AHxrGiSFCt5pT2j5AB4SbaxHI5Ax1GeDX/DDn7W3/AEQfxN/35T/4qtf/AGR/2oPDW06p8AvHJUxmQva6JNdIijqWaFWC/iRWraNrGg3Z0/XNKvNOulUMYLuBoXAPQ7WAODXws/bH/ae+DM9u/gH41eJba1tuE069uzfWeMYI+zz74xkcZCgjggggEfB3/gtl4ksfs+mfHn4T2mqRDYkmq+GZzbzBRwzm2mLJI5HPEkS5B4APy/BD9rP9nv8AaKtkPwp+JmmanflQZNJnLWt9Edu5gbeUK7AAHLIGTg4Yjmqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv+CtP7XUvwo+H8P7P/AIC1d4PFfja1Z9Xnt3KvYaWTtK5xw1wQ6DByESTONyk/s3fAbxL+0h8XNG+GHhxmgjuWM+qX/lF10+yQjzZ2A7gEKoJAZ3RSRuyPA3gjwx8NvB2j+AvBmlRadomhWiWtlbRj7qKOpPVmY5ZmPLMSxJJJqqqqqqqqqqqqqqqqqqqqqqqqqqqq/a4/Yr+HX7U2gtfTpFofjmxg8vS/EEcZJ2g58i4QECWIknBPzITlTgsrfF34RePPgb481D4dfEXRm0/VtPbIIy0V1ESdk0L4G+NgDhvYggMCB/wT3/4Ke3QutH+BH7S2uRG2MaWmg+L7qQh1cEhIb52OCCCqrPxgqPMzuMiwzQ3EKT28qSxyKGR0bIYEZBBHUEVVVVVVVVVVVVVVVVVVfts/8FXtE+HsupfC79ma7sdc8SRqYrrxX8txZadJyGW3UgrcSrgHccxAkcScgSSfED4w+OjJK+s+LvF3iS7ySxe7u76dvzZjx+AHYCv2UP8AglbZ2S2Xjv8AaZCXVwVSa28J20vyRNnP+lyqfnPAzGh2+rNytaZpemaJp1vpGi6da2FjaRiO3tbWFYooUHRVRQAoHoBVUo4zVVVUBmqqgMCqqqqqqgMCqqqqqqgM81VVVVVVjPAoL2HWsADAqqqqqqoDiqqqqqqqxngUFAHvVVVVVVVVjPAoKAPeqqqrGeBQUDvVVVVQGelAADiqqqqqqqqqqqrBPSto7msDGBVapq+kaJa/bda1S0sLcHHnXU6xJnBOMsQOgJ/A14n/AGsfgF4WZ4bj4gWuozp0i0yJ7vdyAcOgMffPLDpxmvE//BQzw/Erx+DPh3qF2x4jl1S6S328DkpHvzzkY3DjBz2rxX+2/wDHTxHG0GmajpXh6JgVI02yBcryPvzGQg4I5XaflBGOc+J/iR8QPGrOfFnjXWtVWTrFdXrvGOQcBCdoGQDgDGea0Lwn4p8UzfZ/DHhrVdXlyR5dhZSXDZAyRhAT0BP0rwz+xt8ffEjK0vhSDR4SwXztTvEixzgkopZ8Dr936Zrwx/wTx1aUrL40+I1pbgD5oNLs2m3HBziSQpjB2/wHPPTrXh39hj4E6Mo/tW01rXm5z9u1BoxkgDgQCMgZBI5PXkmvC3wn+GXglxN4U8B6Hps6nInhsk83/v4QW/DPFVVVVVVVVVVVVVVVVVV7Y2WpWz2WoWkF1byY3wzRh1bByMqeDyAa8YfskfAbxhHKz+CYtHuZAQtxo8htDHnusYzF+aGvHn/BPnVrcS3fw28bQXiDcUstXj8qQAdAJYwVZj05RBnuM8eO/gh8VvhsZH8YeCNStLaM4N4kfn2/TI/ex7kHHOCQevHBrwd8QPG3w+vxqfgvxRqGkT/xfZpiqSezp91xwOGBHA9K+Gf7fusWflab8VvDaajHuwdT0zEUqr6vCflc+6lMAdCa8BfE7wH8TtMGq+B/E1nqcagGWNG2yw57PG2GT8Rz2qqqqqqqqqiARiioPQ0QRVVVVVVVVVVVUQCMUVx0qqqqoqD3ojHFVVfFD4ieHfhJ8O/EPxK8V3HlaV4dsJLufBw0m0fLGv8AtO21FHdmAr4meP8AXfir8QfEPxH8Sujan4j1Ga9uAgwqF2JCL/sqMKPYCv2GPgnd/AH9lzwP4C1i1lt9ckszqWsxTAh4ry5PmvEwyQDEGWI44Jjz1JJqqqqqqqqqqqqqqqqqqqqqqr/gt98PWuvB/wAMvitbxIq6bqV3ot24UbnNxGs0IJ3Zwv2W46KR85yRwD/wSN+IQ8MftI3/AIFuLjbb+M9BmiijzjddWxE6H3xEtxx70R2qqqqqqphkZFVVVVVVEAjFEdqqviNePLqsFkG+SCHdj/aY8/oBVVVVVVpetajo8vm2FyyjPzRnlW+orQvF2n60Vt5MW91j/VseHP8Asn+lVVVTAEcVVVVVVVVVVf8ABT348z/FX4/XHw/0u73aB8Ot+nxojgrJfHH2qQ47qyrFg9PKPTJr/glJ+zDZ+LNfu/2jvGenJPYeHLk2fhuCaPKy3wUGS5weCIlZVQ4I3uxBVohXjX4V/DH4kxLD8Qvh54b8Sqi7Yzq2lQ3RjxuxtMikqRvfBGCNxx1rxL/wTf8A2OfEv2mX/hU39l3Vxt/f6bq13B5e3H3IvNMQyFwfk5yT1Oa/4dWfsk/9AbxN/wCDp/8ACv8Ah1Z+yT/0BvE3/g6f/Cvh9/wT9/ZL+HTGey+Eena5clsmbxCzan2IA8uYmIABj0TOcE5KgjTdL0zRbGLTNG061sbSHPl29tCsUaZJJwqgAZJJ+pNVVeIvC/hnxhpj6J4t8O6Zreny532mo2aXMTZUqco4IOVZh06MR3r4of8ABNf9lH4krdXNj4Jn8HalcsW+1+HLk2yoc5wLdt8Cr2wsa8HAI4I+Nf8AwSe+Nfgj7Zq/wm1iw8eaVFueO04stQVBg48tz5chAz91wW28Lkha1nQvGHw+8Q/2d4g0bWPDWu6dIsggvLeSzurZwQVbawV0IIBB49a/Zg/4K1/Gr4TS23hr41m4+JPhZFEa3NxKF1a0Ax8y3B/4+BjcSs2XJI/eKBg/AL9pn4MftMeGf+El+E3jC31BoFU32mTERX2nkkgCeAksgJBAblGwdrHBqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr4v/FPwn8Evhn4h+Kvje8+z6N4cszcTkfekYkLHEv+3JIyRqP7zivi/wDFPxX8bviZ4h+Kvje8+0az4jvDcTkfdiUALHEv+xHGqRqP7qCv+Ce/7MK/s8fBmHVPEenND418ZLFfawJoyktnHt/c2hB6GMMzMMA75GByFXFVVVVVVVVVVVVVVVVVVVVVVVVVVVVftIfs3/D79pv4fTeCPG1r5N1Dul0jV4YwbjTLgjG9M/eU4AeMnDgDoQrL+0h+zf8AED9mT4gTeCPG1r51tNul0jV4YyLfU7cHG9M52sMgPGTlCR1BVm/4Jxf8FHZfhTLpvwF+PWtvL4KkZYNC125cltCY8LDMx62p6Bj/AKn/AK5/6uGaG4hSe3lSWORQyOjZDAjIII6giqqqqqqqqqqqqqqqrxV4r8MeBvD174s8Z+IdP0TRdNj8y7v9QuFghgXIALOxAGSQB6kgDk1+3d/wUy8TfH43/wAK/gtc3vh/4cusltf3LKI7rX139WON0MBVRiIYZgzeZkHYv7P37NXxV/aV8Vjwz8OdCZ7a3Zf7R1e5Vks9OQ9DLIAfmIB2xrl2wcDAJH7Lv7G3wp/Zb0MDw9aprXiu5hCaj4lvLdVuJv7yRLk+RESAfLUknC7mcqCKqgM8ZrAxgVVVWM8CgABgVVAZ5qqqqqqoDPNVVVVVVAZ4rAxgVVVVVVUq0AAMZqqqqqqoKMZqqqqqqqsZ4FAADrVVVVVVVUBnpQAAwKqqqgue9YAGBVVVVjPAoKMc1gYwKqqqqqqqqqqxngVtJ71PNb2sElzczJFFCheSSRgqooGSSTwAB3rxJ8d/g14SEi678S9AiliBLwQ3i3Eq/Lu5jj3PyMYGOcjGc14l/bv+Cmj7o9Dh13XpBuCNbWYhjJGMZMrKwByeinocgcV4m/4KF+LrtXj8IfD/AErTsjCyX90923U/NhBGBxjjnBB65rxR+1d8e/FRZbj4g3enwkELFpcaWe3IGcPGA/bPLHGTjAOK1TV9W1y6+261qt3qFzjHnXU7SvjJOMsSepJ/E14V+Cnxa8bui+Gfh7rd2jkATtaNDDk7eDK+1B95Tyehz05rwv8AsGfGLWYxN4hvtD8PpjmKa5NxKD9IgU/8f/OvDH/BPr4d6eqyeLPGWt6zKpzttY47KJuTwVPmN029GHIPrgeFf2a/gZ4NdJtG+G+lSToQyzX6teurDb8ymYvtOVBG3GOcYyatrS1sreO0sraKCGIYjiiQIqj0AHAqqqqqqqqqqqqqqqqqqqqqqqviJ+yn8FviLHJNceFotF1ByzfbtHUWrlj1LIBsfJwSWUn3GTn4pfsP/E7wXu1DwXInjDTR1W2j8m7j+sJJ3DoMozE8kqBVjqHifwRroutOu9S0PWLCQrujZ7eeFh1B6Ee4NfBX9uy4tlt/D3xotzPGCqJrlnD86jPJniX72P70YzgfdY81pOr6Vr+mW2s6JqNvf2N3GJLe5t5A6SKe6sODVVVVVVVVVUQDRXjg0R2qqqqqqqqqqqioPQ0R2qqqqIBGKK46VVf8FLv207b44+Jl+DXwz1NpPA/hm5LX12gZBq1+hZSRz80EY4XIG5tzcjYa/wCCXP7JE/x7+MMHxQ8VWG7wP8PbyK5l3423+oqRJBb4IIZFIEkg6bQqn/WVVVVVVVVVVVVVVVVVVVVVVVVf8FFvhnL8U/2OfiJo9jbPNf6PYrrVoEBLK1nIs0mF3KGJhSZcHP3shWYAV+z/APEh/hB8bfBPxKEm2HQNat57rlhuttwWdflBPMTOOATz0PSo5IriJJ4JUkjkUMjochgehBHUEVVVVVVVTDHSqqqqqqmHcVVfEO3ePXFmPKywKVP0JBH+fWqqqqqqgSCCpwR0rw743kgK2WtSNJH0Wc8sv+96j360jxyoskTq6sMqynIP0qqqmGDwaqqqqqqqq+MXj+D4VfCnxd8SJ1if/hG9Fur6KKXO2WWOMmOM4IPzOFXqPvdRX/E38Ua5/wAtb7VNWu/q888j/wA2Zv1r4I/DHTPgz8I/Cfwv0qKBY/DulxW8zQAhZrjG6eYZwcyStJIeBy54HSqqqqqqqqvi38CvhN8dND/sD4peCNO1yFFIgmlj2XFsT3imXDxn/dIB75r9pj/glT458DpdeLP2f76fxdokS75NEuiBqcPPIi2qEuFA542v0AV+teHPE/jz4TeMotc8Mavq/hbxNoVyyiWB3tri1lRsMjDgjBBVkYYPII6iv2Uv+Cw+h6vDD4P/AGq7JNKvwypb+KtLtWa2mBGD9pt0y0TZGd8YZTu+5GFy3hrxN4d8Z6BYeKfCWuWOsaPqcIms76ynWaGeM9GR1JBFVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV/wWQ/aXl8ReNdL/AGafCuqv/ZnhtVv/ABGsUhCz3sigwwtjqIoyHx03TDulf8E0P2a/+F0/GQfEDxJp0U/hPwBJHc3Ec6ZS8vWDG3iAIwwUqZGHbagIw4qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr44/Ar4dftD+A7n4ffEnSWubGVvMtrqAhLmwmAws0DkHY4BI5BBBIYFSQf2qf2VvHn7LPjxvDniJG1DQdQZ30PXI4isV9EDyCOdkqggPGTxkEEqVY/8ABOL/AIKOy/CmXTfgL8etbeXwVIywaFrty5LaEx4WGZj1tT0DH/U/9c/9XDNDcQpPbypLHIoZHRshgRkEEdQRVVVVVVVVVVVVVVXxN+Jngf4O+B9V+I3xG8QW+j6Do0JlubmY/gqIo5d2OFVFBLEgAZNftr/t4/EL9rvXv7ISNvD/AMPtKumk0jQo3IeYjIW4uyCRJNtJwB8kYYhcks7/ALHP7Dfjf9qDWl1rWFvvDvw/tMm51s2+GvGBK+TabhtkbcpDPyqYOcthW+Gvww8B/B/whZ+Bvhz4as9F0eyUbYbeMAyvtCmWRuskjBV3OxLHAyaqqoDAqqqqUA81VYzwKwMYFVVVVVQGeAawMYFVVVVVWM8CgABgGqqqqqqgM0BxVVVVVVQXPJqqqqqqqsZ4FAADrVVVVVVVUFzWBjAqqqsZ4FBR3qqqqoLmsADAqqqqqqqqqqrxP478FeCoDP4u8W6Ro67Nyi9vEhZxgn5VY5Yna2AAScHFeL/24vgn4ceW20WbVfEdxGSoNja+XFuHXLylcj3UMD2yOa8Xf8FAfiDqLyQ+DPCWj6NAT8sl2z3kwAxgg/Igzhs5U8N6jJ1/9qj4/wDiOIwXvxL1C3j+bAsIorMqDjjdCqt2GCTkevJrW/FHibxLJ53iPxHqeqyFi++9vJJzu6E5cnnHetB8GeMPFZK+F/CmsawQSCLCwkuMEAEj5AegIP4ivDv7HXx/8QsGfwamlQnP77UbyKLBBAxsDM/fOduMA89K8Mf8E8vEEzRyeM/iJp9qB9+HTLR7gnk8B5NmOMHO088Y714Z/YW+Buiqra1DrXiGTYA4vL4wpuxyVEAQgZ5wWOPU14Y+FXw08GNHJ4V8B6Fps0f3biCwjE3UnmTG89e54qqqqqqqqqqqqqqqqqqqqqqqqqqqq+K/wF+GvxjtGHizQkXUliMdvqtt+6uYfT5h99Rk4VwyjJ4BOa+Of7Lnjr4MSPqqD+3PDTMBHqlvHgxZz8s0eSYzwfm5U8cgnaPgd+0H4z+COsrJpk7X+hXEi/btJmc7JADy0Z/5ZyYJ+YcHjcCAMeAfH3hf4m+F7Txd4Q1Fbuwuhgg8SQSDG6ORf4XXPI+hBIIJqqqqqqqqqqIBGKKg9DRGOKqqqqqqqqqqIBGKK+lVVVVTyQwRPPNKkccalnd22hQBySewxX/BRH/goh/b39p/AH4A69/xK/ntvEfiS0l/4/Oz2ts4/wCWXUPIPv8AKr8mS/7Mv7N/jv8Aam+Klh8MfA6pbh1NxqmpzRlodMtFIDzOB1PIVVyNzMoyASw+C3wc8D/AT4a6L8Lfh7paWek6PDtLYzJczHmSeRjks7tlie2QBgAAVVVquraVodhLqmt6naafZQY825uplhjTJCjLMQBkkDnuRWu/tC/AHwt5H/CTfHL4f6R9p3eR9v8AE1pb+btxu275BnG5c46ZHrWsfty/sf6Hemwvf2jPA8soUNus9US7jwf+mkO5c+2citc/4KO/sS+HoYpr/wCP+jyrKxVRY2V3esCB3WCFyo9yAK/4ej/sJf8ARdf/AC2dW/8AkWv+Ho/7CX/Rdf8Ay2dW/wDkWv8Ah6P+wl/0XX/y2dW/+Ra/4ej/ALCX/Rdf/LZ1b/5Fq0/4KC/sYX1pBeQftCeGVjnjWRFmM0TgEZG5HQMp55VgCDwQDWh/tpfskeIYZJtP/aQ+HkSwsFYX2vwWTEkdlnZCw9wCK0T49fAvxNaNf+G/jT4E1W2jkMTzWXiO0nRXABKlkkIBwVOPcetWl3aahaQ31hdRXNtcxrJDNC4dJEYZVlYcEEEEEcHNVVVVVVVVVVVVVeWdpqFpNYX1tFc21zG0U0MyB0kRhhlZTwQQSCDwc18dvhdf/BT4yeMvhTqAlL+GdYuLSGSUgtNAGJhlOOPniMb9B97oOg/4J+/F5PjN+yv4P1S5u2n1bw5CdB1Qu5dvOtVVUZmPLM8DQSE+sh5PWmXuKqqqqqqIBGKI7VVVVVVEAjFEY4qvF2gnXNMxAB9ptyXiz/Fxyv48fiBTo8bmORSrKSGVhggjrxVVVVVVVeHPFNzokiwTFprMk7o+657r/h0/nVneW2oWyXdpKJIpBlSP88GqqiARjNEdqqqqqqqqv+CqPjKfwr+ybfaTbvKp8V67ZaWzRqOFBe6OTkEAi1xxnOcYwSR+wH4EX4gftcfDvTp4ma20nUG1eZwpPlmzjaeMnDA4MscS+mWGQRkGqqqqqqqqqr9pf9in4M/tNWE95r+lJoniwqog8S6dAq3PyjCrMOBOmABhuQAArLX7R/7Dnxy/ZtluNV1/RU13wmspWDxDpQMkO3nb50f34GxjO4bNxwrvjNfAD9qT42/sy682t/CbxpcWEFw2b3SbjM9hfdP9bATtLYAAkXa4GQGAJr9lH/gqP8Fv2gp7Dwb49WL4e+N7uQQQWd7db7K/kwABBckKFdzwIpMHJVVaQnmqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/aC+Mmgfs/fBzxT8XPERR4PD9i0kFuzbTdXLYSCEe7yMi57Ak9BXijxH4o+JnjbU/FWuTz6r4g8TalJc3LIhd7i4mcsQqjJyWbAUewFfsjfAa1/Zy+A/h74dvDF/bLR/btemjIPm6hMAZeRwwQBIlbukSnrmqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq+Lvwi8B/HLwHqHw6+IujJqGk6guQRhZbWUA7JoXwdki5OG9yCCpIP7Uv7NHi79l74mT+CPEEjX+mXKmfRdYWBo47+3z154Ei5AdATtJHJDKT/wTK/4KEXfgPUtM/Z1+OPiSH/hEbjEHhzW7+Yg6RJ0S1kc5H2djwrMQIjwTsI2VVVVVVVVVVVVVXxN+Jngf4O+B9V+I3xG8QW+j6Do0JlubmY/gqIo5d2OFVFBLEgAZNftr/treOP2vfHHnTfaNH8C6NM39gaB5n3Oo+03GDh52X6hASq9WZ/2Hv+Cfus/tCPbfEz4mNc6N8PbecGCEIUn14qfmSJsgxwgjDSjJJyqc5ZNC0HRfDGi2Phzw5pdrpul6bAlvZ2drEI4oIkGFRVHAAAAxVVVAA0eaqqoDPArAxgVVADGaqqqqqqAwKqqqqqqA4zVVVVVVQGeM0qg1VVVVVUFzWBjAqqqqqqrGeBQAA61VVVVVVVBfWqqqrGeBQAA4qqqqxngUFHeqqqqqqqqq8R+MfCPhC3+1eKvFGlaPERlWvrxIN303EZ+grxZ+2v8AArw1vi0/WNQ8QTpx5em2bbc/78uxSPdSa8Wf8FCPElzvh8EeAdPsR0WfU7l7lj77E2BT7bmH8q8X/tOfHLxtHLb6p8QL62tZQVNtpwWzTaeqkxAMwPoxNJHqGr322NLi9vLqQnADSSSueSe5JPJrwh+yz8dvGirNYeAL2wtj1n1UiyA4JBCyEOwOMZVSORnAOa8If8E89QkVJvH3xCt4SCd9tpFsZcjPGJZduOP+mZ6+3Oh/sP8AwC0hFS/0jVtbIQqWvtSdCSTnd+48vkDj0x1BPNaD8E/hD4YJfQ/ht4dt5SSfNOnxySDIAIDsCwHA4zj8zSRxxRrFEiqiDCqowAB2AqqqqqqqqqqqqqqqsZ4FYPpW0kcVtNbT6itp9RW0+oraTxmtvvRXsDW0+orafUVtPqK2mipxWD6UR2qqqqqru0tb61msb22iuLe4jaOaGVA6SIwwysp4IIJBBr9q39lJ/Bb3fxK+Gunl/D7EyalpsS5OnE9ZIwP+WPqP4P8Ad+7+z98b9a+CXjSHU4ppp9CvWEeraer4WaPkCQDBw6Z3AjBPK5AY1pGraZr+lWmt6NexXljfQrNbTxHKyIwyrD6g1VVVVVVVVVVVFQelFTiqqqqqqqqqqioI4ojHFVVeNfG/hH4c+GL/AMZ+OfENloui6ZC0t1eXcmxEUDOB3ZjjAVQWY4ABJxX7b/8AwUj8RfHaLUPhZ8HZLrQvh/JvgvrtgY7vXFDHr3it2AX93wzAkOcEoP2Zf2YPib+1X8Qo/Afw7sUjht1E2ravdKwtdLgJxvkYDlmwQkY+ZiD0AZl+AvwL+Bv7D3wiTw/aa5pOkQNiXXfE2tXEVm+pXKozGSSR2CoiqHKR5wihupLs3xA/4KS/sYfDt5rW9+NWn63eRfdt/D9vNqQk+YA7ZoVMPAOeZBkA4yeK8a/8Ft/g9pplT4e/BvxbrzIo8ttVvINMSRt+G5TzyF2YYEjJPBVeteMv+C1X7QmrJJb+Cvhz4H8PxyKwEtylxfzxnPBVvMjTIHB3IwPXjpXjD/gpv+2v4wkYP8ZZdHtjIsiW2j6Xa2gjIXbxII/NIOSSrORk9OBjxN+01+0b4z8xPFPx6+IGpxSXBuPs9x4kumhSQ5+ZIt+xMBmACgAA4GBWoajqGr3suparf3F7d3DbpZ7iVpJJG9WZiST9a0Xw14i8SyyQeHNA1HVZIVDSJZWjzlAeASEBwK0X9n349eJYpJ/DnwQ8farHC22R7Lw1dzhCRwCUjODitG/Y/wD2qdeums7H9nj4gxyJGXJvfD9xZpgEDh5lVSeR8oOepxgGv+GE/wBr3/ogXij/AL8p/wDFV/wwn+17/wBEC8Uf9+U/+Kr/AIYT/a9/6IF4o/78p/8AFV/wwn+17/0QLxR/35T/AOKqb9lT9p+GV4X/AGc/iaWjYqSnhK9YZHoREQR7jitX+CXxn0C8Ona78IvGmnXSqGMF3oFzC4B6HayA4NappWqaJfSaXrOm3VheQ48y3uoWikTIBGVYAjIIPPYg1aXd3YXcF/YXUttc20iyQzQuUeJ1OVZWHIIIBBHQ1oX7Qvx+8L+f/wAIz8cviBpH2nb5/wBh8TXlv5u3O3dskGcbjjPTJ9a8P/8ABRT9tXwzKJtN/aB12ZhD5IGoW9tfjbxztuInG75R82N3Xnk58M/8Fhf2wtBEY1WbwT4j2W4ib+09EaPe4x+9P2aSH5zg8DC/MflHGPC//BcPxVb/ACeNP2fNKv8Ad5YEml+IJLPZ18w7JIZd2eMDcuMEEnOR4Q/4LS/s26xItt4u8C+O/D0jyMBMltb3kCIFyCzLKsgJbK4EbAcHOCceDv8AgpX+xT41SNLX422OlXLKpe31mxubExknGDJJGIyfXa5AHJNeEPiz8K/iEF/4QD4meFPEu/dt/sfWbe8zt+9jynbp39Kqqqqqqqqr/gtR8E/7C+InhH486Ta4tfFFm2k6syjhby2G6F2yc5khYqABgC255PP/AASI+NieEPi9rXwZ1e52WPje08/T9zABb62Vm24P9+EydD1jQYOchlBGRTDHSqqqqqqmAI4qqqqqqogEYojtVeJvB1vrW68tGWG9A5JHyyf73v7/AORf6deaZcNa3sDRSL2I4PuD3FVVVVVVVoPiG80KfMJ8yBz+8hY8H3HofetP1C01S0S8spQ8bj8VPcEdjVVTAdRVVVVVVVV/wWV177P4P+GHhfbKf7Q1LULzIbCDyI4U5Hc/6Rwe2D61/wAEg/Df9pftC+JPEk1tFJDo/hOVI3Y/NFPNcQBSo940mBPv71VVVVVVVVVVUsUU8TwTxpJHIpV0dchgeCCO4xX7T3/BML4X/FxrvxZ8H3s/AfiqRWd7aOEjTL6THG+Jc/ZycDLRLjkkxsxJPxh+BvxS+A3iU+Fvij4RvNHuXZ/ss7put71UIBeCUfLIvK5wcjcMgHiv2Rv+Cnfxh/Z2GneC/HBl8deAYZNjWd3Mft9hEeD9mnYnIXqIpMqcbVMedw+BX7Q/wj/aR8HR+NPhL4tt9VtlVPttm37u706Rs/u7iE/NG2VYA8q20lGZcMaqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv+C03x+F9q3hb9m3Qr2JodNxruvqhywnZWS1iP8Ad2xtLIV5z5sZwNoz/wAEuPgLB8U/jufiLr1p5uifDqOO+jV4yVl1FyRbDPT5CrzZByGij4IJqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr47fAn4fftEfD68+HnxC0zzrab95Z3cQAuNPuACFnhYg7WGenRgSrAgkV+0J8BPGv7OPxM1H4ceM7dnNuxk07UFhKRalak/JPHnPBAwy5O1gyk5Ff8Etv2+/7Yj0f9ln4yatK+ormDwhrd1NuE6BcrYTM3IcAERMThhiPghA9VVVVVVVVVVVXibxLoHgzw7qXizxTq1vpmj6PayXV9eXDbY4IY1LO7H0ABr9vL9tnXP2u/HsUGjw3ek/D/w7I66HpkspD3DHg3dwoJXzWGQAP9Wp2gklmb/gn9+xBL+0Jrw+JHxM0y8g+Hejy/uoyvl/25cKeYVbIYQqR87r1PyAg7ilhYWGlWFtpel2UFnZ2cKw21tBEI44Y1AVURVwFUAAADgAVVVWM8CgABgVVVVAcVVAZ4qqqqqqoDPNVVVVVVAZqqqqqqrGeBSqCMA1gYwKqqqqqgM8ZrAxgVVVVVVVjPAoAAdaqqqqqqrGeBQUVVVVAZrAAwKqqqgue9YAGBVVVVVVVeJPHHgvwdGJPFni3R9GDDKC/vo4C3XG0MQTna3A64PpXiz9tf4FeGt8Wn6xqHiCdOPL02zbbn/fl2KR7qTXiz/goR4kud8PgjwDp9iOiz6ncvcsffYmwKfbcw/lXiz9p/46+Md8eofEPULOB+BBpuLJQPTMQViP94mnfUNXvg0j3F7eXUgALFpJJXPAHckngV4R/Zp+OPjXypNI+HWpwW8mCLjUEFlHtO35wZSpZcMD8oORnGcGvCP/AAT38X3vlT+N/HWl6WhwzQafA93Jj5cqWbYqtywyNwBA+9njwj+w/wDA3w55U2r2Op+I7hMMW1C8KR7xt5CQ7BtyD8rbuGIOeMeHfBvhHwhb/ZfCnhjStHiIwVsbNIN312gZ/Gqqqqqqqqqqq1rxZ4V8NqzeIvE2laWEj8xje3kcGEzjcdxHGQRn2rUf2hfgdpbpHc/Fbw05cZH2fUEuAPqYywH0NX/7U37P2nW5ubj4naa6AgYgimnbn/ZRC36Vp37Vn7PeqRvJa/E2wQRnBFzbz25/ASIpP4Va/tIfAi8uI7WH4p6AryHCmW58pR9WbAH4mtO+Kfww1l3i0j4keFr54xl1t9YgkKj1IVzirW5tr63ju7K5iuIZRlJInDqw9QRwa2j1raK2jsKwO1VVVVVVVVVVVRANEA9qKg9KKg9DRU9jRU46UR2qru0tb61msb22iuLe4jaOaGVA6SIwwysp4IIJBBr9pn4OS/Bz4lXWnWNu40DVc3WkSMc/uzjfH9UYlfXbtJ61+wd8YH1PS7z4Pa3cqZtMja70dmPLwlv3sWT/AHWZWA64Zuy1VVVVVVVVVVVVRUGipFVVVVVVVVVRAIxRXHQ1X7UX7eXwT/ZhtZ9J1LUf+Em8aeWTb+GtMmHmKcHBuJcFbdMgA5y+GBVGGcftH/tW/GD9qPxIms/EfXEXTrNj/Zuh2KmKysARglEJJZzzmRyzHOMhQFFaf+21+0J4U8DxfDL4WeKrf4c+FIWkddN8K2otHaWTdvla7bfdO5Vgu5piQI48coCPE3izxV411WTXfGXibVde1KXPmXmp3sl1M2WLHLyEscszHr1Ynua8B/svftE/E8B/A3wX8W6lbnpd/wBmSQ2+fl486QLHnDqcbs4OemTXgf8A4JJftXeJzDJ4mj8KeEImYecmpasLiVF34O1bVZVZto3AFwDkAsDnHg3/AIItaWkST/EL463U8h+/a6NoyxBMMek0sjbsrj/lmuCe4HPhf/gkr+yP4fZG1a18X+JQsxcrqmteXuXAGw/ZUh+XjPGGyTzjAHhn9hL9kHwkLYaV8AfC8/2Tf5f9pwvqWd2c7/tLSb/vHG7OOMYwMaD8IfhN4WitIfDHwv8ACWkJYSeZapYaJb24gfdu3IEQbTuO7IxzzRUHoaI7VVVVVVaz8HPhF4itp7LxB8K/B+p29026eK80O3mWUhg2WVkIY7gDz3Ga1/8AYp/ZM8SS+dqH7PvgqJvJ8rFhpi2I2884g2Ddyfm+9054GNc/4JbfsZasIRYfDzVNG8rdu+xa/eP5ucY3edJJ0wcbcdTnPGPEP/BGj4H3MSL4V+KnjnTpQr721AWt6GJA2kBIocAc5GTnI5XHPij/AIIveP7RHPgv44+H9VYQ5QappE2ngyZOVPlvPhcYO7k5JG3jJ8Wf8Epv2wPDkjR6P4b8OeKQsiqH0nXIowwK5LD7V5JwD8pyM56AjmvFP7JX7TfgyA3XiH4DeN4LdYzI9xDo0txFGoOMu8QZU/4ERV3aXVjdTWN9bS29zbyNHNDKhR43U4ZWU8ggggg9DXg39qj9pP4fMf8AhD/jt440+NmZmtxrc8kDMwALGJ2KFsKo3Fc8DmvBv/BXb9szwuxbW/EXhfxeCzELrOgxxAAgAAfYzBwMEjvljkkYA8D/APBcG0d1t/iT8ApY05Ju9D1sOfvDAEEsa/w7jnzeSAMDOR8P/wDgrD+xp45eG31Lxhrfg+4nyFi8QaQ6AHcRhpLczRrkDdksBgjnOQPAPxt+DnxVjRvhr8VPCnid2jEhh0vV4biWMFQ3zxoxdDg8qwBHcCqqqqqr9tb4DQ/tHfs2+L/h1BbvLrUdqdR0Ep94ahbgvCgycYkw0JJ6LKxHIFeCfGHib4XeOtG8ceGrh7HXfDWox3dqZFI2SxODtdeMqcFWU9QSDwTXww+Ifh34tfDzw98SfClx5ul+IrCO7t8nLR7h80bY6MjBkYdmUimUYxRGOKqqqqqqYdxVVVVVVTDuKqr7T7LUrc2t9bpNGezDp9D1Fax8O5U3TaJP5g/54SnB/Buh/HH1q9sLzT5jb3ts8Mg/hdcZ+nqKqqqqqq0LXbrQroTQnfE/EsRPDD+h96s7y21C2S7tJRJFIMqR/ng1VEAjGaI7VVVVVVVX/BZjWvtHiX4W+Hfs2PsNhqd152/O/wA57dNu3HGPIznPO7oMc/8ABGnRpZ/FXxQ8QrOgjstP061aMjljNJOwI9gICP8AgQqqqqqqqqqqqqrx98PPBHxS8MXXgz4heGLDXtFvMGW0vI9y7h0ZT1Rh2ZSCOxFftTf8EsvGXgKK78a/s+TXni7REaSSfQJE3ajZpjI8nH/H0oAI2gCT7oCyZJHw3+J/xJ+CXjO38a/DTxXqXhnxDp7FBcWzbSQGBaKWNgVkQlV3RyKynAyDX7G3/BVXwD8ans/h98dE0/wT40f93b6iJfL0zVHLKFVS5Jt5Tuxscsp2khwWCCqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqvEWv6L4T8P6n4p8R6jDYaTo1nLeX93McJbwRIXkkb2VVJP0r46/FfWfjp8YfF3xa155ftPibVJbmOKV95toM7YIAe4ihWOMeyCv2BfgsnwU/Zn8M6beW/l614kj/trVSyAMJbhVMaHH9yERIck/MGPGcCqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv2pf2aPCH7UXwzn8EeIJEsNUtWM+i6wsCySWFxjrzyY3wA6AjcAOQVUj4k/Drxn8GPiDqvgDxlYy6drugXZjcpuUPtOUmiYgFo3GHR8DIINf8E1v27rX9oXwlb/CL4n6xbxfEjw7arHbyzTsZPENrGmDOC33p1A/erkluZAMbglVVVVVVVVVVV/wU8/bnPx18Uy/BD4Xas58AeG7rF/eQu6DW71DySMgNBEwwgIwzgvkjYR+xD+x9rH7Unjw3OsGew8CeHpUfWr4RsDdHIItIWHHmMMbjn5FO7klQ2haDovhjRbHw54c0u103S9NgS3s7O1iEcUESDCoqjgAAAYqqqqUZ59KqqqgM8VVUAAMVVVVVVYzwKwMYFVVVVVWM8CsDGBVVVVVVSr270AAMA1VVVVVAZ4FYAGBVVVVVVWM8CgAB71VVVVVVWM8CgoHeqqqoLkdawMYFVVVjPAoL61gYwKqqqqrxP8VPhp4MZ4vFPjzQtNmj+9bz30Ym6gcR53nkjOBxXij9uv4KaKWi0Jdb8QyYOx7Wz8iLOB1MxRh1I4U8qe2CfFn/AAUD8c6hvh8GeC9I0iM5AlvZXvJB7jGxQfYhh/OvF37Svxx8a+ZHq/xF1OC3kyDb6ewsk2nd8hEQUsuGI+YnIxnOBSR6hq99tjS4vby6kJwA0kkrnknuST1rwr+zP8dPGDoNM+HGq20TEZm1KMWKBfl+b98VLDDA/KCTzjODXhH/AIJ8eL73yp/G3jnTNMQ4ZoNPge7kx8uVLNsVW5YZG4Agfezx4R/Yf+B3hwRTavY6n4juEwxbULspHvG3kJFsG3IPytu4Yg54x4Y8B+CfBUAt/CHhPSNHXZtY2VmkLOMAHcyjLE7VySSTgZqqqqqqqqqp54LaCS5uZkiiiQvJJIwVUUDJJJ6ADvV38Q/Adkp8/wAY6OXEfmCKO8SSRlzjKopLNyD0B6VffGDwDZqdmsPcuMfJBbuTyM9SAPbr1/Gr79oDSYzjTfDt3OMjHnzLDxjnoG71dfHzxNJgWek6ZCMYO9Xc59R8wq5+M3j6dVWLU4ICDyY7VDn/AL6BrXbrUvEwkTXtc1e+ilbc0E2pTmIHBGRHv2rwSOAOtf8ACr/h9/0Klj/3yf8AGv8AhV/w+/6FSx/75P8AjX/Cr/h9/wBCpY/98n/Grn4SfDm7VVl8L242njy5JIz/AOOsM1P8E/htNEY49BeFj0dLuYkfTLEfpVx8AvAU8Xlxf2lA2fvx3AJ/8eUj9KT4BaHYXMV7oXifW7G4i3YlWRd4yMfKVVSOCQfXNaf4Y+KmgvAPDnx88WWcFup2W8s8k0YJzk7DKExznBXrz1rTPHP7TejyLFH8X7HUraJSqJf6NCS3uzKu8n6sa0n4/ftEWkWzWdF+H+o7IgFaGS6t3dh1ZvvKSfQBRn2rS/2nPFsmV1v4RxwYQYe119Jdzd/laJdo/E1pv7SegXsqQ3ngTxVYZTLSTCzdFPp8lwzH67fypf2g/htHC8moXGsWbxvt8o6PcXDNwDuBgSQEc465BBziof2nPgTLGXl+IVraEMV2X1pPaOSMZwssasRz1xjOfQ1oPxT+Gfip4ofDnxB8O6lNMFKQW+pxPL833QUDbgSSBggHPHWqqqqqqqqqqqqqIBGKIB7UVBHBr9rD4S/8LT+FF7/Z1t5ut+H919p21cs+0fvYh/voDgd2VK+H/jXVvh1400fxvojf6XpF0syoW2iVejxk9ldCyH2Y1omsaf4i0Ww1/SZ/OstTtY7m2kxjfHIoZTj3BFVVVVVVVVVVVVVRUHoaIIqqqqqqqqrxT4v8JeB9JfXvGvinSPD+mRtte91S9jtIVOCcF5CFBwCevY18W/8Agq/+zB8PkmsvBV1q/wAQNTRfkTSrc29qHzyr3EwXjHO6NJByB64+N3/BWH9oT4n6Zd+HfAdjpvw60y7V0efTJXn1EISw2i6baEO0qN8caOCCQwyALu6ur+6mvr65luLm4kaSaaVy7yOxyzMx5JJJJJ6mvhx8APjb8XpYU+Gnwr8TeIIpm2i7tNOkNsh+YfPOQIkGUcZZgMqR14r4a/8ABIP9pDxVLbz/ABB1vwz4IsmYCdJLn+0buMHqVjg/dMQM8GZecc9x8Nf+CQf7N/hWK3n+IOt+JvHF6igTpJc/2daSEDkrHB+9UE54Mzdue5+Hf7OPwG+EvlSfDr4R+F9EuYcbLyDTka646ZnYGU/i1UV9KqqqiAR1ojHFVVVVVVUQCMA0R2qq8TeC/B3jS1Fj4x8J6NrtuI5IxDqdhHdJscAOu2RSMMAAR0OBmvGf/BP39j7xyk39o/BHRtOml3lZtGeXTTEzLjcqQOqcdQpUrnsea8c/8Eb/AIJawJJvAHxK8W+HJpNxEd9HDqUEZ427VAifAw2QZCTkcjHPjj/gjp8eNGM0/gXx/wCEPEtvGxCR3Jm0+4kG/CkIVkjB2/MQZBjBALcZ8c/sJftb/DxJJdd+BviC6giXcZtIRNUXbkjd/ozSED5SSCAQMEgZFappWqaJfSaXrOm3VheQ48y3uoWikTIBGVYAjIIPPYg18P8A9sr9qj4XvCfBnx68YW8NvnyrS81Fr+2TLFjiC48yMZJJPy8k818N/wDgs1+0p4Xa0tPiF4Y8I+NrOJl+0ytbPp17MAFBAkhJhQnDHPkHljxgBR8D/wDgqt+yp8W4YLDxP4iuPh1rkjJG1p4jUJbMxHJS7TMQQE43SmI99uMmtP1HT9WsodS0q/t720uF3Qz28okjkHqrKSCPpVVVV/wVF/Zzl+Bf7Smo+J9JtGXwv8SWm1rTnwdsV0zj7bAD6rK4kAAwqXEa9jX/AASD/aG/5DX7NniO/wD7+reG/Mb6fardf0lCj/pqaIBGKZc8ZojtVVVVVUQCMURjjNVVVVVRAIxRGOM1VVdWlrfQm3u4I5o26q65Far8PLG4zLpVw1s5P+rf5k/DuP1rVPDur6Qc3lm3l9pU+ZT+I6fjiqqqqqrw34hn0O7AZma1kP72Pr/wIe/86t7iC8gS5tpVkjkXKsveqphxkVVVVVVVX/BZK1uU+IXw4vXt5Rby6NdxxzFCEd1mQsoPQkB0JHbcPUV/wRjvrOO++LmmPcILqeHRpooj1ZEN4HYewMiD/gQqqqqqqqqqqqqqq/ax/YF+F37SyTeKNNEXhTx0sZ26xaW42XxC4VbuMY8zsPMGHAAGWCha+NXwG+KX7Pvit/CHxQ8Lz6ZcFn+x3SjzLW/RcfvIJR8si4ZSRwy7gGCtkD9i7/gqP49+BEen/Dn4ztqHjLwFDiK3uS/m6lpEYUhViZ2AmiB2gRucovCMAoQ/D74i+B/iv4RsPHfw58T2Ov6DqSk219aPuR9pKsCDgqwIIKsAQQQQDVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV/wVi+MDfDH9k3U/DWn3Txap8QL6LRYSgBZbc5luWOf4THGYj1P74Yx1H7H/AMH/APheX7RfgvwFcxRPpjXwvdVE0e9Gs7cebKhH/TRU8oZ4zIM8Zqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr9uz9jvTv2nvAn9s+GbaKH4ieHYD/AGLcPN5SXcW7c9rKTkYb5ijHG1yMsFZ8+GfE3jn4SeObPxL4Z1LUvDXirw1eloZlUwz2c8ZKsrKw4/iVkYYILKwIJFfsX/tYeHP2ufhHB4ztIbfT/E2lstr4k0iJmIs7nnayFhkxSKu9eTjJQsWRjVVVVVVVVVVX/BV39tJvhb4Wk/Zw+G2rIPFfimxY+ILqCQFtL0+QbfJxggSzqW7hkj+bA8xGH7PnwJ8YftG/FDS/hl4Nj8uW6zNf3zxl49OtEIElw4BHC7lAGRuZkXILCvhB8JvBnwQ+HmkfDXwJpqWmmaVDt3Yy9xKeZJpD1Lu2SfTgDAAAqqqsZ4FYAGAaqqqgABiqoAGqqqqqqgBjNVVVVVVAYFVVVVVVAZ5oDAqqqqqqxngUAAMZqqqqqqrGeBQUAe9VVVVVVVjPAoAAdaqqrGeBQUd6qqqsZ4FAADiqqqkeOKNpJHVUQEszHAAHUmvE/wC0L8E/CDPHrnxK0VZYvvw2sxu5FOQMFIQ7A8jjHTnpzXif9v8A+GmnK8XhXwrruszJ91pylnC/APDZd/Y5Tt3rxT+3r8XNXWWDw3pOhaDE2fLkSBrmdOCPvSHYeoP+r6gdsivFPxt+LnjRpR4j+Iuu3UU2d9ul20MB5J/1Ue1O5/h6YHQCtM0fVtcuRY6Lpd3qFxjIhtYGlfGQM7VBPUgfiK8J/sl/Hnxbski8DzaVbt1m1aRbTb9Y2Pmf+OV4V/4J6azLtl8b/ESytsZ3QaVaNPu69JJNmP4T9w5yRxgE+Gf2K/gL4daOS70LUNdlj+6+p3zNk5JyUi2IeDjBXHA7814d8G+EvCFv9k8KeGNK0iIjBSxs0g3fXaBn8aCjuawMYFVVVVVVa98Wfhf4YVv+Eg+Ifh2xdAT5U2pRCQ4ODhN2489gKm/al+DbsYtD1rUtdkWURsumaPczKuf4vMKBNucDIbv9cXP7Tj3MIfQPhfrRLKMf2teW9mMk9/KaZgAOvy59Aau/j78SLvc1honhrS8lSqzGe/x/eBIMGc84OBj0PWrr4q/FG8kEp8ZtZkSMxjs9Pt1Qqei4lSRsDt82eOSauNc8TXiqt34x8RS4i8tsavcIHHfcquFJP0/SpNM02W6W9k0+2e5UKFmaFS4CkkANjPBJI9MmoopZ5FhhjaR2OFVVySfYVaeCfF98gktfDOpOhAIY2zKDk44JHNQfCb4m3MhhXwl9mO9VEl1fwCPBOCxMbu2AOfuk+gq0+AfxIuGVb7UvDVgBPhmimnuyYuPmAMcXzdflzjp83PGl/s43yuV8QePxcRbsj+ztJFq4XBwN0ksw64529PTOat/gx8P4N3m6VNcZxjzLqQbfptI/Wv8AhUHw6/6F3/ybm/8Ai6/4VB8Ov+hd/wDJub/4uv8AhUHw6/6F3/ybm/8Ai6ufgx8P59vlaVNb7c58u6kO767iauPgZ4InjCRNqMBBzujuASfb5lIq5+AHhh1UWms6nGwPJkKPn6YUVc/s92jlfsnimWMAciS0D5+mGFTfs+akshW38S2zxj7rPbshP4An+dS/ATxgiu0eoaRJtB2qJpAW/NMZ/Gpfgp49jjLpZWshA4RblQT9M4FN8IfiIqlj4dPA7XUJ/TfX/CtfHf8A0LF5/wB8j/Gr/wAMeI9L3HUdBv7cLuy0lsyr8vUg4wR79KngguYjBcQxyxt1R1DA49jWpfDnwLqwYXnhbT9zElnih8liT1JZMEn3/wAa0rwXf+FRjwH8QPFnhpFbK29lqb+R94n5o2yGHzNwe5J55qw+J/7THhlVFl470HxPFCMJDrOmCFmUKQAXh2sxzg5LZJ5J6g2X7YPjLRpRH49+CN+LdSS95ol8LrC56iIrwNoY5Zx2BAzkeHf2xvgJr7eRdeKbjRLkD5rfVbKSFlOOVLKGQEHgjd16ZrQvFnhXxTD9o8MeJtK1eLBO+wvI7hcA4JyhPcEfWqqqqqqqqqqqqv2lfh9H8NfjN4h0CztzFp9xML2wBOR5Mw34HfCsXTnn5O/U/sM+Mj4l+CqaDczvJc+Gr+W1AfJPkv8AvY+fT53UDsEHQYoqD0NFT2NEVVVVVVVVVVVVVRUHoaIxxVVVVVV8bf8Agop+y18EluLG58dp4t1yBth0nwxtvpFbJUh5twhjKkYZWkDjsp6V8av+Cunx68eC70n4VaNpfw+0mePy0nT/AE/UR2Y+c4EaZHTbEGXJw5OCPGfj7xz8RtYfxB4/8Y6z4j1N+t1ql9JdSAcDAZySBgAADgAAdqihluJUggjaSSRgqIiklieAAB1Nfs0f8Ep/jB8VfI8SfGd7r4deHW2ulrPAG1O7U9QISf8AR+mMzDcD/wAsyOa+F/7C/wCyp8IxDP4X+D2jXd/Dki/1lDqU+47cspnLBD8o+4Fxk4xubMUMVvElvBEkccahURF2hQBgAAdBjtVVVUVBHFEdqqqogEYojHFVVVVVVUQCODRHaqqqqvFPgvwd45046P418J6N4gsTnNrqlhHdxc9fkkUj9K8f/wDBOD9j74gPPdSfCqLw/ez5/wBJ0C8lsRHksx2wqxgHLH/lnwAB0AFfFr/gjQ0VnNf/AAN+LDT3CN+70zxRAFDr3/0qBeGx0Hk4JPLLXxf/AGUv2gvgXPc/8LH+F+s2VhbSFP7Wt4DdWL8ZBFxHuQZAyFYhuuQCCB8L/jt8ZPgpfjUPhT8TPEHhlzIZJIbG9ZYJmIxmWEkxy8AffU9B6DHwi/4LRfG3wvDFp3xf8B6D45gjVFN7ZyHSbxjjDO+xXhYng7VjjGc84Ix8LP8AgrF+yH8RY7e21/xLqvgbUZI8vBr9gwiDhcsFnh8xMcHBcpu44BIWvB3xC8AfEWwbVfh9458P+J7JMbrnRtThvYxksoy0TMOqOPqp9DVV/wAFAf2aW/ad/Z11jwzodgk/i7w+39reGzwrSXMSkPbgnHE0ZeMAkKHMbH7lfDbx94m+DvxH0L4heG3e21nwzqKXMSSArlkOHicddrLuRh6MRXwa+K3hn43/AAx8O/FHwlOjWGvWSTmESB2tZcYlgcj+ONwyN7qccYogEYFMO4qqqqqqphkcVVVVVVUwyOKqqqiAw2kAgjoa1jwNpOpbpbVfsc57xr8pPuv+GK1jwprGjAyTQCaED/WxfMo+vcfjxVVVVVeDvEZ0y5Gn3cn+izNwT/yzb1+h6frVUQCMURjjNVVVVVVf8Fn/APmjv/cc/wDbCv8AgjldWqfFTx/YvcxC4l8PQyRwlwHdFuAGYL1IBdAT23L6iqqqqqqqqqqqqqqvih8J/h58ZvCd14K+JPhez1vS7lSAk6YeFuzxSD5o3HZlIP4V+2H/AME+/Hf7N73XjfwfJc+KPh0JP+QgyqbrS9xwqXSKACCSAJkAQn7wQsoP7Ln7XXxc/ZP8YR694B1d7nRLuZDrPh25fNpqUYxnIwfKlwPllQbl6HcpZG/ZZ/a9+Ev7WfhD+3PAepfZNasY0/tjw/dyAXWnuVGTjjzItxwsqja2MEK2VFVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVf8Fk/i2/jL9pDTPhZay5svh3o6LKhVgVvL1UnkIJ4IMAtOncHJ7D/gjp8LUFr47+Nd3Hl3kj8PWDhhwAEuLkEdec2mDwOD17VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV/wU4/YyfxPZXP7SPwu0eWXWbOPPiuwgVnN3Aiqq3aKOjRquJABgph+CjFv2Vv2lfGf7K3xc034l+E2+02hxa63pjEBdSsWdWlhyQdj/KGRx91lBIK7lPw1+InhP4ueAdC+JXgbUft2h+IbNLqymK7W2t1Vl/hdWBVl7MpHaqqqqqqqqq/as/aG0D9l/4I678V9ZFvcXdqottG0+aXZ/aF/ID5UI7kcM7Y5CRue1eLPFPjf4weP77xT4ivL/xB4o8UX++RsNNNczyMFSNFGSf4URB0AVQMACv2H/2VbD9mD4UxWmrQQTeNfEKx3XiC6VQTE+35LRWBIKRZYZBwzFm6EAVVVVKAeaPPeqqqAzVVjPAoAAYFVVVVVQAPFVVVVVVAA1VVVX9/YaXZy6hqd9b2lrAu6WeeQRpGPVmJAA+teKf26/2RvB2uReHNa+Ovh972Wbyf9AE1/Ej/AC/fmt0eOMDcMlmAGGyRtbGoft4fsg6bpz6ncfHnw48Me7KW5lnl4xnEUaM56jGF55x0ONW/4Kl/sd6deG1s/Get6pGFBFxaaDcKhJ7YlVGyP93FeHP+Cmn7GviDy47j4n3WjTyziFItS0S8TOcYcukbxqmTjLMMYJOBzXw9+Mfwm+LEBn+GvxI8OeJfLj8ySLTdRjnlhXIGZI1O9OSPvAdR61VVVVVUAMVVVVVVVQHeqqqqqqqxngUAAMCqqqxngUFA71VVVAZrAAwKrV9b0Xw/ZNqOvavZabaJndPeXCwxrgEnLMQBwCfoDXiz9rP4C+Et8UvjmDVbhOkOkxtd7vpIv7v82rxV/wAFC9Fi3ReB/h5e3Ofuz6rdLBt+7nMce/P8Q++McHnJA8U/ts/HbxEXj03V9N8PwuCpj06xUnaRj78u9gfdSDnkYrxL4+8ceM5Xm8V+MNY1cuclby9klUckgKpOFAJOAAAM8Yrw54G8aeMXMfhPwlrGslThvsNjJOExjOSoIGNy5z0yPWvC/wCxR8ePEkYlvNG03QY2GVbVL0KT/wABiEjD6ECvDH/BPPSI1WXxn8Rru4JPzQaXZrDtAJziSQvnI2/wDHPXrXhb9kj4CeFVjePwPFqs6EEzarM9zuwcjKE+X25G3noeK0jRNF8P2S6boOkWWm2iY229pbrDGuAAMKoAHAA+gFVjPAoLkcmsDGBVVVVXib46fB3werHxB8SNCgdULGGK7E8uAM58uPc3IHHHPQZrxJ+3n8GtJMkWg2Ov67KpIR4bVYImw2MlpGDjIyR8noDjtqP7fHj/AF+6+xfD34V2Ky/wrdTS3zHk5JWMR4GMcZOMHk9m+JX7XPjAZ1Dxta+G7VgD5cFtAj8DggorOM9wWH0q4+GV74gVD4/+IninxQyADZe6hI0ZAJIGGZjwST97jJrS/h94J0UKNO8MWCsn3ZJIRK44x958t096hgmuZFht4nlkb7qIpYn6AVp/w58caoAbXw1eAHoZ1EIPH+2RVj8CPFlwc3t7p9qoJGC7O31wBjH41Zfs/aemDqPiS4l5GRDbiPjuMkt+ePwqy+CngS1Cie1u7zaDkz3JGfrs21afDvwRZ4EHhewbCbR50Xm8f8Dzzx161a2FnZR+VZWkNun92KMIOPYVtFYGMCqqqqqqqqqqqvtC0PVCW1LRrG7JIJM9ssnIGAeQe3FX/wAIfAN+Qw0U275+9BM6fhjJH6ZrU/2ftMkBOj+IbmEgcLcxLLk49V24/I1q3wQ8aWGWsVtNRQdBDLsbHuHx+hNap4d13RTt1bR7y0wcBpYWVT9D0Nav4Z8Pa+mzWdFs7w9mlhBYY9G6itV+A+gtOt/4U1e+0S7jOYikhkVD6gkhwe2d1J4o/aq+GCltB8faxqdlEm1D5w1BY0AwAIpwxXAHRVwPWtE/b6+LulM9t4i8OeHtT2bgSYJLaVWyOCQ5XAwRjaD78V4f/wCChngq7eNPFHw+1jTQxIZrK6jvAvofmEZPv6e9eBf2i/gz8RTHb+HfHVgt5Idq2V8xtJy2M4VZMb+P7m4dfQ1VVVVVVVVVf8FEdEig8TeDPEahPMvrC5tGIHzYhdHGfb9+cfjX/BO3VJo9Z8a6JjMU9raXGSx+VkaReB7iTn/dFVRAIxRUGivoaI7VVVVVVVVVVVVEAjFFeODRHaqqppYoInnnlSOONSzu52hQBySewxX7R/8AwVT+CHwhlvfDHwxgb4i+J7ZmidrSXydNtZBwd9zg+aRwcQqytggyIa+O37b37R37Q3m2Pjbx3LY6JLvH9h6KGsrPa2cq6qxaUYOP3rPx+Oar9nb/AIJo/tD/AB2+y65r2l/8K+8KzYb+09dt2W4mQ94LTIkfggguY0YHKuelfs9/sQ/s+fs3RQXvgzwimpeIo1w/iHWAtzeE45MZwFhHtGq5HUnrVUQCMUVwODVVVUQCODRHaqqqIBGKIxxVVVVVVVMoI460R2qqqqqqmhiuIngnjSSORSro65DAjBBHcYr4uf8ABP39lX4xCe71b4ZWugarPvJ1Pw4f7OmDv95yiDypGzzmSNuc+pz8Uf8AgjT4ktd158GPi1YagvzH7B4jtmtnUDpieEOHJ54MaAEDnnj4mfsR/tT/AAneR/FPwZ164s49x+3aRENTg2AkB2e3L+WDjOJApwRkDNWGo614b1Rb7Sr+90vUbRmVZreVoJoSQVYBlIZTgkH6kV8Pv+CjH7Znw4SO30z446xrFqrAvB4hSPVTIAWODLcK8oGWP3XBwFGcKAPA/wDwW4+LemxrH8Rfgx4U8QbcjfpN9PpTMNoAJ8z7QM5DEkAA5AAGMnwb/wAFnf2ZNcQxeLvCfjjw1Oqs25rOG8gbBACho5N+4gk8xgDB+bpn9vJv2f8AxB8brv4n/s4eOrTX/DfjbzL+9tktZrWTTtR3n7QvkzRo6xuWWRWIwS8gHCCv+CVP7Uv/AArzx6f2fPF17FH4f8aXfmaRPNLtFrqZUKsYzxicKqAdfMCAffNUwyMimGOlVVVVVVMO4qqqqqqph6VVVVVWt+CtJ1UGW3QWlwf441+VvqvT8Rg1rGg6lokvl3kB2E4SVRlW+h/pVVVVXgnxF9tgGkXkn7+Ff3LH+NR2+o/z0qqYZGRVVVVVVX/BYvSbub4N+BdcjVPstp4me2kJbkPLbSMmB6Yhf9PWv+CSesxaX+1Je2MkLO2r+Er21RgceWRNbTZPtiEj6kVVVVVVVVVVVVVVVUsUU8TwTxpJHIpV0dchgeCCO4xX7aH/AATHiuRe/E/9mbSUikVWl1HwhEMKQqEs9l/tHH+o7ljsI4Q+AvH/AMQ/gp48s/GngPXb/wAM+KdBuGEU8aBZIXGVeN43BDKeVaN1KkZDAjIr9hz/AIKK+BP2otPtvA3jaWx8L/Ey2hUPYSTBINZIHzSWZY5LcFmgOWUcguqsy1VVVVVVVVVVVVVVVVVVVVVVVVVVVVV5d2lhaTX19dRW1tbRtJNNM4RIkUZZmY8AAAkk8Cvjl8Q5fi38ZfG3xMlkdl8S69eX0IcEGOGSVjEnPOFj2KM9gK/Ye+Gy/C39lj4feH5LZoby+0tdVvVkBDia7PnlXBVSGVZFQgjjZjJxk1VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVLFFPE8E8aSRyKVdHXIYHggjuMV/wUR/ZFb9nv4hf8J14K0po/h/4tuXa1EaAR6ZeNud7TCgBEwGaIf3VZefLJr/glx+2jH8CPHo+DHxF1nyfAXjK8X7NcTMqx6RqT7UWZmIysUgVUck7UIVztAcmqqqqqqqqv+Clv7Vsv7SHxwn8O+HrlG8F+AJrjT9IMUhZb2bcBcXZ7EOyBUx/Ainqxr/glV+yxF4l1uX9pPxxpaS6bosrW/haCZNyzXgyJbrBPSIfKmQRvZmGGiBqqqqrGeBWBjANVVVjPAoAAYFVSgHmqqqqqrGeBQGBiqqqqqrGeBWABgGqmligieeeVI441LO7nAUDqSewxXxL/bZ/ZZ+E6SJ4p+Mugz3ke5fsOkSnU7jeFJCMluH8snGMybRkjJGa+I3/AAWQ+G+mefafCv4Ua7r0q7kjutZu49Pi3fMBIqJ5rumQh2ny2IYg7SOfiR/wVJ/au8dS3EPh/wAQ6T4LsJWYJBouno0gTsDNP5j7sYyybMnoADivGvxM+IvxIvPt/wAQfHniDxJcBiyvqupS3RQ/7O9jtGOMDAArwz8JPip4205tX8HfDbxRrlinW507SJ7iPOdpAZFIJz2HPB9DUfwP+M8urpoCfCTxkNSdQwtG0K5WTaf4ipTIX3PFaX+xB+1PrFu1zafCO8jRXKEXWoWls2QAeFllViORzjHXng14h/ZG/aV8MNIupfBrxFMYiob+z4Bf5LAEY+zl9w55IzjocUV17wrq6ki/0jVLNgy5D288JIyCOjKcEH6Gv2fv+CoP7QHwilg0jx/fN8R/DwbDx6vct9viHcx3eGZj7Sh/QFa+Af7Rfwq/aV8Hnxj8Ltda6jt2SO/sLqPybvTpWXcI5o8nBxnDKWRirbWbBxVVVUBnmqqqqqqqAzzVVVVVVVYzwKAAGBVVVYzwKAAHWqqqrxJ498D+DYXm8WeL9H0gIM7by9SJjwSAFJyxIBwACTjivFP7bPwI8PK8em6vqPiCeMlfL06xYDcDj78uxSPdSRjkZrxR/wAFC9cmVovBfw6sLUgnbPql49xuGR1jjEeONw++eoPbB8T/ALWHx88Uq8Nx8QLnT4H6RaZEloU4AOHQCTnGeW6njFapq+ra5c/bda1S71C4xjzrqdpXxknGWJPUk/ia8L/BH4ueNGi/4Rv4da7dRTf6u4ezaGA8gf62Tancd+nPQE14W/YL+LusGOTxJqeh6BCQC6PObmZcjkBYxsOOh+fr0yOa8K/8E/vh1pu2Xxd4v1rW5Vz8ltGllE3Xqvzt0K9HHI7g4Hhf9nf4I+DXEug/DbR1lU5SW7iN5Ih9VaYuy/gagggtoI7a2hSKKJAkcaKFVFAwAAOgA4xVVVAZ6UAAOKqqkeOKNpJHVUQEszHAAHUmvFH7QfwT8HKw134l6GsiEh4bW4+1yocgYMcO9h94dR6noDjxZ/wUF8C2G+HwZ4L1fWJF4Et7KlnGT6jG9iPYhfw614s/bn+NviDfDocukeHYTwps7QSyY92mLjPuFWvE/wASPiB41Zz4s8a61qqydYrq9d4xyDgITtAyAcAYzzXhj4PeNfEoSc2A061YAia9ymR/srjceORwAfWvDvwG8IaQFm1eSfV5x18393F+CKc/mTVlp9hpluLTTbG3tYVORFBEI1B+g4rRfht4014B7PQ5ooj/AMtbkeSvTtuwT+ANaR8AHJV9f19QO8VnHn64dvb/AGa0n4S+BdK2sNI+2Srj57tzJnGOq8L29O57cVZ6fYadD9n06xt7WMdEhiCD8hVSyxQxmSWRUVerMcAVceKPDdqds2uWQYEgqswYgj1AzipfiN4PjTcmpvKf7q27g/qBWlanaa1p8WpWLMYZs7Cy7TwSDx9QarUvE2haRdpYalqKQTvH5iq6tjbkjOcY6g8Vaa3o1/gWWrWkxOMKkyk89OM5B9qqqqqqqqqqqqqqmVXUowBBGCCOta38M/BOvKxudDhglb/ltajyWznJPy8E/UGtd+AN1GGl8N60kwGSIbtdrew3rwT+AH9Nf8IeJPDMmzWtInt1zgS7d0bfRxkZ9s5rVPhx4d8f3kGm6hokNxd3MixRTKCkgY8D5l5wOODkcdK8dfsRfFLwdazaxBqvh/UNNjmC5ju2SZELAbmWRFUkZ6KxJxwM4FeLvBut+CtT/s3WYANwJhmTlJlHdT/MHkfiK+Hn7Q3xe+GAht/DHjG6awiwBp97/pNvtGPlCPnYMAD5Cp96+GH7evgrXlTTvibpEvh28JAF5ahri1fnqQP3kfXphxwTuHSvD3ivwv4ushqHhXxFpur22AfNsrpJlGcgZKk45UjB7g+hq51LT7OeG1ur2GKW4z5SO4UvjGcfmKqqqqqqr/goF4ytNY+IOg+C7SdJW8O2Dy3IXrFNcFTsPv5ccTfRxX/BO7Q7h9Q8aeJGYrBHDa2iApw7MZHbnPG0KvGP4x6VVVVEAjFFQelFT2NEdqqqqqqqqqqqogEYrV/FGg6JlL7UYhKv/LJDuf8A75HT8a1b4rTSAxaLp4jHaW4OT/3yOB+Zr44ftXfCH9nn4eW3jn4i+II1ub+AnTtGtGV73UJQPmWKMkHarcNI2EXIy2SoP7Uf7efxs/afup9J1LUf+EZ8GeYTb+GtMmPlsATg3EuA1w+CAc7UyoKopzmq/Z2/4Jo/tD/Hb7LrmvaX/wAK+8KzYb+09dt2W4mQ94LTIkfggguY0YHKuelfs7fsD/s7/s4/ZdX0Dwx/wkPiq3w3/CQ66q3FxG47wJgR2+DnBRQ+DhnaqqqqivpVVVUQCMURjiqqqIBGKIxxVVVVVVVMM8iqqqqqqqYZGRVeP/gr8IfipE0XxG+GfhrxGxXaJtR0yKaVBjHySEb0OOMqQa8ef8Epf2SvF+ZNA0fxH4Nm650bVmkRm+bql0JgASwyF2/cAGOc+OP+CMPieDzJ/hr8btLvt24x22uaVJabOm0GaFpd2fmyfLXGBwc8eOP+CXv7X/g3zJbDwTpfim3i3bp9D1aJ+BjBEcxikbOTgKhPynIHGfG/wB+OPw1imufHvwi8X6FawMVe7vdGnjt8gc4m27GHuGIqKWWCVJ4JWjkjYMjoSpUjoQR0NfsA/tWxftN/CCODxHfI3jvwksdnr6FNhugQRDeKBxiUKdwGMSK/yqpTJAIxTKOlEdqqqqqqiARiiMcZqqqqqqIBGKI7VVVVVVzbW95C1vcwpLG4wyuMg14o8GS6WHv9MDS2gGXUnLRf4r79u/rVVVVbzzWsyXFvIUljbKMOoIrQdZh1zT0uo8LIvyyp/dbv+HpVYBGKIxxmqqqqqr/gqt4eTWv2S73UmsWuDoOv2F6sgYj7OWZrfecHkEXGzByMuD1AI/4J06+vh79sf4ezTXc8UF7LeWUqxE/vTLZzJGrAdV80xnngFQe1VVVVVVVVVVVVVVVVftn/APBP7wf+0Zb3PjvwGtn4d+I6qC924ZbbV1VcLHcKudrgAATKC2BhgwC7PE3hb4gfB/xtJoPibStW8LeKdAulcRyBre4tpUIaOWNxz1AZJEOCMMpIINfsBf8ABT6x+JQ074NftIa3Fa+Mri4W30bxE8aQwatuztin2gJFPnCqwAWTKjh/v1VVVVVVVVVVVVVVVVVVVVVVVVVVVV+3X8RE+F37IvxR8ULdPBczaDLpto8chSRZ7wi1RkIIIZTMHBHTbntXwd8DP8Tfiz4N+HixysviLXbOwlMasxSKWZVkf5eQFQsxPYAntUUUUESQQRpHHGoVEVcBQOAAB0GKqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqvix8L/Cfxm+Hmt/DXxrYJdaXrVs0T5XLQv1SVD2dGCsp9QO1fHT4LeMP2f/idrHwv8a2+LzTZA1vdIhEV9btzFPEe6sv/AHywZThlIH/BLf8AbNl+PPw8Pwe+IutJN488F2qLbTzyEzavpygKkzE/flj+VHbktlHJJZjVVVVVVVV/wVE/ag/4UD8Abjwb4cu9ni/4jRz6ZYskm17O02gXVyMc5COI1IIIeUOD8hB+BHwf8QfHj4r+Hfhf4cil83V7tVurhIyws7ZTmadvQIgJ56nA6kV4B8C+Gvhl4K0T4f8Ag7T0stG0CyjtLOEAZ2oMbmIA3OxyzN1ZmLHkmqqqqlHeqqqoDPNVWM8CsDGBVVVVVUADzVVVVVVXjn4lfDz4X6Sdc+IvjfRPDdltcpNqd9HbiXaASEDEF25HyqCSSABkivir/wAFYP2bPAssmn+B4tb8e30bMpfT7f7LaKy8EGabDEE9GRHU4JzjGfiX/wAFc/2h/FTyW/w70Lw54ItDu8uRIP7RulyTjMkw8o4XaP8AVDJBPQgD4h/HL4x/FpgPiV8T/EviOJW3R21/qMkkERwoykOdiE7VztUZIyeea8F/DL4h/Ea5Fn4E8E61rsgJDGxsnlSMjGdzgbVA3LyxH3h6ivBP/BOL9obxP5c3iKHQ/CsDcsNQvhNLj2SAOM+zMv58V4F/4JgfDLSUWf4g+Otc8Q3A5ENjGlhByCCGB8x2wSCCGXpyMHFeC/2Yf2fvh9tbwt8JvD8UyZ2XF3b/AG2ZM7s7ZZy7jIdgcHkYHQACqqviD8LPh18VdIfQ/iF4P03XLVlwv2mH95F7xyrh425PzIwPJ55r9pD/AIJ26x4Lsbzxp8Erq813SbZDLc6Lc4e8gUZLGFlAEygfwY34/vmvhB8X/HvwK8e6d8R/hvrTafq+ntgggtFdREjfBMmRvjYAZX2BBDAMP2WP2mPBv7UnwvtPHXhxorLVYMQa5opnDy6Zc8/KehMbhSyPjDL6MrKtVVAZ4zWBjAqqqqqqoDNVVVVVVVjPAoAAYFVVVjPAoAAYFVXij4wfCzwW0sXif4gaFYzw532z3qNOMEg/ulJfqpHTqMV4s/bw+EGjb4fDVhrPiKYfceOD7NC31aTDj/vivFX7f/xE1LdF4R8I6Lokbfx3Lveyr93o3yL2bqh4YdCMnxR+0P8AG3xlGYdd+JOsNEww0VpILNHHoyQhAfxFQwXupXawW0M11dTt8qIpd5GPoBkk14W/Zh+O3i9Y5dN+HOo20LkZm1HbZBVJ+9iUqxHf5QTjpmvCv/BPjxlfRpN4y8d6VpWQG8mxtnvHH3flJYxgHlgSNwGBjOePDP7CvwS0VY21wa14glH3xdXhgjJwRwsIRgOh+8eR1xxXhj4XfDfwWq/8Ir4G0TTJFORNBYospIJIJkxuONxxk8ZwOKqqqqrGeBQUdzWBjArxJ8RfAHg4SDxV410PSXiBLR3d/HFJwu7AQncTjBAAycjHWvFH7anwD8OKy2fiG+16ZCQ0Wl2LtjkdHl2IepPDHoe+AfE3/BQ6MK8Pg74bMTj5LjU7/GDk9Yo154wfvjuPevE/7a3x68Rq8Vprun6FFJ1TTLFVIGAMBpS7jkZyDnJ644rxN4/8ceM5Xm8WeMNY1cuclby9klUckgBScKAScAAAZ4xXhvwN408ZOY/CXhLWNZKnDfYbGScJjGdxUEDG5c56ZHrXg79iT44+KGSXVtN0/wAN2xwTJqV0C5XIztji3tuxk4bb06im/ZL+FngcrF408eap4k1JDl7DSIktI0cbcpJI3mHbkMDgBuRwuOdP8BeA9EuYrnQPCVlZPD/q3ZnnkDYALb5GYhjtH3cAEnAXJFaJ4a13xJObbQ9Mmu3X7xUYVfqxwB+Jrw/8BCypP4n1YgkZNvaDp7F2H06D8e9aH4L8LeHAp0jRbaKRRgTMu+T/AL7bJ/WqrVfEmh6LldR1KGKQDPlA7n9vlGTWp/Fm2QmPR9MeXGR5k7bR+CjJP5itQ+IHinUMr/aP2dDn5LdAmOvQ/e7+vpVzd3d7J5t5dSzv/elcsfzNW2l6negGy065n3A48qFmzjr0FQ+CfFc+wR6HcDeON4CY+uSMfjXgrT7/AErw3badqVr5E8DOCu8NkFiwPBI71XxF0TW7/wAQvdWelXlxCIkRXjiLjIGTjGeOamt57dhHPC8bEZCupU49easfEGuabgWOrXMQUYCCQleP9k8VZfFbXoIxHeW1rdEDhypRiffHH6CrX4u6c/F9o9xF6eVIJP57asfHvhS/IRNVSFv7s6mP9Tx+tQzwXMYlt5klQ9GRgwP4iqqqqqqqqqqqqpI45Y2ikRXVgQysMgg9QRVp8O/CWn69D4i07S0tbqENtSLiMlhjOzoCATjGK+OOja9qWgwXmnSh7GwZpLuBeGPTD+4UZ47Zzz28Larp2maokeuaba6jpF3+5v7O6gE0csR4JKMCCQORke3c18Sf2HvhP4ySXUPBrz+FNRdSy/ZD51o7HnLRMcqOwEbKB6GviZ+yl8ZPhrcu7+Gptf0wbimoaPE9yoUAkmRAN8eAOSw2+jGra61DSbwXFlc3FndwEgPE7RyRnBBGRgjgkGvhh4gj1/wVpbv44ufFN5BbgXV5d3jXEyyMS5Ri5LKAWIAbsB9a0jxt4j0VVittQM0K4xFOPMXjsM8gewIqT41aZY2az6j4d1SVhnzBYIk+0AZztLKxyRjChjyPrWmftUfAXU5Gtn8fwaddRbRNb6lazWbxM38LeYgGRjBwSB68itI+Ifw/8Qhf7A8deHtS3yiJfsepwzZkOMINrH5uRx15HrVVVeKPHfgnwVAbjxd4t0jR12blF7eJCzjBPyqxyxO1sAAk4OK+Mn7dnhfSrKfRfg/G+r6lIhQarcQGO2tyQRuRHAaRgezKF6HLDis+IfG3iMBpLrVta1m7xukcvLcTSN3J6kk9TX7P3wrh+D3wv0zwk08Nxfvm71GeEfJJcSAbtp/iVQFQMeSEBwM4FVVVVUQCMUVBHBoggVVVVVVVVVdXdpZQm4vLmKGNeryMFH61rPxQ0ay3Q6VE99KMgN9xM/U8n8vxrV/HXiPVyUe+NtEQR5VvlBj3PU/iatLG81CUQWNrLcSHokaFj+lftN/FLRf2Xfhi/jvxs0T6lfyNbaFokcoM+oXO0nnH3IlABeTnaCowWZVb45fGvxT8ffHT+PfFtpYWl39ljtYrexEgiiiQsVADuxzliTyASScDNV+zh/wTx/aC/aJisvEEGjJ4S8I3arImu60jRieIjIe3hHzzAg5DfKh/v1+zR/wT/wDgL+zWINbsNI/4SrxdHtb+39agSSS3cd7aPBW36nlcvg4LkVVVVVVUVBHFEdqqqogEYojHFVVUQCMURjiqqqqqqqYdxVVVVVVVMMdKqqrUNPsNWsLnStUsre8sryF4bi2uIhJHNGwKsjqchlIJBBGCDX/BQb9i27/Zs8af8Jv4Isml+HXia5f7Esccjf2NPwTayOc/KQSY2JywVgRlMt+wL+0L4T/Zw+Pln4o8a6W0mj63anSbzUY7iVH0yOWRCZjGh2zRgopdGViAoZPmUBrC/sNWsLbVNKvre8sryFZra5t5BJHNGwBV0ZSQykEEEcEEGiARjNMuRgdRVVVVVVTAEcVVVVVVUwyMiqqqqqogMNpAII5BFeM/DA0qb+0LCPFpKfmQD/VN/QH/AOt6VVVVeHtal0PUFuFJaFxtmQHqPX6jr/8ArqGaK4hSeCQPHIoZWHcGqYZGQaqqqqqv25/C7eL/ANkf4n6Slu0xt9EbUNqyBCPskiXJbJPQeTkjqQMDk1+zj4mbwb+0B8N/E4lnSPT/ABVp0k4gALtD9oQSqoJAyyFlwSOvUVVVVVVVVVVVVVVVVVX7Vn7IPw4/ao8MC21+L+y/FOmwOmja/bpmW3J5Ecq/8tYS3JQ8jLFSpYk/GT4KfEf4C+M7jwL8S/D8um6hFue3lHzQ3sIYqJoX6PGxU4PBHQgEED/gnZ/wUyvbS90/4E/tL+KGuLS4ZIPD/iy/ly1u3AW2vJD95DwFmY5UnDkqQyQzQ3EKT28qSxyKGR0bIYEZBBHUEVVVVVVVVVVVVVVVVVVVVVVVVVVVVf8ABanx2NE/Z98JeALe6SO58T+JhPJGWQtLbWsLlwFPzYEs1uSy4xgAn5sH/gl74JHi/wDa40PUpLZJ4fCul3uqyo6qyj939nRiG7rJcxsMZIYA9iRVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX/BQ/9lBf2h/hYfFfhDS0k8e+DYZJ9OEURMupWv3pbP5eWY43xgg4cFRtErNXwd+LHjD4G/ErQfil4E1F7XV9BulmjAbCzx9JIXHdJELIw9GPfmvgd8YfCfx9+FHhv4teC592neILNZTAzZe0mHyzW78D545A6EjglcjIIJqqqqqqmmht4XnnlSOOJSzu7bQoAySSegxX7dX7Rkv7TX7RXiDxrY3byeG9Mb+y/DiEnAsoWIEgHYyuXlPcbwOwr/gk5+z5/wAIZ8NtR+PPiGx2av41za6T5i4aHTYn+Zh3HnTLkg8FYYmBw1VVVVYzwKwMYFVVVjPArAAwDVUo4zVVVVVVjPArAxgVVVVVqmqaXomnXGr61qVrYWNpGZLi6upliihQdWZ2ICgepNfG3/gqL+zd8LftmleDb66+Imu2+5Eh0YhLISDGA94w2FCD9+FZhXxi/wCCpf7SvxLiuNK8I3eneANKlYbV0ZC94VA+61zJkg55zGsZwAOmc65r/ibxnrMmseJNb1PXdWvX/eXV9cyXU8zk93clmJJ7nqa+Hn7Gv7RnxKSC60n4c3mmWE20i+1lhYxhWBKuFkxI6nHVEbqOxzXw/wD+CW1moS4+KnxNmkJX57Pw/bhMHb1E8wORuzx5QyB2zgeAf2O/2c/h00Nzo/w00+/vYX3reavm+kDA5DASkqpHGCqjGAevNW9tb2dvFaWkEcMECBIoo0CqigYCqBwAAAMVVVVVVV+3/wDsnaZpNndfHn4b6X9nRpt3iTT7eP5AXP8Ax9ooHy/McSDplg3Hzk/sYftMar+y/wDGjTfFj3M7+F9VZLHxNZISRLaM3+tC4OZISfMXGCcMmQJGq1urW/tYb2xuYri3uI1khmicOkiMMqysOCCCCCODVVjPAoAAYFVVVVVVAZ71gYwKqqqqqqxngUAAMCqqqAzwK8V/HH4QeBw6eJfiHottNH9+3juPtEy/WKLc/wCleLP2+vhlpW+Hwn4b1nXpl+68u2zhb6M25/zSvFH7efxc1dZYPDelaFoMTZ8uRIGuZ04I+9Idh6g/6vqB2yK8XfGb4q+O/MTxV4/1m+hlzvtvtJigOd2f3KYTozD7vQ46cVovh3X/ABLdiw8OaHqGq3J6QWVq87/98oCa8Jfsh/Hnxb5Uo8H/ANjW0mP3+rzrbbc7esfMo4bJ+TsR1GK8Lf8ABPO5YxzeNviNEgAG+30uzLZOOcSyEYwcj7nPXjpXhn9jP4B+HFja48MXOtzx9JtTvXcngg5RCkZ6914OCMVoPhfwz4WtjZeGPDumaRbnrDYWaW6nkn7qADqzH8T61VVVVVVPPBbQSXNzMkUUSF5JJGCqigZJJPQAd68TftF/A7wizR618TdF8xGKPFZym9dGBwQywBypBHIIGK8T/wDBQH4baaWh8KeEtb1qRQcPcMlnExwMYPztjkg5QYI4yDmvFf7e/wAWtYja38MaPoegRsDtlWJrqZTyOGc7O6nlOq+hxXin47/GPxo5bxD8R9bnQnJhhuTbRZ9fLi2p+nFWdje6lcpZadaTXVxJnZDDGXdsDJwo5PAJrwx+zR8dvF2G0r4aatDH/wA9NQQWK445HnlNw+YdM55xnBx4Y/4J/fEvUmjk8U+K9C0aFvvLAJLyZOSOVwidADw/ftXhn9gD4X6aqv4n8Ta7rUwQArEUtIiccnaAzdeQN/HfPWvC37O3wR8GuJdB+GujrKpykt3EbyRD6q0xdl/A1qOpaL4X0n7TezwWNlaoERQAoAA4RFHfA4UeleOPjDquvmTTtAMmn2BJBcHEsw9yPuj2H51oug6v4ivV0/RrGS5mPUKvCD1Y9FHua8I/A/TbDbeeK5lv5xgrbRErEp9zwW7eg+tWtnaWMC2tlbRW8KDCxxIEVfoBwKqqvbQX1lPZmRoxNGyb1JBXIxkYptN1B7+XT0tZZrqJ2V440LtuBweB71pfww8RXyrJdmGyQ9RK25/wUf1IrTvhXoFsA1/PcXrjqC3lr+Q5/WrLw5oOnAfYtItI2AwG8oFv++jz2qprq1tyFnuYoyRwHcLn86/tLTf+ghbf9/V/xqOWKaMSQyK6noynINVLDDOnlzRJIv8Addcj8q1HwF4V1EZbS44H7Nb/ALvH4Djt6Vd/CGBmZrHXHRcHaksAY57ZYEe3ar34V+IrcFrSa0ugBwFcox49CMfrWoeG9e0vJv8ASbmJVzl/LLKOv8QyOx718KrN59flugzCO2gJOD1ZjgZ/DdVa9rtj4dsRqF+JDGZAgEYBYk+gJHYGrLx74UvgAmrRxNnG2dTHj8Tx+vFW+pabeBfseoW0+/O3y5VbOOuMH2qqqqqqqqqqqrm3hu7eS1uEDxTIUdT/ABKRgj8q8R6PJoGu32jS5zaTsilurLn5W/FcH8a+GOtpr3gnTpzP5k1tH9nnyckMnAz7ldp/Gq8U/DnwD43jKeLvBujauSMCS7skkkX/AHXI3L+BFan+xv8ABmbUP7W8NRa74UvCc+boupvGQd2TgSCQAHgbQAAAMAVY/Avxtoqxww/EK1123XCldR0/7POBg5PnRMVY528GPJ5y2a1zwlrvh0K2pWqmMj/XQkvHn0zgEfiBmvid8IPDvxJsmkkRLHWEA8nUEiy2B/C4yNy4/EcYPUHX9A1bwxq1xoeuWT2t5attkjcfkQe4I5BHBFadr+u6PG8Wka3f2SSHLrb3LxBj6kKRmrX4xfFyyt47Sy+Kni+CCIYjii1y5RVHoAHwK/4XZ8Zv+iu+NP8Awf3P/wAXWpfFH4m6z5f9r/EbxPfeTny/tOrzy7M4zjc5xnA/IVDBe6ndrBbQzXV1O3yoil3kY+gGSTXw/wD2Tvjb8QJlaPwnNoVlwWvNaVrVQCeqoR5jcZPC49xkZtfC0Xg1P+EdEVsJrJVhmlghEYmZQBvwPXGcEkjNeB75dQ8K6fICN0UXksPTZ8o/QA/jVVVVVVVUVBHFFT2qqqqqq1jxToWhqRf36CUDiFDuc/gOn44rWfirfT7odDtFtk5Almw7n329B+tX2pX+py+fqF5LcP2Mjk4+npWj+CfEWtASW9iYYT0luP3a/UdyPcA1pHwt0e02y6rcSX0gH3B+7QflyfzH0qzsLLT4BbWFrFBGo4WNAtf8FAPj7c/H/wDaU8SanZ6i0/hvwxM2i6BGHVo/IgYrJMpXhhNL5kgY/NsZFJwoA/Z9/ZT+Nf7TWtDTPhh4WaSwhm8q81y+3Q6fZNtBIkmCnLYKnYgZ8MDtxzX7NP8AwTM+BPwIEHiHxfZxfEPxYm1lvtXtF+yWbjnMFqSyg5wd8hdgVBUpyDVVVVVVVUQCOtEY4qqqiARiiMd6qqogEYojHFVVVVVVUwx0NVVVVVVEAjFEY4zVVVfEDwF4V+KPgvV/h9430tNR0PXLVre9tmYruU8ghgQVYEBgw5BAI6V+1z+yx4v/AGVviZN4X1eKW68OapJNN4a1ZiGF7bKw+VyAAJowyCRcDBYEfKyk/wDBN39u+XwDf6d+z58Y9dt4/Cdwxj8P6xeSFf7KlY5FvI2MeQ7E7WYjyycE7D8lMMjIpgB0qqqqqqph6VVVVVVEAjFEY4zVVVVVVPDFcQtDNGro6kMrDIIrxX4YbQ5hc2uXs5WwueTGf7p/oaqqqvBHiIW0g0W8kAikY+QzH7rH+H6H+Z96rAIxRGOM1VVVVXxL8Lp44+HHirwU9ulwuv6JeaeYXkMYkE0Dx7SwIKg7sZHIq1urmxuYb2xuZbe4t5Fkhmico8bqchlI5BBAII6V4S1+LxZ4U0XxTBEkcesafBeIiS+YFEsauAGAG4YbrgZ61VVVVVVVVVVVVVVVVX7Q37OPw1/aY8Ef8IX8Q7CUNbyGXTtTtCqXWny8ZaJ2BGGAAZCCrDGRkKR+0h+zf8QP2ZPiBN4I8bWvnW026XSNXhjIt9Ttwcb0znawyA8ZOUJHUFWb/gnF/wAFHZfhTLpvwF+PWtvL4KkZYNC125cltCY8LDMx62p6Bj/qf+uf+rhmhuIUnt5UljkUMjo2QwIyCCOoIqqqqqqqqqqqqqqqqqqqqqqqqqqqv+C3fi+S++L3w48AGSUx6N4bn1NVMahAbu5MRIYfMSfsQyDwMLjktX/BGvweHvviZ4+uIGBhhsNMtJdy4bcZZZ1I+8CNluc8A5PUj5aqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr/gp5+yz/AMKq+I//AAufwVpEsfhTxpOz6gsYzHYao2WkHqqzAGQA8bvMAwNqj/glH+1mPgt8Vj8FvF99L/wiPxDvIorVnlxHp+qnCRy4PAEw2RORzlYieEqqqqqqr/gqT+0CnwS/Zj1Pw5pVw6+IviKz6JYbGKmG3Zc3c2RjgRfuxg5DTIcEA1+zt8GNX/aA+Mnhn4V6SZYk1a7Bv7qNcm0s0G+eXnjKxq20HguVX+IVoOhaR4Y0LTvDPh+wisdL0m0itLG1iGEghiQJGi+yqoA+lVVVVKO9VVVSjPNVQGeBWBjAqqqqqqUDGaqqqvG3jvwX8N/D1x4r8feKtL8P6Pa8SXmo3KwRhiDhQWPzMcHCjJJ4AJr48/8ABXzwxpEt3oH7PXg5telVWRPEGth7e2DdmithiWRcd5DEQRypFfF/9oX4zfHrUEv/AIq/EDU9cWCRntrOSQR2tsxzkxwJiNDg43BdxGAScV8Lf2a/jV8ZGWXwN4Fvp7AnB1K6X7NajjPEr4DHGOF3EZHHIr4Yf8EvvD1jsvvi/wCPLjU5di/8S7Q08iJH77ppAXkXtwkZ757V8O/gT8HvhOif8K++Hmj6TOmcXaw+bc4IwQZ5C0hGOxbHJ9TVVVVVVVVVajp9jq+n3Olalax3NpeQvDcQSLlZI3BVlI7ggkH61+0j8IJPgb8Ytf8Ah/E80unQSC40qeUfNLaSjdHk4G4rkxswABZGxX/BK7463XxW/Z6PgLXbzz9a+G08emhmJLPp7qWtGbsNoSWEAdFgUnk1VAYFVVVVVVAZ4BrAAwKqqqqqqxngUAAMCqrxb8ZfhV4F8xPFXj/RbKeHO+2+0iScY3Z/dJl+qsPu9RjrxXjH9vT4YaMzweENC1bxHKudsjAWcLYJAwzgvzjPKdCO+QPFf7d3xd1rfD4asNF8PRH7jxwG5mX6tJlD/wB8CvFfxZ+JnjkOnivx1rOoxP8Aet5LthD+ESkIPwFaF4X8S+Krk2Xhjw7qer3C9YrCze4ccE9EBPRWP4H0rwr+xh8efE217nw/ZaDC2dsuq3ip0znKR73HK45UfeB6ZI8K/wDBPbRotsvjf4h3tznO6DSrRYNvXpJJvz/CfuDHI5zkeFP2TPgN4S2SReB4NVuE6zatI11u+sbHy/8Ax2tN0rTNHtE0/R9OtbK2j+5BbQrEi/RVAAoAAcVVVVVVa/4v8J+FIlm8U+KNI0eN/uvqF7Hbg/QuRXif9sz4B+HFdLfxRc63PH1h0yydyeAeHfZGevZuD1xXif8A4KGDmHwZ8OPpPql79ePKjH+6c7/UY714o/bO+PfiRWitvElnokLkkx6XZIncEAO+9xjHZhnJByK8ReNfGPi+Qy+KvFerauxbd/p17JNgjIBAYnGASBjpmvDXgDxx4zlSHwn4Q1jVy5wGs7KSVRyASWAwoBIBJIAzzivDP7Enx68Qqsl7o2l6DG6BkbU79ckEZGVhEjKe2GAIPXFeGP8AgnjpEarL40+I13cEn5oNLs1h2gE5xJIXzkbf4Bjnr1rwr+yF8A/CrpcL4LXVrhCCJdVne5BxtODGSIzyueU7kdDitF8O+H/DVoNP8OaFp+lWw6QWVqkCcf7KACqqvGHjbRfBlj9o1GXfcSA+RbIfnkP9F9W7fpXinxdrXi+/N9q1xkLxFAnCRD0A/qeTXgH4Taj4o8rVNXL2WlnBU4xJOP8AZHZf9o/gD1GjaHpPh+xTTtHso7eFB0UcsfVj1J9zVMVVckgADkntXiH4j6PpObfTiL+5GRhGwiH3bv8AQfmK1vxXrmvvm+vSIgcrBF8iL6cd/qcmvCGsnXtAtr6Q5mA8ub/fXgn8eD+NUsNrbebOkUURf5pXChc47k/41qfxA8LaYWja/wDtMq9Y7dd/6/d7etaj8W7t8ppWlRRjHDzsXP5DGPzNfD7xNf8AiO2vTqUqPNBKuAqbQqsOB+an8/pVfF+FVn0ucNy6Srj0wVP/ALNVQXFxbP5ttPJE+MbkYqfzFWPjzxVY4CatJMoH3ZwJM/ief1qL4q+JY02vDYSnPV4mBH5MKT4uaoEAk0q0Z8clWYD8ua8OfEg69qdvpL6MY5JsgOs+4cAknBAwMA9zVXl5aafbPeXtwkMMY+Z3OAMnA/UirXxF4QLyT22r6ZHJJgSMZFjZ8ZIznBPU/mah1HTrjYLe/t5d4+TZKDu+mDXxF0LW9fs7S30iFZkidnlQyBSTgBcZIHQtV54Y8Q2BxdaNdqB/EIiy/mMinRkYxupVlOCpGCDVKzIyvG5VlOVIOCD6iodc1q3z9n1i9j3ddlwy5/I1a+N/F8JWK31meRiAqh0WQn0+8DzSqwQKzbiBycYya2kjgVa+JdAvbh7O31e2M0blDGz7SWGQcA9eh5GRW0+orb71t962j1raPWto9a2j1raPWto9a2j1r43+Dry41uy1vR9OmuZL6MxTJBEXYumNrEDJOVIH/AK+DPhXxb4akvxrenm1s7uNGjV5VLb1P90E4yCeuOgrafUUVPaipx0ojipoYbiJoJ4kljdSGR1yGHoQe1ePvCK+HbxLywVvsFySFB58p/7ufTHI/H0zXw71K1nuZfC2rwQXVhfjcsNwgdPMXkcHjnHfuBV/8JPhTqtwbvU/hj4TvJyADLPotvI2B0GShNSfs0fAWR2kb4W6HliSQsJUfgAcD6V/wzN8BP8Aolui/wDftv8AGtP/AGdvgZpkxntvhT4bdmQqRcWK3C4yDwsm4A8dcZ/M1ovhbwx4aiEHhzw7pelRhNgSys0gG0dsIBx7VXxT082viNb1V+W8gVif9pflI/IL+dfCTUQ9rfaQ7DMbiaME8kEYb8BhfzqqqqqqqqqiAaK+hojtVVI8caGSR1VVHLMcAVrPxJ8P6Zuis3OoTLkYiOEB/wB/p+Wa1rx/4h1hWhW5FpAwwY4PlJ+rdfw6Va2d5qM4gs7eW4mbnZGhZj6nitH+Fmq3YEurXKWaf881+d/8B+ZrR/B3h7RNj2dgrzp0nm+d8+oPQH6AVVftl/EnxF8NfgDr8ngWxur/AMY+J9ug+GrO0hM0819dBlBiQcs6RiWUDB/1WSCAa/ZR/wCCSA/0Hx1+1Hd/3Zrfwhp0/wDukC8nX/gQMUR/unzOqV4c8N+H/B+hWPhfwpotlpOkabCIbOxs4FhigQdFVFwAKoqCOKI7VVVVVVVVEAjFEY4qqqiARiiMdDVVVEAjGaIxxVVVVVVRAIxRGOKqqqqqqIBGAaI7VVVXx6+Avw8/aM+Hl78OviJpnn2s37yzvIgFuNPuACFnhYg7WGcY6MCVYEEiv2iv2dviB+zV8RL7wH4306drdZn/ALK1cWzR2+qwDBEsTHIJwyb0BJRjtJ7n9hf/AIKVTfDy10v4OftB6jLceF7aMw6X4lcPNPpqgDy4J1UFpIRgqrAFkyoOUA2aXqul65p1vq+i6la6hY3cYkt7q1mWWKZD0ZHUkMD6g4plHSiO1VVVVVRAIxRHaqqqqqpgCOKqqqqqqurWC7ge1uYlkikXDIw6ivEfh+fQbzySxkgkGYZCOo9D7j/CqqqBIIKnBHSvCOv/ANs2PkXMgN3bjD56uvZv6H3+tUwyMg1VVVVVfHzwtL4I+OHxA8ISxso0jxNqFvGWt/I3xrcOEcJ/CrJtYDkYIwSOa/Yd8VDxn+yT8L9XEm77PoSadny9n/Hm7WuMe3kYz3696qqqqqqqqqqqqqqqqqvjt8Cfh9+0R8Prz4efELTPOtpv3lndxAC40+4AIWeFiDtYZ6dGBKsCCRX7VP7K3jz9lnx43hzxEjahoOoM76HrkcRWK+iB5BHOyVQQHjJ4yCCVKsf+CcX/AAUdl+FMum/AX49a28vgqRlg0LXblyW0JjwsMzHranoGP+p/65/6uGaG4hSe3lSWORQyOjZDAjIII6giqqqqqqqqqqqqqqqqqqqqqqqqqqv+CsfiKXXP22fFmmSQsi+H9N0yxjZpS4cNaR3GQMfIM3BG3nkE5+bA/wCCR3hhdI/Zq1bxBJHAZ9e8U3MiyISX8mKGCNVfI4IdZSAM8OOecCqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq+MPwo8J/G/wCG2u/C/wAaWzSaZrlsYmkjOJLaQENHNGezo4VxnIJXBBBIPxV+Gnif4O/ETXvhl4xtfI1bQLswTAfdkUgNHIv+y8bI6+zCv+CcX7VLftO/AeBfEupNc+OfBbR6d4haUjfcgqTb3fHaVFYEnBMkUvGME1VVVVX/AAVk+NUvxS/anv8AwbY3yTaJ8ObVdJtlRSAbpgJLtjnncJGER7fuBgdSf+CQPwWgtPD3ir496tbxNc6hP/YmjlkO+KGMLJcOD0w7tEo7jyX7NVVVVWM8CsADAqqqsZ4FYAGAaqgOKqqqqqgM8CsDGBVVWoX9hpVhc6pql9b2dnZwvNcXFxII44Y0BLO7EgKoAJJPAAzX7S//AAVY+G/gGK58L/AS2g8ba/teNtWkZo9Osn7MvG65OeylU6HeelfF344/Fb48eIV8TfFXxpf69dxbxbRzMEgtFbG5YYVASMHaudoGcDOTzXwT/Ym+Nfxn+z6p/ZH/AAjHh6XDf2pq8bR+YnrDD9+TjoflQ/3q+D/7B3wI+FTR6lqejt4y1lVH+la7GksMbY58u3x5YGRkFw7KejCo4o4Y1hhRURFCoijAUDoAOwqqqqqqqqqqqv8AgqD8N11Dwn4W+KtlAvnaTdPpl8wJ3NDMN8RPbaro49czDr2/4JUfEpvBH7VNl4WuLpI7HxxpVzpriQgKJkX7REclhhiYWjX72TLjGTkUBnmqqqqqqsZ4FAADFVVVVVVYzwK8TeOfBXgqAz+LfFmk6QuzcovbxImcYJ+VWOWJ2tgAEnBxXi79uP4L6ArRaBJqniOcHA+yWphi685eXaemeinJx2Oa8X/t8/EbVlMPg/wxpOgRn/lpM7Xsy8DoSETrk8oeo9Dnxf8AG74tePI5YPFPxA1i8t5gRJapP5EDg9QYo9qH8q0Xw9r/AIluxYeHdD1DVbntDZWrzv8A98oCa8I/scfHfxYUebw1b6DbuCRPq9yIccdDGm+QenKd/rXhX/gnto0W2Xxv8Q725zndBpVosG3r0kk35/hP3Bjkc5yPCv7K3wG8JbXtfh/ZajOM7pdVZr3d16pISg4bHCjoCeRmrOxstOtkstOtILa3jzsihjCKuTk4A4HJJ/GqoDPAoAAYFVVVXin42/CLwYso8R/EXQraWHO+3S7WadcAn/VR7n7H+HrgdSK8U/t6/CLSGlg8N6Vruvypny5UgW2gfkj70h3joD/q+hHfIrxV/wAFAfiJqW6Lwj4R0XRI2/juXe9lX7vRvkXs3VDww6EZPin9pD44+MFli1f4k6skMuQ8NlILNCuCNpEIXK4JBBznvmv9N1O9/wCW13d3UvvJJLIx/EsxJ+pJrw1+zn8cfFyq+i/DPWvLZA6SXkQskdSMgq05QMCO4NeGP+Cf/wAS9SaOTxT4r0LRoW+8sAku5k5I5XCL0APD9+1eFf2BfhTpMaSeKdd1vXrgABlV1tIT93JCKC45Dfx9Gx1Ga8MfAL4MeDir6B8NtDilQYSa4tvtMq8EcSS7mGQxB5570kccUaxRIqogwqqMAAdgKqqqq+InxMsfB0R06yVbrVpEysWflhz0Z/5hep9hg1qWpap4g1J7/UbiS7vLlhliMlj0AAHT0AFfDj4Prb+Xrni+2DSgh7eybovfMg7n/Z7d/QVXiHxdo3hxCl3P5lzjK28fLn6+g+v614h8ba34gLRPP9mtSMfZ4iQCP9o9W/Hj2rRvD2r6/MIdMs3kAOHkIwifVun4da0H4YaTYbZ9Xk+3TjnZjbGp+nVvx49qvtR0jQbNZL25gtIEGEU8dOyqOT9AK1v4sIFaDQbHccYE9xwB9FH9T+Farr+sa2+/U9QlmA6IThV+ijitM8O63rJH9m6ZPMp6OF2p/wB9HA/WtO+EupTENqmpQW6/3YlMjfTnAH614a8HaZ4YMkllLPLLMoV3lYdvQADAzVVV7oGiajk3uk2kzHqzQjd1z161efDDwtdPuhjubX1EMvH/AI8GqX4R6Qzkw6rdqvYMqsR+OB/Km+D6k/L4i47A2n/2deFvh5e6BrsGqXF5bzxxI4AXcrBiMcDoRgnvVeP4bq68LXVpZWk1zLM0YCRIWIw4JOBz2qXRtXgfy5tKvI2x914GB/LFVb6hf2gUWl9cQ7TlfLlK4PqMGrbxv4rtQBFrk7YGB5oEn/oQNQ/FDV2URanpthexg5AaMqc/mR0z271YfEXwlcYXUNE+yMepWFZFH4jB/So9N0a+tUkXTraSGZAwD24GQRkZBGR9DU3g7wvOwd9BswQMYSIIPyGK/wCFb+FFniuILGSFomDALOxBIOeck1tyOtam13Fp1zJp8RlulibyUGBl8cdeOuKv9O1HTpjFqVnPBISeJUKk+pGev1rTvE2v6SAun6tcRqOiFtyj/gJyP0rT/i1rdvhdQsbW7UdSuY2P4jI/StP+K3hy5wt7Dc2bdyyb1H4rz+lWPiXw/qXFlrFpI39zzArdv4Tz3FVVVVEAjFYHoKKg9qKgjAoqCMA0V44NFcjHFeLtKXVvDl9ZkDd5ReMkdGX5h9OmPxqwu5NPvre+hPz28qyLzjkHNIySxrJG4ZWAKkdwaKnoKIqqr4o6X9t8PrfRx7pLGUMSBkhG4b8M7T+FeB9VGkeJbSZ3xFMfJk5xw3A/AHafwqqqqqqqqqqogEYrUtT03SIPtGpXsVuh4BdsEn2HU/hWtfFaBMw6DZGU8jzrgbR+Cjk/jj6Vq/iHWdckL6lfySrnIiB2ov0Ucfj1rSPDGt66QdOsJHj5/esNqDHX5jx+ArR/hTYw7JdbvWuXHJhh+RPoW6kfTbVjp1hpsAttPtIreP8AuxrjJ9T61VVVVVUQCMURjiqqqqqqqogEYojHFVVUQCMURjoaqqogEYojHFVVVVVVEAjGaI7VVVVVVUwyMiqqqr49fAX4eftGfDy9+HXxE0zz7Wb95Z3kQC3Gn3ABCzwsQdrDOMdGBKsCCRX7VP7E/wAWv2XdaubrVtOl1rwW8+zT/EtpETC6sTsSdRkwS4ABVvlJ+6zV+zr+2V8dP2ZroW/gPxN9s0CSTfceHtU3XFk5ySWRMgwscnLRlS3G7dgCvBn/AAWZ+GN7bqvxE+DvifSZhGcnRbyDUFZwRjAlMBAI3HvjAHOci2/4KwfskXNrDcS6h4stpJI1ZoJdFJeIkZKsVdlyOhwSM9CRzVr/AMFKv2KbvyVHxn8qSbaNkvh/Ul2E9mb7PtGO5zj3xX/Dc/7Iv/Re/DH/AH+f/wCJr/huf9kX/ovfhj/v8/8A8TX/AA3P+yL/ANF78Mf9/n/+Jr/huf8AZF/6L34Y/wC/z/8AxNXf/BSP9imzuprOb42xNJBI0bGLQdSlQkHBKutuVYccEEg9QSK/4eWfsS/9Fr/8tzVP/kav+Hln7Ev/AEWv/wAtzVP/AJGr/h5Z+xL/ANFr/wDLc1T/AORq0v8Ab8/Y71uzF9ZfHbRI4yxULdwXFq+R/sSxq2PfGDXhH4rfC34gsU8BfErwr4kZWZSNI1mC8IKgFh+6duQGUn0BHrVVVVVURjgVVVVVVMMjitY0q21qwexuRw3KNjJRh0Iq/sLnTbuSyu02yxHBHY+hHsaqqrStSuNIvor62PzIfmXOA69was7uC/tIry2bMcq7lz1/H3ogEYojHGaqqqqr/gpZ4Oj8Jfte+LpraPy7fX7ez1OJPmOC8CpIctnOZY5G44GcDGMV/wAEk/Fi65+zNqHhySZPP8N+JrmFYg5JEMscUysQegLvKMD+6T3qqqqqqqqqqqqqqqqqqr4u/CLwH8cvAeofDr4i6MmoaTqC5BGFltZQDsmhfB2SLk4b3IIKkg/tU/srePP2WfHjeHPESNqGg6gzvoeuRxFYr6IHkEc7JVBAeMnjIIJUqx/4Jxf8FHZfhTLpvwF+PWtvL4KkZYNC125cltCY8LDMx62p6Bj/AKn/AK5/6uGaG4hSe3lSWORQyOjZDAjIII6giqqqqqqqqqqqqqqqqqqqqqqqqqr9tzXIfEX7Xfxev4IHiWLxdf2hVzklreUwM3HYmIkexFf8E6fD7eHv2N/h5BNawRXF7DeXkrRAZlEt5M8bMR1byjGOeQFA7VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV/wAFV/2Zp/HPgq0/aB8I2Xmav4QgNvrkKIN0+nZLCbOeTC5ORjlJGJIEeD+xT+0pqH7Lfx70P4gNK7eH7xhp3iS3Cs3mafK6+Y6qOrx7VkUdym3oxrTtR0/WNPtdW0m/t72xvYUntrm3lEkc8bgMroykhlZSCCOCCDVVVVXj3xhpXw88C+I/H+uFv7O8M6TdanebFJbybeJpXwACSdqHoDXiLXtc8ceKdT8T61PLfaxr+oTXl3Icu89xNIXdu5JZ2J7nJr9nn4aJ8Hvgd4I+GptYre40LRYIr1IsbTdsu+5YbeDumeRs85znJ61VVVUoB5o896qqpRnmqoDPArAxgVVVVVVADFVVfGr44/DT9n7wTP49+J/iBNN0+NvLt4kQyT3sxBKwwxjl3OD6KBlmKqCw/as/bi+LH7UOpz6ZeXsvh7wPHJm08N2c58t8MCsly4AM8mVUjd8ikDaqkkt8EP2avir8ftTFv4K0PytLifbd6zekxWlv1/iwS7cY2IGPIJAGSPgD+wx8J/gu1tr+tQr4u8Uwnemo30O2G2bH/LGDJUY6h23MDyCvSqqqqqqqqqqqqq/a78Kw+MP2a/iBpkqxk2mjyaghckbWtSLjggE5xER75wSASa+Bfi6XwD8avAfjWK8S2GieI7C6kkknMKeWk6GQO+RhCm5W7bSc8UBngGsDGBVVVVVVQAxVVVVVXV1a2Vu91eXMUEMQy8krhFUepJ4FeMv2p/gb4KLwXfja31O6QkfZ9KU3ZyAMjenyDqByw5yOxx4w/wCCgkxZ4PAHgFFAzsudYnJJ5OMxREY4wf8AWdSR2yfGX7Ufxy8bF473x1d6dasTtttJAs1UEAEbkw7DjozHqfWv+Jnrd/8A8vN/e3Lf7UskrfqSa8H/ALLHxz8aCGWz8DXOnWsuP9J1VhaKgOcEo58wj5f4VPUHoQa8I/8ABPi5Z4p/HvxBiRAR5trpFsWLDvtmlxj8YzXhL9kT4D+E/KlHg/8Atm5jx+/1edrjdjb1j4iOSufudyOhxWm6VpmkWiWGkada2VtH9yG2hWJF+iqABWABgVVVQGeBWABgVU88FtBJc3MyRRRIXkkkYKqKBkkk9AB3rxT+0h8DvB7Sxav8SdIkmhyHhspDeOGBI2kQhsNkEEHGO+M14s/4KB+BbDfD4M8GatrEi8CW9lSzjJ9RjexHsQv4da8Uft2fGvW1aLQk0Pw+mTse1s/OlAyOpmLqehHCD7x9iPFHxR+I/jUsPFXjrW9TRgR5M965iAIAIEedoztGcDkjJrQfC3ibxVcmy8L+HdT1i4XrFYWclw44J6ICeisfwPpXhb9jr49+KFjlfwnFo0EhAEuq3SQ4yeSYxukGOp+X6ZPFeF/+CeczLFN41+I6I3/LW20uxLdhnbNIR3yOY/Q98Dwx+xj8BPDZWW48N3etzIMLJql679iCSibEOc91OMAjBrw94K8HeEYhD4V8K6RpChCmLKyjgJBxnJUAnJAJz1IoLnvWBjAqqqqqq+J/xOi8NRPoehyrJq0i4dxyLUHufVvQdup7Aww6p4h1RYYVmvb+9l4BO5pGPUk/qSelfDr4XWXhSKPVNWSO51gjhgcrbg9l9T6t+A4zmr29tNOtnvL65SGGMZZ3OAP/AK/tXib4m3N3vsvD4a3izj7S3DsP9kfwj9fpUFve6ndiG3ilubiZugBZmJ6k/wCNeG/haibLvxI4dsZFrG3A/wB5h1+g/M1bWttZwJa2kEcMUYwiIu0L9AK+IfinW9AMFtpscccdzGT9pI3MGB5UDoOMcn19qvLy91O5Nxe3MtxM/G52LH6D29q0T4ca/q6pNcothAf4pgd5HqE6/nitG+HvhvSNsj2v22cf8tLgbgPovT9M+9KiIgRFCqowFAwAKqqqqqqqsZ4FbSe9S28M6eXPEki5+665H5Vc+F/D17k3GiWTsx5byFDHHuOau/hj4TuQfKtZrYkdYpm/PDZFX3wet2BbTNZkQjos8QbP4jHv2pfhd4khv7eGeGGa2aVVllhlB2KSMnDYJ4J7dqVERFjjUKqjCqBgAe1XVxDZ2st3cPtigjZ3OOgAyT+Qq1+ML7tt9oalf70U2CPwI5/MVYfE/wAK3pCTzT2bHgefFxnjuuePrjpVjrGk6ooOnalbXBx92OQEj6jqKmhhuImhniSWNhhkdcg/UGtS+HnhPUssdNFtIRjdbMY8fRfu/pWo/B8HLaTrPrtjuI/r/Ev4fw/4VqHw68WWBONOFyg6PbuGz+HDdvTvV1ZXdlJ5N5azQSf3ZYyh/I1Y67rWmYGn6rdQADG1JSF/Lp3rwTeanqPhu1vtWuPOnm3EMVCnbkgdPYZ/Gq1X4g6Do2qz6PfpdLLbhdziMFTlQRjBz0PpUXxO8ISPse9mjGPvPA2P0ya07X9E1bC6bqltO5HCLIN3/fPX9KYhVLHPA7DJ/Krn4rfD212+b4mgbdnHlxSSdPXapx+NRSxzxJNC4ZJFDKw7g9DVVVVWmAHTbQY/5YJ/6CKKg9KK8cGime1FR0Boqexq+s4r+ynsZxmOeNo2HsRir6zlsLyexnGJLeRo2HuDivC2rjXNBtNQYjzGTbKP9teD+eM/jVVVVVVVVVaz418O6HlLm+E0y/8ALGD52/HsPxIrWvilrF6Gh0mFLGMgjeTvkP4ngce341NPd38/mXE0txPIcbnYuzHt15NaL8OfEOrbZbiEWMBGQ84+Y/ROv54rRvh34e0jEk0Jvpx/HOMgH2Xp+efrSoiIERQqqMBQMACqqqqqqqqiARiivpVVVVVVVUQCMURjoaqqogEYojHANVVUQCODRHaqqqqqqmAI460R2qqqqqqIBGKYY6VVVVappWl65p1xpGtaba6hY3cZjuLW6hWWKZD1VkYEMD6EYr/gop+z1+xd8I7u7v8AwX4xv/Dnj+9w8fg3RoY721T5NweSMshskfchyXPDZjiYBsV4c8LeJ/GOpx6J4R8Oaprmoy48uz06zkuZmywUYRAWOWZR06sB3q//AGI/2tNNs5b64+APi944V3MsFj57n/dSMszH2AJr/hlz9pr/AKN0+J3/AISF9/8AGq/4Zc/aa/6N0+J3/hIX3/xqv+GXP2mv+jdPid/4SF9/8ar/AIZc/aa/6N0+J3/hIX3/AMaqx/Yk/a01Kzivrf4A+LkjmXcqz2PkOB/tI5DKfYgGv+GF/wBrr/ognif/AL8p/wDFV/wwv+11/wBEE8T/APflP/iq/wCGF/2uv+iCeJ/+/Kf/ABVa7+yr+0x4bkuk1j4A/ECNLKPzJ7iPw7czQIgXcW82NGTAHU7uMHOMGr7T9Y8PakbLUrK80zULVlYxTxNDLEcBlODgqcEEfUGvg9+35+1H8GJI4NJ+JF14j0tNo/srxOz6lBtVdqqjOwliQAD5Y5EX1Br9m7/gp/8ABj4xy23hn4jxp8PPEzqAGv7pW066fgYjuDt8tjydkiqBkAO5qqqqphkcVVVVVVURg4FeNfDw1Sy+3Wkebq2UnCjmRO4+o6j8R3qqqq8Ba15FwdGuH+SYloc/wtjkfiB+f1qmGRkVVVVVX/BZDwY9p8Qvh58QlXK6ro1zpbkFjtNrMJVyMYGftbY5ydp4GOf+CN/jU23jT4i/DiW5Zv7R0u11WCFmchPs8rRSMo+6CftMQPQnavUA4qqqqqqqqqqqqqqqqqqr4xfCHwR8c/h9qfw48faTFe6bqEZ8t2X95aTgHy54m6rIhOQR1BIOVYg/tIfs3/ED9mT4gTeCPG1r51tNul0jV4YyLfU7cHG9M52sMgPGTlCR1BVm/wCCY3/BQi08CfYv2dPjv4klTQZpFh8La5ezAx6WTwLSZjysBONjk4jPynCEFKqqqqqqqqqqqqqqqqqqqqqqqqr44apf678afH+t6pcefe6h4o1K4uZdgXfI91IzNhQAMkk4AAr9kfR7XQ/2XPhRY2ckrpJ4Q065JkIJ3z26TOOAOA0hA9sZJ61VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVqml6dremXei6tZxXdjfwPb3NvKu5JYnUq6MO4Kkgj3r9rb9n7UP2bfjdrnw+MVy+iNJ9r0C8nXJubGTlPmwAzIcxOcDLxsQACK/4JC/tPw/En4U3H7Pvia+ZvEnw/h83TDK2TdaSz4XHOcwSMIzkABJIQM84qqqq/4Ko+Ox4I/Yu8X2kN0kF34ourLR7YsyZbzJ1klUK33iYIZhgcgEsMbcj9jjwKnxI/ai+GvhSa38+B9divLiIkAPDag3MinJHBSFge+M4ycCqqqqsZ4FYAGAaqqrGeBWABgGqoDiqqqqqoDJxVVXxL+JPg34ReB9W+Ifj3WItN0XRrcyzyuRuc/wxoM/NI5wqqOWYgd6/ao/aZ8X/tS/E2fxx4gRrDS7VTb6Jo6ztJHp9vn34Mj4BdwBuIHACqB+x5+w9cfFCGy+KHxZt7iz8KF1l07S8FJdWUc73PVIDwBj5nGSCowzaPo2k+HtJtNC0LTrew0+whWG1tbeMRxwxqMKqqOAAB0qsZ4FBCeleXkYzWwetbARwK2DGMVs9qKAjAFFBjpRQHoaMfHBooRRFVVVVeItJ/t/w/qehfaPI/tGzltvN2bvL3oV3YyM4znGRVeC9YuvEXg7QvEF7HElzqemW91MsQIRXkiVmCgkkDJOMk/WqqqqqqgM81VVVeJfGfhHwbaG98V+JtM0mFULBry6SIsB/dBOWPsMkmvGX7c3wg8Pl7fw1BqniWdSQrW8P2eHgDq8mG6nHCHoe2CfGn7dPxa8QLNa+FrLSvDNs5PlvDF9puEXPQvJ8h44yIwep44x4p8eeNfG8/2nxd4r1TV3BBUXd08ipjONqk4X7zcADqfU14O+E/xK+IDRf8Id4J1bU4pW2rcx25WAHOOZmwi8+rDofSvCP7BHxN1iOK48W+IdH8PRuAWhTdeTx+oKrhPykNeDv2H/AIMeHGiuNdj1PxJcRtuxe3HlRZB4/dxbcjpkMWB57HFeGPBHg3wVa/Y/CPhbStHiIAcWVokJkwAMsVGWOAOTknHNVWM8CgABgGqqqAzwK8W/tAfBjwRG/wDb3xF0cTRjLW1rP9qmHTAMcW5hncCMgcc9ASPFn/BQPwLYb4fBngzVtYkXgS3sqWcZPqMb2I9iF/DrXjH9uD42eJVe30S50zw1btkf6BaiSUqQRgvLvweeqhSMDpXifx3418azGfxd4t1bWG3blF7ePKqHJPyqxIUDc2AAAMnFeGfh7478aMq+EvButauCwUyWdjJKiknHzMo2qM9SSAO9eEv2Ifjh4k8qbV7HTPDtu+GLahdh5Np28hIt53YJ+VtvKkHHGfCv/BPfwjaxpJ418e6rqMuATFpsCWiKfl+Xc/mFhwwzhScjpjnwn+y/8CvB2yTTvh7p95On/LbUw16xPriUsqn/AHQKs7Gy062Sy060gtrePOyKGMIq5OTgDgckn8aqqC561gYwKqqqqqvib8Q4vB1iLLTnjk1a6U+Wp58lf77D+QPU+wIqC31PX9UEFukt5fXspOOrSOxyST+ZJP1r4d/Dqy8F2X2i42T6rOmJpgMiMf3E9vU9/wAgKxngV8VtGvnSDWYp5JLaIeXJEScREnhwOnPQ++PXjwz4B1bX9l1OptLI8iV1+Zx/sr9O/T61DB4U8CWJHmRWocfMzndLKR+p+g4HtWifEWw1vXV0mK0eGGVSIZZG5dxzgjtkZ79cetV4l8PWviXTf7PuZDGQ4dJFXJQjrj6jI/GtD8KaH4eQHT7MGbGDPJ80h/Ht9BgVVVVVVVVYJ6Vtz3oKD0Fbc962isDGBVVVVrulDXNIuNKNy0AuFCmRVyRgg9PfGPxq9+EGqxKxsNVtrjHQSIYyePx5zxV/4E8V6ecSaNNKOzQDzc/guT+Yp0ntZtrq8UsZBAIKsp6g+orTvG3ijS9ot9YmkRRgJOfMGOOPmzgcdsVpvxfuUwmr6THIO8lu5Qj/AICc57dxWm/EXwpqRCHUDauf4bldn68r+tRSxTxJNBKskcihkdTkMD0II6ipYop4zFNGsiHqrLkH8KvvA3hO/UrLotvGSODAPKI46/LirGyt9OsoLC0TbFbxhEHsB/Oq8Z+B/FGr+ILvVLKwjminZQgWZVICqFydxHpVz4B8X2sfmS6JKwAPEbrIePZSTU0F1Yz+XPFLbzxn7rqUZSPbqK+GmqeItU0+V9WfzrWP5YJ5M+Yx78/xD3POfXtWi/8AIGsP+vWP/wBBFVVV4nvk03w9qF4xGUt2C7um4jC/qRVlavfXlvZRn5p5VjXAzyxA6fjSoiIscahVUYVQMAD2qqogEYoqDRX0NfFbRPserxaxEmI71MPz/wAtFGP1XH5GvhRrPk30+hSyfLcr5kIJ6Oo+bH1X/wBBoqaqqqqqquru0sYTcXlzFBGvV5GCj9a1n4paNZb4dKie+lGQG+5Hn6nk/l+PetZ8b+I9bBjuL8wwn/ljbjy1/HuR7EmtP0rUdWm+z6bZS3DjqI0zt9yegHua0b4T3c6pNrl6LcHrDDhnH1boD9M1pHhnQ9DUDTdPjjfHMjfM5/4EefwHFVRUHpRGOKqqqqqqqqoqD0ojtVVVVVVVEAjFEY6GqqqIBGKIxxVVVMMjg0R2qqqqqqpgCMiqqqqqqiARjNEdqqqr9vr/AIKJp8IJ7v4N/ArUrW68aLuj1jWAomi0XjBijByr3PrnKx9CCxITR9G8c/Ffxpb6Po9pq3ijxRr9ztjjUvc3N3KeSSTlmOASWJ4AJJABNfsz/wDBJLwzo1rZ+Kv2l9R/tnUztkXw3plyyWkGQDtnmXDysDwVjKoCOGcHNeCPh74F+GmhQ+Gfh94R0nw9pcChUtdOtEgU47ttA3McklmySSSSSSaZR0ojtVVVVVV40+HvgP4jaYdG8feDNE8R2W0qLfVLCO6VQSpOA4O05VTkc5VT1Ar48/8ABJj4QeN4rvWvgrq0/gbWpGaRLGZnutOkY/w7WJkhBPOVZlXOAmMAfHL9nf4sfs6+Jz4W+KHhiWxd+bS/hzLaXq/3oZgMN7qcMvcCv2Ov+Cifj39nyWx8C/EJ7zxX8PgyxJDJMWu9IT5Fzbs2d0aopxASFz91k5z8PviH4K+K3hDTvHnw88RWut6Fqke+1vLfIDYOCrKwDIwIIZGAZSCCARVVVUwA6VVVVVVMMjiq8Z6Euk6j59tGRbXOWXjhW7r/AF/H2qqqkd4nWSNyrocqR1BHetC1SPWdMhvUYbyNsqj+Fx1H9foRRAIxRHaqqqqv+CtHgV/Ev7M9p4ttrfdN4R8QW9xNJlvkt51e3YYzjmWS35I4xgYya/4Jv+N18E/td+DVuJ0itPEC3Ok3BbOSZoWMQGCBkzpCOcjBPGcEVVVVVVVVVVVVVVVVVVVXx2+BPw+/aI+H158PPiFpnnW037yzu4gBcafcAELPCxB2sM9OjAlWBBIr47fAn4gfs7/EC8+HnxC0zybqH95Z3cQJt9QtySFnhYj5lOOnVSCrAEED/gl9/wAFA4tUt9D/AGWvjLfynUY8WvhHXJpGk+1LklLGckkh1HyxN90qFj4KrvqqqqqqqqqqqqqqqqqqqqqqqqrV9Vv9d1W91vVbjz73ULiS4uZdgXfI7FmbCgAZJJwABXwS0a18O/BfwD4fsZJXt9M8L6bawtKQXZI7WNQWIAGcAZwBVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVf8FM/2fP+Fw/AebxvoVj5viT4feZqEGxcvNZED7VF74VVlH/XIgfer9mb45az+zj8cPCvxd0cSzR6NeAajaRtg3llINlxDzxlo2baTwrhG6qK8M+JdA8Z+HdN8WeFdWt9T0fWLWO6sby3bdHPC6hldT6EEVVVVf8ABcDxU1n8Ovhd4HEtwF1bWr7USigeWTawxx5bnO4fbDjAxgtnHGf+CRvhSTWf2j9Y8SyJL9n8P+F52V0dQPPmmhjRWB5IKGY8dCoyexqqqqAzR5qqqlGaqgM8CsDGBVVVVVUBgVVV/wAFO/2rZ/i38SJPgt4P1CUeEvBF28d6UkGzUdSTKu/HVIvmjX33noRX7Dv7LTfGvxZ/wnfjOyb/AIQrw7OpaN0BXVLpSrC3wQQYgOZPUFV/iJWOKOGNIYY1REUKiKMBQOgA7CgpPAoIO5oJkYA/OhGTxmhFkUIuwoRDpXlAjgV5XtXle1GIHgUYs8AUY/SjGRwDRTtiigI4NFSOKI7VVVWq2UenapeadDew3iWs7xJcQtlJgrEB1PdTjI9jXwO1G61j4KfD7Vr7TZdOub3wtps81nLnfbO9rGzRtkA5Ukg5A5HQVVVVVVAZ4zWBjAqq8Y/Fj4a/D9ZB4w8baTps0SbmtpLgNORjORCuZG4x0U9R6145/b68G6aZLTwB4VvtalU4F1ev9kh6feVfmduwwQnf8fG37X/xw8ZPJFb+JE8P2j5At9Hi8gjjH+tJMmcEnhgM8gDAxc3Wra/qRuLy5u9R1C8kALyu00sznAAyclj0H5V4J/ZY+N/jp0a18GXGk2rEA3WsZs0UHo21h5jDvlUP6jPgv/gn5YxGC5+IXjuWc4zLZ6RB5YBx0E0mSRn/AKZg49M8eDf2b/gr4F8uXRvAWnz3MecXWoKbuTJxyDJkKeB90DHPqcgZ5qqqsZ4FAADAqq8TeNPCHgy0a+8WeJ9M0iFULBry6SIsB/dBOWPsMknivF/7c3wY8Psbfw//AGt4kmHRrS28mIEEZBeXa3TOCqsDjqM5rxj+3t8T9aV4PCGhaR4cibO2RgbydeCBhnATjOeY+oHbIPi/4ufE7x68p8W+OtY1GOYktbPcskHPXEK4RfoFFeHfBvi3xhcfZfCnhjVdYlBwVsbN59v12g4/GvCX7EPxw8SeVNq9jpnh23fDFtQuw8m07eQkW87sE/K23lSDjjPg/wD4J9+DrAx3HjjxtqWrOBlrewgW0jBwPlLMXZgDu5G0nI6YOfCf7OvwS8F7H0P4c6QZo+VnvYjeSA+oaYsVP0xSRxxRrFEiqiDCqowAB2AqqqqxngUFGKqqqqqq8Y+KbLwfoc2rXW15B8sEJbBlkPQD+Z9hWqanqGu6lNqeoztcXd0+XYjknoAAO2AAB2GK+Fvw7TwnYjVdTjzq91HhwTkQITnYMd+ASfwHqbu7tbG3a6vLhIYoxlnc4Aq2+JunXXiCLT0hCWEnyfaZCQd5xg47LnI59QeMGsDGBWq32l2Fk82rzwx25UhhLyG9gO/0rxD8U7qdntvD0PkR9PtEq5c/Reg/HP4VPcXeo3JmuJpbieQ43OSzMa8NfDnX7qeHULuQ6Ykbh0LLmXIOQQvbp3/I1VVVQ+NfCt1rUfh201uCfUJCwWKIFxlVLMCwBUEAHgntVeM/G+l+CLOG61K3uZjcFliWFQcsAOCSRjr/ADrWPjx4kup/+JNYWljAOgkBlc/U8D8Mfia/4Xb47/5+bL/wGFW/x28awqVkg0u4JPBkgYEfTawq2/aD1FG/0zw1bSjb0iuGjOfXkNx7frVp+0DojlBfaBexZHz+VIsm0+2duf0rQPi54Q8RX9rpdm15DdXbFI45oMYPPUqSOcfrW0etYHaqqqqqqqqr3TtP1KMQ6hZQXKDossYbH0z0rUvhh4WvsvbxTWbkcGGTjP0bPHsMdK1H4RarCC2malb3IHRZFMbf1H8quPB3iW1uI7a50e4QyOEDhC6gk4+8uRVpaxWVpDZwDEcEaxoD2CjA/QUxVVLMwAA5J7VB8YU8xlutEJTcdjRzYOO2VI6/jVr8UfCdyB509zbdf9bATj/vndUHi/wtcqTFr9iMAf6yYR9f97FLqemyBfL1G2bd93EqnP05qtd0/wAOaxEbHWTasyjgtIFeP3BzkVfXdnpnhu6vtPlijt7OzkeJ4gGVAik5AGQQMUqs7BEBLE4AA5Jq1t47S1htImJSGNUUt1wBjn8qqqr4seIVPleHLWUEjEtyAen91T/P8q+GWjNqXiNLx490Gnr5jE9N54QfXOT/AMBqqqqqvGGiL4g8P3NiqBplXzIPaRemPqMj8a0ddVTVIJNIt5pbyBw6JHGWOVPcDt61bu89vFNLC0LugZo2IJQkcg49OlFcjpmioPQ0QQKqqrWPGfhzRdyXWoLJMo/1MPztn044B+pFav8AFfUbjdFo1nHar0Ekvzv9cdB9Oav9S1DVJfP1C9luH7GRycfT0rR/BHiPWwJLexMMJ6TXH7tfqO5HuAa0b4W6PZFZdVne+lHOzGxB+AOT+f4Va2dpYwi3sraKCNeiRoFH5CqqqogEYorxwaI7VVVVVVVVUVBHHWiO1VVVVVVUQCMURjoaqqogEYojHFVVUygjIqqqqqqqmGOlVVVVVVMARkVVV/wUN/bMH7NXgePwT4GvE/4WL4qtWbT5BscaTa7trXToerNh0iBG0srMciMq3w9+Hvjj4xeONO8C+BdGutb8Q63PthhQ5LE8vI7HhUUZZnYgAAkmv2Of2OPBf7KHgvyYTb6v421eFf7c1zy/vdD9ngzykCkD0LkBm/hVKogEYzTDI4qqqqqqpgB0r4ifDfwL8WvCV74F+I3hmz17Q79cTWlypxns6MpDRuM8OhDKeQRX7b37BniP9mK+/wCE08GzXmvfDq+l2JeSIGn0mVjhYbnbwVPAWUAAn5SFO3d+x3+2J4z/AGU/Gfmw+fq/grVpl/tzQ/M+90H2iDPCTqAPQOAFb+Fk+HPxG8GfFrwZpnxA+H+u2+r6Hq0PmW1zEfwZGU8o6kFWQ4KkEEVVVRGRgVVVVVVURg4Fa9pMWtaZLZOBvxuiY/wuOh/p+NSRvE7RSKVdCQykcgjrVVVeBdWNlqf2CRv3V4No9nHQ/jyPxFUwHUVVVVV+1R4Bb4nfs5/ETwTDbpPc32gXD2cTRh91zCvnQjBzg+bGmD1B5HIFeA/Fl74B8c+HfHWmJvvPDmrWupW65AzJBKsijkMOqDqCPY9K0vUrHWtMtNY0yfzrS+gSe3k2ld8bqGVsEAjIIOCAaqqqqqqqqqqqqqqqqqqqv2pf2aPCH7UXwzn8EeIJEsNUtWM+i6wsCySWFxjrzyY3wA6AjcAOQVUj4nfDbxr8E/iHqvw+8aWTafrug3Ox2iY7X6NHNE2ASjKVZWwDgjoeB/wTO/b2/wCF8aFb/A74tazLN8R9Gt2aw1CcZ/t20jGdzMOtxGv388uoD5Y+Ziqqqqqqqqqqqqqqqqqqqqqqqqvhh/yTTwl/2ArL/wBEJVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV+3Z+z5/wzx+0BrOhaRY+R4X1/Oq+H9i4SOCVjvgHYeVIHQDJOwRsfvV/wAEbP2h7fxf8LtX/Z41y9zrPgqRtQ0hXkJM+mzv84UEn/UzscngYuIwBwTVVVV/wW38QQ3Pxu+H/hRTcebp3hV71g3+rxcXUiDbz97/AEY7uBxt5Pb/AII0eG9tr8UvF01tEfMk0yxtps/Ou0XDyrjsDuhPvj2qqqqoAAYBqqqsZ4FAADAqqUcZqqqqqqAyaqq/bv8A2hf+Gdv2fNZ13SL/AMjxT4gzpPh/Y2HjuJVO+4HceVGHcHBG8RqfvV8PPAmv/E7xvovgHwxbNPqWtXSwQgKWEY6vI2OiIgZ2PZVJ7V8Mfhz4d+FHgTRvh/4Vt/L0/R7ZYldlAed+ryvjALu2WY9Mk4wOKCAcUEJFLGO1CLIwKEYxigg7CgmRgCv+ClXxM+JPhD41aR4c8KfELxLoulXnhCB7mw0/Vp7aCZmurtGZ40YKxKqqkkcgAdBUf/BOr9odokM37R6rIVG5VuLxgD3wdwyPfArXPiJ+2x+xD4j0yH4i+JpvFXhrULgbBe37aja3qxhd8UU8o863YKeBhRnnawBr4VfEbw/8YPh5ofxJ8Lecum65bmWKOdQJImVikkbYJG5XRlOCRleK/az+IPx1n/bK1z4XfDr4reLdJXUtQ0mw0yws/EFzaW0c1xaWoACo4VAZJCSQOrEmj+zF/wAFHv8Aor/iH/wvrn/4uvhn+zt+31onxJ8J6z4y+KeuXegWGuWVxqtvL42uJ1mtUnRpkMZchwUDAqeDnHejGCMCv2lf2/r7T/EFx8K/2b7NdZ1oSm2k1yOAXaibIylpFhhMwORvYFSc4Vhhqsf2Yv2/PjPbL4h8Z/EW80UXH76K013xBcW+0kAfLb26usJwOVKofUZqX9jn9u74eqNZ8G/FVdQuwwJt9I8V3MTybSCAwuFijZc/wsSD3FfDuy8S2XgDw5aeM9Tm1DxBHpdv/adzNGiNJc+WDKdqBQBuyAMdAM5OSWUjpVVVfC3/AJJj4Q/7ANl/6ISqqqqqxngUAAMA1PPBbQSXNzMkUUSF5JJGCqigZJJPQAd6+K37a3w38E+dpfgof8JbqyZXfbybLOJvUzYPmdjiMEHpuWvHP7V/xv8AHSyW83iw6LZyDBtdGQ2o68/vATLz0IL4x25OdO0vWfEWopYaRp15qd/cN8kNtC00sjE9lUEkkn9a8F/sY/G3xYsNzqOk2nh20lPMmqT7ZAueT5SbnB64DBc+oBzXgn9gz4b6Mkc/jbXdT8RXIxviiP2O3PIOMKTJ0GM7x1JwDjHhL4beAfAdulv4O8IaVpWwEeZBbKJWz/ekOXb8SaqgAe9YGMCqqsZ4FAADrXjr42fCr4brIni/xvptpcxjJs0k86464H7qPc454yRjrzwa8b/8FAfDFkzWvw/8GXuqMMD7VqUotowc87UXczDHqV5PTjnxn+138dvGe+H/AIS7+w7V8f6Po0X2XaRnkS5Mo69N+OBU0+p63fiSea5v725ZUDOzSyStwqjnJJ6AD6V4M/ZZ+OfjcpJY+BLvTrViA1zqxFmqggkNtfDsOOqq3UeteDP+CeoxHcfEP4geoe00a3+mCJ5R9RjyvQ57V4Q/ZR+BHg5VaDwLa6tOPvTawTeFuCPuP+7HU9FHOD2GLa1tbK3S0sreKCGIYSKJAiqPQAcCqqqqqrGeBQAA4NVVVVVVMyqpZmAAGSTxiviV4xfxj4heaCQ/YLPMVoh9P4nx6sR+QUdq+C3gT7RKvjHVYcxxEixjdfvN0Mn4cge/PYV4h8QWHhywN5ePuY5EUQPzSN6D29T2rX/E+reI5/Mv58Rqf3cCcIn0Hc+55rwt4F1PxGy3Ega1se8zry+OoUd/r0/lXiHxlp3hCzTRrCRr2+hjCKsjFvLAHBc9z7Dn6VqusalrVybvU7x55P4QxwFHoo6AfSvDnw51fWglzfBrG1IyGdPnf6L2+p/WtD8KaH4eQHT7MGbGDPJ80h/Ht9BgVVVV3d2thbSXl7cRwQQrueSRtqqPUmviF8WNR8RzzaXoM8lrpQO3cvyyXGOpJ6hT/d/P0Hwaivl8bWt3Bps1xAqOk0qR5EO5Thieg5wPoTVfE/wveeLfCzadp0ayXcVxHLCrNtGc7WJJIAwrMe/TpnFaH8AJGCyeI9cC8cw2a5P/AH2w/wDZfxr/AIUD4OI/5CWs/wDf6P8A+N1c/s+aA7E2eu6hGuOBIqOQfqAKuf2d7tVX7J4qhkbPIksymPphjV18BfGkC7oLrS7j5sBUmZTj1O5QP1r4c/DHxfoHjax1TWdLSK0tTLmUTI4YmNlXABJHJzkgVXxsvFtvh/dwEpm7niiG44OQ4fj1PyflmrPxZ4osGDWXiPUofmDFVunAJHqM4P41Y/F/4gWOF/t37Qgz8s8CP+uM/rVl+0B4ni2rf6TptwFAGUDxsfUk5I5+gqx/aF0mQgal4cu4Bk5ME6zcY46he9WXxu8AXYBnvrqzyucT2zHB9Pk3c/pWn+NvCGqyrb6f4k0+WZ2CJF54VnY9AqnBP4VVVVVVVVX9s17Y3FmkxiaeJoxIBkoSMZA9qvvhFrMQJsNStLgDPDgxsfoOR+tXXgDxdaHD6NJIOcGJ1fOO+Ac/nU2iazbsI59IvY2IyFe3ZTj15FVVazrZ0/4JEyTYnuUNpGCdpO6QjH/fsE+/514H0xtY8X6Pp6KWD3aM4Gc7FO5un+yDVVVeNPGdr4YtfIgKzahKv7qI9EH95vb27/maY3uq3pJ8y5urmTsMs7E14P8ADaeGdGSyYq1xId9w45BYjoPYAY/M96qqqqqrXT7Gx8z7FaRQmZy8hRAC7E5JPr1qqIBGKKg9DRUelanrmj6Om7UtSggOMhGb5j9FHJ/AVq3xZtI1Meiae8rY4kuPlUH/AHQckdO4rWPGHiHW9yXmousLjBhi+RMehA6/jmrDS9R1WUwabZTXLqORGhbb7n0H1rSPhRqNxtl1m8jtV7xRje/0z0H61o3g3w9oex7OwV506TzfO+fUE8A/QCqqqqqqqIBGKIxVVVVVVVVRAIxRGOKqqqqqqogEYojHQ1VVRAIxRHaqqqYDtVVVVVVRAIxRGOKqqqqqqYY6VXxU+JHhn4P/AA68QfEzxheJb6T4esnuZyzhTIRwkS56vI5RFHdnUd6+NPxa8UfHT4oeIfin4vm3ahrt20ohVspawj5YoE4HyRxhUBPJ25OSST/wTn/Y+074BfDW1+I3jDTYpPiD4vtEmnaSLL6VaPho7VCeVYja0vT5sLyIwxqqpgDyKIxwDVVVVVUQCMA0R2rUNPsNWsLnS9Usre8s7yF4bi2uIhJHNGwKsjqchlIJBB4INft7/sQ6l+zT4jPj3wZuvfh1r96UtWYjzdJuHDOLaQfxIQrGNx2Xa3IBf9gP9szUP2bPHcfhHxnqV1P8OPEc6x30JfcukzsygXqLgnAAxIq4LLzhmRQbW6tb+1hvbG5iuLe4jWSGaJw6SIwyGUjgggggjrVVVMOeKqqqqqmGRxVePtH+x6gupQp+6u/vYHRx1/Mc/nVVVRyPDIksTbXRgyn0IrTb6PU9PgvoiMTICQOx7j8DkURkYzRHaqqqqvjr4A/4VX8ZvG3w6SPZB4f127tbXnO63WRvJb7zEbo9hwWJGcE5zX7CPxB/4WT+yf8AD3WJZ/Mu9N03+yboE5ZXtGMC7j6tHHG//Axnmqqqqqqqqqqqqqqqqqqqqv22f2PtD/ao8Bo2lGz0zx3oSl9F1SWPAmXBJtJmHPkuTkHko3zDgurMPHHwm8dlVk1Lw14s8K6iRujcw3FjdQv1DA5VlZeor9gj9tbRP2t/h41prX2fTviF4bhjTXdPWRQLsYAF5AgOfKc4DDH7tztyQVZqqqqqqqqqqqqqqqqqqqqqqqqvhh/yTTwl/wBgKy/9EJVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV/wUt+BEXxh/Z1vvE+l6e0/iT4fs2q2BjTLvbcC7i/3TEBLgclrdB3NfszfHLWf2cfjh4V+LujiWaPRrwDUbSNsG8spBsuIeeMtGzbSeFcI3VRXh7X9F8WeH9M8U+G9Rh1DSdZs4rywu4TlLiCVA8ci+zKykfWqqq/4LGa3dat+17HYXEcSx6N4TsLSAopBZDJPMS2ScndMw4wMAcZyT/wR50aKD4E+MvEKzsZL3xa1q0ZHCiG0t2BHufPI/wCAiqqqoDJ60cHpVVVAZNVWM8A1gYwKqqqqqgMCqqv+CnXxzl+Lf7R194R02/aXQfh2r6RbRYwv2wMPtkmD/F5irETwCIF47n/gmJ8GxDZ698b9a0/57n/iV6G7jogObmRQfVhGgYcjbIO5pUyMCljGOlaz+2D4S0D9pIfs4X/hXUhffaIIDqwnjECmW1SdWKkghQHCk9sE89K+Kv8AwU3+GHgvW59C+Hvg+88am0mMU18L5bG1fGQTC+yRpBkDB2hSDkEjGf2bv25vhv8AtB60PBtzpVx4U8UOhe2sLu5WaK8AGWWGYBSzgAkoUU45GcNt+Of/AAUP+FHwi8RXvg3w7ot54y1rTXaK7+yXKW9rDKOsZnIYlgeG2oQCCCcggfBj/gpZ8NfiP4otPCfjrwlceCJtQm8m1vpdRW7tAxA2iWQpGYtx+XO0qOCWAJI2D1r/AIKoKB+0J4fA/wChMtv/AEsvK2AjgV/wUWtdFm/ZR8TS6okZube9sH00uSCs/wBqjViuO/ktMOexPtX/AAS7bUpP2ctRW+WbyY/Fd2tl5ikDyvs9sTs9V8wyc+u4dq+OwH/Dz7TR/wBTn4X/APRVjRQHoaMYI4r9vH4wXvwe+AOpSaDf/Zdd8TzDSbCVJdskKyKxmlTHzArErKGGNrOhznAP/BNf9n7RNB8AJ8d9bsY7nXvELyxaS8qZNhaRu0TsgI+V5GV8t/cCgYDNuZARjFNGCOKaMYxTIQMUyA9KIxxXxj1m38O/CPxrrt0u6LT/AA9fTsm4KX2wOdoJ7nGB7kVouk3mv6xYaFpyo13qNzHbQB22gu7BVyewyRVraWtjaw2NjbRW9vbxrHDDEgRI0UYVVUcAAAAAcCqqqqq1LU9M0XT5tV1nUbWwsrZN01zdTLFHEvqzMQAPc18U/wBuLwF4Xjk074d2p8T6jjAuG3Q2kR92I3SfRQAf71fE/wCPPxO+LszL4s8QuNP3bo9LswYbWPByPkBJcg9Gcsw9a+G3wV+JHxZuhD4N8OTT2yttlv5x5VtD67pDwSP7q5b2r4cfsJeA9CigvviJqtz4hv1wXtYGNvaKeDjj94/TGdygg/drw14P8KeDLEab4S8OadpFthQ0dnbLFvx0LFRljyeTk8n1qqqgM8CsDGBVVXj/AOOHwq+Gcbr4u8Z2NvdoDixhbz7knnH7pMsMkEZYBc9SK8c/8FA7eMyWvw28DmUg/Jea1JtB45/cxHPXv5g4HTnjxv8AtGfGfx+zJrvjzUIrY4xaWD/ZIsA5AKx43c85bJ4HPArQvDXiLxTef2d4Z0DUdXu8E+RY2j3D4AyTtQE9Aa8E/sRfGfxSq3Ou2+n+GbY5x9vn3zMMcERx7sc8YYqRyfTPgz9gv4V6J5c/i/WNW8SzrkNHv+xW7dP4IyZAeD/y0xz0714S+HfgTwJCYPBvhLStI3gh3tbVUdwTkhnxubnHUnoPQVgAYFVVVVVVUBngUAAMCqqqqqqq+M/is6H4dGj2cxW81UlMqcFIh98/jwv0J9K8H+Gp/FviC10WFiiyHdNIBny4x94/0HuRV3caV4X0bzWVLe0s4wkcajHQYVQP0rxBr154j1F9QuyF42xxg8RqOgH+PrXgXwAt0iazr0GYSMwWzj7/APtMPT0Hfr06+OvGyaBEdJ0sg37py2OIFPQ/X0H4/W0s7/Wb5bW0ikubqdiQM5LHqSSf1Jrwp8PtP0RUvNRCXd9jIJGUiPoo7n3P6VVVVeP/AIt6d4YaXSdGCXuqLlXyf3du3H3v7x9h0xyRWveJtd8TXP2rW9SluWB+RWOFT/dUcCvAnw21fxdfwyXVrcWulD5pbkrs3L6R5ByT64wP0OkaPpug6fFpek2iW9vCMKijqe5J7k9yaxngUFHc1rmrWvh/SLrWbyOR4LSPe6xAFiPYEgfrX/C/vBn/AEDdZ/78xf8Axyrf43+ApzGJLq8g3gZ8y2J2fXbn9M1b/Fj4eXTFYvEsIKjnzIZI/wAtyioPHngm5jEkfizSQM4xJdoh/JiDUOr6TcskdtqlpKz/AHFSdWLfTB5qryxstQgNtf2kFzEesc0YdTxjofYkfjV98NfAeoBhP4XsV3AA+Qhh6emwjFXnwK8C3TZgOo2nzE4huAfw+cNx+tXv7PERBbTvFDKQvCT2ucn/AHgwwPwNX/wE8YW5Jsr3TbtMgACRkbpySCuOvHWr74V+P7DJl8NXEgAJzAyy5A9ApJ/DrXgLwrraeO9Gg1HRL638q6WZhNA0W0J8+TuA/u/jVfEPVrjQvBeq6nZ3DQzxQhYpFGSrMwUEfi1W/wAZ/iHC5aTWYrgYwFktIwB7/KoNad+0D4ihKrqejWF0qgDMRaFjjqScsMnjsK0v4++F7sqmqade2LEDLACVAe/Iwf8Ax2tI8b+EdeZI9K8Q2c0jnCRNJskJ9kbDfpVVVVVVTokiGN1DKwwVIyCK/szTf+gfbf8Aflf8KihhgTy4IkjX+6i4H5Cvjr4rj1TWYfDVlKrwaZlpypzmYjp/wFePqzA9K+AXh9rrWLzxFKh8uyj8mIkdZH64+ij/AMeFVVeMfiLZ6OsmnaM6XN9gq0gOUgP/ALM3t27+lSPf6xfb3aW6u7l8c5ZnY9K8B+BU0JF1bVEDai6/InUQAjkf7xHU/h65qqqqqqrxJ420Xw2hjmmFxd4+W2iOW/4Eeij6/ka8M6/beJNJi1GAqsmNs0YP+rcdR9O49jWp69o2jJu1PUoIDjIRm+Y/RRyfwFar8XNMgzHpFhLdMOBJKfLX6gck/jitV+IHinVgUbUPs0ZH+rth5f6/e/Wo4rm9uBFDHJPPK3CqpZnP06k1pHwy8SakVe7iSwhI+9Mct+Cjn88VpPwu8PWAD3xkv5QP+Wh2ID6hR/UmoLa3tIlgtYI4Y1GFSNAoH0AqqqqqqqqqiARiioPQ0R2qqqqqqqqIBGKIx0NVVVVVVUQCMURjiqqqIBHBojtVVRAIxRGOlVVVVVVMARx1qqqqqqogEYzRHav+Cvv7Q39pa3ov7Nnhu/zBpOzVvEnlt1uHT/Rbdsc/LGxlKnIPnRHqlf8ABMj9nOL41/HZPGPiTTEuvCvgBUvrpJo90dzeMSLaEg8EblaUjpiIA/eqqqqIBGKZcjFVVVVVVMPSq8beC/DPxF8Jar4H8Y6Tb6no2s2zW95azoGV1PcZ6MCAyt1VgCMEA1+1j+zb4h/Zf+Ld/wCA9SaW70e53XWg6iwJ+12bMQm47VHmrja4AwGGRwVJ/wCCVH7WTeIdJH7MfjzVGk1LSoXn8J3NxKCZ7VQWls+fmLRDLoPm/d7x8qxKDVVRGRgVVVVVVUQAcCvEemjVtGuLQLmQJviwMncvIx9en41VVVfDzUg0c+kyNyp82PPp0Yfy/OvpTAZ4qqqqr/grX8OJfC/7ROnePoLRls/GuiRO85TAku7X9zIoPcrELU/8CHtn/gjl8Rkm0Lx/8IriTD2t3DrlmuR84kQQTnpnjyrfuR83Qc7qqqqqqqqqq8vbSwhM95cpEg7ucZ+nrU3j3Qon2RrdTAfxJGAP1INWXjHQb1gn2owMRwJ12/r0/WlZXUMjAgjII71VVVVVVVVX/BRT9ikfHHw5L8XvhnpTP4/0K2AuLG1gXdrlsuMqehaeNclDkllXy8ElNvwj+K/j34B/EvSPiZ4Av/7P8Q+H7hjGs8W5HBBSSGVDjKOpZGHBweCCAR+y5+0n4H/ao+E9h8SvBkrQzKwtdY06VcSadeqitJEf7y4YMrjhlYdDuUVVVVVVVVVVVVVVVVVVVVVVVeWd3YXc1hf2sttc20jRTQzIUeJ1OGVlPIIIIIPQ18Ib6z1T4S+CdT0+4Se1u/DthNBKvR0a3Qqw9iCDVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVLFFPE8E8aSRyKVdHXIYHggjuMV+2Z8Df+GfP2hPEvgSxs5YdDuJF1HQmdcB7KfLKF7kRuJIcnqYia/4JBftCS/E34FXnwg8Qakk2t/DeZIrRXc+ZJpk24w9eojdZI+M7VEQOMjNVVf8ABXn/AJPK1L/sX9O/9Aav+CQH/JtHiT/serv/ANIbGqqqoAYqqqqAAGKqlHGaqqqqqgMmqqvj/wDFC1+C/wAFvGXxQuXiD+H9JmmtUlcKstyRst4yT03zNGnQ/e6HpUMOt+MPESQQiTUNY1u9CqCRvuLiV8Dk4GWZv1r4S/DrTPhT8NvDnw70hY/I0KwjgeRF2iaXGZZcdi8hdz7saWMUEGMCv2wvDN940/bv1zwVpl39muvEOp6NpsUp6I1xZWkWTyMj5+RkZFfDD4PfD/4Q+D7bwT4H8PW1nYQwCOZ3jVprwjOXnfGZGJZjzwNxAAGAP+Cgfws0n4I/HnSvFXw2VdAh120TU7ePTmNu1jexSkNJDsx5Yysbgqchi2MYFfsifALwn8E/hFoI07T7KbxDrOnxXer6ssC+dO8qq5iEmAxiTIVVPHG7ALGv+Co/wT8K6Jp3h/41+H9Ht7DUdS1NtN1iSBQgvHeJ5IpGUcFwIpQX6kFQScDH7IfjXUPiR+zZ4C8W6rczXF7NpptriaYAPLJbSvbs5wTkkwk56nOTgnFf8FVV2/tDeHh/1Jdt/wCll5X/AA9V+AH2bcPBPxA8/Zny/sVnt3Y6bvtOcZ74/DtXxX+L/wAdv+Cg/ifT/AHww+H93p3hTTrsOYRK0kSSH5RcXs+0IoVSSqAZG5gPMOK+BXwf0b4G/CrQPhno0iT/ANlW/wDpd0E2m7uXO6aUjkgM5OASdqhVyQor9r+78Uab/wAFANWv/A1p9q8R2utaFLo8Hlh/NvFtLMwrtPBzIFGO+aPxg/4KhHj/AIVT/wCUKL/4uv2fviP+3j4h+L2gaP8AGf4f/wBm+Dp/tP8AaV1/ZUcPl7baVovnDEjMojHTvjvX/BWu/vIdO+GGkxzYtbmbVZpI9o+Z41tVQ56jAlcY6HPPQY/ZpsLSx/Z1+GMFnAsSN4R0yUqo4LyW0bu31LMx+ppowelNH6UyA8GnjBHFPH6UyAjBr9vTxQvhb9mLxTGrYuNae206DIBGZJlL5yR/yySTGMnOOMZI/Yw8GJ4//aq+F/huW1+0w/8ACRQXk8JiEiyRWubmRWVuChWFgwP8Oaqqqq8UeL/C3grTTq/izxBY6TZg7RLdzCMMfRc8sfYZNfE39vPRrB59L+FXh86o4BVdU1ENFCD2KQ8O4x/eKEHsRXj34q/EL4nXa3XjfxVeakI2LQ27NshhJ7pEuFU4OMgZxjJr4Z/s7/Fb4sKl14Z8OmDTWIH9pX7GC364yrEEvjvsDYr4VfsUfDfwT5OqeNW/4S3Vkw2y5i2WcTe0OT5ncfvCQeDtWrOystOtIrDTrSG1toECRQwxhEjUdAqjgD2FVVVWM8CgABgVXjDx/wCCvAFh/aXjPxPp+kwfwfaZgryH0RB8zng8KD0PpXxD/b48I6Z59h8NvDlzrVwjbUvr/Ntbkf3lT/WMO2GEZ79ufH/7Tfxn+IjvHqfjC402zcnFjpLG0iA54O073HPR2boPSvD/AIX8SeLb9dK8L6DqGrXjdILK2eZ/qQoOBweenFeBP2Gfi54n8q58Uzaf4WtH2ki5f7RcbT3EUZxkD+FnU5IHrjwN+xD8G/Cyx3HiGG+8UXij5jeymKENnqsUeO3GHZxyfbGjaDoXhyxXS/DujWGl2aHK29lbJBGDgDIVAB0A/KqxngUAAMCqqqqqqqAz3rAAwKqqqqqqqvif4gbxF4xvZo5S1vaN9ngGeAqcEj6tuP418DfDC6doMniK4iIuNSbbEWGCIlOBj6tk+4Ar4g+JzruqmztZc2VmSqYPDv8AxN/Qe3Pc18PPCC6zc/2tqMebO3bCIePNcfzUd/fHvVeLvh9aa8z6hpzrb35GWz9yU+/oeOo/GvB3hODwzYfvAkl9MP30oHT/AGR7D9f5VVVXxc8fSeGNOXRtIuQmp3q8up+aCPuw9Ceg/E9hWk6TqXiHUotM0yBri7uGO1c/iSSeg7kmvBnwh8P+G447vVIo9U1EAEvKmY4z/sIfw+Y88ZGOlAZ4FBQeM0F4xmgABgV400m71zwrqWkWCobi6gKRh22jOR3r/hR3jz/n3sv/AAJFT/CP4iQRGV/DbkL1CXETn8ArE1c+APG1qyrL4U1RiRx5dq0n/oIOKn0HXLZmjuNGvomj++r2zqV+uRxVW17eWe77HeTQb8bvLkK5x0zirbxv4xsyht/FWqgRjCI127KBjGNpJHT2qz+MHxBs9i/2956Jn5JreNs59Tt3frVj8ffFcChL7TtNugAfmCNGxOe+GI6cdBWn/tC6VIFGq+HbqHn5jbzLL26gNt79v1rTPi94B1PC/wBtfZJP7l1E0fp/Fyvf17HtVlqOn6nD5+m39vdR/wB+GUOPzBNVWvaFpviXS5dH1aJ5LabG5UkZDkHIOQexAODxkCrn4D+B59vlTanb7c58udTu+u5TV/8As8KQG0vxOc4+5cW3fudyt9OMfjWq/Bfx5pmWi0+C/QdWtZgfT+FtrHr6djV/pep6VIINT065tJD0S4haM9uxA9R+daN458XaAFTSfEF3FGgwsTP5kajGOEbKj8q8IfHHW9Q1G00bVtEgu5buZYo5bZvKYFiACVOQefcDH0qqqqqqr4pfFKLwxE+haFMkmrSLiSQci1B7n1fHQdup7A2NjqXiDVYrG0SS6vb2XC5OWdieSSfxJJ9ya8G+GLfwj4dtdEgZXeIFppAMeZIfvN/QewFVrGt6XoVt9q1O8SFSDsUnLOR2UdTXin4j6nritZaaGsrNgQwDfvJB7kdB7D3yTWjaHqev3gstLtjK/wDEx4VB6sewrwl4H0/wzELh9txfsvzTEfc45Ceg9+p/SqqqqqqqllihjaWaVURBlmc4AHqTXxHn0C81sXeiXiTySLi58tfk3DoQ3Q5HXHHHXmrLVtU06KWHT9Qnt0nAEgikK7sfSooLq+n8uCKW4nkP3UUuzE+w5NaZ8M/FGohHmt47ONu874bH+6Mn8DitK+E2iWoV9Uupr58cqP3SfkOf1qw0rTNLj8rTbCC3U9RHGFJ+p71RAIxRXjg0VB6URjiqqqqqqqqqIBFEY4qqqqqqqqIBGKK46GqqqqqqqIBGKIxxmqqqYZHFVVUQCMURjiqqqqqqmA6iqqqqqqvGvizRPAXhHWfG/iO58jS9BsJr68kyARFEhdsZIGcDAGRk4r4q/ETXPi58R/EfxL8SSs2oeI9Rlu5VZ9/lBj8kYOB8qIFQcDhR0r9hz4CRfs9/s6+G/C97p7W3iHV4V1XxAJU2yLeTopMTe8SBIsdMoT3NVVVVMARxTD0qqqqqqIBGM0R2qv20f2adO/ad+C2oeFLaztR4q0rN74avJTsMN0o5iL9o5VyjA/LkoxGUUjTNT8XfDDxvb6rps91onibwtqQeJyoEtndwSd1IIyrrgqQRwQQRX7MHx+0H9pb4OaP8TdGWK3u5c2usWMbFvsN9GqmWLJ5Iwyup7o6k8k1VVTDniqqqqqphkcVXiSwGna3d2qptTzCyDH8LcjH54/Cqqq0bUG0rU7e+U8Rv84x1U8N+hNIyyIrowKsMg+tEZGBVVVVX/BWT4Xt4x/Z1svH1jYJLfeBdXjnll2gvHZ3H7mUDvgym2Y47JkjAyP8Agnz8T1+Fn7Vvgy9u79rbTfEMr6Hf4JAkW6G2INj+EXAt2OeBtycYyKqqqqqqqq1nV7TRbM3VycnpHGOrt6D/ABrVdVvNYumuruTJ/gQdEHoBVVofibUNEcRo/m22fmhc8D1K+h/StL1S01i0W7s5Mg8Mp6ofQiqqqqqqqqq/4KXfsSR6S9/+0h8JNGiisX/eeKtIs7dsxyEktfoq5AU5HmgBQpHmc7nI/Y6/aq8V/sl/Fy08caQ0t54f1HZaeJNJHzC9s94LFFLACdOWjckYJKk7XcH4f+PfCfxS8E6L8Q/A2sRanoPiCzS7sbqM/eRh0YdVdTlWQ4ZWUqQCCKqqqqqqqqqqqqqqqqqqqqqq+L8Mtv8AFnxtbzxNHLH4i1BXR1KsrC4cEEdj7V+yvdWt5+zJ8JZbO5inRfBOkRlo3DAOlnErrkd1ZSpHYgg9Kqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/4Kv/BFfHXwSsvi1pNkj6t4Auc3LJGTJJYXDIkg46hJPKfnhV8w8ZOf2Dvj0n7O37TnhLxrqN8lroOpTf2Pr8kkhSNLG5ZVeRyP4YnEcxGD/qvWqqq/4K8/8nlal/2L+nf+gNX/AASA/wCTaPEn/Y9Xf/pDY1VVQGeBRweBVVVAAnFVWM8CsDGBVVVVVUAMVVV/wWE+LDaF8MfCfwd065ZbjxTqL6hfqoBH2W1ACI2eQGlkRhjvAeR0P/BP34Z/8LD/AGjNI1K7tvN07whC+rzloty+amFgGegYSujjv+7OOmQqADFKmaWMYr46gf8ADz/TAP8Aoc/C/wD6KsaEea/4KxLt+IXgMf8AUFuP/Rwr4boP+FeeFv8AsC2n/olK/wCCrC4/Z38Pcf8AM6W3/pHeV/wT3Td+x/4AIP8A0Ev/AE5XNf8ABVpdv7RHh4f9SXbf+ll5Q/Z1+ABuPtg+Bnw++0b9/m/8IxZ7t2c7s+XnOec1ZaZZabaRWGnWcFrbQrtihhjCIg9AoGAKMQxjFfHpcf8ABUfTFH/Q6eFv/RVhRjPQGih6Yr/gqJ8PL7xP8DNM8aabD5jeENXWW6UJkrbTr5bMCATxJ5ORwMEk9BX/AATq+L+mfEb4A6f4Nmu1/t7wMxsLqBnG97Yktbygdk2Hy/8AehboCKaP0pkyMGnjBHFMmOBTICMCnjBr/gqf48RbfwV8LrebLM82sXce8cAAwwHGc85uOSAOOCecf8EffhzP4j+P2v8AxEmizZeDtBaNXBGVurt9kY5B4MUdz0IOQOoyKqqr4gfFf4efC6zW78ceKbTTjKMw25JknmHTKRIC7DPBbGB3Ir4nft5a/fvPpnwq0OLTLYgqupaigluP95IslE/4Fvz6CvEvizxP4y1E6t4r8QX+r3hyBLeXDSlQTnC5Pyr/ALIwBXwx/ZG+LfxGjg1G60xfDmkSkEXeqKY3df7yQ/fbjGCQqnIwetfC39kP4T/Dd49SvrE+J9XjO5brVI1aOI+scP3B2OW3MCMgjpSRxxxrFEiqqDCqowAB2AqqqqqnaOKNpZXVVRSWZjgKB1Jr4i/tf/BvwCz2VnrD+JNRTINvpIEiKQcfNKSEHfgFjx05GfiF+238XPF/m2fhl7XwpYSZAWzHm3BU54Mzjg8jlFQ5Harq61zxRq73V5c32rapfSZeSV3nnnf1JOWY8V8P/wBjn40eN/LutR0VPDNg/wDy21cmOTvnEIzIDwPvBRz1r4f/ALDPwp8Mbbrxfc3viy7VgwE5NrbjHpEjZPPUM7A8DHXPh/wv4b8J6euleF9B0/SbRekFlbLCufUhQMnk8+9VVYzwKAAGM1VVVVVVQGe9YGMCqqqqqqqrxRqjaJ4c1LVoz+8tbV3j4z8+Pl/XFWVpNqF5BYWy7pbiVY4xgnLMQB056mvFN3D4P8GR2GntsZIktLf1Hy4J+u0E59a0nTJ9Y1K30y1H7y4cKDjO0dz9AAT+FWFha6ZZQ6fZx7IYE2oP896qqqqqvGOuv4m8TahrLPuSeYiHjGI1+VOP90D8c18IfB0Ph7w3Dqk8KnUNUjErsQMpGeUQHtxgkep9qCiguR1rAAwKqqqqq4tre6j8m5gjmTOdsiBhn6Grjwp4WvHEl14a0qZwMBpLKNiB6cirj4V/D66jEcnhm3AByPLd4z+asD+FXnwK8C3OfIXULXLZHk3GcD0+cNx+vHWr79ni0ZSdN8TyowBws9sHyewyGGPc4Nal8BvGVoGawuLC+UD5VSUxuePRgAOeOv5VqngHxno4LX/hu9VVOC8cfmqOnVkyO4q1vLuxnW6sbqW3mQ5WSJyjKfYjkVovxi8c6OUSTUk1CFBjy7xN5P8AwMYbP1Jrw98efD+oMtvr9lLpshAHmqfNjJ79BuH5H61puq6ZrNqt5pV/BdwsOHhkDD8cdD7Gqqrm1tb2E293bxTxt1SVAwP4Gtc+D3gbWlZo9NOnzHpJZtsxzn7hyv6f0rwr8GF8L+LbXXV1hLy0tVdkjkh2uHKlRnqCBnOeDkCq+LHiW58MeEJbnT7kwXlzKkMDg8qScsR/wFSPxrSPjx4tsmRNVt7PUowfmLJ5Uh+hX5R/3zWmfH/w1chU1TSr6zc9Sm2ZBx65B/8AHas/ip8P75QYvEsCHIBEyPEQT/vAfn0r/hNfBv8A0N2i/wDgfF/8VUvjnwXDGZH8W6QQo5C3qMfwAJJrR/H/AIS8Qar/AGLo2rLdXIjZyFjYLhcZwxAB69s9DVfFf4ZyeLY49Y0GGP8AtWIhJELBBOnTkn+Icck9Mj0r4d/DbT/A9qZ5nS61SYYluAvCL/cTPQep6k/gBLLFDG0s0qoiDLM5wAPUmvE3xTtrcPZ+HFE8o4NzIvyL/ujqT7nj6ir6/vtWujdX1zJcTycFmOT7Aeg9q8MfDDUNRKXeul7O2xkRYxK/1B+6Prz7d60zSdP0e0Sy021SGJR0Uck+pPUn3NVVVVVVOyIhd3CqoySTgAeteIPilpOn7rfRk+3zjjfnbGp+vVvw4961vxPrXiGTdqV6zoDlYV+VF+i/1PNaJ4T17xCQ2m2LNFnBmf5EHryev0GTVn8JdRGqxQ315E1iEDSyxHDE90APOffpj34rT9L07SoBbabZxW8YHRFxn6nqfxqqqqogEYoqCODRUdBRGOKqqqqqqqqIBGKK+lVVVVVVVRAIxTLjpVVVVVVVEAjFEY4qqqmHcVVVRAIwKI7VVVVVVEAjFEY4zVVVVVV/wVm+Lf8Awgf7N8Pw/sLzy9S+IGppaFVm2P8AY7crNOwxyRuEEbDgETHJ5wf2F/hAnxr/AGoPBXha+svtOkafd/2tqysAUNtajzNrg5ykkgjiIAz+87dRVVVVVMo6CiO1VVVVVTDuKqv+CsH7LX9ga9D+0x4L0/Gn61Ilr4phjXAguwAsNzjPAlUBGwAA6KTlpSa/4J0/tOj9n340w+H/ABHd7PBvjmSKw1RmPFnPki3ueSAArvtc9kdjyVAqqqiMjAqqqqqqIBGKIxxmviPYYktNSReGUxOcdxyP/Zvyqqqq8D6n9v0cWsj5lszsOTzt/h/Tj8K+lMADgVVVVfF3wBZ/FT4W+K/hxehPL8R6Pc2KuzbfLeSMhHzhsFW2sDg4I6HpX/E38L65/wAtbHVNIu/o8E8T/wA1Zf0r4R+PrP4qfC3wn8R7Ep5fiPR7a9ZFXb5byRgumMtgq25SMnBHU9aqqqqqqqq8Uaw2sapI6ODBCSkIB4IHVvx6/lVVVaXqt5pF0t1ZyYI+8h6OPQitK1S01izS7tHBBGGQ9UbuDVVVVVVVVV1a2t9ay2N7bxXFvcRtHLDKgdJEYYZWU8EEEgg1+37+xtefs5eNz4y8FadLJ8OvEc7GyMaMw0mc8m0kYknGMtGzHLDI5KEn/gmj+3FJ+zl42X4U/EXUf+LbeK7wZmkKqNEv5CiC6LsRiAqoWUE/KAHH3WV4ZobiFJ7eVJY5FDI6NkMCMggjqCKqqqqqqqqqqqqqqqqqqqqqv2ydHvdD/az+MNlfqglk8barcqFbI8ue5kmT8dki5HY1+wJrFrrv7HvwzvbNJUSLTZbUiQAHfBcywueCeC0ZI9iM46VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVr2haR4o0LUfDPiCwivtL1a0ltL21lGUnhkQpIjezKxB+tfGn4Yav8GPiv4p+FutrL9o8O6lLbJJIm03EOd0M2OwkiaOQeziv+CcPx4/4X1+yt4Z1DUbzzvEHhPOgaxlNpMluq+S/+1vt2gYt03lx2NVVf8Fef+TytS/7F/Tv/AEBq/wCCQH/JtHiT/serv/0hsaqqpRxmqqqoAYqqUA1VVVVVAZOKqqr/AIKe/EWXx5+1pr+kx3LS2Pg+yttHtc8AFU86bC7QQRLNIuTnOwEHbtA/4JcfD/8Asb4VeI/iHdWmy48SasLa3dl5e2tkwGBycAyyzLjAOU5yMYWPNKgApUzXx2UD/gqHpi/9Tn4W/wDRVhQQHgV/wVnUj4h+Ah/1Bbj/ANHCvhtGD8O/C3/YFs//AESlf8FXU2/s6+HT/wBTra/+kV7X/BPNAf2PPh+c/wDQS/8ATldV/wAFX1A/aK8Ogf8AQlWv/pbe1s9q2cYxWz2r4+Af8PTdMH/U6+Ff/RVhRQHoaaMYwK8S+GdE8XeH9R8LeJNOjv8AStWtZLW8tpMhZYnUqykggjIJ5GCOoINfFb9mX9oD9iv4nf8AC1fgY+rav4bgd3ttRs7YztbW7Elra+hXO5AqgGQgI2Fb5Hwq+Df+CssP9mCH4ifCJm1BFX9/ouobYpTj5j5UoLRjOMDe/XqMc+JP+CsuiLYFfCPwbvZb1sgNqWqrHHH6HCIxf6ZX618O/El341+H3hrxhf6dJp91rWkW15cWkkTRtbySRKzoVbkYJI59Op60yAjAp4+wpkBGK/a8+KMXxd/aA8VeJrC7afS7S4GnaYSuB9ngGzcvfa7h5Bnn9526D/glB8J5fAH7M58ZalaJHqPj3VZNQR8EP9jjAhgU8kEbkmkBwDibnoKqvif8ePhj8I4GXxZ4hRtQCbo9LswJrqTjI+QH5AR0Zyqn1r4pftvePvFscml+AbRfC1g42tcK4lu3HfD4xH/wEbh/eqR/EXjDW2kkfUdb1e/clmYvc3Fw+OSerMcD36V8Mv2IfiP4t+z6l42uofC2mv8AM0Mi+beMO2Ix8qZ5HzMCv909K+GX7PHwq+FBS68NeHVn1JQB/aV+RPcdMZViAEz32Bc5qqqqqq8R+KfDXhDTZNY8U69Y6VZxAlprudYl47DJ5PQADkkgDrXxI/b18L6Usth8MPD8mt3GGCX+oBre3Vs8MI+JJB7ExnkV8RPjf8UPim7J4w8W3dxZsQRp8LeTbKR0PlLhSR/ebJ968A/CP4j/ABQu/svgjwne6gq/fudoigj/AN6VyEB4PGcnBwDXw7/YBt0WK++KXi4yuDk6do4wvBGN07jJyOCAgx2bvXgf4T/Dj4bQGHwR4Q0/S2b78yIXmfjGGlclyOvBOOT6mgM8ZrAxgVVVjPAoDA61VVVVVVQGe9YGMCqqqqqqqxngUFHevjbei18BzQFwPtlzFEARnODvx7fcr4V2C6h4+0mJ0LJFI0xO3O0ohYE+nzBefUiviB4V1jxL9kbTZYNlsHzHIxUksRyOMdB3x+tfDrwjqejajd32sWXkSLEI4QWVs7jliCCemAPxqvHfi3xDoviJ7TTNSMMPkoQnlIwzz6g14B13UPEOiy3mpSI8sdy0YKpt4Cqeg9yaqqqri3jureS1lBKSoUYA4OCMGrz4BeFZiWstT1K3Yt91mSRQPQDaD+ZNQwRwxJBEgVI1Cqo7AdKwAMCqqqqqqqqqqq1jwr4b18EaxotpdN/feIbxxjhhyOAO/YVr3wC8P3m6XQNSuNPkOSIpR50fsBnDD6kmvEfwn8aeHS0j6ab62UZ8+zzIAO+V+8MdyRj3rS9Y1XRLkXmkahPaTAY3xOVJHofUexrwj8ebhGjsfGNssidPtsC4Ye7IOD9Vx9DWlaxpeu2S6ho9/Dd27HAeNs4Poe4PI4PNVjPArBPAFbSeK2n1FeKPB2heMLSOz1y3eRYW3RMkjIUPcjB5445zWq/s8WTgvofiKaPHSO7hD56fxLjHft6fWtR+CPj6yJFvY2t8B/Fb3Kjt6PtPt0q88D+MbAMbrwvqaqgyzi1dlA/3gCK/sTWT00i9/wDAdv8ACl0LW3YImjXxYnAAt3yT+VfDbwb44sPF2mawvhq9hhtp8TPcJ5G2NhtcgPgnCsTgDJ7e20etbR614l8VaT4Yt997NvndSYrdPvP/AID3NeJPGOs+JZGS6m8q13ZS2jOFGOmf7x9z+leHfB2teJXBsrfZbg4a4kGEHrj+8fYfjivDfgbRvDaiWGL7Rd45uJRkj/dHRR+vvRB6YojtVVVVVV4k+IGiaArQRSC9vACBFEwIU/7TdvpyfavEHi7W/Ej4vrnZAPu28RKoPqO59zWg+E9b8Rv/AMS20/dA4aeQ7Y1P17/QZNaB8MNE0vbPqZ/tG4HOHXEan2Xv+OR7ClRERY41CqowqgYAHtVVVVVVVEAjFFeODRUHpRHaqqqqqqqqioPSiO1VVVVVVUQCMUy46VVVVVVVRAI4NEdqqqIBGKYY6VVVTDIyKqqqqqqIBGBRHaqqqqq/4K0fE5vGn7TieB7W4Y2PgTRoLNo9wZftU4+0SuuBkEpJAhGTgxdjkV/wRq+FEVv4f8dfG29gQz3tzHoOnO0eGSONVnuMN1Kuz244wMwnr2qqqqqiARgUw7iqqqqqogEYzRHaq+IHgTwz8T/BOt/D7xjp6Xuja/ZSWl5CwGdrjG5SQdrqcMrdVZVYcgV8fvgz4j+AHxa8Q/CvxLGxm0i5P2W5I+W8tX+aGdccYdCpI/hbcpwVIH/BND9pf/hd/wAFo/AnifVvtHjDwBHFZ3Hm8SXVjjFtPn+MhVMTnk5RWY5kBNVVMOeKqqqqqmGRkV4wsRfeH7lQoLwL5q57bev/AI7mqqqrwdqf9na1EjtiK5/dPx3P3T+ePzqmGRwaqqqq/wCChnw0f4Z/tZeNYIrWWKx8Szrr1m8m796Lob5mBPb7QLheOPlxxjA/4JMfEpPFf7Od78Pri7ia88DazLFFAuAyWl0TPGzY5+aZroAn+714wKqqqqqqrxHdGy0O8uFPIiKqfQt8oP61VVVVoetXGiXouYRvQjEkZOAw/wAfSrS6gvraO7tZA8cq5U4xVVVVVVVVXxP+GnhD4weA9Y+HPjnTUvdH1q2aCZdql4iR8ssZYELIjYZWwcMAa/aV/Z98V/s1fFXU/hz4lVri2RjPpGoiPamo2bMRHKB2bAKuuTtdWGSACf8AglD+2/L4w0+D9mD4sa60ut6bD/xSGo3UpZr22QEvZOzcl4lGY+uYwy8eWu6qqqqqqqqqqqqqqqqqqqqq/wCCmeg2/h/9uD4m2tpDLHDc3FleKZMne01hbySEHuPMZx7Yx2r/AIJYeIW1r9kfS9NN+lx/YOt39ksYUA2+6QXGw4HJP2jfk5OHHYACqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv+Cv3wWe11fwp8e9Jt8w3sZ0TWCqN8sqbpLdyeR8y+ap6Y8tOueP+CPPxzb4fftCX3wj1W9SPR/iRY+XCHwAuo2qvJAdxPyhozcJgcs7RDsKqq/4LE6Pe6Z+1/8AbbpUEereFbC5ttrZJjDzQnPod8L8emD3r/gj1rFrP8BPGHh9El+0WXi97mRiBsKTWluqgHOcgwPnjuOTziqrGeBWBjANVVUBngVgYwKqgABgVVVVVVAcVVVqup2Oi6Xd6xqc/k2dhbvPcSbS2yNFLM2ACTgAnAGa8e+Lb34geOvEfjvU023niPVrrUrhcg4knlaRhwFHVz0AHsOlfs0eA/8AhXHwE8C+D3h8qe00aGS6jxjbcTDzph/38kelUDgUseaVAOKm8H+FbzVBrl14Z0mbUldXF5JZRtMHTG1t5G7IwMHPGB6UsYI4rVvCfhjxBJHNr3hzS9SkhUrG13ZxzFAeoBYHAqGCOGJIYI1jjjUKiKuAoHAAHYVqvh/RfEFstnruj2WpQI+9Yru3WZVcAgMAwIBwSM+5rTdI03SLKPTtI0+2srSHPlwW0KxImSScKoAGSSfqa1bwb4T1+4W813wxpOo3CRiNZbuyjmdVBJCgsCcZJOPc0IxjGa8setGMEYBq58G+E7vVRrt14Y0mbUldJBeSWUbTB0xtbeRuyNowc8YFNFmmj44pk7YpoweBXiL4KfB3xZenUvFPwn8G6zdlmY3GoaDbXEhLHLHc6E5J5Pqa8NfCf4X+CLj7X4L+G3hbQJ9xbzNM0eC1bJUqTmNAc7SR9DimQEYp4+OKZAeDX7avxytPgb8FNUubO8VPEniOJ9O0SLGWEjjEk3sIo2LAkY3bAfvV8JfhvrXxg+Jnhj4YeHTtv/Eupw2UcpjLrbq7DfMyjkpGm52x/Cprwt4a0fwZ4Y0fwd4etvs+l6FYQWFjDnPlwQxrHGue+FUCvHfxF8F/DTR/7c8a6/b6bbElYg+WeZgM7URcsx9gOO+K+MH7afjvxpNPpHw7ebwtohyomRh9unGerSDPlcY4j5Bz85BrS9I8QeLdYXT9H0++1fVL2QkRQRtNLKxOScDJPqT+Jr4YfsI+JNaii1P4oa1/YcDYI0+yKy3JHfc/KRnHpv8AcDpXw9+Evw9+Flh9h8EeGrWxZ02zXRHmXE/+/K2WIzztztHYCgM81VVVVXjv4meBPhnpv9qeNvEtppsZBMUbtulmx2SNcs/4Djvivit+3lqN352kfCHRvsMfK/2vqUavKfeOHlV6cF92QeVBrxX4y8VeOtVfW/F+v3mrXr8ebcyltg9FHRV/2VAFfDX9k34w/EjZd/2H/wAI/prbT9s1hXg3qcHMceC78HIOAp/vCvhv+xX8JPBJhvvEcEvi3UoiGD6gmy3VgOcQAlSDzxIXHSra1tbK3S0sreKCGIYSKJAiqPQAcCqxngUAAMA1VVQAxmqqqqqqqAz3rAxgVVVVVVVWM8CgoA96r4+f8idZ/wDYTT/0VJXwS/5HuH/r1l/kKqq+KtpJD4ijuSCUuLZdpxgZBII/kfxr4T61Ai3Hh+UBZHczwt/e4AYfkAfz9KqqoLkdaC5GBQAHSqqqqqqqqqqqqqrGeleKfht4V8WIz32nrBdHOLq3ASTPqezfiDXjT4QeI/CqPfWg/tPT1yTNCmHjH+2nXHuMj1xXh/xJrXhe+GoaJfPbycB1HKyAdmXoRXhb466Bqhis/EVs2mXLnaZh88BPueq59wQO5qGeC5hS4tpklikGUdGDBh6gjrVVVVVVVV4y+JNtpJk0zQmS4vVyrykZSE98f3m/Qd88ippr7VbxppnlurqduScszn/PavCfwtRQl/4mG5uq2ingf75HX6D/AOtUUUUEawwRpGiDCoq4AHsBVVRAPaioIwKKg9DRU9qIqtd8UaL4dhL6jdjzMZWBMNI30H9TgV4k+I2s63vtrJzY2hPCxsQ7D/ab+g4+taXo+pa1ci00yzknk77V4T3Y9APrXhv4W2Njsu9fkW7nHIgX/Vr9e7foPrUUUUMawwRqiIMKirgAewqqqqqqqqqqIBGKZePWivHBojtVVVVVVVUQCOtEY4qqqqqqqiARimXHSqqqqqqqYAjiqqqIBGKIxxmqqqYY6VVVVVVUwHUVVVVVXxq8fS/FX4veM/iPI7MviPXLu9hDAgpE8rGNPXCptUZ7Cv2Fvhonwq/ZS+Hnh6S2aG9vtKXVb5ZAQ4muz55VwVUhkWRYyCONmMnGSw7iqqqqqpgKI7VVVVVVMO4qq/4Kn/s0f8LQ+FKfGrwvp0T+JPh/bu9+VTEl1pWS0oyBz5JLTAEgBTMRyQD+yx8dtV/Zy+N3h74lWM8/9nwzC11q2iYj7XYSECaMj+IgBZFB43xoe1aDruj+KdC07xN4e1CK+0vV7SK7sbqI5SeCVA8br7MrAj61VUwyMCqqqqqogEYp0WRDG4BDAgg1f2j2N7PZOctBIyZIxnB6/jVVVAkEFTgjpWgaiNW0i3vCRvK7ZB/tDg/4/jWARiiMcZqqqq/4LF/DNrrQPAfxfs4GJsbmbRb9woPyyr50GTnIAMc46EZccg4Df8En/ig/g39ou58AXV55dh460mSBYmkZVa7tgZomwAQSIxcKM4/1nBz8rVVVVVVVXjlmXw9Kqtw0iAj15z/QVVVVVXg3xANNuf7Pu5MW07fKT0jc9/oelVVVVVVVVV+2B+zHoP7UXwoufCs5itfEml7rrw7qLqT9muMDKMARmOVV2NnIGQ2CUFXlp40+E3j17S4+2+HvFnhHVescmyawvLeTIZWU8MjoCGU44BB71+wd+1/o/wC1r8I4dR1Ka3tvHfh1UtvE2nx4UGQ5CXUa/wDPKUKTj+Bw6chQzVVVVVVVVVVVVVVVVVVVVV/wWY8MS6N+1ZpmvKlw0HiDwjaTiR4iIxLHNPC0at0JCxxsR1HmDsRX/BHHxOl58LviD4KFyjNpWv2+oGHyyCguYPL3FuhB+yEY6jac9RVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVftRfB+L47/Abxh8NUs4J9R1DTnk0nzm2BL6L95bnf/ADIqqT/AHWYHIJB8JeJdc+HvjTRfGOin7PrHhnVIL+085CPLuLeVZE3DIPDoMjIP0r4U/ETRfi58NPDHxO8Pcaf4m0uC/hjLbjF5iAtGTgZKtlTwOVNVX/BayGVf2ovCk5icRv4BtVVypwWF/fEgH1AZcj3HrX/AARtvrOTwD8SNMjuEa6t9Ys5pIu6o8LhGPsTG4/4CaqqUAnrR5qqqlHGaqgATVVVVVUBngVgYwKqq/bp8cN8Pv2SviXrsM7Rz3OjnTYSmN+68dbb5ckcgTFsjkAEgHGK+DvgtfiN8V/B/gWSNnh1zW7W0uAoJKwvKolbAIOAm48EHjqKVRjApI8jilUYwKSPPFLGMUEyMCljzQjAFBM8AUIyeM0IsivKBHAryvajEMdKMXpRjPQGinbFNGCMCmj7UydqaMEcU8fpTKDwa8ZeKfD3gTwxqXjDxXqkGn6TpNu091cTOFCKOwz1YnCgdSSAOTX7Tvx+1j9ov4o3njS6jmtNJt1+zaLp8j5+y2yngsASPMc/M+O5xkhRX/BKj9lyLwZ4Zuv2pPiPZLZ3ep2zx+GheDyvslgAfOvG3H5RLjapIGI0ZgSsoNfGf9t3w94dM/h/4UwRa5qADI+qy/8AHrCccGMdZSOeeF4GCwJrxX4x8XfEHW31rxZrl5rGo3DYDzNnGT91EHCLnoqgAdhXwT/Yx8XeOPsviL4h+d4d0Nm3C0ddt7coPRSP3Sn1f5u4XBBrwJ8M/Avw003+y/BPhu002MgCWRF3SzY7vIcs34njtiqxngVgYwKqqqvHfxM8CfDPTf7U8beJbTTYyCYo3bdLNjska5Z/wHHfFfFb9u3xBq/naT8KNK/se1OV/tO9RZLlh6pHykf47j9K1TVvEHi7WG1DWNRvtX1S9kCmWeRppZWJwACck+gFfC/9ij4neN4otT8WsnhLTnwVS8iL3bj2hBGzv/rCpBx8pBzXwx/Zt+E/wrjguNF8OxX+qwkMNU1FVnuA4/iQkYj/AOAAe5NVVUBgVVVQGaqqqqqqoDPesDGBVVVVVVVAZ4FAADAqq+PUUkngy2dFJEeoxs59BskGfzIH418Fp44vH1pG5O6aCVE477C38lNav4h0bQkVtUvkhLglEwWZsegGTXh/xdpPiaa4g04Tg2wBJlQLuBzyOSe3fHUVXjTw6niPRJLeNR9qg/eW7Y53Dqv0I4+uD2q0urvS72O7tnaG4tnypIwVI7EfoRWga1a+INKh1K1ON4w6HqjjqP8APtVAZ4FBR0HNBRiq+LvxG8VaNqMnhrTbc6bE0YYXanMkykdUP8I6jjnI6ivh98aH0SzXR/FS3N3BGQILlPmkjHo2T8wHY9R054xoni/wx4kAGi63a3LnpEG2ydP7jYb9KqqqqqqqqqsZ4FbSRW0etYHaqqvGnwd8PeKXa+09l0q/Y5eSKLKSHuWQEcn+8MH1zXi34d+J/BrGTU7MSWm7at1Cd0ZPbPdT7ECvAnxJ1rwRcCGNjd6a5Jks3bAz/eQ87T+h79iNA1/SvE+lQ6vo9yJYJR0PDI3dWHYj0/oc1VVVVVahqFjpdq15qF1HBCnV3OPwHqfavF/xIvNZ36foxe1siCrPnDzDvn+6Pb8/StF0LU/EF4tjpdsZX/iY8LGPVj2H+RXhTwPpfhiMSqBc3xHzXDrgj2UfwjBx71VVVVVVq+uaTocHn6pfRwDHyqTlm+ijk14j+Kt9eFrbw9H9khxgzSKDI30HIUfmfcUFvdSu8KJrq5nboAXd2P5kmvDnwoubkJdeIpTBGRkW8Z+c/wC8eg+g5+lWGmafpVuLXTrOK3iH8KLjPufU+5qqqqqqqqqqqqoqD0NMuaIxVVVVVVVUQCMURjvVVVVVVVEAjFMuOlVVVVVVUw7iqqqIBGAaI7VVUQCMURjiqqqqqogEYojHGaqqqv2uPG4+HX7MnxM8WrdJbz23hu6gtZWZAEuJ08iE/P8AKT5kqYU53HjBzivh54SufH/j/wAM+A7Ld9o8R6xaabFsKg755ljGCxAByw6kD1qwsLPS7C20vTrdILWzhWGCJBxGigBVHsAAKIBGM0R2qqqqqiARimGRxVVVVVUQCMZojtVXdpa39rNY3ttFcW9xG0c0MqB0kRhhlZTwQQSCD1r9tf8AZ6uv2cfjzrfhSzsZYvDWqSNqHh6YqdjWshz5QbuYmzGe/wAqk/eFf8Emv2kv+Ev8DXn7O3ia63ar4Rja80OWSTLT6e7/ADxc8kwyPwc/clVQAI+aqqYAHAqqqqqqYelePbEWuumdB8t1GH6Y5HB/kD+NVVVXw81Hy7ifSnfiVfMjB/vDg/mMflVMB1FVVVX7bnwy/wCFsfsu+PvDMFv5t9aaa2p2ADYbz7UicBeDyyxsmMc78ZXO4fCPx9d/Cv4o+E/iPYljJ4c1e2vWRV3GRI5AXTGVyGQMpGRkHqOtWt1a31rDe2NzFcW9xGskM0Th0kRhkMpHBBBBBFVVVVVVV4vhM3h28VcZVVYZ9mBP6CqqqqqrwX4iN9CNJvJM3ES/umPV1Hb6j9aqqqqqqqqr/gpv+x7/AMLG8OT/ALQXw803d4n8O2hOv2sQ+bUbGNc+aMsB5kKq3AGXQ45KKD+zb+0H42/Zj+LOk/FbwPL5k1pmDULB5CkWpWblTLbyEA4DbVIODtdUbBKivhR8U/Bfxr+Hmh/E74f6ql/omu2qzwNuUvCxHzxShSQkqNlHXJ2spFVVVVVVVVVVVVVVVVVVVVf8FxPB+R8KPH1un/QS0y6Yyf8AXCWEBf8Av/k/7tf8EdPE89p8XvHfgtWl8nVfDceoOARs3W1yka5HXOLtsfj61VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVf8FE/g8PhD+1D4j+xW3laV4v8A+J/YYHA+0M3nKOTjE6zfLxgFeACK/wCCMnxqHi74Ja/8FNRmiF54B1D7RYL5mGeyvGeQgKeuydZiWHA81AQDy1V/wW9/5Kn8NP8AsX7r/wBKBX/BGGWIS/F+AyoJGXRGVCeSB9uBIHoMj8x61VUBxgGqqqxngVgAYBqqA4qqqqqqUcVVVX/BYTxsNG+BHhbwNBdJHceJPEYnkQsmZLe1hcuAp+bAklgO5emACfmwf+CbnhJPE37UGm6m67l8M6TeaiVJAGSgtwSCDnBuQRjBBAOeMFE4xSqDwKSMdBSoMYpUzSoMUEzwBSxg0sfGBQjGMUEHYVs9BWz6UUz2ooCMYoxgjANNHxTR+lMnGKZARxTpjpTICMCtVvrDR9NutX1S7jtbKxgee4nlbasUaKWZmPYAAk1+2n+2XqX7Q2st4M8GSTWPw/0q43wI6lJdVlXIE8o6qgydkZ6Z3N82An7PHwv0fxL4usvEnxH0jUZfCOnA3L20GIm1aRD8luHYHZGzffkCthQwA3EY+KHx28efFQR6bql7Hp2gWuFstD09fJtYEXAQFB98qAAC2cc7QoOK+E/wF+IvxjuseF9K8nTo22zapeBo7aMjGVDYO9hkfKoJGRnAOa+Dn7Lvw4+Eawamtquu+IYxzqt7CP3bZ6wx5Ii+oJbqN2DiqqgOKqqq9vbLTrSW/wBQvIbW2gQvLNNIESNR1LMeAPc18c/23bHSjceGPg0Yr28UtHLrk0e6GI4x+4Q/6wg/xsNny8Bw2RqmreIPF2sNqGsajfavql7IFMs8jTSysTgAE5J9AK+FX7FHxI8beTqnjVv+ES0psNsuYt95KPaHI8vuP3hUjg7SK+GHwF+GPwjgRvCXh5G1AJtk1S8xNdScYPzkfICOqoFU+lAcZqqqgM81VVQGeM1gYwKqqqqqqAzxmsDGBVVVVVVVAZ71gYwKqq+KumHVPAeqRou6S3jE6deNjBmP/fIavB+qLovinStUdgqQXSGQk4whOG7j+En2r4o6Sb7QU1CJcyWEm48fwNgN+u0/QGvCGuDw/rsF9Ix8g5jnxn7h6nA64ODj2pWSRA6MGVhkEHIIqvif4b+w3667aR/uLs4mA/gk9f8AgQ/UH1r4X66bHVm0aZ/3N8PkH92QDj8xx+VBcjk0FyKwMYFVXivwhovjLTf7O1eAnYSYZk4eFvVT/MHg14j+CPi3SGebSVj1W2UkjyjtkA90PU9OFJ61c2t5p1wbe7t5ra4iPKSoUZSPY8itH+JHjbQwqWPiG5aNT/qrgiZcccYfOBx2x39a039oLX4Aq6roljdhRgmF2hY8dTncM556VY/tBeHJVA1HRdRt2IH+qKSgHvySpwPpz7V4W8XaP4xtJr7RTO0MEnls0sRT5sA4HrwR+YqtX8Y+F9BuhY6xrdtazlA4jkY52nIB/Q1/wsvwF/0NFl/30f8ACrj4s/Dy2cRyeJYmJGR5cEkg/NVI/Crj42fD+GTZHqNzOMffjtXA/wDHgD+lXP7QPhdVX7Jo2qStnkSCNMfTDNV1+0PctgWPhaJME8y3ZfI+gUY/WvhV4+1Lx3b6idUt7WKayePaLeNlBVg2M7mPOVNbcjrXxu8WeIvD2paXa6Jqs1mkkDu4jwCx3Ac/l+tQfFP4gW0gki8T3JIGMSKjj8mBFRfGb4jRyK76+koU8o9pEAfrhQf1r/heXj3/AJ+LL/wGH+NaT8WPinr12tjo9vBdzn+CK0Bx7k5wB7nivCtp4+ZVuvF+r2YJHFpawDI/3n/oB+NVVXVrbX1tJZ3kCTQzIUkjdchgeoIr4p/CyXwxK+u6DE0mkyN+8jHJtSex9U9D26HsT8OfHVz4I1kTOWk065IW7hAySB0ZRn7w/UEj6W9xBeW8V3azJLDMgeORDkMpGQQfQiqqqqvE3xI0bRA1tYst/dgY2xv8iH/aYfyH6VrniDVfEVz9q1O6MhH3IxwifQdv514T+HmpeIdt5eh7OwIyJGX5pf8AdB7e549M1pWj6bolotlpdokEQ5IA5Y+pPUn61VVVVVa1438N6HujutQWWZR/qYPnb6HHA/Eitc+K+r3ytBo8CWMRGPMPzyfgeg/LPvTyXupXO6SSa6uJTgFiXdj/ADNeH/hXq2o7bjWZPsEB58vGZW/Dov48+1aH4a0bw7CYtLtAjMMPKx3O/wBT/TpVEAjFFQelFeODRHaqqqqqqqqqqqIBGKK8cUV9PyojtVVVVVVVEAjFFcdDVVVVVVVEAjFEY6VVVVVVUQCMUwx0qqqmAIyKqqpgCOKqqqqqqYZHFVVVX/BWvxc3h39k86FFMwbxR4jsrF0VwNyRiS5JI6lQ1unToStf8E3/AAWnjX9sTwHFcJuttFkudVmwQCDBA7REAg5/feVnvjJBBGaqmAIyKqqqqqmA7URjjNVVVVVTDuKqv+Ckn7PX/C8v2fL3XdEsfP8AFPgDzNW03YmXmtwo+1wDv80ahwoGWeCNR1NfAn4ua58Cfi14Z+KmgFnn0K9WSaANt+027fLNCfZ42dc9iQe1eEvFOh+OfC2keMvDN8l5pOt2UV5ZTr/y0ikQMpx2OCMjseKqqYZHFVVVVVRGRgV8RrPzdPtr1RkwylD9GH+Kj86qqqtLvn03ULe+jJzC4JA7juPxGRSMsiK6MCrDIPrRAIwDVVVVLFFPE8E8aSRyKVdHXIYHggjuMV8efh3L8JfjR41+HD27QxaBrdzb2gbq1sHJgflmIDRGNsEk4bk5zX7BXxGf4nfsn+AdWuZM3uk2B0e6BLEhrRjChLEDcWiSJyRnlyMkg1VVVVVVV5brd2c9oxwJomQ/iMU6MjGNxhlOCD2NVVVVVBNLbTJcQSFJI2DIw7EV4e1yHXbETLhZ48LMmeh9foaqqqqqqqqr/go1+yBL8CfHjfE/wLpzf8IH4suWYoi8aTfNlngPpG4BeM9vmTA2At/wTG/bUl/Z8+ISfCb4gam5+HnjG6VA8jHGj6g5VUuB6ROAEkHb5HyNjBqqqqqqqqqqqqqqqqqqqqv+CxfgdvE/7JMfie3t0Mvg/wATWd7LN5AdlglWS1Zd2QUUyXEJOM5KKCOhH/BODxV/wiv7YPgcSSbbfWPtenTYj3E+bbSeWB6fvViyfTNVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX/BXD4QReKfgxovxfsLdPt/gjUBBeOFwWsrtljOSBklZxBtBIAEj9zz/AMEzvjYPgt+1n4XF9d+Vo/jTPh3UNx+UfaXTyGPBxi4SHLcYUtkgE1Vf8Fz/APmiX/cwf+46v+COH/JS/iH/ANgK2/8AR5qqAycUfTNVVUACaqgMnGaqqqqqsZ4FYGMCqqq/4LI+MZNR+LvgTwEsu6HQ/D0uoYG0hJLucow4+bO20jOCcYIIxk5/4JMeFWaX4h+NZWYBVstPgAYYJPmySbhjORiLBzjlvwVeMCkjpUAGBSJmlQDpSx5pY6CDoKEZPFCIHpQiHSvLGMZryx60YwRgGjECMCmjzTIR0pkBGMU8eRxTx+lMgIx3rXtY0fw1o974g8QanbadpunQPPd3dzII44I1GWZmPAAAr9rT9svxx+0x4hufhd8JFvrbwIr7BbxR7J9ZKHPnTE8pFkArGSB0Z/mwE+H3wC0nRlj1PxiIdSveCtrjdBFx0YH75+vHseteFfCPiLxnq9v4b8I6JcalfzD91bW0eSAOpPZVHcnAHrXwW/Yh0TRFtPEXxcli1bUABIujwsfs0DZyBI4OZSOMrwmcj5xybOystOtIrDTrSG1toECRQwxhEjUdAqjgD2FVVAZqqqvjD+1F8NfhGJ9LlvP7c8QRodul2MgJjfsJpORFyOQcsAQdpBFfFv8AaB+I/wAY7lo/EWrG20hZC0Gk2hMdunPBYdZGGB8z5xzjGSK+EX7PnxF+Mt0r+HtN+x6QjYm1e9Vkt0xnIQ4zI3GNq5wSNxUHNfBr9nnwD8F7ENo9muo63IMT6xdxKZjkcrH/AM8k/wBlevG4tgVQGaqqoDPFYGMCqqsZ4FAADANVVVVVVAZ4FYAGBVVVVVVVBc96wMYFVVVdW0N5ay2c67o5o2Rx6gjB/StV0+bSdTu9LuAfMtJnibIxypxnv6V4N1SHxR4N0+9uCJ/tFqIrkOM7nA2PnPqQfzrxToE3hzV5bBxmI/PA/wDeQ9PxHQ+4r4b+MEkij8OalKqugxaSMcbh/c+vp+XpVeI9ITXNEu9NZAzyRkxZ4w45U/mB+GahludOvI5kDRXFrKGAZcFXU9x7EV4Z8QWfiXS0v7YhZB8s0WcmN/T6ehqqqqrUdL03V7c2mqWFvdwn/lnPEHH5GtU+CngPUm3w2VzYMTybWcjP4NuA/AVqH7PEoBfSvE6Nz8sdxbbccf3lY9/9mr34F+OrUEwJYXmFyBDcYyfT5wvP6V8I/C+reFPC0thrVv5FzNePMY96ttBVVHKkg/dz+NV8Y/DXiLVPGst1pmgajdwm2jAkgtHkXIHIyARX/CFeMv8AoUtZ/wDACX/4mrf4e+OLpS0XhXUhtOD5lu0Z/ANirf4S/EO6jMkXhqUAHGJJo4z+TMDVt8CfHVwwWVdPtwRktLcZA9vlBNWf7PGrSbRqHiSzh4+byYGlwfbJXP6V4A+G1v4CluZrfWJ7truJFkVowi5XPzAAn1PXpmq+Mcvg/TrWy1LxH4Zn1a4cPFbFZ5IkTGCQ7KwxnOQMEnB9KvriC6uXmtrGGzjY/LDEzsq/i7Mf1rQfCniHxPN5Oh6VPdYba0irhEP+054H4mvC/wAAIEWO68W6kZHzk2lqcKPZnPJ/AD6mtJ0XSdCtFsdH0+C0gX+CJMZPqT1J9zzVVVVdWttfW0lneQJNDMhSSN1yGB6giviH4Ol8FeI5dNQO1nKPMtJHOSyHsSO4OQfwPcV8C/Fy3ug3Hh2+m/eaSDJEzd4SeR/wFs/gwHarDxBoeqhRp+rWs7MOEWUbv++eo/Kqq/1rR9MB/tDVLW3K9VklAP5da1f4r6BZho9Mimv5B0YDy0/M8/pXiDxzr/iHdDPdfZ7VuPs8GVUj/aPVvx49hWkaDquvXAttLs3mOcMwGFT3J6CvC/wx0zSdl3rPl312OQhGYkPsD978fyqqqqqtW8UeH9EBGo6rBG6j/VA7n/75GTWrfGBBmPRNK3Y6S3TY/wDHV/xrWPGHiLXFMd/qchiIx5UfyJj3A6/jmtN0fVNYm8jTLGa5fvsXIX6noPxrRfhFfT7JtdvktkPJhh+d/oW6A/TdWjeGND0BMaXYRxuRhpD8zn/gR5/DpVVVVRAIxRUHpRU9qI7VVVVVVVVVVUQCKZc8GiMcVVVVVVVRAIxRXjg1VVVVVVRAIxRGOAaqqqqqqIBGKIxxmqqqYY6VVVTDuKqqqqqqYDtVVVf8FpfFKiH4WeCbe5QsW1LULqExnK4EEcLBumDmcYHPAzjjP/BGrwr9v+NHjjxk0Erro/hlLJX8nMaPc3CMCW/hYrbMAO43+hqqIBGKIxxVVVVVEAjGaYcVVVVVVEAjGaI7VVft4fs9f8M7ftB6zoWkWPkeF/EGdW8P7EwkdvKx3wDsPKkDxhck7BGx+9X/AASP/aF/4SPwZqv7PPiK/wB1/wCGd2oaF5jcyWUj/voh/wBc5WDfSb0WqqiARiiMcVVVVVVeKLP7boN7Aq7mERdRjnK88flVVVVXgq/+3aFFGzZktSYmyecDp+GCB+FcUwwcZqqqq/4K3fDZfC/7QWk/EGztmS28a6IjTyMD+8u7U+S+DtAwIfsvGSc5zgFc/wDBHH4jRPYfEH4RXV4gkjlt9csLcvy4YeRcuB6DbajP+0M44zVVVVVVVeMtLGnaw8kY/dXQMq8dCT8w/Pn8RVVVVVVpeqXekXa3dnJhhwyno49CK0vVLTWLRbuzkyDwynqh9CKqqqqqqqq+J/w08IfGDwHrHw58c6al7o+tWzQTLtUvESPlljLAhZEbDK2DhgDX7QHwO8V/s7fFLV/hf4uKTT2DCSzvY02pfWr58qdBk4DAYK5O1gy5OM1/wSp/bPb4w+B0+AXxG1h5/GvhC1zpd3cON2q6auAoJJy80Odp4BMYRiWYO1VVVVVVVVVVVVVVVVVVVX7avgL/AIWX+yf8U/CKQedPJ4bubu2i8veZJ7UfaYlAyOTJCgBzwSDg4wfhf4qHgT4l+EfHHmbP+Ed12y1Hd5e/HkTpJnb3+7071FLFPEk8EqSRyKGR1OQwPQgjqMVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVXxY8Aad8Vvhl4o+G2qPst/Emk3FiZO8LSIVWQcHlW2sODyo4NX1lrvg/xFcaddC40zWdEvWikCSbZLW4hfBwynhldeoPUcGv2dfivafHP4GeCPixavEX8R6PDPdpE4dYroDZcRgjrsmSROg+70HSq/wCC3nhprv4S/DbxgNPR10zxFc2BuiwDRG5t/M2AZyQ32UkkDA8sZIyM/wDBI7xF/ZP7TOqaJLNc+VrfhO6hSKNv3ZljnglV3GR0RJQDgkF8dCTVUo4zVVVUBxVUo4zVVVVVX9/YaXZy6hqd7BaWsC7pZ55BGkY9WYnAH1rx5/wUN/ZE+H8rWt18W7PW7pWx5OgwSaiD6kSxgxYH+/k9s1rP/BZD4KwXSp4f+FXja9t/LBaS8e1tXD5OQFWSQEYxzuHU8cZP/D5b4Zf9EZ8T/wDgwg/wrRf+Cx3wLuPO/wCEj+GHjux27fJ+xLaXW/ru3b5otuOMYznJ6Y58Gf8ABSL9j7xnOtmvxT/sS6eQqkWtadPaqQAPm80oYlHOPmcHg8Y5PhPxx4K8faZ/bXgXxhoniPT9xX7XpOoRXkWQSCN8bMM5BHXsa/4KQeLm8Xfth+OiistvozWumwK6BWAht4w+cE5BlMhB67SMgdK/4JleExoH7MkGttAyv4l1u8vQ7Mp3qhW2GMchQbduG5zuPQikTNKoApEzSqAMDrSR54pUAHNKmeBSx8UEHQV4n+Nfwn8HTNa+IPHWmw3EZw8EDNcyIf8AaSIMy/iKsv2mPgXfzC3g8fwK+CQZrK4hXj/aeMD9a0rU9J12xj1LQ9UtNQtJc+XcWsyyxtjrhlJBry+OlGLNGLI4oocYFMgPGKaMHpTJgYp0BHFeM/F3hbwB4cvvFvjPXrTR9I06Jpbm7upNqoAM4HdmOMBQCWOAAScV8fv2h/id+2d4huvDnhea58N/CexuVEcEihXvWjORLNjl3OQwiB2Jhc5YBj4S8FeHvBWnLp2g2QTj97O+GlmPqzd/pwB2Ar4G/sneNviwbbXtcWXw/wCF3KsLuaP99doRn9xGeoIx+8b5ecjfgivh18MPBPwr0JNA8FaLFZxEL585G6a6YZ+eV+rHk8dBnAAHFVVVjPAoAAYBqq+Jfxg+H3wk00ah401+K2klGbeyi/eXNx1+5GOccY3HCg4yRkV8Yv2y/iB8QHm0jwY8vhXQ8so+zy/6XcL0BeUfc4GdqYxkgswwa8I+CvFfj7WItA8HaDd6rfSkARQJkICcbnY4VF9WYgDua+DH7DuhaC1t4g+LVxDrN+uHXSYCfssTA5HmNwZTwMrgJ1B3jk2dlZadaRWGnWkNrbQIEihhjCJGo6BVHAHsKrGeBQAAwKqqxngUAAMA1VVQGBVVVVVVWM8CgABgVVVVVVVQXNYGMCqqqqvjh4ebTPFCazCgEGqR7jhcYkQBW/MbT9Sa+BHipLe5ufCV5KqrcEz2hY4+cAB1/EAED/ZPrXivw1beJtMa0kIS4jy1vKf4G9/Y9DV3aXukX72twrQ3Ns+Dg4KsOhBH4HNeBfHEeuImlapIqagi/Ix4E4Hcf7XqPxHfAXI4r4l+DDmTxLpkQxjN3Go/DeB/P8/U18MtP8QPqy3+nMYbEHbcu4ykgH8IHdueD2/Q1VVVVVVVVjPArbnqawB0qq8ReOPCvhYFdZ1mCKYDIt1O+U8cfIuSAfU4HvWn39rqthbalYy+Zb3USyxNjGVYZHHb6V4t8M2Pi7QbjRL0YEgzFJ3jkH3WH+ehI714Q+BejaWouvFEyanc44hTKwxn9C/44HtVtaWtlbx2llbRQQxDEcUSBFUegA4FVVVVVXx80uC68JW+ptgTWN0Ah25JVxhh7chT+FeBLvVLfWZbPSIGmm1CzmtmjQZZkK7mwO5wucVcW1xav5VzBJE+PuyIVP5Go7++hjEUN7OiL0VZCAPwp9R1CRDG9/cMrDBUysQR+dRxSzuIoY2kc9FVck/hWm/D7xXqbALpb2yHq9z+6x+B+bt2FaJ8JdLtds2tXb3kg58qMbIx7Huf0+lWtpa2MC2tlbxwxIMKkahQPwFVVVV/qenaXD5+o3sNunYyOFz9PWtZ+LmmW+6LRLJ7t+gll/dp9cdT9OK1jx34n1rKT6k0MWSRFb/u1+nHJH1JqzsbzUJxa2FrLcSkcJEhY/XA7VpHwq8Q3+19QeLT4iP4zvf/AL5Bx+ZFaV8MfC+nbZLiGS+lHedvlH/ARgY+uahght4lgt4kjjQYVEXaB9AKqqqqqqqiARiivHBojHFVVVVVVVVVRAIxTKCOKK+lVVVVVVVTL3FVVVVVVUQCMURjjNVVVVVVEAjANEdqqqIBGKIxxVVRAIxRGOM1VVVVVEAjGaI7VVV/wWC8Sf2t+0vonh+G6laHQ/CVsjwsuFSeW4nkYr65jMOT/s+1f8EYfCzWnw1+I/jY27BdV1y108SmQEMbWBpCoXOQR9rByRg7hjocVVMMjiqqqqqpgDRGOM1VVVVVMPSqr/gpd8A4PjN+zrf+JtNtN/iT4e+Zq2nusZZpLcAC7h4yQGjUScDloEHAJNfs5/GjV/2ffjN4Y+KukiWVNIuwL+1jbBu7Nxsni54y0bNtJ4VwrfwitL1PTtc0uz1rSL2K7sb+3S4tbiJtyTROoZHU9wVIIPvVVTDIyKqqqqqpEV0KsAwIwQR1q/tjZ3s9mc5hlZOfY4qqqq+Ht+IdRmsHYYuY8rk/xL/9Yn8qpgCKqqqv+Ctfw4i8Ufs66d4+gtEe88Fa3E7zlMmO1uv3Mig9QGlNqfT5R7Y/4JyfET/hXn7W3g8T3fk2PibztEuvn27/ALQn7lfvKDm4SDg59gW21VVVVVVVeNtN+3aK08aFpbQ+YMf3f4v05/CqqqqqqtA1yfQ70ToS0L4E0Y/iH+I7f/Xq1uYL22juraQPHKu5SKqqqqqqqq/b3/ZXi/aW+Eck/hyyRvHXhNZLvQX37DdAgedaMTxiQKNpOMSKmWVS+fh/468a/Bb4j6R498J3EuleJfC2oCaDzoiDHKhIeORDglSNyMp6gkHrX7L37Q3hb9qH4MaJ8V/DAS3lulNvq2neaJG02/jA86BiOw3KykgFo3jYgbsCqqqqqqqqqqqqqqqqqqryztNQtJrC+torm2uY2imhmQOkiMMMrKeCCCQQeDmvid4Ln+G/xJ8WfDu6m82fwtrl7pUj5B3NbzvETxxyUr9krx0nxJ/Zn+G3i4XHnzT+Hre3upcAb7i3X7POcKAB+9ifgAAdKqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv+Cl/wm/4Vl+1JrerWVp5Wl+N4E1u2KQ7UEsmUuRuHDMZo3kPQjzVz1yf+CLfxdh8R/BjxV8G724T7d4O1YX1mjSYZrO7BJCr1ISaKUsRwPOUHGRmv+CrHgdvGf7F3iu9gt0mufC99Y6tChgEjALOsMjKSfkKxTyMWGTtDL/Ea/Yc8VDwb+1t8L9X8zb9o11NOz5e//j8RrXGPfz8Z7de1VjPAoAAYBqqqgMnGaqsZ4FYAGAaqqqq/ax/4KXfDv4GXl74E+F9tZ+N/GdurxzyLcZ0/TJhxtmdDmV1P3okIwQVZ0YED4y/tIfGr4/aq+p/FHx9qOqxBiYNOR/IsrYZ4EdumEB4A3YLHaNzEjNfD/wCBnxf+KhRvAHw71rV4HbaLuO2MdsDu24M77Ywc9ctxgnoCRoX/AATa/aP1eMPqB8K6IxiVyl/qjOQT1T9xHIMjvzj0Jr/h2B8ff+hu+H//AIH3n/yLWu/8E3P2kNJ3/wBnx+F9a2wmQfYdUKbmGf3Y89I/mOB1wvI5648W/swftB+B97eIvhF4kSKLd5k9pZm8iQLjJaSDeoHIwScHnHQ1oev+JfBusxax4a1vU9D1ayf91dWNy9rPA4PZ0IZSCB36ita1nVfEesX3iDXdRnv9T1O5kury7uHLyXE0jFnkdjyWZiSSepNfsM+K/hzrP7Ovg3w14G8U6dqV3oWlxpq1pBH5EtpdPmSZXiOG/wBY7jzMbZCCwJzSIMUi5pVAGBSJmlQAUqZpUGKbYiGSR1VVBLFjgADqTX7QH7TGp+Mr2fwp8P8AUrix8PRbo5rqJjHJqB6HnqsWMgLxuBJbrtEFtcXcghtYJJpCOEjQsfyFXOj6rZx+bdabcxIAPmeJgOR614D+JPjL4a6smr+EdaltWDAy27HdDcDuskZ4YY4z1HUEHmvg58V9B+MXhNdf0tPs15bsItQsWfLW0uM8eqN1Vu+COCCAYgRgU0QIpoq8TeMfCHg+Dz/FHiXTdLXAKi6uVjZs5xtUnLdD0HY+laZ+0h4A8T+MNP8ABPgqLUddvL6fYZ4bcxQwoBl5C0mGIC7ui87TzyCWQEYFftH/ALV3wm/Zp0J7rxfq6XuvTxM2naBZyBrm5YL8pYc+VGTgGRhjrgMRivHnxF+LX7YniO38X/Fif+yvBNpIZdH8NWcjpDjkK553OxUnMrYJyQgRWAHh7w7dX9xY+GfC+jyTzyFYLOytISzHsFVV9q+AP7GukeEvs/iv4qwWurayPmh0sqJLa0PIy/UTNgg9Nqnpu4IqqqqAzzVazrWkeHtLuNa17U7XT7C0TdPc3MojjjGcZLHgckD6kV8av25HJuvDXwctlABMba7dJnIxyYIj05xh39/l6Gr298R+NNee7vbjUNa1nUpRudy089w+MAd2Y4AAHoAK+D37DfiPxFHDrnxWvJdBsHCvHpluVa7kB5+c8rDxjj5m6ghSK8GeBPB/w80ZNA8FeH7TSbJTuaOBPmkbGNzucs7YAG5iTgDmqqgOM1VVQAxmqqqAzzVVVVVVWM8CgAB1qqqqqqqguawMYFVVVVV4+8Jx+MvDk+lqyJcofNtXYcLIOgPsRkH0zntUUmoaHqayRmS1vrGbjIw0ciHuD3BHQ14E8aWPjbRlvISsd5AAt3bj/lm57j/ZOCR+XUGvGXgu28TWvnQ7YtQiX91IRww/ut7e/b861DT73SL2SxvYWhuIWwVP6EH0968K/FGSBUsfEpaVBhUulXLKP9sDr25HP1q3ubS/t1ntJ4riGQcMjBgatrW2s7dLW0gjhijGEjRdoX6AVVtr2kXmoz6Tb30bXdscPFnB4HOPXHfHSqqqqqqpZIoY2lmkVEQZZnOAB6k1r/xP0LSiYNNH9pTjgmNtsa/8C5z+AI961zxv4h15mW4vmhgPSCAlFx79z+NeD9cTxD4ftb4vumVfLnB6iRRz+fB+hFatrui6FCJ9Y1W1s0P3fOlClvoOp/CvEHx88PWOYfD1jNqcmDiV8wxg9uo3H6YH19PEPxc8beIA0Q1L+z4D/wAsrIGP82yW/XFWVhqOr3QtdPtJ7u4c5CRIXY++B9etfCfTPEmi+EYtK8SWggeGRjbIXBYRtzhsE4O4t+BA7VVVVVVVVfHjxhYy2cPhHT7lJZxMJbwIc+WFHyofQknOOowPWvgfpc9948t72PIj06CWWQ7ePmUoBnscvn8DU0ENwnlzxJIv911yPyp/DXhyRi76BprMxySbRCSfXpQ8M+G1IZfD+mgjoRaJx+lQ21tbKVtoI4geoRAufyqqqqrWdf0jQLf7Rql4kQP3ExlnPso5/pWu/FnULrfBoVqLSM8CaXDSfXHQfrV3eXupXHn3t1NczNxukcsx9Bz/ACrRfh54l1rZJ9k+yQNz5tx8nHqF6n8se9aN8KNCsdkuqTS38qnJU/u4/wDvkcn8Tz6VZ2Fjp8It7C0ht4x0SJAo/SqqqqqqqqqqqIBGKKgjg0RjiqqqqqqqqqoqD0plz060R2qqqqqqqmXuKqqqqqqogEYojtVVVVVVTDIyKqqpgCOKqqpgCOKqqqqqqYAjIqqr/gpR4iXxH+2d8QGgv2ubbTmsrGEMpURGKygEqAEA4E3m89CSSCQRX/BJvw0NC/ZGtdUFtLF/wkXiK/v9znIl2lLbcvoP9G2/VTVVVEYOBVVVVVEAjGaYcVVVVVVEZGBVVNDFcRPBPGkkcilXR1yGB4II7jFftifBCX9n79oXxX4AggSPSHuTf6Hs+79gnJeJRk5zH80RJ6tET0Ir/glX8b0+JH7Pp+G+rXqSa38PLn7GqtIWkksJcvbuc9lPmwgDgLCnrVVRAIxRHaqqqqqIBGK8cWgtfEMzAYE6LIBj2wf1Bqqqq0e8On6pa3gbAilG49Pl6N+maogEYojtmqqq/aC+HLfFz4IeN/hvBCkt1ruiXMFkrkAC6CFoCSeABKsZ5x06jrXhfxFqfhDxNpHi3RJvL1HRL6G9tH3Mu2WJw6HKkMPmUdCD6EV4N8T6d438H6F4z0hs2Ov6bb6hanOf3U0ayJz/ALrCqqqqqqqljjmjaGRQyupVge4PWr61eyvJrNwd0MjJyPQ9aqqqqqq8G+IBptz/AGfdyYtp2+UnpG57/Q9Kqqqqqqqqr/gqX+ybL4Y16T9pPwHpTHRtYlVPFMUQJFpeOQqXOP4UlJVWPTzCO8lf8E7P2u5f2WPjNHb+Jr91+H/jBorLxChQuLRgSIb1QOcxFm3AZzG74VmCYhmhuIUnt5UljkUMjo2QwIyCCOoIqqqqqqqqqqqqqqqqqqqv+Cqnw/fwJ+2f4rvY7bybPxZZ2es2i5Y5DwiGVsknrPBOfQZwAAK/4JHfET/hJf2ftZ+H91d+ZdeDddfyYt+fLtLpRKnBYkZmW6PCqp7ZO41VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVf8FevhXF4g+Efhn4s2UCfbPCeqGzumEeC1rdADLN1IWWOMAHj96xGOc/8Eqviv8A8Ky/a/8AD+lXT4sPHNnPoNwS+AjybZYGA7kzQxp2wJDz2NftB+Ah8UvgV8QPh2kPmz+IPDd9Z2w8vzCs7wMImC5G4rJsYDIyQORWjard6DrFjrmnsgutOuY7iAuu4B0YMuR3GQOK8Oa7YeKfD2l+JtLLGz1eyhu7YvjPlyoHXO0kZww6Ej3oAE4zR571VVSjvmqoDJqqqqr/AIKHf8FA5fDcurfs/fAvWWj1iNntPEviC3fBsSPlktLdh0l6q8g/1fKr8+Snwv8AhT45+Mfiy18G+AdDm1C+nYGWQKRDaR5wZZnAIjjGep6nAAJIB+A/7APwn+GFrZ6z49tofGnidFDyPdx5srd+uIoDw4HTdJknG4KmcCOKOGNYYUVERQqIowFA6ADsKrGelYJ4Araa8f8AwT+E3xSgki8ffD7RdYkkUqbma1C3C5UKds6YkU4VRlWH3R6Cv2if+Cdfi3wb9q8V/BNrjxLoqbpJNHcbr+2HJxGAMXCgADAxJyAFfk14H8eeOPhV4rtvFvgXX73Qtd05yI54eGHPzI6MCrqcYZHBU9CDX7Jn/BQXwd8afsngb4oPZeGPGz/JA4JjstUb5j+6ZifKfAAKO3zEjaSTtCKMUi5pEGKRM0qADilTNftbeMrnwl8JZtO0+48q68Q3AsCR94QlWaXHsVXYfaT8Ro+mS6xqUGnQEBpW5Y/wgck/kK8PeF7LT7ZbawtwgwNxxlnPqT3NXOhlIvmj4I6EV8QfCUOkv/a1ggSKV9skQHCMe49j6V+yX43uvCfxe0/SfNP2DxGDY3MeeC5BMTAEjkOAPo7Y614w/aG+DvgmSa01Xxpa3F5A5je1sAbmRWBwynZlVI5yGIxgjrxXiz9uiAK0PgbwQ7HnbcatOBjg4zFGT3wfv9sd8jxb+0p8ZPGCvDdeMJ9Pt3BHkaYotQASM/MvznoByx4yO5zc3NxeTvc3dxJPNIcvJIxZmPqSeTX7IHgfQPDXhq58b6nqlgdZ1qIiKL7Sha2s0OSSAcruYbmz0Cp05r9sD/gp2dJvLz4c/sz3tncyIoS78XkCZFbqyWkbDaxHyjzmyv3gqn5ZK+HPgPW/ihrlx8T/AImaleas17cNMz30rSy6hLnl5GbkoCMY74x0GD4R8Ia/431+x8J+E9LkvdQvXEcEEYwAB1JPRVA5JPAAya+An7OvhX4KaQl0I4tR8UXUO2+1Qg8A8mKEH7kYIHOAzYyeyrVVVVjPArAxgV8aP2g/AnwVsANauft+szputtItZB5zjs7/APPNP9o9cHAODj4s/G/x/wDGXUxeeLNU22ULbrXTLbKW1vgEZC5O5uT87EtyRkDAHwZ/Zl+IXxkePULSBdH0AOFk1W8jO1h38lODKQPQhc8FhXwl+AHw5+DtlH/wjukpc6tsxNq12ge5ckYYK2P3an+6uB6561VVQGaqqoDNVVUBnisDGBVVVVVVQHFVVVVVVUFzWBjAqqqqqqr4w/DltRjk8XaHCDcxIWvogf8AWIoHzr7gDkdx7jnwp4o1HwjrMOr6c5Ow4lhLYWZO6n/HscGtD1nTvEOlW+saXN5lvcJlSRgqe6kdiDwRXiDw1pXiS1+z6jBl1B8qVeHjJ9D/AEPFa18OPEGmXiQ2kBvYJpAkU0Y6Z6bx/D9envWhaRb6FpVvpdsBiFMMwGN7d2/E1WrXt1e6Tep4Xu7a4v4gUULKp8tu4PUBsZwDjnrU0d9p14yTLNb3ULchso6n19RXhj4qPGFs/EqlxkBbqNOV/wB5R1+o/I1aXlrf26XVlcRzROMq6NkGqqqrUdV03SYPtGpX0NunODI2C2OwHUn2Fa18W7SHMOg2RnYdJrgFU/BRyf0rWPEmt69IX1PUJJVzkRA7UX6KOPx61onhPXvEJDabYM0WcGaT5EHryev0GTWreBrTwX4cufEl9bjWrizQMbXzDDGMkAngEtjIOOMgGtV+LfjPUIvstjexaTaKMJBp8QiCjGOG5YfgRz+FSPe6ldF5HmurmY8liXdz+pNaF8HvHOuFXfTBp0JPMl6fLI/4By36V4f+AXh6xxL4gvptTkwMxJmGMHv0O4/XI+nppmjaTosH2XSNNtrOPABWGIJnHc46n3NVVVI8cUbSSOqogJZmOAAOpNXHi3wpayeTc+J9JhkAzskvY1P5E1/wm3gz/obtF/8AA+L/AOKr/hNvBn/Q3aL/AOB8X/xVf8Jt4M/6G7Rf/A+L/wCKqX4oeAIY2lfxRaEKOQoZj+AAJNat8ePBtkjLpsV7qMgzt2ReUh6dS2CP++T0rxP8ZfGHiDdb2lyulWrDBitThz9XPP5YFabarf3yxzMWBBZznk/5OK+HXinRvA9nNYvpMsn2mTfLco4MjY4VcYHAGcc9SfWtJ8Z+GtaCrZ6pEsrdIZj5b59AD1/DNVVVVU7IiF3cKqjJJOAB614r+KUduz2HhkpLIMq1065VT/sDv9Tx9RVzc3mp3RnuZpbi4lblmJZmJ7f/AFq0D4V6vqKrPq8osISOEI3SH8Og/Hn2rQ/B/h/w+imwsUaZf+W8o3yE/Xt+GBVVVVVVVVVVVVVVRAIxRUEcGiMcVVVVVVVVVRAIxTKDwaIxxVVVVVVVMvcVVVVVVVTDI4ojtVVVVVVTDHSqqqYdxVVVMB2qqqqqqIBGKIxxVV+2BrN1r37VHxavryOJJIvGGpWqiMEDZBcPCh5J5KxqT2yTjA4r/gntosWgfsafDGwhnaVZdOnuizDBBnuppiPoDIQPYVVVTDI4qqqqqqYA0RjjNVVVVVTDniqr/grz8EIvEnww0P47aVAx1Dwfcrp+plejWFw+EdsnqlwUUADn7QxPQV/wTn+M8vwe/af8OwXd80Oi+MmGhaihY7CZ2At3IwQCs4j+bjCs/IBaqqqYelVVVVVXxKtAJLK+UDJVo2454wR/M1VVVVoV39u0azuSwLNCAxz/ABDg/qDX0NMBng1VVVftdeBF+G37TPxI8IwxNHBBr89zaxspBSC4IuIl5ZiQI5UG4nJAycZxX/BOHx/F4+/ZH8Ho0yPeeGmn0a7CtnYYZCYh95iD5DwEg45PAClaqqqqqqqvHNn9m11plXC3MauOOMjg/wAs/jVVVVVVVeD9bOraf9nuJAbm1AVs9WXsf6H/AOvVVVVVVVVXijwzoPjTw5qfhHxRpsWoaRrNpJaX1pLkLNDIpV1JBBGQTyCCOoINftVfs767+zL8YNU+HmpNLc6W/wDpehag5BN7YuT5bMQAPMUhkcYHzoxA2lSf+CRP7XUPjXwc37MfjvV0/wCEg8MQmbwvJM+De6cMb7ZePmeA5IGSTE3AxCxqqqqqqqqqqqqqqqqqqqv+C3vw283Sfhp8YLW3x9nuLrQr6Xb97eont1z2x5d0fx9q/wCCR3xE/wCEb/aA1n4f3V3stvGOhP5MW/HmXdqwlTgsAcQm6PCsw7YG41VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVftDfDRPjF8DvG/wANRaxXFxrmizxWKS42i7Vd9sxzwNsyRtnjGM5HWvCfibVfBXirRvGWgzeVqWg6hBf2cm5l2TQyLIhypDDDKOQQfQjrXgjxXpvj3wV4f8c6M2bDxFpdtqNoc5/dTxLInPf5WFV+1P4Bb4YftIfErwIIGhg0vxNeizV8bvsrytJATgAZMTxngAc8cV+wJ46b4gfsj/DvUZ5Ua50nTm0iZAQTH9jkaCMHAAyYo4m6ZwwySckqOODVVVUBgYzVUBxVVVV/wUM/amn/AGcPhB/ZnhK/8nxx4x8y00eRcE2US7fPusZ6qrBU/wBt1OCFIrwH4G8V/FfxtpvgvwtaSX+sa1chFL7mC5OXlkYAkIoyzNg4AJr4AfAjwj+z/wCAbXwj4ct45b+VEk1fUymJNQuQvzOe4QEkImcKD3JZiFJ4FCMnvQhyOBQhJ/8A1UICR60YCRgCjAcYzTQnGMfpX7Tn7FfgT4+W8/iLRhb+G/GyplNTig/dXpA4S5RcFuAFEg+dRj7wUJXxN+Ffjz4P+KJ/B/xA8P3GmX8WWjLrmK5jzgSRP0kQ46joQQcEED9i/wD4KKX3h+fTvhR+0Fqsl5pUrrDp3im5mzJY8ALHdEjLxZ/5bE7kz825eUt3iniSaCRJI5FDI6nIYHoQe4pVB4FIoApEzSJniv27Ibv/AIR/wlKlwotlvLlZIivLOUTY2e2AHGO+72r4cNCviPbLjc0DCPP97I/oDXh94A6eYRitcl097ZRABkLz9a+JskK+HLxWYZcoFHqd4P8ASvg3G8vxe8EJGhY/8JFYHAGeBcISfyFfFj9mn4c/FUSahcWX9j602T/aVhGqNIx7yrjEv1OG/wBoV8Vf2f8A4i/CSaW41zTPtmkKwEeq2YLwHOAN/eM5IGGAGehPWqpWV1DowZSMgg5zV/4Z8N6ru/tPw/pt5vbc32i0STcfU5B5qOxs4YUt4LaOKKNQqJGu0KAMAADoMdq+F/xf8VfB/wC2S+C7bSorm/wJrq4sxNLsGMIGJ+VcjOB1OM5wMQ/t0/F+KGOJ9D8JTMigGR7OcM5A6nEwGT7AD2FWn7fHiNLhGv8A4dabNAM70hv3iY8cYYqwHOOx/rWn/t96PL5n9q/DO8t8Y8v7Pqizbuuc7o0x29evatO/bp+E915MeoaD4mspH4kb7NDJHGfqJdxH/Afwqw/a/wDgHeQmW48X3NkwbAjuNLuCxGByPLRhjt1zx0qw+PPwX1N2jtvif4dQoMn7RfJAPwMhUH6CtN1/QtaB/sbW7C/woY/ZrlJcKeh+Unj3pRxmv2k/2q9O+FqyeD/A8lrqXimRGWeQkPFpmQQC46NLnBCHgdW4wGmn8UePPEgknm1DXNc1adUBdmnnuJDhVHck9AB6Yr4C/sU2Gkrb+K/jFFBf3jIrwaGvzRQEjOZ2/wCWjDONg+UY5L5wIIILaCO2toUiiiQJHGihVRQMAADoAOMVVVWM8CgABgGqqsZ4FAADAqqrGeBQAAwDVVVVVVQAPOaqqqqqqoLmsDGBVVVVVVQGeBQXIwK+Lvw7fw5qDa/pFsx0u7bLqo4tpD1B5+6TyD0BOPTPw8+It94Fu3jaE3WnXLAzwA4YHpvQ+uO3Q+3BGkavp2vadDqulXSXFtOuUdf1BHYjoQelVWt3c9hpF3d2kMks8ULGJI03Etjjj0zjPtSXF7aXRnjuJoblWOXDFXDZ5565zV94i1HVYFg1Z47zywBHLIgEiDPZxgnqeGyPaq+E1hqE2qzX8c8sdlAhWRQfllc9AR3wOfbj1qqrGeBXjvxx4k0rVp9FszDaxxgFZUXc7qRnqeB1xwOo61dXVxdytc3lzJNIR80krlifqTV3rdjbDaj+c/onQfjV3rl7c5WNvJT+6vX868I6vFr3hjTNWhK4uLZSwXGFcDDDjjhgR0HStU0631fTbrSrpcw3cLxOPZhg/wA60T4A+HLMpLrepXWouuMxoPJjPqDjLeg6j9eNH8N6B4fi8rRdItbQYwWjjAZh7t1P4mqqq1Xxx4Q0UsupeI7GJ0BzGJQ7jH+yuT+lan8evB1oSmn299fsDgMsYjXHrliD+n5VrP7QGv3lu8Oi6RbaczDAleTz2T1IyAM9OoNatruta7KJ9Y1W6vHH3fOlLBfoOg/CtT8Q+H9FdItY13T7F5Fyi3NykRYeoDEZFf8ACdeB/wDoctD/APBjF/8AFV/wnXgf/octD/8ABjF/8VX/AAnXgf8A6HLQ/wDwYxf/ABVXnxx+CthJNDefF7wXFLblllibXrbejL1UrvzkEYxjOa8Tftpfs5+G4iR48/tWfZuW30yzlnLDkff2iMHjoWB5HrXiv/gpTeR6xanwD8MrSXTYLkNcHXLlzJdQg8oEhZfJYj+LfIBnocc+E/8AgrV4BvNHtPDnjv4BXuiW8MqvLd+GtWScyncRuMM8aklUPG6Y7j3UHj4f/tf/ALMHxQe2tfDnxcsdJ1G5UEab4niOkyxkjO0yyE2xbOVwsxycYzkVLazwwwXEkR8m6iEtvKOUmQjIdGHDKQQQRkc1pHjDxFoaiOw1OTygMCKT50x7A9OnbFaT8X7eTbHrWmNGeAZbc7h7naeQPxNaV4p8P60FGnarBI56Rs2x/T7pwetbTW01tNXE0NrA9zcypHFEpZ3c4CgdSa8b+PLnxDM+n6bI8WmrxjoZ/dvbjgfn7eGfCWq+Kbjy7KMJAhxLcOPlT29z7fy614b8GaN4ZjVrSDzbrbh7mQZY+uP7o9h+OaI7VVVVVVVVVVVVVVVVUQCMUVB6GiO1VVVVVVVVEAjFMvGKIx3qqqqqqqmXuKqqqtb8X+EvDUsUHiPxTpGlSTKWiS9vo4C4HBIDkZFRfE/4aXEqQQfETwzJLIwVETV4CzMeAAA3JPpVVTAEZFVVVVVVEAjFEdqqqIBGKIxxVVRAIxmiO1VVVVVTAEcVWoX9npdhc6nqFwkFrZwvNPK/SNFBLMfYAE14l1qXxL4i1TxFPAkMmqXs108SEkIZHLkD2GcV+z/4al8HfAj4c+E7hUE+j+FNMtZtsZjBkjtY1dtp5BLAk55yeaqqqiMHAqqqqq8v7HT4vNvruKBMHBkcLnHp60/jnwopKHVgSDjiGQ/qFrT9Z0vV0L6bfRz7R8yg4YfVTyKqqqqqmAI4qq+J/gDR/ir8OvEfw4144sfEemTWMr7dxi8xCA4GRkqcMORyBXinw7rXgLxhq/hLVz9n1fw5qU9jdeUxHlzwSFH2ng8MhwcA1+y98XB8dfgD4K+J09xFLf6rpirqZjTy1F7ETFcYX+EeajkD+6RjIIJqqIBGBVVVVVV4+thP4dkkyP8AR5UcZPvt/wDZqqqqq8A3Pn6EYCwzbzMoGexwf5k1TAEcVVVVf8Fefh7/AMI/8c/DfxCtoNlv4t0LypWx9+5tH2uc/wDXKW3GPb3r/gjf4/laL4i/Cy5mYxo1rrNlHu4BIaG4JBbqQtsMhex3H7oqqqqqqqq+ItsGhsrsD7rNGTj1AI5/A1VVVVVVWh6pJo+pQ3iMdgO2RR/Ep6j+v1ApHSWNZI2DKwypHcGqqqqqqqqv23v2X7L9p/4PXGj6bBEnjHw95l74buWCqTNtHmWzM2Nscyqqk5ADrG5yEwfB3i7xz8GfiJpvi/wzd3mgeKvCeoiWEyRmOS2nibDRyRt24ZHjYYILKwIJFfsvftDeFv2ofgxonxX8MBLeW6U2+rad5okbTb+MDzoGI7DcrKSAWjeNiBuwKqqqqqqqqqqqqqqqqqv+CkXwwj+KX7HPxAs40zeeG7Ma/aPhTsNmfNlPJHWATr1z83AY/Kf2ffiKvwj+N/gf4jzTtFa6DrdvPesqkn7LvCzgAZOTE0g6Hr0NVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVfto/DiX4W/tQfELwwLRoLSfV5NRsQUwpt7r9+gQ91XzCmfVCDyDX/BKH4nR/ET9jzQNGmk3X/gfULrRLkkqCyqwnhIUAYAhuI0zzkxkkk5xX/BYn4ep4U/ayTxhbQv5Pjbw7aXsspUgG4g3WrKMsckRwQE4Cj5xxnJP/BHTx4l98P/AB98M57v97pGrQapbwuRkpcxeW5TncQGtlzwADIvOWrAxgVVVQGT1qqxngGsDGBVVVeKPE2g+C/Dep+LvFGpxafpGjWkl3fXUoJWGGNSzsQAScAHgAk9ACa/af8Aj9r37Svxi1f4mawstvaSYtdHsZGDGxsYyxiiyOCcszse7ux6Gv8AgnX+z5D4J8DH4y+JNOkXX/FcO3TVmUg22nZBVlHrMQHyc5QR4xubKx5680sOeKWEYxSw5GAKEBIxXkAjGaMAxwaMGeAaaE00IPGK+NXwK+H/AMePCUnhTx1pKzbFc2N9EAtxYSsMeZE/Y8DKnKtgBgRX7Q/7N/jv9nLxWmg+KEW9029UvpmsW8ZWC8UY3DBzskXIDITkZBGQQT+wp+3Xe/A++tfhX8U7+e7+H93LttbtsySaFIx5ZRyWtyTlkHKkll53K1lcW19aw3tlcxXFvPGskMsTh1kQjIZSOCCCCCKVQeB2qNB2pEGK/aW+HM3xG+FGo2enwebqWksL+yUA5dowd6DHUsjOAPXbVjez6deQ31qwEsLBlz/I+3avDXjzStRgQtdx29xj54ZH2kH2J6itS8T2lpDvvb6KFQOPMcLn6V428WjxHcJb2ZYWcJyNwwXb+9jt3Ar9iv4Z3nir4kf8Jzd2sg0nwxGzJKV+SW7ddqR89dqsznHQqmfvDJiBGBVxaw3ML29xCksUqlHjddyspGCCD1BHavi5+xh4S8V+frHw5mh8Oaq7FzaMpNlKT2Crkw84+6CoAwE5yLxrS013VvDS6pp91f6FdvaX8dndx3AgmUkMpZCRnII+oI6g1onwE8c+Nfi/4o0TwtLdaToWj51HWdaw3k6fZOdyknI8yRifLjj3Au4xkAMy6XZpo1hDplpe308NuuyOS9uDPMw7b3wNx9wAPQAcV8Df2PPDWqfD211r4rWd8dY1Yrcw28czQPZQkHbGwBwWYEM24ZXheCGzP+wt8H5Znlj1zxZCrsSsaXkBVAewzCTgdOST71dfsEeE3SUWXxA1aJyG8oy2kcgQ9twBXdjjOMZ9qvf2Ar6ODdpvxSt55sjCT6OYlx3+YTMf0q//AGD/AIlxzKul+LfDFxFt5a4eeFg3oAsb8Yxzn8K1H9jX462UTPbaJpt+ytgJb6jGpYeo8wqMfUg+1ap+zX8dNIMi3Xw31OQxRl2+ymO5yMZ48pm3H2GT7Vqfw7+IGigf2x4F8QWOVLD7Tpc0XyjqfmUce9VpnxE+IGigjRvHfiGwyoQ/ZtTmiyo6D5WHHtUsEc7tJJlnYksxYkk9ya+Bnxmtvgjf3Wp23gPStavLplAvLiV47iGL+OONvmVQ3qFz67gABon7efgO6VF8ReC9c09yBn7JJFdKpzzyxjOMd8Z9q0P9q/4Ea6pVfGy2EoBJjvrSWHgED723aevQHPtxWi+MfCHiUD/hHPFWj6ruzj7FfRz5x1+4x6VVUoB5qqqgBjNVVUBxmqqqqqqgM8VgYwKqqqqqqAzWBjAqqqqqqoDPegoI4rAxgVdWttfW0lndwJNDMhSSN1yGB6giviV8K73wpNLq2jxPcaMxznOWtiT91u5X0b8DzyfBPjzWPBF+J7JzPaOf39m7kJJ7jrtbgfNj8xxXhbxfofjDTxfaPdBmUASwPxJE2OhH9RwfWqrU9B0bWU2anpsFwcYDMmGH0Ycj8DWp/CPSLjL6XqE9qx/gkAlUew6EfiTVz8KfEcEqrDJbXEZYAsj4IGeThsducZq3tNP0u3ZLWCG1hzuYKAi56En8hWsfEnwzpf7uC5a/lHG22G5R9W6Y+ma1P4s63dApplpb2S44J/euOPU4H6VpPxfttC0iQeJTd3t20rGDykHzDA4J4AGf59OK1z4/eIbxTFoOm22nKQR5kh8+QehGQFH0INXXjDXNRnku9VvZL24cAK8zZ2AZ4AHAHPQYHX1rT9F8W+LGI0zTLy9UHBMcZEa/U/dHTua0T4A+JLwpJreo2mnxkZZEzNIvtgYX/wAeNaL8EfA+lqGvbefU5Qc7rmQhQfZVwMfXNWlnZ2FslnYWsNvBEMJFFGEVfoBwKqqrXPFHh7w3GJNc1i2tMqWVHf52A6lUGWP4CvEn7QFlCWt/CulG5YZH2i7yiZ9kByRj1K/SvEHjbxT4nkZtX1meWNicQK2yNR6BRx+fNeIfFnhXwjai+8V+JtK0W3b7s2o3sdsh5A+85A6so+pHrXi79tn9nfwoZIY/GMuuXEasTDpFo8+cDgCQ7YznGB8/1wOa8U/8FKNDi3xeCvhjfXWdwSfVL9Lfb0wTHGr5zzkbxjA5OePFOuftxftK3Ml74f8AAnjaz0CRd9vaaRY3FnZFFK4zM20TNna2GY85KqoGB4Z/4JiftU6+F/tbR/DfhvO7P9qawkm3HTP2YTde364rw/8A8EivFlzbbvFnxs0nTrjy0Ij0/RZL1d5B3jc8sJwDjBxz3C9K0v8A4JM/CKC4Ztc+JnjC8g2ELHaJbWzBsjBLNHICMZ4wOo5450r/AIJe/s06ZFJHeXXjHVWdsq95qkalPYeVEgx9Qaf/AIJpfsvr00nxF/4N3/wrWf8AgmL+zbqEUaWNz4w0tkYkvaapGxf2PmxOMfQCvEn/AASm8B3LS/8ACJfFnX9NBK+UNRsIb0qMDdnYYc5OcHjHv1rxj/wTD+NuirJP4T8TeGPEUSL8sRlks53OT0V1KYwF5Mg5JHbJ+IXwX+K3wpuDb/ELwDrGiqHKLcT25a3kYHBCTLmN+o+6x6j1FfBv9qT47/AVxD8NviFfWmmFv3uj3areWEw3bmBt5QyDdk5ZQrfMcMCc1+zP/wAFAfgX8er2w8EfFSwT4aeNbxVjhu4pi2kahccfKjOS1uzHO2OQ7eAokZiA2rfCvxLYEvZLDfxjoYm2vj3U/wBCaurK8sZPJvbSa3k/uSxlD+RrTvFniTSlCWGs3KKOiM29R/wFsirX4t+JoVVJ4LG4APLPEVYj0+UgD8qb4x6oVIXR7UHHGXY14g8feIfEVsbK6mhht2+/FAhUPj1JJOPbOK8FeEpvFWpeW5ZLK3wbiQcH2Ue5x+HX2NjYWem2sdlYW0cEMQwqIuAP/r+9VRAIxRAIxiioPSioPQ0VOODRB6YqqqqqqqqqqqqqqIBGKK+lVVVVVVVVEAjFMoxxTLjpVVVVVVVMPSvil8afhR8E9Bk8R/FTx9pHhyyRcoLuf97P7RQrmSVuD8qKx4PHBr4wf8FmfCGmS/2d8Dfhje62wU79T8QzfY4lbtsgjLPIpHdnjI6YPWviH/wUv/bC+IMsip8TE8MWbsWFn4esY7QITu6SkNPgB8YMhHyqfvDdXif4qfFDxv8Aaf8AhM/iR4o1/wC27PtP9qaxPdedsxt3+Y53Y2rjPTaPSqrSda1nQLwajoOr3um3aqVE9pcNC4B6jcpBwfSvA/7bH7WHw6EMfhj49eK/KtlCQwaldjU4o1CbAqx3QkUKF6KBgYBAyAa8C/8ABY/9oLQ7iKPx34H8H+KLJWzIYI5dPuWG4ZAkVnjA2hgP3ROSCc4wfAv/AAWP/Z91y3ij8d+B/GHhe9ZcyGCOLULZDtGQJFZJCdxYD90BgAnGcD4c/thfswfFqWG18CfG3wzd3lw22Cyu7k2F1K3zcJBciORj8jHhTgYPQgmqqqqqphkcVVVTDI4qqqmGRkVVVVVVUwA4Fftp+Nz8PP2Uvif4mjumt5f+EfmsYJkZ1aOW7ItY2UpyGDzqQexwSQMmvgp4HPxL+MPgj4fG2aePxD4gsrGdFVziKSZFkY7PmCqhZiRjABORjNVVVVMMjiqqqrxdr7eH9K+0QKrXEz7IgwyAepJ+gH5kVe3t3qNw11e3DzSt1Zz/AJwPaqtLy6sLhLqznaKVPusp5FeFvFNv4gt/KlKx3sS/vI+zD+8vt7dqqqqqqiMHFVX/AAVg+Ev/AAgn7RsPj6wtPL03x/piXRZYdifbIAIZ1GOCdogkY9SZiSOcn/gjh8WGms/HPwPv7lmNu0ev6ZGQDhW2wXXPUDP2XA5GWY8E81VURg4FVVVVVrdqb3R7y1UZZ4GC/XHH64qqqqr4cT831qW7I6jH1B/pVYB4ojtVVVf8FdPAEXiH4AaF48hhQ3fhLX0VnZeVtrpDHIAdpIJlS27quAc5IWv+CZfjpfBX7XHhuznlaO28U2V3pEzAnq8fmxggAkgywRL2AJBJwDVVVVVVVV46gEvh93258mVGznGP4c/r+tVVVVVVVXgbVBe6UbGRsy2Z2jPdT0/qPwFVVVVVVVVV/wAFTv2Uh4U8QH9pDwPp23SNdnEXiaFDxbXrcJcBeyy9G9JMHrJX/BNj9rMfsz/GuPRPE99FD4G8dSQWOtSXEuxLCQMRBeZPACF2Dk8bHY/wiqqqqqqqqqqqqqqqqqqtR0+y1bT7nStSt0uLS8heGeFxxJGwKsp9iCRXxd+HuofCb4p+LvhjqjO9z4W1q605pWQr5whlZFlAIB2uoDg4GQwPev2KviSnxW/Zd+Hvih5/MvINJTTb7c4Z/PtCbd2cAnBfyhJg4OJFOBmqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr/gsP8Nm074g+CPivaWyrDrely6VdugA/fWz+YjOAoJZknKgknIiAwNoz/wRO+K0OkfEXx38Gb+ZwviPTYdW08MflWa1YpKo5zueOdW6YxAeR3r/AILYfDb+2vg/4G+Kdrb75/DGuS6dcMo5WC8i3bm9hJbRqPQycdTX/BLL4iDwX+1TYeHbm78uz8aaTdaYyu+2PzVUXETEFgNxMBjU4Y5lKgfNkHmqqqUcVVKM1VVVf8Fbv2kmRdP/AGZ/CerIRIseoeK1icEjlXtbZ/Q8CcqeceSehr9jP9nz/hfnxWhg1qzkl8J+HNl3rbK20SAk+Vbk9R5rIQcYOxXwQcGre2jijSGGNURFCqqjAUDoAOwpIQBjFJDkdKWEYwBSwkihAOlCEdAP0owjHT9KMIPb9KaEEcGnh7Yp4Qegr4pfCvwZ8YfBl94E8d6St9pl6uQR8stvIAdssTc7JFzwfcgggkH9pL9nLxf+zf43/wCEc18/bdJ1DfLourIm1L2JSM5H8MiblDp2JBBKspP/AATj/baufDOp6d+z38W9ckl0W+kjtfC2oTqXaxmZgqWkj5z5LEgISDsJC5CEbI1B4qNRjFIuelImeK/aE/ZDvte1O88dfCuNHu7yQy3ujuwQPIcl5IXY4BJwShwMkkHota5oGueGtQfSfEOkXmm3sX3re7gaJxyRnDAHGQeelV8IP2YfiT8VryC6fS5tE0AsDNqd9EYwy5GfJQ4aVsZwR8uRgsK8CeAPDHw38MWnhPwnp62tjajJJ5eZz96SRv4nbHJ+gGAAAY+OKZOMda/ar+JLfB39nX4gfEW3n8q70rRZlsZN7JtupsQ25ypBH72SPoQfQg818L9M8U6/8RfDXhjwVqMllrevarbabZSpJs/ezyrGgYkgFSzDIPBHXivjzo+mfBbRbT9n/wAM+JrjWWEsOreJL2a3jhe4vTAkaKRGBhNi+YIzuwJE+YkZr9kP4PwfE74ijWNbtPO0LwzsurhHUFJ58/uomB4KkqWYdCFwfvVVVVVVftH+OvDXw1+G974p1bQtL1TU5R9j0qC9t0lDzuDjIIJKKAzsOMhcZGc1Ywaz4h1eLT9LtZbvUNQnCQ21tCMySMeFRFGByeABgV4W/YX8FnwbZWvjTWtT/wCEjK77q602ZUjViDiNVdWDKuQN2AWIzxnFa3+wGDmTw58SvQCG9038zvR/02/jWufsS/GnS1Z9NGhawACVW0vjGx54GJVQAke+PevEHwL+MPhhXfWPhzriRxlg8sFqbiNdpAJLxblA54OcHtmrm1ubOd7W8t5IJojh45UKMp9CDyK0H4q/Evww6toHj7XrNV6RJfyGM8Y5QkqcDpkcVof7YXx10bYlz4js9Wjj2BUvrCM8L1BaMIx3DqSSfcHJrQf2+fEcCKnif4e6bet0Z7C8e1A56hXEnbHGevcZwND/AG6PhPqClNZ0fX9KkAJyYEnjPIwAVfdk9eVxx1rQ/wBpD4GeIiV0/wCJekREEj/Tmay6AH/luE9fx/A1puraVrVsLzR9TtL63zgS20yyrnAOMqSOhB/EVQGeKwMYFVVAZ4rAxgVVVVVVYzwKAAGBVVVVVVQGeM1gYwKqqqqqqC5oKCKwMYFVUsUU8TwTxpJHIpV0dchgeCCO4xXxF+C8tsZda8GwNLFktLYKMsnvH3Ydfl6+meg0zVNU0K+S/wBLvJbS6iJAdDgj1BHcexrwj8d7C6SOy8XW/wBlm6fa4VJjb3ZeSv4ZH0rTdV0zWbVbzSr+C7hYcPDIGH446H2NVNNDbxNNPMkcaD5ndtoH1JrxB8UtJ0/db6Mn2+ccb87Y1P16t+HHvWt+J9a8QybtSvWdAcrCvyov0X+p5p7q0jlSGe7iiLsFy7YC57n0HvWl/B6Rgr61qwX1itlz/wCPN7e1eNPhLpd74WltPDdkTqcbo8TyzHMmDggk8D5WPQDkCtB/Z7u5dkviTW0hHBMFmu5vcb24B/AitD+FngfQWWW20SO5mUgiW7PnEEdCAeAfcAUqqqhEAAAwABjFVVVrfirw54cj363rNranBIR3y7AeiDLH8BWvfH7w/Z7odA0641GQZAll/cx+xGcsfoQK174w+OdcLRx6mNOhJ4jsl8sj/gfLfrWravZadaz6xruqQWtvEN091dzhETtlnY4HOBya+Jf7dPwR8Ci4sdA1CfxfqcL7PJ0sYt89ybhvkK47x7+SPci58eft5ftOlI/hJ8ONf8P6BdxboJ9Oh+wxzLjcG/tC4KAnA48t0BBAwSRnwd/wSY+MXim6GrfFr4r6NpLzYMotEl1S5IAA2szmNQcADIZwBjr0rwD/AMEtf2ZfCQin8TweIPGNyuCw1LUDBDuH92O3EZx7Mze+RxXgr4K/CT4bJEvgH4a+G9BeHO2ax0yKKUnBUkyAb2O0kZJJwcU8II4FPGRwKeIEYFPHgYNSRjHFOnavGPjfwl4Fsft/inXLexQg+XG7Zklx/cQZZvwFan+114Khumi0zw5q93CuR5shji3HJ5AyTjGDzg89BWl/tYeBL+48jVdG1XTkYgLMFWZV65LBTuHboD17Vpms+GfGmkPcaTf2WrafcIY5ApEisCMFHU9Mg4KsPrXxt/4J+fBz4lW9xqfgeyj8Ea9sPlNp0IWylYDgPbjhR7x7cdSG6V8Yvgl8Q/gV4obwr8QNH+zyPuazvISXtr5AQC8MhA3DkZBAZcjcATiv2C/+CkviD4O3+mfCT46avcat8P5W8mz1WbdLdaEWPykty0tsCTlTlkBBThdht5tG8R6Xb39pNZanp19Cs1vNGyzQzxuuVdGGVZWU5BHBB961D4c+EdQJc6YLdz/FbuU9f4fu9/T0rWvh34M0l1iu/GUemvIuYlvZogTjqQCVyK1LR/hpo8aS6v8AGnwzYpIcI1zdQRBj6AtMM1HrX7OemQPean8afDN4sS5ZItctzn6IjFj9BWnftFfs46TbCz0z4keH7eIHO2NmGTjGTxycAcmtT/a0/Z20ry/tXxPsZPNzt+zWtxcYxjOfLjbb174zzjoa/wCGzP2a/wDopP8A5R77/wCM1/w2Z+zX/wBFJ/8AKPff/Ga1L9t39nexdEtfFV/qAZclrbSp1Cex8xUP5Zr/AIbr+AH/AEEta/8ABY3+Nf8ADdfwA/6CWtf+Cxv8a079tf8AZzvoDLc+NLqwcMVEVzpNyzEYHzDy43GOcdc8dK/4bM/Zr/6KT/5R77/4zX/DZn7Nf/RSf/KPff8AxmtO/ax/Z31WN5LX4n2CCM4Iubee3P4CRFJH0o/tOfAA9Pipon/fxv8ACoPjn8E7mCO4i+L3g0LIgZRJrlujAEZ5VnBU+xAI71pvxS+GOsiT+x/iP4XvvJx5n2bWIJdmc4ztc4zg9fQ1p3iLw9rExtdJ17Tr2ZULtHb3SSMFBAJwpJxkgZ9xVVVVVVVVVVVUVBHFEY4qqqqqqqqoqCOPyorxxVVVVVVqGoWGk2Fzquq31vZ2VlC81zc3EojjhjQFmd2YgKoAJJPAAzX7Wn/BWq00x7vwL+y15V3dLujuPFt5bh4omDY/0SFwRJ0/1kg2+isMNXizxh45+KfiyfxL4y8Qat4m8Q6rKqvc3kz3E8zcKiLnJwOFVBwBgAAACvhR/wAE+vj38RWhvfEWmQ+CtKkXcZtYBFwRgYC2y/OG5HEmzGDzkYPgT/gmd8DPD0cE3jTVde8W3aqPOSSf7DbOe5VIv3ig+nmnt9a8M/svfs9+EY4U0T4N+FQ9uyNFNd6cl5MjJjawkmDuG4BznJPJ55rSvCnhzQpHm0Pw/punySrtka0tEhLAdiVAyKvNMs9RtZLHULWG5t5l2yQzRh0cehU5BFa18FvhH4itxaa98LvCeoRLnYtzosEmwkYJXKZU47jBrxp/wT+/Zm8Xxk23g268O3BXb9o0a+eIj0xG++PPvs+ua8af8ErLRlkn+HfxXmjYFvKtdasA+Rn5czREYwMAny+evHSvF3/BPj9pzwtvks/CWn+IoI926XSNSjbgYwQkpjkbOeAFJ4OQOM+L/h746+H962m+N/B+saFcK5QLf2TwhyP7pYAMOMggkEcg4r4N/tcftEfAQR23w0+J+qWWmrIrtpN0VvLNsdQIZQypuHBKbWwByMAj9n//AILA+BPFU9voH7QXhX/hEbyTao1rShJc2LNgZLxHdLCM9MGXryRjNeHPEnh/xhoVj4n8K61ZavpGpQiazvbOZZYp0PRlZSQRVVVVVMAOlVVUwx0qqogEYojHANVVVVVTDI4r/gsd8Qf7D+Cvg/4cW8uyfxVrz3cny5329nF86/dIH7y4gOdyn5cYILY/4JZ/Dv8A4Tn9rXR9Zng8y08G6Zd6vKGHy79ogi59RJcK4H+x6A1VVVVRGDgVVVXxMtfN0SG5U8wXAyM9iCP54qqq2uZ7OeO6tZmjliOUZeoNeGvENv4hsfOULHcRcTRZ6H1HsaqqqqphkZFVX/BVT4Tr8QP2ZZvGdlbI+p+AdRj1BG53m1kIhnQDpj545DnHEPB7H9i74sL8F/2mfAvjO7uUh02TURp+ps4JVbW5HkyOwHOE3iTjvGOD0NVVMARxVVVVVVf24tL64tAP9TKyYznocf0qqqq+HbsNXuIg3ytbEkY7hlx/M1VMB2qqqv2v/BA+In7MPxL8LJD500nh64ureLy95knth9oiUDI5MkKAHsSDg4wfhh4yn+HXxJ8K+PrZ5Q/hzWbTUB5SgsRDKrkAEgHIUjBIBzg8GopYp4knglSSORQyOpyGB6EEdRiqqqqqqq8VRrL4evUboIwR9QQR/KqqqqqqqrwhqA0/W4QzYS4/dNn36friqqqqqqqqq8deCPDHxJ8Hav4C8Z6VFqWia5aPa3ltIPvIw6qeqsDhlYYKsAwIIBr9oX4H+J/2ePivrPww8Trvexk82xuwPlvbRyTFMv1UYI7MGXtX/BKP9qwfGv4PH4P+LLzd4u+HNvFBC7nm+0vASCTJOS8ZHlPxgDyjklziqqqqqqqqqqqqqqqqqr/gsT8JX8EftN2nxGs7Ly9O+IOjx3DSrGyq15agQTKCSQSIxbMcY/1nIySzf8Edfie1xpXjz4NX1+hNpLDrmmW7EbirjybkjvtBW19gXPQnmqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/4KD/BdfjR+zH4ltrK0afW/Ci/27pQQEsXt1bzUAAJcvA0yhe7lD2FfAD4s6l8CvjR4O+LelmVpPDWqR3E0cRw01ucpPEOR9+FpExkD5ueK0nVdN17SrLXNGvoryw1G3juLS5hbck0TqGR1PcFSCD71+3N8Nv+Fsfsl/E3wjFb+ddLocmoWaAZZp7Mi5jVfQs0IT/gRB4Jr4T+Orn4YfFDwn8RbNpfM8N6za35SMnMiRSqzpgMuQygqV3DIYgkA1a3VrfWsN7Y3MVxb3EayQzROHSRGGQykcEEEEEVVVjPArAxgGqoAAYqqqvix8RtF+EPw08S/E3xAN9j4a0ya8eISBGnZF+SJSeA0j7UXP8AEwrxv4x8SfE7xxrPjjxNcte634j1CW7umRT80srk7UXnCjIVVHQAAcCv2SPgVbfAn4O6X4eubRU1/VFF/rkh5Y3LqP3ef7sahUAHGQzdWOY4h0ApIgBjFJDkc0kWRgClgzQgGKEII6ZryR/dpoRjGKaAHpTwkcAU8IIxipIuMYzXxy+CfhD47/D/AFDwJ4ttIyJ0Z7C98rdJp9yFISePkHKk8rkBhlTwTXxX+GHir4OePdW+HnjG08rUNLmKrIoPl3MR+5NGT1R1wR3GcEAggf8ABOH9rk/HLwP/AMKu8eamj+OPCdsoSaVxv1WyB2rNgAfPGCkb9Sflcklmwi8YFRqCOKjTIpEGKvdJ03V4Vt9U0+2vIlbcsdxCsihgMZAIPOCefetJ8C+C9EdZNF8IaLYOjmRWtdPiiKuerDaowT60seRxXlU0XtTxjFf8FjfGaaJ+zz4c8Fwvi58S+Jo3YFVINvbQyM+Ocg+Y8HIBGM5IyM/8EwfBMXi39rfQtZuzD9h8G6Ze63dmbaFVUi8hGJLDG2W4ibIz93pjJHj/AMUyeOPG+veL5A6/2vqE1yiP1jRnJVf+ArgfhX7Mfw7t/hz8HdDsjAqahq0K6jft3MsqghT/ALqbEx0yp9c1VVVVQGTiv2vPir/wsn4qXGmabdeZovhffY2e05WSUH9/KPq6hQRwVjU96/YV+D0FzJdfGHXbTf5DNaaKkijAfpLOPcf6tT7yd8YqqrVtD0TX7Y2WuaPZajbkEGK7t1mUgjBGGBHI4Na3+zb8DPEDF774baVESwYiyD2YyBjpCycY7dCeTzzWt/sM/CXUELaRq/iHS5QgVQtxHNHnPJKsm4kjjhgOhx1zrf7AniCFWbw38RNOuzglEvrF7fBzwCyNJ274/Ctd/Y7+O2jb2tvDlnq0ce8s9jfxnhe4WQox3DoACfUA8Vr/AML/AIj+Flkk8Q+BNesIot26afT5Fi+UZJD42kAc5BxVlqF/pk4utNvp7WYAgSQSGNgD1GQc1oX7Qvxs8NhF0z4lay6xklVvJheAZXbjEwcYx0HQHkc814f/AG5/ixpiiHW9J0HWE5y7wPBLnAA5RtuOM/d79elaH+354SuCR4l+H2r2AycfYbuO74wMffEXfP046540D9q/4D+IWESeN00+Y5/d6hbSwYGQM7yuzv03Z6nsa0TxT4Y8Tw/aPDfiPS9WixnfY3iTjAOM5QnuCPrWM8CgABgGqqqqqqgMCqqqqqqgM8A1gAYFVVVVVVBc0FBFVVVVeM/hd4b8Yl7uSM2WosuBdwryx7b16N+hxxmvEvwm8ZeHC8p086hbKTiazBkwPUr94cdeMD1q1vL7Tbjz7K6ntZ0ON8TmNlIPqOeoH5V4b+InjgiSL/hKbpvLVQBKiSkge7qT9fWtV17WNbcPqmozXGDlVY4VT6hRwPwFXepWdmCJZgWH8C8t+Xb8au9cvLpvKtVMQY4AXlj+P+FeFPgx4p8RFLvVF/sq0bkvOpMrD2Tr/wB9Y/GtOtGsdPtbF7l7hreFIzLJ96QqoG4+5xmqqqrXPGHhfw2CNa1y1tnH/LIvuk6f3Fy36Vr37QWmwhofDmjTXL44mum8tQfZRkkfita58VfHOvMVl1uS0iJOIbP9yBntkfMR9Sa1XVrDTLSfV9b1OC0tovnnuruYRouT1Z2OBknqT3rxp+2r8C/Ct6uj6PrN74u1SSXyY7Tw/bfad0hICgSEqj7icDYzH25GdM1L9uj44Lj4Y/BnT/hro06v5eseL5Ss4GcKwhZd4yMt/qJF6fMRjdoP/BNG98b3cGv/ALT3x38SeNLxAWXTdNb7LaW78DCs+4lSBzsSI5Psc/DL9lP9nr4QLayeA/hPoNpe2jBodRubb7XeKwOdwuJt0inPOAwA4wAAMeXkYAoxEjGBRjyOlNECOKaLA4p4weCKki9KeMEYqRMcYr4v/EfTfhb4QuNfuQk15LmLT7Yn/XTEHGRkHYOrEdvcivE3ijXvGOsTa74k1Oa+vJicvI2QgySFUdFUZOFGAKi066mUMseAehbip7G5gG54+B3HNeCfHXiLwBrMeseH71kIYefbsx8q4UfwuvccnB6jORg18SP2hdcsLLRNS8EjTfsWs2PmsbiJpJ7eZXIdDhgvHA5X1PcV8TPiN4j+L3huXwh46XT77SJmV3tvsaLh1OVdXxvRhyNyMpwSOhNW/wAJ/hzbeV5XhGwbysbfMUyZx67id345z3rTPGnjLRfDtn4R0bxbrNjoWnk/Y9Ltr+WK2t8kk+XErBE5ZjwByx9TWp65rWteX/bGsXt95OfL+03DS7M4zjcTjOB+QqtO0rVNYnNrpGm3V7MqF2jt4WkYKCAThQTjJAz7ioPAHju6mjtbXwVr000rBI4002VmdicAABckk8Yr/hSHxp/6JB41/wDCfuv/AIitO/Z7+OWqu8dr8JfFKGNcn7TpctuPwMgUH6Cv+GYfj9/0SzWv+/a/41/wzD8fv+iWa1/37X/GtO/ZN/aI1V3jtfhhfoY1yftNxBbj8DJIoP0Ff8MZ/tKf9E2/8rFj/wDHq/4Yz/aU/wCibf8AlYsf/j1aj+yh+0Npcy29z8MNQd2TcDbzw3C4yR96N2APHTOenrX/AAzD8fv+iWa1/wB+1/xr/hmH4/f9Es1r/v2v+Naj+z98cdLnW3ufhL4rd2TcDb6VLcLjJH3o1YA8dM56etf8KQ+NP/RIPGv/AIT91/8AEVJ4D8cRO0Ung3XEdCQytp0oII6gjbWo6VqmkTra6tpt1ZTMm9Y7mFomK5IyAwBxkEZ9qrTtV1PSJjc6TqV1ZTMhRpLeZomK5BIypBxkDj2Fad8T/iXo7vJpHxE8TWTyDDtbavPEWHoSrDNad+0X8ddLgNva/FjxI6sxYm5vmuGzgDhpNxA46Zx+Zqy/bE/aNsYoYI/iO8scOMCfTbSQsB2ZjEWP1zn3rTv29fjrZTGW5j8NagpQqIrnT2VQcj5h5cinPGOuOelab/wUR+IMUSLrHgHw9dSBvna2lmgBX0AZnwcd8n6Vp3/BRnSJXcav8KLu2QD5Dbaws5J9w0SY/WtK/wCCgHwWvjFFqOjeKdOcx5keSzikjVscqCkpY89DtHvitJ/bK/Z11ZYEbx6bKaYkeTd6dcx7OT95whQZxn73cd+K0f45fBrX/LXSvil4XlklYpHC+qRRysR6Rswb9KsNR07VbcXel39veQEkCWCUSLkdRkEiqqqqqIBGKK+lVVVVVVVVMueRTL3FVVVVeM/GXhX4eeFdT8beNtdtdH0PR7dp729un2pCg/UkkgBQCWJAAJIFftt/t7eN/wBqLW7zwl4aubrQ/hja3Cmz0nhJdQaMnbcXTDkkkhhFkomF+8y7z+z5+y98Sf2iNZ8jw3Z/2foVs4W+1y7jPkQcE7UGQZX4xtXpkbioOa+BX7Kfwk+AdvFc+FNE+2a+YfLuNcvv3lzJkfNs7RKf7qAcYyT1pYQaEQxihGDwBQiJGAK8onjH6UYj0A/SjESMECmiB4xTRAjitW0TS9csJtK1nTbW/srhds1tdQrLHIM5wysCCMgHkV8Xf+CenwK+Iyz3/heym8D6u6sUl0mMG1Lk5y9scLtHPyxmPtzxXx0/ZC+MnwG8zU/EGjx6t4eD7Y9a0smWEZ6eapAeI4xncNuTgM1fs7/tTfGD9mPxIutfDbxEy2E8yvqOiXZMllqAAx+8jyMNjgOpVh644r9kn9uf4VftWaculae//CPeNrS3El94evJgWbrue2fjz4xjJwAy5G5QCCaqqqiARiiO1VVEAjGaI7VVUwyOKqqqqqq/4Kw/E2Pxv+1DJ4RsbuKay8DaRBp5EeCBcyDz5juHJIEkaEZwpjIwDur/AII1fDN9N8AeOvi3eQ4fXNTh0myLoysIrZPMldSRgo73CrkE/NCRxjmqqqqmGRxVVVeKLM3/AIev7ZV3N5JZR6lfmH6gVVVWj6rc6LfxX9qcshwyno69wfrWm6hbarYxX9o+6OVcjPUHuD9DVVVVRAIxRHaq8XeGNJ8b+E9a8F67G0mm69p1xp94iEAtDNG0bgZBGdrHqDXjPwvqHgfxhrvgrVxi+8P6lcafdDbjEsMjRvx25U1+zB8T5fjN+z74D+JN1Mkt7q+jx/b3TOGu4sw3BGckDzY5OCTj1NVVEAjFEdqqqqqrxpbm38R3YAOJCrrk+qjP65qqqq8Cf8jAn/XJ/wCVVTAEYFVVVdWtrfWstje28Vxb3EbRywyoHSRGGGVlPBBBIINfEbwhP8PviF4o8BXMvmzeGtZu9MkfIO5oJmiJ445KV+yd41HxE/Zp+GvitrlLie48OWsF1KrKd9xAnkzH5OAfMifKjG08YGKqqqqqqrXVVtEvwwB/0aQ4I7hTVVVVVVVUrMrBkYhlOQRwQa0u8XUNOtr0EZliDMB2OOR+eRVVVVVVVVVf8FKv2Yk+Nnwhf4ieF9L87xl4CgluYREoEl7YcNPCe7lApkReTkOqjMhz+zt8b/E37Onxj8NfFvwtK5m0W6H2u1BG29tHG2eBgeMPGWAJ+621hgqCPAnjfwz8S/BeifEDwZqaahofiGxivbC4UFd8UihhlTyrDOGU4KkEEAgiqqqqqqqqqqqqqqqqqr/gsB8IpfH/AOy9H49023eS/wDh7q0d84SIuzWc+IJgAOQAzwyE8gLE2eOR+wz8Vm+D/wC1B4I8Qz3LRabql6NI1IADBguv3QLZ6KkhjkOOf3ffoaqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqWKKeJ4J40kjkUq6OuQwPBBHcYr9rL4Nt8B/2gPF/w6gjYabb3pudJZgBmzmHmQjjglVbYTxkoeB0H/BJz47/8Lb/ZhtfBGr6p9p1/4a3A0mZJJN0hsWBeycjHCBA8Cj0tTU0MNxC9vPEkscilXR1yGBGCCD1BHavjl8O5fhJ8ZfG3wylRwvhnXryxhL5y8McrCJ+ecNHsYZ7MK/Yd+JC/FP8AZX+H3iCS6aa9sdLXSr1pCS/nWh8gs5LMSzrGrkk8784GcCqoDJqqABOKqqqv+CwPxvax0bwv8ANFvWWTUm/tnW1SQD9yhKW0bAckM4kcg4wYkPOeP2APgzJ8U/jrZ+INQslm0HwSq6jemRMq9xyLWP8A3jIDJzwRCw71HGOgqOLsKjiGOBUcOetJEMcClhBHNLCOwoQ8cCjCcY4ow8dKaEEYxTw8YFSQgjGKkiwMYqSIdq/bu/ZkX46fDg+JvC2nLJ418KRPNYCOImS/t+str8vLMcb4xg/OCowJGNfCf4n+K/gz8QtE+JPgq/a21XRLkSoAcLMnSSJx3R0LKw9Gr4RfE3w18Z/ht4d+KHhCZn0rxDZLcRI5G+B8lZInxkb45FeNsEjchwSOajXPSo1B4pVBwBUaA8CkQEYpYwRjFeWMYBpoxjAqRBiv+C2GsXU/jP4WeHXjiFtZaZqN1GwU7y80sCMCc4wBAmOOpbk8Y/4JzSDw58PP2g/iFDdCK507w7p+kw4iJcG/mmTerggqVMS/iQf4cH4eeF38a+O/D/hJFJGralBbyEfwozgO34Lk/hSRxxxrFEiqqDCqowAB2Aqqqqqvjr48Pw0+E3iLxZBOsd7DaGGxJYA/aJSI4yB32swYj0U9Oo8OaFqXjHxNpvhzT8y32sXsdtEWOcvI4XJJ9zkk14N8KaT4G8K6V4Q0OLZZaTapBFkYLYHLH/aY5Y+5NVVVVAZ4qqrxF8PvAni4P/wk/g7RdUeQEGS7sY5HHy7chiNwOMDIIIwK8Q/sg/AnxAJHh8Lz6TNICDLp15JHj5cAhGLIMcH7vJ655rxF+wH4emEj+EviDqFoQCY4tRtEuNx28Auhjxlu+04B6HHPiP8AYf8AjHpO6TRZtE1xOdi292YZDjGMiUKoJyf4iOOSOK8RfA/4veFTIdb+HWuxRxEh5orRp4l+bbzJHuXk4A55yMUj3NlcrJE8kFxbvlWUlHjdT1HcEEfhXhr49/GXwiV/sP4j60qIMJDdT/a40GCMCObeo6noPQ9hjw5+3V8VtMKReINI0LWogcuxhe3mPAGAyNtHQn7h5PpxXhv9vbwFfskXinwdrGkM3Be2lS8jQ5PUnY2MY6KeTjHevDn7S3wN8TxI9l8RtLtHYfNHqTmyKHAJBMoUHrjIJBOcE4qzvbPUbZLzT7uG5gkzslhkDq2Dg4I4PIIqqqqAzzVVVVVVWM8CgABgVVVVVVUFzQUYqqqqrxZ8TPCnhFmtr28Nzer1tbYB3X/e5AXtwTnB4BrXPj34kvS0eh2Nrp0f8LuPOkHGOpwvv930/HVvFvibXcrq2vXtyjdY2lIT/vkcfpVlp2oanL9n02xuLqX+5DEXb8hmvC/wn8eXFwLl9MisYmXaWvJdhx/ujLdh1A6/XGgfCXTLFluNeuzqUo5ESp5cSn6ZJb8Tg+lWX7Pfh+NlbUddv7jC/MIlWLLdzyG468frXh74e+EPC8guNI0eNbgD/XykyOPoWzt/DFVVVPPBbQtcXEyRRRjLu7BQo9ST0rxN8cfCujhrfRg+r3AyMxnZEp93I5/4CCPevEfxd8aeIWeNNROnW7HiGzzGce7/AHj+eOvFa3ruk6JZT614j1m0sLWPLz3d7cLEi9yWdyAO5yTXxA/b2+CHhFZLbw1PqHi2+VflWwhMUAbPRpZMcY5yiuOR748L/FP9uL9ppmj+CvwwtfCug3BBj1y6hxHHGWwSLm4/dzY6EQxFxycDjHgj/gmUviS/tfFP7T/xi13xzqYVmk0yzuJI7aJ26oJpCZGj46IsPRRjAwfhj8BPg58HrZbb4Z/DjQ9BcRiNrm3tg1zIoGAHnfMr8E/eY9T6mljGMAUsWaWEEcChDxgCjDkYxTQgjgU8IIxinix0p4wRgU8eBipYwRxUiZGK/bC8Sy6r8S4PDiy5g0KxRdgYnEsuHY47Er5Q/Ac9ho9mLmbzGXIU4APrWk+HHvEGxM8elax4ea0UhkwQOhFanai1uNqDCtyB6etfBf4e6T8UNRutA1jVrm0TT4/tUaW5XdIGKq+AwOOic44yM5yKtP2XPhtbMxmudbu8jAWa6QBfcbEX9an/AGbvhPNpc2nro13HNLE8a3iX0omiLAgOozs3LkYypHAyDzn9oPRPjP8AAz4g6j8P/Efi/VJbYN52mX8bmIX1qWPlyqV5VuNrKDwwYZIwT/wSk/aH8B+Nln/Z/wDiX4e0KfxhEktzoOr3OnwGfVLdfnlt5ZSN8ssfzOuckxq2SPL507QNB0d3l0jRbCyeRcO1tbJEWHoSoGRVVVVVVVVVVValo2kazGkWr6VZ3yRnKLcwLKFPqAwOK1L4X/DTWY0i1f4d+Gb5IzlFudIglCn1AZTitS/Z2+BWquklz8J/DCGNcAW2npbj8RGFB/Gr/wDZB/Zy1K4NzcfDO2RyoGIL+6gXj/ZjlA/Sr79hD4CXcUsdva69ZtJna8OpEmL/AHd6sPzBrU/+CeHwxl8saP428UW2M+Z9pME+7pjG2NMd/XOR0xzqn/BOQHz5NF+LP/XCG60X9GkWb9Qn4VqX/BPH4oxOg0jxp4VuUI+c3LXEBB9gsb5/StV/Yf8A2hNPj32nh3TdTPmbdtrqkKnHPzfvSgxwPfkcdcat+zX8edGWRrz4Va/J5Mhjb7JbfasnOMjyi24cdRkd81q/gvxjoCzNr3hPWdNFsQJjd2EsPlEkABtyjHJA59RVrdXVjcR3dlcy288RzHLE5RkPqCORWifGj4u+HDGNE+Jvia2SIsVhGqStFkjBJjZip/Edea0X9s39ofRlWKTxtFqMSR7FjvdOgfHI+YuqBye3LHr9MaN/wUN+Jlt8uveC/DV+o2hTbedbNx97JLuCTx0AA54PbRP+Cing64MY8SfDjWbEEt5hsbyO72jHGN4izk4z0x71of7bv7PmsbFu/EmoaQ7opC32mynDH+EmIOAR3Oce5rQvjX8IPEwVdD+Jnhq5kbOIf7SjSXjqfLYhse+MUjxyxrJE6ujjKspyCD0INVRUHpRHaqqqqqqqplzyKZe4qqqppYYIXnnlSOONSzu7bQoA5JPYYr/god+2xd/tL+Nf+EF8DXzRfDjwxdP9iaOSRf7Zn4BupEOPlABEalcqGZicvhf2S/2SvEP7RXiEarqouNM8D6ZMBqGoqu17lxgm3gzwXIxubkIDk5JVW8KeEvD3grw9YeFPCmkW+maTpkIhtLSBdqxqP1JJySTkkkkkkk0sY6AZpYieKWIYxQjB4AoR5GAK8onjH6V5R9P0ox+1GIEYxTRAjiniI6c08YIwB+FT20c0bQyxq6OpVkZchgeoI9K/aY/4J4eG/Gcdz4v+BlvY+HdcVWkm0Qr5VlenjAixxbvgHAA2EkZ2DLUU8cfCrxnG7Jq3hfxT4eu0ljLK9rdWUy4ZGHRlPQg9wQehr9gz/gozonxntdK+EHxq1KLTviGMW9hqTqI7fXsD5eRxHcnGCnCucbOW2CqqqphkZFVVUwyMiqqqYc8VVVVVWoX9npdhc6nqFwkFrZwvNPK/AjRQSzH2ABNfFfx7f/FT4m+K/iTqQZbjxNrF1qBiZy3kiWVnWMZJO1FIUDJwFA7V+xZ8MU+En7L3w98IvaRQXr6Ql/qATBJubrM8gZhwxUyBM5IwgAOAKqqqqqiMcVVUQGG0gEEcgitasDpmq3VhjiGVlX/d7fpiqqq8B6+dN1D+zbh/9Gu2AXP8EnQH8eh/D0qqqqqmHPBqq/4Kl/DR/An7VepeIbeHbY+NtMttVhKIwRZQpgmXJGC2+DzCATjzV6ZxX/BHj4mJrXwq8X/Cm7u4jdeGtWS/tIzhX+z3SYIAHLBZIWJPOPNUE4KiqqqYDPFVVVVV8SLbZqltdAYEsG0/LjJUnv34I/SqqqrwJ/yMCf8AXJ/5VVYzwDRHaqqq/wCCjfhFvCX7X/jlUZmg1hrXUoC7hmxLbxl84AwBKJAB12gZJ61/wSd8ZN4j/Zck8OTToZPCniK7s44jMGZYZRHcq23AKKXnlAznJVjnqBVVVVVVWuf8gW//AOvWX/0E1VVVVVVVV4Cu/P0ZrUt81tKQB7Hn+eaqqqqqqqqqv+Cgv7Nf/DPHxwuZ9A06K38HeMvM1HQlgTbHbHcPPtQMADynZSAOBHJFznIH/BG79qFI21H9ljxhrLEyNJqXg9ZnJAOGku7VPQYBnCjAz5x6mqqqqqqqqqqqqqqqqqr4heCdF+JXgPxF8PPEUXmaZ4l0u4067XHIjmjZGIwQQQGyCCCCAQRXjLwpq/gTxfrngfxBEkeqeHdSuNOvUQkhZ4JGjkAyAcblPUCv2XPisvxt/Z/8EfEeS5Sa91HS0j1IqCMXkOYrjg8gGWNyM9iDkg5NVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV/wV8+C7ax4P8MfHbSbRGuNAl/sjVnAAb7LMxaBjxkqkpdcZ6zjjqR/wSz+PSfBf9qPS9B1i+SDw98Qof7EvjLIVSK4Y7rSXA4LecBFk8BbhzVf8Fivhz/wiX7VsXja3j/ceOfD9rdySZ63NuDauuCxPEUNuc4UHfjBIYn/gjl8Qvtvgjx98LLm4+fSdSg1a0RjklLiMxS7fZTbxkj1k9zVVSgYqqUZqqqq/ap+Ls/xy+P8A40+IovpbiwvdSkh0neR8ljD+7twAOFzGisQP4mY5JJJ/4J7fCeP4dfs+abr93bsmreNZTqtwZI9rLARtt0B6lPLAkGe8zY4xUcfAA61HHjoKjiGMmo4sjpSRZGAKSAY5pYRjAFCEkYA/SjASMAU0J9KaEHjFPDjoKkiBGMVJF2NSxYqWPuK/4KGfs+R/CP4rL448OWaxeGvG7SXEccUW1LO9XHnxYHAVtyyL0+86gYTNf8Ej/wBoO50fxnqX7OniC/lksPEEcmoeH0diwguokLTxKOweJTJ6AwnuxqMA8VGBwBSAE8Go1FRqCMUq54FeUMcCnUDipFGK/wCCzgx+1F4YA/6EG0/9L7+v2QdRh0v9kL4nQq3lyax4x0u0YqnMgSGSYKx9B5ZI9/rX7JGkf2x+0B4WR4XeK0a4uZCpxs2QSFSfbfsH41VVVVQGTiv+Cgfi6SDSPC3gSB3C3c8uoXAHAPljZGOmTy8h64GBweMfsLeC18QfFq48U3NsZLbwzp7yRvg7UuJv3aA9uUMxGe657ZFVVVVAYFVVYzwKwMYFVVa54P8ACXikBPEvhfSNXAAAF9Yx3GACSB84PQk/nXiP9kT4EeIYnEXhOTSZ2GBPp13JGV4I4Ri0fcH7vUDPGQfEf7AOjy7pPCPxEvLfGdkOo2SzbumAXQpjvk7T1HA7+I/2KfjdoivJptppGuooyPsN6EbGT/DME5wAcDPUYzXiX4ZfEPwcW/4SjwTrWmooJ82eydYyAASQ+NpxuGcHjODWja/r3hy6F94e1u/0y5HSazuXgfj/AGlINeF/2tfjp4YdFbxcusQKQTBqlus4bp1cYk6LjG7uT1Oa8M/t+36IIvGXw+t5nxzPpl2Yhn/rnIG/9D4968L/ALYnwL8S7Y7nX7zQ5mzti1SzZOmc5ePeg4XPLD7wHXIGieKfDHieH7R4a8R6Xq0WCd9jeJOuAcE5QnuCPrQGeAawMYFVVVVVVAYHWqqqqqqguaCjGaqqqq+K3xYuIribwv4WujH5RMd3doeScYKIe2O7dc9MYydM0rVNevksNLtJbu6lJIRBkn1JPYe5rw9+z/fTqs/ifV0thkH7Paje+O+XPAP0DCtJ+FPgPR1Xy9BhupAOZLsmYt16q3y9+wHaoLa3tYxDawRxIDkLGgUZ+goDPAoAAcVVVU88FtC1xcTJFFGMu7sFCj1JPSvFnx30bTt9n4Wtv7SuBkefKCkKn2H3n/QehNeIvGHiPxVOZtb1WWdc/LCDtjT6IOB256+pr4p/tFfCL4PRFPGHiyA6h/Bpll/pF031jU/IODy5UHBAOeK+JP8AwUN+IHiKY6V8J/Ddt4fhkbZHd3Si9u5Dn5SqEeWhI4KlZOehr4d/sTftXftNavB4l+Kup6l4d0lsH+0vEzPJcbGAyILTIbptOG8pD2bIxXwX/wCCf37Onwfa31JvDDeLdbgZXGo+IQtz5bgdY4QoiQA8glWZTj5iQDUcYx0qOPNJGCMCvh3+1l+z18VPiBN8LPAfxB/tTxRbef5th/ZV5Bt8k4l/eSQrGcEdm57ZpYwBjFLGSMCvjV+0h8Ev2dho3/C4/G3/AAj/APwkHn/2d/xLbq78/wAjy/N/1ET7cebH97Gd3GcHHhfxBofjXwxpHjHwze/bNH16wgv9PufKaPzreaNZI32uAy5RlOGAIzyAaMPHFfGv9pD4J/s8f2MPjF41/sD/AISDz/7O/wCJbdXXn+T5fm/6iN9uPNj+9jO7jODjw14h0Txp4Y0jxj4Zvftmka7YQ3+n3PlPH51vMiyRvtcBlyjKcMARnkA18Rv2/B4C/a0tf2W/+FS/bvtOuaTpf9u/295W37clu3mfZ/s7Z2faMbfM+bZ1XPEkeeKkTtipY/Sv2rdOuLL426zcTAbL+C1mhxn7ogSM5/4FG3SvD0qKWQnkNn/P5V4S1m1sdryKrY7GvF2r214zvGFGewrXpFe4RFPKgk/j/wDqr9k2wnl8a6vqSgeTb6UYn653PKhX26RtTgA8VX7V37Pem/tCfDKfR4LeFfE+kK9xoF25C7JiBuiZu0cgVVPYEI38IrRdY8W/DHxraa5o11c6L4l8MakssEoUCS0uoJMjKkEZV1wQQQcEEEV+yz+0FoP7TfwV0L4qaOLe3u7lTbaxYRS7/wCz76MDzYT3A5V1zyUkQ96qqqqqqqqqqqqqqIBGKKg9DRBFVVaz4B8CeI/M/wCEg8FaDqfnNvk+2abFNvYdzuU5PvWt/stfs++ICzX3wu0mItJvP2IyWfOMceSyYHP3enfFaz+wP8DtS+bTrnxJpLDdgW18jqSemRKjnAx2IPJyT21v/gnNasrSeHPipKhWP5Yb3Sg+5893SQbRjH8J6e/Gt/sAfGbTyz6TrHhnVY/M2oqXUkMm3H3mV4woHGMBieR741v9lr9oHQFZr74XarLtj3n7EY7zjOOPJZ8nj7vX25rXPCvifwzL5HiTw5qmlSBihS+s5IDuHUYcDnHatB8W+KvCs32jwv4n1XSJck77C9kt2zjBOUIPQkfQ14c/az/aB8NFFtviLd30SqqmLUoY7veFGBlpFL9+SGBPcmvDX/BQr4kWGyPxV4O0HWI0wC1s0lnK3XOTl1yeOigDB4OePDX/AAUK+G2obIvFXg7XtHkbALWzR3kaHnOTlGwOOiknJ4GOfDf7VH7P/ioRpZfEvTbOVwMx6mHsth25ILShV4xjIJGeATkVpmr6RrlqL3RNVs9Qt87RNazrKmcA43KSOhB/EVVVVVVVVMueRTL3FVVf8FP/ANusTvqv7MHwg1eJ4V8y18Z6nENx3hsNp8bdBggiUjv+7yMSA/s5/AvXf2hPidYeBNKka2slU3OrX+MiztVI3sB3ckqijuzDOFDEeDPBvh7wD4W0vwZ4U02Ow0nR7ZLe0t0H3VUdSerMTksx5ZiSSSSaSPPAFLEAMCliJ4ApYhjFCIYwBQjJGMCvKOMA/pRiJH/1qMeRjApogRjFNCCOKaPjFPECMAU8eBg1JECOBX7T/wCyh4M/aI8PTXHkW+leMrSHbputCPk7ckQz45eIlj7qTkZ5VvGvgrxf8LvF974R8W6ZcaTrekTYkjY4Kkcq6MOGUjDKwOCCCDX/AATn/brt/j7olt8GfiXd+V8Q9BsAbe8mlJGvW0QCmTLEk3KjBkXksMyDgOEqqqiARiiMcCqqiARiiMcZqqogEYzRHaqqqqv+Ci3xNT4YfsjeOJ4buKK/8TW66BZJJj98bs7JlAPf7MLhuOflzxjI/Z0+GL/Gb46+BvhkbSW4t9d1qCK+SLduForb7lht5G2FJGzxjGcjrTDniqqqqqpgCOKqqr4maf5GrQagiALdRbWOOrrx/Ir+VVVVXg/WzrekI00m65tz5cuep9G/EfrmqqqqmAIwKqv+Cxfw8/tj4T+C/iZbQb5vDetS2E7KORBdx7tzewkt0Uehk9zX/BK34i/8IV+1XY+HLm78uz8a6TdaWyu+2PzlUXETHLAbiYDGpwxzKVA+bIqqpgCOKqqqqr4k24fT7O6wMxzFBzz8wz/7LVVVV4E/5GBP+uT/AMqqqYDtVVVf8FjvCkln8VfAHjgpLs1bw9PpysXUoTa3BkICj5gf9MGSeDlcdGr/AII1+LGj1f4meBZpmZbi2sNSt4y4AQxtLHKQvUlvMhGeg2j1qqqqqqqtc/5At/8A9esv/oJqqqqqqqqr4eXITULm0LAebEGAPcqf8GNVVVVVVVVVX7afwBi/aK+Aeu+DrK1STxBpy/2loDkci8iU4jB7CRC8foN4PavBHjDxb8IviFo3jfw5LLpviLwpqkd1biaMqYp4XBKSJwSpKlWQ9QSp6mvgH8ZvDf7Qfwh8M/F3wqvk2fiCzEslqZRI1nOpKzQMwAyUkVlzgZwDgA1VVVVVVVVVVVVVVVVVf8FfPgwnw5/aZh+IWmWjx6X8RtNW9L+WQn26DEVwqnJBO37PIcY5m6dz/wAEefi3PdWPjX4HaleyyCz2a5pMTEERoxEN0BnkDcbYgDjLueCTmqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq+Lfw10P4xfDPxJ8MPETMlh4j0+S0eVBloHIykqjPLI4VwDwSozxXiHQfE3w48Zah4c1aO40vX/AA3qL284RyklvcQvglWHIIZchh7EV+yP8bY/2iP2dvBXxUmuIZNU1HTxDrCxqqbL+EmK4+QE7A0iM6qf4HQ9DX/BaT4bf8JJ+z94X+JNrb+Zc+DfEHkyvj/V2t5Hsc5/67RWwx7+1f8ABMv4iReAP2tfD1nd3KQWni6yudEnkb1kUSwr90nLTwQpxt5bk4yDVYzwKwMYBqqA4xVVVftyfE6X4S/ss+PvE1lMiahd6d/Zdjk8iW7YQbl/2kSR5B/ud+h+Ffgib4lfErwv4AhaRP7f1a3s5JI/vRRvIA7jOfupuboelabYWunWVvp9jAsNvaxLFDEo4RFACgewAAqKMYzUUYIzUcYPJqOLOKSIYwBSQjvSw5GAKEBIowDHFNAelPEDwRTw4HFSRAjipYgRg/hUsfGDUsYHFftY/Bq1+N/wN8R+Dvswk1S3gbUNFfA3JewqzRgE8DeC0RJ6LKxr4c6H8avAnjDQ/iJ4K8G+IIdU0C6i1KxnOmyhGaMhwDwA6sBgoD8wJHOa+HPxC8OfErwtpfifQbyPOoWMFzLZPKpuLJpI1cxTIDmORdwVlIBBBBqPGOtR9ajIxgGo8dM1GR0rcNuBUhB4BqTGMV/wWehlH7TvhacxsI38B2qq5U4JF9fEgH1AZcj3HrX7M3iC1T4K6z4US9JuZvFCXslusgwqJa7Fd1zkZLuFOMcPzwa/YK0f7b8W9U1aS23x6bocmyTfjy5ZJYlXjPOUEg6ED64qqqqqgOK/bW8QPrfx61KxL7k0SxtrKM8YwU84gY/2pmHPOc+1fsC+HBp/wy1rxLJCqy6vqxjRsLl4oY1CnI5+/JKMHp1HWqqqqgAaqqpQMZqqqgMCqqq8S/B34V+MMt4i8AaHdyn/AJb/AGNY5e/HmJhsfMTjOMnPWvEv7Dfwf1dnm0K71vQpD9yOC5E8S8jOVlDOeMj746968S/sFeP7As/hXxhourxKCdt1HJZytwOAo8xeu7qw6D1wPFX7Pnxm8G731r4e6qYU+9PZxC7jA9S0RYKPrike/wBKvd0bz2d5auRkExyROOCOxBHSvCv7THxv8IbI9P8AiBf3cCceTqWLxSPTMoZgP90ivCv7fniK32Q+NPAmn3w6NPptw1sw99j7wx9sqK8L/tjfAvxLtjufEF5oczZ2xapZsnTOcvHvQcLnlh94DrkDQ/Efh7xPZ/2h4a17TtWtckefY3STpkcEbkJHY1VVVUBnmqqqqqqgM96Cgiqqqqvip4nl8LeELi5tJjHeXbC3t2BwVZgcsMdCFBIPritB0W88RaxaaJp6gz3cgRSeijqWPsACT7CvCnhHRvB2mLp2kwAMwBnmb78zgY3E/wBOgzxVUFzWBjAqqqvG/wASdA8EReTcubvUGXMdnE2G9ix/gX3PPoDzXi3x54j8Z3G/VbzbbqcxWsQ2xp+HUn3OT+FfGL9pT4VfBKNrXxVrTXOsGPfFpFivm3Lg9CwyFjB9XIz2zXxh/bi+LHxI8/SvC0//AAh+iSZXyrCUm6lX/bn4I+iBfQ5r4A/sI/Gv49SR67qNu3hHw5MQ7atrFu/mXKt1aCDhpfXLFEOeGJ4r4A/sefBT9n60t7rw54eXVfEUa/vdf1RFmuix6+XxthXnACAHGNxY5JiUYqJM1GgPApEBwAKjjGBgV/wT6XP/AAUC10A/9Bz/ANGGo4sgUkII4Ff8FrU2D4NY/wCo7/7j6/ZaiB/Zi+EJH/Qh6J/6Qw00PHAr/gtUm0/BsD/qO/8AuPr9mCPP7M3wjI/6ETRf/SGGv2iVH/D2vS1H/Q8+Ev8A0Tp9SxjtUqAipEyMV+2F8L7rxDoVr8QtHgElzoUTR3ygfM1sTkMO52MWOPR2PaoJ5LeQSxNyO3rVn4jjRQGcofQ1feIlkUrG5du3GBUkjyuZHbLE8k1+z78OrnwJ4J+06rCI9T1pluJ0xzHGB+7Q+4BYkdixFSDjjtVV/wAFIfgJb+FfFNp8bPDVj5dh4lm+z6zHGgCRXoXKyYA481VYn1dCScvX/BJ79o3/AIVP8dj8J/EOoeX4c+JXl2kXmN8tvqiZ+ysPTzNzwEAZZpIsnCVVVVVVVVVVVVVVVVUVBoqRVVVVVTwQXUElrcwpNDKhSSORQyupGCCDwQRxite+Bvwc8TOZda+GPhqeZmDNMunRxSsQNoy6AMRjjBOOB6Cte/YV+AOsAjT9O1rRM4x9g1JnxjrjzxJ17/pivEH/AATojKiTwt8UGBCgGHUNNzk5OTvR+BjHG09+fTXv2Ffj7pAP9n6domt4xj7BqSpnPXHniPp3/TNeI/g18WPCSmTxF8OfENlEOs76fI0WcE48xQVzgE4zng1YajqGl3Au9Lv7i0nAIEsErRsAeoyCDXhn9qX4++FGUWPxL1S8jDgtHqZW+3jOSC0wZgD0+Ug46EV4Y/4KFfEHT1ji8WeCtE1lU4L2ssllI/B6k+Yuc46KBjjHevDP7fPwb1hlh8Qadr2guWAMktsLiIAnGcxktwOSNn0zXhX45/B/xsY4/DXxG0O6mlUFLd7oQTNkZ4ik2v3GRjg8HBqqqqqqZc8imA7VX/BRr9r7/hmr4WL4X8FapEnxD8YxvDpm18vptqOJb0qOhH3I84y5LDcInWvAPgXxZ8WfHGm+CvCtpJf6zrd0EUvuYLk5eWRgCQijLM2DgAmv2evgF4Q/Z88AWnhHw5bxy38qJJrGplMSahchfmc91QEkImcKD3JZikWetJHngUkQApYyRgClhyOlCIYxQjBHAzXl8YAox5HSjECMEU0QIwKeH0FPHxginiBHFPH2xUkZHSpIwRxX7Xv7Lml/tEeDRPpfk2XjHRInbSbwooFwMZNtK3Xy2PQ5+Rju5BYG0u/GXwu8aw3tpJf+HvFHhnUA8bFTFPZXMT5HB5DKw6Gv2Lf2ptH/AGq/hBa+KSkVp4p0bZZ+JbCPhYrrbxLGOoilALqOdvzJklCTVVVEAjAqqqmGRxVVVMPSqqqqv+CzvxMdrr4e/Bu0nwqRz69fR725JJt7dtoOONt0MkE88Ec5/wCCPfw2i8S/HrxF8R7yzSaDwXoey2kZMmG7u2MaMD2Jhjul+jH3ywyOKqqqqqiARiiO2aqq8e6aNQ8OyyIuZLQiVeOcDhvwwSfwqqqq8FaudK1uJHkAhuv3UmTwCfun8/5mqqqqqIGcCq/bS+Hn/C0P2WviP4Vig866XRZL+0QDLNPaEXCKvoWaEL/wLHQmvhL47uvhf8UfCXxGs2l8zw1rVrflIycyJFKrOmAy5DKGUruAIYgkA1a3Vrf2sN7Y3MVxb3EayQzROHSRGGQykcEEEEEdaqqIBGKI7Zqqqqrxzb+f4buGAJMLI4AGf4gD+hNVVVXgz/kZbP8A4H/6A1VVMMjg9Kqqr/gsF4X/ALR+BXhLxbFBLJLo3igW7lIdwjiuLeUszN1Ub4Yl9CWHfFf8ErfFQ8O/tbado/mY/wCEn0K/07Hl7t21FusZ/h/49c59sd6qqqqqqrXP+QLf/wDXrL/6Caqqqqqqqq8Hz+R4htfmwH3IRjrlTj9cVVVVVVVVVVV/wVH/AGfP+FW/G5fijoFj5fh74i+ZdSbF+WDUkx9pU46eZuWYEnLNJLgYSv8AgjZ+0j/wjPjfV/2bPEuobdO8U7tS8P8AmNxHfxp++iH/AF0hQN9YfV6qqqqqqqqqqqqqqqqqv+Cr/wAE2+K/7KmpeKdK0x7rW/h3dLrUBiQGT7IBsvFyeiCJvObGP+PcHnGD+yh8Xx8Cv2gvBvxGuJpV06zvhb6oqSbd1nOpimJHRtquXCnjci8jGRVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV/wAFZPgdF4J+MGmfGHQ7FIdM8dW2y/2HgajAArttHCh4vKPuySMeTX/BGL4/f8Iz8RvEH7O2t3spsvGMZ1PQ4icpHf28RM6geslugYk/8+qjvX7Vfwz/AOFxfs4fEb4cRWn2q71fw/c/YIvL37ryJfNtuNrE/vo4j8qlv7uGwa8G+J9Q8EeL9C8aaQ2L7QNSt9QtTnGJYZFkTntyorwv4i0zxh4Z0jxbok3m6frdhDfWj7lbdFLGHQ5UkHKsOhI9CaoAE1VAAnFVVVX/AAWV8f8AkeGPh38K7eTd9uv7nWbpMY2eTGIYTnbznz5+jDG3kHKkf8EyfAS+Jfj7c+MbiJjD4Q0iaaJwSNtxP+4UHHUGJ7jr6D6iJAcAVEgOAO1RRg4AqOMHgVHGDwKjiHAFRw5GMUsIxjFCHI4FGE47flTQ8YxUkII4FSQ46VLECOKljBHFSxgjHeviB400jwDoEmuaqWc7vLt4E4aeQgkKPQcEknoB34B8afEnxV47uS+sX5S1B/d2cGUiQcdurHjOWyfp0pUZ/uKTj0FWd7e6ddR3unXk1rcxHMc0MhR0PTIYciv2f/2nL7UNRs/A3xIuhNJcFYNP1Urh2kJwqTY654AfGc/ezksEIzjNRsMcGo2GODSsCODW84xTMMcmpGBHBr/gtfYXifEL4Y6o9uwtLjRr2GKUjhpI5oy6j3AkjJ/3hX7NFns8L6rf5X99fiLGOfkjU/l8/wDOv+Ce2kYg8a69JCh3NZ20MmfmGBK0gx6HMf5VVVVUBngVgYwK+PmoPqnxs8cXMkaoU167gABzxFIYwfxCA/jX7J2kR6N+z/4ThURGS5hmuZHRNu8yTyMM+pClVz/sjtVVVVQGBiqqgM8A1gYwKqqAzVVVVVVXiTwN4M8ZwGDxZ4V0rV12bVN5ZpKyDBHysRlSNzYIIIycV4s/Yr+CfiLzJdJstS8PXD5YNYXZZNx3clJd425I+VdvCgDHOfF37A/i6xWSfwT4z07VVBytvfQNaSAZPAZS6sQNvJ2g5PTAz4q/Z7+M/g3e+tfDzVTCnLT2cQu4wPUtEWCj64qx1DVdDvRdabfXdheQkgSQStFIhHUZBBHSvCP7WPxz8IrHAvi86zbRnPkavELndyOshxL0GMb+57814S/b902RVh8d+A7mFh1n0mdZd3B6RSFcc4/jPU+nPhD9pL4J+NmSDSvH2n21y2ALfUSbNyxIAVfNCh2JI4UmkdJUWSJ1dWGVZTkEeoqgM1gYwKqqqqqgM96VQRVVVVVftD3krXei6eCRGkcspGepJUcj2A/U1+z5pcdxrWqauwUmztkiUHsZGJyOPSMjr396qguRzWBjAqqqviX8YYdGMmheFJ4578FkuLnGVtz0wvZn/MD3OQPEniXTdEsLzxP4s12CztIAZbu+vrgIq5ONzux6kkDnqSPWv2gP29tY8QC48K/BRp9K04/LJrkq7Lqb1ESn/VKf7x+f/dr4TfA74uftFeJ7my8DaJc6rP5nm6lqt5IVggZzkvNO2fmY5O3l2wxAODX7O37APwm+DMtl4m8TxL4x8VwKri5voFNraSjB3QQkEBlIGHfLAgEbTUagnFRrwAKjXgAVEoxxUSjGaiTNRoAABX/BPVQ3/BQbXF/7Dv8A6MNRxA4AFLGOmK/4LaLtHwY/7j3/ALj6/Z3/AGzv2VvC37P3wx8MeIfjj4bsdV0jwbpNnfWksrh7eeKziSSNsL1VlIPuKb9uv9j8jA+P/hY/9tn/APia/wCCsHx1+D/xrPwt/wCFUfEDS/E39jf2x9v+wuzfZ/N+xeXuyB97y3x/umv2X4wf2Y/hCf8AqQ9E/wDSGGv2vfFOkeA/+CoFx438QSSppfh3xR4Z1G9aKPe4hgtbCSQqvc7VOB3qW8/4KM/tr2k3jT4ea9a/CH4f3uX0SGTUJdOmuoQ4KSefDG9y5ZSvzjZEwU7Ryd3jL4h/8FBv2EvEun3XxG8bXni3w9qFywjn1O/k1nT9QYJzEJpcTwNj5guYydpIDANn4AfHPwl+0X8L9N+JnhENBHcs0F9YSyK8un3SY8yFyvcZVlOBuR0bA3Yq4iSRGjdFZWBBVhkEV8W/2Qkv7y58QfDC6gtmmJd9IuDsjDHr5L/wg9kbgf3gMAan8GfixpN01nd/DvXnkXOTbWL3CcEjh4wynp6+h6EVo/wS+LGuXItbTwFq8DZAL3tsbVB153S7R2PT29RXwn/Zk07wfeQeJPGd3DqmqQEPb2sS5gt3/vEkZkYcY4AB5wTgiUAjFSAHiqr4w/DTSPjB8Nde+HWshVi1e0ZIpiuTbzj5opR7q4Vsd8Y71rWkeIfAXiu80TUVn03W9Avnhl8tyjwTxPjKsOQQy5DD2Ir9kr43w/tD/s+eD/ifJOj6pe2K2+sqsflhL+H93cYUfdVnUuo/uutVVVVVVVVVVVVVVVVUVB6URjiqqqqqqqrxT8Mfh143LSeLvA2h6vKykedd2Eckq8AcSEbl4VRwR0HpXif9h/4A+ImaSx0bVNAkdizNpl+3JJycLMJFUdsKAAOmK8T/APBOq9RGl8GfEqCVth2wanYmP5gOP3kbNwTj+DgevSvFX7Gvx+8LrJLH4Si1qCMkGXSrpJs4PBEZ2yHIGfu/XB4rX/Cvifwpciy8UeHNU0e4b7sN/ZyW7ngH7rgHoyn8R614X+KfxJ8FFR4T8d65piIMCGC+dYiACADHnacbjjI4JyK8Kft5fGrQ9sXiGLRPEcXO57m0+zy/xYw0JVR1HVDwo6Ek14U/4KD/AA51PbF4v8Ia1ocrZ+e2dL2JcburfI/QL0Q8segGT4U/aC+Cnjbavh34laJLK+dkFzP9klbG7OI5gjnhSeBwMHoQaqqqmXPIrxh4q0HwJ4V1fxp4nv0s9I0Kxlvb2d/+WcUSFmOO5wDgdzxX7RXxy8T/ALRnxe174reKJGR9Tm2WNnvJSxtEG2GBBnAAUZOMBnZ26sa/4J9fs2RfC34fL8TPFmlqvizxZEskAmixJYWJAKRjIyryH5355HlggFTmOMHrSR54ApIx0A5pIs9KWMYwBmliJoRDGKEYPQZry+OBRjyOlGIdMU0QI4p4iOB+tPEDxTxY6U8YPHepI8cVJHjpUsYI4r/gon+zCut6ZJ8ffBFhGl/pkO3xJbRRhTcwDhboY5LoPlfOcptPHlnP7H/7Sut/sufGXTfHdqZbnQ7vFn4g09XIFzZsw3MAODJGfnT3GOjGtF1nSfEmjWHiHQNRt9Q0zVLWO6sru3cPHcQyKGSRGHBVlIIPcEVVVVMPSqqqIwcCqqiARjNEdqqqqv8Agox8RP8AhY37XvjqeC48yz8PTx6JbLnPl/ZUCTL/AN//ADz+Nf8ABJD4e/8ACK/suyeMp4/3/jXXrq7jkz/y7wYtlXAYjiSGc5wpO7kEBSaYYPFVVVVVTAdRVVUsUc8TwSqGSRSrKe4PBrVbCTS9RuNPkOTBIVBI6jsfxGDVVVV4b1QaxotteM2ZNuyXP99eD+fX8aqqqqYAjiqmhiuIngnjSSORSro65DA8EEdxivit4Jl+GvxP8W/DyZ2dvDOuXmmh2OTIIZmjD52rkMFBB2jIOcDNfsa+On+JP7LXw08VzXPnzvoMVncSkMC81qTbSMcgHJeFie2c4yMGqqqYDtVVVVXihGk8PX6oMkQE49hyf0FVVVXgz/kZbP8A4H/6A1VVYBGKI7Zqqr/go54XXxR+x348WO3SW50pbTULctIU8sxXURkbg4J8oyjByMn1wR+xj4jl8LftWfCvU4XnDTeJrWxPkybGxdN9nOT/AHcTHcO65Heqqqqqqq1z/kC3/wD16y/+gmqqqqqqqqtFmEGsWUpfaFuE3HHbIz+lVVVVVVVVVVftc/Aa1/aN+A/iH4dpDEdZWP7doM0hA8nUIQTFyeFDgvEzdklY9cV4Y8R+KPhp420zxVoU8+leIPDOpR3NszoUe3uIXDAMp5yGXBU+4NfBD4raB8cvhH4U+LPhmZHsfEumx3OxT/qJvuzQnk/NHKskbcnlDyetVVVVVVVVVVVVVVVVVrmi6V4l0TUPDmuWaXenapay2t5buSBNDIpR0OMHBUkcetftAfCXUvgV8aPGHwk1TzWk8NapJbwSSjDTW5w8Ep4H34WjfoB83HFf8E+fjOvxo/Zj8NXN7eNPrfhRf7C1UvksXt1XynJJJcvA0LFj1cuOxqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/bH+BsX7Qf7PnibwJbWKXOt28P9o6CScMl9ACYwpOQC6l4if7srdOo8B+NfFPwp8faJ4+8LXLWOveGNSivLRpFI2TRODtdeMqcFWU8EEg8E18M/H2hfFX4d+GviV4Zl36Z4m0u31C2BYM0ayxhtjY4DqSVYdmUg9K/a/+G3/Co/2nviX4Bjt/ItrDxBcTWUW3Gy1uCJ7cf9+ZY/rX/BOH4kL8R/2SvCSTXTTX/hVptCvNxJ2GBswqCWJwLaS39ADkAAAVSjiqpRmqqqr/AIKveMD4k/avuNBW4R08KaBZWBjVXXY8ga6Oc8FiLleV4xtHUGv+CVPg86f8L/F/jaS3aOTW9bjs0ZlYeZHbRbgwzwV3XEgyO6sCeBiFeM1CgxxUSDAxUSDHFRR8ACooge1RxZGBUcAIxilgyMAUbfjgU8HHSpIcdKliBGMVLHjpU0YHSpkA5FfH/wAVyeJfiHe2UcpNnopNpCvTDr/rTj135H0UVomkC+bzphmNTgL/AHjQ0ItCpiiACjAAGMVqGgRzZR12vj5XxyDU0LwStDIMMhwRX7PHjy5+IPwt0zVNRuWn1KxZrK9kc5LyR4wxPUlkaNiT3JpGGOtI4x1pZOOteaMYzTSAjANSP2Br/gtjooufDHwo8SC6x9gv9UtPI2Z3+fHbvu3Z4x9nxjHO7qMc/ACzNt8NrWcxov2u5mlBUcthtmT7/Jj6AV+wTpK2nwl1bVXtikt/rsgWQk/vIkhiC4HThjIP/wBQqqqqpQMVXi6eC68V61c20ySxS6jcPHIjBldTIxBBHUEd6+CMEFv8GfAkdvCkanw3YOVRQoLNboWOB3JJJPckmqqqoDJqqqgOKqqxngUAAMZqqqqqqAzwDWBjAqqrxZ8Mvh546jdfFvgzSNUeQYM89qpmXp92UDevCqOCOABXi/8AYX+FWtK8vhTUtV8OTnOxFk+1wrwcfLId55x/H0yO+R4w/YZ+LOiNJL4VvdJ8SQBsRpHN9knYZ6lJSEHHOPMPf8fFPgDxv4JkEXi7wnqukbm2I93aPGjn/Zcja3Q9CeleEviV4/8AAciv4P8AGOq6UobcYYLlhE555aM5Rup+8D1rwh+3Z8TNHVIPFuh6T4hjXGZFBs5mwADlkBTnGfudSe2APBv7anwY8TOlrrF1qPhu4I/5iFvuiLegkjLY+rBRx9M6H4j8O+J7P+0PDWvadq1rkjz7G6SdMjgjchI6g1VVVVQGeAaVQeBWBjAqqqqqv2hf+QzpP/Xq/wD6FX7Ozxh9fiLrvYWxCk8kDzMnH4j8xVBRjmqqqr4ufFN7Ay+FfDNyBOy4u7uN8+WD/AhHRvU9unXp8RfiJ4T+FfhO88Z+M9TW00+zXAA5knkP3Yo1/idscD6kkAEj47ftE+Pv2h/EUNtcRyWejxTGPS9DsyzAlm+VnA5llPyjOMf3QMnP7NX/AATj1rxObXxf8excaPpZy0Xh6Jil3P6GZx/qVP8AdHz+uyvCfhTw34L0O18NeEtDstJ0uyQJBaWkIjRAOOg6k9yeSeSaiANRAGoxkgVGASBUaggCo14AFQr0AqNQTxX/AATyUH/goTroHprv/ow0icYFRxAgAV/wW8TaPgt/3Hv/AHH18FP+CSf7OXxJ+DPgL4i6540+JEGpeKfDGnareRWmo2SQxzXFtHK6xhrRmCBnIALEgYyT1p/+CMf7L6jjx38Uv/BpYf8AyHX/AAUU/Yz+F/7Ip+H4+G+veKNS/wCEr/tP7Z/bl1BN5f2b7Ls8vyoYsZ8985znC4xzn9l1Af2YfhAR/wBCFon/AKQQ1+1/4T0jx5/wVEuPAviBJX0vxF4o8M6derE+xzDPaWEcgVux2scHsahs7WxtYrGytore3t4xHDDEgRI0UYCqo4AAAAA6V+1H8PdI+J/7P/j3wfq8ETrcaHczWzyDIhuYUMsEnHPyyIh46gEd6/4I++L9R+2fEbwBKzSWBis9TgBYAQygvFJgYyS6mLnPHlDjkmpVyODUqgjAqUZGBUoHIqUA5FSgHIqUAmiOcVVf8FJvhVH4Q+Llh8RNNtFisfGloWn2LgfbIAqSHAPBZGiboMnceTk1/wAEXPjI1nrvjf4CaldoINQhXX9JjdwuJo9kNyqjqzOht2xngQscEZIqqqqqqqqqqqqqqqqqiARiioPQ0R2qqqqqqqqoqD0q9sbLUrZ7LUbOC6t5Mb4Z4w6tg5GVPB5ANeLf2W/gN4y3yah8O9Os52zifTA1kwPriIqpP+8DXi3/AIJ3+GrnfN4G+IGoWDclYNTtluVJ9N6bCo99rH+deLf2Ivjx4a3y6do+neIYEyd+mXi7sf7kuxifZQf614k8GeL/AAfcfZPFfhfVdHlJwq31m8G76bgM/hXhL4ufE/wJsTwj491rTYo/u28d2xh/GJiUP4ivB37f3xV0UxweL9D0bxJbqvzuFNlcOcAZ3pmMdCf9X1bsABXgv9vH4OeIjHb+JrfVfDNwxALXEH2iAZB6PFlsZAHKDqO2SPCvj/wP46t/tPg7xbpOsIASws7pJGTGM7lB3L95eCB1HrVfHn47+OPgD+0hLd2MS32gazpVnNdaZNJ8kyKzIZYyD+7l/dsuSMHAyCAuP+Cn37Z3hjxh8HvD3wm+FmsyyDxi323XgymOS3tYJMR28i9meZdxAP3Yh1VwT+xZ8CLf47/Gmx03XIPM8OeH0GpauhxidEYBIDntI5AYddgfBBwajQGo488CkjGMCo4wRSR54ApYwBiliJGKWEY6UIhjFeWMYxRjBGAKMQIwKaHI6U0ZAxjNPECOKeMYwali9KkQEY71IgxipExxWpafZ6lZz6fqFpDdWt1E0U8E0YdJUYYZWU8FSCQQeCDX7VPwLuPgF8XNS8KWsFwdBu/9L0O4lyfMtm/g3d2jbKHvwD/EK/4JG/tLS+MPBN9+zn4pvUbUvCEJvNAd2O+eweQ+bGSTyYpHXH+xIoAwlVVVRAIxmiO1VVMMjiqqqYdxVVVa1q1noGjX+u6gzC1021kuZyi7iEjUs2B3OAeK8VeI9U8ZeJ9X8Xa3N5uo65fz312+5m3TTSF3OWJY5ZjyST6k1+z54Df4YfAvwD8P57fybrQvDtlbXiZY/wCkiFfPPJJGZS5x0GcDAFUwyOKqqqqqiARiiO1VVfEzS/I1GDVIwNtymx8D+Je5+oIH4VVVVfDTUCs13pTNwyiZAT0I4b88r+VVVVVEZGAaqv8Agp94H/4Q79r3xHfxQ+Vb+KbCz1aBBHtHzRCGQg5O7MtvKxPHLEY4yf8AgkR4zfXv2b9X8JXD5l8MeJZ0hAVsCCeOOVeSSM+YZ+BgYxxkkmqqmAIwKqqqq17/AJAeof8AXpL/AOgGqqqrwZ/yMtn/AMD/APQGqqr6GmAzxVVX7QvhyLxf8BfiL4YkWAnUvCupQxtPHvWOQ2z7Hx6q21geoIBHNeHNZl8OeIdL8QwQpNJpd7DdJG5wHMbhgDjscYqKWKeJJ4JUkjkUMjqchgehBHUYqqqqqqrXP+QLf/8AXrL/AOgmqqqqqqqqreQQzxTEZEbhsfQ1VVVVVVVVVVX/AAU++BTfCn9oKbxzpkCJonxHWXUoQqhdl4hUXaYHUlnSUnuZvY1/wRT+OMsqeM/2dtYv2dYlHiDRI5GJCDKQ3SLkcDJt3C56mRgOWNVVVVVVVVVVVVVVVVVX/Bar4FR6V4o8IftEaPbbYdcjOg60VRVX7TErSW0hPVneITIc8BbZPWv+CUPxtbwL8bbz4S6resulePrbFsryAJHf26s8Z56F4/NTjlm8sc4GKqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqpQMV/wUM+DX/Cmv2nvElvp+nxWuieKsa5pSxfdCTk+coHRNtwswCDouzAAIFf8ABGT41Dxd8Etf+CmozRC88A6h9osF8zDPZXjPIQFPXZOsxLDgeagIB5b/AILWfDn+wfjj4L+JlvHsg8W+H3s5Oc77iyl+duWJH7q5t1xtUfLnJJbH/BG74heXqfxD+FFzcZ8+C21qyiz93YxhuGx7+ZbD8PeqAGMCqoDjFVVVX7aviL/hKf2sPipqfnXUnkeJbmxzctlh9lP2fA5PyDycKOyheB0H7APh4aB+yp4M8y08i51L7XeT/vN3meZdS+W/UgZiEXAxjuM5qJcgCol4AFRKCQKiQEgVCgPSooxgAVDECOKjiAAGOaWEkYxRg44p4sDkVLECOKmjx0qZARxUyDBFTJkY9K8Wi4/4SrWRd+Z5/wDaE/m+bndu8xs7s85z1zXhlYzZwAEdP1zzWiQadJakXDAYXjivES28c5MRGAa8QhP7Tk2EZ2jP1x/+qvDHxC8beC7KXTvCviW8022ml814YHwpfABbHrhQPwHpVv8AtLfGq3eInxiJVjIykmn25DgdiRHnnuc596sv2ufizaSmSZdEvFK4Ec9mwAPr8jqc/jjmrL9s/wAbxxFdQ8KaHPLu+VoTLEAMdMFmOevOfwqy/bZUmGPUfh0QMASyQ6rnnHJVDF69i341Z/tneApUY6j4Y1+BgflEKwygj3JdcVZ/tV/B68dUn1e/swVyWnsJCFPodgY5/T3r/gp/rHhn46/s9aLbfC+4h8Q61o3imG8e3htnW6W1+zXCSbFZQ5Bd4SVHXaDzgV8L/DGsaB8PtE06/wBFv7OZIWZ4bm3eN1ZnZmBDAEct0r9jPTo7L9n3QblJSxv7i8mYEfdIuHjwPwjB/GqqqrGeAawMYFVXgv8A5E3Qf+wZb/8AopaqqqgMDFVVAZ4FYGMCqqgM1VVVVVUBgVVVVVU8ENzC9tcRJLFKhR43XcrKRggg9QR2rxj+zD8EPGivJe+B7TTrpgQtzpWbNlJIy21MIx46sp6n1rxf+wBMC03gHx6jDnZbaxbkEcnGZYge2B/q+oJ74HjH9nH40eB98mseBL+e2jxm5sFF3Hg55JjyVHB+8Bjj1GdG13W/Dt6up+HtZvtMvFGFuLO4eCQDjgMpB7D8q8F/tn/Gjwsyw6vqNl4ktRkeXqMAEi5OeJI9rZ/3twAOMdMeDf26vhZrcaxeLtL1Tw3cE/MxT7ZAB/vxgOe//LP0/Dwv448HeN7Q3vhDxPpmrxL982lyshjPoyg5U8jggdRVVWM8Cgo6CsDGBVVVVVV+0Hpc1xo+l6vGCUsp5I3Hp5gXB/NMfjXwm8YWnhHxIW1I7bO+jEMsmP8AVHIKsfYd/Y57Um1lDqwYEZBFVVV4m8beGvCUO/WdSRJSMpbp88r/AEUfzOB7140+M2v+I/MsdH3aXp5ypCNmWUf7TduOy+pyTXxE+JXgr4VeG5vFPjnXYNOsovljDnMlw+OI4k6u59B0GScAEjxNrnxs/bl+KiaX4W0SaSztCRZWAk22mk25ODLNIeNxAyzY3MQFUHCrX7M37GXw++AtrbeINTgt/EPjXZmTVZ4spZsVAZLZT9wD5h5mA5DEZAO0QjtUIGOKhAxwahGelRDJ4NRDJ61GASADUYBIFRAY4qFQQMV/wTwAP/BQvXQP+o7/AOjDUagniokBAxX/AAXDUKPgqAf+g/8A+46v2Uo8/ss/Bw4/5kDQ/wD0ghqWMY4r/gt6oB+C+P8AqPf+4+v2W1B/Zg+EAH/QhaJ/6QQ1+2X4v074e/8ABTy/8e6vFLLY+GvEvhrU7pIhl2it7OwlcL7lUOPetG13RfFGi2PiPw3qtrqel6lbrcWd5ayiSKeJhlXVhwQQetftp/FzR/g3+zn4z1681OC21PU9Ln0zRonkUSTXdwhjTYrA7ygYylcEbYznA5r/AIJB/Di+sfDPjz4qahYMkGrXNtpemTNld4hDvcYGcMpaWEbsdUYA/eFSgY4qUcHFSgYIFSgZwKlAzgVKBnANSgZxTDnFVX7e3w9Xx7+zfr13b2bT3/haSPV7XaDlVjO2cnH8IgeVj2+UHtkfsc/FFvg3+058O/Hj3DQ2dtrMVrqDBgALW5zBMTkEECOVmx6qMFThhgegrA9BRUGioIwK2j1oqD0NFTjg0VOOlEdqqqqqqqqqqqqqogEYorjpVVVVVVVVTKCOOtEdqqrq0tb63e0vbaK4glGHilQOrD0IPBrxf+yd8A/GMchuPAFnpdwy4SfSCbIx9OQiYjPCj7yHqT1JNeM/+Cd0DK8/w9+ITqwzstdZtwc8HGZogMc4H+r6Entg+MP2Sfj34OV5pvAlxq1upwJtHcXm7nHEafvPTqg/Q1/xM9E1D/l6sL61f/ailib9CprwR+1j8dfArQxWvjWbV7SLA+yawv2tWAJON7fvAOSPlccYHYY+OPxpv/jprmleJ9a0C103U7DTVsp2tZmaKcK7uGVGyU5kfjc3Uc8V4q8J6H4z0ltH12182EncjKdrxMOjKex//Ua/4J/eA/Bnw48Ca3o1lqcNz4o1LUjNqDMuyR7dAVt1Ud1VS7HGcNI/OCtRoMACo0AAA60qpGhkkcKqjLMxwAB3pPFHhY/8zLpX/gZH/jSR54FJEPSljB4ApYcjk0IhjGKEYx0oxAjpTRAjAposdKeMHjFSRelSRgjGKkQDipY/SpUBHFSoCMVImRgV+358F4fil8DrzX7C23674J36lZMqEs8GB9pi4ycGNQ/A5aFBwCa/Zv8AjZrH7PHxq8L/ABY0gSypo92BqFpG2DeWcg2XEPPGWjZtpPCuFb+EVpWqadrul2etaPexXlhqFulxa3ETbkmidQyOp7gqQQfeqqqph3FVVUwweKqqIBGKI7VVV+3Z4sHgv9kL4p6x5m37RoL6bny9/wDx+OlrjHv5+M9uvavgB4Kb4jfHLwB4FECTR634ksbadXgEyiFp081mQkB1Ee4lSeQCKqqIwcVVVVVVMBniqqvGml/2r4euI0BMkA86MAZ5Uc/mMj8aqqqvC96dP1+yn34UyhH+jcHP55/CqqqqqYc4FV/wWa8GND4h+GvxDhgRlu7K90u4lWAAoYnjliVnzlgfOmIXGF2sf4jX/BGnxWLP4m/ETwN5n/IX0G21Hb5ec/ZZzHnd2/4/OnfPtVVVUR2qqqq1JEk066jcAq0Dgg9xtNVVVXhKXyvEVk23OXK4zjqpH9aqqpgCOKqqliinieCeNJI5FKujrkMDwQR3GK8R6LL4c8Q6p4ennSaTS72a1eRBgOY3KEgehxXwD1mXxJ8Cfhx4hnhSGTVPCWl3TxochDJaRMQPYZxVVVVVVWuf8gW//wCvWX/0E1VVVVVVVVVVVVVVVVVVVf8ABTP4P/8AC0f2YdW12xilfVfAc663brHHuLwoClyreiiF2lJ9YV6DNfsb/GdPgD+0t4E+JV5ePBpVpqS2usMHKr9huAYZ2YDO4IjmQKRy0a9CARVVVVVVVVVVVVVVVVVftZ/BC2/aK/Z78ZfCkxob/UrEzaTIQuYr6EiWAhm+6C6KjHI+R2GcE1oWseIfh94w07xBp3m6drvhrUorqATRFXtrqCQMu5TyCroMg9xXwk+JOh/GL4Z+Gvid4cVksPEenx3aQuctA5GJImPdkcMhI4JU44qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqgOAK/4Kz/AAZi8ZfBHTvi3ptkjan4EvQt1Kq/O1jcssbA85YLL5JHB2hnPALGv+CdnxrHwM/aw8H65e3fk6N4ikOg6vk4BguiqoxODgJOsEh9RGRkZzX/AAWK+HP/AAlv7KUXja3j/wBI8DeILW7kkz0t7jNq64LAcyzW5zhiNuMAFiP+CePxC/4V5+1t4GnmuPLs/EE8mi3K5xv+1IUiX/v/AOSfwoDJwDVUBniqqqqvjVqt5r3xk8ea5qDI11qPibUbicou0F3uZGbA7DJPFfstaTbaP+zl8M7O1eR0k8LWFwTIQTumgWVhwBwGcge2OvWogCQBX7X/AO3L4p8OeMP+Gf8A9m61bUPGctwLO/1KK1+0PbXDEAW1rGQQ82ThmIZVzgAtkpoH/BNb9oX4zafH4m+P3x9msNSvFW4WzuVm1ueFmRRtlLyxojqPlwhdQFABx08bfsRftefsr2b+OPgB8WtV8RadpamWa30ZpbO6C/Kzk2W+SOZNw5QM5O0Er6fsF/t2w/tGxt8NPiSlnp/xB0+2M0MsC+VBrMCYDOik/LMoOXjHBGXUBQyp8R3lg+G/iueCVo5I9DvWR0O0qRA+CCOhr9mL4Q/tJ/tYa9rXh34c/GCTTrnQbNLq5bWvEF9CjI77QEMSSEnPqB9aT/gl3+3g/C/tBeFv/Cs1b/5Fr9gv9kL9on9nHxd4o1v41fErR/E9lrGmxW9jDY6ze3rQyLJuZitxDGFBHGQSa+JPj/wZ8JPAusfEf4ga1DpPh/Qrfz726kBO0EhVVVHLOzMqKo5ZmUDkivHX7Zv7Z37cXju58Cfso+Htf8LeHLOTkaPOIboRtlUmvb/Ki3ztcqiMgGSMylQ1J/wR8/aD8URJrHj742eGTq8igOHe7vyqgcAyuqkkdMAYHYmvhN/wTq/aw+BPxX8I6n4c+NUT+DYNWt21qPRNWubSVrXzFedGt32xujhQD8zH+LaSADMowRUyjPBqZRyBXx+8IzeF/iNqFwsLi01lzewSEHDM5zIM+ofPHYEdiK0HV0sm+zzthCcqx/hNW2tMkW1ZOCOMVrOtoqnzHy3OFzyammknmaeRsuxyTXwn/Zz8ET+A9KvPH/hprrWruMzTl7uaIxqxyiFY3AyFxnjOSc1cfspfCe5gaGJdYt3bGJYrwFl57blI9uRV5+x34ClRRp3ifXoGB+YzNFKCPYBFx+tXf7F9lJNusPiJNFFjhJdLEjZ78iVf5Vd/sb+Mkh3WHizRZpc8JMksa478hWP6Vd/smfFe2m8qE6LdKB/rIrwhfp86qf0q9/Z4+MthEJp/BE7KW2gQ3UExz9EcnHHXpWv+A/GvhayutT8R+E9W06xshm4vLmzdIIhkDJkI2gZIGc45q3uba7hWe1uI5o2GVeNgwII7Ee1Weo6hpzM+nX9xbM4wxhlZCR74Nad8VviXpQVbLx7riqkflpHJfPIiqMYAViQOg6CtO/aV+MNgFWTxNFeIkexUubKE4xjksqhieOpJ681p37Xvj6AKmpaBoV2qx7cpHLE7Nx8xO8j14AHXt0qw/bJsXBXU/AU8WFGGg1ASbj34KLgfia079q74VXzBbk6zp43hd1zZhgAf4v3bOcD8/atK+OXwj1coLTx5psZckL9qZrbGBznzQuPx61p2t6LrKB9I1iyvlKBwba4WUbT0Pyk8H1qq8F/8iboP/YMt/wD0UtVVUBk1VVQHFVVYzwKAAGBVVVVVUBmqqqqqqgM81VeMvhB8MfiDGy+LvBOl6hIxybgw+VP/AN/U2uPpu9PSvGP7A3hO+33HgXxlf6XIcFbfUIluY885AZdrKPu9d3Q9c8eOP2Uvjb4GV55vCp1q0jGTc6Mxuh15/dgCUY65KYx34OLa61XQdSFxZ3N3p2oWbkB43aGWFxwRkYKnqDXgT9sP4zeDTHbalrMXiWxQBfJ1VN8gAGMiZcOT05Yt09zXgj9uX4WeITHbeLtP1HwxcucF5F+12444+eMb+vHMYA4OeuPDvijw34v02PV/C2u2Oq2cgBWa0nWRR7HB4PYg4IIINKvYdaAAGAaqqqqqq1fSrLXNMuNI1GLzLe6jKSL0OPUehHBHvXjnwNqngfVPsd4DLaykm1uQuFlX+jDuP6EGvhx8WL3whs0jVle70gk7QvMluT/cycFc9VP1GOQYPin8PrmPzY/E9sBnGJFeM/kwBq5+K3w9tVVpfE9u244HlxvIfyVTitU+P/he2jK6Vpl9fSjpvCwofxyT/wCO14i+NXjLW98FhcJpVu4xsth+8x/10PIPuu2ta1vTtJsrnXfEWsW9laW6+Zc3l7cCOOMf3ndiAB7k18av+CgfhXw75uh/Byyj8Q6gNyvql2jx2cJ6fIvDynOeflXoQXB4+DfwB+Nf7ZvjAeOviBr2oR+HEkH2zXLxNvnIGIaGyQLsLZUg7QETknJwrfC34TeBPg14Ug8H+ANEjsLKPDTSH5pbqUKFMsr9WchRk8AdAAMAJjGKi6VF0qLpUPaosEgCouoqLGQKixnGaiwRx6VDg9Ku/wDgnT+3FoXxN17x98M9VsvD13fajdy21/pnilrG4EMsrNt3x4YAgjIzS/sg/wDBV9uF+O3ij/w5l5/8cr9nL9mL/gpF4P8Ajf4O8T/Ff4xeIdU8IadqSy6vZ3Hj+6vUmh2kFWhZysgyRwRX/BTf9kX4x/tVD4bD4SWWlXH/AAi/9q/2h9vvlttv2j7J5e3IO7/USZ9OPWvgR4R1f4ffAr4deAfEaQpq3hrwnpemX6wyb0E9vaRxSBW/iG5Dg96kwRwa/wCCmv7Jfxg/alPw3/4VPZaVcf8ACMf2r9v+3Xwttv2j7J5e3IO7/USZ9OPWvgf4S1jwD8Evh74E8QpEmq+G/CumabfLFJvRZ4LWOOQK3cbkOD3r4rfsC+MfjF+3lq3xa8daLpt58KNaWNbwJqCrcuqaQluAI8FlP2hF+YEMB8wIIBF1+yr/AMFC/wBmCa+8Pfsp/FP/AISDwXNO72Gn3FxZ+dbIzlsGG+XyUbnLNEQHOTgZwLD9gz9sj9pzxpa+KP2uviE2j2Fk2PKe7gvLkISN6WsFsTbQBgoy2Rg7TsfBFeAvAHhL4W+C9J+H/gXR4tM0PRYPJs7aMk7RkszEnlmZmZmY8szEnkmpMY614t8aeFPBNj/aPirXrTTYSGKCZ/nlxjIRBlnIyOFBPNar+1p8KrOcw2kWuaiuSBLbWaqpweuJHQ89en5VoP7Rfwn8SSR248QnTJ5BxHqMRgA4yQX5QY6ct9M07JIoeNwysMqQcgj1FS9TU3X8ab7xqq8RaFp/ijw/qfhnVo/MstWs5bS5TaDujkQo4wwIPyseoI9q13Rrzw7reoeH9QC/atMupbWcLnG+NirYyAcZB6gGvgV47k+KHwU8B/EW4nilufEnhyxv7oxSK4SeWBGlTKgDKyFlIAGCCMDGBVVVVRAIxRAPaioPQ0VPaiO1VVVVVVVVVVVRUHpRHaqqqqqqqogEcUR2qqqq8WeA/BXjuzFj4x8K6XrMK/cF5arIYz6qxGVPJ5BB5NfFX9ir4ERaHqXiiz1m/wDBUFjC8806zG5tolCjJMcmXIyMhVcEliB2A1KKwh1C6g0u8lu7OOZ1tp5YfJeWME7XZAzbCRgldxwTjJ61WlarqWh6jb6to99NaXlq++GaFirIfY/TgjuDivgh+0PpfjoQeGfFskNh4gAVIpC22O/PqvZHP9zuT8vXaPjX8ctL+E9ilhZQR3/iC8iLW1szfu4QCAHmAIYKecAfe2kZHWvF3xF8beO7hp/FPiS9vlLFlgaQrDH/ALsYwq9B0Haq8BfF34gfDW4STwt4gmjt1Pz2Mx823fnJzGeBnJ5XB5618EPjZ4f+Mmis9uqWOuWSj7dpzPkqOgkjP8UZOOeqk4PYlYxjAFLETSwgjAFeSMcCjCCMYpoRjAp4iBwKeMEYxTx4GDUseRxUiZGMVIgIxUqYqVAKlQDkVdQRzRPDLGro6lWVhkMD1BHpX7Sfwv8A+FO/G3xV4DgjkWxtbwzacXTbutZgJIgD0barBCRxuQ8DoP8Aglr8YJ/ij+yxpnh/Vb6W51XwDdvokrTEbjbKBJakY/gWJ1hHQ/uT9TVVVEAjFEdqqqYZHFVVUw9Kqq/4K0+IE0X9kifTWu54jr3iSwslSMkLMV8y42vjquLfdzxuVe+K/wCCanhj/hJv2zfAPm2P2m10r7bf3H7zZ5Xl2c3lP1BOJjDwM5zyMZqqphkcVVVVVVMMjiqqiAw2kAgjkEVrunHStXu9PA+WGQ7P908r+hFVVUCQQVOCOhrTbtb/AE62vFYHzolf8SOaqqqqYZHFV/wV18Lwaz+zNpniECJbjw/4otpA7KSxilimidF9Ms0bH/cr/gmV4tXwt+2H4Rtppkig1+2vdNldnK8tbvJGMDqWlijUA8ZbPaqqqph6VVVVX/8Ax43H/XFv5VVVVeF/+Rgsf+uv9KqqogEYzVVVftT6FJ4b/aV+KWjvYRWSReL9Te3t4goRIZLl3i2heAPLZCB2zjAxiv2BNYtdd/Y9+Gd7ZpKiRabLakSAA74LmWFzwTwWjJHsRnHSqqqqqqtc/wCQLf8A/XrL/wCgmqqqqqqqqqqqqqqqqqqqrWdH0zxDo99oGs2iXWn6lbSW13A5IEsUilXU45wVJHHrXxc+HWq/CT4neKPhprKt9q8OapPZl2UjzkRj5coyAdrptcHAyGBr/gnr8c7L48fsseD9Wa9SXW/DVqmha1H0Zbi2RUV25PMkXlSZ6Zc4xjAqqqqqqqqqqqqqqqqqv+Cp3wAHwV/ae1LxJo9lDB4c+IkZ1mwWJcLFckhbyIj187MvAACzqB0OP+CQvxwF/oPiX9n7WbjM2lyHWdFLyZJhkKpcRAE8BZPLkAAOTNITjAzVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVUBngUfSvHng7SPiH4J17wHr8Xmad4h0240+6XuI5oyjEYwQQGyCMEEAgiviB4J1v4beONe8AeJLZoNT8PahNZXKMuMvG5XcPVWADAjIIIIJBzXwx8Ut+3H/wAE+Jkui8uv+KfCN3o99vYBhq0MbQ+ccMgw0yJMFLAbXUMcZrQda1Dw1rmneI9Im8q+0q7iurWT+5LG4dD+BUV4L8T6d438H6H400hs2Ov6bb39qc5/dTRrIvPf5WFVSgGqqqqvij/yUzxd/wBh29/9HvX7N4H/AAz58MB/1Jmk/wDpHFX7RHxOk+DXwO8ZfEi1kVbzSNMb7Azx+You5WWG3LL3XzZI8jpjOSBzX/BKr4P2nifX/FXx88VWq6jc6ZcDTtJnuR5rLduvmXM2WBxIEeIBs5xLJnrzAB2PaoQCo5r9tLwZffsf/ti6R8VPhfaWumWWoSQ6/pVtbkJFFKrFLm3KLjbGxDZUDbsm2g8EDxvqVjrXwf8AEWsaZP51pfeGrqe3k2ld8b2zMrYIBGQRwQDX/BFz/kqHxG/7ANt/6PNQAdKiUEAV/wAFm/jNqut+M/Bn7Nfhl7ieK2gTWNStrdCzXN3MzRWsWByzKgkbbjnz16kcfsw/s7+Fv2Yvg1ofwx8Pw28t9BCs2t6jHGFbUb5gDLKTgErn5UDZKxqi5OM1OBnirgDoDUwBqcAmpgM4HpXxN+H2lfEbw9Jo98whuYiXtLoIGaB//iTwCOM/UA141+H/AIn8Bag1lr2nusRYiC6QFopx6q3rgZ2nBHcUs0qDbHK6j0DEVPNHFG9xcTKiIpZ3dsBQByST0GK/ZB8EeDPjIbv4kQa1a6to/hzWJNO+zRx70nuo44pCSxG14wJk+7kMe+ByjjHWlk7A0sgxwaEnGM15gxjNGTjANNICODX7QOhReLPgT8RfDE0yRLqnhTU7YStF5giL2sgD7cjJUkMBkcjqOteCoJrHwfoNtIdstvptsjbT0ZY1BwfqK+Bnw/8AC2ofBfwVP4m8HaNfXkmkRS+bd2MU7lZBvU7iCeVZT+PNXv7P3wd1DZ5/gi1Xy848meaHrjrscZ6d+n41e/smfC272eRca5Z7M58m7Q7unXejdMdsdav/ANjaxcA6X49niwpytxp4k3Htyrrgfga1H9kHx9AGbTNf0K7VYydsjyxOzc/KBsI9OSR17da1T9m/4waYGceFheRoBl7W7ifOeMBdwY/lWqfDrx7or+XqngzWrf5toZ7GTax9mAwfwNI7RsrxuVZTlSDgg+orSviT8QdDYHSvG2tW6hi3li9kKEkYJKElScY6jsPSp/B2gTZK2rREk8pIf5HIrwt+1vr+h6ZZ6Vqfg/T72KygWFDBcPbsVVQqk5D84HPHPtWlftefD67YR6routWDFjhxGkyAAcEkMGyTxgKe3Ppo/wAfPhFraoLfxrZ27tgFLxXtthIzglwBx0yDj3rTdZ0fWYvP0fVrO+j2q2+2nWUYb7pypPBA49aoDAqqoDPArAxgVVUozzVVVVVVjPAoAAYFVVVVVQGeBWBjAqqqq8Z/C/4e/EKAw+M/CGmao2wos0sIEyAjB2yrh16DoR0HpXjj9grwdqZkuvAPiq+0WUnItb1PtUPTorZV17HJL9/Xjxz+y/8AGrwF5s994Qm1Oyi3E3mkn7Wm1erFVG9VxzllXj6HGla1r3hjUPt2hatf6TfxZTzrSd7eVOeRuUgjkDj2rwH+278WfCyx2niaKx8U2akbjdL5FxtB5AlTjnPVkY8D8fAP7Y/wY8a7bXU9Wm8MXxYKIdWQJGxPcTKSgUerlPpjmra6tb23S6srmKeGQZSSJwysPUEcGqqqqqxngUFAHvWsaNpmv6dLpWrWiXFvMMMjDofUHsR2I6V4u+CvibS9Rlbw5ZPqWnH5o2DqJEyfuspIJI9QMY545Al8C+NYZGifwlq5ZTglbKRh+BAINQeAvG1zIIo/CerBsdZLR4x+bACtL+Cvj7UiDNp9vYIRw91OB69l3MOncdxQ+C3hXwpo194n8f8AiwQ6bpVrLd3swIt4beGNSzyO7Z+VUUsTxjHtX7Tfxyv/AI8fFfW/E9rJcWvhlLt4/D+ltI2y0tF+WM7ST+8ZQGdjklmPOAMfsi/sJ3/xFj0/4m/GC2msfC0qx3OmaWGCy6tGfmV3IOY4CMY6M4ORtXazaXpWl6Hptto+iaba6fYWcQitrW1hWKKFFGAqIoAVQOAAMCqTB4BqHB71CcjrUJBGAahPHBqLrgGouuAajIyMGoyMjFREEYzUJHAzULDAwajYZwDUTjGAaSQEdaMgI+9UsgIwDUzgjANSMMdamIIxmpSCeDUxB6GpSCMA18d/jBafCbwys1ssVxrmo7k0+2kzjjG6RsfwrkcZGSQPUjxH4l17xfq8+veJNUmv7+4OXmlP5AAcKo7KAAOwq18I+I72ETW+lybCOC7KmfwYg1qGlajpcnlahZyQsem4cH6Hoa+D3xr1n4f6jb6Rqt2914cmk2zQyZY2ucAyRnBPGMlOh54BOaiube9t4ru1mWWGeMSRyKchlIyCPYipeTim5PWqqv2ufDzeGP2lfiHprQNEZtZkvdrOHJ+0qtxuyOx83OO2cHpX/BMTxW3ir9ivwEs9y091ozX2nTlowgUR3cpiUYwCBC0Qz1JBzk5NVVVVVVVRAIxRUHpRXjg0R2qqqqqqqqqqogEYojHeqqqqqqqpgCOOtEdqqqqv2p/B/wC0b8ZdVg8FeCfBUtr4Ns5h5tzLqltF/aMuR+8kQybxEhB2rtyeWwTtC/HvXbX9nLx/J8NPHsMtxrkFpFcXEemFJ0gEgyiszMvzFcNgZ4Yc9h+y74VuP2nLXXdXsHm8PaTossdut1PELhrmZgWKKisuNq7SxJ/jUDPJGm/sceHEEQ1XxnqVwQT5n2e2SHcO2NxfHGOuau/2YPhF4T0TUNf1G4166i022ku3eS8VWRY0LHb5aL2HoTXiDXL/AMSazda3qdxNNcXL5LTTPMwUAKql3JZtqhVyxJwBk1pejCcB5U3MeikdKPh5jDgwgjHQrWr6P9mBlhXG37y/1FfCzx/qHwx8daV4wsGkZLSULdwI2PtFu3EkZ7HK8jPAYKe1WUkF5bQ3drKskM6B43XoykZBH1Fftd/HrUv2aPgtefE/R/Dlrrd5DfW1pHa3M7RRjzXwWJUEnAB445IOeMHRf+CkH7b3xHik1n4Sfsx6drWjxN5bS2XhrVdVCSDqDLBKq5/2cZFX/wDwUY/bf8AWcusfFb9lSDTdN24juJ/Deq6UiEdS0kzupGO3GPWvgJ/wVK+B3xb1Wz8LeO9JvPh5rd9KY4XvrpLnT2OfkBugEKMf9uNVBwNxzTR8YxX7df7YniD9kmLwUvh7wVp2vyeK2v8AzGvbp4hALb7PwAo5LfaOuRjb3zxpP7eX/BRHxHpVprugfsr2mp6bfwrNa3ln4H1maGeNhlXR1nKspHIIODXhb/gq1418MeJo/C/7QvwNbRpYmEd82nrNaXFq27DMbW5ySAM/KXByOvp4A+IHg74q+DtN8eeAtcg1bRNWi8y2uIj07MjKeUdSCrKcFSCCK/bY/bQ8T/steKfDfh/QPBWla5Hrmny3Mkl5cSRmMpJtwAvUV+yv+0dof7TXwwi8Y2sdtY65Yztba1pUUhY2cuSUIzyUdMMrdCdy5yjASoCOK/bU/aw8Q/su/wDCHDQfCWna3/wk327zftk7x+T9n+z427eufPOc+gr4aeKrjx/8M/CXjq8tI7WfxHoVlqMtvGxZYmngSQqpPJALEc+lSrkY9K/4Kn/D2MJ4M+KttE3mBpdFvHI4I+aaAD0P/Hz+Y6YOf+CQHxRl8J/tDar8M55kFl470Z9iEkM13ZhpoyO2BCbrIx3ByMHNVVVTAEZFVVUwweKqqIBGKqq/4LLa1FB8BPBfhwwMZL7xet0sgPCiG0uEII9T54/75Nf8EjtGutU/axe+t5IlTSPC19dThyQWQvDCAuAcndMp5wMA89AaqiARjNEdqqqqqogEYojtVVXxO04R3lpqaD/XIY3+q8g/kf0qqqq8DXDXHhq1DHJiLJk+zHH6ECqqqqiARjNEdq/b08MN4v8A2QPifpKRzuYNHF/iEgMPsk0dyScg/KPJy3faDjB5r9mXxQ3gv9on4aeJhctDFY+KtPNy6Rhz5DXCLMADnkxs49eeMHBqqqqYZHFVVVUsaTRNC/3XUqfoaqqqvDTpHr1gztgecAOO54H6mqqq/GmHOKqq/wCChOjRaF+2P8SrGGdpVkvbW6LMMEGezgmI+gMhH0Ff8Eu9Vu9R/Y/8PWdyyGPS9U1G2twFwQhuGlOfU7pX/DFVVVVVVa5/yBb/AP69Zf8A0E1VVVVVVVVVVVVVVVVVVVV/wVs/Zzggk0z9pPwvp217iSPTfE5jBO5goW1uG9PlXySfaIV/wTA/abn+Af7Qtj4R17VPJ8GfESSLS9SSQgJbXZJFpc5/h2yN5bEkKEmdjnYuKqqqqqqqqqqqqqqqqr/gqH+z63xy/Zg1XWdHgRvEPw7Z9esTtG6W3jjIu4QTyAYcyBRyzwRjvX7PPxh1L4B/Gbwt8VtNiaddEvQby2XGbm1kUx3EYzxuaJ3Ck8Btp7Vpep6dremWes6ReRXdjfwJcWtxE25JYnUMjqe4KkEH3qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqUAmqr/grX8F18G/GfSfi/pNoy6f48svLvmAJVb+1VUY9MIHhMBA6sySH1r/AIIrfG7+yfGXi/8AZ+1a+2W2vW/9t6PG74X7VCFjuEUZ++8Jjbgfdtzk8Cv24Phinwh/av8AiV4Lt4His11qS/sVZiwFvdqtzGoY5LBVmCZJJypyc5r/AIJk/EkfEH9k7w/plze/aNQ8HXdxo1yWb5giN5kAxtGAIJYkGM52dc5AqgBjg1VVVV+1ToUnhv8AaX+KejvYRWSReL9Te3t4goRIJLl3i2heAPLZCF7ZxgYxX7Eesxa9+yx8O76IzERac9qfO+9mCeSE45Py5jOPbHTpX/BSm5uIP2V9ViguJI0uNWsY5lRyokUSbtrAdRuVTg91B7V/wS+traD9l22lgt4o3uNevZJmRApkcFFDMR1O1VXJ7KB2qA9gahIxgHpX/BZY2pn+EWwxfaPL1rzMY37M2W3PfGd+Px96+Ghuv+GJPDAvTL9p/wCFV2nnebnfv/stN27POc5zmv2Lj+2GPFHiMfseH/ic/YIv7Y/5Bv8Ax7+Z8n/H/wDL9/8Auc+vFI//AAXE/gb/ANRWv2JJP2yP+EW8SD9sk/8AE5/tCL+xf+Qb/wAe/l/P/wAeHy/f/v8APpxX7VEza7/wV70PStYCXtlD418IWqW86B08lo7B2jKngqWkkJB67jU0gIwDUzAnrVwwPepiDnmpyCetTHOcHpUxBzzV5DDcQtDcRJLG4wyOu4H6g1e/C34dXEjTP4O0xWbqI4RGOOOFXAH5V/wUy0yz8L/G3RfD3h8S2elz+F7e7eySdzD5xublGcISQCVijBwB90V/wRyvLlvhV4/sXuZWt4vEEMkcJclEdrcBmC9ASEQE99o9BStxwaWTtmhJ2BoSDHWvMyODRkBHWmkB714m0mLxJ4d1Tw7PO0MeqWU1q8iDJQSIUJA9QDmvAjM/gbw67sWY6TakknJJ8pa/Zs/5IR4L/wCwYv8A6EaqqoDJqtU0HQtcj8rWtGsNQTbt23Vskox6YYHivih8J/hBYeC/EHia98GadaHTNOnuxJaK1vtaOJiMLGVB6D5ehOOCa0/xjr11dQ2UVpBdSzyBI0CkMzNwAMHHXFav+xtqce59A8cWs+d5SO8s2i2/3QWVmz7naPXHatY/Zb+Lelhjaabp+qBD1s7xRkYySBJsPHTGM56A9a1f4afELQiBqvgrWYFJAEhs3ZCSM4DAEZwDxmoJ57WZLi1neKWM5R42Ksp9QRyK0b4w/FHQCv8AZvjvV9qBQkdxcG4RQowAFk3ADHbGOnpWiftbfEvTwkerWmkaqgGGaSAxSNgYyChCjJ5Py/TFaN+2R4dmUDxD4N1G1I4JsrhLjPA5w3l45zxnjjk1o37Rnwf1oiNPFiWcv9y9gkhwMgZ3Ebe/TOa0vW9F12H7RomsWWoRAAl7W4WVcHODlSeuD+VAcVVVjPArAAwDVVVVVUozzVVVVVVQAAqqqqqqAzXjr4L/AAv+JYeTxl4N0++unQL9sVDDcADoPNQh8D0Jx7YJrx/+wFGY5bz4ZeMjvBBSw1lOCO+J4x+QMfcZbvXj/wCCvxP+GTyt4w8IX1raROE+3xp5tsxP3cSplRnsCQfYEEV4K+JvxA+HU5n8E+LtR0ndIJJIYZcwysOheJso/p8wNeAP2+9TtvLs/iZ4RS9jHDX2ksI5B16wudrHp0dQPSvh/wDGz4X/ABQVI/Bvi+yurtk3GwlYw3K4GW/dPhiB3ZQV96qqqgvc1VVVV/wV7/abbwr4LsP2b/B+sIup+KFF34k8lwXgsUYGKBiOVMsi7j0OyPB+WTn9lL4F3Hx8+Lum+Gbq1kfw/p2L3XZVYoFtkI/dhgQQ0jFUGPmAZmHCki3trezt4rS0gjhggQJFFGgVUUDAVQOAAABiqpOuAahI4FREZxURGcVERnFRkZHNRkZHNRnIGDUZyBg1EwwOajYZ4NROMcGo3BAANRyds1cX9rY2st7e3MUFvbxtJLLK4RI0UZZmJ4AAGST0r4rft66ZpN1JpHwl0OHV3iYBtU1JXW3bHURxKVdh23MUwR0YYNX37ZH7Q93cvPD44htEbGIYdKtSq4GOC8bNz15J614M/br+JekXEcXjTSdM8RWYQK7Rp9juMgY3BlBT1JGznsVr4bfFzwT8XND/ALb8IakXMZ23FncAJcWzejoCfX7wJU9jwamcEYBqRuOtTMDwDUx4wDX7SPi648W/FzWw0xa20eU6fbIScIIuH6gdZN5/HqQAa+HPhiG6Ua3dRh2DkW6sOBjq3uc8D0wfwtNDaaPKx549K17w9bXVrJZ3kAeNxgqR+o9DWu6U+i6rcacxLCJvkYjqp5B/L9a/Zs8TTa78Oxpt3KXm0W5a2TcST5RAZOcdssuOwUVKeeDRwTwaqq/4KE6VHp/7UXiG7jmZ21Oxsbh1IxsItkiwPbEQP4mv+CO+uDV/2SrvT/svlf2L4vvrTfv3ebuht592MDb/AK/bjn7uc84BAIxitooqD0oqCMA0V44NFT2og9KqqqqogEYoqD0NFTVVVVVVVVVUQCMUV44NVVVVVVVTKCOOtEdqqqq7u7SwtJr6+uore2to2kmmmcIkaKMszMeAAASSeBX7QvxUuvjd8bvGvxUuXlK+IdXmntUlcs0VqDst4yT/AHIUjToPu9B0r9iD4bRfDT9m7wlZPCq32u2/9s3rCPYWe5AdNw67lh8lDnn5O3QRKDXx9a6j+DXih7OASyG0UFS2MIZEDt+CknHtUKq00aseCwBzXh2KJ5VD4/Gm0vSv7HEwkXzMfdrxDDCsjKuMciq+EEk1z8KPBVxPK8s0vh6wd5HYszMbdCSSeSSe9f8ABVaPb+yLqJ/6jth/6G1f8Eo4w/7IunED/mPah/6GtCHjFf8ABRD9gfwJ4g+H+tfHH4PeGbXQfE/h23e91XTdMtVig1a2X5pn8pcKkyLuk3qPnAYMGYqw/wCCU/7RWs/FX4X6r8I/GN9Leav4A8n+z7uaQvJcadLuCIxJJJhZCmeAEeJQPlJP/BaZNp+Dg/7Dv/thX7MUYb9mf4SEf9CLov8A6Qw1+3n8GPCHxU/Zv8a6lrelWp1fwno1zrGlaiYwJrZ7aNpmRXxnbIiMhU8HdnghWH/BH3xbq2peCfiP4FuJd2naJqNjf2oPVZLqOZJAO+MWkZx0ySe5z/wV/GPiR8Px/wBQO5/9HitG1/Vf+CfX7Yd3atc+b8NPFjpMs0Q86O50edy0M8ezO57cll4GW2PgbZAaiuLa9toryyuIp7eeMSRSxOGWRSMhlI4IIIIIr/grwoB+FAH/AFGv/bGv2dl/4x5+F2P+hL0j/wBI4qlUZwK/bU8Dw+Ov2avG9ky/vtJsDqsD7iuxrU+a3scxo64P971wa/Z68ep8L/jr4A+IM9z5NroXiKyubx8A/wCjCZROOQQMxFxnqM5GCM1VVVUQCMURjjNVVMARxVVVMOeKqv8Agsvon2j4E+CfEf2rb9h8Wi08nZnf51pO+7dnjH2fGMc7uoxz/wAEjtZutL/axext44mTV/C19azlwSVQPDMCuDwd0KjnIwT7EVVUwHaqqqqqph6VVV46sDf+G7gqMvbETLx/d6/+Ok1VVVfDSUtpV1BgYS43A/VQP6VVVVVTDnAr4ueGx4z+FHjTwf8AZpbn+3PD1/Y+TEcPJ51u8e1T2J3YH1qwvrzS7621PTrhoLqzmWaCVODG6kFWHuCAa0bVbPXtHsNc09ma11G2juIC67SUdQy5HY4I4qqqiARiiO2aqqqqqqrRmVNYsHdgAtzGST2+YVVVVMM81VV/wVW0ZtM/a0vb06elv/a2gWF0sgQKbnCtD5hI+8R5OzJ5wgHQCv8Agj3qtnN+z/4u0ONm+1WnjGW4kBXgJLZ2ypg+uYX4+nrVVVVVVXiYkaBfYP8AyxNVVVVVVVVIjSMsaDLMcAepqqqqqqqqqqqq+Jvw98PfFj4fa/8ADfxTB5ul+IbCS0nwMtHuHyyL6MjBXU9mUGvib8PfEHwo+IGv/DfxTB5WqeHr6S0nwCFk2n5ZF/2XXa6nuGBr/gnz+0j/AMNL/s36H4g1nUPtHizw5jR/Ee9sySXMSjZcHufOiKSFsAbzIo+4aqqqqqqqqqqqqqqqqqaGG4he3niSWORSro65DAjBBB6gjtX7bXwAuv2bf2j/ABX8PksvI0S4uDqfh1lUhX064ZmiVc8nyyHgJ7tCx6Yr/glb8doviL8D5vhVrGoNJr3w+lEMSyvlpdOlJMDL3IRhJERztVY+fmAFVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVUoGKqv+Cgvwb/4XL+zB4os7Cz8/WvDEf8AbmlhU3MXtwTKigAkl4DMoUYyxX6V8APizqXwK+NHg74t6WZWk8NapHcTRxHDTW5yk8Q5H34WkTGQPm54r/gsr4E0q+8YfDb9obwq6XWj+N9B+yNdREhZWhxNBIVKggyQ3AAOeRF0XGW/4I6fEpNN8e+OfhLeT4XXNNh1WyDuFUS2z+XIqgnJZknVsAHiEnjHNYzwKwMYBqqqqr/gp14Jl8H/ALX3ie+FskNr4nsrLVbVVXGQ0Ihkb3JmgmJ9zX/BLjxvFrnwR1nwVLMpuvDGtsyoG5W3uEDoSNxIJkWfsBwMZO6v2rfh3qnxY/Z08ceBdCgkuNSvNOE9nBG2Gnmt5UuEjXg8u0QUDjO7GVzkf8Em/jBpdovin4GapdwQXV3cDW9IVsK1y3lrFcoCTyyrHAwUc4Eh6KahYZxmoWGcZr9vDxXP+0/+2Ronwg+Ht7FfR6TJbeG7WaNA8f2ySXNxIWXLFY2cIxPC+QxHGSfGemWeg/BvX9D08MLXT/DFzbQB23EIlsyrk9zgDmv+CMTBfif8Ref+YDbf+jzUcgxkGknAGAa/4K9+A/EPgj9oHwf8c9Fa4W21vTYoorpkWRbe/spMgYxgAxvCwVwdxWTGQCF+AXxv8MftB/CXw/8AFHwvdW7Jqlqn261jlDtYXYUedbvjkMjHvjKlW6MDUsgAwDU8gPQ1MwAxmpmB6GpmGOtTMMHmpyMYzU54wDX/AAVas7VPif4K1BIVFzNoMsUkmOWRJ2Kg+wLuf+BGv+CM08vlfF6AysY1bRGVCeAT9uBIHqcD8hSyDHBoOCOtBgRgGg5PQ1vNFz60WGMZpnAHBr4a3X2z4f8Ah6Xy9m3ToY8Zz9xQufx25/Gv2UdQm1P9nzwfczqiskE8ICAgbY7mWNT1POEGffPSqqqAGKqv25vi3FoPhS3+FWjXmNS17E2ohDzFaK3yqT2MjqOn8KMCMMM/stfD+T4g/GfQraW283T9Hl/tK+LJuUJCQUVgeCGk8tcejHg4NVVat4X8M69n+3PDumajnGftdmk2cdPvA1q37Nvwf1fDjwubKTIy1pcyR5AHTbkr+OM8da1r9jbw7MpPh3xnqNqR0W9t0uM8HjKeXjnHOOPQ1rv7JXxM01Wk0i50nV0H3EinMMh45yJAFHOf4vyrW/hL8S/Du86t4I1eNIyQ0sdsZoxzj76ZXk9Oee1RS3VjcCWCWWCeFuGUlGQj3HINaF8cviz4dAXT/HOoyoONl4wuhjjgeaGwOB0xjnGMmtB/bE8Z2ZVPEPhnStTjUYzbu9rI3B5J+deuOijofXI0D9rj4cakEj1ux1XSJSQGZohPGue+5DuOOv3fzrw78Tvh74t8tfD/AIw0u7lkxsg88RzHOMfu2w/VgOnU460ozzVVVVVVjPArAxgVVVVVVAA1VVVVVYzwKVQeBWBjAqpI45Y2ikRXVgQysMgg9QRXxC/ZR+DHxC8+6l8Of2JqU7bze6Qwt2Ld8x4MZyepK5PPIJJr4hfsMfEnw1HNfeCdStPFNpGCwhVfstzjjOEYlW79GyccDJxWs6F4g8Kao2ma9pN9pWoQHJhuYWhkXBwCAcHqDz7V8PP2uPjL4AWKzm15fEWnRn/j11gGdgMjO2bIkHAwAWKj+7Xw5/bS+E/jWSDTvEUk3hPUJcDF+4a23ccCcYAHXlwg498VZ3tlqVpFf6deQ3VtOgeKaGQOjqehVhwR7iqA71VVVVX/AAUB/ZO/ae8L/FPxb8c/HWl/8JXoXiG/lu31vRI5J4dPgGFiinQjfAkcQjjDNlMKBvJr4U/Gr4l/BPW5Nc+G/im40uS42C7t8CS3u1XO0SxNlWwGYA43LubaQTmvgn/wUb+Gfjj7PonxTtP+EN1h8L9r3GWwlb13/ehz6OCo7vWkazo/iHTLfWtA1az1PT7tN9vd2c6zRTL/AHkdSQw9wapcAjmoj2BqMjI5qMjPWoyMjBqM5AwajIIGDUTAjGaiYEYzUTAcZqNgRgGo5B0zUcnQA0kgIxmv21Pjff6jrjfCHw3qLxadYIray0T4+0zkBliJHVUG0kZwWbBGUFaF4e1DxBcGCyQBUx5kjcKn/wBf2q3+FOnGEefe3jvjlk2qPyIP868SfD290eF7yxlNzAgy6FcOg9eOo/L6V8N/iFr/AML/ABdZeL/Dsq+fbHbLC/3LiI43xt7EDr1BwRyBXh3xHpvi7w9pvifR5C9lqlrHcQFsbgrqDhgCQGGcEZ4IIqVx0BqRgec1IwJwDXxbilh+KfjBJo2Rjrt6wDKQcNMxB+hBBHsa+HFzby+H7RYwAYiyOPRtxP65B/GtBv7KGHEyq3HANeIbiCV2MeMV8RLiGfxI6RBcwwqjkd25P8iB+FfsrapplvpOs6ZPqNrHeT3SPFbvMqyOoXBKrnJGSBmpCOcGqqq/4KD6t/aX7UniSz+z+X/ZdnY227fnzM20cu7GOP8AW4xz93PfA/4I46JdaV+ydqV9cSRNHrHjO9uoAhJKotvawkNkDB3QseMjBHOcgVVVVUQDRUEYFFfQ0QcVVVVUQDRXjg0R2qqqqqqqqqiuelEdqqqqqqqphkcVVVX/AAUK+KKfCj9kXx/qsFwkeoa5Y/2HYIWKs8l4fJk2kA4ZYWmkHT/V9QcV8GPh/J8VPiv4U+HqMyprmqQwXDqMmODdmVwPVYw57dOoqxtbaztorOzgjgggQRxRRoFVFAwFUDgADAxUSjHFeMfDMXjDwhrHheVgv9pWUkCMxICOVO1uPRsH8O9ahYXmk39zpeo27wXdnM8M8TjDRyISrKfcEEVomsgBQXAkXqOn4ij4mlMHlCQ9K17WFkVo0cGRhjA7V4P8Lap438UaX4S0WPfe6pcrBFkcLk8sf9lRlj7A1oWjWehaPY6Hp0ey1062jt4FwBtRFCqMAAdAOgAr/gq2mP2QtRP/AFHtP/8AQ2r/AIJOoG/ZB049f+J9qH/oa0IiRgCrywtb+1msb61iuLe4jaOaGVA6SIwwysp4IIJBB4Nf8EwlHgr9uLxD4P0n99ZvpGr6WXuPmk8mG4idWyuBvJgTJxjBbgcY/wCC1yBT8G8f9R3/ANx9fAj/AIKBfsheDPgZ8OvB3iX4vfY9Y0HwnpdhqFt/YGoyeTcQ2kccibktyrYZWGVJBxkEiv2tf2+bL9oDw5efs8fsseFvEPiS78T5tr7VI7B1aa33fNFb25UyESD7zuE2LkYJYlf2Gv2Y9Q/Zk+Dp0PxRcRTeKfEN3/aGsLDJvjtW2KsdupHDbFByw6s7YJUKa/4LBLj4lfD8D/oB3P8A6PFftdfs2f8AC+P2V/DOu+HrXf4u8F6BBe6Yqjm6h8iM3FvwCSWRAyDu6KOAxNf8EyP2lf8AhMPCkvwA8XX+7WfDUBn0GWQ4NxYAgND05aFjxzko4AGIia/4K9gBvhOB/wBRr/2xr9nZR/wzx8LiP+hL0j/0jiqZRjitb0u11nS7zR70Mbe+t3glCnBKOpU4PY4JrWtKuND1i+0S7eN59PuZLeVoySpZGKkqSAcZHcCv2efGj/EX4D/DzxxPd/aLnWvDNhc3UhkaQ/aGgTzgWbliJN4JPJINVVVVMMjIqqqiMHAqqpgCOKqv+CrvhVPEP7IGqas1ukp8M65p+oKzSFTEWkNruAB+Y4utuDxhieoBr/gnf4vi8Gftj/De+urxobbUL2fTJVE/lLKbq2lhiVsnDDzXiIU9WVcc4qqqiMjAqqqqqqIyMCqqrmCO6t5LaUZSVCjDHYjBq4he3nkt5Mb4nKt9QcGqqq+GEiCTUYi3zMsZAx2G7P8AMVVVVVTDI4qvGejWvh3xhrvh6xkle30vUri1haUguyRyMqliABnAGcAV+zTrsnif9nX4X69cahFe3N74P0t7meMrhp/ssYl4XgEOGBUYwQRgYxVVVUw54qqqmZUUszABRkknGKqqqrD/AI/rf/rqv8xRHOBVVVMAV4qqr/gsRonkfHHwX4i+1bvt3hQWnk7MbPJupn3bs858/GMDG3qc8f8ABGbWLmfQvit4fdIvs9ld6VdRsAd5eZLpWBOcYAgTHHc8njFVVVVVXif/AJAF9/1yNVVVVVVVVadG0uoWsUYyzTIFHvkVVVVVVVVVVVVV/wAFcf2fP+QN+0d4csf7mk+I/LX/AMBp2x+MRJ/6ZCv+CX/7SP8AwoP9o+x8P67qHkeE/iJ5Wj6pvbCQXJY/Y7g9vllYxliQFSeRj0FVVVVVVVVVVVVVVVVVX/BYL9nj/hY3wSsvjZocOdZ+HMjfbFBx52nTsqycdzHJ5bj0Uye1fsY/HRf2ev2gvDvjm/nZNEumOm64AxH+hzlQzkDqI2WOXHcxiopYp4knglSSORQyOpyGB6EEdRiqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqxwBVVX7WHwf/4UV+0F4y+HFtFKunWd8bjSmePbus51EsIB6NtVwhI43I3AwQLHXE/aj/4JManoV5dNfeLfgFqUMq+ehMn2GMkRFWUgBFs5pYwCD/x68r916/ZH+JjfCH9pHwB44e4aK0t9Yjtr9gwGLW4zBMTkEEBJGbHqowVOGFAZNVVVVV/wWM+FUt94Z8D/ABn0+BmOlXMui6iVj3YjmHmwMSPuqrxyrzwTKoyD1/4Jq/E2HwT8fW8I6hNts/GuntZJkkAXUZ82InrnIWVAOOZBz2MLZGM1+1d+xV8SfBPxHb9on9mCC7knS7/tK50nTYx9qsLoMCZLWID97GxJJiAJGSMMrYX4e/8ABV/xB4b0tfD/AMa/hNPd67YKI5b3TZ/sjTEAD95byL8jnqSrbSTwijiviD/wUZ+O/wAey/w1/Zg+FuqaRd6goie/tQ1/qIVh8xjCIEth94GQ7ioG4MhGR+wN+wtdfs/u/wAU/iolnP45vbYRWVlC/mrokTA+YpkVikkrghWKghQpCswYmviXID8M/Foz/wAwK9/9EPX/AARpYL8TfiHz/wAwK2/9Hmo5hjg0swI61+0B8DvAn7R/wx1L4Y+PbPzLW6/fWV2g/e6ddqrCO5iP99dzDHRlZlOQxFN4G/ba/wCCa/jfVte8FWU+q+EZm3XV9DYte6TqUCFijXMandbuACSdyOuSA5ViW0n/AILTazFpkEPiD9nmyvNQVSJ57PxQ1tE5z1WNraRlGMcF2+vavhr/AMFPfi/8b/i54U8H+CPgHFa6Bf6zZ22tyW8lzq1zaW0jgSSh440VAqB3O6M/Kjcj7wmkB4BqZxjANTMCMA1MwPANTsCcZqZgT1r/AIKparcT/GvwtojJH5Fp4XS4RgDuLS3U6sCc4xiFccdz14x/wRy0b7P4a+KHiEXW77bfaba+Tsxs8lLht27POfPxjAxt6nPCyds0smeM0JPQ15nGM15mR1oyZGAaMgI600nbNfCGG7sPBaeH9RIN7od7dWFxiTeA8czZAPoAQB7AV+wjrMWofB280rKCXS9amQqDyUdI3Vj9SXH/AAGqqgMnGaqvjR8XvD3wZ8Gz+JNXlimvZAyabp5k2veTY+6MAkKMgs2MAe5APi/xVr3xB8V6h4r1+c3WqavcGSTaCQCcBUQEkhVUKqjnAAFfsq/BM/CLwELzWYCniPxAEn1BXUA2ygHy4P8AgIYlv9pj6CqqsZ4BrAxgVVVrHhHwr4jUr4g8N6XqWe91aJKRgEAgsCQcE89s1r37L/wh1sE22j3ekyscmSwu2Hp0WTeo6Y4A6n2xr/7Gk4LyeFvGsbjB2Q6hbFcHHGZEJ74/g49+la/+zX8XtCMjJ4bTU4Y/+WthcLLu5PRCQ56Z+73HfitU0XWNDuPsetaTeWE4/wCWV1A0TfkwBrQPiP498LFBoHi/VbREACwrcsY8AYH7skqcAY6V4e/a4+JGl7ItctNL1qMfeaSHyJT9DHhR/wB8V4a/a9+H+qBIvEelaloszH5nCi5hUcD7y4c9SfudB68V4a+Jnw/8YFI/Dfi7TLyaQZW3EwSYjA58tsP3Hbrx1qqqqUDGaqqqqqsZ4FAADAqqqqqqVe3egABgVVVVeKPBnhPxtYf2Z4t8O6fq9sM7Y7uBZNhOMlSeVPA5GOlfEb9grwxqfn6h8MvEc2j3DZaPT9QzPb54wqyD94i+7eYa+InwJ+KXwukdvFfhW5WyRiq6hbL59s2O/mL93I6Btp4PHBrwF8VfiD8MrprrwR4pvNNEjBpbdW3wzEY5eJsqxwMZIzjvzXwz/b2065Cad8WPDjWkuQF1HSULxderwsxZQBySrNk9FFeD/Hfg34gaadX8F+JLHV7VTtdraXLRHsHX7yHjOGAOOaqqqqqaGG4heCeJJI5FKujrkMCMEEdwR2r9o3/glr+z/wDGn7Vr3gS2/wCFc+J5ct5+k24axnf/AKa2mQo+sRj65O6v2h/2Hf2hf2a5Z73xn4QbUvDkbEJ4h0cNc2ZHYyEANCfaRVyehPWvhd8cPir8GL9774b+NL7SRM2Z7UES205xjLwuCjHHAYjcOxFfCL/gp1o175elfGvwjJp8p2qNW0VTLETwCZIGO9AOTlGcnOAoxmvAfxQ+HnxQ0wav8P8Axjpeu2w+/wDZJwzxEdnjPzoeRwwBwQe4qM8jBqM5AwajIIGDUZBHWomBGM1Ew6ZqJscZqNhwQaRsgYNRSA96R8jAPNJJxjNLKMYzXi7XZfFPirWPEk8skj6pfzXJaUAN87luQOB16Dgdq+H+kQ2mh2YVV3zoJXYD7xbkZ+gwPwrTfDv2q3MirnaMmtb05YCy4HFeKNPh0vX7yxgAEaPlFH8IYBsfhnFfsaa1PqXwZ+wyg7dJ1a4tostn5SEm4GOOZm459e/Er54BqVxjANSMAMZr9rf4fTaJ4wj8d2NufsGugJOyphY7lFxg44G9FDepIc14V8Uz+HLkhlMlrKf3sY6g/wB5ff8An+tWHjzQp4A8Ws2yD0lkEZH4Ng14m+JGn28DxaZOt3dHIUryiH1J7/h+lO91qF2XbfPcXEnQLlnYnsB3J7Vq+h6voF22n6zptzZzoT8lxC0ZPuAwBx0/OvDnxf8AiH4Y2R2PiOe4t1P/AB73h89MY6Ddyo/3SP514a/ae0udI4PFuhTWsu3DXFkfMQt67GIZR+LV4e8deD/FigeH/EVndyHOIQ+yXgZz5bYbGPaqr9qbxSnjL9oj4ga7Fc+fF/bc1rDKHDh47fECFSOCpWIY9sV/wTD8KN4V/Yq8BNPbNDday19qMytIHDCS7lETDBIAMKxHHUEnODkVVVVVVVVEA0VB6GiDVVVUQCMUV9KqqqqqqqqoqD0NEdqqqqqqqph3FVVf8Fqviawh+HPwZs7pNrNca/qMBI3ZA+z2rABsgc3gOVweMHhgP+CYHgBPEPxn1nx5cw74fCWklYW2A7Lm6JjU5IOP3SXA4weeuMgxjgCowMAVEvAr9o79nS+8a3T+PPAlvE2rCP8A4mFiSVa82gBXjPTeFABXjcB1zw11aXen3Ulne201tcwMVkilQo6MOoIPING7uiuw3MmP941pGj6t4h1K30bQ9NudQv7ptsNtbxGSSQ4ycKOTwCfYDNfswfs1j4WQ/wDCZeMY4pfFd3CY0hVldNOjbqqsMhpCBhmBwBlQcEloowQAK/4KwqB+x/qOP+g9p/8A6G1f8Elo9/7H+mkf9B/UP/Q1oQ5GBXxG8aaD8MfAfiD4h+J7pINL8OabPfXTs2MpEhbaPVmwFAAJJIABJAr/AIJEeGNU8eftJeOvi7qtrFMmk6JKZplLJ5V9fXCshVRwQY4boYPTjA4yP+C2agH4M/8Ace/9x9fAf/gnt+yD40+BXw58ZeJfhF9s1jXvCWl3+oXP9v6lH51xNaRSSPtS4CrlmY4UADOAAK/a9/Zs139gTxX4V+OP7OfjTXdP0a+vvsrx3E4ka0u1VnSJyABNDLGsvyOpxsYEsGGP2f8A4v6Z8fPgz4W+K2mxJA2t2QN5bLnFtdxsY7iMZ52rKjhSeq7T3r/gsKP+LlfD4f8AUDuf/R4r4cqD8NvCn/YDs/8A0QlftqfCPxN+yZ+0VpX7QvwttorPRtZ1T+0LHaC0VrqA+a4tnUEERSAuwUEAq7oMBK/4KQ+NNL+Mvwj+B3xk8Llf7H1WLUtyGQM9vPKttuhOOrRvBMjEcbk96/Z1AP7O3wux/wBCXpH/AKRxVMMjAqYAj8K/aw0L/hHf2kviLp/l3Cebr1xd4nXDf6QfPyOB8p8zK/7JHJ61/wAE0/ETeI/2Lfh609+l1daat9YzAKFMQivZxEhAAGRD5XPUggkkk1VVVUQCMUR2qqphkcVVVRABwKr4/fDtvi38E/G/w2hZhceINDurW1KkArOYyYjyVBAkCZBIBHBI61oGt614L8T6b4k0eaWx1fQb+K7tZOUeC4hkDoexBVlHoQRXwz8d6T8Ufh34a+I2hsn2HxLpVvfwosgk8oSxhjGSMfMpJU8AgqQQDxVVVMBnAqqqqqqYDOBVVXjO1Np4lvowuA8nmDjGdwBP6k1VVXwy/wCP69/64r/OqqqqqIBGKr9rjSbzRf2o/izZ3yoJJPGWp3KhWz8k1y8qfjsdcjsa/wCCf+sWuvfsdfDG9s45USLTJbUiUAHfBczQuRgngtGxHsRnHSqqqpgCOKqqq/8A+PG4/wCuLfyqqqqsP+P63/66r/MUw96qqrAIxRHbNVX/AAWa0Zmi+E/iCHT02q2rWtxdBADk/ZGijZupHExA6D5vU1/wRz1e2g+LPj3QGu2W4vPDsVzHBhsOsNwqsx7ZUzoOefnOO9VVVVVVa4A2i34IB/0WTr/umqqqqqqqqvD8Rm1ywQMBi4RufY5/pVVVVVVVVVVVVXxN+Hvh74sfD7X/AIb+KYPN0vxDYSWk+Blo9w+WRfRkYK6nsyg18Tfh74g+FHxA1/4b+KYPK1Tw9fSWk+AQsm0/LIv+y67XU9wwNf8ABP79oef9pL9mjw74p1y9+0eJ9Dzo+vu0gZ5bmBVAnbvmWIxyHgDe7gcAGqqqqqqqqqqqqqqqqq8ReH9F8WeH9T8K+I9Oh1DSdZs5bK/tJhlLiCVCkkbezKxB+tftM/A3Wf2cfjh4q+EWrtLNHo14Tp13IuDeWUg3283HGWjZdwHCuHXqpr/gmn8dovjD+zrY+GNU1Bp/Enw/ZdKvxK+Xe25NpL/umIGLnktbue4qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqoDJxR54Bqqr/gsH8G2vtG8I/HjTY2aTTWOhaoMA4hctLbv6gK5mU9cmVemDn/gl38TdH8O/HnUPgx41naTwf8Y9Fn8PajZuoaGa4ZG+zlxzyQ00I4IzcnIxkj4heCdZ+GvjzxF8PPEUXl6p4a1S4067XHHmQyMjEYJBBK5BBIIIIJFfsmfFF/jL+zl4D8f3V59qv7zSUg1GUyM7NdwEwzs24A5aSNm5/vDBYYYqKqqqqr9pP4QWfx5+Bvi/4WXCJ9o1jTm/s+Rn2CK9jIktnJ6hRKibvVdw71o2q674G8VWOt2AlsNa8P6hHcQiWMq9vcQSBl3KeQVdRkH0r4R/EbR/iz8N/DvxG0GVWtNdsUn2j/ljL92WI9fmSRXQ8nlTyetQvjAzWqeHPDfiLyR4g0DTdT+z7vJ+2WiTeXuxu27gcZwM49BWk6fpujWUWm6RYW1laQ58u3toViRMkk4VQAMkk/U1HIBjBqOTHQ1HKCMZpJeOGoTYGM0Z8jGaaYkda1LwF4B1S7l1DU/BHh+8up23Szz6ZDI7n1ZipJP1q1tbDS7OLT9MsoLS1gXbFBBGI0jHoqjAA+lSyDHXmpZBjrUr5Gc1K4x1qVxgnNTNx1r/AIKYazJqv7Sn2FzDjSPD1nap5fXBaWb5+T82Zj6cbfqf+CQ+kxW/wO8Ya+s7GS98VtatGRwohtYGBHufPI/AUJARjNBgRwaDnHBoOa8w+tFyelFyR1osMYzXw01VJfGXxF0MuA8HiW6uFU4BIeV1JHc48sZ9Mj1r/gn74oW28TeKfBssx/0+yhvYUZjgGFyj4HTJEy57kIPSqqlHfNV8Xvi94U+DXhSTxJ4kn8yaTKWFhGwEt5KB91fRRkFmPCg9yQD8Tvib4q+LPiq48V+K73zJpPkt7dMiK1iB+WONeyjP1JJJJJJr9j79mp7Y2nxe8fadLHOpWXQbGUbdoxxcuvXnPyA/72OVIqqpQDzVVWM8CsDGAaqqurGy1CH7PfWkNzFnPlzRh1yO+DXiH9nz4ReI8vP4OtbKU9JNPLWu3p0VCE6DHKnqT1Oa8Q/sZ6XJmTwp40uoMfdh1C3WXPT+NNuP4j909hx1rxF+zP8AF3w/uePQItVhTrLp04kz9Eba5/75rVNF1jQ7j7HrWk3mnzj/AJZXVu0TfkwBrw58WviT4SKjQfGepQxoMLDLL58Q4xxHJuX9K8M/tieKrIxw+K/DlhqcQbDS2rm3kwT1wdykj0wucDnvXhn9qL4UeIFjivtRutFuGXmO+gO3IHIDpuXHoTjPpk4rS9Z0fXLYXeiatZ6hB2ltZ1lX81JFYzwDWBjAqqqqqqUcZqqqqqqAzzQHGaqqqqqpI45Y2ikRXVgQysMgg9QRXxK/ZD+EPxC829sdJ/4RnVHQhbnSkEcZbsXg+4e+doUnPJ6V8S/2Ovi74BEt9pOnp4o0uNCxudMBMqD/AGoD8+ep+TeMckjpWh+IPEfhDVV1Tw5rOoaPqMGU8+0naCRf7ykqQcHGCDwe9fDH9u/xdorwab8TtHi16xUBWvrNRDdr6sVyI5OMDH7v1LGvh58Zvhp8VIN/grxVa3lwibpLOQGG4j45zG4DEDpuGVz3NVVVVYzwKlghnheCeJJY5FKujrkMCMEEdwRXx+/4Jjfsy/G5pdX0bQG+H3iBlOL3w1FHBBK2MAzWmPKYZOSUEbsermvjt/wS2/ae+EEsuoeFdAT4j6Ei7hd+Hoy90nQbXsz+9LE5/wBUJRjkkdKsNS8TeDNb+1aXqGp6FrGnylDJbyyWtxbyKeRlSGRgR04IIr4Vf8FGPjn4Dji03xgLHxxp0bKP+JgPIu1QcFVnjHOefmkSRs9+1fCr/gob+z58QWtdO8QatdeC9VnYIYdZTFvvwCcXKZRU6jdJ5fToMjOjavpWuafBquianaahZXC7obm1mWWOQZxlWUkEZBHFRN2zUbAcZpG6EGkYEDBqNwRjNI+OAaR8jINJL6GlkBGM0JOODXj3w7J4S8a674ZkhWL+zdQmgRFYsNiuQpBPJBXBBPODXw61yG80aCHzQZrNRFIvQgDhT9MAfka0/wAQtb2+xHxkVruqxOkk0sgVVUliewHU14h1JNX1m71CMfJK/wAmRj5QAAfyAr9kK50XRvh1F4dk1WBda1WebVTYPKPNEJKwq4XqFIiB78MD0YVI4xgGpHHUmpXB4BrxR4e0Xxbot14e1+xjvLG7TbJE4/Ig9QwPII5B5r4k/sxeMvCl1Jd+EoJvEGlYLL5Sj7RH7NGPvfVc59BV9p2oaZP9l1KxuLSYDPlzxNG2M4zg89QR+FeH/APjTxXIkfh/wzf3gfGJVhKxjPQl2woHB6nsa+EnwAsvBVxD4k8Uyw3+sRgNBCgzFaN6jP3nHr0HbnBrWdJ0rXLNtP1jTra+tmOTDcRCRcjocHv714m/Zx8C6wXm0V7rRZ2yQIm82LJ7lGOcewYD+nij9nrx7oO6fTIYdatl6NanEgHvG3OfZd1Xdlf6VdG1vrW4tLmE/NHNGY3Qj2OCK8O/G34jeHFSGLXmv4E6Q36+cOufvH58dsbuh+mIv2prhdFvjdeEw2qR2shsvImzFLOFOxXViCqltoJDE4J9K134Z/Eqz1hbfV/DepT3moXQjWcDzxPNI3H7wEgsxPc5z1718L/A9l8Mvhr4U+HWnFDb+GNFtNNjZRgOIIlj3fU7cknkk5NVVVVVVVVVRAIor6GiO1VVUVBojFVVVVVVVVEAjg0R2qqqqqqqmGOhqq/4KZ/EOX4hftieM0SdpLLwutvolmGYkoIIwZR95gB9oknIAxwRkBi1f8Ex/Aw8P/Aa88Xzw4n8V6zLJG+MbreACFB+Eiz/AJ1EOlRgcD2qMdMGohnArxV8MvAHj1QPF3hPTtSk27RPJFtmVcEYEi4cDnoD1weoFJ+yj8AWwD4Bz/3FLz/47Xg74feCPAtuLbwh4W03Sxs2s9vABI4zn5n+83QfeJ6D0qFQcVCoIAr/AIKxr/xh7qR/6j2n/wDobV/wSn8c+BtA/ZJ07Ttf8Z6Fpt2uu37GC81GKGQAuuDtZgcGvG/7UH7OHw3she+NPjd4N09SpZIf7XimnkA6lIYy0jj/AHVNftuft3+IP2wL+z/Z0/Zo8N6/d+Hr68Bme3gk+2+IZEBKxrAnK264MhVgSxRWIQJg/sMfstp+yx8DLLwvq7eb4s12Qal4jkDKyxXLIALdCpIKRIAmQSGbewwGCj/gtwoB+DOP+o9/7j6/ZfUH9mH4Qc/8yHon/pDDX/BQ7wI/j39kXx/Z21v5l3o1pHq8By37sWsqyytgEZ/cLMOcgZz1Ar/gkL4+/tz4MeLvh3PJvn8L66l1HxjZb3cfyrwoB/eW85zuJ+bGAAuf+Cww/wCLlfD4Z/5gdz/6PFfDcA/Dbwn/ANgOz/8ARCV8ffg34c+PXws1v4aeI0VU1CLfZ3WwF7K6TmKZD1BDcHGMqzL0Y1471Lx/4G8PX37OvjvT5rWXwt4jlu1gmkJNrMYzHKqDkNHIBG6suAeWG4OCP2dAD+zt8LgD/wAyXpH/AKRRVKOKlGRX/BQPT5LL9qvxbcvPDIL+3sJkWNsmMCzhjw47NmMnHoVPev8AgkHrf9rfsoXWn/ZfK/sXxbfWm/fu83dDbzbsYG3/AF+3HP3c55wKqqqqYdxVVRAIxRGOM1VUwBHFV8Uvit8P/gv4MvPH3xL8TWuiaLZfK00xJaWQglY40GWkkbBwigk4J6Akft0eBvDOk/GR/iv8NpvtfgH4sW58R6BeKrBWeVv9LiIIBRkuPMJjwNiui9q/4JFftM2sunX37L/izUtt1BJLqXhMyuAJIzl7q1XJ6q26dQASQ85JAUZqqpgCOKqqqqqmGRxVVXxLt/L12KZVOJbZck9yCQf0xVVVfDL/AI/r3/riv86qqqqqiOcCv+Cguiy6D+2P8TbGadJWl1CC6DKMYE9rDMB9QJAD7iv+CXesy6r+xx4WsZIURdH1DUbWNgeXBupJsn0OZiPoBVVVURkYFVVVrMjRaPfSoRuS2kIyO4U1VVVWqu1zCsThXMihWIzg54NMMjrVVVcUw5xVV/wWJ0aWf4JeCfEKzoI7LxV9laMjljNazMCPYCAj/gQr/gkzrMul/tUT2McKOur+Fb21dmODGBJBNke+YQPoTVVVVVVV9/x5T/8AXJv5VVVVVVVVV4PiEviK0BQsFLMfbCnB/PFVVVVVVVVVVVVV/wAFefgT9nvfDf7QmgaZ8l1/xJ/EMkY6SKN1rKwA7qJYyx4GyJe4r/gk/wDtDz/CH9oy2+HGrXu3w58TPL02VHkCrFfqSbSQZPVmZocd/OHXAFVVVVVVVVVVVVVVVVVX/BZj9nz/AIS34baD+0H4f0rzNU8Gyf2drUsY+Z9Omf8AdMwxyIp2wMYx9ocnI6fsKftBf8M8ftAaNrmrX/keF9fxpOv72wkcErDZOew8qQI5OCdgkUfeqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqpRk1VVX7QPwss/jb8FfGPwtu1Tdr+lyRWrv0iukxJbyEZGQsyRtjIztqyvNf8AA/iiC/s5ZdN1zw/frLE+BvtrmGQEHByMq6jrxkV/wUMg03xh8SfCX7R/hrTYbXRfjP4TstaZIeEh1GNBb3sHYl43jTc2MMzk5JJNf8Edviz9t8PeNfgjqF5mXTZ01vTI3m3MYpAIrgKp5VFdYDxxumJ4J5A4xVVVVVAZ4r/gqF8A5/hT+0BcfEDS7Tb4f+IvmahG6RhVivhgXUZx3ZmWXJ6+ceuCa/4JifH2PRtav/gD4iu2EGsytf6AzniO4VP38OSejoiuoHAKP3eonBAwaikBAANRSAjGail7ZqOTA4NJKCODSzY4zQn44NGcEcNTT8YBp5SR1qSUYwDUknvUsme9SyA96mfPANTODwDUz5PXpX7amvR+Jf2o/iFqMTwsItRSzPlPvGbeCOA8/wB7MRyOxyO1f8EzNEGj/sj+HL/fEf7a1HULvCJgjbcvB8x7n9x19MDtQYEcGg5HehJkYzXmZ4BrzBjGaMnbNGQdAaLk9K8JeIF0X9ofxJaTSbYdV1a/tWyTgOZ2ZDgd9yhf+BGv2e/GyfD/AOMfhjxFcT+XafbBbXhJIUQzAxMzAdQu/fjnlQcHFVVAcYr42ftIeAvgxZy2V3dLqniNoybfSLZwXUlcq0zdIkOV6/MQcqpwcfEr4meLfiz4nm8VeML9ZrlxshgiBWG1jHSOJSTtUfUkkkkkkmv2VP2VP7T+x/E/4n6b/ofyzaTpE6f6/us8yn+DoVQ/e6n5cBqqqxngUAMYFVVKM81VVQGBiqqtS0rS9ZtWsdX061vrds7obmFZUPBByrAjoSPxrxJ+zT8I/EbSTJoEmlTyEkyabMYQPpGcxj8FrxJ+xrq8KmXwj4wtbr/phqEDQkYA6Om4Ek5/hXHHPevE3wS+KfhMsdU8G38kK5Pn2ifaY8A43Ex7toOP4sHkcVp2qano90t7pGpXVjcr92a2maJxgg8MpB6gH8BXhj9p/wCLPhwJDdatba1AgAEeowBmA5z86bXJ56sT0FeFv2xPCF+scHi3w/f6VMcBprYi5hHHLH7rqM4wAG69eM14Z+IvgXxmsY8MeK9Nv5JF3LAkwWYDGeYmw449RVVVVQGeAawMYFVVVVVjPApVB4HaqqqqqqsZ4FYAGBVfEz4D/C/4tR+Z4u8NxNfgAJqVqfIuVA4ALj74A4CuGAzwAea+Jv7C3jvw59o1P4d6jD4lsI/mW0lIgvFHfAPySYA7FSey1PBr/hTWTBcQ6ho+rafLyjq9vPbyD2OGRvyNfC39tf4l+CI49L8Yovi/TEG1WupjHdx/SfB3+p8wMx/vCvhb+0V8Lfi3HHbeH9dW01Vh82lX+IbgHvtGcSf8AJ98VVVjPAoAAdaqq+MP7M/wI+Pdo0HxV+GWi63cmPy49RaDyb2FcEAJcx7ZVAznbu25wSDivjX/AMEXtNn+06v+z98TZbZ/mZNF8TR+YmTk7UuogCoHyqFaNj3L8c/GD9jf9pb4F/aLn4hfCXWoNMt95bVrGIX1kET+NpoSyxgjkCTacZyBggeCPiZ8QvhrfHUvAHjXWdAnf/WGwvHhWUccOoO1xwOGBHA9K+GX/BUT4xeGmitPiV4d0jxjaK3z3EajTrs8Y+9GpiIB5x5YzyMjIx8NP+Cjf7Nnj14bHWdb1HwdfyYXy9cttsJbaC2J4i6Bc5AaQpnHQZArwx4r8MeMNMTWvCXiPS9b0922rd6deJcxMcA4DoSM4I79xUb44BpH4yDUcgIxmkkx3pZAehpZCBjNLMMda/a6+CN/rkp+KnhOyM88NuE1m3jGXZEGFnUdTtUbW9FVTjAY1pup32k3K3en3DRSL1x0YehHcVbfFW9jiCXOlRyP/ejmKD8iD/Otf8Z6xr8f2eV1gtyOYo8/N9T3r4ZfCnxV8UdSuLfw/pkk9vp8RlupN4iHAJWJXb5RI5G1c8AnJwoJGuXfinSPFUt1qUd5o2tWUq7Y1VreS0KKBGqDhkCqFC+gAxXwc/azFyYfDfxUmRZWYJBrCIFVsnAE6jhf98ADHUDBYx3lvd28d1aTxzQzIHikjYMrqRkMCOCCMHNSPgdakfvmpXzwDUrjGAalYYwDUrDGM1IwPOaPPeq1jQNE8QW32PXNJtL+LBAW4hD7c9SM9D7jmvEP7NvgbVDJNotxe6RMw+RY386JT67W+b8Awr4reNPAfwl+JN38M9c8WQ3F7ZRRNNcQ27mKJ5BkROcHa4Xax6qA6/NncF0vV9K1yzXUNG1K2vrZiQstvKJFyOoyO/tXw/8A2pfjV8OvLt9O8WPqthH/AMuOrqbqPvgBiRIo56K4FfDf9u74d+JpINM8e6Vc+F7yTCm6DfabQtwMllAdMnsVIA6t3rRNd0TxLpkOs+HtXtNSsbgZiubWZZUb6MpI/DtVVVVVVVVVUQCMUV44NEdqqqIBGKKg9DRHaqqqqqqqogEYojHFVVVVVVEAjFEY4q7u7SwtJr6+uore2to2kmmmcIkaKMszMeAAASSeBXjvxXeePfG/iHxzqKst34i1W61GcPJvYSTytI2WwNxyx5wM+gr9m/wongn4D+AfDIXbJa6DatOMggTSRiSXBAAI8x2x7Yzk81q+u6N4b0ubWte1KCxsrZcyTTPtA9APUnoAOScAc14x/bFsbSeSz8B+G/toXIW91BzGjHB5ES/MRnB5ZT1GBnIj/bN+JyMpfQPC7KDyBbTgkex87ivAH7ZPhPWrqHTvG2iy6FLI20XcUnn2+TjluAyD8GA6k+llcW15bQ3dncRzwTRh4pY2DK6kZDKRwQRg5qIA45qHp+FQgdqiAx17V+0P+z74R/ab+GFz8LPGmravp2nXFzFdC40uSNJlkiJKf6xHBXJ5GAT2I60n/BE74WuOfjb4q/8ABdB/jXhL/gi3+zhprJP4s+IHj3XJY5i3lQ3FtZwumBhGUQs+c5OVdeoGBgk/Bn9mr4G/s7aXJpfwf+HemaE1wu25vQGnvLkZziS4kLSuueQhbapPAFTgdBX7VX7Gvwv/AGuP+EYPxI13xRpv/CJ/a/sX9iXUEPmfafJ3+Z5sMuceQmMbcZbOcjHgbwZpfw58B+HPh5odxdT6d4X0i10uzlu2Vpnit4liRpCoVS5VASQoGc4A6VremWOtaZeaNqUHnWd9A9vcRbiu+N1KsMggjIJGQQa/Z1/Yb+F37LfivUPFvw78X+NbybVLA2d1aatd2ssEib1dWxHbxsGUrwd3Rjwc1+0r+xb8LP2pNb0fX/iBr3irT7jQ7R7a3TR7qCJGRm3EsJYZCTn0IrR9HtfD+hafoFlJI9vptpHbQtKQXKxqFBYgAZwBnAFS9/rXx/8A2GPgt+0P4xTx94tvPEelawLVbaeXRrqGEXSp9xpBLFJllHygjHAAOcDHg3wnpvgLwZoPgXR57maw8O6XbadayXLK0rxQRLGhcqFBYqgyQAM54HSpun4VL0r9vC4trr9rDx5Ja3EcyK9ihaNgwDLYW6suR3DAgjsQRX/BJv8A5M/03/sPah/6GtVVVVUQCMUR2qqpgOoqqogEYzX7VX7aHwm/ZV0I/wDCRXaa34tuoS+m+GbO4VbibsskrYb7PCSMeYwJOG2q5UgftD/tN/Fj9pvxafE3xI15ntrdmGm6PbMyWWmoeoijJPzEAbpGy7YGTgAD9mT9iT47/Hv9nfxPBrkCaT4YlhGreBYtUUpLcaoBjzIM8xW00WUkY4WQ+Q4DeUCthfeNPhP49ivrN7zw/wCK/COq5XfHsmsby3k5DKwPzK6EFWGOCCOtfsW/tieEf2q/AMPmXFvYePNFtY18QaPnblgApuoBnLQO3bkxkhWz8rPVUQCMUR2qqqqqiARiqqvilCRcafPuGGSRQPTBH+NVVV8MgDfXuP8Aniv86qqqqqmHpX/BTnSbzTv2z/G15cqgj1S2025t8NklBYQRHPod8T8emPWv+CS2sWuqfsovY28cqyaR4ovrWcuAAzlIZgVwTkbZlHOOQe2CaqqqmHOM1VV4mkaLw/fshGTAy8jseD+hqqqqsP8Aj+t/+uq/zFEZGM1VVVMMgYqq/wCCsOk2eo/spfbLlWMml+JrG4t8NjDlZYjn1GyV+PXFf8E0b67tP2z/AAHb21w0cd7DqcNwo/5aINOuHCn23Ih+qiqqqqqqr7/jyn/65N/KqqqqqqqqvAMQk1xmLf6u3ZgPxUf1qqqqqqqqqqqqqvjT8MNI+NHwo8U/C3W1i+z+ItNltkkkTcLefG6GbHcxyrHIPdBXiHQfE3w48Zah4c1aO40vX/DeovbzhHKSW9xC+CVYcghlyGHsRX7JPxztP2jf2ffCHxTjk/4mF9Zi31eMqF8u+h/dzjAAABdS64H3XXpVVVVVVVVVVVVVVVVVeMvCWgePvCWseCPFNgl7o+vWM1jfW79JIZUKOPY4Jwexwa/aC+DWv/s/fGPxT8I/EQd5/D980cFwybRdWzAPBMB6PGyNjsSR2r/gnB+0F/wvD9n6y0LWr/zvFHgHy9J1Le2XmgCn7JOe/wA0alCScs8MjHrVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVUo4qqqgM8V/wUv8Ag7/wqv8Aaf1rWbC28vSfHUY1u2wOBPISLoZyckzK8nbAlA7Zrwxc/wDC3v2H/E/gmVfP174JeIIvEWlfufMkbRtRK297EmBlUjuRbTsxJ4c8YXK/sL/Fdvg9+1D4I8Qz3LRabql6NH1MADBguv3QZs9FSQxyHHP7vv0OBjAqqqqqlGTX7WfwA079pX4H658NZ3ig1M7b3RLuRc/Zb6LJjb2DKXiY9klbHOKli8V/DTxq0Mq3Oi+JvCuqFWU4EtleW0vPqAySJ+Yr9lz4/aT+0N8K7DxjbmODWLXFrrVkrjNvdKBuYAchHHzr7HHVTUcnGQajkBAwajlBGM0kuO9JKCODSzHHBzQmBGKMwIxkGjNkYFNKSPvVJMAMA1JJnqakkzwDUsgIwDUsnGAa8S/FbwJ4bla1v9fhluV6wWwMzA5wQduQp9mINf8ADQfgWWdoDBqqKCcStbrtOPQByefp+VfHPwx45tfiH4m8XeKfD9zbR67rV3eLciMGJzNK0nDKSoJ3fdzkdO1fsr+GG8Ffs4fDfw7JE0U0Phu0mnjdApjmmjEsikDuHkYZ74zSydgaEmRgGhICMZreMYreKLgjAoyDGAaaTPGa8f3lxp/xU8R6hZyFJ7bxBdSxMOCrLcMQfzArTbmW/wBJ07U5rOW1/tGxgvI4pRhljmiWRD+Kupz3Br9nn44eF/HPww0WTXPFelwa9YW4ttQt7i7jilLR/KJShOcOoVsgYySBjGBNe2VvaG+uLuGK2VNxmeQKgX13HjHvXjb9qX4IeBkdbrxnb6tdKGK2uj4vHYjqNyny1PbDMP0OPin+3B4+8WxyaX4BtF8LWDja1wriW7f1w+MR/wDARuH96tL0nxL4215dP0myvtZ1jUZi2xFaaWZ2OWZjyTyclj9Sa/Z8/Y20/wAHT23jL4ppb6lrUJ32ulriS2tD2aQ4xLIO38Knn5jgiqqqAyaqqoAAYqqoAE4qqqqqq8T/AA48CeNAT4m8K6ffSsMGd4tsuMY4kXDjj37D0FeKv2O/CF+sk/hHxBf6VMclYblRcwjgYA+66jPUkt16cYrxX+zD8VvDTNJZaVDrlsCcS6fJuYDtmNsNkjsAwHr0zfafqei3rWepWV1Y3kDfNFPE0UkZB7g4IORXhb4+/FfwkscFj4snvLWPAFtqAFyuAMBQWy6geisOgrwl+2Tp0/l23jjwtLbP8oa601/MTPcmNyCoHXhmP5c+Evip8PfHConhrxXY3M7jItXfypvQ/u3w3XuBjp61VUBxVVVVVUo44oAAYFVVVVVWM8CgABiqqq8efDPwJ8TNN/svxt4btNTjUERSOu2WHPeORcMn4HnvXxT/AGC9TsY5NU+EeunUUUZOlao6xzH/AK5zABGyezhMAfeNeJfCXijwbqJ0nxX4f1DSLxc4hvLdoiwBxuXI+ZfcZB9a+FX7XnxU+G3k6bqd7/wlGix4H2PUpSZY1HaOfll4AADb1A6KK+FX7TXwq+LPk2Gmaz/ZWtSYH9lakRFK7ekbZ2y98BTuwMlRWM8CgAB1qqqqqvjH+wZ+yv8AHB7m/wDFXwr0/TtYucltX0LOnXJctkyN5WElc5IJlR+D7DHxe/4Ir3kKTah8CPi2lwFX93pfimDYxIPP+lQLgkjoPJAyOWAPy/Fb9ij9qL4MJdXfjb4O65/Ztmx8zU9NiGoWoTOA7SQFwinjG/aeQCAeK8N+LfFfg2/GqeEPE+raHejGLnTb2S1kGOnzRkGvh9/wUn/ac8FyQQ63r2l+L7KJgGh1exVZCncCWHY27GcM+7B6gjivhz/wVo+HOppHa/E/4ca3oFx8qm50qdL+AnAy7K3lugJz8oEhAxye3gD9sL9mj4kiKPwx8YvD63MuAtpqMx0+Yt/dVLgIWP8Au59enNW9zFPEk0MqyRyKGR1OQwPQg9xSyAjg0svGM0JAR1oSDHBr4j/sn/DzxtdSaroMsnhnUJTmQ2kIe3c+phyAp6fcKjrkEnNX37FHxDjlC6b4p8OTx7eWneaJg3pgRtxjHOfwrwr+xRp1vdLceNPGMl3Cj5+y6fB5W8Ad5GJIyeCAvToeePDXhnw74M0eLQfC2kW+nWMP3YoVxk4ALMTyzEAZZiSccmvij8IPBvxT05odbs1g1GNCLbUYVAliOOM/31z/AAn8MHmvih8G/F/wrv8Ay9Yt1utOlci21G3BMUo44YdUbBHynvnBYDNfCb47eK/hdKlgjnUtCZyZNOlfGwnOTE2DsOTkjGDzkZOR4K+Ifhb4i6T/AGv4X1ITohCzQuNssDHoHXqOhwehxwTUsmRgGpHwOtSMMZzUjZ71K+eAacgnANVVftZ/treHPgxY33gbwBeW+q+PHTYSoEkGk7sgvL2aUY4i7ZBbj5W8JeFPGnxb8d6f4T8NWV9r3ibxLfeXCg3TS3E0hJZ3bk4+87uegDMTgE1+zP8AshfDv4CfAfS/hDquhaV4gupv9M8QXV1bidL3UHC+Y6h14RdqpHwpCxqT8xYn4ifsHfDbxFHNeeAtSu/DF6VJSBi11aluOCrneucHkMQM8DjFfEr9l74xfDEy3WpeGJdU0uMtjUdLBuYtoGSzqBvjGO7qBnua8F+P/Gnw61Ua34I8SXukXfAdoH+WUDoJEOVkX/ZYEe1fC79vpJZItL+Lnh9IQcKNU0pCQO2ZISSfclD9Frwn4z8KeO9JTXPB/iCy1ayfjzbaUNsP91h1Rv8AZYAj0qqqqqqqqqqIBGKK8cGiO1VVEAjFEY71VVVVVVVEAjFEdqqq1LX9B0d0i1fW7CyeQZRbm5SIsPUBiM1qXxS+GOjeX/bHxH8L2PnZ8v7TrEEW/GM43OM4yPzFaj8f/gfpcC3Fz8WvCjqzbQLfVYrhs4J+7GzEDjrjHT1q6/al/Z9s7d7mX4paSyRrkiJZJW/BVUsfoBX/AA2V+zZ/0Un/AMo99/8AGak/bo+ACOyLqusuFJAZdMfDe4yQcfWtS/b6+Btk6Ja2PinUAy5LW1hGoQ+h8yVD+Wa+Ln7cfw58Z/Crxj4P8I6R4ssdb13Qb2w0+7uba3SO2nmheNJWKzMcKzBuFPSrL9mbUReQHUfEts1qJV89YY2VymRu2k5AbGcEgjNXX7YOmw7P7O8CXM/Xf52oLFj0xhGz39K+Nvxavfib4ieKzuJU0CwcrY25JAkI4MzDAO5h0B+6Dj1za2M12f3Y4Hc02gS7MiXn3Wrm1mtW2Sr9COhr9kr40Xfh3xLB8NvEN68mk6zIEsHlct9luTwqDJ4R/u4A++V6ZY1GQcYqEgjANQkHABqEggDNQsMAZqJxwAaWTA4NO4IyTUzgjg1MQTgGpyD3qYg8VOR0zUp44NS4IwDUuCOtS4xipcYJBqXkVKQRX7T+tR+IP2iPiNqMMaog8R3kCbZN4YQyGIODjo2zd7Zxk9a/4J06JLoH7FvwxsJp0laWxuroMoIAE97PMB9QJAD7iqqqqqpgM8VVUQCMUR2qvi/8efhF8BtC/wCEg+K3jrTdCgZSYIJX33F0R2ihXMkh/wB1SB3xX7Qv/BX3xZ4hgv8Awz+zx4W/4Ry0l/dx+IdWCzXu3PLRQDMcRPABcycE8K2Ctta+OPin4yW2tk1nxV4q8Q3ZwCZLy8v7hzkkk5d2JySTk9Sa/Y2/4Ja6T4HlsfiR+0rZ2Wsa/EyzWPhhJRNaWLDYytcMp2zyAhgYwWi95MjEUMUESQQRpHHGoVERdoUAYAAHQYr/AIKNfsHf8Lm0ub41fBzQIj4806Mtq+n267W123VeGVRw1ygAA6GRflySqCvh78QvHPwc8c6d468Ca1daJ4h0SfdDMgwVI4eN0PDIwyrIwwQSCK/Y3/bI8FftY+CvOh+z6R420iFf7c0LzPudB9ogzy8DHHqUJCt1VnqqYc8VVVVVVMOcCqr4qf8AML/7bf8AslVVV8LogRqUpT/nkqsR/vZAP5fpRHbNVVVVUwBGBX/BVn/k7nUP+wDYf+gNX/BHn/k2XxL/ANj3d/8ApDY1VVVUw9Kqq8cSLH4ZulL4LlFX3O8H+QNVVVVh/wAf1v8A9dV/mKojnGaqqphleKqv+Cllra3H7F3j+We3ilktpNMkgZ0BMTnUbZSyk9DtZlyOzEdCa/Yhlih/a0+FzzSoinX41BY45KsAPqSQB7mqqqqqqr7/AI8p/wDrk38qqqqqqqqq+HURae+lDDCoi4+pP+FVVVVVVVVVVVVVX/BWT4HReCfjBpnxh0OxSHTPHVtsv9h4GowAK7bRwoeLyj7skjHk1/wRc+O//CP/ABB8Ufs963qmyy8VW/8Aa2hwySYUX9uuJ0jXHLyW4Dkn+G0qqqqqqqqqqqqqqqqqq/4LEfswS+M/Aun/ALSnhOxRtT8Gwiy8QIinfPp7yARSgAcmGSQ5/wBiRiThK/Ye/aKH7N3x30zxNq1zKPC+tRnTNfjXnFvIQUmA6ZikCPnrsDqPvVFLFPEk8EqSRyKGR1OQwPQgjqMVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVWOAKqqpRk1/wVW+D3/Cwv2cl8e6fFEdT+Ht+t7kx5d7OYrDcIp7cmGU9sQnviv2W/HWi+CPjFplr4vvYbbwh4ut5/DXih5h8iaZfoYJpWIIK+VvWYFSCGhB9j4y8Kav4E8X654H8QRJHqnh3UrjTr1EJIWeCRo5AMgHG5T1Ar9ln4sL8bv2ffA/xIkuUmvtS0tI9SKgjF7DmK44PIBljcjPYg5IOTVVVVSjiq/4Ko/shXF+0n7T3w60ppJIoVj8X2cCs7MqALHfKoBwFQbJTkAKqNj/WNX7MH7Q2vfs4fEu38WWImu9FvVFtrmmo3/H1b5zlQTjzUPzIeO65Cu2fBvjDw/468M6b4w8KapDqGk6tbrPa3EThgynscdGBypHUEEHkGo5PQ1HKCMZpJcDGaWUEcGlmOMA0JzjrRnBGAaaYkYBppcjrTzDoDUkuepqSXIwDUkgxgGvjR8Xr6e/n8JeFb5obe3JjvLqJiGlfo0anso6EjqfbrHFJM4jhQux6ADNPpOoRpva2OB6EH9M1d2lteW8llfW0c8MqlZIpkDKw7gg8Eexr4B/FtPFFkng3WhbQalp8AFp5UYiSeFABgKOAyjsABjkDg0HBHWgwIwDQcnoa3k8VvouT0ouSOtFu2a+JeD8RvFWD/wAxu8/9HPXjP4Ga5rHwA+CPxM8EeHb7U5r3wBolrq8FhavO4aPT4THOVQE42KUZjgDZGP4q1HSNW0h0i1bTLuyeQZRbiBoyw9QGAzVeDvhV8R/iDKkfg3wZqmpo5AE8cBWFckj5pWwi8gjlh0Poa+Hv7A3iXUHhvfiZ4nt9KtsgvY6aBPcMOcgyH5Iz0OQJBXw7+FHgD4Vaa2meB/DtvYeaALi4OZJ7gjoZJGyzckkDO0ZOAM1VVVUAMVVUBk4qqqgOKqqqqqlHeqrW/Dfh7xNbCz8Q6HYanCudqXdusoXPUjcDg8DkV4u/ZN+G+veZP4flvPD9y24gQP50O49zG5zjPZWUfpjxb+yn8TvD5ebRY7TX7ZTwbWTy5QMdTG+O/ZS3b3xqmj6tol2bDWtLu7C6UZMF1A0Tgeu1gD2NeE/jt8UvBpSPTvFdzdWyH/j1vz9pjIx0G7LKPZSP1NeDv2x9Kudtt478NS2b7R/pWnN5qFu+Y2IZR9GY/wA68KfEnwH43Cjwv4qsL6Q5xAJNk3AyT5TYcDHfHr6VVVVVVAZoDjNVVVVVVAd6qqqqrxX4M8KeOtJfQ/F+gWWrWT/8srmINtP95T1VvdSCPWvin+wWrvJqnwj11UBOTpWqOcL/ANc5gCfwcf8AAq8aeAPGfw71U6J418OXmk3fOxZ0+WUDq0bjKyL2ypI96+FP7XvxU+Gvk6dqV7/wlOix4X7HqUpMsa+kc/LLwAAG3qB0UV8Kf2m/hV8WfJsNM1n+ytakwP7K1IiKV29I2ztl74CndgZKiqqqqqqq+J37J/7OHxmeS4+JHwc8N6rdzbvMvktfst225ixzcQFJeWZj97qxPUmviZ/wRe+D+utJd/Cv4n+IvCszKNttqVump24YADC4MUig4ySWfBJwMYWviV/wSX/a28Defc+GtI0DxxZxbnV9F1NY5vLG85MVyIiXwq5SMucuoUtzjx/8H/it8Kro2XxK+G/iTwxJ5nlodV0uW2SU5YAo7qFcHY+GUkHaSCQK8E/FP4lfDW5F38PvH+v+HX372Gm6jLbrIcqTvRWCuDsTIYEHaAQcV4L/AOCk/wC1h4Q8uO98Z6b4mt4toSHWtLifgZyDJD5cjZyMlmJ4GCOc+DP+Cv8Aq8Qht/iF8GLS5JUebdaNqrQ4IXkrDKj5BbHBkG0HqxHPhX/gqh+y9r7Rx623ivw0xYK76hpQlRcjlgbd5WKg8fdB9q8Nftlfss+LIBPpXx38JQKYw4Gp3405sE4+7c+Wc+2M+1aR4l8P+IopJtA17T9TjhbbI9ncpMEPoSpODRkHrTSjHBppcjANPJkda1bT9M1rT5tK1ext72zuF2ywTxh0cdeQeOoB+tfGH9mK+0Mz+IvhxE95pqhnm01mLTW4AyfLJ5kXjpnf0+9kkeG/E3iDwXrUeteH9Qmsb63JUle4zyjqeGU4GVPHFfCX4+6J8RUi0bV1i0zxDt/49wT5VzgcmInOD32E5HYtgkSOD3qR88A1K4xgVI3HWjz3qviZ+2D+z58LIZ01n4gWeqajCm5dN0VhfTu39zKHy4246SOnb1Gfjv8A8FDviV8SrOfw38ObNvBWizqUmnim8y+uFIwV80ACJTn+ABuPv4JB+GXwt+Ivxv8AG1p4G+HHhq/8Qa9qL58qBC3lqWCtNM/SOMFhukchRnk81+wt+wt4U/ZN8KDXNcFprPxJ1m3C6rqyruSzQ4JtbUkZEYIG5+DIwBOAFVaqvih+yv8ACD4oRz3V1oK6Lq8pLDUtLAhkLnu6Y2SZIGcjd1wwyTXxY/Y++K3w087UtLsv+Eq0SPJ+2abETLGo7yQcsvAJJXeoHVhXhPxn4r8C6smueD/EF7pN8nHm20pXcP7rDoy/7LAj2r4P/t5wzPDonxm05YflCrrWnwkrn1mhGSBjOWjzzgbO48OeJvDfjDSYtd8La3ZarYTcJcWkwkXPdSR0YZ5U8juBRUEcGip7GiDVVVVVVVUQCMUVx3qqogEYor6VVX/ibw3pdwbTU/EOm2k4AJiuLtI2APQ4JBq/+PvwP063NzcfFzwk6AgYg1eGduf9mNmbH4VqP7XX7Oelzrb3PxNtXdk3A29jc3C4yR96ONgDx0znp61qP7dvwCslnNrfa5qBhJCC300r52DjK+YVxnr8233xWpf8FD/hbFGh0jwX4quXJ+dblIIAB7FZHz+QrUf+CjmlxTqukfCa6uYdnzPc60sDBsnICrC4xjHOe/T1u/8Agot4xd5jY/DfRokJbyVlvJZCg/h3EBd2OM4Az7Vdf8FBPjbcW7wQ6H4OtncYWWKxnLJ7gNOw/MGtR/bf/aGvoFhtfE2nWDh8mW30qFmIweP3iuMc56Z4HPWtS/a3/aK1WNI7r4nXaCM5BtrO2tz+JjjUkexrUv2gvjjqrpJc/FrxShjGALbU5bcfiIyoP41qPxM+I+sTLc6t8QfEl7MqbFkudWnkYKCTgFmJxknj3rUdV1PWJludW1K6vZlTYslzM0jBQScAsScZJ496rTtG1fWXeLSNKu754xl1toGlKj1IUHFWHw2+Imq3AtNL8A+I7ycqSIoNKmkbA6nAUnFQfAv413M0dvH8IvGQeRgqmTQ7hFBJxyzIAo9yQB3r/hmL4+/9Es1r/v2v+NQfsdftIXMEdxH8NXCyIGUSapZowBGeVaYFT7EAjvWm/sS/tEX/AJn2rwlZadsxt+06rbtvznOPLd+mO+OoxnnHx9+C/jX9mrwNH8QfiilhbaVPfx2EAtLoTyyzujuqKox/DG569FNfCDx9pHxs+IWmfDbwnZX9vqWqLM0U1/EqQRiKJ5WLsjOwBCED5T8xA4zmvGH7O3i7wP4U1DxZrWsaPJBYCP8AdWryO7l5FT+JFAxuB/Co08yRYwfvECtC0xJikaKMDgDFS+D2Wx+0GPjHXFeINMVVeLaMgfL9atrie0uIru1lMc0Dh43HVWByCPxFeFdXXX/Dek66rxsNRsYbkGMEKfMQNlc845qFhgc1EwBxmonGMZpHxyDUcoxjNLMMcGnlBHWpZAR1qRxjrUzgjrUrA9+lfFH4ueCPhPpS6j4u1MpJMG+zWUCh57kjGdi5A4yMkkAZ614s/bf8c6hcTReEPDelaXaE4je6DXM4AI5yCqDOORtOM9eM1a/tkfGGCYSznQrtQCPKmsSF+vyOp/Wvht+1p4Q8Y3EOjeLbT/hHtRmbbHK8u+1kPb5zgoT/ALQx/tVMeMZqU/LjNTEY4NatqFnpOnXOqX83lW1nA808m0tsRAWY4GScAHgDNa1qlxrusX2t3aRpPqFzJcSrGCFVnYsQoJJxk9ya+BHhOXwJ8D/h94KuI2SfQvC+nWU4e3MDGSK2jRy0ZyUYsCSDyCTmqqqqqphkYFVVV4l8R+H/AAfod74n8Va1ZaTpOmwma7vbyZYYoEHVmZiAB/jX7VP/AAVrlLaj4F/ZhsUChmhk8X38RJYYILWkDAYOcYklB4B/d8hhreveOPih4r/tLX9W1nxT4k1i4WJZbiWS7urqVjtRFzlmJJAVR6gAV8BP+CTfxy+JH2LXfivf2vw90Kb53tp0+06m68EAQAhItwyMyOGU8mM9K/Z8/ZF+B37Munyw/DTwyzancqFuta1Jxc304AAx5m0BFOMlI1RSTnHpVV/wUK/4J6xfFGLUPjl8DdGSPxlGrTa3odum0a0AMtNCo6XOOSv/AC1/66f6z4SfFv4h/AL4h2PxB+H2rS6XrelSFJI5FJjuI8jzLeePjfG2MFTgggEEMoI/ZU/ac8HftUfDC38c+HUSw1S1YW+t6M86yS6fcY9uTG+CUcgbgDwCrAVTDI4qqqqqqYZHFVXxSkZrrT4SRtWN2HHckZ/kKqqr4ZwbdGuZ8nL3JUAj0Uc/qaYc9aqqqqogEYr/AIK7aNa6X+1Lp17byStJq3g+zupw5BCuLi5hAXA4G2FTznknnGAP+CN2q3k3wa8daE7J9ltPE6XEQC8h5baNXyfTEKcfX1qqqqphkcVVV8RJTHoUcakfvLhVIPXABPH5CqqqrSIlm1ayhYnD3EanHXlhVMPeqqqIBGKI7VX7cf8AyaT8UP8AsBP/AOhrX7MH/Jy/wk/7HrRf/S6Gqqqqqqr7/jyn/wCuTfyqqqqqqqqr4cf8xD/tl/7PVVVVVVVVVVVVVV+3F8F2+On7NnirwtYWiXGs6XD/AGvo6kDP2q3BbapIOGeMyxA8f6zkgZNfCn4iaz8I/iX4Y+J3h75tQ8MapBfwxltol8twWjJwcBlBU8HhjXhPxPovjfwro3jTw3d/adJ1/T4NQsJ9u3zYJo1kjbHbKsp/Gqqqqqqqqqqqqqqqqq8ReH9F8WeH9T8K+I9Oh1DSdZs5bK/tJhlLiCVCkkbezKxB+tftM/A3Wf2cfjh4q+EWrtLNHo14Tp13IuDeWUg3283HGWjZdwHCuHXqpr/gmZ+0F/wuH4Dw+CNcv/N8SfD7y9Pn3tl5rIg/ZZffCq0R/wCuQJ+9VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVQGTiiB0FVVUo4614k8P6V4s8O6p4V121S503WbKayvIHAIlhlQo6kHIwVYjn1r4qfD7VfhR8SPE3w01uZJr3w1qk9jJMiFVnEblVlUHkK6gMAecMK/aJvh4+tPA3xzWaGa58ZaGtlrzxQeWE1nTQttcBsfKZJLcWN0xGObzOBnn/AII6fF1GtfGvwM1K+w6SJrukxMRyCBDdAZ5422xAGRy545zVVVVQHGKrUNPsNWsLnS9Usre8s7yF4bi2uIhJHNGwKsjqchlIJBB4INft+fsZah+zZ47k8W+DNNup/hx4jnZ7GYJuXSZ2ZibJ2yTgAZjZsFl4yzIzH9i/9r69/Z415/CviwzXngTWbgPdIgLyabMQF+0RKPvKQAHQckKCOVw2ha9pPiLSbPXtA1O31DTr+FZ7W6t5A8c0bDKsrDggiklB70kxAxmlmBHWhKMcNQmOMA0ZjjhqaYnvTSjpup5h0Bp5cjk1JKDwDXxE8RSeGPBmra1bsfOgt9sJH8MjkIp/BmB/Cgsk8oUEs8jYyTySa0XRo4o1ijXJP3iepNXWgSC3DBOvtWt6OCjyRr+8jUngdQK8Ma7c+GfEOneILRj5lhcJKFBxuAPzKfYjIPsaguIriGO4gkDxyoGRh3BGQfyoOa8ztmvNGMA15mRwaMoIxmjIOmaMhPANfEG5tr7x74kvLO4ingn1e7kilicMsimZiGUjgggggiv2X4ZYP2aPhJBPG0ckfgXRVdHUqVIsYQQR2NTwQXMD21xEksUqFJI3UMrKRggg9QR2pPAPgWO/XVYvBegrfJH5a3K6bEJAgO7aG25xnnHrzQAxgVVVVVWM8CsDGBVVSjiqqsZ4BrAxgVVVVVWM8CsDGAaqqqtZ0DQ/EdmdP1/R7LUbZusV1Asq/UBgcHgc14u/ZN+G+veZP4flvPD9y24gQP50O49zG5zjPZWUfpjxl+yv8S/DKSXWjxW3iC1Q8GyJWYD1MTc9eylvX1xd2WraDqH2a+tLvTr62fPlzRtDLEwPXBwQQRXg39o34qeD2jiGvnWLNAR9l1QGcfg+RIMdvmx7HFeDv2wPBmrbbbxlpN1ocwUZnizcwk9/ujevqBtb6+vh7xX4Z8XWjX3hjXrHU4UIDtbTB/LJ6BgOVPscVVVWM8ClXjANYGMCqqqqqoDPFYGMCqqqqqqvEXhfw74v0mXQ/FOiWWq2E337e7hEi57MAejDPBHI7EV8X/2ELabzNZ+DV/5Dcs+jahOWX2EMpyR9JCf94dK8V+DPFXgbVX0PxfoF7pN6n/LK5iK7h/eU9GX3Uke9fCD9sD4lfDUR6Vr1w/irQ1wq219OfPgA/wCeUxy2MYG1tygAAbea+FH7Qnwz+MUa23hnWDBqojLyaVer5VwoHUqMkOB6qTjviqqqqsZ4FBR3qqqaGG4he3niSSORSro65DAjBBHcEdq8ffsTfsn/ABNeWbxd8B/CslxP/rbjT7U6bNIQwOWltTG5OQBknOOOhIrxv/wRm/Z01tGm8E+N/Gnhm5OAEknhv7cAKedjIsmSdp/1mMAjAzkeNf8Agin8Y9Okc/D74v8AhDXoU5X+1bW40yRhtB4VBcLndkAFgMYJIzgeMv8AgmX+2h4OYM3wibWrdmVFn0bUra6G4gnHlhxIAMcsUC5wM8ivFnwI+N3gQSnxr8H/ABpoSQxvK8moaDc26CNPvuGZACgwcsDjvmopZIJEmglaOSNgyOpwVI6EEdDWh/Hn44+Gcjw78ZPG2mqzh3S21+5jV2HQsofDfiDWkft4ftb6JJJLZfGvVJWlXawvLS1uwPoJomCn3GK07/gpl+1jZWUdrc+KtE1CVM7rm50SBZHySeRGEXgHHCjgDOTk1H/wVg/aOSJI38I/DqRlUAu2m3gLH1OLoDJ9gBVn/wAFa/iklpEmo/CvwrPchcSSQ3FxEjH1VCzFR7Fj9ab/AIK1fEVh/wAkl8Of+B03+FeMP24oviFr8Graz8KNK0WSWT/TbnS7t90oJOXKMMM+TnPBPQnpjQfEGk+ItOg1rw/qUd3bScxzREjBHYjqrD0OCKg/a1+IHgrwiySeB4/G95Z5K79VNlO8Y6jPkyCRx2+6SO5PW4/4KvTwTSW1x+z68UsTFJI38UlWRgcEEG0yCDxitS/4KqeMJ7t30r4Q6Pb2px5cVxqkkzrwM5dUQHnJ+6MDA561q3/BUT4zT3CvovgPwXawBMMl1Fc3DFsnJDLMgAxjjHY888a9+3/+1BrduLW38cWelIUdJDYaVbqzhgB951ZlI5wUKkZ69MeMfi78U/iErxeN/iL4i1uFyT9nvdRlkhUkqTtjJ2KMqpwAOQD2r4d/Br4s/Fy8Fh8Mfhx4i8TyhirtpunSTpERtzvdRtQDemSxAG5fUV8Cf+CNHxH8RT2mr/tA+MrTwpppw0+j6M6Xl+wwMoZuYIjyfmHnD5ehzkfBb4BfCX9nvwsvhL4TeDbLRLV1T7XcIm65vnQECSeY/NI3LYycDcQAAcVVVVV8WP2YPhR8W/Ov9T0b+ytblyf7V00CGV29ZFxtlzxksN2BgMK+MH7JnxP+FLy6ha2Z8SaCgLDUtPhbdEv/AE2h5aPpnILLgj5s5A8BfEvxz8MNWOs+BvEd1pk77RMkZDRzgZwJIzlXAycZHGTjFfBz9uXwj4pFvoXxRtk8OansCjUUYtZztxkt/FCTnody8ElhwKtbq1v7WG9sbmK4t7iNZIZonDpIjDIZSOCCCCCOtUQDRUEcGiCBVVVVVSPHFG0srqiIpLMxwAB1JPpWpfFf4WaM6Rav8S/Cti8i5RbnWbeIsPUBnGRWp/tb/s6aT5f2r4oWMnm52/ZrW4ucYxnPlxtt698Z5x0Nat+3t8BdPj32Z8R6mfM27LXTlU45+b966DHA9+Rx1xq//BRjwbCs39g/DfWbwqR5AvLyO23jIzu2iTbxnpu7euRqf/BRrxPNK50b4Y6XbRlMItzqMk5DepKomR7YH1rU/wBvz46X/l/ZbTwvp3l53fZtPkbfnHXzJX6Y7Y6nOeMap+2L+0bqqTwv8RXtoZnyI7XTraIxjOQFcR7wBwPvZI6k5Naj+0B8cdUnW4ufi34rR1TaBb6rLbrjJP3Y2UE89cZ6elX/AIm8Sarbmz1PxDqV5ASCYri7eRcjocEkVVl4A8d6nLDBpvgrXruSfHkpBpsrmTPTaApz+Fab+zx8c9W8z7L8JvFCeVjd9p02S2znPTzAu7p2zjjPUVZfsc/tHX8UM8fw4eKOXGDPqVpGVHqymXcPpjNWH7BXx3vLgQXB8OWSYJ8241Fio9sRozfpWnf8E7viXLG51fxz4ZtnB+RbYTzgj3LImPyNWX/BOG9ktUfUfi9BBcHO+OHQjKi88YYzqTxj+Ef1q1/4J1eB0eE33xE1yZFK+csVtFGXH8W0ndtzzjIOPerL/gn58EbW5SefWfF94iZzBPfQBG4xyUhVuOvBHT0rTv2H/wBniymMtz4Z1DUFKFRFc6pMqg5HzDy2U54x1xz0rTv2R/2ddLd5LT4ZWbs4wRc3lzcD8BJIwH4Vpv7PPwM0rzBa/Cbwu/m43fadNjuMYz08wNt69sZ4z0Fab8LPhjo3mf2P8OfC9j52PM+zaPBFvxnGdqjOMn8zWm6HoujeZ/Y+kWVj52PM+zW6xb8ZxnaBnGT+Zqqqqr/gth4pa0+Hvwx8EiW4C6rrN9qBRQPLJtYY48tzncPtZxxjBbOOM/8ABMbw82p/HfV9ekgZodG8OTFZA4GyaWaJFBHUgp5vsMDPbPxx0yXWPhH4ktIGO6O1W4OFzxFIsrfoh+lRuI5EfGdrA4+leH9QjhZHDAjqKm8YK+mC0LjAHArxFqKMJJdw4HH1qGGW4lS3t4nllkYKiIpLMx4AAHUn0rwZpA8O+E9F8PBdv9madBa43bseXGq9e/TrUTDpmo3BAIPSonHABqOTtmlcY4P4UJTjFNKSOKeQY61LIMYzUrg9DXxD8baX8O/Buq+M9YV3tdLg3tGnDSMSFRB6FmZVz71488c+IfiN4nvPFfia+e4u7pvlUn5YIwfljQdlUHp9SeSTVpo89yodzsHYY5qXw/IqZSU57ZWpoJIHMcq4YV+yp8W73xBYz/DvxHfNPeadCJdNmlbLPAMBoie5TII6naSOAlTEYwDUxGMA1+1/4vXwT+zb4/1gTLHLcaRJYQk5zvuiLcbcEHI83IPbGeQDXwS8DH4m/GPwP8PTbNPH4i8QWVjOiq5xFJMiyMdnzBVQsxIxgAnIxmiARiiMcZqqqqqqIAOBVV+0n+1J8Kv2XfB7eJPiBqvmahdRv/ZOiWzA3epOpAIjU8KgLLudsBR6khT+09+2H8Xv2p/ETXvjHU/7N8PW8hOm+HLCVltLVezOCczS4xmRu5O0IpCj9lj9hD4y/tSvFrmjwxeHPBi3HlXHiLUYmKPg4cW0QwbhlwRgFUBGC6mv2df2N/gX+zLYIfAfhhLzXmUifxFqiJPfyZGGVZNo8pCOqRhQe+TzVUQM4FVVf8FM/wBhX+3INW/aZ+Emn/8AEwto/O8U6LbW3/H0gOGvYggz5gB3Sg8MoL5BVt/7O3x/8b/s2fE/TfiT4KuWc27CPUdOaYpDqVqSPMgkxnggZVsHawVgCRXwV+Mngj4+fDfSPib4A1JLnTtThBlgLAy2M4UGS3mUE7ZEJwR0PDAlWUmiARiiO2aqqqqojIwKqviTOJvECRKf9TbKpGe5JP8AIiqqq8DWwt/DFoCPml3OefVjj9MUw4+lVVVVVX/BYgf8ZNeGuf8AmRLT/wBLr6v+CNP/ACTL4if9h22/9EGqqqqiARiiO1VXxLlxBYQbfvO7Zz0wFH9aqqqvD0TTa7p6IRkXCNz7EE/yqmGR1qqquKYc4FV+1ZYWeo/sxfFm3vYEmjTwVq0yqw6PHaSOjfUMqn6ivhLdXNj8VfBl7Y3MtvcW/iGwkhmico8brcIQykcgggEEdKqqqqqq1SQQ6ZdzEZEcDtj6Kaqqqqqqqq+Hlwqahc2pfBliDAepU/z5/nVVVVVVVVVVVVVVX7bnwfi+CX7S3jHwlp1m1vo95cjU9IUtkfZrkeYFXvtRzJEM8/u+/U/8Eg/jO3xG/Zml+HupXiSap8OdSaxVN4L/AGGfMtu7AYIG7z4xnPEPXqBVVVVVVVVVVVVVVVVVf8Fe/wBmiH4k/B+3+O/hrS0fxJ8Pl26i8cf7y50l2y4JHJEMjeYAeFV5j3r9kb483P7OXx38PfER5pf7GaT7Dr0MYJMunzECX5RyxQhJVXu8SjpmtL1PTtb0yz1nSLyK7sb+BLi1uIm3JLE6hkdT3BUgg+9VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVSgE9aPPeqqqA4AzVf8Fefg1L4e+KHh7426ZbILDxbZfYNRZIsFb22ACM7dCZICiqOuLdu2K+Gk8vjj4O+PfhHJdJ9p0pV8Y6FCVO+SazjaO/iTAJw1lI9w2egsAcj5g37KXxfl+BX7QPgv4jvcNHp9lqCwaqAeGsph5U+RkBiqOXUHjcintmopYriJJ4JUkjkUMjochgehBHUYqqqqAycVVV478D+F/iX4N1jwD400qLUtE120e1vbaQfeRh1U9VZThlYYKsAwIIBr9sT9jvxn+yn4z8mUz6v4K1aZv7D1zy/vd/s8+OEnUD2Dgbl/iVP2Rv20PEX7PF2PCfiSG41zwNeTBntFk/e6Y7MN8tvnggjJaI4DHkFSWLeDfG/hX4geHbPxZ4L1+z1jSL9N9vd2kodW9Qe6sDwVOCpBBAIIpZTjg5pZgRjNCUdmoS5GAc0ZTjqKMueN34U0ox96mmzwDTyg9Wp5cjANfG1Wn+G2qhHfKGFiq/xDzUyD7c5/AVYOkd5C74wHGc9q0OWJJFLYq91TTn00RKi7wOta1JEsM8pIxtPX0xVaIstvothbzIUkitY0dT1UhQCKEmehrzBjrW8EVvFFxjijICODV/qNpptlcajfXCw21rE0s0jdERRlmPsACar4ZQywfDbwnBPG0ckeh2aujqVKkQICCOxqgMnFVVVVVSgGqqsZ4FYGMA1VUozzVVVVVUozVVVVVUBk4rxB4W8OeLLP+z/EmiWWpQDO1LmEPtzjJUnlTwORivGP7H/gnVUe48G6rd6HcY+WGUm5gPXj5jvGeBncceleM/2cPin4O3zf2H/bNon/AC8aXmfjjqmA468/LgYPOOatL3VtB1AXFheXenX1s5AeGRoZYmB55GCCCK8G/tWfEvw2YbbXJLbxDZxkBlu18ubbjoJV7+7q55/LwX+1J8MfFPk2urXkvh69kbb5d8P3WfUTD5QPd9v8s2d7Zajax3un3kN1byjMc0MgdHHTII4NUBnmgMCqqqqqrGeBQAAwDVVVVVVQGeaqvFfgzwp460l9D8X6BZatZP8A8srmINtP95T1VvdSCPWvi9+wheQvLrXwb1FZ4sFm0XUJtrg+kMx4PYbZMYwSXOcVrGi+IvButyaXrWn3uk6rYS/NFKjRSROp4Yd+oyCPqDXwj/bb8c+DIrfRPH9u/inS4/lF08uL2MevmHIl+j8n+9Xw8+LXw8+Kth9u8EeJrW/ZE3TWpPl3EHb54mwwGeN2Np7E1VVjPAoKAPeqqgM8CgABxVVVV4n+Gfw38bCYeM/h94a177TGI5v7T0mC68xB0VvMU5HseK8RfsLfsf8Ailp21L9nnwbCbiEwuNOsP7PAUjGVFuU2Ng/eXDZ5znmtc/4JSfsVatLFJYfD7V9GWNSGjsfEF24kPqfOeQ5+hArUP+CM/wCyre3kt1beLPiZYxyNlLa31azKRD0UyWjNj6sT70P+CJfwtP8AzW7xV/4LoP8AGpv+CHcLSubf9p1kjLHYr+C9zKOwJF6MnHfA+gr/AIcdf9XQf+WT/wDd1ah/wQ9v47KV9K/aXt7i7Vf3MVx4QMKM3ozreOVHuFP0rwP/AMEivjr8P9T/ALQ0L48eFTG/E9pLp9yYpx/tKCOR2I5H0JB+MvwF8d/BLU47bxLapc6dc4FrqlqC1vM23JTJGVcc/KwBOCRkc18SPgn4b+IUyahHN/ZOqbgJLuGAOJVyMl0yN7BQcHIPQE4Ax8Of+COel/EbwjZeKtK/as0ydLoHeth4Ua5SFuuxma7jYOFKllZFKk49zon/AARD8IW8Mi+JP2hNXv5Sw8prHw9HaBRjkEPPLk++R9K8Nf8ABGz9ljR/Ll13xF4/16X7OElS41OCCEycZkRYoFdeQcKXYANzkgGvBH7BH7Hfw9nW68OfAHw1LKkhkR9WWXVijEAZH2t5cY2gjHQ5IwSTVhp2n6TZQ6bpVjb2dpbrtht7eIRpGPRVXAA9hVVVVVVVfGX9kT4a/FQXOsaXB/wjfiOX5vt1mmYpm/6aw5Ctnn5l2tk5JOMV8WvgH8SPg3eMvirRjJprSbLfVrQGS1mJHA3YyjcH5XCk4JAI5r4QftE/Ej4M3CQeHtTW80YyFptIvcvA+epXBzG3fKkc4yGHFfBr9pv4bfGWNLDT706TrwHz6TfMFkbpzE/3ZR7DDcZKgYzVVrvjvwL4YMi+JfGehaS0RAkF9qMVvsLDIB3sMZBBH1rWv2rf2eNCcx3vxS0yUiRo/wDQ4prsZHU5hRxj0boexrVv2/PgRpzqlnB4n1UMWBa009FC4xgnzZEOD2x6c4rVv+Cj2lx4XQ/hRdXGUOXu9XWHa3b5VifcPxFax/wUO+LN3uj0Twn4WsEaIrulimnkRzn5lPmKvHGAVPI5yDitY/bU/aL1bckPjaDToniMbR2emW65znLBmRnVsHGQwxgYwea1j49/GzXty6l8VvFDo8RieKLU5YY3Q5yGRCFOckHI5HHStR1XVNYnW51bU7q+mVNiyXMzSMFBJwCxJxknj3rTdG1fWXeLR9KvL541y620DSlR6kKDgVo/wE+NmvbDpnwp8UOjxCVJZdLlhjdDjBV3AU5yCADyOelaR+xj+0ZqxgZvAS2MMyk+beajbR7OD95BIXGcYxt7jtzWk/8ABPn41XyxSajrXhXTkaTEiSXk0kiLnlgEiKnjoNw98Vp3/BOHV5Y3Or/FmztnB+QW2jNOCPctKmPpg1pf/BOv4cwuDrfj3xHdp5eNtrHDbnfxzllk468Y7jnjnS/2DvgFp6QJd2uvamYXy7XWpFTMM5w3lKgAxx8uDjvnmtN/ZJ/Z10rzPsvwwsZPNxu+03VxcYxnp5kjbevbGeM9BWmfBf4QaK8EulfC7wnbTWybYpk0eDzVG3b9/buJI4JJycnPWtO0bSNGR4tI0qzsUkOXW2gWIMfUhQM1VVVVVVTDIyDVVVVVV/wWwv7yX4hfDHSnuWa0t9GvpooT0SSSaNXYe5EaA/7or/glPYWcmo/ErVHizdW8OmQxSbj8qSG6LjGccmNOSMjHuaEcU8TQTRrJG6lXR1yGB4II7g18Yfhjf/DXxTPZrbyNpF27PptwRkNH12E8/MmdpzyeDjBFWeoz2eFU7k/unjH0pvEZMe1Ucn0JxV5ezXj7pG4HRR2r9lX4SXXjLxjB411W0xoegTCRTIpAubkcoq8YIQ4ZvT5R/FUbAYINRt0INRScDBpHxyDSS9s0svGAaEwxgGmmBGM08oIxmpJAeAakcY61+3XrN7bfD7RNGgdVgv8AVd0/HLeXGSo+mWyfcD8dOgW4u0RhkLyRivD+kLeMqgDn2rWfCRsrYSPHjK5GRXiOyVELAfNGeOK+DGqXGj/Ffwpd2p+d9Uhtzzj5Zm8pv/HXNStk9alOTjPSv+Cpfjj+zfhv4T+H9vPtl13VZLyZVPJito8YPsXnQ/VPY1/wSg+G/wDwnH7WVh4kubTzbPwRpF1qjmSPdH5zKLaJTlSN+ZzIoypzEWB+XBph3FVVVVVTDI4qv2xf22Ph9+yp4WntBcWuteP723DaR4eEhyN24LcXGOUhBU8cM5G1ccsvxJ+JvxD+NnjW68a/ELxHf+INe1GTaHmYvsUsSsMKDiOMFjtjQBRngc1+xN/wS3+0xaf8Vf2ntKli2yebp/gyaPaTtZSkl6c9Dhv9HxyCN5+9HVhp9hpNhbaVpVlb2dlZQrDbW1vEI44Y0AVURVwFUAAAAYAAqqph3Bqqqv8AgpZ+xEnwd124+O3wv0zZ4J167J1exiwF0a9lfIKKMbbeRj8oAwjZXhWRR+wt+11qv7LXxPiXWLuefwF4imjh8QWQJYQc7VvEUKzb4gSSqjLrleu0rpWqadrml2etaPexXdjf26XFrcRNuSaJ1DI6nuCpBB96phzxVVVVVUQM4BqvFN2L/wAQ39wrbh5xVT6hflH6Cqqq020Fjp1rZD/lhCqfkAKYZ4FVVVVVX/BYj/k5rwz/ANiJaf8ApdfV/wAEaf8AkmXxE/7Dtt/6INVVVVUwGcCqr4kTB9VtoAB8lvknPqx/w/WqqqrwnG0viKxRMZDk8+ykn9BVEZGBVVVUwHY1X7UH/JtHxb/7EXWv/SGavhf/AMlM8I/9h2y/9HpVVVVVVWuf8gW//wCvWX/0E1VVVVVVVVpGoyaTqMF9Hk+W3zqP4lPBH5Va3MF7bR3VtIHjlXcpFVVVVVVVVVVVVVX/AAWD+FC6n4K8G/GfT7ZPtGiXr6RqLjO5oJwZIWI6bUkjkHY5nHXt/wAElPitL8Pf2ttN8KTzIunePtNuNJn8wnCTKv2iBhg/eLw+WOv+uP1FVVVVVVVVVVVVVVVVVqOnafrGn3Ok6tY297Y3sLwXNrcxCSOeNwVdHRshlZSQQRgg4Nftp/s3al+y78fdf+Hv2WUeHrqQ6h4auXbf5+nysfLBbOS8ZDRMTgloy2MMpP8AwSq/aP8A+FhfDa5+BPiO63a54FgEumPJJlrrTWcjAHX9w7Ih7bZIgOhqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqUccGqqqxngGsDGBVft0/Br/hd/wCzN4t8N2dn9p1jSLc6xo6Km5zc2ys2xBgnc8ZliGMf6zGQCa+F/jaX4b/ELw/43jtEvI9Jvo5bqzkzsvbYnbPbuARlJYmkjYZGVcjPNfHP4Xt8IfiXqng+2vX1DR2WK/0DUiARqOl3MazWdyCvykvDIhbaSFfcuflNf8E0/j3B8Zf2drDwzqV3v8SfD3y9Jv0ZyzSW+D9km5yQGjUx8nloHPAIqqqqUZNVVV408E+EviL4Zv8Awb448P2etaNqULRXNndx7ldSMZHdWGeGUhlOCCCM1+2h+wF42/Zsvrjxn4NjvPEvw6uJmKXyQl59IBPyx3YUYC8hVmGFY8EIWUH9nz9pn4kfs6eIRqXhO9+26PcvnUdCu5W+zXQOAWAB/dy4UASAZGACGXKn4CftQfC39oTSftHg/V/s2rwIDeaLesI7qA45IGf3iZ/jXI9cHilm45NCYEYBrzhjG6vOH96jMCMZzTTDsaaYkdaeUeuaebPANa7p1vrukXmj3R/d3kLRMcfdyOv4Hmta0i+0HVLnRtSi2XNpIUcc4PoR7EYIPoRWleIhDEIbhirqMK/r9afxFD5eDdx9OgbNazrRvh9mtyfKHViMFv8A61fCrwZN408X2lm8DPYWria9cplQg5Cnt8xG3H1PY0HB6Gt3YGt5PQ1vNbzW80XPrXxn1iLQPg94612aJpo9N8NajctGpwXEdtIxAPqcYrRdJvNf1iw0LTlRrvUbmO2gDttBd2Crk9hkiqpRxVVVVVUBxVVQGTVVVAYGKqqqqqgMDFVVVVVQHFVVeLPhv4F8cpt8VeF7G/kAwJmTZMo9BIuHA56ZxXjL9jWzm3XHgHxO0DlifsuqDcgB9JEGQB0wVY+/HPjT4RfELwCS/iPw3cx2wOBdwjzYTxn765C/RsHg+leF/HPjDwVcrd+FfEd9pzK+8pFKfLc/7cZyr9BwwPQV4J/bG1O1VLP4geHkvUGB9s07EcmAOrRsdrEn0ZAPSvBPxc+HfxACx+G/EttJdYybOY+TOO3CNgn6jI5HrVVVVVVQHGaqqqqqqAzWBjAqqqvHnwy8CfE3TP7K8beG7TU41BEUjrtlhz3jkXDJ+B5718X/ANh7xf4UEmr/AAxupvE2mjLNZSqqXkAHTGMLN/wEKewU9as73xJ4L15LuxuNQ0XWdNlO10L289u4GCOzKcEgj0JBr4Qft2atpnl6L8X9PfU7fhU1axjVZ07fvYuFcdOV2kAHhia8I+NfCfj7Rotf8Ha/aarYygESQPkoSM7XU4ZG9VYAjuKxngUAAOKqqC571gYwKqqqqrGeBQXPegABgVVVVeIvDeg+LtGuvD3iXSrfUdOvIyk1vOm5WBGMjuD6EYIPIINftG/sk658LHn8W+CUudY8KszNIgQyT6aoGT5uB80fDfvOwADY4J+EPxp8bfBfXxq/hW+32szL9u02YkwXigEAMB0YZOGHIPqMg/Bz42+CvjZ4eGreGbvyr62RRqOmSn99ZuR0PA3IcHDjg+xBUVVVVVVVVVqOq6Xo8C3Orana2ULPsWS5mWJS2CcAsQM4B49q8UftFfs62NlPY+IPiZ4Yv7WWDM0EEo1BJEJwVKxBw3T7uM+3NfHub9ma5uZdY+Ces6wlzJOFk002Li0KkfNJG8pV4xkfdIbk4AUDhdRaJ1kgDK6HKsGwVI6EYrQf23/j3oOgQaAutadqBtyPLvtQtPOuSgGNjPuAcdyzAsSPvda1r9rf9onXfluvibewKN21bK3htcBuozGik4wMEkkdjya1v4h+P/EysviPx14g1UNH5bC+1OafKZztO9jxkk46c1a2l1f3EdnY20txPKcRxRIXZj6ADk1ofwN+MniRY5NF+F3ie4ilVjHOdLljibacHEjAL1BGM9RWifsWftFa0qySeCItOiePesl9qMCZ5A2lFdnU9+VHT6Z0b/gnf8VruMSa34u8MafujDLHE81w6seqsNiqMeoLDP51pP8AwTh0eONjrvxVvLh2Vdi2mkrCEP8AFktI+4dMcL09+NN/YB+BVh5n2q78Uajvxt+06hGvl4znHlxJ1z3z0GMc1pv7JH7Omk+Z9k+GFjJ5uN32m6uLnGM9PMkbb17YzxnoK034V/C/RfM/sb4b+FrHzseZ9m0eCLfjOM7UGcZP5mkjjijWKJFREXCqowAB0AHpVVRAIxRXjg1VUQCMURjiqqqqqqqYelVVVVV/wWt/5Kj8N/8AsAXX/o8V/wAEpuD8UMH/AKBH/t7UTc4BrxF4Y0Hxjo8uheI9OivbObqjjBRh0ZSOVYZPI5rxd+yFq8Ust14H8Q211CXJS0v1MTovoJFyGPTqF/xH7KXxcYcWel/+Bo/wrwH+xtbwXCXnxE8QR3Soc/YtMLKj8/xSsA2MY4Cg+9aPpemaJp0Gk6NYW9lZ2y7Ybe3jEaIM5OAOBySfqc1E4Ixmo3xxmkfHQ1HLwMGlkB6GhKQOtCbjGaM2eM00hI4P508gHQ5NSSg96/bl0m6vfh5ousQjdFp+q7JlxyBJGwDfTKgf8CFadOtvdI7HAPBrw9q62jK24ZHrWteLTfWwjeTO1cDNeJL1XQxq3zSHj6d6+CmjXOufFbwvaWoO6DUYrpyFzhYT5rdx2TH49+lSMMdalbjrX/BQ34jR+PP2i9Q0iyufMsfCFnFpUe1m2mYZkmOCcBg8hjJAGfKHXANf8EaPhaNE+FnjP4uXkKfaPFGqx6dZlgjFbe0UlmUj5lDyTsCpPPkqcdCaIBGM0R2qqqqqiARjNftlftk+Cv2TfBXmzfZ9X8b6vC39haF5mN3UfaJ8cpApHsXI2rjDMnijxL41+Lvjy98S69dX/iDxP4nv9zkK001zPIwVI0UZJ/hREHQBVAwAK/YM/wCCdWhfBiw0z4t/GrSLfUviIzLc6fYSN5kPh/g7RgHbJcc5LnIRgNnK72ph6VVURkYFVVV4p8MaB428Nap4P8U6ZFqOj61aS2d9aS5CzQyKVdSQQRkE8ggjqCDX7ZH7Luvfsr/Fq68JzmW78NaruuvDmpOoH2m2yMowBOJImbY2cE4DYAcV/wAEn/2tmVk/ZY8dXrPuaW48IXLgYQYaSezZs9OGkj4JyZFJxsUUwyOKqqqqqmHpV9crZWNxeMMiCJpCP90Z/pTszsXdizE5JJySaqq8PWhvdcsLYDIadSw9gcn9AaphkdaIGcA1VVVVX/BY7RrqD4++DvEDyRG3vfB6WsaAneHhu7lmJGMYInTHPY9OM/8ABGLWYrjwn8UfDqwsJLHUdNumkJ4YTRzoAB6jyD/30Kqqqqph6VVeOJhN4kuVUgiJUQEHP8IJ/U4qqqq8DxNJ4ktmUj92rsc/7pH9RVURziqqqYZHBqv2oP8Ak2j4t/8AYi61/wCkM1fC/wD5KZ4R/wCw7Zf+j0qqqqqqrxI7R6DfMjYPksOnY8H9Kqqqqqqqqrw74ludClMbKZrVz88WcYPqvv8Az/WtP1Kx1W3FxY3CyL3HQqfQjtVVVVVVVVVVVVVftM/DIfGP4A+O/hxFb+fd6vo032CMttBvIgJbbJwePOjizgZxnHNeDfFWr+BPF+heOPD0qR6p4d1K31GxdwSFngkWSMkAg43KOhFeDfFWj+PPB+heOPD0rSaX4h0231GxdwAzQTxrJGSASMlWHQmqqqqqqqqqqqqqqqqqr/gqJ+y//wAL++ANx4y8OWm/xf8ADiOfU7FUj3PeWm0G6txjnJRBIoAJLxBQPnJHwI+L/iD4D/Ffw78T/Dk0vm6Pdq11bpIVF5bE4mgb2dMjnocHqBXgrxj4c+IfhDR/HPhHUUvtG12yjvLK4UEb43UMMqeVYZwVOCpBBAINVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVUB8tVVUACcVVAZIGa/bK+DX/Cif2jfGHgW0s/s+ktd/b9HCptQ2dwPMjVOAMIWaLgYzGR2zXiLw4Pjp+xbonxT0xftPir4G3g8O+IwDull0O5kaTT535JxDK0tuOB8mOyE1+wj+0J/wzt+0Fo2u6tf+R4X8QY0nxBvbCR28rDZOew8qQI5OCdgkUfeqqqqUcVVVVahp9hq1hc6Xqllb3lneQvDcW1xEJI5o2BVkdTkMpBIIPBBr9r7/glel+958Q/2YLWKK5eR5r3wjNKI0fcSSbJ2wqYz/qXIXGdrDCoY5PGXw28WblGreGvEug3RBV1ktLqynQ4IIOGRgcgg4PY1+z9/wUw1HT1tvDHx/wBPa+gVdieI9PhAmXnjz4BgOME/PHggKPkYktXgf4j+CPiVocfiPwH4p0/XNOk4860mD7D/AHXX7yN/ssAR6UJgelGYEYyKMwximnzwDTTEjrTSZ4BzTMW4zVfGHw74U1OytrvVb5NO1FmMdtc+UX3ADO1wOdnv1Un3INn4/wDCF5qMmjNrKWWoxn/jy1CN7OdhkgMscwVmU4yCB3HrUUMtxIsMETSSMcKiKSSfYCvCXwe8Z+KZ4zJp0mmWTAFrq7jKcdtqHDNn249xXgnwXongXSF0vR4yWc7ri4cDfO3qT6DsOg/EkhzjAOaEhxjNeYMYBoSDHBrzfejICOtGTtmv20PEX/CN/st/EXUPtMkPnaR9i3RjJP2mRINv0bzdp9ia/Zi8Or4s/aO+F/h6Wwa8t7zxfpi3UCsVLQC5jaXkEEARhySCCAOKqgOMVVVVVWM8A1gYwKqqA4qqoDJxVVVVVUACcVVVVVVAZOM1gYwKqqqq8afs9fC3xqjyzeH49LvWLH7VpoEDFj1LKBsbnnJXPvya8b/sjeONDE154QvbfX7RMssPEFwF9NpO1iOejZOOBzitV0fWvDmoGw1jTrvTr2E5MVxE0Trg9QDg9R1rwN+0l8UPBEcNidWTWtPi4FtqQMpVeOFkBDjgYAJKj0rwN+1b8OfFPl2viAy+G71s/LdN5kBPtKAMcf3go/SrW7tb+2jvLG5iuIJl3RyxOHVwe4I4IqqqqAzxWBjAqqqqqqAzwKwMYFVVVVV8UvgR8M/jBb48X6Cv29I9kOp2h8m6iHYBwCGAycK4ZRknGa+MH7HfxH+HC3Os+HIj4n0GIlvNtIybmBM9ZIRycDqyZGASdorwd448W/D7WovEHgzXrnS7+LgSQkEOPR0OVdf9lgRXwc/bo0LWRbaD8XLNNIvW+QavaozW0h7GROWjJ4BI3Lk5OwdNM1TTNb0+DVdG1G1v7K5TfBc20yyxyr6qykgj3FVQXPWqqqqqoLkdaAAGBVVVVVVJHHLG0UiK6OCGVhkEHqCK/aS/YyFwbrx18GNMUSEmS88PQqqLjHzPbDgDpnyu+Tt7IfCPjHxX8OvEUHiLwpq11pep2jYLRsV3AEExyL0ZCVGUYEHHIr9n39pnwt8bdPXTbz7Po3iqAYn0xpsi4wuTJBnllwCSvLJ3yMMaqqqr2+sdNtXvdRvYLW3jxvmnkEaLkgDJPAySB+NeIP2kfgP4ZVjqnxV8PMY1JZLO6F4wwcEbYd5z7YzXiH9v34G6T5kej2/iLXJB5gja2sVijYr90kyujBW9dpIGcgdD4i/4KPXTh4fCfwuijII8u41HUy+Rt5zEiDHzf7Z4HbPGvft3/tAawCNO1LRNDzjH2DTFfGOuPPMnXv8Apitd/aB+N/iPeuq/FXxKySIySRQag9vG6t1BSMqpBzjBHTjpV7fXup3L3uo3s91cSY3zTyF3bAAGSck8AD8K0P4Y/EfxPsPhzwB4i1NZEV1a00uaVdrfdbKqQFORz0rQ/wBjb9orXDGy+AGsIXLAy319BDswO6by+D0GFPX05rQ/+Cd3xTvNj+IPGPhnTUdFJWBprmRCeqkbEXI9mIJ745r4z/sYeP8A4UWja7pN4fFWhxIDPdWlqYprc9y8O5yFH94Ej121aQ2ltcwzzWkd2kTqzQTMwSUA5KsUKtg9Dgg88Eda/Zw8XfsqePFttAvfhV4V8P8AitxGq21/aJcxXUinANtLNuIYkg7CQ5JABfaWrTtJ0vR4Da6RptrZQs5do7eFYlLYAJwoAzgAZ9hVVRAIxRX0qqqqqqZc9KI7VVEAjFEY4qqqqqqogEYojHFVVVVf8FsdLvofHvwv1qSDFndaRf28Mu4fNJFNEzrjORgTRnJGDu4zg4/4JWapbw6v8R9EZJDPdW2nXCMANoWJrhWBOc5zMuOOx6d424HPSomBGAaibIxmtW+Ifgnw7I0GseJrGCaMNuiEm91x1yq5IPtjmtP+NfwvvZPKh8XW6sMf66GWEfm6gVp9/Z6hbRXun3cNzbzLujmhkDo49QRwRSP0INI4IxmkkGME0r8cGllGMZpZuODXmgjrRlGODTTAjrTy5GAad89TXjfwtpXjzwrqfhDW0LWepwGJ2X70bdVdc8blYKwzxkCvHvgbXfh14ovPCniCDbc2jfJIqkJPGfuyISBlT6+uR1Bq01a4tVCH51HQE4IqbxBK6bY4znH8RqaWSdzJK+5j61+zB8Jbjw3p7fEDxBb+Xf6nb7LCFhzDA2CXII4Z8DHov+8RUzAjANfEfxtpfw58Da7461g5s9CsJbuRN2DJsUkIDg8scKOOprU9Q1vxp4mu9VuxPqGsa7fPNIEVpJLi4mcscDkszM3Tkkmv2afhSnwP+Angb4WGLZc6FpEa3w3Kw+2SZluSCvGDPJKR14I5PWqphzxVVVVVftFfHvwZ+zX8KtU+KPjSTzIrTEGn2CSBJNRvHBMVvGTnBbaxJwdqK7YIU18Z/jF46+PPxE1T4lfELWJb7U9RkIjRm/d2cAJ8uCJeixoDgAdSSxyzEn/gm7+wj/wqDS7f45/F7R7WTxpqtuj6HYSpvbQ7d1OXbnaLiRWGeMxr8uQXdRVMARgVVVRHOBVVVftffs2aJ+1D8GtS8C3XlW+uWmbzQL9kBNteIp2qSeRHIPkcehz1UVdWvjL4W+OGtrlL/wAPeKvCupjjJiuLC8gkyCCOVZHUEEdwCK/ZA/aK0z9pz4H6L8QY5LePXYF+w+IrOJSotr+MDzNqnOEcFZUGThZACSytggEYzRHaqqqqqYZHFeP74WfhuaIMA9y6xL+eT+gP51VVXw5tPtHiETleLaFnBweCfl/kxqiMjANMPeqqqqq/4LMf8lN+HX/YBuf/AEeK/wCCLP8AzWP/ALgX/t/VVVVVMAR1qq16cXWtX04YENcPtOMcA4H6AVVVVfD6LzNeZt2PLt2bGOvIH9aqmHPFVVUQCMCq/ag/5No+Lf8A2Iutf+kM1fC//kpnhH/sO2X/AKPSqqqqqqvGUgj8OXQD7S2xRg4z8wyPyzVVVVVVVVVVZ313p8wuLK4eKRe6nr9R3Hsa0jx9FIBDrEIjIH+ujBIP1XqPw/Kra6tryET2k6TRnoyNkVVVVVVVVVVVVV+2V8OV+Ff7TvxD8IQQtHaDV3vbNWJbEF0ouI1DHJYKsoXJJOVOTnNf8EqviXL8Rv2NvDVleXTXF74MvrrQbiR85xEwlhXG0DC29xAgxu4UEnJIFVVVVVVVVVVVVVVVVVX/AAUo/Zgl/Zx/aCvb/RLFIvBvjxptV0IxqQlu+4fabXGBgxO6kAZAjli5zkD/AIJL/tIqjX/7NPivVmJkaTUPCiyuTg4Z7q2T0GAZgvTPnHqaqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqrAxgGqqqUA96qlGehr/gsD8Fm1jwf4X+PGk2iNceH5f7I1dwAG+yzMWgY8ZKpKXXGes446kf8ABNf4haBoXx8m+EPjwJceDfjFpM3hbVbSV8RvLMP9HbqBvL5iU8keecDJBH7QXwa1/wDZ++Mfin4R+Ig7z+H75o4Lhk2i6tmAeCYD0eNkbHYkjtX/AATc/aE/4Xj+z5ZaFrV/5/ijwD5ek6lvbLzW4U/ZJz3+aNTGSTlngkY9RVVVADAANVVVVV+0P+yR8Fv2m9I+yfEHw75OsQRlbLXtPIhvbXpwHwRInGNkgZRkkAHDD9o//gmz8evgbPea34V0yXx/4Rhy66jpFuWurdMZPn2oJdcYYl03oFGWZc7R4M8e+NPh3q6694F8U6lod+F2meyuGiLr/dYDhl9jkV8Kf+CnPjvRGh074u+F7XxJahdr6hpwW0ugcD5jH/qnOQeAIxznPGD4B/bh/Zs+ICQxQ/EK20C8kTc1rr6GwMXGSGlb9zngjAkPP1GbDVLDVrSO/wBKv7e8tZc+XNbyiRGwSDhlJBwQR9RRYkYzR571Va5rmi+GdIutf8Q6ra6bptjEZbm7upRHHCg6lmPAFeMviFpnxJurXXfD80kujGD/AEGV49nnoTkSgHna42kZwdu3IByK8V/CXV/2l/ivoXwqsImt/DvhxRqfibVhHu8hX4jtkPGJXVWOPRg+CEIPhzQtF8J6Bp3hfw7p8djpelWsdrZ20edsUSKFVQTknAA5PJ6k0GBHBoOfWhJ2zQkyOtCQEda8wHgVvHqKMgoyAjg1/wAFOPFf9jfs5w6FFcYk8Q6/bWzxLcbC0UayTsxT+NQ8UYI6AspzkDP/AATP8Jx+K/2yPBL3CRPb6JHe6jKjuyEmO1kEZXb1IleJsEgEKc56GgMnFVVVVVSgGqqsZ4BrAxgVVUo4qqqqqqUcdaqqqqqlHGaqqqqrGeAaUA8DtXiHwt4b8WWf9n+JdDstSgGdqXMIfbnGSpPKngcjFeOv2OvDmoRy3fgDWZdKueSlneEzQH/ZD8ug6cnf06c5rxz8HfiH8PHL+I/DswtMkLe2/wC+gbH+2v3c4PDbT7V4O+IvjXwBcm48JeIrqwDsGkhVt0UpHdo2ypPGM4zjvXgP9si0n8ux+IuhfZ3LYN/pwLIB6tExLDHcqTnPAFeFvGXhXxvpw1XwprtrqVvxuML/ADRk9A6HDIfZgDVVQGBiqqqqqrGeBQAAxVVVVVVWM8CvjB+yv8M/i2bnVZLRtD8QzqSNUsk+++ODLFkLIOmfusf7wr4tfs4/E74QXE02taOdQ0dD+71ewVpICD038ZjPYhgBnoWHNfC/43/Eb4Q36XXhDXXFpu3TabdFpLWf13R5GD/tKVb3r4NftdfDr4qSQ6Jqzf8ACNeIJMKlpdygxXDd/KlwAT/ssFJyMbuaC+tVVVVVjPAoKB3qqqqqqqqr9pL9kvR/ijFP4w8CRWuleK40d5YVjCQ6qeuHIwElJJxIeucNxhlePxL4G8RhZEvtF1zR7gEBg0M9tKpyD2KkcGv2av2ttM+JMdt4L+Il3b2HixpPLtZwnlw6lnOMY4STjG04DEjbydo8QeNfBvhJQ3irxdoujAqCDqF/FbZBJAPzsOCQR+BrxJ+2L+zx4bYwv4+j1Kcf8s9NtZbjIyRneF2dum7OMHuK17/go14Jtwf+EX+HGt6hjG37fdxWefX7nm//AF/ate/4KE/GHUQYtD0DwzpMZxhvs8s8oI68s+3B/wB3PvXiD9qP9oLxMoXUfirrMOFAH9nslh0JP/LuqevXvxnoK1XWdX126+3a3q15qFztx513O0r4yTjcxJxkk/UmvDvw5+IPjBFl8KeBte1iNukljpssydduSyqQBkEZPAxXh/8AYy/aH8QKZD4HXTYgpxJqF7DDkggY2Bi/fOSuODzWg/8ABObx5cuo8T/ETQNPXeQTYW814Qu3ggOIuSeCOw5yeleH/wDgnZ8M7Ly38S+NPEWqSJsJW2WK0jcj74IKu21uOAwIGeT1Gg/sc/s7aCi7Ph/HfzBCrTX95POWBOful9gI4GQoOPqc6D8P/AfhV1k8L+CdA0h1curWGmxW5DFdpI2KOSvBPpxVVVfHb9jLwl8QVufEnw+Fv4e8QkM7QKm20vHJz8yj/VsefmUY9QeteMvA/jD4beIJfD/i3R7nS9RtmyocYD46PG44ZfRlJFfAP9tXWvCP2bwp8WHutZ0ZdscGqL891ZryPnHWZOnOd4AON/C1oPiHQvFWk2+u+HNWtdSsLpA0NxbSB1YfUdD6g8jvVVVFQe9EY4qqqqqmUEcdaI7VVEAjFEY4zVVVVVVRAIxmiO1VVVX/AAWu8KNefDP4a+OBbMy6Rrt3pxmEgAQ3UCybSuckn7GTnGBtOeoz/wAE0fEraR8fr3QZJW8rXfD9xEse4AGWN45VbHfCJIMD+8TSnB60kiopZmACjJJOMV8UPjNqOrXdxoHhS8a202ImOS5ibD3J6HDDonpjk9e+KRHkYJGhZj0AGTUltcRLult5EHqyEV4Q8ceJfA2oDUfD2otCTxJC/wA0Uo9GXofr1HYivht8QtL+Inh9NXsQIbmM7Lu1LgtC/wD8SeoPGfqDSOAODSSAjGaWTA4OaWUHvQk7A0JMjGaMmeAaaQetNKMYBp5Mjk1JIMYBr4hfDbwd8TdKGl+LdLW4EZzBcRny5oGx1Rxz36HIPGQcV4m/Ys8QRXhbwd4v065tWJITU0eF4x6ZjVw3bnC/StM/ZB+J12//ABMtQ0PT4w2GL3DyMR6qFUg/iRXi79m+D4ZeALnxno+vXWo+JNFmiu1l8lEhRFb5tsbbslQd2ST9wYA5B8C/tcusaWHxE0hpGGFF/p6gE+7xEge5Kn6LXh7xh4a8ZaeNU8Ma1bahb8bjE3zRk9A6nDIfZgDX/BT34uw6P4J0X4N6be/6d4hmF/qUSkgraQt+7DDPR5hkcHmA9MCv+CavwRT4zftS+H7nU7NptD8EL/b9+cEK0kDL9mTOCMm4aJih+8kcg9aqqYAjiqqqqtQ1Cw0mwudV1W+t7OysoXmubm4kEccMaAlndmICqACSTwAM1+3V+1lqn7U/xbuLzS7uWPwL4ckktfDVmQy748gPdurAHzJiobBAKoETqpZv+CW/7F3/AAmer237SnxO0bdoWkXBHhewuoflv7peDdMrrhooyfkIPMqE8eX81VVEdqqqYAjiqqqr/grt+zQuka1pn7S3hDSWFrqzLp3ikQoSsdyqgW1ywGdodAYmbhQ0cXVpDn/gmB+0R/wpv4+Q+Btdv/K8M/ETy9On3thIL4E/ZZfbLM0R/wCuoJ+7VMBniqqqqqIBGK+J9/5l9a6YjcQRmRxju3A/QfrVVVfC60Agvr0j7zrGpK9MAk8/iP0qqYcYB6VVVVVX/BZj/kpvw6/7ANz/AOjxX/BFn/msf/cC/wDb+qqqqqIyMZqppFhheZgSI1LEDrwKd2kcu7ZZiSSe5qqqq+HEaNqV1KfvLBtH0LDP8hVUwyOtVVVweM0R2zX7Z2q3ejfsp/FO7sWQSSeGbq2bcufkmXyn/HY7fQ1+z7o9r4i+PXw18P3skqW+p+L9KtZWiIDqkl3ErFSQQDgnGQaqqqqqq+IE4j0aKAMMyzjIx2AJP64qqqqqqqqqqqrO/vdPk82yupIX7lGxn6jv+NaX8QHBWHV7YMMY82IYP4r/AIflVhqmn6pF5tjdJKAOQDgr9R1FVVVVVVVVVVV/wWE8CppHxf8ABfxBgt/LTxJoUlnIwIxJNaS5LYznOy5iHIAwoxnnH/BEDx9HHqvxP+FlxLDvuLey1m0TcofEbPDOcdWH7y35HCnr94VVVVVVVVVVVVVVVVVVV+2/+zXD+1L+z/rXw/sY7dfElky6j4cnmO0R3sQOELfwrIjPGT0G8N/CK8NeIvFfwy8aWHiXQrm60fxD4cv1mhZlaOS3niblWU4I5BVlPUZB6mv2ffjX4a/aE+E2hfFHwzIiJqUOy9tN4L2N2nyzQuAcghuRnG5GRujCqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqoDJABojtmqqqUcdaqlAxXxw+GGmfGj4ReLfhdqkUDx+ItLlt4WnB2wXGN0E3GTmOVY5BweUHB6Vc2+v+CPFEtpcJLput6BflHXI321zDJgjIyMq6/TIr/goH8ObL9qr9lP4fft4+CLK3TWLLRbeLxRZ2Y8xfs7Psl2sCSTa3ZkQ5GdjszFfLxX7E37RF5+zf8d9H8UXV80fhrWGXTfEUJGVNpIw/e4/vROFkBHOFZc4cgxSxXESTwSpJHIoZHQ5DA9CCOoxVVjPANVVVVVSgHvVfHD9ir9nP9oBLi78beAbe01udWxrujn7Feq7HJdmUbJWzn/XJIOTxXxe/wCCPXxI0R7nUvgt4/0vxNZrlo9N1dPsN2BnARZBuikIGCWYxA88cDPxI/Zw+PHwiluE+I3wm8S6NBbMyyXsli0lpkddtxHuiYe6sR0PevD3i7xX4QuTe+E/E+raLcEqxm069ktnJXODlCDxk49Mn1rwX+2t+0r4JuYJbf4mX2sQQrta11pRepKOeGZ/3nfqHB4HOOK8I/8ABU68SCODx58JYZpgvz3WkakY1J2jpDIrEAtuOfM4BA5xk2//AAVA+BL28TXfg3x5HOUBlSOztHVWxyFY3AJGe5Az6CvGf/BUyI2skHw8+FLfaGVvLutav/kQ/wAJMMQyw6EjzF9Perv4ofGH9rP4laT4b8d+K7y4024uvMk0+0/cWlpCnzOyxg4yFBAZ8tkgZOasLAyyW2mafbqC7LDBEi4AzgKoA6DoMV4T8L6N4Q0hdK0ayhtw7ebcuiYa4mIAaRz1ZjtAyegAAwAAA5HehJngGhIOgNCQEYzW8Hit4PANbhjAP6Vv9/0osCMZ/Si46Zr/AIKs+MBceIPAXgGGbH2OyutSuED53ea6xRErjjHky4Oedx4GOf8AgjJ4LnvPid8QfiIVlEOk6DBpQbICM11OJTxjJIFmOh43cg7hilGaqqqqqgOKqqUZNVVUAMYFVVVVVjPANYGMA1VVVVWM8A1gYwKqqqqpRnmgMDFVTokiGN1DKwwVYZBFePP2aPhj41SW5s9KGg6i4+W505RGmR/eh+4ffABPrXj/APZe+I/g3zrzSrZPEWmxciayQ+aF9Wh5YHj+EsPfrjT9S1rw3qa3ul313puoWrkCSJ2ikjYcEcYI9CK+H/7X3irRttj4+09Ndtu11AFguE+oACOO2MKe5JrwN8WvAHxFij/4RnxDbyXbqSbGZhFcJgc/uzyQO5XI96ABqqqqqqoAEZqqqqqqrGeBQAAxmqngguoJLa5hSWKVCkkcihldSMEEHqCOMV8Yv2IPBvjDz9a+Gc8HhnV3YubN1JsZiewVcmHnH3AVAGAnOR49+HHjX4Zaz/YPjfQLjTLpgWiLgMkyg43RuuVcfQ8d8V8Fv2u/iF8LFt9D1qQ+JPDkZVRa3T/v7Zc8+TL16fwtleABtyTXwv8AjP8AD34waZ9v8F62s08Uavc2E48u5tc9nTJ78blJXPQmqqqoLkdawMYFVVVVVVVWu+J/DXhe2F34l8RaZpMBUkS312luuBjJy5AwMjP1FeJP2xf2ePDbGF/H0epTj/lnptrLcZGSM7wuzt03Zxg9xXij/go14WgRo/BXw41S+ZkIWTVLuO1CNjglI/N3AHHG5cjuK+N/x+1z456taatrXhXQNKuLFGjinsIZBNJGcEJK7OQ4UgkfKMbm9aW6uFdZEndXQ5VlbBBHQjFSPJNI0kjs7uSWZjkknqTXhn9nj44eMNjaF8L9eeOTGya5tTaRODnBDzbVI4PIOBxnqK8M/sB/HDWtkmuz6B4fj43pc3hnlGc5wsKupIwOrAcjBPOPDP8AwTj0SLZJ4z+Jl7dZxvg0yxWDb1yBJIz57YOwY54OePDn7D/7PWgKjXnhq/1yWMqVl1LUZDypzkrEURgeMgqRx0658OfCX4XeEWik8M/Dzw7p00O3ZPBpsSy5U5BMm3cSDzknNVVVVVVVXxH+Fvgf4saC3h/xtokV7EA32ecDZPascfPFIOVPAyOhwAwI4r49fsr+M/g3LNrmnrLrfhUyHZfxRkvaKT8q3Cj7vYbx8pOOhIWvhD8c/H3wX1db3wtqZk0+WTdeaVcMWt7kcAkr/C+AMOuG4AORkH4MfHrwL8btI+1eHbo2uq20Ya+0m5YCe36AsMffjyRhx6jIUnFVVUQCMUV9KqqqqogEcUR2qqIBGKIxxVVVVVVTAEZFVVVX/BTbwG3jr9jXxs1tA0t54da11i3VcYAhmUTMcgnAgeY8YOQOcZB/Zt8bx/Dn47+CPF88jJb2mrRxXLq+0rDNmGU9DwEkY479MjORXxj8Ry6D4JmhtZdtxqTi1Ug8hSCXP02gjPbcKtrdrqdLdDy5xn09a0jRIo41jhj5xyccn61c6IyR5aPjHQitf0hbJ/tECgIxwy46H29q+AXiqbw58QbOyabFprH+izKTgFj/AKs899+B9GPrSSY4zSPnoa8a/G74afD12tfEvim3S8TrZ24M8wPuqZ29OrYHvzTftr/CdLgwDSPE7oHKiZbSEKRnG4Azbsd+mfavBHx7+FfxAuItP0DxXAt/KSEsrtTbyuc4wobAcnrhSTj6HHmHHBFGQ4xkUZAf4qaUYwDTy54Bp3HrmpJMjrUsgAwDUj8ZJrVbK01bT7rS7+IS215C8M0bDIZGUqw/EE14r0C48LeJdU8N3LbpNNu5IC398KxAYexGD+NaPreseHr+PVNC1O5sLuP7s1vIUbHcHHUHuDwe9fHX4c+IPjH4pu/iBd+KJJ9euoo0kS6QLCyxoEUIEAEYwucAEEkngkmv+CVnwf8AD/wt+Cep6rLrGnXXjHxVfCfWrW3ulkexhhLpbQuFPHDSSZ/6bY/hqqqiO1VVVX/BWj9q+XwzoUX7MngPVmi1PW4RceK54HKtBZnBitcgdZvvOAQfLVVOVlIr9jX9mrUf2o/jZpfgV/tVv4csv9N8RX0CcwWaHlFboskpxGpOcFi2GCkV4b8N6F4Q8P6d4U8MaXb6bpGkWsdrZWcC7UgiRQqqo9AAKI7Zqqph6VVURkYFVVVXxa+G+ifGH4ZeJvhf4hO2x8S6ZNZPKIw7W7OvyTKp4LRvtdc/xKK8Z+E/EPw08ca14J8QJ9l1vwzqc1jdCFyQk8MhRijDGRuXIYdRgjrX7JXxj/4X1+zx4L+JNxNE2pXlgLfVVSTdtvIGMUxI6ruZC4U8hXXk5yWAI4NVVVVVXiLURq2t3l+pyskhCHGPlHC/oBVVVfDy3EPhqOQKB58rv164O3/2WqphngHtVVVVVf8ABZj/AJKb8Ov+wDc/+jxX/BFkZHxj5/6AX/t/VVVVVURzgV4mmFv4fv3JHMDJycfe+X+tVVVVfDVARqMhUZHlAMR/vZ/pVUwyMCqqqphziv8AgpFrn9h/sa+PvK1D7Lcah9htIcHBl33sHmIPrEJM+2a/Yt0SDxB+1d8LLC4hllWLxJbXQWPOQ0B85Tx2BjBPsDVVVVVVXxFuQ09lZq33EZyM+pAHH/ATVVVVVVVVVVVVUFxPbSrNbTPHIvRkbBFaP49liCwaxEZR0E0YAYfUd6tL20v4RPZXKTIe6nOPr6VVVVVVVVVVf8Fd/Cn9r/s7aD4nhTM2geKId5L4AhmhlRsDufMEP0Ga/wCCT/jhvB/7aPhnTGuEht/Fem3+kzu84iXBgNxGpyPmLS20ShcglmXBOMGqqqqqqqqqqqqqqqqqqv8Agr3+ys3w9+I9v+0Z4O05x4e8cTeTrqoBts9VC5D4GCFnjUt3/eRykkb1Ff8ABMf9pofCD4rn4UeJ7vb4W8fzxQxMx4s9SyFhkyTgLID5TcZz5RyAhzVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVKATVVVUAMACqoDgDNV/wVA+DP/Cr/ANpa+8V6Zp8VvovxAtxq0HlfdF1wl2COu5pB5x7fvxg9QP8Agjn8S9F8f/DT4kfsreNrS0v7ARvqMFpO+Td2N2gt72Erkfu1byuR3uWyRxX7WnwD1L9mn49eJ/hVdJK9haXH2jRrmQc3NhLloHz3IX5GP99HHav+CWn7SP8AwtL4RH4N+I7rd4i+HlvHFavJJlrvTCSISB/0x4hIHAURdyaqgMmqqqqqoDjrVVVeNP2av2fPiG5m8Z/Bfwdqly2M3UmjwrcYBLY81VD4yzHG7BJPrXxO/wCCTf7MvjNZbrwRJ4g8C3xU+WtheG7td5IyzxXG9yMA4VJEA3egArxT/wAEbPjLaXgj8E/FnwXq1ruYGXVYrnT3A42nZGk4yfmyN3GByc8S/wDBKf8Aa8jieRNB8NSsqkiNNbjBYjsMgDJ9yBU0MsErwTxtHJGxV0dSpUjggjsa+EvwG+F3w08O+HvGvg7X77xBqfibRIrqa+uVSNYY5QrCJIkJ8tgyncGZiCAM9c/BzQDqfiRtVlXMOmJuGR1kbhf03H8BX0NBiOM0HB4oMCODQc44NCQ4xXmDGM15gI615gxjNGQY4NGTIwDX7cfjj/hOv2mfF88U2+10SZNJtxnOz7OgSUf9/vNP41/wSP8AAKeGP2X5/GEqZuPGOv3Nyj5/5YQbbdFxuI4kimOcKTvwcgAmgOKqqrX/ABJ4c8K2H9qeKNf03R7INt+0390lvHnBONzkDOFY/QE9q8SfttfsleFd/wDaf7QXg2fy7czn+zdQGofKM8D7P5mW+U/IPmPHHIzq3/BUD9ivTrM3Nn8U7zVZAwAt7Tw9fq5B75lhRcD/AHs1a/8ABVr9j65uobebxL4jtUkkVWnl0KUpECcFmCbmwOp2gn0B6Vpn/BSH9ijV76LTrT45WqSy52tdaLqFtGMAk5klgVF4HcjJwByQK8HftJ/s+fEExReDPjX4K1W4mUslpDrcAuAA+zJhZhIo3EDlRncMdRmqUDFVVAZOKqqqqqlGT1qqqqqqUZqqqqqsZ4FKAeB2qqqqrx18Ivh98RUZ/E3h+CS8KbFvoR5Vwvp845bGOA2QPTmviB+yB4q0bdfeAdQTXbbvaTlYLhPoSQjjv1U9gDWo6ZrXhvU2stTsrvTdQtXBMcyNFJGw6HBwR6g14A/as+IfhQQWPiJ08SafHwRdNtuAvtKMlj7uGPPX0+Hnx9+HHxIaOy0zVjYam5wun6gBFK5/2Dkq+fRSTjkgVVVVVQGeM1gYwKqqqqqsZ4FAYHWqqgoA968VeEvDXjfRZ/DvizRbXVNOuMb7e4TcMjowPVWHYjBHY18af2F9Y0lrnxB8HZ31OyGXbRrqQC4i55ETnAkUDorENgdXJqzvfEngvXkvLG41DRdZ02U7XQvbz27gYI7MpwSCPQkGvgj+3QD9m8NfGeD0ji121i+gzcRD8SXQenydWrRta0fxHpdvregapa6jYXaboLm2lEkcgzgkMODyCPYgiqxngUFHeqqqqqqR44o2lldURFJZmOAAOpJ9K8VftFfA7wYrDXvifoSyRkh4bS4+2SoQQMGOAOw+8Oo6ZPQHHiv/AIKE/CbSllh8K+Hde164TPlu6JaQPwcfOxZxyAP9X0JPbB8U/wDBQ74n6kzxeE/CWgaJC33WuDJeTJyDw2UToMHKd+1eK/2mPjv4zWWHWfibrEcM2Q8FjILJCuCNpEIXK4JBBznjOcVPPe6ndme5nnu7q4fl3YyPIx46nJJrwt+z98a/GbRjw98Mteljl+5PcWhtYWwSDiWXanBBB54rwz/wT++NWsCOXX9Q8PaDGVBeOa6aeZcrnAWJShwcA/OO5Ge/hz/gnJ4Qt1B8X/EjWL9jnK6baR2mOBgZfzc4OecDIxwK1j9hL4EXnhy40nR9M1DTtTk5g1U3ss8kRyDgxs3lsvGCNoOD1B5r4nfC3xT8IvFdx4R8V2Aimj+a3njB8q6iJ+WSNu6nH1ByCAQRX7LP7XEOgW9t8N/izqpGnRgRaVrExLfZh0EMx5Pl9lc/c6H5cFUeOWNZInV0YZVlOQQe4rGelYPpRHFVVVVVVVVTwQXUEltcwpLFKhSSORQyupGCCD1BHGK/aC/YmttQ+0eLvgvapb3X3rjQCypHJ0GbcnAQ/eJRjg/w44U6fqHiXwP4iiv9PuL7Rda0qbKOA0M1vIOCCDgjjIIPUEg8Gv2d/wBsbRvHy2ng74mTW+leJWIit70KI7a/OQFHX93KxP3fukjgjIUVVVRUHpRHaqqqqmAI460R2qqIBGAaI7VVVVVVUwx0qqqvFfhrSvGfhbWPB+uRebpuu6fPYXke1W3QzRmNxhgVOVY8EEeoNfEDwXrHw48deIfh94hi8vU/Dep3GnXa448yGRkYjBIIJXIIJBBBBNfsyfE1fi58DfCfjGW5aa/axW11Jncsxuof3crEnnLMu/6OOT1r9oDVNPntdO0qC7SS6gmZ5Y15KArgZPQHjp1rQDGNQAc8lCF+v/6s14daDenmEYrXJNOa1UQAAhefrXilohZTA98AD3zxXw/OPHvho56ava/+jlpJQRjNftLftAX/AIZml+Hfge9eDUjGDqOoRMVe1DDIjjI5DlcEsPugjHOSp82eQklpJHOSTySe5r+zrwruEB6eor97BICC0ciHIPQg9iK/Zk/aJ1LUb+3+G/j3UjcyTDZpN/MxMjOMAQOf4sjJVjzkYJORgykjAoyEjrTSDHXNPLkYBp5PU/hUkuRgGncdzUkmeAalfsDX7V/hc6b42s/FEEZ8rWrXbK3/AE2iwp+nyGPr6H8K8EeC9b+IXiex8IeHBanUdQYrAtzcLApIBONzEc4HAHJ7A18Mv2C7vQdRtNf8Y/Eq7tb60k3xx+HC0DxsBwVuXAYcnsgOO/PFla/YrK3svtM1x9niWPzZ33SSbQBuY92OMk+tVVMPSqqq+M/xX8MfA74X+Ivil4uuEWw0CxecQmQI11LjEUCE/wAcjlUX3YZ4zXxB8d+Lvi94/wBY8f8Ai68bUfEHiO9a4uWRMbnY4VEUdFACqqjoAB2r9g39mGL9mL4G2Wi6vAh8X+I2XUfEUo52SkYjt1OM7YkIU9cuZGBwwAphzxVVTAEYFVVURzgVVVVf8Fc/hTF4M/aKsPiJp9r5dn490hJpmCsA15a4hlxxt/1X2YkA5yxJAyC3/BGP4kXl34f+IXwjvJ2a3065ttZ09CuQpmUxXAz2H7qAgdMljxzkjIwKqqqqrxlqn9leHrmZGHmSr5MeTjluMj6DJ/CqqqrwrB9n8OadHhRmBXwo/vfN/WqphkdaI5xmqqqqv+CzH/JTfh1/2Abn/wBHiv8AgiwMj4yAH/oBf+39VVVVVTAZ4rx/P5Ph5ow2PPmRMY64+b/2WqqqqvhvG4027kI+VpgoPuFGf5iqogEYzVVVUwHGDX/BXbxOuk/s46H4bhuUW413xTAHiaMkvBDBM7kHopEnk+5BOO+P+CXfhyXXP2v/AA9qcazlfD2l6jfP5ce5QGt2t8uf4VzcDn+9tHeqqqqqqvGN59s1+4CtlYMRLz0wOf1zVVVVVVVVVVVVVVZX95p0wuLG5eGQDGVPX6joR7GtE8cWl4Ft9W2W82cCQfcb6/3f5UCGAKkEEcVVVVVVVVV+3x4cl8Vfsf8AxN0uFZ2aDS474iGPe2LW4iuCSP7uITuPZcntX7JnixPA/wC098KvFE1ylvb2fi7ThdSvGZAlu9wkcx2jJJ8t3xgE5xiqqqqqqqqqqqqqqqqqqq+OXwd8J/H34UeI/hL4zh3ad4gs2hEyrl7SYfNDcJyPnjkCOAeCVwcgkH4q/DXxd8Efidr/AMM/F1vLZa54X1BreUhWj3bcNHMnfY6FJEburqe9fsC/tPxftH/Bm2g8QX6P428JLHYa4jt890AuIrzGSSJVB3Hj94kmAF25qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqpRxwaqqrGeAaqsZ4FYGMCq/4Kj/B3/hZf7NF14u0628zV/h/drqkRUfM1q37u5TkgABWWUnr+4wOvP7Gnxs/4Z9/aT8EfEi6u/I0mDUFs9ZYngWNx+6nYjBzsVvMA7lByOtf8FbP2bf+FufARfi14d0/zfEvwy8y7l8tPnuNLfH2pTjr5W1JwScKscuBl6/ZX+N19+z38dPDHxJguGXT7e5FtrEQyRNYykLOCBkkhfnUYPzIp7Va3Vrf2sN7Y3MVxb3EayQzROHSRGGQykcEEEEEdapQD3qqqqqqA4xVVVVVAZPWv+ChHwPn+Dn7UviLT9K0/bpPjCQa5o0cKg5W5dvNjVVGF23CzKqDomz1FfDvwqvgjwNonhQTNK+nWaRyuxzukPLkeg3E4HYYr4YaCuieErVnjAnvh9olOOfmHyj/AL5xx6k1VV9DWT2Nbz61vNb631vr4oeP9M+F/wAPPEHxA1c5ttCsJLjZjJlcDEcY93cqo6DLDJHWj/afiHWOsl5qOp3P1aaWRv5lj+tfszeD7f4efBTwx4At3jc+HbGKylkjB2ySpGu9xnH3mJboOtYzwKwMYFV8Vf27P2Vvg+72fiX4t6XqGopjOn6GTqUwJOCG8nckZAySJGU49cgH4jf8Fn9Bh8+0+EfwXv7vdGPIv/EeoJb+W/Gd1tAJNw+8OJl7H2r4k/8ABRr9rv4lS3CS/FOfw1YzMxSy8Nwrp4hB7LMuZ8DgDdKTx65Na5r+u+J9Rk1fxJrd/q1/L/rLq+uXnlf6u5JP514d+EHxY8XxrN4V+GXirV4mVGEtlo88yBX+4xZVICkAkEnGATVh+xr+07qd3HZW3we1hJJM7WuJIYEGATy8jqo4HcjJwOpq8/YV/assbWS7m+E0zJEuWWHVrGVz9ESYsx9gDWqfsr/tG6Pbrc3fwW8WSIz7ALXTnuWzgnlYtzAcdSMdOeRWs+Hdf8OXAtPEOh6hpk5ziK9tXgY4ODwwB4PFfDj4/fGz4RSwv8NPip4m8PxQtuFpaajILZj8x+e3JMTjLucMpGWJ6818JP8AgsF8bfDNza2Xxd8KaH4z00MonurWP+zr7G0KWBTMLHI3bfLXJJG5QRt+BP7fv7M/x7EOnaJ45i8O6/JsUaL4iZbKd3bACxMWMcxySNsbluMlQMVVUo4qqqqqqA4qqqqqqAAGKqqqqpQDQAxVVVVVV4q8CeD/ABzaCy8WeH7PUo1BCGVMPHnrscYZfwIr4gfsb3UTS6h8N9bE0eCw07UDtcH0SUcH2DAYxyxzmvEvhDxP4Ovv7O8UaDe6ZcHOxbiEqJADglG6MM91JFfD79or4k/D+OOxg1NNW01MAWeo7pAi+iOCGXjoMlR6V8Ov2mfh1488qxv7v/hHtVfA+y38gEbt6RzcK3YANtYnoDSOkiLJG4ZWGVYHII9RVVVAADFVVVVVVADGaqqC9zVVXxe+APw7+M9iV8SaWtvqsce221a1UJcRYzgFv40BJOxsjk4wTmvjR+zF8RPg076hdwLrOgGQrFqtnGdqjt5ycmIkepK54DGvhT8bfiB8HdUW88I6y4s3l33WmTkvbXPAB3JnhsADcuG4HOOK+Cf7Vfw9+L6W2kXMy6B4mkOz+y7qTKzt6wyYAfI/hOGznggbiAAOtVVVWp6rpei2b6hrGpWtjax/fnuZliRfqzEAV4r/AGvP2ffCTm3n+IFtqc4OPL0qJ7wH1PmIDHj/AIFXi3/go1okW6HwJ8OL26z92fVrtYNv3c5ij35/iH3xjg85IHi/9uP4+eJ98Wm6zp3h2B8jy9Lsl3Y/35d7A+6kfh0rxP8AELx341lebxd4y1nWC5yVvb6SVRySAqk4UAk4AAAzxivDngfxn4xlEPhLwnrGsuXKYsbGScAjGclQQMAgnPQEV4W/Yg/aC8SlWvPDthoELrlZdVv0XseqReY46AYKj7w7ZI8Mf8E4owqTeM/icxO357bTNPxg5HSWRuRjI/1Y7H2rwt+xL+z74ZKy3Hhm812ZFwsmq3rv2IJKR7EOc91OMAjBrw14D8EeC4zH4Q8IaLooZcN9gsI7cv0zuKAE52rnPXA9KxngVtJ6Vtz3raPWvin8JfBfxg8Nv4c8X6asu1WNpdxjbPZyEY3xt26DIPytgZBr4xfB3xX8FvFcnhrxLD5kMm59Pv40IivYgfvL6MMgMh5UnuCCf2Uv2rX8FvafDT4laiX8PsRHpupTNk6cT0jkP/PH0P8AB/u/dR45Y1kidXRhlWU5BB7iqqiAaIB7UVB6UV44NFTRHaqqqqq+Pf7M3g342WTagoTR/E8KH7PqkMQ/fYAAS4A5kTAABzuXjBxlT8Q/hv4w+FviKXwx4z0l7O7TLRP1juI8kCSNv4kODz17EA5Ffs4/tkaj4Saz8EfFa7mv9DG2G01ViXmsRnpJ3kjGRz95QONwwBpeqabrem22r6PfQXlleRLLb3ELh0lRhkMpHUEVVVRAIxRGOKqqqqZQRx1ojtVUwBHFVVVVVVEAjFEdqqqr/grh+znqui/HLRfi94Q0K5urT4iQJaXiW6GQ/wBqQIEwFA+XzLdYiB3aKVj3r9lDQPiX8EfAOsaJqmuJCviG5hu0sIhuOnuEKyHf03uPLVsAgeSuGOTiKxvNZna2t4JbqeUMxVVLMcDJP9aila3mWWFvmQ5UkVpHiGGVFAkCSd0Jwc+3rV3ryrDmacKoH8RxWt6udRk8uI/ulOQTxuPrXwA8LS6140TXJEP2TRV8xicjdKwIRf5t/wAB961/XIPD3h/UvEFwhki0yzluXQHBYRoWIH4CtT1C71fUbrVb+Yy3V7O80zsclndizH8STWiaUGVRsBdvvHFJ4Wla38wR8Y9K1zRgFKFcOv3T6Va3VzYXcN7ZzPDcW0iyRSKcMjqcgj0IIFeBvEp8W+DND8SnZv1LT4Z5FU5CyMgLr+DZH4UZCe9NICPvZppRjANPJnoaeQAdeaeTPepJB0FahqFnptnPqOo3kNra2sTSzzzyBEiRRlmZjwqgAkk8AV+0J+2d4C+I/jrSPhR4Fs4dS0uC+YXHiCRioacB0VLcfxRk7QXPDZGBgBj4O8EeLfiH4hs/CngnRJdT1O8kVQinakKbgHlkfokaA5JP0ALEA6ppviDwJ4pn02+jm07WdDvNrAZV4Zo24ZT9QCD34Nfs1fGu2+Nfw+h1C8lRfEOk7bbWIRtGZMfLMFGMJIASOAAwdRkLk1VURkYFVVV/wWJ+Pg1jxb4f/Z30DVN1roEa6rr8UZ4+1yr/AKPE/PVIWMmMYxOhySOP+CWP7Nlv8Y/jZJ8TPFOm/aPC/wAOvKulSQEJdamxJtk7bhHtaY4PBSIMCr81TAEcVVUQCMUR2zVVTD0qqqq/4LJeEItT+BXg7xrHaNLc6F4n+ymRYN3kwXNvIXYvjKKXt4V7AkrnkCv+CS/iBtF/a3g01bu3iGveG7+yKSkBpgvl3G1M9W/0cNxztVu2aojtVVVVXxR1DzLu00tH4ijMrgHjLcDPuAD/AN9VVVVvBJdXEVtCMvK4RRjuTgVBClvBHbx/diQKufQDFVRGRjNMPeqqqqv+CyerWc/xl8CaFGzfarPww9zKCvASW5kVMH1zC/Hbj1r/AIIt6ZexaX8XNYkgxZ3U+j28Um4fNJEt4zjGcjAljOSMHdx0OCO1VVVVUwyOtfEufEFhbD+J3c89MADp+NVVVVfDyJk0F2JB8y5Zhj6KP6UQM4FVRHOKqqphkcV/wWR8am68afDr4cQ3LD+zdLutVnhDOA32iVYo2YfdJH2aUDqRuboCM/8ABG/webvx78RfHzQJ/wASvR7XTI5WZwf9JmaVlUfdI/0RCSeR8uOGNVVVVVUsixRtK54RSTx6VcTNczyXDn5pXLHJzyTmqqqqqqqqqqqqqqq0DxXeaMVt5cz2nTyyeU91P9On061ZX1pqVst1ZTrLG3QjsfQjsfaqqqqqqqr456LF4k+CXxB8OzztDHqnhXU7V5FGSgktZEJA9QDWh6xe+Hta0/X9NZBd6ZdR3NuXXcBJGwZcjuMgcVVVVVVVVVVVVVVVVVVVVf8ABW/9kaH4i/D9v2kPBOmu3iXwZa7dcgt4Mm+0wHJmbHJa3yWJP/LLfk4QV+zD+0D4i/Zr+LmlfEXRmnnsFb7PrOnRvtF/ZsR5kfPG4YDKT0ZV7Zrwf4t8PePvCuk+NPCmpRX+ka1aR3VncRtkPG4yPoR0I6ggg8iqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqgBt4NVVUACQM1VAZIGaqta0fTPEOj33h/WbRLrT9StpLW7gckCWKRSrqcc4Kkjj1r44fDDUvgx8XfFvwu1SKdZPDuqS28LTgb5rfO6CbjAxJC0cg4HDjgdK/4J3/HPSv2jP2U9Bh1iSK91nwzb/8ACO+IbW4VX8wxRhY3ZSPmSWAxkkjBbzF52k1+1t8C7v8AZy/aB8XfCx0/0CxvDPpEm4t5ljN+8gOSSSQjBGyfvI31r/gmB8b1+K37ONr4O1K9STXPh3KulTRmQs5siu60kIPRdgeED/p3PQYqlHHWqqqqqABOKqqqqqlHHWv27vg/4N8dWfw+8f6xdNBrfg3XGewRFB+1RSR7njYZGAskMDhsHG0rgeZuHiTxh4Y8BaUfFPjKeaLR7SeEXZhjLybHkVDtUEEn5qtZ7W6tYbmymimt5Y1eGSJgyOhGQVI4IIxgiqqqqqqq/wCCnHxbbSPCWh/BnS7tkuNelGo6oqOBm1iYiJGXrteYbgfWD61+wV8LG+JH7QOk6pdIx03wcv8Aa9wQBzLGwEC5PQ+aVboeEPTqPhh/yAJ/+vxv/QErU9U0vRNPuNX1rUrWwsbOMyXF1dTLFFCg6szsQFA9ScV+0R/wVi+EHw6+1eHvgtY/8J/rseU+3b2h0yBvXzPvz4PaMBSOklfG39s39oz4/TzR+OviPfwaTLG0f9iaS7WNjsIwVaJD+9zzzKXIyQCBxXgv4beP/iPenTvAfgzWNenT/WCxs3lWIccuwGEHI5Ygcj1rwF/wTR+N3iJo5vG+saH4StjjejS/brheRnCRHyzgZP8ArByAO5I8A/8ABNT4F+G0hn8bajrni+6VMSxy3BsbZ2xjKpCRIvPODKe3XnPgv4PfCr4dbW8DfDzw/osy5/0i00+NJm+996XG9uHYDJOASOnFVVVqGnafq1m+n6pY295bS43wXEQkRsEEZU5BwQD9QK8f/sY/s3/ERZJdQ+G1jpN46kLd6ITYOhJBLbI8Ruxx1dG6nuc18VP+CYPiLTo5dR+DvjiHV0UsRpmtILecKPuhZ0Gx2PA+ZYx3z2r4g/DD4gfCnWz4d+IfhLUNDvufLW5j+SYDgtFIuUlXPG5CRnvX7NX/AAUW+Pf7PMttot9qbeOfCEShDoms3TF4EGP+Pa4wzwkAYCkPGAT8mcEfs4ftffBX9qHSXuPh7rzW2sWqg3mg6kFhvbfjlggJEif7aFgO+DxVAcYqqqqqsZ4BrAxgVVVVVQAJxVVVVVUoyAKqqqqqsZ4FYAGAarXNA0PxNpsmkeIdJtdRs5fvQXMQdc9iAehHYjkdq+If7Hmh36Pf/DfUjpk4GRYXjtLC3sshy6/ju/CvGnw38bfD26W18XeHrmxEhxFMQHhlPXCyKSpOOcZyO4FfDz44fEL4bPHBousG505Tzp15mWHH+yM5T/gJHvmvh1+1J8P/ABp5Wn67L/wjmpvgeXdyAwOf9mXgD6MF/GkdJEWSNwysMqwOQR6iqAzzVVVVVVQGaqqAzzmqqqqSOOWNopEV0YEMrDIIPUEV8a/2IPDHi3zde+FLWvh7VWZnlsJMiznzz8gAJhOeyjb22jrXizwf4s+HuvyeH/FmjXekapand5Uw2nAJAdGHDLkHDqSDjg18Dv21/Ffgk2/hz4m/aPEWhIuyO8BBvbQADHJwJl46Md3OdxACnwh4z8K+PtDh8SeDtdtdV06fhZoG+62ASrKcMjAEZVgCM8gVrGuaJ4dsW1LxBrNjplomd1xeXCwRrgEnLMQBwCfoDXjD9sj9n7wh5sP/AAmn9t3UWf8AR9Ht2ud2N33ZOIjkrj7/APED0Oa8Yf8ABRr/AFtv8P8A4cevlXWsXf8AvAZhi/4AT+89R6NXjD9sj9oHxh5sP/Caf2JayZ/0fR7dbbbnd92TmUcNj7/YHqM1rPiDXvEd0b7xBrl/qlySxM17cvO5LHLHcxJ5PJ968H/BL4t+PvKfwl8PNbv4JsbLr7KYrc524/fPtj6Mp+90OenNeFP+Cfvxj1kxy+JtU0Lw9CVBdHnN1MuRkgLGNhx0Pz9emRzXhb/gnh8M9NKy+LPF+u63Iq/6u3VLKJjggkj52xyCAHGCOcg4rwp+zf8AAvwU4m8P/DLRVlU5SW8ia9kQ+qvOXZfwIqC3gtYI7W1hjihhQJHHGoVUUDAAA4AA4xVYzwK2kitox1rA7VVVXxG+Gng34reG5fC3jXSlvLRjvidW2S20gGBJG45Vhn6HoQQSD8c/gZ4o+B/ig6Rq6m70u7LNpmpomEukHY/3ZFyNyduCMggn9mD9rDUfh1d2vgb4iahNeeFJNsVtcyZeTSj0GO7Q9ivO0YK9Cptbq1v7WG9sbmK4t7iNZIZonDpIjDIZSOCCCCCOtVVVVUQCMUVBGBRU9qI7VVVVfEb4aeDvit4bl8LeNNKW8tGO+J1bZLbSAYEkbjlWGfoehBBIPx4/Zy8YfA/VDLch9V8OzsFtNXjh2KWIP7uRcnY/B4zgjkHqB8Av2mPF3wU1CLTZpJdW8KzSD7TpkshJgBJLSW+ThHySSPut3wcMvgTx54V+JXhq28WeDtVS+0+5yu4Da8Tj7yOp5VhkZB9QRkEE1VUQCMURjiqqqqmAIyKqqYdxVVVVVVTDuKqq+IvxP8I/DDRzqniXUAJZAfstlEQ09yw7IuenTLHAGRk8jPxa+Mvij4uaij6qwtNKtZTJZaZE5aOFsFQ7HjfJtYjcQMBjgAEivi58ePB/wqtHtJp01LXmX9zpkL/MuRkNK3Plrgjryc8AjJH7KPxL8C/Fj4VWPinwrbQ2upqBBrloW3S290o+YE9SjffQ9NrdmDAfEr4OXF9dz+IfCSK0kxMlxZE4JbqWQ+/9316elXdnd2M7Wt9ay28yHDRyxlGU+4PIqvBnwv8AE/jSdGt7VrOxIy15cIQmP9kdXP049SK8JeG9K8HaJBoWkRsIYuWdzlpHPV2PqfyHFfEstJ8NvFccalmbQ70KoGST5D9KUgMCRnB6V4fniWRGJBHarbWdMXRzA0aGTHDeleI54ZJWKEd6lIeRmXoScV8EEmtfhH4UinQox02NwD/db5lP4gg/jRlJ4zTSEjrTSDpmnlyMA07jHWnkzwDXx0/ab+E/7P8ApZuPGuu+bqkqFrTRrICW7uenO3IEa853uVXggEnAP7Rv7Y/xO/aCuptJluW0DwkspNvotnKR5g7G4kGDK3fBAUHouea/ZW/ZC+M37Tniu3HgOwOk6Hp86PfeJr6JltbPac4Tj99LxxGvfG4opLD9kv8AZ8u/hG/ifxD4jt/+Jrd382n2LsuCLGGQjzB0IEzqGwRyqRkda/bh+BX9vaSfjD4Zs86hpcQTWY1P+utl4WUDuydD/s8/w18Afi9ffBj4iWPiVXlk0q4It9WtkyfNt2I3MFyAXT7y57jGQCasr201Kyt9RsLhJ7a6iWWCVDlZEYAqwPoQQaqqojnAqq+JHjvRPhf8P/EXxG8RybdN8NaZPf3IDBWdYkLbFzwXYgKo7swHeviH44134m+O9f8AiF4mumn1TxFqM19dOzZw8jlto9FUEKAAAAAAABiv2KfgP/wzr+zn4X8A31nFDr1xG2pa8yLgvfT4Zw3qY0EcOR1EINVRAIxmiO1VVMOeKqqYZHWqqqr/AIK2anY2H7JT2l3Psl1LxLY29qu0nzJAJZCOBx8kTnJwOMdSBX/BMWwvLz9tfwDcWtu0kdjDqk1wwHEaHTrmMMfbe6L9WFUw54qqqqrxXfHUfEN7cB9yrKY05/hX5Rj8s/jVVVeBrD7f4ktsqClsDM2e23p/48VqqogEYphxj0qqqqr/AIK76xa6p+1Np1jbxyrJpHg+ztZy4ADObi5mBXBORtmUc45B7YJ/4I2aTeQ/Brx3rrqn2W78TpbRENzvito2fI7DEyfr6UwGcA1VVVVTDIwK+I04k1mGBWBEVuMjHQkn+mKqqqq8CxeX4at33Z8x3bGOnzEf0phz1qqYDPBqqqiMrjNft9fESL4lftY+PdVs7lJbHSb1dItdvIAtEWGTB2gsDKsrZOfvYBKgGv8AglR8Pf8AhEf2XYvFNxBtufGes3V+GYYbyYiLeNfpmGRh/wBdM9CKqqqqqvEMqw6FfO3e3dfxIx/WqqqqqqqqqqqqqqqqtC1260O6E0J3xPxLETww/oferK9t9RtI7y1kDRyLkeo9j71VVVVVVV/YWeqWFxpmoQJPa3cLQzxOOHRhhlPsQSKr4WSy3Hww8ITzytJLJoNkzu7EszGBMknuTVVVVVVVVVVVVVVVVVVVTQw3EL288SSxyKVdHXIYEYIIPUEdq/4KJ/siS/ssfGaSfwxYuvw/8YNLeeHX3lxaMCDNZMTzmIsu0nOY3jyzMHx/wS2/ayi8Ha8v7OPj3Vki0bXrnf4Ynm4FvfSH5rYsTwsxxtH/AD04HMlVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVAcAVVVSjPSqpRk1VV/wAFcP2cYLrR9M/aV8L6bi6spI9N8UGMH54WwtrcN2G1/wByT1PmxDotf8E3P2kf+Gdv2j9K/tvUPI8J+NNmj63vbCQ72/cXB7Dy5SMseiPJ61/wWi+Ara/4C8M/tEaJYtJd+Fpho+tPHEDixncmCR26hUuG2Ac83Q6c1/wTY+N7fB/9pTSdH1K9aLQfHSjRr5TIFRZnObaQg8ZEuEzkYWVzz0NADAAqqqqqgMnrVVVVVUo4Ar9rDxf/AGz44tvCtvLmDQYP3gH/AD2lCs31wgjHsc+9ftp+M3itdD8AWspBnJ1C7AOMqMpEOvIJ8wkHuqmv+Cd37QSePPAjfCDxNqO7XvCUKjTjKwBudOGFRV7kwkBDxwrR9ecVVVVVVeKPEuieDPDmpeK/Ed/HZ6ZpNs9xdTyHARFGT9SegHUkgDrXxu+K2r/Gz4n678RtXRoTqc+LS1L7ha2yDbFED0yqAZIA3MWbA3Gv2EvgVf8Awa+C1tq/iXTPsniHxsU1S5jkjKSw2u3FtE4PopaTBAIM5B6V8Vf2zfhH+yt4IlHim9/tjxRd3DPYeG7CZftMqlUAkkJyIYsgjew+bDBQxUgftJfthfGn9p/VWfxx4gaz8Pwzb7Lw7p7tHZW+CdrMmf3sgBI8x8kZONoOK+CP7L/xc+Pd4p8H6A1to6kibW9QVobNMHDBXwfMcf3UBI74HNfCL/gnT8GPAnl6l49kuPHGqrtIW7UwWcTDB+WBDl+cg+YzKRj5R1rS9J0zRLCHStF021sLK3XbDbWsKxRxjOcKqgADJPQd62k9BWwnivLJHWvL96MeRgGvLPqKMZ6A0Ubpiip6YqvFng7wp470Sbw34z8O6frWmXH37W+t1lTI6MARwwzwwwQeQRX7Q3/BNyS3juvFX7Ptw0qq25vDN5Nlgvf7PO5+Yjg7JDnGfnJwpsL/AMZ/DPxdFf6fc6t4Z8TaDdZR132l1ZTocEdmRhyCDjuDX7FX/BUTSfHD6d8Lv2kb6y0fXiqw2HihsQ2t+QFVVue0MzHcfM+WM9MIcbqqqqqpQDVVVVVUo4zVVVVVAZOKUcZqqqqqqUA1VVWoafp+rWUunapY295azrtlguIhIkg9GU5BHsa+I37IXhjXPN1H4fX39h3rZP2OctJaufY8vHz6bh2CivG/w18a/Du8Fn4s0G4s1ZisVwBvhm/3JFypOOcZyO4FfDz44fEL4bPHBousG505Tzp15mWHH+yM5T/gJHvmvhv+1D4C8cPBpmtOfD2rSkIsV0+YZGPGEl4H4MF9s1gYwKqqqqqgM8CsDGBVUBmsDGBVVVUBngVPPBawSXNzNHFDCheSSRgqooGSSTwAB3r43/F/9k/XdDl8N/EfxXo+uhN3kx6buvJoJMYJjkhBEbfVgOMHPSvFF34Xj1y7TwVLqk+jbh9kfVIY4rjbtGQ6xsy5ByMg8gA4GcDwr8RfHHgW7nvvBnijUNFnuYjFM9nMYzIvocdcdR6HkVrniPxD4nu/7Q8Sa/qOrXRz+/vrp5356/M5Jrwj8BfjJ472N4X+HGt3UMn3LiW2NvC30ll2p+teEf8Agnp8UdX2TeLvE+ieH4W+9HFuvZ09cqu1PyevCP8AwT/+DWibJvE+oa34kmH345bj7LC30WLDj/vs14V+DHwm8ENFL4V+HegWFxBjy7lLFGnGCCP3rAv1UHr1ANVVVjPAoLkcmsDGBVVVVVV4/wDAHhb4m+Frvwh4v05buwuxkEcSQSD7skbfwuueD9QcgkH45/AzxR8D/FB0jV1N3pd2WbTNTRMJdIOx/uyLkbk7cEZBBP7LP7U118MLqDwH47u5bjwlcSYt52y76U7HkgdTEScso6feXuGtbq1v7WG9sbmK4t7iNZIZonDpIjDIZSOCCCCCOtVVVVVVEAjFFQehojHFVVVq+kaVr+mXGja3p1vf2N3GY57a4jDpIp6gqeDX7SX7Imp/DVbrxr8PI7jUvCqAyXNu7b59NGeSe7xDj5uWUfezgufhH8Y/GPwZ8Sx6/wCFrzfC/wAt5p8zEwXadwyjow7MOQfYkH4PfGrwZ8afDqax4avFjvYY1OoaZK+ZrJzkYb+8pIO1xwR6HIFVVEAjFEY4qqqqph6VVEAjFEY4qqqqqqIBGKIxxVfGz9ofQfhpBNoWiNFqfiVlKiANmOzJAIaUjvyCEHJ7kDBPiXxP4i8b63LrfiLUrjUL+5bAZyTtBJIRF6KoJOFHAzXwY/ZTnvfI8S/FK3aGD70Oj7isj9RmYjBUdCFBye+OQf8Agqz+xYhg/wCGmvhToCRpaW6weLtOs4TgRoAsV6karhQqAJLyAFWNsf6xq/Z0/aA8Ufs7ePY/Fmhx/bdOu0Fvq+lvIVS8gyDwf4ZF6o+DgkjlWYH4ZfE3wb8X/Btj458C6qt7pt6uCCNslvIAN0Uq5Ox1yMj3BBIIJvtM03U4xDqWn212gBAWeJZBz14IPoKtPC/hewl8+y8N6XBJjG+KzjRvzA9hSSY6HNJICMZp1iniaCaNZI5FKujrkMDwQR3GK+J/ge6+HvjS/wDDkyP9nR/NspHH+tgYnYwPfoVP+0prTdWa0xHITtH3SB0oeJUEW37SnTpurVNZa7BjhY4PVj/SvAPgvU/iB4rsPC+lxsWuZMzygZEEQ+/IfQAfmSB1IqygttPs4NPsolit7WJYoYxnCIowAPoABRlJ4BoyZ700gxjNNLkYBr4vftK/Bn4I28w8e+NrOHUY0DLpFq4nvZMjK4hU7lB7M+1fevjj/wAFLfiB4zt7jw/8INH/AOEP02ZDG+pTuJtQkBGDsI+SDgkZG9hgMrqa0zS/GHxH8VwaVpNnqviTxHrdzsihjV7q5vJm/NnY9Sfxr9lX/gkbfaiLbxl+1NPLYwLIkkPhPTroeZMo5IuriMnYD02RHdj+NTxXhnwr4Z8E+H7Lwr4O0Cw0XR9Nj8u0sLC3WCGBckkKigAZJJPqSSeTVXdpa31rNY3ttFcW9xG0c0MqB0kRhhlZTwQQSCDX7Q/wlf4OfE2/8MWySnSbhRdaVLIclrd84UnuVYMh9due9fsJ/GD+3/DNx8JdZnzfaCjXGmOz8y2rN8yeuY3b1+66gABKqqph6VVf8Fh/jN/wjHwf0D4K6Zdbbzxrfi71BAc/6DakMFIx/FOYWBz/AMsWGDnj/gnJ8FIvjX+1J4cg1SxS50Xwip17UkkXKOIGUQowJAYNO8OV5yobIIzVVVMBniqqmAI4qqogEYzRHaqqq/aJ/Zu+Hn7T/g618DfEqfWY9Osbs3ludMuxA8dx5TxrJyrKxVZHwGBGTyD0r9mv/gnJ4H/Zk+M7/Ffwx8Q9Z1i3TTLmztdN1G0jEkTSumJDNGVDbY1ZSvljJbdlcbTTDI4qqqq1W7Gn6ZdX2RmCFnGe5A4H50xLHJOSepNVVV8MdPCWt3qbpzI4iQkdhycfiR+VVVUwB4FVVVVf8FK/ELeIP2zPHaJfpdW2lrY2VvtUARBLKEyJkDJImaXJOTnjOAK/4JL6DFpH7J/9oRifdrfie+vH80fLlVhg+TgfLiAevzbuewYe9VVVVUQCMZrxnP8AaPEl2QThCqAHthRn9c1VVVV4Pi8nw1YpuzlGbpjqxP8AWmA4xVUw6YqqqvEg15vDuqL4VFmdbNlMNNF7I0cH2nYfK8xlVyqb9u4hWIGeD0r4ofsW/tUfD3XEj8YfDDWNQk1W7VE1OykW/hnllcgF5kLbCzZyZdp5yeDmvhZ4EsPhf8NPC3w501keDw1o9tp6yKgXzjFGqNIQABudgWJwMlie9VVVVVXjy7WDRBahvmuZAMY7Dk/qB+dVVVVVVVVVVVVVVVVV4K1ttPvhp8z/AOj3TYGT9x+x/Hofw9Kqqqqqqqq+E/8AySvwb/2L9j/6TpVVVVVVVVVVVVVVVVVVVV+0p8A/Cf7S/wAHtc+E/ixNiX8fm6feKPnsbxATDOv+6xwR/ErMv8VfEr4c+Nfgt8RNZ+HHjjTpdK8R+G7zybmMMRtYAMkkbcZR1KOjD7ysrDrX7A/7W1v+0r8M00fxTfQL4/8AC0KQ6tEXVX1CIAKl6qDs3AfAwsnZQyiqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqxngGiO2aqqpRx1qqUcdaqq8d+CvD3xI8F634B8V2f2rSPEFhLZXkQ4by5FKkqedrDOVbqrAEcivjj8IvEPwI+KviL4VeJX8270G7McdyIjGt3AwDRTquTgPGytjJxnGcg1+xj8UNP/AG8P2I/EnwZ+Imopd+KNK02Tw9rM0oBkZHjY2F9jB+YBR8xyTLbO2ORWu6N4h8A+LtQ8Pamk+m674b1KW1uFVmjktrmCQqwB4KsrofQgiv2Z/i5a/HX4E+DPidDcebdatpqLqIKBNl7FmK5XaOg85JMcDKlSAAaqqqqqlHFVVVVVWoX9ppWn3OqX0ojtrKB5pnJwFRFJY/kDXiTXLrxN4g1HxDek+fqN1JO6lt23cxO0H0AIA9gK/az/AGKpviz+yvotx4U0oN8QvBtq+p20aw7ptQSUeZcWWV5LYIMYwcvGF48wsPh/488W/CTxxp3jXwpdtY6zotxuVZFOGxw8Ui8EqwyrLxwT0PNfBL4y+E/jt4AsPHXhSdV85fLvrFpA8thcAfPC+McjqDgblKsBg1VVVVVf8FBf2oY/GWpy/A3wHqjPoulT/wDE/uIj8l5dRsCsIIPzRxsMnPBkA/uAn/gn3+yrP+0r8ZIL3xDY7/Avg6SK915nxtu2yTDZgYOfNZTv6YjV/mVimf2+P269M+GPijW/hr8J72HUPGduEgudRVEmttIO351wciSdTxsIKqT82SpSpJPF3xH8WBpJNS8Q+I9duwoLFri4u53OAO5Yk4Ffs0/8E6NN0pLLxt8fFj1C9+Sa38NRNmCA4yPtTj/WMCRmNfk+XkyBio07StP0mxg0zSrG3s7S2QJBb28QjjiUdFVVwAPYUIsjAFCImhDkYoQDpivJHQCvJFGEHoKMIPAowDHAow+lGE4wDTRHpijGOgooR05r9oz9ln4eftD6I51a0j0zxPbQlNP123iHmxkfdSX/AJ6xZ/hPIydpUkk/GT4K+PfgZ4tn8JeOdJkhIdvsV8iMbe/jGP3kLkYYYZcjqpOGAPFfsB/8FIdT8CXmmfBX9oTxC134TZY7XRPEF0Rv0bHCQ3D9Xt+gDtkxcAny/wDVxSxXESTwSpJHIoZHQ5DAjggjqMVVVVUBx1qqqqqqAGMCqqqqqUZHFYGMA1VVVVWM8CgABgVVVVVqGn6fq1lLp2qWNveWs67ZYLiISJIPRlOQR7GviN+yF4Y1zzdR+H19/Yd62T9jnLSWrn2PLx8+m4dgorxv8NfG3w7vBZ+LNBuLNXYrFcAb4Zv9yRcqTjnGcjuBXw5+P3xF+G3lWen6r/aOlR4H9nX5Mkar6RnO6PvgKduTkg18Nv2ivh58Rng06O9OkaxMQq2F6wUyOeMRv918ngDhj6VVVVVQGBVUBngVgYwKqqrxr8Xvhh8O1lHjTx1o+lzQpva1luQ1wRjORCuZG4x0U9R6140/4KC/CvRGe38G6Bq/iWVc7ZWAsoGwSBhnBfnGeY+hHfIHjL9vX42+IkNv4dGj+GYezWdr58xBAyC8xZeoOCqqRnqcZrxZ8QfHPjy4+0+MvF+ray4IKi8u3kVMZxtUnao+ZuAB94+prwn8LPiR482t4O8Da3q8TZ/f21k7RL97rJjaPusOSMkEda8I/wDBP/4ya55c3ifUNE8OQn78cs5up1+ixZQ/99itT/4JzaJF4Vkj0X4iXlx4lQ7o5bu2WKzlGPuGNd0iZP8AHubA/hNeOPhx4t+GOuv4c8Z+H59LvkXcgkXKzJ2eNxlXXtlSRkEHkED4FftmeLPh8bbw38QGuPEXh5SqJOz7ruyQDHyMf9Yo4+Vjn0YdK8FeO/CXxF0GDxH4N1y31KxmH3o2w0bf3XU/MjezAGqqqxngUFyOTWBjAqqqqqqqq8f+APC3xN8LXfhDxfpy3dhdjII4kgkH3ZI2/hdc8H6g5BIPxz+Bnij4H+KDpGrqbvS7ss2mamiYS6Qdj/dkXI3J24IyCCf2T/2qh4Ha0+GXxGvv+Kedtmm6jIedOYn7kh/54knr/AT/AHfuo8csayROrowyrKcgg9xVVVVVVVEAjFFQRwaI7VVVJHHLG0UiK6MCGVhkEHqCK/ac/Y7EovfiJ8INOdpmYy3/AIft4s7s/ee2Uc5zyYh1ydvZT4L8b+Lvhp4lh8S+EdVm0zVLXdGWCghlPDI6MCGU45UjqAeoBHwB/aL8K/G7RIoPNg07xRbRE32lF+TtwDLFnloySPdScHsxqqogEYojHFVVVRAIxRGOKqiARiiO1VVVVVTDI4r46/tQR2IufB/wyvEkuPmivNXXkRdisBzy3X5+g/hySGGhaD4h8aa7Do2h2VxqeqX8h2xg5Z2PLMzE4A6ksxAHUmvgr+znoHw3jh17xAsGq+JCAyzFCYrI4GViB6tnP7wjPoF5zV3aWl/aTWF9bRXNtcRtHNDMgdJEYYZWU8EEEgg8HNf8FCf+Ce1/8Ar+7+L3wh064vfhveTbryzXMknh6RzwrHktbkkBJDypIRjnaz/AL9onx/8As8+Jm1rwjdLcafeMi6npNwx8m9jU5/4BIBkLIBlcnqCQfgb+1B8J/j5p8X/CKa7Haa2EzcaFfSLHdxkDLFFz+9Qf30yBxnaeKoEjoaVweAeaWUjgmvjB4M8D+MvDWfGWp22k/ZWza6nK6o0DH+HLEBg2MFe/GOQDV38NvFLW9zqvh/TrjXtHt7mSBNR0+3d45CgBZtuNwUA/eI25BAJxTWd2l19he1mFzvCCExkPuPQbeufavBvwP+I3jS4RbXQJ9PtDgteahG0EYHquRuf/AICD+FfCn4UeH/hXpTwae7XepXar9svZBguQPuoP4UzkgcnnknAwZuODRlJ6V8Svjr8JPhBbLP8AEfx/pWis/wDq7aSQy3DjGSVgjDSMOnIUjkeor4jf8FSPhlo6T2fwz8F6z4kul3Kl1fsthbE4G11HzyOMk5VljPHXnI+Kf7d/7RnxQin08eLl8L6XMwzZ+H0NocDjBnyZiD3G8KcnjGAPC3g/xn8Rdfj0DwZ4b1bxFrN225bWwtXuZnywBYqoJxlhljxzya/Z6/4JBfE7xjLBrv7Qeup4I0jdk6Rp8kd3qU6+hcFoYAQQQ2ZW6govWvgn+zb8FP2d9GOkfCbwHYaPJLGEutQKma9vMYP724fMjDIyEyEUk7VUcVRAIxmiO1V+2D8J/wDhZfwouNT0228zW/Cu++s9q5aSID9/EP8AeRQwA5LRoO9fDHx7qXwx8eaN430olpNMuA0kQOPOiPyyRn/eQsPbOe1aFremeJdEsPEOjXK3FjqVslxbSj+JHUMp9uD07VVURkYFVX/BRr4tp8Xv2svF11ZXST6Z4VZfD+nsoP3bUsJuT1BuHuCCOCCMZ6n/AII+fCRPCnwJ1v4s3tsgvfHeqmO1kySTZWZaJevAJna5zjqFXJOMCqqmGRxVVRGRgVVVTDnAqqqqqiMjAqqqq+Il2bXw1JGrYNzKsWR/30f0WqqqrQtPGl6NaWAGDFENwP8AePLfqTVVVMB2NEds1VVVfte6xda9+1N8Wb27jiSSLxhqVqojBA2QXDwoeSeSsak+5PQcV/wT10T+wP2NfhlYfavtHm2Fxdb9mzHn3c823GT93zNue+M4GcBgCOKqqqqqtTuRe6jdXanImmdx9CciqqqqvDUTReH9PRiCTbq3HuM/1phkcVVMMjg1VVX0ojnFVVVVVVeOdRF7q/2WNspZrs68bjy39B+FVVVVVVVVVVVVVVVVUrMrBkYhlOQRwQa0i8Oo6Xa3hPzSxAt/vd/1zVVVVVVWqalY6Lpl3rGpT+TaWEDz3Em0tsjRSzNgAk4AJwBmq+GVndWHw18J2F9bS21zbaHZRTQzIUeJ1gQMrKeQQQQQehqqqqqqqqqqqqqqqqqqqqr/AIKm/sXv8c/Ai/G74d6XLN478GWbJdWdtE0j6xp4O4xqigkyxEu6Y5ZWdTuOzb8C/jR4w/Z/+J2j/E/wTc4vNNkK3Fq7ERX1u3EsEg7qy/8AfLBWGGUEfCP4qeEfjX8OtF+Jnge++06VrUG9AwKvDIpKyRODyGR1ZT2OMjIIJqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqABIGaIHQGqqqAGOKqgBjg1VVX/BWv9nWXxT4P0r9oLwvpjS6j4ZX7Drohjyz2LMTHMcdopGIJ/uy5PCV+wT+01L+y5+0FpPirU5nPhXXVGleJIt5AW1kZcTgcjdC4STpkqHQEbya/4K6fBqH4fftKxfEvRg76N8TdNTUUkC/uheQhYbhI2HDAqIJSc5zOe2K/4I7/ABmEsPjD4CavqMrPFt13RYX5VUysV2oJ6cm3YJ33SNgfMTVVVVVAcdaqqqqoDJABr9pfxOvh74WXtlFNsudalSziAYZ2k7pDjuNilT6bxz0r4S+E/wDhNfiJomgyReZbvciW6BTcPJj+dwfQELt57sPpVf8ABTb9h650HU779pH4Q6E0uk3rNN4t0u0hUCwkwubyNFAzG/zNL3VsucqzbPgB8f8Axp+zv40PivwmIbu3u4hBqemXLMIbyHcDg4PyyLyUfkqSeCrMrfBr42+APjt4Ti8VeBNWWbYqi9sZSFuLCVhny5U7Hg4YZVsEgkVVVVXNxb2dvLd3dxHDBAheWWRgqooGSxJ4AAGSTX7XH7fFnf2N/wDDH4DarIyzboNR8TQsUG3o8doevPQzccZ2ZyHHwJ+B/jn9of4k6b8MPh/aRSahfZkmuJ22Q2VuuPMnlPUIoI4AJJIABJAr9of9pLwN+xD8KY/2P/2V9Qik8WQRkeK/FFuMNbXboqzupy3+lvtAPJEChUB3KPL8EeBvGfxU8W2vhPwdpF1rOtanL8qICx5PzSSMeFUZyzsQB1Jr9mD9kzwV+zvoUV4IYdV8aXlvs1PWWBOAcEwwKfuRAgc4DPgFv4VVYs9aWEYxihEOwoRE9BQhJGM0IQeK8kdK8lcYwK8lfQUYRjjFGEEcUYTjg0YjjGKaIYximiBGBTQ+lNEcYxXxj+DXgn44+CbrwR420/zbeX57W5jAE1lMAQs0THowz9CCQQQSK+PHwH8bfs/eNpfCPi63823l3SaZqcSEQ6hCD99PRhkBkJypPcFWP/BMr9u3/hHJ9L/Zn+MGqyyabeXAg8KazczZ+xOwASxkJ/5ZMwxEf4GbZ9wr5dVVVjPArAxgVVVVVQGTiqqqqqxngUoB/CqqqqqoDJqqqqqqxngGtW0fSdesJNK1rTba/s5hiSC4iEiN9Qa+Jf7H2kahHNqnwzvv7PuvvDTbty0L+oR+WQ+gO4ZwMqOa8VeDPFXgjUTpPizQrrTbnnasyfLIB1KOMq491JFfDL9pzx54CEenaxO3iLSVwBBeSnzoQP8AnnLycYx8rbgMDGOa+HXxh8CfE+2U+HNYQX6x75tOuP3c8Xr8p+8BkfMuRyOaqqoAHmqrGeBQGBVXV1a2Nu93e3MVvBEuXllcIqD1JPAFeNP2wfgF4MV0bxrHrdymcW+ixm7LYBPEgxFyQBy/Ujtkjxt/wUV1y4L23w78AWdmgJC3Wr3DTswwOfKj2hCDn+Nh0+leNP2lvjl49Z11z4japFbtkfZdPcWUW0knaVhC7wM4+fceBknFaTo2r6/fxaVoWlXmpXsxxFbWkDTSOemFVQSeo7V4K/Yu+P3jNIp5fC8Hh+1lTcs+t3AtyOM4MShpVPQcoOT9ceCv+CdXhq0aK5+IHjy91FlfLWumW62yEA8KZH3MQRjOAp5IB714L/Zo+BngJUbQvhzpUtwmD9q1CM3su4ADcGmLbCcfwbRycAZpI44o1iiRURFwqqMAAdAB6VVeO/h74O+JegyeG/GuhW+p2TncgkBDwv2eNxhkbtlSOCQeCRXx2/Y48Y/DJbnxH4KNx4l8NRK0khWPN3ZID/y1RR86gYJkQdmLKgGT8P8A4k+NPhfri+IPBOuzafc8CVFO6O4UHOyRDwy/Xp2wea+Bf7X3gr4p/Z/D/igweHfEz/KsMkmLa6P/AEykbox/uNz6FqqguaAAGBVVVVVVVVVV4/8AAHhb4m+Frvwh4v05buwuxkEcSQSD7skbfwuueD9QcgkH40fB3xL8FvGVx4a1yF5LORnfTL8JhL2AHhh6MAQGTqpPcEE/shftQP4bubb4V/EfWFGiSAR6PqFw3/HlJkBYHY9IjztJ+4cD7p+SqqqqqqqogEYorgcGqqq/ab/ZJ0/4hR3/AI/+Hdutr4qI8y5sVISHUiM7iP7kx4+bO1iOcElq0zUvE/gPxLBqemz3uja5pE+5GKmKWCQdQVI+oKkYIJBBBNfs2/tQaR8aLX/hHPEKW+l+LraPc1ujYjvkA+aSHPII5LJkkDkEjO2qqiARiiMcVVVVEAjFEdqqmHcVVVVVV3d2ljbSXl9dRW8EKlpJZXCKgHUkngCvj1+0pe+MWufB/gW4ktNCV9s96pKS32OoHdIye3VgBnAJWvh18NfFHxQ1waJ4ZtA3lgNc3MpKxWyE43OffsBknBwODj4W/CPwr8KNH+waHEbi9nA+16hOo82c+nH3UB6IOnck5JqqmhhuIXgniSSORSro65DAjBBHcEdq/bG/4JOWWsS6h8R/2W4LewuWVp7rwdK+2KZ+pNnIxxETyfJYhB0UoAErXNA8dfC3xW+k+INK1rwt4j0mUEw3MUlnc2zjowzhl9QR9Qa+Fn/BQr48fD63ttJ8R3Vn4102D5cavuF3tx0Fyp3Mc87pBIeSPTHw6/4KRfArxZHHb+NYNW8GXpVd/wBpha8ttx6hZYQWIB7tGg5B9cap+0L8KNP0yHU7TxMuqrcJvhisIzI7D3zgKfZiDXiz9qTxZqiPa+FdOt9GiJI85z58xHQEZAVfXoe3Prf6p4j8ZatEdT1K81S/uZBFEZ5i7FmOAq5OAMnoMAV4E8M2/grwjpfhm2MbGytwsroMCSU8yP68sSaEvavEfxA8D+DY2k8X+M9D0NFRXZtS1GK1AVm2hj5jDgtwD3PFeM/27/2Y/BbTW0nxGj1q7hz+40a1kuw+CR8sqjyTyv8Af6EHoQa8ef8ABVW0WKa2+GHwsmeQqfJvNevAgU9i0EOdw9hKP1yPiL+27+0n8SWnivPiHcaHYzbgLLQV+woikglRIp80jgD5nPGR/E2dG0LxX4715dL8PaPq3iHWtQlJW3s7eS7ubiRjyQqhmZiT7kk18Kv+CWn7WfxIe3utc8LWHgbS5trG68QXipLs3ENi3i3yhwB92RYwcjnByPg7/wAEdPgz4WaDUvjJ4y1fxxdqql9PswdMsskHcrFGMzgHGGWSPpkrzgfD74W/Df4T6N/YHw18EaL4asDjfDp1mkPmkZwXIGXPzHliTyaqqphzxVV+058LYvhP8W9U0bToPL0jUf8AT9MULhUhkJzGPZHDoOvCqTya/YQ+KD+JPAl78ONUu1e98MyeZZBm+ZrSQk499km4Z7B0GBgVVVRHavjP8QoPhL8JPGHxMnWKT/hGdEur+KKXO2aWONjHGcEH53Cr1H3uoq0ttY8Va9DZwvLf6rq92saGaXLzzyvgFnY9WZuWY9TkmvhH8PNO+Evwu8KfDPSn8y28M6Rb2CynrM0cYV5DwOXYMx4HLHgVVVRGRjNVVURzgVVUw9KqqqqqiB0BqqqvindAtYWKnkB5GGPoB/Wqqq8M2X9oa/YWpGQZgzD1C/Mf0BqqqqYZHBphzxVVVV451az1/wAa+INc05ma11HVLm5gLrtJR5WZcjscEcV+zHozeH/2b/hZo0unpYz2vg3SluLdUCbJjaRmXIH8RcsSe5JPemGRgGqqqqq8Q3YstDvbnIysLBc+p4H6kVVVVVWhf8gPTv8Ar0i/9AFEZGM1VEAjGaqqr6GmHOM1VVVVVaxqMek6bNfSYJjX5FP8THgD86mlknleaVy8kjFmY9ST1NVVVVVVVVVVVVVVVVVXgl2fw7ArAYR3C8dRuJ/mTVVVVVVXx91mXw58CfiP4hghSaTS/CWqXSRucBzHaSMAcdjjFeGNDl8UeJdJ8NW9wkMurX0NokrgkI0jhAxA7Ddmqqqqqqqqqqqqqqqqqqqqqr/gqL+xL/wozxs/xu+Guj7PAXi68c31rbx4TRNQc7igUcJBKSzR4+VGDJhR5YP7AH7Xkv7NvxCPhnxffN/wr7xTKq6mCMjTrjgJeKApYgAbHUYypB5KKKiliniSeCVJI5FDI6nIYHoQR1GKqqqqqqqqqqqqqqqqqqqqqqqqqqqqqlAJqqqqA4AqqAGABVVVa/oWj+KdC1Hwz4gsIr7S9WtJbO9tZRlJ4JUKSI3sysQfrX7RfwX1f9n34zeJvhVqpllTSLsnT7qRcG7s3G+CXjjLRsu4DhXDL/CavvG8f7XP/BM278P6jd/aviD+zzeW90IzKomu9HAMIlw3VEhkIbHObVCTlwG/ZI+LyfAz9onwT8RLu7aDTLXURbasQ5VfsU4MMzMBncERzIFxyyL0OCKqqqqoDgCqqqqqUAnrX7W3iddS8Zad4Xgm3R6PaGSVQw+WWXBwR1zsWM8/3ug7/sf+Fmlv9b8aTIdkES2NuSvBZiHkwfUBY/8Avv8AOpoYriJ4J40kjkUq6OuQwIwQR3GK/bc/4JiavpF9qHxW/Zn0WXUdNuJHm1DwfaQ5msjgs0lmM5ljJH+oUb1JAQMp2p4P8beNvhj4jTxD4L8QahoGsWuYzLbuY3xkbo3U8MuVGUYEHAyOK+EP/BTq5tbeDSfjZ4RkvGTCnV9EVVdhjq9u5Ck5xkqyjGcLxg+H/wBuL9l7xFCHi+KNvYShAzw6hZXFsye25k2sfZWNf8Nefs0f9Fk8P/8Af1v/AImtS/bN/Zg0q1N3dfGDSXQEDbbQz3D5P+xGjNj3xXxC/wCCm/wr0a1uLb4c+Fda8R34UiGa7VbK1z2bJLSEDrt2LnpkZyPjT+1d8Z/jnJLaeKPErWOiOfk0XTMwWoHbeMlpf+BlsHpiv2aP2QPi/wDtRa+lp4M0drHw/bzKupeIb2Mra2qlgG2njzpAMkRoc9MlQd1fH34l/DL/AIJvfCKD9nn9ntrW/wDij4htGbXPEThftVkrqv8ApEuMlZGB/cQ52xqN7ZyDL4K8E+L/AIpeL7Lwh4Q0u41fXNXm2xxqclieWd2PCqBlmZjgAEk1+yz+zD4c/Zv8Gvp8NwupeJdXWN9a1IAhZHUHbFED92JNzYyMsSWOOFVYh0pYieAKWEY5oRDGAKEWegoQkihCCMV5Q6ZryR2/lRiBGP6UYVxRhB6GjDxxTRHpimiBHSmiBGBTREcCmjBGMU8IIwBXx0+CXhX48/D6+8BeKE8rzsSWN+kSvLYXC/dlTP8A3ywyNysy5Gcj4qfDLxT8HvHmq/D7xfaeVf6XMVWRQfLuYj9yaMnqjrgjuM4IBBA/4Jn/ALZUvx38FN8I/iJqyS+PPCdqDbzOCH1XT02qspIGDJGWVH7sCr8kuRVVQGTVVVVVUoGOtVVVVUACaA4qqqqqqAGKqqqqqlHGaqvEnhXw54v01tI8TaNa6lat/wAs54920+qnqp9xg18UP2P7y1E+s/DC8+1RD5v7IumxIo7+XKThv91sHA+8x4q6tNd8K6w1rdw3ulapYSDKOGhmhfqD2IOCCD7g18MP2uvEGhiHSPiNbtrNiihFvoVAukxwN/IWQY78N3JY14W8YeF/G+lrrPhTW7bUrQnBeFuY2xna6nDI2MHDAHkVWM8CsDGBVV4n8Z+EfBNh/anjDxNpmjW2GKyXtysO/aOQoY5Y8jgZPI9a8d/t5fBnwu72nhiPUvFV0hZd1pF5EAI45kkwSD2KqwOM56Z8cft9fGDxGs9r4TstJ8L20hPlyQxfarlFz0MknyHjjIjB6kY4x4t8e+NvHl2L7xn4r1TWZl+4by6aQR+yKThRyeAAOTXgz4NfFT4hsn/CG+A9X1GJ8bblbcxwckAZlfCDr3bpk9ATXgr/AIJ5/ELVljufHPi7SdAiZQfItY2vZ15OVblEHQYIZuvtivBP7DvwK8JhJ9V0u/8AE12BzJqlz+7ViuDtjjCLjqQG3EZ68AjQPC/hrwpY/wBm+F/D2maPaZJ+z2FolvHnOSdqADqT+dVVVVVXx2/Y48HfE03PiPwUbfw34mkLSSFYyLS9cj/lqij5GJwTIg7sWVycjx38PfGPw012Tw3410K40y9RdyCQZSZOzxuMq69sqTyCDgggfAf9tHxN4BjtfC3xHS48QeH4hshulIN5Zr2AJIEqDptYhgDw2FC14R8X+GPHmg23ibwhrVvqmm3S5jnhJ691ZTgqw7qwBHcCqqqqqqqqqqqr4ofC/wAJ/FzwnceEvFtl5kMnz29wgAltJQPlljbswz9CCQQQSK+LXws8SfB/xneeD/EURYxHfaXYjKx3kB+7Kn16EZO1gRniv2Pf2n8/ZvhL8Sda/uRaBf3B/D7NI5P+6Iyfdc/cFVVVVVVVVUVB6GiO1VVftM/sw6Z8ZbE+JvDC29h4wtIwqyv8keoIBxHKR0YD7r9vunjBW9svFfw68VvZ3cd7omv6HdDIBMctvKpyGUj8CGHBBBBINfsu/tUW3xTgi8D+PbqC38XQofIn2rHHqigZJUDAWUDkoAAR8yjGQtVVEAjFEdqqqqmGRkVVEAjFEY4qqqq1rWtK8O6Vc63rd/FZ2NlGZJ55ThUUfzJ4AA5JIA5NfHP9oHVfipP/AGHpEcuneHLeQlYC/wA92wPyvLjoMYITkA8kk4x8Hfgt4g+LurPFayGw0i0OLzUXj3KhxkIgyNznjjPA5PYHwd4J8OeAdAt/DvhnT0traBcMxGZJm7u7fxMfX8BgACqqqqvi58Bfg78eNGTQvi38PtK8SW0X+pe4Ro57f18qeMrLHnvsYZ718R/+CLngO+1B9R+FXxW1fTLYl2OlazAlwCTyqpcoA0ajkfNHIcEZJIOfEP7IfiD9nuQz6z8NZbWNGXGshTdxEngAT8hCf7vyk+lVXxh+JWsfDrSrGfwrqcljrlzchrW5iID26xkMzrnvnavTufStV/ak/aM1mOOK7+NXi6MRklTa6nJanPuYipI9jWq/GD4ta7JHLrfxS8XajJEpWNrvW7iUoO4BZzgVFDLcSpBBE0ksjBURFJZmPAAA6k+leBv2Qf2oPiQYW8IfAnxjcwXDBYrq50x7O3cl9nE0+yPAYEE7sLg5xg18Nv8Agjp+0T4n8i6+Ininwt4KtZNvmw+c2pXkWdhP7uLERwGcf67lkx0IavhV/wAEhv2avBUdvd/ELUNf8f6jHtMq3U5sLNmCkHbDARIASQcNK/3VHTdu8CfDD4c/CzSRoXw38DaH4asdqK8Ol2EduJdowGcqAXbk/M2SSSSck1VVVVTAEcVVftz/AA2j8WfC+HxtZxE6h4SmMjY/itZSqygj2YRtnsFbjmv2bPiI/wAMvjHoGuy3LRafdzfYNSGcKbeYhSWxkkI2yTA7xiqqqYDtX/BXX4hf8Ir+y3F4Mt5f3/jXXrW0kjx/y7wZuXbO0jiSGAYypO7OSAwP/BOT4df8LH/a/wDAlvPB5ln4euJNbuTjPl/ZULwt/wB//IH41VVVUR2zVVTD0qqpgCOtVVVVVTDnrVVVfEW5+0eJpIf+faFI+R6jd/7NVVVfDS2EutzXBHENucc9yQP5ZqqqqIBGM0wHY1VVVV8Lv+SZeEP+wDZf+iEogEYqqqqqviLdmHRY7VW5uJgCPVVGT+u2qqqqqsABYWyqAAIVwPwFEAjFVRAIxVVVUwHGDVVVVVV4x8RpqswsLJw1rA2S4/5aN6/QVVVVVVVVVVVVVVVVVVV4atWs9BsoHBDeXuIbqNxLf1qqqqqqr/gop4rtvCf7H/j2SUo0+qw2+nW0bBvnaa4jVuQOCI/MYZwMrjuBX7GfhL/hN/2r/hN4eaCGeJvFljc3EM0PmpLDbyieVGXnIKRMDnjnnjNVVVVVVVVVVVVVVVVVVVVVXxA8BeE/il4J1r4eeOdIi1PQfEFm9pfWsg+8jDqp6q6nDK4wysqsCCAa/a5/Zh8V/so/GHUfh1rnm3mkzZufD+rsmF1GzY/K3HAkQ5SRezKSMqysf+CYv7ZqXcFn+zR8UdZijlt4wng+/uGCCRQQPsDN3bnMWeo3JnIjU1VVVVVVVVVVVVVVVVVVVVVVVVVVVVS4qqqsZ4FVWM8Cqqqqv+CrX7PX/CxPhFa/Gbw/Y+ZrvgHd9t2L80+mSEeZnufKfEg7BWlNfsmfGy1+A3xq0rxTr8LXXhLV4ZdF8W2I3EXmkXQEdypC/MxUbZVUEZeJRnrXxX8Bz/C/4l+J/h7cXkV5/wAI/qk9pFdwuHjuokciOZGHVJE2up7hga/Yf+J3/C2v2WvAHiae586+tNMXS78ldrefaEwFm5PLLGr5zzvBwudoqqqqxngGqqqqqpRx1r4g+JD4w8ba14kEzSR3147QF85EQO2Mc88IFH4V+z/4YPhb4VaNBNAI7nUEN7PxgsZTlCffy/LHPpVVX7Rf7CP7P/7STXOteI/DraF4rmXjxFou2C4kYDAM64KTjhQS6l9qgK618VP+CRX7Q3hF3ufhlreg+P7MYCRJKul3ZJOCTHO3lAAYP+uJ68dM6/8AscftV+G7pbPUf2e/Hc0jZwbDRJr5ODg/PAHUfnz1HFf8MtftOf8ARuXxP/8ACQvv/jVWH7Jv7UWpXkVjb/s6/EhJJm2q1x4Xu4EB/wBp3jCqPckCvhz/AMEvP2t/HssMmq+ENN8HWMjY+1a9qKIQBuDHyYfMlByuAGVc7lIO0lh8Dv8AgkZ8HPAt5ba78YPEl54/voVVhpyxGw09JOp3KrGSYA4xl1VgDuQg4H7Tnx78AfsY/A1tW0bRNGtLpIzZeFfDlrGltFLP22xR7cQx5DvtAwMDILrXiHxD42+LPji51/xFqV94h8UeJL4GWaU75bqeQhVUAcD+FVUAAABQAABX7HH7K+n/ALO/go6jrkMdx431+FG1efIcWqjlbWIjoqkjcQfnYZyQqYWMngCliAGAKWPIwBSw8c0sQ6AZoRE9KEOaEAPavJBHWvJHYijEDxkUYQehFGEHpTQ+1NEen8xTRgjkU0QIwKaIjinjB7U8RHSnjB4r9tn9mCH49+Af7a8NWCnxv4bid9MZNqNexHl7ViSBg43IT91s4wHbPw0+IvjD4M/EXRPiP4MvGsdf8N3ontzIGAJGVeKRQQSjoWjdcjKsw718B/jL4Y/aA+FHh/4q+FHRLbWrUNPaecJHsbgcS27kAfMjgjOBkYbGGFVVKOOtVVVVVQHGKqqqqpRkYFYGMCqqqqqAycZqqqqqqxngVgYwDVVVePPhl4L+JOnmw8VaNFcOq7YbtFCTwd/kkxkDPboe4NfFP9lnxl4Mkk1PwkkviLRxk4iTNzAB/fjH3h7pnocha8M+K/Efg3VI9Z8L6zc6deREYkhbAbHZlPDL/ssCD3FfCr9rfRNdMOifEeGPSb4gKmox/wDHtKcclx1iJ49V5PK1Z3Vpf2sd7Y3UVxBMu6OWJw6uD0II4Irxp8VPhx8O4w3jXxrpOkuykpDcXA85wOpWMZdhyOgPUetePv8AgoN8PdH82z+H3hvUfEVwu5Vurk/Y7fPRWAIMjDvgqh6cjPHjv9tP47eNY3tbTX7bw3aOrKYtFhMLkHgHzWLSAgd1ZeT9MXl9rXiTUhNf3l7qmoXLhFeaRp5ZWJwBk5JJJ6e9eBv2VPjt4+WG50zwJdafZS4IvNWYWaBSCQwV8O6nHVFbqOxzXgf/AIJ02ERguviP8QJbg7QZbLR7fywGx0E8mSRn/pmpx6Z48E/sz/A7wBGh0T4e6ZcXKnd9r1GP7ZNu9Q0u7Z0/hCj8zVUBxmqqqqqqq8dfD3wd8TNBk8N+NdCt9TsmO5BICHhfs8bjDI3bKkcEg8Eivjz+yH4x+E8dz4k8NPL4g8LQje9wqf6RZr3MqDqox99eO5C18NPi147+EetDW/BWtPbM3E9rJl4LkekkecH68EdiK+Bv7U/gD4zRw6PJIuheJyDu0q6lBE5HUwScCQY524DjDcEDcaqqqqqqqqqqr4y/B7wt8aPCE3hvxBbqtzEGfTr5RiSzmxwynupwAy9CPcAjx/4A8UfDLxRd+EPF+nG1v7Q5BHKTIfuyRt/EjY4P1BwQQP2Q/wBp3/hN7az+FXju5/4n9nb7NMv5H/5CEaL9xyeTMqjOf4wCT8wJaqqqqqqqqiARiiMcVVV+0h+zfonxu0T+0dNEFh4t0+IiyvWXCzqOfJmxyUJ6N1QnIyCQde8P+K/h74lfSNdsL3Rdb0yVX2MTHJC4wyOrA/QhlPoQa/Za/altfifaweBPHd3Fb+LbePFvO2ETVEUckDoJQBll7/eXuFqqphkcVVVVEAjFEY4FVTDI4qqqvEviXRPCGiXXiLxFqEdnYWabpJH/AEAHUsTgADkk18aPjp4g+LV/9kCmw0C1lLWlip5c9BJKc4Z8dhwucDux+CHwB1v4p3cWs6mslh4ZhlxNc9HuSvVIR354LHgc9SMVoui6V4d0q10PQ7GKzsbKMRwQRLhUUfzJ6knkkknk0QCMURjiqqqqq/an/aZ0b4SaNP4N8P8A2XUvFupW5UwSqJYrCJxjzJlOQzEH5Yz16t8uAyx32rXcrxQy3Nw4knkEcZY4VS7vgdAFDMT0ABPQVXw+/wCCe37O/jf4deG9Y+Nvw1fW/FE1n51xLJqt7ZtCsh3rFsgmQfKpUHIJ3bumcDRf+CcH7E+gXTXlh8CdPlkeMxlb3Vb68QAkHISad1B4HzAZ6jOCa0T9iz9kvw/LJNYfs7+BJWlUKwvtGivAAPQTBwp9xg14a8CeB/BcK2/g3wboehRrCsCppmnRWoEajCoBGo+UADA6CqqmAxgVVVVVRGRjNEdqrW9H0/xFot/4f1WHzrLU7WS1uY843xyKVYfiCRXjLwxfeC/FmseEdSBNzo97LayMUKb9jFQwHowAI9iK/Z18dH4i/Bnwz4inmMl4loLS9ZgAxnhPlsxxx820P9GHTpVVTDI4r/gs544Gq/GLwL8PYbpJY/Dvh+W+dFZD5Ut3NtZTj5g2y1ibDdmUgDJJ/wCCLPgJJ9e+JXxQubfD2dnZ6PZy5B3CV3mnXGcjHk2x5GDnrwaqqqqYc9aqqYZHFVVEZGM1VVVVVMMjiqqq8RXIvNev7hSCGuHCkDGQDgH8gKqqr4XQbbXULjJ+eRE6f3QT/wCzVVVVUw4xVVVVXwu/5Jl4Q/7ANl/6ISqI5xVVVVXxJu/N1O1sgQRBCW+hY8/ooqqqqqrD/jxt/wDriv8AIVRHOBVURziqqqYAgYNVVVVV418RC3jbRrGX964/fsD90f3R7nv7fWqqqqlVnYIgLMTgADkmtK8CajeKJb+QWkZ6KRuc/h2/H8qh8AaJGP3stzKcc7nAGfbApPB3hyMqw04Er/ekc/pnFf8ACMaB/wBAqD8q/wCEY0D/AKBUH5V/wjGgf9AqD8qk8G+HJNx/s/aWzysrjH0GcVL4B0KTGx7qPH92Qc/mDU/w5iIzb6qwIHR4s5P1B4q58AazEcwS2847YYqfyI/rV14c1y0BM2lz4A5KLvA/Fc0ysrFGBDKcEEYINVVVVVXhrRJNa1BEZD9niIaZiOMen1NAAAKoAAHAqqqqqqr/AIK/+LG0v4EeFfB8EzJJrviZZpAHA3w28Em5SOpG+WJsjoVHrX/BInwm3iP9szSdYW2eUeFtB1HUSyyBREGjFruIJ+Yf6UFwMnLA9ATVVVVVVVVVVVVVVVVVVVVVVX7Yf7LXhb9rH4QX3gTVUt7TX7JXufDmrOgDWN4F+UM21mELkKsiqMlcEfMqkeMfCPjn4M/ETUvB/iW1vNA8VeE9RMUypIY5LaeJsrJG6npwrpIpwQVZSQQa/YN/bG0z9pjwMvhnxRdwQfETw3bD+07YZX7fbqVRbyMHPBLKsgB+VyDgK6CqqqqqqqqqqqqqqqqqqqqqqqqqqqqqXpiqqqAyQAaqgMkAGqqqqtV0vTtc0u70XV7OK7sb+3e3ureVdySxOpV0YdwVJBHvX7W/wCvP2bPjpr/w2xPJpG4Xug3M3LXFhKSYiTxuZCHiZsAF4mIAGK1vXtV8R3Nve61ey3dzb2dvZrNNIzuYYIlhhTJJwEhjjjUDACxqMcZP/BGv4nAj4gfBi8uf+eOv6dCF+lvdMTn/AK8wAB65PSqqqqgASBmqqqqqr4yeIk8L/DDxBqYmCTPaNbwEnB8yX92MepG7P4E14W0KfxR4k0vw5bEiTUruO3DBSdgZgCxxngAkn0AqCCG1gjtbeMJFCgREHRVAwAPwqqqqAyQM1VVquqadoel3mtavexWljp9u9xdXErbUhiRSzux7AKCSfav20P2mdV/ah+NOo+LEuZ08LaSz2PhmyckCG0Vv9aVwMSTEeY2ckZVMkRrX/BN79mRILeP9oXxpp+Z5d8Xhi3lVlMa4KSXRHQ7gWRM543N3UhIh3pY88CliGOaWPIwBSw57UsQ6UIsjgfnQiJGBQh4ryR2xXlAjA/lRiGP/AK1GIHgGjCDwKaI44pou2M00QNPDxwKaLjA5p4geBTx44p4wRgU8eOCK/wCCjH7O8Pw38dW/xa8K6dHB4e8XzMl5HEpC22o4LNx0AlUM4A/iSXoMZ/4JPftKf8K0+LM/wR8Uax5Phvx9Iv8AZyzy7Y7fVgAse0E4BnQCI45Z1hHaqqgOMZqqqqqgATiqqqqqxngGlA7dqqqqqqlAxVVVVVUoB5qqqqqv2h/Bv7PkkFxq/jfxZpfhLXZUMi3UcgaabAGS1sp3TcY+6N3I5rW/GumWV3Na6M41GONysdyFaJJB2YKwDY9iAas/jJ8TtJ0650jQfGuq6RY3Y/e21hdPAp56jacgnocHkcHgmj9t1O8LEzXd3cyEknMjyuTknuSSa8BfsXfHTxzDFez6Ha+HbKVVdJtYn8pmUjPESBpAcY4ZV69euPAP/BPf4e6OIrv4g+JNR8RXC7Wa1th9it8/xKcEyMO2QyHrwM8eCPhP8NvhvAIPA/gvS9JYIEaeGAGdwBgbpWy79/vMep9TVVVAZqqqqqqqgM9KwAMCq+PP7FfhbxvHdeJ/hjHb+H9fxvewVdlneEdcKP8AUueOV+UkcqCxevE/hXxR4D16XQfFGkXekapaEM0My7GX0ZSOCDjhgSD2NfAX9t3VvDi2/hb4wzXWracvyw6yB5lzAAAAJR1lXjJbl+Tnd20LXdG8T6Pa6/4e1O31DTr2PzLe5t3DpIvTgj3BBHUEEHkVVVVVVVVVVVX7QfwG0T46eE106WZLHXdO3SaVflciNiOY3xyY2wM45BAIzjB1rR/EfgPxPcaPqcN1petaNdbWALRyQyIcq6ngjsysOoII6iv2Vf2kLX4v6GnhPxLOU8YaTbbp2YALfxKQPOXAADAlQy+pBHBIWqqqqqqqqiARiiMdDVVX7QX7Pvh345eHfLcxWHiOwjP9mans+738qXHLRE/ipOR3DeJfDXiv4beK59C120utI1vSJweGKPG6kFJEcdR0ZWU88EGv2Wv2pbX4n2sHgTx3dxW/i23jxbzthE1RFHJA6CUAZZe/3l7haqqYDtVVVUQCMCqqmGDxVV4l8S6J4Q0S68ReItQjs7CzTdJI/wCgA6licAAckmvjR8Z9b+Lmt738yz0Ozc/YLDd07eZJjgyEfgoOB3J+AfwCvvidfJ4g8QRy2vhi1kwzDKvfODzHGey9mft0HOSun6fY6VYwaZplpFa2lrGI4YYkCrGoGAAB0GKqmGRxVVVVX7UH7UGm/BvTX8LeFpoL3xneRZRDh001GHEso7sRyqHr94/LgNZWXiz4j+LEs7OK913xBrl0cAkyS3ErHJYk/iSx4ABJIAr9nv8AZe8M/CHwtOfENta6v4k1q1aHVLhk3RxxOMNbxZ/g7MeC55OAAB8M/wBne7v/ANp2b4W6zZPc6V4Zv2udRMowJbOMhot3A4lDRDAAyHJGOtVVVVUw9KI7VVVVVTDniqr9vHwIvh34rWnjC1hVLbxTYh3IzzcQbY5OOgGwwnjqSfqf+CenjPI8VfDu4m/556paRgfSKck5/wCvcDj156VVURkYzX/BRLxq3jn9sX4jXiXCSW+lXsWlQIk4lWMWsKROoIAwfNSRivVWYqelf8El/BSeGP2RrPxDt/eeL9evtRLEqTtjZbQAYAIANqxwc8sTnBwKqqqmGR1qqojIwKqqojnAqqqqqmAI61VVcSiCCScjIjQsR64FSO0rtI7bmYkknuaqqr4axNH4edyQRJcuy4/3VH9KqqqqcA5ANVVVWk/8EzP2L9MUed8J59QkWYyLJd69fkjnIXakyqVHTBBJHUmtL0yx0XS7PRtMg8mzsLdLe3i3FtkaKFUZJJOAAMk5qmHPWqqqqvFd0L3xDfSqwIWTYMf7I2/0qqqqqtFdpdGsJHbLNaxknGOSoqmHNVTDmqqqYAriqqqqvEuux6HYF0Ia5lBEKH17sfYf4VLJJNI0srl3dizMx5JPU1VVVAFiFUZJ6CvC3haHS4EvL2INesMgHnyvYe+OpqqqqqqqqubKzvF2XdrFMMcCRA2Pzq88EaDdZMUMlux7xP8A0ORV58O7lctYahHJ6LKpU/mM1c+EPEFuedPMg7GNg36ZzU9hfWwJubOeLA58yMrj86qoLO7uzttbWWY+kaFv5VpXgXUrt1fUSLWHuM5c/QdB+NWGn2mmWqWdlEEjXt3J7knuaqqqqqqr/gsJ45TWPi/4L+H0Fx5ieG9CkvJFAGI5ruXBXOM52W0R5OMMMYya/wCCH/gZn1D4o/EuYoFihsdItgACxLGSabPcAbIPY5P90VVVVVVVVVVVVVVVVVVVVVVVX/BS/wDYdi/aO8Et8Vfh1p3/ABcnwpZtthjVmOtWMYdzahFzmcMxaIgcklD95WT4bfEXxn8GPiDpPj/wbfS6drugXYkRX3KH2nDwyqCC0bjKOmRkEiv2Yf2kPBn7TnwztPG/hueC31SBVi1vRxNvk0y5I5Q5AJRsEo+MMM91YCqqqqqqqqqqqqqqqqqqqqqqqqqqqofdxVVVAZPWqoDJ4NVVVVV/wU+/Zul+MfwYT4j+GbJH8TfDxZrtgq/PdacVzcRjH3mTYkq56BZAOXqv+CfPxFl+G/7W/gG8a5ZLTXr06Hdxj/lsLtTFEpwrHAnMD9uUGSBkiqqqpQCaqqqqqv2vvEP2bw5ofheKXD3109zKqtztiXaAwz0JkzyOSnUYr9lnw4mtfE5NTmhDxaLZyXALDI8xsRr+PzsR/u5qqqqpQCaqq/4Kz/tNS+CvA9n+zv4Sv2i1jxfCLrXZoZSrQaarELDxyDO6kHn7kbqQRJx+yr8Crj9oL4w6V4MmEiaLa/6brcyA5S0jI3ICCMNISsYPYvuwdpFaXpdho+nWuk6XaR21nZQpBbwRrtWKNFCqqjsAABSR54FLGAMAc0kWaWIDiljJ4pYh0pY8jAFCEkYoQgjpQhHTFeSPavJHtRhBGBimgBGAKaH0NNEcYxmmiB4FNFgYFPGDwRTxCnj4waeL0p0BGK+M/wALtH+Mfwy8QfDjWgqw6xaMkMxXJt51+aKUe6uqtjvgjvWp6f4g8AeL7rSrstY654b1J4ZNjZaC5gkKnB9VdP0r9lj44Wf7RXwJ8LfFKFUjvb+18nVIFGBDexHZOoH90upZf9llqsZ4zVVVVVUozVVVVVAZNAcVVVVVVAcYqqqqqsZ4FADGBVVVeOPif8PvhtZ/bfHHi7TtIUpuSOaXMsg55SJcu/Q/dB6V48/4KE+CtLMln8PPCV/rkqnAur5xaQdPvKo3O3YYITv6c+PP2uvjr4+WS3n8XHRLORcG10RDaDrz+8BMvPAIL4x25ObDTde8UaqLTS7C/wBX1O8kJEUET3E0zseThcsxJP5mvAf7EPxw8ZLHdavptn4XsnXcJNUmxKRnHEKbmB9n2dPplv8Agnd4ZtdCxbePL291tCSJLi2WK1kGOF8tSXXnPzb2/wB2vGnwh8U/Ce8GneIPDosopDiG6gTdBP8A7sg4JwPunDAYyBXwy+Pfj74YPFZ6dfLqGjq+ZNNvPmTHfY33ozjPT5c8lTXwx/aE8AfE0R2Nvef2TrDYH9n3siq0jHtE2cSfQYbvgVVVWM8CgABgVVVVVVUBnvWBjAqqr4mfCXwJ8XNEOieNdFS6VeYLqP5Li2b1jk6j3HIPcGvjl+yx4++DTzavFGde8MAjbqltEQYM8Ynj5MfPG7JQ5X5gTtHwg+Onj74Laut74W1MyafLJuvNKuGLW9z0BJX+F8AYdcNwAcjIPwW/aF8AfG3Th/Yd6tjrcUe+60a5kAniAOCy9BInT5l6bhuCk4qqqqqsZ6VtJ4rbkda2jGM1tHrW0UVB6GioIwDRXjg1+1F+zfbfGnQV1rw5Hb2/i7Soz9mlcBBex9fIdu3PKseFJIOAxI0nVvEngbxJBquk3V3pGt6Pcko4Bjlt5VJDKyn8VZSMHkEYJFfs+/HbRvjn4Q/tSKKKy1vTyItU08SA+W+OJEGd3lNzgnoQy5O3JqqqqqqqqogEYplx0qqr9oL9n3w78cvDvluYrDxHYRn+zNT2fd7+VLjloifxUnI7hvEvhrxX8NvFc+ha7aXWka3pE4PDFHjdSCkiOOo6MrKeeCDX7LX7Utr8T7WDwJ47u4rfxbbx4t52wiaoijkgdBKAMsvf7y9wtVRAIxRGOKqqqmHpVUQCMCq1/XdK8MaLeeINbvEtrGwhaWeVuyjsB3J4AA5JIA5NfGj4z638XNb3v5lnodm5+wWG7p28yTHBkI/BQcDuT8A/gFffE6+TxB4gjltfDFrJhmGVe+cHmOM9l7M/boOcldP0+x0qxg0zTLSK1tLWMRwwxIFWNQMAADoMVVUQCMZojtmqqq/aU/aT0T4H6J/ZummDUPF2oRE2Nixytup48+bHIQHOF4LkYGACRI/iXx34maRhd6zrut3ZJCqZJbmeRuwHUknpX7L/AOy/pvwb02PxT4pigvfGd5Fh3GHTTUYcxRHuxHDOOv3R8uSxAIxmrPwf4c0/xTqXjOz0uKPWdXtoLe8ugPmkSHdsHt9/BI6hVz90YqqqqpgCMZpgOxqqqqqmGRxVV+3R4QXxD8FW1+KMtceG9Qiucgn/AFUh8pxjvy6HPbb6Zr9lLxc/g/48+FrgzFYNUuDps6D/AJaCcbEBwCeJTG3b7vJAyaqqr4n+Km8d/Evxb43e5a4bxDrt7qJmeMRmQzzvJuKjAUndnAGBX7JvhZvBn7Mfws8OyxzxzweE9PkuI5yC8c0sCySpwBwHdgB6AcnrVVVVTDIwDVVVSyRQxtJLIqIgyzMcAD3NX/xF8M2TmKOea6IOD5EeQPxJAP4etWvxJ8M3LhJXubbJxmWLj/x0mrS9s9QhFxY3UU8ZHDRsDTAZ4qqqqqiMjANVVeK5xb+G9RkJXm3ZPmOPvDb/AFqqqq+Hf/ItJ/12f+dVVVVMAePaqqqqqpgOxqqqqup47W2lupD8sMbO30Ayf5VJI8sjSyNlnJJOMcmqqqqq8ISrP4bsmX+FCpHuGI/pVMKqmHvVVVHBXFVVVV7eW+n2kt5cvtjiXLH+g9z0rWNUn1m/e9nOM8Iv9xR0A/z1JqqqqrwNpC3uoNqEygx2mCoI6uen5dfriqqqqqqqqqqqqS1tpW3y28TtjqyAmvsNl/z6Qf8AfsVVVVVVVVVX7ZXxFX4q/tO/EPxfBM0lodXeys2ZSuYLVRbxsFPKhliDYIByxzzmv+CUvw5/4QD9jTw3qE0fl3fjLULzXLhM5xvcQRHIZhzBbQtxtxuwRuBJqqqqqqqqqqqqqqqqqqqqqqqv+Cqn7B//ACE/2qPhDpP9+58b6ckv0/0+JD+PnAH+64X/AFjV+zD+0h4y/Zj+Jlp438Nzz3OmTMsWt6OJtkepWwPKHIIV1ySj4ypz2ZgfhR8U/Bnxq+H+kfErwDqLXejazCXhMibJInUlXjkX+F0ZWUjkZGQSCCaqqqqqqqqqqqqqqqqqqqqqqqqqqodAKqqpRnvVUoz3qqqqqpoYriJ4J40kjkUq6OuQwPBBHcYr9tb9n+X9nP4/a74OsrVo/D2ot/aXh9yODZyscRg+sbh4vU7Ae9aXqd9oup2es6ZP5N5YTpPbybQ2yRGDK2CCDggHBGK8I+I7Dxl4U0Xxfpbo1nrmnQX1sUkEgMc0aupDLwwww5HBqqqqUcVVVVVWM8Cv2ofEP9tfFKfTopd0WjWkVsArZXeR5jHqRnLhT0+7jtmv2QfD/wBk8K6z4lkTD6jeLBGSOqRLnI9i0jD/AIDVVVVSjjrVV4q8S6P4M8Mav4w8Q3X2fS9CsJ7++m258uCGMySNjvhVJr48/F3XPjx8XPE3xV8QFln129aSC3ZtwtbdcLDCPZI1Rc9yCe9fsB/Av/hT/wAD7TWdXsvK8R+NfL1O/wB64eKAr/o0J7/LGxcgjKvM4PSkjzSIDwKSIEcUseeBSRDFLHkYFJCDX7YP/BQi+8A+KLn4I/s82EWseMknNje6t5P2lbG6J2fZreHBE1wHIBzlFYbCrksE0T9hX9tz9oy0XxL8aPiPPo8F0oKWniTU57m4CjJTFsgKRLkn5WKFST8vNS/8Em/jz4aibVvA3xj8ODVY1IQK91YllPUCVFYgn0Iwe5FfszeCviR4F+Cnh7w38X/Euoa94vgWZtTvL69N25ZpnKKJSzFlWPYAScnGSATga74A8U/tKfty+LfgPefELUNOsdT8da+IJbkyXkNmLd7uUFIDIozsjKAArjd1xVz/AMEffE2jQNqng/8AaMiOs22GsxL4dksl357zR3MjJxnkIf1zXw7/AGoP2jv2JfjJbfB79pLVb3xD4Xgjijninuftj29q5/d3dpORvdVAI8tjjCsmEYArGIp4kmglWSORQyOpyGB6EEdRimiOOOaaMHginiBHSnixwKeMEYA/CnjxwKeMEcU8eOBUkYI4p0B4r/gpv8H4fB3xX034n6PZeVYeNLVheFFO1b6AKrscDC74jEcdWZZG9a/4I1fGOXTvF3jD4Eajct9l1e1GuaYrH5VuItkU6gZ+88bRHp0gOTwBVAZPWqqqqqoDjrVVVVVKMjisDGAaqqqqoDJxVVVVVUACaqvFHjPwj4JsP7T8X+JdM0e2wxWS9uVh37RyFDHLHkcDJ5HrXxD/AG+/hj4ceWx8CaRf+KrlFG24ObO1z3+ZwZDjn+DB4w3OR8QP2yPjl46lmhs/Ev8Awjdg7ZS20ZfIdR0GZuZScdcMATzgcAQwa14l1RYLaG91XUrxjtRFaeaZsc4AyzHArwF+xZ8cvG3lXN/ocHhmxk2kzaxJ5cm09cQqGkDAdnC8nGRzjwB+wP8ACnw6sN3421DUfFN4nLxM5tbbOOyId5wcnl8HjI658L+C/CPgqxGmeEPDWmaPbYUNHZWqw79o4LFRljyeTk8nnmqrUNO0/VrKXTdUsbe8tJ12ywXEQkSQejKcgj2NfE79jzStQSTVPhjeLYXWSzabeSFoW9o3wWU+zZHuorxD4X8TeDNUOl+I9IvNLvYjlUmQoTj+JT0Ye4JHvXwr/ar8X+CxHpPi8S+I9JHAeWT/AEqAcfdc/fGM/K3PTDADFeBfiP4O+JGlDVfCesR3SqB50DDZNA2Puuh5HpnkHsSOaqgBjNVVVVVVAZ4FYGMCqqqqeCC6gktrmFJopUKSRyKGV1IwQQeoI4xXx9/YgsNVF14t+DSxWN4A0k2gv8sMuF/5d2/5ZsSPuN8pLcFAMF4/E3gXxIFkS+0TXNHuQQGDQz20qnIPYqRwa+AX7b9pqH2bwl8aJ0tbk7YrfXkjxHIef+PhR9w/dG9Rt5ywUAtUE8F1BHc20ySxSoHjkRgyupGQQR1BHeqrGeBW3I5NbRjAqqqqqqqv2xf2aD4zsrj4reBLEtr1lCP7S0+CEE6hEv8Ay0UDkyqp5HO5VAHKgN8NPiN4k+FPjGx8a+FrhVu7MlXhkyY7mJuHikAPKkfkQCMEAj4W/Ezwz8XfBln4z8LXO6Gb93cQMfntJwAXif8A2huHsQQRwRRBFVVVVVVVVEAjFEY6Gqqv2kv2fdM+OPhdTYm3svFGmKW06+dMCRecwSEc+WxOQeSrcjgsG1bSfEngfxJNpOq2t3pGtaRcAOhJjlt5VIKsrA/RgwODwQcEGv2V/wBpWy+LGiQeD/Ft+kfjPT4juLgINSjX/lqgHG8Ljeo9Cw4JC1VMMjiqqqogEYojtVVqN/Y6TYz6nqV3Fa2lrGZJppWCrGoGSST0GK+Ofxz1X4sar9hsTLZ+HLKQm0tCcNMw482XHViOg6KDjqST8A/gFffE6+TxB4gjltfDFrJhmGVe+cHmOM9l7M/boOcldP0+x0qxg0zTLSK1tLWMRwwxIFWNQMAADoMVVVVMPSqqv2hvj3onwJ8JLqMsSX2u6juj0rTy2BIwHMkmORGuRnHJJAGM5HiHXvE3xF8WXWvavLNqmua3dAv5cWXmlYhVVEUfRVUD0Ar9k39mNfhJpv8AwmnjW2gm8XajEBHEQH/sqIg5RW7yMD87DpjaDjcWqmHPFVVVVVTDjA7VVVVVTDIwKqviL4Wj8b+AvEPhB1UnV9Nntoy38MjIQjfg20/hVleXOnXtvqFlL5dxayrLE+0Ha6kEHB4PIHWvDWt2/ibw5pXiO0CiHVbGG7iCtuAWRA4wcDPBHOBVV+0P40f4dfAb4h+OYLv7Pc6J4Zv7m0k8xoz9oWB/JAZeVJk2AEcgkV4f0W68Sa9pvh2xkiS51S7itYWmJCK8jhFLEAkDJGcA/StK0ux0TS7PRdLg8mzsLdLe3i3FtkaKFUZJJOAAMk5qqqqogEYojtVVXj3xVPqt9LpNpLiytn2naf8AWuOpPsD0/P6VWn6lf6VcLd6ddPBKvde/1HQj2NeGfiBaauUsdVVLa7PCuDhJD2x6H2/XtVVVVVUQOgNVXxFuBD4Zki3AefMiAEdcHd/7LVVVV8O/+RaT/rs/86qqqqYZAxRHOKqqqqpgCKqqqvGV19k8N3jK2GkURr1GdxAP6Zqqqqqq+HN0JdKuLMtloJsgegYcfqDVMMjg1VMMjg1VVXBGKqqqvGPiA6pdfYbSTNrbnqP437n6DoPz71VVVVXhGzFnoFsNoDTAytx13dP0xVVVVVVVVVVVVVVVVVVVVVftSfFZfgl+z/42+I8dykN7pulvHppYE/6ZNiK34HJHmyITjHAJyACR4N8Kav488X6F4H8PRJJqniHUrfTrFHJAaeeRY4wcAnG5h0Brwb4U0fwJ4P0LwP4eiaPS/Dum2+nWKOQWWCCNY4wcADO1R0AqqqqqqqqqqqqqqqqqqqqqqqqmhhuIXt54kljkUq6OuQwIwQQeoI7V/wAFLv2Dn/Z+8ST/ABn+Fum5+HXiG8P2ixt7dgPD9y//ACzOBtW3dj+7PAUny8fcLfsVftfa7+y149A1OW+1HwJrUgXW9JhcHY2ABdQq3AlQAZAK71G0nhSvhTxV4c8deGtM8YeEdXg1TRtYtkubK7gJKzRsMgjPIPYggEHIIBBFVVVVVVVVVVVVVVVVVVVVVVVVVVUeRiqqqUcdaqlHHWqqqqqABIFf8FJP2ev+F5fs+Xuu6JY+f4p8AeZq2m7Ey81uFH2uAd/mjUOFAyzwRqOpqv2EPEg8V/shfC/VBdSz+Rov2HdKuCPssr220ey+TtHsBVVVUo4qqqqqgMkDNeMtdbxR4s1jxCzEjUL2WZAf4VZiVH4LgfhXwW0E+HPhZ4c014vLlayW4kUgghpSZCDkA5G/BHbGO1VVVVAcdaqv+Ct3xu/4Qb4Iad8I9H1PytW8fXf+lxxvh10+3KvJnAyoeUxL1G4CQcjcK/Y7+DkXxv8Aj34f8L6jZLdaLpzHU9Zjc/K9rCy5Rh3V3aOMjriQ8jrSJmkTPAqOMEUiZ4FJGAMUkWetLGAMV+1v8W5vgb+z34u+IGnz+XqsNoLTSiACRdzsIo2AJGdhYyH2Q9elf8EoPgBpOsQ65+0f4rtftuoW98+l6CtzFu8lwivcXSlh8zN5ixq6njEwOd3CxZ4AoQ5oQA8V+z0gP/BWfVU/6nnxb/6J1CjAMcD9K/4LKHRl8Q/CyGAL/ay2WqNdfIQfIL23k5PQjcJ+ByOc9RXwj1DVfC37LngrXdct57nUtI8A2FzeRXEpEks0VgjOrOcncWUgtzyc81+x5+3EP2tPEviHw6Phd/wi39g2Md1539t/bvO3vs27fIj2465yfpTxg8Yr4/8A7ew+Bnx9sfgcPhP/AG39t+xf8TT+3vs2z7SwH+q+zvnbn+/z7U8YIwRUkQxTp2NPHjpUkYI4qRMVJGMcV+3r8Nbb4ifs0+J5ltt+oeF0Gs2bhQSnkcze+DAZemOcHtiv2XviP/wqP9of4e/EKS7+zWuk69b/AG6XzNm20kbyrnncoH7mSQfMwXn5uMiqUcdaqqqqqscYzVVVVVKBwPSqqqqqpQMVVVVVVXl7Y6ZZy6hqV5Ba20CF5Z55BGkajqWY8Ae5r4hftr/BLwQ72Wl6pceKb1CV8vSFDxA88mZiEI46pu6j618Q/wBuv4weLt1p4UWx8JWRckfZF8+4ZT0VpZBjjjlEQ59uK1fXPEPirUvt+vaxqOsahLhPPvLh7iV+eBuYknknj3rwB+yR8cviHFDeWnhT+xrCdNyXmsyfZVI7EJgykEcghCCOc8jPgL/gn38PdH8q7+IHiTUfEVwu0ta2w+x2+f4lOCZGHbIZD14GePBvw88D/DzTxpfgnwtp2jwfxfZoQryH1d/vOeByxJ4HpVUBxVUBk1Xi7wT4V8d6U2j+K9EttQtyDs81PnhJGC0bfeRvcEGvip+yP4k8PSSat8OpJdc07ljZSYF1APQdBKOvQBugwetaTrGveE9YTUtG1C80vUrNyokidopIyOGU9/UFTweQa+E/7Xtlf+VonxShS0uCwWLVbaPET54/eoPuH/aXI56LjJsb2y1O0hv9NvILu1uEDwzwSB0kU9GVhkEH1FYGMCqqqqqrGeBWABgVVVVVVfGf9n7wD8bdM8rxBaGz1eCNls9WtQFmhJxgMOkiZAyrdidpUnNfGH4E+Pfgrqws/FFh52nzsRZ6pbKzW8/XA3Y+V8KSUPPfkc18A/2o/GPwYuotHvnl1rwq8n77TpHy9vkjc8DHlSAD8hOwknoTuHw7+I/g/wCKnhyHxP4L1ZL21fCyoflktpNoJjkX+FhkcdO4JGDQUetYGMCqqqqqqqqq/bK/ZrXwreXHxb8CWCpot3JnWLKIH/RJ3Y/vlHaJyQCP4WPHDYX9n746618DfGC6rCJrzQ77Eeq6csmPOTtIgPAkTqCeoyuQGJHh/wAQaJ4s0Sz8R+HNTg1DTdQiEttcwtlXU/qCDkEHBBBBAINFQelEEVVVVVVVVEAjFEY4qqr9pr9nTSvjR4dk1fRrWKDxhpsJ+wXAIT7Uo58iUngg87SfuseuCwP/ABUXgrxF/wAvuj63o91/tQzWsyH81YEV+zV+0VpPxu0D+zNRZLTxbpduG1C1HAnQYX7RF/skkZX+AsB0Kk1VMAOlVVVTD0qppYbeF555UjjjUs7u20KAOST2GK/aM+PDfEjUP+EY8K3c0fhqyf5mzt+3yA8OR12D+FT/ALxAOAPgV8Fr74t66z3by2mgaewN7cqpBkP/ADyjOMbyOp/hBzzkA6LouleHdKtdD0Oxis7GyjEcEES4VFH8yepJ5JJJ5NVVVVMMjAqq+Mfxf8K/BXwfN4q8SyGWRiYrCxjbEl5NjIRfQcZZjwo9SQD8QPiB4p+J/im78YeMNSN3f3ZwAOEgQfdjjX+FFzwPqSSSSf2QP2Xh4ItrP4rePLbOv3lvv0ywkTH9nxuv33B5EzKcbf4ASD8xIWqpgCOKqqqqqvj9+23+zn+zi8unePfG6XeuxqT/AGDo6C8vcgZAZAQsRPGPNZAc+mTXxZ/4LJ/FXXftOnfB34faL4WtXwsV/qjnULtRx8yr8sSk9MMsgwfXkeOv21P2r/iPJI/if49+Ltk0bRy2+m339mQSIyhWVobURowKqBgqRyx/ibOteINe8SXSX3iLXL/VLmOMRpNe3LzuqAkhQzEkDJJx0yT61Wh+KvFHhjzv+Ea8S6ppP2nb532G8kg83bnbu2EZxuOM9Mn1rwf+3Z+1/wCBmLaJ+0D4ruSzM2NYuV1bkgA8Xay4GAMDoDkjBJrwJ/wWR+PuhukPj3wJ4R8U2wkDM0Cy6dcFd3zLvVnQfLkA+XweTu6V4B/4LLfA3XGjt/iH8OfFfheZ1GZbR4tTgQ4YkMwMUmOFAIjOS3IUDNfCX9qT9nv46yrafCv4saHrd86uy6cZGtbwqn3mFtOEm2jH3tmMYOcEVVfHXw8nhb4x+MtDiTZFDrFw8Kc/LHIxkQc8nCuoyevWv2S/Eh8TfADwrNJMZJ9PhksZQxY7PJkZEGT/ANMxGeOBnHbFVX/BTTxU3hb9jDx2tvctDdaw1lp0BEYcMJLuIyqc5ABhWUZ6gkYwcGv2HvCzeMP2ufhRpKRzubfxJb6hiAgMPsmbkk5B+UCHLd9oOMHmqqqqqpgM4Bqq1q9OnaReXyMA8EDsmf7wHH64piWOSck9Saqqrwv4/udLRLDVg1xaqAqOPvxD+o/X+VWGo2OqW63Wn3Uc8bDqp6fUdQfY1VVVUw54qq+Kd0FtbCxDcvI0hGem0ADj/gR/Wqqqr4YzB9GuYMkmO5J56YKr0/I1VVVUwyOtMOaqqqqpulVVVXxLuwljZ2IPMspc/wDARj/2b9KqqqqqvAF+LXWjaOwC3cZUZOPmHI/qPxqmGRwaqmGRwaqqriiBnAqqrxt4gFnbnSbST9/Mv70j+BD2+p/l9RVVVVVVbw+RbxQZH7tAvAx0GKqqqqqqqqqqqqqqqqqqqqr/AIK/fGrjwp8AtG1L11vW4on/AN6O2jfA/wCuzlSf+ebEfdNf8EgvgmPiR+0nP8SNStfM0n4baf8AawxGQb+43RWykZH8IuJAeeYhxzkVVVVVVVVVVVVVVVVVVVVVVVVV4h8PaF4t0K/8MeJtItNU0nVLd7a9sruISxXETjDI6nggg4xX7e/7DPiT9krxn/b2hrLqXw18Q3jpomonLPZyEM4srg/89FVWKt0kVSwwVdV/YJ/be1j9nvxLa/Djx9qbXPw11e5xIZiznQ5XPNxEACfLJ5kjA771w24PYX9hqthbappd7BeWd5Cs1tcQSCSOaNgCroy5DKQQQRwQc1VVVVVVVVVVVVVVVVVVVVVVVVVDqKPNVVUo461VKOOtVVVVUACcVX7VH/BOD4z+GvjFqs/wQ+Ht14h8G69dyXWlrYNGv9nByGa2dWYbEQvtRjwUAOchsfsffCTxH8C/2bfBHwu8XSwPrWkW08l6sDBliluLmW4MQYEhinnbCw4JUkcEVVVVAcYqqqqqvinra+Hfhz4i1fcoaLT5UjLEAeY42J1/2mHHfpXh/SZNe17TdDhz5mo3cVsmCAcu4UYzx371BBDawR2tvGEihQIiDoqgYAH4VVVVUAMACqr/AIKNfGOX4w/tUeKDbXqT6P4PYaBpgRSABblvPP8AtE3DTkN3XYMkAGv+CXXwrj8O/CbVvinewKb3xhfGG0cjJW0tiycHGQWmM2QDg+WnccIgAwKjjBwBSJngUiYFRx5pEHQdaSMYr/grbeXdn+zLosFtO0cd74ztIbhR/wAtEFpduFPtuRD9VFf8E17G1h/Yy8AzQW0Ub3MmpyTMiBTI41G5UMxHU7VVcnsoHQUsQxSxDoBmhESMAD8qHxlsP2fP+Ch3jP4u6noc+sWugePPEglsoJhE8gmku4MqxBGV83dg9duMjORrf/BY/wAMSadJB4O+Bus3Wry/Jax3uqxpFvPAJ8tGZucfKAM9MjrXwq/Zm/aO/bn+Otv8cv2jfD8+geEhLFJcfarNrRbm2hYFLG0gc+YI2yQZGyAC7bnc4b4m20UHwu8VwQRJHHHoF4qIi4CgQPgADoMV/wAEckD/ABL+IYHbQrb/ANHmniGMYr9v9dv7fmhr/wBgT/0YKeMEU6Y4NSRgjFOnY1ImOlSICMinQDite0Wx8QaNf6DqkXm2epWsltcR/wB6ORSrD8QTXijQLzwp4l1bwtqKst3o99PZzhoyhEkTlGyp5Byp4PIr9mvx1cfE39n74d+O72dpr3WPDdlLeyNuy1yIlWY5YliPMV8Ekkjkk5oDiqqqqqAyQAaqqqqoDJxSjiqqqqqoDjFVVVVfEX9on4P/AAv8238UeMrVr+LI/s6yzc3G4Z+Uomdh4I+cqM96+JH/AAUG8Q6isth8LfC8WlRE4XUNUxNPjJ5WJTsQ9OpcdfY142+JnxA+I9419438X6lq7tJ5ixzzHyomxj93EMJHwTwqgcn1r4e/AX4tfFLZN4O8F3tzZlwpvpwLe3XPUiSQgNgckLk9OORn4ef8E8raM29/8UPGjSkENJpukR7QeOhnfnGeCAg4HDc5Hw/+Cnws+F8a/wDCEeDLCwuFXBu2UzXDZznMrkvzk8ZxzjGKqqrGeAawMYFVQGBiqqvin8CvA/xUt5J9RtFsNYwPL1S2jAl4GAHHSRcADB5A6EV8Uvgb43+FVw0urWgvdJZ9sOp2qkxHJ+UOOsbEdjxnoT1r4ZfGfxx8K7wNoGomfTnfdPptwxaCTpkgfwMQANy4PTORxXwq+PXgf4rxC0065Onayo+fTLtgJDgZLRnpIvB6cjGSACM1VVVVjPAoAAYqqqqqqqtd0HRvE+kXWgeINMt9Q069j8u4triMOki+4PuAQeoIBHIr9oX9i/W/A4ufF/wshudY8PrmS408nzLqwXuV7yxj2y6jGdwBevh38SfGHws8Rw+J/BervZ3aYWVDzHcxggmORf4kOBx17gg4Nfs//tQeEPjZbRaLdFNI8WRxFptNdjtuNqjdJAx+8vJOwncAD1A3VVVVVVVVVVXdpa39rNY3ttFcW9xG0c0MqB0kRhhlZTwQQSCD1r9qP9ni4+CfiddS0KOWXwlrEh+wSu+9raTGWt3PU4HKk9V7kqxr9kP9pBvhlrSeAfGmpY8J6rL+5nnkO3S5zn5h1xG5IDDgKcPkfNmiARiioPQ0R2qqqqqqqiARiiO1VVftXfszr8WdNHjPwXbQReLdOiIkiACf2pEAMIzdpFA+Rj1ztJxtK+FPFfiXwB4ltPE3hnUZtO1XTZcxyKMEEcMrKeqkZBU8EEg18CvjVoHxv8Gpr+mL9m1K02xarYnObaYj+E90bBKn04OCCBVEAjANEdqqqogEYojtX7UHx5OpzXPwz8G30i2sDmPV7uJwBOw4MCkc7QeGPc5HQHPwk+FOt/FrxKNF0xxb2dsBJf3jjIt4yccD+JjggL3PXABI8N+GdE8H6Ja+HfDthHZ2Fmm2KJB+bE9SxPJJ5JOaqqqqqiBnArx1428O/DrwpqHjLxTe/Z9O02IvIQMs56KiDIyzEhQPUjp1r42/GTxH8bPGlx4m1mWWGyiLR6Zpxk3JZQ5+6MAAscAs2Mk+wAH7Gn7MX2lrL4yfEHT4JLYoJdB0+ZdxZsgrdOM4AGDsUg5J3cYUmqqiARjNEdqqqqvF3i7wv4C8Nah4w8aa9ZaNomlQma8vryURxQpkDJJ7kkADqSQBkkCv2u/+CrPjn4h3F94F/Zzur3wl4ZVnhl19WMWo6gA3DwsMNaoQMjH7znkpyteGPCfjD4h+II9C8JaFqev6xeMXEFpC08rc/M7YzgDOSx4HUkV8Kf8AgmB431uO31P4u+L7Xw7C6hm0zTFF3dDrlXlz5UbA4OV80EenbwZ/wT//AGZPCVuiXHgm48Q3SZH2vWb6SVmBAGDGhSLtnOzPJ5rSPgn8INBEv9hfCnwfp3n4837JoVtD5m3ON21BnGTjPqa/4Vr8Pjx/wgvh7/wWQ/8AxNa98CPgx4njMfiD4T+EL8mJolefRYGdFbrsfbuQ+6kEHmvGX/BPX9mXxazzWnhK+8O3EikNLo2oPHzhQCI5PMjGAvZRnJJyea8Y/wDBKiwd2m8AfFq4hXedttrGnCU7SeMyxMvIGB/q+Tzx0rx1/wAE6v2kfB8L3WlaTo/iqBOT/Y99+8VcnrHMsbE8DIXd94Yzg41vw94p8F6sNN8R6HquhapBiQW99bSWs0eDw21wGHIPPqK/Z6/4KaftG/BGWDSvEust8RfDSN8+n6/cu91Gvfybz5pEPAAEglRR0Qda/Zl/bO+Cn7U+l7fA+tNp3iS2h8y+8N6mVivIADgugBImizj50JwGXeELBa/bk0IaR8eru/C4/trS7W7+9nO1TBnHb/UdPx71/wAE/NXe6+Fuu6LI0rGw1xpELPlVSSGP5VHb5kY/VvrVV/wWL1m5079lzRNNtbxIxqvjO0huIiFJliW1upOM8gCRIjkY7DOCQf8AglZotrqv7Znhm+uJJVk0fTNSuoAhADObV4SGyDkbZmPGOQOcZBqqqqqmHvVV4y/5FfUf+uB/mKqqqqsNRvtMnFzp908EgGMqev1HetI+J8qBYdasxIAMedDwfxU8flitN8T6Dq+1bLUomkOMROdj59MHr+FVVUw96qviBqS6j4hkijbKWaiEYPG4ct+px+FVVVXwvugt5fWJ5MkSyD22nB/9CFVVVVEAjGaYe9VVVVURkYFVVVXxBvPtOvG3VsraxKmPc/Mf5j8qqqqqqrW5ls7mK7gYCSFw6EjuDWlalb6vYRX1swxIvzLnJRu4NMMjAqqYZGBVVVcUwGcCqrxDrcOh2JmJDTyAiGM/xHufoP8APWp5pbmZ7ieQvJIxZmPcnrVVVVVVBKLi3jnUACRAwGc9RVVVVVVVVVVVVVVVVVVVVWva5pHhjQtR8S+INQisdL0m0lu726lOEghiQvI7eyqpJ+lfHr4s6v8AHP4veKPijrEku/W753topODb2y/LBFgEgbI1ReD1BPfNf8EyfgDL8C/2W9DuNb097XxJ44b+3dUSQndEkoAtoyDgqRAI2ZSMq7uD0qqqqqqqqqqqqqqqqqqqqqqqqqq+Jvwy8D/GLwPqvw5+I3h+31nQdYhMVzazDGO6ujDlHUgMrrgqQCCCK/bU/ZA8V/sifE9/Dly11qnhHVt0/hzW5IdouYh96GQj5RPHkBgMZBVwAHAH/BPv9uvUPgzrVh8G/ipq/nfD/Up/Lsb25l50CZz13H/l3Zj86nAQkuCPnDRSxTxJPBKkkcihkdTkMD0II6jFVVVVVVVVVVVVVVVVVVVVVVVVUACcUcdqqqoDjFVQHGKqqqqpQCaqqqqqgOMVVVVVX7VetDTvhkmlpJ8+q6hFEy7sZRAZCcdwGRB26iv2d9GXWfi7oaSDMdm0l0/IBGyNivXP8e38KqqqqrAxgVXxl+IMHwn+E3jD4lzrFJ/wjWi3V9FFLnbNLHGxjjOCD87hV6j73UU7at4m1suzSXup6rdZJPLzzSN/Nmb9a+F3gWw+Gnw88N+ANNbfB4f0yCzEh6ysiAM56cswLHgcntUa5pFwABUaDFRpnmkTjA61GnFImeBX7fXwu1L4p/sq+M9I0OGebUtIhj1e1ghAJm+zOJJE29WJiEuFHJbbjPQ/8Eifjro9x4Y139nzXNQt4NVtL19V0NJXVGuoZFAniQcFmjZPMxydsjH7qcLFmlhBHShDkcCvgHpthq3/AAVr1rTNTsre8tZ/HXi9ZYJ4hIkg8nUeGU5BH1rSvAHgrQbwahoXg/RNOulUqJ7TTooXAPUblUHBpocjAFfE+0ln+G/iuCCJpJZNDvVREXJZjA4AAHU+1f8ABG68s1+LXj7S3uEF1P4cimiiPVkjuFDsPYGRAf8AeFPHxg1+39c2s/8AwUE0mC3uYpXtn0OOZEcExOWRgrAdDtZWwezA9CKkj9KdARinTHFSxjGBTrke4p1HSpUHQVIoIr9uLwd/whf7Unjuyji2walepqcLCPYH+0xrK5AyekjyKT3Kk4GcV/wSb8WjxJ+x/pujCTd/wiuvahpuPL27dzrd4z/F/wAfec++O1YGMA1VVVVSjJqqqqqlGaA4xVVVVVAZOKqqrxL4s8MeDdMk1nxXr9hpNlECWmvJ1iU47DJ5PQADJJIA618SP2+/AGgNJp/w60W68S3S5H2qYm1tlOccZG9+h/hUdOTzj4i/tT/Gz4lFoNS8XS6VYE5Fho260i6YIYgmRx7OzDPTFeGPB/irxrqS6R4R8O6hrF4cZhs7ZpSoJxubaPlX1Y4A7mvhz+wF8QvEKR3/AMQNds/DFsSp+yxKLu5ZepBCsET0B3MQc5Xjn4e/snfA/wCHWyey8Jx6zfI4Zb3Wtt3IpHQqpAjUg85VAc9+BiqqqqlAPNVQGTiqqqq4tre8t5LS7gjngmQpLFIgZXUjBUg8EEcEGvi5+yHpuqfaNe+F7pY3jtvbSZXxA/r5TH/VnvtPy84G0YFajpmveEtZay1K0vNK1SwlB2uGikidTkMD1HIyCPqDXwk/a71LS/s2gfFBXvrNF2JqsSZnTHTzVH+sGONw+budxJNaRrGleINMt9Z0TUIL2xu03wzwvuVx7Ee4II7EEGqqqoDjNVVVVVVUBngUAAMCq/aG/Y48PfEf7T4t+HaWuh+JmzJNbBNlrqDd9wH+rkPXeBhj94ZJcazovifwH4jk0vWLO90bWtKnyVbMckMiHhlYe4yGU4PBB71+zf8Atowau1n4F+MV7HBenbDZ66+FSdicBbjHCN90B+h6tjqUeOWNZInV0cZVlOQQehBqqqqqqqqqvHPgnw98RfCuoeDfFFn9o07UYikgHDRnqroecMpAIPYgda+MXwn8Q/Bvxvd+ENeTei/vbG7UHZdQEna49+CGHZgRyME/sW/tGnXbW2+Dfja+Lajax7dCu5GZjcxKGZoHJzhkUfIehUbeCo3VRAIxRGO9VVVVVVUwyMiqqq/bH/ZpXV7e7+L3gHT4kvbaNpdcsYYyDcqOWuEA43gcuMDcAW6g7vhT8UPEvwi8ZWfi7w3cuDCwW7td+1LyDILxPweCBwcEqcEcivhv8Q/DnxT8H2HjPwxc77W8T54mYGS2kH3opACcMPT0II4INVTD0qqqq/ac+Oq+DrGb4f8AhO9X+3b2LbfTITusYXXoCOkjAgjuoOeCVNeB/BWu/EHxLZ+FvD1sZbm6cb3IOyCPI3SOR0VQcn8AMkgH4cfDzQvhl4WtfDGhxhvLG65uSgV7qU/ekb69AMnAAHamGRxVVVVVVqeoWGkadc6tql5Fa2dlC81xPK21Io0UszsT0AAJJ9q/ad/aDv8A43+LDa6ZI8HhXR5XTTIAzD7QckfaZFOPmYdAQCqnHUsT+yH+zW/xR1lPHvjOxYeE9JmBhgmi+XVZlJ+QZ6xKQN56E/Lz821I44o1iiRURBhVUYAA6ACqqqphzgVVVXiDX9D8K6HfeJfEurWumaVpdu9xeXl1KIoreJBlndjwAAOtft1ftw+J/wBqrxfJ4f0Ce40z4baJdN/ZOmhirXzrlReXA7uwztQ8Rq2BlizN+y1+yN4v/aP1Z9Rkmk0bwfp0ypfaq0eWmbILQW4PDSbTkk/KmQTnIVvhP8Fvh38FvDkfhr4feHLfT4diC5udgNxeugwJJpMZduSfQbjgAcUsORgD86WDIwTQgB6CvJBHSjCMcCmgHpTQcYBp4eORXjr4ceCPiVoknhzx34X0/XNPkz+5u4Q+w4+8jfeRv9pSCPWvj3/wTOeKO58RfALU2kKqCPDuozAFiMAiG4cgZ77ZMd/n6CpoPG3ww8XhJV1nwt4n0K5V0IMlnd2Uy4ZWUjDo3IIYY6gg14e/ae8c/tJ6NY/8LMjtrzxJ4SthZy61HEsT6jBI7vF5qoAA6ESDIABDA4zuLf8ABO3UfK1nxvpHk5+02tnP5m7G3y2lXGMc583rnjb78VX/AAWm/wCSX/Dj/sPXP/ogV/wRq/5Of8T/APYhXf8A6X2FEds1VVVVTAEdaqvE9v8AavDupQgMSbWQqFHJIXIH5gVVVVVVVp3irxBpYVLTVJdi9I5DvX6YOcfhWn/FKdcJqumo/wDtwNtP/fJzn8xVh458M34UDUVgcj7lwNmPx6frUU0NxGJbeZJEPRkbI/MUwBHWq8WeNbTSbd7PTZo571wV+U5EPucd/b8/d2Z2LuxZickk5JNVVVXhbURpevWd0zAIX2SZ/utwT+Gc/hVVVVRAIxmmGR1qqqqqqI7VVUzKilmYAKMkk4xWqXZ1DUbm9Jz50rMM9gTx+mKqqqqqq8I+Im0S98mdybO4IEg/uHsw/r7fQUrpJGHjcMrDKkHII9RVURkYFVVVTAZwDValqFtpdnJe3TgIg4Hdj2A9zWr6rc6zeveXLdeETOQi9gKqqqqqq8E6sl/pYsnf99ZgKQe6fwn+n4e9VVVVVVVVVVVVVVVVVVVVf8FWf2mLDwz4Gj/Z18J6qj674jaOfxB5EgJtLFSHSF8HKPK4jbH/ADzVgRiQV+wh+zvP+0n+0f4b8H3dn5vh3SJBq3iFmjDJ9jgZSY2ByD5rlIsf9NCegNVVVVVVVVVVVVVVVVVVVVVVVVVVV8Z/gx8PPj/8PNS+GXxN0NNS0fUVyCMLNaTAHZPC+D5cqZOG9yCCrMp/a8/ZD+IX7I3xCbwz4mjfUvD2os8mga/FCVh1CEHlSOfLmQEB4yeMgglWVj/wT/8A+CgEvwyl0/4I/G7WGk8HyMsOi61cPk6KTwsMrHrbdgx/1X+59yKWKeJJ4JUkjkUMjqchgehBHUYqqqqqqqqqqqqqqqqqqqqqqqqpQCcZqqqqHQfSqoDgCqqqqqUA1VVVVWM8Cqqqqqr9sLWC+oeHPD6SACGGa6kUPkncVVSV6jGx8Hvk+lfseaV53iXxDre3/j0sY7fO7p5r7un/AGx/zmqqqqh1FVX/AAVi8ft4Q/ZSn8NW1yi3HjPXLTTmQSAP5MZa5dgM5K5t40JH/PQA/er9h7wP/wAJ5+1D4GspUzb6TenVpmzjZ9lQyxnG4E5lWJeM/eyQQCKRRgAVGueBSKCeKRQQAKjQYqNM80iA8Co0B4FInGBX7W3/AATo+IfgDxz/AMLv/ZJsbo28d2t6dC0ljFd6PcBlIkswDl4y2W2Lgx9ACv3fA3/BW346/DfzPCnxx+Edh4j1XToxFK7SPoV75nB3Tp5UiZweiRRjkH66h/wWd8Tarbrpng39nC1h1e5kWO1Nz4jkvVZ2OAohjto2YkkAAMP1r9lv4i+O/i98CfC/xC+JnhOXw34m1aOd77TnsprVY9s8ixtGko3iN4hG65LfK4+Zup/ZxXP/AAV+1ZR/0PvjD/0TqNGIkYBpojjpTxA8Cviz4A+Mn/BNn9pz/hO/AdjKPDM93MdAvZ0aa01DT5G3NYzt/wA9FUBWGVfKLIuMqaj/AOCtnxU+I8SeDfhD+zOs3jC/UJaFNWl1Qb/4mFtHbxsR3/1mB3yK+M3wY+K/wj/aC+G+ofGvU59Q8ZeOJbPXdalcrIsF1JqMiNAJUyjskccLME+VTJtXKhWMsYxUiY6VIgI4p1yMVIoB4qVewqRQRxTqM4Ff8FU/Di6d8cvDviOGOBE1fw1HHIFJ3vLDPKCzDGMbHiUc/wAJ6d/+CKnib7R4X+KXg1xj7Bf6dfx5mzv86OaNsJ2x9nTLd9yjjAzVVVVVKBiqvL2y0+H7Rf3kNtEDjzJpAi5+pq9+Ifw/06UQah468PWshXcEm1OFCR0zgt04NXHx4+D9skjyePtMYRAlhGWkJx1wFBLfhnParz9qD4K20PmweKprtgceXDp1wG+vzoo/Wr39rn4TWsQkgTW7xi2PLhs1BAx1+d1GPxzzV5+2V8PUh3WHhvxFNLnhJo4Ylx35EjH9Kl/bR8OLC5g8D6i8oU7Fe6RQTjgEgHA98H6V/wANtjGB8Mv/ACtf/aKvP21dckm3WHgKxhixwk188jZ78hV/lX/Dafij/oStK/8AAiSr79s7x9IynTfDHh+BQPmE6zSkn2IdcfrX/DZfxQ/6AXhb/wABZ/8A49X/AA2X8UP+gF4W/wDAWf8A+PV/w2X8UP8AoBeFv/AWf/49X/DZfxQ/6AXhb/wFn/8Aj1D9sv4oDpoXhb/wFn/+PVa/tnfEZJ1a98OeG5YRnckUM0bHjjDGVgOcdj/Wv+G1fFH/AEJOlf8AgRJV5+31eaXMq3vg/TZ9rAvDb3T7yO4BwQpx6/ka+IX7fXxL8SQNY+BNFsfCcLAbrguL2599rOoRQR/sEjswrxD4l8ReLdTfWvFGu3+r38gCtc3tw00hA6DcxJwOw6CtM8K6tqToWga3hJ5klGMD1C9Tx+HvXws8Hfst6FcRXfj+Xxbr91EN+y4sktrQsDwvlxTO7dR95tp28jnFeFP2iP2eNBsF0XQZ18P2MAzHDFpEkcZOAOFiU84A5I7dasf2h/gvfwmaDx9ZKA20ieKWFs4HZ1Bxz16Va/Fr4W3scclv8RvDZ837iPqkSP8ATazAg+xFWWt6Lqcpt9N1iyupVXcUhuFdgOBnAJ45H51VVVUAAMVVKOKqqqgBiq+Ifwv8GfFDShpnivSxK0YP2e7iwk9sSOSj4OOx2nKkgZBwK+Lv7O/jL4WvLqcaHWNABG3UbeMgw57TJyU543cqcjkE4Hw1+LvjX4V6h9q8NajutJWzc6fcZe3n4AyVz8rcD5lweMZxkH4TfHXwb8WbRYbCX+ztZRczaXcSAyDAGWjPHmJz1wDxyBVVQGeKwMYFVVVVVVBc96wMYFVVfGn4B+Bfjho4tPEVsbXVbaMrY6tbKBPb9wpz9+PJOUPqcFSdw+LfwX8c/BjX/wCxPF2n5hl5s9RgVmtrsY52OQPmGRlTgjjjBBP7N37XOs/DB7XwZ48luNV8KErFBMWLTaUvTKcEvEBjMfUAZXptbw/4g0TxXoln4j8OanBqGm38QltrmFsq6n9QQcgg4IIIIBBqqqqqqqqqv2gPgnpPxx8DSeH55IrXV7MmbSb6RSRby8ZDAclHA2t1xw2CVFahp/iLwN4ml0/UILjStb0S7w6H5ZLeaNsggj0IBBHB4IOK/Zh+Pdr8bfBYTVJoo/FOjhYtUgAC+cMfLcIBxtfnIH3WBGACpNUQCMUV9KqqqqqqphjpVVVftgfs1HwVfT/FHwNZE6DfTE6lZQwgLp0rdHUL0iY+2FYgZwwA/Z6+O2s/A/xcL6NTd6FqRSLVbIsRuQHiVPSRMnHqCRxnI8P+INE8V6JZ+IvDmpwahpt/EJba4hbKup/UEHIIOCCCCAQaojIwKqqr46fF+y+EvhQ3Vv5U+uX+Y9NtX5BYY3SOBzsUH8SQOMkg/wBueK9c63Wqatqlx7yS3Ern8ySTXwM+Den/AAl8NCOdYrjX79Q2o3aZI4JKxJnoi5/4Eck9gKIBGM0R2qqqqqv2z/2km8XalP8ACbwNqWdD0+XGq3cEhAvp1P8Aqh0zGhHPUM2D0UE/s4fAfU/jl40WylWWDw7phWTV7xDtIQniKM4I8x8HGegBPYA6JouleHNGsvD+h2SWmn6dbpb2sCZxHGgAUDOSeAOTye9VVVVMPSqqppYYIXnnlSOONSzu7bQoA5JPYYr/AIKJft/an8cdZ1H4K/CfU2tfh3pd00V9e28oLeIZo2+/uUkfZQwyig/PgO38Kp+yX+y7r37RvjaMXkF1Z+C9KlB1nU4xtJOMi3hJ4MrcZ67FO4/wq3hPwj4e8FeHrDwn4T0i30zSdLhENpaQLhIlH6kkkkk5JJJJJJNJCPTNLDkdKWAEYAzQgJ6DmhATxj9KMGOMfpTQH0FNACMYp4OOBmpIQRxUkPYiv2i/2WPh1+0RoTQa7arpniG3iK6dr1vFma2PHDrkCaPjGxjwCdpUnNeB/hH44+BfxP8AE3w98b2nkzLaxzW08eTDfQhyEniY/eXkj1UkggEEV+xh8SPCHw7+MkcXjLXo9Lg8TWLaTYSTkrFJePLE8SM33ULCN1UtgFmVQcsAar/gtpf3kdh8HtLjuGFrcTa1NLEOjSRiyVGPuBI4/wCBGv8Agi7pdhN8Y/H+tSQZvLXwzHbwybiNsctyjOMZwcmGM5IyNvGMnLDniqqqqqYZHFVUsSTRNDIMq6lWHTIPWrm3ktbiW1mGJIXKMMYwQcH+VVVVVVVVUM89u/mW87xPjG5GKn9Ki8U+I4VKprd2c/35S3881d65rN+pS71W6lVuqNKdvTHTp0qqqqqq8Ha0mtaLCWfNxbKI5geuR0b8QM/XNVVVVMBjAPSqqqqqo9aqq8WX39n+H7uZT87p5aYz1bjP4Ak/hVVVVVVVV4M8WJbBNH1OX92TiCVj9z/ZPt6Ht9OlVVVVUw569q1nxFpuipi4l3zEfLCnLH6+g+ta3rt7rs4luSFjTPlxL0T/ABPvVVVVVVVaRqc2j38d9Bg7eGU/xKeoqxvbbUrSO8tJN0cgyM9R6g+9VVVVVVVVVVVVVVVVVVfEz9oH4J/By3lm+JfxP0DQpIlLG0nuw902FDHZbpulc7WU4VSfmX1FftGf8FapNS0+78K/s4+H7mxeeMxnxLq8aiWLPUwW/wAwB9HkJ/3OhrTdM8ffFvxqmn6Zaa54v8WeIblmCIJL28vpjlmY/ed2wCxY54BJOATX/BPb9j9v2TvhA9t4rhsZvHnimZbvXZ4MP9mUKBFZrIPvrGNzEjgvI+CVCmqqqqqqqqqqqqqqqqqqqqqqqqqqqr4z/Bj4efH/AOHmpfDL4m6GmpaPqK5BGFmtJgDsnhfB8uVMnDe5BBVmU/tefsh/EL9kb4hN4Z8TRvqXh7UWeTQNfihKw6hCDypHPlzICA8ZPGQQSrKx/YE/4KCP8LPsXwW+OGryy+DeItG1qbdI+jHtDJjJa2PRT1i6fc+5YX9hqthbappd7BeWd5Cs1tcQSCSOaNgCroy5DKQQQRwQc1VVVVVVVVVVVVVVVVVVVVVVVKMnj0qqqqqqqqqqqlAxwaqqqqodRVVVVVV+1Bqjaj8WbuzLMRplnBbqGUDGV83jHUfve/Oc9sV+yNpTWvw/1HVJFYNfamwQFgQUSNACAOQdxcc+g/Gqqqodaqv+C0PjRp/EXw0+HUNwgWzsr3VbiJZwS5leOKJmTGVA8mYBs4bcw/hNf8EnfDIu/iJ478Y7M/2XotvYbt+MfaZjJjb3/wCPXr2x70gBIFRjiolyAKjUE0i8AVGoxxUag1Goxmo0yKjQEYrV/DHhzxJDFB4i0DTdVjhYtEl7aJOEYjBIDg4OK0LwZ4S8NyyT+HfC+kaXJMoWR7KxjgLgdASgGR9aSHIpYcjAFGDjAFPCMcCpIhjGOavbK1v7WawvraK4t7iNo5oZUDpIjDDKyngggkEHg5rSPCfhjw0Jv+Eb8OaXpX2nb532KzSDzNudu7YBnG44z0yfWpUFOoIxUqjGKdR0qRRnFSqMYqQZFOMgipQK/wCCtPhe6ufDXw78aQxL9m0++vdPnfZyXnjikjBPpi3l49zX/BHPxemjftGeIfCM6p5fiLwvKYm2Et50E0TqM5wFMbTE5B5C9Oc1V7f2GnRCfUL63tYy20PNIEBOOmSeuAa1L41fCbSkL3XxB0VwEL4trkXBwOvEe459uprVf2rfhBp4c2eoalqZUDAtbFl3Z6gebs6e+PxrU/2z/DsW/wDsfwRqN1hsJ9pukgyPU7Q+D7c/WtS/bM8YS7v7I8I6NbfvCV+0ySz4Tng7SnPTnp7c8an+1Z8YL/f9l1LTdO3Nlfs1ijbPYeZv4+uTWo/Hf4v6oZTc+PtTTzRhvs7Lb4wMfL5YXb07Yq98c+NtSlE+o+MdbupFXaHn1CVyB6ZLdOT+dVZaPq2pIz6dpd3dKhwxhgZwp98A4q1+GPxIvo45bP4f+I5o5fuSLpcxU++7bjHvUHwD+MNzMlvF4C1AM5wDIUjUfVmYAfiatv2YPjXcTrDL4TitlbOZZdRtyq8d9rk+3A71a/sk/Fq4nWGVNGtlbOZZb0lV477VJ9uB3pP2NfiazqH1/wAMBc8kXM5IH08mv+GLPFH/AEOulf8AgPJVl+xNfyQk6j8RbeCUNgLDpZlGMDnJkXnrxj8a/wCGI/8Aqp3/AJRf/t9WP7E+lx7/AO0viFdT5xs8jTVh2+ucyNnt6V/wxZ4X/wCh21T/AMBo6/4Ys8L/APQ7ap/4DR1/wxZ4X/6HbVP/AAGjr/hizwv/ANDtqn/gNHWu/smfDjwvpkmseJfihPpdlF964vBDCgOCcZYgZwDx7V8UfEHwZ8NSSaZ8OPEuteJ72JwGupbVILPH8W1id7EEdlCnqGNaj4j1fUwUmuikZ/5Zx/KP8T+NeD/AfjP4gal/ZHgrwzqGs3S43paQFxECcBnb7qLnjcxA968Jf8E+/iLqmmQ3/izxLpWi3EnLWCZuJIx6M6/IG68KWHTnsIf2FPGWkzsNH1nwxIgA2zTyzrIeOcjymxz6H0q+/Y++LFps+z3OgXu7OfIvHGzGOu9F6+2elX37K3xptNn2fw7a3u7OfI1CEbMY672Xr7Z6Vffs7fGjT3WOfwFduXGR5E0Uw/Eo5x+NXnwe+K1lN9nm+HPiNmAzmHTZZV/76QEfrV34U8UWHnC+8N6pb/Zt3nedZyJ5e3727I4xg5z0qrHxBr2lxrFpmuX9oiNuVYLl4wG9QAetWXxl+LGnuz2/xG8QOWGCJ7+SYfgHJx+Fad+1D8a7Axq/iyO8jjTaI7mwgOcDAJZUDE+5PPfNad+2T8TbURR3+keHr1EJ8xzbyxyOM+qybR6fd7Vpn7bco2R6z8PUb5vnlttSK4HsjRnJ/wCBCtL/AGyvhnd+VHqeja/YyNne3kRyxpjOOQ+45wP4ep9Oa0z9pj4KapsRPGiW0rrkx3VpNFt9ixTbn6GtJ+Ivw/17YujeN9CvHZVYRxahGzgHpld2Qe2CM5rGeBWBjANVVUACcVVU6JIhjkVWVgQysMgjvxXxi/ZJ07W3uPEXwwWDT75zuk0lsJbyHv5R/wCWZ/2fu+m2ru013wprLWt3Be6TqthIMo4aGaF+oPYg4IIPuDXwZ/a2DLb+GfirIBtASHWkU88gATqPx+cegyOrVb3FveW8V3aXEc8EyB4pY2DK6kZDKRwQRg5qgABiqqqqqqguawMYFVVVXjbwN4V+Ivh648LeMdHg1LTrjkxyDBjYAgOjDlGGThhgjJ9TX7Q37L3ij4KXcms6b52r+EZGVYtS2jfbs3AjmUfdORgOBtOV6E7a+BX7Q3jL4HawX01/7R0K6Zft2lTudjAHl4j/AMs5MZG7BBz8wOBj4bfE3wd8WfDMXirwXqf2m1ZjHLFIuyW3kHVJE/hbofQgggkEGqqsZ4FYPpRFVVVVV+2T+zv/AMLA0JviT4O07f4k0iL/AEyCGP5tQtlHYKCXlTjb6rkc4UV8MPiP4h+E/jXT/G3hqbFxZNiWBmIS5hbh4nHdWH5EBhyAR8OPiB4f+KPg3TvGvhqZms7+PJicjzIJBw8bgE4ZTx+R6EVVUy9xVVVVVVRAIxmiO1VVajp1jq+n3Ok6naRXVnewvBcQSruSWN1KsrDuCCQR71+0/wDs93fwV8Uf2josLyeEtXlP9nStJva3fGWgcnnI5Kk5yuOSQ2P2TP2kJvhfrcXgfxffSyeFNUmCxPJIAulzM3+t56REn5xkAfeHOQyPHLGskTq6OMqynIIPQg1TDniqrxv4y0TwB4YvvFWvz7LWyjJCD78z/wAMaDuzHAHbucAEj4iePNZ+JPiy98V60+JLhtsEIOVt4QTsjXgZAB645OSeTX7LnwOXwtp0PxG8U2a/2zfxZ0+CRCGsoWBBY+jyKfTKqcdWYAjtVUw54qqqqr9s39o1vh/pD/DDwZf7fEerW+b65hkIbTrdugBUjbK/buq84G5TXw1+HXiP4q+MbDwV4Xt993ev88rqxjtox96WQgHCKO/qQByQK+Gfw38NfCfwbY+CPCtuy2lmCXlkwZLmVuXlkI6sT+AAAGAABVVVVTDI4qqr/grH+11P4H8OL+zT8P8AWPK1rxJZ+b4ongcFrbT5AQtrkHKvMAS6nnysZG2UV+zz8C/Ef7QnxJsvAmhM1vbBftGqX+wMLG1UgNJgkbmyyqq5ySw6AEj4Y/DTwp8JvBOl/D/wVYta6TpURSISPvkkYks8jt3dmJYngZPAAAAjiyMAVHCMYApIMilhGMAfnQgJGBXkAjrRgGODTQHHHNPCCOlPAO1SQgjBFSRdsVLDgYr9rbT7NvA2k6i9pC11DqyxRTmMF0R4pC6q3UKxjQkdCUXPQV8fZZYPh888ErRyx3sDI6MVZWDcEEdDX/BNr9sRv2ifh23w78d6s1x8QvB1qv2qadhv1Wy3bEuRyWd1JRJSR95kbOZMCv8AgtXpFlP4I+F2uurfa7PVdQtoiGwAksULPkeuYU+nPrX/AARnv7yP9pDxbpcdwwtbjwRPNLEOjSR3tmqMfcCRx/wI0w96qqqqojIwDVVXjzTzYeJ7sBcJcETISMZ3dT/31uqqqqqqqqqqqqqqqvCevtoGqpNIx+zS/JOoHbs34dfz9aR0lRZI3VlYZVgcgj1FVVVTjqAaqqqqqm61VV8StQDNaaUjD5QZnH6L/wCzVVVVVVVVV4Z8atYomn6uzPbqMRygZZPY+o/Ue/aC4t7qJZ7aZJY2HDI2QaogZwKqiQBkkAAd61LxboenZRrsTyj/AJZw/Mfz6D861fxzqmoAxWQFnFjHyHLn/gXb8MfWmZnYszFmJySepNVVVVVVVVeHvENzoVzkZktpD+9iz+o9D/OtP1Gz1W2W6spg6Ecjup9COxqqqqqqqqqqqvif8AtF+J1zNqH/AAsn4n+E7+dQrXHhjxpfWCrtUKNtv5jW6nA5IiBJyTkmviF+wf8AtUx+bc/Cn9u34hz5z5Vl4h17UItvpm4glbP/AH6FfEL4I/8ABUz4eebNP40+K3iCzjzi58PeNLvUN+P7sSTed+cYrW/2gf2s/DWoSaR4i+N/xb0q+h/1lre+JdQglT6o8gI6elf8NP8A7S//AEcT8Tf/AArr7/47Wm/tpftXaTYxafafH3xhJFFna1zfm5kOSScySbnbk9ycDAHAFf8ADcf7W3/RefE3/f1P/ia/4bj/AGtv+i8+Jv8Av6n/AMTX/Dcf7W3/AEXnxN/39T/4mvEfx8+OfjCI2/ir4zeN9XhLM3k3viC5mjUtw2EZyoBHGAMY4rw54Y8S+MdXt/D3hHw9qeuapdMFgsdOtHuZ5WJAAWNAWY5IGAO9fA7/AIJLftQfFSW11Hx1Y2Pw20KVgZJ9ZPnXpj5BMdnGd24EfdmaHIOQfX9mH9i74I/spaO0XgDQ3vfEF3Ckeo+IdRPm3dyVByF/hhQkn5IwAeNxYgGqqqqqqqqqqqqqqqqqqqqqqqqqqqqq+M/wY+Hnx/8Ah5qXwy+JuhpqWj6iuQRhZrSYA7J4XwfLlTJw3uQQVZlP7Xn7IfxC/ZG+ITeGfE0b6l4e1Fnk0DX4oSsOoQg8qRz5cyAgPGTxkEEqysf2Cf29r74FX1r8J/ivqE958PLyXbaXbZkk0GRjyyjktbknLIOVJLqM7lfQtd0XxRotj4i8Oara6lpepQJcWd5ayiSKeJhlXVhwQQRzVVVVVVVVVVVVVVVVVVVVVVVL7VVVQAPFVQAPFVVVVVSjiqqqqqHWqqqqqr4raimq/ErxNextGyHVJ0RkbcGVHKhgfcKD+Nfs96amm/B/w9EBHvnikndlXG4vK7DPqQpUZ9qqqqqXr+FVX/BWXxKNd/a4utLF1FL/AMI74dsLDYi4MW4Pc7W9T/pO76MK/wCCU2ifZvgt4p19mkDah4maBUZcLthtoSGU98mVgf8Ad+tIMnrSAYxmowMcelRAYxSAEgCkAwADUQGAKQDAGaiUdqiUYApFBOBUaA1HGCKWMHgCmiIFPGCOBUqDGKlUY4qVQRUqjGM1IBnrUqjpUgGcVKB2qUA1IMggGpBzgHtWr+O/Bmkg/b/FGmxsP4FuFd+n91cnHHpX7ZT+F/j98Hrj4f8AhXVIm1WPUra8s7i6tGESlCQ53EbkPlvIAQpJPy8Biw/Zp+BOrfs9/Fzw58Y4/F1vqWo+H2nKWC2JSKTzYZIWBkLk4McrfwggkenOrftXfF7Us/Y73S9LzjH2SxDYx/11L9f84rV/i58TtdVk1Lx5rTo4IaOO7aJGB9VQgH8RVxcT3czXF1PJNK5y7yMWZj6knk1pfgvxhrsYl0TwnrOoJt3BrWwklGPXKqeK039nT4y6owEXguaBd4VnubiKELnvhmBIHsDWm/sf/Eu72Pf6poNihbDq1xJI6j1AVNp/76FaZ+xb/qpNZ+IHr5kNtp31xh2f6HlfUe9ad+x38OLcRPqGt6/eSITvUTRRRv6DAQsOMfxVYfsv/BeyTEvhia8cPuD3F/NkdOMKygjjuO9WPwa+FGn7/s/w80F/Mxnz7JJsY9N4OOvarDw14c0pon0vQNNtGgXbEYLRIzGMYwu0DAxxx2qqqsZ4zSjPeqqqqviL8aPhl8KrdpPG3i2zsrjy98dkpMtzKO22JctgnjJAHqQATXxO/wCCgOvalHNpvwo8OLpETjCalqarLcL6lYgTGp6feLjrxzx4w8d+M/iBqI1bxp4n1DWbpdwja7nLiIE5Kov3UXP8KgD2r4afstfGX4oGG60zwvLpelyEZ1LVQbaLaRkMikb5BjuikZ6kV8NP2Dfhn4XWG/8AH17ceK79QCYTutrVHBzwinc+OB8zYPdecDR9D0Xw7p8ek+H9IstMsYc+XbWdusEaeuEUAD8qqqoDJqqvtM03U0WPUtPtrtUOVWeFZAp9RkHFXvws+GeomZr34feHJZJwRJIdLhDnIwTuC5B985q+/Zx+CmoOrz+A7ZSgwBBczwj8QjjP41qH7IPwivU2W39t2J37t1vegnHPy/vFbj9eBz1rUf2J/D0quNI8d6jbEvmP7TZpPtX0O1kycd+PpWqfsU+LYg/9i+M9IusNhPtUMlvkep278H25+tar+yZ8ZdPJFnpOnanh9oNrfouRj7w83Zx29eelaj8Evi5pmPtPw71x8sVH2e0a45H/AFz3ce/Sr7TNS0x1j1LT7m0ZxlVnhaMsPUZFaR4m8SeH2D6B4h1LTWViwNndvCQxGCflI5I4+laP+0N8ZtEVUtPHt9OqgDbeKl1kA9zIrH8c5960j9sz4l2eI9W0fQtRQZy3kyQyH05V9uP+A/jWj/tt6FMVXxB4EvrUZAZrO8SfjHJCsqd+2fxrRP2qfgxrJSObxBc6ZI4XCX1m68kcgsgZRjoSTj0JrQfiB4G8UbV8O+MNH1B2xiKC8RpBnOAUzuBODwR2pQMVVVVfE/4P+DfivpotfEFn5V7CuLXUIFCzw9cDP8SZJJQ8d+DzXxU+C3jL4T3/AJetWy3WmyuRa6lbgmKUDHDDrG2CPlbvnBYDNfCD4/eLfhRNHYIx1TQGctLpssmNhOctE2DsOTkjGD3GTkeAfiJ4S+JWjLrfhTVEuI1wJoGws1uxzhZEzlScHHY4yCRVVVVVVQGawMYFVVVVVfWNlqdpLp+o2cF1bToUlgnjDpIp6hlOQR7Gv2lf2OLvwcl348+FVtNeaFGjzX+mFt8tiM5LRd3iAPIOWULklhkr8Mvif4v+EfimDxZ4Nv1huoxsmglBaG6jPWOVQRuU4HcEEAgggGvgX8fvBnxy0NZ9IuFs9dtYVfUtIkY+ZbknBZDgeZHnow6ZUMFJxW0dqwO1VVEAjFFQaKg9DRBFVVV+2T+zv/wr/XG+JPg7TtvhvV5f9Mghjwun3LHsFACRPxt9GJHAKiv2Vfj/ADfBnxh/ZWuXLt4U12VE1BDlhaSdFuVHsDhwOWXsSqioJ4LqCO5tpklilQPHIjBldSMggjqCO9VVMuOlVVVVVVMB1FVVV418GeH/AIg+FtQ8IeJrNbnT9ShMcgwN0Z7OhIO11OGU44IBr41fB3xF8FvGdx4a1iKWaykLPpmoGPal5Dn7wwSAwyAy5yD7EE/sYftHLItn8F/G17hx+78P3cmACACfszn14+Qnrnbn7gNMMjiqd0jQySOFVQSWY4AA71+0X8ZJPih4o/s7R7onw5pDstkq5UXD9GmYHB56LnoPQs1fswfBBfG2rJ458UWRbQtMlBtYZFIF7OpBGQRho1wd3YnA5G4VTD0qqYZHFVVVX7Q3xs0r4H+Aptdl2T6zfboNIs2JHnTYGWOAcIgO45xnhcgsK1HUPEfjvxPLqGoXFxqut63d5dyN0lxNI2AAB6kgADgcAYFfsx/AKw+CPgpDqEEUvinV40k1W5AyY+Mi3UgkbUJIJH3jk9MAMOeKqqqqogEYojtVfGH4naD8F/hd4n+KniVWk0/wzpst28KHDXDqMRxKecNI5RATwCwzxmviT8QPFfxi+ImufEXxdOLvXfE1+9zcCFDtDucLHGpJIRRtRVycKqjJxX7Gv7O9v+z/APCe2sNStY/+Eq1/Zea9MACQ+D5duDjO2JWIxyN7SMOGwIos4AqOIEAAVHCMYApIR360kBIwBSwChApGMfpRgXoB+lNbgjingPTH508IPQc1JCCMYqWLHFSxY4r9rpNvw403H/Qci/8ARE9ePvht4l+KXg3VtC8JQLcajY2zaglseGnSH5nRP9vbkgdyMd6+B/xf8T/AX4qeHviv4QKPqGg3PmG3kYql1EwKywvjna6Myk9s5HIFeAvG/h34l+CdC+IPhG9+1aP4hsIb6ykIw3lyKGAYZO1xnDL1VgQeRX/BZ7Q/tHwH8EeJPtW37B4uFp5Hl53+faTvu3Z4x9nxjHO7qMc/8EiNZutL/a0ext44mTV/Ct9azlwSVRZIJgVweDuhUc5GCeOhDAEdaqqqqqiO2aqvitpYe3s9YjXmNjDIQOx5X8AQfzqqqqqqqqqqqqqqqq8AeKgAmgahKeTi1c8/8AJ/l+XpVVVUwzwD2qqqqqqYDPFVRIAySAAOSa1/Ujq+r3N8HLI7kR5GMIOBx24qqqqqqqqqrDVNQ0uTzdPu3hY9QDkH6g8GrL4jXsYCX9jFMO7RsUP5c+/pUfxE0llzLZXatngKFb+oqf4i2C5+zadO/HG9gnP4Zq9+IOqTgpZ28NuCOCfnYfnx+lXur6pqWRe380oJzsLYX/vkcVVVVVVVVVVVWn6ne6XOLixuDG3cdmHoR3rSPHVheBYdSUWs2Mb+qE/zH4/nSPHKgkidXVhlWU5Bqqqqqqqqqqqq8UeDfB/jfTjpHjPwpo+v2Jzm11SxjuouevySAj9K8ff8E2f2R/HbS3EXw9n8NXcqsDcaBfyW20kEArExeEEE5GExnGcjivG3/BGzw7PLNcfDj43ajZRhT5NprekpdFj2DTRPHgY7iI14u/4JM/tQaB5svh678H+J4h5phSy1N4JWVfuBluI40VnHQByoOcsBgnxX+xJ+1l4NkaLV/gL4ruGWRUJ0q0GqDLLuGDamQEY6noDwSDxXiDwl4q8KSpB4p8M6to8kjOsaahZSW5YpgMAHAyRkZ9Miq+GP7Wf7SHwbtE0/4bfGHxBpFnHJE62vnLcQjywAi+XKHXYAANmNpHBGK8If8Fef2yvDQUa1rnhTxZt3Z/tjQUi3Z6Z+yNB07Y/HNeDP+C4Piq38mH4h/ALSr/d5KzXGi63JZ7P+errFLHLuz1VC64xgsc5Hgj/gsl+yn4keC28VaV418JyvsE015piXVvGS2CQ1vI8jBRyT5YOOgJ4r4f8A7ZX7K3xRjh/4Qz49eD7ie4z5VpeaithcvhSxxBceXKcKpJ+XgDmqqqqqqqqqqqqqqqqqqqqqqqqqqqqvjF8G/h18evAN/wDDb4oeHodX0W/w2xiUkt5VzsmhccxyLk4YdiQcqxB/bX/Yo8cfsheOPJmNxrPgXWJm/sDX/Lxu6n7PcY4SdVHsHALLjDKn7Fn7d3iv9mS+j8GeJ4p9f+Hd7ch57EPmfS2Y/PNa5OMHO5ojhWIyChLE+DfGnhP4ieGrDxh4I8QWes6NqUKy213aybldSAcHurDOCrAMp4IBBFVVVVVVVVVVVVVVVVVVVVVVKARVVVAZIAqqAyQAaqqqqqh0qqqqqlAJqqqqqrq5gs7WW8uZNkUEbPI2CdqgZJwOegq9upb+7nvbggy3EjSPgYG5jk4H414M01tH8HaFpDhw1jplvARIu1gUjVeR2PHSqqqqlANVX/BRLWotf/bO+Jt9DC0SxXtralWOSTBZQQk/QmMkexr/AIJwaVcad+yr4evJ5I2TU9Qv7iEKSSqi4eIhuODuiY8Z4I9wEweBSYIGDUfT8Ki6AVHjI57UnOBUeCODSY4qLGMVERwM1HjOM1ERjGajIIAB7VZeINCvbo2NlrVhcXK5zDFcI7jHX5Qc8U7qRgGnII61KQRwalIxipMYxXxl/aWtNHa48M/DyeG7vgCk+pD54oDyCI+zuODu5Ue/bwFBeW/gbQI9RnnmvG06B7mSeUyu0rIGcliTk7iec1Lg8A1rXiDQdBi87W9asbBMZBubhY8/TJ5rXv2g/hlpOUg1W41OReqWVuW9P4n2qeD2PY961v8AakkcPH4e8KKp/gmvLjPbuigd8fxf41rHx5+JGrApFqsGnox5Sztwvp0ZtzDp2Pc5zWqeItf1s51jXL6+9ri4aQDvwCeByeKsNH1bVn8rStLu7x84228DSHOM9FB7VffDrxvp+g3/AIju/DV3FZ6bZy3U2/asmyOMuQqEhixC4Ax1IHFXf7aHgFb23t9P8N61LA8ypNPcCOIRoWAZwqs5bC5YDgkgDjORov7HHga0KPrniTV9RZQuVhCW6McfNkYZsHjADDHqa0j9nT4OaOqmLwbDdSAAF7yeSbdjuVZto/ACtJ8IeE9BYPoXhjSdOZWLA2llHCQSME/KByRx9KqqoDJAFVVVVUBk0o4qqqviX+138GPhuZrEa63iHVIiyGy0fbNscDo8uQi88HBLDn5eMV8S/wBtj4weOjNZeHrqLwjpjlgkemuTclSMYa4PzZHPKCOrHTdd8UaqLTS7C/1bUryQkRQRPPNM7Hk4GWYkn8zXw0/YO+Jfihob7x9e2/hSwYgmE7bm6dcZ4RTtTPA+ZsjuvGD8Mf2Zfg98KXgvdA8NLfarbnKapqZFxcK395TgLGe2UVT19TVVVVVKKqqqqqgOMZqri2t7uB7a6gjmikGHjkQMrD3B61q3wk+GGvEyar4B0KaVmDNKtikcjEDAy6gMRjsTjp6CtZ/ZO+DGqq32TRr7S2YH5rO+c4JPUCTeB9MY9q1n9iPQpizeH/Hl9ajJKpeWaT8Y4BKsnfvj8K1j9jL4l2eX0nWdC1FBjC+dJDIfXhl24/4FWs/AH4yaEoN78PtTlyAR9jVbvqcf8sS/p+H41fadqGmyi31GxuLWVl3Kk8RjYjpnB7cH8q0H4geOvC4VfDvjDWNPRMYigvHVDjOAUztIGTwR3rQv2sfjLowRLrWLDV0jGAl/ZL0Axy0exj65JyT1PWvD/wC27wIvFXgX6zafd/Tjy3H+8c7/AEGO9eH/ANq34M66qrc63eaPKxwseoWbL3PVo96DoDyw6j3xoXi7wp4pQv4a8S6XqgUZYWd2kxXpnIUkj7y9emRVVquk6XrunT6TrOn299ZXK7Zre4jDo468qeOoB+oFfG79lS/8OLP4o+GkE9/pYO6bSxuluLYE9Y+pkQeh+Yf7QyR4M8b+J/h/rcWv+FNWlsrqPhwpyky90kXoyn0PfBGCAR8Fv2gPDXxZtU024MemeI4k/e2DvxNhcs8JP3l6nb95QOcjk1VVVUBngGsDGBVVVVVUFyOKCgV+03+xxbeJPtvxB+EtikGrnEl5okSqkd313SQ9AsnQleA2CRhj83h7xF4r+HPimLWtDvLvR9a0qZlPBjkjYZV43U4PqrKfcGv2e/2hPDnx18OeZH5Vh4ksIx/aembvu9vNizy0RP4qTg9i2Cegraa2mtpog9MVVUQCKK4HBqq8SeHNF8XaDfeGfENhFe6dqMLQ3EEqghlPcZ6EHBB6ggEcivjv8HdW+Cnj268L3jPPp82Z9LvGH/Hxbknbk4A3r0YAYB6cEV+w/wDH7+0LSP4LeLL7NzaozaDcTScyRjlrbp1X5mXJ+7lRgKoqqogEYojHFVVVVVRAIxRHaqqq+Nvwb8PfGzwVP4Z1YJb30WZNM1ERbns5uOR6o2NrL3HoQpHi/wAJeI/h74ovfC3iSylsdU0yba68jpyroe6kYZWHUEGv2Uv2i4fi94fHhbxRdKPGGkQ5nYgKL+EHAmUDowyocdMkMOGwpAIxRHbNftZ/GZdNsn+Fvhu6Rrq8jzq8yMD5MR6Q8dGbq2cYXA53HHwl+Gmp/FXxla+GbFzDbKPOvrrH/HvApG5h6sSQoHqwzxkjQNB0nwvoll4e0O0S2sdPhWKCJR0Udye5PUk8kkk8mqYAjAqqIBGM1VVXijxNongzw7qHirxHfpZ6bpdu09zM/wDCqjoB1LE4AUckkAZJr44fF3WfjV4+vPF2pF4rRcwaZaMR/otsCSqcdWOSzHuSewAH7EP7O/8AY1jD8Z/Gdh/p19FnQLaVMGCFhzckH+J1OE9FJPO8YphkcVVVVVVEc4zVf8Fmfjg+leGPCX7Pej3O2bW5DrmshXZW+zRM0dtGR0ZHlErnuGt0Pev+Cfnwc/4Wn8e7HW9StY5tE8FINTu1kGRJODi2Qe/mYk5GMQsD1FeI9Sk0Dwzq2uwwrLJpthNcrGxwHMcZYA+xxX7GP7YviX9p7xP4h0DXfBmmaLHolhHcxyWlxJIZCz7SCG6CoohwAKjiGMCv2zP26vE/7LvxM0n4f6B4A0rW0v8AQY9Tkury7kjKl554tgVR0HkZznnd0GObT9t3/goze2sN7Y/snR3FvPGJIZYvAmtOkiMMhlInwQQQQRQ/4KjfHX4caha6X8cv2a4tPuW3CaLbd6NK3QgpHcrIeEdCQSc5ByoYV+zl+1/8Fv2nIJbXwNqlzYa9axtJcaDqqLFdpGpAMihWZZI8sPmViRkbgucV+158dNX/AGbPg1c/E7RPD9nrF1BqFvai1upWjQiViCcrzkYo/wDBYH4jnj/hTvhn/wAD5/8ACj/wV9+I5GP+FPeGv/A+f/Cm/wCCvHxGYY/4U/4a/wDA+f8Awr9lD446r+0b8Hrb4l61oNppFzNf3Fsba1kaRAI2ABy3OTmpYgeDUsQ6Gv2wE2/DbTf+w5F/6Inr9mJQ3xUgH/TjP/IV+3N8Crf4KfGe4m0KDy/DvitG1LTkGMW7lsTwDpwrncoxgJIgySCa/wCCOv7QE+seG/EH7OPiC83yaDu1fw8XcZFtI+LmED0WVlkGASTPITgAV/wVg8M/29+x1rGqfZYpf+Ec1vT7/c5wYt0v2bcvqf8ASdv0Y1/wTp8SxeFf2zvhnfXDt5V3e3FgyrJsDG5tJoUznqA8iHHcgY5xTAEVVVVVVMBnANVXiPS/7Z0S705VBeSM+Xk4+ccrz9QKZSp2kYI4II6VVVVVVVVVVVVVVVQJBBU4I6Edq8GeMxqATSdWlAugMRSsf9b7H/a/n9etVVMPeqqqqqpgOxqq8a6qdL0OVYn2zXP7qMjtn7x/LP4kVVVVVVVVVVVVVVVVVVVVVVVVVVWGrajpbFrC8kiz1UHIP1B4qw+IUq4TUrFXHd4Tg/kev5irDxPoeogLDfIjkfcl+Q/rwfwoEMAVIII4qqqqqqqqqqqqqpYop4ngnjSSORSro65DA8EEdxivE/7OP7P3jNXHif4J+CNRkaHyRPLoNv5yICSFWUIHUZJPBHU+teKP+Cbn7HfidpZ1+FbaRczMpM2l6tdQBdoAwsZkMSggDOEyTz1JNeKP+CPvwK1HzJfCXxE8a6NLJI7BLpre9ijB+6qqI43wpx95ySB1zzXiv/gjf8S7MN/wg/xl8Mavjbt/tXT59Oz/AHs+WZ8Y7evtXjP/AIJtfte+D3LRfDWLX7ZcD7Ro2pQTjJJGBGzLL0AOdmAGHOcgeL/hz8Qvh9Olr498B+IvDc0n3I9X0uazZuM8CVVJ4r4c/tBfHL4RbI/hl8W/FfhyBM/6JY6rLHbtnd96DPltzI5GVOCxIwea+FX/AAWZ/aM8JCGz+J/hfwz4+s0yZJ/LOlXsh+XH7yEGEAANx5Gct1wMV8FP+Cpv7J/xdEGna54sl+H+sy8fZPEyiCAnnJF2pMIGAOZGQnIwDzi0u7TULSG+sLqK5trmNZIZoXDpIjDKsrDgggggjg5qqqqqqqqqqqqqqqqqqqqqqqqqqrxr4K8J/EfwnqngbxzoNprWg6zbtb31jdJuSZD+oIIBDAhlYBgQQDX7df8AwT18Yfsp6hL448JzXHiL4Z310Ut78pm40lmPyQ3YHGDkKswwrEYIRmVT+yp+198R/wBlfxObjQJf7V8LajOj6xoFw+IrgDgyRN/yym28BxwcKGDAAD4J/HL4cftB+CIPHvwz1v7dYPIYp4JV8u4s5h1imjySjYIPcEEEEgg1VVVVVVVVVVVVVVVVVVVVVKOKqqodaqh1qqqqqqAGABVVVVVKM9DVVVVVfE7Uzo/w78SaikhSSLS7gRMF3YcoVU4P+0RXh/TDrevaboyq7G/u4rcLHwx3uF4z35qqqqqlAxxVV+3X/wAnefFP/sPP/wCgJX/BPkj/AIZE8Aj/ALCP/pxuaTBOAaQgrgGoyD0NRHjANIRwQaQggYNREYxmkIIGD0qNh2NRsMYzSMMAg18eP2idM+FFt/YWhJBqPie4TKwO2Y7NSOHlxySeMJwSOSQMZ8Z/Ezx58Qblrjxb4ovr9SxZbdpSsMZ/2Ilwq9B0HYVXw5/aG+J/w3uIY9P8Qz6jpkZAbTdQkaaIrnkJk5j7/dI5PINfC/4seGfi14Zj8QeH5vLmjwl7ZSMDJaSY+63qpwSrdCPQggSSA8A1ruu6R4e0yfWNb1CGzsrVC0s0rYCgencn0A5PavjT+0ZqnjwT+G/CfnaboDAxzMxAmvhx97+4nGNoPIJ3HnaK139rDVzbrY+DvClnp8MSeXHJdytOwUDAwq7QpHHUsOK174z/ABO8RMxvfGF9ChJxFZsLZQOeP3eCRzjnOeM5xTNdX9zl2luLiZgMkl3djwPcmtG+EvxG15Uew8JXyxyfdkuUFupGAcgyEZGCOR17Vo/7Mfim6AfXNc0/T1P8MStcOOvUfKPT+I9fbFaV+zd4LsgH1XUdR1F/7u8Qp27KN3Y/xdD+NaZ8NPAOiFW07wrYBk+680ZmYc9QzljRSOKNYokVERQqqowFA6AD0rxRpH/CQeHdU0AXPkf2lZTW3m7N3l+YhXdtyM4znGR9arwVrN14k8G6D4hvo4kudU0y2upkiBCK8kSswUEkgZJxkn61VVVUBk9aqqqqpRnoaA4AzVapqml6Jp8+razqVrYWVqm+e5uplijiX1ZmICj3NfE79u34a+FPN07wHZzeLL8IQsysbe0Rvd2G58dcKuD03DOR8Tf2kfi78WN9t4j8Tvbac+4f2ZpwNtb4OchlBLOMHHzlq8C/C34hfEy8+w+BfCd/qzqSHkiQJFGQAcPKxCJ1H3mHUV8Mv+CfdukcGp/FnxSZJCMtpWk8Kvpunbk+4VRyOGIrwT8N/Afw408aZ4H8KadpEWwK7QQ/vZQOnmSHLyH3Yk0Bk8GqqqqqoAYqqqqqgM8VVYzwKwMYBqqqruxstRh+zX9nBcxZyY5ow65HfBrXP2fvg34gUi88AaZA3ZrFWtCCAQDiIqO/Q8HjNa9+xb8P74PJoHiLWdMkYHaspS5jU4wMLhW68nLflXiH9izx3Yq0nhzxLpGqop4SZXtZGGRjA+ZfUnLDp3zXiD4B/GDw0N2o+AtSlQfx2SC7GOeT5RbA+U9cY4zjIqaG6sLkwzxS29xC3KupR0Ye3UGvD/xr+LHhcqNH8fasEQYSK5m+0xqMEcJLuUdT0Hoewrw/+2l49sWSPxF4c0fVYl6tDvtZW57tll6ccL2714b/AGx/hjq2yLX7PVdElP3nkh+0RD6NHlj/AN8V4a+JXw/8YFF8N+MdKvpX+7Alyol/79nDD8q+OX7MOj+O1ufFHglItN8Q7S8luFCwXxA6HoI3P97oT94clhqema74R12XTdRt7nTNW0yfDISUkhkU5BBH4EMPYg96+Bn7Vdvq/keE/ijdx298SEttYYBY5yTgJKAMI3P3+FI64Iy1VVVjPAoAAYqqqqqsZ4FBRjmqoKSOK/a7/Zf/AOFh2MnxG+HulIfFFohN/aRLtbU4gOqjvMoHHdhxyQorwn4s8TfD7xNaeJ/DGozabq2my5jkUYII4ZWU8FSMgqeCCQa/Z6/aF8OfHXw5vj8qw8SWEY/tPTN33e3mxZ5aIn8VJwexaqqiAaKgjg0VPaqoqD0NEdqr4/8AwY0n42+Abjw/cKkWrWe640i8I5hnC/dJ/uP91h06HqoxdWviLwL4oe1nW40rXNBvsHa22S2uIn6hgeqsuQQewIr9nT43af8AG/wHFq0kkEWv6bth1i0jG0RynO2RQSTscKSPcMP4TVVRAIxmiO1VVVVVTDuKqqqv2qv2d7b4v+Gj4i8N2aL4u0eIm2Kqqm/iHJgYkjnuhPQ5HRiR4c8Q+IvAPiiz8Q6JcS6frGj3O+Jmj+aOReCrK34qVI6Eg18D/jFoPxr8D23iXTJIodRiAi1TTw+WtJ+4x12Ngsrdx7hgPjN8UNO+FXg241uZ45NRnBi021LAGWUjrjqVXILfgOpFTS614s10yytcalq2rXXpvkuJpG4AHcljjHvXwS+FNl8JvB0Wklkm1W92zancLyGlx9xf9heQPXk4GcVVUR2zVUw5wKqq/bW/aBXx74jb4YeE79ZPD2hTZvJ4mOL27XIIBBw0cedo45bceQFNfslfAJ/jH41/trXrQnwr4ekSS9Lp8l5LnKWwORwR8zYzhcA43qaSOOKNYokVERcKqjAAHQAelUQCMZqqqqqqYVX7XfxmHx+/aL8a/Eu2mlbTL2/NvpKySbttlAoihIHRdyIJCo4DO3JySf8AgnV8Jv8AhXn7P9n4l1Cxkg1bxtMdSm83732b7tsAOm0xjzR3/fdegHxCQD4d+KD/ANQW8/8ARL1/wSRUN8RvHoH/AEBLf/0caij4AFRRDAAFf8Fck2/tI+Gx/wBSPa/+lt7UcGccV4m8GeF/G+h3Phrxj4e07W9KvF2z2V/bLPFIPdWBGfQ9q/ba/Zs1P9ij4u+G/jL8EdSvtN0DU7559LKM2dIvE+ZrbfnLxOhbaG5Kb0bdtJb9n/4p2Xx1+C/hH4sWVqlufEOnLLcQIcrBcoxjnjU5JKrKkignkgDODxTwAjAp0x/wVmKD/oef/aNSxAjA61LECMYqaMEYqaMEYr9sm7EXgfRdO8rPn6t5u/d02ROMY9/M/T3r9layluvii00bIFtNLnlcMcEgsice+XH61/wUI+FEfxE/Z/1HXrS3Z9V8FSjVbYpHuZoANtwhPUJ5ZMhx3hXPGa/ZP+MM/wACP2hfBPxI+3S2+n2OpxwatsI+exm/d3AIPDYjdmAP8SqcggEfti+Cm+IX7LXxQ8LRQJPPL4burm2iaAS+ZNbr58ShSRhi8SgN/CSG7V8K/Fo8AfFDwf478zZ/wjmv2OpbvL37fInSTO3v9zp3ojIwKqqqqqmAzxVVXj/Rv7J1+WWJSIL3MyZHRj94fnz+IqqqqqqqqqqqqqqqqBIIKnBHQjtXhv4hvbRpY66HlRRhbkcsP94d/r1+tWt3a30C3NncRzROPldGyKqm7UetVVVVUwyBiq1fVrLRbJry8kwo4VR1c+gFat4jutamle+hjkQgiBCSPI6cgjr05zwfyqqqqqqqqqqqqqqqqqqqqqqqqqqqqstY1TTsCyvpYgOihsr+R4qw+IN7EAmoWkc4A++h2N+XIP6VYeLdC1DCreCBz0ScbP16frSukih43DAjgg5qqqqqqqqqqqqqqqr+wsNUs5dP1Oygu7WddssE8YkRx6FTkEfWviF+wh+yf8SfMl1j4OaPpt3Jki60MNpjqx6ttgKox/3lb1618V/+CO13Es198Efiqk+1f3emeJoNjEg8/wCkwLgnHQeSBkckA8fGT9mf44fAO7eD4n/D/UdMtPNMUOpxqJ7Oc9tk6ZTJGDtJDdiAQQPgN+1z8f8A9m7UIZ/hf8Qb2205GzLol4xudPmBJJDQMdqkknLJtfk/MM1+yn/wVc+D3xrFv4T+MC2nw58XNhUmubkf2XfHgDy53IMLk5/dy8AYxIxOBVVVVVVVVVVVVVVVVVVVVVVVVVVqOnafrGn3Ok6tY297Y3sLwXNrcxCSOeNwVdHRshlZSQQRgg4Nft6f8EutS+F0WpfGL9nSwutT8HwRy3esaAz+bc6Qu7Je343S26qeQcvGEJJdclfgn8cviP8As++N4PHvwz1v7FqCRmKeCVfMt7yE9Ypo8gOuQD2IIBBBAI/ZV/bK+GH7UugBNGuU0XxdZQq2peHbuZfOXCjdLBzmaDccbwAV43Ku5c1VVVVVVVVVVVVVVVVVVVUMbcZqqqgMnrVUoBNVVVVVQHGKqqqqpaqqqqr9o3UBYfB/XFWcxyXJghjwD82Zk3D2ygbrXwW08an8VvC9sYnk2ajHNtQHI8v95njsNmT7A1VVVVSjjrVV+3X/AMnefFP/ALDz/wDoCV/wT6OP2RfAQB/6CP8A6cbmkPAINREEYBqM9s0hAPBpDkYBqNgRjNRtjjNRuOmaRscZqOTgc1418W23gnwfq3iu6TzE0y1eUR5x5j9FXPbLED8a1vWdS8R6vd65q909ze38zSzSMclmY5P4dgOwwK0jw3GUWS6jEkjDO0jhf8al0CBotrWsZGOhQVrmhixH2i3B8vPzKf4f/rV8APiHcfDn4l6XftclNN1CVbPUULEKYnIG8gf3CQ3rwR3r4j/FLwp8M9KGo+IrwmaUH7NZw4aa4IIB2gkcDIyTgD64FfE74teKvilqn2rWbjyLGJv9F06Fz5MOM4OD958E5c888YGAKr4SfBO0+J1jNqUni+3tVtJNlxaQwGSdM/dJzgANhiCNw49cgaL+zj8MNFCvdWF3q0q/xXtwcZ/3U2gj2INaX4c8O+H0KaFoWn6eCMN9mtliJ6dSoGeg6+gpyDzmpWyMA1IRjANSEY61IcnANSkHoa8Y6Za6L4u1vRrEMLaw1G4t4QxyQiSMq5Pc4Ar9ma/vNV/Zu+FGqajcNPd3ngjRpp5X6ySNZQlmPuSSaqqqqUA96qqqqpQMcGvFXjHwp4H0w6x4v8Q2GkWYO0S3c4jDN6KDyx9hk18VP2/9IsJZdK+Eegf2m65X+1dTVooc9jHCMOw6HLlCMY2nOa8ffFb4h/FC8+2eOPFl/qgWQvFbySbYIWIxmOJcInHGQMnvXwx/Zz+Lfxa8q58LeGHi0yRtp1S/b7PbKP7wY/NIB38tWI9K+GP7CPw18KeVqHjy9m8WX4T5oHU29ojeyKdz46ZZsHrtGcDTNK0vRNPg0nRdOtbCytU2QW1rCsUcS+iooAUewFVSgGqqqqqrHGM1VVVVSjJqqAyaqqqlHFVVVrPh7w/4jtxaeIND0/U4V6RXlqkyjkHowI6gH8BXiL9ln4M6+h8nw9PpMxGPN065aMj/AIC25P8Ax2vE37Ef+sm8G+OPXy7fU7b64zLH/wABH3PU+1eJv2Y/jJ4a8yT/AIRf+1YI8/vtMmWfdjPSPiQ8Ln7vcDqcVqek6rot01hrOmXVhcpndDdQNE4wSDlWAPUEfUGvDHxh+KHg4qPD3jjVII0BCwSy+fEvGOI5NyfpXjT423HxO0xbT4g+FdMu9Qtoyllq9jutriDLZIcZZJU/2CF74IJLVXwJ/aa1LwClt4T8ZtLqHh1cRwTAbprAZHTu8YGfl6jjbwNp0rVtL13ToNW0XUbe+srld0NxbyB0cdOCOOoI+oqqpQDzVVVVVYzwKAAHFUBngUFAqq/ai/ZDg+Ict18QvhrDBa+JNjSX2n4CR6mw53KeiTHoc/K5wSQcsfD3iPxd8N/FMWt+H7+70XXdJmZQ2zbJC4yro6MMHurIwIPIIr9mz9pPRPjjog03UjBp/i7T4gb2xBws6jjz4c8lCcZXqhODkEE1VVVEAiiMVRAIxRGOKr9t/wCAQ1/S3+MXhLTd2pabFjXI4hzPbIuBPgdWjAAY/wBwA9Er4H/F3V/gr4/s/F2nI1xaMDBqVnnAubdiNyj0YEBlP95RnIJB8Pa/pPirQrDxHoV4l1p+pW6z28yHIZGGR9D2I7HIqqpgCMiqqqqqogEYqqqqr9tL9nMTR3Xxm8E2Sh4wX1+zjVVBUf8AL0oGMn+/1J4bsxr4IfGHxF8F/G1t4j0XzLqzmIi1HTfMKpexHIwfR1JyrY4PqCwPxr+Kd78WPGk+tbpYtLtcwaXbSHHlQg/eIHAdz8zdccLkhRX7I/wfWCD/AIWn4gtAZJQ0ejxSIcovR5+fXlV9tx7iqqqYc8VVMPSqr9sX4+r8KvB//CHeHLzb4p8R27CJkIJs7Yna8x5yGb5lQ46hj/Dz8PPAevfE3xnpfgfw1Cr32pzBA78JCgGXkc/3VUFjjnjABJAr4dfD/wAO/C/wbpvgnwxbeXZ6fEFLsPnnkP35XI6sxyT2GcDAAAqqI7ZqqqqqmAI61/wUF+MbfBT9lTxlrtjdpDq+uQromlZcKxmuso5XOcskImkAx/yz7dR8JPAF58VPid4Y+HlkJN2vanDbSPGQGiiLZlkGePkjDt0P3eh6VomkafoelWWiaTbLb2Wn28dvbQqTiONFCqozzgAAc18Q0z8O/FOO2iXn/ol6/wCCRihviP4+H/UEt/8A0cahjB5qKMEA4r/grwu39pPw0P8AqRrX/wBLr6o4gcAVHACMYr/gqL4W0vWf2MvFuq38O+fw9f6bfWJ2qdkrXkVuTyCR+7uJRxg89cEg/wDBIbVLnV/2V9Rsp44lTSPGF7awFFILIbe1mJbJOTumYcYGAOOpMkAx0p0H/D24p/1PX/tGpofSpowRwKnjGMipkHav2171kTwnpcdwmGa7mliGM5HlBGPcDlx6Hn0r9j/T0m8X67qR8zdb6YIRj7uHkUnPv+7GPx/DXNJ0/XNKvNF1W2W4sr+3kt7mFiQJI3UqynHOCCR+NeP/AAndeBPHPiHwTeljPoOqXFi7MMFjFIybvoduc+9fsjePYvjT+yt8PPFmsbNRfU/DyWWpm4TeLqe33WtyXUqAQ8kMhIxt5wMjk/E/wRefDX4keKfh3qBY3HhnWbvTnZhguYZWj3fQhcg9CDmv2NviF/wtP9lv4Z+M3l8yebQIbS6k27d9xa5tpmxtUDMkLnAGBnAJGCSO1VVVVUwHY1VV450E67obrBHuurU+ZCAOTjqo+o/UCqqqqqqqqqqqqqqqqq0zWNS0eYT6ddvEc/Mucq31HQ1ovxHsLvbBrEX2WXp5iAtGT/Nf1+tQXFvdRCa1njmjbo8bBgfxFN0pgM4BqqqqqtR1vSdLXF/qEMTD+AnLf98jn9Kv/iTZxnZpthJNx9+Vtg/IZJ/StX1m/wBbuftN9NuI+4g4VB6AVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVaajf2BzZXk0POSEcgH6jvVn491iABbpIbkdyy7G/Tj9KtPiBpc2Fu7aaBvUfOB/X9KtNd0a/wLXUoGY9FLbW/I81VVVVVVVVVVVVVVV1a2t9ay2N7bxXFvcRtHLDKgdJEYYZWU8EEEgg1+0R/wS5+C3xRivPEHwnCfD3xK6s6QWsW7S7iTqA9uP8AUA4C5hKqoJPlua+OP7P/AMUv2dvFY8I/FDw61jPMrPZXkTeba3yDGXhlHDAZGRwy5G4DNf8ABMb/AIKDf8IJd2X7O/x38VbPC822DwtrF6cjS5CcC1mlJ+W3PARiMRngkRkbKqqqqqqqqqqqqqqqqqqqqqqqqqq/bx/4Jc6P8Um1b4xfs8WdvpfjOZpbvVPD+RHbay+AS0HRYZ2IJIPySM2SUOWaKfx/8IPHTm3n1nwl4u8NXjxOUZ7S7sZ1yjqcYZT95SO4JB4Nfsb/APBS3w18T1sfhx8fL6z0DxhI3lWeslRDZaqxPyq2Plt5SMDBwjEcFSyoaqqqqqqqqqqqqqqqqqqqgOBVVVL1/Cqpev4VVVVVVVVVVVSgY4qqqqqv2sr17b4aWltG6D7Xq0SOp6lRHI3H4qv+TX7Mdkl18YNLnaQg2dvcSqAPvExMmPycn8KqqqqodKqv+CiWixaD+2d8TbCGdpVlvbW6LMMEGeygmI+gMhA9hX/BNzXG1b9lvSLA3MMo0XVb60CRkbog0pm2vjo377dz2Ze2KjIIxmo2BxzSMOCDSMCMA9KjfIGDSMBjBpGBAwajkHTNI4IwTSPjjNftVXs8Pwa1CCGTatzd20cowDuUSBse3zKp/CtNRXv7dWGRvHBrRIYpHUPj8avNMsE05ZEdSxHSvEMMRhnQ42lGz+VVqvwh+HvxKs7XxP4p0eabVr+ygaW6jvJkI+UHCru2AdRjb3rVf2S/h5cl5NM1jW7J2I2oZY5UUAc4BTdz7tWpfsg3Cbm0rx3FJlztjuNPKbV5/iVzk9Ow/DpWofssfEi0wba80S8BYgCK5dSB6kMgH5E14O8D/Gr4XeJbbX7TwfdyBQRcQQzJMs8X8SsI3PPcZ6EA4rT9RTVdPt9RS2ubcXEYfybqExSx5/hZTyCOhqR88A1IwxgGnbPOakbPANSsCMA05GM5qQgnrXjXULPV/GOvarp83m2t7qdzNBJtK70eRmU4OCMgjgjNfszWF5pX7N3wo0vUbdoLuz8EaNDPE45jkWyhVlPuCCKqqqqUDHBqqqqrx18TfAPwz09dS8c+KbLSYn/1aysWkl9dkagu/wDwEHFfFj9vvUrsTaP8HtF+wx8r/bGpxq8p94oeVXpwX3ZB5RTXi3xp4s8eas+ueMfEN7q98+R5tzKW2D+6o6Iv+yoAHpXwu/Y9+MHxKjg1K60tfDWjzEEXeqqY5HX+8kP324xgkKpyMN1x8LP2O/hB8NJYtTu9OfxPq8eCt3qyq8cTDvHCBsXkAgtuYHowqqqqUcVVVVVQGTgVVVVVVKBjg1VKAaqqqgOMVVVVVQAJxWp6RpWt2jWGs6ZaX9s2d0N1AsqHIIOVYEdCR+JrxT+y/wDBzxQzSr4cfR52zmXS5fIxkk8Icxjr/d9B0ArxP+xFqCMZfBfjW3mB6QapA0ZXkf8ALSPdnjP8A6D148T/ALPvxe8JpJPqPgq8uLeIEmexxdLtHViIyWUf7wFTQy28r288TRyxsVdHUqysOCCOx9q+Gvxt8ffCrzIPDWoRS2MrF3sLxDJAXOMsACCpwAMqRnv2rwn+2zotwsNv428JXNnITte506UTJyfveW2GUDuAWP8AKvCPxl+GHjp0g8N+MrCa5chVtZmNvMxPZY5ArN6fKCOnqKxngVgYwKqqqqxngUAAMCqC5rAxgVVVX7UP7J1n8Wkfxr4ES1sfF0Sfv45D5cWpqo4DHosoAAVzweAxAwyo/ibwL4k3Rvf6Jruj3JGQWgntpUOCOxUjkV+zP+1dovxatbTwd4wnisPGccZA+UJFqQUD54+yyEZJj9iV4yFqqqqIBGKK+lUQCMURjipI45Y2ikRXRwQysMgg9QRX7VHwHl+DPjhrzRrdv+EW12R5dMYZb7M3Be3Ykfwk/LkklMZJIav2Ifj0/hvXI/g/4ovcaVq8v/EmlkYAW12x5iyedspOBzw+AB85IqqIBGKIxxVVVVVUw5wKqqqpI45Y2ikRXRwQysMgg9QRXx5+EXhP4YfFy/sPCt/FJYSQpcRWAJZtOdxkxMT1GMMvcK65zjcfBx8IS+MdDsfHWq/2fot3epFdzg42oTzk/wAK9AW/hBz2q0tLawtIbCyhSGC2jWOKNBgIqjAAHoAAKqqphkcVVMMjrVfETx5oHwy8Gap438STFLLS4C5RBl5nPCRoP7zMQozxk5JABI+InjzXvib4z1Txv4kmV77U5i5RBhIUAwkaD+6qgKM88ZJJJNfsd/AcfCrwT/wlXiGz2eJ/EkSvMrHJtbb70cOOzHhm98D+GqqqYDOBVVVVVMMjiv8AgtH8Vk1DxZ4C+CunXuV0izm1rU4kkVlMs58q3DDGVdEimOM8rODjoT/wS6+GQ1/4pa78T7623W3hXTxbWbvFx9qucqWVj3WFJVIHOJRkjPMKjNfENR/wrnxT/wBgS8/9EvX/AASIGfiR4+A/6Alv/wCjjUKAgAVCgJAr/gr8u39pXw0P+pFtP/S6+qGIHAqOEYwBX/BXH4l6b4O/ZgX4fNdxf2n491eC3itj99ra1dbmWUeyyJbqe+ZV9TX/AASu+H1z4M/Y/wBE1K7s1t5vFuq3msFQ7MzqzLBG7A8KTHboQF42lT1JqSEYxipV/wCNurL/ANT3/wC0amjGMflUyAdKmQcgVr/iTw34cjEniHxDpmlo33WvbtIAfoXIr9qvWrPxt440n/hEdV0/XLS300Rr/Z1zHcN5xeRnG1CWICKhzjA9a/ZE8O6jpOkeJdR1Gxu7Vru4t4lWeBowRGrnIyOf9Zz+HrU6jmv2/vCsfhj9qDxPLBHsg1qG11CNfm4LwqshyeuZI5DxwM4GMYr/AII/+L0179l7UPDUkyef4Z8UXUCxBySIZY4plYg9AXeYYH90nvX/AAVV+GkvgT9rTVvEENm0Vh420621WBghCGQJ5EwB/vb4S7DqPNB4BFf8EafiemtfCfxj8Jry7iN14Z1dNQtIjtVzbXSYYADlgssLknkgyqCcFRR61VVVVTDI61VUwyMA18QfCh065bWrCIC0mb96o/5Zue+PQ/z/AAqqqqqqqqqqqqqqqqqqsNT1DTJfO0+8lgbvsbAP1HQ/jWn/ABMv4gI9TsorgD+OM7G+pHIP6VaePfDl2B5lzJbOf4ZYz/MZFW+saTdgG21O1l4HCygkZ6ZGeKqmZUUu7ABRkknGKu9f0SxQtc6pbLgZ2iQM2PYDk1qPxI02DMem2slywyA7/u1+vqfyFal4y8Qampje88iNhgxwDYPz6/rRJY5JyT1Jqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq11bU7IBbTUJ4wOirIcfl0q08ea1BhbgQXI7lk2n/x3A/SrL4gaXPhb22mtmJ6j51H4jn9KstX0vUQBZX0MrEZCBsN/wB89aqqqqqqqqqqqqqvjt8Cfh9+0R8Prz4efELTPOtpv3lndxAC40+4AIWeFiDtYZ6dGBKsCCRXxy+Cfjf9n34j6j8M/HtrEmoWOJIZ4G3Q3lu2fLnjPUqwB4IBBBBAIIH/AASy/bfl+MnhmP8AZ++KmuvceOfDlqz6RqF3KTJrVih5VmP354QQD/E8YD/MUkaqqqqqqqqqqqqqqqqqqqqqqqqqqq/bF/YJ+FX7W+lHVbop4a8d2cPl2PiO1tg7OoK4juYwV89MAhckMmcqcZVvjz+z18VP2bPHEvgL4reHX0+9C+ZaXURMlrfxcfvIJcASLyAehU8MAQRX7H//AAUo8ZfBxrD4ffGV7zxT4JjVLe2vA26+0dFAVdhP+vhAAHlsQyjlW+XY3gnxz4O+JPhix8Z+AvElhruiajHvtr2zlDo3qp7qwPDIwDKQQQCCKqqqqqqqqqqqqqqqqqqoYKgA1VVSjPeqpQDVVVVVYzwDVVVVVUvSqqqqq/bDvIk0jwzp5VvMmubiVTjgBFQHPv8AOP1r9kK2nf4gareKmYYtGeN2yOGaaIqMdeQjflVVVVVADAAqq/4K7aJ/ZX7Vlpf/AGrzf7Z8JWV3s2bfK2zXEO3OTu/1O7PH3sY4yf8AglJ4iW5+GPjXwmJIS2na9HelAD5g+0QKmW5xtP2bjjqG9sRtjgGkYcYNRsCMZqN8cZpGHUGkcEYzSPjjNK4PQ0knbNJKCMZr4weFpfG/w013w7aAtczW3m26gZLSRsJFUcH7xTb+NAyW8wOCkkTdCMFSPatD1qKeJZI3wwHzLnkGpNekeHyzIcY6V4m1lGia0ikDSyDDYP3R3r4eeErjx1410fwtboxW9uVE7KP9XCOZG/BAxppsjANNJkdaeQYwDUko7GnfPepHBGAadgBjPNSODwDUjDGAakbA61I2eAalYYwDXibWP+Ef8Papr32fz/7Nsprnyt+zzPLQttzg4zjGcGq8FaLdeHPBug+Hb6SKS50vTLa1maIkozxxKjFSQDjKnGQPpVVVVQAxwaqqr4jfF34d/CnTxfeOPE9rYM6FoLQHzLift8kS5YjPG7G0HqRXxR/b38X628+mfC7RotBsWBVb+9UTXjf7Srkxx8cY/eeoYVqur+IfF+stqGsajfaxql7IFMs8jTyysTgAE5J9APyr4WfsQfFHxzFFqni9k8Iaa+Cq3kRe7kHtACNnGf8AWFSDj5SDmvhZ+zX8JvhJFFPoPh5L7VUwTqmogT3G4c5QkbY+f7gHvmqqqqqA4AzVVVVVAZPWqqqqqoDgDNVQAxwaqqoDJxVVVVVUozVVVeJPAfgvxhGY/E/hfTNSPGHuLZWdcej43Dp2PTivjv8AsvXvg8XHiz4eQTXuiAs9xYAF5bBAMlgSSZIxg8n5lGM7uWFV4O+M/wATfAflxeHPF96ltHtC2dw3nwBR/CEfIUY4+XB9+BXg79trUoPLtfHvhOG6QbQ13pj+W4A6kxOSGJ68Moz9ePB/7QHwl8bKkemeLrW0unH/AB6agfssgOeg3/Kx74Un9DVVVYzwKAAGM1QXPJqqqqqv2jv2XvDXxq02413R4oNN8ZwxAW99yqXQQHEU4HBBHAfG5cL1Uba1/wAPeLPh34mfR9dsL3RNc0uVX2MTHLC4wyOrKfoQyn0INfsv/thWviqOy+HnxX1FYteaRYdP1WQBUvs/dSU9FlzgBuj5A4b71VVVVMueRVEAjFEdq+Lfwz0T4ueA9R8E62u0XKb7WcD5ra4UHy5B9DwR3BI714r8Ma94C8U3/hbXoDaappFyYplVujLyGU9wRhge4INfsr/HKP4z+AEi1e5U+J9BCW+qKV2mcY/d3AA4+cA7sYw6twAVzVUwBHFVVVVVTDI4qqqq+KvxC0/4YeCL7xXeCOSaICKzt3bb9onb7iDv2LHH8Kse1X15rHi3xBNe3BkvdU1e7LMFGWllkboB7k4Ar4zfD/xl8NvGB8OeMrNYJfs6S2zRPviljYdVbAzhgVPoVI6YNfsXftCf8JBp8Pwh8ZajnU7CI/2Nczyc3MK9YCWbJdAflAH3Fx/DzVVRGRjNVRGRgVX7a3xzj+I/jNPAXh28Z9A8LzOsjpJlLu8xtaQY4IQZRT7uRw1fsWfAU/EPxcPiH4lsWbw74cmVoEkjBS9uxgqnPVU4ZvfaO5qqqqYe9VVVVURkYFftyfE5/i5+1b8RvFaT+ZZ2+rvplhtdmT7PZgW6MgJOA/lGTAwN0jHAzX/BOj4fJ4L/AGcNM1ie2jS98WXk2pzMMFvLz5UQLDtsiDAdvMPQk1AARjNfEQf8W48Uj/qCXn/ol6/4Jm/F34Z/CHx34z1L4l+MtP8AD1tf6TBFay3jFRK4lyVGAecc1F+2z+yeOT8dPDX/AH9f/wCJr4c/tL/AT4p+Io/CXw9+KWi67rEkTyrZ2sjFyiDLNyBwK/4LAgf8NL+GgD/zItp/6XX1Q/FH4YAD/i4/hf8A8HEH/wAXXxd/b3/ZX+DOl38+qfFXSPEGrWasI9G8PXC6hcyzAsPKJiJSJsowPmMgXjPLKCLf42/8FVv2n7e+Oj6lpHgfTZlhlkg3S2nhzTs5PzthGupQvQAGRhnaEj+Xwv4S0XwT4V0fwZ4ctfs2laDp8FhYwZz5UEMaxxrnvhVAqeIAYFSjP/BXph/1Pn/tGp1APFXCjoK/aF/arvdP1O58E/Cy9jRrYtFfauEDkSA4ZIM8cYILkHk/LjAY6hqOoavey6lqt/cXt3cNuluLiVpJJG9WZiST9aKkcEY44yK+Hfxj8a/Di8jOmanLdabuHnabcSFonXjO0HPltgY3L7ZyOK8FeOtA+Ivh+HxF4fnJjclJYJCBJbuOqOATg9/cEHvX/BU3SPI+MPhTXftG77Z4aFt5WzGzyrmVt2c8587GMcbepzx/wRP1m6uNB+Lfh144hbWN3pN1GwB3l5kukYE5xgCBMcdS3J4x/wAFjfhPL4n+C/hn4tafAzz+CdVa2vNkRIW0vAil2YdAs0UCgHjMp5HQ/wDBNT4wp8Iv2rvDceoX32fSPGMcmg35YrtzPgwE7uB/pCQgnIIVm65ILdaqqqqphkcVVUen4VPBDcwvbzxrJHIpVkYZBB6givGXg2XQJTe2StJp8jcE8mEnsfb0P4H3qqqqqqqqqqqqqqqqqqqo5ZYW3wysjY6qcGvt99/z/T/9/D/jTMzsXdizE5JJySaqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqrLxLrdgQINQkZQPuSHeP16fhVh8Q2ACanYg8cvCcf+On/GrDxPoeogLDfIjkfcl+Q/rwfwqqqqqqqqqqqqr9uv9lax/aZ+FEp0a1iXxv4YjludAuGyDLkAy2rYOMShFAJ+64U8AtnwJ428ZfBz4iaP468LXU+k+JPCupJcQeYrI0c0TfNHInBKnDI6HhlLKeCRX7OPxy8N/tH/AAZ8N/F3wwPKi1m3Iu7Rmy9ndRsUmhbv8rq2CfvKVbowqqqqqqqqqqqqqqqqqqqqqqqqqqqvjD8E/hb8fPB03gT4seD7HX9JkbfGs6lZbWUcCWGVcPE+MjchBIJU5UkH9sz/AIJsfFH9mRr3xx4OFx4x+G8bZOpxIDd6YuB/x+RKBhc7gJUBTgbvLLKp/Z9/aZ+K/wCzT4n/AOEg+HGubLW5kQ6lpF0C9pqCrnAkTIIIDHDqQwzwcEg/stftu/CP9p3TrfS9Nv00HxrHCDeeHL6QLI7BNzvbN0njGH6YdQuWVQQTVVVVVVVVVVVVVVVVVUOg+lVVUtVS1VVVVQ6iqqqqqodKqqqqr9sS8aTVPDGnlBtht7iUN3JdkBH/AI4Pzr9jiyaTVfE+oCQbYLe3iK45JdnIP/kM/nVVVVVAcAVVf8FpPCci3Xwu8dQJKyPHqOm3LF12IQYZIQF+9lt0+TyPkXp3/wCCVPioWHxM8Z+DGnZRq+iRXqoVXa7W02wcn5t2LlsAcEbiegpGBAwajcEYzSPjjNI4IwDzSP2zSvjoaSQEYzSvjoaWQEdaWTHQ0sox1r9oD4A351G68d+A7A3EFwTJqGnwhmlWUklpY1Gcqc5KjkHJHB+UGa3kIBeORCQeqkHvT6nqDqUa8kwfQ4rSdH1XX9Qh0rRbCe9vLhtscMKFmY/h29+gr4D/AAVtvhjpraxrPlz+I7+ILMy4K2idfKQ9znG5uhIAHAyWmGODTTA8A08pPANO4x1p5M8A07gDg1I+eAadxjANOwPJNSOD0NSNgdeads96/bA8Tx+Ff2ZviNqckkaifQ5bAGRGYZuiLYDC85JmAB6A4J4Br4EeD5fiB8bfAPgiK0S5Gt+JLC0kjkgMyeW86CRnTBygTcWzxtBzxVVVVVAcAZqq+Jnxt+GnwktDN4z8SwQXRTdFp8P725m9NsY5AP8AebC+9fFH9vHx14kjn0r4caXF4ZsnJUXsuJ7xl9sjZHnnOAxHGGGM1PP4g8W60ZribUNa1fUZeXdnuLi4kPHU5Z2P4mvhP+wx498XeRqvxEuv+EW0xsN9m2iS8kH+592L/gWSP7tfDb4F/C74UQRf8If4VtYr6NCralcIJrt8jDZlPKg91XC+1KB0HpVVVVVVjjGaqqqqpQDVVVVVVgYwKqscYzVVVKMmqqqqqoAYxVVVVXxx/ZX0/wAU/avFfw4hhsdZbMk2mgBIbxuPuEkCJzgn+6xPO3k1fWN/o2ozafqVnJbXlnKUmgnjw0bqeVZT7joa+HEHwT+KyweFfG2nnwp4mkHl2uqabJ5UF/IT8oeIgxxN04UKGJIBU7RXi39i7x5pZln8Ja5puuQICUilzaztx0AOU9slx26dvFPgPxl4Jn+zeK/DOoaY27arzwERucZ+V/utxnoT0PpXg/4t/EfwEUTwv4uv7WCM5Fq7+bB0wf3T5Xp3Az09BXg79trUoPLtfHvhOG6QbQ13pj+W4A6kxOSGJ68Moz9ePBnx1+FnjxYo9E8W2kV3JwLK9b7PNuxkqFfG84/ulh154NUoGM1QGe9VVVVVVfHf9nzwb8dNEEOqp9h12ziZdO1WJfnhJ5COP+WkeeSp5GTtKkkn4i/DXxj8K/EcvhfxppL2d2mWifrHcx5IEkbfxIcHnr2IByK/Zh/bJfSlg8A/GTV5ZrQlY9O1yc7mgyQPLuG6lO4kPK87sjlUeOWNZInV0cZVlOQQehBqqqqpgOoNUwyMiq/ba+BB8ZeHP+FpeGLJn1rQoduoQxRjN1aDkv6lo+T/ALpPoK+DnxS1n4O+PtO8a6ODKkB8q+td5UXdsxHmRk/gCCcgMqnBxivDHiXRPGXh7T/FPhy/S803U7dZ7aZO6kdCOoYHIKnkEEHBBqqIBGKI7VVVVVRGRgVVVVftNfFdfiL42bSdIui+haCWhtyrHbPL/wAtJcfX5R14XP8AERX7IXwoXWtYk+Jmt2m6z0qQxaYrgESXGPmkweoQEYP945BylftD/BHTPjf4Fl0nEUOu6aHm0a7cY8uUgZjY9RHJtUN6YVsHaBUieI/A3iVo2N3pGuaJdkHaxjltp427EdCCK/Z1+OOnfG7wSmoSvBB4g07EOrWaELtfHEqLkny37E9CGHbJqqIBGKI7VVfte/HN/hD8PzpPh+8VPE/iMPb2TLJte0ixiS4AHIIB2qePmYHnaQfh54D174m+M9L8D+GoVe+1OYIHfhIUAy8jn+6qgscc8YAJIFfD/wAC6F8NfBuleCfDkGyy0u3WNWZQGmf+OV8cF3bLH3PYVVVVMMjrVVVVVfFHxgnw7+Gfi74gOqMvhnQr3UyroXBFvA8vKggkfJ0BBPqKdtS1vUi7tc3+oX8+SSWlluJXb8SzMx9ySa+HvheHwR4I8OeC7eXzItA0m10+N8k7lhiWMHnnotQ4IAHWtX0i28QaJfaDeSSJBqNrLbStEQHVZEKkqSCAcE4yDUf/AASW/Zxc4PjT4kf+DGy/+RKj/wCCRv7NrnB8bfEr/wAGVl/8iV+z3/wT7+DH7OnxFg+JngjxN40vtUgtJrVYdVvbaWEpIAGJWO3Rs8cfNX7Uv7Anw7/au8X6d488QeNfEWhazp2mRaYos1hkt2t0kmlGY2Td5hec/NvxhQNueaj/AOCL3wvcf8lq8Uf+C6D/ABr4f/8ABIz9krwpfxah4gj8XeMCm0m21jVVjg3AEHC2scLYJIOGZvugdNwbwd4P8JeA9BtfC3grw3pmhaRZLtt7HTrVLeGMd8IgAye56k9amKk8VcYOeam/YE+Dg/aOP7T48S+Mv+Eq/tf+1Psf2y2+w+dt27dn2fzNmO3mZ96uMEEDrX7T/wAQbj4dfCbUtQ06Ux6hqki6dZyDOY3kVizAjoRGkhB9QKjjaV1jQZZjgVpehKwARNzd2Iq68PuIsPECMdCK1XTvsb74x8hOMehr9mvxpdeGviFb6G85/s/XgbeWMtwsoBMbgeuRt+jn2r/gqLrD3nx08P6LHexywaf4XicwqVJhmkuJ927HIJRYjg9sEdef+CKmkWUPgj4o67Grfa7zVdPtpSW42RRSsmB2OZn/AE9K+Lvw8074tfC7xX8MtVfZbeJtIuLAy94WkjKpIODyjFWHB5UcGtb0jxB4C8W3+g6is+m654d1GS2nCM0cltcwSFWweCrK6H0IIr9mX40ab+0F8C/CPxTsriJrrVNPRNUijG37Pfx/Jcx7ckqBKrFc9UKt0YVVVVVTdKqqqpoYbiJoJ4kkjcYZHXIYehFeK/h5NaF9Q0FDLByXt+rx/wC76j26/WqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqtP1zVdMKizvpFVT/AKtjuX8jxWn/ABCBATVLP/tpB/8AEk/1rT9a0rVAPsN7HI2PuH5WH4Hmqqqqqqqqqqq/4Ko/s1RfDj4jW3xz8LWKRaD47uXTU44otqWuqBdzMccZuFDyepeOUnqK/wCCPf7Rt74C+Ml18Atc1F/+Ee8fq82nxyy4jttUhjLBgCcAzRRmM45ZkhHYVVVVVVVVVVVVVVVVVVVVVVVVVVVVTQw3EL288SSxyKVdHXIYEYIIPUEdq/bN/wCCTPhb4hHUfiT+zTHb+HfFErPNdeGZGCafqMjPlmgYn/RXwW+XmI4UARcsfEnhjx58JvGUmh+JtI1fwt4m0K5DGKdHtri1lRsq6HgjBAKupweCD0Nfsof8FUdV0BbLwH+0yZ9U02NUhtvFdtEXuoADj/S4lGZlCkZkQeZ8vKysxYeEvGHhXx94es/FfgvxDYa1pF/GHt7yymEsbgj1HQjuDgg8EA1VVVVVVVVVVVVVVVVRAwAKqqpaqlHHWqqqqqHUVVVVVVDpVVVVVfteXnmeNtG07y8eRpXm789d8rjGPby/19q/Y3smTTvFOoGQFZprWILjkFFkJP47x+VVVVVVVV/wVp8DjxP+yk/iaK2RpfCHiCzvmm2puSKUtasuTztL3ERIXqVUkYGR+wv4xTwb+0/4MnnkjW31aaXTJd7BcmeNkjAJ7+b5fHU9ByajcDgmkYdjSOCMZpHxxmkcEcHmkk7ZpX9D+FJKOmaWQHoaWQjjNLKCOtCQEcGvFPwm+HPjSU3XiDwraTXLEk3ERaCRjzyWjILdT97NQfsxfCOKVJH0y/nVTzHJeuA3sduD+RFeF/B3hTwXaGz8LaDaafGwAcxJl3x03Ocs34k0ZSehppM96aUYxmmlzwKaQHqaeUYwDTyZHWnkGMA07g9TUkme9O/cmpHz0NSOMYBr/gqN49XRPhB4f8AwTMtz4o1czSLg4e3tVDOM9M+ZLbnn0P4f8Esfh3P45/a98P6w1r51j4OsLvVrrcgKg+UYIucjBEs8bDGT8nTGSKqqqqHQfSvid8avhv8ACGyFx428RRW9zJHvt7CEebczjkZWMc4yCNzYXI618Wf25/Hvi7ztJ+HVt/wi2mNlftO4SXkg9d33YvooJH96oYPEHi3WlgtodQ1rV9Rl+VEV7i4uJD6AZZ2P4mvhP+wd4x8R+Rq3xS1L/hHNPbDfYLYrLeSD3PKRZGDzvI5BUV8PPg98N/hXYLZeCfC1nYyYxJdsvmXExxyXlbLHqeM7Rk4AFVgHjNKAaqqqqrGeAaqqqqqlAxxVVVVVAZIAqqwDxmqqqpQMcGqqqqqxngGsDGBVVVVVfGD4EeFPi7Zefdr/AGfrtvHstdSiXLAdkkXOHTJ+o7EZIPjrwD4p+HGvSeHfFemm1uVG6J1O6OdM4DxsPvKcfUdCAQQPgL+1Fe+Ffsvg74iXMt5oq7YrS/I3SWK9Ar93jHHqygcZGALe40jxFpUV1bS2uo6ffRB43GJI5kIyCOoINeNP2XfhH4ujlkttB/sK9ZcJPpZ8lVPUfuuY8euFBI7jjHjH9jHx9pBefwfq1hr8APyxOfsk/T0YlPbO8duPTxH4T8S+Eb3+zvE+hXumXBztS5hKbsYyVJ4YcjketeCfjl8UPACwW+g+K7l7KABVsbs/aIAoP3VVs7B/uFT155rwT+2zpN0FtfiB4Xlsn2gfa9MbzY2bvmNyGQfRnPt3rwj8Q/A/j2EzeEPFFhqZVA7xRS4ljU9C8Zw6/iBWBjAqqqqqqqvi18H/AAV8Z/DR8OeMLFmMRL2d5CQs9nIRgsjeh4ypypwMjgEfGb4H+Nfgj4hOj+J7TzbG4Z/7O1OIfubxB3HJ2MMjch5Ge4IY/sxfta6h8MJo/BnxEvbvUfCbgLbzEtNLpZAwAg6tFgAFB93qvcNpeqabrmm22saPfQXtjeRLLb3EDh0lRhkMpHBBFVVVRAIxRGOM1TADgVJHHLG0UiK6OCGVhkEHqCK/ax+BbfB/x22o6Ja7fDGvs82nhEO21f8AjtyfbOV/2SP7pr9hz47DQtWHwc8TXmNP1SUvosjD/U3LctFnsr9R/tf79VVMB2qqqqqqI5wKqq/aZ+KZ+HXgV9N0q5Met68GgtGXGYYxjzZPb5TtB/vMD2NeBfBuqfEDxZp3hHR8C4v5dvmMCViQDLucdlUE/h714Y8N6V4Q8Paf4Y0SIx2WmwLDCGOWIHVmIxlicknHJJqv20/2ex4o0yX4u+D7HdrGnRD+2LeNebu2RcCYY6vGAAfVB1+QA/Bz4ra78HPHNl4v0ZjLCpEV/Zltq3duSN8Z9DxlT2YA4IyD4Q8WaD478M6d4u8M3q3Wm6nCJYJBweuGVh2ZWBUjsQR2qqojnGarXNa0rw3o174g1y9jtNP063e4up3BxHGgyzYGSeB0HJr42fFTU/jL8RdS8bX4ligmIh0+1kfd9ltkzsjHYdSxxwWZj3r9iD4Gf8IP4PPxM8R2W3XfEsQ+xxyphrSyzleoyGlOHPJ+UR9DuFVVVVMAR1qqqqqv+CkPi9PBv7GPxGuUmRbjVLW30yBGcqZDcXMUcgGO4iMjY6EKQa/Zr8Ow+Kv2gPh7odyI2gl8RWckySAlXjjkEjocf3lQr+NQkEg57VARjrUBGBioT0APaoSM4BqBhwAahYA4zUDgjGahlA4zSTjHWnnBGAamlBGM1O4xgGp2BOM1+3qc+E/C/P8AzEZv/RdaUFN6u49jjivCsVs8iCUjGe9eIrXSY7FTbMC23njvXiZYhFKAfp+deADdL488NtYrG1yNXtfJWT7pfzV259s4r9vrxJH4j/ak8WpA8bwaSlrYxsqMpJS3QyBs9SJGkXIwCAOvU/8ABIjwzdaD+yW+q3BlKeI/FV9fwB4CgCLHBbHaSfnG62b5hgZyv8JNV/wVz+BbeAvjnZfF/SLJk0j4hWubpkjAji1C3VI5Bx0Lx+U/PLN5h5wcf8Egv2if+Eb8aat+zp4jv9un+J92o6D5jcR30afvoh/10iQN9YfVqqqqqogEYzVVVEdqqvE3gXTddD3VqFtb0gnzFX5ZD/tD+o5571q+ianolwbbUbZoznCuBlX91Peqqqqqqqqq8F+ELHX9OurvUDIPn8uFkOCpAyT79QPzrX/AeraOrXFv/plsvJaNTuUepX09xVVVVVVVVVVVVVVVVVVVVVVVVVVVVVWlheX8nlWVrJMwHIRScfX0qDwLr82BJFDDn/npKDj/AL5zUXw81Ns+dfWq+m3c39BUvw71EKPJv7Zmz0YMv9DU3gLXY/uNbS8fwSEfzAq48LeILYZk0uY4H/LPD/8AoJNSwTQN5c8Txtj7rqQaqqqqqqqqqqqqqqqqqqpWZWDIxDKcgjgg1pnjTWLAqlxILyIdVlPzfg3X881pXizR9VIiSYwTHpHN8uT7Hoaqqqqqqqqqv2lPg5p3x7+CPiv4YXkETXOp2Dtpksh2+RfR/PbybuoAkVQ2OqFl6Ma03UfEvw/8XWmr6XdXGk+IPDeopPbzJgSWl3BIGVh1+ZHQH6ivgV8V9F+Onwe8I/FrQXh+zeJtLiuniifeLafG2eDPcxTLJGfdDVVVVVVVVVVVVVVVVVVVVVVVVVVVVX7SH7J/wY/am8MnQ/ib4bRtRt4Wj03XLQCO+04k5zHJg5XPJjcMh9M81+1v/wAE5vjX+y/PdeI7G0l8aeAU+dPEGm2rbrNcHIvIAWMGMH95lojlfnDNsHwH/aa+MX7N+tnVvhj4oa3tZ5VkvtJulM1lfbe0sWRyRxvQq4HRhX7MP/BRj4OfH1rTwr4nmTwV40lVUWwv5f8ARb6THzfZ5+mcjiOTa3zALvwTVVVVVVVVVVVVVVVVRwQADVVVL0qqXpVVVVVAAnFVVVVVQHAGaqqqqr9rdkb4k6equCV0SIMAeh86Y/yIr9jz/kWvEP8A1/R/+gVVVVVVVX7QXw8n+LPwO8dfDiyXN5r+g3VtZAuEH2kxkw5JIGPMCZyQMZ5FaNqur+E/ENjrmmyyWeqaNex3Fu5BV4Z4nDKexBDKPfivh94z034h+B9B8c6Qy/ZNe06G9iUSB/L8xAxQkfxKSVPAwQcgUj44zSP6GkkBAGaSTHGaSQEYzSyccHNLICMZpZD2NLL2zQlB70JARwaEp6Bq804xn9a80+o/OjKcY3UZAR96mlHrTTDGAaaQkYBppB65p5M8A08gHepJM8A07gDk80754zUjgDANO3vX/BQj4ox/EX9oXUNHsJ1k0/wbbjSY9pyGnVi056n5hIxjPT/VDjOa/wCCNnwjbQfhf4t+Muo2zLceKtRTTtPLkEfZbUEu645AaaR1Oe8A4HU1VVVfEX4ufDv4U6eL7xx4ntbBnQtBag+ZcT9vkiXLEZ43Y2g9SK+LP7eHjHxH52kfC3Tf+Ec09sr9vuQst5IPUDlIsjjA3kcEMK/4qbxtr/8AzE9e1vU5f+ml1c3MmPxZ2wPc18J/2DvGPiPyNW+KWpf8I5p7Yb7BbFZbyQe55SLIwed5HIKivh18Ivh38KdP+weBvDNrYM6BZ7ojzLif/flbLEZ5252g9AKqqAyQBSjjrVVVVVDqKqqqqqgBjiqqqqqAyeKqgATiqqqoDjFVVVVUACcVVVVVVKOK8deAfC3xI0GTw74r01bq2Y7onU7ZIJMYDxsOVYZ+h6EEEivjJ8CvFHwi1AyTh9R0KZgttqaRbVLEH5HXJ2PweM4I5B6gfBX4/wDiT4TXi2E7S6p4dmYCaweQ5gy2WeHPCtycjgNnnnBHhHxj4b8eaFD4i8K6pHfWMxKh1BUow6qynBVhxwR3B6EGq1LS9M1mzk07V9Otb61lGJILmFZUce6sCDXjb9kX4XeKFmudBS58N30gJRrRvMg3Z6mJj0/2UZB0rxz+yb8VPCfnXWj2cPiSxiXd5lgf32PeE/MT7Jv7e+JodV0LUTDPFd6df2rcq6tDLE2PTgqcGvA37V3xX8IvHb6pqUfiSxUKph1IZkAHUrMuH3EcZfePavAv7XHwt8VpFba7cT+Gr9gA0d6N8BbuFmUYxx1cJVlfWOp2kd9pt7Bd20wzHNBIJEcdMhhkHpVVVVVVXjnwF4T+JPhyfwp4z0eLUdOuCGMbkqyOOjoykMjDnkEcEjoSK/aC/Zw8VfA7W5ZxFNqPha5lxYaqE6bskRS44WQAH2YDI7qv7OP7UHiT4Lalb6DrE02p+DZpSbiw4Z7UsRmWAnkEHkpna2T0Y7h4c8SaB4v0W18Q+GdWt9S069jDwXED7lYEZ57g+qnBB4IBqqqqYAjiqYZHFV8Y/hbovxi8Aah4J1crE8w82xuigY2lyoPlygfiVYDBKswyM5rV9K8QeB/E1zpGoJcabrGi3ZR9rGOSGWNuGVhgjkAhh7EV+zX8Z4PjV8OLbVryZBr+mYttYiUAfvQPllAAACyKN2AMA7lH3c1VMMjiqqqqqpgM8Gqq8u7awtJr68nSG3to2klkc4CKoyWPsAM18aPiPc/FDx9f+IPNY6fExt9MiYY8u3Unacdi3LnPQsR0Ar9kP4Wjw94Zl+IWr2yf2hrqhbEMvzQ2oPX2Mjc/7qoc8kVVSxxyxtFIiujKQysMgjuCK/a2+AZ+E/iz/hJ/DdiU8K69MxgCqNtlcHLNBgABVwCUH90Efwk1+x38fl+HHiUeAfFV/wCX4a16ceTLIQFsbtsKHJ7I4Cq2TheG4AY1VUwGeDVft9fGh7W3tfgtoF0yyXCpea1JHJj93z5UBHoTiQj2TsTX7LXwYX4zfE23sdUhZtA0VVvNV4OJUDDbBkEY8w8cc7QxHIpI44o1iiRURFwqqMAAdAB6VVVVUQCMZqqqqqv+Cz/ie60/4A+DvCluJUj1jxWJ55EmKhkgtpcRso+8C0qtycAxjgnBH/BODwyuvftMWeqllB8N6LeX4BYgksq23AA5/wCPnocdz2AMJ6VCRnGagbIxmoWBAwahYEAg1C44wahcEDB6VDL0ANeL/iN4N+H9guoeLdet7BHz5SNlpJSB0VFyx6emK1L9tvwLbXTQ6V4X1q8hXI86Qxw7iCeQu5jgjBycHnoK8PftnfDPVpvs+u2GraKSTiaSETxAcdTGS+eTxtI4684rSPEWi+JNMh1nQNVttQspxmOe3kDqfUZHQg8EHkHg1PKCMZqZxjINftVeA7nx98Jb9NNi8y/0OQajBGFyZBGrCRR3z5bOQO5UDvkRStDIsqH5lPFaPriqoaKTkdVPUVd+I5JYdrS8AetaxqQvH8uNsqDkn1PtX7Pnhldb+IFvrV80cWmeHEN/eTykKkewHZkngfNhsnoEJyMZr4ieK5vHfj7xH41neQvruq3N9+8GGAllZwCASBgEDAJAxiv2NvAv/CuP2WPhh4UeHyZo/Dlvd3MRj2GOe5H2mVSMnkSTOCc8kE4GcCv2xP2f7X9pX4A+I/hukMR1pY/t/h+aQgeTqMIJi+Y8KHBeFm7JMx64rw34g8T/AA28aad4m0WafS/EHhrUY7i3Z0KPb3ELggMp5BDLgqfcGvgL8X9C+PXwh8MfFfw8UWDXrFZJ7cNuNrcL8s0J90kV1z3AB71VVVUen4VVVR61VVf2FnqVu1pfW6TRN1Rx+vtXiP4cXFrvu9BZp4s5+zt99fof4h+v1qSN43aKRCroSGVhggjqCKqqqqqqqvCVj/Z/hyxtyuHaLzGBGDlvm5/PH4VXiXwLYayr3ViEtb0jqBhHP+0B0+o/WtQ0680u6ayv4GilTse49Qe496qqqqqqqqqqqqqqqqqqqqqqqqqqrw14KN8iX+rBkgYZSEcM49T6A/nUNvBaxCC2hSONeiouAKqqqpYYZ08uaJJF/uuuRV54Q0C9BzYiFiOGhOzH4dP0q++Hbfe03UAfRJ1x+o9var7wzrlhkzafIyj+KMbx+nT8aqqqqqqqqqqqqqqqqqqrSvFOr6SQkdx50I/5ZS/MPwPUfyrS/GmkaiVinY2kxH3ZT8ufZv8AHFAhgCpBBHFVVVVVVVVf8FNvhBF8MP2nNT1zTbdI9M8d2y61EEXAW4dmS5U8D5jKhkPX/XDnOcf8EV/jA3iD4VeL/gpqV073PhLUk1LTlcAAWl2CHRcckJNE7HI6zjk9BVVVVVVVVVVVVVVVVVVVVVVVVVVVVU0MNxC9vPEkscilXR1yGBGCCD1BHav2uP8Agkz8M/i39u8cfAWS08CeLpN802meWf7L1KQ7ifkHNq5JX5owY8L/AKvLF6+MXwN+K3wC8VyeDPi14K1Dw/qK5MJnj3Q3aDH7yCZcpMnIBZCQDwcEED9mv/go98bPgPFbeGfE07ePPCMCpHFp+p3JFzZRrwBb3OCwUDACOHUBQFCcmvgH+1f8Ef2k9OM/w28VZ1OCMPd6JqEf2a+ts+sZJDgd3jZ0Gcbs8VVVVVVVVVVVVVVVVVVUvSqpelVVVVUOtVVVVVQ6D6VVVVVX7Uf/ACVe4/68YP5Gv2Q0QfDfU3CjcdblBOOSBBDj+Z/OqqqqsA8Zqqqv+Cgvwfb4NftUeL9KtrVINK8Rzf29pQRAi+TdFmdVUHCqk6zxgccRjgA4H/BM34st4p+FOpfC/UrpWvfBt3vs1OAWsrgs4A5yxWUSgnoA8Y9KRwQBmkkxgZpXBHBpJOMZpXBHBpJe2aWQEdaWQ9jmllBGM/nQkB4BoSdg1eafUV5pI/8ArV5p/wAijKemRRkJGN1NIMfeppQBgGmkJHBp5B3NPKMYBp5MjrTyADANO+e9SSDGAadx1zzX7Rfxhsfgf8Ite8fzvG15bQ+RpkD/APLe8k+WJcdwD8zD+6relaZpviLx34qtNH0q1n1XXvEOoJBbQoMyXV1PIFVR/tM7AfU18EPhZo3wS+EfhP4U6EkX2bw3pkVs8kabBcT43Tz47GSVpJD7uaqqrxr8QPBXw60k63438S2WkWnIRrh/nlI6rGgy0jY/hUE+1fFj9vPxDq/naR8JtK/se1OV/tO+RZLlh6pHykf47z9DV/qHifxxrv2rULvUtd1i/kChpGe4nmY9AOpPsBXwn/YY8e+LvI1X4iXX/CLaY2G+zbRJeSD/AHPuxf8AAskf3a+Gnwe+Hvwk0sab4J8PwW0jJtnvZFD3Nzzn95LjJGedvCjsBVVVUOtL0qqqqqh1qqqqqqA4AzVVVVVL1/CqpQCaqqqgOMVVVVVSjPeqqqqqoDjFVquk6XrunT6RrOn299ZXK7Zre4jEiOOvKnjqAfqBXx4/Zl1PwB9o8VeCop9Q8OKGkuIid0unjPQ93jAI+bkgA7um4/DL4q+LPhTri6t4cvC1vIQLuwlY+TdL/tKP4h2Ycj6Eg/C74seFPixoSapoF0qXcSL9t0+Rv3tq5yMH1UkHDDgj0OQMZ4BrAxgUBngV4s8AeCvHVo1n4t8M2GpqU2K80I82Mf7Egw6d+VI6145/Ym0W88y7+HviWWwlOStnqQ82LPOAJFG5R0HIc9ea8c/Br4kfDsmTxP4XuYrUHAvIB50B4z99chfo2DwfQ14P+IfjbwBdfbPB/iW801ySWjjYNG5xjLRtlG6DqD0rwJ+21OhjsviP4YWRQMG+0o4bgcboXODnqSGHsteDPib4D+IVsJ/CPiezv3ABe3D7Jo+M/NG2GHfnGODzxVVVVVV4h8PaJ4s0S88N+I9Mg1DTdQiMVzbTLlXU/qCDggjBBAIIIzX7S/7LWsfBW6/4STw7Jcar4QuZNq3DrmWwcniObHBB4CvgAnggHG79nr9oXxH8CvEe+Pzb/wAN38g/tPTN3Xt5sWeFlA/BgMHsV8HeM/DPxA8O2ninwjq8Go6beJmOWJuVOOVYdVYZwVOCD1qqqqIwcCqI7VX7dnwQ/tXSo/jN4ctM3enIsGtxoOZYMhY5sZ6oTtbAJ2sDwENfs6fGG4+C/wASbLxDPNKdGvB9l1eFF3b4GI+cD+8jAMMc8EdGNWN7aanZW+o6fcpcW11EssEqHKyIwBVgfQgg1RAIxRHaqqqqqYZHBqq/a7+JsfhzwkngDTLrGp6+u65CnBjtASD/AN9su33Aevgz8PJfid8QtN8MlGNkG+0ai65Gy2QjfyOhbKoD2LirS0trC0hsbKBIbe2jWOKNBgIqjAAHoAAKqq8d+CdC+InhLUvBviO1Wax1KExtkfNG3VZF9GVgGB9RXxR+HGu/CjxvqXgnX4yZbKTMFxs2rdQnmOVOTww7ZODlTypr9jj4+t8RfDh+Hvii8eTxHoMAaGeZyzX9qDgMWPJdCVVs9QVPPzYqmA7Gvil4/wBL+F3gDWvHOqlWj0u2LxRE486Y/LHGP95yo9s57V4i17XfHniq/wDEWqu95q2t3rzSiNCS8sjZ2ovJxkgKo6DAFfs0/B6D4NfC6w0S4hxrWpYvdXdsEid1A8sH+6igLjpncf4jVVVVVEZGAaqqqqr/AILeyyiH4MW6ysI3bXWZAxwWAsACR6gMfzPrX/BLOztX+JnjO/eFTcQ6FHFHIRyqPOpYD2JRP++RULAjANQsCAQahcZGDULj1qFx0zUUgxjNRS4wM18XPitYfCrwsdXeJLnUbpjFp9qxIEkmOWbHOxRyfXgcZzXiTxJrXi7WrrxD4hv5Ly/u33SSOfyAHQKBgADgCoNE1G4TesG0dt5x+lXel3tmu+aH5f7wORXw0+KPif4Xa4mq6FdO9s7AXlg7kRXSjsw7MBnDYyPcEg+HfFGk+L/D9j4l0S486y1CESRE4yOxVsE4ZSCpHYgippc8A1M4IwDX7Qn7MOo6bqNz4z+GemyXen3BaW90yLmS1bqWiXq6H+6MlT0BX7rLJE5RgyOhIIIwQe9PLLIMPKzf7xzXgP4beLPiNqK2Ph3TXeFXAuLyRSsNuOMlm6Zwc7Rlj2FftHyaN+zV+yR4m03TLhJNV16D+yhcsfLe6uLoFJGHUjZD5rKo6CP1y1fAX4aT/GP40+CfhhCkrJ4j1u2tbpogC0VsXBnkAJGdkQkfGQfl4qKGKCJIII0jjjUKiIu0KAMAADoMVVf8FXP2Yx8LPiynxu8K6bKnhr4hXDvqBRP3dpq2C0oyBx56hpgCSS4nI4AA/wCCU/7UsHwo+JFz8EPGeqxW3hbx1cK+nyzsQtpquAiYPQCdAsZJ/iSHkDdmqqqogEYzVVVHrVVTdfwqvEXhDSvESGSVPIugPluEXnp/EP4h09/cVrvhvVPD83l30OY2OI5k5R/oex9jzVVVVVVWk2D6pqVtp6ZzPIFJHYdz+AyaVFRQiKAFGAAMYqq1vQ9P161NpfR52kmOReGQ+oP9K17w/f6BdG3u03RsT5UwHyyD29D7VVVVVVVVVVVVVVVVVVVVVVVVVSI0jLGgyzHAHqa0TwhpmnWyNd20dzdFQXaRdwU+ig9veuKPWqqqqqqr/Q9J1ME3ljE7EffA2t+Y5q++Hlq/zaffSRn+5Ku4fgRjH61e+DNeswWW1W4UD70Lbv04P6VNBNbv5VxC8bjqrqVP5GqqqqqqqqqqqqqqqqqrTPEGq6ScWl2dneJ/mX8u34YrTfH9hOBHqcDWzgffQF1P4dR+tWt5aX0fm2d1FMuOqMDj6+lVVVVVVV/wVw+GMvir4C6L8SLOJGn8D6uPtDMDlLW72xOR7+ctsMHAwTzwAf8Agl98UY/hh+2N4Qju7jyrHxfHN4euTvVcm4AMC/MQDm4jtxjOTngE4Bqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq+JHwv+Hnxg8LT+Cfid4P0zxJolwwdrS/gDhHAIEiH70bgMwDqQwycEZr9qb/AII5+I9EluvF/wCy1qz63p7M8knhbVZ1S6tx1C21w2FmXkgLJtcBR88hJxqWmeNPhl4vm0vU7XV/DHibQbrbJFIJLS6spl/JkYcEHjsa/Zx/4KsfEb4fxW/hj466ZP450VGCpq0DrHqlsmB1zhLkDHAco+WJMh4A+DP7RPwa/aA0j+1fhX45sNXkijD3VgWMN5adB+9gfDqM8b8bGIO1iOaqqqqqqqqqqqqqqqqpelVS9Kqqqqodaqqqqqh0H0qqqqqv2o/+Sr3H/XjB/I1+yJ/yTXUv+w5L/wCiIKqqqqHUVVVX/BWL4CS/Ef4JWfxY0DT0m1n4eTPLdFEPmSabLtE3TqI3WOTn7qiQjGTn9mT4y3HwJ+Mmh+OWlk/stn+x6zEgJ82ylIEnA5YoQsqju0ajpVpeW93bxXdncRzwToHiljYMrqRkMCOCCMHNJICMZpXx0NJICMZpZD2NLKCME0sgxgH86WXAxmhMDxnNCVTwDQkGOG/OhIexrzT6j8680+o/OjISMA0ZMjG6mlHrTSjGAaaXIwDTSE96aQdjTyk9DTyADrTyZ4Bp5ABwaeTPANf8FA/2ho/iv8RU+Hnhi9WXwz4NleNpI2yt5fH5ZZAQcFEAEa+/mHJDDH/BIz9mufxj8Rb39ojxPpudD8HbrTQ/NAxcanImHcDnIhic9QPnmjKnKHFVXiTxP4c8H6TLrvinXLLSrCDh7i7mES57KCerHHCjk9hXxc/b2VHuNF+Dukq4UlBrWoxkA/7UUBwfcGT8UrxB4k8S+NdZfWPEus32sanckKZrmVpZG54UZzgDOAo4HQCvhD+xT8RfiBFb614wk/4RPRpfmVbmIteTL/sw8bAcEZkII4IVga+FfwM+G3wesRbeDtDUXjjE+pXZEt1N25fA2jgfKgVe+MkmqqqqqAyeKAGOKqqqqoDJ4qqqqqqHQfSqqqqqWqpQD3qqqsZ4FVVVVVSgY4NVVVVVgHjNVVOiSIY5FVlYEMrDII78V8fv2Wf+Pvxr8LbD/butCt4f++ntwPwPlAeu3steFPF3ibwBr8Wv+GdSl0/UbbchYKCGB+8jqeGHHQjqAeoBr4JfHnw78XrD7GVXT/ENpEGu7Bjw/YyQnOWTOMg8qTg5GGNKAaqnRJEMciqysCGVhkEd+K8e/sy/Cnx3510NF/sXUZcn7XpmIcse7R42NyOeATk8gnNfED9kD4jeFg954VeHxPYrklYB5VwgHrEx+bjH3CxJ7Cpodb8NasYZ4r3StTsZOUdXgmgce3DKfyr4f/tcfErwgkNj4haPxRYRDAW8cpcAe0wBJ+rhjz16Y+H/AO0p8LfiB5VpFrP9j6i682epkQknuFfOxunHOSOw6CguR1raMcVgegoqCMUV44NEdq1TS9N1zTbnR9YsYL2xvImiuLedA6SowwVYHggiv2p/2Urj4VTy+OPANrNc+EJn/fwFmkk0picAMTktETwHOSCdrHOC3wL+Onin4G+KV1jR3N3pV2VXU9Ld8JdIO4/uyLk7X7dDkEg/D/4geFfif4Vs/GHg/Ulu7C7GCDxJBIMbo5F/hdcjI+hBIIJqqphkcVTAdjVX1jaanZXGm39ulxbXUTRTRSDKyIwwykehBIr9oP4PXfwW+I174ZXzpdJuP9I0m5lXmWBjwpPQshyjHjJGcAMBX7C/xwGu6M3wc8R32b/SYml0aSV+ZrYHLQ5JyWjJyoH8GcYCVVEc4zVVVVVMMjrVajqFlpOn3OqajcpBaWcLzTyvwI0QEsx9gATXxM8cXnxH8b6r4uvAyLeTEW0LH/Uwr8saemQoGcdTk96/ZW+G/wDwhXw9TXtQt9mqeJNtzJuGGjgA/dL+IJf/AIHjtVVVHrX7U/wJX4y+CftmiQoPE+hI82nnABulxl7cn/awCueA2MkAk14T8U+IPAHimw8VeHbp7LVdJn3xMQRgjIZGHdWBZWXuCR3r4TfErRfi34E07xpojbRcpsuYD962nXHmRn6Hoe4IPeqYZHBr9vX4vx+KPGNn8LNEvVl07wyTNqBjfKyXzLjaccHykOPUNJIp5Wv2Gfg3H448fP8AETWrZn0nwjIj2ysPlmvjzH+EY/ecYIby+xILAEVVVVVVVVat4o8M6ErnXPEemaeIwC5u7tIdoPAJ3EYycVqf7QnwZ0lzHdePrGQhyn+jRyXIyOvMatx79DV9+2B8JrTZ9nt9fvd2c+RZoNmMdd8i9fbPStQ/bX0ONM6V4Cvrht+MXF8sI2885VX56cfrX7Z5sv2x5vCTatBN4Yi8Ii98hLeYXLTm58jduLKuMfZ1xgfxGv2ffhzpP7OOp6jrPg++u9QvNWtI7e6/tIq0YCtuyixhCuT6luPzp/jV42MheOSxjBPCLb5A+mSTS/G/x4owtzZ/+Awpfjv8QFGFurL/AMBh/jS/H74iKMLdWP8A4Cj/ABq0/aM+IdtJvkXSrkbcBJbYgD3+Vgf/ANdWv7UHjWOQG70PRJYwPuxpLGc/Uu38qs/2sL+JCL7wVBM2flMV+Y8D3BRs18bPiXL8TvEdpqKWslpaWdmIobZ33bWJJds9Mk4GcDhRXhvT0uJDdSLnYcICO/c1p+iNcoNqZ/CtU0fyUKOgIxggitZsFsLsxx/ccblHp7V+yH4okm0fXPCU8pIs5ku7cEk8SAq4HYAFFOM8lzx1qWUdAalkGMZqaTjrzXin4d+AfF0/2vxJ4R0q/ueM3EtsvmkAEAFx8xHPTOKX4KfCW1lSaHwBpBdDkCSDev4q2QfxFQ2tnp9tHY6faw21vEMRxQxhFQewHAr/AIKh/FePW/G/h/4QaXeK8Ph23N/qaI2cXU4HlIwxkMkI3DHBFx7cf8EdPg63iT4veIvjPqNoj2Xg7TvsVg7oCRe3WQWU5yCsCyqcDpMOexqq+PXwZ8MftAfCfxB8KfFaoltrVqVguzCJHsbgcxXCAkfMjgHGRkZUnDGviX8O/GPwX+I2tfDnxjatYeIPDV6YJ/LLAEjDJLGxAJR1KSI2BlWU96/4J7ftaWv7S/wjg0fxHfZ8e+D7eK11xZGAa+QArHeKNxLbwo8w4AEm7gKy5qqqqqqputVVN1/Cqqe3guoXt7iJJY3GGR1yCPpXiD4aRybrrw/IEbqbaQ8f8Bbt9D+Yq+sLzTbg2l/bPBKo+64xx6j1HvVVVVVfDPRjNdS63KvyQAxw5HViOSPoOPxqqputanplnq9m9jfRB43H0KnsQexrxFoFz4fvzaTHfGwzFLtwHH+PqKqqqqqqqqqqqqqqqqqqqqqqqq8NRpJr1grjIEwIHuOR+oqqPWqqqqqqqqp7e3uo/KuIY5UP8LqGH5Grvwd4fuxxZ+S2PvRMVx+HT9Kuvh1AwzZam6+iyxhv1GP5VP4B1uNsRSW0oycEOQfxyKl8JeIosbtMc56bXVv5GrjRNXtv9fplyoHfyiR+Y4ogg7SMEdjVVVVVVVVVVVVVVVRySQuJIZGR16MpwRVj4z12yAR7hblAMATLk/mMH86sPiBp0+E1C2kt27svzr9fUfkas9QsdQj8yyu4plxzsbJH1HUVVVVVXxz+GcHxl+D3i/4XzTxQP4i0ma1t55VLJBOVzDIwHJCyBGIHPFXFv4p+HXjGS1nW60XxJ4Y1Mo6g7JrK8t5MHkdHSRPzFfsr/G+y/aK+AXg/4sQS2/2zVbEJqkMJ4t76ImO4TB5UeYrEA/wspGQQTVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVXxy/Zm+B/7R+ijR/i74A0/WZIYylpqIUw3tn3/dXCYkUbjkpkoxA3Kw4r9pf/AIJCfGD4bS3viX4D3r/EHw2jM6ac4WLVraPsCgwlwQMDMe1mPSMVBceMfh14pE9rPrPhjxJotwQHjaWyvLKZeDyNrxuOnY18Cv8AgrH8W/BX2LQvjRotr440iL5H1GBRa6ki8AEkYil2gdGVWY8s5PNfBD9rP4CftDItv8NvHlrPqwj3y6LfKbW+jwuWxE+DIFH3njLoP71VVVVVVVVVVVQAJAojtmqqqXpVUo4qqqqqh1qqqqqqHQfSqqqqq/aSnlm+MeuxySFlgS2SMH+EfZ42wPxYn8a/ZRt7eH4VtJA2Xn1Od5RuzhgqL+HyqvH+NVVVVDqKqqrVdL07XNLvNE1eziu7G/t3t7q3lXck0TqVdGHcFSQR71+1v+zzqv7M3xt1r4dXCzzaOzC70K+ljKi7spOUIPdkO6Jj/ejY9CK/4J1ftHReKfDI+BXiu9jXWPD8Jk0OSRyDeWYOWi56vETwAcmMjAxGxKuD0P4UkvbNLICOtLIR3zSyg8ZoSehoS8YzQlB70JARw1CQ9jQlJ6GvNOMCvNPqaMpI4oyEjAIoyZH3qaUdAc002eAaaQnjNNIMdaaXIwDTyDHJp5RjGaeTPU1+3h+1fH8L9AuPhF4E1Fh4v1y0/wBNuoW50q0kBBIYHKzOPujqqnfwShPwP+C3jb9oH4l6R8L/AAFZebqGpyfvbiRGMNjAMeZPMVBKxqCMnHJKgZLAH4OfCjwn8EPhpoHwt8FWvlaXoNmsCyMoElzJ1knk2gAySOWdiABljgAYAq9vbLTbSW/1G8gtbaBC8s08gjSNR1LMeAPc18ZP26fC/hzz9C+E9qmv6lGxR9SuEK2cZHBKAENKcjr8qnghmFePfiP41+J2s/27438QXGp3SgrEHwqQqTnbGi4VB7Ac96+D/wCyV8Tviv5ep3Vv/wAI1oTYP2/UYGDyqe8MXBk9ckqp/vdq+En7N/ww+D9vDNomjrqGsoP3mr36iSck9dnGIh2woBxjJY81Q6iqqqqqXr+FDpVVVVVL1/CqqqqqrAxgVVVVVSjjrVUtVVUOoqqqqqqAGODVVVVVDrVVVV8f/wBmiw8dw3Hi3wJZw2viYsZLi3DhI9R4Gc5O1JOM7uAxJ3ddwim8ReDNfLwS3uj6xpkzKSrNDNBIMhhxgg9QRXwC/aQ0v4lQ2/hTxQ6WXimOPAbAWK/2jlo/R8DJTHqV4yFAGKqqqvGHw78EeP7T7H4v8NWepKBhHkQrKg/2ZFIdep6Eda8f/sTPmW++G3iNSMlhYaocYGOiSqOeeAGX0y3U14u8A+MvAV4LHxf4cvNMkYkI0yZSTHXY4yrfgTXw7+PvxM+GzwwaRr0l7psICjTb9jNAFHZATmP/AIAR75r4d/tg/D/xQkNj4wik8NaixCl5D5tq59RIBlOnRgAMj5jyatLu01C1ivbC6hubeZd0UsTh0cHoQRwRVVTAEcVV3aWt/azWN9bRXFvcRtHNDKgdJEYYZWU8EEEgg8Gv2qv2Vbr4XXU/jzwFaS3HhG4kzPbjLvpTseh7mIk4Vj0OFbsW/Z8+O2t/Avxiuqwie80O+xHqunJJjzk7SIDx5idQT1GVyAxI8IeL/Dnj3w5ZeK/CeqRX+mX8e+GaM/mrDqrA5BU8ggg1VUQCMZqmGRwaqv2o/guvxl+G01rplsjeItE33ekttG6RtvzwAnGBIAB1A3KhPArwr4n17wF4p0/xToNwbTVNIuRLCzr0ZeCrL3BGVI7gkV8M/H2k/FDwLpHjnRRsg1S3DtCX3GCQHbJGTxkqwYZwM4z3qmAzxVVVVVMMjANV+2J8SP7D8L2vw802423uu/vrzacFLVG4HqN7jH0RwetfAz4dP8TPiJp+iTxFtOtj9q1FscCFCMr/AMCJVf8AgWe1IiRoscaqqqMKoGAAPQVVVTAZwKr9tr4DL4d1V/jB4XtMadq0+NZhQACC5bpKAB92Q53E/wAZz/Hx+y18cH+Dfj1ItXuSPDOulLfVFK7vIIP7u4AHPyEnOM5Rm4JC4R45Y1kidXRhlWU5BB7ivjn8TrP4Q/DHWfGszRtdQReVp8L/APLa6fiNcdwD8x/2VaoINb8X+IY7aBZ9R1jWr0Ii53SXNxK+B9WZm/M18G/hjpXwh+Hek+CNMWNpLWIPfToD/pN0wHmyc84LDAB6KFHam6VVVVVqeq6Voto19rGp2ljbJndNczLEgwCTlmIHQE/ga8S/tMfB3w3vjHif+1biPP7nTYWn3Yz0fiM8rj73cHoc1r/7a2nozxeFvA1xMP4JtQuxERz3jQNnjP8AHx71rf7WHxh1dSlnqOm6QD1FlZA8YIPMpcjOc8c8DGK1r4l/EPxEGTWvG+t3cbnJhe+kEeQMZCA7RxnoO59TUUMtxKkEETSSyMFREUlmY8AADqT6VpXwp+Jetqj6X4C12aOQEpKbCRIzjg4dgF6jHWtN/Ze+NOo7Wk8LRWSNHvV7m+hHXHBVWLA89CBjBzVh+xn8SJ2hbUNf8PWsbrlws0skkZx0x5YBOeD82PrX7Wfw11v9lj4PD4rz3Nr4m2avBYz2ETG0EcUqviUSEOWIdVXZs6NndxivgN+0nJ8X/jHoXw51jw7baLp2uyTRJdJdmWSF1hd0HKgNuZAnQfeB7cwfA7wv5YW41TVHk7sjxqD+BQ/zqP4EeDnHOpax/wB/o/8A43SfAHwW4/5Cetf9/ov/AI3Sfs8+CHHOqa3/AN/4v/jdT/s3+EpUUWWu6tE2eTKY5AfoAq4q4/Zj02VFFl4wuImB+Yy2Ykz9AGXFXP7L2rByLHxdZyqF+UzWrRkn6Atge9ePvCuqeC/E9x4c1d4XuLZEPmQ7jG4ZQwKlgCRzjp1BHavCk8ZgMeRujfkex7/zrw7qltaqGkVWwOhrxFfwXJZkA57V4olSS9SNDkonPtmv2RreU6v4jvVx5cVtBG3PO5mYj9ENSTDGAall9+allHrUsgPOamkGM5r4leP9A+GHgbWvH3ie6WDTdEtGnmJYKXI4SNc/xu5VFHdmA718QfG+t/ErxtrXjzxHP5mo65eSXM2DkJuPyov+yq4UDsFFfsLfAyf9n79mnwr4N1bT/sev6hGdW1yJlAdLy4AJR8AfPHGIoj15j6kYqqpulf8ABUf9kOX4w+A0+N/w/wBIa48ZeDLUrqFtACX1PTASzAL/ABSQktIoGCUaQfMRGo/Z9+OfjD9nP4q6N8UvBku640+TZeWbMFS/tGI863ckNgOoxuwSpCsOVFfB74seD/jj8N9D+KHgW/8AtOk63b+YgYFXgkUlZInBwQ6OrKexIyMggmqqqqqputVVN1/Cqqq1HS9O1aD7NqNpHOnYOOV9weoP0rWvhh1m0K7/AO2M5/kw/kfzrUdJ1LSZfJ1GylgbOAXXhvoeh/Cqqq0bRrzXb5LGyTJPLuRxGvdjWl6bbaRYQ6daLiOFcAnqx7k+5PNVVN1qta0ez1ywexu0HIzG+MmNuzCtW0q60a+k0+8XDxnhgDhh2I9qqqqqqqqqqqqqqqqqqqqqqqqtDuVs9Ys7h8bUmXdnsM4J/Kqo9aqqqqqqqqqqqquLOzuxtubWGYYxiSMN/Orjwn4euTl9MjU8/wCrJTr7A1P8P9GkBME9zEccDcGA/AjP61efD/VIQWs7mG5AHCn5GP4Hj9avdI1PTs/bbGaIKcbiuV/McVVVVVVVVVVVVVVVFNNBIJbeZ43XoyMVI/EVpnjvU7QrHfot3GOpPyuPx6H8R+NaXrWm6xHvsbgMyj5kYYZfqKqqqq/4Kv8AwRbwL8bbL4taTZsulePrbNyyRgJHf26okg46F4/Kfnlm8w84OP8Agit8dI9K8UeL/wBnbWLrbDrkf9u6IGdVX7TEqx3MYHVneIQuOwW2c96qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq+N/7K3wC/aKsng+LHw30zVrzyTFDqsam3vrcdRsuIyHwCAQpJXsQQSD+0R/wRs+KHhCe4139njxJF410YbmXSNUkS01KAAZCh/lhuOB9790ckAIeTXifwl47+F/ib+xfF3h7WvC+v6fIJFt762ktLiFlY7XUMAw+ZThh3HBr4G/8ABSv9o74QfZ9K8QazF4+0CGMRrZa6xNxEB3jul/eZ6D955igDAA618Kf+Cp37NHj9YbLxlc6r4D1J1+ZNUtzPal88BJ4d3GOcyJGOCPTPhnxb4U8baUmu+DPE+k69psjFUvNMvY7qFiACQHjJUnBB69xVVVVVVVVVUOoo/wBKqqpelVQ6VVVVVQGTxVVVVVUOg+lVVVVV+1H/AMlXuP8Arxg/ka/ZE/5JrqX/AGHJf/REFVVVVDqKqqqv27f2ULX9qP4SSWmhWtqnjnw5vuvD11LhDIcfvLVnJACSgAZPCuqE4ANaPrHi/wCGvi+HVtJur/QPEegXh2sUMM9pOhKsjKwyCDuVkYYPKkYJFfsyftGeHf2ifAUOt2z2tn4jsFEet6THISbaTkB1B5MTgblPOOVJJU0HI4NK/oaWXjGaWQHvSyccNQlI4oSg96EgPANCTI4NeaSMA15pxjP60ZewNGUnoaMme9GQEdaaUetNLkYBppCRjNNIMdc08oxgGnkJGAaZ88A1+1l+3FoPwfju/AXw2mtda8asrxXE4cPBozdMvwRJKDnEXRSMv02Noui+PvjJ4+t9G0a01LxP4t8T3xEcYJlnu53JLMzE/VmZiAoBJIAJr9hv9jTQv2T/AAAzaqbLVPH2vKH1vVYo8iFMAizgY8+ShGSeDI3zEABFRRxXxg/bF+Gnw0eXR9BlXxVraAgwWM6/Z4W6YkmGQCCOVUMR0OK+Knx2+JXxjuxJ4x1zNlExaDTbRPJtofomSWP+05Zh64r4S/s9/Ez4yzCXwxo/2fS0bbLqt7mK2U55CtgmRhjlUBxxnGRXwe/ZF+GXwsNrrN/bN4j8RQgN9tvlzFC+OTDD91fYtuYHoR0qqodRVVVVVL1/Ch0qqqqql6/hVVVVVVVVVVVL0qqXpVVVAZIAqqqqqqHSqqqqqXr+FVVVVfHT9n7Qvivpsuq6XFBYeKYU/cXmNq3O0YEc2OoxgBuq8dRkHXNC8S+BPEUmka1Z3Ok6xpsoYru2vE4wVdWU89iGU4PBBr9nj9py28SwweCviRqUcOsqVjsdRlwq3uSAEfsJOQAejf733qqqqq1HStM1qzfTtX061vrWUYkguYVlRx0wVYEGviJ+xr4J19Jr7wHeyeH75iWW3kJmtWPpg/MnPoSBnhcYr4h/BL4j/DKdh4k0CR7IEhNRtAZrZ/feBlOh4cKeM4xXgb4o+PPhxeLd+EfEdzaJn95as3mQS/70bZU9+cZGTgivh3+2l4c1d4dN+ImjnRbhgFa+tC0tuzdyU5eMfi/uRWkazpHiDT4tV0LU7XULOYZjntpRIjfQiqpgO1Vd2lrf2s1jfW0Vxb3EbRzQyoHSRGGGVlPBBBIIPBr9q/8AZnuvhLrMnjDwdp0sngy/kHCsZDpsrH/Vv3EZP3GOf7pOcFv2af2h9S+BfiZ476Oa+8Maq6jUbNHO6I9BPEucbwOCD94DGRgEaXqmm65pttrGj30F7Y3kSy29xA4dJUYZDKRwQRVVRHaiARjNVVftufBRfA3jNPiN4fsfK0PxNKRcqigJb32CWAA6CQKX/wB4SdBgV+xB8Zz4L8bf8K01u7K6N4pmAtN2NsF+QFQ+v7wKsff5vL6DJqmA4xVVVVVHp+FXl3bWFpNfXk6Q29tG0ksjnARVGSx9gBmviv46m+JHj3VfFbgrBcS7LSMjBSBPlQH3wAT7k1+yj8PI/CHw5j8R3cONS8S7bliy4KW4z5Sj2IJf33j0qqqqbrVeIdA0nxTod94c1yzS6sNRgeC4hcZDKwwfoe4PY4NfG/4R6v8ABjx7d+EdQc3Fow8/TbwjH2m3YnaxHZgQVYf3lOMggn9iP46r4m0EfCLxNfL/AGrosO7SJJHO65tF6xc9Wi7AH7mMDEbGv28Pi5/wl/xAg+HGkXW/S/CmftO1vllvXHz/AF2LhPZi4r9gf4R/8JJ4yvPipq9ru0/w1mDT968SXrryR2PlxtnB6NKhHSqbpVVXiz4meAPAykeKvFmn2Mq4zbtJvm5GciJMvjHfHceteKv2z/CliGg8H+GL7VJMYE13ILaMEjqANzNg4BBC5556GvFP7Ufxd8Sl47bWoNFt2JxFpsIjOMAD523P27Eck+2NV1jV9duzqGt6rd6hdMMGe6naVyP95iT3P514c+EfxM8W7G0DwTqk8b/dmkg8mI/SSTav614f/Y3+IeoNHJ4g1rSNJib76qzXEqcjPyqAp4z/AB9R75rQP2MvA1iVk8Q+JNW1V1H3IVS1jY4Ocj5m9CMMOnfNaL+z98HNCZXs/AdhO46m9L3YJxjO2UsPwxjPNaZo+kaLbi00bS7OwhAAEVtAsSgAYHCgDgcVVV+158Lbj40fsz/EP4c2Fu1xfajo0k1hCiMzS3duy3ECAKQctLDGO/XkMPlPg7xRqfgjxbovjLRZNl/od/De25PTfE4cA9cg4wQeCDXhbxJpXjHw1pXizQrjztP1iziu7V8jJjkUMucEjODyM8GopMc5qKTuDUUvQg1HKOxqOUEdeaSbjBNftO+BJL61tvH+mwhmtEFvfqq8+WT8kh9cE7T3+Zew4sr2axnE0J9mU9xVn4ps2QBpjG2OVcf1rUPFUBQrbsZX6DjAFSPNdTl2JkkkboBkkn0FfBLwPL4C8DwWt/EY9S1BvtN4p6oSMKn/AAFQM+5NPMCMA1JKMcGpZcjOalkB5zUsgPf6V/wUG/afh+Jnic/B/wAE6h5vhrw3ck6hdRSHZqF6owQMfejiOVB6F9xGQFY/8Exf2VZfjj8Xofid4r0tJvBPgG6jnlWUkLfagMPBDj+JUO2VweMBVIIkqqqm6VX/AAUw/Yrl+C3jGb41/DXRG/4QPxPcu+oW9tEfL0O9dgSpA4SCVmymPlVtyfKPLDfsDftlX/7LHxDbSvE93PP8OvE0yLrlskZmazkClUu4VB4ZcgSAAl4x0ZkjxpWqadrul2etaPexXlhqFulxa3ETbkmidQyOp7gqQQfeqqqqqputVVMPeqqqqpoIbmJoLiJJY3GGR13A/UGtT+Hnh2/Jkgiks5P+mLfKf+AnI/LFXvwx1eE5sr22uF9GzG35cj9am8E+KYcb9IkOemx1b+ROK0n4d63fuGvlWyh7l8Mx+ig/zxWjaHp2g2v2XTodoPLuxyzn1J/p0qqqm61VeLfDcfiHT9se1buAEwOf1U+xwPpU0UkErwTIUkjYqysMFSOCDVVVVVVVVVVVVVVVVVVVVVVVV4duzfaHZXLPuYxBWJ7lflJ/MVRHOM1VVVVVVVVVVVVVVVXvhrQ78Ez6dEG/vRjYc+vHX8avvh5EwLabfsp7JMuR+Y/wq/8ACmuaflpLIyoM/PD84474HI/EUysrFGBDKcEEYINVVVVVVVVVVVVVb3E9rKs9tM0ciH5WU4Irwv4mTW4fstzhbyJcsAMBx/eH9R/kVVVX/BQT4N/8Ll/Zh8UWdhZ+frXhiP8AtzSwqbmL24JlRQASS8BmUKMZYr9K/Z1+K918Dfjn4I+LFq8oTw5rEM92kTlGltSdlxGCM43wvInQ/e6HpVpd2moWkN9YXUVzbXMayQzQuHSRGGVZWHBBBBBHBzVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVfFL4NfCr426APDHxX8BaP4n0+Ni0Md/bB3t2OMtFIMPExAALIVJHGcGvjr/wRV8Las82sfs7fEeXQ523MNE8Shri23FuBHcxr5sSBeMOkzE/xCviv+wd+1l8HJZD4q+C+u3tjGu/+0dEh/tS2Ccjcz2+8xjjpIFPTjkZ8M+LfFXgrVU13wZ4n1bQdSjUql5pl7JazKMgkB4yGAyAevYV8Nf8Agp9+1R4CeC31zxDpnjSwjky8Gt2S+aUJywE0Ox88nBcvjjggYr4df8FgfhPrbR2vxN+G+v8AhiVlINxp86alACBnJyI3UHGAArYJGTjJr4fftV/s4/FPyo/A/wAZfDF9cTY8uznvBaXLfSCfZJ/47xVVVVVVVUOoo/0qqql6VVDpVVVVVL1/CqqqqqodB9Kqqqqr9qP/AJKvcf8AXjB/I1+yJ/yTXUv+w5L/AOiIKqqqqHUVVVVV/wAFK/2FZfiJaXv7Qvwd0Se58W2yq3iPSLcFjqduke37RCnUzoqoCi/6xRkDeuJPhl8TfGPwh8Y2PjnwLqrWWpWRwQQWjuIzjdFKufnjbAyPoQQQCP2dP2l/An7RPhmO90S5jsfENpCG1XQ5ZMy2rZwXTp5kRJGHH94Btp4r6Gg5Ax1oSDoDQlPTOaEwxjNCUEYzQkB4BoSA8BqEvYMK80n+KjIccNRlOOWoyZ6NRkGOtGUdjTTDGM00pI4NNJnq1F89K+K3xz+FnwW0wal8RvGFnpryIWt7IEy3Vz2/dwrl2GeN2NoJGSOtftB/8FB/iF8TFufDXwvW68G+HXLI1wkoGoXkeeN0i/6gHAysZzyQXYEivhJ8Gfib8dfF0Pgf4V+EbzXtVkXfIsKhYraMdZJpWwkSZwNzkAkgDJIB/Yt/YZ8Ffsm6FJqt5dW/iLx9qsITUdb8jYlvGcE21sDkrECOWOGkIBIUBUX4tftB/DP4OQeV4n1n7Rqrpui0qyxLctxwWXIEanPBcgHnGcGvjB+1p8TfisJNMtbn/hGtCbcPsGnzMHmU9ppeDJ6YAVT3XvXw/wDhd47+KOrLpHgnw7dag+4LLOqbYbcH+KSQ/Ko+pyewNfBz9h3wZ4PFvrfxMng8T6vGwdbNVIsYSMcFWwZuR/GApBwU4yYIILaCO2toUiiiQJHGihVRQMAADoAOMVVVQGSAKqqqqpev4UOlVVVVUvX8KqqqqqqqqqqpelVS9KqqodaqqqqqgBgAVVVVVSjPSqqqqgBgAGvjF8EvDHxg0pI9RJstXs0YWWoxKCyZBwjj+OPPO3gjnBGTnxr4K8RfD/xFc+F/E9iba9tjwRykqH7siN/EpxwfqDgggfs8ftRQmGz8BfE2/fzQyw2GsStwRjASdieD0Ak75+bGCxqqqqUDHBqnRJEMciqysCGVhkEd+K+In7KHwy8cPNf6RBJ4b1OQEiWwQeQzerwnj8EKZ7mviX+zx8Sfhl5t7qGlf2lpEeT/AGlYAyxqvrIuN0fbJYbcnAY14R8feM/AV4b7wf4jvNMkYgusT5STHTehyrf8CBr4aftp6fdCHS/ijpP2OThf7UsELxn3ki5ZfcpuyTwoFaD4h0HxTpkWseG9YtNSspfuT20okXPcEjoR3B5HemGR1qq1jSNM8QaVd6HrVlFeWF/C0FzbyrlZEYYZSPQg1+0t+z1qfwM8UqbE3F74W1QltNvnTJjbnNvKRx5igZB4DLyOQwX9jz9pJvhvq0Xw28ZXv/FL6pOTa3EmT/Z1w+B17ROfvDorHdxlyaqj1ojIwKqq+J3w90X4p+BtV8D64FWHUYSsU/lh2tpRykqjjlWwcZGRkZwTXijw3rngbxPqHhfXLd7TVNHumhmUZGHU8Mp4yp4ZW7ggjg1+zR8YoPjL8MrPVrqdP7d0vFnq8QcFvNUfLLjrtkXDdMbt6gnaTTdqqqqqo9Pwr9rT4hyeEvh6PDOnTbL/AMTM1uxDYKWygeaf+BZVPo7elfBzwCfiV8RNJ8LyBvsbyGa+dcjbBGNz8joWwEB7FhUUMNvClvbxJHHGoVERdoUAYAAHQY7VVVVMOetVX7SvwUtvjV8PpdPs4kXxDpO+50eYhRmTHzQknGFkAAPIAYIxyFwdE8Q+IPAPiO08RaNczadrGjXIkhYqVaORTgqynqOqsp4IJBGCagg1vxf4hjtoFn1LWNavQiLndJc3Er4H1Zmb8zXwf+G+n/Cb4caJ4EsDHI+n24N3Ogx9ouG+aWT1wXJwD0XaO1U3SvEfizwx4Rs1vvE+v2OlwuSI2up1j8wjqFB5Y+wya8aftk+EdM3WvgjQ7rWpgxH2i5P2aHHZlBBdvoQn17V4y/aH+K/jUvFd+JpNNtHBBtNMBtkwRggsCXYH0ZiOvrVjp2qa3fLZaZY3V/eTt8sUETSyOxPYDJJya8J/sqfFjxMkdxf6fa6DbNg7tRlxIRnBxGgZgepw23OOvNeF/wBjTwRpypL4q1/UdYlAGY4QLWLPOeBuY8YH3h0J74Hhj4Y/D7wYq/8ACM+EdNspExicQh5eAAP3jZc9PXrk9Sao9aqqqqr/AIKEfs+T/s+ftKeIdNsbPy/DfiqR9b0FkjCokM7sZIABwvlSiRAvXYI2P3hX/BN347R+JvBlx8Edeu2bVfDSvc6Uzn/W2LMNyA9SY5HP/AHUDhDQODkGopcYwajlHY1HKMAg0koIxmkm4xmpRBdQPbXESSxSoUkjdQyspGCCD1BHaviB+zXM002qfD+5jZGJY6bcPtKnPSNzxjoAGxjH3jWp/D7xzo7umo+EdWiEZ2tILR2TJ9HUFT+BrRvhV8RNekRLDwjqKq4BWS5hNuhBGQQ0mARgdvb1FfCn4EWPgu6j8Q+JZ4NQ1aPmCNBmK2P94Z5Z/Q8AdhnBp5gRwc08xIwDUko9cmpZcjrXivxd4b8GaHdeJfFmuWek6XZIXnuruURogA9T1J7Ack8Cv2pP+ChuteOI7rwN8DJ7rRtCkUpda46mK8vB0KwjOYIzzz988fcGQ3wG+A/xC/aM+I2nfDb4daU093dtuu7ySNvs2nQD7887gHYgHAzyzFVXLMoPwR+Dfg74BfDLRPhZ4Gt2XTdHh2meVV867lY5kmlKgAu7EknHHAGAABVVTdKrxl4N8LfEPwtqfgnxtodrrGh6xbtBe2Vym5JUP6gggEMMFSAQQQDX7aP7I3if9k/4knSZXbUfCOutJP4e1QIRvjB+aCX0mjBUHnDAqwxuKr/wTc/bvHwg1S3+Bfxf1m6k8F6rcImh38z710O4djlG43C3kZhnnEbfNgB3YVVUR2zVVTdaqqbtVVVVVHp+FVVVVVTdaqq+IXhkyKdfsY8so/0lQOSOgf8ADofz9aqqqqqqqqqqqqrw34RvNf8A9Id/ItFODIVyW9Qo/r/OrbwT4ctVC/YfOYDBaVyxP4dP0q78EeHLpSFsjC2PvRORj8DkfpWp/Dm5jBk0m7EwHSOX5W/Pof0q9sLzT5jb3ts8Mg/hdcZ+nqKqqqqqqqvh5erLp9xYMw3QSb1B/usP8QfzoAEgU3Wqqqqqqqqqqqqqqqqqq+0nTdSXbe2UUxxgMy4YfQjkVf8Aw+0+YFtPupIGxwrjev8AiP1q/wDBuu2OWW2Fyg6NCd36df0p0ZGKOpVh1BGCKqqqqqqqqqqqtOvZdOvoL6E/NC4OPUdx+IyKVldQyMCCMgjvVVVSxRTxPBPGkkcilXR1yGB4II7jFfH34cP8IvjV41+G2zbDoOs3EFrww3W+7dA2GJPMTIeSevU9a/4Jx/FRvi1+x94C1K7uXl1Hw/atoV6XYMwa0Plx5IAyTAIW5Gfm5LH5jVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVXxV/ZV/Zz+Nwml+J/wc8M61eT48zUfsYtr0gbsD7VDsmA+ZjjfjJz1r4p/wDBFP4Ra5HcXvwh+JviDwvdGPMVnqsKalbbwuAAw8uRAxHJLPgkkAjC18T/APgkx+194BluJvDnhvR/HOnwsxSfQ9RRZTH2Jhn8t92MZVN+D0JAzXjL4eeP/h1frpXxB8DeIPDF6+dttrOmTWUhwFJwsqqejoenRge4r4d/tM/tA/CfyI/h/wDF/wATaXbW23yrH7c09ou3bj/R5d0XSNF5X7o29CRXgD/grh+0L4cijtPHPhzwt4viRQGuHt2sLlyNoJLRHysEBjgRD5nznAC14D/4K9/A3XIli8f+BfFPhe6K5LW3l6jbgjtvBjfPp+7x64r4W/tS/s9/GqcWPw0+K2iateldy2Lu9pdMNxXIgnVJCMjsvdT0Zc1VVVUOoo/0qqql6VVDpVVVVVL1/CqqqqqodB9Kqqqqr9rK0ht/ibazRA7rnR4ZJMn+ISSrx+CCv2Prmd/CGu2bSZhi1JXRcDhmjAY568hF/Kqqqqh1FVVVVV/wUB/4J26P4pF58afgTpwtPFF9d7tW8PoyRW2oMwJaeHOBHMSMspO18kja+d+ha/4w+G3iqLWNB1DUNA1/RrkgOoMUsEqNhkdT7jDIwweQR1Ffs5f8FD/CPjC3s/Cnxunt/DuujZDHrGNtleNgDfKeluxPJJ/d8k5UcVb3FveW8V3aXEc8EyB4pY2DK6kZDKRwQRg5qskdDQc9M0JOMUJB0DUJT0DCvNPrXmnGARXmn1FGUnuKMp6bqMgxw1GQHgHNFyeBRJPGa+K/7TvwR+DCSQeNfHNmNTjyBpNiftV2WAyA0aZMeR0MmxT618af+Ck/xA8WRy6J8INHXwjp7rtbUbkrcX0g77RgxwjBI4DtwCGU8VJJ4r8e+JN8kmreI9f1ecDLGS8uryU8D+87sePU1+zR/wAEmPiZ8RRB4l+Pl/deAtCba6aXAiSandqeoIJKWvGOXV2zwYx1rwp4L/Z6/ZB+H7WHh3T9H8F6EXBmkZmee+lC9WY7pZ5MA4HzEDOAAK+M37cnibxMs+g/Cm2l0DTXG1tSnA+2yc/wYJWIEf7zdwV6VY6d4n8b66LXTrTUtc1i/kJ2xq9xPMx6k9Sfcmvg3+wbJIINd+M99sAYMNDsZskgY+WaZTxnkFYz0wQ4PA8OeF/Dng/SYtC8K6JZaVYQfct7SERLnuxA6sccseT1JqqqqoYz1qqqqql6/hQ6VVVVVS9fwqqqqqqqqqqql6VVL0qqqh1qqqqqqHQfSqqqqqXHTNVVVVDoPpVfFH4VeF/ix4dl0PXrdY7hQTZ38aAy2snYg919VPBHocEfEP4eeJPhl4kn8M+JrTZNH80EyAmO5jzxIh7g4+oOQcEGv2d/2n00GC18BfEm8kewUrFp2qOc/ZR0EcxJz5Y6BudvQ/LyqOkiLJG6srDKsDkEeoqqql6VVVXxL/Zp+GvxH82+/s/+xdXkyft1ggTe3rJH91/c8Mf71fEv9mn4k/Dnzb7+z/7b0iPJ+3WCF9i+skf3k9zyo/vV4S8beLPAmpDV/CGv3emXXG4wv8sgHQOhyrj2YEV8Mv20LG+aDR/ihpS2crEINUsVJi+skRJZfcqW5P3QK0jWdI8QafFquh6na39nMMxz20okRvoRVV8QvAPhz4m+EdQ8GeKbNbiyvo8A9GhkHKSIezKcEfkcgkV8W/hV4k+DvjS88HeI4mYxHfZ3YjKx3kBPyyp7HoRk7WBGeK/ZT/a8NmdP+FvxVv4ks0jS20jWJMJ5O0bUhnPTbgACTtj5sg7lqj1o4x1qqqv28vgub2ytvjP4ftGaa0C2muIgHMXSKfH+yT5bdeGTgBSa/Zc+ML/CD4oWd1f3KpoOtFbLVg5wsaMfkm9vLYhieflLgcnIV45Y0kidXRxlWU5BB6EGqqqqqPT8K/aF8fD4g/E/VL62l3afprGxscNuDRxkguO2Gfcw9iPSv2OPh8dF8I3nj2+jX7Tr7+VaAgZS3iYgn1G992R6Ih71VVVU3X8Kqq/bftfBlj8cb6DwmGW7kto5taRTmNbtxk7fQlNjN/tMe+a/Zl8VeF/BHxo8O+KPGEKtp1rM8Zmc/LbPIhRZmGDkIWz2x94cikeOWNZInV0YZVlOQQe4r4g/Hz4afDp5rLVdcF5qcQOdPsR50oYfwsR8sZ9mIOOcGvHf7Xvj3xA8tn4Ptrfw9YtlVkCie5YdDl2G1c9flXI/vHrV9qGs+ItR+1alfXup39ywXzJ5WmlkboBkkkntivA/7M/xU8aCO6l0caHYuf8Aj41TMLEA87YsFzxyMgKfWvBn7H/w/wBESO48W3154hulJLR5Ntb+3yKd5x6l8H0HStA8L+HPCtn9g8NaFY6ZBgBktbdY92OhbAyx9zk81VVR61VVVVX/AAUK/ZVb9qH4JSQeG7VH8b+EGl1Dw8SwU3JKjz7TJ4AmVVx90eZHFlgu6vBXjLxT8L/Gmm+MvC95Jp2t6Hc+ZCzKQVYZV0de6spZGU9QxB61+z58ePCv7QXgC18XaDNDBqESrHq+liXdJp9xjlTkAlDglXxhh7hgASOhqOXHQ1HKD0NJMDwTSTccGlmHTNCYHgNQnx0ajPnuKaYdC1NMCODTzEjrWua/o3h3S7nW9f1ey0zTrNN9xeXk6wQwr/ed2IVR05Jr43f8FKvhh4Mjm0j4TWbeNNX24F226CwhPqWIDykeigA/36+Lvx0+KHxz1z+2/iN4puNQETu1pYofLtbIN/DFEPlXgAFjlmCjczEZr9lX/gnh8aP2lZbHxJf2zeDvAc6+Yde1CAl7pBxi1gyGlJP8ZKxgBjuJAU/Av9nz4Vfs5eD08GfCzw1Fp9u2DeXkgD3d+4LEPPNgGQjc2AeFBwoUcVVVVN0qq+NnwZ8D/H74bav8MfH+mpc6dqkLCKYIDLYzhSI7mFjnbIhOQeh5UgqzA/tJfs7eOP2Y/ihffDXxqqXARRPpmpQxlYdStWJCTID0PBVlydrKwyQAx/4Jpft8QmHT/wBnL44+JW80Mlv4R1q9k4IxtWxmkPQ9BEzdc+XkYjBqqPWqqm61VU3aqqqqqPT8Kqqqqqm61VU6JKjRyIrKwIZWGQR3BFeLfDzeH9TMUYJtZ8vbsc9O6/Ufywe9VVVVVVVVVVVXh/R5Nc1SKxThPvSt/dQdf6D6kVBbw2sEdtboEjiUKijsBTdfwqqvbCy1GA217bpNGR0YdPcHqD7itd+H9xbhrnRXaeMcmFvvr9D3+nX606PG5jkUqykhlYYII68VVVVVVV4S1P8AszWoWdsRT/upOOx6H88UOoputVVVVVVVVVVVVVVVVVVVV/pOm6mu2+s4peOGIww+hHIrUfh7E/z6VebD/wA85uR+Y5/StR8P6vpeWu7Jwg/5aKNy/mOn41VVVVVVVVVVVmrLaQIwIYRqCCMEHFVVVX/BXH4d/wDCN/tAaL4/tbXZbeMdCTzpfLx5l3at5T8hQDiE2o5LMO+BtFf8EQPiA8/h/wCJ/wAKrm5wlneWWs2UOGO4yo8Nw2cYGPIthycndwODiqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqta0PRPEulXGh+ItHsdV067XbcWd7brPDMAQcMjgqwyAcEdq+JP/AATU/Y0+Jfmz3HwhtPDl7JnbdeG5303y8/3YYz5H5xnFfEH/AIIg+Gp0e4+FPx01OyZWPl2niHS0ug4JXAM0LR7SBvOfLO47RheTXxA/4JO/tleBo5p9N8IaJ4wt4PvS+H9XRyRtBysdwIZG5O3AQnIPGOa8YeB/Hfww8Rt4c8d+Fda8L65abZDZ6nZyWk6A/dcK4B2nGQw4PUGv2WP+CnPxJ+FMtj4O+NE15428JIyxreyOH1LT055WQ/8AHwoz92Q7gMAOAAteBPH3gz4neFrLxr4A8R2euaJqCk215avuVsHDAg4KsCCCpAIIwQKqqqh1FH+lVVUvSqodKqqqqpcZxmqqqqqodB9Kqqqqr9r2zRPGmi6gHJabS/KK44ASVyD+O8/lX7G95LJY+KtPKp5cMtrKpxzlxKDn2wg/Wqqqqh1FVVVVV8UpWGmWUAcYacsV9SFxn9f1r4w/sRfDH9qaa9ub128PeL4bBjZ67aRAl2UqqLcR8ecgHHUMBjDADB/aG/ZM+Nf7M+tPY/ETwtK2kySFbLXrEGaxuxkgYkA/dscH93IFfvjBBPwY/ap+M/wJVLHwb4jW50ZSzf2NqaG4tMkkkquQ0eSST5bLknnNfCf/AIKSfCLxdHaab8SNOvvB2qysEknKm6sc9AwkUb0BPZkwuRlyMkeF/GPhLxxpiaz4N8TaXrli/S40+7S4TPoShODwRg8gg1VVVZOMA1k+tHnvVV45/ag/Z/8Ah3HIfE3xV0ETxLlrSyufts4Py4Bih3suQykZA4JPQEj4kf8ABUHwnYLLZfCnwDfatNtwt9rEotYVbPURJud1x6tGcn25+JX7Z37RHxPE1tqXj640bT5t4NhoY+wxhW6oWU+Y644w7tx9TnwP8PPHfxN12Hwx8PfCGr+ItUnYKlrpto87jP8AE20HaoAJLNgAAkkAE18Bf+CQPxR8XNFrXx58S2/grTCoZdL08peahLkdGYExQjkc5kOQQVHDV8FP2X/gF+zJokj/AA88H2Gmzw25N9r1+wlvJUAy7SXD8qmBkqu1B1wK+M37cvhzwzJNoHwotINf1BMrJqc5Is4j/sAENKevPyr0wWzx4y8c+LviFrUniHxnr9zqt/LwZJiAEH91EXCov+yoA9q+DP7GHj/4gSQ6v43jm8K6CcNiePF5cA/3Ij9wf7T46ghW5x8Nfg/8PvhLpY03wVoEFtIybZ72RQ9zc85/eS4yRnnbwo7AUuCMA1VVVVQ6iqqqqqXr+FDpVVVVVL1/CqqqqqqqqqqqXpVUvSqqqGM9aqqqqqhyOtVVVVUvX8KqqqqHI61VfEP4eeGvid4an8M+JrTzIn+aCdABJbSAcSIexH5EZByCa+KXwt8SfCfxI+ga/F5kT5eyvUUiO7jB+8voRwGXqD6ggn9m79pF/Cb23gDx9fFtEYiOwv5WybA9kc/88vQ/wf7v3UdJEWSN1ZWGVYHII9RVVS+maqqqq+Jf7NPw1+I/m339n/2Lq8mT9usECb29ZI/uv7nhj/er4nfs2/Eb4aJPqMlkusaNCCzahYqWEaDqZE+8nHU8qP71eDPiF4y+Huof2j4Q8QXOnyEgyRo26OXHZ4zlW/EV8Lv2x/DmtJFpXxLtl0a+J2rf28bPayf7wyWjJP1XuStafqOnavZQ6lpN/b3tpcLuhuLeUSRyD1VlJBH0qvjJ8G/CXxr8JSeGfEsHlzxbn0/UI0BlspSPvL6qcAMh4YehAI+KPwu8WfCHxZceEfF1l5c8Xz29wgJiu4iflljbupx9QcggEEV+x1+1HAYLP4Q/EnVnEwcRaFqVzICpXAC2rseQQfuMSc52cYUGjgng0en4VVVWvaHpnibRL/w7rNstxYalbSW9zE38SOpVh7cHr2r4xfDPVPhH8QtV8E6krtHayl7GdwP9JtmJ8qTjjJXgjswYdq/Yt+MX/Cw/hwPB2rz51rwikduSz5NxaEEQv9VCmM9cbVJPz4qqqqqvj945Pw/+Fur6tbzrHfXafYrEnGfNlBGR7qodv+A9K8H+Gb7xn4o0rwrppxcapdJArlSwjBPzOQOcKuWPsDWiaPYeHtGsdB0yLy7TT7dIIFwMhUUAZx3wKqqqqbk4zVV4z8VaX4G8J6t4w1lmFlo9nJczBfvMFXO0e5OAPcivE3iHWvG/ifUfE2szNdanrN29xMVU/NI7Zwo5wBnAXsAAK1H4ba14c1ebQNRurBpLZYzNLa3AnQMyKxQFeCy7tjDsysM8VN8Z/ifceDtL8DyeL7tNO0q2FuhgPkyTIv3RI64Ztq4UDOMKM5OSfDfhLxL4wv10zwvod5qVySAUt4iwTPQseijg8kgV4F/Yy1W8jS9+IfiBdPQn/jx07EkuP9qU5VT7BXGO4rwX8K/AHw/iRfC3hq0tp0Xabt18yduuSZGy3OTwMDnoBxVVVVVHrVVVVVN2r/gpP/wT4vvFF3f/ALRHwH8P3V9rVzI8/inQbUBjcAIzNewKSCZPlAeJAxcsGUbg274U/Fzx58FfFkPjL4fa21jeovlzRuu+G6iyC0UqHhkOB6EHBUqQCP2fP23vhX8arW10bW72Hwp4sKqkmnX8ypDcyH/n2lJw4PHyNtcE4AYDcQcdDSyEd6SfAwTSTDGA1LMcYzmhMOma89cYzRnUjANa54k0Lw1ps2seI9asdK0+3UtNd31ysEUYAySzuQAMA9TXxA/4KHfs0+CRPbaf4pvPFV7BkG30SzaRS3y4xNJsiYHdnKs2Nrd8A/Ez/gqV8QtaWWx+FvgjTPDsLLtF7qMhvrgHP3lUBY0OMDDCQdeeRj4ifFr4l/FnVDq/xG8bapr0+8vGl1OfJgJ6+VCuI4gcdEVRXwI/Ye/aP/aI8q98EeBJbHRJdhGua2WsrLa2MMjEFpRg5/dK/H4Z/Zo/4JbfBL4L+R4j+JYi+I/ildrK2oWgTT7Nhz+6tiWEhycb5S2dqlVjOc1VVVU3Sqqv2pv2ZvBX7UnwvvPAviaC3ttVgV5dC1kw75dLuSBh1wQWRsAOmcMMd1Uj4xfB3x/8CPH+o/Db4k6K2n6vpzZBGWiuoiTsnhfA3xMAcN7EEBgwH/BPL/gobF8UotO+Bfxz1pI/GcarBoeuXL7RrYAwsMzHpc44DH/W/wDXT/WVR571VU3Wqqm7VVVVVR6fhVVVVVU3Wqqq8RaHDr+lyWMhCyD5oXP8Djofp2PsauLea1nktbiMpLExV1PYjrVVVVVVVVVVVfDnTRDp02pOvzXL7UyP4V/+vn8qpuv4VVVWveFtN11C8i+TcgfLOg5/4EO46f4itZ0HUNDn8q8iyhPySryr/Q/0qqqqqqrwprP9s6WjyNm4gwkuTySBw34/zzTdaqqqqqqqqqqqqqqqqqqqqqtQ8L6JqSky2SRyEcSRDYw9+OD+INX/AMPblMvpt6ko7JKNp/McH9KvtC1fTiftdhKqj+MDcv5jiqqqqqqqqtB01tV1W3swuU3bpPZR1/w/Gqqqqv8AgsL4MXVPgx4M8dRQNJPoHiJrNisBby4bqBmZiwPyrvtoV5HJZeR0P/BIPxknhf8AbIsNDZUJ8X+HdR0tSyFiCipeZBBG04szycjBIxkgiqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq+MXwN+FHx98JyeC/i14K0/xBpxyYTPHtmtHOP3kEy4eF+ACyEZHByCQf23v+Cc/j39lm7m8aeC21Dxb8NZpDt1PyN9zpGSNqXoRQoByFEwCox4IQsqn9kz9r/wCIH7K/ioy6Q/8AanhPVZ4zrWhzklJFBAaaDkeXOEBUN908BgwC7fh18RfBnxY8GaZ4+8Aa7Bq2iatD5ltcRH8GRlPKOpBVlOCpBBFVVUOoo/0qqql6VVDBGAaqqqqodaqqqqqh0H0qqqqqv2wP+Rk8P/8AXjJ/6HX7Gn/M3/8Abj/7XqqqqodRVVVVVfFSUNPp0AHKJI2fqVH/ALLXwrhLXl/OAMJEik9+Sf8A4mtU0rS9c0640fWtNtdQsbuMx3FrdQrLFMh6qyMCGB9CMV8f/wDgk18FviTPPr/wj1WX4d6vJudrSKA3WnTNjIAiLBocnAyjbVHSM18Y/wBgb9qT4J77vXvhtda5pKyMg1Xw5nUYCF/jZUHmxIRjDSxoOQOvFaH4h1/wxqCat4Z13UNJvo8bLmxuXt5VwQwwyEEcqp+oB7V4L/bo/aY8Fxpap8QW1y1jUAQ61bJdkkbRkykCU8Ljl/4iepzXhz/gqb43tY1Hi74T6HqbhGDHTdQlsQW3cEB1mwAvBHc85HStG/4KjfCmfP8AwkPw68WWXyAj7G1vdfN3HzPHx79/QVpf/BSf9nHULdpruPxZprq5UQ3WlozMMD5h5UjjHOOTng8dM/8ADxn9mj/oK+IP/BS3+Nar/wAFKf2c9Pjjezt/F2ps7EFLXTI1Ke582VBj6E1qn/BUb4Uw3Cponw68WXcGzLPdNb27BsnICq8gIxjnI6njjJ13/gqf4ruYyvhj4QaTYSeUwDX+rSXYD/wthI4vlHdc5PqK8Uf8FEf2mPEIkXTte0Xw8sjD5dL0pDtG3BUGfzSAT82c5B6EDivGnxh+KvxG3J45+IniDWoW/wCXe7v5HhXG37sWdi8opOAMkAnnmvBPwx+I/wASrz+z/h54C8Q+JbgNtaPSdMluih/2vLU7RjnJwAOa+F3/AASj/an8fxR3/iew0PwLZM3TWr3zLhkwPmWGASYPbbIUPB9s/CT/AIJD/s8eCmhv/iXrWufEG+jbLQzMdNsm44/cwsZMg5PMxB4BGM58GeAfA3w50dPD3gDwfo3hzTY/u2ml2MdrHnnkqgAJJJJJ5JJJ618ZP2mvhv8ABuN7DUL06vr2Pk0mxYGRevMr/diHA6/NyCFIzj4v/tEfEj4zXDweINTWz0YSBodIs8pAmOhbnMjd8sTgk4CjivhL8AviP8ZrvHhbSfJ02Nts+q3gaO2jIxlQ2DvYZHyqCRkZwDmvgt+yp8OvhEttq8tv/b3iWLDHU7tBiB8YPkR9Ix7nLcn5scCqWqqqqqHUVVVVVS9fwodKqqqqpev4VVVVVVVVVVVS9Kql5GKqqodRVVVVVUOlVVVVUuM4qqqqpeRiqGCBg1XxD+Hnhr4neGp/DPia08yJ/mgnQASW0gHEiHsR+RGQcgmvjB8H/EXwh8RHS9TU3OnXJZtP1BUwlwg7H+64yNy9uoyCCf2bv2kX8JvbeAPH18W0RiI7C/lbJsD2Rz/zy9D/AAf7v3UdJEWSN1ZWGVYHII9RVV06VwRwaqqqjjHWic18S/2XPht8QPNv9OtP+Ec1d8n7VYRARu3rJDwrdySu1ierGviR8B/iN8MXkn1vRjdaap+XUrLMsGP9o4yn/AgPbNeAPi34++GdyJfCmvyw25bdJZTfvLeX1zGeAfcYPvXwu/ay8C+OHi0rxQq+GtVYYBuJgbWU9PllONpPXawAHQM1I6SIskbqysMqwOQR6ivjJ8G/CXxr8JSeGfEsHlzxbn0/UI0BlspSPvL6qcAMh4YehAI+KPwu8WfCHxZceEfF1l5c8Xz29wgJiu4iflljbupx9QcggEEV+yX+1oNUFl8LPinqn+nfLDo+sXD/APHx2WCZj/H0CufvdD82C1N04NVVVX7b/wAHf+E38Aj4h6PDu1fwlE7zqqZaezJBkGf+mfMnsN/c18EviXd/CT4laP4zgZ2toJfKv4l/5bWz8SLjuQPmH+0q1p2oWGr6fa6rpd3FdWd7Ck1vPE25JY3UMrKR1BBBB96qqqq/bK8dLrXjOx8EWUu630GHzLjBGDcSgHHH91Av4sePX9jDwJ/aHiDU/iDe2u6HSo/sli7Lx58g/eMp9VjwD7TVVVVVTdaqv24/H0eseHLL4T+GNat2lub3ztcEUu7yY4sGOFwO7OwfGcgxDOMjOleGdL0oK8cQmnA/1sgyQfYdv8814f8ABvivxYLk+GvD99qQs4mlna3hLLGqgk5PQHA4HU9Bml8apb6hbTxaXDdW0EyvLBcMyidQQShKEMoYDBIIPPBBr4Val4J1vwBo+t/D2wtLPRL+ASwQ28QjCN911YDq4ZSrE8kqeTTYxiqqqqqo9aqqqqpu1V+2P/wS88E/G64v/iL8Friy8IeN52M11ZSJt07VpCzM7uFBaCZi2TIoZWK4KAsXHxa+C3xR+BnieXwh8VPBepaBqCs4hNzCRFdqpwZIJR8kyZx8yEjt14r4b/tZftAfCu1i03wr8Rb59NibK2Goqt7Co7qolDGNc84QryT6mvBv/BUzU441g+IXwptbiTbzc6NftCM+0Uofg+vmce9aF/wUj/Zv1bZ9vl8UaLumEZ+3aWH2rx+8PkPJ8vJ4GW4PHTI/b4/ZPHH/AAtf/wAoWof/ABih+31+ycB/yVj/AMoWof8AxitY/wCCl37N+lxb7H/hK9XPm7Nlnpaqcc/P++kjG3gd88jjrjxT/wAFWrBI2h8E/CC4mc/cuNV1URhcN3ijRt2Vz/GME98c+NP+Cif7Tfi6I29h4h0jwxE67XTRdOVSfo8xldT7qwrxN4v8W+Nb/wDtbxj4p1fXr7aF+06neyXUmAAAN0hJxgAfgK+Fn7HH7Tfxn2zeAvg7r1xZnbi/voRp9qQe6zXBRHwBkhCx6ccjPwt/4IxfELV4o774wfFXSPDylsmw0W0bUJiuBw0rmNI2znoJBwPXj4Lf8E9v2WfgeyX2i+AE8R6ujArqviZl1CZcDA2oVWJDyTlI1PPJ4GKbrVVVVVN0qqqv2wf2TvB/7Vvw0n8O6hDa2XirTI3l8Oa26ENZznB2OVyTDJgK64OOGA3KtfEP4eeOvg3461HwJ470a60TxDodxtmhc4KkcrJGw4ZGGGV1OCCCDX/BPL/gobF8UotO+Bfxz1pI/GcarBoeuXL7RrYAwsMzHpc44DH/AFv/AF0/1lVVU3Wqqm6/hVVVVVHGOtVVVVVTdaqqqviN4dDINfs4xkYW5AHXsG/kD+FVVVVVVVVVVV4chSHQNPSPobZG59SMn9TVN1/Cqqqq6tbe9ge2uoVljcYZWFeI/AtxZF7zR1ae3HJi6un09R+v86qqqqqvDusvoepJc8tC42TIO6+o9x1//XUcsVxGs0MiujjKspyCPaqqqqqqqqqqqqqqqqqqqqqqqq98P6LqAJudOhLN1dV2N+Y57VefD2wkybK+mhPYOocf0NXXgLW4Bm3MFwOwR9p/8ewP1q50LWbXPn6ZcAAcsIywH4jiiCDtIwR2NVVVVQW811MlvbRNJI5wqqMkmvC3h5dEtC84VruYfvCOdo7KP6+/0FVVVVf8FJfDy+IP2N/HTJYtc3OltY3tuFYgxFLyESPgHkCJpcg5GOeoFfsI+I08LftifCPU3vLi3E3ia3sQ8BIYm6zbhTj+FvN2t22sc8ZqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqrUdO0/WNPudJ1axt72xvYXgubW5iEkc8bgq6OjZDKykggjBBwa/4KQfsCP+zZrg+KnwpsLu5+GuuXDCa3CtIfD1wxG2F35JgctiNzyCNjHOxn/Yq/a+139lrx6Bqct9qPgTWpAut6TC4OxsAC6hVuBKgAyAV3qNpPClfCnirw5468NaZ4w8I6vBqmjaxbJc2V3ASVmjYZBGeQexBAIOQQCCKqqo896qqpelVS9Kqqqqodaqqqqqh0H0qqqqqv2xLLZqXhfUPNz50FzFs29NjRnOfff+nvX7HN5Kmr+JtPVU8ua2t5WJHIKM4GPbDn9KqqqqHUVVVVVXxOnEmvQwq+RFarlcdGLMT+mK+FcAWyv7kKcvKiZI4O0E/+zfqKqvEPiTw54S0mfXvFev6bo2mWqlp73ULpLaGIAZJZ3IVRgE8ntX7Uf7W//BNvxIb6z8aeDrD4qa3HGU+0aLpO2Ut5e1VGoExHZgj5o3fbgEAsoA+K/iX4ZeKPE0l/8KfhnP4I0Xc/l6fPrcmpvgnI/eSKpAHIA5IGAWYgsa0Pw7r/AIo1BNJ8M6HqGrX0mNltY2rzytkhRhEBJ5ZR06keteHP2PP2mvFUayaX8HdchDIzgakI9OOA205FwyEHPQdSORxzX/DBf7WP/RKP/K5p/wD8frS/+Cfv7U+oXDQXfgGz01FQsJbrWrRlY5HygRSO2ec8jHB56Z17/gn/APtQaJbi5t/BFnqyKjvILDVbdmjCgH7rsrMTzgIGJx9M+Nfhd8R/hxKIfHngXXNB3vsjkv7GSKORsZwjkbX4B+6T0NV+w78Xv+CfOtT6b4R+JfwH8I+DfGZ5j1TWWk1LTL2bAHySXskptnIXIVzsBB2vuYKbDT7DSrOLTtLsreztYF2xQQRCNIx6Kq4AH0qvGPjnwj8PtEk8Q+M9ftdKsIhgyTEkuf7qIuWdv9lQT7V8av23vFXi0z6B8LEuPDmksGRtQdh9tuAR1UjIg742kt0O5eg8P+G/FXj3X00fw7pV9rWrX0hby4UMsjkn5nY9hk5LsQB1JHWvgp+wxpGlx23iH4xyLqN8DuXRbeX/AEeP08115kPfapC8YJYE1ZWNlptpFYadaQWttAgSKCGMRpGo6BVHAHsKqqWqqqqqHUVVVVVS4zjNDpVVVX2saRpjLHqWq2dqzjKrPOsZYeoyavfi38LbCKSW5+Inh390cMkepRSODnH3VYsfy4q+/aJ+C2n7PP8AHlq/mZ2+RbzTYxjrsQ4696u/2rPgzaz+VDrd9drj/WQ6fIF+nzhT+lXX7Y3wqt52hh0zxJcquMSxWkQVuO26UH25Aq9/bR8DxyhdO8J65PHt5acxRMG9MBm4xjnP4VcftreHlgZrXwLqEkoHyJJeIik+5Ckj8jX/AA27/wBUx/8AK1/9or/ht3/qmP8A5Wv/ALRV5+2vrkk26w8BWMMWOEmvnkbPfkKv8q/4bV8Uf9CTpX/gRJVv+2t4hWZGuvAunSRA/Okd46MR7Eg4/I1/w2//ANUx/wDK3/8AaK/4bf8A+qY/+Vv/AO0VZfttaRIjHUvh9dwMD8og1BZQR7kouP1qL9tXwi0qLP4M1dIywDsk0bFR3IGRk47ZH1Ff8NnfC7/oA+Kf/AWD/wCPUn7WnwcdlU6jqagnBZrBsD345q1/aV+CF5OltD47iV3zgy2VxEvAzyzRgDp3NWPxn+Euo7/s/wARdAXy8Z8++SDr6byM9O3T8a07xh4R1ZIn0vxVpF4s7bYTb30cgkbOMLtY5OeMDvVUtVVVxVVVVVUvIxmqqqqqHUVVVVUvpmqXkYzVV418FeHfiD4dufC/iexW5s7kcEcPC4+7IjfwsM8H6g5BIPxg+D/iL4Q+IjpepqbnTrks2n6gqYS4Qdj/AHXGRuXt1GQQT+zd+0i/hN7bwB4+vi2iMRHYX8rZNgeyOf8Anl6H+D/d+6jpIiyRurKwyrA5BHqKqgccVkEcGqqjjHWic8Cqp0SRDHIqsrAhlYZBHfivij+yN4I8XJLqfgjy/DWqkljFGhNpKfQxj/V9uUwBz8pzmviB8LfG/wAMtRNh4t0SW3Rm2w3aKXt5+M/JJjBOP4eCO4FfC39oT4g/C54bGy1A6nokZO7S7xt0ag9fLblo+SThflyckGvhZ+0D4A+KkcVnYX39na0Vy+l3bbX4/wCebfdkH05x1Ar46/A7wz8dPCX9hawfsupWW+TStSRMvaSsBnj+JG2qGXvgHggEfED4f+Kfhj4pu/B/jDTjaX9ocgjlJ0P3ZI2/iRscH6g4IIH7Jn7VkHjGCy+F/wAR75ItehRYdL1CQhV1BQMLE57TAcA/x/733mxjFVVVU8EF1BJbXMKTRSoUkjkUMrqRggg9QRxiv2kPhS/wh+KmqeHraArpN2ftmlMRx9nkJwn/AABgyf8AAQe9fsI/F6LxB4SuPhPrN5nUtA3T6aHPMtmzfMoPcxu3f+GRQBhTiqqq1/WtP8N6Hf8AiDVZSlnpttJcTsoyQiKWOB3OBwO5rxNr994r8Q6l4l1N83Wp3UlxKMkhS7E7RnsM4A7ACvgp4HT4ffDPRPD7xFLwwC4vtybW8+T5nB/3chPogqqqqqjya8f/ABQ8E/DPTxe+LNZjgkkGYLSP555/9yMc44xuOFBxkjNfFL9qPxx488/SvD7t4e0aT5fKt5D9omX/AG5RggH+6uBg4Jbqby9tLCA3F5cLEg7t3+g6mv2d/wBmvw/4w8K6b8RfHZnnt9TTzrHS1JjBiyQrysDk7wAwUEYBGSSSq6Xo2k6Dp8WlaJpttYWcAxHBbxCNF+gFftlfBY/Dvx2fGeh2ip4f8TyNIFjQKtrd9ZI8DoG++OnVgB8ua/Yi+NJ8J+Km+FuvXirpPiKYNp7SOFFve4wFH/XUBVx/eVMfeOW6VVVVVVHnvVVVVVNzxVV4m8KeFvGujTeHfGXhvSte0q5x51hqdnHdQSY6bo5AVP4ivix/wSb/AGWfiC733hCz1rwDftg/8Se7M1s53cloJ9+MrwBGyAEA4PIPjn/gi78ZNKE0vw8+LHhPxFHEpZI9Stp9MmlwmdqqvnoGLZUbnA6EkZIHij/gmV+2n4Y+0yj4Qf2ta22z/SNL1izn83dj7kXmiU4LYPycYJ6DNf8ADBn7YX/Rv3in/vyn/wAVR/YM/bCx/wAm/eKf+/Kf/FV4d/4JkftqeIWtXf4RJpVtdKWFxqWtWUQiABI3xiUyqSRjGzOSM4HNeC/+CM/x51d4JfHPxG8GeHraTaZEtDPqFxEC3zZTZGhIXkASEE8ZHWvAP/BGj4G6I0dx8Q/iP4r8UzRqAYrRItMgdsMCWUCWTHKkASDBXksDgfDD9kr9mv4NPBcfDr4NeG9OvbWTzINQntje3kTbt2VubgvKvPo3GB6DFVVN1qqqqqm6VVVVftl/saeCv2s/BXkym30jxvpELf2Frvl/d6n7PPjl4GP1KE7l6sr/ABD+Hnjr4N+OtR8CeO9GutE8Q6HcbZoXOCpHKyRsOGRhhldTgggg1/wTy/4KGxfFKLTvgX8c9aSPxnGqwaHrly+0a2AMLDMx6XOOAx/1v/XT/WVVU3Wqqm6/hVVVVVN0qqqqqqbrVVVVLFFPE8EyK6SKVZWGQQeCDXiXQpdA1N7NstE3zQuR95f8R0NVVVVVVVVVVWH/AB42/wD1xX+Qqm6/hVVVVVeIPBlhrDNdWzi1uics4XKv9R6+/wDOtV8NaxpGWurUtEDxLH8y/wCI/HFVVVVV4N8SrZsNJ1CUCBz+6kY/cb0PoD+h+vFVVVVVVVVVVVVVVVVVVVVVVVVVVxZWd2MXVpDNx/y0jDfzq48H+Hrgf8eAjIHWNyv6ZxU3w80xzm3vrhOOjAN/QVL8OZwQIdVjYY53RFf6mm+HusbiEu7MjPBLMD/6DS/D3VywD3dmF7kMxI/DbVt8OkBBvNTJHdYo8fqT/StM0TTNIQrY2yoxGGc8sfx/pVVVVVftpaZe6t+yj8U7XT4PNlj8N3M7LuC4jiHmSNyR0RGOOpxgZPFfs363aeGv2iPhb4jvo5ZLbSvGmkXcyQqC7JHeROwUEgE4U4yQM9xVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVeIfD2heLdCv/AAx4m0i01TSdUt3tr2yu4hLFcROMMjqeCCDjFft8/sL+J/2U/G0/iXw5ZS3/AMMdevHOj36Bn/s1mJYWVwSSQ6jIRyf3irnO4Oq/sG/tw6z+zx4jtPh147vpb34bavdhZBIxZtDlkbm5i6nysnMkY68uo3ZD2F/YarYW2qaXewXlneQrNbXEEgkjmjYAq6MuQykEEEcEHNVVVVVS9Kql6VVVVVQ61VVVVVDoPpVVVVVftiWXmaZ4X1DzceRPcxbNvXesZzn28v8AX2r9kG8lTx5q+nqqeXNpDSsSOQUljAx7Yc/pVVVVQ6iqqqqq8eXH2jxReAPuWPYi8YxhRkfnmvB2p6N4Y8Dz694g1a00zT45nlnu72dYIYh8qZLsQAMgDJPWvit/wUo/ZN+GFjM9p8QP+Ez1KOQxpp3hmL7WXIJBbzyVgCAjr5mSDlQ1fGP/AIK//GXxXv074N+FNL8DWgkbbfXRXU7x1HCkCRBCmRyVKPg4AbAO74hfFv4n/FnUV1X4l/EDXvE1zH/qjqV9JOsI54jQnag5PCgDk+tfDb9mX45/FkQz+DPh1qk1jNsK6jdxi0tSrfxrLLtVwByQm44xxyM/D3/glvqM62958VPiXDbfMDNp+hWxkbbnoLiXAU4/6ZMAT3A58C/sPfs0+BI4Wj+HsOvXcSgNda7K16ZSO7RNiHP0jHWtJ0bR9BsU0zQtKs9OtIvuW9pAsMa/RVAAqqqryys9RtZLG/tIbm3mXbJDNGHRx6FTwRXxp/4J+fBj4lxy6l4MtF8Da2V+R9MgH2OQ9t9twqj3jKepzXxN+C9z8L/DqzarqlnfaimryWcstlIzwMgDbTGWVSwPls24gcMBjrX7CX/BR3xB8IL/AE34T/HHV59W8Ayt5Nnqs26W50MseCW5aW3BJypyyAgrwuw/Gn9tfwX4Oin0P4Zy23ifWTF8t9FIHsbckcHep/fHGDhDt5+9kEV47+InjT4l602veNvEFzqd0ciMSNiOBePljjGFReBwoGTyckk18E/2NvHXxI8jXfGSzeGPDz4ZTPFi7ul5/wBXEw+VTj7744IKhga+Hvws8B/CzSF0bwR4et7CMj99PjfPcH1kkOWbnsTgdAAOKqqqlqqqqqhj1qqqqrWPHvgfw/vXW/GGjWLpvzHPfRo5KfeAUnJI9AM5rVv2mfg1pLmNfFLX0isAUtLSV8AjOdxUKR0HBJyfrjVf2zvBVuGXRfCms3hVW2m4aO3BYdOhc4PrjI9K1X9tPxLMHGh+CNMtCQNn2u5e4we+dojzx9Me9an+1h8Yr/f9k1PTdO3Nlfs1gjbB6DzN/H1ya1P45/F7V9/2r4g6unmNuP2ab7Ng+3lBcD2HFal4t8VayZW1fxPqt8ZxiU3N7JLvAGBncTngDrVWXhfxLqQhOneHdTuhcECEw2jv5hPA24Bzk+lWXwd+K1/KYYPh14hVlXdmfT5IRj6uAM89OtWP7Ofxp1FGe38CXKhDgie5hhP4B3BP4VZfsp/Ge7hMk+hWVmwbAjn1CIsRgc/IWGPxzxVl+x78V7tGae78P2ZU4Cz3khLe42Rt+uKi/Yx+JbSos/iHwwkZYB2S4nZlHcgeUMnHbI+or/hirxR/0O2lf+A0lf8ADFXij/odtK/8BpKsv2JdSkRjqXxEtoGB+UQaY0oI9yZFx+tf8MQ/9VO/8on/ANvq4/YjuFgZrX4lRySgfIkmkFFJ9yJiR+Rr/hijxSRx420r/wAB5K/4Yo8U/wDQ7aV/4DyVe/sW/EKOULp3ifw7PHt5ad5omDemBG3GMc5/Crr9jf4q29u00Op+G7l1xiKK7lDNz23RAe/JFTfsmfGSOF5E03TZWRSRGl+gZiB0GcDJ9yB71cfs0fG+2gaeTwLIyoMkR31tI34KshJ/AVefBD4uWMHnzfDzW2UHGIbUyt/3ymT+lX3gHx1pjrHqPgrXrVnGVWfTZYyR6jK1NDLbyvbzxNHLGxV0dSrKw4II7H2qx1bVNL3/ANmandWnmY3+RM0e7HTODz1P51p/xh+KumPutPiL4g4TYFl1CSVQOOiuSB0HOK079qP42aeY1fxbHeRxpsEdzYQHOBgEsEDE+5PPfNaZ+2j8RbfamqeHfD94qx7d0ccsLswx8xO9h68BRye3StL/AG3dKk+XWvh/dQY2gNa36zZ/vHDIuO2Bk/UVpH7Xnwd1Ngt7c6vpQLEZvLEsAAM5/cl+D09c+g5rR/jh8IteVTp/xC0ZSwBVbq4+zMcnAG2Xac+3WrS9stRgFzp95BcxEkCSGQOuR15FcEcGqqqqqXkYzVVVVVfQ1wRwaqqqh161Q64zVVXjXwV4d+IPh258L+J7FbmzuRwRw8Lj7siN/CwzwfqDkEg/GD4P+IvhD4iOl6mpudOuSzafqCphLhB2P91xkbl7dRkEE/s3ftIv4Te28AePr4tojER2F/K2TYHsjn/nl6H+D/d+6jpIiyRurKwyrA5BHqKqvoa3HpW72otngUee9VVVWraRpevadPpGtafb31lcrtmt7iMSI4znlTx1AP1Ar4r/ALG1jeLNrXwquhaTAZOkXTkxuc8+XKTlOP4WyP8AaUV4h8OeIvBusyaN4i0u50zUbYgmKVdrD0ZSOo9COD2NfCn9rjxb4PS30XxvFJ4h0mMhROWAu4E4HDHiXHXD8n+8B0+KHgj4b/tdfDsP4V1+z/tfTf3thebB5tpIyn9zOuN6xvjkeqhhu24Pinwt4p+HPim58O+IrG40vWdKmG5c7WRhyrow6g8MrDgjBBr9lH9pwfFqwHgjxpPFF4s0+DKTllQanGvV1XtKAMsoGCPmGBkLVVVV+2H8Jh8TPhRc6nptt5ut+Fd9/Z7Vy0kQH7+Id/mRQwA5LRoO9fDDx7qfwx8eaN440olpdMuQ0kQOPOiPyyRn/eQsPbOe1aDremeJtEsPEOi3S3FjqVslxbSj+JHUMp9uD07VVVX7YfjR/D3w4g8MWspS58SXPlsQcHyIsPJgg55YxLjoQzV+z94Lfxz8VtF094y9rYSi+u8DgRxENg+zNsX/AIFVVVVVXd3aWFtJeX11Fb28KlpJZnCKgHUkngD618W/2vILN5tC+FaR3EoDJJq9xHlEbpmFD9713MMexHNX2oa74q1f7VqF5e6rqd7IFDSO00srk4CjqSckAAV4/wDg54g+GXwlufiP43uE0u7uJorbStKZd8s0snOZCDiPEayPtOT8mDtNfD3wtq/xY+I+g+Dkknnn1i+jhkdMbooc5lkA6AJGHfp0U8Vp2nWWkada6TptusFpZQJBbxLnEcaKFVRnnAAApu1fFf4b6N8WPAmp+CdZVVW8j3W0+0FradeY5F9MHg46qWHQmtd0TXfBPiW80HVYpbHVtGu2ilVXw0UqN1Vh7gEMOCMEGv2bvi/B8YvhpZ6rdTp/bmmYtNWiDAt5qj5Zcddsi4bpjduUE7Saqqqqo4x1qqqqqqbr+FVVVVU3Sm6/hVVVVVHk1VVVVTYxjNVVVVftl/saeCv2s/BXkym30jxvpELf2Frvl/d6n7PPjl4GP1KE7l6sr/Eb4beP/gv44vfA/j/Qb3QfEGkTfNFKCpOD8ssTjh0OMq6kgjkGv2Cf+Ck1r44/sX4H/tAan5PiZsWukeJ7iQCPUzwI4bkn7tweQJDxIQAfnOXqqbBPFVVN1/CqqqqqbpVVVVX7S3xu079nb4LeIvixf20V3JpMcaWlnJJs+03EsixooHVsFtxA52q3IAJHgP8A4KV/tneHprbSIPH8XinzJFjt7TVNHhuZJHZ8hQ8arK5YnaAWPBAGMCrEXwsbZdUe3e9EKi5a3UrGZMDcUBJIXOcAknGOaqqqq8W6AviDSmgjwLmHLwE/3u6n2PT64PanR43aORCrqSGVhggjqCKqqqqqqqqqvDsy3Gg6fIrE/wCjopJ9QMH9RVN1/CqqqqqbpVat4M0bVCZUjNrMf44RgE+69P5H3rWPCGr6SGk8r7RAP+WkQzj6jqP5e9VVVVeE/FnlbNK1WX5PuwzMfu/7Le3oe38qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr9sHWv7A/ZY+Kt99m8/wA3wnf2uzfsx58LQ7s4P3fM3Y74xkZyP2f9Dl8T/Hn4beGredIZdW8XaVaJK4JCNJdxIGIHYbs1VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVfEDwF4T+KXgnWvh5450iLU9B8QWb2l9ayD7yMOqnqrqcMrjDKyqwIIBr9tH9knxR+yL8VpfCd411qHhXVt9x4Z1qaNV+3QLt3o204EsRdUccZyrhQrqK/4JmftpN4evrX9nD4ra6g0i7YJ4T1C6kCizmJJNm7nqjk/u8n5W+TkOoSqojiqqqXpVUvSqqqqqHWqqqqqodB9Kqqqqr9rixSf4e6bfLAzyW2ropcAnYjRSZz2wWCDJ749a/ZcvltPi3Z27XDRm9sriFVGf3hC79px2wmeeMqO+Kqqqq+hqqqqqv2uv+CgXhX4a+Ita8I/C6O18TeKkupI5rqRs2WnMrspVtpBllXaAUUgDOS2QUPxL+M3xQ+MN+l/8RvG2p615Du1rbTznyLXcxLCKIfJGMk/dAr4f/s+/Gr4pxpceA/htrWp2sh2peGDyLYn0E0pWPoR/FxketfDH/gl/4kvpI734veObXTLfILWGiDz52XjIM0ihEbr0WQdD6ivhf+yZ8A/hJ5Vz4Z8A2d1qcXI1PVR9suQ3PzKz5WM4OP3apkdaqqqqqq/aI+KI8KaL/wAIho85GratCfNdSQbaA8E5/vNggegBPHGfD9h4A1W/fS/idoiar4curaZLqDYTID5bGNoiCCkm8KA2RjJ5Aya8I/B3wV4L1CXUdOhuL64EzNbT3xV3gTJ2gBQF3BcAtjk5IwDgaUdN1TxXoXgyXXtL07UfEd6llYfb7kQo8rnC5POBkgZx1IHUgH4J/sj/AA++FQh1vWY08SeIgi/6VdxAw2rdT5MRyAc4+dstxxtyQaqqqqX0BqqqqqqrX/H/AIH8Lqw8Q+LdKsXU48qa6USEjAOEzuOMjOBxmtd/aw+Euklo7C61PWHU4H2O0KrnB7ylOMjGRnrxmtd/bQ1STdH4Z8EWsGMhZb66aXPTBKIFx343HtyK1r9p/wCMWsF1h8Q2+mxSZzFZWca4BPZnDOMdPvZrWvHHjPxGXOveLNWvxJnclxeO64JyQFJwB7AYqz0+/wBSlMGnWU91Iq7ikMRcgdM4HbkfnWk/A34ua2wSy+H+rRksVH2uH7KMgZ6yleMd+hPHWtK/ZG+Lmoxh7tdF0w7c7Lu9LHPp+6Vxn9K0z9ii9dg2sfECCNVcZS208uWXv8zONp/A1pn7G3w4tgj6nruvXrq2WVZY4kYehAQt+TVpn7MfwW00IzeE2vJY2yHur2Zs+xUMFI+oqw+D/wAK9MTZafDzw/kPvDS6fHKwPHRnBI6DjNWOk6Vpm/8AszTbW08zG/yIVj3Y6ZwOep/OqoYyKqqqqqhyOtVVVVVc2lpfQNaXttFcQvjdFKgdWwcjIPHUA1qHw2+HerP5mp+BPD11Js2B5dMiZgvPAYrkdT+dX/7OXwU1HZ9o8BWi+Xnb5FxNB1x12OM9O/T8avf2PPhHeKiW82vWZU5LQXikt7HejfpitR/Yh02RFOkfEK5hYA7hc6csgY9sFXXb3z16+3Oq/sX/ABGtQX0nxBoN8qpna8kkLs2egGwr0xyWFax+zN8atIY7vBzXkYYKslndRShiRn7obcB2yVAz9RWreCPGegx+brnhDWtOQLu3XenywjHrllHFWd9e6fMLnT7ya2mAIEkMhRsHryOa0b49fGPQTmx+IerS5JP+myC86gD/AJbB/T8PxrQ/2y/ibp6iLWNM0TVVGcu0LQydAOqNtxxn7vfrWh/tteEbohfEXgzVbDOebSdLoDkYzu8s4xnPHbvmvD/7SPwY8RMkUHjS3spm6x6hG9tt5xy7gJ2z97vWmazo+t24u9F1azv4CARLazrKuCMg5UkcjmqqlxnGaqqqqqhyOtVVVVDBHWvoa4I4qqrx74G0L4jeFrzwp4hg3210vySKoLwSD7siEg4YevoSOhNfFL4W+JPhP4kfQNfi8yJ8vZXqKRHdxg/eX0I4DL1B9QQT+zB+0L/YMsHw58e6tjS3xHpV5OeLVu0TsTxGf4SfunjIX7tVVVVVVVVV40+H/g/4h6YNK8X6Fb6jCmfKZwVkhJxko4wyk4GcHnAzXxV/ZD8WeFPtGr+AZJfEOlJyLXbm9jH+6oAl/wCAgH/Zrw/4j8Q+DdYj1nw7qlzpmo2xKrLE21h6qR3HqpGD3rx7418J/tC6DbWfxDjt9B8cabD5Wm+IoYz9mukGSIbpBlkVmJIdQQjMThVLA2l3rHhjWorywurjT9T0y43RyxOUkglQ8EEdCCK/Zl/aK0r42+HF0rU5Vt/F+k2ynUbcgAXKjCm5jAAG0sRuUD5GYDoVJqqo9ODVftXfCOP4T/FS7i0qzWDQddBvdMVE2pECf3kKgcDY3QDorJX7BHxcfUNO1D4Qa3el5rAG90cyNn9ySBLCM/3WKuB1O9z0Wqqq/aw8YL4o+LN1ptvKr2vh+BbJNoIBk+/Iee4Zth/3B9T+xd4J+w+HdW8eXcWJdUmFpaEjkQx8uw56M5A5/wCefvVVVVXxP+MHg34UaaLrxBe+bezLm106BgZ5uuDtz8qZBBc8fU8V8Vfjn41+K9y0Op3X2HSFYGHS7ZyIlI6M56u3ueB2Ar4W/BTxr8V7wf2LZ/ZdLjk2XOp3KkQx9yF7yPj+FemRkqDmvhf8D/A3wrto5NIsvter+Xtn1S5GZXz94KOka9sL2xksea/4KEfEP+2fHej/AA3s5IzB4dtTc3e18n7ROAQrDttiVGH/AF1PtX/BPT4d/wBp+K9d+Jt9DmHRrcWNiWHWeXmRhg9VjAXnjE3txTc8Zqv26vgx9usYfjLoMH76zRLXWo1H348hYpgAOqk7GPoU6bTX7NPxhk+D3xItdRvbhxoOqYtdWjUZ/dnOyX6oxDeu3cO9I8csayROrqwyrKcgg9xVVVVR6fhVVVVVU3JxVVVVVNjGM03JxVVVVVHrVVVVVN2qqqqqv2sv2TvAH7V/gBvDfiRE0/xBpyvJoOvRxBpbCUjkEcb4WwA8ZPOAQQwVh8aPgf8AEr9n/wAa3HgL4n+HZdM1CLc9vKPmgvYQxUTwP0eNipweCOhAIIH7CH/BTGXw8lr8If2mPEc9zpxaOHRfFV0xd7XJCiG8fq0fORM2SnIclcFIpYriJJ4JUkjkUMjochgRwQR1GKo9aqqbBOBVVVVVN0qqqqr/AILHfGxJZ/CX7Pmk3O7yf+J/rARgQGIeG2jOOQQPOcqccNGcHgj9nnRrXxH8f/hl4evZJUt9T8Y6TazNEQHVJLyJWKkggHBOMgjNHrVVVVRxjrXxD8OCGT+3rOMBJDi4UDo3Zvx6H3x61VVVVVVVVVfDvU1udLfS2Pz2jEqMdVY5/nn8xVN1/CqqqqqbpVU3StW8K6Pq4LyQCGbHEsQ2n8R0P41rPhHVdJLSJGbm3HSWNen1XqP5e9VVVXhzxlLpwWy1NnmtxwsnVo/8R/L9Kt7i3u4VntpkljYfKynIqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr/AIKOeJ18L/sd+PGjuUjudVW00+3Voy3mGW6iEi8cA+UJTk4GR64B/Yf8OjxR+178IdM3yr5PiyyvcxJuP+jSC46en7rBPYZPaqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv2mP2c/A37UXwo1L4XeNk8jz8TabqkUCyT6ZdLyk0e78VZQRuRmXIzkfFj4WeOvgR8SdY+GvjywbTvEHh662SGJjtccNHNE+AWR1KurYBwRwDkD/gnf8Atcr+0J8PP+EE8a6qknxA8I2yLdGRj5mp2a7US7yxJZ8lVlP95lbjzABVHoPpVVVLyMVVL0qqqqqh1FVVVVVQ5HWqqqqq/aVsPtvwg1eUFt1nLbzKqjOf3qqc+2HJ/CvgZqP9l/Fvwxc+aib7zyMsM/61Wjx9Tvx9TVVVVVVVVVf8FCf+Cicury6h8Cf2e/ETR6fGzQeIPE1jMVN0Rw1tayL0j7PKD8/3VOzJf4U/CLx58avFkPg34faK1/euu+aR28uG0iBAaWVzwqDI9STgKCSAfgB+w78J/g3Ywan4h0+38XeKWQGa+1G3WSC3fuLeFgQo6Dc25jjOVB2iqxngVtJHArYa8skda8e/G7wP4Clk0+5u31HU4zhrO0AZoz/tsflX6dfar79rHxJIxOm+E9NgXcNonmeUhe4yNvPv+laH+1lE0/l+JfCTJCekljPuZfbY+AfruFeFfGHhzxtpi6r4b1OO7h4Eijh4mI+669VP+RVePPGWm+A/DF34j1JtwhXbDEOs0p+6g+p6+gBPavEGvan4o1m71/Wbkz3l7IXlc/kAB2AAAA7AAVXwq+Fuh/EtNQsLvxvFpWom3lSzto4t0wkKHZNh8K6qfmKAknbztByfir4a8b+APiXrXhjxxdXJ8Q6NebJZ3kJZsANHIhzkKyFGX0DDpX/BPL9p9v2lPgbAviPUWuPGvg5o9P18yEb7kEEwXXHaVFYEnBMkUvGME1VVVUvX8KqqqvEnxD8DeEC0fiXxZplhKgyYJbgebjGciMZY8Y7d68Tfte/DvSy8Ph3TtS1yVR8sgT7NC3B43P8AOOQP4Oh9sV4j/a++IWp74vD2l6Xo0TZ2PsNzMvXHzN8h4I/g6j0OK8QfFX4keKDINb8batcRy53wLctFEeSf9WmF7nt046CrDTNR1Wf7Jpdhc3k+MiKCJpGxkDOFBPUgfjWh/s9fGDX9rW/gu6tIzjL3zpbbQc8lXIbt0AJHHrWi/sY+K7lQ3iDxjpdhn+G1ge5I4GM7vLGc5H9TWi/sf/DawIk1fU9Z1Rx1Rplhj6jsi7s8Y+93rRvgh8JdBKHT/AWlMyAhWuojdEc5zmUsc579R06VaWNlp8ItrC0htogSRHDGEXPc4FDqKXkYzVVVVVDqKqqqqql5GKqqqqoYz1qqHUYqqqta+H3gTxFvOu+DdFv3ffmSewjdwX+8QxGQT6g5zzWtfsr/AAW1lHMHh240yWTOZbK8kXBIxkK5ZBjrjbj61rf7EWiyq7eG/Hd7bkZMcd9aLNnjgFkKY56nB+leIf2PfixpStJo76RraZ+RLe68qTGR1EoVR1J4Y9D7Z1/4T/EvwwrPrngbWbeJBlphaNJEvJHMi5Xse/v3qx1C/wBLuVvNMvri0uE+7LBKY3X6EHNeHf2jPjJ4a2pb+Nrq+iHWPUVW63df4nBccnsw6AdBivDf7bOvQbIvFvgyyvB0aawnaBh77H3gn8RXhv8Aax+D2v7I73VL3RZm4CX9qcZ/3496ge5IrQvFPhnxPD9o8N+IdO1SMDJa0ukl2/XaTj8aqqqqql9M1VVVUp7ZqgR0zVVVfEP4eeGvid4an8M+JrTzIn+aCdABJbSAcSIexH5EZByCa+Jvw4134W+LLnwtri+Z5fz210sZVLqI/ddc/kRzhgRk4zX7Lv7Q8oltfhl481JDFtEej387kMG4C27nuCPuEkYxt5yoFVVVVVVVVUx7A18Vf2ffAXxVRry+tTpms4wmpWigO3tIvSQdOvzcYDAZr4o/A7x38Kbktrmn/atMZsQ6paKXgbOMBjjMbc42tjJBwWAzWpaPZamv7+PEgGFkXgj/ABFaHqfi/wCGPiOy8W+F9RltL6wk3wXUPI9Crr0KkcFTwQSOa+BHxv8ADnxx8ILrelstvqtkEj1bTyfmtZSDgj1jbaxVu4BBwQQKqm6YBqv2sfhN/wALU+FF6NOtfN1vw/uv9O2rln2j97EP99AcDuypXw/8a6t8OfGuj+N9Eb/S9IulmVC20Sp0eMnsroWQ+zGvDHiPSfGHhzTfFOhXHn6fqtrHc27kYOx1BAI7MM4I7EEdqqvFPiCz8KeGtU8S35XyNLtJLh1Lbd2xSdoPqSAB7kVfXmo+INXuL+5Ml1f6lctLIQCzSyyNk4HUksf1r4d+E4fA3gbRPCcQTOm2aRylFwGlI3SNgerlj+NVVVXxt/ao0zwk8/hj4dyW2qawoZJ74nfBZt0wo6SuOe+1TjO7lRe3uveLtckvb2a81XVtRlyzENLLM54AAGSegAA6AACvg9+yKHjt/EfxWzhhvi0WJyD7GdxyO52L7ZbqtWVjZabZw6fptnBa2tugSGCCMRpGo6BVHAA9BV3dW9jazXt3MsUFvG0krt0RVGST7ACviX4zufiJ4/1/xtdF92sX8s8aOADHETiNOP7qBV/DqetfsoeAf+Fe/Avw5p89v5V9qsR1O9Bj2N5k+GUMOu5Y/LQ55+Tt0FNycZqtX0jTNf0q70TWbKK8sb6Fobi3lXKyIwwyke4NfGX4ZX/wi+IeqeCbyR5orZhJZXDrt+0W78o/1xwccblYdq/Yr+MT+OvAz+A9bujJrPhaNVikkfc1xaEkIf8AtnxGfbZ3JqqqqpunBqqqqqqbBPBqqqqqm6U3WqqqqqPNVVVVU2DgA1VVVVR6fhX7RH7Nvwv/AGnPA58FfEnSmc27NJpup2xCXWmzEYLxOQeCANyMCrYGQSAR+09+yl8T/wBlbxgvh3x1ZpdaXqDSNo2t2qn7PqEatjjr5cgG0tETldw5IIY/sT/8FEPGH7Nr2fw88fR3XiP4bmRtluhDXekFyCXtyxw0edxMJIGWLKVOQ3gL4g+Cvil4UsfHHw98SWWu6FqKk217aPuRsEhgQcFWBBBVgCCMECjgng1VU3WqqqqqbpgGqqqr4t/FLwl8Fvh1rnxM8b6glrpWh2rSuC2GmfpHEg7u7lVUerDtXxe+KHib41fEvxF8U/GEyPqviK9a4lRCdkC4CxwpnJ2Rxqka5JO1BknrX7IH9j/8NUfCX+3PN+z/APCY6bs8vr5/2hPJ/DzfLz7Zo9aqqqqPT8KuYIbmF7eeNXjkUqysOCD1rxJoU2gai1qwLQvloHx95f8AEdD/APXqqqqqqqqqvD+ryaJqkV6hyn3ZV/vIev8AQ/UCo5I5o1licMrqCpHcHvTdaqqqqpumKqm6VVaz4Q0rVwZEQW1wf+Wsa9T7r3/Q+9ax4Z1TRmLTw+ZCOk0Yyv4+n41VVWk65qOiy77Ob5CcvE3Kt+HY+45rRvF2masFikcW1xjBjkOAx/2T3/nVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV/wWH+IsWmfDTwR8LLa4YXOvavLqdwscuMQWsexVkUHO13uQy5BBMJOcrX/BH7wA/iz9ruDxW8coh8E+H77UBKEbZ5sqraKhI4BKXMrAHr5bEdKqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr/gpr+xg37SHw4T4j+AdIa4+I3gu1cWsMCDfq1ju3vakAFndSXeJQfvM64JkyPg98VvFfwQ+JOhfE/wVcpHqmh3IkWOQZjuIyNskMg6lHQshxggNkEEAj4JfF/wr8ePhjofxQ8Hzf6FrEG6S3Zw0lnOvEkEmOjIwI9xgjgiq6gc1VVS1VLVVVVVDqKqqqqqh0qqqqqvijph1j4b+JtPRXZ30qdo1TqzqhZR+LKBXhrUzoniPStZWQobC9huA4XcV2OGyAevTpVVVVVcEcVVVX/BTr9sa8+DvhRfgh8NdXa28ZeKLXzNRv7abbLpNiTjCkHKTTYKg9VTcwwSjD4Q/CXxb8bfHdh4A8GW8b3t3l5Zpm2xWsK43zSHqFUEcDJJIABJAr4M/BjwT8DPBNp4M8GadHGI0U3t60YE1/MB80srdSSc4GcKOBgCsZ4FBSeBQjGOTQQYwBQQkYFeUSK/aF+Kc/gTR4vDugXfla3qkZbzFPzW0GSC49GYgqD2wT1ArEk8nUu7nkk5JNRaLcSKCzheOmM1c6bcWw3EblHUjtXgjxrrfgHX4Ne0S4IdDtmhJ+S4jzyjD0Pr2OCORXhvxLpPinwzZeKtPnC2V3b+bmRgPKx99WPQFSCD2yDXxs+Jj/EXxPtsJGGjaZuiskIxvJ+/KfdiBj0UDgEmq0/SNV1eTydJ0y7vZNwXZbwNIcnoMKDye1aJ8H/i3dyxXmk+EtTtZo/njklYWjoQcZBkZSD+vfpX7bPwE+J+u/D+3+MXie202fU/Cka2moS2zlp7myZgEkcKu0mN3PIOdrsTwmT+wv8AH2X9nj9ozw14pvdRS18O6vMuk+IDK+2NbOd1Blb2icJL64jI7mqqqqqHXANVcXFvawtcXU8cMUYyzyMFVR6knpXir9o34S+FVKHxKmrzjGIdKUXORjOQ4Ij9ON2f1x4p/bJ1i4Mtv4N8K21ohGEuNQkMr9PvbFwFI9CWH8q8T/Gn4oeLi66v4yv1hds/Z7V/s8YGcgbY8ZA465PGevNadpWp6zdLY6Rpt1fXLfdhtoWlc8gcKoJ6kD6kV4Z/Zq+LniVVlPh9NJhbGJNTl8nGQDygy46j+H26g14d/YxtV2S+LPGksn96DTrcJj6SPn/0CvDv7Ofwg8OBJI/CcWozL1l1F2uN31Rvk/8AHa03StL0a1Wx0fTbWxt0+7DbQrEgwAOFUAdAB+Aqqqq+hpfSqqqqqqqqqqqXkYqqqqquKqvoa4I4NVVUvIxmqqvEPgHwR4sLP4k8JaRqUjDHm3NmjyDgDhyNw4UDg9hXiH9kT4RaysjaXb6losrZKm0uy6hsHqsu/jJBIBHTgivE37E/ia18yXwl4v0/UFGWWG9ha2fHPyhl3hjwoydoJJ6YrxN8BPi74U8x9T8D6hNDHkmeyUXSbRn5iYixUYUn5gMcZxkUr3um3eY3mtbq3cjIJjeNhwfcEV4Y/aQ+MfhZgsPjG41KEdYtUH2rdyT99vnHU9GHb0GPCv7bn3IPHHgr133OlTfTgQyH68+Z6cd68JftD/CHxiYoLDxfb2d3JgC21EG1cMTgKGf5GJx0Vj1HrUM0NxCk9vKkscihkdGyGBGQQR1BFVVUOD1qqqqr6GgQRwa+hoEEdaqqr4pfC3w18WPDT6Br8WyaPL2V6igyWkmPvL6g4AZehHoQCPHfgjXvh54nvPCviK1MVzaOdjhTsnjydsqE9VYDI/EHBBA/Zg/aGTX7e3+HHjzVCdXjITS76dv+PtMACJ2P/LQc4J+8MD7w+aqqqqqqqokAdaPPejjHWry0tNQtpLK+tobm3mUrJDMgdHB6gqeCPY18Xf2QNN1X7Rr/AMLXSxvHbe2kyviB/XymP+rPfafl5wNowK8QeHda8MarcaB4j0qewvrZtstvcR7SPf3UjkEZBBBBINfD3xT4g+E/jG08a+CblopYWC3lgZNsV9BkF4m4OMgcHB2nBGCM18O/H2g/E3wlZeL/AA87i3ugVkglAEttKvDxuAThgfwIIIyCCapsE4Bqq/ax+E3/AAqz4r3n9nWvl6Jr+6/07auFj3H97EPTY+cDsrJX7AvxY+2abqPwe1e6zLY7r/R97dYmb99EPo5DgdT5kh6LVV+2F4uTQvhhH4bikX7T4iu1i27sHyYiJHYdz8wjU/7/AOB/Z18HDxr8W9DspofMtNOk+33QKbgEhwyhh0wz+Wpzx83fpR5qqrVdW0vQ9On1bWdQt7Kytl3TXFxII0QdOSeByQPqRXxy/af1Pxt9o8LeAprjTdBPyTXXMdxejkMOD8kR4+X7xH3sAla8B/DvxZ8StZXRPCmmPcyDBmmYFYbdTn5pH6KDg47nGACa+D/wC8JfCe2W8RU1TXnXEupTRAFOoKxLz5YwcHBy3c44DYxgGq/bG8bDwV8AvEHkzbLvXtmk2+RkN5xPmg8j/lis3rzjjGa+DfgVviX8UvDPgjZuh1O/RbkZIP2dMvMQRzny0fHTnuOtJHHFGsUSKiIuFVRgADoAPSqPJ4NVX7Z3wg/4WF8Oz4v0mHOteEkknCqmTPakAzJ9V2hx1xtYAfNmvhH8R9S+FHxB0jxrpzyslnMFvIEbH2m2biWMjocryM8Bgp7CtC1rTPEui2HiDRrlbix1K2S4tpR/EjqCp9uD0qqqqbpgGqrxd8Vvhd8Pw3/CefErwt4b2bd39r6xBZ7d33c+Y64z29a8R/t8fsc+Fkum1P8AaB8MTizYLINOeTUCxJA+QW6SeYORymQBkngGta/4Ks/sX6XdJb2HjvWtYjaMMZ7Lw/dIinJG0iZI2zwDwMcjnOQNQ/4LJ/srWd5La23hT4l30cbYW4t9JtFSQeqiS6VsfVQa/wCHzn7Lv/QifFL/AMFdh/8AJtaH/wAFpPgLced/wkvwr8fWG3b5H2FbS739d27fNFtx8uMbs5PTHJ/4LNfsunp4E+KX/grsP/k2tF/4K2/sfaqlm19q3ivRzcsolW90NmNqCcEv5LSZAHJ2bjjpk8V4S/by/Y88bBf7G/aB8LW+7dj+15n0r7vXP2tY8e2evbNaXqula5YRapoup2uoWc2fKuLWZZY3wSpwykg4IIPuDVVVN0ABo9aqqqqOMdaqqqqqm6/hVVVVVN04NV8SPhv4K+LngrVPh78QdBt9X0LV4THc20w6d1dGHKOpAZXGCpAIIxX7Z3/BOz4gfs5XV/468CQ3Xij4b+Yz/bETddaQpPCXSDkoBx56jacfMEJUH9nL9qP4s/sv+Kz4j+G+sIbO6Zf7T0a83PZ6iqhgvmICCGXcSrqQwPcgkH9ln9tv4P8A7U2kwW2g36aF4wSEve+GL6cGePaBuaF8ATxj+8oBA5ZU6VVU2CeDVVVVVN0qqqtf17RPC2iX3iTxJqtrpml6ZbvcXl5dSiOKCJBlnZjwAAOtft7/ALbV/wDtQeLY/C3gqe8svhz4fmb7DBITG2qTjIN3KnGBjiNG5VSScM7KPgL+yZ4o+L/wj+J3xtulls/DngHQbu6tXZNo1G8ihMjRq2eBHGPMY4OTsX+Ikfs/a1F4b+PXw18RTwtNHpfi/Srp4kOC4ju4mIB9SFxR5qqqqo9PwpsHABrxFokGvaZJaOo81Ruhf+646fgehqWKSGV4JkKSRsVZWGCpHBFVVVVVVVVV8Ptd+02p0W5kzLbjMJJ5ZPT8P5H2putVVVVU3SqpulVVMqupRgGBGCCOta34Fsr7dPpbLazH+DH7s/gPu/h+Vahpl9pc32e+t2ifHGeje4Peqqq0rxZq+lARLMLiEDAjl5x9D1H8q03xto19tjuHNpKeMS/dz7N/jio5I5oxJDIrqw4ZTkGqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr/AIKHfGWL4y/tOeIZtMvkudG8KKNC0542yjCBm851IABDTvLhucqFwSMV/wAEYPg7P4T+B/iT4xalDsn8e6otvYZUHNnZF494OeN08lwpGB/qlPPGKqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr/grB+x9/wqH4if8AC/fAembPB/jm8b+1IYzldN1ZgXfA6iOcBpByQHEo+VTGp/4Js/tTxfA34pH4d+M9VS38FeN5UhklmfbHp1/wsNwTj5UYfunJwAGR2IEZqgOBzVVVLVUtVVVVUOoqqqqqqHIxVVVVVTQQ3UElrcRiSKVCjoejKRgj8q1bTp9H1S80i5BE1lcSQSBlKkMjFTweRyOlfDzWF8QeA/D+sK+5rrTYGk68PsAcc8nDAjPeqqqqh0H0qqr4x/FHw78FPhd4l+Knil/+Jd4bsHuWiD7GuJOFihUngNJIyRrnjc4zXxG8feKvi/8AELW/iH4snF3rniS/e5uBEp2h3OFjjUkkIo2oq5OFUDmv2Nf2dofgL8MYZNbsI4/GHiJEuNacOWMIGTFbA9P3asQ2OC5bkgKaClulKgHA5NCMkYFCIdBQi44FCMdM0EHQV8ctbk174qeIZ3fK2l0bSNcY2iH92R+ak/U1oVksoEhGSxwPpWjeFWvYcxpnj0rX9CFmWUr07YrULdbe6aNRweQK/Z9sX8f/AA48QfD661e6srWG9jkme2UCRoZVOYwzAgAtEc8dyO/Gn/sy/CqwAFzpt9qGE25ub11yf737vZz+nPStN+GXw+0dg+neDNHikViVkNojupIwcMwJAx2zjr60kEcEaQwxqkcahURVwFA4AAHQVXiHQdM8U+H9T8Ma1b+fp+r2ctndxZxvilQo657ZViK+LHw+1P4U/EjxF8O9WWTz9Cv5IEkkXaZousUuOweMo49mFf8ABOv42r8bf2WvDFzfXjT654SX+wNWMmSxkt1XyXJJJcvbtCzMerlx2NVVVVxcW9rC1xczxxRRjLPIwVVHqSeleL/2lPhV4USSO21v+27xFykGmjzVY9B+9+5j1wSQOx4z4r/a/wDHOqM8PhTSbDRIDkLJIPtMw4xnLAJ15xsPbk858TeNvF3jKf7R4o8R3+pENuVJ5iUQ4x8qfdX8AOp9a8K/DXx543K/8It4Wv76M5xOI9kPBwR5jYQHPbPr6V4W/Y68WX4iuPFviGx0qNjl4LdDcSgdwTwoJ9QWAz+FeGP2XPhP4fjRr/S7nW7kAZlv5ztBwQcIm1cHPRg2OOe9aXoujaHbCz0TSbLT4B0itbdYl/JQBVUOlVVVVUvP5VVVVVVDoPpVVVVVL6A1VVVVVdRVUMEDFVVUOuKqqqqrxH4J8H+L4xH4o8M6Zqm1dqNdWyu6D/ZYjK9T0I614p/Y++Feuq8ugvqOgXGDtFvMZoskjlkky3rwrKOa8V/sZ/EbSN83hfVNM1+FeFj3fZZ26/wuSgHA/j79O9eKPh/438FyGPxT4V1LTVDbRLPbsInPH3ZB8rdR0J614Y+IHjfwXIH8K+K9S01Q24xQXDCJzz96P7rdT1B614T/AGz/AB5pZhg8W6JpuuQIMPLEDaztx1JXKe+Ag/Dt4O/ao+Efivy7e81eXQbt9o8rU4/LTceuJVJQAHuxX1x1xZX1jqdpHfabeQXVtKMxzQSB0cdMhhkHpVUDkdaqqqqBx3rgjg106GuCODVVVfGb4QaF8XfC8mm3kUcOrWqO2mXxGDBJjhWIBJjYgBhg+o5ANeJfDWv+CdfufD/iCxlsNSsJMOjcEHqGUjqCMEMOCMEV+zX+0CnxHs08F+KplTxLYwZjmY4GoRrgFh/00A5YdxlhxuC1VVVVVUcY60TnvVMQeAaqvH/w08HfEzSTpHizSkuAvMNwnyTQH1R+o+nQ9wa+Mf7N3i34WQy67ayrrHh5XwbyJdsluCcKJU7ZyBuGVz12kgHwD8SvGPwz1Yat4T1Z7dm4mt3+eGcejp0P14I7EV8IP2kfB3xPWHSL500XxA2FFlPJ8tw2OTC/8X+6cN9etEgDrVVX7WPwm/4Wp8KL0ada+brfh/df6dtXLPtH72If76A4HdlSvh/411b4c+NNH8b6G3+l6RdLMqFtolXo8ZPZXQsh9mNaDremeJtEsPEOi3S3FjqVslxbSj+JHUMp9uD07VX7ZHiltY+JsHhuKZzD4fsURoznCzS4kYj6oYR+H5fsTeEljsPEHjmeH55pE0+2cqBhVAeXB64JMXYD5e/aqrxp4z8PeAPD1z4m8TXwt7O2GABy8rn7qIv8THHA+pOACR8Zfjl4k+LeptFI8lhoMDg2mmq+RkZAkkI+85yfYDgdyfgv8BfEfxavlvX3ad4et5ALm/dSDJzykIxhn9T0XqecKfBXgbwz8PtBh8PeFtNjtbaIDe+0eZO+MGSRurMfU+wGAABTc8A1X/BRPxsLzxP4Y+HltIpTTbN9Qutu0/vJW2Ip7gqsbHHHEgPPGP8Agnh4IOpeN/EXj64jbytEsUs7ckMAZp2JJBHB2pEQQTx5inHQij0qqOMdaPPev2n/AIQv8I/ibd2thbqmha0WvNJKDCxox+eH28tjtA5+UoTyeP2Efi5JqWm33wh1u83S6chvNHLnkwlv3sQP+yzBwOTh37KMVVV8Wv2xP2Z/ghLLZfEX4waFZ6jCzpJplpI19do68FXgtw7xnPHzhRkHng4+Jv8AwWn8BadLcWPwj+EOr64UZkjv9bvUsIzj+MRRiRmU44BZDg84PFfEH/grD+1541S4ttD1/QPBttPlduiaUrSBCrKQJLgysCcg7l2kFQVK8ivGv7RPx7+I6TweOvjN4z1u2uN/mWl3rc72+GXawEO7YoK8EBQCDVvbXF5cRWlpBJPPM4SKKNCzOxOAqgckk4GK0L4CfG7xP5TaD8IvGF5FM5RJ00WcRbh1BkKhR+JFaV+xH+1NrMby2fwivoxG20i7vrW1OfYSyqSPcVpv/BPH9p++tRPdeFdJ0+QkjyLnWIGcY75jZ1wfrX/DuX9pf/oFeH//AAbL/hWq/wDBNX9ovTpI47O58I6mHUkva6nIoQ+h82JDn6A1/wAO5v2l/wDoFeH/APwbL/hWsfsKftT6NHcTyfC2S7gt32iSz1O0mMg3YDJGsvmEHg/dBA5IGDjxb8Gfi34Ejln8Y/DTxNpFvCSGubvS5UhyOuJduwj3BNeCviJ4/wDhtqY1r4e+Ntc8N3wYMbjStQktXYgEDJRhuGGYYPGGYdCa+Cn/AAVz/aG8Aywad8UrPTfiJpCKiF7hFsb6NV4ys0S7HO3OfMjZiQDuHOf2ef2+v2cP2j5YNF8M+K20DxNM2xPD/iAJaXUzdAISGaOcnkhY3L4GSq1VNzxR571VVVUen4VVVVVVNycZqqqqqmxjGaqpoYriJ4J40kjkUq6OuQwIwQR3GK/bD/4JVaN4va++In7MdpZaNrUjGW58KO629lc8MT9lY4WBydoEbFYueDGBg6ppfjj4XeL5NM1Wz1nwt4n0O4G+KVZLO7s5QAVI6MhwQQR2II4Nfsuf8FavE3hx7Hwb+0xaS69pa4iTxPYQKL2AEgA3ES4WZFB5ZAJMLnEjHn4d/FD4d/Fvw7F4r+GnjLS/EWlS4H2ixnD+W2M7XX70beqsAw7iqPWqqqqqboADVVXxs/aC+Ev7PXhabxV8UvF1rpkYjZrWxVhJeXzDgJBCDuck4GeFXOWZVBI/bI/b4+IH7U11J4U0q3l8M/D23nD2+jpLumvmQ/JLduOGOfmEQ+RDj77KHr9jv9jLxz+1Z4yRFjutG8D6bIrazrzRYG3P+ot8jDzNgjuqD5m/hVk+DvgvTvgvcfArw1pdvpnhl/D82iQ24j3qkMkLRsWGQZGO5mYk7nYsSckmtQsLzS7+50vULdoLqzmeGeJxgxupIZT7ggivhd40g+JHw08KfEC2eJk8SaLaagPKUqqmaFXIAJJGCxGCSRjBqqqqpunBpuv4VXxB8O7W/t2zj4bi4UDoezf0P4VVVVVVVVVVje3GnXkV9aPtlhbKn+YPsRxWmalb6vYxX9sflkXlSeVPcGqqqqqbpVU2MYzVVVYBFXllaahbm1vIEljbqrDp7j0PvWu+Brm1DXOkk3EQ5MR++v0/vfz+tEEHaRgjsaqqq0v72wffZXcsLd9jEZ+o71Z+PNatwEuBDcgDq67W/Mf4VbfESyZcXenzIcf8s2D8/jio/Gvh2QEtePHjoGhb+gNQ+INDuHEUWq25Y9AX25+maBDAFSCCOKqqqqqqqqqqqqqqqqqqqqqqqqqqqv8AgoB+1DF+zl8HJrHw7qbQ+OfFyyWehCIZe1UAeddnPAEasAvcyOnBAYj4QfCzxX8bviZ4d+FXgi0+0az4jvBbwA/diUAtJK3+xHGru3+yhr4c+A9A+F/gHw98OfC1qkGleG9NgsLRFXGUiQLuPqzYLMTkkkkkkk1VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVfFX4Y+EPjP8Otf+F3jvT/tmh+I7Nra6jAG9OhSSMsCFkR1V0bB2uintXx6+Cvi79nr4ra/8JvGsP8Ap+i3BWK5WNkjvYG5iuI8/wADqQfY5B5Br/gnJ+1DL8ffhGfCXi3U0n8aeCFjtbwuCHvLMjEFx/tNhWRyOdyhjjzBkYKgA1VVS9cA1VL6CqqqqqqqqqqqXpVVVVVDrXx80T+w/i14ggRMR3U4ukO3bu81Q7Hqf4iwz3Ir9lzWxq3wotrEyBn0m7mtiOc4LeYM8+kmB0HGO1VVVVDkCqqv+CxHx1updR8Mfs76JebbaCMa3rqoSDJIdyW0R9lXzXK8gl4zwVGf2APgl/wtH4xxeLdXs/N0HwTsvZd65WW7JP2eP8GUyH/rmAfvUqAjJpUJGBSxDoKEYAxQTPAFCMmhEMV5YxjFfGnSZNF+K3iiylzufUZLgZI6TfvR09nH9a8PXKKqoTyh5FeG/FCadD8rLnbjmvFGsRXrM4Iya1SVZrxipztG2v2P9Kmj0XxJrTEmK6uoLdRt43RKzHnPpKvH+PDRgjAp4wRjFMhHApkB6UR2r/gp78L49K8X+G/izp1qwj123bT9ScH5RPCAYmI67mjLLxxiEdD1/wCCOvxd/wCEa+MXiT4PajebbTxnpgu7CN5sD7babmKoh7tA8zMRziBcggZFVXjT42fDXwG0ltrXiSGW+iJVrKz/AH8wYdQwXhDz/GVrxl+2Fr99vtfA+gW+mxHgXV6fOlPTkIMKp6jnf+FeKfHXjDxtcfafFXiK81Fg25UlfEaHn7qDCr1PQCvCPwt8f+Owj+F/C95d27MVF0y+VACOv7x8Lkemc14S/Y21Gfy7jxx4pitl+Uta6anmPjuDI4AUjpwrD+vhL4EfCzwaqSad4Vtrq6QY+1X4+0yE+o3ZVT2yoH6mkRI0WONQqqMKoGAAPSqqqpeRjNVVVVUuDiqqqqqoYIwDVVVVVDr1qqqqqqGCMCqpeRjNVVVVVVVVUvXGaqaGG4he3niSWORSro65DAjBBB6gjtXi39nj4Q+MTJPfeEbezu5Mn7TpxNq4YkEsVT5GJx1ZT1PrXi/9iXUYnafwJ4vgnjySttqsZjZR2HmRghj/AMAUfnx4x+DHxN8CeZJ4i8I3sdtHuJu4E8+AKP4i6ZCjHPzYPtwa8OeMPFXhC5N34W8RahpcrfeNrcNGH9mAOGHA4IPSvB/7Znj7SCkHi/SrDX4AfmlQfZJ+nqoKe+Ng78+ngv8Aai+Eni9EhutbOgXhHzQaqBCueM4lBKYye5BPXAqCaG4iSe3lSSKRQyOjZDAjggjqCKqqqqoHtmqBx0NAgjg1VVXx8+Bun/GDQo5rOSO08RaYjfYLlhhZAeTDJjnaSOD/AAkkjgsD/wAT3wlrv/L3per6Vce8UtvKh/MEEV+z98cLD4t+HxY37eT4l0uFft8RAAnHA89MADDHqv8ACTjoQTVVVVVHAHWic8CqY9gaqqrUdOsNX0650nVLSK6s72F4LiCVdySxupVlYdwQSCPev2ivhFqnwI8VLNbxS3ng7WJXOmXBYu9qw5NvIx6kZ+Uk5ZRnJKsBaXkVwiXVlcBgDlXQ4KkfqDXwe/ay1zwutv4f+Ifn6xpSAJFeqd1zbj/aJ/1qgevze56VoPiDRPFOk2+ueHdUt9QsLpd0U8D7lPqD6EdCDgg8EA1VV+1T8Ln+F/xf1S1tLRYtI1onUNMKLhQkhO+MdhscMuP7u08ZFfsEfFV9a8Oah8J9UnL3Ohg3mnFjnNq7gSL/AMAkYH/trjtTukaGSR1VVBLMxwAB1NeOfEL+LvGWt+JmYkanfzToD/CjOSo/BcD8K+APhZvCPwh8N6ZKm24uLX7ZOCpVg8xMmGBwQVVlU/7tVXxI+JnhX4W6Cdc8TXhBkJS1tYhuluXAztRfyyTgDIyeRn4n/FHxN8VfEMmta7cGOBSVs7GNyYrVOwA7t6seSfQYA+Af7NN746e18X+ObaS28Nkb4LYsUlv/AE6cpGf73BI+7wdw0/TrDSbC30zTLSK1tLWMRwQRIFWNQMAADoMVVHBPFV+0d4xXx58bvF/iGKVngOotbWxIIzFABChAPIBEYbHHXnnNfsS+CT4Q+Aul31xEyXfiS5l1KVWUZCsRHFggnIMcaOOmN5461TdMA1VHpVftQfCAfF34Z3Nrp0O7XdF33ul7Uy0rqp3wj/roBgf7QQnpXgrxdrHgLxZpfjHQLgxX2lXCzREMQHHRkbH8LKWUjuGI714N8V6T468KaV4v0OXfZatapPFk8rkcqf8AaU5UjsQa1XVdK0LTbjWNb1O00+ws4zJcXV3MsMUKDqzuxAUD1JxX7RH/AAVx+C/w2ln8PfBnTW+IutRMUe8SU22mwkdcS4LTH/cXYez18cP28/2nvj6lxp/iz4i3GlaJcNltE0AGwtSMkhW2kySqMjiV3+6p6gGvC/g3xb441NNF8G+GdU1y/fpb6faPcPj1IQHAwCcngAE14B/4JyftA+LkhuvEkWj+ErWRN5GoXXnT4IyuIod2D0yGZSOc8jFeCv8Agl78K9JCzeOfHXiDxDMmf3dpHHp8LZ3feX94/AK4w45U5yDgeFP2N/2aPCCBdM+D+h3bckvqqNqJYkAE4uGcDoOAAAckAZNaL4X0Dw7bG08PaFp+mQnGYrO1SBTgYHCgDgcV9nJHH8qFvkYz+dfZ/ejb8cUbc/5FNAeuBTQkDAFGMjgGviJ+zJ8CvihDMPFvw10d7uZGU39nALS5UnB3ebFtZiCARuyOoxhiD8Xv+CYl7ax3GrfBPxi16FXKaNrYVZCe4S5QBTngBWRcd3NeMfAnjr4X+IDofjTw5qWgarbtvRLmIxk4PDxsOGGRwykj0Nfskf8ABU74k/CSex8FfHW5v/G/g1P3aX7ES6ppy4ONsjEfaE3EZWVtwH3WwoQ+APiF4J+KnhOx8c/DzxLZa9oWpKTbXto+5GwSGBBwVYEEFWAIIIIFN7dhVVVVVN0qqqqqqbBPFVVVVU3Sqqj0/Cv2gv2W/g1+01oC6N8T/Dfm3dvGUsdZsiIb6wycnypSCMZ/gcMh6lc1+0t/wTP+OvwJ87xB4Qs5fiF4TTczXuk2jfa7RRzme1BZgMZJePeoCksU4B+Fvxj+KPwT8Qr4q+FXjnVPDeo/KJGtJf3dwq8hZomzHMgJztkVlzzivgV/wWNB8rRv2ivAf9xF1vwzH9AWmtpX+rM0b+u2PoK+FX7QvwS+ONmLz4VfEzQ/EDbdz2sM/l3UQ55e2kCyxg4ONyDODVVVVVNg4ANanqul6LYyanrOpWthZw48y4uplijTJAGWYgDJIHPcivip/wAFC/2TfhQlxBffFG18R6jBuA0/w2n9oyOyqCVEiHyVPIHzyLzkfwtj42f8FhPiJ4iSbSPgT4KtfCdudyrq2rFL68YFeGSHHlREEnhvNBwOmSK8WeMvH/xY8Vtr/jLxFrPinxDqUixLPeTvdTyEnCRoDkgZOFReBnAA6V+yf/wSt8c+PLqx8bftE2914U8Njy54tBzs1C/XIOyYdbVCOCD+9GSMIfmrwd4N8LfD7wtpngrwVodro+iaPbiCysrZNqRIP1JJJJY5LEkkkkmq/b9+GM/ws/ax8e6WLTybLW7863YMMbXiu/3rbR2CytKmOPuccYr/AIJPfFqLx3+za3gC7v2m1TwBqL2rJI+51tJ2aaA9c7cmZFzwBFgcDiqqqpulNgnAqpoYriJoJ41kjdSrKwyCK8VeG5dAvSYlZrOU/uZCOn+yfcfrVVVVVVVVVeCdfXSr77DcuBbXRALE42P2P0PQ/ge1VVVVU3aqpu1VVUenFVWu+FdP1oGZQILrHEqr97/eHf69a1XRtQ0ebyb2HAP3HXlX+h/p1qqqqqqsdY1TTcCxvpYwP4M5X/vk8Vp/xCmTCapZrIM/fhO0gfQ8H8xWm67pWrDbZXiNJj/Vt8rfkaqqqqqqqqqqqqqqqqqqqqqqqqqv2m/2p/hr+y14OTxD41nlvtUv96aRodo6/aL+QDk8nCRKSN8hyFyMBmKqfjl8bPG/7QfxH1H4l+PbuJ9QvcRw28C7YbO3XPlwRjqFUE8nJJJJJJJr/glR+xfP8HvBg+P3xI0vyfGPi+zC6TY3EQ8zSdPY5DNkZWWYBWI6qm1Tgs6iqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv+CsP7J7fGb4TL8avBumJL4v+HlrI90kUQ83UNKzulTPGTBl5lHoZgAWcA/s2fHLW/2dfjFoHxQ0dGnhspfJ1OzU4+2WUmBNF1A3FfmUngOqEg4xXhLxRofjjwtpPjHwzfJeaVrdlFeWU69JIpFDKfY4IyOx4qqql6/hVUvX8KqqqqquCODVVVVVLyMVVVVVQ6iv2vtDW18V6H4gRUUahYvAwAAJaF8kn1OJVHPYD0r9jvXSmoeIvDLuCJYYruJc4I2Eo56ZOd6d+Me5qqqqodKqru6tbG1mvb25it7e3jaSaaVwiRooyzMx4AABJJ4FftC/FW8+OHxt8ZfFS7ZtviDVZJbVH6xWqYjt4zwMlIUjXOBnbmv2GPhRF8MP2fdDmubRY9W8VKNYvWK/NiVR5Kk5PAhEfy8YLNwCTSx5pYxilTIwKWLNCMdBQTPQUIyRQiyOBX7V3wsutUs4fiRoluZJdOg8nU4kXkwg5WUADJ2liGPPy7TwFJqCeS3kEkTYI7HvUHiIomHVwfbmr3XJbhSkQK5/ibrWh6JqniXWLPQdFtWub6+lEUMa92PcnsB1JPAAJPAr4c+CLb4f+C9N8KwSrM9pGTPMq7fNlYlnb1xk4GewA7U8QPSmTHFPGCOKeP0p0BGO9ftz/D6Px/8As2+JgsTPd+G1XWrUgfdMGfNJ9vIaYduoPsf2fPiU/wAHfjf4H+Jgk2w+H9bt7i75YbrbcFnX5QTzEzjgE89D0rW9e0Tw3p0mra/qtrp9nF96a5lEa57AZ6k44A5PavG37XXhDSC9n4M0yfXJ1yPtEuYIAc9sje3fsB059PGvxz+JfjtZLfVfEMlrZSHmysQYIsehwdzD2YnoK0Dwv4i8V3n2Dw3ol5qU4+8lvCX2+7EcKODycV4O/ZB8XamEufGer2uiwnrbwEXE/wBCQdg7chm+leDv2ffhb4MCy2vh5NTulYMLnU8XDgjoQpARSDzkKD78DCIkaLHGoVVGFUDAA9q4I4NVVVVUvIxmqqqqqXp9KqqqqqbxL4bgleCfxBpscsbFXR7tFZWHBBGeD7U3xS+GMbNHJ8RvC6spwVbV4AQf++6f47fB9GKH4g6SSpwcSEj8CBzV5+0j8ErKb7PN47gZgM5hs7iVf++kjI/Wv+GnPgb/ANDx/wCUy7/+NVcftYfBuGZ4otW1GdVOBJHYOFb3G7B/MCv+GtPg9/z/AGqf+ALf41/w1p8Hv+f7VP8AwBb/ABr/AIa0+D3/AD/ap/4At/jX/DWnwe/5/tU/8AW/xqx/ar+C127LP4gvLMKMhp9PmIb2GxWP54r/AIae+Bn/AEPP/lMu/wD41Vl+0V8FtQRnt/HtmoQ4IngmhP4B0BP4VafG74RX0wt4PiJoisQSDNdCJeP9p8D9atfiX8OL6dLSx+IHhu4mfOyOLVYXZsDJwA2egNWWt6LqUpg07V7K6lVdxSG4V2A4GcAnjkfnVUvpmqpfTNVVVQwRxVVVVVfQ1wRxVVVeMPgZ8K/HJebXPCFmt05ybu0U20pOMZLJjdx/eyOnoK8afsT38Jkufh/4rjuUAJW01Rdj8DoJUGGJ56qoHHPceM/hZ8QfAErp4r8LXtnEmP8ASQnmQHPAxKmUz04zkZGQK8I/Efx14DmE3hHxTfacM5MKSbomPq0TZRup6g9a8E/tsahb+XafEHwul2g4a80xvLkHXrE52senRl+leB/jH8N/iIqx+GPFFrLdEZNnOfJnHOOEbBb6rkcj1qqqqoHPBNV06GgQRjNVVV+0t8AU+JGmnxd4Vto4/EunxHzI1TH9oxgDCE/89FA+U987Txgr4b8Sa/4J1+28QeH76Ww1KwkyjqMEHoVYHqCMgqeCMg18H/i1oXxf8LjW9LQ217bER6hZOcm3kI7H+JTgkN374IIFVVVRIAxmic96piAMA80ee9VVVXjnwT4e+IvhXUPBviiz+0adqMRSQDhoz1V0POGUgEHsQOtfFf4Z+LPgT47uPC+quChHnWNyhzHeW5YhXx2PBBU8gg44wTo+vQaqRblDHc7SdnUNgZJH4DOK+HnxQ8Z/C/VP7T8J6qYlkI+0WkoLwXIHZ0z9RuGGAJwRk18Ivj/4O+LEMenwyf2br6x7pdNnblsDkxN/y0GBnsQOoxzVV+2T8LIviJ8I7rWbODdrHhPff2jBcs8OB58f0KAP05aJRwM18IviLf8Awp+Imi+OLAPIthOBdQKcefbt8ssfpkoTjPAbae1fGjxxZ6N8FNa8VaTepImoacsdhKODJ9owiMoJByFfd7AZwcYrwP4dfxb4y0TwygJGp38MDkfwozgM34Lk/hUUMMESQW8SRxxqFREXaFAGAAB0GO1V8UPif4b+FXht9e16bzJnyllZI2JLqQD7q+gHG5ugHqSAfiD8QfEfxL8Rz+JfEt35kz/LDCmRHbRjpGg7AfmTknJJNfs7/s0Ra1Bb+OviTpxNi4EmnaXKCvnjqJZR12eiH73U/LwyIkaLHGqqqjCqBgAD0FVR6cGq+KXiz/hBPhv4m8YK+2XSdKuJ4OM5lCHyx0bq+0ZIIGcnjNaVpl3rWqWejafH5l1fXCQQJ/ed2CqPzIrw5odn4Z8O6X4a08YtdJsobSAYx8kaBF4+iiqb0BqqbpgGqr9r74Qv8NfiXPrumW6poXimSS7tAg4hm4M0XoPmbcAOArgD7pr4Y/tu2P7L3wl1+38U+FdV8SW9pJ5+jW9nKsYSaQ7THK7Z8qJm2neFcgs3ytmv2kP2wfjf+1HrBuviL4l8nRoJC1j4f04GCxtOnITJMj8Z3yFmGSAQuFHwb/Zl+Mfx1uEbwN4WkXS9+2XWb8m3sosHB/eEEyEHqsYdhnkYr4Sf8E2/hD4RhttR+JN7eeM9VTDPCzNaWKNwQBGh3vg5GXfaw6oOleHfCnhzwppqaP4V8P6bo9gh3La6faJbRKcAZCIAOgHbsKWAkc0sAxwKWAngChB2zQgGMYr7OOw/SvIHoPyowA9v0poBjAFNBngGmgPpTwA8Yp4OOKeHHQV8SvhR8P8A4u+Hn8M/EPwvZ6xZHJiMqYlt2OPnikGHjbgDKkZHByCRX7UX7E/jH4Dtc+LfC73HiHwMH/4/SF+0afuOFW4VcZGSAJVAUnqEJUH9kz9rn4i/sn+O4td8N3MupeGr6QLrnh6WcrBfR8AuvURzKB8sgGR0OVJU/B74x/D/AOPXw/074lfDXW01HSNRXBBAWa1lAG+CZMnZKuRlfcEEqysaqqqqbpgGqqqqqo4PSqqqqqboADVVTdKqv2if2Cf2eP2jftWr694Y/wCEe8VT5b/hIdDVbe4kc950wY7jJxkupfAwrrXxs/4JU/tH/DFLjVPAsVl8RdHhbh9KXyb0JkgM1q5JJ+78sbSEbvQEi6tPE/gnX2tb221TQNb02Qbo5Uktbm2fGRkHDIcEenBr4W/8FEP2tPhSlrZ2PxQn8R6ZasD9g8SRDUVkAOdpmbE4XthZVwOBjAx4I/4LReIoIobf4k/A7Tb6QsBNd6Hq72oUY5KwSpLk9ODKPrXh3/gsN+zTqjWtvr3hLx9o0sqnz5WsLa4ghIBONyT+YwOAARH1PIAya/4ewfshf9BrxR/4I3/xq7/4KzfsjW9rNcQ6h4tupIo2ZIItEIeUgZCqWdVyeg3EDJ5IHNeKf+CzfwdtIC3gn4ReMtWm8skJqs9tp6788DdG85xjvj8K+IP/AAWK+O2vpcWnw88BeFPCdvMuI5rjzdSuoDkEFXYxxE4BHzREc9q8Vft3/tfeMVlTV/j54lg85VVv7LePTCApBG02qR7TxyRgkcHIJrxV458beO7wah438Y634hulZmE+q6hLduC2Nx3SMxydq59cD0r4bfs+/G74wuo+Gfwt8R6/C3/L1bWLi2Xkr807YjXlWHLDlW9DXwf/AOCPnxc8RtBqPxl8Z6R4PsmVWew08/2heg4OVYgiFCDgZV5B146Z+AX7H/wD/ZttY3+HHguJtaEeybxBqZF1qE2Rhv3pAEQYYBSJUQ4BK55qqr/gsN8EP7e8BeGvj1oum77zwxP/AGVrM0ceW+xTtmB3bPCRzkoB/eu6/wCCbnx6i+CP7SGlafreotb+G/HCro2okviOOV2H2aZgeMLLhSxI2pK5zgEGqqqpulN1qq1jSrbWdPl0+5Aw4+VsZKMOjD/P861DT7nTLySxu02yxHBHY+hHsaqqqqqqqqrwXr/9r6f9luZAbq1ADZ6uvZv6H8PWqqqqm7VVNg4ANVVURkdaqqu7O1voGtruBJY26qwrxD4MudNDXenF7i2Ayy4y8f1x1Hv/APrqqqqqqoEggqcEdK0rxlq+nERzSfa4R/BKfmH0br+ea0nxZpOq7YvN+zzn/lnKcZPsehqqqqqqqqqqqqqqqqqq1nxH4e8ORRz+Ide07TI5mKxve3SQByOoBYjJxX/Cz/hp/wBFE8M/+DeD/wCKr/hZ/wANP+iieGf/AAbwf/FVVVVeOfjF8JvhmjN8QviX4Y8OusZcRalqsNvK4C5+SNmDOcDgKCT2Br9pD/gq38OfDGkTaD+ztH/wlmu3EbINYu7eW3srA9NwjkVXnbrgYVOh3MMqfGHjLx78XvGc/ifxhrOo+JfEuszIjTSgyTTvwqIiqOB0VUUADgAV/wAE/P8Agl/H4XktfjN+0/4ZtLvU2jWTQ/CV7Es0doGUHz7yNgQ0ozhYWBEZBLZfASqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqmhhuIXt54kljkUq6OuQwIwQQeoI7V/wAFFP2Wv+GYvj7e2/h3TJYPBPi/zNT8Ot5eI4AWHn2ikAD9y7ABeSI3iySTk/8ABJf9pS5vrfUP2afFmo+YbSN9R8LySuSwjzm4tRk4wCfNUAZ5mycAAVVUOuAaqh1wKqqqqqh0H0qqqqqlqqqqqq/at0JdS+GkerIp8zSL+OQsAThHzGR14+Zk556Y7mv2cNYOkfF3R0LYjv1ltZOCT80ZK9D/AH1Tr2qqqqpelVX7f3xJf4X/ALJPxA1e1kxfatYDRrQZYEteMIHIYA7SsTyuCccoBkEivg14Bk+KXxV8K/D5GZU1vVIYJ3UZMcG7Mrgf7MYc9unUVZ2dvZ28VpaW8cMECBIoo0CqigYAAHAAAAx2pUAAFLHnrSxgDApUyMCliyKEYxigmeAK2EjrTwpIhjkUMrDBUjIIr4mfsmaJ4ju5tZ8B31volzLlnsZIz9mZvVdvMQz2AIHYCr79l340Wlw0Fv4Yt71F6TQahAFb6B3VvzFeHP2S/ihq0y/26NP0ODJ3ma4E8mP9lYyQfxYV8MPgv4R+FtmRpcH2zVJV2z6lcIPNYf3V/uJ/sjrgZJwKaLjimj4xTICMCnjx0pkB4FSR56V4y13wlp+mXOl+KL6Pyb2B4pLYMxkkRlIIAX5hkEjPH1FeC/2M/DGm3DXXjjXptaCv+7trZTbRlQf4yCWOR2BXHqetXuqanqa26ajqV1di0hWG3E8zSeVGowqLknaoAAAHArwR+z38TfHAjuLfRf7LsXz/AKXqRMK8eiYLtnsQuPcV4K/ZM8BaCI7rxVdXHiG7U5KNm3tweo+RTuOPdiD6VpulaXo1mmnaPp1rY2sf3ILaFYkX6KoAFVVDBAwaqqqqpfQGqqqqr3ULDTYhPqN9b2sbNtDzyBATjpknrgGtT+N3wk0aMyXnxB0aQBd2LW4Fyfyi3HPt1rVf2tPhBp5cWd3quqbSADa2JXdkckeaU6dOcfjWpftq6DFuGj+BL+6xIQv2m9SDKc8nar4PTjpyeeOdS/bR8ZS7v7H8IaLbfvCV+0ySz4Tng7SmT056deOeNS/ax+MN8JRa6hpmn+YflNtYq3l+y+Zv+nOetX37QHxk1GRpLjx9fqWXaRAscAx7BFAB9+tXvxE+IOpQiDUfHfiG6jVtwSfVJnAbGM4LHnBP51e6hf6lKJ9Rvp7qRV2h55C5A9Mk9OT+dVF4a8R3EST2+galJHIoZHS0chgRwQQORirf4ZfEm7gS4tfh74lmikGUkj0mdlYeoIXmrL4LfFrUEZ7f4d66oQ4InsmhP4B8E/hX/ChvjD/0T/VP++B/jVv+zT8brmBJ4vA0gVxkCS+to2H1VpAR+Ir/AIZi+OP/AEI//lStf/jtf8MxfHH/AKEf/wAqVr/8dr/hmL44/wDQj/8AlStf/jtf8MxfHH/oR/8AypWv/wAdq+/Zz+NWniPz/Ad0/mZ2+RcQz4xjrsc469+tf8KF+MP/AET7Vf8Avgf41efB74rWU32eb4c+I2YDOYdNllX/AL6QEfrV78OfiFpsIn1HwH4htY2baHn0uaMFsdMleuAePar3RNZ02ET6jpF7axs20PPbtGC2OmSOuAePaqsdY1fTEaPTdVvLRXOWWCdowx9Tg1ZfFL4l6cIUsviF4jiSAgxxDU5igwc427sEexGKsf2ivjTp7s9v4+u2LjBE8EUw/AOhx+Fad+158X7Ixm5m0bUNibSLmx27zj7x8tl578YHtWm/tseK4liGr+CtJuSpPm/Zp5INwzxt3b9vGOua0v8Aba8Ny+X/AGz4G1K1y2H+y3aXG0eo3BMn24+taT+1v8GtSCG81HVNLLAki7sGbbg9D5W/r14z74rTPjh8IdXQPafEXQ0BQOBdXQtjg+0m059uo71Zahp+pwmfTb63uo1baXhlDgNgcZBPOCOPeqpcEYzVVVVVUuCMZqqqqp0SRDHIoZWBBVhkEV41/Zo+EnjRJJf+EeXRr1zkXWlEQEEeseDGc9/lz7jOa8bfsceP9DD3PhDULTxFbryIuLWft/CzFT36Nk46c4rWND17wvqR07XdKvdLvocN5NzC0Mi+jAHBxxwa8B/tL/FTwN5Fr/bX9tafDhfsmp5lwo7LJnevHTkgYHGBivAX7Xvw78TmKx8URT+G75yF3TnzbdiR2kAyvP8AeUAcc9cafqOn6vZQ6lpV/b3tpcLuhnt5RJHIPVWUkEfSqqqB7E1wRwaoHtmsg8A1VV+1V8AV1CK6+Kfgy0VbmFN+r2MMRzOM83CgfxAHL9iBu6g7vhp8Rde+F/iu18UaFKWMZ2XNqXKpdRH70bfXqDg4IB7V4F8b6B8RPC9l4q8O3Sy210g3xkjfbyYG6JwOjKTg/gRkEE1VEgDrROeBR571RIxgGjz3qqqqqvj18GNE+N/gebw5fBINTtN02k3p4NvcbSAGIBPltwHXHIweqqR4g0HxD4D8UXnh/WbefTtY0a6McihirRSIeGVh26MrDgggg4INeAfCOgftHeFbi+8IPZ6N8QNGi3appW0x22ppwFni7RM3IZR8u8jhAwNappXiDwdrj6fqdrdaXqunTcq2Y5InU8MpHuMhgcdCDXwY/a3a3WDw18V5neJVCQayiFmXAwBOo5bp98AnPUHJYWl3aX9tFeWF1Dc28y7o5YnDq4PQgjgipI45Y2ikRXRgQysMgg9QRX7Q/wAMn+E/xY1rwvDamHTZZPtelknIa1kJKgHuFIaM+6Hr1rTvjVe+I/2ftL+FN9M73ei6rlZCD81mqExIT04d2AHYIv4/sieHm1n4uw6m0O6LRLGa5LMDgOwESjpjP7wkA4+6T2qviP8AELQPhn4WuvE2u3CYjUra22/a91Ng7Y14PJxyccDJPAr4g/EHxH8S/Ec/iXxLd+ZM/wAsMKZEdtGOkaDsB+ZOSckk1+zf+zf/AGj9k+IXxC0//ROJdM0yZP8AXdxLKp/g7qp+91Py4DVVU3TANV+3x4pOifA5dAhkXzPEWrQWzoSAfKjzMW6HgPFGOMfeHOMg/sheEf8AhMP2gPC8UkW+30mV9SmOM7fIUtGf+/vlD8aqm5PBqqbk4Bqq/wCCk/7Unwe8BaJpnwZ1ErrHjK8vra7dLeQY0KDPzTTHn5njZgsXBIbecALv1fS9P8Q6PdaRfoJrS/gaKQA9VYYyD2PcHscGv2Xf2AfBUVlZfET4q6ja+KGmZ2stJgU/ZFAYqGmzgyNkHKcKDwd9av4k8N+Bm8P+G1hgtzqNzFYafZwIEWJOEG1VGFRflUAADkDgdEhzSQ5GAKWAHrSw5GAKWAYxQgBGAKEBIyB+lfZyRwP0r7OfT9KNucdP0owE9v0poBjpTQDoKeA4xingB6daeH1FSQccVeWVvd28tpd28c0MyFJYpEDK6kYKkHggg4INftv/ALGsPwye5+LvwvsdvhW4mH9qaXGp/wCJVI5wJI/+mDMQNv8AAxAHykBf2Mf2s/E/7KPxQh12CS4vvCWsMlv4i0gSkLNFnidByBNFkspxyNyZAckeFvFHh/xv4a0rxh4U1SLUdG1qzjvLC7iBCzQyKGRgCARkEcEAjoQDVVVVTYOADVVVVVHGKqqqqqm54zVVTdMA1VUcY618QvhJ8L/izpy6V8S/AGg+JreP/VLqVgk7QnnmNyNyHk8qQeT618R/+CSf7L/jCWa98HzeJfBNy7bki0++FzbAnbnMdwHfGFbAWRQC56gBR4s/4IwfE6zDf8IL8afC+r427f7W0+fTc/3s+WbjGO3r7V4h/wCCVH7YeipO2m+FvD2vGGYxoun67ChmXOPMX7QYsLjnDYbB6Z4r/h2b+27/ANES/wDLk0v/AOSa/wCHZv7bv/REv/Lk0v8A+Sa0X/gkV+1fqsskd9d+BdHWNQVe91iVhIfQeTDIc/UAV4L/AOCLlqrrP8RfjrLIvIa00XRgh6jBE0sh/h3DHl8Eg5OMHwb/AMEuv2OvCUEC6h4E1TxRdW8iut1rWszlmKnPzRwNFEwPdShB6YxkV4L/AGaP2e/h26z+C/gt4N0u5XOLqPR4WuMEhsecyl8ZVTjdgECqqqqviZ4A0L4qfD3xD8OPEqM2meI9OmsrgpwyB1IDr/tKcMPcCvi78L/E3wW+JfiL4WeMIkTVfDl61tKyAhJlwGjmTODskjZJFyAdrjIHSv8Agn3+09F+0d8ErODxFrSXXjnwoq2WvI7YlnHIhuiO4kQcn++r9OKqqqmxjFHk9aqq8c+Gzqtn/aNnGDdWynIA5kTuPqOo/Gqqqqqqqqq0jU59H1CG+t2OUPzL/eXuD+FWl1Bf2sV5bOGjlQMp/wA96qqqm6/hVU3X8Kqqo/dwKqqqvEfguC+DXmkqsVweWj6K/wDgf0/nU0MtvK0E8bRyIcMrDBBqqqqqqqtI8W6rpRWMy/aYB/yykPQex6j+XtWkeJtK1kCOCUxz45hkGD+HY1VVVVVVVVVeOv2tf2Z/hukv/CXfG7wpBNB/rbW1v1vbhMqGGYLffIMggj5ec8V4y/4K0/syeH3mt/DOmeMPFEqbhDLa6cltA5BwCWnkSRQRyP3ZOOoB4rxL/wAFlx/pNv4O+An937Ndan4h+hbfDHB/vAYk9D/s1rP/AAWF+PU9yr+H/hv4AsrcRgNHeQ3d05fJyQyzxgDGBjaehOecDU/+Cr37V1/eyXVrL4P06J8bba20YtHHgAHBkkd+SM8seScYGAP+Hqf7W3/QZ8Mf+CRP8a1r/gp/+2HqkUcdj480nSGjYlnstBtWL+x85JBj6AGtd/ba/ay8RR3UV/8AHvxXELyTfIbG7FkVO7d8hgCGMZ7JgY4xjita+PHxx8S+T/wkXxn8dar9m3eT9t8RXU/l7sbtu+Q4ztGcdcD0rWde1zxFcre+INavtTuEjEaTXly8zqgJIUFiSBkk49z61VVWlfGn4x6FdjUND+LXjLT7pVKie0165hcA9RuVwcGvDP7d/wC154SRE0r47a/OEhMQ/tNYdSO3IOSblJMtwPmPzYyM4JrX/wDgpr+2LrW1bT4j2Gjp5ZR0sNCtPnz33SxuwPupFeK/2nP2ifG+5PE/xv8AGt7C23Nv/bU0cOV6HykYJn3xmtK0jVddv4tK0PTLvUL2fPlW1rA0sj4BJ2qoJOACTx0BNfCn/gnF+2D8XJbKXTfhJe+HdMu2XdqfiZxpkcCNuAkaKT9+y/Kf9XE5wVOMMCf2RP8AgnZ8GP2WIbXxNPCnjH4gKrb/ABFqFuALQsMFbOEkiAbeN+WkO5xuCtsFVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVft0/syWn7U3wB1fwbY2UUnivSM6l4YndxGVvY1I8osSAElQtGdx2gsrnlAR4Q8U+K/hR4+0rxfoTy6b4h8LamlxCs8bKYp4XyUkQ4OMgqyHqCQepr4F/FrQ/jp8JfDXxT0AosOuWSyTQBsm2uF+WaE+6SKy57gA96qqGM9aqh1qqqqqqHIFVVVVUvX8Kqqqqqviroo8Q/DbxJpKxiR5NOleJCBzIg3p14HzKvPbrXh3V38P+INL16Jdz6beRXKrjqY3DD+VRSxTxJNBKkkcihkdTkMD0II6jFVVVS8jFVX/BZ/x41t4T+HHwvt5kZdQ1G71i6QEZQ28awwkjbnDfaZ+jAfIcg8Ef8ABMvwN/wkXx5vPF88O6Dwpo0skb4+7cTkQqPbMbT/AJUqADApI80qDoKWPNKgHAoR54NLFkYAoRZGKEY9a8taMakYFGMEYFNECOKaLjGKaMjgUyAjGKePjinTjFeKviP4S8LeZBeaks95HwbW3+dwfQ9l/EivE/xn8Ta2r22lbNKtmJAMJJlI937f8BAPvUNve6lc+VbQTXVxK2QqKXdyfYck1+0F8cm+BWtp4Nu/Ctzc6/Lax3QjmcRwxxPnaSwyWPyngDjuQQRX/BLzxjpfxk+E+v8AjTxRomjv4r0jxLLa+bFCS0VsYYZISoYnZ8zSqCDk7OSaqqqqGCMA1VVVVQ69arW/F3hTw2pPiDxNpem47Xd2kRPBIADEEkgHj2rXf2ovg9o25LfXLrVZEzlLG0c8jHRn2qc57HHBrXP20bJd8fhrwNNLydk19diPAz3RA3Uf7XB9a1z9rX4r6pvTS30rR1JOw21p5jgZyMmUuCccZwB7CtY+MfxT17euo+PtZKyb98cFyYEYN94FY9oI9sYHarq6ur6drq8uZbiZ8bpJXLMcDAyTz0AFaV4G8aa7t/sXwjrN8HUMrW9jJINp6NkDGD69K0v9m34z6sVMfg2S2j37Ge6uYotvHXaW3Ec9QDWmfscfEq7EUmpazoFijZ3r50ksiYzjgJtOcD+LofwrTP2J8iKTWviF6+ZDa6d9cYdpPoTlfUe9ad+xt8NrZYn1HXPEF5IhO9RNFFG/PAwELDjH8X+FWX7LPwVtEZZ/DVzeljkNPqE4K+w2Mv61Z/A34RWEscsHw+0dmiGFE0HnA8Y5D5B/H61ZeAvAumo0em+C9CtVc5ZYNNijBPqcLVraWljbra2VtFbwpnbHEgRVycnAHA5JoYyKqqqqql5GM1VVV9pmm6mix6lp9tdqhyqzwrIFPqMg4q9+HPw91OUT6j4E8O3UqrtDz6XC7AemSp45PHvV98A/g5qCzLP8P9LXzyS3kK0OMnPy7CNv4YxV9+yn8Frt1a38P3dmFGCsGoTEN7nezH8q1D9i34dzJnTPEviC2kL5PmvFKoXngAIp9OST/WtR/YiuFV30j4ixuS/7uK50wphc93WQ5OP9kZ9q1T9jX4n2m99O1XQL9A2EVbiSORh6kMm0f99GtT/Zs+NWlsRJ4ImuE3lVe1uYpg2O+FcsAfcCtT8C+NtEQyaz4O1uwUIXLXWnSxAKOrfMo4HrVvcXFpMlzaTyQzRnKSRsVZT6gjkVo/xh+KeghU0v4ga2iIAEilvGmRQDnhHJA/KtI/a5+MemgC9vtK1bGf8Aj8sFXOf+uJTpWjftusG2eIPAAIySJLO+xgYGBsZeec87vwrRv2vPg5qhxfXeraRgkZvbEt0A5/cmTr0/CtD+KHw48SkR6H440S7lOcQreoJOoGdhIbGSBnHeqqqqhgHrVVVVVVWveGfDviuwOl+JdEsdUtTnEV1AsgUkYyufun3GCK8d/sX+EdVV7vwDrNxolxyRaXRNzbnjgBifMTnqSX+lePfgZ8TPh1JM+u+HZZrGLJ/tCyBntyoz8xYDKDg/fCn2rwl498ZeA7w33hDxHeaZIxBdYnykmOm9DlW/EGvAX7ar5isfiR4dUjIU3+mDGBjGWiY888kq3rhe1eEvHvg3x5Zm+8IeI7PU41ALrE+Hjz03ocMv4gVVUDjgGgQeAaqsnoDW4HisgjrWR6iv2oPgCfB99L8QfBtlI2iXsrPqFtGg26fKx+8oHSJiTgYwp4zgqB8CfjLqPwh8UCeRnm0LUWRNTtlQMxUZxInIw65PfBBIPYjSdW03XdLtNa0i7S5sr6FZreZAQHRhkHnkcHoeaLDtRJIwDR571RIA60TkYFVVVVVTEYwDVftefs7t8VNAHjbwlZ7vFWiW5HkopLahbrk+UBn765Yrxk5K9xjwJ448RfDfxXp/jLwreLBqOnSFoy67kdSCGR17qykgjg88EHBFlZfCv9rX4a2HieSzMc2DF50LgXWmXAALwlsfMOQcMMMrK2ASCPix8EPGXwlvR/a0H23Spm22+p26HynP91v+eb/7J64OCcGvhF8e/GHwnuo7SCdtS0Fm/faXPJ8q5OS0Tc+W3J6cHPIJwR8PviT4S+J2iDW/Cmo+cq4FxbSgLNbMf4ZEycHg8jIODgmv26Phc/i/4b2/jnS7RZNR8JyM85Vfme0fAk6ddrBH56DeeMnPhO3EWmGYjmaQkfQcf0NfsSaBFF4d8SeKGjBkur2OzRjglREm9gOMjPmrnnBwPSvF3ivRPBHh298UeIbrybKxj3OQMs56KijuzHAA9TXxZ+KOt/FjxXNr+pFoLSPMdhYiTclrFxwPVmxlm7n0AAH7Mn7PkOveR8RPHenM1gjB9LsZ0+W6PaZweqDspGG68r96qqqbBOAar/got4nN3428K+Do5WKabpkl66jGN08mwZ5zkCDoRwGGOpr/AIJzeFVm17xh42ljYG0s4NOgYg4bzXMkgHOOPJi6g/eHI5zRxijz3qqPPQ1X7Xn7SeifssfBXVfiTex295rEjCz0DTZmYC9vXB2KdvOxFDSPyPlQgEMy51rWvG3xa8d3GtazeX3iLxV4p1HdJIV3zXl1M4AVVUdSxCqqgAcAAAAUfhB4z+EPgnwnpPja6judQm00faHiO5YZVJzBuBIYohjUsOGIJHFfs+fEzTvDVrq3h/xJe+TYRRNe28jAnYygCRABySw2kAdw3dqtvGmo+P8A4z6Br1+WRH1u0S1tyxIgiEy7VHv3J4ySTxmkhyOaSHIwBXjL44/BX4c3Vxp3jj4q+FtGv7NUaawutUiW6QPjafI3eZghgfu9OenNQftifstzSJCvxv8ADIZ2CgvcMoBPqSMAe54rwl4w8FePLB9U8D+L9E8RWUb+W9zpWoRXcat/dLRswB9qWAkcCvFnxX+E/gDUo9G8efFDwl4cv5oBPFaavrdvZyvEWZRIEkdSVLIw3YxlSOxrw5r/AIa8YaNb+I/CXiDTdc0m73fZ7/TrpLmCbaxRtkiEq2GVlODwQR1FfZwRwP0q7e1sraW8vLiKC3t42klllYIsaqMlmJ4AAGST0rw98afgr4x1m38OeEfjB4J1vVrvd9nsNO8QWtzPNtUu2yNHLNhVZjgcAE9BXjH4m/DD4fXNvZePfiP4W8N3F3GZLeHWNYgs3mUHBZRI6lgDxkU37Rv7OZGB8ffhx/4VVl/8dp/2i/2dcYHx9+HH/hVWX/x2n/aK/Z2PH/C+vhz/AOFTZ/8AxyoXtry2ivLO4juIJ0DxSxsGV1IyGUjggggg1JCO1SRAjGOa1HTrPUbOfTtQtIbq1uomingmjDpKjDDKyngqQSCDwQa/bG/Zsk/Z5+IqpoUMz+D9fVp9Gmkk8xoSuPNt3Y85QkEE5yjLyWDY/wCCSX7Vcvh/xJJ+zF401RF0rWmkuvC0koP7i8wXltt3QLIoZ1BwN6kDLSAVVVVU2CcA1VVVVRxiqqqqqpuTjNVVN0ABqqpsYxmqqqqqo8mqqqqqbpgGv+Cq/wCypL4/8Fx/tDeCNOR9c8IWrJ4gghhO+804c+flerQck5H+qZyWAjAP7Kv7RniP9l/4vab8RtFja809lNpremggC+snI3oD2dSFdG7MgzlSwPgnxp4Z+I3hLSfHHg3V7fU9F1q1W4s7qFwyup7HHRgQVZeqsCDgg1VVTYOADRwelVVV458NDSrr+0rJALW5Y7lA/wBW/XH0POPTn2qqqqqqqqqvh9rhjlbQ7h/kfLQE9m7j8ev4H1qqqqbBOAaq2urW9gW5srmK4hfO2SJwynBwcEcdRimwTgVVVTAbeDVVVVWveG7LXIsuBFcqMJMBz9D6itS0y80m5NpexbHAyp6hh6g+lVVVVVVUCQQVOCOlaN43v7ALBfg3cAGATw6/j3/H8607VtP1aHzrC5WQD7y9GX6jrVVVVVVVV8UPCg8efDPxb4H2b/8AhIdCvdO279mfPgePG7t97r2qtD+EvxV8USWsPhn4ZeK9We+j8y1Wx0W4nM6bd25AiHcNo3ZHbnpWgfsU/tYeJUgbTvgJ4thFxKIkGoWX2AhiQMsJymxefvNhcc5xWg/8Evf2wdYz/aHgjR9DxIFH2/XbZ8g9WHkPLwO/f0BrRf8Agjv8cbjzv+Ei+J3gWx27fJ+xG6ut/Xdu3xRbccYxnOT0xzo3/BGfw7BLIfEPx/1G9jK4jWy8OJalT3JLXEmRjtgfWrX/AII+/s4pawpe+PfiRLcLGolkivrKNHfHzFVNqxUE5wCxx6nrX/Dn/wDZo/6Hf4m/+DKx/wDkOtY/4I6/Bee5V/D/AMVPGtnbiMBo7xLa5cvk5IZY4wBjHG09Cc84H/Dm/wCGf/RZfE3/AIL4P8a/4cwf9XJ/+Wb/APdtf8OYP+rk/wDyzf8A7tq6/wCCNvxCS5lSx+Nfh2W3WRhDJLpc0bumeCyhmCkjGQGOPU9a03/gjX44mvYo9Y+OOhW1oc+ZLbaPLPIvBxhGkQHnA5YYBJ5xg/8ADmD/AKuT/wDLN/8Au2tN/wCCNngaKxij1j44a7dXgz5ktto8UEbcnGEaRyOMDljk5PGcDx3/AMEbvDs6vP8ADL40ajaMrHZa67pqXO4ErgGaEx7SBvOfLOTgfLya+Jn/AATV/aw+G9vLfW/guz8X2cClnm8M3f2phhQflgdUmc8kYWMnKn1UnVNH8V+BNf8AsGtaXq3h7XNNmDeTdQSWlzbSI3B2sA6MGHXggivgZ/wVA/aq+DDW2m6l4uTx5oMC7BpviQGeRRgAFLoYmBAAwGZlHPy5JNfs3f8ABT/9m/4+/ZNB1/Vv+Fd+LJ8L/ZevXCrbzyHtBeYWN+SFCyCJ2JwqHrVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV/wVz/Zjt/hT8Ybb41+E9M8jw78RZJZNQEanbBq4O6Yn085T5o9WEp6Cv8AglN+0fF8PviNe/AnxPesmi+OZVm0lmb5LbVFXbt6jAnjUJnk74oQB8xIqqHUVVDqKqqqqqhgjANVVVVUOuBVVVVVUUSRDG6hlYYKkZBFeLtEk8NeKdX8PyIVOnXssABzyquQCMgEggAg45BBr4Ma8PEnwt8N6iZC8iWS28pZiWLw/u2JJ5ySmfxqqqqX0qq/4K0eMovEv7WMvh+GXI8JeHbLT5EG4BZJN90Tg8ZK3MfK5GAOcggf8Er/AAgdO+F/i/xtJbskmt61HZozKw8yO2iDBhngruuJBkd1YE8DCR560qA8CkjGOaVMjApY88CljAGKCE9BQjyKEWegryq8qjFntRjGOK8XeLtJ8HWltPqTF5b24WC2gTG+RmIBI9AByT9O5FNGCMCnj4xXjH4peEPB2+2ur77ZfJkfY7bDup/2j0T6E59Aa8XfGPxZ4oL21rN/ZVi2R5Fs5DsP9qTqfTA2g9xWk6Lq2vXa2Oj6fNdzn+CJM49yegHueK8M/AK5dlufFuoLGnBFranLH2ZyMD6AH6itG8MaF4ctzbaJpkNqp+8VBLN9WOSfxNf8FVfC62fjzwL4yDLu1TSLmwI3HI+zSiTpjAH+l9c889MDP/BFfxh9n8T/ABO+H8tzu+3WFjqdvC1zjZ5EksUrJF33faIQzjGNqA5yMVVVVLgjAqqqq8TfErwD4OVh4k8W6bZyJnMLTB5eBk/u1y56enXA6kV4j/a++Hmmb4vD2l6prUq52PsFtE3XHzNlxkgfwdD6jFeIP2wvH2oBovD+i6VpKEna7hriRRnjkkL0/wBnn26Vr/xg+J/idWTWPHOqyRsctFDN5EbdDyke1TyARxx2qC3vNRuRBawTXVxKSQiKXdz1JwOT3NaL8Dvi34gUNp3gLVFU9Gu4xag8A5BlK5GCOe9aL+x38Qr0h9a1rRtNj/uq7zyDkdgoXGM/xfzrRv2MvB9uqnX/ABdq1846/ZYo7ZTz6N5hxjA6++a0n9m34NaSg2+EEu5ApUyXdzLKWBOfultoPQZABx9TWk+EPCWgMH0LwvpGnMrFgbSxjhIJGCflA5I4+lDqMUvIxVVVVVfQ1wRwaqqqqpfTNVVVVVQ5GM1VLgjFVq3hDwnr5c654Y0nUTIQXN3ZRzbiBgE7gc4AArWf2bPgtrQYy+CoLWQghZLOeSDbk9Qqtt/MGtc/Yt8B3hZ9B8TazpzMGwkwS4RTj5cDCtgHqCxJ9RWt/sVeNbVXfw/4s0fUAmSq3Ecls7gDjAAcZJ4wSB71r37OXxm8Phnn8EXV5Gudr2DpdbgMchUJYdehAJ59K1TRtX0S4NprOlXlhOCQYrqBomBBweGAPB4rQfiB468LhV8O+MNY09ExiKC8dUOM4BTO0gZPBHevD/7XHxg0YBNQvdM1tOmL2zCsBx0MRTng8nPU5zxjw/8AtuaPKVj8VeBry2AHzS6fdLPuOD0RwmOcfxHqfTB8O/tGfBvxLtS38bWljKeseoq1pt6/xOAh4Xsx6gdTirW7tL+3S6sbqK4hfOySJw6tg4OCOOoIqqByMZqqqqqq+hoEEYzzVePv2avhX48WW5OirouoODi70xRDlic5aPGxuepIycnkda+IH7JfxK8I+Zd+Ho08UafGpYvZpsnUe8BJJPshc1Z32seH9RFzp95eabf2rkCSGRoZYmB55GCpBH6V8Pf2xvGeg+Rp3jqxh1+yT5WuowIbpR7kfI+B6gE92rwD8ZPh18S1EXhbxFDJehdz2FwDDcLxk4RvvAdyu4D1qq+hrJxisgjANVVVqOnWGr2E+l6naQ3VpdRtHNBMgZJEIwQQeoIr9oD4KXnwl8SGewR5vDupyM1hMFY+Qc58hyc/Mo6HPzAZ65A/Zk+PB8AamngrxVeKPDmoSkxXE0hxp8p7jqBGx+8OACd2R82aokdM0TngUee9VVVVVRIA60ee9VX7af7Op0G+uPjD4L09Rpl5IDrVrEoAtpmwBOqgcI5xu7h2z/Ecfs5/HPUfgh42TUJnmn8P6jiHVrNCWymeJUXIHmJ2J6gsOM5APh3xp4eVwbLWNG1a3DKflmhuImGQR1BBGDXxu/ZRvdE+0+KfhjBLeaeN0lxpRO6W3HJJi7ugGBt5f/e7eFvFfiXwJrsWu+GtSn0/ULYlSy/xDPKOp4ZTgZUjHHtXwq/aA8GfGjSZfBHjO1tbHWNQtntriwkJFvqCOpVxESe4JBjJ3YPBbBI1rQ9P8M61qXh3Sbx7uy029nt7e4dNjTIkjBXK9iQAce9fsieMfC938HbiyguEt7jw3eXA1YyP93JMiy+y+XhfrG3vX7QHxoufix4l+z6bLJH4d0x2WwhJZfOPQzOp/iPOOAVU46k5/Zs+A8nxD1NPGHie2ZPDmmzKY4pI+NRlU8oM8GMEfMe+do/iKoiRoscaqqqMKoGAAPQVVVRwBR571X7XniUeKP2hPFk0U6yQadNHYRBSp2eTGqOMj/poJDzyM47Yr9g7wxDonwHg1oRL5viHU7m6ZzgsVjbyAuQM4BhYgHPLE96psYwDVUenBqq/4K1/HKb4i/tHJ8MtPnkbRPhxaLatEz/u5r+ZVluJBjB4UwxYPIMTkcNX7NGjeE/C0uj/ABz8KeD5/tcIuLS2GriSW3iuNqiR4iCu9lViobJAEjZG4Db45+NWo/EDw2NB1nw7p8cqSrLHcwM4MbjqQpJ4KkjGe9V4YZk8S6S8bFWW+hKkHBB3io4gQCajhzxiv2q/Cel+Of8Ago7ceBtaaZdP8Q+I/DunXZgYJIIp7WyjfaSCA21jg4ODVx/wS9/ZbubaWCOy8UW7yRsqzRauS8ZIwGUMpXI6jII9Qa/aG+AXxJ/4J/8AxE8PfEP4W/EO9k0zVHkXT78L5UyNGys9rcoPklRhsPI2vhsoNoz8DPiNZ/Gb4Q+E/ifaLAn/AAkGmRz3EUD70guB8s8SnqQkqyJzz8vPNf8ABSvw+3xE/bL8C/D7RLpDf6roWl6TgAExT3F/chAQSoziVDyQORyK/wCCSXxPOvfDfxT8JL+4Z7jwxfpf2IYAYtrkEMi45wssbMc95hyeg8g+lf8ABR/4pn4X/sx61ptjcrHqnjWVdDthvG4RSgtctt6lTCjxk8YMq89Af2SdC1z4Yfto/DHS7n5L24+xzsssYBWLUdMEu0gE4IiuseueSAcgftTfsS+F/wBqTX9D1/XvG2qaHJodnJaxx2dtHIJAz7iTu6HtXiz/AIJO/D7w74X1jX4Pix4imk0ywnuUjexhAcxxlgDz0OMV+xd+y5oP7UnifxHoGveKb/RI9DsIrmOSzgSQyFpNpBDdBTf8Ejvh2OP+FueI/wDwAh/xrQ9Dj8P6DpugwTNNHptnFbJIwwXEaBQSB3OKkiB6CpIgeMVLEDxjmv2kPgtpnx2+E2seBLyKP7cUNzpE78fZr1Fbynz2B3Mjf7LtVhfeJvh/4uttSsJ7nR/EPhrUlmhkA2y2d3BICpwejI6D8RX7N3xs0b9oj4KeF/ixpAiik1izA1C0jbP2O9jOy4h55wsittJ5ZCrdGFVVVR5NVVVVU3TFVVVVVR5PWqqm54zVVTYxjNVVVVVR5qqqqqm6YFTQxXETwTxpJHIpV0dchgRggjuMV/wUB/Y/uv2ZfiT/AG94VspX+HviyeWXR5ACw06b7z2Tt1G0HMZblkHVmRzX/BOL9tl/gV4oj+EXxO1vb8Pdfn/0a7u5m2aBdHcd68HbDKxAkBwqnEmV/ebopYriJJ4JUkjkUMjochgRwQR1GKqqbGevaqqqq8s7e/tZLO6jDxSrtZTxXiHw/d+H7z7POd8T5MMoGA4H8iOMiqqqqqqqqoZpbeZJ4JCkkbBkYdQRWjanFrGmQX8eAZF+dR/Cw4I/OqqqbrX/AAVT+Pf7QHw0l0T4f+ELlPD3gzxPZO0msafK4vL2VMrNbM/AhQK8Z2rksDndjcg/4JOftNT6Vr1z+zR4svd1hqvmX3hiSRwPIuVBee3HqJFBkXkAMj8EycN1qqqm6YzVVVVVarpNlrFqbW8iB4OxwPmQ+oNazo13ot2bW5XIPMcgHDj1H+FVVVVVVVVvc3FpMtxazvFIn3WU4IrRfHkcgFvrShG7ToOD9R2+o/KopYp41lglSRGGVZTkH6EVVVVVVVWj6DoXh61ax8P6NYaZbvIZGhs7ZIUZyACxVQBnAAz7Cqqqqqqqqqqqr4i/CP4YfFzSjovxL8B6J4jtdpCC/s1keHIIzHJjfG2CfmQgjJ5r47/8Ei/B+rxXWufs+eK59CvmZnTQ9ala4sz6JFOAZYxx/wAtPNJJ6gV8ZPgH8WfgF4ifw18UvB15pExYi2utvmWt4uM7oZlyjjBBIB3L0YKQQP2W/wDgop8ff2YvsXh231T/AIS/wTBIu7w7rE7MIIxgFbWfl7bgcKA0YJLeWSTn9mT9sX4J/tX6LNefDXXJYNYsIw+o6BqSrDe2inA37ASHjyQPMQsuSAcE4qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr9pn4G6L+0f8AA/xV8ItYaKKTWbMnTruRc/Y72M77ebjnCyKu4DBZC69GNa7ovi/4T+P73QdTSfR/E3hLVWhl2Nte1ureTG5T6h1yD9DX7J/x9079pP4IaH8SYFig1M7rLWrSNs/Zr6LAkX2DKUlUdklXPOaqvpVV9Kqqqqql6VVVVVQxkVVVVVVL0HNftR+Hf7F+KU+oxRbYdZtIrkFVwu8Dy2HQDOYwx6/eznmv2QfEH2zwrrPhqSTL6deLPGCeiSrjA9g0bH/gVVVVS4ziqr9szxVL40/ar+KmtyyNIE8T3dlG7XBm3R2r/Z0Ib+6UhXA6KMAZAzX7BfhiPw3+yx4LQpH5+pJc307RuzBzLcSFDz0IjEakDjKnr1KoOgpIxjJpUzxSRgilTPApYwegoRjp1oJnoKEZIryuMUYsjAoxnHBrWtV0/QNLudZ1a4ENpaRl5XIJwPYDqScDHqa8d+O9U8ceJH16dmt0iIWzgVz/AKOg6YP97PJPr6AADxT8VvBXha0Sa81IT3MsYeOztwHlwRkbhnCdR94ivGXxr8YeK99raT/2RYNkeRauQ7D/AGpOCfTA2g9xWj6FrHiG8XT9E02e8uG/giTOB6k9APc8V4S/Z5IEd54zvvf7Fat+jP8A0X860vQdI0GzWw0XToLSBf4Ikxk+pPUn3PNOnapE9K/4KkeHWv8A4I+HvEUMczyaT4kRJApGxIpoJQWYdfvpEBz/ABHrnj/gk94qbw9+2DpWkLctGPE+hahpxURhhLtjF1tJP3R/oobIwcqB0JqqqqpfQVWo6lpuk2j3+q6hbWVtEMvNcSrGij1LMQBXin9p74TeGwYrXVp9bnGf3emw7wPTLsVXH0JrxN+2R4nvC0PhPwvYadHyBLeSNcyHnhgBtVTjHB3d+TXiX4u/Evxcpj17xnqM8R+9DFIIIzwBykYVT09PX1NadpWp6zdLY6Rpt1fXLfdhtoWlc8gcKoJ6kD6kV4d/Zm+L3iEI8mgRaVC3SXUZxFj6ou5x/wB81oH7GFuFSTxT42kc5G+HT7YLgd8SOT2/2OPfpXh/9mj4PaCsbN4abU54/wDltqFw8u7gdUBCHpn7vc9uK0rQtE0KH7Noej2OnxEAGO1t1hXAzjhQOmT+dVVVX0NLg9DVVVVVUOR1qqqqqhjPWqqqqqpcdM1VDg9aqqqlx0zV3aWl/btaXttFcQvjdFKgdWwcjIPHUA1r/wAAfg74kLPqHgLTopGOS9mGtDnB5/dFc9e/BPXNeIf2KvBF6rP4a8U6tpcjHO25RLqNeRwANjdM9WPUemD4i/Y4+KOloZdDu9I1pQOI4pzBIT9JAF/8e/KvEXwy+IXhMv8A8JD4M1azSMEtM9qzRYAyT5i5U4Ayea0fxDr/AIdnN14e13UNMnPWSzuXgY8EdVIPQkfifWvDv7Vfxk0DalxrdprMS/dj1G1Vsdf4k2OeT3Y9AOnFeGP22NAunjg8X+D7yxJIVrixnE6j/aKMFIHsCx+vSvCvxx+FPjBY10bxrp6zuufs12/2aUHGSNsmMkc525HGc45rgjg1VVVVVfQ0CCODVV46+FHw/wDiRCE8W+HLe7nVSsd0mYp4+OMSKQcDrg5HtXxD/Yz8T6S81/8ADzVY9Zs1BZbO7YRXS+wbhJOO/wAnpg1qOl634Y1VrHVLG80zUbRwTHNG0MkbDoQDgj1Br4d/tb/EPwjHHp3iYJ4nsEwA11IUuUHtKM7u/wB8Mc45Ar4c/HX4c/E5I4NE1lbbUmHzabe4inB77RnD/wDASffFVVZ9DW44xW4HitwrI9a8ZeEtC8eeG73wr4jtmnsb5NrhW2shBBV1PZgQCO3HIIyK+J3w2174WeKp/DGuBZNo8y1ukUhLmI9HXPToQR2IIyep/ZV+PC6pa2/wu8X33+m267NHupWJNwgyTCxPRlGNvYr8vBA3EkjGaqqqqqqokAUee9VVajp1hq+n3Ok6naRXVnewvBcQSruSWN1KsrDuCCQR71+0n8Cr74I+OHtLRJZvDeqFpdIunySF43Quf76Ege6lW4yQP2K/2hv+Efv4fg/4z1HGmX8p/sW6nk4tZm6wEs2AjkfKAPvtj+PivjZ+zR4e+JCT+IPDQg0jxHgszrHiG9bk4kA6MSR+8GT6g8Y8T+FvEngbXJdC8R6bcadqFs2QrqV3AEgOjdGUkHDDg4607vI5kkcszEksxySfWvB+qeJNG0vWbO31S5tbLxCY/ttop2icRsWTeOvBY4+p9a+CHwh1H4t+K0sSJINGsSsupXQUjCZH7tDjHmNyBnoATzjB0fR9M8P6Va6Jo1nHaWNlEIoIIxwigYA9T9TyTyaqqqm6YBqpHjijaWV1REUlmY4AA6kn0rxdr03ivxXrXii4LebrGo3F5JuAB3SyM5yBx1btxXwI8Ojwr8GPBWhGIxSwaJbPOjFTtmkQPIMrwfnZv8T1qm5OAaqm9AaqviN4vufiF8QvFHj69LfaPEus3epy7wobfPM0rZCgKDlz0AHpXwG+Evh7Rf2c/BngDV9KtriJtGhnu02Di5mTzJXVhzuDyMAw5xXxT+FGsfDfVCCsl3pFwSbS9C9v7j44Vx+R6juBWn3jafqFrfxoHa2mSVVPAJUg4/So4s1HFkYAr49qB/wVT0pQf+Z28Kf+irCkhGK/4K4fE/wofA/hX4NWl/BceIDrS6zdwRy5e0gjgmiTzFA4MhnJGSDiMnGCDX7F3w61b4ZfsvfD3wfr1vLDqEOmvdXEMow8L3U0lyUYdivnbSO2K/aUk/4SH/gqp4I0nSIZZ7jSfEHhpLlAuNuxobhyPUCJwxPsfSvD9xL+yX/wU/vNNupktdA8Ua3JbuVl8qL7FqmJIgSeAkUzxZzx+4PP8VfZjX7bd7c/tPftzeBv2atHfzNM8Ozw2N3JbkysjzhLi+lGDj93bogK9mhbJHRfjat34Q/4KwaEdM09bKCXxR4YtrRPI2Rm3ktbO3byxwNoG9BjgFSO1PADxivifDj4b+K8c/8AEjvf/RD1/wAEhkD/ABG+IA9NDt//AEcaki4wRXxU+JHhD4O+BNW+Injm/a10fSIg8pjTfJIzEKkca/xOzFVA4GTkkAEhf2xv21f2m/Et1p37MvgVdF06wbc7W9tb3JjGGwtxdXY8hWYAkKAhO3jODnS/2qf2xf2ddVsD+1t8NJtS8LXsqwS6xa21ustu7NkMJbQm3Zthb90wVm2DDDDZ/ZL/AGtfiL8ff2h/GXgvWbrTG8KWOnX19pEdvbASIiXkEcOZeC48qUjOBk4OBUseRjvX/BR/4Sw+AvjXB430yCOLTvHNs1yURSu28i2rPx0+YNE5PdpG47n/AIIxfGF1uvHHwH1O+yrxx69pELFuCCsN0BnjndbEKMHhzzziqqjjHWjz3qqqqqm6AA1VVVVVHnvVVTcnGaqqbBwAaqqqqo9Kqqqqqm54qvi18KfBfxt+HusfDLx/pzXmi61CEmWN/LkidWDJLG38Lo6qwOCMjBBBIP7TP7Nnj39mH4j3PgjxjZNJYzs8ui6tGpMOpWwPDqezqCodDyrHupVm/wCCcf7fsukS6H+zd8aNQaSwkaKw8J63Jkm1JwkVlOe8f3Vjf+DhT8mClVT88Z7VVVVVq+kWWt2TWV7HlTyrD7yN6itX0i80W8ayvY8MOVYdHHqPaqqqqqqqqvh3qphvJdIkf5Lgb4wezgc/mB+lVVUeT1r9sX4GWv7Qv7PvifwIln52s29udR0BgoLJqECs0QXPA8wF4SeyzMeuK8E+MPEvww8caN438NXDWWueG9Qju7UyKRslicHa68ZU42sp6gkHgmvhv4+0D4p+APD/AMRfC9yk2meItPivLfEgcxh1BMbFSQHRsow6qykHBBqqqmwQAKqqqqqtV0q01i0azu0yDyrDqh9RWr6Vc6PevZ3IzjlHxgOvYiqqqqqqqqtJ13UtGkDWc52Z+aJuVb8O31FaJ4t03WAsLsLe5PHlueGP+ye/061VVVVVVVVVVVVVVVVVVVeKfCXhfxxodz4Z8Y+HtO1vSrtds9lf2yzxSfVWBGfQ9RX7UP8AwShltYrzxn+zHdNMitvfwjf3GXVe/wBluJG+Yjg7JTnGf3hOFOm6n4++EnjVNQ0y71zwj4s8PXLKHQyWV5YzLlWUj5XRsEqVOOCQRgmv+CfH/BRrT/2jYYvhR8Ybux0z4lwKxs7hFEEGvxgEkxr0SdVB3RjhgN6cblSqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr/gsf+y//wAI/wCJdN/ah8KWn+g+IJI9M8SxRx/6m8VP9HuTjtJGhjYkABok5Jl4/wCCZf7Rk/we+N0Pw713UvL8K/EGSOzlWQjZb34yLaUE/d3FjE2MA+YpOdgxVVwRxVVwRwaqqqqpeRjNVVVVUMetVVVVVS9ODX7YHh37V4c0PxRFFl7G7e2lZV52SruBbjoDHgZIwX75r9lvxIuifE+PS5pMQ63aSW3JAAkUeYpOe/yFR7vVVVUMZFVd3VrY2s17e3MVvb28bSTTSuESNFGWZmPAAAJJPArxBrN14j13UvEN9HElzql3LdTJECEV5HLMFBJIGScZJ+tfBHQovDXwa8CeH4pVlGneG9PgMix+X5hW3QF9uTgsQT1PXqajQY60iZ6UkYApUzxSxjHNKhPApYhivFPxc+HXg2eSy1rxJB9si4a1t1aaRT6MEBCn2bH60n7S/wAKWYKb7UFBONxsmwPfivC3jjwf41haXwtr9rfmMZeNCVkQerIwDAe5FFAe1OERS7sFCjJJ4xXxy+J48Z6v/YOiXO7RdOc4dTxdSjgv7qOi/ie4xWlaPqmu3qado2n3F7cyfdihjLtjueOg9SeBXgn9nB3jiv8Axzdsh6/2fbMMj2eQZ/JfzrSfD+j+HbJdP0PTYLOBf4Ikxk+pPUn3PNOgIyKdMdKkQEcU65GK/b38Nr4h/ZX8ZqkCyXGmra3sBZyuwx3MZduOp8oyDB9fXBH7E/iWXwn+1r8J9UhedWn8UWlgTBIUbF032Y5P93Ex3DuuR3qqqq1TV9J0S0N/rWqWlhbKcGa6nWJAf95iB2NeLf2sfhvoO+Dw/HeeILlNwHkJ5MO4djI4zgnuqsO/pnxd+1l8SNe8y38Px2Xh+2bcAYE86bB7GRxjOO6qp7+mNa8Q694juftmv61e6lPkkSXVw0pGcDjcTjgAfgPSvC/we+JfjIB9B8H38kJxiedBbxHPcPIVB+gyfzrwz+xrr90qzeLvFllYjg+RYwtcNjA4LNtCnqOAw4968M/szfCPw0Y5X0KXWJ42ysupzebnnODGoWMj6r/WtO0rS9HtVsdI021sbdPuw20KxIMADhVAHQAfgKqhgjAqqqqqlPA5qqqqqqXBGKqqqqqhyOtVVVVV9K4I4NVQwR1qqqvoa4I4NVVV4n+Efwz8ZPJN4i8FaXdTykmS4WHyZmJ6kyR7XP514n/Yv8BakrSeF/EGqaLMx4WUC7iUZ7KdrdPV68T/ALIPxW0UtJoqadr0IGR9muBFJ05ysm0evQnP6V4h8I+KfCdwLXxN4d1HS5CSEF3bNEHwSMqSMMODyMg14W+J/wAQvBTKPDHjDUrGNMYgWYvFwQR+7bKHp6dMjoTXhL9tPxlp5it/GPh3T9XhXAae2JtZuvLH7yMcYwAq9OvOa8HftUfCPxX5dveavLoN2+0eVqcflpuPXEqkoAD3Yr6464sNR0/VbZbzS7+3vLd/uywSiRT9CCRVVVVXToaBBHBqqrxd4C8G+PLMWPi7w7Z6nGoIRpUw8eeuxxhl/AiviT+xhf23m6n8L9W+2R8t/Zd+4SQe0cvCt7B9uAOWJrW9B1/wnqr6Vr+l3ml39uQTFcRNE49GGeo44I4PUGvht+1X8Q/BPlafr0v/AAkulpgeXdyETxj/AGZeSfowb2xXw3+NXgD4pWyf8I9rCRajtJk0y6IjuI8dcLn5xjnchIGRnB4qqqqqvjP8J9K+LnhKTRblkg1K13S6Zdtn9zLjo2OqNgBhg9iOQK1XS9c8HeIJ9L1CK407VdKuMMATG8UinIZTwR2IYdsEV+z38a7f4s+GzaarNGniXSkAv4goQTKSQsyDPQ4AYD7rHoAVzVVVVVUSAOtE571VVVfFn4X+HvjB4IvfBXiJdqT/ALy1uVTc9pcKDslUeoyQRkZVmGea8a+DfEnw28W33hLxJbG01TS5trFCcN3WRG4JVhhgeOCOlfsj/H5fi14S/wCEX8SXyv4r0CFROXY7723GFW45JLNkhXP94g8bwKr9q/R/Dd18ItT1rVdMtJtR09oRp074WWJ3mRWCN1I2liV6HbkjgEVX7MCeGG+DWhXXhyxjtpZoz/aa7laQ3inbKXI55IBUHohTtVVVUcY60eTxVfGvXF8N/CDxprYmWOS10K8MLMhYeaYmWMEDsXKj0554rw5os/iTxDpfh21L+dql7Dax7IzI26Rwgwo5Y5PTvSRxxRrFEioiLhVUYAA6AD0o4xR571VHk9ar4rf8ku8Y/wDYAvv/AEQ9VY2dvZ28VpaQRwwQoEiijQKsagYCqBwAAAMCtR0fTdd0240jV7OO6s7qMpLDIMhh/Q9weoOCK+L/AME9Z+HN3LqunxyXvh2RwIrrgtAW6JIB0PbdjByOhOKqxmgvrWG8tX3wzxq8bYI3KRkHB56Go4weBX7ZWn+K9S/4KI6tpfgK8+y+JrrXNAh0WfzBH5V41nZCB9xyFxIUOe2K1/4K/wDBWG30S8lj+JOoah+5Km20/wARwR3DqeD5bHZhgCTkMG445wK/YOsvg437Sp8P/tKaD4hb4mx6kV0mXXbky27akrEGOeFk3i4BGUaR2UsOisE3JAMdK8U6ra67/wAFgILqzjlRIvFFjakSgA74NPihc8E8Fo2I9iM46V/wWM+GV1a3Hw++N2nLs2eboV5MpKujgtcW20j/ALej1GMDGcnHwq+PuieKf2UdH/aN1+6/0WHws+pawwUL+/tUZbpVC5/5bRSqoAyeOATiv+CWHgrXfi18c/iH+0740i+03Ft5kUNw4cK2oXrmSZo85+5EpUgk4E69eo/bo1W68Of8FH/COvWUcT3GmXfh65hSVSUZ451YBgCDjIGcEfWngBGAK+KUOPhr4sIH/MDvf/RD1/wR+QP8SPiCPTQ7b/0eakjB4I5r/grt8Qb1vEHgb4TW1+y2kFnLrd3agHEkju0EDtxjKiO4AwcjzGyORXwK+EWh/BL4T+Hvh5ounwW7WFnGdQkjUBrm7ZR50zkfeZmB5PQBR0Ar4heB9C+I3gvWvAviWziudN1yyktbhJE3YDLgMPRlOGUjBDKCCCAa/wCCVw3ftCeIAP8AoTLn/wBLLOpkB5Ff8FGPh3F4y/Z0vvEMMTNe+EL6HUYtg5aNm8mVT/s7Zd56f6sfQ/sHfEOX4Z/tcfDTXROyW9/rKaRdLuIRo70G2y/zKCqtKr8kgFA2DjBqqo4xVVVVVU2CcA1VVVVRxjrVVVHk8GqqmwTgGqqqqqPSqqqqqpuTiqr9pT9mv4eftQfDybwP44tfJuoN0uj6vDGDcaXcEY3pnG5DgB4yQHAHQhWX49fAX4hfs5/EK9+HXxE0zybqD95Z3kQJt9QtySFnhYgbkOMY4KkFWAIIH7AP/BRuzu7Ox+CX7RniVILuBVg0HxTfS4W4UYC293IfuuBjbM3DAYchgGeqfkHmqqqqq8Q6Ba+ILH7LO2yRMmGQDlG/qPUVqWnXWlXkljeR7ZYz+BHYj2qqqqqqqqsrqSxvILyI/PBIHAzjOD0qCaO5gjuIjlJUDKfYjIqqo81X/BR/9lrUvgf8XLz4i6FZIfBfju9lurV4YwiWN65Z5rUruOB1kQ4VdrFQP3Zr/gmd+103wk8ar8E/H2qsPB3iy5UabNMw2aVqDEAHcSNkMowrdQHCMNoLk1VU4HABqqqqqqrXtEt9csTbSkLInzRSY+63+B7iru1nsrmS0uU2SxNhlPaqqqqqqqqq0TxnqGm7YLwtd244wx+dfoe/0P6Vpms6drEXmWNwGIHzRnhl+o/r0qqqqqqqqqqqqqqqqqqqq/bT/Yk8J/tP+GpNd0GGz0b4iabCf7O1Ux7FvVA4trogZZDjCvy0Z5GVLK2v6B42+FPja40HXbLUfDfinw3ejfGWMNxZ3EZDI6OpyCDtZXU4I2spIINf8E5f247P9p/wP/wgfju+SL4l+FrVPtzSyRr/AG1B0F3Ei4+YYAlULhSysDh8LVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVfGX4U+Fvjl8LfEvwm8aQs+keJbFraZ4wN8D5DRTJnI3xyKki5BG5BkEcV8Ufh34p+C/wATPEHw28Txtba14W1KS1ldMruKH5JUPXa67XU9wwNfsMftDt+0d8AtJ8RazfJN4q0Jv7L8QjAVpLiNRtnwMcSxlHJAC7zIo+7VUMEDBqqGCBg1VVVVS+gqqqqqq4I4NVVVVUvIxmvi94aPi74a6/oscZedrNprdVGS0sWJEA5GMsgXPvXhnW7jw14i0zxBaswl067jnUKcE7WBI+hAII9DVnd22oWcF/ZTCW3uYlkicDhlYZBH1BFVVVxVftA61L4b+AvxJ8RQQJNJpXhDVbpInJAcx2krgHHY7cVoGkya/rum6FDMsUmo3cVskjDIQu4UEj0GajTNIoPAqNARSJnpSoAKSPPNLGMV8evjjqFxqV14H8GXz21raM0V9eRNtedwMNGp6hAcgnuR6fejikmYRwxs7dgoyafSdRRN5tHx6Dk/kOasL+/0m+h1DTbua1u7Z90UsTlHjYdwRyK+Avxjb4jWMmheIZYl8QWMe8sq7RdxDAMgAGFYEgEDjkEADIH7QvxUXS7eXwF4euiL2dANRmQ4MMZ58sEH7zDr/snHfiq0jWdU0DUItV0a+ltLuAny5YzgjIwR7gjgg8V8LPjPZeOWTRNZhSz1oLldgPlXGM5K9dpAxkE/T0DoCPenTHSpEBGRTrjipE9K/aA0JvEvwN+IOhRW0M8174Y1GOBJgCvm/Z38s89CH2kHsQD2rwhe3HhnxBofjS4025msNL1aCVnVCFkaJ1kMYc8btoHHuDVVNLFBE888qRxxqWd3O0KAOST2GK8Y/tLfC3wnugtdWbXboKSItMAlQHtmQkJgn0JI9OmfGP7WfxB19GtfDdraeHrdhgtF/pE/fI3sMDjHIUEHvWs69rfiK8Ooa/rF5qNy3WW6naVvoCxOBwOK8J/An4peMikmneFbi1tmOPtN+Ps0YGOo3fMw7ZUH+deEv2NtOh8u48ceKZblxtLWump5aAjqDI4JYHpwqn8+PCnwh+G3gpkm8PeEbGG4QgrczKZ5lI7h5CzL+BHb0qqqqpeRjNVVVVUvIxXUVVVVVQ64zVVVVVUvIxmqqqqqocjGaql9M1VVVKcjGaqqqqq4tre7ge2uoI5opBh45EDKw9weteK/2avg94sDSP4Vj0m4IwJtKb7Lt4xwg/d+h+71Huc+LP2JtXgWS58EeL7e8AyVtdRhMLgADAEibgxJz1VR05714v8Ag18TvAyNP4k8HX8NsgJa5hUTwqB3LxllXj+8R+hrw/4o8SeFLwX/AIZ16+0y4yCXtZ2i3Y6BgD8w68HI5PrXg/8AbG+I+hqlv4nsbDxFAowXdfs0554+dBt6ccoT05658F/tWfCjxa6WuoahP4eu2OAmpKEjJ46SqSoHPVtvQ1ZX1jqdpHfabeQXVtKMxzQSB0cdMhhkHpVVVfQ0CDxVVVeLPBPhPx1pp0jxboNpqdtzsWZPmjJ6lHGGQ+6kGviT+xhf23m6n8L9W+2R8t/Zd+4SQe0cvCt7B9uAOWJrUdL1vwxqrWOqWN5pmo2jgmOaNoZI2HQgHBHqDXwv/a38Z+EvL0vxuJfEumjCrLJIBdQjvhz/AKz/AIHz/tCvh/8AFLwR8TtPN74S1qO4kjGZ7ST93PB0+/Gecc43DKk9CcVVVVV+058Df+FhaKfF/hew3+JNMjw0US/NfQjqnX76gkr1J+7zkY8GeL9Z8B+JrDxVoFx5V5YSblBGQ6kYZGHoykg/Xjnmvht8QtE+J/hK08V6G+1Jf3dxbs25raYAb429xkEHjIIPeqqqqqJAGKPPeqqqqq/a5/Z/Hxa8KjxV4atN3irw/bv5CIvzX1uMs0HAJZgclB03Mw/iyPA/jLxD8NfGGn+LfD8722o6VPu2tld46PG4/usMqR6GvhZ8StA+LXgjT/Gvh6TEV0m24tySWtZwB5kTHAyVJ64wRgjgiv20NWNp8NdN0qOUq9/q6F12gho0jckZ7fMYz+H1qvH/AMMNf+G+g+Ftb8SIYP8AhKIJJI4WjKm2ZSCI3PZmRlYA4P3hjKmvh18S/Ffwv1wa34XvvL8zat1bSDdFcoDna6/nhhgjJwRk18JvjL4V+LmkC50mVbTVIEzeaZLJukg5xkHA3pnGGA7jIB4qqqmPGAaqv20tbOjfs6+JI4nlSXUZLa0jZAON06M4b2KK69+o+o/Zj0M+Ifj94G09YWkMWrJd7VcLj7ODPnJ7Dys474x3qmxjANVRwBwaqru0tL+0msL62iuba4jaOaGZA6SIwwysp4IIJBB4Oar4bqP+FfeGAP8AoDWn/olKjUYAFXWnWWp2U2m6haxXNtcxmOaGVAyupGCCD1GK+Nv7P+ofD8zeJvDYe88PPJ8yYLSWOcYDnumTgN9AeSCfCSf8UxpGP+fCD/0WtRRgDivj+uP+Cr+kr/1PHhP/ANFafUUQOOK/4K0eBf8AhX/xy8EfGXww39m6j4jsG8y4t22yG90949s+eoYRy26g+kS+hr4V+LYPiP8ADXwp4/tniZPEei2moDylKqDNCrkAEkjBYjBJIxg18FdYHiH/AILD394LXyPK8W+I7TZv3Z+z2F7Duzgfe8vdjtnGTjNftq/B9vjT+zF478GWds02px6c2oaWkeA7XVqRNGik8AvsMfPaQ8jqPAv7Scnhn/gn/wCP/gXDep/amq+L7OKBWbDx2FzG0sxQZ+6HsSjdcfafcEfsD/BYfBX9lrwboN5DEura5b/25qpSPYfPugHVW9Wjh8mInnmPjjFf8FbdRl8Lftd+C/EWm29ubqx8IafeKJE+WSSPULwrvwQSPkUHkHAxkVLAMcCvipGP+FaeLc/9AK9/9EPX/BHhd3xJ+IWO2hW3/o81LGCK/wCCvfgi/tPiR4G+Iwjkay1LQ5NKLBcpHLbzPKAT2LLdHAPXYcdDXwe+JWifGP4XeG/iPoV5BPDrWnxyzLFx5E+0CaIjLbWSQMpGTgjqep8deLdB+H/hDWfG/ie7+z6VoVlLeXcgGW8uNSxCjPzMcYC9SSAOTX/BKoBv2hvEI/6ku6/9LLOpVHSvjD4TTxz8K/F/g5rfz21jQ7u1iQRhyJHhYIVB43BipHoQDWlapfaJqlnrWlz+Te6fcJcW0uwNskRgytggg4IBwQRWi6tZ+INGsNd05ma11K1juYC67WKSKGXI7HBHFVVN0wDVVVVVUeTVVVVVN0xVVRxiqqqPJqqqqqm6YFVVVVVR5NVVftKfs1/Dz9qD4eTeB/HFr5N1Bul0fV4YwbjS7gjG9M43IcAPGSA4A6EKy/Hr4C/EL9nP4hXvw6+ImmeTdQfvLO8iBNvqFuSQs8LEDchxjHBUgqwBBA/YF/4KPQeC7Gw+Cf7Q2tytosPl2+geJJyXNipIVbe6Y8+SAfllP+rAw3yYKRSxXESTwSpJHIoZHQ5DAjggjqMU3I61VVVVVeJfDdr4htcHEd1GD5UuOnsfb+VXtlc6dcvZ3kRjljOGU/zHtVVVVVVVV4HvDd+HYFZtzW7NETn05H6EVVHGOtVXxj+FHhP44fDXXfhf41tmk0vXLYxNJGcSW0gIaOaM9nRwrjOQSuCCCQfjF8KPFnwQ+JOu/C/xrbJHqmh3JiaSM5juYyA0c0Z6lJEKuM4IDYIBBA/4JzftPN8ffg5H4X8VakkvjXwSqWd+XkHmX1rjEFzt652jy3PPzpuJG8CqqmHOM1VVVVVVXjHw9/alt9vtEzdQLyB/y0X0+o5NVVVVVVVVVUM0tvKs0ErRyIcqynBBrRvHs0WINZjMq9BNGMMPqO/4frVlqFjqUPn2N0kyd9p5H1HUfjVVVVVVVVVVVVVVVVVVVf8ABQz9jy1/aC8Ay/EHwTp3/FwvClo72scEALaxbDDPatgFmkADNF1+YlcfvNy/CL4p+NfgP8TtE+Jfgq9lsdb8O3gkCNlRKo+WWCQd0dSyMPQnvXwJ+MnhX9oH4TeHPi54OLpp+v2vmG3kYNJaTKSksL443I6spPfGRwRVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV/wWU/ZghutK0z9qjwnYv9psmi0rxWiLw0LHba3R44KuRAxJOfMgAA2kn/gn9+0RF+z58fNPm1/U0tfCfitV0vW3lk2xwBm/c3DE8ARyYyx6I8nrVUOlVQ6VVVVVQ64qqqqqqGCBg1VVVVS+gNDkCvib4ZTwd4+13w5EhWG0u28hSORE2Hj/APHGWv2c/FB8T/CnTFlfdPpDNp8pwR/qwCnX/pm0f4/lVVVV1Fft+a3B4f8A2O/ihf3E0say6QtoGjzktcTxQqOOxaQA+xNfs4aS2uftBfDXTBYreJL4r00zQOgdXiW5RpNwPBXYrEg9gaRQBgVGmeBSKDwKRABSR5pUAFfFrxJN4Q+HGua7aytHcx23lW7oQGSSRhGrD3UsG/Co42lkWJPvMQB9TWiaLHFEIolyx+8xHJNSaC6xb9nb0rxLoy+U10iYljGT/tAV4Q8Q33hTxLp/iDTbv7NPZzZEuzftBBVvlyNw2k8d+lD9layu7ia913xzfXt1cTNLNKlqsZcscsTlmO4nJz79Ktf2YPhxbbvOn1m73Yx510o2/TYi9ff0q2+BfwqsmZoPCEDFhgiaeWX8gznFWfgLwVpmDp/hLR4GVNm9LGMMV44LYyeg61HbQQRLBbwpFGgwqIu0D6AVJHkcU6Aj3p1xxUqDpTrkYr4oeLNH8L+G7iDUYIryXUYngispDxMGGG3Ac7ADz9QMjNftZ+GdD8IfDvwh4d8O2EdnYWd7KsUSD/pnyxPUsTySeSTmvAnxh8BaX8FPAfivX/FMMEer+GNPvIEnYtcTLJaxuD5Yy5JDDJx1PJ5rxv8Ath5E1l8P/D/qqX2o/wDoSxKfrjLemR1FeL/iL428dz+d4p8R3d6oJKQF9kSc5+WNcKO3OM8D0rwh8MPHnjxl/wCEX8M3d3ETj7QVEcIxnP7xiF7HjOa8HfscMWjuvHnigbcHdZ6YvOe2ZXH5gJ9D3rwh8I/h14FMc3hzwtaRXUYGLuZTNNnHJDvkrnuFwPaqGCODVVVVVDriqqqqqHXFDkYqqqqqvpXBHBqqqqqh161VVVVVDGcZqq+hrgjg1VVXTpXBHBqqqqqqBx3rgjg14w+Bnwr8cl5tb8IWa3TnJu7RTbSk4xksmN3H97I6egrxh+xLMivceAvF6yYHy2mqx7Seef3sYx07bBz3548afBv4leAFkm8S+E7yK1jJzeQqJoMA8EumQueMBsH2614X8a+LPBV6L/wp4hvdMm/i8iUhX9mX7rDpwQegrwV+2h4v0wLa+ONDtNbhCgC4tj9lnB7swAKN9AE+vavBX7Qvwn8dBYdP8URWF4VBNpqeLaTJ7Asdjn2Vm/KqqqB4wa4I4NVVV4z+H3g34g6f/Zvi7QLXUEUERyOu2SL3Rxhl/A/Wvij+x74j0I3GsfDi6OtaegL/AGCYhbuMAchTwsvQ9NrdAFY81b3Gt+F9YS4tpbzS9U0+XKsN0M0Dj8ipr4X/ALZGp6f5ek/FGyfUIOFTU7ONVmT/AK6R8K46crtIweGJrw34r8N+MdNTV/C+t2mp2jf8tLeQNtPow6qfYgGqqqr9rD4If2JeyfE/wrY40+8kH9q28ScW8p/5bAKMBGI+Yn+Ns/xcfAz4v33wk8WpeyvLNol9iLUrVSTlc8SIuQN69s9iw75Gn6hYavYW+p6XeQ3VpdRiSCaJwyyKRkMCOoIqqqjjHWiR0FHnvVVVVVVX7a/7P6+FNYb4teEbDZo+qy41eGMALa3THiQDrskJ54wHB5+cAfsu/Ha4+DPjdYNWuZX8L60RFqUIG7yW6JcKMjBU8N6oW4JC4/a1+J2neNPFdl4V0G6iubDw8r+ZcQzb0nnkC7gMcEIFC59S4r9lv4W2fxC8cSavrMUc2k+GxHPNA4yJ5mJ8pCO65RmPUHaFIw1ftO/DV/ij8HNZ0SyiV9T08DUdOB7zQgkqPd4zIgzwC4J6VoviUwhbXUnLJ/DKeSvsfUe9aFr+reHdSttd8PanNZ3lsweGeB8EEc/Qg9weCOvFfAv9o7SPiYkPhvxJ5GneJ8NtjQFYbwDnMeScNjqhPYkcZAqjjHWic9Kqv+CiGqRwfCfw/o4MolvPECygrwpSOCUEHn1kQgYxwemBX7B+kHU/j9bXgtVl/srSbq5LkgeTkLFuHPP+t28f3vxqic96qmweAaqvEOm3Ws6BqWkWGpy6dc3tnLBDeRA77Z3Qqsi4IOVJBGCOR1HWq8By29z4L8Pz2lotrDJpVs0durlhEpiUhQWySAMDJ545qJckAV8Uvjj4X+GERsWH9p626gx2ETgeWDyGlbnYMdBgk5HGOR4q/aR+KXihZrYapa6bZTqUe0tLVdrKRghmfcxBHUZwcnjHFaL8e/i7oCQxaf42uzFbxLEkVxHHOoRcYGHU9gBnr7818K/2v7HVLq10L4lafFYTSsI11W2OIMngGVCcoPVgSMnooBI+Mv7O3xt8Tf8ABR7S/i3oHw31a+8HJ4t8NXjazCimAQQRWQmkznO1TG+eP4TUSDGfyr/gpN8APFHx1+AlpH4B8PXWs+JvDetQ3dpZ2x/eTQyAxTKFJAP30c9wIzjgnP8AwT20n4l+GP2atD8AfFjwbq3h3XfClzcWSRajEFNxbNIZYZEIJBUCUx44I8rkYIJ/ZE+APx90H9v7xb8ZPHvwo1fQfD+uXuvXS3syDyQbmdnQKck4O7jiktwRgCrv/gmj8ULv9tObwsPh9qEPwkm8Vtd/2zFAi2yaWT9o8hfm4IT/AEcHGN/O3+GntwB0r/gqr+zN8YPjH8UPBnif4T/C/VPEf2XQJLO/urGMNs2zs8aNkjp5jkf7xrw/Nqt/4c0q+16zS11O4sYZL23RSohmZAZEAJJADEjBJ6dTXxH0271TwD4l0zTrdp7q70e7hgiQcu7QsFUe5JAr/gl9+z78Z/g5488b6l8UPh3q3hy11DR4IbWW9jVRK6zElRgnkDmpUAOBX7Q3wK8JftE/DHUvhp4tLQR3LLPY30cavLYXSZ8uZAe4yysMjcjuuRuyPD3wM/4KJ/sf6neaT8Iv+Km8MXM+7y7CSG9tJnZWw32aYiWF8AbmRVBIRS7gCtE+An7aH7UepaaP2sPE0XhfwBZTw3M/hqzaKOTVQGR9jpbNlQQuC0z742+6gJJH/BKWxupPj14m1FIGNtB4QmikkA4V3u7UqD7kRuf+AmplGalXng1r2kyaDrmo6FNMssmnXcts0ijAYo5UkD0OK/Z71uXxN8Avhp4jngSGXVfB+lXbxISQjSWcTkA+gziqqm54BqqqqqjjHWjz3qqqqqm6YBqqpunFVVUee9VVVVU3QAGqqqqqo81VVX7QP7Ofwv8A2lvBTeDPiVovn+R5j6bqMDeXdabMy7fMicfgSjZRtq7lOBj9pX9lz4nfsv8AjSTw14501p9LuZpBo+uwRkW2pxrg7kPOxwGXdGTlSe4wx/YV/wCCiutfBS50/wCFfxr1a61P4diMW9jemIzXGgYJxjaC8tvzgx/MyALs4XY2h69oninRLHxH4b1W11PS9St0uLO8tZRJFPEwyrow4IIPWqqqqqOMdarxL4atfEFrtJWO6jH7qXHT2Pt/Kr2yudOuXs7yIxyxnDKf5j2qqqqqqqvhrMGs723DE7JFbB6DII/9lqqOMdaqjjHWv+CpP7MyfE74Xj43eF9PR/EvgK2Y3+0HfdaSCWkHpmFmaXnGEMvU4FfstfHXVf2dPjZ4e+JNlPP/AGfDL9l1q2iYj7XYSECZCP4iAFkUHjfGh7VoOuaR4o0LTvEvh/UIr7S9WtIruxuojlJ4ZUDxuvsysCPrVYzwKbk9aqqqqqqqvGugDT7n+0rVP9HuGO8D+B/8D/j7VVVVVVVVVVVbXVzZyie0neKRejI2D/8AqrSvH8qAQ6xB5gA/1sQAb8V6flj6Vp+uaTqiqLO+jZyP9Wx2sPwPNVVVVVVVVVVVVVVVVVVf8FQf2Z7P4R/FKD4seENPaHw34+llkuolGUtNTHzSgf3VlB80A/xebjCgAf8ABHT9pH/hBvihqf7PHiTUNmi+Os3ejeY2Fg1SKP5lHYedAmCTyWgiUDLVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVePPA/hn4l+C9b+H/jLTU1DQ/ENjLZX9sxK74pFKthhyrDOQwwVIBBBANftBfBrX/wBn74x+KfhH4iDvP4fvmjguGTaLq2YB4JgPR42RsdiSO1f8E3v2j4vjj8DLTwlrl60vizwBDFp2oGRvmubbBFrOOckmNNjE8l42Y/eFUvSqpelVVVVUOoqqqqqqGCMA1VVVVQ64BpeRiv2vvChtta0fxlbxny72FrS4I6CRDuQ+uSrMP+Adu/7IvioWPiXVfCFxKBHqduLi3BB/1sXUD6oxJ/3PzqqqhyBX/BUrWpdL/Y08U2EcKSLrGo6bauzEgxhbuObI98wgfQmv2F9H/tz9rD4eWIufJ8u9nud2zdnyLWaXbjI+95e3PbOecYpFz0pFAGBUaDFImaRABSJnk1+03aTTfCS+lhj3Lb3Vu8hzjapcLn35ZR+Naa6pfwMx4DitCkiV13EHmry/099OEaIu8Dk14hliWGaQkYCN/Kq0WwmsdF0+xuAPNt7WONwDkblUA/ypoxjGKeP0qXYiF5GCqoySTgAVdfEH4f20zW9x468PRSxnDI+pwqyn3BbirLUtL1iAXOk6la3sRAIkt5lkXBGQcqSORUiY5FSICOKdcjFfEDx1pPgPSvtl4wmu5QRa2obDSt6+yjjJ/qQK8Q+INV8UarNrGsXJmuJTwOiovZVHZR6f1JNeJ/A/hLxo1gfFWh2+prpsxmto7jLIrkYJZM7XGOzAj2rS9KubyW10XRNNknk2rDa2lrCWOFGFREUdAAAABwBXgr9lP4ieJVS68QPb+HLRsY+0jzZyCOoiU8ehDsp9q8Ffs3fC/wAHGO5l0g63fIP9fqeJVBxztiwEHPIyCR61FDFBEkEEaRxxqFREXaFAGAAB0GKqqXkYqqqqqqqqqqqlweBVVVVVUORjNVVVVVQwRwaqqqqr6GhyKqlORjNVVVSnIxmqqqqqqge2aqq8a/s+fCjxyjvqHheCwu2BAu9NUW0gPPJCjax56sp6CvHH7F3ibT/OvPAOvW+rQqu5LO8xBOT/AHVb7jHvklB+XPijwV4s8FXv9n+K/D17pk38PnxEK/urfdYdeQT0NeCPjd8Tfh95UPh/xRcGzjwBY3X7+DaMfKFbOwcAfKVPvXgT9tHw9qHkWPxC0GXSpzhXvrHM0Ge7GP76DpwPMNeGfGHhbxpYf2l4U1+y1S3GN7W0wYxkjIDr1Q47MAaqgcdDQIPFVVVXxF+D/gL4o2fkeKNHU3SDEN/bERXMX0fByOfusGXvjOK+Kf7LfjvwD52qaFE/iLRlYkSWsRM8K+skQycAdWXI7nFeEfG/irwHqqax4T1y50+4BG8ROdkoH8MidHX2YEV8Kv2vvDfiJYdG+I0Meh6jjAvo8m1mOe/Vojz3yvBO4cCoZobiFJ7eVJY5FDI6NkMCMggjqCKqqvrGz1OxuNN1C2juLW6iaKeGRdyyIwwykdwQSCK+PXwfvPhL4ue2tkkl0HUS0mmXDckLxuiY/wB5Cce42nuQP2UPjf8A2JexfDDxVfY0+8kP9lXEr8W8p6xEk4CMR8oA++3+1xVUSBxmic8Zqqqqqqqq1/Q9I8T6NeeHdesIr3TtQhaG5t5R8siMMEccj6jkHBGDXxb+B2r/AAn+Jc/hK933Glvm50+8Yf6+2LHbk4A3jhWAHB5HBFBVVQqgAAYAA6V+xJppi8J+JNY+zgC51GOAS5GW8uPdtx1483P/AAKq/aY+HK/DH4xa5odrbmLTr2T7fpwOceTMSdoz1CuHTv8Ac6k5r4CfD/QPjz8Jb3QbSdLDxp4NkKW0xGI7y1lJeJZjyWIcTIGHKLszkYFeI/DmveC9eudA1+ylsdRsZMOjcEHqGUjqCMEMOCMEV8Bf2pItQW38HfFC/jhuVCxWWryZAnOQAs57N0/ecAj72Dy1MeMA1VV/wUg1TjwFokV3/wA/9xNAD/1wWNj/AORAPxr/AIJ06ZFN8QvFWsGNzJa6KkCuPugSTKxB9z5Qx9DTHjANVRxjrR5qqr44+D0+H3xo8eeBI4Uij8P+JdQsIlRCi7Irh0QqDztKqCPYiv2SvEy+Lv2bvh1rCyrIyaDBZu4Ytl7YfZ2JJ53bojn3zXxO8cw/DvwVf+JCgkuEAitIj/HM/C59hyx9lNahf6jrupz6lfzvdX19M0krkfNJIxyTgepPQVoPwzNxElxrEzhmGfIi42/U/wBB+dXvwt0p4CLaW4gkx8rFtw/Ef/XFa3od9oN4bO+QZIyjryrj1FfsbfFi+1Nbj4Xa9dvObOD7RpMkhJZYlwHhz6LkMo7AsOgAEYGABUIBAxUYGAKhUYFRoMAA0IVK5zUiDGBUyjGBUqjkVOoxxUoByB6VOB2r46/GPwp8A/htqfxQ8ZJczabpkkKNb2mwzztLKsYWJXZQzfMWxkcKSSACR4f/AG+v2S/FOni+g+L1jp7DG+31O1ntZEJ7YZMN9VLD3r9rT/gof8JIPhvrfgf4K+Im8SeI9btpLFr2C3kit9PjkUrJKHkUCR9pwoTIyclhtwf+CZ3wD1n4b/DjVvih4t06ay1TxqYRYW08e14rCIEpIQeQZWdmwf4UjYfeqYA96mGT1rVtSuta1S81i+ZTc31xJPMVXA3uxZsDsMk18C9FtPDfwR+Hvh2xklkttL8K6ZawtMQXZI7WNFLEADOFGcADPaqqjyaqqqqpunBqqqqqqbBOBVVTYxgGqqjjHWqqqqqpsE4Bqqqqqj04NVVVVfFv4RfD/wCOPga++HfxK0CLVdHvsNsYlJLeVfuSxOOY5FycMOxIOQSD+2H+xL8Qf2VvEcuoeRPrXgC/uTHpGvqqkqSCRBcqv+rmAB5wFkAJXkMqfsbft2+Ov2XdVj8Pay154j+HlwzG40QyjfZuxyZrVm4Rt2SyZCPk5wxDj4c/EbwX8WvBel/ED4f69b6voerQ+ZbXMR/BkZTyjqQVZGAKkEEVVVVUcYqq8S+GrXxBa7SVjuox+6lx09j7fyrUtNu9Ku3sr2LZIn5MOxB7iqqqqqqvhpGwj1CQj5WMYB9xuz/MVVNjGM1VNjGM1LFFPE8E8aSRyKVdHXIYHggjuMV+2p8AJf2dfj5rvg6ytWj8P6i39paA5HBs5WOIwfWNw8fqdgPev+CTf7Qf/CafDbUfgL4hv9+seCs3Wk+Y2Wm02V/mUdSfJmbBJ4CzRKBhar6UeTVVVVVVVVe2dvqFpLZXK7o5V2sPT3HuOtapp02lX0tjPy0bcNjG4diPrVVVVVVVVVVVVYeJ9b0/Cw3zyIP4JfnH68j8KsPiFC+E1KyKHu8JyPyPI/M1Ya5pOpgCyvonYj7hO1vyPNVVVVVVVVVVVVVVVVftT/BKw/aD+Bnib4bzwI+oXFsbnR5DgGG+iBaFgSQAC3yMcj5Xb1rw7rviP4deM9L8T6S0una/4Y1SG8tTLEQ9tdW8oddynoVdBkH0r4P/ABN0L4z/AAt8LfFXw2Nmn+KNLhvo4TIHa3Z1G+FmHBeN90bY/iQ1VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVf8ABX/9l1viD8NbH9oTwbo6Sa/4HVotd8pAHudKbnzGPVjBJyB2SaUk4UV+yD+0FqH7Nvxx0Px2LmUaFcyCx8RW6Lu86wkYCQhepaMhZVAwS0YXOGYHS9T07W9Ms9Z0i8iu7G/gS4tbiJtySxOoZHU9wVIIPvS8jGaql5GM1VVVVVVVVVVS4IwDVVVVVDGRzS8jGa+PXhP/AIS/4XaxaRReZdWEf222ATcd8XJCjrkpvXj+9+FeBfFE3gvxhpHiiEO39nXSySIjYZ4+kig/7SFh+NQTw3UEdzbyCSKVA6MOjKRkEfhVVVLyMZr/AILBanfWH7Lej2lpPsi1LxpZ290u0HzIxa3cgHIyPniQ5GDxjoSK/wCCbml2moftWaDdXAYyabpt/PBg4w5gaI59Rtlb8cVEgOBSKCeKRcgAVGgxSJmlTsK8XeGLbxh4V1TwzdEKmo2rxK5GfLcj5Xx/ssAfwrXdE1Tw1rF3oOs2rW19YTGKaNuzDuD3B4II4III4NaN4kREWK7l2OowGPRv8Kl8RW4iBa8j2kcHf1rXNcOo/wCj25PlZ+YkYLf/AFq+AXw+uviB8Q7CIwMdO0mRby/kyVAVGBVMjuzADHBxuI6Zox+lMgIwRXxX+Jmi/Czw4dX1FDcXdwTHY2anBnkA7n+FRwSe3uSBXjr4peNPiFeSza9rE32V3JjsYmKQRDOQAg6445bJ461Wj63q/h6/j1TQtTubG7j+7NBIUbHcHHUHuDwe9fBj49r4wlj8LeM5YIdYbC2t0qiNLs4+6R0WQ+2Ac4ABwC6jpXjrxpo3gTR21XVZN0jZW2tlOHnf0HoOmT0H1IB8S+I9U8V6xPrWr3LSzzH5QTxGvZFHYD/6/UmqdS6FVcqSCAwAyPcZr9iOPSdO+D9hoOqa7p+r+NLKe9OrXgt0hubiN7uR4WZQAdoheFePlUrtHSqqqql9KqqqqqhyOtVVVVUp4HNVVVVVS46ZqtS1jSNHjSXV9Vs7JJDhGuZ1iDH0BYjNf8Jv4L/6G/RP/BhF/wDFUjxyxrJE6ujjKspyCD0INVVUvIxmqqqqqpT2zVUDjoa4I4NVVV9DQwRwaqqqqqqBBGM1VVWoadp+rWUum6rY297aXC7Zbe4iEkcg9GVsgj2NeN/2R/hj4n8y58PrceG7x8kG1PmQbueTEx4HI4UqOK8e/swfFPwQj3ltpS6/Yqf9fpYaV192ixvHfkAgY5IrStZ1vw1qK3+i6neabewnAlt5WidcHkZGD1A49q8A/ti+NdBSHT/G2nweILVBj7QuILoemSPkfHuoJ6ljXgL46fDL4jLFBoPiKOG/kAzp96PInBPG0A8Oen3Cw5qqBIGAayCMZrgjg1VVXxW/Zo8B/EiOXULCBNB1w5YXlpCAkrf9NYxgN35BDZPJPSviX8GvHXwrvfJ8SaZvsnbFvqNtl4Jvx6qf9lgDxxkc18Lfjv47+FU8cGlX327SA37zS7ti0WCcsU7xseeRxk5IavhV8evAvxXhS1028+wa0FzJpd02JOAcmM9JBgE/LyByQKqq+JXw90T4n+Ervwnri7Ul+e3uAu5raYA7ZF9xkgjjIJHevFnhXXvAfiW88Na7AbbUNOl2sVJw3dXQ8ZUjBB9COlfsz/GhfiV4a/4R7XbxW8SaLEBMWJ3XUAwqzckktkgOfUg/xAUSMYBokngGqqqqqqqqmIAwDzR571+098SP+E++I8+n2Fx5mk+Hd1na4OVeQH99IPqwCgjgrGp71X7Gd3osnwvvbHT72OS+g1aR7+Df88TMiBCV6hWRBg9CQ3OQQK/b1+HDa/4C074hWEW668Nz+VdYUkm2mIGeP7sgT8HY54wf2T/iKfhz8adGmubhY9N10/2XfbsABZSPLbJ4XbKIyW7Lu9a+Kvwg8J/FrRxYa5Cbe+gB+x6jCo82A+nP3kJ6oeD2IOCPiJ8NvFXwx119C8T2JjJLG2uUGYrpAfvo3fqMg8jIyBXwD/aauvCPkeD/AIg3VxeaMWVLW/djJJYjptbu0Q491HTIwBb3NveW8V3aTxzwTIHiljYMrqRkMCOCCMHNVVf8FFNSjl+JnhnRxGwe20Lzy5PBEk8igfh5R/MV/wAE37C4j07x9qjBfIuJtPhQ553RrOW49MSLROTwaqmPGAaqqr/gqh8NpfAH7X/iHVorZorDxnY2us2uQSNzR+TNhizEkzQSMQduN4AAXaT/AMEt/iDDrfwl1/4d3F1uvPDOrefFGcjbbXK5XHr+9jmJx03DOMjP7Xuozx6V4Z0lWPk3FxcTONxxujVFXjp0lbnt+NfD+xhvfESNMAfs0TSqCO4IA/8AQs/hWhaalwypgVrGgJaQBmAyVzXxQ02KXRHucAPaSKyn2JCkfqD+FfAbV5NE+Mng+8iL5k1WK2O04OJj5R/DEhz7VEQQMGoWGBg1EwIGDUMgwMGo5RjGa87jAapJQRgHNTSAjrUrAjrU7A9DUpBB5qYg8V4t8K+GPG2i3HhvxfoGn61pd2u2ezv7ZZ4n+qsCM+h6ivE3/BNr9knXb5r618C6ho7OzNJFp+r3CxszHOQrs4UDoFXaoHavh1+xD+zH8Kr+21nw78MrS91W0YNFfavNJfujh9yuqSsY0dTjDIisMDnPNTkdBU2DwDX7RfiyHwR8DPHnieV4w1noF2IBKCVaZ4ykSkAg4MjIDgjrXg/wzf8AjTxbong7SlZr3XdRt7C2VIzITJNIsa4VeWOWHA5NWlpa2FrDY2NtFb21vGscMMSBEjRRhVVRwAAAABwKqj0qqqqqqbpiqqqqqo8nrVVTc8VVU3TAqqqqqqPJqqqqqmxjGaqqqqrxd4S8M+O/Dd/4Q8Y6HZazo2qQmG8sbyISRzL15B7ggEHqCAQQQDX7cf8AwT58R/s83V/8TPhtHLrHw1mnyyZaS50MueEm4+aHcQqS5J5Cvg4Z/wBmH9qv4l/sr+MX8ReCZ4r7StQ2prGh3jt9nv41PB4OY5VBOyQZKknIZSyn9n/9ov4X/tK+Cl8ZfDXWvP8AI2JqWnTr5dzpszLu8uVPzAdco21trHBxVVVNjGKqq1zQbDXrUwXSASKD5UoHzRn29vatb0K/0K5Nvdx5U/6uVR8rj2Pr7VVVVVVfD+yNvoRuGX5rmUsM/wB0cD9QfzqqbGMZqqbpVf8ABTr4AxfFv4BXHj3SbVD4h+HKy6lG+MGSx2j7XGT6BFWX/tkQPvV+zH8Z7n9n/wCOXhT4oxiWSz0y78vUoIyczWcqmOdcD7xCMWUHjeqntVrdWt9aw3tjcxXFvcRrJDNE4dJEYZDKRwQQQQRVHmqqqqqqqqvGeh/2nYfbbdM3NqCQAOWTuP6//rqqqqqqqqqqqqqq0/xRrenFRDfNIgP+rm+cH8+R+BFWHxCtZAE1GyeM4+/Edw/I8j9asdY0vUgBY30Urf3M4b/vk81VVVVVVVVVVVVVVX/BSn4Cz/Bz9oe/8S6babPDnxB8zVrBlQKsdxkC7h4wMrIwk4GAs6Dkgmv+CLXx9Go+HvFP7N+t3OZ9JkOuaEXlyTBIVS5hUE8BJPLkAAOTPITjAzVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVavpOm69pV5oWs2UV5YajbyW11bTLuSaJ1KujDuCpII96/bK/Zw1L9lz49eIPhs0V3JoLyfbPDl7cLk3VhLzH82AGeM7onIAy8bEAAiv8AglV+0wvjz4fz/AHxXqCHXvBkPm6Kzk7rrTCcFM85aB2C9vkkjAB2MaXrgGqpeuAaqqqqquoqqqqqlx0qqqqqqXB6HtTokiGN1DKwwVIyCK+JnhJvA3jrWPDAUiK0uCbcnnMLANGc5P8ACw/HNfs3+L/+Eq+GFjbTOn2rQ2+wSANzsQAxnHUDYVHuVP0FVVLjpmv+Czt0E+GHw6sftOPN165k8nfjfsgA3be+N+M9t3vX/BK2xtLv9o3WJ7iBZJLPwhdSwMRzG5urVCw99rsP+BGo1wOKjUYqNQaRc4ApF44qNM96jjBFfGf4BaR8Uof7X0uaLTfEUMe1Lhl/d3IHRZcAnjoGGSB2IAA8YfCzx94FuJIfEnhi9gij5F0kZkgYeokXK/gSCO4FIjyOscalmY4VQMkn2r4ffs8fErx/cRuuiy6Ppu4eZfajG0She5RD80hxnGBtzwWFfDf4ZeHfhf4cTw/4fjZyzeZc3Uv+suJMY3NjoMcADgD3JJaIEYxTx8V8dvG8/jr4k6re+YTZ6fK1lZIcgCONiN2D03Nubnn5sdq0PRVvALm4XcmfkT19z7VBoreUAkYAA4AGMVrPh2KVGZIwkwHykDGT71BPc2N1Hc20rQ3FvIHjdThkdTkEe4Ir4deKD428DaR4kkIM9zBtuMDH75CUkwOwLKSB6EV4y+BeseNdbm1vV/H5ZnJEMQ035YE7Io83oP1PJ6037MeBx43zj/qGf/bab9mrbx/wmuf+4b/9tr9pL433H7O3xVvfhpP4Tj15Le1guIb5NRNsZFkQEho/KfaQ24cM2QAeMkD9hKx1r9ozwTc/Gjw/4gk8E32heIZdLitkh+3+Z5cEEpcvmMbWFxtKFSML1OcCJZREizujyBQHZFKgnHJAycD2yfrVVVUOvWqqqqqpeRjNVVXV1a2Vu93e3MUEEQy8srhFUepJ4Fav8Z/hDoRmj1b4n+FreW3AMkDatCZVyAR+7DFjkEHAHetW/a8/Z40kyRSfESK5ljjLiO0sbibfxwoZY9mT05YY74rUf29vgfZOiWtl4o1AMMlrewjUL7HzJUP5ZrUP+Cifh+KBW0n4X6hczb8MlxqaQKFwckMqOc5xxj8fW/8A+CiviOS43aZ8MNNgh2jCT6k8zZ7ncEQfpV7+3/8AG26tXt4NH8I2jtjE0NjOXTB7B5mXnpyD1/GtS/bb/aGvo0S18U2GnlTktbaVAxb2PmK4/LFal+1l+0Pqvl/avidep5Wdv2a1t7fOcZz5ca7unfOOfU1qX7Qnxy1by/tXxY8Tp5Wdv2bUZLbOcZz5ZXd075xzjqavfjB8WtStXstQ+KXi66t5Mb4ZtbuHRsHIypfB5ANf8Jv40/6G/Wv/AAYS/wDxVX+o6hqtwbvU7+4vJyADLcStI2B0GSSaqqqqqw8TeJNLtxaaX4h1K0gBJEUF28agnqcAgVB4+8dWs0dza+NdeimhYPHImpSqyMDkEENkEHvX/C7vjR2+L/jX/wAH91/8XUH7THx8t4I7eP4q68ViQKpknDsQBjlmBJPuSSe9QftjftI28EdvH8SnKxIFUyaVZuxAGOWaEkn3JJPerX9u34+27wvNfaFdCIqWWXTQBLjqG2FTz3xjrxitO/4KHfFWKctq3g3wncw7CFS3juIGDZHO5pXGMZ4x3HPrYf8ABR3UI7cJqfwjt7ifJy8GttCuO3ymFz+tad/wUR+G0sxXVvAviW2hCHa9u0E7FsjjazoAMZ5z6cc8aR+3T+z/AKkYBearrOleapLm80x28rAPDeUX9MfLnqPetG/aj/Z+14R/Yfino8fmuVX7aXs8EevnKu0e5wK0bx94E8ReV/wj3jbQdU81ykf2PUopt7DqBtY5PtXBHBqqU5GM1VVVAgcZqqqqqqr6GgcjGaqqqq8c/CH4d/EZGbxT4atp7ogAXsS+VcDHQeYuGIHocj2rx7+xdr1gsl78O9dj1SJQStjfkQznnhVcfIxx3bYK8QeGfEPhW+OmeJdEvdMuhnEV1A0ZYA4yufvD3HBr4f8A7S3xP8BlLZ9WOu6eoAFpqbtLsAGBskzvXAGAMlfavh9+1R8M/GogstVum8OanLwYL5h5Jb0WYYUj/eCHPb1R0kRZI3DKwyrA5BHqKr6Gt3GK3A8VkHgGqq+sbHU7ObTtSs4Lu1uEKTQTxiRJFPVWU5BB9DXxZ/Y80/UFm1v4WXC2d0SWfSbh/wB05J/5ZOeY+/ytkcjlQK1fRtf8Jaw+mazp95pWp2bgmKZGikjI5Vh39CCODwQa+Ev7Xus6F5Oh/E2KXVrAfKmpRDNzF/vgkCUdOeGAyfmOBXh3xLoHi7SYdc8Natb6jYz52TQtkZHUEdQR3BwRRx0zRI7Gv2mPgwvxK8Nf8JDoNmreJNFiYwhQd11AMs0OACS2SSg9SR/ETXhTxTrvgPxLZ+JdBuDbahp0uVDg4bsyOOMqRkEeh7V8OvHui/ErwlZeK9EfEdwu2eEnLW8wA3xngZIJ645GCODVVVVVVVVRIAwKPPev2gviR/wrX4cX2oWVx5eraj/oenYOGWRwcyD02KGYHpuCg9a8F+E9S8c+K9L8JaSP9J1O4ESsVyI16u5HoqhmPsDXxP0/SNI+IWv6JoNu0Fhpd9JZwo4G7EJ8sk46klCc9TnJ5NfB340X3wX+LQ8QK88ui3hS21i0Q/62HA+cA8F0PzL0J5XIDmtO1Cx1fT7XVdLu4rqzvYUmt54m3JLG6hlZSOoIIIPvXivw3pnjHwzqnhXWIVlstWtJLaZSM4V1IyPQjOQeoIBGK8S+H9S8J+IdS8L6zEsd/pN3La3CqcgSRsVbB7jI4PcYNfAj4hr8UfhT4f8AF0jqbyW2EN8ATxcR/JIefUjcBzww5PWvG/gTwv8AETQpPD/irTUurdsmN+kkD4IDxt/Cwz9PUEcV8YPgr4m+EWqLHqBW90m7dhZahEpCvjOEcfwSYGdvIPOCcHHwI/aC1P4V3H9ha2k2oeGp3y0CtmSzYnl4s9j3TgE8gg5zoutaT4j0m11zQ7+K9sbyMSQTxHKup/kexB5BBBwRVV+39qX2/wCOsFr5Oz+ztAtoN27O/MksmcY4/wBZjHPTPfA/4J26c8Xwy8TauZVKXOu+QqY5UxwRsT9D5o/Kqo4x1o81VVX/AAWX+Ds/if4SeFvjNplrul8E6hJZ6iyRjP2S8Maq7sBnCTRxqoJABnbu1fsCfFeP4X/tEaRa6jcNHpfi2I6PdZk2qkkjKYHIPBIlVFzwQJGx6H9rLQZr7wro3iCGMuumXbxSkZ+RZVHJHpujUZ9SPWvCGrx6LrkVzMwWGRTHIT2Bxz+YFaPqqwBXRx04Oa1HX2uYdrPnAr4oeIIWt10WCRWllYNMAfuKOQD7k4P4e4r9mvw5J4l+M/h2IRb4dOmN9MxXIQQqWU4yP+WgjHsSDUUgI61FJjjNRSjHWklGODSzkDrX2kYwDTzkjANSSgjk1LKCMZqWQHgGppBjANSuDzmpWB5zUrAng9KlbJ69KlYE4BqVgTwa/wCCnvxLtfDnwa034bwXDf2h4w1FXeMDj7JalZHJPY+abfAPX5j/AA1/wTQ+GTfEv9r/AMHtNapPYeElm168DAEoLdcQMAVYZFzJb+hAyQQQKqqbpiqqqqqpuTiqqqqqPTiqqqbBPBqqpsHgGqqqqqOMdaPPeqqqqqbGMVVVVVTYPAq7tbW/tZrG9tori3uI2jmhlQOkiMMMrKeCCCQQetftx/8ABM650Jr74t/s06HPd6e7S3GseFYdpayABYy2S8Fo+DmAZZSRsyp2p8H/AIyfEL4FeOLD4gfDfXpdO1KxkBeMktBdx/xQzR5AkjYEgg8jOQQwBH7Iv7anw7/ao8OR20EtvofjmzhZtT8OyTbm2qQDPATjzITuX/aQnDdmaqqm6Yqqqr/T7PU7VrO9gWWNux7H1B7GvEng290UvdWoa4sgeH6tGP8AaH9Rx9KqqqqsbKbUbyGxtxmSZwq+3v8ASrO0isbSGygHyQxhF49B1qqbBwAaqmwcAGqu7S1vrWaxvbaK4t7iNo5oZUDpIjDDKyngggkEGv2p/g4/wF+PnjD4ZwwSpp2n3xl0lpCzF7GYCSD5z98rG6ozf30Ydq/4Jj/Ghfin+zXYeFtQvGl1n4fyjSJ1fJJtQN1qwOT8vl5iA4x5J4AwSen4VVVVVVVVVVeMNCOlX/2m3TFrcklMDhG7r/Uf/WqqqqqqqqqqqqqqoEggqcEdK0/xbrmnqsaXfnRqMBJhuH59f1rT/iBYzFY9RtXgJON6Heo9yOo/WrPUtP1FN9jeRTcZIVuR9R1FVVVVVVVVVVVVV/wUw+CyfFf9mfVfENhb79a8ASHWrVlQbmt0XbdoWPRfJJlOOrQIK/ZR+Nl1+z1+0F4M+Kkd3LFY6ZqCxaskZJ82xl/d3ClQDu/dszAYPzKpHIBEM0NxCk9vKkscihkdGyGBGQQR1BFVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX/BTb9lZv2i/gVN4k8Lac1x438ArJfaTHEAGvLc7ftVue5JjTegHJeNVH3zXwZ+LHif4HfE7w/8AFHwhNjUNCu1lMLNhLqE/LLA/B+WSMshPUbsjBAI+GPxB8PfFj4f+H/iP4VuPN0vxDYR3cGTlo9w+aNsdGRgyMOzKRVUOtVVVVVQ5HWqqqqqHXAqqqqqqXBxVfte+Ddr6R47tLdAGzY3rjgk8tESO/AkBPso9K/ZX8Yjw/wCPpPDtwwFt4gh8oE5GJo8tGfxBdfqw59aqqHXAr/gtXf2ct/8ACDS47hGureHWppYh1RHNkEY+xMbj/gJr/gkraiT4i+PLwW+7ytFt4/N2fc3TE7c9s7M477fakUHApF4AqNRio1zzSLngVGmeKVAeAajjBFJGCMUsWRxQjGMZoxgjANNGCKePjindpGLu5ZmOSWOST614eSIW1sB08te2O1aTbac9kzTMAwXgYrxBHCrsEIrWFQalOEPG79cDP61+zH57fC9RNGqouoTCIg5LLhck+h3bh+AqRRjFSLzipV44r/gqHo40z9pW2vRc+Z/a3hi0udmzb5WJZ4ducnP+p3Z4+9jtk/8ABGHWbqf4I+OvDrxxC3svFS3UbAHeXmtYkYE5xgCBMcdSeTxiqqqr6GuCOKqqqte8WeFfC0P2jxN4m0rSIsZ3395HbrjIGcuR3IH1NeIv2r/gB4bV1uPiHZ3sql1WLToZLreVIBw0alO/BLAHsTWu/wDBQL4XWLNH4f8ACviLVHUkB5Vito244IO5m6+qitb/AOCh/jC4WQeG/hzo1iSB5ZvryS72nPOdgizkZx0x71rH7bn7QOpuWsvEWm6SDIzBbPS4mAB6L++EhwO3f1JrWPjz8adejMOpfFLxK0bRtG8cWoyQo6nqGVCA2RxyDV/qOoarcG71O/uLycgAyzytI2B0GSSa0vwz4j1yPzdF8P6lqCeZ5Ya1tHlG/j5cqDzyOPcVpf7O3xz1kH7H8KvEkeGC/wClWLWvJ/667ePfoK079ir9oe+d0ufCFnp4UZDXOq27BvYeW7n88Vpn/BPz4uXLwNqnifwpZRSJmQJPPNJEcZxt8oKTnAOGx1IJ76d/wTn1OaBm1b4s2ttMGwqW+jNOpXA5y0yEHOeMenPPFj/wTt8Fxi3GpfEXWp9u37R5FpFD5mMbtud2zPOM7se9WH7APwRs7gTXGq+LL5ACPJuL6FV+v7uFW/WtO/Yk/Z5snd7nwtf6gGGAtzqk6hfceWyH881p37I/7OulzG4tfhnau7IVIub25uFxkHhZJGAPHXGfzNWv7NnwGtLiO5h+FXh9njOVEtt5i/irEg/Qiv8AhSPwX/6JF4K/8EFt/wDEVp3ww+GmjxvFpHw88M2SSHLrbaRBEGPqQqjNf8IR4L/6FHRf/BfF/wDE0PBHgsj/AJFDRP8AwXxf/E1p3hzw9pExudJ0LTrKZkKNJb2qRMVyCRlQDjIHHsKqTwX4OldpZfCejO7klmawjJJPUk7a/wCEI8F/9Chon/gvi/8Aia/4QjwX/wBChon/AIL4v/iauvg38IL64e7vfhV4PuJ5Tl5ZdCtnZj6klMmrr4EfBO8t3tZfhH4PVJFwTFokETD6Mqhh+BFf8MxfAH/olmif9+2/xofsZ/s1kf8AJN//ACsX3/x6p/2FPgBMJBHp2tQ787dmpMdmfTcD098++avf+CeXwjktXTTvF/i+C4ONkk89vKi8jOVEKk8ZHUf0rU/+CcmiTeX/AGP8Vr22xnzPtOkLPu6YxtlTHf1zkdMc6j/wTp8YxGcaT8SNGuQqnyPtNlLBvOON20vtGeuN2PetS/YE+Odj5f2W78L6jvzu+zahIvl4x18yJOue2ehzjitV/ZC/aK0mPzZ/htcTJ5mxTa3ttOT152pIWA46kDt61rPwY+Lnh/zG1n4Y+KLWOJ9jTPpMxjyemHClT+Bq6tLqxuHs722lt54jh4pUKMh9CDyK0Pxp4x8MGNvDXi3WdJMRYxmxv5YNhIwSNjDGQSDWiftT/tB+H1VbD4p6tLtj8sfbljvOMg5PnK+Tx1PPvzWh/t+/G3Tdkeq2HhrV03rvaeyeKQqOoUxyKoJ9Spwe2OK0H/go3AUWLxP8LXV1Q5msNUDBmzwBG8Y2jb1O88jpzxoP7eXwB1h1TULvXtDBcqWv9NLgALkN+4aQ4J49c9QBzXh39oD4JeK1iOifFHw7JJNt8uG4vVtpWJGQBHLtfOOoxkdDioJ4LqCO5tZo5oZUDxyIwZXUjIII4II71VUDkYzVVVVVVX0NAg8VVVVVWveHNA8U6e+leI9Gs9StJAQYrmESAe4z0PTkcg4NfEH9jDQtQVr34b6wdLnJJ+xX7tLAeeiyAF1x778145+GXjj4cXos/F2gXFmrsVhuQN8E3+5IuVJxzjOR3ArwB8bPiP8ADaWNfD3iGWSxj4OnXhM1sR1wEJ+TnuhU+9fD39rvwB4nSGx8XxSeG9RYhS8h822c+okAyn0YADI+Y8mrS7tL+2ivLG6iubeZQ0UsTh1cHoQRwRVVX0NZPY1k+tZPrXj/AOGfgz4m6X/Zni3SI7goCILpAEntyepR+o6DI6HAyDXxc/Zl8Z/Dgyato6ya/oQJP2m3hPm24xn97GMkDr84yvHO3IFeA/iR4x+G2qDVfCWsSWrEjzrdhvhnA/hdDwfTPBGeCOtfB79pbwj8S1g0XWGj0TxEwVfssr/urp84/cuepJx8jfNzgbsE1VftYfBZfDmpn4k+GbPbpeoyY1OJAALa4Y8SAf3ZM88cMDz8wA/Z6+MM3wr8XLDqU8jeHtVIjv4gM+U38EyjPBU8H1UngkLiKWG4hSe3lSSORQyOjZDAjggjqCKqqqqqqokAYFHnvVftaePn8VfEZvDVpcFtP8NoYAoGAbhsGVvw+VPqh9cn9jH4epLLqXxK1C2J8kmy04uvGSAZXH4FVz05YeuPiLfJqnxB8T6lGhRbvWbuVUY5KhpmOP1r4s/DHxD8PrnRNV1UJLYeKtLh1LT7mNSFYOis8Zz/ABoWGQM8Mp/iwP2Ivj59gu4/gv4svsW90zNoVxNJgRyHlrbp0b5mXJHzZUZLAVX7ePw+/wCEd+Jll45tIdtr4ptf3pA4FzAFRu/GUMR6DJ3Hk5r/AIJ+fEH7LqniD4YXk/7u9QalYqeMSIAkw6cll8o9ePLPrRIA4Na94f0TxRpM+h+IdMt9QsbldssE6blPoR6EdQRgg4IINfHH9n7W/hXdy6zpayX/AIYmlxDc9Xti3RJQOnPAfoeOhOK+DHxu8QfCXWEQSzX2gXD/AOmacX4GeskQPCyD8AwGD2I8KeLfDvjfRIfEHhfVIr6ym4DocFGGMqynlWGRkHB5HrVft2yRv+0DfKkisU0y0VgDnadhOD6cEH8RX/BPP/kius/9jRP/AOktrVUx4wDVVVV8Xvhpofxk+F/ij4WeIzssPE+lzWUkwjDtbs6/JMqngvG+11z/ABKK8T+HPE3w38Z6n4V12CfTNf8ADeoyWtyisVe3uIXKkqw54Zchh7EV+z58TNF/aj/Z70/WdZKSXV7bNp2vwxYUw3sYAkZR0XdlJlHO0SJ6GviH8PNe+G+vPousx743y1pdopCXKf3h6EcZXqD7EE6L4z1vRY1t4ZlngXpHMCdo9Aeo/lV98StduoTDbRwW2RguoLMPpnj9KVLzU7xY40muru6kCqqqXeR2OAABySSelfs2fBt/hf4ek1fX7dB4i1dR9oAbd9miHKxA9M55bHGcDnANRyADg1HMCMZpJcDg0k+BwaW4HTPSvtPvTXAI4NPcZGM1JLkcGpZRjANSybu9SyA8A1NIMYBqVwBgGpXAGM1K4x1qVhjrX7bfxm/4XN8etZvdN1H7ToHh7/iV6RtPyFIj+9kGDg75TIQ3UpsHYV/wR5+CE/hH4S+IPjXrNtLHe+OLtbXTUkQACwtSwMinr+8meQEHtAhHWqqm54BqqqqqqPPQ1VVVVTdMVVUcY60ee9VVNycVVVVVRxiqqqqqpueM1VVVVTe3YVVftz/8E5dH+LsWo/Fr4IWNvpfjeOHzL3RYkWK21or1K9BFcEd/uuQN20kvXh7xD42+FPja18QeH7/UfDnijw3ekxSBTDPaTxkqyMrDg/eVkYYILKwIJFfsS/tteF/2p/C40TWmtdI+Imj24bVNLVtqXaDAN1bAnJjJI3JyYyQDkFWaqpueM1VVVEBhtIBBHIIrxB4Ctb8tdaQyW05OWjIwjfTH3f5VqGmX2lzm2v7Z4nGcbhw3uD3FVVAFiFUZJ6DFeCPDTafD/at9FtuZVxGjDmNfX6n9B9TVVTYJwDVU2CcA1Vf8Fg/g2uoeHPCPx402NPtGlSnQtUGCWeCTdLbv6BUkEynuTOvoa/4JffGCX4bftL2PhG9vEi0fx9bNpdwrrkC5UGS2YY53FwYh2/fHPqD93FVVVVVVVVVWoafaapaPZ3kYeNx+Kn1Hoa13QrrQ7ryZhvifPlSgcMP6H2qqqqqqqqqqqqqqqqV2Rg8blWHQg4Iqz8W6/ZgKt+0yj+GYb/1PP61a/EScALe6bG/q0Tlcfgc+3erbx7okwAnW4gPGdybh+hJ/SoPEugXIBj1W3HH/AC0by/8A0LFRyxTLuhlRwDjKnPNVVVVVVVVVV/YWeqWFxpmoQJPa3cLQzxOOHRhhlPsQSK+NXw2vfg/8WvFvwxvxKX8O6tPawySkFpoQ2YZTjj54yj9B97oOlf8ABOr4xf8AC6v2R/BGsXd79o1bw9bnQdULTea4mtAERnbrveAwSHPOZOp6mqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr/AIKffsqf8M9/HOfxl4T0vyPA/j+SS/sFht9kOn3hObi1GPlUbj5iKAoCSBQP3ZNf8Eqf2oovBnimf9nbxnqbLpPie58/w68gytvqBGHh3dQsyquB0DrwMyEmqGPWqqqqqpeRjNVVVVUOowaqqqqql6cHpVfEfwjD468Eav4XkA8y8tz9nYnGyZfmjJPpvVc+2R3qzur/AEPVYL63L297p9wsiZBVo5EbI46ggivCHiOz8X+F9M8TWJ/dajbLLtxjYxHzKfcMCPwqqoYz1r/gsxq1nP8AGbwHoUbN9qs/DD3MoK8BJbmRUwfXML8duPWv+CQ9jdJZ/FPUXgYW00ukxRyEcM6C7Lge4EiH/gQqMZqNcnANIoxgVGvHFRKD0qNQBxUaZwKjQGkQYpYyaEQxxRiBGKaMgcU6Ajj8q+NXgybwL8TNc0Noilu9y1zZkrgNBISyY9cZK59VPSvDusx26raTybSp/dsTx9Kh1t449gk7eta7r8UKMzOGkI+VM8n/AAFMZbmbOC8kjdAMkk+lfCvwg/gn4f6ToFwm27SIy3QJyRK53MPwzt/4CKlUZqVamUdq/wCCsGkRQfGHwfrqzMZLzw0bZoyOFEVzKwI9z5x/IV/wRQ1a8n0b4uaC7J9ls7rSLmIBcESSrdq+T6YhTjtz61VVVVQ5GM1q+t6L4fsm1LXtXstNtEzunvLhYY1wCTlmIA4BP0BrxN+1r8AvDDNDL48g1KZWK+XpkD3WcHBIdRsx/wAC56jNeJv+ChnhiBWTwb8O9TvWKELJqd0lrsbHBKR+ZuGccbhn1FeJf27/AI1axuj0OHQtBj+YI1tZmaQZxjJlZlJGD0UDnkHivEnx3+Mni4yLrvxL1+aOUkvBDeNbxN827mOPavBAwMcYGMVI91f3TSSPLcXNxISzMS7yOx5J7kkn8a0P4IfGDxKiy6L8M/Ec8TIWWZtOkjjYA7Th3AUnPGAc8H0NaD+w78e9ZJXUdL0fQ8EgG/1JHzgDn9x5vXOPwPTitA/4J3azMI5PFPxMsrbGwvFp+nNPu/vqHd0x6BtpznJAxg6J+wF8H7BY5NY1zxLqkqhg6m4ihibJ4IVY9wwMfxnn8q0f9kj9nrRowsPw6t7qTywjyXl3POXx/FhnKgnvtA/KtL+Gnw50MEaJ4A8N6flg5+y6VDDlh0PyqOfeqqqUg45qqqqqqXrjNVVVVVQxnGaqqqqq/wBN07Vbc2mp2FveQEgmK4iEi5HQ4ORWt/An4MeIiz6t8LfDEsryeY8yaZHFI7Yx8zoAx69CcdPQVrP7Ef7O+rANaeFb3Sn+bc1lqc3JPfEjOBjsAAOec8Vrn/BOjwPcLIPDXxF1yxJUeWb61iu9pzznZ5WQRnHTHv0rxD/wTw+Jdn5j+GvGnh3VETzCq3Ky2kjgfcAAV13N6FgAcckcjxD+x9+0N4dDySfD+XUIEIAl066huN/y54RW8zjBHKjn6jOveBvGvhUH/hJ/B+t6Rtxn7fp8tvjPT76jrWh+J/Enhi4N34Z8Q6lpM5IJlsbt7diRnByhB4yfzNeGv2vP2hPDAjig+IVxqMEagGLU4I7rfhcDLupk9DncMkZOea8Of8FFvHNmgTxZ8PtE1TGRusbmSyJGBjO7zRnOScADkcDFeGv2/vgrrBjh16w8QaDKSA7zWqzxLlsZDRMXOBgn5B3Az38KfHb4OeN2jj8M/EjQrqeVQUt3uhBM2RniKTa/cZGODwcGhgjg1VVVVVV9DQIPGaqqqqqruztL+2ksr62iubeZSskMyB1cHqCp4I9jXxE/ZA8D+J5JNR8G3b+G71skwpH5ts5/3Mgpk/3Tgf3a+IfwY+IXwxlY+JtDY2O7amo2uZbZ+w+cD5SewcKT6V4B+LPj34aXIl8K69LFblt0llN+8t5fXKHgE+owfevhv+174M8TeVpvji3/AOEd1BsL5+S9rIf97rH/AMC4H96rS7tL+2ivLG6iubeZQ0UsTh1cHoQRwRVVVVVfGH9lbw142E+u+ChBoetkMzQqu22umPPzKPuMf7yjHqD1rxb4O8S+BtYl0HxTpM9heRH7si/K4/vIw4ZfcZFfB/8Aav8AEXhDydB8fG413R1yqXW7dd2w7ck/vV68Mdwzw2AFrwz4q8OeM9Ii17wtrFvqNjNwssLfdOASrA8qwBGVYAjPIrWdH0zxBpV1oms2cd3Y3sTRTwSDh1IwR6j6jkHkV8YfhfqXwo8ZXHh66LTWUuZtOuSP9dASduTgDeOjAd/Yiv2SfjMNRsx8LPEt6PtNom7RpZGA8yIfeg7Esv3l6kruHAQZqqqqqqJA4FHnvVeP/Flt4F8F6x4tuihXTbVpEVzgPIeI16j7zlR+Nf8AE08Sa1/Fd6jql17BpZpG/AZLH9a+H3hC18BeCtH8IWZBXTbYI7A5DyElpGH+87MfxrxPLDceJNWnt5UkjkvpmR0bIYFzggjqCK1L4SaP8Yf2dPDfgzWVW3uv7AsZLG7eLc1ncLbphwMg46qw7gmtd0TxH4A8VXWh6rFcaZrWiXexwrlHhlQ5Dow+gZWHBBBBwQa/Zi+N9v8AGbwDE+o3EQ8SaMFg1SEHl+MJOB6OAc+jBh0xX7XPw+/4WB8E9Y+zQ777w/8A8TW1wOf3St5gHIzmJpOOckDjOK+FPje4+G/xG8PeN7dmxpV6jzqvV4W+SVR1xujZ1zjvVtdW99bQ3tpKssM8ayROvRlIyCPqCKrUNPsdWsZ9M1O0iurS6jaOaCVAyyKRggg9QRX7QP7Olx8OnbxX4PimuvDTkCaNiXksGPHzHqYyejdicHsT8I/i54h+EviEanpjG50+4IW/093wlwg7j+645w3bocgkHwF8QPDPxK8PReI/C94ZYHO2WKQBZbdx1SRQThuncggggkHNftn/APJy3jH/ALcf/SGCv2E/+SAWf/YUu/8A0IVRxjrR5qqqqr/gsP8As5f8I1410j9o7w1YbdP8UbdO1/y04jvo0/cynH/PSJCv1h9Wr/gnr+0DH8I/is3gfxFerF4a8btHbySSy7Us7xc+RLk8BW3NG3T7yMThMV4g8M+HvF+mPo/iTSoL+0k/gkBBU+qsMFT7gg1rn7IWg3k5m8N+L7uwjO4+TdWwucHsAwZCB9QT/X4hfDDS/hdqkWjeI9fub2+lgFxHDZWarG8ZLKMys+VO5CMbG4yfQH9kWz8E39nrGqWXh2GHWtPuERbqWQzSrC8eBgnAUlllyVVcjAOcVHKCME1HLgcGklBHBpJ8d6W4GOtCcYwDXnDGA1GcdN1NcDGAaknzxmpJcjk1JLkcGpZR0BqSQDvzUsgxnNSuD3qV896/b5/aOh+Dfwvm8G+HNT8vxh4whe3tPJlKyWNqflmueOVOCY0OVO5iyk+Wwr4CfB/Xfj38XvDHwn8PBln169WOecLuFrbr800xHokau2O5AHevCHhTQfAnhTR/BXhexSz0jQrGKysbdekcUSBVHucAZPc81VUeTwaqqqqo9ODVVVVVU3PANVVN04qqqjzVVVVVN0wDVVVVVU2CeKqqqqp+R1qqr9uL/gnz4b/aGtb/AOJnw2ii0j4lQW+WTKx22uBBwk3HyzbQFSXIHRXyMMkcvjn4UeNt8MureF/Ffhu9ZSVL2t1Y3EZwR2ZWByDX7CX7duiftK6JD4C8e3NrpvxL0y3zLEMRxa1Eg+aeBegkAGZIh05ZflyEqm5PBqqqqqru1tbyEwXdvHNGf4JFDDP0Nah8PNJuSXsbiW0Y/wAON6j8Dz+tSfDfVw2Ib2zZcdWLKfywai+G+rM4E19ZqvcqWY/lgVong3StHZLhgbm5TkSyDAU/7K9vxyfeqqqbBPFVTYJ4NVX7QPwss/jb8FfGPwtu1Tdr+lyRWrv0iukxJbyEZGQsyRtjIztrS9S8QeCfE1nrGmT3Ok65oN8k9vJtKS2lzC4ZWwRlWV1BwRwRXwp+IGn/ABW+GPhb4kaWuy38SaTb3yxHrC0iAvGeTyrblPJ5U8mqqqqqqqqqqtQ0+01S0ezvIw8bj8VPqPQ1r3h680KfbKPMgckRygcH2Pofaqqqqqqqqqqqqqqqqqqkd42DxOysOhU4Iq08T67ZkeVqUrgfwynePpznHSrP4h3SAJf2Mcv+1ExQ/kc5/StP8V6HqWFjuxDITgJP8h/Dsfzqqqqqqqqr/grx8M7fw18bPDfxLsoEij8aaO0VyQqgy3VmVRnJByT5MtsvI6IME8hf+CIvxTePVfiN8Eby8lZJ7eDxDp1v82yMowt7p/QFvMsx2J2d8cVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVftT/ALPXhv8Aaf8Agnr3wp19Yorm5j+06NfumTp+oRq3kTjgkDJKOBgtG8igjdmvE/hvxl8JvHl/4Y1yC80PxP4W1FopQjtFLa3EL8OjDBGCAyuO2CDyDX7Fv7TenftP/B208R3M0SeK9F2WfiS1RNgS528Sov8AzzlUbh2B3L/DVVwRxVVVVVLjpmqqqqqrgjg1VVVVS46UMEDBqv2lfBLeFPiRc6nbxgWXiAG8iI7SE4lB99/zemHHoa/ZG8b/AGiw1L4f3kmXtWN5ZFm/gbAkQDsA21uP77VVVX/BXbW/7W/artLD7N5X9jeErK03793m7pribdjA2/67bjn7uc84H/BJjSJIfg74x14zKY7zxN9mWPHKmK2iYn6Hzh+RqIDjFRDPSkAOAKQcAColBAApFBIHpUS5HFRoMYFImaSMEdKEWRwKaPjpUkfHFSJjkV8cPgvpfxd0FIVmistascmxvWjLYB6xuAeUbA55KnkA8g+L/BHirwHqraN4r0a4sLkZKGRcpKP7yOPlcc9QTg8HmkurmNPLjuZVX+6HIFW9td39zHa2kEtzcTNtSONC7ux7ADkmvgZ+z3fabf23jXx7arFJB89lpki5ZX5AeUEYBHBVeucE4xipBzipQMcVKARwalGRxX/BW/SIoNe+GmurMxku7TUrZoyOFET27Aj3PnH8hX/BFbVryHxt8UNBjZPst5pWn3MoK874pZVTB7DEz8fT0qqqtb8Q+H/DVp9v8Ra7p+l2wzma9uUgTjr8zkCvFX7Z/wABfDO6O18Q3uvTp96LSrNnxnb0eTYh4bPDHG0g88HxV/wUL1mXdD4H+HllbY+7Pqt00+77ucxx7MfxD75zkHjBB8T/ALVvx78VFkuPiDd6fCQQsWlolntyBnDoA/bPLHGTjAOK1LVNT1m7a/1fUrq+un+9NczNK7fVmJJrwv8ABH4u+NGi/wCEb+HWu3UU3+ruHs2hgPIH+tk2p3Hfpk9ATXhj9g34zaysc2v3mhaBGfvxz3RnmXg4wsQZDzgH5x1714d/4J5eErdQfFnxE1e+Y5yunWkdpjgcZfzc4OecDPHArw7+yH+z/wCHWEqeBU1GYZ/eajdS3GRkHGwts7dducZGeTWheE/CvhaH7N4Y8NaVpEWCPLsLKO3XGckYQDuSfqaqhyMZqqqqqlPGAaGCODVVVVVfQ1wRwaqqqqqhyOtVVVVVQPbNVX0NAgjrXiD4HfB3xSQ+u/DLw3cygg+cNOjjkOAQAXQBiOehOPyrxJ+wd8A9bQ/2TZ63oD/w/YdRaQZAI5E4kJBJBPIPAwRzXif/AIJy61EHk8GfEuxus7tkGp2LQbemAZI2fPfJ2DHHBzx4r/Yy/aC8LNK8fg2PWraLP+kaVdpNu5I+WMlZTwAfudCO+QPEHhTxR4TuRZeKfDeqaPcN92HULOS3c8A/dcA9GU/iPWvCfxS+JHgTavg7x1rekxLn9xbXrrEfvdY87T95jyOCSevNeEf28/jl4e2Q67No/iWFeGN7ZiGXHs8JQZ92Vv614R/4KJeAtR2Q+NfBOsaNIeDLZSpeRA+pz5bAewDH69a8G/tF/BHx6qr4d+JGjmc9La8m+xzE4JIEc21mwAclQQMdaqqqqqgexNcEcGqqqqqqaGG4he3niSWORSro65DAjBBB6gjtXxJ/ZJ8BeLvN1Hwk3/CM6k2Tsgj3Wsh94sjZ2HyEAddpr4h/Bj4hfDGVj4m0NzZBtqaja5ltn7D5wPlJ7BwpPpXw8+M/xC+GMqr4Z1xjYhsvp10DLbP3PyZ+UnuUKk+tfDD9qzwL43EemeJynhrVjgAXEubaY/7EvG08fdcDqAC3NI6SIskbhlYZVgcgj1FVVVVeNfAXhT4h6O2ieLNIivYOTEzDEkDEY3Iw5U/T8civjB+zF4r+HYm1rw75uu6AnJlRP39uP+miDqB/fXj1C14B+JXjD4aasNW8J6s0Bbia3f54Zx6OnQ/XgjsRXwe/aG8I/FWCLTJXXSvEKpmWwmb5ZcYy0L/xDn7pww54IG4/G74U2PxZ8GT6RiOPVrMNNpdy+QI5cfdOP4XA2nrjIOCVFRS634T14SxNcabq2kXXGRskt5o25BHYhhjFfBn4nWHxW8E2uvwlI7+H9xqNuGGYpgOSB/db7w9jjqDVVVVVEjGAaPPeqr9tLxq9j4f0fwHZzqH1OVru8UDnyo8BAfZnJPrmPt3/AGUvBS+K/ipbapdWyy2Xh2I3sm9Tt837sI/3g53j/rmaqvCf/Iq6N/2D4P8A0Wtftq/AOPxdoL/FfwrYFtc0eEDUooU5u7Vf+WhHd4x36lMj+FRXwe+KOs/B7x9p3jXRwZUgPlXtrvKi7tmI8yMn8AQTkBlU4OMV4c8Q6D428N2PiTQbyK/0rVrYSwSAZDow5DA9COQVPIIIPINfGXwFN8Mvid4h8FvFshsLxjZ/MW3Wz/PCcnqfLZc+hyO1fsdfED/hOfgnplncyIb3wyx0uZQ3JSNQYWx1A8tlX0JRsegqpoYbiF7eeJJI5FKujrkMCMEEdwR2r9oz9nN/Cr3PjzwHZFtFYl76wjXJsT3dB/zy9R/B/u/d+GPxO8R/CvxHHr2gzeZE+EvLKRiI7qMfwt6Ec4bqD6gkH9pnxXp3jz4z674z0eKdLHVYrN4RMm1gVtIUdT7q6suRxxxxX7Cf/JALP/sKXf8A6EKpiDwDVVVVVfGH4V+F/jd8MPEfwo8ZQs+k+JLFraZ4wN8D5DRzJnI3xyKki5BG5BkEcV8ZfhN4t+BvxM8QfCvxta+VqmgXjwNIqsI7mPrHPHkAmORCrqSAcMMgHIH7Bv7T8Pxr8BJ4G8U38f8AwmfhS1RJlJIa+s12olyMklmBKrIf7xU8bwBHJjjNfte6Ik2i6B4mQANb3Mlm57sJF3r+Xlt/31X7JmvLpfxMl0mSXCavp8kaLgcyIRID6/dWTj39qSUYwTSTYGAaSYEdaWY9jmlnHQmhODyGrz/9qmnBGd1NOD0NPMSMA08wx1qSbI4NSSjoDUko9akkz3qWTPANfF34r+Efgz4D1Px/4z1BYLHT4j5cIYCW7mwdkEQP3pHIwB0HJJCgkfGn4t+Jfjh8R9X+IviiVhPqEuLa18zellbrxFAnA+VV74G5izHlia/4Jdfslv8ABX4bN8Y/Gmn+V4y8dWaeRFLGySadphKukTA9HlZUkYY4AjHBU1VUenBqqqqqqbpiqqqqqpuTwaqqbpiqqj04NVVVVVTc8VVVVVVHmqqqqjjFN0xVVVftv/sF+HP2nrD/AITTwbNZ6D8RrGHYl5IhWDVY1GFhudvIYYAWUAkD5SGXbt1PTPiF8FviFJp2owan4V8YeFb8EgMYbiyuIyGVlZT9GVlJDAggkEGv2Cf+CgFl8frW3+Ffxcv7Wy+I9tGfst1sWCLXkUZLIqgKk4AJaNQAwBZABuVKOD0qqqqqmxjFVVVVVR5qqOD0qqr/AIKG/ChfhN+1V4stbO2SHTfFDLr9gqk/duSxm4PQC4S4AA4AAxjoP+CSfxOi8Vfs/ap8Np5ma88Daw+xCeFtbvdNGR3yZRdcYxwOTk4qqqqqqqqqqq6tLa+ga1u4VljccqwrxH4TudHLXVqTNZ5+9jmP2P8Aj/KqqqqqqqqqqqqqqqqqqqqtM8RatpJVbW6JjX/llJ8y/l2/DFaJ4x07VdtvcEWtyeArt8rH2P8AQ/rVVVVVVVf8FV/h7/wl/wCy7L4pt4N1z4L1m2vyyjLeTKTbyL9MzRsf+ueegNf8E5PiN/wrT9sr4b6hNLi01zUG0O4TbnzPtqNBEMhWIxO8LcY+7gkKSaqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr/gsB+yNDrOjx/tV+BNNc6npyx2fi63ggz9otgAkN6dvO6PCxOTnKGP7oiOf2O/2k9S/Zg+M2n+NSJ7jw7qCix8RWMWCZrN2BLoDx5kbASL0J2lNyh2NaNrGleItHsfEGg6jBf6bqdtHdWd3buHjnhkUMkiMOCrKQQe4NUOQKqqqqodcVVVVVVDkCqqqqqHXANLyMVX7RngN/G3w7uLixi36jobG8twFyzooPmoO/K/NgdWRRXw/8W3HgXxjpXim3VnFjODNGpwZIjw6j3KkgZ4zirC+s9UsbfUtPuEntbuFZYJV6OjAFWHsQQaqq/4KX6y2sfto+P0TUUu7fT10+1g2OGWILYW5eMEdCJWkyOobINf8EvNLtLD9mL7XbBhJqXiK9nn3NnLhYohj0G2JPxz61EBjAqIAjg1GATgVGAcYNRAcCoxmolBwKRRgAVEgIwKjjBFKgxgUyAjFSIMVKg6VIvGK1rRtI1y0NhrWl2eoWzHJguoFlQn12sCOh/Wr/wDZ5+DV3LHNL4FtVaL7oinmiB+oVwD+NaF4E8G+EQx8NeGdO053+9JBAFdvYt94jnpnvUg5xmpQM8VMMjAqUAj8KlHy4r/grdpEU/hr4b66ZmElnf6hbLHjhhKkLEn3Hkj8zX/BGXWbqD9oHxl4dSOI2174Ne6kYg7w8N5bIoBzjBE7546heRzmvGn7RfwW8BBo9f8AiBpr3Kji1smN3LnbuAKxBtmRjBbaORzzXjP/AIKD6JAr2/w/8B3d45BC3WrTrAqsCOfKj3FgRn+NT0+leMf2v/jx4vMka+Ll0O2kXH2fR4Bb7eCMiQ7pRwf7/YHqM1qWqanrN21/q+pXN9dP96a5maV2+rMSTXhP9nX42eNdj6H8OdWEL/dnvYhZxkeoaYqGH0zXhP8A4J9+OdQ2TeMvGmkaRGeTFZxPeSD2Odig/QsP5V4W/YX+COiIp16PWPEUpHzfa70wJn1VYNhH4sa8K/DD4deB9reEfBGi6VKuf39tZIsp69ZMbjwzDk8AkdKqqqqpcdKqqqqodetLyMZqqqqqqU5GM1VVVVVA9s1VVVVVUCCOtVQOO9cEcGqqrq0tb63ezvbaK4glXDxSoHVh6EHgivFf7LPwC8YmSXUvhtplrO6tibTd1iysR97EJVWPf5gRnqDXjD/gnX4NvvNn8C+PNU0qQ5ZINRt0u48/NhQy+Wyryoyd5ABzuzx4w/YU+O/hrzZtHsNK8S26ZYNp14Ek2DdyY5th3YUfKu7lgBnnHiXwV4w8G3H2Txb4W1bRpScKl/ZSQbvpuAyPcV4O+LnxO+HypF4M8eazpUCEkW0N2xgyTkkxHKHnPVe59a8F/wDBQb4qaIqW/jLQNH8SxLjMqg2U7YAByyApzjPEfUntgDwV+3h8EfEzRW2vyap4ZuZH2/6dbebDknA/eRbsDGMllUDnsM14X8aeEfG9gNT8H+JtM1m22qWksrpZtm4cBgpJU8Hg4IwfSqqqoHHANAgjg1VVVVVVNDDcQvbzxJLHIpV0dchgRggg9QR2r4k/sg+DPEvm6l4GuP8AhHtQbJ+z7S9rIf8Ad6x/8ByB/drx98L/ABr8NNQNh4r0WW3Rm2w3aKXt5+M/JJjBOO3BHcCvhh8ffH/wueKz02/GoaOHzJpl58yY7+W33ozjPQ7c8lTXwv8A2h/h/wDE7y7C3vP7J1hsD+zr6RVaRj2ibOJPoMN3wKqqqqr4y/so6B4tWbX/AIeRW+jawfmkswNlrc4B6KB+7c8cj5T3GSWrW9C8Q+DNck0jW7G50zVLFwWjf5XjPBVgR+BBBx3Br4M/tb3Nn9n8NfFWZ7iAbY4NYRMyRjn/AF4H3x0G8Ddxk7iSa/at+FOn6xp0fxm8FtBdRSon9ptav5qTxkAJcKVJBAACkjjGD6mvgn8U734T+NbfWg0smlXWINUtkP8ArISfvAHjeh+YdO65AY1Y3tnqdjb6lp1zHcWt1EssE0bblkRgCrKe4IIOaqqokAdaJzwKqqr9pXxV/wAJZ8YdceNy0GkuNOhBz8vk5Djn/poZOmBz+J/Y88JnRPhnP4iuLdUn8QXrSI+BloIvkQHv98SkZ/ve+TVeE/8AkVdG/wCwfB/6LWpI45Y2ikRXRwQysMgg9QRX7W3wJPwl8bnXfD9kU8LeIZGltAkYCWc/WS344A/iQYHynAzsY1+xT8dR4O8Q/wDCrfE18qaNrs2dPllcgWt2eAg7BZOB/vY/vE1/wUE8ARwXnh34nWcSr9pDaXfEDGXUNJCeBySvmgknoijtx+wh8QH8O/E+78D3Vwq2Xim0Plow/wCXqAF0we2YzMD6nb6AV8W/2spNS8eaR8Jvg3Otxc3Ws2tneawjKUctKqmGAng5J2mQ8ddvUMKp0SRDHIoZWBBVhkEV+0f+ziNPN18Qfh7Yf6LzLqWmQp/qe7SxKP4O7KPu9R8uQup6ZaataNaXSZB5Vh1Q+or9iFLex+DH9hi9gmurHVLgzRo43IrkFCV6gEA4+h9KJAFHnvVVVVVV/wAFLP2MW/aI+H6fEj4d6OknxE8IwuyQwQgyazZAZe2OBuaVD88XXkugGZAV+HHxE8VfCjxrpXj7wXqH2XVdJmEkRYEpIvRo3UEbkZSVYZGQTyOtfs+/Hrwh+0F4AtfGfhe48u4jxDqmnycSWNyFBZCO6nqrDhh6EED9oHTRq/wl1pQwD2gjuE3EgfJIu7/x0t+OK+Eer/2H8TfDOoGTYo1GKJ3MnlhVkPlsSfTDHPqMjvSTDGM0k3HBpZgRyaWb0ahOcYBr7RxivtFG4yMU05IxmmmyPvU8w7GnmyOTUkwxgGpJR3P4VJLkda8f+P8Awp8NfCmoeNfGusw6ZpGmRb555T+AVQOWdjgKoySSAK/an/ah8T/tKeL1upo5NN8LaU7ro2klhlAeDNKRw0rADPZR8o7s3/BNb9hS8+LXiOw+O/xa8PsvgTR5hNotncjaNbvI3G1mQg77VCrbs4DsAvzKJBVVVNjGAaqqqqqm5OAaqqqqo4xR571VU3JxVVTdMA1VVVVVNgniqqqqqOMVVVVVTdODTcnANVVVX7bf7Enhf9qfwuda0QWukfETR7crpeqMu1LtBki1uSOTGSTtfkxk5GQWVvG/gj4hfBD4hXXhPxZp9/4d8U+HLtW+WQxyQyKQ0c0UinkHCukinBGCDX7An/BQ4fFuS1+DXx11e3h8as2zRtadUhj1nPSGQKAqXA6LgASDA4f79VVVVU3PFVVVVUenFVR6cVVVX/BYb4WvqngXwX8YbCPMmg30mlX+GYkw3C74mx0AV4mXIwSZh1A4/wCCV/xJTwT+1FbeF7yfZaeN9JuNN+ZwqLOgFxExyRyfJeNQMkmUDHPFVVVVVVVVVVUQGG0gEEdDXiDwPHKGvNFAR+rW54B/3fT6fyqaGW3laCeNo5EOGVhgg1VVVVVVVVVVVVVVVVVVVV4S8VzLNHpWpzb42+WGVuqnsCe47e1VVVVVVfHrwIvxO+CnjnwAImkl1zQLu2tgqklZzE3lMAGXcRIEO3IBxg8E14e13UvC2v6Z4m0afydQ0i8ivLSTH3JYnDofwZRXhjxBp/i3w1pPirSSxsdZsYb22L43eXKgdc7SRnaw6Ej3NVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVavpOm69pV5oWs2UV5YajbyW11bTLuSaJ1KujDuCpII96/by/ZK1D9kv4zTaBpy3Fx4K8RK974YvpVJJhBAktnY8GWFmVTgnKNE5wX2j/gll+1zFpN3F+zL8Q9YYW99Kz+Ebq4k+SGU5aSyJPQOctHk/fLKOXQVS8jGaqqqqodeDVVVVVUuCMZqqqqquKXHSqdEkQxuoZWGCpGQRXxk8D/APCv/iDqWhQQsljIwuLEsOsL8gD2U7kz6oa/ZY+IH/CQ+EpPBt/PuvtBx5G48vbMfl/75OV9gVqqr9sPWpdf/ap+LN9NCkTReLtRtQqkkEQTtCD9SIwT7mv2DNIk0T9kn4eWUsyytJZ3NyGUYwJruaUD6gSAH3FRgY49KiGQBmowCcVGBkCohnoaiAOMGohxSAcCohUajFRoCKkjAGBUqgipgMVIOePSpgOgqYDoDXxw/as8J/C+6k8N+H7dPEHiGJis8KS7YLMjqJHGSX/2B05yV4B8RftWfG/xDO8i+LV0uEvvW30+1jiVOvAYguRz0LHtWiftQ/G3RbhZf+EzfUIg254L63jlV+MYJ2hwP91hXwg/av0Hx7dweG/GlpBoWszlUgmRyba6kP8ACM8xMeysSD/eyQDKBt4q5lhgheeeVI441LO7ttCgdST2GK/4KLW/h/4yeEPDPh3wN4w0q91fR9ZaS4t0cvGsLwsGcyoCuVZUXaMk7z02mv2WYPE37LHi7UvHvhfV7K/1rVNHk0pxc2hMUMTywyllG7JfdAoBPGCfl6EeM/jN8VPiEHTxf481bUIJMbrXz/KtyRnnyY9seeTztrwx4H8ZeNrn7H4Q8LarrMqkBxZWjzCPJAyxUEKMkcnAGea8F/sLfGHxFifxNLpnhi34+W5mFxMQe4SIlfwZ1PI98eD/ANgr4T6KY5/Fmrax4kmVcPE0gtIG4GTtj/eDkH/lp0OOSM14Q+Ffw48Aqo8HeCtI0uQf8t4bZfOPBHMhy54JHJ7n1NVVDBHBqqqqqh7GqqqqqpTnHNVVVVVQOOhqqqqqq+hoEEcVVVVVVA46VwRwaqge2aqqqqBBGM1dWlrfWz2d7bRXEEq7XilQOrD0IPBFeNv2SvgJ45Lz3ngW20u7ck/adIY2bAkAE7E/dnoDyp5ye5z42/4JzA+ZcfDj4iegjs9bt/rkmeIf7ox5Xqc9q8c/sq/HbwCZ5dS8B3moWcCljeaV/pkZUDJbCZdQOc7lXGM9Oas77W/DepGfT7290vULZyheGR4JYmBwRkYIII6e1eB/20fj14MMEFz4mi8R2UKhfs+swCZmGMZMy7ZSfcueeuec+Af+ChPw81kRWfxB8N6j4duDtVrq2P2y3z0ZiABIo74CuevJxz4I+LHw2+JMAm8D+NdK1ZtgdoIZws6AjILxNh06H7yjofQ1VV9DQYHg1wRwaqqqqqq1PTNO1rT59K1axgvLO5QpNBPGHSRT2IPWvif+xxpWoJJqvwvvBp9zks2m3krNC3tG+Cyn2bI91FeJfCniTwfqTaR4n0W6027T/lncRldw9VPRh7jIr4VftV+NPBUkWl+LpJfEejDA/fPm6gA/uSH7w/2Xz0GCteAfib4M+Jmmf2l4S1iO4KAefauQk9uT03x5yM4OD0ODgmqqqqviR8LPB/xT0Y6V4n08NLGD9lvYgFntmPdG9DxlTlTgZHAI+LnwM8X/AAkvPM1FF1DR5X22+pwRkIxOcK687HwM7ckehODXwr+NviX4aGTSJFXV/DV6GS90e5b5HVhhjGedjEEg44OeQeCPGdl4bttbluPB989xot5++s0mOJ7dT1ilXJw6nIzyGABBINfsgfF0srfCjX7tflDTaNI5A75kgz36l14/vjP3RVUcY60SOgo896qqrxTrkHhjwzqviO42mPS7Ka5YHvsQtj8cY/Gv9M1a/wD47i7vJv8AgUkjH+ZJrQbTQvh74U0Hw1d6pZ2kVpBDYW7zypF9olVOgzjc7bWbA5PJ9aqvDMsNv4Q0meeVI449NhZ3dtoUCMZJPYYrwt8RfCHjjUtT03wnqyan/ZHli6uYATCGfdtVX6OcKTlcgcc18U/hzoPxX8D6l4I1+NRFex5guNm5rWdeY5U5HKntkZBKnhjXjDwpr/w98Waj4S1+A2uqaPcGOTaSBkYKuhIBKspVlOBkEHvXin9rK2+I3wUufhh4/wDCl1c6uLWJYNZiu1bzZ42ysrxlV2k4UNhmzuY8dKtrq5sphcWdzLBKoIDxuUYAggjI55BIPsa+B8E1x8Z/AcdvC8rDxJYOVRSSFW4RmOB2ABJPYAmqpiMYBqv2m4PDfg745aj4a0mwi0+2uraC5jSPiMSSLlgB/CCRkAcc4GOK+H/xA8R/DbxHB4k8N3eyWP5ZoXyY7iPvG47g4+oOCMEZr4Z/E7w58VPDkeu6DNsljwl5ZOwMlrJj7reoODhuhHoQQKqqqqo4Ao896/4KffsGf2S+r/tP/B7S4k08/v8AxbotrDtMDlsNfxKvBQkgyqB8pzJyC5X4CfHvxv8As9eN4fF/hC5823l2x6npcrkQ6hCD9x8dGGSVcDKk9wWU+Dvi74I/aE+CuqeKPA2q7re/0u4t7q3lA8+wnMJDRSoDwwyD1wwwQSCDVjdyafe29/AqtJbSrIgccEqQRnHbikm4xnNJMDjDUs5x1pZwRjNCcHo1CfP8VefxgNTTjHDCmnBH3qacdAfzppyRwaeYEfep5sjANSTD+9Xxl+OPw8+BfhSXxZ8QdbW2i2t9ksoSrXV84xlIIyRvb5lychVyCxUc1+0p+1H49/aR8QifW5f7O8OWEztpWiQPmOAHgPIf+Wku3gseBkhQoJB/YO/YD8QftKa7D48+Itle6R8M9NmBkkKtDNrcgwfIgPBEXTfKOn3VO7LJoGgaH4V0Ow8NeGtJtdM0rTLdLezsrWIRxW8SDCoijgAAAYqqqjyeDVVVVVHGKPPeqqqqqbpgGqqqPNVVNycA1VVVVRxjrR571VVVVTdMA1VVVVTYxijyaqqqq/bA/Y58DftX+EEgvJItG8Y6TGRo2vLDuaMZyYJgMGSFiScdUY7l6srfFr4S/EL4CfEK++H/AMQNKl0vW9LkDxyRsTHcR5/d3EEnG+NsZDDBBBBAZSB+wX/wUfs3s9N+Cn7RevtHcxsltofim7fKyqflWG8c/dYcBZjwR98gje1VVVVNgng1VVVVTdODVU3Tg1VVX7XHwzX4vfs3eP8AwNHAk13caPJc2CspJN1b4nhAwQQS8Srn0Y5DDKn4f+MdS+Hfjrw7490dmF74d1S31CAKwUs8MiuBkgjB24OQRg8gjitG1fTPEOj2Ov6LdpdafqVtHc2k6AgSxSKGRhnnBUg8+tVVVVVVVVVVVVavoOm61Fsu4QJAPllQYZfx7j2Na14R1PSSZY0+024ziSNTlR/tDt/KqqqqqqqqqqqqqqqqqqqqvD96+o6LaXcjFnaPDE9SynBP5iqqqqqqv2hPBI+HHx08feB47ZIINH8RXsFqiKqj7P5zGEhU+VQYyh2jpnGBjFf8E9/Gq+Pf2MvhXq5uXlksdFGlOHVFZPsUj2oUhOMbYVwTyVKk8k1VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX7Wf7NXhb9qn4N6p8NNeENvqK5utB1Jwx/s++VWEchCnLIQzIy85VjxkKR4x8I+Ofgz8RNS8H+JbW80DxV4T1IxTBJDHJbTxNlZI3U9OFdJFOCCrKSCDX7Bn7XNh+018NU0vxHeQReP/DMKxazbAhDeRg7Uu413ElWG0OeAshPADLlcdM1VVVVVcEcVVVVVS+lVVVVVSnOOaqv2oPh//wAJT4JHiewg3ah4d3SttHL2xx5g/wCA4D89AretfDDxtcfD3xtpviWJmMEUnl3ca/8ALSBuHGO5xyPcCrW5t761hvLSZJYJ41kikU5DqRkEexBFVNLFBE888qRxxqWd3O0KAOST2GK8Va3/AMJN4n1fxH9m+zf2rfz3fk79/leZIX27sDOM4zgZ9BX7P+gDwt8C/h74daK2SXT/AAxp0U32dcI0ot08xhwM7n3NkgEk5PJqPBGAe1RdBX7Qep6lonwA+Jus6LqNzYahYeDtWuLS7tZmilt5Us5WSRHUgqysAQwIIIBFfszftufGX4UfFbRPF3xO+IvjPxT4LvZH0/VLfVtUub+NYiULywrK5Xzosxtx821ivAetLv7HVbG21PS72C8s7yFZre4gkEkc0bKCrowyGUgggjgg18Q7u5sfh34ovbG5lt7m30W8khmico8brCxVlYcgggEEdDX/AASU+LnxX+JHxF8eWPxD+J/izxTb2ei28ltDrWtXF8kLmYgsiyuwUkcZFJgkc1+1X8XU+Bf7Ovjr4mRXSQ3+maTJHphYE5vZsQ23A5IEsiE4xwCcgAkfB/8Aa0/aZ+FPxL+GnxK+Ivxl+Imr+Db/AFBLm4t9V8QX15a3tilw1vdL5buVd1CyEDDbW2HHQVbXEM8KT28qSRyKGR0bIYHkEEdRUjgjANSEYxmpiCMA1JgnAPapsHgGv2pvi5P8KPh076PMU1zXHa0091ODB8uZJh/ujAH+06noDR825lJLNJJI2SSckk9SatPD5kUGVix9F4Aq68OhEzEzKQOAeRUsTwuYpFwy9RXwU+Lvivxr8HvEGj2N0JPFvhawJs55iJGuk2sYyQ33mBQoSeOVJ614r8e+MvHNz9q8WeI73UmDblSWTEaHn7sYwq9T0ArXGfw34Xm8a63a3VroUDrG+oNbSGEOx2hQwByxOeBzwfQ1+zn4r8KftA/Hrwt8GNPOpWUOvvdebqLRqPKWC0muPlTJ3bvJC84xuzzjB8Gfsh/AjwaI5R4R/t26jz/pGtSm63DjgxcRduuzPJ5xVnY2WnWyWWnWkFtbx52RQxhFXJycAcDkk/jVVVVS8jFVVVVVQ5FVVVVUvpmhgjrVVVVVUDkYzVVVVVUDzjNVVVVVVAjpmqr6GgcjGaqqqqBzwTVVXjT4VfDf4ixhPG3grSdXdVKpNcWw85B3CyDDqOB0I6D0rx3/AME9fhzrTvdeAvE+p+G5WLEW1wn26AeiruKyAdsl27enPj/9ij46eCRLc6foUHiexj3ETaPJ5km0dMwsFkLEdkDc8ZPGbq01fw9qZtr22vNN1GykBMcqNDNA4wQcHDKehH4V8O/2xfjn8PY4bI+Jl8Q6fETi11tTcnB7CXIlAHYbyBgcYyD8Pv8AgoP8O9cMVl8Q/D1/4anKgNd25N7bZA5JCgSKCcYAV+vJ4yfCXj7wT49szfeDPFmla1Cv3zZ3SyGM8cMoOVPI4IB5FVWcdDQIPFcEcGqqqqqqvFfg3wt450ptF8WaJbanaE5VJl5jbGNyMMMjYyMqQeetfFX9kHxBoCzaz8OLiTWrBBuawlP+lx8/wYAWQY+jegPWrHUNe8K6v9q068vdK1OykKlo3aGWFwcMpxgg5GCD+NfCb9sJJng0L4rRKjEbV1m3jwCe3mxKOP8AeQYzj5QMkaZqena1p8Gq6RfQXlncoHhnhcOkinuCOtVVVWo6dYatYT6XqdpFdWl1G0c0EyBlkUjBBB6givjf+yhdaIlx4q+GEU97ZAvJc6Sfmkt15JMR6uoHGzlhxgtnh0eNzHIpVlJBVhggitM1K/0bUbbVtKu5La8s5VlgmjOGjdTkEfiK+DXxPsPit4JtdfiKR38P7jUbcMMxTAckD+633h7HHY0SMYBonPeqqqqv2sfEL6F8HL61iYrJrN3DZKw7AkyN+axMPxr9n7w6vif4w+GbCSMvFb3f2uX5SQBCpkG72LIq88ZYDvXxM8DWXxH8D6r4RvCiNeQn7NKw/wBTOvMb+uAwGcdRkd68N/Fr4s/CvUJ9F0/xHe2xsZGtp9OvCJ4omQ7WQI+QhBGMrg8darxl8WfHnju0g0zXtelOm2saRw6fB+6gUIAFJUffIx95skZ61+zL4Rt/BHwgsNQvDHFca0DqVzK5ACxsB5eWOPlEaq3PALN6146/bU/ZR+G8s1t4s+PXhKO4tlJmtrK8/tCaPHUGO2EjBufu4z3xX7Vf7Y37JHxu1zRH+G3iPV7zxQl0NPN42ky21rdWzBmUsZdrDbJhVO0H96cjABWq8CfE7QPgv4v0r4p+KbTULrSvDc4uruGwjR53QAjCK7IpPI6sB714V/4Kj/sWeKJxaTfFC60Od5Aka6rot1Er8dfMRHRR2y7LXgP41/B74pxo3w3+KXhXxM7RhzDperQ3EsYK7vnjVi6HB5VgCO4FHnvVft/6bHY/HO1uklZjqHh+2nYEfcIkmjwPbEYP418L/Ani3xx8ONY8Z6Yv26Pw7dLDdQKpMoiKbvNH94LznuBg8jJHgDx/4j+G3iODxJ4bu9ksfyzQvkx3EfdHHcHH1BwRgjNfDT4i6F8UPC0HiXRCYyT5dzauwL20o+8jY69QQe4IPHQVVVVUxB4BqpoYbiF4J4kkjkUq6OuQwIwQR3BHav2/v+CaV/4Gl1j44/s9aSk/hVVa61nwzbqTNpfd5rZAPmtwMsyZzFyQDHny/hJ8X/G/wT8XQ+MvA1/HHcBDDdWlyhktr6E/ehmjBG9Dx3BBwQQQCK8OXDP4f0x2kLMbOIkscknYKWcdCaWYdmoTnHWhOMYFeeMYDV54/vUZwRjdRnHSmnJGAaabPG6mnB6HNT3UcMbTTSqiIpZmY4CgdST2FftF/wDBRPwD8PFvvCnwk8nxZ4ljVo2vgSdPs5M4OXGDOwxnCfJyPnyCK8f/ABG8ffF/xVL4m8ca9e63q14+1A5yEyeI4oxwi9AFUAfjX7E//BLjW/HD2HxR/aU0660fw58k+neGCxhvNQwwYNdDGYYSMjYCJWyf9WAC+n6fYaTYW2laVY29nZWUKQ21tbxCOOGNAFVEVcBVAAAAGAAKqqo9ODVVVVVU3TANVVVVVTYPAqqpsYqqqjyeDVVVVVN04NVVVVVTc8A1VVVVTY9aPPeqqqqq/aY/Za+GH7UXg0+HPHOn+RqtlHIdG1y3XFzpsrAcryBJGSBuib5Wxn5WCsvx6/Z9+Jf7OXji58EfEbQ5bdhI/wBg1GONja6nEuP3sEhADjDLkfeQthgDxX7Cv/BSC++HTWfwk/aF12e88IxQrDo+vSRmWbSAowsM2wF5YMcK2GaPgcpjy7C/sNWsLbVNLvre8s7yFZra4t5BJHNGwBV0ZSQykEEEcEEGqqqo81VVVVTYxiqpsYxmqqq4r9p74bN8Iv2g/H3w9W2S3ttL1udrGJQBttJT51twFUA+TJHkAAA9OMV/wTp+JDfEn9kvwc11dJNf+GFl0K6CkfJ9mbEC4DEgi2a3znBycgYIzVVVVVVVVVVVVVav4Q0nVWMyqbac9XjHB+q9P5VqvhDWNNLOkP2mEDO+IZIHuOoqqqqqqqqqqqqqqqqqqqvD9k+n6LaWkilXSPLKeoYnJH5mqqqqqqv+CnXhH/hF/wBr3xNfR2/kweI7Cx1KJBbeUvMCwuyno+6SCRi46sWB5BJ/4IweLE1n9mHXvC81yjXHh7xdcBIljKlLea3gkQlujEyef3yABnHGaqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr/gqB+xA3x78Hf8Lo+GWko3j/AMKWrC8togFbWrBeSnPWaIbmTuylk+Y+WF+DXxc8Y/Av4j6L8TfA175OpaPOGaJyfKu4TxJBKB1jdcqe4zkEMAR+zz8fPBP7R/wz034j+C7lEE6+XqOnNKHm026AG+CTGOR1DYG5SrAAGqqqqqocjrVVVVVDGetVVVVVS9PpVVLFFPE8E0aSRyKVdHXIYHggjuMV8W/Akvw68dah4dVG+xlvPsHbnfbuTt57lcFCe5U1+yx8QP8AhIfCUng2/n3X2g48jceXtmPy/wDfJyvsCtV+0v4si8Dfs8fEnxW8kSyaf4W1BrcSxs6NOYHWJWC84aRkU9MZ6gcjwl4euPF3irRfClmZBPrOoQWUXlwmVt0sioNqDBY5YYUdelW0UUMKQQRqkcahURVwFA4AAHQVFgjioiMYBr9pfH/DN3xYAP8AzI+s/wDpFNX7L/wLi/aR/Zl+LnhDRNPZ/F/gvULLXdEkVNzXLPBcK9oo65lW3I9N/lk8A1/wSp/ahTxV4Yk/Zz8aayh1jw9CZ/DTTud91ZZJkgDMfmaEnKr18s4A2xHHxNI/4Vl4uH/UCvf/AEQ9f8EYiB8T/iLz/wAwG2/9HmkIIGD0r/gst8WZYtA8CfATRrhnutXun1vUYIZDvMceYbZSg5ZXkecgHjdCuASOP2+f2T5Ph3+wz8HNTt9Lij1r4W28Nhr/ANmjVlH24K1y5kQAMi32ApI588nO5ju/4J3/ABiHxn/ZM8E6vd3vn6t4etzoOqFpvNcTWgCIzt13vB5EhzzmTqepZxjrX7QP7UHwY/Zo0GHW/iv4sSwlvVc6fp1vGZ7u+KY3CKJew3KCzFUBIywzXif/AILZeF7fVJIfB/7Pmqanpoz5dxqXiOOxmPzHGYo4JlHy7T984JI5xk/sift5+Df2udT1Pw1pPgDXvDmtaNYLe3i3EsdzaBCyJtSZSrFt7NgGNQVXOc5USkHgGv2+NWvJ/iR4f0N5M2tpon2iJcnh5ZpFc4zjpCnbPHOeMaFAskzSEZKgAZH514Y0KO/ZYwBk+teJ/DSacpRlGQOleIrZI3WRRyDtr9kjV2074v29gLoxrqlhcQGPbkSlV8wD2x5ZOeOmO+DoPwE+E3hm7k1Gx8H2c908pkEl4PPEZzkbEbKpjPG0Dt6Cv+Cjeipf/sm+JbmOYQro95p9wsYjyHBuo4do5G3/AFuc8/dxjnI/4J76zdaD+2X8L76zjieSTU5rUiUEjZPazQueCOQsjEds4zkcVwRwaqqqqoYzjNVVVVVS46ZqqqqqvpSkHgGqqqqqoYz1qqqqqqoHI61VVVVVfQ0CCMZ5qq+hoEEcVVVVUDngmqqqrxb4B8E+PbMWHjPwppWtQr9wXtqshjPqjEZU8nkEHk18Qf8Agn38M9cimuvh9rWo+GrzZ+6t5nN5a5HPIc+YM9M7zjg4PIPxE/Y5+Ofw9jlvR4aXxDp8RA+1aIxuTg9zDgSgDudhAx1xgmw1LXvC2qi70vUL/SNTs5Colgle3mhdTyMqQykEfgRXw4/bn+Mngv7PY+JJrXxdp0b/ADrqAMdzs9FnXnOe7q/XHpj4c/tt/BPx15NnrOpT+FNRfCtFqwCw7uM7Z1JTbk9X2dDwKtbu1vraO7sbmK4glXMcsTh1YeoI4IqunQ1u7Gsgjg1VVVVVV8T/AIG+A/irA0ut2BtdVCBYtTtcJMuOgbs69sMDxnBHWvin+z/48+FjteX1qup6NnCalaKSi+0i/ejPTr8vPDHmvht8XPG/wsv/ALT4Y1Q/ZJH3XOnz/PBP0zlf4WwANy4bAxnHFfCf9ovwN8UEh02S4XRteIAbT7qQATNx/qX6Sdfu8NwflwM1VVVV8bP2afDvxHSfxB4bEGj+I8FmdY8Q3rcnEgHRiSP3gyfUHjHiXwxr/g/WJ9A8TaVNp9/bHDwyj8ipGQynswJB7GvgF8Vrj4V+N4bq4lJ0XVCtvqcRYgBM/LKB/eQknp0LDjdkW9xb3lvFd2lxHPBMgeKWNgyupGQykcEEYOaqqqqr9t/WSX8K+HowcAXF1ISowfuKmD17Pke4r9ijQRd+M9e8RPErrpunLApYA7HmfII7g7YnGR2Jz1qv2wfh5JoHjSHx1ZQ/6D4gULPtXAjuY1APT+8oDe5Dmq+I3i2DwL4G1rxZMRnTrRmhVujSn5Y179XZR+NfEH43fGD4rso+I3xL8ReIIo1RYrW9v3a3hCgBQkOfLQDA4VRzz1JNeEvht8Q/H8hi8DeBdf8AEDIxVzpumy3IQjbncUUhQNy5JxjcM9a8O/sFftUeI2iZfhk2mwSMymfUdRtoNhA7p5hkwTwCEPJ9MkRfCv4ufDbwppcXxY022S+YtAt1a3X2hJ9gGGZh91yDyD1KsR3Arxn8PfFnxV8L3/gLwLZpd65qUYNpbvOsIl8phKy72IUEpGwGSBnGSOteJf2PP2nPCm7+1Pgv4hn27c/2bEuo9emPszSZ9/Tviruz1XQtQ+zX1rd6ff2rKxjljaGWJsBlODgg4II/A18K/wBur9q34Pyxr4W+M2uXllGuwadrc39p24Tj5VSff5Y4/wCWZU9eeTn4J/8ABaHRL9k0v9oL4atpkrMMat4ZLSwgY53W0rF1AI6rI5O77oxk/td/Fv4RfG/xR4a8f/CPx5YeJbSTRRbXJtnIa2Kyu6LJE4WSJyJW4cA8DgYNf8E4r25ksfH2nPLm3gl0+WNNo+V3FwGOevIjT8vc1+0b+zl/Z/2r4g/D6w/0XmXUtMhT/U92liUfwd2Ufd6j5chfhl8TfEfws8Rx67oM3mRPhLyzdiI7qMfwt6Ec4bqD6gkHwB4/8OfEnw5B4k8N3XmRP8s0L8SW0gHKOOxH5EYIyDmqqqokAUee9VVftsf8EuND+KU958Tv2drXT9B8WTyNNqOhORBZamxAy8RHy28vBJGNjlsnYcs1fBb/AIKP6l4Q1CTwV8adGk1PS7KY21prOmxgXMCISoE0RIWUYC/MpVgAeHJr4efFb4f/ABW0RfEPw78XafrtjwJGtpPngJHCyxnDxNjna6qcdqWcEYzzQnHZqE/+1Xnk8BhXnEjAIoznpuFNPkcNTTg87q1zxJoXhrTZtY8Ra1Y6XYW6lpbq9uFgijAGSWdyABgHqa+Ln/BSP4L+CY2sfh9HdeONTKn5rYNa2kLc8NLIuW5wfkVhjPzAjFfGv9rL40/HaSW08U+JmsdEdvk0TTM29qB2DjJaX6yFsHpivgL+y98av2ldYk0v4U+D5b62tZFS91S4YQWVkTz+8lbjOOdi7nI5Cmv2S/8Agm98Iv2b3tPGHicxeOPHsW149UvLYLbaa4bcDaQEkK4IX985L5XK+WGZaqqqqb0FVVVVVTcnANVVVVVRwTwaqqbpgGqqj04NVVVVVTdMA1VVVVVHk1VVVVT9OtVVVVVTYJwK+OfwI+HH7RPgK6+HnxK0lrmxlbzLa6gYJc2EwGFmgkIOxwCRyCpBIYMpIP7Vf7IvxI/ZY8XSaf4htJdS8LXs7LoviKGLEF4uCQj4z5UwUHMZP8JKll+av2K/+CgPi79m+8t/BHjx7/xH8OZPkSzVw0+jksWMltu4KEsxaIkAnBUqc7vBHjrwb8S/C9j408A+JLDXdE1KPfbXtnKJEb1U91ZTwyMAykEEAgiqqj04NVVVVVTc8VVNzxVVVHpxX/BXv4cJ4e+NXhf4k2ybYfGGjNBNwPmuLNlVm65/1U1uOQB8vBPIX/gjh8QfL1P4h/Cm5uM+fBba1ZRZ+7sYw3DY9/Mth+HvVVVVVVVVVVVVVVVaj4f0jVAWu7JDIf8Aloo2t+Y6/jWofDyVcvpd8HGOI5hg/mP8BV94d1nTyTc6fLtX+NBvX65HT8aqqqqqqqqqqqqqqqq8JeE2Zo9W1SMqFO6GFhgk9mPt6D/Jqqqqqqr/AILH+F2tPih8PvGht2A1XQLjTxKZAQxtp/MKhc5BH2sHJ4O4Y6HH/BDvxUyah8WfA80twyzQ6ZqNsgA8uMobiOYnnIZt8HYjCHpgZqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/4Kn/sGt4S1DUP2nfg7oztomoTPP4w0uBQBp87lf9MiUcmKRiTIP4HO77rN5f7JX7Ufi79ln4lw+J9IlluvDuqSQw+JNJADC9tlY/MgJAE0YZzG2Rgkg/KzA/Dr4i+DPiz4M0zx/wDD/XYNX0PVofMtriI9OzIynlHUgqyHBUggiqqqqqXBGBVVVVVfSuCODVVVVVL6UMEDFVX7THw4/wCEw8Gf8JHp0WdT8PK8wAXJltzjzF+owHHXow/ir4c+Nr74e+L7DxNZMzJBIFuol/5bQEjenPcjp6EA9q07ULHV7C31TTLqO5tbqJZIZUOVdWGQR+Ff8FUPGUXhb9j/AFzSTLsl8VatYaXCRuByJftLAFemUtXB3YBBI6kA/sR+Dz42/al+H2nPbtJDp+pf2nKwVsR/ZY2nRmK9B5kaAZ4yyg5zgxEEYBqI9ADUR7A1+0uR/wAM3fFfH/Qj6z/6RS1/wRq4Pxewf+gH/wC39ft4/CTxP+yf+0Zon7THwll+xWHiDUmv4XCZSz1QEtPCy4A8qZSWAz826ZcAKM+EfjJ4c+Pv7LV78UvDUqCLV/DF59qtgTus7pIHWaFgecq4YAn7y7WHDAn/AIIyED4nfETn/mA23/o81GwIAzXhOR/20v8AgqU+tyP/AGh4V8Lau11H+6aWEadpZCwnY3SOadYywbAzctxztr43/DSw+NHwe8YfCzUBEF8S6RPaQySkhYZyuYZTjn5JQj9D93oelf8ABHT4tT+DfiZ43/Z38Tyy20utR/b9PtpFGI721ylzHkc72iKnnjFueQfvavrFhomlXmtapceTZ6fbyXFxLsLbI0UszYUEnABOACa+BfgnWf8Agpl+2N4h8c/FbUrqLwvp0Zv7yzjn2tFYpII7XToiOUBB+d1AztlbIkkDHwf4D8E/DnRIPDXgLwppWgabbxrGltYWqwrhRgFtoyxx3OSepNWHhfwvomp6hrei+HNLsNR1bZ/aF5bWccU13sZ2XzXUBnw0spG4nBkc/wARzKwI61+3z4Zni8T+GfGa5aG7sH09+eEaJzIvGOMiZu5+70GOdHult7nYxwH4B9xWg6+bHayvgitd8StfIS8mTjua128W4mESNnact9a/Y60C71D4j3uvR5FrpOnMJG2ZBeUhUXPbIVz/AMB/ESnjAr9uTSrbWv2U/iJZ3UkipHp8VwpjIB3Q3EUqjkHgsgB9s9OtfshateaL+1R8I7ywZBJJ4z0u2bcuRsmuUif8dkjY9DzQ5GM1VVVVX0NcEcVVVVVX0rgjg1VVVVSnjA7VwRwaqqqqqgcjrVVVVVUDiuCODVVVVVV06GuCODVUDjoa4I4NVVVVAgjBNVVVVV49+EPwz+J8CweO/BmnasyfcneMxzoMYwsqFXUdOA2OB6CviJ/wTs0643Xnwr8aPayFyfsOtrvjAPQLNGu5QOmCjEg9eOfiN8Cfiv8AClnk8a+DL20s1faNQiUT2rZOF/eplVJxwrEN7CvAfxX+I/wwuWuvAXjHUdI3vvkhik3QytjGXhbMbnHdlOK+GX/BQ6cSwaZ8WvCUbRnCtqmjkgr0ALwOTnuSVcY7IelfD/4wfDT4pWv2nwN4wsNScKDJbCTy54sjPzRNhwOvOMcHng1VZPQGtwIxWQehqqqqqpoYbiF7eeJJY5FKujrkMCMEEHqCO1fFz9kjw/4iSbW/hsING1MlnewbItpz1wg/5ZHPQD5e2B1rxJ4X8S+Cdak0TxLpVzpmo25z5cowepAdGHDLkHDKSDjg18If2s9d8KRw6D8QUuNb0uNQkV2hBuoAPUkjzRj1Ib3PSvDPirw54z0eLXvC2sW+o2M3Cywt904BKsDyrAEZVgCM8iqJHTNFh0FEkjANHnvXxN+FHhL4q6MdM8RWm25iU/Y76IATWrHGSp7g4GVPB9jgj4nfDXXvhZ4pl8M66EkG3zbS5jGEuYSSA4HY8EFT0IPUYJ+CH7R2u/DF4fD2vCXVPDJbCwbh5lllvmaInqOp8snGeQVyc+HPEmheL9GtvEHhvU4r/T7td0U0eQD2IIOCpB4IIBB4Iqqqqr9r/VV1H4wvZh4ydL0y3tiFByM7pfm9/wB7+RFfsVaL9l8B63rrx7Xv9U8lSYtpZIo1IIb+IbpHGOgIPqa/aY/bX+BP7LenyQeOPEqah4meFntPDWmsJr2U4BUyKDiBGyMPJtBGdoYgivjD/wAFOPjL8a/Hel3GpWdl4c8BWF4rt4bsQJ2lTkF5rhlDSyhXONojTgfJnJNpdW1/aQ31lOk0FxGskUiHIdWGQR7EEVcfs02v7R3h8aTr3ii+0bQ7HUI5LlbG3VprwqpzGsjkrHjcDko/JXpjn4b/ALGH7OPwxjhk0f4a6dql9Fg/b9bT+0JiwYMHAkykbAgYMar09zm2sbazt4rOzt44IIEEcUUaBFRQMBVA4AAAGK8oYwK8d+C9N8eeFr3w1qKqBcJmGXbkwyjlXH0PX1BI71r+hal4a1m80DV4PKvLGUxypnIyO4PcEYIPcEV+zVbwz/FWzaRAzQ2k7xk/wnZjI/AkfjTReleJ/BnhPxpp/wDZPjHwtpGvWW7d9m1Oxjuo8ggg7JARnIHbsK+JH/BOX9nfxpbyyeGdN1DwbqLZZJ9MuWlhLYON0MpZdvT5UKdByOc/Fz/gnb8evhz9q1HwrZ2/jnR4NzLLpQK3YQYwWtWO4sf7sRlNfs5QX+j6j4o8O6xbXVlfW7QF7O5iaN4mXzA+5WHynlQQcHp6V+zv+1D4F/Ze8YHWPiTa3a+HvEzR6bcalbKZW05vmdZWhVS0kfyndsyw4wr9K8N+JfD3jHQbHxR4T1uy1fSNShE1nfWUyzRToejKykgiv2kf2fJ/D9zd/ELwVZ79JlYyahZRRgfYmPWRFH/LI9SP4ev3fu/CX4p618J/FC65pi/aLSdRFf2TNhbiPOfwcdVbtkjoSD4N8ZeHvH3h628S+Gb4XNncDBB4eJx95HX+Fh3H0IyCCaqqY54BqqqvFGoyaP4a1bV4ZESSxsZp0aT7qlELAn24qv2m/wDgl3+0F8L9Y1LxP8ONMk+Ivhq4uZJYv7JgLahbqzEhZLUZZyM4zFvzjJC9K0bXvF/w/wBfN/oGrar4e1qxkaJ5LeZ7aeFlb5kbGCMMoyp7jkV8Nv8AgpN8bvCFvb6d4007SfGVpAqqZbhTaXbAEdZYxsJ25GWjJzgknnPhD/gpp8DdahVPFWieJPDtzxv3W63cIz6PG28468oPx6Vov7Zf7MmvrC1h8YtGiE0nloL1ZbMg5xlhMibV5+8cDvnFD9or4BAf8ly8Af8AhTWn/wAcrVf2o/2d9Gjjlu/jb4NkEjFVFrq8V0QfcRMxA9zivE//AAUI/Zg8PxTCz8a32uzwF1MGmaVOSzLnhXlVI2BIwCGwcg5xzXir/gql4ZgSWPwT8J9UvWO4RS6rqMdqF4+VmSNZM89VDDj+KvHH/BRv9orxZDPZ6HdaH4Vt5dyq2mWO+YIc4BkmaTDYONyhTnkYNeMPHvjf4haj/a/jnxbq2vXYZikuoXbzmPceQgYkIvA+VcAYAxxXwY/YY/af+OwgvPB3wwv7LR5uRrOtj+z7Tbz8yNJhphkY/dK+D1xzX7P/APwSD+FHgWe38Q/HPxFL491OPa6aVbI1np0LYGQ+G824ww4JMakcNG1aD4f0Hwro1p4d8L6JYaPpVhH5dpY2FslvBboP4UjQBVHsABVVVVVHk8VVVVVRxjrR5qqqqqj04NVVU3J4NVVN0wDVVVVVU3J4NVVVVUelVVVVUelN0xVVVVVUeT1qvHngDwZ8UPCt74I+IHhyz1zQ9RULcWV2m5WwQVIIwVYEAhgQQQCCK/bQ/YC8bfs2X1x4z8Gx3niX4dXEzFL5IS8+kAn5Y7sKMBeQqzDCseCELKD+y1+138Tf2V/FI1Hwvctqvhu8mD6v4buZylve8YLqcN5MwAGJFBPygMGUba+AH7RXww/aU8Fr4x+Gutef5GxNS06dfLudNmZd3lyp+YDrlG2ttY4OKpumKqqqqqmwTwaqm5PBqqqmxt4Nf8FV/h7/AMJf+y7L4pt4N1z4L1m2vyyjLeTKTbyL9MzRsf8ArnnoDX7AvxEi+Gv7WPgHVby5SKx1W9bSLrfwCLtGhjydrFQJWibPH3cEhSTVVVVVVVVVVVVVVVVVXmkaXf5N3YQSsf4inzfn1q68BaJOMwNPAewV9w/XJ/Wr74e3sfzWF7FMP7sg2H+oP6Ve6Hq+ngm70+ZFHVtu5R+IyKqqqqqqqqqqqq0rw/qmsMPsdufLzgyv8qD8e/4ZrQ/B1hpRW4uCLq5HRmXCr9B/X+VVVVVVVVX/AAWR8PJdfDj4deKzYs7adrd1ZC53ECIXECuUIzjLfZgckZGw4xk5/wCCMHif+yP2o9d8OzXcqw694PuUjhVcq88VxbyKzemIxNg/7WO9VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVajp2n6xp9zpOrWNve2N7C8Fza3MQkjnjcFXR0bIZWUkEEYIODX/BRr9grUP2bvFM/wATvhholxN8LdYmGAjmZtCuXP8AqJCRkQsTiNyT12Md20v+xH+2dr37Kvi6TTtVgl1TwFr06NrOnxgGW3cAKLqAn/loq4BQnDqMHBCsvg/xh4X+IHhfTfGngvXLXV9E1e3E9le2z7klQ/qCCCCpAKkEEAgiqqqql64zVVVVVUOQKqqqqodetLgjGaqnRJEMbqGVhgqRkEV8b/hy3w48cXGn2kLjSr4faNPdhxsP3kz6o2RjrjaT1r9lP4lx3dhJ8NdVnxcWu6fTGY/fjOTJH9VPzD1DHptr/gs946STU/ht8MreSLfb295q90m4F8OyQwnHVR+7n5PBPT7pr/glJ4LTVfir4u8dTQeYnh/RY7WNiRiOW6lyGxnOdlvIOBjBOccVERxzUZ7ZqMjNftLHP7NvxXAP/Mkaz/6RS1/wRrPPxeAP/QE/9v6+OPwd8MfH34V678LPFY8u21eDEF0qbnsrhTuinQZGSjhTtyAwyp4Y1+wb4z1j4D/HPxn+x38ZLhtJs/GbSaUy+cAtvqewpG0TMNuJ422qcHe3kcc1/wAEaCB8TfiJz/zArb/0ea/a7+L3/CkP2b/HPj+2vPs+pQaY9ppTLN5bi8uCIYWTuSjOJMDnEZORgkfsaf8ABPTV/wBrbwPrPj65+JH/AAh2m6bqf9n2hOi/bzeSLGrykfv4igUSRjOCCWIyNpof8ESweP8Ahpr/AMsv/wC7q8ZfD3xB/wAE5f2y/CEk/iZtetdCmstXi1GG1aye+0+bdFcp5YaTYSFuYeGcEDPcqP2gLGfxt+zn8SNH8MyRX02veCdVg09onDJO01lKsZVhkEMWXBHrX/BFPxLpdp4n+K3g+WbGo6nYaXfW8e5RuhtpLiOU4zu4a6h6AjnkjIy7gDANSuCMA1KwPQ18YfhnpPxb8DXnhDU5FhkciayuSm421woIWQDIzwWUjuGNeLPCXiHwPrtz4a8UaXNYahaMQ8Ui/eHZlPRlOOGHBq31e7t12bg4HTd1q41m7nXapCcdR1rRdF1bxJq1roeh2Mt7f3sgjggiXLOx/kMckngAEnAFfBX4WW3wo8FQ6NIIpdVu28/UrhOQ8uMBVOAdijgD13HjcalI6A1+1Jplpq/7OHxOtLwMUTwpqNwoU4O+K3eRPw3Iua+Dl/eaV8XvA2qadcNBd2fiTT5oJU4MbrcxlWHuCAaX0zVVVVVUuCMZqqqqqqXBGM1VVVVQOD1pcEYzVVVVVUDjvVVVVVVQORjNVVVVVVA44rgjg1VA44rgjg1VVVUD2zXBHBqqqqqpI45Y2ilRXR1IZWGQQeoI9K+Jn7H/AMFPiSLi8/4R7/hH9Vny327R8QZc4+ZosGNskc/KCcnkE5r4nfsJfFrwaZr/AMGm28X6YhYqLX9zdooGctCxw3oBGzk+gqeDxD4Q1swXEOo6LrGmy8o6vbXFtIPY4ZG/I18L/wBuX4ueBY4NN8UtF4x02FdoW/kMd0B2xcAEt9ZFc89emPhf+1v8F/ifHBaQ+I10HV5F+bTtXxA2R12SZ8twecANuIGSoqqr6GsnGAa3VuB4xW4VuFbhW4VuFeNvAPhH4iaUdI8W6LBfRKG8mRhiSAkctG45U8DpwcDOa+MH7Mfiv4d+drXh7zdc0BOTKifv7cf9NEHUD++vHqFrwL8Q/F3w31hda8JavJaSkjzoT80Vwoz8sidGHJ9xnIIPNfB39o/wp8ToYNI1R4tG8RnCGzkk/d3LY6wseucfcPzDp82M1VVVftDfCZPij4IlGmWsb6/pQabTnPDSf34c/wC2BxnjcFyQMmnR43McilWUkFWGCCK+Ffxf8V/CXWPt+hTC4spyBeadOx8qceuB91wOjjp3yMg/Dj4meFfiloI1zwxeFjGQl1ayjbLbORna6+/OCMg4ODwcVVVXx/1I6t8ZfFl0bgTGO/MG4ADHlKse3j02Y/CvjR/wUX8YfCHwNH8APgXaHSdbtTMde8S3GyVkklJYR2aAkKVjaMNI43BwwCjaHNnZeM/iV4rW0s4dX8TeJNanJCjzLu6u5SCWJPLOcAkk9gSelfs+/wDBMcsLPxP+0JqH+0fDFhN/47Pcxt+axH0+fqK+Kfw60vwBd6fB4a02Oy0N7VILW2iXCW/lKF8seg2hSM+p9K0HxB4O+FPhDTtI8R6rFa6g8PnT2qqZJvMcbiGVQSOygtgcDmm/aM8AC48gWmssm7HnC3Tbj1xv3Y/DNeGfif4E8WzJaaTr0Qu5Dhba4UwyMfRQ3DHvgE/zryge9eUPWv2nvhcNY0gfEHR4h9t0uMJfIAcy2+eH+qE8+qk8/KBXw68b3Xw88W2fia1tluRBuSaBjjzI2GGAPY9wfUDORkVqP7W3jKWfdpPhnRbaHJwlx5szYzx8ysg6e35VoH7W90JY4vFXhGF4yQJJtPmKsg5yRG+d3bguO/NeEfGvhbx7pp1Pwvqsd3HGQsqYKSQsezqeR0PPQ4OCaeLA4r9rXw3p0/hDTvE6aZC2o2d8tu12Ih5i27o5Kluu3eE4zjJr9oCx+1/De5n2K32K6hlyf4ctsyP++8fjX7If7bfxP/ZM8R7NKml1/wAF30mdT8NXVyywsTjM1ueRDNgAbgCGGAwOFK/BL46/C39o3wHB49+F3iCPVdMmPk3UEsZjnsptoLQTxHlHAI9VYEMpZSGP7RnwBufBF9ceNvCNnv8ADl1JungiT/kHOx6YH/LInoei52nHBPwX+MusfCPXTNGrXmi3rKNQsc8sB0kTPRwPwI4PYjQNd0rxRotl4g0O8S6sb+FZYJV7qexHYjoQeQQQeRVUx4wDVVVV+0frieHvgV41v3dAJdJktBvUsM3GIBwOc5kGD0BwTxmvhrpD+IPiL4W0OMKTf6zaQfMm4ANKoJI7gA5PtVfGj9lf4BftA2k0fxR+GulalfSx7E1WKP7NfRcAKVuI8PxtXCsSvGCCMg/FL/gix4eunlvPgx8Yb2wO4mPT/EdmtwvIGB9oh2FQCG6xscEd1Jbx3/wSx/bG8F+ZLp/gfSvFdtFu3z6Fq8T8DGCI5zFK2cnAVCflOQOM6/8Asm/tP+GZLtNZ/Z7+IcSWMfmXFxF4cuZoI0C7i3nRoyYC9SGwMHOCDX/Cqfij/wBE28U/+Caf/wCIrRfgT8b/ABJdNY+Hfg3451S5jjMjQ2Xh26ndUBALFUjJAyQM9MketeFf2CP2xPGaQvpH7P8A4mtxOrMg1WOPSyApIO4XTx7TxwGwSMEZBrwT/wAEdP2ltfRbjxf4l8F+FojkGGS8lvLgHaCDtijMeMkj/WZBU8EYJ+H/APwRf+D2iyrcfEn4reJvFDI2RDp1pFpUTj0cEzORj+66n+VfCb9jz9mj4IyxXvw6+EGh2eowsrx6ldxtfXaOvIZJ5y7xnPOEKjOOOBhjxgGmwTgGqqqqqbpxVVVVVU3TANVVVVVTYxgGqqjjFHnvVVTYJwKqqqqo4x1o896qqqqpsYxVVVVVN0wDTcnFVVVVUenFVVahp9hq1hc6Xqllb3lneQvDcW1xEJI5o2BVkdTkMpBIIPBBr9tn/gmTqHhuTUPit+zZpUt9o53T6l4TiXdNYAKWeW1JbMsZwf3IBdSRs3A7U+FfxY+IPwU8Z2nj/wCGfiSfRtas1ZFmjUOssbfejkRgVkQ4GVYEZAPBAI/ZC/b++G37TNrbeFdeNr4U+IKR4k0ea4/dagVHzPZu2C/ALGI/Ooz99VL1TYPANVVVVVR5qqPNVVU3QCvix4Ftfif8L/Fnw6u1i2eJNGurBWkAxG8sTKjglWwVYqwO04Kg4OKik1Xw7rCTIJ7DUtMuQwDoUkt5o27g8hlYdD0Ir4c+LoPiD8PfC/j21i8uHxJo1pqUaYI2rPCkgHPPR6qqqqqqqqqqqqqqqqqqq80PSL8lrrToHZs5bbhj+I5q78AaRMd1rNPbnsudyj8+f1q6+HmoICbS/gmwOA6lCf51ceEfENv105nA6GNg2fwBzVxY3tqP9Js5ocf89Iyv86qqqqqqVWdgiAsxOAAOSatPD2t32Ps2mzEdmddg/M4HarL4e38pDX15FCv91AXb+gH61p/g7Q7AKxt/tEqj78xzz9OlAAAKoAAHAqqqqqqqqr/grNosWqfsrQX0k7I2j+KbK6RQPvkxzw4PoMTE/UCv+CVOsXumftxeBLK1ZBHq1rqlrchlyTGNPnmGPQ74U59M+tVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV4h8PaF4t0K/8ADHibSLTVNJ1S3e2vbK7iEsVxE4wyOp4IIOMV/wAFA/8Agn5rn7LuuTfEL4eW13qnwu1S4xFK2ZZdDlc/Lbzt1MZJxHKevCMd2C/7HP7bnjj9lXWn0maCXX/AuqTq+o6M0uGt2yA1xbE8LLtGCpwr4AbBCsvw6+Ivgz4s+DNM8f8Aw/12DV9D1aHzLa4iPTsyMp5R1IKshwVIIIqqqqHUVVVVVVLyMZqqqqqvoaXB4Bqqr44/DVPiV4Kms7RB/a2nE3GnseMuB80Z9nXj/eCntWg65qvhTXbTXdJmNvf6dMHjLL0YdQR6EZBHoTX/AAUm+KEXxW/aTbVbWK5jttM8O6fZwxzRqAmUad1Rhy6rJcSLuPO4MOgFf8ErPCo0v4IeIPFU0DJPrviJ40YspDwQQxhSMcg+Y8wwT2BAGcmIjpmkIyOaQ8DBr9pRs/s3fFYZ/wCZI1n/ANIpa/4I2kA/F0Z/6An/ALf0jDsa/wCCnn7OGoahpth+1T8NYJbbxJ4P8n+2ms1xK9vG4MN2AoyZIWI3MekYBJCxV/wRsOPiZ8Q8H/mBW3/o81/wWO+LTR2Xgb4HWF0wM7Sa/qceAAVXdBa89SM/asjgZVTyRx+x98Jx8EP2b/Avw/uLT7PqUGmLd6qrQ+W4vLgmaZX7ko0hjyecIvAxgLJxjNf8Fj/hMdf+GHhL4yabbM9x4V1F9O1AqQALW6AKO2eSFmjRRjvOeD1H/BOj4uH4ufsneELm8uWn1PwqreH79mA+9ahRDyOpNs9uSTySTnPU/tK/AD40/sGftBN+0B8CbKceDZr2W80+5tLVpbbTo5G/e6feIPuw4bYpYgMm0hhIp2+FP+Cy3wVu9Ehl8dfC/wAbabq/SWDSVtb6DoOVkkmhbk54KcDHJzx+yx+3Z4b/AGsfiB4n8J+FfAmp6Jp3h+wW7t7vUJleW7VpdnzogKQkDB2733bjgjYdztnvUrg96+Inww8D/E/Tl07xjocN55X+ouFGyeDkE7JB8yg4GR0OBkGvEf7C1qZWl8KfECRI8/JBqFkHIGT1kRh2wPuckE8ZwNH/AGGmWVJPEfxCXy1c74bKw5ZccYkd/lOf9g9Pfj4efCTwL8LLFrbwppW24lXbPfXB8y4nHHDPgYHA+VQq5GcZ5qQ5PXpUpz0NfF3T7PV/hX4y0nUIfNtb3w/fQTx7iu9Ht3VhkEEZBIyOatbq6sLqG+sbmW3ubeRZIZonKPG6nKsrDkEEAgjoaHsaqqqqqh161VVVVVDGetVVVVVSnI4PSuCODVVVVVUDkYzVVVVVV9DQII4qqqqqqoHtmqqgccVwRwaqqqqBxwKBBHBqqqqqqrx58K/h38TrL7B478I2GroqhUklQpNGAScJKhDp1P3WHWvid/wTxcGbUfhJ4rUjLMNM1k4wMZ2pMi888AMo7Zbqa8cfDTx98NtQOmeOfCeo6PNvKI1xCfKlI6+XIMpIPdSRXwr/AGovjD8JI7fTtD8SNqGjQEY0rU18+FVH8KE/PEOvCMozyQa+FP7c/wAKvHcsWleMA/g7U3wFa9lD2cjd8TgDZ3/1gVQMfMScVBPBdQR3NrPHNDKgeOSNgyupGQQRwQR3qqqqqqqq+MP7K/hrxqs+u+ChBomtkMzQqu22umJz8wH3GP8AeUY9QeteJ/C3iPwPrkuh+I9NuNO1C2bIV1K7gCQHRujKSDhhwcda+DX7WGqeGktvDXxH87VNLUhItSXLXNsvT5x/y1Ucc/eAz97gDRtb0jxFpsGr6FqVvf2Vwu6KeCQOrD6jofUdRVVVftZfChfCPilfHGi2nl6Rr8mJ1UALBd4JYADoHAL/AF39sCooZbiVLe3iaSWRgqIikszHgAAdSfSvBnjbxH4A1638ReGNRa2uoG5UklJl7o6/xKR2/EYOCPg38cvDXxb0xYo3jsNfgQm601nycDAMkZ/iQ5HuDwexNVXxL+Jfgj4P+CNU+InxE1+30fQdHhMlzczH8FRFHLuxwqooJYkADJrxF4otPHOv6n41sLaW3tvEF5LqEMM2N8aTOZFVscZAYA44zXhz9mL4h/tQfHfxcfD1sdH8O2WtTQahrt3EWih2OUIjXI86Q7CQgIA43MuQa+BH7NHwr/Z60X+z/Aui79SnTbfazeYkvLvJBIZ8DamVXEaBV4BILZYrHngV+0J49sY44vBOnJFPdxyLPczHObVgPkC4/jIJz6Aj14YzXMxZmeWWRiSTlmYnqfUmho+pFdwtTjH94fyzTJNbyAMGjkQ5GRgg+tfA74wXl7eweCfFl0Z3m+SwvJDl93aJz3z2J5zxzkY8njGP0r9pH4rS+BNDj8MaDcBNb1eIkuPvW1vypce7EFQe2GPUCra1uL2dLW0haWVzhVUc1Y/C/UbiEPeahHA5AwiRmTH1OR+ma1zwDq+kRNcwst3AgyxRdrKPUr6fQmvB/i3WPBHiC08RaHcGO4tnG5CTtmTI3RuB1UgYP4EYIBHg/wAT6b438M2HifSsiC9j3bGOWiYHDIfcEEV+0fpJ1L4Qa20cZeWzMNwgDYxtlXcT9ELcV8Lo9LuPiFoOn61p8F/YaheLZ3NrPAs0c8c4MTI6NlWUh8EHsTX7X/7BWpfDtrv4kfBexutT8MFpJtQ0hQZJtJUAsXQ9ZIAAevzIAM7hlh+zf+0z8T/2W/HY8cfDbUYiLiPydS0q8DPaajEM4WVFIOVJJVwQyknBwzA/s3/tIfDH9rb4Xjxb4QkQOY/s2u6FcuGn06ZlO6KQcbkYbtkgADrnoQyr+0D8Crj4WakutaH5lx4b1CUrAzZZrSTr5TnuMZ2t1IBB5GT8Afjfd/C3Wxpms3E03hm/f/SYQN5tnPAmQdewDAdR2JArT9QsdWsYNT0y7iurS6jEkM0TBlkUjIII6gijjHWjz3qqqq/by8Wf2P8ACrTvCsE+2bxBqa+Ym7G+CAb24BBOJDD2I+hxX7G/hlvEfx60WdovMg0WCe/mGSMbUKIeD2kkjPp271RIA4NVVVVVVMQeQfpVVVVVU2CcA1VVVVVNycA1VVVVVHk8GqqmxjANVVHGOtHnvVVVVVN0wDVVVVVU3PAqqqqqm54Bo89DVVVVVN0xVVVV+2V/wTh8HfHuW/8AiL8LGs/DHxAnYzXQk3JZaw525MwUHypMBj5iL8zE7wSdw8U+E/H/AMIPG0vh/wAU6Rq3hbxRoNyr+VKGt57aVSGSWNh24DJIhwRhlJBBr9j3/gqZLpsVn8Ov2ob+e5gRRFY+L0hMkiAD5VvEQbnGBjzlBbON4bLSDTdV0vXNPt9X0XUrXULG7jElvdWsyyxTIejK6khgfUGqqqqqOMVVHpwaqqqm5OAar9rjw3a+Ev2nfihodiYvs6eKL2aFIoREkSTSmURqoOAEEgUY/u5wOg/4J4a7L4h/Y3+HF1OYPNtra7s2WH+EQXk8SZBJIYoiE+5yAAQKqqqqqqqqqqqqqqqqqqqqqqn07T7r/j5sbeXnPzxBufXmpfDGgTBg+lQDd12gr+WOlS+CfDsgASzePHdZW/qTUvgLQZFCobmM56rJz+oNf8K90X/n6vf++1/+Jr/hXui/8/V7/wB9r/8AE1H4A0NGy0t1Jx0aQD+QFR+CPDqElrV5OOA0rDH5EVD4Z0CDbs0qA7RxvXf/ADzmoLS1tl221tFEMYwiBf5VVVVVVVVVVVX/AAUstbW4/Yu8fyz28UsltJpkkDOgJic6jbKWUnodrMuR2YjoTX7A+ow6X+2R8Jbma/S0V/EUUAkeXywWlVowmc9XLhAO5YDnNVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVXiHw9oXi3Qr/wx4m0i01TSdUt3tr2yu4hLFcROMMjqeCCDjFft7f8ABNrxF+znLd/FL4SJe+IPhvcTO9xbiJpLnw6DyFlOSZIOyzHBXhX5w7/sx/tVfEv9ljxi/iLwTPFfaXqG1NX0O8dvs9/GDweDlJVBOyQZKknIZSyn9nj9pn4VftNeEB4n+HWs4urf5dR0a7ZEvdObJA82NWPyNglXBKtyM5DAVVVXBHFVVVVUvXGaqqqqqlPA5qqqv2ofhkfDniNfHGkWuNN1qQ/aggOIbrqxPoHGWHuG6cV+1N8Jf+E38Kjxfotrv1vQIyWCLlri15LJwMkqfmX6uOrV+x5c2Xwq/Yy8Iatdpbm6vIbm7VYyf9ImmuJWiBzg5EewNjoEOM4yfhx4im8U+D9O1m6mWS5lRlnIAX51YqeB0zgH6EVGcjg1GwIxmpre2vraWyvbeK4t542jlilQOkikYKsp4IIOCD61onhrw34d80+HvD+maZ9o2+d9jtEg8zbnbu2AZxk4z0yfWkYEcGpre1vrWWxvbeK4t7iNo5YZUDpIjDDKyngggkEGtD8K+FPDkkk3h7wzpOmSTLtkezso4C4HQEoBkVqngnwVr14dQ1zwhomo3RUKZ7vT4pnwOg3MpOBSSDGM0sgI4Nanp2l63YSaXrOnWt/ZzY8y3uoVljfBBGVYEHBAPI6gGtG8P+HPDcMkHhzQdN0uOZg0iWVqkAcjgEhAMnHrTSA961b9nT9njW76TU9Y+Avw6v7ybHmXFz4Ws5ZHwABlmjJOAAPoBWmaRo+hWY07QtKs9OtQxYQWkCwoCep2qAMmpHGMA1K4IwDUjDHWnbPepWHQGpCMYBqQjHWpCCcZrVrCz1fT7rSdQh821vYXgnj3Fd6OpVhkEEZBIyOarRviFoPhH4SeF/E3jbX/AC2uNEtHaSdzJNdSmBGbA5Z2JOSffJPevhv45j+I/hhfFVrpU1hZ3FxKloszhnljRtu9gOFJIYbQWxjqaqqqqqhgjrVVVVVUDkdaqqqqoHB60DkYzVVVVVUDjoa4I4NVVVVVQOO9cEcGqqqqqqge2aqqBwMVwRwaqqqq+hoEHgmqqqqqqq1nQtE8R6dJpHiHR7HVLGb/AFlre26zxPj1RgQfxFfFT9gPwD4mefVfhpqsvhi+dSwsZQZ7Nn9snfGDznBYDjCgDB+JvwF+Knwknl/4TLwndRWMbhV1O3QzWj5OFxKvCk9lba3tXww+OXxO+EF2JvBPiaeC1L7pdPn/AHtrN67ozwCf7y4b3r4R/t4/D7xh5GkfEe1/4RTVGwv2ncZLKVvXf96LPowKju1WV9Y6nZw6hpt5Bd2s6B4Z4JBIkinoVYZBHuKqqqqqqrx38PPCPxI0dtF8WaTHdxgN5Ew+WW2Y4+aN+qngexwAQRxXxi/Z18WfCxn1a13ax4fLHF7DEd1uCeBMo+72G77pOOhIFfDL4veMvhVqYu/DuoF7KR83WnTEtBcdASV/hbAGHXB4A5GQfhT8avB/xZ07zNIuPsmqQrm50ydv3sXXlTwHXjqvTIyATiqqvHfg3SviB4T1HwjrGRb38W0SKMtE4OVdfdWAP6d68S+HtX8GeI77w5q6eTqGl3Bik2E43KeGU8HBGCDxwQatvhxZftAeAj428INBbeNdFiWDWdOGAupOqjZOOmySRVY5xtZlI4wWNleeIvBXiGO8tJLzSNZ0qfjchilgkXghlIyO4KkcgkEYNfAX9orSfiRaW/hnxPcRWfiqNdoXbtjvwBy6dg+Byn4rxkLX7Q3xM1P4R+CrXxnpmqaNAyanDaG01M4F8ZsosUWGVjJnDALzhWJGFNftyW/7RX7S/jdvGFxqNlqHhzSoyui+HLOUw/YEON5COdssrYBaTdubAAVQFUaTYf2XpVnpnm+b9kt0h37du7aoGcc4zjpUHxf0Hwf4asPDng7Rop5La2AeZovIh84/NI+wcsWcszHjLMTk5zXwnn8Qa1oEniXxJezTT6lMTDG3ypHCvC7UHAyd3PUjbnPBrUr2DSdLu9VuD+5srd5pD0+VFJP6CtU1G61jUrrVb6TfcXczSynn7zHJx7c14f0dEhRmQGVxliR09qTQHaHeI+Mela9oiTRMhUB1B2NjkH/CkeW3lWSJ2jkjbKspwVI7g9jXgvWG8T+EtI1+QASX1nHJKAMAOV+YD23Zr9oHXZde+LniGVp/MjspxZxKGJCCJQpA9PnDEj1Jr4Y6JGLJtUdAZbhiiEjogPb6kfoK0vQDdR/ImePStY0gW2VK8jtivG+kxaRr8sVuAI51EyIBjbkkEfmD+BFfshazPeeHdf8AD8rM0en3UU8RY52+crAgegzFnHqx9TXxF0Ua74E8Q6QEDNdaZcJHlScP5Z2nA9Gwa068bTtQtdQjQO1rMkqqeASpBx+lRyQ3MEd1byCSKZA6MOjKRkEfhX7cP7D39jf2h8Z/gvo//Eu+afXdCto/+PXu1xbqP+WfUvGPucsPlyE/Z2/aB8d/s1fE/S/iV4FvpQ1tIqalp3m7ItUtCwMlvLwRhgOGwSjbWHKivg18ZfhJ+1r8JI/FnhOaLU9G1OP7PqemXQAnsZ8AvBOgPyOuQQwOCNrqSCDXxu+Dmq/CbxG0aq1xoV/I7abdnk7Rz5b+jqCPZhyO4H7N3x7fwXex+CfGWpH/AIR64OLSeXn7BKT3OeIm5z1CnB4BY0JElRXjcMrDKsDkEeoqqqqr9unxoviD4tW3ha1ujJbeGdPWJ48nalxN+8cjtyhhBx3XHbA/4J+eDpIdO8UePrmDC3MsWnWjng/IPMm98EtDz0yD+FMQehqqqqqqY8YBpjngGqqqqqOAKPPeqqqqqOMUee9VVVVUenBqqqm5OAaqqbpgGqqqqqpueBVVVVVUeTVVVVVMfQ1VVVVVNzwDVVVVX7RH7Lfwj/ab8Nf2H8RNE26hbxldO1uzVY72w+YMRHIQQUJHKMCpyeAcEftQ/safFv8AZa1dT4osv7Y8MXUmyx8SWELfZpWPIjlByYZcA/Ix+bB2lgpI/Za/bb+Lv7Ld8NP0O4TX/B88oe88N38pERJOWeCTBNvIRkbgCpzlkchcfs7ftWfB39pzQTqXw71/ZqttHu1DQb7EV9ZYIBZo8kPHllxIhZPmAJDZUVVVVN0wDVU3TFVVVgGjyar/AIKT2Etj+2f8QHaya3iul02aEmLYsoOnWwZ1/vDergkfxBu4Nf8ABKS+s7z9k+C3trhJJLLxFfQ3CjrG58twp99jofowqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/bj/AOTSfih/2An/APQ1r9lOaG3/AGovg7PPKkccXj7Q2d3baFAv4SSSegxVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVU0MNxC9vPEkscilXR1yGBGCCD1BHav24/8Agk/PG9/8Vv2UtH82M77jU/BiEBlJYsz2GSAQAf8Aj36/LiPJKx14H8e/ET4L+N4PFXgfXdR8M+JdHmZPMjXZJGynDxSxuCGXIw0bqVOMEGv2Sv8Agpn4A+MMdl4I+M81h4P8ZtiOK7LGPTtScthRGzEmGQgr8jnBP3W5CiqqhgjGaqqqqodetVVVVVS+maHI61VV4u8LaV408OX3hnWIt9tfQlCwALRt/C65/iU4I9xXhzxN4f8AGGjweIfC+ppf6bdAmGdVKE4OCCp5UgjBB5FXGrK3hrQvCWnWFvp+laBa+RbWtsNsY5J3bexxgfgcYziv2dtVFx4c1HR2Yl7K7EgyeiyLwPzRj+NRMOOajYA4zSMOCDUbAjGajYDjNK3HBpHGMZpJBjGaVvQ0JOMZrzBjANGUEYBppMjANM2epp5BjANSPxgGnYHvUjZ4BrxT8afht4Umey1LxLDLdRnDW9qjTsCCMglQVUjPRiOhqP8Aaj+HM9wIXstciTJHmvbRlRjvgSFufp3rwz8Q/B3jZD/wjevQXUijLQkGOVffYwDY9wMe9SsOgNSNxnNTyxxo0sjqiIpLMxwAB1JNfCb9kvXvEPka58Rml0fTjhk09eLqYcH5wf8AVAjjB+brwvBq/wBT1HVDA2o3890bW3S3g86Qt5USDCIueigdAOK+HPh4eE/Amg+HWj2SWVhEkwxj96VzIfxYtV1r+g2U7Wt5rdhBMmN0ctyiMuRkZBORwRX/AAlPhj/oZNL/APAyP/Gr34geAtNZY9R8b6Bas4yqz6lFGSPUZav+Fo/DL/oo3hj/AMG8H/xdf8LR+GX/AEUbwx/4N4P/AIuv+Fo/DL/oo3hj/wAG8H/xdQfEr4c3cyW1r8QPDc0rnCRx6rCzMfYBsmv+Eo8Mf9DHpf8A4GJ/jS+KfDHT/hI9L/8AAxP8aqqqqqlPbNVVVVVSnjrXBHBqqqqqqUgjGaqq8Y/tCfAb4fS3Fr43+M/gnRLq2WQyWl9r1vFP+7yHAiL72YEEbQCc8YzxX/Def7Hf/RwXhb/v8/8A8TWiftDfADxNLJB4b+Ofw+1WWFQ0iWXiaznZAeASEkOB9aqqBGMZqqqqqqqgc8E1VUDjiuCODVVVVVA4GKBBHBqqqqqqqri3guoJLW5hjmhlQpJHIoZXUjBBB4II4xXxd/YX+GPjvztW8Ct/wh2rvltltFvspW94cjy+wzGVUcnaxr4s/s1fFj4PPJc+ItAa80hD8urafma3x2LHG6P/AIGB7Zr4YfHL4nfCC7E3gnxNPBal90unz/vbWb13RngE/wB5cN718I/28fh94w8jSPiPa/8ACKao2F+07jJZSt67/vRZ9GBUd2qyvrHU7OHUNNvILu1nQPDPBIJEkU9CrDII9xVVVVVVTokiGORVZWBDKwyCO/FfGj9ku0v1uPE3wsjS2uuWl0Y4WKTjnyW/gPH3Dwc8FcYMFx4g8Ia4JreW+0fV9OlIyN0E1u44IPRlPUEH3FfBT9qzT/E0kPhj4lS2+nao3EGpcRW9x7SZOI39D90/7JwDVV+2F8Lf7V0eD4m6NabrvTFEGphOr25PySY7lGOCRztbnhePgf8AE6b4V+O7XW5ZJDpd0Ps+pxKu7dCSPmA/vKQGGOeCOjGvjb8AfD/xdsT4j0SaK08RJbj7NdoR5V4uAVWXGcjHAccgEfeAArVtH8ReDNdfTdWs7zSdW0+UHY4MckTA5VlI/AhhweCD3r4Y/ti+HdN8JanL8aNUWxk0Kykuv7SjgZvtcUa5ZSiAnzsDgKPn6AA/e/a//bK8b/tRfF2HxtbT3ui+HfDswHhbSTNk2QVgfPcD5TO5UMxGcAKgLBAT8PfFQ8deB9E8Xize2Oq2glaJ0KhXBKPtzyV3qwB7gA1Wk6Zda1qlppFkhee7mWKMAdycZ+g6mtK0u10nTbXSrJCtvZwrFECcnaoAGT3PFeOrWW48DeIreBC8kmk3Soo6kmJgBVaJNE2xwwKkAg+1W2pWC6cYmRS+ODWuyxOzlSBV06yXMsidGdiPzr4JWU9r8KvDsU87ys1s0gZwQQruzKPoAQB7AV8e9GbQ/i/4otDB5YmvTcqMkhvOUSbgT6lz9Dkdq+F+qRS6X9gLDzLRzx/ssSQfzzWi66lnH8rDOK13VEuizAjmviDqEOoeI5FgIItoxCxBzlgST+WcfUV+xxpNwmjeJ9aYfuLq5t7dOP4oldm/SVaePHQV4u0U+HPFWseHyMf2bfzW45JyEcqCCcEggDmvhBq6+IPhd4Y1LzDI39nRwu7OXLPEPLYknuWQ59/XrUiY6V+3p+xx/wAIZd3fxs+FOixx+G5vn13S7dcf2fMzc3EaDpCxK7lH3G5HyHCfsdfta+Mf2TviZb+ItOnur7wpqciReI9ERxtvIBx5iBjgTR5LI2R3UnazVBJ8N/2hvhhY6tYXdtrnhjxHaLc2V3A4JXI4dGGdkiHKkdVYMrDqK+Jvw08QfC3xLJ4e1xFkRgXtLtBhLmLOAw9D2KnkH1GCf2Yvjw05tvhl4zvpJJSRHo13J82RjiBj17fIT/u5+6KqqqvFviXTvBvhfVfFerPi00izluZQCAWCKTtGf4jjAHckCvE/iC/8WeI9U8T6o+681a8lupz/ALcjFiB7c4Ar4E+Aj8NfhP4c8JzwLHewWgmvhtAP2iU75ASOu1mKZ9FHTpTHjANVVVVVVMQeQfpR571VVVVTEHgGqqqqqpumAaqqqqqmxjANVVHGKPPeqqm5OAaqqqqqjgng1VVVVRxiqqqqqOMU3TGaqqqqqmwTwaqqqqrWtF0fxFpN3oPiDSbPU9Nv4WhurK8gWaG4jYYZHRgVZSOCCCDX7W3/AASsuraS88ffsww+dbnMtz4RnmJeIBeTaSuSXyQT5TnPPyseFFjfeO/hT4zW7sbjW/Cfirw/csu5fMsryxmXKspHDo2CVKnHBIPU1+yt/wAFXbOWKx8DftORNDKqrFD4tsoNyuRnm7hTlTwBviUgkjKKMtXh3xHoHi7Q7LxN4W1mz1bSdRhE1pe2cyyxToejKykgiqqqbniqpueAaqqo4x1o896r/gqn/wAnbah/2ArD/wBAav8AgkB/ybR4k/7Hq7/9IbGqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr9qD/k2j4t/wDYi61/6QzV8Jv+SqeDP+xgsf8A0oSqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr9sX/gnT8Jf2p45/FmliHwf8QVjO3W7O2Hl6gQuFW8jGPM7DzBiQAAZZVC18fP2a/jD+zT4rbwn8V/Cktg8nNnqEOZbO+X+9DMBhvdThl/iUV+zH/wUV+NH7PvkeHNfuZfHPg6PCrpWpXJE9mOB/o9wQzIAAAI23IOcBSSa/Z8/ay+Cn7S2krcfDzxQi6vDCr3uhXy+Re2hI5yh4kUf34yy9MkHiqpeRjNVVVVVQ5HWqqqqqHXrS8jGaqpfN8p/I2eZtOzf0z2zjtXxb+Lfx+8FahJaXmnado9m8my1v7O18+OXuMPJuG7APylQcZ4714b8K+HfB+m/wBkeGNIt9Os/MMnkwrgFzjJ+vA/Kq/Z+1YWXjGfTJJMLqFowVc/ekQhh3/u76jbtmkbIGDUbAjGajbHGaRgRgGkftmlYdjSScYzSycYBoSAjrXmKeM0ZF6A00gxgGmk4xmnkBGAaZs96kcEYBr49/HLUbnU7rwR4NvntrW0dor68hba87gYaNT1CA5BPUken3o4pJnEcKF2PQAZptH1JU3m1OAOxBP5ZqGe6sbhJ7aaWCeFso6MUZCO4I5Br4K/FyfxlC/hvxLco2s26l4ZioT7VGOvA43r3A6jnHBNeNfiF4c8FwH+0rsSXbLmKziIMjehI/hXjqf1rxr8TvEfjR2gmm+x6fn5LOFuP+Bt1c/kPQCqqqqy0rU9T3/2bp11d+Xjf5ELPtz0zgHGcH8q/wCEW8Tf9C7qf/gHJ/hX/CLeJv8AoXdT/wDAOT/Cv+EW8Tf9C7qf/gHJ/hV1oGu2UDXV5ot/BCmN0kts6KuTgZJGByQKqqstX1bTUaPTtUu7VXOWWGdowT6nBqy8feOtM3/2b411618zG/yNSlj3Y6Zw3PU/nVl8ZvivYIyW/wAQtccMcnz7xpj+Bckj8KtP2jPjTZQiCHxzMygk5mtIJW/76dCf1qz/AGs/i3ayRvPNo92EGGSaywH4xk7GU+/BH5cVZ/tl+NUiK6h4U0SeTdw0JliAGOmCzc9ec/hVl+2lpsjMNR+H1zAoHymHUllJPuDGuP1rT/2v/hjdGOO903X7JimXZ7aN0VscgFZCx+u0fhWl/tIfBnVVVU8ZR20mzc0d1ayxbe2NxXaTz0BNab8Rfh/rKB9L8b6Fc/IHKx6hGWUHpuXdlfoQKqqqqBwetA5GM1VVVVVfQ1qGoafpOn3Oq6rfW9nZWULzXNzcSiOOCNAWZ3ZiAqgAkk8ADNftNf8ABYS707Xbnwn+zFoGm3tlasUl8T61DI4uG5z9mtwU2qMDEkhO7JHljAY/Ef8Aa4/aY+LPmx+PPjb4pvrabPmWUF6bO1f628GyI/8AfPFaVoWt67I8Oh6NfajJEu50tbZ5Sg9SFBwKl/Z++PMMbSy/BLx8iIpZmbw1dgKB1JPl8CtV0LW9Ckjh1vRr7T5JVLRpd2zwlgOpAYDIrwf8UPiZ8O2L/D/4i+J/DLMzMTo+rz2RLMAGP7pl5IVQfUAeleDP+ClH7Z3gsRQQfGO41i1jYs1vrWn2175hKbRulePzsDAOA4GRk5y2dL/4LIftVafYRWd34Z+G+pyx53XV1pN0skmSSMiK6ROAQOFHAGcnJPw0/wCC1Xi2C8it/jF8HNIvbV2PmXfhq6ktZIh2IhnaQSHoMGRPXPavAP8AwVJ/Y38cxwx3vxAv/Cl5Nnba6/pUsJXCkndLEJIV4GOZBkkAZJrw1+0r+zv4x8tPC/x28A6nK9uLj7Pb+I7VpkjOPmeLfvTBZQQwBBIBwa0jW9F8QWQ1HQdXstStSxUT2lwsyFh1G5SRkelVVVVVQOeCaqqBx0NAgjg1VVVVX0NBuxrgjg1VVVVVVUkccsbRSoro6kMrDIIPUEelfGD9ib4XfESO51Xwnbr4T111Zlks48Wsr9vMh6KCe6beuSDXxW/Z8+KnwbnZvGHhx204Ptj1ayzPaSZOB84AKEnosgVj6V8KPjz8TPgzf/afBmvsLNzmfS7sGa0n9cx5G08D5kKt2zgkV8Gf23Pht8RvK0fxkY/CGuHCqLqcNaXB6fJMQNh4+64GMgBm5pHjljWWJ1dHXKspyCD0IPpVVVVVVXxc+BXhD4sWTzXcIsNcjTFvqcKDfkAhVkH8aZI44IxwRXxE+G3ir4Y66+heJ7ExkljbXKDMV0gP30bv1GQeRkZAr4JftN654De38N+MZZtV8PAhEkPzT2Q55U9XXJHyk8AfL0wdB8QaJ4p0m31zw7qlvqFhcruingfcp9QfQjoQcEHggGqvLS2v7SaxvIEmt7iNo5Y3GQ6sMEEehBxXxh+G938LvHV94akEj2ZPnafOy486BuV+pU5Q+6mv2Q/if/wknheX4fanNnUPD8Ye1ZmyZbUtjGP+mbEL9GT0NfGv4M+GPivoe/UbiPTNU09C1tqmwHykHLLIMjdH1PJG08g9QdI8QeHPEkE174W8QWGtWMNzLbi8spC8btGxUlcgHBwCMgZBB6EVo/7H3wy8XfGzSfEGsatFpHhd5vtGp6SIyqXUysCsSMMCKOTnf0wAQpG4FPjZ4HtYfD2n6xolhDbQaNEtobeCIIkVvwIwqjhVU8ADgBvaq/Z88M/2n4nn8QTws0OlRYiboPOfgfXC7/oSD6UkYAryVkUo6hgRgqRkEV8TPBN34C8XXuiTQstsXMllIc4khJO0g+o+6fcGtF10WSi3uSdg+6wGdv19qXxLaeVxexYx0Lj+Va34hW5Rra0YtuGGfp+Arwb4T1Pxx4ksvDWkr++u3w0hUlYkH3nbHYD/AA71pul2mk6ba6TYReXbWUCQQpnO1EUKo/ICv2tPhFd+I9Nh+Ivh61knvtKg8q/hjGS9sCWEgA6lCzZ/2Tn+GtM1O80e8S+sJdkqccjII7gj0qx+K9qIcXthcRyAf8sSHB9epGP1rX/iZdX0LW2kwPAHBBmkPzj6AcA++T/WtC0PVfFGtWegaLavdX9/MIoY17se5PYDqSeAASeBXw28B23w68F6d4Ut5RNJbIWuJgCPNlY5dgPTJwPYCvEeu6J4W0mfXPEOpQWFjbDMs0xwBk4A9SScAAcmvi14k0Pxf8Rdb8SeG0uV0+/nV4vtChXJCKHOATgFgxHfBGcdK8B/tAePvh5osHh7Rzp1zp9uztHDeQM+0udxwVZSBkk4zjJPrXgn9rPw9q0kdh440htIlbj7ZbsZoSfUrjcg6f3vwqeHSdf0uSCVLTUdNv4GR0YLNDcROMMCOVZSCQRyCDX7a/7Lc37P/jca94Zt5H8E+I5nfTmPP2CYks1oTkkhV5Rj1XjJKMT/AMExf20/+FH+NE+CnxH1fb4E8W3iCxubiTCaLfudocseFglJVXz8qsFfKjzCfir8MtE+KnhabQtTSOO7jBewvSm5rWX1GCCVOAGXoR7gEeKPDWt+CfEV34d1u3a2v9PmKtjIDY+66E4JUjBB7gg1+zj8coviBpUfhLxNeRp4j0+ILG7ud2oRKP8AWc9ZAB8wzk/e6ZxVVX7eXxV+yabp3wh0i6/e322+1faekSt+5iP1dS5HBGyM9Gr9kv4Yv8R/i5p9zd2qy6R4cK6hel1yrMp/dRnsdzgHB4Kq3WjjHWjzVVVVVUx4wDTHPANVVVVUcY60eaqqqqqjgng1VVVVVHk9aqqbGMA1VUcY60eaqqqqo9ODVVVVVU2MYBqqqqqm6YBpuTgGqqqqqOMdaPPeqqqqqbpgGq/aQ/Y8+C37Tunb/HGhfY/EVvbtFY+IdPAiu4B/CrnpNGDyEkBAy23aWJr9pj9hb42/szyyatrGmp4i8JbsReIdKQtEmc4WeP78DYA5bKHIAdjkD4DftQ/Gj9m/VxqHwx8Wy29jJOJrzRbsGawvSMA+ZDkYJVQpdCr4AAYV+zl/wU3+B/xigs9B+IlzF8PvFcmEaHUJ82Fw+cAxXJAC54O2ULgnaC+MmKWK4iSeCVJI5FDI6HIYHoQR1GKqmwTxVU3JwDVVVN93iqr/AIKdazLqn7Y/i6xkhRF0ey021jYHlwbOKbJ9DmYj6AV/wSf0mz079lL7ZbKwk1TxNfXFxubOXCxRDHoNkSceufWqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr48aJ/wkvwO+Inhz7V9n/tTwpqlp52zf5fmWsibtuRnGc4yM+orwxrkvhjxLpPiW3gSeXSb6G7SJyQHaJw4UkdjtxVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVXxA+HPgT4reFrzwT8R/CemeItDvlImsr+3EqZwQHXPKOMkq6kMp5BB5r9qn/gjrrekNd+Mv2WdTfVrMtubwlqUqpcQLjn7PcuwWUZ6JJtYD+NycVaXnj74TeNRcWk2ueEfFnh66ZDgSWN5YTLlXVgdrxtglSpxwSCOTX7J3/BVbSfEDWXgL9pswaXqTskNt4stoglrOSMf6XEoxAxYDMiDy/n5WJVLGwv7DVrC21TS72C8s7yFZra4t5BJHNGwBV0ZSQykEEEcEEGh1xmqqqqqpeRjNVVVVV9DSkHoaqq+K0UMvwx8WCaGKUR6LdyKskYcBkiZlODkZDAEe4FfCj9r1CttoPxRgIIARdYt0zk56zRjpx1ZPb5epq7tLmxuZLO9t5IJ4W2vHIpVlPuK8A6odG8aaNqAfaqXaI7c8I52N0/2WNI2QMHpXiHxr4X8HwJceJNbt7ISZ8tXJZ3x1KouWI+g7j1qT9p/wCHsMrRx6drsyqcB0towG9xmQH8xWgftGfDTWp1tri9vNKdm2ob+DapPH8SFgBz1JA4PtmzvLa8gjubS4jnhlXdHJG4ZWHqCOCKVgRjPNK+OCaVwR1oPxwaEnbNCXIxmjJxgGmkyODTPnv+dM4x1p5MjANfE/xNJ4R8A634gt5Nk9talYG/uyuQiHoejMp/w60BJcSgbi8kjdSckk1omixxRrHGuSfvEjkmn0CQQ7zHxj0rxHoytE0yLiWIZBA6gdq8N63ceG9f0/XbQnzLG4WXaDjcAfmX6EZB+ta58JvAviiRtV8i5hmu/wB61xb3DZkzg7sPuHI9u5rVf2fbiMFtG8RxSH+GO6gKY4/vKT/6DXhz9pH4OeJdkcfi6LTpm6xalG1tt+rt8n/j1PFJGsbSRsoljWSMsuN6MMqw9QRyD0NfsreMNA1iGb4deItJsZ72ANPp1xNAjmSP+OIkjOV+8OvBbptGbOys7CAW1haQ28QJIjijCLnucCqqqqqdEkQxuoZWGCrDIIoeFvDBGP8AhHNL/wDANP8ACrv4c/D3UJvtF94E8O3MpGC82lwu2B7lc1e/Bn4UagyvP8PdCUoMAQ2awj8QmM/jV7+zp8F7+UTT+BrdWVdoEN1PCMfRHAzz161dfsp/B+4eR4dN1G2D52pFfOQn03bj+eavf2Nvh/JEF07xN4ghk3ctM8MoxjpgRrz05z+Faj+xZbsXfSfiFIgCfu47jTQ2Wx3dZBgE/wCyce9X37GvjqPZ/ZvinQZ853+eZYtvTGMI2e/pV7+y18ZbVFa30C0vCxwVhv4gV9zvZf0rUPgz8V9MdY7n4e645YkD7PZtOOOvMe7H9e1XumajprLHqNhc2rOMqs8TRkj1GRWl6/ruhuJdE1u/09w24Na3LwnPrlSOa0n49fGDRVCWfj7UZAFKj7XsujgnPWUNz79QOOlaT+2B8S7Jgup6Zoeox7iWLQPE+McAFWwBnnlSev4aN+2hoMzKviHwRf2qjhnsrpLgnjqFYJ37Z6d60X9pj4OayoVvE7afKf8Alle2skfGBzuAK98Y3Z4rSPE/hrxCM6B4i0zUwASTZ3aTYAwCflJ7kfnVKcjg9K4I4NVVVVVX/BTf9uS/+J3iW/8A2e/hZrUsPg3Q7hodeu7eQAa1do3MYZSd1vGw4GQHcEkEKhr9nP8AZz8Y/tFeMV0LQkay0ayZX1fV5Iy0dnGewH8cjYIVM88k4UEj4d/sGfs4eAI4prrwjJ4pv48E3WvTfaAxDBv9SoWLGQByhOOCTk50XQtD8N6bDo/h3RrHS7C3XbDa2VssEUYAwAqIAAMADgVVzbW95by2l3BHPBMhSWKRAyupGCpB4II4INeKf2Z/2f8Axmko1/4QeF5JJ93mz22npaTOWGCTLDtfOOhzkHkYNeJP+CcP7NeuOz6XZ+JPDwLhgum6qXAAXBUfaFlOCfmPfPQgcVef8EtvhpJcyPp/xN8TQ25P7uOa3gldR7sAoP8A3yK8e/8ABL3x/prPcfDj4gaPrkA3MLfU4HsZgMnCqV8xHOMfMSgJzwK8VfsiftJ+D5jFqXwf8QXYDhVfSoBqKtkEg/6OXIGB3xjIBwSBWs+FfE/hzH/CQ+HNU0zLlB9ts5IMsOq/MByPSq0v4g+PdDsItK0Txxr+n2cGfKtrXUpoo0ySTtVWAGSSTx1JNaJ+2Z+1j4flkmsP2jPiDK0qhWF9r096AB6LMzhT7jBrwJ/wVy/a48K3ER8U6h4Y8Z24b96mp6OltIylgTta0MQVsAqCVYDcSVbAr4Mf8Fivgl40uo9J+LvhDVfAFy+1UvYpTqdkTnB3MiLLH1B/1bADOWGBnwZ468FfEfQYvFHw/wDFukeI9ImYpHfaXepdRFh95dyEgMMjKnkdwKqqoHsTVVX0NAg8VVVVVVfQ1u4wa4I4NVVVVVVVcW8F1BJa3MMc0MqFJI5FDK6kYIIPBBHGK+Mf7CfgHxvLca58O7mPwnq0mWNqkObGRvaNeYs+qfKP7tfE/wCDPxE+D+qnTPG/h6e1jZ9tvfRqXtbnjP7uXGCccleGHcCvgz+1J8Ufg09vp2n6o2seHoid2jX7lo1B6+U/LQ8knC/Lk5Kmvgv+1N8L/jQsOm2Gof2N4hfg6PqDhZJCBkmF/uyjrwPmAUkqBzVVVVVVXizwj4d8caJN4e8UaXFfWU3JRxgow6MrDlWGTgjnn3r40/s6eI/hbI+s6WZdX8OM3F2sZMlr04mAGFBJwGHB4ztJAr4V/GHxd8JtVF3odz9osJW/0vTZ3byZwcAsAPuvgDDjkYGcjIPw1+KfhH4qaMNV8M3376Pi5sZiFntj23qCeDjhhwfXIIFftPfC1viF4DbVdKgMmteHg9xbIibmniwPNiGOckAMAMklQB97NeAvGWpfD/xdpni7Shvm0+YM0RbaJkIw6E84DKSM44zntX/BUP8Abn0+28Ewfs//AAi15zf+K9NiufE15CwBttPnj3LZEhspLKhBkUgYjYDkSHH7Fug/FHxh8YLPwr8Pr3yNOmAn8QNNGJIIrRCA0jKSCXywVNpDZb+7urxP4Y1Xwlqsmk6tDtdeY5F+7KvZlPcfy6Gvhp8SbS7sW8B+Np91ldRG3t7uQ/6sMMbHJ7ejHp346eItDvPDWt3mhXw/fWcpQnGN46qw9ipBHsa+DfhceG/Atis0LJdahm6nDdcvjaMdsIF49c0keaVMjAr4h/DLQPiVo66ZrAeGe3LNaXcQG+BiBnr1U4GV74HIIBHjL4HfETwbLI82hzajYq5CXdipmUgd2UfMn/AgBnuae3nSc2rwusyvsMZUhg2cYx1znjFeEvgt8R/Gc8aad4cuLW3cnN5fIYIlAOCckZbnjCgn24NfCn4QaH8LtLaO2cXuq3I/0q/aPaWHZEGTtQccZ5PJ7AeWPSjGD2/Svil+yH4a8V3U2t+Br9NA1CZi8tq8Ze1lYnJIA5iPPbK8ABRya1T9k742afcGC08PWepICQJbXUIVU4PXErI3PXp9cV4b/Y8+J+qzRHxDc6XoduSPNMk32iVQc52pHlWIwOC4HPWvhj8EfBvwqs/+JPbm81SVAtxqVyoMr+qpj/Vpn+Ee2S2AauHht4XnuJUjijUs7u20KAMkk9hivjp8XL34r+LHuIJWTQ9PZo9MgIK/LnmVh/fbA+gAHYk2thPd8oAF/vGm8PuEyJTnHdaubWW1fZKv0I71+zP8VLrQNfh8AaxdF9K1aTbZmRuLW4OcBf8AZkOFx/eKnjLZ+KXw18MfFzwJq3w+8X2f2jTtVh2MQSGicEMkiEdGVgrDtxg5BIr4wfCnxP8ABb4gar8PfFduwutPlPkXAjZI7yAk7J489UYD8CCDyDX/AATD/a7b49/DA/CvxxqzT+PPAtqiNNcShpNV04EJFcf3meMlYpCc5JjcsWlIHx8+DFr8VvDwutOjSPxFpcbGwlyFEw6mFyexx8pP3WOeAWzYX+u+D9fjvtPubjTdW0u4O11+V4ZFOCCPzBB4PIPBr4LfFvTPiz4XW+UJb6vY4i1C03glWxxIo67G5xkcEEc4yar4jeO9G+GfgrVPG2ukta6ZDuESkBpnJCpGue7MQPbOe1eMvFet/ELxdqXizWpXuNR1e5MjgEttzwka99qqFVR2AAr9l34Qv8JPhlbW2qW6x69rRF5qYI+aJiPkhP8AuLwR03F8ZBzTEHgGqqqqqo4x1pjkZH4Uee9VVVVUx4wDVVVVVUxwODVVVVVUcAcGqqqbk4BqqpumAaqqqqqm6YBqqqqqqbk4Bqqqqqm54FHmqqqqqbpwaqqqqqm5OAaqpoYriJ4J40kjkUq6OuQwPBBHcYr9pH/glj8JPijLceJvg5cQfD3xA6ktZQwbtLuX5/5Yrzbk8DMXyAD/AFZJJPxo/Z0+M37PurjSfip4GvtJSWQpa34UTWd2ev7qdMoxwMlMh1BG5V6V+z3+3d+0F+zt9l0jQvE3/CQeF4ML/wAI/rha4t40HGIHyJIMDOAjBMnLI1fAT/gpZ+zt8ZYINN8Tav8A8K+8SPtV9P1ycC3kYkD9zdgCNhkgfvBExOcKQCailiniSeCVJI5FDI6HIYEcEEdRijz3qqPNVVU2MAVVftSeI/E/iv8AaN+JGreMdZtdU1WPxJeWct1axeXC6W0hgiES5JEaxxIq5LHaBlmOSf8AglL+1FYaNcy/sy+Lmggj1K5lvvDd6xCZnZQZbVj3LBd6HrneuTlAKqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr4of8k08W/wDYCvf/AEQ9V/wlvhT/AKGfSf8AwNj/AMaqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr9rD9iD4N/ta6Kp8V2H9i+K7OPZp/ifT4FF1Eo5EcoOBPFnPyOflyxUoWJP7T37Gfxr/ZT1z7J4+0L7ZoNzIy6f4i09Wks7kA4AZsZikIwfLfB9Nw5r9l39uH4xfsv3kWm6Pe/wDCReDnkzc+GtRmYQrltzNbyYJtpDluVBQlssjkDH7O37Vnwd/ac0H+0vh3r+zVbaPdqGg32Ir6ywQCzR5IePLLiRCyfMASGyo4I4NVVVVUvXGaqqqqqlPHHaqqvin/AMkx8X/9gG9/9EPVeKPBXh/xja+Tq1oPPRcRXMfyyR/Q9xyeDkV4y+F/iPwezXsSG+09TlbqBTmMf7a9V6denTnPFeMvi3beFvA+m63aiO51PWLZWtIW6KSoLO4HZSQMZ5Jx6katq2qa9qEuqavfTXl3O2XllbcT7D0A6ADgDgVDo+oToHWDaMcbjj9KubK6tcC4hZM9CeR+dfCf4uaz8PNThtbi6luNAlk/0q0I3bM9Xjz91h6Zw3fsRY31tf2kF9ZXCTW9xGskUqHKurDIYHuCCDSuCMZoNxwc0sg6ZoSAjrW8YxRcEYFGQYwDTSA8A0zk96ZxjANftBlpPhFr6ICxxbnAGeBcRk/pVi6R3kLvjAcZz2960OWJXTcRxVzqVg2miJUXeBya8QTQiOVyRtVST+VVpCvDothDKjI6WsasrDBUhRkEdqdhjOar4f6Lo+ufCfwhZ61pNnqEH9g2X7q6gWVf9QnZgRUnwE+GEerW2vaRoT6PqVpMs0Nxp9y8W1l6YTJTHqNvPTpkUMZxmqqqqqoYz1qqqqqq+hqWKG4heCeNJI5FKujrkMCMEEdxjtWq/Cv4a62XfUvAmhTSSEFpRYokhwMDLqA3QAda1f8AZX+DuqBvsuj32mMwPzWl65wT3Ak3gfTGPatX/Yu0SUs2geOb62GSVS8s1n4xwCVZO/fH4VrX7IPxNsGLaVfaNqkf8ISdon6dw6gDnj7x/Ctc+CvxW8Oh21PwHquyMEvJbQ/aUUAZJLRFgBjvnAqWG6sbgxTRy288Lcq6lGQj26g1ofxj+KXhzauk+O9WVExsinnNwi4zwEk3KBz0xzXh/wDbD+IOntHHr2jaRq0S/fZUa3lbkZ+ZSVHGf4Opz2xXhr9sL4d6mI4fEWl6posrY3PsFzCvTPzJ855J/g6D1OK8N/FH4d+L/LTw74y0u7lkxsg88RzHOMfu2w/VgOnU461VVVV/wUt/ajn/AGefgg3hzwnfeT4z8eebYabIjgPZWwA+03IGOoVhGvQhpQwJ2EH4V/DTxF8X/H2j/Dvwqkf9oavMUWSXIjgRQWeRyASFVVJPGeMDk18Kvhf4T+DvgXTPAPg2xWCy0+ICSUqBJdzYAeeUj7zuRknoOAAFAAqqqqqqq88E+DNRuZL2/wDCOi3NxMd0k02nxO7n1JK5NXn7OH7Pt9ayWc3wQ8Cqkq7WMPh+2hcfR0QMp9wRXiD9hn9lzxCrNJ8MIbGYxeWkun39zblOSchFk2FuerKe3oK8d/8ABL74d6lHNcfDvx9rWiXBUmO31KJL2Hd2UFfLdVPAyS5HXnpXxf8A2K/jx8HrebVdR8OR69o0GS+paIzXKRrjO50KiRAO5K7QQeehPwY/aA+L37P3iOPxL8KfG9/o0vmK1zaLIXtb0L/DPCfkkGOORkZ4IPNfsef8FJ/hl+0g9n4H8cJa+C/iBL8kdlLPiz1N9wVRayOc+YxYYhb5v7pfBIqqoHtmuCODVVQPYmuCODVVVVVV9DWTjFZB4zXBHBqqqqqqrW9D0bxLpVxoXiDS7XUtPu02T2t1CJI5BnPKnIPIB9iAa+NP7Aemal9o1/4L6hHp04QsdDvXZopW5OIpiSUJ6BXyuT95RXirwf4u+HuutoPi3RL3RtUt8OIrhCjYycOh6MuQcMpIOODXwU/bg8f/AA+W30Hx8svizQo12JJLJi9txn+GU/60Dn5ZOegDqBivhz8V/AHxZ0Zda8C+I7bUIwv76DOye3PTEkR+Zee5GD1BIwaqqqqqrm2t7y3ltLuCOeCZCksUiBldSMFSDwQRwQa+O37K0lv5/i74V2JkiAaS70aMFnBznNuoHIwT+76jHy5yFHhfxX4k8Ca7FrvhrUptP1C2JUsv8QzyjqeGU4GVIxx7V8FP2gvD/wAV7ZNKvjDpniWNWMljk7ZlXq8THqMclc7hg9QM0ee9ftJ/DAfDnx/JcafFt0jXt91ZYXCxNu/eRD/dJBH+y6+9ftafCK+1fUNL8deFdJkury9mi0++ht4stLI5CQOQOpJIjyfWMV+yP+zxY/s8/Cy10G4ijfxJq226165Rt4afHESn+5GDtHYnc38VeKfBui+NNN/s3V4CdhJhmTh4W9VP8weD+FeM/Bmr+CNXbS9TTcjZa3uFXCTp6j0PqO30wTpN1D4g8QaPa+KtS22UckUE1xISCsAb7pYdgMgE9MjnAGPif+0N/ZFzP4c8ALBJLAPLfUiA6RsOoiXGGx03HI9ARzWq+PfGuuXH2nVPFeqTvklQbllVc4ztUEBeg6AVpvjnxno84udM8V6rbyd9l2+G+ozgj2NfC/8Aablubu30L4jLAglYJHqsSiMKfWZRwATj5lAA7jGTUSo6K8bBlYZUg5BHqKWPI4pYsihCCMAV5PGMUYRjGKaLjimjwKeMEYFPHgYr9onWbrw78F/FWpWRIla0W2BBwQJpEhYg4PaQ/wCI6iKPzZEjB+8QK0LTEnKRKOAMAVN4OZLAXBj+XHBxXiHTUVZIgvOOPY1BPNazx3NvIUlhcOjDqrA5BH41pF42qaLYam5jLXdrHMfL+7llB468c1+3x+zl/wALk+Gp8Z+GdP8AN8W+DoZZ7dYo8yXtpjdLBgAs7DbvjX+9uAGZDXwK+M3i39n/AOKeg/FXwZNm+0W43SWzyMkd5A3EsEmP4HUkexwRyBXw4+IXhX4seBND+I/gjUPtuieILNLqzlK7W2nqrL/C6sCrL2ZSO1ftQfA1vEdrJ8RvCNlu1S0j/wCJlaxqB9piUH96McmRRgEfxKBjlQD8PPHut/DbxVaeKNDlO+E7LiDdhbmEkF4m4PBwOccEAjkCvBHjPQviB4as/E/h+6EtvdIN8ZI3wSYG6NwOjKTg/gRkEEyPHHG0kjqqoCWZjgADqTX7V3x6/wCFueLRoHh253eFtBlZbRh/y+TYw85/2eqp/s5PG4gfsYfA3/hOPFH/AAsrxHabtD8Ozj7HG5wLq9GGXjuseQx6ZYoORuFMcDg1VVVVVUxGMA0xB4BqqqqqjjHWjzVVVVVU3JwDVVVVVU3PANVVHAFHnvVVTcnANVVVVVTcng1VVVVRxjrR5qqqqqnII4NVVVVVTc8A1VVVVVHk8Gqqq8TeF/DfjTQbzwt4u0Kw1nSNQj8u6sb63WaGZcggMjAg4IBHoQCORX7Qv/BJDwb4j+0+Iv2efEP/AAjV+2X/ALC1R3mspD6RzfNJD9G8wf7or4y/s3fGr4A6q+mfFHwDqOlxbiINRRPOsrkZ4MdwmUJ5B25DDcNygnFfBz9qf4+fAR4ofhl8SdT0/TUkLtpE7C6sXJIL/wCjyBkUtjBdAr+jDrXwO/4K8eBtf8rSPj74Rl8MXZ2qdX0ZJLqzJ4DM8JzNGBycL5px79fA/wARfAXxN0VPEXw88ZaP4j05sA3Gm3iXCoxGdr7SSjYPKtgjuBR6VVVVNgnANV8SvHOlfDH4e+I/iHrZQ2XhvS7i/lRpBH5gijLCME5+ZiAo4JJYAAk4rUL3UvEOsXGpXjtdahqVy8srLGAZZZGySFUYyWPQDvxX/FW/DDxz/wAvOh+KPCWrewlsb22l/EBkkT8xX7MPxqs/2g/gf4Y+J0KpHeX9t5OpwKMCG8iOyZQP7pZSy/7LLVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVXibwz4d8Z6BfeFfFmiWOsaPqcJhvLG9gWaGeM9VZGyCK/a9/4JBX8E9348/ZRP2qGWRpLjwfe3ARoBjP+iTyHDDP/ACzkII7OeFqCfxz8LPGYlhbWfCnirw7dkEMslneWFwhwQQcPG4OQQcHqDX7K3/BV+CQaf4F/abt/KKx+VH4vs4iwZh0N3AgyMjgyRA84ygBZx4d8R6B4v0Oy8TeFtZs9W0nUYRNaXtnMssU6HoyspIIqqqqoexqqqqqqXHTNDkdaqtQsLPVbC50vULdJ7W7heGeJxxIjAhlPsQSKrwBd3F54H8O3d3cSTzzaTavLLKxZnYxKSxJ5JJOSTS7WUowDAjBBGc18XdYTVfHOoW1siRWelubO2hjXasYQ4bAwOr7z+PfrXhvTEm/0qRQSThMjp6mrLQnnjBVM8elatoypG0UsYKkYIIrULQ2d29vnIU/KT6V+zP4jn1fwFJpF07u2j3TRRM3P7thuUZ68EsPYY/BXI70sgI4NCQEdaDgjANbhjAP6Vv8Af9KLjGM00gxgU0gxjNM5PQ14i0m18RaFf6DeY8q/tngYld23cpG4D1HBHuK8Q6DqPhjWrzQNWh8u7spTHIBnB9GGeqkEEHuCDWj+IlhRYbt9rKMBz3+tSeJbURc3sZAHQOCfyrWtca/zBbkiL+IkYLf/AFq+F3g2bxn4ttLR4GawtXE165TKhByFPb5iNuPr6GpGzwDXju+utO8F6/qFjO0Nxa6XcywyL1R1iYgj3BAqtF0iz0DRrDQdOVha6bax20AdtxCRqFXJ7nAFV9DQwRwaqqqqqhyOtVVVVVSntmqqqrV/Dvh/xBF5GvaFp+pR4xsvLVJhjOejA9wDXiD9mf4Pa+sjJ4abTJ3z++0+4eLbweiElB1z93sO3FeIP2L7dleTwr42kU5OyHULYNx2zIhHbH8HPt0rxF+zJ8X/AA/uePw/Fq0KdZdOnEmfojbXP/fNaroms6FcfY9b0m80+4H/ACyurdoW/JgDXhr4u/EzwiVGgeNdTgjQELBLL58S8Y4jk3L+leGP2y/FlkY4fFnhuw1OINhpbVzbS4J64O5SR6YXOBz3rwt+1F8JPEuIrrV7jRLjgCPUodgPrh1LIB9SOv1xp+pabq9pHf6TqFte20ozHNbyrIjD1DKSDVTSwwQvPPKkccalnd22hQBySewxX7aPx5uv2jf2ifFPj5L3z9Ft7g6b4eUElU06BmWJlB5HmEvMR2aZh0xX/BOb4Fw+Cvh3J8XdcsduueLkKWTOCGh04MCoA7ea6B/dVjNVVVVVVVVVVV+0t+wp4G+MyXPinwJ9j8K+MW+ZpViK2l+QDxMifcYnGZVBPXKvxjx58P8Axx8JPFs3hTxro11o2s2LB1VjjcM/LLE44dSQcOpIyDzkGv2B/wDgp3LcS6P8EP2ltaaSWRvs+k+NL26JZmOBHDfM3UnkfaC393ePvSVVVQOOBWQRwaqqBx0NAgjANVVVVVVV9DW7jFZB4zXBHBqqqqqq8ffDbwR8UNDfw9458O2uq2hyYzKuJIG4+aOQYaNuBypGRwcgkV8bf2EvGHhD7R4g+Fc0/ibSV3yNp5QC9tlHICgHE/H90BjwAp614e8S+LPh/wCIY9Z8N6tf6JrFhIV8yFzFIhB+ZHHcZGGRgQehBr4Ift7aXqi2/hz40Wy2F4SqR65ap+4kJOMzRjmPt8y5U5PCAc2V9Y6nZw6hpt5Bd2s6B4Z4JBIkinoVYZBHuKqqo4x1okdAaLZ4FE5718df2bdJ+IUU/ibwjDBp/iREZnjVQkWoNnPz+kh5w/ckbuORqGn6/wCDtel0++iutK1bTJtrruMckLjoQR+BBBwQQQcGvgN+07a+KTB4Q+It3BaawxWKyvtuyO9OANr9kkJ78K2cDBwD+2F+3PL42/a4h0zw/rST/DfwVNJoyG3kLRXruVF1d8Eq4WVAEYZBSIEH94a0rU7jTL611fTZwJbeRZYXByCRyPqD+orwxrll4m0S01uwJEVymdhPKMOGU+4IIrTvGmlaj4yufBlirSz2No01xMCNiMGQbB6n5+fQ8dc48T+EdH8ZaPJo2sQbkbmORRh4X7Mp7H9D0PBr4h+CtZ+Ht6LHUpIHE7uLWVHGZUUD59vVR8wHPfOM4Jq2tpbydYIhlm7nt71YeGLZUG+PzmxyWH8hV94YtXjJWHym7Mgxj8KvLSWynNvMOR0PqPWv2VfH0viHw7c+CtTnL3ehhWtWbq1s3AX/AIA3HPZlHakiyOBSxgDAGaEZIwBXl/T8qMZxjFNGCMU8XHSpIvSnQEYr9pPRrrWvgj4ss7QZeO0S5PGfkhlSV/8Ax2M1E/lSpIP4WBxWgajHCySBgR2qfxgH00WpkG0DgV4i1FCsku4Z7D3qKKWeVIII2kkkYKiKuSxPAAA6mtG0waRoWnaTsiX7FaRQbYhhBsQL8o44444qRcjHpX7dnwA/4Ut8XZtY0DTvI8KeLd15puzlLebj7RB7YYh1GMBJFAztOP8Agj1+0zLbanqX7Lniq+T7PeLLqnhV5GIZZlG66tRzyGQGZQAMeXMSTuAFftH/AA/8OaB421e+8E3EUkVr5Ems2EKgDS5bjeY+AMBHCE4/hLLnAdAfgJ8Y7r4V+Jlh1GeWTw7qLBL6AZYRHgCdFH8S4wQPvLkckLj9rD9qW18SWt38LfhtfiXTJBs1bVImI+04IJhiP/PPjDN/Fyo+XO74K/B7xB8afGcHhrSA9vZRYk1LUDGWSzi55PqzY2qvc+gDEeF/DGieDPDth4W8OWS2mm6bCIreEEnao7knkkkkknkkk0ee9VVVVVRxjrTHjrVVVVVUx4wDVVVVVUSAOtHnvVVVVVHGKPNVVN0wDVVRxijzVVVVVHGOtVVVVVTdODVVVVVHGOtN6A1VVVVVNyeDVVVVVHpwaqqqqq/0+w1Wzl0/U7KC7tZ12ywTxCRHHoynII+tfHD/AIJgfs4/FVbjUvB1hP8ADvXJFYxzaJGpsy5OQZLRsLtHPyxNF25wMV8c/wDgnV+0l8FftGp2/hj/AITPQIIzI2p+Hka4MajGTJBjzUwOSQrKACd3Bx4G+Ivj34Za0niL4eeMtY8OakuAbjTbx7dnUHO19pAdMjlWyp7g18Ev+CvHj3QWttH+PPg6DxTZ7lWTWNHCWd6igfMzQcQysSOgMI5PPQV8Gv2w/wBnX47xQw+BPiTpy6rKyJ/Y2pv9hvRI3RFikx5p5AJiLrk4zVVX0NHk9ar/AIKyftD2fhf4d2X7Pfh++RtZ8WtHeayqjJttPik3RqT2aWZFxg/dhcEYcE/sGfBaf42/tL+FdKuLeV9G8OzjWtXkVAQsNswdEbPaSbyoz3w7EdK/4Kx/s+f8IZ8SNO+PPh6x2aR40xaatsXCw6lEnysew86FcgDq0MrE5av+CXv7TVt8J/iZcfCHxjrLW/hjxzLGLEyFjHa6qSqRnAyFEy4jZvVYskBSRVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVWvaxa+HtC1HX71JXt9MtJbmVIgC7JGhZgoJAzgHGSPrVf8Mh/GX/nx0r/AMD1/wAKqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr9p79in4GftWabv8e+H/sXiW2t2hsPEmnARXluP4Vc9J4weRHICBltpUsTX7VH7Afx2/ZXum1LW9K/4Sbwe+TD4l0iB3gjwcbbhOWtn5X73yHOFdiGA/Z8/av+NP7NWtJffDzxTK2lSSBr3Qb4maxuxkE5jJ/dscD95GVftnBIP7MH/BQP4LftHJY+G7i9/wCES8cT4jOhajINt1JtBP2Wb7soPICnbJwfkxhjVVVUOR1qqqqqh7Gl9M1VV8Q9GtvDvj/xN4fsrRrW30zWLu1igcsTEsczKFO7LZAAHPPHNfs6asus/AT4c6iL5rx5PC2nCaZ3LM8q26LJuJ5LbwwJPfNIwIxmvH8MkPjnxAkqFWOp3DAH0MhIP4gg14WljNnGoIypII9Dn/69aBqdnbw4lRW+Xoa8RXcEzs0ePYV4hlSXUSqkfIgU/wA/61+yrbTR6Lr18XlMU11FGqkHYGRSSR74dc/RaWQdAaDg9DQc4wDmhIema80dKMoPAoyZ4BpnJHWmcetPIMYzTPkda+Knwn0f4jWi3KOtnrFshEF0F4cdkk9Vz36jnHUg+JPhd468LTmLUfD1zJGDhbi2QzRt77l6Z9Dg1FDLcSLBbxPJIxwqIpJJ9gK8J/Bzxr4pmjaTTZNMsmALXV5GU4PTahwzZ9hj3FeDvBmieBdIGlaPEWZjunuHHzzt6k+g7DoPxJLsAMA18edRu9K+CPxD1TT5vKurPwpqc0Em0NsdLWQqcHIOCBwRivBei2viTxjoPh2+klS31TU7e1maIgOqSSKjFSQQDgnGQRmqqlI6ZqqqqqqB5xmqqqqqr6GuCOKqqqqr6Gr/AE3TdXtH0/VtPtr21lGHguYlkRh7qwINeJ/2ZvhD4mLSr4ebSZ2zmXTJfIxkk8IcoOv930HQCvFH7GGt26vP4P8AFtregLlbe/hMLEgcgOu4Ek9MhRzz614s+D3xK8EI03iLwhfQ26AlrmFRPEoHcvGWVeP7xH6GtB8T+IvC139u8N67faZPkEvaztHux0DAH5h7HI5rwh+1t8StA8u38QR2XiG2TaCZ08mbaOwkQYyR3ZWPf1z+1d+1bL4p/Zh8Y+H/AIV+H9X/AOEx1+zGmC0MKyeTbzkJcyIwb5v3JkRSAHDMrbcKTXwk+D2v/Ez4vaB8KbqyvdNuNSvQl75tuyS2sCZaZ9rDgqivgEYzgHGa0rS7DRNLs9F0q2W3srC3S3toVJIjjRQqqM84AAFVVYzwKCE0I88ZryweMV5Wf4a8v0FGPjpRiGPu0Yh0xRjGOKMZxwaKkdqI7VXx3/Z/8A/tA+EZfDni2wjiv4kb+zNXihBudPkPdG6lCQN0ZOGA7EKw+MXwd8a/A7xrdeCPG9h5VxF89rcxgmG9hJwssTHqpx06ggggEEV/wS7/AG6ZfHFnYfszfFvU0Ou6darD4R1FkIN/bxRsTaykcCWONBsY/fUEH51Bkqqr6GgQeKqqoHsa4I4NVVVVVVVX0NZPY1uPStwIrII61kEcGuCODVVVfGn9mH4ZfGq2kutT05dJ1/afK1mxjCy54/1q8LMOB97kdmXJr40fs2/En4JXcs2vad9v0LzdltrNopaCTP3Q46xOem1uM5wWHJ+Dn7QvxH+Cd+reGNWa50mSTdc6PdsXtpc43FVz+7cgAb1weBnIGK+CX7Tvw3+N8IsdJu20rxAi5k0a9cCVgBktEw4lUAHOPmGMsqgjJYY4otnpRJIxmjz3qqqvjB8EvDPxc0oJdgWGsWwP2TUYowWXj7jj+NOnGcjsRznxv4E8T/DzXJPD/inTWtblMmN+qTpkgPG38SnH19QDxXxT/ZJ0/wAZeI7LVPAF3Y6E1/dxRajBMu2CNXkVWuEA+7tUlmQYBC8YPB8c/C7SPhhbaNpPhaKVdCisIra0EsplZPKRUwWPJyoVs+pOMAV4T+JWs+ENA1LRtOIL3bBreVufs7Hh2A7kgDHYEZr4A3Msfj9kBDfabGVHLcnqrZHvlRUa4HFftQXEsnj6xtmEqxw6VHtDH5STJISyjP0BPHK+wrwrCjSSyEgtkKPavDumwXTKjlfxrxFpVvagojKcDqK8WwooikBG4MR+H+RX7KE0sfxZjiikKrNp06SAfxKNrY/NQfwpY8iliyKEIPQV5IxjFNCPSni44p48DipIwRxUkY7VdW0NzC9vcRJLHIpV0dchgRggg9QR2r47/B/UvhH4xnsVgkfQ753l0q5Kkgx5/wBWx/vpkA+ow3G7FWeoT2Zwh3L/AHScY+lN4jcptWNie2Wq7vJ7xt0rcDoo7V+zJ8I77xN4jtvHmsWkkWjaRJ5lo7cfablCNu31VCMk/wB4Ac/NiRewqVRnAr9rb4KRfHT4L6x4WtLdX1uwX7forEci6iBwgPpIpaP0G8HtXgXxp4m+FvjvRPHnha5ay13w1qMV5aM6n5ZYnB2uvGVONrKeCCQeCa8ZftYeCrH4BaF8YvCVzFeT+NNOEvh+0kYE+aVG/wAwDtCxw4H8QC5BYGv2MNYHi74meNdN8ZypqK+JNBnn1GW6kw0xE0YfJ46iViSMY28YrxenhG18XaxpngnxAmsaTaXTpbXIBBKZ4GSBux03jhsZHXAsPh5eePfFelaDo11aWt5qt0lsZLqXy4wWOAxPr2wOWOAASefhF8J/Dfwc8HQeEvDoeUlvOvLuUYe7nIAZyP4RgABRwAB1OSWIPANVVVVVVMQeAelE571VVVVRxjrR5qqqqqqY54BqqqqqqbGMA1VVR5PWqqm6YFVVVVVTYxgGqqqqqpueAaqqqqpumAabk8GqqqqqPSqqqqqpumAaqqqqqm5OAar45/sY/s8/tC/aL7x34Git9cmjKrrukv8AY71D/eLKCkpAGB5yyAelfGr/AIJI/Fvwik2rfBvxNYeNrFMn7BcgWN8oC5+XcTFJ0I+8h5XCnJx42+H/AI4+G2tzeGvH/hLVvD+pwMQ1rqNo8DnHddwG5SCCGGQQQQSCDXwo/bn/AGofg80Nv4e+Keo6ppsTZOma6f7RgIxjaDLl414BxG6c/U5+G/8AwWQlCw2nxf8Ag6jNu/fah4bvtvGVBC20+ecbzkzcnaOOWrwl/wAFQf2QPE0CvqfjPWPDMzbAtvq+iTlssSCC1uJkGMDJLAfMOTzg/tx/skY4+PPhj/v8/wD8TWv/APBQL9j3w7arc3fxt0y535CR2Fnc3bsQM4xFG23PTLYHvXxh/wCCv/hCys7vSvgd8PdR1S+eEpBq2uuttBDJ2cQIWeVQMcM0Rz7DnxV4p8c/F/x5d+J/El7eeIfFXiW9UyukAMt3O5CqiRxqBk/KqoigDgAdBX7BX7KH/DMnwukn8SeVL418WeVc6yyDItEUHyrVTgE7AzFj3dmxkBTXxr+Enhr46fC/X/hZ4sDLYa5beWJ41Be2lUhopkz/ABI6q2O+MHgmvjP8HvGvwI+IurfDXx5pzW+oaZKwimCERXsBJEdxCT96NwMg9RypAZWA/YX/AOCj+katp2j/AAW/aB1SLT9RsoEtdI8VXVxiK9VcKkV2x+5KFwPOY7Xx821vmeKWKeJJ4JUkjkUMjqchgehBHUYqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq8R6NF4j8Pap4ennaGPU7Ka1eRRkoJEKkgeoBqv+Gu/jL/z/AOlf+AC/41VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVXlnaahaTWF9bRXNtcxtFNDMgdJEYYZWU8EEEgg8HNftZf8ABIr4f/EiW78bfs5XNj4I8QzM0k+hTArpNyTknygilrViSBhQYgAAETkn4lfCr4nfBDxdL4R+JnhDVfDGuWUhKxXUezdtOPMhkUlJUyOJI2ZT1BNfst/8FRPiV8JIrLwb8Zorzx14Vhbal6ZA2q2cfosjsBcKD0WUhucCQKAo+Efxs+F3x28MjxZ8K/GFnrtgjBJxFlJbVyM7JYmAeNsdmAz1GRzVVVLgjFVVVVVSkYGDVVX7aGgf8I1+1f8AFfTt8red4pvLzMqbT/pLmfgen73g9xg96/YT1/8At/8AZb8FNJcRyT2KXVnKqDHl+XcyhFPv5fln8c1G/bNfH/wfJZaxF4ws4ibbUAI7kj+CZRgH6MoH4qc9a03UpNOl3KNyN95f6j3q18T2bRjbdBTjox2kVqniiIqUtm81yOD2FJHcXtwsUSPNPM4VUVSzOxPAAHUk9q+FXhE+BvBdlos6qLx8z3m3B/etjIyOuAFXP+zSuCODQc44OaEnvQkyODXmDGMijICMAijJxgGmkB6GmckcGmcDvTyZHWmfPANO4AwDTtnvUj8YBp2755qR8jANftsatHo37Lnj+7liaQSWMNuFU4IM1xFED9AXB/Cv2TPDzeKf2nvhTootLe6jk8X6bLPDcKCkkMVwkkqsDkEGNG4PXp3qqrp0rgjg1VVVVVAgjg1VVVVVKR0zVVVVVUD2zQORjNVXi/4O/DXxz5kniDwnZSXMm4m7gXyJtx/iLpgsc8/Nke3Jrxd+xfC5e48CeLTGSfltdUjyBxz+9jGevbYeD1458XXmkeCPiHqXwq8ReINKt/FGktCtzp32tPMzLGske0Z+bKOh4zjcM4qxmfTdVt9csNsOoWoIhuVQb0BGCAfQjqOhrQvjZq1rth1+wjvUHBli/dv9cfdP04rQPHvhbxJ5cVhqaR3D8C2n/dyZ9ADwx/3SaCEihGDwOaERPQUIc0IRjGKEQxihEMcfyryxjGK8sehoxg9BRiHT+lGIHpimhHpTRccU0Z6Ypox2NFSK/aZ/Z48O/tFeAJPD18sNrr2nK8uham4ObScgZVscmJ9qq4wegYDKrV3aeKvhx4xktLlLnRvEfhrUcEK2JbS6hfghgSMq6ggg44BBr9h/9qHT/wBqn4JWPiy7nt08WaMy2HiWzjATZdKoImVAciKVfmU9Mh1GdhqqqqB7E1wRwaqq+hoN2NcEcGqqqqqqqqqqqr6Gsn1o896q7tLW/tZrG+tori2uI2jmhmQOkiMMMrKeCCCQQeDmvjv+wjo+trceJvguItL1BiXk0SaTFtMSSW8pyf3R54U/J2GwVrmgeJ/A2vyaRr+mX+i6xp8gYwzxtDLEw5Vh0OOhDDg8EHvXwG/bq1fQRb+F/jM0+q6ePlj1uNd9zD6CVR/rV/2h8/8AvV4f8RaD4r0i317w1q9rqWn3SBobm2lDow+o6H1B5Heqqqqq/bx/bI8SeKP2tdSu/h54gkl8NeBh/YltbtJutr543JupCo4IaXciuCSVjRgRnA+H/ij/AIWB4D0nx/aaPe2Nhq4kEJuI8BnjYpIEbo4Vwy5746DpU+uazdabDo9xqlzJY25zFbtISin6fy9Mn1qvgrOIfiXo4M/lrJ5yNlsBswvgH1ycceuKRQcCv2q/Ds6X+i+Ko42aGSBrKV+yMpLoPxDSf98mtDv0sbrEjARyDBJ7EdDWmawbdQUftxWpa4bhDuk7dzXiDUFvboRxPuSLPI6E96/Y58LyX/i7VvFcsZ8jTLMW8ZIIBllbOQc4OERgRz98Hjio4wegpIsjGKEQxg15Yx1pogRgU8Q6CpIx6VImOlSoCOKlQDpXjDwf4c8b6JP4e8UaVBf2U4OY5VyUbBAdD1VgCcMMEZrxv+xFepNJdfD7xZBJCxJWz1VCjIPQSoDu79VXtz3qP9kH4xSSpFJb6PErMAZHvshB6nAJwPYE14A/Y403TbiPUPiJraak0bArYWG5IWI/vyMAzA+gC/U5xVpp1jpVlBpum2kVta2sYjhhiQKsagYAAHQVKo7VMoxxUy5HFftn/s+6h4M/aNuNP8H6RIdN8bsdS05FTEcUjt/pKAjgKkmXxgBUdR0GT4O0Ofwn4O0fwe2rXl9b6PA0cInmZ0iLu0kgiUkiNDI7ttXAyxJySSYbq5tllW2uZYhNGY5Qjld6EglTjqMgHB9BXwP+D/j34r+Ihb+ErQR2VqQL/ULgFYIVJGQSPvPg5CDnjPABNeKPDWt+CfEV34d1u3a2v9PmKtjIDY+66E4JUjBB7gg1+zn8Zk+JXhz+wtbuQfEmkRD7QWODdQg4WYD15Ct7kHjcAKqqqqqiQBwaY54BqqqqqqY8YBqqqqqqOAKPPeqqqqqo4J4NVVMRjANVVU3J4NVVVVVR5PBqqqqqqPPeqqqqqYjselVVVVVTdMA1VVVVVNycA1VVVVRxjrR5qqPPQ14s8G+EvHmiy+HPGvhrTNd0uf8A1lnqNolxETgjO1wQDgkZ6jNfEv8A4JVfsx+NnnvfCUOu+CL2WTeBpl4Z7fJOWzDPvIBGcKjIFOMDA2146/4I/wDxw0V5ZvAPxA8J+JraP7iXYl064kyQOExJGMAknMg6cZPFeKP2B/2vPCcs0eofA/WrtYYWl36XJDfh0Gfu+Q7ksdpwmN/I+XkUf2X/ANpfH/Ju3xN/8JG+/wDjVaV+yV+1BrV2LGz/AGfPiDHIVLBrvw9c2qcf7cqKufbOTXw3/wCCWP7UnjZobjxPpuieCbKRd5fVr9ZZtpCkYit/MIY7j8rlCNrZwcZ/Ze/YN+D37MuzX7VZfFXjJoykmvajCF8kHGRbQAlYAdvXLycsN+07RVftSfsq/D79qjwQPDfin/iW61Ybn0bXoIBJPp8hxkbcjzImwN0ZIBwCCrBWHx+/Zt+K/wCzd4qbw38SPDssFvPI403VoFL2eoopxuikHGcYJjbDqGXcoyM/sz/8FAPjZ+zktp4da6Txf4Mt12LoOpzFTbpnP+jTgFoT22kPGAT8mcEfs+/t1/s/ftD/AGXSNC8Tf2B4onwv/CP62Vt7iRzxiB8mOfJzgIxfAyyLVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX99Z6XYXGp6hcJBa2kLTTyv0jRRlmPsACar/hE/FP8A0LWq/wDgFJ/hVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVfGX4EfCb9oHwr/wh3xc8F2PiDT0YvbmYFJrSQjBeGVCHjbHdSM9DkcV+1r/AMEn/in8GxeeMvgd/aHxB8HxZeS0WNW1awTgZeJABcjJ+9Cu4DJKKoLV8Pfib8Sfgz4ri8UfDvxXqnhrW7KTDSW77NxU8xyxtlZFyOY5FKnoQa/Zf/4KteC/GzWfgz9oa0t/CmtMqxxeILf/AJB90wHJmU/NbMTjn5kySSYwADYX9hq1hbappd9b3lneQrNbXFvIJI5o2AKujKSGUgggjggg1VDGcZqqqqqqX0zQ5HWqr/gq74T/AOEc/a71HWNmP+Eo0Gw1HO/du2I1rnH8P/HrjHtnvX/BMDxSmpfCDxJ4SkufMn0TXvPVC4JjhuIU2jb1ALxSnJ4JJx0NK2RweavrGx1exm0zUraO4trhCkkTjIYf579jzXjL4C67ps0l34Ub+0rMnKwMwWaMenOAw9xg+1T+GvEVtKYLnQNRikXqj2jqR+BHpWi/Dnxv4hkWPTPDV6yk482WPyYxjr8z4H4V8LPgxYeC5Y9d1yWO91lcmLZkR2uQQdv94kHqRx2Hcq4PQ0r44JpZOODQkBGM1vU8ZrePWt49aLr0BoyDGAaaTjrTSZ4BpmGOTV7r+i2M32a91iyt5QMmOW4VGA9cE5pmyOtSPxgGnbuTUj5HWnbjrzX/AAUy8SQaX8BtO8P5VrnW9fhVVIPEcUcjswPThhGMH+/x0Nf8EwfCKeKv2yPCFzPCktv4ftb7UpUZCwyts8cZ46FZZY2BPGVx3qqqgcjGaqqqqqge2aqqqqqr/ULDSrOXUdUvreztbdd0s9xKI0jHqzEgAfWvGv7eH7IPw/nNr4h+PXhuWVJBG6aU0uqlWIPB+yJLjG0g56HAOCQK1v8A4K+fsmaVDFJYWnjrWWdiGjstGiRoxjqfOmjGPoSasP8Agsd+yzeXkNrceFfiVYxyNhri40q0KRD1YR3TNj6KTXhb/gp/+xj4nnFpN8TrrRJnkCRrqujXUSvx18xEZFHbLsteBPjR8IfiiiN8OPih4W8Su0Ycw6Zq0NxLGCu7541YshweVYAjuBVVVKcjg1wRwaqppYYIXnnlSOONSzu7bQoA5JPYYr48fEZvi98aPG/xNAZYvEmu3V5bRuAGigaQ+ShwBkrHsXOATjJ5Jr9mr4MaZafs8eDLLxZb3j6zd6Yt3PcTTv58XnkyImGJC7EZE2kYG3pnNa58ENXtg02gX8d6gyRDMPKk+gP3T254rU9H1TRp/s2q6fPaydlljK5+nr+FeHfiR4r8ObYrfUDdWwx/o91mRQB2U5yv0BxXhr4z+GdW2W+sI+lXBOMud8R/4GBx+IAHrVtLb3cKXFpPHNE4yskbBlP0IoREihCCKEQxgV5Q7D9K8oY6V5Y9KMQPajEMcD9KMIPSjDxxTRHHTNNGDTwgjgU0RA4powenWv8AgpR+z/HcWdp8ffC+msbmFksvEawx53x42wXLYH8OBExJOQ0Q4Cmv+Cc37Rv/AAz1+0Xpf9tah5HhTxls0jWt7YSHe37i4PYeXIRknojyetVVVVA46GgQeKqqr6Gg3Y1wRwaqqqqqqqqqqqqqq+KvwZ+H3xm0UaN430VZ3hVvsl9CRHc2hI5McmDgdCVIKkgZU4FfHb9lH4gfBZ5tYijbX/C4I26raxEGDPAE8fJj543ZKHK/MCdo+Enxv+IHwX1gal4O1dhayyBrvTZyXtrsDj50zwcfxLhh69q+BH7T3gP44Wsen28q6R4ljjBn0i5lG5yASzQNx5qgKScAFR1AHJqqqv2wfjYn7Pn7OnjP4lW8qDVLaxNro6sSN19OfLhPHJCs28jjIQ8jqPC/hzXPH/i/SvCmjL9q1fxBqEVpb+a5+eaVwoLtzgZbJY9BkmvC/wAMPCnhr4daP8MbexE+kaNYxWkAl+8di48zPUOTkkjuTXj74d6r4KvGk2PcaXI2ILrb69Ff0bj6HqPQV8NLmO18f+H5ZAxDX8cYCjnLHaPwyRUYyeDXivwnpXjXw/d+HNYQm3ukwHUDdEw5V1znBB/zzXjn4feJvh7qp0zxBYlFYk29ynzRTqCRuVvw+6cEdwKttTvrRQkFwdo6KRkVdarf3alJbg7T1VRgV4C+H/iP4j6/DoHhyzMjsQZ52BEVsnd3bsPbqTwMmvhv4B0n4b+E7PwrpDtKkGXmndQGnlblnIHT0A7AAZOM1HGMcUqZGBSxAjgV5PHSmiGOKdMDBqSMEVKnHFSKOn5VMnHFSKCMVIuRipVGa+Knxk8I/C20A1Sf7Zqkq5g02Bx5rD+839xPc9cHAODXiP8Aas+JWrSyDRU07RYT/qxFAJpF6dWkyCeo+6OD681YftL/ABctJxLda7a6goI/dXFjEq8H/pmFPPTr+VfD39pjQPFFxFo/i2zj0W+kO2O4EmbaQ+hJ5jP1yOPvDgVOVRC7sFCgkknAAr48+PNM8f8Aiy3TTra2ls9AE0NneGNTI5kK+aVfkhGMaYAxnaCe2Kr9nP8Aar1P4RQw+D/FFmdR8Kb2KCCNRcWRYszMnTzAWbkMcjsRjB+NfgHw38d/Alv8RfhveW2p6hZwu1tNajP26JfvQkAbt6kHapwQSQQM8eFvE+ueBvEdp4j0Oc29/p8uVDg4PZkccZUjII9DXw78d6R8SfCdn4q0bKJcArNAxy1vKv30J74PfuCD3qqqqqjjHWmIPP5Uee9VVVVUcAUee9VVVVVTHPANVVVVVTEYwDVVVNycA1VUcAdaqqqqqjwODVVVVVU2McGqqqqqOMUxyMZqqqqqqbk4BqqqqqqPPQ1VVVVTdODVUenBqqqqbniqqqqvGXgnwh8Q/Dl34R8c+G9O17Rr5cT2V/brNG+OQcHowIBDDBUgEEEZr9oj/gkiskt54m/Zu8QpGrM0g8M6zMcL32W90cnHQBZcn1kr4hfDL4gfCjxBL4V+I/hDU/D2qRZPkX0BTzFHG5G+7Iv+0pKnsa/Zx/4KTfG/4HxW/hzxZO3j/wALxMALXVbt/tlsnA2w3J3EKABhHDqAMLtr4Dftz/s7/tBzwaL4Z8Xf2N4jn2hNC1xRa3ErED5YTkxzHOfljYtwSVA5qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq+MmpWOi/CHxxrGpz+TZ2HhvUJ7iTaW2RpbSMzYAJOACcAZrTtPvdX1C10rTbd7i7vJkht4UHzSSOQqqPckgVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX7V/8AwTr+Bn7Ttvf+IodLi8IePZoyYvEOmQBRcSZyDdwjCz5zgucSYwN+ABX7Sv7G/wAc/wBlfWTafEfw15+izyFbDxDpxM9jd9OA+AYn5xskCMcEgFcMf2b/ANs/43fsyXkVr4N1xNS8MmYyXPhzU8y2khb7zJghoX77kIBIG4MMg/s1ft3/AAM/aVS30bStX/4RvxhJkHw5q0qpNKQuSbeT7lwMBuFw4CksijBNfQ1wRxVVVVVDGaXBGKqv+Cz3gP8A5Jt8T7Zf+fzR7skf7k0GML/18Zy393A+8a/4JheMG0r4ueI/Bcs+yDX9E89FwTvntpBsHAwP3csxycdMdTQOOhpWz0NJJnjNK4I60snYmlcHoaV8dDSyZGM0HB4BoOcYBoOa8w+teYfWi5PSi5PGaZx61488feH/AIeeHpvEPiC62RJ8sMKcyXEmOEQdyfyAyTgCviB8f/iH48uXQavNo+m5/d2NhKYxj/bcYZ/oePQCq8NeNfFvg65W68MeIb3T2D7ikUp8tz/tRnKt0HDA9q+C/wAfbT4gEeHfEqwWWvKv7oodsd4AOSoPR+pK+nI7gSOMdadu5NE7u9f8FRPHEOqeP/CPw+tpd39g6bNe3G0EASXLqoU88kLAD0GBJ1OcD/gi74BS58WfEj4oTxMradp1ro1q5U4cTyNNMAd2Mr9mg6qT84wRyCMEdaqqBwetVVVVVX0NAgjg1VVVeKPFPhrwToF74q8Ya/YaLo+nR+Zd39/cLBDAuQAWdiAMkgD1JAHJr9o//gsHBZ3lz4Z/Zm8OW99HG23/AISbW4XVJB3MFqdrY9GlIP8Asd6+Kvx5+Mfxw1H+0/it8RtZ8RyLIZI4bqfbbwtzzHAmI4/vMPkUYBx0rwX8Hvir8RtreBvh54g1qFs/6RaafI8K43felxsXlGAyRkgjrxWi/sDftS6w0LS/DuHTYZovMWW91a1XbkZAZFkZ1PsVGO+K1L/gnf8AtQWVqZ7XwrpOoyAgCC21iBXPvmRkXH414o/ZF/aU8HxyS6z8HtedIlDO2nxpfgA9827PwMc+nfFSwa14b1VFmivdL1OxkSRA6tBNA4wyMOjKehB47Gvg9/wUd/au+ED21pH8RJfFukQcHTfEy/bgy7s4E5InXAyABJtAPTgY/Z6/4KufAX4sywaB8T4m+GevytsR9RuRPpsx7Yuwq+UcAk+cqKOAHY1aXdrf2sN9Y3MVxbXEayQzROHSRGGVZWHBBBBBHBzVA46UpHQGqr9szx0Phv8AsrfFDxWk/kzR+HLm0tpfM2GOe6H2aJgcHkSTIQMckAZGcj4e+Fv+E58f+GfBQm8r+39YtNP8zdt2edMsec4OMbuuD9DVrZ29nbxWlpBHDDAgSKKNAqooGAoA4AAAGKWLPWrzS7HUrY2moWcNzC3WOVAw/I14j+BOiagGuPDl0+nTY4hcmSIn8fmX8z9K8TfD7xX4Udjqelu9uvS6gBkiI9dw+79GANeHfFviHwrcefoepyQAnLxH5o5P95TweOM9R2Irwl8edNvNln4stPsUp4+1QAtGfcryy/hn8KsLqx1S1S9068huoJB8kkThlP4ihEMYzXlD0/SvLHYV5Y9KMQxjFGIY/wDrUYQRwaaI9qaPjGKaIHimi9KaMHginiBFeMPCmkeNvC2r+DtfgaXTdbsZrO7RW2sY5EKNtPZsHg9jg18SfAuq/DHx9r/w/wBbO680G/ltXkCFBMFb5ZVB5Cuu1xnswr9gz43t8ff2YfCPiy/vEuNb0qH+xtaYY3G7tgql2AJw0kZilI4/1nAAxVVVVVA4GKBBHBqqqvoaDcYrIPGaqqqqqqqqqqqqqpI45Y2ilRXR1IZWGQQeoI9K+Pf7C+geKFufFHwdW20TVtrPJozfJaXTZz+6P/LBjyAv+r+6MIMtWs6J4p8A+JJNK1myvdF1rSrjJVsxSwyI3Dqw9xkMpweCD3r9n79uiW0Fp4Q+NcrSwKojg8QIpZ0AXA+0IAS+cAeYvOTlgeWrTtR07WLC31TSL+2vrK6jElvc28okjlQjIZWUkMCO44qqr/gtR8YPt3iPwL8CdNvMxaZbya9qsaTblM0uYbYMo+66Is5552zg8A5P/BM34XReK/i/qXxG1G1aS08GWOLVicAXlyGjQkfxARCc47EoeOMxjgDNXunWOrWM2m6jbJcW1whWSJxkMP8APfsea+JHws1HwVK2pWRa60eSTEcuMvDnosn8g3Q+xOK8LXY0/wAT6Rf4U/Zr+CTDHA+V1PPtxV38UPh1pc32e88aaSsgJDKlyJCpHBB25wc9jXh/x74J8SSi30PxTpt5PkgQpcKJDjHIQ/MRyOcYrVdC0fxFYvpWuaZbX9pJ96G4iDrn1APQjsRyK1H9lT4XajcedavrOnJ/zytrtWX/AMiK5/WtL/ZK+FlpcrcXVxruoIvWC5u0VDyDyY0Vu2Ovf8a8OeGPD/haxGm+HNHtNOtgcmO3iCAn1OOSfc1EgPSkXIwKjjBGKSLIwK8nini4xUsYxgVIoxipkFSqO1TKMcVIvOKlUZxXxU8d2fw28FX/AIqukSWSBQlrAzY86ZuEX6dz7A1res6p4k1e61vWLuS6vr6UySyOSSzH09gMADsABWhfDW5vokuNVnaAOMiGMfOPqT0Ptg1f/Cq2EJNhfzpIAcCYBgT26AY/WtS0280m8exvYtkqfkw9Qe4rwT40+Ivj3wRP8K9AuIXvYkz9qnuTHJ9jGAY1Pc5IH+4cVbfss+MG3i/8Q6NFjGzyTJJn1zlVx29a+Ivx/wBd8DeNdb8Ev4Ot1utA1K4sZ3lu2YStFIU3KAowDtz1OQRz6/slfs9fBP48fALwl8W9Rl1m5vtescahBHfhIra7hdop1jURqyqZEYhXLYG3DMPmbT/2Vf2fdKEn2X4a2T+Zjd9puZ7jGM9PMkbb17YzxnpXh/wj4U8JQy23hTwxpOixXDB5Y9Pso7ZZGAwCwQDJxxk1+1T8Gl0K+f4leGrLbp97IBqkUagLbzMQBIABwrnqf75/2q+BfxeuPhP4mae7E0+h6gAmoW8QBbjOyRc/xKSeMjIJHpjxl/wWQ+AWgX8umeHvhx481ae1naKdrq3gsUG3g7Q0jPnOQVZFIxV3/wAFurRLqZLD9mmWa2WRhDJN4wEbumflLKLNgpIxkBiAe561/wAPvP8Aq2P/AMvX/wC4a0X/AILbaBcXTJ4i/Z1v7K2EZKSWXidLpy+RgFXtogBjPO48gcc5HhX/AILH/sz6wsMPibwn470C4ZWMrtYwXUCEE7QHjm3sSMf8swAcjtk+Af28P2Q/iSAnh747+G7Wbp5GtStpL7vl+VRdLGHOXA+XOSDjO04sNQ0/VrKHUdLvre8tZ13RT28okSQeqspII+lMRjANE56GqqqqqpjngGqqqqqo4Ao896qqqqqm5OAaqqOMdaPPeqqmweAaqqqqqm54BqqqqqqbngGqqqqqbGMA02CcA1VVVVRxjrR5qqqqqmxiqqqqqpvQGqpumAaqqrGaPJqqqqqq+IXwy+H3xY8PS+FfiP4Q0zxDpcuT5F9AH8tsY3I33o2/2lII7Gv2g/8Agkd/x8+Iv2cPE3q//COa3L/47Bc/oFlH1kr4hfDL4gfCjxBL4V+I/hDU/D2qRZPkX0BTzFHG5G+7Iv8AtKSp7Gvgb/wUF/aR+CH2fTYPFn/CV6DFIC2leIS10AvcRzZEsfB4AYqDg7TyD8F/+Cq3wD+ICQ6b8S7W++H+rNgF7kG7sXO3nbNGu5eQfvooGV+Y5ONA8R+HvFmlQa74W17TtZ025UNBeafdJcQyggEFXQlSMEHg96qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv2wda/sD9lj4q332bz/N8J39rs37MefC0O7OD93zN2O+MZGcj9n/Q5fE/x5+G3hq3nSGXVvF2lWiSuCQjSXcSBiB2G7NVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVWt6HonibR7zw94j0ex1XS9QhaC8sb63WeC4jYYZJI3BVlI4IIINftX/wDBHzwt4mjv/Gv7Ll5D4f1Y5lfwrqE7GxnIXLC2lbc0LsRwjkx5bGYlHHxD+GnxH+DPi6Twh8R/Cmq+F9fs8Si2vIjE+3JCyxsOHQlTh0JUkHB4r9mj/gqP8WPhNFa+Ffi7DP4/8NQqkUM8kqx6jZRrx8spH78Y7SncePnA4r4L/tA/CL9oPw9/wkfwq8ZWurxRYF1akNDc2jYHyywuA688bsbWwdpI5pcEYzVVVVVUpzjmqr/gpT8M3+JX7I/it7WDzb3wnJFr1sNjNgW5Imb5QSMW8k5zjA7kDJH7OPj5Phh8c/BXjaa5jt7Wx1WOO8lkxtjtpgYZ2OeBiKRznt6jrVA46GlYHoeaSTtmlcEYzSuQODSyDHWg4PQ0H4wDQk7ZoS8YzXmHpRlJGBRkyMA0ZMjg0z5HWv2hvHknjj4iXkNvNu07RGaytAGyrFT+8cY4+Zgee4C1p2nG8be4O0HAA71B4fDRgpAuMf3a1PQhGpMabHA4AGAatLq6068hvbKd4Lm1lWSKRDtaN1OQwPYggGvh94tj8beC9K8SgqJbuD9+q8BZVJVwB6blOPbFE7u9SyRwxtLLIqIilmZjgKB1JPYV+0R8SG+Lfxq8W+O47hpbS+1B47AsAMWsWI4OBwP3aKTjuScnOT/wTH+Fb/DL9kzw7fXtn5Go+NJ5dduQ0bKxjlwlucknIMEcTjAA+foclmGM4zVVVA5GM1VVVVVA84zVVVftE/tJfC/9mLwOfGvxK1ZkNwzRaZplsA91qUwGSkSEjgDG52IVcjJBYA/tO/tc/Fr9qjxO2qeN9Vaz0K0mZtK8O2krC0sly21ivAkm2uVMrDceQNq4Ufs9/sefFD4/yx6paw/8I94XD7ZdbvoCQ/H/ACwjypmP0IUd2B4r4SfsR/AL4Uw21ynhOPxNrMOGbU9dUXLb+DlIiPKjwRlSF3Dux61DbRxRpDDGqIihURRgKB0AHpS25PFC3GPpX2cEcD9K8YfDbwJ8QbFtN8ceDdH123ZCgXULJJigP90sCVPcEEEHkEGvi9/wTF+G3iVJ9U+EuvXXhPUCrFbC6LXdk7ZyACT5kY6jOXAGMLwc/GD9nT4v/Au+Nt8QfCNxbWbPsg1S3HnWc/ptlXgE/wB1trewr9l/9uP41/suX8Fl4d1dtd8Ihm8/wxqU7G2+Y5ZoTyYHJJOU4JJLK1fszfte/B39qjQXvfh/q7W2t2MKvqegX2I7uzzgFgucSRbiB5iZGSAdpOKoHtmgcjGar/grn4pbw/8AshT6Qstwo8TeJbDTyIgCrbfMusPk/d/0UHjJ3Be2SP2EPDjeI/2pfBqNA0kGmtdXsxVwpTy7aQo3uPNMYwPX0yaSLjFLGOgFJED1pYxjAFeQrqVdQwIwQR1rxV8FPCHiNGmsYP7JvTnEtqnyE/7UfQj6bT714r+EnjPwqZJpdON9ZoCftVoC6gf7S/eXtnIx7mvD/inX/C119r0HVJrVyfmRTlH/AN5TwfxFeDv2gtLvfLsvGFn9hlPH2qBS0R92Xll/DP4V4t+I/hDwbosetX+qRXKXKlrSK1dZHucEAlMHGBnk5x/KvEf7SPjXUpnTw9BaaRb5/dkRieXt1Lgr27L3Psah+PfxUilWR/EqTKp5R7GABvrhAfyNeCv2loLu5i0/xvpkVqHO37dabtin1aM5IHTkE/SraW0v7aK8sbmK4gmQNFLE4dXU9CCOCPemhyMAU0R7U0fHSmiB4FPFgYxTxginix0p4wRwK/4Kf/DL+wPihoXxNsbXbbeKbA2126xcfarbChmYd2ieJQDziI9ccf8ABFf4oT2njDx98Gbu8zbalp8WuWMLYASWFxDPtPcss0OR6RZGOaqqqqqvoaB7UCCODVVVVn0Nbu1ZB4zXBHBqqqqqqqqqqqqr4vfA74f/ABr0X+zPF+mYu4k22ep24CXNryD8jEHKnHKkEHPY4I+OX7N/j34G6g0mrQf2loEsvl2ms28ZWOQnJCuuSYnwD8pJHXBbGa+BX7Svjz4H6hHb2Fy2qeHJJM3Wj3Mh8vBILPEf+WcmAeeQc8g8Y+Enxo8B/GnQBrXg3VVaaJR9s06YhbmzY9nTPQ84YZU4ODkEAkdM1JLHGoZ3VRkAFjjknAH4kgV+178Vm+Nn7SvxB+IceoJe2N7rMtvpcyMCjWVv+4tiuOMGKNG46lickkk/sB/DxfAX7N+g3dxZtBf+KZZNXud4ILLIdsBGf4TAkTDt8xPfJjAIAqPoK8V6z4f0Hw5eah4nMZ05Yyssbpu83PAQL3J6Y/kOa8U+MT4gvIUs9LtdO0yxRorOziXlIyc/M5+Z2J5JJ6k9MmljeT7iM2OuBmmVlO1gQR2IxXwk/aH1zwnd2+ieMLuXU9DYhPOky89oOxVuroO6nJAA24xg2c8F3DHc2s0c0MyB45I2DK6kZBBHBBHeoxx1r9u34rfFj4G/AWb4p/CO+soL3RtUtl1EXlmtxGbSYmLODyGErwYOcYJBByCP+Cff7S+vftO/BW48ReNZLU+KdC1eax1M20AhSRSFkhkVATgFH2dsmNuO5iXJ4r9pD4sw/Aj4C+NviwwRp/D+kyPZK65V7uQiK2VhkfKZpIgfYnr0r4eftv8A/BRL4o/CLXviX8P9Ai8TTaT4jsdIis9F8IG+d1ltrqaeRo4gzgRmK1BY4XNwo5J4/wCGwf8Agrbj/kgniv8A8Nhef/G607/gpd/wUD1vxi/w50fQrS/8WR3E1s+hW3g95b5ZoQxmjNuuZA6CNyy7crsbOMGn/a+/4KyEYb4D+Kf/AA2N5/8AG6/ZU8XfFjx98APCvi3446DdaN43vvtn9qWN1pj6dJDsvJkizA4DJmFIm5HIIPQ14g1fTPDujX/iDWrtLXT9MtpLm7ncEiKKNSzuQMnAUE8elS/8FW/2nR4mj197XQV8Lzas5jszo+N9urqzW4l3ZLCN0BOcjcpzyDWl6vpviHRrDX9Fu0utP1K2jubSdQQJYpFDIwB5wVIPPrUo54qUV+23qU8WleFdGQnybq5uZ3G4gboljVeOh4lbnt+Jr4f6bHf6+skybktYzIARkbsgD+efwrRNLW4KqAMn2rV/DwtIQWXGR3FfFDSYpNK+3Ko8y0cYP+yxAI/PFfBC+ubH4paA9rKV86doZBnhkZGBBHf1+oB7VKATiv8Agor4G/4RP9oy81y3t/LtvFem2+oKUTCeaoMEgGFA3ZhDsMk5kBJ+bA/4Iv8AxGGs/CLxv8Lrm43TeGdbi1C3VjysF3Ft2qPQSWzk+hk56ivjx+1b8Cf2cNOa6+KPjq1tL9o99to1qftF/c+myBTkA9N77U9WFfHX/gsZ8UvE0suk/APwnZeDdO24Gq6rGl/fs3B3LGcwRDqCrLLng7h0rxf8cvjF8QPEln4u8cfEzxDr2radMs1nPqF+84tmU5HloxKoMknaoA5PHJr4SePH+JHgTT/E8+n3FncyL5VzHLEyK0igbmjJHzI2QQRkc4zkGvid+yDrPxo+INre/DjUdD0m/wBTjc3yalI8ELyKMh1Mcbkuwzn5eSuc5JrT/wDglP8AFOWzjfVfib4Wt7o58yK3huJkXk4w7KhPGD90YJxz1o/8EpPiHj/kq3h7/wAAZv8AGtX/AOCVnxegjjOh/EXwhdyEnzFuxc2wUeoKxyZPtgV4o/4J5ftReHBI9n4O0/XoYfMLS6XqsJ+VOjBJTG7bhyqhS3GCAcCvGXwf+Kvw8V5PHHw58R6JCjFftF7pssULEFQdshGxhllGQTyw9a+Hvxj+K/wlvBffDP4jeIfDMu4s66bqEkCSE7c70U7XB2JkMCDtX0FfB7/gsP8AHLwtPbaf8YfDOi+OdOEn7+8t4xpt/tIwSDGPJbH3gvlLnkbhkFfgP/wUH/Zl+Pot9N0jxn/wjPiGeQRLoniMLZzu56CN9zRS5wcBHLdMquQKqqqqokAcGjz3qqqqqqY54BqqqqqqJwODVVVMc8A1VUcYo896qqqqo4x1o81VVVVRxjrR5qqqqqmI9elHnvVVVVVNjHBqqqqqqbGMA1VVVVVNyeDVU3JxVVVHGOtVVVVVVVXjXwJ4M+I/h648KePfC2meINIueZLPULZZ49wBwwDA7WGThhgqeQQa+OX/AASR+GnioXGtfA7xRc+D9SkkL/2XqJa709gf4Ub/AF0Pc5JlHQBR1Hxh/Yn/AGlPgj5134t+Gt9e6TDydX0Yfb7ULx8ztHlohkgfvVTJ6ZrwF8TfiH8LNX/t34c+NdY8O3xxvl0+7eHzAM4DgHDjk8MCOTXwp/4K4fHDws0Nl8VPDGieOLNW/eXMSjTLw5GPvRqYSAeceUCeRkZGPhd/wVC/Zd+IKJbeI9a1PwPqJyPJ1u0LQsQuSVnh3oBjIG/YSRjHIz4Y8ZeD/G+nDVvBfivR9fsTjFzpd9HdRc9PnjYiqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr/gpFrn9h/sa+PvK1D7Lcah9htIcHBl33sHmIPrEJM+2a/YM8PxeJv2xvhJpsunS3oh8SQXoiiDEqbYNOJPl5whiDnthTnjNVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV8ZfgR8Jv2gfCv/CHfFzwXY+INPRi9uZgUmtJCMF4ZUIeNsd1Iz0ORxX7U3/BIb4nfDuW68V/s8XVx478PFnkbRpUWPU7FOyrztuhjugV+gCHrWj6544+GHiz+0NC1TWfC/iTRrh4mkgkks7q0lQ7XRsYZSCCrKfQgiv2bf8AgrjqVtLbeF/2mtGS7t9oRPE+j2wSVTxzcWq4Vgckl4duMACNskj4efE74e/Fnw9F4q+G3jHS/EWly4Hn2M4fy2xna6/ejb1VgGHcVVVVVS4IxmhyOtVr+iad4m0HUfDerw+dY6raS2l1H/fikQo4/FSRXxM8Car8L/iH4k+HOuBjfeGtUuLCV2iMfmmKQqJADn5WADDkghgQSDmv2UviYnxY+AfhLxPNd+fqMFmLDUixG/7Tb/u3ZgOAXCrJj0kHA6VX0NK4PBNK5HGaWQdjQcEYzQc44NCTsTQkBHBrzAeM1vHrRcEYFGQdARRkGOtNJngGriea6nkubiQySyuXdj1Zick/nXh6KNkhUgY2itB0fTLjT2eaRFZVyAe9eKrW3ildY8cGtQVVvJVXGM/0r9lyeeb4bTxzD5YNVmSL5cZXZGx+vzM1V+3d8cI/hJ8GLzQtI1BY/EvjFX0+yRHxJDAw/wBInA6gBDsBHIeVCOhr9nf4O6j8ffjT4T+E+nyywLrt+qXlzGm5ra1QF55QMEZWJXIzwWwCRnNaVpenaHpdnomj2cVpY6fbpb2tvEu1IYkUKiKOwCgAD2r6GhyKqqBwetVVVVVVAgjrVV8R/iD4W+FPgXW/iN411D7Honh+ze6vJQu5tq9FVf4nZiFVe7MB3r9qH9o3xh+1D8WNR+I/ilvs9qM22i6apBXTrFXZo4sgDe/zFnc/eZiQANqj9jb9hmHxVZ2fxX+NmlSDS5dk2jaFMCpu1BDCe4X/AJ5ED5Yz98HJ+XAa1s4LW3itLSCOGCBAkUUaBVRQMAADgAAAYqODI5pIMjAFLACMUsGeAKEBIxX2f3FNATwBWr6FpWvabc6Nrml2mo6fexNFc2l3AssUyEYZXRgQykdQRiv2n/8AgmzHOtx40/Z0t1Sbc0l14Ynn2q+TybWRjhSM/wCrcgYztYYCHw54m8cfCzxdFrnhnV9V8M+JNFuSqzQO9tcW0qNhkYcEcghkYYPII6iv2MP+Cpeg/Eme0+Gv7R1zYeHvE0ny2XiMFLbT78gfcnyQLeU9j/q3OR8h2q1Kc9+RQwRwa/4LYa5b2/gL4X+GmnlE1/q9/dpGAdjLBDEjE9sg3Cgf7ze9f8EvNF+3/HrW9WlsvNi03wvPsmI4hmkuLdV/EoJR9M0qZGBSRgDikjzxSxgcAZNLETxSwjFLGOgFeNPgn4T8YNJewRtpWovybi3XKufV4+h/DaT3NeNvhP4v8Dhrm/s1urAHC3lsdyD/AHh1T8RjPQmpoIp12uucdD3FWWiLdzmMzNgDIAXn/wCtUvhOHZlJJQQOCSDV/p89hL5cwyD91h0Nfs2/EKez1YeANUuAbK9DPYFuPLmHJQH0YAn/AHgMfeNNF6U0fGMA00QPAp4eOlPHxjrTxAjinj7U8YPSv+CkfgePxR+zbeeIEiU3PhPVLW/RgvzlHb7O6g7Sdv79WIyo/dgk/Lg/8E6/Hf8AwgH7ZHw3v5Z9ltq1++jzoZNgk+1xPBGCcHOJXiYDuVAyM5FVVVVVVfQ0D2oEHgGqqqqvoayema3A8VkEdayDwDVVVVVVVVVHHTNEjHBrUtP07WLCfStWsLa+srqMxz21zEJI5UIwVZWyGBHUHiv2hP2G73TGuvGHwVhe7s+Xn8PtlpouRk27HPmLznY3zDHBbIA8KeLvFfw98RQeI/CesXWkatYuQksXBHqrKQQynGCrAg9xX7O37YPhj4sfZfCXjMQaH4tbCRjO221Bug8ok5Vyf+WbdcjaWyQP2+fHt14W+GWjeHdI1S6sdS1zV0ljmtp3hkSK2xIXR0YMrrKbcgjoRkYIBq/+FeqJ8WNN+GdqWWTXNTt7TTpWAO9J5RHG3JAOCcHJAyDyOtaFpNjoGkWOhaZF5dnp1tHbW6f3Y41CqPyAqP8AhqPoK/aQ8Uy3/iO28KQSn7PpcQkmXpmZxnn6IVx/vGtG0wahMTJ/q06jpk+lWWiAxBIogABwAMVqmhRSRGOeLPHBxyPpV9ZvY3LW7nOPun1Ffso+MLnX/BV14cvrhpptAnVIi2SVgkBKLk9cMsgHoAo6AVHgrgGvir8PtO+LHww8U/DTVX2W/iXSLixMneFpIyqyDg8q21hweVHBr/gkx44vfhn+0v4p+CniaKWzuvE9hLbNbEglNQ09nfY2DjiI3fIzyB2JIix61/wWd+M/9m+E/BfwD0yaLztZuG13VQJMSJBDuit12/3JJGmbJ724x3r/AIJ/fBy3+CP7Kfgfw9JYxW+ra1ZjW9YZAQ0lzdgSDf8A7aReTCccfuh16ncu3HtX7N+P+HxusYP/ADP/AIy/9E6lUxByAamwehr/AIKffFsfDD9lXXNIsbzytV8c3Eeh2wSba4iky9ydo5ZDDG8Z6AGVc9cHXP2XZYf+CYOifFSLSGi1+38RyeI7nNvmZ9PnYWYGeuzakE4PICljgbiR/wAE0/iyPid+yxoek3t55uqeCJ5NDuQ825/KjAe2O08qohdIx2/dNjpgTYJwDU2D0NftsaLPc+HvDXiCNWMVheT28hGMAzKrAnv/AMsT7c/SvBGrQ6Trsb3DhYp0MTMei5IIP5gDPvWjaotuVYMMitU8QG6h2s+cDjNfFHXIRZrpMUgaW4YM6g/dUHIz9Tj8jXwJ0m61b4o6KLZfls3e4mY/woinn8SVH1IqXrX/AAVL8IwXPgzwT48SJRNp+qT6bJINoLrPF5ig8ZIBt3xzgbm4+av+CcH7TXgf9mP4veIPEHxM1O7tfDeteHJLVxa2huHa6SaJ4SAoLfdEy9Qvz5PQV8YfCviTXfi9dSaDL4q8Xp4zlOpeG7zUElu9S1eyldhC7jLu8oEbI3JO6Nu1fBH/AIJlfEPxhHDrfxh1lfCGnOu5NNtgtxfyA9NxyY4Rgg8l24IKqea+F37HX7PfwnjSTw98O7G/1BVw2pawv264J5BKmTKxkgkHy1XI655rx54GtfGnh+XTnVEu4gXtJivKOO2fQ9D+fapodR0LVGhkElpf2E+Dg4aKRD2I7gjqK+HHjG18d+H0vgAl9bYjvIgOFfH3h/st1HpyO1G3GMZ/MU9uSOBningHYVc2kU0TwzRq6OpVlZchgeoI9K+In7IP7PHxI8ybXvhjpdteSYJvNLQ2Eu4Z+YmHaHPzH74bPGegx8Wf+CYGvacsupfBnxrHqsS5I0vXAIZ8AcBJ0Gx2J7MkYH96viD8MPiB8KdbPh34h+EtQ0O+58tbmP5JgOC0Ui5SVc8bkJGe9f8ABPX/AIKK3Xw0urH4I/HvXpbnwjcSCLRtfu5S76M7HiKdjybYk8Mf9UTz+7/1dVVUcY60TnoaqqqqqiQB1o896qqqqqmOTgGqqjjHWjz3qqpjxgGqqqqqpsYwDVVVVVU3TANVVVVUcYpjkYzVVVVVU3PANVVVVVTcng1VVVVRxijz3qjjHWjzVVVNjGAaqqqqqqqqqviv+xj+zR8Zmmu/GXwq0mPUpm3NqelobC6ZwMBnkh2+YccYkDDgccDHxX/4I7XcSzX3wR+KqT7V/d6Z4mg2MSDz/pMC4Jx0HkgZHJAPHxO/Yu/ad+EheXxb8IdakskbAv8AS4xqNuV3bVZnty/lhjjAk2t8y5AJArw74o8TeENTj1rwl4j1PRNRixsu9Ou5LaVcMGGHQhh8yqevUA9q+HP/AAUk/a0+HcVvZyePoPFVlbKFS28R2a3ZbHHzTqUnYnHJaQnv1JrwF/wWQsHWG2+KHwYnjYMvm3mgakHyMjcVgmC4IGSAZTk4GR1r4d/8FGv2SfiJ5FuPiT/wjN9Nt/0TxFaPZbM7R803zQDBfB/efwsfujdXh3xR4Z8YaZHrXhLxFpmt6fLjZd6ddpcxNlQww6Eg5VlPXowPeqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/4K4+Iv7J/Zm0rRIprXzdc8V2sLxSN+8MUcE8rOgyDw6RAnBAD46kGv+CT+hf2v+254P1D7X5X9iafqd3s8vd526ylg25yNv8Ar92efu4xzkVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVftKfsU/AT9qXT3PxA8LJZ+IFUC38SaUiW+oR4XCq0m0+cgHRJAwHbaea/af/AOCX3x/+AUtzr/g3Tbj4jeDolMh1LSLQm6tV54ntAWkAABJdN6ADLFelfC/4w/E/4K+IV8U/CzxvqfhzUeBI1pL+7uFXkLNE2Y5kBOdsisuecV+z1/wV50PUfsvhz9pHwx/ZU/Cf8JJocLy25/2p7XmROBktEXyTxGorwV478FfEnw9b+LPAHirS/EGj3PEV5p1ys8e4AZUlSdrDIypwynggGqqqoYz1pcEYzVV/wV3+CaeEPi9ovxm0i22WPje0Nvf7UAC31sqru4/vwmPqOsbHJzgf8Eyfi22jeMtb+Dep3bfZfEMX9oaXGzgKl1Cn70KOpZ4QCfa3HvVVQYjjNBgehoORxmlkHY0JAR1oOCMVvB4BrcMYB/St/v8ApRcdN1Fx0FGTjANfFbwrP4O8f6zoskbCL7Q01qxXAeFzuQjgA4B2nHGVI7VomorCVhdsFT8ue9WnieSGHy1kPT1rW9cWQF5HyT0Hc1I5ldpGPLHNfBfwxP4T+HGkabeIy3UyG5nRhgo0h3BcdiFKg+4NeM/GPh34f+FdT8aeLNRWx0jSLcz3U7KW2qOMADkkkgADkkgDrX7RPxx1z9oH4mX3jnVEa2slUW2k2BORZ2qk7VJ7sSS7HuzHGFAA/wCCUX7LEvw88By/tCeMtPaLX/Glr5WiQSpg22lkqwlwQCGnZQw6jy1jIPzkCqU9s1VVQOR1qqqqqqBwetVX/BXr9paXXvFFh+zP4XvkOnaC0WpeIXjY5kvGQ+TASD91I3DkdC0i946/Ye/Z1j+OHxMGteJrFpPCPhVkub9Hi3R3s+cxWxzwVJG5xg/IpXjeCLe3jjjSKKNURFCoirgKB0AHYVFCMdKjhyBkUkGRSQAjAFLb5FC3Hb+VG3GOn6UbcEf/AFqa3OOKeAEdK/aT/Y4+F/7RFhNqN9ZLovi5IiLXXrOMLIzBdqLcL0njGF4PzKBhWUEg/Gn4HfEH4C+MJ/CHj3R5ICHf7FfojG21CMY/eQuRhhhlyOqk4YA8V/wT+/4KP6j4AvrD4L/tB+IpbvwnN5dtoviC7k3PoxACJDOx5NtgABzzEevyfchminiSe3lSSORQyOhyGBHBBHUEUpHY1/wW01wXHij4T+Gha7fsGn6pd+dvzv8APktk27ccY+z5znnf0GOf+CUGkXFx4p+Ieto8fkWmn2Nu6kncWlklZSBjGMQtnnuOvOEQYwKSPPWlTIwKSIEUsYxgClizSxjGKERPAFftPX0+n/DNbaE/LqGpQwS8/wAIV5P/AEKNa02Wa4uEtC4O7OC3bAz/AErwxowHEgUyOeSOwq/8Ni3tBKVHK8V4nsI2tJgQMxqWU+hFeHtVk0PXtO1qKR1axuo5gUALfKwPAPB6dDxTREdDTR8YIpogRxTR44H5U8YIwOtPF6U8YIwetPHjiv2m9Fh1z9nj4k2E0Ukv/FLahNGkedxkigeRMY6/Oi8d6+EetN4b+K3gvxEmpJp7aV4hsLoXbuEFuY7hH8wseFC7c5PAxVVVVVVVVVZOMA1uB4rgjg1VVVVVfQ1k9jWT61uPtW6t3tW72rd7UWz0rcfaiSehoknvR571VVX7Qn7JnhD4zxzeIdFaLQvFu3IvVT91eEbQBcKBk4VcB15GedwAFeOvAXiz4a+I5/CfjTR5dO1K3UMY3IZXQ/ddGUkMpweQSMgjqCB41+J/jj4i2Wi2PjPXZdUHh+2a2spJgPMEZx95hy5+UfM2WOOSa+GPh3wxefG7wJ4q8QFIv7EvpWSVmKgl4JFjBII4EpibngYPYnMfJqPoPao+gr4zfaj8Tdd+2H955qben3PKTZ0/2dv9ea8KeWbYAdfMO768f0xXhpbBlAuCMY5rxKtqGYQkY7V4p2G7iCkZ2H+dfseIx8R+IXCkqLKIE44BLnH8jURBGAaiI45r9tbSp/2Wv+CgWn/FrQofLs9Rv7HxZDFFCMMTKVu4zhlDGSSGdiMqcTDJGdx0zULLVbG21PTrhJ7W8hSaCVOkiMAVYexBBrxejfttf8FP10Bla98M2PiFbFkeASRDS9MBM4PYJM0UpBbvcKMchaRwRjNb+Otfs4ED/gsVrBB/5n/xl/6J1KpXBGAakYE4z0r/AIKU+Kda/aF/a+8E/sx+FLzfDokltp5EcJfyr+/aNppGxjciQfZyecJtkyRlsa78L/CWr/Ce7+DJsEh8N3GgNoaW6LgRWvkeSoXGMFUxjGMEDGK/4Jh+NdS+Df7S3jP9nvxfcpbya0s9k8IYFRqmnyONoLYIBj+0jjkkJkdMSkE9alwTgGvHvhDS/HvhXUPCmr5FvfxbRIoy0Tggq6+4YA1428G634B8SXfhjX7Yx3Nq52OAdk8eflkQ91YDI/I4IIGh+PtX0eJbWVVu4EGEVztZR6BvT6g1f/FHUriHy7Kwjt3IxveTzMfQYH9annudQuWnnkeaeZuSeSxP+elfAD4X3XgbQptb1228rV9WVcxMPmtoRyEPoxPJHb5QeQal68dq/b18Nv4i/Zf8WPAkjz6S9rfRqrqoIS4QSFs9QI2kbAwcgdehr/gnjFo2tfsy+Eb59HtjqOhzalYpePCrShXu5JmCPjcqnzFBGeStRwZHSo4MjAFLbivj98OWurX/AITnSLcma3UJqCIvLRjhZfXK8A9flweApr4f+OZfAPiKDVyxa0k/dXcIJ/eRnrgeo6j/AOua1b9qTRIbgx6L4Ru7yEEjzLi6W3PbkKFf37+nrxpn7UOhXEoTWPCl5ZoTjfb3KzkDHXBCd/8APavDnifw54xsTqHhzVYb2JceYqnDxE9AynlTweo7VJBxxUsIxwPwqaEYyBXxk+Dfgr43+CrrwP430/zbeX57W5jAE1lMBhZYmPRhnp0IJBBBIr46fBPxZ8BPH934F8VKs2xfOsL+JCsd9bsTtlUHoeCGXnawIyRgn/glV+2LcePtBT9m34jau0/iDQLUyeGrudmZ72yTJeB3YnLwgjaOP3Qxj92SaqqY8YBqqqqqqpjngGqqqqqonA4NVVU3TANVVHGOtE571VVVVVMcnANVVVVVTcnANVVVVU3TApsE4BqqqqqqPPeqqqqqPTg1VVVVVNjGKqmxjg1VVVNgnANVVVVVVVVVVVXxG/Z9+CHxcaSb4j/Czw3r11IoVr25sEF0ABgATqBIBgAcMOg9K+In/BI79n7xL5918P8AxN4m8HXT7vKh81dRtI87iP3cuJTglBzNyqY6ktXj7/gkP8eNBWW58BeM/C3iuGNm2QytJp9zIMnaQjhowSMZBl4Jxk9a+IX7Kf7R3ws81/HHwa8TWVvDnzLyCzN3bJj1ng3xj/vqvDHjLxf4I1Aat4L8V6xoF8MYudLvpLWUY6fPGwNfDT/gpf8AtW/DxorfUPGFn4xsI2H+jeIbMTNjBBHnRlJSTkHLO2Co7bgfhn/wWE+Heq+TZfFn4ZaxoExyrXmj3C38OeMMUfy3QfeyBvIwOu47fhd+1D+z78aXS3+GvxX0LVryTOyweVrW7YA7SRbzhJcZxztxyPUVVVVVVVVVVVVVVVVVVVVVVVVVVVX/AAWW8VRra/DDwPBJEzvJqGo3KmNt6ACGOIhvu4bdNkcn5V6Dr/wRK8OS3Xx48eeLVmYR6b4R+wtGIiQzXF3C4JfPBAtT8uOck5G3mqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/ac/4Jz/ALPX7TE974lvtJl8J+M7vLN4g0ZQjTyY4a4hPyTdsn5XPTeK/aS/4JuftH/s7fatb/sD/hNPCdvlv7b0GJpfJQfxTwcyQ4HJOGQf3zXwv+MPxP8Agr4hXxT8LPG+p+HNR4EjWkv7u4VeQs0TZjmQE52yKy55xXwD/wCCv2j3UFvoP7Rng6WyuhtQa/4fi8yF+QN01szbkwOS0bPkniNRXw8+J3w9+LPh6LxV8NvGOl+ItLlwPPsZw/ltjO11+9G3qrAMO4qqqlPA5qq/a/8AgFa/tJfATxF8OUhiOtLH9v0CaQgeTqEIJi+Y8KHBeFm7JKx64rwz4h8SfDbxpp3iXSTLp+ueHdQSeJZoyrRTRPyjocHGQVZT1GQepr4R/EzRPjD8OdD+IugL5dtrFsHe3L72tpQdskTHAyUcMucDOM45qqr6GgxHGaDgjrQfsDQkPrQk4xmvM968z3oyZGAaMhPAosTxmvjT8Jbf4maTHcae8UGuWIxbTSkhZEz80b4zx3BxwfYmta0PWPDuoS6Trmmz2V3CcNFMhU/UeoOOCOD2NLd3KrtWd8Dtmnd5G3O5Y+pNeB9T0DRvFmmap4n017/TracPNAhHOOhIPDAHB2nAbGDwa8a/Fb4efDzwe3jvxd4rsbHRfK8yG4Mm43GRkLEoy0jHsqgmv2qf2tPE37RmsR6bYxXGjeDdOfdZaU0gLzSYwZ5yvDPyQF6IDgZJZj/wT+/YX1j9orxPafEn4gaW1t8MtHucyCZWQ67Kh5t4sEHywRiSQHsUXLbikUMUESQQRpHHGoVERdoUAYAAHQYqq+hrgjiqqgcHrVVVVVVQIIxmviR470T4XfD/AMRfEbxHLt03w1pk9/cgMFZ1iQtsXPBdiAqjuzAd68a+LvEPxL8ca3448RSi61rxLqc19dmJMB55pC7BF5wMtgKOgwBX7L3wctvgh8GdB8GCFRqckX23WJAuDJeygGTPqEAWIHAysa5Gc1FEMCry907SLN9R1a/trO1ix5k9xKI0TJAGWYgDJIH1Io/tC/s9wSNDP8dvh5HJGxVkfxRZgqRwQR5nBrQda0DxLZDUvDut2Gq2m7aLiyuUnjzgHG5CRnBB+hHrWq+JPC/h6SODXvEel6ZJKpaNLy8jhLgdSAxGRS/ET4b/APRQPDf/AINYf/iqHxF+HBGP+FgeGv8AwbQ//FVEkc8STQOkkcihkdTkMD0II6img7YrWfFHhLQLlbLXfFGkabcPGJEivL2OFypJAYBiDjIIz7GoXtr21ivLO4iuLeeMSRSxMGWRSMhlI4IIOQRUsGBwK+NnwS8DfHfwNd+BfHOnebby/Pa3UYAnsZwCFmiY/dYZ6dCCQQQSK+P/AMAPHP7O/jmbwd4wtvOt5d0ml6pFGRBqEAON6f3WGQGQnKk9wVY/8E7/APgohL8LZdO+BXx01ppPBkjLBoeuXLktojHhYZmPW1PQMf8AVf8AXP8A1cM0U8STwSpJHIoZHRshgRwQR1GK/wCC1XPxQ+G4B/5gF1/6PFf8Ekxk/FXH/UG/9vaSMEUiZ4FJGMAmljz2pYwBjFLFkYpYhjpSxjGAM1+0xoMurfCy4u4QxbSbuG6KqOSOYzxg8ASknpjGc8YqwuBaXkVweiNz9Dwa0bUki2SI4IIyCO9XniNp7YRF84GAK8Uaki2zxhhvlG0DPbua8E6HL4l8YaNoMUPm/bb2KN1K5GzcN5IHYKCT7A00Z6EU0YIwKeIHtTx8YIp4gRxTpng1JH2FOgIwa+JUePh74o/7At3/AOiWqqqqqqqqqqqvoaycYzW4Hisgjg1VVVVVVVVVVVVVVXj/AOKPgD4XaX/avjnxPZ6XGykxRSNummx2jjXLP+A474r9pj9qi3+OdvB4c0TwbbWGj2MxkgvL5FlvWPHKkcQggAMqls4GW7VVfAz4gt4/8FwTX0wfVNNItr3nlyB8sh/3hyf9oNUfIABqM5A5r9o/wtLZa/a+LIImNvqMQhncJwsyDjJHqgGM/wBw/hpGpnTp/nyY3+9jt71Ya4piDQzBh6qa1XX4Y4y08w6cDPJ+gq+u3vrl7h+Nx+Uegr9lnwZN4d8Ez+Ir1Clx4hkWRFOciCPIjOPcs5+hFRMOOajbBwDX/BX/AOEo8TfB3w38X9OtN934N1I2l86Q8izu9qhncdlnSFQDxmZsEE4PwA/amfQP+CX178Tft2dc+HekXHh2N/LYrFeIyQaeMAkkBLiy3H5f4vujBr/gjD8IJrOy8cfHfU7KWP7b5eg6RKwCiSNSs10QDyRuFsARxlHHJB2rKMYzXmjHWv2cm/43Cau2f+Z+8Y/+idRqSQEYBrxR4i0nwl4c1XxVr12ltpujWU17ezuQBFDEhd2JPAAVSea+DHhb9pf9qX9oLxZ8YPglcJY+Lba9uNZnv579YxYG6d1WNHcHJ2M6KAuAqHGMAU/wP/4K3nhvjDn/ALmCH/4ivi74V/aG/ZZ/aD8NfEr4xTpeeL5r2DX47yG/EovjFNh0eRRwWEe1hjG1xkEHFaB4g0rxX4e0zxRoV2lzpusWUV5ZzKQRJDKgdGBHGCrA/jUhyetTHPArx54A8KfETSv7I8U6Wl0ibjBKp2S27EY3Iw5B6cdDgZBFeJf2OruOaSXwl4xhkiJJig1GAqyDjAMiZ3d+dg7cVYfsieMXkxq/ijRraPcuGthLOcfxHDKnI7DPPqK+H3wI8F/D2RdRRH1XVUOVu7pR+6P/AEzQcL9Tk+/apORgGpDk9e1ftFaPHr3wD+ImltZyXbyeGNQeGGMMWeVIHePAXkneq8d+nOcVX/BLfUrjU/2cL+0nSMJpfiq7toSoILKYLeUlueu6VhxjgD6mKAYBIqOAEAYpLcngCvip4y0zwB4OutW1C3jupJ1NvbWsihlnlZThWGeUwCW9gfWppWmkMjYGTwB0HsKttC1C5QOEWMHoHOD+VXWh39qm9o1kAHJQ5x+FeGfE2s+ENZg13Qrsw3MBxg8rIvdGHdT6fjwQDXgPxfp/xA8MW3iGxURuxMdzAG3GGVQNy578EEexFTQ56CpoRjIFTxAciv2wf2frf4+/Cq50/TraP/hKdB33mhTMACXwPMgJxnbKqgY4G9YyeFxXw88e+LfhF4+0b4g+Dr1tP1/w5ercWrumdrrwyOp6qw3KynqCR3r4NfFPw58bvhZ4Z+K3hR/+Jd4k09LlIi+5reTlZYWI4LxyK8bY43IaqiQBR571VVVVVMcDg1VVVVVUxycA1VUcY60eaqqY8YBqqqqqqOAOtVVVVVRxijz3qqqqqmI7HpVVVVVU2McGqqqqqpumAaqqqqqm5OAaqm54Bqqqseho896qqqqqqqqqqqqqq+JP7NfwD+LzTT/EX4S+G9Yu51KyX7WSw3ZGGGPtEe2UD52Iw3BOeoBHxG/4JC/BDxCslx8N/HPiTwhdOw2x3KpqlqgzyAjGOTOMjJlPb0Ofib/wS2/ai8CebdeGdM0fxvYx4IfR70RzhTnOYZ9hJGBkIX+8MZ+bb4o8G+L/AARqB0jxp4U1jQL4Zza6pYyWsox1+SQA/pXwu/bT/ab+EDpH4T+LWsXFiuR/Z2ryf2jb4J3EKk27y8nklCpyTzyc/CH/AILC6Zdyxab8dPhm1kXYBtV8NSGSNc4HzW0zblUcksJXOOApPX4SftPfAT45iKD4ZfE7R9Vv5Y2caY8htr0Kv3j9nlCyEDHLBSvQ5wQTVVVVVVVVVVVVVVVVVVVVVVVVX/BWbxd/wkH7UEHh6K5zF4Y8N2lq8K3PmKssjSTsxT/lmxSaIEdSFQ5wQB/wRD8Hiy+FfxK8fBP+Qz4gtdM3eZnP2S3MuNvb/j9698+1VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV+0l/wAE3P2b/wBor7Vrf9gf8IX4sny39t6DEsXmyH+KeD/VzZPJOFc/3xX7Rn/BLr9pT4Fyz6t4Y0VviT4XRjs1Hw9au91EvYzWQ3SIeCSY/NRQMs46V4P8beOfhf4mi8SeB/EureGtdsWKC5sbh7eVOfmRsEZU4wyNkHoQRXwG/wCCvnjDSJbTQv2hfCcGvWSqsb67okS294D3eWDIikP/AFz8oADoTXwk+Pnwf+OujprHws8e6Xri+XvltY5dlzbjgHzYGxJHyQMsoHIwTmqpfTNDBHWqr/gql+zE3wv+KQ+OPhbT2Twz4+uWOobQNlrqxDNIPUCZVabnOXE3QYA/YL/aVj+EPjU/DjxbfLF4T8V3S4nlk2pp16QEWY8cI+EjckgKArEgKc1VVVZ7A1uOMA1vOMZreaLk9KLkjAosSMZonPGarxD4W8O+LLIaf4j0e11CFTlFmjyUPqrdVPuCKuf2bfhVPM0sWl3sCnpHHeuVXjtuJP5mtT8I/AX4S6Hca14ltPD+j2AjYS3erzhsgrtYK0pJyQcbU5JbAHNXPjD4Y+JPE+r2Hww8UPrOm2EuI5ZYHhcqe4DAFlByN2BnGcDIr49+BtW8Y6La61p97eXNxoULpFZPK8ieSWLMIkyQrbiWIUDdk5ycV+xd/wAE0vGHxol034k/Gq2vPDPgTck9vp0kbRXuuR9QFBIaCBhz5p+ZlxsGGEi6BoOieFtFsfDfhvSrXTNL0y3S3s7O1iEcUESDCoijgAAAYrgjg1VUp7ZqqqgcjGaqqqqqgcdK/wCCv3xtbwX8EtI+DekXiLqPj+98y+UYLLp9qySMDzlC85gAPRljlX1r9ib4V/8AC1f2hPDtreWvnaV4ff8AtfUNyblKwEGNSCCCGmMSkHqpaoYwea/a2/au0T9mfwtbw2Frbat4y1qNzpOmysTHEikA3E4UhhGCcBQQXYEAgKzL4E/Zd/al/bXurf4ofFfxvNpHh/Ul8yyvNTUyFo+QptbJCqpH3BJjDZ3AtuJMH/BJD4fmJFm+L3iFpAo3FdPhUE9yBk4Htk/Wv2T/ANg64/Zl+Kmq+OJPiAviSxu9Jezs4/sjWjxO0iMWePc6uQqlQ24Y3N8vPy/8Fd02fEb4fjP/ADBLj/0cK8Mf8Eh28S+G9J8Qj9oP7P8A2pYw3Xk/8Ilv8vzEDbd32wZxnGcDPoKH/BGtjx/w0b/5Z/8A92V4Z8Pjw54a0nw99p+0f2XYw2vnbNnmeWgXdtycZxnGTj1poMjiv2jLnXv2of2gPjF4/wDCx+1aX4MsZbqF40PlnT7OWG2DjccjeGabGCck8AZK/wDBMf4nL49/Zwi8I3dwj6j4Gv5LBkIO420hM0Dntj55IxjtDyO5lhyOKmhBHFfF34O+APjX4Sn8G/ELQo9QspNzQSD5ZrSUqVEsL9UcAnB5B6EEEg/tJ/s7+Kv2bviC/hDXZPt2nXiG40fVEjKpewbscj+GRTgOmTgkHlWUn/gmX+3onhaXSv2afjHqk8um3t0lt4T1mdy/2KR9qpYy9xCW4jb+AttPyFTH/wAFqOfih8OBn/mAXX/o8V/wSOXcfit7f2N/7e0iA8Co4xjNImaROw60kQPFLGMYA5pYs9aWMY6Ve6baanY3Gm39us9tdRNFNE44dGGGU+xBIr4r/C3Wfhf4hfT7uJ5dNuHZtOvDyJowehIAAcDAYceo4INWOr3lgAkbh0H8Ddvp6VL4qunTbHAqtjqzE/pxVxcTXUpmnkLue5r9mT4QXWkRD4i+I7MxXVzEU0yCWPDRxsBmb1BYEqOB8pJ6MKaPjGKaIEYFPGQMDmnjBHH5U8eOlPGCOKdBjFSR18SkB+Hnigf9QW8/9EvVAgjg1VVVVVVVVVVVVk9Aa3HpW4EVkHgGuCODVVVVVVVVVHGOtEj1osMYFFs8CvG3wv8Ah58R4kj8ceDtL1hokKRS3NuDLEp6hJBh1GewIrxl+wB8JNcMlx4R1fWfDUzLhIhILy3Tg87ZP3h5K/8ALToMcE5r4pfEPwL8Lfi14q+FOpa3Ndv4X1WbT31KK3DQzNG21jtRmZWBBVl52sCMnFabFcax4dtfFum2lzPot7v+zagsDiGXY5RtrkYOHBU+/FfBzx/J8PPGtrqUsn/Evu8W1+p6CJiPnx6qQD9MjvRurWG2N1NcxJCq7jIzgKB656YrT9Z0jUZDBYarZ3MiruKQzq5A9cA9OR+da94f0vxXotzoOswGW1ukwwU4ZSDkMp7EEAivH/wi8U+Brh5mtXv9LJYx3sCFgqj/AJ6AcocevHoTg4V2Q5RyDjscUA8rhVDO7HAAGSTXwf8A2etU1+9h17x1YS2OkxEMlnKCkt0ewI6onrnBPQdci2SKGJIYI1RI1CoirgKB0AA6DFRMOADSNkDB6V8afhxY/GP4ReLvhffiLb4j0me0hklJCwzlcwynHPySBH6H7vQ9KtviV4t8P/CDxJ+zrLaTpHqfiuz1KaEfeSa3inhkhKjOSzvAeO8A9a/ZS+E6fA79nnwN8NJLZIb7TNKSTUlUk5vZiZrjk8kCWRwM44AGABgCQ44NGXIwK/ZzP/G37VmB/wCZ98Yf+idRp3AHBr/gqV8XG+HH7LepeHNPumi1Px3exaPEUAJWDmW4Y5/hMcZiPU/vhjHUf8EovhOPAf7OEvj6+tPL1Lx/qT3YZ4dj/Y4CYYFPcjcJ5FPAImGBzkyMAOtf8FY/hW3jP4Bad8RbG3aS98BaoJJSCPls7rbFLx1J80Wx46BWJGOR/wAEzviq3xE/Zi07w/fXLS6l4HvJdIlLgAtAMSW5GP4RHIIx0P7o59TIRjrUhBPXpUhyetTEHjNTEYwDUpwMCpSMYzUhBBr4h/8AIgeJf+wPd/8Aolqr/glBGG/Z38Qn/qdLr/0isqih6cc1FAOOOaSAEAEfhX7WeuTXXjbTvDqSn7PptgJCmCP3sjHcff5VT9a8O2CXU7TSKGEWNoPrWmaI1yoCrk/StT0Q2y4ZO3pXiGwS0uhLGMLLnIx0I61+ytq06+INa8PbmMNxZLdAE8K0bheB2JEn6D0qeLI4HNTxDqBU8Y6CpkweO1ft8/Cyz+Gn7QF/eaRa+Tpviy2XV4VVCESV3ZZ1BwBnzEL4GcCRemcV/wAEavifP4g+D/i/4VX135knhHV47yyRsApbXitlF9QJYZWPUgy+hAqjjHWic9DVVVVVUcY60TnoaqqqqqicDg1VVTHjANVVUTk8GqqqqqpsHgGqqqqqpsYwDVVVVVHGOtN0wDVVVVVU3PANVVVVVTYJwDVVVVVUeaqjyeDVVVHpxVVVVVVVVVVVVVVVVVeJ/CXhTxtpT6F4y8M6Tr2myNuey1OyjuoWIBGSkgKk4JHTua+K/wDwSx/Zo8frNeeDbbVfAepOvyNpdwZ7UvnlngmLcY42xvGOAfXPxd/4JU/tE+AFm1HwHcaT4/02Nm2LYP8AZbzaCMM1vKduSDnakjng+2fEXhfxf4D1n+yPFnh3WPDurW+JPsuo2clnPHzw2xwGHIPOOor4N/8ABQv9p74Ni00+38b/APCVaJax+WuleJEN4gXttmyJ12jhQJNo4G0gAV8GP+Cr3wI8drb6Z8UtM1H4f6tI20yyg31gxJAXE0ah0Jzk74wqjq5614c8UeGfGWkQ+IPCHiPTNc0u4z5N7pt2lzDJjrtkjJU/gaqqqqqqqqqqqqqqqqqqqqqqr9rLxqfiH+0r8SfFa3LXENx4iuoLWVmcl7eB/IhPz/MB5cSfKcbRxgYr/gmT4HfwN+xZ8P4rlMXWuR3OrTYZiCLi4kaIgEDH7kQ5HTOSCQc1VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV8fv2If2bf2kYbu5+IHw/t7fXrlcDxDpGLPUI2AwGMigrKQCRiZZF6ccDH7Rn/BI74+fCqWfXPhFInxM8OBiVis4hBqdsp7PbFiJQMgbomZjySiCre48ZfDnxSs9pcaz4Y8SaLcHa8bS2V5ZTLwcEbXjcdOxFfAf/grV8XfBP2LQfjVotr450eL5H1GBRa6mi8AEkYim2gdGVWY8tITzXwQ/az+Af7QsUUPw28f2U+rPD5r6JeH7NfxgZ3fuX5cLjlo9yjjnkZ+lKQeAaqvi38LfCXxq+HWt/DPxtYJdaVrdq0T5XLQP1jmQ9nRwrKfUDtX7QnwD8cfs4fEzUvhx43spQ1vIz6bqHlbItTtNxEdxHyRhgOVySjZU8qa/Ye/bQt7230v4IfFnUY4bmBEtPD2sSkKsygBUtZj0DgAKj/xcKfmwXqqqqqqqqrx3+0V8DfhrJNbeNPijoNjdWxKzWaXP2i5jI6gwRbpB0x92vHf/BT/AOGWko1v8PfA2ueIbgcebfSJYQcgYKkeY7YJIIKr04ODmviD/wAFCv2ivGpe30bWNN8JWZbiPR7T94yhsrullLtnGASmwHHQAkHxB4l8R+LdRbWPFXiDUtZv3Xa11qF29xKRknBdyTjJPfvX7NX7DH7Wnj7xFY+JfD/gmXwjpaIJG1TxQsljBPEwyFWLaZpQ4GQVQr90llyDXxO+Gnib4T+Lrrwd4ptglxCPMgmT/V3UJJCyoe6naR6ggg4INfsd/tFv4rsofhT431DfrNlF/wASm8mcA3cCgDyWyctKoyQerKDnlSWoHIxmqqvoa4I4qqoHFcEcGqqqqqr/AIKkfEeXx9+114g0iO5aWx8G2Nro9qDkAFY/Omwu0EETTyKSd2dgIO3aB/wTD+Gf9h/DHXPibe22258U34trN3i5+y22VLKx7NK8qkDjMQyTjiGMHAFWNhF+2H/wUAn07xQs0/hq11W4R7dnxjTrAPsjxlSFlZFDBfmXz2IPGatLaKGJIYY1RI1CoijAUDoAOwqKIYHHNRQjjiv+CwCbfiR8Ph/1A7n/ANHivhdAW+GvhLH/AEA7L/0QlLbAjGKNuCMcV+118UB8FP2dfG/j23u/I1GDTWtdLZZfLcXlwRDCydyUZxJgc4QnIwSP+CVvwHstR+DHj7xx4t09Z7Dx+zaHFBKgPmWMSOs5HX5ZHmZCCOsHoa/4J+a3qn7Pv7Yvij4A+KLpo01lrrR5DITAkl5Zu7wS7TkEOizKgzz564JzgzQjGRU0XGQKnjA6Cvjr8E/B/wAePh/qHgLxdaRkToz2F75W6TT7kKQk8fIOVJ5XIDDKngmvi38LfFHwY8f6t8OvF8Crf6XLgSxhvLuYyMpLGSASjDBHHqDyCK+OH7RPiT49eD/hxpfjV57rXvAmlTaTNqUsgc6hBvVoJHPUyBcoxOS2wOWLO2P+CR+pWserfE7RXZvtN1b6XPGAvGyJrlW59cyp+vpUaAjikQGkQYwOtRxjHFKmeBSRgCljzwKSIHtSw8Vr3hnQvFOly6L4h0uC/spvvwzJkZHQg9QR6jBFeKP2NtLuZTP4O8Vz2iliTb38QmAz/dddpAHoQT059Yv2MfEbSos/jbTljLDeyWrsQO5AyMnHbI+or4ffszeA/A9xBqmoCXXdUgZXjnulCxxuMfMkQJAORn5i2OMU0QIwKeHjGKeLjinj7U8Y7U6AjHepI8dKkTI4p0BGK+JiY+Hfin20W8/9EtVfQ0G4wa4I4NVVVVVVVVVVVVVVZ4wDWTjANZPrW4+1buMYrd7Vu9q3e1FsjAFbj7UWJGAaJJGM0SSMZqqqqr4w+Jh42+LfjfxkLqK5/t7xHqF/50S4STzrh5NyjsDuyPrX7NOhpon7PHw70uTe+7w1ZzSLKmCGliWRlI9i5H4V8ffFHw1S+uPDHh3wbpU+rQMVu9RiQwiCQZBUCMr5jjuWyoIwQTnF7quqalHbxajqV1dJaRiK3WeZnEKAABVBJ2gAAYHHApWZGV43KspypBwQfUV8Nv2i/HfgS6jt9SvZtf0kDa9peTFnQdjHIclcYHByuOw6jwd4u0PxxoFr4k8O3fn2d0uRuG1o2HDIw7MDwR+IyCDV/wDDvwFrB8zUPCGkyyFgWkFoqOcDAyygEjHbPpWh+DfCPh50l0Lw1pllMi7RLDaosmPd8bj17momBA5qNgOM0jDsajfOMGlbjINXH7C/7KV/8Q5fitffCaK48T3GtNrUt5Lq986SXrTec0jQmfyiDISShTYem3HFLKMYBoSjGM15vGM14d/ZE/Z08JfF+b49eHvh39l8dz395fyar/a17Jm4u1kW4fyWmMQ3iaTjZgbuAMDDygjGa+Nv7NPwQ/aIl0ib4xeC28QNoKzLpwOqXdqIPN2eZgQSoCW8tOTk/KK8K+F/D/gXwtpPgvwppyWGjaFZRWWn2quziGGJQiLuYlmIAGWYkk8kkkmpXyOteM/Cfhzx94W1XwX4v0qLUtF1q0ktL60kLKJYnGGG5SGU9wykMpwQQQDXwf8A2bvgr+z/AC6tL8IfBzaA2uLCuoD+07u6E4i3+XkTyuAV8x+Rg/MalYY4NSMMYzUjDGc1IQTjNSkE4zUpB4FTEY4NSHIxmr28tdPs57+9nWG3tomlmkboiKMsT7AA1X/BJy3kX9nPX3kjZRJ40uihZcbh9jsxkeoyCPqDUMQwOM1FCCBgc1FbgjNftZafcWfxUW4lTEd3pkLxN6gFlP45U8fT1rwpNGrywk/NkMB+n+H514e1WG0Ks2049a8RarDd7nUKM9hXiu4R5YoFIJGWPt6f1r9lrSze/EG7vmV9ljpkjBl6b2dFAP1BY/hU8Qx0q4jHSp48jFXCDqK/4Kl+Dre48E+C/H6CNZ9O1WXTHIADSLPEZVzxkhTbNjkY3ng7iR/wSG8df8Iz+1PN4Tmn/c+L/Dl3aRxGTaDPCUuVYDB3ERwzDGRwxOeMGmPGAaqqqqqqYjGAaqqqqqqY54Bqqo4x1o81VUx4wDVVVVVUcY60ee9VVVVVR5PWqqqqqbpgGjyetVVVVUcY60ee9VVVVUcYo896qqqqpsY4NVRxjrVVVU2CAAaqqqqqqqqqqqqqqqqqqrx18Nfh78TtK/sT4h+CtE8SWQVwkOp2KXAi3AAlCwJRuB8y4IIBByBXxk/4JH/CLxQrah8GfE+o+CrwKcWF4z6jZsc8YLt5yHGQSXcdOBzn42/sRftHfAWC61Xxf4El1DQrTJk1vRWN5aIgJ+dyoDxLx1lROo7nFfDf4wfFD4P6uNc+GPjvWPDt2c+Z9iuSscw9JIzlJBwOHUjIB6gV8H/+CvfxC0WW30342eBNO8R2SqVfUtH/ANDvAeoZo2JikPbC+UMYOcg5+GH/AAUG/ZR+KbWtnY/E2Dw9qVyoP2DxHEdPaMkZ2mVswFu2FkOTwM5GbW6tb61hvbG5iuLe4jWSGaJw6SIwyGUjgggggiqqqqqqqqqqqqqqqqqqqqv2hviUnwe+B3jf4lC7it7jQtFnlsXlxtN2y7LZTng7pnjXHOc9D0rwx4c1fxj4l0nwj4etGutU1u+hsbG3RSWlnlcJGoAyclmA49a8EeE9N8BeCvD/AIF0YYsPDml22nWg27cRQRLGnHb5UFVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVXxy/ZR+AH7R1oIvi18ONP1W9jjKW+qQlrW9g4wNs8RVyATkIxZM9VNftEf8EbPih4QnuNd/Z48SReNdGG5l0jVJEtNSgAGQof5Ybjgfe/dHJACHk14j8MeNvhn4ok0PxToer+GfEOlTBmtryCS0ubd1OVYBgGU5AIYexBr4Ef8ABUT9oX4TtFpnjq5T4k6GihRDrFwY71MDA23YVmJOBkyrIT7ZzXwE/wCCgH7N/wAeobTT9P8AF6eGPElwyxnQ9fZbaVpD2ikyY5gSDgK27plVJArgjg1VftXfso+Af2q/ALeG/EaJp+v6ervoWuxxBpbCUjkEcb4WwA8ZPOAQQwVh8Z/gj8SfgD40uPAfxO8PS6ZqEW57eUfNBewhiomgfo8bFTg8EdCAQQP2a/8AgoZ4g8CW9r4L+NUV54h0OLEdtrER331ouMASBj+/Qcckh1G7l/lUfD74n/D74raIviH4d+LdP12y4DtbSfPCT0WWM4eJsDO11U47VVVVarruh6FHHLres2OnpKxWNru4WEMR1ALEZNa/+0F8C/DHnprnxg8H201tt823/tmB5l3Yx+6Vi54YHgdDnpzXiD9vf9l3QWaKL4gzarMkmxo9P0u5kxwTuDsioy8AfKx6j3x4q/4Kl+CLZCvgj4Wa5qTHADarexWQXg5OI/Ozg44yMgnkY58Yf8FKvj7rySW/hmx8N+GYj/q5bayNzOvAzlpmZDzkj92OuOeteNfj58afiJG9v4y+J/iLUraRSHtGvnjt2zuBzChCdHYZ29DjpxXhHwB47+IN82l+AvBWveJbxPvW+kabLeSDIY8rErHojH6KT2NfDL/glp+1f8QHguNd8O6X4J0+STDz67fKJQgOGIgh3vng4DhAeOQDur4Zf8EcPhRoqQXnxX+JGveJroR5ktNLiTTrYOVwQSfMkcKTwQUyQCQBla+GH7MH7PnwZ8mX4a/CPw5o93Bny9Q+yfaLxQduR9pl3zY+VTjfjIz1qv2gPgho3xs8GyaZIsVvrlgGl0q+KAmN8cxsevlvgAjscNztxWraT4j8EeI5tK1W2u9I1rSLgB0JMcsEqkEMpH4EMDg8EHBBr9mT9pXTfi5pcXhTxLNFaeL7GHDIThdRRRzLHn+PAyyf8CHGQoOOhrgjg1VUpHTNVVUCCMZqqqqqq/aN16LxV+0H8TfEluZ/I1PxfqtxALj76xvdyFFbBIBCkDAJAxgV+x34eHhv9mf4ead9k+z+do6XmzzN+ftLNPuzk/e83djtuxgYwIEGOK/4JbvJZ/tM+IrPWJGgvpPCd5EYrk7JWmF5aM6YbkuArkjr8pPY1DGDgelQxg81FFkCv+CxCBfiT8PB/wBQK5/9HivCv/BM79o/X/DOka3YftP/AGW31CwhuIYPPvR5SOgYLw+OAQOKH/BLT9ppun7VY/7/AN9/8XX2X2r/AILI/FRHvfA3wPsLlG8hZNf1OPBJDNugteegOPtWRycMp4B5+A37SP7HvwZ+C/g34X23x38J7/D+kxQXTxOyrLckb7iQAKcb5mkbqfvdT1r9tz4k/Dix/az0P9oD4A+ONJ115FsdTuXsZGIi1C0cLhwy42vHFDwM5O/I5y3gvxXpHxB8FaD470CTzNO8Q6bBf2pPXy5Yw4B6EEBsEEAg5yBU8YFTxjBFTpkYHav+CjXwCt/iP8K2+J+i2i/8JD4HiaeZwpLT6dyZkwP7hxKCeAFk/vVX/BJi8tF+KXjfTXnUXM+gRSxxHqyJOodh7AyIP+BCkXgAVGgA4qNAaRAeBSRgDFJHmo4sjFLGOgFLFnjrSw8UIQRwKMAxwKaEY6U8PHSmjxwKeMEcdakiGMCnTHFSJnoKkTI4qRMdKlTjirmCKeJ4Zo1kR1KujrkMD1BHpVfCn/kl3g7/ALAFj/6ISqBx0NAgjGaqqqqqqqqqqqqqqqqqqqqqqqqqvG2tXXhvwZr3iKwjikudK0u5uoUmBKM8cTMoYAg4yozgj61XxG16X4UfCGK2s7sy30FpDplnOF2EyeXt8wDJwQqswHPIA96xJPJyxd3PJJySTWm6GrqCyB39SOn0q58PARfPAMY9MVqFg1m/ByhPGe1fso+Prjw945/4Q65mY6d4gUhELcRXCKSrAdtygocdSUz92omBGM1G3bNRsAcA0jZAweajcEYzSPjoaSQYABpXBHBpZMcZoS8YBrzfcU0uRjNNJngGncY5NSSAjGakfPepHGMA1IwAxnmpGBOM9KlbPANSsMYBqRhjrUh461IwPOalOeAacgng18Q/+RA8S/8AYHu//RLVX/BMvSLux/ZN0C7uVTy9S1O/uLfDZJQXDRHPod0TcfT1qCMEZFQQjAyKhgzjiv2pPhNe+OfB8HiPQLVrjVvD+9xbxR7nuIG271HcldoYD03ADJq3uJbWZZ4Ww6HjNWXiu32ATlomA54yP0q/8WQFCtsGlbHBIIA/rU80tzK00zlnY8k1+z58N7nwL4Ka91a3MOq62yzzxseY4wD5aHHfBZj6F8HpVxGO1XCAgirhAf8A9VXCDBGf0r/goLpVvqH7LPim7mkkD6bc2FxCFIAZjdxRENx02yseMcgfQ/sDa9J4b/bF+FeoRahFZtLrf2MySlQGFxFJAUG7jLiUoO+WGOcUSAODR571VVVVVHAHWjz3qqqqqqY4GAaqqpj2Bqqqicng1VVVVVMeMA1VVVVVNjGAaqqqqqmORkGqqqqqpumAaqqqqqm6YBqqqqqqbngGqpumBVVVYzxTcnrVVVVVVVVVVVVVVVVVVVVVfGX9hT9mX43yzaj4k+HkGkazMrhtX0Bhp9wWbkyOEHlyvnndKjnjHTIr4r/8EgfiZoazah8H/iDpPiiBF3LYapEdPus5xtRwXic4wcsYx144GfiZ+z78bPg5PLD8S/hjr+hRwsVN3PaF7ViGCnZcJuicZZRlWI+ZfUV4A+MPxW+FU5n+G3xG8ReG98nmSRadqMkMUrYAy8anY/AH3geg9K+Hf/BV39pnwjFDZeLofDfjS3RdrS6hYm2uTjdjEluUTOSuS0bEhB0JLH4ff8FePgX4g8q2+IXgrxN4SuHxulh2albR+uXXZJ+UR/Cvhz+1N+zr8Wpbe18AfGHw3qV7dMFgsJLv7LdyE8ALbzbJSc4H3epFVVVVVVVVVVVVVVVVVVf8Fe/iz/YPwx8L/B2wk/0jxXfNf32P4ba127VPP8csikcH/Unpxn/gld8HP+Frftb6BrF9Z+dpPgK3k166Lw7k86PCWo3HhXE8kcg6k+S2OhIqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqvit8D/hH8ctAk8M/Fn4f6P4lsXXan2yD99B1+aGZcSQtyRujZTyeeTX7RH/BF6S3t7jxB+zN42lunXc58O+I5VDNznbBdKqj0AWVR7yV8WPgr8VvgZ4kbwl8WvAeq+GdS+YxJeQ/u7hV4LwyqTHMgJxujZlzxmvgF+35+0d8AZ7ex07xhL4o8OR7VbQvEMj3cKIAABDIT5kGBnARgmTko3SvgL/wVK/Z7+KkVppHxDvG+HPiGZljMeqSGSwkcjqt2AFjXjkzCMDpk1a3Vrf2sN7Y3MVxb3EayQzROHSRGGVZWHBBBBBHBqvjj+z/APCz9orwg/g34oeG4tQgXJs7yMBLuwclSXglwTGTtXI6MBhgRxX7SH/BML44/Bye8174dW0vxD8KR5dZtPt8X9smMkS2oJLY5G6IsCBuITOBoniDxR4M1Y6j4c1zVNC1ODMZnsrmS1mjweV3IQw5A49RXhb9vv8Aac8M24tZvGtrrcSsCg1XTopWXkkjegV2Bzj5mOAABiv+Hn3x9/6FH4f/APgvvP8A5KrxJ/wUO/aa15JI9P8AEWj6Csj5xpulRkquCCgM/mEDnOc7sgc14h+NXxf8V3v9oeIvif4ovp+dpl1WbagPUKobCj2AArVfEviPXY44db8QalqKRMWjS7u3lCHuQGJwarw/8O/H/i5IH8KeBvEGtLczCGA6fpk1yJZCQAi7FOWyQMDnJrwt+wx+114xaFNI+APiqAzMyp/atsumYKgk7jdNHtGBwTgE4AyTXgv/AIJGftQ+Imik8UX/AIQ8KwFY2lW71JrmZdwJZVWBHRmUgA5cDngmvAv/AARh8CWqJL8TPjTr2qO0YLQaFp8VgI328gSTGfeA2edi5HYV4C/4J5/sg/D9Y3svg5putXKsGa41+WTUi5BYjMcrGIABsYCAEBc5IzWh6BoPhrTY9I8N6LYaVYw/6u1sbZIIk+iIAB09Kqqqv2jP2c9F+Nei/wBo6cIbDxZYREWV6y4WdRz5M2OSpPRuqk5GQSDe2Xir4eeKntLuO80TX9EuRkAmOW3lU5DAj8CCOCCCCQa/Zu/ak0X4rWVr4T8XXUNh4yjTaEK7I9SCjJkj7K+Blo/qV4yFBBGM1VV9DQII4NVVdOlcEcGqqqqq+Kv/ACVDxh/2Hr7/ANHvX7OSj/hn/wCGX/YnaT/6RxVAucAH868bT6r+xX+3dL4qNnMuiNqr6lFGjsRdaVelxIq5OXKbpUG7P7yEHnANeDfFPhvx34a07xf4P1m21bR9VgE1neW7ZSVD+oIIIKnBUgggEEVCgOAKgjziv+CyChfiV8OwP+gFc/8Ao8V8KIQfhl4QP/UBsv8A0QlJACMAUYCV6fpVroVr+39/wUW1a1vp7q58IXOp3DSzWl0Pk0ixj8uJkbkKJikX3QcNcE56tUv/AASg/ZFUfLo3if8AHW3/AMK/bV/4J0/B34S/ADW/iV8IbLWIdW8OTwXF0l5qDXAltGfy5AqkcFTIr544jb1wf+CWHxVHjr9nWXwHfXfm6j4C1F7UK0u9/sk5M0DHuBuM8ajoBEMHjAnQEY9KnXjNTqM4FahaW95bTWd3BHNBMjJLFIgZXUjBUg8EEZBBr49/DhvhJ8ZPF3w8EeyDSNTkWzBYsfsz4kgJJ5yYnjJ68nqetf8ABKrULKy/aR1W1uptkl/4Ru4LZdpO+QXNrIRx0+SNzk4HGOpFRrmkXJAFIoxxUaDFRoD0qOPI4pEBGBSRZFJFkYFLCMcChEO38qMYIwBRiBGBTxDFSRDpipI6dARjvUsYI6U6cYqRADkVKgIwKdcjHpTqM4FV8Kf+SXeDv+wBY/8AohKqqBI4zQIPGaqqqqqqqqqqqqqqqqqqqqqqqq/aM1q68N/s9/E/xFYRxSXOleDNXuoUmUlGeOzldQwBBxlRnBH1FeHtJ/t/X9M0L7R5H9o3kVt5uzd5e9wu7GRnGc4yK/ayZh4b0JAx2m+clc8EhP8A65/OtLVWu13HoDivClpbzSIshGK8R6RptvZK0EqsSvOO1eJYoxFIARweOPevhY7J8TvCLI5B/tyzGQccGZAf0qNsHGaRsgEGo3BGM1G/bNI+RkGkkBGM0ZljQu7hVUZJJwAK+Jv7ViabdyaL8OLe3vGiJWTUrgFo9wJBEaDG4cD5icHsCOa1T4zfFbV7j7TdfEDW439LW7a2X/vmLaP0rQvjt8WvD0oktPHGo3K7stHfv9rDdMj97uIHA6EEZOMZNfCD9pXSPH93B4b8TWcWk63LxCyP/o903Hyrk5RjzhSTnHDEkCjIMcU0uRgGnlzwDTvkdakkGMA078ZzUj54BqRgBgGpGGM5qRs8ZqVs8A1KwxgGpWGMZqRgec0ee9ftO6rHo37O3xHu5YmkEnhm9twqnBBmiaIH6AuD+FV+xx4UXwb+zB8NNFWJY2fw/b3roqFdr3Q+0MCDzu3THPvmoEBwKgjBwBUEQwOKhgDAV8af2RdN8b31x4q8BXsGk6vcEvc2cy4trl+7AqMxsec8EE84ByTr37O3xo8PSzR3fgDUrlYTjzLFBdBxnAKiMkn1xjIHUCtE/Z3+Meuyxxw+B720R13GS/xbBB7hyDn2xn2r4V/su6J4Iuodf8X3cOtavCQ8ESIRb2rj+IZ5kI7FgAP7uQCLhADx/KrhRgjNXCg9+1XC8EA1cD+VfthAf8M0/EMf9Qd//Qlr9lbH/DUHwf8A+x90T/0vhonNVVVVVVMc8A1VVVVVHGOtE5qqo4x1o896qqY8YBqqqqqqOMdaPNVVVVVTYJwDVVVVVMRjANNycCqqqqqpuTgGqqqqqpsE4BqqqqqqOCeDVU2CcA1VVRxjrVVVVVVVVVVVVVVVVVVVVVVVSxRTxPBPGkkcilXR1yGB4II7jFfE39iP9l34sebP4l+EWj2l9Lgm/wBHQ6dPuGfmJgKhz8xzvDZ4znauPiN/wR08G3iyXPwn+LWraZJuBW0160S7Q5PIEsXllQBkjKMeAD13Dx//AMEyP2tvA26Ww8HaZ4tto8bp9A1JJMZ24xFN5UrcsR8qHG1icDBPi3wH458A3q6Z468Ga74cvGztt9W06WzkOApOFkVT0ZT9GHqK+G/7Tv7QfwiaFfh78XvEmmW1uu2Kxa8NzaKAFA/0abfFkKigHZkAY6V8NP8Agr78XtBSKy+KPw/0DxZBGoU3VjK+mXTHJyznEkTHBGAsaDjrzkfCf/gpf+y18TfIstW8VXPgjVJdim28RQ+TFvPDYuULRBQf4pGTgg4HONI1nR/EOmQazoGrWep6fdLugu7OdZopRnGVdSQRkEcHtVVVVVVVVVVVVVVVV+2z8bV+Pn7Rnijxhp96lzomny/2VockchdHs7dmVZEJ6LIxklxx/ra/4I8/AtPh9+z3ffFvVbJ49Y+JF95kJcEFdOtS8cA2kfKWkNw+RwyNEewqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqvGHgjwb8QtCm8MeO/C2k+IdJuP9ZZanZpcxE4IztcEA4JGeozX7RH/AARs+F3i+3uNc/Z48SS+CtZ+Zl0jVJJLvTZySSFD/NNb9fvfvRgABBya+PP7I3x//Zu1CW3+KHw/vbbTkbEWt2am50+YEgArOo2qSSMK+1+RlRmvgt+1B8df2fLrzPhZ8Qr/AEyzkkDz6ZKFubOc5Gd0EgZASBguoV8HhhXwG/4K8/DvxT9i0H49eFpfCOpP8kusaYHudPZuPmaPmaEHJGP3uMZLc8eCfHngr4leHbfxZ8P/ABVpfiHR7riK8066WePcAMqSpO1hkZU4ZTwQDVfFT9mz4D/G1HPxQ+Fmg65cvjN89v5F2AF2gC5iKzAYwMB8cD0GPFv/AASF/Zk1x5rjw3r3jbw5K0e2KG31CK5gRsk7is0TSNwQMeYOAO+Sf+HMfwx/6LR4n/8ABfB/jXhX/gjz+znpbQ3Hifxn4512aNm3xC6t7SCQEEAFViMgIznIkGSBxjIPgj9hH9kXwBE0ejfAjwzfM6kO+uQNq5YkLkj7WZAp+QfdAAy2MbjnSf2c/wBnvQLwahoXwI+HmnXaqVE9p4Xs4XAPUbljBwa0nwN4J0G8GoaF4P0TTrpVKie00+KFwD1G5VBwaqqqlI6Zqqqqr9oz9nPRfjXov9o6cIbDxZYREWV6y4WdRz5M2OSpPRuqk5GQSDq2k+I/BHiObStVtrvSNa0i4AdCTHLBKpBDKR+BDA4PBBwQa/Zn/at034g21n4G+IV/FaeK1Ihtbl12R6pxxyOEl4wVOAxI28naOCODVVQPbNVVUp7Zqqqqqv2jtIs9A/aG+KGg6crC103xnq9tAHbcQkd5Kq5Pc4Ar9ke/vNT/AGa/hxcXs3myJoNvCp2hcJGPLQceiIoz1OMnnmoADX7S37MHgr9pvwQuga6/9na3p2+TRtYiiDSWkhXG1s8vCx27kyM4BBBANaX8Of8AgoR+xbf3Nl8PNP1vVvD0s7MU0e0/tmwnxnMpt9rPASqAl9sbY2gt0FW//BR/9t6WVILf4CeHJZJGCoi+FdUYsTwAALnk1+xX8RP24PiL8TNW1n9orwRfaH4Kk0aQWENxpEemfZr3z02gROROwaNZslw+MIQVD5b/AIKwfCH4r/Ej4g+A734dfDDxZ4pt7PRbiO4m0XRLi+SFzMCFcxIwUkc4NaH+1H/wVQ0DRrDQdM+A/isWunWsdtAH+Gt4zBI1Crk+XycAUv7Xn/BWQD5PgL4qP/dMbz/43Xij4jfHhv2BLrx3qPgPxLdfFnWPDDW50bTNAm+2wX1wxgEgtApdPKD+aQRwEPB6H/gkh+zP4z+Gdl47+KXxO8C614a1u/kh0bTbTWtJlsblbZAJp5FWVVYxyO0ABAAzAeSR8s8YxivGvhbSvGvhTWvBuuxtJpuvafPYXiKQC0M0bRuBkEZ2seoNf8E+fh38e/2ev2ndY8G+MPhV40tvDOtwXelXerNoFwLDz7dmeC4+0bNnlt5ciKwYqfPB5BBE688elTqMEVcDIqdRkjNf8FS/BI0j4seGPHUKbYfEWjPbPzndPayfM33iR+7nhGMAfLnkk4/4JtavFpv7WfhqyeFnbVbDULZGBxsItnlyfbEJH1IqNRiolBGM0i5IApF4wKjTjApFGABUaA8Co48jpSIMY70sWetLDkdKMPGKeEYxinjwOKeMEcVLGOlSp2FSICOKlQdqkUEVIuRj0qRQDUq1Xwp/5Jd4O/7AFj/6ISqqqrJ6ZoEHjNVVVVVVVVVVVVVVVVVVVVVVV+05p95q37NfxZ0rTbdri7vPA2swwQoPmkkaxmVVHuSQK8Lapb6H4n0fWrtJHg0+/guJVjALFUkViFBIGcDuRX7R/hafxF8Opb2yh8yfRZhdkAcmIArJj2AO4+yVBKYJVlXqp6Vo2tiJVeKT/wCtV74mknh2NITx3Na1qIun8qNsgH5jX7OfhK48U/FLS7gQ7rTRW+3XDkcLs/1ePcyFOPQE9qjYEYB5rxl8RfB/w804al4r1qK0VsiKIAvLMfREGSfr0Hcitc/bYgSRovDHgWSRAflmv7wIcZPWNFPbH8fBJ69Tof7b0onWPxH4CQwlvmlsr0hkGOyOuGOcfxLXw3+MngL4o22/wvrH+lxqDNYXK+VcRfVckMPdSw96VwRwea/aj+Ilxo+jW3gTS7lorjVo/NvWU4P2cEgJn0dgc+ykHgmrS0mvZhBAuSepPQD1NWvha0MYEkbSt6kkfyq/8LQiMm1BjcDgEkg/1p0mtptpLJJG3GOCCO4r4EfEWf4g+Are51Gcy6pprfZb1m6yMBlZP+BKRn3BppM96ZxjGaeTPANO4x15qSTPANO4xjPNO2e9SuD0NSMAMZ5qRgR1qRgT1qVsjANSEHgGq/4KH+KI/D37MuraaZNkniLUrPT4iNwORJ9oYDHqluwOcDBI6kCvCHhq/wDGnizRPB2lDN7ruo29hbDGf3k0ixrx9WFaVZaboGkW2nWapbWGm2yxRK7kiKKNcDLMc4Cgck/Wtb/a8/Zh8Kam+j638dPCEd3FkSRwaitx5bBipVjFuCsCpBUkEdxyK+HPxr+D3xUPl/Dn4n+GPEcyxiR7bT9UimniU5wXiDb0+6fvAdDV9qmlaDpV3rmuala6fp2nW8lxeXl3MsMNtCilnkkdiFRFUEliQAASa8B/Gr4MfEjVJNC+Hfxb8F+KdSgtzcy2ei6/bX00cKsqmQpE7MEDOiliMAso7ihGhTOea1O5s7C0mvr66it7a3jaSaaVwiRooyzMx4AABJJ4FeEvi98JPiVdXNj8Ofil4R8VXFnGJLmHRdbt754UJwGdYnYqCeMmrkDPBq5AJxVwAc1cAY4NXHerj+lft3a0dE/Za8byxXMMU93Ha2sayEZk8y6iV1UHq3llzx0wT2r9irRbXX/2tPhNY3kkqJF4qs7pTGQDvgkEyDkHgtGoPsTgjrVVVVVVTHjANVVVVVVMeMA1VVTEHgGqqqJyeDVVVVVUx4wDVVVVVUSAOtVVVVVTEevSjz3qqqqqjjFHnvVVVVVHGKPPeqqqqqPTg1VHGKPPeqqqbpgVVVVVVVVVVVVVVVVVVVVVVVVVVa3oOheJdOk0jxFo1hqtjL/rLW9tknif6o4IP5V8Sf8Agnb+yb8Skkkf4aReGL192Lvw3KbAplSOIRmDgkMMxnlR2yD8T/8Agjr4jtc3fwb+LFhqC/MfsPiK2a2dQOmJ4Q4cnngxoBgc88fFn9kb9ov4KefcePfhXrEOnW+8tqllGL2zCL/G00JZYwRyBJtOM8DBA/4JO/DLxJ4t+OGpeOoNc1XT/D3hCyEl7BaXUsMeo3UodIIZQhAdFHmyFSTyigghjXinxZ4X8EaHc+JfGPiLTtE0qzXdPe39ysEUf1ZiBk9h1NaV/wAFNPgN4n+OHh34Q+EYNR1HTtbvfsL+Jph9mtYp2BEISNxvdXk2oWYIBuB+YVVVVVVVVVVVVVVf8FIv2lIvgh8E7nwX4evkXxd49hlsLQJLiSzsyMXFzgcg7T5aHghpNwPyEV+z38FfEn7Q3xh8NfCbwxDL5utXird3KRlhZWqnM1w3oEQE89TgdSK8PeH9F8J+H9M8K+G9OhsNJ0azisrC0hGEt4IkCRxr7KqqB9KqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqvLO01C0msL62iuba5jaKaGZA6SIwwysp4IIJBB4Oa+Pv/AASf/Zo+L/m6r4Fspfhnrr7yJtDgV7J2OSC9mxCAAn7sTRccemP2g/8AgnJ+09+z59u1fUPB3/CV+F7T5v7e8PE3Uapyd0sOBNFgAbiybATgO3Wvh38VPiP8JNcTxJ8NPGureHNQVlLS2Ny0Yl25wJE+7IvJ+VwRyeOa+A//AAWF1mxaLRf2i/BKalbhQq654cjEU4IGMy2zsEck8lkaMAdEavhF+0F8Gfjxpg1L4VfELSddZIRLPZRTbLu1UnGZbdsSRjPGWUAnoTVUDzjNVVVVVX0NcEcGqqqqvj3+zv4W+NeiyTCKHT/E9vFix1QJyduSIpcctGST7qTkdwfFXhXxJ4B8SXXhrxLp82narp0uJI24IPVWVh1UjBDDgggiv2Y/2uR4jmsfhx8UrxI9SKLDp2sSvgXj5wI5iekhGAG/iPB+YjdVVQOR1qqr6GgQRxVVVVX7XWl32kftT/F201GDypX8a6rcKu4NmOW6kljbgnqjqcdRnBwciv2ItVttZ/Zb+Ht3apIqR2EtuRIADuhuJYmPBPBZCR7Y6dKgx29Kg6CoMdAag6Vb429at8HAFW+M9ag2lQKYxlMDrVxtxip8c49Kusc1c4xgVcf0qfqauMFeDVx1P0r/AIKq6ELn4X+DPEwjtz9g1+Sz3sv70efAz4U4+6fs/wA3I5CcHHH7E2tPoH7Vnw2vo76K0Mur/ZDJIVAInieEp83GXEhQd8sMc4pACAKiXpzUQz0qNQTgGolBxio1zX7Zfxv/AGmdU/bb1f8AZ0+Gfxn1vw1p2r6pomk6TBa3z2MVtLd21oQzSwr5qqZpizEZOCRgj5aX9iL/AIKhaWP7Usv2uxdz2f76K3/4T7WJPOZeQm2WDy2yQBh/lOeeM18L/wBv39pf9lz4vp8G/wBtiwn1LSgyC4v5bWM3tjE/EdzFJBhLuDIbdwz8HDbkMZsZ7S/tYL6wuYrm3uI1khmicOkiMMqysOCCCCCODS/tLfHo/wDBUAfBz/haOtf8IX/wmf2L+xfMXyPI8rPl4xnGfejECMCniGMAVLGMcV+2j8Yrn4Dfs1+NviDpV61trMdl9i0eVFVnjvLhhFFIobgmMuZcHIxGeD0P7M37bv7SOifHn4av8YPifr2reD/FV2sMttevG8U1vPLLaCfoCojuELEgg/ujwQcGVRinUcipV4xTgEYqQDOBUoGOKlGR+Fa5pNzoGtX+hXkkT3Gm3UttK0RJUvGxUlSQDjI4yBXwE1uHxN8Cvhz4jt4Ghj1Xwlpd2kTnLIJLSNwCR3AOKqqqqrJHANZB4rgjg1VVVVVVVVVVVVVVVVVVVVXjbRbrxL4M17w7YSRJc6ppdzawtMSEV5ImRSxAJAyRnAP0qtC1GLxL4b0/V5rRUTVLGKdrdj5gUSIGKk4G4c46c+lfGT4Har4PvrnxB4aspLvw/IzSERjc1j3KsOuwDo/YcNzyUkeM7o3Kn2OKkubiVdjzMR6E14U8H+IfG+rx6J4a02S7uX5YgYSJe7u3RV9z7AckCvg/8MdO+FvhsaZE8dxqV2RJqF2q481xnCr32LkgZ9SeMkV8R/H+n/Dbwhd+J75VleLEdrbl9huJm+6gP4En0Ck9q8V+K9c8a65c+IfEN9Jc3dy5PzMSsSkkhEBJ2oMnC9qt9LurhdwUKO27rU+kXUS7lAfA6DrWi61q3hvVrXXNDv5bK/spBJBPEcMjD+YxwQeCCQeDXwJ+LkPxa8GJqlxHHBq9i/kalBHwofGVdR12sORnoQw5xk/tE3U1z8XNZjlk3Lbx26RjAG1fIRsfmxP414VtkMLSkDc74/Af5NeH9GivVVSVyR3rxDpcNnuUEcV4ngRLmKZAP3ikH8P/ANdfsh3c0WteI7NZP3UtrC7rjqyswBz16M3500hIwDTOCOTTyDGAaeTPANO4A4NO+R1qSQEYBp3GMk1I+eM1K+RgGpGAGAakYHnNHnvVf8FTfGHHgT4fwN/z86ncg/8AAYosYb/rvnK+mD94V/wTl+HLeP8A9qLQb+eBpLHwlbzaxcfKMBkAjhySRg+dLG3AJ+U8YyR+3j8Y/iX8bfj/AGP7H/wv1WS005bq2sb6OK72Q6neTKkn79kyfJhDgFDnDo7FSVXHgL/gk/8As76T4btLLxzqfiPxFre0Nd3sV79kiLkDKxRKDtQY43MzcnnoB8JP+Ca3wo+C3x48O/GTwR4n1i6stF+0v/YusrHchJpI3SOSKVVQgRh+FdXbOG3grz+1CwP7L/xfAP8AzIWt/wDpBNXwL8dfEj9nzxF4d/aT8EpvttI1ttPnTe6xznyld7abAxsliZwOScxscAqDXwt+KXhH4x/DvQfid4F1D7XoniGzW5tXYjenZ43CkhZEdWR1ydrIw7V8XJg3wu8YDP8AzAL7/wBEPX/BGJgPiZ8Ruf8AmA23/o81cMCcA1csCeCKuGBB5q4I6Z7VcEYPNXB7D0r/AIKieMW0r4T+GvBUMqpJ4g1pp5ASMtDbRncACDxvmiOQQRgDkMa/4JOeD28TfthaVrSuwHhLQtQ1MgOFBDxi05BByP8ASxwMHODnAINVVVVUcAdaPPeqqqqqo4Ao896qqJAFHnvVVTHjANVVVVVRxjrR5qqqqqqbB4BqqqqqjgCmPGAaqqqqqmxjANVVVVVTdMA1VVVVVNg8A1VN0wDVVVYB4zTYJwKqqqqqqqqqqqqqqqqqqqqqqqqqqqq0Xwv4Z8NyX0vh3w7pmlyanP8AaL1rKzSA3Uu0L5khQDe21VXccnCgdBXxt+F2lfGv4TeKfhZrDpHB4i057dJmQP5Eww0MuD1KSqjj3XtXiPQNb8D+KtU8L6zE1nrHh/UJrO7RX5huIZCjgEdwynkelfscf8FI/h1468NaH8N/jj4ibQPG1rCtr/bGosFstWKjCu03SGVgPmEgVS2NrEvsEUsU8STwSpJHIoZHU5DA9CCOoxVVVVVVVVVVVV8R/iF4T+E/gbWfiL441RLDRNCtjPdTNyeoVUUfxO7FUVerMygda/aR+PHiX9pD4uax8T/EQaCO5byNLsDIXXT7JCfKgUnuASzEABnd2wN2B/wSc/ZMl+Dnwtf44eNtKSDxf4/tVNgjkmSx0glXjVh0VpmVZSOcKIQcMGUVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV+0J+wB+zN+0fLea14r8Dpo3ii6Vs+IdCb7HdM553yqAY524A3SozY4BFftD/8ABI39oH4U+ZrXwpnh+J+gryy2MH2XUoPXdaszCQZIAMTux5JRQKt7nxl8OfFK3Fpcaz4X8SaJcHa8bS2V5ZTLwcEbXjcdOxFfAX/grB8bPhxFaaB8V9Ot/iFo0TKpuppfs2pRx4x/rgCspHB/eLubGC4zkfAf9tX9nj9opotP8CeNUtdcdQf7C1hBZ3uSMkKhJWUjv5TOBj05qgQRwaqqqqqlPbNVVVVV8b/gh4X+Nvhc6Rqyra6pahm0zUkTL2rnsf70bYG5e/BGCAR8Svhl4t+E/iebwp4wsVhuYxvhmiJaG5jPSSJiBuU49AQQQQCCK/Zn/a8l8MR2Hw8+KV2ZdHjHlWOsSEs9mONkcvUtGOQG6rkA/KPltbq1vrWG9srmK4t7iNZIZYnDpIjDIZSOCCCCCKqgcVwRwaqqBA71VVVV/wAFJ/CUnhL9svx/GElFvq8lrqVs8rqxcTWsRkI29AJRKoBGcKOvU/8ABNHxKmt/s3RaP9okd/D2uXdpscj5A+ycbQDnaTMeTj5t3pmoCOmagOQMVARkc9qgPYVbsCAM9qt3AwAa+I/7Qfwv+EzNaeJ9e83UlTcNNsk8645GRlcgJkYxvKg5Fah/wUP0q3udmjfCy7u7fH+sutXW3fOT/CsTjpjv3Ppk+DP29vhdr88Vl4r0fVfDcsgXMzgXUCscZBZMP1J52YwMnHStN1/SPEWl2+taDqltqFhdpugubaUSRyDODhhweQR7EVO4I61cODwDVywPAqdsnAPapyCDVweMA1OQc1/wVLwfgB4fwf8Amcbb/wBI7yvghrH/AAjvxp8AeIPs/wBo/szxRpt15O/Z5nl3Ubbc4OM4xnBxSDgDNRgYwPSohwMVGAaiAI61EBxiv2iB/wAbdtIGf+Z88If+idOpAMACv+CzviPwlrfxG+G/g3QporvxRo+n3rarFAwd447h4DaxMFJIf5JmCkA4kUjIYV8CvDGreDPgp8P/AAdr8jPqmheFtNsL1nzuM0NrHHITnnO5T1pQD/wWOCj/AKH/AP8AaNNENvFOoxipVHQV/wAFkviZda3rnw7/AGd/D7fabqSQ61e2yAlmmkLW1moPQHm6yOvzKeAef+CmP7MqfC/4KfBHxXpdn8/hTRIPCWsyoQQzrF50T4HPzSi8JPTLr0yM/st/FhPjj+z14H+JMlyk19qWlJHqTKCMXsOYbjg8gGWNyM54IOSDkyDkc1KBjj0qQAnGakAzwamAI4qXp+FfEr/ko3ir/sN3n/o56/ZU/wCTXfg7/wBiDof/AKQQ1VVVVVV9DW7tWQeAaqqqqqqqqqqqqqqqqqqqq/aE8Et8N/jt8QfAf2dIY9D8S39tbqkAhUwrO/lMsYJCK0exgoPAIFfsg+LI/Gf7NPw/1ONVRrTSE010AwQbQm2yRk9REG99wOBnArXvgt8MfEcr3F/4TtY53DZltWa3O49WIjIDHPOWB/U1Yfs5fCe0l8ybRLi76bVnvJMKR7KRn8citD0TRfDtkmm6FpdrYWyDAjt4gg+px1PueTSNkcHmv2wPEM82saD4WSUiGC1e8dACAzOxRSexwI2x6bj61pVstxcZYZCdj61oegG9CqqZOPSta8MtZRksmOO4rXbNYJRKoxuOG+tfsfeI5tJ+KT6H56iDWrCSNo2/ikjHmKR7hVk/Amv2ldCn034jyauVYw6xaxyox6bkURso46gIpxz94euB4a1BLeQ2sjY3tlCfX0/lWl641qg2vjA9a13xB9rUqHzWr3q3twNhyka7QfX1NfsmaHPDBr3iaVWWKYx2sB6BiuWft2ymOe5ppQRjNNITwKZwepp5BjANO+RyfwqSQEYzTv3J+gqR88A1I4AwDTtx15qR88A05ycA1Vfto/Fm3+MHx+1vV9LuY59I0NF0jTJUUDfFCWLvkfeDTPMyn+6y+lf8Er/hKPCnwl1f4qalabb/AMZXvlWjOnK2VtlVIJGRvlaXOOCEjPbj9r208V/sy/tyQfG220f7Zp+oajb67pQnlG28Coi3UJYL8hD+Yo4JVXjbnNfAn9pf4PftC6FFrHw48WQTXQUfatIu2WG/s225KyQ5Jx1G9SyEhsMcGoJAQADX7T0oP7MfxdXP/Mh63/6QzV/wTo+EWl/tJ/AH4+fA7xB9nSN20m+0W6kU5sdSaO8WOckclQYYwyjqpcfxV/wTF/aF8QfA34taz+yH8W4brT01XU5IdOiuZQRpmrRkrJbkZI2TbcAqSPMVMA+aWHxXmB+GHi8Fs/8AEhvv/RD1/wAEaXCfEv4inP8AzAbb/wBHmvGXi7w74H8Nan4v8W6vBpmj6RbPc3t3OSFhjUZJ4ySewABJJAAJIFeMP+CkXx++NnjGfwP+yP8AChjCiyFLy6szfXsqLkiUoCIbdcD7r+Zk4+YE7ak8U/8ABXzwf/xPde8Pf25aQffsvsejXPmf8AsyJj0/hNfD7VfF+tfD7w5rHxA0u20zxLfaXbz6rY2yMiWty8YaSMKzMRtYlcFjgjqetXDAjGauG681/wAFCPibD8Qf2hb/AEbTruOfT/B1sulRtGTtM6kvcZ/2lkcxnp/qh9T/AMEZPhbLpPw98cfGC+hQN4h1GHSrAsPmEVqpeVh/su86r16wnp3qqqqqmOeAaqqqqqqY54BqqqmOeAaqqOMdaJzVVVVVUx4wDVVVVVUcAUee9VVVVUxB4BonPeqqqqqpsE8Gqqqqqo8ng1VVVVVHnvVU2CcA1VVRxjrR571VVVVVVVVVVVVVVVVVVVVVVVVVVVVVf8FY/gT/AMIV8W9O+NGhaZ5WkeN4BFqLoPlTUoVwSQBhfMhEbAdWaOVjyTVfs8ftzfHz9nJbbRvDviFNc8LQtz4e1ndPbxqTk+Q2Q8B5Y4Rgm5ssjV+z7/wUf/Z++OH2XRNc1T/hAvFE2F/s3W51EEzntBd8RvyQAHEbsTgIetVVVVVVVVVVeIvEWgeEtDvfEvijWbPStK06EzXd7eTCKKBB1ZmY4Ar9uv8AbP1X9prxk3hzwhf3lp8ONDl/4l1o4MR1GYZBu5k65OSEVvurzhWZhX/BNT9h66/aN8cQ/FT4g6WjfDTwrfASxSlT/bN7HtcW2wg7oQCplJwCGCDJLFaqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq+N/7MHwK/aK0d9K+LPw70zV5gpEGpJH5F9anGAY7hMSKOFO0ko20blYDFftCf8EYfGujS3mv/ALNvjC38Q2Cq0iaBrsq216D2jiuABDKf+unkgAckmvH/AMM/iH8Ktcfwz8SvBGt+GdUTJFtqli9s0ig43puADoSOGXKnqCRXwF/4KR/tIfBGK00O/wBfTxx4btmUDTtfdpZY4wMFIrr/AFqDGAAxdF2jCgZB/Z8/4KT/ALOvxylg0LV9XbwF4lmbYmna/MscM7dhDdDEbE5UBX8tyxwqt1qKWK4iSeCVJI5FDI6HIYEcEEdRiqqqqq+hrgjg1VVVVXxO+F3hH4teGJ/C/iywWVGBNtcooE1pJ2kjbseBkdCOCCDivjd8B/F3wT15rXVYHvNFuJSunatHHiOcYyFbrskAByp9CRkc1+zz+1J4j+EFzD4d8QPcax4SlkAa2aQtJYAnl4MnAHJJj4BPIKkknwl4t8O+OvDtl4p8K6nFf6bfR74ZkP5qw6qwOQVPIIwaqgcjGaqqoHIxmqqqr/gs58Pm0v4seBfiZb26JB4g0KXTpmSMDdNaS79zkclil0igntGAPu1/wSq8dRWviPxt8Nrq4XdqFpBqlmh4OYWMc2OOSRLFwW42HAOWIgYDGKgYdM1A3AwelQuODmoHAIANftM/Hy4+FWiweHfC8sf/AAkurws0cjDP2KHJXzsdCxIYKDxlSTnGC732rXzyyyTXd5dSFndyXeV2OSSTySTkkmrb4c+IrmISOLaA4+7JIc/+Og1rHhjWdDAe+tf3ROBKh3Ln69vxr4B/HjX/AILeIlKSPd+Hb+VRqWnsxx6edH/dkA/BgMHsVs9XsdZ0211fS7pLmzvYEnt5k+7JG6hlYexBB/GppBjOancHkGp3GOtTsMYBqdgTwanYYODX/BU7U7WL4K+GNHdm+03PimOeMbeNkdrcK3PrmVP1qvDmrxeIdA0zX4YWij1KziuUjc5KCRAwB9xnFR4I/CouQMGoyOmaixjAqIjA5r9tr/hNz/wUo1j/AIVkf+Kx/t7w7/wj3+q/5CH2Kx+zf6793/rdn+s+T+9xmtdP/BbH+xro35v/ALN5Z8z7B/wjvn4/2PI/e5/3Oa/ZJ8ffCj4H/tJN4g/bQ8EeNR44h1Hz4Nd1u5lcaZdNgi5ubVkE0r7tx84vIBuDCPKhxYXtpf2kN9Y3UVxbXEayQzROHSRGGVZWHBBBBBHBzSkD/gscCD/zP/8A7RppcjGadhjrUrA9DXwCnuf2zf8AgpvqHxTuZZ73w54Y1GfWrRmKqIrOyKxaeAOVUmX7M5VcknzDnOWr9sT4SD44fs3eOfh/bWn2jUp9Ma70pVh8xzeW5E0Kp3BdoxHkc4kYYOSD/wAEc/i+t5oPjT4Fajds0+nzLr2lo7lv3L7YblVH8Kq4gOM8mZjgckyYJxmpTkYFSY7VJgng1NgjANS9Pwr4lYPxG8VYP/MbvP8A0c9fsqf8mu/B3/sQdD/9IIaqqqqqqqqycYBrd2rIPGa4I4NVVVVVVVVVVVVVVVVVX/BW34TSeAv2pJfHFpa+XpnxA0uG/RlVVQXUKi3nQAAHOI4pWJzlpycnkD/gl78SLe88MeKvhNeXi/atOul1axidyWaGVVjmCjOAiOkRIAHM+ec8VVI2eM1G46Zr9rrT5k8Z6LqzA+Tc6X5CHacbo5XZuenSVeO341osqx3DIx5YDH4V4V1qGyZHJHHrXinxFDqKFhtyR2rxFcJIyxqQSWz9K/ZR0t9Q+Mun3ib8aXZ3Nw20djGYufbMo/HFfFT4fWvxG8MPpwZY9Qtd0thM3AWTH3W/2W6H04POMVq+j6roGoy6VrNjNZ3lu2HilXaR6EeoPUEcEciotb1CKPyvODj/AGxk/nVxqF1cja8mFPVVGK8GeCNf8eaxHpGg2jOdw86dlPl26n+J27Dg4HU4wM14V8OaX4O8PWXhzR4yLazj2hm+9I3Vnb3Ykk9ueMCjIMcU0ueAaZ896eQAYBp5M8A07gck1I+ehqRwBgGnfPJNSuDwDTt2Bqq/bF+OkPwN+Dmo32nX/k+JteR7DQ1UkOkrDDzjHTykJcHpv2A/er4X/D7WPiv8Q/D/AMOdAZUvdfvo7VJXGVgUn55WHdUQM5A5wpxzXgjwvo3gTwno3grw7C0WmaFYQ2VmjtuYRRIEXce7YAye5ye9fFP4QfDr44eE5fBfxK8OQatp7MXhLZSW1l2lRLE45RwCeR64IIJFfFz/AIJa/EXwJO3jT9nLx5PrEmnMZ7fT7yYWWpRkN8vkzptjdwDnJ8rocZJAr9jH9vr4laT8SrX9nf8AaelnkuZ7w6dZaxqEJivbG9B2rbXfTerMAgcjersNxZTlP2nJg37M/wAWxn/mRda/9IZq/wCCLMnln4xYPX+wv/b+v+Crv7N02m32n/tVfD60ltZoZIbXxO9oRGYpQyra3oIwwcsViZs9RDgA7if2cf2nLX9pf9kXXda1C5X/AIS3QNAutP8AEcGeTOtu4S4HA+WZAH6YDb152ZP/AARycJ8SviGc/wDMCtv/AEea/wCCv3xWvtL8HeDPhBpd2yR+ILmbUtURSV3RW+xYUPGGVpHdsZ4MKkjkGv2L/gl4c+B/wA8L2On6XFFrmv6bb6nr12yqZp7mVBJ5bMpIKxB/LUA7cKW6sxM8mQTmrhwflBq4cHODX7Xv7R+n/s8fDWbUbGa3n8Waxut9DsnZTh8fNcOvUxxjk8YLFFON2R4Y8N+Lvin440/wxoVvd614j8S6gsMKszSS3E8rcs7HJPJLMx6DJJ6mvgP8KtN+CHwc8I/CnS/KKeHNMjt5pIhhZrg/PPKOB9+VpH6A/NzVVVVVMeMA1VVVVVUxAGAaqqpjgYBqqqmPGAaqqqqqjjHWjzVVVVVUx7A1VVVVRxjrTHI61VVVVVHAHBqqqqqqPA4NVVVVVTYxwaqjgCjz3qqqm6Yqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr9p34GaZ+0X8FvEPwvvHggvbyHztJu5gdtpex/NDISASFz8rYBOx2A61rOj6z4R8RX3h/XdPay1bRb2S1vLS4jDGCeJyrxupyCQykEHI4qD/gm78Lv2tfgf4f8A2gv2VfEcXhHVdbs92o+FNSlkuLC3v4yUuIIpjumhAkVtu8SblKN8oYV8Zf2ffjH+z9r/APwjvxc8BanoE7sRbzzR77a6A6mGdMxyD/dYkd8V8D/23/2jPgFFa6X4S8bNqmg2ilYtC1xWvLNFwMKg3B4lGM7Y3QZJ9TXwa/4Ky/BLxlFbab8W9G1HwJqbKFlulRr+xZ+eQ0a+agJA4MZC7uWIBavB/jvwT8Q9ITX/AAH4u0fxDpz9LrTL2O5jz0wWQkA5BBB5BBB6VVVVVVVVfHD9oX4Ufs8eGD4n+J3ieKxV+LSwhxLd3rf3YYQct7scKO5Fftaftr/Ef9qjVE069j/sDwZYT+bp+gQTbxv2hfNnkwPNk+9jgBAxAHJZv2FP2C/GH7WPimLxB4hhvdD+GelTD+0tXEex79lPNraEjDOcYZ+VjHJyxVW8FeCvCfw58J6X4G8DaDaaNoOi2629jY2qbUhQfqSSSSxJZmJYkkk1VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV8RPhd8Ofi3oDeF/iZ4J0fxNpZbcttqVoswjb+8hIyjf7SkH3r9oT/gi/4U1qW88Rfs2+Nn8PTMrOnhvXS9zaluyxXQJliXA6SLMSTy4FfGj9m344fs96kmnfF34c6roCTyNHbXskYltLphnIiuELRucDO0NuAIJAyK+BP7af7RP7O/2ew8BeO5bjQoJAzaDqyfbLJx3UKxDxAnk+S0ZPc18Bf+CsPwN+I/2LQvitY3Xw912b5HuJ3+0aY7cAETjDxbjk4kQKo4Mh61peq6XrmnW+r6JqVrqFjdxiS3urWZZYpkPRkdSQwPqDiqqqqlPbNVVVVVXiTwz4f8YaLc+HfE+kWup6ddptmtriPcp9CO4YHkMMEHBBBGa/aF/ZR8Q/Ch7jxR4UFxrHhPOWkK7p7DOeJQOqDA/eAAc4IHGfhD8avG3wX106r4VvVe0uSov9OnG6G7UdMjqrDJw64I9wSD8Hfjn4G+NOjC88OXqwalBGDe6VO4E9ucDJA/jjywAcDB74PAr6Ghgjg1VV9DQwRwaqqr/gqf8JG+Jf7K2peItPtml1PwFexaxCEIBaAZiuAc/wAIjkMp6H9yMeh/Zb+KX/Cnfjv4S8aTyxpp63gtNSMr7FW1nHlSuT/sBvMHbKDPGagYYwDULjAINQuOOahkHAzUMgwBmvjp4pm8Y/FnxLq8k2+KO+e1tsOWXyof3aFfQEJux6sfrXw08PxR2A1WRAZ7okKSvKIDjj6kZ/KrHQGni3LHnA9K1rRYjFJBPCro6lWRlyCK8R6T/YmsXGnKxZEbMZPUqeR/h+Ffsb+LJ9c+EP8AZF5cmSXQdQktYw77iIWCyJ24GXdQDn7vpgCaXPANTSAjANTyA9+lTvxnNTuCOtTsDwD0r/gqv4rSbWvAPgiC5w1rbXmoXEIkByJGjjiYr1GPKmAPQ5Poar9kHxFbeKP2YPhjqVoItkHhq0sj5cokG62QW7cjod0Ryv8ACcjtURHHNRntmoyMjBqJhxg1GQDjNftDkH/grppBz/zPfhD/ANE6dUbAjGa/4K+/DHwnrnwD074pz6fFH4k8N6vBaQXqRgSS2s+4PA7dSgbDgfwndjG5q/4Jf+N9Y8Zfsc+EV1ubzZdBuLvSoJT1aCGU+UCB/dRlQeyAnkmgw/4fEBgf+Z+/9o0ZgRgGnlBGM1+3v8X0+DX7KvjrxFb3SRanqtidG0sEHcbi7/dFlx0ZI2llGeP3ffof2Pf2EPjJ+0B8Pb34p+Bvivb+CLJtRk06NS1wst2IlRmfMRA2bn2jJJyrcDAJf/glr+04o5/awz/28X3/AMXXwv0rxF+wh+3h4f8ADPjPU7eWDTdQgsdQv0idYLiwvolUzqDyVQSh+M4eIjnBFOQRnNSsMdakIxwakIOealIPANSkYxmv2jooof2hvihDDGqRx+M9XVEVcBQLyXAA7Cv2DP8Akzv4U/8AYAT/ANDeqqqqqqqqqqvoayema3A8VkEYzXB6GqqqqqqqqqqqqqOMdaJHTNFh2r/gq58C5/ix+zbJ440TTvtOt/Da4fVF2KC5sGULeAcdFVY5m5HEB6kAV+zz8XL34IfFvQfH9u0jWdvN5GpwJ/y3tJPllXHcgfMo/vKtaVqmm65pdnrWj30N5YahbpcWtzC4ZJonUMjqR1UqQQfQ1VfQ0jZ7818cvh7L8RfBT22nRb9W01/tFkowDIcYaPJ6Bh9PmVe1Ok1rM0bo8UsTFWVhgqRwQR2+lWevtEoWUEEd1q68RGRNsYZjjjPAqWV55DJI2WPWv2Z/h1J8O/B954v8Uqtje6sgldbhvL+yWqAkF88KTyxz0G3OCCBf/F74V6PZSajq3xN8KWVrDjzJ7nWbeJEyQBlmcAZJA+pFfG74heBvGWm2Nj4Yl0jXfORZRrFq6TqkZwVWGVCQwbqcEjGOpPD6ah+5KR9Rmk06IYLuW9ulfs8/Y0+HwW2WMOl7Ksu1cHPBGfwI/OjICOtGRfWml4wDTS54zTPkcmnkAGAad896eQYwDTvxyfwqSTsDTt6Hmjz3qr28s9Os59Q1C7htbW1iaWeeaQIkSKMszMeFUAEkngCv2tfj9cftA/FW71qwnkHhnSN1noMLgr+5BG6YggENKw34IyBtU525r/gmL8Bn0bSL/wCPniK1X7RrETWGgI6/NHbq/wC/mwe7uiopHICP2eoZOAAeRUMoIGDzUMvAINf8FLYNPk/bD0tfh+kSeI59H037a0B2O2pebIsJZmwu7yRaAHoAFz0NftMTBv2a/iwAf+ZG1oY/7cZq/wCCM0nl/wDC4MHr/Yf/ALf14o0LQfGnhzU/CPijTotQ0jWbSS0vrSUkLNDIpV1JBBGQTyCCOoINfCObVP2Av20734UeNnlvfA/i7ZptxPdxKsV7p9y+La8ZW+QiN9ySHkACdRk8V/wR9cJ8SPiDz/zA7b/0ea/4LB+DL+Vvh38R4Ekeyj+16Tctt+WKQ7ZYRn1YLPwf+efHev2W/ijovxa/Z+8E+K9IvYJZU0iCy1GKL5fs95BGsc8ZXJKjepKhjkoyN0YGp5RjOank4wDXxi+LHhP4L+AtT+IHjG78uy09MRwIyiW7mP3IYgxG52I6dgCTgAkfHH40+Lfj18Qb/wAeeLJ2UzMY7GxWUvFYW4PyQx5xwOpOBuYliOa/4JbfsYz/AA30GH9o34k6d5fibxDZ48OWTkE2GnyoD9ob0lmU4C9Uj6nMjKtVVVVEgDg0ee9VVVVVRxjrR571VUcY60ee9VVHAHWjz3qqqqqqY8YBqqqqqqOMdaPPeqqqqqYjGAaY5OAaqqqqqm54BqqqqqqbngGqqqqqpsHgGqpumAaqqrAPGabBOBVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVf8ABWX4CweCfijpXxt0C08vTvHMbQaoqRkLFqMCqN5I4HmxbSB1LQysfvV/wRv/AGjJfBnxS1T9nXXrtv7H8cq99o6sTtg1OCImQDsBNbxkEn+K3iUfeNeI/DHhrxjo8/h7xd4e0zXNLulKz2OpWiXMEoIwQ0bgqwwSMEd6+PX/AAR0+Bvj9rnWvgxr178O9WlZ5PsTI1/pzsR90Ru4khBbursqgnCYAA+Nn/BOz9rD4GCe91z4by+ItGh/5i/hljqMBAxklFAmjHzAZkjUHnGcGvB/jvxt8PdXTX/Afi7WPD2pR9LrTL2S2kx0wWQgkEEgg8EEg9a+GX/BVH9p3wOI7Pxdd6J45skXaBqlkILhFCAKFlt9mTkAlpFdjk5OSCPh9/wV4+BfiDyrb4heCvE3hK4fG6WHZqVtH65ddkn5RH8K8FftmfssfEIwp4a+OfhbzbhgsMGo3R02WRi2wKsdyI2LFuigZOQRwQa03VNM1qxi1LRtRtb+0mz5dxbTLLG+CQcMpIOCCPqDVVVeM/if8Nvh1Abjx94/8O+HEWMSZ1TU4rUlSSAQHYE5KsBjqQQK+JP/AAVF/ZW8CpJb+Htc1jxtfLuUQ6JYMsavtJG6acxqVJABaPfjPQ4NfGj/AIKvfHbx20+mfCzTNO+H+lSLtEsYF9fsCCGzNIoRAc5GyMMp6OetXV742+JXidZL271vxV4i1WbYrSyS313dyMegzud2JJ45JJr9jn/gkf4i8WS6f8Qv2pILjRNDDeZF4RjkaK9vAMY+0yKQbdDzlFPm47xmvD/h7QvCehWHhjwxpFppek6XbpbWVlaRCKK3iQYVEUcAAADFVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVeIPDvh/xZot34b8U6Hp+s6TfxmK7sNQtUuILhP7rxuCrDgcEEV+0P/wSB+B/xOnuPEPwc1aX4bazLudrOK3N3psz4yAISytBk4GUbao6Rmv2gv2K/wBor9mm9lHxF8B3E2jKxEPiHSQ15p0wHfzVAMRODhZljcgE7cc18GP2k/jb+z/qqan8LPiBqOlRbgZ9Od/PsrkZ5Elu+UJ5I3YDLuO1lJzXwE/4K9+APFUtpoHx88Jt4QvnVUfWtLL3Vg792aHBlgXoAAZvUkCvCvi7wt460K28TeDPEem65pN4u6C90+5WeKQd8MpIyO46jvVVVfQ0CCOKqqqqqnSORGilRXRwQysMgg9QRX7RH7F8Vwt141+DdmkTohkuvD6jAfA5Ntj+LjPlng87SOFOi654p8A+I49V0S/vtF1rTJiu9CYpYXU4ZGU/QqyMMHkEYyK+AP7Yfh34hLa+FPiG1voniRisUNxnbbagxIA2/wDPOQk42ngnoedooEDjNVVUDjjNVVV4g0HR/FWg6l4X8Q6fFf6Vq9nLZ31pMMpPBKhSRG9mViD9a+PHwl1j4E/GDxV8Jdck8258OX7QxzYx58DKJIJcdt8TxvjtuxX7Cfxqh+L3wK0q0vbrfrvhFI9J1JWcs7rGoEExJ5O+MDJPV1eoZBwAahkGME1FJxjNRSgjGea+IemXGjePfEWlXIPmW2q3KElSu4CRsMAexGCPYivhzfQz6BabGG6HMbrnOCD/AIEH8a0XWoLWAqQpJXHNa/fRTszKRzXju7hvPEtyYG3CICMn/aA5/I8fhX7E+nXFp8P9b1WXKx3uq7IgykbhHGuWB7jL4+qmpZgRgGppQRgGppAeAankB4zU0gPOancY61+3b8QP+FgftK+JvIn82z8N7NFtcrgr5APnA8n/AJeHnx04xxnNTWl1bxQTXFtLEl1GZIGdColQMyFlJ6jcjLkcZUjsa/4Jh+KBr/7K9hpG/d/wjet32n42bdu51ucZ/i/4+evvjtUTDGM0jDgg0jAjg1GwIxmkYEDB6V+1x4s0fwH/AMFQZvHXiCSVNL8O+KfDOo3zRR73EMFrYSSFV7nahwO9XH/BWT9km1tZriLUPFl1JFGzJBFopDykDIVSzquT0G4gZPJA5r9pD9pP4xf8FG/HGhfCj4PfDjVIvDmm3YmtNMQ+dJLO2YxeXkgHlwoqOyjJ2oHcl23DH7OfwhsfgB8EfCHwhsLv7V/wj1hsubgElZrqR2luJFzyEaaSRlU9FIHavjR8UbD4Lf8ABS3xJ8VtT0u41K18NeMGu5bS3cJJMBEBhSeAee9H/gtV8MCMf8KV8U/+DCD/AArw3/wWO+GvibxFpXhyD4OeJoZdVvYbRJHv4CEMjhAxAHQZr/gsr8WZtW8T+BfgNo9w0osIX1rUYIZC++eUmG2UoOjqizkA84nGAAef2bfhbH8EPgL4H+Fvl7LnQtIjW+XcGH2yTMtyQV4wZpJCOvBHJ61K4xwa/wCCx/wpSO98DfG+wt0HnrJoGpyZIJZd09tx0Jx9qBPBwqjkDj9kr4tf8Lr/AGc/A/j24vPtGpTaatrqjNL5jm8tyYZmfuC7IZMHnDqcnOTK2eAalYYwDUhGMZqQgn6VKc8A1+0n/wAnFfFP/sddY/8AS2Wv2DP+TO/hT/2AE/8AQ3qqqqqqqqqqqqqqyegNZPrW6twPFbhWR61kHgGsj1FZHqKyPUVkeorI9RRI6ZrI9aLDoKLDoKLZ6UST0NEk96qqu7S0v7SawvraK5triNo5oZkDpIjDDKyngggkEHg5r9ub9ly//Za+Nt/4dsrRh4Q15pL7wxOZDJ/oxbmBmbJLxEhDkkkbGP3q/wCCf/7V1ro62/wH+JGsLFayyY8NX9y+BE7HmzZjwFJyYyehJTPKAVVdOhpHzxnmv2pPDPgfTIbLX7a2Nt4h1GZlKW+1EnQcvLIuOWBIG4YJ3c5xxo3h7VvELXCaRbJO1pCZplMyIVjXlmwxGQAMkjoOTgVrnxk+HOhPNBL4jhvJoWZfLsQbgOQcHa65Qj0O7BHIJrxb+0b4kvLpB4GM2iRwOHjuywNwSDkMCMiMj2JOec14k8Y+LvGV0L7xf4q1fXLlWZhNqV9JcuC2Nx3OxOTtGfXA9K8MSRxeItOd9BOt4uUCaaN3+luThY8KCTlsDA5PTvV38e/2jv3aaT8C57aNFwUm0S9l+mNpXAx2xXgTUPEer+DtJ1Xxda21tq95bLNcwW8bRpEW5VdrMxBClQcnqD06DTvE2gatq+p6Bp2qwT6jozol9bKSHhLoGXIPUEEcjIzkZyCB8fv2ifjv8D/HGmW3wx8e3eiaXeaZ5rW4gguIpLjzHV3McqON2wRDOOwx3rwP/wAFW/i7o+yHx94B8OeJIUz+8s5JNOnbO77zfvU4JQDEY4U5yTkfD3/gpl+zr4xaGz8Tza14OvJX2E6jaedBknC4lh3YGMZLqoHOTgZrwd8SfAHxEsF1LwJ400XX7dkDltPvUnKA/wB5VJKHnBDAEHg4NNKMcGmkJHBpnAHWnkzwDTuB35qSTPANO/YHmjz3qq/4KEftSx+Xc/ADwBqTGTOPEt9bzcAf8+YKnr/z1B6YCc5cD9mT4Ba1+0J8S7XwxapJDolhtudcvguBb2wYDaD/AM9H+6o69WxhGx4f0vS/D2j2Hh7RLOO007TLaO1s7aMYWGKNQqIvsFAH4VDLkAZ5r4lx/wDBRf4WfGPxR4z+H1vN4j8L61qUl1a6fayrqdokGCI4vJlImidY1Td5YRWYYUkcVdftYf8ABSfxVA2g6J+ztNoV5cqwjv4fBd/E0JCk5DXcjwg8cb1IJwOc4P7Kn7DPxEh+KY/aE/am1X+0PE9tdLe2Gmvei7l+2KflnuZVJT93tUxojEAheQECn486Vqvif4FfEbw1oVjJe6nqvhLVLSytohl5ppLSRERfcswA+tf8Eu/gx8Vfg0PiZ/ws7wPqXh3+1/7J+w/bEC+f5X2zzNuCfu+Ymf8AeFfaDjhq/b9/Zj/4aH+FR1jwrp6TeOfCatcaUFUK97Ccedak4ySygsg/vqBwHY1/wTN+Bfxg+D3jnxpqXxK8Aap4fttQ0mCG2lvEVRK6y5IGCecc18afhT4R+Onw51f4Z+NYZG0/VIxtmhIWW1mRg0c0ZOcMrAH0IypBDEHR/gV+3T+xT4s1PU/hDpsninQrviZtLgF9b6gik7PNs8+ckig9UGQSVV2UnP8Aw01/wUp8eQy6FoP7PkXh+7mwiXzeF7qzaIt8oIa+mMPBIPzAgYyeM1q3xksPgh8D9B8YftCatHpmswaZbwajDEwnlvNQEQ3pCqcOzsrHj5VBySFBYftJftI+M/2kPGja9rztZaLZMyaPo8chaOzjPUn+/K2BucjngDCgAf8ABNr9gkfFW9039oT4u2OPCGm3Yk0LSJo8/wBszxN/rZQRg2yuuNvPmMpB+QEPVVVVRwB1o896qqqqqqY54BqqqmOeAaqqpjngGqqqqqo4x1o896qqqqqmPYGqqqqqpj3B+lVVVVVRxjrR571VVVVRxjrR5qqqqqjjHWjz3qqPJ4NVVUcAdaPPeqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/as+Cdr+0F8B/FPw4+yxS6pPaNc6I8jBPK1CIFoDuP3QzDYx/uO1eGPEfij4aeNtM8VaFPPpXiDwzqUdzbM6FHt7iFwwDKechlwVPuDXwa+KOgfG34VeFviv4YV49P8AE+mxXkcLnLW7sMSQsehaNw6EjglTjiqr4w/sk/s4fHlJpPif8JNB1S/nXa2qQwG0vhg5H+kwlZSAcnBYjk5BBIPxP/4Ik+ANTluL74QfGLWNCLMzxWGuWKX8Yz/AJYzEyKOxKyHA5yea+IX/AASg/bM8Cs76Z4K0fxjaxKWe48PavG4AAU8R3HkyscsRhUJyrdsE+OPg38W/hk7R/EX4X+K/DO3Hzavo89orAsVBDSKAQSrAEHBwcVo+va54duWvfD+tX2mXDxmNprO5eB2QkEqSpBxkA49hWj/tRftJ6A1p/ZXx9+IMUdiqrbwN4jupIUVRhV8pnKFQMDaRiv8AhuP9rb/ovPib/v6n/wATV3+21+1leWs1nL8e/FapPGyMYroROARg7XUBlPoQQR1BFaz8ffjt4jijg8Q/Gvx5qkcLFo0vfEl3OEJ4JAaQ4OK0/TtQ1e9h03SrG4vbu4bbDb28TSSSN6Kq5JPsK+Gf7A/7XnxY+zzeGfgd4gtLKfay3utxrpUPlts/eqbkoZEw6sDGGLANtB2nHwf/AOCJV7OkGofHr4vJbhl/eaV4Ut97A54/0q4XAO3qPJIyeGIHzfBD9lb4Bfs62SQfCf4b6ZpV55Iim1WRTcX1wOp33EhL4JJJUEL2AAAAqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqaGG4he3niSWORSro65DAjBBB6gjtX7Qn/AASq/Zq+Nkt5r3hOyuPhx4kuFYrdaFChspJT/HLZnCkdeImiJJySa/aM/wCCef7Sn7OMs+o6x4RfxR4XjY7PEHh5HuoFXsZowPMtzyoJdQm44V2r4X/Gf4q/BXXI/EXws8eat4dvUbLC0n/dTdOJYWzHKvA+V1YcDjgV+zx/wV+0PUvsvhv9pPwx/ZM/Cf8ACSaHC8tuf9qe1+aROBktEZMk8RqK8E+PPBXxK8O2/iz4f+KtL8Q6PdcRXunXSzx7gBlSVJ2sMjKnDKeCAaqqBHTNVVVVVVA5GM18ef2YPB3xltZdXski0bxTHGfJ1CNMJcEA4SdR94EkfOBuGB1A218Rfhl4z+FXiB/DnjTR3s7gZMEoO+K5T+/E44ZenuM4IByB8BP2xfEXw7W38L/EEXOveHI0WOCZWBurBQMAKTjzEAAG1iCOx42nwv4s8NeNtFg8Q+E9atdU064yI57d8jI6gjqrDupAI70DkYzVVVAgjGaqqr/grp+zH/wknhmx/aU8JabLLqmgRpp/iKOFM+ZY7mMVwQBnMbtsZv7jrnAjr9k74+XH7PXxbsvFFy0kmgain2HXIEUsTbMwPmKueXRlDDuRuX+I1pGqWOr6da6rpl3Hc2d7Ck1vPG2VljdQysp7ggg/jUUgPGailBAGajlHQmv2t/hndWGvr8StLtmey1JUi1AopPkzqAquewVlCj/eB7sK8N+Jbzw5dGWBRJDJgSxE4zjuD2NWfxO8PyQgy3ckDY5R4mJ/8dBFeJPiak0T22hhmdgR57rgL7qOufrWiaLqniXWLTQ9GtJLu+vpRHDEiklmPf6AZJPYAk18NvBtj8OPA+leEbNkdrOHNxKox507cyP64LE4z0GB2qSbIxmpZQRgGppc8A1NICMA1NIDwDXxv+J9h8IPhb4k+Il8YWbR7F3tYZWIW4uT8sMRxzhpCinHQEntVjZa7408S2+nWMUmoazrt8sUMYID3FxM4AGTgZZ2HXA5r/gpH8ALH4A+JvhRoGkW1ulqvgC10+5mieRvtV/ayyC5m+YYAfzY2wuOScquef8AgkR4qWTSviN4JlmUNb3FjqFuhc5fesscpA6ADy4uf9oelIw4wajYEYzUb9s0jAdDUcnHBrx7+zP+z78VNbbxN4/+EHhjWdXlx51/NYKlxPhFRfMkTDSbVRFXeTtAwMCtO/Yo/ZL068hvrf4A+EHkhbcq3Fj56E/7SOWVh7EEV4V8K+E/BGkpoPgvwxpGgaZGxZLLS7KO0hU4AJCRgKDgAdOwpZRjGa139nz9n7xVrF14h8T/AAL+HusarfSGS6vr/wAMWdxPO/8AeeR4yzHgckmj+y5+zBjj9nD4Xf8AhH2P/wAaq2/Zn/ZqsLmG+sP2evhnb3NvIskM0XhKxR43U5VlYRZBBAII6V4h+C/wX8V+I/8AhMfFHwi8Faxr4aNhquoaBbXF1mMARnznQvlQoxzxgY6VJKCMZqSTPANeLvB/g/x3o7eH/HHhXR/EWmPIsjWOq2Ed3CXX7rGOQMuR2OOK8LeC/BfgDSjoXgPwjonhzTWlaY2ek6fFZwmRgAz7IlVdxCqCcZOBTtx1qRgT1qVhjrUjDGM07Z5zX7SXP7RPxS/7HXWP/S2Wv2DCP+GPPhSM/wDMAT/0N64I4NVVVVVVVVVVVVVVVVVVVVVVVVVVVVftPfs4eCf2ovhVffDfxjG0UysbrSNQiOJNPvVRljlH95cMVZDwyseh2sPjL8FviL8BPHeofDv4maBLpup2MhCSAFoLyP8AhmgkwBJGwIII5GcMFYED9lj/AIKCXHhy3svh98d7u4vdPj2Q2PiIKZJbZAMbbkD5pFHy/vAC453bs5GgeINC8V6Na+IfDOsWeq6Zepvtry0mWWKVc4JVlJBwQQfQgjtVVq+s6VoGny6prOoQ2drApZ5ZWwBgZwO5OAeBya+I/ji++Ifiu68RXYZI2/dWkLHPkwqTtX68kn3Y18e/iHPq+sv4P0i+ddPsMpeCNyBPMCCVYdwhUYH97J7DHwR/ZX+PX7RN0YvhR8O7/VLOOQJPqkxW2soDkA7p5CqEgHJRSz46Ka+CX/BGfwDosNrq3x88f3viS/ChpdI0IGzs0bBBRp2HnTL0IKiE57Edfhx+zJ+z58I7e2h+Hfwd8LaRLabPKvF05JrvKEFS1zJulYggEFnJzz15qvjv48f4efDTU9YtLgw6hdD7HYOo5WaQHDD0KqHYe614U8N3/jDxLpvhjTFJudSuUhQhc7ATyxHooyT7A1P8A/g3d6fa2V58N/D9xPZWRtLfUJNOiN5GhRUYrcbfMVmCLlgQSQM9K8V/8E0/hB8UZr6L4nXmsTPauyaLqWk3/wBnnjgfBKyI6NGXVh12kHdn2X4jf8EVNUj826+EfxstZ858qx8R6c0O303XEBbP4Qj8a+Jn/BPj9rv4WI11rHwb1TWLFc4u/D5XVFICgsSkBaVAM4JdFHBxkDNWt3r/AIT1r7RY3WoaNq+nSsm+J3t7i2kGVYZGGRhyCOD1FfDz9vn9pv4exJaHxyvia0RsiDxFD9sJPvNlZiOBx5mPzNeAf+Cq3gzUGjtPiZ8NtV0djhTd6RcpexkkgFmjfy2RQCScFzxwDnj4VfHb4U/G/TZ9T+GfjK11gWbBbq32PDPbkjjfFIFcKeQGxtJBwTg08gxgGnkJ6GnfPANHnvVahqGn6TYzalqt/b2dpbIXmuLiURxxKOrMzEAD3NftR/8ABQy2+z3fgL9n2981pkaK88TlGTywRhltFODu7eaRgclAflceCvBXi74n+LrLwh4Q0u41bWtVm2xRKckk8s7seFUDLMx4ABJNfs8/ATwv+z38PrbwhoYjutRmxLq+qmEJJfz88nrhFB2omeB6sWYo+04zUM2cDPNRTZwM81HMOBmknGACaScEYDUJ+K88YwDTTgjinnyMbqknBGAalmHrk1+0n+2f8MP2fbWfSZLpfEHi9oibfRbOUZjbsbiQZEK+xBY9lxzXxk+N3xE+PHiyXxb8Qtba5l3N9ksoSyWtghxlIIyTsX5VyclmIBYsea/YJ/4JsXvxMktvi5+0ToF1ZeEPLWXR9AmZ4JtY3KGWeUqQ0dvggqAQ0h54QfPp+n2Gk2FtpWlWVvZ2VlCsNtbW8QjjhjQBVRFXAVQAAABgACqqqqjjHWic8A1VVVVVUxAGAaqqokAcGjz3qqpiAMA1VVVVVUxzwDVVVVVUcY60ee9VVVVUx4wDTYJwDVVVVVU2MYBqqqqqqbpgGqqqqqpumAaqjgDrVVVU2MYBqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr/gpX8EV+EP7SWpa3pNkkOg+O4f7YshHGVSOdjtuo89C3mgy4HAWdBX/AARZ+Oba34K8V/s96xeo1z4cm/tnRo2wG+yTMEuEHOSqTFGzjg3ByegFVVVXiP8AZ/8AgN4xljn8W/BLwDrcsTO0b6j4atLlkZ8byC8ZwWwM+uBmtV/4J4fsWazYS6bd/s++H44psbmtZbi1kGCCNskUiuvIGcEZGQeCRX/Drj9hL/ohf/lzat/8lV/w64/YS/6IX/5c2rf/ACVWkfsNfsf6Hei/sv2c/A8soUqFvNKS7jwev7ubcuffGRXhfwP4K8D2n2DwV4Q0TQLby44vJ0vT4rRNkYIRdsagYUEgDoMnFVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV+0l/wTc/Zv8A2ivtWt/2B/whfiyfLf23oMSxebIf4p4P9XNk8k4Vz/fFftB/8Ex/2nvgT9u1nT/DH/Cd+F7T5v7W8PIZpETn5pbT/XJgAliquijq9fDj4tfEz4P61/wkPww8daz4av22iV9PumiWdVOQsqfdlTPO1wy+1fs8f8Ffv+PXw3+0n4Y9E/4STQ4f/Hp7X9S0R+kdfDv4n/Dz4t+HYvFfw18Y6X4i0uXA8+xnD+W2M7XX70beqsAw7iqoEEdaqqqqqvoa4I4NeOfh/wCDviVoUnhzxpoVvqdk53IJBh4X7PG4wyN2ypHBIPBIr48fsleL/hOs/iLw403iDwwnzNcJFmezGCSZkX+EAf6wfL6heK+G3xY8c/CbWhrXgvWntmbie1k+eC4HpJH0P14I7EV8EP2o/AXxjSHSJJF0LxMQd2l3MoInI6mCTgScc7cBxhuCBuPBHBqqr6GgQRxVVWt6LpPiTRr/AMO69p1vqGmapayWt7aXEYeO4hkUpJG6ngqykgg9Qa/bP/Zc1r9lf4wXvhQpPceFtWZ7vwzqExBNxa5GY3IP+siLCNshd2A4ADgV/wAE9v2todEe1+AXxK1eOKxlcjw1qN1KR5MjEf6ExxjaxLFCSMMSnO5AsUucAnmo5QRgmo5uxNXlrY6rYz6bqNrFdWtzGY5oZVDK6kYIIPUV8Rf2SNTguZtS+G19HdWzksum3kgSWP8A2UkPysM8DdtIGMk8mr/4PfFTTp/s1x8Pded8Zzb2Lzr1x96MMO3r/OvC/wCzd8WfE00Yk8PHSLdgC0+pOIdoOP4OXzg9NvGMHBr4UfBLQfhJp1xdadKmp+IbmEq99cJsUeiIBkomQM9SevYAaD+1h4VN1Jovj3Rb7w9qdpI0N0Qv2iFJEbaw+X5wcg8bTjkZPU6H4x8M+K7Y3XhzX7HUo1+8badXKH/aAOVPI4OKlmyMA1LKCMA1LKCMA1NLngGv+CmPxxm1LXNO+A2iTbbXS/L1HWnWTmSd0/cQkDkBY28wg/eMiHA2An/glB+zzL8Ufjs3xY1q1Y+Hvhuq3MZYELcalIGFug9QgDzEg8MkQIw9f8FrdIsp/A/wu151b7XZ6rqFtEQ2AElihZ8j1zCnPbn1r/gk7dXKftBeJbNLiQQS+DbiR4g5Cs63loFYjoSA7AHtuPqajYEYzUb9s0jAjg80jgjGaR8YANJIMcGlkB6HmlkIHWll4wDQmyMA0ZRjg002eAaaXPANM+R1p5ABgGpJB0Bp3z3qR88A1I4AwDTsD3qRs8A1IwxgGvilc3F78TfF15eXEk88+u3skssrlndjO5LMTySSckmv2R9PstN/ZX+EFtYW6Qxv4H0iZlUcGSSzikdvqzsxPuaBx0NAgjGa4I4NVVVVVVVXxT/ao/Z1+Ckslp8Tfi/4d0e9iXc+n/aDc3ajkZNvCHlxkEfd5IPpXjP/AILF/sxaCZbfwp4d8beKJ1UGKWGwitLdzvwQXlkEinaNw/dkHKjjnGq/8FubRJ7iLQ/2bpZYRkW8934sEbHjgvGtqwHPYOfrX/D634o/9ET8Lf8Agxn/AMK0P/gtr4ot/O/4SX9nvSr/AHbfI+w+I5LTZ13bt8Eu7Py4xtxg9c8aJ/wW08I3EMh8R/s/avYyhgIlsvEMd2GGOSS8MW0+2D9a8P8A/BYj9lPV8pq2jePtDdIwxa70mGVGbuqmGdycerKv9K+G37cX7J3xc1ZdB8D/ABu0KfUXYJDa6gk2mSTsSAFiF0kfmMcj5UyevHBqqqqqqqqqqqqqqqv2i/2avhb+1B4GPgn4l6Szm3ZpdM1O2IS702YjBeJyDwQBuRgVbAyCQpH7UH/BPn47/szy3WtXGlt4u8GRM7J4i0i1dkgiXo11EMtbHGMklkycB2NfDb41fFX4QXMtz8N/HOpaJ553TQROJIJWGMM8Lho2YAYyVJAyOhNaL/wUp/aL0uN47638I6wzKoD3umSIVIzkjyZYxznnIPQYxzXij/goj+0x4hEi6dr2i+HlkYfLpelIdo24Kgz+aQCfmznIPQgcV8LYPG/jW5/4Wj8T/Eur65qU4ddMOpXLzGCN/vyIGJ2BsbQFAAUcDBGPBvwO+MPxutdQ0n4Q6ZZm8hVI5dS1C48i2sPMJAkdsMzEAFgqKzHHQivgH/wSS+Bfw4az8QfFzUbv4jeIIj5klvcJ9n0xH4IxACXlwcjMjlW6mNelaXpOlaFptto+iaba6fYWcYjt7W1hWGKFB0VEUAKB6AYqqr9rP4gv4n8eL4Rsps6f4bUxsFbIkuXAMjcf3RtTB6FX9a+JX7Tl/wDsv6THr/g7T7G78ca1FLbaLLep5kemRY2zXnl8b5MMI4w3yEtIWDBCjeNP2nP2ifiJfPqHjH42+M9Rd5PMWE6zNFBE2AMxwoyxx8D+FR+teDP2v/2o/h/cW0/hf4+eNoks1CwWt3q8l7bIAwYAW85eLGVH8PQkdCQfhP8A8Fjfjr4Xkgs/ix4Q0DxxYrgSXFuP7LvW+UKTujDQnkbsCIZJIyARt+C//BTr9lb4vz2uj33im68Ea3c7EWz8SQi3ieQj5gtyhaHAIwC7IWyMLnIHxF+B3wV+NWmmH4kfDfw34niuI12XV1ZI8yrwQY51xInGOUYccdDXxT/4I6/s++LJZL34Y+KvEPgOdlwtsW/tS0Q88hJmE2ecHM2OB05J+LH/AASg/ap+HNpPqvhnT9F8e2EOWI0K6IulQAnJgmVCx4xtiMhORgHnGgeI/GPw38TprHhzVtT8Pa9pcrIJYJHt54HGVdGHB9VZDwRkEYyK+EX/AAU/vraK30j42+DmvQq7W1nRNqSE9i9sxCknuyuuOyGvCf7Y37NXjWCOTT/i3otg7L80Oru2nNGdoJUmcKpIzjKkgkHBOKt/in8MbuxXUrT4j+F5rRkLrcR6xA0ZUdWDB8Y4614i/aq/Zw8MWou9S+NHhWZCrELp1+t+/GM/Jb72zyMDHPOOhx8Tf+CnfgDSYJbH4U+DtS8QXoZlW91PFnaj5eHVQWlkGeCrCLp1r4z/ALSvxf8Ajzc/8V34mb+zEk8yDRrFTBZwN2IjBJcjJw0hdhkjODipPAnjePwhH8QZPBuuL4Wluvskettp0osmn5/dCfb5Zfg/LnPB4r9lv9oif9nDx9N4lbw3a6vp+qW62eoqUAuY4PMVmMEh6H5QShO18LnBCsvwj+O3wv8Ajjow1b4e+J7e8lRN1zp0rCO7tOcfvYc7lGeA3KnsTVI5U4zUU4OAT9KjmGMZpJ+ODSzjGM0LjsGrzz/eFNcZGN1PODwDmvi9+23+z58IYJbfUfHFtrurKhMel6EwvJSRxtd1PlxHpxIynHIBr49/8FF/i38VIbrw54Aj/wCEG8PTZRmtJy9/cJn+O448sHAO2MKRkqXYGvAXw88ffF3xbb+D/h94a1HxHr+oMzJa2qGR27s7seFUZyXYgDuRX7G//BLTwx8MWsfiL+0PDpvijxQqrLa6AEE9hpjENnzd3y3MgBXt5aMDjfhXFVVVVUxB4Bqqqqqqo4A60ee9VVHGOtE5qqo4x1onPeqqqqqokAdaPPeqqqqqpj2BqqqqqqYg8g/Sjz3qqqqqjjHWjyeKqqqqqmwTwaqqqqqm5OAaqmweAaqqrGeAabk9aqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/4KifBmL4mfs33HjSwskl1v4e3I1KBwvzmzfEd1GCTgLt8uY9z9nAHWv2KvjV/woD9pvwL8Q7qaJNKXUBYawZpNiLZXI8maRj/0zV/NGcDdGM8Zqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr9pL/gnz+zf+0v8AatZ8QeFf+Ec8WXGW/wCEj0FVtrmSQ/xTpgx3GTjJkUvgYV161+0l/wAEv/2j/gH9q13w/pX/AAsXwnBlv7U0G2Y3MCD+Kezy0icAkmMyooGWcdK+F/xd+JfwV8TR+MPhZ401Hw7qqLsaW0kG2ZMg7JY2BSVMgHY6suQDjgV+zx/wV+0PUvsvhv8AaT8Mf2TPwn/CSaHC8tuf9qe1+aROBktEZMk8RqK8E+PPBXxK8O2/iz4f+KtL8Q6PdcRXunXSzx7gBlSVJ2sMjKnDKeCAaBx3qqqqqqoEdM1VfHL9i7wt448/xH8NPsvh3XW+aSz27bK6Pf5VH7ljxyo2nHK5YtXivwh4s+H2vyaB4r0a70jVLU7vKmGDgEgOjDhlyDhlJBxwa+B37a3iLwXFB4b+J0dzr+jxIEhvYyDeW4H94sR5q49SGHqeleEfGXhbx7ocPiPwdrlrqunT8LNA33WwCVZThkYAjKsARnkCqqgcd64I4NVVftK/s5eBf2oPhjd/DjxsJbdhJ9p0vUoBmXTrtVZUmUZAcYZlZDwysRkHDD4xfB3x/wDAjx/qPw2+JOitp+r6c2QRlorqIk7J4XwN8TAHDexBAYMB+xP+3Tb3lvZ/CP45eII4bmBBHo3iK+mCrMoHEFzIxwHAGFkY/N0Y7sF45hjGaSbjAOaSYEYDUs5x1oXAx1r7QPWmuAehr9qzwQNI8W2/jOwtttprce24KjgXKDBOO25Ap9yrGrDUdQ0q6S+0u/uLO5iOUmglMbqfUMpBFeFv2lfiT4fMcGp3sOuWqcFL1MSYzziRcHPu27r06V4b/ad8B64Eg1uO70O4bAPnL5sWT2DpzjPdlUfriw13Stcs1v8ARdTtr62YkLLbyiRcjqMg9favjT+0j8J/gbpVxd+MvE1u+pRpmDRrSRZby4YgEAR5+UEEHc+1cHrXjnxfqvxA8Za3431yTdf67fzXk4DEhGkcttXPIVQQoHYADtX7EXwLX9nr9m7wp4Hu7JINcu4Tqmunyyjte3GGZXB53RoIoeg4hFf8FkdBGo/sz+Hdcg0/zptJ8ZW/mTgcwQSWtyrfg0ghH1xX/BNXWYdL/at0SylmkRtW0u/tY1XOHYQmbDe2ISfqBSNjoaRgRgHmkcYxmkfPQ80kgIwTSv6Gll7ZpZewNCUEV5ikYzRkXHDUZAR1ppR0Bp5QRjNPJngGncY608meAadxjg1I+eAakcYwDTsAOvNSNngGtb1a41/Wb/XLxI0uNRupLmVYwQqs7FiFBJOMnjJNfAXRIfDPwL+HPhuCdpo9K8JaXaJI4wziO0jQMQO5C5qqBx0NBgRg1wRwaqqqqr9p/wD4KP8AwG/ZuurzwpbXEvjfxra7kk0TSJlEdpICRsurkgrCcqQUUSSKcbkAINfHr/goj+038ebq4tbnxrL4S8PSZWPRPDjtaR7cj/WSg+bKSAudzbcg4VQSK8P+GfEfi3UV0fwr4f1LWb913La6faPcSkZAyEQE4yQOncV4R/YN/aa8XQrdN4Fj0SB8bZNXvY7dueuYwWkGOvKj2zWk/wDBLj4tzxudb+IfhG0kDfu1tRcXAYdyS0cePpg1p/8AwSm1CWzjfVPjjb290c74rfw4ZkXnjDm4QnjB+6MHjnrWqf8ABKfxLFJGuh/GXTbtCp3td6K9uVPsFlkyPfIrWP8Aglr8a7eTOg+OfBl/EItx+0S3Ns5fn5QoicYxjksOT2xk69/wT4/ao0SIzW/gG11aNImkdrDVrZiuO2x3VmYjoFBz068V46+GHxE+GN+ul/EDwXq+g3DqGjF7atGsoIyCj/dcdR8pOCCOoNfB39tj9pz4FW9tpngL4r6p/Y9pH5UOj6mRf2ccec7I4pgwiGR/yz2Hrzyc/Db/AILVeK7OGGz+LfwY03VGC4kv9A1B7NgQG58iUSBifkB/eKBhiM5CjwT/AMFcP2QvFCKPEOp+KfB8nIYatorzrkKDkG0MxIJJAyAflOQBjPhL9rH9mTx3Itv4W+PXga7uXkaOO1fW4YJ5Cq7mKxSMrsAvO4Ajg88HFhqGn6tZQ6jpd9b3lrOu6Ke3lEiSD1VlJBH0qqqqqqqqqqqqqaGG4heCeJJI5FKujrkMCMEEdwR2r4s/8E6v2Svi/fT6zq3wyi0LVbmQyTX3h6dtPaRiCCWiT9ySSQxYx7iwyTy2da/4IsfA+4ukfw78WvHNlbCMBo71LW6cvk5IZI4gBjAxtPIJzzgfHf8A4J8/ss/s5+HbAT+J/GfirxjqaEW1jd30EVqqgjdcPHFCsiqPuqpk+Zs9QrAeEfCWs+Mdf03wd4V077TqGoSrBaW6YQZx6nhVABJPQAE18GfhLoHwZ8CWXg7RQssygS6heFNrXdyQN8hHYcYVeygDJOSaqjjHWvit48tvhx4E1PxRIyG4hj8uzjb/AJaTtwgx3GeT7Kaii1TxHrKQRLLe6lql0FUZy880jYH1LMfzNf8ABTfwzL4K/aI03wcWZoNH8IafDAxB2tuMskrKcDIaZ5T9SR2xX/BIX9nj9n74n/Cjxf44+Inw78P+LvE1n4kNisWt2Ud7Fa2gtoXjKwyqUBd5J8tgk7F6Y58f/wDBMr9jDx/burfCOHw9dHOy70C9msmTJycRhjEf+BIcdBgcV8XP+CJOpQRT3/wK+MMV2VyYtL8UWvlMRuOP9KgBBOzAx5KgsM5Ab5fjb+zT8cv2ddUj0v4wfDrU9BW4bba3rKs9ndHGcRXEZaJ2xyUDblHUCv2fv24P2if2bfLsPAnjL7foKf8AMA1tWu7Ef7ibleLnn906Z75r9nP/AIKvfAn4sfZfD3xWX/hWviOTCebfT+bpc7dMrdYHk5wSRMqqowPMY1aXdpf2kN9Y3UVzbXEayQzROHSRGGVZWHBBBBBHBzX7Sf7C/wAA/wBpu0mvPFHh7+w/E55j8R6NGkF2WA4E2VKzp0yHBbAwrL1r4x/8EhP2hvBNxPefCvU9I+IGlqrPHGkq6fejDD5TFK3lk7TnKyEnawwDtDeKv2bP2hPBE5t/FfwQ8cab+8MayzaDceVIwGSEkCFH4/uk1d2l1YXU1hfW0tvc20jRzQzIUeN1OGVlPIIIIIPQ14f/AGf/AI7eLLo2Xhn4L+OdUmTG9LTw9dS+WCcAsQmFGe5wK+Gn/BKb9rbx5LbzeIfDukeCdPlYFp9b1FGkCdyIYPMfdjOFfZk9SAc18CP+CTH7PfwyEWsfE+W6+JGtpsZUvlNrYQsMHK26NmTnIIld1Ix8gPJ1/wABeCPE/guf4c6/4U0q98L3Fmtm+kSWq/ZvIUAJGsYGFC7V24xtKqRggV+1r/wSd8YeCp7vxv8As0LdeKNAeRnl8NTOG1CwUjP7ljgXMYOQF4lA2DEp3OEk8V+AvEm+KTVvDviDSJyMqZLO6s5Rwf7rxsPwNfDv/gop+0D4NRLPxJdaX4wtFyANTt/LnUY4Cyxbc885dXJyRkcY8Of8FS/AF0F/4S74WeINN+9u/s29hvsemN/k9e/p71p3/BST9m+7tI7m4l8UWMj5zb3GlgumDjkxuy84zwx4PY5FJ/wUg/ZoUY/tXxB/4KW/xq+/4KXfs2WVm9zbyeKb+RMbbe30oK75IHBkdV4HPLDgevFal/wVT+CcNqX0fwJ44urncMR3MFrAmO53LO5z7bfyrUv+CsXhOG1L6N8G9XurncMR3Orx26Y7ncsbn8Nv5V/w9tP/AEQD/wAuv/7krxf/AMFTvjNqxlh8HeC/C+gQux2PcCW+njG7IwxZEJ2jaSY+ckjbxj4kftG/HL4uI0HxB+Jus6pavjdZLKLa1JAwCYIQkecZ5255Pqa+GfwZ+Kvxm1gaD8LfAOs+JLsZ8wWVqWjhA7ySnCRjkDLsBkgdSK/Z6/4I7a1fSweIP2k/FyadbK24eHdAlEk8gHaa6IKIMjlY1clW4dDXws+DXwt+CXh1fCnwp8DaX4b075TIlpF+8uGUYDzSsTJM4BxukZmxxmqqqqqpiMYBqqqqqqo4x1pjngGqqqY54BqqqmIPANVVVVVVMc8A1VVVVVHAFHnvVVVVVMRjANMc8A1VVVVVN0wDVVVVVUTgcGqqqqqo8Diqo4x1o81VVR4HBqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqte0LSPFGhaj4Z8QWEV9perWktpe2soyk8MiFJEb2ZWIP1r4sfD/UPhV8TPFHw21R99x4b1a4sTJ2lWNyFkHA4ZQrDgcMOBX7Avxmf45/speBfFl/f/AGrWdNs/7H1hmLFzc2n7re5OcvJGIpSQcZl7dBVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX7Tn/BPf8AZ6/aat73V9S8OQ+F/Gdxlk8TaNCIpnkzktcRAhLnOACXG/AwrrX7SX/BNz9o/wDZ2+1a3/YH/CaeE7fLf23oMTS+Sg/ing5khwOScMg/vmvhD8c/iv8AAbxCfE3wo8a3+g3cu0XMcTB4LtVztWaFgUkAy2NwOMnGCc1+zZ/wVs8E+MJbbwt+0TpNv4R1JlCpr1iskthO/AAkjAaSAnP3sunUkoK0DxBoPirR7TxB4Y1uw1fS76PzLW+sLlLiCdP7ySISrDjqDiqqqqqqgQRjNVXxG+F/gj4raE2geNdFivYgG+zzj5JrVjj54pByp4GR0OAGBHFfG79j7xv8NPtWv+Ekm8SeG4E8x5Y0/wBJtV7+ZGPvKOpdRgDJIUCvAfxG8a/DLWBrvgjX7jTLogLKEwyTKDna6HKuPYjjtivgt+2p4M8crb6F8Rfs/hnXXIRJst9juWJ/hY5MX0kOP9o5xSPHLGskTq6MMqynIIPcVVA9s1VVX7WX7JvgD9q/wA3hvxIiaf4g05XfQdejiDS2EpHII43wtgB4yecAghgrD4/fs8fE79m3x1c+B/iRoUtuwkf+z9SjjY2mpxLj97byEAOMMuR95C2GAPFfsuft+eK/hFHa+CfiiL7xP4SjUR21wJN97pijoELH97EOnlsQVGNrAKEPw2+KngP4teG4fFXw98T2esafJgOYX+eByAfLlQ/NG4BB2sAcEHoQaWfjBNLOCMBq88ngNRmOPvUZyRjdXxT8Jx+PfA+peHlWNrp4/Ms2fjbOnKc9s/dJ9GNSxSQyNDNGySRsVZGGCpHUEdjVaHo/hSXS7/xH40+IOjeF9I0oKbmS8cmVgTgeXH1fnjAJOSBg5r4wftgaTpdtceD/ANm2DUtHhkVor3xXcyFb69QjBWFMf6Mued4xJ0xswd3h3wx4y+I3iWLQvCuh6t4j17U5SUtrOB7q4ncnLNhQWPJySfqTX7Dv/BL3WvCview+LX7S+nWQm0xo7jR/CyTifbcA5WW7ZcoQhAKxKWBP3yApRq/4KWeFV8VfsYeP1jt0lutJWz1G3LSFBGYruIyNwcE+SZQAcjJHfBH7K/i1fA37Rnw78RyTtDDHr9vbzyLMYtkU7eTIxIB+UJIxK/xDI71G4IxmkfHQ0rgjGeaSTsTSyAjrSyEDGc0sgPGaEnoaEhxgGvNOOtX3iHRdMlFvqWsWNrKy7lSe4WNiOmcE9OD+VQXtveQJc2lxHPFIMpJGwZWHqCOtNJkYzTSDHWnlHQGnfPU08gAwDTvngGpHGMA07jGSakkz3rxxqd3pHgzXtV06fyruy0y5mgk2htjpEzKcHIOCBwRiq8P6LaeG9A03w7YySyW2l2cVrC0xBdkjQIpYgAZwozgAZ7VVVXToa3cYNZB4BqqrWtb0Xw3pF3r/AIi1ey0vTNPhaa7vb24WCG3jUZZ3kchVUDkkkAV+23/wVG8S/EG61P4W/s5aldaD4WjkaC68SwStFeaqAV5gIAa2iJDDOd7qRnYCVPw3+F3jz4u+JIvCfw98N3Wr6g673WJdscCDrJLIcLGnQZYgZIAySAfgn/wTX8B+Hbe31r40anJ4n1NkDNpVpI9vZQMRyGdSJJiD0OUXqCrDmvDHhLwt4J0iPQPB3hzTdE02JiyWmn2qQRhj1bagAyccnqe9KmetLHngCkiGMAUsY6DmljJ4FCIEVrfhzQfE2mzaL4j0Ww1awuFKzWl9bJPFICMEMjggjBI5HeviR/wTn/Zv8evLe6Pouo+D7+TLeZolzthLbSBmCQOgXOCVjCZx1GSa8ef8EpfH2nia5+G/xL0bWoxudLbVbaSxlx8xCBk81XbAQZOwEkk7QOfFv7DH7U3hBpGuvhPf6lCiuyzaTPFfeYEAJISJjICc8AqCxzgGvE3gHx34LdovGPgrXtCdHWNl1PTZbUhmXcqkSKOSvIHcc1pGt6z4fvBqOg6ve6bdqpUT2lw0MgB6jcpBwfSvDf7Xn7UnhLy00L9oPx9HFDbi3it7jXp7mGKMYwqRysyLgKACACBwOCa8M/8ABUf9tLw7Ctvc/E2y1uKOFYol1PQ7RmQKMbt8caO7EDkuzE9TzzXhj/gtF8drL5PF/wALvA2rqvlhTY/arFyBneWLSSgluMEKoBzwc4HhL/gtb8Lr0L/wnnwT8U6P97d/ZGowaljH3ceYLfOe/THvXgT/AIKa/sa+Omitm+KTeHbyVsLb69p09qAMA5MwVoVGTjmQHKntgnwf8QPAXxDsG1TwB430DxNZpjdcaPqUN7GMllGWiZh1Rh9VPoaqqqqqqqqqvjL8W/DnwX8EXPi7X3aSQnybC0QZe6uCpKoPQcElj0APU4B8c+NvE3xO8X33i/xNc/a9V1SUFhEm1VAAVI0UdFAAUDk8ckkkn9kD9nRvhT4f/wCE38W2e3xXrduAIZFIbTrdsHyiM/fbCluARgL2Oao4A60WHQUTnvX7VvxLfxX4y/4QzTZwdM8OOUkKn/W3RGJCf9z7gGOCH6g1+yN8MxrOvTfEfVIs2mjsYbBGXiS4Zfmf6Irf99ODkba/4LN/BAX2k+EP2htLSVptO/4p/VxuyogZpJrZwM/LiRp1Jwc+anTbz/wSj+PM/wAIv2nrDwVqGpeT4f8AiVGukXUTsAn2wEtZSdR8/mFoh1/4+G4JIxVa1oeieJdKuND8RaPY6rp12u24s723WeGYAg4ZHBVhkA4I7V+0l/wSD+DPxL+1eI/gdqH/AArvX5Mv9g2NPpc7+nl532+T3jJUDpHXx8/ZL+PX7NOpNafFTwJd2lg0my21m0H2iwuT22TrwCf7j7X9VFfAv9rz9oL9nSWKD4Z/EK9g0hJvNfQ70/arCUnO79y/CFs/M0exjxzwMfs8/wDBXH4N/EKGDQ/jfp7fD/XNuDfJvudNnbHUMAZISf7rgqP75rQfEGgeK9GtPEXhfXNP1jSr+PzLS+sLlLiCdP7ySISrDjqDiqqiQBgGmb35o896r4yfsy/Aj4/WTW3xW+Gmka1cbQseoiI297EF6BLmIrKFyc7N209wa+JX/BF74c6pLLefCf4va54fJV3Wy1qyj1GMudxVFkjMLRpyq5YSMACTuJrxL/wR6/ak0eUnQtc8Ca9A0zLGbfU5oJAg+67rLCqqSMfKrNg9z1q7/wCCYP7b1vdTW8PwbiukikZUni8R6YElAOAyhrhWAPUbgDzyB0r/AIdj/txf9EQ/8uTSv/kqrD/gl9+25eXkNrcfCC3sY5Gw1xceI9OKRD1YRzs2PopNWH/BI79rq8vIrW4tfB1jHI2GuLjWspGPVhGjNj6KTWl/8EZP2jJr6KPWviP8N7WzOfMmtbu9uJF4OMI1sgOTgHLDAJPOMH/hyv8AFD/otfhb/wAF0/8AjXgr/ginaI63HxG+O8sq8g2miaKEPUYImlkb+HcMeXwSDk4wfhl/wTO/ZD+GjxXT/D2XxbexbcXPie6N6Dhg3MACQHJUZzH0yOhIOjaJo3h3TLfRPD2k2WmafaKVt7Szt1giiGckKigBRkk8DvVVVVVVTEAYB5o896qqqqqqY9gaqqpiAMA81VVRIA4NHnvVVVVVUxwMA1VVVVVUxzwDVVVVVUxHY0ee9VVVVVROTwaqqqqqmOeAaqqqqqmweAaqmxjANVVVTegNVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX/BXD4UN4W+OGi/FSytmFn440sR3MhIOby0Cxt05AMDW2M9SrYPBA/4Ik/FyKDUviB8Cr+7wbqOLxBpcRZsFkxBdYBO3JDWvQBiFOdwX5aqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv2nP+CZX7PX7Rc974n0yzl8B+M7rLNrGjRAw3EmMBri0JCSdyShidj95z0r9pL9hT9oX9mS7urzxd4Sl1bwvFIRD4l0lDPaMmflMuPmtyeOJABngFutfAv8Aan+OX7OmordfDDxxdWti0m+40e6/0ixuT33QtwCem9Nr+jCv2av+CofwU+M/keHPiUYvhz4obairf3QbT7xjxmK5IURnIzslC43KFZznEUsVxEk8EqSRyKGR0OQwI4II6jFVVVVV06GuCODVVXxq/ZE8BfFN7jXdBZfDfiKQZNxbxA29ww/56xDHJ6b1IPOTuxiviZ8HfiD8JNSGn+NdBlto5Ti3vYv3ltcdfuSDjOBnacMBgkDIr4O/tPfEn4PLHpljeLrOgqf+QVfuzJEM5PktnMWck4GVyclSa+EP7Rnw1+MsSWuhakbDWcHfpF8VSfgZYx4JEi4BOVOQBkhaqgQRjNVVV8WPg98N/jj4PufAvxQ8LWut6Tc4ISXKSQOCCHikUh43BA5UjIyDkEg/tU/8Euviz8G7q48T/ByC/wDiD4O+d/LhhDanp6g8LLCgHnjbj95EOSG3IgAJ8EfEHxt8Ndbj8R+AvFGoaHqMfHnWkxTeP7rr0df9lgQfSvhH/wAFRdc06NdN+NXgxdVRFAXU9CCwznGM74HYIzHk5VowOm3nI+Hv7Xn7PHxLjRfD/wAT9JtrpjtFlqsn2Cfd6Ks23eef4Cw688HEV3FPEk0MqujqGRlbIIPQg+leeP71NOB0Na94p8P+F9PfVvEuu6fpNjHndc31ylvGuFLHLuQBwpPXoCe1fE3xn8MPF/xI1qf4aeMNM1yLKy3v2Bi8cUzZDYfG2QMyltyEj5vpXjj4n+HPh/f6VZa+0oTUy+ZIhvMAXb8zKOdp3dsng4Bq3uNI8Q6Ws9tLa6jYXaHDKRLHKvQ+oI4xitU/Zj8Ca/4ksp4NevPDelyzqNQW2tPthSPPzNEjSJ83+yWx6Yxg/sl+AP2Zvht4Fi8Ofs9TadcYhV9Ru5pFfVLpunmXRIVxk9BtVAc7VAqq+JXg63+Ivw58VfD66l8qHxNol5pcj5I2LcQvETxzwHqeDVPD2ryW8vn2OpaZclG2sUkgmjbBwRyGVh1HQivhd42tviN8OPDHj20aPZ4g0i2vSsZGI3kjVmTAZsFWJUjJwQRnio5AR1pX44NLICMZpX9DmllHQGlkBHWhJkYDV4y8b+H/AAF4eufEviW+W3s7YYAHLyufuoi/xMccD6k4AJHxL/aX+Ivj66nttO1SfQNHYkR2djKUdlIx+8kGGbI6jhfand5HMkjlmYkszHJJ7mvD/inxJ4UvBf8AhnXr7TLjILPaztHux0DAH5h14ORyfWvgV+1H/wAJdeWngz4gmKHWLhhFaahGoSO7fgKjqOFkY5xjCk4AAOAWkB/ippRjANPLngGncHvTyZ4Bp3AHJ5qSTIxmncAdea/al1e20f8AZx+Jl3dJIySeFr+2AjAJ3TQNEp5I4DOCfbPXpXwa0a58R/GDwL4esrNLu41TxLp9rDbuVAleS5jQId3GCSBzxzzVVVVVVk4xmtwIxWoahp+k6fc6rqt9b2dlZQvNc3NxKI44I0BZndmICqACSTwAM1/wUN/bt1T9o3xVdfDL4baxLB8LtGuB5bRhom12dMZnlBwfKVs+XGfQSMNxVY/2X/2X/FX7RviryojNpvhTTJV/tfV9n3e/kw54aVh+CA7m/hVvhb8JvAnwa8KQeD/AGiR2FlHhppD80t1KFCmWV+rOQoyeAOgAGAAM8ClQdOppIs0sYAx+lLHkUsQAxQjzwBQizQiGMZoRjGAK8sEcLXlkjpRjyPu1rnwV+D3ieRJvEnwo8HatJGzMjX2hW05UtjcQXQ4JwM+uBWufsH/soa9hrn4Q2Vu6IURrK+urXbnvtjlVWPuwNa5/wS5/Zz1LDaZqvjPSGVCALbUYpFZuzMJYXPHoCP614j/4JMWbK0vhP41TRssXywajogffJk8mRJV2rjHGxjke/Hij/gl/+0NozPJoGq+E9fiDHy1gvpIJSAOCyyxqoJ6YDmvGn7Kv7RfgBnHib4PeI0jjUtJPZWv2+FAApy0tuXRR8w6nrkdQcWGo6x4d1Jb7Sr+80zULVmVZreVoJYiQVYBlIKnBIP1Ir4Wf8FKf2vfhbLGg+JbeLrFFwbLxVD/aAY8cmbKz54I/1uDnnnBHwP8A+Cw/wd8YvDo/xq8L3/gW/fC/2halr+wYlsHdtAlj4IP3HH3ssMDPgzx14K+I2gxeKPAHi3SPEekTMUjvtLvUuoiw+8u5CQGGRlTyO4FVVVVVVV8QPiB4V+GPhW78YeMNSW0sLRcADmSaQ/djjX+J2xwPqSQASPjP8YvFPxq8YTeJfENyy20RaPTbBTiOzhzwqjuxwCzdSfYAD9jT9mL7S1l8ZPiDp8ElsUEug6fMu4s2QVunGcADB2KQck7uMKScY60SMYBokngGjz3qvjp8S7f4ZeBLrUIZwNWvw1tpsYI3eaVOZMf3UHJPTO0fxCtD0bU/FWu2WhaXGZ7/AFO5WGIMTy7HGWPYc5J7DJrwJ4O07wD4S03wlpbb4tPhCtKV2maQ8u5HOCzEnHbOO1fHH4V6N8b/AIReLPhRrqRfZvEmly20csqbhbz43QTgdzHKscg90FahYeJfh/4uudL1C3uNJ8Q+GtSeGeJwBJZ3cEhDKevzI6EfUV+zD8b9H/aK+BXhL4s6VKnnatYqupW4Iza30fyXERHGAJFYqSBuQq2AGFVVXlnaahaTWF9bRXNtcxtFNDMgdJEYYZWU8EEEgg8HNftJf8Ek/gJ8XPtXiL4Sv/wrLxLJl/KsbfzdLuH64a1yPJzgKDCyKoyfLc1+0P8AsLftH/sz+Zf+PfBf9oaBH/zMGhs15Yj/AH32q8PPH71Ez2zXwX/aQ+Nv7PWpyan8IviHqWhLcNuubIFZ7S5OMZkt5A0bNjgOV3KOhFfs+/8ABYrwVrdvb6F+0b4Wl8O342qdc0WF7mzk5ALSQcyxYHPyebnnAHArwP4+8EfEzw5b+Lvh74s0rxFo11xFe6ddLPHuAGVJUna4yAVOGU8EA0zZGAaJzwDVVVVVVVUSAOtHnvVVVVVVVEgDAo896qqqqqqYgDAPNHnvVVRxjrR571VUcY60Tmqqqqqo4x1onPeqqqqqokAdaPPeqqqqqY4GAaY54BqqqqqqYjGAaqqqqqjgCjz3qqqqqjjHWjzVU3JwDVVVcetHnvVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV/wAFRvhjF48/Zb1DxNBEzah4H1CDU4dg5aJm8iZT/shJfMPT/VDnsf2D/itL8HP2svhz4qMyJY3mrJpOo+YTsFtef6O7Ng9EMiyfWMcHoaqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqmhhuIXt54kljkUq6OuQwIwQQeoI7V+1B/wAEk/gz8WvP8TfBJ7T4beJW3O9pBbs2l3jnoDAD/o3TGYQFA/5Zk818eP2Y/jd+zZrzaH8WvAt7pcLzNFZ6rGhmsL/GcGG4X5GJUbthw6gjcqniv2av2/Pjx+zb5GiWGr/8JV4Sj2r/AGBrM7yR26DtbSZLW/U8LlMnJQmv2b/22fgV+03a21l4Q8Q/2V4oeNmm8Naqyw3ilQSxj5KzoAC26MkheWCHIFVVVVAgcZqqqtY0XR/EWl3Gia9plrqNhdpsntbmISRyDOeVPB5APsQDXxj/AGEre6ebXfg3frA7FnfRr+X5PYQS4yv+6+f94Ditb0LxL4J119J13Tb7RtWsJAxinjaGWJhyrDv6EMODwQa+Dv7cHi/ws8Oi/FCGXxJpeVQX6ELd269yeMTcdmw3fceleA/iN4K+JujDXfBGv2+p2qkLKEyrwsRna6HDIfYjntX0NAgjrVVVV+0B+wX+zh+0VPPrXifwf/YviWfcX17QiLS5lcj70wAMc5zj5pFZsKAGAyK+Mv8AwSH+PngqW4v/AIT63pHxA0uNQ0cG4abf/wC0DFKxiIA7iXLYPyg4B8e/CL4p/Cy6Nn8SPhz4j8MyeZ5aHVNMltkkOWAKOyhXB2PgqSDtJBIFeH/G3jPwkyt4U8X61ozJL5qnT9Qltir4A3DYww2ABnrwK0r9qX9o3Ro3is/jX4ukEjZJu9TkuiD7GUsQPYYrVf2m/wBojWLhbm7+N3jSN1TYBa61NargEnlYmUE89SM9OeBWq6zrGu3C3etateahOibFlup2lYKCSFBYk4ySce5r9lv9nj9pXX/FUPiXwf8ABrxNeaE0JjvLue2+xwvEwzujecospUqG2pubHAGWGdF/4Jb+JPjrqul/Ef4m/FCHQvD95YQvZaVpdiZrvyGXeN8shCROS5Y/LJgYU88jwD/wT8/Zk+HHg6XwloHhTUZJbhF+0avdanLJeTSL0kOCIlbBxhI1U91r4qfsVfELwncTah4AH/CUaQMssasqXkIAyd0ZwH9BsJJ/uioZte8K6ys1vNf6Pq2ny/KyM9vPbyD0Iwyt+RrwB+2n8WfCQitPEr23iqxjwNt4PKuAoxwJkHJ4PLq55rwf+298H/ECpD4iXVPDdyQNxubczw7iTwrxbmPQZLIo5/GvDXjjwZ4zgE/hPxXpOrrt3EWd2krIMD7yqSVI3LkEAjIzX/BQ34Ty/CP9rPxvp0cDJp/iS6/t/TmMRQPHdkvJtHTas/nxjHH7vt0H/BMX4mx+LPgRdeArm6Z7/wAE6k8QjY5K2twWliOcDjf9oXBJI2dcYAVwehpJOwNLID3pZCO9LICMZoSA8A0JDjGa/av+IV54m8eN4Qt7qT+y/DwCeVjar3LKC7n1wCEGemGx94k+DfCZ8RTtPclltIWAO3gu393Pbjr9RVj4Q0uCER2+mW6KB/zyBJ+p6mvE3w902+heS1t0tbrBKui7VJ/2gP59anhubC6aCUNFPA+D2KsPSvgd4/uPiJ8NtM1vUJ0k1GDda3zAjJlj43EDoWUo5HGN3HFNIMdc00oxgGmfI5NPIMYBp5M8A08gA4NO+e9f8FA/EI0P9lTxhFFcyQz6m9nZwlBnO66iLqfQGNZB+PvX7C3hkeLf2vvhRpX2WW48jxHDf7Ijgj7IDc7vovk7j7A1VVVVVVX/AAVV/bXlnubz9lv4Va40cdu2PGWo2kpHmMV/5BysP4QCDNjuBGSMSqf2Z/2fNe/aJ+IcPhmyeS10Ww2z63qC4BtbfPRc9ZHwVUYPOSRhTXgbwN4V+G3hXT/BfgvR4dN0jTIvLggjH5sxPLMxySx5JJJoDPApE7DrSRe1Kg6CkiGKWPPAFLFkUsYxgChESMUIc9BQhHrXlL0zXlL6/pRiUjANGIEYBpofamix0powRginiBHFPHxjFeO/g/8AC/4lwmHx54A0LXTxiW8sUeVMYxtkxvXpjgjjjoSK+KH/AATB+EfiGGe8+GOv6r4Rvtn7m1nc31nuByAQ580Z6Z8xscHacEH4xfsVfHz4Nedfan4W/t/RYULtq2h7rmFFHUumBJHjIyWQL1wxwTXwr+M3xT+CHiNfFfwn8dap4a1L5RK9nL+7uFXkLNE2Y5kBOdsisuecV+yp/wAFZ/APxGltPBf7RFvZeCvEErCODXISV0q5JwB5pZi1qxJzliY8AkunAMUsNxCk8EqSRyKGR0bIYEcEEdQRVVVVVfEH4h+Evhh4ZuvFfjDVUtLO2QlUyDLO/aONM5ZicDHbqSACR8dfjp4p+Ofik6vq7taaVaFl0vS0fKWqHuf70jYG5u/AGAAB+yL+y5B8Qnh+JnxCs2bw7azY0/TpYyBqUi4y756wg8YH32BB4BDIqRRrFEioiDCqowAB2A9KPPeqqppYYIXnnlSOONSzu7bQoA5JPYYr47fE6b4n+Obi+t5T/ZGnFrfTI8/8swfmk+rkbvYbR2zX7IfwuNtaz/FDWLciW4DW+kqwHEfSSXHuRsHTgP1DA1Vf8Fbf2c/+FcfGG1+Nnh2w8vQfiDu+3eWmFg1SNR5mfTzUxIO5ZZTX/BIL9qYeAPiJefs6+MtX8vw/41kE+g+byttqwAUxg9hPGMcnG+KMAAuxNVVVNDDcQvbzxJLHIpV0dchgRggg9QR2r9oz/gl1+zV8dYZ9W8MaKnw28UMp2aj4etUS1kbHBmshtjcdSTH5TsTlnPSv2h/+Ca/7TP7P/maqPDH/AAmvhxf+Yt4bje58r/rrBjzY/dtpT/ar4cfFf4k/CHXU8S/DHxvq/hvUVZS0thctGJducCRPuyLyflcEcnjmv2f/APgsdrFtPb6B+0l4NivbU7UHiDw9F5cycAbprVm2vk5JaNkwBxGxr4P/AB7+EHx70EeIPhP4703XoEUGeCKTZcWpPaWBsSRn/eUA9s1VVVVVVRIAwDR571VVVVVVRIA60ee9VVVVVVEgCjz3qqqmOeAaqqpjngGqqqqqqmPYGqqqqqqmOeAaqqqqqmIPIP0o896qqqqqmOTwaqqqqqmPYGqqqqqpumAaqjwOKqqqmxjANVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVeN/CemePfBev+BtZ3f2f4i0u50672kg+VPE0b4IIIO1j0IrxH4f1Xwp4h1Twtrtq1tqWjXs1leQOCDFNE5R1IPIIZSOa/Z0+J0fxo+BPgT4oiTdP4h0O2uLsZU7bnYFnX5QBxKsg4AHHQdKqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqvFvg3wl4+0C68K+N/DWma9o96u24sdRtUuIZB7o4IyOx6jtX7Vf/BHS0vPtvjT9lXUvIuD5k0vhHVroCNz1EdncNjZ6BJ2IyeZFHFeLfB3j34TeMJvDXjLw/rHhbxLo8yu9tdwvbXEDjDJIucHB4ZXXgjDAkEGv2Yf+CsPxC8BNaeEv2grOfxpoAbautwbV1S0XHG8HCXKggfe2P8zEu+AtfC34t/Dr41eEbXxv8MvFVlrmlXSj54Hw8Dd45Yz80bjurAH8KqqqqBBGM1VVQI6V8RPhV4C+KukjSPG/h+C/RMmCYExzQN6pIuGH06HuDXxg/Ym8eeCvM1f4fNN4s0lcs0EcQW8gHbMYP730+QZ/2QK0DxH4l8F6wmseGtZvtI1K2JUTW0rROvPKnHUHGCp4PQivhF+3ikslvonxi0xIQflOtWEZIHvLAoJ+pT/vmvDnifw74v0qLXfC2t2Wq2E3CXFpMJFz3UkdGGeVPI7gVwRwaqqqqmhiuIngnjSSORSro65DAjBBHcYrxV+yt+zV42aabxN8B/At3cTsrS3Q0KCKdyoAXMyKrkAADG7GABWr/wDBOD9inW7w3158DLKOUqF22mr39omB/sRTquffGTWif8E7f2L/AA9539n/AAJ0uXz9u/7df3l7jbnG3z5n29TnbjPGc4GPBPwI+CXw4aKXwD8I/B/h+eFY1Fxp+iQQzN5YIVmlVd7MAT8zEn5ic8mqqq8efCv4f/Ey0W18a+F7PUmjQrDOybJoQeySLhlHOcA4zjivHP7A2lXAluvhz4zmtHO4pZ6tH5qZPQCVAGVR05Rzjvxz4x/Zg+N3gsvJeeB7vUbZSQtzpWLxWAAy21Muo56sq9D6VNBeaddGG4imtrmBuUdSjxsPY8g1430TTfiUbZvHsMmuyWUDQW0t7M8kkEZYsUR87lXczNgEDLE9TmvgjBYfs+eKNQ8U/Dqwjil1Sz+y3FrdyyTQFQysCBuDbgV4O7+I+vGn/td67EU/tPwbYXACYcQXTw5bHUZD4Htz9as/2wdJeIm/8EXcUgbhYb1ZBj1yVXnrxj8at/2u/h8YUNzoPiFJcfMqQwuAfYmUZ/IVZ/tXfC24hEk39sWzZI8uWzBYe/ysw/Wh+1T8Jsf8fmp/+AR/xq1/aa+DU8Cyy+K5bdmzmKXT7gsvPfahHvwe9aF8e/hT4k1e10LRfFf2m+vJPLgi+w3Cb29MtGAOnc18SJJ5/iH4omubfyJn1q8aSLeG8tjM+VyODg8Zr4dwW6eH7IREHcpZj/tFjn/D8K0Kwsp4CZXVSF4zXiG3gjdljxgV8Q4408SymPGXiQtx3xj+QFfsbzy/8In4gtzKxjTUUZULcKTGASB6kAfkKaQY5amlGODTy54Bp3GOTTyAjANO4xyaeTPANf8ABUjxE1j8EfD3h2GSZZNW8SI8m0DY0UMEpKtzn77xEcfwnkd/+CTHhRvEX7Ymlauts0o8L6FqGollkCiLdGLXcQT8w/0oLgZOWB6Amqqqqqqv25P2nbH9lz4G6j4nsb2IeLdb36f4Zt2CuxumQkzlG6xwr87Egru2KfvitH0nxV8SvGVtpFgLzWfEPiK/2hpXaWW5uJWyzuxySSSWZj7knrX7PXwQ0H4A/DSw8D6QI5r04uNWvVBH2y7ZQHcA8hQFCqOMBR3JJxngUicY71HGMcV+zl+2Z8Ufi/8AtF3/AMIfEmg+FrbR7T7fsnsbWdLg+QxCZZ5mXnHPy/TFJGAMUkeelftz/tU/EL9mEeCP+EC0bw5qH/CS/wBofa/7XtppdnkfZtmzypY8Z85s5z0GMc5+EfifUfHvwq8GeOdYhtor/wAReHrDUbqO2VliSWe3SRwgYsQoZiACScYyT1pYhjmv27v2rviJ+y6PA/8AwgGi+HNQ/wCEm/tD7X/bFtNLs+z/AGbZs8qWPGfObOc9BjHOfhF4o1Lx/wDCjwV461iG2hv/ABH4esNSuorZWWJJZ7dJHCBixChmIAJJxjJPWhDntX7cn7Z3jL9mvxP4V8EfDXQtC1XWdXs5L2+j1a2mmEcRkEcAjEUsZ3M6TZzn7q46nH7DX7U+uftReDPEV54y03SLDxF4d1GOOaHS4ZI4DbTR5hfEkkjbi0c4POMKMdTgxA9xRhBHApoiOhpo8jBFNECOKeL0p4weCKePHSnjBGKdMcV+0D+w18IfjXHcazplkvhPxQyuyalpkCrHcSHobiHGJBn+JSr+rHpXxu/Z5+JvwC106T460Rls5pWSx1W3Be1vQBn5Hxw2OqMAw9Mc1+xP/wAFD/Hf7M97ZeBvGz3Xib4aySKjWLPuuNHUsS0loTwRlizQt8rEcFCST4F8feC/if4VsPG/w+8Tafr+hanHvtr6ymEiN6qe6upyGRgGVgQwBBFVVHHTNEjsa+Mfxr8GfBXwzJrniW8jkvJEYafpccoE9647KOoQEjc5GFB7kgH4x/GPxZ8a/FkniXxNP5cMW5NP0+NyYrKIn7q+rHALOeWPoAAP2Vv2ST4+jg+IfxPsJY/DjLu07TmLRvqPpIxBDLF6YwXPP3fvWtpa2NrDY2NtFb29vGscMMSBEjRRhVVRwAAAABwKqqqv2sviwmjaQPhroV2Pt+pIH1NkbJhtz0jPoznk85CjkYcGvhH8Ob74oeNbPw7bqVtEPnahPnHk26kbj/vHIUe7DsCRpum2Gj6fbaTpdrHbWlnEsUEMYwsaKMAD8BVV+0p8BvDX7Sfwc174U+IykLX8Jk029KFjYXqAmCcAEEhWOGXI3KWXI3ZrX9E8W/C3x1faBqX2rRvEvhXVHhkaGVo5bS6gkxuR1wQVdcqw9AQa/Yc/aesv2qvgPpfja7uLceKdLb+zvE1rEnliK8RQfMVeySoVkXGVBZlzlGxVVVVX7RH/AAT9/Zo/aSnuNc8U+Df7D8Tz7i/iDQSLO5lcj70ygGOc5x80iM2FADAZFftCf8ElP2ivhJDeeIPhsLf4meH4GZlTSomj1OOMDq1oc+YecAQtIx67QKtrrxn8N/FS3Nnc614X8S6JcHbJG8tjeWMy8HBG143HI7EV8CP+Cuvxu8A+To/xj0i1+ImlLsQXW5LDUIVGBneiFJsKM4dAzHrJzmvgR+3B+zf+0R5Vj4I8eRWOty7ANC1sCyvdzYwqKWKynJx+6Z+fwzVVVVRI6Cjz3qqqqqqqiQB1o896qqqqqqOMdaJz3qqqmPGAaqqpjgYBqqqqqqokAUee9VVVVVTEAYBqqqqqqYjGAaY54BqqqqqqY4HBqqqqqqOMdaPNVVVVVTYJ4NVTc8A1VVWAeM0xycA1VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX/AAUb+Hf/AArz9rbxh9ntfJsfE3k63afu9u/7Qn75uFUHNwk/Iz7ktur/AII2/En/AISv9mHUPANzcbrnwP4gnhhiznZa3IE6H2zM1zx7e9VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV8eP2bvg5+0n4Wbwp8WvB9vqiIrfY76MmG8sHIIDwzL8ykE52nKMQNysOK/aw/4JWfGP4GteeLvhStx8QfBMKvK7W8X/Ey0+NQCTPAo/eKBn54s/dJZUGM/Cr4wfEr4I+K4PGnwu8X3+g6pFhZGt5P3dzGCD5U0Z+WWMkAlHBGQD1AI/ZU/wCCpvw4+Kv2LwV8cY7XwR4rk2xRajvP9mahJwM7zzasST8shKYX/WZYLUUsVxEk8EqSRyKGR0OQwI4II6jFVVV9DQII4qqqgQRjNV8XP2b/AIZfGGB59Z0v+ztYPKatp6LHOSBwJOCJF6cMM46Eda+L/wCyd8TvhU8uoWtmfEmgoCw1HT4W3RL/ANNYuWj6ZyNy4I+bOQPBfj/xn8OtVGteCfEl7pF3wHaB/llA6CRDlZF/2WBHtXwo/bx8Pat5OkfFfSv7Hujhf7TslaS2Y+rx8vH+G4fQVomu6J4l0yHWfD2r2mpWNwMxXNrMsqN9GUkfh2qqqqqqr6GgQRiqqqqtd8M+G/FFsLPxL4f03VoF6RX1ok6jkHo4I6qp/AeleKf2S/gZ4ltblYfBsWk3s0RSK6sJ5YvJbJIYRBvLPJ7r047DHxD+LnxV+DvxC8QfDHx54c0iXVfDd69lO0aSReaUPyyjLEFXXa64AyGBr9nfwbpf7Rvw6j8a+GfFVvY39tdPa6ppNxblmspByuJARvVkKsG2gckdVOLj9lDx2JmWz1/QJIh91pJZkY8c5AjbHPvTfsx/FIMVW20xgDgMLwYPv0zVz+zl8X4Zmih8MRXCr0kjv4Arcdtzg+3Iofs6/GQ8Dwd/5ULb/wCOVP8AA34s28rQSeB79mQ4JQo6/gysQfwr4S/Cj4jaH8SPD+r6r4Rv7a0tbwPNNIoAQYPJ5r9ojwjN4W+KGqTiIi01pzf275J3GQ5k57ESb+PQr0yK+HviuGwA0a/lCKXJgdjgAnqp9OeR7k+1WmuNFHtWTHHrXiPxNZ2FrJd3twFRRwM8sfQDua1jUpdY1KfUZhhpmyF/ujoB+WK/Zn8MSeGfhfa3dzAY7nW5mvWDKAwjICx8/wB0oocf759aaQkYzTSDuaaXjANNJnjNPIAODTvkdaeQAYBr/gqr4nW68WeAvBySrnT9Ou7+RAxyfPkSNSR0x/o74Pu1f8EUfB6Xfjf4nePnhTdpmlWWmRSMh3EXMskrhT0x/oqZHXlaqqqqqq/4KCftM/8ADS/x7v7/AEO78zwj4U8zSvD+yTclxGjnzLsf9dmAYd9ixg8iv+Ca3wDjjtbn9oDxFZq0krS2Hh5JIslFHyT3Kn1J3wgjkYlB4NUi55qNM0iA8Cv2G1z+3DrI/wCwz/6GaSPNInYV/wAFd02j4T4/6jX/ALY1+zWgP7Ovwsx/0JWj/wDpFFSxZr/gsEoUfCQA/wDQa/8AbGv2aYyf2cvhVj/oSdH/APSKKhFkV4Cay/ay/wCCl97r93bRan4Z8PX09wieWWje009PJt3wysNrziFyGwD5hHBIFfscXd3+zH+3brXwV1ydksdWubvw/wCZccmQhvNspvl/ikCRqOMfvzkDsYR0x+lGLjimjPSmjB4xTxelOnY1JEMcU6cYNSR8cVJGCMU6AcGvGXgvwt4+8PXnhTxloNnrGk3ybJ7S7iDq3oR3VgcEMMFSAQQRmv2t/wBjbXf2fbz/AISvwtLdaz4Gu5dqXUiZm02RjhYp8cFTwFkwAT8pAON37EP7ZPir9lD4hRC6vLq++H+t3CjxDo6r5mBjb9pgUkBZlAHcB1G1uilfBnjXwr8RPCumeNvBGu2ms6HrFuJ7G+tn3JKh/UEEEFSAVYEEAgiiSehokkYzVftB/tVeEfgzby6Fo72+ueLSo26ekmY7TP8AFcMpyvHIQfMePughq8Z+NfFvxK8STeJPFurXGq6pdkIGf+EZ+WONBwqgk4VQBz7mv2ZP2NHL2fxB+MulLsA8yy8O3UPJPZ7pW/PyiO43YwUKRxxRrFEioiDCqowAB0AFVVVXxV+I+mfC/wAH3PiO+KSXB/dWNs2f9InIJVeOg4JJ9Ae+K1TU9b8X69NqV/LPqGqancZYgF3lkY4CqOT6AKPYCvgV8KoPhZ4MjsrmNG1rUds+pygA4fHyxAjqqAkdSCSxGN2BVVX/AAV4/ZWa/srf9qXwXpqGezWGw8WxQxAM8X3Le8Y/xFfkgbqdpi6KjEfsL/tV6l+yh8a7PxPdTyyeENc2WPie0RN5e23fLMi/89ImO4dyN6/xVp2o6frGn2uraTf297Y3sKT21zbyiSOeNwGV0ZSQyspBBHBBBqqqqqq+N37LfwE/aJ02Wx+K/wANtJ1a6aMJFqqRfZ7+3xnb5dzHiQAE52ElDgblI4r4/wD/AARe8b6F9o1v9nDxtD4msooyy6H4glS1v2Ix8sc6qsEhPX5xCB6mviJ8LviL8JNfbwv8TPBOseGtUVdy22pWjQs6/wB5CRh1/wBpSR718C/+CjX7T3wPa205fGbeMdBhZQdK8Su95tjA27Ipy3mxALjaAxRSB8pGQfgb/wAFXP2cviiINL+ID3Xw21qXI8vVJPtFiSOgW7RQBxnmVIxxjJJGdL1XS9d0231jRNTtdQsLuMSW91azLLFMh6MjqSGB9QcVVHGOtE5GAaqqqqqqqiQB1o896qqqqqqpjngVVVTEAYB5o896qqOAOtHnvVVVVVUcY60TnoaqqqqqjjHWic9Kqqqqo4x1pj7/AEqqqqqqpjk8GqqqqqpumAaqqqqqjgDg1VHGOtHmqqqOAKPPeqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/wCCyHw7yPh58WbS1/5+dEv5/L+k9su4L/19HDN/uj7xr/gi58Sf+Ec/aA8UfDW5uNlt4y8P+dCmf9ZdWcm9Bj/rjLcn8Peqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv2t/wDgmp8Fv2lft3i/w7bxeCPiBPvkbWLGD9xqEh3H/S4AQrFmYkyriT1LgBa/aF/ZY+Nf7MHiQ6B8VvCUttbSyMlhrNoGm0/UAM8wz4AJwu7y3CyKCCyLkV+yx/wUK+M37Nstl4bvrpvGHgSBfLOg385V7VOubWfBaIg/wENGQSNoJDL8Av2lvg/+0t4YPiT4W+JkupLdU+36VdAQ32nMwyFmhycDqA6lo2KsFZsHFVVA471wRwaqq+hoEEdaqviv+yj8Kfil52of2Z/YGtyZP9oaagTe3rJH91/c8Mf71fFf9lH4q/C3ztQ/sz+39Fiyf7Q01C+xfWSP7ycdTyo/vV4L8f8AjT4daqNb8EeJL3SLvgO0D/LKB0EiHKyL/ssCPavhd+31p88UWl/Fzw+9vMMKNU0pN8bdsyQk7l45JQtknhRXhPxn4U8d6SmueD/EFlq1k/Hm20obYf7rDqrf7LAEelVVVVVX0NAgjFVVVVV/wV7/AGY7q/t9O/ad8Iab5ps449N8UxwoSwjzi3uzgYwpPkuSc8w8YBI/Y3/aGk/Z8+K8N/qk8x8L+IFSy1uFX+VF3fu7jb0LREsfXa0gH3qtby2vLeK7s7iOeCdA8UkbhldSMhlI4IIwc0svbNLICODQk9DQlPrQlyOtfF34Zad8U/DY06SZLbUbNmksLornY5GCjd9jYGcdwp5xg+KfCPiLwXqj6N4l0uWyuVGVDjKyL/eRhwy+4PXI6g1a+INbsoRBa6pcJGowqb8hfoD0q7vbu/l869upJ5P70jE4+noK+C/wH1bxdqNr4h8V2D2nh+F9/lTqyPe4wQqjghDnlu+MD1CuqII41CqowqgYAHtTSE8ZppB600oAwDTSEjANNIOxp5c8A0756mv+CgXitfE/7TniC1hmaWHQbS106NjKXA2xCRwoP3AJJXBUdwT3r/gjl4OXRP2atc8WSujT+JPFM5Xa5O2GCGKNQwIGG3+ceM5BXnsKqqqqr/go38fG+Av7M2tzaVcIviDxix0LShuG6MzRt50wHXCQq+D0DtHnrXww+H2t/Fb4gaD8O/Dq/wCna5eLAshXcIU6ySsByVRFdzjnCmvB/hPRPAnhTSPBnhu1+z6ZotnHaWsZ5OxFCgse7HGSepJJPWlXccZqNMmkTAwOtRoMV+wyob9uTWQP+oz/AOhmkTPAqOMEV/wV9UAfCXH/AFGv/bGvh9/wVD8efD7wF4a8BWXwt0C7g8N6Pa6bFcS3syvKsEKxB2A4BIQEgetD/grr8RhwPhD4b/8AA+f/AAr9qv8Aa78R/tWDwuPEHg/TdC/4Rf7Z5P2Od5PO+0eTu3bumPIGMf3jX7MyA/s3/Cg9f+KI0b/0ihr9rz4sS/BL9nfxj46sZ2i1RLP7HpbIcMt1cMIo3XkcpvMnXoh+lf8ABJH4Uvo3w18U/Fy/t2WfxNqC2FgXI/49rYEu645AaWRlOe8I4HU/8FSPA2qfDL48+C/j/wCFA1pPq8UbG5ijKmPUbB0McjOv8RjMIUHBxCcE44+HPjPS/iZ8P/DnxB0Rl+xeItMgvoUWQSeV5qBihI/iUkqeBggggdKaM9MU0YIxiniBHSnjI6U8YIwKdMcCpIwRxTpjipI8dKkQEcU6ZGO9a7omk+IdKu9C13Tre/06/haG6tbiMPHNGwwysp4IIr9sj9lW9/Z78WDW/DdvcT+Bdbm26dcSNvNnMQWNrI3XICsUJ+8o6kqxr/gmV+2vL8FPGMPwS+JeuN/wgXie6WPTri6lPl6FfOxAYE8JBKzASZ+VW2yfKPMLVq+r6VoGmXOs63qNvYWFnGZJ7m4kEaRKOpLHgCv2gf24L3U2uvB/wXme0s+Un19gVml5Gfs6nHlrgY3t8xzwFwCfD3hvxT4/8RRaH4d0271nWdRkZlij+eSRuWZmJ/ElicDkk1+zt+yF4c+FS23izxsLfWvFqHfER89tp5B+UxAgFpOhLsODjaBgs1VVVWp6lYaPp9zquqXcdtaWcTSzzSHCxooySfwFfGr4rX3xX8WyajmSLSbEtFpdsxxsjJ5dh/ffAJ9MAZO0Gv2Uvg2t1LH8UvElnmKFiNGhkAIZxlWnI/2SCFz3y3ZTVVVVrei6R4k0W/8ADuv6db6hpmqWslre2lxGHjuIZFKSRup4KspIIPUGv20v2XtY/ZV+M9/4N8u6uPC+qbr3wzqMy5+0Whb/AFbMODLEf3b9CcK+1VkUV/wSM/bIfVbQfsr/ABI1vddWcbS+Dru7nZnliUZksQWz9xQXjH90Oo4VRVVVVVVVePPhx4B+KOgTeFviL4N0fxJpU6lXtNTs0uEGf4l3A7WGAQy4IIBBBANfH7/gjH8M/E0N3rP7PHi648HaltzBousSSXunuwGNomO6eIHg7j53Qjbzx8eP2Mf2jv2cZWm+Jfw4vU0jcwi1vTsXthIBnkzR58okAkLKEcgE7cCvg/8AtDfGj4CamNS+E/xE1fQVaYSz2UU5e0umAxmW3bMchxxllJA6EV8Dv+CyoP2LRP2hfh5/djl13w4fqN8lrI3+6WKP/eKp0WvhL+0H8FPjtYNf/CX4k6L4j8qPzJra3mMd1AmcBpbeQLLGCeAXQA9qJzxmqqqqqqqokAdaPPeqqqqqqqY54BqqqiQBR571VUcY60xzwDVVVVVVTHjANVVVVVVMeMA1VVVVVMeMA0xyeDVVVVVUxwODVVVVVUcY60TnvVVVVVU2CcA1VMRjANVVVTEHgGqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr/gpN4Bbx3+yP4slt7dJrvw1Lb6xbhowxXypAsrAnlSIJJjkc4yOhNfsTfEFvhf+1l8LPGHnJFCniKCxuZXIAjt7vNrM5yrcCOdz0zxwQcEVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV4z8E+EPiJ4bvfB/jrw3p+u6LqEZjubG+gEscgIxnB6EdmGCDyCDX7XP8AwSE8SeF/7R8e/suyy67o8MZml8J3cpa/hA+99mkPE4A5EbEScYBkYgV4X8V/EL4OeOItf8Lavq/hTxVoF00fmRlre4tpUJWSKRDzjIKvG4wRlWBBIr9lP/gq94W8Zf2f4G/aQhtfDeteX5a+J4PksLtxwDPH1t2YYywzGWyf3YwosL+w1awttU0q+t7yyvIVmtrm3kEkc0bAFXRlJDKQQQRwQQaqqB7Zqqqhx0NcEcGqqviv+yj8Kfil52of2Z/YGtyZP9oaagTe3rJH91/c8Mf71fFT9j34sfDp5L3R9PPizSFPy3Wlws0yDt5lvy69ySu9QOrCvD/iTxL4L1lNY8NazfaPqVsSomtpWideeVOOoOMFTwehFfDP9vjxVpHlad8UtCTXrfdh9RsQlvcovcmIARyH0A8v6mvh38cPhd8VI0Twd4ttLi8YEnT5m8m6UDqfKbDED+8uR71VVVVX0NAgjGaqqqq8VeF9A8b+GtU8HeKtMi1HR9as5bO/tJchZoZFKupIIIyCeQQR1BBr9rj9mfxF+yz8X9Q8Aak8t3o1zuu9A1JwT9ss2YhNx2qPNTG2QAYDDI+UqT/wT2/awhuLWz/Z/wDiHqaxzwLt8MX08iqsicAWR4Hzjkxkk5GU4IQMsoIwDQkBHWhJ6NQlOKEo6ZoSg8ZrWtD0LxJZnT9f0iz1C3OcR3MIcKcYyM9D7jkVffs4/CS8kEkOh3Fp1ysF7Jgn6MWx+GBXhz4O/DLwtNFdaV4VtmuYsFZ7lmnYMB94byQp78AfhRlHY0Zcjg00hI4NNIDxuppB2NPKDwDTyEjk0z54FHnvXxS8XN8QPiV4p8bmSRl13WLq8iEjElI5JWZF55AVSqgdgAO1fsQeBH+G37Jfwu8KzRyxzf2BFfzxSoyvFLeM13IjK3IKtOykHoRiuCODVVVVVf8ABWP41D4l/tKv4B0y+87R/hzZjT0CvlDeyhZLphyRkHyom4B3QEHOAa/4JhfCXz7vxH8adTtvktv+JRpLFv4yA9w+32UwqD0O5x2rGeBSLwAKjQYqNAeaRM4Ar9hYZ/bm1kA9tZ/9GGo4wRikQHgV/wAFglCj4SD/ALDX/tjX7M0ef2cfhUf+pJ0f/wBIoqWMY6V/wWOXA+EP/cb/APbCv2Y4wf2bfhOf+pH0b/0ihr/gr98U0ku/BHwTsLlD5Cya9qUeCSGbdBbc9Acfacjk4ZTwDz8O/AH/AAVv+GXgrSvA3w90FdH8P6XCVsrJT4cPlK7F2yZCXLFnZiWJYknPNftHfDj/AIKY+PvhtdXf7QPh5tT8KeFmbV52jbRFa18qNw8v+iESsqxu5IGRjkj5QR/wSY+KS+KvglrPwsvLlGvPBGpmS2jwQRZ3ZaRevBPnLcZx0DLkDOS0fGMU0YIxiniwOKeMHipIuOOtOgPHepEA6VJGCOBToCMH8KkTHFSp6VKgI4r4mfDvw18VfBGreAfFlmtxp2rW5jbK/NE/VJE9GRgGB9QK+L/wu1/4NfEXWfh34iVnuNKuCsNx5RjW7hPMcygk/K64OMnByM5Br9kf/goKLb9mjTrH4o6XrWseNPDx+wWbyoYxrFsoHkzvOQeVTEbuQzMUDfMWYj4t/Hj4i/GfU2uvFesGOwQ/6NpVqTHawAdCEydzdfnYlucZAAA+B37L3j/40XEWoxwHRvDayATardRn5x3EKcGQ+/Cjue1fCb4M+BPgzoI0XwfparNKB9s1CYBrm7Yd3fHQc4UYUZOBkkmqqqqv2m/jcnjC9fwD4Wud2jWEx+2XCOrLezKRgKRn5EIPOfmPOOAT8CPhBd/FTxQhvIZE0DTXV9QmAxv7iFSCDubBGR90c+mbS0trG1hsbOFIYLeNY4o0GAiqMAAegAAqqqqOMda/bI/Zf8OftV/B+98GXscVv4i0zfeeG9SK4a1uwpGwnr5Ugwjr0+62NyKRfWPjb4R+P5LG6W98O+LfB+rYJSTZPYXtvJwyupOGR0BDKccAg96/YT/a+0L9rP4Swahd3MMHjnw7HFbeJ7AKI/3pHy3MSgn91LtYjptYMvQAmqqqqqqqmhhuIXt54kljkUq6OuQwIwQQeoI7V8ef+CZX7K3x0nu9bTwlL4J8RXWWbU/DLLarJJg4aS2IMLZY7mIRXbu/evjp/wAEkv2mfhYlxqvgGKx+JWjQtw+kL5F8EyQGa0ckk/dysTyEbvQEiaDxd8PvEvlXEOr+G/EOkTBgrrJZ3dnIACDg7XjbBBHQ8ivgr/wVN/ad+Fn2PS/Fmr2vxD0S2j8v7Prq4uwvYi7QeYz5/il83gkehHwV/wCCpv7MXxU+yaX4s1e6+Hmt3Mnl/Z9dXNpu7EXaDy1TH8UvlYII9CdL1XS9d0631fRNStdQsLuMSW91azLLFMh6MjqSGB9QcVVVVVVEgDFHnvVVVVVVVMewNVVUcY60TnvVVVMc8A1VVVVVUSAOtHnvVVVVVUcAdaPPeqqqqqOMdaY8daqqqqqqY5PBqqqqqqY8YBqqqqqqOMdaPPeqo4J4NVVUenWjzVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV488JWPj7wN4i8Cam+2z8R6TdabcNgnEc8TRscAqejHoQfcdaurbUdD1OWzuUltL/T52jkXO14ZUbBGR0IYfmK+EXjuH4pfCnwb8SYERF8U6DZamY1GBG08CSMmAzYKlipG44Ixk9aqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv2sv2CPgn+1hYy6prVl/wAI143SPba+KdNgXzmITaq3MeQLmMYThiHAXCOgJz+0v+xf8dv2VtSC/ETw19q0GeTZaeI9LDz2EzHO1DJtBikIUny5ArHBIyBmv2bP21Pjh+zDeRWngzXU1PwyZjJc+G9UzLaSFvvMmCGhfvuQgEgbg4yD+zN+3P8AA/8Aabs4dP0PWk8P+Lto8/w3qkoSfPPMD8LcLwT8nzAYLKuRmqoHIxmqqqBxXBHBqqqviV+z38J/ivHNL4n8MxRalKMDVLHEF0pHQlgMPx2cMPbgV8TP2EfiB4bjm1L4fapb+J7KMZ+yuBb3YHfCk7HxjswY9AprVdI8Q+EdZbT9Y0++0jVLKQExTxtBLEwOQQDgjpkH8RXw5/bC+MvgFY7K71hPEunJgC31YGR1AGPlmBDjt94sBjpyc/Dr9tr4R+MzFZeJZLjwnqDjlb4+ZblsDIEyjA78uqDj1IFafqOn6tZRajpV/b3lrMMxT28okRx0yrKSDyKqqqvoaBBGM81VVVV+1p+zD4T/AGqfhVd+BtbMVlrNpuuNB1fygz2F1jjJwT5T4CyKOoweqqR498BfEH4I/EG98G+MtMvfD3ifw9dAsoco8TqQ0csUinlT8rJIpwQQQa/Yy/bPs/jDZ23w2+JN7DbeOLWLFtctiNNZRRyyjoswAyyDhhll43KokyMA0JTjrQmHQGhMCODXmgjAIrzAR1rzB6ijIOgoyjHBFGYY4NNKCMA00hIxmmkB4zmi5PAo896r9p7xp/wr79n7x34oSTZNFo8ttbvjOya4xBE2NrZw8qnBGDjnAyR8MPBF58S/iR4V+HenlhceJtZtdNjZRkoZpVj3ewAbJJ4AGTVpa2thaw2NhbRW9tbxrHDDEgRI0UYVVUcAAAAAcCgw7Gg2RWQRwaqqr4sfETSPhF8MvFHxO1yJprLwvpVxfyQIwVpzGhZYlJ4DOwCDPGWFa9rOveO/FeoeIdUkm1HWvEOoy3VwwBeS4uZ5CzEDklmdj6kk18A/hnb/AAg+D3hb4fRQ+XcabYKb7Lht13J+8nOR1Hmu+OuBgZwKUZOM1GuTSLkACo1GABUaDGa/Zh+KfgL4Oftf6741+JGvf2Ro0U2rQPc/ZZrjEjyEKNkSO3OOuMUv/BQz9jsDH/C4f/Lf1L/5HrwP+25+y78RPFWmeCPB3xP/ALQ1vWJxBZWv9i38XmyEZxvkgVV4B5JAr/gsMuB8I/8AuN/+2Nfsypn9m/4UY6/8IRo3/pFDSRiv+CyigD4QAf8AUc/9sK/ZhiB/Zr+Ex/6kbRv/AEhhrwgG/bK/4KUHW0Zr3w5Za816rpMHjGmaaAISOwSZoogQvecnPJahCMVrOh6Z4h0e+0DWbVLqw1K2ktrqByQJYpFKupxzgqSOPWv2KtX1D9l79vG9+EHiG+22mpX934WupGl2pLJ5mbSUKQAxeSOJVyAcTnHUqWi46U8QxxTx44p4wRxTp2NSR56VIgI96dB0qVOwqDxV4e1DxHeeE7HUo59U0+BZruBAT5CscKGPQMeu3qBycZGZFBGKkXtXjz4p+CPh9Gy6/rCG825Sxt/3k7eny5+UHsWKg+tfHPXvDXxw8W6Z4n1fwbZRtokDwWZlPmuyMwb950VsEEquMLubk5zWt6/ovhuwfUtc1KCzt0B+eRsE47KByx9gCa/Z+/Yq8F2+maX49+IWq2vilr+2iu7KxtW3WISRAyszdZ8gqR0XrwwIIgggtoI7a2hSKKFAkcaKFVFAwAAOgA4xVVVVVftMfHpNHt7j4d+CtSB1GUNHqt1Fz9mQjBiRs8OejH+EccN934efD/XfiV4ntfDWhQMTIwa5uNmUtYsjdI3I4A7ZGTgDk14H8FaJ8PvDNn4X0CHbb2q/NIygPPIfvSOQBlj/AIDoBVVVHAHWi3YUWB75oknjNf8ABUj9i9viV4euf2jvhzYs/ifw7Y/8VBYxqP8AiYWMS588ZI/eQoDkAEugAHKKG/Zt/aD8bfsx/FnSfit4Hl8ya0zBqFg8hSLUrNyplt5CAcBtqkHB2uqNglRXwW+Mfgf4+fDXRfij8PdUS80nV4dxXpJazDiSCRTyro2VI78EZBBNVVVVVVVVfFr4AfBb47ab/ZXxb+G2ieJY1jMcc11b7biBeeIp0Kyx/eb7jDBOetfG/wD4IpeGdSR9U/Z5+JtxpFztOdI8UKbi3dif4LmFRJEoHZo5ST/EOlfGr9jz9pD9n3zLn4nfCvVbLTEkZF1e0VbyybHQmeEsqbhyFk2tgHgYIHwh/aA+MfwG1hNZ+FPj/VNCfzN8trFLvtbg8A+bA+Y5MhQMspIwMEYr9jT9vr4d/tUWEHhjVUt/DPxGghZrnQ2lZo7xUALT2rsBuUgljESZEAb7yr5hqqqqJAGBR571VVVVVVTEAYB5o896qqOMdaYg8CqqqY4GBVVVVVVRxjrROe9VVVVVVMc8A1VVVVVMQeAaJz3qqqqqqY4GAaqqqqqjjHWic96qqqqqmOeAaqmxjANVVVTdMA1VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX7avge3+Hn7VXxL8M2kCQwf2299DEm0LGl2i3SqoUAKoE4AXHAAHOM1/wSk8ef8Jv+xh4Y0+WfzrnwlqF9o87mTcflmM8YIwNu2G4iUDnhQc84FVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVWt6HonibR7zw94j0ex1XS9QhaC8sb63WeC4jYYZJI3BVlI4IIINftYf8Ee9A19L3xv+y3fpo+pszyy+E9Ql/wBEnJIOLadjmAjnCPuQ5ADRgc+NvAfxF+DfjObwt468Oax4V8S6TMH8i6ia3mjZW+WWNv4lJXKyISrYBUkYNfswf8FW/iP8OfsXhD49W91448PJ5cKausoGp2SDgs7EYu+MffKuTkl26V8JPjV8Lvjr4Wj8YfCvxlYa9p5wJhC+2a1c5+SaJsPE3BO1wMjBGQQTX0NAgjFVVUDjjNVVVQIIxmvF/gXwb4+03+yfGfhrT9Ytlz5a3cAcxEjBZG+8jY/iUg+9fEn9gPw7qck+o/C7xI+jyPkpp2o7poAeeFlGZFXp94SHrz2r4jfAD4sfCxpJfFXhG5+wJkjUbQfaLYgHGTImdmewfafavB3xD8c/D68N94K8V6jpErHMi205VJcDA3p91+vRga+H/wC374s054rP4keGLTWLYAhrzT/9HuP94oSUf0wNnr25+Hv7TXwZ+JXk2+j+LoLDUJcBdP1Ui1mLHHyruO125xhGboaqqr6Ggc8E1VVVV+3F+w74V/as8KnW9DFro/xG0e3K6VqrLtS7QZItbkgZMZJO1+TGSSMgsreJvDPjX4V+Nbzwz4m03UPDvifw7ebJoHJintJ0IZWVlP8AusrqcEFWUkEGv2Sv27dJ+Icdj8OPjDeW+meJ1SOGy1eRwkOrOW2hWGAIpjlePuuc42nCnJ7Gt56ZrfntXmAjGa8wdjRkBGM/pRkGODRkB460XyMAUXJ6GiSeM1VVX/BTf4y2I0/RPgbo9zHLdvMuq6yUfJgVQywRHHdizSEHBAWM9Hr/AIJP/B+8+If7Ttv47uNOaXRvh7YyX1xMThFupkaG2Q8HLHMsgHH+oJzxtaqyRwDQYUGyMA5rIIxmuCODVf8ABWPxs/hL9jzVtHifa/i7W7DSQwVsgBzdNggjGVtCDnIIJGOcj9jXwbb+OP2lvA2lXlu0traXrahN+6Lqv2aJ5k3YIwpkjRcnjLAEHODSLkcVEowBUa55qNQeKRQcAdq1T/gm5+zJrmq3us3+leITc39xJPMV1Z1Bd2LNgY4GSaT/AIJifsrsOdI8R/8Ag4f/AAr4c/8ABPv9nX4Y+NtH8f8AhXTNdTVtDuRcWjT6o0iBwCOVI5GD0r48/st/Cj9pQaEPidZ6lP8A8I79o+w/Y702+3z/ACvM3YBz/qUx6c+teDPCekeBvCOieCdBSRNM8P6bb6fZLK+9xDDGsaBm7naoye9ImeBXx9/ZT+Ev7TI0EfFGy1Of/hG/tP2H7Fetb7fP8rzN2Ad3+pTHpz61ovw+0Pw98OLH4XaNLeWukabokej2jpNmeK3jhEKEOQfnCAfMR15r4B/sZfA/9m3xDqPin4ZaRqMWpapZfYppr68Nwyw71cqhIyoLIhOOu1c9BQiyOleTxjAr4lfsM/Aj4p/FY/GrxDY63b+LPNtZhd2GomECW2VFhk2YI3KI0H/ARTRnGBTxgjGKnVI0aSR1VUBLMxwAB1Jqfx/4Bivo9Mk8ceH1vJV3R27anCJHXnkLuyR8p59jUU1te26XNncRzwyDKSRsGVh6gjg06ZGKlTHIqVARxXx8/aWttHW78E/Du8WbUceXdarE4KW2c7kiI6v0BbovOPmHy/sbQ3F3J4x1q9VppZ5LVftUp3O7nzmk+Y8kklCfU4rx18fPhp4IVoZ9bXVL1Tj7HppEzg/7TZCL1HVs+gNeO/2nPHnikzWegOnh+wfgC2bdcEe8vVT/ALoUj1qx0/V/EOorZ6daXWoX1y5IjiRpJHY9Txkn3NeAf2XNUvGi1Hx/d/Ybcc/YLZw0zezPyqD2GTjuDzX7XHw98SfDH44674Y12/uryzdvteizzsTusZSTGF7fIQ8ZwAC0bHHNf8ElPjyPiZ8Arj4Waxeyy658N7hLdDKc79On3NbEH/YZJotvO1Uj5+YAVVVVV+0N8fU+Hlu/hDwrIJPElzFmSYrlbBGHDc8FyOg6DqewOgaBr/jbX4NE0S0m1DU9QlO1c5LE8szMegHJLHpyTXwf+E2j/CjwzHp1skU+q3KhtRvVXmaT+6pPIRc4A49SMk1VUSOmaLZGAaLA9OaJJ71VV/wUx/Yil+C/ie6+O3w00pB4D8R3v/EwsrdDjRb6UkkBQMJbyNnYR8qO3l4UGMH9gn9tPXP2SfiQINamu9Q+HniKRYtd0xXJ+zMSoF9CvP71FGCo/wBYmVPIRk8P+INC8W6FYeJvDGr2mqaTqlulxZXtpKJYriJxlXRhwQQRzVVVVVVVVVVX7bn/AATA8A/GzS734gfAzR9M8JePraGSQ6faQpbWOuPkHbIowsMx+bEoADM37zrvXUdN+InwV+ILWGo22r+EfGPha+Vtjhra6sbhCGVgeCD91gw4IIIJBBr9gD/goRZftBWlv8KPi/qFrZfEm2jP2W72LBDr8ajJZFUBUuAAS0agBgCyADcqVVUSMYBo896qqqqqqpjgYBo896qqpjngGqqqJAHWjz3qqqqqqpiDwDVVVVVVTHAwDVVVVVUSAOKY5GAaqqqqqjjHWicng1VVVVVMeMA1VVVVVHGOtHnvVUxyeDVVVYBonJ4NVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV/wV+8Cvo3xv8KePYbfZbeJvDxt3bLHzLi1lYOeTgfu57cYGBxnqSa/4IfeOHe0+Kfw1uH+SKSw1a0UK3VhLDOSc4H3LfAABPzZJwMVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVXxv8A2dvg3+0X4Zfwt8W/BFjrUIUi1uypiu7Js5DQTrh4zkAkA7W6MGBIP7Vn/BKP4w/BUXHiz4PtdfEbwiuWeG2tj/atiOSfMgQETIAB+8i5yTmNAMn4efE74lfBbxXF4p+HPizVPDWt2UgDSWz7NxU8xyxtlZEyOY5FZT0INfswf8FZfBHjL7F4O/aLsovCmsv5cMXiG1Utp9054zMvLWxJ2/N80fLEmMCtL1XS9c0631fRNStdQsbuMSW91azLLFMh6MjqSGB9QcV06GgQRxVVVA9iaqqr6GgQRjPNVXxB/Za+CvxGElxqPhRNLv5P+X7SCLWQHjJKgGNjwOWQ/rXj/wDYE8a6RG958PPEtp4gjUEizu0FpP3+VWJKN0HJKdfbNeLvAPjTwFfHTfGXhfUtInyQourdkWTGRlG+644PKkjivAH7Qfxe+Gkcdr4X8aXYsYxtFjd4uYAoxwqPnYOB93aa8Cf8FBNJufKtPiR4Kms3O1WvdIk82PJ6sYpCGVR14dzjtxz4F+Nvwp+JKxp4P8b6beXMgyLOSTyLjg4P7qTD9eMgY5HPIqqoEYwTXBHBqqqq/a9/Yn+G37WPhoG+S38P+NbFQNN8SwWYeVUB/wBROAVM0JycKWBRjuUjLBvjf8BPil+zv4zm8DfFTwzLpl6NzWtwp8y2v4gcCWCUcOh49GUnDBWBUfsy/t9eKvhXBZ+CfijDdeJfCsKiK3ukfdfacgUBVXcQJYgFxsYhhnIbChD8O/ib4E+LHhyLxV8P/EtnrGnyYDtC/wA8DkA+XKh+aN8EHawBwQehBqqqqqqqqr9ov9s34ZfBDSLvTdG1ax8SeMWiZbTSrWcSpBJxhrl0P7tRndsyHYYxgHcPEGu+L/in43utd1WS71vxH4kv9zLFGZJbieRgFjjRcnqVRUUcDaoGABX7Bf7MEP7MHwMstF1eBD4w8Rsuo+I5RzsmIxHbqcZ2xIQp65cyMDhgBVVX0NZOMUGHQGt3GDX/AAWx8ViLQPhX4GhuIm+13mo6jcRCb54/KSCOJino3nTAMe6MB3r/AIJh+H1v/jVr/iGVIXTSfDjpHuJ3pLLPEAyjpjYkgPP8Q69sZ4BpFyQBUa8cVGuBxUSjGaiXNIoJAFIvAAqNBio0zzSJngVHGCMCkQYwKSLI4pYgRgChDxgV5I9KaHjFPFxjFfHr47eG/gnoSyXCpqGvXyn+z9MWTaWHQyyHqsYPfqx4Hcr8Q/i/8QfijfSXfi3xHczwMwaOwido7WHHTZEDtB/2jlj3JqvB3j/xl4A1BNS8I+Ir3TpA4d44pT5U2O0kf3XHswNfAL9obTfi9atoetQw6f4ntIt8kKEiO7QYzJHnkEd0OSOoJGcXs1taW0t5dzxwQQIzyyyMFWNQMlmJ4AAGSTXx+/ael8SrdeCvhvdmPR5E2XmphWSS6z1SPOCsfYkjLdOF+9Vp4j1/T9JudCsNavLbT71t1zbQzMiTnGPnAPzDHY8V4Y8GeKfGl8uneF9Cu9RmJwfKT5E93c4VB05YgcivA/7I107RX/xB1lYkwCbCwOXJ9GlPA9woOc8MOteHPBPhTwZaGz8L6Fa6ehADmJMu+Om5zlm/EmpF5wDX/BTX4UJr3w80f4t6dB/pnha5+yX5VQN1pcMArMevyTbAB/03Y1/wTc+M0vwb/au8Km5vUg0fxix8P6mHUkEXDL5BH90i5WAluy7wSASaqqqq+Pfx7sfhlYv4f8PyxXXie5j+RDhlsVI4kcd2I5VD9TxgNa2viLxv4iW2tkutW1nVrg4BJeSaRuSST+JJPAGScAV8Dfghpnwn0k3l40d34ivogt5dKPliXIPkx/7IIBJ6sQDwAAKJHTNFuwosCMZotngUee9VVVXiXw3oPjHw/qPhTxPpVvqWkavayWt7Z3CbknidSrKw9CCa/bg/Y38Sfso/EOQadb3uofD7WpifD+sSgMVO3c1rOQMLMnzY6CRRvGDvVP8Agmj+3vL8B9es/gd8VtVt4/hvrN05tNRuSV/sC6kyd5YA/uJHwHDYCFvMyo37oZobiFJ7eVJY5FDI6NkMCMggjqCKqqqqqqqqqq/bu/YR8J/ta+EzruhLaaN8StFtyuk6sy7UvEGSLS6IGTGSTtfBaNiSMgsrXtp48+DnxCnsLhr/AMNeL/CGqNG5il8uexu4HwdroeqsvDKSD1BINfsPftR2f7VHwXtPEuoPbw+LtEYWPiSzhG1UuADsmQcYSVBvA6K29MnZk1RIxgGqqqqqqqokAYFHnvVVVMewNVVUcY60TmqqqqqqmOBgGqqqqqqjjHWjz3qqqqqqYg96PPeqqqqqpjxgGqqqqqo4x1onPeqqqqqpj2BqqJwODVVVUxAHBqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/4K++CRrPwK8LeOILZJLjw34iEEjlVBjt7mFw5DH5sGSKAbV65BI+XI/4JE+Mm8L/ALZmk6GquR4v0HUdLJVAwASMXmSSRtH+hjkZOSBjBJFVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV8TfjRo/wm1jSbfxXo19/ZGrKwTU7bEghlU/Mrp1AwVIIJJ+bA+U14T8c+D/HenrqfhHxFZanARk+TJ88fs6HDIeRwwB5FVVVVftJft3/AAC/ZW8S23g34nXmuya5eaT/AGpb2WmaaZ2kgLyRph2ZUDM8MigFhjGSQCDX/D6v9lf/AKEL4q/+Cqw/+Ta+DH/BRb9lD45a3b+FvDXxCbSNbu2VLaw161axadipO1HOY2bgjbvyTgAHIzVVVVVVVVVVVV/wUE/be8Vfsaf8IF/wjPgfSvEX/CX/ANpef9vuZIvI+y/Zdu3Z13faDnP90V8G/HF18TvhB4G+JN9YxWVz4s8N6fq01tCxZIHubaOVkUnkgFyAT6VVVVVVVVVVVVVX7VP/AATq+A/7T6XfiFtOTwd45nX5fEelWwzO+c7rqAFVuDzgsSsmMDfgAV+0p+xT8e/2W9QdviB4We98PswFv4k0pHuNPkycKrSbR5LnskgUntkc1+zp+2Z8df2ZLoW/gPxN9s8PySb7jw9qm64snOSSyJkGFzk5aMqW43bsAV+zD/wUC+B/7SiWmgLqCeEfGsy4Ph7VLkZmfONttOQq3B5HygLJjJ2YBNA46GuCODVVVAgjGaqqr6GgQRjPNVVaro+k67YyaXrel2mo2cwIkt7uBZY3GMEFWBB49a8ffsRfBvxc0154fhvfC17JyDYSb4N2eSYXzgY42oUA498+Ov2IPjJ4WMlx4dhsPFNmp+U2UoinC46tFJjvxhWc8j3xrWga54bvm0vxFot9pd4gy1ve2zwSAdMlWAPY/lXgX9pr41/D8xxaT43u72zQ/wDHlqh+1xEYxgb8sg74Rl5+pz4L/wCChMZjSD4ieAjvB+a60Wbgj2hlP1/5ad68HftM/A7xwyQaT8QdPtrpsAW2pE2TliQAq+aFV2JI4UmoJ4LqCO5tpklilQPHIjBldSMggjqCO9A44rgjg1VVVfFz4MfDL47eEZvA/wAVPCNlr2lSNvjWZSsttIOkkMq4eJ8ZG5SCQSDkEg/tU/8ABLX4r/ByW68V/BwXvxA8HhmYW8MO7VbBeTiWJBiZQAv7yIAk5JjQDJ8E/EDxr8N9bj8ReA/FGoaJqMfHnWkxTeP7rr0df9lgQfSvg1/wU3eGGHR/jl4XknZcINZ0WNQx7bpbdiB7lkP0Svh78X/hj8WLH7f8O/G+l62gTe8UE2Johxy8TYkT7w+8o5NVVVVVXxV+OXwr+CumDUfiL4ws9NeRC1vZAmW6ue37uFcuwzxuxtBPJHWv2hf2/wD4ifFWO48MfDyKbwd4alUpI0cub68U5BEki8RoQR8ic9cuwOB8N/hR8SfjD4gXwt8L/BGr+JdT2h3g0+2aQQoWC+ZK33YkyQN7lVBIyea/YU/4Jtab8BL6w+LnxjntdY8fRRl7DTrdvMtNEZhjdux++uApI3fcQk7dxCyVVVVVVZ9DX/BZvX5bz9oTwf4aBt2g0zwfHcApkyCSa7uAytzjG2GMgYB+YnnIx/wSu0WOLQ/iF4iMqtJc3Vjaqhj5jEaTMSGz0bzRxgfcHXPCgEgVGMnNIAcAUi8ACo14AFRLUSg4qNQTxSLwAKRQAAKiQEcUiDAAqOPPAqOMEUsYxihHkYAryjjAH6U0fGMVcvBbQSXNxMkUUSFpHkbaqKBkkk9AB3r4v/EPUPin8Q9Y8ZX0zmO6nKWUTMSILZTiJAO2FwTgDLFjjJNaVpAuQssy7t33VPp60mgOYfliG3HQCtX0MRAyQJtcdVA614Y8Sat4Q8QWHibQbnyL/TphLA5GRkdQR3UjII7gmvjh+0n4g+KoGiaLFPo3h3YBLaeaDJdPkHMhH8IIGFHHc54xVaTfRaZqdrqE2n21/Hbyh3tblSY5gOqsAQcH2NfDPxV4P8Y+ELTVPBcFraWYUJJZQRrH9kkAGY2VQACOO2CMEcGpFHSpF5xUyjtXxL8FWPxH8AeIvAeotsg17TZ7Myd4mdCFcdeVbDDg8gVd22s+Fdems5zPp2raPdtG+yTbJbTxPg4ZTwysvUHqOK/Z/wDidD8aPgl4J+KUaokviLRoLm5jQ5EVxt2zoCMZCyq65wOnQdKqqr4+ftFWPgm2uPCXgq9iuvET5jnnTDpp/Y57GTsF/hPLdMGwsNe8Ya7Fp9jFdapq2pTbUUsZJJnPUkn8yScAAknAr4H/AAP0n4U6UL6+8q88R3keLq6AyIVPPlRZ6L6nqxGTgAAFuwot2zRbPAokkYzVVVVVVXxg+EXgT46/D3Vfhl8RdJ+36NqseG2Nslt5F5SaJ+dkiNgg8jsQVJB/ao/Zl8a/stfFC88CeJoLi50qdnl0LWWh2RapbAjDrgkK65AdM5U47MpP/BMb/goRaeBPsX7Onx38SSpoM0iw+FtcvZgY9LJ4FpMx5WAnGxycRn5ThCClVVVVVVVVVVf8FmP2abKfR9G/ah8L2CR3ltNHpPigLwZomGLW4I6ZRgYmPJIki7ITX/BL34zz/Cv9qTRvDV5qHk6J4/jOjXiPKFT7QwLWjYPV/OCxDof3zYznBOMdaJzwDVVVVVVVUSAMCjz3qqqmIxgGqqqOMdaJzwKqqqqqqOAOtHnvVVVVVVTEHgGqqqqqpjxgGmOeAaqqqqqjjHWjzVVVVVUx4wDVVVVVUcY60ee9VTHJwDVVVUxycA1VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVf8FAvCP/AAmf7IPxGsI7fzJ9PsItSicW3nNF9lnjmdlHVf3aSKXH3VZicjIP7JHi+LwH+1B8K/FNxJCltaeLNPS6kljZxHBJMscrhU+YkRuxGM8gcHoaqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr7UdO0yET6lf29pEzbVeeURgtjOMk9cA8e1f8Jb4U/6GfSf/A2P/Gv+Et8Kf9DPpP8A4Gx/41/wlvhT/oZ9J/8AA2P/ABr/AIS3wp/0M+k/+Bsf+NftAaX4W+JHwv1TRbTX9Km1G0H23T0W9jy08YOFHPVlLoPdu3WtN1TU9GvI9R0fUrmxu4jmOe2maJ0PqGUgjpXgD9sjx94eMVl40tIPElmpAMpxb3KLjHDKNrevzLk925yPh9+0L8LfiMIrXTPECWGpOuTp+o/uJM+ik/K54PCknHJAqqqv+C1f/J1Hhb/sn9n/AOl9/UP7JH7KsEKQR/s0/CwrGoVS/g+xdsAY5JiJJ9zzX/BTb/gn18KfhT8MH+P/AMDtEbw9Fpd9DDr+jRyvJamGdhEk8W4kxsJWjUoDtIkyNpXDf8ExvjZrnxw/ZP0W/wDFWoXeo654U1C40G+vrqUySXRhCSxOzHkkQTwqSSSSpJOSQKqqqqqqqqqqr/guf/zRL/uYP/cdX7J3/Jq/wa/7J/oX/pBDVVVVVVVVVVVVVVq2kaVr2m3Oja5plpqNheRmK4tLuBZopkPVXRgQwPoRiv2qv+CP3grxrLf+Nv2a9Rt/CWruryv4autzafcycsRDISWtiScBcNGOABGor4k/Cr4n/BDxdL4R+JfhDVfC+u2UhKxXUezdtOPMhkUlJUyOJI2ZT1BNfsv/APBUX4tfBz7P4W+LQuviF4UXCpLcXB/tOyHAHlzMSJUAB+STnOMOoGD8F/2gfhF+0D4bTxN8KvGllrEIUfabTPlXVm3QrNA2HjOQQCRtbqpYEE8EcGqqqBzwTVVVfQ0CCMZ5qqqq8Q+F/Dfi3T20rxRoGn6tZt1gvbZZlz6gMDg8Dn2FeN/2Hfgz4nSSfw5FqHhe8bJVrOczQlicktFKW45xtRkAwPx8afsF/FXQ3aXwfquleJrcD5V3/Ypyf9yQlB2/5aV4s+G/j7wLIYvF/g7VtJAxiS5tGSNs9Nr42t1xwevHWvC/xE8e+CXD+EfGWsaSAcmO0vHjRv8AeQHa30INeFf27PjPom2LxBFoviKPnc9zafZ5e+MNCVUdR1Q8KOhJNeFf+CgPw71LbF4t8I6zokrZ+e2dL2JcZ6t8jdAvRDyx6AZPhn9oz4G+L2WPRfibookdgiRXkpsndicAKs4QsSewFQTwXUEdzbTJLFKgeORGDK6kZBBHUEd6qqqv2iv2Bv2d/wBo/wC1avr3hj/hHvFU+W/4SHQ1W2uJHP8AFOmDHcZOMl1L4GFda+On/BKz9pH4Upc6t4HtLf4kaHArOZdGj8u9VAcDdZsxdmIx8sJlPX0zV7Y+JfBmuvY6jZ6noWs6e4Dwzxva3Fu+MjKnDIcEHt1rwH+3P+0r4DWK2Xx83iCziOfs2vQC8LnK53THExGFxjzP4ieDyPCP/BU68SCODx58JYZpgvz3WkakY1J2jpDIrEAtuOfM4BA5xk+H/wDgpB+zbrTKuo3PibQt0mwnUNK37RgHefs7y/L24546Y5rQf20f2XvElx9l074v6XE4dEzf29xYrljgfNPGgxxyc4XuRmv+GhPgF/0XH4f/APhTWn/xyvFv7aP7M3g+OU3fxV03UZoyVWHSUe+LkdlaIMmPcsB7146/4Kk6Lb3L2vw2+GF1fQqw23ms3gty2OuIYw/B5wS47ZHOB8Qf+Cg37Rnjm3ex03W9N8KWrrtZdDtTHIwzkEyys7qegyhTp05OZJPFfj7xJvkk1bxH4g1ecDLGS8uryU8D+88jHj1Nfsw/8EmPiV8QZbXxT+0BdXHgfQAySLo8arJqV6ndW5K2oxjlwz9QUHWvhH8GPhj8CfCEPgf4VeEbLQdKjbfIsKlpbmQ9ZJpWy8rkYG5ySAABgAAVVVVVVV/wVW1q01X9s3xNY28cqyaPpmm2s5cABnNqkwK4JyNsyjnHIPGME/8ABMzSZrD9n3Ur+aKMf2l4nuZYnXlmjWC3j5+jI/HvnvSDJ4qIZHBqMAnFRgEgCkAJAFRr8tRgY4qJRgCo1BNRrnAFRKDio1z0qJAcAUiDH8qSPNLFkV5II/8ArU0XHFfGSaex+Enje9tJTFPb+HNQkideqMtu5BH4iq0GKJpEU4x2qy07SX0lpHkUSAcDFeJYYUlYR49quAonkVMYDnH51+zp4Y8PeIPgR4al8Q+GdKvpJobhJPtNhG/mItxIqbgV5+VV5PXrWp/BT4RagipP8NvDqBDkeRp8cB/EoAT+NXv7M/wPu52uZfAkSu2MiK9uIl4GOFWQAdOwq9/ZF+EU9u8MKazbOcYlivQWXntuUj25Hevh98CNF+FmtnVPCnijWzb3MZS+srx4pI7kAHYflRSpUkkHk8kcAnMgGcVKBniphkYFSAcYr9vDwEngT9pPxE1vF5dr4jSPWIBsC5MwIlPAAOZkmOevqSck/wDBHP4mp4p/Z6134a3V00l54H11mijJJEdneL5seMscEzJdkgBR0PJLGqp3jjQvI6qqgkknAAHevjp+0+to1z4P+GV5uuEZorzVlwVXjBWA55OSfn7Y+XOQw8OeG9f8aa5BoegWE1/qF2/CKM9+WY9lGcljwO9fBv4H+H/hPYfay4v9fuogt3esvCescQxlUz3PLYyccAFsjAoknjNVVVVVVVVV8ffgF8Ov2kfh1e/Dj4jaX59rN+8s7yIBbjTrgAhZ4XIO1xkjHRgSrAgkV8ffgF8RP2b/AIiXvw4+I2l+TdQfvLO8iBNvqNuSQs8LkDchwRjgqQVYAggf8Es/2921uHS/2XPjHq7PqMK+V4R1u8uR+/jA+WwkZzkuOkRydwxHgFU3VVVVVVVVVVftY+AbL4n/ALM/xN8EXsFvKb/wzePai4/1aXUMZmt3bgkBZo42yASNuRzXhDxLf+C/FuieMdKdlvdC1G3v7YpIY2EkMiyKQy8qcqORyKjmiuYknglSSORQyOjZDAjggjqMVVVVVVVVEgDAo896qqpiAMA80ee9VVUxzwDVVVVVVRxjrROehqqqqqqpjgYBqqqqqqOMdaY8deaqqqqqqYg8A1VVVVVHGOtHnvVVVVVUx7A1VE4HBqqqqJwOKqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq+LXhseMvhV4z8IfZpbj+3PD1/Y+TEcPJ51u8e1T2J3YFadqF7pOoWuq6bcvb3dlMk1vKnDRyIQysPcEA1oer2XiHRNP1/TWdrTUrWO5ty67WMcihlyOxwRxVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV+25/ySjSv+xhh/8ASe4qqqqqkR5HWONSzscKoGST7CvgFof7VNqtvJBevYeH1YD7P4mDOGUDGI4z++QYxjBRTkHmkDhFEjKzgfMQMAnvgc4qq/4LV/8AJ1Hhb/sn9n/6X39V/wAFWf2wvhhqnwjuv2avhv4m0/xV4j8T3lqdWXS5vtEenwQzJMimRAUaV5Yox5atuUAlgMqG/wCCbvwD8Rfs9/st6N4c8Zaa2neI/EN9PrmqWMgYPayTBEjjcEnDiCGHcuF2tlSMgsf27f29vHHgT4hWX7LP7MFjb6r8SdaaKzvL4L5j6ZcXBTyYIVcCNpmVwxdyUjDDIznbH/wTi/bS8eaPaeMPiT+3V4jsvGNuwubTToJ725t7GUhGwk63MYiYMoz5UWMopBPb4PftjftPfsdftC2P7NX7aeuw694ev5IIofEWoXJke0gkJjivo7tgrT2zMpEhmG9SrElGR0b/AIKxL8V/Dv7PunfFf4TfEfxX4UufCOsRrqp0PXLnTxPZ3WIsyCF1EhWf7OF3/dDvgjJDf8E1fjRrHxv/AGTvDuteKNautX8Q6DeXWj6tfXd1JcTTyxPvR5HkZmZzBNCSSTk5IwCAP+Cqn7QOu/A79nCLSvBHia90PxX401aKysbzT7mS3ura3iPnXEsUqEFD8kcROc4nOPUf8Ejf2g/Hvj3T/iN8Hfi/4y13XvFfhzUk1G3m17Up767EDqIJ4RJK7YSKSGM7ePmuGIzk4/ag+KMfwV/Z68f/ABOFx5VxoehztYtvVP8AS5B5VsMkjrNJGPXngE4B/wCCPWtfGn4l6T8Qfin8U/it418VabBcW2jaRBrXiG6voY5lUzXTeXMzKHCvahWByAzg9a/a0+FX/BRD4yfGi68HfBD4pWngX4YR6fa3EGqQX7aZL5x4liaaDfcySB42YBRHHsdVPOS3iz/gmz+3d8PdPn8f/Db9p698ReJNNZp47O11y+s7q4yrb/KkdirO2cbXKhgzZbsf+CYX7dfjf47X2rfAn44X/wBu8Y6JZm80rVpY0glv7eNwksMyjaDPGWQhguXTeW5jLP8Atu+Idf8ACf7J3xO8R+Fdd1DRtW0/Q3ktL/T7l7ee3cOvzJIhDKeeoIr4Tft1/tSeL/gHpf7N3wf1bxX4p+MHiHxBfLNrVxcSahfRaX5MTL5U87N5bljODITiJItwKswZf+Cd/wCzX+1d8FPHPjLxV+0f4guNSj8QabEtsbnxE2py/afNLyM+WbDEHlsnJ7mv20/+CgHxl8efGR/2Vf2PF1CLUIdQbS73VdNVWvNQvVb54rZ84hijKsHlyCcOcoilm0D/AIJn/t5WtvB45j/a3/szxo+65aNvEGpOUmcncHulBLEhjvIRgSWHzDk/txfGP9ojxgPB3wX/AGpPDqWnj34ZTaiJdTQRr/adreJaNC7CLMbsPs7/ALyPCsGUFQ6OW/ZO/wCTV/g1/wBk/wBC/wDSCGqqqqqqqqqqqqqqq+MXwN+FHx98JyeC/i14K0/xBpxyYTPHtmtHOP3kEy4eF+ACyEZHByCQf2sP+CRnxH+GzXnjL9nWa98deGkV5pNFkRTqtkoAJVAuBdjg4CKsnIUI5+Y+F/Fnjv4VeLovEPhHXdX8MeI9HmZFuLWV7a4gcZV42xg4PKsjcEZBBBIr9l3/AIK36Vqv2fwh+09YRaZc8LD4p023YwSdABcW6AmM9SZI8qcgbEAJOg6/oPirR7TxD4Y1uw1fSr6PzLW+sLlLiCdP7ySISrDjqDiqqvoaBBGM81VVX0NAgjGaqqqqqeCC6gktrmFJYpUKSRyKGV1IwQQeoI4xXjH9mj4H+N1kfVfh9pttcSHcbnTk+xybskliYtoYksc7gc8Z6DHiz/gnt4bud83gjx9qFi3JWDU7ZblSfTemwqP+At/WvFP7Evx28PF303SNN8QQoCxfTr5QdoGfuS7GJ9lBOema8S+APHHg2V4fFfhDWNJKHBa8spIlPJAKsRhgSDggkHHFaB4v8WeFJGm8LeKdX0eR/vPp97Jbk9uShFeHf2xfj/4fYK/jNNVhGf3Oo2UUvOQc7wA/bGN2ME+xHhv/AIKG+I4THH4v+HOm3mSBJLpt49ttG7khJBJnC9twyR1GePD37evwb1RFTW9N8QaNN/EZLVZ4xzjho2LHjBOVHtmtD/aT+A/iJFfT/inoUQKFgL6c2RwDg8ThCDnt1I5HHNafqmmavAbnSdStb2JWKNJbzLIoYAEjKk84I49xVV8SPgn8IvjBara/E/4b+HvEojULFLqFgkssIDbgI5cb0Gc/dIzkg8E5+J3/AAR//Zz8VpJcfDnXvEfga7O7y40n/tK1XKnGY5j5pw20/wCuGQCOpBHjr/gjP8b9H8yb4f8AxM8I+JIY9xEd9HNps8gGNu1QJUyfmyGkAGByc8eKv+Caf7Z3hVpnb4QNq1tEyqtxpWq2lyJCwB+WMSCXAPBJQDI9ME+Jv2UP2m/CHnP4g+AXj2CG2jEk1zHoFxNBGp7mWNWQfnxX/Cqvih/0TfxR/wCCef8A+Irwl+y1+0j46Cv4V+Bfji+hfdtuf7DnigyvUea6hM+2c18L/wDgkN+0r4vuIpfiLqPh7wFY7gJhPdLqN0o3EEpFbsY2IA3YMy/eUZzu2+Av+CMnwZ0d4Z/iJ8UvFPiaSPJaKwt4tLhkO443KfOfG3AwHByM5AOB8Iv2d/gl8BbKay+Efw30jw6blQlxcwxtLczqMYWS4kLSuoIyFZiAcnGSaqqqqqqqq/4KIa1L4g/bP+KF9NAkTRajBaBVJIIgtIYQ31IjBPua/wCCe+lSaf8AsueHrt5lcane31wigYKAXDxYP4xE/iKQDsatNU0u4vptKt9StZb21AM9skytJECARuUHK5DKeR3HrUQBPWowCeKjAJwDUYBAArXfEWh+FNHn17xHqcFhYWwBlnmbAGTgD1JJIAA5Jrxt+21NDdm1+HPheCSCM4N3q+4+Z/uxRsCo9CWycjIGMVF+2N8aI3VvtmjuAclWsBg+3BzXw8/bjtri5Sx+JvhuO0Rzj7fpQZkTn+KJiWxjHIYn2rQdZ0jxHpNtrmg6jBf2F5HvguIHDI49iPcEEdiCDUSg4FRLkYqNQTgVGgIpIxivLBGAP0p4wRxWrabb6pp91pl2CYbuF4ZApwdrKQcHtwa8ceEtT8B+L9X8G6wjC70i7e3cldu8KflcDJ4ZcMPYitC1dUVYnfEiDAz/ABUniWRYPLEnatd1sMGAcGQ9B6e5rTtOvtZ1G10nTbd7m8vp0gt4U5aSR2Cqo9ySB+NeBfCqeCfA+h+ElkSRtKsIreSRM4kkVRvcZ5AZtxx2zUoBFSAYIqQA5AqUe9SgZwKmAxwamA6CpBxjPSv+CpngjzNP8D/Ee3hx5M0+k3UmOu9RLCPw2T/nX/BHHxzLoP7RXiDwQ6M1t4o8NSMCoJ2z20qOhb5gAvlvOOhOSuMAtRbtmtd8QaL4Z0qfWtf1OCxsbZcyTzvtA9APUnoAOScAZNfGr9pHWPiCs3hzwqJ9L8PtlZMnbNeAgZD4J2p1+UHkHnPQfDr4ZeKfifrS6P4ctQEQbri7mBWG3X1ZgDyegUcn6AkfDH4U+F/hXo507QoTLdXAU3l9KP3lwwH/AI6uc4UcDPckk1VVVVVVVVVVX7V/7L/g79qz4XXHgTxFImn6rasbjQ9ZW3WSXT7jHXnkxPgB0BG4AcgqpHxE+Hfjr4NeOtR8CeO9GutD8RaHcbZoXOCpHKSRuOGRhhldTgggg1/wTf8A26of2mvCP/CtviPqSf8ACz/DlqZLh/I8pNXs1KqLlcfKJQWUSLhRlgyjBISqqqqqqqqqmhhuIXt54kljkUq6OuQwIwQQeoI7VXwr/wCSYeD/APsA2P8A6ISqqqqqqqJA4FHnvVVVMcDANHnvVVVMc8A1VVVVVVTEHgVVVVVVUcAdaPPeqqqqqpiDxnpR571VVVVVEgCjz3qqqqqqYjGAaqqqqqjjHWjz3qqY54BqqqqY54BqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqvGejWvh3xfrnh+xkle30zUri1haUguyRyMoLEADOAM4Ar9mTVb/Xf2bPhNreq3Hn3uoeB9GuLmXYF3yPYwszYUADJJOAAKqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr9o74gj4efCzUry1nePUtU/0CwKHBWSQHc+e21A5z6hRxnNeH/C/iPxZfrpfhnQ77VLo4/dWsDSFQTjLYHyj3OAK+H/7FfibVHhvfiLrEejW2cvZWbLPcMPTfzGn1G/6c8eAvgr8Nvhsgfwx4agW8wA19c/v7hiO4ds7M4BITaM9qqqr/gtX/wAnUeFv+yf2f/pff1ef8EM7pLSZ7D9p2Ka5WNjDHN4MMSO+PlDML1ioJwCQrY64PSvh3pt7/wAE0P2s9K0n9or4W+GfFNg/lXFvrkcT3T29sz4S/sS2AskTo2VaMPlWAIyj1aXdpqFpDfWF1Fc21zGskM0Lh0kRhlWVhwQQQQRwc1+zl+0/q/gf9rTxL+0m3wf1P4m65qjaneW1hBdG3mspbuYbrjdHBNgLHJJFtVFH73AIA2n/AIe6/FT/AKMQ8V/+D64/+V1ftyfH74kftn6h4P1WP9lnxR4Mu/C0N5DLKGuNQa9jmMTIp/0WLYIzHIf4s+aemOY/A19+0L+w7pfgnxlYeXq3jT4b2a3EMqvGbe/ksY3RiHO5THPtbDE8r82ea/4InePbvTNf+KPwQ1lZbe5EdvrFvaTKUeF4nNvdblPIOXtgRxgrz7ftizS/tO/8FNvhV+z5pkrX2jeB2tW1a2LHy42J+3XxBPygtax26HGfmULyflC3En7JX/BYKeW/drTw98Ub5iZW81BPHq3zbvvEFV1FcEnKDyycJgbP+C0vxUTw58DvCvwms7lFu/GWtG6uUDHcbSzUEggDGDNLAckj7hwDyV/Ya+DzfA79lfwB4HvLV4NUk01dS1VJCC63d0TPIjEcZj3iLjtGOT1P7Wn/AAUJ+Cv7J16nhTV4r3xT4zlhWYaFpboptkblWuZWOIQw5CgO5BU7QrBqtP2/f+ChHxTsv7e+Cf7EqW+juweG51iyvbuOaM5A8uXdarKQytlkBA4BAyM/sF6t4un/AOCnml3HirR4vDeuanrniT+3NGsY2t4LSc2d7JLarHubEccygKhLY8teSRmv2/P+TNPiz/2L8n/oaV/wRC8B6A2n/E34nTWqS62s1po9vMy/Nb2xVppVU+kjiEsP+mK+9fGrxhc/Dz4N+PPiBYl/tHhnwzqOpw7FUtvt7aSUYDAqTlB1BHrX/BEzwNZa/wDFP4lfFHUilzf+HtJtLGBphvcNfSyu8gJzhsWZUt1xIR0Jqv8AguR4f0W21/4P+KbfToU1bULPWLK6uwPnlgt3tHhjPsjXM5HvI1fsnf8AJq/wa/7J/oX/AKQQ1VVVVVVVVVVVVVVVV+0/+wZ8A/2o9PubvxHoCeH/ABayk2/ifSIUiut4BC+eMbbhM4yr/NgYV0zmv2pf2Bvjx+y1ezahruiN4j8Hbj9n8T6TCz24HHE6DLWzcgfP8pOQrNg4/Z+/ay+OH7NGrC9+Gfi1hpsjZutC1EG40+65BO6HI2McD54yj443YJB/ZY/4KKfBr9o8W3hrWJovBPjaWRIY9G1G7DR3ztwPssxCiQk8eWQr5PAYc1VV9DQIIxnmqqqoHPBNVVVVVVVJHHLG0UiK6MCGVhkEHqCK8T/AX4NeMVca98NtClkk+/PBai2mbAAGZItr9AB14rxJ+wb8GtXMkuhX+vaHISSiQ3SzxLls4KyKXOBkD5x2Jz38Qf8ABPLxTBvbwp8RtKvuAUXULOS153cglDJ0XnOOTxgda8Q/sa/H/QEaaLwjBqsSfefT76KQ9cDCMVc5znhTgdcVr/wv+JHhXefEfgHxBpqR78y3OmypGQn32DldpUcHcCRgg55q1urqyuI7uyuZYJ4jmOWJyjKfUEcitA/aB+Nnhny10n4oeIAkWwRxXN41zGgT7qhJdyhe23GCMAg4rRP25fjzpSxrfX2h6yUDAm+00JvyeCfJMfToMY981o//AAUQ1mL5fEHwwsrnO0BrPU2g2/3jtZHz2wMjHqc8aR/wUC+FV1tj1nwr4nsHaULuiihnjRTj5mPmK3HOQFPA4znFaP8Atjfs9av5aP44ewmkYqIrzTrhMe5cIUA+rVpfx5+CusRh7H4q+FsmTy1SfVIoHZuOiuQx6jkDFadrei6z5n9kavZX3k48z7NcLLsznGdpOM4P5GqqvoaB4wayCODVVVVVVVV+3j/yeD8Vf+w8/wD6AlfsEyRyfsoeCEjkVmjOoK4ByVP2+4OD6cEH8RSDI61+1N4Nkig034kaQzw3Ns4tLx4sq205McmR0wcqT1O5R2rwj+0h8TvCwS3udUj1u1UjMepKZHx3xICHyfViwHHFeDv2sPh/rckdr4jtLzQJ5Gxvk/fwD0+dQGHPHKADrnrjQte0PxFaLqGgaxZajbN0ltZ1lX6ZUnB4PFRjoM18ffjDf/FLxXNb2V5IPDmmylNOtxwHIGGmYd2Y5xn7qkDqWJtNPmvDlBtX+8R/Kv8AhHWKZErZx/dq8sZrNgsgyD0YCv2TvjVffD7xvaeDtY1Bz4a8QTiFo5H+S0uWwI5hnoCcK3IGDuP3BUQB4BqIA4FRAEVEoOBmkUHAFCMFeMVIo6CpgOlftW/szH4w2cHivwclvB4s06Py2SQhE1GHjCM3RXXnax45KnjBXX/D2ueFtVuNC8RaVc6dqFqxWW3uYyjqR7HqPQjg9q+03ONouZMYxjeatLO81K6isrC1murmdtsUMMZd5GPYKOSfav2aP2ZLrwRcwfED4g20J1gxZ0/T2G42JOQXcg4MhXGBztz/AHvuygEYqQZUAGpAOak6/hUoBqapQCKm6fhUnSv+CkGl2+ofs2y3c1s0r6brlncRMCf3THfFuOO22Vl54+Yd8V/wTP1pdF/bX+HTTak1pb3jahaygOVWYyWFwI42A+8DL5eAeNwU9q+Kfxt8H/C20eK/uBf6wUzBpcEgEjZ6Fzz5a+5GfQGviN8UfFnxQ1Uaj4jvB5MRP2WyhysNuCADtUknJwMk5J+mAPg7+zx4i+JM0Gsa0s2k+HMhmuGXbLdLzxCCCO2Nx4GeN2MV4W8J+H/Beiw6B4Z0yKysoeQiDlm7sxPLMcDJPPSqqqqqqqqqqqo4A60WzwK/4KDfsVad+0z4Ck8X+CtNtYPiV4bt2ksJwm19Wt0VibF2yBkk5jZshW4yquzDwf4t8dfBj4i6d4u8N3V74f8AFfhPUhLEZIzHLbXETYaOSNu3DI8bDBBZWBBIr9kL9qbwh+1n8JbPx5oQisdas9tt4g0fzQz6fdY5wMk+U+C0bHG5QQfmVgKqqqqqqqq1zV7Lw9omoa/qLOLTTLWW5nKLuYRxqWbA7nAPFV4A0u+0TwH4b0XU4PJvLDSLW3uItwbZIkSqwypIOCCMg4qqqqqqokYwDR571VVRIAwKPPeqqqY9gaqqqqqqpjxgGqqqqqqjjHWmOehqqqqqqYgDANMc8A1VVVVVUxzwDVVVVVUcY60ee9VVVVVTHsDVUSAOtHnvVVVHAHWjz3qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv2m4ooP2kvixBBGkccfjjWVRFXAUC9mAAA6DFfsB/8AJmnwl/7F+P8A9Deqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq+Ifwc8K/FPVdMvPGct5dWOkI4t9Oil8mN3fG93ZfnPCoAAy4weu7jQfDXh/wtp8eleG9FstNtIxhYbWERj6nHU+pPJJzVVVVX/Bav/k6jwt/2T+z/wDS+/qv+C13w4t9Z+Cngr4o29rvvfDPiBtPldUOVtruFmYsd2MCW2hABBOZOCvO7/gm/wDEFPiN+xl8OL15na60SxfRbhHYsYzaSNDGMlQCDCsLADIAYLkkGv2evEWn/sE/8FIPEHhP4hzJovhm7ur3Q5r10EcUWn3TpPZXBB3bYiUtSxDDYpbLEKwMM0NxCk9vKkscihkdGyGBGQQR1BFVWoW1v+yN/wAFe49TvjDpvhjxpqEl4JJFMUclvqkLhiCwcgJelx8uATFtGwEqP+CUnhvUvjH8cfjL+2F4pt5ZJdT1Cax0x7iPfslupftE6ox4UxRC2jAXokuOBwf+C0nw11CHQPht+0F4cieG+8NalJpV7dQxlnQSYntXJzhVjkhmAOOWnUE9Afit4ntP2+P2+/gj4TVIpdBj8N6LeapaiIPEEe1/tW8jZdzBS0ciwEFmCsoB3EEGv+Cd1roHx+/b+1zxf8e7JLzxC0Opa7Y2OojA/tdbiIhDEw5MMbTsqYGwwqQBsArXNb0fw1omoeI/EGpW+n6XpVrLd315cyBI7eCNS8kjseFVVUkk9AK/Yy8X6L8Qv+CsFv4+8NySvpPiXxZ4q1OwaaPy3ME9pqEsZZf4Ttdcjsa/b8/5M0+LP/Yvyf8AoaV/wRD/AOSVfEr/ALGC1/8ASc18TvBdv8SPhr4s+Hd1N5UPijQ73SpHyRsW4geInjngPX/BJb4oj4GftL+LvgH8RVbQ73xgo00w3RRPK1ewklC27kjhiJLlBhuX2rhiy4r/AILP/Gbwn45+LPg34U+G7+K+u/h9Z3j6tNCcrDc3hgP2cnu6JbIzY4HmgE7gwX9k7/k1f4Nf9k/0L/0ghqqqqqqqqqqqqqqqqqryztNQtJrC+torm2uY2imhmQOkiMMMrKeCCCQQeDmv2sP+CRnw4+JSXvjL9nWax8C+JXZ5pNFkRhpV6xIJVAuTaHrgIrR9FCIDuHxc+C/xP+BPi6fwP8VvB2oeH9Vhy0a3MX7u6jBK+bBIPkljJBAdCRkEZyCB+y3/AMFPfi78FGs/CfxQefx74MiVYkW4k/4mNgijA8mYn94oGPklz90BWQZz8Ev2g/hJ+0P4Wh8VfCzxfa6nGYw11YuwjvLBjwUnhJ3IQcjPKtjKsykE1X0NAgjGeaqqqgexNVVVVVVVVA46GgQRgGq13wF4G8UOZPEvgzQtXZmDlr7TorglgNoPzqeQOM+la7+yH+z5rqHd4CSxm2BVlsbyaAqAc8KH2EnkZKk4PsMa5/wT7+Gd45k0Dxh4i04sXJSfyrlFz90LhUOBz1JJ457nWv8Agnh4pgVv+Ed+JWlXxEeVF7YSWuXz0OxpcDGOf071rf7Dvx70outhpej6zsICmy1JE35HJHneX06HOPbNax+zZ8eND8z7b8Lddl8ptrfY4Bd5Pt5JbI9xkVq3gbxtoJmXXfB2t6cbYAzC706WHygQCC25Rjgg8+tVp/jnxtpBgbSfGOt2RtQBAbfUJY/KAGAF2sNuBxxWnftBfHDSvM+y/FfxO/m43fadRkuMYz08wtt69sZ4z0FWP7W/7ROnW4tbf4lXDoCSDPYWs7c/7TxFvwzUH7cHx9igjik1jSJmRArSSaYgZyByx24GT14AHsKg/b++NEUEcUmg+DpmRArSSWVwGcgcsds4GT14AHsKs/8AgoV4/jtkTUPAfh+e4Gd8kMs0SnnjClmI4x3P4dKsP+CiPiOO4Dan8MtNuIMHKQak8LZ7fMUcfpX/AA8Z/wCqOf8Alxf/AHNUH/BReBpo1ufhA6RFgJHTXw7KueSFNuMnHbIz6iv+Hh3gv/onetf+BUVf8PDvBf8A0TvWv/AqKv8Ah4d4L/6J3rX/AIFRV/w8O8F/9E71r/wKir/h4d4L/wCid61/4FRVe/8ABRayS5dNO+Ek09uMbJJtcETnjnKiBgOc9z+HSv8Ah4z/ANUc/wDLi/8Auar/AP4KLahJb7NM+E1vBPkYefWmmXHf5RCp/Wv2hPBVh+0H8YfEPxg1S4bRbvxC1u0tjZxhoomit44cgnklvK3HPdjXwO+IviX4DeAofh94bi0zULKG6luBNfW7mQtIQSPkkUY49Kuf2pviRPD5UNloVs2f9ZFayFv/AB6Qj9K8QfH/AOIfifQ7zw9q0+nSWd9EY5VFmoOPY9jkDnsaqtG1zWvDt8mp6Bq13p12gIWe1maJwD1GVIOD3HQ1ofxa/aD1f4Za/cX+kR6hoa6ZLG2s3UX2aWJWXZujdSokYAnkKTnBJ7FEMjrGp5Y4FaDpqSlIlAwBgCj4Pb+z/tOz5cdcVr+mIoeEgcjHSiCDgjkdjXw11m68R/D7wx4gvjm51PRrS6m5z88kKs36k1ERxg1ER0qJhgYNIwwMGvN+XGadge9SsD0NTEEYBrxd4G8GeOLQWXi/wvpmsRJ9wXlsshjPqrEZU8nkEdTVx+yP+zw7s/8AwrxQWJJC6peAfgBLgfSvCvw08AeA4yng/wAIaXpbkbWmgtx5rAdmkOWbqep7n1qYg96l6VLgrgGpO/NSYJ4PapfQVNg8A1N04qbGMZ7VJ0r9trRNW8R/sx+MdD0HTri/1C7awWC2gjLu5F/bscAdgqliegAJOACa/Z3/AGdNa+GHibS/iZr+utbeI9Jm87TraxYMts+0jc7kHc2GPC8DGcnPFra654p1hbW0hvNV1S+kO1FDTSzN1J7knAJJr4R/sqafozW/iD4lCHUL1Duj0tcPBGe3mH/lof8AZ+767qREjRY41CqowqgYAA9KqqqqqqqqqqokY60WzwKPPeqr/gqj+xVFPa3f7UXwr0RI5bdc+MtOtIseYC3/ACEFUfxDIE2OwEhAxIx/Y4/am8Sfsm/GGx8d2Au77w9eYtfEWkRSlRfWpP3gpIUyx5Lxlu+RkB2rwp4q8PeOvDGk+M/CWqw6lout2cV5p93ECFnhkUMjAEAjII4IBHQgGqqqqqqqqv2gNch8MfAb4keJZ4Hnj0nwjqt28SHDOIrSVyoJ7kLivD+i3XiTXtN8O2MkSXOqXcVrC0xIRXkcIpYgEgZIzgH6VVVVVVUxGMA1VVVEgcCjz3qqqmPYGqqqqqqqJAHFHnvVVVVVVTHsDVVVVVUcY60x461VVVVVUxAGAaqqqqqqYg8A1VVVVVHGOtHnvVUxzwDVVVYzwDTEHgHpVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV+2/DFD+1p8UUhjRFOvysQq45KqSfqSST7mv+CXH/ACYl8Mf+4t/6dryqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv8AgoX/AME9PjT+1r8adF+I3w48T+CdO03TfC8GlSxa3e3UMxmjurqUsoit5V2bZ0AJYHIbjoTX7VXwSP7RX7P3jP4QW81jBf65Yj+zZ73eIobyJ1lgZymWVfMjUEgMQpPysMqf+Cdf7LXxt/ZJ8FeKvh98UNc8H6ppOq6omqaY2hXc8zxTtEsU4kEtvFwVhgxgnBVuBnJ/bM/YL+GX7X2lQapeXb+GfHOlwmLTvENrbrJvjznyLmPjzouSV+ZWQnIOCytoH7Df/BUL4HRSeEPg98drSXw4Pmto7LxLLHBEoZsAQXEYELkYZhGCpyPmbHHwm/Yl/b/1j4q+DfiX8fv2pYX07w3qlrqTaPHql3qQkMMgcwvb4ht13hQpdWYgNnkgCq/4KL/sCeMv2u9a8HeMPhfrXhfSde0K1uLDUn1uSaEXNqWWSEK8MUjZjcz/ACkY/ekgjBz+xj+zzN+y/wDs+eH/AIU6nPp91rlvJPd6zd2EjvDc3UshJZC6qxCxiOMZVeIwcZya/am+CcX7RPwA8Z/CFXtI73XNPP8AZk12zJFDexMJbZ3ZAWVBKibioJ2luGzg/wDBPP8A4J1ePf2U/iR4h+JfxY17wjq1/caSNO0VNFmnuPIEkgaeRjNDFsbEcartDEh5ASo4av2pv+CU3jrU/ivdfG/9k3xtY+H9Qvr59Tm0m5vJbCWwvS+8y2NxEDtDOSwRtgjI+VipCpoX/BP79uD9oGW10X9sP9pK9h8E2l0JJtH07VGu7i8xtOQoRYFPULJJ5hQhiEwefhb/AME3PiL8Gf24NL+P3gO/8CW3w50e8lFnpH2q5jvYrWWwe1YeWLcxmQGRmyZPmIySCSK/ae+F+v8Axr+AHjj4VeFbzT7XVvEultaWk2oSPHAjllOXZFdgOOyn6V/wTp/ZE+JP7Ifgrxd4b+JOt+GdSudf1SG7tn0O5nmRESLYQ5lhiIOfQEe9V+27/wAEwPDn7SXiO4+Kfws16x8JeOLxf+JlFeRMbDVXVcLI+wFoZThQzqrBgMlN2WPhj9kf/grTJFB4Gv8A9paXRdBTMT6i/i2eeSOLdkmN0Qzk4ztBZcABcoOnxY/4I9zn4NaH4M+DPjPR7/xp/bTX/iTxD4oea1GoJ5TKiRLCkxiVWYkIdxJdiznCgfBDwTqvw0+C3gD4ca7cWk+peFfC+m6VeS2js0Mk1taxxO0ZZVYoWQkEqpxjIHSqqqqqqqqqqqqqqqqqqvin8IPhn8bvCc/gj4q+DdP8R6NccmC6Qhom/vxSKQ8T/wC0jK3vX7Wf/BI34g/DYXPjL9nF9Q8ceHI43luNGmCtqloBziNVAF0MdAgEmcAI3WvB3jb4hfB/xjF4k8FeINX8LeI9JmKGa2ka3mjZW+eKRf4lyuGjcFTghgRkV+yt/wAFZvDfiltO8D/tK2tvoGrSssEXieziK2MxwArXMeSYGY9XXMYLZIiUHGl6rpeuadb6vompWuoWN3GJLe6tZllimQ9GR1JDA+oOKqgc8E1VVVA9s1wRwaqqqqqqqvoaDdia4I4NVVVVaj4d8P6vMtzquhafezKmxZLi1SRgoJOAWBOMk8e9X/wF+Ceo25tbj4T+E1QkHMGkQwNx/tIob8M1qP7JX7O+qTLPc/DW1RlTaBb3tzbrjJP3Y5FBPPXGenpV1+xD8ALi4eaHRNVtkY5WGLU5Cqew3Zb8yak/4J//AAYd2dfEXjJASSFW9tsL7DMBP51/w7y8Gf8ARRNa/wDASKrr/gnTavcO1l8XZYoCf3aS6EJGUe7CdQfyFaj/AME69TjgVtJ+K9rczbsMlxozQKFwckFZnOc44x361/w7y8Z/9FE0X/wElq9/4J6+P0tnfTvHnh6e4GNkc0U0SnnnLBWI4z2P4da/4d+fGU8f8JN4L/8AAy5/+R6/4d9/Gb/oZvBf/gbc/wDyPX/Dvv4zf9DN4L/8Dbn/AOR6/wCHffxm/wChm8F/+Btz/wDI9f8ADvv4zf8AQzeC/wDwNuf/AJHrT/8Agnp8RpYC2q+OfDdvMGIVLdZplK4HO5kQg5zxj0554/4d4eM/+ii6L/4CS1Z/8E7vE0lyiah8TNLhtznfJDp0krjjjCl1B5x3H9K/bO066/ZC+I2j+BHgl8VW2r6ImpR6mYzYIHM0sbQhf3oYqIkYnd/y1XgcE/sl6foX7TOn+Jbi8uLnQ7rw9Nbo1tC3n7kmWQq5Yqo5MbjAzjbz1FWv7JvhQTKbvxTqskQzuSNI0Y8diQ2Ocdq+If7PXw38EeCdV8Uf2zr7TWcH+jpJcRFXmYhYwQIgSNxGcEcZ5HWq+HvwJ+IHxFMV1Yab/Z+mPg/2hfAxxsvqgxuk78qNueCRXgD9mX4ceD1hutUtD4h1KMhvtF8v7pW/2YQduOh+bcQe9fFzR5tY+FPifSrBAJDpcpiRV67F3BQPfbgfWkYo6up5U5FaBqMcZSVW4PSj4wJ077L5ny46V4h1KNleYkHA4HvSI8rrHGrO7nCqBkkn0FfDzRbjwx4E8N+Grtt0+k6Ra2kpwBloolQ9CR1X1P1NRMMYzSMOCDUbjGM0knHBrzRjGaeTIxmpHBHBqRgT16VK2RjNTMCMZqU8YzUxB4BqYjGAalIxgHtUh461IRzzUuCcA1KcnGamIPANXMiRo0kjqqoCWZjgADua8W/GDRNJBtNB2andDILq2IkP+9/F/wAB4968QeJ9b8T3P2nWL55tpPlxD5Ujz2Veg7c9Tjkmovip4CPxT8J/CV9Xa51nxP4hsdHaKzUSfYjczpF5khJAG3fnbnceOADkfDz4WeD/AIZaaLLw3pwFw6Bbi9lAae4I/vN2Gf4RgD0qqqqqqqqqqqiR0zRJIwKPPeqqq1DT7DVrC50rVbK3vbK8heG5triISRzRsCrI6tkMpBIIIwQa/b7/AGSbz9lv4tM+hxNL4G8WNLd6BP1+zEN+9s3/ANqLcuD0ZGQ5LBwv/BIP9r19D1n/AIZR8eal/wAS3VZJbrwfcTSNi2uTmSez5yAkmGlT7oEgkHzNMoFVVVVVVVX7cGt3Wgfsg/F6+so4nkl8J3toRKpICTxmFyMEchZGI7ZAyCOK+AOh/wDCT/Hf4b+G/tX2b+1fF2l2nnbN/leZdxJu25Gcbs4yM+oqqqqqjjHWmIIwDVVVUxAGAeaPPeqqqYgDAPNHnvVVVVVVRwB1o896qqqqqqYgDAPNHnvVVVVVUxB4BonNVVVVVRxjrROaqqqqqiQBR571VVVVVMc8A1VE4HBqqqqJAHWjz3qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/bj/5O2+KH/Ydf/wBAWv8Aglx/yYl8Mf8AuLf+na8qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv2p/+Cf/AMB/2p4ZdY1vS28NeMguIvEukRhJnwDhbiL7lwuSOWw42gK6jIP7T37FPxz/AGU9S3+PfD/23w1cXDQ2HiTTgZbO4P8ACrnrBIRyI5ACcNtLBSa/Zm/bW+Nn7L+o29t4T1r+1vCnnmS88M6ixe1mDffMZ+9BIeodONwBZXAKn9mP9tn4K/tR2EVp4U1VtH8VRw77vw3qTqlym0De0RBxPGCT8y84wWVM4qqBBGCaqqqgccCuCODVVVVVVVVQOOAaBBGAaqqqqqqqqqqrJHQ1uBGDXBHBqqqv+C13hKJrT4WeO4EiWRJNR025Yu29wRBJCAv3cLtnyeDl16jp/wAEwPFK6b8W/E3hGWZkTWtCE8amUhXlt5VwNnQtsmkIPYBv71J1xXxn8BeKviZBpHhfRp4LPTI52ub+7nYEBgNqKqj5mOGc44HTn0+H37O3gHwUYr2+tf7e1NMH7Rexgxo3qkXKjsctuIPQikPTmkI45qMgjGa/aE+EN38M/Fs19p9oT4d1eVpbGVEwkDEkm3PoV/h9Vx1IbFpfT2Z/dkFSeVNHxE/l4ETbsdC1Xd7PeNmVuB0UdBX7J/wOvPHviu18c69ZFfDmhXCyp5i4F7cIcqigjDIpAL9ui85OEYA4BqNxgDNRyAjGaR8Dg0soxwa83IwDTSgjrTvuGAakcYwDUrgjrU8qIhd3CqoJYscAAd61347/AAj0O5NnfeOLJ5QSCLVJLkAjGctErAdfX19DVl8fvhBqtytpa+NrZHbobiCW3XqB96RFUdfX37Goru2vYI7qzuYp4ZV3RyRuGVh6gjgipm461IeMA1IRjg1IQTwau7iC2ie4uJkiijBLO7BQo9ST0rxd8avDejF7XRP+Jtdrkbo2xCp93/i/4DkH1FeJ/HXiXxbIRquoHyM5W1hGyJf+A9/q2T714f8ACHiDxPKE0mwd4wcNO/yxr9WP8hk1+0B8OrTwR+zv4716HUriTWLbSGaC5hdofs7blGU2nOcEjJP4Cv2Vv+ToPg//ANj7of8A6Xw1VVVVVVVVVHGOtFhjAoknjNVVVVVX7U37Pfh/9pz4M6x8L9ZeK3u5cXWjX0ilvsF/GrCKXA5IwzIw7o7Dqa1TTPGPwr8dXGlajDdaF4o8KamUkQMBLZXcEnUMpIyrrkMCRwCCRzX7GX7SelftT/AfQ/iNHLbx6/Av2HxJZQqVFrqEajzNqnOEkBWVBlsLIFJLK2Kqqqqqqr9vz/kzT4s/9i/J/wChpX7K/wDyc98H/wDsfdE/9LoaqqqqOMdaJzwKqqqiR0Bo896qqpiAMA80ee9VVVVVVHGOtE571VVVVVUcAdaPPeqqqqqpjgYBpjngGqqqqqqmIPANVVVVVVMQeAaqqqqqiQBR571VMcnANVVVjPANMQeAelVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVftx/8AJ23xQ/7Dr/8AoC1/wS4/5MS+GP8A3Fv/AE7XlVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVWraRpWvabc6NrmmWmo2F5GYri0u4FmimQ9VdGBDA+hGK/a4/4I/abrIvvHn7KtzFp98d8s3g6+m2wTt8xItLhziIk4Aik+Tn/WRgAHWdC+IHwm8Xtpmu6ZrvhHxPo8wbyp45bK7tXHRhnaynuGH1Br9k7/AIKyaxoj2ngb9qLzdU075Y7fxbZ2+bm3Gcf6VDGP3yBSP3kY8wBOVlZiw8MeKfDXjbQLLxV4P1+w1rR9Rj8y0vrG4WeGZckEq6kg4IIPoQQeRVA9ia4I4NVVVnHQ0CCMCqqqqqqqqvoaB4wa4I4NVVVVVVVVVVV9DWTjGa3AjFcEcGq/4KqfD+Xxz+yBrepWyNJP4O1Wz1lI1zlgrNbyH6LHcyMc9lr9kPxsvgH9o3wPrc96tta3GoCwumkYrH5dyph+fBA2gyK2TwCoY9KHUYNRkZwDUZyBg1GQQMVGcjANRMOOa1jQ9H8S6XcaJr2nQX1jdIUlgmXIIIxx3B9CMEdQRXjj9iuyvJpLz4d+JVsw5ytjqSs6L9JVywHTgqx96T9jD4uvfNafbPDoiUZF0byTyzwOAPL3+3K9vxr4e/sS6Fpl5FqXxC8RHV/KIYafZxmGFiOzuTuZT6AIfftWk2Gn6PYW+laVZw2lnaRCKCCFAiRoowFUDoAKjcEYzUb44zSSDsaSUHgGhICK8wEcGmkGOD+dPKMYzUkme9a/rumeHdJu9c1i8jtrOyiMk0rtgKB/UnAA7kgd6+Lfxz8T/E3UZraC6n0/w+rEW+no+3ev96Uj77HAODlV7dyYree4O2CJ3P8AsrnFTWtzb/66B09CVwK+HvxS8U/DnUFn0i8aayZv9IsJnJilGRnA/hbj7w5+o4rwv4q0nxp4fs/EWjTb7e6jyVb70T/xIw7MDwfzGQQalbPANeI/GHhnwxHv13W7W0OMiNnzI30QZY9ugrxP+0PbKGt/CektK3I+0XnygcdkByfxI6dOa8QeMfE3imQvrmsT3C5yIc7I1x6IuFz74zXhzwN4n8VfPpGmO0AOGuJPkjHr8x649Bk+1eGvgvoek7LnXpf7TuRz5eNsSn6dW/Hj2qOGGCNYLeJI40GERF2hR7AdK/bY1b+xf2W/H159n87zLOC2279uPOuYot2cHpvzjvjHGc1+xvokviD9q74R2EM6RNF4w067LMCQRbzrMR9SIyB7mqqqqqqqo4x1okdAaJJGBR571VVVVVVVf8Ff/wBmuDRNc0j9pbwnpuy31uRdM8TrEDhbpU/0a5I5xvjRo2PCgxRdWkJP/BKn9pH/AIUn+0LD4C8Qah5Phf4keXps/mNhIL8E/ZJfbLM0R/67An7tVVVVVVVX7fn/ACZp8Wf+xfk/9DSv2XJYoP2mvhFPPKkccfjvRWd3O0KBfQ5JPYYqqqqJAHWic96qqqiRjANVVVTEAYB5o896qqqqqqOMdaJzwDVVVVVVRxjrROeAaqqqqqjgDrTH35qqqqqqokAdaPPeqqqqqpjgYBqqqqqqpjngGqpjgcGqqqonA4NHnvVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX7WOpXurftPfFi61CfzZY/GWqQK20LiOK6kjjGAB0RFGepxk5OTX/BOfRLTw/+xP8ACqwspJXjl0ua7JlIJDz3U0zgYA4DSMB7AZJPNVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV+0h+yf8ABj9qbwydD+JvhtG1G3haPTdctAI77TiTnMcmDlc8mNwyH0zzX7W//BO341/stPdeJ1tf+Eu8ArJ+78Q6bCxNopYhReQ8mA8Ab8tESyjfubaP2ef2qvjJ+zNr8ep/DjxNKNLkuBLf6Dds0ljfHABLxZ+VyoA8xNrjA5wMV+yf+3P8JP2pdLg0uyvYvDvjmOPN34avJx5jkKS0ls5AFxHhWJ2/OoBLKoILUDjjNcEcGqqq+hoEHiqqqqqqqqq+hoEEYNcEcGqqqqqqqqqqqr6Gsnpmvif4KsviX8N/FPw71AoLfxNo13psjMMhBNE0e72I3AgjkEZrU9O1vwj4hutJ1CObTtY0W9eCdA+2S2uInKsAynhlZTyD1FfCPx1a/E34YeF/HtpMr/21pcNxLtGNkxUCVMeqyB19OOOKQ9CDUZBGM1EwIxmom7ZqNsHANIwwCDUbAjANRuMdaRgOM0jccGo34xmkcEYzSyY4zSy8YzQlBHBrzQRjdRlyOtPLkcGnkzwDX7Yni24tdI0bwZayMqahI91dYOAyx4CL7gsSfqorTLA6hciInCKMuR1x7Vp2jK0axwxAKo4AFX2imOMh48gjoRWs6V9hulWEfu5T8oPY+lfCv4pat8M7PUbFLBL+C+ZZI4nnKLBIBgsAAc7hgHp91eeK8RfGb4geIw0cmsmwgYYMNivkj/vrJf8ADdj9ahgvdUu1gtoZ7u6uHO1EUyPIx64AySa8PfBDxjrJSXUoo9JtieWuPmkx7RjnPsxWvD3wc8HeHwk9xbNql0uD5l1yqn2QcY+uT70VjijWKJFVVGFVRgADsBUhBOc0ea/4KF6pcaf+y9r1pAkbJqV/Y28pcElVFwkuVweDuiUc54J+o/4Jz6JF4g/bT+GFhNO0SxXt1dBlGSTBZTzAfQmMA+xqqqqqqOPWiw6CixPAo896qqqqqqqqqvjL8LPDvxu+Fvib4U+Kk/4l3iSwe2eQJua3k4aKdQeC8cipIueNyDNeKfDniX4Y+OdU8K6sZdP17wvqclrM0LsjQ3EEhUsjcMMMuVbg9DX7I/xtj/aI/Z28FfFSa4hk1TUdPEOsLGqpsv4SYrj5ATsDSIzqp/gdD0NVVVVVVV+03oM/ij9m/wCKnhyzhhludR8F6vBbpLgL5rWcoQknphtpz2xmvCut/wDCM+KNH8SfZftP9k38F35G/Z5nlyK+3dg4ztxnBxnpUUsVxEk8EqSRyKGR0OQwI4II6jFVVEgcCjz3qqqjjHWicjANVVVTHsDR571VVVVVVTHPANVVVVVVUxB4FVVVVVRxjrTEHvR571VVVVVHGOtE571VVVVVHGOtE5qqqqqqJAHBqqonPQ1VVWM8A0xzwDVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVfG7WLnxF8aPH2v3qRJcan4o1K5lSIEIryXMjMFBJIGScZJ+tfsUaF/wAI7+yN8INP+1/aPN8H6fd7/L2Y+0QrPtxk/d83bnvjOBnAqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq8S+INP8ACmgX3iTVVuGs9Ngaa4+zwNM6ov3mCKCTgZJwOgNeD/Hngv4g6b/a3grxPp2s2q4EjWk4cxEjIV1+8jY/hYA+1VVVVVX7ev7ZT/sc+A/Dut6N4d0/X9f8S6o9ta6fe3TQqLeKMtNN8oLNtZoExwP3oJPAB/YE/wCCgV7+2Fr/AIs8IeK/B+k+GtZ0GzgvrGKyvHl+2W7OY5mKuAR5bmAZGQfNGcYGaqqqqqqqq8ZeKtK8C+D9c8b64Lg6d4e0241G8FvCZZTDBG0j7EHLNtU4UdTgV+xP/wAFMrj9q34v3/wp8Q/C638Myy2M99pNza6k10CsTLmGUNGuW2MT5gwCVxtGRVVVVVVVVVVVVVVVVVVVVVVVVVVNDDcQvbzxJLHIpV0dchgRggg9QR2r9sP/AIJIeDvHyX3xA/ZkSx8KeItryz+GJP3Wm3753Ytz/wAujkFgF/1JOwYiG5j4u8HfET4MeOpvDXi/RdX8J+K/D90rmGZWt7i2lUho5Y2HbgMkiEgjDKSCDX7H/wDwVZ1bSJdP+HP7T9w2oacFWC18YRxk3FvjgC8jUfvlxgeaoDjblhIWLjw54k8P+MNCsfE/hXWrLVtJ1KETWd7ZzLLFOh6MrKSCKBxwK4I4NVVVQPbNcEcGqqqqqqqqqBI4BoEEYzVVVVVVVVVVVVVf8FQfg63wt/ap1vXbG0SLSPHkK63a7ECgTP8AJdA4PLGZHkJwP9aOvU/8ExPipHqvg/xB8H9RvFN3oc/9pabGzfM1rKQsoUAfdSUKST3uB6cIe2aiYEDmo2wcZpG6c0jAgEGo2BGM1G2OAaRgMEGo3BGM0j44JpX44NJIMYzSy5GM0HBHBoSE8ZBrzDRkJ4yKZxj71PKMYBr9r+GX/hYmlXRQ+U+ioisehZZpSR+AZfzrwqyCaZSRuIXA/PP9K8Ny2qsvnYIHUV4kmsXU/ZwAMcCvFsiCONVI3eZkfgD/APWrwR4dufHfiK38O6ZPHFLMjO8sgO2NVXJJ7nsOO5HbJrRP2d/CenESa5qF1qzj+AD7PGfwUlv/AB6tL0HQ9AhMGi6TaWSHqIYgpb3JHJ7dakbPepWycA1KwxwakORjNV/wVH163t/hT4R8LsI/P1DxCbxMygNtgt5EbCdSM3C5PbgfxV/wSJ0CXWf2tv7RjFuV0Lwvf3j+aDuAZoYPk4PzZnHp8u7nsaqqOMdaJHTNFuwoknjNHnvVVVVVVVVVVVVX/BXX4Kf8IH8e9O+Kuk6dFBpPxBsN07xcZ1G22pNlRwu6Jrds8bmMh6hif+CJXxgSG98ffATULpFFwsfiLSoiCCWXbb3XPQkr9kwODhWPIHy1VVVVVVNDDcQvbzxJLHIpV0dchgRggg9QR2r4k+Drn4efETxT8P73f9o8M61eaZNvKlt9vM8TZKkqTlD0JHpX7IvjgfEf9mL4ZeLmukuJ7jw3awXUqshD3ECCCY/J8oPmxPlRjaeMDGKo4x1onPANVVVRxjrROeAaqqqmIxgGqqqqqqqpjngGqqqqqqqY8YBqqqqqqpiMYBpjngGqqqqqqmPYGqqqqqqmOeAaqqqqqqYg9DVUxGMA1VVVEgDiqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq1nVbPQtHvtc1BmFrp1tJcTlV3EIilmwO5wDxV3dXN9czXt7cy3FxcSNJNNK5d5HY5ZmJ5JJJJJr4Q+Gk8GfCbwT4Pj097FdC8O2Fgtq7FmgENukewkkklduMkk8daqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqkjjljaKVFdHUhlYZBB6gj0r9ov4aa5+zx8XprjwbqF7pOm6nuvNDurK5eGSCNj88IdSGBjY7epJUoSck18M/28/iX4XMNh4+srfxXYKQpmO22u0UDHDqNr44PzLk925yPhn+1V8GfigYbLTvEq6TqspCjTtVAt5GYjojZKPzkYVieOgyM1VVVftx6n/wANV/8ABSLwP+zxA8txoHhy8sdGvIkO0HzGW51CVCp3ZEJCHkYMHGPvHT7qf9jH/grJcwzyfZvDviLxI8cgiUeWdP1fDIOcEJBLMhOOc25+8OteIfjp8EfCPiYeCvFfxk8D6L4h3RqNI1HxDa213ukAMY8l5A+WDLtGOcjHWq1fVtK0DSrzXdd1O007TdNt5Lm8vLuZYYbaGNSzySOxCoiqCSxIAAJNeBvi98JfihLd2/w0+KPhHxbLp6o12mh63b37W4bIUuIXbYDtbGcZwfSte8Q+H/C2myaz4m13T9IsIf8AWXd/cpbxJ9XcgDp3Nad+2F+ypq/iQ+EtM/aI+H9xqXyhEXX4PLmZioVI5d3lyOS6jYjFs5GODiGaG4hSe3lSWORQyOjZDAjIII6givFPizwr4G0G68U+NvE2k+H9Fsdn2nUdVvY7S3g3uqJvlkIVdzsqjJGSwHU1Y/Ej4dap4Kb4k6Z4+8OXfhFIZZ21+DVYZLBY4mZZHNwGMYVGRlY7sKVIOMGvhd8efg58bZdXg+E3xF0fxU2gNEuonTpvMFuZd4jycYIbypMEZB2mviH8V/hj8JNKTW/if8QfD/hWyl3iCXVtRjtftDIu5kiDkGR8DOxAWPYc14I/a7/Zg+I+qDQ/Bvx48GX+pOwWKzbVY4Jp2IY4iSQq0hwpJCA474yKmhhuIXt54kljkUq6OuQwIwQQeoI7V+zj4V/Yd8K+LNeP7L2ofDGbxHrELXWpx+Gtdg1C4W3DoG2okrtDAHaPKIFj3FeMkVXjr4m/Dj4X6amsfEjx74f8L2Um7yp9Y1KK0WQrjIQyMNx5HAyeR614I/a7/Zg+I+qDQ/Bvx48GX+pOwWKzbVY4Jp2IY4iSQq0hwpJCA474yKrxT4s8K+BtBuvFPjbxNpPh/RbHZ9p1HVb2O0t4N7qib5ZCFXc7KoyRksB1NaX8Q/AGueDx8QtF8c+H9Q8LGOSQa5a6nDLZFI2KSN56sY8KyspO7AKkHkGrX9tb9km+8Qp4Xtf2ifAkl+8hjQ/2xGIWYDOBOT5RzjAw3JwBkkCoZobiFJ7eVJY5FDI6NkMCMggjqCKqqqqqqqqqqqqqqqqqqqqqv2kP2T/gx+1N4ZOh/E3w2jajbwtHpuuWgEd9pxJzmOTByueTG4ZD6Z5r9rz9gj4w/sn6rPqt7ZS+JPAUkmLPxPZQHy48sFWO6QEm3kyygbvkcsNrMdwX9mH9sf4wfssa6J/BupJqfhy6mVtS8O6gxa2uQMglD96CTDHDp1IXcHA21+zV+1r8If2pfDp1LwDrH2bWbSMNqWgXrql5Z8LltgPzxbnAEq/KScHByo6dDQIIwKqqqgcdDQII4NVVVVVVVVVQJHGaBBGAaqqqqqqqqqqqv+Cov7P0/wAZ/wBnebxboNp5/iD4dSSarbKsYLy2ZUC7jU9R8ipLgdTABzkV8DfirqfwV+Kfh/4i6aJJE0y5AvbdDj7TauNs0fPGShbaTwGCt2rQtc0jxLo1j4i0DUYb7TdSt0uLS5hbKSxuAVYH0IIqNgDwaRgRjPSo2BGM1G46ZqNscA0jZAweajcEYzSPjgGkcEYzSSY4zSuCODSyEDrSyjGM0JQRgGvN4wDRlGMZppQRwaaTIwDX7U/gi48Q+FLTxTp0AkuNBdzcALhjbuBuOe+1lU47AsfXNndyWVwtxEckcEHuPStM8Q28qL5VwA2PuE4I/CtR8QwRxnzrhQQPu55/KtT1B9RuPNYYVRhF9BX7MfhCaFdQ8cXce1JUNpZ5/iGQZGx6ZVVB/wB4VI2eAalYYwDUjADrUjDGc1KwJwDTHJ61X/BUXxYNR+JvhHwXFcLImiaLJduqlT5clzLtIOOQ223jOD2ZSOpz/wAEUfCUjXXxT8dzpKsaR6dptqwkXY5JmkmBX72V2wYPA+duvYketFhjAotnpRJIxmjz3qqqqqqqqqqqqqqokDjNf8FWPhdF8QP2UtR8TwQs2o+BNRt9Vg2D5miZvImU/wCyEm8w9P8AVDnsf2BPig3wk/a5+G/iOW7SCx1DVk0i/aQgRiC8H2cs5LKAqNIkhYnjZnBxtNVVVVVVV/wU48HyeD/21/iHGElFtrElpqds8sisZBNaxGQjb0AmEqgEA4Udc5P/AASQ8cDxR+yenhiS6RpvB/iC8sUhLJuSKUrdK2BztL3EoBbqVYA4GASAOtE54qqqqJAHWic96qqo4x1pjngGqqqqqqqpjngGqqqqqqqJAGBR571VVVVVMQBgHmmOeAaqqqqqqJAHWjz3qqqqqqJAHFHnvVVVVVUxGMA1VHGOtE5qqqsA8ZpiDwDVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVftb+KP8AhDv2Yvihrqzywyp4XvbeCWKXynjlniMMbK3UEPIpGOeOOa+FnhNvH3xP8H+BUtnuG8R69Y6aIUkEbSGedI9oYkBSd2Mngdaqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/a3+LHi/wCDnw303xP4KntY7651yKzkNzAJV8toJ3OAe+Y15r/huv4/f9BHRP8AwWL/AI1/w3X8fv8AoI6J/wCCxf8AGv8Ahuv4/f8AQR0T/wAFi/41/wAN1/H7/oI6J/4LF/xr4p/tHfEf4yaHb6B44bSbiCzuRcW7wWQikjcKVOGBzghjkdDweoBFV8Nf2lPjF8K/s9r4c8XT3OmW+Aul6lm5ttg/gVWO6Nef+WbJXwp/bx8G+LZ7bQ/Hnh690PVJtqLPZRtd28r5wcKoMidjjDd8txyjrLGsihsMuQGUqfxB5H0NVXxS8f6P8Kfht4n+JevMhsPC+k3OoTI0oj80RRlhGCc/M5AUDBJLAAEnFf8ABInwTqvxb/aD+Jn7UfjKKKe8tPNCSYZQdR1GV5Z5EGT92NZFIYni4HU8j/gtr8HmnsvAHx6061ZjbtJ4d1STIICtuuLXjqBn7WCeRllHBPP7JPxgT48/s4eAvifJdJPf6npMcWqMoIxfQEw3PB5AMsbkZzwQckEE/wDBa74H+VP4K/aH0bTYVSXd4f1yZOGZwGltGKjr8ouVLnptjXJ+UD9jf4zt8fv2afAnxKvLxLjVbvTVtdYYOGb7dbkwzswGNpd4zIFI4WReoIJ/4K6fGBPh1+yvN4HsbpE1T4h6lFpqJg7xaRET3DqemPkiiOe0/A7j/gmR8L9F/Zu/Y3v/AI1eOrb+zbnxXbzeJtVuZIsSRaTbxMbYHB+ZPJWS4Xvi5NfC/wAN/FP/AIK2fHrXvFHxL8Sa34e+D/g+4WaDSbWZWS2MnyxWkJICmd40d5JyjFemAHQV42/4JGfse+I/CD6F4U8Oa34U1lbfZBrdrrFxdSGUKMPLFO7xuCRllRY8gkKU4I/4J7fGn4n/ALNX7UWr/sL/ABi1+71LSGvLjT9GVpvNg0+8jVpY3gL/ADJb3EYJCAcPJGdqlpDX/BUf/kxL4m/9wn/07WdfsxfDv49/t0+DPDX7LHh/xOnhz4a/DJp9R1bUZQ80azXlxI8eYVZRK/8ArfKjJUD9+xcbgo/ZW/Y78FfsH+H/AIheI9H8a634mttas7e7vUubWKN4ksUuXxGFIBLCdupAyo55NfssfDO5/wCCmn7UXirxh+0L461M2+mWI1CTT7GRUkeDzdkVpASCIYI93zFVLHPUPIZB4v8A+CNX7LutW9sfCet+MPDtzbybnP25buK4XIyro6huxAKOv3jnPGLSztLC0hsLC2itra2jWKGGFAiRIowqqo4AAAAA4Ff8EVP+TqPFP/ZP7z/0vsK/bO/aTsv2VfgPq3xNNml7q80yaboVo7bVmvplYpuOD8qIkkpH8QjK5BOR+yL+xZ46/b81XUP2nf2qPH3iC60O+vGhs/LIiudYaJsP5bFPLgtEbdGFiX7wkVfL2ZPxt/4I1fBbWfBN0/wK1vW/D/iy0jd7NNUvvtVnetjIilyu+PJGBIp+XcSVfgD/AIJZftWfEjSviRqH7GPxtvLuW8037XFoP28l7qxurUsbmwd8nciqkrLn7nlsoO0qF/4Kj/8AJiXxN/7hP/p2s6/Yq+CHxj/bW8KWfwMuPGV74X+C/gK6e91x7Q4a/vbl2dFVTxLLhFA3ZSFVLY3SBX+NP/BFv4c2Pw+1TVfgX4/8YTeKrC3kntbDX5ba5hvyq5ECmGGIxu2MBjuGSMgDkf8ABGb9o/xTrt34k/Zw8W65cajZaZpq6t4b+1TF2s40kWK4t0LZ+Q+bC6oCAuJCAdxxVVVVVVVVVVVVVVVVVVVVVVqOnafrGn3Ok6tY297Y3sLwXNrcxCSOeNwVdHRshlZSQQRgg4Nftl/8Ei4bpLr4h/snWqRz7mlvfB1zc7VfJyWspXOFIz/qZGAxnawwqHSNX8d/CbxrBq+kXer+FvFXh66JjcB7W6s5l4KlThlOCVKkcgkEEEiv2Ov+Co3hj4mtZfDv9oabTfDHigqsVrr4cQWGpMA2fN3fLbSEBe/luzHHl5VK+hoEHjNVVVX0NAg8GuCODVVVVVVVVVUCemaBB4zVVVVVVVVVVU0MVxE8E8aSRyKVdHXIYEYII7jFft4fsyT/ALMfx01DRNJs5V8H+Id2oeG5nIP7lseZBx3hkJTB5KeWx+9X/BOf9pSOJR+z7411JUBLTeGJ5eOSS0toW9SSZEyO8i5+4tI3GM1E/A5pGAxg0jAjg1G4Ixmo37ZpHz0PNI4IxmkfA4NJICMZpX44NLL2zQkB70JBjrXmD1oyAjGaaQetNKDwDU6xzxPDOiyRupV0YZDA9QR6V8Vf2cNV069n1r4fWxvdPkJdtPDfvbf1CZ++voPvDpz1q7tLqxuHtL22lt54jh4pUKMp9weRUcUk0iQwxs8jsFRFGSxPQAetfDv4AeIvEssWo+KIpdI0sENskXE849FU/cGP4m9RgHtZWVnpVhb6Xp0CwWtpEsUMS9EVRgD8hUjgDAPNO2ec1I2eAalcY4NSNx15qq/a88cx/EP9o3xxrtrO0lpbagdPtf3u9NlqqwbkIJGx2jZxjg789zX/AASk+H7+C/2RdK1u5tPIufGOr3mqtvVg5jDC3jJBAwCtuGXGQQ4YH5jVVVVVVVVVVVVVVVVRIHGaJJ4Bqvi94KT4k/Cjxl8PXXd/wkmgXunLyqlWmgdFILAgEFgQSDggHtWnahe6TqFrqum3L293ZTJNbypw0ciEMrD3BANeCPFFp448FeH/ABpYCH7Nr+l22oQiGcTJsmiWRdrgAOMMMMAAeveqqqqqqq/4Lc/Dx7D4jfDr4qW9n+61rR7jSbmZFbAktZRKgfjaCy3TbecsI24ASv8AgjB8RfsPjj4g/Ci5uPl1fTLfV7RGOAr28hil2+7LcRkj0j9jR571VVVEgcCjz3qqqjjHWmOeAaqqqqqqqmIxgGqqqqqqqJAHWjz3qqqqqqOAOtMeOvNHnvVVVVVUcY60Tmqqqqqo4x1onPQ1VVVVVHGOtHnvVUx4wDVVVUxwMA1VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVf8FV/GSeGv2T7vw+H/eeLddsdPChQTtjY3RPJBABtlGQDywHQ5H/AATd8Dt48/bR+Gti1uklvpN9Lq07PAJViFrBJNGxBIwfNSJQ3VWZWGcVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV8ZPg34X+OPhe18J+LL/VLS0tL9L1H06WOOQyLHIgBLo424kbjGcgc+v/DvT4Lf9DP41/8AA21/+R6/4d6fBb/oZ/Gv/gba/wDyPX/DvT4Lf9DP41/8DbX/AOR6/wCHenwW/wChn8a/+Btr/wDI9ftU/Az4P/Au30nRvCmveI7/AMSakfPe3v7qF44bUbl3kJCpyzjC8/wt6Cq8CfCn4jfE65Nr4E8H6jq+x9kk0Ue2GJsZw8rEIhI7Mwr4af8ABPWYywal8WPFsYjGGbS9HBJboQHnccdwQqH2cda8AfCH4bfC+1Ft4H8I2GmuVAe5Ee+eTAx80rZc9+M45PHJqqr/AILHfGBvA37OGm/C/T7po7/4iasIpVAHzWNoUmm56g+abUcdQzAnHB/Za1v/AIKW/Bj4SWWkfAT4J3EnhLxBMdatrqfQo5mvDPHGFmDO4JVo44tpxgqAR1zXx417/gqh8dfhdq/w6+KfwIln8O3Xl3Fw0Hh6GOaEwOsoeNw5Kn5MEjkqzDoxr/giV8YGnsvH3wE1C6ZjbtH4i0qIgEBW2291z1Az9kwORlmPBPP7WnwUh/aE/Z48bfCyOyt7jU9S015dG89tgj1CL95bHf8AwAyKqs391mByCQf+CKnxlEA8efs6a5cyxXMUi6/pNvLHjGNsF4pJ5BBFqQnvIcDDZ/b61HUv2vv+Cgfg39mDw1Nd/wBneGpIdImeN9wSWYC51C5QYO3ZAqK2R1tiTkYx/wAFELdPAv7AnxC0bwdYJb2mnaTp2m21siFxDaG8toGUZycLCW5PTGe1fsHeGv8Agobqnwl1nUP2RfGeg6L4UPiKWK/jvYNPaWS+W3tyxzcQSPt8poQAG2g7sAEkn/hDP+C2/wD0Vrwp/wCA2jf/ACJXg39hH9vfW/2s/B37SHxjv/D99qdp4o0m+17UrXU4IJJbW2eGNwsUCIn/AB7xbCqgbhkHO45/4Kj/APJiXxN/7hP/AKdrOv8Agijp9lH+zP4v1WO3QXdx46uIZZQPmeOOxs2RT7AySH/gRqvjb/wTa/aQ/Z4+J918av2KPEN3c2S3Dy2mnWN2sGpacj8vDtfEdzADwFJJK4DI2Cx/Z/8A+Ctvjzwx42g+Ff7ZPgj+yJobj7Le65Dp8ljd6fISMG8s2HQZO4xhCFxhGOc2l3aahaQ31hdRXNtcxrJDNC4dJEYZVlYcEEEEEcHNf8EVP+TqPFP/AGT+8/8AS+wr/guRrupW/h/4P+GIp8WF/eaveTx4+9LAlokZ/BbiUf8AAq/Yy0HTfDv7JXwe0/SYfKhl8F6ZeMvrLcW6Tyn8ZJXP41Xxetk8I/8ABZCwi8GHYJ/iJ4febylWXm7jtGvOMEDJmnyeq5JyCM1/wVH/AOTEvib/ANwn/wBO1nX/AARU/wCTV/FP/ZQLz/0gsKr/AIJv/wDKR7xB/wBx/wD9Gmqqqqqqqqqqqqqqqqqqqqqqqr9rz9gj4PftYaVPqt7ZReG/HsceLPxPZQDzHIUKsd0gIFxHhVA3fOgA2soLBv2g/wBm34s/sx+Nn8D/ABW8P/ZJpN7WGoWxMtnqUSkAyW8pA3DlcqQrruG5VJxX7G3/AAUq8bfAj7F8P/i29/4s8BR4jgkBEl/pC/Mf3TMR50e4gGN2+UAbCANp8DePPBnxN8LWHjXwB4lsNe0PUo99te2UwkRvVT3V1PDIwDKQQQCCKB7Zrgjg1VVVA44oEEcGqqqqqqqqqqgSOM0CDxmqqqqqqqqqr9rj9mfw5+1P8INQ8Aam0VprNtuu9A1J1x9jvVUhNx2sfKfO2QAZKnI5CkeLPC3jP4T+Or/wr4js73QvEvhq+8uVDuhlt5oyCro3Bx911cdQVYHBBr9jr9r3SPjno0HgnxfcQ2PjzTbfDxltqarGg5miz/GAMun1YfLkIjDgg1G4Ixmo37E0j46GkcEcHmkk7ZpXBHB5pJO2aVwRwaWTHBNLKCMA0JARwaEgxw1bzjG6jIDxuoyDGM00oxgGmlyMA0zgjk1fWOnahsF/Y29z5edgmiD7c9cZ6dBVvpmlWEhmstNtbeQrtLRQqhx6ZA6cCpJM8Zp3AHWpHB71I/YGpGAGAakYHvTHcetV8Z/iDb/Cr4U+KfiFO8e7RdNllt1kYKslwRthQk/3pWRe/wB7oelaJo+q+LfEWn6BpiNc6lrN7Ha26sSTJNK4VQT1yWYfnXw28Ead8M/h54Z+HWkSeZZ+GNItdNgkKbDIsESxhyMnBbbk8nknk1VVVVVVVVVVVVVVRIHGaJJGAaqqr48+HofCXxy+IvhSCZJY9F8WapZI6ReUGEV1IgIQE7QQvTJxX7D+t3XiD9kH4Q315HEkkXhOytFESkDZbxiFDyTyVjUntknAA4FVVVVVVf8ABW34XN8Qf2RdQ8R2Vqkl94E1a21dGAAk8gk28yg7Sdu2cSMuVz5QOTtCn9hn4of8Kj/ap+H3iee58qwu9TXS78lcr5F2DAWYZHCtIr5zxsBw2Npqqo4x1onPANVVVRwB1onPFVVVVVVVTHsDVVVVVVVHAHWic96qqqqqjjHWmIIo896qqqqqqY54BqqqqqqpiMYBqqqqqqpjngGqo4A60ee9VVUcY60Tk8Gqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr/gsh4/+0eJfh58LLeXH2GwudZukxnf5ziGE528Y8ifo3O7kDCk/8ETPh22rfF3x78Ubi2R4PDmgxabA0kYO2e7mD7kJGQwjtXUkdpSDw1VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV4l8Q6Z4T8Paj4m1mZo7LTLZ55iq7mKqM4UfxMegXqSQB1qX9n39oP9pjxzqHxE8TaI3hey1SZmik1h2Q28K8JFHDjzCAuAGKqrcnPJr4b/sN/B/wWYr3xMlx4u1CNeWvx5dsGwMkQKcEdeHZxz6gGrKxsdMs4dP02zgtbW3QJDBBGI0jUdAqjgD2FVVVX/BQHWtQ/au/4KD6L8CvDF6jWmj3Vj4WgmQELHNJIJLyZs5OY2ldGIHS34B6nQ9F0rw3omn+HNDs0tNO0q1jtbO3QkiGGNQiIM5OAqgc+lVo6p+wt/wAFQhZOiWHhdPExhXdAY4U0fU1+QgL95YEnXleN1ueOCtV8dnT9g3/gp3bfFVIriHwr4ium1ySOAnElpfiWK8QqCchJ/OcJjjbGQv3a/wCCQnw61X4l/Ev4n/te+NDFNqF7qE9hbMqtg3t04ur2QZPGFeFRkscStnGPm+NnwzsPjN8IvF/wq1GfyIvFGj3Fis/P+jyOhEcuAQTsfa2Oh24OQSK/4J+/tNxfsM/F7xp8B/2itOu/D+k6reJHeXUlqzNpN/CGCu6qu94JUYfMA3SNlG1matF+OfwU8SaVb63oHxe8GX9hdKWhuINdt3RwDg4IfqCCCOoIIPIrwX8e/gr8SfFl94G+HnxQ8OeJtb0y1N1eW2kX6XYgiDqhLPGSgIZlBXOQT061/wAFR/8AkxL4m/8AcJ/9O1nX/BFT/k1fxT/2UC8/9ILCtR1Cy0nT7nVdSuUt7SyheaeV+BHGgLMx9gATXw6/ap/Z0+K2hR6/4J+MvhS8hMcbywTanHb3FtvGVEsMhV4zwRhgOVPoa/4LKfFD4E+Pb/wHovgbxBoniDxtokl2mp3mlTJP9ktGEZSCaVDgkyEsqfMUxIfk3/P+x/4a8T+D/wBlv4WeGvGJlGr2PheyW4imZme3BjDJC27kGNCsZXopTA4Ar/gip/ydR4p/7J/ef+l9hX/BUj9nvWvjx+zVNfeD9NuL/wASeBr4avZ2lujSS3cARo7iFEX7zbGEgGCxMIUfeOf+CW/7dPw3X4XaX+zj8WvFOn+HNd8OSPD4fvdRmFvb6layOXSEyu20To7sgQ7dyeWF3MGr42ftlfs7fAnwde+KvFHxN0G/uYYWez0bS9Rhur6/fnakUSMTgkYLthFz8zCv+Cb/AMOfH/7UH7Yus/tceNLJP7H0LUrvULucgeXLqVxGyw20YznESS+ZkcKIowfvjP8AwVH/AOTEvib/ANwn/wBO1nX/AARU/wCTV/FP/ZQLz/0gsKr/AIJv/wDKR7xB/wBx/wD9Gmqqqqqqqqqqqqqqqqqqqqqqqqvix8Ivh18cPBN78PPih4XtNd0S9+ZoJ1IaGQAhZYnGGjkXJw6kEZIzgkH9tv8A4JxeP/2Xbi68beCnvfFvw0LFhqRjButJBZVVLxUAXBLACZQFY9VQkA/szftXfFb9lrxamueBdWa40a6mU6x4fuXza6jGMZyMHy5MD5ZVG4dDuUsp/Zl/ap+F37U3gweI/Amo+RqtlHGNa0K4bFzpkrDoeAJIyQdsq/KwH8LBlUHHQ0CCODVVVV9DQIPBrgjg1VVVVVVVVVUDjoaDA8GuCODVVVVVVVVX7ff7Cmm/tReH18c+CDFY/ErQrMxWbSPsh1a3UswtZSeEcFmKScAFirfKQyPH4y+F/jQxzRah4e8T+Gr/AOZJEMNxZXMTdCp5DKw6Gv2VP2mvD37RPgpJXkjs/FmkQxprenAbRvIx58Q7xOQSB1Q/Kc8M0b44BpGBGM80j8YzSP6GlcHjPNJJjvSyAjGaV+ODmllB4zQkBHBoSHHDV5h9a80/5FGQnpRkJ700gx1zTSjGAaZye9PIAMA08meAadwO/NSPnvUjjGAadsDrzUj54Bp2B4Bqq/4KdfF63jsdA+CWk3W64lcatq4UjCIMpbxn3J8xyOCAqHkMK/4JWfBS6+J/7TVl42vNO8/Qvh1bnUrqR8hBduGjtE453798oHA/0ds+hqqqqqqqqqqqqqjjHWiwxgUSTxmqqqqv25rS1s/2u/itDZ20UCN4imkKRoEBdwrO2B3ZmZie5JJ61/wS4/5MS+GP/cW/9O15VVVVVVVePfB2lfETwL4j+H+ubv7O8TaTdaZebCQ3k3ETRPgggg7XPQivGXhTV/Ani/XPA/iCJI9U8O6lcadeohJCzwSNHIBkA43KeoFfsqfFtPjl+zz4G+JUl0k19qWlJHqbKCMXsOYbjg8gGWNyM9iDkg5NUSAOtE54zVVVUSAOtE571VVVVVVVEjGAaqqqqqqo4x1onPAqqqqqqpiCMA0TnpVVVVVVTEAYBqqqqqqokAUee9VVVVVRIAo896qmOeAaqqqmPGAaqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/bx+Jdv8Vf2qvHeu6fcJLp2m3o0myZGV1KWiCF2VlA3K8qSODzw4AJAFf8EhvhhL4F/ZNh8V39k8N5461q51NTIhVzbR7beIY/unyZHU9xLkcEVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVXwi/4J2fBb4O/HeT9ovS/FvjvXvF8txfXLvrl/azxGe7DiaUiO3jfeRLIM7v4znNVX7UH/BPT4GftZeN9P+IPxB1fxbpOsafpq6d5mhXlvAtxCsjunmLLBLllMjgMMcEA5wMaHpjaLomn6M+p3upNYWsduby+kElxclFC+ZKwADO2MsQACSeBX7VX7FXwh/bAh8Oj4mX/AIj0258LtcfYrrQ7qGCVkmCb0fzYpVZcxIRwCCDg8kH4B/AzwR+zh8LNI+Efw+N7JpGkNM63F+8b3NzJLK0jvK8aIrNltoO0YVVHQCq/aO/Yu/Z//alhhuPid4TZNbtYWhtte0qUWt/EhHCmTBWVVJJVZVdVJJAG5szf8ERfg22oPJb/ABn8ZpZGYlIXtbZpVjzwpkCgFtvG7YBnnaOlfsx/sC/AT9k/X7zxd8OT4l1DX72zayfUtZ1PzHFuzq7RCOFIoiC0cZyyEgoMEc5+PfwU8K/tFfCbXfg3421DVbLRfEH2f7TPpcscVynkXEc6bGkR1GXhUHKngnoeR+y/+y94A/ZL8AX/AMOPhxq/iDUtN1HWJNVll1u4hmmWaSGGIqpiijUJtgQgFScluegHjLwj4f8AH/hLWPA3iyye90TX7Gaw1G2W4kgM9vKhSRN8bK6hlYglSDg9a8W/8EVP2etVvRdeEPiN440GNmYyW07297Go42iMmNGUDDZ3M5ORyMc/AH/glr+zP8CvEVh40ng1jxr4g05lltZ9fkje3tp1YMssUEaKoZSoILl8Hkc4Ir9l/wD4J6fBb9kvx/f/ABG+HHifxrqWpajo8mlSxa3e2s0KwyTQyllEVvGwfdAgBLEYLcdCK+PX/BL39l346azf+LV0rVfBniLUZJJ7m98PXCxR3M7biXlgkV4zlm3NsEbMR94ZJrwB/wAEYf2cfDmrjUvHPjDxb4wt42Qx6e8yWEDgH5hIYh5jA8D5HQjnk5BHhDwd4W+H/hnTvBngnQLHRdD0mEQ2VhZQiKKFM5wFHckkknkkkkkkmvj38FPCv7RXwm134N+NtQ1Wy0XxB9n+0z6XLHFcp5FxHOmxpEdRl4VByp4J6Hkfsv8A7L3gD9kvwBf/AA4+HGr+INS03UdYk1WWXW7iGaZZpIYYiqmKKNQm2BCAVJyW56AV8E/+CenwW+AvxrvPjx4P8T+NbzX777Z5ltqV7ayWo+0tukwsduj8E8fPx3zVVVVVVVVVVVVVVVVVVVVVVVVVTQw3EL288SSxyKVdHXIYEYIIPUEdq/bj/wCCT9vLHf8AxW/ZS0fypBvuNT8GISVYBSzPYZJIJI/49+nzYj2gLHXgjx18Qfgx46tPF/gnWr/w14o0G4YRzKm2SBxlXjkjcEMDyrRupBGQwIyK/Yq/4KE+Cv2lbS28D+OPsXhn4jRKsa2XnYg1nEeXltifut8rEwkkqMEM4zt+hoEEYJqqqqoHHANAgjANVVVVVVVVVVV9DW49KyCMA1VVVVVVVf8ABR/9gaf4z2s3xx+DOjxHxvp9uf7Y0mCIK2uwoOJEx965RRjB5kUBQdyqreAPiH46+EHi+HxZ4I1m40bWrHdEzbAQynh45I2BDKcDKsOoB4IBH7Mn7XngT9oLSbfSp7m30bxpDCPtmjySbfOYAlpLYscyJhSxXlkBwcjDFHxwTSOCOtJJ2JpX44NJICME0r8cGll7A0JAeDQcdmoSHsa8yvNyKMmRgUZCR1pnzwWppB0Bp5c8ZpnyOtPIAMA07571JIMYBp3GOvNSSZ4Bp27A0ee9V488a6B8OPBuseOvFFy0Ol6JaPc3DIAWYKOFQEgFmOFUZGSwHevif8Qdb+K3xA174h+IX/07XLxp2jDbhCn3Y4lPUqiKiDPOFFf8E7P2df8Ahn39nfSxrVh5Hirxjs1fWt6YeLev7i3PceXGRlT0d5PWqqqqqqqqqqqqiQBjNFsjAo896qqqqqv2jtWs/EH7Q3xQ13TmZrXUfGer3MBddpKSXkrLkdjgjiv2DNJn0b9jn4SWdzZfZXk8NwXAj2hcrMTKr4H95XDZ77s1VVVVVVVf8FfvgmPhx+0nb/EjTbXy9J+JGn/aywGAL+32xXKgZP8ACbeQnjmY8cZP/BG342pFP4u/Z71a5x52df0cO4ALARw3MYzySQIHCjPCyHA5JokDgUee9VVVRIxgGjz3qqqqqqqqY54BqqqqqqqjjHWmOeBVVVVVVTHsDTHPANVVVVVVHAHWjz3qqqqqqpjnoaqqqqqqYg9DVUxAGAaqqqjjHWjzVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV8fPida/Bj4MeMfifcvEH8P6TNNapK4VZbkjZBGSem+Zo16H73Q9K0LRvEPj/xdp3h7S0n1LXfEepRWturs0klzczyBVBPJZmdh6kk18Lvh/o/wp+G3hf4Z6CENh4X0m206F1iEfmiKNUMhUZ+ZyCzckksSSSc1VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV+3F/wTf8CftOwXfxB8BNaeFviakZZ70o32bWtiEJFcqDhHyFAnUFgOGDgKF8ZeCfiP8E/Hk3hjxjo2q+FPFmgXEcpikJhntpBh45Y3U/7rK6HB4IPev2Df+ClUXjGXTPgt+0Tq9va6ysK2+j+KbqbYuouMgRXbHhZiNoWQ8SEYbDkF6Bxxmsgjg1VVVUD2NcEcGqqqqqqqqqqqrJxgGt3asg8A1VVVVVV+3L/wTX0T46z6l8WPguLXRfiDN+9vtPlcQ2WtMAdzHA/dXLcfPnY5Hz7SzSDxJ4Y8cfC3xdLoXifSNV8NeI9FuAWhnR7ee3lRsq6ngjkAq68Hgg96/Zr/AOCi+r6FJa+D/wBoC4m1PTAqxweIoYN9zb/dA+0IvMqAZJdQZOOQ5OR4d8SaF4r0a08Q+GdZs9V0y9TfbXlnMssUq5wSrKSDggg+hBHakcEYzSyY4zSuD0NLJgcGlkBGCaD+jUJDihLxjNCXIwDXm+9GXtmjLkcGjISMCmfI600gHANPJnoadxjrTyZ4Bp3AGAaeTPANO3YGjz3qpZI4Y2llkVERSzMxwFA6knsK/bo/arX4xeIv+FbeA9SWTwXoU5aS6gkJXV7kADzOgzFGdyoBwxy+SCm3/gmf+yY3x8+K8fxF8X6Yk3gbwLdRz3CTKCmoXww8NsVIIdBgPIDwV2qeJKqqqqqqqqqqokdM0WGMCiSRjNVVVVVVfHf9pH4Pfs3+Gx4j+Kni23sGmVjZabCRLe35BAIhgB3MASAW4VcjcwzX7Tn/AAVD+NPxlurvw98Lrm6+HnhBt0apZzj+0bxMkbpbhQDFuGP3cRAGSC7jmvhj8JfiV8aPFNt4K+FngzU/EmsXLKFt7OLIiBON8shwkSZ6u7Ko7kV8MfB8Xw9+GvhPwBAkSR+GtDstMRYZGdFFvAkQCs/zMPk4LcnvzVVVVVVVV/wUp+AqfHb9ljxGum2KT+IPBatr+kkRFpGMCMZ4k28kyQmQBeQziPjIBHwN+KurfA/4u+FPivoqNJceHNRS4eFXKGeE5SaLI6B4mdD7NXhLxXoXjvwrpHjPwvfJeaRrllFe2U6/8tIpEDKcdjgjI7Hiqqqo4x1onIwDVVVVVVVUcY60xBGAaqqqqqqqmOeAaqqqqqqJAGAaYgjANVVVVVVHGOtE56GqqqqqqmPGAaqqqqqqYjGAaqjjHWic1VVVMc8A1VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV/wWB+Ma2Hhzwj8B9NlT7Rqsp1zVOSGSCPdFbr6FXkMzHuDAvqa/4JN/BSX4pftT2HjK+skm0T4c2ratctIxAN0wMdqoxzuEjGUdv3ByegNVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV+1p+xv8AC39rjwY+jeKrWLSfE1pGBo/ii2tFkurAgkhG5UywEs26EsAdxIKthh+0H+z58SP2aPiRffDX4laV5F3B+8sr2IE2+o25JCTwOQNyHBGOCpBVgCCB+wL/AMFJIvBNnpnwQ/aG1aeXRI2jttB8TTNuOnJ0WC7YnPkDgLLyYxw3yYMdrdWt/aw31jcxXFvcRrJDNE4dJEYZVlYcEEEEEcGvoaBB4Jqqqqr6GgwIwa4I4NVVVVVVVVVVVVZOMA1u7VkHgGqqqqq+P/7LnwX/AGmNBGj/ABS8Kpc3dvC8dhq9o3kX1hu7xS4PAPOxwyE9VNftXfsEfGH9l+8uNaltG8T+B2mK23iCwhJ8pewuoxkwN2ySUJ6NngfB74//ABU+BWpvf/DvxNJaQXDhrvT51821uscfPGeM443LhgOhFfCn/gp94G1j7Npnxd8JXnh66d9kmpabm7tFH99o/wDWoO2FEp798DwD8efg58TYoG8C/ErQNVmnUFLRLxY7kZ6ZgfEin2KilcHoaWXsTSyAjg0H4wDQlPrQlB4zXmgivNH+RRlGOKMo6A0ZSRgGmkJ4zTSDpmmlB4Bp3z1NPIMYBp3zwDTPngGjz3qvHXj/AMGfDPw5ceLPHniOz0XSrbh7i5fG5sEhEUZaRyFOEUFjjgGv2pf26fE/xkjvPA3w7W68P+C5lMVwXAW71NDtyJSCfLjyCNin5gTuJB2j9l79mLx/+1F8SbDwZ4UsJ4NIjmR9c1toSYNMtgRvck4DSEHCR5yzY6KGZfhH8J/BXwQ+HmjfDD4e6c1nomiQlIVkffJK7MWklkb+J3dmYnAGTgAAACqqqqqqqqiR0zRYYwKJJ4zVVVVVVVarqul6Hp1xq+tala6fYWcZkuLq6mWKKFB1ZnYgKB6k4r9q7/grXp2kfbfA37L9tFqF8N0Uvi6+h3QQMMgm1gYYlOcESSfJx9xwcjxB4i8d/Fjxk+r+IdU1fxT4m126VA8rPc3N1M7YSNFGSSSQqoo7gAdBX7KH/BITxx47Sy8b/tJXtx4R0NmSSLw5bj/iZXSg8idj8tspAHHzSEEgiMgE/Cz4QfDP4JeE7fwR8KvBun+HNGt+RBaoS0jf35JGJeV/9p2Zveqqqqqqqqq/4KDfs2/8M0/tH634f0bT/s/hPxHnWPDmxMJFbysd9uMcDyZQ8YXJOwRsfv1/wSI/aNi8QeDdT/Zz8T6on9peHWa+8PCWTDTWcjEzQrnqY5CXx12ynslVVUSAOtE54qqqqqqqqOMdaY54BqqqqqqqqY54BqqqqqqokAdaY+h5o896qqqqqqY54BqqqqqqokAdaPPeqqqqqo4A60ee9VTHsDVVVUSAODR571VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVapqenaJpl3rOrXkVpY2ED3FzcSttSGJFLO7HsAoJJ9q/aY+Md38fPjh4r+J80s/2TU70ppkMx5gs4wEgTA4U7FUkD+JmJySSf+CV/wBl+C37Mdj4j1vT3t/EPxDmGs3YkJ3R2pXbaR44wPLzJjqDOwJ4AFVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVa74s8K+F/I/4SbxNpWkfad3kfb72O383bjdt3kZxuXOOmR61aXdpqFpDfWF1Fc21zGskM0Lh0kRhlWVhwQQQQRwc1VVVVVVVVVVVVVVVVVVVV/wAJv4K/tn/hHP8AhMNE/tbzPK+wf2hF5+/+75e7dn2xVVVVVVVVVVVVVVVV+0Z+zP8ACj9qLwM3gj4oaH9o+z+ZJpepQN5d1pk7Jt82Fx+BKNlG2ruU4GP2qv2Ofi5+yX4rOkeONP8A7R8P3cmNJ8S2ML/Y70HcVQsR+6n2qS0LHIwSC64c/sef8FB/iR+zFLB4Q1+Ofxd8PpJhv0qe5bz9MU53NZOxIQZO4xMNjEHBjLM9fBz45fC74++E4/GPws8WWus2XyrcRKdk9nIRny5oj80b+xGDjIJHNUDjjNAgjg1VVVUDjoaBB4rgjg1VVVVVVVVVVVVX0Nbj0rcDxWQeAaqqqaGK4ieCeNJI5FKujrkMCMEEdxiv2gv+CWv7PXxlnutf8FRS/DjxHdSeZJc6TB5tlK2ACXsyyovA/wCWTRZJJOSTXxe/4Jf/ALV3wuFxfaP4UtfHWlQ72Fz4bn8+YIOVzbOFmLkfwxrIAQRk8E+JPCnijwbqj6H4v8N6poepRZ32epWclrMmGKnKSAMMMrDp1UjtXhT4+fG3wOiw+FPiv4p0+Bc4tk1SVockAZ8piUzhQM4yMCvDn/BRP9qPQhGt94s0rXUjdSE1HSIRlV6oTCI2IOOTnd7ivh9/wVX090htfip8LriFlT99e+H7oSB2x/DbzFdoz6ynAPfHPhD9un9l7xgYobf4o2ml3MjAGHV7aWy2EttGZHUR4PByHOAcnHOPDnxF8BeMFibwl430DWxMrNEdO1KK53hThiNjHIBBB9MUJQe9eYDxkV5i4xmtR1XTdJs5NQ1W/trO1ix5k9xKI0TJAGWYgDJIH1IrxH+0P8CvCayDxB8YfB9pLCqs1udYhebDEAERKxcjnsOmT0BNH9sj9mIjj40eHv8Av6//AMTTftjfsyHgfGjw9/39b/4mm/bE/ZjP/NaPDx/7av8A/E037Yn7MpGB8Z/D3/f1v/iaf9sL9mY8D4zeH/8Av63/AMTVj+1l+zdqd3HZW/xp8LrJJnabi9ECDAzy8m1R07kZPHU1pHi/wp4jkEPh/wAT6Tqb+V5oSzvY5js4+bCk/LyOenI9a1TWdH0O3W71rVrPT4HfYst1OsSliCdoLEDOATj2NeM/2r/2dPAayLrvxb0GWaJmV7fTpzfyq4OCpSAOVORjDYx3xXxb/wCCnlmLWfSvgp4Nma4JZBq2ugKiAcbo7dGJbPUF2GMDKHJA+IPxP+IHxW1s+IfiH4t1DXb7kRtcyfJCD1WKNcJEuRnaiqM9q/ZI/wCCaHxS+PU+n+MPiZb3/gjwDLtl86aLy9Q1KMgEfZonB2IynImcbcEFVkFfCz4T/D74K+DLPwB8MvDNromi2XKwwglpZCAGlkc5aSRsDLsSTgDOAAKqqqqqqJHTNFhjAosTwKJz3qqqqqqqqr9p/wDbO+DP7LGjt/wmOr/2p4ouLcy6d4Z091a7uOytJ2giJ/5aP1CvsDspWv2nP20/jV+1Jq0yeLdbbSfCyTFrHwxp0rJaQgEbTL3uJBgHfJnBLbFQNtr9lz9hr45/tWalbXPhDRP7I8Ii4MV74o1JClpCF++Ih96eQdAicbiAzICWH7LP7C/wO/ZR0+K78JaS+teLXh2XnifVEV7p9wAdYQBtgiJBwi84IDM+M1VVVVVVVVVV/wAFGf2XW/aa/Z/vIPDejpeeN/CDNqXh0qg82Y4AntVJ5xLGowvQyRxZ6V8Jvid4o+DPxH8P/E/wXd+Rq/h67E8BP3ZFIKyRN/sPGzo3+y5r4NfFrwj8dPhloPxT8EXLSaXrtqJVjkGJLaUHbLDIOzxuGQ4yCVyCQQTVEgcA0ee9VVVVVVVRxjrROeBVVVVVVVUxzwDVVVVVVRwB1pjR571VVVVVUxwMCqqqqqqjjHWic96qqqqqqYg8A1VEgCjz3qqqsA8ZpiD0NVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVf8FVP2h4vh98KLf4J+HtTRPEHjpd1+kcn7y30tGw5IHIEzr5Y7MqzDtX7GP7PF1+07+0L4a+GbwynREk/tDxFNGSph06EqZvmHKmQlIVbs8yE8ZqGGG3hS3t4kjjiUKiIu0KAMAADoMdqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/ay+KXxY+CEGgfEXwPPaX2gmV7HVtLvbRZIhIw3RS702yqSFdfvbM7cjJ5+Fv7enwl8aiPT/GyT+DtSbaA10TNaSMcDiZRlOc/6xVUDHzGtK1bStd06DV9D1O01Cxuk3wXVrMssUq+qupIYe4NVVVX/BVv9p74/wDwV/aI8PeFfhV8VNa8NaTdeC7a9mtLKRVR52vLxGkOQeSsaD/gIqq/4Kt/tPfH/wCCv7RHh7wr8KviprXhrSbrwXbXs1pZSKqPO15eI0hyDyVjQf8AARVVVVVf8Fz/APmiX/cwf+46v2Tv+TV/g1/2T/Qv/SCGqqq+B/7dH7P37R3xTv8A4UfB7V9Y1y703RZNWl1N9Me0szEksETIvnbJd+64XgxhcK3zdM1VVVVVVVV/wUG/aM/bh+Hv7V+h+Bvgw+u6XoM1rar4ZtdM0xbuPX7h13Tlw0beY4fdGYskKkauApck6HPql3omn3WuWKWWpTWsb3lqjhxBMVBdAwJBAbIyCc4qqqqqqqv+cyv/AHUD/wBoVVVVVVVVVVVVVVVVV448CeDviX4W1DwT498OWOuaHqkLQ3VleRb0dSCMjurDOQykMpwQQRmv2yP+CTvjb4ZPqXxE/Z1iu/FfhNPMuLjQSDJqOmJuztiHJuo1B7fvAByH5avh38T/AIlfBbxZF4q+HPizVPDOt2UgDSWz7NxU8xyxtlZEyOY5FZT0INfsw/8ABWTwH47a08IftC2Nv4P12VtkeuWoP9lzccGXcxe3Ynjncncsg4rS9V0vXNOt9X0TUrXULG7jElvdWsyyxTIejI6khgfUHFfQ0GB4NcEcGqqqqgcdDQIPGaqqqqqqqqqqqqqqrJxgGtxxjNbgayDwDXB6GqrX/DXhzxXYf2V4o0DTdYsi277NqFolxHuwVztcEZwzDp0J9a8Z/wDBP/8AY68dyPPq3wI0CylbkNoxl0oIdoUEJavGnQA4KkZ5IJJz4y/4I5fs561FLL4O8ZeNfDl2ygRK9zDe26EdSUeMSEnj/loBXxL/AOCQH7SPhWW4n+Hut+GfHFkjMIEjuf7Ou5AOhaOf90pIxwJm5zz3PjX9lH9pX4duw8XfA3xnZxLjNzHpMtxbglioHnRBo8kqeN2cYPQgm7tLqwuprG+tpbe5t5GjmhmQo8bqcMrKeQQQQQehrTfF/i3RbUWOjeKdXsbcMWENtfSRICep2qQM1/wsX4g/9D34h/8ABpN/8VX/AAsX4g/9D34h/wDBpN/8VUskk8jTTSNJI7FmZjksT1JPrXhrwL438aSrB4O8Ha5rsjTLCqabp0t0TI3CoBGp+Y5GB1Nf8MrftP8A/Rt/xR/8I+//APjVf8MrftP/APRt/wAUf/CPv/8A41X/AAyt+0//ANG3/FH/AMI+/wD/AI1X/DK37T//AEbf8Uf/AAj7/wD+NV/wyt+0/wD9G3/FH/wj7/8A+NV4g+AHx38J2q33ir4J+PdGtmztm1Dw3d26HAycM8YHA5NVXgj4L/F74lzW8Hw++GHijxF9q5ik07SZp49ocIXLqpVUDkKWJCqepFfCD/gkv+0v4/nhuviEul/DzSn2s0l/Ml7dlCFOVt4HIBwT8sjxkFSCBX7Ov/BOv9nf9n37LrQ0H/hMfFUGG/trXYll8px/FBB/q4eeQcM4/vmqqqqqOMdaJHTNFh2osSMCiSeM1VVVVVVVVVa1rWjeHNJu9e8Q6vZaXpmnwtNd3t7OsENvGoyzu7EKqgckkgCv2tv+Cs//AB+eAv2V/ZLjxjeW/wD30LOCQfQebKP721PuyVp+m/ED4u+N0sdNtdd8YeLfEV0zBEEt9e38zZZ2J+Z3bALFjngEk4BNfsj/APBH+y0eWz8dftVzW+oXSrHNb+EbC43QwvwSLuZDiUjoY4zsP99wcVpWkaVoOm22jaFplpp1hZxiK2tLSBYYoUHRURQAoHoBiqqqqqqqqqqqr/gq7+yLL8H/AIoyfHTwRpDr4N8d3TPfJDGfL03VGG6RTycLPhpV7BjIowAor/gmr+16n7P/AMRX+Gvje/ZfAnjW5jV5Hc7dL1A4SO5x0COAscnAOBG2cRkE4x1onPANVVVVVVVVEgDrROe9VVVVVVVTHPANVVVVVVHGOtMQRR571VVVVVUSAOtHnvVVVVVVTHPANVVVVVVMcDANVVE56GqqqpiMYFVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVeP8Axz4a+GXgrWviB4w1BLLRtAspLu7lYjO1BnaoJG52OFVerMygckV8fPjH4i+PvxZ8Q/FLxJKwm1e5P2W2J+WztU+WGFQOMKgUE/xNuY8sSf8Aglr+y63wH+A8PjrxXo6W/jP4hLHf3HmoPNtLArm2tyeqkqfNZeCGkCsMpxVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVfGn40+FfgT4VtfF3i6w1W8s7vUEsY49OijkkEjRyOCQ7oNuIm75yRx6f8PGPgj/ANCv44/8AbX/AOSa/wCHjHwR/wChX8cf+ANr/wDJNf8ADxj4I/8AQr+OP/AG1/8Akmv+HjHwR/6Ffxx/4A2v/wAk18V/22vgJ8Ufhzr/AICv/C/jYJq9m0cbtZWwEcykPE5xcHhZFRunaq8BfFb4jfC+9+3+AvGGoaQ5JLxROHhkJAGXicFH4A+8p6Cvhj/wUXB+zaZ8W/CPoj6po5/Dc0Dn6E7W9SF6LXgL4rfDn4o2P2/wF4v0/WEVSXiicpNGAQMvE4DoOR95R1FVVf8ABav/AJOo8Lf9k/s//S+/qq/4LV/8nUeFv+yf2f8A6X39V8H/APgoV8APjL448a+BNOHiDw3c+AdPutR1a98Q28FtaC2tplhmlWVJpMBWdD84UkNkdDj4hf8ABZz9nbwvr8+ieC/Bfi3xdBaTPG+pRrDZ20wGNrw72MjKfm++kZ4GAQeP2Yf23/gT+1fby2nw/wBYutO8RWkbSXPh3WI0hvEjUgGVArMkkeWA3IxIyNwXIFV+0N+1N8Fv2XvD0GvfFnxR9kmvt39n6Xax+feX20qG8qLI4XcMsxVRkc5IBsP+C2fwJuNeWzv/AIUeOLXR2mZftytbSTLHzscw+YBk/LuUOcZOC2AD/wAFb/jN8M/j14J+BXxC+FHiy017RZ5PEMLSw5V7eZRppaGaNgHikAZTsYA7WVhlWUn9k7/k1f4Nf9k/0L/0ghr41fHn4Ufs9eEJPGvxZ8X2mi2PzLbROd897Iq58qCIfNI/sBgZySBzWsf8Fq/2crPXI7HSfh18QNR0xZGSe+NvawtgHAeKIzEuCMEbzGeeQK/Z4/a2+BX7UWlTXnwn8Ypc39kpa90W9j+zX9ooIG9oSfmT5lHmIWTLbd27IH7U/wC3h8IP2Q9f0Pw58SfDvjDUrnX7OS7tn0OztpkREfYQ5lniIOfQEe9f8E9P2oPAH7Jfxp1n4jfEbSPEGpabqPhefSootEt4ZplmkurWUMwlljUJtgcEhicleOpH7LH7eHwg/a81/XPDnw18O+MNNufD9nHd3L65Z20KOjvsAQwzyknPqAPevjV8efhR+z14Qk8a/FnxfaaLY/MttE53z3sirnyoIh80j+wGBnJIHNax/wAFq/2crPXI7HSfh18QNR0xZGSe+NvawtgHAeKIzEuCMEbzGeeQK/Z4/a2+BX7UWlTXnwn8Ypc39kpa90W9j+zX9ooIG9oSfmT5lHmIWTLbd27IHxD8a6V8NPAHib4ja7b3c+m+FdHu9VvIrRFaaSG2haV1jDMqlyqEAFlGcZI618Ov+Cg37OnxB+Buu/tCXeq6r4T8K+HtYOkXK6/bxJdS3PlRyKsUNvJMZN4lAUDn5JCQFUtX7Kn/AAUS+HX7XHxW1f4ZeB/AfiPSF0vRZdWS+1WSFTLHHNBEVMcbPtYm4B+8RhTzzXxH/bt+D3wu/aN0D9mLxJoXi2TxR4iurC2tL21s4HsUkvXCQ73M4kA3MAxEZxnvX7Qn/BVP9nL4FeKbzwJpcOseOtf02ZoNQTRVRbWzlU4eJ7iRgGcdCI1cAgqzKwIr9l3/AIKR/Ab9qDxJF4B02HVfCni2aN3ttM1kRhL3bklbeZGIkcJhihCtw2AwUtX7TP8AwUb/AGd/2Ytem8Ea9ear4n8XQR7p9G0KBJDZFk3Ri4lkdEj3cfKpeRQwYpgjPwU/4K7fs7fFXxhZeCfFGia74EudTmENnqGqvDJYlzgKssqNmIknAZl2DqzLVftQ/wDBQv4Lfsl+P7D4cfEbwz411LUtR0ePVYpdEsrWaFYZJpogrGW4jYPugckBSMFeeoFfD/8Ab6+BnxD+O/jD9n23tvEeia54JbU11DUtYt7eDT2/s+Ux3DJMszsqgK8gaREGxCSVOFPxP/4LIfs3eC9fuNA8F+GvFHjUWczRyajaJFa2cmOCYnkbe4yOpRQRyCRX7Lv7ffwD/asuv+Ed8I6je6F4sjh8x9A1pUinmAXLtbsrMs6rhicEOFG4ooqv2gP2qPgf+zJpFvqXxa8ZQ2Fxe82WmW6Ge8ul3BWdIV52LnljheMZzgGH/gtd+zpLryWb/DT4hxaOyjN88FoZkb3hE5G0eocn/Z7V8D/2g/hD+0b4THjD4ReM7TW7SPat3bjMVzYuc4SeFsPGSVbBI2sFJUsvNV+0j/wUv/Zy/Zy1+88EXd3qfi/xVYN5d1pmhIkiWcnOUnndlRGBXDIu91JAKjnHwk/4LC/s2fEHxJB4a8a6P4g8BG7kEdvqWpCO4sgxIAEskRLRZJ+8U2AZLMoGatLu01D/AILFQ31hdRXNtc+PFkhmhcOkiNb5VlYcEEEEEcHNVVVVVVVVVVVVVVVVVX7V/wDwTj+Bn7Tkd/4mtrL/AIQ7x7PmRdf0yIbbuTbgfa4PuzA8ZYbZOB8+Mqf2lP2Kfj3+y3qDt8QPCz3vh9mAt/EmlI9xp8mThVaTaPJc9kkCk9sjmvgX+1R8cv2c9RW6+GHji6tbFpN9xo90ftFjcnvuhbgE9N6bX9GFfs7/APBWT4QfEb7L4e+NNh/wgGuyYT7dvabTJ29fM+/Bk9pAVA6yVo+taN4i0y31rw/q9lqen3S7oLuznWaKUA4yrqSGGQRwe1A46GgQeKqqqqoHHQ0CDxXBHBqqqqqqqqqqqqqqqqrJ6A1k+tbuMVuBFZBHWsgjg1wRwa1vw14c8SwxQeI9A03VY4WLRJe2iThGIwSA4ODir/8AZm/Zu1W8l1HVP2ffhreXc7bpZ7jwnZSSSH1ZjEST9a/4ZW/Zf/6Nv+F3/hH2H/xqv+GVv2X/APo2/wCF3/hH2H/xqtK+FXwv0L7H/Yfw38Laf/Z2z7H9k0eCH7Nsxs8vag2bcDGMYwMVVVVVWueFPC/ifyP+Ek8N6Xq32bd5H26zjn8rdjdt3g4ztXOOuB6Vpnw/8B6JfRapovgnQLC8gz5Vxa6bFFImQVOGVQRkEg+xNVVHGOtEjHWiRjg0WGOKLZGAKJJ6GiSe9VVVVVVVVVVVVftP/tnfBn9ljR2/4THV/wC1PFFxbmXTvDOnurXdx2VpO0ERP/LR+oV9gdlK1+0/+2d8Zf2qNYb/AITHV/7M8L29wZdO8M6e7JaW+OFaTvPKB/y0foWfYEVitfsl/sJfGT9rPVY7vQbX/hHfBkMhW98UahbM0A2sA8dugwbiUAk7AVUYwzpkZ/Zo/ZB+Cf7Knh46X8NPDvmavdRlNR8Q6htlv77JBKtIAAkeVXESBU+UEgtljVVVVVVVVVVVVXxY+Fngv41/DzW/hj8QNKS/0TXbVoJ12qXhYj5ZoiwISVGw6Ng7WUGv2k/2dvHf7MPxU1P4YeObZnNuxl0zUlhMcOqWhJ8u4iznggYZcnYwZSSVr9jf/gqLqPwu0fR/hV8e7G61jwxp8aWthr9ovmXlhEMKiTRk/vokXuv7xVGAJDgV4E+IPgj4oeGLTxl8PPFWm+INFvVzDeWM4kXOASjY5RxkBkYBlPBAPFVVVVVVVRIAwDR571VVVVVVVMc8A1VVVVVUcY60xB70TnvVVVVVVRxjrROe9VVVVVVMcDANVVVVVVHAHWjz3qqYjGAaqqqjgDrR571VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV/wVA/a4i+IniUfs+/DzV2k8OeG7kv4guYJMJqF+hIEIx96ODnPOGkJ4/dIx/wCCZn7Jn/DR3xri8VeLLKU+BvAckV9qOYspqF0GzBabjxgkb5Bg/IhXjzAwqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/bB+EXjL40/DPTPC3geC0lv7XXYryRbmcQqIlgnQkE98yLxX/DBH7Qn/AEDdD/8ABmv+Ff8ADBH7Qn/QN0P/AMGa/wCFf8MEftCf9A3Q/wDwZr/hX/DBH7Qn/QN0P/wZr/hWrfsOfHTQtKvNb1eDw/a2On28lxdTyaooWKJFLOx46BQT+FVXw0+AnxY+Ljo/gnwdeXNkxYHUZ18i1Ur94ea2FJH91cn2r4Pf8E/9I8J39p4j+IfjW+vdTtXDx2uhzvZxIcYYGcYlYckZTyz+eKjjWONYlLEIoA3MWPHqTyfqaqv+C1f/ACdR4W/7J/Z/+l9/VV/wWr/5Oo8Lf9k/s/8A0vv6r4KfBHW/2jv2xde+DGna/caPpPiHWtR/4SG6gZQw06C6NxIoB+8xeGIKMEB9jEbVNaL+xL+yRoXha38IW37O3gG5tLe1NsLm90KC5vHUjBZrqRTMz8k79+4HoRgY/aB8BW//AATy/b68P638P7i6svDdneWfiHSoTMZpI9MnkeK5tSxJdkwl1ANx3lMEkk7jq+qWGh6Vea3qlx5Nlp9vJcXMuwtsjRSzNhQScAE4AJr9n/wJqv8AwU4/bS8Q+MfivqWoR+FrGOTVLyzSdsxWKTKlrpsT/wDLMHf8xXGQsrAh23Vp/wCzt8AtL8Mw+DLL4KeB10OCHyVsH0C2kiKZyQysh3EnJJbJJJJJJJr/AIKd/sm+HP2Yfipod98OLZ7LwN41tZ7nTtNa5aUWF5CY1uo03ktsKvbuCxJy7L0QV+yd/wAmr/Br/sn+hf8ApBDXwr024/4Kf/t6av4o+IF29z8OPB0Mlxa6eUaNX0yKfba2gK7WVpmkMrsfmwJBkYUCx+G/w+0zwcnw8sPA+gw+GEhWEaMunxfZDGuMKYtu0gbV6jsK/bE8GWX/AAT0/bc8IfFz4IWiafouqwjV49IX/UwkyPDeWiKCCIXjIKjICmUhcBFx4t+E3wW+L39n6747+Gfgnxr5Vv8A8S+81fRbXUtsL4YeU8iNhG4PynB61/wSK+HngD4l/tJ+JNB+I3gbw/4q02DwPdXMVlremQ30Mcy3tkokVJVZQ4V3UMBkBmHc14H+Dfwg+GN1c33w1+FXg/wnc3sYiuZtD0K2sHnQHIV2hRSwB5weM18K9NuP+Cn/AO3pq/ij4gXb3Pw48HQyXFrp5Ro1fTIp9traArtZWmaQyux+bAkGRhQLH4b/AA+0zwcnw8sPA+gw+GEhWEaMunxfZDGuMKYtu0gbV6jsK/bE8GWX/BPT9tzwh8XPghaJp+i6rCNXj0hf9TCTI8N5aIoIIheMgqMgKZSFwEXH7U93aah+yZ8X76wuorm2ufh3rckM0Lh0kRtPmKsrDgggggjg5r/gn5+zbqn7W/j+X4ZeKNf1Oz+GfhZv7e1y1s5RH9puWCwwxg8/O4DYcg7ESYKVZ8n4Sfssfs+fAjVTrvwk+Fuk+HNSfT/7PkvLdpHmktyyMUd5GZmy0UZJYkkqCT1r/gqnHr837d8cPhSe4h1t9N0hdNlt5/IlS5P+qZJMjYwfaQ2Rg4ORX7O/7FPwI/Z9+HmneEtP8BaDresrCDq2u6lpsVxdX85ALne6krECPkjGFUAdWLMf+Cp/7PHhj9mf43eDvid8HLOHwvY+Lo5bu3sNOjWCLTtRsniLyW6pgRIRLAwUDAcOQcEKv/BPn9kT4d+HPgnofxl+I/hrSfF3xB+JNuPEF9rGrWqXckMV4oljijLg7coyu5xuLyOCSoUD/gr5+yh8NvDXgHSf2hfh34TstC1aHVotM12DTYI7aC6glSQpcPGuB5qyIqZUbmEvzZCAj/gnF8UtX+LX7HvgPW/Ed+95q+kwzaPdzuhBcWsrRQkkkl28gQ7mPLNuJ5r/AILV/wDJ1Hhb/sn9n/6X39Vq/wALPE3x1/4KJfEL4K+GNbfSm8afEbXrPULlSPksY76e4uWwSN5WOBnCZG4qFyM14N/Ya/ZK8FeELLwda/ALwTq0VnbiFr/WdDt769uDtw0klxIhcuxyflIAJ+UKMAftxfBu3/YS/a58M+Nvgskum6NP9m8Q6Fb/AGgv9jmimImtssSzR5QHDAgpNsO7DV4i+JHh7w58K9T+L100p0PS/D82uyHYQ5to7czn5QCc7B0wTmv2EPhFL+35+0X47/aX/aMgt/EGl6LNCBpNxEfs91cyqwgh2rgGG3iiXKH7xaPduy+fEPw3+H3izwsfA/ibwPoOqeHtu0aVd6fFJbKMEfLGV2qcM2CACMnFeHLL/h35/wAFPLTwH4J1Hy/BXibULLTnjvZ8qmmai0XEjFhj7PMch2OSsIJzuIP/AAUW+P2q/s7/ALL2veJfDN7LZ+IvENxHoWjXUTMr2086Ozyqy/ddIYpmVuMOq89Af+CV/wCw/wDDzxx4FH7TXxl0O38UXOrX08Xh7TNRiE9rGkMmyS7kVsiWUzJIgDZVQhPLMNnxa/ZG/Z0+NXhO48JeNfhR4f8ALljZbe/0+wjtLyxc4PmQTxqGQ5VSRyrbQGVlyD+zR8ONf+EH/BR/wd8LfE14l3qPhbxkdPe5RsrOkSusci8khWjCMFPKggHBBAqqqqqqqqqqqqqqqqqqvLO01C0msL62iuba5jaKaGZA6SIwwysp4IIJBB4Oa/aS/wCCSfwE+Ln2rxF8JX/4Vl4lky/lWNv5ul3D9cNa5Hk5wFBhZFUZPlua+P37EP7SP7N8t1c/ED4f3FzoVs2B4h0jN5p7qeAxkUBogSCMTLG3TjkZ+EP7Qnxn+A2pjUvhT8Q9W0JWmEs9lFMXtLpgMZlt2zHIccZZSR2Ir4Hf8Fj7VoLTRP2hfh7Ksy4R9e8NkMrcgBpLSRgVwMszJIc/wxjgV8Jv2ivgf8dLVrn4UfEzRvEDx/6y1ilMN1GMZBa3lCyqCM8lQDg+hwCRxmgQeAaqqqqrPoa3AjBrgjg1VVVVVVVVVVVVVVVVVVVfQ1k+tZOMA1k+tbjjFbj7VuPtW4+1bj7VuPtW4+1bj7VuPtWT61k+tZPrR571VVVVVVVVVVVVVVVeKPFPhrwToF74q8Ya/YaLo+nR+Zd39/cLBDAuQAWdiAMkgD1JAHJr9rj/AIKx3l8194A/ZeLWtuGeG58XXMPzyrjB+yRMPkHXErjd6KvDVp+m/ED4u+N0sdNtdd8YeLfEV0zBEEt9e38zZZ2J+Z3bALFjngEk4BNfshf8EgrTSns/Hn7VphvLpdslv4Ps7gPFC4bP+lzxkiXoP3cZ2+rsCVrStI0rQdNttG0LTLTTrCzjEVtaWkCwxQoOioigBQPQDFVVVVVVVVVVVVVVftDfsu/Bj9qHwsPDHxX8LJdy2yv/AGdq1qRDf6azDBaCbBwOhKMGjYqpZW2jH7WX/BM/42/s5S3fijwrZXHjvwIjMw1TTrctc2KckC6t1yygKOZUBj6ZKEha+EHx0+LHwF8QnxN8KPGt/oN3LtFzHEweC7Vc7VmhYFJAMtjcDjccYJzX7NX/AAVi+GnxE8jw18erG18B66dqJqcDvJpl0x6k5y9rzjhy645Mg6Vpeq6XrmnW+r6JqVrqFjdxiS3urWZZYpkPRkdSQwPqDiqqqqqokdAaPPeqqqqqqqOMdaYgjANVVVVVVHGOtMQRgGic9KqqqqqqOMdaYg8CqqqqqqjgDrR571VVVVVUxzwDVUSAKPPeqqqwDxmmOeAaqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/4KI/tmwfAfwdJ8Lvh3rOPiJ4it8NLAxDaLZuCDPuHSZsbY16jJk42qH+D3wm8b/H74o6L8MfBFo19rniC6Kh5H+WJAC8s8jE/dRAzk9TjAySAf2ePgT4O/Zu+Eeg/CXwXCjW2lQ5vL0xbJNRu25muZOSdzt0BJ2qFQHaigVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX7eHxIuPC/wttvAGiPK2seNbn7KscJy5tkIMoAGSdzGOPAHIduex+GP7D/xp+IAivdc06Pwhpj5zLqyMtwcf3bcfPnP9/YMA89M/C39ij4K/DgR32paW/izVV2n7TrCq8UbDGfLgHyAZAPz72H97FQW8FrBHa2sMcMMKBI441CqigYAAHAAAxiqqq/4LaeHbq2+P3gXxY75ttR8HiyjXYRh7e7ndznoeLlOO2Oeorwb4n0/xv4P0LxnpLo1jr2m29/bMkgkBjmjWRSGXhhtYcjg1X/BVhYfiv+3h4Y+GuiSMb+LRdH8Py+Qn2iRbi4uppUxGpyW2XceEOCcjswNV/wAE3/8AlI94g/7j/wD6NNV/wWr/AOTqPC3/AGT+z/8AS+/r9py01K//AGbPixY6NHLJqFx4H1mO0SE4dpWsZggU9juIx71/wQ2v9Kj1r4xaXM6f2lcWujTW6mMljDG14spDYwBukhyMjORwcHFf8FzZoWl+CtusqGVF15mQN8yg/wBngEj0O0/kfSvgLpupa1+xH8OdH0Z9moX3wq0uC0bfs2yvpcaod38PzEc9q/YJ/ZQ8F/tffETxD8P/ABT451Pw7c6Tov8Aadp9htVladFmjikyW+UBTLH3yd3GcHH/AA5C+FX/AEXDxX/4Lrf/ABr/AIchfCr/AKLh4r/8F1v/AI14Y0OHwx4a0nw1bztPHpNjDaJK4wziNAgYgdCQua/4Iqf8nUeKf+yf3n/pfYV4403Uta8Fa/o+jPs1C+0u5gtG37NsrxMqHd/D8xHPav2Cf2UPBf7X3xE8Q/D/AMU+OdT8O3Ok6L/adp9htVladFmjikyW+UBTLH3yd3GcHH/DkL4Vf9Fw8V/+C63/AMa/4chfCr/ouHiv/wAF1v8A41+0XocPhj9jL4n+Gredp49J+GGr2iSuMM4j0yVAxA6Ehc1/wQx/5rZ/3AP/AHI1X/BSD/lI94e/7gH/AKNFV/wXP/5ol/3MH/uOr9k7/k1f4Nf9k/0L/wBIIa/4K8/8maal/wBjBp3/AKG1f8Ehv+TNNM/7GDUf/Q1r/gtX/wAnUeFv+yf2f/pff1X7NX/KZXV/+ygeM/8A0RqVV/wW9/5Kn8NP+xfuv/SgV+0Rpupat/wTu8V2ulPtnj+GYnY79n7qK0SSUZ940cY79O9fsA/sM+AP2ydE8ZXXiH4j6xoGpeE7q0Q2tjZpIGhuFkKOWfjJaGQYB4284yM/8OQvhV/0XDxX/wCC63/xrwx/wRe+GHhjxLpPiW3+NPiieXSb6G7SJ9PgAdonDgEg9DtxX/Bbi01J/gt8Pb2KOU2EPiiWOdgfkErWrmMEepVJcfRq/wCCdl/pWpfsU/Cm40d0aBNJkhcrGUHnR3MscwwQOfMVwT3POSDmqimhn/4LJl4JUkUfEJlJVsgMsJDDjuCCCOxFVVVVVVVVVVVVVVVVVVVU0MNxC9vPEkscilXR1yGBGCCD1BHavj9/wTJ/Zb+Ost1rdt4VfwP4kum3vqnhvFukjdSZLYgwsWJJZgqux5LGvjp/wSS/aZ+FiXGq+AYrH4laNC3D6QvkXwTJAZrRyST93KxPIRu9ASNR03xd8PvEs2matYav4b8Q6RNtmt7mKSzu7OQDoysFeNsHocHmvg5/wUz/AGqPhN5djqfi2Lx3pQkUvbeJw91MF6NsuQyzZIx99nUEZ28nPwb/AOCuvwD8axW+n/FfRdW+H+qOxWSYo2o2H+yRLEolBJ7GLC5HzEZI8B/E74d/FHSRrnw58caH4lsQql5tMvo7jytwyA4Uko3B+VgCCCCAQaBBHBqqqqqvoa3dqBB4Bqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr9qb/gor8FP2cEvPDWlXKeNfHEDPEdF025URWUqkAi7n5ERHPyKGfIwVUHcP2iP2q/jL+03r76n8R/E8p0uK4MthoNozR2NgcEApFk7nCkjzH3OcnnBxX7Jv/BPn41/tVzxa1ZW/wDwifglZNs/iTUrZmWTjkW0OVNwfcFUHQuDxX7NH7IPwT/ZU8PHS/hp4d8zV7qMpqPiHUNst/fZIJVpAAEjyq4iQKnygkFssaqqqqqqqqqqqqqqqq/am/4JgfAf9oaW68VeFLdPh740nZ5JdR0q1Btb6RuS1zagqpYtkmSMo5LEsX4A/aR/Ys+Pv7LupTD4g+EJbrw8JAtt4l0tWuNPn3YCgyAAwuSSNkoRiQdoYYY/Av8Aao+OX7OeordfDDxxdWtg0m+40e6P2ixuT33QtwCem9Nr+jCvgL/wVx+EHjf7FoHxr0S68C6xL8j6lArXWmO3ABJGZYdxPRlZVHLSAc14f8SeHPF2kwa94U1/Tda0y5UNBe6fdJcwyggEFXQlSMEHg96qqo4x1onPANVVVVVVVRxjrTHPANVVVVVVUxGMA0TngVVVVVVVTHPANVVVVVVHGOtE56VVVVVVUx4wDVUcY60TnoaqqqmOBgGqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv2y/wBr3wn+y34Dl8q4gv8Ax1rNtINA0jO7DYKi5nAOVgRu3BkI2rj5mTX9f8bfFfxtca7rt7qPiTxT4kvRvcoZri8uJCFRERRkknaqoowBtVQAAK/4J7fsR2X7J/w/fXfF0Nvc/EnxPCv9s3EUvmJYQZDJZxMPlIUgM7D7z9CyohqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqXQtDm1eLxBLo1i+qwQ+TFfNbKZ44+fkWTG4L8zcZxyfWqqqqq/wCCkn7JurftS/BOBvBFvFN438F3El9osDhVN9G6hbi1Dt9wuqoynoXiRTgHcv7FH/BSW9/Zd8Pn9nj9pPwf4jOmeHLprbT7iK023+jZkYyW1zBKUZkRmJH8aDKhWARV+Kv/AAWX/Z/0Lwtcv8JfD/iPxP4kmhkFlHfWQs7OCXA2tOxfeVyc7UUkhSCyZBr/AIJyfsu/E/46/HOT9tr48W92NPXUJtV0p7pPJfWdSkJKzpHji2i3FlK7V3rGq5VHAr/gm/8A8pHvEH/cf/8ARpqv+C1f/J1Hhb/sn9n/AOl9/U0MNxC9vPEkscilXR1yGBGCCD1BHavGfhb4r/8ABKr9rw/Ebw14XuNV8BahNcw6UZJnS31bS5juNjJOA2yeLbGeQTvhSTayHBtf+CyH7JM3h5NXurHx3Bf+WWfSv7IjeYMDjaHE3lHOMglxwRnByB+2145+Nv7SK6b+1V8QvCtx4X8Favdf2L4J0qdic2saNLJKmQpcFmBaYqA5bap2xAD9k7/k1f4Nf9k/0L/0ghr9qr4N/Gn9gD9qqT9q34N6K914K1HUpb6KdYnmtbU3J23NhegY8tHeRhGScEMm1vMT5fAn/BZX9mPX9Agu/HOg+LfCusBR9psVs1v4Q3fypoyC6jjlkjP+z3rwB/wVW+AHxV+NnhP4NeA/D3iiceJr42Z1rUIYrS3ikaItEETe0khaQCIhljwTkbh1r9mz4jT/APBOf9tLXdO+Kvh7VRpltHd+H9TZYB5/2OSaOSG7jU4EiMYYZPlI3IxK54B/Zu/a/wDg3+1bL4ki+EdzrE6+FVtDfyX9ibYZufO8sJkksf8AR3zwAMrycnH7VXwb+NP7AH7VUn7Vvwb0V7rwVqOpS30U6xPNa2puTtubC9Ax5aO8jCMk4IZNreYny+BP+Cyv7Mev6BBd+OdB8W+FdYCj7TYrZrfwhu/lTRkF1HHLJGf9nvXgD/gqt8APir8bPCfwa8B+HvFE48TXxszrWoQxWlvFI0RaIIm9pJC0gERDLHgnI3Dr+1l/yav8Zf8Asn+u/wDpBNX/AAQx/wCa2f8AcA/9yNV/wUg/5SPeHv8AuAf+jRVf8Fz/APmiX/cwf+46v2Tv+TV/g1/2T/Qv/SCGv+CvP/Jmmpf9jBp3/obV/wAEhv8AkzTTP+xg1H/0Na/4LV/8nUeFv+yf2f8A6X39V+zV/wApldX/AOygeM//AERqVV/wW9/5Kn8NP+xfuv8A0oFfDfT7LVvg34W0rUrdLi0vPDNnDPC44kja2RWU+xBIrV9F+O3/AASa/aZvPGei+GZde+HmtySWttLK7i01jT3cyR27zhSIbyMR5yVLAqxAeNyG8Of8Fiv2SdW0e3vdbh8Z6HfMo8+yn0lZzG2Bna8TsrLnODwSOSq9K/Z+/wCCk/wa/aV+OCfBn4f+G/Edqsum3Vzb6rqyRQC5lhYZjjiR3ba0W+QMxVhswUHUftjfs92/7Tv7PviX4WRyxQaxLGt5oVzKxVIb+H5otxGcI/zRMcHCysQMgV+xj+2z4o/YI1bXv2ev2iPh74gXQ01R52iiVftmj3JXZIUjchJoJCkZ+V1A+Z1378H4n/8ABX74Qy+Hm0T9nrwj4r8Z+OdVzBpVpLpZhhjlJUAuNxkc4JKoiNuK4JUEGvgH4H+I/wAPP+ClPgbw58Xp3l8azeIrTU9caR1d/tV9aLeOHZcqXBuMMRxuBxxVVVVVVVVVVVVVVVVVVVVVV8Ufgp8JfjXop0D4rfD3RPE1mMeWL+1DSQEd4pRh4jyRlGBwSOhNfGn/AIIufCbxPLNqnwP8e6n4KnZiy6ZqUZ1Ozxz8iOWWaMcr8zNKcL0Ocj41f8E4/wBrP4ICS71T4cS+KdJjjaR9U8K79ShRV+8XQKJowBglnjVcd+Gxomv+KPBWtx6v4b1vVNB1ixk/d3VlcyWtxA6ns6EMpBA7jBFfCX/gqZ+1P8NlhsPEWuab480yJdgh162zOoznIuIikjN1GZPM4OMcDHwk/wCCu37PXjVobD4l6Lrfw/vpGw0syHUbJeOP3sKiTJORzCAOCTjOPAPxX+GHxVsDqXw1+IPh7xPbooMh0vUYrlocgHEioxaNsMuVYAjPIFVVVVVWSOAa3AjFcEcGqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/aC/a0+Bv7M+k/bfiZ4tQalIubTQtPAuNQuuCRthyNikA/PIUTPG7JAP7Tv/BTn42fHP7d4X8BTS/D7wdcfu/s2n3JN/dxjGfOuVwVDYOY4wowxVjIMk/CP4L/E/wCO3i6DwP8ACnwdqHiDVZcNIttF+7tYyQvmzSH5IYwSAXcgZIGckA/sof8ABIz4cfDZLLxl+0TNY+OvEqMk0eixox0qyYHIVw2DdngZDqsfJUo4+Ywww28KW9vEkccShURF2hQBgAAdBjtVVVVVVVVVVVVVVVVVVV5Z2moWk1hfW0VzbXMbRTQzIHSRGGGVlPBBBIIPBzX7Rn/BJL9n/wCLss/iD4WTN8MPEErFnTTrUT6ZMe+bTcvlHAAHksiLySjGv2hP2Cf2lP2cZby+8V+BbjWfDdorSHxHoUb3lksY/jlIXdbjkf61VGTwT1r4XfG34tfBTWF1z4VfEHWfDlz5ivJHaXB8i4K9POgbMcwGfuyKw9q+Cf8AwWN1e1eHSPj/APD2K+h4U6z4dPlSrzyXtpG2twequuNv3Tnj4OftUfAH4+RW8fwy+JmkajqU8Jl/seaX7NfxhR8+beTEhC92UFehBIIJokAdaJz3qqqqqqqqOMdaJzwKqqqqqqpiCMA0xzwKqqqqqqpjxgGqqqqqqqY54BqqqqqqokAUee9VTHjANVVVRxjrROaqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv2vP2zvAP7Lnhie1Fxa6z48vYA2k6AJDkbtwWe4xykIKn0ZyNq45ZfH/AI/8b/GDxvqHjfxvrF1rfiDWp90srjJYnhY0QcKoGFVFAAAAAr/gmj/wT9HwT0qD44/Grw7F/wALA1KMNo2nXK7n0C3ZeWZTwtzICQ3Uxr8uQzSKKqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq+JPwD+CXxiQf8LQ+FPhfxNMq7Y7nUNMjkuIhlThJseYgOxchWGQMHI4rwp+xp+yn4I1Iax4b+AHgmC9XHlzzaUly0RHdDKG2H3XBqq8N/BD4LeDfEknjLwf8ACDwToevzeZ5mq6b4ftbW6fzDmTM0aBzuPXnnvVeNvgh8FviXqsWu/Eb4QeCfFWpQW4torzW/D9rfTRwqzMI1eVGYIGdyFBwCzHuarxH4Y8NeMdHn8PeLvD2ma5pd0pWex1K0S5glBGCGjcFWGCRgjvWhfsVfsmeGtci8SaN+z54Ji1CCTzIZJNLSVYnzkMiPlFIIBBAG3tivG/w2+HXxN0+DSfiR4B8OeK7G1m8+C11vSob6OKXBXeqSqwVtpIyBnBI71pOkaVoGlWWg6Dplpp2m6bbx21nZWkCww20MahUjjRQFRFUBQoAAAAFXlnaahaTWF9bRXNtcxtFNDMgdJEYYZWU8EEEgg8HNa9+w/wDsieJNSk1bVP2ePBP2mb/WG20xbVWPclItq5Pc4ye9eAPgZ8F/hSwl+Gfwn8I+GJ/JSF7nStGgtp5UUEDzJUUPIcE8sSTkknk1XxQ+AfwV+NSW4+K3wv8ADniiWzXZbXGoWCSTwLnJVJcb1UnkqCAe4r4XfA74QfBKyvLD4TfDnQfC0epMjXradZrHJdFN2zzH+84Xe+0MSF3tjGTV5Z2moWk1hfW0VzbXMbRTQzIHSRGGGVlPBBBIIPBzWvfsP/sieJNSk1bVP2ePBP2mb/WG20xbVWPclItq5Pc4ye9eAPgZ8F/hSwl+Gfwn8I+GJ/JSF7nStGgtp5UUEDzJUUPIcE8sSTkknk1q+kaVr+lXmg67plpqWm6jbyW15ZXcCzQ3MMilXjkRgVdGUlSpBBBINeBfhN8K/hd9t/4Vl8M/CnhH+0/L+2/2FotvYfavL3eX5nkou/bvfGc43tjqarxJ8EPgt4z8SR+MvF/wg8E67r8Pl+XqupeH7W6uk8s5jxNIhcbT0547VXjr4TfCv4o/Yf8AhZnwz8KeLv7M8z7D/bui29/9l8zb5nl+cjbN2xM4xnauegrSdI0rQNKstB0HTLTTtN023jtrOytIFhhtoY1CpHGigKiKoChQAAAAK8YeB/BXxC0VvDfj7whoniXSXkWRrDWNPivIC6/dYxyqy5HY44rwh4H8FfD3RV8N+APCGieGtJSRpFsNH0+KygV2+8wjiVVBPc45rxt8EPgt8S9Vi134jfCDwT4q1KC3FtFea34ftb6aOFWZhGryozBAzuQoOAWY9zVaZ8EPgtovjN/iNo3wg8E2HiyS4muX1628P2sV+00wYTSG4VBIXkEjhm3Zbe2c5NV44+Dfwg+J11a33xJ+FXg/xZc2UZitptc0K2v3gQnJVGlRioJ5wOM1aWdpYWkNhYW0VtbW0axQwwoESJFGFVVHAAAAAHArWtD0TxLpVxofiLR7HVdOu123Fne26zwzAEHDI4KsMgHBHatQ/YR/Y71O9lv7n9nbwYkszbmW3sPIjB/2Y4yqqPYACvBPwr+GHwzhnt/hv8OPC/hSO6bdOmiaPBYCVsAZYRIu44UdfQVXxL+B3wd+Mtqlp8U/hn4c8TiJQsUuo6eks0IDbgI5cb0Gc8Kwzkg8E5+GP7OnwJ+DE8178LPhP4Z8OXlxkS3lnYILhlIAKeccvs4HybtucnGSavfhB8JdS8axfEnUPhf4RuvF0DI0WvzaJbvfxsi7UK3BTzAVUbRhuBxVVVVVVVVVVVVVVVVVVVVVVVV8W/2YP2ffjtG5+K/wl8P69cvjN+9t5F4AF2gC6iKTAYwMB8cD0GPjD/wRQ8B6qk2o/Az4p6noNztymmeIYRe27NnkLPGEkiXGOqynI688fFn/AIJt/tg/CNLu9v8A4VXHiXS7ViP7Q8MSjUlkAON4hTFwF6HLRLgcnGDi3ute8K6w0tpc3+j6tYSPEzRu9vPbuMo6kjDKfvKRx3Br4Zf8FH/2uvhk8USfE+XxTZR7c2fieEagHwQeZiRPyAVOJBwx74I+G3/BaHRpmjtfjB8Gby2UKfMvvDd8s5Y44Atp9mBnHJmPB6cc/D3/AIKF/sgfEfZDYfGbS9Fujnfb+IUfS/L+9jMswWI5CE/K5xlQcFgKsNQsNVs4tR0u+t7y1nXdFPBIJEkHqrKSCPpVVVVX0NZPTNZB4zXBHBqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq+PH7Zv7O/wCzra3CePfH1rca1BkL4f0hlvNRdsA7TErAQ5HRpmjU/wB7NftFf8FZ/jB8R/tXh34K2H/Cv9Ckyn27eJtTnX18z7kGR2jBYHpJWk6R47+K3jKLStGsda8WeKdduD5cMSSXl3eSkFmPd3OASSegBJ4Br9mD/gjj4q8QS23in9qDWH8P6YVDp4b0i5R72U8ECeYBo4l9VTcxB+8hFfDH4S/DX4L+FrbwV8LPBmmeHNHtlULb2cWGkIGN8shy8rkDl3ZmPcmqqqqqqqqqqqqqqqqqqqqqr48f8E4P2Vvj15+o6h4G/wCET8QTY/4nPhhlspCR/fh2tBJnjLNHvwOGFfHT/gjz+0J8PmuNV+Eeq6Z8SNHjXcsMWNP1FQAS2YJGMbgYAGyUux6IOleK/Bvjb4beIX8P+NPDWseG9bs23G11C1ktZ4yGIDAMAcZU4YcccGvg5/wUI/an+C/l2mmfEOXxNpccaxppnicPqEKKv3QjlhNGAONqSKuO3Ax8K/8Agsd8MtaRLL4wfDjWfDV1yPtmkSrqFs2FzllbZJHk5XAEnOCTgnHwv/aY+APxoijb4afFjw9rM8jbVshc/Z7vPHW2lCygHPUpjOR2NVVVVVVUcAdaJzxVVVVVVVMQRgGmOeAaqqqqqqpjgYBo896qqqqqqYjGAaqqqqqqOMdaJz3qqJAHWjz3qqqqY54Bqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/bT/4KH+FfgXBqPw2+FV1a698QjG8M06ES22hSZAJl7STD5sRchWX5/wC62t6z48+Lvjm51rV7jVPFHivxHd7pGWNp7i7mbgKqIMngBVRRgAAAAACv+Cf/APwTS0T4Jw6N8aPjhZJqPxFC+fYaQ5WS10EsBsYgA+ZdKM/Pkohb5QWUSVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVfEf4JfB/4v262/wAUPhl4Z8UeXH5cUuqaZFPLCuScRyMN8fJP3SOp9a+Kf/BG79mTxhHPc/DfV/EvgC9aPbbxQXR1Kzjbbjc0dwTM3IyQJ16kccY+KX/BGr9pPwk8tx8NfEHhnx7Zru8uNJ/7Mu2wxxmOc+UMrtP+uOCSOgDH4nfs3fHv4MpJP8T/AIReKPD1pEwVr6605zaFiQABcKDExyQMBj1HqK8JfET4geALn7b4D8deIfDdxuLebpGpzWb5KlScxMpztJH0JFfD3/gqL+194D2Q3/jTS/F9rHnZb+IdLSXGd2cywGKVuWB+ZzjaoGBkHwB/wWktXe3tPin8EJY148++0DVQ56rnbbTKP4dx5m5IA4ySPAf/AAU//Y88cJDHeePr/wAK3c2dtrr2lSxFcKSd0sQkhXgY5kGSQBkmvBfxd+FPxJRX+HvxL8L+Jc5+XStXgumUhQxBVGJBAZSQQCMjOKqqqvoayema3AjFcEcGqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqrx38Uvhr8L9ObVfiN490Dw3arGXD6nqEduXA/uKxBc8YCqCSeACa+MP/BXf9n3wWlzp3wt0jWfH2px5EU6RHT7HO3vJKPNOGwOIsHkhuhPxy/4KSftP/G2C70VPFcXgzw/dZVtN8Nq1q0iZOBJcEmZsqdrAOqN3TtXgrwH4++K/imHwt4B8Lax4n16+YutpYWz3Mzc/NI20HCjOWdsAdSQOa/Zz/wCCNPjnxXDB4h/aP8VP4RsJF3LoejPHPqBBH/LSYhoYT7ASnsdpr4G/szfA/wDZx0U6P8IvAGn6NJLGEu9RZTNe3mMH97cPmRhuGQmQiknaqjiqqqqqqqqqqqqqqqqqqqqqqqqvH/wz+HfxV0J/DPxK8EaJ4n0xskW2qWKXCxsRjem4Eo+Dwy4YdQRXxh/4I4/s4eOWm1D4X65rvw7v5GysMLnU7FeOf3MzCXJODxMAOQBjGPjD/wAEnf2svhik2oeGtC0z4gaXEu8zeH7nNwozjBtpQkjN04iEnBzng48V+DfF/gTV38PeOPCmseHtUjXc9jqthJaTqMkZMcgDAZBHTsa+G/7aH7UfwoSO38HfGjxALSJiVstRmGowAE5YLHcBwgJJJ24OST15rwJ/wWR+MWjpFB8Q/hf4X8SpHkNLYXEumTSDacFifOTO7B4QDAxgZyPCf/BYP9nHWNsHinwj438PzHcTJ9kgu4AB0+ZJQ+T6eXgeteE/28/2P/GkixaP8e/DluzSNGDqxl0oZVdxJN0kYAx0J4J4BJ4rwx8Q/h/44RH8F+OvD+vrJCZkOl6nDdBowQC48tjlckDPTJFVVUSAOtE571VVVVVVTEEYBpjngGqqqqqqqJAHFHnvVVVVVVRIA60ee9VVVVVVMc8A1VHGOtE571VVVMcDANVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV4y8a+EPh54cu/FvjnxLp2haNYrme9v7hYY0zwBknlicAKOWJAAJOK/a4/4KgeJfiJFf8Aw8/Z9F54c8OOzRXPiCQmK/1BMYPkgH/RozzzzIw28x/Mh+EXwY+Jnx48Z2vgH4VeE7vXdYuskpFhI4EAJMksrEJEgAPzMQCcAZJAP7En/BPX4f8A7J9lH4u12a38T/Em4heO41koRBYI/DRWiNyoK4VpG+dvmHyqxSqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq+In7HX7LnxVhaPxv8CvCN3Ky7TdWtgLG5IxgDz7fZJgdhu47V8Q/+CL37OniKO4uPh7428YeELyTPkxyyxalaRfK2P3bqsp+YqeZugI4J3Dx//wAEWf2g9Cd5vh94/wDB/iu2EhVUuGl024Zd3ytsZXjHy4JHmcHgbutePv2Ev2vvhpbS3nir4A+KDbwLumm0uFNVjiXaWLM1o0oVQqkliQF4yRkVqGnahpN7Lpuq2NxZ3du22W3uImjkjb0ZWwQfY14T/aj/AGj/AANtTwr8dPG9lCm7bbf23PJAC3U+U7FM++3NeEP+CsH7YHhpi2teIPDPiwFmIXWNDjiABAAH+iGDgYJHfJOcjAHhH/gtPrMTRwePfgPZXKsyiS50jW2gKDHzERSxPuOeg8xcDueteFv+CwH7MOswAeItB8b+H7lYwXWbToriItn7qPFKWPrlkWvC/wC31+x14wZE0n4+eHoC8xiH9qLNpgDAA5JukjwuCPmPy5yM5Brwt8Tvhr46SF/BHxD8M+IVuFZoTpWrQXYkCkhivls2QCCDjoRVVWT0Brd2rIPGa4I4NVVVVVVVVVVVVVVVVVVVVVVVVVVVVeMPif8ADT4eor+P/iJ4Z8NKyqynWNWgsgQxIUjzWXglSB6kH0rx7/wVB/Y78DpLHZ+Pr/xVdw9bXQdKllJyoI2yyiOFuDjiTg5BwRXxI/4LSD9/afCH4K/3vI1DxJqH++F3W0A/65scTf3l9Hr4m/8ABSD9rv4nPJE/xPl8LWT7sWfhmEaeEyxPEwJn4BCjMh4UdySbm617xXrKy3dzf6xq1/IkStI73E9w5wiKCcsx4VQOewFfB/8A4JqftefGNIL20+GreE9LnXcuoeK5Dpy9cD9yVa4IPJBERBAznkZ+Cv8AwRe+EPhjy9T+OPjrVfGt4sisNO01TplkFH3kdgWmkzj7yvFgEjGea+Hnwu+HPwl0BfC/wy8E6P4a0tW3G2020WEO395yBl2/2mJPvVVVVVVVVVVVVVVVVVVVVVVVVVVVV4r8G+D/AB5o7+HvHHhTR/EWlyNuex1Wxju4GYAjJjkDKTgkdO5r4mf8Eqv2NviNLNeWXgjU/Bl7cNukuPDOpNbjPy/dgmEsCDCkYSNR87Hrgjx//wAEQNVjje4+Fnx3tLh/LOy01/SWhG8LxmeFn4LYB/dfKOfm6V44/wCCT/7aPg9pW0zwNo/iy3gWRnn0PW4CNqAHKx3BhkYtk4VULEqRgZGfGH7Lf7SPgKRo/F3wI8d6dGsixC4fQbh4Hdl3BVmVTGx2g8BjjBHUHE0MtvK9vPE8csTFXR1KsrDggg9D7V4c+M/xi8HCMeEPiz4y0PyrcW8f9m67c22yIYxGPLcYQbV+XpwPSvDv7d/7X3hdrVtM+PniWY2ilYxqLx6huBBB3i4R/MOD1fJBweoFaN/wVN/bI0u1a3vvHOjavI0hYT3ugWyuowBtAhSNccE8jOSecYA0n/gsl+0HDeB9d+G/w8vLXaQYrS3vLZ89jva4kGPbb+IrR/8AgtNrsFqyeIf2erC9uTISkln4me1QJgYBVreQk5zzuA5HHGTpP/BZb4Mz3gj134TeNLS12nMtpLbXLg9hsZ4xj33cehrQv+Csf7I+sRWj6hqHizRGuJNsiX+ilzbDdjc/kPICMfN8hY47Z4rw5+33+x34q8v+y/jxoMPmTiBf7Sin0/DHHJ+0RphPmHznCjnng48MfGL4ReN2RPBfxU8Ia+0kxhQaXrlvdFpAASg8tzlsEHHXBFVVVVMQRgGmOeAaqqqqqqokAdaPPeqqqqqqjgDrR571VVVVVUxwMA1VUxzwDVVVUcY60ee9VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVftH/APBTT4K/B6K40H4cTQfELxSjGMxWNztsbU8/NJcgMHIOPkj3Z5BZOtfHT9o74vftGeII9f8Aij4qlvltdwsdOgXybOxB6iKEcAngFzl2AUMxwMfshf8ABN/4w/tOvZ+LNbjl8GfD+TbINZvbcmXUE3ci0iOC+QD+8bCDsWI218Bf2dfhL+zX4MTwT8KPDEWnW7Ya8vZQJLzUHBYh7ibAMhG5sA/KoOFCjiqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqvFHgfwV44tBYeNPCGieILYRyRCHVNPiu02SACRdsikYYAAjocDNeOP+Ccv7F3j5pZ9S+BWj6bcusgSbQ5p9LETOANyx27pGSMAgMhUHPHJz4y/4Iqfs9as0k/gr4jeOPD8sjMRFcvb38EYxwFXy0fAPJ3OxPTI614w/wCCInxespGXwB8avB+sxiRQraxZXOmEpt+YkRC4wQ3AGSCOcg/LXjL/AIJW/tq+EkkntfhpY+IreJWZ5dG1m2kIAOOI5HSRieoCoTjrivFn7K37S3gZph4q+Afj2wigZVe5bw/cPb7mAIAmRDGx56BjzkdQRV5Z3dhdzWF/ay21zbSNFNDMhR4nU4ZWU8ggggg9DXhv40fGPwb5f/CIfFrxloflW4t4/wCzdeubXZCMYjHluMINq/L04HpXhv8Ab/8A2x/Cvl/2X8e9en8u3EC/2lFBqPyjHJ+0RyZf5R85+Y888nPh7/grX+1xorQNqV34P18QwiN11HRdgmbAHmN9neLDcZwuFyemOK0H/gtB8WbeK0XxP8HPCOoSJJm6awvLizEqbuiBzLsO3AyS4zzjHy1on/Bavwvced/wkn7P2qWO3b5H2HxFHd7+u7dvgi24+XGN2cnpjnRv+Cxf7Ml+1pBq3g/4iaZJMq/aJG061lhgbHzfMlzvZQeAQmT1wOlaL/wVO/Yy1WWSO+8f6vo6xrlXvdAu2Eh9B5KSHP1AFaT/AMFBP2N9asxfWXx60OOMsVC3cFzavkf7Esatj3xg1aftZfsu3trDeQ/tG/DNUnjV0WXxXZxOARkbkaQMp9VYAg8ECovih8NLiVIIPiJ4ZklkYKiJq0BZieAAA3JNcHgGqqqqqqqqqqqqqqqqqqqqq1fxz4I0C8Ona74y0PTrtVDGC71CKFwD0O1mBwa1v9sb9lLQIopr79onwBKsrFVFlr0F4QQO4hZyo9zgV4h/4Kd/sW6Cs6QfFefV7i3mMTQafod65Yg4LK7xLGy8dQ5BHIzXin/gsj+zvpjPB4V8B+Otbkjm2+ZLb29pC6YOXVjKz5zgYZF6k5GMHxN/wWq8UXACeDfgHpdjt8weZqevSXe/psO2OKLbjnI3NnIwRjJ8W/8ABWT9r/xIGGja34W8LZ24/snQ0k2464+1mfr3z+GK8ZftZftM/EAyr4q+O/jW6gnULLaQ6xLbW7gPvGYYisZIbkHbkYHoMQw3+r38dvbxXF7fXswWONFMkk0jnAAAyWYk9OpJrwJ+w3+1z8SRA/hb9n/xb5NyoaGfU7MaXDIpTeGWS6MaFSvRgcHIAOSBXw//AOCKvx8114Z/iJ8R/B/hW1kzvSzE2p3MeGI5QLFGcgAjEp4PODkV8Mv+COH7L3hForv4g6p4o8eXIUiSC6vP7PtGOMZCW22UEE5/1xHA98/Df4EfBj4P24t/hh8LvDPhojdumsNNjjmfcTnfLje/XHzMcDA6ACqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq13wl4V8U+R/wk3hnStX+y7vI+32Udx5W7G7bvBxnaucdcD0rXf2KP2RvEfkf2h+zh8P4vs+7Z9h0KGxzuxnd5Cpv6DG7OOcYyc+If+CVX7EOuQ7bP4V3uizNN5jT6f4gvtzDBym2WV0C5IOAoIwMEDIPiD/gi9+yzqkl3c6L4u+IujSSxkW8MWpWs0EDhcA7ZLcyMNw3EGQZyQCoxjXP+CHXg+4hiXw1+0NrFhKGPmtfeHY7sMMcBQk8W0+5J+leIP+CIHxJtpLtfCvx38M6jGkZNo2oaTPZGV9vRwjTeWN3GQX45xn5a1z/gjV+1vpMMUmn6v8PNaaRiGjsdYnRoxjqfPt4xj6EmvEH/AATE/bg8PRXdzL8EZb62tJCgl0/WrC4Mw3bQ6RLP5pByD9wEDkgYOPFP7KH7Tfgr7U/ib9n/AOIFnBZbPPu/+EeuZLdN23b++RDGcllHDdTjrxWq6RquhX8ula3pl3p97Bjzba7gaGSPIBGVYAjIIPI6EGvCXxr+MfgFI4/A/wAV/F+gRxKqrFput3Fum1TkKURwCuf4SMV4L/4KW/th+DnVJPibF4gtlz/o2taZBOCSQSfMVVl6AjG/ADHjOCPh9/wWb8TwbLf4q/BfS77dndd+H9QktNn3iMQTCXdn5B/rFxhjzkKPhz/wU8/ZF+IMsNneeNNR8IXk7bY4PEenNAM/N96aIyQoMKDlpAPmUdcgeE/iR8O/HsSzeBfH3hzxGjxtIraTqsN2CittZgY2bgN8pPQHimYEYBonPAqqqqqqqjgDrROaqqqqqqOMdaJzwKqqqqqqJAFHnvVUxwMA1VVVUxB4Bqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqrq6tbG1lvb25it7e3jaSWaVwiRooyzMTwAACST0r45f8FN/2dfhN9o0rwjqEvxE1xIyY4dDmQ2QfsJLw5XBHeJZcdwK/aD/br/aA/aH+06Rrnib+wPC8+V/4R/RC1vBIh4xO+TJPkYyHYpkZCLXwN/Zm+OH7R+snR/hF4A1DWY4ZAl3qLKIbKzPB/e3D4jU7TuCZLsAdqseK/ZV/4JO/CH4MT2vjD4xzWnxG8VQ7Xht57Yrpli+DkrCx/wBIPPDSjbwCEVgDUMMNvClvbxJHHEoVERdoUAYAAHQY7VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVWueGPDXieGK38SeHtM1aOBi0SX1ok4RiMEgODg4rXf2KP2RvEfkf2h+zh8P4vs+7Z9h0KGxzuxnd5Cpv6DG7OOcYyc67/wSf8A2I9X8j+z/hvquieVu3/YfEV6/nZxjd58kmMYONuOpznjGuf8ETP2e7iGJfDXxU+IdjKGPmtfSWd2GGOAAkEW0+5J+leIP+CG90kV3P4V/aQilk8wm0tdQ8LGMBC3R5kuW5C9xHgkdFB41v8A4Im/tEQXaJ4b+KPw5vrYxgvJfTXto4fJyAiW8oIxt53DkkY4ydY/4JK/tq6ZZG6svBGg6tKGAFtZ+ILZZCD1OZmjXA/3s1rn/BOL9tnw9LFBf/AHWJWlUspsb20vQAOOTBM4U+xwa1D9j/8Aau0y9lsLn9mv4mvLA21mt/Cl5PGT/syRxsrD3BIqb4V/E+3heef4ceKI44lLO76POoUAZJJKcDFVX/C1PifjA+JHij/wcT//ABdWH7TH7SGlWcWnaX+0F8SbO1gXbFBB4rvY0jHoqrKAB9K0T9sv9q/w9539n/tD+PJfP27/ALdrc17jbnG3z2fb1OduM8ZzgY/4bv8A2v8A/ov/AIo/7/J/8TUX/BTD9t6GJIU+ODFY1Cgv4d0tjgDuTbEk+55r/h5r+3B/0W//AMtvSv8A5Gr/AIeyftf/APQa8Lf+CNP/AIqv+Hsn7X//AEGvC3/gjT/4qtG/4K7ftZ6XdNcX0PgfWEaMqIL3RpFRTkHcDDNG2eCOTjk8dCP+Hyf7T3/Qi/C7/wAFd/8A/Jlf8Pk/2nv+hF+F3/grv/8A5Mr/AIfJ/tPf9CL8Lv8AwV3/AP8AJlf8Pk/2nv8AoRfhd/4K7/8A+TK0b/gs1+0DBdM/iH4Z/D2+tjGQkdlDeWrh8jBLNPICMZGNo5IOeMH/AIfSfE//AKIr4X/8GM/+Ff8AD6T4n/8ARFfC/wD4MZ/8K/4fSfE//oivhf8A8GM/+Ff8PpPif/0RXwv/AODGf/Cv+H0nxP8A+iK+F/8AwYz/AOFf8PpPif8A9EV8L/8Agxn/AMK/4fSfE/8A6Ir4X/8ABjP/AIV/w+k+J/8A0RXwv/4MZ/8ACta/4LNftA3F0j+Hfhn8PbK2EYDR3sN5dOXyckMk8YAxgY2nkE55wP8Ah8n+09/0Ivwu/wDBXf8A/wAmV/w+T/ae/wChF+F3/grv/wD5Mq7/AOCtX7XdzdTXEOoeEbVJJGZIItEBSIE5CqWdmwOg3En1J61qn/BVj9sfULCSztPF+gabK+Nt1a6DA0keCCcCUOnIGDlTwTjBwRN/wUw/benieF/jgwV1Kkp4d0tCAR2ItgQfccipv26v2vZ4Xhf9oDxWFdSpKXCowBHYhQQfccirv9p39pW/tZrG+/aH+Jlxb3EbRzQy+Lb50kRhhlZTLgggkEHg1rXjXxl4ktUsfEPi3WdUto5BIkN7fyzorgEBgrMQDgkZ9z61Xhz4K/GTxi1unhH4S+M9ba6hE8A07Qbm5MsZAIddiHcuCDkcYNeG/wBgf9sjxWlo+l/s8eLYBeMVjGpWy6cVIJB3i5aMxjg8vgEYPQg14T/4JE/tmeI2hXWPD3hfwsJWYMdW16OQRAAkFvsgn4bGBjJyRnAya8K/8EP/AIiXiIfHHx68OaSxhJcaVo02ohZcj5QZHgyuMndwcgDbzkeDv+CKn7PWktHP40+I3jjxBLGykxWz29hBIMfMGURyPgnn5XUjpk9a8Ef8E2P2LPAjwXNl8EdP1a6h2Fp9cu7jURKVbILRTSNF9QEAI4IIrwp8PPAHgK3Fp4F8DeH/AA7CIxEItJ0yGzUIDkLiNVGM846VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVaxoeieIbI6br+j2Op2hYMbe8t1mjLDodrAjIrxl+w3+yF48SRdf/Z58GRtKrLJJpmnjTJG3HJYvamNt2f4s5968e/8ABG/9lbxNFJJ4O1Pxh4PuTIzRLa6it5AoZgdrJcKzkKoKriQH5ssWxXxD/wCCJfxY0qO4ufhf8X/DPiJY8tHbavZy6ZK6hWO0FPOQvkKoyVU5JJXGK+J37D/7V/whSS48afA/xGLOJgGvdNhXUrcAkBS0lsZFQEkAbsHJA68VPBf6Rfvb3EVxZX1lMUeN1MckEiHBBBwVYEdOoIrwJ+11+038Nni/4RD44eLIIYM+VaXd+19bJkljiC43xjJJJ+XknmvAX/BXf9o3w4lvaeNvD3hPxfBHjzZ5LV7G6lwqj78LeUucE/6rqx6DAHw8/wCCwHwJ8Q+VbfEPwR4n8I3L43SwbNSto/XLrskP4RH8K+Hn7V/7N3xV8qPwL8Z/DF9czY8uznvBZ3LfSCfZJ/47xVVVVVHAHWic96qqqqqqpjngGqqqqqqjjHWic9KqiQBR571VVVMeMA1VVVVVVVVVVVVVVf39hpdnLqGp3sFpawLulnnkEaRj1LHgD6146/b3/ZJ8AJKuofGfR9Vnj4WDQg+pmQ7QcK8AaPoepYDORnIxXij/AIK//AjTbsW3hXwB4y1uNWYSXEyQWaHGNpQGRmYHnO5UIwOueNZ/4LMeIZ4o18PfAHTrKQMfMa98RvdBh6AJBHg++TWtf8FhvjtPLGfD3w08B2UYUiRb2O7uix9QUmjwMdsH618Of+CxHgfUZbey+Kvwn1bRSzBJL7RrxL6MZ6uYpBGyqO4DOcDjJ4r4d/tvfsrfE/yIfDvxm0K1vJ9qrZ6xI2mS+Ydv7sC4CB2y4GELAkHBODi1urW+tYb2xuYri3uI1khmicOkiMMhlI4IIIIIqqqqqqqqqqqqqqqqqqqviv8AtR/s/wDwSWaP4j/FLRNOvYVy2mxTG6vOTgf6PEGkAJyMlQODkgAkfGT/AILBadD9o0n4D/DiW5cblTWPEb+WgIyMpaxElgflYFpFPYpzx8Yf2nPjp8eLmaT4l/EXU9RspZNy6XE/2eyi5BULBHhONq4LAtxkknJPwQ/ZW+Pn7RV6lv8ACf4b6nqtn5wim1WRRb2Nue++4kITIAJKgluwBJAP7Pn/AARn+G3hP7D4g/aD8Wy+MtUj+eXRdKL2unI3HytLxNOBgnP7rOcFcDnwl4N8JeAdAtfCvgjw1pmg6PZLtt7HTrVLeGMd8IgAye56nvVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVXlnaahaTWF9bRXNtcxtFNDMgdJEYYZWU8EEEgg8HNTfCH4TXEL28/wv8ACMscilXR9EtyGBGCCCnII7V/wyb+yv8A9Gz/AAp/8Iyw/wDjVXn7Bn7HN9dTXk37O/g9ZJ5GkdYbMxICTk7UQhVHPCqAB0AArXP+CbH7EPiKWKa/+AemRNCpVRY6nfWQI91gnQMfcgmv+HXH7CX/AEQv/wAubVv/AJKr/h0N+xp/0A/FX/g9f/Cpv+CQf7G0sLxR6T4tiZlIWRNdYshx1GVIyPcEV/w5U/ZX/wCh++Kv/g1sP/kKv+HKn7K//Q/fFX/wa2H/AMhVrn/BEz9nu4hiXw18VPiHYyhj5rX0lndhhjgAJBFtPuSfpX/DkL4Vf9Fw8V/+C63/AMa/4chfCr/ouHiv/wAF1v8A41/w5C+FX/RcPFf/AILrf/Gv+HIXwq/6Lh4r/wDBdb/41rv/AAQ58KXPkf8ACMftFarYbd3n/b/Dcd3v6bduyeLbj5s53ZyOmOf+HGH/AFdH/wCWR/8Ad1f8OMP+ro//ACyP/u6v+HGH/V0f/lkf/d1f8OMP+ro//LI/+7q/4cYf9XR/+WR/93V/w4w/6uj/APLI/wDu6v8Ahxh/1dH/AOWR/wDd1f8ADjD/AKuj/wDLI/8Au6tE/wCCHngqC0ZPEf7Qet31yZCUksvD8VogTAwCjzSknOedwHIGOMn/AIchfCr/AKLh4r/8F1v/AI1/w5C+FX/RcPFf/gut/wDGtP8A+CKH7M8dlDHqvxH+JtxdquJZbe9sYY3b1VGtXKj2LH61p/8AwRd/ZQsr2G6ufFvxNv44my1tcatZrHKPRjHaK2PowPvX/Dob9jT/AKAfir/wev8A4VD/AMEtf2FI4Uif4IPKyKAZH8TaqGcgdTi5AyfYAVafsB/saWQhEP7PPhRvs+3Z50Dy529N29ju6c7s575rSv2Y/wBmzQr+LVdD/Z6+Gmn3sGfKubXwlZQyR5BU7WWIEZBIPsTWh+GPDXhiGS38NeHtM0mOZg0qWNokAdgMAkIBk49aqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqvGPw2+HXxEtfsPj/wAA+HPE1vtC+TrGlQ3qYDBgNsqsMbgDj1ANePv+CY37F3j5JZG+EieHryRdq3WgX89kYxkn5YgxhzljyYycADoAK+IP/BEHw1Oj3Hwp+Omp2TKx8u08Q6Wl0HBK4BmhaPaQN5z5Z3HaMLya+I3/AASl/bK8Ab5tP8E6T4ytY8b7jw5qiS4zsxiKcRTNy5HyocbGJwuCfGXw88f/AA6v10r4g+BvEHhi9fO221nTJrKQ4Ck4WVVPR0PTowPcV8NP2kvj18HnjHw3+LHiLRoItu2yS8M1r8pBGbeTdEegHK9CR0JB+Gn/AAV7+PXhhI7T4keEvDnjaBNu6dAdMunwoBy8YaIZxniIcse2APA//BXr9nTX1ig8Z+GfF/hW5K5kka1jvbZTkDAeN/MJ5J/1QGFPcgHwH+2P+y38SljHhP45eFXllYLFbahef2dPISWGFhuRHIx+U9F6YPRgTa3Vrf2sN7Y3MVxb3EayQzROHSRGGVZWHBBBBBHWqOMdaJz3qqqqqqqmPYGqqqqqqqY54BqqOMdaJz3qqqqJAHWjz3qqqqqqqqqqqqrxB4j8PeFNKn13xTr2naPptspaa81C6S3hiAGSWdyFAwCeT2r4o/8ABTf9lb4dpcWuieKL7xtqcO5RbaDaM8QfaCubiXZEVOcbo2fGDxkYPxh/4KzfHLxmJtO+FWh6Z4CsH4W4+XUb0jjPzyII1zg9I8jdw2QGrx38U/iV8Ub4al8RvH2v+JbhGYxnVNQkuBDkkkRqxIjXLHCqABngCvh5+zb8fvizCt18OPg54t160Zdy3lrpUv2bGP8AnsQI8nsN2T2rwh/wSG/bK8TBf7a0Pwp4T3bs/wBsa6km3HTP2QT9e2PxxXh//gh745uZQvir9oDQdOj8nJbT9ClvW83j5cPLD8uN3zZzwPl54g/4IaaetrcpdftNXElyyqLWSPweESJtw3F1N4S4K5AAZcEg5OMH4n/8Eff2rfBMbXngo+GfHtqMkR6Zf/ZLlQFUktHciNOpYAJI5IXoCQK+InwN+MvwklaL4mfC3xR4aVW2ibUtLlhhc9PklK7HGeMqSK8FfFT4m/DaVpfh78RPEnhpnYs40rVJrQSE7c7hGwDA7EyDkHaM9K8C/wDBTv8Aa48Fsiaj4w0nxZbRKFS313So2xgMOZIPKlY5YHLOTlR2yD4N/wCCydyqLb/EL4HxSPyTdaNrBQfdGAIZYz33HPmcAgYOMnwb/wAFS/2SfFTrHq2v+IfCjtkKNZ0Z2BOQAM2pmAznOSQAAckcZ8K/tN/s7eN9qeF/jd4KvZm3bbb+2oY58L1PlOwfHvjFRSxTxJPBKkkcihkdTkMD0II6jFVVVVVVVVVVVVV44+Lvwr+GlvJcfEH4j+G/DqxLkpqWpxQO3ylgFRmDMxCsQqgk44Br4hf8FQf2T/BHmwaP4j1jxhdx5Bi0PTW2Bv8ArrOYkI91LfieK+IX/BYvxte+bbfCz4RaPpS8ql1rl7JeuR/e8qLygp9izj69K+J37aH7TvxcLw+Lfi/rUdkzcWGluNOtwu7cqsluE8wKcYMm5uFySQDXhXwb4v8AHmrp4e8D+FNY8RapIpZLHSrCS7nYAgEiOMMxGSB07ivgr/wSG/ab+JPl6j8Q30n4baUZFB/tNxeXroerpbwsRxyNsskTZxxg5r4F/wDBK39lb4Pxw6j4l8OS/EfXUwzXfiULLbI23DBLNcRbD1xKJWB6NVpZ2lhaQ2FhbRW1tbRrFDDCgRIkUYVVUcAAAAAcCqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq1XSNK12wl0rW9MtNQspsebbXUCyxvggjKsCDggHkdQDXxJ/wCCan7GnxL82e4+ENp4cvZM7brw3O+m+Xn+7DGfI/OM4r4o/wDBEOFmnvPgr8bXVdpEGm+KLDcd2GILXVvjgnYMCDgbjk8LXxA/4JV/tn+BHmey+H2n+LbOH7114f1aGUHLBRtimMczdc8R8DJOAK8cfBv4t/DJ2j+Ivwv8V+GduPm1fR57RWBYqCGkUAglWAIODg4rwn8QvH/gG5+2eBPHPiDw5cbi3m6Tqc1m+SpUnMbKc7SR9CRXgz/go5+2F4LRbeL4sy61bDP7nWrCC8JO0DPmsnm8BRxvxkkkZJrwr/wWX+JtoFHjn4M+GNXxu3f2TqE+nZ/u48wXGMd/X2rwj/wV6/Zx1pTF4q8L+M/Ds6qzbmtIruE4IAUNHJv3EEnlABg89M+FP25v2RvGgX+x/j34Yt927H9rTPpf3euftSx49s9e2a0fXtC8RWrXvh/WrDU7dJDG01ncpMiuACVJUkA4IOPcetVVVVUx4wDVVVVVVVMewNVVMc8A1VVVHGOtE571VVVVVVVVVVXxE/aZ/Z++E/nx+P8A4v8AhjS7q23ebY/blnu127s/6PFulPMbr937w29SBXxG/wCCvPwQ8PLJb/DfwP4k8X3SMNslyyaXauM8kOwkkzjJwYh255OPiT/wVQ/ai8bpJZ+F77QvBFo+5f8AiU2AmnZCpBDS3Bkwec7o1QggYI7+MviB46+ImpHWPHvjPWvEV6WLCfVL+S5dSQAcFydowqjAwMKB0Ar4O/sY/tN/Hj7Pc/Dr4Ra3PplxsKavfxCxsijfxrPMVWQAckR7mxjg5APwf/4IlXs6Qah8evi8luGX95pXhS33sDnj/SrhcA7eo8kjJ4YgfN8JP2FP2UvgrGkvg74OaJc6gmT/AGlrMR1O5yV2sVefd5eV4IjCjBPHJzVVVTQw3EL288SSxyKVdHXIYEYIIPUEdq+IX7E37JvxRST/AITD4B+EZJpWLS3WnWX9mXEjEqctNamORj8g6seMjoxB8f8A/BFn9nvXo3m+H/j7xh4VuTGVVLhotSt1bb8rbGVJD82CR5nI4G3rXj3/AIIo/HTRhJN8O/ih4P8AE0Me4iO/jn0yeQDG3aoEyZOWyGkAGByc8eN/+CdP7Z/gN5zqXwJ1vU4Yd5WbRJIdTEqq2Nypbu789QpUNg/dHNeKfA/jXwPdCw8a+ENb0C5MkkQh1TT5bR98ZAkXbIoOVJAI7EjNeGPHnjnwROLnwX4013QJhIJBJpeoy2jBwMBsxsDnHGa8Mftz/tceEEhTSfjx4kuBCrKv9qPHqRO4kncblZNx54JyQMAYArw5/wAFYP2rNDiWPU/+EM8QsIVQvqWjshLDq/8Ao8sQ3Hvxt9AK0L/gsp40t5QfE3wM0S/j8nBWx1qWzPmcfNl45flxu+XGeR83HOj/APBZD4cTpaHxB8GPElkzsv2oWepQ3QiGfmKbhHvIHIB2ZPGR1rRv+Ctn7LWqSyR32l+PNIVFyr3ukwsHPoPJnkOfqAK0f/gpt+xrqdq0978S77SZFkKiC88P3rOwwPmBhikXHJHJB4PGME6P+33+x7rtq13Y/HPR40SQoVvLa5tHyADwk0SsRyOQMdRng1a/ttfsm3lzDZw/Hvwmsk0iohluzEgJOBudgFUepJAHUkCv+Gn/ANmj/o4n4Zf+FdY//Ha/4af/AGaP+jifhl/4V1j/APHa/wCGn/2aP+jifhl/4V1j/wDHa1n9sX9lfQpY4b79oDwPK0qllNnq8d4Bj1MJcKfY4NX/AO3f+yHp1nLe3Hx28PvHCu5lgWadz/uoiFmPsATWp/8ABTH9jGxsZLq1+KtzqMseNttbeHtQWR8kA4MkKLwDnlhwDjJwK1v/AIK0/ssaV5P2Cw8c6x5u7d9i0iJPKxjG7zp4+uTjbnoc44z4h/4LIfDi2S5PhT4MeJNRZGH2YajqUNkJBkZL7BNsOMnA3cgDIzkeKP8Agsd8ULxpT4K+DvhbSgWXyhql7PqBQADcG8vyNxJyQRjAwMHrXij/AIKgfte+IvMXTvGej+Hkkkdiml6JAdqt/AGnErADsc7vVjXi79pf9oTx2w/4Sv41eMr+NWVlgbWZo4VZQQGESMEDYJG7GeTzWlaRquu38WlaHpl3qF7Pnyra1gaWR8Ak7VUEnABJ46Amvhz+wF+2D8UJYhoHwJ8R6fbuw3XWvQjSI1Q7MyD7UUZ1w4PyBiQG2g7SB8M/+CJfxO1X7Pd/Fv4v+H/D8TbXks9Es5NSm2/ITGzyeSiPgyAsPMUFVI3gnHwu/wCCSv7Ifw+jiuPEmg63481BNrGbXtRZYlfaA22G3ESFCQSFk8zG7qcA14V8G+D/AAJo6eHvA/hTR/D2lxsWSx0qxjtIFbABIjjAUHAA6dhVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVXjj9kj9mL4kRsPGPwH8E3szYzdR6PFbXBAUqB50QWTADHjdjOD1AI8ef8Eev2RfFQml8LReLfBkzKfJTTNXNzCjbMDct0srsu4biA6k5IDAYx49/wCCIHiq38yf4X/HfSr/AHbjHa69pMlns6bQZoWl3Z+bJ8tcYHBycePf+CUf7Z/gkSS6f4E0nxbbQ7t8+g6xE/AxgiOcxStnJwFQn5TkDjPjj9nb49fDVpR49+DXjPQ44VkZri80SdICsYBkZZduxlUEEsCQM8mtJ1nWNBvBqGhateaddKpUT2k7QuAeo3KQcGvB37av7V3gSRJNA+PXiyQR/cj1S8/tONRt24CXQkUAA8DGAcHqAa8If8FbP2o/D/kxeIbTwd4niHlLM97pb28zKv3yrW8kaKzjqShUHGFAyD4U/wCCzlq0aweOfgPKkixsWudJ10MHfd8oEMkQ2jb1PmNyOmDx4X/4KyfsneIJxDq03jHw0vmBfN1TRhIoGPvf6LJMcdumfavCn7b37JXjOJZdH+PvhO3DRs4GrXZ0s4VtpyLoRkHPRTyRyARzWgeLvCniyJ5/CvifSdZjiVGd9PvY7gKHB2klCcAgHHrg0xwMCjz3qqqqqqokAYBo896qmPGAaqqqqY54BqqqqqqqqrWNe0Lw9are6/rVhplu8gjSa8uEhRnIJCgsQM4BOPY144/bs/ZL8BW8kup/G7QNSkRfkh0ORtUaRtpYKDbh1BO3GWIUEgEjIrx3/wAFg/g1o6vD8Pfhr4p8S3EbEB7+aLTIH5XBVwZnxgt1jByAO+R49/4K3/tFeI0ntPBXh/wp4Sgkz5U8dq97dRfKR9+VjEcZB/1XVR1BIPxE/aZ/aB+LHnx+P/i/4m1W2uN3m2P25oLRt27P+jxbYhw7r937p29ABXhPwP408e6kNG8C+ENb8R6gcYtNJ0+W7l56fJGrH9K+GP8AwSu/bG+JUa3N34H0/wAGWb/cuPE9+LYn5SeYYhJMvQD5oxyw7BiPhd/wRL+HelvFe/GL4v63r7rtY2Gg2aadEGDAlWlk815EIBB2iNueCMc/CX9jn9mT4IPbXXw5+Dfh+y1G02mLU7qA3t4jKxYMs85eRDuOflI6KOiqBVVVVVVVVV5Z2moWk1hfW0VzbXMbRTQzIHSRGGGVlPBBBIIPBzXiz9kz9mHxy003ij4AeAby5uGVpbsaBbw3DlQAuZo1WQgAAY3YwAO1eI/+CUf7EuvSxzWXw21LQ2VnaQadr93iUtjqs0kgUDBwECjk9eMa5/wRK+A1xLEfDXxa8e2MYUiVb4Wl2zN2IKRRbRjsQfrWu/8ABDnxVb+R/wAIz+0VpV/u3ef9v8NSWezpt27LiXdnnOduMDrnjxB/wRe/am0yK7udF8XfDnWY4ZCLeGLUrqGedN2AdsluI1O07iDIQMEAscZ1z/glF+29pMsUWn/DLTNaWRSWksfEVigjPofPljOfoCPetV/4J3/tp6Pfy6bd/s++IJJYcbmtZbe5jOQCNskUjI3BGcE4OQeQRWsfsbftZ6Hemwvf2bfiRLKFDbrPw1dXceD/ANNIUZc+2citQ/Ze/aY0myl1LVf2dvibZ2luu6We48I30ccY9WZogAPrX/Cpfip/0TPxX/4Jbj/4iv8AhUvxU/6Jn4r/APBLcf8AxFf8Kl+Kn/RM/Ff/AIJbj/4itC/Z5+P3inz/APhGfgb8QNX+y7fP+w+Gby48rdnbu2RnGdpxnrg+lQ/skftVTypBH+zT8Uw0jBVL+D75ACeOSYgAPc8CrT/gnz+2ffXUNlD+z34lWSeRY0MwhiQEnALO7hVHPLMQB1JArRP+CUv7cGrXbW1/8LNP0aNYywnvvEdgyMQQNoEM0jZOSeQBwec4B8N/8EXf2oNVS0uPEHjH4eaJDKxE8L6hc3FxAASM7Ut/LYnAIAk6EZIORXhX/gh1qDsk3jj9oe3hVZiHttK8OmQyR4HIlknXY2c8eWwwByc4HhP/AIIv/sw6OkM3ijxd498Q3CqwlRr63tbdySdpCRw+YpAwP9YQTk98Dwd/wTt/Yu8DtHJpXwD0G9ljZWL6zJPqgdgMZK3Mki4PUqAFz2rwx4N8H+CdPXSfBfhTR9BsVVVW10yxjtYwFG1QEjAGAoAAxwOKqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq8b/Ab4I/EqOdPH/wj8H+IXuN5ebUNFgmlDMu1nWRl3q+P4gQw4wRivGH/BLj9ifxdI1zH8J5dDuXkVnl0fWLqAEKu3aIjI0Sg8E7UBJGc8nPjD/giJ8IL6Jl8AfGrxho0nlqFbWLK21MBw3zEiIW+QV4AyMHnJHy140/4In/AB30vzpfAnxS8E+IIovOZUv1uNOmlVf9WFUJKm9xnIZwqnHzEEkeLP8AgmF+2z4TaZ2+DbaxaxMqrc6Tq1pciQsAfljEolwDkElAMj0wT4o/Z6+PfgjB8YfBPx3oqHzNkl/4duoEcR43lWZAGAyMkEjkHvVrdXVjdQ3tjcy29zbyLJDNE5R43U5VlYcgggEEdK8M/tIftBeDGT/hF/jd440+NJTN5EWvXHks+ACzRFyjHAA+YHoPSvC//BSf9sbwwsMDfFVNYtoVYLDqmk2s5JYk5aQRiViCeMvgDA6ACvDf/BYH9ofTvJh8SeCPAmswxxlXeO1ubWeVuzFhMyD6CMfhXhv/AILOWreTB4v+AUseIz51zpviEPufttheAYH1kOPevDX/AAV9/Zx1T7NB4j8IeOtEnl3+dJ9jt7m3hxnb8yTeY2QAOI+C3oN1aL/wUz/Yz1eCN7n4pXWlTSSbBb3ug3wYc4BLRxOgB9S3HfFaN+2R+ypr0skNj+0D4HiaJdzG91eOzB+hmKBj7DJrQvij8MvFP2X/AIRn4i+GNX+3f8ev2HV4Ljz/APc2Od3TtVUSAKPPeqokAcUee9VVVTEAYFVVVVXV1a2NrLe3tzFb29vG0ks0rhEjRRlmYngAAEknpWtfHz4E+HJY4PEPxr8B6XJMpaNL3xJaQFwOCQGkGRWt/wDBQT9jnQPJ+3fHHTJfP3bPsVjd3mNuM7vJifb1GN2M84zg48R/8Fbv2ZNIiK6JovjfXZ2hZoxb6bDDGHH3VdpZlZQTjlVbA7HpXij/AILLRBZYPBXwHZmKr5V1qmvY2nI3BoY4TkYyARIOSD2wfF3/AAVl/ag8QebF4etPB/hmI+asL2WmPPKqt9ws1xJIjMgHBCKpJOVIwB4r/ba/ay8ZyNJq/wAe/FduzSK5GlXY0sZVdowLURgDHVehPJBPNatrOsa9dnUNc1a81G6KhTPdztM5A6DcxJxXgT9mz9oH4niCTwB8FvGet29woaO7ttFmNtgpvUmcqI1DKMjLDORjORXw/wD+CRP7YfjMCXxDo3hnwTD1zresLK7L8vKpaCYgkMcK+3lGB28Z+HP/AARC8KW+y4+Lnxw1W/3Z32fhzTY7PZ94DE85l3Z/dn/VLjDDnIYfDb/gmp+xp8NPKnt/hDaeI72PGbrxLO+peZj+9C58j8oxWheHfD/hbTY9G8MaHp+kWEP+rtLC1S3iT6IgAHQdqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqvFnwr+GHj5Jo/HXw48L+I1uFVZl1bR4LwSBSCobzFbIBAIB6ECvFH/AAT5/Yw8Xicar+z34Zg+0SCR/wCyxNpuCP7v2Z49g/2VwPavE/8AwR2/Y/1+UyaSfHHhtTM0gj0zW1kAU9Ix9pimO1e3O71JrxF/wQ88FXQUeE/2g9b07EjFv7R8PxXuU7AbJocEdz0PoK8Rf8ESfjnbBf8AhE/i74E1L94wb+0Uu7LCdiNkc3J7joPU14m/4JJftp6CJP7K8G+H/Eey4MS/2Z4gt496jP70faTD8hwODhvmHyjnHiX/AIJ/ftm+FGuk1T9nrxRObNQ0n9mxx6iGBAPyG2eTzDgjhMkHIPIONc/Z/wDjz4Ylit/EnwS8e6TLMpaJL7w1dwM4HBIDxjIqaGW3le3nieOWJiro6lWVhwQQeh9q0bxH4h8OSyT+Hdf1HS5Jl2yPZXTwFwOgJQjIrSv2iP2gdBsxp+hfHX4hadaqxYQWnie8hQE9TtWQDJq1/bd/a0s7WGzh+PnixkgjVEMt2JXIAwNzsCzHjksST1JNf8PKP21/+i1f+W5pf/yNX/D1b9rf/oM+GP8AwSJ/jVr/AMFff2nLe1ht5fCXw2uZIo1Vp5dLvA8pAwWYLdhcnqcADngAcVo3/BYj47QyyN4h+GngK9jK4jWyju7Uqe5JaaTIx2wPrX/D5H4mf9Ea8Mf+DCf/AAq1/wCCyfxCS5he9+Cnh2W3WRTNHFqk0bumfmCsVYKSMgEqcdcHpX/D6H/q2z/y8v8A7ir/AIfQ/wDVtn/l5f8A3FX/AA+h/wCrbP8Ay8v/ALirWP8Ags3rk9qqeH/2fbCyuBIC0l54le6Qpg5AVbeMg5xzuPQ8c5B/4LH/ABLP/NGvDP8A4MJ/8Kv/APgsZ8WZLOWPTPhH4Rguiv7qSe5uJkU+rIGUsPYMPrX/AA9//aX/AOhI+GX/AILL7/5Mq6/4KsftZXFzLPDf+E7ZJJGZIItFBSIE8KpZ2bA6DJJ9SetX/wDwUu/bPvLuW5t/i1BZRyNlbeDw9p5SP2UvAzY+rE1f/tvftaalFcQ3Hx68Uot0rq5guFgK7uuwxqCh54K4I7YxWsftBfHrxFarY6/8b/H2p26SCRIbzxLdzIrgEBgrSEA4JGfc+tanqmp61eyanrGpXN/eTY8y4uZmlkfAAGWYknAAHPYCtD8MeJfFEstv4a8Panq0sChpUsbR52QHgEhAcD614Z/Y8/aq8YeW2g/s7/ECSKW3FxFcXGgT2sMsZxhkllVUbIYEAEkjkcAmvDH/AASr/bZ8SQrcXPwvsdDheFZYm1TXbRGcMM7dkcjujAdQ6qR0PPFeGP8AgiR8bruVV8Z/GHwPpcfnKC2mQ3V+wj/iYCSOD5gM4XOD/eFeE/8AgiH8JbNIR46+Nvi3V2VWEx0mwt9NDtk7Sok+0bQBgEHOTzkZwPBP/BLH9izwZJHcz/DK68R3MX3Jdb1e4mA+UqcxIyRNnJPzIcHBGMDHgf4A/Az4aNFL8Pvg94M8PTwrGoudO0O3gnbywQjNKqb3YAn5mJPzE55NVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVarpGla7YS6VremWmoWU2PNtrqBZY3wQRlWBBwQDyOoBrXP2dP2fPFEsU/iX4E/DzVpYFKxPfeF7OdkB5IUvGcD2rUP2Ef2O9TvZb+5/Z28GJLM25lt7DyIwf8AZjjKqo9gAKm/4Ja/sKSwvFH8EHiZlIWRPE2qlkOOozckZHuCKm/4JB/sbSwvFHpPi2JmUhZE11iyHHUZUjI9wRX/AA5U/ZX/AOh++Kv/AINbD/5Cqb/giH8JWlc2/wAbfFqRljsV7C3ZlHYEjGTjvgfQVrv/AAQ58KXPkf8ACMftFarYbd3n/b/Dcd3v6bduyeLbj5s53ZyOmOf+HGH/AFdH/wCWR/8Ad1ar/wAENdVhsJZNE/aXtLu9GPKhuvCLW8bcjOXW7kK8ZP3TkgDjOR/w5C+Kf/RcfCn/AILbj/Gv+HIXxT/6Lj4U/wDBbcf41/w5C+Kf/RcfCn/gtuP8a0j/AIIe+OZ70R6/+0BoNnabTmWz0KW5kDdhsaWMY993Hoa/4cYf9XR/+WR/93Vaf8EM7RLuB7/9p2Wa2WRTNHD4METumfmCsb1gpIyASrAHnB6V/wAOQvhV/wBFw8V/+C63/wAa0v8A4In/ALNkNhFFrfxK+Jd3ejPmzWt3ZW8bcnG2NrWQrxgHLHJBPGcDR/8Agjt+x/plkLW9/wCE41aUMT9pvNbVZCD2xDFGuB/u5960/wD4JXfsMWdlDa3PwbuL+WJcNc3HiXUlklPqwjuFXP0UCtI/Ya/Y/wBDvRf2X7OfgeWUKVC3mlJdx4PX93NuXPvjIrRPgL8C/DVo1h4b+C/gTSrZ5DI8Nl4ctIEZyACxVIwCcKoz7D0qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/4KVfEf44eGviT+z38Nfgp8UdQ8FXPxH1y90m6ubYjYzvPp0MLyDBJCG4c4H941/wx9/wUe/6P3/8k5P/AImv+GPv+Cj3/R+//knJ/wDE1/wx9/wUe/6P3/8AJOT/AOJr/hj7/go9/wBH7/8AknJ/8TX/AAx9/wAFHv8Ao/f/AMk5P/ia/a+sP29P2QPAGk+O/E37a11r39sawumwafZqYpuYZZGlAccovlqpx0Mi+teCP2bv+CgvxC8FeH/H3hv9vqV9J8S6XbanYNNYSxuYJ4lljLLj5Ttdcjsa/wCGPv8Ago9/0fv/AOScn/xNf8Mff8FHv+j9/wDyTk/+Jr/hj7/go9/0fv8A+Scn/wATX/DH3/BR7/o/f/yTk/8Aia/4Y+/4KPf9H7/+Scn/AMTX/DH3/BR7/o/f/wAk5P8A4mv+GPv+Cj3/AEfv/wCScn/xNf8ADH3/AAUe/wCj9/8AyTk/+Jr4Yav+1v8ABX/goV8Lv2e/jJ+0fqHjzSfEuh3urXMKL5cDp9k1ARoysMkrJaB89OlVVeOPjJ8IPhjdWth8Sfit4P8ACdzexmS2h1zXbawedAcFkWV1LAHjI4zX/DWP7K//AEcx8Kf/AAs7D/47X/DWP7K//RzHwp/8LOw/+O14K+N/wW+Jeqy6D8OPi/4J8ValBbtcy2WieILW+mjhVlUyMkTswQM6KWIwCyjuKrV/2m/2bNA1W80HXf2hfhpp2pabcSW15ZXfiyyhmtpo2KvHIjShkdWBUqQCCCDX/DWP7K//AEcx8Kf/AAs7D/47X/DWP7K//RzHwp/8LOw/+O14Q8b+CviFoq+I/AHjDRPEukvI0S3+j6hFeQF14ZRJEzLkdxniqqqqqrxT+0L8AfA2vXXhXxt8cvh/4f1qx2fadN1XxNaWlzBvRXTfFJIGXcjKwyBkMD0Nf8NY/sr/APRzHwp/8LOw/wDjtf8ADWP7K/8A0cx8Kf8Aws7D/wCO14H+J3w1+J1pc3/w1+IfhnxZbWUgiuZtD1aC/SByMhXaFmCkjnBq8u7SwtJr6+uora2to2kmmmcIkSKMszMeAAASSeBX/DWP7K//AEcx8Kf/AAs7D/47X/DWP7K//RzHwp/8LOw/+O1/w1j+yv8A9HMfCn/ws7D/AOO1pOraVr+lWWu6DqdpqWm6jbx3NneWkyzQ3MMihkkjdSVdGUghgSCCCKqqqq8UeLPCvgjRZ/EfjTxNpOgaTbY86/1S9jtIIs9N0khCjPua1j/gpf8AsOaHemwvfj1YyyhQ26z0fULuPB/6aQ27Ln2zkV8Kf2mv2f8A44iOP4VfFvw5r91Iu5bCK7EV4BsD5NtJtmACnnKDBDA8qQKqtW1bStB0251nXNTtNOsLOMyXF3dzLDFCg6s7sQFA9ScVr3/BSH9iPw5rknh3UPj9pM11FJ5bSWGn3t9bk5xkXEELxMOPvByMc5xzXw8+KPw5+LegL4o+GXjbR/E2llthudNu1mEbf3XAOUb/AGWAPtVVVeNv2mPgL8O/G3h/4a+Lfiho9r4q8T6lDpum6NAz3V09xNII4lkjhV2hVnIUPLsTOfm4NVVVWveIfD/hbTZNZ8Ta7p+kWEP+su7+5S3iT6u5AHTua8U/8FGP2J/CF0LLVv2gNEuJDJJHnS7S61NMoQCd9rFIuDnhs4bkqSAa8BeO/DHxN8JWHjfwbd3dzpGpeZ9mlu9PnsZG8uRo2zDcIkifMjD5lGRgjIIJqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/wCCjv8AydR+xb/2UB//AEv0iqqqr/gsv8Xh4z/aJ0b4T2U++y+HWjj7QjW+xkv74JNLh/40Nutlj0bePWv+CQfxii+IX7Ln/Cvb69ifVvhxqktiYjM0kxspyZ7eV933Rue4hUDgLbDpVVVVVV8Zf+UyvwI/7J/df+iNbqqrxF4H8FeL9v8AwlnhDRNa2xtEv9o6fFc4Q9VG9Twe46V/wVv/AGZ/hr8CfiL4N8YfC7w1Y+HdO8c2t79q0uxHl28VzatDueKEALCrLcRjah25U4VeS3/BKz9nL4f/ALQHx31e4+KGh2+t6B4Q0U3q6ZO58u5u3lRIvNTGJIlXzWKkjLCPIZdwrwz8PPAHgoRjwb4G8P6D5VuLaP8AszTIbXZCMYjHlqMINq/L0+UelV48/Z9+BvxP0280nx78JfCmtQ38jSztc6VF5plbOZVlADpJ8zfOrBvmPPNftI/DKy+DPx78ffC7S5ml0/w5r11bWDySeY5tQ5MG9tq5fyygYgAbs44xX/BKj9kz4Nz/ALPWkfHXxj4C0TxH4n8U3l21rdarbLeCwgt7h7dVijkXbE5eGRi65Yhh82PlGkaHonh6yGm6Bo9jptorFhb2dusMYY9TtUAZNVVVVVXi/wCEPwp+IEN7B45+GnhfX11FVW7OpaRDcNMFA27i6kkrtXB6jaMYwK/4KAfA3wt+z5+1H4p8BeBbX7L4bmjttQ0u0MzStaxzxKzxEsAcLKJQoyxCbMsTmv8AgkH+yv8ACX4keDvFHxv+JXgzSfFF5p+uHR9JttUiFzBamOCKaWXyHUoXPnxhXbcV2HAU8tofhjw14Yhkt/DXh7TNJjmYNKljaJAHYDAJCAZOPWpoYbiF7eeJJY5FKujrkMCMEEHqCO1fHz9jb4DfHX4fat4W1P4X+GbTVpbORdK1ezsI7O5sbjafLZZ403hA+CyEMrDqrdKr9j/9jv4F/CX4JeDp/wDhWnhzU/FGq6Da3Ot6zfWUd5PcXE0SSSqkkiblhDnCoAo2opILZY2lnaWFpDYWFtFbW1tGsUMMKBEiRRhVVRwAAAABwKqqqvih8SPC3wg+HniD4neNbx7fRPDdjJd3bRgF2CjhEBIBd2KoqkjLMBkZrxP/AMNk/wDBUL4mXniXQPC+oXnh/Trh4rC0+0mDRtBTAPliWTCNOV2F2AMj5B2hQqrqv/BGb9rXT9Fl1Sz1z4c6ncxxhl0211i4WeQnGVUy2yRZGecuBwcE8Z8Z+Bfiv8AvHa6F4y0LXfBfivS2WeASb7aePkhZoZFPzKSrYkQkHBweK/4Jlft9al8d7RPgR8YNQ8/x1pFmZNK1eRvn1u2jHzrL63Ea4JbrIuWI3KzNXibxLoHgzw7qXizxTq1vpmj6PayXV9eXDbY4IY1LO7H0ABrx58bPjR/wVJ/aWsPgX4O1u78PfDA6g80FiqbVg0+Fsvf3YX/WzlQNkbNsR3RFILNI3gn9gf8AZB8DeEI/B1n8BvCmrRfZ/Jmv9b06PUL2clSGkNxKC6OSSf3ZQKSNoUAAftp/A3xL/wAE5/jB4V/aO/ZZ1C90Pwxrl01tPpryvPa2t2q7jayhmzLBPGJGVGJZTHIQy4Tb+yl+0t4R/ar+EGnfEzw0qWl6rfZdb0veXbTb1VUvFuIG9SGVlcDBVh0IYCvFmr6noHhjVdb0Tw5d+INQsLOWa10q0ljjlvZFUlYkaQqiliAMsQBmv2x9R/4Kf698G9e+Icdv4U+HHhHTLd7rUvDnhfWXuNct7NV3SPPdhPLcRqGLm3dPlB+VgCa/ZTmluP2rfg7PPK8ssnxC0Nnd2JZmOoQkkk9Saqqr9uf/AIKSeDv2Y2uvhp8PLa38TfEtoQXidt1looYZVrkqQzSkEMsK4ODuZlBUP+z1+xH8ev22tZtfj9+2x458Sp4XvZEu9J0KS48ubUYGyw8uMHbY2xyMBUDurMV2blkb4ZfAP4K/Biyisfhb8L/DnhsRKV8+ysEW4kB675yDJIecZZicYHQVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX/BR3/k6j9i3/soD/wDpfpFVVV4i1/RfCfh/U/FPiPUYbDSdGs5by/u5jhLeCJC8kjeyqpJ+lfFj4h6r8W/id4r+J+txeVe+KtYutSlgErSLb+dIziJWbkogIRc9FUCv+CSvx3tPhL+0yngjX9UlttE+JVmNJCGQLENSVw9m7jGSSTNAoH8V0M8VVVVVVfGX/lMr8CP+yf3X/ojW6qqr/guf/wA0S/7mD/3HV/wRD/5Kn8S/+xftf/Sg1VV+37/yeV8Wf+xgk/8AQEr/AIJcf8mJfDH/ALi3/p2vKqqqqqqr/grz/wAnlal/2L+nf+gNX/BFT/k1fxT/ANlAvP8A0gsKqqr4T/8AJK/Bv/Yv2P8A6TpVVVVX7fnwx8b/ALQVv8K/2dfDtjrsHh7xh4qN34s1qwtJHh0/TrKEyMk0oVo42kZ1MQk4aSJQM4IrwV4K8J/DnwnpfgbwNoNpo2g6Lbrb2NjaptSFB+pJJJLElmYliSSTVf8ABR79mzw98ff2cPEmrppMTeL/AANp8+saFepAXnxCvmT2w2gswmjRlCdPM8tv4a/Yo8Tar4S/a5+EGq6NN5dxP4w0+wc7mXMN3MttMPlIPMUzjHQ55BGQa/4LN/HO98H/AAm8N/BDQr14rnx1dSXWq7Ov2G1KFYyc5AkmZDwDkQsDjPP/AARE+GsMeifEj4w3ETtLcXVvoNm5jwEEa+fOA2eSfMtsjAxtHJzxX/BQb4c2XxM/Y7+J2lXQRZdG0WXXLaRm2mOSx/0k4OG5ZInTGOQ5GVzuH/BJ/wCOd78Kf2oLHwPdXrroPxJh/sq7hP3fta7ntJeo+YPviHXi4bjOCKr4yWdpqHwg8c2F/aw3Ntc+G9QimhmQOkiNbSBlZTwQQSCDwc1+yd/ydR8Gv+ygaF/6Xw1VV/wUo/bhn/Zf8GWvw/8AhxeQ/wDCyfFduZLaV0DjSLElkN3tPDSM6lI1IK5V2OdgV/8Agml+yCf2pfijqfxg+Llvcav4N8L33m3q6grzDX9TkBk8qSQt84Tcs0ud27eisCJCaqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/4KO/8nUfsW/9lAf/ANL9Iqqqv+CnHxD17wv+zPP8OfBMV3ceLPixrFp4U0i2s5UWaX7Q2ZlCtyySRI1ucdDcrkjNXlnd2F3NYX9rLbXNtI0U0MyFHidThlZTyCCCCD0NeHvEGs+E9f0zxV4b1GWw1bRryK9sLuE4e3nicPHIvurKpHuK+FHxD0r4t/DHwp8T9Ei8qy8VaPbalFAZVka386NXMTMvBdCSjejKRVVVVVfGX/lMr8CP+yf3X/ojW6qqr/guf/zRL/uYP/cdX/BEP/kqfxL/AOxftf8A0oNVVft+/wDJ5XxZ/wCxgk/9ASv+CXH/ACYl8Mf+4t/6dryvip4Z+LHibT7GD4TfFTTPBF3DMzXdxfeGhrIuIyMBAhnh2EHndk56Yr9qj9tn/goL+yZ8TR8N/HHifwNqH2qxjv8ATtSsdCXyby3dmXcAx3IyujqVbkbQeVZSf+CbX7RvxM/ag+BeuePvird6fPq2n+LLjTIWsrQW6CBLS0lUFQTk7pn5+npXj7SfGeueEr7S/h94wtPC+vzeX9j1a70oalHbYkUvm3Mke/cgdPvjBYNzjB/a9/a9/wCCgv7H/jux8J+K/GPgbWtO1q1a50nVrPw+sa3KqQsitGzExuhK5GSMMpBOSB/wTO/al+LH7U3gXxnr/wAWLzTLi70PVoLW0NjZC2AjaLccgE5Oe9VVV/wV5/5PK1L/ALF/Tv8A0Bq/4Iqf8mr+Kf8AsoF5/wCkFhVVVfCf/klfg3/sX7H/ANJ0qqqqqqr9onxPp/gv4BfEfxXqjoLfS/CupTsrSCPzGFs+2ME8bnbCgdywHev+CWnwjv8A4oftd+GdXFpK+k+Bo5dc1CZVG1GjXZbrlgRkzvGcfeKq5GNpZa/4K4eMLnxP+2jruiTl9nhLRdN0yDcqgbHgF2cYGSN1233snOe2AP8Agj1oNvpH7HseoQwyo+t+KNQvJWfOHZRFBlfbbAo47g1Xi3Qv+Ep8K6z4Z+1/Zv7X0+ez8/y9/lebGybtuRnG7OMjPqK8G+J9Q8E+L9C8Z6S7LfaBqVvf2zJIY2EsMiyKQy8qcqORyKhmhuIUnt5UljkUMjo2QwIyCCOoIqviz/ySvxl/2L99/wCk71+yd/ydR8Gv+ygaF/6Xw1VeJ/Eej+D/AA1q3i7xBeJa6XodjNfXs7kARQQoXkck8ABVJ59K/aC+Mmv/ALQPxj8U/FzxEXSfX75pILdm3C0tlASCEeyRqi57kE96+DGr/Ar9gT9mbwR4A+LvxG0HwtqcWkjUNStru633V1eS/vLlooFBmlVZCYxtQnCqMZr/AIe2fsV/2r/Z3/CZ+IPI+0eV9u/4R+48nbux5uMeZsx82Nm7H8OeK+F3xl+FXxt0A+JvhP490fxPp8bBJnsLgO9uxzhZYzh4mIGQrqpI5xg5qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv8Ago7/AMnUfsW/9lAf/wBL9IqqqvibqA+P3/BVP4c/C9LK01HQPgR4fuNf1aC6i8toL+4jR45Y2zmXa8mkOo/hZXPODX/BSX4MX/wY/a58ZxT3f2qy8bXEninT5WYb/LvZpWkVgPu7LhbhADyVRWP3qr/gjR8Xh4z/AGdtZ+E97Pvvfh1rB+zotvsVLG+LzxZf+NzcLe59F2D0qqqqqvjL/wAplfgR/wBk/uv/AERrdVVV/wAFz/8AmiX/AHMH/uOr9gX4mftRfDLxf4qvv2XfhVpnjnVL3TYY9Ut76zluBbQCTKuojniIJbjkn6V/w1d/wVy/6M88Kf8AgkvP/k+v+Grv+CuX/RnnhT/wSXn/AMn1/wANXf8ABXL/AKM88Kf+CS8/+T6/aP8AEPxF8WfHLxl4j+Lnhq38P+Mb/Umk1jS7eJoo7WfaoKKrO7AYA4LN161/wS4/5MS+GP8A3Fv/AE7XlV/wW9/5Kn8NP+xfuv8A0oFf8EVP+TV/FP8A2UC8/wDSCwqv+C5//NEv+5g/9x1f8EQ/+SVfEr/sYLX/ANJzVVVf8Fef+TytS/7F/Tv/AEBq/YP+Nv7c/wANfhBq+hfsx/AXRPG/hefxJPc3eoX2nXE7xXptrZXiDR3UQAEaQtjaTlzzzgf8NXf8Fcv+jPPCn/gkvP8A5Pr/AIau/wCCuX/RnnhT/wAEl5/8n1/w1d/wVy/6M88Kf+CS8/8Ak+q+E/8AySvwb/2L9j/6TpVVVVVVX/BXz9oGXSPh9ov7LngZm1DxR8QbqKXUrO0QzTpZRyqYYgi/NvnuAgXGSVhkUj5xX/BPr9k23/ZV+CcNlrVvC3jfxZ5V94knQHMbBT5NqD3EKu4z3d5COCKr9v3/AJPK+LP/AGMEn/oCV/wS4/5MS+GP/cW/9O15VVXwn/5JX4N/7F+x/wDSdKr4s/8AJK/GX/Yv33/pO9fsnf8AJ1Hwa/7KBoX/AKXw1Vf8FSfHd14G/Yu8aRafdNb3fiSa00eKRWUHZLOrTLg9Q0EcyEDnDZ4xmv2d/hZ4u+NXxt8IfDPwHqUWna7q+oA2l9KGK2nkq0zzEKCTsSJnx329utfDX/gmH+z5oM934p+M76r8YfG+rSGbU9e8S3MgWWUjDGO3R8AHj/WNKwxwwHFftc/8Eovg/wCNfA2o+Jf2dPDUPhHxtp8Znh0+G4lay1VUTmDy2ZhDIwA2smFLfeHzF1+AXx9+Jv7MHxNtPiF8PNQe0v7NjBqGn3AbyL+DcPMt7iPjKnHsysAwIYAj4GfF/wAN/H34SeGfi/4Sjli03xJZ+ckExBe3kV2jliYjglJUdCRwSuR1qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr/go7/ydR+xb/wBlAf8A9L9IqqrxFr+i+E/D+p+KfEeow2Gk6NZy3l/dzHCW8ESF5JG9lVST9K/4JY+H9Z8baX8V/wBrjxfp0trq3xf8WTyWUUo8xIbKCR2/cStlzH500sO04H+iJxxX/Bab4OSeJvg94T+NOl2cstz4K1R7HUTFCuFsrwKBLK/3sJPFCijkZum9ar/glN8YovhX+1vouh6nexQaT4/s5dBuGnmZUSd9stqwUcNI08UcC56faG5Gaqqqqr4y/wDKZX4Ef9k/uv8A0RrdVVV/wXP/AOaJf9zB/wC46v8AgiH/AMlT+Jf/AGL9r/6UGqqv2/f+Tyviz/2MEn/oCV/wS4/5MS+GP/cW/wDTteVX/Bb3/kqfw0/7F+6/9KBX/BFT/k1fxT/2UC8/9ILCq/4Ln/8ANEv+5g/9x1f8EQ/+SVfEr/sYLX/0nNVVV/wV5/5PK1L/ALF/Tv8A0Bq/4Iqf8mr+Kf8AsoF5/wCkFhVVVfCf/klfg3/sX7H/ANJ0qqqqqq/aO/aF8A/sy/C3VPib47v4gLaNo9M07zdk2qXhUmO2i4JyxHLYIRcseFNfsU/smePfHHxPvv24f2sbLz/GniKQ3Xhrw9exZ/saM48md0b7kiRhUijIzGvzN+8I8uq/4Ka+FG8J/ts/EeFIrgW+pzWmo27zEEyCe0heQrgD5RL5qjvhec9T/wAEqdXstT/Yc8CWVqzmXSbrVLa5DLgCQ6hcTDHqNkyc+ufSq8Za5L4Y8H654kt4Enl0nTbi7SJyQHaONnAJHY7cVp2n3ur6ha6Vptu9xd3kyQ28KD5pJHIVVHuSQK0PR7Lw9omn6BpquLTTLWO2tw7bmEcahVye5wBzVfFn/klfjL/sX77/ANJ3r9k7/k6j4Nf9lA0L/wBL4aqv+C1f/Jq/hb/soFn/AOkF/X/BJbR7LU/21fDV7dK5l0nSdSubba2AJDbNCc+o2TPx64Paqr/gpd8LrL4V/ti+N7LSLVLfTPELQ65aRqMYNzGGn42gAfaBPgDIAxznOP8Agip4r1LVf2f/ABh4Ru2322heKDLaMTyiz28ZZAOwDRs31kNVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVf8FHf+TqP2Lf+ygP/AOl+kVVV/wAFTvivf/Dr9lLVPCnh17ttf+JGoQeHLGOzcGYxy5e4Aj5Z0eGJ4CFGc3C+tfs//Cew+BnwT8FfCTT0tP8AimdHgtbqW1QpHcXW3dczqDyPNmaWQ57ua/aA+E9h8dPgn40+El+lp/xU2jz2trLdIXjtrrbutpyF5PlTLFJx3QVeWd3YXc1hf2sttc20jRTQzIUeJ1OGVlPIIIIIPQ1aXd3YXcF/YXUttc20iyQzQuUeJ1OVZWHIIIBBHQ1+z/8AFew+OnwT8FfFrT3tP+Km0eC6uorVy8dtdbdtzApPJ8qZZYznuhqqqqr4y/8AKZX4Ef8AZP7r/wBEa3VVVf8ABc//AJol/wBzB/7jq/4Ih/8AJU/iX/2L9r/6UGqqv2/f+Tyviz/2MEn/AKAlf8EuP+TEvhj/ANxb/wBO15Vf8Fvf+Sp/DT/sX7r/ANKBX/BFT/k1fxT/ANlAvP8A0gsKr/guf/zRL/uYP/cdX/BEP/klXxK/7GC1/wDSc1VVX/BXn/k8rUv+xf07/wBAav8Agip/yav4p/7KBef+kFhVVVfCf/klfg3/ALF+x/8ASdKqqqqr4z/Gf4efAH4eal8Tfibriado+mrgAANNdzEHZBCmRvlfBwvsSSFVmHwZ+CXj39tv4kaV+1n+1b4e/s/wbpW6T4cfDy4G6OKByrC8u1IG/fsjbDAeaVUkCJURqqz1HT9SSV9Ov7e6WCZ4JTDKHEcqHDo2DwykYIPIPWv+C2PwdurDxn4G+O+nWD/YdWsX0HU5o41VI7mFnmty5AyXkjkmAJJ+W2A4wM/8EU/HcOs/AXxn8PZbppbvwz4mF2qM3EVtdwIEAHYGS2uD9Sar9tHxlD4C/ZO+LHiSS9a1kXwre2ltOk/ktHcXMZt4WV8jDCSZMY5JwByRX/BPb4O3Xxp/a18B6GbB7nS9Avl17Vz5avHHbWbCQCRWBBSSYQwkY584DjORVfFn/klfjL/sX77/ANJ3r9k7/k6j4Nf9lA0L/wBL4aqv+ClXw01L4ofsb+O9P0SKWbUdDjh1mCGM/wCsW1lV5gQFJP7jziAMfMF5xmv+Ca3j+2+Hv7aPw5vNQvGgsdbuptGnCxqxka7geKBPm6A3DQEkEHA75INV/wAFqJoZf2qvDUcUqO0XgGzWRVbJRvt18cH0OCDj0I9a/wCCNfw9u/C/7LupeNr+Ha/jTxJcT2jByQ9rbolupwRwfOjuRxnI2/QVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVf8FHf+TqP2Lf+ygP/AOl+kVVV8YMftIf8FSvht8J0/f6B8BdHbxNqyH/R5Ir9/JmjZH5Mqb20jK8ceaPWqr/gpx8IB8Iv2wvGH2WDy9N8a7PE1jun812+1lvtDN/dzdx3WF7Ltqv+CLXxik8TfB7xZ8FtUvZZbnwVqiX2mrNMuFsrwMTFEn3sJPFM7HkZul9aqqqq+Mv/ACmV+BH/AGT+6/8ARGt1VVX/AAXI13TbnX/g/wCGIp86hp9nrF5PH/dine0SM/i1vKP+A1/wRL13TbT44ePPDlxPsvdR8LpPbIRgOsNygcA+v71Tj0BPaqqv22vEej+LP2tfivreg3iXVjJ4muoop0IKyGJvKZlIyCpZDgjgjB71/wAEpdd03WP2H/A+n2E++fRLzVLO8X/nnK1/POB/37njP41X/Bbz4c3t94S+GnxXsbJDb6TfXmkahOoy2bhI5bcHn7o+z3HOMZcAnkZ/4JUftpfDD4BWfij4R/GLxA2g6Prl8mqaVqcsbPbRXPliKaOXYCULokJViNv7tgSPlz8Uv+Cg/wCyD8KdCl1i/wDjZ4f8RziNmt9O8LXker3Fy4BIRfIYpGTtwDK6LkjLDIr/AIKXeJ/j/wDEqy+F/wAWvjTodv4R0nxOurf8In4NCFrnSbOP7ITPdyEAmefzYyUwoRYU4DMwH/BI/wDal+EPwVsfiD4D+LfjDT/C8epyW2p6bfXzlIpzGjpNEW6BwPKKr1bL45AB+GvxO8BfGHwha+Pfhp4lt9d0C8mnhgvYEdVd4ZWikADgNgPGwBxhhhgSrAmqr/grLrum61+2n4ltdOn819I0vTrO5IHCy/Z1kIB74WVQfQ5Hav8Agidrum3P7O3jXwxFPnUNP8aSXk8f92KeytkjP4tbyj/gNVXifxHo/g/w1q3i7xBeJa6XodjNfXs7kARQQoXkck8ABVJ59Kr4Da7pvij4H/D3xFo8/m2Wo+F9OngYjB2tbRkAjsR0I7EEVX7dX/BUb4pfCX406t8GvgbpuiWsHhaRIdT1bUrNriW6uTGGeONCVVI03hSSpLMhIIX737Bn7XM37X/wiuvFeu6HaaP4n0DUDYaxa2Qf7O7FFeOaHeWKowYjYWZlKNk4Kk1VfGn42/Df9n/wDqHxF+J3iGHTdLsIyUjBDT3kn8MMEeQZJGJAAHAzliqgsNc/b68H/Gj9oqL4xftLeANY8TeE/CzE+DPA1jdRixs33f6268z/AF74VWbjDtgHEaCM/wDD7z4Vf9EP8V/+DG3/AMK/4fefCr/oh/iv/wAGNv8A4V8af+Cz3j/xjo914V+BPwwTwpdXy+TDrV/ei+u0LBeYYFjEaODuUFjKDkHAPFfsx/CSP4G/AXwV8MpItuoaVpcbas5ZXMt/L+9unLL97MzyYOTxjk9a/aU+BmgftIfBTxP8INflS3GtWv8AoN6Y95sbyMh4JwOCQsiruUEFkLLkBjX7CHxF8SfsM/thX/wt+OtvL4Xsdfj/ALG1tLqMskE+d9nchl6xljtEgymyct0G4V/wWD/ah0OXwzY/sreBdZTUNb1G+iu/FENo7ObaKMq9vavt4LySFJNnJURISBvU1/wTA/ZF1T9m/wCEd34w+IGkNZePPHbRzXlpPGBLpllHnyLZuTtc7mkccHLojDMWaqvjVqFlpPwb8earqVylvaWXhnUZp5X4EcaW0jMx9gATX7L2oWWk/tMfCPVdSuUt7Sy8daLNPK/AjjS+hZmPsACaqpoYbiF7eeJJY5FKujrkMCMEEHqCO1ftzfsm+Nv2OvjWfFXhK3u4PBOqap9u8Ja1bAj7DKG80WrMM7JYWB2En50QOOQ6p+x1+0/4W/aq+DGk+ONLvrdPEVnDHbeJtLVgJLC9Awx2Z4ik2l426FTjhldV8a+NfCfw58J6p458c69aaNoOi27XF9fXT7UhQfqSSQAoBZmIABJArxRY+Nv+CmX7b+rXfgPS7u20bVryNFuZYzt0jRrcLELibqEdkXfsyQZZdgJyK+H3gTw38MPA2g/DrwfZfZdF8OafDYWMROWEcaBQWPG5zjLN1ZiSeTX7WX7Qdh+y/wDAvxB8XLrSv7Vu7DyrfTbAsUW5upnCRqzAHagyXY8fKhA+YgV+zl/wV8+M+ufGfRfDfxp0Xwzd+FfE+qQ2Uj6ZYyW82kmaTYskZ3uZI1Ljcrh3Krw2c7qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/4KO/8AJ1H7Fv8A2UB//S/SKqvEWv6L4T8P6n4p8R6jDYaTo1nLeX93McJbwRIXkkb2VVJP0r/glZ4f8XeNm+L/AO1x4106K1k+L/iRpNKhcNI8MEE07S+VK2SYPMmEKrxzZ8jAWqr/AILTfByTxN8HvCfxp0uzllufBWqPY6iYoVwtleBQJZX+9hJ4oUUcjN03rVf8E0/jTafBL9rfwrfaxqMNloniyOXw9qk0kYbalztMB3EgRj7VHbFn6KgfPGaqqqq+Mv8AymV+BH/ZP7r/ANEa3VVXxU+HnxW8aahY3Xw8/aB1j4e29tCyXFrY6Bp2oC6cnIctdxOykDjCkCvHH/BH3w98S/E154z+IH7TPj7xDrl+wNxf6jbQzSvgYUZY8KoACqMBQAAAABXhX/gjd4R8DeIbHxZ4M/aQ8d6JrWmyeZZ39hbRQTQNgglXUgjIJB9QSDwTXwk+Fnxb8AaqZ/HX7SPiD4haWmn/AGWDTtT0LT7XypAybZzPbxLLI4VGX52IPmEnJANV8T/2bfjf8Szrumt+2Z440LQdYuppIdN0nQ9OtHs4GclYEuoo1uCqqQm4vuYD5iSTX/DkL4Vf9Fw8V/8Agut/8a+Dn/BMrWP2fdSuNU+Dv7W/xA8NSXm37VBBYWstvclfumSCQNFIRkgFlJAYgcE5+HHhrxh4T8Mx6P45+JF7441RZndtXvNNtrGRkJ+VPLtkSMBemcZPeviV8OPBnxe8Caz8NviDokWreH9et/IvbSQkbgCGVlI5V1dVdWGCrKpBBAr4g/8ABES9m8SXV18LvjdaQaHPIzW9nremO09qvZDLE2Jf97Ynbg9a/Zb/AOCUXwe+AviSy8f+P9fl+IvibTZFm04XNkLSxsZgQVlWDe5lkUg7WkYqOGCBgrD9vX9h3Wv20L34eR6b8QrHwraeEW1IXks+ntdySC6+zbTGgdASv2c5BYfeHPFfAz/gmJ+yj8FGt9UuvBr+OtehXnUPFJW7jViBuKWoUQKMjKlkZ17P3qGGG3hS3t4kjjiUKiIu0KAMAADoMdqqvin8FPjV8QNa1ebwz+1h4r8FaDqcaRxaPpWgacxswI1VzHdPF9oBZlZ92/KlyAQABWo/8EU/h1rGoXOrat8fvGd7fXszz3Nzc2UMks8rks7uzElmYkkk8knJr4X/APBKKH4KeJB4t+FP7VHxA8M6qYzFJNY2kCiaM87JUOUlTIB2uCMgHGQCPhZ4L+IfgvT761+Ifxk1P4h3NxMr21zfaPZae1qgGCgW0jRWBPOWBNajb3V3p9za2N+1lczQukNykau0DkEBwrAqSDg4Iwcc18XP2Ifit8dfDJ8GfE79tPx9qWhu2ZrC10PTtOiueQQJhaxx+coKghZNwBAIAPNf8OQvhV/0XDxX/wCC63/xr4O/sEfEX4A6QfD3wn/bN+IGjaT8+zTpdIsb62iLNuYxw3KSJGS3JKAEkn1OfD1hqeleH9M0vW9dl1vUbOzihu9Tmgjge9mVAHmaOMBELsCxVAFBOAAMV+2V/wAErLD9or4oXfxg+HHxAtPCusa15f8AbVhe2Blt7iRIwnnxtGwKOQiblKkMSX3Akhv2RP2U/CH7InwsPw88N6tLrd/fXjXur6xPbCB72cqqDCAt5caqihU3Ngljkliaqq/ay/4J26D+1v8AEK18deLfjJ4o0qDT7FbSx0i2gjltrUZy7oHPDOcFjjJ2rngDH/DkL4Vf9Fw8V/8Agut/8a/4chfCr/ouHiv/AMF1v/jX/DkL4Vf9Fw8V/wDgut/8a+FP/BHv4TfDD4l+GPiNJ8T/ABBrp8M6pBqMWn3dlEkU8kLh0DlTnAdVOOhxgggkVVftVfsXfBv9rjRbW38f2d3p2vaXGyaZ4g0wql1bqefLbcCssW7nYw4y20oWJOhf8E8v20fBWkxfDzwT+35rdj4Mg8uGAJbXUM9rAFRWWBBOxiChSFjSVV46ruOP2YP+CbXwH/Zx1C28Z6glx498dwsJRr+txDZbTZDGS2tsssTbgCHYySKc4cAkVVV8WP2Ifit8cPDk/hD4mftp+PtT0S6VVubC30PTrCG4CsGAkW2jjEgDAHDZ6D0r/hyF8Kv+i4eK/wDwXW/+NfCX9nT4zfC280e2uv2vvGvijw/pkiGXSNY0awnN1GqBBG1y0bTqMKvKuDnJ6kk1XxH+GvgT4veDNS+HvxJ8NWmveH9Wj2XdlchgGwchlZSGR1IBV1IZSAQQRmtX/wCCSnir4c+Nn8b/ALJ37TniDwG8ki4srtJC8aZLMpuYHTzY9wUCKSIgrnczd9Q/4Jl/Hb4zXdtF+1J+2f4l8T6JZyKV0jTIHVHAJJcGV/Kjk5I3GFz05wAK+B/7Pfwh/Zz8Jjwf8IvBtpotpJta7uBmW5vnXOHnmbLyEFmwCdqhiFCrxVftB/A3wh+0f8JNd+EXjVpYrHWY1MV3AoM1nPG4eKaPPdWUZH8SllPDGv2ff+COdh8NPi5pfj/4mfFS08UaR4a1CO903S7TSTB9ukifdE1wXdgqAqhaIBw2SpYAfNVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX/BR3/k6j9i3/soD/wDpfpFVX/BVn4xSfCv9kfWtD0y9lg1bx/eRaBbtBMqukDgy3TFTy0bQRSQNjobheRmv2Vv+CjX7GnwF/Z28A/CeXVfFUN7oWjx/2kg0l5lF/MTNd7Xzyn2iWXb6LgV/w95/Y0/6Dnir/wAEL/8AxVf8Pef2NP8AoOeKv/BC/wD8VX7QH/BSL9iX46fBPxp8JL/WvEH/ABU2jz2trLdeHpXjtrrbutpyFbJ8qZYpOO6Cv+FS/FT/AKJn4r/8Etx/8RV58MfiVYWk1/f/AA88S21tbRtJNNNpE6JEijLMzFcAAAkk8Cvh5/wV5/ZnPgDwz/wsbXPEH/CWf2Paf299k0F/J+3+Sv2jy8NjZ5u/GO2K8Ff8FSv2NPGuqy6UPiJdaD5Vu032nW9Pe1hbDKuxW5y53ZAx0Vj2r4efGD4W/FrS4NY+Gvj7RPEVtcxySRmyu1dyiSeW7FPvABxtyQBkj1FVVfGX/lMr8CP+yf3X/ojW6qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/4KO/8nUfsW/9lAf/ANL9Iqq/aKjk/ad/4KgfCv4FwxS3Phz4N2a69rw+wqwhnYR3bCQtxJBKE0uE8cGZx61/wqb4V/8ARNPCn/gmt/8A4iv+FTfCv/omnhT/AME1v/8AEVVVeWdpqFpNYX1tFc21zG0U0MyB0kRhhlZTwQQSCDwc142/Zp/Z/wDiLpUWieMvg94U1CyguBcRxf2bHFtkCsobMYU9HYYzjmv2r/8Agj5f6HZSeL/2W7y61aKLe914a1KcGZI0gBBt5TzK7OjjY2DmRQDgGtE8Q+IPDV21/wCG9d1DSrl4zE81ldPA7ISCVLIQSMqpx0yB6V+xZ/wVj8SaNquk/DL9p7UP7U0i7uJUXxjKx+0WjSMpjFwoGGiU7wXA3KGXqENWl3aahaQ31hdRXNtcxrJDNC4dJEYZVlYcEEEEEcHNV8Zf+UyvwI/7J/df+iNbqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr/AIKO/wDJ1H7Fv/ZQH/8AS/SKrxFr+i+E/D+p+KfEeow2Gk6NZy3l/dzHCW8ESF5JG9lVST9K/wCCUnh/UfiZr/xl/bI8W6dFFq3xB8STWWnI4kd7OAP9puI4pH+/AWlto19DZ44xiqqqqqr/AIKyfs26V8Efj1Z+PfCOn2lh4b+JVvLdxWVuixrbX8HlrdqiDojeZDLnu00gAAUVX/BHD9pHVfHXgPW/2ePFWoWklx4Et47vw9udjcz2Mssnnq2cgpDK8IB7CdVxhRVfGX/lMr8CP+yf3X/ojW6qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/4KO/8AJ1H7Fv8A2UB//S/SKr/gqz8YpPhX+yPrWh6Zeywat4/vItAt2gmVXSBwZbpip5aNoIpIGx0NwvIzX7K3wgHwF/Z28A/CeWDyr3QtHj/tJBP5yi/mJmu9r90+0Sy7fRcCqqqqqr/gtl4f0W5/Z28E+KZ9OhfVtO8aR2VrdkfPFBPZXLzRj2draAn3jWq/4JM+L9Z8Nftr+FtF0uSFbbxZpep6ZqIePcWgS1e7AU/wnzrSE59AR3qvjL/ymV+BH/ZP7r/0RrdVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVf8ABR3/AJOo/Yt/7KA//pfpFV+2J+0F4V+NX/BSL4R+FtO1201LwR8PfFGi6bPJLPHLYy3MmoRSX0oJ+XZt8qF9xIP2YnpzVVVVVVX/AAWr/wCTV/C3/ZQLP/0gv6r/AIJS+H9Z1r9uDwPqOl6dLc22g2eqXuoyIOLaBrCe3EjexmnhT6yCq+Mv/KZX4Ef9k/uv/RGt1VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV/wVJ0e/8AEXx+/ZH8P6Vr93od7qfjC6tbbVLRQZrCSS70lVnjDcF0JDDPGQK8bfsA/Er4i6VFonjL9uD4rahZQXAuI4vLgh2yBWUNmPaejsMZxzX/AA5C+FX/AEXDxX/4Lrf/ABrwL/wTt8d/DT7b/wAIT+2z8VtO/tHy/tPyxTeZ5e7b/rC2Mb26Y6818Lf2b/iz4B8d6X4t8Tftc/EDxrpth53n6Fqttbpb3e+F0XeUG75WZXGO6Cqqqqq/4LIfH2w8f/GXQ/gn4eufNsvhzbyvqcscp2yahdLGzRkA7W8qJIwG6q0sqnGDVf8ABJ79kW6+C/w5uPjj45sJbbxd49s1itLK6tjFNpenLIzBTnnM5WKUgjhUi6HdVfGX/lMr8CP+yf3X/ojW6qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/4KO/8nUfsW/9lAf/ANL9IqqqqqqrW/EPh/w1aJfeI9d0/SrZ5BEk19cpAjOQSFDOQCcKTj2PpX7V/wDwVz+HvgOyk8Lfs3NaeMfEEm9JtYmRhY2atACkkXQzuHdfl4UeW4PUV8KP2QP2mv2g/FF1YeF/h1rb3JkMl/qetRvaRRu6yOGlllGSXMbjPOWIz1r9lH/glP8ACj4H3kfi/wCK91afEHxPHsa2jntdtjYSRzmRZYo25dyFiGX4GHABDVVfGX/lMr8CP+yf3X/ojW6qqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/4KO/8AJ1H7Fv8A2UB//S/SKqqqqr/hj7/go9/0fv8A+Scn/wATWr/sVf8ABQ/XdKvNE1X9u/z7LULeS3uYvs0y743Uqy5UAjIJGQQa+PP/AATD/bS0DS2ew8XS/FPSbWOK4MEGpzNMJ2kMe2O3mY7iqsGLgj5Wb0NfDL9t/wAB/BLxJYRa3+w78P8AT9Vgt4LPXZxbSw3U0J8t5WWCYFI3cKHUEYGRzivgX/wVG/Zh+NF5p3h/UdauvBWv6hlVs9bULD5hnEUcS3A+RncMrAcADdk/KatLu01C0hvrC6iuba5jWSGaFw6SIwyrKw4IIIII4OaqvjL/AMplfgR/2T+6/wDRGt1VVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV/wUd/5Oo/Yt/wCygP8A+l+kVVVVVVVX/BYX9mK08Y/DOz/aM8JaNENe8HSCDxA8EIEl5p0pVEkcjlzBJsx6JLIScIMV/wAEtP26PEnhLxloH7L/AMStT+3eFdfuPsnhu+upT5mk3TA+XbAnJaKV8Rqv8LuuMKTiq+Mv/KZX4Ef9k/uv/RGt1VVVVWgf8FGdd1v/AIKPP8E7PxFYz/Ca6um8M2kbW8Sf8TRVwLpJtu9y90rW6rv8tkdWC7sGqqqqqqqqqqqqqqqqqr4r3Px+t/7L/wCFG6N8P7/d539qf8JZqd5Z7PueV5P2aCXdn97u3bcYTGcnH7QP/BSn9rH9mPxdH4O+LX7NXhTT57uNpdPvYNWnmtb+JTgvDKBg4OMqQHXcu5RuGdF/4LQ/G7xLqlvofhz9nfQtV1K7Yrb2dldXU80rAZIVEUsxwCcAdqsP21/+CiepaUusW37Ad6kDKzBLhLqCbCkg5hkAkB4OBtyeCMgim/4LJ+L/AAJq8Hhb43fsl6x4e1SNWa8U6tJaTqMtsK2lxbKwGQFOZOzEf3a/Zv8A2/8A9m/9pyeDQfCPimXQ/FU2QnhzX0W1u5SASfJIZo5+Ax2xuXCgllUVVVVVVVVVVVVV8efiB+2X4P168k+BvwI8I+N/DttaxPE934g+yXs0p/1irGxVcL1HzAkA4ycA+Lf+Cxf7QHgHX7rwr43/AGatH0HWLJttxY6jcXVvNGfdHUHB7Hoe1fsK/wDBRPw9+1rf6r4H8X6Lp/hLxvY/v7CwiuzJFqttgl2hLgHzI8EunXaQwyA+yqr4o/FH/goH8MfB1/4yi+B3wn8XQ6ZC01xY+HPEGoT3flryzJFNbxeYQMnahLHBwCcA/Cb/AIKw/tK/HLx1p/w3+GH7NvhrWte1LcYrdNUmjVERdzyO74VEUDlmIHQckgH4VX37T95rUh+Nnhr4Y6ZozWJaD/hGtYvbu7F1uTajrPbxoE2eZllYncFwCCSKqqqqqqqqqqqqqqqqqqqv+Ct/xo+KXwh+AWh2vwz1nUNCHinXPsGp6tYs0U0USwvIIUlXmMyFCcghisbAcFq/4JSftCfGxv2oNJ+FV34x13XvCnim1vX1Syv7qW7jtHhtZJY7lN5bymLxxxEggMJFByQmKqqqq/aM/aX+FH7LvgZvG3xQ1zyPP8yPS9NgXzLrU51Td5UKD8AXbCLuXcwyM/Ev/gtD+0B4g1J4/hj4K8KeE9KWTdF9rik1G7YDcMPIzJHggqcCMEFfvYOK8A/8Fnf2kvD+pI3j3wl4P8WaaZA0sMdvJp9wFGMrHKjMi5GeWifnH0r9mr9p34W/tU+AR46+GeoyhraQQ6npV4FS802Y5wsqKSMMASrglWGcHKsFqv2rP2z/AIPfskeHkuvHN/LqPiTULd5dH8OWGDc3uDtDsT8sMW7gyP2V9gdlK14Q/ac/4Kc/tPWC+MvgX8D/AAf4J8GT7pdPvtbfe92mcBVkmdTMMHIkSBUJVvm6LXiT/gob+23+yd4usfDf7XfwI0TUNJvpHNvqel5tTdxgkM0FxG8lu5UbW8oqjgFd+zeCPgP8f/hh+0l4Bt/iJ8Kte+36c8hhuIJl8u5sZl+9FPHklHwQe4IIYEggmqqq/a6/4KvfD/4Ga9f/AA4+EOg2/jnxZpszQahdzXJj07T5VxuTcmWuHByrKpQKcgvlStar/wAFhf2wtRv5byzm8E6ZFJjba2uiM0ceAAdplkd+SMnLHknGBgD4Hf8ABazU31S10f8AaI+GdiLCVgkmt+Fy6PbjBG57WZm8wE7dxWRSBuIRjha8FeNfCfxH8J6X458Da9aa1oOs263FjfWr7kmQ/qCCCCpAZWBBAIIqppobeF555UjjiUs7u20KAMkknoMV8Zv+Cq1tcePV+D/7H3w0l+KviWaR7ePUyJfsRnGAPJijAkuIwxO6TdEmFyrMrbxrPjT/AILL6LpUvjeX4c/Dy7jt2a6m8OWawTzLGpLNCqrOWkG0bQscrSNkBSWNfszf8FaPDPjzxdD8LP2kPB8Pw78TNcfY/wC0Ud0sftYIRop45f3lod+R87Oq9GZcZNV+3b/wUi1D9k/4l6R8MfBfgfSvEuoS6WL/AFR767kiFt5jkRRqEHUqjMeTwy9O/wDw+8+Kf/RD/Cn/AIMbj/Cv+H3nxT/6If4U/wDBjcf4V/w+8+Kf/RD/AAp/4Mbj/Cv2Af2xPH37YmieMvEXiv4d6Z4b07w7dWlrYXFjdPKLuZ1kedCH5BRRAfQ+b7VVVVVNNDbwvPPKkccSlnd22hQBkkk9Biv2n/8AgsX4X8C69c+Df2b/AA3pnjOeyYpceItTklXTzIMgpBEmx51B/wCWu9FODt3qQ9Q/8Fgv2xI9QS8e78GSwpMJDaPoeI3UHPlkiQPtI44YNjvnmv2Pf+CsHhH44eJrD4Y/Grw/Y+C/FGpskOm6laTk6bfzkY8o+YS9u7NgIrM6sTt3htoeq+PHx/8Ahh+zd4Bn+InxU177Bp0cght4Il8y5vZm6RQR5BdyAT2AALEgAkfFD/gtR8bNb1C7tvhL8O/C/hnSmbFvPqiy6jegAj5shkiUsAcqY2wGwGJG6vCn/BZL9qvRdSFx4k0rwT4ismwJLabTJLZgO5R4pBg+7Bh7V+yD/wAFDPhB+1rL/wAIta2dx4R8dRQtNJ4fv7hZhcIuSzWs4CicKuCylEcfMdpVS1VVV+2F+0/4b/ZQ+DeofELU1ivNaud1p4e0xyD9svWUlNy7lPlJjdIQchRgfMyg/wDBNr9uz4/ftQfHPXPAHxV1LRJ9J0/wncanCtlpi27idLu0iUlgTkbZn49celVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVf8FHf+TqP2Lf8AsoD/APpfpFVVVVVVV8TvA9p8Tvhr4s+Gt/fS2Vt4s0O90ma5hUM8CXMDws6g8EgOSAeMiqtLu7sLuC/sLqW2ubaRZIZoXKPE6nKsrDkEEAgjoa/Zp+J//C6P2fvh98T5dVtNQvdf8P2s2pT2qbI/t4jCXaKv8Oy4WVCOxUiq+Mv/ACmV+BH/AGT+6/8ARGt1VVVV+218frT9m39m/wAV/EFL3yNcuLc6Z4dVWAd9SuFZYmUHg+WA8xHdYWHXFWmq6lY6rBrlpfSpqFvcLcRXO7LrKrbg+T33AHJ71+zT8ZNP/aA+BXg34s2JRZdc01GvoVYN5F4n7u4j47LKjgcAkYOBnFVVVVVVVVVVVVVVVVVX/BcDT7KX4dfC7VZLdGu7bWr6GKUj5kjkhjZ1HsTFGT/uiv8Agip/ydR4p/7J/ef+l9hVftDfs4/C39pvwDc+A/iboMVyDG/9nanHGovNKmbH723kIJQ5Vcr91wuGBHFfEDwZ4u+BvxV1rwRqV5LYeIvBmsPbi7tJWjZJoZPkmidSCucK6sMEZB4r/gnF+1FrP7UHwBj1LxrP5/jHwpeHS9auRD5a3uFDw3IAG0F42CsB/HG5wqsoqqqqqqqqqqqqr/gqJ+z58N/if+zj4j+KWuaV5PizwBp5utK1W3AWUx7xvtpePniO4kA8q3KkZYN4P8X+Jvh/4p0rxt4M1q40nXNEukurC9gIDwyocgjOQR2KkEEEgggkV+xH+1/4Y/a5+Fqa7EsWneL9D2W3iLSDMpaOXaMXEajnyJeSpIGGV052bjVV/wAEifDWh6P+1F8aoNN09Il0axlsLElmZobf7fgoCSSQfJjyTknaOaqqqqqqqqqqqqqqqqqqqqr4hfDrwP8AFfwjf+A/iN4Zsdf0HUlC3NjeJuR9pBVgRgqwIBDKQQQCCDXwR/ZU/Z9/Zxl1C5+DHw0sfD11qqhLu7NzPeXEiDB8sS3EkjqmQCUUhSQCRkZqqqqqaaG3heeeVI44lLO7ttCgDJJJ6DFeAP2eJP8Agoj8VLz9qf4+PqY+FVjfTWXw88Iu8tuuo6fGSovZgGDRLKwVyq4Z2XBPlom/Qv2e/gP4Z0KPwzoPwZ8E2emRR+WLaPQbfawxglsoS5I6s2S3Uk1/wUX/AOCcnwtf4Ya/8d/gZ4ZtPCmteFLNr7VdF0yBYbG/s4+ZnSIYWCSOMF8phWVGBXcQw/4JdfFHXPhz+2B4S0iwuJW0zxl5uj6naq5Cyq8bNE+M4JSVEOSD8pcDG7NV8aviv4b+Bnwo8UfFrxY//Et8M6e908QcI1xJwsUCk8B5ZWSNc8bnGa/Zt0LXP2+P25NKv/jXqH9pRazeS6tr0akojWltGXW0jXdlYjtigwG3KjEg5GTDDDbwpb28SRxxKFREXaFAGAAB0GO1ftEfA/wt+0V8IPEPwp8VWtuyarav9gupYQ7afeBT5NymeQyMe2CVLL0Yiv2Xvj/4+/Yi/aJkuL5LiOzstSbR/GWiuXCzQxTGOb5Mj99EQ7Rkjhsg8MwOk6rpuvaVZa5o19FeWGo28dxaXMLbkmidQyOp7gqQQfeqqv8Ago58e/EnwV+Ag8P/AA7S6k8c/EfUE8PaB9ljLSRNL/rpF/29n7tMHcHlRhkKcfswf8Edvh3oWiW3iT9qC5uPEuuXUIZvDmn3z21lY7lGUkmhKyzSqf4kdUyCMOMNXiP/AIJy/sXeJdHn0ef4E6PYiZSFudOmntZ4WxwyujjkdcHKnHII4r9vT/gn7r37I1/a+L/Cmq3fiH4c6vcC2tr67CC60+5IZhBcBAFbcqsVlVVBwwKqQu7/AII/ftG+IvBvxsP7Pmqai8/hfxzDPLZW80rbbLUYIml3xgnaolijdGGAWYRc/Lhq/wCCwn7V2peD9BsP2YvAmr+Re+JrP7Z4qmhbEkdiXxDahh93zWR2kXg7FQHKSkH/AII7fAbQPB3wEl+O9xbpN4h8fXVxDDcHrb6fbTNCIVGTgtPFK7HjIEYx8oJr/gs1+zZpumSaB+0z4U0mK3e/uBo/iYW8G3zZSrNbXT7R1Ko8TO3pCtf8Ejv2t9V+KHg+9/Z5+IGrfatd8GWaz6DdTyM015p27a0bsxO4wM0arjH7tkGPkJOr6rpug6Vea5rN9FZ2GnW8lxdXMzbUhiRSzux7AKCSfavhZ+wr41/4KBfE3xX+1Z8ZtX1jwV4J8ZatJd6HZBFfU72xDBIApcFIolgVEWQq27ZlU2bSbT/gkB+xxbWkFtNp3jC6kijVHnm1wh5SBguwRFUE9TtAGTwAOK/4dDfsaf8AQD8Vf+D1/wDCv+ClX7IX7MP7KPw28LyfDbSddj8W+KtWkjt2vdUaeNLOCPNw204+bfLbqO2HY8YGf+CdXwc/4Ut+yP4I0a7s/s+reIbc69qgaHynM12A6B16h0gEEZzzmPt0FVVVX7cniH4l/EW98L/scfBC/l0/xL8So5bvxJrMYfGh+HoXVJ5mKlSPNdhEBnDAPGcGVTXwF/YF/Zl+AWhWFppHw40nxFr1vGftPiHXrNLy7nkYYdl3grACCVCxhRt67iWZvjB+xn+zT8b/AA9caD4x+Evh+CeaNUi1bStPisr+12klfLuI1DAAsx2HKHJypr9qD4B6z+zN8bfEXwg1i+/tBNKkSSwv/L8v7ZayqHil25O0lThhk4ZWGTjNfsNfFHXPjP8AsmfDb4g+JriW41a70t7S9uZnLyXMtpPJatM7Ekl5DBvYnqXPTpVePPhP8Sv+Cpn7V3iq/sNebw98IfhhfNodrqM0O5iUYCYQRnBaaZlaTcwAjj8oNyFVvAf/AATK/Yw8CaLFpZ+EcPiO5EapPqWvXs13PcFc4cqGWJCd3PlRoDgZHAr9pf8A4JF/BD4gaBe638BLVvAfi2GFntrNbmSXTL6QchZEcs0JOAoaIhVzkxtV5a+Pfg18Q5rKZ9Q8M+MPB+qNG5hm8uewvIHwSroeqsvDKSD1BINfsl/GyX9on9nfwV8XL2G3h1LWbEpqcVvkIl5DI0M+1TyqtJGzKpyQrLyepqvE3iXQPBnh3UvFninVrfTNH0e1kur68uG2xwQxqWd2PoADX7X3j74m/tnXHjb9qiKK40/4V+BNSt/D3hu1u9ylvObOEUDZ5zLiaY5JQSQpllCEf8EVP+TqPFP/AGT+8/8AS+wqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr/go7/wAnUfsW/wDZQH/9L9Iqqqqqqqq+J3ge6+GPxJ8WfDW+v4r258J65e6TNcwqVSd7ad4WdQeQGKEgH1qv+CRnji08WfsYaLoFtYywSeC9c1HSZ5HYEXDvN9tDqB0G29RMHuhPQiq+Mv8AymV+BH/ZP7r/ANEa3VVVVX/BRjxJq/7Vn7Yvw7/Yr8EahcJaaPdR/wBryIpZI7u4jEssu0A7xb2Y3ZzwXlU4wSf+Cq/7NGlfAj4yaD4m8F6Wln4S8WaLBDawxRlUtrmyjSB4hj5QDEsDjGOWfgYyf+CK3x3yPF/7Oeuap/1HtBhlk/3Y7uNMj/ri4QH/AJ6MB941VV48+I3gH4X6BN4o+IvjLR/DelW6kvdaneJbocdl3EbmOQAq5JJAAJIFfBr47fCj9oTw1e+MPg94tTxDo+nalJptxdJZz24W5jSORkAmRCw2SxncoKndwTg4+MP7SfwI+ANvHL8X/ijonhyWaMSw2U0plu5oyWG9LaINM6ZRhuCEZGM5r4U/tv8A7KXxt16Pwr8N/jTo9/rE7bLexvIZ9OmuW4+WFLqOMytz91Nx4PHBxVfGH9qj9nn4BDyvi18WNE0K64/0De91eYO3B+zQK8uMMpzsxg56c1pOqWGu6VZa3pVx59lqFvHcW0uwrvjdQytggEZBBwQDXxG/b5/Y9+FWuSeGvGXx10RNSgkkint9NguNUNvIhw6Sm0jlEbg8FXIOQRjg4+Evx5+Dnx30dtb+EfxF0fxNbxKrTJazFZ7cMMjzYHAkiJz0dVP5Gry7tLC0mvr66itra2jaSaaZwiRIoyzMx4AABJJ4Ff8ADWP7K/8A0cx8Kf8Aws7D/wCO14W/aF+APjrXrXwr4J+OXw/8Qa1e7/sum6X4mtLu5n2IXfZFHIWbCKzHA4CkngGq1f8Aab/Zs0DVbzQdd/aF+GmnalptxJbXlld+LLKGa2mjYq8ciNKGR1YFSpAIIINeH/2kP2dvFus2nhvwr8fPhzrOrX8nlWlhp/imzuJ7h/7qRpIWY8dADVVVVVf8Fvf+SVfDT/sYLr/0nFf8EVP+TqPFP/ZP7z/0vsKqv+Ck+r6Prn7b3xTvdDZDbxX1pbPtUKPPgsbeGbp382OTJ7nmv+CHGg6lb+H/AIweJpYcWF/eaRZwSf3pYEu3kH4LcRf99VVatq2laDptzrOuanaadYWcZkuLu7mWGKFB1Z3YgKB6k4rxN/wUh/Yj8JarJo2q/H7SZ7iPO59M0+91KE4YrxNbQyRnlT0bkYPQgn4SftU/s7fHadLD4U/F7w/ruoSRmRNNWc294UAyzfZpgkuAOp2YHfFVq/7Tf7NmgareaDrv7Qvw007UtNuJLa8srvxZZQzW00bFXjkRpQyOrAqVIBBBBr/hrH9lf/o5j4U/+FnYf/Ha8D/GT4QfE66ubD4a/Fbwf4subKMSXMOh67bX7wITgM6wuxUE8ZPGaq8/am/ZhsLuawv/ANo/4X21zbSNFNDN4wsUeJ1OGVlMuQQQQQehrwh8evgX8QtZXw34A+NPgTxLqzxtItho/iO0vJyi/eYRxSM2B3OMCqqqqv2/P+TNPiz/ANi/J/6GlfDz9haT9oL9gzT/AIy/C/TZZviJ4Z1TUI5dOtoVJ120WVSU7EzxgsycksAY8ElNvwA+Ofj39mT4s6X8TvBMnlajpchhvbG4XCXtuSBNbSgjIDBcZ6qwDDBUV8BPjn4E/aO+Fuj/ABX+Hl68mm6mrJLbTYE9jcIcSW8ygna6n8GUqwJVlJqv+CTn/J1Hx9/7a/8ApfJVV8SPi58L/g/o8eu/FLx/oPhaxmZkgk1S+S3M7AZKxKx3SMBztUE45xUP/BTn9hifUE0yP48W4mkmEKs+g6kkYYnGTIbcIFz/ABE7QOc45rwD8TPh38VdCTxL8NfG+ieJ9MbANzpd6lwsbEA7H2ElHweUbDDoQK8QeIfD/hPRbvxH4p13T9G0nT4zJd3+oXKW8Fug/ieRyFUcjkkCv+Gsf2V/+jmPhT/4Wdh/8drwV8b/AILfEvVZdB+HHxf8E+KtSgt2uZbLRPEFrfTRwqyqZGSJ2YIGdFLEYBZR3FV/w1j+yv8A9HMfCn/ws7D/AOO14Q+PXwL+IWsr4b8AfGnwJ4l1Z42kWw0fxHaXk5RfvMI4pGbA7nGBVfEX4rfDT4R6L/wkHxO8d6J4Z0858ubU7xIPNIxkIpOXPzLwoJ5FfCH4zfDX49eDIviD8J/Ev9u6BNcSW6Xf2Oe1zJGcOuydEfgkc7cV4w/b5/Y98B+Npvh54n+OuiQa5bSeVcRwQXF1DBICQySXEMbwo6lSGVnBUjBANadqOn6xp9rq2k39ve2N7Ck9tc28okjnjcBldGUkMrKQQRwQQar4t/tU/s7fAqd7D4rfF7w/oWoRxiR9Nac3F4EIyrfZoQ8uCOh2YPavD/8AwUm/Yh8T6gNM034+6ZDNt3btQ0y+sI8ZA/1lxAiZyw4znGT0BI0DxD4f8WaLaeI/Cuu6frOk38fmWl/p9ylxBcJ/eSRCVYcdQSKqvjD+1l+zj8A7r+zfiz8XNE0PURGJDpwZ7u8VGKhWNvArygHcCCVwQGI4ViPCX/BRP9inxremw0b9oHQbeVWVd2r29zpUeWzj57uKNcfKcnOBxnGRnT/j/wDBbWfHuk/DDQviTomreJtb0+XUbKx02f7Xvto8bpWki3JGOfl3spbB252nFV8Wv2lPgJ8C48fFj4reH/D1x5ZkWxnuvMu3QKTuW2j3SsMKQCEIJwByQDoX/BSv9h/xHqUek6f8fNPinl+61/pN/YxD6yzwIi/iwrQPEPh/xZotp4j8K67p+s6Tfx+ZaX+n3KXEFwn95JEJVhx1BIqtQ1HT9JspdS1W/t7O0t13Sz3EojjjHqzMQAPrXw3/AGrP2evjD8QdR+Fvwu+KOmeJ/EWk2Ml9dQadFNLAtukqRNItyE8iQB5YxhHJO7IBGTVVVVVVX7QnibSvBnwH+IninW5vLstM8L6jNLhlDPi3fCLuIBdjhVBIyzAd6/4JAfs0ap43+Lsn7Q3iHS3Xw14IWWHSpZowUu9TkjKYUHqIY5Gcn+F2iIOQcV/wWj8bX+h/s4+GvBlhLLEnifxRGbwrjbJBbwyP5bZ55laFwRj/AFfvX/BFT/k6jxT/ANk/vP8A0vsKqv8Agp34Kh8Fftq+P0s7dIbTW2tNVgVWyWae2jMzH0JnExx6Eetf8EuviXe/En9jbwimp3aXF54TmuNBkcXHmMI7dgYFYdUKwSQqFP8ACqkYDACaaG3heeeVI44lLO7ttCgDJJJ6DFeH/wBuv9kTxV8Qh8LdA+PHhy68QNN5EKhpUtbiUkAJDeMgt5WYsAoSRixOBk8VXjj4CeH/AIg/Gz4e/GfxFqctxJ8Nre/XSNKe3Qwi5u1RHuC33i6rGgUHIBAYAMM1VftyeDdH8cfsh/FrStZhSSOy8K32qQlkDFJ7OJrmIjPQ74VGfQmv+Ccnh7UPEn7avwus9OCbrXUpr2VnB2rHBbSyvkgHBITaM8bmUEjOar9vLxtf+Pv2wvixq9/LK7WPiS50mJZMfJHZH7KoAXAAxDn15yckkn9gP/kzT4S/9i/H/wChvVf8FAPBUPj39jb4q6RJbpK1joMmqx7m2lGsmW63A+uITx3BI71+wt8S734U/ta/DHxLbXaQW93r0GlX5luPJi+y3jfZpWkJ42oJfMweMxqcjAI8ZeEfD/j/AMJax4G8WWT3uia/YzWGo2y3EkBnt5UKSJvjZXUMrEEqQcHrWnadp+kafa6TpNjb2VjZQpBbWtvEIo4I0AVURFwFUKAAAMAACqr9sSY/tif8FM/C3wD0+VLnQvDt1a6HciNnQNHFuu9ScEc71Tzo8jAzAvI+8YYYbeFLe3iSOOJQqIi7QoAwAAOgx2qqqqqqr/gqlpGrfHD9u/wz8J/hrp0ureJo/Dem6KbSMqcXMk9xcDcQTsQQ3Mbsz7Qq5Y4Ubj8BvhJo/wACPg54R+Eehuktv4Z01Ld5ljCC4nOXmmwOhklaRz7tWo291d6fc2tjftZXM0LpDcpGrtA5BAcKwKkg4OCMHHNfs8fAzwt+zf8ACLQvhD4Qle4s9GWQyXksYSW8mkkZ3lkxnLEtj0ACgYAAFV/wWH8G6P4W/a7TVdKhSOXxT4VstUvQqBQZxLPbE8dSUtUJPrX/AASe8Pahof7FPhW7vgirrWpaje26AEMsf2l4huBAwSYWYYyCrKc84FV+3R8V/GP7Wfxz0P8AYA+AGqSrai883x7q8Acx24jOXikwVVooF+d1LESTNDGCrrhv+Ci/wm8HfA7/AIJ1Wnwt8CaelrpGg6tpsMeFw08m9jJM57vI5Z2Pqx7V/wAEVP8Ak6jxT/2T+8/9L7Cqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqv+Cjv/J1H7Fv/AGUB/wD0v0iqqqqqqqr9vPwPdfD39sb4t6BeX8V3JdeJJ9WWSJSoVNQC3qJg90W5VCehKkjiq/4IgeOLrUPhr8T/AIavYxLbaFrllq0dyGO+R72B4XQjoAo09CD3MjZ6Cq+Mv/KZX4Ef9k/uv/RGt1VVVfGX4o6B8E/hV4o+K/iZXk0/wxpst48KHDXDqMRwqeQGkcogJ4BYZ4zX/BIj4ZeIfH/jH4i/tjfEQy3mra5eT6bYXU8YxNNK6z3s6Z5GD5UasuAAZVzwQP8AgpJ8C0+On7KPim1sLJ59e8Hr/wAJDpAQEs0lsredGFUEuXt2nVUHVyh7Cv2aPjJqH7P/AMdfBvxZsS7R6FqSNfQqzL59m/7u4j47tEzgcEA4ODjFadqFlq2n2uq6bcpcWl5Ck1vKnSSNgGVh7EEGq+NvhL4weNvC9toPwb+K1j8PdRmuj9u1mfQ11SVLbypBthid0UOZDH8zHgZI5GD/AMFCvgh8WPgZ8boNH+Kvxc1X4lz67paanZeIL+KWJnVneN4djvIsZRovuI5UI0R+XO0fsE/H74veGP2ctQ+AX7L/AIQ/4SH4r+J/Gl5ftcTxA2nh3Tfsunxfbbgv8h3OHRASRlGJB+RHtP8AgjLqXjT7d4s+Nn7Suq6t4z1eRp7y6tNN89PNOeWlnk3zfwnOI8dMcA1+1V+zP4z/AGRfi/N8OfEOrJqKCFL7RtZtoWgW9tmYhJApJMbhkZWTc21l4Zhhj/wTq+P2q/tE/svaD4l8S3st54i8PXEmhazdSszPcz26Iyyszfed4ZYWZucuzc9QP2kv2fv2mfjhfX+m+Bf2rm+GnhRoVW30zSfDm+6uHMe1/OvFuI5FUtuwI8DDchiorxxomv8Ahrxrr/hvxZJLJrmlapc2mpPMXLtcxyskpYuA5O9WzuAOeoBq+1n9pn9vTwlpXwk/Z/1D/hAfgro+l2una142ukdJdeuYYVWaK2UbXeBZEMZCFQ21t74by68Y/wDBEKxt/Bs0vgP46Xd34pt7fdHFqejpFaXkoQ/JlJC8AZ8YY+ZtHBDda+B3xa8d/ssfHTSvHWmpe2Op+GdSNtrOmGQxG5gWTbc2ko9CFIwc4ZVbqoI06/0rxLolrqli6XenapapNCzxkCaGRQQSrAHBUjgjvyK/4KSfsx/BjxF+y34y8dWXw/8AD+j+KPCVnHfadq9hpkcE6rHKu+F2j2l42R5BtYlVZw4GRX/BLj/k+34Y/wDcW/8ATTeVX7e37NPwd+I/7N/xI8Val4E0G18TeHdBv9dsdct9OSO8Se2ie4ZTKm1mWTaysrEqS+4gsox+wH/yeV8Jv+xgT/0B6qqqqr/gt7/ySr4af9jBdf8ApOK/4Iqf8nUeKf8Asn95/wCl9hXifxl4P8E6e2reM/Fej6DYqrs1zql9HaxgKNzEtIwGAoJPoOa/a/8A+Cr3wq+G+gap4L/Z21y38ZeN5VaCPV7aMTaXpZPBlEp+W5cDBQRh4iTlm+Uo3wb+Afxx/an8dvpHw88Oan4i1K+ujLqmsXbt9ntmclnnurl8hScsxyS7nIUMxAP7MP7P3hv9mP4LaB8JPDk/2t9PjM2p35j2Nf3smDNMR2Bb5VUklUVFJO3Jr4l/ETwn8JPAOufErxxqP2LQ/D1m91eTBdzbV6Kq/wATsxVVXuzAd6+DHhn4i/8ABV34na78VfjRrGp6F8DvCd8LLTPCOn37JHeXAXeImKkZdUkjeWcgMfMRI9oz5d3+w9+yDe+E28GTfs5eBFsHtxbmaLSI4rwKO4vFAuA/H3xJu96/b6/ZKl/Yu+MWi3vw+13U/wDhGfEKvqHhu8eYpdafcQOvmQiVSCWiLwusg2nEi9WUsf8AgmJ+2hrP7SngTUPh58TNT+2ePvB0ayPesm1tUsWO1Jm2gL5iMRG/97KNyWY1+2V+wf8ACf8AaO+HniO+8OeBdB0f4mGGW70rXrS0S1mu7wBSI7p02+csgQR75d5j3ll5BB0lNNs/EFlH4otbs6fBeRjUYIRtm8oOPNVQSMPtDAZI59K8B+Avhn4L0e0T4aeEfDmj6dJaotu+j2UUMckGMphowNykEEHJznPeq/4K7+B/htqmn/Cn4a+BfBGgxfFDxn4qEWnSWVrHBcS2zqYWjkZAPlkuJ7fBfuj46NX7N37LPwn/AGafAmjeG/BvhHR11y1sY4tU19bJPtmpT4JkkeYguVLs5VM7UDbQAKqqqq/b8/5M0+LP/Yvyf+hpX/BIb/kzTTP+xg1H/wBDWv8Agqn+walmdV/ap+D2jIsDM1x410q3UlhI7/NqMaDPBLZmAxj/AFpGDIw/YX/bG8Q/sj/E8X1w8t54G8QyRQ+JtNjiEjtGm7ZcQ5IxLHvbAzhlLKf4WXwp4q8PeOvDGk+M/CWqw6lout2cV5p93ECFnhkUMjAEAjII4IBHQgGq/wCCTn/J1Hx9/wC2v/pfJVft6ftl6V+yF8MoLzS7W31Txz4maS38P6dMT5ce1R5l1MB1ij3INoILs6qMDcy/slfsEWHj7TdP/aU/bQnuviV8QfFMaahbafrlwbi00y3f54keLOyUlWz5TDyowwRYwV3H44f8E5v2VfjN4Wn0ay+F+g+B9XSFxp+seGNNi097aUj5WkihCRzqDjKuM4yAyk5Gg+NPjd+w78ftbsvCXiJ9J8TeFNSk07UY1y9rqUUcoJjlTP7yGQKrY4OCCCrAEfA74teCf2nvgl4f+J2j6bFNpHiezJutNvEEwt5kYpPbyBlAfZKjru2gMAGHDCv+Cu37N3wi034E2vxl8K+CdE8PeJ9J1y2t7i70zT0tjf28yNGY5hHtDlSsTK7BioQqMBjj/gip/wAnUeKf+yf3n/pfYVX/AAVV/Zp+Dsn7L+u/F7Q/Amg6H4o8H3VlJFf6bpyW0t1DPdQ2zwymPbvUCZXBcNtKYXG5q/4JDf8AJ5Wm/wDYv6j/AOgLXx38BfG34h6LbaD8HfjfafDISRzLqWo/8I2urXUm7y/LEBeaNYcAS7mwzfMpUqRmv26/gx8TvgX8fbzwj8U/iXqHj/ULzT4dQsvEF+ZBLeW0rOOUkdzHtlSZdodl4yDzgfst/FT4v+M/2XfCv7HH7L1y8HjvxZq2o3nifXo2ZE8NaMZAhkeVR+6eU5wQfMVVAVd00bD9pP8A4J6ftF/s36lC194Yl8X+H7zi21zw7bS3MQfvHNGF3wv0xuG1s/KzEMF/4JvfD/4m/DP9kfwl4X+K2m3umass13cW+mXwZbiwtZJmaOORGGY2OS+w8qHAO0gqv/BTX/gobr/w51S9/Z0+A+uPYa8sO3xL4htJcTafvClba1kVspNtzvfGU3AKQ+Sn7I//AATM+EPgDwZp3jP4/wDhC08d/EfWYxeaoNczeWtjJId5hED5SRxwHkkDkvu2kKcH9oT/AIJo/sx/GrwreWvhfwBo/wAPvE6Qt/Zur+HbJbOOKTHyia2i2xSoSBu+UPjO1lJJPwq+OP7Qv/BPj44ar4VjuJYm0TVPI8SeF57gmy1MKMZ74LIQ0cyjOCp+ZcqfgX8afBf7Qnwt0P4r+A7l30zWYdzQSsvnWcynEkEoUkK6MCCM88EZBBP/AAUQ/aW1H9mT9nXUPEPhS/S28XeIrpNJ0GTCs1vK6s0lwEbORHGj4OCA7Rg9a/4Jp/sXfCb4kfCC0/aZ+OWiW/xD8T+Mr68e2XXib2G3ghuGgJkilysszSwSsXfd8rKAAdxPxc/4J5/sk/Fzw3Pocvwc8P8AhW8MZFpqnhewj0ue1fBw+IVVJcZ+7IrqfTIBH7B3wM8U/s3/APBSq++EHi2VLm60XRL8297HGUjvraSJHhnQHOAyMMrk7WDKSSpqv+CkP7fut/Be9X9nj4EyPJ8RdVhiOo6lbqsz6RHN/q4Yoxkm6kUqwyPkR0YAs6lP2W/+CaPwo8D+G7Pxv+0V4dh+I3xO1mNbrWZfEMn2+3s5nAZoFjZmjmKnCtLJvLEEqVVttft8/sJfADXf2dfGPjv4efDDw54N8U+C9Jl1e1utB06KwjmhtlaWeKWKIKjhohJhsbgwTkgFT+xL+194z/ZW+Kmk3Y1+9fwDqd8kfibRi7SQPA5VZLmOLoJ41AZWXDNsCE7WIrxja+Kb7wlrFn4H1Wx03xDNYzJpd5fW5ngt7koRG8kYILKGwSM/n0r/AIKefs6/tGfD/wAG6N8VPih+07qvxP0m/wBYOnS6c2kHSrbTJJEeWNo7eOeSLDeU6ltqkFYwS2Rj/gip/wAnUeKf+yf3n/pfYVVVVVVfED4o/Db4U6Ode+JfjzQfDFhtcpNqt/HbCXaAWEYcgyNyPlUEkkADJFfte/8ABR34SftIeJdH+DEf/CW2HwSF9HP4q1DTrWNNS1ryn3RxwxyuBFArKkhL/OzKvyrtw37NXjf4BeNvhHorfs26no83gzSoRaWtnp4MbWBXkxTRN+8jlOdx8wbm3byW37jX/BaPwTf67+zj4a8Z2EUsqeGPFEYvFUDbFBcQyJ5jZOeJVhQAf89Pav8Agmr8S9N+GH7Y/gTUNamhh07XJJ9FnlkH+ra5iZISCWAGZ/JBJzhS3GcVVf8ABWfW7TVv21/FFhbRyrJo2l6ZaTl1ADObVJgVwTkbZkHOOQeMYJ/4Irwyx/sq+JXkidVl8fXjRllIDj7DYDI9RkEfUGv2nPCPjDx9+zz8RPBfw/ung8Q6x4du7bTyjbWkkaMjygexcZTPbfmvCnwa+Lvif4mwfCnw74B8R/8ACapdLE+lHT5orqycMuXmTbuhVNwZnYAKOSQOa8PWepad4f0zT9Z1D7dqFrZxRXV1jHnyqgDv/wACYE/jVVX/AAU6+K9l8Lv2PvGNqbxotT8Yqmg6fGkuxpDOf3+cEHb9nWbIGQcgEYY1/wAEYf2cr2yg8QftOeJNOeOO/hfRfDXnRcSxhwbq4QkdN8awhl7rMvrVft5eCb/wD+2F8WNHv4pVa+8SXOqxGQD5470/alIKkgjE2PUYwcEED/gkF8S9N8Y/sk2vgWGaJdQ8Baxd2U8AGH8q4la7jlI3HIZp5UBwvMTDHG41+2drdpoH7JXxhvryOV45fBeqWiiJQSHnt3hQ8kcBpFJ9gcAnivhBDLcfFnwTb28TSSyeIrBURFJZmNwgAAHUn0qqr4m+OtN+F/w48UfEjWI/MsvC2j3Wpzx79hkWCJpCgODgtt2jg8kcGv8Agj14H1r4mfHf4k/tK+Lp3u7qzheHz5HbdPqGoSmWaX/aKpG4O4/8twcE8iqqqqqmmht4XnnlSOOJSzu7bQoAySSegxX7Vf8AwU0+BPwB0W90bwLr2n+P/HLRyR2mn6VcpcWlnOOM3k6NhQpzmNCZCV2kIDvH/BM742fsteIfGOueJ/Fniy7b9oXx9ePNql94lgjt1umlcE22mOpMaxlioERKSPgAKUjULVVVfttXOuftg/8ABQq9+Gvw0EupSx3lv4X01txlSMW4JupcAkLFHKbqRiONqM5wSa+GfgDQvhV8O/DXw18Mx7dM8MaXb6fbEoFaRYowu9scF2ILMe7MSetV/wAFB/2tf+GafhR/YPgi6+0fE7xtusPDNlBF50tvu+V7zZ32ZCxgg7pWQbWUPj/gnx+yM37NHwtbxD43je5+JnjhUvfEtzPIJpLUsSyWokxkld2ZDlg0pYhmUKa/4K8/8maal/2MGnf+htX/AARU/wCTqPFP/ZP7z/0vsKqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqq/4KO/8nUfsW/8AZQH/APS/SKqqqqqqqv8AgsZ4P0Xw1+17HrWlpKtz4s8J2Gp6iXk3Bp0kntAVH8I8m0hGPUE96r/giT4v1my+Ofj7wBbyQ/2TrPhNdTulMeXM9pdxRQkN2G29nyO5K+lV8Zf+UyvwI/7J/df+iNbqqqq/4LB/GLV9ek8C/sf/AA/tri+1/wAWX1vqV/bW5IacNI0FjajnDGSbe5U4IMUJ6NXwG+Emj/Aj4OeEfhHobpLb+GdNS3eZYwguJzl5psDoZJWkc+7VX7afwKf9nX9pPxn8OLa28rRheG/0IqjKpsLj95CqluW8sMYS3domr/glL8c0+Ln7LeneEtSvWl1z4cTDRrhZMljaAbrRwST8ojzEBxjyDwBgmq/4Le/8lT+Gn/Yv3X/pQK/4Io6dp8X7M/i/VorG3W+ufHVxBNcrEBLJHHY2bRozdSqmWQgHgGRiPvHNf8FytKsIdV+DetxQYvbu31q3ml3H5o4msmRcZwMGaQ5Ayd3OcDH/AAREllb4S/Ei3MrGNPEVuyoWOFY2+CQPUhVz9B6VX7WX/J1Hxl/7KBrv/pfNXwX07T9J+D3gfTdKsbeztLfw7YLDb28QjjjH2dOFVcAD2FV+3VpVho/7YXxcs9Ng8mKTxRdXDLuLZkmPmyNkknl3Y46DOBgYFfspzS3H7LnwdnnleWWTwDobO7sSzMbCEkknqTX7fn/JmnxZ/wCxfk/9DSv+CXH/ACfb8Mf+4t/6abyq/ay/5NX+Mv8A2T/Xf/SCav2A/wDk8r4Tf9jAn/oD1VVVVX/Bb3/klXw0/wCxguv/AEnFfsH/ALLHh/8Aa8+L2r/DXxH4q1DQLbTfDc+rJc2UCSu7x3NtCEIfjBE5OfVRX7Z//BLK/wD2dvhgvxU+F3ivVfGOnaVI3/CQW1zZhZrSA7dlwixggxqQ3mEkbQVboGI+B3jLwL4A+Kfh/wAVfEz4f2njXwvaXBXVNGuCw86B1KOyYZQZEDb0DHaWUZx1HwsT4aP8PtD1D4Padoll4P1KzjvNJj0ayS0tmhlUOrJEiqEyCCRtBB6gGqr/AILYfEvUvD/wi8B/C2wmlih8Yaxc3t4yDAlisUjxEx3dDJdRvjacmIHIxhv2Jvj5+2R8Mf2c/Dvh34L/ALH/APwmPhiS4vLiHXvtco+3yNcyLI23dhdhXysAAfu89SSf+GwP+Cj3/RhP/k3J/wDFV+2LY/t3/tj6J4a0jxP+xhe6BL4Yup57a7sj5srCZVV48u3yqdiEgdSq56Cv+Cbv7M/7VvwQ/av8O+JvGPwg8QaB4Y1DT7+w1q+ukCxpC1u8sYO1+c3ENuMEEZxxnBFf8FTvgAPgr+09qXiTR7KGDw58RIzrNgsS4WK5JC3kRHr52ZeAAFnUDocf8Em/j03xb/Zmt/Amr3zXGvfDOZdKmMkgZ2sW3NZtgcqqxq0CjHS36nmq/Z/dv2vv+CjXj39oC5he98D/AAbhOieF3lIkga6+eISxkHa4Yi7uAcEr5sOcEKaqqqqr9vz/AJM0+LP/AGL8n/oaV/wSG/5M00z/ALGDUf8A0NamhhuIXt54kljkUq6OuQwIwQQeoI7V/wAFJv2E5/2cPFz/ABS+GWlSyfDPxFccxqAf7CvHLE2xA58hgMxOenKNyFZ/+CZf7eKfATXovgj8WNaSL4c67dNJZX9yxI0K8fHzFv4baQj5xjajnzPlBkJr/gk5/wAnUfH3/tr/AOl8lV/wUn+IWq/E/wDbn1vQ306XVbHwtcWOhaZpcjtH54RUaWPKtx5k8soDKVO0r0Ir/hsD/go9/wBGE/8Ak3J/8VX/AA2B/wAFHv8Aown/AMm5P/iq/aG/Zm/bV/aD+MfiP4xX37KmtaBc+JJIZJtPsIlMUbxwRxMwJbJLmMuxPVnJr/glH8Nvjb8IPgv4o8A/GTwVqfh1YfERvNIh1AHe0csEayBfmICB4gQAB8zuTnPH/BXn/kzTUv8AsYNO/wDQ2r/gip/ydR4p/wCyf3n/AKX2FV/wVH/5MS+Jv/cJ/wDTtZ1/wSG/5PK03/sX9R/9AWq/4LV/8nUeFv8Asn9n/wCl9/X/AARu+H/hXQ/2Zr7x/Y6an9v+J9euI7+9cAuYbfakUKnHCLl2xz80jHOMAV8cfiXa/Bv4O+M/indRpKPDGi3N9FE6MwmmSM+VGQvOHk2LnIxuySBkj9lqxvfjL+2T8PJfGupvf3fiDxtb6lq1xcp5jXsgn+0TbwCOZSrAntvzg4xVV/wWv+E1lonxC8C/GTS9PSNvE1jPpuqTRrjfPalGhZ/VmjlKg+kIHYV/wRL+Ll/aeMfHnwKvbuV7DUdPTxBp8TMBHDPC6QXG0ZzvkSWDPB4t+2Of+Cy3w11/xd+zjonjjRInng8Fa8s2pQrHkpbToYvOznosnlKRg8SZyAvP/BPz/goq37LVlJ8K/iRolxq3gG/vvtMNzZAfatIlkwJHCn/WxEAEpkMCCVzkqfhR8dfg98dNFXXvhJ8RtE8TW3lrJLHZ3I8+2DdPOgbEsJOPuyKp9q8Q/s/aPrn7SnhP9pSDWmtdU8OeHbzQrmyNqHF7BK++IiTcDGY2eYkYbd5g+7tO7xP4g0/wl4a1bxVqxYWOi2M17clMbvLiQu+NxAztU9SB6kV+xlf6l+0N/wAFE/BXi3xveedqOueKLvxDcvIvmATW8M96iqMjADQoq44UBcDA21Wo6fZatp9zpWpW6XFpeQvDPC44kjYFWU+xBIrR/wDgjd+zdpPxNi8ZyeJfFF74dt74XcXhi5eJ4SAwYQSTbd7w5BGDhyuAXJyxr/grz/yZpqX/AGMGnf8AobV/wRU/5Oo8U/8AZP7z/wBL7Cqqqqqr4nf8Eyf2R/ipPfarrHhHxBY63qEjyS6xa+Jr2a4BYN90XMk0QALbguzAKgY25B/bI/4JefET9nHRb74k/DzV5fG/gay3SXr/AGfy7/Sov780a5EkYzzKmMclkVQTX7H/AO034r/ZZ+Mmk+NtI1S7Hh67uIrfxNpkZ3Jf2W758oeDIgLMjcENkZwzA2l3aahaQ31hdRXNtcxrJDNC4dJEYZVlYcEEEEEcHNfF/wCFnhP43fDPxD8KvG9p9o0bxHZm3nA+9EwIaOVf9uORUdf9pBX7SP7NfxR/ZT+JEvg3x1p1xDGZpZdC1mJQsOqW8chVZ4irNsbAVmjJ3oWXPUE/smf8FePh3feC7Hwn+1RqN7o/iXTl8pvEdtpz3NrqEar8skscAaSOY42sFjKE4YFQSq6//wAFR/hP4m1mPwH+zD4C8YfGLxjf/LYWmnabJZWv8GZJpJ1Ekcal/mcxbRsYsyrhj8dPiT42+L/xe8VfEX4jG0HiPWNQc38dm5aCFowIliiJd/3aKiovzt8qjk9a/wCCaHw5vfhx+xl4BttTDC88QQza5IpbKrHdSNJBt4BAMHksQc/MzYJGKqqqq+Mj69/wVP8A2xbb4Y/DzUZYPhF8Md0V/rkEqMkiM+J7qLqrPM0Yih+8CsYkIALqPCnhXw94G8MaT4M8JaXDpui6HZxWen2cWSsEMahUUEkk4AHJJJ6kknNV/wAFRf2GNb+PWlWfxt+D+hvf+OdAtfsup6Xbqok1eyUllZMsAZosvhQC8isFGSiKf2Sf2q/Hv7H/AMUj4t0C0+26Xe7bTxFoVx+7F9ArHgEjMcqEsUfHByCCrMp0z/grN+xPf+G11y68e63p16Y9x0a68PXTXSnA+UtEjwZ5x/rccHnpn9u79vjx/wDG34BXunfC34Q674b+E/iW+g0y58W+IALa51WXJnENpCj/AOqxbOJJAZQVYIwiLAN/wT5+HN78TP2xPhjpVqHWPRtaj1y5kVtojjsf9JGThuGeJExjkuBlc7hVV/wWB+MX/Cv/ANmSH4dafe+VqnxF1RLMok3lv9itys1wwxyRvFvGw4BWYgnBwf8Agmr8Hm+Dn7IXg60vbVoNU8WK3iLUFYj710FMPA6EWyW4IPIIOcdBVVVVVfGz9nH4O/tFWGn6V8YfDF3rllpnm/Z7aPWb2xjPmGMt5i200Yl5hjI3htpXK4yc/Hr/AIIw/DXWtHudW/Z58Yan4d1uJXeLSdal+2WNyQPljWXAlgOf42Mo6DaOTXxH+HHj34LePdS8AePtEu9C8R6FcbZoXOCpHKSRuOGRhhldSQQQQa/4JiftQ63+0h8B5dO8day2peM/A10thqd3M6mW8t3UtbXEmOdxVXjLHlmhZiSSaqq/b9/at0/9lb4HXmqadco3jXxQsun+GLYSAMs20eZdkHnZArKxwDl2iQ4D7h/wTC/YrvfgR4Rn+NnxQsm/4WF40tQIbedtz6TYOQ+xuT++mIR3zkqFRflPmA18Xfiv4J+B/wAOta+KHxC1T7FomhW5lmZQDJKx4SKNSRukdiqquRksMkDJH7D/AMJvGH7Wfxp1L/goB+0Dp7rALpofAOjSr+7to4nIjmQjbuSAb40LL88heQ4ZQTX/AAV5/wCTNNS/7GDTv/Q2r/gip/ydR4p/7J/ef+l9hVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVX/BR3/k6j9i3/soD/8ApfpFVVVVVVVX/BbL4Y2mrfCbwJ8XraKU3/h3XJNJnEVsCGtruJpA8rgZASS1RVB+XNw3cjNf8Er/ABXf+Gf23PA1nb63/Z9lr9vqOnagrSBFuozZTSxwsT1zcQ25A6llUe1V8Zf+UyvwI/7J/df+iNbqqqtc1rSvDeiah4i1y9S007SrWW6vLhwSIYY1Lu5AycBVJ49K+EPwO+IH/BTz9oz4ofHGDx7qHw803T7yOSw1IWDXkkAYlLW0TE8WHS3iy7IxAOOB5gr/AIdFfFT/AKPv8V/+CG4/+WNf8Oivip/0ff4r/wDBDcf/ACxr9t//AIJw+Pv2evhanxqvvjfqfxMjsb6Cw1EXejvbvp9tKWCSmR7mYlPOKR7cD5pgfWv+CUnxzb4SftR6d4R1K9SLQ/iPCNGuFkwALvO60cEkfMZMxAc5888E4Iqv+C3v/JU/hp/2L91/6UCv+CKn/Jq/in/soF5/6QWFV/wXP/5ol/3MH/uOr/giH/ySr4lf9jBa/wDpOar9rL/k6j4y/wDZQNd/9L5q+E//ACSvwb/2L9j/AOk6VX7fv/J5XxZ/7GCT/wBASv2Tv+TV/g1/2T/Qv/SCGv2/P+TNPiz/ANi/J/6Glf8ABLj/AJPt+GP/AHFv/TTeVX7WX/Jq/wAZf+yf67/6QTV+wH/yeV8Jv+xgT/0B6qqqqr/gt7/ySr4af9jBdf8ApOK/4Iqf8nUeKf8Asn95/wCl9hWo6dp+safc6Tq1jb3tjewvBc2tzEJI543BV0dGyGVlJBBGCDg1/wAFCP2OtS/ZU+LMl1oFrLP8PvFcklzoF2FyLVskyWUh7PHkFT/FGVOSwcL/AMEmv20I/h94ki/Zl+JGqQw+GfEFw8nhu+uJVRbC/c5NuxYgCOc5288S4AB80lar/gt7/wAlT+Gn/Yv3X/pQK/4JCeOrTxT+x/Y+FkvYpLnwbrl9YSQABXiSWT7UpIzkhjcPhuhKsP4TVVVf8FSPgEPjZ+y9quuaTb7/ABD8PJDrljti3PLAiFbqHIBIBiJkwOrwRg4GTX/BM/49J8C/2ptAXVb9Lfw/42X+wNUMshWOMzOpglPYFZljG5uFR5OQCTX7f3x4H7Pf7L3ivxXYXnka7rUf9iaE2zcReXKMN47ZjiWaUZyMxjIOcH/gnv8AAp/gB+yt4R8NajbeTruvRnXdaUoyMtzdKrCNlPR44VghbtuiJ71VVVVX7fn/ACZp8Wf+xfk/9DSv+CQ3/Jmmmf8AYwaj/wChrVeNfBXhP4j+E9U8DeOdBtNa0HWbdre+sbpNyTIf1BBAIYEMrAMCCAa/bX/Y98U/shfE3/hHri4uNX8JayrXHh7WngK+dGGIaCUgBRPHxuC8EMjgLv2r/wAEsv284vENhpv7MHxj1yGPVLGMQeEdXvLli9+mTtsZGbI8xAcRHIDKFjA3Ku//AIJOf8nUfH3/ALa/+l8lV+0trF74e/bC+Kuv6ayC70z4laxc25ddwEkepSsuR3GQOK8MeI9H8Y+GtJ8XeHrxLrS9bsYb6xnQgiWCVA8bgjIIKsDx61VV/wAFef8AkzTUv+xg07/0Nq/4Iqf8nUeKf+yf3n/pfYVX/BUf/kxL4m/9wn/07Wdf8Ehv+TytN/7F/Uf/AEBar/gtX/ydR4W/7J/Z/wDpff1/wSG/5M00z/sYNR/9DWq/4Kffa/8Ahhb4n/YvN8zy9Mz5Wc7P7TtN/Ttt3Z7Yzniv2H9btPD/AO178Ib+8jleOXxZZWiiJQSHnkEKHkjgNIpPsDgE8VVf8Fr9Pspf2Z/CGqyW6Nd2/jq3hilI+ZI5LG8Z1HsTHGT/ALor/gj1pV/qP7YUd3ZweZFpnhfULi6bcB5cZMUQbBOT88sYwMn5s9ASNR07T9Y0+50nVrG3vbG9heC5tbmISRzxuCro6NkMrKSCCMEHBr9qn/gjna6pe3fjP9lfVrfTzIu+Twjq07eVvHX7LctkqDxiOXIBJ/eKuFXXPDXx0/Zg+IUUWsaf4o+Hvi7TWLW8ytJZzYBwWilQ4kQ9Nykq3qa/YC/4Kia98SvFeh/Af9odbSXW9Wk+y6L4riKW/wBrnP8Aqre6iGE8xzlEkjxuYxqU3MXP7VkMtx+y58Yre3ieWWTwDrioiKSzMbCYAADqT6V+wT49svhp+2F8LfFWo3FvBa/21/Z80txxHEt5FJaF2ORtAE+dxOFxk5AIqqrStW0rXtNttZ0LU7TUbC8jEltd2kyzRTIejI6khgfUHFf8Fef+TNNS/wCxg07/ANDav+CKn/J1Hin/ALJ/ef8ApfYVVVVVVVeWdpqFpNYX1tFc21zG0U0MyB0kRhhlZTwQQSCDwc1+1T8JLf4FftE+PvhTYRSx6foWsSLpqTMWcWcoE1tuY8sfJkjye5571+y9NqFx+zP8I59WluJb6TwLorXL3LMZWlNjCXLluSxOc55z1qvGvgLwR8SdAl8K/EHwjo/iTR52DPY6rZR3UJYfdbY4IDDPDDkHkEVL/wAEz/2HJtXGtv8AAWxFyJlmCLrGoLBuBBA8gXHl7eOV27T3HJr9qHxd8L/2H/2TvFmrfDXwto/hGa6tTpfh+10e2SzabU542jikygBd41DzszHcywtzk1+yZ8A9S/aW+PXhf4VWiSrYXdx9o1m5jHNtYRYad89iVwin++6DvWnafZaTp9rpWm26W9pZQpDbwoMCONAFVR7AACqqqr9sH46fED45/EA/sMfsrXUNzrerRmLx94lgdmi8OWJYLNCzjChyp2yYYkbvKA8xzs/Z8/Z8+G/7NHw3sfhr8NdK8i1g/eXt7MAbnUbkgB553AG5zgDHAUAKoCgAVVfFn9kH9mf45XUupfE/4NeH9W1Gf/XajFE1leS8qRvuLdo5WxtA5Y4GR0Yg+Af2BP2OvhnqSav4U+Avh83kUgkil1Rp9VMTjGGT7XJKEIIBBXGDz1r/AILH/Hqy8ffGrRfgv4fv3msPh3au2pFJP3b6jchGZMDhjHEsQ3dVaSVcAhs/8Eaf2c5fCvgXW/2j/ENo8d/4uVtL0NXBBXT4pAZpPpJPGoHtACOGqqr9vPUrr9q7/got4N/Z00ua4l0fQLqx0O48pVIjaZluL+4XkElIWCkEjm2OPU2lnaWFpDYWFtFbW1tGsUMMKBEiRRhVVRwAAAABwKr/AIdFfFT/AKPv8V/+CG4/+WNf8Oivip/0ff4r/wDBDcf/ACxr/h0V8VP+j7/Ff/ghuP8A5Y1/w6K+Kn/R9/iv/wAENx/8sa/4dFfFT/o+/wAV/wDghuP/AJY1/wAOivip/wBH3+K//BDcf/LGqqv+C1nwZ03U/h94O+PWn2ONW0TUP7D1GZBzJZzLJLEX9o5kcLjHNy3XPH/BD+bUF+InxRt4pbgWT6LYtMisfLMomkEZYdNwVpMZ5wWx1NVXxt+NHgH9n/4b6t8TviLrMNjpemRkRozfvLycg+Xbwr1aRyMADoMscKrEfst/Ajxt+1N8T1/bi/aw8N+RJJ5f/Cu/CFxlrfS7NfmiuXRuWOTvQsBuYtLtAMW2q/4Li+JdfsvDvwj8JWerXEWj6tdavdXtmjYjuJrdbNYXYdygnmx2+c+1aH+3F+1n4a0TT/Dfh746+ItP0vSrWO0sbO3eNI7eCNQkcaKFwqqqgADoAK/4b8/bK/6OG8Vf9/k/+Jr4j/tXftF/F7wy/gz4l/F3XfEOiSTJM9leSKYzIhyrYCjkV8L/AIv/ABM+Cuvz+KvhV4z1Dw1q11ZtZTXdkwV3gZ0dozkHgtGh+qiv+CUn7T3x/wDjX+0R4h8K/FX4qa14l0m18F3V7DaX0isiTreWSLIMAchZHH/AjVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV/wUd/5Oo/Yt/wCygP8A+l+kVVV8Q/8AgmR/wn/j/wATeO/+GpfiXpP/AAkesXepf2faT4htPPmaTyoxv4Rd20ewFf8ADpv/AKu8+K3/AIEf/Z1/w5C+FX/RcPFf/gut/wDGv+HIXwq/6Lh4r/8ABdb/AONf8Om/+rvPit/4Ef8A2df8Om/+rvPit/4Ef/Z1VftJ/Biw/aG+BfjH4OX939l/4SPTylrcEkLBdRus1tI23kos0cTMo6qCO9eK/CniTwN4k1Hwf4w0W70jWtIuGt72xuoykkEi8FSP69CCCMg1+zfr+i+E/wBoj4W+KfEeow2Gk6N400i8v7uY4S3givInkkb2VVJP0qvjL/ymV+BH/ZP7r/0RrdVVV/wV2+Po+GH7OyfC3RrnZrnxLuDZuUl2vBp8JV7lhg5+cmKHBGCssnOQM/sHfAVv2dv2Y/CXgnUbFrXX9Rh/tfX45IgkiX1yqs8bgdWiQRwk8/6r0qq+MPwy0L4z/C3xT8KvEh2af4o0uaxkmEYdrdnX5JlU8F432uuf4kFavpniT4ceNbzRr5JdN8QeF9Ukt5lwVe2ureUqwwRkFXQ9R1Ffsw/G/R/2ivgV4S+LOlSp52rWKrqVuCM2t9H8lxERxgCRWKkgbkKtgBhVf8Fvf+Sp/DT/ALF+6/8ASgV/wRU/5NX8U/8AZQLz/wBILCq/4Ln/APNEv+5g/wDcdX/BEP8A5JV8Sv8AsYLX/wBJzVftZf8AJ1Hxl/7KBrv/AKXzV8J/+SV+Df8AsX7H/wBJ0qv2/f8Ak8r4s/8AYwSf+gJX7J3/ACav8Gv+yf6F/wCkENft1aVf6z+x78XLPTYPOlj8L3Vwy7guI4V82RskjoiMcdTjAySBX/BMvVLDR/25Phdd6jceVFJcahbq2wtmSbTrmKNcAE8u6jPQZycDJqv23PGvhPwP+yd8Vbvxbr1ppkereE9T0qw898G5vLm0liggjUcs7Ow4A4AZjhVYj9gP/k8r4Tf9jAn/AKA9Vq2raVoOm3Os65qdpp1hZxmS4u7uZYYoUHVndiAoHqTivBnxD8AfEjTX1n4d+OfD/imwik8qS70XU4b6JHHVS8TMAfbNVVV/wW9/5JV8NP8AsYLr/wBJxX/BFT/k6jxT/wBk/vP/AEvsKr9oH4FeCf2kPhRrXwm8d22bLVIw1tdogM2n3KcxXER7OjduAylkOVZgfjV8HvHf7O/xU1n4YeN7V7TWdAusRXMJIjuos5iuYW4JR1wyngjOCAwIH/BNT9s21/aT+FsPgHxtrSSfEvwbaiPUFlkZpdVskKpHfZb7zfMscvJPmYY7RKqiv+C1Hws1LxR8E/B/xV02zluF8DaxLb35Tpb218sa+a3t50FvH65lHqa/4J9/tfS/slfF97vX983gbxasNn4kiSMu8ARm8m7QKNxaEySZUZ3JJIMFthHgfx14O+JfhbT/ABt4C8R2OuaHqkKy2l7aSb0dSAcHurDOCrAMpyCAQRU00NvC888qRxxKWd3baFAGSST0GK8G/GG1/af/AGz4rT4dagl/8PPgXpt3JeaxayM0Gp67ep9mWJCCA6RQfasMNylixwf3bipoYbiF7eeJJY5FKujrkMCMEEHqCO1ftifAaX9m79ofxb8L4Le4TR7e6N3oUk+SZdPm+eH5sneVBMZbu0bcDoPBPxV1z/gpb8ef2fvhzrsEs2i/DfQ/7b8bqzGIXl3DIiTOyjIZJDHaqu3lRdSYKkkCqvLu0sLSa+vrqK2traNpJppnCJEijLMzHgAAEkngV4H+J3w1+J1pc3/w1+IfhnxZbWUgiuZtD1aC/SByMhXaFmCkjnBqqqv2/P8AkzT4s/8AYvyf+hpX/BIb/kzTTP8AsYNR/wDQ1qq+P/wH8A/tJfDDVPhV8RLSWTTr/EkFxbtsmsblM+XPEegdSTwQQQSpBBIPxv8Agv8AE39lf4v3XgDxc7adr+hzR3en6lp87Ks8e7dBd28gwwBK5B4ZWUg4ZSB/wRT1C91f4x/FTVdSuXuLu80WCaeV/vSSPclmY+5JJqv+Cqvws1L4dftgeJNcktJU0rxvb2+safM3zK5MaxTru9RNFIdvBCunGCCf+CW37fPhPSfC1l+zR8bPEdpo0mnyLF4P1W6/dwzxuzFrOaTG2N1Y5jdyAwbZkMqB6+KHxS8A/BnwTqPxD+JXiW00TQ9LjLTXFw2C7YJWONeskjYwqKCzHgCv2HNV8XfEXwB4n/aG8b6fLp178W/Ekus6bp8jsfsOkxQxWtjFjO0nyrfeZAqmTzAxHSv+CtulX+o/sV+Ibuzg8yLTNY024um3AeXGbhYg2Ccn55YxgZPzZ6Akf8EYdUsNP/aw1qzvLjy5dT8D3tvarsJ8yQXdnKVyBx8kTnJwOMdSAa/4KweNfCfh79jDxb4V1rXrS01jxVcWFvo1i7/vb14b+2nl2KOcJFGzMxwo+UE5ZQf+CQ3/ACeVpv8A2L+o/wDoC1X/AAWr/wCTqPC3/ZP7P/0vv6/4JDf8maaZ/wBjBqP/AKGtV8cfhpa/GX4O+M/hZdSJGPE+i3NjFK7sohmeMiKQlecLJsbGDnbgggkHUNO8W/DDxvLpuq2NxoviXwvqW2W3uIh5lpdQSdGVsglWXocg+4NfBD4raB8cvhH4U+LPhmZHsfEumx3OxT/qJvuzQnk/NHKskbcnlDyetV/wW68a2tn8Jfh38ORc2/2nVvEU2q+SVYyhLW3aLcCPlVc3mCDyTjH3Wr/gi38C73w/4C8VfH/WrN4m8VzDStELjHmWlu5M8q8fdafCdetu3HQn9q39pK//AGdfGnwYNxcWkXhnxj4sbSPEEk8YPlQSRbUlDkjYI5HWRj/dU8HpVfGH4J/C34+eDpvAnxY8H2Ov6TI2+NZ1Ky2so4EsMq4eJ8ZG5CCQSpypIP7Rvwi1T9mn9oHxV8KodYuJJ/CepIbC/VhHK0Lok9tLlD8rmOSNjg8N6Y40OGXx18MNPt/FUTiXxDoMa6kiKYmDTwASgD+E/M3Havij8P8AxF8Gvih4k+HGui4g1bwpq01m0rRNAzmJyEmQE5CuoWRSCQVZSCQQa/YH/bN8LftR/CrTdN1jWre3+JHh+1S31/TJZAsl0yDaLyEcb0kC7mAH7tyynjazV+3z+1dpvwu8Ht8CPhtq/wBv+MHxG8vR9E06xbfNp/2pli+0S7eYyQ5EQxuZypAKqxHwj+Hun/Cb4WeEfhjpbI9v4W0W105ZVQL5zQxKjSkKANzsC7HAyWJ71/wV5/5M01L/ALGDTv8A0Nq/4Iqf8nUeKf8Asn95/wCl9hVatq2laDptzrOuanaadYWcZkuLu7mWGKFB1Z3YgKB6k4rwZ8Q/AHxI019Z+Hfjnw/4psIpPKku9F1OG+iRx1UvEzAH2zVVVVVfGvw7dftwf8FGvE3h74bv9osvEPiCGxOoWyGSOGysoIraa8z/AHAluzg9GJUD7wrTtPstJ0+10rTbdLe0soUht4UGBHGgCqo9gABVVqOo6fpGn3Oratf29lY2ULz3NzcyiKOCNAWd3ZiAqhQSSeABk1+3D+0h4u/bu/aE0v4ZfBrTNQ1jw5pF4+neFtPtQ27VpycSXrISAoYLlS+NkS7m2ZfH7Bv7F2i/sh/DeWHVZbTVPHviPZLr2qRJlYgANlpATz5SHJ3EAuzFjgBFSq0/4s/CvVvGdz8ONK+JnhS88WWW/wC06Db6zbyX8OwAtvt1cyLgEE5XjIqq/ab/AGmviF8QfiFJ+x3+x3Ml58QLtWTxV4qRz9l8I2uQJCZFzicA4JGTGSFUGUgJ+zL+zJ8PP2W/h5H4J8EwveX92wn1zXLpB9q1e6xzLIecKMsEjyQgJ5LFmaviz8W/h98EPA2o/ET4l+JLTR9G06NiXmlVXuJAjMsMKkgySsFO1F5JH1r4K+PtZ+Kvwo8L/EnXvCP/AAjFz4m09NQj0o332treCXLQbpNifO0Jjdl2jYzFcnbk1X7b/wC114b/AGSvhLda6s9peeNNajktvDGkyPkyz4wbiRAc+RFkM3TcdqZUuCP2J/2L/iB+2v8AEu7+InxAu9Qj8Dwao114k165ZjNq9y7+ZLbwueXlcsS8nSMNuOWKq2h6Jo/hrRNP8N+HtNt9P0vSrWO0sbO3jCR28EahI40UcKqqoAA6ACqrxl4q0fwJ4P1zxx4glaPS/Dum3Go3zoAWWCCNpJCASBnap6kV/wAElfBGpfGT9p34jftQeKrTzJNL+0TRSPFuH9panLI0jpIe6widWA5xOuSAcNVVVVVVVf8ABZ34k6LoH7O+g/DQ3EL6z4s8QRzx25PzLa2qM0kox6SPAnPXefSv+CQ3wF1f4Vfs+X3xH8SWLWmpfEy6hvraGSIo406FWW1ds9n8yaRexSRCOtV8Z/jP8PPgD8PNS+JvxN1xNO0fTVwAAGmu5iDsghTI3yvg4X2JJCqzD4MfBj4hfts/ELTP2rP2rNDfTvBGmt53w6+HNxloUhJBW9u0IG8uArYYfvcAkCJURqqH9pjwx4g/aPtf2cvh/a2nibUNP0u71LxbqVtqSiLw8kTLFHCyqrebcPMyq0O5GjXLnONtVVf8Fef+TNNS/wCxg07/ANDav+CKn/J1Hin/ALJ/ef8ApfYVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVV/wAFPtf0Xwn+0R+yB4p8R6jDYaTo3jS4vL+7mOEt4IrzSXkkb2VVJP0r/hvz9jT/AKOH8Kf9/n/+Jr/hvz9jT/o4fwp/3+f/AOJr/hvz9jT/AKOH8Kf9/n/+Jr/hvz9jT/o4fwp/3+f/AOJr/hvz9jT/AKOH8Kf9/n/+Jr/hvz9jT/o4fwp/3+f/AOJr/hvz9jT/AKOH8Kf9/n/+Jr/hvz9jT/o4fwp/3+f/AOJr/hvz9jT/AKOH8Kf9/n/+Jr/hvz9jT/o4fwp/3+f/AOJr/hvz9jT/AKOH8Kf9/n/+Jr/hvz9jT/o4fwp/3+f/AOJr/goX4e/Y0/aQv/8Ahb/wd/aH8E2PxEPkw6na3V48VvrEKhY1cuVPlyxoqjPRkUKeQDVfsmf8FW/gz4k8AeF/Bvx88Q3Xh7xpZ2/2K+1W7iaS0vfJhBF3JMPuPLtIKkf6wnHDCtV+L3wz+Nf/AAV1+CHin4VeM9P8S6Ta+C7yymu7JiyJOttrDtGcgchZEP8AwIVVV+0L+138Bv2X7e1Hxc8XS2OoajbyT6dplpYy3NxeKhwdgRdi88ZkZFz3r9pH9qvxD+0r+0Anxl8Y6JE+k6fcQR6X4ckkDxW9hDJuFuzFSGMhLs7FTlpGwAoVR8Dv+Cg37Lfx8n0fQvCvxA+weKdX8tE8PapZy29ykzhv3QYr5UpG05MbsAMZIyBVV8cf2qPgJ+zglqvxi+INvoVzfwtNZ2gtZ7me5UHBKRxIxxnjJwB3Ir9pb4laF8Y/j749+J/hjTpbHSfEeuXF3ZQzALJ5RbCu4HCu4AdhzgsRlsZP/BO79vmH9k/VNT8DfEe01PVPh94gmSc/Y/3k2kXYG0zxxswDI67RIud3yIy5IKvof7Zf7MHiT4eXnxW0j4xaO/hXTrqK0u9RmjmgFvPKCY43SRFdWYA4BUV/wU0/af8ABX7TXxzsLr4Z3r33hbwnpI0+1v2haIX07SNJNKiuAwT5kQBgMmNmHDCv+CUn7aHwc+B/g7xN8GfjB4kh8MxajrDazpeqXEDtBK7wRRSwyOufLIW3jZdyhTlxuyVU658Xvhn4a+GcXxk17xnp9l4Kms7a8j1qViIGguSggkBxnDmWMDj+IV/wVT/ax+G37SnxA8JeG/hRqf8AbGheBLe8V9XFu8SXdzctD5gi34LRoLeMBtoBYsQWXax/4JR/tffCv9n2/wDGfw9+MPiH+wdM8TyWt5pmpyW7PBFcRCRJY5WQFl3q0ZUkbR5bZIyM/Eb9un9lj4XeGbXxN4l+LGnyR6no41fSrSyiknuNTtmd442gQL82+SJ0BYqoKksVUFh8SfGl78SviJ4p+IupW6W934p1q81WeFGyI5LiZ5WUHAyAXI6D6Cv2Nv28PgH8X/hx4B8DXPjS00j4gf2fa6Vc+H7mGSJ5buKIIxgPzK8b7Cy4YkKQGw2RXxp/af8AgR+zykS/F74h2OgXNzavc2tpJHJLPcxqcHy441Ysc8Yr9ob4ow/Gz44+N/ita2DWVr4m1q4u7S3kXDxQFsRBwGYb/LCbsEjdnHGK/YE/bt+Avir4I/Dn4QeJ/HVvonjvRbWx8NRaTfQvEb11CwWpgcApIHURrjIYNncACpbxP4c0fxj4a1bwj4gtEutL1uxmsb23dQRLBKhSRSDkEFWI59a+OvwJ+L/7Hnxi/sTXodT0y90fUvtXhrxHBE0Ed+sLq8V3bSAkBhmNioYtGxCtgivBP/BZD9qC60e28Ir8L/CPivxXOwgsruKyuRJcuQAN1tC/7xyd3EZQHIAUY51L9mj4t+N/gF8U/wBqP9tnUv7b+INt8Otd/wCEW8MuiLZ+Fozp8v70Qr8guW2qeMlMAkmQDy/gd4Q+Mnib4h6NqfwO8H63rvibw9qFvqFmdNsWuPss0TiSKRyBtQBo85YgfKa/Z7vP21fEGpR+IP2ldN+GnhjSTp7ImgeHYp57z7S3lEPNO00kShdsoCxlh8/JbAYf8Ft/+E9/4VN8PP7I83/hD/7cuf7b2dPtnlJ9j3Y7bftntnHfFf8ABIT/AIT3/hsCx/4RTzf7E/sO+/4SfH3Psnl/us9s/avs2O/3u2aqq+K/x++DPwM/sv8A4W58RNJ8L/23539n/b3ZftHk7PM24B+75sef94V/wVF/bO+H/wC0zr3hbwP8ItQuNQ8M+EmuZ7nUpLUwpfXcu1QYg4D7ERDgkLuMh4IVTX7AP7SmhfstftEWPj3xbFdt4c1XT5tH1iS1jEjwQTPG4l2YJYJJDExVcMVBxn7rfC39rf8AZv8AjXrdr4Z+F3xa0fX9XvLU3UVhCJY5/KVQzFkkVWUgEZVgCOhANV/wUd/Yth/aj+Gq+KfBVgn/AAsjwjCz6UR8p1O25Z7JiWVQSx3ozZ2sCOBIxr4cfEfx78FvHum+PvAGt3eheI9CuN0MyDBUjh45EPDIwyrIwIIJBFfAX/gsL8AfHOj2tj8bbW9+HviFFVLiZLaW+06d8YLxvErSxgkE7ZEwgYDzHwWqOX4cfHj4ZSCGWx8UeC/GOmyQsyM3k31rKpRsEYYAjIyMEdQQea/bD/4J2/F/9mHV77xBoemXvi/4dFnltddsrdpZLCEH7t8ir+5ZRgeZ/q2yCCpJRfhx8Z/i18H72S/+FvxJ8R+FpZmDTrpeoyQRzkdPMjU7ZAPRgRXg7Wv27v26NXj+Guk+OfHHi7T5WWO/FxqMkGlWqZz5l2VxFgAEgMGc4wgYkKf2Uv2afCP7Kvwg074Z+GmS7vS32rW9U8so2pXrKoeXaWbYuFVVQHAVR1JYmq/4LLfs9w+Lfhhof7Qugaa76x4NmXTtYeJMmTTJmJR3PXEVwwAA7XMhPQV/wR1+AreAPgbqnxm1qxeLVviJdAWXnRANHp1szpGV/iUSStKx6BlWI8jBqq/4LM/8J7/wzLo//CN+b/wjn/CSQ/8ACReV12bG+z7/APpn523Pbf5ftX/BLv8A4T3/AIbP8Ff8IL5vl+Xdf25t/wBX/Z3kt5vme27ytv8A008vvVVXj7x/4M+F3hK+8d/EHxFaaHoGmeX9rv7tiI4fMkWNMkAnl3Rfqwr/AIKLf8FDfgb4o+BGufBb4K+Mk8U654sWGC7vbK3Y2tlaCRXlzJIoDO6psCqDgOTlSBn/AIJb/t2fB/4NfDnUvgb8a/EjeHVTVpdQ0TVp4GktXSZYw9s5iQtGwdHkDvlSJCNy7VDfC/4vfDP41+H5/FPwq8Z6f4l0m1vGspruyYsiTqiO0ZyByFkQ/wDAhVV+25+yB4Y/a5+Fr6FI0WneL9D33Ph3VvJUtHLtObeQnnyJeAwBGGVH52bT/wAEd/BXiv4c/H/4veBvHOg3eja9ouj29vfWN0m14XE/5EEEEMCVZSGBIINV+2n+yX4e/a7+Er+Dbm8i0zxLpEjXfhzVZELLbXGMMkgHJikACtjJGFYAlAD8cP2d/i/+zt4pn8KfFbwbe6VIszx2l95TNZ6gqnHmW82NsikYOOGAI3BTkDwH+1t+018MdFi8N+Bfjn4w0zSbeNYrewGpvLBbIucLFHIWWIfMchAoPGc4GP2Xv2Sf2k/26/H2ifE/9ozX/Ft58ObGYzyan4ivZ3fUoS4drWwR2DLFITgyRhY0AbaSyha0nSdN0HSrLQtGsorOw063jtrS2hXakMSKFRFHYBQAB7V8f/hHpvx5+C3jD4Q6pNDBH4m0uS2guJYfNW2uBh4JimRu8uZY3xkH5eCDzV1p3x1/Y3+N1pc3ljqfgvx34TujPaSSxcOvzRl0Jyk8Ei+YmRuR1LDkE14V/wCCuf7XHxKt7H4d/D74MeFNW8cal+4guLCyupy7EkCRbbzSFIBUlmYoCCSNvA/ad/ZZ8UeDf2K/ir+0J+0l4g/4TH42+IbfSUnvJWWSDw7bNqtp/oVmq/ImAzB2QAcsq/KWaT9j7w5+0tH8YvD/AMSP2afhzqHiTX/Dd4TFIbNmsVLoY5IriYsiIjxylWzIh2vnI614b+MfxV+DPha++JH7dXjb4Y+DtK1GaCy0ew8PwXbi2nxPIyyzSM5lleNB8qDb+6OMEnP/AAUX/aS8H/tQftFy+M/h80s3hvQ9Hg0XTbya3eB71IpZpWm2P8yhpJ3C7grbQuVU5Ff8Eu/28vhT8DfBeqfA3426xcaFYz6s2oaJrbwvPbIZljSS2lEYLRAMgkVyCh8yTcybV3eAvH/gz4peErDx38PvEVprugan5n2O/tWJjm8uRo32kgHh0dT7g1X/AAUf/wCCdF58dZ5vjj8DdPhPjyONE1jRt6QprcagKsqMxCrcKoAO4gOqgZDKN/7EX7cPjT9h7X9T+EPxs8F+I5PCF7dCWfTbiBre/wBBueQ8kUMwXcr4AeJivKhlIIZX17/grJ+xVpfhiTXNK+IGq6zqK2/mR6LbaBeRXDPtz5ZeWNIQc/KT5hXPIJHNaV+z9+0J/wAFOfj7bfGn40eF9V+H3wktYwmmwzkpK9irZWC1STlpJc7nuNgj6kZ2oleFPCvh7wN4Y0nwZ4S0uHTdF0Ozis9Ps4slYIY1CooJJJwAOSST1JJOa/4LZWd3qHw1+F1hYWstzc3PiS4ihhhQu8jtAAqqo5JJIAA6k1+yH/wVavfhH4dsfg1+1J4c128TQVFrZ6/BFuvLeFVAjhuoH2s5UDHmA7iNuVYgsfFf/BWf9izQPDr6xofjrWPEt+F+TSdP0C7hnLFScF7iOOIAEAE7z14BFfs9fsnfFT9vH9pHVP2q/j94VuPDngO+1ZNR+w3kJ3axHEQkFjEGALQJHEkckxA3KuF+ZiyV/wAFI/8AgnxqH7Rgi+Mfwdt7KLx9plq0ep2EzmP+3YI0/dKjZ2rOm3Yu4AOHAZ12LV/p/wAQfhJ4wa0vrfXfCHijRpmUhhLY3drIMq2CNrKfvDI96s/2yv20vFNva+CNI+PXxFv57ry7e2gsNQmN5MykbVWSP98znaMkMWbnOcnP/BOP/gnn4r+FPij/AIaL/aGgz4yljdtE0iabz5LBpl/eXdy+Tm4ZWdAmTsDsWy5GzUdQstJ0+51XUrlLe0soXmnlfgRxoCzMfYAE1/wVN/bd+CXxZ+EWmfBf4N+MofFF5e6xDf6reWkDfZre3hR9sfmOBmRpHjb5MgCNgxBIB/4J2ftHeEf2Yv2jbbxr4/uLi28NazpM+j6ndQWxna1jleKRZSi5YqskEZbYC23OAx4Pwv8Ai98M/jX4fn8U/Crxnp/iXSbW8aymu7JiyJOqI7RnIHIWRD/wIV/wW3/4T3/hU3w8/sjzf+EP/ty5/tvZ0+2eUn2Pdjtt+2e2cd8V/wAEhP8AhPf+GwLH/hFPN/sT+w77/hJ8fc+yeX+6z2z9q+zY7/e7Zqqr476r8RNC+DHjbWfhHpv2/wAZ2Wh3UuiW4i81nuVjJTbGQRI4PKoQQzAL3rwZ/wAFNP21/hr4pubvV/iO3iBkmZL3RvEumpJCJFLAqVQRywkEkFY3ToAQcADwB/wW3+Et5o4PxU+D/i3SdURUBHh+W31CCZsfO2ZpIGjGcEL8/XG7jJ+IP7ZX7UH7cGjSfCr9jf4F+I/D3hzxCstpqfjXWR5Ijty+xwsy/ubc7A2/a8spyyxqGQM37EH7Dng79j3wteTjUE1/xv4hhhXWdYMW1I1UZ+zWwI3LCHJJJw0hClgNqKlarq2laHYS6prep2mn2UGPNubqZYY0yQoyzEAZJA57kV8T/wBvH9kr4TWtxJ4i+N/hzULu2ZkfT9Dul1S5EittKFIC2xgQchyuMc18WviD+2P/AMFMZYfh58Ffhjqfw++C91N5k+ta6WtRqka/MklxIM+YmDGVt7cSDedzO4UMn7Hv7Cfwo/ZH0X7fpMf/AAkHjm+t/K1PxLdR7XZTgmG3jyRBFkA4GWY43MwChar9qL/hPf8AhnT4j/8ACr/N/wCEp/4Ru8/s3yf9bv8AKOfL/wCmm3dt/wBrbXhP/hJP+Eq0b/hDftX9v/2hB/ZX2TPnfavMXyfLxzv37ce+K8Pf2z/wj+mf8JH5X9rfY4vt/lfc8/YPM247bs4r/gpJ/wAFGtX+Dmq6z+zh8GIbi08YLaxDWvEbMUOlLPEsqx2y4+aZopEbzc4jDDbl+Y/Bnxk+L3w3+3f8K7+K3jDwv/akiyX39i67c2P2p1ztaTynXeRubBbONx9a/wCGsf2qP+jmPir/AOFnf/8Ax2v+Gsf2qP8Ao5j4q/8AhZ3/AP8AHa+C/hfx/wDtc/tE+Cvh34w8aa3r174j1RY7u/1a/mvJo7VAZblwzsWJEMcjAZGSBkjqLSztLC0hsLC2itra2jWKGGFAiRIowqqo4AAAAA4FeMPiz8K/h6G/4T74meFPDWzbu/tjWbey27vu5811xnt618Zf+CpPwj0TzfBn7N2i6r8YfH13GqadZaNYTvaLK/ALuF3y7cqdkKtuJCb0OSvw1/4J3ftA/tY/FKf4/ftza5Lo1vd3CsnhuCX/AEqWBGBS2UIStnbYLKAGMp+YkKzeY3grwV4T+HPhPS/A3gbQbTRtB0W3W3sbG1TakKD9SSSSWJLMxLEkkmqqv+CunxgT4dfsrzeB7G6RNU+IepRaaiYO8WkRE9w6npj5IojntPwO4/4Jf/B5PhL+yF4Vurq1SPVPHDP4ivWBJ3LcBRbcnt9mSA4HALNjOSTVVVVVf8FNf2ov2xfgv+0R/Yvg3xfrHhHwLHY20mgTWNrH5OoO0amdpJGQ73WUOnlkkKqqdo3kt8Dv+C0nxH8PQ2ui/HzwBY+LbeNQj6zozLYXpwDl5ISDDKxOBhPIAHPPe/8A+Cx3w18TbNC+CHwE+JfjDxTd5S00ye0gh8xzgLgW8lxI3zEDAT+dfCb9hj46/tbfF+P9pv8AbxKaXbI0X9meC4U2M1vGxeO3kjJb7Nags2YyzTOWkLlCdzwww28KW9vEkccShURF2hQBgAAdBjtXxz+MXhP9n/4T+IvjB42W7fSPDlusksVpF5kszySJFFGg6ZeWSNMkhRuySACR+1j+2F8Tf2s/iFH4v8TTPo2j6UxGgaBa3LPDpaEjLB8L5kzYBaUgE4AAVVVR/wANY/tUf9HMfFX/AMLO/wD/AI7X/DWP7VH/AEcx8Vf/AAs7/wD+O1r37SH7RHinTZNG8TfHz4i6vYTf6y0v/FN7cRP9UeQg/iK/4IyfBUeEvglr/wAa9RhiN5491D7NYN5eWSys2eMkMem+dpgVHH7pCSTwtVX/AAV5/wCTNNS/7GDTv/Q2r/gip/ydR4p/7J/ef+l9hVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVVXxX+APwZ+Of9l/8Lc+Hmk+Kf7E87+z/ALejN9m83Z5m3BH3vKjz/uiv+GA/2NP+jefCn/fh/wD4qv8AhgP9jT/o3nwp/wB+H/8Aiq/4YD/Y0/6N58Kf9+H/APiq/wCGA/2NP+jefCn/AH4f/wCKr/hgP9jT/o3nwp/34f8A+Kr/AIYD/Y0/6N58Kf8Afh//AIqv+GA/2NP+jefCn/fh/wD4qv8AhgP9jT/o3nwp/wB+H/8Aiq/4YD/Y0/6N58Kf9+H/APiq/wCGA/2NP+jefCn/AH4f/wCKr/hgP9jT/o3nwp/34f8A+Kr/AIYD/Y0/6N58Kf8Afh//AIqv+GA/2NP+jefCn/fh/wD4qv8AhgP9jT/o3nwp/wB+H/8Aiq/4YD/Y0/6N58Kf9+H/APiq8Bfshfsz/C7xZYeO/h98GvD+h6/pnmfY7+1iYSQ+ZG0b7SWPVHdT7Maqqqqqqqqqqqqqqqq8V+DfB/jzR38PeOPCmj+ItLkbc9jqtjHdwMwBGTHIGUnBI6dzXgz4N/CD4bzvdfDv4VeD/C88v+sk0XQraxZuMHJiRSeOK1fSNK1/SrzQdd0y01LTdRt5La8sruBZobmGRSrxyIwKujKSpUgggkGvCvg3wf4E0dPD3gfwpo/h7S42LJY6VYx2kCtgAkRxgKDgAdOwqtW0jSte0250bXNMtNRsLyMxXFpdwLNFMh6q6MCGB9CMV4M+HngD4b6a+jfDvwN4f8L2EsnmSWmi6ZDYxM/dikSqCffFVVVVVVVVVr37N37O3inUpNZ8TfAT4c6vqE3+su7/AMLWdxK/1d4yT+daF4d8P+FtNj0bwxoen6RYQ/6u0sLVLeJPoiAAdB2qqrxB4d8P+LNFu/DfinQ9P1nSb+MxXdhqFqlxBcJ/deNwVYcDggitE0PRPDWj2fh7w3o9jpWl6dCsFnY2VusEFvGowqRxoAqKBwAAAKqryztNQtJrC+torm2uY2imhmQOkiMMMrKeCCCQQeDmvA/wx+GvwxtLmw+Gvw98M+E7a9kElzDoekwWCTuBgM6wqoYgcZNVVVVVVVb+EvCtn4ku/GVn4Z0mDX9Qt0trvVY7KNbq4hQ5SN5gN7Ip6KSQO1VWq6RpWu2Eula3plpqFlNjzba6gWWN8EEZVgQcEA8jqAa0r9mP9mzQr+LVdD/Z6+Gmn3sGfKubXwlZQyR5BU7WWIEZBIPsTVVXjX4bfDr4lWUOm/EXwD4c8VWluxaG31rSob6ONjjJVZVYAnaOnoPSvBnwx+Gvw3t3tPh38PfDPheGXPmRaLpMFirc5OREqg8814p8JeFfHWg3XhXxt4Z0nxBot7s+06bqtlHd28+x1dN8UgKth1VhkcFQeorRdD0Tw3pVvofhzR7HS9OtFK29nZW6wQwjOSERAFUZJOAO9VVVVeMvh54A+ItgulfEHwN4f8T2SZ222s6ZDexjJUnCyqw6oh6dVB7CvDH7P/wG8E6gureC/gl4B0C+VlZbnTPDVpayhlO5SGjjByCAR6HmqrVPDHhrXNR0vWNa8PaZqF/oczT6ZdXVokstjKyFGeF2BMbFCVJUgkEjoa8UeB/BXji0Fh408IaJ4gthHJEIdU0+K7TZIAJF2yKRhgACOhwM1oX7N37O3hbUo9Z8MfAT4c6RqEP+ru7DwtZ28qfR0jBHQd6qq8afDH4a/Ei3jtPiJ8PfDPimGL/Vxa1pMF8qYOeBKrAc814K+E3wr+GglHw4+GfhTwr5+7zf7E0W3sfMztzu8pFznYmc/wB1fQVVVVatpGla9ptzo2uaZaajYXkZiuLS7gWaKZD1V0YEMD6EYrwZ8PPAHw3019G+Hfgbw/4XsJZPMktNF0yGxiZ+7FIlUE++Kqqrxj8Ffg38RLn7b8QPhL4M8TXG4N52saDbXr7goUHMqMc7QB9ABWj/ALNP7OPh69GpeH/2f/hvpl2qlRPZ+FLKGQA9RuWIHB9Khhht4Ut7eJI44lCoiLtCgDAAA6DHaqrxT4S8K+OtBuvCvjbwzpPiDRb3Z9p03VbKO7t59jq6b4pAVbDqrDI4Kg9RXhj9n/4DeCdQXVvBfwS8A6BfKystzpnhq0tZQyncpDRxg5BAI9DzVVVVp/wm+Fek+M7n4jaV8M/Cln4svN/2nXrfRbeO/m3ABt9wqCRsgAHLc4FVVVVV4w+AvwL+IWst4j8ffBfwJ4l1Z41ia/1jw5aXk5RfuqZJY2bA7DOBXhjwb4P8E6euk+C/Cmj6DYqqqtrpljHaxgKNqgJGAMBQABjgcVVVVeOvhN8K/ij9h/4WZ8M/Cni7+zPM+w/27otvf/ZfM2+Z5fnI2zdsTOMZ2rnoKtLO0sLSGwsLaK2traNYoYYUCJEijCqqjgAAAADgVVVVVVaxoeieIbI6br+j2Op2hYMbe8t1mjLDodrAjIr/AIZN/ZX/AOjZ/hT/AOEZYf8AxqvC/gfwV4HtPsHgrwhomgW3lxxeTpenxWibIwQi7Y1AwoJAHQZOKqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqqr//2gAIAQIRAT8AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHYc49x1Ydx1ojrrsOu+w60mOrN9R1Jvqe0l3OeXccpPqXfcu+5dvqa9y7OaXc5pdzml3OeXc55dznl3OeXc9rLue1n3Paz7ntpHtpHt5Ht32Pb+R7ddj28T20D2sH1OeL6l09gAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHYdSC6jrx6DxD6Idab6jnJ9TcAAAAtcAALouAAAAAXa2OeS6irSFXfVCrRZ7SL6l09mAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAOUVux14LYeIfRDqzfU3GBYAAAALAADZuAAAAAAAFzcUmtmKrIVVPcTT2YAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAANpIlWgupLEN7IlUlLqAJXGAkAAAAFgAGx6gAAAAAAAAXAFUkhVYs0YAAAAAAAAAAAANpIdamuo8RAeKj2PrULbCxFNiqQezAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAlOK3ZKvFbEq8nsNt7gAFh6ha4AAABa5YABsAACxYsWRZFgLIsiyLFgAAC4ApNbEavcTTWgAAAAAAAAAABOtCPmSxEntoNt7sB6oEEZyjsyOJfVEKsJ7MAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAG0lqSrwQ8Q3siU5S3YAAAkMCwAAAFrgANm4ABa5YsgAAAALIsAAFwATa2I1e5dNaAAAAAAAAAATqRgtSpWlP0AAGG4BCvKO+pCpGa0AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAJ1YR6k8RJ7aDbe49QAAA3B6gkAAABa4ADYABYsgAAAAAALAABuACk47EKqlp1AAAAAAAAAK1Z7RNwAAewIANtiniOkjRoAAAAAAAAAAAAAAAAAG0j2kO6PaQfVGjQAAAAAAAAAAAAAEpRitWSxK+yidWct2CQIAAAEhha4AAAG4ADdwALXLAAAAAAAADQABcAGiFVrRiaa0AAAAAAAAK1X7KGrgAAAAA9ilWcNOhGSkroAAAAAAAAAAAAAAAAKlaMPUniJy20G29wE2tmRxE11uQxEJaPQ3AAAAAAAAAAAAbSRLEQW2pKvOW2g9QBLQYAAAJXGBYAAACwAPUACwAAAAAAAABuNAAJgA0Rm4vQjJSWgAAAAAAAVZ8sRgwAAGHQAAp1HB3RCcZxugAAAAAAAAAAAAAAAK2I+zEeoAAQqyhsynXjPTZgAAAAAAAAAATqRjuyeJb+ElJy3YAG4AAAAkPUEgAAALAA9QAtcAAAAAAAAAAGrgAJgA0Rk4u6ITUloAAAAAAAV5XlbsCHsAAA9gQADCnUcHdEZKaugAAAAAAAAAAAAAADEVvsxAAAApYhx0lsJpq6AAAAAAAAACpWhD1JV5y20AAAQ9gAAASHsAAAACQAPUALAAAAAAAAAAAFrjQAJgA0Rk4u6IyUldAAAAAAA9EN3YMGAAAwAB7AUqrg/ITTV0AAAAAAAAAAAAAAFepyR03AAAAClVcH5CakroAAAAAAAAJ1IwWpUrylotAAAAN2AAAFh6gkAAACQAPUALAAAAAAAAAAAADQAIA3CnPlYAAAAAAVXaDA3FsMAABgtgAAMPV5XyvYAAAAAAAAAAAAAAbSVypPnlcAAAAClVcH5CaaugAAAAAAAdrFTEdIjdwQAACGdAAABIewWuAAAFrgA9QC1ywAAAAAAAAAAAADQAIAauFKV1YAAAAAAr/AAdQAAB7AgAGAUKnPGz3QAAAAAAAAAAAAAGKnaPKgAAAAAoVeV8r2AAAAAAAJyUVdlSq5+gPYEAAB0HqAAAFh6gkAAAAA9QC1wAAAAAAAAAAAAAGgAWoA0U5WkgAAAAADEfAgB7G4wAAGAAPYAhJxkmiMlJJoAAAAAAAAAAAAAKkueTYAAAAAGHq3XKwAAAAAAqVFBak5ubuwGAAAAgAABIewWuAAACQAwC1wAAAALXFRkxUI9Weyh2PZw7Hs4dh0YMdBdGOhLoxwkt0AAAAAADQAbgDRF3SYAAAAAGI+FAAPYAABgtgAADD1LPlYAAAAAAAAAAAABiJ8sLdwAAAAABNp3RSqKcbgAAAAAVJqCuTk5O7ABAAAIYAAAW0HqCWgAAAkAMA3AAAAC1yNFvcjCMVoSqRXUdfsh1pM9rPue1n3PbSFX7oVaDFJPZkoRluiVDsyUWtGAAAAAA0AADVyn8CAAAAAAxC9wAGDAAjBzdkRw0EtdSeFg17uhUpSg7MQADAChV542e6AAAAAAAAAAAAAxMuaduwAAAAAAUqnJLyNGgAAAACclFXZObm7sAewIAAA3YAAAkPYLXAAANwBgBYAAABRctiNDuyMIx2JVIxJ1JSAAAFUkupGtF7jSkidH+UAAAAABoAQBBe6gAAAAAK69wANxbDAAwsLRcgJRUlZlai6b8gAGAFO/OrAEpxjuxVab6m4AAAAAAAAAASajFsbu7gAAAAAAYapdcr6AAAAAO1itU535AAwAABAtgAABLQYJaAAACQA9QCwAAARg5bEaKW5ZJEq0VsSqSkABYsWLFixYsRk47EKqlp1KlNSXmNNOzAAAAABoAQWvoW0AAAAAAqK8GAB1HsABRVqaAJRUlZlSm4SswAewAYaN537FebdTR7CxFRLceIqNWuBCpKD0ZTqxqLTcAAAAAAAAAAxUrQt3AAAAAAAIycWmiMlKKaAAAADEVNOVAAMQAAC2GAAAIewWuAAAJXAGAJAAAEaUpEaMVvqNpIlWX2SUnLdgFixZAAABZEKltGVIKS8xoAAAAAGgDcpRvIAAAAAAaurAAMGAFLWmgArU1OPmAAwDCrRsk7tsABNxd0UqiqR8wAAAAAAAAAMTK87dgAAAAAAAw1T7LAAAAKk1CNxtvVgA9gQAAAAAALYYJAAAFgGAJAAAk3oiNC+4oRWyJTjHdkqzew23uAWLIAAAAApz6MrQW40AAAAANXAEQjaIAAAAAATVpNAAIYAYaV4W7AAYmnZ8y6gAwCjpRb9QAAhNwd0QmpxugAAAAAAAAAm+aTYAAAAAAAJtO6IyUopoAAADET5pcq6AAMOgAAIex0AAA3HsG4AAAkAPUC1wAC19iNFvcUVFaE6sVtqSqSfUALAABoXRdFy6LougAE1OANIaAAAAAauBShd3ABtLqOce57SHc54vqXT2AAAArq0wAOowApz5JXE01dABOCnGzJJp2YA9gCOmHAAApVHBiakroAAAAAAAACq7QbAAAAAAAAMLP7IAAAVZ8kGxasABgAAC2GAAAJDBIAADcAYBYAEm9iNFvcUYxWhKrFbakpykAFrlgFFvZHs5dhwkug2XAAAC5dBSetiorSAaAAAAAaIxcnZCikrIJVIodVvYcm+o0AClJbMVaSFWi9xNPZgABiFswAHsDACjW5HZ7CaaugAxULNSQAwDbD/ANdwAAChV5HZ7AAAAAAAAAYp2gkAAAAAAAAQlyyTE01dAAAGKneXL2FsAAIAAAAAAOgAAACQDAEgBJvYjQvuKKitCVWK21JVJSAAsCpyfQVBdWKnFdCyLIshwT3HQi9h4d9GOlJdBqwAWRZFiwEZWaKy2YA1caAAAAIwciEFFaE6kYk6kpCAGgAAjVkvMjViwAKyvBgACGABSquD8iMlJXQBUgpxaGraADAeuH/ruAAAGHq8y5XuAAAAAAAAGKd5JAAAAAAAABhp3jbsAABJqKuSd3c6AA9gWwAAIex0AAAQwQAAFrgD1ASuAouWxGguokktCVaK21JTlLcABJvYjQk9xUY9RRitkOSW7JV4rYlXk9j20+4qs+4q0hV/I9tFntIPqJprQcU0Sop7EoNAAFkywVNaaZcAGgAAAhSvqyyKlRvRAC1AaugAACM5R2ZTqqWj3AGrqw1Z2AA6gABTqODuiE4zV0AYmFp37gDAX+7/ANdwAAATad0U5qcbgAAAAAAAGId6jAAAAAAAAChPlmgAAMVK0bdwABgAAC2GAeyn2PYTsNNaMAWw9Q6AAAJaAMAsRg5bEaKW5okSrJbEpuW7AAjSlIjQivMUUloSlFbslXXQdWT6jdwtcsgAE2tiNaS3I1Iy2GkyVKLJUmgAAavSAvcAaAAAhTtqwqVL6IGgABoAABhTrNaMUlJaAV42kAA9gYABTqODuiMlJXQGIhzQ9AB7AQ1oMAAACjU5JeQAAAAAAABN3k2AAAAAAAABTlzQTAADES5p+ggAGIAAACFGUtyNOMdixYnTU1YlFp2YD2BIAAC1wB6gQpykRoxW+o2kiVZfZJSct2ABGlKRGnGJYlJRWpOs3sPUUW9iUJJXYouWiOWwAAAQqtaMumrkWpK6J009UWd7AANWpA0FwBoACFJy1ZCnrdhUnfRADQCAaugAAGroEXa1RGu1uRqRlszERvG/YAAQwAAw9TllZ7MBpNWJx5ZNAAUtaLXqAAAAYepzR5XugAAAAAAAbsgAAAAAAAADCy3iAASaSuN3dxbAA9gQAAIexClKWxClGPqWuWQDVzEQuuYEMFsAAAloAwjSlIjRjHzJSjFakq1/hG29wASb0RGi3uRpxjsSnGJC7V2VKijotxtyeok27IjSS3NEirKMlZMpWTGk0SoroyUWtwAABSaWhTtKnZEKl9GWROknqhwkugEIOTKz0SAauNAA0Ba5CklqyUknbqFSp0QANXAAGgAAGG6Avz0wADqPYAAClPngmBioWaYAww+tNoAAAAhJxkmhNSV0AAAAAAAVdIMAAAAAAAAApS5ZpgAGJlamHQAGHQAAEinQvrIS7FgAJx5k0HQAAALXAI0ZSI0oxJTjHdkqzew9QAUXLYjQ7iSS0JVYolVlIo0+Z3ZVqcqstx6hRa1J1ukR3e5ZgpNbCqp7kmnB2Eruw4NAAAUHuiatJojNrYjUUtAcUzlj2JSSWpV1imADQbgNCi5OyIU1FeZUqpaIpaybKk7aIAAaAQDV0AAAwQwoS15StDlkAA9gYAAYadpcvcCtDmg0AMMLs0NAAAAGGqfZAAAAAAAMR/DYAAAAAAAAAUpc0EwAMXLVIAAYAACV9EUqSirvcsAADKkbTaHsCAAAS0CNKUiNKMSdSMd2TryexuAEYSlsRopbmiRKslsSnKW7CMXJ2RJqEdBu7IwcmVYpRVghRb3JpKVin8CJRUtx0Yk48rsKLa0I6SVxNPYHCL6A01uAFJ2kVGnK6AfvQOeS6nPLuPUlrTAAaDcIwcthRjFaFSrfRBGXLHzNwABq4BuA0AAAD2BNp3ROKqQ0AAFsMAANtUQkpxTQFWHLNoB7Bhd2iatJgAAAEW4tNEJKUU0AAAAAABif4YAAAAAAAAAYWW8QAK7vUYgABAABRpcqu9xIAAB7FZe8MOgAAWuRpykRpRiSnGK1J15PbQCwEaUpEaUUSkktSVbsSk5bgFGFlcqy5mRi5OyLRhElJ1HZEKaivMnUUST5ncFOSPbPsTkpMpNco0mOnF7E4yj1FUkuorDSkiVHsNNbgAFLWLQBDWFgABoIQ5jSKJzchoAAAGgEA1oAAAwWwwoTt7rKlFSd0OhM9lPseyn2PYz7HsZ9h05R3Q9gAAexhJ3TiBio7SAYYX4ysrVGAAAAGGnrygAAAAAAYr4EAAAAAAAAAFGXLNAASd22IAHsCAAMPTu+ZiQAAAV+gAABGDlsQoqO+pKUYrUlWb2JO4CQRoyZGnGJKcY7slWb2G2wApw5pFaVo2Ra5GKiipPnemxSilG6KlTogAATa2Yqr6lSSlaxCHMiUGiMnHYjVT3Gk0SSu7HspNDTWhKDS1Ao9R7gUXuiSs7AAEKV9WSkoolJyAaAAANwABoAAHsCGBTcnHUcordnPDuc8O57SHc9rDuKUZbMq0U1eIAAFKXJNMCvHmpsAZh3+8RiF+8YAAAAJtNNEZKUU0AAAAAAGL+FAAAAAAAAAARfNFMAm7RbDoADDoAAld2RCKSSAsWLFgAMRsgQACTeiIUOsj3YonW6RG2w3AhRbRGnGOxKcYkq0nsAAJXKUUo3Jy5ncptKSKknJ8sSUOV2OZ8tgAAAITUVYUlJaEqSexKDjuKTWw9SM42SuVUnEr7ICl8LAKT94qr3gAtchTtqyc1H1JXeoA1cAAAGgEAwAAGD1QU4c0rFWpyqy3HqAHoUavMrPcxFPld0AADKUuaCYE48smgGUXaojFL30AAAABhZ7xAAAAAADF7IAAAAAAAAADDSvC3YAraU2AAMAAMPG879hbG5ZIuBc0Y0AYjZAAEKTl6EYRitCdZLRajk5O7AYQpSnsQpRj6kpxjuTquWwAAbkY3aRVajGyIwckxJt6EIKK8yrJSegDi1uU6Skrs9jAdDsyUHF2YABtsRq9yq00rEIcw6ckNNGrSRX6ALSmARdmitbQAUXJ2RGCiTqJaIeoNCAaAAANwAAAAGC2GUYqMLsnLmdwABOz0NKtMas7AAD2MJK8WgMTG07gMi7SRi1sAAAABCXLJMTTV0AAAAAAYvZAAAAAAAAAAYWVpNAGI/hsQADEAAYaPu3LXANw3DViTsBiNkIAKdFJXkTnGK1J1ZSDoAouTsiFBLclKMVqTrN6IAAASKMbu5U96dkNqESlC2r3Kk76IClDmdyvbmIVFGNup7eQq6e5VkpPQpRThqOjF7DoSWw4tbgAU5qIpRezCavb1K+6CMeZ2KrSVgAeoEKbkJKKJ1L6IAGhANAAADQCAYAAPYEWu0is+WCSAADqUJWlbuYiFpX7gAD2MLK1S3cDFrRMB7BidaaYAAAABhp3jy9gAAAAADF7IAAAAAAAAACk7TTAMT/DEAD2BAAFKNopD0Rcvobh1A6A0GJ6CAtcpUbavcq1eXRbjdwQEKTluKMYrQnXW0S7buwAABIKfuwuUld3H787dEVZWVkBCHM7EmoRG7iV3YdGQ6cl0GrCk1sxVn1FViys02rFKEZR1JUOzJRa0YAXZSV5FZ3kFONlcnLmYARi5OyHF3sRpJbkpKKJTcgAAWoNXAAANwAHqgAAGFJXkiu7ysAAD2EVUp07gABB8skwMRG9NgBL3sOgAAp0J1FdFSlKDswAClPlmmAAAAAAYvZAAAAAAAAAAEHzRTAxP8MWwAPYFsAAldpEdADqAPYBDRiN0gEm9EU6Sjq9yrVtpHcAWwk3oiFFLVk6kYonUcwtoAAABV92nY+CBCXKyT5ncCklGNycnJgtGj2sH1FKL2Za46cX0JUF0ZKEo7hGo4qyI1k9xpSRUpqL0LFiyLIpRUY3Jau5ThfVlWVlZAAUk7lkTqrZDdwAAauANAAADQC2AYAAMMOt2Td5NgAALYoNOLiSjZtAADIO8UwkuaLQwGUveoteoABGLlJJEYqKSRUgpxsyUXF2YABRnzQQAAAAAGL+FAAAAAAAAAAGGd6duwGJ/hnQAGAAFFXmhbAHUAex0AexW1mCi5OyKdJRXmVa3SIARg5OyIwUVoVK1laI9QQAAAkCV2kVtXFFZ7IAKukEgAE2tmKrJCrJ7iaktCdFP4TlZZEZOOxOXMyFNSiSpSWxyS7Di1oxz9yxGPMyTUUPUsWAhByZpFFSfNogAAAaEA0AAA9UAgGAAD2ClpTuMAAAoytNGIjaVwABmHd6aAqxtNoBmFd00NWdgAMJG8m+wGLp7SQABhp2lbuAAAAABivgQAAAAAAAAABhXq0Bif4YADEAAYdXmdADqAGxuD2J6yZGLk7IhTUEVat9IgBTpuR7sYlSo5aLYYC2AAAtcCGs0VGlNFRqUtCUFyJgE5uSVwAAFdbEKiejJwUl5jVgI1HFWFOL6l13KrTegld2NIok22ANJig5OxpGJOTkwAAABoWoNXAAAGgAAAAZLSih7AAAM2K6UqakgAB7GEd4NAYqNp3AexhX77RWVqjAAwi9xsCcVKLQ1Z2AATs7oi1KKaAAAAADFK8AAAAAAAAAADDu1RAYj+GAAIAAwy3AAAAJOyEnJ2IQUIlWrzaLYAKdJy1exJxgic3JgFrgAAFgZS+NFf4gavTAAAC1ywFOp0ZUhzaoAAhGyJy5mACV3ZCSiib5gAAAAauANAAANaAIBgAAyvpBI3QAAD2Cl71NxGAAYN6tAYqN4pgPYoO1RGJVpgAYX+GAYqFp37gAGFlePL2AAAAADEK9NgAAAAAAAAAEXaSYFf+GwAewLYAAwy91gAAAFZ2iUqfKrvcrVOZ2WwAUqXNq9iclBEpOTuwewCAAASA3KfxIrJuaSJRadik7xsNWdgFSThpuNNOzACwAFOd9GVY21QAQjd3KktLIAEr6IjFRROV9EG40AAAANCBq4AAAwAewAAJXaRiOiEMAABbGHdpWK0bTYABhnapYCur02AQdpIxa1TAAwnwP1AMTDmp+gABRlyzQAAAAAV1emwAAAAAAAAAApu8Ewr/wANgAw6AAGHXuIAAAB7HJeSbK1S3uoAKdPnfkTmoIbcndgMAAACwMSI6SRU0mmVlqmUnaRVXvAUqiWjHFSWpKh2ZJWdhU5NaIVKQqPdk4pOxCEXFDpRZOMIrzAXvwAIx5mNqMR6gCi3sRioonO+iABoAAAANwaAAAYC2AAAIK8kYj4kIYAABB2kmYmOqYABRdqiAkrqwwHuYnWmmABg3o0ANJoqQ5ZOIABSlzQTAAAAAKivBgAAAAAAAAABh3emFb+GwAYAAFBe4gAAANyrU5VZbgBCDk7I92nEnJyd2dAAQAACQM3AraxTJrmhcTs0VldJgEZyjsKtF7k7Sk7EZRUVqOcV1HVj0JPmdylfl3JzUdFuPUCk9bE4vmYRg2JKKJS5mARg2WSROd9EAANAAAANCDcAABgIBgAFL40V/jDdAAAMexU9+lcAAi7STAKqtNoBkvew4AGFladgAxdPaSAAMLLeIAAAAA1dAAAAAAAAAABhXo0FVXgwAYgAjh5yXYeGl3J05R3RSVooAMHgMTjKns8PByf9bvZfMx+T47ApPEU3FPro196bQAMqTUI3G23dgCi5OyIQUIlSpzvyAGAtgAALAxIB+9SKTurE42dhSTp6gBa4AOLSuRm1GyACHxIdrlkNpEpOQFrkaaW5KSRKTkAAA0AAAA0boGgAAGAD2AAKPxFbWTBbDAAB7BR96m4gAD2E7oDEq1QBlL3qDXqABSdppgBOCnFxGrOwAFOXLJMAAAAAJq0mgAAAAAAAAADDO07BNXiwAEAUaSS5nuWLDQrAFrmQ5XDLsFGm17z1k/P/AIG3/DmJw9HF0ZUqivF6MzDCTweJnQnvF2/yfzWoYHIsZi6Mq6jy04ptyei0V9O/y07sOhUnzyALXKdNQXmVanM7LYEAwAAASBm4BRacWj4JE4qUdB32LNgblghC2rGtbIhSS3KrVrAAU1eSKvxHM+4AlfREYJInPoh6gAAA0AAADV0INwAAHsAtgAAKHxFTWTBDAAAWxh37zRVVptAAFJ3ggMWtUwHsYXWLQ1Z2AATugAxULTv3AAKMuaCAAAAAKytUYAAAAAAAAABRdqiDcaAewLYCjDmkJADVjcDI6Cr5lQg9uZfhr+hjKnKrGDrXlY4nyTE4zNorDQu5RTfZWbV2/RIybgjCYW1TFfvJ9vsr5dfnp5HGdeVHKZU6a+JpaLZbv0WlvmbmInZcqEAUKf2mVql/dQHQGAgAALaAxLQApytIrx2ZTmloxu7KVrNdSpDlegWLXIwSJSV7IUVFaE6qWiHqByy7HLLsKnJkYqK0JwbdwAhGyJzvogAAAAaAAABo3QNAAAAgGABQ3ZP4mMHsAAAU3aaMSveuAAUNaaAxa91MAwr95oqK02gAKTvTQAYqN4X7AAGFlq4gAAAAGJVqgAAAAAAAAABF2aYFRWk0Aw6AYeNo37hY0BpM2YHC1lm9G/d/+kszJO2hlqlzK5xRmlfLsyoVqL1UXddGr7P+t9TLcxoZhho16Lun96fVPzX/AAdjQxeSZdir+2oxbfW1n96s/wATE+H2T1pXXNH0f+aZiPDONv3OI+Tj+qf6GK8P84pfw1Gfo7f+lWMZlGPwivXoyiu7Tt9+xThzSsVZqEbIABgdAAAQMWoACtOA1bQSuyyZUtysSCEUkWHoyc3IBU5MjTUQ0G0SbewnclTT2Gmgpxu7lSXRAAAAADQAAA1dCB6otcaAAGAD2AAodR6sYboBJvREqU4q7QAdTYxKvFMAAw2tMDEq9MAw7tUMQrVGABh3ekgAnFSi0MACEuWSYAAAABi1qmAAAAAAAAAAEHeKYVlaowGAWuQikkgAHqLUMpxCw+Oo1XspK/p1/AxVLnjdGEoWdzjXEqtmbgn8CS+e/wCpw/ntXKsRzLWD+Jd/Nea6fccb1oYjLaGMw8rrm0a80/u2t66FPizN8JFcldvyl7353f4lDxJzCL/e0oS9Lr9X+Rh/ErCy/jUJR9GpfnymF41yTEae25X/AHk1+O34mHxVDER5qM1Jd00/yMbw7leLT9pRV+6Vn96tf53Mz8OOdueDrfKX+a/y+ZmXDuZ5fd16TS7rVfetvnYEAwAAAWwMS0AAoys7FaHVBGo4k5KSViNO+4qSbNEiVW+iNwSb0RCmlvuNpDd9ixZFkctxo5raMaTQ6SLKMR6gAAAAD1AAABoWpGmluWQSpxZKm0AD2AWwAAUdItg9gWwww8Fy8w0mrFSHLKwAwl71AAAwvwtAYhXpsApaVEYpe+ABhdaYAFaPLUaAAKMuaCAAAAAxS91MAAAAAAAAAAKLvTQYhe+AxAUY800JaAAHUDhjO6eOwsaUn+8grNd0uv8An5maZlQy/DutVfourfZf1oYivOvVlVqO8pNt/MaIZhWjhpYO94NqVuzXVeuzMRK8rdhAUZzhUThJp91ozD8ZZtgrJVOddpa/jv8AiZf4jYOraOKpuD7r3l+jX3MwWZ4PHQ5sNVUl5PVeq3XzMx4VynH3dSklLvH3X+Gj+aZmPhxWheWDqqS7S0f3rR/cjH5Pj8A7Ymk4+fT71dfiAIAANwNwAAhJTiVKbi/ISuQhbVk5cqFVsvMlJy3AjTciMVFaDfYsAA0SjdWFJrYVRvoS5pdDkkezkezkezkckjlfYaYAAANXAAAGrohDlXmKJoXQ0mNNE6alqtwABAMAI6UgYIYYfWAYmF0mAPYKXvUmgADCbsCtrTYAtJJmLWqAAwnwMADFxtNMAAwstXEAAAADEK9NgAAAAAAAAABhneNgxS1TAEBho7sfYAAZughKUXzRdmVq9WtLmqScn5u4bojrNk3eTYgKEbyuVneYU6k6clODaa2a0Zl/HGb4S0ZTVSPaWr+9Wf3tmXeIOXV7RxEXTf8A4EvvWv4FDE4XGUualNTi+zTXz/yMz4JynGpuMPZy7x0/Db7kvUzPgDMsKuag1Vj5aP7n+jZWo1KM3CpFxkt01Zr5M6AAAgYkAAEZOLuiMlJaHJFPRCRVknogEm9ERpJbjaQ9QAALFkF0OUe5zx7nPHuc8e5zR7l0+oNJjhF9B0kOk+g4SQAA0AABTjd3EtLj1ABx7FWF1dADABgBLSkAwewYZ+60EkpKw1Z2AFsYd7okrNoADC/G0BV+BgD2MVrBMADCfCwAMYtEwAClLlmmAAAABNXi0AAAAAAAAAAIwz1aDEr3UD2BAUI2ggAAYh7HQAWnMw6AUElFserAQYfE1sNUVSjNxkuqdn+BlviDmOHtHEpVI/dL71p96u+5lnGOU4+0VU5JPpLT7ns/LW/kY7LMHj6fJiaamvPdej3XyZmvhzCSc8DUs/5Zfo1+qfqZjk+Oy+XLiabj2fR+j2YAAG4AAEKV1dlkloTqLZEqjasBGDkRiorQb7FgNAAtoBK9gCyLIsAcz7nPI9oe0ixNPZjSY6cWOm+g00AAABCNlYeoABJE48rAewC2B7ABW0ikAweqDDPVoDERtO4AUH75WVpsADDfxAKvwMAexV1or5AAYT4WABilemAAFOXNBMAAAACatJoAAAAAAAAABFF2mgrq9Ng9gWwEFZJAAAdRj2DcHpCX9dAAjpRAEAGV8RZllzSoVXy/yvVfc9vlZmV+ImFq2hjYcj7rWP3br8SlWwmPoXhKNSD9GvR/5MzbgDAYm88K/Zy7bx+7dfJ28jNuHMxyx/v6fu/zLWP39PR2YAgewgAApR5pE5qPqSk5bgEIczLJIbuAaAAHUGhoABq40AAKTXUVRinFmjQ6cWOm0DQABBXYtgAA3VirG8fQABAMCKvJFfdIB7Atgw7tMDExvG/YA6lLSaMSvfAAofxEBV+BgA9cOABhPhYAFdXpsAAMLK8WgAAAAMQrVGAAAAAAAAAAhOzuboqK8GDAIK8khAAAV8RRopOpNL1diNSFSKlBpryHsdAKmlNiAJ6UUAdAewC2DCY7E4Op7TD1HF+T/q/zMp8RJxtDHwuv5o7/ADW33W9GYHMsFmNLmw9RTXVdfmt180ZxwLl+NvPD/up+Xwv5dPlb0Zm3DuYZW/8AaIe7/MtYv59PR2fkC2BnQAAKKSi2Ntu4BGLk7ISUVYeoAthlwQFwY9iS1AABq40AACbWwqj6inFjimSpdiUWmABTWgAABJElZ2AYIBgUleaK2sgHsCGU3aaAqR5oNADI6SRilqmABT0mgKnwP0ACGtBr1AAwnwsACqrwfoAAGGladu4AAAAGLXvJgAAAAAAAAALYKTvBDV0NDAKKvNC2AADEV40KUqktkjFYqpiarq1Hq/w8jJsfLDYhRb92Wj/z/roPYNAr/wANiAK38NAAMAAoYirh6iqUZOMl1TszJ/EKvTtTx8eZfzLR/NbP8PmYLMMDmVFyoTU4vdf5p6r5ozvgHCYq9TBP2c+32X+sflp5GY5VjMvqezxMHF9Oz9Hs/wCrjFqAAAtKQAU42Q3cAOhbQ0YgAGE1oAAANAAACk0KomNJolR7DTW4G5FWAAAHqistUwGAD2Aor3iprJgAhgndJgTVpNAPYMTrBMACHxICp8D9AAo60mgAMJ8LAAkrxYAAQfLJMAAAADFRvBMAAAAAAAAADoGGd4tBUVpNDEBh1eZ0AGy4HEtXlwigvtP8tf8AIqycp2XQoyb91mBquthac3u0vv6gW0MR/DFsAYj4UAgGCAAMLi6+FqqrQm4yXVaf0vLYyTxCTtSzCP8A28l+cf8AL7iUcFmWGs+WpTl81/w6+9Mz3w/nBOrl7uv5Xv8AJ9fR/eypSqUpuFSLUlumrNfIAAD/AJdAEI8zJPSxYANxgHUAYSWgAAANAAAAm1sKquo0pIlS7DVhQvZi2AAAEVY+6A9gFsDAoLcerAGD2Ci7wQFdWmwBbFXWigAI/EgKnwP0ADDaxaAAwnwsAB6oAACk+aCYAAAAVo3ptAAAAAAAAAABhnaVgrq02CAwy95gAaMQcU39nS9X+hSs5u5H+LoYbPMww8VCnU0XRpP80UuL8bH44xl96f5/oUeMqL/i0mvRp/nYo8SZbV09pZ+aa/Hb8SeIo1qd6U1JeTv+QAYjZAIAEAAC2MszfGZdU9php27ro/VbfqujRkfHGDx1qWJ/d1PP4X6Pp6P72Zzw9gc1hatG0ltJaNf5ryZnnC2OypuUlzU/5lt810/Ls2AAJXIq9OwCTbsiMVFG7GACA6C2OoA9gkAAAA0AAADV0KTWxGqnuSipLUSsrAAAATV0AMEAwo6RbABhugwzvGwGJXvIAHrQAAW6AqawfoAGG6klqwAwnwsAAAADDO8LAAAAA1dDVnYAAAAAAAAAApO00GJXvIEBht2G4MYW1Diak5YRTX2X+en+RVThO/cpRurshwtgJ0Ic0WpW1s3vbzuirwbQf8Oq16pP8rFbhDGx+CUZfev0/Ur5JmFFXnSfy1/K579OXZr5MoZ1jqPw1G156/mUOKqi0q00/TT8Hcw+f4Ctpz8r89Px2/EjOM43i7oxGyA6A9gOgAAD2EZFxhjcttTn+8p9m9V6P9Hp6GW5vgc1oOVCXMuqe68mv6T7nEPAcal6+XaPrDo/8L6ej080VqNSjUdOpG0lo090ACWhSl0KlN3uhUpMjFRWg3fQGAFgFsW0LFDD1cRVjSpRvJ7Ir8HZpSo+05VJ9k7v8rP5NjQ9gYwAAAGgAAAYRm1ogAAAB7E1aTAYAMIfw2AD2BbDMM9WgMUtEAdSGtFgALdAT+BgBht2VPjYAYT4WAAAABhXq0AAAABXjaowAAAAAAAABALQWqMUtEwWwGG3YLYewzqHYMVh44ijKlLqjE4adCo6VRaoyfAPFYhXXux1f+XzBosWMbh6NWKVSKl6q5X4dwFXaLi/J/53X4FfhSotaNRP1Vvyv+RiMoxtD4qbt3Wv5fqUcRVou9OTT8mUOKcZFJVbTX3P8NPwMLxDgq2knyvz/wA/87EJRnFOLugGAAAIGLYMLi6+FqqrQm4yXVf1+Bw9xzRxVqGNtCfSX2X6/wAr/D02M74dwWb0/wB4rTW0luv815P5WM5yLGZVV5K8dHtJbP8Arqn+WoFrhsKcWtxziuopuQkDADqB0OgcE5ZGnhni5L3pXS9F/m/yFUi3ZPU40yyOGxca9NWVS9/8S3+/T53HsD2HuAAAANAAADV0ELJ3YmmgGgYFx7APYqr3gHsAD2CP8JgACGUHaaAxK9xADKOsGgAAJ6xYAYb4mVfjYAYT4WAAAABQlaogAAAAMXHVMAAAAAAAAAQBSd4IxSvAOgGG3Z0Og9hgdQMRhKGIt7WCdu5So06UVCmrJdgAxHwoAxGX4XEv97BN/j961MfwlRWuHm15PX8f+HMVkuNw+soXXda/8H70YfF18PK9KbX9dtjCcVTj7uIhfzWj+7b8jCZlhcWv3U7vts/uGCAABbA9WAHDvGOJy61Gv79Lt1j6eXk9OzRCpgM4wd1apTl/WvVNfJo4l4NrZdevhrzpfjH17rz+/uxbABBaAMAGcrk0luZRwXhYUFLGrmm+l2kvus2++tu3c4m4YjgYrEYbWn1T3X/A/J976BgKaw+W0oLpFffbUhX9/RnGlJVcrVT+Vp/fp+owexK9zUuXQAAANAAADQRWgIewdRtM6B0AKy1QDAWwPYI60gAGIZTdpIDEfwwBmH1TQAAEtmAGH+Mr/wARgBhPgYAAAACdncTTQAAAAYmN6foAAAAAAAAAIAwzvGxXV6bADD/GdBbD2GjdAgDqBdG5iPhQBSjeaMQ7ySDFZXhMV/Egr99n+H6mM4VnFXw87+T0f37fkV8NXw07VIuL/rYwfEOLoWU3zx89/v8A87mCzzCYq0VLll2f6PYQAAD2EABlOcYvLK3tcPK3dPZ+q/prozIeJMJm9L3PdqLeL39V3X9NI4n4IjPmxOXxs93Dv/h7en3W2couLcWrNABFprQ3BgA1c4RwSxOZxcldQTl92i/Fp/IrV1B2RWpU8Zh50p7STT+ZVpyp1JQlum19wL3sFTt2X5FKElWbOKKc55NNRV3o/wAUMHsPcLIsaovcAABq4AAA0JXaEA9gerLILj2OoFZaIAYIAp602gAGD2DdBX/hsAexh+pJasAAHsAFD40Yj+IwAwnwMAB7AAAFCXNTQAAAATjzRaAAAAAAAAAFsAYZ+80TV4tABRdpoWxsbnUaNWXAOoAV17gBQjpcqS5pNgtgq0qdSHLNJrszG8MUKl5UHyvtuv8ANf1oYzLMThH+9jp36ff/AJmBzzF4WyUuaPZ/p1X5GAzzCYu0b8suz/R9fz8gAEDFsAAUK9WhUjUpScZLZrdHDPGVLHqOHxVo1ej2Uv8AJ+Wz6djifhKlmUXXoWjW/CXr59n8n5YjD1cPVlSqx5ZLRpgCdhO60NxgAcA006lefZRX33/yMfW5ZNmXz5kZ/BQzOul/M39+oZBXWLymk76pW+cdP0IYWXPqilVpxqqm5Lma0XXTfQz7gyhi71cJaFTt9l/5P008upi8HXwlV0q8HGS6P+tV57D2HuADRsAAA0AAAQXvC2AGbFzoW0HsMCstABggGUNU0NW0ABh0CGsUFb+GwAw27Ki95gAt0A9joAUP4iMT/EYAYT4WABP4WAAAYWW6AAAAAqxtNoAAAAAAAABbAFF2mgkrSaAIuzTEAAAdQAqq8GBGPM0kVGoQsgAYOKkrMx/DeGr3lS9yX4fd0+X3GNyzFYR/vY6d1t9/+Zl+f4nC2jJ80ez3+T/pGBzTDYyP7uWvZ7/16ACB6gAALQ4X40ty4XHy8lN/lL/P7+5xFw1h83pXXu1UtJfo+6/LddU8dgcRgq7oV48sl/V13QAm0JpovoABwFUSrV6fdJ/c3/mY6hzSaMBT5UziGM/7TrOStdu3mtk/R2DhLPYYKo8PXdoS1v2f+T6/LzK+Nw9Gj7apNKPe/wCXf5GcZ1Vx2P8ArFNuKj8PRpLr6vcyHjdScaGP+U//AJJfqvmt2ZhleCzSgo1kpLo1uvNP+kzPuEcVl6dWl79Pv1Xqv1WnewAAWsbgAA0AABBanQAeww6HQ6AgrfCAPYAGUbplVWkAD2BbBS1ggrfw2AGH+JlVe+wAjq0A9hbAFH40Yn4wAwezAAq/A/QAAAw7tUQAAAAGKjaSYAAAAAAAAAAnZphWVqjACk7wQAAB1ACSvFoChTsuZlaXNL0ABgglGMo8sldGYcM0at5Yd8su3T/gfl5GIwuIwdRKonF9P+AzLuJpwtDE6rv1+ff8/Uo16VeCnTkmg6AtQAAQHC3GEsFy4XFu9LZPrH/OP4rppoZ3keEznDJSa5rXjJa2/wA0+366mZ5ZiMuxDoV1Zr7mu68gBbiaZc3AOGMfHB5lCUnaMvdfz2/GxVoRqGKxFHA4aVWbsonEeRUszwEZUNZwXuvuu3ndbefqxpp2e4HQMm4jxuWPlpvmh/K9vl2f4d0zJuIcFmkbUnaa3i9/l3XmvnY4k4Kp4hSxGBXLPrHo/Ts/wflu6lOdObhNWa0afQAAaNwABq4AAEAAYwQti2gWCt8IA9gFsCi5OyJcsIorRuroAHsCGUdaaCt/DYAYf42Vl7zACGskA9hAFL40Yr40AGD2YAFb+GwAABOzTN0AAAAGKjeF+wAAAAAAAAAAU3zQTMSveTADDS0aAAAxGfYKjU5G231t0/ryuYevTrwVSm7pgAicWpNFOi3rIrT5Y2W4CAYIAr4elXg4VI3XmZlwzOF54Z3Xbr8u/wCfqYbGYnBVL03Z9V/mjLM+oYu0J+7Pt0fo/wBPzB7CAABbAHC/FdTLZKhXd6L++PmvLuvmtd80yvBZ3g1FtNPWMlrZ915d119dVmuVYnLcS6FdarZ9Gu6/r1ASBSNy4BlPGqo0VSxcXJrZq136ptfffUz/AIjq5naEVy0106t93+iOCc/2y+u/8D/OP+X3djjfIfY1Pr1Fe7L4vJ9/n18/UB7AyFapSmp05NSWzWjRw1xpDE2w2OaU+ktk/Xs/wfl14n4WpZpB1qXu1kt+kvJ/o/v02r0KlCpKlUjaS0aYAA0bgADQBucrfQ5JHs5EYWQAMZoIWwAVtgABCTbsiMFFFSfMyk1KNmSjyuwACGUP4aCv/DYAUPjK3xMAKavNAPYQBT+NGK+JABg9mABiP4TAAACi700AAAAFSPNFoAAAAAAAAAAexhnenYxK91MAKMuWaEAAZ7jHhsK+V+9LRfqSnGLs2cP410cSqTfuz0+fT/IAJSSkjRlSrGOnUk3J3YCABbAAZjlGGxsfeVpd1v8A8EzDKsRgpe+rx6Nbf8D0Mr4jq0LU6/vR79V/mv68ijiKWIgp0pXR0AAAAOGOKKmVVPZ1PeovddV5r9V19TMcuwOe4Fa3T1jJbp91+q++zWma5VictxLoV1qtn0a7r+vUSuACkFy6AhOUJKUHZrVPszI80o53l7hVV5W5Zr9fR/g/QzvKamWYyVCWq3i+66P9H5gwbG7sOE+L3BxweNlptGT6eT8uz6ddNuKOGaeaUnVpK1aK08/J/o+noVaU6U3TqK0lo090wABo3ALNnI2ezueyR7OJyoG0c0e5zxE09gWwwexugex0ACtsgAYIhBRRUqX0QU5csivG6uADEMo/w0GI/hgBQ+MrfGwAoq9RAS+FiAIfEjFboAMHswAMT/CYAAAYV3i0AAAABVjyzaAAAAAAAAEAD2MK90V1emwAKc+aKYABxTVvWpw7K/3v/gCTm7mFquElJbxYmmtADEbIu1sAC2B7AAAEqcKkHGSun0M04ZsnUwn/AID/AJf5MwmOxOBq3g7Pqn+q/pmWZzQx0VFaT7f5dwABAAHDHEtTKq3JN3oy3XbzX69152M0yzB55gUrpp6xkunmvLuv12zLLa+X4iWHrq0l9zXdeTAAvYUu5dMLlzI83qZZjI1o6x2ku6/rVHEWVU85y5VKDvJLmi+67fNfjYd4uzWpcZJ3AOEOLHFxwOMlptGT6f3X5dn022tbi7hdZhTeJw6/er/yZLp6rp93a0otOz3AAtcUGezORFkaDku45odQdSTHOT6jbYBT2QLYYGxobgAVugAMKMPtFWfRAwg1OFhqzsAMHsUl7iDE/AgAofGVfjYAUNaiAn8LACO6MV0ADB/CwAK6vTYAAAYV+80AAAABio6pgAAAAAAACAB7GHdqiJK8WgAMPOz5RAAcUp/Wl/h/VlFrlZR3ZBWikAV9kAB0BgAAHQMyyahjld6T6P8Az7mMwOJwNW01Z9Gtvk/6ZlPEl7UsU/SX+f8An9/cTUkmnoAC2AADhXiaeV1fZVXejLdfyvuv1X6md5Lhs7was1zbxl6/o+v3mMwdfCV5UK0bSj0/roAG4F2czOY5jgTPrS/s+q9HrD9Y/qvn3Rxxkn1XEfXKS9ye/lL/AIO/rc5hu4AHBvFHt1HA4qXvL4W+q7Pz7d/XfjXhnmUsfho6/bX/ALcv1+/uFrnI2ezRypA5IdRDmxybAGroAAhshgA1csWBDB7FX4gAexGPM7FSShHQBhSlaVivGzuAMHqiKtFIMVskAFD4yr8bGAYZfvAKnwMAI7oxXQAMJ8LAAlFSi0NWdgAACg7VEAAAABiY3p+gAAAAAAAAgAZB2kmFSNptABtsU5qUUwAscU0W/Z1V5r9V+pKnKLsjLsK6taFLu9f1/AAK/QAAGCAAAewivh6Vem6dSN0zNuH6mFvUo+9D8V6+Xn95lOeVcE1CXvQ7dV6f5bem5hsVRxNNVKUroAAABbHCXFDwE1hsQ/3T2f8AK/8AJ9e2/e/EnD1HOMOpQaVRL3ZdGuz8n07b9716FShVlSqxtKOjTAeoAAUqk6U4zg7SWqfmjL8Vh+Icocau8laS7SXVfg0ZjgauBxU8PVXvRf39n81qAARck047owvH8oYOMKtLmqrRu9k+z6691a336YiUKtaVSMFFN3stl5IshySHNdBybHqDRuAAwAIbIYABuCGD2KmsmAD2KMbK5UlzM6AwJ+/TuAPYEr6AYndIAKHxlb42MAwq98Cr/DfoAEd0YrdABhPgYABiqVnzIAAAi7STAAAAAkk1Yas7AAAAAAAAIAGEXeKZiY2kmABSnyvyEyxYsPUzeFGWCn7Z2Xfs+jKtCpTkotb7dn5p9Th/Kp0n9Yqqz6L9QAr7oAEAwQAAgZ0AzjhyNVOrhlaXVdH6dn+Bg8bicBWbho+qf6oy3NKGOp80HaS3XVf8DzBAABuBwfxT9XccFipe49It/Z8n5fl6bcWcMxzKl7egv30V/wCBLt69n8nptOEoScZKzXQZYAADhTO/7Mxy53+7npLy7P5fk2cd5IsRh1jaS96G/nHv8vyb7ACTewqYopGg5o4Z4aynMcIq85OUtmr2s/lr6O/yM8yitlmLlQqareL7rv8Ao/MAA2AAewAENkdCviKVCHPUkkjDY/DYr+DO9vv+56gFkABLYerABK7sVWowsgWwPYCi7qxJWdgHsFJXkgMRrMAKPxorfGxgGFWrArfw2HQCO6MVuhgGEX7t+oAA0mrMrUXB6bAAAEXeKYAAAAGIjaowAAAAAAAFsADEUHemjExvC/YAAo1be6xMvcDi6vyYBQX2mvw1/wAjh2g6WW0k+uv3u/5AA9iv8QAIAFsAAIHqwAzXJaOOjzLSa2f+f9aE4YrLsRr7s1/XzRlGd08dFQlpUXTv5r/IQAAIA4N4o51HAYqWq0g31/uvz7fd2vxnwwsRF47Cr318S7ruvNde6899wAAA4LziOPwLwtZ3nBW16xe33bP5dziLJ5ZZj5UUvcesfT/gbP7+ooNigkaDmhzbHqHD+dVMqxaqrWL0ku6/zXT/AIJnmVUM9y5SpNOVrwf6ej2fZ69CrTnTm4TVmtGuzAAauIAAAIbHQzbHPF4hyv7q0Xp/wTD4iVGoqlN6owteOIoxqx2aAAAqOyYAFJXdys7yBABSdpFZWkAGHV5gV9ZsAKPxorfxGMAwi3Ar/wANgtgI6yRit0MAwq/dgAA0mrMr0HDVbAAAUXemgAAAAMVG6TAAAAAAAAWwAMRhno0TjzRaAAFsU63LoxNNXRcucYVXUxFKgt0r/e7foUKcadKNNbJJfcNAD2K3xgAgewHQAAWwLVgAY/L6GNpclRa9H1X9djHZfiMvrJS+TX9aPyMkz+OJSo13afR9/wDg+XXoAAHQATad0cIcTrMKawuIf72K0f8AMu/quvffvbjPhn6vJ47DR9x/EuzfVeT/AAflsAAAotmXY3EYCuq9CVpK6+/y/rUxeNxWNqc9ebnLz/RdPRDkkObew9QAOB+IPYVVgK0vck/d8n29H+fqcd5Amv7QoLymvyl+j+T7gADAAYAU9jMarpYSpNb2f+RiG3aKKV4TS7nDVVzwbg/st/5/5gAAVvhAAh7sLkgQDBaMrq8UwAwy1bAq6zYAUfjRW/iMewBhV7rAxH8JgtgKavJGJ+JDAMOrUkAAA0mrFehyO62AADDa0wAAAAKseaDQAAAAAAAHQAGIw7tOwVY2m0AD2OgJtbMjX/mITjPYxC+t8RKK2i1/5Krv8UwuaMsWJLQqayYFripSfQ9lJIaaHsAAAdB7CAAMThaWKpOlVV0/6ujNcorYCpfeD2f6Ps/zMj4gVW1DEv3uj7+T8/Pr67gAgAKNapRqRqU3aUXdPszh3PaOc4NxqJc6VpR6O/VLs/w27X4q4dlleI56avSls+z/AJX+ndejAAFBsUEhtIc10PD/ABeGlVnQnBe03jK2tuqv+P39jjDJngMwc4L93Uu169V8n+DQAAJtO6OFs6hm+BdGvrUirST6p6X+ez8/VHEeSyyvGul9h6xfl29Vs/v6gAbiAB7ABAz2rCGAmpO17Jetyp/FKtnNWOF/4NT1/QAACpUUVYAEruxVdopDABgS1pADMMvdbAnrJgBR+NFb+Ix7AGGX7sDE/wANggKWs0Yn4xgFNWgkAAANKSsytSdOVugABhfgYAAAABUjyyaAAAAAAAAABFN2mgxMdUwBD2ACg0mzhhfWczq4h+b+bf8AlcABlT42EaLluRhGOxYsNJqzKlKyugAAAewtgABFahTrU3TqK6fQznJKmBlzw1pvr28n/n1Mi4gtbD4l6bJ/o/8AP7zcAQAMsZdmFfAYmOIovVfiuqfkyhWwXEGWaq8ZKzXWL/zW6fzM5yitlmKlQq/J910f+a6MAUGxRSG0hzb2HqGCxdXB4iFek7Si7r+uz2ZnGFpZ/kqqUdW1zR9VuvzXqNAABk+aVctxkMRT6bruuq/rrZmeZdQz7K1Oi7u3NB+fb57Ps/QnCUJOMlZoABgAMAUoxTbdkjOMzqZji0qesU7RX6+rK8G0pIoxcpX7HDlFwwXN/M2/0/QGgAG1YrK8bgBTV5FV3lYewAMCOtJgDKKtTQPYeoAUPjK38Rj2AMOv3aAxP8MYgKCvNFfWbGBa5bQAAAKlNTjYkmnZgAYXZgAAAAGKjqpAAAAAAAAAAtgTukzEK8ABDADMK3scJUmuif4qxwZRtQq1O7S+5f8ABAA6lX42U6VtXuWuWQG40VafVCAAEDOgAAgqU41IuEldPoZ3kUsI3Voq9P8AL/gef3+eQ597K2HxD93o+3k/L8vTbcAAN2Bw7ntTKcUp7wlpJd13Xmv+B1M7ynDZ5gE6bTdrwl/XR9fv3RXoVKFWVKorSi7NeYoNiikOSQ5NgAcAZw41JYCo9HrH16r5rX5PucbZOsFjvbU1aFTX0l1Xz3+b7AAAcB557Kq8BVfuy1j5Pqvn+fqcd5GqFZY6kvdnpLyl3+f5+oAD1QtgAYBxPmnLH6pTer+L/L59fL1OFco5I/W6q1fw+S7/AD6eXqZ1l0sNXc0vclqvLyMHg6mKqqnTX/A8yjRjRpxpx2SsIewAMnJWaI+9CwAUVuyTux7AdAYEP4TAGRVopBUdoMACh8ZW+NgBRVqaAxXwDEBhl7zKrvNjAgryQAAABi6dmpIADCdQAAAACtHmg0AAAAAAAAA9gWwUHeCKivBoAQwA4jq8mBcf5ml+v6HDNH2eW0+7u/x/ysABfUVP33JiQWLFgGicOVgACB6sAAOgEoqSaaumZ7kTwrdair03/wCS/wDA/pmQ597O2HxD93o+3k/L8vTYQAMWwBw/xVXyuhOi488XrFN2s/8ALrbv2u2ZlmNbMMQ8RWtzPsrbf11uxySHJsAAMNiKmHrQrU3aUWmvVGYUKXEOSKVLdrmj5SXT84slFp2e4AAQnKElKLs1qjLMXQ4hyhxq7tcsl2fdfg0ZhgauCxM8PVXvRdvXs/RrUADqAD2AzPHwwWGlVe+yXdmU4CeZYtzq6xWsn38vn+RTsopLoThGceWSuilRpUly04pLyVgGwBsnLlRuUnrYqq0mAR0pgAtgewCqNR5QC12kBXdqbAAw694q/EwApr3EBivgQxAYeUU2mN3Y9gKKvUQAAABWhzQaAAwnUAAAAAKkOWTQAAAAAAAAPYFsGGlugkrNoBAAcV1PdpU/V/l/wTLIezwsKf8AKkvwAB7Cd6luyErljQuFy5a+w0VI8ysAAIOoAAAEoqSaaumZ7kTwrdair03/AOS/8D+mcP55a2GxD/wv9H+n3CAB6gKLYopDkkcCf2bWqShUgvbLVN63Xktk19/VdTjPh36jW+tUF+7k9V/K/wDJ9PPTsAAAeH+b8lWWBm9Jax9Vuvmtfk+5xrlH1LMHVgvcqXa9ftL79fmAABwnnX9m49c7/dz0l5dn8vyuce5Mq9COOpL3oaS849/k/wAH5AAMAAM7x0swxip0tYrSPm+/z/IyzAQwWGjSW/V93/WhB2Yg3ABslJJEtREXaSK62YBPSnYBggewABBXkgMS/dSAAw+7KmsmDASsrAYr4UMQAPYDCxvUv2AAAACStJoAMJ1AAAAADFQ2kAAAAAAAgAewdAou00FdWmwFsdQAzxe3zOnS9F97/wAijK0hAEmkinHS/VmiNwADckipG0gAFsCAAEABOEZxcZK6ZnuSvBz9rSV6b/B9vTs/l68PZ57VLDV373R9/J+f5+u4AJN7CgkNpDm2GGxFXDVo1qUrSi7pmV5hhc/y186WqtKPZ/5dU/1TM9yarleLdGesd4vuv8+4AAWMNXqYetCtTdpRaa9UZ1h6ee5H7WkrytzR9VuvzXqWAAA4PzWOZZc8LX1lBcrXeLVl/k/+CZ7lUssx08O9Vun3T2/yfmgAEADOI8x+q4b2cH709PRdX+hwtlt28XNbaR/V/p94EZXRcNCw2hkqi6D1BBU96ncBK7K2yQDBAAAUVeaAxL95IADD9SWrZ0GEVeSQBi/hQwAHsBhI6NgAAABVX7xgBhVowAAAAAqQ54NAAAAAAAIAHsAEXdJmJXvJgLYACn+/z5vs3/5KrfmgpT5o+Zc0Gyb5pKKEAAAb6FWN4+gAA9hAACAAKlKFWDhNXT6Gc5RPL63ND4Hs+3l6/mZBnSxkPZVX+8X4rv69/v8AQCMNNTRIlO+wAZDnVXKsWq0dYvSS7r/NdP8AJsznLMNn2XJ05Ju14S8/8ns109UV6NSjUlSqK0ouzXZoALAHh/mjvPAzf96P6r8n95xdlf1HMp8qtCfvL57r5Pp2sNIaAAMhzSWW4+GIW20l3T3/AM15pHGeV08wy5YujrKCumusXq/8/v7gAdQAqSjCDlJ2SKsqmcZiktE9F5RX9X9SjRhRpxpwVklZAEJJgwlNIlJyAOoQ96FgIK8kVtwGCAAAoL3wMRrMADD9R7i2GFFXqIAxeyGAA9gKMOSCQAAAAVHebADCr3GAAAAABXhyzAAAAAABAwGAGHd4W7GJXupgdAAOH/32Pq1vJ/iwjJxd0Rqxkjnh3J1ltEox0uAAABJXGrOwADFsAALYAARicNSxFJ0qiumZhga+WYpWfnF/117r9DJ81hj6N9prdfqvJii2KKRKaWw22AAcE8Q/VK31Ou/cm9PKX+T/AAevVnHHD3t6f16gveiveXdd/VdfL0A3LABgMZUweJp4im9Yu/8AmvmtDizB080ydYqjq4rnXpbVfdr6oBoAA4EzeOJwjwVV3lDa/WL/AMnp6NHEeUvLcwnRS916x9H/AJbfIAHsAHFWY+yorDQest/T/gnDGX+xoOvNe9Pb0/4P+QARjd2CU3fcbb6gAPcKPVElZtBSXvFXWQDBAMADDrVgV/4jAAw/Ue4MMOr1EAYvZD2AAChDnmgCrVVNEsTUezse2qdxV6i6kcXJbq5DEU5dbABUkowbAAw6/doAAAAADEwvC/YAAAAAAEADADDPVorq9NgdAAxlT2eHqT7J/kcK0/cqz9F91/8AMDoCRCNkkAAAA9iqrSAAZ0AAOgAAgzDL6WOoOlP5Ps+5kvDawU1WqyvPolsv8/yHOKdr6jk2AAAcHcQ/2hh/q1d/vYL/AMCj39Vs/k+rOL+H/wCzcT7akv3U9vJ9V+q8vQ3LAAHAOZKvhZ4Kerhqv8L3+5/mZ/lry/MKlDpe69Ht923yAaAAynMZ5fjKeJh9l6ruuq+aOMcup5llccXQ1cFzLzi9/wANfk+4ACAq1I04OcnZJXZShPN8zvLZu78or/gaeooqKUVsgAgrK4pXuMAAYUviKq99hSWpPWTAewAMADDrcCt8bAAjOUdgGGFXvMAxfwoewdAYBhYWjzdwCtPnm2ABCtOGzIYmEt9BNNaMbSWpiKyn7sdgAKKtTQAAAAADSasTjyyaAAAAAAEADACm+WaZNXi0ALYAzufJgKj8rfe0jhqny4K/dv8ARfoB0CkryQtgAAACstEAIGAAAAAthRbEkkYr2k6Uo0pWlbR72Z9axuAxznUb51vfqv8AJ9DAY6ljKCq036rqn2AAAMHi62Erxr0XaUXdf12fUw9bB8Q5W7rSSs11jL/NbrurdzM8trZfip4eqtV+K6NeT/4AAAZHmTy7H08R0Ts/R6P/AD9Tj7LVWwsMbDeGj84vb7n+YA0ABwFmqr4aWBqO7hqvOL3Xyf4NHEeUvLcwnRS916x9H/lt8gA6gcV4506McNF6y1fov83+Rwvl/sMN7aS96f5dPv3+4YBFXdio7KwMAAYtiHxIrr3kFHqS1YD2AjBy2J05R3GABh/hYFX42AAAwwi0bAMUvcQ9g6AAld2RGKjFJAVpctNsAAC7Ww22AARVopAAAAAAGKhqpAAAAAAC2ABiACL5opjQC2AOJp8uCUe7X6v9DJafJgKS8r/e7gPYMOt2AG5YtcAAqq8WAIOoAAAAWuRh3G0kSk2GdZRDH0tNJrZ/o/L8vvMvx1fLMS7rylH+uvb/ACMPiKeIpRqU3dMAAA4Xz2WVYtOWtOWkl+T9V+VzinI4Ztg1Voa1Iq8X3Xb57rz8mxpp2YAAcK4qnmuSywtbVxTg+9mtH92i80YzC1MLiJ0KnxRbX3ADQAZNmMsvx1PELaL181s/w/E42y6OOyyOLo6uGt+8Xv8Ao/RMAGEpKKbb0RFTzjNNfhb+6K/r72RioxSWyGAU1pcm7sHsAAPYWwtyutEFLZgD2Apx5YpDSasVYcrsABQXuAVfiYAAdR7BhV7jAMSr0x7AtgewGGhzTv2AMW/cSAAAAAIq8kgAAAAAAqQU4tDVtAAAAAAFsADEAGHd4W7E17zB7HQA4rnanSh3bf3W/wAzBw5MNTj2S/IB7BQXuIAFsAbgBJXQAgW4AAgAIxctiMUkSmug9QOpxBkqxcPbUl+8X4r/AD7fcZFm7wNX2dT4Hv5Pv/n/AMATUkmndAABa4cDZ/dLL6z/AMD/ADj+q+a7HHGQ+wq/XqK92XxeUu/o/wA/UAAODMy+p5nGEn7tT3X69Px0+Z4gZZ7LEwxkFpPR+q2+9fkANXAA4FzKOLwEsHVd3Do+sX/k7r0sZ3lssux1TDvZPTzT2/DfzAHsdDifHewwnsov3p6fLr/l8zhXAKnh3iJby29F/m/0B7AJXdibSjYB7AEad9xRS2CVOLHTcQrawQU/gYACV2kBUgpxsNNOwAUV7iArL32AAHUewYdfu0AVVem0PYFsAGHhyw9QDGbRAAAAAKEb1EAAAAAABiYWlddQAAAAAFsADEAFCai9SbTk2D2ADiZ8+JpU/L83b9BJJWQD2CmrRSAHsdA0LC2ABjWoC2BAACACNO+5okOd9gADiXJt8XRX+Jfr/n9/c4azizWErP8Awv8AT/L7uwAAJaBTqSpzU4OzWqfZoybMaGfZY41Vd25Zrz7r13XZ+hnOV1MtxksPPpqn3XR/11AAE2mmnqYhLiDh7mSvNq6/xx3t66r0YANABw9mf9nZjTrt+7tL0e/3b+qPEHK1UoQxsFrHR+j2+5/mACMyqSzTNfZQel+Vei3f5sp0404KEFZJWQAU11KjuwAIRuJXNAaQ0Th1RPWkgj/DYAMh8SAKlJTXmSi4uzAIK0UgMQvfYAAD2CkrU0ANXViUbNoFsB0IR5pJFlYAxi0iAAAAAYWOrYAAAAAAFWHPBoAAAAABbAAxAACHsAGa/vM5pQ7cq/G4CGJXdhADEMuXudBAD2Jr3mAth7CAAFsBa5GmluSkkSbYgAWwNJqzM+yl4GsqlL4JbeT7f5GQZv8AXaPJN/vI7+a7/wCf/BAASuAcP5zPK8ZGqtYPSS7r/Nbr7upxTlEM2y9VqGs4q8Wuq6r5rVefqxoAA8Psy5alTBSe/vL1Wj/C33M4ry36jmlSMV7sveXo9/ud16ADV0ABw1iaecZI8NW1cU4P7tH91vmjF4aeGrzoz+KLafyAM6xv1TBzmnq9F6v/ACWpwlgrueJkttF+v6L7wBg/diADCMbKwADRNL2dgX8MAGR3QASipKzJ0GvhGinDmlYAxK1TAABj2BKySADEwtK/cFsAYWG8gAxUb079gAAAADDxtTQAAAAAABiKfLK66gAAAAB0ABiAAEPYAJfvM/Xk/wAogIZTV5oWwAbFrlkNAgB7FT4mB0HsXSV2yrmmCpaSqr77/kVOJcvjtJy9E/1sT4twy+Cm362X+ZPi6TXu0dfX/gIlxbiGvdpq/wA/+Af6143+WP3P/M/1sxv8sfuf/wAkLi3GX1hH7n/mYHP8bjaypUqKb9XZLu2RjZeZiuKMBRqukm5W6pXX5/kU+Icuqf8ALyz80/8AhiljMNW/h1E/RoWwA9gDE4aniaMqVRXTK1LEZRjVZ6rVPo1/wdmYHGU8ZQjVp9fwfYAFsAHAmecy/s+s9VrD9V+q+fkca5H9Txf1mkv3dT8JdV89/v7AAGWY2WBxlPER+y7+q6r5rQ46wMMVl8MZT15Ov92Vv1t+IAMAOCcz+p5kqcn7tT3fn0f36fM8QMr9lioYyC0no/VbfevyAOKsU62Jhhoa8v5v/gW+8y/CRwuFhRXRa+vX8QBkFdlR3dgAZTjdiQABNe60C/hAAw3AAcYy3QopLQAxCvC4AAMexBXkkABWhzwt1BAQg5uyIxUYpIAJRUotEk07MAAAAKNBzd3sAAAAAAAFanzwt1AAAAADoAAgABDADAfvM7qPs5f5AIZRV5oWwAPUYI6gA9ir8RJqKu2YjPsBR+3zPy1/Hb8TEcWvajT+bf6L/Mr8QZhW057LyVvx3/Eq1qlV3nJt+buUsJiKq/dwb9E2U8gzGe1O3q0v1IcK46S1cV83+iZHhGo371Vfd/wUR4Qjf3q11/h/4LP9UqH/AD8f3I/1Rof8/H9yFwdRktKr+5GCwOHwVJU6KsvxfmzOsNj8ZD2eHmox67pv5pPTy+8qcL5hC9kpej/zsVclzCnvSfy1/K5UpTpu000/PQo4/FUf4dRr5u33FDijH0/ial6r/Kxh+L6MtK1Nr01/y/Uw2cYHEaQqK/Z6P8RgBnWVxx+H5V8a2/y9GZJmUsvxLhU0i9Guz7/LqJpq6AAAoV6lCrGrTdpRd0/NFKeG4iyiz05lZ/3ZL/J/evUxeFq4WvOhVVpRdmAAcJ4iGZ5LLCVXdxTg/RrR/ovQxWHnh686M94tp/LQAewARk4yUovVFeMeIeHuZazav6Tjv97uvRjQV6saNKVSWyTf3GR0ZY7MnXn0vJ+vT/P5AAyCtG49WADKa0AAB7E42lYX8MAGEHeKAAACceaLQAAMexh1eogADEUuWV1sxBCnKbsinSjBWQABiqdnzIAACFWK0lFMgqU1okOlB9D2NPsKlBbIAAAAAAAAxFPllddQAAAADoAAgABDADIbTzKrPyf4yQCGUPjOgAdRmgMYBjeIMBhdHPmfZa/8D8TG8V4iq7UYqK89X/l+DMRi6+Id6s3L1MNlWMxFvZ03bu9F97MPwnWlrWqJemv+X6lHhjA00uZOXq/8rFHAYWjb2dNJ+iv9+4ABGFxtRRKTkASjGSs1dFbJsBW+Kkvlp+VivwlhJ/w5uL+9f5/iYjhPGQ1ptS/B/jp+JiMDicP/ABYNeq0+/YwuZYvDW9lUaXbdfc9DC8XVFZV4X81p+D/zRhM5wWKsqc9ez0f/AAfkAcUZTdfW6S/xf5/5/wDDnDGa+0h9VqPVfD5rt8vy9AEAAcHZ39QxqpVH+7qaPyfR/o/LXoce5L7SmsfSWsdJenR/LZ+VuiAAOCMx+q5mqUn7tRW+e6/y+Zx7l3sMfHERWlRfitH+FvxAAAPD3M1CrUwUnpL3o+q3XzWvyOL8s+pZpPlXuz95fPf8b/KwcVYv2WEVJPWb/Bav9DhnB+wwSm1rPX5dP8/mdAC12kVHZWAAauJAABuVlsyP8MAGFF3ggAAAK8OWYAA9gwqu2wACUVJWZLCa+6yOFX2mKKirIAAJRUlZlai6b8gAAL22IYmS31I16cupo1oAAAAAAAAVIKcbDVnYAAAAAABbAACGANpK5wsnKrVl5L8bgIZh/iZ0ABHUjKMl7ruMexl+Z/XcwqqD9yCsvNt6v8NP+CVKkKcHOckkur0MfxbhqV44dc777L/N/h6mOznG4xtVJ6dlov8Ag/O5g8pxmLt7Gm2u+y+9mH4RUbPEVPkv83/kYbK8Hhrezpq/fd/ewGdAAIU+rJSUUNtsAABpNWZisiwGIu5U7PutP+B96MXwhUWuHqX8np+K/wAkYrL8VhXatBr8vvWhg86xuEsoTuuz1X/A+VjBcV4eraNdcj77r/Nf1qU6tOrFShJNd0SjGcXGSumZpgqmV41Spuy3i/0+Wz8vUy7HQxuHjVj13XZ9UC2AADhXNYZtlrw9fWUVyyT6q1k/ns/P1M2y6eX4yphp/Zenmuj+aAApVJUpxnB2ad16o4gpxzjh9YmC1SU18viXyV/uABgBl2NngsXTxEN4tP17r5rQ41wcMwyqGMo68nvL/DK1/wBH8mGcTeYZsqEHomo/5v8AP7iEIwgoR2WiEAQXUm7sACKu0LY3GXOhcvcCqrxZDWDABhh37rQAAAFeHNH0AAHsGGjaFwAAAAACUVJWZVwzjrHVAAAApNbMjiZrfUjiYPfQTTWgAAAAAAAYmnrzIAAAAAAewLYAAQwDEy5aM5dk/wAjhSPuVX6fqAhmH+JnQAOpxBmUp1Xh6b91b+b/AMl+ZhMXVwtRVKb1/P1MNWhiKUakdmjijNfq1D2FN+/P8F1+/b7zKs4ll0KnJG8pWs3srX+/cxePxOMnetNy8unyWxgOGMbirOa5I93v8lv99jA8NYHC2k488u7/AMtiySsitsgQMACFO2rJzS0W43cABAAtglFSVmY3hrA4jWC5H5bfdt91jHcN43D3cFzx8t/u3+65h8XiMLPmpScX/W66/MwHFkXaOKjbzX6r/L7jMMNQzXBtUpJvdNd/+DszIswlgMW6VXSL0fk11/R/8AAAA4azT+zsxp1ZO0XpL0f+Ts/kce5Sq2Hjjaa1ho/8L2fyf5gAHAGOVbCVMHPXl1S8nv8Aj+ZmuBeBxtXDv7L09N0/mrAD2AA4JxsMdlc8HV15Lq392X9NfcZ9TeVTrwqb07/Pt9+n3nCuGdXEzxEteX83/wAD8wAPhiAAQWoLYewDAJq6KWsWgAYUH71gAAACtS5XdbAAPYUXJpIjBRVkAAAAAABVw8Z+TKlOUHZgAAARnKOzKVeM9HuAAAAAAASipKzJwcZWYAAAAAD2BbAACGAZjLlwdV/3X+RwtH/Zpvz/AEQCGYf4mdAArTVOnKfZN/cYmrJ3d9WUpuMkm9zJMwp4fAVJVXpB3+/p82Y3F1MXXlWqbv8ADy+RlWAWOxUaDny3/TovOxgMlweCS9nG8u71f/A+VgCtsgQMAtchTSWu5UqW0R1AAEAM6ABjspwmMX72GvdaP7/8zMOFsRRvKg+dduv/AAf60KOIxGEq3pycZL+tV+jMdjHi6ntZRSk97bPzt0fc4azP6zh/Yzfvw/FdH8tn8u4AABwlmVPM8teFr6yguVp9YvRfho/v6mc5ZLLsbPDvZPTzT2f3b+ZZDQBwrj/qWa0pN2jL3X6P/g2Z4hZfyV6eLjtJcr9Vt96/IAewAHCOZfUs0hd+7P3X89vxt8rnjhGnQnQcH71VPmXlC1n87pf9unD+E+r4CCe8tX89vwsHUCKuyp2AAILQFsPYLjAHsU92iS1YA9gg7STAAAAJRUlZlWk4PyABRctEUqPJq9wAAAAAAAaTVmVMInrBkoSi7NAAAAUK3MuV7gAAAAAAGIpc8brdAAAAAAPYFsAALYAM3dsDV9DhhWwT/wAT/JALYZh/iZ0ADM3bBVbfyv8AIrazSK0UmrGR4KhjIVaVZXWj3a117epieDaMtaFRryav/l+ouH8zwNeFanFS5XfR/o7PUhJSipd+4BW2QIHuAk3oiEFFeZUqdECAAEAPYAAMdleFxsbVY69+q+ZmPDOJw95Uffj+K+XX5fcjB4utg6yq03Zr+rMyziPDYu0KnuT7PZ+j/R/iAAbljh/NXluPhWfw7S9H/lv8jjTKVjsCsXR1lBX9Y7v7t18+4DQBj7Z3w17Vaz5eb/t6O/32aXqADAA4kx2L4u4lpUIK8kowS/wq839/M/SwkkrIGBBEndgAEFogWw9h6mgAPYjpUZUXvMAewFOXNFAAAADSasyph2vhHFrcIUZSIQjFWQAAAAAAAASipKzKmET1gSi4uzAAAItxaaE1JJoAAAAAAAxFLld1sAAAAAD2BbAAC2OoBnrSy+p8vzRw4msAvVgLYZQ+M6ABi6Tq0JwXVNfgYiLVn2I3nJI4WpNUqk+7S+7/AIc3AArbIEHUEm3ZEIKKKlS+iAQAAtgBgAAHEOTQxFB1qUf3kddOq6/P/hgyviPEYS1Op78PxXo/0f4GDx2HxlPnoyuvxXqgASA4IzdYrCvBVXeUNvOP/A29GjibJ3luPlCK9yWsfTt8tvSz6gNAeHuOUqVXCSe3vL0ej/T7zO8B9RzCrh+ieno9V+DQAwAxVeNChOq/sps8FsLSr5/UxNX4owfL/ibV368t/kzivLvqOaVIpe7L3l6P/J3QMDaIMACK0BbD2A6AD2HpUKy1AHsBRlZ2AAAAA3LJbAAAAAAAAAAThGas0VcNKOsdUAAAYaV4W7AAAAAAAEoqUbMqQcJWYAAAAA9gWwAAtjqAcQO2XzXp+aOH1bL4fP8ANgLYZR+NC2AAzrAvDYmWnuy1X6r5FOleSjBasy7BrC4aNLr19eoABW6Ag6iTbsiMFFFSpfRACAAFsAMAAAOI8r+qYj2kF7kvwfVfqvu6FLJ6eZYVV8K7TWko9L912v226aWITxWAr6XhNf181+BlPElHFWp1/dn+D/yfkAlcAyzMKuAxcMRT3j+K6r5oz/L6Wd5UqlDWSXNB/mvntbvbsNA1dAcMY/6lmlKo3o3yv0en4Oz+R4h4Dlq0sXFbrlfqtV96v9wAwA4qxPs8Eqa3m/wWr/Q4FqyyuNDFLfm5n6PS3zj+Zx9gI4jA08bT15Ov92XX77fewewJXZUfQGAG4gsi2g1oHQLXGh7E375WV0mAPYAhPmiAAAAAAAAAAAAAAAFbDxnqtyUXF2YAAYeVp27gAAAAAABWpKcfMatoAAAAA9gWwAAtjqAcRO2AkvNfmZErZfTXr+bAWwUviQtgAMThaOIhyVY3RhMpwmFlz04693qAAPYrfECBJt2RGKiipPm0QAIAAWwAwAAAx+Cp4zDyoz6/g+jMsxdTKse41NFtJfr8t/8AhzHZdhcfSSqK/Zrdej/pGaZJiMA+Z+9Duv17fl5mUcSVcNalX96H4r/NeX/DFCvSr01UpSun1FsAHAec2bwFR+cf1X6r5nGuTfU8Z7emvcqfhLqvnv8Af2AYGP8A+FrhlVVrJR5v+3o7/fr94A9gA4knLFZjDDQ6WXzf/AsU6cacIwjslb7jhmtDNshlhKj1inB+lvdfyVrehXozo1ZUpq0otp+q0Y9ggiTuwYALdC2AA6Ah7D2J/Ex+9TAHsAQk4u4mmroAA9slNxYAAAAAAA2krsliYLbUWKh1I1IT+FgAAAAVKUaisypTlB2YABF2kmAAAAAAABiKPMuZbgAAAHQHsHQAAWwAcSf7i/VGSf8AIvpej/NgLYI6MQAAAAPYqayYISbdkQgooqT5tEAAtgADoAdQAAEBxVlftKaxVNax0fp3+X5ehwtmftaX1Wb96O3mv+B+XoSjGUXGSumZxwva9XCL1j/l/l93Yy/MsTl9W8H6p7P/ACZlma4fH0703aS3XVf5rzADD16mHqxq03aUXdPzRUjQ4iybTeS0/uzX/B/B+ZVpTpTcJqzWjXmDA8PsaqlCrhJa2d16PR/LRfeZtgngsdVw/wDK3b06fhYB7ADaSuzJk8bm8q72V5fovz/AOA8x+r5i6En7tRW+a1X6r5nHWXfVsz9rFe7UV/mtH+j+Y9joLSIDACO6FsAV60KMHUm7JEeJsK6lnFpd/wDgEJRnFSi7p7APUZLVspO6sNWdgHsAFOpyuz2AAJu8mUKnMuV7gAAAAAAYipzSstkD2NtinipLSWpGcZq8WAAAAFSnGcbMq0pU3ZgAFN3gmAAAAAAABiKNveiAAAB0B7B0AAFsAHE3+5L/ABL9TJ/9wpegC2BFN3SAAAACQ9WEVfREIKKKlTm0QAAtgADoAdQAAEBOMZxcZK6ZjcPVyjME6ey1j5rt+jMJiaeKoRrU3o1/S+QZtkFDHJzj7tTv39f89ydPF5didbxnH+vmjJuIKWNSp1fdqfg/T/IAOB84+rYv6pN+7U28pdPv29bHHmUeyxEcbTXuz0l/iWz+a/FeYPYDhPHfVM2pNvSXuv56L8bHiDgfZ4yniUtJqz9V/wABr7gHsAZ3iFQwFSV9WrL56HCOGUcPOs927fJf8OGHrzoVoVYPWLTXqji6hDM8khjKX2bSXo91+r9AWpPawDACO4tgDibGOVSOHWy1fr/w35irQcrXOGcW5Qlh5PbVenX+vMAlsFJ2kVVaQD2ABlGpf3WADdkMg2ndEWpK6AAAAAAMTUcYpLqA9gIycXdMp4qMtJaGjQAAABOEZqzKtCUH5AAYZ3pgAAAAAAAFejy6rYAAAOgPYOgAAtgA4m/3Jf4l+pk/+4UvQBbAii7xHqAAAAkVXowtcp01FeZUnfRCAAFsAAdADqAAAgDP8s+u4V8q9+Oq/VfP87HCuZeyqvCzektvJ/8AB/P1BGYZdh8dS9nVXo+q9DM8pxGXVfe1j0kv60ZkfEiqWoYp69Jd/Xz8+vXzAjKUJKUXZowFelxBkrjU3a5ZeUl1/KSMTh6mHrSo1FaUW0/kARbi01ujiGKzbh2OKitUlP8ASS+Wv3AAHF+ItSp0V1bf3afqZTh/YYGlT62/F6sDgfFQxuVVMFU15br/ALdlf/goxuFnhcTUoT3i2vuKauyerAYFSpCnHmnJJeehiOI8vo6KXM/Jfq7L8SpxhZ/u6X3v9Ev1J8X4+StFRXyf6s/1rzHuvuI8WZine8X8v+CUuM8Qv4lJP0bX+ZmGM+sznXStzf1+Q6fu3OHarWNh5pr8P+AAVH7rBOzKqvG6AewHQGFKopKz3AKrtBjEUJ68oAAAAABiviQD2ACFWUNmQxcX8SFWpy2YAAADVyrhU9YEotOzAMK9GgAAAAAAABpNFahy6x2AAAAewdAABbABxQ/9jj/iX5MypJYGlbsgFsCKD1aExoAAAtcexWeyCnT5Vd7lSpfRAgABbAAAAAAAgA4jy+WDxar0tIy1XlLr/mv+AZTj447CxqrfZ+q3/wAxbBXoUq9N06ivF9DOsgqYFupT96n36r1/zMi4jdG1DEu8ej7eT8vy9Nk1JJpgcFZt9Ux3sJv3KmnpLp9+3zXY4+yt08RDGwWktH6rb71+QMDgbEQxeWVcHU1UW1b+7L/g3MVh5YevOjLeLa+52AAzj/bc5jQWytH9X+YBwVmH1XNYQb92p7vz6firfM8QMB7HMI4hLSovxjo/wsU1oPUBmNzPC4ON6ste3X7jG8VV6nu4ePKu71f+S/H1K1etiJc1STk/PUw+SY+uk402l56fmUOEK8v4lRL0Tf8AkUuDcL9upJ+ll/mf6nYD+eX3r/5ElwbgmvdnK/y/yK3Bb/5dVvvX6p/oYvDyoJ0pbxdn8tBzXs7HD0W8ZSXr+TAKz0Ap+9Cw1Z2B7AdAYJtO6KdVTVuoGIltEYi9mmhNNJoAAAAADFrVAPYAAI1Zx2ZTxfSRGSkrpgAABVoxqLzJxcXZgYZ2nYAAAAAAAACth76xGraAAAD2DoAALYAOJv8Acl/iX6mT/wC4UvQBbAiLtJMQNAABa4SZJ8zuUqdtWValtELYEAALYAAAAAAEAGZ4GGNwsqL36eT6P+uhkGNlgMa6NXRS0fk1t/kLYCUYyi4yV0zPeHXhr18Orw6rqv8AgfkZFxBLCNUazvT/APSf+B5fd5wnGcVKLumCbTTT1KEocQZDyt+81Z+U1/wbP0ZOEoScZKzQwOBsb9XzRU29Kia+e6/K3zOOcF7DNXUS0qJP57P8r/MBg2krsyBfWs1niH0u/vdvyYBTqSpzjOLs07r1RxNTjm3D8cXTWqSn+kl8ru/obQAxGIpYem6lWVku5mfFFSreGFXKu/X5dvz9Cjh8Ri6lqacpP+tX/mYLhNu0sTP5L/P/AIHzMLl2Fwq/cwSffr9+4LcQwOIMNyYyfaWv9fMVGbdjhnD82JdTpFfi/wDgXAexVd5WApOzsVY2dwewHQGAQr2VpDrQS0ZJuTuxiGYeV427AAAAAAYpe6mA9gAACE5Qd4spYmMtJaMAAAMVT5o8y3QFN2mmAAAAAAAAAVKUZrUqYeUdtUAAA9g6AAC2ADib/cl/iX6mT/7hS9AFsCClK8fQLlr7AAWGxlWVlYpU76sq1OVWW4LYEAALYAAAAAAFsABxZlqhUWKgtJaP16P5/wBbmQZksbhFzP346P8AR/P87gG5xBw77NPEYZadY9vNeXddPTbI8+ngpKlVd6b/AA815d193nTqQqQUoO6ewcB5n7HFywknpU2/xL/NfkjjbLfquZOrFe7U1+fX9H8xgYXESw1eFaG8Wn9zOO8PHFZbSxlPVRf/AJLJb/fb7wGGcV/YYCrNb2t9+n6nCNBRw9Sr3dvuX/BADhHMqcskxNCrqqak7f3ZJu33p/eVNrBmud0MDHl+Kfb/AD7GLxuJx9XmqO76JdPRGW8LTmlPFOy/lW/z7f1sUMPSoQUKUUkAEXdAGc5Z9dpLk+OO3n5Ectxcqns1Td/T9drGWYBYPDqnu936gSaJasAkueAPYDoDAAGIZRlyzQAABUmoRue3qXvcpVlNW6gAYlXpgPYAAAClXlDTdEKkZrRgAASSasAJ3SYAAAAAAAAAToQn6lShKPmgAB7B0AAFsAHEiTwPzRkrbwFL0/VgLYEFJtSsIC5oaGhcGyclFFOHO7sqTUVpuPUFsPYQAAtgAAAAAAWwAGLw0MTQlRns1b/gmVYqeVZg4Vdvhl/n+vobgBxDw9vicMvVfqv1RkWezwU1SqO9N/h5ry7r5+sJxnFSi7pmGrzw9aFWm/ei018jPsFTzvJ41qOskuaP3ar9PUYBkrWa8Myw71kk4/Nax/QaBhxbX5cJCmvtP8F/SMjoexy+lHur/fqAGExlWhCpCD0mrP0un+lvmY3LqtDD0MRLaonb5O3+T+ZnfESot0MM7y6vt6ef4L8sFgMTmFa0NX1b6er/AKZlmS4fAxTirz7v9Oy/psYAEHoDAAbKkr6IYBSfQqRtIewHQGAAMQwhLmimAABipXaiCbTuilUVSPmAFSPNBoB7AAAAEZOLumUsQp6PcAAB7gUXemgAAAAAAAAADEUuV3WwAD2DoAALYAOIlfAS8mvzMid8up/P82AtgQUvjQ5Lm5RMAAbJzUURi6j1JSUIjbbuwFsCAAFsAAAAAACAAOLsvvGOLgttH+j/AE+44azD61g1CT96Gj9Oj/T5AAcQ8Pb4nDL1X6r9UZBnrwklRrP92/8AyX/gd/vIyjKKcXdM4Bza8ZYCb296P6r9fvOLsreBzKXKvcn7y+e6+T/CwB4eY3lrVcK3ulJfLR/mvuOIsH9UzStSW17r0eq/OwMOKZOvjqVCPb8W/wDgIhBQiorZABBHHvHWFxWCoZVlusaaXNPZ3trGP/tz6vbTV5PkdXHy5paU117+S/rQw2GpYakqdKNkgYAEXZiBoBjdyc1sgYAnZ3KiUo3Q9gOgMAAYhhhpXTQAAFd3qMCE3CV0RkpRTQAVI8s2gewAAAAFLEuOktURlGSumABN2i2AYfWmgAAAAAAAAACcFONhpp2AB7B0AAFsAHEP/Ivn6r8zIP8AkXU/n/6UwFsCCj8aK3xlOqnowuXRdFxslVW0SMXN2PdhEnJydwBbAgABbAAAAAAAgADEYeGIoypT2krGBqzyjNHCo9L2fo9n+TNwAOIsg9nfE4ZafaXbzXl3XT024dz32DWGrv3Hs+3/AAPy9NsBjKmDxMMRTesXf/gfNaHEmCp5zk6xFDWSXNHva2q+7p3SAOG8Z9UzWjU6Xs/SWn4XueIeD5cTRxCXxJp+q1/J/gDCP+18Q36Rf/pK/wA0ABxDn2jwuHf+J/ov1+7ueHPh/wD6x1pV8VLloQ3t8Um72S7LTV/JdWquF+qzlh7W5G427W0AYABGV9C5dF0XG0lqSqX0QgYAUpXVipGzA6AwABiGFGVpoAACesmAUKvK7PYAMVC0kwewAAAABGcou6ZDFp6SIyjJaMAxU0o8vcAwvwAAAAAAAAAABiYWkpAdAewdAABbABxBG+Xz8rfmjh93y+C7X/NgLYEFD4yt8bLXKcZJasLFgqqSerIq7siEFCJUnzMAFsCAAlKMVeTsipnOApL3qq+Wv5XKnFGXx2vL0X+bRPi6gr8tNv7l/mf64f8ATn/yb/7UlxfWb0pL72Q4vml71FP52/RkeMIt+9R0/wAX/ARDi7CNe9CS+5/qinxJls3bnt6plLMcJW/h1Yt+qv8AduAAACAAFscXZdz044qC1Wj9Oj+/T5nDWYfWsGoSfvQ0fp0f6fIADc4iyL6s3iKC9x7rs/8AL8jhvPL2wld/4X+n+X3djgHNlOlLAzesdY+j3XyevzZxPlLy7MZ04r3Jax9H0+T0ATad0cRL+0+G4YlatKMv0l9139wE5qEHJ7I4Vg62Mq13vb8W7/oAHEWdewi8NQfvvd9l/m/wMiySWNn7Wov3a/F9l5d38vTgDFxwuYqgtIzjZLpdar8mvmccYH6vmsppaVEpfPZ/ir/MBgAD3FU7nPF9Rzj3HU7DbYIGAEXZ3KkeaOgHQGAAMQwIvmimAAEtwAoV00ovcArQ54NA9gAAAAAE2tiOKmt9RYuPVEsXG3uolJyd2AYX4WAAAAAAAAAAFSCnGwHQHsHQAAWwAZ7G+X1F6fmjh13wEV2bAWwIMOveJQcpuxCmo+pYAauVIKUbFKnyq73KtS+iAAWwIDFZvgsNpOor9lq/w2+ZieLY7UKfzf8Akv8AMxGf4+tpz2Xlp+O/4lSrOo7zk2/PUpYDFVv4dNv5Mp8O5jP/AJd29Wv87/gR4TxrSvKK+b/yP9UK/wDz9X3Mjwe2ta2v+H/gk+EKifu1U/VW/Vk+EcSl7tSLfzX+ZPhXHx25X6P/ADSKmRZjTWtJ/Kz/ACbKuHq0napFr1TRQxuJofwqjXo3+Rh+KMdT+NqXqrflYw3FmFqWVWLi/vX+f4GGxuGxKvRmpfn924AAgABFejCtTlTmrpqzMDVnlGaOFR6Xs/R7P8mbgAThGcXGSumZ/kssDU9pT/hvbyfb/I4J4qlSxVJ1H+8g/wDwJdV62/z7nGWAp5hlccZR1cFzJ94u1/0fyYBwZOONyWrg5vbmj8pL/NsqQlTm4S3Wn3BnVX2WX1ZeVvv0/U4So8uEnPu/yQBneaxwND3fjey/X+uplGV1cyxF5P3VrJ/11ZGlClTjTpqyWyMFiZYXEU68N4tP7mce4aOJy6ljKevK/wDyWS/zt94DAAGAA9hAwAKUrqxUhyyAGAAJXdirCMbWGBhpXjbsAAE17zABlDEX92QBXp8s/UewAAAAAAAAYX4WAAAAAAAAAABXhyzDoD2DoAALYAM4jzYCqvI4Zd8E12k/yQC2BBQ3ZYsABYrTsrIAAWwVa9KjFzqSSXmY3iqjC8cPHmfd6L/N/gYvN8ZitJz07LRf8H5mGwOJxLtSg3+X37GG4UxE7OrNR/F/5fiUOGcDT+JOT83/AJWKODw9D+FBL0QtgADoDSkrMr5PgK/x0l8tPysYjhHDT/hTcX56r9H+JieF8dS1glNeW/3O36lSlVoTtNOMl3umYTiPHYfRy515/wCe5guJ8HXtGp7j89vv/wA0iM4zipRd0AIAAQcX4ByjHFRW2j/R/p9xw3jvrWCUZO8oaP06fh+QABiKFOvSdOorxfQzPLq2V4lcr03i/wCuq6nhXxRTzrK5YOtb2lLRrvF/pe68rpGdZbLL8dUw72T09Ht+H4gcAYv2WYToN6Tj+MdfyucXYNYbN6qS0l7y+e/43DiuryYFR/ma/DUyGj7LLqS7q/3u/wCQGLxVPC0ZVaj0X9WG8Tm+N85fcl/kv61Zl+CpYSjGlTWi/F9ye50MjazbhmWGesknD5rWP/to0DAAGAA9hAwAE7O5OKnHQAYABRV5or7oYFCVpoAAK0bVGADChW+zICrTU426klbQAAAAAAAAMKvcYAAAAAAAAAAGIjeF+wdAewdAABbHUAzKPNg6q/uv8jhaV8LOP979EAtgQU4csQAAKyurgAFavSoQc6kkl5mP4q3hhY/N/ov8/uK1eviql5ycpf1sjB8NYuuk6nuLz3+7/OxhOHsDh9XHmfn/AJbCikrIAQAAtgDqFahSrR5akVJeauYzhXCVdaLcH96+7f8AExuQY3C3bjzR7rX8N191jB5jicJK9GbXl0+4wHFdGpaOJXK+62/zX4lOpCpFSg00+qAAADE4eGIoypT2krGUV55ZmbpVHo3yv9H/AF0AAGY/A0sbQdKp128n3MuzDMOGc2jiKLtOH3Si90+6a+59mtM8xuD4iyijnWC6e7NdY+T9H96knswMlxf1TMaNa+ikr+j0f4HiNhPeoYlecX+a/UOMKl5UqS83+SX6lCkqVGFNdEl9yA4hzN4zEexpO8I7eb7/AKL/AIJkWUrA0Ly+OW/l5f5+fyIIlqdDw7xvLiKuFb+JKS9Vo/vT/A4lwSwma1qS2vdej1/C9gYAAwAHsIGAA9ijL7JWhZ3QDAAMPHdlfdDAIyUopgAGKjqmADEFCtf3ZAYmjzLmiAAAAAAAAGHVqaAAAAAAAAAAAaTViSs7APYFsAALY6gGJjzUJx7p/kcKSvCrH0/X/IBbAinHmkkILGg9QsBNXVgAbSV2ZlxJRo3hh/el36L/AD/LzMRicRjKl5tyl0/4CMBwxXrJSrvlXbr/AMD+tDB5dhsIrUo2ffr94AAgABbADADHZJg8ZdzjaXdaP/g/MzDhrF4a8qfvx8t/u/yuYPMMTgp3pSt3XR+qMt4mw+ItCt7kvwfz6fP7xNNaAAAg4vwCUo4qK30f6P8AT7jIsf8AXMFGTd5LR+q6/NAAbhneUwx9Cy+NbP8AT0f4HCXElXIsZOhXTdCp7tSP4cyX80Xqu+2l7puD+CSkujWzXdeTAzr/AIU+Fo195RUZfNaS/UM5/wBozqnS7cq/G7/BgcSZn9Vw/soP35/gur/Rf8A4Xyr2k/rVRaL4fN9/l+foEdIj2Dh3GfVM0oVel7P0ej/O54i4LlrUcSlunF/LVfm/uBgWbORns2cjHFoAB7CAAB7C0E1OJKLTsDAA3ElCCRX3QwDDSvFoAAxEean6AAxAUK3OuV7gV8Nf3ojQAAAAAAAFNWgkAAAAAAAAAAAYiNp37gPYFsAALY6gDSascLvlr1ab7fk/+CAthiKC3YWHqBYLJoGSVm0Bjsww+DhzVZa9F1f9fcZlnWIxt435Ydl+vf8AIy7IsRjLSfuw7vr6L+kYDK8Ng4/u4693v/wPkHUAAQAAtgBgAGY5JhMam5K0u63+ff5mZZFisFeTXNDuv1XT8vMyzPcTgrRvzQ7P9O35eRl+aYbHQvSlr1T3X9dwAAMdhIYrDToy6r8ej+TOG8VLB494eppzaPya2/y+YAMA4lyV14/WaK95bruu/qvyOGc49lNYWq/de3k+3o/z9QOCakcZlFXBzeza+Ul/ncqU5U5uEt1p9xhl9Y4hlL+Vv8FYK1WFGnKpN6JXEq2cZh/if3RX/A/H1KNGFGnGnBWSVkG0BgZ5bNeGI4haySUvmtJfqDFBsUUixYsWGidO2qAB7C2AAB7BTnysnBSQ1bQYAFKF3dleeqS6FdaIYBRlaaAAGk1YkrOwAxAJtO6KVRTj5gVKMKm+5PCTW2pKEo7oAAAAAACnHmmkAAAAAAAAAAAGIjeF+wD2BbAAC2OoAZR+7zerD/EvxuAthiKStFCHqAMB6j2Kq94M24gp4e9Kh70+/Rf5v+n2IwxOOr6XlN/18l+BlnDtHD2nX96Xbov8/mCDqAAIAAQA9gAAaTVjNOGaOIvPD+7Lt0f+Xy+4q0cVgK9pXjJbf8BmU8UQqWpYvR/zdPn2/L0NGtAAEHFeCdDFRxMPtfg1/mv1MsxixmEhWW7WvqtwAAjJxkpReqOOeFlhYUs3wcf3FbdL7FTqvKLd3HttorX4dzf65R9lUfvx/Fd/8/8Agh4fYpU8wnRb0nH8V/wLnFuEWGzetFbN8336v8bnC69rj6tbyf4v/gBxXmOiwkH5y/Rfr9xw3lv1bDe1mvfn+C6L9f8AhgJfCMDgatDF5VVwc9VFtfKS/wA7lejKjVlSlvFtP1WhGJa5ymgGg0NFSNtUAPYWwAAPYCnO2jJ01L1JJp2ACFFvcnJQiPUqa00xgBCXNFMAAxMbTv3AGIApzcJXRGSkroAlFSVmVsO4ax2AAAAAADDU7LmAAAAAAAAAAACS5otFgewLYAAQAEf3Wftd3+cQFsMRBaWGAACGjEyjCPPJ2SM34glXvSw7tHq+r/yRlmTV8c77Q7/5d/yMHgaGEp8lJW8+r9QED3AAEVMbhqfx1Ir1aRPPcugruqvxf5InxJlsVpO/yf6pH+s+X939x/rPl/d/cR4jy2Su6lvk/wBEynneXztaqte+n5lPF4er/DqJ+jTAex0AAAxWDoYun7OtG6/L07GbcPV8HepT96HfqvX/AD29DKM/rYJqnP3qfbqvT/Lb0MLi6OKpKpSldMAC5nGB+uYOdJb7r1X+e3zOEcdyVZYWT31Xqt/vX5APUADhH6tmGHxGT4xc1Oor2/O3Z7NPo1fc4lyDG8K5y6De2sJdJRez/SS736WZl+Op43DxrQ67rs+qMgxbwuZ0KvaST9Ho/wAGeKdD2MY4tfySX/gOq/M4PppU6s+7S+7/AIcxWIhh6Eqs9or+vvMqw0s0zBzq6r4pf5fp6AblTodQOAcYqOZui3pUi181qvwucYYH6tm9RradpL57/imJGwAA0SimrDVtAHsLYewAA9gCFS2jJRjJakqD6M9jIjQ7sUIrZE6kY+pKTlqwh71KwwAw0tHEAAxMbwv2AGIAKFXlfK9mABXw/wBqIAAAAcrY00BSoym/ISSVkAAAAAAAAAAABWjabB7AtgABDADNF7POaUu/L+dv0AQyCvJIWwAABisVRw1F1asrJGc53VzCfKtILZfq/wCtDKOHXVtVxKtHour9ey8t/QjGMYqMVZIAQMCTUVdsxOf4Chpz8z8tfx2/ExHFrelGn97/AEX+ZX4hzCr9uy8lb/g/iVcTWq/xJuXq2yFOc9Iq5DK8bO3LSl9zIZDmM3ZUn87L82f6u5n/AM+vxj/mf6u5n/z6/GP+ZLI8xi7Ok/w/Rk8txkL81KWnkycJQdpKzKWMxFH+HUa9G0UeJMxpbz5l5r/Kz/EocXvatS+af6P/ADMNxBl9dJe05X2lp+O34ikpK6d0AACNzOOGIVb1cIrS6x6P07em3oYPG4nLq7cNGt0/yaMrzahj6d4O0luuq/zXmAAZ1Rll2aKtT2b5l+q/royjXjWpRqQ2auAAGW42eCxdPER3i7/LqvmtDxR4epZ3kkcVS+OnrF/3ZW/B6em5k2ZTy7FOFTSL0ku3n8vyE00mmeI9WOM4LeLvqlB/+BWi198vwOFafLgL95N/kv0OLcd8OFi/N/p/n9xw/l/1TBpyXvS1f6L7vxbAjuie51AyrFfVcdSr/wAsk36X1/A8QsInChiV5xf5r8mVsLOlRp1ZbTvb5OwAADRVWtwGLYewAA9gQEako7CrRe57WHclWXQlUlLqD2DDvdE42k0AFKXLNMAAklJWGrOwDEADMPV542e6AAxFD7UQAAhQnIWEXVn1amKhTXQUYrZASpQluiNGnHZAAAAAAAAAAAABiY6Jg9gWwAAhgBxMuTE0qnl+T/4Jo1oCGUVeaFsAABjsdQwdF1arsvxb7IzPM8RmVdNrTpFdP82zKOHI4eKrYlXn0XRf5v8ABAAIGYzN8HhFapPXstX/AMD52MZxVWneOHjyru9X/l+ZiMZiMQ71ZuXr/lsYbKcbiP4dN27vRfiYfhKtKzq1EvTX/L9SjwxgIL3ry9Xb8rFLLMHS+Ckvu1EklZAAIlGMlZq6K2VYGr8dKP3Wf3or8K4Cp8F4+jv+d/zMRwhXjrSqKXrp/n+hicpxuH/iU3butV96uYfF18O70puPozCcWYiGleKku60f+X4Iwee4HFaRnZ9no/8AL7mAABm2SUMfG+01s/0fdfkVaOKy7E2fuyWz/Vd1/TMlz6njo+zqaVO3R+n+QAHE2B+sYJzivehr8uv4a/I4TxvtMPLDyesdvR/8H8wAAODsVTzHKJYOrry3i/8ADK9v1XyONeH54avOol70G1Lztopf10s+5wvmvtIfVKj1Xw+nb5dPL0OLMzvwF7G+vtVD85/p+Bkrhh8qhOWyTb+9sy6lLNM0dSotL8z9Oi/JegBDcnuARWhiKUs34ZhyazsresXZ/fqca4WGF+q0IbRhb7uvzAANAKiumAzoPYAAYIAAB7BSdpIxEdUwAKUuaCYABiI2mAxAAynNwkmiLUkmgAMRQ5fejsAAtGNpLVkq9NdSWKXRH1qfZH1qXY+t+R9aXYeLfREMTF6S0NGgAAAAAAAAAAACtHmg0ALYAAQwA4rhenSl2bX32/yMJP2mHpy7pfkCGUFeZ0AAIYiE606S3ja/zv8A18zH4+jgqLq1Xp0XVvsjHY3E5piU2rt6Riunp+r/AEMjyCngYqpV1qP8PJfq/wCnV1iwADH57hMJeN+aXZfq+n5+RjuIMXirxT5Y9l/nv+RhsFiMU7UYOX5ffsYThNvXETt5L/N/5fMwuVYPDW9nTV+71f3sAAAAA6mKyrB4r+JTTffZ/ejGcIp3eGn8n/mv8jGZbisI/wB7Bpd+n37GCzvG4SyhO8ez1X+a+RgOJ8LiLRq+5Lz2+/p8/vE01dAC2Ax+X0MbS9nVXo+q9DMctxGXVkpbdJLr/k/IyPiCOJSoYh2n0ff/AIP59AAlFSi09Uyg3lOb8r+FO3/br6/LR+qAAA4MzH6pmkYyfu1Pdfq9vx0+bPEPLIwxUcSl7tRWfqu/qrfczNcDUyzGKVN2W8X+nyM7z365k1OlF6SmpNdpRi1+PN/WpxHz5fl1HBS0lKMb+nKn+N0cMYL2GD9o956/Lp/n8wCG5LcWwHC+X4DMMQ8LibptXi07ardaprVeXRmT5VDLMN9XhNyV21fpfpoeIiXtcP6S/QY0PY6DLXLXQEtRqzsDB7AADBAAAPYFsP8AeUwAMNKz5QADFRukwKMYyqJMrxtUYAMMJU+wwAGk1Yr0fZvTYADcAAClWcHboJqSugAAAAAAAAAAACUbNoBAACGAHE1PmwSfZr9UZLPnwFJ+VvudgQzDrVgAFSpGnBzk9Erv5GSZxThLFYmu7Xadv/AtF+CMdjcTmmJTau3pGK6en6syPIoYCHPPWo9328l+rCp8LADHZnhsHG9WWvRLd/15mYZ/icVeMHyx7Lf5v/hjBZZicY7Uo6d3ojBcMYailKs+d/cv+D8/uIQjCKjFWSAAAAAA6gNKSszHcNYLEJuC5JeW33f5WMfkOMwmrjzR7r9VuvyMDm2LwT/dS07PVfd0+VjLuI8LirQqe5Lz2fo/8wEAYnC0sTSdKqrpmb5NVy+pdawez/R+f5/lkPEPtrYfEP3uj7+T8/z9dwDi/Bc0IYmPTR/p+v3oyHG/WsDBt+9HR/L/ADVgAAjJxalF6ozOEc84dVaKvK3Mv8UfiS/FIzTL4Y7DOk9+j7P+tyrTnSm6c1ZrocW4qnnXE04Yezpx5aaa2aglFyXk7NrysRhGEVGOyAIEjoBhq06FWNSm7SjqmZVmFPMMHDEQ6rVdn1X3niJFc2Gl/i/9tGPYYAthg9ioveBg9gABggYAA9g6GHl9krQ5ZAAm000JqSTQAFeN6bAi7STMTHZgAwjJxaaIyUopoACcFONmVKbhKzAAAAAo1eR2exuAAAAAAAAAAABiI2ncBAACGAGd0+fAVF5X+5pnDVTmwXL2bX5P9QQzD9QAr55gqNRwcrtdlco1qdamp03dM4rxvscF7JPWenyW/wCi+ZTpyqSUIK7eyMiyOGBpqc9aj38vJfq/0AmroCcoxi5SdkjM+Jt6eF/8Cf6L9X9xSoYnG1bQTlJ/1q/8zL+GaVK08Q+Z9un/AAfyIxjGKUVZIewtgADoAAdABgAZhw/hMXeSXLLuv1XX8H5mYZLisE7zV491t8+3zMtz/E4O0W+aHZ9PR9PyMBmmGx0L0pa9U90ABXoU69N06iun0M6yWpgKnNHWm9n28n/Wpw/n3trYbEP3uj7+T8/z9dwMbhY4rDzov7S/Hp+JwviZYfGSw09ObT5r+n+AAAM8PMxUqdXBSe3vL02f6feziDL/AKhmVWglondej1X+RxZCEccuVauKb9btfkkcIU8O6tScpfvElyrunfmd/KyXz8mAENhgC2OB83WGxTwlR+7U2/xf8FaeqR4ixbWGf+P/ANtGDDoC2Oox7FVag9wYAAwQPYAAewdCLs00VIqcLoADDzuuUABq6sNW0Af7yiADAwk7xcewABVpqpGzJRcXZgAAAAYafNHl7AAAAAAAAAAAAYmN4pgxAACHsAGLp+0w84d01+BwrUvGrD0f5ghmHWjAM9xrw2FtB+9LRfq/67kpxi7NnDuNdLEexb92X5/1ocTY36zj5RT0hp8+v46fIp1JU5qcHZrVGUZjHH4WNXZ7Nea/q4E2la7DH5jQwdPmqvXourMyzevjpe87R6Jbf8EyzhyriLTr+7Ht1f8Al8zDYWjhqahSjZB0HsIAAWwAAtgB7AADSaszM+F6Na88N7su3R/5fl5FahicDWSknGS2/wCAzKuKIztSxej/AJunz7eu3oRkpJNO6ACtRp1qbp1FdPdGdZNPL6nNHWD2fbyfn+Zw/nqxMVh6799bPv8A8H89wDiOhLB5jHEU9ObVeq3/AEfzMNXjXoxqx2krgAG5w/mH1DMqVdvROz9Ho/u3+R4h5fzQpYyPT3X+a/X7zimXNmDXZL/MxOAxuT1aNaStzLmi+/Rr1TTTXz2aMFi6eLoRrQ2f4PqvkAR+E6gCIylCSlF2aOJc1jmeWYWt9q8lJdmkr/fuvUY9hgwWx1GFXoHUGAAMED2AAGD2ChL7JWhyyACMuWSaE01dAAVVabQGFlo4lSPLJoAYFGfLNMAAK1FVF5kotOzAAAAClLlmmAAAAAAAAAAABUjzRaBiAAEPYADIf3OY1KPqvuYDMP8ACAcU1L14Q7K/3v8A4BZzuzB1ZQcZJ6xf5FHg/Cp81ao5P7v83+JQyXL6HwUlfz1/O4kkrIDHY/nzihhYvSN2/Vp2+5fmZvnlLB3p0/eqfgvX/ISxOPxGl5Tf9fJfgZXw/RwtqlX3p/gvT/P8gBbD2EAAIAAQALYAAMXg6GLp+zrRuvy9OxmvDtfCJ1KXvQ/Feq/VfgZVnlfAtR+KHb/Lt+RgsdQxlL2lGV1+K9UAFehTr03TqK6Zm2VVctrpxfuvWL/rqv8AgmQ51HG0/Z1H+8X4+a/X+rBxJg/rGBlJLWGq/X8PyOEsZ7TDyoN6xeno/wDg/mAAwMC1nnDfsnrPl5f+3o7P56N+pVisfnri9ua3yitfvsPhyPEvDNbCwX7+jJypv1WsfSVmu17N7HDeYSwuJeGq6KTtr0l/wdn52AF8B1AEZVltXMMVGhT67vsur/rrYpcL5XDD+xdPmXdt3v38vlY4iyOWWYhKLvCWqf5p+aAepuhHQYFXZAgYAAwWwPYAAYPYE2ndEkqkNBoAZh53XKABXX7xgUZcskYmOqYAwCjLngmAAFaiqi8yUHF2YAAAAUpc0EwAAAAAAAAAAAKkeWTQxAAA9joAD/cZ95N/+lL/ADYD2MP8IBxQn9b/AO3f8yi4qDKHUgrRSCniaFSXLCab7JphjsXDC4edae0V/wAMvmz65X+sPEKVpu7v6/18jL8sxGPqe7t1b/rVmBy+hg6fJSXq+rABbAgABAACAAAAAzfhqnXTqYb3Z9uj/wAn+H5lKtisvxF43jJbr/PujKM8o4+Ki/dqdu/mv8t1+IAYvCUsVRdKqrp/1dGNweIyrFKz1WsX3/rqv0MnzWnmFDm2mt1+q8mEoqSszBN5XnHs2/dvy/J7P8mAA9QMh4kWUYDGucrWg5R/xJWS+ba+44bTnmcZPV2b/A8PMTyY6pRv8Ub/ADT/AMmzxOyT6hntetTVozlf0cve/G/33MjzJY3Cpyfvx0f6P5/ncD7AtgBHAuDUcPUxD3bsvRa/i3+BPFWlZHFeFjisqnJbx95fLf8AC4dAOoG4VdkPYQMAABbA9gABg9gKE7PlK8LO6AOpGXK0xNNXQAYj+Ix7Atir79JMAYBhJbxAAAnCM1ZlXDyhqtUAAAAYV3i0AAAAAAAAAAABiY6pjEAAB0ADiJexx1Ksuy+9P/hhNNaA9ih8ABxTQd6dVea/VfqTpyi7JGWYZ1K9Ol3ev6/gHEeYyUvq0Hbv/kQnKElKLs0ZVjvrmGU38S0fr/wTjDHqUoYWL21f6L9fmgy6pQqYWEqCtG23bugAEHUAAQFgAAGAAAGZZTh8fC01aS2fVf5ryMbgMTl1ZKWj6NdfRmScRRxFqOIdp9H0f+T/AAYAZll9LHUHSnv0fZ/1uRlispxvaUfua/yf9amBxtLGUI1qb0f4PsHF+DtKniY9dH+a/UynF/W8HTq31tZ+q0YAwDivEcmCVNbyf4LX87HCdKU8dJpX5Ytvy1S/NpfM4WxP1fOKEu7t/wCBK36nibllOvikprSpH8U9/loZfiamVY9xqbbSXl3/AFXl6iaaugfwABE4Vio5LTa683/pTX6FWtaqolePtMvqxfWMl+AdA3OoWTAq9B7CAAADoDAAGD2AE1UgSi07AdR7GGqfYYAVnebHsC2KPvU3EYDAKMuWaYAAAVcMpax0ZKLi7MAAAMM7TsAAAAAAAAAAABiI3h6DEAAAtgA4qpXo06nZtfev+AZdV9rg6cvJf5A9ih8CAMwwccXh5Unv08mV8NVoVHTqRs0cP5ZKk/rFVWfRfqGZ4h1K1Sp3b/r7iLcHc4XrNVZ0+6v93/DmJ4RniK0qs8RrJ3+H/wC2HwV/0/8A/Jf/ALYyvKpZfGVP2nMn5Ws/ve4ACB7gk3sKlJnsfM9lE9nHscq7ABZHLF9B04sdNDgxxaB7AAAAYnC0cTSdKrG6f9XRnGSVcBLmXvQez7eT/rUyLiPltQxT06S/R/5/ebgGd5RDH0fd0mtn+j8n+BlOZVcsxLU17u0l/XVFOpCpBTg7p7GbYP63g6lJb209VqjhDF2lUw0n5r8n+gDADiebxGOpYaPSy+cn/wAMeFeWxxmcY6gl7rpTS8nzw5fusQlOhWUlpKL/ABTPEClGvgKGJjsn+Elf9EcVZbz01ioLVaP06P5f1scL5j7fD/V5P3oben/A2+4JfAAEdjhGaqZPGC+y5L8b/qVKF6lzGy9hllWfaMvy0B6FkzRHQAexV3HsLYAAHsADAAGIYBSnyyK8LrmQAxNp3RTmpxugG0lcbu7j2BbGGerRVjabQDAClLngmAAAE6cZqzKtCUPNAAAFJ2mmAAAAAAAAAAAA0mrElZ2AAAEAGfUvaYCfdWf3P/I4bq8+BUf5W1+v6h0KHwIAGWEYnVIqqPIrHDF/ra/wv9AexU+JgAIUG9hUL7sVOKNEh1IrqOsug6rHUk+pzSfUu2XYXZzPuc8u57RiqXOdMumOKY6aexyNIAAACcIVIOE1dPozPMglg261HWn+K/4Hn95kPELoWoYl3h0fbyfl+XpsmpJNMA4lyX28HiaK95bruu/qvy9DhrOPYzWFqv3Xt5Pt6P8AP1DFL+y85518N7/J7/qjRrQHqAGG/wBsz9y3UW3/AOA6L8bHgxiIR4ixCf24NL15k/yTOM8v+qZrNpe7P3l89/xv95iP9u4PjLrGK/8AJHZ/gmVKcakHCSunoTjVyfMdNk/vi/6+8pVYVaanB3T1Q/gACDOBcwjGpUwkn8Wq9Vv+FvuZLDwlK5xpj40MCsPF+9N/gtX+iAsWAB7E9ZDOg9gAB7AAwABiGAFKXNGzJx5ZWAYUqjhITTV0GInywt3B7B0KLtNGJVpgABhalnyAAAAFTCwltoTw1SPS40AACd0mAAAAAAAAAAABiY2mAAB0AAxFJVaMqfdNfejharadSk/J/do/0BbFFe4gB7CRoNGY0HCrOHVNivNqKOF6N6859lb7/wDhgexU+JgFrkaMnuRpRQ2kiVZLYdWTG2wAFsAAIDmaFMTTHFMcOw1YAAAaUlZmfcPPD3r4dXh1Xb/gfl6GQZ+8M1Qrv3Oj7f8AA/ITUkmmAcSZP9Wqe3pL3Jfg/wDJnDucfW6Xsar9+P4rv6rr9/c4vwfPRhiFvHR+j/4P5mQYv6zgINvWOj+X/AsPUAK9VUqUqj6Jv7kcI0nOrVrPeyX36v8AJHAuMlhMzliIbxafrrt81ocd4SGLy2njKWvLZ3/uy/4NvvOCmsXktbCvvJfKS/zuSTTszibLliMN7aK96H5dfu3+84VzFSg8LN6rVenVfr94/gQARepQrVKNSNSm7SWqZkXFFbMqqwzppTtvfTTd2s38r690cVe2Wa1Y1ZXasl6WT0XRagAASY9Rg9gAB7AAwAAQwApy5ZXK8OaN0AwKdWUNtj61C2xUm5u7B7AJ2aZildJgAAm000QkpxTQAAAATpxmtUVcK4q8dQACi700AAAAAAAAAAABio3in2AAAWwABg/9mzqUOjbX36r9AWxSXuoAY9gWqOI8BKNT6xFaPR+T/wCD/W5GCb0WrMmwLwmFSl8T1f8Al8vzB7FX4gtcjRb3IwjFaEqkYkq0nsPUewtgABAACABik0KaZo0OC6DVgAANGrMz/IPYXxGHXudV2815fl6bcP586LWGxD93o+3k/L8vTbcCvQp16bp1FdMxmGxGU41cr21i+6/rRojWpZtl0uX7Stbs/wDgPU4TxLp4ieHl11+a3/D8gAM/reyy6q11VvvdvyOFKPJgXP8Amb/DT/M4P+Ot6L9TharHNchlhKj1inD5W0fy6eh4fzlQxuIws9JW284uz/Mz7D/V8zr0+0n9zd1+DGk1YzGhPKsxU6e17r07fmvQoV4YjDwqwejVwB7AjhOr7POKD7tr700cdUuTNm/5op/mv0AAAqPQOoMAAYIBgAAhgAUZc0eVlSHLKwhgADAKvvUUwAAMNV5XyvqAAAAAYigmuaO4AGGd6YAAAAAAAAAAAE480WhoAAFsdQAM/i8PmEK68n80/wDhhNSSaEU/hQA9xlhEoxknGSumUcswdGpz06aT/rbsA9ir8RGi5bkYRitCVWK8yVSUgOg9hAACAAAAFsCbWwprqaNDh2LAABurGf5F9Wbr0F7j3Xb/AIH5HDme2tha7/wv9H+n3dgLozjLYY/DuH2lqn5/5PqZPmE8txThU0i9JLt5/L8jM/8AYM3VeHwtqXye/wB+v3iakk0AHFtTlwcYd5fkn/wDJqXs8vpR8r/fr+pwf8db0X6nAWYewzF0G9Ki/Far8Lr5lSh9Q4uhNfDWT++zv+KT+ZnHD6xfEE6k1+7UFOXyTSXz5fuuHEGX/W8I3Fe9DVfqv662OE8wtzYST84/qv1+8AYQd0ZLU9nmWHl2nH80eIVO2Moz7xt9z/4IAAMqO7BbgwABggAAHsC2GABCXLJMqxU4XQthgADAI+9RaB7AABQq88bPdAAAAAGIpqE9OoAYV6NAAAAAAAAAAAAGIhyz9ToAALY6gAcU0ebDwqdnb7/+GRlFf22Cpy62t92giGyAOo9WHU6gByK9yc4xJ1JSABbAgAAAAFsAHUATa2FNMaTJRaAACUYyi4tXTM+yV4KftKS/dv8AB9v8jh7O/rEVh6z99bPuv81+K+YBxRlXPH63TWq+L/P5dfL0KuK9vhI05v3obf4X0+TtbyfkcPYr6xgIXesfdfy2/CwAcYTcp0aa8399iioQgqcX8KSOD7Kdb0X6mWZgoVKeKoO/K7rzs/y0+ZmOWxzRYXFUZ2cGpp23Ts2vK+ncx6isLVl/df5MFsZthZ5ZmKqUtE/ej+q/T0MLiIYmhGrDZoBhF2ZhJcteD7NfmeIsVzYZ/wCL/wBtAACbsrgxAwABggewAA9gWwAAUJ3XKVYcshgADAMPrFoHsAAEJOLuinUU43QAAAAGMXupgBhXq0AAAAAAAAAAAAYmF4X7C2AAEMADNqHtsFUh1tf7tf0OFq/NRnS7O/3/APDCEDBD3LHU6hYCrUadkAAIOoAAAACABgAKTQmmShfYaAAK1KFam6c1dPdGa5ZVy3EJxfu7xf8AXVGSZvHHUbS/iR38/P8ArZgSipJp7MzvLHgcQ1H4Hqv8vkcJYrlrToN/Err1X/A/IAM9/fZzRp/4V97OJalbCYmliqMrNpp/LXX7yliatKnOnB2UrX+XT/Myuj7LBUodor792cD476xlSpyfvU218t1/l8jNdcBX/wAEvyY4sOIsv+t4FuK96Gq/VfNfikcJ4/WWFk/Nfqv1+8ACEno1ueIiXssP6y/QAAZOV2D2EAAAwWwPYAAewdAAAUnF3ROKqQ0GAAMQGGfvNFRWk0PYAACnUcHdFOpGaugAAAAxj91IAML8YAAAAAAAAAAAA0mrDi4uwAAhgANJqxk/+y5pKg9ndfdqvy/Ejq0IGAwN2HQHsVviQAAgYAALYAAAAQAAp33HFMcWgADG4Oli6LpVFo/wfdFSnicpxi196O3Zr/JmX4+ljaCq0/muz7AZpl8MdhnSe/R9n/W5g6s8FjYSkrOL1/J/gJpq6AKq9rxGl2a/CNziih7XL3JfZaf6fqYel7atCmvtNL73YSSVkcCY72GYug3pUVvmtV+F18zOv+RZiLfyT/8ASWWHFMfwGaUJ5XmSqUtr8y/Vfp6GHrwr0o1YPRq49gCLszxEdqWH9ZfoW7AFhsqStogGLYAAAFsD2AAGD2AABlCf2TEU7O6AAGIDDu0yurTY9gAACM3F3TKWIjPR6MAAAAxFTnnp0ADD/wARAAAAAAAAAAAABiYWfMAAIewABnkXhcyjXj1s/mtH+X4lFqTi1sLYGdToboFsIOgFbdAAAMWwAAgAAWwAMAAIysaNEoW2AAM4yuGPocv21s/09GZZj6uWYpqa02kv66op1IVYKcHdPYDivLkrYuHo/wBH+n3HDOZ/WKP1eb96G3mv+Bt9wBgF7TiCpLs5f5GJw6r4edN/aTX4HD2HdTM6cX0bf3L/ADDB4mWGxEK0N4tP7jOa0auTVqkHpKDa9Gv8gaGrxOJMA8Tg3KK96Gvy6/5/I4Tx6lTlhZPVar06/j+YwClFTqRj3aPEedo4aPfm/Dl/zEy5dF0N3JzS0QAwewAA9gAYAAwewAAMvbYTVWBKLi7MABiApO00Yle8hgAAAQrzhpcp4iE9NmAAAYiukuWO4AFD+IgAAAAAAAAAAAAqQUotDVtAAEPYAA4nw/Pho1VvF/g/+DY4er+3wlN9Y6fd/wACwtgYzoWuWOh0ACvugAAHsIAAQAAIANxAAAm0JpolDsAAcSZP7en9YpL347+a/wA1+Rw1m3sZrC1X7r28n29H+fqBmWF+s4SpS6taevT8TCYqpha8asN1/TRhsRTxFGNWm9GgOH17TOK8v8X/AKUhK5gKNLA5riatR2jHb/t53VvkYbPcHXqezTafS/X+vMOG6scyyCWGm9UnD8NH8k/wK3AeH9janVfP52t9y1X3v5mKw1XDVpUaqtKOjQ9iSMVCeUZpzRWid15xfT80QnGpBSi9HqAZXT9pj6EO8or8UeJE06+Hh2Un97X+QpNHMmXQ5xRKbYAbsGAAPYAGAAMGAAA9ijU5ZeRiKd1zIAAQCdmYpaJjAAAACGInDTchiYS30E01oAEpxirtlXFN6RAAKXxoAAAAAAAAAAAADEQs+ZAAB0AAx2H+sYadLuvx6ficH17VKlB9rr8n+h0Bj2Og0XDoAFfoAAC2BAAAAAAAMAADYjJMlG+wABxHlH1Wr7ekvcl+D/yZw9m31yj7Oo/fj+K7/wCf39QM5wv1fH1IJaN3Xz1/4BwtmXs6jws3pLVevb5/n6hucKrmx1aXl+bDPa8auNm47LT7v6sRqRk/dZkuMeKwicn70dH/AJ/ccG4JYfLvavebv8lov1fzI4iLlY47wKtTxUV/df5r9fwBo4qwPtsMq0d4fk9/8/vOF8d7bDOhJ+9D8n/l/kAcL0fbZxh49pX+7X9DxCq8+aQivswX5t/5AwAAQMAAYIBgADEMAAOo9goVOaPK90VqfI9NgAFsAV9aaGAAAAAJtbMWIqLqfW526Dxc/IliKj6jbe4AAU/jQAAAAAAAAAAAAEoqUbMlFxdmAALYAAjL+z87T2i3+Ev8r/gfXU8wWFT2jzfO6X5AAWRZAAY2rCjSdSbskYasq1GNRK3Mr/eAAIOoAALYAAFsADVxAAAEZX0JRvsABiKFOvSlSqK6ZWpYjKMcrPVap91/wdmYLF08XQjWp7P8H1QcX4a0qddej/NfqY+jKn7LGUtp6+kluvv1X/AMqx8cbhY1Vvs/Xr/mYrOY4LGqlXXuSSafbo7+X5fllGbQwDrVbXlJaL59fJfiZdUqPAwqVXeTV389f67GLqOWr3bLODTOFqn7ypDuk/u/4cwcFRy2jBdIx/JEa96ljiyCqZNOT6cr/FL9QauitTjOLjJXT0ZhpyyrNeST929n/hez/Jm4HAVD2mbqX8sW/wBP1OMayq51Wa2Vl9yV/wAbgwAB7CBgADBAAAAhgAAMIycXdHu1YEouLswHsC2AKutFfIYAAAAAAAAAQ+JAAAAAAAAAAAAAFelzK63AA6AABxThdIV16P8ANfqZDjqmIziNSo9ZJr7l/wAAAAAxGfYKjU5LtvrZaL+vK5QxFKvTVSm7pjskcRZz9cq+ypP93H8X39O33lHko0YRk7WSX4EsXh4q8qiS9Uf2jgv+f0f/AAJf5izHBt2VWP8A4Ev8z6xR/nX3oBAwAAQFrnKwBAAMAAAIyvoSjfYADO8rWOw9o/HHVf5fP87HD+ZvBYj2NTSEnZ36Pv8Ao/8AgBn2G9vl9RLdK6+X/AuZLSjmGW1cJJ6xd15X2/FO/qcP42eCxjoVdFJ2d+j6f5M4xw16NKsuja+/X9PxKdN1JqEd27FSmo4dwj0Vl9xiN4laScVY4Wi/rMv8P6ow0va5dSl3jF/giFG1bmOKpKGSzi+vKv8AyZP9AJK5xZgbxjiYrbR/p/l80ZDjfrWCjd+9HR/Lb8APDjDaV678or8W/wBDMcR9ZxlWt/NJv72AAAxbAAAMED2AAHsCGAAAwKNTkfkVqanG63GgewLYAqfwUMAAAAAAAAAI/EgAAAAAAAAAAAAAr0re8gAWx1AAexnGOxtSpKhX0Sey28n3ZkdTkzGi/O336fqAFrliwZ7jXhsLaD96Wi/V/wBdyVSMXZs4exro4lU2/dnp8+n+RxRnfKnhKL1+0/0/z+7uBGMpO0Vdn1at/I/uY8PWSu4P7mEZSi7xdmU8xxlO3LVl97/IpcRZhTteal6pfpZlHi2a0q00/R2/B3/Mo8SYCro24vzX+VyjXpVo3pyUl5O4AAEY3Ektiw4pjpocWgAAXYAAAIyvoSjfYADijKlCX1qmtH8Xr3+fXz9ThvNPrND2FR+/H8V/wNn8hpNWZkjeCzeWHezvH9U/nb8TirLuSosVDaWj9ej+a/rU+srM8imm7zgtflrf5pffcyKh7bMaMezv92v6Bj8M6dWdLrFshTlKSujheg1CpVfWyXy3/NHCWMjicsjTb1h7r/T8PyI4RKV29DjvGJUqWGW7fM/RaL77v7gHsY7DRxFGVKW0l/X3HD2IlhMe8PU05tPmtv8AL5gZOllvCs6z0lKMpfN6R/QHsAADB7AAAC2B7AAD2BbAAALYYBQq/ZZXo83vLcHsHQAqfwUAAAAAAAAAAt0AAAAAAAAAAAAADSaKtPkfkALY6gAGbZOsfQcofxI7efl/l5mFcqOKg3o4yX4MrVqdGm51HZIp8R4Kc1HVebWn5/oJpq6AA4pq3rU4dlf73/wCzndmDquElJbxZmXClDEN1MPLlk9bPVP9V+PoY7LMVg5WrQt59H8zIalOrg4SSXMtH8v+BYAJ4PDTVpU0/kipkuX1N6S+Wn5WKvC+Cn8Dcfn/AJlbhKov4VRP1Vvyv+RXyLH0Vd07ry1/Ba/gJ1KU7ptSXyZheIsdQsnLmXn/AJ7/AH3MJxPhK2lVcj+9ff8A8Ap1IVIqUGmvIAOhFaFgsixYtc5UNWABo3AAACMr6E49UAFWlCrTdOaunoyvSr5Rj/d6arzX9aPzMLiaeJoxq03o/wCvwOI4SwuZQxEetn84/wBIrYeljcK6ctYyX/BT/UyutLLswlRrfC7xl/n/AF0OFsHKGY1Ob7Ca+d7f5hxBlU6kvrFJXfVfr/mYbB1sTU5Kcbv8vUweFhhqEaUen59WZFnNTLMRzpXi9JL/AC80VuM8sjR54Nyl/LZp/NvT8WZjj6uOxMq9V6v8F0SAJrQ4nwssPi44iGnN+a/pGBxUcVhoVl1X49fxMLh5YmvCjDeTSXzdjjqvDCZVSwVPq0rf3Yr/ADsDAADdgwAB7AdAYAAw6AAAzoMAKNXmXK9ytRv70RgAVIp0rLoAAAAAAAAAAtwAAAAAAAAAAAAAJwU42ZKLi7MBDAAMPszifJfaJ4ugveXxLv5+q6+Rn+YOvKFKL0STfq1f8vzFVhJ2TOG8Y6tB0ZPWO3p/wP8AIADilXxUf8P6sotKLKHUgvcRioRnHlkrowuCo4bm9krJ626AAAtgrYWhXXLVgpeqMVwthamtJuL+9fjr+JjOHsbh7tR5l3Wv4bmHxWIws70pOL/rdf5mB4qTtHEx+a/Vf5fcUMRRxEOelJNeQEVd2EAAWQ0NWADYAAACMrk421QAZ/lixuGbgvfjqvPuvn+Zwxmfsav1ao/dlt5P/g/mcV4b2mDVRbxf4PT87HDeI9vl0NdY+79234WOL8tVli4Lyl+j/T7jJ8yWGwtare9STS+5PX+uo8diXPn9o7+rMnzH67Q974o7/wCfzAAQ1oElczzBfWsFOCXvLVeq/wA1ocJ4y6nhpPzX6/ocA5d9YzF4hr3aa/F6L8Lv5HHWP+s5q6cXpTXL893+dvkAAA9hAwABgAwABg9gAAewAAUqykrPcrUFNXW4007MASu7GmxONpNAAAAAAAAAAAAAAAAAAAAAAAAVqXOtNxoEMAAw/UaM0qc1ao13aXp/ww4NK5w1VaxiX8yf+f6AAcU0HanVXmv1X6k6coyskZdhXVrQpd3r+v4BX6B0AAEDAxmV4TFr97DXvs/v/wAzH8L16V5UHzrts/8Ag/1oUa+IwlW8G4yX9ar/ADMu4np1LQxK5X36fPt+XoRlGUU4u6Ka1OgAANXJK6AA2AAAATUkSjYAOJMteFxCr09Iy/CX/B3+8wWIjm2WTi/js0/Xo/1ODcTaVWg/Jr5aP9BSo5hQqU7e7dx+7S6+e33lXBywdSdOpuj2s27nDmI5MZFdJJr9f0CwAh7A9ia1MdQqZLnC5lZaSX+GSv8AkzIqFPI8hdepu1zv1a0X5L1ZWqzrVJVJu8pNt+r3HsAADFsAAAwQDAAGD2AABgAAUq9/dkVaMZrzJRcXZgUY800SnaqkYmNp37gAAAAAAAABuAAAAAAAAAAAAAAFelf3kCHsAAYfqMzFNVJf4n+pOSdNI4bi3jKfkn+QAGOwkcVh5UpddvJmIwlbD1PZ1I2f5+nc4fyqdJ/WKqs+i/URX6AtgABAxbAGNy3DYyNqsde/VfP+kZlw9iMKnOHvQ/Feq/VfgZdnGJwTSg7x7Pb5djKs0w2Nhem/e6p7r/geYBYsiyLANElZgA1cAAABOxpJDVmAY7BwxeHlRn1/B9GZRjKmV49wq6LaX+fy/IxVaeV5tVlT87f9vK6+66+44Sk5Zd6Sf6GfNvFVfUUF7O5kjf1ql6gAAGW4CWOxtLDx+07P03b+SuzxeyGrUxM8ZRh7lJQUuyT91fivxMu4vnmXDGGwDfv0/dl5qPwf+S2v5oGAAD1B7AAAwQAAAxDAAAewAAMo12tJbEoxmieGkvh1PYzfQpUuRXe5Od58yK0FUhdAAAAAAAAAAAAAAAAAAAAAAAABXpcrutgOgAGH3YtUZ5hvZ4ypHu7/AH6/mKlNuzOGKHNXnU6JW+//AIYAAB7FfdAgAAHsIADNOHqGKvOl7s/wfqv1X4lWjisvrpSvGS2f+T6oyXiWGJtRxHuz6Po/8n+H5FrmgAXLdgktAAGrgAAARdiSUkAHFmWXisXBbaS/R/p9xVrVKrTnK7Str2Rwc74Gav8Aaf5I4gotYyou9n+COd8tjIaLljKUfO/3ajLXNgADgDLearUxklovdXq9X9yt95xRSwdTJ8TTxXwSi0+92rK3ne1uzOHK8sLj5YeenNdfNf00AAALUGAAALYHsAAAhgAB1HsAAAiFSUNiNeEt9D2kO5Wrpq0RmGqacrMRQ+1EAAAAAAAAAAAAAAAAAAAAAAAAaTVmVabg/IEABQ+MRnuVyxcFOn8cfxXb/Ip4DE1J8kabv6W/4Yy3AxweHVPru/X+tAAA3HqV/iBAAAtgVOTHTkNWADF4DD4yl7OtG6/FeaM4yKvgHzfFDo/8+35MyPiaVK1DFO8ekuq9fLz6emyakk07oAA3JIAAauXAAACEuhOPUDcqUYVqcqc1dPRmY4KeDxMqMum3mujODWnhai/vfojibBuSjiIrbR/p/XoeyhzXscM4N+9iJei/V/p95uXRo0IDLsBVx2JjQpbv8F1bKXBmVxo8koty/mu0/klp+DM8yWplmJ9m3eL1i+/l6rr8n1Mhy9YHLqVFr3rXfq9X923yOIMojmmBlh72lvF9pLa/k9n633OKMFVy/M/aW5ZX+6UdH+hQrxr0Y1I7SVwAAewgYAA9gOgPYAAewLYYAADAAAWwAMTtsUaymrPcr0Le9EAAAAAAAABatAAAAAAAAAAAAAAAE4KSsyUHF2YABR+NC2N0AAAdB7FbWQAAAtSMEixZkopolFpgLVoSuVIRnFwkrp9DPeG5Ya9fDK8Oq6r/ADX5fiZDxBLBtUazvT/9J/4HdfNEJxnFSi7pgAEh7gADVzY3AAAIu6JRswjuLY4ry32+HWIgvehv6f8AA39LnCeLo0KFX2sktVuQqUMTTfK1KL0Hw3gufm1t2vp+V/xIU4U4KMFZI6AjqBwJglGjUxL3b5V6LV/fdfcSxSUrIznD4SrQhXxHw02pbX26ej6mL8QqS0w9Fvzk7fgr/mYzjbOK91Gagv7q/V3f4nFNKpjcJOrOTlOOt27vTR6vXb8jhTEe2w0qLesH+D/4Nz2XmOk+jHFoAGdAAAGADAAGHQAABDAAHsC2ABgm07oo1lNWe5Ww99YjVgAAAAAAACGskAAAAAAAAAAAAAAAFSmpqxJNOzAAg7SQtgaAALXB6jJ6tgLYAApR6iQXGrkopqw1Z2CO6FsG5xBw5y3xGFjp1iunmvLuv02yDP5YOSo1nem//Jf+B3XzXnnGeewtSw7Tk1e+9r7W/qxgeIq8aiWIfNF+VmvuE1JJpgD2JbgAAWLgAAJ2NJIad7EEFeUIUpSn8KTv6FaUZ1ZTSsn07JaL8DLMfLCV1NP3XuvL/gdC6auhjAYtQ4VioZLTa683/pTX6FStaqomLSqZZWi/5ZfkS2JK8h04yi4vW5lMpZdnDoSejbj9+z/L7yxYauTp22AN2D2AABggGAAMHsAAB0GAAPYOgAMBO2xSrqWktyrRjP1KlKUNwAAAAAAAKavNAAAAAAAAAAAAAAAAVaSmvMatoABHVBvoAALYYSdkAIAC19CKSVgAauTjdBD4kLYA4j4f5ObFYdabyXbzXl3XT02pV3FWKVZydmZBiHWwUU946f5fgAPYnubgAANXLFy5dGhoXRc1ZB2JxuiK90ZxDV5MC0vtNL9f0K7cpcvYoNp8plNV1cDTk+1vu0/QYxgwOE5Kpk0IrpzL8W/1KlC9VMxrVLK60n/LL8h7FtSxxfhXTxFPEw66fNbfh+RgMUsVhYVl9pfj1/ENBqxUjZg9hAwAABAAADB7AAA9gAAGAAABTxDjo9URlGa01KmGjL4dCdOUN0AAAAAAAUVeogAAAAAAAAAAAAAAACrSUldbjVtAAou8EBuNAAh6hVfugwQAFNXkJD1ABoqRsyHxC2AM1k44KrbsypTSqWROHJJHC0rwqx7W/X/IAKnQAAAAsiyLFiyLICDurFkM4ov9Vh/i/Rkbe1dxW9rocP8A/Ivh8/zY9h7BuMYcCY5WqYRv+8vyf6EsNGUrnGeNjQy72CfvTdvktX+i+YxBxDhPrOXzS3j7y+X/AALmR559TwbpyjzO+nTR7/j5dTLc6o42XJbll27+gPVFRXQMWwMAAewC2B7AAAIYAAMAABgAMAAUnF3TIYrpITjNaalTDJ6xJRcXZgAAAAABQV6iAAAAAAAAAAAAAAAACrSU1dbjTTsAYd6NAFyyZYshsBlWV3YGAAFFbnQAAnG6sQ+IWwBmcOfBVV5P8CvpJMcnOaRwtC1KpLu0vu/4cAKnQEAAAAAAQj1BnENH2mBk19lp/p+pXi4y5kUItvmMpo+ywVOL7X+/X9R7G40JG7NQwuJq4atGtSdpR2KXHlD2N6lJ8/ZWt9+6+5ma5nWzHEOtV9EuiXb+uoxBi60aNCdSWqSZOUacTD4hxmqlN6pmHrRr0Y1F1Vwkho3B7AAAwAYAA9gQwAA6j2AABiAB7AAARk4u6ZTxKekiUYzWupVoOGq1QAAAAABhleYAAAAAAAAAAAAAAAAFSkprzGmnZgUHaVhbAAAFWSirm4PYBbABSXujLXHsWAGjltMWwBKKkrMx2DdGrKjPo/8Ahn9xToqHqZNhXh8HGL3er+f/AALABU6AJgAAAAAKTQnc6FWnGrTcJbNWMdg6mFrOlNenmu5lWXyxddRt7q39P+CWSVkBoFjodABiDiKbjgWl1aX6/oVbzqWKa5aljh+blgILtdfiEipo2IGAADBAMAAewLYAAAYAACAB7AAABTqyhsU6sZrQrYdPWI1bQAAAAAMKveYAAAAAAAAAAAAAAAABUpqa8yUXF2YJ2dyLTV0AADZKSirsqpThdCABABTWiGD2AAa1FsAGaZRTxqUk7SXX/MwPDcaVRTrS5rbL/MACp0AEwAAAAAEBXw1GvHlqRTXmUqNOlHlpxSXkBZFiwAAxBxIm8Fp0aG+WqxPmqpo4cjbAp92wexV+IWwMAAAQAAAwewAACGAAAgAYAAACbTuiliFLSW5Voxn6k6coOzAAAAAMKvdbAAAAAAAAAAAAAAAAACdNTVicHF2YUJ3XKLUA0GxsnWW0RtvcoSuuUnHllYBggAjsA9iw9h7AMWwAAAFTZABcAAAAAEbjQAAAAgQZrh3Xwc6a3tp6rUrUnLVblGk46vcy3DuhhKdN7pfi9WD2Kq1QPYAAHsAtgewAAMHsAADOgwAB7AtgAYAAAAUsQ46SPdnHuiphU9YkouLswAAAAw6tTQAAAAAAAAAAAAAAAAAE4KSsydNxYm07ohNTWhcuDlFbslWXQlJy3YEJcskytHmjdAMAAjsMHsbjSLKx0Bi2AGblhABU2QAFwAAAABCHsAGaZ5WrVHCjLlgu2jfncyfOqqqqjXldPRN7p+vYsAB0BAZtklWNV1KEbxetluvl2MoyOrKqquIjaK6Pd/8AAA6FXdAwAAYAMAABDAAB7AAAMAAYAAAAEZyi9GQxMXpIajNdyphmtYjQAAAEI2ikAAAAAAAAAAAAAAAAAASipKzKlJw9BNp3Qq8rE60nocze7AAKErxsThyuwPYBAEdg1HsALYHsLYAerGXGjcAqbIAALgAAAAhDAM5rujgZtbvT79PyK1Vxdo7lGo5aPdGAr+3wtOo92vx6gAdB7CDcAOhV3QMAAGCAYAA9gQwAAY9gAAYAAAAAAAEKkobMp14z0ejMR71SyKlNwtcAAApR5ppAAAAAAAAAAAAAAAAAAANJqxVpOOq2EMAAIS5XcqRU43QDBAEdgHsHQAewtgHsG4boWwBU2AAAuXQAAAXEABxO2sHH/EvyZy89VoiuWrYyBt5fD5/mwAGPY6FkM3FsB0Km49gAAewCAAAGHQAADqPYAAGIAGAAAAAACbTTK9PnjddAAADCw1cgAAAAAAAAAAAAAAAAAADcqUbaoYLYAAoTuuUqw5WDAAhqkAZRwhicfRVapLki9tLt+drrT5mc5Bicsmvae9B7Nfk+zAHsLYB7HQAQBU2AAAAuXLoui6LouXAQdAQ0cSU3LBX7NP8ANfqT9ypco3nUbMjpuGAp363f4sAED2OgM6C2NWD2J6yGAADAB7AAAwewAAAwAAEAD2AAAAAAAovmpor0L+9EAALXKcOWKQAAAAAAAAAAAAAAAAAAAFWldXiCAAE2ndDSqQGraD2AWwFPWKAyPL1jsfTova936LV/ft8yU4U0kZlg6eYYOdB9Vp5Po/vJQcW4vdAPYQPYex0AOoBU2AAAAAAAI7IBiGYzDrEYedJ9V+PQrUNXCas0YXCupUjSprVlKnGlTjCOyVvuADoDDYZ1OgEhsAABgAwAAEMAADoMAABAAwAAAAAAMLK8WgrYdS1juNNOzAAw1K75mAAAAAAAAAAAAAAAAAAAAFWjfWO51AAChOzsVofaQAgKL90DgSipYurU7Rt97/4Bja1pMwNTmVjP6SpZnXiv5m/v1/UB7CAex0AsAFTYAAAAAAAhsBugDPsfCpmE/ZpWWnq1u/09Ejhas55hZ/yv9AAtceobsHsMZ0AqOyGAADAQDAAHsCGAAD2AAAewLYAGAAAAAABh3adgKlKM1ruTw847agJNuyKeG6yLKwAAAAAAAAAAAAAAAAAAAAFWkparcaAAEU5KcbMqQ5XYYAUXugOApJV60erS/Bv/ADMxpOTaRl0GkcSyU81rtd/ySQD2EAxjtY0Hqx7AFTYAAAAAAAg9bC2AY2VIuUGk7Pv2M9yXDZfRg4TblJ9bWt6W9OpwzCVLM4xmrNxf4q6/AuAdAewloM3BltDUZUd2MAAGB0B7AAAwWwAADHsAADAAGAAAAAABF8skwAlCMt0exp9hRS2QAAAAAAAAAAAAAAAAAAAAAFSmpLTcasAARlyu6JRU46EkAtgpu0kLYOEMWsPmkU3pNOP6r8UkYjDubuiEY4ek5Sdkld+iMXiHiMROs95Nv72BIg7pAMsho3EgAqbAAAAAAAC0EwHsSlbRD1OKJutjaVCPRfi3/wABEsvisVSrw3gmvlbT7v1FNML3Eh6g9QYAE5WQwAAGADAABg9gAAOo9gAAYAAAAAAAAAU3zQTAAAAAAAAAAAAAAAAAAAAAAAAKlNSWm41bQABbFKdnZlandXQCAi7pMIycZKUXZoyfinB4uilXmoVFvfRPzTemvbocT8TUK1J4TDSun8Ul27Lvfr06ddM54bxWXfvF79J7SX4X7fl2YFR2TKXwoAAACpsAACi2ez8z2fmODQAAAEJdC5uVJW0QGHX13P23sm//ACVafikcqsSh2FNojJS2L3BsDdAA2TlzMeoAAPYBAMAABDAAAQwAAYgAewAAAAAAAYZ3hbsAAAAAAAAAAAAAAAAAAAAAAAAFSmpLTcasACClPmVnuVqfK7rYAKT0sbgHDleOMyejz6+7ytPy01+SM+4IUr1sv0fWH/yL6ej07PoVac6UnCatJbp7oq7FL4QAsAAVAACMerLXOW44jRKCY4tAAAAptEXdXY3dhiqyoUJ1X9lN/ccH0XKtVrPtb79X+SAnC+qE2noKaY2iVRbIhKzLlxtF0XG9Cc+bRD2AAAYHQAAAewIYAAPY6DAAAQAPYAAAAAAAMK90AAAAAAAAAAAAAAAAAAAAAAAABUpqSutwABNp3RFqcSpBwdgQJ2dxNNXQB4fYpTwtWg3rFp/Jr/gfiZrj/qODniORy5Vsv627vojMsxr4/ESr1nq/wXb5E5czKWwAEqU4xUnF2fWwAFXoXACMbsSuDAaLDSY4JjgwAASu7E3aNgOJ8R7LA8i3k0v1/T8ThjD+xwMX1k2/0X4IAnC+qACM2j2kWc8e46kR1V0RKTYPUAAB6huAwAAYdAAAGAAA9gWwAMAAAAAAAMM7VAAAAAAAAAAAAAAAAAAAAAAAAACrTvqgAWwQk4u41GpElFxdmAU520YmbljgXEqlmns2/ji181r+SZRzHDYjFVsGn70LXXdNJ39NbPt8zi7hepgJvEYaN6L3/u+T8uz+T8yl1ADg/I6eKk8VXjeMdEns339F+foVaNOrTdOcU4vo9jiHKf7Nxjpx+B6x9O3yACr0BMAjGysGyAAsiyLDSZKNgAKcepUd2BxTVdbGU8PHdL8X/SKFNUaUacdopL7gAnBPYaAAYkD2AAAewCAYAAMHsAAB1HsAADDoADAAAAAAACk7TQAAAAAAAAAAAAAAAAAAAAAAAAAVafVACAhU5H5FSCnEaadmboCFS2jE77F2ZVi1hMdRrt6Rkr+l9fwMfnVennNXF0JWkpO3pe1vS25kWe4TOsLpbmtaUX+Pqn/w5n/ATk3Wy7/wB/8Atr/R/f0J4erh6rp1YuMl0aszoAZJQjhcqoxS15U/m9f1IYl8+5x1hozwVOst4yt8mv8ANIAZV3AuEVdiEh6gAANXGiUbMASvoO0YgNpIy2+Pzh1nsm5fJaL9Ai7xQATgpDVtABm4D1AAAYAAAAwQwAABgAAwABgAAAAAAAtGboAAAAAAAAAAAAAAAAAAAAAAAAAKtPl1WwAD1RSqW0exVpqa03NnYATa2PaS7jbYYPGV8JWjWoS5ZLZ/1+RkPHWFxaVLGWp1O/2X/wDI/PTz6GLwOEx1NRrwU10v+j6fIxfAOAq3dCcoP71+Ov4lfw9xaf7qtF+qa/LmJcDZsnZKL+f/AACvwlnFGEpype7G7b5o7Lrvf8B2WDgk76L8ilOTrNHFiTyaX/bv5oAexU3ACmtA6A9gHsABuSjdWAKcepUd3YGzPMT9XwM5X1ei+f8AwLnCmF5aM6z6uy9F/wAH8gpvSwAEoqSGmtAHqJAwAAAEAwAAYLYYAAM6DAABiAAAAAAAAAApu8EAAAAAAAAAAAAAAAAAAAAAAAAADSasVIcrAWwDKNX7LKtLmV1uAAAZZn+Y5fph6rS7br7npr5WZg/EOqkliKKfnF2/B3/NFHjzK5r31KL80n91m/0I8Y5K0n7a3/bsv8jH8R5VVwVWMK6bcXZa9UZdUWKyylNdYr70rP8AEp0Lz0ONqqp5ZGmnrKSX3Jv/ACAGtCp8QARVkkDB7AMAAauiaCMeZ2JtRjoMbuHFeJc6sMPHpq/V7f15mAwyw2FhS7L8ev4hB2kAASimiUWhiVwHqAAAwFsAwAAYdAAAGAAACAB7AAAAAAAAFB3poAAAAAAAAAAAAAAAAAAAAAAAAACcVJWJJp2YIBhSq/ZZUpqS03GraAAAU3eKAOCc0hOi8FN+9HWPmnv9z1+fkxQindLU4wzSGMxipU3eNO6+b3/RfIC1yWxP4mARV2hCAexcWp1AAETjdWCMVFE5czAlJRi23ojL4vMc29rLa/N8lt+iAE7oAAaTROm1sAwAABgA9gAABDAAA6j2AAB7AtgAYAAAAAAABhXeAAAAAAAAAAAAAAAAAAAAAAAAAABVp8yutwAApVukidNTXmSi07MAAKT1sLVAUqs6U1OnK0ls1uV+KM0r0fZSqaPeySb+a/SwFrjGS1YBTV2LY6APYB7gABJEYWdyrO+iAOJMZ7DBOCes9Pl1/DT5nC2E9nhnWe8np6L/AINwCm7qwAAE6fYYAAA9gEAwAAewIYAAAwAAYAAwAAAAAAADCPRoAAAAAAAAAAAAAAAAAAAAAAAAAAK1PqgQDQFKrZWZKKkicHEAATs7kWaMsBYshsJABSQMNB6oDQYAA9ipLlWgAZzWlmGYqjTd0nyr16v+uiKNGNGlGnHZKwBB2YAABVhdXQAAA9QOgG4AAMOgAAD2OgwAAYAAwAAAAAAADCv3mgAAAAAAAAAAAAAAAAAAAAAAAAAAKtPl1QA1cAhUcfQTjJE6LWqAAKUrqxuFy5cAm9GAFJaAwsiyAsWuAAFVXQBm2N+p4SVRb7L1f9XOFsE51ZYmS0Wi9Xv+H5gARd0AAATjyuwAAPYBIB7AAAA9gAAGAAACAAAAAAAAAAMM/wB4AAAAAAAAAAAAAAAAAAAAAAAAAAA0mrFSHKxbANALYjJxehCopepOkparccWnZgBF2dxNNaAADKr0sAFP4UIAAbLgACJq6AOI8ZLE4pYenqo6esn/AFb7zLsHHCYaFFdN/XqABTetgAACtG6uAAD1AS0AYAAPYEMAADqPYAAHsC2AB7AAAAAAAABRdqiAAAAAAAAAAAAAAAAAADHYuOEoutJXStf0btco1qdamqlN3T2YAAAAAAABKKkrMlFxdgGroACFbpIlGMkTotbABTnbRiYADZN8zACmtEdAAGWuWQi10ACJDWoZvj1g8LKd/eei9f8AgbnDOBdbEPET1UdvV/5fqgADYTugAAJq8WAAPYDcB7AAAMOgAAAhgAAw6AAwAAAAAAAAp6TQAAAAAAAAAAAAAAAAAE69KDtKST9T61Q/nX3ox86FfC1KSmrtNbre2hlWcV8vqe7rF7r/AC7P+mYHHUMbSVSi7r8U+zAAAAAAAAJwUlYaa0AaAWwEZuOxGpGROnGRKlJbagEKnRhdGg2hk5X0QAENkdAAHqMuHQYASJr3mGc42WYYxU6WqTsvN9X8/wAjAYOGEw8aMem/m+oAAU30AAAAAHqAloAwAADcewAAD2BgAAwABgAAAAAAAAtGgAAAAAAAAAAAAAAAAAOK/wDkYv0QBkEMdLFJ4V2tvfa3n+nXsAAAAAAAAFSHMtNwGtAAAjVa31IzjLYlCMtyVB9GSi1uEZuIqkWXT6kppE5uWgmABDZHQADqPUsIQ9QAZU+I4jzP6vQ9hB+9L8F/wdvvOF8tu3ipryj+r/T7wAAFKzuaNAAAPcAGBuAAAAwQwAAGAAACAAAAAAAAAAA3QAAAAAAAAAAAAAAAAABmVf2GDqVOyf39PxMnyOtj5820Fu/0Xn+CMJg6OFpKlRjZL+rsAAAAAAAACpC+qAaAQAEaskKtF7iaa0JUosdB9GezkuhYGrgmAEdkdACwIBAADMyxdPCUXVnsvxfYw9Gtm2O97rq32X9aIpUoUoKEFZLRAAA9RK5Td1YAABgA9QEgGAAAw6AAAA9gAAewIAHsAAAAAAAAAR+FAAAAAAAAAAAAAAAAAAGMwlPFUXSqfC7X87O9inThTgoQVktkAAAAAAAAAFSFtUDWgAAAJtbCrSQq0XuKUXsxpMcIvoOlFjop9T2Hmexfc9i+57F9z2PmJWQAhoEAhAAVJxhFyk7JGdZpPMsQowXurSK7+fqzJ8sjgcPyv43v/l6IAAY9SwQdmAAAAMC1wDcAAHsCHsAAAzoMAAGHQAGAAAAAAAAAQ+FAAAAAAAAAAAAAAAAAAAAAAAAAAAADSaJw5WDQCAAABSa2Yqsu57ZntvI9sh1ont4nto20PbLsRlzItoAh7B1GHQWwAaI4jz761J4eg/cW7/mf+S/HfscO5M6UViqy1fwrsu/z/IAAbCwBF3SYAA9gA3AS0AYAADBbDAAAYAAAwABgAAAAAAAAEPgQAAAAAAAAAAAAAAAAAAAAAAAAAAAASipKxKLTsNXANwAAAEwBoL3Kfwo6AAM3RYfY6DznAKfJ7RX+dvvtb8RNNXQNpK7ZxDxD9YvhsM/c6vv5Ly/P034cyH6xJYmuvcWy7+fp+foVd0AA2FgB6lJ3VgAB7ADAtcA3AAADoAAAdR7AAACAAYAAAAAAAABD4EAAAAAAAAAAAAAAAAAAAAAAAAAAAABOHMhoaAQAAAAmANBR+EAANi4kcR46VKkqEHrLf0/4JzRva+pw1jpTi8PJ7ar06oxGIpYem6lWVoruZ3xFUxt6VH3af4v18vL7zh/h2WJar4he50Xf/gfn6CSSsirugBs3LXLAD1Cm7SAAHsAwBIBgAAPYEMAAAWwwAAewLYAHsAAAAAAAAAEfhQAAAAAAAAAAAAAAAAAAAAAAAAAAAABVhdXQ1cAQAAAAmANXKXVCAACyDiKs3jZ/3Ul+H+bOVtXMFmbwdSNZK71028jG5hiswqp1Hd9Etl6L+mZJwvy2rYta9I/5/wCX39hJJaD2Ku4NoeoWAGPUBaM3AAAeoCQBuAAA9Q6D1AAAYMAAGAAMAAAAAAAAAFsgAAAAAAAAAAAAAAAAABYStLDvERjeCdm+z8+1+gAAAAAAAAAABUhyvQaANwAAAC4BT3EMAAugz5P63V9f8hOPszD4SeJnTpw3bt95leRYXAK696fd/oun9ah0HsVX7wFiwA2ABB3igACS1GAAMAAB7Ah7AAAHUewAADEAAwAAAAAAAADcAAAAAAAAAAAAAAAAAAOBMTBYyphaivGpHZ7Nr/gNnEnBsqHNiMErw6x6r07ry3Xn0AAAAAAAAAACSUlYlGzsNAIAAAAC9wjo0IYAAHEmH5cY30kk/wBP0OWafLY4fw/PjYLpG7/r52Aew9iprIsAA2AAUtYgAFTSTAEgAAABgkMAAAWwwAAewIAAAAAAAAAAAirySAAAAAAAAAAAAAAAAAA4Y4Yy/MMvVeunzXa0dtj/AFGyj+WX3mD4Ry3CV416SalHbUOJuD6WKUsThEo1N2tk+78n+D69xoAAAAAAAAAAKkOZaDVwBAAAAAXCLukdAAAM8y6WLoJwXvR28+6HFp8rWpkOWyw1J1Ki96XTsv8AggMexLVsAGx6gABR2YABWXvAWuAMAAANzoAAAMGAADBbAA9gAAAAAAAAApK80AAAAAAAAAAAAAAAAABlXFmNy3DLD0YxcU29U76+kkf8RBzP+SH3S/8AkjIuLsyzHH08PKEFF3vZO6SX+JmKxVDC0XVrSUYrdv8Ar8DiPiytmTdGjeNLt1l6+Xl9/kAAAAAAAAAABVhZ3Q0AbgAAAAFKV1YWoAAA6UHLmsrgIGPUB2G7gAAFHqAAV1qgLAPYAAAYJDAAAB7AAAMAAYAAAAAAAAAYdXqIAAAAAAAAAAAAAAAAAAOE8ZhcthWx2IetuWK6t7u34a7amdZ7is1q81V2itorZf5vzAAAAAAAAAAACSTViUbOwwAAAAACMrO4hgAAAPYJvRhdDdwAAAKO7AAK60QJAD1AAAA6AAADOgwAABAAMAAAAAAAAAMKrybAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAqwurjVwBAAAAAFOV4i1QAAAJD1Cq9AAAAAKO7AAKqvBlrgDAAAGCQwAAB6gAAD2BbAAAAAAAAAAABhVo2AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAVI8rGgBMAAAACEuViY0AAAPYHsVXd2AAAABJt2RGKirAADV1YtYAAAANw6D1AAAB7AAAMAAewAAAAAAAAAGHjamgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAACpHmiDQAmAAAABSn0Yn0AAAQEmkrj1YAAAAQhyrzAAAqK0gHqAAAMEh7AAAD2BgAACAAYAAAAAAAAAbkVaKQAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAVYW1Q9QAQAAAAEKi2YXANDQ0G7g3cqyvogAAAApw6sAAArLZgwAAA3C2g9QAADdgAAD2BAAMAAAAAAAAAKMeaaQAAAAAAAAAAAAAAAAAAA0AAAAAAAAAAAADSaJw5WNXABO4AAAACk1syNXuJp7ANpdTmj3OaPcunsOyROpfRAAAABCHMwAAAmrxsMAAAGCQ9gAAAQwAAGAAPYAAAAAAAAADCx1bAAAAAAAAAAAAAAAAAALXLFgaAAAAAAAAAAAABpNWJwcWNABcAAAAAOaXccm+oAPUAAAAFFydkKKSsgAAAKsLO4AAABbQeoAAAweoAAAgAGAAAAAAAAABRhywSAAAAAAAAAAAAAAAAAANLF0XQNgAAAAAAAAAAAANJqxOm47DQAXAAAAAAAAAAAC1yEOVeYAAAA0mrFSDiwAAGCQ9gAAA3HsAAA9gQADAAAAAAAAAClDnmkAAAAAAAAAAAAAAAAAAAIbAAAAAAAAAAAAAAnSvsNWAC4AAAAAAAAAABThbVgAAAASipKzJ03FgADAsPUAAAewMAABgADAAAAAAAAAAw0LR5u4AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAEoKSJ03EADcAAAAAAAAAAp076sAAAAAGk1YqUnHVAAPUEhgAABuwAAAEAAwAAAAAAAAAhBzkkhJJWQAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAATop7EoOO4AXAAAAAAAAACFO+rAAAAAACdFPVDi07MBgD1AAAGdBgAAMEAAwAAAAAAAAAw9Pljd7sAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAaTRKinsSpyiAXNwAAAAAAAC1yFK2rAAAAAAAJRUlZkqHZkoSW6HqCQwAAAeoPUAABgAAAAAAAAAAAUKXO7vZAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABKnGRKg+jHCS3QFzcAAAAAAC1yNJvcUUloAAAAAAAADhF7odCD2HQ7MlRmug00AAAIewAAD2BAAMAAAAAAAAAp03N2RGKirIAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHCL3Q6EXsOg+jHSmuhaS6F0AAAAAKLewqUmKiuoklsAAAAAAAAAADSa1JUIvYlTlHcAAYPUAABgtgAGAAAAAAAABTpubsiEIwjZAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA0mOEX0PZxfQdKLPZRPYxPYxHRXc9iu57Fdz2K7nsV3PYxPZRPZx7HKlsAAAAAAAAAAAABOjGROjKPmACHsAAAPU3AAAAAjTlPZEcI3uyWFVtHqThKLs0AAAABSoOer2IxUVZAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABOlGXqSoyiDBgAAPYEAAwAC1ynhm9ZCSSsgGk1Zk8LF/DoToTj0AAACFGc9iGHjHfVgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAASpxluiWH/lZKMo7oAAAYdAAGABToSn5IhSjDYAAJU4S3RLCxezHhZdGOhUXQdKa6Hs59mexm+gsNUZHCd2RowjsgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADclQhIlhpLZkoSjugAAQAD2ALXIYect9CFCEfNgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAASpQl0JYZPZksPNbajhJboGC2AAYot7Ijh5sjhYrdkYRjsgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHGL3Q6FN9B4Wmx4aPRjwq6M+q+Z9VXc+rR7iw9MVKC6AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAH/9oACAEDEQE/AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA5WcjORns/M9mjkRyROWJyo5UWRZFkWRZFkcqOVHKjlTOWJyxOWJyROSJyRORHIj2aPZruez8z2Z7NnIzkZyssAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABucrZyM9mjkRyoAAAAAAAAAAAACyY4pjgh00ezY4MaAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAALNnI2ezRyoAAAAAAAAAAAAAAAAAcUxwTHB9BpoAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAALXORsVNChFDAAAAAAAAAAAAA0NC6Lo0AAAAAHBMcGAAAAAAAAAAAABa5ySZ7Nnsn3PZMdOQ4tAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAcrZyMUEWQW0GAAAAAAAAAAABdFwAAC5cugAAABpMcOw1YAAAAAAAAAAAUGxU4osgABxTHSXQlBoAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAALXFTbFSRypLQAEgeoAAAAAAAAAAFy4AAAAAXLoAAABpMcOw0AAAAAAAAAAouRGCQAAAC2JUovYlBxAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAFBsVNLcsgHqAFhgAAAAAAAAAA2AAAAAAAXLoAAABpMlBoAAAAAAAAAjT7gAAAAgaTJ0+wAAAAAAAAAAAAAAAAAFrnK+xyvsAAAAAAAAAAAAAAJN7CpPqKCQAPYAEg3AAAAAAAAAAbAAAAAAAAC5uAAABKCew1YAAAAAAAAhDqwAAAAAHsSgpDTWgAAAAAAAAAAAAAAAARg5CpxRYBpMdOLHTa2AAAAAAAAAAAALXFTbFTigAAYAWGAAAAAAAAADHqAAAAAAAAAFwAAAHFNDVgAAAAAAAhHmYAAAAALYAlFSRKLTsAAAAAAAAAAAAAAABGn1YAAEopkoNAAAAAAAAAAAKLewqS6iSWwAAD2ABIAAAAAAAAABsAAALFixZFkWRZFkWRYsWAAADcAAAGk0SjYAAAAAAAgrIAAAAAEAwlFSQ1Z2AAAAAAAAAAAAAAAIQ6sAAACVNPYasAAAAAAAAACg2KmkAAAAwAsMAAAAAAAAAeoAAFiyAAAACyLFgAALgAAA0mhqwAAAAAAbhugAAAAAB7ASipIaAAAAAAAAAAAAAACEbsAAAAJRUkNW0AAAAAAAABRcthU0gAAAAAEgAAAAAAAABm4AAWLAAAAAAFkNAABcAAAJRugAAAAAAirtAgAAAAAWwAFSN1dAAAAAAAAAAAAAAFriSSsAAAABOKkhoAAAAAAANyNPuAAAAAwA6DAAAAAAAABgABYAA0Lly5cC5cuXQAA0mNAAFwAAAmrMAAAAAAhuAAAAAAIBgFSNmAAAAAAAAAAAAABTjfUAAAAAJwutAAAAAAABJvREYJAAAAAD1AEG4AAAAAAAA2AAWuWAC6LgAAABcubgA0AAAAAEldAAAAAAENwEAAAAAAPYAaTVhqzsAAAAAAAAAAAAAEVZWAAAAAAqR6oAAAAAAIwcmKKSsgAAAAB7AAloMAAAAAAAAbAALAA2bgOSHIuy7Ls5mcxzIumAAFzcAauNAAAAADWoAAAAAENwAAAAAAWwABUjdXAAAAAAAAAAAAAKauwAAAAAB6ko2YAAAAAEYuTEklZAAAAAAwDcHqAAAAAAAAwANywA2PUByQ3c5WzlOVHKjlRyo5BxY0XZzGjAALm4A0AAAABPcAAAAACG4AtgAAJNJXHVk9hVX1IyUloAIBgBONmAAAAAAAAAAAABBWiAAAAAAE48yAAAAABRcnZEYqKsh7AAAAAAAIewAAAAAAADAAtcAG7g7Dl2G2xRbFFIAAAcUxxBSAAC4A0AAgAAlqwAAAAAIbgCAAArO7sAm09CE1JAAMAJWsAKLexyS7AAAAAAAAAAAJXdgAAAAAACpHqAAAABuQhyrzBgAAAADAAYAAAAAAAD1AC1wBsG0hy7G5ylkgAAAAaTHGwnY3AALm4DVwAEAAPUAAAAACO6AAAAKmsmAJtPQjJSVwOgPYAKj90glyjpxZ7OIEopkoNAAAAAAAAAAFNXYAAAAAAA0mrDVnYAAAAKUOoDAAAAAHqAJDAAAAAAAAeoAJXAGxtDk2WuKJZAAAWLFixYAHHsJ2AAA3AauAAtQAm9AAAAAAAAEAAE17zACEuVgIBgFXoi2gADSaJxswAAAAAAAAAKa90AAAAAAAKkdLgAAAEY8zsJWQPYAAAAAewAdBgAAAAAAAwASuA7Ep9htss2cqLAAFixZAABZFkWLEkRfQAABO4DVwANwCTuwAAAAAA3SAEAAFVa3AApSurAgGAT1mgAAaTViUWnYAAAAAAAAAErKwAAAAAAANXQ1Z2AAAApxsgGAAAAAMAQAAAAAAADAC1wHNdBtsUbnKgACxYANC6LouXRdF0ADSY1ZgAACYDVwAAm7KwAWbOVnKzlZYAAACGsQAAAJx5lYatoAEXZ3NGgB7AD1mAABKKkhq2gAAAAAAAAEVdoAAAAAAAAqx6gAABFXdhAMAAAAAAFsPYAAAAAAAYAWuaJDmug22KNxJIAA3LDaRzRLply4AAAXLoJrQjqhoAABMBoAENpIbuCi2KCOVIABxTHBMcGNNAABS7DAEAAFSnzLTcasAFKV1YFsDAN6gAABOF0AAAAAAAABTWoAAAAAAAA1dWGrAAAFNWVxbAAAAAAPYABgAAAAAAD1ABzXQbbFBsskAADcV1HU7Dm2BcuznZzo5kzcAAAHqrEAauNAAAmA0ANpDdxRbFFIAAAAcExwaAAg7SGAAAATgpIcWnZgEXZ3NxAMD/l4AAAFSPVAAAAAAAABTWgAAAAAAAAVFrcAABK7sW0OgPYAAAAAYAh7AAAAAAADAdhy7G4oNiikPUANBySHN9BtsSb2PZs5EckTkiOCHT8zkZyMaLtCmXTAAAFoy4DQAAJ3AaBy7G4ooAAAAAHFMlCwAboAOgAAEoqSJRadgCk7xsAMB/xAAAAaTRKNnYAAAAAAAAgvdQAAAAAAAATV4gAAU1d3AGAAAAABzLuc0RWsMAAAAAABg2kOXY3FC4opbAwBtDkN3Em9hUu4oRRfsAABa44Jji0XFITTAAD7QF7g1cAANwJPogUQQAAAAAEoJ7DTQEHeIwBAAAEoqSsNNOwFN2kAPYB/GAAABOPMgAAAAAAABLRAAAAAAAAASVnYAAIK0RAMAAAAB7DkkOTYaEZNF0wAAAAAAJNDlctcURJAMByQ22NiTZGmuoNpHMmOyNGAAAEoJ7FmmNW0FI0AA+0AXBq40AASmlsSlpoCiAbgAAAAAbjgnsOLRTeth7AAAAAVY3VwBO6uLYAJ/GgAAACpGzuAAAAAAAG4AAAAAAAABVXUAASu7AtgAAAAAlJIlNyGNgIg+gAAAAAA5JDk2WucqLADByQ22JNkrbIjG4opIbSRKo3sashFpk1dGqFMTT2AAAGkyWkhx7D2Iz7l0wG0iOoABuNABKfYUbhGPUaAAAAAAAAPhkAB0AAACcbSsBSd1YQDCp8SAAAAGk1YatoAAAAAAAR1aAAAAAAAAAJK6sAAU1eQdAewAAAASqdEMbAAiwAAAAAckhybFFs5UAJA2kOVx6nK2KKJysrEY3Egq62FDuWQDSY6fYSdx7CaYAAFRaC1Q0mShbYQXYlcjuwADcGgbSRKVyML7ktiMUwGgBAAAAAABNaEHeIwBAAABVjdXApu0gBhV6AAAABUj1AAAAAAAIfEgAAAAAAAAAmrMACmtABgAAAMnO+iGwAAQtUAAAADkkOTYotigkMASuNpDm3sbiiWSBtJCXMxIlJJEZNsJTS2E7onuJtbHOxO6G0S1Wg00F2GjACauiKsgLWkWRZAtJAABubkmkNuTIwsDV2C1Bq4AAAAAAADV0RfKx6oAQAAAbko2dgIu8UwHsFXZC1SAAAAGk1YkrOwAAAAAAFP4gAAAAAAAAAqrZgAQXuoQDAAAAnO+iGwAAEQ2AAAAckhybFFsUEgNwG0hybEmzkLIAm76EI2Q2khtyYkkiUr7Cg2JJKwOKY6aIqyJq7BTYmmOKYaoU+5dMAAnugCWkgAAJTSRrJiikAADQAgAAAAAAnG+pGdlZnPE5l3OZdzmic8e4pJ7MAAArR2YFF6WFsDCp8JH4UAAAAFSN1cAAAAAACnuAAAAAAAAAE1eIACWggAAAAqS0sMAAAIbAAADaQ5t7CTYoIAYOSG2xRbFBGiQASdkQjd3CUuZkY2RNtuxGHcAAGkxwT2IxsSlYUkxpMcH0NULY5kaMTT2AqdDoBURugACUraISciMUloNXAAQDANwAAAAAAkknoWbOVnKzlZyMaaIT6MAACavGwFN2khAMqfCQ+EAAAAGroas7AAAAAABT3AAAAAAAAAAas7AC1aDoD2AAAB7End3AAAAgAABKfY1bFDuWAByQ22KLYooB6gBN3diKsrFTWJGKSuxNNHKr3AAAAlG400KbQmnsNJg4u5C9yAE9WgCauiGqAAlK+iIxuWSQDQAC1BoAQAAAAABJ2RCPMxrQARa5OFtUQldWAAAmrSsBF3SYDJq8WU/hAAAAAqx6gAAAAABS6gAAAAAAAAAVFqAR+JADAAACo7RGAAABAAAlJIbbFC+4kktAAckhybEmxRQJAwAeiIK7uSkkMlK5FWQGjJStojnZziaaAANxwXQgrEpWFJM0ZoQAesgBq6IbADaSJSuRhfcAQNAAA0AbgAAAAAT1diKtoAANXQ/dkbgABWWqYFJ3iAxq6KXUAAAABq6sNWAAAAAAKXUAAAAAAAAACqroAh8SEAwAAAqvWwAaWGaWHY0Q7AQAAlLohRbFFIAHZIlO+wk2KKAtcB6gBU2I2jE1kyb6EYgTdkQ2JRuz2aHTZFWRN6imznTLpgASjccWgj1IbA3ZEFd3AACUkh3kyMEgAEDQAIGroAQAAAAAEVd3AYAtgqRuim7qwAAVVeIFF7oWwPYKe7QAAAABUWtwAAAAACl1AAAAAAAAAAkroAp/EIAAAAJu7YWLDQ9joPYGAQACUr6IjG4AEpJDbYodx9gBLQGACVyesrFR9D4YkI31Ak7Is5MGc6OZPqbjSY6a6DgyC0Jtp6Cn3Lp7ABZMnsQWgTd3YirIAG0kXVrkp32FFsUUkAAC1GrgAbg0AAAAAAE9iC0BDAEBH3ZWAABq6sBTdpCAFpMAAJTUSMlJaAAEldWAAAAAAKXUAAAAAAAAAAkrOwFP4hbA9gAAAewwew9h7FroexoaXGAQ2AJSvoRhfcAGSnfYUWxRSDcBK4AAC1kfFIkroSsrAT1dhJJA9UcjGmHMxTE0wcUxwfQ1QpXRcuXLknditYlLSyIRvqABO1jcUO5YAAAGgAQNXQAgAAAACp2ErIEPYAAqKzuJ3QAASVmwTsxAMlpNAADaSuN3dyMuV3E00AATVmAAAAABT3AAAAAAAAAAKi1Ap/EdAewAAAT+EYPYtdHQYPY0GAiGwOyRKVyMO4ANpIbbFDuAwBLQGACQ9ER2ZBABHV3AAaTHBMdPsNNCn3LouNJiVkN2ZzJl0aMtqN2Qldhc0YEmkjWTIxsAAAAgaADcGgAAAAACWsgAYAFRXiU3eIAAVF7wEHeKAZVWxugAKz92wFKXQAAqK6AAAAAAp7gAAAAAAAAAFVaICn8QDAAACp8Iwew9joPYHsG40CEtENpIk7kY2ACUkjWTIxSAHqAlcAASuEtiKvEirITblYAUUgAAHZjj2E7ADjccWWIrQexuy1kAXG0kayYopIAAAAFqNXABA9UAIAAAABayAFsMAWwPUhpKwAAVlqBSd42OgPYqrQjrFAAVt0AnZ3N0AA1dDVnYAAAAAKe4AAAAAAAAABUV4gUviAYAAAVegy1xgABuCVzSw3zMjGwASlYScmKKSAHsAJaAwAtoMlsQ2DaQAAANj1AlETsAASd9BKyABj1ZFJAAAAAA0AG4NAAAAAAQ1YAh7ACAnpK4AAVlogKL1aFsD2Jq8SnrEACt8QBSd4gAFVa3AAAAACn8QAAAAAAAAAA1dWAp/EgAAAAq7h0AAAGiC1JSvoiMbIAJSsJOTEkkADA3AAEg3JbEbcommie9zdAOVpGjQANgASiRfQAJOyIoABu4lYDcAAAAEbgAgewBugAAAB7EAAAAKquiDvFAAFVXiBTdpIQEldWKWzAArbgFN2kAATV4gAAAABD4kAAAAAAAAAAElZhT+JA9gAAAKu4AAACL6WIx6gBJ2Qk5MSSQAACBgAMHsR1i0Q2Jq6IaoCcb7CbQp9zdDkhyQ5C1Q27nMxNsDZgDdkWuwAdhu4o2AC4AAAAbjQAbgwEAAAAS2IbAAMACSuim9LAAFRXiwFoxAMp6SaAArLYAE7q4ABNWYAAAABHdAAAAAAAAAABUXvBT+JA9gAAAKm7OgAAAEY3ABtJGsmJJIABgAACQwCG9haSsNXRB9ABxTHB9BaIkm2crOViVkStcUbgEloLYG0jVsSsgBtI1Yo2AALgAAACAAQMAAAAAn8JD4QAWwwBbAR0nYAAkroAi7xQDFpMACqrxAClLoAAVVpcAAAAAAAAAAAAAAACqtgh8SBgAAOoke0QpJ7E9WxgGNzDC4Kn7TETUV5/ot38jAZzgMe2sNUUmumqf3OzABISuy2gA7JDbkyMUkAAPUBAwAGAbSJrqRd0Ne9oAA9QDQaTYAPY6F2JXEkgBy7CVxJIAABMAAAA3Q0AAALYAAACexH4UACHsAICppK4AANagUtYnQGS0mgAJq8WAEXZ3NwAJK6sAAAAAC1QAAAAAAAAABUXuhHdAwAAnK+iHYugewA7WOIc2lmWNlUv7i0ivLv892YTF1cNWjWoytKL0/rt3Rl+LhjcLTxENpK/p3XyegZhxFgcHXhhnLmqyaSitXdu2vRL117I3LEVZABJ3IxsgAB7AHQHqAJDACa1uaSiJ2YaAD1CUux0HLsRQAEtiOxZAA3cUQAAAEwAAAEAAgYCAAAAnsR2QAAABVWhHWKAAJr3mBR2aFsD2Ku6AAGgApO8QACaswAAAACOsUAAAAAAAAAATV4gAABN2QwDcaAz6u6GWV5p68rX36fqQp+0momIw7otHC+f4TAZPJ4ufKoyaXd6J2S3erfkZ94gYzGJ0sGvZw7/afz6fLXzOA8NHEZ1GpUl8Cb1e7ei9Xrf5FiCvqAE30Ix6gAAwED2AAYASV0QfQnG+xbQmRldA9QbuKOg3cUQC6LockN3E1awASdxRsAAAAC1AAAA3Q0AA9UAAAABU2FsAAMACavEp7AAFRe8wKO7EBVWiI6pAATXvMAKTs7AAFVaXAAAAAKfwgAAAAAAAAADV0AtYoAAKju7G4APVAcWJvJq9uy/wDSkZfZV9TOXBtcpwvk2HzXK8Rh660clZ9U7br+tVoZtlWIyzFSw1darbs10a8n/wAB6ltDB5/mmDt7CvJJdL3X3O66djD+JGdUlaXJP1j+Putf5GG8VJXtXw3zUv0a/VGE8R8krL945U35xv8A+k835GCznLsbph68ZPsmr/dv+BJ2RGN2AAAwFsDAEMAAfuyNx7FyO4wk7huhRSAbSG7gWElYaFI3CT0IoAAAAA3AAABAALYGAgAAAqdDoABdWOaLejAFsD1RT3aAAKvxAUviEBUXukNYoACqveABOzTAAJK6sAAAAAU9gAAAAAAAAAAktQhrBAABJ3AAQ0GcYX6zl9ait3F29bafiRm4SUkVa7rNHA+FdHKlN/bk38tv0OJuHKGc4XkelSPwy7Ps/J9fvPD/AA9TDZpicBiYWly6p+TX36O67rUxXBuSYu/PQSfePu/lZfejEeF2WyX7qtOL87P9F+ZifCzFx/gYiMv8Scfy5jF8CZ7h1f2PMv7rT/C9/wADE4PEYaXJXpuD7NNP8TAcT5tgWvY15W7N8y+53t8rGV+KDSUcdR+cP/kW/wBfkZZxLlWZWWHrJy/lekvue/yuAAPYAAAsMAAmrog+gSimJWHLsc2huKIA3ctcsXRe5cuDjfY2OY3YAAAAAAAAAXViUr7AKTFJMGAAAAE9WkAAVHrYE7oBALSYABWWqAp/EgCorplP4QAKvxAAQd4oAAmrMAAAACnuAAAAAAAAAAFRWkFN3iAATdojAsWA3QM4qyKpgMVKrBfu5u6fZvdf5eXzMpyqvmWJjQpL1fRLu/61Zh8PTw9GNGmrRirL5BPLKE8ZDGpWqRTjfun0fo9UQWgBWpU6kHGpFNdmrox3A2S4279lyPvD3fw+H8DMvDHG0rywdVVF2fuv9U/m0Y/KsbgJ8mJpOD81p8ns/kZZxdnGX2jSrNxXSXvL8dV8mjLPFChNKOOouL7x1X3PVfezLs7y7MVfC1lJ9tn9zs19wAwEDAEAAASVmRkmhkpXErjiWSAbSG7lhsALAhpM5UJJF0XRdF0XRoAAAAgAAAk7jYFzRkZW0N0AdAAAB6yAAKnxBTetgEBPSSAAKy0QEPiQBJXRS2YAFbdAAUneIABVWlwAAAAIfEAAAAAAAAAAFZahS2YABV6IYAAi2puyUIzjyyV0UqFKjHlpxUV2St+QD+ES0AJ7EVoFWlTqwcJpNPo9V9xmXAGS4y8oQ9nLvHRf+Au6t6JGZ+GuZ4e8sLJVV/4DL7m7fiYjCYvA1eWtCUJrumn6r/MyvjzOMDaMp+0j2lq/v3+9teRlXiNleLtHEJ0peesfvX6pLzKNelXpqpSkpRezTun80MBbA9QBbDAAAaTVhpobbQRVgGOV9i1zRG4ALYuhoAsWLFixYAuXLo0AAFqAABJ6DAATIPoMBAAAC1kAAVVqgTszdABVWwtUABW+ECHxIFsD2Ke7QAFbdAAUeqAAJK8QAAAAFo0AAAAAAAAAAIrLRBT3AAKjuwAABAA9UkAE9WkABisJQxVN0q8FKL6NXX4ma+G2WYm8sLJ0pffH7nqvk7LsZtwRnGX3k6fPBdY6/et156W8zAZpjcvqc+FquD8tn6rZ/NGUeJs1aGYU7/3o7/OL/Rr0MtzrAZlDmwtVS7rqvVbr7gHsAbgwAAJStoh6iiKKQDaQ3csNhuWAOgdC4gAACyLIsWLBcugAAACTuAACZe6AQAABDVgAFVaICm7xA6BUXukNYoACr8IEPiQLYHsR+NgAVt0ABS+IAAJKzsAAAAAtUAAAAAAAAAAiorxYQ+IAAlqAAB0EAG7QAPWQAAGbcM5XmabxFJc38y0l963+d0Zv4Z4qleeBqc6/lekvv2f/AJKVqGMy/EctSMqdSPqmvNf5oybxGzDC2hi17WPfaX37P5q/mZPxPlmbRX1ep738r0kvl19VdeYwBD2AAAk7IUblkgBuxuaIeoWPQuNDQPYOgIAAAACxZFguXQIAAk7IYGhoaAEHrYAAAB7ENgACorxApvWwC2CavFlP4QAKivFgQ+JAgPtgAVt0ABT+JAABVWoAAAAENYoAAAAAAAAABDV0MjugAHsM6AABl+U4/MG44SjKo1vyxbt622+ZiMLXw1V0q8HCa3Uk018nqAC1aABayAAAYY3L8Ljafs8RTU4+a/Lt8jOfDKnNOpl1Sz/llt8pbr0afqjMMqx2W1eTE03CS2fT5NaP5MyXxBzLA2p4j97Dz+Jekuvzv6oybiXLc2j/ALNU97+V6SXy6+quvMAYAAE9XY0tYAeiNzYeoDEho6DBaosPYELYAAAAACyLBc3AAkMAABG4C2AACfwkNgACSvECLs0AgauilswAJ/CwI/EgQEvjAArboACPxIAAKivEAAAACntYAAAAAAAAABbBUVpMAAn8IxgWRoxoMqy2rmWOpYOj8U2kvLu/RLVmT5RhMowUMJho2hH72+rb7vr9ysrI8Q+FqWc5XOrCP7+km4vq0tXF+TW3Z287gEdwAjuwAAYAYnDUcTSdKtBSi+jV0Z34a4eqnUy+XJL+V6r5PdfO/wAjHZZj8rrKOIg4SWz/AFTWj+TMi8RcZhbUscvaQ7/aX6S+dn5mV5vgszpe0wtRSXVdV6rdf1YYAAA/iACTubI3AQ1dgPY6DEaD1AQAAAAAAWTLApGjAJAABuCI6oBAABPYjsgAAaAWqBAU92gAJbMCPxIAKnxoACtugAFugAAaurAAAAAU9wAAAAAAAAAOgVlqgjqkAFT4R7gItcaN0Hg/gVWzyddr+HBteraX5Nma4r3uW+iMqxDqU+VvY4jwMcDm+Jw0VZRnJL0vp+FgCG4AQ3AABgIGGLweHxlF0cRBSi+jV/6fnujPfDXerlkv+3JP8pfpL7yMsdleKuuanVj6p/8ADP7mvIyDxIhUtRzJWf8AOtv+3l09Vp5IpVadamqlOSlF7NO6fo0AAA/iAJOyEh6nQBbAB0OgACAAAAAAADl7GwpA30GAAAwjuAAAFTodAAAmveAhrEAI/GAA9gI/EgArboACtugAAAAJqzAAAACOkkAAAAAAAAAAFZXQQ1igAq7HUDyA6jPBXl+s4zvyx/N3/T8DO3NSlbe5kDb37GZ8F5FmVWVbE4dOb3ack2+7s1f5mK8JMmqa0ak4P1TX4q/4mM8HsXH/AHXExl/iTj+XN+RjfDriLCq/sOdd4tP8L834GIwGKwk+TEUpQfaSaf4+qACAAAMBbAwDNckwOaUvZ4qmpdns16Pdfk+qZxBwBjcBerhf3tPyXvL1XX1XzSMj4lzDKJ3oTvDrF6xf+T81bzMg4uy/OEoQfLU/le/yfX8+6QAA9h6MBjdx7B0A6AAdAAEAAAAAAACGkxxL2HqM6AAB0CIwFsAAT1aAAAqrUCnsAH2wAHsBHdAdArdBbABW3QAAAAFRagAAAAboAAAAAAAAAAqK6YU9gAq7INjcAGeD2NjRzupQb/iQdvWLT/K5mmEvUv0ZlGGdODk+pjvE7OqOZV/YVIypKb5U4q3Km7aqz1W93fzRhfGDGxX+0YaMv8Lcfz5jCeLWTVdK1OcH6Jr8Hf8AD5mC4zyDGtKjio37N8r+SlYtRxFPpKL9Gn+hj+B8gxt3PDKL7x93/wBJsvvTMw8I8NLXB4hx8pJP8Va33MzHw7z/AAabVL2iXWDv+GkvwK1CrRm4VIuMl0as/uZAAAAB7ACHsHEPBGAzW9WH7ur3S0f+JdfVWfqZrkuPyfEKFePK+kls7dU/+Ga6pHDXiHKko4fM3zLpPqv8S6+q17p7lGtTr041KUlKL1TWzABkkJqw5IbubAdAOgADMRiKWHpSq1ZWit2yhxplNWt7LmcfNqy/O6+aRo0AgAAAAAAAFsDSYNXLAwAARukAgACXxIAACqtEBS6gLYJfGgAHsBHdALYK2yFsgArboAAAACqtEAAAABDWKAAAAAAAAAEA9RlLqAFXZAxAD2GZLmlTK8wo4ynvBp27rqvmrr5mV5lhc0wdPFYeXNCauvLun5p6Ndzj7ialkmVTUZfvqicYLrqrOXpH87ILmhe5gcwxeElfD1ZQf92TX5MwPiRxBhdJVFUXaST/ABVn976LoZf4uYaVljMO4+cWn+Dtb72ZbxlkeYWVHERUu0vdf/k1r/K5jctweOhyYmlGa80n93b5GZ+FeT4i8sK5UpeXvR+5u/3SRmvhtnmCTlTiqsf7u/8A4C7P7rlajUozdOpFxkt01Zr5AA9gAYAthgYzBYfGUXRxEFKL6P8Ar8ehxL4f18JevgLzh1j9qPp/MvxXnqzIOJ8dk1X907we8Xs/Ts/NfO5kfEOCzij7TDy95bxe6/4HZrT5gMBxZytlkkMDoB0OgB1DjvNJVMSsHB+7DV/4n/kvzFKLdkzgjNJYrByw9R3lT2/wvb7tvuAQAAAAAAACB6oehe49hDLBZMsiwNCI6xAQAA/iAAAqK8QKe4CCp8SAAAjugEFb4SPwoAK26AAAAAmrxAAAAAp7WAAAAAAAAAEATVpFPcAKuyOoxAD2GGWZ7mWWc31KvKHNuk9H8tr+e5jsfisdWdfE1HOb3bd3/wAN2WyGrgJENwDLeIs1y231TESil0veP/gLuvwMp8W8VG0cfQUl3jo/ud0/viZTxxkeZWVOsoy/ln7r/HR/JszLJsvzKHJi6MZrzWq9Huvk0Zx4TYepeeXVeV/yy1XyktUvVSM44YzXKX/tVFqP8y1j960Xo7MBgIGADADifgfC5opV8PaFbv0l/iXf+8te6ZUp5jkmNs706sdv810afzTOFuOKGZ8uHxVoVunaXp2fl93ZDABgHQBCG0ldsznjjF1K7hgXywXWybfnrdJdtL9+y4X4qlmEvq2JsqnRrTm76d/z8ra9Azms8RmFad95P7r6fgVcFy0eZHAtdwzPk/mi191n+gCOgAAAAAAAG4SBgbosdTqdRj2CGwAAA/iAAAkrpgU/iAQVegAAC3QCCr8JD4UAFbdAAAAAPUaAAAACm9QAAAAAAAABA9gqrUh8SACp8J1GI6APUYdB7BoyyGyG4BLREFoGU8V5vldlhq75f5XrH7ne3ysZN4sYeo1DMaXI/wCaOq+cXql6OT8jAZngMzoueGqxqR62d/k1uvRozvw4yfME50Y+xn3jt847fdy+pnvAeb5UnNw9pTX2o6/et1+XmMBbAAh7AAGc5Hgs2oexxMb9mt0+6f6bPqjiLhbGZLV9/wB6m9pLb0fZ+X3NnCfH0ocuEzKV1spvdf4u6/vdOvdRlGUVKLumADQ0AAjZnGWOeFyqSi7Ob5fv3/BNfM95u0TB4mphq8K0Pii0/uKdSNWnGcdmrr5hjFbGTv8AzP8ANmJqweESW5wdUhTzek5ySvda6atOy+YCOgAAAAAAACHsMHsD2OgBuzdHQIDAWwAEviQAAAEPiQCCr0FsAAC3AQVfhIfCgArboABbgAAFRWkAAAAC0aAAAAAAAAAFsD2CqtELcAJq8R7m5shbCZoWA6HQAIbgE30ErIAw2Kr4aoqtCbjJdU2n96Mj8U8fhrU8fH2se60l/k/uT7syXifKs4j/ALLVTl/K9JL5Pf1V15mfcCZRmyc5Q9nU/mjo/mtn+fmjiDgLNso5qnL7SkvtR6LzW68915gPYAQwAAxGHpYilKlVipRejT2ZxXwNVy7mxODvKl1W7j/nHz3XXucKcZ1srksPiLyo/jHzXl3XzXnhsTRxNGNajJSjLVNADVxoaAA3R4h1GqWHh3cn91v8zK8MqtzF01Cs0cPTc8qw7f8AKl92gcTYR4fNa0O75l89f1sSxE5Q5TF0ajoc6i+VNJu2ive2pw5x5iMFahjb1KfR/aj/AJryevZ9DBY7DY2iq2HmpRfVf1o/J6iAAAAAAAACWwwWwPVgAtxD2CG4wEABPoAAAS3CHxIAKuyFsgAewC3ACp8DKfwgBW3QAC3QAABVWwAAAAEXdIAAAAAAAABbAwmrxBaoAaurDGBa5YeoDHsAEdwHoiKuwACM5QalF2aMg8TczwNqeL/fQ83aa/7e6/8AbybfdGScU5XnMP8AZavvdYvSS+XX1V15nEXh7lea3qUl7Kr3itH/AIo7P1Vn3bM/4UzPJZ2xMLw6SWsX8+j8nZjAOgwAAHZnF3Al+bF5bHzlBfnH/wCR+7scM8U4nJa3L8VJv3o/rHs/wez6NYDH4bH4eNfDz5ov+rPs/IAauNDQMA8Q6V6FCp2cl96X+RluJ9lcxVT2lVs4blTeV0YwleySfk92vVXDjDIJ4+lHEUFepHS3df5rp6vyKGBxNet7CnBufa2vz7fMyXIqOBy/6tUSk5fFdXTb6eltPx6nEfh84qWIy3XvD/5F/o/k9kZZm+PyfEOVCTi9nF7O3Rr+mjh3jbB5najW/d1ez2fo/wBHr2uAAAAAAAAD2GMDqAdTqD2CG49gEABOzRF3QAAE/iYQ+JABV2I/CgAewAAVNYsp/CAFboABH4kAAATV4gAAAAU3dWAAAAAAAAABg1dBDWKACas2dAAAHsAC3Am+hFWQAA9gp1Z0pqdOTUls1o16M4d8UMZhbUcxXtYfzL41+kvnZ9W2ZfmmW5zhnPDzVSD0a7eUovVejRxL4XYfEKVfK3yT/kfwv0e8X5ar/CjH5fisDXdDE03Ca6Nfiu681owAAAHsBxfwTDH82LwStV3a6S/yl+D666mQ8QYzI8U3G7je0oPS/wDlJd7abPTQyrNcLmeFjiMPK6f3p9n2f/D7AA0NXGtADirL5Y3K5xgryj7y+W/4XLO90zAYKrjMRGhSV5S/p/gcL8RVspzKUMTpCbamn9mV97dLPR+XohNSSad0AGfcK4DN481RctTpJb/PuvXXs0Z7wxj8olesrwe0lt8+z8n8mzhbjyphnHC5g+aGyl1j691+K89lTqQqwU4O6eqa2aAAAAAAAAYWGhCAOpcOgQ3AAAbSQryZB62AAAn8QQ+JAHQq/CR+FABLZgLcAJ/Cyn8IAVugAEPiQAAA1dDAAAACm9QAAAAAAAAAYElZlPYAKq1uAAHQyrwy4gzDCrERhGCaulN2b9Ek7X6c1u+1mZplmLy3Eyw2Lg4Tj0f5p7NPo1oAAmmiUuxGN2AADAWxgcwxWArqvhajhNdV/Wq8nocNeKNDEctDNEoS/nXwv1W8X57eiM1ybLc7wqhiIKcXqmt1frGS/wCGfW6OKfD7HZRzV8PerR7pe9H/ABLt5rTvYQ9gAAGAcXcHUs1g8Rh/drpfKXk/Ps/k9NVlObY7Icc5RTTWkovS67Ps+z6el08ozfC5rhY4jDyuuq6p9n/Wu60ABq40NFgM44F9tWdXBTUb/Zd7L0aT08raem3D3DNHKk6kpc1R6X7Lsv1Z4gcNXTzLDr/Gvyl+j+/uzw/4j9vS/s6u/eivc80uny6eXodAC1yrRp1qbp1IqUXo01dP5HFfAc8KpYrL05U+sd3HzXVr8V5rbhPi+tlM1Qre9Rb26x81+q+7XfD4iliKUatKSlGSumuoAAAAAAF0XQ9Sw2PVB1AOowCG4MDoAyT5mRjZE1Zid0AAFT4gh8SAFsVPhI/CgAn8LAW4Atifwsp/CAFboABT+JAAABNWkAAAACdmgAAAAAAAAAYFRalPcAJq8RgAHhrw/TzfOoutG9OkuZro3tFP56+aTRUr06bSk7Hinw/TzDJnjIL95R1T7x+0vRL3vKz7sAErpgo3LJIAAGAPYOHOMcyySaVKXNT6we3y/lfmvmmcO8WZdnlK9CVpreD+Jendea+dtjirw3wmPUsRgEqdXttGX/yL81p3WtzMcuxWAruhiqbhNdH+a7rzWgAAAAcWcJUs4pe1pWjWjs+j8n+j6ehlmZ5hw9mD0tJaSi9muz/R/NXT1yjN8LmuFjiMPK66rqn2f9a7rQABosWRYCpThUg4TV01Zp9UcRZPXyDM1Og7RvzQfp09Vt5qz6mQZzSzbAwxENHtJdpLdenVeTAtcsWDjPglTUsbl8dd5QXXziu/ddd1rvwjxZUymsqNZ3oSeq/lfdfquvqUa1OtTjUpyTi1dNbNAAAAF0XLlwLFiwzcEHUAErjR0CG4MBbASlcjGwSV0QfQAAJ/EFNe8AIqfCQ+FABP4WAt0ALYl8LKXwsAK3QACn8SAAACqtQAAAAIu6QAAAAAAAAgGBVWxD4gAJxs7AAHgxhIwy3E4nrKaj/4DFP/ANuMZi71G31KEI43L6lCe0k4/JqxKLi3F7oAgWQAAAAMChXq0KkatKTjKOqadmn5M4V8TlLlw2a77Kov/bl09Vp3W7M5yLLc9wqhXipJr3ZLdX6xf6ap9UzingrH5HNza56PSaX4SXR/g+j6AAD2AAOLeFaWcUPaU0lWitH3/uvy7dn5XMozXG5BmDdmmtJRel/J+a6P9G75XmmGzPCxxOHd4v70+qfmv60AALFgsiyOIcjpZtgZUJaS3i+z/wAnszhfOKuRZm6WIVoN8s12ae/yf4XFaSTTuiyLFgDjfg1TUsfgY67yiuveS8+66773vwXxa8tqLCYmX7mWz/lb6+j6rpv3umpJNO6AALouXAsWLFkWQASFsCB7G4PY6DL3GEQewCCb6EI9QCSszdAAE/iYUviAEVPhIfCgAqfAwFugBD2KWzACt0AAp/EgAAAqrQAAAACm9LAAAAAAAACAAqK8RaMACpG6uNBoaAeD75uH60Vv7SX/AKRAzGnOVWNjJotUWYmSlWm1s2/zAIAAAD2AQMAOF+NMfkc1CL56LesH+cX0f4PqjJs9y3PsI50JKSatKLtdeUlrp96fc4u8M7KWKylebp//ACP/AMj9z2ROEoScZKzWjT3TABgABxfwnTzej7aikq8dn/Muz/R9PQyDPcVkONd0+XacXpt+TXT7upgcdh8bh418PLmjLZ/1s+66AAAWRYseIvDvNFZlQWq0mvLpL5bPyt2Z4fcQfW8N9RrS9+mvd84//a7elvMsgADjrhH2DlmGDj7j+OK6P+ZeT6ro9dtuAuK+Vxy3FS02g3/6S/8A237uwF0XLhYsWRZAAABIQ1c6AjYujc6jBiI7C2GAhuyIrmYATV0Qd1YAAerCj1GAip8JD4UAFX4QI7oAQ9ilswArboABOzNwAACavEAAAACm7MAAAAAAAAQMAkrqwRd0gAJRs7DADwXzGP8AtWBb10mv/SZf+2mNy+bq+7G6M3xccoyWviG7OEW/+3rWS+bsg0LI0IAAAD2AQMBD2DAZhisBXjXws3Ca6r+tV3T0OEfELC5ry4bF2p1+n8svTs/J/JvZcW8C4PO4OtStCutpdJeUkvz3XmlYzTKsXlmJlh8VDlkvx80+q8wBgAAca8IrMqbxeFj++juv51/muj67PpbhbiavkmJcKl3Sb96PVPa6811XVaPo1h8RSxFKNWlLmjJXTQAAAVqVOtTlTmrxas13TMzweJ4ZzpSpPSL5ovvF9H8rxZlmYUcwwlPE0X7slf0fVfJ6AAEoxlFxkrpmL8NI1MdKdGsoUnqlZtrultp2d799tcLSnRoQpym5tK13u/N2CxYsgAAAACQXuPUDRljYQwYhbCBgTd9CKsgAFpKwADAorRjAKnwkPhQAVvhAj8SAEPYo7MAK26AAKUrqwAAA1dAAAAALRm4AAAAAAACB7ABJWdim7qwAE48yGgA4JxmNwue0J4OPNO7XLe3NGz5l222vomkzA5phMbRdWlPRfEno4tbqSesWuqf5HilxrQxsVleBmpQTvOS2bW0U+qW7e17dmAENgAABgLYAEMA4O8R6mFccJmcnKnsp7uP+Lq157rzW2c5HlvEGCUKtpResZLdeafby2ZxLwtjcixHJWV4P4ZLZ/wCT7r81qD2AAADjfg9YqMsfg4/vFrKK+0u6X83ddfXfg3iyWVVlh8Q70ZP/AMBfdeXdfNa7wnGcVKLunqmuoAAAcZZAs2y9+zX72GsfPvH5/mkeHmffVsS8vrP3Z/DfpLt/29+aXcALlwscW8WZzlmMeHhCMI7xla7kvnp6q113OH87oZvgo4ino9pLs+q9Oq8gAAAAACQZZlOOzOuqGDpOcuyW3m3sl5uxnPDObZPb69QcE9E9Gr9rptX8rgFwN0JDEdBAwZFXYABNa3FqgAl8LApfCMAqfCQ+FABW2QEPiQAh7FHZjAK3xAAC02ITUl5gAADWoAAAAEHeIAAAAAAAC2B7ABUXvFN6gAE4X1Q1caA8J8F7bPJVntTg383aP5NniFjI4riLEuO0Wo/+ApJ/jcAEQ2AAAGAPYAWwwA4T41xmR1FTfv0G9Y9u7j2fls+vdUa2V8RZbpapSnunun5rdNf8FdzjHgfE5JN1qV50G9H1jfpL9Hs/JuwwAAAOOuEORyzDBx03nFdP7y8u/bfa9uBeLfq0o5fi5e4/hb+y+z8n07PyegAAAceZHLLswWMoaQqO+n2ZLf7918+xwxncc3y6Fa/vrSS7SX6PdetuhdFzcsWQHEuQUs4wTovSa1i+z/yez+/ocP5viOHszcayajflnH9V5rdd1p1KVWFWnGdN3i1dPugAAAAAJCV9EcE8N0siymFO37ySUpvrft6R2XzfVmY4DC5ng54WuuaE1Z/5rs1un0ZnWV1MrzCtgqm8G16ro/mrP5gAXLjRFADCWxBaAAE1dENUAFT4QKfwjAJ/CQ+FABW6AU/iQAS2ZR2YwCr8QAAXtsQmpadQAAJ/EAAAABTfQAAAAAAABbA9gAqrYTs0AAE4J7DVhpFjwkwsaGX4rGz0TaXygrv/ANK/AxmJlisTUry3m236t3ABEPhAAAABgAABw/xFjckxKrYaWj+KL2kvP9HuvvMi4gy7iHBOVOz0tOD1avumuqfR7P1ujjjgCplrljMAnKh1W7h/nHz3XXuAAAA0mrM424S/s2o8Xhl+5k9V/K+3o+nbbtfgTiv61COX4qX7xL3W/tJdH5pfevNagAAGZ5bhcxw7w+JjzRdn2281qv8AIwWX4PAUvZ4emoR8vzb3fqyxZAAHiBw0sTReYYePvxXvLvFdfWP5eiPDviNp/wBmV5ecH+cf1XzXYAAAAABnCWCjjc9wlCSunON/RO7/AARmlfkiooyzFXqct9GeMGCjRz2FaK/iQTfqm1+SRuWABhHcAewPViAAHqQ3sAFXZAQ2Q9gCfwkPhQAVt0BT+JABL4WUdhgFTWTAAAIT5l5gABU+IAAAACLs0AAAAAAAB0BgBUV4hF3igABjSY4dhxaMA/7K8PZ1HpKcJffN8q/BoABEdkAOSOaJowAEDAHsAAGV5ri8sxMcThZ8s1+K7NdU+xwpxfg8/wAPy6RrJe9D9V3i/vWz6N8c+Hjwynj8tj7m8oL7Pdx/u9106abAAABXoU69KVKorxkrNPqmcT8PV8ixqnSb9m3eEuqa6N91+O/e3CHE8M4wvJUdq0F7y7r+Zfr2fk0AANoubljxLweLVGniKdR+y2lG7sn0dvPbXbTucEZ4syy2MJu9SnaL9Psv5pW9UwAAaTVmcYZDPJcwVbD6U5O8Wvsta2+W68vRnC+ewzjARq/bWkl59/R7r7ugAAAAAM8NsNVrcTYeVON1G7fkuVq/3tIz1O+nYyNT5433ueNP+/4X/A/zEtAYAdBK4C2HsEdWIAAFpIAKu4EdkPYAn8JD4UAFX4gKfxABP4WUvhGAT1kwAABO2xCSkgACruAAAAARd0AAAAAAAAMAJK8QpvSwAABKLlZLc8SpLL+GsNgIvrGPyhH/ADsAAiPwoHNIbbNDQIyvuD2AQMBD2AAB7GCxuIwVeNfDzcZx2a/r71szg3jfD55SVGraGIS1XSXnH9VuvNanHnh8pqeYZbH3t5QXXu4rv3XXpro2raAAABmmWYfMsJPDV1eL+9Po15r+tDEUMfw1mqs7Tg7p9JL/ACa0a+Rkec4fNsHHEUfRrqn1X+T6oAbRctcsgMfgqONw08NWV4yVn/n6rdeZkmLq8OZ86Vd2inyS9HtL8penqaNaAABnmUUc1wM8LU67Ps1s/wDPurrqZBmdfh3N3CurRvyzXl3+W67r1ITjOKlF3T2AAAAAIU51ZqEFeTdklu2zg7hyhw5ljnXaVRrmnLtbpftFfe7szLDOrBSS2Mswlp89tEeLeOjiOIPYx/5dwjF+rvL8pIewMAEiO9gFsEtiK0EAAD+IAJ6yAACfwkPhQAVfiApfEAFT4Cn8KGAPUAAAIy5Xc0aAAq9AAAAACm9LAAAAAAAADAAaKe4AABwxg/recYaj0c439E7v8EeMOLU8bhsMn8MXL/wJ2/8AbQAOhH4USlfRDG7gG5GV9BgLYAEMAAGFCvVoVY1aUnGUXdNaNNdUcEcd0s3hHCYtqOIXyU/NdE+6+a0ulx9wCsUpZhl0f3m8or7Xmv73ddfXdq2gAAAcUcO0s5wfs9qkdYvs+z8n1+T6GQ5ziuH8xaqJpXtOPp+q6d9r2Zh69LEUo1aUrxkrp90NoepYsgAPEnI1OlHMaS1jpL06P5PT5rscA528fl3sKjvUpaesej/T5K+4AAHiLw/7aisxor3o6S849H8uvk/I8O+IHicO8vrP3oK8fOPb/t38rdgAAAADwv4V9pP+1sRHRaQT6vZy+Wy87vojxQ4s9tUeU4WXux+Nrq19n5bvz06Hh5xXRznLIUZy/f0klJdWlopLvdb9n6oz/PcJkuBni8TLRbLrJ9EvN/hu9DMMdVx+LqYqs/em3J+rf5DB7ACRGPUlowEEy2gAAEviQAS1YRV2gAJ/CQ+FABU+JgUviACr8KIL3UMB7AAAAFKV1YACr0AAAAAIOzAAAAAAAAGABUVpEdGgAHsAeGOE9vn0Z/8APuMpfhy/+3HiRi/rPEVddIcsV8kr/i2AA1oX91IbAAExO6AWw9gBbDAABgQnOnNTg7NaprRprqjgTjuGawWCxjtXWz2U0v8A27uuu66pcfcBLEqeY5fH95vKK+13kv73ddd99wAAA4l4Mw+cYinXU+SS0k0r3X+a2v272SMqyrD5ZhY4bD35V3d3d7/8MrLyLFkAAGKw1LFUJ0KqvGSafozLcRW4az9wqv3Yvll5xfX7rSQmpJNO6AACpThUg4SV09GvIzbBYjhrOlKi9E+aD7x7P8U/v6mW5hRzDCU8TRfuyV/TuvVPRgAAABwxkNXO8yhhYfDvJ9ord/ovNo4uz6hw1lEaWGSU2uWmu1lrL0ivvbV92VJSnJyk7tlGvVoVFUpScZLZptNejWpjMwxmNmp4qrKpJdZScn+LYPVFhgWEri2JoWqBBLWQAAA4pu4A9gIK8g6DAnsR+FABLWTApbgBUi2lYtoPYCfwsAAAAg7SQAFXoAAAAAEXdAAAAAAAAMACqtEwWqAB7AHhDhbzxWIfRRivndv8kZ9ifrWZ4iuvtzk/vb/QAENe6MAABMAewAAAAAQnOnNTg7NaprRprqjgTjuGawWCxjtXWz2U0v8A27uuu66peIXAikpZnl8NdXOK/wDSku/8y6773uAADG7lrniM81o04zpVH7B6NLSz83u0/uvo90cC8T/2hQ+p4iX72C0b+1H/ADXXy17gAAHiVkvtKMMwprWPuy9Hs/k9PmuxwHnSx+WKjN+/StF+n2X92nqgAAOM8hWa5c/Zq9SGse77x+f52PDrPXQxEsvrS92esb9JdV81+K8wAAAA4IyKlw/k7xOK92clzTb+ykrpfJavzb8jijP6ud5jPEy0jtFdorb5vd+bGtBoBbA9QsJXBEldEQDeQC2AAACXwsCktQWwwKmwtkAD1YFHcAFsD2Aqv3QAAAAWqACr0AAAAACm+gAAAAAACAYAE1eIQ1igAewBwC/qHCmJxj3vOS/7dikvxTJq6GgBLoSfQeoAACFqgGAAAA9gAKVWpSqRqU5NSTumtGmuqOBeNoZzRWGxLSxEflzpdV59181pe3iHwOsK5ZlgI/u3rOK+z/eX919V0322AAbLXLBisLRxVCVCtG8ZKzX9f0jOMsxfDmaL2cno7wl3X+fSS/Ro4ezyjnGCjXhpJaSXZ/5dv+HAAAxWGpYqhOhVV4yTT9GZFianD3EDo1naN+SXo9pfk/QAAA44yaeVZnHF4fSM3zJ9pJ3f6Nf8A4eziGbZfTxK+LaS7SW/+a8mgAAAPDjhtZpmX1iqv3VGz8nL7K/V+iT3PFTiXkispoPV2c/TeMfnu/l3YDQ0WGFiwogIewtGB0IoBbAAABU+ECmtAWwwKnQWwA9gCjuwAQPYCs9kAAAAEfhQAVegAAAAARdmAAAAAAAgAABqzsU9gAewBiP+E/w9jHrKK/8AJ5X/ACYyUbMaLFhKyuMAsWLABFiBgIewAAPYAAw2JrYatGtRk4zi7prdM4N4tocQYN06tlWirSj3W3Ml2fVdHptZvj7gt5RW+t4WN6En/wCAPt6P7L+T6XAbNywAcR5DRzjBOhLSS1i+z/yez/zSMjzXFcO5o1Vi0k+WcfLuvNbp9e9mUK9LEUo1aUrxkrp90wAAPEvJ1anmFNa/DL/21/mvuOCs3/tHKYc0rzp+7L5bP5q2ve4AAHEWTxzXLqmG+1vF9pLb/J+TZwLm9TLczeCr6Rm+Vp9JrRf/ACL9V2AAACjRnWqRpU1eUmkl3b0SMLRw3B/Djc9XBXfTmm+n32XklfoY3F1sZiJ4is7zm236v+tOwA0AKLYkkAIew9wH8JFe6AgAAAqfCBT+EEMCp0OgBP4WAUeoAIHsBN80gAAAAS0QAVdwAAAAAIO8QAAAAAAQAABUWpT3ABgHiPbB8O4XBrR3ireUYtfnYGk0ODRysUO5N9B6gABYBCBgIYAAMAAMuzHE5fioYnDS5Zxd1/k+6ezXVGQZ3gOJ8rfNFO6tOD6N/o94v9U7cY8KVshxnKrujL4Zf+2vzX4rXyTHqWAADj7hn67Q+vYeP7yC95fzRX6r8Vp0R4f8TLD1Fl2Il7kn7jfR9vR9PP1AAAzLA0sdhKmGqbSTX+T+T1ODMdVyfO3g6+im+R+Uk9H9+no7gAAHiJkssLjI4+irRnvbpJf5rX1TOF85Wa5ZTrt++tJf4lv9+j+YAAB4WcOrFYuWZVl7tLSPnK2//bq/Fp9DxR4hWNx0cBRfuUt/OfX/AMBWnq2ADDlViyAACZugl8JFe6AgAAAq7ICHwoEMCp0OgBV+EAo9QAQBUdogEYOTFSijkj2HTi+g6KexKnJABFXaQAFT4gAAAAAKb1sAAAAAACB7AABVWhD4gAYGS4f6zmWHo/zTivvaPF3EXrYWguik/vaS/JgASdwAAAEQ1QMBDAABgAAcP59islxscVh35NdJLqn+j6PU4y8Q3nNB4PC0+Wk93JJybXbdR+Wr7rVHs5cvNbTa/mWAAAOOeGf7NxP1rDr91N/+Ay7ej3XzXRHBPEqzXC+xrP8AfU1r/eXSX6Pz16gAAeJGUuhjKePp7T0flJbP5r8jhzNVmeWUsRf3rWl/iWj+/f5gAAZ1llPM8DUws/tLR9n0fyZwPmdTKs2lgsRopvlafSa0X46fNdgAAMNh6uJrQo0leUmkl3b0Ri6tDhHhlRhbmgrL+9N9fvu7dlYqTlUm5yd29W/MAHqWFsAABJaC2CWxHZAIAAAKvQCPwoEPYBpM6AFb4UAUd2ACAKzu7AEY2jYACUIy3JUmthqxa5Thy6sACesgAAAAANhO6AAAAAAED2AACSvEW6ABgcB4f2/EWFj2bf8A4DFv9DxSxHtc85L/AAQivvu/1ACWiGAAADCHYAFsMAAAAAZuZfUw1LFU54mnz00/eje112uf2Xkuf5HGhQivYyXu8qtyvuuzT3XXVPcz7IsVk2NlhcQtVs+kl0a/Xs9AAADHYGhjcNPD11eMlZ/8DzXQxOHx3DObqz96Lun0lH/JrRro79rmU5pQzPBwxNF6S6dU+qfmv+DswAA4hypZpltXDfaavHyktV/k/JnhvmroYypgKm09V5SW/wB639EAAAeI+TPD4qGYUlZT0flJbP5pfevM4XzlZrllOu3760l/iW/36P5gAB4U5CsRjJ5jUXu09I/4mtX8l+afQ8UM/wDruZLBUn7lHR+c3v8Adt5O4AC1BAAAtiWxHYJnQYASkkhSUtgACrugI/CgQ9gBbAFboAUtwAQDshu7uBBXkgAANyyAAHqwAAAAACm9LAAAAAAC2B7AAASVmADA8LMP7TPXP+SEn97S/U47r+34hxUuzS/8BSX6ABU2sMYGyHZjGABDdD2AFsMAAYAANlrlg4M4urZDirS96jN+9Ht/eXmvxWj6NcQZFgeJ8sXJJNtXhNa2f+T2a/VGYZfiMBiZ4bER5Zxdmv8ALyfRgAAHF3Dsc4wVoaVYaxf5x9H+Dszg/iCeTY50MRpTk7ST+y1pf5bPy80hNNXQAAcY4Opk+fRxlDRSamu3Mn7y+/V+UjBYunjMNTxFN+7JJr5gAAZ5lkczy+rhXvJaeTWqf3/gcA5nPL81lgq+iqe7Z9Jrb9V6tAAEKcqk1CCu3ol5laVHg/hi0bc8VZf3qkuvpfX/AAoqTlUm5yd29W/MAGWBAAAthkQkDACbuzYjLmQAFTcCPwoEPYAWwBW3AKXxAAgKrsrAFJagAAAAD0QAAAAAARdmAAAAAALYHsAABUWotkAMDwhoKWJxVXtGK+9t/wDtpndf2+ZYirf4pyf3yYAT3ABgzoNDQBEewAAAD2AAY9SwAcAcaPKayweLl+4l1/kff/C+q+fe/HfCEM8wnt8Ol7eC93+8v5W/xj5+TbJwlCTjJWa0ae6YAAB4h8NWf9p4df41+Uv0fyfc8PuI/rNH+z68vfgvd849vVfl6AABx5lP17KZVIr36XvL0+0vu1+SPDXNvbYSeBm9Yar/AAvf7n/6UgAADxDyqWDzGGOoqyqdV0mvyurP1uzIc0hmeXUsTHdrXyktH+O3kAB4YZH9ezb61NXhR1/7efw/dq/JpdzxVz14nHxy+m/cpavzk1+i09WwAFqAAN2G7hzMjJMYtwe6BgD2AjKzN0AFT4gIfCgHsALYAq/EAQ0kgAArO8gCj1AAAAAJ/CAAAAAABTd1YAAAAABbA9gAAKkbrQSsrADA8KoqhleLxL/m/wDSY3/Um3J3e4AT1Z0ARuWSDRjHsAIewAAADABs3LAAHhrxlflynGS8qbf/AKR/8j93Y8S+DueMs2wkdV/ES/8AS16fa8te7AAAKtOFWDhNXTVmu6e5nuV4jhzNlKi7JPmg/Ls/TZ91r1MizelmuBhiafXRrtJbr+ulgACUVJWeqMM3w1xNyN2pqVn/AIJbN+is/VAAAHE+Uf2pllTDpe9vH/Etvv29GeGubOniKmAm9Je9H1W/3r8gAOHMPS4Y4WeJrr3rc8u7b+GP5R7XuYnEVcTWnWqu8pNtvu3qwAYtgAJOyGXC4KV9BfED+JAwOhLYAjOwmmtACWrAh8IAAgCesmACd0ABJ2jceoBR6gAAAAFV6JAAAAAAARdmAAAAAAtgYAAAAAcIr6twRiqq3aqv58vKvyGAMYwBlrjVywMAR0QA9gAAYA2WuWAACMnFpxdmjgPiyOeYN4fEte2grP8AvLbm/SXnrs7HH3CP9i4v29BfuKjdv7r3cfT+Xy9LgAAHE2RU84wEqO01rF9n/k9n9/Q4PzupkuZOhiNISfLJP7LWifyej8n5I0aAAPE3KualSx8FrH3Zej1X43XzRwbmv9o5RSlJ3lD3X6rZ/NWb82AAAcV4SpkmfrFUNFJqcfW/vL77/JmDxVPFYeFem/dkk180AcF5L/a2c0qEleEfel/hj/m7L5ni3nVoUstpvf35flFfm/kmABuwACTuABuLVg/iBgLYewAJtbEaie4SdkAU9gABAD1YAUneNgAK8tkAFJ+9YAAAAAqO8gAAAAAACDugAAAAA6AwAAAAApL6t4dN94v/AMmqW/J/qMAlsPcARowBq49gAWyAQ9iEJTkoxV2+hheFM7xSvSws2u7i0vvdkYfwx4hq254Rh6yX3+7zFHwizGS/e4iCfkpP81H+upS8HqSl+8xja8oW/wDbn+RT8IsuUv3mIm15KKf32f5H/EJMk/5+1fvj/wDID8JMkf8Ay9q/fH/5An4R5K4vlrVL+sf/AJBfmjPPDvJcnwjxGKxkkui5U232Svq/uXVtLUla+mxl/hnnuMwixCUYX2jJtSfnazSv5tPyMV4d8R0P+YfmXeMov8L3/AxeS5lhE/rGHnBLq4tLTztYAADLcxxGXYuGKw8rTg7r9U/JrRrsYTE5dxbkj5l7s1aS6wkv1T1T6qztrYz3JcRlGOnhK+8dn0a6Nev4PQAAA8ReHeVrMqC0ek/0l+j87d2cA8QfX8F9Uqv95TX3x6P5bP5PqAAZvl8MwwNXCy+2mvR9H8nZnh3mE8HmdTA1dOe+naUf+Bf8AAADj7Kfr2VOrBe/S95en2l92vyPDbNvbYSeBm9aeq/wvf7n+aAPCvKoYTLKuY1dHUejfSMd383e/ojiHNp5rmdbGS2k9PKK0ivkkr+YAPYWwALYk9BgAC3D7R0HsAtgAATa2G2wCm9bAACAlomABCVpABKSirsk3J3ABOzuboAAAAJzSWgAAAAAAAQdmAAAAAHQGAAAAAHENqHAWGh/Mqf4+8MAn8IwDoLYAewtUAIjsQpzqSUYK7fRbmXeH+f42zVDkXefu/h8X4GX+EMFZ43E38oL/wBud/8A0kwPh7w9hLP2HO11k2/w+H8PwMNgsNhY8uHpxgu0Ul+Ris3y/CaYivCH+KSX5sxHiBw5QWuJTdtkpP8AJW+9oreK2RQdoxqS9Ir9ZJ/gVfF/CqP7vCyb85Jfik/yKnjDNwahg0n5zuvu5F+Z/wARexv/AEDR+9j8X8b/ANA0fvYvGDGJ64aP/gTM5zvG5viXiMXPml0XRLsl0X9M4OzjIsprfWMdRnUqL4WlFqPom1r5vbouphvFHh6rbnlKF/5ovT15ebfyv5mG404fxCXJi4L/ABPl/wDSrFDE0MRHmozUl5NP8jG5DleNv9Yw8JN9XFX+/f8AEx3hdkGIu6UZU3/dldfdLm/C3lYzHwhxkLvB4iM/KScX6XXMm9uy9DMuDs8y9N18NKy6r3l63je3zsABwZxRPIsepy1pTspry6SXmvyuupxtw1S4gy1VcPZ1Yrmg19pPW1+z3Xnbo2Si4txas0AABicPSxNGVGqrxkmmvJleniuF8793Xld1/eg/81p5NeRg8XRxmHhiKLvGSuv6/PzAAONMLUynPoY2irczU1/iT95fPd+phMTTxWHhXp/DJJr5q4AAE4RnFxkrplCUuGuJeVu0FK3rCW33Kz9UaNaBgsJUxmJp4ekrym0l6t2OOsXTyPhmOAoOzmlTX+FL3n81o/8AEAA9QAFsS3AA3BbC1Q/iOg9gEElqwAAATswOgwEBU+EAApy5lYCUklqTm5MAApSurAAASi3sx8yOZ9zml3HJvqAAAAAAAAQd1YAAAAA6AwAAAB7AHiHejwxg6PnD8IMYBPYYA9joACAMn4Bz3M0pQpckP5p+6vkvifyVvMyjwmy6glLG1HVfZe7H8HzP1uvTtl+UYDL4cmEoxgvJWb9Xu/mZnxXkuXXWIxEVJdF7z+6N2vnYzDxbwULxwdCU33k1FeunM3000+Rj/FDPq91Scaa8o3f3y5vyXyMbn+a426xGInJPo5O21tr29dAABs3LAFOpOnJSg2muq0Zg+Ms+wlvZYuX/AG8+ZfdK6MD4tZrSssTSjUXleL+/Vf8Akpl/ivk1eyxMZUn6cy+9a/8Akpl+eZbmC/2WvGb7Jq/3br7jNOGcozO/1rDxk31taX/gSs/xM18IsPO8sBXcX2mrr71ZpfJmccGZ1lacq9FuC+1H3o+umq+aQB4YcV2aynEy/wCvbf3uP6x+a7I8T+Ffq9b+1cNH3Jv312k9pekuv97/ABAAAHHOQf2lgHWpL97Tu15rqv1Xnp1PDjPlTqPLar0lrD16r5rVed+rAAPEHK/rmUurFe9SfN8tpfhq/Q8OMz+sZbLCyfvUnp/heq/G6+4AAA8TMp56NPHwWsfdl6PZ/J6fM4JzX+0Mop8z96n7r+Wz+at87h4V5QsVm0sXJe7RWn+KV0vw5n5Ox4m5x9ezp0Iu8KK5f+3t5fjZP0AAQACGAAMWxEfxC2HsAgqL3gAAAIO8Q6DAQFZ6JAACbWwqytqSq9htvcAAE7PQjJSQAAA6aew6bQAAAAAAAARdmAAAAAAwAAAB7ARi5SUVuzxacYYTCUl3l+CS/UYBU2GAPYCrQq0ZctSLi/NWAz7htZNkOGlWX76tJyemqSWkfLe7Xf0MPhq2JqqlRg5SeySbb+SMi8KMwxSVTHz9lH+Vaz/yX3trqjJeDsmyhJ4einNfalrL73t8kkZvxRlGVJ/Wq6Ul9lay/wDAVd693ZeZmvi65Xhl1C396f8A8iv/AJL5GacVZxmV1icRJxf2VpH7lZP53BDAAHLsWuAAARk4tOLs0ZXx1n2X2UK7lFdJ+8vTXVL0aMq8XMPO0cfQcX3hqvudml82ZXn+WZpG+Drxn5bP5xdmvmjOeCslzW8q1FKb+1H3Zeumjf8AiTM78KcwwydTATVWPZ+7L/J/en2RicLXw1R0q8HGS6NNP7mUqs6U41KbtJO6a3TWzRwznOG4oyWVPEpOVuWpHztuuye6fRp22OIsjrZNmM8JU1trF94vZ/o/NNAAAHGOT1MmzSOJw+kJPmi10kndr5PVeT8jJszp5lgKeKh9pars9mvk/wDMui6AK1KFWnKnNXTTTXk9GcM1JZJxK8JUejbg/n8L+bt94AABmeAp4/B1MLPaaa9H0fydmcB46eW5xPA1tOe8Wu0o3t+q9Wg4PowyDhOWNqr3pJ1Gvl7q+aS+bK9aderKrUd5Sbbfm9WADOgAHQYy1yyLFkNaB0I7j3FsPYBBVWqAAAAg7MFsMBAVnd2AAAAAAE2tiNRPRgAAA0mOnFjptbDQAAAAAAAU5XVgAAAAAYAAAA9gMspe1x1Gn3lFfe0eL1S9XCU+ym/v5f8AIYBU2AB7C2PC/g+hh8FDNMTC9WesLr4Y9GvOW9+1rbsz3IsFnODlhcVC6ez6xfdPo/z2ehm2WVssx1XB1vig2vXs15NWa8meGvC/9pY767XX7qk9P70ui9Fu/ktmzinhGlxBVw/tqjjTp810t5X5ba9LWfR7/MyrI8tymlyYSkoLq+r9ZPV/Nme+JOTZanCjL21RdI7fOW33cz8jO/EbO8yvCM/ZQfSGj+ct/usvIbbd29SIC2GAA5dhRuAAAAEJyhJSi7NGTeJOd4C0as/bQ7S3+Ut7+tzJPEjJsxShVl7GfaW3ylt9/L6GY5Tl+a0eTFUo1Iva/S/VNar1TRn3hNOKdTK6l/7kt/lLb5O3mzIMyx3CucReJpuKek4tWvF9V3tumt9r2bOO+HqefZVHE4X3qkFzQa+1Fq7XzVmvNW6sAAA4ryj+08rqUYq8170fVf5q6+Z4c5z7DEyy+o9J6x8pLdfNfil3ANwPEnLnQxtLHU9OZWb/AL0dn935GTZhHMMvpYpfaSv67NfJ3QAABx/gJ5fm8MdR057ST7Sja/6P1ucJQXEFTCRo/wDL63y/m/8AAdb+h4rZlDC5ZRy6lpzu7XaMLWX32t6AAbsAAexuxiLDsywHQRLcWwAgqK8QAAACEroWwwEDaSJO7uAAAAAABGo0KSktAAAAHFMlC2wAAAAAAAnYTTQAAAAAwAAAB7AcNU/aZ1hI2v8AvIf+lK/4Hi3O+ZUIX2hf75P/ACGAVNjqALVGAwssViqWHjvOSj97SK7hhMPGlSVklZeSSsYHGOVTlbumeJ3DtbG8RYWOGj71eNvnF6t+Si1fyRk+U4fK8DTwlBe7FfNvq35t6nFOeyyXLJ4yNJzs0rXslfRN+V7LTW7XqZ7xjm+ctrEVbQ/kjpH7uv8A282B0IgLYYAN3FEewAAAAAZJxXmuTtLC1fd/lesfu6X6tWfmZB4o5djLUscvZT77wfz3j89F3Mbl+X5vhlCvCNSm9uvzTWq9U0ZJk8cqw7wtOo5U0/dT3invG/VX2urq9rux4k8M/wBm4/63RX7qq2/JS3a+e6+dtEAADDjXKamU5rHGYd2jN80Wuklq/wAdV626GRZrDNMvp4mO7Wq7SW6+/bysAbhxlln1/J6sEryj7y9Y/wCauvmeGWZe0w1XBSesXzL0ej+5r8QAADjbKv7Qyioor3oe8vlv96v87H0WKFXFTxdSorwoWUX51L3S9FF/+BHiBmv9oZ9Wad40/cX/AG7v/wCTXAB7CAAGDEPsW0EB0ES1SI7ACCSurAAAACdtiElJDAQ2kidTm0QAAAAAAAJtbEavcTTWgAAAE4W1QAAAAAABCVnYAAAAAYAAAA9gODIKef4Rf30/u1PFWfNncF2pxX/k0n+owCpsPcYBwhCMuIMEn/z8h+DTM6k1FW7GUVZynr3PFLOMZlE8FisJLln762T0fI2tU97Laz8zLfF/FwSjjcOp+cW4v1s7pv5pE+POGs7wVXB16jp88WvfVt09brmjpurtdCtTdOpKF07Nq6d07dn1XZgMiAthgDdxRB7AAAAAAGR8T5nk0+bCVLR6xesX6r9VZ+Zw74l5bmPLSxf7mp5/C/R9PSVvVmb5ThM3wUsLiFeEu26fRp9/62OJvDrMcqUq1D97RXVL3kvNfqtOrsAAwOJsmWbZdPDr494vtJf57P1OAs5eX5g8FXdo1Haz6TWi+/Z+duwAgaTRl18g4r9i9IOXL/27P4fuum/QAAAaTVjgXLcF4feH9fG1JKKlz1W3/eajSXnpyL1b7k5OcnKTu2ADAACQMQgGD2EP4SOwMEBNWYAAAAnbYVRPcunsEppEpOW4AAAAAAAAJtbEavcTTWgAAA0mrDVnYAAAAAAAhK6sAAAAAwAAAB7AcAQc+I8Kl3l+EJM8TZKXEE0ukY/kMAnsMYBkmMWDzLD4h7QnGX3STMzo+0pKSMuwdqqsjxpxcZ43C4ZbxjKX/gTS/wDbR6AARAWwwG7iiA9gAAAAAAPD7i+tl+Nhg8TUboT0V3pB9GuyezWi1v0Dijw6y/Nuavh/3VZ9V8LfmvPuterTM4yPHZTiPYYunyvo+jXdPqvy62YAwDxByN4TGRx9FWjU3t0n3+e/qmcJ54s2y6M5P95H3Zeq6/Na+t10AFqg8Tsu5atHGx6+6/Var8L/AHGQ5iswyyjib6yWvqtH+KYAAGV4GWPx1HCx3nJL0u9X8lqfSdzGpg+E8PluH0jOceb/AAQWiflzcvzRwdmn9oZPSm3eUfdfqv8ANWfzADdgABI6DVxIA6FgW5vEhsDBAVI3QAAAABuAAAAAAAAAApNbEaiejAAAKi1uAAAAAAAJ2YmmgAAAAYAAAA9gPDiClxJQfZT/APSJL9TxHmpcR112UV/5JF/qMAn8IzoAHh/xFTznJqalK9WmlGa66LR+kl173XQnKhhqUqkmoxSu2+iXVnFmeyzrN62M+y3aK7RWi+/d+bYAdAjsB0GDdxRsAMAAAAAAA8POJ/7Xy/6vWl++pJJ95R6S9ej89epiuLsRw7mTwWZpzoy1hUXxJdpLry7N/FZJtO5WoZXn2BSko1aUtv8AgPdNfJo4s8N8VlqliMDepS6r7UfW2681t1XUHsAGb5ZRzLBVMLV2kt+z6P5M4bzKtw/nDpYnSLfLNfk/lvfs33NGgEHF2W/X8nrU0ryS5l6x1/FXXzPDHMeehWwUnrF8y9Ho/uaX3gAAeFeWfWc5eJa0pRb/AO3paL8OZ/I8YpRzzH4nBX92MeReUl71/lJ/geHGYSw2YVMBV059l2lHdfNXv6IAYgAAYXL6AHQYjqLYhuDAAnGzAAAAAAAAAAAAAAACFRrRiaa0AACorxAAAAAAACErMAAAAGAAAAPYDwzhzcQ03faMn+Fv1PECanxJiX/hX3QihgEthgwDKc5x2VYhYjBVHCf5rs07pr1RnnHOd5zQ9hiavudYxSSfrbV+l7dbXAAYiOwHQY3cUbAAwAAAAAADIc5r5Rj6eLo7x3XdPdfNfc7PocS5VhuKciU8M7ytz0359n67NdHr0Mk4hzLIcS5YeTWvvRezt0a79L6NdzhjjXL89ioQfJWS1g/zi/tL8V1SOLfDnC5nzYnA2p1t2vsyfn2fmt3utbmPwOJwNeWHxMHCcd0/627PZgAeI+Q80Y5lSWq0n+Sf6P5HAWe/XsB9WqSvUpaesej+Wz+XcAGk0Zd/wg8WOjtBycf+3Z/D911f0AAA8N8PTyzh2tmNVfFzS/7dgmvzUvvMTiJ4itOtUfvSbb9W7s4soTybiKOMpLSTU16395fN3v6lCtTr0YVabvGSTXo1dAM6AADGdA6AAwFsLSQMACUVJWGraAAHJeN0AAAAAAAJN6IVCTHRkhxa3AAAAAjJxehGSktAABq6sAAAAAAABCVtGAAAAMAAAAewHhgr8QR/wyOPP+SixXqv/SUMAeww3AAAAQtgNEhu4o2AAYAAAAAAAeFvE3sK7yuvL3Z6w8pdV/29uvNeZ4o8M/VcUszoR9yo7S8pd/8At7r5p31ZTqTpzU4NqS1TWjT8jhHxNUuXC5s9dlU/+S/+S+9bsz/hvLc/wqjXV3b3ZrdX6p9U+2z9bM4k4Vx+RV+TERvB/DNbP/J90/ldagBicNSxNGVGqrxkmmvJlKWI4Xz2z2i7P+9B/wCa18mvIpVYVqcalN3i1dPyYC2DxNwDp4ihjYaXXK/Vap+tm/uMlx6x+X0cT1lFX9dn+NwACMHOSjFXbONJRyXhCngYaSkow+7WT+dnf1DxFyz6zlaxEV71J3+T0f6P5Hh7mf1rKVRk/epO3yeq/VfIB6sAAHsM6AjL8vxOPxMMNhoc05aJL+tu76FbwdziGG9pCrCU7fDqvkm1a/rZeZiKFWhVlRqxcZRbTT0aa3TOoxIOhI3QwACcLoAAS0RUjbVAAAAAAAUoJK/UHsEqSexKLW4AAAARk4vQjJSWgABJWYAAAAAAAEJ30YAAADAAAAHsB4V/8j1/4JfnE4114gxX+L9EABJAbgAAAgCUrijYAAYAAAAAAAUqk6U41KbtJO6a3TWzRk2OwvFuQOFdayXLNdpLqvwku23RmbZZXy3G1MJXXvQdvXs15NWa9Q4S47xmSSVGpepQ/l6x84vp6bPybuUa+V8Q5deNqtKejX6Pqmvk1uu5xlwBiMmbxGGvOh36x/xeXaX320uFho8QsjWLwSxtNe/S3849fu39LnhznSrYaWAqP3oax84vdfJ/g12AQcaZf9dyWtFL3oLmX/bur/C6PDPMFVwNTCN6wd16S/4Kf3gAHBOXPHZ9hqdrpS5n6R1/GyXzPF3MHUx1DBp6Qi5P1k7fkvxDE4eniKE6M1eMk0/RqxwViJ5Vn88DVfxXg/8AEtn87NL1AQAAPYYB4P5DGlg6maTXvVHyx8op6/fLT/t0WJpOfInqeMWRU6NajmdKNuf3ZeqV4v1aTXpFBuC1CS0IO6GAAFSPVAAlcGk1Yas7AAAAAAFKKbuLYHsA0mrMlSfQsAAAAJtPQjNSQAFRe8AAAAAAAAQnfQAAAGAAAAPYDwr/AOR6/wDBL84nGn/I/wAV/i/RAATWoBuABYAigG7kY21AABgAAAAAAAcB8Sf2NmaVSX7qpaMuy7S/7d/Js8UuG1isKszoL36atLzj0f8A2639zd9kPYMh4hx2S4lV8LL1i9pLs1+u66M4a4qy/iDDP2ek0veg910+cX387Oz0ON/Dh0ebHZXG8d5QW684+X93p000WyC5UhGcXGSunoZlh6vDWfKVL4Yvmj5xfT7rxf3mFxNLFUIVqTvGSTXowQSipRcXszhiTybiiWDk/dblD9Yv52X3gAHhDl/PisRjGvhSiv8At53f3cq+84tx/wBfzvE107rmaXpH3V+CA8QcHPAZvSx9LTns1/ihb9OVmBxcMZhaeIhtNJ/eh7C2AAw+GrYioqdGDlJ9Em39yMv8NuIMYk3TVNd5u34K8l811ML4PXV8TivlGP6t/oUPCbIoO85VJeskvyin+J/xCzh3+WX/AIEyp4VcPyi0lNPupa/imvwMX4O4CS/2fEzj/iSl+XKZRlyyjJKWDTu4Rtfu27t/NtkcWnV5bniVQWK4Vqz6x5Zf+TJP8GwHqRQWuQ0dhgAAThYAgryAqR0uAAAAAAUdhbA9gAlFS3JUX0HCS6AAAAEavRmjQBVWwAAAAAAAAQnfRgAADAAAAGB4TxTzupfpTl/6VA4wk5Z9i2/52ABUWgwA0DQAERWgSdxRAAAYAAAAAAAB4d5/DN8qeBxL5p01ytP7UHor99Pdfyb3OLcgnkuZ1MM/h3i+8Xt81s/NAYHHYnA144jDTcZx2a/rVd09H1ODeO8NncFh69oYhLbpLzj+q3XS6vbjnw8jjebHZbG1XeUNlLu12l3Wz9d5wlCTjJWa6Bujj/Jfr2XfWKa9+ld+sftL5b/J9zw3zhVcNPATfvQ1j/he/wBz/MAPEPCzwebUsdS0cknf+9C36cpg8TDFYanXhtNJr5q4AHB6WTcG1Ma9JSU5/P4Y/fZfePUDj3LfrmTTnFe9T95ei0f4O/yR4bZj7fLZYZvWk9PSWq/HmGAGS8MZpnE7YSk3HrJ6RXz/AEV35GSeE+BoJTzCo6kv5VeMfv8Aifr7voYPL8HgKXs8NTjCK7JLbq+/qzMeN8gwDcamJTkukfefp7t0n6tGM8XsBD/dsPKf+JqP5cxiPGDMmv3OHgvXmlp8nHXz/A/4i9nn/Pql/wCAz/8Akyn4vZypJzo02vJST+/mf5GE8Y4uyxOE+cZfo1+pDF08fldLE0vhnGMlfezV9bN66667kKElinI48ap8I4nm7R/GcQCIIlox6gAANJonCwFNdQGrqw1Z2AAAAAAo7C2B7AAA4xe5Kiug01owAAAjNxE01oBUV4gAAAAAAAARqdGAAAwAAABgeFf/ACPX/gl+cTjT/kf4r/F+iAAaurDQAAACRaysSfQjEAAAYAAAAAAABw3ndXJ8yp4uD0Wkl3i91+q80mcfZJSz3JY4zC+9OC5otfai1dr7tV5q3VgFKpOnNTg7SWqa0afkcD+IMMx5cFmErVuktlPyfaX4PprocccA082i8Zg0o11utlP17S7Prs+jVajUo1JU6kWpJ2aejTQiUVJNPVGJhPhniLmivci7rzhL/gXXqiE4zipRd09gDxDy/wCs5O6qXvU2pfLZ/nf5Hh7j/rOTKk3rTbj8t1+dvkAEYOclGKu2cfyWV8K0cBF78kPlFXb+9L7wCrShVpypzV000/RnClSWTcSywVR6Sbh+sX87K3qbsDAZdiswrrD4Wm5zfRfm+y83ojhrwtw2H5a+Zv2k/wCRfCvV7y/BeqMZj8uynDKVecaVNaJbbdElv6JGd+LUVenllK/96f6RX4Xa80ZrxFmmaP8A2uvKS7bR/wDAVZfhcBgHhrj45hw1Spt3dO8H8tV/5K0Ry2ftdVp3PGDMY4fJqWDi9aktv7sdX+PKAiK0BE1dEXdAAABKmnsKm2xJJWAKi1uAAAAABR3aFsD2AAAHFNWZOk1sAAAFKVnYCSugAAAAAAAABSa2IzTAAGAAAAMDwr/5Hr/wS/OJxp/yP8V/i/RAAE1qMAAASIRvqTl0Qo3AAABgAAAAAAAB4VcROvQlldZ6wV4ecb6r5N6eT7I494ceT5rLkX7qpeUfLvH5P8GgOhtscB+IP1hwy7Mpe/tCb6/3ZefZ9dnrvxvwLRzmk8Thko4hfJT8n59n8npqq9CrQqypVYuMouzT0aaGeI+UfWMDHGQXvU9H/hf+Tt97OAM1+uZUqMn71L3fl9n9V8gDGYWGKw1ShPaSa+9WPDrEzwma1sDU0ck9P70Ht9zf3ABwfgFjc9wtF7cyb9I+8/vtY8Xcc6mPoYVPSEXL5ydvyivvADjXKqkc/wALXo6Oq4q/96LSv9zX3CDhXgjHZ7NVPgoreTW/lFdX36Lrro8pyTLcjwjp4eKjFK8pPd26yl/wyXRJHE3inRo81DKlzy2538K/wrr6uy9UY/McXj6zr4qo5zfV/p2XktAAaA3OAOMFw/jXGtrQqWUu6a2kl5X1XVeaSK3F+Q0sL9ali4cnlJN+iivev5WuuqOMuJZ5/mksTa0EuWCfSK7+ber+7oAkAg2YAAAAATV4gAAEY8zsezjaxODiABS+IWwPYAAACVNSJQcdwAAFowBrUAAAAAAAAAFNoU0wAYAAAAwPC+TWfpLrCX6P9DjqKjxDiku6/wDSUAATSsNXAAASErjdlZCjcAewAAMAAAAAAAAMrzKtluNpYui/eg7+vdPya0ZxTldHijh+NbC6yspw+7WPq1dW/mtfYatoHQDgDj/4MtzGflCb/CMn+T+TOOOB6WdUnicOlHERXyml0fn2fyem1alUpVJU6iaknZp6NNGKw1PE0J0KivGSafo1Y4cx9Th/PJYeu7Rb5JffpL06+jYAZ8nk3FkcStIyan8npL/240aAPCXA+1zWriHtCFvnJ/5JnHOO+ucQYma2UuVf9url/NN/MAMXgaOJqUpzWtOXMvWzX639UjB5nRxOKr4aPxUmk/mr/wCa+RwV4dSxqjjcyjam9Yw2cvN9VHy0b30W+dZ9luQYNSrNRSVowVru3SK6Jd9EjifjPMM8m4zfJS6QW3k3/M/XTskAAMANAEhR0uAgmhO6AAAAACSs7AAAUlpcGk1YnDlYARdpIWwPYAAAAaTVidNrVAAAdAJr3mAAAAAAAAAAQldWAGAAAAMDwzny8Q013jJfhf8AQ8QYKPEmJS/u/jCLAAJ/CJXjcaAA3LCVx2SEuZlkkAAAAwAAAAAAAAZ4TZ/aU8qqvf3oev2l92q9GeJHD/8AZmbOtTVqda8l5S+0vv19HYOgB4f8fX5MtzGXlCb/AAjL9H8mce8Dwzek8Xg42xEem3Ol0f8Ae7P5PSzVSnOnNwmrNaNPRprozxJyW0oZjTW/uy/9tf6fccF5usxyqHM7zh7svls/mrfO4B4n4Dnw9HFxXwtxfo9V+T+84Yx313KMPW68tn6x91/fa4B4XU4YLI8TjprRtv5Qjf8ANsrVZVqkqknrJtv1YAM8IfCDF4TH18/zxWlVk+Slukua8Zz87fDHotZau0eMON8LkVP2ULTrvaPRecuy7Ld+S1MyzPFZjiZYjFTcpvq/yS6LsloAADV0NABa5YjG49gEEtRaOwAAAAAVV1AAAgrRQEoqSsNWdgAg7xTB7AAAAATpJ7Di1owAErtIAqfEAAAAAAAAAAJ2YtQGAAAAMDw2/wCSio+kv/SWeIn/ACUuJ/7c/wDSIgAFT4SHwkodjcsWLFkWuKHcdkjdiSSHsAAAAwAAAAAAAAexl+OrYHFU8VRdpQaa+X6PZ+RnmEo8W8Mqth177XPHupLeP5x9bMatoIAR4e8efWFHLcwn7+0JP7X91vv2fXZ67+IXAqx8JZhgY/vV8UV9td1/eX/k3rvmWApY7C1MLVWkk16efqnr6nC2Oq5FnksLiHaMnyS7Xv7svS/Xs2wDivAfXcmr0luo3XrHX8bW+Z4Y41TwlbCt6xakvRqz/FfiAVf+Erw9SvaU4ff7SV3/AOSyf3AAeHvAiXJmePj2cIv8JNf+kr572PGjxijwbh44PL4qeLqbX1jBK12+8nfSP/bz0spRx0sfFYuUnJ1FzXe7vrcAAAaGrlixYtcUUIYCBkkJ3QAAAABNXiAAAtkAVIcyugAou6sD2AAAAAGk1ZkqPYcWtwClG7uAVdwAAAAAAAAAApu6sAwAAABgeHE+XiTDrupr/wAkk/0PEeCjxHXa6qP/AKQl+gABU+Eh8KCbT2C6LoI2a0JbDdxKyFsPYAAAKVGpVlyU4tvsldmF4Mz/ABTXs8JL/t5cv/pVjD+Fmf1bc6hDTrL8PdUtfw8yl4QY9pe0xMF3sm/u2v8Agf8AEHP+o3/yn/8AbkPB/BqKUsVJv/Cl+F3+ZW8HaLa9ljGl5wT/ACkir4O1FH93jE35wt+PM/yK3hFmqa9lXpv15l+UWYnw24joxuqKl6Sj+Tab+VzF8O5thL+3w04pdeV22vva34gAAAAA9g8J8/8AZV55XVek/eh/iS95fNK/yfc8SOH/AOzM2damrU615Lyl9pffr6OwNAPY22PD7jj+0oLAY2X75L3ZP7aX/ty/Fa73PEfgde/muBj51Ir8Zr/25f8Ab3c8Scl5K0MwprSXuy9UtH81p8l3OEs5WaZXCpJ+/H3Zeq6/NWYDSaszhd/2TxVPCPSLcofrH77L7wKNGVarGlDeTSXq9DxVqxwuUYXAw2cvwhG36r8AA8O+CljprMcbH91F+6n9prq/7q7dX5Jp8dcawyai8NhnfESWnaK7vz7L5vTR+JWEqY7LJYqT5pwlzNvVtPR/i036Hh/mH1rJo029abcfluvwdvkAAAdBx7HKzlZylgQwEDGroWjsAAAAAIkrOwAAdAAqU+qAKcuWQPYAAAAABpMdKLHRfRiovqxJJWQBV3AAAAAAAAAACLswGAAAAPYDw/nycSYV+cl98JI8TIKPEE2usY/lb9AACr8ImlFEpNjY9QCLsycr6ISBbD2AADK+Ds7zJJ0MO+V/al7q+Tdr/K5lvhFN2ljsRbygv/bn/wDImX+H3D+Ds1Q52us25fh8P4fgYfC0MPDkowUY9kkl9yMXn+VYS6r4mEWujkr/AHXv+BifEbhyjdfWOZrooyf42t+JV8Wcjg2o06kvO0bP75X/AAP+Iv4H/oGl96KnjFBSahg215zs/u5H+ZR8YcO4/vcI0/KSf5pFLxey1ytUw80vLlb+66/MoeKmQVF73PH1j+Put/5+RhuOuHcQ7QxcV0968f8A0pL79jC4/CYuN8PVjNf3Wn+TZjsly3HX+s0IzfdxTf37ozDwtyLE60VKk/7ruvulf8GjM/CXM6KcsJVjVS6P3ZfjdffJf5ZlkeY5bLlxdGUPNrR+j2fyYAAAA9gweLq4TEQxFF2nBpr1RnmEo8W8Mqth177XPHupLeP5x9bMatoIsaIClVnSnGpTdpJ3TW6a2aOBeMoZ5hvq+IaWIgtf7y/mX6r57PTxa8OIRw1arhl+4q7r+SXR/wCFu1uz07HAuY1MszeWBraKb5Wu0le36r5oA48pywGfUcdDqoy+cXb8kinUjUhGcXdNXXzDgzCLFZ/hKf8AeT/8B979DxbxaqZpRoL7EL/OTf6JAHBPCc89xv7xNUYayfftFeb69l8jizibDcOZfGNKK9o1aEdkrdbdIr8dvNYnFVsXWnXrS5pyd231ZjsJDF4Wph57TTX3qx4c4qeFzStgamjknp/eg9vuv9wAAB0AAEMBAwkiLugAAAexBtgIrLW4AALVIACpT6oApS5oj2AAAAAAAACrugAAAAAAAAAAKbvEGAAAAPYDgmfJxBhHe3vW+9NHipDlzyLtvTi/xkv0AAKmqQxsAEEVcBbD2ADB4DE42qqOGpucn0Sv/S89kZL4UYyvaeYVFTj/ACx1l9/wr/yYyjg7Jcrs6FFOS+1L3pfe9vkkZnnuW5bG+Lrxh5N6/KKu38kZn4s5bRvHB0pVH3fur9X96RmHifnuJuqTjSX91Xf3yv8AekvkY7Osxx1/rNeU12cm192y+4YAAEZSjJSi7NGB4vz3BWVHFSsujfMvuldfgZf4t5lSdsVRjUXleL/Vdui/G5lvifkWLtGrKVKX95affG/42KGJwmNouVGcakHpo1JfhdGb+HGRY9OUKfspd4aL/wAB+H7kvUzrwwzjApzw9q0F/LpL/wABf5JtlWjUpTdOpFxkt01Zr5AAAAHhLnqhOpldR/F70fVL3l9yT+TPEfIv7NzmVSEbU6vvL1+0vv1t0TQABgcbiMFiIYjDy5Zxd0/677NdVocNcQYLifLJKcVzW5akPVf+kvW33bo8d/D6twtnkMdh7+wraxl2lG2jfR2s13s33SyHNIZnl1LErdrXyktH+P4AeJWBVbLIYhLWnL8JaP8AHlOCsb9bySi27uN4v/t3RfhYPCnC+1zyVXpCDfzbS/Js49xf1riLEyT0i1H/AMBST/FMDKMqxGaYynhMOryk/kl1b8kiMcu4SyT+5BfOcn+sn9y8kZ3nGJzfGzxeIfvPZdEuiXkv+C9WLYOIE8m4rjilpFtT+T0l9/vfeaNAAAdAABDAQMGridmAAAE9iGwCKqvEAAIO8UAAVKd9UBCXKx2aAAAAAAAACruAAAAAAAAAABSdnYGAAAAPYDhefJneDf8A08h+Mkjxap2zShPvC33Sl/mMACbux6gblgQyIC2HsBgcvxWOrKhhqbnN9Er/AD8l5vQyDwp0jVzSf/bkf1l+i/8AAjB4DAZXh3ChCNOC32Xzb6+rM68TcnwLcMO3Wmv5dI/+BP8A9tUjOPEXPMwvGFT2UO0NH/4F8X3NLyJzlOTlJ3b7gDAAAAMJjsVg6iqYao4S7xbT/AyjxUzXC2hi4qtHv8MvvSt96u+5kvHuSZolGNX2c39mej+T2flZ38kZxw7lmb0+XF0lJ9HtJeklr8tu6M/8KcXh71ctn7SP8rspL0ez/B+TMRhq2GqulWg4yW6as18mAAAwy/HVsDiqeKou0oNNfL9Hs/I4twNHiThlYqgryUVUj3296P3XTXdLsAAGQ53ismxsMVh3qt10a6p/1o9TNMsyTj3hueGrrmpVV/29Ca2a7Si/k13jLXKskzHg7iDEcOZlu/eg/szX80fKUd+zi09UwM/wX13K69BK7cXb1Wq/FI8L8beGIwre1pL56P8AKIeEGFUaWLxMv7sb+l2/zRmGJeKxdWu95ylL722B4e8MRyfL3jMSrVaiu7/Zjul5X3l8k9jjriuWd47kpP8AcU7qK7vrJ+vTsuzbGB4n4BTw1HFpaxbi/R6r7mvxOFcf9dyehVb1tZ+sdH99r/MAAAAAQwEDAnHqQd1YAAAqdiGwCJElZ2AAKT0sAAFSn1QFOf2WAAAAAAAAFTWQAAAAAAAAAAC0YtUMAAAAewGV1PZY6hPtKL+5o8XqVq2En3U19zj/AJjACTshgC2LoaAQC2HsRi5NRirtnDXhji8Zy18xbpQ/l+2/X+X56+S3MvyzLsnwrhh4KnBat7bdZN7+rZxB4oYHCN0sBH2s++0F+svlZeZnPEmZ5vNvF1W10itIr0S0+b18wABgAAAAGRcbZxlFoUanNTX2Zar5dV8mkZB4k5TmVqeIfsanaT91+ktvvt8zOeH8tzmioYumpdntJejWvy2fVM4k8Msfl6lWwT9tTXT7a+X2vVa/3Uhpp2e4AA9gPCXPXOnUyyo/h96Ho/iX32fzZxzkX9k5zUpwjanP3o+j6fJ3XpbuAAHBfFdXIsanJ3oz0kv/AG5ea/FadmvEbgbDcY5TTxOEa+tUvfoT6N6Plb/kqJWfZ2lZ2ac6dWnJwqwcJrRxe6fVPzWzAyL/AISeL54faMnKPylrH/20ODf+E/grEYrrL2kl6pcq/GIHhxwx/amP+tVo3pUmn5Slul5pbv5LqeKHFX1ej/ZWHl78177XSPSPrLr/AHfKQMDifAfXcor0Vvy3XrHVffax4YY/mw9bCN/C1JfPR/kvvAAui6LoumAAIYCBgbjXKxNNAAAP3mQ2AQyqtbgAFN2kAABUp21QEKnRgAAAAAAAEtWAAAAAAAAAAAUneNhgAAAD2Ai3FprdHivFVsBhK8drv/yZJ/oMAKnYYA9g6FwQtUC2Mi4bzDOq3s8LDRbyfwx9X+i1fY4a4Hy3JUqiXPW/mfT/AAr7P5+ZxLx7luT81KL9pWX2V0f959PTV+XUz/ivM86m/rNT3OkFpFfLr6u7OgwAB7AAAAAAcPca5rkzUaU+emvsS1Xy6x+Wl90zhzjnK86SpxlyVf5Jbv8AwvaX59bI4l4EyzOk6lvZ1v5ord/3l9r8H52OIOGMxyStyYqHuvaS1i/R9/J2fy1AAAyTNKuV5hSxlN6wd/VbNfNXR4i5XSzjIo4/Du7prnT7wklf8LS+QACQHhvxisFUWW4uX7uT91v7LfR+T/B+TbXiXwh9bpPM8LH95Be+l9qK6+sevdegHiBSlgc6o42nu0n/ANvQf+XKU6kakIzi7pq6+Zmb+oeHlOmtHOMP/J5KT+9Nhg8JVxmIhh6KvObSS83oTeD4Q4e015F/4HN/5v7oryMbjK2NxE8RWlec2236/p2XRB1AMgvk/FssLtGTlH5PWP8A7aA2XC6LougTuAAthgIGASjdEXZm4ABN6WILQgAthk1eIABsLVAABZNWKkOVgRm4iqxYmnsAAAAAABJ2QAAAAAAAAAAAUnaVhgAAAD2AOM/9q4MwlbqvZv74NP8AFjACb1AB7A9gER2DhDw6r5io4rH3hR3S2lL/AORi++7W26ZVq5ZkWATk1SpQ2/yXVt/Ns4p8ScXmHNh8BenS7/bl/wDIryWvd62HqC2GAAPYAAAAAATaaaepwx4mYzActDH3q0u/24/P7S9Xfz6GFxeV57gXKm41aUtGnr8mnqn669Tivwwq4dSxOVXlHrDeS/wv7S8t/UaadnuADQHhbnEcdldTLa2rpbJ9YSvp52d0/JpHEuTyynNK2Ee0Xp/heq/DfzAACcIzi4yV0zwi8QnmnteHsynfF4f4W96tL7Mr9ZxTSn30ldtyt4h8Jf2Ti/rWHj+5qP8A8Blu16PePzXQPEvB+1yynXS1hL8JK35pHBmMeLySg+sVy/8AgLsvwseKDWFyLC4NfzL7oxa/VB4U8OtuWa1o6axh/wC3S/8AbV/28eJPEn9pZj9VoyvSo3XrL7T+Wy9G1uAtwDxCw88FnFHHU9HJJ/8Ab0Gv05TD1oV6MKsNpJNejVxsZcADcTuAC2GAgYICcb6ojKxowAlNdBLmYR0kAthhJWdgACk7xsAAC2JxUlYkmnZgCbWxCopaPcAAAAAAKj6AAAAAAAAAAAAnZj1AAAAHsAVf9r8O0+sYr/ySpb8kMAJagAtUWGAYahVr1I0qUXKUnZJbtvscHeHNLA8uLzFKVXdR3UfXu/wXnucU8aYHI4ODfPWe0F085PovxfRW1WdZ9js4xHtsXO76L7MfJLp+b6tgC2GAAPYoZJmeIX7nDTl6Qk/yRR4E4irO0cLL5tL82ij4acSVHaVFR9ZR/Rs/4hdxD/LH/wACR/xC7iH+WP8A4Eir4bcSwlaNBS81OH6yT/Ar8EcQ0b82Ek7drS+7lbv8jE5RmGFX7+hOHX3oyWnfVAAAAGU5xjcqxCr4So4y/Brs1s16nCniBgs55aFe1Ov26Sf919/7r17X1ZxbwFg86i61G1Ov/N0l/iS/9KWq80rGaZVi8sxMsNioOMl+Pmn1XmADQcIZ1/ZGcUcS37l+WX+F6P7tJeqR4tZIq2FpZlTWsPdl/hfwv5PT/t4ASADjCrjsnxuE4gy6fJWoytf8rrqnrGS2alZ6NnBnFWU+IfC6xHL8S5akOsJq10vwlB9VZuzulxBkmIybHzwlbpqn3i9mvXr2d10OJMGsZlOIo9XFteq1X4pHgpU+t4uOXv8A5+Qfyk0n91vxPGHEXxGFoX2Un97S/QyvLq2Y42lhKPxTdvTu/RLV+SOKszpcMcPxo4V2lZQh321l6pXd/wCZq+49QEAeJGB9vlKrrenJP5PR/jY4Gx/1vJKSe8Lxfy2/BobAAATABbDAQAgCUL7CbQpo50OfYbbFFsSSQPSQC2GFVdQACk7OwAALYKsLq63AAhUvowAAAA0NGBKSSHqAAAAAAAAAAABB3igAAAB7AHCb+tcD4ql/Kqq+fLzL8WMAewwDQuXQwMtyzFZliY4bCw5py6fq+y82cIcFYTIqXPK067Wsu3lHsu/V9ey4y8R4YbmweVyUp7Oe6j5R7vz2Xm9qlWdWbnUk3J6tvVt92wYC2GBCnKpJRgrt9EZb4fZ/jrP2Ps495+7+Hxf+SmA8Iaas8ZiW/KCt+Lv/AOkmC8O+HsKv4PO+8m3+Gkfw/AwuWYLC/wC70Yw/wxS/JFbEUaKTqzUb92l+ZX4pySim54unp054t/cm3+BW484cpRTlil8lJ/kmf8RG4Z/6Cf8AySp/8if8RG4Z/wCgn/ySp/8AIlPjrh2pBSji4287p/c0mUeJclrcqp4um29lzxv917/gUq1OtHmpyUl3Tv8AkYvJ8uxd/rFCE/WKf5ox3hvw9ik7UXB94tr8Hdfh6mP8H46vB4n0U1/7cv8A5Hz8jMvD7iDBXboc67w978F73/kpUpzpycZqzXR6MAADbY4Q8S6mG5cLmrcobKe8l/i6yXn8S8+mcZJlnEOCUatpRavGS3V+sX+mz6nE/CeNyGvy1VzU38M1s/J9n5fddADVwODcZS4h4YeDxDu4p05d7W92X3W17pmOwdXB4mph6qtKDafqnYEgAM1wEMfgquFntNNej6P5OzPA3javwpxLLC17+yq+7OPnG+q81r67X1OM+HKPEWVqrhmnUiuaD7p62v2ktuzt0uTg4txkrNaNM8FMNLC+JdLA205pr/wBOaf3R/E8Va6qZ8oX+GEV+Lf6nhNkStVzOotfhh/7c1+Cv6o8Qs//ALVzeUabvTpXjHzf2n83p6JAPYWwBnGDWNy+th+sotL1tp+Njwxxtp4jCPykvlo/zRRxlOtXq0YvWFk/Vq/5AaAWARHYAWwwEAIAHFMcGcjFBiikAVEJ3QLYYSV1YAATszdAAC2BlWFndAAU530YAAOcUOo+h7SQ5yfUbbAUmuo5t9QAAAAAAAAAAAApvoAAAAPYA8K2q+VYrDX+1/6VG36Ek07PcAl8IwsAAzJcjxmcYuOGwsbt7vpFd2+i/wCGWpw5w1l/DmCcYPXec3ZXt+UV0XTrd3Zxr4iTx7lgsuk40tnLZy8l1UfxfWy0YgYCGZLwfnGbtPD0moP7UtI/e9/+3UzKPCbBUbTx9V1H/LH3Y+l/ify5TLsmy/LocuEoxh6LV+r3fzZmfFuS5ddYjER5l0XvP7o3a+djMPFzBQvHB0JTfeTUV11suZtbdn6GN8Uc+rt+ycaa8o3f/k3N6bL0Ri+J85xatWxU2u3M0vuVl+BOUpycpO7YABTqTpyUoNprqtGYTivO8Lb2WKnZdHJtfc7r8DBeKme0LKty1F5qz/8AJbL8DAeLuAqWWLoSh5xakv8A21/mZbxZkmY2WHxMW30b5X90rN/IzDKMBmEOTF0YzXmrtej3XyM38Jsur3lgajpPs/ej+PvL736GccC55lacqlLnh/ND3l8+q+aSAAA4U40xuRVFD46L3g+nnHs/wfXo1hcXlfEWXNxtUpTVmnun2a6Nf8FPqcZ8CYjJZuvh7zoPr1j5S/R/J2e4FrjR4aZ1/Z+cxozdoVvdfr9l/f7v/bx4sZL7DHwx8F7tVWf+KKt+MbfcywAAcdYOplmdwx1HTntJf4o2v+j+Z4Q8b08Vh6eBqy92a5qbfRvVw/y87rqkeKHCqoVf7Vw0fdk7TXaT2l/29s/72u8jwqya/in9YtoqEp/P3ab/AD/H5nGsKuYcV1qNLWUpRgl52jH8ziPF0+F+GFQw7tNRUI9Hd/FL13l6gD2FsAMw1WGScW1FN2heV/8ADJXX3afccA4yeNWLxE95zv8AetvlsWuMAsWuBF6gHQYC2AEAAAASV0QelgWwwJqzAAKbvEAA6AyUVJWGrOwAFOd9HuAA9UWuKEmKl5nsonsl3PZeZ7J9z2XmSpvoAAAAAAAAAAAAEHaSAAAAHsAeENe1fFUb7qL+5tfqZ3Q9hmWIpfyzkvuk0AT2GDACvl9WjhKOJn8NRyt/27ZN/e7fJmRZFjM5xkcLhY3b3fSK6t+X57LUyXJct4Zy1xi0opXnN6NtdX5dl08223xtx5Wzqbw2Gbjh102cn3fl2XzetrR3BAwOH+A82zdKoo+zpv7Uuvot3+C8zI/D3J8stOUPa1F1lql6R2Xzu13MzzvLcrhzYutGHZPd+kVdv5IzjxbpxvDLqN3/ADT2+UU7/e16Ga8V5zmd1ia7cX9le7H0srJ/O4AAAAAZXxVnGWWWGxElFfZesfud0vlYyjxcekMxof8Ab0P/AJF/j73yMo4kyrNop4SspPttJf8Abr1+dreZnfBOTZteVWlyzf2o+6/n0fzTM/8ADHNMAnUwn76HkrTX/but/wDt1tvsicJRk4yVmgADIeIMbkuKVfCy9U9pLs1+T3XQ4e4jy/iLBuVNK9rTg9bX790+jtZ+Tulxx4fzy5yxuAi3R3kt3D9XH8V101ACE5U5qcHZrVPs0Y2EeLOEueCvUceZLT447r5tNejAAA48yv69lE5RXvU/eXovi/DX1SPDHOJywssMpWnSfNHuk9dPSX3XRwrnmH4myeVPEJOaXLUj3ut/Rr7nfscE8HSybivE1Jq/LT5Yy/mhOcZL5pws1te/kcA4annPEuMzZawpzqW2+JylFfck/nZnidnX17N/q0H7lFW/7eesvu0j6pgD2OgBxfmeY5ZhY4vCWcYu0k1fR6J6NPR+fVGe5zPNsUsTUgoysk7Xs7ddTww/g4lecfyYi9xDFsXGzcEbgAC2HsAgAAAA+GQLYYFRXVwACi9WgJtqOhB3iB0BhWj1AAITUkAAAABKCkNW0AAAAAAAAAAAAE7oAAAB7AHhXiPZ544X+OEl9zT/AEOO8P7HiHFR7tP/AMCSf6gFTYYAGHoVMRWhRpq8pNJLzbsjjThDEVqWV5bgY3cFOLeyXwXk+yvdvzdlqzJcly3hnLXGLSilec3o211fl2XTzbbfG/G1bPa3saLccPF6LrJ/zS/RdPUIggZkPC+ZZ1U5cLD3VvJ6RXz6+iuzh3w8yvKrVaq9rVXWS0X+GO3zd32aM84nyzJ4XxVT3ukVrJ/Lp6uy8zPfFHMsXengV7KHfeT+ey+Suu5WrVa9R1KsnKT3bd2/VsAAYAAAARlKMlKLs0ZJ4k5zl7UK0vbQXSXxfKW/383oZDx3k+b2hGfs6n8stH8ns/vv5Ge8J5TnMf8Aaqfv/wAy0kvn19JJryOI/DjM8sTq4f8AfUl1S95esdfvV/OwABluZYrLsTHE4WfLOPX9GuqfZnCXF+E4gw/K1y1or3o/rHvH8Vs+jfHfh/8AU1LH5dG9PeUF9nzj/d7r7PpsAeEmc8tStls3pL34+q0kvmrP5M48yX+y87qwirQn78fSW6+TuvSwAAThGcXGSumZTKXD/E7oSdoc3K/8Mvhb/wDJWzhfiCrkmYwxMNY7SXePX5rdeZhcRRxVGFei1KMldPyZwFg6nCXAccRi01WmpVpJ6NTqtyjFp2aavGMl/NcrVZ1qkqk3eUm233b3YAwAxWHpYijKlVV4yVmvIznLKmW46phZ/Zej7ro/mvx0PC+d4YqPbk/9u/yEI6jAYiyELYAAWw9gEAAAAE1pci7o6DAsmrDVnYACm7SQDV1Yp9UB0Bg0mrDVnYABNp3RGSkroAAAACceZAAAAAAAAAAAAFN3iAAAAAcBYj2HEOGlfdtf+BRa/U8U8P7LPVP+eEX9zcf0AKnQYwMt8N+IcwwscTTpJRlquZpNrvbez6Xt32Mwy7FZfiJYbFQcJx3T/rVPo1ozwtyb65nH1mS9yir/APbz0j+r+RXr0qFKVWrJRjFXbeiSXU4442q55XdCg7YeL0X8z/mf6Lp6gRApUqlaap04uUnokldt+SRwz4XfDiM1f/cNf+3Nfkvv3RjMdluS4NSrSjSpx0S2+UUtX6JHEnijisVejlq9nD+Z/G/TpH8X1TRUqzqzc5ybk929W/mPYAAGAAAAAHD/AIg5tlPLTlL2tJfZk9Uv7st15Xul2OH+M8qzpKNGfLU/klo/l0l8vmkcS8AZZnCdSC9nW/mS0b/vLr66Pz6Gf8L5jklbkxMPde0lrF+j6Pydn8gAMFjcRgq8a+Hm4zjs1/X3rZnBvGWHz7D+zqWjXivej0a/mj5d10flZvxA4EWD5sxy+P7veUV9n+8v7vdfZ9NgMlzOeWZhRxkN4NP1WzXzV0eJ2WU8wyalmNDX2dnfvCdvv15X5K7AAA8Tcs5alHHwW/uv1Wsfwv8AcjhvMv7RyujiG7yas/VaP79/meFFWtUyOSnJtRnJRv0VovTyu387ni7XxscBQpUofuZNucr7NW5Itb2leTvsnFX1aABgAzxCyR4rBrGUledPf/D1+56+jZ4XyV8VHr7n/twjpoJHQLDHsdAjsA9gDoPYBbAAAAA1dEdHY6DARWj1AAAPhmADArR6gAEZcrE01oAAAABUjrcAAAAAAAAAAACm9bAAAAAGT4j6tmFCt/LOL+5pni9hrVMLXS3Uov5Wa/NgFToAeGnD1PN86TrK9OkuZro3tFffr5pNE61ODSk9TxX4ep47Kfr8I/vKPXvBvVfL4l21tueG+Tf2fkcJyXv1fffo/hX3Wfq2YjD0sRRlRqxvGSaafVPRo4s4fnkeZzwrd47xfeL2v5rZ+avswZCEpX5Ve2oZBw3j87xHssLHRbyfwx9X+SWr+84a4Py/I6d6S5qr3m1r6LsvJfNs4p8ScJl6lh8BarV7/Zj93xPyWnd9DM81xmZ13XxdRzl59PJLZLySBD2AABgAAAAARk4tOLs0cNeJ+MwfLQzG9WntzfbX/wAl87Pz6GExuWZ5gnKlKNWlLRrf5NPVPya8zivwvnSUsTlOq3dN7r/C+vo9fN7E4ShJxkrNaNPdMAMJi6+Erwr0JuM4u6a6f11Wz2ehwbxhQz/DOnUSjWivej0a/mj5Pquj0ejTfiBwM8unLH4KP7l7xX2G/wD21v7nptYA8PMdTzjh6eX13dwvB/4JJ2/VL/CZlgamBxdXC1Pig2n8nv8APcAAOJcs/tHK62HS95q69Vqvvtb5nhlmXLOtgZPf3l8tJfp9x4X0uTh6D/mlJ/jb9DA5tlPFmCxWFg+aMJOnNdU9HGS8mrSi++m6ZnWU18qx1TB1t4vfuujXk1r+AA9wAZOEZxcZK6ehwpk0spzbGYf7NouL7xblb7tn5oQjcBDHsLYIgh7AA9gFsAAAABNdSDugBEkpKw1YACOsUBVXUi7pADAnHmjYAAIT5WaNaAAAABJXVgAAAAAAAAAAAIuzQAAAAwEeIi+u8M4XGdbwl8pRf62AKm50A8GcLGGV4nE9ZTt8oxT/APbmYrF3q6vcjThjcBUoVFdSi4td01b9bGL8WsylFQwlCFOK0V7yaS7bLy2+4x/GOfY5WrYqVuyfKvujZEpOTbbu2HQyTIlR4Qx2ZVF71RKMf8KlG7XrLT/t04Q4CxOcuOIxF4UO/WX+Hy/vbdr9JSyvh/L9bUqMP69W382zivxCxea82Hwl6dF6f3perWyf8q+bY9gB7AAAwAAAAAAynOMblWIVfCVHGX4NdmtmvU4V8RMFmzjh8TalWelvsyf919G/5X8mzirgfAZ5F1PgrdJLr5SXVee6720M6yLHZRiHQxcLPo+jXdPr+a62YAYHHYjBYiGIw83GcXdNf1t3WzOFOKMLxFgXGaSqJWnHprpdd4v8Nn3fHfBc8lr/AFjDK+Hk9P7r7Py/lfyequw8OM3+oZ5CEn7lX3H6v4X9+nzZ4sZP7DMKeOgtKis/8Uf842+5gAAZinw/xUqsdIc3N/27L4vu1S9DDTnkPAqqJ+8qd15SqPT7nJfcZRxxLgrjPD42rL/ZcRFQqryTdp211hzJ6K7jzJbniTw7DM8uWY4bWdNX0+1Dd+tt15X8gB7gAzNszpZdhJYip02Xd9F/XTUrcVZrUxHt1U5X2SVrdra3+dzhvPY5rhnKStOOkl+q8n+H4nQAYzQ0CIIewAMBbAAAAANXVhPlkAIKsdbgAU/hQE1eJTelgBgE1aQAAQnyiaa0AAAACaswAAAAAAAAAAAIu6QAAADARH/hR8PWusI/dyS/+RQBU3GB4RyUuHKiW6qSX/ksf8zMKc5Vo2MoTVJmIkp1ZSWzbEr6IxWTZlhaSq4jDzhF7OUZJfe1YMlyqrmmPpYOlvN29F1fyV2f2PgngI4CUE6SSXK9rRta/fbXv13OI+KMv4fwy9prO3uwW76fKPn913oZ/wARY/OsR7bFS0W0V8MfRfm3q+50HsAAAA9gAAAAAADhLxJxGBccNmLc6WylvKPr/Ml966X2MXg8r4gwCU0qlKWqa6eae6a/4D6o4t4IxmRzdSPv0HtLt5S7Pz2fk9EAZVmmKyzFwxWGlacfufdPun1MmzfL+KMqd4ppq04Po/8ALrF/PRrTi/hWvkON5N6UruEvLs/Nde+/WyITlCSlF2a2M7hHifhBYiCvPlU1/ij8SX/kyXyAAAzfg+XEOc5bRhG/NVjCf+Bu8n/26k/vPEqUaXDVWEdE3BJekk7fgeJuE9pl1Kut4St8pL/NI8BeL1m/DGHwNeV6lKFlfdxi+X74tW9GvM454beS5nKNNfup3lD9V/269PSwHUAJHH+NcsRSwyekVzP1en4JfiKM5K62OD8a8PmtNX0n7r+e342OwA9hljoEQQwAYHQAAAAAqRurkHdWAWwSipKw1Z2ACn8CAI6TsAMArLRMAABSa2I1FL1AAAAKq1QAAAAAAAAAAAFN3VgAAAGAjw2ax3D+KwMn1kvlONvzuSi4tp7oCe4B4L5hF0sVgm9U1NL192X3Wj95isBL2uiujO8VHKsjxFeT1jB/e1ZfjZGjPCbhSjKk83xEU3dqnfpbeXrfRdrMxGHpYilKlVipRkrNPVNPozjfhv8AsLNp4eC/dy96F/5X0+TuvufU8JMicKVXM6i+L3Yei+J/N2XyY9UcTYfH0M1rU8dNyqJ7vqujXk1stlsC2HsAdBgAD2AAAAAAADhvivH5FW5qDvB/FB7P/J+a+d1oZLnuW8Q4JypWknpKDtdX6Ndn0ez+8428O6mA5sZlycqW7ju4eneP4rrdXaAOHeIMTkmOjiaDutpR6SXVP9H0f3FWnlnFeTLW9Oa0fWMl+Uo7dn5pme5Licnx08JiFqtn0a6NeT/B3T1TDwkzdTpV8um9vfj6PSX42+9nFeU/2XnFfDJWineP+F6r7k7eqAADwpy/2+cyxDWlKLfzl7q/DmPFevGnkMYt2cqkUvPSUrL5K/ojjDCfWckxEFuo83/gL5v0PBzP6+X0VVov3qU727xaV0/J6o4hy3DcVZAqmGacmuaD8+sX2vrF9nq9iUXFuLVmg6gBI40m5ZxVXZRX/kqf6mHwkZYZyMvl7LH0pLpOP5oewAxhuEQQwEDAAAAAAHsNcrN0C2Aqx6oAIL3UAT0kmAMAmrxaAAACNRrRiaa0AAACorxAAAAAAAAAAAApuzAAAAYCPCTF8uNxGGv8UVL/AMBdv/bjifCfVM5xVHopyt6N3X4MCe4Bwrn9TI80p4yGqWkl3i91+q80jLc1weZYWOJwtRSg+q6eTXRrqnqjxT4yoYyKyrBTUop3nJbNraKfW2787K+jDKMMssyTD0I7xhFfO2r++7MNjHGrdM8Z8BGeAw2LS1jJx+Ulf/238TLvFejgMJTwtHA2jBJL9526/Bu935n/ABGW/wDzBf8AlT/7Q4u4po8QVadZYf2c4ppvm5rrdL4Vtr94LYAFsMNByRzHMy7AAC5cugAAAAMszTF5biY4nCzcZr8V2a6p9UzhHjXCZ9S5JWhXW8e/nHuu63XXSzfHHh0qnNjsrjrvKC6+cV37x69NdG1bQA4L4sq5FjLT1oz+Jdv7y81+K07NcV8OYbiTLYyoyXOlzU5dHfpf+WX4aPyeIw9XD1ZUaseWUW00900cKZt/ZecUMS3aKdpf4Xo/uTv6o8XMpUqdDMYLb3Jej1j+v3oAAPDGjDAZFicxqLRtv/t2C/z5j6QObzwfCeVYibvNV6cnbqlSqKX381vmVIU8RQcd4yVvk0eG1WWHzHEYSe7X4xdv1Z4V8SexrvK60vdnrDyfVfNarzXmeKHDv1LHrH0l7lZ6+U+v/gW/rcOoAM42ouGbzf8AMov8LfoUMZy4fluZTTdfMaMe84/mv0N0IDdjARHYFsMBAwAAAAAEPYkrog+gLYBpNWJx5XYC1y2gBVWiIapAMAJq0rAAABGTjsRmpAAAEldAAAAAAAAAAAALQ3AAABgHh7i/q/ENC70neL+advxSPFHB+xz51P8An5GL+73f/bUBPcAYkDVzNNKKSMHKbxEkzxYs+GY3evPD8mCFsCAC6SHNPYbbNzlZynKjlRZDALIsiyLFiwXNAAAAKFerh6satKTjKLumtGmcE8e0s3isJi2o4hbdFP07PuvmuqXHfh/HHKWOy6KVXeUVopea/vd/5vXecJRk4yVmgDw34x+p1VlmLl+7k/cb+y309Jfg/VteJPB6xlF5lhI/vYL3kvtRXX1j+K9Egytrifg72MnepyuP/b8PhfztFv1GmnZ7gAGZv+yOAYUlpKcIr5zd5fg2fSZws3wZgLa8lSLfpyON/vaOBcy+u5NTTfvU/dfy2/Cy+Rhv+E/jeUekpP8A8njdfi0UK9ShVjVpStKLTTW6a2ZRqYXjDhxp2vNWf92ov8nr5xfmYvC1sLXnQrRtOLaa7NHUAGcf5dKdOni4r4fdfo9n99180cvmcD5c6+PeIa92mvxei/C7+4Gi5uPRAdBCWgdBgIGAAAAAAh7BNWYndHQAnFSQ1bQKcbyACavEpaoAAKsbq4AAABGq1uKpFgAANAAAAAAAAAAAAQd4gAAAwDLcW8JjKOIX2JRl9zTPF3CKVHC4qOyco/ek1+TAnuAjdgGArLMcnoV4680Iy+bSuYbB81WyR4zY2NPLMPhU9ZTb+UVb85IELYEDHJDbZa5yM5UAAMAAAsixYLgAABGUoyUouzRwLx/HMOXAY92rbRl0n5PtL/0r10fHnAccyjLHYGNqy3j/AD//AG35+pOEoycZKzQB4dcX/wBp4b6jiZfvoLRv7UV19Vs+617niJwh/ZeJ+uYaP7mo9l9mXb0e67arTS/hJm3ssXWwEnpNcy9Y7/ev/STj3Kf7Oz2tGKtGfvr/ALe3+6V0ABgcK8ViqWHjvOSj97SPF3FRp4TC4OGzblb/AAqy/wDSmeNOBhjMhoYWppzXXp7u/wAmeHmMng81q4CrpzJq396P/Av9xx7F4LPsPjI9ov5xl/lYTUkmjw04heX5ksHUf7utp6S+y/n8L9V2PFbh1wqxzWitJWjP16P5rR+i7nUAHsYihTr05UqivGSs0cRcLUMqoSxXtW6d7Wtrrsr3S+dtOzOEHRlk1GdKPKpXb9btO76vT/hkBoXABIAAWwAgAAAABD2CSuiDs7HQAJwUkexfcjFRVkADV0Ud2gAAaTViUbOwAAAAKTWxGonowACa95gAAAAAAAAAAAU3rYAAAGAGeJZrwJTrrWUIwfzi+WX/ALcBPdgIQB4ScTU62EeU1pWnC7hfrF6tLzi7v0emiZL2dNObsl1e33niFxJDO84lOi70qa5Y+dnrL5vbySuCI7AhzXQbbOVs5UIewAAMAAAAsiwXAAAE2mmnqcBcerHKOX5hL97tGT+35P8Avf8ApXrvx/wJHGRlmOAj+9Wsor7Xmv73f+b13atoBgcbXwWIhiKEuWcXdP8Ar7n5GU5lgOLMmaqRupLlnHrGXl+cX+qZWwuK4T4hg6mvs5Jp/wA0Hp+Mbp9nc8WMtjiMvoZhT15HZv8Auy2fomv/ACYADgHBrFcRYaLWkW5f+AptfikeK2L9tnkaSelOCXzd3+TR4w/wMJ6z/KJxfRlk/EUcZSWkmpr1T95fO2vqeJVOGIwGGxlPWN2k/KSuv/STh3E/Wcpw9Xq4q/qlZ/ihNppp6nD2No8VcNujiHeTXJPvzLaXrtJefoZhgK2AxdTC1laUG0/l19HuvIABo40oqrkWIXZJ/dJM8PKvtMlUf5ZSX5P9QAA3IoB7AC2HsAgAAAABDAmrO5F8yAAAAIaTsAABVjdXAAAAAKc+jAAqLUAAAAAAAAAAABOzAAAB7AHQ8O5xzHhuvl8ns5R+U1o/vbKkJQk4SVmtGE92B0FsAUq1SjNVKUnGS1TTs0+6a2Mx4xz3MML9WxOJlKHbRX9bJN/O4CI7Dkhu4oNnKkAPYAAHsAAAABuNBcAABNppp6nAXHKzOKwONl++S0f86X/ty691r3PEXgfm580wMdd5xX/pSX/pS+fcA4Q4lq5FmCrJ3py0mu67+q3XzXVnGHD1DiTKo1cM06iXNTl3TV7ekl9zs9rnDX/C9wlLA1vjinTd+jj8La8vd+aJwlCTjJWa0ADwlw3tM4q1X9iD+9tfpc40xP1nP8XO97Sa/wDAfd/Q8Yf4GE9Z/lE8R8t+sZWsRFe9Sd/+3Xo/xs/kUsQsx4KqU27zoNfcmrf+Str5GScSPBcNU6UH+9c3CPldqTfy5vvaDw+4g/srN4wqStTq+7Lsn9l/J6PybPFjh/mjTzWktvdn6fZf6P1iAA0Z/S9plWJh3hL/ANJZ4ZVL4GvT7Sv96X+QAAJCWgIewB0HsAgAAAABbDAaurEHZ2AAAAHpNA9gAAqQ5XoAAAAAU5c0QAqrYAAAAAAAAAAAAg7xAAAHsALY8Jsd7PMa2Gb0nG/zi/8AKT+440wH1LPcTSS0cuZekve/C9gkB0FsAPYtdFkNCL6Ci2JJAwAAAHsAAAAAG40G4AAU6k6c1ODtJO6a3TXU4E40hnVD6viWliIrXpzLuvP+ZfNabeIfBX9n1HmGCj+5l8SX2G+q7Rf4N22aQB4X8Veyn/ZOIlo9YPs+sfnuvO/cw2VfUs1q4iivcrr3l2nHaXpJN380urPELK/qGfVuVe7U99f9vb/+TXADwgoqFDF13teK+5Sb/NGMlVrVZYicWlNtp99dbPrbqeMCcqODS3vL8omeZLUhGrgMXG0pRs11XNG6+aTWnR+ZlmaSyh4vB14XU4yhJXtaSuk/O2umhl3NLF0Yr+ZafNAcKZjR4m4eeHxLvJLkn320l6tWd/5k+xmuXVsuxtXCVvig2vXs/RqzXkwAexjYc+HqR7p/keF8m44qPbk/9u/yNwABK4CGADAWwAAAAAthgFSPUg7oAAACpumD2AABpNWJRcXYAAAACju0AFXZAAAAAAAAAAAAFN62AAAHsALY4Qx/1HPcNWvpzJP0l7r+69zxbwHJjMPi0vii4v1i7/lL8AkCWh0AB7C2LjCKTA6D2AOgwAAYAAAAADVxouAAGExdfCV4V6EuWcXdNdH/AF8n1OFeJcLxJl7VRL2iVpwe2vVLrF+e2z7vjjhGeR4vmpK9Cfwvs/5X5ro+q80wITlTmpwdmtU+zRwVxNHPMuU5v97DSa8+j9Jfg7o8W8q9rgqOOitYPlfpLb7mvxADgb/ZODsXiOr9o/ugkvxX4nhtQwWbZbicrxkFKKakr7q6s2nurcq27+bMVlmFxWIo160eaVK7jfZN21t3VtO2+9muKMX9bzrFVr6Ocreidl+CR4g5d9Vzd1Yr3aiUvns/yv8AMyf/AJGOH/xx/wDSkaMPD3P/AOys4jGbtTq+7Lyf2X8np5Js8WchTjTzSmtfhl/7a/zT+QAEopqzPDD+LiV5R/UAASErIBDABgdAAAAADoMBDSasRfLIAAACqvdRHWKHsAABKKkrEouLsAAAAFFasAKnwgAAAAAAAAAAAC0FqgAAGAIu001ucbJZvwlSx0d1yT+9WkvlfX0HsMFsAA9joHQI7ALYYAthgAAwAAAAAAaDcAAyXOMVlONhi8M7Sj06NdU/J/8ABWph6+WcWZM9LwmrNdYyX5NPVPrvsziHIcTkuOlha621i+kl0a/VdHoBwvxBVyTMYYmGsdpLvHr81uvMzjC0M7yWrSpPmjVheL894v77MlFxbi1ZoAwz+q+HEpX1lF/+TVLfkzwwxyw2fxpt6VIyj8/iX/pNvmZhilhcHVxD2hGUvuTf6Em5O7ep4i5d9YytYiK1pO/yej/Gz+RkCTzbC/8AXyH/AKUgudThjHUeKOG5YbEu8kuSfe6+GXrs/wDEn2MwwNbA4qphaytKDafy6+j3XkADVzwvV6uJ9I/mwAEiKAFsMBAwAAAAAAYCCpG6uU5XVgAAAqK8CnrFD2AAAGk1Zk6TWwAAABTjaIAVPhAAAAAAAAAAAACm7qwAAAAhnANSGbcMVcvqfZ5ofKSun97f3FanKnKUJqzV012aGHQ6AAzoHQRHYBAAhgAAwAAAAAAGrhe4ABwfxRWyLHKd26UtJx7ruvNdPu6nE2Q4TifKoujJOVuanLpr0fWz69tHujE4athq0qNaPLOLs0+jQHhTxFKXNlNZ3teUP/bo/qvn5Hibwx9Rxn9oUF+7qvXynu//AALV+t/IAz9+w8P6EP5o0/xal+hleOeBx1HEx+xJP7nd/ecf46NDhrETi786UV58zS/K7NzHYSGLwtTDz2mmvvVjIsPKlntClUWsaiT9U/8ANBcPDjPVlucRp1HaFX3X5P7L+/TyTbPFnIXCvTzOmtJe7L1Xwv5rT5LuAFaXJTlJdEzwtheWKl25Px5v8hq5YsWLCiAAAgYAAAAAAAIHqNOMhNNAAAEleLKXwjAAAAlCMiVNoAAAp076sACfwgAAAAAAAAAAABF2YAAAwEPY8KMx9jmdXCN6VI3XrHX8nL7jxAy14HPa6S0m+df9vav/AMmuhh0EdQB6sLIYQ2AQMBD2AAB7AAAAAAAG40XAAPDfi/6jXWXYqX7qb91v7Mn08lL8Hr1Z4l8JLGUHmeFj+8gveS+1FdfWP4x9EBw5mf8AZubUMVeyjJX/AML0l+DZm+V4fNcDUwlb4Zrfs+jXmnqjM8ur5di6mErq04Oz/Rrya1XkwOPv3PB+CpLTWmrelN/8AZm9XGcQcM5bg8HHnq1Hqv8Ar0nGTb2Su03fq1bdGceGefZbhXiZRjOKV3yNtxXW6aW3W1++2pY4pw08s4ljiqaum4z+56r5tfiUfEDEe2vVpLk7K9183o/uXyMLiqOKoxrUXeMthbCdtjLK1Hi3hf2VR++1yy8pxtZ/N2l6OxXoToVZUqitKLaa7NaMAzep7LLq8+0JP7os8LqdsPiZ93Ffcn/mNXLMscrYopAAPYAWwAgAAAAAewAgJxuinK2gAABa6KPUYAAAASpxkSpSWw1YAFFvYjSS3AAJ/CwAAAAAAAAAAAAKcrqwAADAQ9jh/Mf7OzShiukJK/ptL8Gzxey1So4fHRWzcH6PWP5S+8YdBFjcCwXAh1AQMAAAAYAAAAAAANFwADw64t/tTC/UcVK9amtG/tR7+q2flZ9zxC4T/sjGfWcOv3NRu392XWPp1j5XXS7Dg3NFj8iw9eT1UeV+sdG362v8zxT4a+sYeOZ0F70NJ+cej/7df4PyA8UbU8jwVLfVfhC36m54a5TLAcPUZVfineevRS2S8mkm/NshWp1LqLueIvD8MnzucaKtTqLniu1918mnbsmjjfHvEZj7JP3aat83q/0XyLvtocAY9v2uEk/7y/J/oLYR4W559TzN4Kb9ytt5SV7ferr1seKeR/VMyjjacfcqrX/Gt/vVn5u4BxdX9jkeJl/dt/4F7v6nhrR5MonP+ab/AAUV/mCAABD2AOg9gEAAAAAIewAtgCpGzuiEuZAAAFP42MAAAAAaTHTi+g6UT2MRU4roWAACXwsAAAAAAAAAAAABOzE00AADAAhFcQ8D8u81C3nzU9vnK33M/sVrIHmbW9VQXpytt/N2V+6AFqguD2AGZdgq+NxEMPQjzTk7Jf1+PYzPBPA42rhXLmcG4t+a0f4gD2ABgAAwAAAAAAAGhMADAY6vgcTDE4eVpwd0/wCuj2a6rQweJy/i7I3zL3Zq0l1jJfqtGu6t3sZ1lGIynHVMHXXvRe/Rro15Nfds9Q8IcyUqeIwEulpr5+7L/wBt+8yHG08QsTlGJ1lRbjZ/apv4X/4C0n8nfU4ryGeS5pUwzXu7xfeL2+7Z+aMp4MlnWSvF4GX76nJqUX9pWTTi+js7WejtuuvFvClXPI4TDc3LTg25S62SSSS7vvsut9nn+Gw7zuthsJFKClyRS/u2j822rt9Xr1MXy4bCxpQ0SSS9EjCYz96mnseNOFjLCYTEdYylH/wJJ/8Atpm9R1swrS7zl+bKuCth+Y4KqOOcU4r7Skvwb/QQihWnRqRqU3aUWmn2a1TM0o0+LOFVVpK83Hmiu046NL11j6O41bQDxFxHsskcL/HKK/8Abv0OCMP7HIaCe7u/vk7fhYFsAACGADAQAAAAAh7AC2AGk1Y1ixNNaAAAQ+NjAAAAAAAAACWwAAAAAAAAAAAABCVtAAAYAHhJmf8AvGAk+01+Uv8A2045yShgOEp0KC92nJSX/b0//trABoyxaw9Qyrww4gzDC/WFGME1dKbab9Ek7X/vW+7UzPK8XlmJlhcXBwnHo/zTWjXmtBJt2W54fcHLKMN9axMf381/4Cu3q/tfd3vjPa47HVqlKLk5SlLRN7vfT1KeT5jVly08PNvsoyb/ACP9Wc7/AOgOr/4Ln/kS4czmMbvCVEl/cl/kPLMbb+DL/wABf+QAthgAA9gAAAAAAABoTAA4I4nlkeYJzf7mdlNdu0vWP4ptHiBwxDOsvWLwyvVpq6trzR3a037x89FuHAeZfUc/w8m7Rk+R/wDb2i/Gz+RxriqmQcR4XNqa92ceWa7pNX+dmrecTxAyOlneTLG4b3p01zRa+1F6teemq81bqeEGYcuKxGDb+JKS/wC3XZ/fzL7jEVoUKMqs3pFNv0SuYPEOpmVOtUermm/nK7M4TdNfMy+lONZ3PGSSWS4eDWvtF+EZf5maQ9lmFWPacvzZWxaeFUTgyDlnFJropP8A8la/UQxHhLnfLUq5ZUekvej6r4l81Z/J9zj/ACRZXnVRQVoVPfj8918nfTtYDxRxWuHw6/vSf4JfqZZhvquBo0P5YxX3JIAAAWwwEDAWwAAAAAhgAATjzIhKzsAABH+IxgAAAAAAAAA9gAAAAAAAAAAAAAhK+jAAGAdDS+pwXkGR4bCwxuAvOUl8Und+astFZ7pL5s42oe34excbbQb/APAfe/QYAXHqHhpw9TzfOk6yvTpLma6N7RX36+aTRUrU4NKT1PFPh+nmGTPGQj+8o6p94/aXol73lbTdnhpwZ7RxzbGR0X8NPq/535fy/f2AqVKdODlOSSXV6I/tLBf8/o/+BL/MjmGDnJRjVi2/7y/zCrSp1Y8tSKa7NXMTw1k2Ju6uFg2+vKk/vSv+Ji/Dbh3EX5aTg3fWMn+TbWnTSxjfCGg7vCYlrykk/wAVb8vuMw8M8+wyvThGov7r1+6Vn91zGZfi8HPkxNKUH2kmvzAAAZuBcugAAAAABoTAA8L+KXXp/wBlYiXvRV4N9Ut4/LdeV+x4k8Lf2bjfruHj+6qvXtGfVej3XzWyRGTjJSi9UcbRjnXCNPHw3ioz+/SS+V3f0PCviL2+HlldV+9DWPnG+q+Tf3PyP7NfDfG9CUVajWb5e3v6OP8A27Jr5WOOMZ9U4exU72bjy/8AgVo/qJ22MBiYZtlVHEwd+eKl87ar77r1MJgJKreSskeNGYxlXwuCi9YqUn87Jfk/vOM8A8Pmcp292orr8n+P5l5tWb0OAMC5VquKeyXKvV6v7rL7xDQjKMxqZdjqWLp7wafquq+auvmeIuXU83yGGPw+rppTXnCSV/uVn8gM7f8AavGEKC1jGUY/KPvS/wDbgQAAAAgYAAAAAAthgAAFSF9UQlbRgAIIfxGAAAAAAAAAA9gAAAAAAAAAAAAAIy5kAA9gBbDOCeLp5Hi/Z1XehP4l2f8AMv17rzSMzhDGZXWjB3jOEkrdU4v87mAy/FZhiIYbCwc6ktkv60XdvRdTFeEfEFDDurFwm0r8sZPm+V4pX+fpcnCUJOMlZrSz6AAeDGEjDLsTiespqP8A4DG//txi8X+9u3uU6ccbgKmHntJOPyascP8AijjcBGOGx9P2kI6XVoySWm3wuy06Pu2ZLxJlmcU+bB1VJ9Y7SXqnr89vM8QsNXwmdVqUptwlaaV3bXy20d0gEbbFPOcyoy5qeInF+UpL8mYbjfiHD25MXJ2/mtL/ANKTMJ4q53SsqsYTXmmn+DS19DBeL2GlZYrDNecWn+D5fz+ZgOPOH8Y7RxCi+07x/F6fiThh8XRtJKcH6NP80zNfDjIsdeUKbpS7w0X/AIC7q3kkvUzjwtzbCXlhJKtHy92X3N2+5t+RiMNWw9R060HGS6NNP7nqADGPULlwLm4AAAAANCYAYXFVsLXhXoytOLTT7NGCxOC4vyBqatzq0l1hNdvR2a7pq+7M0y2vluMqYSurSg7evZrya1Xkzw5r0804crZdUfw80f8At2abT+9y+4wOOxWS5lGtT0qUpO69NGvRq6ZxNg6fEOQwxmD1nG1Sn3utXH1a0t/Ml2PFDNoVeHsP7N6VnGX/AG7y835uIeFvGuHwtL+ycbNRje9OT0Wuri3stdVfq2r7Gc59l+U4V4nF1Eo20XWXlFdW/u6tpanEGc1s4zKrjaujm9F2S0S+S+969TP8jp5rhvZt2nHWL7P/ACfX7yjwRms63JNKMf5rpr5Ja/el8jLcvo4DDRw9JaL72+rfqdC9x7iPDDNIZhlFTLa+vs7qz6wlfT77r0sZ7lU8rzGtg5/Yenmt0/mrGMxMMNh5157QTb+SueHuHnjc4rY6prypv/t6b/y5gWwAAIewAtgAAAAAAFsMAAAJwtqiE+jABBF2mAAAAAAAAAA9gAAAAAAAAAAAAAE7MTTWgAPYAQyp0PDXjJUJRyrGS9x/A30b+y/J9Oz02enhbwzDAYetjakffnKUV5Ri2n56yTv6IjiKUp8iep4u5BTweY08dRjaNZPm/wAa3flzJr1abAA8HteH66W/tZf+kQMxpTlVVjKE1SZimpV5tPq/zMNWqUaiqU5OMls07NejRmueY7NFT+tz55QVk7K9uza3t566sBAwAwOaY3AT5sLWlB+Tav69/mZX4q5rh7RxcI1V3+GX3rT/AMl+Zk/iHkeY2g6ns5vpPT7pfD+Kb7GYZTl+aUVDFUo1I9L/AJp7r1TRn3hPZOpldT/tyX6S/R/NmYZbi8BWdHFU3CXZr8u681oBLYYABcNwAAAAAaEAHAXEzybMlCrK1GppLsn0l8uvlfyPE/hlYzCLMqC9+mve84d/+3fyb7I8KMx9hnE8M3pVi/vjqvw5jxFy76lxBWsrKpaa/wC3t3/4EmeE3ETvLKaz01lD/wBuj/7cv+3jiDhJ51m+DwsouOFpRlJ26tyXuLtey9FexHhzKI4b6ssNDkta3Kv8r3633vre5x/wkuH8wSo3dGpdxv0tvG/W2lvJq+twAGDEcD5ysrzqlUk7Ql7svSXX5Oz+R4t5NaVHMoLf3JfnF/mvuPEbM1hsrWGi/equ3/bq1f42XzPD7LfquTqq171RuXy2X5X+YAAAhgHQewCAAAAAAYCAAAnC2xGdtGaMBD2BaoAAAAAAAAAAAAAAAAAAAAAAACErMAABD2KnQ1T0MopvCZDh4N+9yRu+rbV5P5u7KeK/e2T1PFfDxxHDXtusJRl994/+3AAeC2YR/wBqwTeuk1/6TL/20xWXydX3VdGb4uOU5LXxDdnCLf8A29ayXzdkEAAHsALYYGTcVZtlDX1Ws+X+V6x+57eqs/MyHxSwGLtSx8fZT77xf6x+d13ZjMBl2b4VRrQjVpvZ7/OLW3qmcS+FuIw6dbK3zx/kfxL0e0vTR+rKlOdObhNWktGno0/QlsMAABAAAAAANG4AeG/Ecczy54DEO9Skra680Nl93wvyt3M3yyXCnE1KpC/slJSi/wC7f3l8tV6WfU8Ycu5qWGx0ejcH89Y/lL7yVLF5DjcPXTtUSjUXlfVJ/LdebRkmaUMxy6njqV+Wavrvpo18ndefTQeYz9pe/wAjxSwEcbw3KtFa03GS9L8r/B3+Qlc0LJgMGIWxlGJo8YcHuCkpTtKD8qlN21+aTfqcRVquf8RrDU9k/ZrySfvS/N+iRRowo0o0qatGKSXotEIAABbDABgIAAAAAABAAAE6fVEZWE01oC2KjtFijencpu8QAAAAAAAAAAAAAAAAAAAAAAACEujABgIexU2Qym1PKKDjtyx/9JKFGaxLl0PEmShwlVi93yf+lJ6fcABw3nlXJczpY2nry7rvF6Nfdt52ZlWeZfmmFWJwtVSja711j/iXR+v5HilxrQxsVleBmpQTvOS2bW0U+qW7e17dmEAYHQewAhgBkfE2Z5NU5sJUtHrF6xfqv1Vn5nDXiHl2bONGv+6qvo3o35P9HZ9Fc4j4Oy3O4N1Y8tXpNb/PuvJ/Jo4k4VzHJK3LiI3g9pL4X/k/J6+q1AAAEboAAAAAHqbG4BkWcVspzCnjKW8Xqu66r5r7nqcV5Rh+J8iVbC+9NLnpv84/NaW6SSvsZZhKPE3C2Gp13/JfvenK0vRySa+dzxWhGOf6dYR/VfocIxUOFcKo/wAi283f/h/MlXl9a5TiZKfC+LT/AOfcvyGWB6oN0WGGb5hHL8BVxUvsK69dkvm7I+jdxZRoUo5ViKi5sQ5uCe7lH3pffFt/JHGfh/DJOOMZmUV+7rrngu0p/wAT/wAmTa7KVgWwAAAAgYC2AAAAAEPYAQAAAThfYTcWKonuc8bblSfNojltGxF8rsAAAAAAAAAAAAAAAAAAAAAAAAEJXVgBgBU2GcC42OZ8M4a71jHkfk4aL8En8ylls/a6rQ8ZMxjSyyhgovWcr/KK/wA2vuAAAIbAwEDAQ9gADhbxExuVctDFXq0V/wCBR9G915P5NGFxmV5/gG6bVWlLRp/k1un/AMOujOMvDetlyli8uTnRWrjvKK7+cV33XW6uwAABAAAAAANXAA8KuJOWcsprS0d3D13lH57r59zD4ShhouNGCim23ZWu3q36s8XYWzuk7b01/wClT/4B4f1o4vhTDpPVJx+cZO34W+8eETq8xxxWWD4VxLe7jy/+BNL9RK4NXEMuNgeJmaKNGlgYPWXvS9Fovvd/uOCsZj8JxDgsRgv4lOcZLe1k7tO32WrqXdNo8ScBTzTIaeY0NeS0k/7krX/9tfyYAAAh7ACBgAAAAACHsALYAAACUVIdNo5JdiFO2rGVI9SE+jAAAAAAAAAAAAAAAAAAAAAAAACMlJAMAKi0GeG/GdPJMTLDYt2oVLa/yy2v6NaP0T6GL4lyfC4Z4mriYciV7qSd/RK7d+iV7nGHEk8+zWeKatBe7Bdora/m2232bteyQ0AWAEQ2B7AIGADaOZAAZRnmOyjErEYOfK+q6Ndmuq/FdGnqcJ8b4LPoKn8FZLWLe/nHuvLdde5xv4cQxSljcsjapvKC0UvOPZ+Wz8nvOEoScJKzWjT3TAABAAAAAADQgDDYqtha8K9GVpxaafZo4fzmjnGXUsXS+0tV2l1Xyf4WZ4wwazLDy7wt90n/AJng7n9OnKrldWVnJ80PN2tJfck0vU+rUnPntqeMefwcaWU03dp88vLRqK+d22v8L6gDGGZZhRwGFliKr0XTu+iXr/wSrxvms63PCSUf5bJr5vf8V8jIc7pZrhfaJWlHSS7Pv6P/AIHQ4kzP+0czq107xvaPotF9+/qzhvOpZRmEMQleO0l3i97easmvS2x4ZZph884clg5y5oxTj6wmm0/xaXZJGYYGpgcZVwtT4oNxfydr/MAABDAOgAAAAAACGAdAAAAABm5OFiE76MAAAAAAAAB7AAAAAAAAAAAAAAACdnoJprQGAdCa0GAGhY0NwaBEfhAAGASlfQ0LoTsJpoB7DKNapRqRqUpOMlqmtGn5HBHiJTzHlwWYNRrbKWyn/lJ9tn0s7I454CpZvCWLwa5cQvkp26PtLs/k9NVVo1KVSVOpFqSdmno013LFiwAgAAAAAAaNwewzwt4jWCxzy+s/crbeU+n/AIEtPVRPFDI8wzLG4X6nRlUfLK9ldLVbvZX8zEYTMcnxcVVhKlVjZq9012a/zQvF7iFUPZWpuVrc3K+b1+Llv/27byMVia2KryrVpOU5O7b1bYA9h7B4gY1yrUsKnolzP1ei+5J/ee81dLQyXE4uFSdDCv3qsXFXdtX19V0MF4YVpa4rEJeUU3+Lt5dP8zB8A5Hh9ZU3N/3n+isvwPDDMKWVZxTw8Eo06i5LJWSb1jZLT4tPmzxXyr6tmcMbFaVVr/ijZfjG33M5jmRowAOgwAewCAAAAABbDAAAAAABg0micLEZ9GAAAAAAAAD2AAAAAAAAAAAAAAACMrMTTQ9gBbE1dDAABDFqCEtAGB0HsBJ9BsC4mboHsMNtjgPxCVXly/NJ+9tGb6/3ZPv2fXZ66vjrgWnnFN4rCJLEJeiml0fn2fyelmuAfDiOZweMzOLVNNpQ1Tk07O/VJO6srO6eqtrxL4T5ZWwkp5XF06sVdK7al5Pmbab6O9u66k4SpycJKzWjT3TNwELYAAAAAANgewzLsPiMRi6VHDfxJSSj6t6emvXoZbgVgcLCi5uTW8nu5N3b+bbdtlstDjDhmhn+WToNfvFdwfaXr2ls/v3SHFxk4tWaABj2DjSblnFVdlFf+Sr/ADMPg1LDORlTdPMaPL0nH80I6FOrOnOM4OzTuvVHFVOHEXCCxlJXlGKqLya+NfJc3zRcugUrgAAIGAtgAAAAA6DAAAAABAMBq5OnbYjJoUlLYAAAAAAACWzAAAAAAAAAAAAAAAAjKxugBD2JIAAEMQyKuAMBbAwbABEWEthgHh7x77bkyzMJe9tCT69oy8+z67PXd13Tg3vYwmN9tLllueKOVRwPENSUFaNVKfzd1L8U38wAWwAAAAAANXEMPCrArE8Rwm1f2cZS/Dl/ORmmJ5Zct9EZXiOdOBxzgo4PiPF0oqy5ub/wJKX6iAB7BxrTcc3qPuov/wAlS/QoYxLD8plEHVzKio9Zx/NCOg2eE2ZxxGX1suq68juk/wCWW69L/wDpRnuVyyzMq2Df2JNLzW6fzVmBuJ3QCHsAIGB0AAAAAAAQAAAAIAAJU09hpoVRrcUk9gAAAAAACfwsAAAAAAAAAAAAAAAAhO2gAh7E1qAACGI3IK7AHsAgZLYYACE7olsMA4JpRq8RYOL/AJ4v7tf0M2qqlBpdTKcU51E/M8a6KVfB1erU19zi/wBQN0CAAAAAAAaGHg1yf21X/m9m7enNG/6Ge8/NLl3Mh5rq54pNPiet6Q/9JR1ADoHH+Ab9li4r+6/zX6/gWeyZwRgJV8x9tb3aav8AN6L9X8gewcAZr/Z2f0JSdoz9x+ktv/JrP5HFfhxUz/NY4qnVVOLjaTtd3W1ldX00d2rWW5xd4eY7IKSxCmqtHZySs4vpzK7snsnd66O2lwi9QFsMA6DAAAAAABD2AEAAAACBgADSaJUuw00RqdxNNaAAAAAABU+EAAAAAAAAAAAAAAAAIysbgFRAAAbgkQVkAMAHsTGWuFgBMlsPcYHB1dUOIMHN/wDPyK+92/Uzii5xTMqwnLVSXc8aq6li8JRvrGMn/wCBNL/20AEAAAAAAAwZ4V46OG4kpwbsqkZR/DmX3uKRmuF5pc3RmV4bkTlY44x0cbxFi60dubl/8BSj+h1AAMXhaWKoSo1VeMlZlbw+xHtrUqy5O7vdfJaP70ZTlVDLcMqFL5vq33/roD2DI8vqZjmVDCU3Zzklftrq/ktSrVhRguZmIo4bMcJUoVFeE04teTVv+GM2y6eX46thJ705OPrZ7/Nagg3AAHsAgAAAAAQ9gBbAAAAAtgewAADSaJU+wm0yM0wAAAAACp8IAAAAAAAAAAAAAAAAEZWNwKiuhgABcRFXYADAWwT3LXLm7Lg9gRvEYBTqSpzU4OzWq9UZRmNDOsro4uG00n6PZr5O6MPhIUdU7s8Qs5jmmf1qkHeEPcj6R3t5OV38wAQAAAAAADSYMweLq4TE08RSdpwakvVO6OH89wmd4CGLoPfddYvqn/Wqs9mca8TUsiyudRSXtZJqC63727R3fyXUk3J3b1OgBudRh0AeweFGGjV4kpyf2Iyf4cv6mb4jkm/IyfEc87dzxSoRpcTVpL7Si/8AyVL9AQthD2AEDAWwAAAAALYYB0AAAABbA9gAAAlFSJRaIz6MAAAAACrsgAAAAAAAAAAAAAAAACMrF00LYkiS6AAG5YSvsR0dgAGAgnuIEblgDoMAtc4L47xPDspUpR9pRk7uN7NPvF+el01rZbbnEfi3Vx2DlhsBSdNyVnJtNpdUktr9+nTXUaABAAAAAAAEjoNXMvzXHZdUdTCVpU5PflbV/XuvJmOx+Kx1Z1sVUc5vrJtv016eWyBBdDZ0AGA9g8I5xjxDZ9YSS/B/kjPaLnKSRkVFwlFdjxaq8/Eco/ywivzf6giOwthgC2AAAAAAAFsMAAAAAAGAAAA0mTp22IzaE01oAAAAAVd0AAAAAAAAAAAAAAAAACdiLTQya6jQBYsWFDuWSJrqJ3QAPYBBIWwIualgFsMA2NyxcaAEAAAAAAAMYhgG5YsMAYD2DgvNI5bn2GxM3aKlZ9kpLlbfkr3+RjcJ7ZJx3RgsJ7GN3uzi/M45lnmKxUXeLk0n3UfdT+aSOgQ2AAHsAAAAAAAAIAAAAAGAAAAEqaexqmRqdxNNaAAAABU1kAAAAAAAAAAAAAAAAAAm09BSUkNXRKNiyLILNih3EktgGrqxF2dgAACQgRYBghgI3AaN0AIAAAAAAAYwYCV9EcGeG+X4HBxrZhSVStJXakrqN+iT0v3bvrtY4/8ADvBTwU8dltNQqQTbjFWUoreyWiaWqslfVWvYbuAAD2DocDeI+BrYKng8yqKnVgklKW0ktE2+j7333vulx14kYKng54PK6nPUmmnJbRXWz6trRW0W972AIbD2AEDAQAAAAADAEAAAACAYAAAADSaJU30NUyNRPcAAAAlqwAAAAAAAAAAAAAAAAABO2wpJoaTHBCgkWQABNa3Iu6ABgIkFkzQBjBDAewloAdRgIAAAAAAAYxDLDR4f5bHH8RYanJXjF8z/AO3VdfikjHYt0rRi9TB4n28WpbnE+WrLs4xOFirKMnb0eq/BoA6ADLaAMAhsMAWwMBbAAAAAAh7ACAAAABAAAAAAA4polTa2KfwiaewAABJ2QAAAAAAAAAAAAAAAAAABGVwAABq6sRdnYAGAiR0DsAxghgdQAYwEAAAAAAAM3QbhueDsIvP6rfSlL/0qCM8ruEpMyKq5tPueKUIrieu11UP/AEhL9ABgI6l7gMBEPhAAewAAAAAAC2GALYAAAAEDAAAAAABpNWIOzsAAAVXpYAAAAAAAAAAAAAAAAAAAIyvoAAATXUi7oAYBI6B0M54ywuArOhTj7Sa31sl5Xs9fkZJxDhc1g/Z+7Nbxe/qu6/pjAQwR1AB7D2AQAAAAAABIHqIYjwjxEaXETg/t05JffGX5RM4w3PN+ZlGF9nFeR4lYmNfifEuO0eWP3RSf43AAEdQQMsgQvhQAgYCAAAAAA6DAAAAAAWwPYAAAAAACatIhPowAAJO7AAAAAAAAAAAAAAAAAAAAjLowAAGro+Fm4AwJI6Bn2YvAZfUrx+JKy9Xovu3+ROet3qzKcxngcZTxEPsvXzXVfcRlGcVKL0YCGB1AB7D2AQAAAAAABIBDEZDmssrzShjI/Ykm/NbSXzV0QeHxlGFWDUoySaa6p7NepmGNw2V4KpiqztCCbf8Al6t6JdW0jHYupjMTUxNV+9NuT9W7sNxgC2DdG6LHUZ0EgAWwMBbAAAAAAAAAAAAHQGAAAAAABVWwRnbRm4AFSWlgAAAAAAAAAAAAAAAAAAAAjLowAAJxuiD6ADAktQPECs44KlT7yv8Acv8AgmBw3tpNmIpezqtHD9Z1sqw8m/spfdp+gCHsB1AAYCAAAAAAAGgBq4kPU8O8JicBw/RhXbbleST+ynsl2VtbdG2eLU5z4f02543+5/rYAAHsdBHQR1GEVqAA9gAAAAAAEPYAQAAAAAMAAAAAACorxAUmhTiwHYlU6IAAAAAAAAAAAAAAAAAAAAAjK2gAAElZid0AwJLQe4eIUG8PQl2bX3pf5GUVo05O5j5qde6OGIOOUUE+35tsBD2A3QAIYCAAAAAAAHsMBbBQnGFWMpR5kmm138vmcD8ZZhn+MrQq04xpwinone7eibbtayfRf5+JNanieGqk6T5kppP1jJxf3PQsMAEMVjRHQGaMSIrQAQMBAAAAAAIewAtgAAAAQDAAAAAAAaurABdrY5pdxtsAAAAAAAAAAAAAAAAAAAAACMrG4AA1dWE+Vm4DBq6GHGeDeIyqTjvBqX3aP8G2RlKDvEpwnWqKKV5SdkYPDxw2Hp0Y7RSX3KwCJIBDNwbubIAQAAAAAAANAFg8LqMMFkmJx8+rf/gMI3/NspcRVXleLwNbVVZKa8pcycvk1+KXdjQ1caALaG4bgG4lcAAAWwAAAAAC2GAdAAAAAQAAAAAAABJWYAAAAAAAAAAAAAAAAAAAAAAABGVjcAAnG6IS6AMCSCcIzi4yV0zOuEsbhKzeHg503tZXa8mlrp32ZwrwtXo1VjMVGzXwxffu+3kt769DIuKcHmn7r4Ky3i+63t3X490BFEwDcaCw9QBAAA7Fy5dAAAANDSZZCQGN/wCEfgCMFpKcEvnUd5L5Jv7gTuOKY1YaLFhoANDcSErIAQPYAAAAAAAGAAAAAAgewAAAAAAAVFqAAAAAAAAAAAAAAAAAAAAAAAAEZWNwACcbEZXQASQ0FixxVh5YLPK6g7Pm5k1/e97T5s4c8QXG1DMnddJ//JLr6rXuupSq060FOnJOL1TWqZBak1qAFy49QAQAA2aFy5oy5oAAAFkNAZVgXjsdRwq+3JR+92Z4vYyNPCYXBQ2bcreUVZfm/uATGrocbFriiSV0NFixYsWuKNkACWgMBAAAAAAIYAgAAAAFsD2AAAAAAACqtEAAAAAAAAAAAAAAAAAAAAAAAABGVtAABq6GuViaaBg1dDXQA8TMHyYyjiEtJRa+cX/k/wADJst/tHHU8LzqHM93+nm9kurMryvD5bhY4agvdX3t9W/UirIqLUABVacpOCkm106gARGgAYwAuaBcugAAFqB4X5csVnqrNaUouXzfur82/keJmY/XM/qQT0pJRX5v8W18gBMAHFMcWcrOVnIKKWwwC1wewAtgAAAAAEPYAQAAAAAMAAAAAAAKivEAAAAAAAAAAAAAAAAAAAAAAAAAjLoAANJo1ixNND2AkrjVyweImDdbJ/apa05J/J+7+bRXyrFYbB0Mdb3Kl7NdGm1byel13+RwbxfTzKmsNiZWrr/yfzXn3XzXWyKnQYBxpn1TCQjhMPK05K7a3S7Ls3+XqUq1SjUVSnJqS2a3OHM4/tPBKpL446S9e/zACOwNADAAsWQXLphuAAxbAeFmDjg8nr4+ropN/wDgME9fvcvuMdip4vFVMRPecnJ/N3ABMAAEh7AAkDA3AAAAAAFsMAWwAAAAAMAAAAAAAJK8WAAAAAAAAAAAAAAAAAAAAAAAAAEZdAACUboTszRoYDjfYaLIznBPGZdXw6WsotL1tp+NjAZLh6uRUcFiI3i4RuvO121531RxBw9jMjxavfkveE15bekl/wAFaHDniKoxjQzP5TX/ALcl+a+a6lLE0cTRVWjNSi+qd0MA4jxUsTmded7+80vRaL8iWGlCPMcAYpwxtSjfSUb/ADT/AMmwAjsA0D2GAABuFw3ADcCMXJ2RxNy5DwZHBx0k4xh85az+9c33hJagAnYAC1weoAAAgAAAAAAYAAAAACAYAAAAAAAbgAAAAAAAAAAAAAAAAAAAAAAAAARldAAE432IysPUAaTRyRLWDG4LD42hKhiIqUHun/X4nEXh9i8G3WwN6lPt9pfL7Xy18upg8xxuX1XLD1HCXW2n3rr6NGC8ScypJLEQjU8/hf4afgUPE3AyX76hJejUvzcSPiHksoptyXly/wCTZh+NcjrzhCFb3pWSXLLd9NrfjbzMTd4uV19p/mzFUoLCJrc4Kus3p27S/JgCI7ADVyQwaLXGW0LXGBuAgBiWgWOBsr/tDPaEGrxi+d+kdfxdl8zxbzP2mMo4GL0guZ+stvuS/EJIABOxuAJDABIGAAAAAAAIewAgAAAAEAAAAAAAABJasAAAAAAAAAAAAAAAAAAAAAAAAABO6AAJxvqiLtoAIGgDNOHcszPXE0k5d9pfetdOzujHeGNCTbwtdrykr/irfkyv4cZvB+5KEl5Nr77pfmyXAufptKhf/t6H/wAkZdwrnNLHUZTw8koyjd6dGr9TO8O8PmNaHaTt6XuvwKuM5qSimcB4dzzJz6Ri/vdl/mAIjsAEtQGDOghgDBC2Bm5a5YPCfK40cJXzCppzPlTfaOrfo3v/AITiDM3meZ18X0nJ29FpH8EgkroAATsJpglcHqAbgbgIAAAAAAQwBAAAAALYHsAAAAAAAAVF7wAAAAAAAAAAAAAAAAAAAAAAAAACdmaNAAEo9UJ2AAaAJLUA47ymca6x0FeMtJeTWz+a0+XmiyODMpng8E6tRWlUs/RLb9X8wBC2AHsMBi3LD2Og9gABA9RJICnTlUmoQV23ZLzZxFOPDfCKwtN2k4qmvOUr8z/9KfqANAAAncGACQMAtoAAAAAAdBgC2AAAAABgAAAAAAAFVagAAAAAAAAAAAAAAAAAAAAAAAAAEZWAACUOwnY3BA0ASWgwK1GnWpunUinF7pmH4TymhW9rGlqtk22l8n+twAR0AJbDAYhnQ0sO1gAYIbViKAPDXJvr+dRrSXuUfefrtFffr8jxWzdYnMoYKD0pLX/FLX8Fb72ASQAACn3AC1wHqAlcAAAAAAAAAAAAABgAAAAAAAFVbAAAAAAAAAAAAAAAAAAAAAAAAAABCXQAAJRLtCaYA0A1dDQAACACYwGrltRnQaLaAB0BISuAHBmCpcP8NyxuIVpSTnL0t7q+7p3ZjsZVxmJqYiq/em2383cAaugAACL6ACQMAS0AAAAAAEPYAQAAAACAYAAAAAAABVWgAAAAAAAAAAAAAAAAAAAAAAAAAAEZXVgAB7DSZqhSuIGroAkhoAAIrYAJgFwFqNlzRlgLaCQyO4Bwjkf9sZtSwz+Be9L/AArf79F8zxWzqNDCU8spOzn70l/dWy+b/wDSQAJLUAAATugtcB6gbgAAAAAAthgCAAAAAQAAAAAAAABUXugAAAAAAAAAAAAAAAAAAAAAAAAAACaaAAGhpMcbClYumgaAauhoAAtcigAluB0AHsWuWLag0JA2EQDw2yanlmUzzHEaSqK930gtV9+svSxxHnE83zOri5bSei7RWiX3b+dwAJLQAAAiCQMASAAAAAAAYB0AAAAA6A9gAAAAAAAAnrFgAAAAAAAAAAAAAAAAABkuU1M1xkcJSklOSfLfZtJu1+l7b9zGYPEYOvKhiIOM4uzT/r8dnugAAAAAAABOwmmgAGtAHHsJtCmmPUAkrjVwA3EhKyACQHQBG7AGLVAMEBwhkEs5zSnQa/drWb/urp89l636HihnkcFl8Mtou0qm9ukF08rvT0TQAANAAALcSAAtcAAAAAAEPYAAAAAAAYAAAAAAAAS1QAAAAAAAAAAAAAAAAAFLA4mtHmp05SXdJv8AI/svHf8APmX/AIC/8jIaONwOaYfEujJKE4t+69r69O1zirhDBZ9QtU92qvhmt15PuvLp0sZ3keNyfFPDYqNn0fSS7p9V+K2dmAAAAAAAAJ2NGgAGgGkxxaFJo5kwCUb7FixYsWErAASAYHQQAILj1BC2LX0RwXkdLh7JniMV7s5Lnm30SV0vkt/Ns4hzmrnGY1MXU05tl2itl92/nd9QAAkgAAAGAJAAAAAAAthgCAAAAAQDAAAAAAAAB7AAAAAAAAAAAAAAAAAAeF3/ACT0f8UvzAOO62SQyxxzRXv8KXx37x7W6vbo73s3boAAAAAAAARlYABq4AhwTHFou0cxdMGkxxZYUWzlSAAJAMtcsjodABi2A6AiOx4bcMf2jjfr1eP7qk9P70+i9I7vzt0bPFPiVKKymg97Of5xj+r+XmAAA1dDAAA6DALXAAAAAAAGALYAAAABAAAAAAAAAAAAAAAAAAAAAAAAAAAAHDuB+vZthsPa6lON/S93+Fzi7jXB5FScE1Ou1pHt5y7Ly3fTq1m2bYzNMVLE4qfNJ/cl2S6L/h9wAAAAAAAAIy6AANACBwTHFli7OYumA1cAAkIaGbIvcYADA6AjIcmxGb42GEoLV7vol1b8l+L06mZYzA8J5ElTWkFywT3lJ99t3eUvK9uiMViquKrzr1pXnJtt+bAAASuTWoAAHQeoAkAAAAAACGAAAAAAC2B7AAAAAAAAAD3AAAAAAAAAAAAAAAAAAMozWvleKjiqFueKdm9bNpq9urSbtfS+99jEYiriKsqtWTlKTu29W2AAAAAAAAAEZX0ABq4AA0DimOLGmHMznZz3OY5kcyOY5h6lkDSGDABjA6BRo1K1SNOmryk7JLdtnB3C9Dh3L3Os17WSvOXRJa2T7Jbvq9drW4z4nnnuPc46UoXUF5dZPzl+CsugAAFgnqgAAGAJXAAAAAAAWwwBAAAAAAMAAAAAAAAAluAAAAAAAAAAAAAAAAAAAAAAAAAAAAEXdAA0AIBoAsmcqOVHs/M9mxwZys5Wcg9C4MYPYWwA9xgCTk0lueHvA39mwWPxsf38l7qf2E//AG5rfstO54kcYxxMpZXg5e5F++11a+yvJPfu/TUAALA2DWoAAAFtAAAAAAABgC2AAAAAQDAAAAAAAAAJbgAAAAAAAAAAAAAAAAAAAAAAAAAAAAnYTTQBuAC1AaAANwaAluD1DdFjcQAk29CHh9xNPDrELCPltfeKl/4Dfmv5WuThKEnGSs10CMJTkoxV2+hwDwAsCo5hmEf3u8Yv7Hm/73l9n128QuOlgYSy7AT/AHr+KS+wuy/vP/yVebVo7AABYGwJIAAAEgAAAAAAEPYAAAAAAEAwAAAAAAAACW4AAAAAAAAAAAAAAAAAAAAAAAAAAAAEXYAGgA3Bq4AAbg0E0B0LjYM3BnhJw3TxuMnmNeN40rKN9ud9f+3V9zafQdSHNy31PF/hunQqU81oRtzvln/itdP5pNP0XcwGX4rH4iOHwsHOb2S/rRd29F1ODfD/AA2TpYnFWnX/AAh/h7v+991tb8ecfwy+MsDl8r1tpS6Q9O8v/SeuuhKUpycpO7ZHYAC1ywNgEldAAAG4AAAAAAAthgCAAAAAFsD2AAAAAAAAAB7gAAAAAAAAAAAAAAAAAAAAAAAAAAAAEH0ABoAAauAAF7g1cmgOgBuguJHhrh44ThalUW83KT9eZpfgkSxX73fU4hymlnuUSwdSXLzWd7Xs009PWzXzMnyHLMiwrhhoqKSvKT3duspdvuS6JHGfiZzqWDymWmzqfmof/Jf+A9GNuTu3qIjsAFgG7gAAAAJAAAAAAAAwBbAAAAAAMAAAAAAAAAGAAAAAAAAAAAAAAAAAASxtCOJWGlK02rpd11t3t1XTcAAAAAAAAAAAi7oAauACAaAAC4TWgwADRlkNnBzUuFMLy/yfk9SpGf1pdjE4uOByueLqK6pxlJrq7K9l67HE/HGZZ7Jwk+Sl0gnp/wBvPTmf3LskAiOwFiwNj1AAJbgAHQSuAAAAAAAIewAAAAAAIBgAAAAAAAAAAAAAAAAAAAAAAAAAAAeI2EnLAUsZTdpUpbrRpPqn6qJwrx3DE8uFzB2nspbJ+vZ+ez8nuAAAAAAAAAACdhNNADQAIBoAAB6oYDAAPCvGwxfDcaDetOUov5vmX/pX4EsDL23LbU8RsXHA8L1oX1naC9W9f/JUzoAiK0LADYAAbk1qAALVAAAAAAAB0GAIAAAAAQAAAAAAAAAA9gAAAAAAAAAAAAAAAAAOLuLszyzM3h8O1ypJ6q+5/wARDzv+aP8A4CY/jbNcbhp4es4uMlZ+6HCfG9XCOOExrcqeye8o9l5r8V000N0AAAAAAAAAAEXbQAGgAWoDQAAEkB0AAPDfiunkmYyp4mVqNWyb/la2l6atP1v0I1ISgpxknFq9+lu9+x4n8W0s3xkcJhZXpUr69JSe7Xktk+ur2sAIWyAbAAA3LFToAAQ2AAAAAAAEAAtgAAAADoD2AAAAAAAAACXwsAAAAAAAAAAAAAAAAADOeC8vzXFPE15zUrJaNJaesX+Z/wAQzyj/AJ+VPvj/APIHEXBOU5XltTFRqTclZJNxs23bpFepg8HXxlaNDDwcpvZL+tF3b0XU4X4MoZXFV69p1u/SPp5+f3W6gAAAAAAAAAARd1YAeoAG4DQAASQ0AAALGYhUvZKo+Ttd2+7Y6ACNC4AABa5YKmwABDYAAAAAAAWw9gAAAAAAAYAAAAAAAAAVPhAAAAAAAAAAAAAAAAAADjTA4zNZ0Muwy0bcpPpFLRX9buy30+7IeHcHk9Hkoq8nvJ7v/JeX66gAAAAAAAAAAAnYTTQA1cADcBq4AA1dDQG4AAMAitgAAAtcAnqgACAAAAAAAG4MAQAAAAAgGAAAAAAAAAFXYAAAAAAAAAAAAAAAAAAAAAAAAAAAAAACD6AA0ABuA1caAAktRoAABgBFagAAFgBslsAAR3AAAAAAAQ9gAWwAAAAAtgAAAAAAAAAAq9EAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAARd0ADQAG4A0ADV0NAGjLBYABEVoAABYAbBu4AAAAAAAAAMA3AAAAAAHsAAAAAAAAABU1kAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAARdmAA0ABcAauNAEkNXAAAAS6AABYAHqEncAACOqAAAAAAEAAgAAAABAMAAAAAAAAAB6sAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAIO6sAA0ABcAGgBx7DQ0WAsWLFixYigALXLADHqEn0AAAIdgAAAAADoMAAAAAABAMAAAAAAAAAJu0QAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABNNAANAAG4ANFgaTHHsNAWuWZZli1xRACxZAA2A3ZAAABF2YAAAAACVweoAgAAAAA6A9gAAAAAAAAAKr0sAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAJ2E00AA0AAXAAsixYLIsgACxYsAAx6gPYeoAAAEXdAAAAAAloPYAFsAAAAAAMAAAAAAAAACbuwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA2IyTAAaAAC4AAWRZFkWRZFkWRYAAY2AEncAAAA2E00AAAABa4PUA3AAAAABAMAAAAAAAAAJOyAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABT7m4ANAAAXAAAAAAuhsACT6AAAAAJ2E00AAAACWg9gAQAAAAAdAYAAAAAAAAAFR3dgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAE2hSTABoAAAuXNAAAAui4AASfQAAAAACMkwAAALXB6gAAAAAAAMAAAAAAAAAJNJD1AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABTYmnsANXGgAAAC5cuXLlwAAAbsAAAAAACnbc0YAAAloPYAEAAAAACAYAAAAAAAAAVJXdgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAU2KSYA0AAAAAAAAA5dgAAAAAABO2wp9y6YABa4PUAAAAAAA6AAAAAAAAAAE5WVgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAUmhTRdMNxoAAAAAAAHJDdwAAAAAAAAuznZzo5kzcEh7AAkAAAAAADAAAAAAAAAJSSQ3cAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAALtHOznRzRZp3AAAAADQckOQ9QAAAAAAAAAAFNiknsPUALAAAAAAgGAAAAAAAABKSSJNtgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAXZdnMzmZzM5mcxzHMcxzHMzmZdgAAAAAAAAAAAACk0KSYCAAAAAAAAABtIdRdBVO4mnsAAAABKaQ22AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAACk0KSZ0AAAAABAMAAJVF0HqAKo+opxYAAAOSRKbewAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAACk1sKouomnsAAAAAgGABKaRKTkAACk1sxVGe0RzxfU5ovqc0e5zR7jqRHU7Dm2AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAACm0KonuJp7AAAAA9gAc0iU2wAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAFJrqKoKpFl0+oAAgGNpDnFDqPoNtgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAXaOeXc9oz2jPa+R7XyPavse1Y6khyb6gAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAH/9k=)
Psychopharmacology of pleasure
- Dopamine
- Serotonin, Norepinephrine
- ACh
![[[@cock_sleep_2008]](http://dx.doi.org/10.1038/ncpneuro0775)](data:image/jpeg;base64,/9j/4AAQSkZJRgABAgAAZABkAAD/7AARRHVja3kAAQAEAAAAMgAA/+4ADkFkb2JlAGTAAAAAAf/bAIQACAYGBgYGCAYGCAwIBwgMDgoICAoOEA0NDg0NEBEMDg0NDgwRDxITFBMSDxgYGhoYGCMiIiIjJycnJycnJycnJwEJCAgJCgkLCQkLDgsNCw4RDg4ODhETDQ0ODQ0TGBEPDw8PERgWFxQUFBcWGhoYGBoaISEgISEnJycnJycnJycn/8AAEQgB0QHHAwEiAAIRAQMRAf/EALoAAQACAwEBAAAAAAAAAAAAAAAEBQIDBgEHAQEBAQEBAQAAAAAAAAAAAAAAAQIDBAUQAAIBAwIEAwUFBQQFCQcDBQECAwARBBIFITETBkFRImFxgTIUkUJSYiOhcoIzFbGSQyTBolMWB7LSY4OTo0RUdNHh8XOzlDXTJVU0ZLTUNhEAAgIBAwIEBAMGBQQDAAAAAAERAgMhMRJBUWFxEwSBkSIyobFCwdFScpIz8PFigiPhorIUwkOj/9oADAMBAAIRAxEAPwD7/SlKAUpSgFKUoBSlKAUpSgFKUoBSlKAUpSgFKUoBSq/cd82faVL7lnQY1uGmR1DE+QT5ifcKrV757bdGlinnkiS2uZMTKZFv5uIbUB0VK5WfvnbslosTtwDdtxmJ/QUmJYEX5pcpnW8ai/AabseQrU0fd2S+ufe4cNTzhwsRWAHskymck+3SPdWbXrXdlVW9jrmZUUs5CqOJYmwAqsye5Ngw4xJkbljKrHSoWVXZj5IiFmY+wCqA9s7bkSdbc2yN1kJufr5nljv7IBphHuCVltuzbNj5s254ODjwS2GNFLDGqEJETqtYAD1ki48BWHmXRM16b7lg3eO3MbYeJn5vAkmDDmA4fmnWJTf2GtI7l3vIv9J29LFxADZ2RDCLE8TaE5DcPdU25POvKw81uiRr00Qf6v3eJmb+mbe0Gn0xjMlEmr9442m3wrXJ3H3Hkyx4OHsrYOQTqnzsxkmxI4xz6Zx5A8rnwX0+ZqypT1beA4Irz/vXD64N3x8hrcYsrD0oTw5NBKrD9tYM/eWSdE244WFHb+ZhY7ySk+z6pyg/umrOlT1L9y8K9ir+h7iHFe58q45asXCI+IWBT+0Vkm+9w7Sb71iJuWFyObtiMJowB80uI7OWBPjGxt+GrKlFlsush0RCPesJcywbTuGRtvC24xQ3U3HFlgZlnKj8QT3Vb7dv2zbqCdvzYp2F9UYYCRSDYh42s6/EVXmOWI6scgpxLQtwuSb3Rvu+7l7qjT7Xs26MMnMwMfJlHpLzxI0i2+6xYE8K2s3dGfT8S+yt22rBNs3Ox8Y2vaaVI+B/fYVD/wB7e1v/AOawf/uYv+dVZFsOxQHVDteIjeYgjv8A8mprQwuhjeNGQixQqCpHkQRanreH4j0/EuoMiDKiWfGlSaF+KSxsHU+5luDWyuKl7ejxpTm9vTf0fP8AOFb4svsyMW4ja/4hZh51ZbT3R1ctdn32EbfuzD9IAk42SBzfFlPP2o1mHt51ut1bz7GXVo6OlKVsyKUpQClKUApSlAKUpQClKUApSlAKUpQClKUApSlAKUpQClKUApSlAKUpQClKUApSlAKUql3bunZ9olGJJKcncWF49txB1sl/+qX5R43aw9tAXVU2690bXtU5wm6uXuGkOMDDjaacqeRIX0oPa7Cqdz3Tu7F8zMGy4jD04eDpkyf+typAyA+yNP4qz27Zv6acuSLNyJ582QS5E+TokkZlRYkBYIvpVV4CudstVtqzao+pk+7d27gLYuHjbPCw9MuW5ycgA8v0ICkat75DWo7PmZVzu285uWSLGOFxhxW8tGLob7XNWEUkoYQZFjJpLLIgIVwLX4G+ki/K9bri9r8fKuTyWfWPI2qrsQcHZdo2xmkwMGHHkf55UQdRv3pGux+JqfqbhxPDlXlKwUjzQlZTmQKPqNISQWF5I1JbRfzBYke2socrGyVR4JkkWQak0sCSPdzrKeRoomZBeQ+mNePFzwXl7a9hgSCKOJALRqFBAtyFqAzqNgoYonxyQWilk5eTsZVP2PUq1aJQ0WRFMoJWQiGUX4C9yjW9jcPjSAbq9tXtqiuJpJWki66BLpa6CNreIWS5+PCkCSTalqjxR5rDVPNoJtZFCNbzuxRb/ZWZhyLHTkkHwJRCPiAB/bVgSbbUtUctuCKP0op2+8Vdoh77Msn9tZxnLcqZY44hc6lDmQ29h0oKQJNlKytS1SAY1plDxkzxAsR/MjH31Hl+YeH2VItXlqA0RZmHMiyRZEbxt8rBhx+2o2LuBdbzwyopklHXKgxAK7BfUGJ0kDgbWqTDFoeZSLqX1Jq42DAMQPZqvW6gFR87Bw9yxmw8+FcjHcgmNxw1D5WU81YeBHEVIpQFOg7k2Nte35Db1gD5tvzXAykXx6GUf5h8ll/vVf7P3Btu9q64rtHlQ/8A9Tgzr08iE8rSRNxHvFwfA1oqt3jacLcIxlTF4MvFVmxs/HfpZEVuJCSD7p8VNwfKulcrWltTLoumh1tK5GCDudYY1l3/AFSBV1scOEktbjyK/wBlYtu3cuyFsncOnvW2rcynFhMOZEo49QRa3SYAcwtm8r11WSj0kxwfY7ClasbJgzMeLLxZFlx50WSKVTdWVhdWHvFba2ZFKUoBSlKAUpSgFKUoBSlKAUpSgFKUoBSlKAUpSgFKUoBSlKAUpSgFc/unczY+Y+1bPhNue4xaTkqHEMGOHGpevMwazEcQqqWt4Vjvfchx5jtGxoubvT8DHe8WKp/xstl+UC9wnzN4edQtnwm2nDOPIkk+TJI02ZlnSWyJnN3nPqFtXgv3RwrF78dtzVazvsan2vdtz9W/7rIyEn/I7cWxcbSfuOwJmk9t3A9lWGFt237ajR7fixYysbv0lClifFm5t8a2R5MUjdPikgJHTkBVjbxAPMe0V6JQZmhAJ0KGdvAFj6V99hevO7We7OqSWxspSsXdY1LubKvM1kEfPLJCkqR9WVJY+ko56mbp/ZZuPsrfFEIltfU54vIebN5n/R5VrVWyHSWVCiRnXEjcGLWsHYeHAmw+2pFqoFqGygsxAUcyeA+2vbVi2PDI4keNWcDSGYAkAm9uNWAQzlQS5WOwYmAByk3KJpDZFXUfmNi1rVPtR4llRon4q40n3Goa7hDBhY2RlEoZXjxiDxIlZujY/wAQqhJtpJS3skTLVpyz04RIQSEkjZgouba1BPwvepNqXAYKTZmuQPdz/tpBDzTS1ZWpakAxtS1Z2pVgGFqWrOvLUgGNq8tWdqWqQDC1eWrO1LUgGu1LVkxVFLMQqjiSeQqMJZ5+MCaIuFpZQQTx46Y+B+LWqQDdavK8iieNSJJWmZjcswUW9ihALCs7VIKY1HyB1pIsawKk9WYEXBRD6V8uL2+w1J/ZUbFvJrymv+uR0wTcCNeCW/e+b40BIpy40pUKUkDZPaWXJNiQPkdvZTmXJxIFLy4UpHqmgjW5eF+bIoup4jyrrcDcMHdMWPO27ITJxpRdJojqU/8AvHlVbVNl7LNj5Tbv27KMHdD6pY//AAuXb/DyY+VyBYSLZh7a7Uyxpb5nO1OqO0pVRsPcGJvsD6FONn456eft0pHWgkHNWHip5q44MOVW9dzmKUpQClKUApSlAKUpQClKUApSlAKUpQClKUApSlAKUrXPPDjQyZOQ4jhhVpJZGNgqqNTMT5ACgMpJI4o2llcJGgLO7EBVA4kknkBXI5e953cbHF7dkbF2vlkb5ps0nHimArfNysZWGkfdvUWKGXuz/wDdN6Dja3bXtu0MdMRhHyT5aD+Y0nzBG9Ki3C96vIXikjBhIMY9KaRZbLwsvsHsrjfL0r8zdadWaNv23C2uD6fBiEaMdcjElnkc85JZGuzufEsalUpXE6GMkaSrokGpb3HmD5gjiD7RUfCjlibKSWQyt1tSueegomlT7rVKrVCGMk7FSoLgLfxCqq6vdegNtaI/18iSQ30QHpxqeRewZ5B7r6R8a3llRS7nSigsxPgBxJrRhuvTSNmvM4MzqRYjWdXq4en5rWPGiQJNq15DyRY8ssKCSRFLKhNgSBe1bgKyAqpEMRpYBlOpTxUjxB5GvA6GQxA+tQGYW5A3A4/CtDMMIhQpaF9RjQc1f5tCjybw8q348TRp+obyudcpBJGo+C38F5CtQDZauN7/AIt0li2wbfEWRMlXLalUGdiEgU3IPE348q7OqXfRrzdhx7ai+eJNPhphikdifdwIrN1NWj0+xyen7mmRJW4qzi22lXJcIXZFaRdDsoLpzsxHEX9hrRkFknxJALqXaJ/YHUkH+8gFS7VF3Fmiw5Jk4tCVlAte4RgzC37oNbg8rZJtS1ZEWJFKQQxtS1ZWNLGrAMbUtWVjSkAizZcePMsMgN3UFCnrJJOm2hbt8eVVe99wR7UcOERsJs2eOGMyDQgUuvULFuPynyq6ihWLUR6nckvIbajckgEi3AchXLdw7HDvfcu1xPNInQgkyJNJBCiOROnZSPvOePurN5S032PT7SmK2Zes4pWtrOFP21bR0IXcWc62hhTiAEDSP7CWbQvw0miYjKD1cmaVjzJYIPgIlQVLPEk+deWqnnIEcUi5pglkeaFIxNEZAL6y5Ui6hQ2kAWvxF6mV4IQJmmLEsVCAeAUG/AeZPM1nUYMLV4RWdq8PtqQUh5X6pTEU/wA3jLY2IiHPlx9R9P21vtWvGHVMmVa3Vssd7X6aX0n+Ikt8a3WqNAwpXkvV0HoBTJ90SEhT7ytz+ytAhyJh/m2VVvxhhJKn952CsfcAPjUgpk+Xixv05J41k/2ZdQ39296zilSZOpHxQkhT56SVuPZwokMMX8qNE439KgcT48PGs6ArN12fBzlbLaIrmwpeLKgdoJ7J6hH1YmRtJtaxNqhzLnbLCm+7Pm5WRixgZGZtuXK+SkuMVBcwtMWkSRF9a+qx5W41dZLFMaZwNRVGIXlc2Nh8aY8PRxYcd7N040ja44HSoU3HtqqzWzI0mXmPPDlQRZWO4khmRZIpF5MjjUrD3g1sricPJyuzi0TJJl9tElo+mpebAuSzAqvGTHF/Tp9ScrWrsMXKxs7Hjy8OVJ8eZQ8UsZDKynxBFeqtlZSjk01ubqUpVIKUpQClKUApSlAKUpQClKUApSlAKUqJum44u0bdk7pmsVxsSNpZSBc2UXso8SeQFAat33vbdjgWfcZun1G0QRKC8sr+CRRoCzt7hXL5Em6d15DY+4Q/0/Y4HR3wWa+Tkt/MRMrSSsaDgxjBJbhc+FZbbiyCRu4+4GUbrlLZQ5BTEhY3TFx/LhbWRxZqnY3XlR7xnHWR3eRzwdrtYaRzHpA9R+HnXC+V6qvzOladWZy9XKfRCUECEiUsCQzj7gAtwXx4+zzoMfNJBbOI9iQxqtv4xIf21JVVRQqiygWAHgK9rkbFKVqyZ0xceXJlBMcKNI4UXbSo1Gw8ahUm2ktW9DbXoFYQyxzxJPCweKRQ8brxBVhcEV7JEZQF1lU++F4Fh5auYHnaqiPTR9DTNGM4GC7Lj/4joxQsQeCIykG1+Z+HnUmKGOJdEahV5kDxPmTzJ9prJVCgACwAsAOQArICtEFq9tXtegXqwQh5oJkwbf8AmVPw6UtTLVCny8eLLjWaeOJIlLNrYBi7+hVUePC5Pwrf9VqfRDDLJwuX06E92qTTc+6tQQ32qkzj1u59ngAv0IcrKc+ABCwr+1jVoFz5EOowwOeQAaXT8SY7n4VU7auRlb/uuQZzpw0gwFdUUamAM8vAhrAFwKllsu7/AC1O+DT1L/w47f8Af/x//IvrVryrLizsxsBG9yeA+U1qOAzEl8zJIPEqHVB8DGisPtrN9vxJYzDPGZoyLMsrNIG/eDkg1qDgSFQhVuCOA8K9tUWPa9vibXFjJG/LUl1PH2qQa2/SQeTf9o//ADqQDbYnkKN6Bd/SPM8B+2tL4GLIpSSPWh5q7MwPvDMRWtdo2tTcYcN/ain9hvSASipHMGlRo9swIW1Q46xseBZLobeV1Ir0RT4wvE7Txi5MUh1PY/gkPl5N9tIBvtVDswXO3neN3FzEHTb8ZzyK44vKVHl1WP2VL3jdlwtpny8Y68lrQYsXJjkSHRGhVuRDG5Brfs+3LtW2Y2ADqaJP1XPEtK3rkc+9yay1Nku2v7jvT6MF79cr9Ov8tYtd/wDiiXavLVnStQcDXXlqzIrXLLHAuqQ2BNlHMk/hUDiTUgC1Rsj9dhhqfms2Rbwi/D/HbT7r1nbKnv8A+Gj8LWaUgj2+lP21sjhjhXTGLXN2Y8WY8tTMeJPtNQotblwHlWEjBELkEheJCi5t7q20AuQKgI4ljZwinUSNV14gA8rn2+FZEVH2wI2Ek6rpOQTOwPHi5LWv5AcBUkio0UxrysiKwZlRS7kKqgszHkAOJJrJTRkgSmLG42kYO9jb0R+s39hawqRUfGVm1ZUgtJNbSptdIx8iXHv1H2mpFAOXKqTIwcvZJ33jttPUT1M/aAQsOUtvU0Q5RTjmCvB+TedXdeM6xqZHOlUGpj5AcatbNOUGk9yz2/OxtzwsfcMNteNkxrLExBBKsLi4PEHzFSa4bFPcPb2KJcBY9w28SSzybS0fSyYo5pGnZceRWKuy6+CMPYDXW7XumDvODFuO3SibGmF1YcCCODKynirKeBB4g16a2VlocWmtyZSlK0QUpSgFKUoBSlKAUpSgFKUoBXG71kydx7rHtOEQNs2qdJtzyGXUkuRHZ48SP8RjazyeANh5ipncu6ZkuTD23s0vRzsperm5ai5xcS+lpF8OrIfTHf2nwrPAwMTbMSLBwYhDjwiyIOPtLMTxLMeJJ51yyXhQt2bpWdWZx48cbdQ3kl/2sh1NxN+Hl8K20pXnOgpQ8jxt7fKoUc+ayuY41nRWKpI5MJcA2PDS/wAvn97mKoJteOiyRvG3FXUqw9jCxqOuRl6gJMJwPvMskTAe71Kx+yti5UWvpyBon4WEg0g3/C3FT7r0gScx2PFumAc7ZNyIUYfTeCEnU6rLc3VuI0cOXga7ECqPcE+j37bNyW4TK1bflEcjqBkgLe5wRf21egUooUdj0e8v6uRZ4S9aqs0lCV19Nvm1PxAFJJI4kMkjBEHNmNhWp5nZjFjKHcfM7fIn71uJPsH7KyjxlDiWUmWYC2tuQ/cX5V+HGtweUx6uRLcY8WlbH9WYFRe9haP5iPHjavRh6zqyJZJiRpKX0R+2yJbn7SakVkBWiGuOGKO3TjVNIsCqgWHwraBXtegVQRc/Nh23CyM/INosaNpGvwvYcFHtY8BUPtvAlwdpi+qFszJZsvMN7kyzHWbn8osvwqFkH/ePdlwI7PtG2SLJnOOKz5K+qPHUjmIz6n9vCulsef7ai1c9Fov2nfJ/xYlif3ZGr3XaqX0V89W38DG1e1rlyMeG3Wmjivy1uq3t5aiKjneNnUlW3DGBHAgzR/8AOrWnc4ql39tW/JSTK9qD/Wtm/wD5HF/7aP8A51ervGzsQq7hjFjwAE0d/wDlUld0X0sn8FvkybStcWRjz3MM0cgXnodWt79JNbbHnaqYaa0eh5S1Q5srIhx3nyOhhIOT5MlwB5vp0r8A3xrm3G493j6VJZIdjv8A5jN6ZgOSAQ3TxkYlwv8A0jH3Cstx4vojriwu82b4Ur913sv3vsjaqSdyb0NwiiQ7XthdMOdyQJ8r5GmGizMkfFV4jj41eQYGRDc/1Cd9QtpcRsinzTWrP8CxqXj48OLBHjYyCKGFQkca8AqqLAVsolG+73JmyK9kqrjSi40Xavj4t6shSJucbaoZIZ0uP0pEMbW8bSIxF/etqyilyZlJVYwVOllLMGUjwI01LrTLE5PVhIWUWBv8rAfdb/QfCqczWy5rlQHiiW/qIBkYjyXVpA9/GvYsaOI67mSU8DLIbtx8PAAewVlFkJK3TYGKYc4Xtq8OVuDDjzWtpFQGBFY1stwv4Dmaitlo11xVOTIBf9MjRztZpD6R+0+ypBTaRXM9290ydvpDj4kBlzssH6dmH6a2On+Jrn5as9z3F9sg+pzJQusiPGxMdOpNLKeUaF/mPuUW51E27Y8h3O671J1d0mC6k0o8UKI3UjijVlsGQ8Sw5msWl/TVw+/Y9Xtljo1n9xTnROFSY9S37l1+RL2LHmxtoxIchzJOqHrNfUNZY6tJ4ekHlbwqeRUaCDIwYlhjIyIIwqohGmUKOB4/K/7KkpIkqCSM3U/Ag+IIPEEeVWDhaztZ2cTZt6aLXsaZllBWSK5KXvFcAOD7/EeFR+omcVWI6sdfVM3EXI5Re8Hiw+HjU4iouOCsmTESDaTWoHAhZAG4/wAWrjUIbaV6a8rJRw5k2Hiag4iTZCdfI0iOR2mSJeN1J/SLnl8gU286muodGQ8mBU+4i1egWAA8OFAKpMqOft7OfuDaoWmx5eO9bdEf5ij/AMXBHyM6feH319oFXdOXEVa2dXKDUqC1xMrHzsaHMxJFmx50EkMqG6srC4IrdXIbBMNl3zI7eICYGerZ20DwV73zMdfIBiJFH5j5V19epOUmupxahwKUpVIKUpQClKUApSlAKE2FzyFK5fdN/wA/Lz5dl7bERkx7LuW5TAvDjFhwhRBbqzWOrTfSv3vKo2kpYSnYhdthcqDK35iXl3md8kOeJECsYsaMHh6ViUH3k1dVV7VFDs23rtss66MK6JI5VS0Z/UVio4D5rWHDhU6HJTIIaD1w2N5eIFwRYLcerx415bOW2d1okjdQkC1za5sPaaVFzJulLippL63ZtKi7HRGx9I95FQEqlYxs7IGdOmx5oSCR7LjhWYoD0V7pDAqwBU8CDxB94oKyAqohQdwBo9nz4o2PVwBHn41+YWN+oov5KUZfdVwkj5saSRExQOA2sfMwNjZPJfzfZ51G3LFbcFnxFI6bY8seQpuNXUX9NA4tbiLt7PfXvbs7ZWw7bO5JZseMMW5llXQx+1av6vNfkdnr7Zf6Mj//AErp/wCDLFEVFCIAqrwAHKs6Cou5Zibfgy5sjrGkRTU7mygM6qzfAEmtHBJtpJS3okiYBWXIEnkOJNc+N9z90JXtzC6kN7f1LN1RY5Hj00/mSfYBWX+7LZwDdwZ8+4EgasZD9PigjyiisW/iJpM/ap/BHf0FT+/dY/8ASvryf09P9zRvyu59lxZvplnOXk2J+nw0bIfh59IMB8TVbuMvdu+wfTbVif0fFlFpcnLkC5BU8Cqxxaylx4866XFxMXCj6WHBHjx/ghRUH2KBW+rxb3fy/eK58WJq2LErWX6sr5a91RQvnJzeF29u8GNHiHdhg40Q0x422wqijzJln6jsTzv51vHaW0vxzHys6TxfJyZW/wBVWVefHlVxk5WNhQnIzJkghUEmSRgo4C/jzqsbfIZ/SkeXDA3ETnEyCzqRzi0xmwPm1j5DxqrGu0+epm3vM7bavwb1bolRvzdYb+JjjdsdsWaSLasduLR6nQSX0tpJ9erxFWEe1bXEixx4OOqKLKohQAD+7UZt0MUSJgbXm5VlHTRYhCLeF2y3itW1Zd9kRWTb4IWJOpJ8k3Avw/kwuOPvrSouiSOds+W33ZL287Nm7+nbd/5OD/sk/wCbXj7VtcqNHJg47owsymFCCP7tYI2+hv1cPFK2/wAPJkvf+PHAqP8A1fMhfTnbNlwxgXM8PTyU8PuwOZf9Srx8DPq3/it8zGXtTtqYgvtWMCBb0II//p6a1HtDZV9WMuRiuOT4+TMhHu9ZHH3VZYO6bduS3wslJSCQ0fyyKRzDRvpdT7xU3TWeFf4V8jovde4WizZI7cnHyONfsGMZS58O6ZEmUhDI2eqZaahyJWQD9nGrIP3hihepDgbii8GETSY0hHmBIHT4V0GmvNJqLHVfbK8jdve5bwsyrlVVC5VWi7cqw/xKFu5o8Vym7bfmYFrHqtF1obefVx9Y4e2rLC3LbtyXXgZUWStr/pOrG3tANxUziKqM7tnZNwcTTYix5Cm65OOTBKCfzxaSfjViy2afnoZVvb2+6t8T71fOv9Nof/cWtq8qhbH7l2kA4U671iIAPpsq0WUAOeidbI5t+MfGt+H3LtuTOMLK17dnk2GHmr0nPGw0MfQ9/DS1Tktnp5/vD9veHbG1lqt3TVr+av3LziC0lhinQxzIsiH7rC4rUuHEihEeVVHIdVzb+8xNSSLC54Dzqsz9/wBl20quZmxJI/yRIepIfdHHqb9lVtLdwc6Uvd8aVd32qpf4EhsDGd1klUzMnydVmcD3Kx0/sqHum9Y21mPFjQ5WfN6cbb4LdRr/AHmH3EHix4VEGd3BvBC7djHacM8Wzc1QZ2F/8HHuQPfJ9lT9s2TB2nqPAGlypuOTmzHXPKfzOfD8o4VmW9vm/wBh3WPHi1zNWt0x1c/12W3ktfIh7XtGSuQd43qRZ91cFY1T+Vixnj0YPb+J+Zq4rOsSKJQcsmS2S3K3kktFVLZJdEYEVGlHQlWdR6HIScD28Fk94PA+z3VKrXNEk8TwyC6SKUYexhaoZPSKhZOqDIiyUj16/wBGUL8+kXdGA8dJvw9tSMV2kxoncFXKgOCbkMvpbiPaK9kjLmM3tofWfbwI/wBNQpgjpKgeM6lPIivTTpIsjSgWdxZrcjbxI5X9temsspjSlKyBSlKFK7esCfNxopsEou5YMq5e3vJcJ1kuNDkcdEiko3sNW+w77jb7jO6I2PmYzdLPwZeEsEtr6XHiDzVhwYcq01Sb0JdryIO5sG4nx2ix86EGyZGLJIIyJB4vEX1xnmOI5GuuK8Pi9mYvWdTtqUpXoOQpSlAKUpQClKUBT9y7nkbZthOCFbcct1xNvR/lM8vBWb8qLdz7BVftuBj7JtseHGxdIFLSzP8APK59Uk0h8XduJNR3dd77llzvmwtkD4mGwN1fKkA+pkHtiW0XvLVZSR9XQGPpVtTL+K3IH48a8+W0uOiOtFpPc1YkOiMSyIFyJv1JjzOo8dOr8o9IqRz50pXM0eO6xo0jnSiglmPgBWmFGeQ5Ug0uw0Rofux3vx9rcz8B4VvpQCshXgrIUQPQKwmlaNQIxqlc6Y0PK/m35RzNeySrEoJBZjwRF4sx8hSGJwTNMQZnFjb5UXnoX/SfGtIh4QmJjSPxYIryyMBxYgFmb3moPasbR9t7Wr8zjo3Djwf1j9hrzueYwdv7gUv1JIjBEF+YvMREoFvElqhdbNzhHsWwuMfFw0SDcN0UXC6FCmDF8DJ5t9330bi3w/P/ACPTjxO3t3qqq2SbWf21WOvX+vYm529suSdr2eEZ+5ixkS+mGBT9/Ik8Ofyj1Go8ewNNlRZO7z/1PcUYSeoFcTHX8McF7G/IavV41cbbtuFtWMMXBiEUd9TeLOx5vIx4sx8zUmKFIQwS93Yu7HiSzeJP7PdWlWdba+HQxbMqJ09vNU9Hd/3LfH9K8F8ZMUil6nUmk1W4JGg0ootbzJY++tkssWPG008ixRJxaSRgqj3s1hUHcNzbGkTBwofrNym4x4ymwRT/AI2Q3Hpxjz5nkKzxe3scSxZm7P8A1LPjOpJphaONv/7eDikdvP5vMmuirJ5WzTHvX1vTOz4c2fHISPqbdDHXTwJMk1mYX/2atW0YO/ZMiNk50WFCDdoMOPqOw8jPkXt8Ix76uqVtVRJKzD2Da8OTriI5GTq1/VZTNPKG81eUsV9y2FWdKVSClKUApSlARM3a9u3FQudjRz2+VnUal9qP8yn3GoH9M3bb7f0rN+ox1FhhbgWewvf0ZS3lH8eurqlAUbdwR4isd4xMjbtBsZWQzwm/is2PrAH74U1ubd9vnjEmFnY0pVuKCZPUB8y3vwPlVtWmbFxcldORBHKvO0iqw4fvA1ILJATfNpaVMeTLihyZBdYJJEDH2L6iG/hJqwK1p/pe2CKWAYcAhmGmaMRJpceTrax+NQDsmThFW2TNbGjUjVhZF8jHKgWCIHYPF/A1vy0gSWZFR8zCxM+BsbNgTIhbnHIoYfC/I+6oR3qXCKJvuI2EDwObG3Vw73sNUtlaO/8A0iAeF6tRpZQyEFSLgjiCD4istdy1s006tprZrRnGZ3YGI0iTbbOyrHywM1pMjFawsBp1q6/aa34ubDsBtuWwrtqA6f6hgos2OeHBmZFEqA/mWurpWPTSc10Z6377JeqpnnLVaa2dbL4rf/cmRsTOw9whGRg5EeTEwuHiYN9tuXxrdVVlds7PkznLjhOHmG/+bw2MElz4sY7Bv4gahRDufD1jFyId4iiOh8fLtj5KEcv1YgyNceY40lrdfFfuMeliv/aycX/Dk+n5X+34vidCRXlUq90YUTCPdoMjaZCOeXHaIkcws0etD9tW0GRj5UYlxZknjIuHiYOLH2qTSU9mYvhyU1vVpPZ/pflbZmRFY1mRUfMm+lxZ8i1zEjOB5kDh+2oYNWErLE5Y3DTTMnsUyNZfhW815FD9PDHABp6ahbDlwFZVGU0TSrEBcFma4jReLMQL2FaWjyJ1tKwhQ80j9TkeRc8B/CPjWeNaYvlk6hISsPC1owbC1/xEaq3Gsspga8rI1jWSmpsmFJBG7FSTpUsCFJtewYi16zkkjiv1XVLcTqIHD41qymHS6IUO83oSNhqU+bMPwrzNY4234WIFEECKyiwk0jWfO7WvxoDJZ5ZuOPHaMgETS3AN/wAKcGPxtUbN2bD3PEkxNzL5KSgBjqMekq2sNH09OhgwBBHGrClJ7AjdtbnlrPN27uzmXPwkWXHyz/4rFYlEmbgLSIRokHnx8a6SuM30z4LYfcOJGZZdqdmyIVBLSYco05KIBzZQBIo81rrsbIhy8eLLxnEkE6LLFIOTI41Kw94NenHblXx6nKyhm2lKVsyKUpQCqzuLdDsuy5e4ovUliQLjx/imkYRQr8ZHFWdcn3jmQR5/buHNkRxJLuAkkjZgC3ThlaO4J5dXTx87VG4TfYLc2bRtybTtuPt6nU0K/rScbySsdcshvc3dyWqbQgjnWLyKhUNe7sFUAX4nj9gAryHcypSlQopWLusaNI5sqi5PsrPxoQ9Fe3Cgsxsqi5PkBXgrVlXZFx1JVsg6NSi9l5u3kPTw99aRDLFUuv1MgIkl4qGFikZ4olvDhxPtqTXg9nAeFRtxz4drwMjcMj+VjoXKjmxHyqPax4VrxFau1lWql2aSXdsoe5Vyd63HC7bwZGiCMubuU6cDFEv8qx/Exvp+FdJg4WNt+LFg4cfTx4F0xoOPDzJ8STxNV/b+3yYeI2Vl8dx3Bvqc5j4O4usS/ljX0irgUqv1Pd/kd/cZfpr7ejmmLt+u/wCq/wCyvgZVWT5WbnZL7ds5CmJgmfuDWKwcLmOJTcPPa3A+lb3N/lr2afI3HJfbNskMQiIGfnLY9Ic+jFfgZmH9wcTxsKucTEx8HGjxMVBHDELIoufaSSeJJPEk8Sa7Vr1Z5GzRtu14m1xNHjKS8hDZGRIS80zgW1yyHix/s8OFTaUrZkUpSgFK0ZWZiYUYlzJ48eMnSHldUBPkCxHGqxt/kyQw2fbsjO0sFMzj6WDjzIkyNLMB+RWoC6pVP0O4MqS8+ZDgwEfysSPqygkf7ef0f91Xo2OJhbJzM3IN9V2yZI+NrcsYxC3spILelUTdo9vyMzyYrSOxuzvPOzE+1mkJrbD23teOoTGGRCqnUqx5WSqg3vwXq2pILilVQ2iZJC8G6ZkYvfps8cq+79aN2/bWCx9zYzsRkYmfD9yOSN8aT4yI0yf92KAuKVSjuH6bprvODPt7PfVLp6+OtvxTwago/fC1bQZEGVEs+NKk0Li6SxsGUj2MtwaA2UpSgPCAwKsAQRYg8QQa598LK7dZp9qjbI2ji021oC0kJ4erCF/kABvF/dtyPQ0oCHi5WLn48eXhyrPBKLxyxm6mxsfiDwI8K2VX5OyumRJuGzTjBzJjqnRl6mPOR4zRXWzfnQhvO9a/6w+HIsO+Y/0LMxWPLVupiPYX4y2HSJ8pAPYTWWiyS58Vpn1rkzQcLFYmXSbeNnR/2VoO2XlWY5mSZEFlYtHe3OxtELj2VYe2vKhSHNDnEDpZAZL/AKkTxpdh4gN8o+K1VJ2tsUpfIgjkxsouTJPjuceVWPHTaHQoHs010FamhQzLPykUFSR95T91vMA8RWWk91J0x5cmNzjvas7w4nzOMi7W7l2zccncsDfF6UjGR4slXm1oBw6o4AsFHMWrpY8STLxlbdQsjyoBLjD+SL8bafve3UTUrMYiExrxkn/SjFieLCxJt4KLk1ttYADkOFYVVXadfE6Z/c5M/H1OM1USq1q4W0tbkMYMUSAYn+WZTddF9Fz+NL2IPj41pRsvNhdJYxiAs0cmmTW+kEq2khV0k+fhVhUVdSZkqWHTlRZVI56l9Dg/DTVOJpG3iPT0snJTTYAGUyCw5C0wf9lbVhZJGleV5GI02YgKB7FUAX9tSKxNZZTTIiyLpa9vYSp+1bVEnkycTGkk1JIIx6GfVqJ5KrKvzG/DgRepxqKD9TIrob48Z1Bh/iOORB/Cv7T7qyaNceuLLVJ26ks0ZKuFAAMZGtF5kD1A1LqHkTOuXCETXHEryZGn1OuoaI9Kjib+rw8K2w5kOQ+iESNYXLGN0Uey8irxqA30pSoU9HAg1Sdpbp/Smj7a3CFoVlyMsbRlghoZkWaSToAjijovJSLaRwq1ldmPRha0p+Zxx6Y/Efb5CoG97c2XtEsGIenlYwXIwJbXZJ4P1ImHvK6T7Ca3S3F+ZmylHX0qHtW4RbttmHucPCPLhjmUA3trUNpv7OVTK9RxFKUoCPuGZFt2BlbhN/KxYnnccvTGpc/2Vx20bRHl4T5++48eTuG7EZOYJ0D6Fb1Q4wDg2WFbL7xerXvSTqYGHtIsTuubBjMD/skb6mb/ALuEj41KJuSfOuOa2yXmdKLdkdMOCIaYjJGp+6sj2+F2Nq1YSSOFlyJRNJDrhUhStiraXLC5Go6amVHgKjKy4lFiDHK3kTIun7f064myRSlKhTRmG2LL5ldKjldjwVR7Sa2xSCaNZLaS3zKeJU+Kn2g8KxERMvVkbVp/lLawXhxPtb20aAl+pE5ic8XsAVbhb1L5+2qQ3itMJMmZO5WwhCwxt56gJXI+1R8KxRM8sepLCF+6Uja/x1SEVlho0YmR5Glbqkl3tc3VfBQorSIShVJu/wDn952rZ/8ACVm3HK8brjkLEhH5pGv8KvKp9u0z9w7zkgXMC42Grfuq0zgfGQUfRd2dsD4vJk646Nr+a0UT+HKS751B3XLyIlhwdvsdxzmMeOTxEaC3VyW/LEpvx5tZfGpw51F2aDqZ25bnMp6rzHEgJPAQY/psoPLVIXJ8/hXWqlnmbhFjt+BjbZhxYOIumGIWHiSSSzOx8WZiST51JpSupgUpSgNWRk4+JBJk5UqwwRAtJK5CqoHiSap0zN43kMcJDtWAw/Ty50BypAR80WO/CL2GW5/JXkcf9b3CbJylLbdhSGHCx3HokmjNpMl1PzaX9MfgLFvK11UbLBX4mybdiS/UiM5GWbaszJYzTHSLCzvfT7lsPZVhSvagFe2ryvaoFKgblu0O3lIEjfLzpgTj4MFjI4BALEsQqIL8WYgfHhUYbXuG4Hqb1lFIjqA27Cdo4gC3p6kw0yyNptexVfy0BNyN12zEkMWTmQxygajEzr1Leei+r9lQh3Ltzv08eLMyG5AxYeQUPC/8xo1T/WqfDh7btiSSwQQ4iEappEVYwQOOp2FvtNRm7j2MAFM6OYE2vATNxHHj0Q9udUhlFurzAsm35lgbeuNY/wBkjqaqsvCx1klz8HBzdtzXA6mVhJCS4B/xIdbpJ8UJtyNWS9xbY/FDkOOWpMTJYfasVq9/r+3/AIcr/wCzyv8A9GgK1N27jRS0O3PuKrYaGj+hkN+ZvNI6N+wVYRdxbf1fps7XtuRwAjzV6QYnwiluYpOX3HNbY95xpQWjhyityLnFnXl7HjU1ry9x26WBos7Enlx5PS6SYc0qG/4lEbcPhQFrSuPfMw9vbqbHuBwxGPVtu5JNFiMCb+h50VoTx9Ok6fy1f7LvGLvmEMzGupVjHNExUtHIvzIWQsrc+BUkEcRQFjWLokiNHIodGFmVhcEHwINZUoCgba8zZQZNhXr4fN9okewFzctiSuf0zx+RvQfDTzqZgbji7lE0mMxDRnRPBIpSWJ/wSxt6lP8AbzHCrOq7cdmxs9xkqz4ufGpWHOxyFlUfhNwVdfyuCKjRUyRXlVI3LL2yRcff1RY3OmDdIQRjtc2VJ1JJhc+30HwPhVsaw0UxIHPy5V5WVYmoUxNQ8vQkuNkMSNEnTuOVpv07N7NWmppqNmIXxZlC620FlXzZfUtr+NxwrJTZWJ4c+XnXqsJFWQcnAYe4i9Rp7zyjFt+mAHnPgRf0x8/vW4+z31llMCPrAGNxjHiqjh1R5t+TyHj7q3GszWJrLKYaVBLADUbAm3EgcuNe0pWSisZXEMTzOPRGrO3uUaj/AGVrkxopdWsuA/zqruoNv3SK8bCxGUq0KkEWPC39lUHuKpGPGXFpHAkkvxOtvU1zW8Gxv5VHiZklbHkYsba4mN7lORBPiVP7LVvoCJ2boxot02cNf+n50vTXyiyrZkf2dYr8K6auLwtwx9l7qz0zyYsfd0wxjZNiYhkIJIjDI44I7gKVvYHlzrtK9VHNU/A42UNilKVohyu8yfVd2bbifdwMTIznFgfXMy4sfusOpU+q2F/qO59+mYerG+kwkJt8gh+rNuH4sirKvNlc3fgdqfaK0emPNJtYzxgBvAtGW9JPnZ+HxrfWLxpKuiRQymxsfMcQawUypUf6eVNPQyHABuVl/VBB8Lmzj+9WcMxk1JIuiZLa0vcWN7MrcLg2oDbXoryshRAyFa8dlMmSgPqWS7DxGpEI+2tgrRKwhy8d7cJyYHPt0tIlx48VI+NaRGS6odon+nn3k6dWvd+la9rdRIUv8L1fCuexomj3vcsAEWlzMbcVF+Ogxkuf+0gtV618/wBh1x/2s6/0Vf8A31/edKtY7M6tDlKpuY8vIVx5EyF/7GFZLUPbZPpt53DAKFUygmfBJ4MxAgnUe1TGrfxV1puee2xd0pSuhgV419J087cPfXtKAo+1bntzbC4IkOOnWB59X/F5fnvVxVTtpbC3DM2maTVrds7CvYXhma8ka/8Ay5Sfgwq2rPUor2lBVB7UDdtyO3wosEYnzspujg4t7dSUi/qPGyIoLOfBR51KysqDCxpczKcRwQIZJXPgqi54Dn7qrdoxcnImbfNyQx5U6lMTGb/w2MxDCMj/AGj2DSe2y8loCVtm2jBR5Z3+oz8izZmURYuwHBVH3Y15Ivh77k1U/c+fDu42ddhypJnEksUiy4wV4Y3WNphqmBAu68Dx9lWeTuqRbjj7ZColmkIbIJcIIY2DaGa/zM7JZEHE8TyBrTLt+S/c+JuihfpYcHIxnN/V1JJYJFsvlaM1SGMOZsfcGrHyEgmmilnT6Ofpu/8Al5mgaTpkn0lkuDas23vaMfG1YMsWRHDPBiPFiujdNp5Ux0DBD6QC9UzdubjHiRDCjgizRuG5ZbyvxUrlJlrC0mkam/mx6h7PZVfj9u9wNlY802KkcaxYMD6p42ZRh5sWUxCQwxIFKKdIX40B2n9V2zrTY31sHXxlMmRF1U1xoObOt7qBbxrWd72UAsdxxgokWEnrR26j2Kp83zG/AVxMvbPc06wJNjwXx13GN2WVFjc5sUiK8UaRKQNTDVrbV76stx7Vych80wY8AE2w/wBLh+UWnDOQvLgvEcaA6z63D/U/zEf6UiwS+tfTK+nRG3Hgza1sOfEVhjblt2ZNNj4mXDPPjm08UUiuyHydVJI+Ncvk7Nvq5mZj4+NDJh526Ye5PltNpKRQnG6kYi03Lg4/DjYj21s2DY912/dY5Hi+mwceKaEI04yVKuyMi42qNZYl9N2DMfAcfmoDrCLgg8QeYNVM3bu2mQZGCh23KW5XIwiIbm1v1I1HTlHsdTXuTs0xkfI27ccnCnbjpLfUQEk8bwz6gP4Ctav6xl7c2juCFYYmfRFuOPqbGOogL1g3rhJJ+9dfzVCmzEz83DyItt3so0k3pw8+MaI52AJMboSenLYatN7MOXIgXFQdxwIdzw5MSVimuzRTJ88cinVHLGfBkaxBrzZs2XOwEfJUJlxM0GXGDcLNEdD2t4NbUPYRVIT6UpQGMkcc0bwzIJI5AVdGAKspFiCDzBqiwOtt2dJskzNJAI/qNtmckt0QwR4HY8zEWWxPEqfZV/VPveOxyNqzozZ8XLCv7Y8hGx2X+86n4VGtAibXhrKsTXM2YmsOB4HiPGtlYVllIGPM8GDH1/VKhMAUc2ZWMacvxAA+yt0EJiQl7GWQ65mHIsQBw9gAsPZWrHjd5pZZQAscsgxxcHmTrflwJvp93vqUajKjE8xUaCUNBE8jDUw4kkC58a2TRySELrKR29RQ2Ynyv4D3VoO3YBZnbFidm+ZnUOT8Xuayykggg8a8qOMOOJQMQnH0/Kq3MfE34xk2+y1ZRzEsIpl6cxuQL3VreKN4+7nWSm6lKVCkfLYRdHIIJEUihrfhk/SJI8QNQPwqRWnKilnhaCKTpdQFWlsGK3HDSDw+2soOv0lGRYyr6XdeAcj74Hhq8vCqQi7ntePue35m3yjhmKbtexEgA6bqfAqyqRVr25uMu7bFgbhOujImiX6hPKVfRKPg6mtFRe1Jejk7zs5vfEyzkxHw6WaPqR9khkFdcL1a+JjItmdLSlK7nM42KWPD7l7iiyXETTNi50es6QYPp48UuCeHCSFgfhU4vJlKyQ6oomBByPlYg/7IH/lH4VBSFcnu7esvKi1S4aYuNhO5DaYZIutIUH3dUhN/PSKt68uT72dq/agBYADw4UpQEHiOIrBoUpSgFZCsayFVEMhWjNACwTMwVMeZZZCeQXS0d/hrvW8VlYMCrC4IsQeRBrSIzPxqh3nRtm8bdvzWELA7dmNy0pMwaKQnyWQWPvq2ibpS/Sub3BeAk8SoPqX3pcfAitmViwZ2LNh5S64MhDHKnK6sLGq1K/I3hyKl07JurTrZLfjbRx49iQKgbpizypDnYKq244DGXFVuTgjTLAT4dVOF/BrHwqv2LcJsaZu3d1LLmY11wciQWGXjp8siHkWVeDD410IrVXOpnLieO3F6p61strVe1kZ4GdjbliRZuI2qGUXFxZgQdLI6niGVgQw8DUmuenw8vbsuTdNlQOZjqz9uJCLkG1utGx4JMPM8HHA8bGrPb93wdz1rjyFciK3XxJQY54ifCSJrMPfyPhXZOTi0TqUpVIU3cWOywQ7vBwydqc5IsBd4bWyIePg8f+sAfCrNHSVFkjYPG4DI6m4KkXBB9tbSAwKsLg8CDyIqhhx932JBjYcA3PbVa2PEHWLIgi4kRDq+iVU5LdlNvPnUaBeVlaqn+u40ZK5WPl4un5mlx5CgsL8ZIRJH/rVaRyxyjVE4ceam9AQNyx8fPnw8GWUfpyrmPjgi7rAdSah+ES6T7xUuTKiTqKrh5Y7AxggsGb5QR4Xrn9z3LE2vumCfMlSFX290R5XCKSZkNrn2CtGOu47huu5Z215+OkLR47ITCZ1a8bWZG6kfhwrPPWN3MQt9j1L2rdebfGrxqytZPjLtxiUmTdk2SJMlt5myfrpZbvFMQLF3GmSdSCeagIn4YwB4m/Q1Wdu//gNrv/5SD/6a1Z1pOUn3OGSvG9q78bNfIoQ+XvuRmRwZsmFgYcrYjDHCiaWVApkbqOH0INVhpF/G9SMbbMrbZEeLcsjJgLWmgzCspOqygpJpV1IPtI9lez7PMuTNmbVmNgzZJDTx6FlgdwNPVaNtJD2ABKsLgcaiRbPvjbnFuGfuUMyRLYY0UM8ceqxs+g5bpqF+ZWqYJKb3rmixDFpynyJ8d01GyrApkMoOniCpTh+aqfad4zDDhZsuTlZMTQGfc/qMYRQxIsLSF4ZBFFf12AF2uPtq5fETCy5t1mlXVOscLAhgtywQaFBb1udK+2wqHDk7BmbfBsqTyriAx40TyJJEspha3SEsiKj6unpIB41zczrZdY/Ye7G8Tpbjhtar4q1o5cdLcobT2lRr0K/b+5HycDcGlyosvLjjTNxRCVPTWf0pDJ0v9k/BvG1T4s7Owt1xsHMzmnedzHMkuO0cZJjZ0ONLHHpv6eKu58eNS8mDZH3WHFkCpmvjyjoqthLjvZXSQhbEXFwL3qPBsWE5xdyx8rJy/pm6uHHNMWVBYoyAML8jb1XNRK2imYff5ltf275X9O2NXq3HFPVqK8bRtKnSPwL81U7h3D29gz/07ctxxYZ5AFbGlkXUQ/ABkP4r+NWSTxzozwMH0kqbcbMvNW8iDzFcR2zhYuZ21E+ZCs0u6RvLubSL6pZZWYS9QnjwPpA8LWrV7cVMSeKqku8GE9vZkW1hydpyyw28ObnHlAL/AEik/wCGVBMY8LFeVqlYevG33OxtIEGXFHmRsOZlX/LzA/wpGfjXMJmZeHtmfsOdrys3aETO2nIZS8mRjQurRH800TDpv58D96uk3BJF3rZcxCVTVkY0y25rNF1V1e5oB8a0nMNdSNF1SlKpBVXvU6xjAhtd8jNhRBf8BM7H4LEatK5nIzYs/u/H29F1ptWPLkTSEGy5EvTjjUHlqEUjE/vCo9gi8rE1lWJrmbPK0zSCGN5WFwiliBzNhew99bHdY1LubKOZqNofIdXlXTEhDRxn5mYcnfyt4CoUY0TQ46RuS0nFpGPMux1t+01tNemvDWWUwNYGks0UIXquF1myA82PkoHE1pGTqGpYJreF1C/scqf2VllNprCSNJV0SC45jzBHJlPgR4GsUyI5HMfFJBc9Nxpaw+8PMe0VsrJTQJHhcRz+qNrCOf2/hk8j5Hx/t314QGBVgCDwIPEGtGnIgFoR1o/CNms68eOlje48gftoCRStBy1VwjxSpcX1GNivxZNQrw5sQlWFUld3BK6Y2tw53Zgqj4mgJFVuw5CZfdu8nGlDRYeLi4uUoIN59U0oHmNCPY+0+yp0TTsLzRrHfkobWw9/AD7Kqd0X+n7ttG94qBZmyo9vzXA+fGyzos9rX0yhCpPL41vG4sp8iX1R2tKUr0nE5OciDvPMiXguXt2PkNxPF4ZpYb25fK4v7hVjUHf1GL3LsucQAuXHk7c7cfmKrlRez/Bf7anV5sq+vzOtPtFRoo48SeRVASPJfqA+HVYWcezVpv771JrxkV1KOLqeYrBoO6RqXdgqjmzGwrxHSRA6HUp5H9lRsaN5WaWc9RUdlxlaxsqm3UPm5Pj5fGpdACQOZAubC/mfCshUfJg+oi0A2dWSSNvJo2Dr/ZWyKVZQSPSymzoeanyNEDcKzFYCsxWkQizZATKxkkjdQZGVZNN04oQt2W+m5/FaptYMqsjBwClvXflb21SSdybPgTfSx5RzABxixg+TJHYfeMYb0m33jetSlu4LTFkyNrHS1434qYLPc9rx92xfp5i0bowkx8mPhJDKvyyRt4Eftqvx98yNtyF23uQLE7ELjbooK40/754iKT2E2PhWyLe9xyYxLh7HktGwujTyQwXHgdLMzWPurXPJ3LmwtjzbNgmFwerHkZRkVx+EBIuB9pqT1Uz5OGenHjsq+nm4OnjkpW9H3rr81/mdCOV/A8qjZm24WeY2yog0sJDQzKSkqEfgljKuPtrkIcTvPaZQ+z4KrheO2TZizxKLcoWZY3j9g1EVcRd240TdLe8PJ2iUEKzZEZaC58RPHdLe02rSuus189vmZyeyutcVq5lvFLVtdfzUq3+EosI8HdcYucbdpJEYHpxZkSTBCeVnj6EhH7zGtlu4f/N4X/20v/8As1Kx8jHy4+tiTJPH+OJg6/apNba6Js8brDaahroRUi3kr+rmwa/HRjMF9ltU5Navot716/6uLatXT+lj0afwfNq+OqrIV7VkkFfIvcCMGgyMSdRa8csUkRPnaRJJAP7hqJu+7jClhR4NOQUeQlWAPNYokRuV5JZFHqFrXq8qk3HbvrNxWLIUvDlQSQPKhCulmWaJ19qMDxA/DSzcabnXAsfqL1daw5+X+IMVzN0gzoMfd+h0sgiJXxi6mN2DMmrXfUGKlb8ONuHGoO39yZ0j4qvLh5H1EvROLCSMlR1DGSyhmF0X1tdRwvUqTCy45o9w3XPSUbUboGRYUdWGkSzuWa7AE2sFAbwqHt0OyTfRYuFumA+diTCdHx2jaWU3curaWDEFWI4Vlq/T8dzvW3t0nyVbNxrVPj+rutP07ErH3rc/qDtHRiXNxmc5btqMSQ2Jx2W3+1NrXPCzX4ir3bs1NxwcfOjUos6B9Dc1JHFT7QeFVa7I+G8e5RTtJuCNI+ZJoBOUj3JgK34BbL0+Ppt76yw9naPasfb/AKuTEzIlDyyYjqCHa7N6JBIpXU3itWvKdf8AHYxn9B0nHEylpK1+rnv024+Bd0qoOL3FBHaDcMfJYA2+qxyrE+F2x5EA/uVjE/dak9eHb5VI9OiWeMg+3VFJetnlNu8WEu1tJ/IGYnV95SRYv+9KVz+C0+TssewrhZK5QnJeWWFo4o1GUZuqJJAFay8VC8Sau8zL3BMYnO2pJoQuufpZEZC6LNq/zAgHC1wb1yx7v7i35y/b8MeFgcDDPkJ1J5B+IpcIinwHE1yyOtdbOJ0iJbPd7a17UVKUl0avyb41q6zxb/qZY5OFvUkk++xhSI8tcmHFaN/qOjj6sfprxt+pGXYDT96rrt4g4+Y8YIhfNyWhuCPSZDqNj+fVXN4fde97XlRY/cqRS4UpCHOhQxvG7NZWlTUVKcbEi1qte58l9lj2rIRujs8WR0NwCj0okqNHDI5HEKsum/vualHV/XVtxMrrJPc3yKvpZKJbcGvt4LZSt4/eVm7ZX1O4Zr9v4MkWTETDnbz9T9DAJIx4hklE5j5FjGQOV6qu3NwxNlTOXde49ty455nyUWB1DJJIbzDgeIZvVpVefKrfudMNMPDG4J/+0rmwnc1HCPoksLzW5x9UoX8POuvx4cSOCNcSONMewMQiVQljxBXTwsfZV/uJ6wpPL9rOU27K/rXcOFmbfDMcLb48kT50kbwxu0wRFhi6oVpOK6iQLC1X29wzS4PWxo2myMSSPKhhQgNIYWDmMauF3W6j31ZGvK0lxULoRuXJqwdww9yx1ycKZZYzwOk8VbxR15qw8VPEUytxwMG31uVFj6vlErqhNvwhiL1Hk2japJnyHwoTPLxll6ah3I5F2Au3xrS2Fsm2as9sfFxOn82UyIhW/D+YRfje3OtSSCNLvGfuoaHYoWghIIfd8xCkaW5mCCQK8p8ibJ7TyqL2viRKmTn44cYs7dLCMnF5Io2d3ymJ4lsiWR3v+HTW1mye4wqqsmLspN5WkBjmzF/2YRrNHCfEmzNy4DndKqRqscahEQBURRYADgAAOQFZbKj2tM88UC6pWte+lRcs1vBVW5b4UlmIYQQ2aduNjxCD8b+zyHjXkUCRHVcySkWaZ7FyPhaw9g4VkpELyyZJmlxpTDEqtjiy31nV1GK6r3tYCpEU8U664muPEEFWB8mVgCD763Go2RjLNd0Yw5GkqmQnzL5XB4MB5NUZTaajyTMXMGPYyj53IJSMfm5XbyW/7K0RvJMVXIyulIo/Wxk0xtq5XLambT5abXqQzY+HEAxWJOOkeJPM2HNm/bWWUxjx44SXHqlaweZuLtbzP+gcKyNZBtSq1iNQBswsRfzB5ViayykfLTVAzKbPF+rGx8GXj4W58jW4XIBA4EX+2sJ79CW3PQ1vsNaI8TCkgj0Rr02RShS6+kgEaStiOHlWSkqlaUYxy/TubhgWhZjckD5lN+ZX+yt1QCsJYxKmm+lhxRxzVhyYVnSgNcEwniEg4G5V18mUlWX4EVX9wuItuTIYEx4+XhTykcxHHlRO7fACpIyY4JJVlDhWlNpAjMgOlODMoOn417mQQbrt+VhBlkTIieFipvYspUcuRBrS0afiHqoOlpXNY+7Z+T2Kd2T/APJLt0knhb6mKJgfZ/MWles4Enu3Dly9iyJcVQ2bg6c7Cve/Wxj1VUaePrAKe41qw8uLPw8fOgN4cqJJo/3ZFDD+2uhrlJO2tz2l5ZO2cmL6WRml/pGaG6Ku51P9PPHd4gTx06WXytXPJR2hrdGqWjcsK15EohglmJ06FLA+0Dh+2om27oM558SeFsTcsTSMzCkILJqF1dGXhJG33WHxseFSMnU0kOMDpZ2Eslx/hxkE/a2kV54acM6mePCuPBFjryiRU48+Atc1spSoBRVUMzj5mABP7t7f20rk9+7ry8fOj2zt/H+uyo2/zWlWdQR/g+nx8zfhRtLVnbB7fJnvwxpSk223FapdW+h1c+RBiwvkZMiwwxi7yyEKoHtJqoXe8/dCV7ew9UPEf1LMDRwf9XH/ADJP2CmHsQnePcN9lOfmgBkjkXTBDcfJHDxW4v8AM1zV8Kql+H5lbw4tEvWt3crGvJb2+MeRS/7trmESb7mTbi3C8Gow4w8wIYrXH7xNXOLjY2GnTw4Y8dPwxKEHD90Cs6EkKSq6iOSiwv8AbW0kjlkzZLqLWfFbVWlF5VWiNGPIsOXLg3sNCzwi/IOzKyAexluPfU2oK4aTS/VZkSNKVVEQgOIwCx9LEfMdXE1t+jCazjzSQFvBW1oD5qkgZR8K2ciWKEB1KOAyMLMrC4I8iDUdFzVUBpYnI5t02W/wDmiw5TOWlyjpPyxxIqAe9m1sapCh3TsjByDJk7NLJtGc3q14zMkTt4dSNCPtW1QNvyN/hlG0ZW7Ph7uoJhiz4458bJVebQTqI3ItzBOoV1zY0xK6MuVQPmFo21D3lOHwqNuWxYO7QNBnmSYW/SctZo25iSPSAAw86y6Leuj7LRM9mP3t3VY8/wDyV6WtWuS9P6pleHyghjet9wVDbjtP1kBGoZe1P1Rp8SYZdL/ZerDbu4dm3VjFhZatMoBfHkBjlW/nHIFaoe0bllwZZ2LenDZ6L1MTKA0rlwDh1LclkX76/GrLcNo2vdozFuOJHkAiwZl9Y/dcWZfgaqndOfB7/NGciwq3HLj4N6q+JzW1X+rhbf4OpNqr7giYYH9Qibpz7a4zI5LXOmP+cnI8Hi1KaqW2TeMSSZO2t3kjijAAxc8fUwB/wJI15F4c+dqpt43fuyHDy8ff+ltsLRSqk2NitkxTXQ/ptKZGEerl6lqvJH3Va/L5kp7NZf7WalvB8lk/oht/7ZNG6Yzd0915ceSLYeAVxIYQTpk0fqGSVb2Yh3OnyHvqx3PsnAOJfpAMgujDgykcirDip9oqmx5W7Y3CLJnnXI27cEikxs1HDp1BEiyxO4txDC6nxB86v9z7ywBiEdUEkWVV4sT5KouSfYK81uLtd5J5T9Pl0g9lfVrXFX2/9tJK/T6l9/Of29Cx7J3bJ3Db58PPlM2bts308kp+Z4yA8Tt+bSbE+JF6uM/asLcSjzqUni/k5UTGOZON/TIljYkcV5HxFc/2fsubg4U+5ZKnG3DcpevJjvxCxKoSGKRfBtI1G3EE28K6NMmeNQcyERgX1yo4aNQBfUS2hgOHlXrpy415bwp8z5ebh6t/T+3k+PkQXxu5MZAuHnY+Zx4nOiKPb9/FKKf+zrJ96jwUixM5lyt2ZCz4eArOxI4nSrH0L7ZGAqN9VndxenbZHwto4h88ArPkC9iMUMPRGR/i8z90feq3wdvw9th6GFCsSX1NbizMebuxuzMfFmN62cznt+l7jzdhzymHDhwPiT9WGVzNkkdNvQqw2jDHl87VG7OlwjgxFdOkqCvuI4V2dcBkdqrjbxNg9tbh9LIYvq5MCaNpIYxI5W6SqwKajeycfZYVxy1fKt6w3WVD6yev2168MuK01VotySnjx/ijpqbe+pMI7ZkhgCDGw0jmbgiw9pqx23dduydqxtj7iibDy5MZIMnD3BQgmsgR9ElzHJexPpa48QK07b2XN9ZDuG/ZozXgIkhxIkKQLIvFXcsWZ9PMXsL+FdZNDDkRtFPGssbcGRwGU+8HhVxUsna1tHaNF0gz7jJR1pjxt2WOfqaiXbt4aHF4HU2vNftXc2ORC0ZfacqazfU4oH6kEl76pIb2N/mWx86yHbcOKWl2TLytrlHqhSGZ2xVbmAcWQtFo81AHstWXcHbkEJhOJK+JiTyoqMh4YOXyx8vGD+lFZ/0pEHBg3vvjt24T5caTZydLN215cPdceO7KstlPVUczGwUMvDk3srN6ur5VcJnKrTUMtu39+bcxNgbgi429YVhm4q3CsDwTIg1cWhkt6T4cjxq7rj95x2yo49y2h0O87cDkYLKw/UFvXjSkf4cw4H22PMVbz719RsOPuW1lTNuKxJgahqUS5BCqXHDhHclh7DW625Lx6mbKGZ527yjJbbNohXM3FQDMWbTBjBhdGyWHq9XgijUfYONYw7LG06Zu6TNuGYnyGQBYIzfUDDji6KR+I3f81S9u27H2vFXEx7txLzTNxkllbi8sjc2ZjQzyTHThqGF7NO/yC1wdIHFyCPYPbWgbJshIbA3eR79OJeLsRx4f+08K1dPKmJ6snSU3tHDxa1vvSMOf7oFVUe/bc8suJsjjetzA1TiCRCq8So+on+SNbg2UXPktcfJmd29zbw8UU7R7JH/JmghK4U0sRIljll60WQ8etdIK/Nz02rL01egPoqfR4loVaOIsflZhqYjhx1HUx99buJFxxB5Ecq4zDzNtimgw9+7ah2+bMkaCGdEhyIZ50BfSp0iQa9JZCy8fGxqmhk3juGNszY9pw9vxEj6uFMmVNFJI12AxnGI8SxyDm+oELy41NN5LqfSjyrCuJ2fN71xcxNs3mXGfKyYHlwI2u8UrxfzImnQLJG4UhuIe4vXQw77CMpNv3OF9vzXA6aykNDKf+gnX0ty5Npb2Vl+GpSfPj4+QAMiGOYDkJEV7e7UDWiPDjx5zLCtw/wA2olin/wAstey/lqWa8NZZTE1gayNYmssp54/+3lUXb9H0MHSv09P6d+ei50c/y2pkx5sySJDKkIIZU9BkZgVtxuyBePvrZiyLLjQyIulWRSE/Dw+X4cqhTKWISAcdLodSP5MP/dwNYJkAv0ph0pfugn0va1zGfHny51urF0SRSkih1PEqwuLjlzoDKlRwDijizPjjxYlmT3seLL7+IqRQGuONo3lbVdZGDKtrafSFPHxuResJ8YSB5ICIcuxMeQBxDgekva2sDyNb6UBS7Em4Tdrbp2rKqQbpgYz4v1QbXDI2TE0keQOAYAs3FSOFKm7SNO+7+vEa8bCk9huuRHf/ALu1K9PL6OXgco+qPE6ilKVsyVm59v7TvEsWRnQE5MAKw5MTvDMqnmolhZH0nyvauYm26Ps3cpcvHxpW2HMiiSaZGknfElh1euVXLuYnDXZx8pHGu6pUtVNQyptFBBPBlQpkY0qzQSgNHLGwZWB8QRWyoW4drS48su49rTLgZkhLzYUgLYWQx4sZIlt05G/2icfMGsNr3Vdx6+PNC2HuOG2jOwJCC8RPyuCODxuOKOOBrz3xuuu6OtbJkpoZAxfHl6RY3ZWGtCb8Ta4IJ9hrCKQYimOSFYIhdhJCP0z4ksALqff9tSaCsGvA0R7jgyBminWQLxYpdrD26QbVLjdZEDowZGF1YcQRXmonmTWo4sDFjYoz3JaNmQ3P3vSRx9taRkkishUVFy47qHSZBbS0l1f+IoCrfYKzJzLHSIQ3gSXYA+0ALf7aqISKyLBV1MQqjmSbD7TUQY+TJHonymueZgURcPIE62HvvWYwsQsjvEJHj4o0l5CD5jWWsa0QyTNgll6OO3XdTpk6diEt4u17f6alVokx4JirSINSX0OLqy356WWxFZwxmJAhkaS3JpCC1vaQBf3mtENte15XtUFZvu0/1XEXot0s/EYZG35A5pMvED91+TCm07lJv2Ckqg4rC8WdGLh4519MkCngVKn732VB7q37L2Pbfq4EAmlkGLhxMNTSTvfQW42RBpLNzNh4VyO0dq73Ljmabdszqy+uTozPAhc82CQlBx8zxrFrqllu21svzPVipbLgdbNVVLfTezhKfur1mdH4fE+pRxpEgjjUKi8Ao5Cqvf0OTjY+2qwBzsiKJwfGJG68wA9scZHxrlNk3nd+3t3x9k3vJfO2/Ofo4uZOdU0MxBKRyP8AfSQ+lb8Qa6STIxZ94yNymcDD2GCSN5PBZ5VEs/Ln04VUfxEVullZSjzZMdsduNvNNbNd0cr3ZtGOubljaIJIS0cOPPj7empppsyW8geH5WCY8Zc8vDjWeFuWH2lmRjfu3INvL+jF3bDx4lLg/ddIi+hrfMFY/ZXT4WQmz7XJuubE5zd0nOQcZBeZ5JbLBjorafUkSKp8BYk8KqN+zMfcdqyIu5oYMX6PcMSNGEjMimYxkjqWj/USKVrkcKOmk0fF/h8UdsfuU2q+6r6tNm//ALKrvW++nZ6HVwbpt2ThncIMqJ8RQWbIDjQoHMsfC3tqqhSXukGbLjMewagcXGYMr5gH+LkKbWhJ4on3hYtw4VwyDacoON53CHac7Hsh9JD5zRus0GVm4rCxjKgGzC7G5uOFX0P/ABAl6bwMmPkPE6wvvELv/TwWBYNIVV3Q8hp5X+8KVv8AxKPy+PY1k9p9TWC3qQ2uP6nHWnS68tfA7DdNxh2nBkzJVMmiyxQR21yyN6Y4Yx4s7cBVfjbP9fGMzeppMqWdVc4gkZcaG/qCRxoV1W/G9yfZyquxMvYHyodz3jf8TPzYuOMOrHFBAWBBMMGtrNYka3Ja3C4r3A7pwxt+Nh7VDLu2ckaq0OKt40a3+NO36aD41p3r3Ryr7XO3HpWXdtcarzs9EZ7zj4vbmOu5bZkSYkysEhwAzyw5UjnhD0Gb525BktbmbjhUvtLHvtzbvM/Vzt2b6nLccArfIsAFgQIgNNj43rzbdmypMxd53+RZ9xW/02PGScfFVhYiINxLkfM5+FSNl1w5m74Lfy4srrY4HhHkxrM3/emSok2+T0jZftNZLUx0eHG+TtHqXWzjalf9K6vq/IuKUpWzzEbcMHG3PCnwMtdcGQpRwDY+wqfAg8QfOuD3CfdO3t5TcNxU5HUh6O4TQJ6MjHgJaPMUcAs8IY9WPxT1L8pA+iEgczaq7fME5+3OsIRsmBlycRn+UTRHWh9x+U+w1Gk1DKnGpFheKRUmhZZIpAHV0IKsp4hgRzBqg7YgkEWxYbyWjwZd2fSvyu0OQ2MlwfJZiRVhtez5Gz5mTgY5U7NLbJ2xCfVAWP62MotwjUkMnlcivNo23MwdwUSlGgjyM8J6vUqZrx5UfDSL+tXFcqVdW0bs00mWG97ljYKQwzlycpiFihGqWXQNRhiTmzPwHuuSQONcfuvdm3Z2FmxZG9R4QjilWPbNuZjNqVWEaZGWiMqajZdKAC/AM1djk7Y2buc80jhY0wmxIWXi8bzkmV18jpVK5TtvtKXGwcnF3yPCycXMEBfHije7GBEjjMrs9mACA6Qtga23CkylJB/pePjdu7VvMW5Y+Hn7PjR5H10cMY1I8OmLGm0G/SbVY34tz51YdgPNl7U2TDO77OpEG2Y82h5YxGLS65I1S667hAeOkXPOrTA7R2TAx9xwosWM4O5yCSXEKjQtkVNCnnpuuoDwPKuJ2/C2Lsz/AIhQ7dC0ox5sMDGhhaaZ/qG5tkRx89S6iPTpAseFc9018djWzk7HuvYH3zAiTHhikzsaTq4ck8skKxOylOqGhVm1LcMo8xVX2Fuy5zbvgzIozYcyXIyMhTpXIM7G00MThXEelRxNdxpr500WZt3/ABOiysiaWHHzseaM5G4pE0bpCeqYcKWJh010m/rsR43olKaDesnQd44OHPtD5U+NNlZOJc7fHiu6S/USjox6TEVNtTC/hwvXMbZufdu99u4265uPHuWJK647bdFjq80iKxiyJRIZdOmyng/zHyuK+lEqYzIPUmktdPVcW+7p539lcn2DgZeHs2XhTw5OLgjLnG2w5YCZC47nVdgouvqY240WiD3Mts3psHExciZ5MnZMpBJj5kgZp4QxJ0zeLRpaxPzJ43HqrpkkSVFliYPG4DI6kFWU8QQRzBrle0ehjz7xsf0s+DDHMuRg7dlA6lxWVY+sh1N6XmRja/A1NG2Ze0z69oyAuPMxLYWRqMBYm5CleMTsT8yjSfFb8altyovDWJqtTeguR9JnYWTiTadQbpmaFvPRLj6+R/EBU2DIhyolnx31xNfS1iORseDAHmKwzSNlOXKlKyUxaSNGRGYBn4IDzNqyrGWNZY2ja4DCxKmxHkQfMVoOS2Oq/WLpHJsheMV/NvFL+0W9tUhJIBFjxB5itEA6D/ScdAXVAT+G/FL/AJP7LVuZ0RDI7BUHHUTw4e2oxj+sZXkDJDGSY14o5axXWeTKOPAfb5UBKpUZY5cUXjaTIj+8kja3HtRm4n90n3VtM8AhOSZFWBVLNMxAQKOZZjwFvG9AaNt//P71/wCgwf8A6mdSo+wZcM8W990zHobdMoixp5OTYuEkhOSPyO0jlfZx8aV6Ifpx1g5T9c+J11KUroZFKUoBVRu/buHu0sWYHkw9yxwVx9wxiFlVTzRtQZZEP4XBFW9KA5Ex927dpTIw4d5S/HIw3XFlA/NBkMUJ/dk+FYPv6xHRkbXucT8yowpZR/eg6i/trsaVzeKr8PI0rs5TC3dNzjaTb4ZG6cjQzjIBx2jdbFldHHUvZhw01NjGZ1dUskYi42jRG1fF2bw/drzO7akm3GXc9t3KbbpsjQcqJEjlhleMaFkZJVJDaAFOki4ArRLsvc8SFsTeYJpLcI8rDAQn97HkRh+2sPE50iDXNdSxFZVQne83bCsHcO2z48puFycGOTMxZLeKtCpkjv8AhdR8a2Y/c+3ZeXFgYqznKn1GJJ4JsZSEGprPkIgJA42FzU42W6LKLsVlUTp5jx2edYZDxJiQNb2Ay3v79NZJiAKVeeeW/MtJY2Ph+mEFCG7JyExseSZzxVWZV5sxAJCqviT4CtMWcyY0MmXFMrsi9VhExAbSNRKpqZRfzFZ4+JjYxvDEFY8S5uzk8uLuS37akitA9RldQ6HUp4gjxrKtGRlY2GnVzJ48dDwDzOsYPxcitEG97VPMYIJxPIF12iVnBBtxVgNJ5+BrSIzm/wDiNAwwNt3UKzRbbmLJk25LFKjQtKw/IxXj4C9XOzbth/Rr6hyr196M8fq21xgzTnCM8zRlCxlON6odWsqW4cqq9v2Xamllxdu2mSLEgnfGeVHjWNTGdLlY3cvprnar5K9GpiIaex7KVaxWx5avircuVbV0ezq5e/huUXdRm37dsDZ9mkWPcJJkyIpiNQhXHIladh5KQLeZ4Vf7RjzbqV2hsQYm1bTOfrmEnXXMykYTAJJ6WKhzrk1Di3p8DUiTZ8XCzG2zt/HGLm56CTctzAJkjxg2j0uTfqPxEYHBfmtwrLdc3CwsJtg2x0xcPEjCbllAnRiY1raLg6jPKPSgHqudR8L9MdOKcuW3L8zzZ8qyWXFRWtVVTvC7nk2frkye43Tqpjs2DsGODqE0rt0mlsBcGSX9MHwQFuRNUHe+E21drbZt31ER3DIz455xINZmyGLTSzIrHkkh1i4tYAV0eBFaIb/nwth7ft2Ox2rb7eqKBY+M8yeEzILBb+leHzE1TdwbPm7psePkllXedzyseVWkvpiUB5osQeIVR6T+Ylq1aeLjeHHmYx8fUpz+3kuX8s6mXb/aWI2L1HXXI/qkkclnZjxLOx4kmqzd9rh7e3ODcIEHSeWOLNhsCJYWcKykfiW91P8AoqXtHeWPgI2FuN8PLh9M2NkeiRSOfBuY8iOBrDqTd67nBDhKW22CWObOyyD0yiNr6Ubj5ncrb08hxPhXjql9PFPnP1ftk+re2T/keWyeF1fGH9O308PHsWmT2hJtIaftoK8RfqZG1ZARlkFwSsE0qsYiRw/9lXGw7/tG5wjHxLYmTF6JdukAjljYDiNHDUB+IVc+2uL33b8PuF8nbcHCfKykfqQbmCY4cV3Hr/XPF/ULlEDc/u16eLrrT5dPh2PGs9c0U907T0yrWy/nr+v8ztao8vcDte8ZuRMrHG/pn1V78L4jvrUe0rKtc3i4E2yzQ7XvO5Zu3NKdGLn42QWxJ/HSVyRL0ZLnlyPgamdw7Bk4uDJmDc8zLtHNiyLkujIseVGYdZ0InpSQozE8lBNWt2/069ddiZvbY8evrTVqatVf1rwe346FJibZuXeMg3PeZXOu7QYasRDArckVFIBIHNjxNZ5GBuHZcw3PapZOihDZeEzFopo1+ZdLEhXtfSw8edxU/tPfIsaH6TMXoZeOelkwPwaOReDK3+jz507s3qPNiXAwl6+ZlHpY0KcWZ24ch4Dmx8BxryKzjlyfqTtPXtB7nX/k9L06/wDrxvH6f4uXfxL7fTHuUWyvBBFmR5OSJI4ck6Y3VsWdxq9EnIcR6arIJ5NtTKxkK7dLNndI4uKY2jRVx1kJhfJ6calwysbr7At+NdEips+1YOPIonlx1gxYQLAtJpEIKluXiT7K0Ym44u65k2J9Ejxo7a5XeFixiYxq5i1GSxI9JI5V67Jct4bPJhy2WGHj5UqnLb68uX2vT8JOcxd5ycpsJJd0ixUghn1sOkxYw5Jx7hzdD+nYtb5vC1asbP3CVM/fWlWMomF9RjhB05lJKF/VdlVl9S24jzrpMrGwI9+hWeBXGfA8YQqnTBiPVd2BHFjwFRM3cWx8Lc40wxlLgao8pw6KTGkSzo9mHq9LWt7Kw01q7befY6rJSzjHgX/Ik4fCqVfU1rLXXbUrpt83Zd0fCxU6atmHG6Zkh/lhtHU0G8+th67k20+HjUHbt8fb+4tzxt53Po7ZE0keMuQqJCJ1e7qMiwtaPSyoxv6jbgK6TA3E5e5Q/U4aYmVLASpZkkdljtcKyC6n13seYrnu/u3MrIgn3PZRFjSlXl3qSRmCZGPjp1FhliX0vrKAarXA8bVpbNNzL69Dy52prGOuOKx9LlW8ZUpnZxMkqLLEweNwGR1NwykXBBHMGuE79hzMHfNh7gwsrGGVBKuHh4GRfVK+S5jkk+dPSitc11nbGdm7psGBuW4QRY0+XCs3QgJKKjC8dtXK62NvCvO59tO5bLmY8OBDn5ksTQ4yT6QFaT06+o/FQvzcOPCiUM4tluUI518+/wCK2Msm3bflR4/1uVt8r5Yw5F1YzQRqPqGyRdfSBpA9XEm1Tf8Ah7LiY8edsiwZK7liTyjcHlLzRIVe0MYyCWQt02B0r7a6HuPZpt+2ttoWUQ4+U6JnPx1nGB1SJF5M9tNzyBNaShkk87czYdz2DbdwgWNI8jHjfRCoWNTazIignSFa4tVhK8cMbSzOscaC7u5CqB5ljwFZwY0ONDHj48axQxgJHGosqgcABXAbpvk/e+TP2lsJmxIRLPib7kzY5ITHWw/TduCvJYqoPHxqQWSpXuDcX7h3j/diCDKzdwzFxIJrF8VlxIWlZzOXUamDAFV9K6SRcmulxd/mbLm2TuGERTdVMaPPxklGHJNInUEKySj0SLyHME8jfhXVY2BiYcMUGPCiJAoWOygEWUJfgOZA41z3f/0jdsZmNl5xwXmA+laOxlkmjYSxxQpzZmZR8vEVGk+gTJ8mpikUjBMtLnHlPJzaxt7x8y1uim68ay2sTwZTzVhwZT7jXLt3l27g7fi428ZvVyFghGVJErzqJ9AYqZoVZRJcE2verjDzY5JIZYplycbNXVDkxm6yEcFbhcA6Rpb22+HFp9jaZZ0pWEwkaJhCQsnDSTyvfxteslM6VDbcY4HEechxiRcTNxgNv+lHBT7HtUmKaGcFoJUlUcCY2DAH+EmqCHh42MmVmgQRh0lFnCgGzoktvgWNT6xVEQsVUAudTEeJ5XP2VlQCqHurCxW2bOyTCGkCxu9iRqCSITqW4VuA+9V9VR3Mde1HCU/q7hPBhQj800qg+B4BAxNWv3LzD2Z2OldOiw02tptwt5WpXtK9ZwFKUoBSlKAUpSgFKUoBSlKAVA3bZ8De8UYm4RlkVhLE6M0ckcighZIpEIZWF+YqfSgOabYu4sRbbfvS5KqLJFuWOshsOV5scwvf2kGqrcNx7z2zMwNuOPtuZkbg7iLpnIjCRRANLNJq1AKmoeNySBXdVzecwj7wxhLw+o26ZMYkji0c0byge3Syn/4ViySTcGk3O4iTuR3/AMxPgQoLfyYppGPn/MkjA/bWR2vImV/rt1ynjI9SQmPFUDx9UKiQcPz1lte8jcEw3iW0eYJ5l1+kpjwt01f3uxU8fA1og37rxZk2dGI4IYxk4kKH1ZGO7NHEzavxMtrcuIrGi3cfA61w5LLlWsrTr/E+K/EgR4mLnusWw4UPQjNn3vMT6j3/AEvWLPK9x85Oj96uh2ra8Xby8iF5smTjNl5DGSZ7m9tR+VfJFAUeAqO+fvOINWXiQFDFJIFx3YmHpoXAl1qoKkjTdfHwqTte7QbjGmkNHO0Mc5jdGS6SAHXHrA1LfhWk1MT8yWw3VXaFZLd1asl8iiTYMo5BP0CRZC5r5SbmcgnSrTtKCsIvdum2mx4VZbZsMGNkZefmRL9S+ZPkQyh24Ru10JAIXl7Kn7jnS4aQpBGJcnKlEECM2lAxVpGd2sTpVUJNqgYe55mZLNFkvjJDjIJWy4JNcbcwwZGIZdFuN+BqKtE/+h3tm9xfG3KSa1iz5NTGkuYnpsat5SNnbKnyclIZ0GPBgYjdOXJkBZhpZbSC4NuBAA9RNq2bX26FaHK3JIlaBjJibdjgjGx3bnIb2M03nI45/KBzMPH7hyTtmfuWXFGmdhqjwwyXCrBPZopXPMagfVb8NTNr358nMmgyJsWeCOESnJw3LRIbtdJC3IlRfn4VpXrK13OVva5q1tZ1UUcPWe370WbHF3rb8mBGY484nxHcXU8C0Emm/tBsaoIsl8jbdlEpY5GPnx4mSX4Ey44lgkb+IpqHsNbMTuLIGBlO+MqTQBcqGIk/qYUn6izcBwITVceYtW/Pgixxtoi4iXcY5nccdTyLI7N8b8KvJPY55MV8bi6jWN57P9p73PhLkbZNnXj6mFDLKqywQzhiq6lB6yNbiPComXl7lsmbtOKmQs2C4lkzA0UcemFTDGNAiVQOmZdXuq/y8eLMwMjEnYpDPG8cjqQCFYEMQTfwqDOu0bjH/VWyNWPiwT48jg2QRyqhl13F7gIKzZPo4269tzvgvRKqvjVkndP6E39dYp9UTM7FJum87mZ94aCRRjYeLI+KGTh1YX6buR98aiV48OFSBumbLkbVhyMyZXVlh3LEThdoota6LabI/wAynyqY+y7Y+HPiiaaQR439OlIYGQ6yJb3K8ZGLg35ca3Hbdvfcot5d2OfgBopRGeB6iiyMpHHSH9PvqJXmZ6rr46nS2T23F1VNVW0W46/Zxo35vXzKDddrX1Y+ZFjq+QZBi7bhRLl5syH5mM+Z6U58W0hV/Fyqsz8Tunt3EZsmSTK2iYGJ4zP13xoSttE3VhKOpHAmwA/EBxrutq2x8aSfPzisu55Z/WlUcEiUnpY8d+OhB9rEnxqyZVdSjgMrAhlIuCDzBFbdZ1Wj7nmxZuKdLrnjtvXx/iq+lvE+Z9uRbL3G/wDR+4IQu84aj6TMiZ8ebIxgo6eso+pnjX0sGZrgalJBvXbbT2xsmyO02BjWyHFmyZWaaW3iBJKWZQfIWFczueyY3bu4Yu6zY31eyY0okidbrkbaxPFo3Qh2xjqOqO9hflau7R0kRZI2Do4DIym4IPEEEVKw25SVlv8A59jWalqVrxu74ry6udJ6q1ell1RX7yrBMTJAJTFyo5ZQLk6CGiJsAfl6mr4VXwbDknJhORHi48cAmU5WEDDPMJQyWIVVEY9Wo2Y+oAi1dFSq6puWZp7i9Kca6bqfB/5s5/I7exWytujlBy8WA5DyJmu2QxMiKFsZtXK1RsjYTjY2642EkMEW6dSKFEGlE146RJqCjh+orcvOrjc55YMjbBGbLNktFIvmpgmf9hQVF3pMzO2nPx9qnMe4Ih+mdTYrOgEsYNxyJsD7DWXWvbf/ACNL3ObT6pjv3Vuc+ckWfEY9wbKUNjjwytOR4/piJV5cdXH7Kn7xg4m44GXh56SSYkqMJ44S6uyAaiq9Ihje3Ic+Vctv+4zbv29tO8bbmyYS5E0cbTIxQxtlI2MC9uP6WQV1D2Gui7czJtw2PAzMiNo5ZYVLB36haw0iTUeJEgGoX42PGnj4mLXduKf6VC+Ldv2nIf8ADvP3GDOyu3N0XOjOPAsu2QZiIiR4SSNHFy9ZdtQBvflbwr6KULKQGKsQQGFiQT48eHCuD3vB3Lt/uxe8482OPacx8TC3WN1u6Qk9JSCQQF6hUlgRzru4pYMqPXBKssbXXXGwYeR4rcVqDmfPOypUwu6t/wAU7plZ2JLOBiO0ROPNkuobLk6kMYiDrIui1wK+j2qPtu3Ye04UO3bfCIMWBdMca+A8ST4kniSedS7VQYVVbRsG2dvxZgwAyLlzyZmU8rl2Mj/MdTcbC1T8/LGBiSZbQzZCxC5ixkMspH5UHE/CuCy+9d57lnO1dj4k0UydGXJ3HJQRdGMyGOVGgylGptNiNJqQCx7k74wdsxpV2uSPKzDjHKxZQDNisBJ0tDyQN6CTwuxCjzrj/wCnHf5WTb8OTcs3OlMuTvEjmHboP5a5qYCozDX6tGoD1Et6jarPC/4Pw4OdDuJ3E5ORFnw5jSSx8Xhju8kZAOnVJJxvavomPi4+FAuNiRLDAhYrGnBQWYu32sxNRlRV7f29tm2xZeOkMTY2TlPl9Exr00uqoqonFRpVPCuS7Nz8Tftr3HBxo5Ijj5k2RHNp0Rf5iaSeEwfetp9g511+/dw7P27itlbtkLEukskIIMsgBCnpRkgta/G1fPto7rmhXdd0xdtfJycy24TYao2KsOLGVw4AgdX6sjrZm0ekedYsm0zSaTO+w8o5EaiYdPIsS0Z4E2Niyi54X5+RqTXLbZvsG5ZWZiznHTJw9EmT9JOXjKyEqZIndI3Dxkevhb21eq+ak745KTaArqzgxlka4IBXUupSOPAc65NQbTJhAYFWF1YWZTyIPMGoEEOOyfTT2GRETGHUGNyqnUhVl0n5bcRUyKVZVJF1ZeDo3zKfJhWdQClCbAm17C9hzNRI2zNWtccIjkl45JF1A/iXphxx8RegJdVQDbh3dgYgGvH2vHkz8geCzzf5fG1e3R1WH21LlzkxUkmzl+mgjGp8hjqiC+LMy/Lbx1AV52dCZoc7f3DBt5n60AYaSMWIdHGFj+JF1/xV0xVm09jN3p5nTUpSvQchSlKAUpSgFKUoBSlKAUpSgFKUoBXO944M0u1tu2C6xbltCyZeJI66lICETQv+WVOHsNj4V0VRtwxfrsDKwrgfUwyQ3PEDqKUv+2gOd/pTy4+3/Skwibbzt8kqqG6SMqSB9N7D5WX3kVul7Y2uEq+KPpIRE+PkqCX1wso0gF2OlkdVZSKw7bzWz9iwMh1KSiIQzoeaywfoSqb+Toay3VXyMva8IfyZJzkZItcMmMhkVT7DKUrhpOq/wj1L3GWta1pd1STWnWe/eOnY2RbVueSt9x3EyAxvGiRxdItrQxq09nIbSGJ0gAaq3TbblYyfUYOQq5MOCuHCWQFdSMG6jamUW4cql3rRm4ePuGO2Jlr1IHKmSO5AbQwcK1uakrxHjVldn8zPrZNNawunGqr8kjLOwn3WKMrKcdoHE2NKFDXkAK6mVrXQhitvEGq9u2JJnypJ8uNWzYVxp0x4FhTpaw8gUambU41KWYnn7KuAa9o4e6/EUzZcailo+Cb3nd+JTZfaeK2v+lyfQdaIwzrYyq+lleJiGf7hB9962S7DuGX1mzdwQyzQ/S6oIBEOi7q0qtd3LFlGlTf03JFW169vSK9i/wDs59JvLWzaVrf1PXoUWb23i4Ucsu0RGN8jHfAlhXVIHWayI7FmJURE6r+V6lbpEkC7RBH8kWbjxrfyWORR/ZVnVL3Lmw4EW3TzB3IzounDEpklkISQ6Io14s1VJTooM5M2S9Ur2do6vVvzfgSu5GkXYMsxkjggcjmIzIok/wBS9UGbi7rNgdwPiZciY7S5R+mEKMrroF9LsC51flrpcf6nMgvucCwh7j6MP1AFP+1YAAtbmBwHmamRlYo0iQHSgCrqYsbAWF2a5NHVN6vpBvD7l4qKqqm1flqk9Pp01n+E5PcmzjNkrLPIMeHLVm4TLEF+mi0erGPVRdZY3B0351K2Eyz7onXDiSPBjOSsjFizLNKsEjFlRuKXI1KDa16ucXOWbKzxpKriyJCzHxtEsxI9n6te7Npmw03Iw9GfcAuRMpJLeoehTfyW3Cqq/VMlfuU8TxukaJJz28Px+RY0pSuh5Dx0SRGjkUMjAqysLgg8CCK5ZNXZ+asLMf8AdvLYJDfiMGZjfSWPKFzy/CeFdVWnKxcfNxpcTKjEsEylJY2FwVPMGs2rOq0a2Z2w5VSaXXLHfS1fytX/AFLobqVzWzZkmzTZGwbtKelhR9fC3CY2V8QHTaWQ+kPFcKbnjzrene3acjALu2PZjYOWsh/jI0/torJqXp0h9yZMNqXdV9ajkrJaOj2se9yllOzspKsNzxxcG3Bg6MPirEVJmDR5cE6WAkvDN5kWLxn+FgftqL3H+r/RekNercsdwV4jSqySM1/LSpNSc4XxmfQXMRWZVXi142D+n28OFS+5ipy2dt0ME+4dsZT9Lbe4Nc+0zMLJj5xOt4FI+X9QCaPzOoVX/wDD6fP2/dMrYcxJZGeJZXMzktiHFVMY45DoNSMTqiZW4r7Qa7bPwcLdsNsTMjE2NLpYC5BBB1JIjCxVlPFSOVVCP3LsI0gN3BtiABAWCbkijnqLWjyP9VveaiZWjppYIMmFoMmJJoXFnikUMrD2q1waxwNvwdsxUwtux48XFiv04IVCILnUbBfMmo207ztu9QtLt8wdozpyIGBSaF/FJom9SN7xVkK0jJkKUpWiCsTWVeGoDA1R9wZvcOFEjdv7VHuchv1VlyFg0+A0hh6j4niOFXcj9NGexbSC2lRdjYXso8TXJv8A8RO0kiynyM76XIw1Zp8DJRoMkFRcxrFKF1N7AahSuxuwTn7e792Zsm4brMwlilLFo8I6hJ0sVHLLYMLFjzHDlXXPBC2QMsxqclEMSzaRrEbEMUDcwpIBtXIr/wAS9tzsiKPY9uzd3i6ZkzDjQsJMa5GjqRyBQdQ4izVU5/d3cO8bdDuGxYckeHPuMce2ZkbKJZ04o0T4rdTUAwbUxYLYXrDTZpQae5dyycrvfC2yLUzwsccQiKPSsOSlp5nnSVpOAswRkUcBXf5EbyWkiIEyEmMnkb80bgfS1clsWbJkHd8fdJpIe4HlT6lzGscTJA6wK+H6bPEhNiTc35+FdImZNC4g3BAJLEieIHpuALl9PFl/bbzrlfdLsbX5kiIxT6ckIVexX1CzjjxRvcRW2o6kR5F1N4cka0YchIBx4/nXiPcak6W4cDx5Vkp5SqXOzM993g2fYpoXz5bz5sWQrSR48CLYM3SZWQyPZVF/M2qT/uvu+53j7h3NfoyfVg7ajY6yDxSeZ3eVkI5hdN63XHa2vQjskRcTHi7v3Kbq3k7e21xHoB/SzctTdtX44oCLW5M/urtq1Y2Nj4cEeLixLDjwqEiijAVVUcAqqOAFba9FaqqhHJuXIpSlUgpSlAKUpQClKUApSlAKUpQClKUApSlAcpuWBn7HuM28bXC+Zt2a2vdNuiF5Ultp+rxgSNVwAJIxxbmOPPEbthZmTs+bhTLPiZb5GMsq34SGPq6CDYqw6BBBHCutrk+5e1NvkTI7g27FCb7i6cuCWMuOpJAQ5BjU6GeRFMeorfjWLUT1W5pWjQua9BqLg52PuWHBn4raoMhFkj8wGHyt5MvIjzqTXI2ZVkDWF69HGqQzvXtUz9xbeZhj4SzbhKbAHEjMkWokgKcg6YQeH4+HjXvQ3jcFX62YbdAbF8bDbXMw8VkyWA0/9Wt/zVQbszeFjmbb9tjGdua21Y6sAkIb7+TJyRfG3zHwFV8+2HHn23Ozpfq9zfNhWTJtpRFKSXix4+PTjv8AxH7xNXWHiYmBjri4USwwpyRfE/iYnizHxJ41F3f5ts/9fD/yZKskgtL0rGvRzAoDl3TJbYe42iJE2Zl5cWO9yW9ZXDS1uPpIsBXYxRrDEkSCyxqFUewCwrl45FGywvAQ4y89Dra9rTZ9yw+HKurrdeplilKVogpSlAfPO7ZpN77ki7eLn6HDiSfIh+7JNKW0a/MIq8ByufZVs/aeEcIgxrbTysKh93bXlYG6p3VgxNNEY1g3GKMFpAEJ6c6qvFgNWlrceR860f7/AO2PihEyI3dhpVFYMxPkFHEmvHkVfUt6qblfSfUw2yPBiXtrJcW/UScPlO7+BF7bE2JvEfb7szYONKc7CW5/TYxTRPELn5PUGVfA3ruy1cTs+Nl42Zi9xbpG+MdxyVwsTGkFnWJ4pSssg5q0j2Gk8hzrtL10ryVKq28dfwPJ7h0ea7xxDa22mPqj4kdIp4B04CjQD5EkLArxvYML3Xy8qhZm7S48/wBDBCuZuBXWuFC51BT8rSsyhY1NuBY8fC9Z7tnS44gwsMr/AFDPYxYuriECjVLOw8RGvG3ibDxrXt+3QY+M2NCzHGDN9RMzAy5UnESzTSD81/8A3LwqnIpt22jM3iZJm+lwtxQAHJwlyJcqIlb6GyYZMYWH5uHsqZ2zj75tGS57h3M5ccy9PHEsjvISDqBEIDKp087Mx9tVmf3dktLHhdv7dK2C87YjbysTPjxSRsEk/ShVpNKk21EAE8uV6sxs3duRDuGMcvBwteqGLIjglkkkSRF1TCRsjUpDMQAQeK+VaXL4EcHVR5M0kiKuNIsLC5mcqtvZoJ1/sqVUbEx4sPFgxILiLHjWKMMSx0oAouTxPKt966GT29eGvL1zXd3dq9rrgxpiNl5W4y9HGTV04w11X1vZje7iygXPHyoC73HKlwsGfMhxpMySFC4xYbdSS3NU1WBa3IVxcffvYe/52LtU8ayS5TsoGdjBFSVCCqyGccGY/LbxHGu6YnkeY52865zuXtjD3uKZxg4mRnTxfT/UZusiOMHWNAj4htRvdSD7azJSZJumy4b5n6kcL4z48Wayrps8+mPHV2AseDAewVKKw4sdgqQQxgmwARFHifACvlGd2xn7Qu5Ys5hx9m2/Bx8nK3CNZ8aLJmh/l6oopX1yoVvrHja4qZswxu4cOVtxzt3wMrcMXrZsWU8QTLxVHTvFeLRoHEelVazcb3BrFtpNIkq+fJ3jmS5eXJPjQxdAyZEKJDjJLLHNCIBGz60kWPjI3C5HjXViFwwhsI2vqELEmJtP34GHqjbjy8PLxqL2/tu27diyQ4uEuC+T68jGvqX03j0KbnUqfL/8akBPpHOJKScOwkx5CfXFY2Ivz0pcWPhfjwrnZyzSRjFK0cn0uVE0azHQmu2kyEApoZPSSePLjfj41q7X7d7f3bt7b9xy8Qz5c8K/VTzySPKZkukpLlyR61PI1YgCdXxsgXdbEkcNQ5rItuR93I1q7OaeB952mcALh5rSY5HAGLKVckED99nreFqWjN9kXm3bTtu0xtFtuLHjI51SdNQC7ctTtzY+01MpSu5zFKUoBSlKAUpSgFKUoBSlKAUpSgFKUoBSlKAUpSgFKUoDjMxf90M+XIKkdt58hlkdRdcDKc3d38RBMxuTyV/YavlZWVXUhkYBlYG4IPEEEcwatGVXUo4DKwIZSLgg8wRXMN2vmbW7SdsZawY7HU205YaTFuT/AIDA9SD3Ldfy1zvSdUaVujLWou4YEW5QpjZDuMfWrzRIdIlVb/pSEcdBNiw8eXKoDbru2GdO6bFlKFHqyMEpmRG3iFUpN/3dMburt/Kl+nTPjjn5dHIDY73HhpyFjN65xZdDcplmsCJLCEjVIYI2WJVAVUJ0iyqOA9PCpF6xBuAw4g8QRyNKSDK/x9lU+6ZuM39MPVVT9bCxRjZlGiS+oHlbxq3vVXvSCVtsD8U+ui1IRfUNEnD3VUyE4ZySgHFRsgNydeEfK9+o3Aj929ZR/XNxeSJHv6URGYDyuzMt/sFbb16pswPkaSDn9vRn7Y23Ww6kUmNISBwLJlITwvwBrsK4+Bxi9soWIk+kk0yaPEwZdmAv4+muwrrTr5mbClKVoyKhYu4fU5+fhdPT9E0S67319WMScrcLXtU2qX6LeMXdM/MwhjSQ5pia0zyIymOMRn5EYcbVLNqI76nXFWtlkVmk+P0y4+rkv2SP63lyJJmYu3PPgRuydRXHWcI2hpIobepQQfvAnwFSMxNvwHiykw4jmTyrFC4jVXaSS9yXCkiygknyqGu2b1jwPtuHlQw4bOxiyQrfURRu5cxqttBIuVDX+F6l7wkinAy0XUmLkh5h4iOSOTHZuHPT1b1lO0OZ6f8AWDramF3qq8YbtEO0tR9HKdm32KuLuSHdNjXcGw9E6TY4OJIQSpllRYplNuKkNqU29lbMXecnMnZIYcMR9aWFVfKtMek7R36XSPzab2vWC9sTx4W0xxzIuVhfTxZj2OiaGGRZtPncMl0J5XPnWrbdqnwXM8uHguqTzTtmMG+pCtI73/ln1KDb5qx9ek9tTtavtIyPHDbtbim3ounVT4Etc3Hysc7vFHd4YXVVI9SsPVJF79SAH3VQbtJkZu4bV2xjxzyxAJkb59I4h0QtfQJJTYqrSKSVU62Hxq2woZYtlmadOnLkS5GQyXvp6zmVV+CsK5iDdpdg7p7ln3XPgx9sh0ZvRABlyGyY1hx47sNf6YhsAvi1Fq/gjy5Eq3tWuys0uuifc7HA27t3YJSmFDjYM+SryO1wssqoQ0js7nWwUtcknhVb2l3RuHcsuZKMOOPa8abIgiz1c/rFJNMQjT2Jxdr2va3jVZhbFmd3zYO/d44mMMVcdxi7IVkJiMxRurMzsLvZbadNgPbUfvXcRiLg9rbTtglxMqSGaWHDdIjLGk/6mJEijg7sgueVtV7WraOZ9GDVymzblLune2/kMVxdrgx9vWMEsrSMzzvICPTex0kcxVvtOZm5WD1c5YFz0ZxPjYsgkSJgdSQNJyLqpXUeV64nsrJhHem/Ys+2Libif1GXFYPiY8QCrputh15W+dtIvb2UXUH0vUK5rvna9x3rtzIxNpVDuCPFkYpfgweFxIOk5+VzpsDXQ3ry9SRByfZ/ei9yJFizxadyWKWfM6Q/Rj6c5x0UFjqOu11PjY11RNfM58WLav8AiYfo5f6Bg5mMk2bIxiSPMeOVfRj6r6dbsqvyJ9XnX0hm/wDhUsyo15EMOTE0GTGs0L8HikAZWsb8VbgeVfPO58fOwd8wcqLJzc+ebIWGOF444tvgxpWDvjK+m+tlhBFieXq51a947r3Bt+ZhjaIWXHmRsWfOkUNDDNlSxxY8gQHVI6EH02+9WWJsudnbw26b50XyMaJIMZIHkaJJAjo8nqCKXfXcnT6RZR41mY1ZouTENeQsIIkDjKgJ5FnWxA9jFTce2sDkxZOPDnKAqoR1o5hZlSQaJEfyIv7uFeY8jHGwc4lmJiSKfiBcPpGtv3X4+4mvXxQZciFU9TqXQt8rq5IliY87auPs1cK5GjJg8bvGql5cYdXH/NG3paK5PEi3j+Wmzui90ZxjN0zMDFnBF/UYpJo9Xl8rKK148xmgw806laNjDMo48z0m1e51U+yte3KIO74YFAQfQ5RAU8DH1sd0uvhpeSQV0x/ejN/tOwpSleg5ClKUApSlAKUpQClKUApSlAKUpQClKUApSlAKUpQClKUApSlAK05OJi5sfRzII8iI8THKiut/cwIrdSgOZk7MxMZ3n7fy59mmY3McB6mKfMHEm1Rgfuaa0Lkd07e+jdNrTPgHLN2txqNvx4uQysP4HautpWXVPdFTaOQTu/t4v0cjMGFNa7Q5qSYrjhex66oL/Gvdw3Pbcg7a2Pm48o+the8cqN6dMnq9Lcq60qG4MAR7a5nf9g2KV9s6m2YjXzY0N4IzdWVyyn08jYVn013LyLISIQCHUg8QQQRUXM3jatujMufnQY6LwJkkUG/kFvcn2AU/3H7Q5/0XE/7Jal4Pbfb+2uJcDa8XHlHKWOFA/wDftqp6fiOXgcnt02Pve29x7ZgvKyTNLNiySRvGSmbHrRoxMFOkSq9ja1dvt2SubgYuWpBE8SSAjl6lDVz/AHIDtO77b3EptjyEbXuXgBHO98eZvD9Ob08fBzU3t2bofV7LLpSTAkLQIt+OLMWeBuPkdScPw1a6NrvqHqpLyuT33uPI27d58Iblt+3xQYkWUi5yMzzvI8ysiFJoyAOkOSseNdZUKPbkTdcjdC+psiCDH6ZAsvQeZ9QPt637K2ZKmPu/EXGilysXIR1x4cnchHGZEwxMgk0ztwPpHEgAkDiQK3xd1bdLnHCWLI0rknBOWY/0Ovp6iprvx1A8CBasNw7afLyM5sbPfExt1VU3PHWNHMlk6JaN24xs0YCE2PLhxrKPtmGKNIo5isce4DcUUKLDSNIhHHkB40Bs2vubB3bIXHxop1EqNNjzSIOnLGhUFlZWbT8w4PpPsq5rmtq7Ri23dl3Xro8iLKt4seOCWXqkEtlyx/zmFuZA48a6WgFc7gSOs25Y7GxgzZQAPwyKmQPE/wC1roq5ss8XcW5YzLZJYcbKjbzJ6kEnh4dJfGsZPt8jVNyNjSM+Pu4LFtOXOOJvb0R8K5buzCw5O6MOTLyjhCR4QcxQupFeOeNo0JVjrkMYVGHFb8ONdBh263cdv/Nr/wD4mNVP3b9THuse44cirLgDHmbVEZwqB5IppigIsYY8kSC9cU9Tb2O5x4lxooseMsViVY0MjF2sosNTtcsfMmvjpi2zI3OFuhmbSm47nI2PlRvLJOiIhjzIYBBrNpJ2Zb2A4k+FfRR3ZsUuDNmw7lFDjqzwQ50wtE8qLqJh16erp/Lz5V8/7cztx2nelwIsjGTdd26E2TJkwSyTwrkNJO2NBDFqstm6ja2AW/Gt1nUjPrW3YGDtGFFt22wrj4kAtHEtyBc3JJNySTxJPE1yG4rm4P8AxP2vNx4epBuOFJjShCEJEWp2kcsEVtHpsNRNr2Fdpqr5f37n5I7m2KQR5OEcbJMUMmXIIcGbVpUtHLA5kW4f1k29ItalXLDR9VLgAkngOJPurVDkwZUKZGNIs0MoDRyoQysp5FSOBrFJdaK4ZX1AHUnFDcc1/LXBdr7tJsPcGZ2TnrCRefcsafGkJhhjkPXbHZJACgUEsONSZnwBh/xCzcjB3nZJnnxciLrA4m3ZMdjFOw6IzWk5SLGXvoOmxtXa7fuu37rHLJt2UuYmPI2PNNH8vVQDVx5Hn4cK5bvCcbjuGzbUEmONJOssjmFGxZlaNtOiVrkyJqBsvAjnWU/b0E5i3HDzsjALacx8eAh8X6nGVI9ZxyBr4Lp03sffUbUKSpbmPeUqf1LY8mKFJXw83TJkGQXiZomdY+iWGokevVb0gV1kSJFGscfyKPSfPxufaedcth9v7TJuBfJkbO3oKJtzlnf1KJRYfpJZAdN41sPSlxeukxNaxGJ7Xhdo1I/AOKf6hFYs9vAqMYUBE+JNZlBJAta8ctyB8DcVpZ2WFJ5G/UwnKTu621R8mbh+JSr/AArfOVinx5iSNbGBrcjrF11fxLw9/to6L9VoYEx5MbI45rdPMeZVj77VCmEsKLLJCRaHMB1FeFpgPm1DxZRz8xWnZmOT3RNIwtJi7bFFKCBqDyTyk3Pt6Vx51tjWTK25VICZCiwuLhZYjYHj+Zb+6vOzF+rXc+4LgpumT/lrcxBjKMZPtdHYe+umJfV5Gb7eZ1FKUr0HIUpSgFKUoBSlKAUpSgFKUoBSlKAUpSgFKUoBSlKAUpSgFKUoBSlKAUpSgFVO9/Ptf/rov+RJVtVTvfz7X/66L/kSUBbUpSgIm57fj7tt+VtmXfoZcbQyFTZgGFtSnwI5iuQ2+fPysOLMhUf7xbIz4eZA7aBMUsJInbj6ZlCyRseRseV67qua3vZ9wh3D/eDYUWXLZBDuO3u2hcuJL9Mo59KSx3OkngQbHwrNk3qt0VPvsy32zdsTdYmaAlJoiFycWT0zQuRfRKngfLwPhU6uJgzNj33IUqz4e7wXXpOWxM+LhxBS4Zl/vIas9O8x2WPdWKgW/Wx43e/7ydIfsrKyr9WjLwfTU6Olcxk5G7YuLNlT7uI1hBkaQ4yFAqjiNGrUT7mq52abcMjbMafdUSPMlQPLHGGVRq4qNMl2VtNtQvwNarZW2I01uTqUpWiCuf3ljBve2SBPTlRZGM8luTKEyIwT/A9q6CqbuQaMTGywATi5cDkkDgsjfTyG55WSU1m6mrXgWu6KHGuh36d+CTZoRPE3EOPj/wDLrRmYY3DO3LDmaRVyoJYz9OVErxvohI1OCEC9I/bUpFUQzgNq6u62lHDhadF08PYgrbEXfIzROoXFSXTcG7SNbqaNPP7/AMftrzHU5/G2zesiTE27cMvGyNrw8nGnhKIEmZYLLGkapGqaep873N7W4XqPtG4bZgb1nyT71JHr3KeLF2RAhkeSR9DvMqRmVw0jEpc+lfOuxlhWQLNmsI1jYNCoOkRnkDq8W/Z7Kps7FXIGS+SssUbTY2RDuOKmmQPFwMguGIsqi+pdJ41UxB01yOdfPe+9+bdcfK7Y2SAZufDkRJm4xRnLRgLMGiK+jTqGiTUw8vGrCXbO4MfMzIId2GRjbquPBJlZUYaSPUsqM+OsOiLio08V5kHjVvte07fteHjbdhwLHiNHJjMOAZtJJDObetiFJJ86Jpa7jfwNvb26Q7htOLOgMakdFC0P0yuyD1dCK5snA6fdVFum24vc3c8UeXisMLatePO/V6ZmmlSOZQiwkPpVDZyxsQbWrd/uviBcaTDycvGysbKIhCTEx47SMfqGgimDIBIlzxBHHhU/C2zH2qeYQO0s0jo5ed9c00pDtI8jG3D1rx8ALUmJaYjuRsfZtpwpZH2/HtPgGXpGR5JEgFtcUcCuzLGCXU2W3AVMy8WIFNvjB65x20uGbjISkQd1VlvyLHzsa17nnY+JjxrjRHPmyshCVhICyzXuilwCLXQXtfSoJPKke6rjdxf0rcUUZWTAr4mWt9DAk3x7HlZlYg+PvpruNDWsEu4Q6VWPD7h2g2RowREdYuvlqgyFHFT8p9q1YbZlwZxfLhuhnRTNCx4xzRFoZYz+ZCuk+4VhubHCzMPdVsIgwxM4sbWhmb9N/ek2n4Ma1Pjrt+/x5YJEG5BoJFvwXJCq6sBb/FSKx/Mo86m5SxzSq4zyOLiIrJwFyNDBrgefCmW5ikxWtcGYRk+ADo4B+2w+NbpEMkbxjm6lR8RaoJd8rE25rWeSSGSTwC9MdV+fhdLVAQd4ysn6Z9owXMO57nkth4UgvdBIollyRbwiiZm94rscHDx9uw8fAxV0QY0axRL5Kg0iub7Wx491zsnu1gTFMDibSDyGNGbSTgHkZpF/uqtdZXpx1418WcrOX5ClKVsyKUpQClKUApSlAKUpQClKUApSlAKUpQClKUApSlAKUpQClKUApWrJnjxceXJlv04UaR7C50oCxsPcKjNu2GseBKzEJuRC4xItzifJ9f4fRGaAnUrFpI1tqYC/K5AvyH+mgkjZDIrqUF7uCCOHPjQGVVO9/Ptf/rov+RJUufc8DGmxoJp1WXMJGMt76yqlza3sFa4JcDe8XDzoyXj/AEsyAH0sNSakLr4eluRoCfSomTueHiiBnfUMidcWMx+odV72Bty5VI60V3HUW8f8wXHpv+LyoDOlYdaG4HUW5YoPUOLDmvv9le60NrMOJKjjzIvcfsoCHuey7VvMPQ3TEjyU4WLr6lsdQKOLMvHyNc3tUMu37pu+0Nky5GPitjSYQyHMkiQyxWKan4soeNrGuwSSOTV03V9J0tpINiPA2rls/qQd6ITwhztsKrx4GTFn1cvMLkVjKpq/A1R6mbxLue8Y22MW6GKq5+WoHpfS2nGiZv8A5imS35BXUVR7KS25btc30nHRR5Dpa/7XNXlMaiq8dRZ6sUpStmRULd8E7lteZgK2h8iF40f8Lkehvg1jU2lAfPtsyWzIc1wwJlkx90x2tbg2hZl96ywSLx9lToEaLedwxowRLOyZccjAsqoY0x3Kjle8YFvjUafElxN7kgIEOIciTH0KP5mPuMfWS1uRTKRwPfUl42bK2ySSW8c8Zx5k4qZHRTIp1KRz0nUPGvLZQ2jsnKJafSpIWiVsvKAs0nBiNJtYubInHwFvdWxsWTLBXM4x31dCMnT6SGUu3Aty4jl76wytxwNuMePI4ErnTDiQqXlYkXAWKME/HlXu2f1LLyGyM+MYmHo/Swbh5SeBL5DrdfciH3k1Epa6BuEVG5PmMs2TteLJjxxSIJclo2czlG4Ji4o+clvvnQPG5FTMfD3toD9VFjxM05yUBaQmG7awraRpYjjezAcbcuNeYGRve6YUXccObDDDL+pibfNEFhGOWsOrL/MEhT7w4A8LWrSO+dpyMt4jpOMDKuPaeIySyQBmIkgVjJGraDpLD3+FdfTRnkeL9XO0m8SpkxYOMHcRqrLNlMgZF0Q80Tj6PvsefC19Y2rLw8OTJMBk37d3SKWYAyCDVe2lnvZMeO5v4t7xXs/fmHH0XysY446KZrY088MUohkBMbhXYa2OkkRqb+drgUj7py3yNyyPpZJ9rxMgIM+PpqIsZ40dnEbESPouWY24Dzp6fiTkZ7djyf1w4y4skO27ZirBtV1ujNq6c8oNvmsFQceI1HxqTvW1S58kYiiP1Agn6EnEBZUaGaG58P1EH7aiyd77ZJPLjYLx2EeRDhzrNEzNNEpYXxweoqkxmzMOPuNYbF3TlTzwRbj+u+XPi4sBQKgQzbfHmyO1uYL6vtp6esyXkXJgbd9rMeTAYxmQaZoWF9DOtmU+1GquzDny9tpmdJjnYyRZDqQQerjOrS+A56G+FZxd2GSfJxsLbJciSFst2AkjQBcSboSNdyPma5UVI2vel3lp9vkwpcRMnE+sx3laNtcGR6OKxs2hhqvY1PTjr4jmWFwTdeR4j3Vzu5Qybw+H23jOVbLaWXcXU2MeCrskvtDTFumvx8q2Ym8Jt/b0GRnt18vGtgyQw2Mk2ZGegIYx4vIw4e+9XHa2yZG2wz7hudm3fcmEuXpOpYVF+nixt4pECePi1z41MdJcvZFtaF5l7FFHBEkMKhIo1CRovABVFgB7qzpSvQchSlKAUpSgFKUoBSlKAUpSgFKUoBSlKAUpSgFKUoBSlKAUpSgFKUoCJukUk+2ZsMK65ZIJUjXzZkIA4+2uWxO0vpG7flBy5zC2nPgycqSeFEfDmif9KV2T52C+kePlXaU58DQHzHPwp37azHyictcafB2jFUsAk0OPnRXkVr85dQRj+WrOfZZ5ppcvD2ZsfaepjNlbITFEcow9YPKI43MQ064zYkdTRY8LV2Y2/BGKmEMaIYkekx44RempRg6aUtYaWFx7akUBwmPsE8efi7k20hMUbjJLi4VomkxIJMYRBratKKcgdUqhOm9+d6wxe287bdrwU23b0TMOyZGNmAto15UhxiqTSIwZj/M0nVw8xXfUoD51gbFu0OZHLHgPHC82BMCYsfGUHGbJ6paHGZgnCVPxEgewCoCdt78YbnbXSSXBy8TLVBBGDNL0nT1rK8sy3je0kjFuPtNfVKUBxG6drI0u8SYO2xhnXCfb2RUW0scjNM8XLQ1guphYms1wt3OXiYH9OkEOPuW45kuYZESMx5SZhg6bIzSXPXUE6bqa7SlAcV2ds247ZuEjy4rQ45xhDI00WPC/URgUVThuVmFi15HUN9pAsO8CuGm2b23ybflos7eUOUDiOf4WkVvhXS1HzsLH3HCyMDLTXj5UbQzJ5o4Kn+2o1Ka7hPWTn0zfoc3eEQBcgYCZkZtfUYurGb+B02XxqobvDd17Hjz5OlH3E6lCAuqLWkP1rS6L/K2MNdvMitEeVJFA2FmyFcvZg+27uxA1vg5CiNM1bn5bCORj7GFX47MwTDIiZEg622DamYBSCBGIfqbcupoAX3Vmn2x1Whq289yTP3btWLmnCnbSsbpBPlF4RHHK+kKjK0ol5uASEIHiartz7xeMbfkYOLOuDkZYjOa6IYpoFjlkfpWk1jV0/SWUXqfN2lhybmc9JTHHLIMjKxulC3UlULZuq8ZlQHQNQVrH4m8eTsuJ/psY7jkf0vDm62LtpEfTQFXjMXU0CUqBIdN29NbMmzeO5hBgM2CrLlNiQ50TSKCojllSLS3q+b1V7H3JJj4pkycaXKlbKz4l6AiRVixcl4V1vPJEl9AHC9zx4VpXsu6Spk7pPka8aPCiLpGOnDDKJowBGqgt6bMfGts3aMUmRj5MeY8bwS5chPTiclcyZsl1j6qt02DNYOvqtQFXu2XDm7rte44tzjZ0eDNE7KRqUTsy8/ECbiPbWG+4IyO358ZMiTF6WSrpkwm0qf5kEshBHH1mt+fsqxY+DsEOT1J8DDD4ssoAIOPPA+OZAg5XjCk++oO5bqpxBg5kEuDmT5cKrFIt43MmQg/RmF45AL387cdNcL/fp4HWv2knbjLs/UxpNsL6GEb7hgL1OrpA9UyO3W1C/H5x7a8w+6cPP3mTb8d9GLDEWlmnBx+pKWUCNDNoOlRfVYc+FWZ6egoh6eHGSHcc5GJ4qtuPFjxI4k8Kykxhmx9LKQDFIsMYqOI/PzsPyisJw5grWkEf+gbVc9fPlOB1fqf6b1VGIr6xNwBXXo1C+jVp9lQ8eHt/cMvJw8XcJpoF67f0+Fg0MbTao5JrhOIvIdKaioPEC/LOfY+2Ntx5M5tpxQuOmsaYELHT8qrwPqJ4D21li6dnwfrM+Mncc5lMmPFZ3aQj9PEg+X0xL6R4Dix8a36jM8DLM2jbcifHw48qfDnmxlxJDE4WSfFx7X6npbTp121izeqwrXjbRsudkTPj7hOcXPP1kuGjgQyxrpx0vdNWg9LjZhq8eFaMjGyCzY07k7vvQ6c7wk2xcKP+YsTW4aQxAY/M7X90mIx4W6bnmONOFgYWNCouTpWITTuF/gKU9RjijKLbtjl3LKwoM2ZVUTE4CMBCjyr+sy+i/p64OjUVDG9r1jF29suPkLhQZkseWiw5kMoZDLEceJMBJE1IUN0WzBgedRtnhkgzNvXIYNmT4eTmZfgepkTQyG49nyj3VZxRat7ysg29GNBCvmNTzSN8DwqPIy8UY7fse0bblvDFkyyTyY+Tq6rhmKZU/VlckKOIc8KwjbZNnzrJkmTMhxYNrxsQEGSWymaNQoAu7BefIDnavMvJx8HdZM7MlWDEgwf1Z5CAoLTekeZJ0HhWfaW0JNl5nd2dhmDcNxbTiJMP1IcNFWOIEfceULrYc+IHhW6N2eu0GbJIy7e7XdMxu499RDu85Lx4iHVBiBgFtGOTzFVAeW1zyHDn1dKV1SjRGBSlKAUpSgFKUoBSlKAUpSgFKUoBSlKAUpSgFKUoBSlKAUpSgFKUoBSlKAUpSgFKUoBSlKAUpSgFKUoBSlKA5zujZJsvpbztiK26YKsphb5cvGb+bhSex+aX5Nb21E7V3qFYcfCaRn2/IuNpyZTZ1Kk69vnB+WWEgqvmB5jj11cT3Ht0WzZU+7GLqbDuBA33GHKGT0qmfGF4jiAJNPHk/gTWbSvqXxXcq7M7alc7t+8S4OTBtW7OZUyDp27c+GiU81gmI4LLb5Tyk8OPCuiqppqUGo0YpSlUhz2b/wD9Gf8A0K//AFjVdvUX1WPkq7KcfDEeTJHo1OXiP1AIfUNJAQeF+NWmfEy78k1xpkw2QDxBjlU/t6lR8YK2ZuCuLqZIwwPIgwJcV5sn3v4HWv2o9xhHkBMpRaEC2InIBANIkA/MPl/L76Zu4Ym3hPqZLSSm0ECgtLKw+7FGt2Y+6ua7e7b3Kbt94Nn3jJwsrBy8nG6Ex6mOVhmcJGAy9SNTHp4o1enIz9njZY+3srD3LIfTPm6H3JACCTIJYS8knKwRtNV43vuvAKy8iZlZM7ZUD7hFrnYiTatjiIZ9S8PqcxxdQEPj8qfma1TI8dNuE++7zMJMwRWldbmKGP8A2OKh42Y2BPzOefgBBwdy2/A6jpgbtLkzkNk5cu35JllYCw1ERgADwVQFHgK8yMndd1zsdsbYM6fCxVXJhScR4gkydRCNJ9Q6sFiX1AaT6jxHCpxs9IZZXcsdvgbGTK3ndCI8rIXqT35QY8QLRwX/ACAksfFiagCHJm2uFJQIsrfctZZ4z6rQt+q8QtzIxoQnvqdJsHcO/RtBvM8W2bbMpWbCwz1shwR8smTIoRRfmEjPvqY3Y2xZLNLuYn3HIICpkZMz9SNRyWHo9IR+9QCfGtLFbd6GXddCp3HcsDau4zk7lkJixf02wMhsWY5PBUXmx9gFZ4w7i3XJny9oxlwcKeOJEytzRlkJjMl3ixVKuVYOLdQr7qvtt7U7e2iY5ODgRpkkAfUSFppQB4CSZnYfA1c1tYkt9SO76aHN7f2ZgQ567zusr7tuyhQmRkBVjiKXt9PAg0JbUbE3b210lKV022MClKUApSlAKUpQClKUApSlAKUpQClKUApSlAKUpQClKUApSlAKUpQClKUApSlAKUpQClKUApSlAKUpQClKUApSlAKxdEkRo5FDo4KujC4IPAgg+FZUoDgpNvTYph2zuAabYNwJj2mdr3gc+oYUjj5bH1QPz4aeYFdJ29nZEsc227g/UztvKo8xsDNC1+jkELwBcKQw/EDU7c9swt4wZdv3CIS48wsynmCOKup8GU8QfA1xWM+64WYYMh+vvuy3OrgG3HbZD81rgdS4/hkXybjhri56Pf8Aea3UdVsfQKVows3G3DFizMOTqQTDUjWI9hBVrEEHgQeIrfWzJRbtJo3nAS4tNj5S2PMlGgcW/bWmFNGVlte/UMb+70CO3+pW7uCD/O7Pm3t0ciSI3IAtPBIvj46lWtSOozJYvvGONx7ruv8AaK8+X7vNHWmxh22wj3fuHF4D9fHydPD/ABsdEvYceJirpa5TbXePvTIiuenk7XFIRwtqgyJFv58pq6uu1PtXkc7bsUpStEFKUoBSlKAUpSgFKUoBSlKAUpSgFKUoBSlKAUpSgFKUoBSlKAUpSgFKUoBSlKAUpSgFKUoBSlKAUpSgFKUoBSlKAUpSgFKUoBSlKAUpSgFc13dgyrBD3FgqWz9n1SlFHGbFYf5nH4eajUv5lFdLXhAIIIuDwINGpUMHIY+dDtEw3nEcSbJuRSXL0W0xvLbTnL+V7gS8fJvxV2HPiK4vBxo9rz83teZA2EVOTtkbAaDiSkrLjAHmIXNrfhZatu3ck47TbDO7NJhgPhvISzSYjn0EseZja8be5Sedc6Npuj6beRuy05LqbO6o3bZpMiIkPhSw5gIIHpglWSTmRzQMKjyiNc2B7+p0kiU+Y9Mlh/cvV7k48eXjTYswvFOjRSDzVwVP7DXKYcrLtWE7Au+I6wTFh6rxMcSVv7WrOZbP4Fo90ZySDF7p2SfhbLjy8FuHmqZScb/9Aa62uM7l1wbem6RKWm2ieLPRV4kpEdM6/GFnrsY3SVFljYOjgMjKbggi4INaxOax2M3Wp7qUGxIv5V7XzTfNvbN7q32RMLbcmSHHxOnNuUzQvExjlIaEoj+VybipWN3BuoxpsmLPaPA2/Z8LLUZcAlnklyI5kV5ipB+ZFYgcz4gV0Mn0GlfPsbe9ylz02zPnTMbHzNukSWWOBZVGV19SMuO8iKU6Q0m9+NasPuPuWPb8LNnzYciTdcPMlhSWNIY4JMYjptrXncN6tXC/kKA+jUr5vJuO85v0mLkbnLDNi7nhl3khx0Zop1fSrtjSSwMNSN8pv5jlWbd3b8Bk5gBWOVc1I8eT6VVgbFjmZXj0zNkSMDENSvGOfh4gfRaV8+y907px8aaVd2jLR7WN5U/SpzUerG+b5G8/m9tfQFOpQfMA0B7SlKAUpSgFKUoBSlKAUpSgFKUoBSlKAUpSgFKUoBSlKAUpSgFKUoBSlKAUpSgFKUoBSlKAUpSgFKUoBSlKAUpSgFKUoBSlKA5/uzb8mfDh3XblZ9x2hzlY8SfNMmkrNi/9anD94CqvJy4JsXb+6tuJlTEH1KlLkviSgDJjsvMhPUB+Ja7SuN26MbVu+49v6dMAP9Q24W9PQyGPViX/AOXNq4eTCueRRF1ujdHvV9TsEdJEWSNgyOAysOIIPEEVzDQSx5+77ezi0xXKxfNVyEKNf2CWJj8aldsu+KuRscrFvom14bMdTNiSkmK5/wCjYNH7lFN4jSDedtywtmyEmwne/s+pjB/7Jre+l/qpK8yV0tHwPIiMnGQzKLTIBKo5eoWce6nZeTJNsMWNMby7dJNt7HlcYsjQIT7SiqTWGKqxGeBQQEkLqDytL+p6fZqJrV27K+Pvu+bbJ6UleHcMUHhqWWMQzFfO0kXq9p9tYwv6mu6NX2TLfL2DYs/J+szttxcnJ9I680MbvZfl9TqTwqU2HiO0zPjxs2QgiyCUU9SNdVke49SjUeB8630rucyHDtG1Y4iEGDjxCEKIQkSLoCEsoWw4aSxIrJdt29FiRMSFVgV0gURqAiyfOqC3AN4251KpQEFNm2iLFODFgY6YhcSHHWJBHrVg4fQBa4YXvWbbXtrTTZLYcBnyIzDkSmNNckZ5xu1rsp8jUulAR2wMF1KvjRMrRfTEFFIMP+y5fJ+XlUjlwFKUApSlAKUpQClKUApSlAKUpQClKUApSlAKUpQClKUApSlAKUpQClKUApSlAKUpQClKUApSlAKUpQClKUApSlAKUpQClKUApSlAK5jvGIYsOH3KoOvZpC+RpBJOHNaLKFhz0raT+GunrXPBDkwyY2QgkhmVo5Y2F1ZWGllI8iDUalR3COYzpDhZeBvUZZo8d+hkqnJsbKKoWPsjfRJ7gan91yHG2pc8EKMLIx55GYhQIhIqTEk24CN2rnotn7oGCe11xUXEXq4v9ayZUkVsI3WMpAjdRphG2n16VuL3NXn+7edPh4+27jvEuTgRII8iIRRxvkqv3MiT1Eqw4OF06qxSrVXVmrNSmikbubBGT9VBBnT4LL0pMuHDneAsCTG6OFu4PEXUEcuNWnb0Wfn7tkb7lYkmFijHXE26GcBZ3Uv1ZZpY7kpdlUKp425106qqqFUBVUWAHAADwFe1a461coOzegpSlbMilKUApSlAKUpQClKUApSlAKUpQClKUApSlAKUpQClKUApSlAKUpQClKUApSlAKUpQClKUApSlAKUpQClKUApSlAKUpQClKUApSlAKUpQClKUApSlAKUpQClKUApSlAKUpQClKUApSlAKUpQClKUApSlAKUpQClKUB/9k=)
- Opioids, endogenous morphine-like NTs (endorphins)
![[[@clapp-niaa]](http://pubs.niaaa.nih.gov/publications/arh314/310-339.htm)](data:image/gif;base64,R0lGODlhwgEsAfcAAAAAAP////Xp6qWipOPF4MulyQwEDCQcJLKxsreLuhQMFZRepGktjN3c3qKcxxwbJAwLFcHA3AQEDRgYGXuBty08lFRppuXr+PD0/L6/wSwwNtLn9arh+IbT7mHA2oG5w6TL0QQMDAwUFBwkJPT8/AQMBAwUDBwkHPT87tXW1Ozt2RQUDPz8xPz8zPHxw/z81PT01Pz83PT03/z85ywsKvz89PT08vz3RPz2Wfz1aPz1ePz1ifzpC+zhZfTsdPTsfPTshPTsjOzkjPz1mPTslPz2qPz2tPjlOfrnSPTsnCYlHPz2vPzhINzEHPTcJOvVKPTcLOvXOvzqWPjnafzsdPTkdPzsfPvYCuzMFPTUHOzMHfTUJPzgLuLGKfTZOOnRTPTeVPTdXPTkfPzshPzsjPzslPz0xPXLC+zEDPrWFPTMFPTULPXWRPTWTM22RPXcZPTcbPTihPTjjOnYifTklPzsnOvemJeRdd7Wsfz0zPLszLy3n+zEFPTMHOzEHPzUJPzTMPzcUPTUVPzcXNnCXLajTenRZvzjdMaxXPTcd/zkfPzkhNnJgfTknvzrpPzsrOPWnrGqjfz01PzMFPTEFOy8FPzMHfTEHPTKKOzEKOK8KNOwLvTLOOvGNvPMReTDSfTTXOnSePzkjMu+jO/gqfvst9PInsvGs/S8DPzEFOy8HOK0I8SfL+a7OPXTZ/zklLWmdO3guO20FPzEHOy0HOy2JLORNPzSVOvFWOPEa/zknDk1KvnrxOjgyPz03tnTwfa6G+SsJOqzLJ+CNoVuNo2AXuqsHPa3JOysJPzXh86zdPnbl21gQvTr1+jj2F5QM6eNW0pBLvSoHPapJOSfJOGjNPzNe5yYke2fJPDGhPLOkOesVO2/evzMhPzKjXFsZt2FJUhFQvWtbfbCkPrMo/m2gPN8IPqJM/mUR/mfWz88OmJfXYyJh8pjJF8zGKRRIYVBHVRQTvz08TUsKXt4dyUcGxQMDDc0NCYkJOno6MzLy/z8/AwMDAQEBP///wAAACH5BAEAAP4ALAAAAADCASwBAAj/AAMIHEiwoMGDCBMqXMiwocOHECNKnEixosWLGDNq3Mixo8ePIEOKHEmypMmTKFOqXMmypcuXMGPKnEmzps2bOHPq3Mmzp8+fQIMKHUq0qNGjSJMqXcq0qdOnUKNKnUq1qtWrWLNq3cq1q9evYMOKHUu2rNmzaNOqXcu2rdu3cOPKnUu3Lsh8DfQhGHDtGjt6gL+tG0yYMGDA7Nj1vTZgAIIM+vQ1aJAv3759djNrbpliAL146mhMUGCgH4DTAEyjXs269erS/BTInjABHz4a93KrUxdu3Td61xDoy7e5uPGJGdbR4HfaAIR68+6FY+a7WDFodyJF2rOdeyRY2q/B/7pzxzq9b+jXMYv3LFruefNu15swH4ICCAZO9+tnoLa6dfQgkMJxBBYo0ADq5KfAf3fsAQkdjrDAwhJFVDjEEEW8MEMML8QwwwwvULhECxOyUIQRLJAo4RImDiFhHYo8IiMppMyRSy7KwAJNMcw8E8499SiAmgI0hBOgDQYmKZc+39CQ2i53kCKhhEY4YqUjRiyRRB260PFICy+E2SGHLTwiihEtLFFlHY6o+YgRcDryphFlLGIlinSOccgUVhSxw58txKAHHrAU88wu9eTXzwHhXEOckpCelQI96jCnQDywRBgDC6UYUcojcjripRkyNqJLm0vkMWYMeagphy4sLv/xSCOPyEpHHZ3KWKUuccj55iN0yAEHKKIsIYooZQQaQwt/7jBEHIgUEw8+pk2wjj6RZutVDQh8cw9zBtxTDCQs0FnKC2Z82si66j5ixruONNLuqqVA2AgducbrbpXyyvqmI0nE8asRoMrhSiJfGkFHEmaE2UIdoixSxhBj6DAhLMwcAAAE8WCr7cdT7aMPO/EcYJoCu3wzyhIvpKmLHKOWYiWEpchcK7qNtCkqvjH07EgicdCRMx1EE1GKGbImUauMMr4854SPJAKKHCPKWmyYZhRRRxyikCEGKCMaUYQYxTipwDqPgqw2UgJkQDI++QFQTzh3mHKiEQ3LTMcyXrb/kO66S8tYit9yytyIHI50+IIjySSii+GmnsoiqI98SjDASWCpagtGSJ0Iyyw4AkcSeaQpqxhiBBExiixQvMMUxeADwAPsrG07UDboc8066jxgWj/1qPPNHqVzzrQZMUB+dB7pCv6p4EjLKWsjyTzCoSSPxBFHKSwwbSWtS3z6aeagysnzC3nU6woo5+aRhCtyTBjhI2LAUceJJy4xBBlDWJGDIcwQ0j0ycLsC1iQfGdidOg4QN3vgIxzF2EMsTgQiTz1PTuiqmSNKsYTw1eyDz+NgKWhVs2CVgkNLoF4jkFYz74FPTevyVajyQKYlOMIVcVhCDJbwikSU4RFEEEW5/xIBhy+VK0sVMgIZ9oQEN6ijHwqgB2YMSEWU5M4z8bjHBBTFD3xE4xvXMAWKwPQCUIFIfJUjxQrfVa8V1qyDIJTREppXORtuEH2P0EUjPPWmTqnLS3B0xCsakYTnsYxDZqgSwjr4CDkEzRFi+NwS4JAINCENiWUwAhAE0QZO/EEHxZgAANQxoCqakiM1aAACrvGNH21RPxOYRzjAaIpHpChNeJtjHu5IqqOxsXItlFe65piuD1bODK2a1dGOxyk5Uq5yoEpCI6oGqmD9anAxkAS6jCCHRJBKFHJIghzqQAQisskVcChCHliANBaUIQxVaAMfMsEJT0xhB6GIBgAmMP+AU/ozIrlb5TeyiA97xK0/RVpHdiDBspaVa3Dp4yAvipmHPAhuCZKIQS+RJiP01ayi4YPmI0rHi1nlDHDAPOanrKQuObUgD5cjGsFqBSYUNvJzRtDeleggCjHIIQiL8GER5LcEMXCCE36oBCcOEYYwWEEHSCAGBPpBj39alSD50Ms1PhMO0dgDNQZQAD7UEY9v3GEUkPDQEdWEzJDWKqLEfBcegSmJGQyTF7woBQ2R9q5kPoIUx2zezGjFNPGlCVi2lNkGW9ACw0HoeC/IKOd0cawNEg1gdQhWHcpABzKIAqhqsl88OZEJTLRBDFWAgyIUMQgkFEKU67jq7WzQgAz/IOAv6/gRPhTwVf0YoB5Figc9zgoJ7jGWc0boGdQ+GL7mmeGl7+qrqor5URC9i5iqquh1P7jB60ITcCGdo7xeoLdzmeGF4RNVztpkhFV1b7NluN9mM6s9OVSJCIuIQx3EQIR3lqEKb2hDGwQBih8oogqoe8MgpEAIan1DJA3wzDriEY91sMNjsp1K7vwyUN3Wwx77QU1sDrCL3gx3D6bgXopApCLGSghpyvoU0pbAi5Aub7p6DRPzWHg0D6GLxn1FHxvf+EEWkgpCbJzjCEc0qy9pVE7FrNJji9DBXuBhD9wRj3hgMQpG0KEIizgEHETxiCIQIQlaqx8Z3hAEILzB/xNvQLAi3hCGQwBYEGHQgTLqAQEMcyQD4RBlavhRGgDwQx0IyLBTdMe7AzBHPxCgDw14s45iZOcUpOhUCz60LDCZbkMh6iCIHLbpGVQJXcUEJgcxyryKZrBmbc3DhtJXx+nyYsdmyCv0XPA3X9LYX++SUcPQVUfvneoRkNDOHRLDDvIMtzzQhgUjQnEIO5GzDuRcBJvhIAZFcKISVajQDwT8BjgE2Kk7KAYAqpqQfKRAH5BJQQNqwJB1mIYGzCgGLJShDGh8Yxf9kMCDFW2UfCCAHuGA22m6OB19R8IUkMiohF5gop5RHG8WZ2wpvhGNSX/DBWVyxEArTOE76EFWGv8FoTFHKmTkMW95MZjorGU0Ue0ik3kueF6mhym+8MFRXsk82kubi8YqjcLZ9Gj2Yvpyh2uQhzzNvoMy7KA1zpYhCPX7LxAUoQpZVKGDZBDEG94gCEGwARSHyIEtBG4QAVwjHjR4gAHmzp8JHCCh7EBAAwoSDwDgAxrJCHw3uhH4wENDY9+oLQIeo48UWIbgOImwtwT9W+E9XE1iGwKcPA2ncj1iTMMc0wtMcQLWTMAU3TNBa06PNQ96sIUuRyaratwwViFvBhGVRAtqXtGXt7COeYV1zSY6Qr2KL7oqzxIkrmFpdjiAAhTAcgZOQf1TeCcSTl+2pUcB3zoEVQxlUET/EMiQiUwIgkVleIMYwtAGT3hC7DlgBgCugdVvPCA1sYxGNMLB/13gYzRDQgPxcGH0AAC7kA3LkAyD1w3WsICEpwxwUw+P5hwK4B/CpXeQ5xL5MADeIiQAYA/zEA/FMAqytiyeN1MyliYclD6Jg0jhEygt4yQAEA93YG8AcAITUg8GOBjhcBrqYHHs9C4rIj6soiq29y4ZlT7Iky63ly42R3wgBIVshHy0Ql2uJyudsgSmEHXsQAEb4AAWkAHOoAJkqAK/8At68At7UAALcA2R8HSwQAbYRgZVcHXg5wgCFgaZBARVsGbtxwZtoAOEYHdpgwCyMwGYMgdk4CzNYgWK/2AIblAo8bALEyABG6MAB6AMydCADMiJDWgNhgcBD8AMpPgMz6AO8aEAlrhP9/ANCEBvGUgS+5ABlCJoCjAPCoV6E9JeM6CCF+Vcl2QGngcm0XVILXAHp3EHLTMDyAgAd8ACpccMFtd3E/AC1zBLLBILFCZGd7QqkhBdGqKEr/Yh6dNqzANkyNRXn7JjzJNXXnKOz/MIxDdHW6gY2wF9FGAB81YDNSADGZCP+tAMCUAACbAAezAAH/AXo4BmZZAIW6MIGBIGoGBuVhAGb5B+nNQGgSAG9yABaCMQCMAPEsAMc+Anq0UGWjMGizAGinAIg2AFnLMDhlAIxBANEBANov+wgJzIgDwJislQKbAgCsmQgILHDYXCDPMgJItyLbH4ERu4DtRiaAJ4B3jQaSTiL3sVTI/gAsDGTryAB3gAcbHQQeykVyTSgydwXL24VabQAjq4DsryDfv0AvHQDxNwIqZwGpEQOtajTRYnCeU4A98oCYD5KXX1jex4jrfGjjWDPtrFQbSiY38lbK1iBnjgdNuBhhlgARbwigJAbxHgABsAfb0wkGyYAR3AAR2gkENwCKAwBqgVBEYAB3VmbgIWBKAgCFVABbhwD/YxARBwD9cAN8WAIVqTCIlAB1USB4sgB2IQYFVgBEMwBYEQCFygCQ9wDwqokz3Zk8kADRKgDoL/N3iFN5TL8AqMUAyBBgAGoA791JQY0QDXKGhzQw+mwCIs0DN89Tcs5y/qNUe6cmXY93RuiAcs1EFF4CThgC6BEigsdgL9EA+P0At7UHr48ALrAABKUAdDYAqmsQdl8nlG6As6NlGOSZgvJ2uD2Wo9Y6KA+S55pU0w+i6A5SERBWvn6HQDkAG90AzOIA++IAA2UAAJkAAzEAEUAIYU4AsFuQAJAAIcsAEd8AHkYQeHIAhiAARwsAhGUAXwNHag4AnPOQVFAAsHIAGx1B5zZwDMUARlEF9EEEmZVARxIAZksAhhGm5k0AZswAbAUAsHgA+f2J2DN6iE15GA54AOmAze/7AMuqAL2lAM95AaiAafE5EP16iU+BAPkQByjIWVaTJRpNKNSwBY97JGRmAHb8hszRZ12RELuGQESjCD5HVCZNQhOtgaxRADGUoDJwIJpjEKjTVSqgJqgLmYeUCY6JNXKkpDrUaOyTp76bKYQ1ZSjbBXt8aELWMKTXcKztAMIDClxFEABUAATloD+RiGNRADBYAAKiADHhCv7PCGkbBmWwoHKCkGYlduXgoEiyAGQ1AMEhAOsLBgRXAHEAABoyBlQZQIb8ClWCcGiwAH7jcGRWAFnsAJnUALx0CJuaCAO9mJhUqe4DkP22moi9qojqALsKAOqREPpWSpDDEA8SBK/f9AA8OTXCDSQaRShcMUbCxHa6WiC8J4meSxdIuxbIqBBx20BLswSiFHah5Sev0gAvxgAvhQDPnZqyiCBx9aq8ikIYKZmMmKokg4A75AmBOVUWN7mLB2c3hlTMuwQR2Fo2aAfXvgDL6QAh0gpR9QA0TKhgnAjyngDKmkDzIwA/zoDCCQAXhwCm9opWGgZv9lbmEAB0Bgp3UwBlaQCIVwIYPQBhgCC7CAfqJQB3JAdmJQB4lQbW7GCWBwIVPgCZ3QCbUADDaJCN25k55InuHQD8WwidypqIzaqI8KC/MAAFEkswnRAE1yGviwDpGgQ+jSIWbQXaBya83VPOeyLKmWM0v/YLTNNgBYdg0JQL7Y53TsgAcoQo28cGp4sEXXMAOzug6lCqtpgqEAQAMS4rUAsAchsjx1xWlmm6y9V1EfkrbtSJgvmoTA9EsmRTRxkAgwsz2ggjSkkB2/IAMxoA+p6QF/KwBN+gv8WAP7wLdTarg1kAII0AsxQKF1M51WYAVAkLktOQiHQAZkQATk1JI7UARDAAqgcD8XUi7Cgrqg4ENBAAdVEAdeygnh9jrthwmZMAvPIAHQkAgja6i9mwzKUIGaaA2eKMaL2g3eYLzLEEoAcA9+xrwIEA9CogDhEAm6hyJCRozC1JgRFV2V0yE79nuQAHWZmQJOSgAFkAG/kAED/8oO5BIJpxEPndeDAIAHL6CDzEBxcDQ4GVoPKdKMAHxzZ4u2kVVR2sTAgEnAklBzd6VkRJMESiNOzBlJwyIIifA1ZVBO2DYH26EHHwIDHxCv+jADeaEPziADKQACidu3HOABGVADDvB8FqAHL3AKXFYElBQHVrAIQOA1YzcGYxAEbpoIdVgEa/YG8lUHBJMIruAIQVUG5MzEExsGYloEYxAGUtBJmJAKxCABxHAIhDqoXcwMEhAN28mTI1vGZ+yojNCDClA7bpwgfvcNVbkhxaRNvTdCyKRXrDJ8RoY1bYXBkaAYe3AKGwwDhZwABRADzqCGTncNLhADknwC6lB6AP/wDZsCWzvEXMfog797GiBqBhYN1BtSc4SZtmx7yhtSmMvkLrIiSETENVhHROYWpngGB3Cgm7QJB1PACA9nBilwCi+gB6fQC74Qrh6QAjMQrlMKrh3QAR4QzBQQARHQmS+AB5EwCkXwtM/gLIuo1zogsWXgNelEzlJDNUUgJ0RASUYgCnEgNnEwdqkFBoLwA1IMBm3gBZwwCcMwkv5s0AC9xYfaD9+wCLtLqIraqGksJPGAJE2JAC67xteAPDX1cu5ie+IzXVjDXLD2otOFNHvQbHuQAjDwITOAAAtw3CpQAzMAb6fgdHuwKX23GuuQIksAW6/ma5P6yCsAAHsJ1LP/J2u+8FGEmU3Jinu8oCHfuFL6MiGoGwfmlghecwhjFwacZH4ANnZ0BgpkFwqjMAcDoKQdIswdcAFT2gsDvsy/0Awf8AEI4CHQbAGn0AKxoAy5kAOzGg06MARlQAUXTjFjQAYfvgNi4zUTKSFAHFR0oCWZVAdVgGeDEGBtYAVUAAZgQAVS4AVr8AescJOH0Lv/bNBePBqwALKebdrk6Q3J8ArK4CTqsHeQlwJ1OUqRoDjVa3uyYgaCeXxC9pgqtzk0JJgS3hd70AsZ4ADBrNy29Zm+AH0OMH1O1wuMxa1mhQcp8i6jMAq9gGoruE5LEAkj2AKmsAcTdWso6iGpXAq8/xADvpC2Qr1LtjorACMHZyI2qVtuTBwHfEibYycI5udmZWfVFkl2yjAKXgiGv6AhDfDBH6AHZu0BvyCYeuALDaAHvQB9A/AusTAKhoAEFy4FY7ADHA4A4eAnQbM/ymAK+6MIcEAFj2AKoxAKO3A/j0BOYEabuSkIntAGPkAFYUAFRQAEXtAHXAAGD0ADivDZRs6JySCw+MANJxuyKOuAjZoNPYgPbWxV7CAk8xAJPQMm1pWsuSZjHPK20YVrBzpHGdUzG9ICe3ANYw6Gz5cCJVzCm0kASKqG7ACiuKRDftNqEpJNM1pRE8UCLaMq6KJdNDTASs0h6JMqyUM0SLMl/f+lPSdSBhTLCRZ5uSB+3wCG8+vHBpwAB1ZgbuZmCKaAjxZAydkEAiB8CjHQAAvu9B8iA2b9C9mkUeHLCPacvM+gAzngLFxfBIlQDyJQDDQdDXMwBlQADYImAc/gCEWQBK8QBHFqbjDOp2EA7M4yBFLQB0eQA+oAAWFc5D1JxjyZDM8AAAR9sj8e74SXDPJ3APduSvnQgytAD6si1MmTjgHfPoPuhAava+mY8D0TAy7QF/qgB9B3Ac9cwoq7DwhgAfjYDL+AfbCaSIPzLh1Pylt+wH0lXQfsCxvC6NOqXXmkQXTQKwrTU9ojBkkwBGLQBpjQCYIABwMmBmOg6bTZBm//wFTZnqUJNgVVMAd7wJkDsAS9AAL6kAe/4MIe4gvZ5P76EKVTiuXDCgmhsGBOgm+kCBDMJgCIx0KIBAAAJuBLGG0HrIQTaCSMN0RUkUWJDlWZIiiTpzBg2FjZ8WPIDyROkOBgJoHYIWvdYsaUWbPbTZrJut2TwEwnTpszg968mWwZMwD19AVg2tTpU6hRpU6lWtXqVaxZtVpFcACAulMxXpTKM+NFnheSluTJY8atmVJu48pl+4gs3FKleKXNE0Psixd4rp1yFmNABQsWItSYIaPGYxkUEqeY0evUNVMs8vBaa2YvW9Bt80hiK4k06dKhJc2Y4SvPEl6l7Mp+RCeO/6MkROIkimNkzJs3YsiIGbMIjiBPmNocD6Poh5g3YcKAEhQmuiA4VoSTif7GUJxly4zg6cDBQwbAevpWLpAgAa8U5T184BXDjF07bwbpmJewX8ISEnpmiUdEAOCeIXRACgAhogFgniKKeEah4hxJZLkqBGkjE07AoAIMKaiYQoqUtoDiCFYgiAYmoISqyRqhrElGmQN6SiYZmmIkaqiiXmGmnwNS2GpIIos08kgkoWKHn37WaeGFFvASazRJzBjNM8/iCi3Lt9rybAkrSfMrhhZa2COSU1RozQEKMqihgQ86WGwfxpyRxxcQOgAhkkj2MsM0t16IAbXNSpEkLUn8Mv8NtSqt/HMGsfIi0BHciKCDtzqIEAMOUA6po4wqDiGiDOKAUKSKMDQMo40qyhguOukEic4TDIEYgjvgOPmktxYy6OCCDj4oJQMKKAirgAIIWAABMz6Yb5RIHyElCEWm2KE/fJh5Jh6BAIhmiFJMAKCYInYwJSFYGPrGiCJIGWUUOhaJI45Qp2ijDU888YJEMHSY4ogssuijEjX6wOcBXGAkKkegFi6Kxn6eufHFhRWmyaabfDzQhiQ79vhjkLFaBwAF7oDyS9n6svKFMN/KK1D72ooLNZd5sbIvX8TSI5I98gQhn8f2ESDODTpooAY66QTBgw04QCCSWFroi1FI2WL/uRRBR4P0NNFeJkssAh8xog466CBClEXegKMORRJ5Q1bhhgji1irIsHuMU4/j5A1Xq2hD1lQ15KQN4MaoY4wwOOIEE1waYeEXDyJH4BQKCJBsCfcKWGCAFkoxBZK4WnBEjiKGMHWHiZ7ZYfVrAXhGh1IgAACWCO3gZ9wTxmU3BhbYNa6MIKp42xM2OPQCEzZ2mAKJI5zwIxhaeHimn2JY1PF6m2akQQKJcWxxR+yLmjCckMs3//wi8wkHAHxMkfotvFYj7awrbY6Ll0dUNk0vtDZzy8u2+EVQvxjAADzAAQ6AYAZMaYYH5HO0CDiAMkQL1gDw4JYZHAowjAGM/wQF/xhI/Q8uZqDUEuhgBJbVwRHsIoK8xiAGMQgCFMOBThjg0KrSAW8MQ+BhGUaVIUwIQgxAEIMnhLgqNniiEzYEg3BQdao2sKENoBDOKT4wgFhQLgKSeUQsFvBFU7hFbGBqQSI8kYQijGEMqPNWDtSog4k4xAUGusMSjDAKdCkBAN8onSm+UYxSkKFTQDhEGNjAiUNiYg1awEQOjoAEEmXBGH64gS0kEI0pMOxiOnJYMrixve5hjGLfq4lO7gEAeqBPlatkJVNS0B919MI+sLFZbAxVtS3BD3+J4kWiRggaz7DlZolazS/2YEAEfgBpj0FA5EBQAwdYgE0NyEDkPGDMPf8soRRjeoEeLHOma0QinDzDQyweEaEl4MYIcZDDEhJRBjP4JglLKMKmqlBDkBhHDIp4Q90i9JsxFKEMQLjnEIoAhDdwQhBVeKEn4DAEOODLE34ITj+FBwcYTicMmFCFIOogtt5JxgIDMAIL9HAKSBghL454xArLgK8qHHSN/YmGDg6hiDGkbggusAcAaOCIIqyPH0EFgBLmUAR1KKQFZaAOHN4QRU54YQ2Z0MIWtnAELxxBB1LIAi0wcYNJ0EABboCRJhs2yph4EpQ/cdFM2po9ZUxAARloZV3tmiR9eCUeeYgSXvLy1xeY5X/C5MVovDQoM8TAF38SjTCFmdhBxWD/BqeYnLM8kAIB5CkFNkhBClBQg2JtwAEOqIE+QOAMGZxiD7JhQSxMEYk7sEO27IhtbGd7hzvAghGOqNAbXGEEGLJLOEUgw0X9pqoYViEINyRD6arAhkQsoQxwQCQOgVCdKhAyDGUoQkQ1lAlBqPE4U9gIP6XDiUpwogqLKMM5GzG5/D2pdy1NghwSAYpFBE86piKDDvQYDSq84RBWqIe3ijBH/wwEAMy4Y0L4oeA7LFUMfmMDG7yQCar6QRVrkAIUoCCFIEgBC6rYghdS0ZKXNKytZs3JJ230PYYNhSaiKMaB7npjHGMlAwVeB5m0+deXPaI+g3pZaKZmM9RUbVA2/0uLWwqbh5wlagaTk4Ez9OGMGuSpaA3YhzzyMQMHVEAyEdhHABiT2j00wg6wta1k6CHOO1zjGnHGLW6LAQtlVGEIghAEC7gTITEEoQzAkU6q+gmHiB6iDNapAkOFF9MhvKETfshEGNj1A7XFUAz0jOh52xCEIhyCVRMWnnQ4pAiEJi4ROCVDGZ5kBEfUAQaPmEKF2SCG0i3Chm9Q4zMmwAwpwGHAvmawC2RXjF0AwATrYBcLIjER9t2BBSwIwg49kQkL90EV265EJ3TgBChYawoB0zYwNgGBebhiky/Cnlu7MSN8vBitKh5lMhzEjhznW99NQYAC9gglr2mzlo4oRf/V8mLkP5EGfpJNlGum1pZergZSjdmDPmwQtKGVR076yJwM2OSmx3A8AaqFxR2KUQzcVi6aA1Ctak2xB3DGlh53nkMbwADrh4aaONZZlYbgAKo3gAHUzHkDGYwwBOLUkw1+oEUmykDPQwCHDYvoXYZm1YanA4FwUUf02zDxhtLxudGC4AR+I0TePJCgAZEYXBusINAaqu0NXqBCgOGQiEWQwQpTYIELbkc7SJDCjmJrwRJIkdIi1GGpei5eVregimD4wcNAkIIXcrADKRwBYKqgRSqiAQFoLEInbz3rW2f0AAlAw3sOi9G6360MBdQjH/um/V0zMIF+0KMF037EEsz/Mu0nEagsYilTkx9rGoj3kjUZpBLy+yLZ5csAAZR5zGOWNp8GLCABAkgAlmUQgZ9lP1kJMDnKwzkACkTQAqfQgwq8SSwKjGIPCWgPPehxBzcEgoeHGEIdDjGFHKCCHLCXQVgOI8gQwiGDILCCRFgbFqinMQiCKQAFjvKDQ2AXhHqqIDCD7lKbMPAETgCCJSCDNig0OJgCOPhAIDg6TggDUzkkPbgAeXgBOciDfdCHdWCGPyAc4dgU4AAOT7iEMiiDQ7g7H1q1FoiFEACAOygpI1Ap2SiF3YM127CjIRgDP8CELXAeWkAGVeACJBgCH+ACa8k8L+iqWbCFeZAABXgG/2WYGHrjpIt5N2hQgAPghtGjmBVrGFFAim+oPUBcJX0osGJgGTOIhVEwhV/AAzwwBVO4IKyJAcG4g1FIlC1BC+QzA1+YAcYoDecbjU2cuGY4Bep7jAAQAH3IABtAAe3TnBTwhThJIARYAGRJgPu7g0gYhV+IBZGiAMd4jBRIPwugAFPYPs2xM0LAqbjRAUIoBmYohkIIBHsBu0MonkwoujGogh8oAiMgFUJCDj+ghDOYAuKqAqdqg0WQLul4myhKhHr6m1SxDqcKgzroxsGJjr9JxQbAABvAgBQIh3AIBC9YKCLaCEKDAzbohDqoAxhKBCJ4BTGQAyOwHQCABiMIG/9tsgu3IBA6wDV6agNjyAItUAWmQwZacIIp0AHNAwLKOwIoSINZYAYFUBEN6Ic2fEO2Yr04vIk+7AefOKtRIiVPmoAHmL1APEqQaQB1YZkleIFvSAioTAgaYIEkXB+oPIFI6KBhSpRBkThJ4IVNpBLQWD7JUgHC4EROrIEAyAcO2IAamEXtq4FeOKCiOYUvWgA5o0Q86AW0iAEEQIBmaIAEILMtshwKiAX3+KI9mLNiMAQrIAMeWhCo3IVcCIQwGIO/Ibt+yhdFqIMisAJFWIRDQI5LYAVigIZIEwQUXA7usKEUjA44QBxPAIXAIbTtOjqyU44wGIQ7iId1GIAGaID/dXiGQAiEwTmEMfgBcwyJ4HAqIhiCRTCOeYmDi3iEUYAFO6iDUmCpsJkNMKkDOKCCIZgCYwgGJGA6DasFY9gCHUAJH1idKeCCI4iCXZCABxgGdSuGeIMAN/yJeZOxm4C9eYCx8HGRZEAKfENKBUWSfTildQC4JYiBpwQAfjCAEPiPe9g9aDsBaAOASIgBIysL1pAyXuAF1mi+0ZAsv5gBGMgAWWIMtUyBA+qAFJDLXqgay/qAZtCDvzyFARiFWFCPFACBDJiBfSAA+lsAfcgHkRqAGPiFLzqFXtgDWCgGaLCVIYCG/zgBdegPn6qwMOAEKcgBJDgkVcAEKtgBHRDA/zV9Ki6YECXIgUDwBLUpQTgIot+ojhtCQA2RImGzjhUqgjfABD/AjjeQAlwohEigpnWQggrjhBaMgzFwKj7brk8ZlSCIg7vbJzl4hQgRKDp4hCRohEdoBEcw1SR4hN6xFynwA3DIhBzABFrIAj/wAz44gh8YAx0ogx0YAiq4Ai/giV2oBlcoCmvQTzYshjx0GGYVimRQBwmAhTxkMVJ6PQO4hwXNVqfIhwH4hnVYh29gh+mLinjoh3A4GbeQUIVQjxKNhb2Y0G/wC1MYiAkAjE9kDV/YxBgorBN1PjNogTGJgZOSJWdoAE5cmqZRoBlwhmJxUQQ4LcmCgV7Qhz3AA/+0iA/zeCZkUZYECIAGGK1mkAQ9EBTG6IVRMDlGKJ2kmoeV4AJiSAhbAAQ2+AM3GIZh2IRJwARM+ARE+AIpKIRh+AQkCIQjQAoaSKQSzMyOUoRCe4NTSZU24IR86acwEIOL7C5OAC9zVAQqQII4uAAEKAYpgFRO6AQ2eAMgMA55rAMrJIM4aBsxWAQxoE45+Cg5kAPcoANSFVXbuIgxAAMkgAJaoIY10IE+OMmAyYItuLzVIYIdQAIe+JFwOII3mEMcKQYFkABlDUpRepFkqLGf1MOddNZ7MAC60laktIFrCAcF64f/SAgFwId4YIcGYAp26AcaiAXBiosZeMoTkCz/wAhePVIH1hCLO3gAfMADxRoUKhkUXzA+SIkLpmyEpjwUFngEUyCFU8ic0rKmZ3qBYokgypAsX6iyBvgF9hMLXwEWZeLYZQkAOqmBfFga9GANFzAFWIAGRFDTZNsFLuACJviDYSAGnMWEZHPdnkiDSXCQaNCjfgiBYeACB0kICbCFS+gESAVBVegEMBgEWIEOPgNBEFwoQRBBegpTheonU2mDUMCADCiGQMCEFswXQSADIuCn5SiCJaCnONiURBADUehh4FmhjFAhOaADltIUVzBHNtDC5yFcHHieE6FVP7ACudkBR0oDPzgYQxgEi8meOpyAN+QR8NmRZIAFBYiG/2n9TxcpihpLJdQFxHygB4boB3xYhzsAp2KIB3WoB9dVgHXoCgUYBa00gwh9ypKxswiLhf+gB0ixj8BKC7/whdOgZHt9gRI9i1AtPEeIA6xhgSVgSEYYBe3jPs/KgA+IAE7Mg2HcIgRYvvJ1BhVIgVPImRhoAGt65RlojwKYgXxwBtYAAQTygFOYAX1gE3aABmiIgiOYEIV4BgLmAR6whGNwEH6IhmQDgGG4ggmeAF8DgH6IAmIYCBHYBVYABlrY4KythUxg52sTBB+IFeJJSBJ+g7khAw2hlcQxDi8IhQ0I2/P0hDFQFQU0R1kZAjAR1Cl6g0RIWzEYwnpsQCKog/9ECILciAPqeIMm1jBaKE9MkAJjoIYPywJVyITV0QEwyAI+kAZbgIBwwAF1o7d3KwZzzUmgDApr4IYJmAebrpjSswll4AdsjeN9qwF6KDAFiIdIWIJpYwEomTYjIIVIWAd8MAAF4IdvoB9g6t2ohMosSohIGNG9yKCGy9d87UoYGAdtEB24SAQ5uA8xcEd6Go4wIARl+CLN+YXqk4EUKIwBSAwLAGazVgFZPl/njYEhRYBZBgEQ0AMUGNLzyIPyMI8MiIH0i6Y7gIZVmIQm6FCpHAZgAIeEYAVpnpB5WGAAEIEo4IFNQAhmiFwA2AUeUANVoIZaWIMtyARVqAVqCIb/TDAGT9g7fEmid84QWlFAfKSVGwIDTgiFBhiAYiieNtgUp4KhAUM07nrHeSw6eomDuQFPUCgDUWgDUQiCRTgVVDkeLfCDWuC8VUCCkMYEHNACY+gDfzlDPqAFbIAHl7iBLw4KHMEHBXjDOdRJjKGBOxTddbsenZiHCahdos6xAZiICVgHPACMKCGQuMhIpu6db+iHedCDvgoNrg4BdTjxe1AHXsCDhLgG5l0PEMqZ5TuUGCCHdGgHRvgoOrg7RyACUPg5RyAoOACDT2CEPfiiBIA+Z3CgDnCGF9AHBAjFR5aBfGiGFJjYj1u+BtC4D9CDDmiaa4Kc+egF8B0tC2CH/0JYBTVggjQYhmdQgiWE2WEA512Yh10oMH4ABga+gRu4gmSLBunBXSboKmpABifYgj5gOtteA2NoJCrABHbOBAyujnvxhEZDNHOEVKnz524FhMFJHKbNNLndoSIIgnvJRj6j1Hu6Jz4TA1DIBCLKriHYATCQKlqlBQ0LBigIBmrwgyNQBWPQgsBVg/U2Bmx4htTLAYUh4wOVNzI+q2dVADwMynbLnukZgAi/MRtYB9eNBzwok7VAC/y5C23KSBaIBgnISiwBjReogaecABaAlBegyli4nW+YgRQ9hW/4hhuVhFAcFEwWB3OAB9QklUT44UKKTSBwm6Ar8iXQg2aIAf8Y8HeiSaAYaAZkAeYYmNhmkIFm6AU8SD/JUAHGiI+i+YBf0Lhr2vj07ZXEoABlKAQtAEMo4IE0sARNIAYDcYc5V4gVmIATUAIaqGbXwYEc4IE/54GXpYE0YAJjAIcvdAItSHRqyAQn4Lw1wIRV2G1jqAUpypd7wZcwmAIyGAMluqFBsANuLQZPV25UETAYEo4nLAM6NQ4RVqiog8eFmii5hQNFOAkLAxhaCIZtI0lj2DY+4Lw+cAJaTfxaAIZngABEsIKygrHXgwB18E8Z06RnhQBlKGPSE91kgIY9yna70oeJoIFIKJPOIYtDyQMtQZlSYIFRgIBd6KuVOYvAogf/hRg+vGCBUzoBE2WN9ekHPYisqjGDFyCFdDAHd2AEt32nRQCFH45bv8nMNmCEWIgBFfAZSWimDvAAEJCEViyAGIA/B2gGFeiFX7hsC9DrGpABy0KAPKgmDxiAIXuBIQWBUgAIPHuEMHJTycsDAMyuWLrCYxeAaLYAmNByQwcXTaumQdy1QweTegCe8WAGYF6aPrRoqYLiJUsWJyyz0Fqlyhi1TGuM+fHCKRMmNmA8ZWrz5scPQWwOiRlkZ8O1Ylw4cRL0BlSYKmSAKNK6hEWdMECGVOFUyU+mN2KqeAoD5w3bTG4PjRkyxYuXI6qCGVPlx49fvaqyaIHpx1gtVapS/0WDoMyKtcjWuk2mbLkbvnrZul2uPPmz5WT3FHBL1tmyZ9SXkymDoC4A7NiyZ9Oubfs27ty6d9seoABAvFIszOTJ86iUGTOS8phZUpx5KeHfAHxrsYT48uczpk/IMyNGnlKPWEQCAIDGtT3xzH+LsVxSjBdmjCzRhq6duyBkyCQqEsQTEGR0FUYmmbDxRhi5kGLGBx1swEEKsXzgAQiS/LJAAQQk0IsFFFzgQAYqxNICBRZ0KIkvKTSjBwgZ9BLfL3jkEYN7+nTAgQcIPFJEHblUg8xDAJRADCusENMPAMQcw89IOfQAUTTSRGNeMTiYBIAtJQFQDyvUtLRGH1xs0f9HFrWw5CUWqoCTCQ5b+NEHYKqsgUQmtHDCRhhTDJXnFKRswE4xbLRRVRhtVLHIWlXAUYURLAwBRxlFjGGWnYtWEcYYZYjxhidVtRHGEFaE4QUUmBgTjF99jImJFLTUcgRMfQRDjZd+TNIYImNIxhlonHWTTGPKJJPaaZUNiw8+vKpW2WmqcYbPATbwNi211Vpb7TX8GEBPCy0U5+1xzzGXXB7eRseCOvaMYh0vL4xbXAzTlaDHDDNIEt2M05m3bzgvuIvcC6Us8UgS3KDzDjGZEvFGGWS0UcYQYoghCFCCvGXIHMs0yEEHGbjXjCQz6LEAyQno0WGJGegxYh4IIMD/C4sdfJACjTXTqE8Cp7xg4wYy2+EIHYQEQ0sabpyw777RpDEJMeYhCUA/tkgJwApI75LGDU2bNwwwSCCBRUx+0EJNMLNygoMWXkLRRx9avLnFGl6oQk3cbCDBhieCBlJKA4Ae0YYnn7axCBBvWAUHHIqQUQQZjNbRRiWZgAHHFHAwZUQZYQgahieekDFFG0hgstebbR/RRyZS8JG6E1n4MSsyb94KASuLDLsraJMl80w/0JjmbK+6K6PAPcMue3yxvzaWwrXNO/88bwMYYM8dL7RAnL+8CEyju8kRV5zApExAwxLO5bGcGbxgf40667T7HS+8zDjDHuHgM8E97Lzg/54ZjxxXBx3qEARxnANhQxjCIgShHzgMoQ5rEcSdFvUGOMyhEQjwgAc+8KJ6cfAXAzDZDE5hIge0AA8LagH3eCazFzjDAQNoRgwKIMMFnEIFGPRAJHSxjFCswktY6wEz5jGBCcyDGDhgAiVmYYtoDDEarJDGNKb0jGEM8RlQOMMRpPCMeeyCFbRggxTysoW9UAMZOGnDENbgl1i1LgtHQIITbrIGKSDBC2vgRNy84Cd6FAMQg+qEIKrwg0OEQRBhmKAgBIG5hxUBDpngRFaqsJZDLKEIoBCExDj3BjB0AhNbqIRfWreFI/DBD0hQVQ5e8jovwekKz5BAIRYhmV0py/8ayfgGPmABPF91JhnFUMjvjpc7ZfEyGSZBAPSSqUzo6WMCBqhecwQmCflE5wXZSR+5mNOCSPQjHi0oxYyKQxzsoTAGxKFRduDDwRnUYAYt8Ff/GkGHOExwCNw4BzygMQQgVEEQZCDCIg74hoF6Ai6EDAIdYmGGX5wiFv6KAQdtBIIXgScWpHgBDOxAB3xJogUZuFEGe0EBB3ToBQnI0AIG0IJYIGAPjdCFGFpBK1qQ6QhcYMhNj4CRPvAhDTkYQg6uIA1kMKYfz8ABF25wAy6owgs4uMIkJgGML3pNCnACDEu+OISvkWlMcFLFFpAAhb3MyQlO+AtQshCLBtBjGFz/6FwmPEGFSyGoUF2xShswUYWIAc4TVahCV94wBDIMtAz8bAPgamGMTBzBD2rwAhzH6IcjOCGLkl1JYfqQBnVAABGyxB3uiMmsyNSSmMWAgO9KO8zggcaYALjGMmMrW96ogzosMIL/HIEc6CiHXLs9H3NeQI9+FMM6wAWu+mqWB/lJornv8UUMfPFQ+ZTLOHGIAxwSmQRxmAMesLDLGz4Fhx8wrlBwAIMk4dCGOhiBFEvwVwt64QAH9GIGKfAABzgAAvmVizikIIIYGvEI77lgQh/Agz48VKKFkmwBe2CBeIbQiESwoRLgoAYtxBamS/ThBn6oBRLG9Jcj7AAHskJG/xogEo4cMIELOWDqmyoRtgzzAYxgy0If/MIHJOwACWr4y1/2QgstHGELfABrFsAqNj9k4QqxiEBb/8AJTGSiE2EIAxgOOai9juENVbbYQAVR0EMgrg1i+AFcyDBYMXNibKrIAdve+NVaODaLKgnyFrJAiU08gAaumCVlSNssz4xWtZzhBjcMvVrRcsaXAGDHbCMtadmwAwDqGEIRdOGIRjRCYGbw9LiSw4vmWnNG6wBAJAKmnOXwjxcQBQ92itPc88161PFjwRKMQAdXvMEVoGCDHLgLD0TooJCDwgQogiCGKyMIDpvCpBGSQIpu8aJD85XEKfDLgQ+4YLnMaYQh1P8Mh0QMWH28wAMezBCLDnXICGY4RQL24D9O75oTeqEV2bykBbDqxRhOUANhmqqD09EiGLcy6g1KSdk4cUIKrSu4McBYyhwHww9QgEKRtbCSjfPlTYDhw9r8ApgsQCENvUgBPQqRkjhBVlBT+AkaizCFTrRlUEYJ3Fs4F4ZErEUMYxDDTzpxE2TkACZQOKsqCk62LUDhyH5RBZmYJgFm3G6Wn7H6MIVZWl8Ra9Gp+UwyoNEPeky67LK1wQEUMIod6IIOm6bDgMXziFF/Gl/+Qt8L4mEAUwSsFOCZ0QvUFzJfODedhZdEc3hBDnFkoxFB4JRVxMCGUKSju8qYgiD62gn/TiBuoIeEi+HYEIQ62MEFJbSAAxpAgV/0YkIeyEBykFMKQ7RhCmM4hCCA4IgklKJbSxDOKSjADkiUwghGKEIRHqELbbjCE3yISdLxndWKJ5kWlS1yJeYYcmRcIgo96AFNoO46nrIhB0dXCTg4gYQgF3xOPI1JwYNhJrK17VSqKHLSUQUFJDDhFxFYByukQZxQA17AjRR0Ai10whRYwVB8SqcUlKDkHM4NwRjAAecoxiNlgjH4gFjhGEuY0azQgusE2ZuogSrggwIIy2XwitUFWmhpXdcNGi/FoO78EqSZHQ5CzzUARxEswivIAR0EYe+Jh24lx+8hB3wwR7mogwLg/0HAuBrinQ9zQZd7EN57XCHilYI4oIM5oEM65AIoVAUckMEhUF53MQIhtQHeINYbTMFaDNSiZJcnwIF/NMILrJuJUIDfxcIenELsCYwQWAzeAAEcWMEYWEwRGIEuNEIi1oEj5JYjFEEZjAEoCIMqYEEftE7+lY1iKMbQAAYS5ACsgFyIZQEWrIQT6AAS8AEt8AHTkVwWIAEQIAFl0QI4CEPTKQZLHEFjDQZNgODYGMMRqNGpOJbYDI0a2NQvZAAApsElaBwBQsFheIHYYELLsYGgBJ3NTRCncB7jXAobqEItcAJeVIIU7MAUQMEY5R9iZJiqAFkfXAExSMAzmEbuFP9aC+ajMH0d8gTP1okCM/TDAOQgQTZPPPQDLOzAKxCBHDSkHJQCNfnPckSHGVDhcryTEI0IcoCH+vjC+UCXL4TkrImLOIlHIcBDO5jDOaTDNnzCQAEBFRgCF8IDI4gK4LCBoiBIEHTZJnVFGwjCFEgiECxBCV0DBUCCEfgLC7DAN4lHI9hBNmzDoAxBV/yAGgoFLniCKwzBDiSCV1IOGGACFsjC0LDNFpyVqyADqrCEStRCFmwBFegAG4wfjm0BF2gBT+jAFJggkZGKFEABGOxADsCRLVJD9YlNWK0jVgHGqVDDqyBBkgXDGjTWSszREbBBCjDjJqwcMtDNGFGDFuT/2BbYkR1hQifUSQLG1adAECe8ARlkF+JwAs2RoypowUccgTSqwiogw2D8hRb42CWIHw10lvEwSz+yoNdxnXIaGgz6SjKwBj4YwDpIS0FWJ2/QwASEgiLEAR2Ighx45yPOmxnMADV5B3kyB3yMBh5806gpoXvAB0g6l7eJS3T4jzLAA0qqJEviQhsEQitw4Tswwg4gVuZNEOaVQRn0EycYhVuIwQEtTiTuiBHIXnQMDB0QQS5sIThUAxgUQRVk2U9kgjaUAzUYgxRIgRo8X+vg301kGJDFyUrESZIxGRTkgBQcgQmyBFilkl/oQNHxgReIYigiwTn6gMPNjZcknZIe/4EX5JgqqCVW1QJv+gEm4GUlTMHAic05ek0KIMA6NMGP7QU4uMlKMB3bqKMTuEn+eYmBYOOCfsrLpQXonKYn0oIUDFxj8SZbopXDHQYwzGM02OPWDSo+NufVLafWOWcycAMzKAAEHMB56IN1Tupt5MMDzMPtucJ1AWEcNCSnOUI5fdqqwdqMhIM98B2+yNoMEF5I+oK9OFf6PEf/7J4jLMEPDAN+tsM5eOE2ZEITAKgyDIGYAU4k5R4FUkxRjAEZ1IV/JMp36oLAPIIj6JYcgAIZGEJKmgM4fEIReFktZIIf5EI54JMt4EAOoEEf8KITeA3UCZkxrEQnKkacuMnbhP8YJr7OSmSCDkgBTGzBmADpwIGBj/7AR+BA03Vm2WTYXtiNGrjN3LiKX8Tfu9bCFuhAj5XSGqwBF7xBCgxAPJQSYAQDOESsxUFBaBqdk66SYlgjlVEF6JhJ7bWB0M0KNYADFKhiiPEFXxTGX9BiHyADNgwnIjCasyCqcfrjcWbdCj7ncyoDMyTEAxQDN/DOAUgqpV5tbDgD8XQZKEhMHYiCV3LnI4aT//SPq/1dDJxaqiHhR9IISEJUSLqH+mDTEsiTKNRBt27BF0ADPLyDOXQhSxJQOxSCh/5ks1VBG4ACxIACJvxFEhiBfvSgVWCemSVBQ8ZBIiwMJxTBDrBCtp7/gxwQCEu0wjgcjDsYAxRIARZggdfExBqIYNrQWdIBmS7SVBbwQduMJkyEppL6QQ6oYpL1ph+E0RZIARDswA4AgRRsQZqILDjEDiiygRo4QY6VUW1CY9n4wfNRwRT0QV/chBMEAVQsRI4p3dNlARcU2ZtogTqWztjQlB+sgca5TRjRChj9xNjULDhggvkx6ZIBBpwYQ2L+qQSEwyIE2goqp3HOYALzowM7C9N2AzfAQjFEgwL0wwMwQ2kkwysE5DxQJ9ZSKtrhwyEEQtcuQh3Iwbglgtg2ihls2nE4hzWFDD0AAD2AR+G5anQ1F3TJJzZFh67RgRzUQR1AEBUUASEQ/0Pf6icXtsMwwOQbAAFPhgFV7NUbmAUf1MESdEXjXNmyzaF6BY5rXrGkvME7pOSubgNR8EE2iEM7zIMtBEYWQJYXhGkwqIpi4JjIiWNi1CaOZRbJleJf8BSPWZW8TiMSVELFJi864pisgAM40FQfYIEUSC/1duYa0QSGfaIbnVIZmQkazMGfMAMToGXFfVUmcoHr1CYXwMlXDRmcBJzbrB+GXWYndiYk14JKqAruns5fvC9uAkM0GAA0iALw9AovJXMy01LWIXOvMOozRIMQIYkE3IMGP2ej7Q4ArEMIY+1oEAIggEIVEAEZiIFXeqUYOAJ9vHAj6Jb2mIF70E83xf+He9CafIaMu4xLhRKhKNDBIgDUTyrCGEhBICCCEvvt35pDO3zCFEyBI2QO4PxEGCRoXj3MEjTOG4JZViCSoQABBDEOFbABMfjtrooDN+TCOOATPFxBJUgyHeVi2cSE68CEHkMcYPhBaGaiH7DuX16cqgBcHeHuxxlDJ+AAH1ByEOyAFfiYmECyvmVBJYCBj7nOTGViH5iJmfDExf1lwWWYLIxC3xBDi1Uf28QrkNECxtFuquDYCLbNWFGDH/grS+yb8w7NJ8ZvDphgYwbDKF2CEOWCPR4qoirzMnOdPy5aaMBCP0gABFxqOBSDMvxKYDdaNhwAP2RAN1PqOvRDISz/xbiV81vAwSKoc661ANAI2BI4gqvNSDPUwwPQSz0b3vnYC3OoDxHW6iPEgSisBRxwgjhn1xtQQSCwQq4mNDgYQhhIIudQBSYIwhgkRRiQQRAcEKKclyFdmeVIiiDQxUcbQRUEQiDgKkl74TmYwztsghpgVh8M5piQ0f5RbyYfhlpmFS1gwSjxAf/CkRPUMWHwAchSL535ASfogBOkLhXswA9ApljVrC9mgTGoHxQkWa0wmesUXFN5wRQcwRQgwcb1wVdfQFvxwFm+iQcmXYzuBS/+mFCLXFjlWI6t0pskHZwggy1eWMHVwpHxr0qUDf0xQR/QwAHYEg0yMGETefI8//BncAM0wIIybLCRG8svhUNmW+c+VBoxIEEjGcWzCUIS3Na0goUc6EISPEIjwHMevMAMrMc11ECtrZq35fPvGUE7JwEdDIwYLIIibMqCDpRBvYEUrIF4g4NKosM2CMEQvAEbUMWC9pPnGAEQMIWiVOCVWYVaYE5WkEEYRIEW05UUIAI8uEN+/m0aowrYIEE64pgxgMMaINU66jS8emIwVEIynoET4MC/ocEZYMEZXIEsyAIapAETpLcqVAKPeQEYneMO/CUOeAGteAG/0gRaL+/Y5MVKwEnB8cHvQuaGd6IayMIpbMA3DEN6x1mOqSU43DFWrUGEzzQAywmcMdmb4P+EyPHshzk12SisMBQdLez1m9QCsiQCoClwAhd5YR8nMa1GoxUa8iTDPFz2lFdnBkhANCABWVBFlw0KELAAjzRKHVxXp8bBI5jTjHCTOtTA36Vq+lyHGViH/wSQHIjB8S0CaBvO5wEWYL2BH1DCKgyDO4x3OnADKFylGHuCIBTBElTBBGUZ48ABKIACELgF5vjToZsZBf5AFRwCV7oBMbgDqKMxrz5BGtysKGKByKauWC3ZKqvCkVWCMSAGKAyBI8QCzPQC3a8MKUBCNiRC0K+BLLT9GvTYYxnvDsylF8QRLQiskYmNKpzVqeTF3HjS3KiC7b3RQB/ZSiBDBjQAAFL/AjVsgRb8efRFcmgCcCfDe20awxrkwJiMoJmwONtkgf5C8oXFTibkgNiopTHwpiocw1/DgaAp7QITvMIXKpIf+fE4Gjc/PEGiXT20ARUcgmyWQXhVxRgUQQPxSBWAQq+Bgi7IR3LEQG1dgznxwnHwQvn4j8CwwCMEoSgkARmIQmpLEu4ZRT9NkKKUhYlTQjBsPUIDBLp0uL6AadPGE6c3RYZUeTNFkBgjDd/8INMmjJEycIoUASWoihgydaqUKbIjEBIum4a5c/cO3Llz6D7d2DFGihpq1JBIcZLFjypVfoL6UVOpFTdTKRpgwJAiQ1SpEahG2JAvXwM8pnIZGiLl/0yWG1NybFHDJxg1PlJ0mA1KzViWLLT8OOkDzpMXP8iMaekpBfAWWmmr6YuwrokaWlvWYgpGayfdPkX7IDkydKgTP0fK8slSyxityUP7aNHiB1xq1eCo0cKEZPJjWquQIUv1TEIhONasdfPtu3e34L+JFy/eOzhy4cuXK3cOvHn0bsm4TcBnI0B27du5d/f+HXx48ePBx8stZYygTkPACML4ZshEhmE4ebIfp4URI49YRDKAj5QiSmmkEUceKTAJR+poRA4xRCGCiDjEKCMIQd4Iw5M2xFDkjTfgOAQOQVShhZZaSJxkE2LgececmQYKow02NCwDCEUUEQMOOIIgKf+MkDwJY4ggFCmijjAEgaOki4YoYgow2FgjizSyYIWYl1pEB5cgdkCCj51yQAKKPvoAKihjKmmDlHxSQIAdetaJJxw4w4kzznjiWecbdgZAIIJ8LrhgHxnsSAQMJ65Agw/IqMGCrTCDCiYYNpDAgg8ooDCGDb2GUuOIH3L4dAtjgsFmmwgy+IYJoFShBgwt4IKMFqGE0qI0P2Idc7I+LjPGD1FVgYISosT0I5jVUqMmLT+gsEvWwYy5YhgJmJGCN+KUi26435IxzlrohvvWW2yhYy64ZMLpZwDy1F2X3XbHYwcAZnLYYYo2rHDPkymWZKghTzrhhI0fiYxjETKseKb/n3B2qGMRMeh4WBQ66ogjx0QmFEMQUOIg4w1B7AuDjDKqCAmOMDAckURV2BhkDTfggaedFtPZxpAqqBhiCDEWCeIQK3Iko4o2kBSaDDGAKGIjI6sYAo5M6ijDk0zXmEwNNa5YZcWYBUpEii5ryeGII2jdwo9KKumEFAw2GGAdde4J55s7YDEFkjnqzoWRUSK5o5hv7FxnHXoG0KeBDS7IxxlIcmlFFtag2MGKI7LoY1VVLOPDDySQqMQLKPhQBQskchjDBx+k6DUYWRiJ4JpveBgNnDVWrWVVuohSBajBSJRrsF/7IPGxysKcPChVVGuHtRGp8WNy0rTgg48+mPji/4EHQDkk227DzZ5b7cV9TrrnnksGmn7WcRf99NUHrwEFaJCC3jbq5SSTNjoawgoqwKC/PvsE8TgMb/CCJzQgAWJwYRAWEwmOcOShNyQCCIk4yBjqIAYMteENJsHYIYLWIX+NSBVvqMLKdlCIl8XsHOkQxxwS4QMrFO0NYhDDFErGiSocokeHEMQYGAIEHMJhCItYiA84UcQ1+GFMk+MDE0zIoplsQxjUsJwTYNMHJ6jhDHHIxwYysA4NxKMYcyiCEVjAgiUU4RH7ecUiOrKDIiyBDEIYxTWK8abAsQMBKfhTPnqhjG1MQQdIUAMWKGeMyhxRV1Log6VsRQsg7eBLUP8YkTFkAYkIfIMYqcAMNbSgCmTsZChIzIJQ/GCiWGlhLlJEgmZERQstfOlzuaNcsVITjFnSShXBUIUWLgEFyxgKN9OyRjK2tT1xMYc4xeyeca4Vru9hKxnKgMA91ldNa6JPHbnBARs40SROGKMWSxpCk7zAiU5kIhP/wgQ6i5gJQCBiAhIYBhKo0CFFVMFCAYRDDIH2vyAMoQxiaANIlhS0hkUECFUAw4gycQhFYJAKUmiDLU4oM3HkIhRx8EEYqACEkoWBDSHJIBz8wEMykAEIFypDkIxABm5igla6sotRbhAKFWltZtXgAxa2MKawlOICp1JHPJSxpCWYIQ95WEL/KUrxiAPRIRFxeIUcXrGEOvCnI0Wwwyju4Ld4CC4FGCABCWoAAyKAIQtnqMQusxA9J2zBCapYwxHQAo42BGkKkvucKmTxiy4OgxKyCkZQFCUXJMZVVkX5yYgyFzbGOuFLdLlMXbKABVoiQ5ZaMAYyYqWKLXypD5fYxAQm4IZBDLN7zUSmcZS5TGYi81rgY04ydqGAFFwTt7kFDzv68QwuoHMKLwWDOKlwhHJiYp1+wIQXkOCFTAgFE1wYBgQgMAwcDAIjIPWYhWL4AxAdIghk4NlBgFiERYRhDAENGRmaRov6waENnDjEG8AghTAMo6IuEkc2smGxkr1BRjwsQtDq/1AEGfoQDjQiw0TYkCnmaWELWziNZ3QwBERUVCDbqAYWsHAGTOjhAggIhzqUUYQWLOERpTADUnnBVKc6gkFycAQd5GCGJNRhP0Qy8RJ2EAdE9A1P7MhAAy7glGbYARdpkQUf1NDWVc0Ven6Ywg52oAMvEI8PnVhTPDbBhN/RAhlEgQxmiUc5lAlrC2TKwhEspQpjeFYKR5gMEuAaNicYo5ZECR4IIYMFNn/uCsSQwDzyxRtDJ+c3sV2ttqbTaNc+U7Ximw4zAJAu3V760vk4wAM0MRgpPIkNO8DZONfgBS/M9Yiq4ISVg0K7LaRhuhAgxhGQcBD7cEIQARyDQ0T4NP/x4hNkE4EDEFC6r0NkghaewEQb8JmjqIFhC1WAhktgdiUVloO/2TDEJ+LQER+AgQxFsMJDxFCFMZBhCEsgQibqiytckogWnNmBD/Bb7QxrIgrOWJs6vkHGFpiBFynmxYrNwNSmwhjGj3CELh5Bh0bUIQlLUNASjPA0KwyhBS6ARR2/eg19bMAp+cADNz6hBbXiORNcOAsSdqAlK/xkcleAQwMGEA8w4MDMtLgEYWnlh9MU7zFByYJm/IAF5ho31X34lGbYfASyXKaW1Nj5LnOZS2pIjjR9SEM0JLCLT0ihQ64QO2qFo2jVEscb3fBGMta+dke/Flzgu1YyigEAemD/Gu+5XQcAiAEMbBjX1DpIAs6msAUobMELUtBLMFbd1tGoAhhMgIYCIPAMLxwBYAnJUEXm26MCjzE9nhgDHN8QMjG8sSGd4AMcOqGhHHUoDJnAxBinQCVqvwOF+i1HOcahjWxklA44EoQPfmCFHRhhCVZQBRg2tyziqQI0WFDDvHKghXrjVBz6YIc6isGCGLxgxQZPavhd3PDyOwLGddAFHYxAY/QrSBSiKCMV3MiIrgKOHghowBbz0QxSJOITqKESpCR0qExLrkwuuIAUNoAengEHuOQ0ZEUVwmYyng/6OmvoJkNZckAKOJAP3owKfEBMwMQLdIAKuMB4gkELwCGm/4hiL8AhE3bAC4whLrSuD7LpAIjBDTzhE2yhEA4hGaqFXBJtXBiNmNaO7djOG9KumBANOCTtmJTBAOIh76qwmvQBAmiARExtDbRACkaNCiJszXAACnrFlzCHrkDoCHDADfBh0GyBC7igiKIGJKrgo35gjIwADgBGDgwMDgos3CaiCjgBE8gA18SAg/YJDCrBE2iPDabgBxBhGJ7BJdwha2LmSgRCHMRh97ih97RhDlIsDPrCDMmwEoaFKCrhC3XgCG4gCloC98wBHdCBGO6ABWbg+5hq4FZs/FxgqZqKDpbqQAzEERIkCaZKDtwvCeigDPYjDnBsX+xg4+7kGpoCA/8ugATkQQ+WAQ7YoA+2wAdETQbTAMKQQA9SYB2I4QaggBYOr5H6QApgIwskzAVTBnfSrGz4QHS2xFe+BAvWIJCOAJIkaXnWgBomp+cugRpihxVJBJW0Tg2YQQEkAAI2TQLwYRBa65jABe2OsO2OUCOdkAjNzjeoQwGoyQpTEn3iAQCGYRKgABOQSAqIYAh+gM1KAzDKUDJUTZGqjtauoAtwAwKiwQ0A4Q+8QEbAABLfICE4wggEIRNKD2isQD+AoA6WYAyaJhMughOQxA6NxA8WgmMGIcBMsAoIoRCIgRIr8WVwrx1ybxZViBPLgRs2cRtwAQjAAAuy4ArSoMnU4Av/qYCKjuAMVgG/MFEgsoEXctFABo4X8uAF8oDgzOBAlkDi6CAJ9uP9jpEOREEOyqAzM5MIXiFB6qDAWGAixkAZIqEBbCAFrmEA8kGsMEAGeIEOyABywKStoAAH8iEC4sEWriAoFM9WVFAKpmZZVEEDTYQyoGCUHqMEvQAutkBysoDlpsAJxmAK1MB4MsEglTMLIjAtjkAKRmkwmId5rmATmGEXaGAeooEY5CsIVyu2uMc3kLDt8DMkj2kkmwM58AEf9kElB1Rd9iEDsvAPuGANJMwLdoAKoODysAATAkkz6EIoEg8LFGUNwoQJmGAYaIAio2EYWoELyNMKxuANOKES/6KSDMDAE0TiB94gCCzz3JYgRf0gE0pGaGrkQnCtI8bNE/zgEgJoCoq0ynJABwwBEdJSLSuR2lbELTNRLjsxF3LhCzpBl5CADGlhcmiBEtLAFJjBSmQiHcrBDlLsEcxgqcxAEiSTFwauqZiqI4ygBcrICNBoxmjsYeRADozAQMpgpeLjTsWxCJyCju6EHq5hyJzCBmTADBrhB9iAEpCADEIsHIKBEmhhLbzAk6RIzhbpdx4ShISCOuNqIY0hzXTpCOyCD3xgnLJAB8BAVMBBNGIlA5UHII8AC1YFHAYLifzgD5jAErYgWP8AE3CBJInQP0symfQz7ZRV7oawmZJhHv8mIB8IFFvFYx9YUh3fapGkAArUAK4oYUKdk5RIhNYSJRjC7K3kIhWOgRhAVAIO4BmGgRDIAge84A8AYQrGAIcUYQwEahFQc0L0sA0ygRMC6CHGwF/JcgeWYAgiigsSdGK5AAkCwQqsYBDIQgfABgeozAcIgRVsgRia1EktMUpnQhM3URxCIfZCQxX44AxC4QIG4B6qpNpmQhy0oRT+TcXG76h+sRSWABbCgRnsJB6+ocQcgU8jZqroQOHqAN2UIRxOYAJOIByKqgxsoGaVwQpM4Q7sSHAy4AKygwRq0wjy4AKugRnO4AP7AAe2oBIghUvtAijCzLBoJzMWKzLmMSj/nNNWjA8JYBUJwAwchAJzsgBwY2XpbGUnPqk1ROMItuASMGEN1kAYPCHunCnSGO0+lylaEU3RaEsBGiBbT/c7MkAB8IENuGByDA9zhmUNcsAuHnInHhALXNAPDK95VOEKmMAWnuEBJIAiaQA+ocENQuFOv4L6vpAFhoDKdCAl2EJ0PkUKqCAHpoAK6sAOGGFJh6EYiIEZniEaouEe2pMGaGAXjNZekZQKpiAQJIWe4mMOJPEdqsRJL1EmVpYbtuETOiELegEdmWEIYqEYLBGF0EEc7EAyJeEF2hRODW4JvgEAKrgfKhgAvmEIFMFiHKEIHMEkQHgH6g6DMbgYdkAP/2wgAkZhB1igCHRADkahjgBHT/ThT/aBBHzTFMjAFSLFJ/igEjBnM/AxVuCKcmLldghLFah3LnpqKHQAXN+KWFyplG6H6irlS+5WimrhcWMFuVQBuYoIWaGQJDmStY5jXIhJWqWVdE0Xdd94O/aOGVKFD16jp8JTrhRvHj3ndjk1lJRzWchGro4gDdJAE4aBPR8AAohXAiaABt6GGZihGKChEPRGGWBBGQpBk6GBk4shkp/hGdShPfFhAqiLePuBkamLtA4AHw6glIlXAdQBGt5XEAJmEA5B1IogByyBFpggDVRhE9yAZNcSZRFTLsUBDxBAHRiBTs0IFp4hZxWYFP9i4PsKjvz0o+5CgBkARwkqeBQW4RB46E7HyBGGABYqmAbuQBmgAR8qWBmMICvyQQ9ioAgOoQ7yYwjmgKv8Bk+uAQEIBxsDQAX0gAXiABQwAQ3MRi6YZYhvB3h0SSiwQNSQQLPEph13IM4EKZe4VFiKJzzXDEycwFUsFFLEzDvRaZ04wRU2V9GMyT5TCzi4gRmUYVtEdwip1VrhWKcDIB/wQQRY4Qp2KQeubAtyyQ/i8RthA1KOwDHEbKOhQM58Tkwu4RJSIRVmoRVYwZPN9wFeWQJQ+ZQZWay/GqwpUgFIS33VwWiZgRgmuRAQQRk+4ROQBA6UFBqYAUT7IRoGYRD/TMZk4msIDoF+MKGo+QAHbuAKrgALVmGYnxQu0aEYisBOzeAFWiAS8gsdtIGa86AUksrFioCCJ6AIKnsUKrgYAKoY2nMXvqEROmIXAGACPFjUIEEE4mUfLoApnmIUDkEU8mNOX8AIQkEZuupOAmcA9AgDAmAfaJMUQgEXhAELDIUPKIFSMEd3SETPEo8VV+U4Q+dLeurwwIl4fvUeLUUusE45p3gyeOXU1gCdPGGlXXpcnBAK4W456K4f8IGm17haqKMeaGCnd/oa+mEeCtk6wUQwWsMy3Ktr9ixWICPowmZyLmEocsdykYsLUiEQdqAKQgEROvlon8FORBxpI7mt/90aEQhhDuTAYaFXBzrWMqxABwCBDc4togIBMKygEKKhGHAIDEDBSNbJEcZgfwhbKOxscijBGJhgErCAJfA3Z89BHFq7jNSUBV4AFlakRaR8MUvBgV1ATutOBPiDBUghBAAgslmyhGmgCBrhzL+BBaTWxBohFGQgANgGT9QBFqggDpAP/caoCE6KysaAEaBhhoMsj/5kj/SgFOTAFdggiNHgDGJFDWxFeSyjc9ZKCu6YuYAiHlEJj381KLYANowhE1ZJiTzpV1YFE4wLE8AYvpuppW0aOWYdtsYFYQ4AGtSYI4cJFiQgHAJ8p8MhXnCACzpQktZ1MUaJE6SgEpjTdv/SYrDYTIopY5csg8IxgRZWjQqwVwcYomNzAGeqbNyP9FOyt0jxhw0K8STGzUg8gUP8gBPGgEM6ZGOnADDoaRACIRD4mhPsp0Utt6iVxbC2wDHKkxKygAmuhkqgOYHFwQWAexkfIQb0oBiiGRJiABcLrhSMILTpBhIoDQBgoREqmBms4JwBIG8qGBbulBRGwRQYgREQgB5kWRmKgR4a4RXioAwUjkjK4BWIgIMXYRHyxwqKwAX22Y7yJNGLjARsoBdIYQ5AQRiggAuugESMgRoQTy6sE1yBojK8IMLiEXCRKALrGAlm5wGhoJPczLOSBfPSCRPg21p6fb6hdY03kpj/mGGRn4GmeT2ZKI0dhF2nG+AA+AEWcOBTskBUimdEFNeT6AIZlLPqdGVqZsoF9bEshOK52M2+wmAKjC9jjY/K3Chix82h4EARwmAQOCEIFsE9fiBnHMI9MCgMVCEqEaJIA4EKDgEXPsENlIEQ3IAQDAEUOOHop8CIGKNSnAAN14AWAqnJgMIYUAEJGoAdXgKF0sEOlqAMoooOWmAGTEHLW4QbamAGCI4FSLiEAWAEjKARMDkUGIGER0EZ3HlJIqGEue+EAWLMjiUsipSRQyTJoyRy4iSKIyZOnEVwEhE0soPKmDnK7nyLt27dN3YIIuTDQOKCjRku5uD6pEoWmjRX/7BggYIESpYsUI4cwTllZx8/WfoYVUVLmBQ+tPwc8VJUlapVqvoEA1cLU1NOnDxZS9bNWrexYsuSPRu2rNiza62ZbUs2WTJoNCRMYAbrq1y5opQpOJAvgODBhAsbPow4seLFjBsPHtBvxBcma6zSMkorWLAta4IhC6YqWK0+tC7z4aNqyxE1fvpc4uNlR44+qvzYVsUJSRs2YH6MqRLmjRgyQMgUOQQHyJsqQKqMEfOGjxwycKq/OSRmDBnoYdr4qcQV0CFDiGDdKYY+fbHz0JTZgQPIyxo/l5EgMYrES6UjUrYMlUoJKCkMsI4t8LxjzjnoiENHHHC44kgMM8AADf887ZhjjjgyxGCGGSx8AwAAIvDDzwThmGKEEbDsEmKLykASIjRDGLGHCEqIAEA4GCmyCBlGLGFEHaKI8koZDr3ikEQNgZIIC0vUoQgcohQxRJVkzBHJHcysE0889LAzgD4XjImBDfnEAokQb3DCRxZHOHEGH1tAAYUX9sm5hR9bZDGnVdRIocVQtmXhhBdQ0BYaNdSsYkwmXnQiSCJquUWpWWzBFVammWJq6VrdgMXMAQBIcIA6zBQDDTTFHMAPO469CmusssIK4i5qpOEHNbU4QRs1xkCxhTHGBEOLVKEV60dtffi0J22xIcFHa7QF4yhXbfhgxRRttFHFD9kpUkX/FVHCIcYhZJDxRiVxlLHcG9dVMUQRY8AhiCeYtMYJInfsu956+/7Lr7+FfPGHGrRsoUMOWWDBRRrF5qSFVFucYUgE7DCjAynEWJggOrnAAUocLLzQQgvKcGwOOrzEkEcLxQAwQS+l2PGIEUUYYUo/AOxyxx46j7IEjvHE+8ISRaiTYxGLHCLKjEGWMWQZ6B7ypESiCOmKHCwYUYYYidSRRNTnxjuEFaHoS09IIdEzQAYpNHABBmU244IjVaxRyRlXEOpEUUW1poVqWQRDzWytSbtFDkhkgdRVvtKCiReBXOeWK5W6pSnmcIGl6aZodYqWXNwUE84BEEjQTz8SjKpA/zwNzAp77LI7Fg4Az1xxiTGqcKEFLbrm4IVU1NDS91Cl2eYHFl7kcEQW9LGxgxd89LFFxMPfm0kmnExBRRucLDcFHMsdAsZycDiHbiZiiNFG+ePXUQQQQAjCCSaVsKLqv9fcAcs117ATiQBmyTzsOE8xbOEHNWRhB1NAAy568QVaGOMI/qFFLaRwBlzogx3F0AELWgCLA3VsG4KAQymc9AgWQIIYCEpZLGbwAhBNQA8FecQjHGGEl53ACDU4RYhgYYR4hKgYO2ABC14GgGgMYQyJmMMoRhEJ8+wLFrBABCNK4YgyLEIUYUsEER5BBFFEpAyOQEgd5LCIMhQhi2SwQv8RXvACFyhDVVwCyTpIkoIx7SMANojFHMIwsTg5rypaIAoFPXMEpCCvD17AwZv8pChqrUEKOZCCFAIRiEFoMpOHOITm2DKWbkCDG5zy3Fs+l6m9WGOOxWCGK5lxDwDUYwCzq6UtZ5cPDfSDGamohSqc4AdkNAUJiayNMSphvFogoza22QIxaUOL5W2hKoM83r38gAkkgIETmfBE+aYgiDCITwyKeEMYxjAEOLThDet8gyDWGYQqgWsKbMiELdCzL//twRQIKAABFnANPJwCAQDdQyT2p6phbEINxOxEAzaQgUzIqQ+9q4UOroALixXDCjYbQhw2diEF4SJFdChDzR7/MYwWnuOF9BBRLJZgwxsa4Q4hukMklPDDHdDhBCFSgjp4GoIc0UGKBaQHwNjBQaNGYhRzSOMi4kAHR5zRIXIw0iuSYDUi1OERRRBFGZbQgiJMYUZUWAQjYNFKtdHjGvrIx5gu4AxShCITZzgDFpwQLT/kpFiKRIoqsoAEKUBlWtQAhzHW4IUj9KAQw3hGNKKhjl1EgxnQIEQnv+IptyRDGRC4RzIshUrOZS6UnfvUXk7LDAMYgB63bK1rHaOPB/SDGLO4BGmK1Ye++UoKU1hconyprCxoYSf0aQ0fiCWo0NRGFWtAAhu8kIladMKbUwhDcIRTBXPGgTqgCIMn3NUd/3IBYV5wmAIuVLWeSHyAAx5AAC8KkAACJGABMSiAfRewBzzsYX/7UmganMCJASAgA5GoBB/qWglaIOEKbYjAAIpBhSGQ4Qc7GAIiOKYgbgThEHGowyscYQZoqNQMLeXHSx9RihvebB4tCscEAEDEIkCCRS3aRSR2BrA7+G/HBdzxHQrIP0aQYYtJcIQuxCiKBr1CDhFRkhHMYIQvGoFeKRwOGQQyo1iY4g5pW8eXMgC3CzQAD6FYwxUoURsKJosWq0AULXiiEz8UMjPUQIYTABGNB5xOAhCAwATwsecJRKMYuVAEWDD3KWhI4BmH9lxawtIeUJrS0aTthiig8eJvvP9205wuTAYm0I9nTGISfnWeH0CjgyEEJVHGSJa0WgPM0hSLWMM1nipOg5MjrCF7XGFDGKYA3vK6SxFiEF8VDuGuN3gCDsjO7hvgkD9YjAIEHLhABz4QA2csYNsF2EcC/LkABOjDARSggHnQ04W6umHACGBELkIBh5eEAQlh0McA2OGc54jBCjvAxcYwdA4ShgGN63KELVQq0FPkwQwpJoUjHMGCUnhkDzPAgzKUsQiuFiESxfgGECFBD3/p+Bp72MMp9pCABJAcAQaNRAHXA4s5QI0IjkhCIuQgJDkQiQ6LiMgjgOSIItTBFXBYwhLicLU6lKEMRihZC0gGCY+IhG3/+nhoAyDxBTQokDVypg0favHmoQiqV9RIDRPyPFlbEGMCCmCGIQrxjLqQihkYF+1cJMCMz4bylN0oxi45B5dTBl5TmxUVazuN+Ndm4AD9iIYmrlAbXikyBzuAlh+GVZplutoJh7qNsZCXrFvbZyeFVEXkOMEG3lg3OGEQQxnGUC/XVwcOnKhCsceQLUTA3BS80IcHOuCBD8xgHw1IQAYEsA99bHsBeKBABCJggWuMwjzQeIIazsCICGQgAm97aD7cKoAM3NsQVPhBFYD9BipQwUDtOMc5qtGJ64pvCiIGuBk4tIQU37ARQDKChGLQAkoXVTWzNfFiCj0WCXuAAB1w/229oALx9U8FoAL6gAAk5z8/dgeMUAd1YASPIAai4AgNQnMhGBEbyEVh4woWYQQPYTNYVQdO5yRFsAM74AhbtjYDkEdjFgZMoAa6o2akEQxQQBTVYxuZsUx98AddkAlMkAaWMAnDoGfMkANMwAlwJ1utowxy8RWKlneoFEqbpQD48CmgI3ioJBabNQH8gACJx4a3lAKxhA/DMAlpkCeZoWDRUwlFKBWg8R9VMT15IhW38WpZwD0+URla4CiYUAuc0D7h5C5kMARi4AmcEATUYQWHoD7PcWW5MEUDQAEZ8AIgEHy9UAMBQHzX9jr6kAC/8ALl5gAWMACQAAvQcAeIwP8DW9ALG9QlIfENX8IO/oMAA6AMOaAI4HQtGSEFrIBh59AKbPBOkygFtnAh5lANN5QEjUAHjaCNpZBiSwBHS9AIujCAKZJFo6BjCXgKv9ABG7ABHqAPNZByBbAAGQCP8jgAJ3dQ+zIKRQBTEvEIRvJwXYQQdKBzEpEIidCBb4CQUZYIjvACecACj0AGVdAGY3AzMpgLxSASAxABcEMKl6cKi+MHTpAZSKAGPSEonoEaI1kVa6AKmQAMk+AG+CABNMAKlgAImLANxYAPANB2WbhZEKAOohV4ZxgOEgANjZZZjoYpmzIX/UADpdiGUxk7+/ANBiAB0cAKPEBqUhEbbFD/CYTyH0fIB8I0FD0hJ1BQCcvVBzoBBTowBZXEK4giFbnBFWCAl2r0Br4mBlUAB8/hB6IwBkBQBodQCPzjAORmAawYC82QAhFQAw2wjtcmD/tQAzMQAw1gARZAAbGgZbMIC0jwCBtwDepADKf5Ss+wi20DC1JABdrCBj9QBFbACZnBQgnCjGDgPZ1wDMYwDNNYDT+QbIlQHQ9HB3TwCC2QYkbWCI6QYnXACPuSgAPQCzHwAQzoASmwD/+0bSggXwIgj73QCwgQQAVkChH5gUbQEAtRB0TwYe1ZBgSJdA3pCCmYIg6RQkbgCEEgBqCQCYtgNEOwCFQgBWygDPTwDQMA/zf6sA1nkAVtAgXGQA338RO1kQXHgwyjEQzCcAR+sGu1wAWecJQQ8AzVgAmDMATLsCowwwzcIAr3YA+w0GibQil3N5SgxHeDZynJ8AwA4CpUCaSzggCxpADPsAlXcGZeoANQwAdOUDyqQA2gAaVS6qSwkQOYEIhYgATzQUyWREF0aXpesIi6yQlAUATeJU7iVAZVoArTkR09QIuj4HzQhwCYmZgW4ACSyQHAJwOXiZkz4JgtYAa8QA7Tl4EX4ADqMAxXkAqpcAY8cAaUkAM3sAkCdg2CoBFh4GtDQAW1cFiBkFK42QryQR/UkAntAJzZ0wna0waHUAUHGQc35Ag2FP+CjXBDWRIJCEBuFNAMzvABHkCPguEMGeAMAZAPCzBfC+AMijkAWXIHdlAHUOUIiSAGj7CBRrA1UiVVE0EEifAKRJCCD3eQNxM2DuGfdFAQSiMI70QliNCLbqMPjIAGlTAnhCMFPME4tcFXUYoF1CAMSIBNmIAJjoIDw0CTD/AMbmAcO2ANK3oAp7JoRJkWiJYM82AAWYhoOYqje/cpsGAA6hCkIRsr+8AOPVmktjCHOOAfexIotaAoUaooTcEFUAA5OuAEw2Kh9NEHWAAGOoAERIgUq4AJu+YFYFBPYlAEytYGwQEHZBAGlVCJrWoIdzAKpIAAm2kBpzADv0AB+fD/ir0giu3lC/pQnS+giqv4ArxACuMwR2elAc/AA4yTQKSBDIpTCXvgYK2ZEc9lBWCgFZhwBEjAfglSDf4BpU7QBe0ADhhSdsOSCannCW0QTtURBElgBFglB0YQdddwCnOKpzXgC70aGDUQAcCXAgGgfAugB4l5AeVmCgPEc3QwdLGaBBwIZbO6EEoSB3LAZFAVBA9RB44gCgbJTqLwI0UQBO7UBo6wRFQADWyzAREgr2kABYqCA1LRh8zkGaGRCZa3BliaTZbQCszwAHx2D6eCCNYwOvgQhvUwATJKo5QyFsngd/EAeJU2WjqaSvMwAYEhsv/bGDZADzQwKvNADIbA/wSpkAa21TsvqxmZEZJIoApacASkMcFS0RROoAaBUHlg2moREzmpNwU7EAZc8QZA0JeCUAlehXqEIG0u0AIZQAGnICHOUG6b2QwxQLa9oJi/0AvxNV9moA8gIIyw4AZYwArBwAS2cQmZARo/IQuIkAHi9wac6gUj/FzZswb+wQrvEFKF2wfIsBOsMI3tEDHB4KHJggmpZ8I/UAfUgbSjQHIusKufWwOJ6Xz50AHW5gECEAA1gAJ3TAGvSAF5cAr7o4Fg8xA193MRabmPIAoPIQffGgdS9YFLlwRdcwiJsMkKWQQFUQfQsbxGQAaHIAWEQBIPhQdrcAbWC6XGAssPDP8590F6kPO9XJAK1EAM86AAEsBnE7AL4fAMBzABD6AAu6AMiqAIlVMpn8IN7ssNeqc5GruUa5EM0SAB+gDA28wY+1CaENAPIhANxLAJo0ZYxILGfsAHgdUHfMAaozFNYLfOUhBYR8AUgQhcL4kJYDAFYKCpq/oGVwYdbRoEnsAGnwALkFAKvxALLxADElIDGbCZGZBtMXDHeVpuvwDEC3AKHcBeA4AIm6AKaxkockZnb8Y4wdA2CGAKOiAFaxCX8nEvSGEMauAGXky4WrBMEkTGGFIN9ux5qlAnFuQFbABskmsFyhAJeJAHekABnAkDNZACFNAAT50CHn1tfrxH+yD/A0+9mDPQC3vQM/r5Cg9BkPl5Q2AUEVUVvJn7CImgEGCzCK5QQtR6kF9lBkUgykTAAuxyCFNABqfQAIPdAKEgoVJwasuEGqVhDMhQZ0QBLMS1CnrCGVB6DMewCbDADNEwD8YMAQqgAH7GZ7tQDLjwBpbTzMnADHg3o6VUhmaRDOrAD6fLzbWtGPrwDXXhkzSwqMFELL7zSxR1KP9hDIHSFM/DQHIZ1HxlG2CQA2DQCbWQCargCT9QiW+ACefTHV8wCpBwXwng0L3qC74QA80gjwkQAzJMyHmQrHtAbRtwbSF9PIICTISDxkVRCYjgAPqQAT1gH/nRXFDQONTQBzjQ/wU4fQ7gMDyKggTgcCHgsAlpYBTS4gRb4CtbsAVeoOGc4AWIEAmkUAoNrQeSMAM1oA+D/NS+ELb6MAM2UAMvXgPO8AskPgMy8AsBBGUESQdl7QgwVQTW2hCvoHQ1Iwo105CXG62gEClPhRxh8IJLUAb0sgikPHtWUAoXkA+DvQGnQAhpQAkSdBn0UQuEMzxaACw6caHYMMHU4zvD8wcN04S1wAq2wFjDYAvDsAvgLGiElgyiwDlo+ADR7IVliBbOXA/4sEe2reiJkQHfoA6kogVpgAzIoAaEcwlOOhTCRR9t1jsTXAnQowM68ANHkIe20YetwVsuGV3aIwhW8APe0/8GnhBOgsAIppAA+SCPsaAHIHBtKlCP/1SnXo0HMeALBaBwv+AByX4HpiAOw8Mr09OhxlAsW0cJsoC3EXAKPtE8WCAFiVQawVAJjvQEi5sg4BAM0q4DPVDGw7UTTkBMg0MLgQssFNUHbvAiKZcAevDQEuLVCHCZzqAHKUBuDlADuJ5yfXqZKmBQemAEsntDcoCcSaALNbfjs4pDjyAHOEQHP1JzYkB0SBcRYiAIL1gE1NF6ZEAE5VIdcfB9KTDFz2cK25BgRLFmmqEoI9kTwOSyzGQV4AAOQi3gl4AJWwAMs8AEljALwMAFSAB3ddEPCqAOxaAM1rAIfc4Mf4e/o6X/9WUxFwAQD4sO9oqxDrbDAxLEBWMOBRVOUSWNFEaxCqChPMT0A0OQSOy+J7ilBfSsBW0gBdljXevaHSZsXWJg6ym3ALHw3h4AAjMAX+eNmb3AC5KQAiAAAzDUAnhAnoxQDglODTiQBWqABTVxa5gQBduCC6AQCqeAB7+gDdoAFuOgDd6QDbNfDtkgF7qQDegAcNXQCq0ABDuQCeeAIayQBozDB4GrBcZQC4HbN+qsBppACP0E7C+gB7/gDDGQAr1g3ixeA873iilgXxE4Aw2QAtUfCS4QJKUAZXVAB0lAB2IgBzakEEDy8DWDQ1Cuc+RSBvy5gQABpIiZMnDCvEm0/6hMnSBi4Lz5lS8DvW/X9GWIYKrVmTR8aKlCFowaOFpZoCDZgkXVSlV+VFGDmenIlo9b1tQKhgmTKlptpEjBMagQMw0GAEig8Qxat2XzIMBK1k3qVKpTrUm9OlVUuH4DAnwFG1bsWLJlzZ5Fm1bt2rH58EGwNUkVEi9+uDjp4ydLXj8utfShFSwYLT9YsuTY4UVVny1OjkBZ2aeSlzVbxgzJxGnIlE6ewggSFMagoCp2Ii1YEGkJiA4cPHyY0Qz1AhUzbMdg3eFDL0l5liTphqidOXPgprBpAyqUtWzavI2DXq6cuHTp0F3Hnl379XPEzaETJ91btnTEwWlSFUxVlv8jKlVByUtzMRZCCQoUWJDADGsP+mI0s28BBGagwAEHLDilgAAHAME1EPDYoxQjHCnFDDNKeYWOVxKJo8IkHlniNyJEocORIpYwQg4x6IhDlDpEgcORJRwpo4g6DoHjoTeAqIMMReAQBJINMogklDvoGUCfBvQxxQtKPgKJGvX8WAOJLFzKQovF/KAFpmC08MMYP7bYQqQ1FMvkp0DCmIIKKoYwpRh1JpBAggmeCceAe17hMxk//awKq6qS4WaCA2xgK1FFF2W0UbKu6YcGLa5AAgksoNhipb786OPKlmgJKZgtHnMCCj+o6UsLKZD4aws/tFBjiiGG4ISNHQ5BThD/T6wYo4oq3nijCjkggaSUFmL54LVefEnhl19gsM22ZjrYwLVTLCzFkVw6seUdc84R57npqkPnnO7aQfedd+Bhl1133oU33nbhURfd4cw1l7hz0pkOF084UYUPLJjgY7GWtmAiilFmg+SXDjDQrQUFBUhggRgysCBjGJyZzeFqPdjDlEckLGUJMx6Rgw6V43jk5EeMeERlOeJIRA6YRWmxxSQWSaQMFooIogwixkjkV1CqiINmVwKZI58UUjBiCEbYQTKCpxlRpRJVjFFviyy2gAKv9bRsqQ+RggmJsC0wMdOLWjChQoo22njjITJYWGKIUKCJhwYIDFBAAXWKgUUb/110WSZxQKOiKqpkdGEGgG8cpbxyyy1fB4BorjgCiUr6cMIllxbzdHRaaOkDdJ66rOUjKXTYgg9P1XhzBza8sOK4NgThJIgirADCoEFAoSPEUiRp5pdenOEvBdt88WUGX3LzwJQLH5mjlUuOAEff4tpRFx5453l3F3eeIeaZeL6hp5hhbLGlEPhtGeYdYtJPP1542X1nuG/RSUd4xCEOQ4AiE6vjwxFmIghGRMIOS/iFB1zzgTz0Ij/5acEM9IEAXkjiBbw4RS/08AEOdMADo5hDHUwGM0ewSA6NQNkjXoCiRzRCQzNLRCKSUAeViUIOoiCCHBIBhzoYoQ5lQGIVHv8CCjiIYRG/gkMp8pEPEuzDBngYwijoQQ8E6CMCp8hFJYzxHinghT1r0FRfLoGFWkgJJoTBRJhqsYY1GKMTOwBDcuAwxEUUoQhlGAMVdmAEU9AjHPUwigEOoA5m3GEUyzgc4hSXOD7pghEHeEADLrdJTnayLDa4BwCekYZJECZ1LqkJfHjSEtFdiRrIyEKXgmGMLeRAClvow0r4gAQiDMGWx0EOGDhRhT/C4RBvmNsb6PAIGc4gBjHQRwcuoBtJSA96L+jFBz4QiZKhrBWq4AQXcFGed7iDfO+63zBYsYlN0OIYOADEEXawBwc0AAO8mMIVcoCDGzCBB9jAxhF4cIb/SeQyC1HYBCuGcT959W841wngALehHsdcCQ6vcATMEPCaU7QgD6dYwDV+8cxn9gIEH9BHC17w0ddEghF7NFYpakiHZJRoCcx8wQvMcFMb+vAVKavDDulAojKIIhFviIMRikAEPzqkCskhAxGOWgoSNOAa7EDANb6xCDIUIRLfwKrVTrENVCAhB3vJghPKZIxO7cUls0RbX3IZDCrV0RM6QEIb9phDOTziiGUQwyEcwQIW6A0WxQjHW/rRDwUskhnFgIYyRsEIZcACGt8IxwEMsA5Pdtazm2wADQCwi7hcwa0sgQJ8ArOYvLDWD3zIS2BqwQcw4DULLQmYFIawAx9M/wEJbAADGDxBmioIwiBt8MweOzSDF7QgBR0wIQLy8ExfrDQPNWSmI0DRCVl0wgtHyAY6ylmIVWjCD8BARSosgYkz0aIWUsjM6+7AjgxcIB+xSIIOjsCFLIzkErlkyRFukAVKnOEKlMgLK2yRv3fR614AHCAuwJAFPmAiEXVwBDNNgQdegMDDHTRpBvIgiQa5hsMy1QYk7MC7MBSRDh9qxAtBJNPptmCnjtCFyhrhiDq8AmZlYFESypDDOPixDEYwwiIUIQZQCIIIQz5EPiJAj3gUYx0aKAZgaahV+qYgA6OYghTOALYwSYk9XsALl0LlB5wMpq7GYIMOwBCGHBIZyP9laGEiRIG3MpBhCEguginusA51vEUC/ZAA4CAAgUMDAAAGgMAE8KGObwzAGZ/FdKbV0gCuSCAarODBFQCsCrDNlZVc0oKW/BA6ltBiDXgtmB9mCYUpjEEHtvaCFzjBCU+AQRB0+/Wuc+QJUJiBBWZwAQheg4cWVDMGecjDyRwRBzh4ohK1yEQgOPGJcpjDHdC4wRow0UZsZwLbnMhEJtjAiU5wQgo5gAVY63sBGbigCluQBTjYCiVjYMJKo0OGKrAADFkAAxh96AIriLE/b50DwuL4hBa2oMwMj6zEHjgFL1rjgQxonAOuiRApGpGEOPyLDcQ17hFJRIcd06EUz77/kCMakePs0sEIvzGqIlS0CN/ZSKljcEjdFuEQKpiC0IwoAhkOMYQyLAJvRTACJO5Q6QxkIAWP0EIaICMlvRyhjF97iWBANctMoDETxvDEGKQQLEXU2Yk+K0IcnN4CR5BhsEgmLAti0AJTHJYZz1CHOsLBjMd+Yx3MKDQ/HD2BcNyhFzXQdOQl/5UB0ADR0bAFKdUg17eKCSSo45JdtJS6ltjSrVxThSzAIAUq6OB26K5VG8Ywhodwwg90awMmnkwGDc1hDhV6xN5L5ohEgALd5s7EIcbghWqYo5xR8EO6kW9ucKK73W1AdxumkINQWJmLKbjABZxhh08YQxa0qIRH/2hRRr4ggxqqqMVKjHE6VaRie8oYxjPcQS/iQHgbnUmExCGHi+siadKNR8gAD3gNbYCkDeGESsgM7WODSuCEOAiCOqszOWiBFjg2G8qxilsmFkgROHCFC7MRqFM5MSg+UFCEMcCRXKgsaIgEWKBBZWCEUGgBIyiDV6gDONmia4iAC0gBQ0CFLtm3TtCBLAC70xEMmPADHGADVcgEaugEKfAEV9irIUoEMZADY6sDIsgwIniFIshBI+BAFmiuFliCIriVMdiBHZgVHTiR3YoD/HsGfDCKCYiHSJiByfNDTLuGe0A00uICHriEvpAtnhgMT+kD/loJMDEGL7AlJlCD0//pEkk8Aic4k3HLNlshgyngHV4LA+wDAoc4uTe4hSpgskcogzAgg+IKA09It10DgjLABGGAh3oghj84nVoALjYARjb4F3bjBGHKBM+4jB3IhWKIh3XAqgYIv14wBVzoBDVAAz6QgjX4Gi55vy0RDDdTAyTYARxIA4VqsAcLoG3AhVAQBVNQwOohBQ8woQ9gpj3Yg0dahjgQhE7RFE6YginMBCoQg88ADU/oBDhYghxkgZhhuQ8xghcLETpYwUdgARMxoj0KLEF4gzEIBVi4g2JghzsQyZAshva5g2uYLBdxBEdogT1gH6vLCO5iKzuSgug7gr6IktbBBB3gBFrIBGT/4IQ2+ARXKEEgEKJE0LklEME6kINF4CFRQJHByoM0/JmiEgOi7Jmmg7qFIAMrCIS1QwRmOICjmIdrgLw/RMtOGoB7qJN7GIY0uAI1gD/C4IlV4wu1Sg+9QBUkwIGE2oRjgAk1+AR24gIvwARauARCcANG2IEpeANPSCZdEYMf2R1caYMwYIPiygQyEANk8gRMyIR22wEy6IR1C4YpQDdagDO5+cVaQbde27U2AAMyUCodkJtCMDxnRIAUgMb7mgNQoIIsSIPQkZKWwAspEYzCOAkukAVjoARgMMf9u5fqSIdcgIMGQjpC+oABmAOlEjJRCIXPFBhP4QMvQIIu8Udh/xJGdKPARViEOKiDF8moJEiCRsCZwaoDJhMEGTqipFMiUaiCQVAERvjIYhBJkbyGk0zQA6WHO4CFUeAxNCwFWKCHa6i6FGAE5zwYaqgFJMgl1aQGruGEHOCEWuAEYfiCQcCFEpSDMggCnBkqM1yCFyEDJFKEImCBGlGpnKpIR0gC/dQhXSCCRygCFyEDOKCbQ3hDQRiGaDAKdUiBtJTSTUIAdaiTXXjLK/gIVFLCvACw1UKCNdALJlACAOgHVgAGVbiCaNAcJlAMVdCECXC0QggE0PgMYGkDRTgE4YKDMQCNNzgIT/CDKnjFgozNIbAC5EA3KsC+TojA4DJGT2CDdv/zBM8IjSFYwyF4AzYIhByAA9xch3VAkhTYAPuagVhIhDYIE1rgAzLhktMxBrZyDCSQtYCbhM/ZhIVzh3foHgBKh22ATLZzhSrYAUV4A97xg/TDgjNQAzUQGMPIgvnTgiPYiShYt7NThS+YG1UUAySTg5lRweJBmRwyghdYqiQggh9xoirwSAO9hmuwxw84KXvcg3dlhwRt0EgwBRY4Q0igmgF4mlP4BEooGA49gjXIBJdIG0lEWE5oBUGYgjBwhTjYofhcCCJQIR0kgjlgBEaABWXYA8kyhV4wgxf4mQlJBEFwBTmgT1HIqKCRA1DwhDcQgzFQhEBAgiMohF0AAHz/OIUpBdrKQYBwuNJhmIVZwEmJ2wstca9ZkkQoUINZKNPRmoUs4AE23Zw+qIVJGAZHE6U/wISgbINBkIJA0AEqEAQ2EA1BmDPQwL43ACxfCQNee4MhKC7k4oQ3AAPkQC7InBt2Gy6DhMyIFQQruLkiEIRMCINDOIQpuDVlsLJQxaoL2AcSyIdegIRE8IQ1SLPToRKvO4IcAIyPMJtauAHTYgXx8Zb/+VX1yARh6IQpEMpPwIXazQVX4IZQyAZuyAZrYA5uuIpsEF7oyIZxKN7jhY5x0IbkZV7lbV5yGAdv2IZcOARFUARlQNBI2AM8MCkO2ACO64VTCBB4tdc7iARe/2ABCWEBLQorfRiFrJm/NThYnWid25MCYhyuKShBDCuDJCgCOrDYMvDYBDXQIwnJ+TpJfX0gI4ADUIBPHluRGSECMRCEo+nTzkQus4qcCdiDoPXgRskAopUA0roCuSAMJdyUVWIPKMiCSZgHrx2GK7hazUmDS6CFNOBZxZsATghbSZUCRBgGaCAEuQEWOx1FgxSEIXjFRXhM2aO9X7PUX3uDKRAuTvi1TugE2ZybUxSEdguCnxGESmiDX9ErMoDDHWAEaGCGZmSHJLmAKrKBXmgEzc0EWfACHaDiO14DKOkDY3gvPqAELEiDJrCF7tEXAOKG5JWORR6gAaLOR6ZOAP/SDnyh5Eq25Es2l3coBJvNBZGMhAFwAAoYENZwkBm4D4pZgF84hUh415OEhCWokBdwgWuoUIwQ2Fj1gmzMtUygBX/TNSz2DBUsMkcYkYp1BAI90ANWUHZg5vlq5kiAhDh4AxnxUZopooJoA6c8ViVatzYYgiIoBgA4gF/44HJmi30IgCrtBwOIhk2YBGSgBSdgtb5Qwr1AmCt44TJVAiZYU82RC+iUAAAgBsWDBkBwTTb12mcIBMa90zfYtUoQBCN4xSO9TDGogjudm4yutWOVzV2r1N2B4kpN1ir4mTBoN1A41tA4hBrtSitIhFEYtPXBKvDDAMvtBTuIgyGgAiT/eB3FGB0OxQFM8ARc4IaoEIfu8I5vyRd7sRd1CZ95aZf905+ppuqqpup6gIVb8eQ9oIAIOBAVSIF3jFL8qJgByYAEuAZWZuZ9hbYXmAE8YGaMyABlgLMc2IJdrgUvYIMoqJVKdYVF8KGgwplXGIJGiIRiAEkFfVd7HICNS+sBQGB2gAVGKNelEgUxoFhRQKYigwNMeANFEASh/rMdiBx8UAFzRu21GAANAIAJYAZkAAYn4ILW6VIt6FInIFMAYAaeJYYZjoZJuARgIIbW5gLRCocjwNlncLRoQGhiuLUwCAMlwoRKqIQ3ABoyCAI4AIOjOQToBhJeC40qGIIf4GhK/63UXcOEdkM3PxCEIhiC7hbFlBYWrlSyIjBXITjsdWDj3cyHC8CAAJgBF3iET5xUYWiFbciGZYDecUGH73nqqjYfeHkG9HkGeCAGdrmfdBqGDa8f+/FwDoeGEKcfaBhxWwjxOwjxFIcF9zaSazCFX+jqA9GHGviFDHAGdK6BikkAFCCABOjxBNiDSAjJX2Aut56BPIiECkUABNiDMIBCugjNWhHGSiU2OWAIUUDXhWiEA01re2wQD0CA7t0A3fgFBNAmkjRfM7BsaoMDMqA2QhUDP3iDIBCmTniDN9wB5Q6Hs0ztPi+LfaAHOZ0HWzAtaigJV3kP+BAw3mYFABABJv9Q7s3JAqnVHB6IHBHwBCkQBBHQ7RvIAZ6NBh0ggzc4mjBIN0yQPaWqgyo4pmB5iENog8z4laWrAsfEvqA8b0cVWy8ABC+otd0KhLEFVFBYiCAIKiaOAxbokagxLMNrRiSxJwzAAHqLBW0gB3KYjnIxB/ARn3c5J/TZcFtYJy14Ai2gBTXIAibg5/2ahDSQSz9IAx6YhCswMHrHBnAYCYByTlQAhBzQgRwIMymYgjCTFaWald36sxkl0D2IhRg4kIwRgBrQoAgQgACogQYAgXwIAAXp8QUI33q9hl6IFtvgO3rAqgzQByAAeDDQNb+FYtotsjqwEZlvBI88yT0IoWj/ugAO+IBmeC4TAgE9oBbXOMn5ugZIOKI4cAVQkHmHIAMy4IQ2/4y5kZXdUgfJ8fOsL4sGiId+gABmSINpqIXWmosj4AMnuIEXfoYZ/rtKZ4JGB4BCyAFGcLRhuIFCcDREQIIpgANDMIQpKINfuejM6AQ/8IQjMoLgCZZegYOnygxFiPpWFA3s67W5+UxMYAM4wAVEQIRCKARYuEdlIIQCCvb4dKKkySEbiQMkqjsr+GaPZYZQ5aJeiAVsF4dy4fZ4QR9xD4Y+uARpOIPgpwQ1SINXbQkmGN204glkgGcoOILU2Qv5A4fpn/63wTY2GQSNxJEpOAQx+GYk+iNA6qMy/1CGSODeZpiBDHAAFaiBGjAQCqCAGmgQ3aiBfJgNBGgAB7iGAXhXgJAxY+CMPEZKXaM3IEMDOlKmtIn45k0YUK4uJqpDp4WRR0Ui3bmz51QvPc0aeOjg4YM8GyBW+hLQgQMHD3v2RLoWEtKQRa7gFCki5hCRImHEjBHUBoyVIUWGDGE0gV+kAFavYs2qdSvXrl6/gg0rFuy+AQcA7GKVqo+qWrSQIFGligkNAMx4sAJQYh6AaDx4PAMAAJ8SJYLV3bAlGJGUQTp07FA05BAcMWE4cfJ0qdMUoJY9DXEEZKIgTm/EmF4U5g0cUGwEhRkUiM0nN4XuFMudOyS7YiGhFf/6VKaIIjHGxSSSUyeOnCKORBlfBHWHKFOjyolDZ67dO3ju3D37di0DqSFT2CC55alVLT+Y+mzRosqPKmNuVWXxEyzYmiNqVAVjjCpe5OCEH330QYsffiBDDTjgUJOJF5l0AgYYnngSBhyHBFFHGXGIQgYRZYgxohxkjJHLSAkkUMAMNexTQz4WEBCBBSl0sEFNKQSQQgII5ENBBBRYsMc1e9RQw0B5lMJCDHsopE8KRFABBi5wDGEFHIlwySUddLBgRBF23MHOHnjAkAIIHeiTwUoN7BPAPr2kIE8AL60UyykgfHANO3fYMQYccRRRRiJVkFFEFYf8EIYnnCwi5hD/OuQwjAT4ODOWppty2qmnYTUQDgD12HKFXLRssQUtqkxiGDNMXBGYYH6lYZhgtwIQAiduCFZIDjgUs8s3Q4jxRhVxvIFJJpkJAkoYY1QhCCZGGEEGaYJYy4kYQbAGhyAUDXIIIYXoFlKZCd2hk7nlmhKHIsVxqQgRi5ThSB1BxFFFIo2QQs446ZyznXffzXPPOtfok49V++xTEBCKgMIJG15wAgUm88lFnxZOJGjMGkgcoaoqWCCRwxq0YLKGKn1kwZaD1GDCxrIXYgYbHHUYUQccpwWxSCKiECGKuyomQAABC/wC4z7yEGmBBc6s6YEHDVgFYwoUbOCABacMcI0K/wNJYoYZvLzAAh7sMJRBKIe4kogRH8oRdyJx0CGKGY/UMYpIvzTjCwg6etCLL77YuU8+a34gQA1t6uHMTB0MABIscigiSh3GBQHEGMaOEUYnsrwhJhlTSHFEYPF8mrrqq7OeFT0SKDDMJJeoQgstwVySBl/M4JDGKiLMipdgxcBSfDGCEcOEYbsYgkg/dg1RBRugVPFGJplgEkYVVohRBRwYGrFEGVUU+4bPnpBxORzewjGFK7DsFtI1ORW9wJF7DMDOn+qGxAgZZCiCOUSogyOSQAQ69Isc5QCYOd5BsGdAgxFAOAQj7vCNeMTjG+wYgJTYMQcyHGIpbwCDsjqBif9VLSgLW+iDH/gAhSNAQVXG4MMRpACFSvSnDy88Qh8cBA5VrCETtYiIJ9pQmjZ0CAjrW1QcxLAIfDkxEaNYQAEKsAB91KABDZjB1SigD8N94AMpgNHCZuQ0CvQiA9c4RQ1ikIc8jO2NLCBFQsJxhzFYQw5LKEIdkkCHZCQnbktYgiNgsUYZ1MAXM9mRCtaERRBwYAMeAIGckoSjC3TgA/i7gzJaUxknVuEyb+gcG7QlKR8EAgle4MsdWufKV8KyK/tgBwQgIDvbuWULV6gHAOKBg0tMghjBi0aufLCDHQxhBxMAgBKOMAxcAeAEcdiBIDrBiTC0wQ+daAOWjAAtOHD/cwhvEwMZjrMzIJRhEUOBgxVygZuQfCCTA/gFAgoggAQsoBf1tOKRIsEOdY2iDnIghzi4QYci8EuBDHRgPdzxDVjU4SkAlMxTdjAHWNxhHfEIRzyCYIXzTMEKl+EEGDiBQj84YQspPAISoAAF+QxIB0iADwxLJoUsOGg/tZjQUjAThky8gURwWFQiDkG+OCyiiT5jxB4W4FRJ4KgDIJiBPHzRgDE2LAMOcMAMYrSPBlAAjVxEAAJm4MY8hE1sZmDBKdYBjTKUQTmDdEQj6BC3V3wpfKSIxB568aLFSe0DvchRTZzBgZlMMklJcobUJtmLU1wDFt/zxFCrAApPVEIQ/51jSlCCgs1AHKEQCnjAL2Jp2tO6cgAKkAAxJlGLYLAsDcxQBzRuYIxLOOEZ0WitbolxzN8WIxrPYAMXhqGOCZzgGYbQwRs8gQlOtMETyxpqEYwwwSpU4YllKCcRyqmI9XEJDnJYhCJC8c6ccACTH2iGM5y6gATUoGhHSwDXiKQTdowiG+g4xznEMY4FBoyhu2XEHodQhjrUgQwhcgoR5sAIZYwCFtDwTfEQgQhDiGE2U6ACG+QyHyfkRxVasGkOvFCfNewACXzwwhGcIIUx3DQY4MhEMJDhBTawwYht6EQmymCEEJoPu4sSAxDUKYdDhAIPvzgFDFRA2KnVoIosCv/AkGzkgH3oo4q+SNIMnIETgUgizGYoBZmbZITQaMQRg3yEI3TRCF3Q4c2NcAQdInGKZmQRBCn4RQb6tkgPpMAXH1hJpv46gwaAAAHNiMEvQBKKHYMChI6ShSc+KggfVDco50llDpgBgHDUALWiHvWm9oGACUhAdshQUBZwQKkjUIMaTJhEKoChilScAQfJPKYOmHADSq0BGJSSQg4G0QZOLIuyFBEE+aoLLWMRoXveK/IbBCGG9YHiDf8z7x1gsQdTIOBxH/DFPgjAIhXU4GhOPcVWbWQmncDiHe0wB38b6A4luIMYkDhzdRMMIg/VIRSMAMk7fbMb39BjN4Vwwxv/csAGTNDHD1loGR/8AAUpPGYNAfKCDqDAh5JdfAc3jTUUarHTHC9FEJXgRBGWIAZuqvNQQ12Ez8iwCDjYQRIDkUE8VeIMZxTNijNwAAUIENZmFI1FipXBKRDgjBm0oAUvWMIjqu4IvFnOj0mgliMKCOc3fwmvo8BDDBqA2DEmSR+BRaQ8nLFlNenjRTMY3ED0AFlGWO8NQ9hZG1TRhipAJAxO0XSO2UCpeUiglaRePOO7ggAF2DIVqqCGKnIwBBxw4Xa28zAtKjGFYwaBDDuo4Q5ygAlqHOI8j8LmNvVOIs71u9o76xZsBHG9b35rEWTAaLoocIoX4CkDVzH7B/Kx/48CJCADQ9+qBSjQ6JDswRbmaGBDiUEIoOxREXVgAZ0TQa856M3g5uKf/NaVG+B0gi2088MWOEYLY/QhBzqYvCpeyLLPewEMo8dCMKgxIf54QUREhN8FBWUIgrUtyqIUWRUEQRAcQhzEAkGo3QeUlQC8l1OhQAM4jQUggLmZ2wLIQD483S/sgR7EgBm0QB48wpwVUCPIQRwkwXiJAh0YkL3c1ZeEnSnggd9EUibNgAqkgAykQAb0Qj6oCbnhyI7MwOGAAAgMDh5Ewih8DxyMTxgIgicIwhQIAhsIXlAEwRSAQf5RSiFAwAP0QuOhYRqWhQHY0iSs2hQMQelkgRocCP8L1ccW6MCu7YBH/cAxZQI1hAFEZAYZVIEnGEtQVEEbnIYRsEARnMYhKAV2tUEm0EcmLEq1BWIFQcM1EJ0F/AKjxYIzUE0+5Egm2UAAKJY+bOAvwMApYNQeZMM5tIM7jMKZQYwoGIEjyIGa1YEoJJUyvJP86M9NnIIpWMdN7EHxhIRuFAMrqAIlzIf9yRg1+MERTF4fHMEcesEOEMERcFwnGIODXIIfoAx6YIg1DQcZJELEANUSvUEcGFVxIEci8EIM2GMv8E0DOIM+OBUWzYA+UEAFulcCQNIHEGEJtkAdlMIj0EEdNEIS4NXP9OI8ikISOEK00c0ryIEozKAdkF3/CjRWBjhOJslAw0CSVKEAJEnSVEFSTShfL+zBKEAiHCgRawQiRVgIGXjTQ5TU55WOp4FaGgol412DAUyALQBDJVieF7iYDamQH1ADLUjBHlLlDoxBDhwTVHqBFLRBFExBEVgBGBiLOMHBNYWOGSjKU8DBUSiCJ9SOKmSCEVlhGwQCRsGCkNjIAAyEVlmAA5DiBvhc1SxWBmzZDMBkTuTCObwDMwSFoYCCKDQiDfJRGVwUPZhLTjiAjSCAC/SCCvRCLMRCL/QCHhhjJGDUhBXDMKzCJeiQE2hBLcRaJbSMKmACFGQBLXDCDnQcx61BOFLegsQMGLQBG3QCHIjJtXmC/zU9ChZaG7JsSPe0gRjkwQvYoyRAkgfoQwx0lZJw2dMBHQLgCOCYwikQEnOE3QH5ohiMVxyMyJYgWBx4X92IAd0QEB7oARdRIA+uZBaVogcIAEpIzSPRhAcgQAzogTFCDGtsjxhMwaKEgXD6wBhYiIVgQhWMARJwASYgnuINZYeiFjv0w1FeARXoQBYgATKp2IJQg+clExAc0xj4QA6IXn1gAsUwxQ5QwWpUgVM0F2uETxkAQREQwSG8wSH4gCcYg30IUSbkGCd0AjTAgikMQFhxYJdRQANojZoE1hIiUg2kAPI5Q5JIAikYyTDAQz34VlLBgSs4Qgs8AlxFFCkEI/9f4QEFOAAB9KWhzZ098oILkKZ1YFRuDAPuHAg5ogzpcEEKkSMbpJhLQQGAqALtiBj2cIKNjkEREGJztUEYEGdmcJNlsMa1OUsc8IIkvMAvdIB6+QIqKskSIg4i/aAHHNYHmAIkCOkLJsIAzQt8zk0QJAJQ7IyBtYYo9Mx8VqRzxIIvmNVArIlKZEAiqQRMeCkI6MPbBVbcHegcUIbsec8bWIEVjFAbgCFm4Bgn+ACHscER2AIEHMAZeui7xhI99EM9bEIOIAEWrAHGSYEW+AEWIAMfTGUQgJ4UEBsQ4IAqzNgRrEEg7AAQTEEYhMEhQAU4WSi1QMUQCMpqHAInBEP/LQiRMQSDhABCK0ApKSwB0Q2AzjVDWBHJJ2aAPuhBJzaALyTdAnQZCGSAKSRjIcCCHfgERhiBm/pRGdRZSNBpL7xABmzgs7IqllJACjAa8iUAJEBChBUDMQyqMayKXHjBpBwBLWSCNQKsDrDUDe2HxM3HJVyMxLDBFIzBGPwAEJSGhhybIirFtnbLUCmCHDxCC/xCSvgglw2OSibW3LmJBwwAJNQN0SpCIvARUq2TIhhKG5DBARIrdm2XcShCEMBVEJACDJiVsjrDoK0XVY0uCIQpw0wrCABhBpzCPZrCr+7Mt3RGSO0ANrXtebBBIHDCGmyYKoHW6cDr8L7SOqAF/xIwweS9RUuxxaogQYvqZh8QiA9IAVSqghWk2A/kqBSAQRjUQfSwxhAsgaQQohVOwRTQGCZgQv+Bg8c4QRdAwygUASnwgtS9AHUqrQWkrB68AArogwOAFRrVrD4gFgLsgTLUwQsYgSgkgisIQhywwB55yCLwXiSYQi8smj0igJ36Qj6cog0ICZEgndEswJkggE5MmC3YToLQwhHoABU4QTAIAw6ogRYQW8i0H4BI3IL0Ab/WKBg8RJFujiKC0zVFy98tylqGKrQkAh08gpt8wC+8gM7t6ZrUxFTFgCTkAWm6IM0lga++gSOQSBP9qhO9ASeAghFtCBCwJyFShhjojP8gFEELZDFazUAz9AIvqIkYzZ2apMBXidtAVCcpzMHsvUF0jdIbAAEZlBQnfB4YIIHM1AIYUO9WIsEa0EA/XAPxcvLqxENf3EAfPMgT8JDm0QKjih4ZIAE1rMFj6AB9qIEOEMGL/QD3RpdVNugU6KSY7EAogQEbcCVsrsH6vozENQEiMIIpsAgCtECY9UIe4KMeSO0MVJnTSAI+vZdKZlIkKAMZTN28gBMcLMFyPIU7iYR9SkIMQE3czR3yLUA+BEkEaE0vsAg+7cHjRFZqbsJ8UIMwqFI2GgMnSIEabAEUgAwfrEEOqNTE0UcfQJx7ADGyue35TkR0sRNmIIVlrIb/9lybsSTCK0ACKYxNKQyEL8RAL5DuL5hVWtEBEWwJECSYt8zLGNzccwZBWWKhszxi91zbt4SQIBjBWpkBHOWBPTbDkzUDAReW2QFm6YaZHtiB7K7PsXmrIQ7BL3OhDkBsh52yDvRdbFjCutZDLHRyWXfKPqiDXcxCrK2BE/BB/wWDKpRMH+6AFlCjFFBBDtAHLQSCQhvTFHiBjYLB551vZ0nocGbCJYcjC8UahOCHFrgBIwTdAuBBHjShYMVAFR0NAvhC07wuL/wIZ6aEB1yDMnzCIZzZzV1WpbEGGfwA/FhwLEznYF3xDGi20jUNBRimUyUAqmKSBxiPLQgINSAD/wxN3BNIARJQnBcgbxawFAyxEA8viB8gQd8ZQxjswBQccnR1wkTA5fa8gbOsxs6AAbNV2yEo5COUQnXO9ingwf3CkRk4ghwvwiGIwYcccjmRAbeAwhjEQR0UImWJJZEtAhAgimVsKjcF9eU4ApnxghnEwuN4AB7MhCQRZmMhwBSXAi88QszNHiL7QFzeLhWQDo5OoluQFBv4wSGAQSZMQmCEwwyY9YyPRT7QgAQwgzQYQ+n0AchSHulQwRDkgPKyVBa8JRb0QSUQmxSsQe8qC1OwARg4BRiCjMqoVP8ByINASC1kASVsgjK0CD6ZAgE3dQu0yHwZ7i+EmZr8wlDjAf8IxCQhrAJJDfaxUQhmtEEOfEExxC/9kg0Bq1dmt4hT1YA8IIAD9E0z8EJl68GgrYQ2jAI0qOYl2E5Bt8wWgAwUpEEacMENdAIrECxDX0LL8Gt9DCd9eAEYTsz1MCkb2M5wVgg2sUZ0mcYb4BhrsOkL5MES1G8LsMBQj9kABUEb5Ex0QKIg+CqigJN0YKrNaEgZDAE6qWVmOIoglAELEMFqxEEjVJ0Z4MkHlIKbqMQpGGYGbCaTPEIMxoG7YKIgDPYUZAYVSAEQDMEPDIEPcAJcmhCyYYIUPJzazkM/FAONEzxY6MMEGAAxXIG9ZkGAUF7ITKWJ2c7GIEipO68X8IH/FqjvGriHF+Rrw0KBJW+BMdBHrCGDjD0INcBW7RBCJDjVNZSCUq9EHvyCe5G7IL+AzJ/C3SwDHQiBG8yHLHCCKgW2odbmJkCpKTzCg/vtaH9AM2Nz0nQnn+hDUc+ACyQaJDwCJMDPJhj5KqBUfqjBEdwAD2gBKxBDNDzAAxACF/CBgVB8DEGlJ6SoQxd0YHvBONIYNbjFKT9K9yLbNUWXIrZBhRbBzS1CHbwCiARF9xgYKERUOe3ManT0aETn24RSt46Bh5DTEpBBNpmGcAKB+GiPIrCGI4zNKZyCSPMCAmiSC1BnDLTAmMnBfIb35MOGFULssQ02MoFhSdFCJ5RS/yaALRh0Qi2owh+wwgRAQFUU/PNvBQJAgD0Ugg54wW0iw6phwYAAARRQQqn7gRZkgYHMR9hPd6pEnBZswUNkAYFIAVssyMnTAjJo+YOArDF0Ajcks8nGAkB88ODhlKQXvU71mjEjRYFeL0Bw2OABQalHuuYYUtMnCy1VUKAckdInGDVVfFhBU2an1JJYvVro+4Dgpb5TeHrl0fdLkqSIHDz8iqEnwwAXLUrZYQTNjRNVyFSluTIpi5ti0Q5IgLB1xKY0fbb08ZOljxcuWsB5QsJHrKo+fZDk8NJHFS1qd43RWuPFyxE2mTJxYsOpk6c2nji9GTKlzZQ3YNi8KVJEEf+cMmPaDBpC5tCbQYoOHRoEBk4YQYeGiBkkSFGVKkQ2wxGzhEwbTm2qiAkzm8ybH0BKLyqSpM6SUnQeJUn2KM8SMzGMlEn0BrSgN6XfvLHOeMoUwVaGUGHjhQ0mWl6QAJoFjJMUTKpq1fozTIKCSAHw59e/n39///8BDFDAAQksMD92+jnBkCOy8EOVYILxg48scmBwCy3qMkaVLMLiiKMtLjnJDz/e4oMPJJDIQoodpnCCD1qQ8Qg+cGik8S5a1GhFlCIeKaWUWE45JY+DENBnoQIKSGCBWFLwoAOCLlomlzWcyAIKDLdwwosKH6SFFltgYeSRRxxwwIJTXjCjhV7/OnjylzxAeDIDXjrgACghy6SAAjNKIYWRQtxIIxVgjtlkmGdo4AcACfBZJxwI8PFqiyxUcasPLZw4wg9qOvHCIwkvRUIKKCgtCRxqaBkLiSPW0MKYTNZYIxNVMOGLEy90CIMNMDxhIxAkihgiDE66YwOQHXTIQQpgk5UiEDagHQRZKdo4BAgyiqhjDOuMcCSMNtoAoozsirACDiAWcYwMIzgD4pBOQEnEEzjqgEMOFsZ4IwzUqAhEBxbhgIMNKXJgYwodkPgjhyFywEGKI95D0Q1WWPH3r0zOu4EYCBSgZx8DQQ5Z5JFJDvAbAOZx4qtNaanSDyTW8KNKLyGEywko/7ZwKwckxBrrCAzH6gML8HJwwo+SaKG0lmDAaYdGCGnpw5hP4qjDkR6XaCGGXsq0AIEZEiCAgAUQYOGUD/ZoxBEiDKmlDwbJ4shKHFKslBZWEJmjiFMoaEDPPGKAiIMLOvjglw4I9yCWDAb64CUKNjDzlG7nQOSJLg6dR4F++plAHXr2CACPAyZghYdLSPRjiy0cXINnYxxUxcFUnViWVD+MoebUpOHygqxaZB/RDy0wGR6TVWPdgotoJogmB0A4QaKQCZQohAYlsKcBn+0nGOZZN2g4YBdfF8F2iCrAzQ3cN1x745Ai4tg3N2uLIAMIMQQRrA0/PKkijDKKUAVBsP9BDEUoRD0mUAwpCGIKbjgAPj5RiAdkT3vawwcrLGELfACAg0ooxg0wwYlKmWUYE+hHOHpRMhWukIUtxE88ABCNNKRhU31wAl2wUKksUAp3qEKCF0hFC2OAIQeksqEX1BC7j+xgB2BwETKosQZNqcIYNaKGg4JhF07FgQ5Xe4SaEOCAC+jpBQlQ0gJGwYJu6WIZyQBFLWBXpbdMCiw4a0vuEGGHIuyBAg6ggAV4ETh9OIkibHrSB3ghiVPQRBLN0JMFLEAKyl3FhP1QwD3WEQlnfAw/4ZAAMXhAFhKBZQt8UMUWjkCXB8muLqoD4ltmVxJVGM0LrBsRLYKxoQaNki3/VkIPF5jBQTcwARNpiAYAJrAJDi6TmcQ4AiCCycFCAKINYZgM+sD1LU8IgptwUIQRxOCJ68wvCPZThGm0wwlPhGFdwvKEJ6ywg2H0AwAh+EQgpIAIDn5iGMz05yZssUxFcZAZ7KHCFCBjCVbMAwAPuIYLIRpRifbHBvfoBzNOFwwtQKEPfECd7MiShVpQIxiTYmUw4EKqHfJhlFncgg90sEO6GAM9LG0Q04LRIC/hxQ+CIEIRGvEIs1ngjxR4AS8GsIBIkOIRjnCEHFzRCmpoSHaT4pBKk8hKWqzCDUK42h+91gI9PAQEH3CcJDJgVjT54gUtSEEGYvELPZ1iCUao/8Mc4gEBGsTjDr+oAX9MoQAazEINWzDiWHa5Q+H1DHdj2ciIxKLFoJGlUpWihha4oCljzEwLR0DREY7gBg4S4wpZSEM9APAMSwxjGLbYBTILMQxifOIITFDCMp8xCcDsoFyHMUwbBGGYMEyhCvkSp/uqgFzXtE9f4hyCETAj3Bz0k4PP4AISRAsAN3yBtba47S5iOwxAMJQGrMiEG14LAGpIYTJT+AswgMEMCABgHX+d6H3xq8IGHKAfwzjdy9bwll2CpVQm8QMyggHFoAlvRCDxEhbiElIHZYELJJqUXVAa2bvY7QiCcMUr7DoAosairY5IQhEc8Yo4gKITL8plz/84lAUn0LFLs1PFJkJhtUbswRRLSEFZM5AHXjwnBi1QgUFe8AKftAmRZjDCEuo6hFDQgAYzANDJSAtZjkCWLUeAAu4wizosBCMTWeDD6lh6NFSRSKZ1wWXLoMCzYACPGlvIARVSqQYmMDQaV0gDKzhoC2BQYhI8eAYA8MGDVEijtoDmxzEncIlL0AIMO/CBFzLhBVUQazBgaAMY3mCbMIThMYIYtakVMYVRh7oNYyjCD6DlqyNQl4Os+DMA+uGGG3CBC0zY4DNu8Ic/AGICAMDoH26A3l2wQgpDGEIbMK0KYPxBgwAIhy/ym21tFwgBCoBAIXhAaU1x6C1aEIsxslj/FwjdJUuMpcVGc6aKShAMCk64IZstvAWmkVQLN80lWdRQCWOswTGDOIQRWNACRyQiEVPIXxbUMCH4sK6XN4uxFnAJlVpAxQ+fmMMySFEKXsSiAxsASi+GgvIYMCRJetBDye/UAj5lawfF6Ec8AqQOCLDiCn3wCEfM/RYv4AAKdulLv/tADU5w9AhGw3gWHXShpM3OJFlAjxZKorsoMBEKfKDGJIgBABFAgQfBrAcwPFKLSRxaCX+wCxIMDYBdKBMAw5hE1ERyhC2YZw0+kBVf2iAFwniaDb0qvGEY04ldDYYNLFpDrdaQhn5O4NDzSAWgtav3I1jittHQLSAAQQMA//DjGbb4xA148IdLeMHZgyFMYCyxiteGw8rbtv3t94MgfGgCGGDwApt32CAqTh2OfsClDUvkoFHK7t1y8UKVKpU6B91Fdza0sWJ1mIaAb2EHzg6DLCphIj7MWHYkTSVkPStT4yfYS7KjxCpyIYdlNKJPH+DAk/CgDz8OYGsJSNICzCBOnsQUzOARSEEXRIEQlEABMgBAZuCBJA0+xsJBzI0PiIgkTqlBsgQcbgWVckYLjqZSNGpT1sCWcokPQEvNdMcLmOj3qAEbAgoAwO21cgs+VOEKDg0fmACKuAAYig2jOu/ubMizOIoWRAUEOQo9RsQLMAETZCUTnBA91sAj9v8CBEXlQsJC8kZvE4ptGIJBmLigD3am8whlDbiA1kZvHogBCjlhDIag0waDCUPotdah9nDvDrftZHbBGKRA05DPy0ypUsQiwWJHLLJAC1BnRFQBaIyvRbKg6dyMUjziLkpiy9qiLoLnUjpqDZhoB6QADQLRD1hlp4KBC77sJNDjEnEpl8xtRCqhFnJBG0DuERjHAxyHAiLATH7hF8JGSVwgFsxqD2LBAJdBFHDhtb4hQJxhAuZhEtxsLIBHp9ogpmCHLvyg3tJC74pwdozPwKiBDdbALeymlqjoLsABE7hOi87htpjhBujJ1uCDD+JuBHAAGIxBC/uhCcpO7FYhaQz/CyR+72H4oLOqxFVQ6hCLp6qkMPoupRLQg1Jk5wr6yQAsIeyUILvcIAxzIAfIkFbM0BaYh5nwgRPWIJ7IAwkggw1qgRb+oBUOwADuAA9lUtvCIbW4IGcQawqkIKsU0SQqS4dgaXYSSxV8YCd/yA9W8pYqJS8UcUO+jBpqYbGi7gh0IAg8MYigwrBA0Eu2gGfcAgrCMHYyTsYOMbJqwRVksRGWwRROoRTMoI/+qCDO6BrMgE8k6RHIYRkWgRDuAQBwLkAaYAJ2IQ3spt9k54aMYRq3IC9YCgX9IBujBkMqa/omEWbYjIouxLLugg+oYAiKjmmwIZjACwCUgAl4SA2u/+CY8AEHKIEWJuGYVuAZiOGY6i4VrCRFRCLOtoASTkQKtkCLtICOhqeUoGBUyMJLUgUkfI4akOEKAkoCVoHzUktRNoEL1KBhOm8WwAITUsHPaoEViCG9iOEPwoAKyCMl2eBBkGEShgECDiAWZjI+JyofaKAfSCtEhIYKpCCHhAeXpK8uzA0EFdGkbGgHqAAL+uJFKmXGWAooK2UNcmAK3SJ1ZAcJnAAJhuAHpEAKnCLr+sAUjW8NcKCUuOD5hEfdkuaGKEvdPiERlmEZjCBGjQABIIkChgQPEiASYsE4xsQIdCEZrKEYDsAvOQlA8oEZZyFEIgTjgoYPtuQ3I6RKKP8lLZAgGKgqdrAIKs5jLjikLmxpU6hvCjxzU8ABG8ABAEyABnmALugiFQ5tBLjgEoBBE0zAnzhoF3rOsMoCJJDgLJxUB44gd1AF6M5sC6ghE3AAQ7RgpFDKZUoiGLQwBIJhFtCQFQoLB3Bgg6LBNrNgE3Zh2SYhs07LJqmACjhBMFA1E9BN7Q4tHuxQPmF1hfRhAiDAFq5AdvhgWbDgo5ABwZQPIhmLlSILpXbACvgAC1gHE1FEscYC404JYbAgzS5FdY5ADbSgQo6ACoxGd1BFxlpHLqzu3tKMAnvmUtwsL4QBF6whCRyhEXTBCBBiCZLMCMakXh2hDoZAEaDhmPj/gR4KZB6SiRIw0fhkJyTsTc1E8XayEd0+BT5w6UY2SkolsUHIgvqMgQ10gHWgCByAIb0AwNbEog9qARiOqTRVYRbCLoaeYWUZih+6YGUqQW4u5C04EQm06LKQoEM2pRaOAD7oIos45Ja8hD3riTlngTYBwBaw4RL6NAidgAn8QALkrhYyyxYUhRiQoFgBIwpSVYgy4RIyoT5jMlbJtmT2YQAkIJmAgQ2OIBXNDZfQTXUuDGdS5Wj84LFwpxLiSQ1+bnaIcJecAOtU4QiQpWctZYeyJImwwCF1wClsBEKMASo6qwQZpBUt5UTBoi1oYSVzRxXaAA6SYRkeoQXaqgVa/6CpdEEX8HUH3mBfDQAA1KEBC2QdAIAYnHElo08Vho4LBA5p4uxnDjVn/HN2kCF3cilL6o0sasZD9s0fK8suriBllWASesYJ5BTSyGISGEoJruAKeEAiR+sKwqJSBFRD+gAKdEAKHvMueJZUQBAqO2JDYsdygccPwBcANoEWgIEV6AkAnAZSR7XPQCt6OUgCsKeDMqHZrCATFC/WOqEWoLAWUsEWICWFyvaCQ2Yf6AEAaIAPdSDO3Bc5K6UPjQ8JoEAccfaGfI4PmEgyf+fduED4Ltchx8ATO5fNZogVpI3McoAkKHFjB5WH9O4tniJ1UAfpTpSKcFfejmcKiiBGWf+AMg4BDm6Bn54BtfrhHgYgZE7BAGjADyhBLOoXBX0T3R71Gk84i4CnLoh4c7OupLaALxqEpEZkjk8Fs4xPeIJBDbjQADBKLFCwD4DhGSRgHv4gDbjwk77CCdCOBiRgU5XID3IIQlTlCLToVOQGKRt2lOjiU/iAkiWSHx4gGEJk7SCgdMABGbJgEnZBAp7hCo4gDIGBGG6Lg0IgGlrhCNzQvXwFWmyDFgIjPo4gmMIhBjD4mA0EhqKBcL2MSjp5+sIACjTkh3bo+ORCDSilEg4mLFIHaiArEp1AiDAUDPYtKq5gE6IBAoYhFZChFpziLmJk3UxCC7SAOZvOlNpPQiz/rFxVQQ3SABguYRImoTXTgAd4AAdy4F/AgBUyZ3P64QHiQXZFBoaeIZQksQ/UAAzWAhOk2RgwWp8ftrJ2yG4gBJMfkUFsLFXs4lRCAgtS5VK6FRhGFmbYAvqAgSNcRhUyixLAslJS0GEZ8s2yaBCzTmbaAupssEHbL0uQIZUlTdJ6JqB3CIoy4Qg8yqrqzfgGhaFZ4QtuABACwdl0AKGgxQugjRbaIxjaAwnmoR+K4VWROa4pqi+ZYQecwi0kky6oYQqQQN6A6Ia4cmduplJWB/seNd0aZI1VYQV3wGaRARuAYROeYQK0onRSQXaMl6QhhJXu4maUaKjfwmiAIQ0E/1oTbOG0UdsWWIsYiOEZ1GEXTIiDBGsdBkAAAqBIQ8YZ8OGTmKDCFtFJveAMVmRW+pApMTGyTIqkScotqBmkIOthq24l+219rZcWLkEuyk2VPvR2NuQtHqYuKEyVCHb6ZAfB6tYuaCGWgQZCjvM/jQETRkWLKgV1IitVKMUYLhYHjA+nsa9SpAEbjoELGK8TWSQQPg0yMs0KOiHwkIAVTnkU7EuuJXw/UmC3SYsSFNEjDMsYkEAHkOhmJsVBvEAKpiAklO/dSiWLVpUWknWxmQYc/KBCupcVnkEBAOAAroEe+uEANmFNoYiV4GiqqpmkyG/ONtctVOYKUqEPNmE2Kf9bAqC8H6Ccg6RcAgxgAvbqGwYgBSCq20SAGG4gDS6hQTii8YYAE/gAYaKPxeWojn+SEMnC3pxiuoNnU0rix2GJOTnKGND3SlTRhozGS9jiWns2aqBvRe3mlgb1ZzFBBypMZjb3dloRtFQBUU/4VGJky1bRI368wzEhGDhaE9vN+KACE6CFWHzA1aaAPKbAB1JSB9qADZCADf6AGAr5ESJ8wnU9Aygbo5jgoz6iZ/1gQ8/3ZvTupC3UlmzqmY8GKuT3KagvlXmACdygxm+cHmw7AE7mBGwVGzJs/V6aLqCCsl4QGf6ACfzMUBBlviSABtbh3eF9HeiBHtjhGjIgBfL/4bbvawAUQAKYYQ2ugAnoQg3EdAegQA2WJc1oYWDa7SdF8EQYZAiVLxOTjvpMokFNYlvB4IPtRqXHIiyWdws8vBIWMQw/whq9RGihzvjO44OxABP0u2XCgswdBnW2FRxyKWoKLME2TsNz9r0jnhI8K+hwSXcDQTDAwAesgFcEgwpScum9ADJALxpcuQVwW9clHAFMyLp6zkFm7C7UAAuwoG3j7EJ/aFVEhERcJmiyCAR1ykq9ZBqQYRjSmYOxfT++QSuYAbLTQIYvF7zRfVCOgRpS4hk0B9c8hx4a8Opl8hREDx+GYSou4USGIAimwLPWgC2kBldSRKfO2xjsrPMf/zEE36JnEOzOIctKx2KxpyAHJDFGVkmPwUFThn1UQhtDrHdg66IW+rGy8oIqD54FU2lae+an9RocagEin2LO3CwpYSlVvIwuho6KxNvUkb5YoM1gBiMK2KATMm08AEETdrsYXoDxsR6Z9UH05sF07g5nMDlGHiZTRGVDf6gSFMuGWIcjVinOjQ8ggKVhsomZkn4SNLCzEaChw4bXJgDAR2xTmitXJqWapBHYpTWfbBF7tksJPwD9JtxbN6DBw5cwY8qcSfOlinUSJESzxfGMlCE7ciCBkqWoKj5QoKjy46dWMFV9/FDzIxRp1GDUVDnhQpQpMmrBaPUJZiyrHyeqjP9NkeLEj6pgcJHRUrUUq1JaUHAc6aMlC12+qmj56UO3sFu41DLpOIIFyQ4pWbTM9UNpqdvCZVshoUsL7lJak/l2pqUGyREtSE8PXlq4E5gpVKaEAcOGE5tObNhkymSbjRdOf2yJmKAsRs3jyJMrX868+ct88QDwY6YpFRcnYP10RuKlj5MpOn4sVqMqKVMnWfxo6UMLGdQ+N67MWjUsmj0ACuIhON4gHgT880TzzIADRrPLPCeIIAFCCuATzjctOSehhPsgcE9OuxBDDRM57KCDDjlEdtkl7BHWWTCDwQUGElh4sUVZyHgnxRFF+WEMVoiBFVVg1EyBwxZuGQMONav/tMcZMm7NtcURW2RBmHp9OKmeW0wx1RkycB2BhBpe7DDFXpMxZYxYW3BGTSdI8BEkktqpskVXtFBDTRZVLUkYFFtMdmMmYIRBxhBUhNGGJ5yokkkbt+GWGyecMEGMBPjYYdyElFZq6aUv7XPNAQBMQEwnPByDDCXsOUEYFlLo4ANkXviBJ2Fa0FVLU6qkkoolX9QnUUL0pBDAPsnp8809E0CQkwHIQqDABDSosw49A+iD6bTJDaCOASlFM8wnOOSgAxdqXAIaXWvs5cdXyEhGDRtHqBHVV1DREoYUamwxljFYyknNFkhoVws4RzwxrirUzIWlrJN15kcWUDDZx8OW/1lGZVSdlWXMGjp4kYUUUiCxRrxn+cEHEv2q8hWafERVC2thefFjlX54QUWabEhRlhdQ1DJXYJgE8toQVoBBWye7gSEFJ5l4sSgngHDxTD/RODIDtVRXbbVD++TzjQKdPjMMNqlcUVlgqhyxwxHozahGFkeIS9okGanCCjG7TIDQAevsN2E+KWSAwN8I6JNCPhJeQwMN7Di3Tzg0xPNQPPhcM+Ep6+AjAQAi7MKMLZs8YQkwwGSExA1pUIIMNk/Vsi8U7M31lBbUrFgJnnRhRc2NR+gAWlll6hslzEnORVhYahwx4xZq0lUlH6rM6tdT7Bnjhw5hIPUl61mM6YUXa/8Ilb2ca3jBxxZbVEKLzmR174UaSXH5YzBeHEGLMd7JGpgftnESxBi1EV1LLW2gwm1y4wU0tQEQgNhFP5hRhBpc7YEQtFQDvsEpAyjBa6w4xixmMQke6IUJR7jBDUCYAx4AQxWaKAQxomE5ABiABnljyKWAhSkaAAAAB3BODU4CAMk1BB8A+MaEgCWPSKxjHlyTAAQOsItnMIMYwygEIbrgh2MAg4OTEAgP0gC6NASGPfaiBs7c8hWwGAMJOZhLnKDgB9udxUlOYs3KCOOekQXlCMz7C1N2xBSC1aItgtGBFCqxhY7haQ1w6R7HTgOX2SmFNSd6yhZysD70uAkHaxD/ixaaBMc2sgZ/mYgDGdrAiU4Exhht0EEbMMEJUq5BPOxqxQMkUAwWODCCuMylcmxwjXBM4HINEhAUbcEKN7ihEKywBTRGwkIF9ONu4aBHBnTpHH3c8IbTjMktG0KCh+xDIjh0iA3pMa0UXOMb4cCHAi63ICVO4AFK2IWAnjgMWxBzFVu4BBavAAxsMIEJabgEMGiBjTF1jDXU2EtaglGLojTpLbUTjGXsCAQWgWYwe4wMxOT0JuZtoTta8EKa3mQM+h0BZ06Ik6uYlCSd6ewoxtsL+egCBVOpAgtTyAGQ0iO9HflmDEMQVCcsBoYdgME2SMgEG4CCBHYVQgEKQEQL/7ZJzapa1SH5GMA31HEAA1wOABIIgQT4wY8FoUSJB7hHPNiRARk6xG/ZvKpMvgGAwwFgHQ7RxzzikYHDBaAG63jACvCxDoZ8EyUAIGcAxlm1feyjARkYADu+sY5w3IMG+JiAV/txQwkYQFlKoEGABkQMKCJzFZpQhRXTMMKMZBGglwiXHofHGmToTEpQsEJQpFILiF3CCfaKzFuMsQUcKKUPaGOYF7CQnlWooihRGktWOMa6JL2FFpfgwg24ANCHMcVJkqHFT7xQCSodgXV82N4UdhCGMHACK8Yo6hSaWpufDGFFIXwUPhgRAxrK9b9VtYE+EMAOelC2sutYxzfocf+NwLnVvw1pwDX1BuCHAJEe9OiUQxCAEonwIwDquCFnAaCOhkhkHShxyTghTDVgObYh+7BBA/QR2QJ/Ix7qUAcNHqAABT0TsfwQwQPwMQ95OrG0w2AFK1axiiwwISNhyyIwJlG6S9AlC5cY0xGGoINKgCMY79KKQ2OlM4yRtw+m0YIfsBC8hfHFSbT4MheG0oeU9XEWl7BFMaBokUkMJgv2IlgbhiA+k9HCNEUhWZ/a8AZPEBUoU6CNF6YwhCEQkA1MeIYEdlGGSVX401V98Yt/dZwM3xCvoA4AhwGQAmsCYAANycANJyDNAdxQcnQFgLQkMgAUO+4eiZXrqF9SAwH/OOMXCBjANQwcDxxjlsc5eeYzlbgsfOzCyE8c5pJXcYluA0Ma2EiDh9IgDfcM9KVZMNVloLADNvShYxq7xGBAIzw4xupgKbtyFtIwCVvsop39EEE0WEFlJ7SlLEtdLsGCQb7z5vSkbBBUWlTBhh0ACglCo/QUtseoS/whGv14xhCmluqSm9wh8+iUhvPakgbQgx2+SsHLXVKDZPvqV8l2SQBkzo4GNCBayUExDRpiQ8ftQ9YA0Ft0hh4AG7Cj5wGQyDVscMMUhFixJ6fJPmaggl+cYg/XsDGO7zGPzEL1ctP2rAgOcIAiC+gZxIBGIYjJCmrUAhjHOAbceMCE9KR3/wdIwEQOnKCmhUmGoX44Ao3SY6WduScLk7gEM3KijmLc4Rvz8Cwz9u2XGy0VE2poY1aQAgUqgOEMU5hCG8LQifPxaQhT4ITMfKADMCjNC5kwBiC+QAMJMJDkWQ++XF3NjhC82iEhVgc/ODsBdlyTHynYRz0AEI5YYzMA17imCJKPnMOq4+nAFgFDZB0CnYe4+lgzMQDYsQ+6huPqwmcOhGvgi2OfIhLXuAM91hGP99PgABOgWWbVWcoyAUQ2WsTACk/wZMCQCjqQJkgQR8+FFjiiJaxDC+VyGSbTQaywCwBQD9fgaTXADpyyC5pwBYXRBzmwBmqCJaGBBMagCjowBf9rcFR0YQyZMAbuxQlj4CXb8xudsQY44AYPYADFsARUFX9KCEF0hQ+LBQCO0xDhcEPxsA4jVoWc5Th0JQLAkmFOmA+cRQP0YEMkhhy2dk1oKDnkp3NTWGK/4jcMIRGJYwMSYXz0wGJLSCGOtQ81UAMz4Au94HX4d3mUFQ4HYnbGkhMPMA9wNwyd4AVwcwVbBAx9wARlghWFJAXIgwSQoQYcMQmb8Awi0A/h8Asx0QtTeAAE5xZbIEgEMxV9xBTg0AlBsQZs4AlEAxo+cFSYsF5SoDRs8DG0wAk5wAS2oCxSlYR5uIyYAkR4tYUyNIXoF2L3IIUAUI2uth8hJkTOxw//OgdE6FcTUzgB4VCO4XAS1SdrJaBzGSYCLnGG0ySHDeF8N4R1zNhixMaHM/ACvIAHpjAKsFAMjPNLYCUC1taIrLAJwXAJV5AGqRA63cIDVyBCV+AHc7ML/4EP7DcTgCUBE8ATWaEDO0Awz+VdhAEObTAGv/gbmUA0h8IGUcAJr3FUP5h7XiAFmDAJj/IAjPACyniPQMkcqzZNruZDU4hqUxiF0eGGNiRExictS+cQS3kcYBhEj3NDNoB0OtcAzBcPJ1GNASAC6ydONyREQWlVe+iHkoAHkVAM8XAP6tRO/KAA9VBkBPJExcAMemkgs+RC90APAnAc+1AMHmkL0hA7/zuQCXbHeG5GDV8gSNyBCbTQkpngCYRCQL/BcZlAcYEnUE9DA6HgaWc5mt2HYp2yAlxDfdYYhSEWhW3YEBmmDhzGdFPIdAGAYuE4E9nXD3EVAGd4DSkAACugc74JTiRGOPsARInTEAjAWfZImgBGf7+wB5P1fma3WdJ2TRBggNF0CssxmBJQD7aADZygA7AjGBDTJPG1GKZhI/9TmaRUG7KHCZjgBfSJCSSzBpggUCAXDa8AfNAZoDBxWGh4Q/xAOEe5msAylQFgTSIQHYq1hYTzhG5IEzaQDzbAYgKAoQGwoS9hA8kmLQ5xof4VYwLQTQIKanvYEPLQdRmwBwOgbP/scA0xegq9QHJ4eBzf0A/4UDDp8RTMlR7kUwlgsBhOcATXVQud4DNIkwmsxAmstAZOKlJrsB6WcAn/JnIAmqIC+psz1gDZN5ZJ2RDnZ41gGQDAVnUN4Wr30Es3lJtcGqctNmyYMgOTNw/B4GeCwQU04gfk0xhswT7X5Qed0AaDshucsAZRsAaMSi4zsj1QMAnB0HsM9JNyCpTANgEvIZZOCGxRCGxuGGK2aWpnug+mdkOgeqmquowxMIXRAAyhFzMsRRRFlQNu8TCCgai+MZm/oZ/cMxnbw4ntkgqsMEvEgISrOpr750MOcQ0Khn0sARHRCq3LGQBcOZYPISzfkA//uJms3ip8vHAPITcLgQEaTrIFakBpUvBFavYe4eMFwYAJjOoWLqIK63FlJIM8qWALxQINLZCj3wqdJUqnDyFr/UCcA0AD4eASNvAAUBiwEFtyeGA5zJAKWIIi81ZUWjAmGOUqDYdIOPMZVeo8C6cKaAQFfDAJwwABEAALohmxEBtiqukQwYkS+FAC1wezOvtfpvBLxAAM7oFRtLAiiVEmIsM2TVImxCUVawAkZFMLZQEO4NCKVOAEanAFjzIBo/CyO7uqQIQPIvpW6sBZ/XAPFNa1aJtL+xAJUDUMqRAlwXUEfgAOmYCkCyNS0PUXTlGvTxEnTyG1tAAeEdgHqfAo/wfACFuatnGaDxM6oBfqVoobuRBUA3fgkazABFwQFVDQL1nBBcHVnsAjGEQBUViBJNQwJGhkLlOmaTRgB5YquaQJsDAGu7T7QDWwo/jwBTeQBcWzGQVDFE7COmQjtH0gvIVxO+nxFYrHPIShBjygabtACq9bu9Rbvdb7EHYKAPPgBWlQvE5QFomXFCZSGM+lHQanHVGSBbXQZE9ygVDhFmmQBiCnDmZwvfZ7v/Y7A65qRdpxO6iSAyhLF0uSJ1sgGXzUJKZCC+nxXD8KZrfKBH2gQOHAtfhbwRass3kQYs8gKjjCB+DBGEtReptRwAw1F2yTbpLxPOlmIygiGQuTBv+1kHKodsE0XMMBGwu99wwD9RXGEAaPkRRFoSWCIRXUgCRoZi/a0b8LExhhcT9ZcAW2sCx7YMNUXMWXagr1IAHEMAsVw4lsQ0lHigS1sB4nMhe/yAdfgRiJFxXZ4ac8wAqadQ2ya8V0XMdKuA97oAAQMAyzkBXowQdgMAYeo3iVsAZtQTaBmya2Iyd48aNZAR9RYDnFMMd2XMmWXHJ3YAAfiQ1+wBVqkCoaQzJ+GhUwA8D9qy9iJhXggAyTkAOeOr2XHMuyfFXgWQ/gkAZ8+h2QkYkUExZJHHjsEQwu+GeipwppEB3hkLizvMzMTE2Apb1akAbxs14ixTHtKie3GhX/6isncvEZc4S1/UADetDM5FzOuqS/ABANuOwEUPCAHYM2hFHED1Mj7HG6q+AenbEvN7CTpmjO/vzPVRMDGvxP8sIdphFo0LMUNWIyWSEXWIENV1AIxbIHlAzQFn3Rx9ELNLBAPIBKZ7bQoBErJeldYbG+cgIMm2A5d0CwGN3SLk0TeGCs1pEngyEZnzQX6CG+Y2I72BDDd/XSQB3UM4HHejwMV0AYnIEiliQYRdEx/tIZ2HAM0QCFsCzUVn3Rd8APHzkJgXGx5fFITFEJomwZl5AKT3MPzXDVam3V4HkAm5AKcCHMtLAF14EefdAiTsBcfXAJPGC4/bzWgP3SzzwP/6sADIjx1crlBKYxF3uxryKgAFMc2JLt0jMQHbswUGWBXEmhJWjkBbdzBGlQrBLArJNd2hYt0FAzC8hgDCK1uTNCFFhAF2mwCvjQD99Q0aad27HcC5n3DNgggzlwBFRgytJzBFwgv1St28r9z3iQxfDABDpgBU4gBQAcJ30AIs9AYsq83Nwty3tQLMPwIVEyI29BCTpQDHXVC9293uR8DZqMCDfQB5WgNsYADNDgkd7J3vo9yzVAmA/wCVeAKkjQB8BgCyvAD5GA2/u94BfcfnXVBcDABjiQBsGQxRvJ4BheyTVg2dQgBcYlwwpecikQoyRO4oGpHPpwDb1ZKVk1AP/KCKZnm+FXrb9QY9zZHQ5V3RxZw7j5cAGM6+MX2uM87uNC/uNDzuMOkWtke01hixxCNy1n2LgO4XwhALkybtXNMA/9UFplm9ZVkwIWEOZiPuZkXuZmLuY0lGv4sOb4cAL1cHNBd43TwmEHChMDMAHqEOJX/s+/YDn8cAB/TTUgcOaEXuhj7hAZdgKZ0hAxmpULBucNimEuJ+exZmAwl1fXoA82QA/7EaP7oFXscOIcJgICAOoy9HN60+gI8OgPgQCWTpx73sx5zA+R3VhgbgEekOsegOu5zuu7ruu/3uvAvusfkOQA8ADahI7X1A96E6aoWoa3WaA+hMw2tA758Kb/1/QAvsJhKiFiNAAszqepNoCzU2ig2RQd11QCsBbr7V2tVrMBwx7v8j7vwP4BKACbAFACOZZjYvgrqfkN32B8JZYPGckOMltiYbpWaSotpgkAJ8AO116WO1qGq0Zr6J442aepOyTxnFV9tsYP7JCwAGAC7M7gKZDrHdABHpDyLN/yLP8BLp/yK6/yuX5Lp4qGJXZYqBaVuxl9FPqE1QcszhjtSbem9dgQdNUPR3dD+3FYt519ir4P93HbATCF1ZhhEyAtWrOtJa/fJADvMR/2Yh/2HvABad4pJa7ivyJ1DfHkKOaEbV+GJ1GtUYliZ+pq2YR0fSMdEwpET3/svyKW/+seHdWIdJhzD+7e9evdACvPAR3g+GMf+SkP+R1AQ4lOQyUq+HE/DwEQlUhfhgbQQw7x9tFeoa52c3ovayLQ91YJ9X91EoNP6foQD8Zpm4q/3nu4ohTSEDVg+YA/oLwGLE/ejhOaqiNwVzTkqaWfVzc0AOCOEktf5wHg99gH+PsA+w1B+L4pTdZqaut+++C/HEm/7/dwWbC2AqIf7UNXs+EgLDdUYrlGD/pgmrCGYqZ/QyeQbPdRjXQOEPkCBBgB4FuAawAmBKjBD8CAgfEA3AsgkR8CGwMAAEAw0ONHkCFFjiRZ0uRJlClVrmTZ0uVLmDFlzqRZ0+S3jTlzHuwH4P9agH0SHwzEmRMfABoD1ekEsC4iAA0e9QHo9yBnvwwBNAIQGEBE0wDsAPALYGMjxADhAOALkG8C03A25c6lW9fuXbx59e4luS/DN3qBBX/TF5bwQATrfgK9dk8dgr+L97ELRyMe2gAIvi0OMLWfvm+WCwdoANjGQHbfsoJmN5DetxQDB2wG6oyeOhrhWvPl3dv3b+DBhfPep3cqgNgoiw9n3tz5c+jRpeM9Pnr6dezZtW/nLlzfCXzJu48nX978efTp1a9n3979e/jx5c+nX9/+ffz59e/n39//fwADFHBAlAYIJx4EEwyngZLyiScc62BCQB2nVBpAnYMC+EYdzAj/9PBD5/JJwIERSyTxRBNTRHHFAjwqiikAsiKpgY06kokehVZaZ62BHMoQRCCD7G2fCBgw8kgkk1RyySNdBECEb6L8Zp1vuhopn55sjEkstlTCKaqw1pFRSDLLrItIJtNUE0mPcDwgpH32WQfCdfChYbcGesoqg3A4bCs0fMKxkU/CwglURrHuyUCdQK270M54ZMRxHtfi6ehCdhC4J1DxBtjUMvHMFHXUj4pc4FRUU1V11VUZONXJCRrIpwFaBdqnp7dyoqctPfdZAamyjtLpp60c2girsJ7UaYLTxGJKn31woiiAEwDYFSdjN1ooWZ36CZVUcMkkwFVWyzUXVVfb/zxWp3iAeusEfVI4YCJeY1xq2h3Ds2HHCfbZ6p4GEPhKHW7XsQHHhwKgAYC4bJj3IBzBPGpXhL+xoaiOqj2ogXrACvdjIcc9d2RzB9oH4QlEEMEhp/Z5a1c5gc0nhIkUOi2Ah/NJAYGN9OEZgNFwEiFZEQRoy6GfUtCH1gHe+uZkAChV2NoAcNw2z4TfuoedFJRmEGSwPzSVZLJPTZcohQTYRx4b2o7zLbR2TCqfbJG1IVemrsnA5oES6mefhLqk2yfSwulpJ6inXZhieknrCSJnN1JgneXCtjxAERPQfHPOO/f88xHVPUGkfb4aoDi566XhHmADcHoAdq4ZYAAbeP8WoSuxyOJyoMF/Whgfdhpg/emIB5q46taxRisfesLJtd3Lo/cvTpOBivN666kvbnvtR7JaVlppLe6txVLHMsa9AWht4QqZp8ffs5QCdvejHzIrRuMNSjx/xpNynCMB0EADKUBdjqR3QARq6Fj9YGBPWvOVxUiEBvvIx0ayspSFJAQA8aBHtRaylX6sYylUE8ub6veTnohphE7Bif+OhyP/pSB+XzlBYKwCvQTmMGwvYsquFEC4AKQOax1JH8QOt5bYaOQBatkI9MQygoE0AGkK3IgG1EIwGPIPeTF8XAAygLeoWUmHYwSXzrp2xq4ZrWs3y8fSBrLGKMaLd9fIlEf/NNIvBNAjA8tp49cCoLSbZYAeELGBHPsYR4EcEiiAHEjt2MGOMZFRkges3EoqqRWFyGM41JtkJz25Er/d7JOjJOV09EElUZZSlatkZStd+UpYxlKWs6RlLW15ywTuDAG7RECEhJOPXV6SNAOIJC6NeR+JMOUAHQKORvohRtQAoASpPGY15SORfhygHtWq0XB4xg9oaoUf9xCmNc2plws4IALrZGc73flOeK7TI3JbjrzohYABiAefDNplZujBDjHqgx3/9GMK8mmDOvYzj9e4Gc8mIICF1oB3xEQMAvxC0I8Ikh7XCOc5PaqSCFRApCMlaUlNetKRLid16nrmPBp3/5wM3C+ZCkkOD/kRQYaxbh0V7AcT18JPhYywcWJZiA1KsMGc8ENGOzocP5j5UaiaJKQopWpVR+oRCWa0Z7kzmljqQcGehHBHG0TIRsJBDw1sJDZjVUjwcrKOscblZ1BiYmsSspCGbASuZsVkPza6sBVEVbAmIZJILVABCxxWsYtFLGMV21jEInaePJJKjSr4kH2o5Wkzo1oAWBeXhcUlf04Zq42OsysF/i19EKmB0wAHgNGVziADUQtFcHQCBjUArh0dbG9DmljgBle4wyUucE220oGkLys69UqMwMoR2jIsAA7ZTUWAtaNpdcaCiFHr3sCZv4MkZCj7cAhaJELO9P8pJByc6W17S1Vc+MY3uMvJKkvzATUa7K1LWJORWuIyxYHkK4gAINhAjpOc9KVgb7cbSEHCC1uglPcp0+LTVzaSXfe697fy5XBwEavS1pFGa1HsBz8kkqEhRjcu86qQZ6W7owJrN2HJ6oc8vNuVozx4KHk1L71gl5UUIOypGYbqPjZAASQnWclLZnKTk4zVJyXIwlpiXc94lyUVU5FrRYEIjCu7lgwgYMTfxLH+7soQCVuXIhKxxx5/piUiD5aTdZlpTiTAXmf5jzTddHFU9lHlnEBPImDSbj/AmJWtdOUrTsldWeKXFh41AIzSjXOl6TIAKkkJoJUcHGrLMqWvXUP/MY28TaA4g2nOeCYDdYJQcVKwDnrYoDj0WEdHTgkzWo8G03j6xqbUwQ5qWlrYeTlwScoJp48Uu3oeKeexgTJsaA+HiRiuSXWifW31/K6YNNHHBCbwLWyHmzv5uK+4zX1udKdb3etmd7vd/W54q+caCNqpixBU3ZogIBwH2ReEVOKgePjRIymIR71bEq1wbJsvBI8Hb+XyDUHF+z1ByYmNZLiRGNcER0O77JBJd/Ft/wzcKZGA+oLzM1/SZR8b8TRv2DEBHEq8JjuiGWoT4hDR2mR30VrHyEdy8ZTr9htGcwn5giN0h9uE1grPSwvZLQ9oRF3qU6d61a0udWdMuAQF/8YupS92D8uMSR/xoIFl4MwOdWjgHvTQZLKgWLWehyUcA7iG2ltDj3ts7Y01WgcNHhNFVCKmMveokkgaEA8NqCMDR7lGcfLRa79jhjKzA/s32h4W3BC+K81DADtW9xN6fH4gzojSaZqXAc+v/SMXWl11vxGPINMgibhZx9gNBncZnQzscx8JO8C+9mbtu026Qc3vY52ZpYwAoKTp+z3W8TUDya7yCAF7yz8qD3hkX/vb5373va/9XxSwMkNT2Akk0jBh5QQi+sjWo32K8Wi+qeMBwKBOhEo4GilL/ZnpCYMitxE984h86JikIpx8sAqdyJD6ywmuY4ouWRi86QdAS/+YBKMWAJiXt3INuBiIr6gyn1mWjTCar9iN++usj6gzekEYBrEBmmkNtgJAF8uJDJAineCHwtgRC8O4BBQsG3gHH/xBIAxCIRxCH4SHXwCKHSk45AgAhtmRuMgXfXCQHJEIgrGBpUgKDaKHBnCWXRELJSgLmukItbAHyOCmAfgOAtsHrKmHDMiAgmCLvXmm6WKYLdwIfBuwO2uAEfoJiZiAAdCt7WKiKuEyJpytn1kuhQizr7iILyKrOGSQhQmBfFqY0bm4WLuGLuKms8qHo7gHn1nE06iW1nCW4BmhVDqOjtCgDDifn/gmG5i/rbAouSmMpVCHBkgB1iGYoriGFFj/GPURngsULHnwwXYoRmM8RmRMRmV8h15AQoZJiDNUHycMgIl5xQSTiAPgmgbIAFoksMlKiicKAAFwCDEkK0grsPPaMyBylgbwjBr4ma+hh8sAiYWBnhToiZ9wCE87Csqpr324IS9axXxwFohYigpZCuhZinDYB8+ARI+xmi2aFSskq3lpF4bkMxwxgdOwitaoxTeCK4FLH0tpo1Wkvw2KFum6uJHUh1VMHAo6i3xAqLFAnmnBiS7BiW2BKhtoB3MwB3DwSaD8SaEMSqIcSqBsh14QPxpZB7FYNbKatI3gxWwxgYYLAKv4kUYLHDCELrWAHv+aMHWUEeWSoRCYDIWQ/6iRsAHq8gi4uSwtuSJIwyGF9KIS7IjhGQjmgrS4eMSpQS2IfMGcmCBR7JuN+BpXDICO8R2PebaQeD8aWAyiCoC0wimjaDwFoojjgBHmaRynQ57Riaqd7EnRHE3SLE3T7EmkBAoqnK57UAvriou3YMpHoqMG2AeEch4Y5Ed1CawSEkdyjMssU7OwLA6R8wyzZBbekZWPsAF76CyXsZ9HM8m4kAgcYp126Yl70JvfNEi8NMe5bMi+1MCFkBs6ih3ZsUDL3ApdAwCNtMD1aYrtoZVgWzVfTJjzGQB+4AdRos/DAb2JuEhreSTzRMmabB0cGQrQ5MnTXFAGNYfUHDCCqf+ypDi/qTkI23wkASA7tNAgG5CILjFJgglHteRKc/xK4cy/CtkRfrjIfrCB9OkIG6gW61uYAtuKn6gW6MmHr9gVtbga6nKoOLk4u/SYvPxKvlwc8UyWB7ieAdAjqzQ5dVRAAFCAjewHdqA4/7k5gbsGfICefTiKhEQKcxyALvWI0NqfET2dP/qnywSKzsSJz9RJchgHcqhTO71TPM3TPB2HcSC680JJsvpKZ4kHdpiXoVmKENgMDJIxGsC7+BGL0TmfcvRKSkvHi9ugsWqXOBSIo5iA21ArkHCWfTMW/2SYb6gWexAIJmpUYQGmqEQAYRnSCrFO+dnLwqTGzoLIEb3/k2SCiLeormkTlvb8VUwiMHoYGJD4Gd2gBwcizEdLn1+jh7XEiX6Ih32hCsBwCF1snNSBmpwssu2picpJx61oDbikoo3Ah8JogPp8EhvBRJ1ArciUVOCUzugip4vzKXwQCJHzIrwJAfaCMrMazAHLCQWwkSvCGxLMiXAoiF3JS5cKU1tdwomZNQBYgeLIAAJkuYEg1kZiIn7AFlnzWB6igXL7CIQJNAH8in5RF50QrZ/BnzpTB6NZKQFToG+VuW2MDYbUh9PYRj/awkFKpTwapJNti2vAKN7JgOTQB30wGqCNohmUWp51Wi/aqJspJGhppCYNHpLII4hQGisRqEEiqDpIYzR6EA/AoYeO2Nk/yoCse9uviVqtdbWSbAtuNJka6Fo/YsmuoCAbuEVM0s+3HduBsqiREFp28CWEBIkGSFpI+ogGGACMMLDDFUCmXdrkyIe8lTm9cDaaEKZKmrO+YImuPBObGMcN4tyj+FCY4DT8hC6QIN3S9dxSmsvt4KHorIlkCkDbtSbKCFjsQIB4UAcMSbmZ8Lx4QN7fbV7nfV7ojV7pHbaAAAA7)
- Cannabinoids = psychoactive compounds found in cannibis
- Endocannabinoids (endogenous cannabinoid system)
- Cannabinoid CB1 receptors in CNS; CB2 in body, immune system
![[[@flores_cannabinoid-hypocretin_2013]](http://dx.doi.org/10.3389/fnins.2013.00256)](data:image/jpeg;base64,/9j/4S9ARXhpZgAASUkqAAgAAAAPAAABAwABAAAA1AUAAAEBAwABAAAACAUAAAIBAwAEAAAAwgAAAAMBAwABAAAAAQAAAAYBAwABAAAAAgAAAA4BAgASAAAAygAAABIBAwABAAAAAQAAABUBAwABAAAABAAAABoBBQABAAAA3AAAABsBBQABAAAA5AAAABwBAwABAAAAAQAAACgBAwABAAAAAgAAADEBAgAeAAAA7AAAADIBAgAUAAAACgEAAGmHBAABAAAAIAEAAEwBAAAIAAgACAAIAENyZWF0ZWQgd2l0aCBHSU1QAID8CgAQJwAAgPwKABAnAABBZG9iZSBQaG90b3Nob3AgQ1M1LjEgV2luZG93cwAyMDEzOjEyOjE3IDE2OjE5OjAzAAAAAwABoAMAAQAAAP//AAACoAQAAQAAAAwDAAADoAQAAQAAAKgCAAAAAAAAAAAGAAMBAwABAAAABgAAABoBBQABAAAAmgEAABsBBQABAAAAogEAACgBAwABAAAAAgAAAAECBAABAAAAqgEAAAICBAABAAAAji0AAAAAAABIAAAAAQAAAEgAAAABAAAA/9j/4gxYSUNDX1BST0ZJTEUAAQEAAAxITGlubwIQAABtbnRyUkdCIFhZWiAHzgACAAkABgAxAABhY3NwTVNGVAAAAABJRUMgc1JHQgAAAAAAAAAAAAAAAQAA9tYAAQAAAADTLUhQICAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABFjcHJ0AAABUAAAADNkZXNjAAABhAAAAGx3dHB0AAAB8AAAABRia3B0AAACBAAAABRyWFlaAAACGAAAABRnWFlaAAACLAAAABRiWFlaAAACQAAAABRkbW5kAAACVAAAAHBkbWRkAAACxAAAAIh2dWVkAAADTAAAAIZ2aWV3AAAD1AAAACRsdW1pAAAD+AAAABRtZWFzAAAEDAAAACR0ZWNoAAAEMAAAAAxyVFJDAAAEPAAACAxnVFJDAAAEPAAACAxiVFJDAAAEPAAACAx0ZXh0AAAAAENvcHlyaWdodCAoYykgMTk5OCBIZXdsZXR0LVBhY2thcmQgQ29tcGFueQAAZGVzYwAAAAAAAAASc1JHQiBJRUM2MTk2Ni0yLjEAAAAAAAAAAAAAABJzUkdCIElFQzYxOTY2LTIuMQAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAWFlaIAAAAAAAAPNRAAEAAAABFsxYWVogAAAAAAAAAAAAAAAAAAAAAFhZWiAAAAAAAABvogAAOPUAAAOQWFlaIAAAAAAAAGKZAAC3hQAAGNpYWVogAAAAAAAAJKAAAA+EAAC2z2Rlc2MAAAAAAAAAFklFQyBodHRwOi8vd3d3LmllYy5jaAAAAAAAAAAAAAAAFklFQyBodHRwOi8vd3d3LmllYy5jaAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABkZXNjAAAAAAAAAC5JRUMgNjE5NjYtMi4xIERlZmF1bHQgUkdCIGNvbG91ciBzcGFjZSAtIHNSR0IAAAAAAAAAAAAAAC5JRUMgNjE5NjYtMi4xIERlZmF1bHQgUkdCIGNvbG91ciBzcGFjZSAtIHNSR0IAAAAAAAAAAAAAAAAAAAAAAAAAAAAAZGVzYwAAAAAAAAAsUmVmZXJlbmNlIFZpZXdpbmcgQ29uZGl0aW9uIGluIElFQzYxOTY2LTIuMQAAAAAAAAAAAAAALFJlZmVyZW5jZSBWaWV3aW5nIENvbmRpdGlvbiBpbiBJRUM2MTk2Ni0yLjEAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHZpZXcAAAAAABOk/gAUXy4AEM8UAAPtzAAEEwsAA1yeAAAAAVhZWiAAAAAAAEwJVgBQAAAAVx/nbWVhcwAAAAAAAAABAAAAAAAAAAAAAAAAAAAAAAAAAo8AAAACc2lnIAAAAABDUlQgY3VydgAAAAAAAAQAAAAABQAKAA8AFAAZAB4AIwAoAC0AMgA3ADsAQABFAEoATwBUAFkAXgBjAGgAbQByAHcAfACBAIYAiwCQAJUAmgCfAKQAqQCuALIAtwC8AMEAxgDLANAA1QDbAOAA5QDrAPAA9gD7AQEBBwENARMBGQEfASUBKwEyATgBPgFFAUwBUgFZAWABZwFuAXUBfAGDAYsBkgGaAaEBqQGxAbkBwQHJAdEB2QHhAekB8gH6AgMCDAIUAh0CJgIvAjgCQQJLAlQCXQJnAnECegKEAo4CmAKiAqwCtgLBAssC1QLgAusC9QMAAwsDFgMhAy0DOANDA08DWgNmA3IDfgOKA5YDogOuA7oDxwPTA+AD7AP5BAYEEwQgBC0EOwRIBFUEYwRxBH4EjASaBKgEtgTEBNME4QTwBP4FDQUcBSsFOgVJBVgFZwV3BYYFlgWmBbUFxQXVBeUF9gYGBhYGJwY3BkgGWQZqBnsGjAadBq8GwAbRBuMG9QcHBxkHKwc9B08HYQd0B4YHmQesB78H0gflB/gICwgfCDIIRghaCG4IggiWCKoIvgjSCOcI+wkQCSUJOglPCWQJeQmPCaQJugnPCeUJ+woRCicKPQpUCmoKgQqYCq4KxQrcCvMLCwsiCzkLUQtpC4ALmAuwC8gL4Qv5DBIMKgxDDFwMdQyODKcMwAzZDPMNDQ0mDUANWg10DY4NqQ3DDd4N+A4TDi4OSQ5kDn8Omw62DtIO7g8JDyUPQQ9eD3oPlg+zD88P7BAJECYQQxBhEH4QmxC5ENcQ9RETETERTxFtEYwRqhHJEegSBxImEkUSZBKEEqMSwxLjEwMTIxNDE2MTgxOkE8UT5RQGFCcUSRRqFIsUrRTOFPAVEhU0FVYVeBWbFb0V4BYDFiYWSRZsFo8WshbWFvoXHRdBF2UXiReuF9IX9xgbGEAYZRiKGK8Y1Rj6GSAZRRlrGZEZtxndGgQaKhpRGncanhrFGuwbFBs7G2MbihuyG9ocAhwqHFIcexyjHMwc9R0eHUcdcB2ZHcMd7B4WHkAeah6UHr4e6R8THz4faR+UH78f6iAVIEEgbCCYIMQg8CEcIUghdSGhIc4h+yInIlUigiKvIt0jCiM4I2YjlCPCI/AkHyRNJHwkqyTaJQklOCVoJZclxyX3JicmVyaHJrcm6CcYJ0kneierJ9woDSg/KHEooijUKQYpOClrKZ0p0CoCKjUqaCqbKs8rAis2K2krnSvRLAUsOSxuLKIs1y0MLUEtdi2rLeEuFi5MLoIuty7uLyQvWi+RL8cv/jA1MGwwpDDbMRIxSjGCMbox8jIqMmMymzLUMw0zRjN/M7gz8TQrNGU0njTYNRM1TTWHNcI1/TY3NnI2rjbpNyQ3YDecN9c4FDhQOIw4yDkFOUI5fzm8Ofk6Njp0OrI67zstO2s7qjvoPCc8ZTykPOM9Ij1hPaE94D4gPmA+oD7gPyE/YT+iP+JAI0BkQKZA50EpQWpBrEHuQjBCckK1QvdDOkN9Q8BEA0RHRIpEzkUSRVVFmkXeRiJGZ0arRvBHNUd7R8BIBUhLSJFI10kdSWNJqUnwSjdKfUrESwxLU0uaS+JMKkxyTLpNAk1KTZNN3E4lTm5Ot08AT0lPk0/dUCdQcVC7UQZRUFGbUeZSMVJ8UsdTE1NfU6pT9lRCVI9U21UoVXVVwlYPVlxWqVb3V0RXklfgWC9YfVjLWRpZaVm4WgdaVlqmWvVbRVuVW+VcNVyGXNZdJ114XcleGl5sXr1fD19hX7NgBWBXYKpg/GFPYaJh9WJJYpxi8GNDY5dj62RAZJRk6WU9ZZJl52Y9ZpJm6Gc9Z5Nn6Wg/aJZo7GlDaZpp8WpIap9q92tPa6dr/2xXbK9tCG1gbbluEm5rbsRvHm94b9FwK3CGcOBxOnGVcfByS3KmcwFzXXO4dBR0cHTMdSh1hXXhdj52m3b4d1Z3s3gReG54zHkqeYl553pGeqV7BHtje8J8IXyBfOF9QX2hfgF+Yn7CfyN/hH/lgEeAqIEKgWuBzYIwgpKC9INXg7qEHYSAhOOFR4Wrhg6GcobXhzuHn4gEiGmIzokziZmJ/opkisqLMIuWi/yMY4zKjTGNmI3/jmaOzo82j56QBpBukNaRP5GokhGSepLjk02TtpQglIqU9JVflcmWNJaflwqXdZfgmEyYuJkkmZCZ/JpomtWbQpuvnByciZz3nWSd0p5Anq6fHZ+Ln/qgaaDYoUehtqImopajBqN2o+akVqTHpTilqaYapoum/adup+CoUqjEqTepqaocqo+rAqt1q+msXKzQrUStuK4trqGvFq+LsACwdbDqsWCx1rJLssKzOLOutCW0nLUTtYq2AbZ5tvC3aLfguFm40blKucK6O7q1uy67p7whvJu9Fb2Pvgq+hL7/v3q/9cBwwOzBZ8Hjwl/C28NYw9TEUcTOxUvFyMZGxsPHQce/yD3IvMk6ybnKOMq3yzbLtsw1zLXNNc21zjbOts83z7jQOdC60TzRvtI/0sHTRNPG1EnUy9VO1dHWVdbY11zX4Nhk2OjZbNnx2nba+9uA3AXcit0Q3ZbeHN6i3ynfr+A24L3hROHM4lPi2+Nj4+vkc+T85YTmDeaW5x/nqegy6LzpRunQ6lvq5etw6/vshu0R7ZzuKO6070DvzPBY8OXxcvH/8ozzGfOn9DT0wvVQ9d72bfb794r4Gfio+Tj5x/pX+uf7d/wH/Jj9Kf26/kv+3P9t////7QAMQWRvYmVfQ00AAv/uAA5BZG9iZQBkgAAAAAH/2wCEAAwICAgJCAwJCQwRCwoLERUPDAwPFRgTExUTExgRDAwMDAwMEQwMDAwMDAwMDAwMDAwMDAwMDAwMDAwMDAwMDAwBDQsLDQ4NEA4OEBQODg4UFA4ODg4UEQwMDAwMEREMDAwMDAwRDAwMDAwMDAwMDAwMDAwMDAwMDAwMDAwMDAwMDP/AABEIAIsAoAMBIgACEQEDEQH/3QAEAAr/xAE/AAABBQEBAQEBAQAAAAAAAAADAAECBAUGBwgJCgsBAAEFAQEBAQEBAAAAAAAAAAEAAgMEBQYHCAkKCxAAAQQBAwIEAgUHBggFAwwzAQACEQMEIRIxBUFRYRMicYEyBhSRobFCIyQVUsFiMzRygtFDByWSU/Dh8WNzNRaisoMmRJNUZEXCo3Q2F9JV4mXys4TD03Xj80YnlKSFtJXE1OT0pbXF1eX1VmZ2hpamtsbW5vY3R1dnd4eXp7fH1+f3EQACAgECBAQDBAUGBwcGBTUBAAIRAyExEgRBUWFxIhMFMoGRFKGxQiPBUtHwMyRi4XKCkkNTFWNzNPElBhaisoMHJjXC0kSTVKMXZEVVNnRl4vKzhMPTdePzRpSkhbSVxNTk9KW1xdXl9VZmdoaWprbG1ub2JzdHV2d3h5ent8f/2gAMAwEAAhEDEQA/APRaqs+jqeVl5WWx2Bc1jaMYt2mos5f6u79J9oc+z1Pb7PToVv7TT2Jd/Va53/UNcoY9YcBkv91lg3AnXaDqK2f6+9WElIftVI+kSwfvPa5o/wA57WtRedQnVPJq9INDNKLXtZbWOAHGNzP3fU/m7G/8J6n0/ppTN+dS0BwILDoLCQ1nyc8t3/8AW96rP6vSJi1gI7Ma+yf7X6Fqxc433WX7XivI3FrXOG7ZB9rdn7rWqmyvrDvaLKHuiIax/P8A0/8AX/wXLn8RmSRCMY60OPilJvx5KAAMzKWl+mhF6B/WhHt3k+TWtH/Sdcgv6xc7hh+dh/8ARLaVm14nVGWj7TkYrah9NpkWag/QZU63b79n0/zFY9PGA1yC4+DKnflsdUop81zX74j4fq4f+lPWyR5fl/3CfH1z/wCh6Ux6neRBZWf64c//AM+WOQzm5B7Vt+FbP+/BygfsoOrrT4e1jf8Av9ibIdTj1ttsx8r03GN2kD+U79F7Gfy0wZeZmaGSRPaJkf8A0kyHHgiLMIgf1gP/AFIz+2ZHiz/ttn/kFNufe0zsqPxrA/6jaqrM/pTzDXa+b3ntv/wWNs+j/LRcfqHQC5pvs9j/AKPsyBOgO7c4V+z3f6NSwxc6SKnMf3zkj/04sUsvKjeMT/dEJf8ARbdXVnNdNocxvd9biYH/ABV3qN/zFqU5Yc5zD+lIDXB1bTBDhubu/Nrs/k71WGP0w1U2Y7Gk5G30nEFxDTE2bLN23bu/Pb/O7FosYytgYwQ0cBaHLwzxBGaYn+7X/felp5pYpEe1Ex73/wB6jOQ1oLnse1o5JbP/AFG5VOo9Zpwoa1pusJIIafa0j817v3/5CvXPqZW42uDWRBJMfisfqduLR0v7PAbZZJpqI1/RlgudDvj/AOCJ+cyjinKMhAxiZcRHFw8Pq+VZiAOSAkOIGQHCDw3xeKI9e6kazeyio0g+4gyW/wDGNa7cz/NVqj6x4bmsF7XVPd9LSWjwJP8AKWAy6zHuFwYGWNAc5g0D63CdpHu+k1Tvrrpydpb6tLWh9YkjdUfcGy385u72PWVj53OAZe4J7DhyR04ifmHtxhOPyZY+3/6U9zG6M+Vwk1wGG5uB1rt65Th+nj9f/pP1vYtcHAOaQWkSCNQQU6wenZzsO6nCcTbjXgHHeYDmh3DXreWriyxyRsbg8Mo/uy7OdkxmBo7HWJ/ei//Q9PxnQw0nR9MNI8W/4Oz+21GQrqi+H1w25n0HH8WO/wCDf+d/5NSqsFrA8AjsWnkEaOaf6qSmTnNY0veQ1rRLnHQADuVl29Wwcxpx6LJuLmupDwWNe5jhZW1r3D857Pai9en9lXR/In4b2blyL7qGuFVljWvfADSYJLtGgfynLN57nsmDLGEIiQ4eOV9deGh+78re5TlMebHKc5EHi4Y100e2FGJkhtz6WOc4fntBcP5LufoLD6x0vOsyS6l1teOBDG44EAHa536Jv5/tczfZVb/wexaXR8z1ceoWH32gkT3ew7L/APO/n/8Arliv2W11MNlrgxjfpOcYA+ZVmeLHnxRmDwCQGTiFbGP6bBGc8WQwI4jEmHCfA/oPH0dC6zbDDk3hwPudsbU2CSY93u/c+h/1tXa+iNY+s2Zt+Ta3aHNqADXbTuex/wCZ+k/O2M9Rn/bfp6d3U8HKZ6VGQJmXNh0uaB7msbt3WO/4Nn01ks6p0jDyvtVmezNyi0jHrb7KamH6bubNrnbPfa+zf/g1FjwctGJyGccsR19MoeXDjZJ5c8pCAicZPQ8Ql/jTdyjEsZ/NsrxR/JAc/wCb3Kt1CyujHdYHmyx8sxzY6d1kHSqoQx+3a5Qx+q1ZfTzfZkM9BkuyrmggNafcyhrfp+o9jmKjgVDrGX+1eot2YNJ2YOM76Jg/T2fnbdv9uz/g8etWhwnFp+qEo/3DDi/7prm/c1/WEH+9x0hxWdSynhjTdtcYNjS5rWx/JZtZ+77FqYWDfTcTfmudtaSWPbqBp+kY+/ft2fvMWh9vo/lfGERj6MgSIftMwRqD/aVfl+XwwIPuHLkGvFKZv/Fj83+Hxs2bNlkD6BjgdOER0+2Xy/4CFjKifWEtoZrveSS8j6LnOsl/o1f4P/hP0n/G0epdfx8QtFr/AEGOJAcQZMRu3O2vbU33fnf+BrSOOXPBseXsa4uDD58B372z8xDf03HfY55LhuJJAIiTqeysZfcoe3V9f7GHHwX67c2jrOEymzNew2VVyPUY19jw4HbtLrN3ptd/pHuqqWLdl3Z1z868QbGlra26iulr/H857rG/pLHLpOoZGF0vHbVsqrpu3B4ePYRG5+/+u38565uo149jTg2NyKJJx3A7w6t4324dv711Hu31/Tsx7PX/ADFU5mWQcvOAkZTjXu+ni/VS+fh/f/r/ANRsYBD3oSMQIyvg14f1kflv91NcwmjGcdXAWUE/1Tur/wCg9PdXa6rFcxrnOsx/TbAn3T7Q7/rTkJmLXdgPrfmX001xNYDTrt9zqnfz9r/pXXP9T1Ge/wDm6UZzL6sg0/tC+tr2NAuivYJDvQa1jW7KmN/0zFWGDFwiZzREJiHAY8UjxaD9Ph/8T5P1k2wc2Ti4RiJlEy4xLQcPq/d/2+P0QZ5TqK7W177W3Y9TW76w0tAaN7Xe8tf/ACt66HpeTZlYFV1oh7hDu0kHbu/tLmGW5dP6tmkXmse5uVq4n9+u0Dc1zv32fo1qfVy94fbRY8kOh1TS7d/X26lSclnBzyiBIDJxemq9uUPl9z+tweni/T/zazmsRGEEmJMBH1XfGJ78H+F6v+7f/9H1VBe19VhurBc10epWOZGgsZ/K2+1//qP9IZJJTXvZVnYltLXaWNLD4tdGm5v0mub+6uKvw623kZNLRfURu3DVrm6sd/5CxdzZSyw7tW2DQWN0cP8AyTf5D/Yq9tNTntdmY7Liz6NwZu8/cz3WM/6bFS53kvvHDKMuCcdL/eg2+U5oYeKMo8UJa+Umn0nFLcHHFoj1XvdB0IDg5zD+8136Nr1m/WTMusyrKaxvbjNGyuYDrCN+7/pbF0TbBfcCwzXTMkd3kRs/62x3vXN9Xps/bFlXDr3M9PTkODWS397a4OUHxCEsXKY8ULIEowP9cCPX+9Jl5KQyczkyS0JjKY/q2f8AvXKqsyHbhfT6cAua8OBBAPs9v0mWbf0n/f12VfTsPJ9PMyKWuvfWzfI0JaNzdzP5G96wcizp7cK23DrpvtrYHY8P3F5c7Yx9vv2X/Rss2bf8D6a2Pq5m5ub042ZrQ25lj2cgktHurL9oDN+x35ij5DFGGWWPJwy4wJxgNY3Dr613OZDLHGcOIcBMTI/N6/7iDq+Bdn5FeE0htL723ZBnaTW1kbWfvO3f+lFLqRyabWMxKWPZWGs9MnYBWAI9M/Rb/mrQootN77sge4GK9dI18E978B7tltlYe3T6QDh+K088eKNWBr+loC0cRqV0Tp+j0cFuZ1MnXprgOdbqwYmO2/8ASfn/ALi0ek2XW2F1lLscwQWOLXHb7djv0ct7qz9lxOfVMf1m/wBys1U11CKxE8nklQ48MuIH0gA3obZJ5Bwkam+4QUdW6ZkZBxacqp+Q0lpp3DfLZ3/oz7/bscrayOodIpqFmfgUNOcwm1jBDQ95O929zW+o31He6z0/pqfT7OrZjLGdQqbTWRAdXuY/dJc72Wbv0LqnVs3f6Rl//BqaGSXGYTHCd41qJRY5QHCJRNjaV/oySZbMbqDm0GkXis7vUd9Fpjbpo7d7US7pHTrcV+IaGMqfE7AGkFv83Y1zf8JX+Y9Wa62VNDKxtaOympBEAmQGp6rLNVejxmN9XOs2ZRc8tb9neQy29xJeGujcytu9/p2/yvTVrL6V1FjmVuqFlLGGttzCPo/SZ6u4td7P39npq+zo2aOoHIdkTWTIEnSdnubr9L2fnqPV+idSzckXUZrqqxumsOc2QQGgez6Ppub6jHtVPPE5Ygzwy9Jr0yr0/NK/3vk/cbOKXtkiOSOo/SH6Xy/905lT8gtGNkYz762CQ2CHsH71dn7iHjvrq6hU+jdsbY2PUADon3B21dB0fp+fhgjKyDcIIa0uc7l25pd6n+jZ+j/lqxf0zDusFxra28EOFgGu4cF4/wAJ/bVeHw+Uo458RjLHMSj7ojLJwQ2/WR4Z/wCOzS5yMTOPCJRnEg+2SIcU+vBLij/iP//S9Pvyq6Ppgn4CVVb1zp+2x73OqbV9Iuaf3bLTt2bvo1UWverr6qrPpsa74gFDGDhgOAorAcQXDYIJb9GdPzZURGX3ARKPt68Ua9XgyA4+Aijx9DfpXxsrHy6hdjv9Ss8OAI7T+d8UZQrqrrBFbGsBMkNAEnx0U1Kxon1vD/VpjcdHsdo10fRM67Ht/e/9R+mHJvyWNa8UgkHX87Tu32+5jlbSQkLBF14hING6vzeXxsbpVfUW0MwNwslrri3ZtD2u37nMad/Hp/T/AJ39J+j/AJ2wv1gt6h02jHxukelRj2Bwe9+7duBY6GWDdtc9nq+936T/ALaXQuY1whwDh4ESg2YdTmuawBgdy0AFhj96pwLFBLl/1cowIjMjhjk4fUI9Y3/WZo5/WDIGUAeKUL9Jl0k53TczqL+gPyc5zHZADmstqkB0fo22bY9jvUWWGQIA0W7n1tu6fbh5LCxtjPT3VhzmGdGgelusZuXMn6vVN3h9N+0aFm55a0CdBH5mv53/AKUVLnsU/wBXxcUhCAiZiNjj/T/utnlckfXw1HikSImX6P6P95sbAfzQfktfoV7wbMZ07QN9YPb817R/0Vzn7IwC76NnYBgsfECRs2B30Pd9BbPROm/Yr33Ma9t9zAxjLHOdtZO51rq3H9HW1382389Q8kP18TC+vFppw/1mXmj+qkJ1/V114v6rs5LXOfU0PLJcdpH7+0ubI/Pbta/2KVNtj3OY9oBZAL2mWkn80Ttd7U9dDGO3uJss43u1P9n81n9hRcHU2eowF1b/AOcaNSD2sa3/AM+f+rFtuUnSQvtNI+kSweL2uYP86wNUvVq/fb94SUzSQ3X0N1dY1vxcAm+01n6G5/htaSP8+Nn/AEklJUkE2ZBBLaYjs9wBP+Z6jf8ApKdVjbWB7eDyDyD3a7+U1JT/AP/T9Lp6ng352R0+q3dl4gab69rhtD2tsZ73N2P9j2fzblaQK8LDrybcuuitmTeALbmtAe8NAa3fZ9J3tYxTdkUN+lYwfFwSUkSQ25FD3bW2NLjw2RP3IiSlJITsisEhsvLdDtEgEfmuf/Nt/tOVW3q+NXy+oeLTYC7/ADKRckiUox+YiPmab6SyLPrBjD6Di7ybW4/jYaECz6wz9Blnx9jPy/aEeE9mI8zhG8x9PV/0XdIDgWuEg6EHghC+yYwMtqaxw4c0bT/nM2uWE7r1hH8ySfE2kf8ARqZUhftrL7NZ/ada78tyPCVh53AP0ifof+6ej+zs/eeQOBvd/wCS/wCqU2V11iGNDQdSAI1XMftrK/cq+6wf+jlJnXLwfdWHeQstH/f3pcEkffsHc/Y9OksGjr7S6Ld1IPDnRawf1topuWqzMrJLLC0PgObsO4Oa6dljI93u2oEEbs2PLjyXwS4q3bKh6Vf7jfuCj9qpH0nFg8XgtH3vDU1mQGuNbBvsA3OaDAaD9Fzz+buhBkSBjAZDQD5BSQfUvmNrA6J27j/5FRZm1kO3gsLDDzBLRoHz6m3bt2O3+9JTYUPSr3+ptG+I3RrHxU0klP8A/9T02qoXtF1/vDxLKz9FrT9H2fRdZ+85Ha1rRDQAPAaIWKR6Ir/Op/RuHm0f9/b70ZJTF7GPaWvaHNPIIkKte2ynaxjnejc4MJklzNx/Nf8AS2v+h/wT1bVTLsZdQ5tJ32Mc17WNOrjW5tu1k+38xNlOESBKQiZaRBNcX91IBOw2eZzPtWV6j2bAWlzceqyfTaGu2tZsZ9H2tVZtfUWj349XHDbYE+Grf9f/AD11x6fgZMX+n/Oe6QXNmdZLQWqhm4WTVePsT241YEbizfJJad24+5/t3s2+rXs+n+kVHm83NYgZGUY4jLhjwC5Rj+jx/wDozBj5LGT6hxz3lKR+Y+TlUYHVXvb6mGxtB+lb6pEAgn2tt2ep7tjP/BFcHSz3NI/tF3/UNemrx+vXQXZFAsn3BlTnaSfz3/ydm3d/wn84jnF6hW+s3Zte4Bnq1117gdf0oaz6VT3t/m3+r/n+n+lqQyc1nP6ozl3Jy5Pb/wDUf/TbH3bBDeMR/gxRjpre76x8GOP5QxDuowcZrXZGQ2sPdtYTXoXc7R71sMpe7WuhrB+/cdx/7bCD1EWtxLjuF9gaW1MeA2pzz/gy0e9//CKxDks1cWbLGAAuXCcktPGc8npQY4R8sL+g/KLmtb0zdt+2Vbjw0GsHx+j6hcp0P6JY4A9Qrg6AttZzG793b/0lmdPdbeDjNZiNyWvIe2vGGwNIbWzb6rd/s9H+c99a2sfpTrLybG4r6mAtbX6Td1cnc3jb7f0bPZZs+gliwYJz0jlyxB4fcPB7Jr/n8ChXDYAhY+U3xpn9N6eaWPa71heWtqLnS07uHN2bN/s/SLSporoYGViB3PcwNuv9kKtTTWXt9MzTSINhgD26iqrYGMZWxzd9vp/8X++g5vW8bHcGeo1m6Q1zyBJA3RW1zmf9J3/bivfqcEP0ccL/AOd/3SKs3Wvd0bACxwcYaQZPgFnsbQymptVjWXWPOx5ILnRt+1E7v532Md9L/g3/AOjVavq3T3Mddmu/RN4se7e0uB2+m2pjWsdZu+hsreg4ByOqZjusx6dVbX04eOYJLZAyLbHf4O236H6P/wAE/wAIcWWGWPHjPFG6ujHb+8oinTPqNe2t5Ja7+asgB4dBdDhX7PoNf+bX/N+lYosfcx86D1Tz2FkbNr/5D/S/Qu/6z+4pWPBYHjs+uyeIBc3e7/MdY5S2l25rf8HY0mTw0OY93/UKRS2NcWACP0RdtDRywk7do/ep3H2/6L/iv5u6qLGutY87CW2PkPa6CPcHNd/0f7atUPc9nv8AptJa6PEfnf2m+5JT/9X0+6t0+tV/OtERwHtH+Df/AOi3/wCD/wC3N5K7G2MD28O+/wCB/lNUkB00WOsAJpfq8NElrv39v7r/AM/b/wClElKzCRjujvAPwJWfImJg8jWD8VpWBt9BFbg4PHtcDIPgsS/p2HkXsyLqy62pzHtMkQ+szW7aPzmrD+LQ/Xwkb4ZQof3oylf/AEmxgPpIG9uth2zNZ/Omxv3/AKX/ADbfd/11itLPqpDm0OdoS92xw5Et+k3/ALbWgtblZyngxSl80oRJ8dPm/wAJhmAJEDu1802CgljgxoM2PJDYYPpO3H6O1YtORXiZfr5eXU+wAtxsDHd+8N3q3ufsc+30x+cz9H710JAIg8Fc5X9UqDc/1KaGVOMOLBq5sl3tbG2vdu9yHMTyxAGKJlKWnF0gtdKrqX2vDFrHsraQ511rXhza2NJ93qfR37Vj0uZ1nNblZBNPScN23Fq1DrHj8/2/pP6//bP/AHJVjqnTX2bOl4NQppvvY7INQDQ2pjB7tv8AWb/256az+rYuLVnsa69+JUwNx6a2Fza9jHN3sft9n6z67WPstd/hP7abnzSw4o68UzUeIj9KvVLhir6W9LX1HAZLGzWJ42ECfkEfbjZLW2DbY3gOHgfpN0/N/eYuUoz+ltrrpry6nbGhjJeJO32bv7e36a2ehXNtdY6l4spcJ3NMt3AlnP8AZVbludyyyxxziKloCAYriHQdRY8hj3D0QT7WiJb+Yw/u7VTyug4+Va51pD2OJdsexro3EOdt3f1f3VoV30WkiqxthaYcGuBgjTWERXcuDHmAGSPFwm46mP8A0UAkOPkUdL6XifZBQ11eVuD2ucG7u7vc7879xrEDoTvs4f0t1m/buuw7QQTZRYZc5sf4Wi/+farnVcTC6lsxbWutewkw0wBI2PFmo3M2pj9XOmtwzi4zDjHcLK7a3EPZY0em29hn2v2ex/8ApGJmKGSGScaAwUPaiBHQ/p/11Gq8WOZR1NzmjGu9B54sawEEBrtv6N/tb7nfn+xVsDC6g0WZWZkutFztceCPTaPb6e/cfV2x+f8Apf8AhLFlY+J9ZMjNZaRbc/Ct+na8srJYdlgr3fTbc1uzfXX9BdHbdXU79I19bt+5oc0wd2jmCyv1Ktz2l3t9Tf8A9tqaUATEkn07D9Hi/eQmtqp9HTaaXaQRuYR/wjIc3b/mJYvpVvFVJaKntLmMaRA2kBxYGf8AGN+j7EwLI9Sh7YP0hI2uP8n03bmbv9WKVrf0YucIc339iQW+/wBr2j85jXNTlP8A/9b1VJZ/Ur+ossqZhsGzm20lh5/Q7Ay2yn+a9X7dv/wleI/F/nMhierMyhgbzV9oy6jsdUx1YL4f6Qv/AJx1VTbqx9p9N1n6Nn6L+cTeOPFw3r2VR3psuxxuL6j6Vh1JHBP/AAjPz/8Az5/wiE8Ul360z03/AL4J2O/t6f8Agn/TQh1LKLHWfs++AGkN9m8zu9T2ue1v6Lb+/wC//B/o/wBKmHVcggEdOyZJ2wQwaxu/0n0P+ESnjhMcM4iY3qQ4gkEjY02twsvY2uCyobnOHEkbGMH9l2//ALbR1UoLywZNVLqxbrZjvhrgZjeNdm/9/wB3v/8APg+o9Suw6m2sxX2jdD2yAQ2DL2xva53/AAacB0WykIgyOw3ocX/Ra1fVsh+c0CPs7n7A2NYJ2Ns3LYXD9MZ0GrrGNRjuLX72+mQXbYO+yup1VjvZ9J/5n89ZYyldH1rqHUsQ1MwcdtwsBL3ue1pBDmfQa76X6M2/S/P9JNz5I4oGctIx/kGDlZSmJeri1bNFdluS6+5pYazDBwD9If2lLK6fjZR3PEWRG4Rx/KDg5qrYefnWdKsycyptN7A4AMIc0kADcz3O9vq/v7P+LWFl2WV1PtaNzwCX2ul2wBrnHIfWP0uT6cf0er9Nd/g1R5znccRjj7YzDLEZNTw1E/Kf02yB+DtjoNIkBzQ06ECpokeCvYuHRiV+nS2ATJMAEn+ztauTxcrLsteywOaxoEwXNLDA9MOlzm3/AGpv6wz0v6J/R8n9OtfED8+l+JZdYyyqX02scQ8bmvq1f+d6bn+oouT5jl/eEBh9uch6ZcXuf9L5Um6Vl9ExMO6zrGJSXZVRddsZO95cS6ytthd/hNzv5z2Kzg5+Z1BtjH47sQAaXBweN247qh9B+70vSf6m3/CWf6FUOgdP6zhNt+25V733PHptvsFo3Na9z4+l6dN276Df9F9CtbtV/qOcwsLHsA3jQgE9tzVfhg9vJKUJVCXzw+b1/vxlxej+6shk44CRiY30l6ZfUL00V0N21iPE9z8URJZlvXK6892F6FjixzWusiG+5r7NzS6N+z09r9n+mo/0iOfmMWACWWXCDpsZf9FdGJloBaF9fU6cuzJyLwzDYCZLoDQA2XP09zPbZ/r9AeeOs5lxd0zKYKGlzLGjaSDDdjYsHtfW/wCnv+mx62MnGoyqH4+QwWVWCHsPBHgVDDwMTBrdViVipjnb3Bvdx/OVIchOM+GOSXsy9c5GX67j9X/U/wBL9OC8zBGo9X4NLpWJnsJOeGOO0tLtrJd7pr/m2j/B/wA5/LVt+C3aRS91YOvpzLD+83a7d6bX/wDA+mrSSvYsXtw4eKU+tzPFJYTb/9f0XqWTXWS0vAcBwZWVRYzLx8+osryWgUfonvNLT73fSug7du3crvVvQ9V3qbJgfS9bw/4D2rMo+zfY+o+l9i4o3fafX9H+cdt+0faf/AvS/wAL/OKrKPLfeIEy/XWeCP8Agy4v0f3GKuZ9wa/qdeKuPb9H+p8y+Rh1DZ6nT6muDHa25YNgHo3M2+72u/Oq3+p/N/aMj+dW3ZldVFjxThNfUQDS91oYZIaXC6vY709ri76HqLnv1bbVP7N2bPzPX9WPQf8AzOz9N9L/AOFfr/4NdgrTK08C7qNpeM3HbjwBs2uDwZdZ+cD/AKJtG72M/Sb/AOcVxJJJSJ2LjOdvNTN/78Dd/nfSQ7sMPbtaQW8hlg3tnxaZ9Rn9lyspJs+HhPHXDXq4vlrx4lDwc7NbXb063DP6o5zNo12gH/grvof2vprmjgWtkOzcpreGtLoLY3e3f9Jzv9f+L7HIj0XTEaTunxHG33b/APR/y1WHpepZs9TfGn854D+d9X9F/nrO53Hy8pRvL7UuAcPp9yBx/oUuFvLHBOhGdkhoAaAHjRo/N37d/wDa+mtbomNdj3PyTbbf6rfSortI2nXe97Xbd2yv8+3/AM+WI7fW9YT6Md49Ld/0fctGjZ9ofs2xt/O3erM9/V93pfuKHlMWKOaBOcTIPohGBjxS8ZFRutkjKTuFlrjY8fR7NHb2M/8AJKBP2e0uP8zaZJ/cd4/8W/8A6D/66sJLZWo2X0vO1j2l3gDqplrSZIBPihZX80P5v6Q/nvo/+dKoNkf4D+zuhCXDpxVvpf7ytei/VekDqQZORbj+mHCKzAO8bdzx/I/MVpt1FTRX6ocWiInc4/2WqnprH2X+1P47leo/mx9D/rf0VXx/d/fycH8/X6z5tv8AorjxcIv5ejE5DollVjx4wG/9GwsciMe2xjXsMtcJB8ipKtj7PVfxv3O/m92yJ/P/AMF63+kVla//2f/tNiZQaG90b3Nob3AgMy4wADhCSU0EBAAAAAAALRwBWgADGyVHHAFaAAMbJUccAgAAAqcEHAJ4ABFDcmVhdGVkIHdpdGggR0lNUAA4QklNBCUAAAAAABCoazx2PW+xlPEIbospFYArOEJJTQQ6AAAAAACTAAAAEAAAAAEAAAAAAAtwcmludE91dHB1dAAAAAUAAAAAQ2xyU2VudW0AAAAAQ2xyUwAAAABSR0JDAAAAAEludGVlbnVtAAAAAEludGUAAAAAQ2xybQAAAABNcEJsYm9vbAEAAAAPcHJpbnRTaXh0ZWVuQml0Ym9vbAAAAAALcHJpbnRlck5hbWVURVhUAAAAAQAAADhCSU0EOwAAAAABsgAAABAAAAABAAAAAAAScHJpbnRPdXRwdXRPcHRpb25zAAAAEgAAAABDcHRuYm9vbAAAAAAAQ2xicmJvb2wAAAAAAFJnc01ib29sAAAAAABDcm5DYm9vbAAAAAAAQ250Q2Jvb2wAAAAAAExibHNib29sAAAAAABOZ3R2Ym9vbAAAAAAARW1sRGJvb2wAAAAAAEludHJib29sAAAAAABCY2tnT2JqYwAAAAEAAAAAAABSR0JDAAAAAwAAAABSZCAgZG91YkBv4AAAAAAAAAAAAEdybiBkb3ViQG/gAAAAAAAAAAAAQmwgIGRvdWJAb+AAAAAAAAAAAABCcmRUVW50RiNSbHQAAAAAAAAAAAAAAABCbGQgVW50RiNSbHQAAAAAAAAAAAAAAABSc2x0VW50RiNQeGxAUgAAAAAAAAAAAAp2ZWN0b3JEYXRhYm9vbAEAAAAAUGdQc2VudW0AAAAAUGdQcwAAAABQZ1BDAAAAAExlZnRVbnRGI1JsdAAAAAAAAAAAAAAAAFRvcCBVbnRGI1JsdAAAAAAAAAAAAAAAAFNjbCBVbnRGI1ByY0BZAAAAAAAAOEJJTQPtAAAAAAAQAEgAAAABAAIASAAAAAEAAjhCSU0EJgAAAAAADgAAAAAAAAAAAAA/gAAAOEJJTQPyAAAAAAAKAAD///////8AADhCSU0EDQAAAAAABAAAAB44QklNBBkAAAAAAAQAAAAeOEJJTQPzAAAAAAAJAAAAAAAAAAABADhCSU0nEAAAAAAACgABAAAAAAAAAAI4QklNA/UAAAAAAEgAL2ZmAAEAbGZmAAYAAAAAAAEAL2ZmAAEAoZmaAAYAAAAAAAEAMgAAAAEAWgAAAAYAAAAAAAEANQAAAAEALQAAAAYAAAAAAAE4QklNA/gAAAAAAHAAAP////////////////////////////8D6AAAAAD/////////////////////////////A+gAAAAA/////////////////////////////wPoAAAAAP////////////////////////////8D6AAAOEJJTQQIAAAAAAAQAAAAAQAAAkAAAAJAAAAAADhCSU0EHgAAAAAABAAAAAA4QklNBBoAAAAAA1sAAAAGAAAAAAAAAAAAAAKoAAADDAAAABMAZgBuAGkAbgBzAC0AMAA3AC0AMAAwADIANQA2AC0AZwAwADAAMQAAAAEAAAAAAAAAAAAAAAAAAAAAAAAAAQAAAAAAAAAAAAADDAAAAqgAAAAAAAAAAAAAAAAAAAAAAQAAAAAAAAAAAAAAAAAAAAAAAAAQAAAAAQAAAAAAAG51bGwAAAACAAAABmJvdW5kc09iamMAAAABAAAAAAAAUmN0MQAAAAQAAAAAVG9wIGxvbmcAAAAAAAAAAExlZnRsb25nAAAAAAAAAABCdG9tbG9uZwAAAqgAAAAAUmdodGxvbmcAAAMMAAAABnNsaWNlc1ZsTHMAAAABT2JqYwAAAAEAAAAAAAVzbGljZQAAABIAAAAHc2xpY2VJRGxvbmcAAAAAAAAAB2dyb3VwSURsb25nAAAAAAAAAAZvcmlnaW5lbnVtAAAADEVTbGljZU9yaWdpbgAAAA1hdXRvR2VuZXJhdGVkAAAAAFR5cGVlbnVtAAAACkVTbGljZVR5cGUAAAAASW1nIAAAAAZib3VuZHNPYmpjAAAAAQAAAAAAAFJjdDEAAAAEAAAAAFRvcCBsb25nAAAAAAAAAABMZWZ0bG9uZwAAAAAAAAAAQnRvbWxvbmcAAAKoAAAAAFJnaHRsb25nAAADDAAAAAN1cmxURVhUAAAAAQAAAAAAAG51bGxURVhUAAAAAQAAAAAAAE1zZ2VURVhUAAAAAQAAAAAABmFsdFRhZ1RFWFQAAAABAAAAAAAOY2VsbFRleHRJc0hUTUxib29sAQAAAAhjZWxsVGV4dFRFWFQAAAABAAAAAAAJaG9yekFsaWduZW51bQAAAA9FU2xpY2VIb3J6QWxpZ24AAAAHZGVmYXVsdAAAAAl2ZXJ0QWxpZ25lbnVtAAAAD0VTbGljZVZlcnRBbGlnbgAAAAdkZWZhdWx0AAAAC2JnQ29sb3JUeXBlZW51bQAAABFFU2xpY2VCR0NvbG9yVHlwZQAAAABOb25lAAAACXRvcE91dHNldGxvbmcAAAAAAAAACmxlZnRPdXRzZXRsb25nAAAAAAAAAAxib3R0b21PdXRzZXRsb25nAAAAAAAAAAtyaWdodE91dHNldGxvbmcAAAAAADhCSU0EKAAAAAAADAAAAAI/8AAAAAAAADhCSU0EEQAAAAAAAQEAOEJJTQQUAAAAAAAEAAAABThCSU0EDAAAAAAtqgAAAAEAAACgAAAAiwAAAeAAAQSgAAAtjgAYAAH/2P/iDFhJQ0NfUFJPRklMRQABAQAADEhMaW5vAhAAAG1udHJSR0IgWFlaIAfOAAIACQAGADEAAGFjc3BNU0ZUAAAAAElFQyBzUkdCAAAAAAAAAAAAAAABAAD21gABAAAAANMtSFAgIAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAEWNwcnQAAAFQAAAAM2Rlc2MAAAGEAAAAbHd0cHQAAAHwAAAAFGJrcHQAAAIEAAAAFHJYWVoAAAIYAAAAFGdYWVoAAAIsAAAAFGJYWVoAAAJAAAAAFGRtbmQAAAJUAAAAcGRtZGQAAALEAAAAiHZ1ZWQAAANMAAAAhnZpZXcAAAPUAAAAJGx1bWkAAAP4AAAAFG1lYXMAAAQMAAAAJHRlY2gAAAQwAAAADHJUUkMAAAQ8AAAIDGdUUkMAAAQ8AAAIDGJUUkMAAAQ8AAAIDHRleHQAAAAAQ29weXJpZ2h0IChjKSAxOTk4IEhld2xldHQtUGFja2FyZCBDb21wYW55AABkZXNjAAAAAAAAABJzUkdCIElFQzYxOTY2LTIuMQAAAAAAAAAAAAAAEnNSR0IgSUVDNjE5NjYtMi4xAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABYWVogAAAAAAAA81EAAQAAAAEWzFhZWiAAAAAAAAAAAAAAAAAAAAAAWFlaIAAAAAAAAG+iAAA49QAAA5BYWVogAAAAAAAAYpkAALeFAAAY2lhZWiAAAAAAAAAkoAAAD4QAALbPZGVzYwAAAAAAAAAWSUVDIGh0dHA6Ly93d3cuaWVjLmNoAAAAAAAAAAAAAAAWSUVDIGh0dHA6Ly93d3cuaWVjLmNoAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAGRlc2MAAAAAAAAALklFQyA2MTk2Ni0yLjEgRGVmYXVsdCBSR0IgY29sb3VyIHNwYWNlIC0gc1JHQgAAAAAAAAAAAAAALklFQyA2MTk2Ni0yLjEgRGVmYXVsdCBSR0IgY29sb3VyIHNwYWNlIC0gc1JHQgAAAAAAAAAAAAAAAAAAAAAAAAAAAABkZXNjAAAAAAAAACxSZWZlcmVuY2UgVmlld2luZyBDb25kaXRpb24gaW4gSUVDNjE5NjYtMi4xAAAAAAAAAAAAAAAsUmVmZXJlbmNlIFZpZXdpbmcgQ29uZGl0aW9uIGluIElFQzYxOTY2LTIuMQAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAdmlldwAAAAAAE6T+ABRfLgAQzxQAA+3MAAQTCwADXJ4AAAABWFlaIAAAAAAATAlWAFAAAABXH+dtZWFzAAAAAAAAAAEAAAAAAAAAAAAAAAAAAAAAAAACjwAAAAJzaWcgAAAAAENSVCBjdXJ2AAAAAAAABAAAAAAFAAoADwAUABkAHgAjACgALQAyADcAOwBAAEUASgBPAFQAWQBeAGMAaABtAHIAdwB8AIEAhgCLAJAAlQCaAJ8ApACpAK4AsgC3ALwAwQDGAMsA0ADVANsA4ADlAOsA8AD2APsBAQEHAQ0BEwEZAR8BJQErATIBOAE+AUUBTAFSAVkBYAFnAW4BdQF8AYMBiwGSAZoBoQGpAbEBuQHBAckB0QHZAeEB6QHyAfoCAwIMAhQCHQImAi8COAJBAksCVAJdAmcCcQJ6AoQCjgKYAqICrAK2AsECywLVAuAC6wL1AwADCwMWAyEDLQM4A0MDTwNaA2YDcgN+A4oDlgOiA64DugPHA9MD4APsA/kEBgQTBCAELQQ7BEgEVQRjBHEEfgSMBJoEqAS2BMQE0wThBPAE/gUNBRwFKwU6BUkFWAVnBXcFhgWWBaYFtQXFBdUF5QX2BgYGFgYnBjcGSAZZBmoGewaMBp0GrwbABtEG4wb1BwcHGQcrBz0HTwdhB3QHhgeZB6wHvwfSB+UH+AgLCB8IMghGCFoIbgiCCJYIqgi+CNII5wj7CRAJJQk6CU8JZAl5CY8JpAm6Cc8J5Qn7ChEKJwo9ClQKagqBCpgKrgrFCtwK8wsLCyILOQtRC2kLgAuYC7ALyAvhC/kMEgwqDEMMXAx1DI4MpwzADNkM8w0NDSYNQA1aDXQNjg2pDcMN3g34DhMOLg5JDmQOfw6bDrYO0g7uDwkPJQ9BD14Peg+WD7MPzw/sEAkQJhBDEGEQfhCbELkQ1xD1ERMRMRFPEW0RjBGqEckR6BIHEiYSRRJkEoQSoxLDEuMTAxMjE0MTYxODE6QTxRPlFAYUJxRJFGoUixStFM4U8BUSFTQVVhV4FZsVvRXgFgMWJhZJFmwWjxayFtYW+hcdF0EXZReJF64X0hf3GBsYQBhlGIoYrxjVGPoZIBlFGWsZkRm3Gd0aBBoqGlEadxqeGsUa7BsUGzsbYxuKG7Ib2hwCHCocUhx7HKMczBz1HR4dRx1wHZkdwx3sHhYeQB5qHpQevh7pHxMfPh9pH5Qfvx/qIBUgQSBsIJggxCDwIRwhSCF1IaEhziH7IiciVSKCIq8i3SMKIzgjZiOUI8Ij8CQfJE0kfCSrJNolCSU4JWgllyXHJfcmJyZXJocmtyboJxgnSSd6J6sn3CgNKD8ocSiiKNQpBik4KWspnSnQKgIqNSpoKpsqzysCKzYraSudK9EsBSw5LG4soizXLQwtQS12Last4S4WLkwugi63Lu4vJC9aL5Evxy/+MDUwbDCkMNsxEjFKMYIxujHyMioyYzKbMtQzDTNGM38zuDPxNCs0ZTSeNNg1EzVNNYc1wjX9Njc2cjauNuk3JDdgN5w31zgUOFA4jDjIOQU5Qjl/Obw5+To2OnQ6sjrvOy07azuqO+g8JzxlPKQ84z0iPWE9oT3gPiA+YD6gPuA/IT9hP6I/4kAjQGRApkDnQSlBakGsQe5CMEJyQrVC90M6Q31DwEQDREdEikTORRJFVUWaRd5GIkZnRqtG8Ec1R3tHwEgFSEtIkUjXSR1JY0mpSfBKN0p9SsRLDEtTS5pL4kwqTHJMuk0CTUpNk03cTiVObk63TwBPSU+TT91QJ1BxULtRBlFQUZtR5lIxUnxSx1MTU19TqlP2VEJUj1TbVShVdVXCVg9WXFapVvdXRFeSV+BYL1h9WMtZGllpWbhaB1pWWqZa9VtFW5Vb5Vw1XIZc1l0nXXhdyV4aXmxevV8PX2Ffs2AFYFdgqmD8YU9homH1YklinGLwY0Njl2PrZEBklGTpZT1lkmXnZj1mkmboZz1nk2fpaD9olmjsaUNpmmnxakhqn2r3a09rp2v/bFdsr20IbWBtuW4SbmtuxG8eb3hv0XArcIZw4HE6cZVx8HJLcqZzAXNdc7h0FHRwdMx1KHWFdeF2Pnabdvh3VnezeBF4bnjMeSp5iXnnekZ6pXsEe2N7wnwhfIF84X1BfaF+AX5ifsJ/I3+Ef+WAR4CogQqBa4HNgjCCkoL0g1eDuoQdhICE44VHhauGDoZyhteHO4efiASIaYjOiTOJmYn+imSKyoswi5aL/IxjjMqNMY2Yjf+OZo7OjzaPnpAGkG6Q1pE/kaiSEZJ6kuOTTZO2lCCUipT0lV+VyZY0lp+XCpd1l+CYTJi4mSSZkJn8mmia1ZtCm6+cHJyJnPedZJ3SnkCerp8dn4uf+qBpoNihR6G2oiailqMGo3aj5qRWpMelOKWpphqmi6b9p26n4KhSqMSpN6mpqhyqj6sCq3Wr6axcrNCtRK24ri2uoa8Wr4uwALB1sOqxYLHWskuywrM4s660JbSctRO1irYBtnm28Ldot+C4WbjRuUq5wro7urW7LrunvCG8m70VvY++Cr6Evv+/er/1wHDA7MFnwePCX8Lbw1jD1MRRxM7FS8XIxkbGw8dBx7/IPci8yTrJuco4yrfLNsu2zDXMtc01zbXONs62zzfPuNA50LrRPNG+0j/SwdNE08bUSdTL1U7V0dZV1tjXXNfg2GTY6Nls2fHadtr724DcBdyK3RDdlt4c3qLfKd+v4DbgveFE4cziU+Lb42Pj6+Rz5PzlhOYN5pbnH+ep6DLovOlG6dDqW+rl63Dr++yG7RHtnO4o7rTvQO/M8Fjw5fFy8f/yjPMZ86f0NPTC9VD13vZt9vv3ivgZ+Kj5OPnH+lf65/t3/Af8mP0p/br+S/7c/23////tAAxBZG9iZV9DTQAC/+4ADkFkb2JlAGSAAAAAAf/bAIQADAgICAkIDAkJDBELCgsRFQ8MDA8VGBMTFRMTGBEMDAwMDAwRDAwMDAwMDAwMDAwMDAwMDAwMDAwMDAwMDAwMDAENCwsNDg0QDg4QFA4ODhQUDg4ODhQRDAwMDAwREQwMDAwMDBEMDAwMDAwMDAwMDAwMDAwMDAwMDAwMDAwMDAwM/8AAEQgAiwCgAwEiAAIRAQMRAf/dAAQACv/EAT8AAAEFAQEBAQEBAAAAAAAAAAMAAQIEBQYHCAkKCwEAAQUBAQEBAQEAAAAAAAAAAQACAwQFBgcICQoLEAABBAEDAgQCBQcGCAUDDDMBAAIRAwQhEjEFQVFhEyJxgTIGFJGhsUIjJBVSwWIzNHKC0UMHJZJT8OHxY3M1FqKygyZEk1RkRcKjdDYX0lXiZfKzhMPTdePzRieUpIW0lcTU5PSltcXV5fVWZnaGlqa2xtbm9jdHV2d3h5ent8fX5/cRAAICAQIEBAMEBQYHBwYFNQEAAhEDITESBEFRYXEiEwUygZEUobFCI8FS0fAzJGLhcoKSQ1MVY3M08SUGFqKygwcmNcLSRJNUoxdkRVU2dGXi8rOEw9N14/NGlKSFtJXE1OT0pbXF1eX1VmZ2hpamtsbW5vYnN0dXZ3eHl6e3x//aAAwDAQACEQMRAD8A9Fqqz6Op5WXlZbHYFzWNoxi3aaizl/q7v0n2hz7PU9vs9OhW/tNPYl39Vrnf9Q1yhj1hwGS/3WWDcCddoOorZ/r71YSUh+1Uj6RLB+89rmj/ADnta1F51CdU8mr0g0M0ote1ltY4AcY3M/d9T+bsb/wnqfT+mlM351LQHAgsOgsJDWfJzy3f/wBb3qs/q9ImLWAjsxr7J/tfoWrFzjfdZfteK8jcWtc4btkH2t2futaqbK+sO9osoe6IhrH8/wDT/wBf/BcufxGZJEIxjrQ4+KUm/HkoAAzMpaX6aEXoH9aEe3eT5Na0f9J1yC/rFzuGH52H/wBEtpWbXidUZaPtORitqH02mRZqD9BlTrdvv2fT/MVj08YDXILj4Mqd+Wx1SinzXNfviPh+rh/6U9bJHl+X/cJ8fXP/AKHpTHqd5EFlZ/rhz/8Az5Y5DObkHtW34Vs/78HKB+yg6utPh7WN/wC/2Jsh1OPW22zHyvTcY3aQP5Tv0XsZ/LTBl5mZoZJE9omR/wDSTIceCIswiB/WA/8AUjP7ZkeLP+22f+QU2597TOyo/GsD/qNqqsz+lPMNdr5vee2//BY2z6P8tFx+odALmm+z2P8Ao+zIE6A7tzhX7Pd/o1LDFzpIqcx/fOSP/TixSy8qN4xP90Ql/wBFt1dWc102hzG931uJgf8AFXeo3/MWpTlhznMP6UgNcHVtMEOG5u782uz+TvVYY/TDVTZjsaTkbfScQXENMTZss3bdu789v87sWixjK2BjBDRwFocvDPEEZpif7tf996WnmlikR7UTHvf/AHqM5DWguex7Wjkls/8AUblU6j1mnChrWm6wkghp9rSPzXu/f/kK9c+plbja4NZEEkx+Kx+p24tHS/s8BtlkmmojX9GWC50O+P8A4In5zKOKcoyEDGJlxEcXDw+r5VmIA5ICQ4gZAcIPDfF4oj17qRrN7KKjSD7iDJb/AMY1rtzP81WqPrHhuawXtdU930tJaPAk/wApYDLrMe4XBgZY0BzmDQPrcJ2ke76TVO+uunJ2lvq0taH1iSN1R9wbLfzm7vY9ZWPnc4Bl7gnsOHJHTiJ+Ye3GE4/Jlj7f/pT3Mboz5XCTXAYbm4HWu3rlOH6eP1/+k/W9i1wcA5pBaRII1BBTrB6dnOw7qcJxNuNeAcd5gOaHcNet5auLLHJGxuDwyj+7Ls52TGYGjsdYn96L/9D0/GdDDSdH0w0jxb/g7P7bUZCuqL4fXDbmfQcfxY7/AIN/53/k1KqwWsDwCOxaeQRo5p/qpKZOc1jS95DWtEucdAAO5WXb1bBzGnHosm4ua6kPBY17mOFlbWvcPzns9qL16f2VdH8ifhvZuXIvuoa4VWWNa98ANJgku0aB/Kcs3nueyYMsYQiJDh45X114aH7vyt7lOUx5scpzkQeLhjXTR7YUYmSG3PpY5zh+e0Fw/ku5+gsPrHS86zJLqXW144EMbjgQAdrnfom/n+1zN9lVv/B7FpdHzPVx6hYffaCRPd7Dsv8A87+f/wCuWK/ZbXUw2WuDGN+k5xgD5lWZ4sefFGYPAJAZOIVsY/psEZzxZDAjiMSYcJ8D+g8fR0LrNsMOTeHA+52xtTYJJj3e79z6H/W1dr6I1j6zZm35Nrdoc2oANdtO57H/AJn6T87Yz1Gf9t+np3dTwcpnpUZAmZc2HS5oHuaxu3dY7/g2fTWSzqnSMPK+1WZ7M3KLSMetvspqYfpu5s2uds99r7N/+DUWPBy0YnIZxyxHX0yh5cONknlzykICJxk9DxCX+NN3KMSxn82yvFH8kBz/AJvcq3ULK6Md1gebLHyzHNjp3WQdKqhDH7drlDH6rVl9PN9mQz0GS7KuaCA1p9zKGt+n6j2OYqOBUOsZf7V6i3Zg0nZg4zvomD9PZ+dt2/27P+Dx61aHCcWn6oSj/cMOL/umub9zX9YQf73HSHFZ1LKeGNN21xg2NLmtbH8lm1n7vsWphYN9NxN+a521pJY9uoGn6Rj79+3Z+8xaH2+j+V8YRGPoyBIh+0zBGoP9pV+X5fDAg+4cuQa8Upm/8WPzf4fGzZs2WQPoGOB04RHT7ZfL/gIWMqJ9YS2hmu95JLyPouc6yX+jV/g/+E/Sf8bR6l1/HxC0Wv8AQY4kBxBkxG7c7a9tTfd+d/4GtI45c8Gx5exri4MPnwHfvbPzEN/Tcd9jnkuG4kkAiJOp7Kxl9yh7dX1/sYcfBfrtzaOs4TKbM17DZVXI9RjX2PDgdu0us3em13+ke6qpYt2XdnXPzrxBsaWtrbqK6Wv8fznusb+kscuk6hkYXS8dtWyqum7cHh49hEbn7/67fznrm6jXj2NODY3IoknHcDvDq3jfbh2/vXUe7fX9OzHs9f8AMVTmZZBy84CRlONe76eL9VL5+H9/+v8A1GxgEPehIxAjK+DXh/WR+W/3U1zCaMZx1cBZQT/VO6v/AKD091drqsVzGuc6zH9NsCfdPtDv+tOQmYtd2A+t+ZfTTXE1gNOu33Oqd/P2v+ldc/1PUZ7/AObpRnMvqyDT+0L62vY0C6K9gkO9BrWNbsqY3/TMVYYMXCJnNEQmIcBjxSPFoP0+H/xPk/WTbBzZOLhGImUTLjEtBw+r93/b4/RBnlOortbXvtbdj1NbvrDS0Bo3td7y1/8AK3roel5NmVgVXWiHuEO7SQdu7+0uYZbl0/q2aReax7m5Wrif367QNzXO/fZ+jWp9XL3h9tFjyQ6HVNLt39fbqVJyWcHPKIEgMnF6ar25Q+X3P63B6eL9P/NrOaxEYQSYkwEfVd8Ynvwf4Xq/7t//0fVUF7X1WG6sFzXR6lY5kaCxn8rb7X/+o/0hkklNe9lWdiW0tdpY0sPi10abm/Sa5v7q4q/DrbeRk0tF9RG7cNWubqx3/kLF3NlLLDu1bYNBY3Rw/wDJN/kP9ir201Oe12ZjsuLPo3Bm7z9zPdYz/psVLneS+8cMoy4Jx0v96Db5Tmhh4oyjxQlr5SafScUtwccWiPVe90HQgODnMP7zXfo2vWb9ZMy6zKsprG9uM0bK5gOsI37v+lsXRNsF9wLDNdMyR3eRGz/rbHe9c31emz9sWVcOvcz09OQ4NZLf3trg5QfEISxcpjxQsgSjA/1wI9f70mXkpDJzOTJLQmMpj+rZ/wC9cqqzIduF9PpwC5rw4EEA+z2/SZZt/Sf9/XZV9Ow8n08zIpa699bN8jQlo3N3M/kb3rByLOntwrbcOum+2tgdjw/cXlztjH2+/Zf9GyzZt/wPprY+rmbm5vTjZmtDbmWPZyCS0e6sv2gM37HfmKPkMUYZZY8nDLjAnGA1jcOvrXc5kMscZw4hwExMj83r/uIOr4F2fkV4TSG0vvbdkGdpNbWRtZ+87d/6UUupHJptYzEpY9lYaz0ydgFYAj0z9Fv+atCii03vuyB7gYr10jXwT3vwHu2W2Vh7dPpAOH4rTzx4o1YGv6WgLRxGpXROn6PRwW5nUydemuA51urBiY7b/wBJ+f8AuLR6TZdbYXWUuxzBBY4tcdvt2O/Ry3urP2XE59Ux/Wb/AHKzVTXUIrETyeSVDjwy4gfSADehtknkHCRqb7hBR1bpmRkHFpyqn5DSWmncN8tnf+jPv9uxytrI6h0imoWZ+BQ05zCbWMEND3k73b3Nb6jfUd7rPT+mp9Ps6tmMsZ1CptNZEB1e5j90lzvZZu/QuqdWzd/pGX/8GpoZJcZhMcJ3jWolFjlAcIlE2NpX+jJJlsxuoObQaReKzu9R30WmNumjt3tRLukdOtxX4hoYyp8TsAaQW/zdjXN/wlf5j1ZrrZU0MrG1o7KakEQCZAanqss1V6PGY31c6zZlFzy1v2d5DLb3El4a6NzK273+nb/K9NWsvpXUWOZW6oWUsYa23MI+j9Jnq7i13s/f2emr7OjZo6gch2RNZMgSdJ2e5uv0vZ+eo9X6J1LNyRdRmuqrG6aw5zZBAaB7Po+m5vqMe1U88TliDPDL0mvTKvT80r/e+T9xs4pe2SI5I6j9IfpfL/3TmVPyC0Y2RjPvrYJDYIewfvV2fuIeO+urqFT6N2xtjY9QAOifcHbV0HR+n5+GCMrINwghrS5zuXbml3qf6Nn6P+WrF/TMO6wXGtrbwQ4WAa7hwXj/AAn9tV4fD5SjjnxGMscxKPuiMsnBDb9ZHhn/AI7NLnIxM48IlGcSD7ZIhxT68EuKP+I//9L0+/Kro+mCfgJVVvXOn7bHvc6ptX0i5p/dstO3Zu+jVRa96uvqqs+mxrviAUMYOGA4CisBxBcNgglv0Z0/NlREZfcBEo+3rxRr1eDIDj4CKPH0N+lfGysfLqF2O/1Kzw4AjtP53xRlCuqusEVsawEyQ0ASfHRTUrGifW8P9WmNx0ex2jXR9Ezrse397/1H6Ycm/JY1rxSCQdfztO7fb7mOVtJCQsEXXiEg0bq/N5fGxulV9RbQzA3CyWuuLdm0Pa7fucxp38en9P8Anf0n6P8AnbC/WC3qHTaMfG6R6VGPYHB737t24FjoZYN21z2er73fpP8AtpdC5jXCHAOHgRKDZh1Oa5rAGB3LQAWGP3qnAsUEuX/VyjAiMyOGOTh9Qj1jf9Zmjn9YMgZQB4pQv0mXSTndNzOov6A/JznMdkAOay2qQHR+jbZtj2O9RZYZAgDRbufW27p9uHksLG2M9PdWHOYZ0aB6W6xm5cyfq9U3eH037RoWbnlrQJ0Efma/nf8ApRUuexT/AFfFxSEICJmI2OP9P+62eVyR9fDUeKRIiZfo/o/3mxsB/NB+S1+hXvBsxnTtA31g9vzXtH/RXOfsjALvo2dgGCx8QJGzYHfQ930Fs9E6b9ivfcxr233MDGMsc521k7nWurcf0dbXfzbfz1DyQ/XxML68WmnD/WZeaP6qQnX9XXXi/quzktc59TQ8slx2kfv7S5sj89u1r/YpU22Pc5j2gFkAvaZaSfzRO13tT10MY7e4myzje7U/2fzWf2FFwdTZ6jAXVv8A5xo1IPaxrf8Az5/6sW25SdJC+00j6RLB4va5g/zrA1S9Wr99v3hJTNJDdfQ3V1jW/FwCb7TWfobn+G1pI/z42f8ASSUlSQTZkEEtpiOz3AE/5nqN/wCkp1WNtYHt4PIPIPdrv5TUlP8A/9P0unqeDfnZHT6rd2XiBpvr2uG0Pa2xnvc3Y/2PZ/NuVpArwsOvJty66K2ZN4Atua0B7w0Brd9n0ne1jFN2RQ36VjB8XBJSRJDbkUPdtbY0uPDZE/ciJKUkhOyKwSGy8t0O0SAR+a5/823+05Vber41fL6h4tNgLv8AMpFySJSjH5iI+ZpvpLIs+sGMPoOLvJtbj+NhoQLPrDP0GWfH2M/L9oR4T2YjzOEbzH09X/Rd0gOBa4SDoQeCEL7JjAy2prHDhzRtP+cza5YTuvWEfzJJ8TaR/wBGplSF+2svs1n9p1rvy3I8JWHncA/SJ+h/7p6P7Oz955A4G93/AJL/AKpTZXXWIY0NB1IAjVcx+2sr9yr7rB/6OUmdcvB91Yd5Cy0f9/elwSR9+wdz9j06SwaOvtLot3Ug8OdFrB/W2im5arMyskssLQ+A5uw7g5rp2WMj3e7agQRuzY8uPJfBLirdsqHpV/uN+4KP2qkfScWDxeC0fe8NTWZAa41sG+wDc5oMBoP0XPP5u6EGRIGMBkNAPkFJB9S+Y2sDonbuP/kVFmbWQ7eCwsMPMEtGgfPqbdu3Y7f70lNhQ9Kvf6m0b4jdGsfFTSSU/wD/1PTaqhe0XX+8PEsrP0WtP0fZ9F1n7zkdrWtENAA8BohYpHoiv86n9G4ebR/39vvRklMXsY9pa9oc08giQq17bKdrGOd6NzgwmSXM3H81/wBLa/6H/BPVtVMuxl1Dm0nfYxzXtY06uNbm27WT7fzE2U4RIEpCJlpEE1xf3UgE7DZ5nM+1ZXqPZsBaXNx6rJ9Noa7a1mxn0fa1Vm19RaPfj1ccNtgT4at/1/8APXXHp+Bkxf6f857pBc2Z1ktBaqGbhZNV4+xPbjVgRuLN8klp3bj7n+3ezb6tez6f6RUebzc1iBkZRjiMuGPALlGP6PH/AOjMGPksZPqHHPeUpH5j5OVRgdVe9vqYbG0H6VvqkQCCfa23Z6nu2M/8EVwdLPc0j+0Xf9Q16avH69dBdkUCyfcGVOdpJ/Pf/J2bd3/CfziOcXqFb6zdm17gGerXXXuB1/ShrPpVPe3+bf6v+f6f6WpDJzWc/qjOXcnLk9v/ANR/9NsfdsEN4xH+DFGOmt7vrHwY4/lDEO6jBxmtdkZDaw921hNehdztHvWwyl7ta6GsH79x3H/tsIPURa3EuO4X2BpbUx4DanPP+DLR73/8IrEOSzVxZssYAC5cJyS08ZzyelBjhHywv6D8oua1vTN237ZVuPDQawfH6PqFynQ/oljgD1CuDoC21nMbv3dv/SWZ091t4OM1mI3Ja8h7a8YbA0htbNvqt3+z0f5z31rax+lOsvJsbivqYC1tfpN3VydzeNvt/Rs9lmz6CWLBgnPSOXLEHh9w8Hsmv+fwKFcNgCFj5TfGmf03p5pY9rvWF5a2oudLTu4c3Zs3+z9ItKmiuhgZWIHc9zA26/2Qq1NNZe30zNNIg2GAPbqKqtgYxlbHN32+n/xf76Dm9bxsdwZ6jWbpDXPIEkDdFbXOZ/0nf9uK9+pwQ/Rxwv8A53/dIqzda93RsALHBxhpBk+AWextDKam1WNZdY87HkgudG37UTu/nfYx30v+Df8A6NVq+rdPcx12a79E3ix7t7S4Hb6bamNax1m76Gyt6DgHI6pmO6zHp1VtfTh45gktkDItsd/g7bfofo//AAT/AAhxZYZY8eM8Ubq6Mdv7yiKdM+o17a3klrv5qyAHh0F0OFfs+g1/5tf836Viix9zHzoPVPPYWRs2v/kP9L9C7/rP7ilY8FgeOz67J4gFzd7v8x1jlLaXbmt/wdjSZPDQ5j3f9QpFLY1xYAI/RF20NHLCTt2j96ncfb/ov+K/m7qosa61jzsJbY+Q9roI9wc13/R/tq1Q9z2e/wCm0lro8R+d/ab7klP/1fT7q3T61X860RHAe0f4N/8A6Lf/AIP/ALc3krsbYwPbw77/AIH+U1SQHTRY6wAml+rw0SWu/f2/uv8Az9v/AKUSUrMJGO6O8A/AlZ8iYmDyNYPxWlYG30EVuDg8e1wMg+CxL+nYeRezIurLranMe0yRD6zNbto/OasP4tD9fCRvhlCh/ejKV/8ASbGA+kgb262HbM1n86bG/f8Apf8ANt93/XWK0s+qkObQ52hL3bHDkS36Tf8AttaC1uVnKeDFKXzShEnx0+b/AAmGYAkQO7XzTYKCWODGgzY8kNhg+k7cfo7Vi05FeJl+vl5dT7AC3GwMd37w3ere5+xz7fTH5zP0fvXQkAiDwVzlf1SoNz/UpoZU4w4sGrmyXe1sba9273IcxPLEAYomUpacXSC10qupfa8MWseytpDnXWteHNrY0n3ep9HftWPS5nWc1uVkE09Jw3bcWrUOsePz/b+k/r/9s/8AclWOqdNfZs6Xg1Cmm+9jsg1ANDamMHu2/wBZv/bnprP6ti4tWexrr34lTA3HprYXNr2Mc3ex+32frPrtY+y13+E/tpufNLDijrxTNR4iP0q9UuGKvpb0tfUcBksbNYnjYQJ+QR9uNktbYNtjeA4eB+k3T8395i5SjP6W2uumvLqdsaGMl4k7fZu/t7fprZ6Fc211jqXiylwnc0y3cCWc/wBlVuW53LLLHHOIqWgIBiuIdB1FjyGPcPRBPtaIlv5jD+7tVPK6Dj5VrnWkPY4l2x7GujcQ523d/V/dWhXfRaSKrG2Fphwa4GCNNYRFdy4MeYAZI8XCbjqY/wDRQCQ4+RR0vpeJ9kFDXV5W4Pa5wbu7u9zvzv3GsQOhO+zh/S3Wb9u67DtBBNlFhlzmx/haL/59qudVxMLqWzFta617CTDTAEjY8WajczamP1c6a3DOLjMOMdwsrtrcQ9ljR6bb2Gfa/Z7H/wCkYmYoZIZJxoDBQ9qIEdD+n/XUarxY5lHU3OaMa70HnixrAQQGu2/o3+1vud+f7FWwMLqDRZlZmS60XO1x4I9No9vp79x9XbH5/wCl/wCEsWVj4n1kyM1lpFtz8K36dryyslh2WCvd9NtzW7N9df0F0dt1dTv0jX1u37mhzTB3aOYLK/Uq3PaXe31N/wD22ppQBMSSfTsP0eL95Ca2qn0dNppdpBG5hH/CMhzdv+Yli+lW8VUloqe0uYxpEDaQHFgZ/wAY36PsTAsj1KHtg/SEja4/yfTduZu/1YpWt/Ri5whzff2JBb7/AGvaPzmNc1OU/wD/1vVUln9Sv6iyypmGwbObbSWHn9DsDLbKf5r1ft2//CV4j8X+cyGJ6szKGBvNX2jLqOx1THVgvh/pC/8AnHVVNurH2n03Wfo2fov5xN448XDevZVHemy7HG4vqPpWHUkcE/8ACM/P/wDPn/CITxSXfrTPTf8AvgnY7+3p/wCCf9NCHUsosdZ+z74AaQ32bzO71Pa57W/otv7/AL/8H+j/AEqYdVyCAR07JknbBDBrG7/SfQ/4RKeOExwziJjepDiCQSNjTa3Cy9ja4LKhuc4cSRsYwf2Xb/8AttHVSgvLBk1UurFutmO+GuBmN412b/3/AHe//wA+D6j1K7DqbazFfaN0PbIBDYMvbG9rnf8ABpwHRbKQiDI7Dehxf9FrV9WyH5zQI+zufsDY1gnY2zcthcP0xnQausY1GO4tfvb6ZBdtg77K6nVWO9n0n/mfz1ljKV0fWuodSxDUzBx23CwEve57WkEOZ9Brvpfozb9L8/0k3PkjigZy0jH+QYOVlKYl6uLVs0V2W5Lr7mlhrMMHAP0h/aUsrp+NlHc8RZEbhHH8oODmqth5+dZ0qzJzKm03sDgAwhzSQANzPc72+r+/s/4tYWXZZXU+1o3PAJfa6XbAGucch9Y/S5Ppx/R6v013+DVHnOdxxGOPtjMMsRk1PDUT8p/TbIH4O2Og0iQHNDToQKmiR4K9i4dGJX6dLYBMkwASf7O1q5PFysuy17LA5rGgTBc0sMD0w6XObf8Aam/rDPS/on9Hyf0618QPz6X4ll1jLKpfTaxxDxua+rV/53puf6ii5PmOX94QGH25yHplxe5/0vlSbpWX0TEw7rOsYlJdlVF12xk73lxLrK22F3+E3O/nPYrODn5nUG2MfjuxABpcHB43bjuqH0H7vS9J/qbf8JZ/oVQ6B0/rOE237blXvfc8em2+wWjc1r3Pj6Xp03bvoN/0X0K1u1X+o5zCwsewDeNCAT23NV+GD28kpQlUJfPD5vX+/GXF6P7qyGTjgJGJjfSXpl9QvTRXQ3bWI8T3PxRElmW9crrz3YXoWOLHNa6yIb7mvs3NLo37PT2v2f6aj/SI5+YxYAJZZcIOmxl/0V0YmWgFoX19Tpy7MnIvDMNgJkugNADZc/T3M9tn+v0B546zmXF3TMpgoaXMsaNpIMN2Niwe19b/AKe/6bHrYycajKofj5DBZVYIew8EeBUMPAxMGt1WJWKmOdvcG93H85UhyE4z4Y5JezL1zkZfruP1f9T/AEv04LzMEaj1fg0ulYmewk54Y47S0u2sl3umv+baP8H/ADn8tW34LdpFL3Vg6+nMsP7zdrt3ptf/AMD6atJK9ixe3Dh4pT63M8UlhNv/1/RepZNdZLS8BwHBlZVFjMvHz6iyvJaBR+ie80tPvd9K6Dt27dyu9W9D1XepsmB9L1vD/gPasyj7N9j6j6X2Lijd9p9f0f5x237R9p/8C9L/AAv84qso8t94gTL9dZ4I/wCDLi/R/cYq5n3Br+p14q49v0f6nzL5GHUNnqdPqa4Mdrblg2Aejczb7va786rf6n839oyP51bdmV1UWPFOE19RANL3WhhkhpcLq9jvT2uLvoeoue/VttU/s3Zs/M9f1Y9B/wDM7P030v8A4V+v/g12CtMrTwLuo2l4zcduPAGza4PBl1n5wP8Aom0bvYz9Jv8A5xXEkklInYuM5281M3/vwN3+d9JDuww9u1pBbyGWDe2fFpn1Gf2XKykmz4eE8dcNeri+WvHiUPBzs1tdvTrcM/qjnM2jXaAf+Cu+h/a+muaOBa2Q7Nymt4a0ugtjd7d/0nO/1/4vsciPRdMRpO6fEcbfdv8A9H/LVYel6lmz1N8afzngP531f0X+es7ncfLylG8vtS4Bw+n3IHH+hS4W8scE6EZ2SGgBoAeNGj83ft3/ANr6a1uiY12Pc/JNtt/qt9Kiu0jadd73tdt3bK/z7f8Az5Yjt9b1hPox3j0t3/R9y0aNn2h+zbG387d6sz39X3el+4oeUxYo5oE5xMg+iEYGPFLxkVG62SMpO4WWuNjx9Hs0dvYz/wAkoE/Z7S4/zNpkn9x3j/xb/wDoP/rqwktlajZfS87WPaXeAOqmWtJkgE+KFlfzQ/m/pD+e+j/50qg2R/gP7O6EJcOnFW+l/vK16L9V6QOpBk5FuP6YcIrMA7xt3PH8j8xWm3UVNFfqhxaIidzj/ZaqemsfZf7U/juV6j+bH0P+t/RVfH939/Jwfz9frPm2/wCiuPFwi/l6MTkOiWVWPHjAb/0bCxyIx7bGNewy1wkHyKkq2Ps9V/G/c7+b3bIn8/8AwXrf6RWVr//ZOEJJTQQhAAAAAABZAAAAAQEAAAAPAEEAZABvAGIAZQAgAFAAaABvAHQAbwBzAGgAbwBwAAAAFQBBAGQAbwBiAGUAIABQAGgAbwB0AG8AcwBoAG8AcAAgAEMAUwA1AC4AMQAAAAEAOEJJTQQGAAAAAAAHAAgAAAABAQD/4ROraHR0cDovL25zLmFkb2JlLmNvbS94YXAvMS4wLwA8P3hwYWNrZXQgYmVnaW49Iu+7vyIgaWQ9Ilc1TTBNcENlaGlIenJlU3pOVGN6a2M5ZCI/PiA8eDp4bXBtZXRhIHhtbG5zOng9ImFkb2JlOm5zOm1ldGEvIiB4OnhtcHRrPSJBZG9iZSBYTVAgQ29yZSA1LjAtYzA2MSA2NC4xNDA5NDksIDIwMTAvMTIvMDctMTA6NTc6MDEgICAgICAgICI+IDxyZGY6UkRGIHhtbG5zOnJkZj0iaHR0cDovL3d3dy53My5vcmcvMTk5OS8wMi8yMi1yZGYtc3ludGF4LW5zIyI+IDxyZGY6RGVzY3JpcHRpb24gcmRmOmFib3V0PSIiIHhtbG5zOmRjPSJodHRwOi8vcHVybC5vcmcvZGMvZWxlbWVudHMvMS4xLyIgeG1sbnM6eG1wPSJodHRwOi8vbnMuYWRvYmUuY29tL3hhcC8xLjAvIiB4bWxuczpwaG90b3Nob3A9Imh0dHA6Ly9ucy5hZG9iZS5jb20vcGhvdG9zaG9wLzEuMC8iIHhtbG5zOnhtcE1NPSJodHRwOi8vbnMuYWRvYmUuY29tL3hhcC8xLjAvbW0vIiB4bWxuczpzdEV2dD0iaHR0cDovL25zLmFkb2JlLmNvbS94YXAvMS4wL3NUeXBlL1Jlc291cmNlRXZlbnQjIiB4bWxuczpzdFJlZj0iaHR0cDovL25zLmFkb2JlLmNvbS94YXAvMS4wL3NUeXBlL1Jlc291cmNlUmVmIyIgZGM6Zm9ybWF0PSJpbWFnZS9qcGVnIiB4bXA6Q3JlYXRlRGF0ZT0iMjAxMy0xMi0xMFQxMzowMTo0NiswNTozMCIgeG1wOk1vZGlmeURhdGU9IjIwMTMtMTItMTdUMTY6MTk6MDMrMDU6MzAiIHhtcDpNZXRhZGF0YURhdGU9IjIwMTMtMTItMTdUMTY6MTk6MDMrMDU6MzAiIHhtcDpDcmVhdG9yVG9vbD0iQWRvYmUgUGhvdG9zaG9wIENTNS4xIFdpbmRvd3MiIHBob3Rvc2hvcDpDb2xvck1vZGU9IjMiIHhtcE1NOkluc3RhbmNlSUQ9InhtcC5paWQ6QThFRjBEQUMwODY3RTMxMUJDOTdEQzMwMTc2MjhERjgiIHhtcE1NOkRvY3VtZW50SUQ9InhtcC5kaWQ6ODlBQjEzMzQ5NTYxRTMxMUFBOThCNjc4NzlFOUVCNDMiIHhtcE1NOk9yaWdpbmFsRG9jdW1lbnRJRD0ieG1wLmRpZDo4OUFCMTMzNDk1NjFFMzExQUE5OEI2Nzg3OUU5RUI0MyI+IDxkYzpkZXNjcmlwdGlvbj4gPHJkZjpBbHQ+IDxyZGY6bGkgeG1sOmxhbmc9IngtZGVmYXVsdCI+Q3JlYXRlZCB3aXRoIEdJTVA8L3JkZjpsaT4gPC9yZGY6QWx0PiA8L2RjOmRlc2NyaXB0aW9uPiA8eG1wTU06SGlzdG9yeT4gPHJkZjpTZXE+IDxyZGY6bGkgc3RFdnQ6YWN0aW9uPSJjcmVhdGVkIiBzdEV2dDppbnN0YW5jZUlEPSJ4bXAuaWlkOjg5QUIxMzM0OTU2MUUzMTFBQTk4QjY3ODc5RTlFQjQzIiBzdEV2dDp3aGVuPSIyMDEzLTEyLTEwVDEzOjAxOjQ2KzA1OjMwIiBzdEV2dDpzb2Z0d2FyZUFnZW50PSJBZG9iZSBQaG90b3Nob3AgQ1M1LjEgV2luZG93cyIvPiA8cmRmOmxpIHN0RXZ0OmFjdGlvbj0ic2F2ZWQiIHN0RXZ0Omluc3RhbmNlSUQ9InhtcC5paWQ6OEFBQjEzMzQ5NTYxRTMxMUFBOThCNjc4NzlFOUVCNDMiIHN0RXZ0OndoZW49IjIwMTMtMTItMTBUMTc6NTQ6MTYrMDU6MzAiIHN0RXZ0OnNvZnR3YXJlQWdlbnQ9IkFkb2JlIFBob3Rvc2hvcCBDUzUuMSBXaW5kb3dzIiBzdEV2dDpjaGFuZ2VkPSIvIi8+IDxyZGY6bGkgc3RFdnQ6YWN0aW9uPSJzYXZlZCIgc3RFdnQ6aW5zdGFuY2VJRD0ieG1wLmlpZDo4QkFCMTMzNDk1NjFFMzExQUE5OEI2Nzg3OUU5RUI0MyIgc3RFdnQ6d2hlbj0iMjAxMy0xMi0xMFQxNzo1NDoyNyswNTozMCIgc3RFdnQ6c29mdHdhcmVBZ2VudD0iQWRvYmUgUGhvdG9zaG9wIENTNS4xIFdpbmRvd3MiIHN0RXZ0OmNoYW5nZWQ9Ii8iLz4gPHJkZjpsaSBzdEV2dDphY3Rpb249ImNvbnZlcnRlZCIgc3RFdnQ6cGFyYW1ldGVycz0iZnJvbSBpbWFnZS90aWZmIHRvIGltYWdlL2Vwc2YiLz4gPHJkZjpsaSBzdEV2dDphY3Rpb249ImRlcml2ZWQiIHN0RXZ0OnBhcmFtZXRlcnM9ImNvbnZlcnRlZCBmcm9tIGltYWdlL3RpZmYgdG8gaW1hZ2UvZXBzZiIvPiA8cmRmOmxpIHN0RXZ0OmFjdGlvbj0ic2F2ZWQiIHN0RXZ0Omluc3RhbmNlSUQ9InhtcC5paWQ6OENBQjEzMzQ5NTYxRTMxMUFBOThCNjc4NzlFOUVCNDMiIHN0RXZ0OndoZW49IjIwMTMtMTItMTBUMTc6NTQ6MjcrMDU6MzAiIHN0RXZ0OnNvZnR3YXJlQWdlbnQ9IkFkb2JlIFBob3Rvc2hvcCBDUzUuMSBXaW5kb3dzIiBzdEV2dDpjaGFuZ2VkPSIvIi8+IDxyZGY6bGkgc3RFdnQ6YWN0aW9uPSJzYXZlZCIgc3RFdnQ6aW5zdGFuY2VJRD0ieG1wLmlpZDpBN0VGMERBQzA4NjdFMzExQkM5N0RDMzAxNzYyOERGOCIgc3RFdnQ6d2hlbj0iMjAxMy0xMi0xN1QxNjoxOTowMyswNTozMCIgc3RFdnQ6c29mdHdhcmVBZ2VudD0iQWRvYmUgUGhvdG9zaG9wIENTNS4xIFdpbmRvd3MiIHN0RXZ0OmNoYW5nZWQ9Ii8iLz4gPHJkZjpsaSBzdEV2dDphY3Rpb249ImNvbnZlcnRlZCIgc3RFdnQ6cGFyYW1ldGVycz0iZnJvbSBpbWFnZS9lcHNmIHRvIGltYWdlL2pwZWciLz4gPHJkZjpsaSBzdEV2dDphY3Rpb249ImRlcml2ZWQiIHN0RXZ0OnBhcmFtZXRlcnM9ImNvbnZlcnRlZCBmcm9tIGltYWdlL2Vwc2YgdG8gaW1hZ2UvanBlZyIvPiA8cmRmOmxpIHN0RXZ0OmFjdGlvbj0ic2F2ZWQiIHN0RXZ0Omluc3RhbmNlSUQ9InhtcC5paWQ6QThFRjBEQUMwODY3RTMxMUJDOTdEQzMwMTc2MjhERjgiIHN0RXZ0OndoZW49IjIwMTMtMTItMTdUMTY6MTk6MDMrMDU6MzAiIHN0RXZ0OnNvZnR3YXJlQWdlbnQ9IkFkb2JlIFBob3Rvc2hvcCBDUzUuMSBXaW5kb3dzIiBzdEV2dDpjaGFuZ2VkPSIvIi8+IDwvcmRmOlNlcT4gPC94bXBNTTpIaXN0b3J5PiA8eG1wTU06RGVyaXZlZEZyb20gc3RSZWY6aW5zdGFuY2VJRD0ieG1wLmlpZDpBN0VGMERBQzA4NjdFMzExQkM5N0RDMzAxNzYyOERGOCIgc3RSZWY6ZG9jdW1lbnRJRD0ieG1wLmRpZDo4OUFCMTMzNDk1NjFFMzExQUE5OEI2Nzg3OUU5RUI0MyIgc3RSZWY6b3JpZ2luYWxEb2N1bWVudElEPSJ4bXAuZGlkOjg5QUIxMzM0OTU2MUUzMTFBQTk4QjY3ODc5RTlFQjQzIi8+IDwvcmRmOkRlc2NyaXB0aW9uPiA8L3JkZjpSREY+IDwveDp4bXBtZXRhPiAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgIDw/eHBhY2tldCBlbmQ9InciPz7/7gAOQWRvYmUAZEAAAAAB/9sAhAABAQEBAQEBAQEBAQEBAQEBAQEBAQEBAQEBAQEBAQEBAQEBAQEBAQEBAQEBAgICAgICAgICAgIDAwMDAwMDAwMDAQEBAQEBAQEBAQECAgECAgMDAwMDAwMDAwMDAwMDAwMDAwMDAwMDAwMDAwMDAwMDAwMDAwMDAwMDAwMDAwMDAwP/wAARCAKoAwwDAREAAhEBAxEB/90ABABi/8QBogAAAAYCAwEAAAAAAAAAAAAABwgGBQQJAwoCAQALAQAABgMBAQEAAAAAAAAAAAAGBQQDBwIIAQkACgsQAAIBAwQBAwMCAwMDAgYJdQECAwQRBRIGIQcTIgAIMRRBMiMVCVFCFmEkMxdScYEYYpElQ6Gx8CY0cgoZwdE1J+FTNoLxkqJEVHNFRjdHYyhVVlcassLS4vJkg3SThGWjs8PT4yk4ZvN1Kjk6SElKWFlaZ2hpanZ3eHl6hYaHiImKlJWWl5iZmqSlpqeoqaq0tba3uLm6xMXGx8jJytTV1tfY2drk5ebn6Onq9PX29/j5+hEAAgEDAgQEAwUEBAQGBgVtAQIDEQQhEgUxBgAiE0FRBzJhFHEIQoEjkRVSoWIWMwmxJMHRQ3LwF+GCNCWSUxhjRPGisiY1GVQ2RWQnCnODk0Z0wtLi8lVldVY3hIWjs8PT4/MpGpSktMTU5PSVpbXF1eX1KEdXZjh2hpamtsbW5vZnd4eXp7fH1+f3SFhoeIiYqLjI2Oj4OUlZaXmJmam5ydnp+So6SlpqeoqaqrrK2ur6/9oADAMBAAIRAxEAPwDf49+690wbm3XtfZeHq9xbx3Hgdp7ex/g+/wA9ubL4/A4WhNTPHS0wq8plaikoaY1FTMkaa3Gt3Ci5IHv3XupGB3Bgd04ig3BtjNYnceAysAqsXnMDkaPL4fJUzMyCpx+Tx81RRVkBdCNcbstwRf37r3Tv7917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de66v/gT7917rFPU09NGZqmaKniX9Us8iQxj83LyMqjj+p9+690F24u+ejdnrI+7e5uqNrpEVWVtxdibQwqxl20oHOSzFMFLtwAeSePfuvdArV/Pn4hxVT0GJ7t29vbIKpdKDrLG7m7Tq6n0yPaki66we5/u2KxNYRluQR9ffutkEZI6cqL5YY/cdM0+w+ifkzvQuqtQlunsv17R1yszBnjyfblV19QQJHpufK6OQRpDfj3XqYrUdJyTuH5j56qnpdpfDvBbUpWsaPOdvfILaGNieJgtjVYTrPBdlZKnmFzePynlSNQ4Pv3XseZ6x1GF/mD7iWZH338UesYpAGglwWxOz+zcpTEmM+OWfPbr2NjKgKNY1ClS5sbDke/deNPKvXX+g/wCYmTpZI8383YsXUNUrNG+xfjj15iFigWMKaZm3Vmt6STBpCSX9JtYACxJ91odSKb4zd3yySHO/OTvzIQyRMgp8TtPo3bYjkPpE8c9F1lNVBghPpLldVjbj37rZNeA6hD4X5SaMx5D5jfNStJPMkPbu3sQ+kf5tQcLsHH6dB+pFi/8Aav7914mo4dOM3wp2pXQ0kWX73+XWY+0C6ZKj5L9k46SWSyiWeoOAyWHWd5wtmUjxgE6VX37rXWOp+CHStbIz1u5vkNVoXMiQS/J3vtaeFyLM8MUO/wCIIzf7Ec+/db1N69S6b4KfHimgjifGdk10iElq3I95901+QmYuXElRW1O/HnmkW9gWJsoA/Hv3Wums/wAvH4gyu7zdV5GaaV3kmnm7Q7eklnlkYtLLLId+3klkcksfyT70DXyPWwx4A466P8u34dj69TVf/oze3f8A7PffiaEDr2oji3Xj/Ls+Hn/Ppqs3HN+zu3bWPH/Pen3vr2o+vTr/ALIh8b0pfs6XAb9oIEg+3ploO6+5qR6GJV0Qihli34JaVqZQBGynUlhY+/deqa18+m5PgX0tTqy4/dnyNxesAs1B8oe+0YyqLJPeff01po/wfp/UH36vXtR6yN8KMHBRpRYX5GfMXb0Uc5qI3ofkbvHJzgsLyRNLuldwtJBJJ6ir6rEWXSLj37rXWNPiV2Bio6o7X+a/ysx9TOsSxS7jzPV++oKUxyBmaKl3J1jUKxmUsramPBFraR7917ri/Rny/wATA67a+bj5SRqgShOx/j111n0EAhKfbLNtDI7BnXVLZy51n6i39Pde6yRY/wDmD7eESLuX4l9m08EaiZ8ptvtXq3L17rCQx+5xeb7DxNI0s9vpSsEW/wBT791bt+fWel7m+XODqxS70+HEGdoo011Gd6f762JuKKRi5ULR4Psai6uysllszamW17C9ufdax69Scz809gbIgmqe1usvkT1VR0gT7zKbi6N3xujAUt2RHeXcnVdB2FgRTwu41y/cCMD1X03I914inn0KXXXyY+PfbUq0nW/dHWu7soQDJgMXu7DHc9NqOlVrtr1FVBuLHuzGwWeljYn8e/da6HC/+B/B+n9f+JHv3Xuu/fuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3X/9Df49+691qW/wDClvJ7x+XvaH8un+TN1RuGXC5/5od2xdjdz5PH0suRq9o9NdZzLHTbhyWNgZfvMTSVEmZzYilaOKSo2zGNQPqT3XuhG/4S9dvbl2h8ffk3/LK7bqayHur+W18j+wOtJ6DLVL/fZDq7d+69x5Xa+YoaKoLSRYmPdVDmki8UkkCUctKVIWRL+691tGe/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917rq4/r/vv8f6e/de67BB+nv3XuscssUKNJNJHDGvLSSusaKB+SzkKBx7917oP8z291PtxmTcPaHXeCdX8bJmd67axbLJ/xzZa3JwEPx9Pr7917oK8p80PiLh5DDX/JromOoEzUxpoO09mVtV9whYNTilocxU1BnDKRoC6i3Fr+/dW0t6dI2f8AmB/D6Keamg7rxGWqIagUjQbc27vbc0jVLelYoht7bOTExZuAU1KTbn37rWk8adRE+d/UeQnek2tsL5L71qxJURQRbb+MfdfgrJKdGe1Plc5s3C4XRMF/bkkqUjckern37rXWX/Zpu0s4inYfwo+SuXckB33w/UvVdLGHRXRgd0dkPkJFuSraaYspH0tY+/dWoP4updNvn5xbkGrH/H/onraIOl27A763Fuyv8R1a2GN2B1U9CZU4shyGk3/WLc+JApU9a7RxPUjJ9efMndCp938lerut4nRlqabrjoKbOViKw5NLnOxOyM7TLMnNpHxpU8Hxj3rUuc9eqvkK9MQ+Huf3BGf9Jvy4+Ve+fPTvT1tBht94HqbBVkcmvUPsOqNq7UyMBs9gUrdYCjm/13x4de1AUAx1Kx38v34nUrQ1Gb60q9/V1O5kGV7R352F2RXMdTuBLJvTdGZiZFMjWXRpAJFrH34nSKk469rPr0JeA+NnxX2arSbc6L6J2/4lVpKig662LSSqtPqdXlqlxKzfs8tqZuOTf2ke/sUV3e9hVFBJJdQABkkmuAPOvDpQLW7kZVW2kZzQABWJzgUAHmehJxu5+scNTLR4jcOw8VRp+ilxuW2/Q0yf8FgpaiKJf9gPZF/Xfkv/AKa/a/8Asqg/62dG39VeaD/zrd//ANk83/QHUXLdu9W4QQfxTsHaNJ9z5PBfPY+XyGIKXt4J5badQ+v9fZTunun7b7P4X7x552uPxK6f8ZiatOPws3D59GNh7f8APG6eL9DyluEmimr9CQUrw+JR0hct8nejsQFMm/cdXOxYCPEU1dlGuo51GlpZEW/4u3PsG7l94z2Z2wAvzvBO3pAskp/PQhH2ZHy6E23+yHuluLEJynNEo85WSMf8bYH9g6SVb8yelqRqcQZHP5Tzh2kagwFValCGwE4qzSnVJfgJq4+tvYVvvvaez1oYPA3C+udYNfDtn7KfxeIY8nypq6EFr93H3PnWUzWNpAV4eJcJ3f6XRr/nTpE5X50bGppdGH2duzLxA2M074vFX/xSOaoqHI/17H2Dtz++jyTbyhNq5U3O6i/iYxQj8lLO38h0J7H7rXNs0ZbceY7C3f0USSn9oVR+wnpHVvzxmE7DHdYs1LpXQ2Q3OkVSXsdeqOlxFRAEB/SQ5JH1A9hS9++03jN+7vbqtvQUMt4FavnUJA4A9O416Edr91RTGPrudqT1P9nbErTyy8qtX1wPl0w1fzs3g8oOP6+29Tw6BqSuzORrJTJdrsslPS0SCO1rDQTf8+yK7++tzS8itYci7fHFpyJJ5ZDq8zqVI8eg0/aeja2+6tsKRkXnOF48tTlIYkFPsLOf59MeR+cHaFSqDHba2djCuryMyZXIGS49JXyVlMIyh/wN/ZNe/fL9xLgKLLl/abYgGuJ5a+lKunRrafde5JhJ+r3vcZweFDFHT9iNX+XSDPy377JJ/vNhlBJsq7WxfAJJsNRJsL2F/wAewS/3pvexmr/WS0A+VnEP8/8An6FQ+757VAUOy3JPr9TJ/k64t8tO+3VkO6cUoYEao9sYlXW4tdSVIBH+t7q33o/et1ZP6z261FKi0gqPsxg/PP2dWX7v/tSrK37iuDQ8DcykfnSn+Hpux3yk75xzTMu9vv8Az6bjK4TD1aw6SzXgVKamMRa9jy1xb6W9orH7yvvTYPMV5xExcD+2t4XC0P4aIlKjHn0pu/Yn2ru1iQ8sGIIf9CnmQmv8RLNWnHgOs9f8qu+chD4DvGChuwbzYzAYelqPT/ZEkkFQArfnj25e/ea96r6LwW5ujhH8UNtCrftKt/IdVtfYb2qtJfFHLjy44STzMv20DKa9S4/lv31HDHF/efEvojVPLLtfGPM+lQNcjqVDSG1ydIuSePaxPvS+9aJHH/WS0IUUqbSIsfm3DPzH7Ok7fd99qmdnOyXAqa0FzLQfIY/ZnA6lJ8v+90qI5mze35Y0eJ3pW2zRrDMsenXG8kcqzqs9jcqwIvxb2pT713vQlxHI28WDxKQShs4wrAcQSG1gN5kEEeRHSd/u7+1rQyRrtt4shBo4uXJWvAgFdJI8qgjGelJ/s7Pb9z/uL2Vz9B/C8nx/62OR7Pv+DE90xX/dfs5/5tTf5Juicfdk9vf+U7c/+csX/WrpQUvzo35HFAlZsfa1VIgQVM0WRytJ57fraKIx1C05cfQFnCn+vs/tfvp85RpAl5yZtsrCgZllnj1ZyQpVglRwFWoc54dEtx91nlh2ma25pv41JOkNHE+n0BNVLU+xenSk+d+4kWX7/rbE1Dl/2DRbkrKRUjsfTMtRiasyyX/KlR/h7M7T77G9qJPrvb61dj8Ph3bpT5ENDJX7QR9nSK4+6rtbGP6XnW4Rad2u2RyT/R0yppHyIP29K+L54YLSxn633AriOMqIc1ipFeU8SKC8cRSNfqrEXP5A9ihPvs7DpPi8g3wfSPhuISNXmKlRj50/LoPP91TeNY8LnOzKVPGGUY8uBOfUVp6E9LLH/NrqioaJchj934rVSRzTSSYmGshhqiqtJRK1HVySzMjEqJAgRtN/yPYvsvvi+11w8S3tluttWMMxMCuoemY+xyxocBgoVuOOg3d/dl9wYVka0utvnIkooEpQla4fvVQKjitdQ+fQk4v5NdHZYN4t/wCKo2SGKokiysVZjGjSWw8Wqspoo5KhSbMiFyLf09yJtn3i/Znc6+Hz1bRNoViJlkhpq/DV0ALA4IUkjoEX/sl7o7ef1eULh1LlQYSkoNPxdjEhT5EgdCVgOxtg7nH+/e3jtvLtrlTx0WYopZi0Sq8toBMJisasCSFsL+x/sfPvJXMi/wC6HmywuzqIpHPGWquSNOrUaA5x0D925Q5q2Kn745dvbZSAavE4FDw7qUzT16RnZHx36F7hik/0m9P9c75ml9a5LPbSwtdmYZCoC1FFnjSLmKCoUAFZYKiORbAhgQPYuBBAINQeg5Whp59AKnwuOx9Mvx3+Q/fHSDwSiSn21U7ul7m6zCarvSvsbuAbpkpKZlAULjshj2RBZWH19+qPXr1a1+3p6qs/82+tVgSv2J1H8msLFCqT5XYm4KzorsUuhu8p2ZvabeOw8rI8Y+se5MYC/AiUc+/de6xYz50dL0GWpdt9y02+vjRuqqnjo4cX39tSq2Rt+srJCVWPEdnwy5XqfNpI62jNNnHZ7qCoY6ffuvU+fRxaWrpa6mp62iqaeso6uKOopaulmjqKapglUPFPBPCzxTQyowKspKsDwffuvdSPfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XVx/vr+/de6xz1EFNE09TNFTwoLvNPIkUSD+rSSFUUf659+4cevdBhme9ekduO0e4O4urMFIkTzvHmOwdpY2RYItfkmZKzLwsIo/G12tYaT/T371P+r0/w9e4cekQPl/8AE08j5O/H0jjkdx9efkAj/mIf6H3okDJOOveg8+lDjvkn8dcxDDUYnvvpfJQVBIgmoe0dkVUcxD+MiJ4M44kIk9PF+ePe+FK+Yr16hyaYHQlYTdu1dzBztvc239wCJQ0hwmZx2WEasSFZzQVNRoUkcE8e/VHr14gilR0/3A/I9+69137917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuv//R39ybf7ce/de61EMv/JD7l/mR/wA1v5rfLf8AmPYntrp7o7BYza3WHwf/ANDvee3Nt7xr9i7fqcth5cvU1ezKncGW2zjsliaabJVNBVpSSyVm45VbWYXv7r3U34ufydvkn/K5/nVbJ7r+GO29+9q/AHvPpduv/kzuXs3uvbW4d/7Z3JXPlp1zWZj3NUYnc+7zgNzbewWQpmo6SqkWirKqBZAbqPde624/fuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+69025XMYnBUUuRzeUx2Hx8IJmrsrW02Po4QAWJlqqyWGBAAL8sOB7917ose6fm58ZNs102HpuzqHfm4IAwO2uosPuHuDPtMoU/amg61xO5jTVRVgfHM0bc+/dboeNOkmnyU7+3yXXp74a9kfYyApS7p7+3ZtPo/DMxk0rVHbscu+exXomiIkUPh4JmB0sqNe3utdZTsb5073jH96O9elukqUvd6DpjqvJ9h58Ra/wDM/wB8+2szFhQ5ibSXXbIIYXU249+63jy6dqP4kSVmh9//ACV+U3Ych1GppqjtSPr/ABFQJIzHJEcV1DgevkFOwYkK0jspPDcC3uvE18usUvwH+KFcqjcHV8285FMx+439v3srflQfuPEJlaXd28My2iXwrdRZSR9OT791rpa4j4efE/BSxz4v42dGU9VFKkyVjdW7Mqq7zRKqxzNXVeHnq3nRUUB2ctx9ffuvdCZS9edW7ZRXoNjdf4CMSKVaj2xtzEoJPT4yphooBrBQWtzwP6e09xd29quu4uY40rxdgo/aSOnIopp20wxO7eigsf2AHqFlO2up9qx+PIb52fjERZW+2hytA7jwMySotNRNI/kR0I0BdWoWt7A27+63ttsNRu3PO2RNnHjxs3bgjShY1BBFKVrjoW7b7e89bzQ7ZynfyqaZ8F1HdkGrAChBBrWlM16CbM/MbpXFSSxUmVy+feNA6HDYOseCZixUxx1VZ9pDqW1ySQtrWJ9xZu33sfZzbHlit90u72RVqPAtpCrf0Q76FqPPI6kLbPu6+5+4LG9xt1taIWofGnQMPmUTWf8AVw6D7JfOzZsOj+E7H3RkLs4l+8qMZjNCgDQ6fvVhkLk8g6SAPz7AW4ffT5Si0ja+TNynya+I8MNB5GlXJJ8xxHQwsvus8zSavr+Z7CLApoWWTPmDhKU9egxyfzt3lM6jDbB23RIJJdbZLJ5LIyPF9Igq0seOWKQf2rlwfxb3He5ffU5qlK/unkiwiUMa+NNLKSv4QNAiAbzbJHp0NrL7rHL6A/vLmy9kNBTw4o4xXzy5kqPTA6DTKfLzvDJErT5vCYVPO8qjF7fpGk8Z1BKVpci9aHijDD1aQ5I5PuOty+9X7zX1RBvFnaIXLDwbaMkA8EJl8TA8jTUfM9DWy+7v7X2ZrNt11cnQB+pcOBUfiAjCUJ8xWnp0ich3/wB2ZM1H3PZe5I0qhaWGgagxsSrpClYPsaGGWnBt/YcG/wCfYLvffj3j3Dxxce4V+qyChEXhwgf6Xw0BT/akdCqz9ofbKxEPgcl2bMhqDIHlJP8AS1uwb8x0h8lvbe+Y0/xXem7MgEBUCq3BlZFCn9S6fugpB/xB9gy/525z3Wn7z5u3SegoNd1MR+zXQ188Z6E9lyryvtxLWHLlhCxNe23iH/Pn+DpLMjNq1Sztrvq1VE7Br3vcNIQb35/r7DZkc6qu1D/SOa8a5zXzrx6PQqimlFFPRVHDhmlceXp1hFJTj/dEP0t/mo/+jfbPhQf8o6f7yOnvGl/3637euS08S/pRF/4Kirz/AF4HuyJGnwRhR8gB1Vndvicn8+sgQDgG3+sB/wAU931D+GvVfXrkBb3osTg9e6796691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XRFxbn/AGHvYJHDr3XAxqbX5A+l+bH+ov8AT3vUfP8A1f6uPXvMnzPXHwR6tYGl7Fdaeh9JFiodLMA355591ooOpVCvQiowaHHEUPDreo00k1HGhyK+tDjpbYLsTsDbDq+3977pxYSSGUQw5mtlpmanuIVlpKqWelliQG2hkKEcEexjsnuFz3y46vsXOG5WtGVqLcSFCV+EGN2ZGUDGkrpI4joMbryXyhviMm78s2NxVSKtDGGo3GjKqsD56g1QfPo0HXfzS3lhKmKj7Fx9Pu3FvNGJctjYIMVnKKA8O/2sKrjskEJvptA5AsGJ95Kchfe/5u2qaK159sE3TbSwrNCqw3KA11HQKQyhcGlI28gxNOoJ5w+7Ty3uMbz8m3j2F8qmkUjNLA58hqY+JHXhXvHmQBnqwnYfZmzOysWcts7OUmWgj0iqp01w5DHysAfBkKCYLU0kov8A2l0tzpJHvO7kv3C5R9wdu/enKW9RXduoGtR2yRE0OmSNqOhz5inoT1iJzVybzJyXfjbuY9rkt5mBKN8UcgHnHIO1gfQEEeYHSqzOFw248XWYTcGJxmcw2SgamyOIzFBSZPGV9NILSU1bj66KekqoJAbMjoykfX2NegwMgHojm6vjDuPoWiqd/fCGSTaOWwdRU53LfGXIZ+qj6F7fon1SZbbeJxGTORp+mN6VlOpOIy2B+xxsdeka5GkqKV5CnuvDjnoy/Rnd2yvkB1/j9/7Kkr6VHqKrDbn2puCkbE70693niHFNuXYO/Nvys1Vt/d22cgGhqqeS6sNM0LS08sUr+690MHv3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917rokj6c/8AIj7917oofZ/zJ692bn8lsDr3Dbg737VxrGHJbJ6zGPmxGz5yrfu9p9mZmrx3XPWFFAy3lXJ5Fch4/VDSTGylPd3dpt9tLe7heQ29igJaSV1jRaCpq7kKMZyen7e2nu547a1geW5c9qIrO7VNMIoLHOOHRJN4/Izee4ahqruL5NUvXuOHmB6W+FuMh3Hl4CSDAm7Pkf2BhCclUQqxWRcBisHEGBAllGl/eMHPH3yvYjkp5baLmZt33JTTwtvjNwK1oAZu2AflIaU6nzlP7sfu7zQsc8nLw22ycVEl64hNP+aQ1TV4cUHRXt6dnfH/ADNYtQnQ1X2bVpMlRUZ35F9v9odq1tbJCnlNSmCqtzzYHGyTIqSMsCrGTcaPp7xb5p/vEd6lmC8ke21tHAD8d9O0hbPERweGFPAisrH5efWQPLv3KdujTxOauepnuCPgs4lQKaf78m1FvMYjWvr0gJOzKelcttfrToHYESUstFDHs3onrfHy01J9x9wyRZHI7eydfrjllUljIdTHUQWufcAb798/7wu9SSm35xg26JwQVtLSBCM1BDyrK+oDAatafPPUx7T91r2W2tIxccuTX0imuq5uJTXGQVjMa6TkkU+VadcD2tvR3U/xjCBuQAuxuuEDEuYjqRNoKr/unT6gbn2EG+8/94hmVj7xb0CPR4APzAgFfz6Ei/d89kQGA9s9tof+a5/YTNUdQ6nfuVydxmsZ1tuaIkMtPubqLqXO0qSlWjWpigq9mhVqdAKhwb6bj+vswsPvY/eNsJGkX3XvpqqRSeK2mUfMK0AAb+l+2vSW5+7j7H3SKh9vLWKjVrFJcRsccCRMcfl0/YHdfTNPX0mS3V8W+ka3KU0P2397uscdXdEb+SAlWJh3J1dW4Nlmjs0gfShDHgqL+5m5N+/97qbMsNtzhsG2bzbLTU6hrOcgUrlA8JY8cxCp8wOou5m+5x7e7j403Lm93+2XDDCtpuYRx8m0zUrQYkOPU9HL68723lhqylyvx+7p3d2V4z5Mj8SvlBksPVbi3JTRqZKjHdI/Il0o8hjt1Q0yacfR7nnzFHkZgIZJaEP9ymcvtV97b2j90rmy2ePc32jmiY6VtL0LGzv/AAwzBjDKT+Ealdv4B1iX7h/dy9x/b6C53OawTcOXohVrm1JcKv8AFLER4sYHFjoKjzbq0fpbubZXfPX2J7F2LPkkxldPkcVlsHuDHTYPd2zd1YKskxe5tkb227VH7zbu79r5enlpa6jluUkTUjSRPHI+T32jPUB44g46Ff37r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3X/0t/j37r3Xvfuvde9+691737r3Xvfuvde9+691737r3WKWaKCN5p5EghjUvJNM6xxRoouXeRyFRR/Un37r3QUbo7/AOidkcby7n6q2q+nUItwdg7TxExXVousFdloJX9fHAPPHv3XugXn/mBfDGGvGLg+RHXuWyJnlplotuVldumoeogv5oUj25QZUvJFYlgL2AJ+gPv3Xum9/wCYT8U1d1G9t5yhHZfLT9Hd71FO4UkGSGph61eCaE2uHRijLyCR791uhpWmOnaj+fXw3qsjHiZ/kJ1/gsnKYwlFu+trdk1B8yGSM6N30ODsrICbnjj37rwBPAdGa2xvLaG9sbFmdmbq25u7ETqrw5TbGcxmex0qPfQ8dbiqqqpnVrGxDG9vfutdKT37r3Xvfuvde9+691737r3Xvfuvde9+6910TYE/W3++/Pv3Xukbv7sXYfVm1Mtvrsnd+3di7OwcIny25d1ZaiwmHokY6Y0lra+aGJqiokISKJS0s0hCIrMQD7r3RXX+Rnb/AGtjcbW/F3oiu3DgMu8oh7U78r8z0psaKmDMsGWxGz67AZHtnd9DUBdcTriMbS1ERDR1RDA+/dboBxPXUnRfyo37Go7Q+Xdbs3HVI1V22PjZ1ntrYIiB0E0lNv3sKbs/dxjT1IZ6cUMrqdQEbAW91o08uHU3E/Az4x02RpM5vPZOU7p3LSFJI9yd+by3d3RkDMjKyTR0W/8AM5vA0UylBY01FCALgCxt7917VTFejUYjEbY2biYcXgsRg9qYKhiIp8diKDH4LEUkSC5ENHRxUtHTxooJ9KgAe2ZriC3jeW4mWOJQSS5CgAZJqTSgHE9WijkndY4ImeQ0oFBJJOAKAHpGZrvDqLb0ksGW7D2tTzwSNFNTx5OGtqIpF06kkgoTUyow1jgj3HW8e8ntXsEkkO6c+7ZHMhoyrMsjA+hWPWQflTob7Z7Z+4G8JHLt/KF+8TioYxMikHzBfSCPsPQRZ75l9L4wTJj6zP7injbQExOEqY4ZD5NBK1eSNDBoC+oNcgj6c+4s3r723tFtyyrY3V9fyqRiG2cKc0w83hr8/PHUgbZ93D3LvzGbq2tbONhxmmUkYr8EfiN8uHHoLM986sWism2Ng5KskAXRPnMnTUEJ/cKyK0FElbNYxAFTq/UeRx7jbe/vq7YisnLnI9xKaCjXMyxite4FYxIeHA148RToc7V91fcX0PvnNsEQqarBEzmlMULlBx444cOPQN5/5mdvZYzriIds7Zp5AFh+zoJclVwKJdYcVORlkhabx2QnxaSASACfcR7997v3U3Rpk2mPb9tgYEApEZpF7qg6pWKk6e09mk8QAepI2j7tPt9YeG24zXt7IMnU4jQmlPhjANK5HdUcCSOgmzXe3c24FnjyPY24EgqFdJKbGTQ4eDRIwZkC42GncWtwQ1wOL+4v3n3v92t+8Zb/AJ93BYJKgpCwgUA+QEKpw8iTWnn1IG2e03tvtPhtacm2ZmU1DSBpTj/mozD7cdBpk8nl81IJczlspmJAFAfK5Ktr2sn6f+BU0q3X8G3A9x1uG6bju7tJu+6XV25pmaaSU44fGxH8uhxZbft22J4e27db28fpFGicf9KB03pEiXKogJNyVUAk3+pP1J9lwRFPYgA6XFmPFj1l976r1737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XvfuvdPW3Nzbh2dm6PcW1stVYXMUTDx1dI+nyxagzUlXC2qCsoZbWeKRWRh/Q8+zzlzmTfeUd3t9+5b3SWz3aIjS8ZoSAalHB7XjbgyOGUjyrQgp3zYtn5l2y42bfdvjudulGUccD5OhHcjrxVlIIPy6tD+Pnybou0alNpbppIMNvlKd5aY0nk/hO4oadHkqZaBZGkloqymiTXLA7MLHUjEekdKvYv7xdl7lzx8scyWsdpzoIyU8OvgXapUu0QNWR1UanjYkU7kYjAwU93PZK65CifmDY7l7nlYuA2sjxbYsQEEhFA6MxojgA1wwHEm4vdb/S4P1/H+295RdQCDUV8uiNbio8ZsH52bFy+yI6bH1Xc3SvYdf8jKSmVabGzYLqep2rT9S9nbkkUpS0ucxuR3FX7fgqZrS12PqGjLNHjo1j91byPSt3B85/jPiM3U7U2/v2q7W3fRSrT1W1ujtqbs7ny1JUsQi02RfrjDbixmHmLkC1bU09j9be/daH29ZU+SPYmegebZHxE+QuVbxLJTNvMdZdX00+pkurJu3sGHNUjhWJtLQq3pIIHv3W6fMdRY+4fllVfceD4bUuPWMEQfx35F9fwPO7K3jIjweD3AkUasAHu97N6Q3Pv3Wum7Fdq/NoyzHO/EbraGmWEGEYT5PUldVyVBkQBZEyPVOKgigWK7M3kZiQAFN7+/dbNPLj06Qd1/J+GpMeW+Fue+0SoSN63b3e/UOXZobkSVUFJla/bU8kajlUbQ7D6gHj37r359ca75bSbaV5N9fGf5T7TpobmoyNF1dR9j42BFuZJpJepty77rVgjUX1GnAI+lzx7914inn0I3Uvyc6J7xrMjiOtexcPmtzYaNZs3sjIw5LavYGDiYAibNbB3ZRYTeGLgubeSaiSMn6MffutdDuDcXHv3Xuu/fuvdVAfJfvXdPamK3Vl6TK7s2j8Vdrb0yfVqQbIy8u0ewPlN2Hg67IYzdeNpd/UM/8AGeuPj7tPJYqooK6uxPjzm5auCeKnnpaFRJVwp79+9G3exnIz82XW1PfbjNcJb21uraFeaQMwMr0OmFFUs5VWY4UCpqJV9nva6+92ebk5cttwS0tI4WnnmKlykSEKQiCmqVmYBFYqvFiaChrqze5qzM47H7cpKLFbW2Hgi6bW612lRphNh7Wpma4ixeApdEE9ax9U9dV+evq5S0k0rOxPviP7o+7vPvvBvM+7c7b5LNB4haG0UlbS2U/CkMAOkaRT9R9UjGpZs9dWvb/2z5Q9tNsjseVtqSK6KgS3LgNdTnzaWamruP8AoaaY14BekoFEk7qwHipxGI4h/mtbDWJChjADxqbLZiNJP59xkMAAYA6HtKkk5J8/PqULAWAtbgWFgoH0AA+gA9+GSAOPW/L5dGb2p8Ut/wC6dobe3tVbq2BtLC7ro1yW3k3Hkcw1XXUDMY1qZVxWJq6ekZmUkRvKZLc2HI95Ze3f3Nfdn3K5Q2nnXatx2W12a/iWWATzSGWSJuDlIomC/wClJLj0Hljfzt96H255G5l3Tla/sdzuNys3McxhijCK4/CrSyLr45IUD8s9cNy/Efurb9BU5bGYvAdhYmk9U1X17nINwV0cQUSCebASR0uZRArfRYpHW3qA49lnOv3Pffjkq0m3CTlyDddtjJq+3y+O4AGW8BlSYqPPQrkeYHRhyr95v2f5quIbJN8l269k+Fb2PwUJ/hEwLxV/0zKD5E9Fmkp/FNJBPTGnqKaQwz01RTGCoppo7hoZoJkSaCaMsRpZQwuePeMckUsMssE8Tx3EbaWR1KuhHk6MAyn+iwB+XU+xyRypHJC6vE6hlZSGVlPAqwqCD5EEg9Y/BBawhi/wHjUj9HjsQAAR4/T/AK3unTlOopiR3q40BijOkEodNqpRraZVMauksRKEOGINha1vdZFDxyAkjz+ymQR/SU0KnyIr1tGYMjgA0/wHiPmCMEeYPR59p/IbfWxu4OwNtdWU+18bvbu/41fHn5Fb/wB2b8GSruu+osdgH3/1B2B3XubEYqSiyu8dzbootkYHH4jBUs9LUZ3Jw65qiGnpZ5T9Aftp7j2P/A/cme5nOl6LSwj2GKa7mkYuB4SCMuTSrPJoBCjLuwAFT1xn565Fux7w8y8h8r2Zubtt4lht4kAXUZG1qgFdKKmohmPaqrU0GQGmf7/3jWVlTVR99/LPcWQlkQnccXYPWvVmM1wyljNhettsdU5nAYegq04NPXzZGeNAB5depjh5zB/eL28G4yxcp+1bXG1BiFlvLpoJXUHtbwYo3EeoZ0sxYfb1kvsf3JbuSxik5i5/SHcioLR21t4sanzXxJJELU4VRQDxGOh26o+X3eWPyFDi8R2Hgu5Ukmp6aPrnvzF7e6x37lYwQjU2zu+Ng0tL1tkc7Kh/ZpNw7cxa1UgC/fR6ricPan76ntX7jX9nsO8wz8vcxTsFjW5ZZLWVzwVLtKIrscKkqxkkgAljTqJvcP7rXuJyPaT7ttph3nZIlLO9sGWdFByz2zd5VRlmiLhQCSABXqzLpj5Idfd05DdG08bDuDZXaWwDSL2F092FjBtvsjZ0dfrGMytXhzUVVJm9qZrxMcfnMVUV+GrwrCCpdldVzBrkf6v+L/LrGuhAr5dD+DcXHvfWuu/fuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3X//T3+Pfuvde9+691xZlUFmYBVBLMTYKALksfooAHJPv3Xuiibq+b3RGKz1ZsvYWU3H352JQtJDUbE6A21kO0snRVKawafcGfwSHY2z3jdCHOYytAEsb/Q+/db0nzwOmal7K+bu/fHJtL46dadL4uUF0yHfHah3PuTxvHqiaXY/T1BmMdTyq/DxvuMEf1B+nuvHB6fZur/lvuWljfcHyl21sWqlQCpouoeitviGnchg60uW7V3D2HNOnqBDtSRMCOB+PfuvAgeXTQvxBzmUMr71+XPy53V9wrCekx/Yu2uuMcdbxykQJ1hsjaGQpVVoyqhKq4RiLke/der6DrpP5fvxarKiGt3dsncvZmRiBVq/tbtPtPseSqUiRStbSbr3jk8XVqVlYEPAwINvpb37repvXoXNs/F7427MiWLavQXTeBVH8ivjettoU8wf9s6/uFxH3Ba8Sm+r6qD9ffutEk8ehppMfQUESQUFDR0UMYCxw0lNDTRRqOAEjhREUAf0Hv3Wupnv3XumzK4XDZ2kmoM3icZmKGoQx1FFlaClyFJPGfrHNTVcU0MqH+jAj37r3RWd2fBX4sbnyZ3FRdUYzrzd66npt7dNZPOdNbupKlyp+8jzXWeS2xNU1EbKCPuRMn1upBN/dbqaUr0zJ018pusaWeTqL5Jv2nS06AUGxvlJtrG52N4wY9UMXa/W2O2lvOknEakJPkaPOHUbujck+69UU4dQ8f8xKjYGTpdtfLXq3N/G7IVs9PRYrsafKw78+PO4qyY+NYqTt7C0VJBsuomkU+On3ZQ4CR7gRNKffuvAE8B0dLH5CgytDSZTF11HksbX08VXQ5DH1MNbQ1tLOgkhqaSrpnkgqaeaNgyOjFWBuD791rqZ7917r3v3XuvE29+690Xb5AfIDH9LY/buGwu3q3snuPsvI1G3um+nMHW09DnN/Z+lp1qcjVVWSqY5qTamxdqUTfeZ7PVifZYujFz5KiWnp5vde6DnYXxeym4d3YXun5WbixfcfbmI8Fds7aFLQunRHRNc8Kio/0R7NyqSVGQ3OrExy7szZqM7UKtqf+H07GlGiwHE9eqR0N3ZXeHXXVQji3Tmz/FqiHz02BxcLZDNVEJ1hJjRxG1LTyOhAkmaNCQbE29xZ7h+8vIXtkix8ybx/u0ZA62sK+LcMprRtAPapIIDOVU+RPUgcl+2XOHPrM+wbZXb1fS1xIQkKnFRrPxMK5VAzAcQOig7l+dmSmili2dsGKlkL6Ya7cuTFSqx8gucdi1iJlP4BqAo/N/p7xU5g++rfSRyxcq8jpFJXtku5tYofPwoQufkZaDgesh9l+6vapJHJzHza0kfnHbRaTX08SXUKfPw8+VOgH3N8pO7tzao13V/d6nOgiHbNFBjXGhtX/AyT7muIc8MNdipt7hHmL7y3vBzDrQcziwhONNnGsVKHj4japDUY+OnUrbJ7Ee2eysjNsRvJRXuuXaQGuPgGmPHHh0CuS3DuLNW/jO4M7lQDIQuRzGQrEBlFpGEc1Q6Ayg2bixHH09w5uXMPMG8mu8b9e3eSR408sgBbjQM5Ar8h1J1jsmy7Ya7bs9pbmgFY4Y0OOGVUHHTIsMSABI0UD6BVC2+v9AP6+yYALkDPRoWZhRmJ/PrnpX+n0+l+bf61/wDW92qfXrRz12AB9P8Aff763vxJPE9e+XXgLcD3rr3Xfv3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917rokD68e9hSeA690v8AqWhy+U7P2HSbefIR5Rtz4mb7nFXFXR0MFUkuRq2YjxpSR0SuJTJ+2VYqbkgex97WWG8bj7j8lWmyG4XcG3CBg0PxpGrgyvX4VURhteqilSQa1A6BnuHebZt/I3NlzuywtZfRSjTL8LyFCI0HmXL6dIXuBAIpTqyfvr5HZDZWboelOkMBie1vlDu/Fvktrdd1eRq6Ha+yNvSyvTP2n3duDG01dPsjrTFzowj9Byedql+yxkM0xdoe1IHpw65aAGg9Oi79a/y19vZDdW8+2fl92rvH5W9rdl1uCyW7sPmXk2T0DhYdtR1Y2vsvZfTe36wUlZsTZv8AEqpaCn3JW56WWWpmq52eqnkkPutk+g6sb2ztTa2y8PS7d2dtvA7TwFCuiiwe2sRj8FiKNT9Vpsbi6elo4Af9pQe/da6frC9/z7917r1h7917ruwP1F/fuvdcdK/0/r/vP19+691yt/r/AO3Pv3Xui7/IL427E7+wdH/Enqtm9l7WqYcx1b3VtKKmo+yurt10LPLi81tzNGPzVOOE7aMhiKlpMXl6N5KarhkikI9+6900dJ99x5z46HtruWuwmzMj17Tb+wvcuXLGh2xis/0/ns7tTfu4qLySVElJt2vrdtz5CkiZ5JYqWdIyXdTf3XqU6JJtjv8A+U/y47SwmN2kMp8YPi9uHce7dtbc3RQ4CirvklviXa20qnPz7gyCbxoMts/qfbRq2pY1onxWRzBllEE8tLMlRBF7rYNPLpp7N+IPZnQ/W24sJtrcHZPyp6LzW8c72NvDZOcptoSfIPpzcOcq58xn99/HCfb239qbc3jiK3LVdVW5bY9fTxz1ZqJziatJZFoZYr94vaPln3o5MveUOZA0dXWWC4jCma2nT4ZY9QKnFVdDQSIWXBIIkH219yd+9ruaLTmbYgslAY5oHLCK4hb4o5NJBGaMjDKMAwrShIru3Yk2Bwu2t8bfzeP331Vv2B6zYHZeAjqIsPuCFGkSoxmRx9YkWT2tuzEzwyU9fiq5IqqlqoZYmUNGwHE73r9h+dvY7e0suYoFn2G4kYWl9ED4FwBTsZTqME4GTE5yAShZc9dU/aj3g5U919pa62SYwbvAgNzaSf2sNcagcCWEnhIoxUBwpx0H0iNqMsRXzBNNnZvHIoDOiPYnxjW19Sgt+Pp7hTqWPs4deE4H+ejaI3sCbujEyCNCHUmxdjwDY25Pv3Xvl0N3WXyM7Q6ppxjNt7kpsptaVrzbL3RTxZ/a8okZmdKajqW+4xfn0vc0skQPJsSPc1+2H3hfdf2jCW/KfMrNsgav0Vyvj2vGp0ISHh1ZqYHQZJIJ6ijn/wBlfbr3H1z8w7EF3cigu7c+FccKd7Dtlp5eKrHAFadHO6w+QHW3ZGTpKGn83TPYs7A4xJcx9xsHN1gAdaXF7jYw123ayqBPhhrLwOfSHNx76U+y332+R/cO9seXOdNvHL/NMtEjZpQ9jO44Ik50GJ24qsygH4RIzUBwT90/uqc2ck2l5vfK93++eXUFXUIVu4k46nhFVkVfxPESRxKAVIVvbHVe2+3618b2DGNi9nY9RSUXYFHQU0YyEpib7Og7BoYFWHL4iRnXxV9OVniB1amUkex17+fda5K964W3e2ddq57SM6LyJF0z0U6UvEAAmiBppkUiSME6WI7SFPZ77wXNPtXN+7Zgdx5Qkk7rWRjqhyCz2rEnwpKVBQgxvTIBowrA3xs/cvXG5MztDd2NbH7iwcgSppVdJoKmGZRJQ5LG1WpIa7GZKnYSwSqQrobGzAgcWOceTeY+QeZt25Q5s21rXf7J9MkddSlTlJYmH9pDKvdHIBQjDUYEDqbyvzTsfOewbdzLy5fi42e6WqNShBGGjdeKyRntdTkHIqCCUxHSzyiChokaWrqWSipYowzSS1lbIsNOkasWJZ6iUAC5+tr+wyIZ7mlpaoWupSEQDJLuQqAfMsQOj8yxwBp7hwtugLsTwCoNTE/IKCehB3c9Zj98fIzIUFTXU2KzPYPW/wAbKen+1aCiqNq/D7rHCUObpMVVTVVTVPh6zuzs3OzVEIEMMlTRJLoZ9bHpT983cL32/wDZ72M9mtvnMFu9kr3iJqCSrZRQosZJrVfqXeRlJyVSuB1gf91qytuc/c73X9z72HxZkuW+mZ6F43vJJWZwPI/ToEBGKFgMk9IG34HIH+P+H+J+vPvmrmpp69Z5UA49cWUMCrAFTcEEXU8g2t9CLj3pgGFGAI62OII4joZ9qdk5uprtkLX70qNmb568qz/oZ7ump2yVd10lWUGR2B2FSqUrd+/Hve3ijp87hZpfNQgR5CgeGrpIXTOX7sf3t929urzbuRPca/lvPb6Z0jiuJHLS7aT2hqmry2nDWjEtEO5CFDL1iP7+fdv27nO1vebeR7JLfnONWeSBAFjv6ZIp8Md1xKsKLMe1xqIbq8343d4nvLr+XL5vb52R2Ps/PV2wu3NgPWDJDZnYmEgo6jI0OOyyxxxZ7a2cx2Qpctg8nEBFksNX0tQArOyJ2IilinhgnglWSCRA6MpDKysKqykVBVloykHIII65lujxs8cikSKxUggghgaMpByCpBBByCKHowftzqvXvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691//U3+PfuvdAT3z3/tDoTbuPrs1QZ3d28d2V8mA6y6p2TRrmOxe0t1eFZUwG0MKZoEaOmSRZchkqqSnxeIpCamtqIIVL+/de6Lxs/wCMvZndq5De/wA2d3ZDLvuZb4/4q9ebuy+M6A63wGpmx+B3RU4ZcHmu795GAq2YyGXmbCS1ZaOhx8VOiPL7r2PLo6+0Nl7Q6/2/j9qbE2tt7Zm18TClPi9u7VwuOwGEx8CIqLHR4zF01LR06hVH6UF/fuvdKYcce/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de6iV1DRZOjqsdkqOlyGPrqeakraCup4quirKWoQxT01XSzpJBUU80bFXR1KspIIt7917okm4vixu/qiYbr+Fe86Dqaro5aitrvj5ueOsyXxk339xPJV11FBtekEuS6YzmQmkLJltreCnWb11WOrQWU+62KefQrdGfJLbncFVl9l5vb2b6l7w2dCj9g9G78ajh3jt6IzCmTcOBrKOWTEdg9eZKcj7DcWHkqcdUqwSRoKkSU0futdGQ9+690G/b3ae0+lOtt39pb3qpabbezsRNk6yKkhNVlMpUlkpsVt/BUKssmT3FuPLVEFBj6VP3KqtqIol9Tj37r3QDfGXpbc2Emy/yB72anzfyW7ZoFkz9QytLj+n9gVVX/ABfbXQHX6SvIMVtbZsbRnKzxWlz+eWevqWe9PHB7r3Cp6dvkj8goeqsWmA261LW79zEEhp4mcPHtzHyI6jN10XqDzM/FLA1vK41N6BzjV94L31g9sdtj2TYHjm53vE7FJqLWJgQLiRRWpJxEh4nuYaFNZx9mvaObn6+fdt4WSLlO2cBmAobiQEHwYzXC+cjj4VwO44qeyWRyeayFTl81kq3L5WufyVuSyM71NZVyf6qaaQliFvZVFkQcKAPfLvct03Ler653TeL+a63OZtUksrF3dvUkk/YAKADAAHXQKwsLHa7O32/bbSO3sIlokcahUUfJRjPmeJOSSeoY4/qf9f2g6V9e9+691379Q9e697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/daqPXr3v3W+ve/UPp17rizon6mVb/AE1EC/8ArX96JC/EQOtgE8AT1iM8Q/3bHz9PWv8AxXge9B4/41/3odb0P/A37Os0KzVCl6eCaoQHSzwQTToGtcqXhjkXUB+L39vRwyzLqhjeRfVEZh9lQCOmpHSJtEsiq3oWUH9jEHp5p9s7rq44paTau5qqKfT4JqbbubnhmDNoUxTR0JjcM/AINr+zi35Z5lukikteWtyliemkpazspqaDSwjo2fTA/n0Wy79sNu0iXG/WMbp8Qa4hUqRk1BevDPS6xvRvcuWkkio+td0q0QiLtX0IxUVpH0ApLkZaeOXSeWCFmUC5FvY0sPZT3a3ORorP283MOtK+LGIBQmlaylQfmASQM8Ogpe+6vtvt8aS3HOtgUatPDcynArSkYYj5E0BOB0OW0/hT2Pl5YZN3ZfB7SoWW80VJIc9llPkC+NYofFjk8iDUG8z6b8rf3NXKv3O+ftynR+at1s9ssc1EbfUTYalKLpiGoZB1tTgR1FnMH3nOTbCOROXNuudwugcM48GLhWtWrIQDgjQtfI9KTK1NTsbc2R+MXw1x1BmO9K2hoZ+4+9d20gzu0fjttjJRrLR5Xdni8FNuTs3L0Uhm2zsqnkhMpK5DJGDHqHqc3vbL2i5O9qtvltuXbRn3CYDxrqYh55acAWoAqA/DGgCjjQnPWKXPnuRzN7h3sNxvlwq2UJPhW8dVhirxIWp1ORguxLHgCBjo4nSPQuweiMBkMbtKHKZTcO562DN9h9j7tyD7g7H7Q3TFTLSSbo39uqpUVmayTRJogiAioqCC1PRwU9OqRLKA6ARPQ1+/da697917r3v3Xuve/de697917r3v3Xuve/de6A75K9hb36o6G7T7E632bNv7fO09p12T23tiKKtqFqsgGipxkqyjxlLW5bI4zbsM75KrpaKGWuqqWkkhplad4wfde6qYxeEzvYvWPX+w9s1m5JfjtsPMYHbm29z5vAZPaWR+ZXzF7E3LJm8r2nVbXzFHS5Sbozqjd2SyO7PBVQx0ucztMCBNj8UrVPutk16srwu1sBtDu7pzrjbmQqafHdedHdi5VcPNURTSZQ5rdGxMM25co0gNXWZiqyENXLPUk3mnq5Xcln9+610ay1wL3/H55/2/B9+691Wx8oOiqfq595d/9cYBc111mZqvcHy2+Pqc7c7I2zIsDbi7x2Fj5P8AJdr/ACB6+oKU5Nqqk8H96KKlkpqrVWijqIw1zfyhy7z7y9uHKvNm1x3myXKFXjcA0J4Oh4pIh7o3UhkbIIPR9y1zLvnKO9WO/wDLu4Pa7tbuGSRTT7VYcGRhh0YEMMEU6rb7C6Uy+y9zZiGkzG3260SDE5zbXbe7dzbd2T19l9p7opFym1shLvXc+RxW23ra3HSgNFFUPK7IWVNJHviXzH91T3YtPc3mX2+5P5Zut0srR1khuyBFbtayqGhaW4ekSy0qsiqWOtW7aHrqpsf3ifbmfkPYucuY9+t7C6ulKSWoLSzLcR4lVIU1SmMnuRmULpYDVUHpI4TaO1t2MlNsT5DfFHsHLGeGlbA7P+R3WWQzIrKhHaCkipJ83SitqZzGyxrCX8jKdN/Yk3D7jn3hLG0W5g2TbbqUipiivY/EXHA61RCfIBXNeiSx+9p7M3ly9vLuV/boDQPJaPoPz7C7AedSuPPpp3ZsbeGw8gMZvTaWa2xWEjxDMYyWlp6pbEpJQV+lqDIRMGurwSyKQbjg+8auauTObeRr5ts5x5avtsvQ1ALiF0Vj/Qkp4cgPloZq8cdTry9zXy1zbaLf8sb9aX9oaEmGVXZf9Mg70PqHVSOklJRwTKVZAnGnVGAjAFQhsQLfoFuQbD6ewyQpBDKCD0IakEFWII6P38Ze55d9fa9Ldn5pP43DElN1bvTKyL51nUaY9hZmrmkM1diMhGNONllJkhmRogWBQe+ln3NfvOXy31p7Re427mazlATa7qZ/1EcCgsJZGNXVxi1Z+5SphJYGPrBD7z3sLZCzuvcvkfaxFKhLbhbRCistf9y40AorKT+uFoCpElAQ9Ry7O6wou4dm5XY2boafH9pbMoMnN19nq2MJXyPj0lra3YGWmlFpcZXCImiMhdYJSrILCzZW/eW9hNp95+ULm5sLGNOftvgkaxnCdzgDV9HIahminIooYkRy6XXgwbHf2J94tx9ruZraC6upH5NvJlF3DXtWpA+pQcEkiwXpmRKq1agivDoCqw692dQVWfSIYYb/ANtPWpWLpiR3rFWi+5EmkKsGUaFmDAC68i3vjt7LSbZZ+8ftk/MkQXbI99tlnSQUCsJNIEgPDw5tDEHzUj166Z+6n1t17Xc/DZJC16+z3BiaPJZdFWKEcdcWoAjiGBHTBjaatq/i70TunIYzHYnOZvtb5lSdh4ygQLV4js+u+Sm9sjuPGbgHlmlGcp6ZYUlMzeZwgYjSV95tf3jVhfLvXtZucqVsjb30AI/36JIZWHpQrlfsPWK/3H7u0O2+4VjGw+rM1pLT1i0SotPOobB+0dB7NrMsamR443RljaPQgWc3AEjuSJS4caE021Le/vmr5U6zs4GvXbTNB/wJ0LFcKJ9VgLkKizKb2cr6mYWQD3uR4gdQGlaAZ9f9ny6rEslCD3NU8PTqQCjorAq6SpdSCrpIjDgqRdXRlP8ArH3XDDjg/wAx/m6sa5zQg/s6M30N2xv7r/dMHYewoc7vLde0sFS4vsfqWgmWpr++uicFrkSPaNHObVvenQcU9RXbciDLNnsBNV4ZmLpQmHqn9xn35O62J9l+btyZt1tUaTa5ZWqZLZQuuyBNO63prgWprCzIopHTrnl97X2gXbL7/XR5bsQu2XThdwSNaCO4YnRdUAppuMLMcAShWP8AadX3bD31tPs3Z23d/wCxc7Qbl2fuzF02Z2/nMc7tT19BVLdWaOVIqikqoJA0U9PMkdRTTo8UqJIjKOj3WD/Su9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3X//1d6DuvuHa/RnXOb7G3TBlMnTY6bF4vC7a29SLkd0713duPJUuC2jsjaOLMsP8S3PuzcFfT0VJEXSMSTa5XjhSSRPdepXoG/jt0hvPF7i3D8hvkBU4/M/IrsegTGS47HVZym1Oi+t1q/v8P0n1rUSRxoaKjkWKp3Fl40jl3Jm0NRJ/k0FDDT+62TwA4dG89+611737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Rc/kN8fcf3ZicHmcFnH667r64rZ9w9LdzYvHU9fn+vdyyQiGrp56SYwpuLYm66RfsdwYKokFHlaCRlPjnSCeH3Xum/wCO/fGT7KbdvW/Z23KbYHyH6jOFoe2djUU1TV7erIc3BVSbb7L61y9ZHDNuLqzfqY+okxtQ6rVUVRT1FBWIlXSyavde6DPtynbub5edI9Lzr59jdH7cl+UnYVG4V6TL7yOYrNldC4esAN3XFZylzm4PG11FViKSQi4Q+/db8q9HUzGTpsHhspma5iKPE46tyVWyglhTUNPJVTEAAknxxH8H2g3XcbfaNs3HdrskWtrBJM/n2xqXbh8lPSrb7Gfc7+x222/3IuJkjTy7pGCr/MjqhXcu5chvLcme3XlZWmr9wZOoyczOblIpnYUdMtvSI6SiWOJQOAE98Q+Z+Yb7m3mHeOZ9ynaS8v7h5iSa0VmPhoPQRxhUA4YwB11d2HZLPlzZNq2GwiC2lpAsQ+ZUd7H5s5ZifOvTN7Iujbr3v3Xuq3v5knafyc642x8ccP8AErO4HEds9n/IRNnwY7cmIx+Ww+9sXherewuwZOvqtq+mqTi03jWbQiohVwaJ4XlUq6i59zB7QbFyZvN3zbPz1aySbFZ7X4paNijws9zbwCcaSNXhCQtoNQaHHUVe6G78y7bBy/Dyvcxx7jPegAOKq4SGeUxk0OnX4emo4EjpO7A+aeT7776/l9y9WZWbFdTfJDp/5P7i7J2DksdQtncP2N1Ni9r0Z2dnKueJ8nhM513u6orqOphjeITMoZwysp9r919uYOWeVvdMb5Dr3vaL/b44J1dgjw3LSMJ0GQUnjCOpNaDFRkdFllz9Pve/cgS2FwE2u9t7lp4yo1LJCIwyseKsjl1ZacRjFOhUp/nxsGaqwWebqTvGDondHcUXRW2vk7Ltraw6dzPYNTuuXYeOlghi3c/YMOxsvveFsTSbhlwqYyestpfxsshIpPavdUjvLU79tv8AWiCw+tk27xJPq0gEfjMD+l4BmWE+K8KzGRVriuOjqP3P2lriEDbrv90vdC3W50oYi5fwwwHieL4Rk7BIYgmqndQg9Q+x/wCYJ19sLM91x4jpv5AdpbC+NeXfb/yC7e602jt3K7B6yzVDR0mT3LjGTMbuwm6d61uxsbWx1GdTA46v/hsROos6lfbm0e028bnb8u+PzBtdluu7prsrS4kdZrhCSI27YnjhEzKVh8aSPxDw6vuPubttjPuog2m9ubCwYLcTRopjjOkMwGqRZJDGp1SCJHKDiK46Wu//AJmbQ2/uza+xOsOsO3fkpurcvUWM78OO6OxO1chTYPqHOztT7X3dmcvvXdmzcUs+8Zo5BiMZDLNka/wuVhAW5Ldp9udwvLG+3Ped6sdosob9rLVdvIpe6QVkjVYYpm0xVHiykLGlRVulm4e4O3wXNpZ7Xtt1uFzLbC4IgVDohaulmMkkYq9DoQEu4BKqQK9Qt2/NLbuK3jt3rLYHSXfvdHa+V6qw/de5OtNhbW21itzdX7B3AzwYOXsybsDdu0MRtzdObrYJqajwiVNRkqmenkCx6AHLm3+3N3Pt13vW6cx7Zt+xJfNZx3EssjR3MyCreB4EUryRKpDNNpWMKwq1cdUvvcO0t7uDbbLZr263RrVbh4kSNWhjbh4viyxKrmhAj1FyVai0Fekjm/5jnRdJ1f0B2jtTa3cPZtL8ld27o666z2TsPY8Vb2N/pK2fQZOozvX+69tZTL4v+7O4MbkcPUUNU08xpKKaJpZ5kplM3sxtvaDmZ965p2W7vrCzfaII7ieaafTAbeUponikCN4iMrq60GplNFUuQvSSX3V5fXa9l3K2trqc38jRxxpGDIJUDlo2QsNLAoymp0gjLBe7pg2780O2s38xcX0DP8U+7sLs7MdC7I7EnmyeN61Tcuz9wbo39kNt5Hce66um7RqKUdeYSmgFFOtJFU5AZKGVo4pICjlRd+3ew2/t7PzTHzvt0u4R7pNANLTmOWOKESKkY+m1ePJXWusqhjIDFWFOkdtz9u0vOcOxvy3crYvZRymoiDozuQzOTPTSoGlguptSmgIz0rs98/eutvyZvc0/Vfdld0BtjtdOldx/KTF7e2xVdL4nfP8Aein2NXSlDu6PsKv2Thd8VAw9dn6bCyY2nr1ddbRoZAhtPard7lba0XfNuXmaay+rj29pJBdPD4ZmAH6XgCV4QZlhaYSFKYDHT0rn90doguJi233R2iO6EDXIWPwxIW0E08TxSiv2PIIigavdQE9Oe4vnh1DtfdvaHWWY272HT9xda9u9d9P0nTgxWDO/+0cn2w6PsHePVdE2fSg3J13l8bFW1s+UkqKZcdS4urNUsTxqr0tPbLfr2w2XeLe7tDsN5Yz3Rutb+DbLbf20VywjJjnVtKiMBtbSR6CwavSmf3L2W3udy2+S0uRukFzFCsOga5fGp4ckY1UaNgWJYldIR9QBWhO+eCVuCVuLghgbcEhh+oX/AD+fcX/PqQ1OpQ1KV69791br3v3Xuve/de697917r3v3XuuiQPr72ATgde661D/fW/4r7tobr3XRcD8H3oqw8utVFaV64tNEltbot721Oi/T/XYe6Eqpo7BTTzIB/ZWtetqGauhCQPQdYzV04veWMWvyXS3/AEN7oZoRT9dM/Pq4jkIwhr6Uz04LR5BigTF5VzIUWMLi68mRpCqxhLU9nMjMALfW/HteLC/ZlVbC4LEgCkUuS2FC9mS3lTj0ja9slDFr6AAA1/VjxTjXuxTzr08z7Q3lTKXqNm7tgRSAzy7bzCKCSFAJ+zsCSfZtLyhzbbqXn5U3REHmbS4H5f2fRdHzLy3KQsXMe3sxHAXMPClf4/TqY2wOwEYq2wt6gg2P+/YzBt/sRSkfT2sPIPPall/qTvBI9LO4P+CM1/LpOOcOUCARzZttP+emH/oLp/o+l+38hSw1tH1lvKamqE1wynFeEulytzFUTQzx8j6MoPs7tfZ33XvbeK7tfbvdnt5BVT4BWo/0rlWH5gHoouPc326tJpLe45325Z0NCPErQ/aoIP5E9KDG/HLvHKRPND11l6ZUk8ZTJ1WJxkpNg2tIquvjd4iD+oC1/Z5Yfd7959xjeWHkG6jUNSkrwwsfmFkkUkfPgfLopvPej2usZEjk5xt3JWtYllkH2EohAPy6doPiz3tPPFAdlLTeVwhqKnO4NaaHV/uyZ4q6aQRj86UY/wCHs0t/uz+9c88ULcnCMMwGp7i30L82KyMaDzopPy6L5vfn2qhikl/rMX0iulYJtR+QBQCp+ZA+fT0fh53n+MVts/625qX/AImlHs9P3UPeYf8ALI27/srT/oHosH3i/aw/8Trz/smb/oLp8Hwp7f0hjX7IViqkocxkgwLAEqWXDlSVPHBsbezUfc892CqsbzZwSOHjy4+X9hTHyx0VN95r26ViPpdypX/fUf5f6N59Zh8I+1mpkmfO7IgnctrpZa7LMIwrWX/KUxjIxdRfhePp7Vj7nPud9MjtvOzLOa1QyT4zjvENDUZ4Y4dMN953kHxSq7VubRClGCRZr/RMnl9uadSIfhTvKKDyZrsDYmGlMjLHCTkaqNouNDieYY/1sSRp0G1vqfaiH7n/ADekXibxzzs1pMWoFJmkqPIlz4QqfMBcep6Zb7zXLkkhTbOU91uYgo7h4amvmCq+Jw8jX8uniP4b4SnoRU5ru7b1HJEheueChx/2UKqxJKT1eajkChLcuBY+zaP7pW0QWgud294rCJ1zJoii8NQD5M9wDw46gBXiD0XSfeP3Sa6MO2+2F3IjGia3k1k081SEj8gfz6UWO+M/x0p5WfJd0rkoTEVEKbq2tj7TXUibywB5GUKCNB4N/wA29nth93j2Bt5S24e731EBSmkXlnHQ/wAQZakAeS/z6Jrz3r95Joglj7beBKG+I2t1Jj00mg/Mfl0qaTqj4aYWtw1DWbjwWWyE8kcVMuT3xNWpk5ixjVa6Ojq4qC0juBpYRoSBx7Eln7YfdJ2i92ewu+YbK73B3ogm3FpPGfh+oscgjycUIVT5DoivOf8A7yG5W25Xdvs93b2iKS3hWQQxDj2FkMmAK1BYjoxeK+P/AEtifLJQ9a7UvUpGshqceuQUonqTxjINVLFyeSgW/wCfc+7d7Ge0O2NK1n7e7YBIADqi8UY4afFLheP4QK+deofvPdn3LvxEt1zruB0VppkMZzxqY9Jb/bVp0rsd1xsHDpLHitk7ToEndZJVpsDjIxI6rpVntS8sq/T2KNv9v+RtpSWPbOT9sgjcgsFtoRUjgT25p0Hr7nDm3cnSS+5mv5XUUBaeU0B4gd2K9KSjxGNx8bQ0GNx9DE7mRoqOjp6aNpCAC7JDFGrOQPra/sQ2u2bbYxmKy2+CGImulEVBXzNFUCvRNPfX11J4l3ezSyUoC7sxp6VJPTgqhQAAFA4AH0A/AFgAB7WhQooOHSYkk1PHrl/vre99e6K58ou3d3bDwW0Ou+oosdV9/d67hqdgdQpmKU12A2zVwYqozO7e0920SSRzVWz+rds00uSqYEu9dV/a0K2arVl917pf9EdKbZ6C66xfX+26vKZuZKvI7g3dvTcdQK/eHY2/Nw1LZHd3YG9MqVEmU3NubKyPNNIfRDGI6eFY6eGKNPde6GT37r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+690WzMo+8/lBtXEShZMN0z1tXb7mjYSgNvPsrIV+z9tzWkpmpZjjdsbezZusokhasUlSHUj3Xunaj+0rPk7n5QIWrcH0ZtimY3jMyU24997nqABYGUQyvt2/NgSnANjb3Xuh9HHHv3XukrvvIYbE7I3jlNxosm3sbtXcOQzyNEk6PhaLEVlTlEaGQhJlahjkBU8N9Dx70TQEk468KkgDj1rC9PvjMF1p0D2H3x13tjt75L4bp7Z+P6+272dRS7q6o+KPUOSxtLkNibO2ZsDIVEmCr+5txbRXH1+6txSw/fx1UwoYZ1pKeOH3gj95n74Uvtbvlz7ee323xXPOFtoN1POC1tal1DiJVVlM05QguNSpECAdTGi5e+w/3Z4/cLa4OdecruWDliUutvFCQs9zoYo0hdlYRQhwQuGaQgkBVAJH7Kd+7T3tiZdudvfFn4u9p7ZqvElZiMx1Zg6VGp4G8kEdMZ6PKwU80EwDRyCO8bC62PPvGnYf7wH3bsJV/f3Ley7hbahq0rNbuV8wpV5EB9CykfLqdt3+5n7cXasdm33dLKamCzRTrX1IKRsR8gw+3rFtPG4DHUKYL4i7zk2fjK2WJch8D/AJVZ/Jb1+L++aW6Cp290p2jWLmt6/HXds8byvjZaWSpwi1jRq+L0lpEys5M+9J7FfeBs05C5+2hLHcL9fD+j3BUe2mfgFguV7RJmqahBICRoqadY681+wHu37N3Z5w5Q3BruytCX+psmZJ41GSZoDVtGBqp4semuqgr0FWToNq7jxW5d59c0W7Nv0Gyt3jYHbvUHZNPT47uT469jyxNUU2y+ysVTVNZSV2Cz9IVqtu7ioZ6rE52hkjlp6iQG/vB77zX3YNx9krr+svL8kl37bXEwRHdtc1nI+UguD+JGNFhlHxEqjjWQzZZew33gLL3TgOw74iW/PUERdlUaYrqNfilhHFXXjLFwABdO3UFQU+uNo6mJpo5IWUl6dninQKwdJ4WhRpvuqWRQ8RUgo3II94lAup1I5WQEEEEhlIOGUjIZeKkHtah8uskGAZSGRWUihDCqkEUIYHBUjBBwQaHq2rrTf8/bfWOD7E+8U9gbGmx22ew/Gx+4mrMfp/unvMpcsy5ajjjWd/oZ0Yc2991Puoe8n+u77X7e26XavzrtOm1vRwZiopDcGpJpPEAS3+/RIPLrkb94n2x/1tef7tbC308rblquLSvwKGP6sFfWFyQB/vsofPoh/f8A17Js/etfuDFUVXSbS3hkZM5hq56rKVTw5/ISVOS3BjZM3mq+orMvuKjzEU9ZOKSNaPH0lVRRrYtYYF/fb9k15E55i9xeXbXweU9/mJkWPAt9yUGSWgH9mtxTxoz5OkoHlXLv7qHumObOVZeRd8ujJv20R0jL5M9kexK/xeFXw3HmjIfWialmk3DHvfdm1ttZbd+Y3zNjd19+dM7ayFPBurde7dqYqLE03yb+M+CyMlPg853bNtemWi3rs2Weh/vtR00NRSTDJRKs0u8g+4XI/wB7v2ztfZb3V3Ubd7pWaq1henSPHliGiO4hBKh5yn6d3bE1lU64+PbGvOnJfNv3aufJvdP29sfq/b24ZlurUAkQpKdclvNpBZYtdHtrgV8NgEk/poytwUDbY27vnb2bxO+OuN5Cqj2xv7bX3i4mrr6ORocntzMUGQp6HO7L33gZwYsjg8rT0uSoKhGjeO6k+8IfeD2O9wPZPd4tv5z29GsJ2It7yDU9rPSpKqxAaKUDLQyAOvlqA1dZae2Pu5yb7rbXJecs3brewgePay0E8NfxEAkSRE4EqErWgbSTToXfjdvHr7aG/Kmk7Owm3q/DbpxP8AxW59yYSk3DjNm5hqtaijy2QxNek9K2MnCCnrZ2HkjjYOnAcETfdn5x9tOUPccH3W5bsb7la/gFv4tzEsyWUhdSsxRwyiJj2TSaS8a0YHSHBIPfvljnzmXkg/63e/Xdpv8AZymYxW8jRPdRhSGiDqQdY+KNCdLt2nuKkGj7h+NWzs/VtJtfG4nqze2qGpqafGSyz9Y7sx9ZKKs5vHUsQnlxuVrqVRFQNTzQYtmlvUGMJx0T90fuUe1PPu3Pu3t54XL/ADHKviRSQVewnBUsqyQ6iqo9dXi25VgKHQ64OE3t/wDeo9xeT7yPbec3l3nZo28N0mot3FQgNomKhnK8DHODmo1Kc9EBnpt39fbpNPW0uZ2TvrZ+WhcqJUjyOEzlDDSVySY7KUclRjskaRa2LVPRzVFK5coHb1D3ym5t5P8AcD2a5yj2rf7W52jmyykWe3lRuIRyIrq0lFBLGWGH4U1K6qSV66Hcs80cl+6fLEt7s9xBuXLt2hhnjcAkagNcFxFWqPTipyKBlJFD0cjoburcW0d4ZzffUWKOQn3TV126u/vivRywU1J2NmHV58/3l8WvuGgoMJ3LkSrVO4tmStBj91SXnp5IMgWmqOtf3a/vWbH7t2tnylzcyWHuZHHTSSFh3DQAGltiTQS41S23xJlkLR/Dzl98/u8bx7b3FxzDy9G93yGz4YCslmSSRHOBkoK6UuPhbCvpamq4XqPuDrrvPY+K7E6u3LSbo2tlGqKX7qGOoo6/FZbHyfb5bbu48JkIqXMba3Rg6sGCuxtfBT1tHOpSWNGFveYvWMvQme/de697917r3v3Xuve/de697917r3v3Xuve/de6/9bbw+zl7/8AnvkYcw1VN1t8H9n7cyOAw6yuuHzPyQ7uwWZln3JlKcxCmylV1n1C8UGNUs4o6rctRN6ZViK+63wHzPVg4+guLf4e/da679+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XiLi39f8AYf717917omHy12PvDCx7Z+UfTlBV5Ht7oCiy9XX7Rx7CMdz9LZGSirezOo8jGiFq3KTUGMXLbZdg7Um4qGAIPHU1Kye690mvizu7aHavyB+VHb2zMsM9t7deA+L8G2cssTRwzbWq+oDvvFNTCQCYLUHfbSSqQPHNqjYCSNwPde+XRgfkjlajEdI9jVVM8kc74MUEbxu0TquUq6XHyssieoFYapj+LgWP19w394Hcpdq9m+f7qB2ErWXhAgkEeNIkRoRnCuepN9mrCPcfc/k22mUGIXfiEEVB8JHkGPtUdUsIAFsBYDgf0IH0IH4498fWxQUFP9Q/yV66YcSWPE565+69b697917oqHyS6e3p2l2D8N9y7TGLOO6M+TdL21vpsjkRQ1Ee0YurOxdoNJhoPBKcplBmdzUv7AMf7Wp9XpsR5ybzDtuybV7g2d+XE25bMbaGi6gZfqYJaOajSuiNjXyIHr0Bubthv943LlK6tFQw2l94klSQdHgTpigzmRcfb0VbAfBrsDYP8ynb3yM2NV7ch+LlbgO7t87j2Q1d9luPaHfndW18LtLf1ZtPGpAUm2x2Au18flagKyimyf3MhH7gDDq49zdp3T2hueVdzWX+uiy2cMctC0ctlaSPJCJWBxJD4jxg/ijKKfh6Bi+3u7WXuLabvYNF/VkieZlNdUdxMEWQj1WXw1dhUd+oimrovvSn8vPP9Qbl2h15uH4SdIdw4PZHbn94cJ8oc38mN8YWSr2RD2FLvXb25tw9DS02Qhm7S2jSzIi0lOy4qrrqWOYPGrN7FPMPuvZb9Zbhu1r7jbjt9zcbf4b7cm3wvSYweC8Ud4CoEErZLsPEWMlaE06D20+3W4bXeW1hc8o211bw3gdLtriQfpiTWrtb6GrIopgEoWAIKjAFbK/Hr5rdWYj5efH7pLYvTu/up/lf2V2z2BtXvDfPZ9VtnO9Lr8gKJKTsyh351pHtzI5DsSfbc0lRUYV8ZW06VYeOKfQFJ9kcPNntxvlxyJzVzHuu4W2+bHZ20MlpFbB0uzYkm3MFwXAgD9olEiNShZKk9HMnLvPG0Rcz8vbPt1pPtm53Msq3DyMpgFwKSh4tDGTRVihV1qKB9PSwo/jv8j/iT2Ptrc/xY692N8hNnZP4qdK/G3dOB7E7SHUe5Nubl+P8OWoNj9hU2Wl27uWhy21dw43PTDK46JVrIJY1aAsDb2gfmzk/nzabyx533e42u9TfLvcI2gt/qY5I70q0sDKXjIlRkXwnNUIajDBPSwcuc2cnbhZ3PLO3w7jbHbYLV1kl8Fka2DrHICUkDKyyHWozqUEGnb08ZTrP5kdN/I7d/wAnOqusOou9cp8hOluotm919c1HbFZ1VFsDtrqODKQ4ncew9z7i27nzuXrHKQ56aKppJ4oMnGYFmS7MV9prfevb3mDlLb+Td73i+2yDatwuprS4FqlyZ7a6Ka4p445I9FwugFWUtGQSDgdXutn522bmG65k2vbLW9k3Cyijni8QxeFLErgNG7RuGQ6yCpCtUAitSOmXpj4RdpdUn4TVWSzW1dyZ/rH5BfJr5C/IfJ4mrqcZhqXdHyJ2NvenrMR1zj6yBq3LYHC7l3NBRReYxTPTRNUuAWKhVzB7k7FvY9xo4beaG0u9q2+xsFYBnMdjNDRpyDpSR4oi/bUBjpHCvSbavb7edtm5Nnd4pJ4b+4ubrSSqK06TkiNSK6VeQKK0JArgmnQ6b02B3ztj5x7F79632BtDsHrfdfRGI6H7MkzHYcex9wdbjC9pVe+03rjMTUYPLrvmjqsVlJoBQQyU04qYkvIFYkBnbt35XvfbPcuWN63a4td5t90e+t9EBmW4L2wh8EtrUwFWQHWdQIJx0I902rmG357tt+26wjuNsuLIW0pMnhvFSUvrAKuHw5GntNRxoeq/dqfy58v1zufP9e5T4T9KfIvZ1d3RubfG3vkLvL5Mb62TImyt59hz7/Wn7D6SpabIw5Td2w5cpNT06Y8fY5Q0cEr+JmkJlO/92rTdrO13aD3H3Hab9NuSCSxhsIph4sUAgrBdlgVimCKzGTvTUw7lp1HNv7b3237hNZTcpW19bNemRblrqRaJJIZT4kOhgzITTtJV6KTpNehv7T+JvyR3x8npPnbTx7Ch7r6K37hdj/GTqaXPYmLaO6vi865PC9o/6TN5Phpa3GdodjUW7shk8U8TNDt96KmpR5BPM3sN7Nz7yZtfJq+2ckt0OXN0tnm3G50MJI9w7HtxbRa6G2tzEkcgI/XDs4poA6P915N5r3DmSTndLeD97WNwI7SAEaXtT2yl2KV1yB3dCcRlFBrqc9W/0GLyuVkgixOIzGSerZEpoqDGVtbJKzr6EjemppIpXGki4OlrG3uEbLa903KaKPbtpurmSQjQI4ZG1kmgAIUjP29TLcbjYWMUku47jbQCMVcySommnGoZgehUwXx87r3JCKnHdf5mCmZIpFmzL0WCEkcrtHqSPJTxTM0ZW7roDKObe5O2T2G939/hFzY8h3ccDAEG4aO3qCadolYNjiagduR0At294PbTZ5DDd83W7zAkEQh56ECuTGpXPD4iK9CTRfDTuupqYIapdpY2nkk0y1cudlqvt4wCfMaeloTJOLgDSpB59yDafdF93bi4t47lNrt4Gbuc3LPoFK1KJHVgTgBTX7OgZdfeT9tYIZZLc7hPKoqFEATUfTU70H206X2N+C27JIwcx2DgKSX7gDxY3D19cjUvo1OJqmehZKj9VlKFRxz7HG3/AHKuZZF1bpzxYxSeJwiglkGioyC7RnVSuCpAxk9BK8+9TsSPSw5Runj0cZJo0OrNBRVcEV4kNWnl0uqb4JbeWdHq+xNwzUqlvNDBh8VSzOuk6QlTI9UsRDWJJjYEXH+PsaQfco5dWZDc8+X722dSrBCjn0o51BT69rAj06Cs33qt7MLiDk+zS48maaR1Gc1UBSf2jrLVfC/qvH6pMj2PuOkhWysanIbapNL3v6pZseFF1/Fgfzf27P8AdA9srHVJfc/bjFCP45LNKH5sYgPy6rD95bn67CpZ8nWMkpP4UuXqPsD9Jpvj/wDFnCmp/jvcNRU6LWjbeG3YHp/Et5Bpx9AXkZ7/AEN/8PYef2J+7Tsvj/vn3WklAp/xOtVK04/2URrX5jo6T3e999zEX7r9u0TjkWlwwavD+0ei/l0nqzZnwjoKaZJOw8/VvE5Jkoczk62ocMABHGKXCtE6gjiy3ufr7JLzkz7nO327CTny8mcGuqOeWRjXyGi3Kmny/b0bW3M/3n7ydSnJ1rGCPheGJFHnU65tQJ+f7Ookea+D+E+0lh29ujck1OFQielz1UlWRGVM1dDWVtFSSFvqfQq6rWHtIm7/AHNdn+lkh2LctwkjFDqS5cOQPikEkkSGvyAFfIdPybb96LdBPHLu9lZRvnte3QoCeClEdxQeVT9vThT96fF7BCogwfRL1MUkiSeSqweAcSOECEqcnV1k8AUC1gApPPtXB71/dt2UTwbN7KGSJm1EyW1samlMGV5XGPSg6Sze1Xvpuvgy7p7pBJFFKLPPgV9I0VT+dT1NPzA2Hj6aKgwXR1HFQ06BKaGafA0cMKEklFpqbFTKi3N/Sfz7Wn72HJFnAlntHs1CtlGoCKz2yAD0CrAwH7eky/d05tupXuN09z5GuXNWYCdyT/pmkWp/LrHVfOfLW04/rHDxaQAhq85M6rpX0WWmxyD0EDgEcf090ufvqbkAq2Pt3aogGA9wzUpwwkIGP+K6ch+6zY1Ju+d7hq8dEIHHj8Uh/Pprm+c/YDIywbI2hE5X0vLV5mdVb8Eok0BYf4XH+v7LJPvp87spEXJ21K54EyTsB9oqtf2jpfH91jlMMPF5o3Bk+SQqT+dDT9nTNL83e2X4XB7Ggta9qDMyD8X/AM5lyfz7KX++R7oMe3ZdlUfKO4P+G46Mk+7ByCB3blurZ/jh/wAkXTZUfNHuaYnwLs+k4YftYOolsSB6gajJvytvzcey+f73nu1LVo02mIDyFu7f8elPSyL7tXtvEKyPuT19ZlH/AB2IdNLfMDvN9QXNYGMkWum26ElP8UBZrn/Xv7Kn+9Z7ztX/AHbWSD5Wsdc/aT0Yp93b2tTSTtl2323D5/ZTpkm+U/e80jyHewh1oU8cODwscaAkHUiGjYhwf7XJ9lE33lveuZ2b+uarUUottAo/6tkg+mejKL2H9rI4wn9WC1DWrTzE/n3UP2dMVd8he7ch6Zuydwwj6aaEUNCCP6saaiRuD/Q+yO899/eS+7ZfcG/UZqIxHH/x2ME/keje09ofbCzzHyVZscf2muT+TOf8HSbrO3O0q1QtX2VvORUvx/eCrpwOOSTTywk/7H2QXfuj7lXahL3n7dzGo/5SXX9pUr/M9HNv7fciW7VtuS9tDH/l3Vv+PBh/l6Tcu5NzZmVIptxbnzFURJ44hmc1kKhgAWfxww1MsjKoFzYcD6+w3JzBzJu88aPzBuN3PnSPHnkY0yaKrkmnnQYHHo6j2XYdqjdotmsbaHFT4MMY+WSoFT5Zz070GwuyNzfanH7P3zm46x2jo5nxeblpZmj16lSqrVWmjCFDcuyi49nVlyN7h8x/TGy5T3m7ExIQtDcFWpx0tLRBT5kdFt5zbyVshuBd8xbVbPGoLjxYAyg8Kqnca/IHpd0Hxi7xycHnh66rKdPK0Jiy2RxGOlJGm7mnqKxpHp/Vw1iDY2+nsa2H3cfeTcovFTkKWNNemk00ETVx3aXkrozgipwcHoJ3fvh7YWUvhPzfG501rFHNIPPAZUABxw+fHoRsZ8I+2q2SdMjNsjCxxxK8U0lfU5ETuW0tEIqPHI0RReSxOk/T/H2Ptt+5z7oXckgvZtns0VQVLSNLqJOVpFEKEcSTUHyz0D777zvINrGjWcW6XUhYgqEWPSONdTyEMDwoMjj0J+z/AIJSpVQzb83fSS0EcoefE7VoZIWrY4pEeOKbJ5BVMUU+kiRUp9enhWF7iSOU/uWPHdpNzpzXE1iGqYbOMgvQgqGllGARUNpj1UNFYcegLzJ96cSW7w8q8uyrdFKLNdSA6CQQWEUZyV/DWSleK0x1YpTww00ENNAgjhp4ooIYxfTHFEipGi6iSQqKB9SfeekMMdvDFBCmmFFCqPQKKAZzgCnWIEkjzSSTStWV2JJ9STUn8z1m9u9U697917r3v3Xuuj9D/rH37r3RI+tJqPt75h929jvSST4b44bexPxw2PXlhNQy7u3bTYXs3urIUSOG8VdSI22sQ8i2ZTRTx3szA+63ig9eju+/da697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuuj/vv9t/rG3v3Xui79WWyHcfyTzkeUmroU3R1/tNKJpIHixEu2+v8bXVVLAI0EsX3NTuMzOrljqe4sD7917rF1p9ruDvf5Cbwp6WldMN/o46khysDOZKuXaeCyG8ctQVKPCgVsXkuxCgKs6eo83BA917ox3v3XuiqfNjcLYP40dlY2E3ruw4du9OY2KzNJLXdz7qwnVyiFUZHMkEG65Jrg+lYy1iAR7sunUuo0Wufs8+tEsASgq3l9vVFnZmTgzHY++q+jXx0H958njcTDeMinwuClGAwlKphVIilPisZCilQAVA9/OZ7rcyTc3+53uFzPO9Xvd4unHA9iTNHGKjiBGigHzAGT1259tdhi5X9veSdgiWi22124PH43jEkhzmpdmJ+fSA8kjyyJEsJSIxh3aQ6vIWvLGYkUldMRBBJsSfYA6GnXhIrkQzxGKR7aVvrR2RTKTDIhF2hAvf0kNa1/fmAZSjfCf9Q4ZqOIPEHh16pWrLggf4fI/I/wDF9CrmNz5HdUNV3DT7Wl3r3v1D1tkcHuXC451pct8qvinjpRleyem9zsiFc52f1biopt2de5CUNUU+ZoGph+3WVIk6f/df967f3n2LePu7+8Uq3hu7CRLO4fMtxAiktFIxx9XbACaCX4mVKmrpU4A/eA9qp/a3d9t96/bRWtFt71GuIU+CCVj2Soo4W87ExTR/Cpeg7GoEXncXj8dVUU+DzFPufZ+5cDg95bB3dRkmg3l19u7HQ5naO56W4BQZTD1SGaI2eCoWSNwGQj3gD7pe3O8+03PfMHIm+1a5s5f05fw3Fu/dBOvlSRKavSQOvkOsyPbvnrbPcjlDZubtposVylJY/OG4SgmhPn2N8JplSrcD0NHxT7Bpuv8At/FYrLTeLZ+/Yf7g7nhaREgXHZ9/BhsjImkRx/wTNtEVc+oIzm/PuUPup+5je2nvLy7c3Vxo2DdXXb7sE0QLOw8CVvL9KfTk8FdvXoB/eI5DHPntjvcMEAbeduVry2xVi0S1mQfKSEMKDBZVxjo7HZHWR3xtTfXXdTRQPvLDUmXyuyqs01I1bFunbsYrpsLT11RTT1VHit3UFAqVMdOYmqDDBdrLY9dPvG8hN7kezPPXLcNuH3b6b6q29Rc2p8WOnzcK0X+lcjz65s+yXOKcke6HKG+TzlNsM4gnNMeBcDQ9a8NOpZB6FQeqoaacOIamBp6eeKSOeMqZaato6iJg0bIysk9JUwSC6kFWUi49/P2juNEo1LKjBgQSHjcUIKEUKOpGCCCCOuzDojB4XVXjYEEEAqynjUHDKw4g1DDjjpQYTsPc+wt1bw3vR7QyHb2G7STDU/yE6hpJ4v4x21jcDTS0cHZ3XsFRJS0VN8ndm4FVWlnDRybxoqOKgqXNfFRT++iP3fvvA7b7lbX/AMD794OQbjsm4r4NluNw1ZhM39lBcStlXqD9Lc18RZAEcsCD1g97zezW4e3t9/rxezIayvbF/GurOIERiOv6kkEa/wChn/iRb08MoSyBeHT7vzZ+JwZ25ntq5ym3v1V2Vtyi3x1RvmnicUu69l5ZUnoWrIJ4opMfuLFeRafI0UyJPTVKkOim6jFT319mt99kOerzlbci02zy6prG4IxcW4YijilBPD8EyZFaOo0uOsj/AGi90Np91+UoN/sqRbrFpju4AcwzaakrkkwyULRN6VU9yno0nxk7qoqukxfQ3Zdcy0NRUSU/U296+ZpJdvZisKiHZWcqpma+3crNq+3NgtNIwQ8FCuU/3N/vLtyvd7Z7Pc9XleXp5RHtt1IxP000hotnIThbZmxA9f0nIjPay6MevvP+xH7+t7/3M5QtB++IY9d/booUXEa/FcxjzmUf2q0/UUax3KdQ+b2612j2DD/dHsakkwuWw5mxGE3xSU0Um5tkvDVTVH8KklnjkbIbIqcnKJqygBUuD5I3Q++gXvV7Kcpe93KsnL3MiGLcYavaXkenxrWYrxBI/UiagEsTHS4BwHCuuGvtd7pcxe1XMI3zZHEtpKNNxbPXwriOuBj4JFyY5ANSE5JTUvVZ26tsbo6y3pktq7iiqcHura+Tjkp62kllp1nEMvlw+5sBXRuT9nlI0Wpo5EcuENidQPvg9zhynzN7a85bpyxvkElnzNtd0KOtUNUbVDcwOKNokFJIXBrQ0rUEddfOWuYuX+e+WbHftpljutj3C3oVYK1QwpLBMvCqEmORDioqBQg9GK6w7bzNRv6r37tHdW1unPkLmaOGiy26M/S1sXQ3yPFBSxwYzE/IvbeImhkwe+qZIRDjN8YkQ5KmQ+KqSspv8kk6Rfdy++om8z2nJHvJeQQbkVCW25mkUMxAAEd7UhYpmpRZ1pC5IDhHI1YN+933WJNrjuea/bG2mnsQxafbx3yxA1Je1xqkjHEwnVIoHaWXC219AfJHG9yTZ7Zm6Nq5bqXvHY1PRVHYHT+6K2ir8jQ47IM0ON3tsjcOP04nsjqncNRHIuN3Djx4JXRoKiOlrI5aaPo6jpLFHPE4aBwCrA1UhhUEEYII4EYPl1g/IjRSPFIpWRSQQQQQRggg5BHmDw6Mz7t1Tr3v3Xuve/de697917r3v3Xuve/de6//19x34TUdRV0Xya39XUsUdX2L8vu86+CsSQyyZDAbJy9B1XtqSWQu4tBiNiJGiiyqq/Qe9nqzeQ9Ojt+9dV697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de69bm/v3XuqqvhFtybpv5lfzC+gCgpNp0OZ6K7n6coGBApOte1NtbtfM4fHjTphw21+0cLm6KkgDEQU3iQBV0j37qx8j0er5C7dyW6end+4fFQyVOQfEitpaaBfJLVPjKmnyDU0aXBaSeOmIFub29xB787BuHM3tJzrtO1RF782okRBkuYXWUoB6lUNBxJoOpF9ot6s9g9yOVdy3CVI7NbjQzsaBBKjR6z8lLDqk+O5BJuCTcqQQVJ+oKnkEH3x3JDUIOKDyp/q/ycD102r5U6ye9db697917roi/+++v+B/w9+69137917rjbm9/6e/fl17rsi/8Ar/g+/de68Rf/AH3/ABX37r3XRHBufoD9bfj+tx79SvDr3T9tzaO7N4VH2u1duZrPzKQsgxdBUVMcRbhTPOiGmp14N9TAD2fbDyvzHzTcG15Z2C8v5gQCIImcLXzZgNCj/TMOiXeeYth5di8bft6tbSP/AIZIFJp/Cp7m/IdGN2p8Oe3twNrzQwezKWysrZOqOTrZNQUkR0eMMiRsoY31yDkEf4+8guWPule6W+EvvAtNptqA1mfxZD60ihrp/wBu44U6hrfvvIe321UTbBc7lP5+GvhoP9vIBX/aqePRiNr/AAk2Dh4DV753Ll9ySqpaRKaSPbWJhs99f7Tz1zgKLnXOFuTxa3ufeW/udcj7TCbjnHmK63CUCpCkWkC544LuccdUlDk09Id377zvNm4yiDljZLeyiJxqBuJW+WQE48NKenS3jxnxH6xJlf8A0Y0lVT3kD1dXR7kyyuCh/aSeXKVwZXjBAQDS30t7Gce3fdd9um1v/V2K6j85ZEu56gg0AczSVBAoF+E8AOgu9994HniiKd8lt3H4Fe3iIoRkqIkNQTWpyPXptzHzF6V2+ppsFFm84YwwSPCYL7GibSLqFnrmoUCMSbHTb6n2X7p97D2f2KM2+ypeXmn4RbW3hxk+gaTwwD/tfU9LNu+7n7lbuyy7oba0Q8TPPrcfaqaz/PoJMx87qxiy7d65iRdXpmz2cLFhx+qnx9NZWv8AjyEe4q3j7693IWXYfb5VUcDc3BP/ABmJP5aj9vUi7b91S3C13jnEljxEEFP+NSNn/eegvy/zO7jyHlGPG18Arn0GjxEtfKi3+gkyNROhPH10e443P73PuzfeILBNtskPDw4DIV/OViD/ALz0Odu+7X7c2ZBvHv7phx1yhAfyjAP8+g4y3yF7szRk+67GztKkq6WixIosRGoP4Q0FLDKh/wAdV/cd7l7/AHvHuhk8fn28iVvKERwgf840B/n0NLD2c9sduCiHk62cjzlLyn/jbEfy6Q2R37v3L/8AF03zu/IXBBFTuLKutj9RoFUqWN/6ewVf8+c8br/yUuct1nxTuupqUPHAcDPQps+UOU9vzY8sbdCfVbeIH9umvSTnMlV/wJnqKm7aj9xPNUam/wBU3mdwT/j9fYZmmmuTW5meQ1r3MzV+0kkno/ijigAEEKR/6VQtPsoB1hWnhT9EaLx9VjQEkfm+n68e2FjiUkrGor8h06ZJGADSE/aesgQD6f776/0t7cLGtR1XHCmOvFbi1yP9bj34N6iv29ex6dc/devde9+691737r3WN5EjF3ZV5A9TBb3NgOSOT70WVfiYD7TTrYVm+FSegV+UtT2Rtf4qfJXsHrui3LRbn2T0V2vunbm5MRhaqq/ged23svK5ahysc8tHUY9JsXNAsx8oKqFBYEe5F9uOUNz5k5v5Utv6tXl5s8u4wLMVglMXh+IhlEjhdKgRkljqHbmo6A3PnMthsfLO+yrvtrb7ilpI0YaWMSa9DaNKFtTVcALjLY6ql7I/mEbe3N8MvjmnTfySxVR8mt57t+Ee3Nwx0tDkqHcu4K7f++NgYTtDHqcxtWHASHJQZGsSuaJkVInZozyvufuVPu6c3XPuDzI28chGPk1Rf/TmeRUjJZJlsjGDJrYBjGfPSoDNjqF+YPezYIuUdhi27mnXzCfpfGMahz2PC1xq8l1J4gHqcY6sJ312VgvjTvr+ZNvvszdXcu58B8Uuw/h1tjsLYWPgxW9cHtmP5DY/BYTbb/HPbmOgxecr1kk3DFW5ygyEhrK+vj0Ud2ZVJvB90Xnvdtp5ds9xvuXttmt/FE0kIuJp5hI+oNM1FjZo/hQAINNamueipPvHcsbXfbxdWlpu994+gxpMYEijKrQqoUllVjUk1ehyARjozXTnYe2Nydlb56o+QHTvyW+MO98F0ru75KdX4DtPFdW0snfPSWxJo6bd9ftqfa+8N2xbJ33tmoraD+KbezUsGSxsWUp3kQgS6BZt33JdojMZ3j3BuZqSVZYLaOMFPNQZDIwYmvcKgCmK9El396reXEoseTbWOqdpkmkcq2ckLoDACmO2prnoPeg/5gPxH7Fr/i1uvdfxR+XXXfx2+Xmf2zsrpH5JdsVuzKXZme7X3bipa/bvXG4tlbV3M++dv4bcuToavGYXcFXjosTnKunLU8xgeOUyNtf3RvaHb/BN3Bf3ro+r9a4YAjyQrEIwVH+9fPoF333kPcq7V1t5rO1DJT9OAEg+bgyFyGP20+XQmby/mkfFjreu7W3v118CO5+2fin8cuwM1sTvT5f4fH7Al2DsfcO0cnFheycjtvae692v2f2btPqrLTrBn6/D42akomjn8flELn3Iu1+xftDs5jay9v8AbtaPqDSR+MwP+mmLmg8hWg9OgTf+7XuVuesXXOd9pZNJCP4Skf6WIIKnzNK/PoZO1v5oZ2B2TUdN/GX4k71+Te8dn9YbU7l7Kh+O6da0FB1Z1r2W1TVdd5Nsj2Bntq0G7t49iYujqa3F7bxQlyFRRRGZymtAZB2/l/YNpWNdq2S0tgldPhQxx6dRq1NCimomppxPHoGXu77tuRc7julzOzUqZJXetOFdTGoHlXh5dMPyZ/mQd6dJ47Nb1j+KHyp7L6i2V1tju2eze0dmZDp/rjC7K21V4eTPbhwuK2rvfduJ3p2Hvfr7BwPPnsTjIHfHyIYVkllt7N/IUAr/AKvy/wCL6LvT/V/qp0FXan8zT+Bdo4XpDqjoj5R/K/sDdXxn2N8udpYzqWr21j8FkuoN8SVbUOSzm4t+7kw2N2Zk6PGQJLHDXEyZKpqoqOkWSctp0RU1p1oDz8/2dILq7+bB132zXfHncOD6Q+T+3Pj98nN7YbqDrP5Ob8xWD2t19P3lnqDLVNJ1fXbRqtxy9ifaR5jAV+Fbc0dEcDLmqR4IpHX9z3vrfSn7J/mq7i653b8ktu7I+L3zL7oxHw13ZLgvkPv7rmt2XR7M2TgqPZO3t+VG49tPundmOyHYeSodt556mfbuKgmytPS0Uk8oWOWDX7r3R2uiv5kW2uwPkHsToTbU2a3/AI7sP4o7L+Y21uxJkx8W3Mn1Jv3etHsrAU1DJqTcA3PJJWx1jxVERgWAmMuJQbe691Gpv5wWw939HdFdn9P9A9qdodhfJXv3uf499M9HpuLrPY24dybo6Jze+cfvnMZPeu+N2YbYOAxCYvYdTW0kM1a+RrPPFDDTvL5AnuvdWh9Tb1zvYvW+zt77n663Z1HuLcmEpsjm+tN9PhJt27Kyj6463AZuo23lM1gKuqop42Ano6uennjKujWb37r3Qh+/de6R+/t/7L6t2fuHsDsPcuI2fsraeOly24dyZyrSjxuNoYiqBpJGu8tRUTyJFBBGrz1E7pFEjyOin3XuiWfy294UW7Oh90mswO5trdiUHendNT2zgd64xMHu2PeO7t9ZTfeOzGWwb1M2TxdNuPY+58RWUMdakFUtFLGkkSFCo917qwP37r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691CyWRoMRj63LZWspcdi8ZSVWRyWQrpoqaioMfRQSVNZW1lTMyQ09LS08bPJI5Coikk2Hv3XuiQ7S7QOwOrdxb7ixcGV7P+Qu/N8766l6/gpGhzu6KLIGDF7DyOYpYW+7pNtYnZWLxuRzeVlWOHG417ykSeON/de6FX4f0NfB8cersrnZMdV7v3jt9N978yuJq5K/H5zfe8p5s/uvMUddOqVFRQ1mYrZDTeQa4qURRm2i3v3XuhQ7W7V2V0tsPP8AY/YOWXEbZ27TiaqmWNqirrJ5XEVHjMXRIfNX5TIVDCOGFLlmPNlBI917qvLtn5Edc/JaT4n/AOjTKVGY29N2f2B27uzF1MDY7LYlPjt1VuTdVNg9xUDs70VT/fjP4F1Ulo5FZZEZlAPsg5rvxtXKvNG5lNX0+23UtK0rogdqV8q049G/L9kdx3/YtuD6TcXsEdfTXKi1/KteqlFmIpROQS0iiUKAZW8kx1Ktrq0hDPzzc29/NVEdcSORRmFT+eaU+VePn13XZdDugPA09OGK08ulPtjZu7t2vU0m0dr7h3VU4+nFXkYtv4qvzElHTs5BqagUyTvBTtLcIXN+LAcexLsHKfNPNc11bcrcs7hudzBHrkW1gkmaNcjVIEU6VqKAnicAE9EW88zcu8txW0/MW/WdhDM+iNriVIQ7/wAKmQrqNKVA4VHSfki1iSGRCpDtHIjqQ8csTlGWRGCukkMqepTYhlsR9fYfIZSyMpDgkEEUIINCCDkEEEEHIIoc9HanWquGBBAII4EHIIPAgjgRgjI6cttbjzG2cvhN14Craiz23clS5THVKOpMVfjpw4jlMJMbQz6DHNHyrRyMhFiR7NNi33deWN62nmPYrlod6sLhJ4JFOkrJGajP8LfA44MjMpwT0W7ztG279tO5bHvFuJdrvIHhlUioKSKQceowyniGUEZ6nvj6HEwdidf7epYaXZvXW6sB2907QRMNOA+Pnysjze4qfYVFGSWiw3UPyF2ju7C0sa/t0tJW00KWUKB0I++NFt3uj7Sezv3hditl0TxfTXRWlUE66kjkI84LyOaEA5XxAPPrCj7sE977fe5fud7M7zcHXHIZoAa0Z7dtLsg/4fbPHL89FTWnSWqg6okqM6SRHhkL3QORaUBZEUSRSqrBmv4wpP1984mDEMI2o3FTWlDxU18iDQ16zpFCe4ArwI4gjzH2EYI869XP4je43Ttzqbuyg8Zq83iMac+kT6/Fu/ajR43P085IXS9XHEWsf1oxJv8AU/Qz7G8/w+5vtVyVziGVrm4s1S4WtStxD+lOrYBB1qTniG9KE8Vvdnk1+RPcTmzljSwt4bpmhNKaoJf1IWB/0jAYwCtPXqtH5I7Aj647o3pjcTTmPB5asTeO3VjjtFVbe3XH/GKaKmAZI3loqmWWnVjYII9Nh74q/eN5BPtv7zc78uwwFNqkuTd2tBRTBd/rAD1CO0kf2oeHDrqf7Hc5Dnr2v5U3uWcPfpALa4zUia3pES3mNaBG+da9Aqkl9M8MrRupV45oXKSQyxNqWSOVCGSaCVQysCCrrcG/PuEPTjXyIww8xQ+RBypGQcgjqWiqsGDKGUihByCDggg4owwwOCDQ9CViBFhsO20JKZIOsu/aTfe/usqYArQ9Y/MHqSiXcfc+ytuwySFMVtf5L9WPPu2GigKQJuTGZV0jBqffUve7uH7yf3LzzPusBl575UikLS6ayNPYaRM6nJpdWhDSZoZKscjrnntFvL7D/ejGwWM3h8n79KgCaqIIL0kxA5pW2uaqmKhMDiegxmjWqgKg8MFeN1Zo2VhZonWSOzoQbG6kG3598stKuhUkUIwc/kRThTiD5ceuhrao6Y7gcg8PmCD5eoPVq/X276/svqPYW/8AKHz5xI63YW7KogiSrz20hFDR5KrUkulTlsDPC7F7PI0Zb83994/uo+4957m+ynLe67tOZd9sWewuXJBaSS2oqSvkktLEUYk0qSWpnrkB94jke05C91N82zbI9G03YS7gQDCJPUui8BSOQOoArQCnl0gvlTtKl3d01gewxETuTrXO0my8tWRRI01Zs3cYkOGnrJixeI4HJjxU8hB0iXSCL8QH9/327srzk7YfdCzt6bxt1ylncME1eJaXBIUymuBBLTQTXMhUUrXqY/ubc63VpzLvPt9czV2y+ga5gUtTRcQULCMUyZYi2sVFfDBNadVyKWYNTzhSxUlTpBSeMfqAVixdolKiQkAEtxx75MkBlowqpH7f29dG1OVNf8n+yPl0bToxqyjquhexnyuSi3Htn5cdbdP7PygqZKqej2H2ds3dKdk9fVckzu9Fsbca0WPrlpGP2rZrH0kkSCoQN76y/wB3puG+3vJfuBb3+7Tz7La7hbRW0UkzyLBSAtIIkdm8JGDR0C0VmBxg9c5fvo7fs9lzTyfPYbbDDutxYzPcSJGEM364WNpGUASuCrCrVdQRU0IpsE++hnWFvXvfuvde9+691737r3Xvfuvde9+691//0Nxb+Xm9RJ8cpHrC5rH7v+TBq9f6vOPkH2QGBvzcKB791ZviPR4vfuq9e9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691WJ2du+g61/mxfF6nmlnif5JfEvv3rGUPUpBjf4t0zvfYPZu2L05F6rKz4/dObjiuSUiDgfUg78urDKnqzhgGjZWGpWDKysLhlNwVYW5DDg+6MoZWVlBUjIOQfkR59aBoQQaH9lOqfPkb0ZX9Ubhq8/jIzUbD3Fk55cXNGsjvgqupLVMuHyL+JYoIBK7faOWOuMaTZlseUP3gPZa+9st+n3nbYjJyTf3DNCwBP07yEu0EuKKuokQsTRlGk0YUPRD2Z91LXnzaINov20c1WcCiRTQCZF7RLGK1JoAZRQUY1FQcFsVg3I/PP+FuPyLj8+8eCPPy/1f4Opt65e9de697917rom3+++v8Ah72BU0691xLgf8bIH+9kW97K44g/n170FD1KoqWsydQtJjaKsyFU4ulNQ00tVO/IHpihV2IuRz9Pb1ra3N/NHb2NvJPcv8KRqXY/koJ6Yurq1sIjPfXMcMA4s7BVH2kkDoYcb0Fvyogir90thOuMO7XOS33lqXDO0YQuXpsaZHyFVcDgKnuW9r9jueJ4E3DmIWWwbQcmXcp0tzppXUkJrM4+xB9vUaX/ALu8qRzPZ7CLredxGPDsYmmANaUeSgjX8z0qcHj/AIz7Gn8u69x7i7ZycDRMtHtfET4/bazpYuPuq6poJK+NWHHrtY/RvYo2Wx+7vydOZeaeY9w5ov4yp8Oyt2is1PnV5Hj8QV4nUKjgD0Q7re+9vM8Hh7Ds9ly/ZtXuuZRJcFTw7EVxGfnTB8x0Lc3zQxeAoFxPW/VlHiKCnj8dN/Fa6npokIPEkmPw8LFyQSeZySfr7lKb73m1bDYfuv2+9s4bO1UUTxpFQA+rRQKa/nJU+Z6j+L7tF/ut4b/nPnuW4umNW8NGYmvkJJiP5JToH8/8r+786CkO4sft2BgB4sBiaaGQWN7isrvvakX+hsR7ijffvR+8e9B0g32CwiI4W0KKR9jyCRx+RHUj7R93/wBsdp0tJtEt7KPO4lYg/wC0TQv+HoE87u/d+55pZtxbq3BmnmJMgyGWrpoTc3Kim8y06rwOAoHuHN75v5q5jkkk33mS/vC5NfGndwfTtqFH2Up1J208tcu7HGkez7FZ2wUY8OJAf96pq/Otek2kUcf6ERf6lVAJ/wBewHsNBVXKqAej0u7fExPWT3bqvXvfuvde9+691737r3Xvfuvde9+691737r3XvfuvdcC4H1BH9fpx/ieeB7tpIUsaAfaOvDJoMnp62/t7cO7KxcftfA5fcFYwv4cTQT1mldRXVJLEpgiQOpGpmAuPZxsPLfMHNN0tjy1st1f3RzpgieSgrpqWA0qK4JYinn0Vbvvuy8v2zXe+btb2luPOV1WuK4BOomnkAT0Y3bHw67g3CkdRlVwWz4Hl0MmZrnrciqC4LrQ4lKmKzW4DzqTcHgX95C8s/dN91t8jjm3IWO0xE0InfxJaDNfDhDDjwDSAmo4DhC++/eP9vNpdodv+q3GWnGFAkfDFXlIP5hCOjGbY+DGy6ERy7t3ZuDccwKs1Lj1p8Bj7gy6lHgFVXskist/3gQVNjY29z7y59zHk+zEcvNHMt9fzAglYgttGSCfNdchBFK94NRg56hzfPvQ80XRZNg2Kzs46U1Sap5OAya6I6g1/BTPRits9E9SbQWJsLsPb0dRDqZa2uo1ytcZJI0ikc1eTNVN+4EFwCFvcgC/ufOXvZj2v5XEJ2fkiwWdCSJJIxNJVhRj4k2thXzzT0A6h3e/dD3B5h1LunNd20Rp2I5iSgJIGiLQMV88+pPQbfNPqzK90/D75N9IbVye2tv7m7e6H7W6z2xkd0VzYfbdJuDeuyczt3Ez5etp6epqKbGxVuQQzPHDK6xglUYgAybGiRLpjQKvoBQfsHQEJZsuxZvU5PVX/AMqPjxuTs/8Alv8Axm+HWzNwbFqe1umcv8FMju3OTVWTpNiVqfGPefWO5N+x4XcCYSStrJ8jR7Kq48WWpEFRLLGJDErsy3611Wb/ADT+uO1Nj9P/AM23vnHZPZWOxHy97z/lj5bqASyZPNZXbdd0x2L1nsieq3/iUo6OGlp5d2tTVCQ0c9S89Cj2eNyB7917o1m8esPlr2t3lvL5SfL7H9LbNPRPwr+TXSfXG0OmOxt4dlxbz3X2xgf4l2T2bla3dOz9mjY+1lxuzaWlwm3xHW1cH3EpqZ2MaFvde6IH8GuiPlt8lvhN/KS2h2RQfH7bPxp6Irfjr8o5uxNp703lk+2u08H1HS5HOdRdVv1dkdpUuH2Nl6TL1iJuLMnOVdLWQ0iyUcKGUxj3XuhxqvhJ80NrfHr5UfBHrxvjnkvj38lOye9M5tf5Ebn39vTE9odMdY/JPeVXvDs7a+Y6TptjZLGdi7328+ayVNgK2nz1FQzCeOSrCeMoffPr3Q1p8Z/kp8afkP2n2v8AEDbnRPaOyO8ehPjv1Nmtp98dobv6wyHV++/jPtWv2BsvsGGu2tsjep35s7cuz6yP+J4iE0FfFW094JdEhYeBByOHXs+nQCfMT+X38gPknv75D5nP9dfF7v4d2fHXY3V3UW7u4+4u4Nnba+IO+MbsDMYDszJbH6SwO3NwY7eOP3jvbJ/xzG5P7qnyyyLHDXMYotLe+eKfb17Pp0bXoL4m9ldV/KHande4cvtCs2pg/wCWT8efhhXUeMyeQqc9L2p1Nk4a3cWbjpJsVSUkuyauOM/Z1fmWpkY+qnj9+p6EEfb175GtegI2X/L+7o278L/5e3x0yO4eupd8/E/5c9b999kZGmzmZl2zk9l7O7M7S3pksds/IS4JKvJ7gkxe9qRIYainpYmnSVTKoVWb3XugowOH+au9O2v5znXPxs278b8ts/tz5P1nXbbw7e31vjZm6+nN4br+LXVOFzu/Bt3AbP3Tiu1NsDaOUiajxUdRi66HLU15JZKeX0e690POA+HPyI+Lnb/xj7R+KKdLdr0nUnwG2r8C96bd7z3lurrF44NhbmxO79l904HI7V2tvI5unXP0k5zG32WmqZ6VxHTVCSHyJ7r3QidCfy8fkDt/+WnsX4q9q/HD4g/N6ep76+RPY3ZWE3j2pv3p3C4jNb87A3Bvbr/sTpPeVD1zu/ce0M7hK3cVUlUsYocpjklApK2RkkMnuvdXP/AHpHuf44fD3ozpH5B9pP3J2717tSfD7s342UzufSq8ubyuQwuAp9y7oSPdG6aPZe3qykw0GUySpX5KKgWonVZJGUe690PnbHbPX/Sexc32N2ZuKl21tXBJCs9XOslRWZHI1kq0uKwGAxdKk2S3BubPV8iUuOx1HFNWV1VKkUMbuwU+690VHaPWfZ3yW37tfuz5Gbfn2D1bsyqod09GfFvKtQV2Wpdywp5cX2/8hpKYVeLrOyMOJNeC23ST1OO2vIwqJpqnKKj0Xut/Z0uesocRF8xPlK234xH9x198dpt8iPwCnm3saTsaDHTyLGBIcqdjw45J2f1tTJSj9IT37rx4L0b337rXXvfuvde9+691737r3Xvfuvde9+6910Tb+v8AsBf37r3Qeb87Z646xhppd9bww23569xHisVUVP3GfzUzOsawYPbtEtTnM1OXYDRS08rD6kAc+/de6BLcXyhGPxlTmaPrfcO3duwSwwR717ryuF6T2jVPUxwyQnHx7nnqd75OR1lIWCHCmaSRDGBqZNXuvdALURd4/JZSKrF0eV2QlRT1NDjcpjty9fdBZB4f3qap3HR5unpO4++qGkqNMq0Rots7brXiTytKtwPde6OD1n0thdh1uT3dmsjV787P3JQQ0G6Ow9wQwjIVGPhYTJtnbOMiDY/ZWxKOpBenw9AEg1WknaoqNUze690H/S7z9Pbxznx5zjSrgqmr3FvnobKyB/tMhsSuyC5HP9dieRmvuDrLNZORY4NRMmBqKR4wfBUeP3XuqkP5sHfY3j2NgOjdv17S4DrZRm94JER4KnfGUgH8PopCrESnb+Fm1W40VFUR9V9+690Wv4W7XUT9/wDcFQ+REPV3Q28NsUEdHVyQwTZfuOow23mE1OZFpZHp6PBCUP8A55QNI4NvcMfeK3tOXvYv3T3V9ZUbRPENJ0sGnUQqQTTgZKkeeaZ6lL2V2ht792PbzbgVod0hlOoVBWAmUjzpUIR6ZqcdNzqoNNEdBGtOCIyP2Y2dSqOC2rUoPo9Qt/T38+qiiqMUAA/YBxPnnrs2T3MfU16On8Kdzmj39ufYkdQaGv39tkz7YyEJZJ6beGy55dwYaFXVh6KylNVGVt6zx+ec4/uFc9Q8ue7W68q3TBYd/wBuMcTYxcWpMqLnyeMzY8yBWlM4k/fD5Sm3v2627mS3FZdnvAzrQkeDcARu2P4XWPzGCes/y96+pvvMF3dgaGGhx2+6l8JvjHUsYiiw/YuOhD1FWsSqFhp9z0URnt9fOjk8vf2Z/fo9moeT+b7P3P2K0WPYd8k8O6RaBYr9VrrxgC6jBLesqMTQv0h+6N7nScyct3fIG73DPu+0Jrt2Y1MlmzU0GuSYJCAPRGUcE6JPFw9QhJOmUMLktZZEVtIuiqqg/QAt/ibm3vArrMUHpcbVqzW7x6+2pIYb9ndafJrp6CWorAjtmNhYzZPyh65pqWiekkFQaWfbG6kQrNFIhr30q6k26H+w1pL7j/c699uQZqvPtF3JdW1e4RnwUvUVF8v1IJKkcS5p1g97w3MfIn3ofafnCPSsW5wRQz0wSNclo5J/5pyR1H9EV6RI0VEFiAyzR8gopUh14BRgVIXji1vfO5WDqrqBQiv7c9ZwsGWorkdWEfEHcwz3UXZ2wahmkqNnZrEdh4ceZG/yPJn+B7jgjUSqAIJ4FmbQhUNISTew99S/7vPnUy7Rz77eTvm2uUv4RqrVJx4cwAZqijojHSuklyWNSB1z3++pyqId15Q50hQUuIXtJSFp3RHXEWPDKOy5INEwONGb5kUNPW7d6O3dqH34xe7Nh1q2Opodu19LlMa4AcxKkdPlXUXAke+o3FvYX/vFeX4LffPa/mpX/wAYubW6s245WFo50PErjxHHkTXzA6P/ALke8zS7Vz7y85Jghmt7leHGVXifhnPhr8sYyckPGpJ3H9iZPJc3OmRNKsCS5RVZCCFVeSCT75v9ZzHPRhthwY/NdW7QxldTS1NZt350fFbI7fmSrnpRQ1nYmQynWeVc+I2mpa3A5urp6mBw0U8E7Kw+hHTv7gcq7xyL728nS1aOTS2ljqX/ABq0lgOPL4FJ+zrAL75cY23m32s5hTEnhupIww+nuY5RQ8eD0HQAvTPQzVFDJpMlDVVVFIVBCmSjqJKZ9P0OnVFx/h75iNC9q8tpJ/aRSPGftRip/mOs/UnW6jiuk+GVFcfYyhhX8j1YX8STVVfRXbNABC9Hge0dsZmnQRwRyUbZrBJS5KcOp8shqmgiU6rGwNhb31f/ALu29uZeSfcjbnl/xSDdoZEWg+KW3Aep4muhaelPn1zq++zZQJzVyRfqn+MS7bKjGtarHOSmOApravSw7sqIKH40dpSVMoT+Nbp66wVAoDM0lfDlJcs6PYWji+zhZgxP1W39Pcrffjv7ez+7zzBbzsRLdbhYxRinxOJ1kpXy7I2OfSnE9R790yznuferZpoVBjt7K7kfPBPBKVHr3OooPWvDqrh3E7xrGGIimDvJ60QMgVgilGHlZ1fjhkNjf3xKpStTw66r10ip4U6EPF5jP0nTXyT+zrVodvbT6rk7ekyMgkU4PsvqfN4zdnTtdiJ4mRoNx5TfOOp8dDGNTVUFRJEqkke86PuC71zdb+6+87DtB18pXO3NLuKkMVjeI0tJEYEaZmdmjoaq8eriQCMRfvj7Zy7L7e7RvG4VXmSC/WOzKkanWQVuEYEHVEFAc6aFX0muSOtmXAVtZksJh8hkKRsfkK7FY2srqBwQ9FWVVHDPU0jAkkNTTuyEf1X32H65ndO/v3Xuve/de697917r3v3Xuve/de6//9Hcl+EdRWwbY7/2pXRwQybF+XXyMwVHDBoGnEZffM2+cLJIiH0PUYzd0b8gEhr/AJv791tuNejp+/da697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917qmH56/eD5Kbf7kxEDzZH4RdM7J7zikjSYn7Df3feIwXYVC8kUisyT9O7EzokiNtQmQ/QWO+rggKerloZI6mOGpgmWWmmiWaJ4yHjnimVXikRwOVZSCCDyD711TrBksVjczQVeLy1DS5HHV0MlPWUNbClRS1UEqlZIp4ZQ0cisp/I9o7/brDdbK623c7OO4sJ0KSRyKHR1OCrKwII+R6UWl3dWF1b31jcyRXkTBkdGKsrA1BBBqCOikbt+FnWedq5a3b1dndmSTB/8hxslPX4dJCp8bR0eQikmhQObsqTBSOBb3i1zP9z/ANt95uZr3Y76+2qVgf04mWWAGn4UlVmUcKhZKeQp1P8Ay/8AeV552mCK23a1tdyRfxyKyS0rkF4yFP2slfM16CKf4IZwTH7XsjHNTh2sajbs6zlNC6LrFkTGH8l7820/4+4rm+5Pu3i/4vz/AG5hqfitWBpTHCUita1+XCnUixfeq2/wv8Y5LmE1B8NwpFa54x1pTpwovghKYqY5LslxOQ33iUG3YzCD6gn2slRXl/6X1j+vtdafcmYpb/Xe4VJTXxPDtRp/o+Hrlr/ptX5dIbn71ffN9HyV+njRruDX569MdPsp0p6D4K7OjSE5XfG662VWQ1Ao6bE4+CcB9UipG9NWTQrJH6eJCynkH8exNY/cs5QiSE7hzjuc0oA16EgjVs5AGmQrUY+IkcQfLoju/vTczu0gsuWLCJDXTqeWRhigJOpAaHNKfb0oNx9EfHDp7bVZuzdGErMhS48QpF/F8pWZCsyNeb/b0dHSrLTwS1ldILaQoSwJOlQT7P8AffZL7vvtPy7d80cx7PLPaWyKP15nleWTOiONAUVpJDxAUDzNFBPRLtHur7z+4u9W/L2ybnHFPMSf0okjWNPxO70YhEHA1rXGTTqsX5efzFYvjN0lvvt6j2tDsTZm0afG0mG2t13jKGbsXem5txZWk23svZmLy1SixLm90bhyNPSRmNUSLWXZmCG+OVh7tc48+cyW3J/tdtW28q7Zc6h4kESLcRwRoZJZZ7rQTSOJWYiFQSRpDGtepq3H245W5M2afmbny/veYd0SlEmdvBeZyFRYoAwB1OwUa2IzUgdEM2Z23/MFy2dXN/Ivofozbm1dw7N3buEbk2931ujeG4+oM/its1ed2/h+4qbc22cXT5rE5CuiWjyFft2aX+HSFmETxjV7jPmWz9vt0kluNv573vct+S4jRjdWwJu0aUJI9s/jStG6g60WcAMvmGx0Nth3LnawgRLjlLbbLZ3iYoIZQogIFQJlEKBgaUJj1UOaMM9KDB/MLpraXT/WO9/kP3h8e9jbm3l1Rh+zslPtnfclR19mcTWVNJjKvcPWNfnYqbcG6tnyZmtigpJRC1VKXF4/qQSXft/zFf7/ALvtvKvLm63Njb3726CWGkyMAzCOdVJSOQICWGrSPXo7teetltNl26+5g3uyjupbVZmMch8Nh2gmPXpZl1MKdtaEY6FbMfI3oPb/AFJju/cz3F15QdJZmnxlThe0ZNyUUmzswman+0xMWIylO0y5KtyFWDFFTQK9S0qsmgMrAEUHJ/NV3v0vKtvy/dvzHEWD2/ht4qaRViykDSoGSxotM16Op+a+XLbZo+YJd3hG0uFKya10tqNAAa9xJxQEmuOPSU2X8yPip2NhJtzbE+QnVm6tv029NpdcVOXw+5Ip6CDsDfk0tNs3Zc0zwxGLcm5aiB46WkI8rOpBCkW9rtx9u+edouFtNz5VvYLpraW4CvGQTBAKyygeaIMs3CnCvSCx5+5R3G3a5s96geETJCSGFBJIaInH4ifLj8upXfPe+G6568+RLbQ3Jtav7l6X6D3d3ONiZGY1lbjaGhwW4KnaeZ3FhYJaaq/u5l83gJYLrKhmMTqGU8+9crcq3W67vymNys505f3DdI7TxgKAsXjEqoxBAdUkByKCoPV+Y+ZYNv27f1sLmNt3tbJpwhyQNEhQkA1oSh/YekdX/NP48dY7I6ZyPyF706o6p3p2j1hsHfSYHce4abDzytunbuKr67JQYt5KusxW2RmK2SGKrqylOunR5WZWIXp7cc171uXMMfKXLN9e7ZZXk0OtEL08OR1ClgAGfStSq1JrqpQjoth5/wCX9v2rYpuYt5t4NwurdH0lgtSQKtQmoWvmccanph7T/mAfGrpr5B9Z/Hvf3ZO0MDm+y9hZzfVNuOu3Nj4cPgoqer23TbKxFdGkc7VlV2dHnnlw0sDtDKlHISbMp9q9j9qeb+YeU935s23aJ5LWzu0hKCNtbAiQyuCaACDQNYNCC4+fTW6e5fLu08ybdy9dXChriDxBJrXTllVFArUltRI+Sn5dCrv75ZfGbqvsTD9Sdld9dYbG7MzoxJxuytyboocfmm/vBN4Nv/fxu322GOfmOmhFbJTmqJHj1XHsg2vkPnPfNpuN92fle9udni1apUjLJ2Cr0Iy+gfHoDafOnR3fc7cq7ZuMO03+9W8W4PporOoI1/Dqqe2vlWlfKvSw7y7j2b8euoeyu7exZ6ym2Z1ZtTJ7s3CuPpxVZOqgx6pFS4rFUzNGKjK5rJTw0lKhIDzzoCQPaDljl3cea+YNo5b2uMHcb2dYk1Gign4mbzCooLMf4QaVJ6W8xb/ZctbFf79ekm1gjLmmS2MAepY4A8yadFR6071+bdTUbQ7E75+OvR/VXx93XjpNxborMb3ZmMn230Hs6XBVG4KPdfa2Ky+1sbs/NwUlHFEmVo8LVtU46SYkCVY3sPN65W9t0F7s3LPNm5XvNUDiOMNaIttey6ghjt2WVpkJJJjeVArAZ01HQE27mrnukG7bzy1axcvSKztS4/WhQKzr4gaNUYimlgrYP8QBPWb46/zKvip8h+r812hQ9obM2RQbb33UbE3Di9ybpoJajB1OR7AyfX/XmXytdBBDRUON7TrqGOow5kKho6lFkYNz7pzd7N88cp7zBssmy3FzPNbCZGjjajhYVnnRVJJLWysRIBXKsRUU6W8ue6fKm/7bPuBv44RHP4bBnXGqZoozx4SEBl86Mvz6OZR9g7JyO+tydY47dmFr+xNn4jAZ/deyqKsjqtxbcw26Xq49t5DM0EAd6GLPGgmNIrkSTJGXVStiY4m2rcoNtsN3msZV226keOGQqaSvHTWsfm5XUtdINCaccdDiHd9tub652yC7Rr2GNXdQcqrV0lv4a0NK0x0YjafRXb29dDYTZGXhpXfR/EM5GMBRKSDdhJk1gnnRRyTHG31Fvcj8reyHunzeQ+08mXUdrqA8W5H00YrxzNoZiPPShGRSvQM5g91vb7lolNy5pt3uAK+HAfHc0+UVVBPCjMBjPRoNqfBjKzqJ9872hogZATjtrUf3LmGx1BsllFjSOU3H0gYCx+twRkfyt9y2/lRZudOcFhbV/ZWaazp9DNMAA3zETAfOtRB/MH3prONjHytyw8i6T+pdPpAP/NOKpI44Mg8uFMmN2v8AE/pTbLLK+2JNyVSMWFVuuumy5uQ62FHaDH6CHI0+E/QfUi/vIDln7sXs9y2ySHltr+5H+iXkjTHhT4O2Kny0dQ1vvv37n74Cg30WUBxotUWHh/S7pK446+jC47F4vE0sVHisfRY2jhUJFS0FLDSU8Y/okVOkaKL/AOHudbLbdv223js9usore0QUCRoqKB8lUAdRJeXt5uE73V/dSTXLGpZ2LsT8yxJ6bc1unbW3UaTM5nHY4gFhHPUR/cNweUpkLTyXK24U3PHtZQVrTPSag9OgdzfyH2zRmWLB47I5yVbhJ3CY6hY86WV5w9VIhNjxGLqfe+vdBDmu9995QSRUcuOwUL3CjHwGaqC34H3dYZQGC/UrGvv3XugnyWUyWXmaqy2RrclMSWM1dUy1FuByolZkiWwH6QB7917qLTQz1sqw0VPUVkxOlYqSnlqZSQCwVUgR2J0rew/p7917p+zPXNeMDWZHfGHwOI2lTiGpr6vsJ8TS4FGpZIquilmpc0s6VE0NTGklOFhaQTorIA4B9k++8wbFyxtd1vXMm8W9htEK1ead1jjUfNm4k8AqgsTQAGvRltOzbvv24W+1bHtk95ucpokUSGRyfkq+Q8ySABkkdABuP5A9NYX7imp8ruHsCc64ZIdt4RKDCVUbGNJkbNboaA1MFTTyOp00bqSCGBU84ec5ff19meX3mg5ast0366UkaoY1trc/DlZbjuYccrEeBxkHrJnln7n/ALp7ysU293G37PbsK0lkM8wyeMUFQKcaGUE1FDgjoJf9mnocTBSY7aPTO18biaBGpqOjyu4MoaamoomtRU9Bitr0228Ti6eGIeuCNGiJ4TSPePO9f3iXPFxOP6u+2+0W1sGb/cie4nYio0H9PwVWg+L4gWrQgDqatr+5NytHHTe+e9xnnNP7CCGFVP4v7QyMwPAcCBxBPTDN8suyZABT7d6tx0gYsk1LsueZ1PJWy5LO11O2gm4DIb25/I9x1c/fx+8JPE0cd7skDkcY9v7hX0Mk0gx66fLob2/3P/ZmJw7wbvMuO17zGP8ASRIc/b1G/wBmu7j/AOVjYwP509b7TUkD8W+yPsgP31fvHAhRznbinrY22f8AjPRsPupex/nytcH/AKjLj/oIdY5PlX3DLG0b1GyNLrpIHXe14yRxxrio43X6fUEH3tfvrfeNVg/9c7Ykf8uNqf8Anzrx+6h7HEU/qvcj7L24/wCgus1J8qOxoYhDV4DrfKKWQyS1G1KihqnVW1SIs+IzNB4TIONSpcD2fWP38fvBWmgXV3sl4FYE+LYAMw8xWGWJQDwqBUcRkdE979z72audX00O72rEGnh3hYA+tJY3r9hOelzh/lhi5KmlTdPVsdPStMErqzZm7cjDUwUxbiaixe46XJ09RVRg8rJUqjgcaT7lPl7+8U5kjntE5t9srCW0Mh8V7O4ljkVPLRHMJFZl9GcB+AK9R7vn3JdmaG5flnnu7S4CfppdQRuhYcQ8kRRgp8iEJHmD0Z/qrcfQHamRmxWyd5ybL3luDJNL/dreG2sXistuKuip44BXTZTESrRbiyUtHAscbGV6rxRrFbSg95g+1v3rfaH3XvLbZdp3WXb+ZppGWKzvlEU0tP8AfTqWhlLDKor+ISCNGOsZ/cL7vPuX7c2txue47dHebDEoZ7q1bxI4wT+NSFljAOCzJopQ16MxF8cMiZI/uN2Uf25ZBMYMZN5xET6/F5KgxiTTe2oEA/Ue8kvPqD+PDqN2R31T9Qbh2f0L0z1rle9e3psEd0V3Wm3N1bb2zV7P6zoag0lZ2BvTce5ZYcNg0y2WkFJiKWpaKfN15lWC0VPUzQe690xzd4/LvcMEmO2Z8LK7amaqT4qfP9y93dY4zZuKZyB9/kYOtMj2NuzJRQA6vt6ajRpSNPljB1j3WyKefT3sX4tPWb/293f8i94y93dzbZjmn2ZE9A2C6c6cra+Iw18vTvWr1WQjxualpz4H3Dl6rKbhmhBVKqnhdqce69X5dDf3H2xszovrHefbG/aySi2vsrDy5WuFNE1TkslVPJFSYjAYSiT97Jbi3LmKqDH4+kjvJVVtTFEgLOPfutdBF8SOsN3bL2LuHsDtWlhpO8e/d11Hb3bdBT1LVlLtbLZjHY/GbY62xtVIoeXE9Y7KxePwqNdlmqKWecH9737r3RrPfuvde9+691737r3Xv99/xPv3Xum7K5fFYPHVeXzWTx+HxVDC9RW5PKVlPj8fRwILvPVVtVJDT08KD6szAD+vv3Xuiybm+Vu1IsZU5LrnC1nYWMpll8++6rJYzr/pnHeMgGXIds70lx+36+nUNq/3DplpCgJCHi/uvdAhH2d3P3HFNT7crt3Z2iqfNAsHQuF/uDsilKSiF48h8hu3KQV2cVFkLNNtnCI9kJiZm06vde6EDrr41b2w81XlKvN7Q6rqsyGbOVHWmLrN5dpZbzovlTcHefaTZnN183oUMaTFUahtZjI1KV917obtmfHzq7ZuX/vMmErN07zsurfPYOYye+93agCC9Jmdy1OQkxIe5ulCtNHyRpA49+690Nlh/vv9gf8AiPfuvdd/T/fX9+690Uf5sbx2D130NnN970hqZcntbJ4vK9bPicjJiNxQdmxySR7Wm2/k4bzUdQskkn3XDRTY41EcyPE7o3utilRXh1qk5/PZndmdze6dx1rZLcO5ctX53OZByQavK5SpeqrZghJ0RtNIQi3sqAKOAPfutdWN/GDER5j4WfIqj2xVUk+7absvaW5d44SOUz5qbr/b1JRtTV1LQK4mho6WsllledUdGjikVrfX3il99PZt/wB69gOaI9hXWtvPbXFygViz2sMoaULp/gOiVjQjw0cHFT1kR91vdNn2r3k5dk3Z9JnjnggYsAq3E0ZWMPX+NQyKKg62Uj0ILSkmMSREMUKSizNpdLer/Nqxe8bEgD6m3vh4MqCDVT/q/wBXyz11mIzwz0pto7syGx907Y3xiG05Da2bxu4KS4B1/YVCSzwOh/FTSmSJh/Rz9PZ/ypzPf8l80cuc3bW+ncNsvYrlPn4banU/J49an5E9EvMewWfNXL298sX4/wAUv7WSBvl4ikK32q2lh8x1bd29gsdv3rvuPb+KT7nG7j2Zj+59iqNRMORx1LT7lhkgCJIf3aKongIC6nUaRb6++333hdhsPdr7uPM13t0XirJtcW6WnqHhQXUZwCasgZCANTVIqK9cnfZfeL72598Ngt71zGybjJt9yPIrI5t5K1IAAYKwrwIr1TVE2uZ3FyklPA68OVU+sGzFjFqIP0Fjb63498IqggEcKD/B5/P16690KsynyNOlrtarhw2d6Q3dNkMVjV2X8zPjKKifM0iVFJJhO5B2L8etxUVPOxD43I1VD2VGYZxYGWNVcFWt76P/AN3fOt5uXvNyvN3Q3W2Wb0+x7mAinnUS/sx6dYK/fagNtae2W/IQJoLu7QHzwsEy59Kofz/Ppiy+N/g+azeG1F/4Pm8ziSz8M38NydVREuPwx8HI/Hvnbulj+7N13faqki1u5oanifCleOp+3T1m/tt5+8ds2zcgKfU20MtBw/UjV8fLOOjK/C7KSQd4ttkiWSm33szee2J444klJZsO+foGJqIwgWCsxBIWMk62W/vKr7kPMMux/eC2GxUt9PutldWr00/hj+pQmue1oT8OanNR1jx97DZo919md2u2p423XltcJWvm/gPw9VlHHFAfOnQv/JqmqarozCVBSCOHb/bNIKguSKppdwbUrqNYowbhlinxw1AW4/rb3l1/eHbbNce2XI24qE8K137Q1Tmk9u6oF/26nVnOKdY1/cpvUh5+5tse7xLjaKrTIrDNGzV9O1hpxxr1XkxVp4FGgskc8lz4jIqsFjvZj5lUsbal4/BP098jh8IPy66S1zUdGC6uyWOw+3tjNlYWki3L85vhlgMYSWSN8lh9y7h3pVyKQDqOJxeCkqpBcAInP9D01/u64Xt7b3j3aeMrZotkC5+HsSd2HzKrQn0B6wF++5Mstz7YWMbBrgi8YKPi7pIUU/YSCB8x0CmVqI6vLZmrhYPDV5vNVcLDkPDU5SqnicW+oaN7++au5TR3O6brcQmsUl5Oyn+i0rkfyI6zw2+F7fb9sgkFJY7aJWHoVjUEftHR+/iO9TB0p3E66kp8h2NsfHs5ClJlp8JLVTRISqkSRMY2axYhSPoD76nf3dcEy8o+51w0TCB9zt1VjhWZbfuAPAldS1pwqPXrnz99uaJuZOQ7dZR4q7fMWUcQHnGk0+dDT7D01/LjdDY/qvrDYFM1RG2690Z/fGWe3jimp8I67fxVMG8TmcpLJLMur9JHH6uC3+8P5uMe2e3nt+hkVbmSXcJSMqwhrBChHmRJIzhge3StRnpZ9ynlpJdy505ycKTBFFZx1+JWm/WkYegKIqn1qfTog4VUWNRGDEjRgwxkRBokYa4o3H+a1xgqGHKk3/HvlyGUEMVJWowDQkVyAfKq1APEVr5ddBGBZSFNCRxpWhIwSPOhoaHjw6Mxvzc/SO8919Z5TZ26uxenNn9eHEZul6fk6g697E6hzXZmHlWpxPaXY1INw4PffYu5NqNEqYeOpyJoKCUfdx0hq9E0XUf22++F92/2+2u12Hlr2y3PZbOQJ47wwW71dEC65XWbxrnTUgOcn4qAmnXPrnr7s/vpznuM+8b7z1YbrcRlvBEs8yEIzE6I0aHwoQ3EqCFHAkgdWH9FfNGrzO8sD1t3HU7ErDvnKz4HqfvDrUZvF9b7/wB1U1FLk5OtN2bU3XLVbj6i7cnxNPLWY/GVdXkcdnqeCY4+ulmhemXOz2+9zuRPdTaJt85C5ih3CwicJIF1LLE5UMFlicK6Eg4qNJodLGh6xG5y5D5t9v8AdI9n5u2Sazvnj1pqoySICQWikQskig8dJqPxAdWIAg8XF7XI/p7HnQR679+691737r3Xvfuvde9+691//9LcQ6nH+jP5sfJbrmeJaPE95bR69+SGzFAijhrs3hKCDqLtqKAKxearopsLtuqnBCsoyCtZg1x7qxyo6PQP8ffuq9e9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+690QPA7Swu/PlR86uv9zxiWh310h0DtyekqlLmq2fuDbvbWAyMtHDN+3NQNXz1Mbsvp86sCbj37rdcU6F74b76n378bOrK3JFl3JtfCT9ZbyppmY1dHvfqnI1nXW7KetDjyJVHNbZmka/BEgYekgn3XiKGnRnQwJZR9Vtfg8XFxzax49+61137917r3v3Xuve/de697917qrX5wZzJ1PY229uyzyfwfGbaTJ0tHciBshkK6ohqK1kvpedYKVY1b6qtwPqffNn75O8bhcc+bDsUkzfuu224SolTo8SWRw8lOBfTGqBqVC1A4nrOb7sG1WUPKO8bwkQ/eM98Y2f8XhxxqVSv8OpixHmaeg6o/wD5knTvY/dHxaymM6iwI3d2T112b073jtTZH3VPRyb7q+nuwMPvGt2ZS1FW6UiZHPYqjnjpBKRG9UI1JFx7g/2e3/aeXudYZd+uvp9nu7K6s5JqE+ALqBoRMQM6UZlLEZC1Pl1KPujs25bvyyW2iDxr+1uIbhY608TwJUkKAnFWCkCvnTrNS/LzAd9YTcWx9l9H/KHGbi3V1h2XLuob86K3lsDB9W5JdiZqeDbu79w7spcZjcvnstmXXHUNNg3yjVUzrJdYvV72/IFzyxeWe6bhzLsr2sN7b+H4N5DM9wpmQF4kjZ2VFWru8wjCr2mrGnSROdI98287TbbBuS3T2ra9dvIixFUJCuzhVLNSgCeJU8MZ6Iz8Nejt1Unc/wDK8y/YXUGZhoOq/wCVvnts5fJbz2RM9H112jJvHY0UGCrpsxQvBtnfLYj7sJC5irRTtLpGkv7k33B5ksH5e96YNq36MzXvOSSKsUo1T2+iYlwFeskJancKoDpHECkc8pbBf/vv28e/2aURwbAVJaNh4cv6GCSO16Bh5NxHDHQET4btrqLrDoPb2z+vM/iOw8T/ADVfmNvvqfZU3VWR7Frv7mUND2RlcZktvdCQZbaWQ3f19X4zMGVspja2kGBhlWupnLlQwmjuNk37euZrq+3aKTbJOStqhuZhcrAPGLWyusl8UmWKdXWnhuj+MR4TClT0RzQ7ns217XDHt7x3Sb/eSRIYi50f4yRpt9UZeIh6l0dfDBDjNB0osps+bLfGLflXs7F9o9ld/dZ/zIOh/mB8v+sY+isz1Jvvar1m6cHurJxdfdI1NZl6+Ta8GwMbHXYtaetyNXlUhqZ2dpmZPaSHcBb867Wm5XFnacrXnKd5tW2XBvEuYpKRPGDPdgKPEM7FJSUjVKoAAig9P/SR3PKl7+70mud5td9t7y8iELJIoMkbN4UBLMVEYqtGdnoxLFiQBb3RWbr7370/mXdo7I6j7nouv9/fy2sb1r1VuLeXWW6tmy9r7lx0nas1VS7NwOfx9FuOac1uZSmipKqmp66d4/IsJhkhkkIrKKy5Y5a9ntk3Pfdubdbbm5ri6jiuI5BbIRbEGV0cp8KFyys0ag0LagwB7M97v+78+bpa7JeJaS7AsUTPE6GRlFzqCKwDcXC5CnhQUIJZO/M130cbT9LzYHszrHZFZ/L/AOr8BsvI9U/FHHd5dhfI/tPJdd1eF3X0pubeue23uDFdaYvYkyRI+Lr1oWY1U1UtTGyAe1PLEHK/jtzCLu0vdzXmid5Rc7k1nDYW6zho7uOJJI2uDMCWEieIAFVPDNcl28nmL6e12eTbrm3tTs8aJ4Nr40txIRIGidyrogj7TpYIX1E+IFFOlB1Zis10xVfyi999qdV9jyYPaPwh310pv2fFdU7q39mNkdn7kxHTU209tb2w2CwWa3Ft+Wsbb1bDFU1kCU9NLC6vJGL+0m9TQcxx+++17HvdmLqfmKK8hDXEcKTW8bXgkeFndI3oJEOlSWYGoB8lu0293sd37Zbluu03Rtk2n6dyI5JGjkK2xUSKoZ0FVkyQACCDSp6DHsbqGvh3z88epu/O1vlzsPbvyI7q3bubG7c6d+Ke3O7do94dWb02ltyi2s+2+w4uot+5uj3RtGGgfEmhrMpQNhKmkjlpFhVvJ7Odn5ggba/bHfuV9l2C5u9p26KMvd7i9pLZXMMjtL4kBuoU8KUkSa1STxgxD1Pb0WbrtEse5857ZvdxukMN9dOwWG1WVJoZEQIFkEErakFV7mBjI7QBQ9WYfNbojd3cnwO7S6M6yFTlN9VHWOy4dg0m6KuKnr9x7i62yW0d1YLCbhq5ylOmW3O+01pKlpCI/vJzqIW9oX9u+abDl33P2XmbeiI9rF7L4zRioRLkSxO6KKnTH4mpAprpAoCcdTBzpy9e7v7fX2x7QDJeC3QxiQ0ZmiZZAGOKElQCTQCtT0Lfxt7y3f8AIvc3U2wMX8UPlxtLfm8ZcdRdk47sn4/b02NsbpP7PHM25Mvv3sbdVBj9j5XbdLkaN6aj/g1Tk6rJmWIxU5Uvpkrb/ute5u83VzJsz2L7Dlre8ecLFcIWorIoDTBtJJZWjUrQiuoCoBvPvAclbbtcVtuCXI3YIUlt0jrJG2k4YkrHQnAZXYEZGOBcvjd8dsH1x/Lh+Y/RPdfxL797k7+2x218iNgn4wU/TG9Ns7W7Yy3fXfG+I+jajC9pxbZXAbg2HWUWXoc7V57G5Jods4yJqmZoHijDZkQ/d32y45z2jnze+Zr9t3tILNY4reTwokktYI4i2uheQSFCxU6AysUYEEjrGub3gvYOV7/lHatjtV264kmLSyqZJGWWWSQALUIjAOF1dxFAVYEClmf8j/44b6+IGB+QXxL+SXV1bkvkx17v/C9hbh+XVZgdwZ7AfLbY++dt45dj7rwnZu44K6d8x1NS419oVmAat8uLTFw1EcKpWMxmu25W5asjZGz5fsojbKyw6YYwYldzK6xnTVA0hLsARUmp8uovuN93u6+p+o3e5fxiDJWR6SFVCqXANGooAFQaAdX7u0cCtLLIkUar6nkZUjQD8lmKgAf6/s9+09FQoBSnQfZvtfYWC8iVO4aSpqYyQ1JjNWSqLgkadNKHjRrj+0y+99e6BzPfI121xbbwCpcELWZqbn6rpdKKkJv9Tw0o5sf8PfuvdAzney99Z8OmQ3FWQ07cmkxbfwumAYAFSaUpPIgF+Gc8Ej37r3SXxuIy+fqGGLx2RzNUx9bUsE9bJzYXeezKurUCdTD37r3QmYjo/sDK6JKihpMLC1iXylWgmVbA3+1pBUyC4NrEqQRyPfuvdCniPjjj49D57cdZWFT66fGU0dDCwVwbeaU1E3rj4NgD+QffuvdCZiOn+vcOUaLb8FbMmkifKyS5GQlGLKStSzQ3F7cKLgC9/fuvdKHOVmG2NtnO59KGjosft/CZLM1MNFSw04aDFUU1W6IkCLdjHEVHB+vsm5h3u15c2Het/va/SWNpNcPTjphjZ2A9SQKAdGWzbXPve77Vs1rT6m7uYoV/00rhBX8z1rc9ndtb77lz9TubfWbrMg080kuMwonlTA7epJJPJBQYjF6zTwpAlgZWDzyN6nc3t7+ej3G91ed/dvfbnmLnTeZZ2eRmht9RFtaxsarHBDXQukUBchpHNSznrtPyN7d8q+2+0QbJyvtkcIVAJZqAz3DgUMkstNRJOQoIRRgKOg69x/0NR14k2Nhc2JAva5twL/i591YkKSBnr3nQ8Ohwq9o9W0+N2fU4LI1fYk+X25FU7sosZ2rsfrTeG3N3POVqMLitodjbdodvZ7E0tN61qX3FSvO40qnIPvNj2+9nfuo8z8sbDJzD94efb+cbmAeLBJ4dvHFOVBaKk0GnShNA7S0kFGDAHrE/nX3P+8Zy9v8Au0WzezEV3ytBK3hTKJZzLED2uTFLUMy5IWMlOBGD0nNz7Gw2Ehxs0uZ3BsGozdYlDgcF33teLq9s9VzkrT4/a/ZVHmNydJ7tylRMNKU9NuCOeQsoWMsbAz52+4lz1t23PvnthzTYc07RkqkbJDcFKVHhkO9vO3GqrIhIoQKmnSPlL733Kd9ertPPvLt3sN+MM5DSwq3nrXQk8Sj1KOPnTpDZnCZnbeWrMDuPE5HA5ygKCsxGWpJaHIU3kGqNpaadVkMUq+pJFvG68qSOfeFO87Lu/Lu53eycwbXcWO9QNSWCeNopYz5akYA54hhVWGVJHWVe07ttm+7dbbvsu4wXe1TCsc0LiSNx50dSRUcCvFTg0PTZ/r8f77/Yeyzhxx0Y/Z1iM0QYx+RTIBfQPU4B+hKKS+kn829t+LGG0lgT6Aiv7OPWyHC6tB0+vAft6UVRt3L4ehx+4txVFF11gJ6mMUO9d+5aDYmEhqQyvDVY7JZiWjyGTqIH0tGmMhqqpn0iNCxUe5g5B9jPeD3HubNuTOStxMHijTeur2trAwyJjcyiOmggEGLVJUDR3UrF3OfvD7ZckQXS8y82WRnVGraRstxPMKUMXgJrqW4Uk0rQmuAerg8L2T8l+59r7b6/6GwWW2Ftml2/hsNu75d9z7dq8ZV5lIsZDS5POdE9N55YN1bvzuQP79HmN0wYjCwswm8GTUGFu+XKW27rs3K/Lu0b5uZvd5tbGCGe4Nf15o4lSWXu7jrcFqtk1qc9ceeYr2w3PmDe9z2qxFrtdxdzSwwin6UckjMkdFwNCECgwKYx0aro/oLr3oPb1diNl0+WyOZ3FXDN797C3flJ9zdkdl7nZGSfc2/t314ORzuUfWywx/t0dDCRBRwU9OqRKIuifoa/fuvdcJHWNGkdlVEVmdnYKiqoLMzMeFVQLkngD37r3VfO0aqs+ZndWI7PCLL8SuiM7NU9Rl1iqcd8he7cc0tFP2/CkivFWdYdRyiak2vMFKZTcBnykTGGioJZPde6sIAt/wAVP1P+v7917rv37r3XvfuvdM+f3Dgtq4ev3DubMYzb+BxVO9Xk8zma6mxuMx9NHbVPV1tZJDTwRgm12YXJsOffuvdEr398rcjlchRbc6rgpsLDmI0qcRvDdO287uXdG8cc0TPJV9P9I4Ewb13dTkkLHmMucLgQw1pNUxcn3Xukhg+guzt91km5ew62fa9FIzTJubuHJYTtXtNkmaOQNg9iwpD8f+n4xqtCsFDna2HSNUgk9Qblmht42luJkSIcSxCgfaTjq6RySuscMbNITwAJJ/IdCzitq/GTr+rhrs1l6XsDdUHiQ7l39mK3svcEEkTuyS0S1i1+J24sUim0eNpqKGMAKEUAD2At091fbzZ2Ed5zVamX+GMmY8SOEQfAIIPpTPQwsPbznbc08S15cuBH6yDwgfs8TTWvlTj0IQ+SHWEfoSbOaI7ohTCziLQp0r4xrUBNI44HH49hI/eD9ug7ILi8NCRUW70NPMfI+XQjX2Z52ZVbwbYEitDMtR8j8x59K/bXcnXm6poaTG7gp6evncpDjsqkmNrJHvwsaVIWOVmH0COx5/r7FHL3uvyJzLLFbbfv0aXzmixTAwuT8g4oa8RQk/LoP737d84bDHLcXuzu9mgqZIiJEA+ZU1WnmSAOhPBvf6ce5FFfMdAnHl1y97690heyOyNm9S7Nze/d/wCbpNv7XwFK9TXV9VIFaR9JFPQUMFxJXZOumtHT08d5JZCFUe/de61W/ll8qd3fKXsD+P5GKowWxsA9TSbB2WZyyYvHyOA+Wy6xt9vVblyqKGmkAIgS0MZ0qS3uvdJbo3oxu0J6/c+78/8A6Pundr1UVNu/f9RTGqnnyMsQqKXZ2y8ZYybi3nko7MsMatHSwnyzWWwYFc++4fKHtjy7c80877zHZbRGQqk1Z5nIqsUEa1eWVgDREVsZYgV6FXJ/JfMnPm+W3LvKu1yXW6SDUQKKqJ5yyuTpjjXzZiKnABJA6M1WVXRmOhXb3VvXGd6oMaNTYvvui3bm8t3tt3Khh/D97U9OtZBsvMUNBVIk1btiso6jFZijMtFOAkxI53Xf94Xd3HOFpDF7e2o9tmm8O4Wd2lvJLd6q8oVaQqdBr9OUkEgrGzgmozUg+5ZDDyxdSPzpMeehHrh8NRHaJMtGWNix8Y1bAnBTwzRwhAoUnJh94ilqm3xj9jYrf2NxS5zPRdbyVS9Y9j7QGSgwlN310fFWGWspevNwZKpgTObbqJGymws5UnH1QkpHoqueJfvK+wGybJYf6+Ps1d2197M7lokZbckjb5JTpOkZItDKdOkgNaO3hOoUAiQvYf3l3jcb1vaT3St7i29zLDVGskwo14sY1UkbANyqZD5W6jAlUlq6k0F8chhsvhqAxVbXVHAvKiqkdgrglyzNyxt7wwKhqqfhJpwB4/bTrKyrDSV+IU6ue6DzaZ/rL44bjq5Pu2josz1VuCSRJpiY6OqrsNS09RJJpT1Y2pQek3C30khbDu191Lek5z+7ryKl3J4rw2cthKDU18B5IArE0/0IJWn+1NOuQ33iNpblf3r5ue3iMcclzHexED/fqpMSoHD9TVSv2ketPW4cFPtjdm4Nt1SeKowGVzODlQwtDY4rKz0o0pM4qEVo1BUFT6SCT9PfDrf9mn5d37feX7kUnsb2e3YaSlDDK0Y7GyoKqCAfw0PE9dZtl3OLe9m2je4W1RXlrDMDUNXxI1b4hgnNDTANeoe5ZsjT9B/JDJY3E0Odq9k7P6w7kp8XXssaVEnSXfvWPYU701S5H2ldBjMXUtDKCCslv8Qc3f7vXcXtvePmTbtVIrjYXZsZLQ3ELjPp3mvWKH31LFLj205evCtZYN4Cg/KWCQH/AI6OhI7pp4aXuPtSGniENP8A6QN0TwRAWCQ1mUnrYwBxwVqL/wCx94pe8Ngu2e7XudYJEESLfr0BRwAM7t/lr1kR7V3h3D2w9vL1nLNJstoSTxJEKr/k6cPj1kjhvkH07kRG8oG+MHSsiM0ZcV9Q+LYGbXYBEyRfTpOsLYm3s09hNxO0+93tRfhC2jfLZaA0qJSYD/1dr6UFDx6Q+8diNy9qPcWyLBdW0ztUitPDAl/aSlK+ROOrAe8MHT1XS3yGxsUsIk27Xbez1KtTG00hpsHvOanneBrfs1ckNUE1/WzH8H31y++htB3T7u3PbCRA1m9tc1ZdR/SuY9WjzV2DFQwyAT69c2vuu7l+7/ezlIFXIulngIVtNfEgemr1UFQSPkPTqpyIapZnPFj4k9RYWX1P6WRdBLt+CymwPvhpkEU+VP8AV/P1662/nX8qdDs4rMJ118UftMVFNLmfkr3z23X5moIdMNh+m/jB2XjsbXwxk+mvfP7hp4YbA2LHi9iOo/3XlHJv3RPeLnZyY5Ln95yq4J+GG2FtEacMSFl+fHjnrnz94KvNH3kvbflVQJIYht8RSlMy3BmlBPzShr0BMKskESH9SRIGJ/1QUar24+vvlpCpWOBCO4AV+2gqT10KmIZ3YcC3Vl/x/oZMP8ddsCZGR949g703ckcvlJkoMfFjts4+oiEqItOkhoZdSDUHYagbH32l+4hsMm0+w6bjPGySbnu11cCrYKKUgRgo4V8HB9c9csPvebvHuXu69nE4ZbDbbeBqYo51zOtfOnig+vAdc+7+ps13btLZa7Pq8T/fvr1s5RxYPNZNcV/enbOUqY8lT0uFrahoccctjq5JB4J5IxKjg6uLe0P3xfu9c0+8NtytzLyIkEvMe1Rzxvbu/htPBLpYCBj+n4qOCdLlQwb4sDpV92X3o5f9s7rmHY+bpJo9k3F45EmRPEEMyalLSqO/w2Q5ZAxUj4SD0R7LdKd0YE1K5nqLsehNHD9xVsNqZKvp4IQjStJ97ioq6hmUIp5SRrH6298uN39lfd7YpJo919sd8hEa6mP0csiBK0rrhEiEfY37Ougm3e63tlu4ibbOf9ol1tpUfUxoxJAoNEhRgfkR9nSBqsPuGiglq6va+6aakhBaSqqdu5amp0UFAGaeopYolVzJwSQOD+R7A8+w79awyXF1sF/FbKKs7206KATQVZowq/mT+XQtj3nZ7iRYIN4s5JWwFWeJmJ+wOSaD0/LqXtzJ4hZK7Bbherm2VuuGlxO86SkkMGSpsbHXLPi934GoK+bF716/zVPFmcDkYtM9HkaJGVgrOrST7G+7u8ezfuHtHNu3XTnaXlSHcIAaJPZswEoZeBeEEzRN8SulAaMQQD7ue2u2e6PJ25cu3kKDdURpbKWlGhugtY9J8o5f7KYfCyNWmpVIvd+I3c25984DPdW9sV+Oru8OmxhcZuzOY2OOmxPa+yc7RyVfWffm1aaJ3hi272ng6V5aiCM2xucpa+hIAp0LfQLt+4WW7WNnum2XKT7bcRJLFIhqrxyKHRwfMMpBHmPPrjVeWlzYXdzYXsDRXsEjRyIwIKSISroa+akUPRwPa3pN1737r3Xvfuvde9+691//09yj5i4er2ljOu/lLt7D5LNbj+LW48nvPN4fCQtUZjcnS258PLtruzb9FRrpOSq6DasybhpKblp8hgadEGtl9+62PPo2u3dxYPdmAwW6Ns5Okzm3NzYjHZ7AZrHSrUY/LYXL0cOQxeSop19M1LXUVQksbjhlcH37rXT37917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917rgHBNrEe9V4EA9ewPMde13NrH/X/AN99Pfuvdc/e+vdFG766j7IXsHZvyR6BODqu3tiYDI7J3RsHc+Snwu2O8epMjXLm6rr+uz8UFaNp7vwOei/iO2sy8E9NSVs1RT1SGkrJpIvde6Lf8Ju9dpbg+Q/yb6vxCZbbUu88pj/kJB1lvCjXB9hdYb1ylPjtk95bB3lt+WVpKOuot4YrH5ykqqVqnF5Wi3AtXRVNRCxf37rZIxTq0csFtc2ubD8XPJsL/Um30+vv3WuuXv3Xuve/de697917r3v3Xuq+PnNsyWam2fv+lhkePHyVO3Mu8cWpIoa0rV42ondRdFFRFJECxteQD+nvBX75/KM89ryrzxaRExwF7SchahUkIkiZj5DUGWp82A8+st/uucyxx3PMXKU7jXMq3EQJySnbIqjzNCrUGcE9V2htV/8AA2/1/wDW/wACPeAPy8+sx+ubPK4UPLI4XlA8jsFP4KhmIFvfvy6oEjWpWMAn064kuwYM7EMdTXZiGe1tTC/Jt+ffurBVBBCioGPs9OgK7z+OfWnyGo9pR9gRbqoc51/mavcGwN87A3tuTr3sDZGWyFA2Kyk+2927YraPI0lNmMU5pqymfy01VDZZIzYWFHLPN+88pvfHa2ha2uowk8M0STwTIDqUSRSKykowDI4oymtCK9BvmHlTaeZFtGvVkS6t3Lxyxu0ckbEUJV0ZWGoYOaEcQes3R/x46x+PWM3JRdcUGe/ie988m6d+703ju3cO/Owd+bkhoosZSZfd29N0V1fmstNjsZClNSRa0p6SBdEMaAm+uZubt55tms5d2ljEVtGY4Ioo0hghjLFisUMaqi6mOpjQsxySercv8rbRy3HdCwhZp7htU0jszyStQLV3cliaADjT0A6HEvKTcyuxvruzsTr/ANWSSTqsBz9fYY/LoQ6EGAgApTh5en2ddBpQGCyOqv8ArCuwDm97vzz/ALH37Hp17QhoSgJHD5fZ1wSSYyBIXnaaoZY1jhMjzTs30jWOK8kzO3AADEn8e7hGLrGFJkY0VVBZmJ4BVUEsT5AAk9VZY9BaUKEXNTQAU4kk4WnqadGl6z+J/Z+94YMhl3GwNv1DJIGy61EmbrIGIvJTYOGSPwEqbqap4r/6kj3kp7efda9wedI7fct7VNl2V6Gtwha5dTShS3BBT1BmZK/wHqCed/f/AJI5Zlls9sU7tvCVH6RHgIR5NOQan1EatTNSOj39cfGTq3rk0dfDiP7x7jpBqXcW4SK2oScgapaOhIGOoLH9GiMuo/tE8+82+QPu7e23ILWt7DtQv9/iz9VdUkfUeLJHQQxU/DpTUv8AEePWK3OXvXz3zktzaTbl9Hs8h/3Ht+xdPo7/ANpJ8yzUP8IGOjBs6ohZyFVFuzMQqqALkktZQAP9t7nYYA6iToMdydwbF240kEmVGUroyVaiwy/fSIw+qyToRSREf0Mlx/T37r3QJ575E5qpDxbcwtJi4yCFqsk/39UL/R0p4fFSxsP6MZBzb/H37r3QKZvdW5dyvqzmbyORFyVgknaKjS/1CUcHiplH+up9+690zUlHVV06UmOo6iuqXIVKWhp5KiZibADxxK1r/wBTYe/de6GLb/RG9susc2RFFt6nYq3+Xv8Ac1ug/UrR0uoRvb8PIhHv3Xuhy270PszDmObJrU7kqksf9yLCOhD8epaCC0ZFhYCRpBb37r3QxUdDR4+BaagpKajp0/TBSwx08Sj/AAjiRV9+691L9+690i9/b/2n1ntXJ7y3plo8PgcUsIlnMU1VVVlXVSpTY/E4nHUiT1+WzWVrJUgpKOnjkqKmd1jjRmNvfuvdAbjtp9kd4yUu6ex63efUuwJKaCbbfT23c++3945BJmEwy/bm5dvv9/RZCSJU8WAxdatPRqWFXPUynxQe690Hna+2oem8njabA7y3I+wu0MD2XtjeXXO59xZTdtNBFj+uM9uCm3jsh89U1mUwFRhpcQYsjDHUmiqYKsMYROqOQZ7jmx/1u+fP3m+nb/3Ne+If6H00ganzpw+fQn5JN0Oc+UDYpqvP3pa6B6uZ49OfSvHqi6H/ADMP9fEl/wDkkf19/N7AKQQjz0j/AAddypP7ST7T1k9u9V697917r1geCAQeCCAb/wCwIN7e/AV7NINcU60Tp7q0p0LnVG+67ryPLZXI4Or371tl2XYWR6Xeiptw47vbe27qWpotl9OYLaGWSqw2a3RuLKlZpJTF48Ri4Kqvq3jpYJGOaP3Gtr9x7/3Ugu+Ub+4g5Ds1c7qSzmzeN0YRQBCfDa7eTS0ega0UM7ELhsVPva7jyLZ8gSWvMVjBNzldMv7voq/UxlXXxJi9A626qCrBiVkcqqgtlTzbG/lx9mYvqbr7bGb+QFJkc1hdvQjI7T7G6zwHbu0Nj5HIZGpzVXsrrPdqZPYfbGP662mtaMTiKKsz1akNBSR6VUEoOq/PvtB7Y+6Cwnn7kix3K6jUBJnVknQA4VZ4mSUJ/RLEceueXKfuRz3yIZf6n803dhFIavGjVhYkULGFw0esjiwWuB06Y7+X/wBr0dWJf76/FeJNEsQnf41dhZl4g4ssqY7NfJGpxckwsD60IX8e4+sPul/dz265F1D7V2LuARSWS4mTP9CSVlr6GlR5HPQzuvvFe9l5CYJvcK8VCQaxiKJsf00jVqeorQ+fQrba+DG5qMtFuL5Kbxx2LYuWwXSHWfUnRtKyTMr1FI+4MXtfdG/2oahwSyjNCQcWcFQfco8ue2XtzyhGIuV+Q9osIxUfpWsQajMGYa2VnoWAJGqmB1H++c8c58yyCTf+bdzvGqD+rcysKgUUhQyqCBUcPtz0PHW3xH+P/VuaG7MD1/SZrfhbySdkdg5PM9l9ivNpRWeHeu/K/cGexsbCNf2qOangBHCD2OSzMFVmqiigHAAfICgH5DoKgAVooBJyfM/MniT9tejJf73711vr3v3XuuibW/x/33+x+vv3Xuq++3t3Z/5Wb03V8VOnMhV4zrPbWSgwHy37qoTLFSY/FzxrNnfjf1hko2X+Idqbuxcq0+4q+FjBtLD1TqWOUqIIoPde6PhgcFhtr4PD7a27i6DCbf29i8fhMFhcXTRUWMxGHxVLFQ4zGY6jgVIKWhoKKBIoo0UKkaAAAD37r3Tr7917rom3v3Xui4b2+RWAxWWzG1uvscnY+69vCU7unpsvSYLrvranp4XqKqv7O7Jro5sFthKOnjZ3ooRW5UgcUun1DRwCetgEkAceicTUm9u+TQ9jb93pi6HZOJyH8Q21vvKbWNNsiiaNJHSs+OnUu4nrKze244wRHDvfecFTGhYT4fEqjeQEe/czbHyzYNue+bjHb2g4FjlzSulF+J2+Sgn8ujTZ9k3Xf71LDaLJ5rk8aDCj1Zj2qvzJA/PpXYbsXE9eQZWj6p22lBV5mo+4z/YW8quq3X2PvOsAVf4tubOZCRqqrqJApMcUsjwUyEJDFEihBivzP94verqSaDlTbI7a1qQs036kpHkwT4ErxAJemK0PWQ+xex22Qxwy8ybhJPcU7o4jojB811/Gw8iQF+WOkHnt0bl3RN5tx57J5c3uIqqpcUqck2io4vFSRKNR4Cf8R7gne+ZeYuZJDJvu9XF1k4kc6BX+FF0ov2UPUu7Vy9smxRiPZ9qgtx6qvcaerNVifnq6YgqLwihR/QAD/evZEihBpVQB8ujokn4jU9d+7de64soYWYXA5/III5DKRYqwP0I5HvRAPEY/Z+w+R+fl148DSlf9XH1Hy6sR+Pm6sjunYMZys0tXW4TIVGFatnYPNVwU6Qz0skr31SSx09QsbMeWKXN/r7z09juZtw5l5IjO6TNLfWdw9uZGNTIqBWRieJYI4Uk8dNesOfdnYbTYubJfoIljtLmJZgiigRmLKwA4AFl1ADAqfs6du5+8OtuhNl12+ey8/Bh8XTJIKGgQx1Gdz9cgBjxW3sSJEqMpkZmYAIllQHVIyICwmPqMutYb5afLXevym3r9/kPusB1zgaiYbF2Is4MVBE3o/jme8LGDIbnro/1yXaOmQ+KHjUze690BnVXW+4u3+xdodZ7VCLm94ZiHFw1U4JpcZSaXqcnmK0KGb7PE46nlnew50AcX91LKtSxooBJPpTOfl6ngOt0rw41pT7eHR7+5s/tGOp211h1bTGm6y6hpKzam1nbU43Nn5qx23d2DkV8peWs3LkojaQuzinRALKbDhh96/wB6U94vchv3RK39UNl8W1tO6qzSByLi70jAErKEjPExxqRTUeutn3dPaxvbPkhX3KMf1n3UJcXJp3RJoBhtif8AhYYs4OPEY/wjoGzFKOY6iQm4JWUI6NYuxHCgpr1AFhcgKLC/vGDqf6YGB0IWwqrcWXqsVtbbi4Wbd2Mz1VufrSh3IY0wtfvCpxU2I3H1tkqyQM+O2X8gdomo21kjwqVk1BX6fNQxkZi/c890bXl7nK49puchFP7ac2B7aaCXMUd3JGVR1DVVBcf2MlKAsY3+JK9Yufed9vJ965Zg9x+WhJDz1y6UnSaIkSNbRuGcEjLG3P6sfogkU1DU6SOYp8dUxUGZ27BkItqbqoF3Ls9cxEUytHj5KuqpKjb2diRkSPdWxtwUNXhMtAT+zkcfKp4IJhL3t9rdz9nfcnmPkm/SQ2Ucni2cpGJ7KUloJNX4mUfpSeayRsCBiss+03uHY+6HI2zc120ii9ZPCu4x/oV1GAJV08VR8Sx+RRxQ4PVgHxK3Ech0tv8AwSnXVbJ7C2vvSmjE8rmCgz1I2OrJJIw5pxBFW43TpiIYu5J/B99A/wC7u5lFxyp7gcpSSVls9xiuo1OonRcxaCafCFEkJ4UyST69Yb/fW2Ewcy8mcyIhCXVlJbuaADVA+pcjuLFJRxxQADouXyywowfyL3+yK6Uu4q+k3bj2mlikeppd04ejyzTIWUyxxNViYLFcNGqeoHj3hP8Aeq5fflv7wPuRbNEyx3V0t4mo6iy3UayFq+QMniAA5WlOsqfu8bym+ezPItwHVpbe2a1fSKBWt5GQLTzOgoS3A1x0HuwdsQ9hR9n9TzxQ1MXc3QveXVUVHV1ElLQ1WU3X1xnYsCldURAz08Eeap4HEiBpI3VWUEj2IvuY76Nj+8PybG8zLFfw3VqaGmppYS0YPqNcYx606JPvR7R+9fZbmSVYg0tnNb3IJyVVJQkhHz0Oa/KvSGj3YN/7a6x7DDF/9IXS3S29ZnaujyjfxDL9abbp82kuUhd4slPBuDH1cckwJLyqxazah7Kvvc7K2yfeM9zIiAI7m5hu0IFAUuLaN6j/AG2oE+ZB6W/dl3Mbp7H8isCddvHNbMCakNDPJx/2pUgelOhB6XuO7+nGAUt/pG2qBcRX5zFEfSWIl5tyEv8A4+429nRX3c9rx5nmCw/7SE6kH3Pofbbn+v8A0Zrz/qw/Vp3ZuKpsxhvkltihmnmjy+zN6z0ApUaprqutwFdFuGOlpkVbSF6ijlVxb/Ng2Nx77ifeL2R+Y/Y33W2pRJ4rbPPIBGKsWhpMFUcTUpSgzQnrkx7Jbsux+7Xt3uBKeGNzhQlzRQspMTMT5UDkgnFadUxUR1UsDkANKnmYAMo1TEyNYO8jKLt9L8fjj38+cbBkifyKg/6v9XDrtA3YWBNaEj9mOh1r48HuPZHTW4Id+4LGt0b1r8m9t5LYFfUVdNufcG/+98xsbE7dy2FoFx8tFl8Fj9kY/MLUVBqY3pZ5EGg6iRn9yp7v+3u3/ci5s9uF5khg5+8KeE2kgKyS/VXgk1RYIkTwS1WB7WBDAdYX8z+2fOt996zlvnZ9ilk5N8WCYXKENHGLa30FZc1R/EpRSO8EEVz0CDx1MoSmooXqq+rlhocdSxqrzVeRrZUpaGmhjZ0Es09VKqql/UTb3gNFDcXU0VtaxNJdyuqIiipZ3YKiilTVmIAp506zNeWGFHnuZVS2jVndiaBUQFnJPkAoJJ+XVw1fgI9lYbY/XyyrI/Xmxdu7XrZELvGcxHTNks2U1XfQmSrXUcAhVsb/AF9/RZ7U8nnkD205G5NYnxtv2yCJyTWsmgNKammPEZqYFBQUx1xD9weZP64c8c2czgfp31/NKnroLkJX56AK/PpEdg9kdW9RT0OK39Wbjz27a6go8q2xdj0tBJkMNjchpNDV7lzWVkjxuLqZ4mEyUoD1AjN2ABB9xP70fer9uPZbdI+XNzivNy5qMayNa2gSsKP8LTySMqRMwqVQkuV7qUIJkf2t+71z17qWDb3tz21jy5qZFuLksBK68REiK0jhThmFFDYrUHpK4n5bdQs5jparvDYCArUffVa47cdA8yxSS3qKPbeWgrojB4QgKxy+RpVAFgfcVcvf3gvtPuF0tpvfL++bZbkgCVkiuEAPEulvIzKB8lJPGh6kDefuZe49hbNPte8bTuE4BPhK7xOacApmRVJP+nHQubZ+Q2P3bWx0O2u+NtZerqYqtP4RujKJt+eY0WJx+dycbYneONpaeYYrHZKBqiVWkgimLwiRpI3VcouWvd/2l5726S85d582i9s+DK08aMCeAeGcxyLXyqtDSvUAb77be4fKd9Hab3yfudtdnKlYZJFanmkkOtTT1U1+VeiD/Kup2HlO2a6p2McTLXRYqg/v3kNulW2zkOw1838bqsAFcU01OaUxR1Dx/wCTvVa2W/JPG772V57bX/vPvk/tmImsDEovZIc20t/qbxngIJQrp0CQpRGlDEZJ66ffdytee7T2u2qLn3Wt14jG0SWvjx2VB4STV7gdWoxq/esRUHFOhN+MnZ1HhN/fH7feRrftKjaWUyHxk3rVvNG38T6x7vqv49009ZOzqk1Nszujby4qFjq8Cbg9J0uQc6/uGe57cze3u4+3O5Ts27cvuGhLGpexuGYxAedIJA8dPJDGPTrET733IP8AV/nSw51s4Au270hEgAoFu4ABLUcAZYyknzYOeNer4h/xPP8Ar/T3nj1iJ137917r3v3Xuve/de6//9Tf1kjSVGjkRZEdWR43UMjoylWR1b0srqSCDcEH37r3VfHR8w+KHd83xHzNXDSdN9mruLsD4eVdVLM38FeikmzXbPxxerqHZWfZM1eM9tWAku+3KmppY7ph29+6scivn59WEgg8g39+6r137917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917rxNhc/Qe/de6L1398jdkdAYWirNwGoyueza1K7e2xjDE2QyDU6EyVtSZGVaHEwSlUkna/LWVWYEe4L98ff3k72M2ezu9+8S53q81/S2cOnxZdA7pHJIEcCkqHkNckBAzVHUte0/s7zP7t7ldW+zaINpttP1F1ID4ceo4RaCskrCpWMeQqxUGvVO++vmZ37vTNz5Kj3lWbKxgqfLj9v7V8dJTUMRp44TBPkHg+/wAqGZC5MxsHY6QB75P86/e798ebt3lv7PmqTZtvEpaG1stKJGNIGlpdHizVyxMhwxNABSnRblP7tPtNy1tkVldcuR7nfGLTJcXVXZzqLahGD4cVPhAUV0gVNa9JLF/Kb5D4jILkabtndVTOqSp4ctUQZagZZhpbVQVtPLStJH9UJW6H6ewrtf3lvfnab9dxg90dzlkUEaZmWaPu41ilUxkjiGpjyz0Idw9hvaDcbU2cvt/YRxEjuiVon7cjvRg1DwND0ZLrr+Yr2dgZ6al7FwOJ3tig1PHUZDGRx4LPpEiuks6JHfF1czuyuwKRAhbCxN/eQft99/r3E2SW3tvcDZLTeNsqqvLCPproKKgvQAwyMaglaJ8JC0J6hfnH7nPJe6xT3HJ27XG2X51FY5SZ7cmoKgk/qqKVXBbjU8KdWr9Xdw9e9xYWTObD3FTZmnpjHHkaNgafKYmomUvHT5THShaijldFJW40uBdSR76be3HutyL7r7TJvPI+/RXdvGQsseVmgYioWaJu9CRkE9rCukmnWBXPHt7zd7dbkNq5t2l7eVgTG/xRzBcFopF7WAPEDIxUDoofzI6ZxO3N4ddfPjYmBqx3N8W6bMtuxtt0UU2Z7Y+NeehSLt/q/LU/o/jU+IxcK7jwGsST0uXxCRQaVq5g0igg8OgYKZqOj4YTM4PeG3sLuTA5Glze3dx4rGZ/B5ahl8lHlMPlqOHIYvJUc6AeSmraKoSRGFtSP791rp9HHA+g9+691737r3Xvfuvde9+690x7j27h92YPKbdz1FHkMTl6SWiraWS4EkUq21Iw9UU0TWZHX1I4BHI9lG/bFtfMuz7jsW82om2y6iaORD5q3oRkEHKsMqQCOjLaN23DYdzst42q4MW4W8gdGHkR/hB4EHBFQcdV972+DWSiklqeud2wTUzMPFhd2xyLPBH9CseboI387D6jyU4JtYn8+8F+cfuYXsc0txyDzRG9vUabe+DAquMC4jU6vOhaKtKefWWvLH3o7Z0jg5x5edJqZmtSCGPqYXIAr5hZD8h0VbdvSXbOyJJBntk5r7aLTfJYiD+O4xw7pGhWqx3mKFpHACuqt/hb3jNzR7Ne5/JsjjeeSrwwKP7WBfqYeIAOuHURk8GUU4mnU88u+6Pt/wA0Kn7q5ot/Hb/Q5j4EmBU9slK48wSOgsM0aNokdY5AbGOX9qUH8Bo5NLqT/Qj3F7kI5jc6ZB5HBFPUGhH59D9VLjUg1J6rkftFR135Y/8Ajon/ACUP+K+660/iH7et6X/gP7D10ZolBJljAH1JdQP6fk+/a0GS60+0da0vUDQan5Hp8wW29zbpqUpNsbfzOfmlcRxjFY2qrI9ZTXZqhE+2jAT1XZwLc+z/AGPlvmHmWaK15c2C7vp2an6MTuK0rlwNAxnuYCnRRu2/7FsED3O97xbWkSip8WRUbjT4SdRzjC8ejVbB+GPYmeqKep3xWUWzMQyiSalpp4sxuGQFbrGkUWrGUbEmzF5JCtj6T9feTfJH3Q+et7nt7nnG8g2jaiKsikT3R8woVT4ScclmYinwHqBObPvK8o7VDPByvbTbnuAwrMphtxn4iT+q9PIKq1/i6PT1p0B1r1cVqcFhPvs0UVZdxZxxksvdQLilllQQ45GNzpp0iB/N/eaPt57He3ftqRc7Hswl3igrdXFJp8fwMRpiX+jEqfOvWLfOnuxzrz1qg3bcvC2ypIt4B4UOf4gDqkPzkLfl0NEskMEUk00kcMUSmSSWZ1jjiVRcvI7lVRQBckkD3L1B1G3QB7s7/wBv4ppqPbNO+4a1dSitJMGGjkFhxPbzVir/AM2l0n8N7317ote5+wt27uYrmMvKaQ6tONoQ1DjlVuCrwRENUf0/eaQ+/de6RagKAqgKB9AAB9PfuvdKDbu1dw7rqhSbfxc9e2rTJUACKhprmxaorJLQxKpP0uW/oPfuvdGU2p8esdSqlVvCvfKznS38MxzSUtBGfyk1VdaurHNjbxD37r3Q+4fAYXAUy0mGxdFjYFFtFJCkbNbi8koHllY2+rMSffuvdO9h/wAj5/3v37r3Xfv3Xuve/de6R2898YLY+Oevy9QDKyMaLGwMjV9fKpA8dPCSLRgn1yMQiD6n37r3QY7a60k3fu7Ddv8AY9dWZnLY2nNX11smXTFtHrWOvpvDLlaPHIAM3viupZWSTLVmuWlgkaCkSnRpTN7r3Q+WAFha/JAv+eT+b/19+691Uj2X2ltb5A9k/LHZOxp33H2R1j1dT7O65gStifG1226nIUFZ3TJtKCML5tzvkqOGhqpVd2mpqaGKPShk8kIfeR2nmTfvY33J2flO2M29T7eQI1JDvGHVpljpxcxK4VfxGg8+pW9jty2PZ/djkbc+Y5xFtMN8rM5AKo+lliZ68EEhXU34Rny6q4VlYXU3Xkf0II4KsDYq6ngg2IPB59/P4rqw1KcVp6Z8x9o8x5cD12YqBgkV+3+Y9R8/PrtiqjUzBV/1TEKo/wBcnge/FlUamYAfMgdeGTQZPS/2f1fv7flNV5LbW3J5dv42GSoy27ctU0u3dmYiniuZanKbszc1FhKSGJVJYmUkAHj3JHIHtH7ke6M/g8jcoXd9EDQzU8K2T/TXMmmP8lLH5dAXnP3L5E9v4vE5t5ntrSbiIa+JcN/pYI9Un5kAfPoPdv8AYHXOd3dUbI6N2/vP529j0E7UVbifjptrddZ8dtj5GFpUkl7W+SUtBjNq5GGlmhZTi9s1FVVVDo0bVNOpMgz19qP7v9obyLdvebfoJ7NKEbfYs9HPmJ7oqrFB5pAqk5Bkp1h77i/fIe4tZtu9sdnkgnaoN7eKhKjyMFsCyhvMPMzUx+nXqyfpDo/5jUe4sF2ZW9O9SYbsHDYKqwe08/3TuyKk2p1Tg86gOaxvTfx+6XpNw4jZldlVVIcrmMnuzJblzVPDHDU1qQD7dejfL/Lmw8p7Ta7Fyzs9vY7PAKJFAgjQeWoqOLHzZizE5JJ6wg3ned25g3G53bfN0mvNymNXllZnc/KrZAHkooo/CAOjfr1v87cpCkuR+UHSe1asPMz0m1PjXk8xjzG5Xwxio3P3A1ZqhAN2sNRP0FvZ10W/l1hqsd/MB2O/32O3P8be/cXTeAz7cy21959G7ryUMSf5UMZufH7g7L2xTZCpf/NrU45adfo0gHI91rpUda/Lbae6d343qrtHaG8fjx3VlFqP4T1v2vTUFLT7yaiYCtk6u7Aw1ZkthdmRQqQ5hxtc2RjiOuakhANvde6Nl7917r3v3XusNRUQUkE9TUzRU9PTQy1E888qQwwQQo0ks000hWOGGJFJZmIVQLn37r3RAK/tvsP5f5iq2Z8X90z7I6BxdfVYnsr5X42lWbKbwmpJXpMxsP4vfxCmfG5SsDK8Fdvx1qcVi5AyYuOurFaWk91sgjj0cbq/rHZPTexNudbdd4OLb+0NrUP2eLx6VFVW1DvLPLWV+RymTr5qnJZrOZjI1EtXX19XLNV11XNJNNI8jsx91rpf+/de69f/AHn37r3RIew+2V7Cot25iLfUvWfxj2DPkMPvvtXD1Ph3N2xuCmqGw1ZsfqnJUhlrcZg6DNn7Cqy9CsmSymU/yDFaGV6o+68BXA6BvJ0tJSYHB43duxsPsvZFAaLKda/FikpqOPD4WCCX7zHb2+QUuPeaLdO7autWOshwAlkoqSoUSVT1lWvmjhr3Q927HkSIbbtqpc8zyrVYyeyFf9+TUof9LGCGbjgZMncge3F3zdIb68LwbDG1C4+KVhxSIHGPxOaqvDJx0ntwbnz28MkMnuLIS19SLR06sFipaGNyFENBRxhYKOFQBwo1EAaiTz7wp5i5l33mvcW3Pf8AcXuLn8NcIi+kaCioPWgqfxFusqtl2LaOW7H6HZ7FYbcDNMs5Hm7nuY/bj0AHVLe/P5kHyA2Fhvkv3nN8Vtjbp+JfxW723n072huvbveNTTd8UeB2DlcHit09m43rDL7Lh2tl8Rimz0VS1BHmkqpadHKeoexzYchbDfSbBtH9Y5YuZdxsknhRoK25aQMUiMofWmrTQNo0g8fLoCX3O+9WUu53o2pX2O1uWjdg3fRKBiF0AGldVDIDpB49DH8pv5hmL+OHffx16joevpd87T7MpNr7x7z7KTNDFUHx56l7G33g+p+r9+ZjHmnmfJru7srccFJ4HaLxU0UstzpJBTy7yNLvey73uMt74N3Azx28enV9TNFG00sangAkSFtXmSBg9Gm8c7Dbtz2m0t7YyWstGleoAjRyERjgk1kIXypQngOhm+Vnyjy/QWY6m6s6z6myvfPyO7+z+5MD1H1RQbioNm4MUGysbDlt99idj74yNNXwbN612RQ1cDVtWtPUVE89RFBBG0jgAo5Y5ch3uPctw3Lc1s9jslUzTFdZq5pHHEgp4kklDpGpQKFmIA6MuY+ZZdqFhabdam43S6r4aA0AAFSzHyVaiuCSSAAScOXRW+PmPlN2Zjavyf8Ajx1VsDELt457bPanSPc1f2PsrJ5JMhT0suxs3t3du1dob0wu4Vo5zVQ1iQ1GOqIo3XWkgCn29WXKsVrHc8vb3cSy69LQ3EIjkApXWGR3QrXFCQw+Y69se5cyT3b2u9bZGiBah0fUPQggojfOtCPKtejV2/F7E/k/i/8AX/W9hc8D0LjWhpx6BXsT539ofHPADYuy+nslt/I5/I5DK0XYnZNPO+385Suq0tPVbNxVA0UGQWOOlDEzVZvYkx2PGdfsBtD7d7e293KrLLfXU09CfwkhEIHkCE4ZrTVWh6xE94twG4c6XEEcitHawRxYGQ1C7qT5kFuOPTy6qs7V7f7J7t3O28O0t2V+7M6sbU9E9SIqfH4ikdtb0WDxVKkVBiaR3sWWFAzkXck8+5s6inoNfr7917o6nxJSfa23fkF21Q1FNS5rbmxcH11taqkd1q6LL9qZtMfla3HBYyor49sYuqSN9SshdiBzxjn97HnXcORPYfnXddou1h3a6WKyiY4YG7cRv4Z/34IvEKcACK1x1Nn3duVrPm/3e5V23crdpdtgaS6kUCoItkLqH/oGTQG9QadRZESM0iqqhI50Rb+I6QI5ETSZSW1EcDR6z/rX98G1XSAi/CBT8hw67Ak1LMeJNepP/EfW30H0+tvp9fe9J8x17UoxXrEJJqWuo6uklenqY5o5YZ42KSw1dJIlXRVMWmMl54JYbglgFtx9fd4pZYJYZ4JWjuI3V0YcVdGDo3+1YA/OlOmpo45o5IZow8Loysp4FWUqyn5MpIP29DFvSMZ3Hbh3dCaaKHcOT213GuIx6JHRYHNdotm9ld0U9NG0vlhpcl2p1zSZvxKmiGfcEvPr5z9+8luz+8v3b/ZP30mjT+sNrPJt24GP4FkkJD1BOpdU8COoOR4p/DTrDH2LsR7Y++ful7SK7DZriFbuzD/EyRgMhDAUYrBM6MfPwx59Df8ACyrnlqu99viSNKCbrvG56ZVQNOarCblpWoFDskYhhMdfKroNV7qb392/u8Nyuovcf3A2hCv0k+yRytUZ1w3OlCD5AeK1V4N+XTv317CCXkjkvc3DfUw7q8S5xolgLPUeZrGtD5Z6dfmhg5qzC9OdmxrdJ8XX9c5yYGQkZHblU9bifKqmV382IqZNJPLmO50qBc1/vDeTpLXmbkHny3hH095bSWMrcT4sJ8WEt9sbSUpUkqa8Oi37lfM6z7HzhydNIfqLadLuMYp4cgEUtPskVK8Pi+fRQNnboq9jbu2rvSi5qtp7iw+4I1sD5VxddDU1EDA31LPTI6MPyG94Dcp8x3XJ/NXLXNtg3+N7ZfQXK+dRFIrMv+2QMv59Zi8ybHb80cub7y3d/wBhuFnNbn5GVGVT+TEEfZ1MyOzsbtPbWV2zhaiCpoem+7u3upk+2hSMw7I3pnG+RnRVTIKeioabwVvV3bsdNG6IVd6B/WxBtnZ9/rlq2v8AePbT3e2UiTZ972wW7OOGpFFzbEmtKtbyOo8yU86HrED7m3ME9rYc9e2u6Epue2X3jqjYNGPgXIp6rNGhOTTX0tfjjixnPkR0xjCs7J/fagyU325HmjgxCyZRptBiciBHox5HBGlCb3v7xV+73tY3n3z9qNvKuVO9QSHRxpAHnJ88DwxqPktesjfencBtftL7h3oZAw2uVBq4Ey6YqfaRIdI82A6tAoMrQw7/AKuvqXD4fI5/PUORfVJFHJhs9NXY+sZmS0qx/b1eo2sbLe4t7+gi+tYNwtL6wuRW2uInjf8A0silTwyONKjI8s9cZLW4lsbi1vIGpNBIsi4rlCGGOByPPHrjqnzeGzsj1zvDc/X2Zplo8jtPLVmM8KgLFLjlmZ8PW02kspoq/GvHJCdRZ0IY8k+/m/525S3TkLm/mPk3ebYw7lt15JCy1wVBJida5KSRFHQ5JUgnPXcflTmXbub+Wtj5n2qcSWN7bJICPJiKSK1ODJIGVhwBBpjpPWBIOm55AOm5/wAQOL3NvYW40avA+tP9X+CnQgIQAg4H+r/UOjf/ABJ64Sr3BJ3VuahaXanX1Rq2dBURSrBu7sZ0K4yOhd0FPkMTteMyVNXIj/tVKxL9QQM7PuT+xl3znzfB7p8wWLLyjs8hNqWHbd3wNAUBpqitKku2VaXQq/A3WIn3q/du35Z5Zn9vdovAeZNzSlxpoWt7QmpDjistwQAi8fC1McMOjt7X8eR3JLmM6fuqTFQZvemfeZ440qYsJS1GYqEmnnOiKKqqUSMu3CBuffXPd90g2TaN03u9NLWzt5ZnNadsSNIcnAwPsHXNrbtvl3TcbDarUHx7mdIlxXukYKKDz48Pl1TzntyV+9c9nd45aaSpym7ctW7iyFRKYjJLUZWZqhQ5gSGA+CBkiXQiKEjFgBx7+b/mrmbcOc+Z+YOb91lL7hud5LctU6qCViyIK+UaURRwVVAHXcblrYLLlXYNn5Z2+NVs7C2jhWgpXQulm+121O1cktU9NdhwT+PoSfpf/H/XPsgy2Dn/AFenR2FVRQAAdcWgFUpDUslUh9NxSS1ERCNfSGWJ4yEY/S/B91MAmAc2xdTwOgsD5YOkg5FOPXhJHHwnRSP6QBz8iQeHUUJQFiiinDD0NGjKhX90SlWjQrpPmQEgi5I91SSEkIjrUYoCKj5U8vs6uyvl2DZzU1z86/5elFTUk8+zO5cimSOIotrdRZzd81eY3eOlze2NwbYzfX0iooINcOxKHGGkF1cyqdBJBBzJ+4pFur/eBsXsJWWxTZr43VK6WiZEWJX8q/UeGyVplWoeI6xb+94+3r7OXC3aqb190tBbk0qJAzNIRmv9gJA1K/EKjIPW0rQSzz0VJNVQ/b1U1LTy1NPcEwTyQo80JIAuYnJW/wDh77V58+uWfHhw6l+/de697917r3v3Xuv/1d/j37r3QF/InoDZvyS6yyXXO76nMYWdchit0bK3xteqXG726x7D21UjIbP7G2JmNDvid07YyaiWJ7NFPC0lNOklPPNE/utg06B74+/InNjdUXxe+S2W2xgPldtzDVOUo1oEfC7c+Q+wcZJ9tT9z9S0lYRHUw1USD+8WCp5Z6zbOTEkcqmiko6qo914jiRw6Or791rr3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917oEfkD3Jjejutc3vSsSCqyYC43a+JmaQDM7jrI3/AIfROYlaRKVShknk4CRIebkXh/3x92tu9mfb/dub7xEl3AfpWkDFv17pwfCQ6QWCChaRvwopyCR1JPtP7c3/ALoc57byxaFo7M/qXMop+lboR4j5wWNQqDNWIxg9a8O9t77q7H3Nk94bzy0+Xz2Wl8lTPI5+3p4lP7NBjqa5iosbSAaYoksABc3JJPBfnLnTmb3B5k3Dm3m/dHvN9uWJZ2+FFBOiOFD/AGUMa0VEAAGSatUnsHytyrsXJuyWPLvLlglvtNuKKoA1MTxkkbi0jcWc5NfIADpK+wv0I+ve/de697917pfdZ9mbv6i3dQbz2XkXo8lSFY6ykleQ4zOUBa82KzFMjKKijnH0b9cL2dCGHI79uPcjmv2q5qsOb+UL8xX0JAkjJPg3EX4oJ1FNUbcR+JGoyUI6BvPXI3LvuFy7d8tcy2YlsZKlHFBLA/4ZYXpVXX04OtVaoPWwT0h3ds7vvZa7h28yrPAsWP3XtyrXXV4HKT0+ubHViuvjqaadCzQyrqjmiN+DqUd1vZj3m5V96uU4+YuXJdN3Fpju7Zj+pazkVMbYAZTlopF7ZFoRQ1A5E+5/thzD7V8xybHvcYa1k1PbTr/Z3EINA6+asMB0OUaoyKEgJ8O2qOqs53T8RsmskNN0fuxN09PCZ2Y1/wAeO2KnJbh2JTUkkhLTw7C3JBmtskA/tQYqnuAHW8v9RyRgdHq9+611737r3Xvfuvde9+691737r3XvfuvdcNH+J5vf6fn3qmSa9aKginSdyWzdp5lZ0y+2sDk1qVVan77EY+pM6oQVErTU7O4UgWueLceyHcOVeWd2SVN05esbhZAA3iQRPqA4V1Ka08ujey3/AH7bWjbb97u4ChquiaRdJPpRhTpg/wBEXVn/AD7vZf8A6DWJ/wDqX2Q/61ftp/0wGz/9kkH/AEB0c/64PPflzluf/ZTN/wBB9SKTq3regqI6ui2HtClqYtQjng27iY5U1qUfS4pLjUhIP+B9qLX229vbGdLmz5H2mO4WtGW1hBFRQ5CenTFzzvzneQvb3XNm4yQNxVriUg04Y1dLGmoaSihWnoqano4EAVIaWCKniRV/SqRwqiKo/wBb2Lba1trOJYLS3SKECgVFCqAOAAUAD8h0G5pprmQy3ErSSniWJYn7SanqUBb839vgAdN/4egx372pgNjo1M5/imcdNUOHpJUEkdwNMtfMNSUMJBuNV3YfpU/X3vr3RON3b93NvSqaXMVrx0Ya8GHpHkixlMLW/wAzfVUy/wBXkLEkcAfT37r3SN/33++H0Hv3XumrOZ3C7Yw+Q3BuPK0OEweKgapyOUyMywUtNCguxJN5JZWH6Y41eRzwqk8e/de6kfDbsTY3yV3x2jS02Eqp9qdcxbaXD11fLNSvuarzMmX+5yM2NXQ9Ni0joIxTxSkStdmkUXVR7r3VptLR0tDBHS0VNBSU0ShYqemijhhRQLALHGqqo/1h7917qR7917r3v3Xuve/de697917oNexeycZsKiVZEFdm62J2xmLRrFwt1NXVv9IKKOSwJ/Ux4UHkj3XuizbFwOY7Z3xJm9wSvWY+hnhrMzOwIpyqkS0WDo4y58FPI6AlB9IlN7lr+/de6PDYKAAAAAAFAsAPoAAB+Px7917qpH+Y/wDNZ+s8TWdEdWZIL2HuXGSR7z3Nja0JNsDBViBRQUstM3li3Zmqdz476ftKc+Q+t47e691QZtDeG6dg7mw28tmZzIbe3Rt+tTIYrM0FRIlVDUIwaUTktaspqtSVqIZNSTxsyuDf37/B17q1rr35G9R/JzPU+J3V8Nd17q7bejjqcvX9I5SChx+aaL0VOUy8E2QwMGFp2NmMlZM92YjWbC8A87fdf9kPcDc23rf+SIV3ZjV5bd5LVpP+aggdFc+ZYrqJ4nqY+VPf33Y5N29dq2bm2U7cgokcypcLGPRPGVyo/og6R5DozUfxK7v3JkTD1l1/0F8U9s+ChK733zi6n5Qd3yrUqZcguO2zmq7E9S7RraIhVp56mbcAD+pobKVa/Kv3YPYfk+aK62v24sZb5K0lug104qa/8SGkWo4A6agYB6pzF79+7vM8bwblzzepascx25W2U4pnwVRjXzGqh6HTZv8ALn6Hpq2j3J3dkN/fK/e1JLHUxZz5GbkbeO1sXUr6tO1emsdTYPpPZ9FE9vDFQbejeMIt5GI1GdoIILaFba1gjitlFFREVEUDgFRQFFPkOojknnmleeeZnuGJJZiWY141Ykk186nPR6sXiMVg8fS4nC42gw+KoYUp6HGYujpsfj6OnQaUgpaKkihpqeFFFgqqAB9PbvTfTj9OB7917r3v3Xuve/de6QvY3WfX/bm1MjsjszaGC3vtTJqfusLuCgiraZZljdIa2ikcCpxmUpPIWp6umeKqppPXFIjgMPde6Jnh8v218NMrDtnd3+kXvr4oVEMr7d7REOU7E7q+P7xetdp9nYvFUdbvHtbq9YATjdy0sFdncUFNPlUqYQlevutk18ul9U/OTpfL001N1DSdhd+bvaljnx2y+qevN15CtqJKhL00eX3LuDGbe2Ls6BpCqyzZjKUKQXOvkaffuvUIz0kaPoLt35I5Ok3L8xKvD4Xrikqocltv4jbBzNTk9l+eF0noa7v3fcKY+fuHLUUouMHTQ0u04pFBlhyTKky+69X5dHroKChxdFSY3GUdLjsdQU0FHQY+hp4aSioaOmiWGmpKOlp0jgpqanhQKkaKqooAAAHv3Wupfv3XuvH37r3RNu3t70/adZunrrBbzp9p9R9eLWTfJvtaiyn2Aw9BjKaOvyPTu385FJGKDcmXoG17grYJDJhcYTAClbVRGH3Xus3VuxaXds2M7l3Xgm29svaOMMfx46jq8UuIxPW2zsbQ/b0W+MxtxAI4+wdz0UXlpxJH5MFipI6WJY6h6xnT3dwtpa3N05AjijZyTwooLGv7PLPTsELXE8FugJeR1UU9WIA/w9FOymUrM9k8hm8jMZ67LVk9fUSXuC87llVPz4ootKIP7KAD3y93Lcrretxvt4vZdd3dStK7epY1H5BaKvooHA9dAdv2+32qws9rtI9Ftbxqij5KKftJqW9STXqIltaX4GtCT/QBhc/6w9oulbiqN606o+6I/l/Y/vzcvy8qfkJv/wCS1L1LuD5/90bpT4vPutdk/H3s3FYvMbXym292blwNPt2l3VvHbOeyFKs88P8AFRisiaZQ0bIpBl/eed5NltOVl2S0sDuSbPAhu9AkuYmKurqrFyqOoNBRNSqSdQPUR7fyim6X2+3O5yXP0f18p8GumORQVIOmgZlY+WqjEZUjHQJ5H4n/ADX+btL/ADEOyMDnejunevfmRnMn0Ls/aPffUXZVX27g+lPjlFVbP6mzm0Mhi9ybax+xcduTedNVbjxq1FDMVqpIqti6FF9nkXMnKfKTckWEqXdzebYizu8E0Qhae6IkmV6q5kKR0jcqw7argV6D8nL3MW/jmK8igSO3uGMaa0IcJHqVSvcv4tUnwn4qipp0JO1eyu+6Zv5cv8wXfvQvbu66vrrozuH4ofM7YO3dk5fMdq9XZupy22qObujB9dzQQbg3ttibePXTSV/8LhqZ58XkYp4VlRT7Lriw2VxztyTY7xaok95BeWEjuqwyqFYiBnyiNol7Q5A1qVNOl0FxukX9X+ZJ9qmcRQvBOoVvEjPZWRQaOw1xGulSQjBgDSnSx+F8UOU/mD9jbs6Bz/y/3R8S8h8WK2Tc+X+QmR+QEmwY/klme4YMoMXsXG980+MrKOtodj61MWLpRSUkIaIMP0+0XNTCLkextd4g2uPmNdwAQWwt/E+lWEirtblgQZKZdtRNT0Z8sRytzfPLtz3p2T6bJlMwXxTIGICzZqFJrigHDq+jr/YmX7Az9Ji6GmqGxa1MYzmVWM/bY2iWz1GudisZq5ovTFGDrLNe1gT7CvIvJO6c8b5abbZ27/u0ODcTAdkMYyQWJprcVVFGSTXABPQw5w5tsOUtouL24njF8YyYIie+RzhaLx0qcsxxQUrU9WD7x6s687F2dN1/vfaOG3JtB6WKjXDZOkSaCmjhh8EEtDKNNRQVcEYsk8LxzL+GHvo3Z2lvYWttZWkQS0hjVEUCgVVAAFPkB1g/cXFxdXE91dSF7mVi7seJZiSST6k5PWr/APNX4sV/xe7SbFYxclX9Y7sjlynX+criKiWONWvktr5KsjRVfKYJ2UIzhXqKV0ksSHsp6apgnonY+o/1/wAe/da6PJ8dAKn42/JCjhCtUY3sDo7cdYGeNPHiY33PiWnS51SuK6sjXQB9GvzY+8Nvv4WV1dfd/vZ7dA0dtvFhLJU0ogkZCR6nU6igzmvl1k990O7ht/eW1hkYiS42y8jjxWr6Feh9BpVjU4xTz6ZKm6xq/IMUsMh9egWEihtRCSErY/QAX+lx74sHiaddT+IFOjxfHPq/q3trqbsHC5igpabsDD5r7ibdjtVtlttYHK0UMO2M/QQxVSwz7fxebp5YstB4nMkMpe4KKfedn3XPaD2s97vbL3H5V3WxWH3Ut5/Ft73U/iRRSRgWzIitQwCZZEuF0nXqyahSMQfvA+5fuH7Uc/8AJHMe33bSe3s8Oia0CpomkSQm4V2KkrMYmRoGqNOnhQkdFI3zs7cnX26cps3dlB/D8/ga6GGshUyTU08My6qXJ46oTQtZichA4lp5rGMobn1AgYc848n8w8gczbtyhzXYm236yk0SJnSw4rLExA8SGUd0TgUKmh7gQMn+WOZ9l5y2HbuZuXbsTbNdJqRuDAjDRyLnRIh7XUmoPyIJXOMxmLfq7Nbmk3dt6jq6Pbu4NgybNkqJV3Zlstl+zdhb427laOgEJp5tvU2JxGUWoqC4kimKKAQxPvJfYucOTx9yvnfka936CLnA80JPDa1/XlUtCwYR/EYxGJGMgpp0gGlaGAt25X5mH3q+VebLbZ5JOWzsbxy3FD4cZWOVCpetBJrKAJ5qxNMCg1/Cm/8Aeju0/hulnP8AbIuu6caLC5MQ4Ivp5/r+PY4/u96D3c5zp/0zp/7TIugz99H/AKdvyqK1/wB3n/atL0Yzsnbkm/8Ao7svZcUQqcriqek7I23CwZnar2tY7ioqfS2pJa7bjy2Atcpc34tnL97jkObn32M5rt7K3Em7bbov4BSprbHVKF+bW/igep+zrEn7t/N0fKHu3y5PdTaNuvtdnLmg/wAYGmJmHmqzeGT1VLTyiaEKWVmj0rIAEFwVDRyeNXk0JNGQygm+k++EisGVW4qR+2vr114IoxBqGB6Wkm6qZtu5XDptPb0Oa3DFsOj3PvlXy0m5NwYvrGHcVJsHHVlPLXPg6d9u47dFXRrVw06Vc9F4oJXZIYws0b575c4cx+z/ACx7LbtYWUvL203CyW9yVb6sKhcxxFySgSMSMgKqHaOiM1B1Fm0e0XLexe5u/wDultt3dR7vuUTJNAGUWxZwniSaQuos5RWILFQ9WAqejAfC3FyP3HlN8PC70PXexdzZzzLLUwCLI11GdvYZQQjQu9VU5OUIBbUq6vVp9y79xrlebfvfey3cQlrTZtvuZ2apCrJKotogTShLeJJ2kioFc06jX72+/wAO0e0N1tfigXO53sEKrQVZI28eTiaimhDqAJ8sV6N/yR+4dTtcyM1iXdzqct+CXYk/7H32s+xvz/y/5euV5pX5dMPYXVXW3dlLQSbxyOW2ZvzEY+HFY7sLGU5ytLk8TTF0oMZujDArJPLj4pikNXEUmWIAFiBY4t+/n3VeTffC5TmD6+XaudUhWL6uMCRJo0JKJcQsQJNNaJKrLIvw1Ze3rIH2e+8NzR7TwSbN9FHuPKrys5tnOho3emqSCUAlNRHcjAoTkKCa9BvtL4m9U7YqhlOwd/5HtEU8/lo9n7Soa/bOAyEaaTHDuLM5H/cmacyR3aOmKMVuNR1G0Ee3P931se27jHuPubzed1gilBW1tY2toZVX8M7szSspPxpGU1AUr1L3O33zd4v7NrLkHltdumeMhrm4dZ5Y2OKwqqiIMBwLhsmoAp0PmUyr5AUkEVJQ4jD4qnShwW3sTBHRYTA45OI6HG0cQSKJT9XkI1yv6mP499DNr2vbdk26y2jZ7GK12u2iWOKKJQkcaIKBUQUAAHy+3rCzcNwvt2vbrctyvJLi/mkLySSMWd3bizMakk/M/Z1LwFVR0MtZ/HMfV1e2c9hMttrPCKnYiTCZ6maiqpaaWVBTtNCWDqNVn0aQefbHMGyWPMmw7zy9ucerbr+1lt5R6xzIUalccDX8unNp3S62Tddt3mwal9aXEc0Z/pRsGUfYSM9V6b5+K/a+xzXZHbGHk7M2FC8smO3Ts4rlZRj08jxPmMLE75fD5Ix6FmheLwxtfQ5X3w69y/um+8PttdX5g5am3jlmN2MV5ZgTaoR8LS26frRPoUagEaPVXS5HXWbkP7x3tlz1bWvi77FtnMMgHiWtyfDpKcMIpmpHKhPwHUHIoCoOei7TyrTyvR1yT4+qVT5KaugmoalVbQobxVMccgRjKoVrANe63942TxPBK9rdRPFPTKuGR8/0WCsPtI+fU6RSLcRrcQMsluThlIdTTy1KSv5VxwPQ2bu7o3Dvzr7YnW1ZuTtrqvB7Bx02Oo9wfGXtmp6c3fmy0dPCtVuSol23uvDZiak+3LqjwKhmd2blz7za9sPvt8w+3nLvL3KF57ZbNf7Ht9skKujPbzsExrZikkJc8W7e9qsck9Yoe4H3Utn513zeuZbPnvcrTd724eUrIizwKWzoQa45Ao4KKnStFFaDoNsRsbtyeeOXrD+Y9ncp/lFUzdT/AMw345dLdvYTJivjRXxdB3/13tXaG7MBrmHFXUxzMurmM2v7ye5c+8j90/3XjSy575P27atzYFCm5WMDR1egbTfQIUWvmzNG1RjhXqAt59jPvD+3JlueUuYL2/sUGrXt95Mr6Uyuq1ldX7f4VEg+3o6PSO0tv1W+uvtofK3I0PT9FVbtwue2T1Dguvtk7b6B787G2vKMts/KRd+7b3FvKl7Xj2/lEXI4naVfW4CqevhinmxdSYQFyy9uuQ/a7kzbZJva/lza7Pbb2jtLZaXWcD4azBnLgZKgOVWpoBU9Y686c28+czXyxc+b7uF3f21VCXRYNET8QETBNBOAx0Amgqerx1vbn68/72fcidAvrl7917r3v3Xuve/de6//1t/j37r3XvfuvdAn3r8fOsfkVtWl2v2Ph6iSowmRj3Bsjem366bb3YfWW76VSMbvbrXemP0ZrZ+68ax9FTTOFmjLQzpNTySQv7rdSOgS6j7p7A6+7Kw/xh+TlTQVu/cxiayt6U7yxlAmE2l8jMRgYJJ81jKrEB5KbZ3eO18XCtVmMHFLJSZGk15LGkU61NLQ+610dgG4v/vf+HB9+69137917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuqbf5kW9srWb92b16VhjweEwbboUqVeaqyuWmnx5Mx0hokpaSmIRQSG8hJ+g98mP7wXnDc7znXlHkQhV2a0sjeCnxNPKzR1Y8QEjSgWtDqJPAddGfuX8sWNvyxzLzfVjudzdC1zgLFEqydvrqds+lKdVt++e3Wa3Xvfuvde9+691737r3Xfv3XujC/GTuut6Q7PxWbkqJP7p5uWDC7yoDLItM+LqZURMsIVdYmr8PIRJG7AkRl1+je56+7l7w3vs57j7Zu8lwf6rXhW3v4iToMLsAJ6VA8SBu5S1aIXX8XUNe+Xtjae53I24besI/rBaqZrOQAahIgJMVSCQkwqrAUzpPl1ah8i8pT9SdodFfKilWlO04K2PofufLKG0UnVXb2WxR2VvCoqRqjix2yO2abFNNKy6Ycdl61yVAJHfC3niuYIp7eQPA6hlYZBVhVSD6EUI+XXHiSOWGSWCdCsyMVYHiGU0IPzB49HeDc25v+T9P8P959vdU68jrINSG4uy3sRypKkcgHgj37r3XL37r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+690AXb3a42wsu2tvvfcc8CmqrRZkwsE1ijKpVllr547lFPEY9TfgH3XuicTTT1M01TUzS1FTUSNNUVE8jyzTytbVJLLIWd3a31J9+691j9+690FnbHcWyunMCcxuuvD19TGxwW2KKWF8/nagiRIjSUjHVT44SraaskAgiH+qayn3Xuqee5u9d5915ha3cUqYvbeOmM+B2bQ1DyYbCkQiGSslmdI5Mplpo7+SplA49KKi8e/de6vO/lRdL5HYXS+d7NztJNRZPt7LUlfiKadGjlTZmBgmpMHWMpclRmaqpqalAVU+Exn8+9EE07j17q1P3vr3XAuBfg8f8R9eTxx7917rwcEiw4Ivfi3P9D+feqjh59ar8uufvfW+kF2Dvug2Jhfv6iL7yvqpDTYrHKwRqqp0li8jcmOkgUapHANhYDkj37r3REMhkM7vPPmpqmlyebzNXFBBDHqsXmbRT0dIhLeClhDWUC4Cgk83Pv3Xuj8bA2jT7L2zQ4aPxvVhfuMpVILfd5GYBqmW59RjRvRGD9EUe/de6Kv83/ltjfjB1yBhmpK/tbecNZQ7Cw9Qn3EFE8SBKzdWYpwy/7icL5QURiv3VRpjW41lfde61ZszmMtuHL5XcGdyNTl87nsjWZfNZatfyVeTymQnaorK2pkFwXmmcniyqLACwHv3Xuhe+PXQe9vkd2XiuutmU8qI7RVu6dyNTtLjNn7dEypV5jIuSkZnYApS0+pZKmayrwGI917ras6J+P/AFr8dtmU2zOuMHDj4GEU+czUypLndz5VYkily2cyBXy1NRJp9KAiKFTpjVR9fde6Gv37r3Xvfuvde9+691737r3Xvfuvde9+691737r3XvfuvddAAfSwuSeBbkkkn/XN/fuvdd+/de6A/dPyS6R2Tvio663Vv/F4XdVBjsXlMvTVdNlDi9v0mdkliwn95tyQ0Eu29tVGZ8DtSw19XTzTxqXRSnq9+690rtxds9Y7SwSbl3Hv7aOJwU1P91SZGpz+NMORhMXmT+FLDUSy5aWaLmOOmWWSS40K1x7917os24e3N+9y73231L1/i919U7K3tt/cWfyHb2Zpf4Nv7IbY29Lh6evh652TkaSTKbZ/jj52KCDP5uOkkp7yPR0U8irNH7r3UPa2xNkdmV+L6n69x9PjvjH0Zn0h3JSU6T1eP7g7UxFacgdvVeUq5J5N07a2ZnR9/uKrqJKh8zuIrDM8n21WJPde6OvlaP8AiGLyFBr8ZrqGroxIALoamnkhDgH8rrvb2i3K2+t2++stenxoXjr6a1K1/KvSmzuPpLyzu9NRFKj09dLBqfnSnVSVVRVOKrKvE10TQVuLqZsfWQvw8VRSyGFlINjZtN1P5BHvl3c2Vxtl1c7XeIUvLaRopFPEMh0kfPhWo8jXh10Ctbu3v7a3vrSTXbToJEI81bP8q56x+2ulHXiWa2pmNgFGoltKj6KL/QC/HvXXgAK0UZ/w9eZma5Zi7HgliWY2HFyf6D6e9Ejz60KLgLQfIdSKGCvydZHS42mr8lkX4ipqCGesrSE4FkgDyIqr+TYD2psbS83O4Wz220luL0/6HGpd/wBi1I+04Hr0murqysbZri9nihsxxZyET/jVKn5Cp6MrsT4557OSQ5DftXUYagUgrhYZlly1WgCt+9UpJJFjInLWIXVNcG+ng+8g+Rfu+7rupi3DnJ2stvDYgQjxpF40ZlxCprwFZKg1px6hLmv3k2vbllsuU4EuLw1/WYUiQ/0UIBmI9TRBUceHRzMLgsTt3H0+KwlBS43H0qBIaaliWNABwXcgapZX+rOxLMeST7y42jZtt2Kwg2zaLKK3sIlAVEWgx5k8STxJNSTkk9Y37juN/u15Nf7ldvNduSSzkk58gOAHkAMAcAB08ezTpH0A/wAjeg9p/I7qzO9b7pjjhlqV/iG2c+IRLWbX3NSxy/wvOUZJVi0LyNHMl9M1O7ofrce691qO792NuTrTeu6Ov930MmP3JtDM1eEy1O0csccslLKVhyFEZURp8ZlKfTUU0oGmSKQEH6+/de6NJ8Ga6hyfaG8OpMpLBBRd69Ybn2DQyT28cO8KBY907MmXlGMwyWKkjQBluZByPce+7HJkPuJ7bc6clTIC24bfKkdRgThS8DcRlZVUinQz9u+aZeSueOV+aYydFlexu4B4xFtMq8D8UbMOH5dcZYJkE1JVJJT1ULTUlXE2qJ4KqnkemqYmK6WQxVETKSLHjj386jwT2zy290hW5ido3B4q6Eq6n5hlI67cLLFMiS27hoHUMhHAowDKw+RUg/n0J3S/aeT6h31ht7UkMtZjxFJid2YK2hdw7Vr9MOaxDxy6gszxL5acnlJ405sWPuTfZ33R3b2e9wNl532zXJbxN4d1AtKXFpIR40WajXgPET8MirwBPQA9z/b7bvc3kzdeVr/Sk0g8S3lP+gXKA+FJXFFyUkFcxs3mB1Zj2V15113DtvbVBnZKit2/nMbS1/UHb2HpYqzcO3cVXzJLBgcnBIwhz+OxlV+1U0UziWEgmMhvp2M9yfaH2w+9HyPsfMcVwFuri0WXb90hVTPEjjV4bioEsVarJC5GlgdJRxUcxuRfcvn/ANgObd22WSEvaxXBjvdvlYiKR1xrRhUxvShjlQdy6SwdDTrXy2Ftj56/LjfPyAqfhtjPjvj+kvjv3XvHoGi7J7bOUzU/fG/Ouqp6DfWQwGOx+79p1vX2wsLldFH93JDk6maYs8YdUK+8Nrj7jW68tW9nDf7buPNG43LyLJLtt3Z2EVnGGHhsyX6SSXUsgJYoGjRKaA7Du6yWX72Vpv8Ac3j2m422xWMMYKJeW1zdvcOQWYB7Ro0iRD2BiHLA6iq/D1YHRfIvob+X5gq7aHyT34Mr8o+z+rcDvDsfrn469e9lfILG9HdXYqp17i3burPbC2jlMhith1WbjljjyuTpsek8NPrSN/FIwyj+7H92eX2LvObN+3feEvN53GkMQEYBt7RHZ1R3qwM0pKtOE/TVkAQtTV1Avvz77j3bteW9n2/bGttssS0sjMxPj3DoFLKlF0xx0YRaquQ7FgtdPQidqfzFvid8Y67qTM777UWrqe49l1PZfTOL2Fs3eHatX3FsWnnxVFkp9j4vYWEz1VudKuh3BHK9NChmOPM1QVEUErLlvJFHNG8MyBoWUqykVVlYUZWB4hhUH5HrHJXdHSSNisisGUg0YMDUEEcCDkfMDoh/zE+RfxY+O3aVZQ02/su21KzB7K35uZ8D172BurC/HfD9pRxV+z9u/IDcmB29kMF1NV5mF2lxozdRS1P8P/cljjVdR44+933OOeuWeb94vvb7bIbzke6dpLRBPFHNG7lnaxSKV1aeRNLGERa2ePStNYoemvtT957lXfeW9rsucbmW35qtkWO4fw3eKRVAUXZdFYRq2pRMX0qslWroNQRvcnzITr/5ndm9Q7xzuYzuxIPjl1R2H1jsbqzrvcPaO/8AcG5c5nN4PvfO7dxXX+Dz+583g8ftnF09TVTPG1BRU4EhKtINUccqewm/+5HtXy1unt9yvJd86vv+4W14zSrFGkFvDC0cchmdI4pPEZwBh3IIOENBfzB7xbXyV7g75b82b9HDyqm1W00KiPWzSSTSq7jQrOy6VTIGkDNR53pYP5J/FX43fEzq7sin7Wx++tufJWv2xl9sZ/qzbm5OxNw9uZvLUdXV7W6w6k2DtLG5Peu7snt2ipKmfJUsNGstBJDO9UsARgvUf7svsSPY7kaax3OeKfnPc3Wa+lQVUECkVvEcVjhWql6DXIZHAAIHWBHvz7uN7tc2RXljDJFyzYxmK0jf4mGr9Sd/R5SAaCulAgqTU9F370+duL3B1J8XO1vif2FS5HCdl/zCvjr8XuzVzezqzGbnweG3bvSuwPanWm79k72xdBuXr7fFNBFEkkdTTU9fSIyyRHS6uckeOeoQ6XfUXzBocbmPkbTfIHelNjqHG/zON1/CLoJaPbUrNPNkdt7IrOtdiVZwVFLJLVVWSydez5av0qAQJpQoQe/de6MFvn5U9DdbZrvnb29N8HDZf4ydQ7c747vpP4Bna0bL6q3Z/GP4BucT0VBPDnDWDAVRalomnqofGNaLqW/vt69w4dFK3f8Azo/glsHG22Zv6t312PmsRTbh68pcx1D2uNt712tW5BMPSbq6UoqnatHJ8iausy7/AG0Ee3fvaaExyyyMUiZveP8A7/8AuPz9yPsdhae2nLMN9zFdsyPcTyRR2e2IsZf6i9LyxhVcV8HxHjibS1XLAI0w+0PJPKXNe63VzzxvctpsdqoKwQoz3N7JUL4NqFjbUVr+qY0eVdS9tKssbqH+Yx2D35tLHdrdX97Z7cG0splsnSJBPt+mw1Hjs3hcjLjMvtfNbMzODparA5LC5GIxT42qhjlhsCQAQTyv5t99PvScjc23NjzP7i7ha73EUm8IfTSWrJMuqN41RGhlgcUMehzGVIoTnroNy37R/d75w5cguNh5IsptsKmMvWdZ1dCVdZGZllWZCDq1KrBvLo4WyfmXTmWhXf8AtOTE5UaEn7D6qqf4HknN1X7zL7PlZcNlUAN5RTzQFuQq82OQPtv/AHg28WYstu91eVEu4FoJL2xIilpWhkktXBRqA1YRSpX8KZp1D3PX3MNtuBc3nt3zG1s5OpbS8BkjJpXQlyveteC+IjU836NdHuDa3c2Enq1p+vPkLtynjVshFX4CKl3vhkQr68lj1ipt1YtlCi06CaM2ADW4Oam17j93v7ymzyyWsGz79AB3pLEq3cPAd6MEuIh6MKKfIkdYs39j7z+xe5xLM+57O9ex45WNtLxPa6loJKVypBPGoB6LbuP4odO7tlkq9h7qynU2QZgwwG6Y590bSaVWd1ips7SyQZnGR+VjdahJlW5+v0940+4P931ytukst77ac1zbTIw/3FuQ11b18tMuoTxj0qZQvEL5dTtyZ983mLbo47Tnzl2LcUX/AIkQEW8/2slDE5+Y8Mnz6J32f032J05k6fG7xxoWhyPkbBbhpZlye2NyU8cRIfEZ2nURySshEkkUqrUKp5S3Pvnd7nez/uB7RbuNp532NoYpGYQXMZ8S1uVXNYphQaqZaNwkg81pnrNjkD3M5N9y9tbceU91WV41BmgfsuICxp+rEc6dWBIpaNjwavSXwu7cjRYjLbTq6fGbn2NnYxDunrHe9DHuTYO5KXyaohl9sVkj0sVQksQlpa2mNPXUsqLLDLHIoPuvtp7we4ntHukG5cj8xzW8Ier2rs8llMDxWW3ZguRWjpocHIbHWuffbHkn3KsHsubNkjln00juEVY7qE+RjmALYNKo4eM8Cp6Pf8cPk5k+mMVR1OSz+5t5fG6DP7e2zvHF763DPujsP4mzbkqFxO29zU298rrznaHxrymblhopajLSPntnNIpnqa2gBNN2c+7z94XYffjl+7mS0Ww5wsdAvLPVqAD1Cz27GjPbuQRkaonrG5JoTy+96fZbefaHeYIprg3fLd2WNrdadJYrQtFMoqqToCCQDpde9PNVuoBB5HI/B4sf8RY/Qj3kR1CfXfv3Xuve/de6/9ff49+691737r3XvfuvdA/3h0xt3vPr+u2Rm6zI4Cvir8duXZW98BIlPuvrnsDb1R/ENpb+2lXOG+zz23cmquoN4aqnaWlqFkpp5on917oM/i93ZubsDGbp6v7fhx2H+R3R+Qotqdv4fHQNRYjciVcDz7R7g2RSysZH6+7Vw8P39GLsaCtWrxspM9FIT7rZ+XDo1nv3Wuve/de697917r3v3Xuve/de697917r3v3Xuve/de9Otfr5wlm+TO+9TOwFDtdVDOzBQMDSkKgJIRbkmwsLm/wBffDD75ZJ+8RzvUk0hsafIfTJgegzwGOutn3XlVfZTlbSoBM10TQUqfHfJ9TT18uine8Xusg+ve/de697917r3v3Xuve/de66YBlZTyGBBHPIIsRxyL+9U1ArTiD/g69UjK/F1d/8AGPI7c+THxSzfVfY9FS53FfwbOdR7yxlnV67bVbifs6OaVjGBT1s+HrFKSRlnjliWS4f6dwvube40nP3sztNrfyod32WT6CSlamOJFNu7gimp4SK0JqVLE1NOuS/3nOSI+TfdDcJ7NGXbN1jF5HXgHkYiZFyTRZQeIHxUGAOhE+HG796ZTq7Idbdp138S7Z+P+7Mv0vvvKyWjn3RFtqGirdib/MJIlCdgdcZXE5R5CArVc86jmNgMr+seTSuOjYs6ILuyoLE3YheFFyebfpUc/wBB791rrtWV1DKQysAykfQg/Qj/AAPv3XuuXv3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de6rn7D+4/v3u/7ryeU52s4mB1eG6/bcH/AHX9uF0f7Tb37r3SP9+690UP5A/LDb3U8tbtHaaUW5ux4VVKuGcmTAbRlbxSqu4dLxS1uSkpphJHRxHgEGV1BCn3W9JPl1VNXZTefae7Jq6plz+/d5Z6pZRHRU9ZnMvVyM7SJQ46hoYqhqejpyx8VPCqQwrwAPr7914gilerbvhz/LKzmYyOL7H+S2FOJ27TLFX4XqqpmH8WzNUHjlpp97fbSFcdjoCtzj1cyymwmKrdD7rXV80FLT0kFPS0sMVJR0kMMFNTUyLBBTwU6COGnhhiCxRU8UShVRQFAFgLe/cOvdI3f3Yu0etsLLnt2ZiDHUqqy01PqWSvyVQFZkpMbRA+aqqJCLCw0i92IHPsG878/cre3m0PvXNO5pb2wqEXjLK1KhIox3O54CmBxYgdCXlXlDmDnTc02nl/b3muDTU2Qka1y8j0oqj9p4KCeqqu3Pktv3sytqKXHVtdtDaALxUuFxlW0FZWQngT5rIQeOaeZ1/3TGyxJ9PUeffMz3U+8Pzn7j3lxa7ddy7VypQqtvE5WSRa4a4lUgliPwIQi8O493Weft57J8q8lW8N1fWsW4cw8WmkUMiH0hjaoAH8TAsflw6Nd8H8xn8ntHeMOVy2TyOOxmfoaTERZCaWqSk8tAaisjpqifXNod3QlNbKh+lr85Ofc33Xe9y5V5rg3PdLi4sba+jSASMziOsRaQIzVNCSCV1EA5xXqAvvPbbtFhzJy9Lt+3ww3k9o7SmNQpekmlCyrQVABoaAn506ODuncuO2lhK7O5N7U9JFdIVI81XUt6aekpwf1TVEnpH4UcngH3mT1jJ1X3undOX3jl58xmJy8rlkpaVWtTY+k1s0dJToLKFQW1P+qRuST+Pde6MP0JsQRQNvfLUrCeoD0+345lKmGksUqMl4yFIkqmukZP0jBYfq9+690u+9+69odAdZbi7L3hVpHRYilaLGY8FjV5/P1KMmHwdDGoZ2qK+qAUkAiOPVI1lUn37r3WpD2z2xvfu3fud7J7ByJyG48/MCYI3kONwmOhuKDA4WGV3FLicbE2lFWxd9Ujet29+690xbI2VuXsfd+3NibNxkuY3RurK02JxFDHq0vUVD+uoqpQGFLQUUIaWeVrJHEhJI49+691tpfGH457Q+NHWGM2Jt2Gnq8xMseQ3puk06RZDdW4pY/wDKK2qcAuKOkDeCjhvphp0AA1Fmb3XujGe/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de68foffuvdEk3Xhts/G/uDt3vLO7fmr+rfkRR7CTuLcdNjq/cU+yd29f7dOycBndxYGmpa6aTrvNbSjpaOqqYYnXGVlKsk6eCpklh917rhgO2fg9jqmfP9fwdc5vOY+sgijXrjrufcu4oMlVOYqSLH0e29uVlbSzzSPoV0EaC/qZV59+63Q0rTpuynXfZXyC7Rx/YUdFu7oPrebrvJdebsGUqKfHdw9lbZyGZpM9S4fDUmNq6mXp3HwVsUwqsg1Q2eqIZ/FDDQuiVK+610czae09t7H23hNn7RwtBt7bG28bS4jBYTGQCnocbjqSIRQ08EYuTYC7OxaR3JZyWJJ917pRWFrfj37j17oJ+wun9q9glamthmxuZhj8cGaxuhKkxllfx1cLqaeujBB0iQalvww9xpzz7Vcs896bm+je33dV0rcRUD0rWkintkHpqBIOQR0OeUfcLf8Ak7XDZus23MamGSpWtKVUjujP+lND5joEZvilMZZPtt7BafV+yKjCBpwlh/nXjr442e/5VQP8PcNS/dll8WQ2/OA8CvaHt6sB5VKyqCfU0HUpQ+/a+Gvj8sVl89M1B+QMZI/MnrnTfFJhMprd7M1PZta0mFSOcnSdGl562aNQG+t1PHu9v92Y+Kv1fOH+L+fh29G+VC0rD7cdVuPftjEfpeWVE1fxzEinnhUU1/PpcYb40bBoFRsq+Y3BOukuausNFTOVLn/gNj/AdLBgCC5/T7Gm0fd55GsND7i11fTildchjSor+GPTxqMajw+3oKbn71833hcWIt7OI1poTxGFafieucGlAKV6GzBbX29tuD7bBYXHYmIAAiipYoXf6/52VVEsx/xYk+5f2bl3YuX4Ft9l2i3tYgOEaKpP2sBqY+pJPUZ7lvW7bzL4267lNcSV/GxIH2KcD8h0/hQOQPZ10W04nrx+h/1vfuvdBD2J3Ntfr2SOhqDLmM1IAzYbGyQfcUkTxs8dRXyyuIqWKRlAUG8hBuFI9xbz77s8t8iOlncB7reG/wBAiK6kBBIaQkhUU+VTqNahadD/AJP9ud95wV7mDTb7Wo/tpA2ljUVVAoqxAyeCjgTXpE7b+TOzsq8UGfo8jteeQqpmnC5DFoxZgC9bTKs0UYFrs8SgE/0F/YQ5e+8RyluTRw73aT7dMaVZv1YQSfORaMAMVJQL8/PoS737Jcz2CvLtVzDewrU0XslIA/gaqk+lGJ/PHRMv5kvxSoe3tiP351zRRVvYOyMOJ85Hi1Sd97bDo0eeo0CA6avL7eivPTPcs9MJYgT6AJ7t7iC7giubaZJLd1DKykMrA8CCMEEenUPywy28skE8TJOhIZWBBBHEEHII619Nsbmy+0Nw7e3ltqrNJndsZjF7jwVbGb+PIYuphraVvqNUcrR6WX6FHIPHt8EggjiDX8+mzkEHgf8AiurLO+48Hm9xbc7e2lDHFtHvTa1H2LQxQHXT4zctQfst9YAFPTHPityRSFo+GUTD8e+Hn3y/bZfb33m3PcLKAx7Dv0X10Qp2rMW0XaCgABEtJAvksmMddYPuvc8nnL2vsrK8n1bvsz/SSV+IxAardz5msdUr5mM9AJGRHNJDwA954rmMagW/eCInrtG/JY8ktx9PeKVKdZFg46NT8fPkXJ1c8Wz98xZLcHUtTXjJvjaFRNmtn5gTCdc/tZnKkxyvf7uiJKTKzOoElw+V/wB2r7z27eyd+uwcwma89spnZpIVGuW0kc6jPbAkEqSSZbetGrqjo9Q2Onvt7Bbd7qWh3jZRFbc+xKAkrHRHcIooIrggGjAYimoStAj1ShUmnS0Hfn8uis7/AOpsZ8RPkD8xvj92p8gu0/kf8TO8firjtlbsweU293llBurMdU9zjc27duVXVu7tn7tklhNfWrJRTUkhe4Men32k5c5l2HnDZdv5k5c3SG92a6jDxSxsGVgR6/hYcGVqMrVBAI65ab5se78t7re7Jvu3y2u6wOVkjkUqykH0OCppUMKqwyCQegm3l1l3v1v8ve/PkT2T0/8AzChsn5g9M/HmYYr+Xr2jgtwZTqvsLrvZNdtPevQXc2NpJcM+4cNDLkTPh89Sk4h3mqQyrq1E8pToqycmvQ+9W/F/K9bfLT+WTnetekuxdhdD/Hb4GfJnrVIuw8tjN3bp6R3Xv3P7ErdobA3nu6lrq2KbflZilyMDTULywiISwrIIwR7917otm++hOwOre+v5hm3uz+p/5lXcXU3za7NrOydmVHwf3vt5utOw9nb46yw+w94dSd24DMVmOOyt27akxL0cWQrnNHPh543idDEw91KIwIaNSta0IBFQag0I4g5B4g0IyOvCopQ+VPypSh9RTFOFOhrwFT3D8G/nvurujrb4Md8difHCs/l9fFP474XL9B4bYW6+ytl7o6zzG6spHsmqxWTzmIye96mWlemxm5czQ1WmkqBBUyieFgyINt2faNmjnh2fara0hlmeZ1hjSJXlkNXlYIAGkc5Zz3N5npbebnuW5PHJuO4TXEiIEUyu0hVF+FFLEkIvkowPIdFTwfwe+SXR/TX8tTszP7R+QNdk/jzvr5jb0786i+He+sXie/uocb8xs3mt14KXqed5oqHdtZ1W9bDidxY+gnSaWkqag0zuqtcx6RdDlWfFTcu/9g9Ybq6A+N3zXoN170/myfEb5I92t8sM5itw9ybm211rNTYjcne24MfBmqqHZ+0Mdt6mp6V0qnirpjTeV4ypX3piqqzMwCjiSRQfafLrdDUAKST6Amv2ev5dOHfvxQ+VG3uvu8uycV8f9+7sy/XH88vBfOTa/XWIGIi3V3L0ZtHH7Fo8tX9ayVGSXE1+aq6UVU1DSVM9PLVtSSIoDWv5GWQao3VlrSoIIr6VB68wKNpcEN6HB/n0HXafW/yj+S+V/nGdnwfEnubq3FfJj+XX1Z0v8adn9gx7cXsbtDcG1v8ASLSPisrtrCZvJ0uz98ZCozKOcFPVSz0lJLA0soebxrb0+dD+XWuFenz5Gdab7b54fEHP1WzaxcD0H0fvjrTP1xoaGLH9W1v+hzp/Dbf2rAkcnjxJOSochBAKONU8iSi/JJ4x/eF5ugn5q+9Tse478W3G43zaYbaFmNXtrNZNSIFAUxw+ICQfNgck166Y+zXKbpyz933cbTY1Fqu1blLcSBVJ8a4kQK0hpXVIEoCScLpFFx0HPw42DvLr+i+TcO8Ns5DbJ3f8x+9N/bWhr4oYlzOzNzy7blw246BYZJVbHZh6eZkY2ZmViVuT7gf3f3zad9n9t32ncEuRacobbbTFSSY7iESiSJyQO+MFRTIAIANOpp9rtn3HaIud03GyaHx+Yb2aIEABopDHodafhahpw88dG/MOm/gfxDgeMrrhNlRF9NwyLGq3UIVFyb+4jBpwPUpFfz6dMBuPPbUzNDuDblfldu57HSCShzOFqpIKqmJZxpNRAUaWFkQGSORWiYMAQ3sy2Xe965a3az3/AJd3a4sd8t2BjnhdkkUg1pqU9y+qNVDwKnov3baNr37brjaN726G82uYUeKZQ6MPsNdJ9GFGXiCOj0dffLjaO64xj+9qOoxecOnwdmbOxMSfdxiISKN37Rg8cU1QFBY1NGgdhYMi2uekXsx9/QRQx7H72WbtKMR7laRDIpwurZODVFfEtxpIIHhqanrBr3R+56XlfdfamdVjPx2FxJwJPG3uGwR/QmNR5SMMdL3tzs7pmo6T31s6LtTb3YK5qjp6nZG38JR5V8piN2U9VBV0GXb7mkSLCUq06ulSvk0EMU5JII7+8x79/d9599l+aOXdt5wtd0364RGsooUkaSO5VlaKViUXwQmRIWI7SyZ1EdA72J9nvejlD3T5f3i+5YuNv2eF3F3JKyLG0DArJGoVj4rNgpp4MA9BQHqsaa6hJwG1Q8sqAnVGbCRQnkjUtp5Ba+m3vkgTUk/6v8vXSenRiPjFjcZuntcdb5+kGR2r2/sbsLqzduNk5pcjg90bUyCTJUx8+aKKSAOoB1BrFSD7yj+5pv8Ac7F94Pk6GB28HcYrm0lUGgZGhaVdXqFkjVvt6x9+9Ds0G7ey3NE8qr41lJb3MZIqVKyiNgPQskhH2dXJ/CjdOT3p8SvjxuPM1E9Zla3qbZ9PX11VIJquuqcVi4sPJXVcwVfLU1hx/lkawLO5J599zOuSnRoffuvde9+691//0N/j37r3Xvfuvde9+691737r3ROflB1/vvG5fZXyZ6Pwpz3cPTS1dFn9lUS0lNW94dG5ieGq371EK6cxqu4Kealizu13lfxQ56gSEhYa2oJ91senRi+seytm9xbA2p2d19mIs9s3emIp81gslHFNTSPTzFo5qSuoqlIqvG5bGVkUlNWUk6JUUlVDJDKiSIyj3Wul37917r3v3Xuve/de697917r3v3Xuve/de697917qgz534jIYz5IbmrK2FIqbOYTbeRxcizRyGopIccuOmd0Q6oGWspXUKwBIF/oR74g/fY2q+277wHMV5eQhbe+s7SWEhg2pEiETEgZQh0YUOSBUYPXV/wC6puNpe+zmyW1tIWntLq5jlFCNLNJ4igV+IFHBqMA46J57xN6yQ697917r3v3Xuve/de697917r3v32jr3VhX8ubeTYjtbc+ypWkNNvLbDV1OqgtGmU25MJvI5aZUiWTH1Uo9KMzsFBIA955/cA5vbavc3mPk+Wpt9224yL/zWtWBqasAAYmcYUkkLWgHWHf3y+Wl3DkLYeZUp4+23wjYniYrhdNBRSSRIinJAAJpk9G+7nq5fj18j+vfkWHkpuq+3aPAfHz5AzGfx43bWZky9TJ8f+08hC1o44KPdWeq9rZGqZgsdPnaSST9qmuvX3rmoBj8ujzOsRkjMhGsh441YizG13AQ3B9IP+w9+691gmqpKepgiaG9PPaMTqblJyRoV0FgqFfz/AF9+691P9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3RX++Ng1tZU0u8MLQy1jCFKPO09JGZajxxH/I8gIUDSzKgYxyaVJC6SeASPde6DnrjqXLbrrkrM3SVmK21TsrzyVEUlLVZTkWpKFJAsywyD9cxUALwt2Nx7r3Q67o+M/x93tko8zuzpzrzP5aOnp6QZLJbZxtRWvT0kaw00U9QYfLUrDEgVfIWIUW+nv3XuhD2xsDY2yYTT7O2dtja0Jvqj2/gsZiA2oKrajQU0BYsEF73vb37r3SpkdIUaWSREjjRmkkkKqiIgLM7uSAqqBySQAPdJJEiR5JGCxqCSSaAAZJJ8gBkk4A62qlmVFUliQABkkngB8z5dEs7g+YW3Nsfebf66WDdW4F1wS5u99t4qXlGaOUHVmamBx+iK0WocueR7xD91vvXcv8ti82TkFU3PfVBU3H/ESFuFQeM7KfJOz1c5HWSft393bet++n3XnJ2sNnJBEP/EmUcaFT/Yqw/E3d6IMHqtrc+5dwbzzNTuDdOYrM1lqonXVVkmvxR6iy09HCLQ0dLHeyxxKqge+efMfM2/c3btPvvMm6S3m6yE1eQ10r5Ii/DGi+SoAPWvWamx7FtHLW3Q7TsW3x223x8FQUqfNnPxO582Yk9MErBELN9F9Tf6wBJt+b/wBPZGDpVj5Af6v9Xnw6NqMaKi1JNP2/5vPq5H4xbEq9gdRYWmy8QpMrnJqndOUhdgGpXyixGmgmJOlJKfGwRBx9A1/fW/7uHJd3yR7W7La7hAYt0vne8mQ8UMwXQhHkwiWMEeRr1zd98ea7fm73C3S5sZA+32qLaxsODCInWw9VMjOR6inQI9rb4k3puSUU07NgcPJLS4iNSRFMwISoyRUGzvVOvoJ5EQH0ufc8dRH099TdWy7vq481nKd02vSSXWN9UZzdVGwAp0/Sxx8bf51xw5GgcaiPde6N5uTce3dj7by+59y5KiwG2NtYuoyWWylWwhosZi6GEyTTSaFLBIoksqKpZjZVBJA9+691qp/Mv5Q5f5P9q1Wbpp6in602tJWYjrPCulRShsQ0gE258pQySELns+y6jqUPBTaIvw1/de6KKSACTfj+gJJP0AAFySSeB+ffuvdbFH8sj4nZDqnatf3R2JhDQb939RQQbXxmQg0ZTamx5As4NTFIgkoMruWfTNNHfXHTRxK1mLr7917q2D37r3XvfuvdRqmqp6SMy1M8FPEt9UtRKkMagfUl5GVePbM9xDbIZJ5kjjHmzBR+046cihlncRwRM7+iqWP7B0y/3u2uU1/3iwegErq/idJbUDYj/PA3v7JhzTy6VDjfrLRUivjJ5cfPo0/q/vmrR+5rrV/zSf8Azddf3w2p+dx4Qf8AkTpP+vvuo5s5ZP8Ay37P/nMn+frf9Xd//wCjNdf843/zdS4dw4OpIFPmcTOWNlWLIUrsx/oFWYkn2sg33ZbkgW+72rseAWVCf2aq9Jpdp3WEFpdtuFUcaxuP8nTtHIki6kZXX8MjBh/twSPZkjrIKowK+oIP+AnpCyspoykH5inXP3fqvXvfuvde9+691737r3XvfuvddWv/AL17917rgkUcd/Gix3/VoRVuf6nSouffuvdZPfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Qcdrbwk2RsfNZumKDIeOOgxOsEr/Eq9vBTuQCL/bgtLb6HR7APuZzXJybydu28W+n6/SI4NXAyyHSpp56cvTzp0MOQ+XRzRzRtu1yg/R1Ly04+Gg1N/vWF/PqsiaWeplmqquearq6qR56qqqJGlqKmeQ6pJppWJZ3dje5986ZJp7mWW4upmkupG1O7GrMx4lmOWJNTU8K0GAOs3oYoreNILeJY4EAVVUAKqjgABgAdYyLi3uvzHEdOEV8+jY/GLdk/3eX2JWSiShemfMYmGUKywyeVYspTRarXhnWZZCnIBDEWBPvKD7uHNNwlzunJlzLW00G4gH8BqBLGPkahwvlkgU449++fL0HgbdzPBHS4LiCY/wAQ01jJ+YoRXzFAa0HVMv8AMR+JM3Q3YcvY2z8cIupOyMtLNSx04Hh2hvKtE9ZkNuPFraSPG5HxSVVC5AjUF4B+hb5bdY49J/4l5rHdn7J3Z8XszV09LumtyM/Y/QGQyNSsFGu96eh8W6tiGeQaIIt54qlEkKEhDUoTbUBfHj7zXsxH70e217tdhEg5v28m5252NB46ijwMx/BcpWMg9qtofGnqafYf3Rf2t56tdyu2c8t3iiC9Vc/pE9swH8cL0kBAqRqUfF0mK2grKSoqcdX08+PyWPrKilq6SqjeOoochSTSQVFNVQhlbyU06FXQmxI98Hbq1urK5ubK9tZIL2GRo5I3FHjkRiro48mVgQR6g9dfbae3ureC7tJ0ltJUDo6GqujgMrKfMMpBB+fUQTqP8+DA4vcNygADt6ZQPG9o4yxsfT9D7Y6er0KHWXdvYXTmQqazr3c9TjYa2QS5bAyU4y23c0yRKAcnh5kamkn8EoAmQxzhW4a3uS/bX3e9w/aPcJr/AJD5jktI5nDTQOBLazEYBkgchSwGA6FJKY1dALnr2y5J9yLOO05u2RLiRFKxzAmO4iByRHKuaVzobUhP4ejYbf8AmJsncdSlP2l1vLtGpm0rLvXrSV6mmilOlHq8psutkZ5aXUGlf7WUyKnAUnj3nl7ef3hEUlxDY+6XJggtzQG7sHaQLgAvJayd4FckRSOwBACHrEDnb7l0sUE957f81CWYVItrwBSQSaKtwlFrwH6iKCc6gOjB1VHDDS4fK4/I47O7e3Fj48xtrceGnM+LzuLlsqVVMzASwSRuSksMqrJE4KsPfRvl/mDZea9k2zmPl3co7vZLyISQzRmqOh4EeYPkykBlYFWyCOsId52bdeXt23DZN7sJLXdbWQxyxOKMjjiD5HyIIJDAgg0IPXHH0FflK2nx2MppqytrZFhgpYBd5nIPFiQgRRyzMQqqLk2Hs4/w9FhNATQ9OdXtvJ0dPX1ST4XKQYeoWkzMm3s/ic8cLUyMyRwZiLF1E82MaRlKr5VUEqRe4t7J9s5h2De5byDZt9tLue2cJMsMscjROa0WQI7FCaHDaTg46M77Zt32pLSTc9ruraKdNUZlheISKOLRlwA65GR69J9m0i44YHgi/DGy3IH+P+x9nHRb0Vv5hdh7hpt5HpDB1VXitp7HxuGlzMFLVTUibs3RmsdS5iry+ZkpfHLkoKaOsENNA7skIUsy3YH3x2+/B7uc0b57l7h7YQXs1tyls0cQeGNygup5ollMs2gjUqK4SJGwCHbSSQR00+6d7b7BtXIllz/NaQzcx7m0hSR0Dm3gjdoxHFqBCF2VnkdRVsKCADUunXHbm+eqc0mU2/lak0M7mHMbbyVXVZHae5sfI5MuPzeNmkeONZYI7/dIqTU7WKSXFjjH7Ze7vPntJzBa7/yhvkyBWHi20ju9tcoDUpNGSRkYEihZEIBVvLqfOfPbblH3G2WfZuZNqjYlT4U6Ii3Fu9KLJDIADjzjYmNwSGXNerKdu5vB792pD2FsWaeXbz1UdJkqKpJbL7J3F4453weaWw88MbuDR1yXhrIdLXD6l990PZ33f5V96uTrbmvlmRklUiK6tn/tLW5CqzxN/EprqjkUaZEIYZqByQ9y/bXmH2s5muOXN+jVoyC9vOn9ncQEkLIn8JxR4ydUbgqcUJLz80dmNkMvtvvnB0hbD7vx+N2pv406amxHYWHp2ho6muhRPKIs7iYgqTn9rVTrchnF+dX39Pai92nnDb/dXabEnYtzijt7x1qdF5EG8NnQcFmgULr83jAahZa5t/c+9x7XcuWLz253O9H73sJHmtVbi9rIR4iKx4mKZi2njpkquAaEeH+PPJsb3/2IPvnzUUFBjrNED5/yp1737q3U/HYvK5eqWhw2MyOXr3SSVaHF0VTkKoww2aaYw0kc0iwQrbW5AVb8+1+2bVuu9XaWGy7Zc3l+wJEUEUk0hA4nRErNQebUoMVOekG4bntu02zXu7bhBa2SkAyTSJEgJ4DVIVWp8hWp8us2b2/n9vOtPubbmdwDzJ5Ejz+FyOJ8sTHR5I/4hSwB42bi/IP0933bZd72GUQ77sl7YyEVAuLeaE0rSv6iKKE4Hqemdt3jaN6jMuzbvaXkYOkmCaOYA0rQ+GzeX7PPpkEJjOuncKGIZo29URDOHkdLEMJGUWXnQv8AT2XVqB6dGNAMDh1xdZ5lMboIVbT5GDpKSrLqZUUoVZdY0MWAuCSB9Peut54dCb1Bu4bC7V6+3ezlKXDboxjZEILscRXy/wALy6KNLEAY2texUahbjnkSB7Uc4NyD7m8ic4gnw7DcopJMVJhc+FOFqDU+E7HAr6Z6BPuTy0OcOQOcOWdIMt3YSrHXH6qr4kXp+NBxx69XZfAzyUvxT6q23VMBlNg0+5+s87BZVemzvXG9Nx7MykUiKqaC9RhfIvA1xur8hgT9GFUbujYGM5BHAg5B/MdcSe4YcUcYI9CMEfkcdG/9+691737r3X//0d/j37r3Xvfuvde9+691737r3XR+h9+690QH7hviF8gBQzU9JRfGf5W79M1DWU6tT0fTXyg3Ko+5ociCWip9k/ImtpxJTSKUjo95eSNl/wBzEZT3W68Oj/X5t/vv+K/X37rXXfv3Xuve/de697917r3v3Xuve/de697917qmH+ZLt9aLs7Y24xVtK+e2pVUBomiVVpRha/WJklDEzfc/f2IIGjR9Tf3yN/vCdkWz9xeS9/8AqizX21vF4dAAn08tdQP4tfi5Bpp04rXrpH9y7dmueSuadmNvpW13BZNda6vGjppp+HTo/OvVc/vAHrMzr3v3XukhvrsHYXV+26zePZO9dq7A2nj3iirdybxz2O27haeec2p6eTIZOengNTUEWjiUmRz+lT7N9k5f37mXcItp5d2W6v8AdHBKw28TyyMBxIVASaeeKDzI6KN537Z+XrKTcd83KG1sEIBkldUUE8BViBUnAFany6btldtdWdk7VqN89d9k7F31sujFU1durae6cPnMDjhQwNU1v8SyNDVywY5qOmUySicxtHGNTALz7f3nlXmfl3dI9k37l69st4cDTBPDJFK2ogDSjqGapIAKggnAJOOmNr5n2De9vfddo3e3uduUEtJHIjoukVbUysVUgZIJqPPpp2B3v0f2vXVmK6u7k6t7HyuOpzWV+L2PvzbO58nR0YcR/eVGPxGSqqyKk8jBfKU8YYgE3PtXvvI3OvK8MVzzNyhue3Wrmivc200KMaVoHkQKTTgK1PkOk+085cq7/M9vsnMVld3CipWKeKRgPMlUdiAPn9nHoVhz7C3Qm6FPo/eMfX/cHXW8J2p0pMRujHrXy1QqGhgx2QZsZXVDJTESu1PT1jOgAN3AuCPcl+zXNsfInuryBzXOyLa2u5RCUsGKrFIfCkYhO40V6gDFQK1HQA90+XJObvbvnHl2AObm4sJPDC6QxkSkiLVsAFkoa+RNM9bE3ZnXW0O4uud49Y75xyZnZnYG2sntnP0LHQ8+LzFHJTSTU0xUtSV1KZFmp51tJBPGkiEMoPv6I1ZXVXU1Uio+w9cUGBVmVhQg9Fe+Gfe7b5pezPjxvzNRV/yA+I258d1R2n5RLFVbuwsuEo8t1l3FRRVQWWXGdnbMqIKmoZC8dPm4a+lBPgBa3XiPP16O9a/1/wBf37rXXvfuvde9+691737r3XFmCi5vb/D/AHv370x17zA8+khvTf2zuvcJUbh3tuLF7aw8CtqrcrVR06yyAXFPSxMTPWVT/wBmKJXkb8D2F+bedeU+RNom37nDf7bbtqTi87hNR/hRfikc+SIrMfIdHvLnK/MXN+5RbPyxs1xe7k/BIVLUH8TMO1F9WYgD16r03n/Mq2lRVjUvX/Xeb3TTQylTls/k4dsUtRGpYGSjo0o8rkmD2BUzJCbXuo94Ic4/3hXKu33j23I3Id5ucCvQz3UwskZfIxpomlIPEa0jIHEDrL3ln7lfMN5bC45t5vtbCUr/AGMEZumU+jPrijqOBCs4rwPQUzfzK+xGqZGpOr9lxUJmVoqepzebqKxIAV1Ry1UUFPBJMwBs4iCi49J59xhP/eI8+NdSPbe2+0LZ6+1WuLlnCYwXCqpale4IBWnbjI/h+5Nygtui3HPW5G605ZYIFQt6hCSwXhUFiT6jpfYD+ZljHqUTdPUeVo6Zp1DVW3tz0eVenpdJLu1LkKDEmonElgEV1BU31C1vY52P+8U215gnMvtdcw25YDXa3kcxC07qxyxwlnr8IDCoPxDoI7r9yPcEi1bD7g28koU9txbPEC1RQBkkl0qRWpIJr5HyMX1Z84+lez8/FtgTZvZeYrZFixEe8aajoKLLzMjN9tS5Kkrq2ggqyV0pHO8TSsQE1Ege589s/vle0HuTvcHLomvNo3adgsAv0SKOds9iSpJJGr4oqSMhckBNRNOof58+7B7l8ibQ++NFbblt0QLSmzZ3eFQQNTRsiOyZqWRWCipagFenD54d/b++Lfw4+R3yH6s61m7c7B6d6s3Hvna3XkaZKRc/kcRArlqyDCwVWYqcXh6ZpK+ripENTLS00iRWdlIywGRXrHTot/wA7m7b7A6gyHyX7W+dPxc+WHxy3P1zTb+xnYHUfU46fpercxjaWXMb6wuay69n75xtVtHa+KVllXLw4/O4+amf7znUqb690pOh/wCaf8bO/OzNldXYvbXf3WeS7e2xubevx93P3f0jvDq7YvyO2ltCijzOeznSu589D9ruZKbbU0eXjpalaCvqMRItZFA8F3GmIVS7miAVJPAAca/ln7OtgFiFUVY+Xn1Vb2h/Pb+MfyBy20Nn9aV3ceA6c3t2BH1MnceY6m3ht/qut7YqspX4TH9a7x7GqoIcRt/J5LJUBjp6dTLTyPLEJ50d1iGDv3id89yuaxvvK3Lf01ty7YwNPLBHdx/vDcLZArNcLApLizUHUEqHmALMNNAMqfZfbOSOWpNp33fIprnermQRxymBvpbOUsQIvEZdBuWIoW+FCQgIJqSxdU/OOn7F+WHfHxjqelO5tvQ9S5PamKw2/avrDctLtmdshsvI7m3BW78zdS/8K2ljKqpxxTbU41Lm6VllULqA94o777ZnZ+ROVucY+Y9uke+WRmgFxH4g0yiKMQIO6QgPW4Wg8Igg8D1kLtHuLJuXNu/cvTbLdLBbSKEl8NytDGXLOdNFDkdlT3VFOhepflz0tWdR9J93wZLcJ2B8gd87G6762q225Vrlazc/YufyG29sQ5jEmQzYailyuLmWaZ2ZYEAYjn2HX5A5hj37mLltki/em12s1xONY0iOBNcmluDMFIoBxrjgej+PnfZZdo2XelaX6O/uY4I+xq65W0qGFKgV4k46YMP/ADGviHsjuffO2OzV7mrNqfGzsTD7d+Tm7dqdL7z3V170hBXzYRNubo7W3fi6SbFbb65z2ZzVPTDIapW0xVDtCsMTye5q9lvu/wC+82b/AMpb/vsFuOS3Rbxv1lLyxpIypH4YOvMqASVFAhpUk06iz3P96tp2HY+YNs2aST+smprZToYBGdFYvqpQkRuSmfiGaUr1bPtn+al8cvkXvfKfHzqH/SlhN7Zzr+r7Q2Dmexuq929bbW7s6Noc0228v2r0Pndy0NHHv/ZSZQpAKuKOBmgmWojR4HjlbqTTT26aU8vTrATNTU1Nf5+fQ19YbCffOfSllVosHjVjqcxKhKl4i2mLGwuOUlrdJBP1WMMfrb37r3R9Ujx+Hx6oopsdjMbSm12jp6Sjo6WLUzO7lY4oYIoyWYkAAXJ/Pv3XutbL+YB82KjvvcdR1l1rl5oumNs1umqrqRpIP9I2fo5CGyc7AI8m18bKpFFF+mpe87i3jVfde6rTt/jb6/X/AA/3n37r3Vq/8ub4XS9uZ/Gd6dkY+nk6t2xkpX2tgq6GVn3xubGTL4MhJExSM7Z2/WIHu2tayqQJbRG9/de62K2kSGNnldY441Z3kdlSNEUXZmZiFVVX6k8Ae6u6xqzyMFQCpJNAB8z5U6sqs7KiKS5NABkk+gHn0DW5u79sYaV6XFRT7hq4yVdqN0hoEYfUGtkDCTSfr41ce4l5g94uW9qkkt9tR7+5WoOjtjB+chHl56QepF2X2y3zckjnvXSzt2FRrGpyPkgOK+WojoGc13VvPKB46GSjwUDGwFDF5qrT/Q1dVrsf8VRfcRbv7wc3blqSzkhsoD/vpdT/AO9vX+Sj7epK232x5bsSsl0sl1MP9+HSv+8r/gJPQX1+RyOUkM2TyFbkJCxbVWVM1RYn6lVkdo0Jt+FHHuOL7cb/AHOTxdxvprh61rI7N+dCSP2AdDm0sLOwUJZWcUKD+BQv+AAn8yeoAjj/ACi/8kr/ANGj2i0R/wAP+DpYXkPn/M9eMaH+wtv+Cr/0b79pj/h63rf1/meveNf9SB/iAAf94A960Rg1CCvr/wAVTr2t/Xpzx+XzGKkWXGZXI0DrbSaatqIlv9b+MP4zf+hU+zOx3fdNrkWTbdxngdeBWRwP95qR0XXe17ffqyXtjDKh46kUn9tK/wAx0KWD7u3djCkeUWkz9KvBFQn2lcVtbirp1KM3+Lxn/X9yRsvvHzTt2iLckjvrYfxdklP9OuCftU9AXdfa/l++DvYl7Sc/wnUlf9I2afYwp0YXZXZe3t56qWkM1DlY0MsmMrdImaMEa5aaVCYqqNC3Ok6h+QPc8cpe4Ww83M1tZu0W5KtTFJTVQcSpGHA8yMjzA6iDmXkzd+WAJroLLYk0EqfDU8Aw4qT6HB8iehEBv7HQz0Evs4dd+99e697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de6Kb8rKqVMPs+iGgwVOYyFVJcesSUdAFi0tewUipa/wDXj3jH95i6lXaeVLIBfBkupXOM6o4wFp5fjav5dT37DW6NufMd0SfEjto1HpR5M/8AHR1Tv8gPnB8efjPvfbHW/aeW7AffO7to5Lf2H2111092Z23lF2ViMxDgMjubKU3XO2dxy4fFU2XnWAyVAjBdgByR7xr2TlHeuYbSa/29IBZxyCMvLNDCviMpYIDK6VYgHArw6nLd+bdr2S6is7rxGuHQsAiSOaAgEkRo9ACRk0GenbF/NL4w7g+OW7/lptztrC7h6B2Di89lN672w9FmKmp2sdreFdxYXcG1WoId1YfdOHepjWfF1NJFWoZF/bswJbk5T5gh3615an25k3mZkCISoD6xVWR66GQ+TqxU+vV4+atll2ife47sGyiBLnNVKgkgimoEUIIIBB4jpRfC7+YP8Ye9u9MBs3rXfefoN/Y/Fw7qrNgdldedh9O7zyHX2XQ0D7yweA7G23t+t3PtaGSqi8lTQrURIxQMVDA+5D5J5d5j5L545P3m9tVG3S3wtnljljkiDSgxmOR4mcK1SCFalWUDoDc2cxbJzVypzJttvIwvFtDOqOjKx8Iq2pBIqE+YqK0BJ6Opi/mH8DP5gvw3797V2d2nj9/fH3rFexcT2zuKlwmfoc/1xm+qMdLuHN5GbbWRxlJuWmyWDoqZMlQMlMy10BRoTKjkHPIgqxU8R1iOCCARwI6oB3tsPPdS13Wu+Nt7jnzey+wts7W7e6I7cxFFX4ODeez8xRUO4Ns7lpKKsWDIbfz0NJWQPW46cLUUzOAQY3VjrzBpWh/1Z8vt62PmMdHwfJUfyt2pkO39nx0kXc+2cXBN3v1nQxSRVmcWhjSlHbWx6MsTX46vpok/ilHEpenmBYX/ALfM37433YNy3C73T3l9vrLxp3j17pZoCXcoBW9gAPcQgAuIgKkKJV1HUDnh92P39s7C0272w50uRFGG0WF07AIoY1+lmYjA118CUmg1GNqDSQAqMHW6nUv0vwQfrcW/BBBBBsR75g1U/CwI6z9INaFSD/q/1V4de4H04H++t791vr34J5tYk2F/oL2t+Sbe/A0IPWm+FurFPinl8jlvjxurFZKQ1NHsrsrEjbRk1ySUMO6MPLW5ekSaVEZqWWqUSCMquh2Jt763f3eu/wC63/t1zpy/dz69r23dU+mB4xi4hEsiA8NHiVdV4qzt6jrm599HZtus+duVd5toBHuF/tzePTAYwyGONiP4tHaW8wo9OhJ7a3Bnuqeht37yosVl48pvkxbGwebpKerji21hq3W24NxVmQgjIxcdZEgo6SVmTyTOCGt7l374PuJu3t97Nbz+4be7/ee6EWYuIVfTZpID408sqgiI+HWOIsVBkkXOD1G/3Z+Sdu5z90drG7TwfQ7epuvBkK6rmRKeFCkbH9TvIeQAGiIcZ6rP6z3zuDq/dGM3Tgq3KUcUVZj33HgcXkf4dS7rwNJXR1tZtjNlIp48hRZFFMcpcMXBsCoZiePftH7n8wezXOe2848sEnw2C3FvrPh3luWrJBJQZJBrHIQWSRVZaVavTD3J9v8AZvdHlW+5Z33AZawTadT20wHbLHWhAB7XQEK6VBrjq0mpSgkocPmcNXjL7c3FhqDcOAzCwrB99ja2ITFJ4Ih4KbIUM6yQTwISIpE039/QTypzNs/OnLOxc2cv3KzbJuFsk8LDjocV0uOIdDVWByGBB4dcZuYdg3Plbft25b3qEx7tZTtDKv8ASQ0qD5qwoyeWkgjj0Sb5kY6Sj+Q+7a9gxpd0YPZW6MZKYHhEtDW7ZoKAC8jP5pYZ8a6uy2UH02uD74q/fQ2efa/vD84TzV0X8FpcpggaWgEJFThiGiaumoFR8+uqH3Wdyg3D2U5YiiI12ktzA4qCdQmaSuOAKyCgOeJ6BTa3Xu8t8UWfrtr4N8tQ7agppM3N93QUixNXif7GgpUr6mnbKZWvWmkMFHTiWomCHShPHuEeT/bbnrn615ivuTOW5r+12i38e7aMoPCjozCgZgZHKozCNAzkKSB1K3NHPvJ/JdxsdpzTvsVncblN4VsH1fqPVRxUEIoLKC7kKCRU9OfUnbW4umdyf3hwipl9v5SBcdu/aNdNKmL3Ng5SzPSVOlZmgqYfIDRzRxh6OT1AlCyk89nveHmj2W5ut+auWJjLbNpW7tGYrDdwcdDYIV1qTFKFLRt5FSyko9zvbPYPdHl2fl3fohHcLqa3uFUGW2l4a04akNKSxkhZF9GCsLMsHntk722Xkq7HUp3j1L2HSPt3c23a6f7TNYesAWobBZgxsTit14J/3aOqH7VXGolQlSdPbHlzmX20+837V3vgILvlrcYTBd2rnRPazUqYpAtTHLE1HikXBosiEg9cqt72Lnz2F9wbXxT9NzBYyCW2nUaoZ460EkZOHikWqOhyp1I4B6IP298d9y9X0k269vT1e+uqXmfwbppaS2W2ol2ZMVvvEU4Z8ZUU0YVFq41emqOGDLfSOSvv1913nj2UurndoEk3b2+ZiUvY074FJ7UvY1/s38vGUeC57qoTpHR32e+8Byp7qQW+3S6Nu5zCgNaO3ZMwHc9pIx7wePhMfFTgAyjV0X9GV1DodSsLhhyCCLi1v6/7f3jJ6HyPU+hqkinDoTurezJeschuaaTaW1t8YbeO08hsrc22920882MyeAyc0EtbSl6V0mjFQIAjj1K6cEcD3MHsp7z8wexnNd1zZy7tVnd3U1mbZ0n1gCMujlkeMh0clACcgjBB6jP3V9rdn92+XbblzetwubW3huhOjQ6SdYRkoyONLLRq+RBrQ56WeN7M2LhYDS9cnuv48U3lWV8DsXf1B3X1FUmyho8j0r3vQZ/AU1KwAHjw9Ti2VFsjoxJ95s7b/eD7Bvcf0HuN7RtNZPhjDcRXSkf04rqKP8gGIp59Yo7j9zDedqkF7yP7jIt4uVWWGS2avylt5H/OoHTRn8JitzT02VwlFszdMUqPNmKnp3KUXXW9mcXkmqa346dx5ykwLVY5Z021vWdJWNoaO5VAG9y9sPuf+980W7+2/upHyfzFOWaSyuURIixyQLa5eLw+4/8AEacoa1VeA6N7P3C+8z7Rxvt3PHIL8y7JDRUuoSXcIMV+pt1kaQEDHjwh/wCJqVPSfGzNi1c8WPou7+v9t7glaKEbM7yx26vjzu9aljaWlWj7OxVBhq6aJbFfs6+pWVTqQlefYT377gPu3aI9xynzJsW92wIFUkeBzXz7vEjx5DxKmop0J9m++X7c3MiQ8x7Bu+1XBrXUiTKKcBgxvngf06D16f6f48drZaCSTGUW0K6i8bl8zjuz+uq/E0sIRnNbPkaPc0kMMEUIMuokHxrf3Ek33QvvG+N9H/rbyh2JUP8AU2uhc01Mwm7QPiAPcRwFepJT7zXsaIjcHnmMhc6Db3Ac4rQKYsny9K8cdWc/BjeuAze/vlttjYe+9sdo9cYPsXrXdOJ3vsrPY3dG2DvPf/UW16zsrbVBnsPV1+NrJ6DcOGXKzokzyRPm/wByzH33K5csrnbOXeX9svZhLe21lBDI4rRpI4lRmySTUrWpJOcknrktvFzBebvu15axlLWa6lkRDQFUeRnVSBQCganAcOrFfZx0Xde9+691/9Lf49+691737r3Xvfuvde9+691737r3SA7R6z2d3H1/uvrHsDEpm9nbzxFRhs3jzJJTz+KUpNTV+PrYStTjcxiK6GKroauFknpKyGOaJlkRSPde6L/8Uu0915Om3l0D3FXmo74+PtZQ7d3Pk6lyJe0evsikzdYd4Y3Usf3NNv3A0hjyvjBSj3JR5CmOkImr3Xujf+/de697917r3v3Xuve/de697917r3v3Xuq1/wCZThKmq672FuCGmganw+85aKuqz4lqYVy+KnjooI7r5pIpqiAlgp0gqCR9PfPn+8L2a4ufb7knfIrZWt7Tdykj9utRPCwjUY1FWde4A0BoSOHWZv3LNzhg515r2mSdxLc7aromdJMMql2PkCFYAYqa06py98leukXXvfuvdVn71wmD7S/mibQ2L2li8fufZ/VnxEre0+odnbjpYMptmbsbcPYzbW3hvmLBVyS4/K7jwO24oaOGSSOVqOKoLxhGOr3kfs15ecsfdn3fe+WLiS23Xc+bFs764hYpKLSK08aC2MqEOkcktXZQR4jIA1QKdY/7rbWfMHvvYbVzEiy7dY7Ebm1hkoYzPJc+FJKFbBkSMKobJVXNKE16D3uHZ21esflR8h8P1bt/C7SxPaP8truXendu3tq4+lw+Iqdx7TydZg+s97ZPDY9afHU25Mhjsnk6EVHijkqqenAYvpv7EPKG7brzL7ZcgXXM9/LdXO2e4thb7dLM5eQRTIsl3bo7ksYEdIZdFSqM3lXoPcz7btuxc983W3LttHDBe8qXUt5HEAqeJG3hwysq0XWytKmo9zBck0wUfoOk3rsqb+U72z2x0P1v091Vi8LtjrfafcHTucxmf3/2Hvbt3q+Hb3Xu3+4KGLAbYrcHsLczJJV1sUbZiRcqsRd0QM7Srz1NtG8p96PlblbnjcN35mlmmvJrDcI3itLS3sb0yXctixklWS6iqEjZvp18DVpUk06jrlRL3arj2P33fuWrbb9jWCKCO6tHDzTTXVsI4VuAI4ykMh73AMv6oXIyTsU2K+k8FfSR/Qjgj/YH3z6qDleB6zkXIB8j17zmlaOrVQ70csVYisTpd6WVKhEe1vQ7RAGxvY+9rMbZ47pV1NEwcA8CUIYA/IsAD8q9VeEXAMBJCyKyE+gYaSftANR8+to/ZmWlz+0dq5yoijgmzG3MJlZaeJmaKCWvxtNVSRRs4DskbykC/Nvf0m8o7lNvXKnLO8XEapNdbfbzMq8A0sSOQPkCcdcLOYtvTaeYN92qOQvHbXk0QY8SI5GUE/M0r1VZvrqjfC/M/wCSfY3R0VJjO+tkbR6F7i2XTVVRFjMR3Ps3N4XdGwu0Oh94V06yU0e3t4J1bjZcdXvGzYDcIp65GCGpjmEXRPXqxXo3vPY/fmy03ds+bIUNZQVk2C3psjclIcPvzrXeWP8A281sbf225mNZgdy4apDI6NqhqI9NRTSTU0sUr+690Mvv3Xuve/de66LAcE2Nr/7b3qorSuevdEW+UnzIxHTX3OytkR0O5ezZI4mnSovPgdpwT6v3s1JTyo9RlvGNUVCpDeoNKVThsMvvJfez2j2jE/KPJ6Rbh7iso1Bu62sVavdcFWBaemUtlIY1DSFVFGyg9i/u5bl7lGLmTmZ5bPkcMwBWiz3RA+GEMDpiJw0xBHFUDNkUv7z31vHsbMy7j31uPJbmzMrMy1ORmvDSK5/zGMoIwlDi6VQBaKCNFH+v75Dc487c2+4G8ycwc6cwXG47sxJ1yt2pXisUS0jhT0WNVA+fHrpXyxyly3yXtUWy8rbLDZbaoHbGO5qfikkNXlb+k5J+wdJT6f77+v19hb7ehHpHGnXvfuvdde9UFa0FevUB4jroqG4Iv9Dz+CpDKR/RlYAg/UEX97ORQ8P9X8x5HyORkdaCgGoGej59GfMTuraPW+79hbW2JH8hOzcXQUFf0v17uPfmJ6+qd7R0tdTjc/XcfYO4oK3F0ucfbpnqcEMgvhqKiH7SWaNHR06b/co+8TuF3uP+tBz/AL687SRg7VcTsGaqA67GSZyHcsAHti2pqh4yxGgDAb71PsjZ2tovuXybtCQqjkbjFECFoxAS7jjAooBxcUouVcD4j0UHZ/8AL67s+TncPzu7A2R8WMh/K76W+UnwR7L+Nu8esM/urYWQrO+vkz2A864XvzcnWPTue3PsHZ1B1ph5Jce+Xp5o81uJK6XyxmMXPTnyB6wL+fSV6y6Z3js2PoGXuf4EfMHrztX4+7eqNvUPefcfzhm7e6B6538Osa3rLMbm6K2XU92bwy+6YN+bcaroqBH2/jP4dja0+ZYjDo9wV95DmqTlX2k5jltL7wNxvjHaREEhiZn/AFNJHAiFZO7yBPUp+zPL6cwe4G0R3Ft4tlarJcSDy/SXs1D08Qqf5UPVF/xo2l8kvkx/L66G+Jm3/j9Jh+s8739Lvar+UD792nUbGw/VfXfyd3R2Bnal9qfdx77pO35c3hHxVLQxUktI8UiVX3SgvGmK/N97yjyb7oczc73PNIfdYtpEA23wpRM1zNt8cEY8Sngm10OJWcsHqCmjgep45ZsuY+ZuTdr5YtNlH0Mm4mVrvXGVWOK78VqqCJPFJTSFCaanVrAFOrcNpYDt3Ynzw+SuVqen9z57p35O4DpPIYnujDbi2mNt9e1nV3W2b2ZuDB732/kMvS7skydXXfbyY80FJUxTLP6mTQ1oK3G92DdfbHlCKPfYYt/2aW7VrR45RJOLi4WZHidVaLSo1BtbBscCCOpZ2/b972nnrmMHZ2l2rco4WWdGjCxtFEyMrKzq4JIGkqrCjZpTojeweiPm/uTq34U/DSi+J+Tp1+NPy06e3FvDt2q7J2PU7Z39sTYXa2azOF3L1Tt3HZKfcWQNZgculdmEycVCcZDA8cYnlIAl6C75O3vnHnDf9h5qN5v3M20XFvZ2EVvMZkurm2WNormRl8ONVZdKMhdSCXcxopPUfNb8y7PsGw7Vu+yC32bZNxhnubp5YwhhhmLB4wGLOSoB0MEYE6QHYjq32r/l0/J/E/DH+fV05j+tIczvr5gdr73zvx6RM1tambtLCZnpLrLbmOqkqp8qIcFSR7mxVfTRR5SSmdPEzlVVwTn37b8nf1F5L5Z5cmnSbcLOySGSZV0liCzlQeJRGdgvlxPn1iFzdzEeZuYd13iOJoraeYukZaukUVK4wGYIC1OOK8OhG7I6e7Szn8wj4N9yUu2RL1h0T8OO7+k9+bijyWJEG2d47hw/VlHt7b6Y9apa+vTJ5PbmRImpopqZDFdnUFbjnhgcOgz1eL0RhI8ZsSmrygFTnqqoyM8lwWeKORqWkQlfoI4ob2PILH37r3VcH81D5P1mzduY34+bHzE9BuTetIuX3/X4+fxVWM2Qxlgp8AJoXWWnqd01APlt6vsoXX6S+/de61/hwNIsFHAAFgFXhQAfoAPfuvdHD+GPxQznyj7IjoqqKroerNqVNNVdi7hp3WCRomBqKTa2JlZW1ZjOiPS7KL0tMXlNm0A+63inz62qsPhcJtPA4/BYKgoMDt/AY6CgxuPookpaDGYyghEcMEMS6UihghT/AHi5JPPukjrFG8kjBY1BJJwABkmvyGetorO6oikuxoB6k4A/b0UXsjsyu3dVTY3GTS0m24JWjWNCUky5RgPuqsix8BI/biva3LXNrYj+4HuLd80XM237dO0XLyMQAMGcg/GxGdB/CoxTLdZIclckW2wQRX19Gsm8uASSKiL+ig/i/ib8h0E4AH0Fv969xdU1BGCPy6kPSONM9d+9db697917r3v3Xuve/de697917r3v3Xuve7LQMCetdSKStq8dV09dQVEtJWUkqTU1RESGilQ3UjmxQ2sy/Rhwfai1vLmwu4L2xuDFdRsGVx5H/KPUeYqPPpi6tba9tp7W8hWS3kUqynzB/wAvofLo9HXm8od6bfiyDKkGRpn+zytKh4irEUMZIx9RBUodaX+nI/HvM/kXm2Lm3ZI71gEv4jonT0kAFWH9FhlfzHl1izzby5Jy1u8lnUtaP3xN6oTwP9JeB/b59Lz2NOgv1737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+690Vn5TYqep21t3MRCQxYfMywVIUKUWPK0vijmdr610zQKgsCPXz+PeN33k9qlueXdg3ZNRjtbsowFKUnWgY+fxIFFPNs9Tj7FbhFBvu8ba9A1xahgfUxPUgfkxP5da/meyUGN/nBdfTTZKnxiH+Wh2aPNU5CHHo1vkzsNwvlnmgVioUm1+LE/j3j3bRvJ7X3YWMsf39FwFf8AiLL6f0qcM16lu9a3T3Bt2udPhfu966uH9rH1W58qqzbef6z/AOFCe7+p6vC1/TOS6c6XwmUy+1JqWp2Xn/kpgeu6sdx5TE1+NkfD5LcFFiazBUuenpmcvWRqsrGVGAHnLizwX3srb7krDdFuJmCuKOts0q+ErA5VSfEMdRShqMdAbcjbywe41xt4/wB1xjHcPgMgjfX6VOY9XnUUOR0YD4wUfcSfzOvjNB83N37H3LvvF/C2trfgtnOmNrV+yOss3tnLU+0qT5A7d3zSbpyec3llO3sFi4MVNS04r3xIxhmqYYkkJCq7B9qaxsDylbSpYtzRaC/WdtcoZZ62zpoCoImJcM2nXqABJAr0ivI7hJd0bfiouv3Ncm28MaUaqnxNVWYlzRGAB06CTQMCABfWmKo/hD/K+6n+aWKqZMB0H85fhX8hfh98x6CggkG3sT3ZL/pvoviN8ksxBTF46XITZRJ9hZ3IFFSSmy2MaUloQ3vNByTJJX+I/tqa/wA+seFwqj5D/B1tEfFXobr35Bfyv/hX1/2Ph0raU/Er4+T4nKU+iPN7XzUfUm2IYc1gq4AvTVlMx5HMUyXSRWQke69b6pO7a6t7l+FXdSUkGZzG3c5iZ6mt6+7Gwamig3NgnfQKqkciSleV4GEWQoJPIqtdWUoQx9kV08f9XHr3oDwr8/8AJn9nQ34v5JdH9wqsfeu0KrqXsCZw1V271Jio6/bOeqDo8lZvPrZ3UwVU5DNNUY9yxZrgW494l+733O/a73Qkut32yI7DzZKxdrm0RfCmc8TcWzUjdicl49EhOSx4dZHe233mvcD2/SHb72Ubxy6gotvcu3iRJ5CG4FXWnkjh0AAAXpe4zouk3VE9f193r0DvPDCnkrPvpd+xbSyNNTK7i+S27uGlgytHOqrdxpKo3FzwTg3vX3B/e6wu3g2i92XcLTOmUTvbk5p3RyRsVNM4JXNK46yy2r74ntVeWol3Oz3Wyuxxj8FJhWn4XR1BAOMqCeNOmSXq/Z22Vr67uTuvrrYe3sZXVdLLjduZWPfu/d1x0Urxk7W25h1ZRjcqia6OvqJI45BZtOm/s65R/u//AHR3Lco05137bNq2UU1tBIbudh5iNNKRq3lWQkAUOknoq5l++XyFY2Dvyrs1/uG7EHSsyi2hU+Rcku7ZyQgyMagekBvT5wZrHbcHWvQG0qPrDrqgrTX0uTzhh3J2BuHLMPHU7m3HUSp/A1yeQhVVWEQypSx+hDwLdPfbP2x5R9pOV7blLk2xaLbUbW7udc08pADTzyGheR6CuAqgBUAUADAbnvnvmT3G5huOZeZ7tZL1wERFGmOGNfhiiSp0otTTJJNSxLEnoEOpfkt2b1Ruunzj7jze7tq1NYTvbr/cWUrcxtfeOCqTLHlcXV4mvmnoo5JKWokNOY0QRzBbCwA9jXcdusN32+82rdbKK42y4jMcsUih0kjbDKysCGBBIoft8h0F7O8utvvLbcbG5khv4HDxyIxV0dcqysO4EfIivDo0vc+0tv7T3qsuyZJp+vd6bewXYPX1ROfI52vuuiFfBjTMrSLJNgq3y0b+pmXxAMdXvgd94z2tg9ofdrmLlPb43Xl+QJd2ZYkhba4DERAn4vAkDxAgntVK8euw/sj7gy+5Ptxs3MN6ytvMZa2utJGZoiB4hA4eNGVlpQdxamOjIfFDcGW3Ftzf2ychkKOXH7AwtFu7adBLS1Jy0MGVzxpt1U9PXNWCh/gkXljnNOKYTiok1eQodHvNj+759x7m72vm/wBrNxuS6WJW9s1rUrHKSt0inzQTaHA4KXb16xS++dyPFably37hWVsFN2GtLphwaSIBrd2/pmLWpPmEX065/MXGfxTZ/Um/4UD1mAfOdZZ2RUYyPRx23Htln0IxfTTy1SQrxqcHkn6E394lylpn9uOfIIwQ3j7fKwBGVpPB8s/rAcCfn5Gv3JuZBo545OlfgYryIVH/ADRl/aPBJpjopuxcht6Wrrto72mrYevd/wAWNw256/FztSZjaeQoclT5XZfZ22KxP3MbvHq3ddPTZfH1CEMr07o10d1OJf3a/eK69lvc7a98mmP9Vr4rbbhH+E27sNE9OBe3c+IPPw9a1AY9ZHe+vtjb+6fIV/tkUAPMNnquLF/MTKvdDXiFuEHhnyD6G8umHP0uYos3nsduimhot34HcGW2pv6kpYY6egTfODMBzGXxEUP7SbY35jq6j3Nhyn7f8NzEcaf5lgDr713tNH7V+61+2zRD+pm+qdwsGUdirKazwKcgiGWpTP8AZPH5U6JPu3+40nP/ALd2tvu01eaNoYWd0G+NtApBKwxl4xpfGJEapqT0qOq+0tzdQ7gmrcAlFk8ZkYFp85tTMBpMDubExSaloa6GIealqMfKwalrYj9zBIx0kqWQxz7S+7/OXstzOvMvJ92pWRQlzayVNvdxA18OVRlWGTHMvfG2RqUsjD73J9s+V/dTYDsXMsDBkJaC4joJraQimuMnBDDEkbdjjjRgGFoWwN443eG3n3/1Pkp/tmplp96bHrZKfI53bEkhZKnG57EmN49wbUlL2p6wxMkkberSwa3bj2f95+R/fTlJdz2dkW9MOi92+Uo09uxw6yJwlhapCSafDlU5o2pRyj9zPa7mr2k5jO3brG5tRJqtL2MMscyjKsj/AIJhTvSoZG+RBIEdh/GPZXYgq851NLSddb8lD1UmxKqoCbC3ZVM8lTOu266oLNtnJ5CeQ6aaQmmY2VGiXkYpe+f3GNm3xdw5o9nXXb97OqR9tc0tJmyWFsxqbaRmwqGsGqi0jUV6yI9pPvbbntDWvL/uaGvdpFEW+QE3EQwAZkFFnRRxfE1KsTIcdEH3Dgdx7My1TgN44DKbczdDqWqoMnQz0ky6CimaOGVAZqeSRiI5IjLEyi4a3Pvl3v8Ay7v3Km7XOw8z7Pc2G9QmjwzoY3FMVAIo6k8HQsh8mNR10C2ffNm5h22DeNh3WC82qUVWWFw6n5ErlWHmrAMPNR01LJFJ+h0bkghWBIKsUNx9eHUi/wDUeybjkcOjWgPHrzxo6hXRXX8K6hl4P9GBAsR70yq66XUMvzAP+HqykqaqSD8sdOUWWysNMaGPJVv2JEoNBLUPU48idDFNqx9WZ6FvLGdLXjJK8exDsPNnNXK0yXHLXNG5bfMtaG3uZoqahQ4RwMjBNOHRDvHK/LPMEbRb7y7ZXkZpXxYI3rTIyVrg8M9NktPjpS2rbexAJEWObx9b9eRfcIiLGBUmDa0ZqrRjSTJrYjgk8+5Wj+9B94iCOOGH3k3xYlUAd8RoBgdxiLH7SSeo5l+7/wCys0jyv7abaZGqSaSjjxoBKAK+goOhP2BuPq/b2Wp8hmuqaDaWUpqX7Kj7M+NlXL8euz8PGqLHHUvB11Jgtjb2FNGoAp8xjZ45ALMGBt7nv2y+/p7icuSpZ+5djHzBsxbM6KlvfRg8SAqrBPTiVZY3NKa89RHz790Dk3e43uuRLx9n3IcIZGea0Y+ncWmhrwqrSKP4cdW5fGburfkG9cR0r2lvej7Zx29evqztL4+95xYfH7azvYmzcBX4rE722Z2ftvDxU+AxnavXddnqAz1OMigoctRVQmFNSzwVER6t8o81bJzvy3s/NvLl6J9lv4FlibANG4owBNJEIKuvFWBU8OudnMnL26cq73ufL+9Wph3W0maKRDnuX8S4FUYUKN+IEEcej8X/AN9/vPsSdEfX/9Pf49+691737r3Xvfuvde9+691737r3XvfuvdE2+VnX24MfPsv5RdW4+urO2vj3/E66vwGHi8lf230flXpKjtnp+amVkGTyNfi8cmX24j6zT7lxtIE0pPPq916uKdGa6/33tPtDY+0+xdiZml3Ds3e2Axm5ttZqiYmDIYfMUsdbRThXCywTeKULLFIFlhlDI6q6sB7r3Sv9+691737r3Xvfuvde9+691737r3RQfnJtT+9Hx03nLHHG9TtWbF7vpmlqWplhGHql+9muqMJ3GNqJgkTcO5HINveK/wB8rlZeZvYPm2RYw1xtjxXyksVC+A41nGGPhO9EPEkeY6yB+7Bv42H3m5aWSQrDfLJaMAoYnxlOgZ+EGRUqw4AdUD3B+n/E/T8fX6H3w2BqAacRX7R5ddbAa9e976t0X/vH4y9ZfICo2dmt2vu/bG+uuqqvrevu0ust2ZHYfZezDl4BS5qiw26cVqlfDZumUJV0NTHPSzWBKBhf2PeSvcfmTkSLeLLaltLrZNxRVurK8hS5s7gJmMyQPjXGalJFKyLWgamOgLzhyBsfOT7fd3zT2+7WjkwXNvIYbiLWNLhZFzpcYZSGU0GKgEMnWnxD6d6uwPamJxqb13bmu7sVUYPtrsbsne2a3x2bvnES4mvwtPjclu/LSGoo8bicfk51oqWljgpqZpWdY9Rv7Xcye7PN/M9/yzeXRs7Sz2aRZLG0tLeO3s7dw6yF0gQaTI7ohkkcs7hQCadItg9seW9gs99t4jcXN3ucZS5uJ5XmuJU0MgVpWNQqqzBVXSq6iQBXpAdY/wAv/oTrDN9cZqHKdz9hJ04kH+iPbnbvcG6OwdldaVdHQ/wygy20NnZBqfBY/MYvGHwUdQ8Ur0kZ/a0nn2e8ye/PPXMljzDYtbbNYfvev18thYw2txeKzF3S4nWsjo790ihgHI7qjHRPsPstyvsd7s9499uF6NvA+ljubiSWK3KrpRoo2bQGRe1WKllHwkdHa/1+f8f6/wCJ/wAT7hc5z1LwFAB1hqQDTzgkKDDICx+ijQ3JtzYe25f7KXPFT05GaSR4/EOtnLp2qqKzqvrqoqsfUYqofZW3Q9BVyU81RAI8XSxxmSSlklgbzRoJBY3CsAbEEe/o19qbm4uvbPkCa62+S1nbZ7SsTsjMumFFFWQlTqADCh4EVz1w99wooYOfOcooLyO4iG53NHQMFP6rE0DANgnSccQaYp0Xmv8ABhv5gmBlZ2SXsD4i7hxkarGuiSbrntvDZBXeUnUXjp+wHCra1iT7H/QP6f8AuL401G5t7UXeXS29B018i8ViIMFNviLCRZ3afZ21aJpJqTYPdey1qcbHvTbEFQ7NQ1kNRSZvCSOz0FXEjzQze690MfUeY7WzOzaabunZ219ldgUlbXY7K4/ZO6ajd+0spFRSiKl3Ht/J1+KwmWpcbnI/3Uo62mSrpOY5GkIEje690JpNgT/T37r3RL/l/wDJyPo3bUGB2u9JVdm7pppThop1E1PtzFamhn3NkYNY8rLIClHC3pnnUlvQjA4kfen+8Ynszy9DsfLbRy+425REwKw1LaQZV7yVTxIIKwRn+0kFW7FYHIz7vXsi/unvcm676rpyRYyL4xUlWuJcMttG3lUUaZx8KEAdzClEFVU1VdWVmRr6qeuyGRq6nIZCtqX8lRXV9bM9TWVlRJYa56mokZ2PAJP9Le+Jt1dXN/dXV9fXDzXs8ryySOavJJIxeSRz5s7GpPXVu2tbezt7e0tIVitIY1jjRcKiIAqIo8lVQFA9B61PWH2x0o697917r3v3Xuve/de697917rPS1dbj6ukyONq5qDJY6rpq/HV9OxSooa+jmSoo6yBh+mWmqI1dTyLjkW9qLO7u7C7tL+wuZIb+CVJYpENHjkjYMkin+JGAYA4JHSa7tra8tp7O9gWWymRo5EYVDxuCroR6MpIPy4Z62Ofjj2x/pm6h2nvWZUTLy08mI3NAiFEh3Jh3+yy3jH6RDUyoJ4wLgRyqPwff0BewPuePdz2q5Y5xmUDdXRoLtQKBbuA+HNQfwuR4i/0XA64z+8fITe2vuHv/ACvHnblcS2xrUtbTd8VT6qCUb5qemL5XbUyW6+nM2mIpqitrsHV0G4Uo6ZDJNUU+PkZa8JELtL4qGeSTSoJOjgH6eyP7znLF/wAz+0u8x7ZbPNe2UsV0EQVZliYiWg4krGzNQZOnHRr7D79ZbD7j7Y24zRx2V3FJblnwoaQDRU+VXULnGckdUk7O2jsnrvbmP2jsHbuA2dtTFvXPi9ubcoabFYfHSZTIVWXyT0mPgCRwSV+TrpqiWwBeaVieT75T7lu91vF5Nf7rub3F++kNJI2qQ6VCKCxoTRQAB8uuiG3bTbbTax2VhZCK2UkhVFBViWJwKZJJ6H7YHS/ZPZrq+1ttVJx3kVJM7lQ2Jw0N7EslZVIr1ZjBuVgSVj/T3IHI3s/7h+4TqeW+XpTZaqNcTfowL/tnAL04lYw5p6dA3m33M5K5LR13ze4/q6VEEVJpT8iimi14VdlHVoXRnx72z1BQ/fyGLOb3rIDFlNxyRFRBHJpd8bhYZLijx6MLareWa13P9kdIvZn2J5f9qbFbyRlvOcZY9M10RhAQNUNup/s4gQM01vSrGlAMGvdD3d3n3GufpE1WvK8T6orcGuojhJMwprkPGnwpwUeZHrKyPDjMjNGxSSKgrJI2H1V0p5GVh/iGF/c8dRH1WIpLqrsSzMC7MTyXkJd2/wACzMTx7917o+eH3Ji9kdMUm7ssRBiNr7AfcmTZQEtSYrDPk6xhZbeSRIWP0uWP5v7917rUL7R7FznbvYm8+zdxOzZbe2erM5LEXd0oaSdtOMxcGsnRTYzGpFCqrZRpNgL+/dbrinXDrPrjdnbu/Ns9b7HohXbn3XkFoKBZLiloo1HlrctkZB/mcZiaRWnnckelNI9RA9+611t0dA9LbX6A6p2t1htVA9NgqJWymVZVFXuDcFUBNmc9XOFDSVGQrSxUG/jhCRj0ovv3XuofeO6Di9uR4GnmMddn3KS+O4ZMVAQaslr3Q1DFYxbkgt7hv3m5jO2bBHsltMVvb4kNTyhX4/s1mifYW6k32v2M7hvTbpPEDa2YqCeBlPwfbpFWPzp0UQCwA/px9LfT/D3it+QA+WAPsHp1kT137917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/faMde6E7qPcUmA3jQ07SaaHOlcXWK3CeZgzUE31srpP6bj8N7kj2s5gk2Tmy2hdz9JekRSDy1H+zf7Q9AT6HoB+4mzx7py5dTha3VrWVPkuBIPsK5p6jo73vMXrGXr3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917pi3Lt3G7qwWU2/l43lx+VpXpqgRt45UBs0c0MliY5oJFDofwwHsn3/Ytv5l2fcNj3SMtY3MZRqYI8wynyZSAQfUdGWzbte7Dullu+3uFvIHDLUVB8iGHmpBII9D1SJ8rfgL8de0d54eq+SPx/wCue463bmPrcNsXd2+NutkJ4ttV1aK+pxdFWwVVKsCTVa+WopHLfujUAVIPvBDmGx539pdzvdmttzuLfa5pNUUyErHOAKKxOFEoWoZKhwa0qCD1lps0nKXuXZWu6TWyNuMSaZIqjXGcVBBBLRE0Kt8JHEVFOsMfQXR0fUFX8f6XqPr2h6Kr8FVbZreosVtjG4jr+pwFdKJq7FS7fxUNHSGCunAeZhaSWT1Oxbn2A/33vDbrHvp3Sdt3RtSzFy0gYYB1NU4BoK4Ax0MxsG1Ltr7Qtoi2DKQUUACh+QFPM+VMnGejL9P/ABh687N351V2JvLrjb+Xpvj7kq3NdR53J429TtDclbgJtozf3Oq9SNTwvt2V6WqClomiVI2BNrTh7Dcsb9unME/MMkk0XLUdDIakLdTq4aJc/GsT1diMBwF4nqJveDedm23aYdmhWNt8aqqBQtDCy0dj/CXB0qDlgWPkOjkP8VvjlJ8fqr4py9M9fz/G+u29X7Tq+l6jb9JUbAn27k8jPmK7EyYKZZKc002WqXqv9UlQfIpDAEZqEliWPxHrF/oXtp7U23sTa+3dlbOwuP23tLaODxW2dsbexNOlJi8FgMHQw43EYjG0sYCU9DjqCnjhiQcKiAe9de6C75DdCbL+RvWma673lCYRUoavb+4KeKFsrtbPwKWoM1jJZEJVopLLNHcLUQM0bEBrj3XutUrvDo3sT4978revux8WKSvhR6zDZikEkmB3XhfK0cGbwVWVCywORaWFrTUspKSKDYn3XugbeKKQq0kUchX6F0ViP9iwJt7917ruNI4v83HGhP1Koqk/65UAnn37r3XL37r3XYuD/vH0J+vH0H+Pv3XurHNm1676+HGxK4Ms2Z6G7K3B19lGIkeoh2b2Bq3PtbW9wgo6fLJNCl9RBsoIuR752f3hfJoveUuR+fYLcGbbr17SVqZ8G7UMlTw0rNEOOQzimCes1vuWcz/S8zc18nzTER3tqtzEtceLbGj0HEs0Uh4YoueHSs+MO6YNqd87MNfPHT4beUeR6+zbPpVfst10s1BTu7eQOwpcq1NKAF9KqxuPr7wl+6xzqORPfbkTdJ5gm3Xk5sJyTjw7seGhPyWYRNXy86A16yt+8LysebfaHm+xigL3trEt5DQVOu2bWwHzaEyD5mnyHVhWc2FFuzaXbXVufSKnyEu2Mnk8TJUhkNBu/Yxly2JyELBHdVkhEgYgeqCRhc399ifvAcgbf7j+0POvLV6EEq2bXFvI3+hXFqplhkBzShBVvVGYefXMr2Z5vvOR/crlPfLVm8I3KQTKP9EgnYRSpTgcHUteDBT5dU4xFBGhaMCkrIw2nSNELypqkjIVRGkDjkkk3dre/nvjYTRJIyDTIoNDwII/kD12gIMTsofuQkV+zFft/wCL6GXdsR3jtbY3Z8NNNVZtY8V8eO0TTEeSbL4Okyme+Nm/K2AgGabJUH8X2bUy3JZ62i1+mFQOjG0zx/eM+59uuy3wM3uJyD+pAwq0slvFHVKmgLCa0V4mHDXbqxzTrBrd4W9kPvNbZu9rSLkrm7slGdCSyuA4oDjwropIKjCTEDj0EExJjjnjGoxskwADkshGl7IroGcxObajYHn8e+dYYMAw4HPWchBGGweB6Vm0N5bq6+3HQ7u2Tnazb24scGFLkqPQ2unksZ6GuppQ1PkcbVAWlgmDI/14NiBHynzdzNyHzBZc08n7xJYb/b1CSpTKmmqORTVXifg8bAqw9CAQRcycsbBzhs91sHM+2Jd7PN8Ub+TD4XjYd0ci/hdSCOGRUdWWdcdt7B73Wnx9KKHYXbk8d63Z9XKtPtbeNUgVZavZ+TlEcFFka2U+Q0ErBwTZdQBc9kPYD72/J3uzHYcs8xum0+4ZUKIXP+L3bAAlrSU0XVJUkW7nxV/CJANXXMP3j+7hzR7cte77sivuXJKtUSqKzW4JrS5jFWCrwM6jwzQFtBNOllufHYfdOO/uJ2/tifcmLxkjJRLVyy43eW0p2Txfc7cz1vuqeMKAfBIZKaYAXFgPc5+6HtFyH7v7KNj542RblIyWhmQlLm3Y41wzDuU+qmsbUAZWoOol5D9yOcPbXdv3tyluzQSPQSxtR4JlGdMsR7X9AwAcV7SM9Es7C+I+6cbHW5/qyobtTalIhq6ilhpY6Lf+30ivKsWV2yXEmWSFSdNTjzKr2J0D3ym94fuVe5Ht/LdbryUrcxcpKC+qMAX0KjiJLYZmAH+iQVJ4mIddDvbL71XI3OMVtt3NLpsnMhIWkhJtJWOKpMQTDU/gmoB5SHoo5pRFJJCfuaWaBmimpnaWKamljR4TFNT1C+WCWK9yrqCHAJHHvDNg0ckkUiMkyEhlYFWUg0IZT3KQeIIFOso0ZZESRXVo2AIYEFWByCrCoYHyIJHXZhYgj7moH6rEOnGpETi6W4035/JPvXV6DrlomFys5IufTJGrAAurFQVKN6YwVXn83N/futHruGUuCrqqSrYyRhi4W9yrK+lS6Ef2rDm4/Hv3XuPVgHxrqsdCnw/3JHXI02yvk721sXcFKHlWWgpO5eld6f3dSVNSxiiyWewNGVvdGnC2Gu3vsf8AcC3ayu/ZTcdoguw1/Zb5deLHmsazaZIuOAJBVxTHxDjXrmJ98bb7y3907TcZrYrZ3W1W/hv5SGLUkn2lTg1zTSeFOruP7P8Aj/vN7f1/r+P9f3nD1ib1/9Tf49+691737r3Xvfuvde9+691737r3XvfuvddEXt/r39+691XfQ1NN8GO1pcNlq2DHfEHvzfNfXbXylSrRYz45d9b2yS1tds3IzrGKLCdOdz5ypqazF1MrR02F3TUS0blYMlSCH3W/IY6sQBv+LG9iPfutdd+/de697917r3v3Xuve/de6Z9wYai3Fg8xgMinkoM1jK7FViem5p6+nkpZSutXUOqSEqSDY829le97Tab9s267JfJqs7y3khcY+GRCh4gitDjBoaHpdte43Oz7nt+7Wb6bu2nSVDn4o2DDhQ0qKHORjrWQ37snK9cb23PsXMwtBX7Zy1RjDrLFailQiTH1kTsAZYayheORXsNQb8fQfOfz3yduft/zjzHyXu8LJfbddPFng8daxSLgVWSMqwNBWtSAajrt1ybzNYc5cr7JzTtsga1voFkxTtc4kQ04FHDKRmn2dJD2E+hP1737r3Xvfuvde9+691737r3WKZHljeCJGklnXwQoou0k0x8cUaj8tJIwA/qT70Y3lVoo1LSONKgZJZjRQB5kkgD7evCRYmEkjARr3EnACjLE/IDJ+XW0N11SVWP2FsjH11PJS11DtHbtJWU0thLT1VPiaOKeCQKWAeKVCp5+o9/SLyHaXVhyPybYXsBiu4dqtEdDxV0gjVlNPMEUPXDHm+6t73m3me8tJA9rLuNwyMODKZnIYfIggjorXeYkwnzC+Eu54oFSnza/Ibq/KZApERpz/AF9it6YbHNI8iSKZ8jsBnRUBLeM34F/Ys6D4Fa9HWiQoiqxBYD1Eajcm5PLEsRc8X9+611k9+69035XJUeIxmRyuQl8FDi6GryFbMbWipKKnkqamXkgWjhiY/wCw9otxv7XarC/3O9k0WdtC8sh/hjjUux/JVPSiztJ9wvLTb7VNVzPKkaD+J3YKq/mSB1rR9wdnZLuPsjdHYeS1xpm6wR4eiYkrjNu0INNhKBBcgFaMCWS36ppXP59/PB7se4+5e7PuBzHz1uWpRdzUgjPCG1j7beIfMJ3ufOR3PXa3225HsfbnkvZOULKha2jrM44yXD90zn/b1RfREUdBr7jvoddNmazeG23iMpuHcWWxmAwGDoKjKZrOZqvpcXiMRjaOMzVeQyWRrZYaWio6aJSzySMqqPaqysbzcru2sNutJbi+mcJHHGrPJI7YCIigszE4oATXpHe39nttrcXu4XKQ2kSlndyFVVGSSxwB558gT0Auw/mB8We0MdvDK9fd/wDV+6qDr/C1e5d6zY7ctPG+2ts0IZqvc2Spq1KSrG24QvNfGj0puAHuRccb77Te5vLM+02u/cibnbT38witw0JpNM3CFGUlTKa18MkP8ugbtHufyHvsW4zbVzPaTR2kTSS0kXsjT4pDU/2Y/j+H59P3VHyY+PXe1Xksd013P112Xk8PSw1+TxW1NyUdflaPHzsEiyUuMYw1zY15CF+4WNoQxALAke0XNPtvz9yRDb3HN/J+4bdbTOUR54WWNnHFPEygemdJYEjgMHpby7z/AMm82TTW/L3MNrdTxrqZY3VmC1pq0gk0r50pXFehvuP6/wCt/j/j/gPYK/wdC8En8PXZFv8Aff7z/re/dbBr11791vq0H+WlvOrTO9k9eTSzSUM+Pxe9KCJtTwUlVBUDC5PSWfTC1Yk9O1lX1mMknj30l/u7+brlN39w+QpZGazeCG/iBqQjqwt5aZoPEDRnC5Kkk4AOCf32OWoH27krnCONVuUmks5GwGdSvjR1xU6CJOJwGAHHq3Nl1cGxBFiDze9wRY8WIPvqURUHFainXPzz6RLda7ANcckdk7TbINKs71jYDGGoaZTdZTIaYnyBub/W/sJHkDkZrxtxbk/bDfM2ov8ATQ6tXrUpx+fHoR/1x5tFotivM1+LMLQJ48mmnpTVw+XS1jjSJFjjRERBZURQiKt+AqqAqgf4exYiJGixxoFQCgAFAPsA4dB0szEs7EueJPE9c/dutdQ8jC1TQV1MhAeoo6mBS17BpYJEUmwJsC3v3Xuqw/G0JaGS2qF5IXtexaF2iPBAaxKf0v7917rL8s+xGxfwB39FTVUkNfUnEddzlP3JhFmdxUMLxPra8cVTh5WW4vpQm309+62Mkda3a8fQGwH4/A/40P8AeffutHj1f9/KW6FpsDsPP/IDO48jP76qarbezpJ4lvR7Lw9SI63IUjMC6tuHNwuGbgmGlS3pY391snA6uJ9+610RPtDNyZze2ZdmBgxkxxFIqm6iGiYrIw/xlqGdj7wt9yt5k3nnDdHYj6e3cwRgcAsZyftZtRP5dZSch7Wm1cs7coH606+M/qS/D9igAdE972+WHxo+MH92f9mJ7x686Z/vn/FTtM79zJw43CMGKQ5hsawp51lTGffw+YnSE8q88+w3tmxbvvPjfurb5Z/DA1aBWmo0FftPp0d7jvm27UyJfXAQtwyB/hI6BlP5mPwOr9g9k9mbU+TXXXYu0uocZtrNdi1PWtVXb0yG28Tu/dGN2Xt7I1GMxlF9zUUmR3LloKZXi1hSxY2UE+155O5jW6tLOba5Ipp2YRiQaNRRS7DupkBSfy6LzzdspguLiO5DLEAWoQcMQB8JbjX06PVIjRO8bizoxVgOeQbH/A29hr/D0JEcSKGX4SOulBY2UXNnP9BZFLMbnjgD3sKSQPPrzusas8hog6Cbo3u7rj5HdW7Y7m6jzNVuHr3eL5+Pb+XrMVXYWqrG2zuXMbQzAkxeRjiraX7fPYCqiXWo8ioHX0sD7Xbltl3tN7Nt96oW6j01FQcMAwyMZBB+Xn0h2zcrfdbRLy2r4TFh+asVPp5joV7j/ff7x/t/aChHEdGHXd/99/tv9697pkAnj16vmevf77/Y+/FCPn1oNUYGeur/AOB/33/GufeqU49b/PHXY596631mpZ2pKukq0bS1NVU1QrflWhnjkBH9SNPt+2lMFza3CNR4po3/AN5cMD+0dJ7qEXEE9uVqHjZfyKkH/COrIYJVnhhmW+maKOVb/XTIgcX/AMbH30CikEsUcq/Cygj8xXrDSRDHI8Z4qSP2Y6y+3Oqde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XvfuvdIne+R2NS437ffMuGbHTuumjy0UdX5pB+l4qPRNMzR/llX0g/Uey7dNp2ze7VrHd9vhubMmuiRQ61HA0INCPIjI6WWG4X+13C3e23ksFyBQMjFTQ+VQRjouVXkfjXV5AmfaRWIxrrrafHZCkolMd1CGlpaqJwQP1MIrEfX2Bbj2d9tLqUzy8o23if0daDHoqOF+3GfPoWwe5fPdvGsScyTlQT8Wlzn+k6lv59GuwcWLhw+NjwkVLDhxRU7YyKijWKkWieNXpvBGoUKjRMD9Aeeefcg2NjZ7ZZ2237fbJDZQoFREAVVUcAAOA/1cegbdXVzfXE13eTtLdSMWd2JZmY8SSenX2q6Y697917r3v3XuisfLr4z4L5PdU1+0ap4MbvDENJmdg7keMFsPuGKFlWmqmA8smFzCfsVcYIujK49ca+/de61M87g8ttjOZrbO4KKbGZ/buVr8HmsbULomospjKl6Wsp3U82SSO6n+0pUjgj37r3TV7917r3v3Xuve/de6OV8MN34OHeG8+md4Zanwu0u/8Aay7Piy9dKsNBgt/4qq/iuwMzUySFYoo2yqmkdmP0nUXA9x57scg2nuh7dc2ciXkgj+vtWWOT/fcy0eGTgfgkVSflUdDb265yuuQeduW+brZS5srhWdP9+RNqWZOI+KNiB8+nvcu2s7tPNV+2N1YvI7d3Fh6x6eqoqhJ6CtpqmjnAWroZGVGkp2kiDw1EJaN1KkE39/PVv+xb/wAqb1fcvcybdNYb/aylXjdTGytG2JIq0DqSAySIWBXSQc9dotn3jaeY9rtN52O9ivNmuYwyuhDoyuMq9K0ahKyRvRgdQI6NNX/NLsOq2rLjq3a+2W3tJts7Uk7UpzXnMy4qeiNBX1tft5/LjJs/VUZ8UdQpdY9Tvo5AXLm7+/H7qXnt1c8jy7bY/vmW0Fs25jUZzGVKSOYGrEZ3Si+JXSDqfRUgDG21+6X7fWvPFvzZHuF3+647kziw7RFrDB0UTAiTwVbOimqlE10qSUIRQtAIQFeEJ4tIIkQBLKFN76nRl/P5HPvC5VVVVUFAABT7MCn5U6ym1MSzOe4kn9p/1f5OhF64qc9lMbv7rfCOzZXsnZWUx+3EDiTT2ZsOZeweq8hGJFhQ1g3ptimpj+kaKtwDax95afcs58fk33v2nZLjS+x8xxPYXEbHtLkGS3anmdatFTiRKR8xjd96nk6Pmf2o3Hd4gw3XZJFu4nX4glQkyg+Q0sJK+TRL+aMz1Rj66b+O4em+xwe7cDg984Oid4m+ww+9sFRblpcbqYmK2IkyT0Z1cXpzcD3D3vZyKntr7s+4HJMETCwstxkNvXytpqT24HyEUgA88dSZ7R83Sc8+2vJnM9w1b24s1Wf/AJ6IT4UtfmWTUf8ATceoCCyJ/wAES/6f9SP9T6R/sOP6e4s6kccOuxqV45EZkkidJYpI3aOWKaNleKaGWMrJFNE6hkdSGVhcEH3tWZCjIxV1YMpBIZWU1VlYEFWU5DDIPAjrTKHVlYAoykEEAggihBBqCCMEGoIOQejkdc/LbL09LQ7Y7poazf8At2jjFPjd2UJhi7F27GpUR2rZSlNuegiS+qGqHmNhaRjx7zm9kvvu858ji22D3Ijl33ldaKtwCP3hAoIAq7ELcooJqJKS+QkPDrEf3V+6fyzzX9RvPIkkez8wsSxhz9FKx9FA1W7E8ClYhU1QcejiYFKLceMi3x1Pub++WCp9DPlMC0tFubbtRpLGj3DglcZXF1US2LHQY2HP099Tvb33L5I909mi37kbf4b2zOHVTpmgaldE0LUkiYcNLAfIkUPXPfnTkTmv2+3aXZ+bdlls7kVKs3dHItaa4pRVJVPqpPoQDjpk3hiOve0InHZuwcVnMs8Rgbe+3yNs76hZFCxTS5GjjNDk54+bpVQsr/kf0B3uX93f2k91kuJ+aeVIV3qRKC9t/wBC7QjgTInxFf4ZVkU+YPQn5E96Pcf26aCLl7mKVtqQ1NpPWa2I8x4bHsr6xlGHr0Trt/4yS7KwFdv7rzccu99j4wxHPUWRoxj967RinmWGOqzFBT6qPJYZJZFV62n0rGTdkABYcxfvAfc+5l9oNru+ceXN1beOR4SPGZkCXlqpYANMifpzRAkapkCFAavGFBbrPj2b+81snuRuFtyzv22jbebZVPhBW1WtyQKlYmbujlIBpG2oNwDVoCVMG/I/r/h+P6fX3hr/AIOsouI6wS/tvHPzpuIpQATqWQhY2ssbu7o/AuQqqxJ9+60ePS923vbJ7J2vv/LUJeQbOXYHeMFOjHW+Q6E7H2zvur8MSkPNLU7OhytOUWxaOQ3OlT7zh+4NzbNs3vHuXKxlpt+87TLqBNB41ofHiZV/E+gypQAHTmpA6xK++Jy7BuPtptvMXh/45tm4IAwH+hXQ8OQMeAXWsZqcAj59bNv8Wx/8I/jn3Mf8K/h/8X+8uPF/D/tvvfudV9Pj+39V/pb32P65kUzTH+r58Ov/1d/j37r3Xvfuvde9+691737r3Xvfuvde9+691737r3Sd3btHa+/Ns57Zm9dvYfde090Yuswm4tt7gx9NlcLm8RXwtBWY7JY+sjlpqukqInIZHUj8/X37r3RGv41vP4LJRYrcj9idwfEusyklPiN6LSV29+xPizjHjiWiwe+kpBXbt7F6RopNSUecWGqy+2aZViyJqqNVrKf3Xuj17b3PtzeWAxO6to5/Dbo2znqKHJYTcO3slR5jCZfH1C6oK7G5THzVFFW0so/TJG7Kf6+/de6fPfuvde9+691737r3XTfQ/wCt70eB69SuOqk/5i3T2RXI4HujDUks9A9HHtrejxeo0UkEg/u9k3iuWMEyyyQSMAFRlQsfUPfL37/XtRei+2T3e2mzZ7ExC0vyufDKn/FZiKatDBnjkbghCFjQ9Z+/c59xbX6Pd/bbcrkJd+KbmzBBo6sP14wf4lorgcWBagx1Vt75pevWeOOHXvfut9e9+691737r3XvfuvdDB0DsSbsbuXr3aoo462jqNw0eRzENRTT1NIcLhpBksj94kBV1p5YoBHquFV3W5t7lj2M5Kl9wPdrkXllLVZrZ79JZ1dGaP6eA+LL4mmhClU0VqBVhXqNvd7mtOTPbfm7ffqGiuUs3jhZWVX8aUeHHo1cWBbVTNQDQE9bKiIqKqqoVVVVUD6BVFgo/wA9/QsqqqqqCiAUH2dcWiSSSTk8eiZ/MGnNLlfiVuqMS+XbPy/6qp7xGFLU++cRu/rioMrzAnw23auoJZm+n9fdutr5/Z0c7/ff8T791Xr3v3XuidfOTfI2X8f8AdNLFPLDkN7VNBsugEVtbJlHafK3ujARfwakqA3KtZuDf3il987nReUPYnma3imZNw3h49vi00qRMS03EHt8BJAeBo3aa9ZC/de5WbmX3f2KZ4laz2xHvJK+RjoIsClT4zJTiMZFOqDgAosFCgf6ngfQfQf0H4/w98PPIDyHXWnJJJJ69791vqtf+ZK1LX4z4gbQ3g0a9K77+Z/Uu3e7Iq6Xw7eye21hyuS2xt7dru6Ur7Yzu9KOjjqYqgiCZlRXuOPeRf3d/Ft5/dndtoQnnKx5Pvptu0isiSkxpNLAACwnjt2kKMg1qCSPPqBffF0kX2+2/cjTlq55gtUu6mkbRnxCkcp4eG8wiDBsHANeHQp/PHZ+yaH4l/KHPHa208buyj+MHcmAw+aiwOGodw0u2pdsyzZLEY2qgpYMiMCJqenaWniJp1ZEJUED2GfY/eN7n90/bWyG63Um0vzLYSSRmWRommWYBZHQsU8Qhn0uRqILZp0de6e17BDyBzpdW9lbJfjZbtFcKusR+CSyhqFtNQpYA0rSuadFx+OfVPc/cnaXwo723l0tgfj3sv469E1GGpMgd67U3d2B3hL2H11tvA42gqIdlUsNPgdgYqmpf4o8GSmlqnrZEVY1Ku3uQvcHmjk/lLlf3k5H2bnGfft55g3oSMv001vbbaLW7mkZgbhiZbl2PgholVBEGLMe0dR5yVy3zRzDvPIHNd9y9FtO3bRtbqriaOSa78eFFCnwqhI0AMhVzUyEUUUJ6R6/IfudP5cfSvbcvZObj7O3N8p9pbAyu8ZJMamYy236v5f5/YuQ2yyvSCnlSTY9AKGWOOLyiljJ4sW9nDchcoP8AeF515VTl2EcuW/LM90kA1GOOVdhiuUlrqqKXDeIrE6S5HEEDpj+ufMQ9ouXN4O9yfveXf0hZ6qHaM7sYTGRSh/SBSlK6QfMHq5eYBZ5lX9KyyKB9QAHYAD/Ye8P1roWvxU/1f5espYW1RqSc0H+Af5a9Y/e+nerC/wCW1/zOXev/AIjd/wDYf7+TFW/1veef93sae7POXz5fP/aXB1h999Gh9ueWPX99D/tGl6ur99feuaPXvfuvde9+691737r3XR/33++59+691Xv2jhf4FvzcVIF0QVFX/FKVRa3gySCoFrM5AScuvNiSPwPfuvdFX+SuEzW7/j12btHDQz1tTNHgN10uPp42mqKut2dk0yTw0sCBnmqanG+ZAigu5CgAm3v3Wxgg9VU/HT439g/JbftBs7ZuPq6TCiVZt270qqKpXB7WwsUscdfJNVtGsFRmXVilNRK3mklILBUVmHutdbb2yNn4Lr7Z+2dj7apFocBtPCY/A4mmULeOixtNHTRM5UANNKI9cjf2nYk/X37r3SklYorEcWRzf+hUEj3RyVUsPIHrYFSo9SB1W/VStUVlbUSMWkqK2rmcn8vLUSOxt+LlvfP27lae6vJ2y7zOTXj8Rr/OvWZttEsFvbQrSiRqB9gUU/Z0B3yPoMfWfH3vt67HY2uan6N7fanevx1DXPTs/Xu4LtTvVwTNTsxUXKFSbC/0Hsw2R3Tc9uCOVDXEdaGle8cfX8+ivmC2gn264aWIM6odPy/1fPqh3tPHRxf8Jn+uJMRTYrFZmt+JfxLiiyseIovImQk7A61+yrMikEUEuUipKt1meKVz5NJBILX9yPYysfdu4Ejkxi9ucVPDRLhfQnIr5cT0ALiCBOQo5o41Fw8EdTSuaR0x9vRxsTuLuT4lfOj/AELdofMbs/uTqDtf4Pd2/IjeO8u9qLZE/wDoV7I6Q3ZgcbuHsDYMO2dtYCgwHXtdhNyTVT7cnWspaUUUarI/JYmnhsd85fF/abHFDfw7jFCixF/1Y5UYqj9zd4ZANQArqNadK7aa92ndWs5dwc2klqZCSFOlgUJIogotH86haVHRfvi58i+06b5rfCzbe2O0/n92l0F8t+uvkVXZbeHzZ2f13tTYvaD9f9f4vsHYvZ/xy21t3F4LduwqSoNRIJaKroaahqcPWU7orNpb2b7xtVm3L++S3Fpt8W5WUsNFtWkLxh3Mbxzksysc8QxIZSOAIBNt263S7lbLFdTPBcK4fxNNDQBgVoqsvngqoKkGlckH/iTv7GRfBH+X305i+4/mvguxt8U3ys3NhehfgVtzZlZ3D2Zt/A/I7sShyG+t1733jiK2h6/2BsOqrUi1y1lBTZCuq0idpCoUK93tJG5i5lv/AN3WD28f04Mt2zCNC0EZ0KqsNTsATQAkAE46bt7xYtl2y2+tljZmnbSmkcHbLMwICgkA1ZMkULcOnnrD5NfLv5C9Cfyp9tP8iuyuot799/Mb5bfHHuvsrEbf2PF2funrvo3A9wQYz+8WMnxuY2diu1JcdseBZshRxSUtHmy1ZGkoRUbV3s2x7ZunOM42uOa3t7K3niQliivK0RYChVigLYQmpU0PGvVo963a9tNitzeMsjTNGxoMgeMh4gitEHcBgioHl0nt5Z75r7Z6k/mo56h/mBd/Sn+WHvzMf7L9LV4nruo3D2XDFsDa3dFRjPk/nn2kG7ZwUOJy529RU1NDifBCZKp3lldFjvbpy/Lecnxvy3b03iMCchnonc0QaBdZ8M1Ac11CppTHV3bdkt96Vd1k/wB19WjwCSAqyAP20etSuAppk5p1aF0p3x2b2D/MnwfXOW3JXR9Vbm/lg9KfINesohT/AN3KHsvePcf8N3FumhU0/wDEHqUwVSKNyZmiipwupQfUQdf7XZ23Kj3UcYN4u8Sw686jGsYIU0xStTwyej+23i9l31w9yViNgzAcBq1HTxzXgAK5PlU9Eewva/zZ7g+D/wAd+29sbv8AkZ2Lseh+UPyvpflpm/itWbJHyyr+jdn9s9q7V6tk6eOcxVZj85g9nVeLoP4tQ4uE5qvxFMiUrOS4YRNYbBZ8x7lZTw20V0bS3NuLgP8AT+I0UTSeJpYEFgW0knSrHupihTPfbtNs1nMk7m2NxIH06dZAklC6aihp24+I4pU1BuR+LO/9ldpfHXp7f3XXbG5O9Nl7j2bRz4HtvelDT4ve29oaOpq8dV5DeuLpcPt+HG7wochRS0eTh+xpWjrKeQNGGv7j3fLaaz3W/trqzW3uEkIMamqr5jSatVSCCvccEZ6kbYLmK62u1liuDLGVHcaV4cDQKK+uBmtQDjoxWNpTXZPGUS/qq8jQ0wv+n96qiTkfkc/7b2j222e93HbrJCKzXESftdRX9hPSzcJxa2V5cngkLt/vKk9WOoqoiooAVFVVA4AVQAAB+AAPef6qEVUUUUCg/LrDcsWJYnJNeuXvfWuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de6acznMTt+glyeZyFPjqKEXaeocKGP4jiT9c0rf2UQFm/A9+690U7eXfmbyjyUe0Y2wmOuyHIzxxyZaqS5AeFG1Q49GXkcNKP6g+/de6AasrKuvqZa3IVdRW1UpLS1VZPJUTsbknVLKzNb88WHv3Xuh8626Tk3FSQ53djVdBjZir0WJRTTVtdAtmSoqpHHkpKWa/pQKJHUXJUED37r3Ru6OkpqCkpqKjhjpqSkgipqaniXRHDBCgjiiRebKiKAPfuvdSffuvde9+691737r3Xvfuvda5P82XqrH7O7v2r2Nh6CKkpu1Nt1LZ14EWOOo3XtWamop6x0BF6itwtZTa2sNRhubnn37reKH16qt/33++/p791roRetupOzu4MumC6x2LuLele8qRSyYihkOKoC17yZXOTiPEYyFByTNMv0+h+nv3WyCOI6tO6t/k/7zy1HT1/cPZ+P2lJNFrfbuyMcmfyNK7rxFVZ3JtBjvJE318MEqH8Nxz7rxNRw6MuP5Qnx4MAjk3l2tJJ4lR5DmsKoeQAXmES4KyfuDUAD6T791rpX77+I/wAgaLruq2ztjs3rX5DNh6BhtTAfKLZtfBXRSQpIIcRT9s9dVlLvHB080WhEqvtqx4HUEo6kr7AvPXtlyB7l7eds555VtNxgp2tIlJYifxRTIVlib5o4+YPQs5U575x5HvPreUuYrmxmJqwRqxv8pIm1RyL8mU/Ijqn/ALB/v51XV1A+Qnxb79+L0KSvHHu+ipZPlh8d6mVTKztB2x0jhKvf2yccYgrRSbm2tTJa4edSvvAP3M/u+K/Ubl7Q82jUSSLHcif95hvIxX7PqEPzk6zD5F++fPGYLH3I5aEiUAN3Y9p/00ls5ox8z4Tr8k6SW0d1bF7EnWLrffOwOyKqR4h9r11vTAbpyodtNQkdVt/FZD+81DIDMNcVVQwyK7aXUPdfeFHNf3fPe3kq6a1372x3XQGoJbeFryFjSvZJbCQEUzUhaedOsp+XPfD2n5ot1m2znuxViK6LiQW0ozTuSbQAa4oGNfKvQ/8AVm29+YzsHYuexWwt+5CPbu+NsVM4xe1dwPLG1Dm6I1NI0/2ISGUwakcOwAUnVxf2W+2+xc7WHO3J+/7dyZvc0djvNpIzQ2lzVTHcJqXUI9KnTqB1EUrnFejPnveOUrzlHmfaL7mnaYjebXcovi3MFCJIH0sBrqVrQigyaUzTpFbgo8X48uNsZHDZzZW1e1+/+n8HmtvTRVtFGnXPcG7hT4HIVdO00EeZwmJ3BBAlOSGSjgiKr4wpOTX94Fyzdbf7x7RzUtiU2vdtniAkCEK1xatIkqOwGkyiMxmldXh08h1Af3Md9hvPbbd+X3ugbzb9yZ9BbKw3CIyMq1qIy6OCaU1kjiekov3CjSRHMosFkEhje2tVAkDKQzJFdiwPqPAA+vvBUgihPWX9eIzTrvXUE/5iNR6eWnLfVnD+lIwbqoUj8G9vx711vPDr1qgi5NOOAdOmVvV4+RquvpE3N/qU/wAffiAQQQCP9R/w9ezio6fNubt3VsrLxZ7a2bzG2sxAVEWX29X1NJUlQ8eiGojhI+6hkdmvHIkkQQeoWJ9nfL3MnMPKW6Q75yrvl1t27x8JbeRo3IBrpbTh19VcMp8x0Ub3sGycybdLtHMG0W97tj8Y5kV1B/iWuUb0ZSG+fR0dm/MiiyNOtF3PsSLclRJZf9IGxDR4Hc7LdgajJ4a0WDzNQD9WTwa7crfn3nt7Y/f/AOZNoig2z3W5cG6wAgfW2fhw3NP+G25pFIR/HG0RI/BU9Yd8+/c22XcpJr/273o7fIc/S3OqSGvpHMKyxj+jIsg/pU6XOc+RXRuJwO7JNt5vce8q3NbSze26LaeS2dW4OGsfcVG9C4z2WqJpMeuOxXkEzCMM8kkY0c2PuYPcv77Xslvvt1zZsmyW9/uG839hJbpaz2jwwkzoUJllftCRatbUqW00SpIPUa8i/dT919q515d3TdpLOx22zvI52uIrhJZFETB6RxrRi0mnSvADVVqUp1W5EpSKONm1GONELG41aVA1WNyNVr++RiKERFBrQfP/AC/4OHp10nLamd6UqSafb1yddaOn+qRl/P5B+tiCRf8AFxf3bqp6FDo/D027Ox9ubQyDRig3/jd19f5MTCNPNQb62VuHa1ZToHZkSoqUyWiPkkOwAufc9fdd3N9o+8F7VXSSoituXgsWpTTPDLEwqeBbXpHnVgB1Df3g9vTcvZb3Dt3iZyliJAFrWsU0cgPnhdOo/IdWT/6YMr/w01/pF8x/vX/sr/8AcvX5v3v7+/wX/RV4PNqv93/fH9rVe/l/x999PLgfsp/Kn8qfl1x4zqrQV/l/mp1//9bf49+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XRF+b2I/Pv3XuiObm+Lm7OstzZjs74c7txXV+fz+Vmz2++jd1w11f8AHbtfKVUzT5LJSYKg8uR6f33lncmTcG241jqZrSZDHZBuR7rYp59KzYny52tWbyxvUnde09wfHTuTKMKfB7T7Elopdodg1aRJLUf6H+1scx2Z2KkesWo1kos6i3abGw2Nvdep5jh0bcEm9xb+n+t/vfv3Wuu/fuvde9+690zbh2/it04TLbdzlKldh83jqvF5KjkuEqKOthaCdNQ9SNof0sCCrWI5Hsq33Zdt5k2bdOX94thNtV7A8MqH8UcilWFfI0OCMg5HRhtO6X2ybpt+87ZOYtxtZlljccVdCCD8xUZHAjB613fkJ0JuboXetThshTS1G08pU1M2ytxRq8tJkcaG1rjqidvVBmsbEwSeJ+XCiRSynjgr78+x/MPslzdPtd7bNJytcyO1hdipjlh4iFifhnhGHQnuHetVbHYL2d92Nl91eWor+1uFXmCBFW8tzh45OBkC8GhkOY3HDKMAwyAvuDupc669+631737r3XiQoLEgAAkk8AAfUk/gD37UFqzGijj9nn1r5dXGfy+ukK/au3sp29uWlSnyG9aGCg2rSzI61dJteKbzz186SRK1PJmKyJWRQTeCNW+j299aPuJ+zV/yvsO4+6fMNp4e4bzEsVmhB8RLMNqaRwQChncAgVNY1U8Hp1ze+917oWvMG8WPt7s1xrstslMl0woUe6K0VFIJ1CFSQeFHYjivVlXvoR1hl0Un5mLDH1lsfKSwJUPg/kX8acrSoxZGSrHd2yaCKWOReEkiFcWBIINiPz791sef2dG1H59+61137917qo3+ZluVZcz1Vs2OWA/Z0e4d0VcKPL9yjVT0WJoWmjDCAwSCGfQSC4ZT9AeeW394nzF4m5+2fKccifpw3V261bXVjHDGSPh0kCQjzqDmnXQH7kuyFLLn3mNkYeJJb2yEgaTpDzSAGmrUKpXNKH16q7982Os7uve/de6Q/ZHWmwu4Nkbi627O2tit67F3XRigz2281AZqGthWRJ4JFaNo6ikraOpiSanqIXjnp5kWSN1ZQfZ1y7zHvnKW9bfzFy3uctnvdq+qKaM0ZSQQwoQQyspKsjAqykgggnom37YNo5m2u72bfLGO422daOjioI8j6gggFWBBUgEEHPRbdq/AX4x7SxG+cRBtbeO5R2H1/mOqdwZXsDtTsff+fp+t89GkWW2btvM7s3Jk6zauHrEij1igaGRzGhZyVHuRt099vcjdbzZLw7laWx2+/jvoktbK0tYzdxf2dxLHBCizuuaeKHABIAFegLt/s7yVYW26W301zP8AV2jWrvPc3E8i27rpaKN5ZXaNCOIQrUgVOB0bTb2Exu1cFgdtYGBqHDbZw2IwGEpfNLO1HisFQwY3F05qJ3knnamo6VE1yMzsVuSSfcU7he3O6X1/uN84ku7qZ5ZCQO55GLyGnAamJNOArQdSNZbfbWFja7dbKRawxrGorWiqoUDPyHRP6/8Al4/E3K7nyG7Mj17mKuqq990/aGNwcnYG+hsraXYUW4KTdFZu/Y2yEzq7Y2hm85m6JJshLQ00Rq1eWN/RLIrS5D7/APupbbbb7Zb8wRIiWLWTSC1tvqJ7UxNCsFzceF41xHHGxWJZHIQhWHcqkRpL7Lcgz7hNuEu2ys7XQuVQzzGGKcSCUyxQ+J4Ubs4qzIgLVYGoJBOuSWZnY3ZmLMfoLkk8D6D6+4Z/wf6v29SsqhBpX4euvfurdWX/AMtDAST727P3SVqBDjduYXbyOvi+0efK5CTJzRzE/vfcxRY2Mpb06JDe5t76J/3d2xST83e5HM5SQQW1hb2gPboLTSmZgfxa1ESFaDTRmrU06wi++xu6w8u8j7CGTXPeTXBBrrAijESkeWkmRga51KKY6uH99XeudvXvfuvde9+691737r3XvfuvdFC+RuM8OewGYUWWuxk+PexJJkoqgzoSNOlR46o83uSB/T37r3RdQSpBUkMCGVgSCpH0IIsQw/B9+690bv4+7rpazEVW0WpqKjrcOGrab7KkpqKOtx88oV5pYqaKGOSrpqhtMjkFnDKWJNz7917oxgFhb/ff8j9+6910Vv8AX6fQjix96p1oitM9V67uxE2B3RncXMhUQZGoeAkaQ9LUuammkQWHoaKWw/xB/p7wU5r2ubZeZN52yVKGO4dl+cchLo32EE/s6y65b3CPdNk2y+Rq64FB+TL2sD8wR+zpC7j2/id27c3DtPP0xrcDurA5nbOdolmmpjW4XP46pxOVpBUU7x1FO1RQVciCSNldNV1IIB9ksFxJbyxTRECVGDA8cg1GOHHo2ubeO6glt5hWNxQ/YegDynxD+P8AmvjFh/hzkdlTz/HjAbU2bsnFbHG49wRVNNtrYGTxWY2pjzuaLIJuOZsdkcJTSNK9QZJtBEhYMwJonMG6x70+/rcf7tGd2LUHxOCrHTSmQxHDorfYNvk2qPaHjJtFVQBU/hpQ8fkPPpX70+PvUHYfaOL7k3vs6l3LvvE9TdhdGQVeTrK+XDVPVfalbj6/f208ntsVC4PJ0u4ZsZCsks8LzRxhkRlVmBT2+8X9rZNt8E2m2M8cxoAD4kYIQ6uIpUkUPH7OrTbDt1xci6lirIIjHxNCp08RWh+AYpnz4nounSn8tj4kdAdg9edpdd7R39JvbqHH7mwHUeS3x3R2r2NR9XbO3bg6jbuY2DsTC7y3TmMPt7YxxVSyw42GERQuqOvqij0m24c5b7ulvc2t1NF4M5UyBIo08RlbUHcqoLPX8RNePqei605N2izminVXLJXTVn7QRQqKsaA+YGD1iy/8tD4gZLZvTuyaDZW99lUnQcO+6DqzcnW3cHZ3XnYGA292huOu3Z2Ns2q3/tPcuM3RnNmbyz2RlnrMbWVE1Pcr4wmlSPJzlvy3N9cyTxym5KGRZIo3RmjGlG0MpUMowGAB61LyftTw2sKIyeEWIKsynubUwqrA0NBUVI+Vel/198Ffi31Vhuhdvdf9ZjbeC+MfZPYvbfSGKptzboqKTZO++18fuPF74yiCty1TLmaXK0G7K9Epa5qinpvMGjVWRSE91zTvl5JuctzeanvIkjl7VGpUpTgMHtGRQ46ei5U2aH6Tw7engyF1yeJLn19XP29KLK/EP4/5vb/yh2tktkzz4P5l19Vk/kVSDce4Ym3zW1mzcbsGompp4sis22NW1sRT04XGtTKGTyAayT7ZTmHdY5NmlS4Afb/7DtXso5f07sn8Va+fTx5c2wrua+EaXXx5P8Oj19B5U6RPc/wD+LvfVd1Vlt/7O3VTZ3pnZj9abK3HsHs/sPrLcknV1RDQQZDq/dWe2LuHB5PeHX+YjxsX3eNyEk8Uza2NjI5ZRt/Ne87at8ltNGY7iTxHV40dfEqSJFVlIVxXDLQ9NXnK223jWzsrCSNQtQzAlf4SVYVB4kGo+XSeyH8t/wCJ0/VPXHS23drdidY7C6iym/8AJdc0vTndXa/VGZ21D2jl6jNb8wEO4dlbrxWVrtqZ6rqSv8NqXlpqWJUSnEQRbOJzfvYvbvcJZIZrqcLr8SGORWKABG0OpUMKDIFT516pJyjtbWlvZxhkjjLadLutAxJIqrKfP1p8ujX9XdYdf9K9d7N6m6q2rjNkdc9f4Sm25tDauHWX7DDYmlaSQQpJUSz1dXU1NRNJNUVE8klRUzyPLK7O7MSO+vbrcrue/vpjJdytqZjSpP5UA+QGAMDHR3Y2Nvt1tFaWyaYUFAPy6MJ1NiP4vvzDhl1Q4wT5af66R9qmmnDfUWNVKn+vb2N/a7ahuvOm0iRKw22qdv8AarRf+NMP2dBX3C3E7dyrf6WpLPpiX/bGrf8AGQf29Hl95l9Yv9NWezNDtzB5ncOUeSPGYHFZHM5GSKJ6iWOhxdJNXVbxQRAyTSLTwMVRQWY8Dn37rx4dU1fFz+bZuz5I1HWvZj/F6PZ/xN7sx+ezPWvbtP3jtLd3Z2KwOOrMjQbeznavSOKwMEu0IN5VONZYqTH5zM5DGF0FZBGS2iH+dPfj2w5C3SXY+YN+YbxGAZIoopJWj1cBJpUhTpo9CcqQfMDqQ+V/avnnnCyO47LsxNhmkkjLGj0FToLEavSuBXFcGjz8Lv5zfRvzC7D+VfU67PzHUu//AI45nflXtXDb13BiRD3x1ZsrdG59kS9m9fVrQ48JBHu7aVTQZHHSpJUYyWWnLu6y+kb2nO3Kd7aWN9Fv9sltcmMRmRxGS0sSzxppcrSR4nV1jPcVyBg0DN3yxzBZXE9rNtMxmi16tCNIAEkaNmBUEFVdShYYDYrkVMb8bv5jPQndPw+6G+X3Ze7dhfHHbfenXOR7FodudodlbVx8mCoMDS1+Q3LSy56vnw1Dkzt7FUDVVXJHEghgOplA9iKG6tbmSeK2uY5JYjRwrBih9GANVI8wQOiaSGWIRtJE6qwqCQRUfKop/Po72zd57T7E2pt7fWxNyYPeOy924ihz+1917ZydHmtvbhweTgSqx+Ww+WoJp6LIY+tp5FeOWJ2R1Nwfb/TfSm9+691737r3Xvfuvde9+691737r3SL3rvrC7Gxv32VkaWom1LQYyn0tW18yj9MSswCRISNcjWVR/UkA+690RTd+8c1vfKHKZmRQIw0dBQQM/wBnj4SSQkKMfXKf7cp9bkfgce/de6S4DMyqqs7uwVURS7uzGyqiKCzsx4AAuT7917ox/VvTWQlrqPce8KRaSgptFTQYSc3q6qpDBoZslBYpDSxAaliYl3a2oKBY+690bcD8/wCH+t/vH+w9+69137917r3v3Xuve/de697917r3v3XuiO/Oj4o5T5VddbexG1s9iNv7z2Xn5M3garPxVJw1dT11IaDK4nI1VDT1WQo0nh0SJLFHIBJEoZCDce690U3pP+UZs/A1lJme9d7S79ngKS/3P2pDU4HaxlR1ZUyOUnk/juWgIWzRqKRW/Nx7917q3Dauz9rbGwtJtzZu38PtfBUCLHSYnB4+mx1FEFUIG8NNHGrykKNTtdntyT7917pSe/de697917r3v3XuurH+p/3j/inv3XuiJfKT+WZ8FPmTS1DfID429ebp3HIqmn7FweOm2F2vjJY5YZ458T2jsSo27vvHzRyU62KV9rXFrEg2R2iOqNirfLH+DqpVWqHUEfP09Ps6JZmPg53R8bospK/eXzL+YvxbgglbIdU1PyP7FxnyE6/2jBS+Kq25tOo29X4ak+Qm1Y8ehT+F11VidzmEeOCsyMrCCT2t611Z9cV+efn1ftpTQKU/4qnpT5dLMdCdGd47NxvcPwEqepN0bKnw+E2N2b0PLlMlsraO4k2JR1WM2/MJMfjqrc/QXyY64p5pKD73JYmSWso2bH5qkdUp56UAe4/tpyd7rctz8rc7bULrbS+tCCUlglAKrNbyjuilUE5HawqrqykjoXclc9cz+32+wcxcq7k1tuKrobAZJYyQzRTIcSxsRUq2QaMpVgD1X12fleheqd40+xeyu35Pjzu2uqYaSm2v37hIa+haecyhY6Ls7pSr7F2ZWQExjxyVyYqV19bxx/p986ecP7vDmG3uZZfb/wBw7O4si2I9xjeGRRk5mt1kjc8BmNCTU4HWbXLH319nlt1TnLkmeK6UZeykSSNmxgQzsjp5nEjgcBU9JeLc3R9bk58Dg/lt8QNzblplhkm2vgu/9rzZ6GOdwkclTR1sNElCObkVDxNb6AkH3El79xX7wlpby3EO17RcutP04r9PEYVpUeLHEmBmmqvlx6km1+917N3M0cUl1ulujD45bM6AaVodEjtngO0ivS+ruvc7h9u028dx5Xr/AGns/IsUw2690do9eYjbu45UbTJT7Xykm45INx1EZ5aOj8roLEgewPtn3SvvE7rNcW8ftndQPEMm4lt4EbNOxnlIb8vLPn0Kr/7yfsnt8EFweeoZ1k8oYbiVl/06rF2/mePUbHYfryqDnJ/IXpfblOscci5bKS9iy7VYOQpQb6o9hPsozRTHxuqV72YHmwPuUdu+4H76XduJr7cOX7OU0pE91LK5qKmpht3VSOBqSPQ06AF798j2nt5dFrt29XUYJq6wRRrjFQJZlYg8eAxxFenrNdL9gYrBpvTAUmH7F2LU6mj3z1Tn8d2Ns6rVdKGU5fbMlc+PlRVsI5oYSWNmsbe4V9wvu4+8vtkHn5j5Mnl2tT/uVZ/43b/aWiGtK/8ADI0+3I6lXkz3y9sOe2SHZeZ4otwNP8Xuh9NPwrRVkOiQ/wCkc/y6DRZA4NrhkZo5Y3XTLDMlvJBNGfVDPE3DowDKeCB7hA1BZTWoNCDWoI4gg8CPMHI6ltSGCsuVIqCMgj1BGCD6jHXfuvVuu7gAk/QXv/gLEk/i1vfutHoR+iwV7f6cJ9VuytkyL6Y7G+4aJ0I8VoiBfgr9R/jf3IntB/09r2uH/hwWHz/4kx+XQG9zBX229whX/liXv/aO46Od91Rf7IZ/o/8A9yX8L/2d3/RN5LQ6vtP9nj89/Pqt/CPJ6PJp1+H06ff0Yaj42v8AFrr+da9cSKDw9NMaKf8AGev/19/j37r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3SI7D612B21tTJ7G7N2dtvfmz8wqrkNu7qxNJmcXM8d/BULTVkcqwVtK7a4KiPRPBJZ43VgCPde6KcvR/yB+P1DJ/stHY0fZ+yKWQT0/QvyQ3Fnsw2NoIRc4TrDveJMrvXa8CwosdLS7ipty0cRsqSUsVyPdbFPPpedS/LjrvsHc8fV288buHovvqKFjW9JdtwUuB3RXmHiprevs5DUVO0O2tuK6OUyG3K7IReMXmWBrxr7rXRrPfuvde9+690geyOt9pdq7Syezd6YuLJ4fJRkA2CVmOq1VhTZPF1RUvRZKjc6o5F/PBBUkEFe4Ht9yt7ncr7hyfzftq3G0XC48pIpBXRNC9CY5YzlHHDgaqSCKOTucuYOQt/seZeWr4wblAfnokQkao5F4PG4wyn7RRgCKH+9/i12N0bW1dXVUU+5djeef8Ah28cZTSSxQUaSIsC7kghQjEVzI4B+sLlSVYDgcTfe37tHuB7MXl1d3Ns+5cl628K+hVmCoCNIu0UfoSEHJzGSCQwGB1W9qPfnkz3Stre3gnWy5p0r4lnIwBZyDqNsxP6yVH+nFQCCcks6FZBdGVhx6gQQb/0IuPeOwoQGVgQfQgj9ox9vU4Gq4ZSD121kGpiFUfViQAP8STYAe9EhRUsB9poP28B+fWq1wASerGvh/8ADx98yYztPtOglg2fFNFWbX2pVxaH3bo9cOVzEUq648Ar2aGKwaqIubR8N0D+6l909+cjtvuX7k2BXlNXElnZSKAb2mVmmBFVtqgFY6VmoCf08Nhj94n7xicsi95F5DulbmJgUurpTX6UHDRQkVBnIqHbhHwHf8NysMKQRpDEkcUUSJFFFEqpHHHGoRI441CqkaKAAALAD31rjjjhRIokVYlACqBQAAUAAGAAMAAUAx1zid3kd5JHLSMSSTkkniSTkknJJJJNes3u/Veik/M7RN1bs3GGWKGfM/Ij40Y6iM7FUkqv9OOxa1YVCgs8skVC4VRyWt+Pfutjz6NqPz/vuObe/da679+691Sd/Mj/AOZ17O/8RvH/AO9FkPfH3+8I/wCnu8nf+K+P+0qbrpj9zD/p23Mn/S5/7V4+q+veCPWX3Xvfuvde9+691737r3Xvfuvde9+691737r3XTMEUsfoLXsCT9R9ALkm/0A5J496qBlq0H2/soKk18h5mnWuJAAyf9Xn5dX7/AAo6jqerulsTUZekkotz76qP7352mqI/FU0UVXEkeDxkylVlV6PEJGzo/KTyyDj6e+5P3Qfa6f229oNsl3S2eLmPeZPrrlXUK6BwBbwsKVUxwBSynKuzjHXJP7zHP8XPnubfrt9wsmx7Wv0kDKSVcoxM0ooaEPLUKRhkVSa9HA95UdY+9e9+691737r3Xvfuvde9+690A/yDxP3uy4MkoJfC5amqD6rAQVatRzErpJchpFtyLe/de6Jf7917oROpq+soOw9sGjvesq5MdVIA7h6Oqgk+41Kv0EfjDBvopW59+691YJ7917r3v3XugU7b67l3PRx5vDxas9jojGYFZU/iVCCXMNyADUwElorn1XK/ke4h90eQpOZLVd42mOu+wJTSCB40fHSa/iU1KZzlfMdSR7f83psVw22bjJ/uqnauo1Phvw1f6U8G/JvXonzBkZ45FeOWORo5IpFZJI5FJDxyIwDI6EWIPvFNkZWZHUq68QQQQeBBBoQa+Roesi0dXVWRgyMKhgagjyIIwevE291p/h6v173rr3Xvfuvde9+691737r3Xvfuvde9+691737r3XJEeWSOGJHlmmdYooolLyyyObJHGigs7u3AAHu6RvK8cUalpHICgCrMSaAKPMn/D028ixrI7kBFFSSaADzJJwAPOvRy+ptgybTxc2QycSLnssqmoTUGNDRr6oKIECwkLHXLYkarAfp95b+1/JMnK+2PfbhHTerqmsVr4cYysdf4s1c+tB5dY28/c1rzBfpaWMpO025Ok8Nb8Gf7PJfl9vQv+5SHAdR/0xbozFPt7bO4s/V4zKZqlweDy2YqcPg8bJmc1lqfGUFRWzY3EYiL93K5SuigMVPTL6p5XVBy3vfHrR4HrQ/2Luvpnb/zy6xpP5ZOd7mput+3u4d77k+XPxn3J1/2TtLo3qHYFdjshmslv6p2v2Tt3Dy9DduQbvmp6WHEYyoMOQd3BphEAzc5vdrbd0ey9z290bSyEtlMf3NfKYkv7hmlrFbMkTs9xbJCzBmmQGPTh+FMyOQb21jl5M/qBPcn6mCu421Ge1ipEQ8gZlCxytKFoImCyksCmKlDL012BjvjFkPlZ1RtjLVHyA+LHyc+c+fotpPj6mjr+4Pj5v3uje9P3B1PNSS06VeR/je1mGb2+wDIuWoo2i1CU+w1+/Nsn5xh5F5guo/6sb3s2zqZdQYWt/BZw/TXINSF0SVhnB4xSNX4ehE+ybknL0vNm2Wsn7323cr9vDAIaa2kupTNEBTUda0eKmBIkbDJPXH4+dV0O7enf5Cu3999eTZ3DbWft3dOX2/u7a0uQx+DzGF6X3nl9o1u7cLk6KXGU8tNm4aWSKGvjME1QiApJ6R7Xb1zVuHLXNf3lN75a302m6FrURTQyKGat3EjmHJDDRqBIBIGqtB0W7dy/bbxsXs7te87QZ7EmTxI5EbCmByNfArk8ajOR1uP/ABX+QVFkY8X1NuLF7a202NxsFDsqTbuModuYOqpaJAhwgw9GKbFYyuCEGnipIooZVDBUVgAZl+7794+9513CPkvn6aFd/Mdba6FIxclcFJlroFw1QV0ALJRgFDDIE95vY635Vsn5o5NilbZlY+PASXaAH4XjNNRhHBtRJTFSVyD8D6D/AFh7zL6xj679+691737r3XRNvxf37r3QfdhdgY3YeK+5nUVeUrdcWKxgcK1TKq+qaZgS0VFBca3tckhRyR7917oie4Nw5bdGVqMzmqo1NbUWAAutPTQrzHS0cJJEFNFc2HJJuWJJ9+69030VDWZOtgx2OpJq+vqm0U9HTRmWaV72J0DgIn1Zj6VHJ9+690cfrLpum2rNBntwPDkdwIpalgT9ygxDPyZINaB5q8KxUyk6V/sD8+/de6Hf37r3Xvfuvde9+691737r3Xvfuvde9+691737r3SD352Ft7r/ABaZHOSytJUSmHH46kVZchkJ1F2WnhZ0AiiB/ckYqiAi5uQCDOdueth5D2wbjvUrl3JEcUYDSyMPJV4UGNTEgLXJrQdCblflPd+br1rPa4l0oKvI1RGgPmxHmfwqMmhoMdFEzXya3vWzzfwXH4XCUbOfAJoJcnXLEVYL9xNJNFSmS5DXSOwItyL+8Wt5+8XzjdTyR7RZWdna6jp1K00gH9IllQMeOFoDg1p1kHtnshyzbRRfvO9ubm6p3aWEcdf6IALU8stUj06S1N352rTzJI+4qerADAwVWIxxhfULXYQQ08pZCbizL/jccewvB74e5ltKssnMEcigU0vBDpNfPtAbHlQj59CCf2l5EmjMa7O8ZNO5Jpaj8yzDPzB6EXbvyhzEVSke69vUdZRu0SGrwRlpaunW2maVqSrmnjqyT6gqvGfwL8e5B2D7yO6wzqnNGxwS2RZQXt6o6rwLGNyyyHz0hk8wKmnQL3n2KsGiMmwbvJHcAE6J6MrH8IDKoK+lWDeuOje7c3Hh904ijzeCrEr8dWqWhnQMrBkYpLDNFIFlgnhcFXRgGUj3lNsO/bTzJtVrvOy3iz7dMKqwqDUGhVlNGVlOGUgEenWPe7bTuOx39xtm62rRXsZoynIzkFSMFSMqRg9P3s56LumjL5/C7fpGr87lcfh6JCQ1Xk6ynoae4F9IlqZI0ZrfgG/sr3be9n2K1a+3rdLezsx+OaRIk4V+JyAcemelu37buO7XAtdrsJ7i5P4I0Z2/MKDT8+gWy3yi6MxEhil33Q1sgLBlxNJkcoFKGxVno6SWPUfxYm49xFuf3kPZjapDFJztBNIOPgJLN/ONGB+0EjqSrD2P90dwTxE5TmiT/hrRxfydwekfN8z+k45GjSq3RUKpsJoduVAjf/FPNNDLb/XUewpJ97j2fWR0jutxkUfiW1cA/ZqKn+XQkj+7d7muis1vZKx8jcLUfbQEfsJ6Kv2D/wAN39mbuye/d4dJx1+9c/8AbruXdOH2vktqZzdC0i6KY7uyO1M1gqndQp4gET+INU6UAQekAe2G+997RKaA7ofstf8Arp08v3a/cyhxt4/6iB/0B0M/V/Y/wi6yx4xPW2ztl9ZUdTaOqhw/WUOAafyH1SZSsx+IZ65mJu8k0srH6kn2d2f3p/Ze80CTmOaDVT+2tp0A+06CKDzyadFN393z3SttbLskU2kV/TniNfsBZTX0xnpZZ3rb4Q/ICdY9xbB6B7DyUsvljmym1tpzbgaYrbyQVc9DT5oSFW5KODz7kzYfdD275nlSDYuc9uubhuCLMgc/IIxDV/LoCbxyFzvy9G028cr3sEI4sYmKD7WXUo/b1m6X+B/w3+PW4sluvpb43dUde7kyyRJWZvC7YpXyQEU9TVXo6mvNY+NkmnrJHlam8TTkjyFtK6R3Unyx0EC5OK/6vn0bJqaBoDTNBC1OVKeBoozAUP8AZMRUoV/wtb3vhw610VHe/wAK+kNy5qo3rsvGZjoftGZ1nXtLoPKt1nuqepiZ3jbcVDiYW2jvukd3/dpc/jMnTTLdXQgn36poQDjr3nUjqvr5E/HvujbuQr91776sru/KSmpKpYe0vjPh8ftnsrcMzU1THjYO2uia6skxNZlUylRFNU7k2lJVyz08TxvhoUZXjxl92vune1Hu1dz73e2U22c0ujg3VmUj8V2ACPcwlDHcFPMnTIy1BfhSdvbr7w/uN7dWsO0WN5FfbAsiEW91qfQoqHSGXUGhVgcAVUMAaceiD4as2tvLA53dnWO7qTsfaW1tyZDZ+7Mpj8LntuZjZe6cdXTY+bB772buWjoNxbTraqqgk+zeeI09aqMYZGKuq8sfeT7sfuX7MOt3uVom58rMG039mrtCuk0pcIQXtiaimvsPk5p10H9r/f3kT3OU2llOdv5hBH+J3Lqsj1FawSYScYNdNGH8NMlvFTFVL46SZJle2uank8kcaEIxvLAzaJpI3vH9QT7x2VlbKMGHyz+2nD8+pvJ4VBp5fP7Ohk6MqsBiu09r7r3XVwY3Z/XAy3aW8cpVTmmpcRtbrjEV26snlKucAmKCD+GoovYSSMsYOphedvuz8szc1+/PtptkcDSRw7gLuQCo0xWitOzkqDQBlQZoGZglat1EPv1zAnL3s/z1es+iWaz+mStDWS6IiCipydDMcVKqC1KA9H4/uF2L/wANefxr+BV3+lb+G/7NR/djx/7lf49/pU/2Yf8Aur9p4v8Ai7fwT/cZ4dGr7j021e+/uoaq0FPsx9tP506440FNNcUp/Lr/0N/j37r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XvfuvddEA/X/fX9+690HXZvUfWfcu3f7q9o7H25vjBrUR1tHS57HxVU2JyUJBpsxgcioTJbfzdE4DQV1DNT1cDgNHIrAH37r3RTJ9u/Jn4rzHJ7Ly28Plt0HFIXyfW+6a+lyXya68oGdTJWddb9ytXjaLufC49CzNhdwSQ7gMYApspVyBaV/de6M71F3n1d3jiMnlut910+alwFdHid27drKWuwW9Ni5x4RP/AHe35srOUuO3Rs3PJFdjS5Glp5WQa0DIQx917oW7g8fX/ff8b9+691gqaanqoJ6epp4qqnqI2inp540mhnjcaWjlhkDRyIy8EEEEe2poIbiGS3uIleBwQysAysDxBU1BB8wQerxSywSpPbyMk6kFWUlWBHAhhkEeRHRec98S/jzuSvbJZLqzb0dU4byfwtanCwSM8rzPJJS4qejpnmeSQ3cpqI4+nuB97+6/7Dcw3p3DcfbWwW6INTCHgViSWLFIGjUsSTViK+VadS5tPv77v7LaCys+eLxrcUp4pWZhgAANKHYAADANOo+J+IXxzwtbHkKPqvATTxq6LHkmrctSESAAlqLJVVVSO62urMhKn6e2Nr+6r7AbReJfWvtrYvMoIAmMk8efMxzO6E+h01Hl0/uP3hfePdLVrS456u0iJBJiCRPj0eJVcA+YBofPoyEEENPDFDBDHBDBGkMMMSLHHDFGoSOKONAEjjRFAUAAAC3vICKKOCKKGKNUiRQFVRRQAKAKBgAAUAHAdQ5I7yyPLK5aViSSSSSSakknJJOT6nrN7c6p1737r3RKvlZWLnezfhp1VTnXXbq+RdJ2LV06+QONtdGbO3FvvIVxZf2xT0u5Ww0TB+C9QoHJ9+62PP7OjqD+trXsf8b/AOP+t791rrv37r3VJ38yP/mdezv/ABG8f/vRZD3x9/vCP+nu8nf+K+P+0qbrpj9zD/p23Mv/AEuf+1ePqvr3gj1l91737r3XvfuvdO+39v53dmbx22tsYquzmfzFQKTGYrGwmerqpiCzFVuEjghjBeWVyI4owXZgov7N9i2De+Z9527l7l3a573e7uTw4YIl1O7DJ+QVRUuxIRFDM7ADoq3retq5e2y93ne9witdqt4y8kshoqjyr5kkkBUHc7EKoJPVi3Xn8uHclfRfxDtffVLtJ2X9rC7Wjp8xURXVNLZDM5FYcfHKrkgxwxSqT9JPefnIf935v17ZvuHufzpFtbEYgstM7LgCslxKFjBrXsRHFciSmOsNOb/vn7RaXS2nIPK8m4L5zXRaFWp/vuKMmSh46nZT6p05b1/lt5CDGVGQ6z7KjztZBAphw26cfTUkddOjOZo487iXkhpWdNIjElMyhr6nA+hhzh/d7XkW2z3vt37hi9vETtgvIkQSMKkhbiE6UJFAA8bLWpZgKUSctffTt5L6G1535Ja1tmfumtZGcopAoTBKKuAaklZQacFPRFNxdG9z7SrPsdw9Wb5pJiXWOSl29XZijn8f6zTV+Fir6OZVBvcP9D9L+8KuYfZT3f5VuzZb37Z7zHLU0KWzzxtTiUktxKjD5hvtA4dZV7J7q+2vMNv9TtHPe1yRgVIa4SJ1rw1JMUdT5UI48MdGO6l632R0bgT3L8m9tyyVVZLFH1T1Tkac/wB4szX0EwqKrceTwlQ8cVPTRN4hAK39uJbyOup4lOQftf7f8l+y2xn3a+8by8xupWA2bZZVrdTyxsHe6lt2KqqIfDEXj9qCsjjU8a9Qt7hc6c0+6W7D219kd7XwIgTuu6RkfTwo60W2imUEsx7tZh7mNEU6VcgSpv5l2+f4zLLTdYbZG3vIfBQT5vJ/xn7cE28tfHB9h9wR/qYNA/x9yBN/eIc5fvd5YPbnbf6v6jSNp5vqKVxWQDw9VONIytfPoGx/cm5Z/dqRzc83/wC99I1SCGLwdXyjLa9Nf6derLul+4tqd27Io967XmljSVzR5fD1ZH8QwGYhjRqvF1qqLExlg0cg9E0TK6/Ww6H+0Xuvyz7x8m2fOHLUjKjHRPA9PFtp1A1wyAcaVqrjtdaMvmBhP7j+3m/e2PNF3yzvyAsvfDKoOieFq6ZEJrStKMvFGqp9SLnuUOgH1737r3XvfuvdMW5NxYza2Hq83lZhFS0icICvmqZ2BENJToSPJUTvwq/7H6An37r3RB9578z29sjNV5Gpmgx5c/ZYWGof7CihX9GtBpjqalhy8rAljwLAD37r3SYjx2QmpJa+ChrJ6KCXwz1cFNNNTwTFBII5pIkdYnZGBAa1x9PfuvdGJ6G2LlI81Lu7LY6eioaahkgw/wB5C0MtXU1pTy1kEUgEgp4qYFQ9gGMht+n37r3Rsh9P98Pz7917rv37r3Xvr7917oMt69W4DeBar0/wrMAHTk6ONAZyRbTXQ+lapP8AE2cD6N+Pcd84e2+xc2Brgobbdaf20YHd5gSLwfPme70PQz5Z543flw+Bq8fbSRWJie35o2dB+Q7fUdFkz/WO9tuSyK2MlytETZazExtWxOn+rkplU1FOw/2pTb8E+8dN89ueb+X3krtr3Nr5SQAyKR6slNa/MEZ8qjqbdo555a3lYyb0W91T4JSEIPoG+Fh9h+2nQeyK0TmOZXhlUkNFMrRSqR9QY5Argj/W9gJ1aJ2ilUpKOKsNJH5GhHQ0jZZUDxMGT1U1H7RUdcffgK163Ude/wCJ+n+PvXXqj16979x69UevXvr/AL1/sfe6H0638/Lrq4LKgN3Y2VBy7E8WVRdmN/6e9fiCDLk0AGST6ADJ61+Ev+Aefl+3pe7e6z3juSWMQYqfHUjldeRyiPR06Rm15IopFFRUEKbgKvP9R7HOw+3fNfMEkQt9se3s2OZpgUUD1VWGtvsAz0EN4535d2YOJNwWa4ANI4qO1fQkEqPzPRntk9VYHZ0iV7GTLZnTp/iFXGgSlvbUKGnF1p9X013ZyPzY+8jeT/bXZOU3F3m53X/frgUXyPhrnRX1qW+fUH8zc9brzGptqCDbq18NSatThrb8X2cPl0KIAH0Fr+5H6BPXfv3XuujyD78evUrg9ES+Vfx+z+8q2Hf+w8fFXZWOjSk3LgqYJBX5lKUn7HJ0lyiVtfSRMYmRjreMLpN1t7wu+8x7E7zzdfW/PvJdilxu6QiO6tlxJOqn9OWOtFZ0U6XQ0ZlC6SSKdZQew3u3tXLFvPypzTdGGweUvbzsKrEXoGjfzVGI1auCmtcHquHIY/I4epNLmMfkMTWRkIafJ0VVj6hXS3pEdVHE+pSv4/p7563+27jtFy1pum3XFrdVIKyxPE1Rg1DKuf8AUOs0rO72/cYBPt13DPbEV1ROrrQ+dUJGeoZfUWYsSXPrJJOoggi/9eR7SUYmlDWnp/qr9vSkLGNNFXtGOGB8uhP6W2hk979obOxWMgklSjzNFm8pPGEC4/F4mqhqqmslMjxgKHjWMAHUXcAe5O9m+U9y5v8AcrlPbdvtmdYbyK4mYAUihgdXaQk8KGigDLFqDPQD9z+Y7DlrkbmG/vJVDS2zwRA175ZlKqgA86EsScALU9XkXAAvx9B77MdcwBgAdeuP6+/de66DKfof8fz7917pp3DllwWDyuZaKSdcZQVNb4IlZnlMETOsdlBIVmA1H+ytz+PfuvdVzZzPZTc+Uqc1mat6utqjybjwU8IZjFSUaAlYaSLVZVH1+puTf37r3T9sjYmb33kPtcZH4aCB1/iOYmVvs6JNXqRTYCprGUHTEpuDy1l59+690d7Z2wtvbJoxTYmkD1UiAVmWqgkmSrG+pEs6qpSK/wBI0sg/pfn37r3S19+691737r3Xvfuvde9+691737r3Xvfuvde9+6910SB9ffuvdVd9o7nqN3b83DkpqlqmipsjU4zEIZTJBT4/HyfbIKVSFVEqJYnlchQWdub2HvnB7j8yXHNHOu+7hJOz2sdw8MAqSqRxHQNAPwhyC5pSrZPWcXIexxcv8rbRZRxBLh4VllNKM0kg1HURx0ghRUmg4dVg5/tTsOk/mndYdJ0+7crF1Pm/gr2f2XldiIab+C12/cJ3DtrA4vdEqmA1ZydFhqySnQiUJ43I0359+g22xb243HdTaKdxTeIohJXu8NoHYqPlqAJ86jpNdX12nPNnZLORaGyZivlq8ZBX14EjoIt77i7m+Xnzh7s+L2ye++y/jd0P8Seuercz2XnujKjAYTt7tjuLuWjr89g9vLvbcWD3Em09hbD2tQLNUQUdKZ8jWzhXkEQsprYwbRyryftfMN5tEN9vO5XEqxLOHaKGCAhXbQjLWSRzgsaKoqM9E24S7vzLzPfbLa7k9pttnGhZkprd5AWUVYGgCippk6hU0FDYF0fsDffWPX1HsvsPuXdPfucxWVzbUPZe+cHt7B70yO2qusNRt/D7qXa1NQ4TN5rAUT/bSZOKmpmrgod4le5IG3ncLTcr97uy2mOxiYCsSM7IHABZkL9yqTQhSTTyNOhxsdhebbZraXu4NcupOlmChqVNKlQAT86DHlx6PT8aNw5Gi3vUbbWaR8TmsbWVktKX/ahr8esbx1kaEHS8kJMbaSNQIJvYe5s+7xvt9Z823HLySE7ZeQPIUrhZIgCJADwJFVYLStQTkdRj72bNaXPLkG9mMC+tpkQMBlkkqCpPmAe4VrTy49HR3fuSi2ftnO7pyRb7DAYqtytSiEK8qUcLSiFGPAkncBF/xb3lzzRzBZ8qcu73zLuFforG1kmcDiRGpbSK+bEaR8yOsa9g2a65i3vadisqfVXdwkSk8AWNNRHmFGSPQdUeb/7C3R2huGr3HuutmqGqJnfH4oyyHGYSjJJp6GgpC7Qp4oiA8gGuVwWYm/HGnnvn7mT3G3u637mO9Z2leqQhm8GCM/BFFGTpXSMM9KuxJY5HXUDk/k7Y+Rtog2fYbZUCikktAJZ283kYAE1NdKk0VaADqnD56/Nnuj4pd+/H3C7F2Vt/enSs/XfY/cnyaoDi6ys37jOpNibr2Ztjce7Nhz01TGiVGxqHdv8AF6unaGXz0lLL9At/cg+13txy7zxypzVPuW4y23MP1UFrt7BgsLXU0c0kcMwI4TGLw1NRRmAp0AvcHnnfeWOZNlisYEl2hIZJ7sEEyCCNoldo85KeLrI46VPDj0MOS7y7a7F+UXbHx86c3v17t/b8Xw16i796m7IyuzJd9UcO5+xOxs3hXzeRx9Nn8Im6dqV20sdE9LSpPTFZJRL5GA0Eji5Z2DZ+TNi5q5g2y6luf6wXNlc26zeCfChgjfQrGOQxyLKxDHQ2FIpXIWvzNvm9c17ly/sd9BHbttcc8MpQyAO8ki1IEialKqPMetadA58YN/fO7s/vr5C7L31370XktlfFfvHA9Z7txW3PjfX4XcHa+HyGwMRvyplwWZl7OrI9kZKYZhaKIvDXqrx+QkhtPsQc57X7Y7NyvytuG18r7ku473tr3ETSX4ZLZ1neHvQWymVQELEAoc0pivRNyru3Pu7b3vdtd75bNa7ZdKkirbsDKpjSQ0JuCEI104OO08es3we+QnyZ+VmSXtvJdzfHnH7Ood6782x2t8PqTrzK0vc3RWO29ms1gtuUW4N7SbqG4P7/AEktFS1dcMjhIMVUU9SRTNwpLfuTynydyLbnYoOXd1a/e3hktt1M6G1vGdEeRki8Lw/ByyJ4cxl1KdY8un+ReZeZ+a7wbo++WS2onlSawMbCW30khAzmTV4mASGjVTXtxnq1dUUOHAs66WVx6ZFYXIZWWzKR+CDx7goYcSDDg1BGCD6g8QfmOpqqSCpyp4jyP2jzHR0Pi13zuHC7tx2wN35yryW1s/px2EnytQZ5sBmLsaKCOtqGM5xuQJMWh2IjlZCtgSDmD92n3v3vZ+Z7HkTmreJLnlu9pHbtMxZra4OUUSOamGXKFGJ0OUK0BI6xl99/afadx2C95w5c2uO3320/UnEahVnh4OxRRTxIx3agBVQ1c56tJUkjn68++kw6wb+zrl7317rqw/p7917oqfaHww6D7Nyo3Wm1JOt+yIHSai7T6iqU6+33DURPUy082Sr8PAuL3alLNWztHT5ukyVMhnkIjBkcnTASI8TgNE3EHII9CDgj5HrYJUggkEGv5jzHofnx6KJuz4Od+4bF0eP2JlviP27/AAijGOw1T3l0puPrbPxUUKBaSDM5r487l25t3NyQKir5V2/Sm17KObw5zR93r2S5z3E7tzJ7a7ZNuJrqkSMwM5NKmTwGj8Q4wzgsPXqTuXvef3T5XtDYbLz3uMVliiNIJlWnkgmEmgeoUgHzHQGdRdYx43svC7F+eu3cP0sazO46n2p1v13s+gxnw27u3PFlErNpUOR7xymQ3FvjsbLRZOiiq6DZm76nbqTV8UZTGZAxRsBDyP7Te2ftsLg8ickbftk0oo8sUf67qTXS8zl5WSuQuoKCBjAoSc1+4nPPO5i/rZzVebhGhqiSOfDQ0pVIV0xqxGC2kkiuc9Xsf2fpz/T/AI1/wX8e5D6BfX//0d/j37r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XvfuvdcWBZSAdJINmABKkggMAQRcHn37r3XUaeNETUz6VClnN2YgAamP5Y29+691yIuLe/de6K/3X8Y8L2VuHH9pbC3ZlukPkJgKEYzb3dezKCgrcnU4UTRSzbS7D2nkdO2+09jzqhtjsukj0TsZqCakqP3ffuvD7Og8w/wAqdwdQZnA9dfM3b+J6yzmYqabD7a782wtdJ8aOxcvWVLwY7GLuDKVFRleoN5ZBQt8NuUx0sk946DJ1919+690eNTqUEEEEXBH0IP0IPNx7917rv37r3Xvfuvde9+691737r3XRv+P6+/de6In0XO/yE+QPYXygVJP9GOxsXmvjr8f3qIXRdyx4jdJn7x7VokcvC+E3TvPA0uBxFQhvUUWAlqVtFVxk+690ey3JPPP/ABF/+K+/de679+691Sv/ADJqOqj7g2NXSU8qUVV17LTU1UVHhnqaTP1UlTBGwP8AnIEqo2YH6Bx/X3yF/vCbW4T3T5KvZIWFrJsRRHp2syXMjOPtUOlf9MOulP3LrqB/b/mm0SVTcpu6sy1yqtAoVj8iVYD7D1Xj7wJ6zG697917r3+vx/j/AE/x+h9+rTNetHgerYviRtzbfSHTSd1bkw8Fdv7siath2fFIymsTasTrHR00Dsh/hlPWPC9XVuAXaMxrc+lR1E+65y9y57N+0w93uYdqSbnjmB3WyBP6gslIEaqSP0UkKtPOR3MpQZNF658/eC3nevc73Hf202bc2j5S2ZUN4R8BuiCXYgf2jrURQg4DayBxPWPdO+N1b0rJavcOarKtHd2ix8Ur0+LpImbUkFPRQssVowB63DO1rk39t8zc5cy83Xcl1vu7SyoSSsQJWGMGnaka0FMCurUTxJ6VbDypsPLVtHBtO3IjgAGRgGlYj8TORXPoKAenUPb+6dw7UrqbI4HL11FLSyLIKdaqdqCpUNqeCro2kaCaCYXD3W9je9wPaTYuZt+5YvYNw2Tc5opY31BAxMbeqtGTpKsMHFaZBqK9P7zy9su/2s9pu23xSxutNRVda4+JXpqBHEZpjh1ZpsfdNDvXbGL3JQgxx5CC81Of1UtZExjrKRwS3MFQjAH8rY++i3J/M1pzfy7tu/2VVSdKsp4pIMSJ89LAgHzFD59YRcy7Dcctb1f7Nd0Z4W7W8nQ9yOPtUg/bXrWE/mQ/PDqPZfyAraPsXcu4anMTy5PbXWPW2ytm7o7G7Ay239pVIpdwZ/H7L2XjczloMK2aeRpq6dIacsVQOWXSOO/vdbc9feF94uft52C1T+qWxTjbknu54bS0g8LUCvj3EiR+LPMJJCikyFdJIAAp0t9qb3lP2U9teUNp3WR35j3eH62SK3ilnmlMgBU+HEjvoijKJqIChi1DU9FHb5r/ABzTpXP9/VO8c3Qdd7U3NQ7J3SMlsLe+N3pt7emTrcfQY/aeV6/rsHTbvp85W1GWpvFEKVhKk6OhZSD7g0ezfuEecrDkSHaIn5gurdrmApcW728tugLNOl0shtzGoR6kyCmkg0OOpbHuxyb/AFau+apL6RNtglEUitDMsySsyqEaEp4oYllouipqKcerKv5RPzi6Y7c7m311f13vGvqcjW4embc+xN07Y3PsDeu2c3Rwz1+ByeW2ZvXFYPPQUGbxSVUUFUsDwStGF13FveZ33P7LnT2g9zzypzXZInLfNdq/0k8M8N1ayXlkpc6J7d5Imk8LxFZNeoChIAz1i595W+5W9zeQjzDy/O533l65T6iKSOSKdLa6OirRyqjhPFCMGpTjT062OR9B76o9c/uve/de697917onXyA3XJkM7S7Th0fZYRYa+rYHU82Tq4XCI34VaWkfgWveQn+nv3Xui+Wvxyb8f48/8T7917oy/wAb55vvt2UhQtSNTY2pdyT4kqVeeJUMf6NUkRvf62X+nv3XujYAW/J+lrf8i9+69137917r3v3Xuve/de697917roi/5I/1v+N39+p1qma16ZsltzBZgf7lcTj8gbW11VJBLIB/hKU8i/7A+yncNi2bdhTc9rgnPq6KT+RpUfkejGz3bdNuINjuM0QHkrED9gNP5dIWs6Y2BVs7pipaFm/5UK2phVT+SqM8sYuf8PYLvPaTka7qf3Y8RP8ABI4p9gJI6FNt7j82240ncFlX+misfzNAf59Mz9CbKbVoqM7Hf6Wr420/62qlN/ZS3snyia6bi+X7JRj9qHoyX3V5lFNUVqf+bZH+Buuk6E2YoAaqzrkfVjXQgn/YLSge/L7J8ogd9zfMfnKP8iDrbe6vMhJ0w2oH+kP+VunGl6Q2DTsGkoq6tIIIFXkJ2Ww+qlYfCpB/N/Zhb+z/ACTAwZ7SeU1/HK//AD7p6RT+5nNcw0pdRRf6SNa/tNT0usVtDbODULicHjaIqbiSOljaa/8AXzyh5r/8hexjtnK3LuzKo2zZreIg1BCAtX11GrV+degvf7/vW5km+3SaQHiCxA/YKD+XSiA5v/h/T/efZ9ToooPIdd+99e697917r3v3Xuve/de64sur8296IJ4HrRFfs6Te5dm7W3lQHGbqwWMz9CTqEGTpIqgI9iuuJ2HlhksT6kZT7D/MPKnLfNlkdu5l2S2vrImumZA9D6qT3KRXBUgjyPRzsvMG+cuXYvdh3We0uv4o3K1+0DBHyII6K5un4VdX5itkq8DX53aEbU5QY/Gyw11AtTrDCpWPJLPOFKXUxiQKfqNJHvHDmP7n/tvvF29xs19e7VGY6COFxJGHrUOPG1vwwU16SOFDnqcdl+8xz1ttssG621ruLK9dcqsjlafCTFpXjkNpr61GOqvfmL8ict8C/jZ3d2X8W5R2b3R1Bk9tb43nPuPY8Fbt3cPVGxN0Y6r7k6626Bm6OuodwT7IiyMtNXRLKiV0KX1KthHXs7zF7P8AtFzvf8i2G5X2682312LR9wNskVrFIp0LbKzSagni1EkiIymQipIUDoZe5+0e5XuPypYc3bhZWm3cu21v9QlmJ2kuHVl1m4YCPTq8PKIzKwUcAW698ov5vW78rN8KuvfiDhsVj93/ADA3titwUe995UVDvCh2z8e9s7M/0idmb2g2tR19DDXVQw1Zj6CkeWsiiWtrQpsbMJQtPvO2Mi863t9yTd2217DGFn1zR+K908/gRW8ShSneQ7andSFjbspQ9R5cext2p5ZtrPma3uNw3VqxKkb6EhEfiNLIxYNQVQBQpqXGcHoF+jvkR/NX7t+cO8fiBhPnB8d6Si2H8athfI/K7yyXwclpslXf3q7Oy2xa7YC4KH5BTUlPHT0OHNTHlY6ubU04Q06lSTMXtp7lbF7pcuDmPYre4giEzQyRTBQ6SIFYrVSVdaMCHU0IPAdRxzxyRu/IW8nZd3lhkmMYdHiJKMhJUHIBU1U9pzj06ML3T/NI7W62/mXbL6cxOJ2fW/B/YfZPTfw/+THYk9FP/ejEfL35P7N3LvrqEYPLef8AhdBs3a1PisHjM5HKzSR126adLEgaJD6B3V9bosiNHIqujqVdGCsrqwsysrAqysDyCLEe/de6DKt6a65rqv7yTbsMMhcvJFSVFVSU0rs+tzJTQzLFZvoQABbj37r3Qh4/G0GJpIaDG0dNQ0VOgjgpaWFIYY14+iIALm3J+pPJ59+691N9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XErcg3It+B+feiK9e6q37MwdTtzsDdeNqIVhR8tU5GjEcfjgkoMnI1ZSyQrcjQokKG39tT75u+4uy3Gwc88zWNxHpVrt5Y8UDRTHxEK/IVKn+kD1nNyNusO8cqbFdwvqYW6xvU1IkiARgf2BvsI6qk+QHRXyrT5sda/Lf44YXoDeVLtj4yb4+P+4dod1dhb669mWs3V2Did60+4cPX7O2Bvj7yCngxYheOUQnU5P4B9q9k3jls8pbhyzv0t7E8l+lwrwRxyAhIzGUIeWI6sk4J8uirf8Aa9+XmO03vZ4IZVW2MZV2ZCDrV6ghHHkAQR+fSUyHx7+ZXXPyPyvzD6Mx/wAdMxvbv/p/r3Yfyz+O++d/77wGxJt+dXivg2X2L0729i9gZXMSS47F5J6CtgyuDh+9plDLok06FSb7ynuGxJyvu8l8lpZXUktpdRxxs5jlprjmiaUDOkEFXwQa1HSJ9k5mst3XmPb0gNxcQqtxCzsF1KCFZGEbcKlSClCADUHo+vSq95jYsM/yLm6sPZ9Zmc1W1eO6ZTc8mwtu4CoqtW3du0OV3glPn9yZLFUI01mSlp6NKqdiY4I0ABBW7/ub6512AXH7uVAAZ9PiMR8TkJVVVj8KhiyjDEnob7IN3Forb34P1pJJEdaAeQq2eFPICtcDo9nxn2zka3d1TuwJ48RhaKrxrTkXFXka9EBpYeR/wFgGuQ/QFlH1vacfu8ct7jd80XPNATTtVpC8Woj45ZQKqp/oKAzelQPPEVe9u/WdtsUHL2qu4XMqyU/gjQnub/Tt2r9hPRq+2dq1W9+uN57SoGiSvzu36+ioTM5jiNY0YkpUkkH6FknjVSfoAb+8mPc7lm55y9v+buVrNlF5fWMkceo0XxKVQE+QLACvlWvl1AvIe/Qcr858tcwXSsbW0u0d6ZIStGIHmQpJp8uqMqyjr8XXVWJy1FUY7KY6aSlyFBVxtDVUlTCxSWKWJrEWYcH9LCxBIPvi5fWN7tl5d7budo9vucDlJInBV0YcQVIB+xhUMKEHrqTZ3lpuNrb3+3XSTbfMgaORDVXU8CD/AIRxBweiM9k9Obv3X89OgO1pNpw5vp7bHxh+R3WO/snWz46TGrmexc9sOXD7Yr8TUTfe19PuHE4usVykLwiNWVyLgGRNl5isLD2u5m2MXpj3+berC4gQA1K26TapFYCitG7IckNwp1HW97Be7h7gbLfvZ69oWwuI3YlSAztBQFSamoU4pQ06Kr8FPib8hPjn8vO9f9INFTZj4/bS6F2r0j8YuxYspR1GQzPWeO7Y3L2HtbYufxrVcmWpc31fh9zPhmlliWGWko4Cjvf2Nfc3nrlXm3kDln91SFOZ59zlvNwgoQqztbRwyTRtQLouGjE3qGd6gHoL8g8pcw8u877oNwiVtjt7NYLWUGpaHxZZI0b+lEHMdaZVV6NB8Yup+yeuO7f5hW89zbZ/h2H7l+Q+A391PUVGToTBvDb+M6c2xt01uujlrKjDwPuLGPSsamJJUALhGUC4M5z3zZt45b9qNts73Xc2G1PBdAKT4TtdSyUzpDHQwYBWzwqDjoS8n7Pu+071z9e3FlRbm7Dw9yjWBDEvkTTuDDh5dFV3V1T233/8rPjV3Vtr4Ybu+I3bXVHZ1Jne+vkXuHevWTYvsDp6hxeSos71FjJOtdx5TL92wb3qqinFJPmcbRLjoozIzo37fscWO+bDyryRzfy/e+4NvvuxX9kVsrFIbjVDdllKXTi4jVLNoQDVYpHMhYChBr0ELjad45h5w5f3Wy5Qk2rcra5rc3BkhpJAAwaEeE7NL4hI+NFCU1Ag46uRWxLleAWNhfhQeQo/wA947f4esgKUoDxoOh5+N2wMlv7tTbzQ0rthdq19LuPP1xV/toIqCTzUNEZVBArMhVogSP8AUUVm+gv7nH7vfIl/zt7mbC8ELfujbJ0urmX8KCM6o49Xk8sgUKvHSGYig6ib3p5utOVOQ93WSZRud/E1vBHUaiXGl3p/BGhOpuAJA4nq55Ddb/4n/e/fXYeZr1zZHAdc/e+t9e9+691737r3XvfuvdI3f/X2ye0tmbi697F2xh957J3ZjpcVuLbOfo4q7F5ShmKt45oZBeOaCdElhmjKTU88aSxukiKy+690Qv8A2Xb5af3c/wBlm/0zr/su/wDer/meX95Mz/sz3+gn+H/c/wCg777+Hfa/3x/iv+4j+/X338R/u1+54P4v/lnv3W8fn1//0t/j37r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+690zbi27gN3YLK7Y3ThMRuTbmdoZ8Zm8BnsdSZbDZfG1aGKqoMnjK+GejrqOojJV45EZGB5Hv3XuiUN0b3V8aMeKn4o7ifsTrrGzeY/FruLclTPQUGJA9eG6N7jyIrdxdeNTIoFFic//G8CoAggOMis6+699vQrdNfKzrLt3NVOw6iPcXVvdGKpBV7g6N7Zxf8AdDsvGQoCs1fisfPPUYrfG3EkU6MvgKvJ4xwV/eDHT791sinn0Zm9zax/x/4p/r+/da679+691737r3RLflpvndWaqdhfFbqjOT4DtP5EPm6bNbqxvilynU3RG3YqZe2e0qdGYijzf2uUpsDt6WQBP49l4JhqWllHv3XqYr0afYmztvdebL2psLaOJhwW1tmbfxO19u4eBtcWPwuCoocbjaZZOWmKUlMl3Yl3a7MSxJ9+690q/fuvde9+691Vx/Mu2fUVOC6237TwzSQYTL5XbOUlWVzDTQ56mp6zHM9OI2UearxrIZWdQCVWxLD3ze/vD+VJ7rYfbznaCFzDZ3c9pM1TpRbhFkiqoFO6SIrrJFCVUV1dZxfco5ihg3rnblOaVRLdW8VzEtBVjAzLL3VrhJFIUA1AJwB1Uh75YddDeve/de6c8JiZNwZvC4CJ/HLnszisGkgDMYmy9dT0Al0orsfEKjVwpPH0Psx2bapN93nZtiiYrJfXcNuDnBmkWOuATjVXAP2dIN13FNn2vdN3kXUlpbSzEevhI0lM0GdNOI+3q3HtqspI9yUmzsOPFt3rjEUGzcNTcBU/htLDDWzWQBfKzxrGT/zb99U/dG8tIt+tOUtnGjl/YLWOwt0/5ooFdj6mvbX+j1zy9vLS4k2a45l3Gjb1vNxJeTN5nxWLIvyFDq/23QX+416kD7ePXiLi39f6+9HHXjwPRtvj5ulqHY3YmPaZY5NvU9ZuGkd2UJFHPi52djrYDSlRQ6ibBeeTf3lJ7E8yvacl8+2PiBZLCN7lCaUCtAxJ4+Tx5PDPWO/u9sa3PNXJ134ZK3jpAwFclZRQYHmr09cdabPW+T3Uvzv/AJiFJ/G9pYP5Db12b0Tm+gcn2LT19dg8h1BT7Zykc4xeOx2TxGay+28T2RNWNm6XGVUUq1csTyEEA++bvM0tluXsx7Ebtd2t5cckpuW6ndvpWVZV3GW5D6pHdJI0mktigt5J1I8MMq+dM0tkS4s/dD3V2yGS3i5pazsRt/1Cs8f0kcBXsRWQtGs+syBGHeQTQkVLT8pu3+5OxviJ8lMJ2JF1Nle0OjvnH0H1TQ5nr2k3FtjZO8K/Gby6l3Nha7KrmsvujO41zW58UlRIJ5nihhuiXWxkH2z5T5P5e91fbu85fO7Q8s75yXud68V20U1xAr29/C6IEjhjYBYy6VUAswzTPQJ5w5n5i3vkPm203aGwO9bbzDYwq0KvFFIwuLJ01hnlZAC4VqM1KGnkOrSfhd1h3htb554r5qfIOs6yxHYdVt7ZfRu2+uunq/c2Y2xiNgpvQ5vM5Xdm9d0UOIy+6dy5SvyBjhSOjp6WjpFKAOzlgEeTvdblPlHdfZ/k727i3GbZLHmyHcZ7q+8OGSWW4RbHwoYIHkjggFu7Fx4jtJJlgBginevbPmHmDZ/cfmTnFrKLcbvl2SzhgtQzRokRN0sjvIqM8hlQaQI1WNa01MSRuf8AvuF1yv697917r3v3XuiSd87fqcZvNs0Uc0O4aWnkhmPKLWUMSU1TS6gAit4lSRRySpJvwffuvdAgWVRdjYXA/wBctwFWx9TH8AfX8e/de6PR0vsyXau1hVV8Riy24HjyFXDIgWSlpgmmhpHBUOsqQnU4J4dyPx7917oYvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XvfuvddHgE/4e/cM9e6or+QGyqXGdm9n7Qz1EK7C5zJZg1lJXKJI8rtzeFPLLUwTLoRJIKuhyE0LAC1ri5N/fG73g2W75Q92+dbRHdZ13BrmJz8RWc+PG1QADpLYoMFSCSR1039uNxtea/bLluXSrI9iIJFGBriXwmBHGvaCc5rXqkH+X58QO6um+1dzZXvaipBtP409d5H4nfDjIJlaTKz7l6QyPYu4excl2PkIYKiokxGZrsPU4LbvjlWCbw4drppIJE3up7gcvcwbHZw8sSk328Xa7nug0sojuxAlusCkga0VxPPXIBmwcHoL+2/Ju8bTvd3PvUAFrtsTWdmag64TJ4niEfhYr4UZpxEWePRuuusB3R1D/M27B+W+2dpvktkQ/CzaXW23an+K42OLeG/sF2JuXcuS6+mx5rYa6nXJYqrhAqplSmjkkVw+pOPcle5w5J5P5MXYtyWPmey5lnuZI3WQxm0mt4oH8UrTUjdw0qdasA6ioB61zZyLJzPzJzSm5bcz7Nc7LGiOpXWJ455JFZK1oy1HlRgSpwT1P66/lD98/Kf+XJ3Nlt6/LnvHYnyM+Ve8+yPlr2F0Xt6bpWp6h2r8vardkO/uvNvVe4q3rPLdijH7CzW09u46SpptyIPHReSneOIqp6i8o848vc97JDzFyxfi42p5HQNRlo8bFHUq/cKEYrxUhvxdYHcxctbzynucmz77aGG+VFehINUcVRgRggj04EEcR1spfF7c/b+8vjt0pub5A7En6y7yy/Wu06jt3YdRkcTlm232JHiKam3bRwZHBVmQxNVQy5qGaanaCaRfBIgJ1XAE3RF0PHv3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de6CLs7qLCdjwRVE874rPUcRhosxBGJiYCzP9nW0zMi1NI0jFuCroTdW/Bi/wBxfa7Zuf4IppZDb73CCI51Grt46JFqNaVzxDDyPl0POSOf905MlkjjjE+1StV4SSO7hrjYZR6Y4EHgR0UHJfH7s/H1j01NiKTMwLfx5CgyNJDBKobSuqGukgqIpCBfSQbD8n3ituPsX7jWF01vbbXFeQCpEscqKpzQYkIYE8aHh69ZEWHu/wAjXdustxfyW0taFHjYkEivFAykeVf5dQh0T2szop2sE1sqa3y2M0JqYAu+ipZgijkkAm349oR7K+5jsq/1a0moyZ4aCpoSaMeHHhWnDpY/utyCisw3zUQCaCKSpp5CqjJ6E/aHxgytTUio3vlqaio0NzjMFK81ZUgEgiTIyxpDTxsACCiO/Nrg8+5J5W+7luk06Tc5bokNoP8AQrZizvTjWUgCMEfwqW4jHQC5h98LBITDyvYPJckf2k6gIteGmMEliPmwH28OjgYPAYrbeNpcRhaKnx+OpF0xU8CBRckGSWRuWlmlbl3Ylmbkn3lVs2y7Zy/t1vtW0WaQWMQoqqKD5k+rE5LHJOT1j3ue53+8X0+5bndNNeSGrMx/YB6ADCqMAYHTz7NOkHQS9h9I9cdoBZN17fhlyMa6Ic3jpXxuaiWxUL9/ShXnRR+lZRIo/A9xjz57Pe3/ALkBZOaNhR79RRbiImK4Xy/tEoWA8lkDqPIdDzk/3L5z5GLLsG7stoxqYZAJIT/tGqBXzK6T8+ivZ74I4GUvJtnfmXoOf26bN42iykagnkGppDjZzx/UG/5944739yzY5Az8uc73kB8luYo5h/vcfgv/ACNfPqctp+9Pu8elN85UtpV82gkkiNf9K/ir+VaelOgpyXwf7OpWlOM3Ls3KICRGsjZXHTSLfgtrpqmNCF5I1EX+nuLtw+5t7iwa22/mDabkA4DGWMkfMFGA+zV0PbP70HJEwRbzZNxgNM08ORQflRlNP9r0mH+G3eCFtNJtSRQTYpuQqZLfQqsmPQLf/Ej2HJPule8qatNntTelLqlfsrF5/Po7T7yHtewUtPfrX1tuH7HPXOn+Gnd0xHmg2nRg/VpNwvMUueQVp6BgSB/Qn3e3+6V7xy6RLBtcIJyTdE0/3mOhI61P95H2yjVjHLuEpHktuBX82cdC1tH4K1bTwT763vB9sro8+N2vRSLJMoa5h/imSv4RIvBZackfj3KfK33L7ozQS86c4xm2BBaGzjNWzlfGlHbXzKx19KYPUfcwfemt/Cli5V5Yfx6ELLdOCASMN4UfEjyBcD1rw6PRsfYG1OucLFgNoYinxGORjLKsZeWorKlgoerrquUtUVdS4Xl3YkDgWHHvM3k3kflnkHZ49j5V2tLawB1NSrPI/m8sjVaRj6sccBQY6xb5m5p3/nDc33jmLcHuL0jSK4VFHBUUUVFHoB9teln7FvQf697917r3v3Xuve/de697917r3v3Xuve/de6//9Pf49+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XvfuvdYkWQSSM76lbSI00hRGAObm93Zm5v9PfuvdZLD37r3QXdqdKdUd2Yilwnamwtvb0pMdM9VhqnKUWnNbcrpAgbJbV3HRvS7g2rl7RLarx1TTVK6RZx791sEjgeizxdK/KjpNwOgu7qHtzYsMkjxdS/KibM5zN4unK6hQ7S7+29FUb1jpo2ASGLcWP3E8aWUVCge/deJr5dOcPya7020/j7T+E/c2MiSR43zXUW6OuO6sO8cQAariocZuHbW9DBK/KRnEGo0nlAbj37r1MVr1g3P8APzorBQQ4HHR7/wA73XmFkpdmfG2Tr3dm1u8t75xaGevGLwezt54rAMMdSUtO82QzMky4TF0qNNVVcca3PutdK/41dQb627U7y7r74bAVvyG7jfHSbop9vyvkNv8AVuxMIJjsfpDZmVqEimyGB2dHWT1ORrljhGZz1bWVhRY2p44vdbJ8qY6Nf791rr3v3Xuve/de6Bfv7q2HuLqnd+xiVir8hQitwNSwjIptxYpxX4aRjKCiRSVkKxStwRFI1iDb3Envj7bQ+7HthzXyS5CXk8PiWz47LqH9SAmvAGRQjHjoY9SJ7T88y+3PP3L3NSgtaQzaJ1zVreXsmApxIQ6lHAso61v8pictgMnksFn8fUYjO4atmxuYxVYuiqx2RpiFqKWdbsNSE3BBKupDAkEe/n53Xat12Lc7/ZN8297TerOUwzwuKPFKmGVvn5j1UgjB67O7duW37xYWe7bTeJc7VcxLJDKhqskbVKsPl5HzBBU5HUH6fXj2hAJNAKnpaSBSp6Mj8Rti5XfnyA6/THxT/Y7Sy0G9c9Wxxh4qPHYJjUU6zM6lFOSyIhp0H6iHYj9J95DfdX5J3PnX3x5GSyif6Pa7lNxuZAtVjitjrjDEig8WUJEF+LuLL8PUI/eG5r2/lb2l5va7kQ3W4W7WUCE5eScBWIAyfDjLyNXAoAfi6VPyN+YXQ/QPZXVu2+9d/U+wty/JfsvO7I6uGRx+TqsTl92w1MJ/heVz1PTyUG3IZJMhTU8NRXPFC9RPHHqBPGYbbRzDznuXOO+WNiZWgmmuJwGBZEaV6UBILkBTVQKgKesY/wB8bJyrZcs7RcXARJIo4ozQgEiNTXAIFSaCpAz0sZO2djQdzRdAS5SpXtObq6q7jXAtjaz7X+4NHu2HZFRlWywjOPSqj3JUpB9qX85Q67aefYcG23h2p960f7rRc+BqqK69HiAafi+DieGOjkbvZfvOPafF/wAbeLxABXK1AqDw8/X8ugzw/wAvfj9n/lJu34aYnfaz/IfY2yaPsHcWyjiclFSQberKXF1/jpNxSQLh67O0OMzdHV1WPilNTT0tTHIy6SbGMvK29xcvW/NEloRsksxjV6ipYEjKfEFqpCtwZgR0ij5m2mbe5uX1uP8AdgiBqd1M0xWmkmjKaA1AIqBXo52z4p67A9u4aGSaFM31BvWmaohpnqBHNR4966EMU5jJVXAFxqLC1z7MuWDJc7L7o7Ok5ja95V3BQQuo6oYjIOGRgMKedR6dJOZZIrfdfbzcnRXNrzFZtpLaaq8gQ4PHyr6U6ow7O6E6T7zosAvbnWG0d/ybfV59vV2dxgfMYBq2JBWLhc5SyUuZxcdWFHlSGdEksNSkge+bvKPPfOXJaTScp8yXdgLlE8VIn/Tl0jtEkbVjfTU6SykrwGOs5uZeS+WOaZY/6wbLBdSQs2hnUF0rg6X+Ja+YUitBXh1ixXxz6EwmwF6qxPUGwsf1wNwY7dr7NpcFTx4ap3XicnS5jHbmro7mfI56kylDDOtXUSSzeSJLkhQApuvcLnm93480XnNt/JzF9O0H1DSnxFgdWR4VINEiZGZSigLpY4z0lt+ROUbTaBsNty/aptQmWXwxGukyK6yK7Cnc4dVbU1WqBnHRitoUk+R3ltGip4nqaqu3btuCKGNijzyyZuiARJFBaNm/1X4+vsu5TtZr7mzlSyt4mklm3S0VVGCx+oioA34TSuejHmO4hseW+YbqaQJDFt9yxJFQAIHyR5genW0kosqj+gA+pP0H9Tyff0ngUAHXC0mpJ6797691737r3TdlMRjM1SPQZWgpMjSOQWp6uCOeLULgOquPQ6gmzCxHv3Xukniesdh4WrFfjttUEdUjBo5ZxNV+JwxcSQpWSTRwyKfoygEfg+/de6Xvv3Xuve/de697917rBNU09NDJUVM8VPTwoZJp55EhhijXlnklkKpGgH1JIHtqaeC3ikuLiZUgVaszEKoHqWNAB8yerxRyTSJFDGzSsaBQCST6ADJPyHQK5z5H9KYB6iGs7CwlRUUzoktNiXnzEwZ30WQY2GqSTxn9eljo/NvcQ7z7/wDs/sTTR3nPlnJcRkApBqnbOMCFXrTzzjqSNr9nPczdlhe15QukhcEhpaRDGf8ARCpz5Yz5dJv/AGbbokcHeM9/pzt/Pf8A1v8AYdP3o/ZQVrzY9P8Anmuf+tXR2Pu/+6tAf6vLX/mvB/1s6dqD5PdGV0Im/wBImJpC0hj8OSgyGPnBDaQ5hqKRHETE3Dn0kfn2b2f3jPZe+jWRefLWMFqUlWWNjmgOl4xg+R9MmnRbdeyXunaSMjcnXEgC1JjZJF4VIDKxqR5jj0I2M7O65zKzPi997Rrlp2RJ2p9w4pxE8i6kV/8AKuCy8j2O9u9xeQN2Ep23nXapghAbTdQmhPAHv8+gje8l837aYxf8r7hEXBK6oJRUDiR2+XSxSrpZNPjqad9dtGiaNter9OmzHVqvxb6+xYl1ayaPDuY21UpRga14Uoc18ug80My6tULCnGoIpTjX06ke3+muve/daqPXr3v3Xqj166uB+ffuvVHr14EH6e/db679+691737r3Xvfuvde9+691737r3XvfuvddH6H/W9+68c46LN8gfjtj+44KLLYutiwO9MTA1LSZOaKSagyWPLtJ/C8tFEVlKRysWgmS7xMW4ZTb3jz75ewtj7sw2W6bffpY83WqFElZNUc0Rz4U4WjUVu6ORTqQkijBiOpo9pveG99uJrmwvLRrvlq4fW8YIDxyAAeJETirDDocMAMgivRB6v4m980lVNTxbToa1Im0rWUe4cQKWoBAbXCKuemqLeqx1xqQQf9f3g/efdg967e4lgTliGdEaniJdwlHFKhlDsjj07lU/b1lja+/wB7WXEEU0nMEsTMMq9vKWX5HQrL+xj+XTLP8ae9oK+GgPXmQlkmCWqKfIYeXHqX1aRNkFrxFDpCeq44uL+yeb7vHvPDeRWTciXDSPSjLLAYQDUnVJ4mlSKZqCQSPXBknvX7WyW0l0Ob4gin4WjmEhpx0x+HU1rjIrQ06su+OXU1d1HsD+C5uamn3BlsnUZvMGjkkkpKeaojhgp6GFnISQ0dLAqO6qqu97XFj76I+wftlfe1vI/7o3eaKTfLq5e4nMZYorOFVI1LU1eGiqGYABmrSop1hT7w8/WvuDzb+8ttikTaLeBYYQ4AdgpLM5pw1sxIUkkClaEnofQAPoPc29RV137917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3XuvEX9+6910FA+g960jGOvdesD79QcaZ6914AD6C3vfXuu/fuvde9+691737r3XvfuvddaQfx7917r1h/T3oKB5de69Yf0Hv1PPr3XgAPp+fe6AcOvdd+/de697917r3v3Xuve/de697917r3v3Xuve/de697917r//1N/j37r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XRF78nn+h+n449+690DHefRWwu/wDZM2zN8UtZDNTVEeY2hvPA1TYjfXW+7qEibCb42BuemAyG3N0YKsRJoZ4mKShTDOksEkkT+690Gnxa7W3ruKn3t0p3XV42f5BdC5HGbe31k8dR/wAJx/Zm1MzQ/fde927exDMwosN2Bi4Jo6yCJmhoNwY/JUaHRAhPuvdGz9+691737r3XvfuvddEXBH9Rb/b+/de6J58i/h7sjvOo/vJRVjbM7BEUEDbmo6Vaqky9NThkSm3BihLTpkHjjYLHUB0qI1ULqKen3in7+fdS5N96ZhzDa3Z2fnsKqG8jjDpOi40XMGpFkIBoklRIoAXUVAUZCez33ieafaqL9yzwDcuUCWb6Z30NCzZLQS0YxgnLRkFGJLUDZ6J3B/LQ3q1bNFVdrbXixQLfbVNNtvKT5GSxQRrUUUtfDSQlo9WrTM9ja1/eJ0P93bzYb6Rbr3O25dsr2MlpM0rUIprRpFQA5qA7eVK5pkbN99rlz6SJ4OQL5r/GpWuYljHGul1RnahpxQYr8urCOkfj9sv4/bRr8XtCOqr85lIY6rcO5chpfKbgyNHTSx0zPGp8FFRQNI3hpogsces/ViWOd/s57Gco+xnLF/tfKiSTbxdKHuruXM1zLGhCEgdsaKSdESAKtTxJJOIXub7scye7e/Wl/wAwukW2QMVt7aPEVvG7AsATRncgAPI9Wag4AAdaw/8AMV6M2T8nvlb8Iehe0oJTtrtXr357bfy1VSr48pt3MydUbYrsNu3BvdWo9xbN3LS02SopEIaKppVsR7xr5J3q/wBh2XnXfYQRfwXdlIwI0hibiTxI29Q3cHU4IJHU1837bBum6cu7WtDbSwTop49vhIEK+hHFG4ggHos/wv8AkTuiv+d+7dufKWs/hHdvwf8A5eG/+pPk5uOrjko8Rumk647z21uzbvd+Gq6kRxT4PtXq2WjzWoN+1WPOjH6En/NWxWw5PtZtgj1bPu29RTWqg9y+Jbsr27UHa0M2qOgB1Ch9eg3y/u00PMRG5yH6+xsJI5SR/DICsnHKuv6mcjK+XRPYu2O8dtdd9DfzJMv8Nu8Nr5PC/MTeXzG7X+QFfnOpzsPN/E35KzU/VGR2w+Bot6z9nRYXA9HQbUkpI58Un21RjXkdY421kTnbNqub7eORE5otJVO1pZxWyrN4i3lr+qGB8PwdRuPGDESZ1AAk46IVvNwhg27mU7dMlL0zNJWOngynS34y5pCI8FRTQCK5PVsGwNjdR/Lr+Yp89Np/Lrvze/V2yvil1703VfHeg2v8h9x/H3Ddf7B3vsOHdub+TOHrdu7l2zR7nz2X3lMKI5LJGvx9HS04pmi0uVJv7Pba9tFytDt1hCba+s9wN5M0Yd/EDCFYCzCqhVfU0YK69VTUL0l9xdyF2+63N5dyC4guLT6eIP26SviO9BiupWQOa006Rk9C7uzaWQ2HuTL7OydSa+pwFQtJHltYljztA0MU+K3DT1ChY6qmz2Mmiq45VGmQTXHvjFz9yju3IXOnNHJ29wCPctvvZI2ABCMuotHJHXOh4yrLxwfl11T5K5k27m/lXYOZdpm12F5bI6kkFlNKOjU/HG4ZGHkR0njx7CP2dCno5nwi6ayvZPbeI3jNBJFs/rXIw5vJVzxnwV2fhR3wuEgJIWWZJiKma1/GkYB5YD3l39zj2i3L3A90dt5smgZeU+Xp1uJZCKrJcqCYLZTwZg1JZPNFUA5YdYzfeg9y7Hk3kDceW4pVbmLeoTBGgI1RwMR407eistY0/iZj5Ker6vfbXrlT1737r3Xvfuvde9+691737r3Xvfuvde9+690HfZ/ZOC6r2nkN2Z8ySwUxjp6HH0zRiuy2SqLrS4+iWV0UyyG7M3IjjVnPC+wL7i+4Gye2nK99zRvpZoI6LHEtPEnlbCRR1NKscknCKGY4HQs5J5O3fnvmGz5e2gKsr1Z5GroijXLSPTNBwA4sxVRx6qA7M7p392xX1Uu4MtPR4OSW9HtPGTywYSjgUnxJPGNMmTqdJGuScsC19KqLAcpPcf3h559zL65k3zdHi2VmrHZRMVt41B7ajBlelNTyVqa6QowOiHI3tjyjyDawLtO3pLuyrR7qRQ07nzoeEa/wolKClSxz0E4jVeFGkWsALAD/AGHuLQaCgrTqQTk1Ykn7eu9Avf8A3j8f7Yfj3vUfInr3XtI/qf8AY8j/AG39PftZ/wBX+r8j16gwaZ6xNTQNYtEjEXtdFNr/AFtccXP+8+6MqNprEuPkP5dXDyDCyED/AA9TYairp5Ipqeur6eeB0kgmhrquKWGRDdJIZEmV42Q/Qgi1uPaqK8vLeSKa3vriOZCCrLLICpHAqQ1QR5U4eXSeS3tpo3imtYniYEMGRCGB4ggihB868enn+9m7v+ev3X/sNx5kf71W+zf+tvNlB/yKtz/7K7j/AK2dFw5f5dHDl6w/7J4f+gOvf3s3d/z1+7P/AEJM1/8AVvvX9bObP+mq3P8A7K7j/rZ1v9wcvf8ATO7f/wBk0P8A0B17+9m7v+ev3Z/6Ema/+rffv62c2f8ATVbn/wBldx/1s69+4OXv+md2/wD7Jof+gOlBgO2Oz9qVAqMFv3c9KTIsskFTk5spSTuqhB56PKGrilXQNNrD63+vs/2H3V9yeWJxcbLzvuMTagzK0zTRvQUo8c3iKR9lOiTevb3kbmCHwN15UsnFKBljETqDntaPSRn7eji9Z/NtlkosT2nhY0Ek0NPJu3AjRTwJIwQ1eWwshLwwxFtUj0rOFUE+Pj3lr7dffEJe0233K2lQGdUN9bDSqgmmue3OQoJ7nhJAAJ0Y6xv52+7IFW5v+RNzYlVZhaT1LMQK6Iphgk8FWQCpNNXVhVFW0uRpKavoqiCro6yGOopaqmkSanqaeVQ8M8EqFkkikQghgbEH3nbaXdtfWtve2dzHNaSoHR0YMjqwqrKwJBBGQQaHrEeeCe1nmtbqFo7mNirKwKsrKaEEGhBBFCOpXtR011737r3Xvfuvde9+691737r3SR35vzZ3V+zN1di9h7kw+ztibIwWS3Nu3defrI8fhdvbfw9LJW5PL5StmIjpqGipYWeRzwFHv3XunvC5nFbixGLz+CyFHl8HnMdQ5jDZbHVMNZj8ricnSxVuOyVBVwO8NVRV1HOksUiEq6MCCQffuvdOfv3Xuurc3ufeuvdd+99e697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuv/1d/j37r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+690Ub5J9Mb4zOR27370DUYrFfJHqrG1lBgabMTNR7Z7i68ra2nym5+jOwKhOY8LuSakE+GyLBpNvZ0RVkYMLVcM/uvdC10d3Vs3vzrvE9hbMkqYYZ6iuwm5dt5WMUm59g74wNQ2P3d17vTFMfPht3bQzEUlJWU7ixZBLE0kMkUj+690L/AL917r3v3Xuve/de697917rjq/wP++/r71Xzpjr3Dj164IP+t79XhjHWqjNM9U7dg9b7Tk7Ti3FnNsYfI7y61zW86LZW5chQw1Od2rTbtRaPcK4PIsnloF3BikiiqvHYTRqAePfM/mOO82PfubdgW4dYPrnEi1w+iRnj1DzADAqfmes7tiFlvW0cuby0StN9IhRvTUgR/wCa0+XQY7j6J6X3hm96bm3V1VsLP7k7H6+XqbsDcOT23QVGb3r1gsz1A6/3Pk/EtZmdpiZywo5naMXt9AB7LLfet3tIbO2ttxlS3t5/GjUMwEcv+/EFTR/Q/wCQ9LZ9g2m6nurmexjM80Rjc0Hchrjh8z0qsrsLY+d2LXdY5naO3cn1zktqtsbIbFrcXS1G1avZrY1cOdrT4Z4xRvgxi0WnWAKEWJQoAsPaaO9vYLwbhDdOt8JPE8StG1g6tdR+Kua+uelU23Wk1g23SwKbIpp0eVKUp+z/AFU6Ffr/APl6/EbvkbV7I75+MnT3aGU67hxO2+pMjvzYmHztRtvbO3fDJSY7GnIUsv3W36Sthj+2pagTUqNDqRQbk5efd02rc02ff+YL2ab6a8uKRIxJRtNTLMoPHxHbSHHELQcOsaPeq72xd02vZrCKMT20RMpUCoLHsjJHDSuSPVqkV6Mt8ivh9s/vaWiz9Hkpdm72xmOixVNmKOljqsbX4ym8jUeOy2K1QK8dG8hEMsTJJFGSvqWwBP8AeB+6tyt743MHMMW6PtfOcMIiFwqCWOaNalI7iIsurQSdEiMrqCV7hQdGvs194PmH2mhm2Z7FNw5WlmMhhZikkbtQO8MlDp1AAMjBkJAbB4lG21/LR3Gc4F3j2Xil25DNDrbbOMq0zORgenLTrG2SeSkxUsVUQiuRUaku1gTYYr8u/wB3hvzbwBzZ7hWn7gR1r9JC/jyrpqR+tVImDYDHxcVagJp1kLvf31do/dZPLnJNx++GU0FzKhhjbVgnwwHlUrkgeH3UGRnq0Xr3r3afV21cds3ZeJgw+DxqWihjLST1NQ4X7ivr6qS81bkKtxqllclmP+AAHSLkXkTlf235a2/lPlDa0tNmtxhRlnc/HLK57pJZDl3Ykk4wAB1g3zZzbzBzvvt7zHzNuLXO6znLHCqo+GONRhI0GFVQAPtJPSjzGaw+3sfNlc9lcbhMXTtEtRksvX0uNoIGnlSCATVlbLBTRGaeRUTUw1OwAuSB7F/Qc6A/5D/IzYfx12RPurd9bRmuqleHbmClyNJj5c1XALzJWVTrT47FU7SKairl/biUgDU7IjQ170+9fLPsty0N23aN7vfbksllYxZnu5QKkKFDMsUeGml0lY14BmKqZL9sPa/fPc7e2sNvkW22e3Aa7vJP7G3jJ88jVK/CKIGrnzChmFJO/wD5R939j59s/V79zWApFqo6vD4TaGTnw2GxUEc8lVjxD9m8b5KemSQA1U7O0wFyAtlHGvn37yXvLz7vz7te863u3QrKGgtbOR7aCDS5aNQEKvJKhNDJLV38wANI6fcoex3tZyhsqWFpytZ3jtERLcXcazyTFkAckvVY0cCojjAVOAJNST+/Cj5mz9lVUHWvY2cx2dz00NbJtHfNHU0E1NuKLGoz1uEy9Tj5GoDnKKKF3SZdInRGVwJV9ed33UPvS7xzbusXtZ7q3BPNboTY3ciiJ7oKuo29wulB9Sq1aKQKonUEU8Qd+IX3ifYHZ+Xdvf3E9uowOWlZVu7ZSXFuzHSJYSST4BYhXSp8JjUHQe2wfanZHXu+58nS7J33szeNThZzTZmm2runBbhnxFQGKGDJw4ivrJKCZXFikoRr++hePLrDP7R0tffuvde9+691WF8490zV29tp7PjdvssBhZc3UxcaTk8zO9LA5t+rxY+jNv6GQ/T885Pvm8yyXvN3LHKcbn6Wxs2uXXgPFuGKKfQ6Y0P2ayMVr1m5913Yo7Xl3mDmN0H1N3ciBT/wuFQzU+13FfWg6JL7wy6yi697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de64sLj8f7H/ev9Y+9ilc/6v8AV5j06959Hp+GHa1fRZ+bqjM1pmw+QpKnI7SSoeR5Mfk6S1RkMVSMzER0FXRl50j+iSxtp/Vb3mz90T3OvrfepPbLdrwttU8Ty2QYlvCmj7pIUP4Ynj1yKnBGRtPxdYpfeT5As5tri5+220CbhDIsd2VAAkjfEcrerq+lCfNWFfh6swBv76HdYXde9+691737r3Xvfuvde9+690R/5Ny4/t7tjo74lvG9fiN15F+9+56FF8lM3UPTmZxNdg9v5mNl8cuN7C7aqMNQywMSKrHUdfGQVuPfutg06S+wMinw27ZxfQW46qop/jd3LnqpvjHuOuRFxHVfYeSmqcpm/jRmcw8zGmxG4J5ZslsE1CRoIRVYRHLU1BHN7r3+Hqwe/v3Wuu/fuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvdf/1t/j37r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3RM+4uhd6YDflR8kvjFPjcJ3IKWjpeyOtsnVLiuuvkztbFqRTbd3nKscsG1+zMTTFo9u7wjieoozajr0qsa3jh917oQunPk51v29lazZKtmOvu48Fj4chu7o3smhXbHZ+2YXJRq3+Dyzz0W6dumQWizOEqclh6gMpSpJbSPde6MTcc8/T6+/de69cf1H9ffuvdeDKfoR7917og/eXaG4q/eNbt7DZTLYTE7blNDJHQ1EtBNkcmnrqK2aSIxTPTxrII4V1aSAWt6uMJfeb3I3675rvNh2ncbmz22wbwyI2aMyyficstCVFdKLWlO7zFMq/azkXZrfl613fc7G3ur+8XWpdRIscRwqAGo1EglzSvBfLLFsDvXd20auKDM1dbujb7MFnpK6bzZKlUiwlx9fMS/oJLNFIXD/QFfr7JOR/enmrla5jh3e6k3LYye6ORqzIKfFFK3oclHqD5FejTm32q2DmCCWTbLaOx3UZV0FImPmskY9fJlpTjQ9DnvTrna/ddDDvXYmaoafMOgp6meSOT7WvaBQBR5aBdNTQ5GlFl1lWbTYEMLH3NPN/IHLvvDZQ84clbrDHuhGl2YNokKinhzqKPHLH5NQmmCCtD1FnLPOm/e2V1LyxzTt0r7eDqVQRrTUfjhY1V4246aip8wcdF0ynSXaGJleNtq1OSiVtKVGJqaSujl4vrVBPHUqtv9Ui8+8f9y9nPcrbZHRuWnuI1IAeF45A2PiA1K1D/AEkBHDqZrD3Q5Hv0Vv38sMhHwyoyEfKukrX7G6XHXvx53Lna+Cu3nSS7ewEMoeXHySocxkwjc0wjidloKdyLM7HWVvpX8+xnyF7E8xbxewXnNts1hsqNVo2Kmeah+EBa+Gh4EsdRHwjoMc4e8Gz7ZbS2vLVwLvdGWgkAPgx1HxVNC7DyA7QeJPDo9lHS09DTQUdJBFTUtLDFBTU8CiOGGGJAkcUSDhERAAAPeaNrbQWdvDa20CxW0ahVRQAqqBQAAYoB1ixNNNcSyT3Epe4dizMclmJqSftPUr2o6b66sPrb3qnn17rv3vr3WtN/P57Z+L+890fFr4O/KrufbPVvRu+qDuX5Kd8U+Y3RNtbIbm231Psys2j0ds3F1lLNBPNXbo733xQ5KCHWEZNszO4KRn2R8y723LnLu9cwJtVzfvZ27SC2t11zzsPhhiX/AH5Ie1ScL8RwD0Z7Lth3rdtu2gXkVuLiVUM0raY4VPxSyMeCRrVm8yBQZI611e4fkjnPmX/KIwm4+2N3jsrc22N69GfGrsnMR1wqqTMDq7u/ZOwKuajmpvEhod27TSlyZmQKatq8zPd3J98u91l5stvvn3O97rd3Bu5tmu7yw8aga2tZNrnmggEfwJ9PMrRuF+KVWdqsa9Z77avLk/3aLbZtutYfAi3OC2vPDJpNKL+CKWYue5/GSjpX4Y3VV7R0tMF25ujA/FndPwokzFfTfJDAd9r8A9pyGplfPTbN3rWyZLY/bCTavuP4ZiPjVUzZJ6wH0z40gsHt7jC95S22/wDc7a/eSK0Rvb242L+s9wNI8MT26BbiyOKB33dREqeaTDio6G9tzXf2fIO4e3DXDDmiLc/3NExqX8CSrQ3FeNE28mUsPxRNwJ6Lb03tTE0/UPx7+J2KyuW2j0d2H/M9+UvXG7aXHZrJ4yTcuxep5N17i2t1TXZmkqqfKJjd85TBUyVcUcyPVwwPFezkGR+at73KHnHnj3dS2guOfdr9t9lvrR5I0dYLy7EEct6IiCjPbLK2hiCE1BhQgEALYLGzm2Dl729ad7flC+5u3C3mRXZTLbwm5kS2aQMGAlaNFYVqwWhqCetrr4r9q/B7AfNP4tdc4H+WZV/FvuTf+1+8cF0N8h9i7W6d2f1xv+j6r2bRT9tUGjq3fX98MhiKnHVMZpId14UTiRo5AqSDye+j3s97hWnun7a8pc828qNc3lqv1CoaiK6TsuIjhcrIDTA7SpGCCcM/cjk6fkHnjmLlOVJBDaznwWcAGSBhrhfFQdUZFSMEg09BsT+5K6A/XB/p+frbj/Hj3o+XHj17qkDvjOPuTuXsPJs8rpFuGXE0/mREdKbBxRYqOMiJmVkV6ZijE3ZSCeb++Nvvfvh5g92efr8sxjW/aFKgAhbcCECgqNPYdJ4kUJz1089ptpXZfbbk6yoodrQTNQkjVOTKTnNe4VGBX5dBV7irqQuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de6dNv53IbV3Bhdz4t5Er8Bk6PK0/ikaJpGpJUkkpi6kNoqoQ0TD6FXI9nXL2+3nLG+bNzHtzkX9jdRzpQ0qY2qU+x1qpHAhiOi3edps9+2fdNjv0DWl3A8TVFaaloGp6q1GHoQD1fHtXcOP3Zt7D7lxM6VGNzeNpMlSSxtrUx1UQkZCwA9cMhKMLAhlIIB99tOXN9sOZ9h2jmDa5VewvLdJUINRR1Bp9qmqn0II8uuU29bRebBu+5bLuEZW8tZmicEUNUNK/YRRgfMGvSg9nXRZ1737r3Xvfuvde9+690SPbxTF/zDezqXJvG9XvD4o9XZTaZkZ3ljxOzuzewMduylg1DRCgyW6MdK6KbsXVj9Bb3W/IdGk7H642R23sjcfXPYu3MfuzZe68bNi87gckjGmq6eRklikimieKqoMjQ1UaVFJV07xVNHUxJNDJHKiuPda6JlsDtzdXxTydJ0h8pc9la/ruOvocH0V8tN0VUdThN6YvIzmnwXXPfWejhgptk9xYEslDBlq4Q4rd0CRTpPHk3qKU+62fXqwNSLDm9xcG4Nx/W44PHv3WuuyQPyPfuvdd+/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r/19/j37r3Sbye5IMZI8ctNNKU+pjMYBuARbWw/r7Idy3+HbXdHt3Yj0K+nzPRvY7PLfIrpMig+tf8nSMru2MZQkh8TkJLAn0SU34+v1kt+PYPvPc/b7IN4m1XBI9Cn+foSWvIV/dCqX0I+0N/m6R9d8iMHQhtW3MzJp+oSahB/wBs0w9hS99+tmsq6+Xrwgf0oh/hboR23s/utzTTvFsPyf8AzdI+u+XO2aG+vZ+43tf9NTixf/kqoHsKXn3peXrOmrlO/b7Hh/yv0I7X7vu93VNPMVmP9rKf8C9I+u+dO0KHVq2FuuTT9dFXhR/0NVfT2Fbz76HKllq18k7mSPSS3/6D6EVt91rmO6pp5qsB9qTf9A9I6t/mNbFob+Trbej6b3012A/3i9YOfYTvPv8AnJdmW1e3u8NT0ktv8sg6Edr9zfmq6pp502wV9Y7j/oDo4HS/a+P7q6+wvYeJxFfhKHNTZGKLG5R6aStg/htfUY+RpJKSSSnYSSU5ZdJPBF+feV3tF7mWHu9yJtHPm2bZPZ2V48qrFMVMimGR4jqKEr3FCRQ8Pn1jn7lciXvtpzjuXJ24bhDdXNssZMkQYIwkjWQUDgNgNQ1HHoWPcmdAXr3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3XuuiLgi9rgi/wDS/wCffuvdA73B0B1F3zi8fi+0tmUO4pMJVfxDbWegqsjgN47RyQ+mS2fvbbtZid2bWr+AGkoayAyKNL6lJU+690BifGLufa8Yoer/AJsd4YPDL/wHxfZm3+te7ZqQAqFip9x7w2xTbwqIEjGlfvMhVy35aRvfut1+Q6yD44fIPLo8e7/nT3DKnoaMbC636N2A0ckbE3aeTYW56iVHVgHQvY24t7916o/hHUSs2t8zunlr87tTsjb3yy27TQ+T/Rb2RgNtdTdmTxxuutNp9s7QpqXZVZk1huYqTN4CnhqpPS+RplOse69g/LoOpMxtX5LUOe7E6np89jt7bVrabbncHS+9MRJtjs/YO5YaYSRUW49tVkhlpK6WhAenmiefHZamC1FDUzxeo4ye9HtJf71ezc4csQma/ZVE9uPik0jSssVcFgtA6fiABB1Chnn2s9yLPaLePlrf5VisQxMMx+GPVkxvQEhK5DDgSQcEEBHPDPTVE1JUwTU1VTu0dRTVMLwVEDqbMk0EoWSNh/Qj3iRNDNbTy211A8dwjEMjgqykeRVgCPzHWSkMsdxBHcQyq8LiqspDKwPmCKgg/b0rNk763DsDKrksDUjRKQMhjKjW+Oycagqoq4UZSJYg37cikOv9SOPYm5P5233kfchuGy3HY39rC1fBmAFAHAIoy1qrCjD1Ix0HOaOU9o5tsWst0i/UUfpyigkiPnpb0b8SmoPpXpU/LD5/YroT4b/JXv7G4CoO/wDpzo/sjsHbe2qynqMjh8ruXbe2q2rwEc9bSxjViZMx4TUBxE4gDi9+feXnJ/vtyxzLNt22X9tPZb5cSiNY9JljZmwCsqCmkn+IKR8+saeaPaXf9gW5vLaWK62mNS2sEI4ABY6o2NagD8JavVJud+JXXMXwPk+W6fJT5cV38yOn+PK/KCT5dUfd/aONxrdtx7F/0rttw9Tzb0XptOkoKu2E/u4uA+zOFTSLzEzF679+OTYd5Gx2ttfT3v1KwkiIIgYv4ZqzsCNJ8tNT5V49NW/tLzPLtZ3eaW1itPCMmZNTUAJ+FVOSB6486dBf3ltL499+7m/lUfIeuxHya2TuD57d8bU3F8ndvbX+V/yP29tncWK3z8X999k5jbO3dr4btyhxGz8NBvjG0VZTQYeHG+KKnMSFUdk92X3r2T6vm61bZrvTs6s0jDQdYSdIW0CuTVqgE5pSvSeX203RINkmXcLet8F0A6xpLRtIA504FFpUVyRjra2+NnX3WHVXTGyevemqncdV11tehqqPANu/eu7+wt0QrU19Tkaumzm7N+ZvcO7cnXQVla4P3tXNJGtkUhFUCTuX+Ydp5o2q13nZboTWEoqDwIPmrKcq6nDKaEdAredm3LYNxudq3W2MV3GcjyI8mU/iVhkMMHodfZ10Wde9+691QR8+Pj7sHZ/yR3h3nUZLPbu7E+RO0uuNuZeg3ZBg8jtfr7rnooZ6Pbe1ev6ZsYKvHxbi3hvevzGVaWWR56oRWIVQo5vf3hPPFzZ7R7f8gWLyIbm5mv5mUkApAoghWqkH45ZH+VBXiOs3fuYcm2u4bpzlzleIr/TQR2kS8aPMS8rkcD2Rqo8qk4x1Tpvf4RdZ71x/fGDl3Du3Abe+QPZPU3bO59v4D+CUmNwG+OqZ9vzplNsQNjJPtF3q216L+LJIHVjEXi0MSfeCGy+9HM2zT8j3gsbW43HYNuvbGCWXxGaS2vRKCk3eNRt/Gk8BhQgGj6gKdZc7j7SbJfLzRCt7cRWe6XdrctGhUKkts0RLJVTTxfAj8Sta0qKE9LCp+JXVNX8ucb8z5Ycoe1Mb16dgw47y0v8AdaVl89HTbxqKL7c1Tbuo9u1cuKjqBJZaGQrYm3spj91eaIvam49n1kj/AKsyX/1WujeMODGDVWn05lCzaKf2ig1pjowk9suXpOfk9wiX/e62Zg01Ggk1Hi6SP7UITGGqOwladImp+CHS2U6a3X0pnaveWUw24+794/IbD7rps2mA37172lu3c1RuuHcewdx4amppsLWbayNS6UTskrNA7RzeRXIJvH74c42vOG085WENpHe22ywbU8LReJa3dlBCITDdQyFlkWZFBkGAGAZNNB0WD2e5Zk5bv+XLme5aGXcpL5Jg5WaC4eRpVkhkQAo0bMQhziobUCa20/ys/wCXlTQ712F8sO4vlB8iPkNvv49ZjtjZHR23exa3YWI2XsOn7O2vtzF77z8uK2Rs/AVe5907hxUcUEtXXVMijxBxGHJb31R+5ZzNtvM/tTvF3tfKNlstom/XC/T2jzvAXMVu7SKJ5ZTGWLZRWCDSCFrUnn596Ll695b9wtrtL/f59ynfaID4s6xCTSJJlVGMUUQfSFwzKXNTVqUA2H/eX3WN/UDKVa4/HV1e5jC0NHU1hMziOK1NBJMfJISBGno5Y/Qc+0e4XS2Njd3rU0wxO5qdI7FLZPkMZPl0/a25u7u1tBXVLKqCgqaswGB5nOB1r91NU2Qq63IOCj19fW1zrrZ9LVdVLUFNb3eRUMlgTyRz74V3l099eXl7ICJJpnkIJJILuWIqcmleJ6632lulnaWlnGaxwxIgNKYRQox5cOHl1j9p+lHXvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XvfuvddH6H3sVqKcf9XzH+HrRwCerTfhPu5sx1hkdtVDu1Rs3PT0sJbUVGLy8f8ToUQni0VQ1QlgeAo+gsPfTX7n/ND7v7c3/L00habab5kStaCGcCWMD/AErmVcGgAHWBn3l9gXbeebPeYVAi3K1Vj6+JEfDcn7VCNXzJPmD0cv3lkDUV6x1697917r3v3Xuve/de6J13PSJt35VfETsMMtNBm37l6Py9SeBUDeOy6bsLblHIVGp3OZ6sYRA3VTKx4ub+62K0I8uji+/da6Zdwbc2/uzB5XbG6cJidybbztDU4vOYDPY6jy2GzGNrI2hqsflMZXwz0ddRVMTFXikRkYGxHv3XuidUvw83D12govjj8me5eldsxp4aHrrLNtzuvrfA0osY6LaeF7Vxuc3Htagp+RFR0eXjoYFIWKBFAX37rYIpSnQH7/pvlrtHuTorqba3y/znYu/t4bwxm8d47dm6P6kw+09r9AbSyVO3Yu6d8th8W+epE3ErDA4AQ1tDJV5qsDxs0VFVBfdeqPTq08fQe/da697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r//Q3+PfutenQXbqF6mp/wAGH+8KvuOeaf8Acqb/AEv+QdDjYMQxf6vPoC8//uwf19wpvlf1M9Sps/BD0B+4ONY/PJ/25HuG964Sfaf8B6k/aPwHy6BLcH1f/XP+9D3Dm9/5/wDB1J208B9n+foEs+f1C3PP1+nBPuH96wXHUobT+HoEM+eZP6DXb/Wt7h3e+MnUn7T+D1x1dz8EBf4y7DH/AE2bo5H1/wCPkyh99i/uU/8AiO3Jf/Na8/7S5euYH3qP+n3c0/8ANK2/7Ro+qqviLsfsH+a9TfI75U9tfKL5TdSbZwPyl716J+LXWXxu7mz3Su2eqeveh901PXdLvXcdBteKKLszszee6cXXZOtm3HHkaGCmaCkiplhV1fK/rHfonvzd79xHUn8wDvjp/wCSPy2/mGY7a3TX8uj4+7s6ib4qZHuCDP7k7mOW7SxW8+xN37V6I2rVbEXde8JsFjahv4/S0m32mDxqiQKyL7r3Rot7b6+YfeP8gzZvyOrvmbW7D7e258JOx+6O4eyfjgmw6zcnae69tdY7gy208Dj+xqOkr8R15UY/NUVOd1yYGijr5a+nqKOmnol1sfde6ur+HO4c5u74i/Fndm58rW53cm5/jl0juHcObyU71WSzGbzXWe2cllcpkKqUtLU1tfXVMkssjEs8jknk+/de6Mf7917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de6Lb3B8ccT2HuXH9q7J3Rl+ou/tt7eyG2trdubWiiq5pMHWyGrXavYW0ayWPbvZ+xosnapTGZRGaknLTUM9JO7Sn3Xui/7hofl9Ki4/sr40dD95VdHEYqfffVvceX6nyWRSNAi1E21d87VykuHq6nlzSrna+mibgTMLH2DeZ+QOUucAp37Zo5bhQQsgJjlFfSRCrY8gSw+XQn2DnHmPlokbRuskUJNSho8Z+1HBWvzAB+fQDbpqt2bZeoqN1fGr5MbEx0LO8mRx23tm96bfghQMzy/cdP7szO9Uo47frlwjPp5Nzx7grfvu0Qs0k3K3MgQcRFdICB8hKma+Q1Kft6lvaPfW4RUj37Y1kHAyQNpP2mJ6g/PSw9KefQX5abpj5CbJ331XVbl2fvrbe+Nsbj2FvzYs2bfa+7ZcDubF1eGzuIyezd1JtvfWIqZKCrcajQq0L2Nwwt7ia99q/c3lG9tdzi2KSV7eQSJLbETqGRjQ0Xu8q5Wmk5zjqQovcXkLmiyuLGbdlg8RSjJOpjORTByvnT4geNPXogNP/Lk+ZdT1DT/E3KfNbL1/w5i23D1vJS0Hxtp6f5NZzo2GEY6PqPJ92fx6fbEUUu2UGIl3DTYOPLS48cFZWMntS/MFnFubczp7aXi80F/EyZ/pRcVr4wh8DUSH7wjPor5Ux0VJsl09mmyHne0OwgUADxeIY6U0a/GP4e3UIw1c4bu6ON2p8S/7+7i+HeTwH8S2Nt/4g9tUfYm3tq0m1chkKTP4XGdT7o6pxOyoKl5IXwkONx24Y5UqiKgsKbQUu+oBDbNw3uxh5njuNju5rjc7YxO5ilUhjMkpegjNSdNGGOPHoU7hte13cuxtBu9tHFZOGA8RD2hHSmXH8ePMAdW0fGfaO68DBuDLZqmqsXicxHQpjsbWeWCeoqKZ5zNk2oXA+3WSGRIwzAO4XngA+8kfu88rcy7Jb73uW628tttd2I/CikqrM6V1TeH+EMulQxozAZGOoc96eYNi3WbaNv22aOe/ttfiSJQqFYLSMOPioQWoMKTg56NX7yV6grr3v3Xuq/P5gnUeR3r1lit84HHyV+Z63rqutyMFJGZaufaWThRMw0UMcTzVb46op4KgIOVjErD8j3g59+j2uv8AnL2623nTY7Ezbzy9M8kioKu1jMoWeigFn8Jljl0LkKJGFessfuj+4FrytzxfcsbrdLHtu9RKkZY0UXcbViBYkBfEVnjqcFtC+nVJakMoZSCCLgggqR/VSOCp/B98csHINV8jxx5U+Xp109yMEEH54P7Ou/fut9eJABJ+gBuT9ALEkn/AD34YzWgH8vn+XHrRrgAVJ4dXvfAjZWU2f0FjKvLwvTz71z+U3nR00o0vDicjDQ0GLd0KK0bVdJjlnsSTplHvtj9yHlDc+VPYvbbvdYylxvN7NfohwVglWOOAkUBBdIg9DmjjrlJ96/mex5j93b+CwIeHa7SKzZxkNKjO8oByDoeTRjzQ9HY95f8AWNnQV94ZMYfqDsfIGAVIh2hmozCX8esVVHJSH1gNbR59X05tb3G3vFuI2r2r5/vjD4mnarhdNaV1oU4/LVX50p0OfbOxO5e4PJ1mJdBbcITWlaaXD/z00/PqjaMeONI+WEaKmq3B0gD/AGFxz/h74wKhCqNQ4AceJ4cPXzpw9OuozEElvXNOsoN/x/xv/W97Ip5j9v8Ah61137117r3v3Xuve/de64gkkAKxJbSAOTc/T6fk/wBPe6D160xCipOOuxz+CPqCD9Rb+v8Aj71ngePXgaivXibf8V/Hv3W+vXNwCpW6hhcWup+hH9Qfe/IHyP8AqP2U/n5daDBuHGv+Drv3rrfXvfuvde9+6910fp7917o5/wAH8s1J2LuvCNNCsWX2pFWLE5ImmqcTkUC/brqCMI4K1jJ6SbW5t7zE+5luvgc+807QZEEV1tiyaSe5nhlAGgVoaK5LYJ4Z6xm+9Ft6z8ocv7p4bmS2vyhIppCSxn4sVyyCmQOPVpI+nvpJ1g11737r3Xvfuvde9+690VT5n7MzG7fj5vnMbRrzh+xep4afu7q7MgROuP7B6o8m7sGlRDUBqWqxucioJsXWwzAxS0VdKradWoe68MkdDz13vCh7D2Dsnf8AjI5IcbvjaW2930EUoIkio9yYejzFPFICSRJHFWBWH9R7917pZe/de6LN8lu2t57Cwu19h9O0GAzXyA7nzNZtHqTGbqNb/dPETY/Htlt29j72GO/y+TZfW+3keuqoIGjnyNU1Lj4njlrEkX3XunToX49YDpOk3FmqrPZnsft/sWbF5TuDufd5jfdvYmcxlK1NRAUkB/he0dmYJJpY8LtzFpBicPTyMsMZlknmm917owvv3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de6//9Hf49+696dBhutX+5qLI59S/pRmFtCfkKR7jvmlHM8pVCceQJ8h0NdgZfBiBYV+0ep6AvPxzEsRDOf9aCY8f7BD7hTe45T4gEMhP+lb/N1Km0PGAtZFH5j/AD9AluCCqPktSVjfU3FHUn/WH+a9w9vlvchZR9LLX/SP/wBA9SbtM1uNFbiMD/Tr/n6BTP0tYS5FDkDyfpQVh/A/pB7h3era7PCzmJJPCNz/AIF6k3abm17R9XDUj/fif9BdAnn6KvJYjG5MgA304yvP4/wpv6+4g3u0vQ0n+6+5/wCcMv8A0B1J203VnQVvYK/81E/6C6BLP0GSIlticwSddrYfJkkWP0tSH3D+92O4apP91t1w/wB8y/8AQHUnbVeWIMddwthw/wBGi/6D6uz+CkcsPxp2LFPBPTzLWbn1Q1VPNTTL/v48mRqhnjjlUMCCLjkG/vsN9y2GaH7vPJkc8DxyeNedrqyN/uXKfhYAio4Yz1zF+9K8cnvXzQ8UqOnhWuVYMP8AcePzUkH9uOiCYn4KfP74n9ifIOi/l397fF/bvx3+SvaW6+9a7YHyd657J3dub499wdkSxTdm5rp+s6+3NhMXu/aG5ctAMvDg84aaOkyEkiLOYZGvlX1j11y3n8J/5j2xfl5un5V/Grvz4nbg3P2j8SOh/jl2ll/kV1l2YmQq949OZLeOWr+1dsbZ6q3JiNv48boym8Z6k4iad6ensIVfQNZ917pmyv8ALF+SfU38s7M/AL4wdvdIZ3P91U3yAh+S3bnf2zt7UFLl8n8lJN1ZvsLcvUuyuscvDQ7QqaXdG7J2xuOq5KqhpqKNEYySamPuvdWGfBLrL5L9L/HbY3UHyezvRm5NzdW4Da3XWy8z0Pit+YjAV/XuyNoYDbGBq9y0m/6/IZD++FW+LllqzSslFZ0Eaix9+690cf37r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xrfn37r3XVhe9ubW/2Hv3XukPvTrHrfsilSi7C2BsvfNLGrLFBu/a+F3HHCGvfwLl6Kr8B5+qWIPv1BWtM9e4dFprP5fvxUab7nb/AF5mOvqgO0qt1X2X2f1fEJXKEyih2LvDBY4uDGtrwkf7c3sXc1Jckn1JPWqLWukV67g+GVFjagVGA+S3zBwapNLPFRL3vk9w46PyKyiH7PeOJ3Es1PCD6Fcm1hzce9VPW8enUan+LvdeKnhnwvzo+QxWF2f7Xc23ekt0U8wa9o6gT9aUM8iIWNrSKfpzx791ao/h64UPSXzQxC0op/m9i9yCmEodN5fGXYMj19/IYfvanaG59padBYBjDHEWVR9DcnXXqj+HpszndXyV+O9DPuf5KbP64390zi545t290dF/3nw2Y6126scgq94b86a3PJuPK121cZOY2r6rAZjJ1NFSeWqkoxBDI6+6r0dvH5LGZ3F47MYeuosviMvQ0mTxWTx1TDXY7J4zIU6VNFX0NZTvJTVdFW0syyRSxsySRsGUkEe6sKilKjr3+r06rm7h/l4YDdecyG5ertzR7IqMnUSVdbtnK0MuT26a2pqZKisrMdUwzpkMWJ2lY+ACWEN+kIvHvnx7tfcL2DmjebzmD235jXZp7mQyS2k0bTWpkdi0jxMrCWHUWJ8Lvj1U06FqOszvbj74G9cv7XabJzzsjbnBAoRLmJ1iuNCqFRZFIMctKDv7XpXVqND0UfJfAD5IUNS0FHitoZuERpItbRbtgo4GdtWqDw5SkpaoSJYXJXQb8E8+8WNy+4194CyuTDZ7ZtV5DSviR3qRr/pdMqI1R5nTT0J49ZDWH3tvZy6gMt3f7jbSaiND2hc08jWN2Wh8s19R0YDpv+XPMtXT5rvHMUtXSRMkq7G2xUztT1Z0hvHnc+VppmhR+GgpFUPbmUrwZ29pPuCSRXUG8+8e7xSwJRht1mzFXIzS4uiEJFaVSFQDwMlK1iD3I++P4kE21+1+2yRzMCPrrpVDJ84IBqFacHmJp/vsGh6tZpKSno6WmpKaCOnpqSGKmpqeJQkVPTwIsUEMSD0pHFEoVQOABb30ztraC0t4LW2hWO2iRURVFAqqAqqB5AAAAeXl1gdLLLcTS3E8heeRizMTUlmJJJPmSSST59Svb/TfQF/Jaup6DozsZ6hmVanAtj4tKFyanIVNPR0ykAGytPMoJPCjn3DP3hb23sfZnn+S5YhXsTGtBWryOqIPkCxFT5DPUn+y9rPd+6HJqW6gsl3rOadsas7H7aA0Hn1qJ9u/NXY/xv8AnzuXZHePaW4tr9U5X4jde7n2ftek2dvbeuIPYU/bG9Mfn86lBsTau5K/H5Kp29R08LzVJihkjjCqSwt752bD7dbnzh7XWe4ctbLFPvce+zxyyGWKFvBFvCyJqmlRSoYsQFyCc9Znbvzvb8te4Vxbb1uMq7WdtjZUWOSQeIZpVLUjRyMBRmgoOgt6M/mE4+PD/LPvPM5fe3bPQcH8wPZfSeyNwCkqtvJ0/wBY9g7Q6+weL3DV7d3VisLuKk2Vh9/ZnTVU8lOlYjVvmA8d/Z3zN7USvPyRy3bW9tY8znlaW8mTUJDdXMMs7mMPG7p4rwodLBtB0BTmnRFy97jvCvMW8XMs11s53qO3U00+BFIkC6yrhW0rK9CCK0cngOjMd9/zBtj9Cbt7v21W9a9gb9o/j2vx8oexM1smTEVTpvL5G72h2ptHrzAYmrmhqszu3HYiePL1ESuqGnmiiBEkq+wdyx7TblzPY8uXsW8WltLupvjAk2ofpWEJllndgCFjZgYgSK6gTkA9C3fPc6w2a73aFdvmnt7JLXxHTSayXUmhI1UkEvpIkIqKAqCc9KHanzThpN79gdffIrp3d/xez+zem9w/IrEnee6dm74oN1dJ7PrpMdvPca1+wMhlabB7m2bVmBclhJ2kqY/uY/E8ouQkv/bZn2zat05S3+33q1uNwjsG8KOaForuQViQicIXjlAPhyr2kg6gOn7L3DQXm4WW/wC0y7fLFaNdIWeOQPAp7nrGXCsuKxnNGBBIPSN2Z8/K3LZfpLI9jfGntTp3p35N57G7X6A7g3VuXYmWg3PuPc+Nqc119ht+7F2/kqrdHVU/ZOJpWlxL16zxhnjjqTA7WC/cPayKC35hj2rnCxv9+2aJpL61jjnUxxxsEuHhmkRYroW7EeIIypoCVDAV6SWPuTNNPs53Dlq5tdq3FwlrM7I3iM4LRh0Vy8RkA7dXn2sVJp0VTAfK35P909CfzRcV2b07mOvsT1FP8udkbR7Dod/deGTrhtlddxT7d6zq6LaNf/eLO7nxUlc9WdxUwaka6r5Ay+xzdci8lcvc0+y82zb+l3cX42uWW3ME9J/FuDruA0wEaRtpCmBzqwcZ6B1jzhzXvvL3uTHuViY4LVb5EmV41KFIxpjARixIqT4gx86jofPid8x81VYX4Y9RdodG9q9a4/vbpHbGP6X7e3zndo5OLs/dWwOrcBnt1Qbh2riMxk94bGnzmKSfI4qoy15MpAhd1idwvsL8+e3tslx7g8wbNzHY3T7ZuMhu7WJJU+njmuHjiKSsiRS6H0pIkX9mTxYAnoQcl88Trb8qbPuu03MUN5ajwJ3ZGMzxxB3DKHLoSNRVnADU8jjoV9xfM3c3XfbnXOy+2PjVvvrnrntnt/FdKbB7UyfYfVubyVbvPcUuQh2ZW7i6m2/nq/em2tob0qcc6UVe/mMavG9RHCJAASWvtxZbrsO57lsXONtd7tZWLXc9ssFxGPCjCmYR3LxiJ5IQwLIKAmuksB0Z3XuNPYbzbbfuvL01vt1xcGCObxImJc1VdUSMzKrspAarUqNQWvQF/Ebs/tHbnW3y/wA1tvrzenyA3VR/zF/kxtPD7Wj31tzbVPt/b9FkcJItXmt5dgZelw+09j7dhjZQsInKPKiRQHWSBJz5seyX28+39te7rb7VZPynt0jy+C8hd2V1okcCM0kr4+KgOSzCnRJyrvu57dtvOFzbWMt9P+/7qNE8RF0qGBy0rAKoANAATw0rnpYVn8yzaWB6m3P2BufpLs6j3t178nNl/FDsTprbmW2ZvbdGI7H34cdLt2s2puTA5Vtpb9weUx2XpaimekqYnkMvjcRsrey+P2av7nfbLarPmOybb7rZ5tygu3WaKN4IaiQSRuglhdSrghlIBWuRSq8e7Vkm0y31xs9wt9Dfx2ssKlHZXkK6CrBgjgh0OGBzkAg9ZT89+023P2X1RSfBjuqu776pwGJ7C3b1hB2P1B/BaLqLcGOyGQ25vdu0v4+dpS7hz0uIq6Gn23CJsg1fSzLr8SNL71/rW7ELPaN7k9y9uXli+kaCK5NvcgtdIwEsP0+jxQsYZHM5YR6HWgLdvXl9zN0eXcNuXk25/ftuqyNCJISvgvXQ/i6wtWKuAgq1VNe3u6GTAfNDr3de4fhnh9tYDcuSx3zX2Nvjfuws7OaKgTaWN2LsrHbzr6DdeMlkeskyc8eQNHppyyxVMLksUsSHLv263Pb7P3CnvbyBLjl24hhmTuYSGaVolMbYWgKh6txU0wcdH9rz/t97ccmw21tIU3iORkJxoEcSyHUD5nUBjH+HoPd8fzCtjbKwfdle/X258jmupflVtj4f4bCzbj2pt3Hb97R3hicJlcBkZN4bhq6HbfX+zZkzBSor8vKqQNDYB3kjQm+3e0+47jdcuwDdYEtr7ZZd0d9ErtDbxM6uoiQNJNKNFRHGDq1YoAT0Xbh7m2djabxN+7pWmttyFki6kAklZY2BLkhUX9ShZiKU8yQDb1/Ldyu+d9byl3T2N1nlOmt14fbO4oK7Y+Q3RtffsZhnydDRUWVxu79n1VRhchiMrTKKimcCOdluskSEczN91zYNti95dxl2jflvtutNnlYSrBJEHM7xoUKyjVE0ZBqKsG8iadRf79b/AHl57VWP1+1G2urvc410mVH0iNHcEFCQ4NfMKVPEDh1dKPoP9b30o6wi679+6911cf7xf/Ye/de6AfuD5KdR9JVmD2/u/cM9fv7dj+HZXVezcXX7z7S3pOSyBdu7F2/DW5uahWRdM2Qnjp8ZSfqqamFAWHuvdF33J158hfl0f7sdzbepvjz8ZKuaOTdPVdBuml3L3j3Xi4ZFmXaPYO6Np1Uu0OseucwAi5bF4auy+Vy1MHpJK6kglmjl91vgcHo+1DR02PpKahoqWmoaKighpKOio4I6akpKSmiSCmpaWnhVIaemp4UVEjRVVFAAAHv3Wupfv3XuiL9/ZqHpX5K9IfIzeIp4enp9kbz+P2+93V0yR0HU2c7D3XsbO9f7xyzvcUO2N07i26Nv5CubTDQ1FZQvO6U5kkj917o8ylTypBHBuPoQeQbj63Hv3XuuXv3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de6//S3+PfuvdcSoN/8f8AY+9UFa0z17HXtA/oP9sP+Ke/UHp17rrQv9B/th/xT36g8wOvfKpp17xr/Qf7Yf8AFPftK/wj9nW6n1P7T1140/1K/wDJK/8AFLe9FFP4R+wdeqeNTX7eveJP9Sv/ACSv/RvvXhp/Av7Ot62/iP7evLGqm4/BuAAAP9sB72qheAAHVTkkkmp6ye7de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3XusNRTwVcMtNUxR1FPPFJBPTzRpLBPDMjRywzRSK0csUsbFWVgVYGxHv3XuiS7P8Ajl278fP4ngfjL2dtSPqCprqvIbb6M7l2xmdwYDrGSvqGq67C9W762vncRubb+x2qpXemwmQpsxS4wP4qFqemVKdPdeFPPp6yvYHza2jGKut+PHTna1DCheqg6u7yym29zyjWb/wzB9mde4fb9TIsQvomztNqbgN+ffut9vlXpedMfJ7rburK57Z2NXcWxu19n09PU716Z7Nwk+zuzdr0lUdFPlpcFVyS0+4tsVUvogzOHqMjiJ39KVJe6j3WujFe/de697917r3v3Xui9fKijqq3ojfiUkJnenpMdXSopAK0tBl6Crq5QCRqENPEzkC5IHHuCvvK2dze+yvOyWsRd44Y5GHokc0bu3+1UFj8h1LnsTdwWfupym9xIFV5ZIwT/HJE6IPzYgfn1rqYjprcuM+Y+9fkeMzif7p7m+NWxelaXBwtWDcMW4drdk7p3nWZWpBpxj/4LPj87FHEVlabzIwKBbMeYNxzDZS+3u38ofTub+Hd5rsuaGMpJbxRBRQ6q6ozxGmh416zuj5cuf663PMcjxGyexSEKa6g6ySOfKnB6faOi8Zj4r4/BdW/zOI+6dy4CXrj5Rbw7H7ijyOO++mrNi7SpentvYmnyOcSqpYE/vJtrN7N/iUQpjLGfFFpfWbAXw88SXe++zr8u2kv732a3t7Uq9As0hu3YqpBP6ciS+GS1KVPl0EZ+TVsdl9xRvFxH9BfSSTKVDVRPBjFaYqVKFhpPkD1WzuPrntXHfyicb2t2HlY2+THyw+SfxS+Qe/91Zfb8060e5d291dWYfrVMttyKaCqlxO2tl4PFvJQ+WEjyzIGjLXEv2e8bFN78XGybVDTk/Y9n3KxgjRxmOK0unn0SUorPLJJ3EGoCkgkZji42rcofaqDdr5q7/uW4Wly7FDhmuLYRnQCDRUWOqihwadWM7h+IXcvyM7I7W3v8tch1RtcZn4vdpfEjZO3uicluvcVIuJ7by0GT7B7YzuS3pi8DUUOWrZ8RQpjcFFHPFSQRyCWrlZgxiO15+5d5S2rZNt5FhvpjHvVtuc0l4sUbF7VSIbaNYmkDKutmeUlCxI0oB1Iqck73zJdX+48zT28Ub7W9nEkGt+2bTrlYuqEGippQA0oaux6TeA+Kvys3/jvit1X8idydEQ9Q/ErfHXXYMW6esKjelbv7v3P9NYWfC9YJm9tbhxGLwfVOJiZ4q3MJT12YkqamHxwGOJiwV3XO3Iu1y8673ylabl+/t9tp4NFwIlhso7ttVzokR2e4YiscQZIdKtV9RHVbXlHnC//AKr7Xv8AdWY2jap4pQ8WsvO0APgkoyhY80ZwHkqwopCnp0pfiX37iMf/ADCeq6DKdQ1/VPzEy/e/ZGzt51uW3bRdh7S393Psai2tHtHcu1IcBVYCfamEyVK075Wnr2qnp2CrS6wSGpOeuVLmT2r3ya3v03zYY7KCaFViaCSCzlMhkjl1hxI6sAIyhUHJenV7XknmKzj572iOW2bad0+oljclxIsk8enQyaCpVSMMHBNcp0I9T8Vd6T5f+WxkBuDa5i+FqSL2LFI+TMm62k6EHVAOzf8AISkoXOL9z/ln216X8a/R7J05729Yvd1DaTa+YiDBQKBH/jpuSJM57AV7NR1H0z0uPJG509vKXcVdpjdZfi7i1u0fbjyZs/Lok+D/AJZncqb56iz25qf4353cvVPzBwnyO3J8pMrkexM58ju79p0O/M1uePZ2UpcpiXwfXr4TbuVgxopaeursdVLj4UhSkjLN7ka495eXF2zfLezbdYrS92FrCPblWBLC0kaBI/FUq3iXAaRDJVo0dTIxYyEDoFD2r5he+tJbiSyeeDc1uWu2Mj3EyeK7BG7OwqjBQBI6UXtVK9Cdur4CdxVOwN3bfpM31jvvHZz5/wDbHy33B0ruzPby2x1j3X1nv6kNPguruytybexVVmsZkts5QxZYRLRZDFVNZSxxzJJGbgmsfdPl5NzsLmW2vbV4+VrbbEu4kiluLW4hYGS5gjdlR1kWseWjkVWJUg9Gd97bb89hfRQ3UEvib3LeNAxdYpo3+GORwjHBo/8AZuhZRVSOkrtL+Wt2jgdj7225TVfQWxW3f8//AI//ADAx+z+r6PdmJ672Zsnquk2zHuLYGBpshizXyZySfDSilnZI6eskk80gpvJ4o124e8OyXW5WF48e63Qt+V73azLcNE880ty0uiZirU0AONS5ZANNXIqUlp7Vb1Ha3cTS2UJm3e2vNEQcIiRGIuoGkVYmM9wVQxNdK8AfvDdEbkxnyt+Svfc2Ywcm1+6eh+nOqtvYeM1v8dxWc64qezJsrkM1GadaFcZWrvan+3MMsspMMmtVsuqLrjmm0m5I5O5YWCYXu3bndXMjnSEZJxAFRBWtR4LHuAFCM9SFb8rX0XNPMG9tcR/TXdnBEo7tQMRnLE4pT9RaefHoqlJ8Lu9Ou+vv5eua6zz/AFRuPuj4PbY3bs7Lbb3tkN1YLrPsvAdjbRbae6Ux+7MNhMpuLbWRxJhgqqKeTGTrLoeN0W4b2Nn9xuWd33X3Vtt6tb6Hl/mOaKVZIRHJcW7wSmWPVCzKkiuCVceIpGGBPDoLf1D5i2/buRZ9suLZ962dXUo5YRSCWIRP3BWdSukOv6bg5BHmJmxvid8iNl9efKmi3zhvih8jtzfJz5H1Pb+7+vd+0G89tdQ5rYmZ2Xt/A5LaMflw278jhdxYXK4SL7GrnpcjFPBD5pPDPKBDTcefeVNw3fkybarjfNqs9n2gWsU8BiluVmWaR1mw8SvE6v3orIVJ0iqqa+tuSOZLTaeZYbuPbr+73G/MzxyB0i8No0XQO2UhlKDSzK2qlSAeFx/8jT4yb3+NHTPa23N557D1UOR7KzWb2rsva+V3bubZHUG1Nx1LZfD9R7F3XvqU7nzu2dmRXCSVMcFpKphHFHEI1OYX3c92teceY+eucdvglW2W0sLF5ZY4o5by4hEry3cvg/pCR9ahkXVgKWYk16xx969tuuWdj5Q5Yvp1NwZ7u6WNZHdLaKQokcCa+9kUqxVzpFSwVQooL1R9B/re8tBw6x449Bp2z3H1l0Zs+r372xvLDbJ2tS1FNQpX5aZzUZPK1rGPH4LAYqkjqcvuLcOTlGilx9DBUVlS/piiY+/de6K/Fur5TfJekqzsPFVvxD6mrbRY/sDsDbtBn/kZu3GyqvlyO0+sMk8u1+oqaojYinq9zfxTKr+qTD050t7916o6HLpf419S9EDL5DZmDqa/e+6jDLvvtbeOQqN29rdhVsQX/Ld5b7y/mzOUGtdUdKjQ4+lvppqeFAEHuvdDyBYfW/8Aj7917rv37r3XvfuvdMe5Nt4Dd+Aze1t04bG7j23uTFV2Ez2AzNHT5DEZnEZKmko8hjMlQVUctNV0VbSytHJHIrKysQR7917oi/V+Xznw/wB/7O+NfYG4cjuTorsWvqcF8V+yNxzVlbl9l5umSatp/jJ2JuKpMwyM1NiI2OxMrVyirylDSS4uoMlZS08tb7r3Vgnv3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de6//9Pf49+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3QCd5/HjZXedDh6vJy5DaPY+yparJ9Wdx7Pemx3Y/V24Z6Z4Blds5WanqYKvH1AbTX4ivjqsPloLw1lNNGbD3XugW63+Ru+eut3Y7pD5i0OB2fvmvmbHda97YONsT0h8gY49Ipo8fNkKqf/Rf23LFzWbSyVQ4qJQ0uJqa2C6Q+63TFa9HjBuL8j/X4Pv3Wuu/fuvdQ8hR02RoqrH1kST0lfTz0dVBIoaOanqYnhmidSCCrxuQb+0t7a219aXNjeRB7OaNkdTkMjgqwPyINOnre5ns7iC7tpClzE6ujDirKQVI+YND1Rh2hsKu6x33ndm1iTGCgqmnw1VIhVa/BVbNLjauNvpJphPikIuFlRlPI98XvcvkW99u+dd95Wu1bwIJNVu7AjxbZ6tC6mlGovY1Dh1YGh66k8ic3WvPHKu1cx2xHizJpmUH+znWgkQ+me9QeKEHoPJ4IqmN4Z4oZ4JUaKaCeJJoJopFKyRTQyq8U0TqbFWBUjggjj2BUcxsHWuoGooaEH1BGQehZIiSxvFIoKMKEHOPs64S0lPPCtPUUtJPAhiZKeelgnp0anZWp2SnliaFDTsimMhR4yoK2IB97WWSNzIjsHzkEg541I41rn16o9vA8fgvGDFjBGBTI/YepIuBYkn/E8kn+p/xPuh8qcKdPdd+9de697917r3v3Xuve/de697917r3v3Xuve/de6979w4de64uQqMxIACkkngAAXJ/2A9+LhBqJwOvBdR0gZPVvvxJ2VVbO6gxs2RppKXJbrr6vdM8MwKyx0tekEGJV0JshbF0sT2sGGux5Hvq/91/k+55S9qtukv7do9w3OZ7xlb4gkgVYQR5VhRGpQU1Edc6/f7maDmP3Ev47OVXs7CJLVSOBZCzSkHz/AFWcV4GlRjpy+RPfUPTGBwGL27gzvzufs/Mvs3pLqylqhTVm9t3vTmomrcpUqkr4Hr3ZtCTktyZl0aHF4yJmtJUSU8E2RPUK9JPqD4vQYHPYnuDvfcjd5/I2OnqpH35mqZ4NoddPlBG1ftjovYc0k2J672tRhPAlUqzZ/IxAvX107OVX3Xujbe/de697917r3v3Xuve/de697917r3v3Xug17d6m2R3j17uPrHsTFHLbV3NSxw1SQzyUWTxtfR1MGQwu4cBlILVeF3LtvMUkFdjq6BlnpKyCOVCGUe/deGDXoF/ih2Lu/MYLeHTPbudp873p8e9wJsXfuZSBaGffe266lXLdXduigRI4YF7H2VJBNWiC9NDnqbI00dhBpHuvdGy9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+6910Ta3+Jt7917pi3DunbW0se2W3XuHB7YxSEiTJ7hy+PwuPjIBYh6zJVFNTqQoJ5b6e/de6L5UfNb4n02RfFv371tNPFMtPNU0OfhyOHgmZmXTPn6BKnBQAFDcvUqF/JHv3XujF4LPYPc+Ix+4NtZnFbhwOXpo63FZvB5CkyuIydHMLxVePyVBNPR1tNIB6ZI3ZD+D7917p29+691737r3Xvfuvde9+691737r3X//U3+Pfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3SV3rsnaHYu1s3snfm18FvPZ+5KKXHZ/bG5cZSZnCZehmt5Kaux1dFPTVCalDLdbo4DLZgD7917h0TOj6o+RnxfNJTfHfJDvvpGGofydDdu71rKbsnYWOPphoel+6c82RTMYGhSy0+A3eZPBGgSDMU8WmFfde4nPQi7O+aHR2f3HDsHeWZy/R/aTgK3WfemGn623HUT6gjR7cymZYbM37AXvoqNv5PKU8i2ZXII9+62ft6NcrJKiurLJG6q6OhDI6sAysrLcMrA3BHBHv3WugK7v6P2n27iY5cvUNg87hKapOF3PCY0NCki62pckspSOsw7zKHkiZlKkakZTc+4a93/AGW5c93NvgXcC1tv9rG4trpACyas+HIv+iQlgGZKqQRVGBJrJntr7o757bX8z2KrPs9wV8e3f4X04Do3GOUAkBhg8GBHVTW5utd07YmrSP4XujEUdTPTnceysrRbpwrGDUWNVLiZ6upw0hRSxiq0ikUX+tr++aHPnsx7i+3k0x37l+STbFZqXVuGmt2UfiLLVoQRnTMqEZyePWdvKHunyRznFH+697jj3AqtbeciKZSfwgMdMpBxqiLA/I9IPUv9fwD/ALA8g/6x/r7ioMrLrVwV+RB6kXzp59d3B/Pu1Dnr3lXy6796691737r3Xvfuvde9+691737r3Xvfuvde9+691x1L/X/eD7tobGOPXqdGT+PHQeU7XzdJn8rHLRde4mtjlra1lGrcNVRyxv8AwTFq3ElO7i1VUWKRqCgu7enIn2D9jtx9zt2td93SJoORbSYGSTH+NujBvp4QeKmn6suVVQUWrnEI+8Xu1Y8hbdcbPt8gl5tuIiEQcLdXBBmlPkwGYo/iZqMaKM2r7y3jtPrLZu49972zWP2rsvZWByG4dx5zIyeDG4XA4Wikqq6rm0KzeOmpoDpSNWdzZUVmIB6toqIixogVFFABgADAA+Q8uud5LMWZ2LMTxPE/M/Mmp6Kr8YNhZ7em6t1/MHtXGVlDv/t7E02E6q2nmaWWjr+mPjnBVDK7N2ZNjah5ZMPvjfUrpuHdousv8Qmp6FxoxsXu3Xujs+/de697917r3v3Xuve/de697917r3v3Xuve/de697917ol/yP607AwW/dj/ACr6IwA3L2j1vi6rZnYnXEFXS46fvTonK16ZLM7Loa2slp6GHf2x8upzm1JKmSKnet+5oJZI4chJJH7rYp59Dt053j1d33tOHePVu7aDcuNEstHlaALNjty7VzFNI8Fftvee1cjHTbh2hubFVMTRVNBkKenqonU3SxBPutdC1f6/4fX+n+39+691DXI495zSpX0b1IVXNMtVA04RiVVzCHMmlmFgbWJ9+691Mv8A7z7917r1wfp7917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917rq4/r7917r1xe359+690h979ndb9Z0DZTsbf+y9h45YmmNbvHc+F23TGJCAzpLmK2jSQBjb035Nvr7917otsPzq6Q3HVNQ9T0Pa3fNWsskTSdNdT733Xt5WidonL77q8VhuvI4hJxrbLBSCCCRyPdb0nieHUev7R+aO8dMHW/xm2R1rST8x7l+QnbdBPVU6epRLJsHp+i3lUTtwCI3zlIxHBKn37rR6xn4+/JPfmmTt/5fbpw2PqAklXs3447H2z1Pi4nPMlIu99xDsLseoplJ0iWnrsdIy8kA8+/dbBp5dKDa3wY+Lu28qu4sj1fR9kbrRvKN3905nP907kWfyCVZ6bI9nZTdBxs0Tj0GlWDxjhbDj37r1SRQ8OjU0uMx1BQpjKDH0VFjYomgix9JSQU9DHCwIMMdJCiU6REEgqFCm/v3Wuq7d67Qf4N7+m7x6ygloPi32FuaEfJrqeiaqmwfVu4dyVdJjMd8kOrcKrS0+2MVFlpoot9YmhSKhqKGY5pYVqaSraq91vjjz6sdR1kVWRg6soZWU3VlIBDKw4ZWBBBHBHv3Wuufv3Xuve/de697917r3v3Xuv/1d/j37r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XvfuvdJHemwdkdj4Gr2t2FtDbO+dtV6FK3AbuwWL3Fh6kEWvLjstS1dKzD6g6bqQCCD7917opOd+CHxowWJyWR21hezuusfjaOtyc+G6k7q7g2JjJ46Knnq5YKXb23N7UeGieYIVVIYI7mw/A9+62DTy6Kl8SvgD0j2/wBZbW+QXyDffveOf7kwWO3zt/YHZHc/Z3YXU/UO0txUiVmE2Jsvamb3hX4fKS0+Jkg/i2Vya19bksiJZA8UPjgj91tuNKdGt3h/L3+MOQFNkeu+r9k9Obtx+Kp8RRZjr7amLwmGy2JoZDPTYDfW0cUmNwm9tvLM1zFVKtXECTT1MD+v3ogFZEIqjijDyYHyI4EcQQagg5HVfNCOKmoPmCOBB4gj5EdF2xHx0xuaqd74XDbbi2l3psi2T3L0tmd9Z/JdZ9iYDIS1C4bfHVu7coa/duz8JuVoJI6eQNUU+JyEBoMhRSJGkjxHzd7Ee0/OoeTduUIIb4j+2tK2snCgr4OlGHDDIeA6kXlv3Y9wuVWQbZzLM9qP9CnpPHTzxJVh9oaoqaUPT/s3407H7b2/WZrr/eO6tp53CV8mF3f152Lh6GfdOxNzQR08lTt7c8OOkpJoZ1gk89PUxGalr6WWKoppJIHV2x65g+5Xs0viPytzxdQHgEuY0mXy4unhN68QTUjIpma9o+9PusZReYOVLaUebwSPETxrRX8RfQDIHEnpL574fd04jySY+j2/uaBAWU4nLrS1DgzmJEFNlUpB5PHaRvXpUG1yR7hLfPume7m1CV7C2sdxiXh4M4Rj3UA0TBM07jRiBwrUV6lXafvHe224+Gt3Nd2Mh4+LEXUdta6oteK9oxU8aUPQMZjqvtLAOEzHXm7qPVNNBHIuErauGZ6f/OmGaiSojeMDkPfQw5BPuId39r/cfY207nyJusVXZVP07urFMtQxhxSmQ1dJ8j1Je28/8jbsC23837dJRQSPGRCA3Cocqa1xppqHmB0gJnamIFTHJTXJUfcxyUxLKSHVROsZYxnhgBcfn2B5kNuSLlGiNaDxAUyPLuAz9nQtiZJxWBw4x8JDYPA9tcfbTrpJ4nGoSxEf4Op/3kH214kZ4OtPtHThjkBoY2r9h65+SP8A46J/yWv/ABX37Wn8a/tHWtD/AMB/Z1iWdHk8UTLLKb6YoQZpWA+to49Tmw+th7shR2CKS0hFaKNRp60GaD8+tMrIniONKepwP2noRNq9Wdlb2kjj2xsjP5FJUjlStmoZMbixFJq0Stk8l9rSGJwhsVZr245t7HnLXth7hc4SpHy5yffXCMARI0bQw0PBjNMETSc6SCa0PQO3/n/kvlhHffOZbSJ1JBRXEstRxXw49T1GOIHHo73WXwoxlJLQ5XtDMnM1MTx1Mu1MOPDhCyi4psnkZB99ko1Y2dYhBG1rXI+uZnt39zvbbI2G6e4+7fV3KkM1lBVbc+YSWU0klH8QQRqeFSOOL3O33mb67F3t/JG3fTQMCq3UtGm+bxRjsjPoX1kcaA9Gt392N1P0JsmTdPYu7todYbEwixUkeQztfQ4HFpIUY0uKxkDGJq/JVRQrT0dLHLU1EnpjjdjY5rWVjZ7bawWG32yQ2MShUjRQqIo4BVAAUD0A6xZurm6vriW7vbh5buRtTO5LMzHiWJJJJ+fRP6bC71+b27trbq3pt3cXXfxA2Lncbuva3V++dt1OB358l95YSoTIba3r2NtzKSpkNodKbVykcNfhtvZCliyWfyMMVZkYqelgp6WoVdMf4erDwB9R+f8Aef8AYe/de679+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XRF/98P8AiffuvdFz7G+I/wAce1t0PvnevVO3qrfUlN9pNvrByZTZu9aqBYvBFHXbr2bkMBncgtNF6YvPPJ4gLJb37r3Qfr/L++KssQhzGw9x7qhSdKiOLefbXcG7oYpEjaJfHDuHfmQiWPQxuunSSSSL8+/de6kN/L7+FzUaUI+OfXMcMU8tVHLBjqqnrknnjEckqZOCtjySsVUWtLZTyLHn37rYJHDqHTfD47Dkeq6E787y6ilWoimpttZTeFb3L1msKI4lxzbF7Yl3JJRY+o1Jq/hmQxtSgjVYpo1uD7r2r1HWKbsD5ndWVch3305sT5C7OijFty/H7NvsnsanSL9c2R6n7SzBweSaVebY3dLyFrhaexAHuvGnl06YX53fGyorIcLvneOS6M3XIwR9o/IPa+f6YzSSHQLQVG+KHF7fy0TF7LLQV1XA9jpcgX9+61x4dGhwG7drbso4cltbcu3ty46cAwZDb+axuZopwQCDDVY6pqIJLqb8MePfuvdKH37r3Xvfuvde9+691737r3XvfuvdeJtyeAOST+PfuvdJ/M7r2vt2metz+5MBg6OPiSrzOYx2MpYyeFDz1tTBEt/8T7917oBN0fND4m7OmelzvyK6gjyCRGY4nF75we4s0Y1Fy0eE29VZTLS/6ywkn+nv3W6H06RI+cvW+aZl6z6w+SfccZRHhyPX/QPYEG3qoSaAv2+7N+Y3ZG1ZQC9mIrbCx/offutEUwesEvdXzE3QY02L8OaDaNNUTPEuY7y702fgDTQt/mq6XbfWWO7PyciKPU8LTwy/gc3t7rYp59P1Ntr5vblgEe4e0vj31hqSQP8A6PetN59h5GKQn9tocrvreG3MY1l/1eKIv/X6e/de7fn0xVHxA3Lu6rhrO2vlr8m98xJUQTTbc2tu3AdK7Oq4InWSXHVmK6j23tfM12Oqiul0qMnNIUJXX7916vy6XWy/hh8Wdg5VNw4Ho/YlVuiOTzpu/duNff28kqCbtUpu3fU+49xR1Ejcsy1ILHk+/da6MxFDHBGkMKJFDEixxxRIkcUcajSqJGihEVRwAAAB7917rIBbge/de679+691737r3XvfuvdMm5NuYTd+3s7tTcuOp8xtzc+Fyu3c/iKtS9JlcJm6CfGZXG1aAqXpq2hqXjcXF1Y+/de4dEq+Me88r03umb4Udt52Ws3RsXC/xH4673zs8gqu8ehMcIaTGqcjUKlPk+0eolMeG3LSxt9xPTJRZbR4q1/H7r3R7xe3P1/339ffuvdd+/de697917r3v3Xuv//W3+Pfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XRBI4Njf/AG/+H0Pv3XuiVT/ETO7IyNbV/Gn5A9g/H3BZLIVeVq+sIMLtXszp6ir8hUz1+Sn2nsje9BPWbDhyeRqpaiopMLkaHHNNIzpTRuzM3uvdR26v+esbVMcHyw6TqIJJHWlmr/i9XCspYAx8RYUXdFNSVFRotrJjVCw4UDj37q3b6HpC5v48fOPI7p2hv4fKLoev3fsaqqWxFTU/GPLYgZPA5NY03FsvL5PG9yz1o23uNaeF5lVHeCqpoKiICSJb+6qKefSb39t7+YLiNxUXZu0+qPjhujsnFQ01JW1Gwe4N57Bw/Ym16OV3k2Pv7bm99gZimrbR1Ez4jJx5EVOFqnuheCWohk91s0rjh0NJ+Xmf2kir3H8VPkt12UWM1+Z2zs7F95bSpWewYpk+mM3u/cLwR2LNLLiIUVBdiDwPU62R/SHQpbJ+Vfxz7DqYMftbuTYs2aqGKx7YzOZh2pvEuEZ2jfZ27BhN0ROiKSwakBW3Pv3Vehnq8HhcqImr8RiciE1PA1Zj6SsEYlALtE00MgXyAC5U8+yy92baNy8M7htNtOVJI8SJHpXiRqU0/Lpba7luVlr+j3G4i1UrokZa04V0kVp8+kvW9T9Y5Gpkrch17sysqpQgkqJtuYmSV1jXRGGdqW7aUFh/h7DF57Y+3W43El5f8i7RLdNSrNaQljTAqdGadH9pz7zvYwrbWfN25RW4Joq3MwArxxr8+ojdM9S6SR1pscf6+2cQOP6/8BRb2lPtJ7X0x7ebL/2SQf8AQHSg+4/uAR/yu26f9lM3/QfQdbj7h+KvUDQ0u4OwOlNlVtBGUpMOMztSDcCRMHBjoMFQSS52cNpYaYoGuePYk27lTljZ1t12rl2xtxCulPDgjTSD5KVUED5Dj0RX3MG/7m0p3He7uYuatrlkbUfIkMxFQeg7HzPxO6pKug6H6J+QPeNRSiGKPLYjrup6w2AZp4I6mCP+/XdVR15iaynaCVSZcdHkES9jZrgH6imPL/B0UnJqTnqJLi/nb228UeSzfUvxN2hUA/ewbR83fHc0kLFlkp6bcGfxe2Ortq1TxkWmXG7g8b303sGO+vGnlx6EDYHxC6e2XuHEb7z0G5u4+0sJ5TjO1u8dyVvZe9sXLOwaWXbb5gLt3ZBbTpC4LH4yNU9IXTx7916ppTy6NAP9e/v3Wuu/fuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3TVl8HhtwUUmMz2Jxecxs6sk+PzGPpMnRTI6MjrLS1sU8MiujEEEG4JH09+690WvJ/B74iZavjykvx36rx+SifyRV23Nr0e06uKXyGYSx1G1xiJElEp1agQ1+b+/dbDEefUZvhJ8dxGsVLt/feLRWZguJ7u7ux6gNe8YWn7DRBESb6QAtx79Xr1T69cP8AZJOg/wDlV7U/9KB74/8Atj+/V69qPXv9kk6D/wCVXtT/ANKB74/+2P79U9e1Hrknwl6BV1Z6Ds6dVNzDUd+d7TQSf7TLE/YxV1/wPv1evaj1Jb4V/HZ1KvtjeTow0uj90d2ujqfqro3YhVlP5B4Pv1evaj69R1+DvxgMc8NR1rUZGnqYXp5qfM7/AOzM5TPDINMkf22Y3jWwIsq+l9KjUvBuOPfuvaj69ZsT8Gvh9hZDNR/G3p+aVhEGkyuy8TnnbwNqh1HOQZEsYzyCbm/Pv3XiW6HTa/WfXOx0ji2XsDZG0Y4VCQx7Y2pgsAkSqCAsYxVDShAAfx791rpce/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917oMe0umerO7MFTbb7X2JtvfmHoK+PLYynz9AlRPhsvCjxw5bBZGMx5LBZWOORkWppJoZ1RioaxIPuvA04dF2r/it2Rsx3rPjr8o+1euUQf5PsXtJo/kJ1gscYTw0dNQb8rIuw8JSjRp00G5YEUH0pf6+69xOeslN2D83tlSwY7ePx86w7lp/LBC28Ol+149kVEkTyeOSsr9gdsY+mTHmKL9x46fP11zcJfj37reOjpKSVBIIJAJBsSDbkEjgke/da679+691/9ff49+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XvfuvdBv2J091R23jmxXaHWuxuwqExtGlPvDa+F3AIVfTqNLJk6OompHuo9UTIwtwffuvAkcOi6L8E+pMCXPVW8u+OixIxY0nU/dm+MdgkZ3EjeHaW6shu3Z9OlxpCpj1VFNlAFh791uua9Zv9lV7IERoR82flN/DDP9wQajpds2DwPt0zp6g8opdItpMRa9zquffut1H8I6jn4I9R5wxN2hvTv/uzQQz0vaXevYNdhp3VxIjVG1NrZbamz5dLqOPsNFuCCDb37rVfQU6HfrnoDo7qJAvWHUXXOwpCG11e1tn4LEZGbU2tjVZOkoo8jVuzclpJXJPPv3WuPHoXvfuvdesB9Bb37r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XvfuvddWH9Pfuvdd+/de697917r/0N/j37r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvdf/0d/j37r3RXt//N74ZdU7tzGwe0Pln8bOud87femjz2zd894da7T3ThZK2jgyFImVwGd3JQZXHvVUNTHNGJYkLxSK4upBPuvdCd1X3l0t3phqvcXSXbnWfcGAoKv7CvzfV++9r79xNDXWYijrchtbKZSlpaohCRHI6uQCbe/de6FP37r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvdf/S3+PfuvdfN3+Vlf8AEzG/8KSP5j03y/8Agp3f8/Ov5Ng7DTB9U9C9d5TsjdO291/6OOj3p98ZPC4nde0KilwtJjUqKN6lqh1SWrjXQSwI917o43/CfKj+PUH88P5zZ343bQ3t8D+pMr8f8Nh9l/y/++03ftju3dFVNNsHN5jsOPZ+any2JpdubSyOMyFXT0jZavrqOl3ApgQUvmdPde63wvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691//9Pf49+691p7du/Af+dZ0B/OZ+ZX8xj4F9S/EvsHanyF2htbr3CQ9+9kZKkT+7VDtHrCDKV7bd25m9q5XG5aLcmxSkXlqpYmgu2glgV917odvgn/ACuv5jHYP81cfzdv5nmd+PXXvZOyuq8h1P1n0d8Z6rP5TEVMFVtnLbOgy+6szmKvJJBi6LC7kyMyxDI5Cqq62WLWaangWGT3XutpP37r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+690jNxdj9e7Pjkl3bvvZm14of89LuPdGEwkcXIH7j5KuplTk/kj37r3QL5n5ofEnACQ5T5K9HwCKdaaUw9lbVrhHO660gkNBk6rRMyAnSebe/deoekp/w4D8OHnmpqTvvZ+XqIY2mki2/T7g3G6xKoYyacFhsjrUXAuL+ohf1ED37q2lvTpsT+YX8WaiLzY/dPYeXXXoAxHx/+QORYkW1gJTdYuT4hy3+pH19+69pb06kVHzn6x8dLNiOtPk7uWKrAeNsD8aO35vHA4BgqZ1r9sY94Iai/o1AN/qgvv3VSKYPXOf5o4qJ2ih+NnzHrpVazx0vx43KNC2v5PLVVdNTuh/2l2N/x7917qbTfLevrIo6in+Kny7FPLzG1X1VhsdPpDFS0tBX71pq+n5BIEkaMRY25Hv3XumKT5f8AYAkkEXwb+X00IkdYp/4D0/D54gxEcwhn7hjnhEqjUFkVXW9mANx791YAEfF1x/2cDsP/ALwX+X//AJ5emv8A7cnvdPn17SP4h17/AGcDsT8fBb5f3/7U3TQ/3n/TJ79T59e0j+IdPw+W2WSkWqq/ib8uqZkpxPVUydabZr6imITXNAiY7f1X99NEbgCn8vkI9Gq49660em4fNfDojPW/Gz5lY8INdqj45bsnZoguppFXHT1xIT6Ff13+i+/da68vzx6apqRa3ce0PkVs2EzGBhuX40d3RyxyD1IJYsTsvLPGs0d3VyNGkG5B49+68ASaDj13H/MK+In+Umv7VqdvLR+E1Mu7uuO1dn08QncJFrqtz7JxVN6mYfR/TqF7XHv3VtLenS2wXzS+JO5DpxPyS6VlbzCn0VXYm2sXJ9wYxL4PFlchRP5vH6tNtVvx791ogjoccBvrZO60il2tvHau5Yp1WSKXAbhxGYjljcaleJ8dWVCyK68gi4I9+610q/fuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvdf/9Tf49+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XvfuvddX/H++5/r7917rFNUQU0TT1E0VPCgLPLPIkMaAfUu8jKqj/XNvfuvdAPu75WfGfYUn2+8O/eoMDW3ZVxlZ2DtdstI6EBo4cRT5KbJzyAm2lIma/49+690iF+a/SGUgml2LH2n2q8UTSrF1j0n2xuyGexdRHDl6faMG3zI5jOkNWKD/X37r3TI/wAqeysvKkWyPhb8mc1HN6Y67dtP1j1lQrJd1/yiPdvYUOZp4QVBL/Zk2a4U2Pv3W6fMdYIe1vnBnA6Y74h9b7MkvD4qvf3yZoK+DS7lZjLR7E623DKjwRgGyyMHJsCPr7914geteswxX8wHcg8dXvD4o9W08sdjJgdn9odsZemYo41x1Gc3N1vip9L6SA1LYi4J9+632/PrOPjh3ruWhko+yvmp27WR1DxSVEHUmzOrunIkaFnZY6HJQba3fu2jgkDWkX+KuzgD1D37quPLrlR/BToqTyybxre4+06qoCfdVHZ3fXcG6YqmRSpM74l940mAjkcryI6RI+SAoB9+62CRw6EHDfEn4u4CUVGL+PXTUdZ4/E2QqeutrZLJyoPGf38nksZV19S5MSkvJKzEqLn37rxJ9ehQxfXHXmBDJhNhbLw6s2tkxW1sFjlZtKrqIpKGEE2UC/JsAPfutfl0pPPh6BzCZcbROoB8RkpaZwrcg+O6MA1v6c+9hWPAE9J5Lu2ibRNcRo/oWAP7CembIb82RiKhaTL7x2riqpohMtNkdw4iinaFmKrKsNRWRyGJmUgNaxIPu4hlb4YmI+QPRbd8x8vWEgivt+soZSAaPPGhofOjMDT58D0lMv3t0vgZYoct2nsKilnhaeFH3RiJGkiRirOvhqpPowtb6k+3Us7t/gtn/YeiS99yOQNudI73nPbI3ZSQPqIiSBxOGPSF/wBm9+Nn/P29t/8AUrLf/W326Ntvzwtm/l0Hv9fH2nH/ADu1p+yT/rX+z18uo1Z8xvjbSU01SO0sNWGFNf2tBR5iqrJuQNFPTpjg0r83sPwL+9jbL8mn0xH5jpmf329p4Inl/rlbuQPhRZWZvkqhKk/LpvxnzV+N+SlmjbsGPF+KNZPLmMLnMfFKWYr44Xkx58kqgXKjm3Puz7Tfp/oNfsI/z9JbT7wHtRdMytzOIaKD+rFMla+QqmSPMdY8l82fjfjpxAu/TkgY1k+4xGCztfSgsWHiM8ePC+Zbcr9Rce9rtN+wr4FPtIH+Xqt194L2ptpAi8y+MCtdUUM0i58tQSlfl07R/MT42yU6VB7TwkZeAT/bS0uXSrS8evwyU5x2tKlf0lPqG490O2X4/wCIzfy6XJ76e07xq/8AXO3BK10lZQ32afDrUeY6dsV8q/jvmZRBR9tbSSUpG+ivq5sV/nXWNYxJk6ekiabyOAUDFh9SLD3R9vvYxVrZqdLbL3m9rdwkEVvztY+JQYdjHxwMyBRWvEVx59LWl7p6ira6uxtL2bsSauxrKldTLurCh6dmOkK5asCEk8cE+2ja3K0Jt3/Yej+Ln/kae4ubWLm/bWuIaa1+piqteHF6fs6Vabv2pJS0VbHubb0lHk546XG1Ueaxr02QqZmZYqeinFSYqqeVkIVELMSDYce2/DkrTw2/Z0dDfNlaG3uBvFqYJWCo3ippdjWio2qjMaGgFT07RVVHWh0hmpqtVsJBFLBUKur9IkVHfTqsbX/p7rQjiKdL4p4JtRhmRwDQ6SDT7aV6TGb692DuQadxbF2fn1BL6c1tnCZVdRBTVauoqj1aTa/9Pev8HTvn0EOZ+IXxY3BJLNkfjz1AlZMulsnjNg7ewWWT9toR4cvg6LHZOncROVDRzIwB49+68GPkekNV/BnpmAibY24e8upapIfDBUdZd/dsYOCnXyGRmiwuQ3Rmdtl2JIOqiYFeLe/deNTx49N9b8f/AJSbTpZU6k+aG4cgsSxmhw3yG6o2P2tQftut6eo3Bsw9SbxeOWLUrSy1lTLqIbmxB91sU8+vVPYvzk2CwO6fj11T3fiYrfc5fo7tSo2XuUQRjVPUR7B7bxdPi56l0/zdNHuVyzC2vke/dbop4Hp3p/m31Ph6WKbuHbvbHx2leSOBpO7uts/trbq1D8NEvYGHi3D1w6q4IDfxcBgLjj37rR49GX2dvzZHYeFh3HsHeG1977fqLeHN7Rz+K3HiZWIDaEyGIqqulLgHkarj8+/da6Vnv3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de6//V3+Pfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XVx/X37r3Xrg/Tn37r3QS9g9+dIdTqx7M7d642JIui1LujeWAw9fIZOI0gx1ZXxV9TJIRZVjjZmPAB9+62ATwHRf2+fXR+WqVpOtcD3n3ZKSUM/UPQvaW6MKkgjaRUl3VVbbxG0oRIAArNXCMlh6rG4914qenCP5M9y5pYpNpfCP5BVMMvlAk3ruDpTr5k0iMxNJS5Tsyvroo5vIf1RB10nUv0v7rdB/F07J2d8uq+cR0fxU2Xh6dpkRarc/wAjcWJI4GClp5qLbXXO4R5IyxBjSZ724b6e/daPyPWOoyXzvq5AaHZ/xPwcDVB4yO/+3NwVMdIx9LGOi67wEElVGp5USKjEcMAbj3XjTyr0zVOC/mCZCok09mfEvbNK1QpSOi6m7Y3LVw0d/WhqMh2nhIZ6nTez+FEuP0+/dexTNa9ZV6g+YeTkY5n5l4LDwSNMXpdi/G/Z1DPFDKjLEtNkd47s3qokp2IKu9M+ojlfx7914H5dRpviPvLPhW358y/ljuB0IPi2vuvYnU9G4MapLG8fWfX23K143K3F6jUpJsefexnh1uv9EdM9X8KvhjioPuuxcSd8PT1UKy5Lu7uPfnYOmtUssccke/N7ZLGQys1/21hQE8aeOLLHI/wIT9g6LLze9o25DJf7tbQRBtNZJUQBv4aswz8uPSkx26fgz0ilVT7Tp+hdjzYySB56TYW1tq09f55REYpY4dq4pqqrnCurF11so5Yix9qEsbxyALdqn1x/h6BW6+7vtttH1AvecrLxIqalR/FbPCixai1flWnn1HzHz1+PeLepjosxuXcUkNP5oWw22MiaerlKFhSw1GQWhjSYHgl9CA/2re1K7RetnSq/ac9Ae/8AvKe19m0qwX13csq1HhwPRj/CC+ihHnWg+fQeV/8AMd6zhhRsVsLf+RqDIoeCqGBxkaRFWLSCpbKVSu6sANGnm978e312O4PxSoP29Bi6+9VynHGDZ8s7lLLUYbwYxT11GRs/Kn59Mc38yTaxjk8HU+6zP438Imz230gMuk+MTvG0sqRFv1FUZgPoCfdxsUlc3K0+yvRe/wB6/aNDGPkq8MlDQGeECvlUipp9gP2dJas/mTZN6Sdcd1JSw15j/wAmlrt2yT0SS3HqqIafDU08sem/Cupvbn24NiUcbnHyX/Z6JJvvYXrQuLXkdFuCMF7klQfmBECfyNekBlv5ifcFX4BidpbDw3j8nnM0eay33GoL49Ouuo/t/GQSf16r/i3t5dktRxlcj9nQavfvSc9ziEWWx7bbsK6qiWWvpxdKftPSPy3z0+Q+QNP9nkNo4EQ+TyDG7YjqhVawunzfxiqyBTxWOnx6L35v7eXZ7IV7WP2t/sdEd595H3SujGbe6sbbTWoS3DBq8K+KzkEfIj59InMfL/5H5qaCaTsmrxhhiaLxYPD4TGU8t31+SeNKGbyyi9g1x6fx7cXbLBRQQV+01PQdvffL3Yv3R35teLSKUhihjDf0mARs+nQZbk7i7a3fL5dy9kbyyf7kcqxDOVlBSpLFF4EkSkxklFTo/iNjZOfqbk39vx21vFiOFR+XQT3bnvnffHD7tzZfS0INPFZFqBQdseheBPlmtTnpAVlVWZGoaryNdX5CrdUR6qvrqutqnSMWjRqipmlmZIwbKC1gPp7eov8ACP2DoMzyz3UpnuriWWcgAs7u7UHAamJOPLPUJqanc3khikNrBpUWVgB9AGkDMAL/AE926Y8GInUYwT88/wCHrtIIYwRHDEgJuQkaLcj6H0qOffiSfM9bEcYrRAK/LrKOPp70RXj1YAAAACnXvr78ABw63+Q6979QZx1qg68OP6f7a/8Avfv3XqAknz6979TrfnXz/n10VUizAMOeGAYcix+t/qPewSMA9VKqeI8qfl1iNPTlVVoIGVf0qYoyq/8ABVKkD3vUxFK46r4MVAPCWnpQU/ZTrsQQgBRGoVSCiDhEIuQ0aA6I2BJsQARf3r8h14RxgABcDh6D7BwH5dOePyuWxPm/hOYzGK+40Co/hmWyOP8AP49Xj8/2lTD5fHrOnVfTc2+vurKrfEgP5DpZa3d5ZeJ9Fezw6/i8OSRNVOFdLCv59KLGdjdi4OeSqw2/97Y2omh8Es1NunNB3h1iTxMZa2Qadag8AH/H3RoYWw0SkfYOjS05q5q2+RpbDmbcYZGWhK3EuRWtO5z59Lan+RnfdLPS1EHbe9BLR0BxlOZcjFVRLSatX7tPUwSwVdVf/d8qvP8A7VYe2PobQ1Bt0z0fx+6fuVC8M0XO24eJHGY1rIGBWoNWDKwLejEE449CZT/N/wCSVPTLT/3ww05RKaNKip2lhZKnTB+tpHWJFmkqVsJGYX/K2J9sHaLEk0Rqfb0Lo/vC+7EUQi/ftu5AADNbRau05JIGk6hgmnzweho2d/MY3pQRZBN97Ew+4pZFDYqfbVZLt4U0ghKiHIwZA5UVEMs9maSNleNbgIxtZNJscTEGOcqPmK/5uh9sX3qOYLVLleZOW4LtyP02t2MAU0wHD+JUE5JUggcFNcD5s/8AmE9P5po4d2Yjdex6lnjjaaoo4s9ilLnTJM1dh3lqI44iLsXgU6bcE3ARS7NcKT4bBx+zqTdj+8/yNuGlN7sbzbpagVZBLHnz1REtT7Ur0a3YXbPV3bWNnl2VuvCbnp7S09bj1fRXRowYOldha+OCujhljvzJCEdfpcey6a2ntzSWMj/B+3qaOV+eeU+c4GuOWt8huQGIKglZFPzjcLIAaYJWhHAnoGd4fCT46bozdTu3AbPreod+1YLT9h9DbizXTO7ppmkMwqMnV7CrMNj9yTGU6v8AcpTVyk/UEce2Pn0LK1wTnpPR9Z/MnrCWn/0cd8bP7z2tSs3+/Q+Ru1f4FvQ0KKClJR9x9WUlDBLWfULPk9rV8jceSRjdh7rZIPl0/wCwvlhjMjv7F9P909dby+O/beflqIdn4PfMmMzGxuzJaSnkq6uDqztTbs9XtDduTpqOFpnxUz4/OpCDI1AI1LD3Wujb3B+hv7917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3XuutQva/Pv3Xui/9tfKTofpOvgwW/uw8ZT7yrYvNi+u9uU2S3r2ZmEZQY3xXXWzqPObxroZdQAlWi8Nzy49+62ATw6DGk+VHZO7HB61+G/yMzlAZUWPNdhQbB6Vx9RE4DJVQY7sDd9Hu9acobkTYqGUfTRfj37rdB/F0oaff/wAvsqB9v8curds66WRw+6PkJVV5hrBIyRQTU+1+qcijRmPTI7pKbC6gE2Pv3WsV446ijN/Or/n3HxWv9eO3+1rf7D/jDZ49+x1vt+fUqTeXzMxtLC1R0V0buSs16KhMB33ujEwaSWYTQNn+mwxAX0lGsdRvfT9Pdax69MFZ378mdtVDTbv+FG7sjgYoTLNk+pO4+suwMpGU1F0TbG4put8pVMVA0LAZmb6WBsD7r1B69LvqP5ZdIdy56fZW39z1u2uz6KGaoyfT/ZmAzPWnbGNipgWq522JvKjxObyVBSry9ZQJV0OkgiYqQT7rxBHHoyNx/t+ffutdd+/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de6//1t/j37r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691xdgiliVVVBZmYgKqgElmJIAVRyffuvdFn3z8yvi913XS4bcXduxajccbyRDaG08k2/d7SVELmKSkh2bseLcO5papZRoKCl1BjyB791uhpWmOkAPld2VvZFXo74id6bvSVGeHcvbNLh/jxs4xsFENQB2HVf6QqiJmcNaLbrEoCVJNh791rrqLb3z237Z892N0D8fMbKWZMb15s3cXdm8YYnjUqk26N+Vmzdpw1cUtwdGCqoiOQT9Pfut48unbI/DrD71SL/AEwd4/IztMSRRDJYibtTJ9abPyEqEtJr2j01B17jftZCSPDK090OlmYe/der5U6Errz4t/HHqiRajrzpDrHa2QTUf41QbPwsm4pC5DOZ9x1lLU52oLMASZKhrnn6+/da6GmryFBj/H99XUdCJA3j+7qoKUOEALePzSRhggIva9r8+9hSfhyek1xd2toAbm5jjB4a2C1px49Axuf5MdE7Q1jN9n7V86Cnc0uLrTnaspUSGKN1pcKlfKyKytrsCYwLtb2qjsbuSmiBq/PHQC3n3b9uNhLLuHN1n4godMbeM9CafDEGODxPBeJp0Au4/wCYV0tjFmTb+N3luupjepiQU2HTE0crRIxp5xV5aop2NLVSWAYRl1HJT8e1qbJdtQyFVX7a9Rpuv3oOQLIOu22t9eSgkYjESGnwsHkI7WOAaVHGnQQZX+ZRKYYf4F1E4qfLeo/jO64BT+DQ3EBx+MllM3kt+oBdP+PtUuwio13WPkp6A1597JjHGNu5JImr3eLcjTTPw6IySa09ARXPQV5H+Yb3XV01RBQ7c67xM8hJgrY6LN10lKnk1C9NV5MU87aPSdQX+oA+nt8bJbClZGI/Z/k6Bt196P3Bmjkjtto2uGUk6W0yuVHlgyUJpxxn5dBfmfmd8j8yrod/RYqJ6hZ1TA7fwmOaLQCBBHUNS1VS1Mb8rI7lvyT7UJtdihqYdR+ZPQRv/fr3W3FWVuZhANer9GGJKf0dRVmK/Ik18+gazPZ/Ze4WRs52JvfKGOomqoRVbny+iCeo/wA7JBHDVRJDqHFlAUDgAe1S29unwwID9g6AV/zfzdumn94c1bjKFYsK3EvaTxK0YUr6DAGBTpETlqqSSWqklq5ZpGlmkq5ZKqSaZiS0srztI0krMblmJYn8+3Rj4QAPkAOg7IDM7vMzOzGp1EtUnzOqufnx+fWNURAFRVQD6BFCgfX6ACw+vvZJ4VOnqoRFwqgfYAP8HH8+uRF/qT/vv8Pp711YivHrv37rw69791vr3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917rjpH+P1/2/+B/qPfutdOuCzmZ2tmaDce3MnWYTO4qYVGOyuPl8FZSyhWQskgBDq6OysjhkdWIYEce9MiyIY5FDRniD5/5elm27hf7NuFvu203bwbnCQUkQ6XUivn6ZIINQRginVjXRvz9ytFPjts910keRo6iqpqKLf+LgSlqMdCyNEanc2JiDLWoJtBeopFjKIWZozb2R3Wyhgz2h7gK6T/k6yo9vPvL3tvJa7V7gQ+NasyqL2MAOinGq4iWoYA0q8eQKsUPVqmKyuOzdDS5TE19DlMbXRCejr8dVQ1lFVRNe0lPU07yQyqbfVSbew8yspKspDA8D1mVZ3truFvFeWN1HNaSLVXQhkYeqspIPSD7g6g2D3rsDO9a9k4Y5nbOcSnlP29VUYzMYXL46ojr8HufbGcoJIcptzde28pBFV47I0kkVVR1USSRsCPdelfQAfHzsDs7aO+M58X+/8rFuffu2cC+7+pu3vHS0P+nrqGlyEOIly+bxdMIafFdsde11XSUO6aaBEpKpqykyVMscVa1PTe690cwe/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3XugO7t+QvXHQmMws+86zJ5Lcu7shLhevettm4qp3X2d2VnokWR8LsXZeM1ZTMz00Th6upIioMdCfNWVFPCDIPde6AzH7H+VHfklRlO3d5T/GfrHJQvHQdM9P5Khre463HTC8bdld4otbSbYyboo8mP2dFG1MGKLmJyNXv3XsdL3Y/WfTHxwqKfYnRnU+Go97blpJMvk6mnimk3Jl8WKz7Ks3j2D2ZuNq/cO6Y6PMVUK1Jqa+vyrCbXFDIqm3uvdCIlN3tkIZJKjMdZ7YlqMXenoqbDbh3TJjcxU7RijImyM+V2/FkqTE75MkqMtNTtWY0LGRDMTJ7917qfVY7uGOOqag3TsCWUw5j7GOv2jm4o0qJnwH8DNVJSbnZ3hoYoMkalUVWqGqKcKYhC/l917rJG3c0VbB5YusqzHGoC1Ahm3RQV6Uj7yceSLyQ11O81L16ysVbSJswCLx07Aj3Xuo1Dne5B9kMp1/s86zihXvi9+VUq0wn/vG2YenWt2xSGoXHijxiQglDUPWTE+NaceX3XuuT7n7W+1Lx9XYsVponlSCXf9CkC1/9z/4rHSyzx4aRvA28f9xDSqjWi/yzSU/b9+690GvYPW/W3ySiyPX3b+w6jCbv2lURbi2RuPE5VqTfe0UDxRYfszrHsvbqw5LZeZGYgnhjSCqhrStMwqoGppvHJ7r3SY+O3a27sMvc/UXyC3bicjvf445PGS5PtbIR4zauL3/0/u3F1Wf677Mz8Aejw2AzyY6grcXuARCGgGVxFRUQrFBOkUfuvdYn/mA/FRmnkxfYeZ3TiaR0Sp3Rsnq/tbe+zU8khhjaHeG1NlZjbldDJKpUSU9TNHxyQOffut6W9Ohp6t+QvSPdq1P+ijtPZG+aqhjEmSxGDz1HNuHEIdNjmttTPBuDDNdgLVVNEQeLX9+68QRx6GQf4/X37rXXvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvdf//X3+Pfuvde9+691737r3XvfuvdRquto8fTTVtfVU1FR06GSerq54qamhjFrvLPM6RRoL/ViB7917ot+8vmb8T+v6h6PdnyH6jx2QikEUuKp97YXMZeORlZlSTEYSqyOTRmVDwYh9Pfut0J8umil+aXSGajLbNj7Y7AcoZKdNk9D90ZmKtUMi3ock2xaXDVat5AVZKkqynUCQCffuvAdN1X8pN71rLBsr4ffKDcs8ysYqjN4TrjrnEg2k0Coq989kYjJ02powOaI21AkWvb3XqfMdNE/aPzlzjIds/FHq3Z8Del5O0/kfE9dH+5H+7/AA/rbrvelI0fiYnT96G1rb6EH37r2PPqXFSfzBchVJ9zm/h/tjHmaQvHQ4DuLd+Qipzcwqs9XnNpUslQlgHPiVTckAWsfY632/PrC+w/nw7u6/Iv47U6u7OtOvxx3dOsKliywid+743lVB6dZVSw5sD7917t9D1lq4P5geGkeTH5H4jb+pI41cUORxnbnXGSq5EjJeCPI0mR7CoKXyy20u1LIEH1B9+692/PrHR/I3v3ajwwdx/DfsmkSxjn3P0Tu3Znd22VZWGuo/hDV2x+yEptBuAmBnmNiAh4v7rVB69CJsP5b/Hbsbc52Jt/s/DUPYKIrv11vOly/XfYPNtQi2Xv3Hbc3HV+MsNTQ08iD63sQT7rXRjvfuvde9+691737r3Xvfuvde9+6914+/de6JhvX5RZ7du5cz1b8SNn4nuvsXbuYfb++975rLVeB6D6cyEIhNbS7531QUddU7r3bQLOL7Z25HXZNX9NbJjkPm9+691govh8m/Wx2d+V3ZW5PkNuSkaWaPa8T5HrfozESTu7/b4vqDbWabHZ5aeN/EKncdXm6qREB1ISyn3XqkcOjDYzb/T/AE1iC+FwPXPV+EhhdpGxeM23s+hEEERaRm+0goI5fHDESfqbL+fd0jeQ0RCT8hXov3Hd9s2mB7vddxgt7ZQSXkdUWg86sR5dBXu35l/HraTSQvvqDcdXGyB6PaFHU7gcByl2+6pkXG2RXDH9++n6Anj2tj2y9kofC0j+ljqLd79+va/ZNSnmIXcoYDTbI03Gn4lHhmlRXv6AHef8xjZNBrh2FsfcW5JjE3jrc7NT7bx6ymINHqpx9/kZkSY6XGiPgXUn2sj2OY/28gX7MnqM+YPvU8vWniR8t8u3V3LSgeYiGOtMYBdiAePaPkei95n+YT3TXrXR4fAbD28k73op/sspmK2gQOhZWNVWU9HWu4Ugs0SCzX03Htcuy2opqdz/AC6jG/8AvQe4F0s62G2bba6iCjaZJWUeh1MFY/Ogp6dF73L8jO9t3K0eb7T3UYHVFalxFXHt+mIjnNRGTHhIqFi8ch4ctqKgKSQLe1sdlZxfDbLX593+HqL9390vcffUMe485XvhHisbCFfiLDEIQ4OBU1AAyRjoIKyuyORkM2RyOQyUxeSQy5HIVtdJ5Jm1zSB6ueYq8rcsRyx+vtQFVRRQAPsA/wAHQGnnu7ptV3dyyvUmruzGp4nuLGp8zXOOoKxBT6VVRe9lFv8AX+gHJ976aVQooB1l9+631737r1Ove/db697917r3v3Xuve/de697917r3v3Xuve/de6973Qnh1UsBxPXveurde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+6111/r8f65H/EX96JA61qrw69f+lj/rH34EE0HXi1OI661AfXg/05/wCIFvdqH068GB/1DrvUPrfj+vFh/S/PF/dtDk6VQk1pSmf9R6r4seTq7R54/wAFa/nTrrUv4IJP4BBP9foLn6D3doJ0BaSFlQebAqP2mgr8q9VFxESAHBY8ACCT+w9ZI0klJWKKWVhyVjid2tcC5CAkDn2hvL2x26JZ9x3C3t7csFDSyxxqWNcAuwBOOHn+R6VW1vd3rmOzspppKVpHG7tT1ogJoPM+WPUdSo8dkZn8cOPrpXILaEpZi1l+p/QBYew9ufPXIuy2ovt2522i3s9YTW95bhdTcFr4nE0wOjmy5W5p3Kf6Ww5Y3GW40k6VtpiaDicoBj7a9SP4HnP+dNlP/OOX/o32Hv8AXn9njke6fL9P+e6D/oPo2/1uPcTz5D3ev/PLL/0D12MDnG4GHyVxa96WRfqQBywA/I/1vdZPer2ciGqT3T2GhrwvIW4Ak/CxpQA5+wCrEA2T209x5SVj5C3Ytj/iNIOJoDUr60H59dHBZwf8ubJn/Wo5j/vIW3u3+vP7PkA/66WwZHnewD+Rev7QOq/63HuISQORN3waf7iy0/I6c9Q5qSspy4no6qExnTJ5aeRBGf6MxXSDz7Fu1808rb4tm+y80bbdrcKWi8G5hkMgFcoqvqbgQaA04nGeg/f7Hvu1yTx7nsV7bvE2l/EglTSaVoxK0B9M0Pr1FDA/TnmxH9Pxz7EDRyIQGQivD54qaeoHmRjPRQsiOaKakcR5jy/w4+Xn1y91/Ppzoxvxu793H0pvTExHJ1UvX2ZytDR7swMgnrqeCjqJnhbK4Sj8pjocnTz1CySNCmqojQoQSQQgvrKO7jLaQJgDQ/Z1KntP7lbr7f7/AGMf1TtyxcTIlzD3OqozEeJClTokBYFyorIBppWnV/rTxRwPPI6RQJG0rzSMI40hCmRpZGcqI41jFyWsAPr7BufPj100FDkefVPE3y/2x8jPlfh92/GTr7sT5LbT+LeG7C2JQ7t6uxuNoust6dy9lPi8LujGwdzbtyeA2A+zesMDtVocjU0VXkDVZfIhIKeZqBm9+6sAKcejYR4P58dkItRmN9dGfGXFTvqTB7J21lu+t901M1wI63d28JtkbJp8ioW9oMHWwoTbXIBc+60dPlx6lp8Pc5moR/pF+W3yv3nUzRBK+LDdh4XqrDTyBkcyUuO6s2rtWuoASn6UrWsCRex9+611Kp/gl0WhqGyGU703BLVEeabcXyT75y8pGkq4jNT2EywLKps4QKGsL+/deqTx69R/Af4zYxpZMVtnfeLmmiWCSooe7+7Yp2gEiyGHyN2C7KjOgLWsWtz791up6wn4LdV0sz1G3+xfk5tSc1MdVG2B+UHd3gheIkxxxY7L7yy2K+1QGwhaBoyOCpHv3Wuos/xa7o2+yT9ZfN7vzFSqXL0PaeF6v7lw0w9TRQvHltnbfz0cIY2YpkRKyiwdT6vfuvdMeX7j+THxtgGe+Ru3NkdudJY9FfdXd/SOIz23d29b49VvWbu7F6Xy9duN6/ZGMW8uRyW3stWVOPpled8cYI5JE91s08q16PNicrjM5isbnMLkKLLYbMUFJlcTlsbUwVuOyeMyFPHWUGRoKymeSnq6KtpJkliljZkkRgykgg+/da6CvtP5BdMdKvhaXs7sLA7Wy25ZJ4ts7akeqy28dztTQTVVX/drZmCpspuvPpSU1NJJM1HRzrEiFnKge/de6JP8Dew+rO5d+d+9lZbPzZ35QT783PitzYPd2Dzm3Nydb9JYvclfjOotu9aYTd+FwWZp+o8ztunp8pLkaKmWHLZ2tqXrHNVGIofde6s6Bulxf9Jt+P8AkXv3Xugbx9fRbX7b3PS5iJaJ+xYNsNtjMNTTUdBl67C4fLQzbTFbWbkrY8nuekpMZVV/ipMbRImPsZHmdSy+690MoN/99/vrjn37r3SP3Yd8xRxz7NG3ql443+4oM2tZHJO9yUakq6eZYozbgrIoB/1Q9+690FNP3bkMJkI8X2Hs6v25JIQqVlIWqacgABpRE1vPCX5Jhkl0g/Q8+/de6G/CZ/DbjokyGDyNNkqNvT5qZw2hx9Y5YyFlhkH+pZVPv3XunGpqIKSmqKuqnhpaWlglqKmpqJUgp6eCGNpJp555CscMMMalmZiFVQSePfuvdBV1dSz5eTcnZdfDPT1e/qqj/hVLUx1NJUUWy8EtVT7Zpq2hh3PuTb09dO9ZV1gr6D7cVtJVQCRNUYt7r3VdVT1ngvkt/M87zxu6o4s7078dOkPjUN47Crg7YPfHfOeyvb279iSbtxt2od07X682Fm/vqfH1SvTnMZKGoljZqWnI91uuKdW109PBSQRU1NDDT08EaxQU8ESQwQRIoWOKKKJUjjjRRYAAAD37rXQKdrfG/pbuiB237sPEVWdRQcXvjCfcbV7H27VREvS5DbPYe2pcXvDb9fSTHyRvS1kfrHqDC4PutgkcOi2YzsPtL4f52i218i97TdofGvNV1Fg9j/JnPQU9Jvjq7OZPIQ47BbJ+SRxtLS4XIbey9RURUmM33BFSRGtZKfMQRSTR103uvY/PqwJWDC45B+h/BH1BBHBBHv3WuuXv3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r//Q3+PfuvddXH0vz/T37r3RRuxfmFsrbm/Mv051ZtLfHyG7vwIok3D1z1RjaaoodjS5Sm+7xb9p9jZ2pxPXvW8NXTlZBDX5D+JtC6vFRyhl1e69001Wy/mh2oKSTdPavX/xp25URh6/avTe3/8ASn2L4plXXRTdsdiUWL2pjaqEcGSi2lLpYnRMwCv791sY8uuqT4E/H+vq6bKdpU2/vkHnaUsY8x372NunseJWkBEz0+06yvo9gY4SiwZKXEQRkAen37r2o+XRk9odU9X9fU6Umw+uNibKpkRY1g2ptHAbeiCILKpXE4+kBC/4+/deJJ4npfe/da66sP6D/X/P+3/w9+6916w/33+vf/e/fuvdd2/Pv3Xuve/de697917roAD6C3v3Xug17P6b6q7p2/LtftjrzaPYODlB00O6cHQ5X7WS4IqcbU1ERrMVWxsAUnppIZkYXVgffuvdFux/xs7c6Wq4Kr4zd2Zf+59OoWbob5AVud7O2AaaKMpT0Ox+xKitm7T60EYJAD1O4Mcg06aAAe/de6UGwflrhKne+O6e742lkfjv3Zlqh6Pbe2N25KHK7D7PlVXnE3TPa8FHjdtb/c0qh3xrx47cFP6hNjkVdZ917o3YIP0P0Nj/AFB/oR+D7917rv37r3XvfuvdV9b43ju35e783r0J1FuDMbL6G67zLbU+Rfem2q+fH5/eG5qcK2e+PXSudoJYZ8Tk6CCVId37np5fNhfIcbQ2yLT1GP8Ade6NnhsJ1b8fesKLB7fxW3OtOrOu8GKehxWMposXgsJiKNLhIoYxqnqqiVizMfJUVdTIWcvK5JukbSOqItXJwOi3d9323YttvN43e8jg223QvJIxFFUf5ScACpJoACT1Vh3l86N871rajD9T1dfsTaEZ8aZjwQx7yzIFtVT5mNRHt+lZhZI4bzsvLyLfQBNabRFEge4o8p8vIfL7esGfcT7xfMm/3MtjyTPJtuxjAloouZvOpJDCBf4QAX/iI+EEcy2WzOfqTV5/LZTOVPkeQTZfIVORKPLNJPK0S1csqQK08zvZAoDObAA+zYKi/AgUfLrHe9vb3dJBPud7NcTVJrLI0hBY6jTUSBUknAGeHUAKALAADngCw5+vA4t730mCqDUAV/1cPT8uvWBtcDj6e/deoPTrv37r3Xvfut9e9+691737r3Xvfuvdev8A77/ePe6cetVGc8Dn5fb14c/77/Yc/wBPfuvEgZPDrq4sTcAD63Nrf69/p7uscjmiRsfsBPVTIihizgAca+X2+nXQdTYAgk/QDkn/AFgLk+9tBMilpImVPUig/MmgH5nrQmiYhVkBY8AMk/YBk+uPLPDrmoZ2VESR2Y2VUjkZmP8ARVVSSefx7Szz21rBJdXd3DFbIpLO8kaqoHEszMAoHmSQB59PRRzXEscNvbSySuaKqxuxYngFAUkk+QFa9Z/s63/lSrf/ADjqf+vXsk/rfyf/ANNjs/8A2XWv/W3o1/q9zH/0zm4/9ks//WvqWmDzbqsiYjJFHUMjCklIYH6EAgH2Grj3h9o7S5ns7v3S2CO7iYqym9iqrAVINDT+dD8z0cQ+3PuLcQR3MHIu7vbuKqwtXoQeB4VA+0V6mrtTcxAYYSpIIDC70w4IuLqZw4+v0PPsHzfea+76sjwv7tberKxBIS4OQaGhEBU/IioPEV6ESeyPu7IqOnIV2VIqO+GufkZQQfkQD1MXZW5rAigp11LyHqabyJfnSQ2rQw/NifYQufvffd6Exik5u3CQxSGjR2V34bEcGVgF8RD5VAqOIHQgh+7x7vPEJI9htEMiCqvdW4dK8VIzpYfaepEewM+6anNDAdRHjkqSzADgG8UTLZv9f2Tbh9972RtLkw2q71eQ6QfEitAqVPFaTTI9R/paf0j0a2n3X/c+4hElxJtlvNUjQ9yS1BwascTrQ/6b8uuY6+zlxeoxwFxciaUkD8kDw82HtA/36PZ0I5i2ff2koaA28IqaYFfqMVOK+XHpUv3WPccsofc9pCVyRPISB5kDwRWnpUV6mHrqu/52tCf+nNSP969hn/g9+TCM+228j/qItT5Dj/sdHn/Al8zkmnOu20/5oz5+eTj7Ouv9HVcP+XpQ/wDUmp/4nj3sff25Krn243mn/Ne161/wJnNAz/XTbj/zZm/z9Sf9HB/53Q/x/wAgJF/8CaoG3+wHsjb7/wBAHcJ7RuUqaE7ioJHlUfS4NOIqQD5nj0cD7pFyQpb3DQNQVAsqgHzofqMj50H2Dh13/o4P/O6H/nB/+N+6/wDB/wAX/hIn/wC5iv8A2y9e/wCBHn/8KIn/AGQ/9vPWdeuaXSPJmKkvb1eKlhVCf9pDyMwH+x9kl19/rmA3ErWHtbYLaau0S3s7SU/pMsap9mlR0ZQfdK2nwUF1z5dm4/EUtolT/ahndvtq3Xf+jqi/521b/wCc1P8A9He2P+D65r/8Jftf/ZXcf9AdO/8AAk7F/wBN3f8A/OCD/P1kj67xykGXJV8i3uyrHTxFhbhQw16Dfm9v8PaG++/pz5LBMu3e32z29wUorPLcTBW1fEVJjDDTVdNRk6q4oVVr907laKRDec3blNGGqQqQxkrSmkEBtJrnVQ4xTz6k/wCj7Cf8rWS/6nRf/U/sk/4On3f/AOjBy9/2Tzf9b+jX/gV/b7/o77v/AM5ov+tHXR69wZFvu8nz/wA3ov8A6n97H36fd8EH+r3Lp+X08+f+q/Wj91b29YEfvjeBX/h0X+WCnUv+4u2gAPBWMQACxrZQSQLEkKoAJPPsLv8AfP8Af93d13na0QsSFFhGQoJJAFWJoAQKnJpnPR4Pu0+0oRFba79yBTUbtwT8yAtAT8sdZItk7ajfV9lNLwRonq5nj54uQpQ3H459odw++H94K/tXtl5rtbUkg+Jb2Nukgp5BmEgofMac+o6U2f3cvaO0nE7cvXE6gHsluZXQ1+SlDUeR1Y9D1J/ultm4/wBxMH4+s1UR/sR5rH2Qt96b7wzK6n3Pu9Br/wAR7MEA/P6fB+Y4dG49hfZ5SP8AkC25IP8Avycg/aDKQfsII+XUxcBgEZXXD44OhDKy0qXBH0PJt7CM3vf71XNtPaXHuxv7W8iFWU3T9ynyJFD+yhHAU6Pova322hlhni5B2lZUIKkQLVSOGDWtPnWvU9KajjbXFR0cbi4Dx0lOjgEWPqWMHkewXdcz827hA1nuXNW63FmaVSW8uJENOBKtIRUeXp0JLfl/YbOYXNly9YQ3IrR47eFWzxFQgND5jrOCFJKgISLEooQkf0JUAkeyaUNcClxI8q1rR2ZxX1oxIrk549GkSeCQYo0Q8KqqqaelVANPl17Wx+rE/wCxPttbe3U1WFQfsHSnxZTjxGp9p678j/6o/wC3928KP/fa/sHTf5nroux+rE/7H+osf949+8KL/fS/sHW6mlKmnXhI44DH/b+9CGEcIl/YOvVPr17W1rFiV/1J5Xg35U3BF/dfCSMmSKPTKfxL2sK4NGFGFRg0ORjrbO7roZqpSlDkU+YNQfz6aK3b2Fyan7rHwF/r5oF+2qBckm0kOm9yfq1/cs8l++nu77fOP6t89XwtBQfT3Dm7tjQAD9G41gBVAA8MpQDqO+Zvaz285tUjfOVbY3FaiaIeBMM6j3xUrU5JYNnyPQdbp2lRYahOTpKyRIVmjjkp6tkYsZmOkwTAIx8YHKkG45v+Peev3b/vR81e6nN45D5t5bt2v2tJJkvLRWRUWFV1fUwEuoDk4mjdQHZUMZ1AjE33o9jOX+QeXX5t5f3ycWa3CRNb3BDMfFZqeDIACWQDKspqgL1qD0JvQnStN2BFm+yt8bnxvX/S3Wa/x3eu+czWRYyjFPho0y9XTUmQqaikgooKWijEtVXmQRUyEadTkKM0Nwvks00gVuGBx5D7egN7P+1F/wC4+6NuE05t+W7KZPEkAOqRxRxFDwBJHxy/gBFAWx0e18Lvr594yIbsw+4+qPhXk3kvsnNR5Tb3bnyr28Bakn3nAHoMt1T0XnbiVcQx/vBuihI+9/h9FM9JVA7rpMBQKATwHR+dq7V21sjbuF2fs3buE2ltPbWOpcPt3bW28XRYTAYPFUUQhpMdiMTjYaehx9DTRKFjiijRFA4Hv3XulBYf77/ffT37r3Xfv3Xuve/de697917r3v3Xuve/de6h5A0CUNY+TNIuNWlqHyL15hFCtCsLmsataotTikFNq8nk9Gi9+PfuvdVQ9YfIuLoT4bdW0W1qKLNbs7Nznb8Hxd2hmXlago+mcbvHd249o9g7ukxyJVYzprqrpxqHLV1RGFkGJFHQwE1lVTI/utmlccOmf4xfGTF7L716j727NzOZ3z8oO5MP2b2Nunf+88ZTLuk7Bo9t4Pb+E2VQUgNXRde4Olk3hT1v8Dxj01PRt46V/O1M00nutdH277+Nmz+9Y9v7g/iea677g6/avrOpu79jyU1F2B11k6+Lx1cdJPURT4/cm0cyFWPLbfycVTiMrANM0XkWKWP3Xq+Xl0iumPkBucb/AJPjb8iMZQbS78xmHqM1tbP4ilnoeuPkVs7ELTx5Tf8A1TJVVFXJjsvinqEO4dq1E82SwMkqyJJV0MkVY/uvdGi3LtrA7xwWU21uPHQZbBZqiqsZlMfM0scdTR1kD01VCZaeSGoh8sErIWjdHs3B9+690jKbZW7cBPJ/dffdZJjJq77n+C7vx8W4qWhgq9wUNfkKXE11NPicnRUlFt6GXHYymMkkFL5BLIJyuk+691Lp6ntlIqZKzDbAqZxLjxWTUm4dw0kDQOmYbKPS00+36uRZaWSOgSAPIRMJZ3Yx+ONJPde6SG8c9lKbCoO0OuIqva7xU7ZvL7Tyc27P7uuMBTVNVW/whcRjd0VYbdU7Y6iONpaqoaILVTpTx61T3XuglzOHz3TmWpt1bOyYyu08pOiNL5FqaOeNXsuNy8kGqKRm9S09UhD6ha2rhvde6FTee/aLc3Sm6dxYzIvgTNjhi66eXLU2GqcFUVtdR4yrtlKjb266SCeKKrLwNJjqqKfUqlAGJX3Xuhyo6aGipqaipY0hpqSCGlp4Yo4oYoYKeNYooo4oFjgiRI0AVUVVUcAAce/de6ID/L5pqvc+G+TPfmVjRch398se5M1jbTYqplg2N1VloOh9iUE1Ri6eIM0OH6yM5SV5pI3qWDNcke/de6OTujs3ZmzslDis7mFhrTj63M10FJT1GRfBYLH43KZSoz+448fFUS4LBSQYaojhq6lY4J6lPDGzSkJ7917pW4rMYvOUMGSxFfTZGhqUjeKopZVlQiWKOdFcD1RS+KVWKOFddQuB7917qLuXbeA3ht7N7U3RhsbuLbW5cTX4LcGCy9JDXYvM4bKUstHkcZkaSoSSGqo62lmeORGBDKxv7917ol/x6z9Z0D2Efhbv/cOYy1NTYbI7w+K+89yzS1NZvfpzFSxRZTqyszdS80mc7B6HM0NLO0khq8htqpx9awaRK10917o9YIP0Pv3Xuu/fuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691//9Hf3JI+n9f8T/vXv3XuiJ7+7J3/APIjszP9AfHfd9XsbZvXuQXGfJP5DYSigq8lt7JywJOvR3SdbkaarwdT23V0colz2YaOqg2dRyRoscmUqIkpPdb4ZPRmenOl+tOhdi0HXHVO16bau1cfV5DJyQR1Nbkspms7map6/O7n3Rn8tU12d3TurcGQkaorsnkKiprauZi0krH37rXQqe/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3XukD2Z1d153Hs7LbA7R2hhN8bOzaItfgc9RrV0zTQkvS11HKClXi8tQTWlpaylkhq6SZVkhkR1DD3Xuiay7s7N+Feax9J2juvLdq/EDKVdDh8d3DuyrFb2f8cMjWzR0GIxfcWaKRv2F1BX1ksNNT7snvmcLPKq5l6qmY5GH3XurAqeeKqhjqKeWKop5445qeogkWWGeCVBJFNDIjMkkUkbAqwJDA3HHv3Xuiq/LTs3ee1Np7S6y6jrIKHvH5Abug6v6zyM0ArE2dBPRVOW3/2rV0JV1qaHq/YtFWZNEcGKbIrSU7giex91sCteht6s6w2f0113tTrLYONXEbV2fio8ZjackPUVMhklqcjmMrVaVkyGdzuTqJq2vq5Ly1dZUSzSEu7E+qBk8OtdUx/LD5C5nuHeOR2zjqt6frbaeZmp8LjI4p6V8xlcfqpKvOZlJljmmlhrFljpYmHiiRfIoLPq9i3a7SOGCO4qrSSKCCCCNJ4Up69c4ver3Q3DnrfrvZrW4K8pWM5WKLSV8WSOqNLMrAMSrhhGjDSoGqhLV6KWBY3/AN9z/vPsz6hTrv37r3SA7C7W6x6kotvZLtLf+0+vMduzdGN2Rtiv3flosPRZzeGYWZ8VtyhqpwYmydclO5jViq2UksB7o8kcYBkkCgmgr5n0Hz6MNt2fd95kuYto22a5khhaWQRjUUjXDSMP4VqKn8unnJ702lht37a6+yu5MTjt87zodxZPaW0qqp0ZzcWN2hDTVG6q/E0mkipptvwVkT1Lal0LIp597Z0UqpbvYdo8+k8VlfTWV1ukFq77bA6LJIB2oZcRAn1kIovSmsbXP0va/wCL/wCv+fd6ZoBnpLXtBPGnXufooZj9TpUtZR9SQoPv0g8FQ0xCKSBViEFTwFWIGfSteqo5lbTEpZvRQzH9ig9OVLhcxWaftcXXy3Krr+2kRAXGpSXkVFVCpBJPA/r7j3mD3a9rOVS68w+4uzWsqo7aPqY5JOw6WASIuxcN26KaieA6GW0e33Pm/FTtHJ25TRlkAbwHRe8VFWk0KoK51ZUA5PT5T7F3HNpL09NTBlLHz1SFo7f2ZI4fKwJ/w9wxvX3zfYbavqhZb1uG5SxsAFtrOQCQHi0ck5hQqv4qlW4hVPUj7b9273X3DwTPt1lZRsCSZ7larT8LpGHZWb8NK+ppXp8h65nN/uMvAg0rp+3pZZSGNrhvI6cf63uIt1+/3syeGNi9r7yTuOo3F5ElU8mURxvQk+RJHEA1HUiWP3TNyOv96c926jSNPg2zt3eYbxJE7QPxDPyp1Mh2Xtda58TNmDUZdaCPJNiBkcfDlosc87Uq5VsXHqr/AOFzVaNElQ0fhMqlA5YEe4u3T783u5dCK52rlbZbK2Ergs0VxcK1aERl3lVUdVoWCZYNWgHEeWX3VuQLcP8AX73uty2kBaNFFpNDq+BG1V8tROOnyPZW24rP/D3lsukmepqZFJtyWAkVdZPPFvcc3/3u/f8Av/EjXnaK2RpNQ8CztUKg/gDtE7aAMUYkkcST0L7L7vPtFZsrf1YaYhAv6txO4OKFiodRqPGoAp5ADpxhwGFp/GIsRQgxkGNmpxI4PNiXk1s55P1v7Au7+9/vHvZvBufuhvTR3Ao6JcNEhGKgJEEVAaCoQLnPQusPa3222v6Q2XI22K8BrGzQrIyn11SaiT/piep6U1JEweOjo43X9LpSwI63/IZYwRcewNdcx8z31vJZ33Ne6z2jgBo5Ly5dGANaMrSlSKiuRx6FUGx7FazLcWuxWMVwK0dLeFGFcGjKgI/I9ZwbG4VAebWRQRf+hAB9krKXVleWRlPEF2IP2gkgj5HoxVVQgrGgI4UVRSnpQAjrl5X/ANUf9u3/ABX2z9Jbf74T/eR/m6e8WX/frftP+fr2tiSb/X/X/pa3+t7uIIRQCMAelBSn2dUqa11NX7TX9vHrrUfrYe7hNNArEDqpFTU9dX/wH+9e96BUmp/b1vr1/ftIODw6rpHHz66v71oUZp1vT8z173frfXvfuvde96691737r3Xvfuvde97691736h9Ovde9669173vr3Xveuvde97691737rxNAT142H5/r71Xj6dVUk1r10Hi/1a/8loP97PtP46g5dQP9Xz62DXrJGnnbTAsszWJ0QxvO4UG2orErMF1cXt7djLzErChkYCtEBY/mBwHz6qWIJHSgx+0N2ZVqZMbtXc1e1aVWjNLgsnJHVFibeGY0qwEG31LheDz7PrDlbmzdTaLtfK+5XHj4jKWsxD+fadIGft61rPp0tKDovuHJGZaXrncsZgEZk+/gp8apEmrT4WraiFZz6eQhOn8+xrYeyXvBuTSrbe3W5KUAJ8VUh48KeI66j6gVp58evB856UeO+Mfd2SMoGzVoPCEJOVzWLpBLrJFoCk9RrKW9X0t7Ee3/AHave3cTKF5OW3CAZnuIU1En8NGetPTrevpVY74fdxViSvVrtbEMjhUirM3LVNMpXUZEfH0NRGqqeCGKtf8AFufYo2/7o/vBeRyPdLtVmwagWW5Zyw9QYY3AH20Pp17X8ulbSfCXessVO9ZvbbNJM4U1NPDjsnWCC7AOkVSZaZZyqcglEBP4tz7Flr9y3naSG3ku+dNrilNNarFPJpzkK2pA2OB0ip69rHp0IGO+EeBi+5XLb9zVWHWEUhx+KoaAwSLJqnabzy16VCzRWULZdB5ufY82/wC5TsaNdfvfnu8lBC6PCgjiK5q1S5lDBhigoRxqeHVTk16ENfjX8fNhYqr3VuzFUNbi9rUVdn8nnt/5YVOEwtBjaaSsr8vkIquSmwVLTY6jgeRp5YrRRqSW4v7yI9r/AGW5K9prm+3HlWK6/e11b+BJNNKZHMWsPoUUCRrqUfAoLDDFh0Q77y1sXMqWUW/7XFdwW8vixpINSCTSV1lD2sQCQNQNK1ABPQB9c7Bl+adbiO2uz8LPhfifhMtjs38b/j3V4mo27Rdix4KqFThe+e7tvyRUUuWoMvXRLWbS2pWQjG0VAtPkq6nmr5oUoJdJZjVmJb16Nre2t7SFLe0t0it1FFVFCqo9AooAPsHVkXvXT3Xvfuvde9+691737r3Xvfuvde9+691737r3XvfuvdAz8huoafv3pLsrpuq3JltpU/Ye2Kzbsu4MKzfd0K1DRS6KimWam/ieGrjD9vkaFpI0r8fLNTsyrKSPde6JQvxzr9t7y2HiN97rxfYfdfdE2M2ru7PYDa8Oydidc/F/qOopd35Pp/qLY8dfmRsfYWWzkeKoazyVNXX5iet11NQyU9LHTe690bupfJVXyjxVMYqR8JhehM3WJIWkFZBmM/v/AAtKqqgXxClnx+Be5J1akFha/v3XujBD6D37r3RePk30vju5usMlR06vjuxdkvL2B0xvOinNFm9hdsbaoaqp2juLFZFLSw071v8AkmRpmvTZLF1FRSVCPBM6n3Xuib7U+ffa3d2J662z8VvjyO5Oysr1ltDeXcu9dzbzh67+OvRe59w4bGZCbr3ce/Uxe6Nx7q7F11jzjAYPFVtRQ0Jjlr5qQyxo/uvdKai7H/ms7Tp6vKbz+M3xD7Xp456SSHbnTnyE37svdL0TtIldFST9q9ZjbFfkKZSjx+asoIpQGUlCQffuvdCd0187+puy96UfT2/8Jvf42fISojJTozvzCx7P3Pm3jA803XW56aryfXvbWNBuVn23lsi2jmSOI3Ue690doi9uSLf09+690AOBwuJ2Tuqp6jkoKX/R7vHC12a2JhYqJVx+DmxLRQ7u2nj8dg9p0OIwOAo1qqTJUs9dkZq6srq+qWMBKdQvuvdArlKKr6w3bndqzVeQi2turG1GOlq6eurMbVHD5aCakp8pTZTHS0tfQZPA1MpvUU0kdQioWVgxUj3XujVdZ7in3RsvCZOuSSPKxUxxeajkxuVw2nMYp2x+Rnp8ZnJp8xTY6tnp/uKNqp2mno5YpSSHBPuvdEO/l6V8fSeP378EN64+t2/2P0Fu3f26tnVeSQrQ9ydGdpdjbq33svtPaGTb9rNyUE+5JcLuOBCanF5qjInVUqaZ5fde6Opv/D5TE5fG9l7Wx0VdlMNRyY3d+JgpIWr9zbFSQ5GtpqNcfgchuDcG5sGYpWwmOFVTUclTWTCT1SKy+690w1GF2/8AwufsnrTO0ezaqolqsnloROkG0c1k6nMQV+46HeeFhNRi8dujL5alXH5PLfbyZigVZI1YFSh917pUbJ7Jiz9aNrbiohgN8UtA1XPjxIs2G3FS0NPg48tubY+RL+fL7OjzObSkgqKiKlqXkRtUC2ufde6Yu+uits997Ng29l8jltrbm23maLePWnY+2Jkpd4dYdhYZJxgt57YqpA0JqaP7h4aujnWSiydBNNR1cctPPIh917oN/j38gcvuXM5bofvCkodm/JzrzHpNuXCJGaHb3a+1opXo8b3d1BLM7Lm9i7p8Qero43krdtZFnx9cqskM1R7r3Rt/fuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691/9Lcp+U3aG6qrN7E+K3TmayWA7s78pMzVVO9cTTJVTdI9KbbmoKfsvuGoaYGnp85DHk4cLteOTio3JkqeQq0FLU6fdbHqeHRjuresNldNdf7U6x66w8WA2ds/FxYvD45JJKmdlVnnq8hkq+dpKvK5vL180tVXVk7yVFZVzSzSu8jsx91roQffuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691FrqGiydFWY3JUlLkMdkKWooa+grqeGroq6iq4np6qjrKWoSSCppamCRkkjdSjoxBBB9+690QTrJJPhr2thPj5lsjVN8a+4MtPT/F3IZD7mpXqXf3gyGYzvxryGYlaS21MvQ08uS2EallaCKCswwZlp8cj+695dKjblMN//AD77Jz9UDLRfHfoLY+wcBBLpeKk3T3huDK733jlqdSDoqpts7IwVLq/UsfkH0kPv3W64p0NvyL3+etult/7ngmEGShwk2LwrAkOc3mrYvHeEgqRJFNVCUH8aL82sVNnbpdXEcEiBoye4HIK+YI8weBHUfe6XMr8pcg8y71DJpvFtzHCakfrS/pxkEcCrMGr8utdwM73aR3lkNi8rsXeRyAWdmPLMx5J/P19jG3trazgitbSBYrWNQqKooqqoACqowqgDAHXLl5JZXaWeZpJmyzNlmOSWJ8yeJ+fXWof7b6k2A/2/tQAWIC5J6a1r5kD7epdNQ1tcdFFSVNUzAW8ELuvqOkFnA0KNRtcm3sM8yc5co8nWsl7zXzRt+3QICT9RPHG2FLUCE+IzFQSFVCTTAPR1svL2/wDMc8dvsOx3d5I3DwoncZIUVYDSASR3MwArkjj1X5/MP6CxHdGQ+DHUHZmHgqNj9r/Lup2Jn6WeWN6mOizXx77gQ5CnSPyPTZHFSItVSyAh46mFCLEe4O3X70Xs5PbcxPsu7XW7ptFn9ZOttbyKrRCRYKRSziJXfxJEOO3RVgxpTrJr2r9n/dTYH37dTZQbbuU9gqWxmmUlZRcQygTJCZGWN40dXBzmhFDXotvxzqt41vz9+Ffxk77zVTku/wD4o9c/NDqnf9dSLGK3sfryHYOwsz0Z3PipJjJ93R9q9f6EmexaLLUVTG9pBp9x7zX96PebT25uPcXlPle0k25Y7NYJbl5CGkuZJILuC4jjK+HPZtGCNDlZVdT8LdSy33fdgltecduk3q4g27ebu3uGjgSMfTtCWcxxM+rVE7s9NSgopoOA6UXQ2Q+aneW3epfk51ztX5e5/cW+PkB93uPbmb3Z8dYPhNTfHmj7fyWyN5dfUmyZ92w9l4nNbI2LjpahMzNAucmz9M/kh8bqhj7n/wC8RzNZ7tzVy3vPuRtdht67UVjFnFeQ7il61oJYpEdYGgMc8zaDHrKLAw7yw1AV7b7I+1W2WtrbJydbzvGf7SfVJI/oXbUoJ+xQKYAp0JuYz3e/eWw/5iHyg2z8r+1fjy3xE7Y792H0j1XsVNnYzqjbGJ+OeCpsvHk+/tv5fbtbX79rO4qhXeqFRV0sdPj6iIUqhgPcF3u4WsG8e0/KXNOwjmOTf7Cwlvbq+nu57mRr2QoEspDPpgFkMR6Yz3K2s0J6kbbtr2nZ4/B2jara0jqSBDDHHxNT8IqAfOh6EvZ28+6fnT21tTZ8ndndPxS29t34H/HD5JZDanQ1fgdt7x3b2r8ihmKiuz+Wy24sBuKqquuetVwi01PilRIJqmp/ypyLD2Eb3auWvbLYr7cI+WNs3u8m5ov9vEl4jSRR29joARRG8YW5udeppSWIRQUBNejB5ZJNOuQmg8+lKuP7n+SXy/7D+J9V8pu6erdmfGL44dD7prdz9GSbR2N2Z372V3A+apK/trdWVGCz1DTbI2quBRUxGNihx8uUqH8raNK+0wuOXuUfb/aeeV5I22+3Ded3vIxHeiWe3sbe0CMlrEmuNvGlL1MshMnhqCM9Up0WTZnePyo+SHVfwE2NT/I7cnVu8OyPlb8vvjh233n1zgtuxZXt7rz497W3xHSb6wOPrsfV7fxO7N1Uu2Uenr4IftqHLa6qONtIT2M7/lnknlHfPdLdJOUIb2wtNj2u/trOeRytrPfSQkwuwIZoo2kIZCavF2Egknr3+HobNkfGbcVJ/NLxtNL8rfldnhtX4Q9Xbor5cpvTalTPvLE4rvnN4CfY+9Pt9mR/f7FzMlL9/XQReCebJzySCZQ4QB3cucrV/ZN5RyRscYn5kuIxohlAiZ7KNxNDWYgTJURoSGUIFBWoJ68RToBdodmfPnv/AG/uH5MdJbd+YOS7Ah+S29cBsvYNDun47434O0nUnV/c9Z1zuTrDd2xtw7touy5ctUbLwVVWZHOVEUWWjzjq1OhpvHqEm4bL7X8p3VnydzHdbAm1nZ4XmnaPcG3hrme0E8dzHMkTWyqJnRI4BWJohRu6tPeR6X3b/fXd2zvnnlPgjhu9s3J118jO2Ood+4jvBa2Q75+JODzuM3RuvcPxPpsymAl2/Tbi72i6/SPZBrp1qqPG5OrGhn8R9lvL3LHLl/7Ywe5k/LEY3TarG6ga0p+hukiNHEm5aTJ4jJZGYm80DSzxx8ACOvdXaysrMxWMRAubRcnxj8JdjqYoOCTzcXP1946RoVoSwOOnusQ5IHtzrfXek8n8AE/n8f0/2/uniL+XXq9etza4/wBv+P68XsD+PfhID9tPn1UMDinXDXF/x1jN/wCjqT/iLAk3Hun1ERpRxxpx8xxHXi4HXeqP/joP9sf+Ke7+Kn8Q/b1rWPTqRTUtVWl1oqWrrWjAaRaOkqqpowxIUyLTwyFFYjgm1z7et4ri7LCztJpmX4hGjOVrwroDUB8iePW9YHEdKGg2PvXKQtUY3Z+6K+BZDC01LgMpLGJQFYxlhSj1hWBt/Q+xBYcm85bpA1zt3J+6T24YqWS1mYBgK6T2caEH59e1g9LHGdE9xZdaZ6Lr3PrHVi8MtdHS42O3q9Uxr6mCSmUlTbWq/j+vsY7Z7H+7+7Lbvae3u4COX4WkEcS0HmxkdSv+2Ar1vUPXpXUXxZ7vqxKz7Xx9AYtNhkNw4lDNqBa0Appqq5W1vUU5PsU2X3XPey9Ex/qxb26LwEt1DV8E9oRnpwp3U/Z02WUj06cF+JndElLR1KYrAeSqp/NNRTZ+nhqsdL5HX7SqPimp5ZwqhtUUjx2YC97+1qfdN96HtbO5Ta9vWSWPU0TXKB4mqR4bkAozedUZloaVrXrQ9ajp/h+GvasiQtPltm03kWJpkeuyUz0+sqZEJjoPHM0NzfSwDEcHm/s/g+5v7qSJCZ912aIvpqPEmYx1pq+GIByoqKA0PkadbJ1Ux0o3+E26lUsewttKo51NhMiq8f6omv8ASD7EEv3LeZoI5JpvcTa0hUEktbyhQPViZaAfb17VqooBLfZ04UXwxpT9z/E+16GO0oFJ/DsRStqg0DUaoVWTGmbyXsE9On/H3e0+6VsLeP8AvH3nsFAfs8JITVKDMmu5+PVU1FRTz6f+ju/K1k/3k/5uudR8PdqUhtU9yxwHjiXFYhDz9ODlQefaDcvu1+1+zzfT7r94bb7eeldMgs1ah86G6B6UQ7Vu1x/YbdcOPlGx/wAnUui+KHV0UL/xTt2pqptZKyUL7foYlhCiyNDKa93kDXOrUODa3592s/Y37vdvbyvu33irKWRSWrFdbfEAtOGkvKxP2HPkvT45e340UbNdf84n/wA3TefjV0zDWTGbt/JmhMMYp6eOHF/dRz3IlkkrVpJIZonH6UESFT9WPsJNyX90+O9nE33jwbLSAqDw9YfzLSiEq6kcFCLTjqPS1OTuaJF1Lsk9PmtP5HPUiD4+/H+injlrux9z5Wm0uGpadoI9TEAIzT0GIM0fjPNrjV+ePdJNp+5ls08Mu4e9t9fWxB7IFdqHhUvBaFlI/hqKjy6Uw8j81yVC7Q60/iKL/wAeYV6douoPjDQzx1D1m7sskZbXQy1uVaCYMjL+6IqOil0qWDDTIvIH1HHu7b99xraZra6XmLftyXUawhL1lOOLgwwmgrUUetQOlUft7zVKaGyRR6tJH/kY9OUmyfjBSmkaDZOWrTTSF9BqMuqSkyiW1YlZlIlqY7iwU3Gjj6e2r33W+5Nt7WLWPt/vd6EJY0jnVeNaSie6j8QH+GjCmKdK4vbPmNyfFa3jHqXr/wAdU/z6lzUvx6geOSi6doKiRZBJ+9BTUwV0YOjWarnEg1DkEW9otx+8Z91q1eCTZfu9XF1IrAnxBBDTSag5llDCvEH9h6Xw+1u6sSJ90t0H9HW3+Renaq3f12DGaHp/afBLSGsxmGSzAgp4xBjnuwbk3/2Htndfvle3iG3fYPu37WKGrm4a1BBFNBj8G2c8eJNOjCD2pc6vqt8FfLRGT+2rDp7j7lNKTJQ7LwlHMwCeaKVY2MWpWkjJho4mIYLx6rA2Nj7Uf8H7u9upfavZ7abe47Rq+oY9uoFlokCN3KCFzRWoSCBQqB7U2le/e5CtM/pj/oL/ACddJ3nuV5Y448LiZNcihKeJq16iVC/+bhCFmMpXgELbV+Le0cX94J7pzXqQW3IWyy6pBohQ3bTOrMAqLRiS7CoUrG1SKhadPH2u2VIWd90uFABqxCADGCa0AFfL06M7RVD1NLSzyQSU0k9NDNJTylTJA8kau0MhUlS8Zaxt+ffV3ZL+63TZtp3K926SzvLi2jkkgcgvC7orNE5GC0ZJUkYqD1Ck8SwzzQpKJERyoYcGoaVHyPHqX7NOmuve/de697917r3v3XuiLfI/FT99d2dP/F2WKWfrKio5PkD8h6Wy/Ybj2htHNQY3qnq/MB1eKqw+/wDsiJ8hkKVhpqsdtqenkvFO6t7r3R6FAVQoAAAAAUWAAFgAPwB7917rv37r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+690Wza/2e4vlF2xmfNFPU9c9a9d9eU0DUk0U+Pn3fW5vfudKVhIiqqfKUkWHJUAmN6X6+oge6905YGatrPk32RqpH/huF6c6uoYMgZleM5LK7p7Ir6+gjpx64HhpKWklcm4kEq2/Sb+690YH37r3QT98bzk676T7d31Ty+Ks2j1tvXcGOb0amyuN27karFQRB/S89TkY4o41PDOwH59+62OI6DL4T/H3A/Fr4q9H9G4KkFM2y9h4f+8k7rGKzM77zkR3Dv7cWTljjj+4ye4N45Stq5nIuXlsLAAD3XjxPRojIo+v5OkE2ALE2Cgn8n8D8+/da6Cjuro3qj5FdfZjq/ufZGF35sjOrE1Vh8zDIJqOtpnWfHZrBZWkkpsvtvceIq0WeiyNBPT11HOiyQyo6g+/de6JX0h2J3B8Xe2NsfEf5N7pyfaGx9/1WSx3xG+Vefmo/wC8e/GxOMrM3P0B3+adKWCLvHbeAoJ58Rn4oo6PeuLpHdlhy0FRHUe690e7sbZx3lt2OnpJqeg3DhMnRbl2jmJsbBmDg9zYvyCgydPja2po6KpqFgqJoUE7+JfNqYen37r3QZbmWm7i6vTcNHjloNy4GbIrXYX76iytThM/haiXH7t2jVZLGSTY2sqcfV00kEjU7yQtNENDEWPv3Xugs6U3Y229ytFKJGwe9q7F4rKpT0ePiixm8/G9NgdwS02I29U7j3Dkt9UBFFkK/IVoosZDhKRboJufde6Er5NfGPaHyW2jisfks5uHrzsfYeWO7emO69i1MeP7D6f33BCI6Tce26qVXo8ljqtEFPlsNXpPis3QF6arhdGVk917oN/j78hd/Lv6s+L3ymx+A218jsDhn3BtHde2Yamg6y+S3X1CIYKzsbrCKvnnqcNuTC1DBNz7UmllrMJM6TwyVWPmhqffuvdDnunZmWwWTqd4bEpFyP8AEI46beXXdRUGLDbyxujJQxfwlq+uGC2flqOszcuRrKuGjlmyywCnm9TJJH7r3RdtwYajTCUe7tkvUZrrmtrqTN0uKzGMyFBlNl5+ENJishJhM5S0+X23ncTJMJ6Cpnpo56WQoZEZdJb3XuhL617kkoKalxG8qmsyOFSpxeFwu85Y5a/L081UKLGYnD9gQ0ImqZ83WNSVVfWbijo6Hb6xOqSNBKrK3uvdLHvLorZPyG2liqLK1ldgtzbar4d3dUdrbSngpt69Y7zigJxG9NlZnTNCWCsFqqOYTY3L0TvS1kM9NK8Z917oHOuPkNvjrzc+3ekvl/Q4Pae/8/k5Nt9Zd1bejeh6X+QFZBA09FTY6WpnqG6w7YyNHE7zbTyk3+VzRytiKmvhVlh917o7YN/fuvde9+691737r3Xvfuvde9+691737r3Xvfuvdf/T3B/jhjn338pvmf3pkFkkXEbr2T8YNj+VldcftfqLbMG6t3GjKSuI0zPY/Ytd5lKo16GO97C3urHAXo+P04Hv3Veve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xugb796YwXf3VG6+r87WVWHObpqWs27urGqpzext7YGtp85snfm35C8bQ53Z+58fS19MQ6a3h0MdDsD7rdadEs/l+dk7m7N7D+W+T7CjoKLtXbm8OpthdqYXHwNS0+I33sPrWn2luB6Onf1/wXc2Qw8mbxrn/l3ZSBLt49Te610OPzst/su24/+1/tK/8A5/KW3s02av1q/wClb/B1BP3jz/zC3c/+em2/6vL1SDicdLl8hTY+mOlp3PklILLBEt2lmdeGbQv055NvaD3L9wNp9r+SN/533qMy21nGvhwqwR7iZzpigVjXSZHoC1DoQM1DTrBbknlLcOeuaNq5W21tE9y51SFSyxRKNUkpApXQowKjUSAOhlx+z8Bjgp+2+/mBJM9dpl+tiNMAAgXTbj0n/X98n+e/vXe9PPTzQxcwDZdoZQv023hocZrruSTcSFq5OtRUAhVp1n5yp7A+23K6xyybP+89xB/trzTJ/vMIAiUenaaDpSqFjUJEqxRqLLHGAiKLAWCqAALD3jvczXF/dSX+43D3F+7FmlkZpJGYkksXcli1STWtcnqZreGK0gjtrWFIbZRQIihFAHkFUAU+VOkFvTq/YvYmY6yz+8sKcvlunN/R9n9cVQr62j/u/viLb+Z2umZaKklijyQGCz9XD4KgPCTLr06lUhbY7xum1W282m3XIjt9xtfprgaQ2uEyJKV7uFWjU1GRTj1fgaj/AFYp/g6TNd8e+msj8gtr/Kir2TSnv/Zmwct1dt7sanyGSpK2m2LmpKmWrwdbjqepjxeWMDVkwpqioiknpUmdY2VWICyPmrmKHle95Kj3EnlW4uhcvblVIM600sGpqStFqoOk0BIqAem9LV6B+l+BPxix/ZE/aGH252Ht7J1fYH+let2TtvurtXb3TNd2W2Viz0+9qvpvE7npdiTZetzkK1lQn2gpampvJJExJufS+53Ok2zrs09zazRJa/TLNLZ2sl2LfSU8EXTo0wQJ2Du1AfCRjr2k1FRjrD2d/L8+JXcHYu5e0N+dcZmuz+/a/DZXtLA4Tsbf21Ote48pt3wHCZLuHrHbmex2zexK2hFNGC9fSuahY1WbyKLe7bN7pc97BtFnsu2btGttaqy27vbwy3FoslS62txIjSwAkn4CKE9tOtsvoOlV3n8Nfj18i9w7Y3b2TtndFHuzaO167YWK3P1j2PvnqDP1HXWTnWoyHWuayXXeZwU2b6/qZxr/AIXUFoIiW8ejU10fLPuBzZyja3ljs93C1hPMszR3FvDcp46ii3CidX8Oceci5NB1srjA6ZOy/gj8Xu1P7gz5nZG4to5XrPYcHVOz9x9Rdl9gdQ7ppeqKeOOKLrHL7j2LnsVltybGRI+KGvknCMzsrBnJ9vbP7l87bJ+9Fg3CG4jvbn6mVLq3guovqakm5SOWNljmNcOgHl6dU0t6dCrh/jt0ntyl6Gx+2evsTtnFfGKTMzdFYbb81bisRsKTcO16/ZubNPQQVAiy7ZLAZWoSVq3zu88rTkmY6/ZJcc2cyXL8zyXm6PNcb1p+tdwHaYJIJU7iBp0uq00gUAC/D1qhBz029g/GXp3sztrrfvTdGH3PT9p9U0X8H2tufae/t4bK+92+ubg3NFtXeuN2zlcfj99bTptxU6VseOyiVFMs4J0kFgX9s5y5g2fYt45Zs5oTsl8weaKWCKYLJoMZliaRHMEjR9mpKGnzp1ulfmekBVfy+/jrvPtHNdmY7ZfaEW4t3b3o9/7v2d132523tjqneXYaVVHVQ7r3V1Ns7cNHsnJ5zIV+Oilqy9OkVfMhadZCTcRbLz/7hXu2WvLu1RJfpDD9LE4sIbu6iiYFRAl0YpJohkqoLal/ARQAWGmmRnoytF/Ku2z2PtHtfauR6ZrKPB969t4vvfszPbo3nnsLvHPdubdyOLzG1t60W5/vn3lt+s2nVYOnhw8VGYIMbTK0EKJDI4aVuUva77xu/XfL+72O1fQrt+3PZWxuxFBElq6MskJtWVjIs2phI0kRaRqO+QD1VqVx0fjDfCveFeEn3JvPCYmSRY3mhx1FV5qoDtIROJKieXHU7ymMahIAdTn1D8+xdsX3L+brqKGTmLnCxsqhdUcMbzuvcarqJiTC00kAj8NKCvVtZ4Y6FCg+E+wooYRkd17urqhJIXqJYWxdBTzojMZoUp1o6h6dJ1IBPkZ103Vhf3Kdl9y7kWK2iXcebN2nuQyl2UwRKwByoTw3K1GCdZYeR69r48OltR/G3oHbfimyGKhrJKSV5jNuPcNTU6hJdUhqoGqoKaaCMN6FaM/439nTex33ZuSIzPzDJZL4Dks19uH8ZwsiNKikD8IZK/MjpVb2N/dMqW1pLIx4BUY/4B1xk2F8YcStTDJgNo1GurkyDRBKrKOk08ccZipWV5/FTaIhphRhGpJIUXPsFbrzB9xzlZL6G7vdhnljm8YpGJbttRCgCPw/EUoAo0xqxRfIVJqc23J/M9wEaLZ5grGlWAUf8aIPTmu6OkMRURVWL2dhzWQKxp6nHbQx1PNC2lo7LPLT07xsyMRcfg8+ym6+9d90/YbqO42PlJrm8SPUklttMcZBP4NcoiZSRSpIp51PR1B7bczTV8VIIs/ikBx69oPXGft/aqq322yTNIL6RNFiYIz/AEJZIpmH+tb2E7/7/Pt8kFdr9nruW5rwle0iUD11KspJHpo/Po2j9qtwJXxt2gVfOisaftp03J3Y9K8jY7Z+KoxIApIqvHIwF9PlNNQxq4UngX9g2T+8E3q3mujsntBtsET/AAlrt9VAO3xPDtkDUP4QR6Ajj0ZR+1NsAvjb3IW89MQA/m9f9XDqDL3hux0dY8fhYWYEK6x1UrRknggPOEYgf1HsJ3n3+/eO4tpYrXYNht52GJBHcOyfPQ82hvz/AGdGEfthsSspe7umAPCqiv7FqOmao7d31OoC11DTNcXkp8bAJD/sZXlA49gjcvvp/eF3CNEi5lsbShqWhsoQzYoAfE8WoHHAH2dGUXt3yvGSWtZXxwaQ0/kB0zz9i75qZBI+5K+IhbaaVaanj+v18aQhb/4+wRuP3m/vAbncm6l90tyiemnTCIYI8eYjSOlT60r0Yw8m8rwLoXZomHq2pj+0nptqN27qqm11G5M3IwFgRkJoR/yTAyJ+P6ewpuHvL7wbtP8AVbj7o7889AKi8ljFBwGmIov/ABnPnXpbFy5sEApFs1sB/pFP+GvTfPmMxVC1VlsnUgDSBPkKqWy3vpGqXhSfx7DW48485by0T7xzhu12yLRTNeXEmkeg1PjpbHtu2w18HboEr6RqP8nUQz1DAhqmpYEWKtUTMpH9CpkII9lL7huMqNHLud08ZFCrTSspHoQXII+RBHSnwYRSkKA/6UD/ACdI7fW89pdabJ3f2NvvL023dk7C21mt47w3BVpPJS4PbW3qCbJ5rLVMVLFPUyQUFDTvIyxo7sFsqk8e6bXs1xvW5bfs+12Im3O6mSKKMAVeSRgiIK0FWYgCpAzkjqlzcw2kElxPIFiQEknyoK+h8h0He7PkV0dsbqbaXeu7ewMPhepd9z9bUu0d71NLkpcbnJ+36/E4zraOmgp6GbII26q/N0scHkhQRmYGUoASDXbuSOYNz37cOV9v2Npd9tEuDLCAupBaBmuK1bTSJUYtRjWh06sVTS7pZQ20F1JcKsEmnSfI6qAUx56h+3p/2l3B15vfsft7qbbWUrKzffRFftPF9n4qp29nMbSYSt3vg33HtyLH5vI46mwu5Vq8QhklfHTVKUr2jlKObe2Nw5W3PZ9o5e5hv7OJNq3RJXtnDRsWWF/ClLIpLxEPw8QKSOFeq2e62t9NPDbyEmOlcNTIqOIHl8+hOBQ6dNjrsECC+q5sAgUHVc/S319kiSROypGQ7EhQF7iSeCilak8AOPkB0YmgqzGlPXH+Hp6pdvZ+tF6TB5eoXyiEtFj6rQJGtZGdkVQTf88AexttHtr7j7+C+xe3+9XKCURlo7KfSrngrMYwFPzJAAySB0W3G87Ramlxuluh01zItaevE9KWj6u31WG64JqYeRYy1bVUtOFBA/cCiR2eNfzYXP8AT3LmzfdH+8LvTVXkBrNNYQm6uIIqf09IdmZfWgrwweiKfnvli3qP3prNK9iO1flU0Fenr/RDkaOPzZ7cm3MJEmp5vNUtLJHTKdPnGv7ZGDNxbULH8+5h2L+7990L4Qyb/wA2bPYL4pDBBNcuEHBlosSMT/CWWnmeiG790NmjMn0tjcSmgpXSgJ86mrGn5HpoqcF1xizpr+yGrpY5XWohweHasAVFD6fNE1QseteA5LLc+5L2z+7qsAIX3r3WuWcSEusFlGisnkFaSZyr04sQwrwUdFE3utMdQttkQCmC0jEg/MBQCP2dNxy/UMEJKQb8y8zSDQh+0oCiabmzoyI9rX/rbn6e5H237gHszaxzruW879dsxFGN0kJQUyAIYk1AnJrw4DHRRL7n8xOymGC2jFMgIW/48x/zdOFJvnqWmPk/0fZicFRETVVlNUx34OopJXFBMbfWwP19yhtX3Qfu87VKs6+3sNw3hFCLiWecZ4nRJIyaj5sAMk9E9xz3zVcLpO6sua9qqtPlUCvS/wAH2v1Bi5YjQYCsxE0hDSVC4JHalLqA5NRHJNOIvTY6OD/T3J/KXtF7YciTm65R5D2ywu9RYSRQIJQWUKdMh1OoKqAQrAY4dE19vm8bkNN/uU0qejMSPzHA/s6Gfbm9tq7qUnBZmkrZVXU9IC0NbELAnyUk4ScaRa5C2HuRuirpV3H++/4p7917ru9/fuvde9+691737r3RI/iEKrf25vkn8jMjNJNH2x3DldkbHhLB4KHqvoKev6y239owdwqZ/dNFncw4U6dWRH5B9+690dz37r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+6910fp/sR/vfv3Xui8dM1UmS3f8ityVL0QpJO3G29RVlPVLMr0OzNj7UxdSaxrKtLNQ5Y1cTqSQvj5IuQPde6ifHeb++D9n90BCuP7Y3s77Ra86rV9e7FoIdmbUy3imldEG4Xx1ZkomRUD01ZESCbsfde6MmSB9f8Aff8AG/fuvdEx+cNVLlOsthdVUkpiq+9e+ul+ry6r5HTAyb0oN6b2bw/qkhOyNm5GOQ/RVkJP09+62KVz0cmWSOCOSaaRIookaSWWV1SKOOMF3lkdyFREQEkkgAD+nv3WugG2rs/DdjZbNdnbswdfJJl6jD4ramKzEu48f/DNubB3HVZbb2UqNs1WbrcBTZut3Q0+QiydDFEa/HGi13VAvv3XuuOQwW5OnaKq3FtPKZ/duy6BKFsxsLcWVpclW4jCUFEaGat2lurcWVxstCKepqpMvl5MxV5F6iGnaKn8J0Kfde6gfJLo3bfys6F3L1sdw1WBm3HRYbdfWvZe2pk/jOwOwdu1lFuzrDs/a1dE4b7/AGvubH0ddHodVqYFeByYpnB917pN/DDv3O/ITpGizm/cTS7b7m693RufprvvadHIJKTbnc3WWRbb+8osedEbfwHPyRw5jFm3qxWSp2ub3PuvdCsGm2p2lLTyLlarBdnUTVazS/xWuxuE3VtujpKJ6E1tdmRhMBR7gwxjalx9HReasrIKuokYnV7917ot9ftnD4DsnLbH3HQpXbK3X5cDk8dPLUU8FdtrcZSWmhaammgniFJXpGNUbIw8dgR7917oxWy9wbn2/lz172LU/wASyKRSVG1d9imWkpN646P+H+ZcnTUGIoNv7W3JR1+Salp8ZHU1M9bSUjVan/OrH7r3SU+TXxt2x8mNhUu2slndwbB3vtLOUW+OoO4dlTQ0u/eoeysOsv8ABN57XqKhJKWqRBK9NksZVLJj8xjZpqOqjeKU2917pHfEn5A7k7axO++tO4MNS7T+Sfx83DSbC7t25RxiHC5qorKAZHZvb2xUM08j9bdwbeUZTGK7NLQz/dY+YtNRSO3uvdCJvjAQbKy9Zv7GYqWs25uWWLE9p7Tw2HkrXzq5f7PEU2+osXgcDVZjO7uxppKCimnqaqKlpcGs8rG8Ce/de6LfurC1XXG7a2gpXhyFBNSv9sldFHV4/O7ay0bxT4rL0rhqXIY+qgL086MCjhQfqRb3XuhswOTzGxdrUu9dnit3P1lJFUZHPbGk++rs/sCjikr6/PVOyXoo8pU5bBbXxtFDj8TtGhx0b6bGnn/TCfde6FfeOzOs+8+vsvs3e+29vdhde7woZKHNbfztFDkcZXwhwWjqKeYeWjyeNrIwyOpiq6KqiDI0c0YK+690U3ZuQ3/8SOx+tult6bu3B2r8fO3dwZHZnTnYe9MhLleyuo99xYisz+3+mt+52SJ6rsLZ25MNha8be3FWSfxilqqZcfkGq/LT1Y917o/Xv3Xuve/de697917r3v3Xuve/de697917r//U3Ifgb/Dq7qTsDddAkiyb7+TPyd3XkZJJDI1RXy91bwwzTaiANApcNCigcKqAfj37qzcadHX9+6r1737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3VavVuFouq/5ovyhwdJaCl+Unxw6S78NOlNOsEu7+m81n+j921QqXP2wqqjbWQ20Xhi9R0eR7lrn3Vvwj7ejhfIHrVO2OpN5bOQzjIVWObI4TwOFY57Dn+IYiMgsiPFPWwLG6sdJVz/AIe1VncG3uIpcUBofsOD0AfczlQc6ckb7sKlhdPEZIqHjNF3xg5yGdQpBxQ9UO9e0j09fmkrYJqXKUUa0NTRVMTQ1NE6TyJWwzxSKksM8NRF43VgGBFj7w5+/wAbjuict+2u1wRk7DPf3MrvXBniiVY0I89Mckj+mT8usaPun2FuN754ubuMrvsFtFB4ZqGRXkJlOcqRIiRtWhpjHQoe+alKY6zbPE149e97611737r3Xvfuvde9+69173rr3Xv99/vv9v735V8uvdTMfj63K1UVFjKOryVZM4jjpMfTy1dQ7kkaVigSRr8fm3HPtXtm27pvd2ljse13F7eMR2W8bzPk0FVjVioJxqag6qWAp69Gm2P8Rt/bip0r9zVVJsyiez+GrQ1uUaIi+pqSJkjpyQRcPID9QbEe8sORfuf827zDFufPu9Jstiyg+DGFmuaGtfEJIhhoPVpCDxUdNktI1FX/AGfs6Mngegugdj0incTYzdGQADS1m5K9KoKyIY5PtsTSyinjiYtcqySMGAIIPuXpORPug+3Fujcy71tE9yiVLXl8LiVs6GbwI305JyFhop9KE9GlpsW9XtBa7XO/qQhA/aaAft6EWLsrrbatM9BtvHKsEYYpTYHEQ4+jLi1hrKUsdnJ/VY/4+yrc/vn/AHeuR7efbuTNvub1YlOlLC0WCB2U0A8STwV7qk66MMVJ6FVp7ccyXJRp44oIyfxuKj/arqP5dJbI9718gdcTgKanvqCy5CreoYcehzBTrENS/kaz7gvmf+8L5luFki5P9uLO1BLhZby4adhw8NjFCsa1GdSeIwyKPSvQptPau2Ug3+7O39GNAv8Axpif8HSJre1N9Vr3GYShQX/aoKOmhClgBbXKk8hsPpzx7gLfPvf/AHhN+laQc9Cwiq1EtLWGIaWAGnVIkrnT+E6tQJOT0KbXkHla2UBtvMresjMf5Agfy6StbuDP5LV9/nMtVB7allrqjQ2m+m8aOkfAP9PcQb97ke4vNCyJzFz7vF6j6dSy3cxU6fhJQOqVHkQoPz6PrbZNns6fS7XbxkeYjWv7SCemVlDNqYlmP1ZyXY/0uzEtx7BMiCaRpZiZJTxZjqY04ElqliPKtejICi6VoF9ABT9lKddhQPp/r/QDn+vA9+0gBQPL/V/qHDrdOB8+u/r9ebG/vYAHAde6797691737r3Xvfuvde9+691737r3XRIAuTYD6n8f7f37yJ8v5ft69xNBx6zU9NV1jrHRUtTWO5AVKWmnqWJLBQLQo45Jt9fr7XbVtO8b5Klvsez3d9OxoBbwyykmtKUjRzxIHzOKjpie6trVS91cRxqPNmVf8J6XON6w3vk1jdcOaKOS37mTnioyAULBjCDJUW4sRpuD+PeQXK33Sff3mtIZ4+Sf3fauAdd9KluQCpYExd0wrwIKBlYgEUz0FbznzlmyJRr/AMWQeUSlwf8AbYH86dFn/mJ9N1mM/l6fOTJZXNwFaX4m9+TyUmPpXZpI4estwzyRiepkRUkPjIDaGHPvKn2z+4fzFy3zTyzzVzL7h2nibfuFvc/T21s7hxC6SFDLI6FWLDTqCMKZp5dAje/cm2v7O4s7PapAJFYanYClQVrpUGuDWhI616PkT0z/ADDMJ/K6+Em5e6e2fh/uf425HfP8r9qXaXXXUvau2+40weT7S6XTZ9N/e/Nb8ye1/wCLUKVtOMlKccI6kJL4hEWT3k1y591T2q5b5y3znq3k3a43rcEvVkWW4AiUX/iLOY0jjRgfDkaNNTsF+KmoAgGXHNu73NrbWbFBDFpoKH8BUioJI4qDgD+fRnPnV8iu6+t3/nhpsrvTcPV8/Q3dP8rPbvRe5RV4uppekMX3LktgxdoV+26DOxSYoYzPx5OrqsrT1Wumq4xIs1oi3s/237snsBtX030ntRtJMQ7TIkkxrXVUtNI5J1ZyaHgRTHSWTmfmCQSBt2mo1K0NK0FBWgHAcPPozua3zvr4efM2PpTr75O9/wDyM2P3l/L1+UfdW8E7i3jit51uxe2Oj323JsnuvrjJ0uExUex8F2Am5q6hagxsK4CSSmpZKGBfCwMp7dyjyns4kXaOVtttA7Bm8G1giqy8GOiNakeRNaeXRS1zcPh7iRh82Y49OPVf3SnyF+WWw/i7/J3+Z3+zu/IPuft35Ud/fGvp7ubZO9t77e3F1J2lsTu5M1t7du0MB1ZjcPHh8FvXqCjxKVzZ2kk/jL1sFTNk3Jl8aiGpyK46Yx1m3D3z8ou3PhF/MA/mKZX5rfIjrzuf4+9x/KSn6k682j2DjtrdC9QYj4zdg1W0di9S7/6fpsE2I35XdpYzEr/GJc3I+QrpMvEaMx6Y/fvy69U9D3tbcPa3zW+Wnf3XW+e7/kJ8aNq9B/Ez4e9ibQ2N0X2KvVuZp+wPkbsfMb93/wBw72yFdiMrUbwwuy8pRR4XH43JB8FSGGYVEbyOxHuvdBd8++1t246j+UFL0XvL+YZ2N2v8Wvi7tjee7t9dDd0dX9OfG3oLc0nXGZ3nsnsDflDmpMJiO2d5dmUeJGbz+KWPJ0Zx7LDRxweRIj7r3Tngsn3l8sPmV171fm/lR8gOnusMx/KI+Ofyc3dtPoHc2M68m3P3Z2PVvS1W7YdwNhclmNqwUmSr/vJ6XH+AZJYIaWob7ZWjb3Xui99EdtfKOj+M38sL5lb5+XPe/Z/ZnyD+ZnW3x97W2Pn8ntrE9G7h6b3Tufs/rGowB6rwWAo8Yu81bYtJm5t0tMcvU5WSUlkhKxj3Xuh63hQdtdqZ7+cdvKt+dHyO6kg+HvZO44Ohto9ddk7c2XsjptdvfG/ZHZ1Hu/e+KbD1eT3ttPP7mklo0xWYlGEVDV+BHqpnZfde6F34UfJ7uztj5l/F+XsDeGVwWH7L/k39I/KHcvX61KYPakHc+e7aw1PvfsOhw0ngjoq5dv1TrUuD46THShZAsa+/de6g9G9s/On5Qfy4fj73JtHeXyC+ReI2t8xvl2nyT2j0H3Xt7qz5Mdu9H7Y7J7W2110OoOyK2bE0+Xw/XNXLiaiTBUOTxk+coIYYIawBRHL7r3WwT8EO3esO9fiR0l2h032D2d2h15uLa08OE3l3W8sncFZPgc1lNvZvF9mvPQ46dt77Zz2KqcZkC8WtqikYl5SfK/uvdG49+690U35Cd/Zza2XxPRfRdLgd5/J/sOhln2ntvKyVUu2etdrh/t8r3R2/LjVeow2wNsar01KWhrdyZER46hu7zTU/uvdBl8HNtbq6Nqu5vilu/c+Y31/on3Nid/7D3xm1oaaq3J1/3ZFktzTQLi8bQUVDhU232Tj9xUcVHEZkp6IUyiRgffut+Q6P/wC/da697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3XugB7U7825sWCrwW2hT787NqKeqGC2PhKuOUUtVHC7plN/ZuMviuvdm48jy1uTyktPEkKMIRNOY4X917otmx8Durs/Y1P07sWsmj6znfOZLvTvdIJ8bB3Ju/cdfV5bfO0upUqYlyFXtncuZr6iCu3MB9vT4sfa45552NRS+690PHw+bDUfx2622jiMbBgH6zw56rzO1YHmb+6Oe68nl2zkdtlKljUxQ49qFTTeSxko3hkF0dSfde6CD+YF8pa744dTwUmza6mpu0+waqbC7PleOnqnwVFTxrJnN1vRTh45f4bBIsUAkUxmqmS9wpB917ogfxE+TfYXye+QPxc2J2ia3cOY6cXvnsWr3bBjIIMdnJv7kYTZOzKjPigjhpaXceMpd65hFcRxw1KsrqPIre/de6ui7e81btWLa9LUtSVW+s1itmiohrsFRVkVBlpzJnZaKPcmIzeHys0O3qSrd6GWnY1cKvGjI5Vh7r3Sjydfhevdt0LQYxaHbeFXF4iOkxVPDBQ4HDoIsfSvDRReOOHGY2MRpojH7cQ4Fl9+690qVeCqp1dWiqaaohDKy6JYJ4JUuCvLJJFIjf4gg+/de6DPqSapj25k8BV1FTVz7O3XuPaq1VXUbor6mehoMg9ViHqcvuvGYyszFX/B6+nE09MJqBZg0UE0iRXHuvdFC64w3+iD+Y333tqhtRbS+VHR2w+/KKgHnSkk7Y6izDdRdn5CkjFKKMV2Y2VmdmvV6ZvJIaVXZOS3v3Xujd9w4Z8ls2oy1FSxVOd2RW0m+duMZNvUU8WV26JKiWKmzO58VmcXt/8AimJkqaGev8Blp6SqlMbKxDD3Xugp76oIMthtp77xUqSJPFTwitpHM0MtFkYBk8TVRzxqA8ay38b8ahILfX37r3QqnDYXtvrzBtmVkWapo6bJY3M0BpVzm19yw0dTj49xbbrqiCtXEbjxUtRMaepVGeF2P4JB917qX17nc1UU2Q2ru56U7x2rUCiyU9FFWwUOYxdT/lGCzmL/AIpM2RyEM2MligraookBysNUkfpS3v3Xuia/Jvb9V1T8rfiJ8nNoUk8M+9N7xfETvOPH08sqbo627Qo81lur6/ORQnQ0vXfcWMpDRVTKWpqfOVseoRzN7917qwXKY2gzOMr8PlaSDIYvK0VVjMlQVcYmpq7H18ElLWUdRE3plgqaeVkdTwykj37r3RHtzYqePrzGUlWobIdX7wzfWUlSa3ak00+BpGSq2vLNjtpBMbgVbBy0jU+PeOOqp6ZozOutrn3XuhT+OGRnei3PiXLNT0tVQZGn9ZKxvWxSw1CqpUWDmkVjz/sB7917pWth16s3Rh59rUksWyd77gOHzO1MXjQ9DgNy5ierr6TcmBoqOXGY/A4/M5WprqncNRMtXPV1UsLoA2st7r3QNfOWpqdu9e9WdoSM52t0x8jemezOw4YYUmqW2JQ7gm21mK+DWbxx7bqdz0+WqGBH+SUEwuAT791sUPR00dXUMrKysAyspDKysLqysCQVZSCPfutdcvfuvde9+691737r3Xvfuvde9+691//V3IPgJQx4bonObcWeKebbff8A8msPWeF9Yhqou+uwKtqd2/46RxVi3H49+6s3xHo7Hv3Veve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de6qK+WW58jsT+Zz/Lz3rTzFdtJ1/8AILrPsNRM0cVHi+49wdQ7Z2HkquFGUPD/AKSsZj6RGk9EbVZP1Pv3VgKqerdLcW+n/I/fuq9V/wDyC+KOU3BvWt7R6zWgfKZykSHeG055FoRl6yBl8e4MTVOwpI8kYYws8Mir9yw1hw5IMK/eJ9u7r3V9tJNn25QeZNsn+rsxgeKQjRy2xY4XxkbtY9okRNdRwjC35Fl2L3Im542AR/SbpALfcLc9tSGDJdxHhrRlXxI6VdSxVgxNSI5PH12FrZ8bmaKrxGQpZpKeoosnTy0dTFNEdMkbRzKoYo3BKll/x98itzsr/ZLybbt52+e0v4mKvHMjRupXiCGABpitCR8+pULjj1EBVv0srW+oVgSP9cA8e0iyowqrA/YQfs4deDitOvHgFmIUD6kkAf7yeB794ij4sfb/AD6tXrh5IywRZEZyVVUVgzszGwVVW5ZifoByT7r9RDqVfFXUSAACCTXhQCpJ+QyfIHqocdLfA9cb+3PUQU2B2duKveoRJI5P4XUUtL4pL6J3rK1aemjhax9TMBx7H3L/ALZe4vNIjfYeSNynhcAh/BaOMg8G8SXQmn+kDT163qHr0LtH8YN9U0a1O9MvtjY1G6qyLXZA5jKT6iLR0+MxCyM8g5B/cNm/r7FG9+y27cmWK7l7o827NyzYsoKLcTfUXMpP4Y7a2DyMR56SQD59GW3bTf7vKItvtJJSD3FRgD5saKPzPQgYTqzqHbPlkyFNnuxq+klRKk5Soj25gKKQrdY6jHUM01fZvqFqXXUv4A9gce4fsJynT6LljeObdyIqHu2/ddj21FUt08S5lWtCPEK6gadpp0M7D24urohrzdIYUAqVQ+IxAya8FwPME06EWi31V4+jig2dj9sbWxbRL9t/djGY8q8JC+OSPJaag1IIHEisQwP19oty+9j7tRwXG18mwbVyrtDgAw7dZRpIAKEapplaQuKEaioqCag8ehztvt5yvbqkzJJckitXYhT/ALVdOPlXpmr85nMo7Pkczk60vqDLPWTmLS5uy+BHSHST+LWHuEOYufufObppZ+Z+dN0vnkrqEtzMU7jUgRq6xhSc6AgUeVOhba7RtdiqrabbBGBkUQVqPmQT/PpqCqv6VVb/AFsByfySbXN/YNjgjiNVRR9igf4OjIljxY9RcjkcfiMfX5bL19DisTiqOpyOUymTq6egxuMx1FC9RW5DI19XJDS0VDR08bSSzSuscaKWYgAn2rhhmuZore2haS4kYKqqCzMxNAqqKkknAAFScDpuSWOFHllYLGoqSeHr1kpaqlr6WlrqGpp62hrqanraGtpJoqmjraKrhSopKyjqoGkgqaWqp5FkikRmSSNgykgg+6OjxySRSIVkRipBBBBBoQQaEEHBBFQQQeHXo5ElQSRsCh4EGtes/NibGwIBNuATewJ+gJtx/X3XiaAf6v8AKPs6tUVAqK9e9+63169iPrz/AE/r/Qf1PvRIHmPzx/Pr3y8+uZRg0yWBkp1jeoiDoZadJReJqmIN5KdZP7JcKD+PexUhSAaNWnzp6Hgf29MLdW7voSZS/oD/AIPX8uuJBHBBB44IINiL3sfeqmo7T08DXhw69731vrr37r3XEuAdP9o8BRyzG9gqqLsxP9Bz7bMia1jBBkJoAMsceSjJPyFTkdaJpUngPPy/bwHStxWx925qzUGCrfEdP+UVarQU9msQ2urMTMtjf0qePczcnfd+96ee0Wbl727v/oyQPGuVFpFnzDXJjLAVBJRWwagEdEF/zXy9tpK3W6R+IPwp+o37Er/MjoRcP0ZmKgxtm8rSY+Igs9PRK1bUjlSAJpBFTre5uQG5HvJ/kn+7+513Jrefn7nCz222Iq0Nov1M44dplcLCDkjUA9CMAg9ArcfdGxiDptdjJK/kz9i/bQVJ+zH29CniepNmYuzPjjlJlZXEuUlaqsVH9mEBIACfxpPvL3kz7mvsRygY55+V23fcFIPi7hIZ6EfwxAJCATkqYz5VJoOgJuHP3M9+GQXogjPlEoXHzOWP7R0IdPS09IgipqeGnjFgI6eJIY1H5ssaqPxz7yY27att2i2S02rb7e1tVGEijWNB60VAAK9BKWaWdzJNKzufNiSf516YM7vfae2kLZrO0FG68fb+ZZ6snkWWkp/LUE3B/s249mHTXRbe4Oyeruz9gb06t3HsuTsDY/Ye187sveOBzbz4nDZ3bG5MdPh83iqpqeVMoaXI46rkicx+JwrHSymx9+690VDsbY/XnanUWz/j/vDr/bNX0h19N1rUbI62gTI0+J25N05W4fJ9YTUmRir485LJs3I7fopqbXUlXemXyBxcH3XuiQfOX4SYf5KdIfIzafW+B2Phu2/k9v344br7h3dvWbK1OK7IoeiexNoZaGl3fFIuYp5TF19hqzF00MNIkFQZwk4IYuPde6E7Yfwk+MXQ+L7yj+NvT22ertzd17F3PsrIZhMruvPVq4yuwO4cftLalDV7mze4a3bHX+3snm2mp8Livt8fTXJjhuqAe699g6B/4C/ykOpPjF118btz1/x62jXfKDqTqbAbOz/a+Al3dVbXj35/BVw29N99f7e3JkqTaeA3HuuliCV+cosRR5KsTU876mdmqzxxxyyyyKsKLVmJAVQOLMTQADzJNBTrxwY1OGf4R5sfQDicdDfvX+Wr8P8Adfcs/eu9Pjt1xlO1K3cuG3buDKruLMJtrde9NutTnAby391tg9zJ1zvjdODkgilhyGUxNXOJIkkYu6KwhLmD7ynsJyy00e6+6u1GeNirJA73LqwpUMLdJKcRkEg+tR0ILXlTmS8CtBs02k8CwCj/AI0R0oO//gV8XPlNuLA7v+QPUuxuyN07cxFRtuiz8+6d5bZyeR2lW1n8QqtlbrqdmZXbz702RNX/AL38Iyxq6FZGYrGutgQv/wAGJ923P/MTYv8Askvf+2fpd/UPm7y2Z/8Ae4/+guk32t/Lg+G3dO6Yt9dpdBdb7xz8GycJ1zVUsWd3fhdvbi2Jtahlxe0dqbs2NtvMYPZ+9cbs3G1Dw4dcpR1LY5NIhZQiBdj74f3bT/4M2L/skvv+2fr39RObBx2dv96T/oLoZtr/AB46v2Zu+DsranX+Bwe9sL05tT4+Ue5KeurDlYOk9mTpUbQ65pqesyU8VVhcFUIrRMInqjb9yVh7N9q+9T93fePqPpvdjbIvCAJ+oE1sCDwKGeJNfz01K+fHpNNydzPBp17NKa8NOlv+Osafn0mqP4g9IYHrvqvqig6TxFH1p0bvfDdqdR7Rpf43UYnYG/cLmM3uDCbsxM8eTqK2TI0Oc3FXVCJVTTxeSpfVGVChZk2TmflnmZI5OW+Zdu3BGFR9PcwzMfsRHLH9lR5jB6Ibi1u7TV9VayRkfxKw/mRT/J0SKm/ldfHfsLvT5Zdw/KXqHrztjLdz/I3Hdp9fTS5TeWNr4uuqHrDrnbUewu18ViMjt3Gbxw8G9doVOShw2UjyeMRpllUK8ki+zwggkHiOmMYIII+XRsu9PiB8XfkxVbAr+9Omtq79r+rEq6bYFe9TndtVu3sNkUpo8ttGOp2bmNvzZLYeago4oqzA1RnxVVHGEeAqSDrr3Rldv/yrfhPu349dcdMbt+P+J2/sXY+6t9dhbF2nsTdO/wDrR9j5PtCqkq9502Gyew91bdzeMxW7BIGr8WlT/DmGhBAqxoq+691YP1f1h130l13tHqjqjZ2A69612Bg6Tbuz9m7YoIcXgdvYSgQiCioaSKwRdTNJJI5aSaV2kkdnZmPuvdFY3p8md0drZjP9R/DTHUG+95Yyvbbu9++cvA8/QnR9U10yc8uZBji7f7HwkbXg2vgpJ446vQuVrMfEG1e690N3Qnx92X0Dt3K0WAqcxuneW8sou5u0u1d5VUeV7E7V3gYBTy7k3hmkigSTwQjwY/H0yQYzE0YWmo4IIFCe/de6Cv4+5+bs/vz5T9qUCyy7Hw2d2b8ftlZNnP2ucr+oabcFZ2Xk8apH7uPo9/bzqMQJlJSSfEzaSbe/dbrgDo5nv3Wuve/de697917r3v3Xuve/de66Jt/X8/T/AA9+690Em+O9Oruva4YXP7nhqd0yKGpNkbapK7du+a8uhaFKPZ+2qbKZ+Qz2sjGBUJt6vfuvdF/3/wDJfddDWxYagx2zepmrxC9BUdrZV90dk5OnnjDq22ehOt6jKbtr6xkJ0pkK7HmNhd4iPSfde6QM2y+4+2aWmFbT9s71pKmSGdq3tzcMfx362hML08sUq9X9Xwv2fnoXMGv7TL1ECSeRkkkVSyj3Xuhf2T8Rdn4+koY+w6jHbwo6KqXIUvX23sBTbE6Zoa5VjEVXN17i6mqG88lT6LCt3JWZioawYaGHv3XujbQwxU8EdPTxRQwQRJDBDEixQxRRqEiiijQBI4o0ACqoAAFgPfuvdFn3bhsr072Jl+49t46uy2wN8QUFP3XtXC0pq8hjMtiaeLH4XuLC0CHz101BhoUx+4KaBWqKjHwU1Uiu9G6S+691rk/NjvwfIjvzcu6sbVfcbL24DszYJDExSbexVRKZ8tGpLAfx/KPLVAj6wmMfge/de6sA/k6bBZ8h3P2lUweiKLb+wMTO6NySZc/mvA9wmkM1Ir8Xvbn8e/dbJwB1cfvVpoNwdZ1a/wARNKu8aqiqkoKjcixk5Ham46elbI0GDpKnHV2PjqdJZsq0NFTyaJQ/3Cwq3utdLTKY+DLYyvxdUoenyFHUUUwI1Ax1MLRMbfkjVf37r3QP9LZWqjxmZ2LlHJyeyMjLjRq1HyY15H+zcMb3VJFZVF+I9P49+69097QpI8b2J2vFHTwRfxaq2huJpIqGpgmqZKjb64aaSpyFTuHJLkZUGDVVEFDjoaeMKlpnLye/de6Jn8upv7mfMP8Alq9os+NgpKztnuz4/wCYnq8jJjqiopO5+ls1ncNRUyiVKXJmXdnVePK00quxk0tHZxz7r3VjxUMjIx4ZWUkEg2YEEg/UHn37r3RbocfW13SG6Ns18Vc2R2VWZzBpUZGTdNXLXQ7Zyhq8TkI83vKnjzG4hVYgxeWvXXBPUeUQuyKD7917p/8Aj5kWrNjyUjFiMXl6yCMu4b9mpSGtVUQAGKNHqGFubkE+/de6c8/NR4bujr+aN6SCt3ptzd2ArIhU7ZoKvIxbbio8/j5pUqcXJurcIxRqKhYoKarjpaNaqWWWNy6ke690B/zchqJ9sfHhKUMZV+aPxRmYI+g+Cn7bwc9WS2pPStLG5Iv6gLWN7e/de6Og3I4/r/tv8ffuvdFT7SljNL21Q1MlQPtdw9f5XGrLX5SeBqjI4WGkkipaOpwtDi8ehGPmd46SrrTI95ZmidhH7917rrpeuodp7H3pvTIxM1PHX01Kv21PW1dVOaZIoKak8VFTVc+ievyCL5FjYRhy72RSR7r3QlYPZW4dz5HEbv7Nni+6x1QuZ2/11BDj6zAbQr6qgwMsH8Wr2WsTdW69q5jH1Zx+ZphjxHDWyIITw/v3XuiVfzDvk71Jhdrbi+Fr752liu9vkf1lnsNh6TeOXods7N2F1/uiY7Oz/afYO8s7NQ7dwuKwcVbUPj8f9xJl83XwpBR00gMssPuvDiOrE9mPh22ltr+AZuk3LhYsHi6XGZ+grabJUeZo6OjipYcjT11FLUUtUlUsOvXHI6kngn37rxya9Kce/de697917r3v3Xuve/de697917r/1txX4eeLbe+fmh1fJCaWt2t8qt1b6ip2qUmeXA927W2p2VjMjHEoDU1LU5TK5GJFI/VTvYn37qx8j0eP37qvXvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XvfuvdVHfJ/YWR7t7T+Z1PhIpf70dNfE/psdfM0UTlewsdv7eXf+DqKKQ63jlq89sDCwSCwBES3uPe+rqQBn16tC6/3dR7/ANh7J33QBVod67S25uyjRW1qlLuLD0eXgQN/aCR1gF/euqdK1lVragDb6f4f63v3XumzI4LCZdSmWw+MyamKSAivoKWr/ZmAEsQM8UhCSAeofQ29lW4bFsm7ro3XZ7W5XSV/ViSTDcQNamgPn69e6D/I9H9R5UQpW9d7XZafX4RT46OhtrVVYH7H7cyCyiwa4H49x/f+yPtJuYiW79vNrKxkldEIipUUP9nprUevXuodJ0B01QVEVXTdc7cE8JJQy08tSgLKVOqGpllgk4J/Upt7TWvsL7O2M8d1b+3e2iZOGpC4FcHtdmU49R1ap9el7RbO2ljYaenoNsYCjhpLGmjp8RQRLAVfyKYtNOCjK/II5B59jux5P5T22G3g2/ljb4YYjVAlvEoU1rVaJg1zUZ6r0meyN5/3PxC/ZhGy+QZqfHROAUiVVHmrZEtZo6YMLKeGcgfS/uAvvR++/wDrJcl26bIY3543VmiskahWFVAMt06H4lhBUIpw0roDVQw6F/J3LR5i3ErOWG3Q5kI4mvwoD5FiMn0r59UN/wA27u/tXqr4g5zLdab0yWzOyO4u5OhPjpRdo0xSXN9dY3vjs7C7E3LvbBzTgw0Gcw23q6pGPmI0U1XLHIBqVffLP2etz7k+7bb77iTS71cW1hf7jKty7SG6eyt3uI4G9ITIBWNAF0BlCgHqXuZSu0bPBYbZGsEcjrH24C6nVainFu6tTU1A6iYL+Wx8Q/ivRbk7T6h2XvnE7x2N1F23iN312R7m7UzUXfmMr+vs3Q52Hvmhze6snQ75rss8RqlrhFTVlBVuZKaSIAIC+696PcHnqe02HmHc7R9sutwtTEFs7VPoWWdShsSsamBUB0aKsjqAJAST0x/Vzb7Hb/3hGrmQRAtVmo1QPizn8wSTkU6Jz8cfl/8AIXL7d/lp/GD4i9F/GvYeP7+/l40/yLMvYm5O1qzY3QuL2pmtubfTb+CxePytdvHsHBiLPR01JS1ORhrmqJRPNWiKJ0eRucvbvlCC895uePcHmrerqXaebjt5W3itVmvTKsr63ZkWKCTsLOyRlNI0pFUg9FNlzDuMcW02G3WsSCS31UJYgaVQ0ya8DTiMefn0K+2/5kfcG7fjt0dn32X0d1h352T80uxfg9v7O9ibk3PN8aOtt/dS1u9odz7xgylPXYTdebx+8qfZywbYxM9bRTVOTr44JqkBCWD157M8u2POHMlou57pfcq2fLdtvUEdvFGNyuIbtYPDhZNLxIYTMWuZlVgscbOqZws/rXuLWELeDGt2ZSjMT2dpetKjidBA8zgV8yg9p/zMvklB1Xujfm9dqfGreD4b+aL1R/L5wO4uoG7JPXe8Np7m3Vitsb57V29nM3ufLfxjI0c+aSOhhppno6GupainqTUFQ3s33D2T5Lbf7Haduvt6gWTke636SO6+m+oimiieSC1dI400KwQs5YBnR0ZAlemYebN2Fq08qxMfqxF2g0pqUHP2H+WDx6GX5k/I/cmfzP8AM++IFTtvb8Ozeq/5Ym4O7MXu2OTJNujK5rsfE9tbUyeEydPLO+HGHoKTa8UsDRRJMZZXDsy6QA17ccmWdpbeyPuHHfTHcr/naOzeLHholu9pKroQNfiFpCrAkrpGM9LN43i4uDum3NGvgi0D186sJa/l29Ibu/5i9n/E743/AB/zG3u1Pgh1zt+i+GHVO89s7W+Su7ux07V7d3Zt7q7EVeQ2js/a+xq7Ex7Z2q+PooaeLPTDKD+I1PjanVIyzmvLHt1sXP8Azjzdb3uyc03d0/MlzDJNt0MH0trE9ywEkskquZJdTFjCDF+kNWqrYL/33f7XtlgtvNAiCCtHNWY1b5gjhk91PNekhH8l/mR378r/AOWrvrorM9PbB6q+TfwZ7N70qOp+zD2blqZKiZuk8ru1d3rs/M4vG53c2113J9vtKsjihSnSWqNWjmRQDE8le3HKft/7zbXzTb393zBsvNEFkLq2NuhpS8SIw+MjMkcvharmNiakRlCCDXw3ndL/AHja5babRHJbqxU8MmMmmc4Iqfm1BTpK/JH+cjlusd9/LCn6/wB2fESiwnxP3xk9gU/SvbWV7YX5C/IrM7DwuIzHZbbDyOz422V1+J6vIzYvasdbR5eTLV9IzVP20Ui2Xcmfdwt972jkNt2tN/a5323Sc3lqlr9DYJO7pbeMs1Zp9IVZLpleERRvRNbA9Wv+cryG4uzE0SxxMQENSzFRVlGcVBoK6s5oK9H1/mIfI7enT/8ALg7u+RfTlbUbV33WdUbFq+vc5lqONqvYmR7ozuy9q4zdGRoahPFHk9kUe+DVlZF0R1lMpYFQR7if2e5O27f/AHn5Z5Q5jjE+1rfz+OiHEws0mlMSt5rMYNNRmjY6EO+7ncLy497bMI5XUDPlVgpz9p1D5Ag9MPUP8rX47dC13X3ZHTcO9k+Tux8c1c3eW+e4e2s7X9271y23KmhyM/yKov71VND2BsjcOayTVlVjYqaAUpWP7JoDGvtZzB76c4c1Juuy8yC2PI91Jp+hitLVFs4UkDAWDCIG3lVV0LIS2savELAnpFb8s2tpZLfQTOdwVGYvrbuIDehz5HOCeAHRN/gR3X/Mirf5eHdHda03TPyb7M2J2t8hq7r7Y9XU9rJvnfOP6z+QW/KHtLYtTuXL5yaho/DtTETU3X6ImmmWCngyAlBPuU/cLkX2t3n3s5Y5H2Pb97sLa9sbHxmt4IZlha42+CS3n8CCPVp8Rtd+W+IGR4tNB0UbXu292uxX1/JPG6pKdOs04SPqUEnJNNK446fn1Zt/Lt+QW+/5i2D7C+QXXuzsTs/4eTZjDbT+PO8d0x5mh7S7Ty2Ex6jt3d2dwEk32GC2Xt7eEkmAxEIgSsrKrG1dTI/hMI9jOP7hfuRc2HK6LudpbbrMZmv5JZQ8Fugk0wR28UUZlnleMGR3aSOMakQCoZumU9zLRJ7l54WaAKNCqMlqVNWY9tDjgeB4Y6tYxfRuFg0vl8nkciw03hpxHQQXFiwJTyzuh5H61Nv8fc7cq/3fft/tzxz84827nuslFJjiEdnDUU1A6fFmZTkf2inSRwYV6Ir73Q3acFbCyigU+bfqNT86L/LoTsRtDbWDAGOw1BTPdf3jCstQxS2ktUT+SYkaQfqOefeWHJnst7V8gRheVOQ9utJMVk8MSzEilCZpdcvEA/Fxzx6BN/v+9bp/uduUzj01UX7NK0FPy6UErwQI8s0kcMaLqkkldY0RR+XdyFVR/ifcoU4fLomoKk0yegyz3cOwcAHU5mPKVaEqKTDA5CQkAEAzRkUkYJI5aS3P59+oK1pnrf5dBFmfkdWyF49vbdggRlYJVZioeWUHkB/s6QogIuPSZTyD7317oI872bvrcBkWu3DWU9Owa9Ji/wDcbTAeq6H7fRO66Wt6nYkfXn37r3SGhjlq6jTTxTVlXKwFoY5Kqokd2ABOgSSMWc/U/U/439+690usT1fv/N+NqTbVbBE5H+UZMx46AAsVLEVDLM2lk5shI+tvfuvdCTjPjxmmjM+4dw4zEwqnlmSkieseCJUZpWkqJ2paZPFa5PK6bni3v3Xuqq/+HLvidlM9U7b6l65+Qnaz1LZp9ndq7r2PlOvvjnvnHYCc02Sz22uw9IrsnhJJIZXx8z0lPFlYo3kpZJYQrsa7dtUt6yeJqjhIJDEcQONP9nqAfd/395e9tLW9tdsa33PmqGREls1nVHgDrrR5QAXYHgViDOpIDhaipPMT/Nz3P3Z1pubsroqkqcXtPCbpy2xZtsdRdVNX9n1+eoaujpY4sLDnarI52rpcvFXQV+OrxPSQy0Ey1JMcf0PLbbNr8IyxRSTENSh+Ko4do9eI8iKdYs8++9vvwN9g5a3DeNs2D6m2WUNCyJEkUitl7uTW6vGVKSBBqSUaFBPRiv5fXcT/ACTj3R21mdy9m5fKYvK7h2EMH2nU5rH7t2lunaOXq8LvjbuZ29W5OtpaHJYzIxGN1iaWJ4rMrsjC3Ln+8m553KG49s/b7arie22SW3ur65WNzGJ38T6aOOZUILLEFk7GLIxcHTqz1lp9xXlTdJOYOf8Am3nTf/3rv0SWkNnKbiS5jFvND9SZrd2OikyumQqyAKRgdK3qndHyC+S3e3yNym2+8Knp7rP4wfJIfH7DdQ7a622HuxOyBtHaeyt2713P25uTdePrN1UC71bd7UuFp8FVYdMdQwJVM9VKzD3gVvm3cn8i8q8mW9zysl/ve+bN9c92880X03iSzxwxWkUJWI+D4WqVp45tchMYEaivWfcF5uG6393MNyWC3gk0hMEmmkk0PEHUQOBx8+gW6F/mBSds/PjsXp4dt9a5TqrfOT7u6g6B6yw0mCrN+7S7G+KJ29/f7fW8JaaePP12I7lfM57+FUtTohji2ovgsamRvYp5q9phsPtRs3Mh2C8TfrZLO8v7l/EEEsG6BzBDF2+GptNMPistSTc9w7AOk1lzBcS7y8E1yBDIzIFoKqUqNX2Ek/Yq1zU9GB+K+7O9Mp8g/lL19ujuKs+RXSvV0HWuCwnaeW2PsbZlZh/kPkBuXJdy9MbZk67x+Nwm4dsddbZfb8lUKhKqvxGWrpKKarmkWVYgnz3t/LEHJ/Im72OwDZ+ZL/6iRrZJ5pw9gpiWzvJPqHZ45bl/HCFdKzRIsixqpBZZsV5fS7luMEtyJbJDlsVDCoxQDBxxFc/LrJ3hvvuXfPyy65+I3VHas/QeOyPx43x8j97dn4DaO0N79iZyDAdg4HrXbfX+xsX2DjM5s/G4+HJ5eTJZ2tmx9ZVmnWngg8HleX21yttXLW2cg7x7gb/sI3aVN3g2+C2eWaC3jMkD3MlxM0LJKzkKIoVV0QEuzayunpzdLu+uN2i2m1uVgTw9bMfPKgAVBFe7OPTIrXorPy9/mD7i+JHyD6A6uyfdHX64nqnA9Uby+XxztHt7am8+2tvd4dh0/Su2m2HtuKsabbWa2awrt8ZODFuU+0pI4SVp3K+x/wC2PtTDzby1zPzRtGyX9tut1NdRbQ9rLNps5rKA3kpkuECO4m/Tso2kIJLs2WBqVbvvc9rc2trLdRSxxqviagp16zo4GuAanzwBwr0f2l797t23/Mc2j0nvLdW3d6/HrenxM7M7OpNgvsPDrnMPurr7tDrbacW5q3sKskrc3uOtyNJuyrC0q/aUEFMsKiN5tcxnf2d+9nzXyN7UfvTm1brfbez5gtrSZ7ibXdLbXEFxM/07aFLPD4QKx3DSmQlwHQFKBve+VLLdN+WDb1S2aaIMoVaR66KQCAcVBoaZBzQjjapuKv6a6/2bX9n7tOydnbL29iP7y5XdmfixuHxeHxkUK1H3tXWVSxJAQrDSv62kIVQWIB6jbFvNhzHsuz8wbTKZNrvraOeJiCpaOVA6EqcglWFQcg46iye3ltrie1mWk8blWAzRlNDn7eik42l7R+btfHubI1PbHx3+LmFqFyPW0G389uHqvvnvHOxh1xvY+5HoGos9111JQJIZ8JgqjRks/KyVmThp6VIaOc26Z4dLmX4MbAzkLY7sft75P9ubbm/4GbN7C773hPtLKICD9vmcNteXa0eZo3I9UFU00EgJV0ZSR791smvl0bja21dsbH27h9pbN29hdqbW2/QxYzBbc29jaPD4TD0EP+ao8bjKCGCjo6dDchY0UXJP1JPv3Wui1fKXtXd2EpNpdG9L5LGU3yE76q6/b+xqmtVqmLrvZmNihl7L7wytDHZZsd1tgasNQQyskWR3DV4+iJKzvp917obuoerdp9J9Z7L6o2PSz0u19jYOlweMNbO1Xkq9otU2QzWZrXAkyOdz2Tmmra6pb11NZUSSt6mPv3XuhH9+691737r3XvfuvdNuWzGJwOOrMxnMpj8NicfE1RX5TK1lNjsdQwL+qerraySGmp4V/LOwA9+690Uzefy4wtPh5ct11gkzeDaQ08HZ2/8AJN1r1C1QX8Sx4jcGZpJdyb+qHlusUO3sXkRORZJBcH37r3QUVNd8h+9fBHjI93SbZnjgNVUR/wAX+OPV1ZHMoWR4MhUR5/5D72oZF1EeBNt0syfUkONOqila4690Luz/AIsHDUlTR5Hesu18TkHSXKbV6UxCdaUWWkuDIdy76kqtwdtbpnkF1eaXOU4lDteMAgLuteHXuh22J1F1h1os39xtkbf29WVY/wAvy1LQpNuDKMTIxky+46z7nOZaZmldi9TUSsWdje5N9VHDr1OhIsP99/h7317rv37r3XX9Rf37r3VTf8y/5dUvXOza7oXYeUc9ib5x3h3VksfVRiXZWz6tbVUEskMgngz25Kc+KBCFKUrvKfql/de613FAjVIwtkUJHHGgNvwkccSLyxJsFUXJ+g9+691cv8Hu2ex/iD1XuvP90dEdiYX4+ZTNRb1zfccWJlZthYyopKSgyO6t1bVlcZ5+vsTSUyVdbkqenY0FKss8qNCjOvuvdXYZzGbZ7a2LTtj8ris5t/cVBht07T3LiqmLNYaaaNqPcezd24uox9ZHR53H09bDS11Ppmamq41UEtG/PuvdRuuN31u4qLKYTcSU1HvbZlcuF3bjYa2nr2WR0NVg81LLQ00GPgbdmBMGTFJCZPshVeBmLxn37r3SFlU7Y75p5VOii31hJI2LaEjNbTRqCgLfrlMlEpsLE+T6n37r3SlpIqSk7wzvjXEw1Wa6w2/V1Qii2hBma44fcmbo456t4WG/MhRUcOQCRtOrYqBnKwkTvKD7r3RSf5mcOQxnSHU/Y2MxmLy9X058vviX2S1LlWEdqCn7t2ptPLz0E5IMGSgxO7Z2iNxqAKchrH3XurFDwDf6cn+vHJ9+690EWLoEo+xeyMMKX7ek3dt/BbnNXDR4+D7iv+1rNr5RnqpNwVmWy1ZHS4+jvfG0lHTR+NVkmkdwvuvdB38cJFpn3pimSUSU9VjnLuNI0RGuogjqfUJtUN2uAOffuvdCPJLkcv3TT08M9TDhNmbGnqK6AT7kooq3ObuyccWPMlLJiBtPPU1HjMFORJFWmuoZpNLReOcN7917osvzknzNXnPhJtPDukQ3P84+n6rMStJodcHsLbu/+yqyKJdaeV6qp2hBER6vQ7G3Fx7r3R82PAPP1vx9bf8AIvfuvdFC7fq5Fh3r5IKyNMlvvbdFSNUpuCnDx4LaFFUVVXQQ5aunxFZRPPW+P7jFQ08BkVknDzo7H3XunDb0FGOteqzJDWZKkPauGq65sfR5d/4VUx5HJTUlRkqnFZ3BrQ4+hyMcAmqKr7ujIYRPTS+RdPuvdGtuo5uL/S/+ufof6En37r3TJmdt7a3FTT0m4Nv4PO0tVH4qilzGJoMnT1MQVlEU8NbTzxyx6XI0sCLE/wBffuvdEM7E6jr/AIh1GV+QPxjwk9F13j3GZ7/+MmD8i7L3NsyKZZ9y9mdN7XitQ7G7d2dixLXNQYxIMbuijp3pZqf79qWqT3Xuj4bU3XtvfO2dvby2fmsfuPau68JjNx7bz+JqFq8ZmsFmaOGvxeUoKlLpPSVtHOkiMPqre/de6UHv3Xuve/de697917r3v3Xuv//X3FN1Ry9P/ODYG91hmi2Z8p+vKjprc1VGJBj6Ht7qdc1vvq+syUrOKeGbdGx6/ceNia2qWegpogbsoPut40/OvR41NwDa1x9Pfutdd+/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de6IrRZak6s+dm9cZu90oMH8puqOvn65zVfGIMTlOwOmp92Y3dvW0dZKTTy7krdobjpMxRUraZaykgrTErillt7rdMV6dvg9karDdd756LzVUJtwfGvtvfXUoWXUlVLsb+I/3x6jyLQyO0i01X1lujGQxvcrI9NIAfSQPda6Op7917r3v3Xuve/de697917r3v3Xui39800v3G263T+wEyFGXAPEz/bzqrG2kBo0Nr/m/vl9/eI7VdjcPa3fBH/uv8O8ty3/AAwmGVVPkKqrEfxEH+HqYvamePTvNpX9UmNwPkKg/wAyOiFfJX469ZfLDpPfXQPb1Dk6vY+/KKhSpqsDkZMPubbmcwuSpc5tXeW08zCGlxG6tobioKevx9QFYJUQAMrIWU4Bcl847zyDzPtPNmwSou5WjNRXUPHIkimOWGVTh45Y2ZJEOGUnINCJM3fa7fd7OS0uMAjB4EHyIIyDUAg+RAOeHRZes/iF8oMNXtSd4/zF+3u/tjY3YW+NhYHZE3T3VHWiZ1d5bVr9oQbo7k3Jtenqsz2ln9r0OQNRRoHxlLLXxrUTpI/sbbz7h8kXMKvyv7O7ftW6vdRTvP8AWXVx4ZilExjtIpSEtY5SND18VhGdKEDoPW/L26dsd5vDPCikKNIzXzOBX7Rp9aHrL8eP5fW2fjzv/wCJO/8AGdn7h3TU/Ez4Y5L4ZYXG5Db+LxtPvbBZLce19xvv3Kz01ZPNiMxFJthIhRQiWArKT5LrY65v93r7m3aef9pm2GKCPf8AmMbu7B2YwyCOVBAtQAyfqk6zTgBTq1nymttPt8xum/Qh0UoOJCj/AJ944+zquz5mfBY9Vdb9IbHp4e+ezOlMt86vk58oO8ewemundsdydl9Wr3ptvdNXg9r0vQebxO99rdgbJrtxbhONq8xV4TK1eKgRaiGCGWTypMPtt7pDft45o3KU7XZcxxcr7btllBd3clpb3RspIlkl+uR4JYZhHHrSFJ4kmJKuzKNJD29bHPt8drGqvJbNOzsygNp+M/CVYEVc17WK4pUmoWnx5+Kvavyk+LndPx87Hz3cvUfSPXHfPT/YP8vHt3fXQvWHQ3f20Mn1dFQ73k3lnej9o4Ha20JNo4Hs+NqfEx5LD4qrzGNlqDPGp8cnst5w575f5G535Z5s2a12/cOZb3aru35gtYL+5vbCVbovB4SXssk03ivb90hillSKRU0tkr05tm0X+7WV3ZzsyWscitCxUI1QVb4QAKD1Kg0NCBUdGf2z/Le3RLuP5Z9i90fLLfXdXavzE+K6/F7sDc1V1ts3ZO29mYinfd0eOzvWeyNv1b0eEx2Pot0kDFTT1Hnq0mqZalnqCsYHv/eeyFnyHs/LnINrtuxcv78dygjW5lmkmc+CSlxNIAzljEP1FClV0oEATJtBypcf45Jd7gzzTQhK0FBQOBTGMPXzz69M3Yf8sDJZ3sB999XfKTdfT9TvH4m9efDnuZk6j667Gz26us+tcDkdv4fMdYbj3p9zL0xurMUmWqFyP2kNdRVAkWXw/cRxzKr2j3zhtdr/AHVvvItvuCW+/wBxu1n/AI3PbxxXE8iu63CRU+riUovhBikgFV1aGK9Um5RmMkb298RWARuSBXBY1XBp8RHAg4NKivSr/wCG7tw7SwnwRHR/ya3F1Pvv4O9P5X4/47etZ1ZtLsGk7U6j3Pi9k4/dmHz+08/kafHbc3BkX2HRz0mQpZpjRSO90kFh7Lh7x2l/ce6X9Z+Sob/auZ9wS+aIXMtu1rdxtM0TxyxqWkjXx2DRsBqAA1A9OnlOWE7bPZXpW4gUISQDVRpHpx7Aa4zXFDTp63D8Ju7dt9o9w70+MHzOy3xz2J39viTtHsrrKv8Aj91j3PDh+zcpiMXhd3736q3PvSqo6zZddvaHD09TXUlVBkaKPIK1RDGpdkKS190OVbnYdg27nn27i3bc9pthbW9yt9c2Re2VnaKG5jhDCZIS5WNlaJylFYtTrdxyteLdTy7fe6IpW1EMK5oATT5geeoVzQcOj39i/GHDfKXpzfXQnZm163d/WPaey6vYe+ZMjM+G/iWMrKWGKfKw5OmjiegzcFdTx19NNSx/5PWxo8agKB7O/Zb2q97eZOa9h5p9tOWJrV7e8Esd7OphsomVtR1M4rOgWqFI1cyKStSSaOcx7py9a7dNt+736O+j4FozknhhaKCK4rT16C/41/y0vkv09vHrFu1v5lvdfe3TPT8wfa/Vdb1T1ZsncO8qLH46bE7ZwfdnbuEoqrdnZeE27SSIxiWLGSZCop4pamR7FW6hbL90j2Yla63fm3kOwuOY7xCbhIZLtbFZHIZ2tYHl1Q6m1Y1kAEhVUEjqGJubN5CR21pfOloldNdJfNa1IAHnxAB+fTNtL+UtvbYPQvyS+J+zPmt2Ltb42999qZ3emE2tt/rTZuH7I6w2H2j2dmeyO/8ArDEdu4+uptxZmn7RTPVWGpsrNDDWYLGTP4BJKVkXJPbdk2bZ217VtVvbPoRCY0VWKxIscalwNRCRqqLUmigAcOg5LPNMSZpWapJzwqSSceWSTj16NH8V/hD178F+w+9sl01vsbE+NXcVTtfdmB+LP8KxeN666a7MxWIhwO9d39Y5M1i1uIxPZWPx1FUZPDCH7VcpBJVxsHqJQTPprozG4O+dlYgSR4xqvcVUtwqY+IxUmqxsGr6kJEVJtzGslgffuvdArnO/t5ZPXHiYKDb8JuA8UYr64Ak2vPUqIEb/ABERH1/qPfuvdBLl9xZ7OsXzebyWS5/TWVkrwi9hZYNSwIDpHAUfT37r3UvB7P3NuKSOLB4LIVasQPOKZqagRTYa3rJxHThBf8E/Xgf0917obtv/AB0ylQEm3PmoKBeCaHExirnsCt1kragRwRki/wClH/1+PfuvdDDielevcWiB8McrKgF58tUzVbOQoBLQho6YBmGq2iwJ49+690ItDhsTi08eNxlBj0F+KKjp6YC51N/mo1+p5/1/fuvdB1vzuPZmwMpj9vVzZ3cW78pCKzH7I2RgMpu/ds2P83gbK1GIw0E74rDJKrD7ytampSyMqyFlIHuvdBH2H3ltncewO4dnLit4bT31T9V78yOO2pvfbddtXK57Gw7arFrMvtSprrYrc9JivuVNSaCpmenItIEuD7et41mmiiZqBmAr9vQe5s3i75e5X5i36wtlmvbKxmnRDwZoo2dQ39Gq586cCOtJLGbQ+X9F8cG+NtPs/rXC1+G6lh6bxPfVD235Is5hFgoNm4/emG6+k27Hltv5yl2JNU1lRS11U0cWRgSKBpFkDKPorfcZLdrOK11aUC6lYtUA8QB504/PrlDdb37aT86pz/d75fzR3G4fXSWMlqAyyZmaCS6aYrLG0ypGjxr/AGRJYDTQumzvj52V0jlu0F2fkE752d2N1x1Zt99r1edwPx/zdLuDq9BsSkhwOT2RR/b0NXlOqvth9/eKomrMXHFJKPIsgubC6slupHjY2vhrVn/RChcEmQ6VFF4sxA9SB0xfc7cv84x8vJdxjad5tL+6dXjR9yV47tvHKvHKdT6byo8JFcaJWZEJFOrIf5dHTfaPx/6jzWO3jRjYy53sre299hdeQbql31lOu9p7ozMmVxmIzXYdTDT5Hd+fklqJ56qqqFklJqhE0sqwq7cWfv3e6Xt3z7zbytsHJhhv932JLiO73CKQyRUleosIzTw7hYHBme5RmRWdokLUJHS37ufL3N2xwbzzZulzcwWW5W1iIYXt1tZZpreHwpNw+mDE2KyxlbeO0KiscQmZIy+no1e5eo+rd2b3z/ZWS2tncD2Du/8AgMm9N29Xdp9r9P5He1XtSOCDamW3tjOud44Pa+8M7tumpYoaOtyePqp0giSF2eJQgx45H9/PcT2/5a/qftB2i75aEjsIdwsIL3SslDJCjzKWWCQ1cx10hmZloT1ktc3a3t0L27jJuqAExu0dSK0btNNQGOFMDHSf2N8aMDtnEdQ7I6rocDvPF9Jbnp93dO9Y9uyrtndGG3XDHnqU1Ow/k/11j8NvSLdeWot15KGQ7wx+boMg9URVSpfypMvLPuv7S+6m77ls/ubyxJypzLvsIsm3XZ5JHsZfElh8CO92u5E0SgMiL9Qh0qFUEL8QtDc38Phtt8njeFV/BnqTRQS2iUFakip0/F5Cox02dbdPfCWq6Pg2N0N3H2D8Z9n7iz+Fm3jsTe3dfYGG3VisZs3taTdnYWytuU3Z++qrbGKk3zvjCVNJW7z2pU1wzFHNO9NVTJU39iT3d9jfvA8pc2y8wHl60532uKGbwLqCxtIriNzbm3R721gjEkn00ZpDEDJA7LGUCU09Huy75y/d2iQy30thdMy6lLMUIrX9OQk4auQwDDgTivRs+4NlfEbvzI7V3H2Hvjrsbr6/GdOyt/bP+QtB1Zv3aOP3VDDT7ow+G31sbf22dyU+3dyUlLElfjmqHpKoQxu8fkjRlx15K9ufvHbWm6bdyt7Zb41jcLE1xb3G3K9vL4ZPguYr6MwvLGSSjKviRktSisai7cdz5NufAe53qESx4DKzE8KGpXOaCuaH8h0hz178Ccdt3tjZNNQbE35Q94YPG7V7Q25tf++/f27t4YXE7Jpeu8Ng3n2rTb93NjoqLaVJHTUzxVVMEmZqjyiplaVpa5f+7h97zmG65a3WSxj2p9vJmtnurqC1EMjzmZ628Adg7yEyaZIiCo0V06B0SXfM3JECXMamW4Z6A6VYg0UAZZgKCnpxJ+fTjiPijn+xO3did4dQ/G3s7a+/dh7RpOttqdufJ/untnrPaW39gY+lxMH93ab497e3lm9271wmUfB0dTXUWYpsCmUrqVJ6uVpf3feUnt/9ze9tNivNi90+fIL/AGK5u2uZrDb7KCCOWYs5Esl9LF9WrDW6qIBEY43MaMB0Cdw5zU3cd1s+3mCZU0q7uWZRShooOnPHIOc9WP7J+IOGnyeC3l8iN6Zf5Ib92/kKLN7ah3ZQUuF6g66y2PUDHz9X9L46SbauCqsUAv22TyRzGeRow4rgeBm3te2bfsu27ftG02iW+12sKRQxJXTHFGoSNFqSaKoCiuaDOegXNNLPLJNLIWlcksTxYk1JPzqejkAW/r/sefa7prr3v3Xugl7v7j2n0R1xnOxd2vUVMWP+3x23ttYwJPuXfm88tIKLafX+z8cSJMtu3eGakio6GnQG8kmtysSSOvuvdBL8ael917an3F3j3kcblfkt29S0L72qcfOcjhOsNn0k0tbtfofryrkAEOzNiipZqupjCPns3JU5Ge5khSH3WzTy6Np791rr3v3XuvH37r3RUu2/lNtnZkNbithtgt7bmo53ocvlavPwYnrTYVXdolXfW84Y65TmTMrCn29iYchuOvdCkVIAfIPde6LTDsrsftw0+7u090Yvb+33qo8rRb27f29TUstM0XphfpX46bmqJts7ApII1tBnN3rmNx1BIm+1hDIqkG/c1cs8rWzXnMvMFlYWwWuq4mjiX8tbAsfQAVrjpyOKSVtMcbM3yFel0d/fHHrfKx53b+BzPbnYtHGkC9gbtqanceeLRqImam3bujzDDQMi2WDE09NSKCBHEq8DEvnr78vtNy20tryrBd7/AH6mlYF8G3+36iYDUPQpG4PkeNDi32C9loZtMa/bU/sHSez/AMvOxsg4GCxO3tvQK1wJYZs1VSqHf0SPUPT06BkK3CpcFTY2PvFjmj7+Huzukqry1su07Vbhq9yvdyMKmgJcpGKjTgKCGB7iDTo1i5fs1H6ru5/YP8HQeN8ie52dn/vzUrqZm0JiMMI11EnSgNASEW9hyTb3Fj/eu+8M7yOPcSVdRJoLazoKmtAPANAOAyaDzPHpZ+59tAp9OP2t/n6Xuzvll2Dg54It0wUO78WHH3L+CLF5tYiWLtTVFMEoZpEvcLJEoIAUMPr7kvkD78funy5dwQ87W1rvu0a+8hBb3YU8TG8YWFyPJXjzhdQ49JLnYLWUE25KP9tR/PP8+rDtp7qwu9cBjty7fqxWYrJwCanl0mORGVmjnpqiFvXDU00yMkiH6MPyOffVfkjnXl33D5Y2nnDlW+Fxsd7HrjalGBBIeORDlJI2BR0PBh5jPQSngkt5GilUhx/qr0oT7FfTXVcfzm+dGB+POErdg7CrKLOd25zHulLTxNFVUPX1HVRske4dxgeRDkQG10OPezzuBJIFhHr917rWlzWay24Mrldxbiytfm87mayoymazeWqZKzI5OvqW8lRW11VIWkmmkc/6yrYKAAB7917q9f8Al6/CnA7M23QfIzvXE0cW4q6FMnsDAbpWCmx+ycI4tTbqy0FcVphuHMLIHpjNxR0zIVAmclfde6tSXsrrbOt/AKrM0Tx5iGjovstwYuvxtBmI9wV2awNLiImz2PpMflKrJ1eErIfslaSZ44yzR+J0Zvde6JXtTZ2M+AXYm0dqbKEWF+EfcW4xtfC7QqK2U7f+Lnc25Kqap21j9jmd5xgej+4ctO9BFiFZMbtzcstKlEIKTImCH3Xujg9iSTbSzO3+y4ZXjxmKtt7fMXlpI4H2llKlRHmaquzm5cLgNv47aOTm/iNdViGorJaSJ4Ywb6T7r3TJ3I0dDVddbwhJb+FbppIXqKdInD0OQ0MSKlrxrG6xELc6W13vfn37r3Sjyuim7m2fUXjX+K7C3ljtRy0EMkz0GX21kIkTCrgZ6rIJHHNK33ByVPFTA6fBM02uP3Xui3fzNdny73+AfysxVLjK3L1+K6h3DvnFUOMnNNlGzHWpp+xMRPi6gFTHkqPJbXimpyOTKiixvY+690bvr7dFFvjYWyN64155Mdu/aG2t0UD1UZiqXotwYaiy1K9REeYp2gq1Lr/Za49+690k87DDR9y9f5ESxwz5baG+MC6Gv2vSS1sVHVbdzMMf2VXi5d4ZkUjRyOq0VZFRUodnqIpGeFk917oNel55IeyOxqGS0f3EtZUNFKNE7S02dqUUjUQ5CxVJJFvoQf8AX917oTuqp1zVDufeYR77v3fmqmmkkpqaleXD4CcbWwcmrHZ/cOKyFPNQYVZaesgeD7qlkjeSGOXWPfuvdFH+TLYjP/OP+XJsuuNdV1OKz/yZ7cgxtPr+whk2d0tNs6gzeWCjSFoK3skRUxJt55/8ffuvdWDVlVTUFHU1tZPDS0dFTTVVXU1U0VPT01NTxNLPPUVEzpDBDDEhZ3dgqKCSQB7917qvbdM0j4zBTzxCKr3XWbl7KrozQbeop0/vhkvHh46yba2UzOEytTBt/GU8f3sNQ4rYhHK1nZgPde6ybV3XLRY+u2rk9vw7w2tk5qevqMBNTLVTUmRo6mnrqLMYvXTVaQV1DXUkVRGzIQk8SSKVdQffuvdCHR7+rsTjZ8a3Y/ZGGpvs4cdhand2x8DuHL45YNryYSPIz5yZIJM9WncMkeXqJKyNzJIhp9Ihf37r3SlTcG1Mu+Sjqu+t/Us+RjyQp56KLEYGjxMVc23ZCmOE21J4fNQy4ab7R5zM8ceRqUbX+0Yvde6XLbr3HsqspavdmVxe6Ouc3UxxUG96SLH4yo21UZTIbgyEcm8Znq8fgINm0WN/h2LoqunM9bU10g86fuax7r3Reem6E/Gz5H7k+NlEk9L0t29tzPd2/H3HNq/hewdy4jL0cHePUG300laPbxrdw0G6MPRIUio1yORhhVYIYkT3W+Ir0fP37rXXvfuvde9+691737r3X//Q3hvkb1FUd3dRbl2NiM++0N5LJiN1da72ij8s2yez9l5aj3T1/uoIEeSWlxW58VTGshWxqqFpoCdMp9+68DQjpr+M/eX+nXrk5bNYf+6fZ2ys7k+uO6dgSOWqNidqbV8MG5cRG0jGSqwGSEsWTwtZ+mvwtdS1It5Co91sinRhvfutde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XB5FjBZyFUAszMQFVVBJZmPAAA/PtuWWOFHllcLEqlmJIAAGSSTwA4knAGT1sCpCj4ieHr9nr0Gua7a2fhar7RqmpyUyl1mOKhjqoqd0CHRLM80MbFg/GguODcj3izz398n2S5F3U7M263W63qMyy/u+NJ0hZdPbJI8sSEsGqvhtIDQ1Ip0Mts5C5h3OH6jwUgjIBHisULA+goTj5gdN9J3Xs2pqYoJBlaJJCwNVWUcaU0WlWYGRoamaWzEWFlPJHsObL9+32O3bc7Xb7r977fbSaq3FzbIsEelWb9RoppXGrTpXTG3eyg0rXpZce2/McEUkkfgSsKdqOSxqfKqqMcTUjpeYvde3M1o/heZx9Y76QsUdQizlmDMF+3kKThiFJtpvx7yF5S93/AGw56WD+qfPW2XsshULGk6CYlgSq+C5WXUQCdOjVg1HQVvtj3fbSfrtuliUeZU6f96GP59I7uDp3rvvnYmU647O2/FuHbOTmo62MJVVmLy+DzeKqErcHujaufxc9Lmtr7s29kI0qKDJUM8FXSToHjcG95GDV8j0V06rK6+m7k+JXzu2btDuvK/6QusvljseTqrZvyGjENBk9y9rdO02W3V1ltfu/BwUtNjMb2tlesKrN46mzWPtQbmiw0BaCkrVeKW3XuIHy6uCkkZE1pGZLEXRbayDx6b2UkXvyfp791rrL7917r3v3Xuve/de697917pD9gbXfde3KqggKLWwvHXY8twGqqYMRCzHhVqEYpf8AF/cAfeU9pbn3i9rt15c210Xf7aVLuz1Uo08Ib9Ik/AJkZotWaFwTQDoS8pb2uwb1b3sn+4zApJT+BuJ+1TQ/l0Sqphno55aSsglpauBik1LUI0U0bqbMrIwBIU/kXBHIJHvhDuNlfbPuNzs+82M1nu8LlZIJkaOaNgaENGwDfYRVWGQSCOsl4biG4hW5t5Ve2YVDKagg8Dj19OI8x1w9pQfUZ6er173vr3XYJU3UlSPoQSCP9iPfuvEAihFR10SSSSSSfqTyT/rn8+/deAAFBw697917riTp+vH45/J/AH9fdWYIuonFf9WP9VeHHr1fTpcba693NujTLS0goaAkH+I5IPDA6n800Wkz1RX/AAUL/tXvIH2r+7L7r+7iR32z7ONv5can+O3weGFgeJhTQZZ6UPwKEOP1BWvQV3vnLY9jJinuPFvKf2cdGYf6Y10r+Zr8ujC7X6m25gPFVViDN5NbP91XIDTwyA3/AMmoeYUsf7T63/xHvpd7Rfc59r/bgWm673bf1g5rSjCe6UeDE3H9C1qY1oeDSa5PPUOHUQb5z7vO764bd/pbI8VT4mH9J+P5Cg6FEaYlNyqIg/4KiKP9sqqAP9Ye8to40iRY40CoBQAYAHoAMAdAY1JJJqSf9Xmegm3V3TsvbTyUsVTLnshHrDUuH8U0UMi8BaiueRKWIk8ekuw/I93690AO4u+t55YyRYhaTblGxIQ0yityJT8FquoXwo1v9RGP9f37r3QN1tbW5KoaqyNZVZGpkYlp62olqpSxNyQ0rPovf8AD37r3WfF4rJ5ysWgw9BVZOta3+T0cLSsgJ/VKwtHBH/tTlV/x9+690N+3/j5uevMc2eyNFg6dwC9NT3yGSC/0IXRRRtb/AGt7e/de6HbbnTmxduNDOuLGWrotLCtzLCtcOLXeOnZVpIjcXFo7j+vv3XuhRRFRQqKqKo0qqDSqqPoAo9It7917rl7917r3v3Xui67/AO09w5rcWT6f6Riosj2VTUlPLuneOWop67YHT9DkLfb1u5poHiTO71rKUtLi9uwSeeeyzVj0tJaWT3Xult1V1PgOqcLV02Pq8luHcmfqYstvnf25J0rt3773AtOsEmY3BXJHFGBGgKUlHTrDQ4+C0NNDHGNPv3Xui2/KLdNHU7pwWFpsU07da7b3ZvbdG6Hx61MWBbfW1dxdc7N2rQyzAQ1OS3fkMvUVFXAGXw43Hs8hHliDxv7te6W0ezXI248+7zZzXMMEsUUcMWGmnlakUbMcRxsw/UlIIRRUAtpUtXeyz8w7bvGz28yxyT2cyGRlDiNWQqzlThtOqun8XDHVVm2euNo7YooqOkxNLWziNUqMlloIMhXVLBUDannjaKKIsl1RFVV/H198dvc37zvvZ7q7u2477zvd2FgrAxWW2yy2dnDQtQqkbiSSSjUaWWRnegJpQDoI8g+wftT7dbZFt+1cp295eBaSXd8kdzcykhagtIhSOMEdsUaKq/M9E+7X7J+QuP7x7m2X0Pi/jrFiehPj51z3nV7e7N2PuKTOdlZHd1X2jPl9q4vsHa+doV67EOH64MdHWNi8kq1VUHmTxKbb2e/g3bkrl2fn7nLnG5Xfd5utvPg7o728EcAtdMslnPrF0C9zWSMyR9qDT3dD+Hljlexu1vNv5T2qG5iAZXS0gR1NGyrrGGU4wRQjy6DPJfODevZG3ty9mdWbm6s6D6T65656M3bvbf3d/WnYPc+5Mnvjvzrah7ZwGyqHZnWWWxMm3tobP2rkYFy24WaqlmqXcwwxwxGVq2/tNtWx3W37Fv8AYbhu/M97ebhDBb2FzbWMawbfdNZvcGe5jk8SaaVCYbYBFCAam1vpB6bh31OSOAJJqcsK+R4dWZ7NqaneGC2jksa1Fn6rdO3MBmqOXasdbWYrMDMYijyYyG3Y6hP4jLhK1KnzUrTKJRTMpls2r3B95ZyQ7tcbVDa3H1S3UkSxMha4Lo5jEZRAdcq00sI9Xd8OOlEaNJTwwWx5f6uH29K/Zmz8/wB21u4OvupNxw4+CmkfA9w/IfGVCTbG6W2tU+aj3dtjrbdwSXbu9/kVl8aZaFPspqjHbKjqGra2f79aWik6Rfda+6buG17lY+5vu3tIhuoGSTbtukNWSRe5Ly8QGgKVBt7cmoeksygqqdF97uKwqYLObVKQQzjgAcFE9Sfxt6YXiSbM5ezPhZ1XszafVOX7Q+O22NmbRweM2ztTaG59/df/AGeNw+3KClx9DRwUedy0jyNRUcUQZ3DSEnUxLG56QDVWpJr/ADr9vQfNa9KXZW3/AIl74aOt66wPx23e5i+8iqtm4rrbOyeGVQ33Ky4WCqkVJFYHX9CD72WZviYnrVKcB0YChoKLG00VJj6Olx9JCNMVJRU0NLTRKOAscMCJEigD8Ae9de6me/de697917r3v3Xugx7e7i676K2LlOxuz9xU+3Ns4ySko43MVRXZXN5vJzrSYTa+2MHQRVGW3NuvcVe602PxtDDPWVlQ6pFGxPv3Xui0dU9S737i7Ow/yk+ReInwVdt6GtPxt6EyJilj6OwmcompK/fm/ooJ6rH5T5B7uxU7U9U8bzUu2MdI+Oo5JJZa2pqPde6PIoIAB+vP+98f7x7917rv37r3Qddkdp7O6rw9Jld1VtQajLZBMLtjbmIpJsvuveW4popJaTbm0dv0gavzeaqkiZvHGoSGJXlmeOFHkX3XuiZdo7i7I37mYNi7yIoa3OY2TMYz409e7rfG5afa92hk3X8k+5sYqf3G2JA4aOoocKQtdIGpoJ8odUQannhtYJrm5mSO3jUszMQqqqipZmNAqgZJJAHmetgEkKBVj0FdFndpdWyY6qxybY7E7NwFLVY/beQxWCp8D0t0vDVLprMR0tstFYRyM66KzL1DPkskYg01SYmECYCe9f339g5f+t5e9pYY903xdSNfSD/E4W4aoV+K7ZTUYKQ1HxuCR0IbDY5Ho94Sifwj4j9voD+3oLc1Vbl3LXVe4c7JlMvWVrtV1OQq455l9fJaP0mGlpkHCIgVEUAf098vuZOZt35t3i933mrepb3ep5CzyTPqJY+SJ8MYGFVEooWlBToVxQpDGsUMaqg8qD/Dx/z9MqwyyK7xxu6RAGV1UlYw30LkXAB9k7UVgrMNXp/xdOrCp8j16KGWeRYoY5JZXNkjiR5JHI+ulEDM1vzYfT3o0ALHgOPD/KR17PkD1zkpKmFUeWCWNJCyxuyMI3ZP1KkljGzJ+QDcX596Dxt8EgJ+X+r/AA9eofTrB9P9792x+L4etHh1Yf8AFXcGO271JuDK7nzFHg8Dj92ZRlyOcrIcZiqSAUNBLUFa2ueGmWMTlyx1frvfm/vrt9waS9f2i3+O4Mn0yb9P4QIogBigZ/DPmDIWLejVB6BvMFBepTj4Yr/Pol/yx/mjbdwdFlNi/Gyoi3JuWeKairez5YNW29us9kaTbFJVR/7+XKopbTM6LRQtZrzfpGcfRH1Q7lMrlM9lcnnc5kazMZvN5CqymYy2Qmapr8nkq2RpqqtraiQlpqieViSeABwoAAA917o2nwU6Ppe9/kbtLb2apBWbS2nFNv7d1PIpMFXjdv1FMuOxc9gVePKZ2pp0dCRrhWT8A+/de62YO3Ex1dF19tXJVlFS4ndfYWFxFfi6uXAik3Jj6LH5TNT7XnxOfweZpc5QZSPFhZqWEU1T4kMkcyGOx917oUcnicdnMdV4vL0VNkcbX01TR1VFVRrLDPTVdNLR1MLBr2WemndCRY6WIvz7917otuS6v25u7b/ZPxe7FpKncfWG99l1393YJUqxLjNmVr/waq2vR1uP2xjsJttdgTPQf3fc19VlpCpqCQacSH3WwaEHpP8AxT3bufePWm8OlO3ckcz230Fn63pTsvNOsLz7xo6bD0OT2D2fHFVQumrsPr/LY7Jy+SIqmUaqjClYufda6UG5RVV/QjUuUmr58ts/KjBVNTWVWVyFRX12187Ngoq2tyuXwWAqMzJX0qLNPUx0sdNNOzGFniCu3uvdLuqyNflanpHdGPWqqKGpyFZQ5pKefdGhIsztHJRpU1GK295cDUx02WoY1M2YApaMOTEy1DRg+690v9+bTxu/dj7x2LmYIqrD712ruHaWVpp2kWGoxu48PWYeuglaJlmWOamrGVipDWPHPv3XuiefyzNy5DPfBn494zNyiTcvW21K7pDdafe1uQkptz9FbkzPUGZp5qrI0tHkHcVezC488SShGAa55PuvdGe3XLKOy+p4I58hGkh31LPDT5HOUlBURQ4CnQDI0NDgq/C5UxzToYUr63HmJ7yQed1MY917ovuzM9P/AKac1M4WKfN1m6cWhhXUyVP7rUfgaUSRxyNLQp6pAyDm4t7917ofukZ8VN1Tsj+D1GPq6SHEfZSz4ptttRtlKCrqaLOIG2fQ4zbBqYc1BUJUfY08NP51fSo59+6909ZHrPY+V7E212xkNv0lV2Fs7bO59nbZ3NJJUisxG2t6V2AyO58XTxLOtI0OXrNr0DyM8bSD7ZQrKC1/de6aO1aiurMLjto4uWupK3fGYi2/NlKCprsZPh8LHBPk9x5WnzMO3dx4ygrqPB0M7UyV8UVJVz6acyK0g9+690ULdFa29t8SRYSFFhq6yh29t2nWNIIkoqNUx9D+zCFjgiZVaXSgCqn0AHv3Xuh4692vnsriaOo2rmqbZuzlqqCemy+Lx1Jkt174mxGeoa2szFTkctHLDiNs7pxkVRj5MbJRLX0g/cjnQ6ffuvdLmTqmrMMEY7M7DqWhpcbTOMxW7fzdJWfwvF57HpNX0Fdt5qaeTIVeairKzSEE9RjqW2lEdZPde6CvMdC7lpnknxU+zd1qmNrRFSZuny+yq2qyVLt7E0uHE+T2m9ZjIlyu54a2syNQuN/bp6iOCmhURam917oS6t9x0e3q3Z8HVlfLiYsNV47HSYLdW3TSmIZqkwmOCzZWsxWRoa84qqfMNIIpBTx0zIsklUY43917ot/yfgz3V/TPS3d+TFLUbt+Me/Ou9z7oqqfL5TNvW7Hzujqzt2li3BmqOLMZhajZO7qmuM1VFHLU1VDFI4VgLe690fZSCLggg8gg3BB+hvc3uPfuvdd+/de697917r3v3Xuv/9Hf4+vv3Xuq8vkoma+K/Zb/ADV2JtnIbi2DmMdgtofM/aG36aors3Uda7eFRDtH5Cbbw1Irz5bdXSCV88eap445ajJ7QmmCK02MpEPurDOPPy6Pnt3cWC3bgcNunbOXxuf23uPF0GcwGdw9ZBkcVmsNlaWKuxuUxldTPJT1lBX0cySRSxsUkRgQeffuq9PXv3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3XusFRUQ00E088qQwwRvLNLIwSOKKNSzyO54VVUEkn2kv76z22yu9x3C6jgsII2kkkdgqRxoNTuzEgKqqCSSaACvVo0eV0ijUtIxoABUk+gHRUewezqrcM743AVNRSYOIPHLURNLBU5UuHjkEnr4x5Q3ReGe9z/T3x5+8p97LefcjcZuVvbjdLmx5Ah1o8yF4ZtwJUpJ4lDUWZU/pJRXkqXkoKJ1PXKHI8G1Rpe7tCku5tQhTRkipkEf8M9TwHl69BEFAFgLAWAA4AA+gAHAHvCdI0RURUUKoAFAMAeX2Dy6kY5NTk9d/wC+/wB9/t/dqD+Efs61pHp12hMUiTRO8M0bao5oXaKaM2IvHKhDxnn8Ee3IJJbW4gvLOZ4b6I1SWMlJEb+JJFo6t81IPz606LIrJIgZDxBAIP2g4PQ77A7aqoJosRu2qElI4SGjzDqFlgkJ0pFkGQfuRPcAS2upA1cer30T+7j98vc7HcLPkv3l3UTbRIFjttzZaSRSatKx3mkfqRyFgEnoGjIAlBBMgibm32+ieKTcOX4KTrUtDXDDiTH6EcdNc+Xp0q/kB1LH3h1PuPY1Nl5NtbmcY/cfXm9KSOKXIbH7K2pkKfcOwt5Y5pIpbS4PclBTySqtvuKUywMdErA9TUdWVGVgVIqCMgg8DXzqPPqGeDUPTf8AGPuSfvbpbaW/8xi49u7yIym1uytqo7Mdo9obKytZtTsHbdnZp0hxu6MTUin1nW9K0Tn9Xu/Wj0P3v3Xuve/de697917r3v3Xuve/de6YsztvB56IxZbE0deCANU8S+VbXtpnXTOtgfw3sBc6+2Pt/wC4lsbTnXlCx3GPHdLGviClaaZRplWlT8LgdGW37xum1P4m338sR9FOM/0eB/Z0F+T6P25VM0mMrcliGbURCGjraVWJBuEqR5wo+lhJ7xN5r+4F7W7rNNdcqb9umzO9SIwyXUCkmuEmAloBUACYeWcZG9h7n73bII722huB/EQUb8ypp/LpA13R+6IXl+yyGKrolUtFraoo55SCbRlDHLErFf7RcLf3jlzB9wD3VsJLtuXuZ9n3C0RGaMOZraWQ1OlKFZI1YihLGXTU0wOhZa+6OzyaRdWc8TE0NArgD1rUH/jNemj/AEPb7sP8goP8bZSn/wCJHsB/8BJ94j/pn9t/7L4f+gejT/XE5W/5S5v+cR65R9Ob5kkRHpcdAjNZpXyUTqg/LMsStIR/rAn2qsvuP/eCuLq3gudo2qC3dgGka+RlQcalY42cjyooJz1ST3G5YRGdLid2H4RHSv5nHSsxnRNa9mzWdig5UmHGU5la2o6h9xVaFHFrHxm3PH59zNyl/d5bzKsc3PXuLDA2KxWEBkP4gQJrgqAT2kN4JAyCpoG6D977qwKCNt2hm+crU/4ylT/xroUcB1ltLb8sVTT401lZF6lrclKauZG5GqONgsETc8FUBHvLj25+6l7L+2t3bbptfLJvd+hyt1fP9RIrUpqRSBCh8wVjBHkegJu3OvMO8I8E954du3FIxpUj0J+I/mehB0qABYWsFAA+gH0HH0A95HBFCqoUBRwHkPy4dBQ56CPe/ce2toyT46mLZvORAhqGidftqSQrdRkK3mOE3YXRNcn9QPduvdFT3V2bvDd5kiyWSakxzkj+E4svSUJX6hZyC1TV2P8Ax0Yj/affuvdIBQFFgAAPwOAP964v7917p1xGEzG4KyPH4TG1WTq5F1eGmTV4476fNPKzJFTQA8a3ZRcGxuPfuvdGR2r8eIysVVvLItIx0ucPiJDHGB9TFWZB1Ekn9GEIQf0c/X37r3Ri8LgMLt6kShwuMpMbTINISmhVC4H0MsnMszn+rEn37r3Tx/vvrf8A3v37r3Xvfuvde9+691xLKL3NgASSeAABckk8AAe/de6LH2F3Bka3Jtszr8rLVVNQmLkzkJ8sj1s8nhamw9gYtMRNnqTqAIbQPTq9+690PO09s0O08FRYajjS8UYlrqrSPPkclKqmtyFZLbyVFVUyi5dyzaQBewHv3XumXs/sraPUGxdx9i75yS4zbO2cdNX18oHkqahgLU9Bj6YEPV5LIVBWKCJeXkYDgXI917rXv2F83xv7ujuBO36p9udYd9zYenx8oEk1P1jmNsRpjthZWtEbyE4yagRIcy6B7SMJgNKWEQe+3tcvvD7Z77yVHdrBujlJ7WRqhFuoGDw+JQH9JzqR8EgMGA7ejHa7xbG7EstTbspSQDiUbjT+kMEDgaUOCeh/3FsDd+1q1aPJ4PISCVFmoq/H0s+TxmTpXAaGsx9dQxz09TSzRsrKysbhhzz74Xcx8qc0cn7vd7BzTy5eWG8wGjxSxPX/AEyMFKyIfwuhKkcD0KWtJGVHtmWeFxUPH3A/lxU+oIFD0THvn4f7b3Nl99dz9kdx9u9A7J7H6y231H27HB2VsXpnrfsTrvZtZuerpMDm9z7x26d1Y+arg3lkKSsOBytFWVNLUGNbN6vcte2EfunvdnsvLvJPtNHvc9lfT3tnczWU7i1ncQiWQTSywWJCNFGwFwJAsgXtPDpJc2v09ZLqTwgRQgsATxoNI78gny6DqT4rw9+ZTKZv4Q4H5WY8b56qwvRm8939D1u2egfi7vrrPaWBqdobSx2f358iNhbk/j8my9q1s+Los3snbGSyQoD4RUTKISMwfbD7q/uff2u3t7v3WwvYW+4yX8EU8Ut9uEF1NIJZWSaCa1gVZplEskMkk8evvCqxYMXTXlnEx8EuXK0OmqqRSnFqnhjAB6sq2F/LB793HsPaXXfc/wAlsN1P1TtrbeA2k3R3xN2fLj4txba2ziqTDYbEdm999s1G5+xew44sfQQrUw0uO2/jKuQOz0ZV9Iy49vfY3219tL+93vl7YFk5qupXlm3C5pNds8rmR/DcgLbqXZiFt0jAB0kkdF9xfTToIj2w/wAIqF/zn8yejq7c/l3fGHFUOHoN07X3J2xDgKaChw1P2vvXcO68BiKKnXTFS4XYUVZjet8DShRYxUOGpo2+rKTc+5dJJLEnJ4/6v8vHpFqNQeh9wnxx+Pe26JMdt/ovp/DUMZJSlxvW2zqSEM1tTFYcMup2IFybk29+69qb1PSF3z8LPix2C4q810dsPHZyKSOej3bsrEr13vfGVcAtTVuL3psR9u7moKylIBjeOqUrb+nHv3XtR9eg4rNlfKL44Y6PJdW7tz/yx66xs0suW6l7ay2Fo+8KDBiN2YdXdwinw2J3hlKFreLFbuiEtag0/wAahcKH91rj0YfpTvXrrv3ac26uvspXSfwvKVG3d2bX3Diq3bW+dgbtoEjbKbO35tHLQ02Y2xubGGVfJBPGFljZZoHlgkjlf3XuhguP6/8AG/8AW/r7917or/dnyf251hn6DrDZe3sr3V8gtyUgq9rdH7FrKBM8mOkdIP729g5ytkGG6v66op5k+4zOVZFcEpRw1lRpp291sDz8uk31r8cdzZne+B75+UOcwPZHdeCpp02JtrbVPXQdNdCxZFAmQg6wweZZ6/N7wrov2a/d2VT+LVUa+Klix1IzUh91ro4gAHNuT/t/9v7917rv37r3Qedn9l7c6p2lV7u3HLUyRR1NHisPhMZAazP7s3Nlp1pMBtHbOMUrLlNw5+vdYaeFSACS8hSJJHX3Xuib0p7BqewEkjpNv7h+XG5tv09RlJa15891Z8Qers1MzQ46ERPSnJ7ozUFOV8ULx1258nE80j02Hp41j917oAd45nHYB9x7C2XW5KsxFTnJazfO+sxW/f727e3XSD7Wr3BvLNxxUwqqCnkVoqGggSGgpYECQxJEqRrxu+9R94/efcff975B5dvBB7fWF00LeEzBr+WE6XkmcEVt1kDLHCBpfTrkLgqqjXatrjto0uJRW4YV/wBKD6fOnQawgeWEWFvJGLWFrFgLW+mmx+n094cfgceQU0+XR2M8etVzsas6z3HkfnDD1TX/ACPzP80mh+dPZeI+M1R1LlPkBO2DpaLc+z12hT7tqYJ5OgKDp/E4lq85mDI2KUd1dQ5T30D2GLfbKD2pbf4Nli9g35TtX3MXibcDIxin8UwqVG4teO2jwGjwXANdIPQalMRN4Iy53HxjopXFCKfKgHRoPn93FufJfIHtHtfana+N21/w0j1x0v2hUde0W+cPt6h7v727Fz+C3n3ps7JbcqsnSV27cTtn48UFTTU1PChMVdkAo1MShAfs9y1t8HJ/L3L9/wAuPP8A6417fWq3BgeRrGwt45IbGcSBGSJ5NxZGZyRVIz5CvSq+lLTzSCWhtVU0r8TE1YfYF4fPo5nyUyw+VHyF+IXxZwHY+8NofHPvPpTsv5U9k5Lrbddbsjc3cWxdqHZVH171ZBvzDyQ5zA7MydXvH+IZ9sdLBV1NNCkXlRdXuL+RrX+oHJfuR7g3Wx211zttO6221Wq3MQmis55RObm68BwUkmUQaLcSKyIXLFSR0rn/AMantLfxCsDqzmmKgAUH2dMHwwHwr2N8luw+pPjn198peh+yIOp6/c+5uou4KLufA9Vbr2Vit90+2z2ptDAdq7j3RRV2dXcQ+0gy2PlpzVUcz6lcWKrfdD/XQ3XkbZOYudN45f3bZG3FYoruzNi93FO9uZfpJpLSOJlTw+5opAQrqKaaZpZ/SLMY4I5Uk010tqoRWlckjj1aoA7kBEaR2YIiICzvI7BEjQfVnd2AA/JPvHxVdmVYoy8hIAABJYkgBQPMkkAD1I6MSQOLU+fWf5Sfy+u7d17Jw27dkb1zW96jG46GvyfSGSrRSYzFVc1N5q6bY8YkgxdXlXkNp4qtVlna5WUkhD3t+7v7ez+1/tBylyneg/vYI9zcg503F03iyJ9kQKx+hKFhx6AO53P1V7PKvwVoPsHVJ1bRVuNrKzG5GjqsdkMZV1GPyGOroJaSux9bSStDU0dZSzKk1PUU8ylWR1BBHuaui/qMOOffuvdXY/ybcDRyZXvrdTxx/wAQpKbZO3IpSG8yUdY2aytRGratIjkqKWMsLXuo5Hv3Xurfu0pqihm60yEOQq6JYO0ds0lTDT1G5IocjT5qHJYdqKrpttYfLvXx+euSRUrvtsckiCWoqIwgv7r3QpmogWWOnaaJZ5kkkigZ0EsscRUSPHHfU6Ra11EAhbi/19+690He88fTrunrLcf2tRPVYvceTwqyQU+QqlpqPc+Br6epknSDOYvHUkJq6CmBqamnyBjH7cUStN5U917oCsRJT7a+c278TQ0yQR9s/Gvbe9M04UL93nuqOwa/aFNVkhFWSokwG/6eCQsxbxUkQFgvv3W8U+fSr3FRyYem+QGMFPWrR5WDD7zpaiejy/2s1XlsPTYjJRU2Xym4MpT10tPPtxGanoaTHUlGkkY0SSySSH3WukrtKrqc70zmqOio5Mnn+u8tS7oweMp6CkydbV1GDrYdy4ykx1BX5vbWNlyNY9NNTUzVVZFTQzuskpKoV9+690ajD5aiz+Ixuaxs8FTRZWipchSzU1VSV0Dw1MKSoYq2hnqaGpVdVtcMkkbEeliOffuvdE8+Mm0d89Wdz/MHrjI7OyeP6py/bOM766h3qyD+B5p+7cEuT7W2lRsaqUxZTavaWAyVfNGsUcfgzsDC7F7e690PlTMa3t0VUkeRiodkdeVlRNVtQ7jp8XNW7qykLeCPJ02ZTa+WqaHH7bZ5qSfHyV1IJ4pYp0jmdH917olUWbqoM+NyUxiFbHmnzMBVWWFpTWvVqNBa4jkV7EG/Bt79wr8v9j/P17o1mCzmdwCVW8NvUOQ3n13uqaq3FkcXRTVFdujYmTixWTq9xGip63LSHL4WapxlJSUGDxdClTHW1Est5EkOjVR17pXt27t+VGjxeH3vnMkj5KGTD43Z+airoqvDvt8ZWhlqMrT43Ew1dHFuSCQK9SqzrFP4DK0Eirvr3RcN+bqy38ZyldlJsUu8a3E1e1ZaPBVb5DH7I21NXxzZXCUOeWnxTZ+t3bNjKKtqWqKNHxxVqeNmBZj7r3TV0/jKbKb1iop5PCzYPOrQsJ5KdhVzUBpE8csDx1MMkMVQ7o8LJKhXUrAge/de6NN1RualrtvUW06+pEG9dl4vH4bdOFq5KyLLRtj5azA0u41oc1lMtuRttbqqcJUVGKra+aSevpbSs7Prt7r3QqXB9+690Wr5I/LDqH4p4XDbg7d/0ipis7LXR0tRsDqLs/td6cY1Kd6qXKU3Wm1N0VOIhValND1CRiQ3CatJt7rYFei3dYfzff5avbW4odm7b+XXVuE3vUTT08Wxezp8z07vM1NMoaanG2e1sRs3LyTqjhgqxMWU3F/fuvUI8unD5fd59d9ndZ5j4zdObx2X2n3X35DS7C23szamZxe7XwO3c1kaKHeXZG848PUVkW29m7G2sauvlq6wxJNURRU0PkqJoo2914U8+rDaKlSio6SijeSSOkpqelSSVi8rpTxJEryOeXkYJdj+T791rqV7917r3v3Xuve/de6//9Lf49+691jkRHR1dQ6OpV0YBldCCGRlIsysDyDwffuvdVw5fbvanwg3bX7g6g6+n7O+FWfnbL7v6W2JS1dT2h8bNzV1XPLuLevR+0YIpYt89P5yaf73LbNovFksPWtPWYaOohmkxye63g0Hn0fbY++dodlbTwW+thbkw27tnbnoI8ngNyYCuhyGJylFIzR+SnqYWZfJFMjRSxtpkhmRo5FV1ZR7rXSs9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XvfuvdF+7v3G8NPRbYppVVq0ityPjlZZRSwsRT07oo5hqZrlvVyEsRz75yff690Jtu2jl/2p2q7VZNxP1V6Fdg/08Z0wwugFDFPLqZqtkRBWQqa9St7ZbMk1zc71cR1WLsiqMazlmB9VGAPU16LeOPfLMClM+QH7PkOpspTh137317r3v3Xuve/de66IBFj9PemUMCrDB/1H9owfljrxFadD31Dvmq+9G1cvVvNTyoz4eoqZGeaKZFXVjjNLJYU/iS8K2J1ekcce+kH3KPvBbsd7T2g513hrjb5o3bbZp3Z5I5EAJsjK70EPhqTbR6SwaqKSKDqIfcTlaBYDvu3wBZAwEyqAFIJ/tKAfFqPceFMn16DbY2PPS/zJ7M2fFpotgfKPasHdW1aZ5Fio6furYSY/aPb2NxkLOVNfuzZ02AzUqKoaWSjrZv8AVn31H9eodOVHr0d8e/dV697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917pvymVx+FoKrKZWsgocfRx+WoqqhgscaXtb+rO7EBVALMxsASffuvdE47E7nyu5Jpcbtqeqw2BRnR6iKR6fJZYcAmV10y0VLwQI0YMwPqP8AZ9+690BoGm9vyST/AFJPLEn6sWP1J5Pv3XuuwCSAB9foOSWJ4CqB9T7917orPfnyj2v1FDWbdwP2e6exyktP/B45teN2rK9OHgrt0SxjS7AuCtBG4nc/rKJYn3XuhQ/lP7o3z2Dn/kNvreeRyWbkrp9jYyHJ1TkUFNNFHuCufC4elB+3x9FRQ1iOIIlCosgJuWudEgUr59e6ue97691737r3Xvfuvde9+6910xCgkkAAEkkgAAC9yT9Bx7917on3cPa0uVqJNs7WyDx4qAyRZjIUj6Gyk4vG9FTVCm5x0QBEhUjyvxcqOfde6UHQewljhbe2UpRqnDU+3I5E5ip+Y6jKKhFlae2iJvqI9RHDD37r3Rj8zmcVt7FZDO53I0WHw2Io58hlMpkqmKjoMfQ0sbS1NXV1U7JDBBBGpZmYgAD37r3Wrn86PlzXfJvsNsftuqrqTqDZdTU0e08Z91KtJumuhmdJd9ZGhCxxmesjBSgSTUYKXniR29+690Rb8erkfkkXJvxawB1XH4/Pv3Xurqfg/wDEn5S57DUO4d7dzdxdIdUtFE2D2Pgd1ZGh3NuCiKLPS1VNRVzV1Ds7CyCclP2BVTfiNFs3vTqkgUSIGoKDUA1B6CtaDqxIpRaj8/8AZ6tNxXxF+PFDuSn3vmetcL2Bv6mjx6Q797T+47N3dBJjP+AlTjsvvaXNvg6hG9ROPWkBf1WuAfdw7rGIlakX8IwPXgMdVOaHzHRjljVVCKNKqAqqAoVVAsFVQAFUDgAe69e6ye/de697917r3v3Xuve/de68RcEf149+690VfuH4zjeO76fuPqHfWS6L7/oKCmxMnYeBxNJntv7/ANv0QlFHs7ufYNXPRYrsjbFH5nNHJJNSZjEOxbH19NrlWT3XukPV9PfMfsXx4bs35PbR6+2e0EMGZg+NXV9ftHf24lCBKtYuwuw92b+l2XBWFSwOLx65Cn1ER1oYB/furVA4DPRh+p+jurOkcXkMb1rtCg2/LnatcnurPSSVWX3hvXMrH42zu+N55mev3TvHOSKSGq8jV1M1jYMFsPfuq9Cz7917r3v3Xukbvvf21OttuVe6d45aHEYilkgpkZkmqa3JZGrfxUGFwmMpI58hms7lKgiKloqWOWoqJSFRCffuvdEuy+4t9y1WG7f3vsiKfuLd9XXbZ+JXx8zFQ0lVsmKupGGQ372IaRqyjxe6Rh5zW7krYFdNu4ZRjYJ3qamX7n3XujMbJ6+TqbrbOwnJybk3pXUGX3RvnfGQpoI8pvbek2Mdq7OZJYEQJTx+BKaipQTHRY6nhpo/REPYf5su5tv5V5lv7en1EG33Ei1yNSQuy1HmKjI6ciAaWNTwLAfz6qQiN4o2P1dFc2/1TgM3+P6j7+cKIloYXPFkBP2kVP8APqS/Mjyr1mjbS6Na+l1a17X0nVa/4vb255MPUU62DToEfj70yvQdP3HSUm6q3dFN3D8huyfkBW09RRLiIsNP2NPiZajaSJT1VSuSpMZ/CgBVNoM2vlFtyLOceaDze3LLvtqwSbbsttt60bWXFuHAmNQNDPr+CmKU1GvSeGFoRLR6lnLfZXy6Lp1b/Lh+Mm3MHvaXvDq3qX5Qdodm9ndn9ndidw9qdSbRye8twV3ZOZnqocDR1eXp85XYXB7L2+afGYyKGp0wx04kVUZrexrzB738+X13tacqcw7jsOwWFja2ttZ2l5KkMa2qBfEYIY1eSeTVLISvF9NTSpaisYEWQSosjuxJJHr/AJukPjP5c02E6D+N/We3fkn2FtLuj4e5LeqfG75M7d23hzuna2w92ZCsCdT732bmarJYTsTr1dqvTYmvoqmaIVsdDBKnhZNJOLj3rjuuceet+vOSLK45W5ljgO57ZLI/gyzxKo+shnQLJbXJl1SRuikqXZTqBJ6bG3nwIIxcN40TEo4FaA/hPy6FPoL4i9wYL5JZb5L95/IWb5E90ZvpmH49bTwezOnMN1Rs7b+wF3hFvio+029iMtuTMZ3c2QzkHked6hIYo2ZVS1iCvfOdNt5m5Z2v2r9uPbmaxsJN2N8Ixczbhe3F0YTAFUeElI1Q00Kpb8Rbj1aOBopmvLq6DSaNOrTQAVr+3q8/pD4y1uJyWJ3r2F4o6uikjrsVtNAlR9tVadVPVZ2ca4XqKViXSnj1Ir6WZyy295q/dw+53uOx7rsfuH7q6Ev7Z1mtdsWj+HIB2SXj5UyRkhlgj1IrhWdyVCgj3PelkV7a0rpOC3r9np0eELb8n/b/APFAPfR/oNdUl/zR/iXUZKM/JXrvDtNVUFLHR9s4nGU15anG0qstDvoRK48kuKiAgyBRC70+iU38TH37r3VF3pJFjdTaxHNwf6EcH37r3VyX8nfeFNR777m2DLNoqM9trbm7qGNnjVaj+7+QqsRXBEb95pYo8zCxsdIQ8/j37r3V1fb1NVT9d7inoYpZ67Ew0W4KSCGhzmVmmn29kaPNLFDitu5zbmUys8goSsdOtZFFNIVWUPEXRvde6ctyUMu4MHQ5vAOseboI4s7typvGTK7wrO+NmkjZg1FmKVvBMqsUIYNclQffuvdBhvveeBz3X+N3mEjSLY+8dobm3JTV0O0/udoQYTNUx3HX5OTeM1PQYWnweKkqZpq2FhWRUqO1JeYqD7r3Qsjamz8hujE9lJiMbWbspds122cPupED18W1M/XYvNV2KpqpW0vjcjX4ikqGXkF4VIPv3XugH3dWY3LZLvPcFLFTtT4na22uvZM1FDtSeKuzdHV5PKZHCrlMfUTbqhq8LLmoUnocgkUET1CyU4JklPv3Xug06Y3FHgN601LVyKMZuOE4WsSSxhM8pLY5pVa6kGobxG/AEp/F/fuvdFgwH82z4H9MZ/s/Y28+5Nw7d6x617e3N1a/b+4+o+56XpnYm+cVnBjdz9X7n7iHWGK6e2pHtLclYcdjII8k9LBQRIJplcc+690fHv75kfGr4y9a7Y7a7g7X29g9n7+rsRiOsmwv3u9M72xndw0X8Q29t/qna2zKTPbk7GzOeobT00GIpatngPlNogXHuvdECb+Z18NtpfFv5D/JXf8A2Fubqmbb259m4zvjG9o9N742B2l1XluzpoNu9Z7Y3bs+p2Ht3dWcx9PhojTYzKpSVKVFJTuTOfE+nagEjUaL5/6vX0Hnw690V/fHzJ2jjK/p6LH7wxWJxPf25cftHpKtx1I244Oxcxldr5DeeINLlaWnqqCjocrtTHPWxSyeOEKoXWXOn3zW51+897v8x7rzUvt/YRbLsGxq73CvHDLdLHFN9O01wZwQC0jJSCNCy8NRoaImnkYsI8U6MR1R8kewNo5mmop8wZcZlKqKGYmni8cE87pHFNLRhRSzxFwFdlVJgp4b8e3/AGe+9rzieZ9r5f8AcmWG+2O+uFi+pWJIZ7aSRgkbFYQqSQFiPEUprQHUGNCOvR3LFgr+vRt8723v/OwtSVOZNBTMpjmiw8K48zDkMHqEZ6oKQ1iFkUEe+kTKVZlPEGnlxHEY9PnQ/Lpb0ldtbWz+7684/A0L1kq2kqpnYRUtHHISPNWVLXVA7XsPU7m9geT7117ob6PoLeWMNNlMXunF0uapHSemEKVsQgmDWbRWDUWXQSCGi0uCQRY+/de6fNxPHV0zL2tszMYDLQxyRU3Zew55Y56OpfE5LDU+ZiyeLZK/H1uOpc1UNTfcJPDTyyF0QMPfuvdOGMz27JagnYfamyd60kstbJFgd9UkmNy1LFV5LbEONpKXO4IxVUlLgsDSZR3+6x9VV5GuqoTJUQxo2r3Xulum/N2UNRTQbj6t3LSrV5CjoEr9s5DE7sxlNHkM5n6GCurzFNi8pS0lBhcVTZCvf7RkpxkI4IjPIklvde6C/e2w/if8m6KlwHbPWPVvYlRkftoqTbvbfX2H/vXTV1bg03DHTUFBvDEQbgx2dpsC4qpFptFXSRDU2gqbe68CR5noEKf48dn/ABJyNdvL4o069o9e1AJ3X8a99ZDEU+6FxsZXxSdId05anTN4+rxCLeHb266rIYmsW8cFbi29be690aLpb5B9ed50mYj2rNmsFu/aclDTb+6u33hKvaHZ/XVfkYXmoqLeOzsnprsfHXRQu1JWwmoxuQRGkpKieMavfuvdDl7917r3v3Xuve/de6//09/j37r3Xvfuvde9+691Xj2TtHJfDXeu5/kn1ZQTy/HzdFZPuH5VdL4Siq6tNv1s0ifxH5M9U4LHwSrQ7kxkbefe+Lp0SHOYqB8iinJ0zfe+690fvDZjFbgxGKz2CyNDmMHm8dRZfDZjF1UFdjMricnTR1uOyWPraZ5Kesoa6jmSWGVGKSI4ZSQb+/de6c/fuvde9+691737r3Xvfuvde9+691737r3XvfuvddE2/wBvb37rRNBXolvaGRTJ74zMkUbxLRNBi3EhW8klAuh5VC3Ajdn4B5sP9h74S/e15nteavf/AJ2uLa0eJbForBtRBLvaIVaRdJ7VYtQAkkqKkA46yV5FsnsuWNuDuGMtZBTyD8B9o8/LpBe8c+hd1737r3Xvfuvde9+691737r3XleSNlljcxyRSRzRSKSGSWJhJG4IsQVdQeD7vFLNbz213aymO7hkSSNwSCkiMHRwVoQVYA4IOOI6q6LIkkbrVGUqR6gihH7Olx8ksDuXsTpjafcvWmMWu7e6B3PQ92bAxCyeObcddtaiyeJ391z91GqlF7H6/yWWw6C4Raqqgd7iK4+hT2Z9xLf3V9teVueYYvDlvIP1Y9QYpNGxjlVqAAHWpNM0BGTx6xW37an2Xd73bWYMsb9pAoCpypH5Ho0HWvYW1+2evtl9m7Krlye1N+bZw268BWAoHkxmboYa6nSoRHk8FbTrN454idUUyMjepT7k/ooODTpbIHA9ZDNc8qLC1/SLXPIHv3XuuXv3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917ojfdG9a7cW56zBJK8WD2/WNSx0ouq1OQgGmprKhePKySkrEDwijUOWv7917oG/fuvdY5ZoaeGWoqZ4KWmp42mqaqrmip6SmgQAyT1NRO8cMMEYN2ZmCj37r3Va3fHzPbIwZLZvTbVVHTSGfH5PsGZPt62eNTNT1VPtKkJLU9LUxkacjJplZDeJEuHHuvdEc2HsXd/Z278NsTYuGrNybs3HU+Ggx8BeV3ckNVZDJVcpkFPRUwJkqaqY6UUEkk2v7r3W118RfjvSfGfpfB9emrpspuOoqqrce9czSKwpsnujKCL7z7JpY46j+GY+ngipqYSerxRaiAWI9+690Z0m34/F/wAf8T71WnXuktnd87N2vNBT7j3Tt/BVFUypTwZbL0FBNMzW0hIqieOQ3uObW9hveeceVOXZYoN/5ksLKeRgFWaeONmJoBRXYGhrxp1UsgwWFelLFNHNHHLE6yxSqrxyRsrxujgMjpIpKOjqbgg2I+nsQxSxzIksThomAKsCCpB4EEVBBGRTq2KVrjrITb/ff8Vt7c690VfujtJJY6nZm26yTyCRodw5KnbSmhQwkxFNOhDMzMbTsp9IGi/Le/de6Bbr/ZNTvjcMGKivDjqcR1WYq1/TT0IYL4k4t9zVsvjjH0+p/Hv3XurCKSlp8fS01DSQrBSUsEVNTQxCyQwwIsccai/Cqii3+t7917qgH+ZH81Yt+1eV+PHV9fK20MNk5KXs3cVO0X2u7MpQSRvFtfE1EMrtUYDGVaaqyXhKqoQRrdEYt7r3VP8A/r/T8/6w/wCNe/de6t3/AJePwUm7Fq9t/ITtaGJOv6GsOT2DtGRS028MljapoqfPZpHXxx7ao6yBjTwepq2RAzWiAEnut1wR1sIAW/1rW/230/wsPfutdd+/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuuj9D/rH37r3RLk7L+Re8PkF3nsHY2O6627s/pSk6sp8XRb+xednrO1puwtsVG5sluKg3Tgc20m1MPg6mCTEQxviK5pq2kqJGYIqofdbFPPoWa3cfyCylHLQ4PrDYm2cuVEf8e3V2DUZzb1NLYhqukxO3dvQZrNRxyC6RTPi2kUjU8ZuB7rXRSd6viuofkdsDe/d+791dk7pw/Vu9Mzs6KPGf5LuHfO4c3tvadH1/0P1VjJjRx7qhojI00zPXZRqavV6qtjpVkdfde6NT1JsbdtZn813L2vR0OP7F3TjosLt/alJNBkqbqXr0TR19PsiHLQ6ocpuTKV6rW7groD9vVVscUMOqmpIHb3XujBSxJLG0UiiSN1aORW5Do6lHVvpwysb+25o0mjeGVQ0TgqwPAgggg/aD1sEjI49VO9vdHbm6yyFXXRUr5bZ9TWVcuOy+PjnnGLpXnZqagzi+PVSVEUciqsvMUtv1Brj3w79+Pu285ezm63u5RWrX3IM08jQXMCu/0yNITHDdrQmNwrBRJmKSmGDVXod7fucF4gUtpnAFQaCp9R6/Z0BZliBsZIwR9QWCkf66kgg/6/vGzXECVaVQw8jjoz68JI2NldGP9FdWPH1sFJP09+DxEgLKpPoDn9nXqHz6Ve39lbw3VIke3Nr53Ma2RVnpMdUfZjXr0NJXTJFRxxt42Gova4t9fY15X9uufudZki5S5M3LcNRXvigfwu6ukmZwkKqdLdxfSCCK1oOmJbq2gFZp1X88/s49GN2p8Q98ZVo5t15jF7WomW8lPSN/GsuRY6VCp4cdD6vqTJJx+PeWvI/3D/cje5Le5543yx2bbuLJFW6u6egA0QIf6RdyP4eii45gtk7YIy7fPA/z9HM656b2V1jC52/QtPk6hdNXncoy1eWqEsl4UqAkaUtJrUsIolVbk/X30E9pfYD279m7V/6sbc8u9SqBLeXBEly4oKqH0gRx1Bbw4woqc1x0G7zcLm9asr9g4KMAfl6/PoVQLC3ubAKCnSLrxNvxf34mnXuo1XS02QpamgraeOpo62Calq6aoRZIKmlqI2hqKeaJwVkimicqykWIPPvwNfLr1MVHWqh83fi1lPjN2vVw4+B5usN71eTzHXuSjik8GOpzP5qzZtZL6o0r8B5wsJZtVRSaHtdXtvr3SO+G3bkXSXyQ603rXTeDBVOVbaO6JGbTHFt/dgTF1NVICyIUx9Y0FQdRsoiJ9+63XBHW2/VU9HkqOopKuCnrsfXU0tNVU86R1FJWUdXE0U0E0Th4p6eogkKspBVlNjwffutdBr1BV00G0ztBK6gq6/rrJVexMjFQRbcpFov4N4zhoZcPtRY8NgBUbcqKOeGhjRGpqeWNXUNf37r3SA7T2TmcDlJ+x9lM6VDx6Nx4mGnFTHWREBZa2SjKNFW00qACqidTwA45De/de6ArFbxo6TGx4wyb2xmLFfBk5cRtzek4xWQmbLbizGZpayTP0uYzkWL3RPuJ46yGGtURwU0EdJ9ssQv7r3UDM7wnyG38PtDFY+Db20cBTUNLjcJTVddkZWTHU0dLSyZPMZOaoyeVniijA8k7u7n1Ozvz7917pM0P/A6iIuCKylIINipE8ZuDwQRa4/x9+691WR/LR+RHxx61+Ev8yH/ZrN37EwHWNL88v5hVZvio7Gy2BODzu3azd7vkcPVYvL1BkyGQyqTNBTUSQSyV0rqkSvI2k+691Xt8G8hgfjR2n/wn97B+XEkuy9kH4OfK3a3W9d2BVVH220u6tw7gxWT6S2rmo8zUNjNsb/rfjRVQYrDRyClqnPkpwdV/ZXve87fy9s+6b7ukxTbrOB5pWCs7BEBJKooLOaDCqCxPl1okAEno9382/wCUHVPyN6Lqdg4/MYDIVuO+TfwkxW49m1dRhMo2BwO4vkRtNYIN/U2Pra6WgTclClUI46lQHhWQILaicdh7/wDIfuXte5co7Kd32zcN02y/W2vL2za3tVaGBmd/GDtQRYYkA6fUdM+KG1BQQ1PMU6o8yWO3B8aPmR8JPgBur76r2r1n86Ml3J8Tdz1K1E65f41bv647cx9Z1tVVcl/9zvRO+al8Z47gNiKyjkX0afeLc0MXNHIfuz7lwxCLeJuW/wB37xDwKbnb3NoRcBaCsd9ABMH4Fw/r0yMq8nmVofy62C6KmnrqyioqRHlqquoggp441kLtI8iqCAis4Vb3JA9Kgn3ils+23287ztOy7VC0u6XVzFFEiVLM7sAKBasAtCzEDtUFvI9JVBbSAM9WABHsiKPJJaOIKpvrkssaqh/OuQgD/X997ER4o4opCPFRFVqGoqFAP8/M5PHo2AwPs6NhtzC7no7dabKrRtY4/D0mX39v9KLEZeuTcWWMdTj8Ht2CorMhTRZOip6Nvv4cnj0RaCsgkpmMjMyW630v/wC63ZWFqXqsJ2Cm4qWbJCeXDb2wmPkWmx9bumpy2UixmX2/DiK5aqj23ULi8Ys4khhESTT+dy5PuvdZMT2ZjJshS7X3pi6jY+666GYQ4fMzRVWHzj0ODxOX3A+2M6qQ02dw+D/jMdLPVSQ0geoVlWP+vuvdNW8OkdpbmMlVjov7t5XllqcdCopJGtcfcY8GOIEkD1RGN/8AX9+690EFVle2+npEp66pXOYC5FPU1f3ORxZW4tF9y+mvxsoAuEkfQPxce/de6FvA5/Y3d+FqsTncHSz1lPRVMGQxdaQ1RT0mUpmoK2pw+Vp2hrYKatp5GheWB4ZdLaW4PPuvdSMPks9sDclFtPdOTrtxba3NWVv90N4ZF6RshQZWorK/JT7W3ZVR0uFxVLAyV9LQ7bjpY6mrq46aVagmRBJJ7r3QOfLPqbMV2Mofkb1FDHjPkX0LicnntoVkMr0cXZWx6O2Y3p0Rvfw2XL7P37RUbrSCcM2IzYpq+maN45PJ7r3Rkutd9YXs/r7ZHZO2/P8AwDf+0tu7ywy1KhKqPGbkxNLl6OGqjUssdVBDVhJFBOl1I9+690t/fuvde9+691//1N/j37r3Xvfuvde9+691jmijnikhmjjmhlR4pYZUWWOWKRSkkckb3V43ViCDwRwffuvdEC6oif4h9y4z4211VBT/AB27hqc7lvi3WV9UEk6637AK7cm9vjUjv+0225MUKjO7JQlHp6Cnr8WupKOkB91vjSg6sABB+nv3Wuu/fuvde9+691737r3Xvfuvde9+691737r3XFrWF/6+/daPA9EY3v8A8fluj/HN1lv+Sl/4ge/nz+8ApHvn7sHSafv65/wr/n6yl5WH/Ia2Ief0ydJj3EvR91737r3Xvfuvde9+691737r3XR+h/P8Ah73/AKuNP59e6MF0fnZAcrtyaSMRR2ylCrNGshd3CViIC2uYcK1gLRj/AF/fSz+789wpgebvay9uI1tU/wAftAWUSFnOm5RQW1y8EeiroiUGuX6hz3R2pV/d+8RxsXP6T0qRQCqHhjzHqT9nQW/HFcb0h3F298T0WSg28r1XyH6Ko5VK0o667I3BWN2Ls/C2vF9v1x2vNUuIAQ1NjtwUCAaQD76c9RB8+jtSySAqsSaiyyHUxIjUqLKCwBHqcjj+nv3Xusq6tI121WGrTfTqsL6b82v9L+/de65e/de697917r3v3Xuve/de697917r3v3Xuve/de697917ogXbm36nAb7zPl1tT5mZs1QzNz5Yqxv30BsLtTVWpD/hb37r3QcQwzVM0NPTRSVFRPKkMEEKNJLNNIdEcUaJ6md24A9+690Vr5Q/Fj5w9j56fbO2dtYKv6paOknx2Pwm78XiJMmZI6aodd7RZWphnrMhj60MqJGDRoqgqGJLe/de6Q3W38o3urO11NL2fvTaGwsKJiK2m2/JPuvcMkCqjL9lqhosLA7lit5Hk06b6Wvz7rYPy6uI+OvxM6d+MmLqIOv8ADVFRuDJQpBm967hniyW6cvGpR/t3rEgggoMd5UDilpY4YQQCVYjV791row2YzWI2/jqjK5vJ0GHxlKheor8jVQ0VHCvNtdROyRqxP0F7k/QH2X7nu22bLYz7nu+4Q2u3xCrSSuqIo+bMQPsHE8BnrTFVBZjRR1XZ3F80qms+6wHUMLU1Owlp6ne2UpmWpe5eNjt7GSgCECwZKqoDXB9MQIDe8Dfdb73FxdfUbJ7Wx+HbEMr38ynxDxB+khagX1WaUEEfDHwbovmvRQrED9vVf1XLPX1M9bkJpq+rqnkkqKyvlerqp3kZ5HaaedpJHJkYnk2uePeD88017dS3t9M095IxZ3kJd2JJJJZqk1JPyz0XEkmpOerY/hJlclkeoq2DIZCqrosPu/J4nFx1U7zjH4yHHYmeDH0uskxUkUtQ7Kn0XUbce+m33Ptyv7/2su4L69kmjtN0lhhDsW8KFY4GWJK8EUsxC+WogYx0b2TFoTU1oehT7k7KbatCMBhZR/eHK07FqhCp/g9E/oNQwvdayoBIgBHFi/4F8rOlfRM6KircpW02PoKeauyFbMsNPTxXeaomck+ok3+pLO5PAuSfz7917o/fW+xqfYm31oNQnydawrMzWgWEtWY1UQwi5IpqRBojF+eW+rH37r3Ve/8AMW+Z46X29N0511WI3am8sPI2WysJinj2JtavVqWaokCyrJDuTMQsy0KsLwx3nIsEv7r3WuCiBFWNAdK8KLlieb3JPqZmJuSeSSSeT7917o53wi+LNR8oO1P4fmFqqbrPZS0eY7AyFORHLWxzSMcXtOinDK8NZnmgcySLqMNLG7AXZffuvdbVeLxlBhsdQ4jF0VPjsXjKSmoMdQUkaQ0tHRUkKwU1NTwoAsUMEKBVUWAA9+691PuAPqLe9FgKiuevddahxza5sL8XP9Of6+/VHmf9R69Tru/+v/tj79Xr3XfvdaCvl17rq4/r70DXh17rv3vr3Xvfuvde9+691737r3Xvfuvde9+690CXZ/VeY3HlaHf/AFvueHYPbeDxE2Exm5a3EtuHbWfwE1Q1adpb+2yldips/t0V7GogaCrpK+gqGaSnnQSTRze690m8Ntz5W1q1UG7ez+lsPG0sX2ldsPqvdL5GOnBvPGybu7Fy2P8AuHXiOQxOiNy0bj0n3XulHszora+2d0x9hbgyW4Oy+y4aWuoaHf3YFXR5TL7dx+UEYymL2XjqGhxu3tkYvIhAKiPF0lM9Siqs7yhVt7r3Q3W9+691737r3XB4o5FZJEWRHBV0dQ6Op+qsrXVgf6H23JDFKjxSxK0bChBAII9CDgj7et1IoQek3PsnZtTNLUVO0ttVFRM2uaebBYuWWVyAC8kj0rO7ED6k39hG59uvb+9uJbu85G2eW6kNWd7O3ZmPqzGMkn5k16dFxcLhZ3A/0x/z9cqXZmz6Gdaqi2rtykqUDBKimwmMgmQOCrhJYqZXXUpsbHke3LL2/wCRNtuUvNu5L2mC7UEB47S3RxUUNGWMEVBoaHIx143Fwwo07kfaf8/ShjiihRYoo0iiQWSONFRFF72VFAVRf+nsUxQQwRLBBCqQqKBVACgfIDA6aJJNSc9cwAPoPbmlc461137317qLW1lLj6Sqra2dKako6eaqqqiUlY4KeCNpZpnb8LHGpJ/1vaPcNwstrsb3ctwukhsLeJ5JJGNFREUs7MfRVBJ62AWZVUVYmgHVanbHyZ3Ru2vmx2ya6r2ztalqG+1rqNnps7m0EckLT1c2phSUE4kLRwIqyW0l2vwOQPvh98PnTnncrjavbncp9m5LikPhyxM0d5dihUvK9SYoXqSkKgOAFZ2JoqjLb9mggRZLlQ85HD8Ir0Hm1O+O1NntCKDdVVlKSLgYzcZfMUToC58ZlmdchEGLE6kmVr2/HHuK+SPvNe9XIUkK7bzrPe7dGf8Ace+rdRFa5FXPjLX+hMD8+lk+12M4o0AFfNcEfPo0m7tu7F+cnQm49mZ6khw+ejOqlcSR1NZs7eVLFJJgs/j5mheQ46rYkMCgaWmeWI+r1e+tXsH737T74coNvltaiz321kEN5a6w5hkIqro1AWhmFWiYgEUZW7lPQP3GxewnEZOqNhg+oH+UefWrlvnZO5evd2bm2BvPHPitz7WylVhM1RHUoSphY+KqpWNi9FX05WoppL+uGRSPr7nTpB1s8/y+O/m72+P2F/jE6y7164eDYu7buWlq3x1JA2BzbFiWIzGG8bOxJvPHL/T37r3Q+7keu2Dv6n3wKqtqNk7qp6Hbm8qN5MnWwbaylFJJJgt3wxz5Wi23tTbUNHLVpnq6SGWeob7IXCxEH3XuhpBVrEEG4uCObqebg/lT/tj7917oAN8dE4nOTT5PbM8WBydRK01RSSIz4eqldizusUYMtDNJe5MYKEm+gG59+690WbcmxN17Tdv43iKiGlUkLkqYGrxsljwRVwgrFqBvaQIbfj37r3SSVmUq6NpYEMrKeQQQQyn/AAI4Pv3Xuidy/wAvb4JVHY9Z2/V/EDoLI9nZPdNVvjIbyy2xKLM5Ku3nW1ZrqvdVVSZWatw0udnrWMxqDSl/L6hY8+/de6d/mH1h1x3V1nRdddvdebT7O2Dm8+lblNvbzwlPmsWcljKRmxNXAJrPj8jT+SQwzwNFPHp9DKAR7xA++JzTzXypyvyBd8sX9xZM27yl7iFmRlZbciOPUDpKSBn1I4YNpHp0muWKKCOiS7M+Kvxn652VVdebG6L622rsit3Nt7e+T2/i8DHHDl947RyFPltr7pzddNLNlM1mtuZKkimo5qqeUwOnpAFwedO78+85b1ffvbeOar243BLaWAO7k6IJkZZo1HwokikiQKBUHPSMySHJY1HVveyvg10/33R9Ldz947B27mN39bwTbg6x3Zk8eU3vtbJ5vHriMjmcVlzJCcUctjIY4/UkvljCNYEKffVD279teUebPabk6Xmvl8ybpuOw2cV+2uWB7tIUXwFujE6GbwkCCNnqQoFDSvRgqKyKzDuIz0Ymv+Je0trUc+R64glizEUbsafLSJWS1iBSTBQ5BlWSgmZQbAehybEi9/Y+5M9o/bf2+vJ9w5Q5QtbPcZOM3fNKBShVJJnd4gRx8PTq4GoJ62kUaZRadAQt6SsjWpV6d6StiFVHKhSSmamqo/Oksf60aEIbj8f7b3IwAFAOHTnR3tiV2Ng7D7SxGvGDK5OfbG9KZ45ttDMZPbWUwVNh6DIVtHhcXQ5ZMfS1+EqKWmmystXVy+JwsiwLGi7690Mfv3Xum7KYjFZygnxWaxtBl8ZVeH7nHZOkgrqGo+3njqYBPS1KSwTCKohR1DKQHUH6j37r3QeUHWtTt+eKTa2+t6YujSXyzYbK5GHd2LqPNmdwZ7InXuWGuzUU2Vrs6qSOlavhpKOCCnEKKb+691GqKXuHFY6QNPsXsP7fFsJqKqx9dsyuzlbRbUkURJMKzcWHomz+9ERj5I/DQ4+Qr+9Iuo+691C2fm9g0e46zFy7ap+vt9NFUvPi8jBS0cuUxgzEGGXL4jIUsslBW4XK5txHRBmhqp7Bvt0vb37r3UzvNsfT9Y7iyFXVRY+txf8AD6/buSeooaCah3YmRpYNttRZfJbe3XT4SrrMvUR0orRQVDU6zs6gEah7r3QlMZp8OzV9IiVEuNY1tDHItXGkz0p+6pI5WSNahFkLIGKqHAvYX9+690VP4BNI/wAMfjgZaj7l/wDRfgl8ut39CfcJFHqk9X7EahLfQabDgD37rx4mnDo4Hv3Xuve/de6//9Xf49+691737r3Xvfuvde9+690EXefTG1O++tdwdb7tevoIMmKTIYHc2En+x3RsTeOFqI8ltLf2z8qlp8RuvaOcp4a2iqEItJHocNE7o3utg06Db4v9ubw3rgdyda9xLjKH5C9JZSn2h2vR41Epcfuennjnl2V3BtnHhzJTbN7ZwVL/ABClSwWjrkrKAnyUUnv3WujTe/de697917r3v3Xuve/de697917r3v3XuuiL/k/7D378+vedeiY9qUFNj985VKbWFqo6XIzB31/5TWRlp9NxdUJQEL+PfDD73/Lu2ctfeA5wg2pWWO8jt72QM2r9e5RjKVrlVYqDo4A1I6yQ5BuprvlixM5BMbMg/wBKhoP8PQfe8aOhl1737r3XvfuvddfX3vhx69173rr3Xfv3XulbsPKjDbuwla6yNE1QaOVY5UivHXIaYtI0pWPwwlxIwP4X+tvc1/d05xXkX3q5A36YSNaNdm2kVJEjql0pgLOzlUMUbMssiswqEx3U6DPN9h+8uXtytgQGCaxUE5TuoKeZ4D7en/5h7E3ZUbW2h3z1RiarN9y/GnP1HY20sDj5WirOwdmVNEcb211KSraald+7IaY0MTq6jO0OOlADRAjv0KEVBx1jH8ujK7F3tt3snZm0ewdmZGHL7T3rgMTufBZJDxVYjNUMWQoZCgLGKoEU6rLGxDRSBlYBlI97690r/fuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Qd9ide0G/sZBTSzmgyNDKZcdkkiWZodYAngliLIZqaoRfUuq4YBh9PfuvdJrZfWG2+tYMhuTM5Slq6qjo6ipqs5lBTY7G4HG0sMk9bUpJUSeGihigQvPUSOCsam5Vb3917pn6R+Vvxk+Sc+5qX49/ITpju+p2XUpSbspuqeydo78qdtzTSSxU/8Zg21lsjLQQVMkLrDLIqxSsjBGbSbe690PU88VNFJPPLFBTwRSTT1EzrHDDFEpaSSWV2VI441BLMSAACT7YnuIraKSe4lWO3RSzMxAVQBUliSAoAySSB178+icdrfMrZO0DPidiQwb9z6en7uCpMO1qKTTcGXKRCR8o8Z/VHSgj8GRSD7xT9zPvXcpcq+NtfJsKb1vQqPEVtNnGfLVKKmU/0Yqj1kHSSa7SPEYLH+XVbe++0d79n5GSr3juafMGOUPDh4Z0gxGK1anhEGEgf7endY29EkqNM686j7wB539xOdfcK9a+5u3ue4jJqkOY7eMVqBHCKJ28NZDOfNj0WySyS/GcdIcW+t/rxfj8fj/YX9gkmvn/q9emuunZVUsSoCgsSTwAouSeR9B78CF1EmgAOfSn+rj5cevUwergvj1j4upvj1hsruDXTz18Ndu6rpZUWCoNRnZRLjMeitHG71E1MsCjVdgWtfSB761/dq5WuOVPaHlqG7Qpd32u8dWABX6g6kXgCT4YTjU5p5Do7tk0QqDx49F4yWRym687VZCZJ63L5uv1JSxKZZXlqHCUtDSoP7MCaY0UcaVH0+vufOn+jmdUdYQ7Lov4nlEjqNz18I88tg6YuncKf4dSPyC1x+7Iv6zwPSBf3Xugh+Yfy12n8XNhT1L1NFlOzdxUdVF19s528s9bVgGH+O5eGJ45KXbWKnYGeUlfK4EUd3J0+691qtbs3TuHfG5s/vPduTmzW6N05WqzWey04s9bkqt9Uroi+iCniUBIYlASGJVRQAPfuvdPnWnWu8O3d87e672FipszubclYtPS00QHipKOMq+SzOQkZkSmxWIpbzVEjEAKth6mAPuvdbV/RPUPWnxE6hw+zIsxi6JI3+93Ru7LNS42r3dumpjUVuSqCSpkIVBFTwJq8NPGqi5DEkHMPNHL3Kdidz5l3m2sdvrTxJpFRSx4KtTVm/orUnqyIzmiqSfl037q+WWyMSzQbax2S3XOBKPuEU4nGpIjaY/8AKK2M1E8ctr6o4j6bfUmwxg5u++NyDszPbcr7Zdbxc941qBbwhl4d8o1sp46kjIpTz4LYtvlfLsFHz49F33J8ou0M4kkGNkxO1aeTSL4qkeqr1C6r6a3INIqeS45EQItYH3jRzR97T3Y39JYNqls9otWx/i8fiTKRx/VmLgVxlUB+zpatjAhqat9v/FdA5lN7b0zVTNVZfdu4q6ado2k8mWrI4bxSCaIRU0EkNPAIpVDJoRdLC4t7g/dufeed+uZrveOcdzuJ3pUtcSgdrB1CojKi6WAZdKjScjpQIolAAiHWGXeO8Z5Ipp937plmgWJIZH3BldUSQG8KrpqlUiIni4J/rf21NzvztcTRXE/Oe7tMiqqk3lx2qnwgfqU7fIkE+pPW9Cf77X9g/wA3TkOyexgQw39vAMhVwf49XEalIYXUyFWBI5B4P59mf+ul7ngqw9x971A1FbuUivzBND9hBB8+q+DCa/pL0JFD8me3qSqpJ6jN47JU9PLG81FU4ahhirYk4aGaemVKiISD6uhDA8j+nuU9u+9V702N5aXV3vlrdW0bAtDJbRqsqilVaSMB1qB8SnUCa9MmytyDRaN9vQ1bW+YFPLK0W9drNQxFXKV+3Z5K5Q/kUJHLQVnhmVFiJLOsreoWC8+5y5S++vbTTGLnnlBoISMS2TmbJYAK0UmlhRaksrtU4C9JJNuI/s5AW9OjS7P7B2hvulWq2xnaHJDQXmpVk8GSpdJVXFVjZ9FZBodwCWXSSeCfeXPJfuRyV7g2i3fKfMNvdClWjDaZ0pg+JA9JEoSBUrSvAnpFJDJEaOpB6Wnsc9Nde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3RXPlnuaTC9Z/weBtM27MvSYmQBwrnHU6tkK/0B1d4pFp0jawKgPZuD7wy+/JzjJy57OrsNtJpud8v4rVgGoTAgaebFQxUiNUalRR6MKNk62GESXpkIwi1/PgOqzPfHSpPHian/P8A6uA6Gvz4nr3vXXujKfFTcxwfacGKkaX7XdeLq8WUjuU++o1OSopJECMTpjgmXVdQurm97e8v/uS84nlr3otdjlkYWe+WU1tQDHjRD6iEkBSSAqSrqJAXVSprTom3yES2JcU1RkH8iaf4emT+Zp8TD2hstO6+vsDJW9k7FpUi3LQ42INWbr2LCJZKt2gV0+8y+2WYVELWaZ6USxC9o1HZroE9VF/Cz5MVHxq7kxm56upqajrvdccO3uwsfTO8iHDzTB8fuWnpv0zZDblS/lH0d6d5UuL29+691tZ0dTt3eW3qeupJsVufbO48ZDV086ilymHzOJr4FlhkVWEtLWUdVA4NiCrA+/de6DbHbM39siKpo9kbooc5t4QyfwjbXYEuZr6rDzih3RXS6N7LU5HP5JczujI48MKtJosdjKZ4qZCTGB7r3Wesw3cOfSrp6reG2djUs9HUQQT7Sw8+dzcNRPjdvS01dHXbmVcXFLjM3T5SIxtQzR1VFPAT4Zka/uvdZcx1xlpqasqML2FvqkzDa5qaOtzNHW4aoUbpl3X/AAauxlbhMhSLiq5X/g8s0cBrIsMfFDIsirIPde6LXv3asKbc2rv/ABmN/hNPuOipUz2HTHZfE0+LzzwEznH4zO0WNy1BjKmpglWOOohik0hW0LqIHuvdNHW+xJt+Z00TSvTYqgiSqy9TGFMvhdykNLTMwKLUVjKQGN9KBm+oHv3XulN2Z1l03u/E5TbW26jLVWVoKumgmOFo9xbjxMWVGfl22+Pqs5QUGRx+KzGJzMDxVimYPjY7yVcaQHWAdz5yHy17k8tXnKnNdo0u1zMjgo3hyxSRmqSRSAEq654gqQSGBBp1VlDqVbh0TyPpjYymKVhmauEvG3hlygamqULgaZDDBG7wv+dLDUPzY+4GtfuaeytrcW9xMN6uI0cMYpbz9OQDikgSJGKHzCsCfUdMi2iqCa46sg29tvB773HuqlzkMGU2rsWLFbBwez5UxJwWOlbBYPNZ6onp8Pnq45QVnkxyw02Zoqarxj0ReBfHUeR8pLW2trK2t7Kzt1itIkVERRRURQAqqPIKAAB6dKBgUHDp/wAn13ldvVU+f6xzlZiK01k2UyezcnV1OQ2luczV2f3BlMfTRVs8ybSze689lkE+YijqmpqeJUWnZAFD3Xugu3LiMH2kuSbG0cW2u39v0Qq9y7GqpUp8nX0sdRNjXycNJULSVeS27X5KiqIsVmvBHT5EQkLwCE917pP7Vz+4crS4emxdQ0HYXXElW2Nxlb9wzb12gGhl3DsSOhnz+CxCbrqYsYlNQV2QM0eNaQyhNJmv7r3Rp9nbwxe8cYayhElLW0c8uPzmGqlK12CzdGwgymIqjbw1L46uV4TUU7S0k7RloZHSzH3Xuld7917pP7ryuYwe2s5l9v7bq945vG4yrrMVtWgyGNxVbuCtp4WkgxNJksxPS4qiqK110JJUSxwqxGplFz7917oq9H81dgYOjml7y2J278aqilmWmqqnt7YtauzBKVDPLTdobIm3h1s1En08s+UpybX0ge/dboTnox2NyHXXbu1sdmsNkto9jbOyMlNlcNmsPX4nc+CnqaWQTUWVxGUoJaukWuoJwHhqIJBLBKAyMrAEe611EwfVezNv5Oky1FSZOorMamMTDnL7hz2chwn8L27JtSGTDwZjI1sdBNVYSVo6qRBrq3YyzF5fX7917oRBYcf659+690Tr4VTUW3ev9+9IxyAZD489ydk9aPSvqWeDbGQz0/YnW0rRMqlaeXrne+MSNhdX8TWJINvde6ON7917r3v3Xuv/1t/j37r3Xvfuvde9+691737r3XR+n/FffuvdEr+UHXe7ds7h2v8ALbpjDVWb7Y6ew2QxO+Ni4siOp736EnmbLbu6xSO4Sq3rt+qj/jez5JTaPMRPRkrBkahh7r3RqNh73212VsvanYGzcjHl9p7029iNz7dycQKrWYjNUUNfRSPE37lPOIJwJInAkikDIwDKR7917pW+/de697917r3v3Xuve/de697917r3v3Xuiq95Uhi3PjKsJGqVmI0M6/5ySalqXBMgFuEimUA88f63vkN/eA7QbP3V5R3lbeJYr3ZSpZQNbvbzuCZPM6UljVCfLt8up19rZxJs+4QFyWjuK0PABl8vtIPQM+8EupN697917oiXyL+WO/Nnd89a/EL44dYbc7X+SfZfXu5u4chN2Fuyv2T09010/tfLU+3Zd/dnZzBYbcW6a+Xce56lcbh8Ti6N6mrqA7ySRRIWMp8ncgbTuXK29e4HOW/T7fydZXkdoot4lnu7y7lQyCC3R3jiURxgySySsFUEAamIHQX3be7iG+h2rbLfxb5wWNT2qBQZNDSpNOB8zgCvQfbU+bfb20d1/Izpb5Q9J7L2L3n0b8ZM/wDK/ZdZ1V2DlN4dO9+dWbfp83SZafaOV3Jt3b+8Nqbg2zurExUGWx9fRyvDHWRTxO6Hk13D2w5e3Cy5Q5k5G5kubnljdN7Ta5lurdYruwuZDGyiVElkiljkjdnikRgGKFWoR0hi5jvYHvrPcrQLeRxa1oaqwyDxCnBoPhzUGpHQc9DfzB+/tyZz4HD5BfHzq3aGx/5im16vLdJbn6Y7Z3PvbcGy87TdZjtmnwvaOy95bG2tOmHqtsI8cuWxFTWU1FWBEmAWRGJ1zX7QcqWVn7pPyhzbfT7nyhPovI7u1jhjmj+p+kLW0sc0vcJMiKRY3dNTL8JHSbb+aLxriwW9tk8C4Wo0sCRVNdCNC+XzNDTGQercwbgH+vP++v7x8FKDTw+2v8+h+MY64uzINaW1RkSLcavXGdaHSeCAyg+6SSvDpmjI8RCGXFaMhDKSDxAYAkHB4HrxAbtPA4P2HB/l1YHjpmqKCinLq7S0dPKzpYqzNEjPbSSv6iff0m8tXzbly/sW4STLJNPZQuzL8LM0aliKYpqrw4dYi3cfg3VzFQgLIwAPHBPVT/x87Uyvxo+Vvfvxi3ukFN0Bu7vjHSfG/cEKtBQ9bb57i2HiO0cl0tnxIfBRYLsHd1fnK7Z9QpETZJK7FWVxRRue9ME1A6tvU3vwRY259+61137917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de6KT88vjLXfMv4c/Iv4tYvfVV1pku8OsM/sXH74paWWvXAV2RjjlpJshj4aqimyOEq56dafIU6SxtUUM00YILe/de6ps67+aVL8CsR8iOsfkv8G+k+ifmf8ZvgFvPvbaG/fj3Q7di6H+WnR3SCrQvDsTc9HgNv7+2TT4XeE9AMjtfN0jyYv7xpqaaeOzFuWaGBPEnlWOKqjUeALMFUeVSzEADiSQBx69WnVfXS/wA5v5jncXbPROH7Dy3yl7N6S7u27lsv8gMZ2j8Rdo/H3rHpfIZLYx3vszcHVHYG0spNnNzderuHRhY8fnpK+pr6OaCrM3kZk98/vfX3NXnXlTnHa7jmfa7K3s7pRZW9juXjTX6rKsU1vfQiNe5aNKuhtEZUxur5PRfPKZEkUOAR6Hj8j0TDZ3ye+dGF+HfUf8wLfPf20N3bSm7qwGxd0fHHHdL7UwOC3r1bnfkBl+k6rI1W/Ka26aTtunlSKro2xopsUsMCwmB5GkkIBveSfbC49wN+9qNs5Vnt9xXbXmivzdySPBcJYLdqi2/9kbXDKwfVLqbVqC0HTZihMjQiKh01rWvRqvj/ANd9zYz+Yp/MAy+Q+Sm589szbVb8f63NddVfWHXdFjt3Y7O9KZyu2PharP0dIM5t2PrWm8dPHNQNHPl/GHri7sSQRzVu/LkvtH7UwRcnwRbjOl6qXAuZy0bJeKszhC2mQ3BqWD1WGpEQoKdNyMDBCAmSDQ/nTpvxfyk7wqfhF8Ae7J9yUMnYnffyF+OvX3aec/u1hEo89tXsXsndW2t0UFJilo1xuFmyWIxMMaTUiRywlC8ZVixLsvJXLS+5Pury0ls37o2raL+e3TxGLJLBBE8ZZi2qQByTpbtNQOvGMeJMoXtVTT7ekHvfuD51ZBf5oHeHXPfOx9t9bfy4O3M9ldqdQZjpzbm4KTs/Z+xtp9e7zz2wO0N41Dx5mk2tlqXPyUeOlxYiy33tTJLPUmGKNFk72u9teReYt29qNluOSXvrzc9iO5X9xJeTRRQRwzyRh4oUwXmlVI3V28MoQFQEsS5BGjCEFKkrqP7erYsD3v8ANLZnzi6A+P3yK+Qe0u9tgfLT4o9gfI6LaW2upcF1nhfjz2XsCbZ+Tp9mdbZDGz1m5d19aS7T3tFSKdx1VdlWqaf7oyxmUwx9JUVFRFiULGAKAcAKYAHkB5Do0/Lq6v4+bOgrJ67edbEJBQzPjcKsiqypUmJGrq1QSbSxpKIkI5F2t7t17oR/kD3ltD48dX57s3eLyyUmLEVJicRSlf4huLcNbrjxOCx6uQvmq5lLSOfTDAjyG6offuvdamXcfb+9O9ew8/2Xv6sjqc9nJFihpaUMuOweGpi4xmAxMTktHQY6F7XJ1SyFpHJZj7917oN6SjrMhVUlBjqSpyGQyFXTUGPoKKGSprK6vrZkp6OjpKeJWknqaqeRURVBJY+/de62OPjN8ftq/Bnqo7+35QUO4u8N7JHRVclCbvjKepC1VPs3DVFUT4KLHBfJkqqNVNRKosGCxgxZ7ve6mz+0nKr8wbnC095JIIre3VgGmlIrxPCNANUjUNFGAWIBUQwtcS6VOPPoPt8bo3nvjKSZ7dsldO2tvs6ZoKiLF4mGVi0dHjqdgIYUUNbW15Htcsb298j+ePcTmb3H3ube+at4+ouFZhHECBDboTXRAlcKOBfLv+JvLo9hhSMFYl+Vf856RrxSxECWOSMlFkAdGUlH5VhqAuGHIP09g8OhqVkU58jn9nV6GmqnZ60P+bqSmOyEih46KrkRlVlZKeV1ZWF1YMqFSCPp/X3UzQrxlX9o/wA/7fTrWfIdYp6SqpdP3NNUU4ckIZoZIg5UAsFLqoYi/wCPe1kjfKOCPln7OHrwr69eofMdR/d+vde9+691737r3XvfuvdSaKtrcbVxV+NrazG10BBhrqCpmpKuIqQylJ4HRwAyg6SSpI5HtTZXt/tl3HuG1301ruKEFZYXaOQUNRR0KnB8sqfMHrzBWUqygjqwj4/97Vu/KptnbsSL+8lNRSVmPy0CpDDmqSkMSTxVFPqvFlYEcSOUHjlS7AKQQelH3b/vA33uDctyTzgEPNEMDSQ3CDStzHHpVxIlaC4UMHYr2yLVgFKkEnu7URL4ifBXo13vMDpB1737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3RHPmpb7Hru/wBf4luD/AkfY0V+frb+vvm//eG0/d3tNU/8TL77f7GEf5ehLy5l7wA92kf4etfrvrsP5Tbg+Z+wPjH8fu4evOlcJX/FPePyC3DuLeXSlH3JWZrObf7YwnX9FgYqSs3XtX+E4xqLMeZ3ikd2eO1vUCuGfJ+y+39l7Ybtz1znyzebrcpzDDt8cUF61kESS1knaTUsUutgUoqsAMno7mkuHukghmCIYy2V1eYFOIxnPQQ7/wDmj8jepvjZ/MOxnZFF1Uvy2+DXXO39843eOycLlW6d7U2f2Vghm+r+zKLZG4K6XLbfqH+0rqLMYSaqmhp62mBjlaN7AR7R7Yckcxc8ezFxsUm4f63PNl7JA0M7r9ZaT20gS7tWmRdEg7keGcIpaNyCtR0291PFDeLIF+phGCB20IqGp/k6EP4dfKv5GYT5y/Hvobf/AHV0F8scH2h0nke+a3eXSuxKbYW5fj7mtujAPQbe7Sptq7y3pteu27v+XOS0mLMs9FXtU0j3jKXBHvKfKnKW17vyl7jcucrbxy7ue0887bYJb31w08O5RT3JieS2LwwTpJCndKF1xFXAJIPSaSaeVJLd50lWS3ZiVA7SorSvDP7cHowfwU/nBfJ/tr4p/Kg/JTEdd4L5O7Y+Ivb3zg+K+6dsbfnpOvO2ug8KN7baonqdvVVaXl3f072btEYvctHHPpelrqKW4ErOeyEqhJpUX4VYgetATx8v2dAsGtD1N7C+Ke7u0fil8f8A5u7BwOMlzHbPQHVfbfevX+0MVJQY+g3XvXY+E3PujeOwdvQGoFFgJK/Iyy1eLidzTKDLDqXWob6tU8K9L34FfP2LpCloeou3aier6mef/fqbnp4mqavr+WsnMk1FWwxBp67aVTUzGQNGDJRMzEBoyQvutdbEOGzmG3Hi6LOYDKY/NYbJQJVUGVxVZT1+PrKeQao5qarpZJYZo2H0Kk+/de6dffuvdIDsXfNJsfBCcKtbuLNzyYTZe3o6nFU+S3XuiWgrK6kweGjzOSxFBV5BqWhmn8T1EIaKBzqFr+/de6LV3XnOvOk+ldo7L3Lujam11xCYYzF6mTGY8NAk5yFbSY3IZLMZimoq/KzyeCEzVTrq0a2CFvfuvdFk+PnzV6c36K/qDZM+a2p2jvet/huHzW/aN6HbGYk8jwzQYjLbazbZOhqkwyzHHkmCZqx0ew5Ue691ajtvb1BtbBY3b+MNVJSYuip6NKnI1c2Qyte1PCkD5DMZOqMlXlctW+PyVNVOzz1EzM8jMxJPuvdFA7221TYndb5HHiOGPMUdPX1sKNAphrWlkplqfB9y9WUqnprl/CsIddIYsxHv3Xuhc66zUNJuSmnaeokou1sDRbgoEEcJp6fdm1cZT4fdlMtNh9rwQ0f3dFT0tVJWZPKzz1U7GGFFSED37r3T5312hk+l+sNydp0Gxsv2HQ7LWizW69vbdqI49yQ7Gpq2A7x3FgKKSnmGeyW1cAZ8iuNVopa+OmeKJxK0at7r3XPcWDxnZGC2t2P13m6AZ1KLGbr2Hu+jkrqnC5/FZXFztj0y9Jisrg13ZtzJYTNzSUtPVTtSw1MsVUq+SNT7917oKsxtum7Txr7/ANiwLhd74jJzYvee14ctj567C7txKQvk8NVZHEVNTjY91YR5kWUxTPDMCPUTYn3XukrQ7rjzORStyeRk2F2tj6YYul3uaIjEZ98ZQ5+lwGB7SxFPNj67L7dw2Sz81X9qlVSxvUkOssRJ1e690YGn7QXb1Vj8P2RjTtmpyM9DQYvdFLIa7Y+4a/K5mvxGAoKDK6IpaLcGbp8ea98e8ckdDTzKrVUttZ917oWoZ4qmKKenkjngnjSaGeGRJYpoZFV45YpEZkkikVgVZSQRz7917rlJEkyPFKqSRSI0ckUih43RxpZXRgVdWUkEEWI9+690T7dPwe6Xrs/Xb36wk3h8cuxa+d6yr3p8f9xz9f8A8VrmPk8+6NkRw1/Wu8llmAaYZXDVTTc6mub+/dbJJ4nrDjdufOnY2mlTsboHvvEQFmSp3vtDdHTG96iJVJjhqctseq3vs+pq3I0mWPDUUZPq0Ace/dex+fT6nZXyygiqFrfi3smpqli/yWTB/IzH1WPml8Rss75nrTA1lOhnst1hkOi5+vpPutdFM68398hfj92z3x3H8lvjHuinw3dtX1nUTZn4w5SPv/aWw8b13spduy1+8MQmP2b21V5WsmqZnq6nG7ZrqWKjpqdAzFHc76saUGerJetu0Ove4tn4nf8A1dvDA762ZnI5Wxu4tuV8OQx80lPIYaujmaM+WiyVBUK0VTSzrHU00ytHKiOpUa6r0vPfuvdf/9ff49+691737r3Xvfuvde9+691737r3XrA+/de6r62Oz/EX5Bx9N1LPTfHH5K7hz+4ujKuVn/h3VnfORkye7Owul2qJbw4/bXZ6fdbj2tAWVIslFlKCJQrUcfv3XurBF+g9+69137917r3v3Xuve/de697917r3v3XugB71xsT4/D5cRytUU1c+PMgZvFHTVULSkOltOt5oVAb/AGHvnR/eEcr2M3LHI3OgtZjutvfNZ+IGPhJBMjSFXT4dTyxoEc0Iyo49Sr7WXki325bcXUQvGJKUyWU0wfQKTXotvvlh1NvXR/1r/wBf9t7917qnv5Iy7r+Jv8x3aXzoy/WXZ3Zfxu7P+KEnxf7eznT+xs52jvDpLee1OxZOwNhbxzPX+1qeu3bk+vt1QZCegqqnG01U9HVqjzIE5ORPJq7dz/7OX/tdbb3ZWPOVhv37ztUu5o7aG9hlthBcRJPIVjWeIorgSsodSQpJx0At1e42bmBN2Ns0tpLFoOkEkEPqBoKk1DMuAc0xSh6Divn3x81e4fk78rNmdL9x7H6Z60/l0d7/ABd6GrO2Ovs11tv35AdoduDIbq3nmdrdabjhp95U2x9v0m3MZjcdNkaWllyGRq5PDGVU+ziJNp9s+XOSeQ9x5k2265ivub7Hc78Ws6XMG321rpiiSW5jJiM0jTSPIsbsscaCpr0XSNcb7d325pZvHawwELUUZic+YBIGkChA4kjAr1XP8KuldvYnKfyw8n8N/j780+vvl/1lD1dtr5jdid47K75250LtP4+z7Qgp/kns2tk79qqnZ8dbunL0dPFgKXZdNT1H3kSFSlOoX3MfuVzLdz23vdD7jc3cuXft/em6k2iCymsZL6W/E1dvlX6ECUiNC5na7JBVqULnoi2yHVcbKNrt5RfxhVcvrKgaRrBDilajt0cDQ1AB625W06m0AhNTaA36glzpB/x0++fYrTu+Lz+3z/n1NS10rq+Kmft8+uveiAeI630dTrIW2Jtf8f7jEP5/46yf7x77y/dWAH3evab1/dSn9skhJ/PrGTnP/lad9/5rn/AOiUbp6t2R2L8uvkd1Fv7GS5TaPeXxY6X3RloIa6bG1dLkuuuxOy9u0WdwOToHgymE3NgKjL0FXRZGnkSooqungmhZZEU+8gugx08bM703r8ctzYrpn5bZ+Cu2xm65MV0h8sq1KLEbS7EE0oTE9edzvCtNiev+74aeyQ1RWnwm7FQy0Zp63yUC+690flXRgpVlYMoZSpBDKRcMpHBUj6H37r3XL37r3Xvfuvde9+691737r3Xvfuvde9+691737r3XvfuvdA/3z0ltD5E9Vbp6g3zkd74bbm7IseKjNdbb63P1rvrC1uIylFm8Tl9sb22dksVuDBZPHZXHQypJDMFcKUkV4mdG917qkXu74Z9JdM9lbsFXuXuH5F9k9n9SQ9Z9odkfJ/svKdvbrk6cq6yrloumsGKuHE4HbOx8rUPNXZSGioIanK1Bilqp5njB94C/e99y90td32D2/wBlvnght/D3C4eMlX8dZAbRdYNQI9LTaeDMUOaU6QXkxUrGhzWv2U4dFR6g+KW1umMrtCbb3cXyd3BtTr3F1uF2B1hv7vDN7s6x2nh6jE1GGo8TBt2agp6zPUODx8whxqZarrxQIiGP1IrDEfmDnm85jt9wW85f2WHcLtg09zBZrHcTNqDFjIGKozsNUvhpHrJNaAkdIvFLkVVR6nz6JN8Nv5Ze2Nh9J9N435Av2hUbx2P2furubP8AR3+mbL7h+PMva0PZO7c3sLsOp2HSy1GDq83j9uV1BUrSw1CY0VqiaWlNQrMZH9w/eW93bmTmGXlQWQ2y5sorSO8+kSO+Fs1tEk8AmIDqjSB1LFTJoqFk0EDp2S472ZKcKV8+rAIfj1s6i+Qmb+S2I3J2bgd9buxG2cT2BtnC74qaXqzso7KxeRwez8pv3YctJPS5nL7bxOVmhp5UmgVlCGVXKKfcVtzZuEnKdtybNa2Um1wSSPBI8ANzbmVleUQThlKI7LqIIJrWhFcJw7aBHjHAny+XRfMX/Lb+OGJyGwpRle9MntvqLtWDubpnrTL9z7mr+r+o98x7ml3ZVTbE2V446SLF5HMVUgalrWrhDTSvDTGBWYkV3PvDzdcRbqDBtiXV/Y/SXdwtnGLm6gKCILLLlvEVBUNGFJYAtrNOnTO9DhdRwT69XT9Lfyw+iaz48/MHafaGE3fTQ/zCcnmt4fIKji3RX4/L09FmttYXbjUO3J0pUn2e8+IwEEssMIl0TH6kiy9CPu58kbvsPJmw71zbaqnMh29bWBdJR7fb1kaaGGVcAzM7tLKxGoVRCezJlbxFI1LDuAp+Xp0lM90R1pn+8urPkNVUWaPYnSnXO9+o+uaiLOVC4Wj2Dvmn25j8vSZfENCUzWTNBtKhEFU7o8RVzYl+MjOlHVsPUmPXG9ebYj0qr1NAMhJpcuGfISPVq1z+TFItx+Dx7917rXZ/mUfI+o7k7oqOvcFXO/XvUNZV4WlhjcGlzG911U+4c6dEkkc6Y8H7Kle/pVZSAC/v3W8U+fVchIW7NYKAWJNgoABJJJ4Ci31PA9+611fB/LL+G1JjMXh/kv2XijJnskGrOpcLVidBg8PPFLSvu7IUUqRq2VzEbE0Gu/gpW8oGuRSuqgUqevdOvzC7gmrcr2DMJRfbe3N6YDadHS1JYLJj8LkzVZwTxABHevUNrHqUxoim4JHKD3n5o3L3Q94L2OVgvL20Xv0UIVgyUjmUSSAjDPcPWpOFRQprpNa7tvVty9t8cjDXezYjThUnGonyVeJ8zwA60nfit2RtVenfgluH4q/ITuTe38wffPaWx6XvjrzEdy9rdlbayPWM+89wp3LUd8bE3Vlsz1/s7bWJ2VDBLTVKrRzR1OnwksTbIfnHYpBvnuBa848qbdB7b29nKbSd7S3gcTiJPpBZyoiyyyNLqVlJdSMmmOgbfXu7mfd472Zk2fw30t8J1UGjQQdRJJNeOPTq33oTufJfHz5CfzAukd67z3D/AHH6tq0+a3UtXuLOV+Rkh6K7J2/U5LsDEUdRlJ6iWfEbM7GwFXTxw+pKWOrWNQi2X3CnMvLdpzTy17c8wWm0w/XXi/uy4CIFrdQuFhaigd8kMikn8RUknolfdN3+m2W8s9wmSY1gIDkjWpopKmoNVya9FYynxno98/yo+7fl93pmu52+W1b0l3p8paPcmF757i2bRbNzHZaZPsTYOCi2XtjeWJ2tNSbHwZoaeOmmo3hQKYyCvHs9fmmXZfe3lbknluDb/wCohv7PbzE1nayGRYNMEzGWSEygysWYsrAk5r1JGy7ou6R7xGxJmtXC6gfiqBVv96rT8qdXA/Bb4zdTdGdQ7C3z19F2G26O4elemc92BkN9dxdo9oRZPL1OzMbuCqrcZj+wd1bjoNtSVOWzdTJIMfHTB1dUa6IgGPnuXzjvnMu/7nt26Namz2/cbtIRDa21tpUStGAxgjjaQBFAAkLUI1VqehRBGkaBlrqKjo7/ALjjp3r3v3Xuve/de697917r3v3XunPC5rJ7cy2Oz2FqWo8riaqOsop1vYSR3BjkAI109RGzRyKeGRiPz7N9h37duWd52zmDYrwwbxZyiSJ/QjirDzRwSrqcFWI8+tOiyIUb4T1b9sPdMO9tobf3TBGYFzOOhqpIDa8FRzFVQ/qb0x1MbBbm5W1/faf295ug585K5c5vt4jHHf2qyFD+B8rIv2LIrAVyRk56DkiGN2Q8Qeld7GfVOve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xui7fJvZNVvHrSsmxtNNWZba9bBuGjpaeN5aiqgp43gyVNBFGC0srUMzuF5uyD8294qffE9ub3n/2fvrjaLWSbe9luEvoo0Uu8iIGSeNVGWYwuzqvmyDiadG+y3ItrxdRARxpNfL0/wA3Wsr3rgfkRsf5ydffJfqT41bi+R+yIPh/vbofM4/aPZ3WHXeXwO8M33DgN+Y+pq/9JOYxSVeKkw+HZWemWVkkcX/SR75hcnX3JO8+028ckb/z3Dsu4vzJBuCPLa3VykkCWckDBPpkYq+twQr6cV6FEwnS6juIbYunhacEDNQfPoEO0fij8nO4vjT/ADKt7b52NtLDfKP5s9ZbZ656/wCjdo73x24cN111x1Xgxhut9i5rsitjw+Bzu+MvU5TJ5LNVsYix6SyxwxMwXV7FWwe4PInLXPXsXtW0brcy8gcq38lxcX0sLRPc3V3J4l1PHbLqdLdAsUcCMWk0qzMM06bkt55bfcJHT9eZaBONAPhFf4q8fLownx6+GW7fj784PjRvj4p9RbY2t1P3R1hR9VfNXrjY8m39qbcw++OvtsUee6v72otuLLS0VZlqHMQZDCZeTGIXqYqtJ5Eb1SexV7Zc2P7u75y5yZzTvlxd84bfzfa3+yXMyyzyG0F2JNxs3loWSIW6JcxmUgIUKBuC9M3UQslkmhQCFoSrgY7tNFb9poQOhD3x/K6+XVR/Ju+Mez+rNpbe2r/MS+LPXvdu1cJtDMZ/b9XiN9db9+1vYO0O7Ohs3uinykW3ZcTv/rzdNLkaOWSsWCkzuKoZDJGyMw7MSuJJZZFFFZiR65Nc/P1+fQJHDh1sA/DHrjdPUPxB+LvUu+sdFit49bfHrp/YO7cSlXS5KLHbi2psDA4HNY8V1G0tHXLS11DJH5YmaKTTdSVIPtvrfVUX8w34HxbLGc7/AOmcYI9rSTT5XsnZNDG5GBmqp1ao3Tt2lhRgmHaaUtW0qqFp7mVLR61X3Xuq2+nvkb3V0LUmo6p7Ay23qGdvNPgJjDmdqVjOoJlm25kxUYwSuLfuQrE5H0b37r3VhW0v5wHbmOx7Um8+rtkbnrUpWSHMYbJZTbsklXc6J6zGzjJUzRAH1LFJFcjiwPHut0xWvQabz/mc9s5fLZHcGzdo7c2vuavomxUO6c5W1e68jhsN9/WZOnxWDwOjF7PpBjKqudYK2aiqci8R0zzSgC3utdV5br3Vubfe4K/dW9NwZfde48nM09dmc9XTZGtmdnZgqPOStNBHf0RRKkSDhVA4HutgkcD0z09VV0dTTVtDVTUVfRVEFZQVtPIYp6OupJVqKOrgkXlJqaojV0P4YD/W9+611t4/EnuCbvX49db9i5CRJM7kcM+L3Ro0j/fzbfqp8LmpGVQqo1XV0LT6QAAJRbj37r3WLsbGUmd3fvvGrTUMmXp+oMNXUc/k2gcr9vSbrzFcsKwwxNv9aE1NJcGZv4S0htEonEp9+690iesMbTdhbPzOxqyslx+RwGTx28tmZqKNamfbmfheR8VnoKKWVaPJnC5dFnWlqVkpZtWiVHU2PuvdGK2HuSt3Jia7H7jxy47de3qt8HuvFGb+I08dU0EdVQVCZGPHUGMrkzuCq6atdKZXipmqTTM3kib37r3RUvjNU1fSPa3ZXw5zEs52zt2hbuL41VdRz5ukd05mSl3B13BMWZ55elt+VLUECuTJFgcli1+iX9+698+jG5aF9odmYbckLV7YfsCFNo7hpw9bUY3G52gSryW1M4fvNw0uC27S1imtoKlqehnrMpXVdChbTEPfuvdS+wOqsJvqJqwAYzcCQiODKxJqWZU1FKfI0+pVqobkgNbyIDwfx7917otNLuTeXWdVVbP3PQR5jAVEc1NW7cy6LWYyvxtXE9PUHE1cqOopqmndlMR1RWYq6C59+690MOz8ftnN0FdJ1Du/I7DyUqQGbbk8MOZwmKFPR7dxFKlPs/JzrDj8bi8HgxTUcOLnpKCOSoeV45JD7917pZR5rt3G1oiyuzts7kxs2RpIY8ntTcE2LqqOgyG58vTyVFbhtxwlZxtvaMVFUVDw1byV1bLLHBBGiKx917qPB3FjI0Q7h2j2PtSTxU0lQmY2bkauCmaba+U3bVRtkdu/xvGznEY7Dyw1TxSyRpXPHTKzzSIh917qXU909dUdNV1lXmcjBTUIrWrJpNrbs0RLjsJhtxVZ0rhGdhHis/SuNIYSSO0S6pY5ET3Xunmj7P68rq+qxUO78EuSo6rJ0VVRVVYmPqY6jD5eh2/kU8Vf9uzpTZvJ09JrW6PUToiFmYD37r3S8AX8AcG3H4/w/wB59+690RHK7axfSHzX6zzOxaGLb+3vlpg+zMD2xt/GRGDCZztPrXblBvbZPZDY6Jo6Gi3XV7Rx2bxuSrIoxNk4VpPuPI1NG6+62KUPr0e6/H+xt9fz/r/X37rXX//Q3+Pfuvde9+691737r3Xvfuvde9+691737r3QQ96dRYfvDqzdnW2VrqnCzZujhqdubqx6I2Z2PvTC1UOZ2XvvAO9hDndnbnoaXIUrXW8sAUnSzA+690Hvxb7tz3am0cvtXs3H0e2/kF07lINh977PpiI4KPdMdN9xiN7bfiNmqOv+0sIsebwVSoKNTVDU7EVFLUInuvdGg9+691737r3Xvfuvde9+691737r3Qadr4yTKbMygiCmTHtBlQHdkAioGMtQQVB1t9vqsp4LW94vffD5PvOcPYbmyPb0ja6254twGtioC2ja5iNIOpxAZNCHDNTINOhjyFfpY8z2JkNElrFwrmQUX7BqpnonAIYAj88++HQINCvDrJAGuRw65e/db67R3ifyRSPE4BGqN2RufrZlIIv8An+v596KqeKj/AFf5fn1V0V1KuoK+hAI/n12zySP5JJZZJODreR2e6/QhixYW9+0jPzrwxx+zz+fn59aWNEXQqgL6AAD9gHXKSaeUBZZ55VHIEk0kigj8hXZlBHv2lQSQMn/D6j0/LrSwxRkmONVPyAH+TrF7305w679+690c7q2FYdibc0u7eWjknOttWlp6mWQon+piQmyj8D33Z+6TYpY/d49sQk0j+NZNMdZrpaWaVyqYGmNSaIv4VoKnrGjniQy817yxUCkunHnpUCp+Z8z69Ft3zUybf+e3QFVI6QUfYXx8722YhLwoa7KbT3R1hvKlpdB/emamx09ZKoHpUaj7yN6CnRvM3gMFujEV+B3LhsTuHA5amNLlMJnMdR5bD5Kkexemr8bXw1FFWU7kC6SIym309+690kerep9idL7VGyOt8TUbf2lDlMnlaDAvmc1l6DDSZao+6qsfgkzeQyL4TAwzkmmx1M0VFSKxSCKNPT7917oRvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+6917/ff77/AGPv3Xuqd/mPSVtN3tl5qt5JIcht3bdVjS8IiWOiippaOSniYG86R10MrFj/AGpLfge+VH3rrW7t/eXdZbpiYprC1eKq0ARUKFVP46OJCWOQWp5dFN6D41Twp0V/3jd0j66tb3atRSmOvdeJABJIAAJJJ4Fv6+9Upk8OvdHk+LPxwyWfyuK7N31jzR7Zx0kdftnCV0ANRuKtT10mWrKaYHw4WkktJAHUPUSKGsIx6syfu3+wW473uW1+4fOFkYeXIGEtpbyKNV24BKTSKw7IEJDoCNUrAGmimoxtbatHkGK1HVmWYFsRlT+f4bXfX/qFl99HQKZ8+jHqseP/ADaf8EX/AKFHvfXujPdm9oQ9V/EXdHYsExhqdvdX1MOJdjokO4ZqP+BYhEs6lmGbnjtYgkC4t7917rUk1TP66iV56h2eWonkcvJPUysZKiaR25eSWZmYk8knn37r3R4fgZ8YH+SXb8D5+keTrDryShz2+ZCh8GXqHkaXCbP1lgCcxLAZKlbH/Io3HBce/de62YeytwUuwuutxZemjjpI8VhHosTS00awQw1M0a4/FU1NFCI0giimlQKq6QqrYfQe4592ubY+SPbrmzmRn0zQWbrF6maUeFCBTP8AaMpwRw6dhTxJUTyrXrX77ZiqMnW5vEeZVkqtq12Jp55dbKs+ZxNdT/dT2JZwKms1yFRqZQfqffJ/k1Ftdss7liS/j63Nak6XUk/MkAmp8yT59Rtz5M78yaStUiij0j5HLdE8+GnRGV+Mfxg6Z6Kz1dtnNbp632hJtvcW5tp0D0mO3BUvn8xl1miqaqio8tWUyw5JEP3IvrQm1re5K545ii5s5r33mG3iljs7mYOiSEFkoirTBKjK1x+fQZ3G7F9fT3aahGxqATwoAP8AJ0Xr53fBzdvyu3T1JunrvsfEdY1uIwu5+ne8ZcjTVs0/Y/xs33ndt7j3TsTGvQ01QVzX8R2+4pWqCkKLWOda8hhL7ee4Fpyha7za7ntb3ivIlza0KgQ30SPHHKdRA06X7qZ7R0q2vdY7COeOaEyAsHT+jIoNG/n0f3t3pLL9+fF35NdH7TrMPtOq7h6c3X1LtPLZmOpXb+Cq83gqnE46Wthx0U9Z/CMTDLGr+GN3CrZVJFvcQQ8yWnLPOfJG/wB4rzrt9/HdyKtNbhXDEAtQampippU9Db27s7hoN5uipEMioisfxMCSfyAP7fPoz3V+1KvYXV3V+wshV0tfkdh9abA2NkK+hEq0FdX7Q2lidvVtbQioVKgUVXU45pIhIqv42GoA3HuNd5vY9z3retzijZIrq8nmVWpVVllaQKaYqA1DTFRjqUFGlVXzAp0uPZd1vr3v3Xuve/de697917r3v3Xuve/VpnrY6P58Qc9HPtfdO3pKmRqjF5uLIwU0soZIqHKUcSXp4i2qKI1lJIXsApd7/Un30Z+5Vv6XHKXNvLb3TNcWl+JkQsSBFPGBWNeKr4kblqY1sT59FG4JSRWpgjo4fvNgcBXou697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de64MByb24/334PupHE+f7OvV/Z0SDu74xV2by1Zu/rlaRqvJT+fL7XqZY6OGSqk1GfI4irltBA87AGSnfShYl1YfpPOT7xP3Odz5h33cOe/aRYfrruXXdbc7LEjSH457aViEQuaGSB9KaizowJKkS7bvSRItveE6QMMBUgehHn0W2q+P3c1JK0DbDyE7BUbyUVdiKmnIYXCrMK5AWX6MLce8RL77rX3gLG4a2f2zu5SADqhmtZYzUVw/jqCa8R5cM8ejld225xUXYUfMH9v+x0dL4/dEt1xG+59ytFPvHJ0a0320WiWn25Ryv5JqGCdCUqayp0p55QLDToQlbk9Dfutfdnb2jt35z5vKSe4F5biPw1oyWMTHU8SODSWWSieLIBQFdEZKVLBvdt0+tIghBFuDX/TEcD9nmOjRe8y+iXr3v3XusM8EFTDLBUxRz088UkFRBNGksM8EqFJYZopFZJYpEYhlIIINj7917rXV/mE/BePpurrO6+pcfJ/oty+QH969rUsTuOvsrXy+jI48KGK7OyVS+nQeKCd1UftOoT3Xuqp/yR/T/kXv3Xuve/de697917r3v3Xur2P5PvaUVTg+0emK+pC1OKydHv8A27TMwHkx2XjjxW4BCGNz9tkaWndgPp57/wBT791sjAPVrG9ocrgt07d3xj8fVZjEpRVe1t5Y6llq6iqiweSqIJcdlsXt/HYiqnzuRos34knM1TBT0WMlqprMwsfda6CnrHbU2O7Cp9w7UqaDcnXe4cZlanD7swtfSZLDVOMqHWanigrqGolp5ZYa2MRKQWV0W6/m3uvdCd2Pt/K0VbR9n7Mpfu94bZx09FXYdJcfRf342kTU1M20qvMZGhyn8DpKfJzpkknpYDVNNSLCG0SMp917ovnywmxmP2v0r8tMJWVMC9Bb52/vrI5JKSupKjJdIdkQQbJ7Yxtfjqynp6xKKDae4Is60FRHG0NXhIHYLJELe63XBHRku36Ojr+st017Vq0hweJfd+OzUJ24ZsVWbZUZ6ky9BW7oxWbwGNngFEStZLTSimRjItmUEe610vsLkYsxh8VloGiaDK42hyMLwTrVQPFX0sVUjQ1Koi1ETLLdXCqHHNhf37r3UPcW2cLunHS4vN0UdXTSKxRyAs9LIR6Z6SoA8lPOn4Kn8WNxx7917one8+pt1bGqzmcDNW5PF05MsWUxheLMYyNDqH3kVPaQpGBzLFdGt61Hv3Xuldsjv+ppI48fvWGWuhUCOLO0MQaqRQApOQpE0rUafzJFZuOUJ59+690ZjCbgwe4qdarCZWjyUBAYmmnV5EJ50zQm00Lj8h1Ug/j37r3T2ALcE/1/3v8Aw/x9+691ArcRiskgjyWNoMhGskEwjrqOmq0E1JUxVtLKFqIpFEtNWwJNG31SVFcWYA+/de6CTY9LU9f7sret4xUNsyrxK57roSyZTKVGKgongpt1beq8g2HkSKlpK6rgrKafJ5WprqySumjjUQ0yhfde6DL5XS0+3Kj449oTr44+ufkn19BkK3SLUeE7WodwdKZGWWUg+ClSp7FppZWNhoisfr791sDy+XRt/wAfX82+ht/S3+tf37rXX//R3+Pfuvde9+691737r3Xvfuvde9+691737r3XvfuvdFH766Z3u29Nv/JH4/pg4e+Nj4Kq2vmtr56pfFbZ776rkqJctUdS7qzMKzf3ezFDm7V+2c+8FT/Bsi80ckbUVdWKfde6ErojvrZPf2z6jcm1o8thM3gcnNtnsTrvddIMRv8A6u3zQRxHL7J3zgWkeXGZnHmRWjkVpKSupnjqaSaemlilb3Xuhu9+691737r3Xvfuvde9+691hqIo54JoJlDwzRPDKh+jxyqUdTaxsysR7SX9na7jY3m3X0IksriJ45EOAySKUdTTNGUkGmc9XjkeJ0ljYiRSCCONRkU/Poje8duHau4a7EBtdMhSpoHuxP2FQ0jU6OzquqSFVKMRcXH19/P/AO+XtfJ7Qe5nMHJSP4m2qVntG7qm0m1GJWLKNUkSqY301FUAqTXrJ/ljehv2zW1+y0nykg/prQMfsPEdJn3En2cOhB1737r3Xvfuvde9+691737r3XE6jZUXVI7Kka8ep3YKg5/1TED37RI7RJCuqZnCqPVmIVR+bED8+tFlUamNFGSfkOP8uj47WxAwOAxGIDySfYUMUJklChyxGtwwSyAq7Ece/oh9pOTB7ee2vJPJQuZJjt23xxF3Chy1Nbagg04ZiMcaCtT1invm4Nuu7bhuBQL4spNBw9BQn7OijfJxIsJ358Ft9tJIslJ3dvfrZo9YSGSl7P6a3sjPL+25kZMjtalCLdQWbk8AGReisef2dHZAt/T6AcD+n+xPv3Wuu/fuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XvfuvdFL+UnQdf25jMZn9rzQJu/bFPWQ0+PqdEUGfx1SY5nx33bFVpKyGeLXA73jJZlbTfUMYPvI+yl97nbbt++ctuv8AWrbY5AkTUUXMT0YxayQEkDCsbP2mrKaA16TXFsJwCCA48+qva3rbsXFVU9DkNibup6umfTPD/AMlP420huJaanmgkUoQQVYgg++a11ylzdYXdxYX3J+7xXsRAdGsriqkitKiMq2M1VmFPPpIduu1z4YI+RB/y9CDsz429yb3liNHtCqwWOkEMjZfdLfwakEMxW0kNPMj5CqIVr6UhvxYkH3JHJ/3f/dbnN4Xs+WJLTb3Ct495+gmhqUYIw8V8Z0qlcZIOOm1s5yaFadHf6n+GO0tn11Nn99ZJN85ikkWeix32ZotsUU8civDUNQyyTVGUqIWW6mdxECf839D7zG9tful8r8rXkG9c5X67zukTBo4tBS0jYGoYxlmaZgRgyHQP999K4bJYzqkOpv5Do6aLoGkABRYKFFgqgABQBwAB9Lfj3lwihAFHAcB5AegA4Dpb11LEk8UkMgDRyxvG6nkMkilGUj8gqfduvdVmZjGy4bMZbEygCTGZKtoytioCwTuI7ISWVTFpIBJNiPfuvdA98v95VY+EfYe1opND0m79iI5sLy4DM7op5Zoxcfpir6cIx4IEq/T37ra8R1QczBFZmBIAJsP1H+gW39pjwP8T791rra2+A3RT9E/HTamMydOYd3b1vv7ePkiEU8GTz9PTyUOJcXZlGFxEcFOVJ4kVzxf37r3S5+WTunVaBXdRJujBpIFYqHQ/dtocA2dNSg2NxcA/j3ir98R2T2hTSxAbdrVSASAQfE+IDiAQDQ4qAfLpdYCs/5dVY7k2nQ7kRJHdqTJRKEhro18n7QbW0M8BdFnTk6eQy34P4980dp3q42o6EXXaE10E8G8yDx/LpHzByxZ78iOzGK+RdKSDIp/C6/jX0qQR0Hb9Y5lGIjyGLkQX0yM9VE7D+pi8Umk/wCxN/YmHNtgw1Nayq58hQ/z6Aj+3O7hj4d7Ayep1An8qGn7T1Moesakyq2UykCwA8xUSPLLKvBYeaYJHCrDgnSxH49s3HN0YjK2loxkPm/AH1oK6vsOOlVn7c3JlU7luEYgByIwSzD01MAF+2hPQsUVFS46lgoaKFaelpk8cMSknStyzFmYlnd2JLE8kn2DLi4mup5Lm4fVM/E9SdaWlvYW0NnaRBLaNQFH+GvUoC/+A/BP5/33+8+2elHXiLfm/wDxX+nNj7917rr37r3Xvfuvde9+691737r3XvejwP8AxfXujufDl6j/AH/yijiNJrwjHIXXy/chKwfY25bxeG0nHFz/AF956/cia4B9xUNiv01bQmbGoORKBF600/qelT0Wbj/oOfXo8Y+g95+9FfXvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+6910Tb37r3TJndyYPbVI9dnMnS42nUHQ1RKqyTMB+ingF5qiS/0VFY+/de6Lnub5EsXan2jiAUV+cnmeBIv9IMdA6yKG/DSSAgf2ffuvdBvT9wdpVuSiNFlDWVkw8NPi6XD009PUOTYWpI4zKXJNtWsAfU29+690eDEy18+Lx02Up46XJS0VNJkKaJxJHT1rwo1TCjjhljmJAP+Hv3XunD37r3Xvfuvde9+690ybk25hN34DMbW3LjaTMbf3BjavEZnFV0SzUtfj66B6epp5o2BBV43PP1BsQQQPfuvdainyh6KyPxz7q3f1lUtNU4aknizOzcpMtjlNoZgyT4mRjdgarHlXo57EgzU7H8j37rdME9F99+6112Bf37r3XEsocR6gZWvpiUF5WA+pWNA0jW/Nh7917oyXx2392n8eO1dsds7d2DvDLQ4oT0uZw429naan3HtjJwhMpjPvDjZYoZXjCzU8pVkSaJGItcj3W6Cla9bNnUHyn6J7wxFDkdk9hYE5CrWCGq2nnK6nwe7sbXToWONrMBk5KauaoUoy3iWSKTSSjsOffutdK3L9XRU+Qrdw9eZmXYG6aynZahqeiTJbTzFTBio8JiJdybV89EtdTbfogzUkNHU0Cid/JKZDwfde6lf3z3riKs0m4Ot8rW08uUoqKizGyshj89QtFls/k6CjlyFFkJ8NmMcuIwlNTVuUlaBqeBqnxwPUFC3v3Xukpj+r6LenQ+7Osd1YZsDh+zNv8AYONye35hUzZHB4zsts1UVmPyctdnd0pNnaD+OSGZ4ap6UVC/5OkcQRF917oIfjnvLcG8/g/tmty1VVtvXa3W25eud11SZHMUeUG7upZsz15uOv8AvsBQZXP02RrMhtaWqjelpp6stKpjjdiAfdbIoejb7QzGOzW2cHkcXWvkKSbFY4rUStUtWajQ08jJkUroabIU+RQOPPFUxQ1KSEiSNHuo91rpSav9pP8AvAsPyTc+/de6gzZTHQSGGavoYZgzKYZqunjlBSnerdTG8gYMtJG0hFuI1LfQX9+690FO8umdrbudslQOcDlJ11/e42KJ6KsLDUslXRXSGZ3v/nI2RmH5PHv3Xui2Z3rbfuwap8nTw1kkFIS8Wf29LMAkY5Ek8ULCtplAHqDq0Yv+o+/de6GDqDtzL7gzFPtLcSx11VNTVclDmI1WOpZqOITSU2QgjURsfEjESgIdQswub+/de6MyOQD/AF9+690FXb2Kqp9tU+5cVTpUZ/YGUpt64ZGpoquSX+FLLFm6Ckgrdw7ZxEeQzO26qso4KitqftqOSoE7K3jA9+690E/zJ21N2b8RO7KPbdVC+SfrPIb62hX0piyEbZ/ZUVN2DtKroJKV5oalnzGApmhkjZ0JKspYfX3Xul9/pjxv+y4/6frJ/Cv9C/8Apb8fkTR9r/cn+932/k06L6PR9Pr+Px7917r/0t/j37r3Xvfuvde9+691737r3Xvfuvde9+691737r3XiARY8g+/de6LD3Z8fMpvPMQdo9Nb8qule/cNQiix++KLGQZzam98XBZ4Nl9z7ImlpaXf+zmYFYJBNS5jDtI0uOraZmkWX3W6/LpMdY/K6CTd1D0t8jttJ0L35MTS4nEZSuaq6u7fMDeJs/wBE9j1MNJjN2UldxKcJV/Z7nxwfRU0OlRPJ7rXRxLi9r8+/de679+691737r3XRFwR70QCKHr3QNdvbObNYpM3QoXyWFikZ47t+/ixqlqY0RQ+uoiIDpxc2IH194RffS9kJOf8Ak+Ln3l+Fn5r2KFiy1P61iNUk6IoVi0yMBJEKEtRoxTV1Ift9zEu1X77bdNSyuWFD/DLwUkngrcG9MHy6KmCCAR+Rcf63vjqrq4VlPaQCPsPU/wDy65e7de697917r3v3XuuvfuvdLDYe223RuWjozxSUhjyOQcqWApqaVXWIj0g/cShU+oNiSPpb3On3cvaub3c909j2Nm07NZOt7eNTV+jBIhEVMD9d6RHuUqrM6VZadBjm3fF2PZp7lczyViQA/iYUJ/2oz8+jwAcD/W+n+2/HvviAAAAMAdYydE/+Y1NP/AehMzTpMz7b+Wvx1ycr08nieKjr99U+2MizkEM8DUGfkV051KxFre99bB4/Z0cAfn+n/E3N/wDbe/da679+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+6911pH9Pr/vv+I96oM469Xrv6e99e660gixF/9fn/AG9/r71Tz8+vdd+99e697917r3v3XuiM96Ypcb2BVVEaFI8zjqLJXAOlp18tFUWNlW5alUkC/wBbk8+/de6Lb2DszHdi7H3TsXLSyU9BufEy49qqFRJJQ1SSR1WPyUcTMiTPj6+njlCFlD6dJPPv3XukX8Pf5Z+Gw+dxfafc25ds9g0uLn+72xsvAQVzYKPL0NWj0uU3TPkEglrKrHtDqXH+LwLKQztIFAPutk18urqxwB/yK3+Fv8PfutdF5+UGGrcv1PkZKJUf+C5PF5usRr6zQ0kksVQ0QH1eIVIcj/UqfeNP3stkv969ntyewjDmxvLe6kFTXwYyyuVpxK6w1PQHpZYuFnFfMU6rHH+Fr/i/0v8A4/4e+UpGk0r/AMV0edVY/NfuX559Cb46qrurN5fEifrHvr5K9W/HPYOC331Z2pld+bLrOyKSsQbp3lnsNvnGYPcNHjMhipnanoqenleGWNQdSsTNHt5y/wC2XNG271Dvdhvi7ztmz3F9PJDcWywSi3IrHEjwtIjMGABZyuoVoBgJZmmVl0FaFgMjqN2j8tu1Pif8gvhz1/8ALbt7qam2p2Z1d8sc72zkeqOst1U2D3lvPYdTtJOlML17tmvO9Oz5dyj+MywyY3HvO2UnLN49C+m+y8jbJzvynz9uvI+w3v19nebYtstzcRF4ophL9W07r4NuI+yokYDwxTNTQ7MjRywrKRpINaDz8qdG6o/mZ8Zch0NQfJnG9pU2X6XymcG06HcmH2xvLL5yfen8Wkwb7EXYGM2/Vb9j3zTZeF6ebEyY5ayFkJdFUavYGf2+5wi5nl5Om2Yx8xJF4pjZ4lj8LSHE3jNIsXg6SDr1lWqKE8OnRIhQOrVXh59Fe+Sfzy27kPh/vPvT4k9jUlbubZHyA6B6h3XFuHZGUxW5tkVu+O4Nj7Y3dtfePXHYuExWc25mMltDcEjUz1dEhMcomp3JUMBlyf7aXcXPthy1z1tJSzuNqvbqMxzI8cohtpZIpYp4GZXVZY6MFZsjS1OmpJh4bPG1RVR9lT0ZTtr5zfE7pvtnOdRb/wC0anau48DuzD7QztdVbA7JrdgbNz+6ZqcbVwG9u1KHatV15tXJ5ePIU3jStyUekzIJShYewhsnttzxv+xW2/bXsqz2csDyIBPAJpUiB8V4rYyCeRFIbKoxOk0rTp1pY0bSxo2PI9MncX8wH4edBbz3Z1z2z3Vjts782HHt6r3ptOk2rvfdGa2vhd0Yxczi915qn2rt3MrQbKjxsiS1mZcjG0GtUnlSRlX2/sPtfz7zPt1hu2ycvNLtt0ZBDIZYo1keNtDxIZXXVNqBCxD9RgCVUjPWmmiVirPRhx49KXrn5pfFnt3to9IdZ9y7e3p2LNjstk8NR4agz77V3jT7fSkl3JD152HUYqHYnYeQ2vDXRPk6bD5CsloVcmVRpfSk3b28512HZP6wb1y9Nb7WHVXLNGJIi5PhmeEN40AlIIRpUUPjTxAPlmidtCuC3+rz6UdZ8pOgaDqjsbvOs7Hx8PU/Um9N19ddi7yOKzrUe2d67I3RQbK3Tgp6Fca2UrJ8TujKU9I0lPDLDI0geNmQFgkTkzmd982rltNpLb3fW8c8EYdayQzRtLG4OrSA0as1GINBkA0HVvEjCF9XbWlfnw6aN3fL7477H7dx3RGf35Wv2jXzbQp6zBbe2Nv3d2M2nUdgMq7Fg7C3dtbbmV2n15Pu0OrUKZitpGljZZCFjIb2oseQ+aty2Kbma221P3KqykO80MRlEH9sYI3dZZRFwcxqwBxxx14yRhtBbu/PoyzoUZo2UhlZkZTywZSVZePyCPYPFSFPmafz4dX6sX+JeCag66r8vKkGrcO4KuohZF/fNJjoocdGlQ/1JWpgmKr/AGQ1/wA++nf3ONgO3e2m473J4ZbctylZCKahFAFgVXI8w6SEDyDfPom3Bj4yL5qo/b0ab3lv0g697917rrULX/B/Nj/xTj3okDj17r2oXt+efwebf0/r79UVA69137317r3v3Xuve/de697917r3v3Xuve/de697917r1/8Aff77/X9+690A3ZnctNtZpsJtwU2Tz4W09S7CXHYlm40y+M3qa0LyIgbIbaz+PfuvdFCy+ZyuerXyOayFTk6173qKl9WhSSfHBGAsVPCt+EQBQPfuvdd4bD5HcGUo8Niac1NfXzCKFLNoQAXknqHUExU0EfqdjwB/iR7917o+uwevsPsTGx09LGlXlJkByWYljUVFVLYXjiPJp6JDcJEpt+TdiT7917oQPfuvde9+691737r3Xvfuvde9+691Rp/ON2PGjdKdmQ06LI0u49h5SsF7tFKtPnsLBLZNJ0yxVeglgbsQBz791sVOB1TTs3Zu7ew85SbZ2HtnN7w3BWyrFBi9v46oyU92YDXUvAjU9DTre7SzvGij6n37rXDq4LoH+UnlcgtHuD5FbobEU7eOb/R9sirjkyDIVuafN7rKPFTMdVmSiRmH4mHv3Xq06tp6v+M3Q3TlGlJ151btTBSKipLlHxsWUztVZQpeqzuV+9ys7uB6ry6T/T37r3Q6BEChQihQAAoUBQBwABawAHv3XugT7X+N3Qvd+3MptTtbqPYm98JmaKXH18OWwNElaaaUliKTM0SUuaxs6SMXjmpqiKaKT1oytz7917okb/y9u1+mseD8JPm53z0stIX+y6y72rn+XHSL0ya5KbE0+H7XyL9nbVohM5vJid1UzhDYo1lt7r3QS7w+V381/wCLFC9X3l8BtmfM3ZuNih+87F+BfYkuP3zLGz0sT1M/x47jjo89U1irJJJJHi85XJcWWyjUfde6HTq7+az8Ye6cXl5urNu/JDeW79uVBxO6OraD419u0fZW091CHyNtDd2CzG28ZRbWzYmvCJchV02NdlLrVND+7791uhHHrD8Id+7t693R2F0J8h9lUPTvbHZvYXZPyS6xwcWVjyW3N4bD7T3PV7mrdp4bcEVsRXdqdTVFWKLdeIpJZhG8kddSNUUM/mT3W2NT0eSp6f6/lytXmaTD1WCyGQqUq6+bbGczu148hWndFJvGqra2kwOSx9HVZDLZ6iR66okjaeugvTztJA7Rn3Veuv8AQ9sZqlqmrpc9lJD/AAm0eV3nu/J0uvCVmdrsfL9jW5yWiWVJdx1SSsI71EJiil1xwQrH7r3Uml6f6uoqE46n2DtcUbY2mw7RS4umqS2Mo9tTbNpqSSapWSd44tp1MmO5Yt9m7RElSR7917puquqaTH1FVkOv87mOvspWVEU9V/DJDlNvVbNUbZjrpanamWefCSVs+B2zHjaeZUQ0ME0jQhXYk+691L2nv2oqsqmyd80dHt3f60888NDT1Kvg920tDS42ry2W2JLWSx5nMYTCfxmmpqyWopaV4qtimgrpdvde6XNPgcHS5B8rS4nG0+SliMMldBR08VU0ROoo0yRq+ljyeeffuvdO/v3Xuo1XTxVVPNTTxxzQVMM1PNDLHHLFLHNG0ckUsUqtFIkiMQVYFSDYgj37r3QXdUS0e5OpdsQyVNLkqKowVVgJpqLK43OU80GPmrcDURLk8Rh9v4qpaIUjRuKehpoY3UoqALz7r3VYH8Uyn/DQn90fuT/Ff4V/st1vHVfcfZ/6ef8AQN9r4/8AgV9z/dv8/X+39PfurU7uGKdf/9Pf49+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XvfuvdIzf3XWxe1Np5fYvY+1MFvbaGdganyu3tx46nyeOqlt+3L4ahGNPWUz+uCoiKTwSgPG6uoYe690Tyn6u+SfxjZH6L3Bkfkl0zTPY9Ddu7rSDtbZuPCKkVP1D3pn5ZF3FjqAACLB7yaVzGuiLNU6hYz7r3Q2dWfKLqntXceS2BTVuZ2L2zg4UnznTvaGFqtjdlUFPJGZBkKDBZUin3dgQFI/iuCqMpi2IstQTx7917oxXv3Xuve/de66IB+v+t/rj+h/wAD70QDxGOvdFd7J6yqMdPUZ7blMZcZKWnr8bAoMmPc3eWopY1tropDdnUcxG5A0/Tk396n7qO48t326e5PtftJuOVZSZbuxiXvsnNWknhQZa1OXkjSrQnUwXw66Zt5L54juY4do3qbTerRY5WOJR5Kx8nHAHg4+fQIKbgG4NxcEfQg/Qj+oI98/QVIqpqPXGfmKVBB4g+nUq5zUZ65e99e697917rPSUlVkKqGhoaaWsrKmQRQU0KGSSRja5sPoiDlieAvJ9mmx7HvPM+87fy9y9tst5vd1KEihiUu7k5OOAVR3OxIVFBZiBnpNc3dvZwS3N3MsdugqWbAH7eJPAepx0cTrvZEW0MSyVIhkzNawkyVVA0rRNpZzTwQ+T6RwRvYkAamuf6e+333aPYa39keTZLfcxBLzxfsHvp4mkZDpZjDDGZKUSFG0kqqiRqsQRSmOXN3Mr8x7gHjLDbosRKQARX4mIHm3z4DHQi+8lOgn0Un5pqP9D2Be3qTvr40Ojj9UbDvzrtQ6MOVazEX/obe/deHHo2tuT/j7917rv37r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3RX/khi9UG2c0if5qesxdRIEc2WeNKmnDyfoVfJC4ANiWb8+/de6Kv7917ofOgNztjNy1W3Z5QtFuCnaamRj6Uy9EoK6L2ANVRhgQPq0Y9+690cz37r3TflMdS5fH1+KrYxLR5KiqaCrjP0enq4nglX/A6HPsu3bbLTett3DZ9wiD2N1BJFIvqkilWH5g06srFSGHEHqnzfGysx19uSv2zmYJYmpZZWxtXIloctivK60eRpZR6JEliADgG8cgKsAffFLn/AJD3r235p3Plbe7Z0aKRjBIR23FvqPhzRngQVoHAqUkBVqHHQghlWZFdeq4/nh1X2H2pF8MV672nk92N1v8APj4+9rb5GM+2/wB+111tI7j/ALybsyP3M9P/ALjMR95F5fHrk9Ysp9nHtpvW1bL/AK4R3S+SAXfK99bw6q/qTymPw4lpXuahpWgxnrUwLeFTycV6du1+st85/wDmMfCTufEbUrcj1x1V0v8AMHB7u3xHHSPjtm7q7Bodlw7FpZpZZVrKbKZ8UNStLLTxllCuC6gm7Oybvttt7Ue4nL1xfou7Xu4bW0UJJrNHA03jMKYKx6lJBNCaYPXnV/qIXXgoPROaTr/5h9P9dfK5+tdsdu7G/wBKv81XsPsrcue6cw+0M93snxD3pT4j+O9k9B7f3JUT4Ku3Dlq+giiJaM5CKhaplggaYD2PJNy5B3/duRxvF3Y3P0XJNvbxpdPKln+9IdfhwXzxDWEVWqM6CwRWIWvTWmVVl0I1PEzTjT5dFryvxQ+TG5eu/wCYe9B1f8n9wzd4/K3+Xp2H1JWfI/PbQ3B3p2J191Dntqvv7de5qnB18NBjP7q0OLlkfH1IirqCiSOORWkBUDGHnjk+z3b2oWXeNmiXbNi3yC5/d6SpZwT3KSCCKPUpLCQsAJAxRnLMMdUaOVlnorEsynu40FOhG/mK9dfNT5DYf53dUpg/mtuz+8u76PHfFfrPpObrPbvw63R0hT1Oy8426u09zZWeDcW7d91eZpMm2Rw1VNFkGqoqeOij8IL+yj2p3X285UuvbTfTNy5CYbdjuM90LiTdY7ykqiO3jRTHFAEMZjlUGPSztIa9XnE0v1C0koSKAUpSopXo1jdG9qnuf+bfulevcmcT8g/jF0d171BmPBjT/pIzW2ug997Z3JtXCzGo+5dMdunJ01NPDUeKBqh0PqtqAJ/rHsn9XfY6zO6r42171ez3UZLHwEkvoZI5G8lBjVmBFWCiny6uEcSXH6eSKD9nn0m+vuge4MBtv+Rhj6jrjMY5vinRZFO9KCGHG06dTNWfFet2VVRZuCnqVhhOR3rIKKb7UTeWqa7lv1e1e7cz7Dd3X3k5Y95iY72y/Rtk/U03JZdUZIzpiGrNKJw6qsTgWa6CAta/s49FX7h6m+UmH+Hvzn+Fu0Pid3F2Rvvt75Y9wdw7K7E29Jsui6dyvTvbHcuzO0KHc9Bu3Kbmpq6v3nh6TFT0E210ohkTUoXLLCCxG2w75yZPz37b+4N/zzYWe22Gx2trLA4mNyt1bWk1uY2jWMgRMWDi41GPSaCrdUZZRDJEIiW11/IkdGk7w2X2vtz5OSdl/E3rn5ddcd9bs3t0ZgO2snj6LaWR+Dfya6m29/AsHuvdfck2R3HUw7dz2x+vqivoaOupaeg3J95RxRJDURsrkF8t3+yXfKCbPzxu2x3nLUFvdvbKxkG8bfdPqeOK0Cx1kSacI7qzPb6HYkowI6u6ssgMSsJCRXGCPU/MeXVurUxq680OISeqerrvs8VCqPJUVDTzmKiURqjSO76lvYEjn3CVhZbhudxYbbt0Bl3O6dIokWlWlkIVVHkDqPDy/LpQ7pEjSOexRUnq4bYe3E2js7bm2kEYbD4mjpZzFq8b1YiD1sql2dyJat3bk/n3269veVo+SeSuWeVUC6rKzjjcrwaUCsrCpJOqQsak1Na9B6V/EkZ80J6V3sZdN9M240zEm3s7Ht6qpaHPvhsomDra6Lz0VHmGop1xlVWQ6W81LT1pR5FsdSAi3tq4mjt4JriUVijQsfsUVP8AIdbVSzBRxJp1pjfy/wDfmP6f+QvV2z/lDnfk/wBQfzTamu7K/vvn+0u3O0dydQ/NFqrJ5dsxn+u3m3Bm+gN7bOxuBqIajD7doqTEZfb324CRNpYnl97t+/8A77XdtzNz37Uc77fd+0itHpks7eA3W25FEv4Jw06SawYzKY3t5RSlOhjBt2228iWd5ZkX4w2okhz/ABIQQKH04jps+Bnzn+RXxm7a7srO7+w93dofHL5+d+/M7ZfW2f3lunMVa9A/LXqfsDsbFbf63wGQrZq3+62y+5+vsTTth6VfHSw5rFmKIo0hBmWT7zfNtjsk0Vns1lufNG0bdtN9fQsWhkudv3C1tmnvIGj1KHsrmRjPF4ZDQyBl06a9Fg2uAy1ldkikd1UihAZWI0mvqOGf29Gy+Dn80Ht3rz+Xr/K46M692RQd1fLT5I9PbhzGL3P3p2VuXGbEwmxurmyOW7K7T7V3bjsHvHf2SGNgqqShpKGipKqur66pRQUjjY+xm33qNj2fmT3U2/nDl2ay5X5Xv4bWS+iY3JlkunVbZEtUjEhcr4kkxD6Y0So1FgvTP7neSGzeCassyk0YUAoKmrVp9g62L/i/2R2p2p05tvdnduxtkdddnTy5Si3JtrrjsSHtDZLmgyVVSY7Pba3T/CcBk2xG5MdDHWQ0mQoaTI0QlMFRGJI2JyI5T5z5V572e15g5Q323v8AaJkVleNsgMKgSIaPE/kUkVXBBBGOi2e3mtpDHPGVb5+f2HzH2dGE9ibpjr3v3Xuve/de697917romwJuBYE3P0Fh9T9OPfuvdFY7W7lLip2zs2rH9uny2epyRp+qy0eJk/LCxWScXA+ic8+/de6K9a3+HJJFz+onk8/2ifz9T7917pXbN2Xmd8ZVcbiYtEMWh8jkplP2eOgdmAZyCvnncr6IlOtvrwAT7917o7mxuu9v7EppFxqSVORqY1Svy9VpNXVBSWEKhbJTUyPyI0t+CxJ59+690v8A37r3Xvfuvde9+691737r3Xvfuvde9+690kN77B2Z2Tt2u2jv7bWH3dtnJNE9bhM7RQ19BPJBIJYJTDMp0TQyC6OpV1/B96qPXr3TNg8N1d1Li48JtvGbN2FioFCJjsVS4vDRtZWlAeGmWGWaQrcgtqZvYH5q9zPb7kcf8jDnbattNaUuLmKJydJegRmDElQSBSp8q16MLPad03HFht003zRGI4040p/PrNTdp9f1UqwQ7rxRkdXYeV5YEsilmvLPHHEpCj6Egn8e4q2z73P3bN3u47Gy94tn+pYEjxHeJKKuo1kmjjjHbnLA+gJ6O5uRub7eIzS7BcaBTgAxzwwpJ/YOlrR5CjyCeSiq6WrjspL008c6jWAyXaNmA1Kbi/49zrsvMOw8x2xu9g3uzvragOu3mjmUBhVamNmAqMivEdBqe2ubV/DubaSN/R1ZTjjggdTfZz0z1737r3XvfuvddWH+x/41b/evfuvdFs73+MOxe7JMbuqGsyvWnde04ZW63762AabGdkbHrLrLHAat4ZKHeG0KqWMLkNvZmKtw+RgZ0khDlJE917oq+40+QG9dk5bpn5k/Dag+RUOHyNJU7Y7a+Pm7NmYvbu8Kyg1nDb4o9r9gb72T2J0L2Lj1e8v8OyOThpJ2c0eSaNtC+62OPHoO9mde/wA03b1VQjqPdnXuzeuKKq0jYPzd3hN8g9/vjw5vT43fvSOP2xkcUkcOhITlM3uaVVBLuzG/v2OtnTXz6VO6Oxf5x209xAUfx1+EvaGz/wBtFqNg9v8AZOD3QzKSZqisxvYmJ2zi6GFwLIkFVXyAm5vax917t+fQlbI73+f3ZFCmHk+Ee1uhdzUqq+X3d3J37tTdfXU8bTTRJ/c7F9R0u4d+Z+seOHySQ5Sl29HTq6gTzG9vdeoo8+l5JhP5gsflykHYnxJqqhkPj2hUdY9sUWIikEn/AD2MXZFXlWEkI/UcXZXN9LAWPuvdvz6R+9+5t+4rauUx3yq+N+79pYTDSUGXXt7o3JS96bAxWTwlXFlcfu+TB4CiwfcOGosBkaOKsliq9t1NEEiKzSSxa7+60BXoeemO8tt9hYXBvT7s21u6izMIi2b2TtnN4/M7S7Ip6daiGGSHK0FDisPjuxK2DFVNdkts0yyz4qJWNzGpK+610YYG4uePfuvdJ7dubj25tvN5uRJpTjcXXVUVPTUldkKqpqI6dzBTUuPxcU+Tr6ieaypDTI9RKTpjUtYe/de6hbDwtXt3Zu1sJXSzTV2NweOpq+WfLZfOu9cKdHrbZjcH+5zIxLVM4jlq/wDKGQDXZr2917qoy1X/ALIt/ev+Lyf3Z/2ev++3n/yjwf3E/wBne8H8O16df2Wv0araLf7Tz7305+L8uv/U3+Pfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+690D3cHQfUffOGpcL2psjE7oXGytV4DNN9zit27SyJsUy+zN5Yaeg3TtDLxOoK1WOq6aYED1Ece/de6LrHt75Z/HDHuNp5qT5jdX4x4Vpdp74yWP2l8ktt7fp0CzQYfsZ1TZfdNdSRDVDDnKfBZSo02lylTM1z7reKfPodulPkb1P33QZJtgZ6ddx7ckSm3r11unGV20OztgZFzY47e+wM/DRbj2/P5NSxyyQmlqdOunlljIc+610OgPv3XuunUOpVgCrAhgRcEEEEEHgg+6SIsiNG4BQihBFQQeIIPkfP1HWwSMg0P+D5j59AHuzpenqpXrdr1EGPkkeaWfG1RlNJJLKwcGllXW1GoJPo0lP6W987PeX7iW2b3eS757Q38G23cjSPLZXGtrd2Zgw+mdam2Udy+GVeOhxpp1Kmwe5U9tGttvsbTKKBZFprAGO4Y1n51B+3oE6nZW8KWVoJttZdnX6tBSNUxMLkKyzQF0YMBe17gHkD3gVuXsD737PeS2N57V700y+cNu08bCpAZZItSkGlaV1AEagCadSdBzVy5cRrLFvMGkjgzBSPkQ1D/AJOuEOzN31Mghi2xmfIwYjzUjU0fA5vNUaIl4P5N/wCnPum3+w3vdul0llZ+0++fUMCR4luYUxk1klKRr8qtk4AJ6tNzPy7AniPvVvp/ouGP7FqejNdedb0u0Y/4hWSR12dqEKvUqjLFQwMAWpaNWu12I/ckNmf6cAW99Wvu2fdf2j2Wt5OY99nj3D3CuU0tMFpHaRMKG3tgc93+jSmjSEaQFQAGEubucp+YnFrbKYtqQ1C/ikYcGc+n8K8Bx4mvQp+8tOgP1737r3RP/mTVqdqdL7cMKVDb0+VPxzwAgcSvqhoexcZu2tm8cfLiiodsyzm/oHjufp791sDj0b8fn/XP9P8AiPfutdd+/de697917r3v3Xuve/de697917r3v3Xuuibe/de+Xn0hcv2f15t+skx+c3ttbFV8QLS0dbnMfDUxAMyfuRNPrjbUpBBAI/p7eSCdxqSFiPs6Dl7zfyvttwbW+5gs4rgVqrTIGFONRXH2Gh6VGOzOJzEH3WIyVBlKb0/v4+sp6yEF1V1Bkp5JFBKMDY82I9tsjqQHUg/MdHNpfWV/GJrG7jmipWqMrD18ifI16c/delXXvfuvde9+691737r3Xvfuvde9+6910Tb37r3Rau+N57bqMVUbNSSWtzkVXRVrGmCvSYyWFywSsm1qPuJIWYeMBmXUCbe/de6Kb7917oSuocTNluwsAIjpXFzS5iocAMFgooyADY8eaadEH+v/AIe/de6P8DcXH0Pv3Xuve/de6DbszrLBdnbfbDZcmkqoJPuMTmaeGJ63E1VxeSESWWWCdRpliJCyL/QhSIu91PanYPdfl1tk3pjDdxvrtrlFVpbeT1XV8aMMSRkgOvEhgrB6KdoWDIPt6rW7H6l3h1lVMM3R/dYV5dFFuOgWSTF1AZ2EUdSzAyY+tZACY5bKTfS7Ae+Wnub7N86+1d3p36y8XY2bTFexAtA+SFDkCsMhx2PjPa70J6OobiOYdpofMdBl/j9R/X3FIIIBBBHy4fken+vA2IP0I5BHBH+sfx7317r3/I/fuvde/Fvx/T8X/rY8XH9ffuvde9+691737r3Xv8Pxe9v8f6/6/v3XuuQDsyxRo8kkjpHFBDG8ss8zkLGkUMYZ5ZnJsoALXP092RHlkjhijZ5nYKqqCzMxNAqqoJZieCgEny68aAVJx0fzoLoCs2tW0e+d6R038YNHrw+AaHyyYKaotqrqydmKfxZae6KiLaESNdiw46Mfd0+7necp31l7gc8rH++vAra2RUE2jOczSsSR9QI6qqqKRB2BJfgU3d34g8OMUX19ejg+81+i7r3v3XumTcuPyeW25n8VhM5NtjM5PC5TH4jclNRUeSqNvZSsoZ6egzkGOyCtQZCbE1ciVCwTgwytGEcFSffu38Sgr5g8CPMH5Hr2fLj1rD9mfBf5OS/JH4+7p+YPyZ697g278Wewqvu7q7D9WfH9+nMn2P2JkcFX7cxm9uyczU7/AN5UOOjw9NWVBmxOBp6Shqqwh5LIFQcYPe1Nj9kOYvdv255V5PvbbceYrYI97c3gniO3zy+ORaW6QRBWaVWjZppJXhKFVqCOh1b+LuCWW4T3KlQSdIWncO2hNT5UIpxB6QWU+BW19z/EPuD4obt3vUV0XZXavbvc21+ycVhWxGa6w7C332nlu2dhbp29RHJzySZvrHcFfGgmSoh+/iSVbRLLpEUW/u9e2HuTy57h7XtCr9Dt9nZS2ruGS6t7e1S0uIpG0CsdzGpopVvDJU0YqOlH0Km2lty9dTMwPmCW1Aj/AEvSN258B979Z9UfBnGdR947bxPfPwY2ZvDr7Adl7z6zyG4uvO19l9l46fHdj7Z3n19jd34rPYnH5mX7esopqTLNUUNXSgkyKxAMtw939q3zmH3Xn5j5Unk5R5suobiS1huhHcWk9q4a1mhuGheN2QBkdXg0SI5GKdUFi8cVmIZtNxCKaiNQKniCCfP7erRviJuzt7pGHFUvaHYWM7k3VnsxXjeOXwGz6frHbFYmZyt8NQbc2nBk8++Opdn0MwpaSWsrKmrq0Baok1NdV3tT71p7Ue5Y5h5J5ali5Rulit59ue7MrzL2x/UPMUjVrsFmdDoEa6jGuhCT05NYHcIEs55QZCahgKaT8gThT5gdXdqbqpsRcA2P1Fx9DYkXHvulC4liikAIDKDQ8RUVzSor65PUekUJHXftzrXXvfuvddE29+690Uvt7twV5qtp7VqiKNWenzGZp5bfdsupJsbQSJyIAVImlBGojSptcn3Xui1+lV4Cqqj6CwAA/oOAAB/sPfuvdDL1/wBN5rd4iyeVeTB4CQh45njvkskgYE/ZwSaRBA4BHmkv9bqp+vv3Xujl4Lb+I21j4sXhaKGhoovUIolN3kNtc00jEvLNIRdma5J9+6908+/de697917r3v3Xuve/de697917r3v3Xuve/de6LX212vWYqsn2rtiY01dAEGWy6hXkpDIgkWioQ6sgqTE4LyH9FwFF+Ry3++d98XfOTd63H2g9qL57XmG3C/vHclCl7cuqutraBgyiXQwaadgTFUIi6yWWZPb/AJCttxgh33fIw9m9fChNaPQ01uRQ6a4C+dCTjiQ7I9z9cp3Vi+iMnu6KXurcnXmY7dx20qqDJ1GTyPX2Cz1JtrL7nOUamfG+KjzlbHAYnnE7FtSoUBI5T3uxc077s+5e5m5xz3m3ncVtLi/ml8WVruSIzLG7SOZWZ0Vn1ZUAUqMDqaI76xtLm32WIpG/h1RFFFABC0AAoBUjoLvkb8x/jb8U02pD3r2RDtjN77kr12TsnB7b3V2D2Ju6HEqr5jI4Dr7YeF3HuuuwuGRgauu+1Wkp/o0mr0+znkX2q569yxuLcocv/VWNppM88skNvaQ66hFluLmSKFZJCOyItrbiBTPSfdOY9s2Ro47yek78FFSafYoLHj5A/OgoehU6B+QfXXd2xML3H8eOyKTd2xs5V19JRbm242Sx6HK4KsehzGEzWHytJj8ris1hK+NoKygr6WGogkuroL+1Mcnuf7C84S21ne7jy3zbb+E7LBL4YkUjXEzCMtb3MLKxKkiSNlJAOTSsb7JzVY65Yo7m1aoo4BKcQQDSqnHqCCPl1ZB1X2R/fSlmoMmIYdxY5EeqWECOHIUzMFFfTRE3j0OQsqXIViCDY8dqPuf/AHpR79bJe8vc1iG39zdrjDzrGNEV5AzaRdwR50aWIjniqQjlWU6XAGPvPfJh5YuYrqzLNs8xIWpqUb+Bj5+qnFQDXI6GBm0/7z/vHvNRm0ivUf8ARPOyvlfjds5qtwGzsLT7mnx0ppqzNVGQMOGFSiss9PRLTRSTVr0s40OwKRkghSfr7wH93/vwbNybzDuHK/IPL8W83FpJolu3n02viAEPHF4YaSYxvRXcFYiQQrNQkCGz2KSaNZbiUoD+EDP+botOb+a29KbcOB27U7q632zuLdMNb/djalU+Mhz26DikM+Um25icvlf4tm2oISGnFKkghUXYD3jbd/fT+8Dukd1ue2Wu3QbXboomMVg8sMZJNHlmkkfwi1QFDnSSBpBJPRkuybchCOzFz6tSvSBz/wAx91bV/jZ3T3ltPa423hKbdG5P7xZXZGE/u7tjIV7Yyg3JnRk1pv4Nt2uyYNLBWVHjp5ageJWL8ewZafeX+85uJtzYc4X84nmaKLwrGCQSSqC7wx6bY65USjFFqQtD8+nztW1LUNFTSKk6uA9TU9IbC/zFtubly+K2/tv5e9J7iz+eq46DA4LA9jdUZfNZyumYpDRYbFUFbPXZOsmcFUihjd3PABPs1uffj73FlBPd326b7BZxLqkkk2pESMebOxtKIB5liMZ6bG3bMxCqUJrwD1P5AcT0ucH8+aqbe2V6lxvfHUW4u08OslVmOuv7wbHyfY2IiSNWlSs2lj8lDmaZaaNC0kbU/kiBJew+i6y+9L96PZthi3m6uJZtinfVHe3O1jwm8tKTrHFEV4Fc1JJAJ6qdp2p3MakeIPIPkfaPL7OjC4n5jbwp5Qc5tLAZWmJiv/DqqtxVSqAESMrTfxCF2kYgi6qq2I/PA82H7/8Az9azg8ycj7Ze250g/TvNbSAAHWav46MSaEAqoGePkzJy9AR+jcMrfMAjo2vVndO0e1aaoTDtUY/N0MSzZDAZLQtdTwPIY0qoZIy0FbSM1gZIidDMFYKSL5zey33h+RPe21uY9heW15jt0Dz2M1PGRCxUSIV7JoiQAZIydJIVwpx0Q3223Fgy+IKxtwYcD/m6F4G4v7nr/D0X9eIJ/Nv99x7917olHavwb643nls/vTq/c+8/jb2Vuf7h9ybx6XrKLC0G7KqpjMM2S3nsato63Z+fzz08jxjMpS0ufjSRljr0UkH3W9R9egQ2z3f80vjItZifl71/je8Ngy7kqpMJ8i/j7gMrPFtTZktbXvHD2/1Ti8fk93Y6sweLjplGQwtJk6OpeRzO0Hi8knut0HEHofOuO1OuvkfFtjeNZ2j03mduYo43O0/XO096ba3pDBumnjw2SpK/ddbk8dQZqh3BsTc+OmfGmnhoLrJqqIfIqBPdV6x97/K7b+FSt6f6DymA7a+UO7aSfDbG6921lY8vT7Rrq6Nqdt/9sZXDrkKTYWwtmxSHIVkte8E9csApKJJqqeJD7r3Tn/spW3f9kx/2UL+8Fb9l/op/uL/ffw/7k/74fb/xD/SH9t5f+Ln/AH6/3MePyf5706/7Xv3Xuv/V3+Pfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XVha1hb37r3Rfe4fjZ1z3Bk8Zu+riy+x+2tt0NVj9l939cVsW1+1doUtYQ0+Potxx09RHmtuVUgDVOFy0GQw1WeZqVyAw91uppTy6ArGd89sfG3JU+0PmTJhsz19VzR0e0vl/s/CPgdizO7pT0uI+QO045ayn6a3PVzyRRQZaCaba2SmcKJMdMVpW91qnR6sdk8bmMfR5bEZCiyuKyNNDWY/J42qgrsfX0dQglp6ujraV5aaqpp42DJJGzIwIINvfuvdTbA88H3ogEU8uvdeAA+g9+oOvcevWB/Hv1B17rsC3A97691737r3XvfuvdEf7dkbsn5j/Gfq/HulVjOm8Pvv5J9hwaA6UFVU4at6p6ihmmVz4KvJ5fcecq4YnA8seKkdb+Mke63kdHg9+611737r3Xvfuvde9+691737r3XvfuvdYppoqeOSaeRIooo3lllkYJHHFGpeSSRzZUREUkk8AD370Hn1V3SNHkkYLGoJJJoABkk/IdVb97/ACf3Bu7LVW3uusxW4TZ1E1RSS5jGytTZDdLkLHLUJUGKOqoMVGyssIidXnB1lgCq+z+zsEjQSTKDKfLyHWHnuP7v7jvd9PtfLF89vsUdVMiGj3FeJBoGRBwUDLZY4oOiglFYs7qHkdzJJJINcryMbtJJI13d2PJJNyfr7M6kcDjqCmAJYkVJNSeJJ9STkk+ZOelnsDfe4ustx0e5tq1Jp6inYLW48lhjsxQs4apx+QplZUlSZF9D8PFIFZSCOW5oY7iNo5OHl8j0IOV+Zd05S3aHd9om0yoRqQk6JUr3I6jBBFaHirUI4dXabS3Njt5bcwm6MRJ5cbncbT5GlJN2RZ0BeCT+k1NKGjcfh1PsKSIYpHib4lND10G2Tdrbftp27ebI1tLmFZF9RqFSD8wag/MdKT3To0697917r3v3Xuve/de697917oIO4d9VGz9uxQ4uZI85mpmpKOTUokoqcIzVWRSMhiWhFkQkW8jg/j37r3RGHd5XeSR3eWV3lllkYySyyyMXklkdzqkkkc3JJuT7917oT+vuuF7Bosx9pl/4XlcTNTBY6ikNTQ1NNVxsY3d0kjmimjlidTp1Ai3v3XujTdc9YYzYEVROtQ+SzVfCkVbkXQRRxwoVf7ShgBJgpvL6iSWZyASeAPfuvdCj7917r3v3Xuve/de6jVdHSV1NNR1tLT1lLUI0c9NVQx1FPNGwsUlhlVo3Q/0It7S3llZ7haz2V/aRz2cqkPHIodGB4hlYEEfIjrYJBBBoei9br+MPWm4jPPjaSt2pXSnWs2DnC0KtpRAP4VUiWjWOyX0xiP1MT7xt5v8Auoe1nMrXFxtlnPtG4Oah7V6RA0Ap9O+qPTxJVNFWJJPSxL2ZAASCv8+i85/4kb8x6yS4DNYLcKhpNFPP5sPVtGunxG8v3NIZpLnUNaqtuCb294z8xfc09w9tWSXl3fdv3NQx0o4a1kK4pXV4kes1Ne4KKevStdwjOGUj58egszXRnbOClVKrZeSrVfSFmwhhzMOpk12Jo3MqFQLEsirfi5PuI9+9gPeLl2ZIrvkS7uFY4a1KXS1pX/Qu4eh1ACoNCelCXVu4/tc/PHSTq9hb6oJ4aat2XuunnqNJhjfA5BjIGfxrZo4XQXfjkj2Dr7289wdsnt7W/wCRd4iuJaaFa0mq1TTGlSOOMkdOCWE8JV/aOnL/AEVdm/8APv8Ad3/nkqf+KezP/Wi91v8Awm2+f9kj/wCbqvjwf7+X9vTniuk+18zU/bUux81SsNOqbLxR4ilUPr0sZ614w3KWOgMV4uOfZvs3sT7v75dG1teQNwiIAq1yq2yCvDulpUeumpHEjrRubdcmUU+XQz7e+Ie7aw08u59y4nCwM5+5pMXBNk69IvGrL4qiU09EJjISpurqLXF729znyz9y7nK9NtNzZzPZWMBPfHbq88oGkcHbTFqrUEUZRSoJGOksm4xCvhxkn59Gu2X0p13saSgrMVt+nnzVBD41z+RLVmTklZSJakNKxgpppCT/AJlE0g2Fh7zC5F9iPbP2/fbr3ZuW433y3Sn1cxMk7PSjSVYlEc5/s1UKDRaDpBLczSijP2ny6Fiw/p/vvrf/AF/cxUHp0nr137917r3v3XuvHng+/de6Cvs/qTavaOPjps1TyQZOkjdMXnKMhK/H6zraMarxT0srj1xuCD+CDz7gj3u+7/yZ727ZHHvCNa8y28ZS2v4sywAtrKMhISWFmy8bCvEoynPRptm6y7c2jSHtGarIfM0pUHipA4EfmD0Rjd/xT7G25E9XhZKLd9Kr6fDjQ1Hklj0avK1HUv45gGGnTG5Ynm1vfNDn37mXvPyZC19tlnbcwWCn/iCWE4FKkm2l72FcfpSSN8s16Fdvu22XJAjuWjfzWWgH+1dcH/bAdF5yuBzuDq5KHNYfJ4ythTyyU9dRVEMqRBinls8dvFquNQJW/wCfeLu57Vu+x3s23b1tVzZbggJaKeJ4nUA0LFZFUhAca/hqPi6M9DaQ6gNGfNSGH7RUV+XHoSunMbsuLceP3b2ZubAbN2Nh87gsTBmt057FbbxGW3vn6+Kh2jtKnr8zPTUtblc3kHCQ0kbmeeTSiKSwHvIj7qvs9/ru+4kM+52sj8m7PouLl1C6HmVg1tbMzKykSMC0iAajGCcA16L9xvjt1qZ1oLlqqgP/ABpqce0cDwrTq0fevYGw+tMIm4uxN6bU2Ft5q/H4hM5vHcGK2zh2ymUmFNjMYmRzNXR0bV+QqDoghDmSVuFB99v1UCiItAMAAfsAH+bqPei9R/Pr4LSvDHF8zfirLJUVEdHBHH8gup3eaqldYoqaJF3YWknkkcKqi7FiAOfdiCMEZ698+jCVPYmwKPeWE65q98bRpewdy4PI7n25sao3Hh4d35/beIlhgyufwu25KxczlMLjZqiNZ6qCF4ImcBmBPvXXunbclDXZTb+Yx+Mqfs8hW46rpqOquAIp5oXRGLaXKKxNiwBKg3HPv3XuiGVfV/YFBUrRPtTJzSavHHJQxrWUbDVpDLUxSGJI7/QuUNuT7917oeutuj1xzx5vesNNV1q6JKLCK3npKF1YuJq5wfFW1CkDSnMSfU6j9Pde6MmAAAAAAOAALAWFuB+Pfuvdd+/de697917r3v3Xuve/de697917r3v3Xuve/de64tq/H++/p79TI60a9Vv7jesk3NuGTIFWrmzeUFUVYuolWrkUqrFmuqqAALmwHHv5dfda43m790Pcu45ikDb+3MF99QVJZPFW4kUhWJbtVQiquohQKYpQZl7GtumybOloKWwto9PrQqOP5nqpjdjqn86/povLFGo/lodzG8sscS/8z+2cTzIyj8e5Q2wE/dN5p7WP/I+seAJ/5Z1x/lP7fn0G7plXnWzZuH0h/wCr0fSS+P8AW4qP+c//ADAB2DVY6Hs8/HD4rR/HBsrUU8eTk6ANBuGfsVOt3qpFlkpD2V6879hf9zxefgW9mfOkF0fuq+y37ljc8v8A7+3f96aQSn7w1RfTfVUxq+l1LbCXI7tGei6wFm/O26C+UNIYo/C1DGnS2qgPlr4+VdP9Hq17b1XtOsiysu0anbFXTnP5Zc7NtWbD1FMd1rLGM+uYlwrPB/eVZ9P3onP3Ye3l5t7xrv490ia2XdY7hJPATwxMHB8Gn6egPQ+FSvh6eynw9D20+i0P9Fo0ajXT6+fQ99HMy9i0QDMofFZZWAJAcCKJgrAGzAMAbH6H3mP/AHfs00f3ldhjSVlik2ncAwBIDUiRgCODAEBqGtCAeIB6BHuiqnlC5YgVFxFT5GpGPy/l0Yru3J1uH6n37kcdUPSVtPtyuEFRHbyRGbx08jRk8K/ilIB+oJv+PfYv7xO77lsPsj7m7vs920G5RbTLokHFS1EJB8m0sQDxBNRkdY47aiyX1qjCoLjqnsKEUIosqgKoH4AFh/vHvgcqhFWNfgUUGa0HUhfZw6pD/mM/H7P/ACP+dHxP2j1/uGfZnduy/iN8pO3/AI773p6uaj/ur3p1r2N1bnth1Fe0TqlRgc7VJJicjDKHiejr5CVNre8rfZPnGz5J9pPcXcd5sluuVLrmParPcoCobxbC5trqOfTUYkjFJYiDUPGtCK9E9/btc3tssbUmETlT/SBBA6nfFHtfYPzO+cPZ+/8APbQ27laLe/8ALK6E2x3B1Xu7DYrPUG1O0dnfIrfWB7R6z3ZtzL09bQzS7T31iZ0SGphYGFYJQPUp909w+Xd49svafYNotN0mjltOfNwks7qKR42ltZtuhktbqGVSrATQOpJQ0DF18ut2siXl5I5QHVAgYUrnV3Ag/McOl38Euh+iMd8sv5neRoOkuosdkOvfmdsmk68yeL6x2LRZbrqCToLaNaF6+rYsEJdnvBkauSqiFAadRVuZf1EsSj3Z5u5un9uvYeCfmvcngveV5zcq11cFbkjcJh/jC6/16ooQ+JqJQaQcdWsoovqdwPgriWgooFO3yoMZz0Enx26/x38vrtDpPoD5SfGjrLc9Pu/vneuK+Lv8xrZmBwOQ3buXtPtGv3JuDGbU+Q6ZKgj7G2R2vuDF5Gpxwyi1tZishGioF0qXURc6bzce8ex81c4cg8839u1ttED7ry3NJIkMVraiOJptu0N9LPZxsiSeEY0mjNTU1FW7dPoniiurdTVyFlABqTmjE51H18+r0GBVihGlkYqy/lWXgg/7UCLH3iXmgPkc9HPS/wCqc7ldvdj7NyOGaX7t85QY6SGIMfu6DJ1UNLX0cqxqzvDLC2pgBe6A/j3KfslzJvnK3u3yDu2wO/17bnDAyqCfFhuHWKaJgoJZWQ1IHmoOKdJL+FJrK4WSgUKTX0I4Hq5Rfp/sSP8AbH39AopnPn1HfXL3vr3XvfuvddAAfQW/1v8Aif6+/de6Lf258PPir3w88/cfx26c7HrKoN9zk90bA23kczOCrKVlzT49ctIrBjwZiPfutgkcD0W+m+L27/hjjf4/8HMXDX9X4smq3R8Nco+Gpdv57Ga2kravpTf+Sp13BsXfdIkjy02Oy1dXbeyTAU2nHFlqo/derjh0Ov8As5XQ3+gX/Zg/7xZP+6X8S/ur/dT+AZL/AEqf6Tfvf4T/AKFf9Gvj/vR/ph/j3+4/+A+H7rz+v/gP+/791rr/1t/j37r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XvfuvdYp4IamGWnqIo56eeOSGeCZElhnhlUpLFNE4ZJYpEYhlIIINj7917oh+X+N3ZvQOVye9vhXl8BjNvZCtqMzun4l76nnx3Su5K2olepyeQ6nzlBTVmR6E3dlJHd2Sjp6vbFZVNrnx0UjyVXv3Xuhi6m+UXXfZ+6a/rLIQZ3q3u3B0hrM70l2dQxbc37DRRBlnzm2VWpqsH2Ls8SowTNberMljSLB5Y3JQe63Q0B8ujJ+/da697917r3v3Xuve/de6DrtvtHZnSnW28e1uwck2K2fsfC1GbzNTFE1TWTJGyQUeLxVEhEuSzebyM8NHQUkf7tVWTxQoC7qD7r3QI/FDrHdu19v7x7a7ZpxD3j8iNyxdjdh0TWb+5GHjo1xvWvTtHKOHoOqdmLDQysCVqMvJX1Y/wCBJ9+63XgOjZ+/da697917r3v3Xuve/de697917r3v3Xuiu/Lfe020+qqnGUNT9tk95VsW3oWT/OnGOpqc2YiCChagiMeoXt5f9j7MNtiElxqb4FFfz4D+fUQe9fML7LybLZ28um8v5BCKcfDoWlofLsGn/bdVMBQoAHAAAA/AAAAA/wBa3sQ/bx6wjHAdcvfut9e9+691Zz8J9znJ9eZvbE0pebae4Jft42KXjxmcj+/plVQNYiWqScAkm5BAsB7I92Sk6yAdrD/B1mD7Abwbvle/2d2q9lcnSK1oko1gD5BtY6Od7Kup5697917r3v3Xuve/de697917oiHd1ZPVdjZWOfhKCkxtHSLZrCnamFSzLq4HkmmYnTwbD8+/de6Cb37r3RoPjfi5vLubONrWmZaPEQ3uFlljL1c5AKi5hDoP+Qj7917o1Hv3Xuve/de697917r3v3Xuve/de697917r3v3XuurfXk8/63/FPeqDr3Xrf4n/b+/U69163+J/2/v359e69p/1/rf6+/U69137317r3v3Xuve/de697917r3v3Xuve/de697917rogH6/6/v3Xuo0tFSzktNTwTMUMRaWGKRjExBaMl0YmNiouPobey652jar2V57zbbeWdo/DLPEjsYyamMllJKE5Kk0rmnVxLKq6VlYLWtKmlfWnCvWv5/Mk2PvH5d/Hv5MfGmlyNNtx8/i86vV9HicfR42k232lsDMxbs6s3NBIlOtZHmKXe23aKRqoOkgDnSQoHvj7f/eT5+5O97NyjS+Tbvb2w5hngn2yzgit7eS0Wdop2kRUDvNIh8YyswYuAV7AB0NP3dBcWKSSBnuWjBDsxLA0qM+g4U4dVObP+Rm4/5l/aX8tDGbjyWZy+J6e6mqfnF8mtv5qvqchR4juXadTmegOstg5eiqpJoPvKbuHGbky/gljuIqBH5VvazmbceeParbPebeL7nndrjd7nc02TZrh7qZ3S0kEe5XF3Cxfs/wAUktYlkWjAykDpuNYbxrJFiUIq+I4oBVuH+GuOjm/F7b3Sld/O/wCzNqb86o6lyGKr/wCXD01FtyKp6v2BDh6Pcg773bLBW1NDDgYKVcxkAywR1ix+dikcbsQEK5Kfdi9/7w+3PKdr7n8wzXV1uHMVztdpeTsnYY7aOeGG5lNGfWxMUUjlm1FEYnVUFm7WCi6lNrEFURhyB6E0qPs8+q3/AJP/AD46LrfmP3J/M8oOwMs/bXw9+cXUvSnSGxsbsLtPI0e4Pgl03Tbh6g+WbY7eOO2hP11TUPY+7O0dxZ8CXJxlF2zSq5VrD3noVZDpZSD/AKh/LoP9b12CzeJ3NhMNuPA11PlMFuDFY/N4XJ0j+SlyOKytJFX46uppBYPT1lHUJIh/KsPeuvdOlv8AEj/Wt/xT37r3Xdvr/j7917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917olXc2yqzBbjrNyU1O/8CzUyTS1CBfDQ5OUaZ4ZwmnwpUtHrRmAVmYi9/fCj793sPvXIHuVvnultu2u3t7vsyTSTqBotb5xpnhlC/wBms5QTQuV0O8joW1jrJL215nt9z2iDZZ5gNztgVCni8QPaynzIrpYcRQHqvTv34SfE75Tbh27u75B9GbS7R3PtPCVm2ts5/N1W5sdlMPt/IVv8SrMPTVu285hZZMfU5A+Zo5C6+TkW94p8me7fuV7cWe4bXyTzfc7dt9zMJZY0WJleRV0q5EscgDqvaCKefQu3Tlvad5mSe/g1TKpWoNDQ0JB+WOmrsj4EfDftvYnWXWvY3x+2TubaPS2MGF6kgmn3FQZ/rvCGMQy4ba+9sXm6He1Hi6uIaaiBsg8VQDeQMefajYvej3T5Z3nmHmDY+dLq33TdpPEvSFiMdzJgh5YGjaAupFVYRhlPAjpuflbZLqG2hltARCKKfxcKfEM8AAaEVoK1p0PHUHTHWXR2ycB1N0b11t7rrY2FepGA2PsnFCjoI6rITtVV9StPF5azI5XJVbtLU1U7zVNRIS0jsefYZ3feucfcbmb63dru/wB65vv3VFw89xOwAVI440BJAAARI1CqOAUdGVpabfstkUiVIbKMEsxoqjzJY4H7f8PGwfqDriq2pDU57PxRxZuvjWCCnV0lGNx/pdlkkAKrV1Di8gDEKqhfrf32e+5P913c/Zqw3T3B9w7NIufdxi8KGDUr/Q2R0uVdgKLcTMAZgrERoix1J19QB7h85Q8wSw7ZtTk7XEdRahHiScAQD+FRXTUVJJPp0LOew+J3Vhspt/KJHWYzL0NTQV0CSrqanqYzGzIykmORDZkb6qwB/HvNrmPZtg555e3vlbdhHc7RfWzwzIrgkpINJIIJKsPiQ8QwBGR1GyGW3dJQCpU1FR5jqrffvx87F2NPVSx4ifcmBSef7PMYOOSun+yRlMEmUx0StVUU3ikAYhXQsjHUB74we5v3Vfdr22uruWLYZd25ZWRxFdWgMreECNLXEI/VhbSQCaOhYM2oAZGlru9pcqNUoSbzBwCT6HomOe6T2RmO+evfkNlmztP2V1Z1x2D1ZtmnGQ+ywQ2v2dkcHmNyHLYaelFTUZcVOApzSzCRBFGWujXBEJxc17ptfK298hyJFHtV/fW11MsiFZ/FtkdYgNekopWUllKkntIIFamHgpJLHOcsqkYNRRuP+DoO+sfh10X098ju/PlP15gstge0vkpisFjO1oYspG2z8jU4StgyTbgxOAFHG+Jz+er6dJ8jKJpIp5gzqiNI5Y8333M5v5m5H5Q9v96uY5uX9jkka0Oj9ZVkUqI3etGSNTpi7QVWgJYAdUjtYobia5QESSUqPLBrX7T0vOv+hOv+td2/Ibee3Bnpct8nt9wdh9oxZTKmoof4/Dsmj2AItrx00FJVYLHPt6hS6+SWZagmRHWygFO88373vu38l7beCIQbDaG2tSqUbwzO1x+qSSJD4jHJABXtIOerJAkZuCK/qmp/ZTHRd9h/y+OrNm7+613zuTt75Qd4UfR+aqdx9E9d99dy1nYfXXUW5KikqMfT7kwOJmxNDk9xbhwmPqpIMXWZuryEtBG37Z1jV7Gu7+8u/wC57Pvm0WPLWw7TJukQjvriwsVtrm7iBDmJ3DskccjgNKsKRiQihxjphNviEiSPJI+k1UM1Qp+Q8/lWtOj2g65EiF3mlYLFEoZ5ppHIAWONdUksjMeAoLE8e4hFZJIoVUmdyFVQCWYk0CqgBZiTgBQTUgU6WnALHgOjq/HToLPLn8fv/etBUYajxEn3e3cNVoYclW5DQRDk66nYB6OjpFctFG4WSSQhiAos3RP7p33YeZYuZ9p90fcLbJtvsNvbxLC1lBS4mnIIWeZOMUUQNY0ajyPRiFRaOGt43WIxtaW51V+I+Q+Q6sAAsLf43/2/vqGBQU6CpNeu/e+vde9+691737r3XvfuvddEX9+690Bn+y0dF/6Yf9Pv+jfAf6W9f3P979FT9x/Ff4D/AHV/vD/DfuP4N/ej+63+4z+J/bff/wAP/wAn8vi9Pv3W6mlPLr//19/j37r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3QOdz9C9Yd+bep9v9kbeNe+Kqxk9rboxGQr9t772JnYwpp9xbC3xgqig3NtDO07ItqihqYWkUeOQPGzI3uvdF2qV+Xfxyp6EUMVR81OrKAyrXR1M+2tj/KPbeKjKCnkpJ3OD6v7rNHSo3kWb+7GWmbnyVszWPuvdDN1F8puku66yswG0N3rjt+4ghNxdUb4x9fsHtnbM+nU0Wc673ZT4rc9PEB+mpjglo5h6opnUhvfuvdGFB/wP+2Pv3Xuui1vwef8Aff6/v3XuiDbhj/2a75L0G0Kd4ch8e/iVujHbk33UxSLU4zsj5Q0dOa3afXr6NdNW4TonGV8GeyqFmA3LV42BlEmPqFHuvdH6AsLXv9ef9c39+69137917r3v3Xuve/de697917r3v3Xuuj9D/re/de6rL+b+dWr3rszbsbRH+DbersnUBJZDIk+XrEhijmgt4lAp8drRhdiHYEW9n20R/pSyEZLUHWJX3hdxWff9g2lTmC1kkbJwZWCgU4ZCE14ivRKwwJt/hf8Aw/2/sz6x6+zh169jb+v++5/p/h7917rl7917o7Pwcyrwby3zhSZvFkNuY3JhFK+ATYzIyUzSS39flaPJqqkcWUgj6eyzdkrHE/mGp+0f7HWRH3eL1k33mOxNdEtqknyrG+kn9kgA+Qz1ZaDcX/1/979kPWWXXfv3Xuve/de697917r3v3Xui+dxdWV+66in3Ht1YZcvDTLR11BJKkH39LEXkp5oZpCIxU0+orpYhWVvqCOfde6AjEdO9gZWsSllwkuHgLL56/KSQxwQRlirukcUkslS6AXCKOf6gc+/de6OvtPbVBtHA0GBxwLQUcf7k7Kqy1lVIddTWTBAAJZ5iSR+BYDgD37r3Sj9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+6914/T3o8D17qr35N7Hrtn9jVO6oIpxhd3zRZOkrgrGGlz0SxrXUEkzEhKiRoVniDW1KxC30ke+Mf3zvbG+5M909w5uS3f+rHMLCdJaExpd6QtxAzgEK7afGQEjUrMFqUI6G+x3QuLRIQR40QpT+j5dVsdC/EnpD41bx753x1Lt6uwuf8Akbv89i9jSV2S++pIsp5spWrhdqUgpoF29tSLL56vrloVMqisrZX1kEAY/wDN3uJzVzxtnKG18yXaS2WyWn01sEXSStFXXMQx8SXRHHHrNKJGo9ejCC1ht2lkjWhkNT/mHy8/tJ6U03x/2DJ3hvL5Dwz7ooe0N8dH0Px8yuUx+c8GPothY3M5PP4+rweONG4xu7KPKZaWSOv1vpKpaO63JcvOO7LyrtXJbx278v2m7ncVVoyWNwyJGQ7ahWLQgBQaanOodbMCGVpxXxCmk/Z1ZH8Mvjp1BR/A2m+GcezZ5+isHsbdvRdVjMzW/wASrd4bO3Pj6+Lc9XncqY4pq/cmabclW+QrfGjTVsrTABjx2Z+6z7s80+7/ALd3W+c32UabtabhNbGaJBHFOqBXQpHU6DGrrEwqQSoapJPQK3W0is7kRwtVSoNPQ9G9+PfSO2Pjb0h1X0FsnMbuz+zen9kYHr3aeU37nTubeE+29s0UeMwkWdz5paI5SrosbDHAJTEhMcag8i5yV6LOhj9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3WCppoKyCSmqYYqiCZdEsM8aSxSIfqskcisjqf6Ee0W5bbt+8WN1tm62MNzt06FJIpUWSN1PFXRwysD5ggjq8UssEiTQSskqmoZSVIPqCMjoJa/ozYFdVPVJRVuP8AISXpsdXy09JdmuzJAyyiM82AWygfj3hzzF9wL7t/MG6TbpFsG4bb4lS0NjeSQ2+pjUssRDhPQBCqAYCjof2nufzdawrCbqKan4pIwzUHzBFfmTUnqIOgdgB1Yrm2CkEoctJocAglXCxKSrDg2INvZHF/d0fd0jkjcpv7qrAlW3FyrAGulgIwSp4GhBoTQjpQ/uvzYylQ1qD6iIVH2VJ/wdNe6qvZvTdDG+2tv0Dbjy3lSg8plnkSKI/vVVVVyvJUpTRNKFCqy6ybXsPYZ94N69j/ALjXL9rN7Ye3G3n3N3kSC08UySusUZHiTzzyM86wRl1RIo3UysQtQqEhVsNvzJ7kXcibzu837otyPE00AqeCKoAXUQK1IOkZPRLO0vkTtrbOc2Vj+1+3dtbKyvaW6I9kdbYHcW56Ha/99d3SwPUQ7U2fjaipp2zGaemu3gj8kjKRfki/MLmz3V9//e+95i5g3PmDe9wtrO3+ouIbLxYrKztlOnWYICqRxCpVnkZ2fOtjk9S9abTypywtraw2sEcsjhVZ6NIzcalm48DSgxTA6d5MzJgoKrKT5eXC0uNpamvrsnNk3x9Lj6KggkqqytrKyWeKKmpKKmheSV5GCRohZiAPcSbRvXMu23kMvL297lBuBkCo1vPMsjNqAQIUcMzeIQEABqTjj0IbmGweB2vIYXtwKnUikU9TUY6UnQfy5212NtLB78677F2r3X1Pmpqqlxu8toZyh3NjakY2slocn/B9xY6SSnyE2Oq4mjkidmsylbg295v+133xvfX2J5htOT/eC0vtz5eh8Nbi2v42j3S1ifIkhlcLJM2lgwjudfiL2q6HPUb7vyHy3zPZy3+wMkF6alTGQYmYYCsmQuogjUpGckHo9kuB2fu+io8lWYTBZ2mq4o62kqK7GUVYXWeEKkweeGRw5hbTe9wOPfYuDZvbf3P2LZ+Zm2Pa932a+hS5gmkt4Zg6SoCrgshIJQhTmoppNKdQDML3brme1kd4543KsKkUINCP29B9kPjn0zkVjWTY2NpfG7SBsdPX493LghlkelqozJHzwp4B5HuN9z+6b933dFjSX21s4dLaqwPNAWJrhjHIpZamoUkgHy6dTdtwj4XTfnQ9M1R8W+mJoJYY9t1dI8i6VqqXOZcVMJuDrhM1XNEH4t6lYWP09kF19y77vtxbzQR8qXEMjrQSJeXOtPmpaR1r9qkfLpwb3uIIJnrT1A6x0vxX6Zp4Ehl29XV7rqvVVudyrVMmpiw1tT1FPF6L2FkHHtuy+5X9320tI7Wble5uXFayS3lyZGr/ABFJI1xwFEGAOttvm4kkiYD5ACnQk7a6q682jJHPt7aGCx1VCUaKuWiSevSRIxEJVranzVKSlB6mVgWPJ59y3yj7J+1PIksdzypyHttpeoQVmEQeYMFC6hLJqdWIGSGyak5PSKa9u5xSWdiPSuOhBH09ymMCnSXr3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuv/9Df49+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvr7917oIu0eg+l+6Y6EdqdZbP3vU4pi2Hyuaw9NJuDBu19UmC3JAsGfwcpJ/VSVMLX/AD7917oGMj8PaCjS/WPf/wAnOo5IF/3H0uC7dyO+NvUbgBYx/druGj7GxktLCq2EACRgfT6D37rdcUp0Wv5E4z50dXbJpYq/uip7J6ITK08nc/ZnUXVtDtr5i7J6zigk/jFdsXCYuryPX+55Q2lspksTiqPcGNxvnnxdBU1qxGP3Wuj8dDbd6f2v1LsbG9BwbdTqSfBw5jZlXtas/ieKzWOzjNlpdwvmXnqqvPZPcFXVyVlZX1M01ZWVU0ks8jyu7H3Xuhg9+691737r3Xvfuvde9+691737r3XvfuvdeP096PDr3WqR/Pd+U3yI+PXyi+ND/HbH4/eeLfbfZfYPfHW38Epctnt+dV9K7dxG797bf2tVS08lfjt0Y/ZuUrclRikZJJpqZVJYXjY5tZZoreHwxVSzEj1A406xt542TlzmHnXmhd3mKTxWVlBFJqKrFPcPKI/EHAozaAxNQFb16TuzfkLVdlfL7YGz+ut24zc/x+7H+CUHyO2hNj6OhnOcz2Z7iw22cJn4s1GrVyodsV7Us1Fr8cdSr6lEicGYkLzJoIMRjB/OtOP8uoDudkj2/le9ub+2ZN7g3j6WSpIoFty7qF+T9wP8NPXosW2vl13fJ8/IsZuHK4N/g12n2z2X8OOnamLD0cdVT/JTpvY23N0ZXclTuxKdZ6vEb53d/H9u0ULTvA1ViiqBWX1UWWQ3FDQ2zEoP9OBX/Y6ENzyvtI5MMkKN/W+3tor6YVx9LPIyBAlcGJfDkJoDR/Tga3tn5rdZdVdlZbqWm2F333JvTZ23cFvDtWj6B6pr+zqXpjau5pXj29leyqikyOPkoq3NQxvVU+LoEr8q9EjTim8YF3GnjRipViwyaCtB6n/Nx6De2cpbhuW3w7m15Z2trMzJCbmUQ+O6fEItSmoWoBc6UqQNXVrfw4qjL3Ji5aWeY0mQ2hmpzxNAKmmlp6GqphU08qxyqy61bxyKGjfggMCPaXcyDak/0hT/AFfZ0PvY1ni9wIIddCbScMAcGmn0wRUVH7erbF+n+3/3v2Hes0h/Lrl791vr3v3Xuve/de697917r1h/T37r3Xvfuvde9+691737r3XVx9Pz/vv9h71UA08+vfPou3Z3yX6864lqcWlVJujctN6HweEeORKaa3EeUyjXo6Ej+0t3lX8p7xh92/vX+2PtbLd7TFdNvHNseDaWjAhG9J5z+lDT8Q75Bw0dLILKaYA00ofM9E+z3zL7UyL1K4Sh2ztyndozTstFPl66nWNyWVpq2dKSc1C8N+yNP9k/n3hDzF9+r3h3R7hNh27adqtnZdBET3MqAHNWkcRtqGGrEKDKkGnRqm2WqjvZmb9g6S3+zY95/wDPTYcf+SviT+f+C+wh/wAGZ94U0I5qsR/1AW/V/wB3WlfgP+9Hp8x3zF7io4Viq/7qZZ1lZ2qavDTU87xkg+Apj6+mp1VQLKwTVzc349iDbfvye+FjbiK9TZ72TUTrktmRiKg6KQzRoBSoB0lvM14dUO22pJyQPtr0ImP+ceXQTfxbr2gnYsn25xmeqIFWOx8gm+8oZizlrFdNhb6+5N2z+8F3xFm/fftlayPUaPp7x0FPPUJYmqa5FCBT59MttSf6HPj5jpd03zf2O8sCVOzt200Lui1E6ti6jwI3+ckWJatZJ/H+ALFvciW394DyBJPAl3yNvUUBYa3Bt30D8RCiQFgPIChYeh6YO1S5pKv8+lEfmd1F/wAq29f9ht+L/ia/2KD9+/2U/wCUTfR/1Br/ANbuq/uu4P4k/af83Xv9nO6i/wCVbe3/AKD8X/1f71/wd/sp/wAou/f9kaf9butfuu4/iX9p/wA3Xv8AZzuov+VXe3/oPxf/AFf79/wd/sp/yi79/wBkaf8AW7rf7ruP4k/af83SgwHyy6azs4ppc3ktvSNIsUT7ixVRRQSEpfWaqD7ynhS/pvIyer/Dn2JuW/vnexPMVybWfmC52yQkAG9t3ijNfPxEMiKK4Jdl/Znqj7dcoNWkMPkf+K6MNi8vis1RRZDD5KgytBOLxVuOq4K2lk4B9E9M8kRPP9feTO0b5s3MFhDumxbrbXu3SfDLBIssbfY6Fl/KtekTI6mjqQfnjpx9moNeq9BN2V350X0xUYaj7h7p6m6oq9xJVSbfpeyuxtn7FqM7HQtCla+Gg3RmcXJk0o3qIxKYA4jMihrahf3Xuk7gPlX8Xt14zdua2v8AJHoTcmH2BjocxvvLYDuHrzMYzZWJqJ2pafKbtr8duKppduY6eqUxpPWPDE0gKhiePfuvdDTjcziM1i6HOYfKY7LYTKUFPlcZmMZW01fisjjKuBKmlyNBkaWSWkrKCpppFkjmjdo3QhgSDf37r3nTz6AR/mF8TI4Hq3+TnQC0UeeTasmQPcHX/wDDY90S1C0ke25Mj/eD7FM89U4jFGZBUeQhdF+PbogmaMTLExiPnTH7eg+/NfLMW9/1ak360XmGlfpjIvj0pWvhV16aZrSnz6GrJ7w2lhchtvE5ndG3sRld5Vk+O2hjMpmsdj8huvIUtBNlKmh21RVdTDU52sp8ZTyVMkVKsrpAjSEBAT7a6PwQQCDjpR+/db6aczg8RuCgmxWbxlDlsbUC09FkKaKqpnIPpYxSqyiRCbqwsy/gj2Scwcu7FzVtk+ycybPb32zyjvhmjWSNqcCVYGhHEMKFTkGvV45HjYOjEP6jHQG1Pxc6cqamaddv11GsrahTUecycFJF6VBWCH7hvGpte1yLk+8br37l/wB3+8uprlOVbiAOa+HDd3CRL8kXWdI+Qx6dGa71uCqB4wP2gdM0/wASOqZa6KpjG5YKVAvkxsWac0s+nVq1zSwvXIXuL6JVtbi3sP3H3FvZCbcILyIbvFapTVbreHw3wa6mZGmFaiuiRSKChGenBv1+FpVdXrTowe29t4TaWIpcFt7G02JxNGpFPR0qaUDOzPLNI1y0s87sWd2JZmJJPvKflLlDlzkXYbLlnlTaIrLY7cHRFGKCrGrM3mzu1Wd2JZmJJPRVNNJO5kmctIfM/wCDp+9iXprr3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de6Jh36Khd60pla9M2Fp/tFswCKKioFQLn0sWm5Nvxa/vhr/AHjo3b/X32dr6fXtZ5fg+mUKQFHjS+N3HBLOAe3gNOrIHWR3tL9OeW7jwlpL9U2s+poNP8v9jrUB/ma7z6m+RHyj+WmzN953fNHuL4YfGrae1vh5W7N6h7f7MocJ83dz1uK79zm9oc51vs7cmG2/ujbFDtPbG2pBV1EM8MGTqQUEbO3s29gNp5l5E9vPbTdtqtrR7Hmrf5pt7Wa9srRpNhiR9ujtzHdTRSSRSNNdXQ0KVZoUpkDok5ovIN03jcgz0NrGBCQrN+oe8MpVSAQFjHlQFvXo43yd+eHTve/RvwZ2RunddbsHpr567WwXaPyM7Djwu863E7c6H2ZR4rKdrdMU+T2Zgs1WUm++3OwUGzpacCN6bEfxR3ZG0Bou9vvZvmbk7nD3g3bb7CO85n5NuZLTbLYyQrJJuEzMtpflJ5EDW9lb1vVOQ0304GoVIMt35jXcNp2W3XUIZhqkIDU0oASmB5khDnhrxUDpWfyLOyupN3fDndGzercthpotgfJT5LyVW2sFgsxgsXtXa+7e7N85br2loYMhiMZQLjqva0MbU8NM0jU0KLHMsUg0eyv74Ww8y7Z7p7bu/MVvKJb3YNqHiySJI80sNhAlwXKu7ahMTqZ6amJZCy56Ovb+4tJNture1FFjlcAUpgyyaQMDgtPs62kOnxKOuNsiUSBvtqrSJAwPiOQqzCVDc+MxEabcabW499cvuULdr92H2oF6sok+knK6wwPhm7uDERqzoMZUx0xo06cU6hT3DMZ5y3wxkFfEXhTjoWvDzrWvz6Ev3lP0C+ve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r//R3+Pfuvde9+691737r3Sdxe7dq5vJ5XC4bc23svmME4jzeJxeaxtfk8NKW0iPK0FJUy1WOcsLATIhv7YiubeaSSKKdHkXiAwJX5EA1HXqjpu312JsPrHBvubsXee19ibdSoho2ze7s5jtv4v7ypJFPSrW5Oppqd6ibSdMYJZgCQLA+27y+tLCMTXt1HFDWmp2CivpnievYHHpQ4fMYrcGMoc3gsnj8zhspSw1uMy2KrKfIY7I0VQgkgq6KtpZJaaqppkIKujMrD6H2/DLHPGk0MivEwqGBBBHqCCa9ex5dOXtzr3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XvfuvddEX/J/wt7917qvrZFRSfDju6t6ky0EeI+OfyP35Xbg6GzUYSn2/1j3dulKjLb66Jr0jiio8DhexsxTVG4dpNdYZspV5LGgK4oUm917qwUXtz9fz7917rv37r3Xvfuvde9+691737r3XvfuvddN9D/re/de614/5g3TG9s3/ADIvin3ThIcTWbJ6m2d3lNv6asrqWmrqCbs/rfF7P2nR0WIm1zZ+nrcliJVqmQBYYW9YsfZ/tsdI7eTigd6/mKDrEP3dvLWw3znnbpZmN3uMO3PEApAAidtYZhWhoO31J9eqj+jPhf8AIf4o/Jr5J7+6nh2nu/oqk+OXae3vhfs6u3RQ4XM7M7A7P7Fx/bUvUGcWuNsRsHbHYa1tTj69gKaChqFiF3uiPRwywzO6U8EIwQehOafZXoKbtzXsnMfL/L9nuQli3c38TX7hGKvFFGYPGU0zK8WkMpyzCvDiGWW/lMdh4z4d7N2btX5Md+535GdXnaPeeytiZ/tPDr8c/wDZp8Huwdm52elwr7WjnxmCy+78tloYaw1upGq/M7lGYCrWbm3jUSMZhmlRTVxPl69GMXuVZy803d3c7DZJsd1qt5JVhY3P0jJ4Qq2ujFVEdRo8qAdGvx+xflZ0V8mflJ2/1Z0dtHubavy/Tp3sBaSr7s291lkuk+4Nj7Dg2VuXb+95MvisjLuXrZ4kE9LX4Baqs8kbIsF3De1AEqTzOqBlkoeNKECmeNR0GZLzlvd9g5d2zcN3ls7nazPGP8XaZZoJJPERoqMAso+FlkoOJrQdXufCLDz5DsXKZmvp0NVhNlA1M1JLM1FSZfMVFJBVQwNUBJ6qnkEFQIWkUOUQMwDG3tHurUt41PxFv8Fehz7A2C3HOO53yqxit7NwCaVBeRVWvzKBuHVpwFhb/fc+yDrMPrv37r3XvfuvddX+trX/AKe9MSASOPXv8HQI7q+QfW20stLhK3JVeRr6WQQ1q4ShkyUNBKJDHNFVVEbpCKinIu8SlpFH4vx7xn56+9v7L8gb/Ny1uW+T3e6wOEnFnC1wkDairLLIpCB46d8alpFqO2uAc22w7ldRLPHCBGeGo0r9nQn7a3Pg934mmzu3cjT5PF1WoRVMBYFZIzplgmikVZaeeJuGRwGH9Pc28lc78r+4fL1lzTyfvEV7sdxXTIlRRlwyOjUeORDh0dVZfShB6Lbi2ntJWguIyso8ulB7FnTHXvfuvde9+690VH5WdtV/X21aHb23al6Tcu8TVQpXwSqlTiMNSLGMhWwcM61NS8yQROANBZmBDKvvDb74/vRuPtrydt/K/LF20HNm+GRRMjUktrWMDxpUwSJHLLDGwppLMwIZR0Y7fbiaQu4rGvl6n0+zqqz6X/xLM1ySWd2LO7sSWd3ckliSWJueffHgKBWnEkkkkkkkkkknJJJJLHLHJJOej8kkAHy6692611737r3XvfuvdcxG5QyBHKA2LhSVB/oW+gPvVR69ep1w97z+fXusmh9Hk0NovbXpbR/yV9L3961GtNZr16nXC/8Avuf+K+91b+I9ap16/wDvuf8Aivv1W/iPXuvX/wBt+R+D/r+/VJFCTT/V/qpw62McOPSw2Tv7d3XmViy20czVYuUTRy1VCsrticosbBjBlMeSaeoilUFS1lkAN1YG3sbcg+43OftlvMO9cl77PaTB1Z4QxNvcBTUpcQ10SKwqpamsA1VgwHVJIopgVlQEevmPnXq3DpzuDB9vbdbJ42Nsflsa8NJuDCTkNLjqyWISK9NKDaqx1RyYZbC4BVgGBHvtT7G+93L3vZyu267XEbbe7RljvbVjmGUrXVGf9Egc18KQDNCpAZSOg5c2z20mljVTwI6An5/dQdS9jfFP5GZzsPq7rnf2a2l8cu9ajauX3rsjbG6sltqdut9wVbTYGuzuLr6nESPU0cUhNO0ZLxIx5UETf0m6o+330z0xtz/hNP17l8L1J1vg67sf4Z/BWl7KyG3tk7bwGV37QZHcvRkmVpt2ZTEYykrs99/9/UM7VUkp8kzvfUxJ917ogXyJ+VXfnxN+OPa/8l3r/cOe233B1l37P8ZejdxUtdNTbg3L8W/krUZPeXQG7KXLU865D+73U3TFVuDH5l1VTHWbepYg3jJBOdshiaKWTDXbMERSOGri/wCQB/PrGv3x5l5g2zfuWbFbiS05DtbG53TcZkYq84s2QQ2KupBVZ5XiV1qDIrUrpqOqVpukOtYPi7u747jEauq4/wCcNjOnZMfCzRVMu0FyOI2lNNFOpEkddJR6p1fVcTH+ns1aCP6R7Sh8P67TSv8Aq/zdQMvNG9ye4e3c7+PTmI+2j3oZshZirygGtRStFI4UPVvfwv8AlJ2Nun5d/wApn4Qd8bhbc3aXwg+Yvcmwf707kq5pc9ufr6r+K3YdL1BvPHVlc7zZbB742lJBpdiZaXJ0k9PcroClW6QRxhaClwjFG+YAqj0+a4J9R1PPsZzfuu7XF4olEnIe5WabhYM7EvZzyTeFebWzVIIhnYtApJIjZVU6aAbnHyh7e3R051JW5rrrB4XdHbm7dwbb606Z2ruOavpduZ7tLfmVhwm1qfc9Vi9WTotqYqWWTI5ienBmgxVFUyICygEm6yU6nfHfvOj7w2dkqnIYsbS7N6/3BWdfd0dcz1AqK7r/ALJwkUD5bFCY2bI7dy9NUQ5PB5EAR5TDVlNUqFLsie690Pvv3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917oIe1evqnetDQ1OLelizWHlmkphVC0NbTzBWkopJQD47yRqykgqGHNr394YffF+7PuHv8A8tbHe8qXdtb887O03g+NVUuYJgPEtWlAPhVZEeORlZQwIYAMSB9yHzdFyzeXMV8rtttyoD6OKMK0cCuaAkEDNOq7+r/j/i/jZBvbbuydv7qwku/e1t990bzqcpk8tn67cPYnYuUiy2585NlZjL91SyNBDBSQKTDSUcEcEYCIB74y+6fKvu/b7/abR7j8k38G7bbYQWEMaWTeCtpapoh8B4UaKQUYu0sbEuzM7HUT1OmzLyvPaTT7dd28kMzMzEsCwLEkghzrU5zwpQeg6T3x1+Ouz/jDsmbq/pHbm68Fs2o35vvsSi27WVmbz0OKz/Ym5Kzd256TBffIz43bzZyummpqCI+GlEjKnB9lnNT+5fujv8G575yxuF7v5s4LasVhMryx2kKxRGQLHR5BEgDSMNb0FSadKLWw2DabSeEXUAtmZidUi0GoliBVjQVY/wDFY6GT4tfCHa/SOyxsLaWIy2zuuX3tv7sObE5jMVec3BX7m7M3bkd67wnjqcg009DQZHcGVndY2P7MThI1AFxmz7afc593/ffmjbeePfmJtj5cS2tYGi0Rxbhdw2kSQQwiFQVtkMSBZJpaTNWqR1OoALdeedh5YtZ7Hlorc7g7MdYJMSFmZq1J7yCxKhcA8fTqyOlp4KSmgpaaNIaamiSCCKMWSKGJQkcaAfRURQB77KbVttjs222G0bXapBtlrCkMUaiixxRqERFHoqqAPs6gaWWSeWSeVy0rsWYniSTUn8z1I9r+m+ve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r//S3+PfuvddE2BP9Pfj17pK72x2ezWzd24nauXXb258rtjPY7be4HRpEweersVV0uIy7oqlnXG5CWOYgAkhPaa7jlmtriK3k0TtGwRv4WKkBvyJ68QaGnHrUc/lafy8v5gHTfz5292J2PsrdfWm0dk1O9D23vvN7gp6vDdqUOUoMnRjE4mWmyFVNvYbiz1RBXrPImmmWPysUlAT3jjyBybzltnOMN9fW0sFrCX8WRm7ZgQQAtCddWoa0BFOPSGGKVZAStB1Y1/PK+EnyV+W21+i8x0Him7Aoutsxuhd0dXfxbH4maafcdLQQ43emNTK1NNj8nV4xKOWklidhLHDUa47jyD2NfdflXfuY4Npl2ZPFWAvqh1AZelHFcEgArTyBqOnbqJ3ClPI9HQ/lRfGzuH4o/DTZXU3eGQgm3tFuHde5ht6jyhzNJsTCbjyC1uL2XDkVZ6ec4tVeWUQE08c1S6RkqoJFXt7sm5cu8s2227q9bsSO+kHUI1c1EYPCi54ep6cgRkjCv8AF1ZJ7HHTvXvfuvdNmVzWIwVPHWZvK43D0ctXSUEdXla6lx9NJXZCojpKCijnrJYYnq66qlWKGMEvJIwVQSQPfuvddVebw+PrcZjchlMbQ5HNzVFPhaCrr6WnrcxPR07VdXDi6SaVJ8hNS0qGWVYVdo4wWaw59+6905g3F/fuvdeJsbWP++/4n37r3Xgb/wCv/T/bf8V9+69137917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xug57a6p2N3d15ufq3sjCx5/Z27qAUOUojLJS1UEkM0VbjcviMhTtHWYjP4LKU0Nbj62ndKiirYIpomV0Uj3XuivdP9z706r7AxXxd+TmSWbeNcJ6foHvGoX7Pb/yS2xiqYuuKytQxFHg/kLtrGQBs9hrxx5hEbKYxGp2qKei91unp0eYG97fg2/23+9e/da679+691737r3XvfuvdFy7w+VfTXx7qcPjexs7XQ5rOwPW4/BYPEVmcyxxscz075WopqRdNJjxUoY1eR1MjghA2lrLrPbrq+DtboCqnJJp1FfuJ7zche18tja81blIt/cKXSGKNpZdANPEZV+FNQ0gsRU4FaGkrp75RdJ97RvF19vKlqcxHxPtjMRS4Lc8V4pJyY8NkhDUVqRwRl3em8yIP1Mp96utvu7Ijx4qL6jI/l0/yJ7we33uMGj5Z3+N78cbeUGG4GCaiJ6M4oKlo9agcSDjorvzf2tLBn9lbzgp2+1yONrdu19RHFGqLWUUv3+PE86nyPLPTTTBAwsFiNj+Pa/aHBSSMt3cR1D/AN4baHi3PYN9CfozRNA54DUhLpX1YgnTX+Ejz6I57Nescs+fHrv37r3XB3VAWbgAEk8cAf6/v3XuAJJwOP8Aq9fl1a/8TOsMhsDYtZl87TtSZ7etTTZWaik0eWgxFPAY8PTTFGIE8sUzzuv1Qy6TyD7D+5XAmmVENUQU/wA/Wavsryhdcs8uT3m4xlNyv3WQqaVSNVpErU4MQS5Hlqocjo1fsu6mXr3v3Xuq8P5lnz4pv5fHQ+H7QxnUG5vkH2NvTfuN6/606R2XlqbDbp3xlRhM9vPddVQVtTQZNIKLZ3X20stlqpvA4MdIEupkDDwyQKHPXvIny6KJ84/5sW3etfgvQ/LLofaWR7e2rvTZ3Te4tu4nB7io9t5jda945Xb2I23trEbilpclSYispZ9xxx18/ilZJlaBNLBmXBD3l97b7nn3Gl+7x7d8yrtAZLhN03cKztB9PDJNNbWygqQyohWeYEFa6YjUMwE+3baLa1G63kHiUoUjqKGpAqT5/Z0Ae3vlZ1tnfiHB8zpUyGP62HTuU7fzWLmeGpz+GfAY+tO5NjVRAWObduJ3Vj58IylRrr1HpGq3vmje+32+WvuO/tnVJN7bc1tFetEkMjDw7gE1PhyRMs5YknQSxPHoax3cTWf1v+ghan5UqCPtB7ft6CL4x/zlMbtn4v4n5Qbo6F7MwO5+we/dz/FnbPxGwOXxe8uzewu98FlpMRiNr7dybw4HBUGU0wS1OQrKvw0uMoaWdpndVQnLz2Yg5n9g/fK85T2rmu13H27k2iLcd1uZC1tZx2LpqW+GtpNM0TMqIRVrhGEZAwVD25mDdNsS4eEpdhykY4sWqe004D5+XVwvx4+TPzv3d2nt/ZHya/l4v0XsbeeOzmQwvbXXnyV67712/tGrxWNXJUW3e0cLjsLtDO4OtzceqClrcZHlqBqsCNnRWEh6uQywzwxT28qvbuoZWGQysKgg+YIIIPp0BSCDQjuHVj3t3rXXRNhf6+9E0GBU9e6qZ+W25l3B3BXUEUgkp9qYjH4NLOHVaqZTk8gAqgaXWaqRHBuQY/6WA4xffS5s/rJ737jtkcha12azhtFyCPEYfUzHAqCDIEIJrVK8KDoRbcoS2HqxJ/ydFk94n9Leve/de697917rmoGsIxS+kOy6lMqxlh+6YwS4QC9mItf3UkgE+XrmnyzSnWwK8GFfSueter5L9s/L3E/L7MYzD5TfOMzWN3XDR9VbSwS5H+7GX200yLhZI8ZCr47NU2Wp2JrpZdZDM+oppFs1uQuXPbO49tbae6gtHtZLdnu5pCvipIK+INR7o9B/swoA4ceghez7iNxMYLB9Q0gcD+fWwdiEyE2PxU+Xp0hyEmOxsmZipwxpafKyUdO+TgicagsEVc0irzwo+v094VzGASzrauTb62EZPxmPUdLEca6aE/PoXprOgle+g/lx/YetdyTtz5kr8y5cfHkd6nff+kYYn+4YGQbZh2l/ExDHTvh7/wAJTbj7cAmNXcH/AHZ5NfvNpOW/atva0SNDaDaPofE8eq+OJtAOrX8fi+LUaPQ0I6B63G4ncgFL6tdAPKnn/LHWxW6gOygpq5fQJELiP/VhCwfxf0YgAj3hGGBAYHs9fL9tB0MQpJ0qO4eXDrgRb3vPmKHqvXXvfXuve/de6Hf4378Owu1cHPPJ48RuYrtbM3BYLHkJUOMqSARY0uVWP1WNo5H4595Efdb9xj7b+8PL1xcS6Nk3Yjb7rBPbOw8B8cNFwIxq4Kjv69Jb6MTWzCncuR/l/aOrVOyNiYbtTrvf3WW45K6HbvY2yt1bCz8uLqEpcnFht34Ov2/lZMdVSRVEdNXR0OQcwyNG4SQBirAWPcUGo6DXRYNw/BDpjcfwi2v8BshX74HSW0uuOp+rsZkKXPUkO/H2505WbTrtqTVefOKejlyc0+zaQVkq0arMrSaVTUNO/l5deqRkDPWvh89eg+t9xfzN96fKPLUeQq+2uvNrwdWbbrnrY2w9PtvI4DG1MctRjXpnmfMYqPI1cFJMk0axQVkoZGYqyjfaLWD6azuwv64Vh+RJ/mPLrlt94Ln3mf8Arn7me3735flufcbaYqwOtWjtoQI42rRYTQOV0mrrUH4gSSyfEjqiTF5DEGp3iKTKfKKH5eVTDN0gmHbEGTp8vHSwv/CrJs77umUGiIM2i489+fZiLOGlCW/thJ/th1Ew9x+YhcQXXh2vjR7AdnH6Zp9IU0Vpq/tqH+0FAf4eh/6q+MHWu/8A+YL8QPk7Nj56Hu3qffuLwu1szDWrSYLL4zLy5KmkoN5Y2OjmqMxHiKDL138OaOaCSCacBi0Y0hLuVnbyxTXcoPixxtSnnUUz+3oa+yvuBzdsu98ve3+zXNv+5Nz3uzYrOjP4bxzI7NGQw0lxGNWCCwXgeOzDmjV9m/O3a+2qyYDZ/wAZ+mv9KEGO8Tf7k+1e6cpuTYeBytQzl4ZoNp7A21mEp7BXWozDtc6FPsAnrrdQ8T1K79657I6/3r/s1Hx4w53Pv3HYCg293N0tC9HQU/yF62wlXLWUSYmrlNNBQ93dfU9VVNtavqJft6yConxVVaGenno/da6Mb1J2xsXvDrzbPaHW+XOa2huuilqsdUy0lTjq+kqKOqnx2WwuaxNdFBkcJuHb+XpJ6LIUNTHHU0dZTyQyKroR7917oSPfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xjxz7917pmy+4MPgKb7zNZGkxtMW0rLVzJHrb/UxpcvI3+Cgn2DOdfcPkj242ob3z1zRZbVtZNFe4kCa2pXSinudqeSBjwxnpft217hu0/0222ck8/oik0+ZPAD5k06DSo736+hfRHV5KrFyPJTYuoaPhivDS+K/Avx+PeK24/3g/wB2ywnMMG+bndgV74LGYphtPGTwya/EKDK5+XQ1h9rebpV1PbQx/JpVB/YK9P2G7X2LnGWOmzkFLOxIWDJq+OkNgDcGpCRG9+LNz7kPkb74P3eefpI7bbPcO2tNwckLDfBrJzQAkgzhUIzQEPk4GeirceROatsGqbaXeL+KKkgH26c/y6ENHWRQyEMpAZWUhlZT9CrDggj8+8lo5EmRJYnDRMAVYGoIOQQRggjzHQRIKkqwoQafOvzHl1z93611737r3Xvfuvde9+691xK3+tj/AEuL2PupUE5p17hnroLY34/23vwUA1AH7Ot1r1z92611737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3X//09/j37r3XRtY3+n59+Pz691pd/NrM/zU4f5lm8IOvp/kZFkV7LxCdEUexI93N1TN1wZcf/AgsVFG2yKjBzUXlOeauJIl8/3NiBbFzmmX3AHPc30RvfE+oAgCeJ4BiqNPDsoVrqqeNa9IJPF8Y6S1er8/5tm4PlNtn4J7mynx0O5KXsMZDZ8PYmS63irp954fY0rk7zrtnDGpLlYWSsWJJpaVWqYqB5WWwDMswe482/2/KU8uyaxehk8Qx11hPxlNOeNOHl0ouNZibw66vl0UL+QpvL5k7t2H3U3yIruz871XR5fbQ6mz/bgzUm4JM1LBkf744/bmQ3Gi5nJbYp0Sldi5khhq3ZYm5cAN+0FzzNcWW6fvszttysvgtNq1lqHWBrzpGPlWvz6btTJRtRNPn0Rf+c73v/MJ2H816fbnVO8e/Ni9bQbS2ZP0tR9SQ7lh2/uzOVEEkmeqZ/4FQz026dzLuM/ay0VV5RFTrEgj0SFmCXuhu3OVrzSkO33F3FYiNDAIdel2I7q6cO2qo0nyp1W4eRZDQkCnl1tQdFZHsPL9LdS5XtqgGL7QyXW2y67sPHeFaU0W86rb2Pm3HTyUkf7dJNHlnlDxL6Ynuo4HvILZ3vJNq22TcV037QRmQejlRqFPIg8R69LFJKrUZp0K/sy631R3/wAKCsjmcP8AA/ZuX23tyXeW5MT81fhFk9vbPgylHg5915yh+ReyKnFbbhzeR/3HYeTN1sSUwqqj9inMmt/Sp9+690Vbt/vf5W9wfzRP5O9F8j/g5m/iPiNu9vfLOv2xuDK/IHqruiLemQk+L+6Ya/DQY3rdTVYZ6GiQVP3FSTDKBoS7e/de6N98A+6vn383aTZ/zch7n6W2P8Vuwe2u0MHtr4mTdOVGU3LF0Psfde7eusLvCu71i3VBn4+58zndsrk5qE4oYKClmNKU8g8o917oiuC7x/me/ML4CfzDvkVlvk/0305111zh/n71B19s7Y/Q8mZ3xm8V0TubsHCwb8zu+azfWJl2luuswm36jC0sOOgkipkSLKFmqmMS+691b7/KdxveVH8BPjDle+e6qbvDce7ek+od27f3DHsSLZGQwO0M31htGqw+08/KNw7iqd557FMZDVZyokhqMjJIXeJDyfde6pwrP5h/8zmk+Ofya+fE/Z3x2j6O+Ifzo7I6Hl+P9P0/kpt1949QbU+SmN6gztZujsuXc8Z693ltvDbgh/gYxNDUQV0lG02Qf98RR+690PPzB/mLfInIfLn5KfHn49dt5ToLHfEnYHV1YlVQfB7uH5jnvjuLtvZWS7HxW2N5ZLrnEVVH0/1xt/A0dDQO0Mi5zIVeQnqIXEFKqv7r3Rh/jB8+vkh3f8wfh71P2V1sOlMF3j/LA3D8rO0+ntzbYyWP35sfvLD9x7E2DU4r7/NrQ5zG7fpaHNVgjoaulWdkkieQhwR7917orR/mLfO3e3xj6qz/AFdmulaDvXtj+b53N8D8Tl9+bIyMmw8L1Tt7fvdW1NvZXK4XC5WmydVuDa+G2XSVpkikRsnU0ohfxrUO6+690Y7Ib+/mF78+RWC/l+df/LTrTZ3ZnQ/xo29318l/lnW/GbBZ3IdoZ3tTf299pdS7L2B0hW73h2fsvF0eO2RW1m4K819bJIwghpVhZ5HHuvdFlyv8yH+YBuPqr4e7A2ZUfHvbXyn3v/Mj7x/l2957uz20dyZjpvOVPT+2Oz55u3NnbdpM3BuLEff0G2qDNw4j751ORD46SoSBmkX3XuhDofmb/MR2L1Z/NK6Okm2t8nvlZ8Gd2dGwdado9ddJ1dBkd9dY9/4Hbm7qzdNf8etvblqYdy9g9ObTqcxVrhMVko48+1DBEhDyMG917qxD+XD3dWd5dO7pzmR+X+y/mNWYLf2QwVRvDC9LN8ed+7CnjxeLqKrrXuHqCfJT5DaO+8FWzSuY6qhxdQ9JNEWgJHlf3XurCffuvdB92d1ZsPuPaFfsXsbbtJuPbtdNS1qQzNNS1+KzGOmWqw+49u5iilp8rtzc+CrkWooMlQzU9bRVCLJDIjgH37rwNOimUNZ8qPjBqxuax25fmR0jTyiLE7lwf8GpvlLsXG6tMNJu7CVtTg9r944yghCqMnj5cduNxzNRV0haf37reD8uhApPnF8Y1hB3T2SnV+SRQKvbvce3N1dTbioZhYSQz4rf2FwEsrRMdLPAZodX6XPv3XtJ9Ossvzj+KPjMuN7p21uYLpLpsei3Dv8AnjV20K81NsnD56ohi1CxZ1Cj8ke/U63pb06Efrn5F9Fdt5Wr2/1x2vsjde5cfAarIbUoM7SR7ux9KujVU1+0616XcdFAhcBnlpUVWNiQePfutEEceqBfnfhd6Yj5PdgybyqsjXxZqaiy2zK2tR0pH2bLSQR47H4cFmhNJhakS00oj/3eGZwGc3H2zPC23wiIAECjDzr8+uSn3kLDmCw93+Zjv00siXDLLas4IX6UqoRIq1BSJtSNp/0TUSAWyUmhra7F19JlMZXVmMymPmWox+Ux1VNRZGgqEN0qKKtp3Sop5lP9pWF/zccezNgGUqygqfI5HUIW9xcWlxBeWdxJDeRNqSSNikiN6o6kMp+YP29XM/H75U7S+R/Wk3QHe+fptu9kPQpj9q75yTRih3RX0MDSYfNvUzvFTUm7qGeMfc08kkUeRBIia8jRqE77bZLC4F5Zrqt61KjiPl9nWevtl7z7N7q8qP7Ze4+5Ja82tEEtrt/guHUVik1NQLdIQPEjLBZxXQe8qAL3vsrsXqSoydL2rtqfD4zD009e3YmHp67JdaZLD0sU082ZOfip5E26Kekgaapp6/xGmRWOt0XX7WJLbToHhlFf4WNGH2jzHzHQA3nlfnHlW+k27mjl+UEMPDubaOWa0uQ2EKSRgmKRjSsUigrXtLDu6rap/wCZR1LWYSfs3H9K/LHJ/GGmr5KSo+YuM6Ny9b8daaggyK4ip3lU5qGtfdtN1tTVzfubgkxS49Yf39XhIcp2vIlozAiMn4qdv+en5dCq19t9/vTLa2lzZSb9GrF7RZS0ylVDPGxCeEsqgisbPrFRWnRxqf5g9e/G/tnZ+E7s+Ifzl3pmt5dg1HXnRp2F0Tt7d/WXcO74du5Dd9Dktg7sXsKGl3nR1m1cNWZOh/ZggMFK8pZmUKCy93BHBSBe6lCfl6D7epr9t/ZaXbZ7XfubjG0698doO4I3FWmY9rug4IBpVssWPV//AEt2ZJ3D1ptfsebrjs3qWbc1LU1UnX3ce2oNodk7bNNXVNCKbc+3abJZeHG1NQKbzRKtRIGgkRrjVYFPp1kd0Kfv3Xuve/de61v/AJ+7p+U/bP8AMg2tSfHHaHSdTt34I9FZGapz3yboe06Hr3Md5fMKlqtuznr2s2Di5RuTd/XfR+z6tKktI0OPg3TIpKyyafeP33lPdLaPa/22updwvtxt7zd5DY28tgYfrIHdC8lxB45EYaKMU1E1QyKygmnRrs9i97eAKqFYxqIatDT8Jp6/s6o2zOyu5+sf5ZfyQ+GW/tv/AMR7G+KvzW+PVNs9OvMVvHMbOzHSHZ/f/X/cfXlX1vVZrGrm8vtDZiZXKYu8itNQw44JPp06jgZbbpy3vfvlyV7nbReMmzb/AMr7iZjcPCkyX1tt9zZ3QutDlFnuCkMpodMjSEqTWnQqaOWDaruzdf1IZ0pStNLOrClfIcPlTodZeru28V81c1/LNx2ydyy/EPsj5Y7e/mE1HYH2k/8AcLE/HMVw7F7O+NMuQ0fZwSbm+U+KpLUAfS2Mq5broYghIb9y/ce19v75T7lAPcWy5ek5eW21Dxn3LSba13OnxkR7U7DX8XiqprXg8Ypvr22tQfo3mEtf6HxFPzfNPToLdj7a7P2B1p1T8naTqXsHsTD/ABd/nH/PTtDtTrrau1Mlld+S9Ndq5fd+wavtXZOzBAmU3ZNshsvBXxR0UUsk+PeZ4NQUkH273mwbzvXMXIcvMNlaTb/7abFaWs80qLb/AFlqkM4tJ5qlYlnCNExcgCQKHpjptFkjSG98JmSK8lLLTOliRqA+VainVpHxU+Qtd27/ADNPiNWfFH5SfMLvPpbd+3vlNmvlz17ueo7VyHxw6khodj4f/QTi6XC7y682zhes6qXcE88VHiPvairiliCsPoT0C+7Fec3S+zvLu087bYbbfNqd7Gpkjl8aGDSbeUSRySI1YmVNQap0GooegnvaQDcppbd6wyKHpQihbiKEDh1tPe8geirro/8AE+9E0p149UW9gZl9w783pm5GnY5LdGanT7oo1QsK1ssFPFIYyUvDTwqgsbaVHv55vcnfDzN7i898wF3b6vd7pwXprC+KyIppjtRVQU8gOhXCmiKJaU7R0kPYL6c697917rkv6uCoNjpZxdFax0s45ugaxP8Ah70eGQSOtg0IPWu5tzYHzWj+YMVbJSdmR7yXsNanNbum/jK7Mm2aMsstTJJk3P8Ad+fa8u3iI44FJFrKEDg+82r/AHn2qb2wkhWWwO2fQ0SIeH44n04qtNYk8TOo09a9A5ItyG4qSr6/E4+WnrYnalglqkrPtY3EMjrBVmmjepp4Gf1iCpdDNF+2TcKw/wAfeEXissLw+MVUrUrqIU4/EtaH8x0MgDXUAKjzx1ryfIrevzOo/lvn6LB5DtGkzUG8kh6uwG3hmRtOt2sKqMYP7WhpUODyOMraU3rpJg/qMnkK2Fs2OStq9qpfbe1mvI7D6VrWt3JL4XjrNpOrubvDg10U9BToI3km5DcHCl/ELdoFaU/wU62EcbTTmnxmQydFRxZhsXj1y1TFSQkx1r0cH8QhSqVGlNDHXl1VdRTQP6e8LbiVdVzb287fSeI2hdRyoZqHTWlStDwrqNT0LxXLMBrI/Yf8nWvFs/r/AOZ8XzFxtdX0HZybtj7JepzG7Kxc02zpNnjLyS1ss2TkJwE215cBdI4ASLFVCavpmxum8+1je2M8UM23/uw7fRIlMfj+LooO2mvxBLktSnz6BscW6Dc6uH1+IM5pSv7OHWxLLp8smgWQu5Qf0QsdI/23vCNa6V1Dupn7ehi/xtw4+XWP3bqvXvfuvddjVxod42BDI8ZKyRupDJIjCxDowBBHNx7qagErIUbFGBoVINQQfIqcg+RFevUBOerpejd8t2F1htjcVS6NkvtDi8yEZm05bFN9nVM5cs+qoEay8/iT+nvvP93z3Bb3M9peUuZrmRTuvg/T3NK4uLc+FITUsQXCrJk/j9KdBi7iMM8kflXoWyAfqL/6/uaek/VK38zL475wbgpPkFtXGT5HDVGKo8F2JT4+jeafDTYzy/wndVWsJkkfGVFLKKSokEYWneKNnazkgV7Ber4ZspHAYGqV8/Vf8o6wB+9v7W7gNzg9z9ls2lsXhSG+CKSYjGG8K4alT4ZU+G5AohClsMSKglngdS6Twugvd0lRlFhc3ZWIFhyfYoofQ06wcWSN1LrIpSlagjqyz+XB8f8AK747Ppe5s5iKmPYnX0dRPtqvq6ULQbj3rUpJRQnGPMumsh23TPJNLNHdUqGiUNqDWD++3yRwG1RgZX408h/s9ZafdT9sr3mDnCLnvcbJ15b2wMYHZeye6aqDQT8awLVmYVCyFADWtLI8jHNs3+YBtXImOKHFd4fGPcW25JAGEtVuzpTfuOz9Csrm6k/3Y7KrNCgglYWNuD7BnXS0VoOjse/de6JN2J0N2P13v3O98/E+uwuN3ZuWpfK9u9C7prp8V1J3zWJAkL7jp62kp6xuru6pKSnjiTcdNTTUmVEccOXp51WOpp/de6csL84Ol6enjpu3/wC+Pxu3bEh/ie0u+drZLZYpZ49Kzpit7LFX9a7to/MWENTiMzWxTKuoW9+63Q+XDpQ03zd+ItZXYfG0fyK6oqqvPZvE7bxKU27cdPFVZ3PVsGMwuKNVFI1LT1mWyVTFT06yunlqJUjW7uqn3XiCOI6NN791rr3v3Xuve/de697917r3v3Xuve/de697917pi3JmoNvYPK5moKiPHUU9QA7FVkmVP8nhuL8zzlU/2PsB+53PNj7a+3/OHPe5Mgtdr2+WcBjQPKqkQxV/4bKUjHzboy2fbZd43Ox22AHxJpQuM0BPcf8AarU/l1VX353jierur+0O+u2s66bV6s2NujsHclXUyhUpsTt3G1GTfG46NgY4pq6WNKWmjRSzzSoLMxsfnD3Df/cj7wHuJskfMm9XO6c4brfJbwiR6xwGeUfpwR/BFDEpPagHahZySC3WVnhbTyhtE30dqsVpCmaUBfSPiZuLMaE5+wdFH+FfzOzfzF+P+9uyF6Ty/UvdfW+693deb6+Ou89z0y5rbHYGHwmO3TtLB5jc7Yunix1BvfbO4MZVxVclJanjqmOlxHybe7XtTae13Ou0bE3N0W5cqX9vBc2+5wQkJNbSO0U7xxBjVreWOVCgerFOILV6S7BzBJvO3XFwtoY76JmBjJFajIrj0KkYpkdANU/O35ibH72+MPSHdvwL2Xsqt+T/AGHktk7fq9k/LTbnZ2ewOG2tgZ9z797AyG08f17jZJNo7HwsIesqWnhj800UIbySoCL19mvaze+TvcLm3lP3iubyDl+xWeVbjZpbWKR5ZFitrZZnunAnnkNEUK3arMRgnos/rNvVvuNhY3u0hGnkAxIGYLQsWp4amgABOfMeo6uy6+7Jy2zclTw1FZU1u3JJVhraCWR5VpopCE+8ofKWMD0/6igIV1uP6ezr7tf3qOcvZDmbabTdd3u7/wBsppRHdWckjSeAjGhubXWT4bw11NGpCSRhlahCsHeb+S9v5isp5YLdIt4Vao4AGoj8D0+INwBOVOej2RSJLHHJG4dJY0kjdeQ6OoZWB/IZTf39CVtc295b293ayiS2lRXRhkMrAMrA+hBBHy6xbZGjZo3FHU0I+Ywesnt/qvXvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3X//U3+PfuvdQKvIQ0ZtKshuL+hQeP9csOfZRuW82u2EC5VzUV7RXH7elMFrLcV8MjHr0wVG78XTBleOuP9dMSE/7C8v59g2890OXbLUJYbrHogP/AD90aQ8v309NDR/71/sdM9R2Zt+C/khynH4WkjNxb6c1Av7Cd794PkixVjNZ7iafwxRk/wA5R0ZR8lbvNTRLB/vR/wCgemKbufaNKulqTOBV4Cx0EAVR/tIFWqgewnd/ez9s7IHxdv3jHpbxn/rP0Yx+2+/y0CT2tf8ATn/oHpkqe/8AYKFGnx+fkaB/JETi6SRon+muItWkxsR+Vsbewnfffj9nbEkTbVvpp6WsRp+24HRlF7Rc0TU0XFln1kYf8+dNU/yj67pr66HdRtf9OLpT/Un65IX9hK8/vGfYmx1GfZuZcelnCf8Ata6M4vYznGX4bqwH2yv/ANa+hL657PwHZ9BkMjt6DJwU+Mr1x9QuUpY6WVpmp46m8KxVFQHj0SDkkc/j3kH7DfeG5H+8Rse+8wci2m4w2G3XotZRdwrC5kMay1QJJICulxkkZ8ugTzjyVu/JF5Z2O7y27zTxeIvhOWGkMVNaqtDUcKcOgk+XHxR2B8x+tNu9V9kZjdWDwO2+3eou56Ks2hWUVDk5dydM77w/YG26Gomr8fkoGxFblsLFHVoI1keAsEdGswnfoIdc+5PipsHu3vL4rd+7ny+6Mfu34jbs7I3h17jsNWUFPgsxkez+vq7rfOQbrp6rH1VVWUlNha95acU81OyVABZmW6n3Xuis9K/yu9pfHvt3+9nU3ya+UW0OgoO2d1d5Yr4dYveu3afofEdlb2rMnltxtSzxbTHY79e1u4MzVZRdqPnGwi5KXy+EqBGPde6G7o74I9M9IfGztj4rY6r3Zu7q7ufdHyF3JviLdOTpHzVUvyW3JujcnYOHpK/EUGLWkxsUu7qqCiKoZoYdGp3Yaj7r3Sn+GfxbPw66Q270RS92dud47a2UlJhNiZjuKr2pXbi2hsHB4nH4DaPX+Pq9pbX2tTVmF2xhsXHFFUVcU9fUOWeaZyQF917ovNb/ACsOiK/4ifIb4Zy7v7PXrf5J9578793pnUy2B/vhi92dh9u4nubL47btadvnGU2CptyYeOCnjmpZ5lpCwaRns4917p27t/l3Qdgd2Z75D9I/KD5C/EPtHsPZO1Ovu6cn0hVdd1+H7jwGwo66m2Hkt1bc7N2LvrEUW+NmY7KVVFj83j46WrWhm8EnlRIwnuvdN3dn8tDbvbG4vj12btr5O/KDp/5AfHvrXN9MUfyH2PvDamR7Q7N6p3acPUby2r2pLvHZm4ds7jnzuYwUGUir4sdS1WPyg+4pTGQqr7r3TV1D/Kc+PXSnVPRHTm0N39v1u0/j38xM1809lVe6d3Uu5tzZHsnOV29shVYXdu48liZcluPbnn33WM8kz/xKdwjy1LvrLe690Inyd+AuO757g2v8ietPkL3l8Tu/cF1/kOos12Z0TWbJafsDqeuzDbip9k782z2Hs/eu1s1HtrcE09dha8UsVfi6mqnMcpSRk9+690mNpfytPjpsTa3xC2ptnMdmRQ/Dzv3dPyZ27nsvuiDcG7O1u4994bfWN33vHuTcWVxc9ZurK7vyPYeQyFZJB9kRVGNYvHBGsI917pV7i+AW3Kne3y77Q6770726W7T+X2Y6O3Bubf8A1tnNsUuV2Hlugtuptnai7No85tXN4iqw+cxitHmqHK0+QpshHIyaYwQR7r3Sz+H/AML9pfEmDt/ORdi9jd09v/ITsCl7M7y7p7UqdvHd2/8AdOL25i9obfQYfZ2A2ts7bWA2ztbDU9FQUOPx8CRxqWkaR2Le/de6OZ7917r3v3XuvW9+691HqKSlq1CVVPBUoPolRDHMoP8AXTKri/v3XuvQUlNSoY6WCGmjJuUp4YoEv9b6YkUXJ9+690CfdHxw6j77xlPTb/21/uexUq1u1ewtsVtVtLs7YuXhB+1zWyewcBJQ7m27kaZyGtDUeCcDRPHLEWjb3XuiBd1bVqMHgsb1N84clkN6dXQ1lZJ1V85MDhcfg8l108cKinwXyUgoKf8Aguza6rjUQf3mhgi2tnLKK2LFVfi86yyvprGXxYjjzB4HqOPcr2s5Y909nj2rfYnjuYW1wXERCzwP5lGNaqww8bAq44ioBBOO0f5fvdWzaaPcPXf8N7u2PXx/e4nN7GmgkzM2Mlj89LUz4IzvDWieDlZMdUVaSEjSBcD2LbTfLO5NJP0n+ZwT9vl+fXPnnr7rnuPyi0k+yWy73s4Jo1t23CqOGu3bz+cTyemkdEbyNHU4uuqcPmKKqxeVop2hrMVlqSox2Ro6mFwGiqKGsihqYZYpAOCtwQD/AE9nKsGXUjAqfTI6xyurea0uJrK9geK6jajxyKUZWHGqsAQR5H+fTT8i+3fk3mPhd8k+hOr+yN0iPevQHaWx9ubUL0uSNQ+f29lfLgsTU5CmqchRVGaSqnpYzFKrjzhFIFgCy/2+2nhuJPplNxpNKYNfyp1NPtZ7zc7cob9yhtc3N10nJcd7Ek0L6ZEW3aWslDIrOqjUSSrVCigouOim5n+aTict/LCptgJ8idvf3arvi3SdH4voLAS7OxnYldvV+tx1dB03J19icfDvObMf3gc0dRSy0+lkQyu2j1+y9I9nSxWZY0abw9NM6tdKUoa0Nepbn5k+8VP7nNy3cbxuMXLse7+M9wkMcNr9CkvjGdrtYlRrc2+TWQlx2Gp673p2f8idi0v8nLZO8e3N7Vu6eoe69iYKD73MrkIsHubbPxZ3xgaqOhSpglhmNFE81GyyCRHj1qQQ3t19utYhtMTWy6ywD+p7DUH889BzaPd3nzdZffXd7DnXcjYW+3yXFiGkJWAHcYljeNSpUfpNpAOoaDpOOtu/4O/LKr+RW3srtzd9AabsrYuPxk24MnRU6w4Tc2OyEtTS0ObpIka2OyEktE61VLYRrJ64iUbShHu23GyaOSNv8XfgPMEcR/m6yn+7x703HujtV7tO+W+nmvbo0M0igCK4jclUmUD4HqhEkfwhu5O00U+nsn6yS697917okHy8gyJm2LUEk4cDN06qAdMWUf7SUtJ6LAzUaWW7c6WsPr75k/3hNru31HtjfO5PLoF5GAOC3RMLEtig1RCiktnS1BxJGnKTR0vlI/V7T/tc9E3FTOujRLInjEgi0sQYxL/nQp4IEl+R9D75t6QpqUUsacQD8xx9MU9Dw6GQNOsflk8Xg8j+HX5DFqOgvbTrK/p1W/Pu2KhtOQKDjgde8qdc/POZEl80vkjCCJw7BoxGP21jYEMoX8Ae9UWjAKADUn5k8agYNetZ4g0PVhHxSlln69y7zSPK/wDfHIku9izn7DFMWZrXcseSTzf310+4IgT2d34KtF/rDcUpUcLe14f4D1H/ADV/yUYzg/pD/CejQe85eg10mt5ZEYjaO58qYWqBjdv5iu8CP43l+1x1RN41ksfGX02vY2+vsK887oNj5L5t3kwtILXbbmXSG0lvDhdqBvwk0pq8uPTkQ1Sxr6sP8PVEEZ1Ro3PqUNySx9fr5J5J9X1/Pv514mLRoxFCRWla0rmlfPjxOTx6FhGklfTrl7c611737r3XveuvDrMsjlViZ3EOtdSam02vc2S+ksRf3RlUVcIusfIfz63qYAAHh1rvfJrJfL+L5h5aPETdjRbjj3XAnUNLtz+MHa8m1/NH/BPsY6QHCy0UtPc5Ezgm/k8n495t8gQe2De2VubtbE2JgJvDL4Ym8XSfEDEnXqB+AKSOFB0EL1tzbcm0ltRftp8NPKo4dbDGJkr/ALHD/wAZEC5QY3GjLCAL9vHk3o4TlUp3XlaY15ktpIBX3hVcCFZrlrQn6cyMUJpUpU6CQcV00PQvBfSgcd4Ar9tM9a6b5P5nf7OjIkcnYX+kf/SQIvs1/jB2V/c3+KKsaSQ2/u5/c/8AuyQSxFvzfye83Ft/aoe1tT9F+4RYipPh+OZtOTq/tPFM1fPAxw6BwO5jcwmphJr/ANrp4kHy4dbG8sj+qMSMYdRtHrbRc8EhSeQPx/h7wcRQQGK93zz/AIa/8X0MdRoaYr1H9u9a697917r3v3Xuu/fh17qwj4Q7lkeh31tGWQstLV4/cVEhK6I1roTQVyopk8h1S0cTGyhV1fW599Lv7v3mpnsPcLkmVu2KaC+jXAA8ZPBmoK14xRsaLpGamp6KN0QFo5aUrg9H5H0Hvo+K0FePRP1hlginSSOZFlilRo5YpEWSOSN10PG6OGR0dCQQQQQfdgaUI4jqjIjq6OtVYUIPAj0I4Efb9nQLZb40/H7OZKny+U6a65q8jSiAQVJ2piYSgpp2qYbxU9NFA+idi3qU3vY3HtUt/eopRbpwv29AC89pvbLcLuK+vOQtqe6SlG+miHA1FQFAOfUHoYsfjcfiaOmx+LoqTG4+iiWno6CgpoKOipIF/RDTUtPHHBBEv4VVAHtKSzElmJY8SeJ6Hdta21lbw2lnbRw2sYoiIoRFXyCqoAAHoAB0T/5bZSh65zPx17/yrfbbe6k7nosNvjLSIzUu39hdzYDK9VZPP5B1ZRTYvDbmz+Hqaudv26aljklbhPeulAFa9HMF/wA/8T/xPv3WuvEX9+6903ZafE0WMr6/O1GPpcPj6OoyGUrMvLTQYuhoaKJ6mqrq+orClLS0dJBE0kkkhVI0UsSACffuvdV8bXqKX5v762NvzC4Gqw/xC6h3gm+dl5PKYyPFr8n+0tuuybM35gsUVSrPR/XVY0mRxFdVLH/ebM/bVdPF9jRQz1nuvdWMj6D37r3Xvfuvde9+691737r3Xvfuvde9+691737r3SN39QT5PZ256GlQSVE+IqzFGy6w7xRGUKqmw8jeOy/0ax9wp94zlvcObfYr3X5f2mFZNxuNkufDUqG1OieJpUH8ZCUjPk5BGR0IOU7uKx5l2S6mYiJblNRBpQE0qfl6/KvWtT/NM2z2p3nsX4//ABB6bx2Gqtx/IvvbbmZ3zkN70G6peq8X0t0BGnb+98d2bktpwSZOiwm9s7hsNhUpopIqiu+6kjjuA9uFf3b9z5b5U3LnL3U5muJU2/ZdllS3WBofq3vtx/xOBrUTEIZIEeeViQVjMdW8ushedVvbz6DZ7KEs8supsHQVQ6iGI8jQAZFdQ+fQc/HeD5R9HfzK+w8Z8i9vdRz4j59dOYzfGO3B8bMB2WnV+E7w+MdNBtmsp95Vm/IZajBbp3z1Jm6ZYpGljpqtMNEi/uobiHnpvbrm/wBg9jn5GvtxW65M3VrZo90ltfq3sN1bxgYVtqeJFb3iPqBXVGZyfhPRZtKbns3M7JfwDw7mH4kDMgMYpk1OdNDxqdB49Cd8aNubi7++dnyk+ZO9cLlsZsfpqGo+D/xKx2extdjZZtu7VyNNn/kd2/iqLIwU7ij7H7F8GKoa5FtVY3EHSxQ8h7n++sOSvZz279q9pu433bdCN+3ho2VwJZ0aPa7J3Ukara2DzSR8VmnFRXgu2eC43XmTcN6uYWW3hPhxBhmoPc3l+IBQfRB1aOYJahkpoI3lnqWWnghjGp5ZpiI4o41HJZ2b6Dn3j3b2N5us8G07ZavPud06xQxICzySyEJGiKAdTMxCgedfSvQ9aWOBWnlcLHGNTE8AqipJ+QHVk2Iinp8XjIKlQlRDQUcMyKdQSWKmjSRQ30YB1+v59/U7yXZX228o8q7bukapuVvtltFKqmoWWOFEkAPoGBA+Q6wsvpIpb68lhNYnlYqfUFiQfzHTn7E3SXr3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r//V3+PfuvdJnOj1L/wT/jf/ABHuP+cvjX/Sj/CejrajQMPn0F+T/P8Arf8AED3j5zGaeN9h/wAJ6G1j5fn0HeS/U/8AyF7gXmL+zk/1evQxseI/1enQc5T6P/wZvcC79wb/AFeXQxs+KdBzkv0t/wAgf8R7gfmfhJ0M7H8PQc5P6N/rn/eVb/inuAuYloZfs6GljxTo53w//wCPS3d/4dEX/urpPfWb+6x/6dl7o/8Aiwp/2hw9Y2feJ/5WHl7/AJ4T/wBXH6Kf/Oo+Su+/iX8ONvd07E7FyPVjYb5SfFPDb33hjKOHITUvVu4u7NqYrsihno3xuWnqKHJ7SqKmGZaeB6kxsRENdvfUbrHnorm7v5t3S/yM/mD/AMszo/4X/JmTd+2t89kfIqP5D7KxGzd4YCk3HtHbvQm4M3swZar3zsvCGSmx28aNJ4loZxN5FBcaPfuvdHF6C/mQbt+TXZ8R6c+HHc26/iHXdsb16VxPzIg3T13T7dy+7Ov6zMYPcO7aHqKqzMHZEvTEW8MDVYiLc4j0TVKCRaUwHyj3Xuii5n+cX3r2/wDHL5kdv/Fn4FdwV+A+Nm2fk7tdu5d7796j27suLuXoPK7jweQXb+3M5m4sz2BtTDY7FLnKmrp4kglkilxSh6tGI917oBPkx/ME+UGZ/le/Aj5C9y53dv8AL9ynefyn+HOxe6e3Np7y6uymQyvRXZ+zpc5vntjbOQpMTvnb/XeF3bLJ9xT0NfRTV+MjjCSpb6+690YP+XN8gfkz33uP57bQ+PHyZPyw+L2waHYWF+GfzS+QezKLLUuU7tzGBzL9u7EyGV64xPVFP3hsPqvLLj3bIUNLQv8Ac1D0X3jhfIvuvdDf/Kc7K+Q26t+fzKesvkN3zuL5C5j4+/Nyt6v2nvPcG2Nr7MWl25J0x1Zu+oxOE2rtKkp8VgNvwZ7clW1JS+SqlhhZRJPK+qRvde6uP9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XvfuvdQ8jj6HLUFZi8pRUmSxmRpaihyOOyFNDW0FfQ1cTwVVHW0dSklPVUtTBIySRyKyOpIIIPv3XuicVf8v74zx1ctXs7b29+pTNJLO1B0x232h1Vt+OeYhmkpNq7L3Zidr4/SR6Vp6OJFufTyffut6j69JndvwLxG46epjTvfuXKvIjpSUfaj7B7qwtEpR0SOJd/bJrd0wxJrPMGYp5bHiQN6g/DdXNuawXDL/g/Z0GeY+SuTecIfB5q5WsdwGaNLEpkFcHTIumQY9G+fHojO+/5b3cO3KuSo21t7rntbGPUMYZdlb43B0ZumjhZpJQW23vij7h2XXtHdVXwZTFppAAjXl/ZrDzDfxadYR186ih/aOsf99+6D7U7nrfaLrctrkauI5VmiOSQPDmQsBwFFkUBRQCpJ6Jfkf5en93N5Sdi5P4qdhbZ7ElqjJUb9xXxx6j7P3SZfDFHPkTvDrDf9bn5KqSV2An8K1DxC7Wa49rE5gttQkfbwJPUFa/tIr1Hm4fdG54bbzslj7zvJsAoBbzpdCPSACF8JJmjADVotCpFK0OBNrvj5sGXJYuLdm9N64LIYfJzVWHrd+fC/wCRVFFgMxHS1FLNk8fmZdpV9LiaxqSaSAVdPMC8cjIrlWI9unmK2NNVo5p8xj5j59ByP7l/OkMdxHb+4u3LHKulwIrkK61DaXAcB1qAdLVWoBpXqyz4m9p/Av434mro6D5CYLK793h9tDubdG6sLurZ33YxxnlpMRisdn8HQxYjDUDVMjKru0k0jl3ckqqkm47jLuDqWGmJfhH+frKX2e9ldk9otuvPp7p7zmC8C/U3LDTqCFikcUYJEcSFmIFSWYlmOQFPDt35mfEfdkVHLt35N9DZMZAP9lFD2tsmOpqDG0iOqUlRmoarWjRNdSgYW+nst6mbowGIzuF3BQRZXAZfGZzF1AvBksPX0mToJwLEmKsopp6eQC/4Y+/de6R/Z/XtB2RtOr27VzGkqRKlbicgFZ/sMpAsgp53iVl80DLIySIfqjH8ge4g97faPafebkO/5R3C5NteBxNaTip8C5RSI3KgjWhDMkiHBRjSjAEL9tv5NuukuFFV4MPVcVHVV24dv5naeXq8FuGimxuSo5HQx1CGNKmISOkVZSSPZaijqVXVG63up5sQR74U84cpcxch8wX/ACxzdtj2W9QMQUkBUSJqKpNCWJ8SCQDUjg/D8QBqOpPt7mG6iE9u2qIipp+E+hHlT16Z/YcoOOafZx+fT4NepmOxuSzFdT4vDUFVlcrVnRS4+hiaepmax/3WgOmNLXZ2sqgEk8ezHZ9m3nmLdbbY+W9rnv8AfZ6rHbwKZJHNM0UcFA7mdqKoBYkU6rJJHBGZZ20RjNTwp/s+nVr3V2yI9gbKwu3VKtVQxGsy0qksJ8vWkTV7qeAI1lOhLAehB771ex3tna+03tpy5ydGdV9FGZbl6113UxDzkH+EN2L/AEEA6izc7z669uLilEJoo9AOHQie5d6L+gf7/kki6X7JeKSSJxtavAeJ3jcBgisA6FWAZWIPPINvcJfeRllh9iPdOSGRkkG0TZVipzQGhGRUEg+oJHSmz/3Kg/0w6olp+ydj1PZ2V6Zp87G/ZmD6/wAN2lldrCkrFko9hbgz1ftjD545AwDGyJWZ3FzwCBJTOmjUyBSD74ZnY90TYbbmY2ZXYJbx7RJarQzxxrK0emuoaY2Vq0pQ8a9CYspkZNXfxp+dOseyOz9idj5ntDb2yc8uczHTG+pes+zaKOhyFM2198Q4XGbilwkslVTQxV2jDZimm89OZILyadeoEe77rsG77JbbBebpa+FbbpaC5tWLKRLCZHiDjSTpq6MtGoccOtBkZnVWqVND0ICxSMNSo7L6vUEYi6jURcAi4Xk/0A9koNW0jJr5eeaV8vPy4+XVgCTQdekili0+SKSPWodDIjoHQ/2k1C7D/WHvykOKoQ32Gv8AsV+VetEEGlM9dtBMiI7xSqknEbtG6o5/2hyAr/7An3oOjFgrqSK+f+EZPXqj16C/dvdGwNndddw9nSZ+n3LtroTH73qez4toTU2dy+2Mp19g5Nw7s2hVUizRLRbxocagZsfNJDJeRNZUNf2f7dyvve573yzsC2bQX28PALTxgY45UuJPDimWgOqFmr+oBTBpU9aLKFdtWADX7R5dKjZO6cbv/YmxuxsFDXx7b7D2dtffO3v4lTfa138E3dg6HcOKjroVeaKHIJj8hGJ41kcJICAxAv7QbtYT7Nu+7bJdujX1lcy28pU1USQyNE1DglSynSacPLqyKXVWHAgH9vSvWOqKRnxyiGVhDHKY2WJgzaVQTFAChP0FyB/T2Wl0DONXcBUjz/NaUp8z1sr5BqnoFumO7tld90vY1bsGDP8Ai6t7j390XuZM3jo6GebfHW9RQ0+45cRDBUVRrMFJJkI/tp28byANqjW3sU8z8p7ryhLskW8PDqv9rt7+Pw21AQXQYxayaBZKKdS+WMnptGEmrQeBI/Z0MJilV2jaKVXQFnRo2V0UckspAKgDkk/j2GVIZdQNRXyyP28Py6vQg08+uf21RrEfgn8hQSCPwy6zGR+sLo1lP8QLe9B0pqLAL9o/Lz8/9nrxUjj1j0MQSFYqv620tpS5sNZsQtz9P6+9lh2/Ovp5CuM5/KvWuuxFKxjURyFpf80oRi0ova8agEuCf6X59+JA1fLj8vtpWn59e9OjVfDurmx/cVTQtCo/iu0cvDMZ1dJoRQVVDVp4lIBu7GzAj9NiPeYv3GdzNr723VjEsbpebHcqWrUr4MkMnbTjUmjV4YpnpBuiH6ZSf4urVPfYsYFOg/1737r3XvfuvddE2F/fuvdE9+cW/uvtufH3f2wt4UVVuzcfde19x9U9adVbfSGt312hvjdmGq8XiMDtHEMTJJJRVNUlXW17qKLD0cMlbVSRwQsw917oxXWGM3RhOtuv8Nvesp8jvPEbJ2pjN3ZCldpKau3PQYKgpM9V08jKrSwVGUildWsuoG9he3v3Xulz7917ojnyPw9J3z3N1d8S9yRCbqjcGzN19292YZpZqcdhbV2HuPaeA2l1nUywTxPNtfPbz3FFW56mKslfj8cKKX9irmVvde6O1R0lNj6WmoaKnp6Oio6eGko6OkgjpqSkpaaNYaempaeFUigp4IUVURQFRQABb37r3Un37r3Xvfuvde9+691737r3Xvfuvde9+691737r3XRFwR/UW5+n+x96IDAgjB69ny49F93z0fSZWWpyO1aqPEVtW6y1OLkDLh6qRSS0kaxKXo5nZrmwZC3Nhf3zd+8B/d+8vc7bhuXOHtJuMOzcyTv4kllICNvlc11tF4Y12jyE6mCBoixY6FLE9Sxyx7nXG3Jb2e+wNcWqAhZB/aqPIGpo6jh608+gfPSnZCrJEuPoTG59aR5mmCS6D6GZGHIH1F+Rf3hFJ9wT7ziMwXlTbHAJAI3GAVHqNQBoeNDn1z1Iw9zeTGKO13L4gHEwtUV4jB6kQdIdh1clqqHG0gRY4xNU5RagCO+nSq08cjARrzbgW+nsy2b+75+8duNy0G4bZs23QhlrJLerKCGJ1UWFHJ0jJBpqrQHpmf3Q5Qt0Jt3nkYkmixFc/OpHE9DpsLpzD7TlhyuQnGaz0VmhqJI/HRUD8+uipyWJnFyPK5LW+gX30N+7t9yPkn2ZvLLm7ma8G++4MJ1RSsmi0s28mtISWYygVH1EjF6GiJH1FfNfuJuXMEcljZp9NtTYK1rI49Hb+GudIFPWvQzj6e84ft49R310fof9Y+/de6Ld2B33RYKWuo9uJRVIxkNVLks7kZDHiaFaNHkq3UFoVmipIYnaSVnWJdJ+tvfM33//ALwKx5S3u95L9mtott23iGRoZr6fWbWOcNoMUESFWuXV+0vrWPX2rroepc5b9sWurRN05juWt7QjUI1prK+rsa6K47aE0OadEn+PXz+2Z8xevW7d+OfcOL7D69h3RuLZ0me2/hp8RBBuTbEsUGYxtRQ5zHU2Si8KzxzQyNH46inlSWNmjcH3ib7p/eb++TyZzJFsHPHMzbJvPgRXAggtrNF8KcUjNQk2oVBFGctG4KtQggDTY+Vfb6/tXuLOwE8akgs7u1CCa8Co/l5inQNdI/zlvjz3d2Nguqequ0txdg7n3Bls7hcJPU9B9xYzZ2Wrtstk480sPZdZsfF7H+0ppsPUxip+/EMksWhGLEAybJ79ffj9sNjk5h5xk2W72W1jhklS7bbmuSkgXQrw2tzHdKzCRCQIi4qCQFr0SLy97d7xci2sormORiQGjMhUEVBpqQrgj1pXq27rvtrGbyk/hlXTjE58RtKKQyeSmrUjGqWShlazEoDcxt6wOefeY/3a/vk8qe+90eU912790e4axM625fXDdogq72sjANqXi8LjWq91WAJAK5v5BveWUa+glM+0aqa6AMleGsDGeGrgT0MI5AP9feaHUf8AXvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvdf/1t/j37r3SV3BLHGw1uq/t39Rtxf2A+bopJGXw0J7R/hPRjY3tpahhc3Coa1yaY6CnK11Gl9VTEPp9W/pa/uBuYNtvpPG02jn8vt6ENrzRy7DiXeIFPzboM8tl8ahctXQLbV/bH0ubH/W9wVzBse8usoXbpT+XR/be4XJEJHi8z2a0Hm46DHLbhwqa9WSpAL8Eyrz9b/7z7g/fuWOYGU6Nonr59vR9B7te2MFPH5429CPWUDoMspunb8YYPlqIfT/AHcv459wdzHybzXJ4nh7Dcn7F6EFt75+ztuoM/uPta/bMP8AN0GuW3ntdA2vN0C8m95kt/a/x/x9wVzByJzmxk08tXZx/B/s9CC3+8b7E2+kz+6uzKPnOP8AN0eH4W5jGZjZ28pcZW09bHDuyGKR4HDqkhxFHIEYj6MUYH31O/ux9j3nYvbn3Nh3nbZbaSTmBCocUqBaQgkfKuOoj91fcTkb3F3Xa9w5F5ps91sre3MUr27iRUkLswRiODaSDT0PSC/mZfFPsT5h9AbI6u6yyO1sZuDbnyb+MncddUbvqq6kxUu2One4dsb+3PRwyY+hyE0mVq8PhpUpY2QRyTMquyqSR0y6izr3yi+J++u5fl7/AC6++NoVW0sfs/4o9kd97u7NosrLV0uczGM7R6PzXW+Ei2zDRUE1NW1cGayKvUiolgVYLspZhb37r3RX/gp8Zf5iPwnqdsfEXDxfF/dfwl2B2z2RuzafeOT3Jv8ATvh+l98bt3Rv7HdRVHUtLt6n2tFvzB5/cv2Z3K24JaF8dBq+wadhb3Xul78WPgD2V1j/AC+flB8PeyN07Wpt09/b2+c9fi9y7XfIZnDbf298ot79j5ralVVxV9JjKmevwmL3nC9ZAihfLGyI7cOfde6AKi+CPzS7N+GX8vLoLujE/Hnae+fhH8tvibvDKy7K3vu/d209/wDRnxl29Ltyq3HG+c2TharG763mrfcJhXinpKZDoasc3Pv3XujIbC+JHf8A8XPlr82+4/jJQdT5Hoz5P9TbR7H250huXcOd2Rh9t/NbZkVXtjLZiGDBbczuN29sLuHaIoarP19FA1Z/GKMzGmlLlz7r3QY/y3fjz/MD6F+Q3y/3t8juu/jDheuvl33VmPkXmK7qjuHsDeO7NjbxfYWw9g4zY1Dh9x9YbXxuewM9Ls9qubJPVUs0ckxjWmZQGHuvdXV+/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3XuvWH9PfuvddWH1tz7917rv37r3WCopaarjMNXTwVUTcNFUQxzRkH6gpIrKb2/p7917pC5HqXqrLljlusuvsoW16jkdmbcrS3kUK4JqcbJfWosb/UfX37r3RbO0Ph3taOjfe/xlgw3x87121Iuc2bntmQV22uu9z5SghkCbS7f682vVYvbW+Ni7mgZqOt89JJkKFZvuqGWKqhjY+69Unj0qOkflfsPs2CDam+paLpvvzC00VN2J0Lv3K0OI3ltrOw64ayo28ldJSpvvYuQliafE57FfcUFfRSRvqjlMkMfiPXrdCOjHZDDbb3PTJ/EcZhs9TEHwvV0lHkoRY3/beRJVWzD8Ec+w7v8AyjytzXCtvzNy5Y7hCOAuII5qeePEViuc4p05FPPAdUEzI3yNOgwrPj11HXTGZtpxUx0CPx0NfkaOD0ljqEMNUqBzq5a1zx/T3BW5fdD+79udybp+QUgbSBpgnuIUx56I5QtfnTPRrHzDu0ahRdVA9QpP8x0IG1tjbT2ZTmm21gMfiVYWlmghDVk9gB/lFbL5Kqa9r2ZyL+5a5H9seQfbi0az5K5UtNvQ/E6IDLJ/p5m1Sv8A7ZqDyHRfc3t3dkG5nZz9uP2dK32PeHDpL1737r3QB/J6sqaLo/fD0zhHnpcfRSEor6qety1DTVMYDggGSGUi45F7jn3jd97i+urD7vnuHLaSBWeGGNiV1fpyXESSD5VQkV8q16W7eoa7hB4Z/wAHWop2tgPkvn/5nW94fjT2j071fl6X4EdSSbsrO4Or892hQZjEP3tv9MdSYKhwO5NtzYavpa28ks0ryrJGQoUEX98zeXrzkSx9hNpPPew7pfwNzheCJbK7jtWR/oLfUXaWKUSArgKAKNVjWvRy3i/V0hZQ3g5r9vRUdrYP5K4Hpz+Z9v6l7DizfyN+OH8xjbnfu5sj09js1sDZ/beH6y606xzHZexV2ZNlsjXNtPcvV8tcrY2rqKlZKylSTkkASDf3nId7zP7CbN+5PC5I3vkeTb4kvXjuJrNrq7uktbnxwip4sV14Z8RVUrGxA49NATeHdvqrIstcedMn+XWD5N/IHenbvWfzN+ZPTnyH3r1x0FS96/DL4xfGvfu0d2z4zalFtXCdtbJyXfPeOMpamVNuVUmUzO8ziKiqqongmo8XLTzloPInt3kPkza+W9+9sfbHmbkm1vucjs+9brultPCHmaWS0nTb7FiP1VCpAJVjRlZXlWRKNQ9akkMqTzI9Ig4UenHJH28OjBb+y+8PhJ8gO79r9B9jdud+bfr/AOW73f8AKLJbZ7Z7QzHeE+J7i6s3NT47rPsnE1uVqK18BjO16DJ1Lz4qh8OLrhQ66WFY0Fgbs1rtfurydylf847Htu0Xyc82O1LLZ2qWIezuoi13bOqAeI1kVUCWTVLGXpI1Tm7N4MjiKQsvhsTU1ow4U6D7D0Of6T2j/LI+RXXHya7s7n7V+WvbHUOyu4sLu3uLM762L8hdodz7Mr8/2jltr9Y1VXU7b2DJ07Vg1VDVYCmokw9PSeGclibnFxLZc1bn79ckb5yJtW18vct7bez2bw2KW8+3TWEyx2qSXQAluBeAUkS5LmdnLJgdVFYxaSrOzFiNQJqDUZp9nr5dILqDoPZOwOhv53G7sRvDt/NZPaPbPzs6zpsRvDubem7Nt5HCf6J8Zk4c1ubamXylRj89vupRzfPVUb5F4AFEmkD2ccyc47pvHOP3U9sutu2yGG523YLtnhsoIZVkF4yFIZkUNHbg8Ldf09eQKk9eVAEv2EhqGbBPy/1Z6FnofZW7fjrv/wDlA5TAd4d3dgSfKnYE3X/dm3uzewMluDr3cOHpPjViOx9j/wB2OvQYtq9d/wCjmrxyUOPbEQQSzUIb7l55HZiHubd223nbZ/vKW97yntNkOXr0XFlJaW6R3Mcjbm9rceLcZluTcBy8njMQJKFAoAAtGpjbbz4jEsuc44YA+zoJ8p3DVx9zfEL5BdA1Hy7k2H3N/MGwHSOR727l+S+LynWPe2zs3vDdey+xtibX+Ksucnmw2ztt5PFSQYTIw4nH1FBHj0mm/wA6ruILXliJ+V/cbkrnGLlz97bZyZJfLYWW2Ot1YTJDDPbXEu7eGFkmlRw08ZlkEhkKJQggUMoDQyRl8y0qWFD6inHpW9bbl2hjuq/kB1znou/dxbm7l/nD/KXYfWXVXxx7CTqbfncm6cdT0e4arZWb7OkyWDGzdg0GEx82TylQmQop5vtY4o2k1eJi3e7HcrjmHk3fLN9mhsdr9stquLq83K2+tt7GEkxrOlpok+ouWkYRRqUYIGLGnHq4ZQro1TquGAAOn+fp0By9n99SfHfubqnF9rd3dP1/X383/wCOnx42Pla3uXG9ydu9T9b9ix7Zrdy9e1ncFJVZPG9jU2FyWYqBBHXSVkUQK09QshiZfYubYOT1515X5iueXNq3GG89tNx3KdBYtZWd3c2xlEVwtmVRrZnVF1FAjNQshUMD014jeFKoJBEqjjUgHyr59Gyx/wAYKvNfLH5f/FqT5Q/Mal6J6q+PvUHeW0Ns0/yD3f8A3up+3+0cZ2LgspuzIdrGVewarB4mq2HFk6bborUwRyNVI705jVEEdSe4EVr7c+2nuEvIPLJ5u3HebywllO3wmA2Vq1tIsK2lPpw7rcNG1zo+oESqNdSW6dEX69xF4smhUBGc1I/wdTepu8e0t67t/kcZvdW/s95O9Oj++l7Rj/i8uMwvam+sP03jW27U7mx8M1Pi87uKorcbLX00UiNIKp5ZIlBJPtjmTlPl/adt+9ja7fs0IG07tt/0lUDSWkD3rCVYmILJGFZUYig06VbrayOwsTq+JWr5VIGK/n0k6HKfID5J7V/mDYLoHtev35Pt3+ZbiMPgtq4bvJ+va7sfo7aOzNoZDsX49dL9w0tRWJ1tk9yzJUeCXHNEizU9RGXi8kkgMri35O5D3H2Zuub+XY7KO45Ekd5X2/6kWt/NPKttuV9ZMF+pWIaQVlDEh0bS2kDrVXlW4EbA0lFBWlR5gHy6vV/ki7l6633tHsWu65o+7tvQdX7r3v1ZvXrv5Hbwrux+2ereysZuCkm3V17V78yGa3I24cDtsmMUFRFkKyKSnqAFk4KLkT91vlberP3u5v3ve7nZZ2bleylgm263Fvb3dveTMYbuOFUiELSCBhOpiRiwFRQAlLeyKbWONC1PEIIbJBAyK+fV+fvor9nRN1737r3Qbdo9v9YdLbdG7O1d97b2HgZKlKClrNwZGKklyuTn/wCA2IwWPBfJZ/N1R4hoqKKeqmPCRsePfuvdFXqt9/Kf5G1kuK6h21W/GHp2SQxVXd3auBgq+6t10AdRLJ1R0pk1ak2XDWxKfBmd4/5REsgdcJIbEe63gZrXoa+pPjB1L09lJ914bFZfdvZWQgkpsz292duDJ9idq5eKcD7qnfeW5Jqytw+KqWUFsdixQYwFRoplsPfutHowoAH0Fv8Aff737917rv37r3RU/kv1PvncU+xu6uk48W/fnRtXmcnszE5zIS4jbnZe0NyUtLRdhdOboyUSSrj6DeeOoYJsbXSRypis/Q0NWymGOZH917oROjO9Nl997Lbde1v4licpiMlUbZ3/ALC3LTDGb66v35jIoHz2wt+YIs8uH3HhnqFuLvT1VO8VTTSTU00Mz+690NHv3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de66IB+vv3XuvWH9B/tveqD063U+vXrD+g/p79QenWuPXfvfXuve/de6hZGOaXH10dMxSoko6lIHA1FJnhdY2C3AJVyD7JOZbe+u+XOYLXbJjHuUtlOkLAaisrRMI2AxUq5BAxWlOlFo0aXVq8orEJFLD1AYV/l1rV/zZex93bD+FXYOxOuqLdWV7h+R+e2x8YOttt7GomynYWVyfa2V/hXYE+zMMtVRSZDcG2uq6bPZGNDNCiPApeRF9Xv50fu3cs2t97ubZe81vBDsnLqz7nfPdfpwRtYhjEt0xB0LJfeDE9QWJYrpJx1lFztuHg7LJDasWkuNKrooTpegxnPYSwJp8PGnRTPgdvHbHSXzz7q6VwPRXefxd6W+SPx/6z7O6a2V8guv8R1etf2n8X9rUnU3db7NocXuHPYuoTJdYy7czWSkMkMrT000roRd/cle8m17hzZ7Ncp82X3Nu0b/AM27FvV3a31xt1y93ptN1ma8sfGZ4437LoXUEYoQAyKp4DoMct3UVtvt3ZCB4LK5iXSGWh1IADQAkfD4deFaE+dOuO2Zvkp/K7qfg70vjfkh1j8pfjD3z8gv9Ae2OpH6rodi9x7Xw/aOZ3dv+k7G693ntje+4absrCbHrK6V8x97RJD9iySCSM2trcv6g/eGh93ubJ+Qtw5e9wNm2T94y3gu2nspXs0htmtbmCaCM2kk4AEOiQkyAoQc9btvruWbraoUvYbm0muNFAtHGvUahg2dIHcCBQVz1sL9Y0tTP2FtpaVJJjS5J6idogxWOkhgnSad2XhYCCAb8G4H59gX7ou17tu33kPaX902c0zW24/UTGME+FbpFL4kkhHwxjWFYk0YsFySB0LOep7eLk/fDO6qZIAqhuJcstFA8zjH7ej/AA+g/wBYf717+jrj1if137917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r//19/j37r3SG3Z+tPz+0BzcjnX+PYX3+uoelB/hPRLumWWvkpp0Bme/wB2fTi/0HuJ93A1OKf5egLuGakjPQKZ4khzzf6f737jLd/xg5H+x0C78Chxw6BXOk3cHnk8/wC3t9LW9xluwGQOHQOvuNPI9AxneRI30sDwP9Y+4w3gDOK/b9vQLv8Ai3/FdAvngLSGw/tf7x7jDdq6nz0DL7i5+XVmv8uDnr3sX6/8f3TN/t9v47/ePedH3KP+VO529f3uP+0eLrJX7uRry9zED/ynD/q0p6Wf8wf5c534W9J7O7W29srF78rt0fIj48dJS4bLZWrw9NSUHdnau3eu8hno6qjpquWSswVLm2qYYSoSWRArMAT7zQ6yL6y/JD5a5zoz5R/Az4+43ZmL3Bi/mF2D3PsvPblrcrV0VdsmDq3qDK9mUVbiqCClmp8tNl6vHClkSZ4hHGxdSSLe/de6dcf/ADDvhRlvkdP8R8X8kOtq/wCRNNl6vbU/W9JkK2eqj3bj8W+artlNuCKhbaX9+qPExvUS4MV5yyRqxNONJ9+690X3uD+c7/Lu6q2Z3Hn6P5I9fb+3V03132Nv3M9ebSrsrXZ6pqOt8xlNp5LadVV0uFrcft7cNbvfGHFRwVzRSq8i1BQ0v7vv3XugU37/ADhMNvz4d/Fr5JfCzbuw+1d0fKD5LfH74vwbZ7J3BuzbW2Oq+w+7MHJmsni965jA7WyGXylVsQmOCpbG0k0E7sWjcqPfuvdCt0R/MI7nz3ZHzS+Nvd3xsp8h8m/hpsbrfsmp2x8Y96jsbZ3de2O3cNnMpsPF7DynYeO62r9r75qKnAS09VjM6KeKFHSpFS0Dah7r3S//AJePzT7d+XWU+X22u6+ldsdFb2+L3yQn6NqNoba7A/0mK1K3XOyOwKWbM7qp8ThsRWZ+lh3itPVigiaiWaJlikkUeRvde6sfqZjT0884UMYYZZQpNgxjjZwpPJFyv+Pv3Xuqh9qfzMt37i/k6bp/meS9Ubfpd37e6N7h7ci6nTcWSk27VVnWW5d44KkxL7ifHpkko8rFtdZZJRAXjMpCggA+/de6Mp2d/MW+KPxy6w6O3/8AKXuXZXSVf3lsfEbv2xt3KyZrNZStSXbWH3BuWfH4jb2Jy+cl27tZcxGtZk5aaOipVdDNJGXF/de6ydp/zMPgV0rJtSDs35UdS7Yqd97O2H2FsakbNz5es3nsbs7K1OG2NuzadHgaTJ1W48Fn6+jlC1NGk0cEUbTTGOEeT37r3RZugf50vxL+S3Y3y26f6xz0NV2V8Z8t2LR7cwFVPk2TuXB9a9eY7euf3ltupXAw0+CwdJk6ufFSRVsgqjNRvLGrxshPuvdAJ8df5sHyd3Bg/wCXR2l8j/j90fg+l/5me49ubG6freku0t77i7M6y3lvTY+e7A2pH2Xs3fOw9uY3MbcqcNtupir67C5GY46XRI0ckTX9+690LHy1+e3za+GG5tpdt9v/ABv6Dn+Hu7fkt158dIaPaHce7M38n6Gj7V3lBsTY/azYObY9B1lkqLIZisgqKjbdLXyZKlpJCTUu6SBPde6uj/1/fuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+690GnY/TPUncFFT47tXrTY3YtHR6/sYt5bXw24TQeVkaX+HzZOjqJ6EytGuoxMmrSL+/de6LxV/y//jLTyffbE2xuzprOQxutBnek+zOw+sazGu7FzLSY7bW5KbbkjM5uyz0M8b/R1ZePfuvdJKs35378QoKjJd3Zyo+RHxvo6yL7zujG7epMV3T0tt5kKyZ3uTam3aemwPZOw8Eqq9fuLAUtFlKCm11FXjJ4o5qqP3Xuj04LO4bc+Hxe4tt5fF7g29nMfSZbCZ3C11LlMRl8XXQx1NFkcZkqGWejrqGsp5FkilidkdCCCQffuvdO3v3Xuve/de6DPuLZkvYHWu7Nq0ziOtyONaTHuxfR/EKGWOvolkEYLmOWoplVgAbhvcT++PIcvuV7Vc58nW0gW+ubUtCTXT40TCWINpBJVnQKwAJoen7aXwZ45KYB6ogj2JtCl37k+zV2vjqXsrJbTx/XeX3aYpFz1Rs3BZqtzmO2nVSeTxfw3GZ6vqKhE8YdZpGubWA4Nybpu/7qi5emuZV2mC7eb6c/AlyVEUjkEV8TSojYk1oKEDoUUSocLkgZ8/XpMz9ZYPbmA7tl6r21tDbG+u513ZuXcWVyVLWvhN19o5zZr7Rx27d9w05qqmupoqWCkirEp47y0kBQIS3Ktd9ub695THMV5c3GzbWYYo0UrrhtEnEzQwVoFqS+gsSA5qTjr2kBZSqgFqn7WIpnokOZ/l6Ukf8ALu6d+CG0c9tCGDrrKdCZXc+f3HhKqfa29avrvtPCdn9py1ODpqapnkj7ArKevipYZ4zGqTxxz6UBtLVr70yt71cze725WtyXvY9wWJI5QJYBc2klpaUckAG3XwyxU1qpKZPTJtR9KsBIpUfnTNcdG/6h+Mvx1+PNbvip6G6c2f1evYWQjq91fwOmq6ioy9JSPV/wnCVNTl6zJzQ7dw8FbJHS4yFo8fTrIwjhAPuM+ZOfOdec4dqj5u5mub/6NKQ+IQBGWprkUIqDxJNI1ysDI1BqYnp1EjhZmiVQT5j/AGekr1f8Mfif0pv6t7R6m6A672Hv+siy1PFuTCY+s82Dp9wSmfcFNtDH11fWYjZNPnZiWq0xFPRCfUQ11JHsx5g90fcXmrZoOX+Y+cr682VCh8KRxRzGKRtMyqrztGMIZmfTQeY6osMKMGSIBvz/ANWfl07VvxQ+NeQ7A7I7VrOmNmzdhdw7Sy2xe09zCLJQz792rn8ZFhM9jdx4+nyMOIrKjNYanjpaqsFOtbNAiq01h7TRe4vPcGzbHy7FzVdDY9suUntIqoRbyxsXRo3KmQBHJZU16FY1Cjq3hxamfwl1Nx6X7dS9ZPUdR1bbIwTVXQZB6UqDFP5esSu2F2Wp2s33H+SkbTUUAMnl/wAnFvrz7JRzHvwTmOMbtNo3gUvhUf41+qZj42O4GU+IRjuz1YBR4dEFU4fsp0DeN+Dvw+w/YMnauL+OnW1F2G296Hsul3NFj68z4XsDH5QZyHd+2Me+RfDbVzVRmh93VyY6mphWz+qoEn09iif3Y9y7nZRy7cc737bL9I1q0WtQHt2XQYZWCh5UCdiLIzeGuE09N+DDr1+EuqtelLu34ofGzfWytzddbs6Z2dl9mbw7LyXc+4cMYslRSVXbuZlWbLdlUeXx2Ro85hd45B1tLWUNTTOyFk/QzAoNt9xOedo3ax3zbuaLqPdLawWyjeqsBZoKJbMjq0ckKDgkisK54562Y42BBQZNfz9ft6xbe+I/xf2jtkbL2t0R13t7aQ7C2d22dvYjFT0eNftHr6npKXZe/wCWKOr1zbowEFDF46l2ZpXUvN5HZmNr73H5/wBzv/3ruHN99Nuf0U1oJXcFxa3BYz24OnEUhZqqB2g0WgAHXhFGq6RGKVB/MdC3BsLZVLvXd3ZFPtjEw7+39tnAbM3ruxIpf4vubam1ny0m3MBlJTKYpMdhnz1aYFVEZfuXuTcWDr7zusm1bdscl/Idns55J4Ia0SKaYIJZEA4O/hpqNfwjq+NTPTuIoegr3p8Uvjb2J1Ts/o3e3TGy9wdS9eSY2br/AGVUU9fS0exp8Ok8ONqNpZPH19JnsDPT09VLCXp6tGlhlaOQspt7P9q9w+eNj5i3Pm3aeaLuDmS91fUzhgTca6ahMjBo5ASFajIQGAIoeqNHG6LGyAqvDpiyPwq+JGT2DmerJfj11xQ9eZ/d+L7ByO1dvY6t2xQDfmEwcO2cTvLFzbdr8bX4PcVBt+BaRKmjlgdoS4fUZJCyyD3T9x4N5teYRznfSb3DbvbrLI4lY28khleFxIrrJGZCZNLq1GpSgAA8IYSNHhDSWr5jNKdW/wD8tPozrTozpzPbc6l2Jt/rvZKbnmpMRt/buPFLRLLDAldl8lNWTTVWSyuTy2VyLyVdTVSyzzTJqZ2NrdKPuPtvvMPLvuD7h8z7nNe7zuW5x2wmloWMdnAo0oQBpjDSlRGFVEK9oz0T7kVVoIkFFVa/meNf9nqx4kgC1v8AY/63vOgcBTor6I7lPkp2B3bltybF+G23sFuOHb9dV4DcvyX7CgyDdA7ZzlHUvRZbC7Hp8RU0ue7x3fg543SeHFy0mBpahDFU5ZJlamO+vdLjqb4nbM2NumPtfsPPZ3vrv96Vqabufs6LH1eUwUE4DVeI6u2rRU8O0+o9sSS6v8iwlNBLMp/yuoqpLyH3XujVhQDf/ff6/wDW/v3Xuu/fuvde9+691737r3Xjzx7917ojfyG2PvLqPfJ+YHR2Ak3DncVgIsP8j+psLSRDL999U7finnxWU28dcUEvc3Ukc1TUYBpbHL46apxEkg8lG9P7r3Rt9ib42p2Xs3a3YOxc3Q7k2bvTBYzcm2c7jnMlJlMNl6SKtoauIkK8ZkhlGqN1SSNwUdVZSB7r3St9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XvfuvddH6H/W9+690VTt74ldYdqbz2F2VX4HEyb+6uz2b3T19mMpSzVsW1tzbkwFXtXO5/ERJPGtDl8jtzIT0bzBXKxTPpClifeCfvh9yvbOfrzmzf8A2z5tflvfN+UDcoPD8Ww3DTKswMyD9SJvFRZG8FtDsAXjNDWQtg58awWxt9828Xttb/2TYEkYoRQE4YUJoG4eR6B7eXwc292Fufr7em+Nt7A3Vurqes3RW9b53LxZOau2jPvbb020t4fwzTCkTU25ts1D0VbBMssE0BAZLgEYn7V/d/feM2Kx3zatl9yOX7fbNyES3KJJdhZ1gkEsGsfTGhilAkjKkMj8DQnocXXuVydfTW1xd7RdNPESVNEJBIofxdAx0Z/J8+Jfx+37H2b1T0j1L1xviCkr8bjtzYHF53N5bbOKylhk8bsgblydbQbHoMil1mhxUVHGyeggodPuVbv7mv3luf8Ab4dh91fvDQPywZVaaG2illaUp8LysUthPIOKtceLQ0PHPREnPHKG2zNdbRy1K12BRTIyKONa0GojOKilR1Z/s/Ye3dmUrRYilP3MyKlXkalhLXVek6j5JSPTHrNwigKPebHst933219itqksuSdpP70nQLc3sx8S7uaHV+pIQNKasrEgVFxg8egJzDzTu/Ms4k3G4rApqka4RPsHmfmc9LX6ce5v6DvDHXvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvdf/9Df49+690h91g6lsL/tD/e29hff2GpB5hf8PRLui1ZD/RPr8/y6AzPfV+D+T9D/AE9xPu1NUhr/AKvy6A9+CKinQKZ4ECQWP+2P0N/8PcZ7uwAcnoFX4ajGnQK50G7XFuf96HuL91ZRU1/wf5+gbekiuOPQMZzgSGx+p/H+HuMd3ZSSAw/1HoF7hkuegXz17PwbnULW+l78+4w3YhdZLDoG334+rNP5cAI697GH/Z90w/P429jv6ge85/uUup5O51oa/wC7df528fWSn3chTYOYs5+uH/VpegI/no9K7k+Qnwm2z1Tt7r3dvZ8G4/lv8PxvDauy8fna/MSde0/fOz5d85OV9tFM1isVittiomq66CSFqKBWl8iadQzS6yL6Kv2F/K46K+LH8yH+VX2j8RugN9YLF03Z/wAnaXuneVHu/uPs7AbT23X/AB03JSbUm3LU793dvDEbSpchumqSCmqLUz1VQRDre4X37r3Uv+VR2DXfGjZ2zv5d3dvw7+RU/wAltofJzvPcu8ez4+icpmekNzx7u7L35v7FfLuT5B5CCPr2Wm3FtnOUtORFkJNyx1ziiSk0rdfde6kfBL4sb+xX8qD+YP1qOlslsvu/vbtP+Z9NjcPuLan9093b/wAp2JvPtbA9Z5mslytNSVeRpdx4CXGQ46slbwyUIhKN4rH37r3Rdeydk5v5k/ytv5TvTNP0B8mcQ3UnzH+A3VHyH2VuHrPtXprf+06LqrZM23+2d00tdSwbe3bi9l7ayRMR3Zi6iKj1qXgqrer37r3RvPhR1pkP5YHyT/mC9FHqbt3efxjy+0NtfNrpfu7GbH3f3H2lun7ulqdods9A7w7GpqXO787h7D2TuHEwVW0sfk6utzZwVeYozIker37r3SF/lV/Iqgx3yL/mkZnefSfyn6p253j8md4fKrr3eHc3xs7W6m2VXdQba6S6w23ksjlt3752/h8BtzOnK7arRFja+enrJIYfKE0XK+691ct8Vfkltn5i/GDqv5M7G2tvLaG0u6tkybx2tt3fuNpcVuylwtZPkKTF1GRoKKsyNIseYpaRaykeKaRJqSoikU2e3v3XutUPa38mLpjL/wAhnsPsTcfxN7aPzrq/j58hc7R7Xm3T35Qb5quzW352BU7SSn6apd50u35chXUi0ckGOGGNPVRyKTC4k5917qyWv3Vn/hN8zOuPkp3t8f8A5Cdr9U9tfyzehvjxsfM9MdGbv7x3B1H2v1nnc7uXsPqPdWzNm4vK7h2f/pdpd1Y2WCsnp4sdVVmIamqpo/CpX3Xuhf6f6myG+f5xW2PkTmPjJubrfr2D+UrsHA7EO+evcXj4eo96bi75yuRznVMVdiBktqbc3/QbNqI4snjMfVu8NLeMkxar+690H3QNRvPqv5U/zmPj/vDpfu6k3T8lext69/8ASG96DqPduX6d3t17U/FnZO0Ymp+2sdiqnZVFug7swNRjhhqmqjyUtWQEhdWLD3XuiL/C34Mbs+D3U/8AJX+Y+zeh+6NydhviNi/Hf5kdRb8fsvtPNdQ7a7923PiKjunYHW++a7cE/Qmf607CpcdFnpdu0+Jp12zW1VPUxeBCR7r3Q49ufKml+QXz5bM/Ln4y/Pak+OPwq7tjh+K/R+xPhd3Zvjanb/ee2ZanCH5fds7825t+t2/mdpbZqslNBsTAwStT0ypJma1nlkghi917raDp5RPBDOqyIJoo5QkqNFKokUOFlicB45Bq9SkAqePfuvdZvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XCRFkUoyo6spV1dQyurDSyMpBBVlJuD9ffuvdEPy3x/7F+N2Rym+PhpS4yv2jmM7W7j7A+JO5cz/AAXr3PVGRZqrMZvojPVMc9N0rvmtq9Uz43T/AHRys8rtNT0M7tXe/de6HPo/5J9bd7Q5TH7fkzW0+wtruKffvTnYeMO0u2NgVx0gw7k2jVyvNJjJmf8AyXLUElbhsgvrpKudPV7917ofwb34IsfyCP8Abe/de68Rcf091YEjBoevdVZ/Knp2r2fuuq3zg8fUSbR3PK9Zk5KaEyU+B3DIS9WlQIgTTUGSt50kYCNZS6XHpvyB++H7HXXIvON57icv7ZIeSt2cyzsikpaXrH9UOFrohnH6qswEaya01DtBEG33HjRiF2/UUYHqPX8uilfgH+v++/23vDAZz5V/b8x6jpfjhXPXve+vde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XvfuvdS6Ghq8nW0OMx0L1WRydZT4+gpUUs9RWVcqwU8Sqt2OqRxfjhQSeB7VWFjfbruG37TtkBm3S7nSGGNRVnkkYIigeZLEVHkKk463wBc/AvE9XbdW7Go+uNjYHaNI3lfG0ofIVJJJq8tVk1OSqf8Ekq5G0AcBAB+Pffn2h9vLH2r9veW+SbI6ntIazP/v24kJknkr/SkZtI8kCjy6CtxM08zyt5n+XRQu3s/uT5NdwZf4n9e5eqwfUuysfR1vy/7DwdVPR5aqptyUJqdufGvZWao2SXE7p3rhpRkN010DpV4jbc0MMDxVWShlgkvpryr0eDa21tt7I23gtn7PwWJ2xtXbGKocHt7buCoafGYbC4fG06UtBjcZj6SOKmpKOkp41RERQoA9+610/e/de697917r3v3Xuve/de697917r3v3XuuiCbc29+690Qjquug+M/yPzvxtraelw3UHek25+3fjPNErQYzD76lnqNwd8dJwEn7emnfKVku8sJTII4zQ1+SghHjoAo917jno/ANxf37r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XvfuvddEgC59+691hnqIaWGWoqZYqenp43lnqJ5UhghiRSzySzSFUjjRRcsSAB9ffuvdFi3n81fi5sfLHbeQ7j2tn93lnij2P12ch2jveWoT/lGXaPXVFufPRTFrD9yBFUn1EAE+/de6RQ+S/eG+hLF0x8O+06iEP4ot0d+5vbfQ22Tdigq4sNVz7u7Lq4ACH0nAQF0+jA8e/dbpjj1hXHfzFq0tlX3R8O9v3eOaDZqbS7g3NTxRKWaSirN8tujbtRUTyiympiw0aobsIW+nv3XhSmRnpcdFfI+o7A3RujpztTZVT1D8hthUEOX3F1/VZAZfbu8doT1CUVJ2r03uxqbH/386yr6+QUsk7U1JkMTkAaTIUtPKYmm9178+jT+/da697917r3v3Xuve/de697917r/0d/j37r3ROvkDnM3jd001Pj8zkqCBsHTTGCkq5oIjKamrVpCkdlLsFAueePb8drbTpWe3Rz8xXqCvc3c9yst4hitL+aKE24NFcqK6jmgPHooub3Xum7D+8ud5Lk2yM/0Fuf18297GybJIreJtFs2fONT/hHUGblv2+jXTerrj/vxv8/QN5zdW6Lyf7+POnkf8vKf6E/T9XPvTcp8rSUMnLdka+sKH/J0Adx5g3+hA3y6r/zVf/P0DWb3Tub133FnCbsOchN+Cbkerj3VuQ+R5KGXk3bT9tvGf+fegJuPMXMIp/u+u6f81X/z9A3nN0bkOu24c39G/wCXhP8A0/4N9fbJ9tfbmQ/qchbQ322sR/596AW5cx8xVem/3f8Azlf/AD9A5ntz7l0yH+8OZuDcn7+c88/nV7o3tL7VyatftnsTY87OE/5OgNe8y8yGUD+sF5pJ/wB+v/n6ud/lEZLI5PqntyTJV9ZXyRdn00UclZO87on91sU2hC5Olbm9h+fdYuVeV+VdVryvy3ZbbbSnW6W0KQq7006mCAVbSAKnNBTrod9y+9vL7k3nGS9upJXXdQAXYsQPp4zSp+ZPR8fkl8mOqPijsTCdk9x5TK4ja24OyutepcZVYfB5DcFXJvPtnd+M2Ps2klosdHLNDQ1Wey0KTVDDx08ZLsbD291mT132d8lOqun+2vj10nvbKZWi398otyb52r1FQUeGrsjQ5XMddbKq9/bojy2Upo2pcFDT7bopJInnKrNKAi3Y+/de6H3jSLn8X/2F/rb+nv3Xuk1vbeO1+utnbs7B3vmKTb2zNi7azu8d27grxM1Hg9s7ZxlVms7l6sQRTTmlxmMo5ZpNCO+lDZSbD37r3RXe+Pnh8evj/wBH9S/IPceW3dvTr3vndXWOy+mB1PsbcfYe5+y9w9x4+bK9c0O1tr4SjbL1Z3LjYTNEXjj0rbVpJA9+690iNg/zLviz2Bs3vvc9Jk+0dsbh+MKYR+8+m969Jdp7b792Kd1IrbN8nTVRteTfGfh3oWC4qfFUldT1j3VZLq+n3Xumrqv5ffGD56ZTuX4kbr637b2jvCj67oc12V8ePlB1BuzqPc+7umd8VVVgqbdlBg88scG6Ng5usppaGqenqHeGVvBVxRM+hvde6sHxmLxmFxuOw2Hx9FisRiKGkxmKxeOpYaLH43G4+COlocfQUdMkdPSUVHTRLHFFGqpGihVAAA9+691P9+691737r3Xv99/vh9Pfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XrX+vv3XugO7j+OnUfe1Pjm7A2x5dwbfl+62jv7beSyW0eydlVya2irtob+23VY3c+DljlcsYoqn7aY8TRSJdffut1P5dAXSbW+bHSf+TbV3fsn5Z9f0MJFDg+1an/RX3zS08JZaeh/0mbdxeS6731UrCAPPksLhZ5mF5qlmLOfdeP2dKGj+V25aOPxb2+JPyr2plo/TNR4vZG0+yMazkkXos51tvfc1HUQnQSGkEBC21KpOn37r1PmOgnovn/T9n5Td2z+j/iN8o+7K/aeVyW0d6Vk+ztkdb9d4Tc9DFF/E9o5LfHam99s4TM5ah8/iroMSmVFDLqhqfFKDH7ZuLa2vLee0vIEltJUKujqGRlYUKspBDKQaEEUI49bGCCGoeiudp47e+3qio3Bvb4pb/wCk8bkaWnr4Mnt7s3q/svrOjyNdUinfC7iyaV+1ZOuKmmlkRmqKgvhCrkRVOpdPvBL3N+4pylzFc3+8e3O9nY9wl1P9JIhlsTJSoWPSRLbKSKEKJI1qNKKB0Z2+5MgVZhqUft6TUu1t2U2Zodu5DaO58PncsalcHjMxiJKF9yfZwtUTvtauE0+I3NpgjeVEoqmaokhXyCLSQfeFnN33Uffnk9p5LjkaTcdvQV8fbpFukIpU1jGmZaZrqiFaYNCB0YrfWsgojhft4/t4dNNbSVmNnakydHV42qQkNTZCkqKCcEKrEGGrjikuqupPHFxf6j3j9f2l3tNybPdrOa0u/wCCeN4X4A4WRVJwQcA0Bz0pHdlcj5Z6wW4v+P6/j/b/AE9sCpGoDt9fL9vXjjjx69/vv+I/3v37r3XXv3Xuve/de697917r3v3Xuve9de6etv7ezu6srDhNtYiuzmWqGCx0VBCZWXV9JKiVilNRwAC5kmdEA/Ps75b5Z5h5y3eDYOVNnuNw3qUgLFCtSK+bseyJQMl5WRR5nrxZI6mVgEHHqzz4+fHWn6xjG590/Z5TfNXE0MZh/wAooNt0chBkpsfJIimWvmA/fqQo49CWS5brf92j7sFt7TRf1u5xEF17hTppATvhsY2oSkBYd0z0AlnAB0/pxnQCXILy+NwfDjFIR/PrN8mPkBUdU43AdfdaUeL3j8le4Rk8J0Z1pUVWgZHI0ccC5vsHdiwk1OM6t6zpKxMjnq8hV8Sx0kLGsqqaN8xekGePl0IHQfTuO6M6zwmxafJTbkzvlr9x7/3zXU0NNmux+ydzVT5ffPYGeEOoHKbn3BUTT+Ms60tP4qaM+GGNV91qtePQze/de697917r3v3Xuve/de697917r3v3Xuve/de697917oIe8OlNm9+df12wN5Lk6OJq7G5/bW6duVpxG8tg71wFQK/a+/dj52NHmwe7Ns5JFmpp1DI41wzJLTyyxP7r3RecD2z8suqY4Nqd0/H/AC/eUeNR6em7u+OmT2ZHTbpoaYMKbJ7r6g33uvau4dm7oqqdVNXTYmpzmNepLtBLFGViT3W6A+fT3P8AKze9VEh238M/llmZzOkEkGQwXUuzhCHjMnmafdnbeLhkgWwVjGXIY2twbe68APXqN/sx3yLjgSsn+BndX2hmeN6ek7S+OVTnEjjQSNUnHN2pBQvDKp0xqtYZWcFSiixPut0H8XTwnzR6nwdJDU9vYbs749mWtpcc0vdfXud2xt+KvqklZIG37jEz/XXhHgcGf+LCG4Hq9Qv7rVPTh0Z7b259t7uw9JuHae4MJufA16eShze3crQ5rEViA2L0uRxs9TR1CA/lHI9+610+Xv8A77/fc+/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3XuvEgfX37r3XVx/X6/T37r3Xrj/AHi9/wAW/wBf6e/de6Cnffe/SfWEU0vYvbfW+yBTxiWWLc+9NvYaqEZbSGWjrshDVy3bgaUJPv3XgK9AxB84ugs88kHXNb2H3FWR1UVH9v1L1J2VvSjaea3htuSj2xDtCKB9V/LJkVhC+osF59+63pPp02HvT5R7xklg6y+H2X23QyFkpd0/IftPZnX9Ip1Dx1X90evm7X3dLTMtyY50oZgbCw59+68RTz6iHqT5o9gC/Yvyg2j1PjpZJPPt344dV0TZT7ZiTFEOxO4K3elQsyKdLywYSlZ7al8Z49+68DTy6n434H9Bz1dNluy4OwO/c/TS+VMx312Tu3saHVYj0bUrslTbCpksxAWHExqBx9AB7914no0u19j7L2RSfYbL2htjaNDpRPs9sYDFYGlKRqFjUwYukpYiEUADjge/deJJ4npUW9+611737r3RVflh05muxNmYbfvWccdF8gOi8s3ZPSeai8UFTW5rHwj+8fWeRq3MWvZ/cG2458DlIJHEAFTDVEeakhZPde6FfpXtzanevWO0O1Nlyz/wTdeMNQ+Or42psxt3NUdRNjdybS3DROqyY7cu0dwUdTjchTOA0FZTSIfpc+69w6FP37r3Xvfuvde9+691737r3X//0t/j37r3RJfkf/x+FL/4b9N/7lVvtfbf2Z+3rHz3Y/5LVv8A80B/x49E7zn6z/wWT/ex7Wx/C329QJufB/t/zdA5myf3z/Qr/vftYKYz0ANwHxdA3nP7X/If+8t7UA1UevQC3L4R69A1m/7X+x/6F9uDj0AdyIBep6B3Pfom/wBYH/eSPaoUp0BLr+1H29XW/wAnr/mU3cH/AIlOm/8AeVxPsNb1/uRF/peukP3Jv+VK5z/6Wo/7R4ug7/4UE0eayXwR2XjNtbjTZ+5cn82PhDjtt7ukxVJnk2tn635GbIp8RuI4HISw0Gc/g1fIlR9nO6RVPj8bkKxPsm6zS6K53L038ueqf5of8nSs+S/zXp/lbis1298taXZ+Aj+N/XHQ7bSy9L8Xt0VFVlTldkZvL1W4Fr6MrTGnqESOG/kBLce/de6x/wAqTbfxm7pwOx/mj8j++Nyj+YzuP5c97bO3vT5r5D7v2zuHb+/sN2F2Ds3D/Eyl6XO7KbasnXeB69xdLJS4AYQmojjXIhy58o917oq/xv8Ahn1V35/LA/mY/JjuTMdy9xdtTZv+aLsfZ8m5O7e2psFsLY3W3Zfalds/YGydo4TeOPwkOCo9y7ciybRz09TNPVTSRMxpSsA917qRv7qrrrZn8pP+TBhviZu/b2zNx9ifOb+WxvCr3tU5vN954TbndG6+vHlyudym3dwb3mMsFNVRq77epq/F0kQ9MaQcn37r3RxvhD2htz4mfLD+bfvr+Y523gD8r+r9tdK9i9l/IJ6Gj2J1NvH4P0O3MzF0fl+qesaBK2u2jT7X3NPl8NnaGWvz2RrNyyRrHVTGeGP37r3Rg/5eO4/9m4+V3af8xzsHL7L2Tluz+ktr9I/Ev44nem1M129sj4tYXc9b2DkO0O4cLg8tXzYLffdW6cvSZP8Agqqx25iYKSmqpWq5J1j917q8H37r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+69035amq6vGZGloKxsdXVVBWU1HkFQSmgq56eWKnrRESolNLM6yaSRfT9ffuvdVp/F75GdL/G7qvYfxm75rIfjh2h1ZiItp5XG9ml8DtTsOup6qoeu7L697Gq6al2hvzD7+yUs2UeWKqGShqaqSOugiqFcH3W9JpXy6OW/yF+N+Yoa2KXvHpHJY2Smlp8lDJ2VsSromo6mNo54MhFJmpITBPCxV0kGllJBFvfutUPRN6bdHxn2/mKboOs7X6n3/wDHntPJmHYGMpu2ts5DNdKdhhmyGJ2lgK2i3C2cxm1svUU7z7XqaSWOpwGThNHEwhmpFh2CVIZTRuvU+XQ/7dyb7Ry+3unu9KrH9l4PcV8d1N3LncTjqtN3yxEyxbB7EnhpTiKDs2npIEekrIhDS7lhjZ0jirI5YXIN85W5b5mgNtzHsFlfW5BGmeGOUUNCR3q1K0Faeg9OrrJInwOQft6V+f8Air0tnS8ke2psDO6lfNt7JVePAZpjM8n27PUUZkbUVuYzZDYWsPePvMn3O/YbmItJHyo+3XBB77OeSGhLFi2gl4ixrpqYzRaAAUFFUe4XUZr4lftFegeynwf2/K+rDb/zdChllZo8liaHJgRH/MRpJDNj3Dx/RnbUXH4HuD92/u++WpZfE2P3Gv7dNbds1vFPRT8CqUaHgfiY11DyXj0pG6MPjiFft6Dyv+Ee+IjH/Dd6bXrgWkEv3dDlMaYlFvEV8ZyHmLi9/wBGm3F/cZ7h9wL3AiMQ2rnzaLgEnV4kNxBppwppM2vVwPw6fU9PrukJ+KJvyIPSTrvh53FTVLw0abWydOqxlK2LOPSxyllBdVgqqJZ08bcG4sTyOPYLvvuP++drdSQWkezXduAKSrdtGGrxoskIYU4ZH2dODcbQgHUwP2f7PUM/EPuz/nW7a/8AQki5/wAP+AftJ/wE/v5/0atp/wCy4f8AWrrf7wtPJ2/3n/Z6VOL+FfY9UtM+V3HtPDrLGXqIkORylRRvY6YSsUFJT1BLCxZZQBe4v9PYs2j7hfupeR2sm780bLZa0JdQZ7h428lIVI0c1wSsgA4ivDps7pbg0CM38uhT218I8BTSRS7u3nlMwulGloMJRRYWBpPVqH3k01fVtFe1tIRvrc/S0wcrfcB5btZYpudefbu9TSA0NpELVSa5XxmeaTTWmQEalc9J5N0YgiOIAfP/AFDoetwbp6D+MWzhX7r3JsPqXa0CW+6zuUx+Inyk0ageOA1Uv8U3FlJmICxRLPUysQFUkge84OTeQeT/AG+2mLZeTuX7awsVABESAO5H4pZDV5XPm8jMT69FrySzH9Rix6Aqi+S/b3dzLRfF3pLNQbYq6eWRO/fkVis91p1xFGSBBW7S63ngo+2+yTOCWiVqbb2OlUX/AIivGoXAAYHVD0KHR/xzouss3uLs3fW7K7uLv7e9HTYvd/cO4sPi8PWQ7boZmqsb1517t3GrLRdedX4iskeeDE0008tTVyNVV9TWVR8w31r5dGW9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+6911b/E/7x/tvp7917r1v8Tz/vr/AOv7917ru3++/wCK/wBffuvdRqujpa+lqKKupoK2jq4Xp6qkq4Y6mmqYJVKSw1FPOskM0MiEhlZSCOCPfuvdFDznwc6PTK1m5upot1fGve2QlM9Xur48Z9+ukyU93kDbg2TBTV/Wu6EeofyP/EcLUs5HLAE391sGnl1zx+zvmrscGnxfcnTfeGMj1tAvaXXOX623gURbRU9VuvrPLZHa9XKwSzSrtmnuzFtNgF9+69UdRanvD5ZYCojhz/wrqNwUoKipy/Vnf/Xe46VV8hRpIcdvrH9Y5qUhBr0CAmxABJv791ug9esCfLzd0CA5f4XfL+hk1PG0dBs7rTPgSKxC2kxXaswaJ1Fw9go4B5Pv3WgK+fXP/Zxcp/3h780P/RV7O/8Atne/da69/s4uV/7w9+aH/oq9m/8A2zvfuvddr8wc1KRHB8O/mW8zcRrP1psekiJ/2upqe0EghFvyxHv3Xuu3+Wu89DmH4XfLiWUKfHE22OrIVkf8IZpO2vHED/qm4Hv3Xuo8Hyn7hyCVC4z4MfJIVUEBkSHO5XpLA09RK11igirZe06qLU7/AKmICxr6jx7917qZQ91/LnMsUofhQu3lKwNHUb9+RvXGNg/cb95ZYtkYfsSsR4E5t4yGJtf8+/dbIA4Hp6kqvm7nVvTYf4xdcRzQm38QzfZva1dRT2NtUFFiOrKGsVWH1E0d/wCnHv3Xu359McnSfyw3LK7bv+ZUm26GpRBLiul+itibVkpnGi/2Wf7GyHbOR8bhTfVHqGrgiw9+69VfTqNF8E+rMtqPZ3YPyJ7r8jvJLTdmd9dhy4WRpbmSM7Z2dltn7YakYn/gOaMwAADRb37rxNfLoU9j/E/4y9bTRVex+hep9vZCGMxR5ek2Pt+bNiMsGZWzlZQ1OXfUwudUxJPJ59+610P0cSQokUSrHFGipHHGqpHGijSqoiAKqqOAALD37r3WT37r3Xvfuvde9+691737r3Xvfuvde9+6910Rf37r3RFdxdf9t/HPs3fva/x92PH2z1p25kV3T2v8f6PP4naO5sN2d4oqXJ9u9S5bcdRSbTrKjeeKpYU3HgKybHisr6ePI09V9zLVQ1HuvVr0pNt/OvoGuzUG0ew8puT49b7qSqQbM+RG2a/qmurJGKqFwu4c2W2LuYNIdKti8tWBiOL+/deoeje0VdR5Kkp6/H1dLX0NXEk9LW0VRFVUlVDILpNT1EDvDNEw+jKSD7917qV7917r3v3Xuv/T3+PfuvdAR2Z09W7/AM3HlYM9S4yOPHw0IgloJapiYpZpTIZFqYRYmX6W4t9faiKYRrShrXqOObuRbjmbcI7yPckhVYwtChbgSf4gKfl0CmQ+IuXrbkb6x8ZN7XwNQ1r244ya3+nt8XgFRpPUe3Xsde3GqnMcQr/wk/8AQfSKr/g1nqwvbsfFoGP523VH/bgZYA+3RuKj/Qug3c/dx3G4rTmyFR/zQY/9ZekTkP5du4q3Vp7Uw8ZN7X2pVtyTfm2bFx7cG6KKfo9B+6+6vutx8POsC/8AUM//AFuHSJr/AOWDuisJt3HhI7g/XZta3JFv+d8Bbn26N3StfA6D139zzebmtOfbcf8AUK5/6zdIjIfyld3VqyBe8Nvx6xbnYtewH1/A3GPbn76j/wCUc/y6Ds33IN8lcN/rjWwFf+UN/wDto6PR8K/iplfijs/em18rvXH71k3Xu2HcsVZQYObCJRRx4mjxhpHhnyGQady9Nr1hlsGtbj2WXt2t26OqFaCnWTvsT7Q3ns/se+bPd77Hftd3gnDJE0WkCJE0kM71NVrggZ6M7vzrfYHaOHpNvdkbK2rvzA0GewO6aLDbvwWN3DjKPcu1cnBmttZ+mocpT1NNDmMBl6WOpo6hVEtPPGroysAfaHqc+vbg652FuzcmyN5bn2ZtfcO7eta/L5TrzcuaweOyWd2PktwYqXB52u2plaunlrcDV5jCzPSVMlM8bTU7GNiVNvfuvdBNR/D/AOKuO7zq/k5j/jl0nRfIivilire7abrTaMHZ1UZ6X7GoqJd4xYpc29dUUH+TyVBm88kH7bOU9Pv3XuhS2Z1d1v11t7IbR2DsPZ+zNq5bMbk3Dldt7Y27isJg8jnN45Gry+7MtX4vH0sFFWZHcuVr5qivmkQvVTSu8hZmJPuvdBVsv4efFPrjauM2LsH44dI7N2XhezoO6sPtTbXWm0cPt7E9vUsjS0nZmMxFFiYaGh3vRM58GSiRaqEcI6gAe/de6UXY/wAafj13DkM5le2Oj+p+yslufYqdYbkyG+dg7Z3RW57rmPcNPuyPYuZqMzjayTJ7Si3RSR5FcfMXpVrUEwQSDV7917pDdP8Awa+GXx93i/YfRXxU+PfTu/JcZXYWTefWnUWxdlbofEZOWGXIYxs5t/CUGRNDWyU0bSxGTRIUW4Nh7917o1Hv3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3XumzK4bE5ylNDmsXjcxRMwZqPKUFLkKUsBYMaeriliLAE8kfn37r3QWVPxy+PVZOams6H6ZqqkzPUGoqer9kTzmokYvJO0suDaQyu5uWvcnkm/v3W6n16hZH4w/G3L0c2Pynx96SrqGoAE1LU9WbHlifSQyMVbB8OjC6sLFTyCD791vUf4j0Cm4f5cvw6z+KyeFp+pZNnY3LpGtbR9Z787I6wo5JKeeKro6sY7YW7tv4wV2PrKeOamn8Plppo1eNlZQffutaj69Kao+NPYOBozD1Z8su+9ozRxpFTU++ajZvd2Ijjj/Sk47I2xkd2VOqwDOcyJSt7MCb+/da6S6Zn58dbLGmY2d0Z8n8NCGRshsnOZboPsSddTkTPtreEm9dgV09mA0pmsallJHJt791vHUn/Z3tsbcIg7e6P+S/Tk8ak1lbuDprcW9NqwFZRBqj3j1KOwMBJA8jegvLE7LyUA9+60ep4/mAfEQ0bVv+l+nCLUfaNStszsRcqJ+PT/AAU7RGXK3NtYg0X4v7917pu/2eHa24GEPVXRnyg7dlmj8tDXbf6R3NtDbFYmsRmQbu7XGwMAkSFuSJmcgHSre/de64Nvb5379LxbW6R6X6FoH1p/Gu4+x6/tHcRRxIkctNsnqqjx+ESZAyyaZ9x2uCjL+ffuvY8+nTEfHfvLcdNVL3T8veyM99y0GjDdK7X2p0PgqJFpY4qymjymNp919iTiepVpFlXOwSRqwVbWufdbNPLpZ9ffEH469a7iTeuB6zxeX7ARVA7I3/X5vs7sZXVi+uDfPYeS3NuWh1yMWKU1TDGCeFA49+69UnFejKj/ABN/8ffutde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3TBuTa22t5YmqwO7tvYLdWCrV0VeE3JiMfnMRVKQVK1OOydPVUk66WPDIRz7917oskPwe+P+DrnyXXOK3v0tPNVJV1NH0n2n2L1ft2qdZfM0cuzdq7lodlrFNJfyCPHIZNR1E+/dbJJ4no3IFgBcmwAuTcmwtcn8k+/da679+691/9k=)
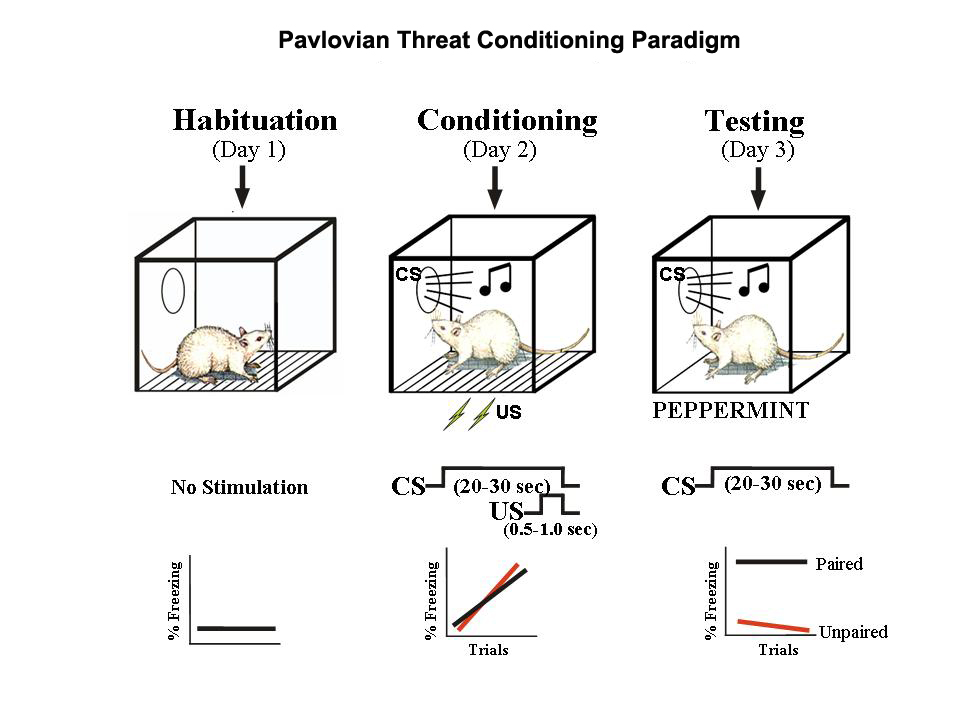{kind=link}
{kind=link}
References
Brandão, M. L., Zanoveli, J. M., Ruiz-Martinez, R. C., Oliveira, L. C., & Landeira-Fernandez, J. (2008). Different patterns of freezing behavior organized in the periaqueductal gray of rats: Association with different types of anxiety. Behavioural Brain Research, 188(1), 1–13. https://doi.org/10.1016/j.bbr.2007.10.018
Clapp, P., Bhave, S. V., & Hoffman, P. L. (n.d.). How Adaptation of the Brain to Alcohol Leads to Dependence. Retrieved from http://pubs.niaaa.nih.gov/publications/arh314/310-339.htm
Cock, V. C. D., Vidailhet, M., & Arnulf, I. (2008). Sleep disturbances in patients with parkinsonism. Nature Clinical Practice Neurology, 4(5), 254–266. https://doi.org/10.1038/ncpneuro0775
Davis, M. (1992). The role of the amygdala in fear-potentiated startle: Implications for animal models of anxiety. Trends in Pharmacological Sciences, 13, 35–41. https://doi.org/10.1016/0165-6147(92)90014-W
Flores, Á., Maldonado, R., & Berrendero, F. (2013). Cannabinoid-hypocretin cross-talk in the central nervous system: What we know so far. Neuropharmacology, 7, 256. https://doi.org/10.3389/fnins.2013.00256
Heath, R. G. (1963). Electrical self-stimulation of the brain in man. American Journal of Psychiatry, 120(6), 571–577. https://doi.org/10.1176/ajp.120.6.571
Hu, H. (2016). Reward and aversion. Annual Review of Neuroscience, 39, 297–324. https://doi.org/10.1146/annurev-neuro-070815-014106
Kadmiel, M., & Cidlowski, J. A. (2013). Glucocorticoid receptor signaling in health and disease. Trends in Pharmacological Sciences, 34(9), 518–530. https://doi.org/10.1016/j.tips.2013.07.003
Kim, E. J., Pellman, B., & Kim, J. J. (2015). Stress effects on the hippocampus: A critical review. Learning & Memory, 22(9), 411–416. https://doi.org/10.1101/lm.037291.114
Kringelbach, M. L., & Berridge, K. C. (2009). Towards a functional neuroanatomy of pleasure and happiness. Trends in Cognitive Sciences, 13(11), 479–487.
Medina, J. F., Repa, J. C., Mauk, M. D., & LeDoux, J. E. (2002). Parallels between cerebellum-and amygdala-dependent conditioning. Nature Reviews Neuroscience, 3(2), 122–131. https://doi.org/10.1038/nrn728
Milner, P. M. (1989). The discovery of self-stimulation and other stories. Neuroscience & Biobehavioral Reviews, 13(2–3), 61–67. https://doi.org/10.1016/S0149-7634(89)80013-2
Musazzi, L., Tornese, P., Sala, N., & Popoli, M. (2017). Acute or chronic? A stressful question. Trends in Neurosciences. https://doi.org/10.1016/j.tins.2017.07.002
Nestler, E. J., & Carlezon, W. A. (2006). The mesolimbic dopamine reward circuit in depression. Biological Psychiatry, 59(12), 1151–1159. https://doi.org/10.1016/j.biopsych.2005.09.018
Pellman, B. A., & Kim, J. J. (2016). What can ethobehavioral studies tell us about the brain’s fear system? Trends in Neurosciences, 39(6), 420–431. https://doi.org/10.1016/j.tins.2016.04.001
Schmaal, L., Veltman, D. J., Erp, T. G. M. van, Sämann, P. G., Frodl, T., Jahanshad, N., … Hibar, D. P. (2016). Subcortical brain alterations in major depressive disorder: Findings from the ENIGMA major depressive disorder working group. Molecular Psychiatry, 21(6), 806–812. https://doi.org/10.1038/mp.2015.69
Watabe-Uchida, M., Eshel, N., & Uchida, N. (2017). Neural circuitry of reward prediction error. Annual Review of Neuroscience, 40, 373–394. https://doi.org/10.1146/annurev-neuro-072116-031109